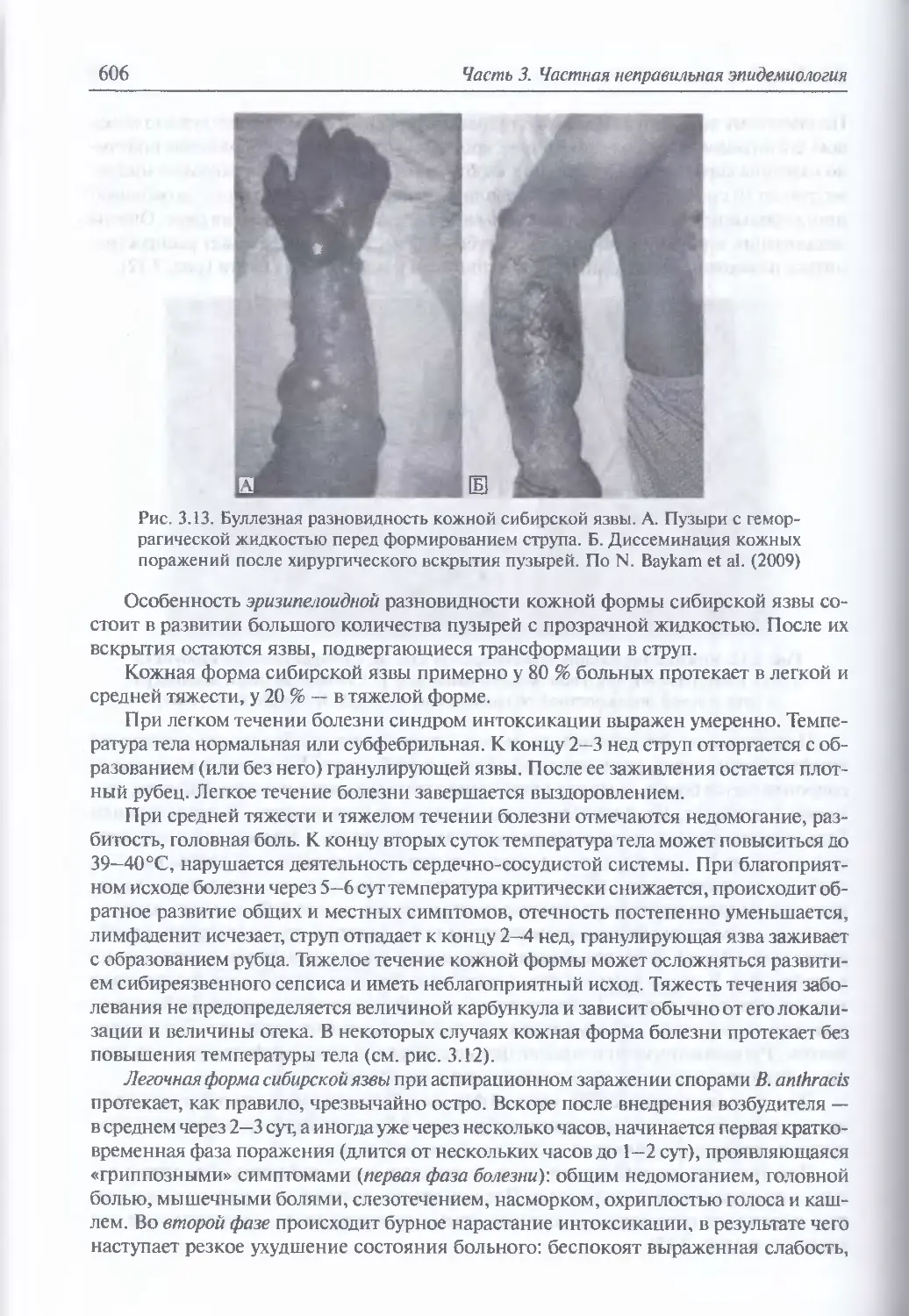
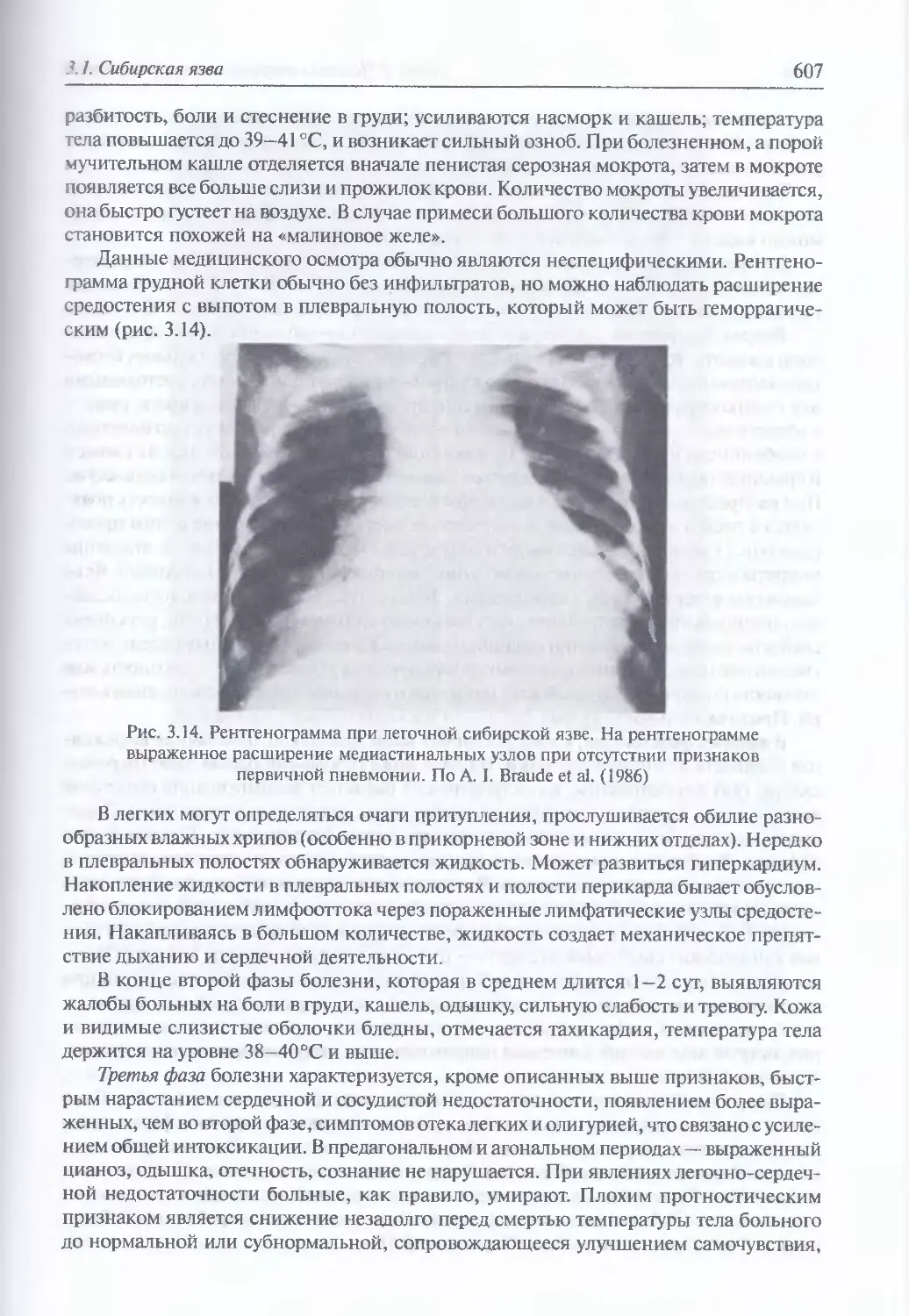
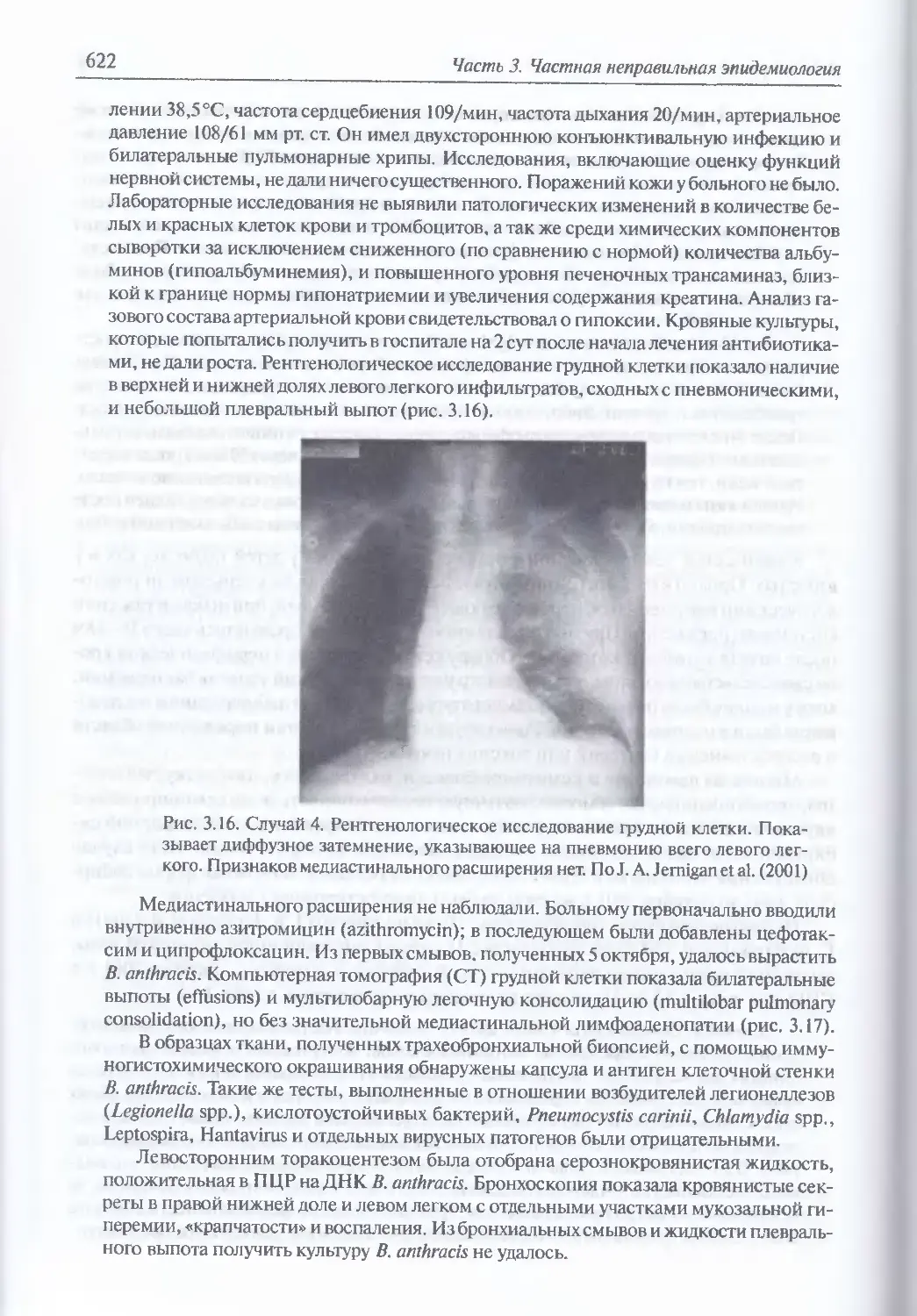
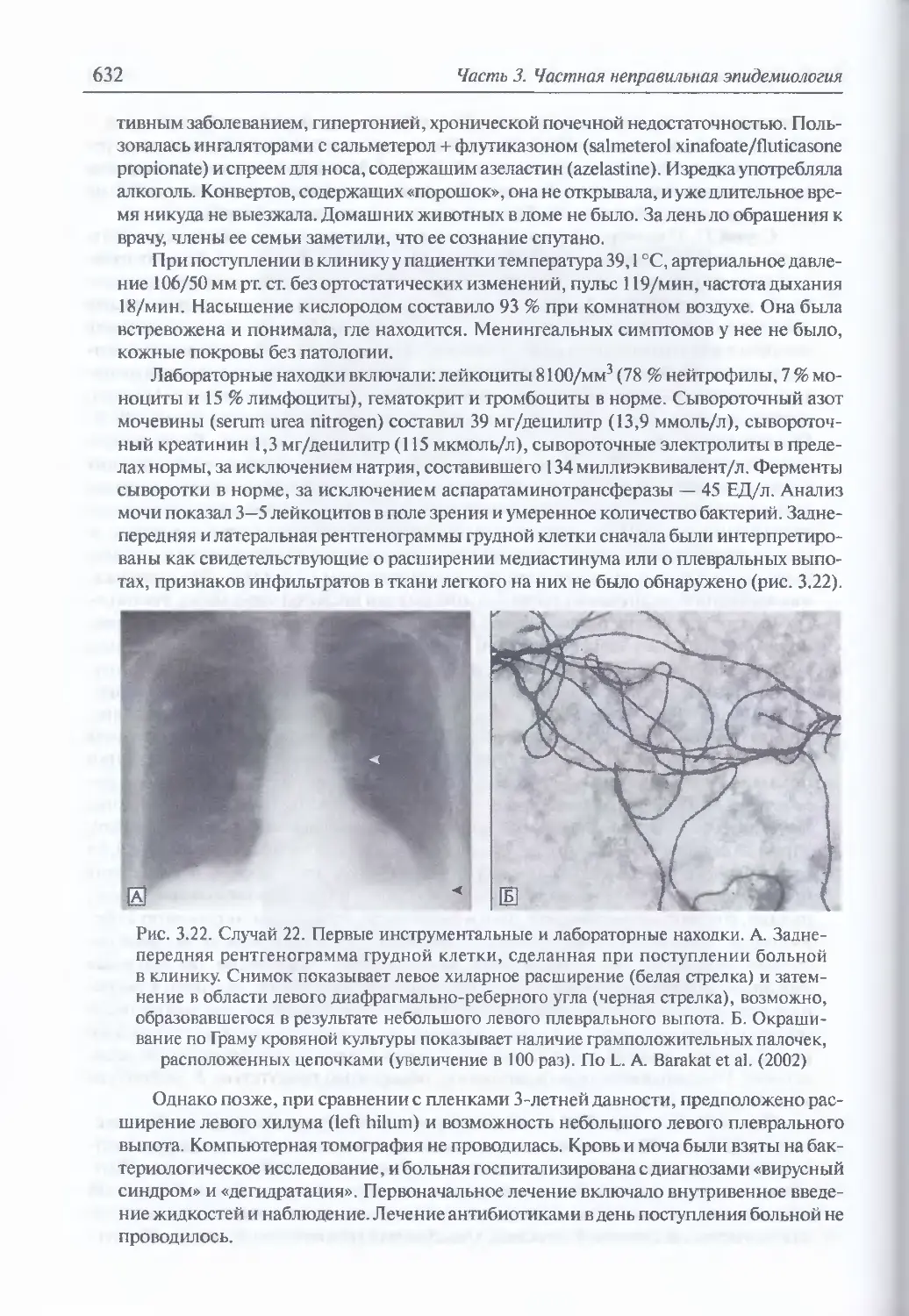
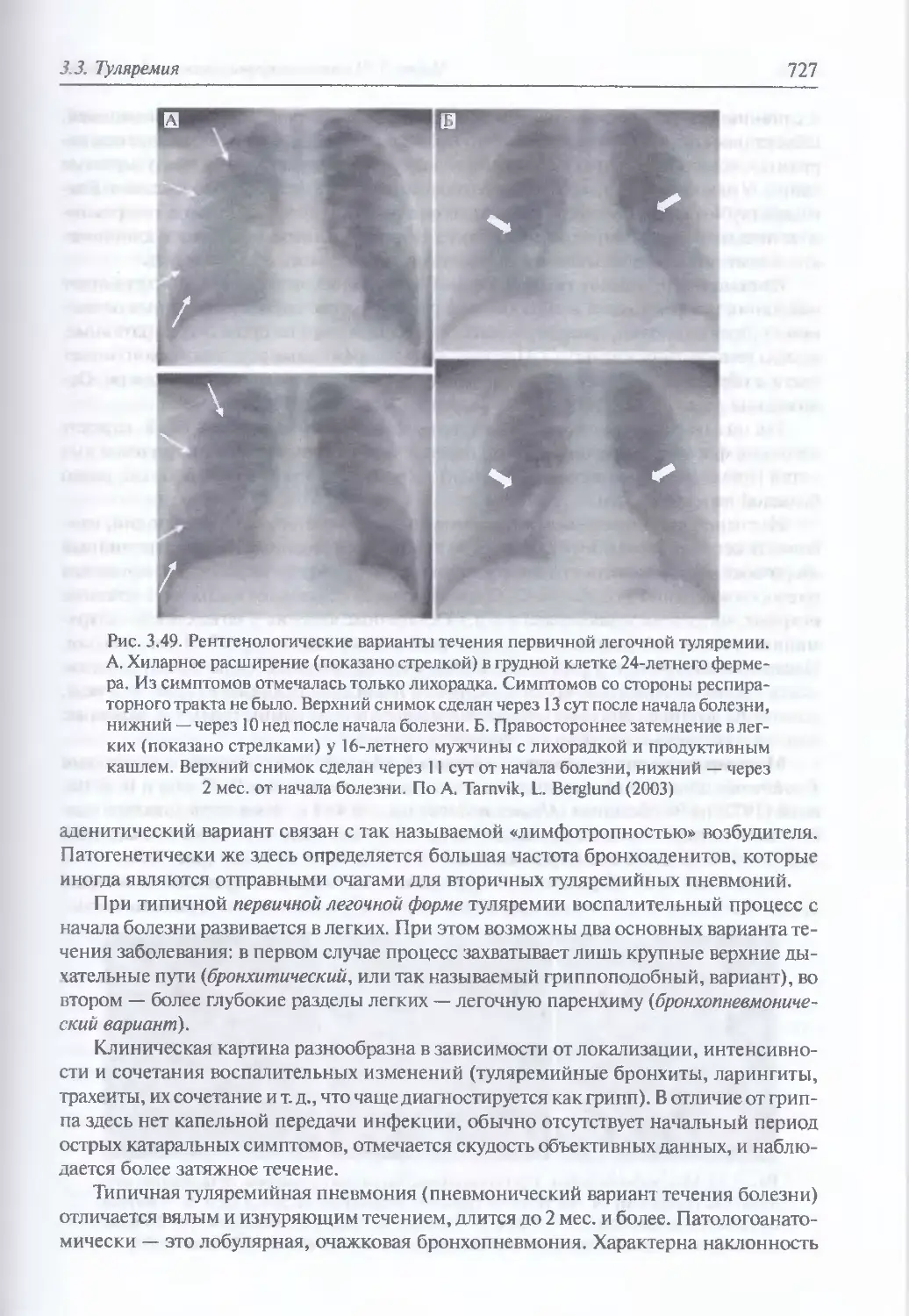
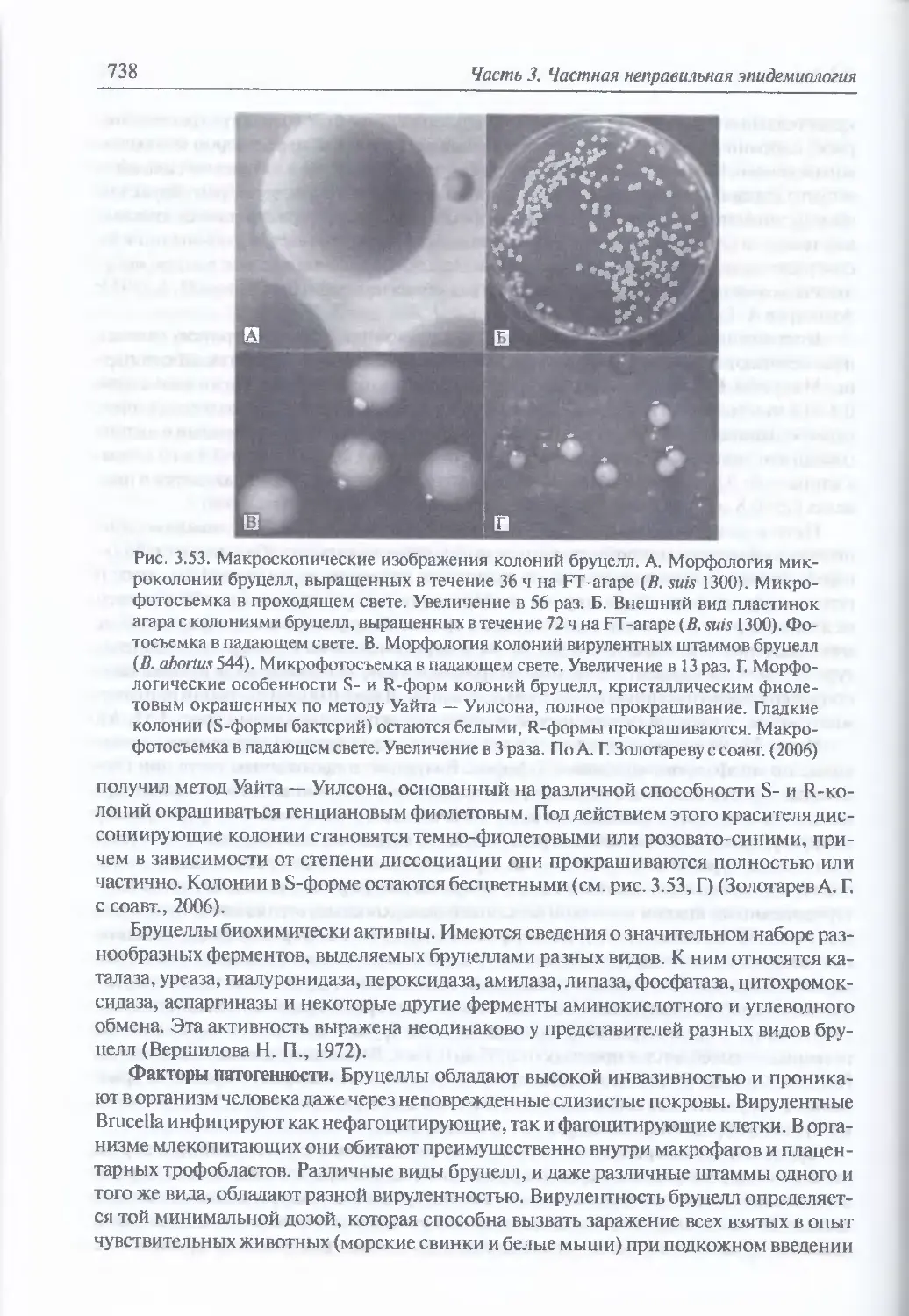
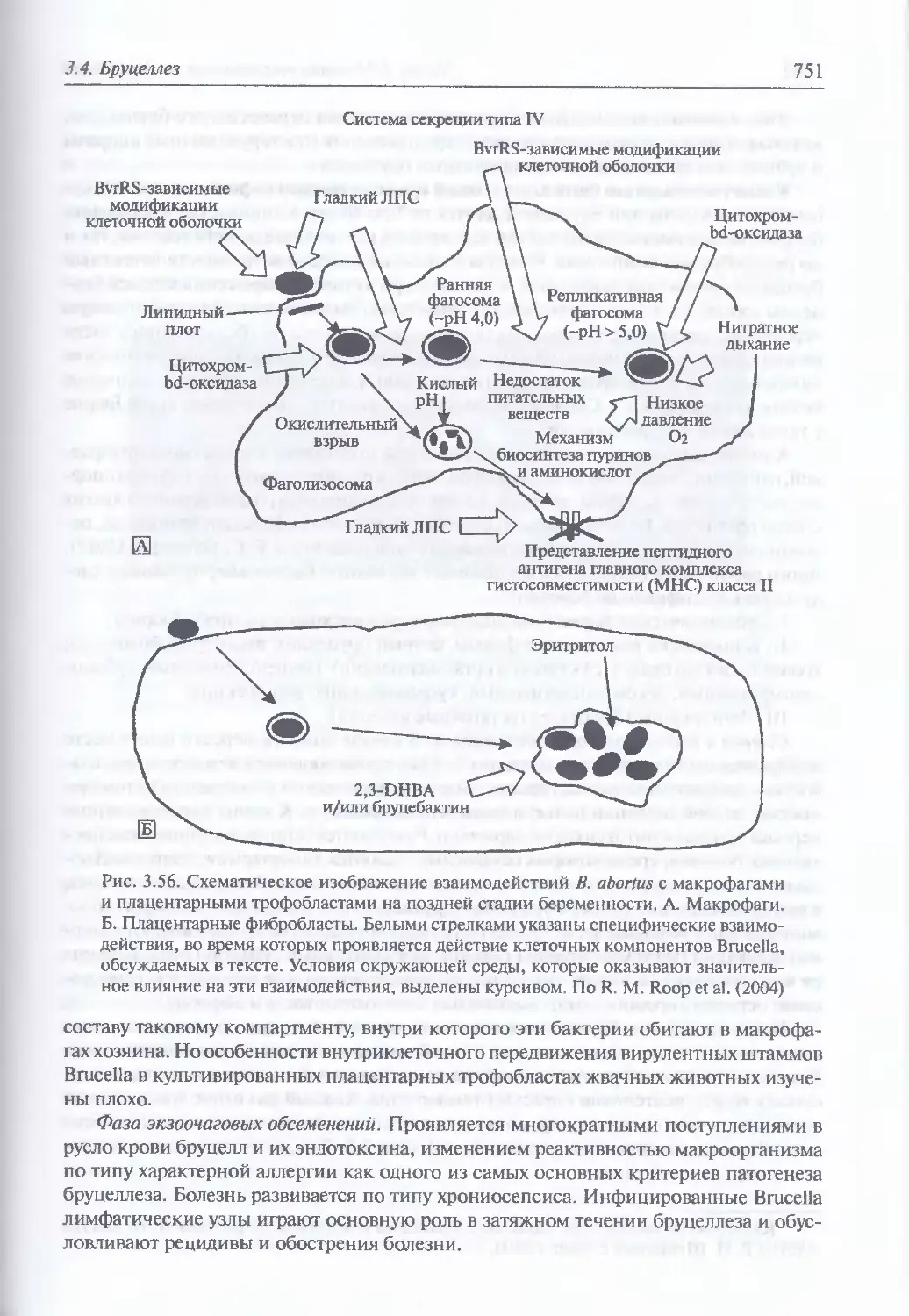
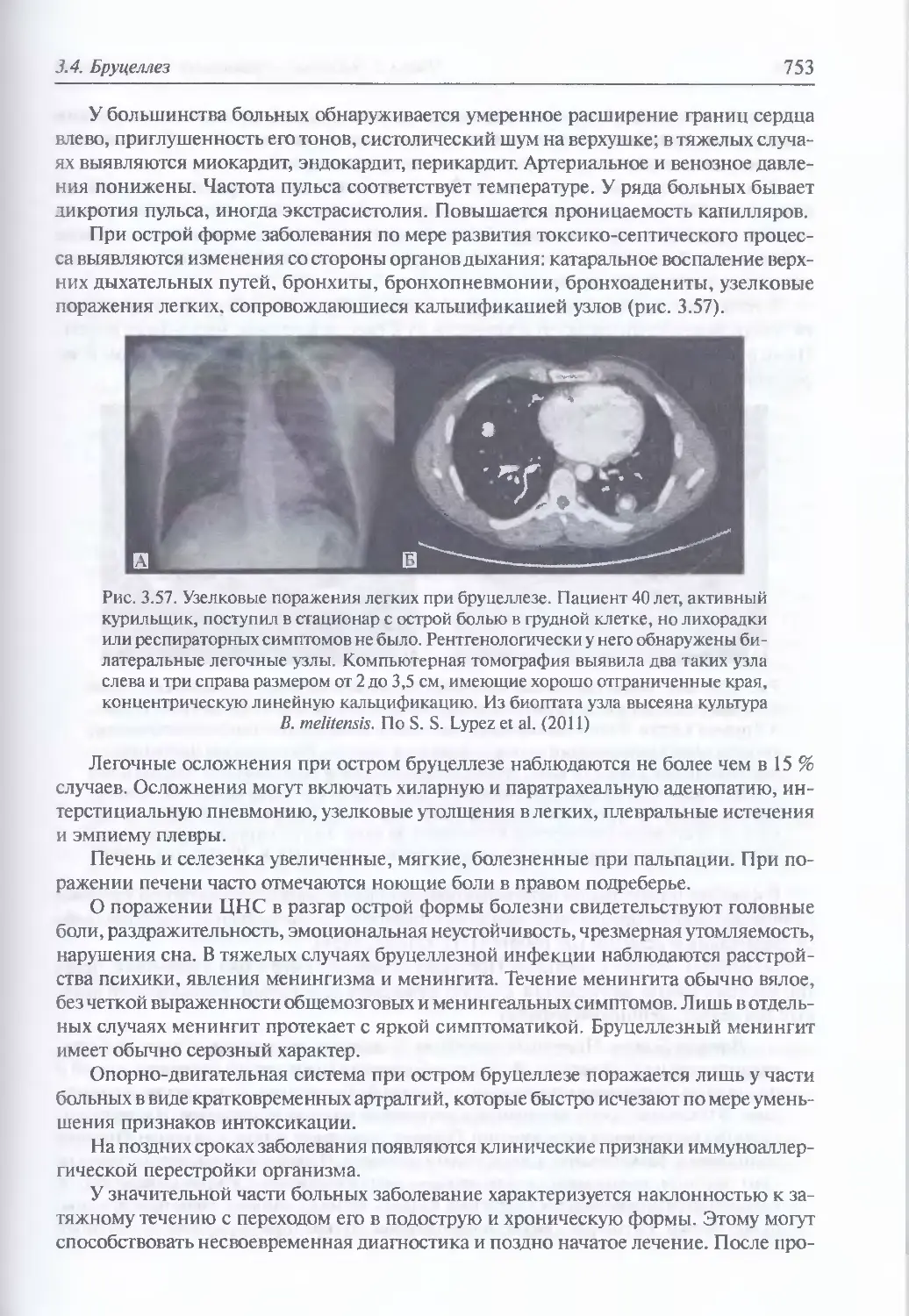
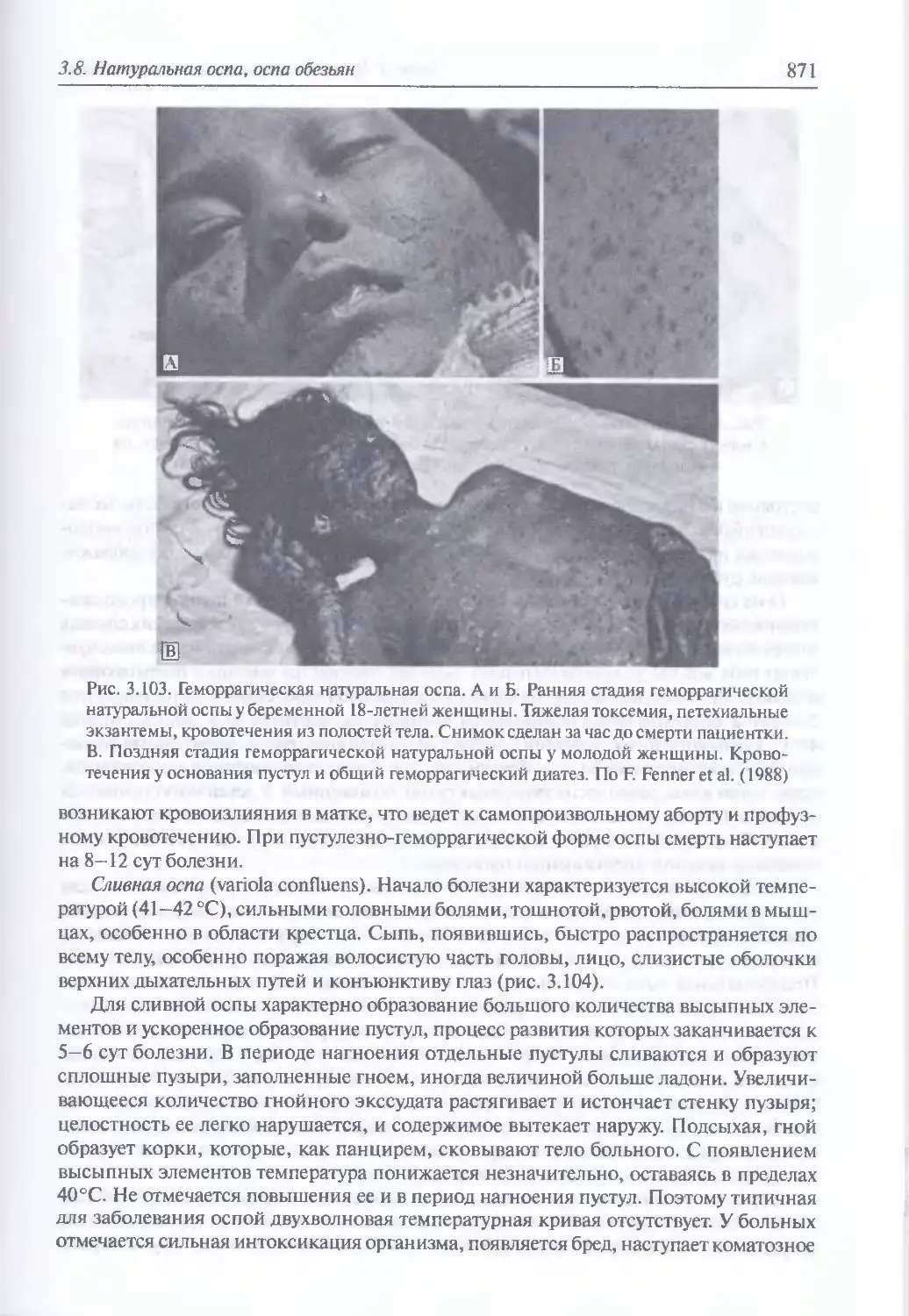
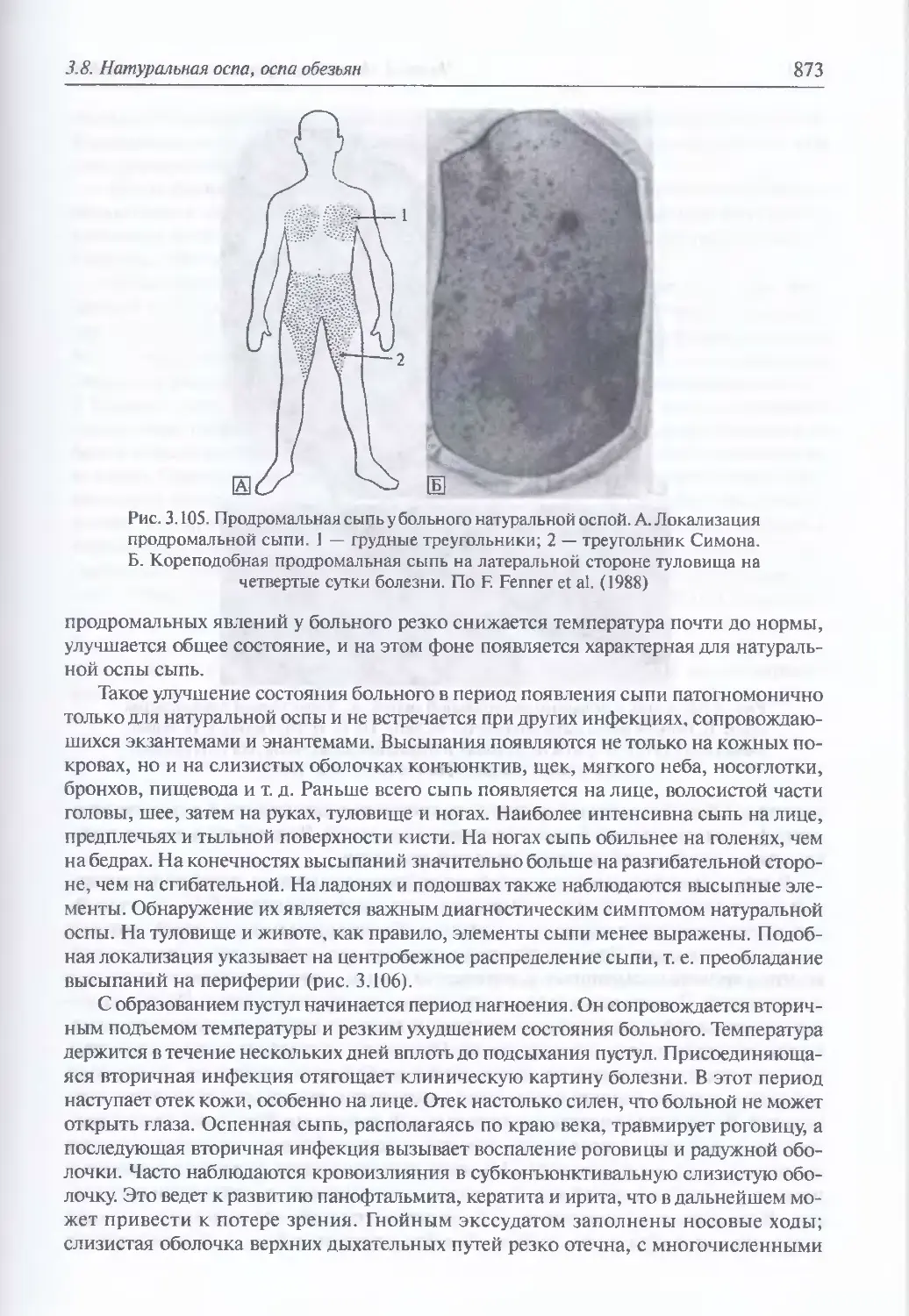
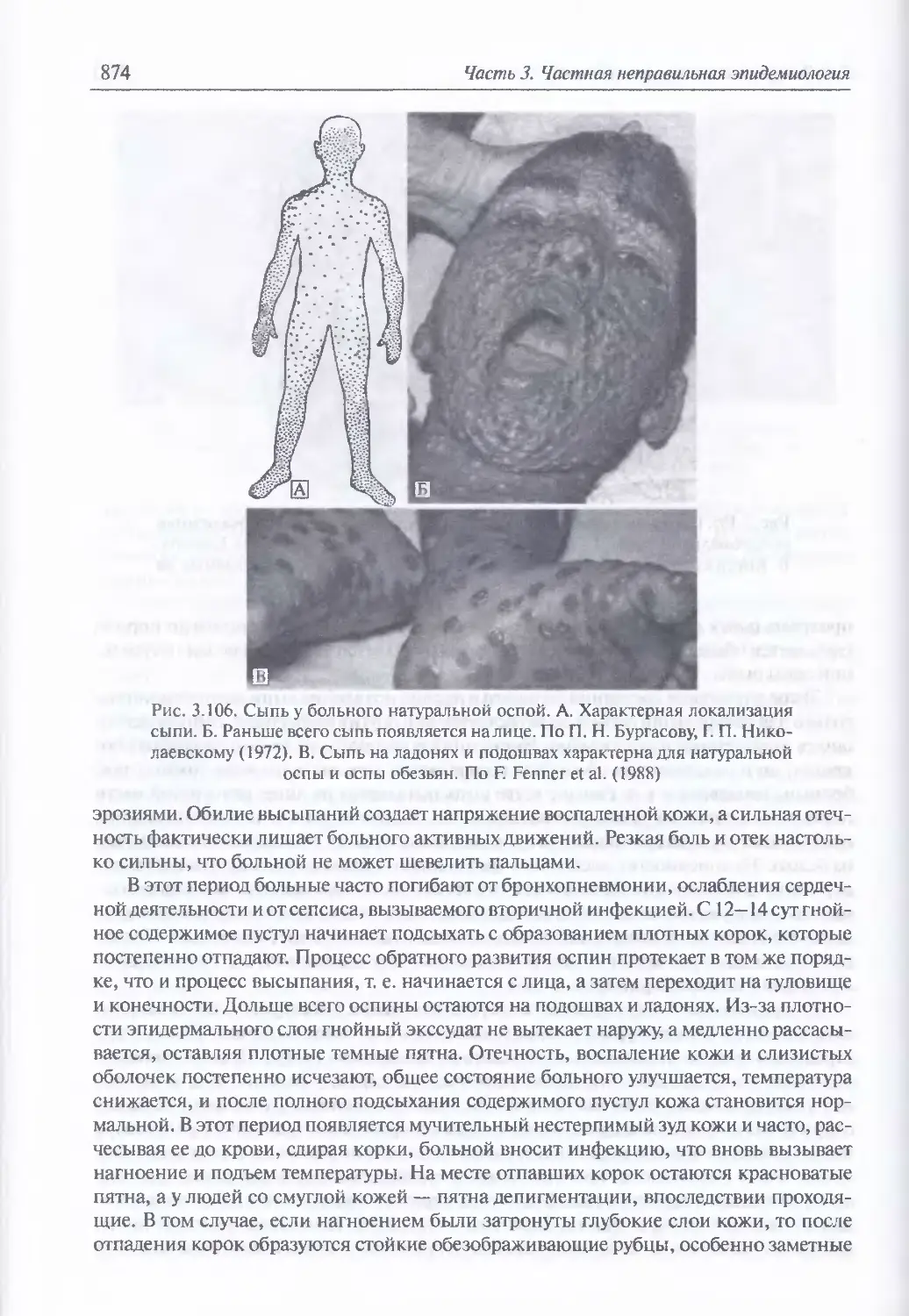
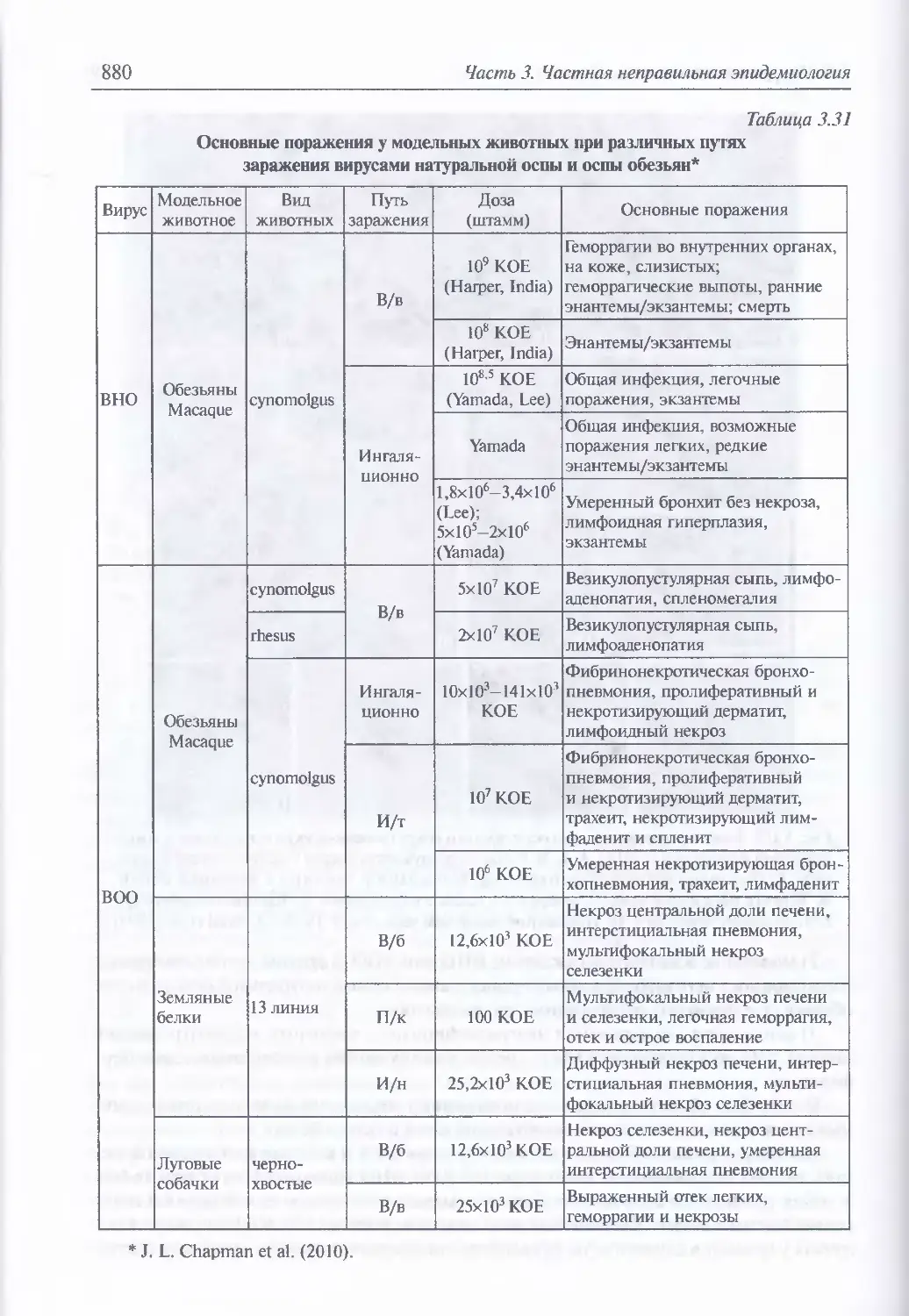
/






















































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































Автор: Супотницкий М.В.
Теги: материальные основы жизни биохимия молекулярная биология биофизика биологические науки эпидемиология биологическое оружие
ISBN: 978-5-93165-328-0
Год: 2013




Текст
М.В.СУПОТНИЦКИЙ
БИОЛОГИЧЕСКАЯ
ВОЙНА
Библиотека БЖД и ЧС
М. В. СУПОТНИЦКИЙ
БИОЛОГИЧЕСКАЯ
ВОЙНА
Введение в эпидемиологию
искусственных эпидемических процессов
и биологических поражений
МОСКВА
«РУССКАЯ ПАНОРАМА»
«КАФЕДРА»
2013
УДК 577
ББК28
С89
СЕРИЯ «БИБЛИОТЕКА БЖД и ЧС»
Стратегический партнер издательства «Кафедра»:
ОАО «Русская фармацевтическая корпорация».
Автор идеи и составитель серии:
Л. В. Примак, доктор технических наук, профессор,
полковник запаса.
Рецензент:
С. В. Петров, доктор технических наук, лауреат Ленинской премии 1991 г.,
генерал-полковник в отставке. В 1989—1991 гг. начальник Химических войск СССР;
в 1991—2001 гг. начальник Войск радиационной, химической и биологической
защиты Российской Федерации
Супотницкий М.В.
С89 Биологическая война. Введение в эпидемиологию искусственных эпидемиче-
ских процессов и биологических поражений; монография / Супотницкий М.В. —
М.: «Кафедра», «Русская панорама», 2013. — 1136 с.: ил.
ISBN 978-5-93165-328-0
Биологические оружие как средство массового поражения запрещено междуна-
родными соглашениями. Наличие химического и биологического оружия послужило
поводом для американского вторжения в Ирак (этого вида оружия там не оказалось) и
свержения режима С.Хусейна. Наличием такого оружия в Сирии и вероятностью его
попадания в руки террористов западные эксперты ныне пытаются обосновать необ-
ходимость проведения операции против правительства Асада в Сирии. О том, какие
угрозы несет биологическое оружие, о принципах его действия и с какими опасностя-
ми столкнется человечество при попадании этого оружия в руки террористов расска-
зывается в этой книге. Монография восполняет пробелы в знаниях о биологических
угрозах, информация о которых широко представлена в западных источниках, но от-
сутствует в отечественных, что до сих пор ставило российских специалистов в неравное
положение с их западными коллегами. Книга предназначена для ученых, аспирантов и
студентов биологических и медицинских специальностей, врачей, организаторов здра-
воохранения, работников правоохранительных органов, но в основном ориентирована
на специалистов их низового звена (которые обычно первыми сталкиваются с послед-
ствиями террористических актов и диверсий).
ISBN 978-5-93165-328-0
© Супотницкий М.В., 2013
© «Русская панорама», 2013
© «Кафедра», 2013
СОДЕРЖАНИЕ
ВВЕДЕНИЕ........................................................... 12
СПИСОК СОКРАЩЕНИЙ...................................................18
ЧАСТЬ 1. КРАТКАЯ ИСТОЕ ИЯ ПРИМЕНЕНИЯ БИОЛОГИЧЕСКИХ
СРЕДСТВ ДЛЯ ПОРАЖЕНИЯ ЛЮДЕЙ.........................................24
1.1. Развитие представлений о природе биологических ядов............24
Античные мифы (24). Ядивитые грибы (26). Ведение войны (27). Змеи (29).
Умопомешательства (29). Средние века (29). Поиски ядов (30)
1.2. Развитие представлений о природе эпидемий .....................33
Античные ми<Ьы (33) Появление миазматического учения (34). Колдуны,
ведьмы, демоны и наказание Божье (35). Антисемитские предрассудки (36)
Появление контагионистического учения (37). Локалисты (40).
«Овеществленный» контагий (40)
1.3. «Биотеррор» миазматиков и колдунов......................................42
Пандемия «черной смерти» (1346—1351 гг.) (43). Флагеллянты (46).
Удобное объяснение причин распространения чумы в Европе (47)
1.4. Контагионисты «сеют чуму»...............................................48
«Сеятели чумы» (48). «Чисто биологическое убийство» Петра II? (53)
Биологические диверсии конца добактериологической эпохи (54)
1.5. Появление военной микробиологии.........................................55
Агробиологические эксперименты на Форту «Александр I» (55)
Бактериологические убийства (61). Способы и устройства для массового
производства патогенных микроорганизмов (61). Технологии и устройства
для длительного хранения микроорганизмов в жизнеспособном состоянии (62)
Взгляды на ведение бактериологической войны (64). Средства защиты
от бактериального нападения и ликвидации его последствий (65). Чума (65).
Сибирская язва (66). Сап (66). Холера (67)
1.6. От бактериальных средств поражения к бактериологическому оружию —
бактериологические диверсии времен Первой мировой войны.......................67
США (68). Россия )68). Румыния (69). Испания (70). Аргентина (70).
Великобритания (70). Италия (70). Франция (70). Появление
биологического оружия? (71)
1.7. Становление бактериологического оружия между мировыми войнами............73
Подведение итогов — Женевский протокол 1925 г. (74). Послевоенные
взгляды на ведение бактериологической войны (78). Микроорганизмы —
потенциальные агенты биологического оружия (80). Способы и устройства
1930-х гг., предназначенные для ведения бактериологической войны (85).
Критические взгляды на биологическое оружие (89). Мистификация
Стида (91). Ошибки профессора Триллья (95). Повышение вирулентности
бактерий (98). Технологии и устройства для производства и длительного
хранения микроорганизмов в жизнеспособном состоянии (98).
Бактериологические диверсии в СССР (102). Военно-биологическая
программа Польши (103). Возобновление бактериологических диверсий
в ходе военных действий (109). А как же немцы? (109)
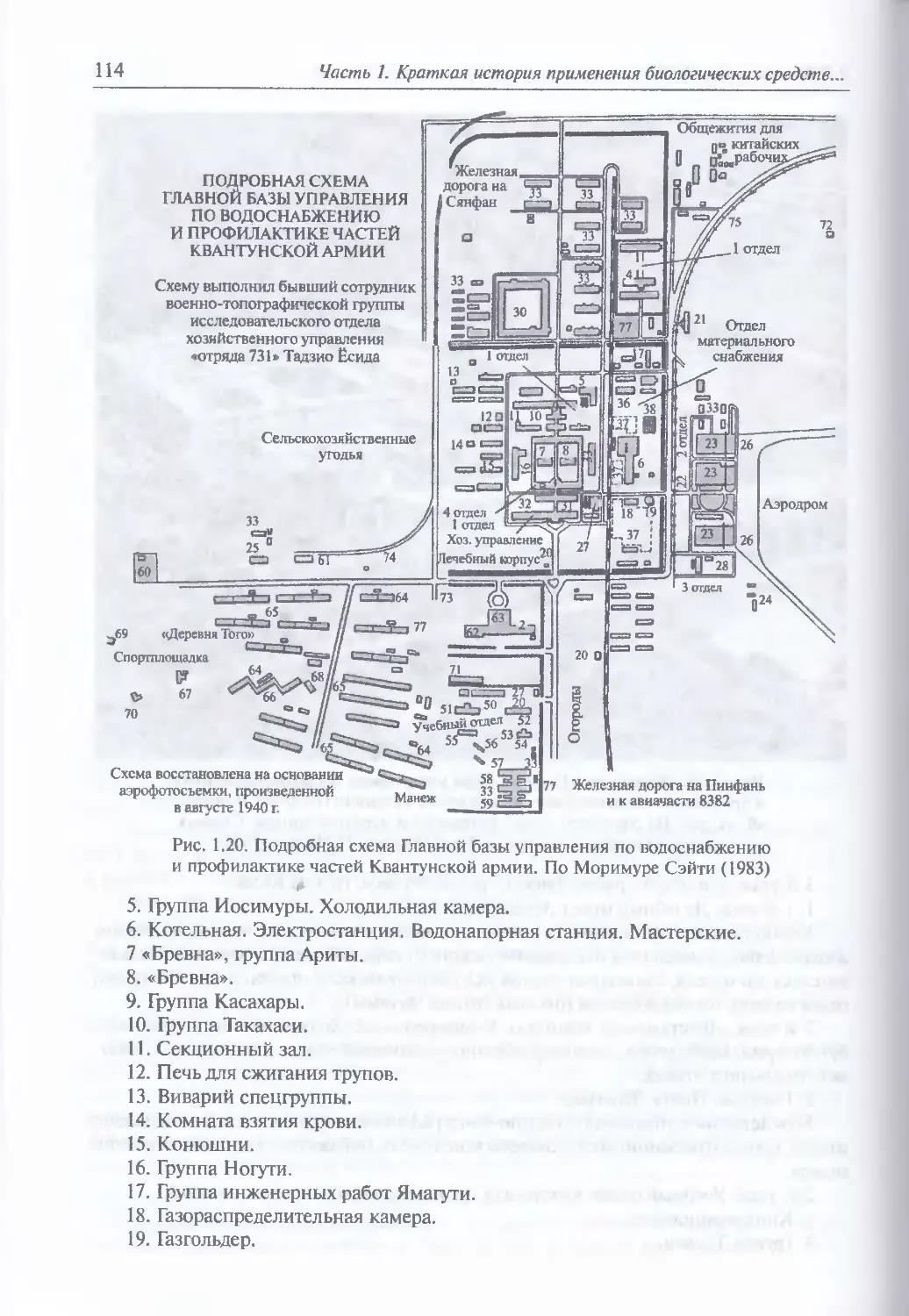
1.8. Крах отряда № 731.......................................................111
Начало японской военно-биологической программы (Ш). Отряд Ns 731 (112).
Отряд № 100 (123). Генерал-лейтенант медицинской службы Исии Сиро (125).
Раоота с «бревнами» (128). Представления японских военных о роли
4
Содержание
биологического оружия в войне (134). Разработка средств бактериологического
нападения (137). Выбор возбудителей инфекционных болезней в качестве
агентов биологического оружия (137). Повышение вирулентности бактерий (139).
Производственные мощности (143). Разработка специальных боеприпасов
и устройств для диспергирования блох и бактерий (150). Лабораторные
и полигонные заражения персонала (1601 Поиск диверсионных отравляющих
веществ (1б0). Возможные масштабы попыток применения биологического
оружия (162). Бактериологические диверсии против СССР (165).
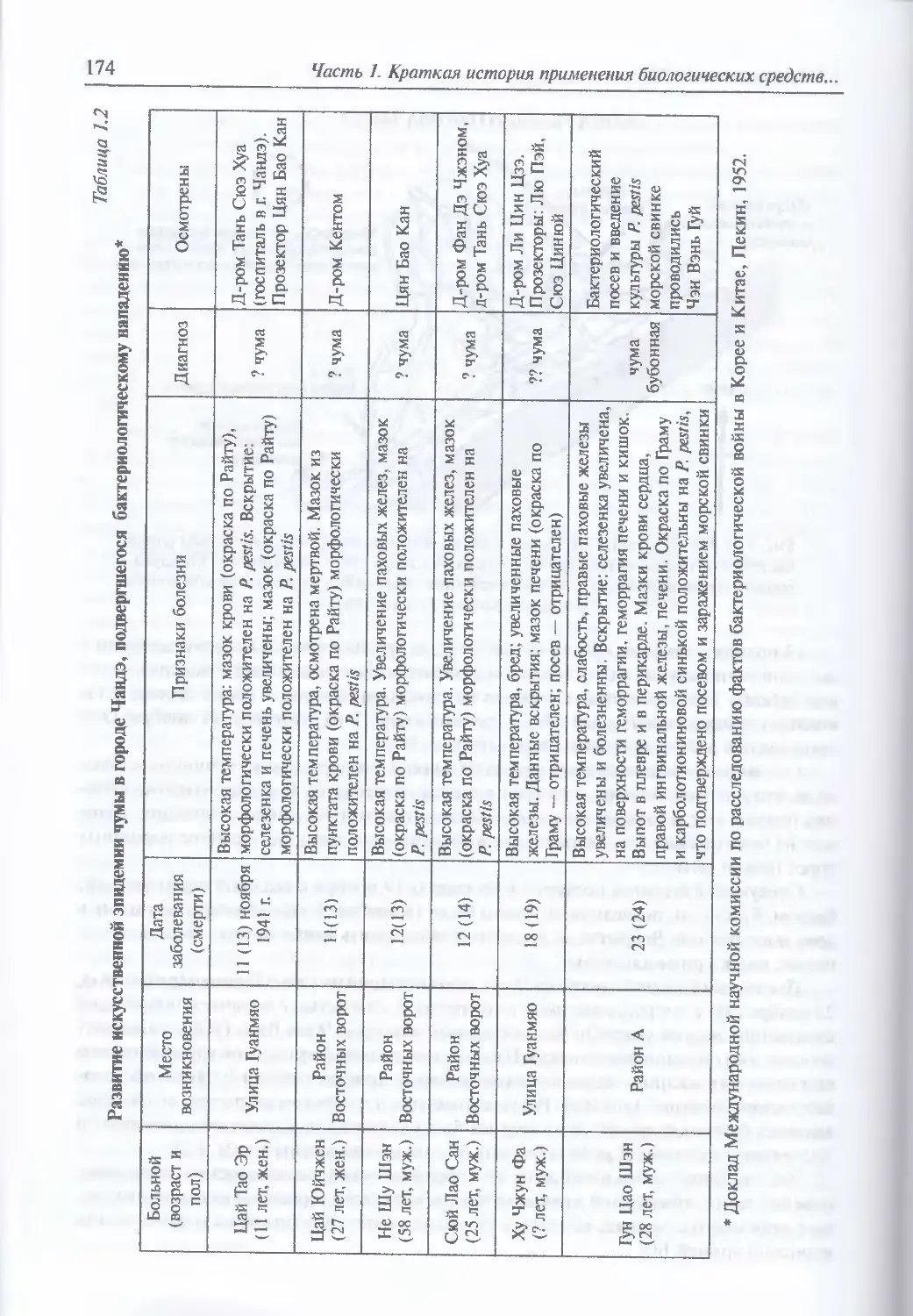
Бактериологическая война против Китая (167). Чума от дьявола в Чандэ (172).
Действия советской разведки (1?6). Конец «кухни дьявола» (177). Судебный
процесс по делу японских военнослужащих, обвиняемых в подготовке
и применении биологического оружия (183). Дальнейшая судьба плененных
сотрудников отряда № 731 (185)
1.9. Биологическое оружие перед Корейской войной.............................186
Взгляды на применение биологического оружия, существовалшие в годы
Второй мировой войны (187). Начало программы по созданию биологического
оружия в США (189). Начало программы по созданию биологического оружия
в Соединенном Королевстве (197). Военно-биологическая программа
Франции (199) Биодиверсии в Европе в годы Второй мировой войны (200).
Несостоявшиеся химические и биологические войны (201). Производство
биоагентов (204). Разработка технических средств для индивидуальных
убийств (207). Разработка технических средств для массового биологического
поражения людей (208). Аэрозоли (211). Моделирование масштабного
применения биологического оружия (214)
1.10. Корейская бактериологическая война.....................................217
Боевые действия в Корее в начале 1952 г (217) От ядерного оружия
к химическому и бактериологическому (218). «Дьявол всегда делает вид, что
он не существует» (219). Почему оказалось невозможным скрыть факты
применения биологического оружия в войне на Корейском
полуострове? (222). Бактериологические боеприпасы (227). Попытки
вызвать эпидемии чумы (231). Попытка вызвать вспышку холеры (234).
Попытки вызвать кишечные инфекции (235). Как по все делалось (235).
Использование аэрозолей (238)
1.11. Биологическое оружие перед подписанием Конвенции 1972 г................239
Масштабирование производства биологических агентов и испытаний
биологического оружия (240). Изучение поражающего действия
мелкодисперсных аэрозолей агентов биологического оружия (242).
Судьба проникших в дыхательные пути человека частиц биологического
аэрозоля (246). Технические средства применения аэрозолей агентов
биологического оружия (247). Аэробиология мелкодисперсного
аэрозоля (258). Отдел специальных операций в Форт-Детрике (263)
Насекомые как переносчики возбудителей опасных инфекционных
болезней (264) Поражение растений (266). Взгляды на ведение
биологической войны (266). Конвенция 1972 г. (268). Уничтожение
биологического оружия (268). Форт-Детрик после уничтожения
биологического оружия (269)
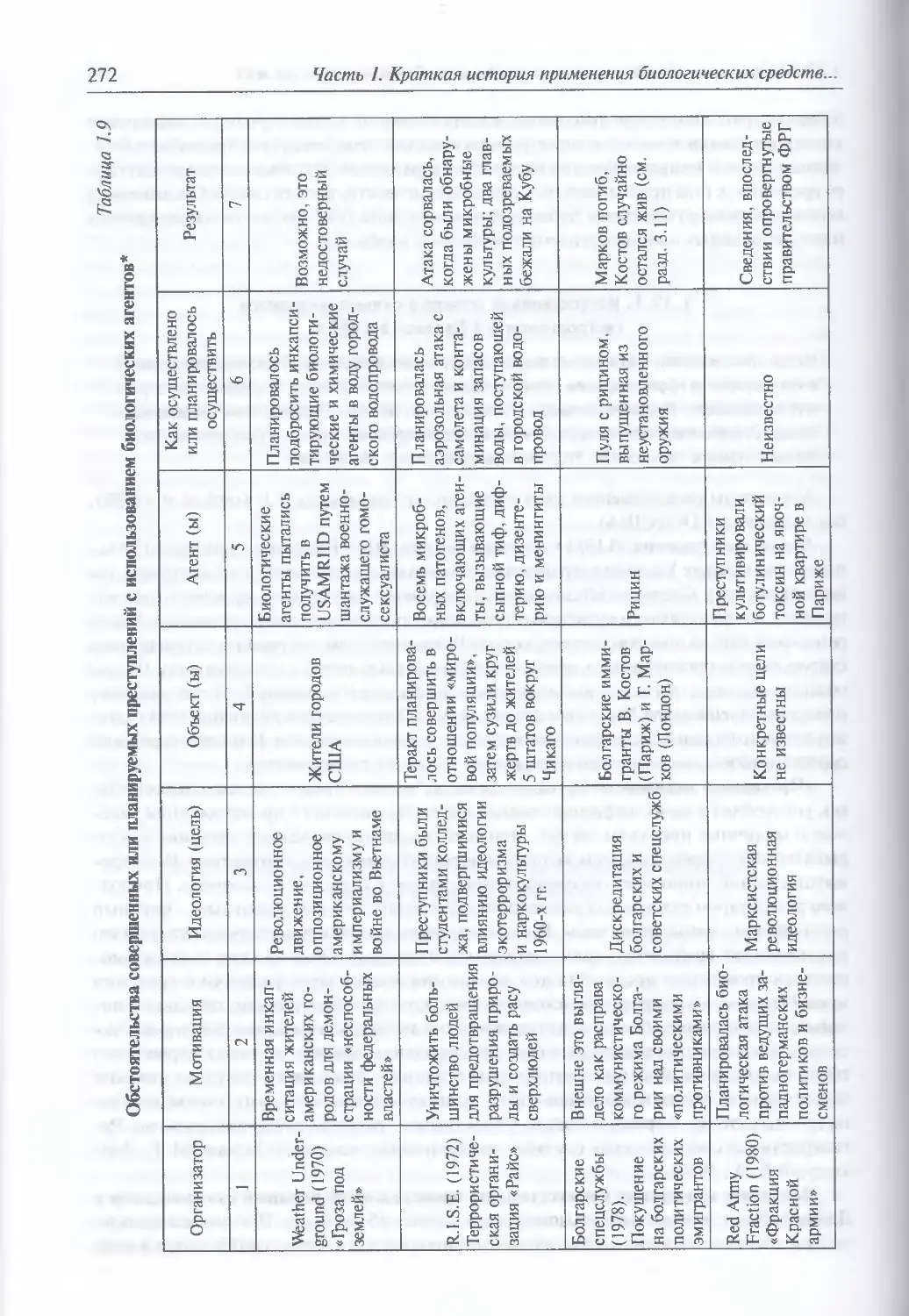
1.12. Биодиверсии и биотеррор времен профанации биологического оружия.........270
1.12.1. Искусственная вспышка сальмонеллезного гастроэнтерита
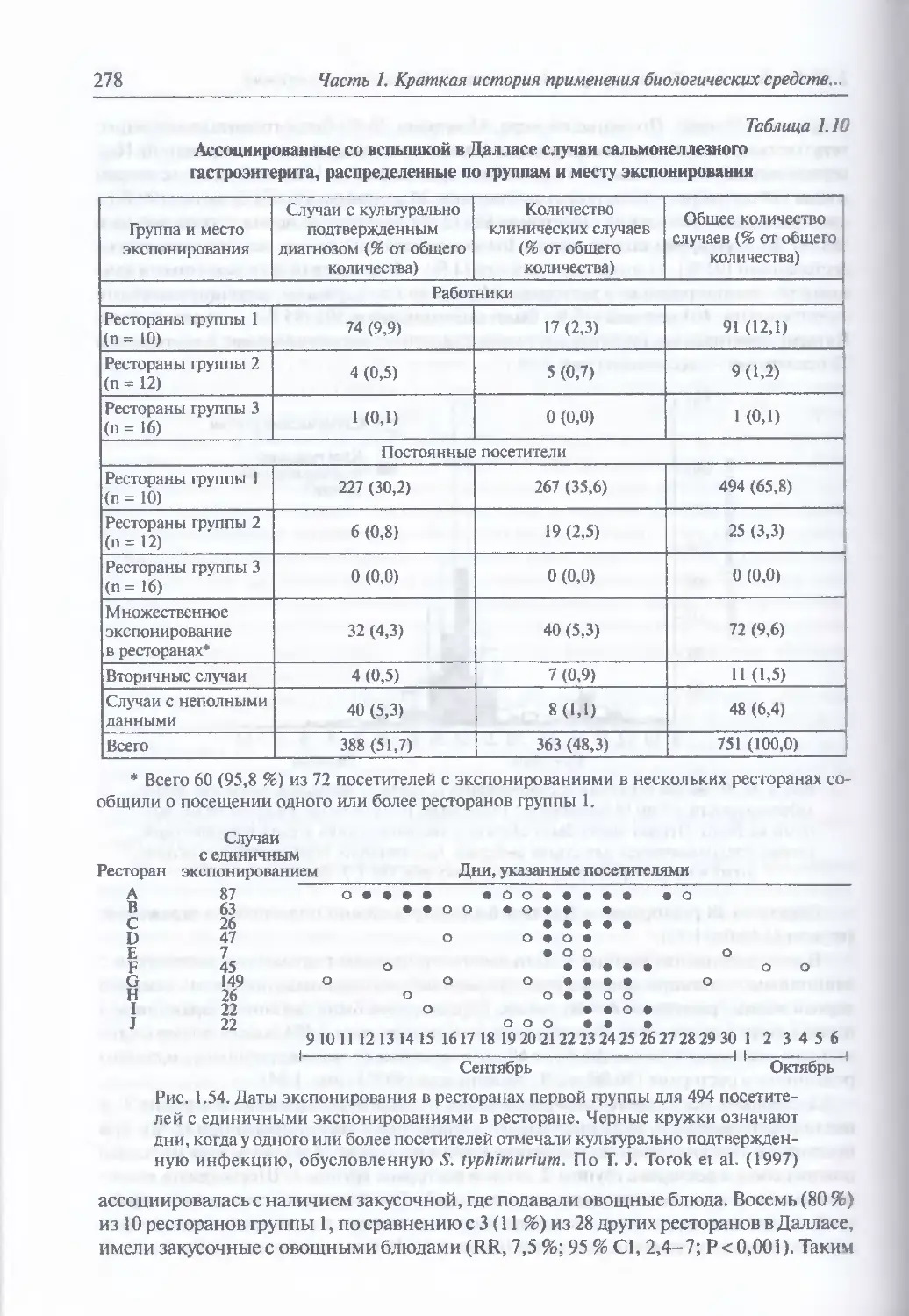
в Далласе в 1984 г...........................................................271
Мотив преступления (271). «Правильная эпидемиология» сальмонеллеза (271).
Появление подозрений в искусственном происхождении вспышки
сальмонеллеза в Далласе (271). Определение «случая инфицирования» (275).
Изучение вспышки (275). Лабораторные методы (276). Исследование
окружающей среды (276) Эпидемиологическое определение места совершения
теракта (276). Исследование выделенных штаммов сальмонечл (279). Уголовное
расследование (280). Наказание (281)
Содержание 5
1.12.2. Искусственная вспышка дизентерии Шмидца среди сотрудников
лаборатории медицинского центра в Техасе в 1996 г.........................281
«Правильная эпидемиология» Shigella dysenteriae 2 типа (281). Мотив
преступления (282). Появление подозрений в искусственном происхождении
вспышки дизентерии (282). Эпидемиологическое расследование (282).
Исследование выделенных штаммов возбудителя дизентерии Шмидца (284)
1.12.3. Инцидент с возбудителем сибирской язвы в районе Kameido
Токио в 1993 г.............................................................285
Мотив организаторов инцидента (285). Обстоятельства инцидента (285).
Эпидемиологическое расследование (287)
1.12.4. Искусственная вспышка сибирской язвы в США в 2001 г. ..............287
Предшествующие вспышке события в области контроля над биологическим
оружием (288). Кошмар из сна (288). Общий ход событий (289).
«Белый порошок» (292). Расследование биологического террористического
акта (293). Поведение людей (295). Эпидемиологическое расследование
вспышки сибирской язвы (298). Два случая ингаляционной сибирской
язвы во Флориде (304). Сортировочный центр в Вашингтоне (305).
Сибирская язва в Капитолии (307). Загадочный случай в Нью-Йорке (308).
Загадочный случай в Оксфорде (309). Исследование окружающей среды (310).
Идентификация (310). Постэкспозиционная неспецифическая
профилактика (311). Брюс Эдвард Айвинс (313). Эпидемиологические
особенности вспышки (314)
ЧАСТЬ 2. ОБЩАЯ НЕПРАВИЛЬНАЯ ЭПИДЕМИОЛОГИЯ .......................316
2.1. Объект, цель и задачи неправильной эпидемиологии............316
Эпидемиология искусственных эпидемий (вспышек) (318). Этиология
искусственных эпидемий и поражений токсинами (320). Клинические
проявления искусственных эпидемий (вспышек) (321). Ограниченность
общей и частной эпидемиологии при описании новых средств
биологического поражения людей (323). Терминологическое определение
неправильной эпидемиологии (324)
2.2. Биологические поражающие агенты......................................325
2.2.1. Классификации биологических поражающих агентов ...................326
Классификация биологических поражающих агентов по степени
важности для национальной безопасности США (326). Российская
классификация биологических поражающих агентов (327).
Классификация биологических поражающих агентов по их
способности к репликации в организме человека (327). Группа
реплицирующихся агентов (328). Группа нереплицирующихся агентов (328)
2.2.2. Реплицирующиеся биологические поражающие агенты.................329
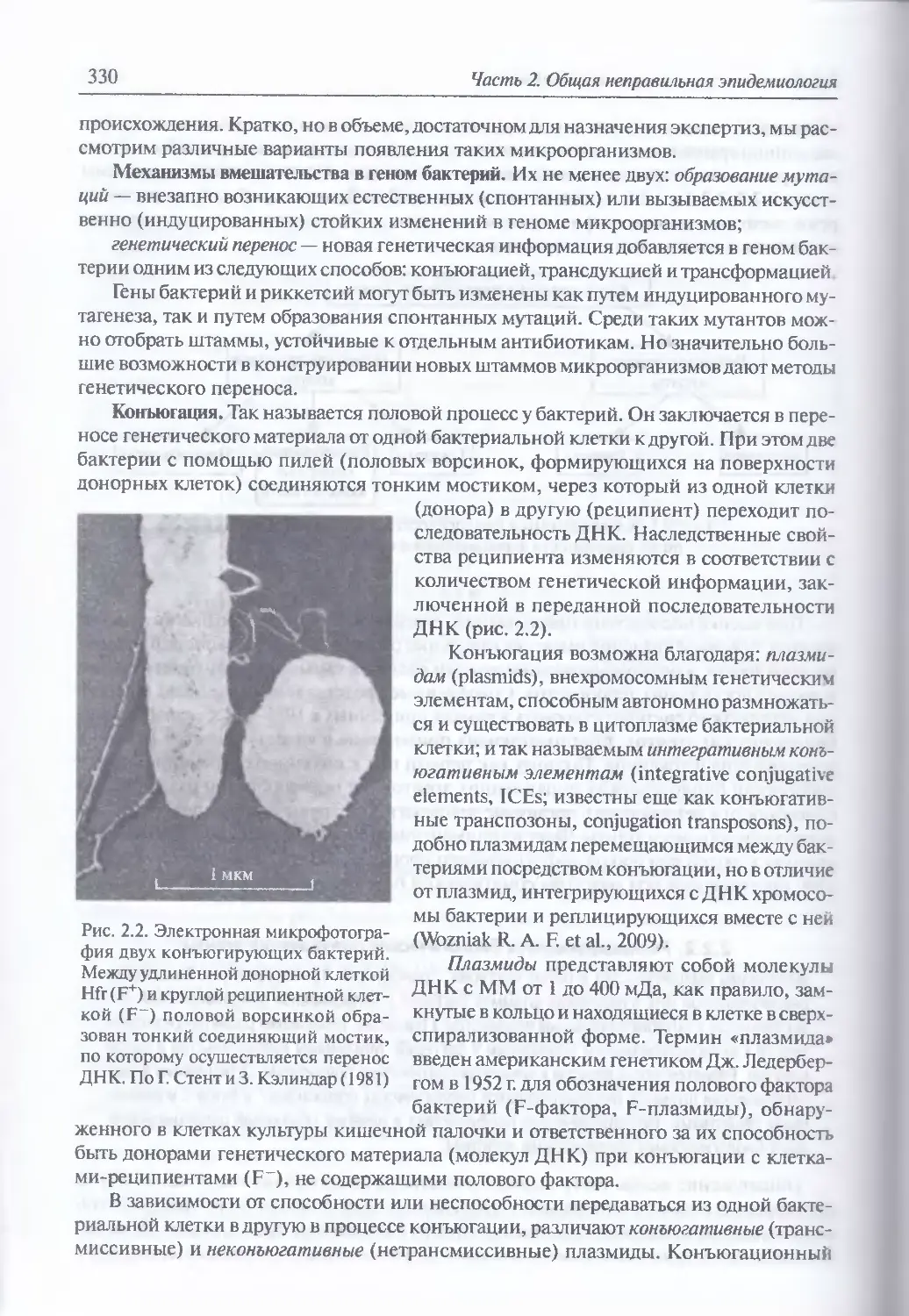
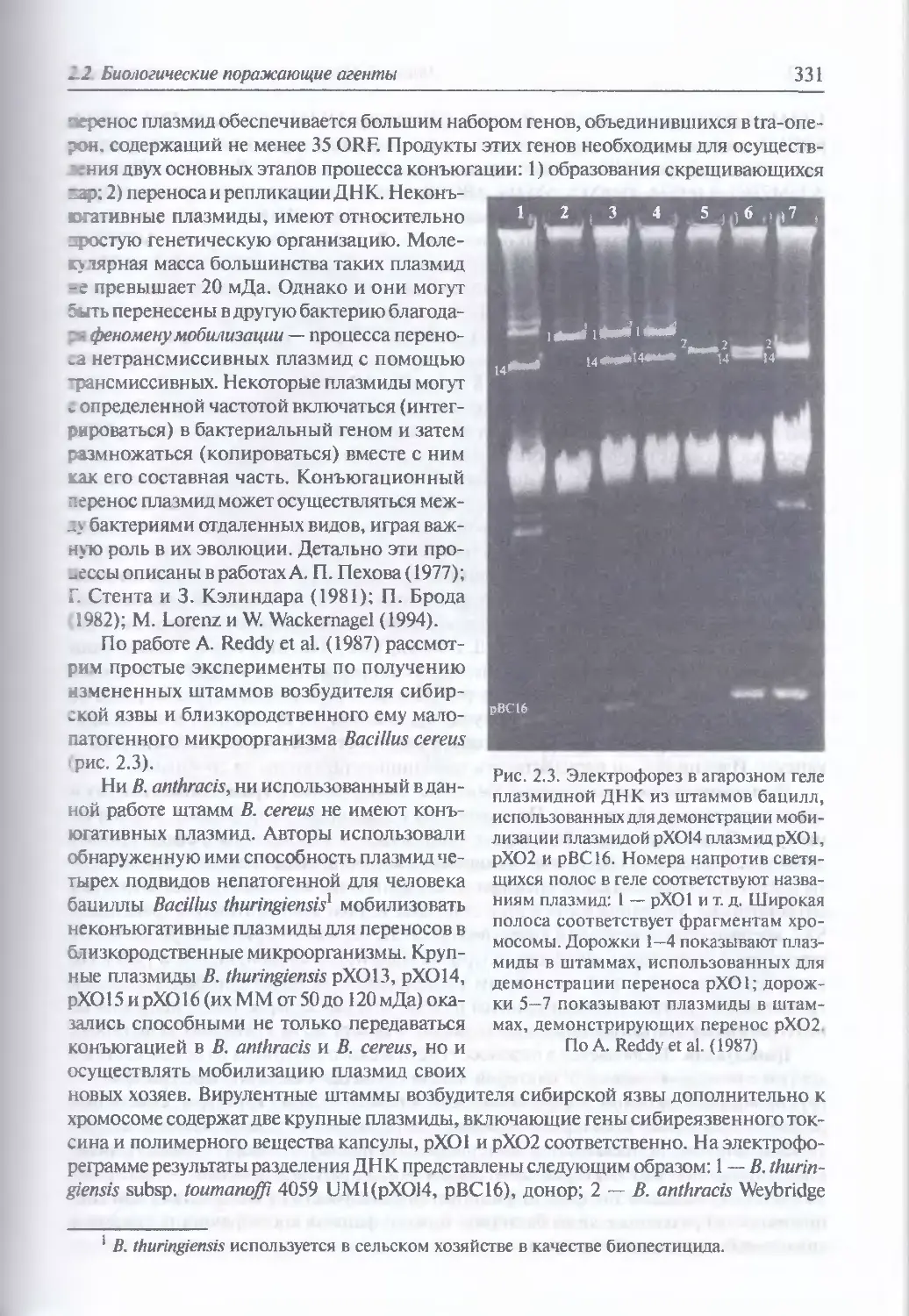
Механизмы вмешательства в геном бактерий (330). Конъюгация (330).
Трансдукция (332). Трансформация (336). Технологии рекомбинантной
ДНК в получении штаммов бактерий с неправильной эпидемиологией (339).
Резистентность к антибактериальным препаратам у бактерий (346).
Механизмы развития резистентности к антибактериальным препаратам
у бактерий (355). Механизмы вмешательства в геном вирусов (361).
Резистентность вирусов к химиотерапевтическим препаратам (367).
Ограничения в конструировании штаммов реплицирующихся биологических
поражающих агентов с измененными свойствами (369). Неспецифическая
профилактика и лечение поражений реплицирующимися биологическими
поражающими агентами (372)
2.2.3. Токсины и яды....................................................... 375
2.2.3.1. Общие свойства токсинов...........................................375
Классификации токсинов (375). Токсикометрия токсинов (378).
Фармакокинетика токсинов (380). Фармакодинамика токсинов (381).
Токсины как поражающие агенты (383)
6
Содержание
2.2.3.2. Токсины высших растений (фитотоксины).............................384
Алкалоиды (385) Гликозиды (385). Эфирные масла (387) Органические
кислоты (387). Лактоны (387). Токсальбумины (387). Закономерности
ядообразования у растений (388). Клиническая классификация
ядовитых растений (389). Роль отдельных токсических веществ
в формировании клинической картины отравления
растительными ядами (391). Патологоанатомическое
исследование (393). Красавка (393). Белена (395). Дурман (Datura) (396)
Болиголов пятнистый (397). Вех ядовитый (399). Аконит (Aconitum) (399).
Безвременник (401). Чемерица (Veratrum) (402). Мак (Papaver) (403).
Строфант (405). Чилибуха (406)
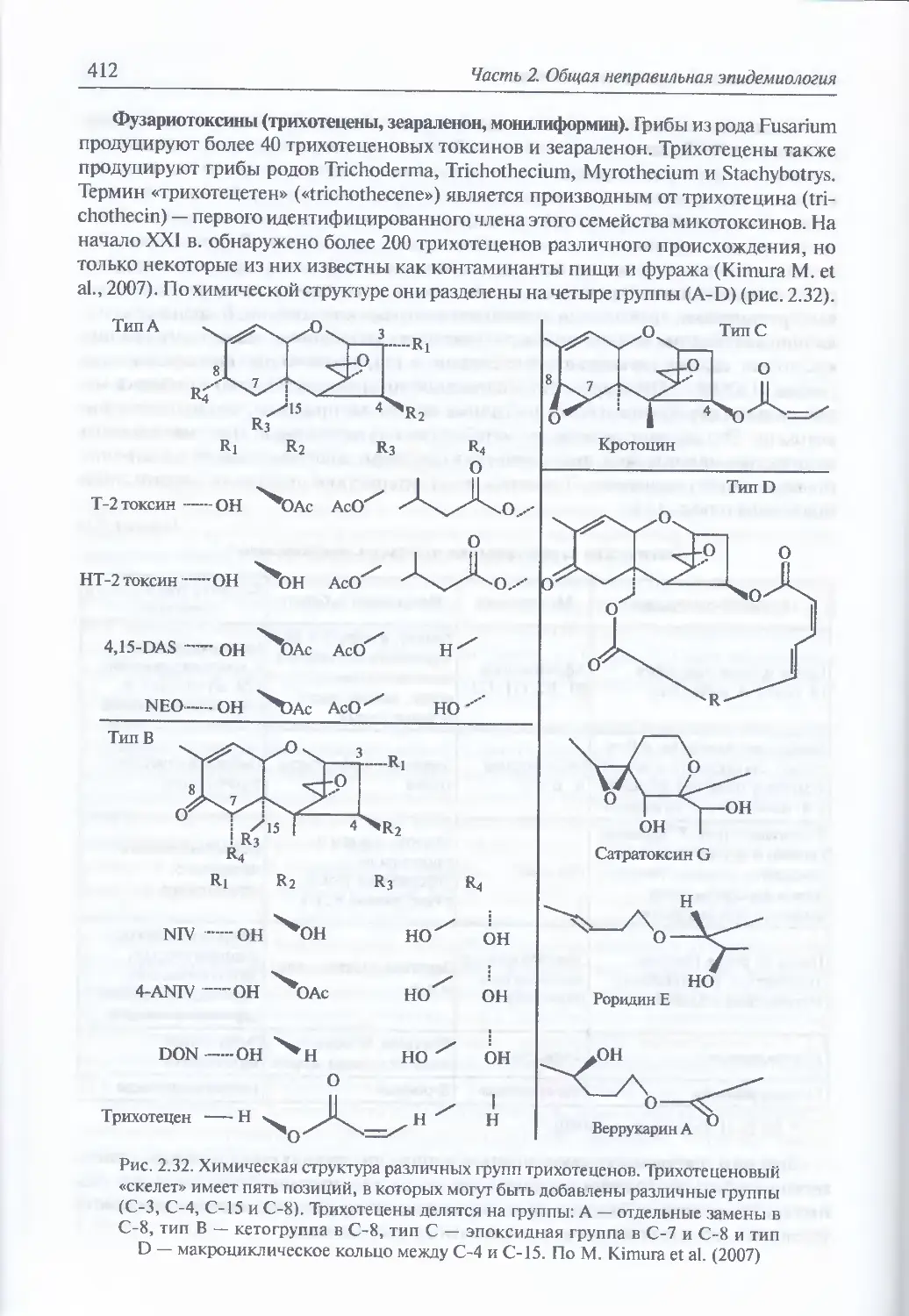
2.2.3.3. Микотоксины.......................................................409
Классификация грибов (410). Роль отдельных токсических
веществ несовершенных грибов (микотоксинов) в формировании
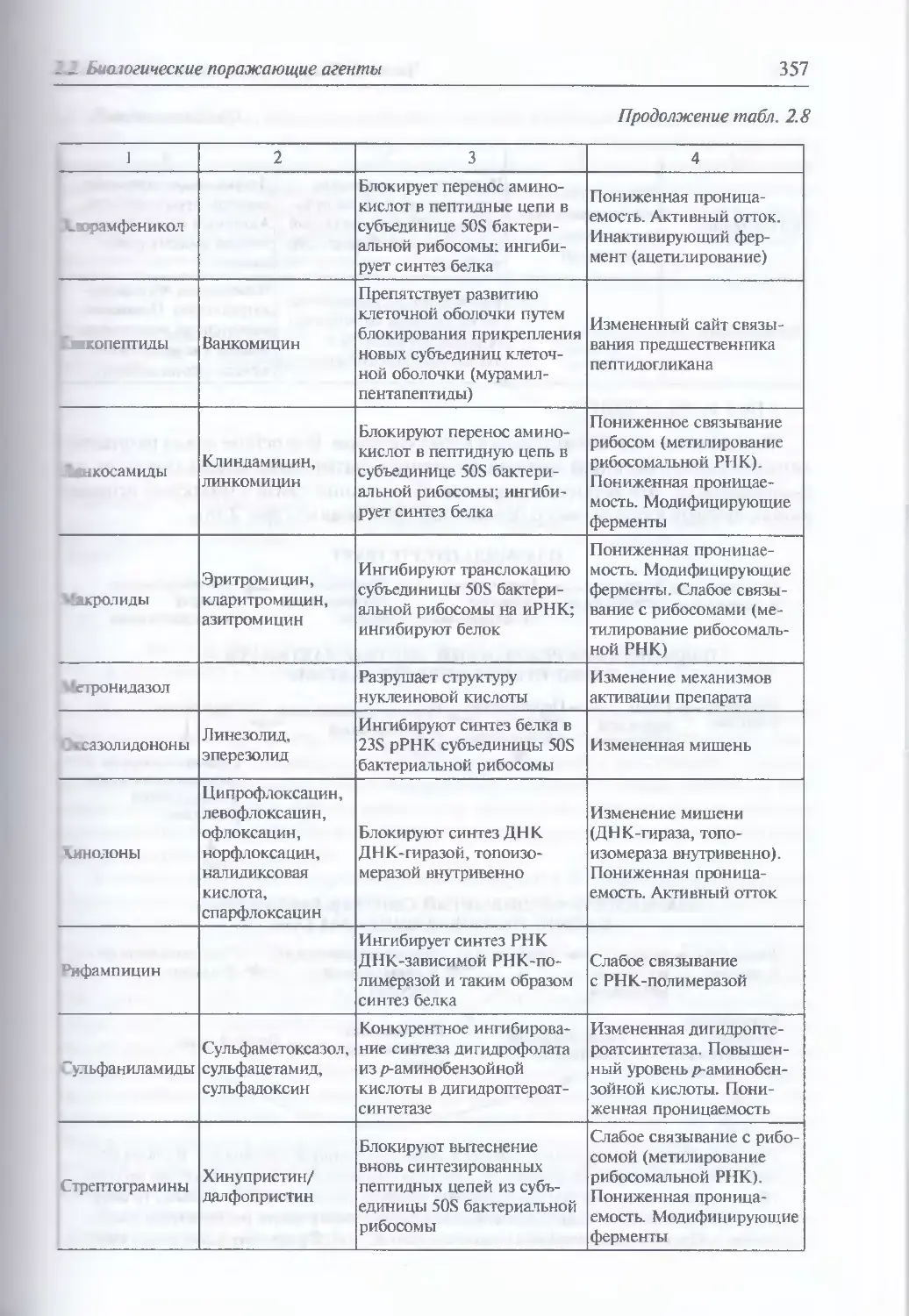
клинической картины отравления (410). Фузариотоксины
(трихотецены, зеараленон, монилиформин) (412). Афлатоксины (416).
Миротециотоксины (дондродохины) <418). Спородесмины (418).
Эрготоксины (418). Охратоксины (419) Микотоксины грибов
рода Penicillium (419). Патогенез и патологическая анатомия
микотоксикозов (420)
2.2.3.4. Яды грибов........................................................423
Роль отдельных токсических веществ грибов-макромипетов
в формировании клинической картины отравления (424).
Фаллоидиновый синдром (424) Гиромитриновый синдром (430).
Паксилюсный синдром (431). Орелланиновый синдром (432)
Проксимановый синдром (434). Судориновый синдром (434)
Психомиметические синдромы (436)
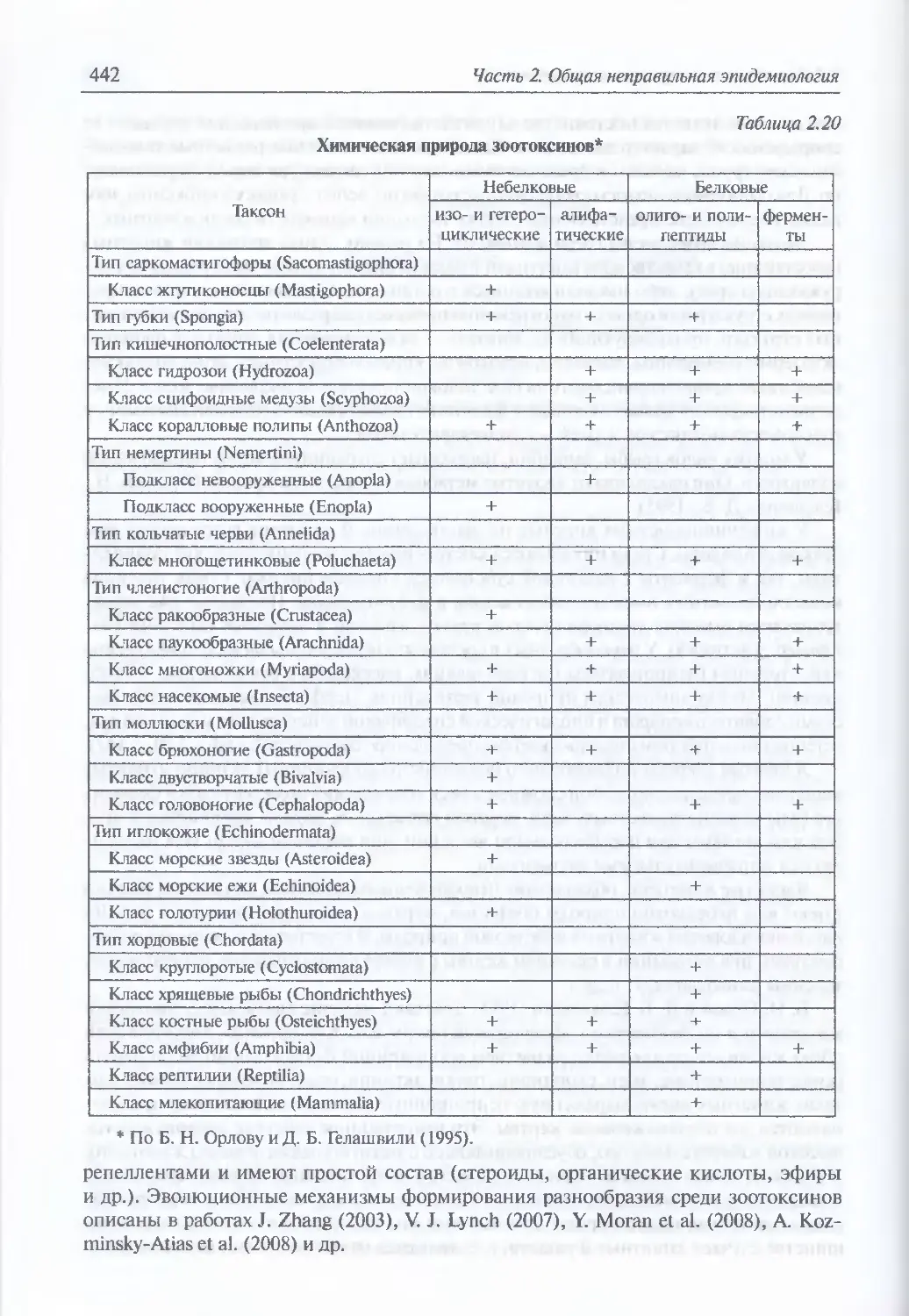
2.2.3.5. Зоотоксины........................................................439
Классификация ядовитых животных (440). Роль отдельны. токсических
веществ в формировании клинической картины отравления ядами
животных (443). Сакситоксин (452). Гимберотоксин (453). Суберитин (453).
Яд гонионем (453). Яд физалий (453). Яд хиронекс (453). Яд хризаоры (454).
Яд цианеи (454). Лофотоксин (454). Яд актиний (454). Эквинотоксин (455).
Теалиатоксин (455) Палитоксин (455) Яд конусов (455). Мурексин (456).
Макулотоксин (456). Яд скорпионов (456). Яд пауков-птицеедов (457). Яд
каракурта (458). Яд тарантула (458). Яд пчел (459). Кантаридин (459).
Яд сколопендры (460). Яд диамфидий (460). Яд мирских ежей (460). Яд
скатов (460). Яд бородавчатки (461). Тетродотоксин (461).
Сигуатоксин (462). Маитоксин (462). Батрахотоксин (462).
Буфотенин (463). Кобротоксин (463) Яд морских змей (465).
Яды гадюк (виперотоксин) и гремучников (кроталотоксин) (465).
Яд гремучей змеи Crotalus durissus terriflcus (466). Мойяветоксин (466).
Яд бумеланга Dispholidus typus (466). Яд древесной змеи Thelotomis
kirtlandi (466). Яд подвязочной змеи (466). Яд ядозубов (467).
Модификация ядов животных (467)
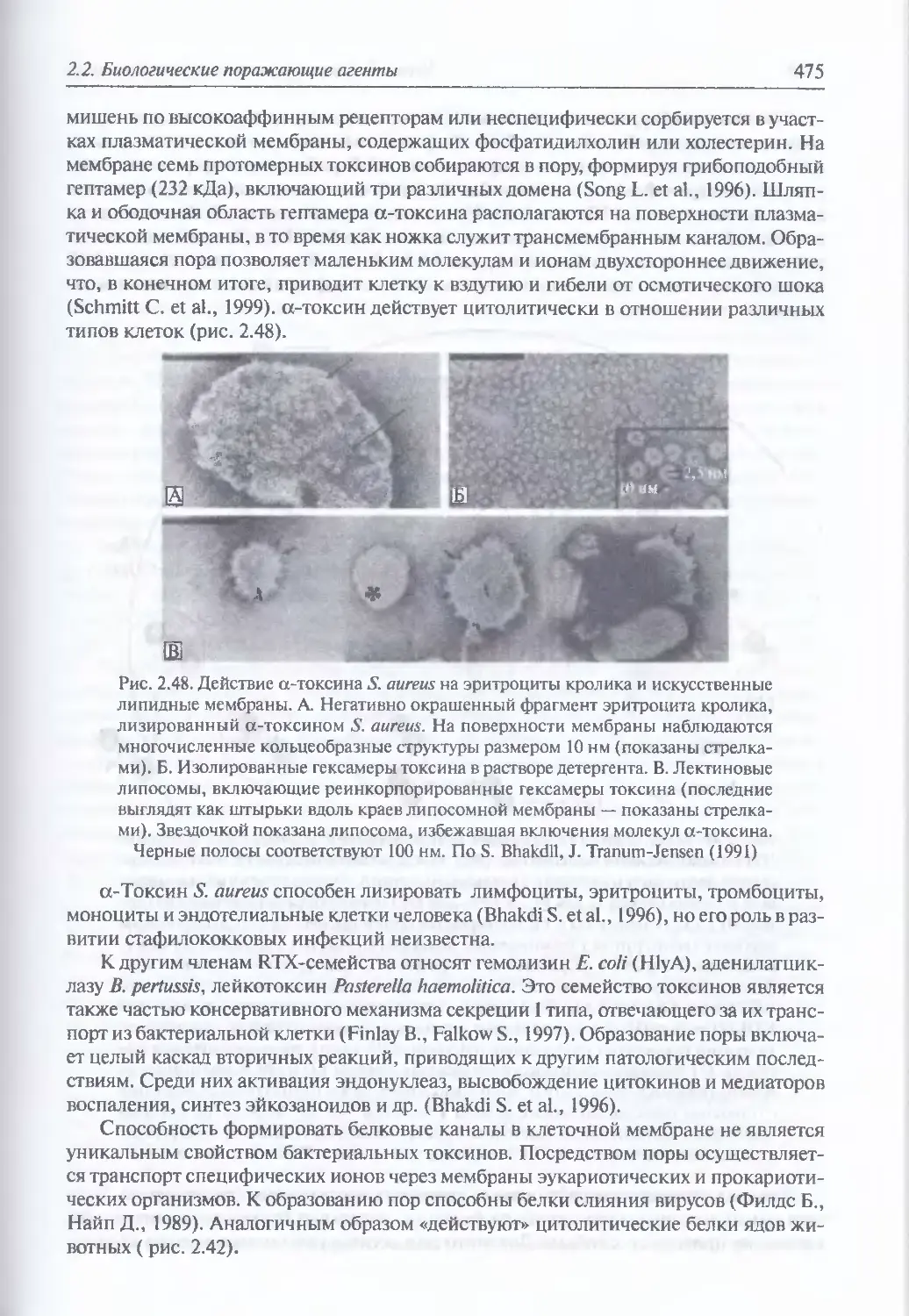
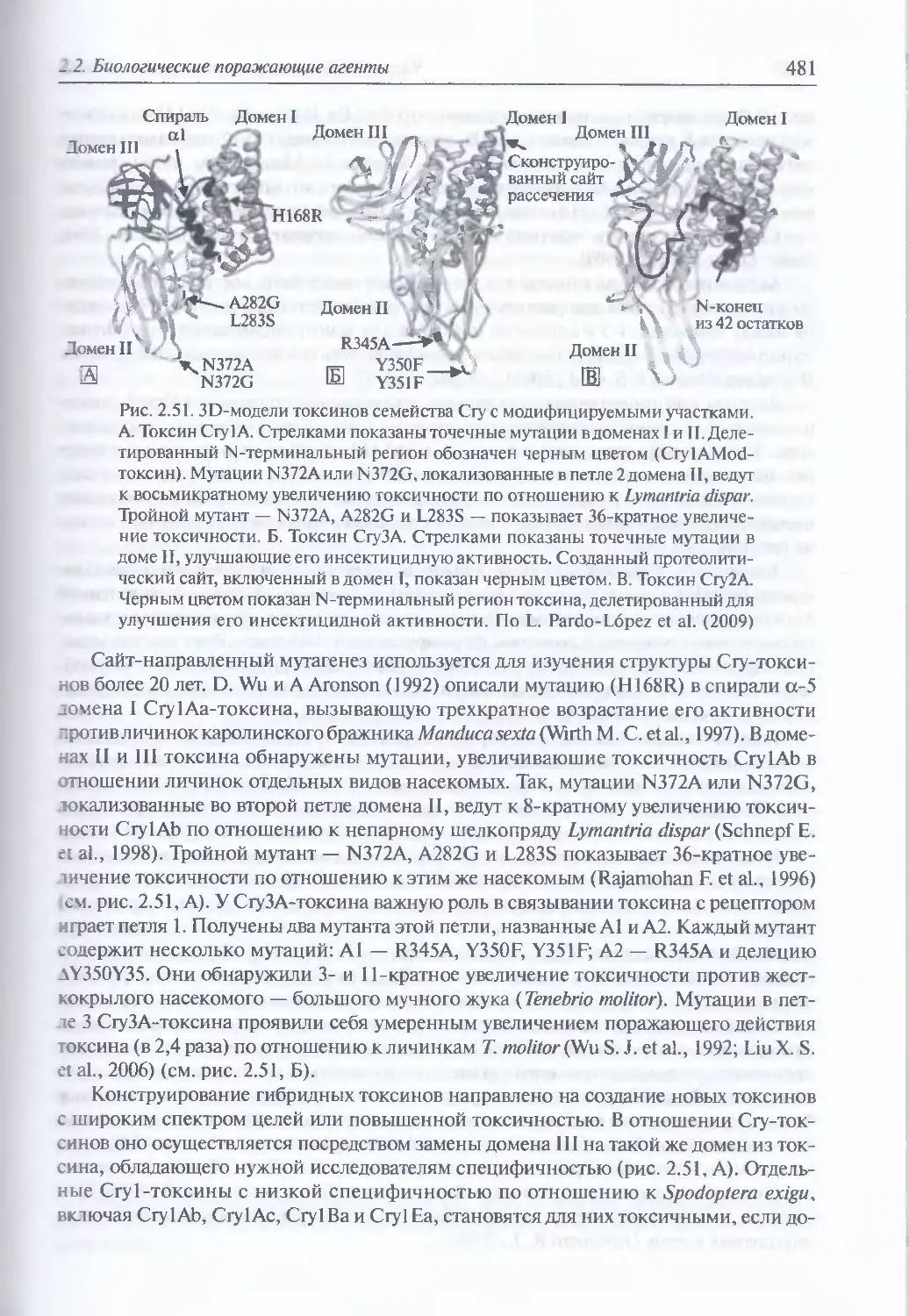
2.2.3.6. Бактериальные токсины.............................................468
Древность бактериальных токсинов (468). Организация и общий
механизм действия молекул бактериальных токсинов (469).
Классификация бактериальных токсинов (471), Порообразующие
токсины (474) Токсины, ингибирующие синтез оелка (476). Токсины,
генерирующие образование вторичных мессенджеров
(посредников) (476). Протеолитические токсины (477). Активаторы
иммунного ответа (478). Гибридные и модифицированные токсины (478).
Повышение поражающего действия токсинов (478). Иммунотоксины (482).
Максимально возможная токсичность (483)
Содержание
7
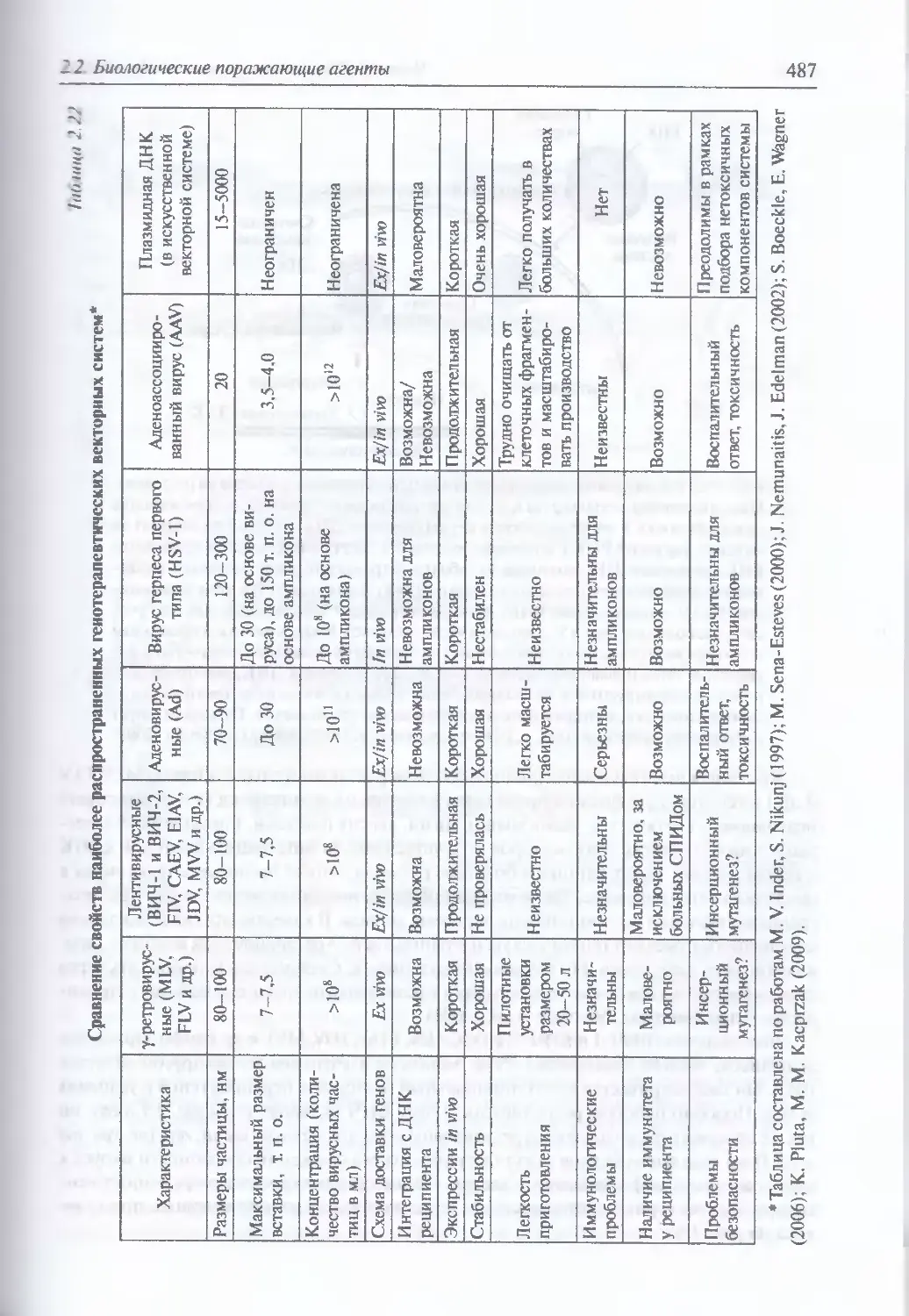
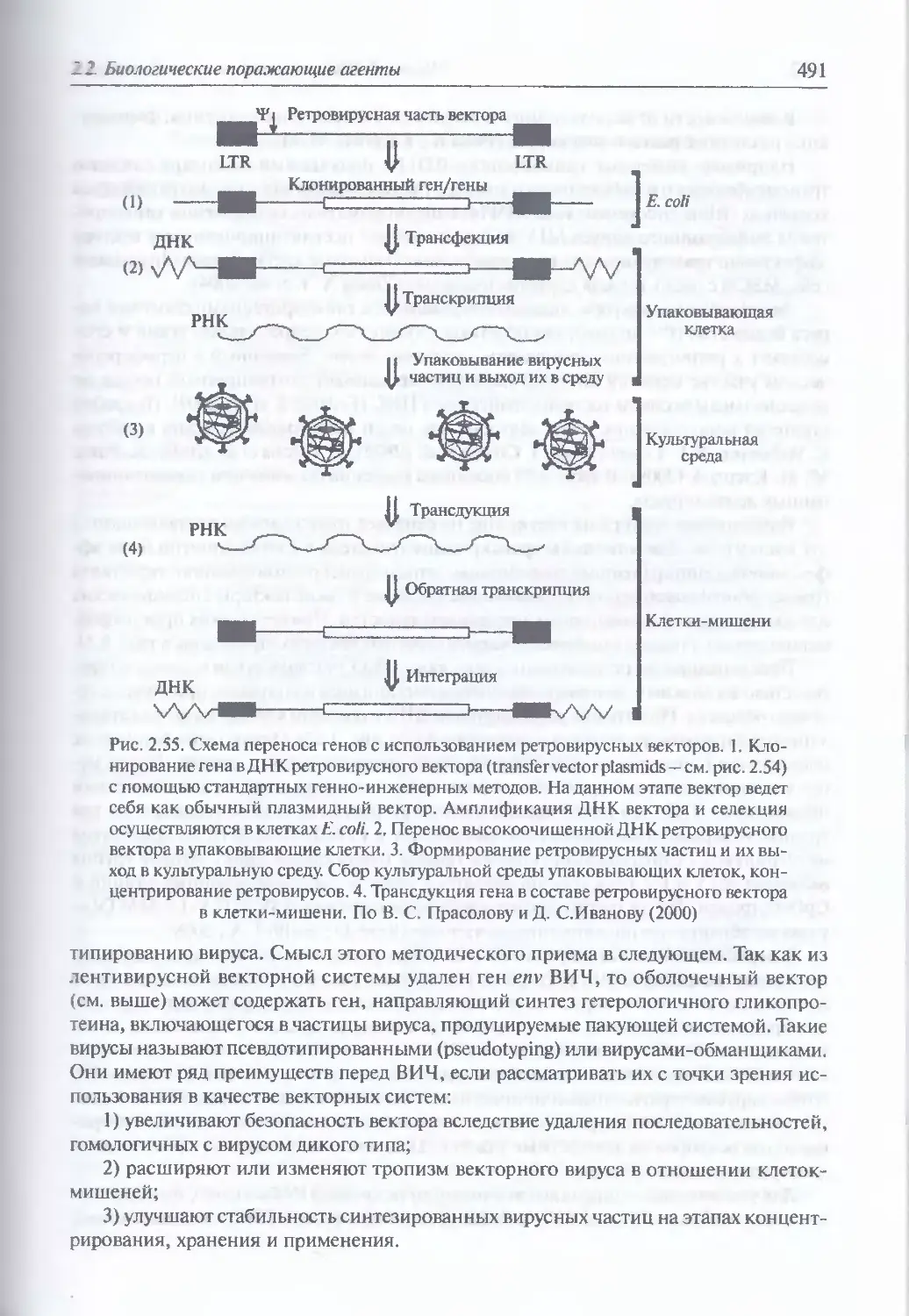
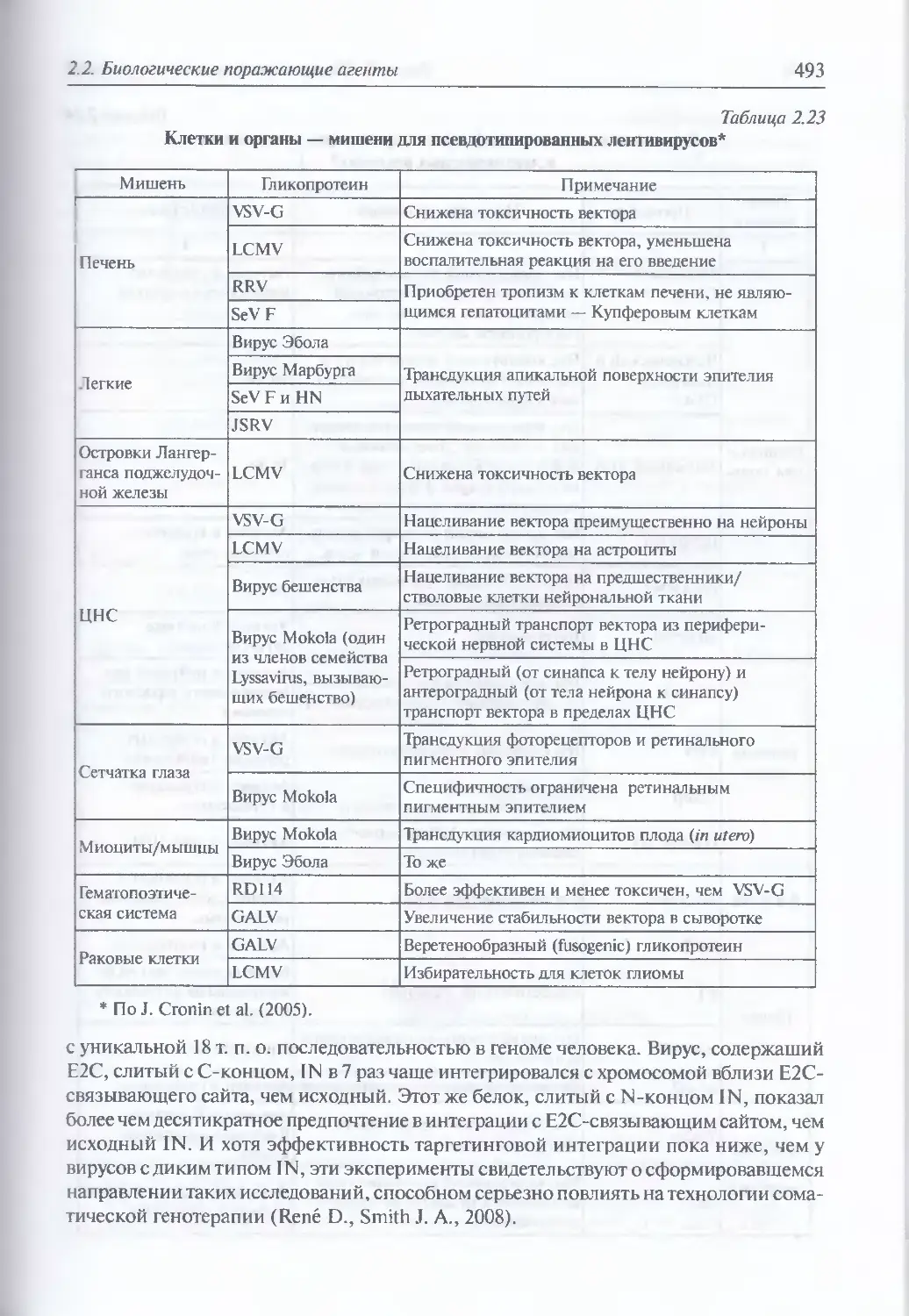
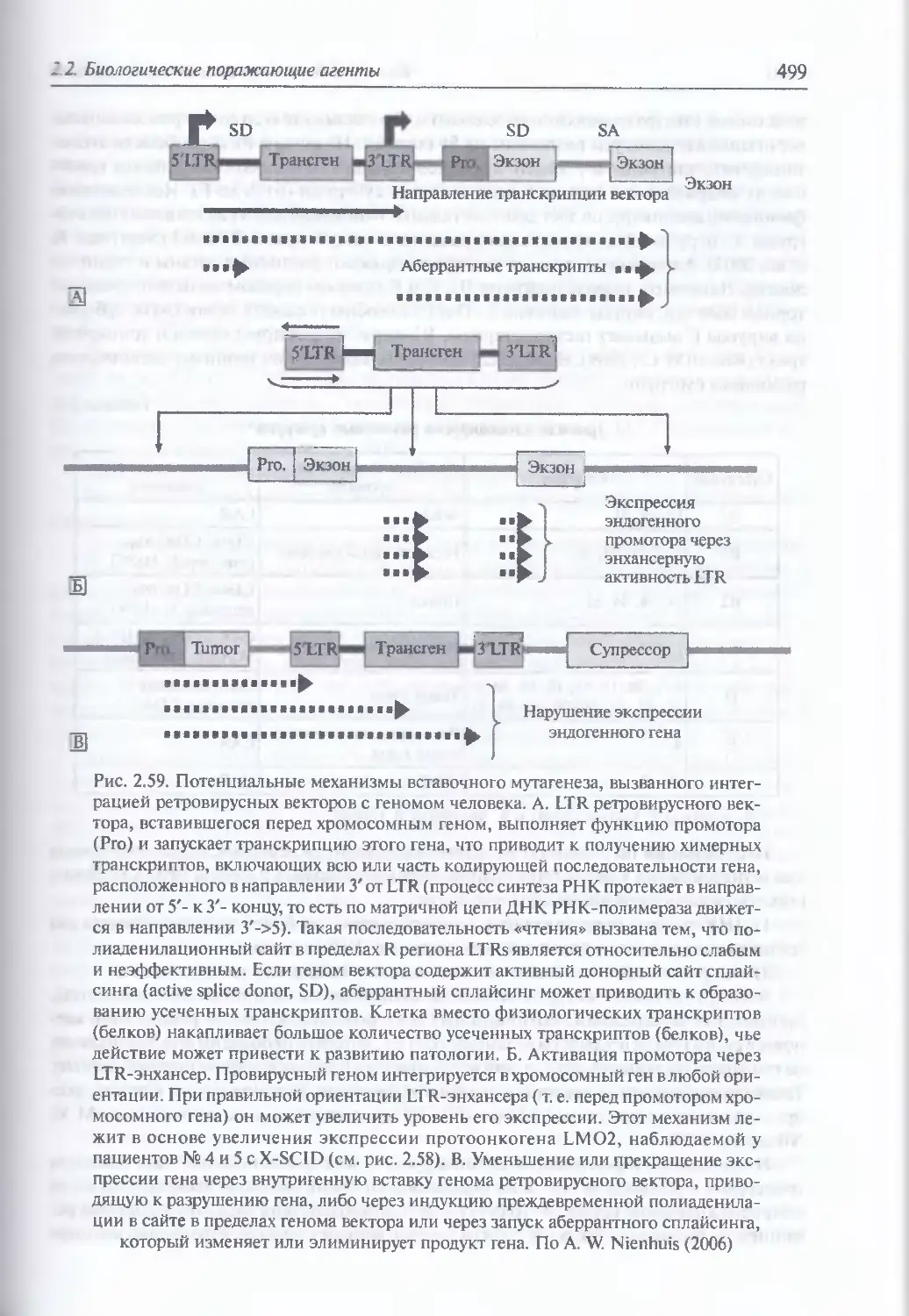
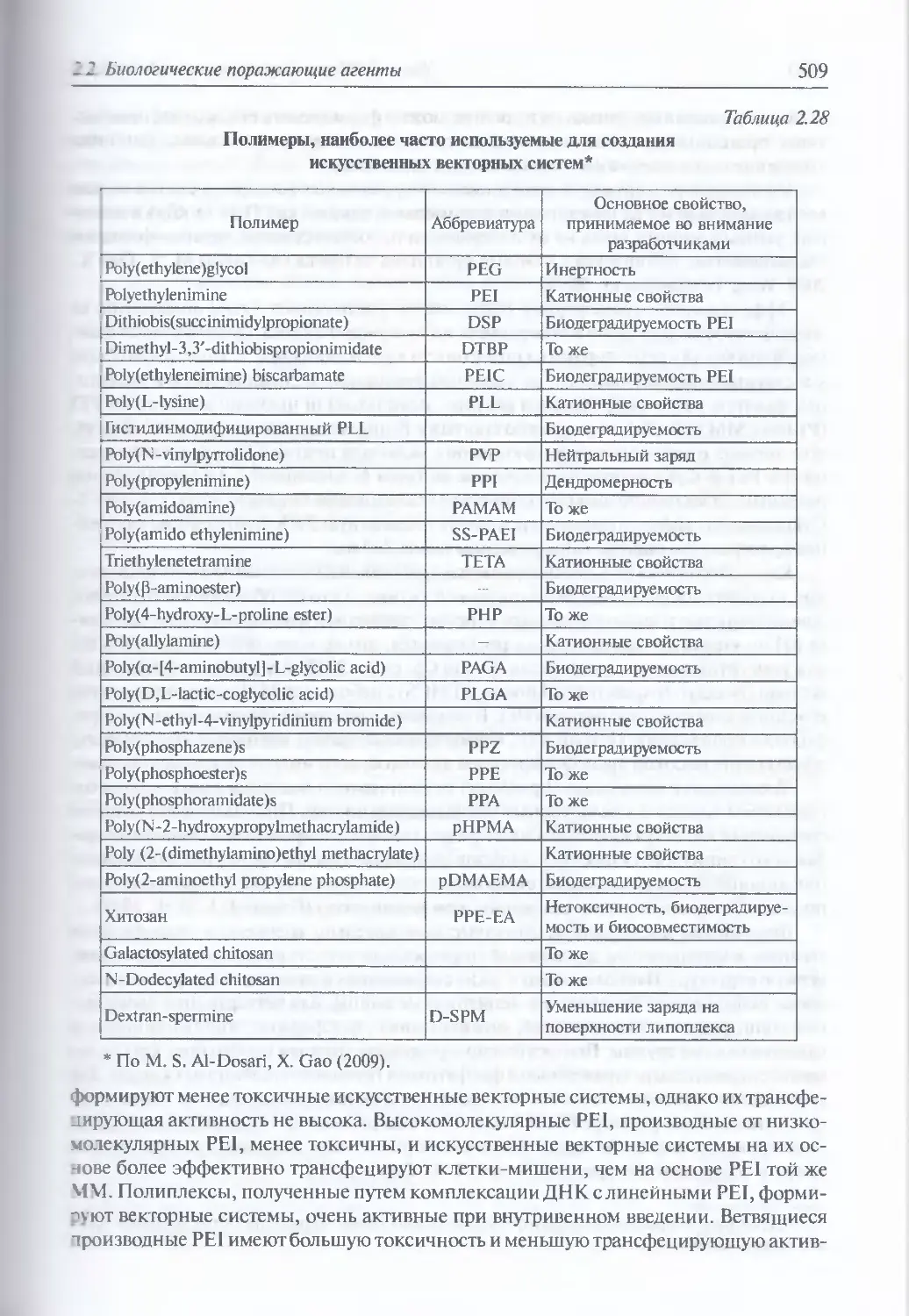
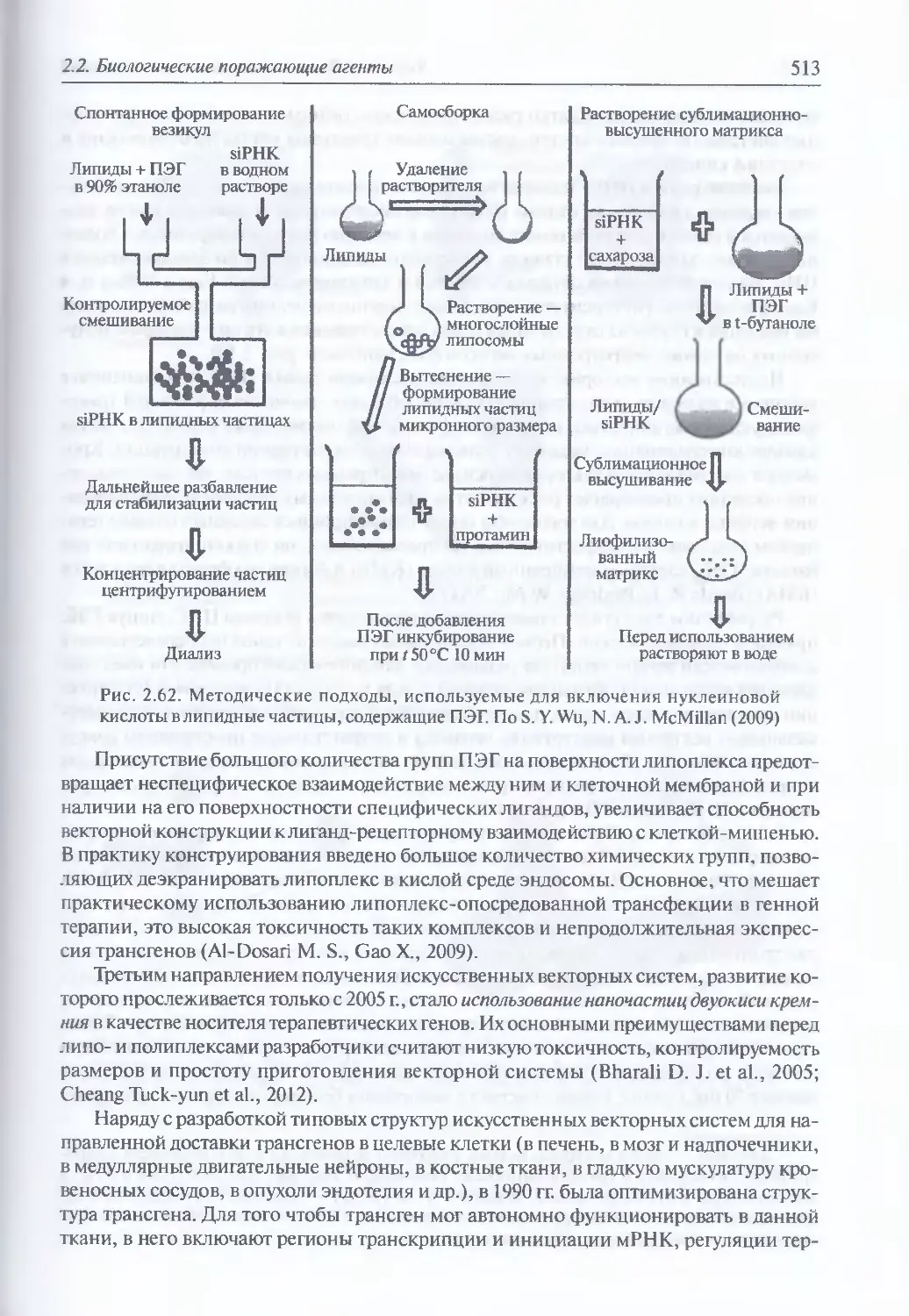
2.2.4. Искусственные генетические конструкции............................485
Искусственные генетические конструкции (486). Ретровирусные
векторные системы (486). Векторные системы на основе аденовирусов (498).
Аденоассоциированные вирусные векторы (503). Векторные системы
на основе вируса герпеса простого (505). Векторные системы на основе
ортопоксвирусов (505). Векторные системы на основе РНК-вирусов,
не принадлежащих семейству ретровирусов (506). Искусственные
векторные системы (506;. Введение генов в ЦНС (514). Введение генов
в легочную ткань в составе аэрозоля (517). Введение генов в клетки
кишечного эпителия (520)
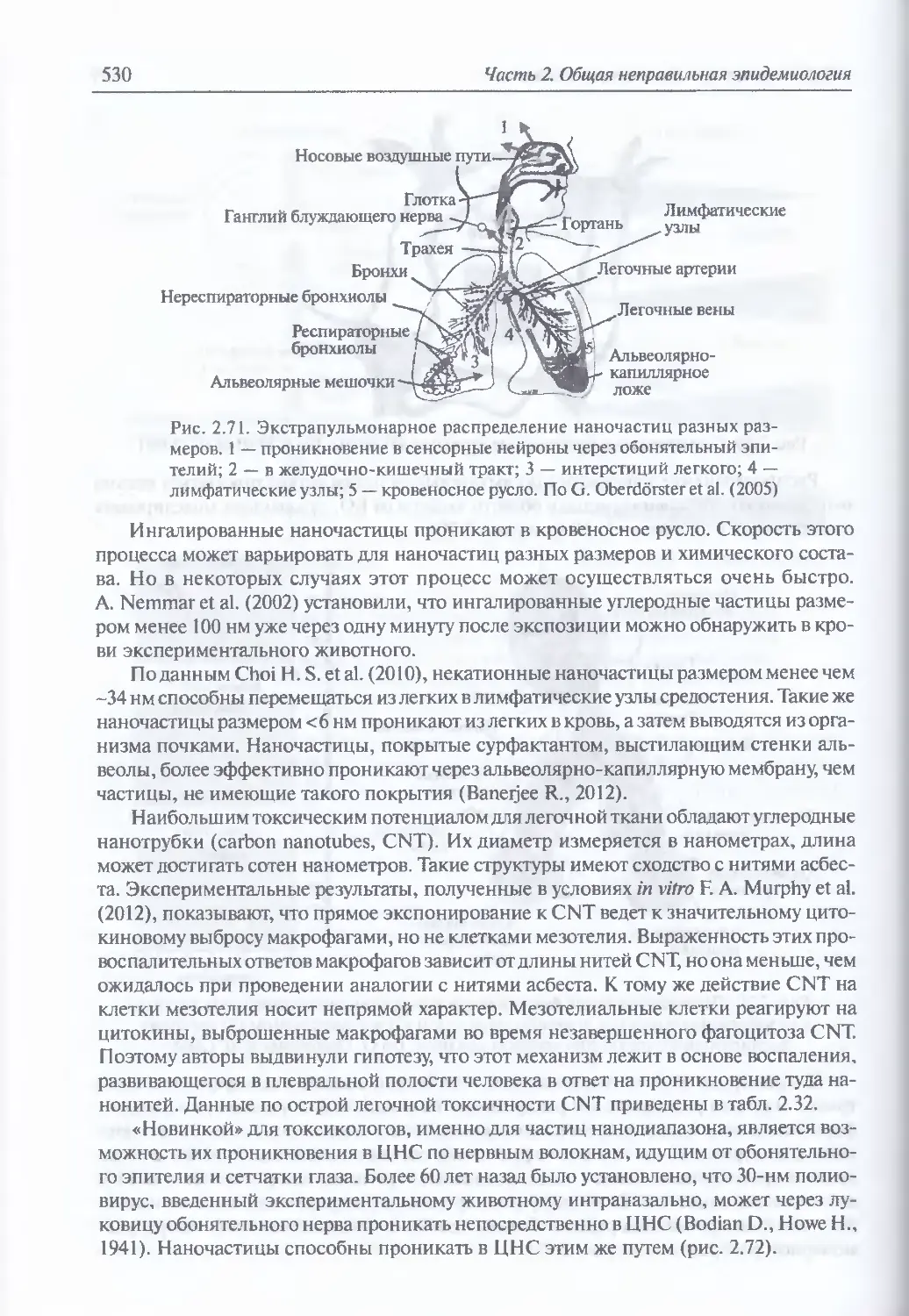
2.2.5. Нанообъекгы ... ..................................................523
Границы наномира (523). Первое боевое применение частиц нано-
и субмикронных размеров (524). Особенности наночастии.
обусловливающие их поражающее действие (525) Пути проникновения
наночастиц в организм человека (528). Случай группового поражения
людей наночастицами двуокиси кремния (533)
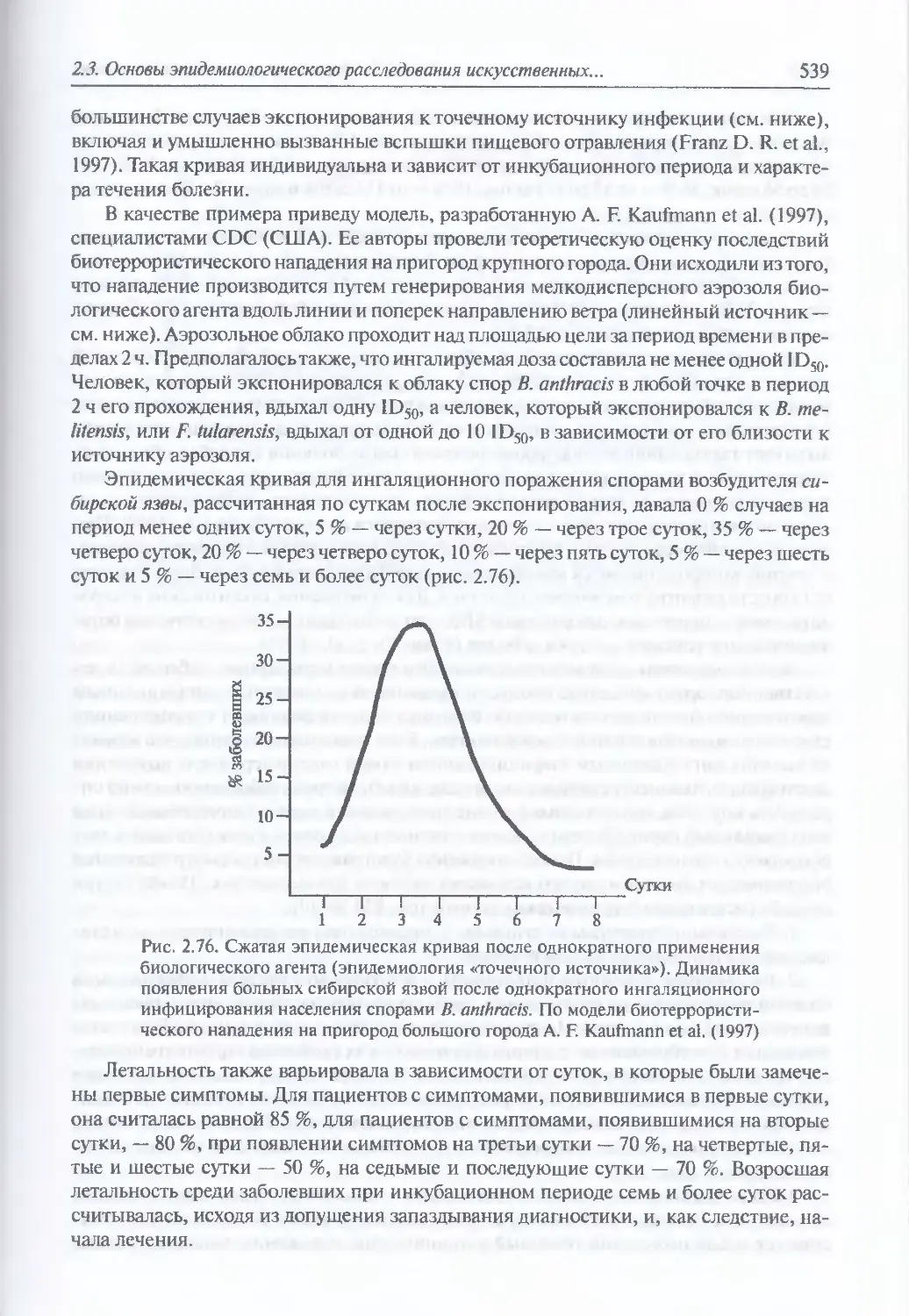
2.3. Основы эпидемиологического расследования искусственных эпидемических
процессов и поражений биологическими токсинами......................537
2.3.1. Характерные особенности эпидемиологии искусственных
эпидемических процессов и поражений токсинами............................537
Эпидемиология искусственного эпидемического процесса, вызванного
однократным применением биологического агента (537). Эпидемиология
искусственного эпидемического процесса, вызванного многократным
применением биологического агента (541). Особенности искусственного
эпидемического процесса, характерные для обоих сценариев
биологического нападения (542) Малораспространенные способы
осуществления преступлений с использованием биологических
агентов (549). Объекты преступления при биологическом нападении (549).
Модели территорий, на которых наиболее вероятно применение
биологических агентов для поражения людей (550). Психосоциальные
реакции у людей, вызванные акцией с применением
биологических агентов (550)
2.3.2. Эпидемиологическое расследование искусственного эпидемического
процесса................................................................553
Подходы к эпидемиологическому расследованию (553;. Текущее
эпидемиологическое расследование в очаге искусственного эпидемического
процесса (553) Ретроспективное эпидемиологическое обследование очага
искусственного эпидемического процесса (555). Распознавание поражения
людей нанообъектами (556)
2.4. Обнаружение биотеррористической организации.............................557
2.4.1. Биотеррористическая организация (сообщество).........................557
Убеждения и мотивы биотеррористов (557). Типовая структура
биотеррористической организации (558). Характерные
признаки биотеррористической организации (564).
Характеристика членов биотеррористической организации (565)
2.4.2. Признаки деятельности биотеррористической организ щии ...............566
Признаки функционирования подпольной биотеррористической
лаборатории (566). Характеристика следов, связанных с деятельностью
биотеррористической организации (570). Распознавание примененного
биологического агента по косвенным признакам (573). Поиск запасов
произведенных биологических агентов (574). Техника специальной
безопасности при осуществлении следственных и иных действий
в лабораториях биотеррористов (574). Назначение экспертиз
и выбор экспертов (577)
8
Содержание
Часть 3. ЧАСТНАЯ НЕПРАВИЛЬНАЯ ЭПИДЕМИОЛОГИЯ.................................579
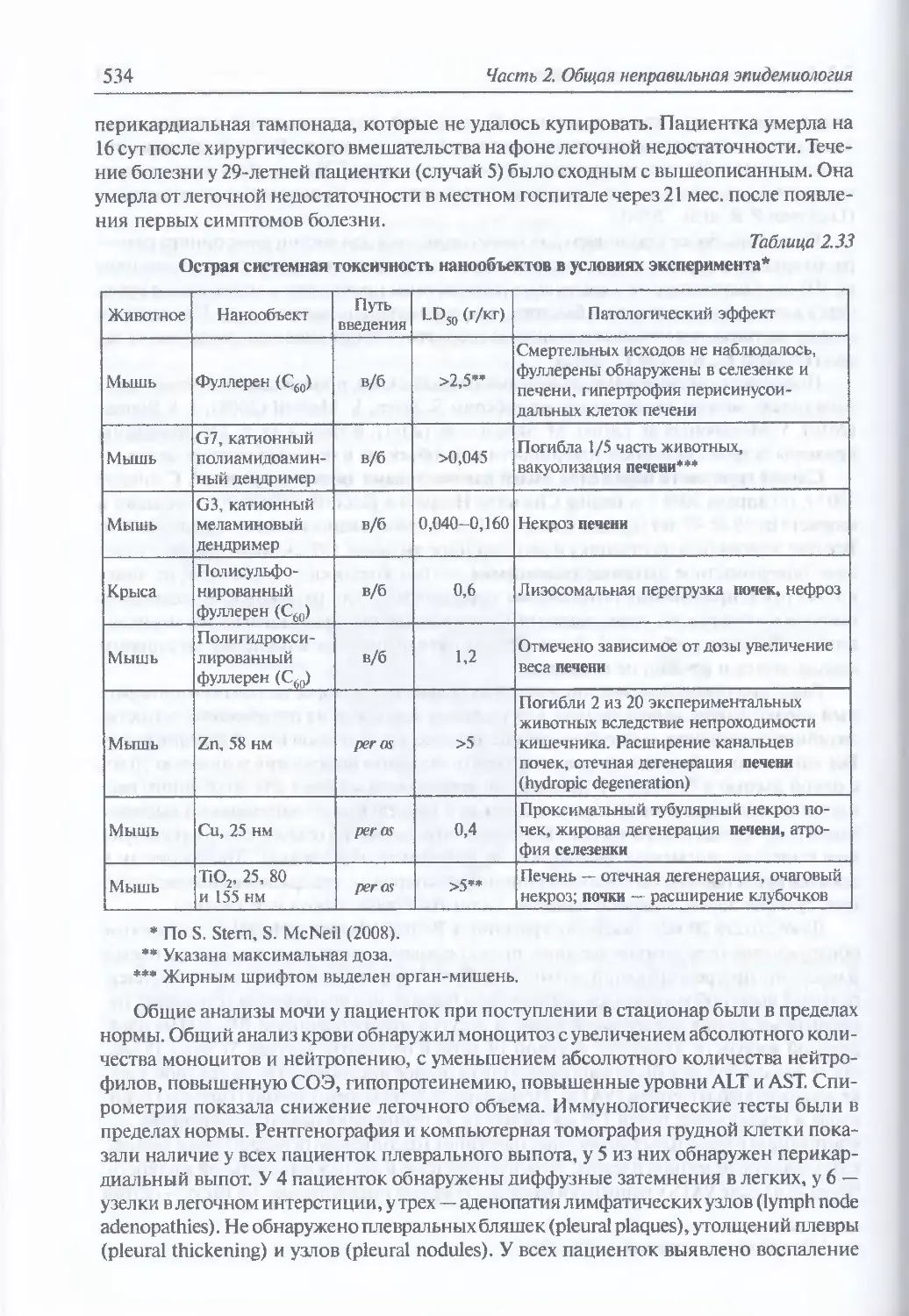
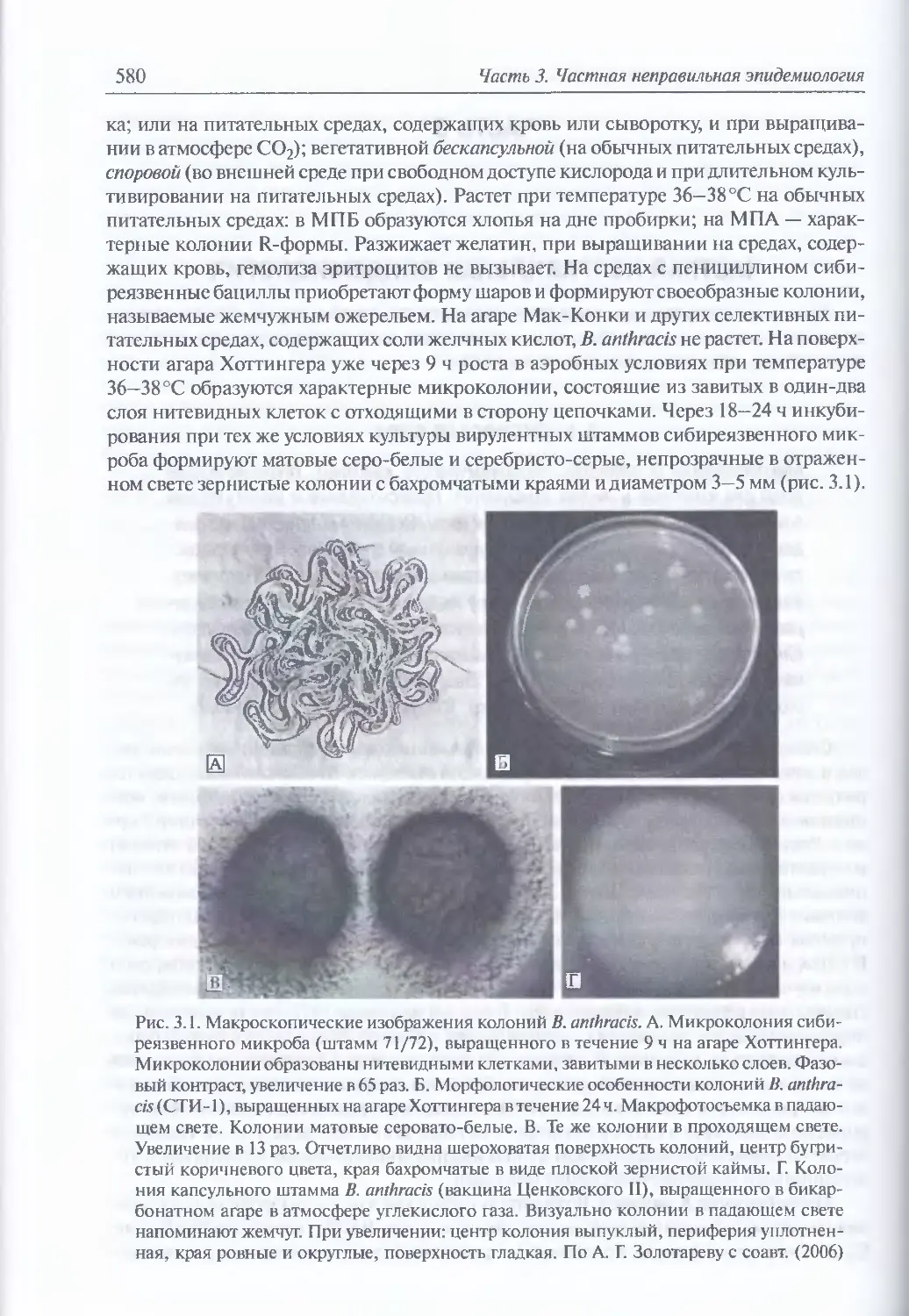
3.1. Сибирская язва.........................................................579
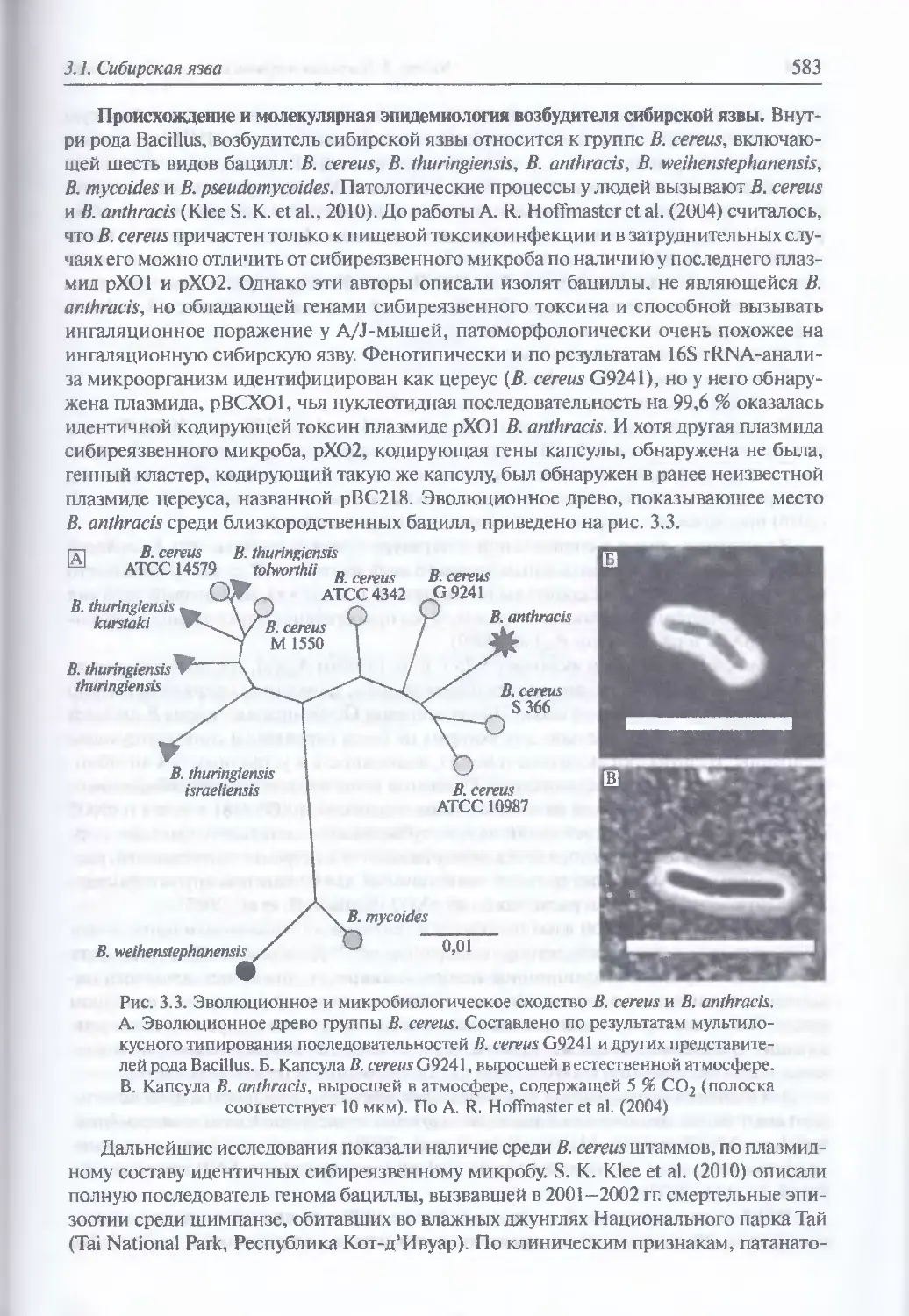
Микробиология В. anthracis (579). Восприимчивость животных (581).
Инфицирующие дозы для животных и людей (581). Иммунитет (582).
Происхождение и молекулярная эпидемиология возбудителя сибирской
язвы (583). Экология и естественная эпидемиология В. anthracis (589).
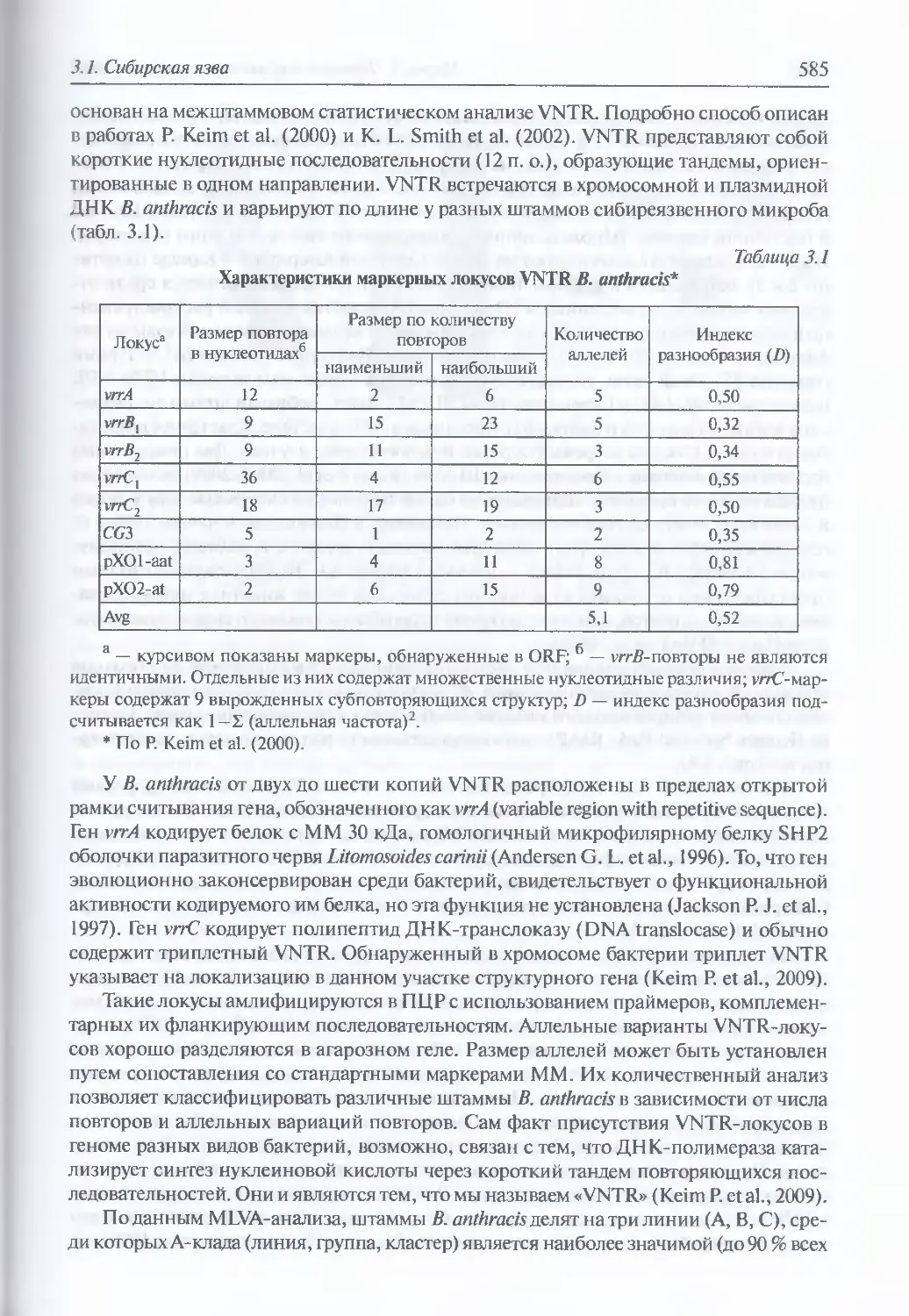
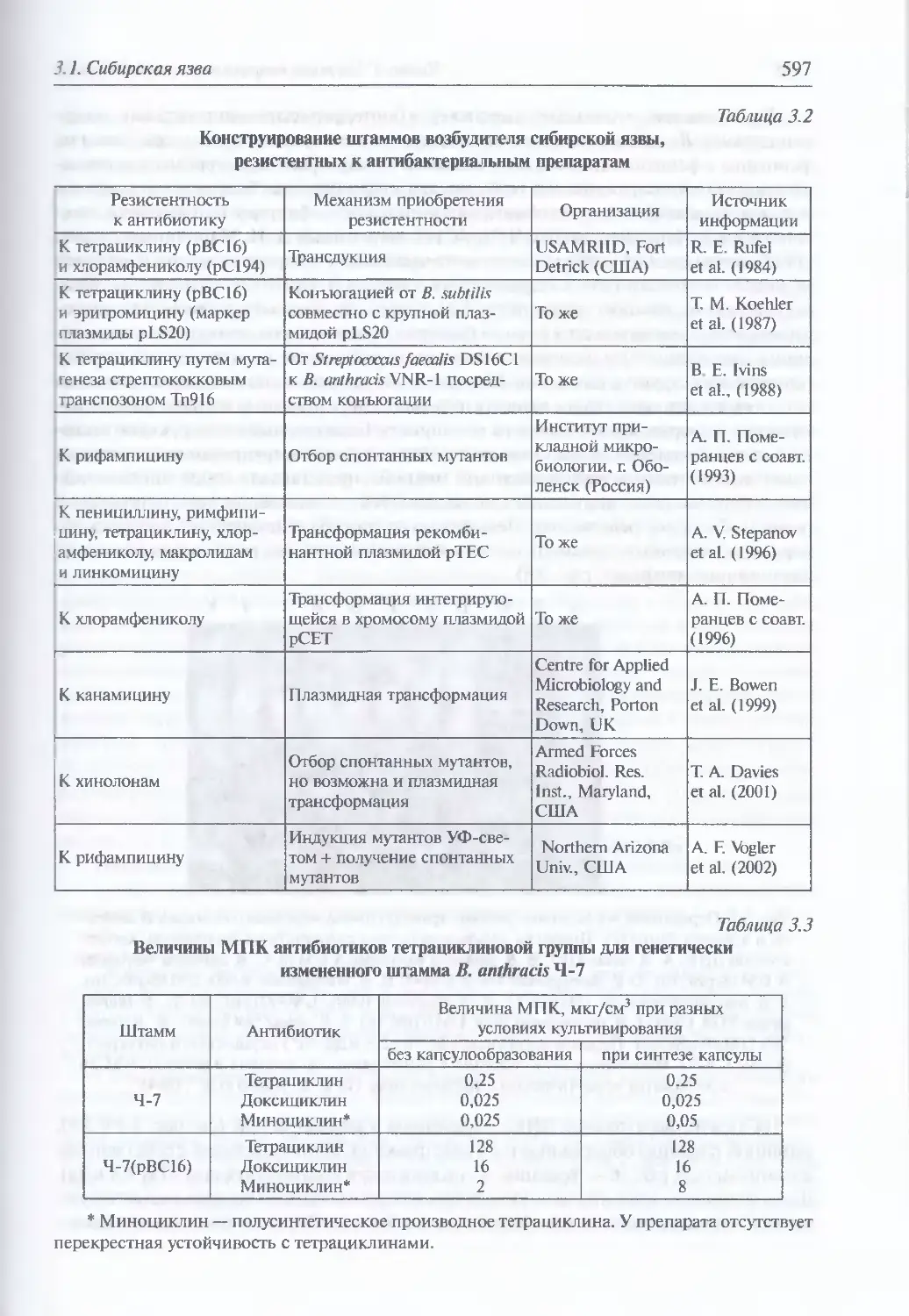
Устойчивость во внешней среде (594). Антибиотикорезистентность (595).
Генетически измененные штаммы и их обнаружение (596). Патогенез (599).
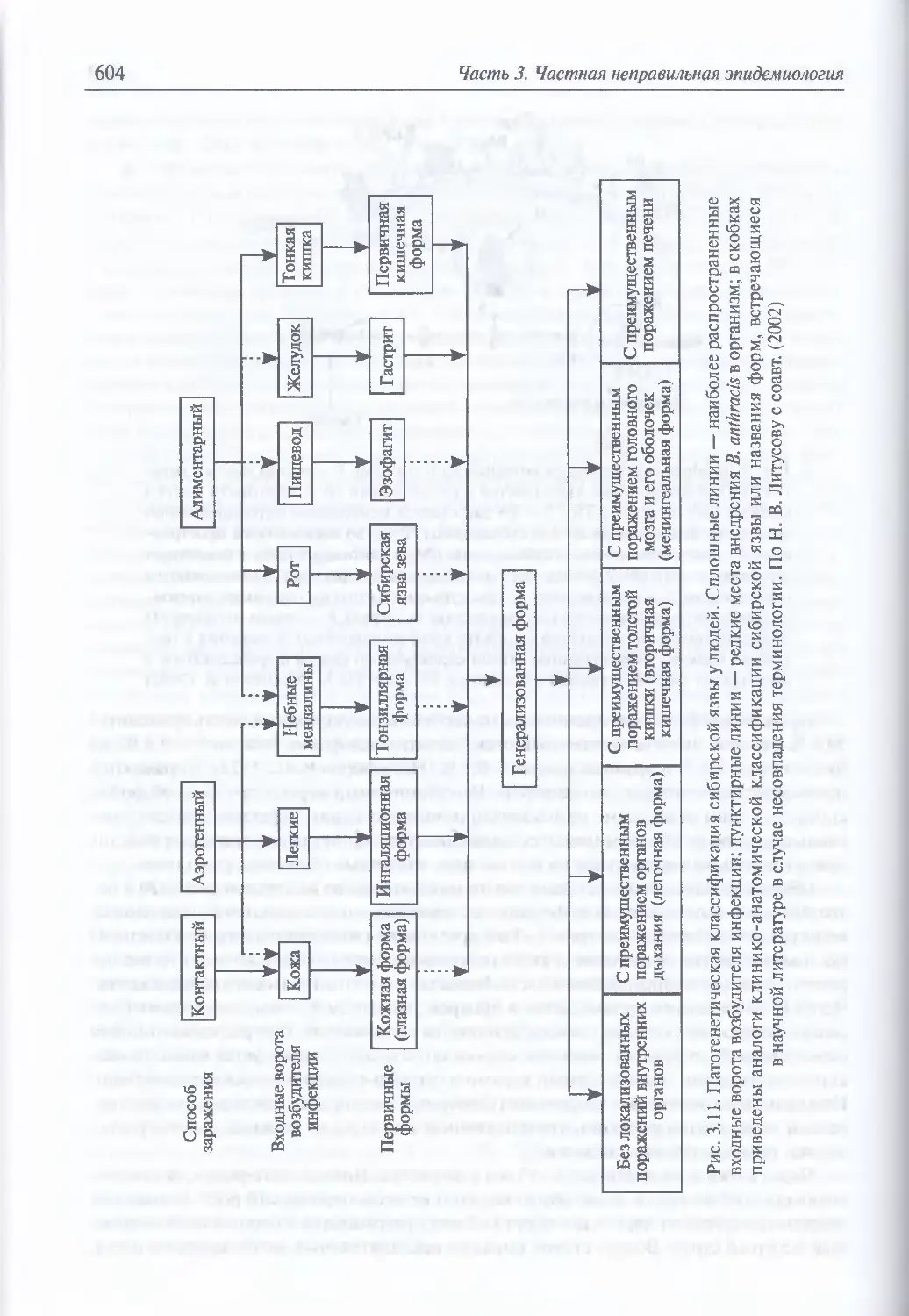
Клиническая картина сибирской язвы у людей при естественном инфицировании
В. anthracis (602). Моделирование искусственного заражения В. anthracis (612).
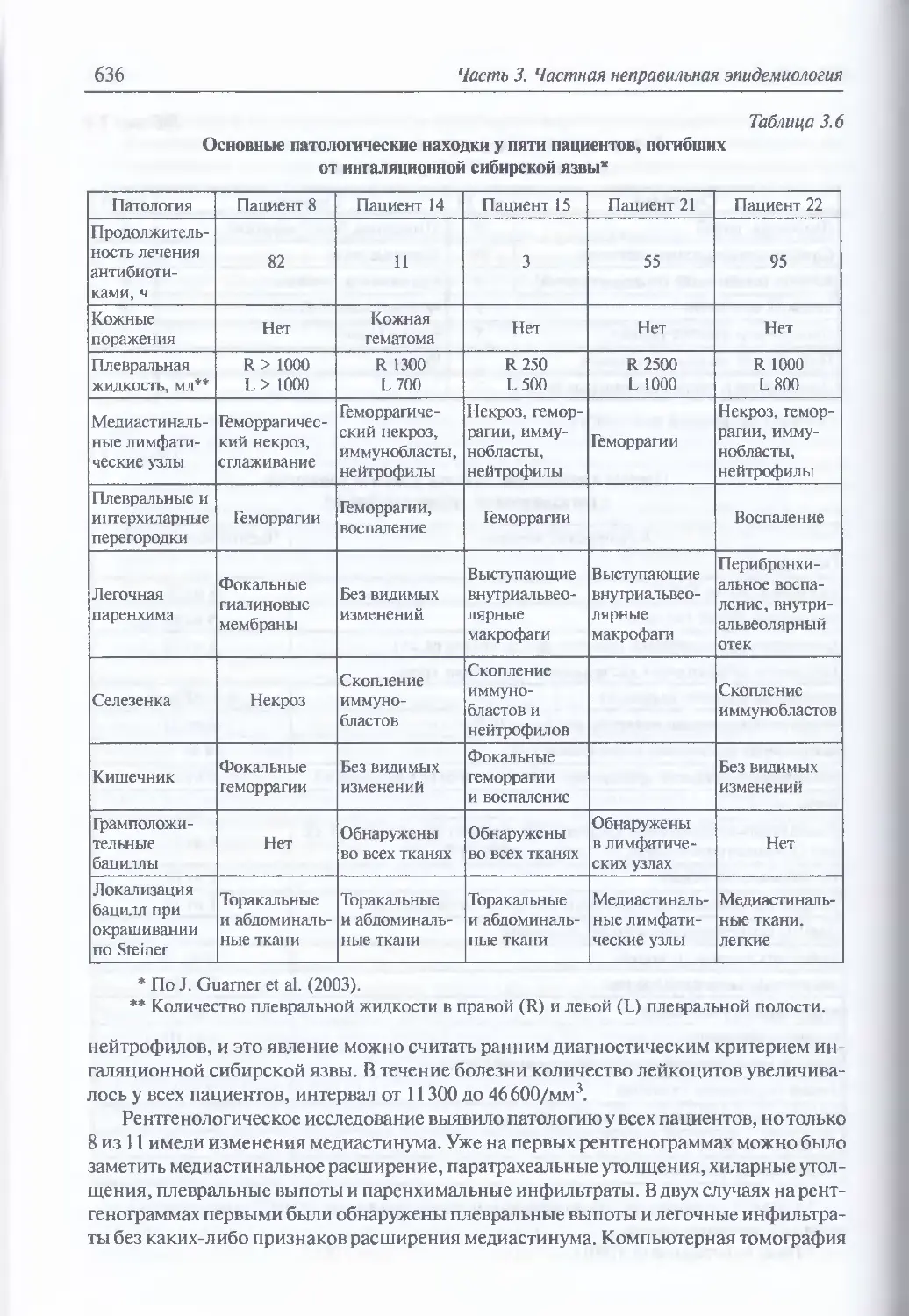
Клинические примеры искусственно вызванной сибирской язвы (617).
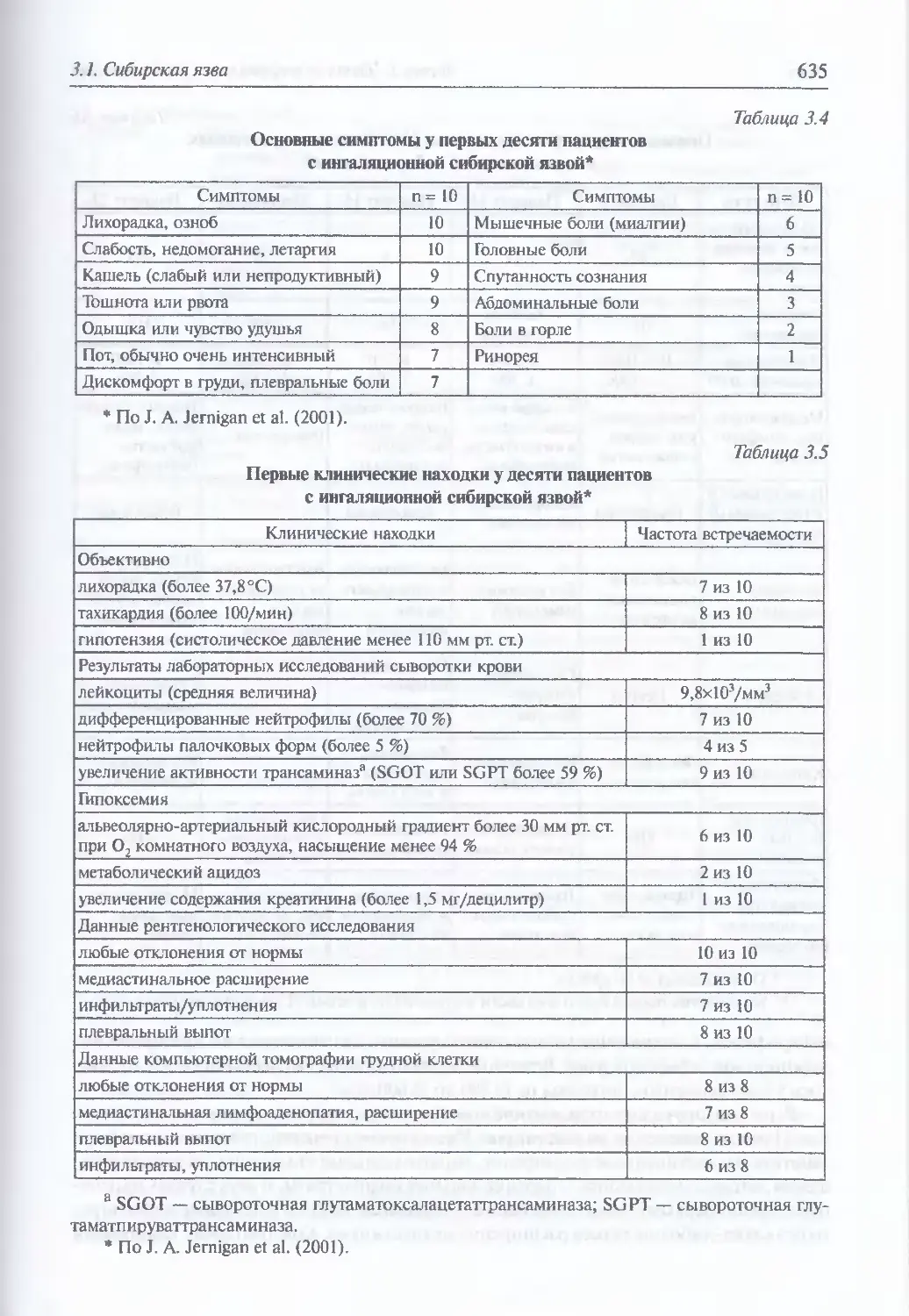
Обобщенная клиника ингаляционной сибирской язвы (634). Диагностика
искусственного поражения возбудителем сибирской язвы (640).
Иммунопрофилактика (643). Лечение (643)
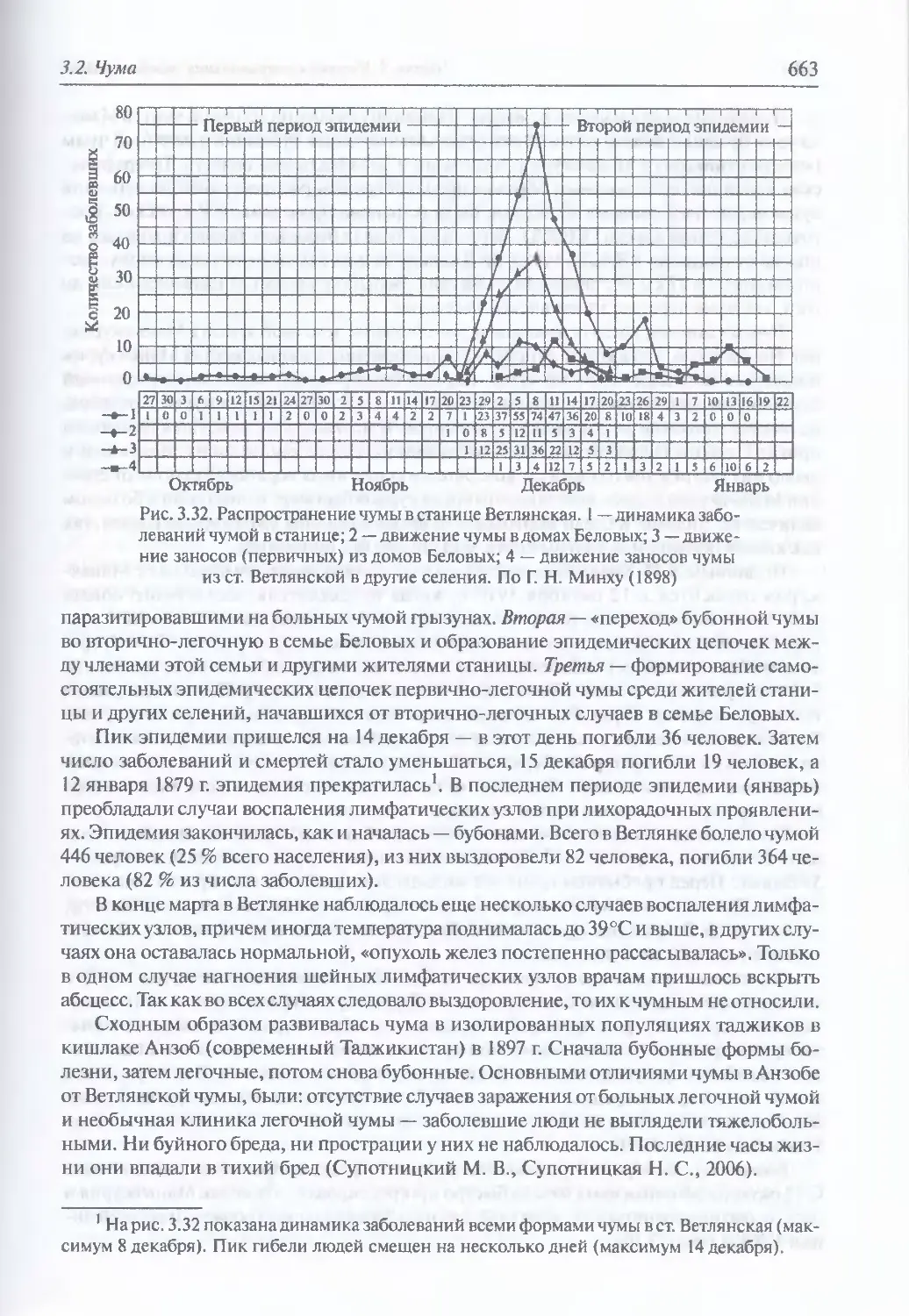
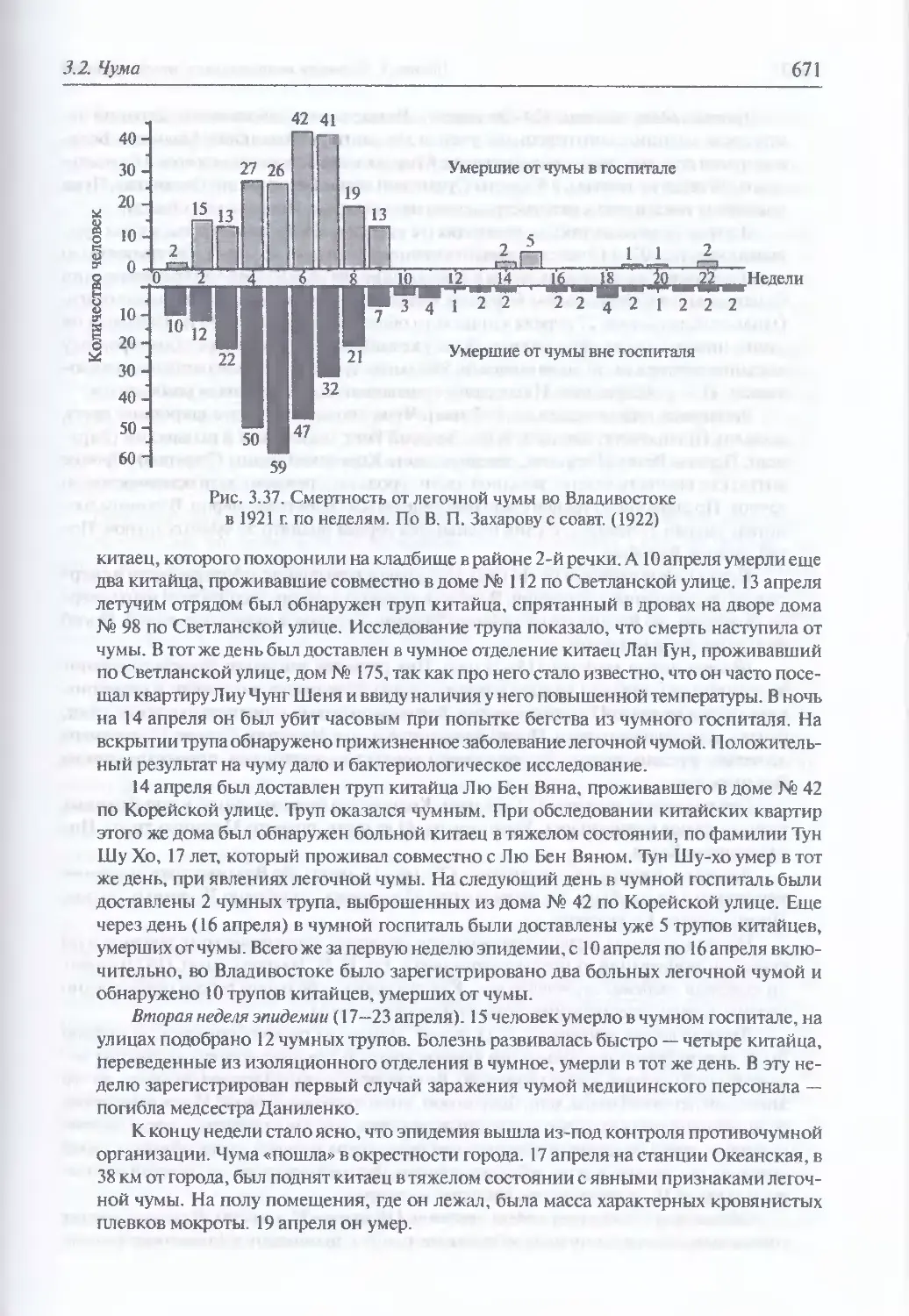
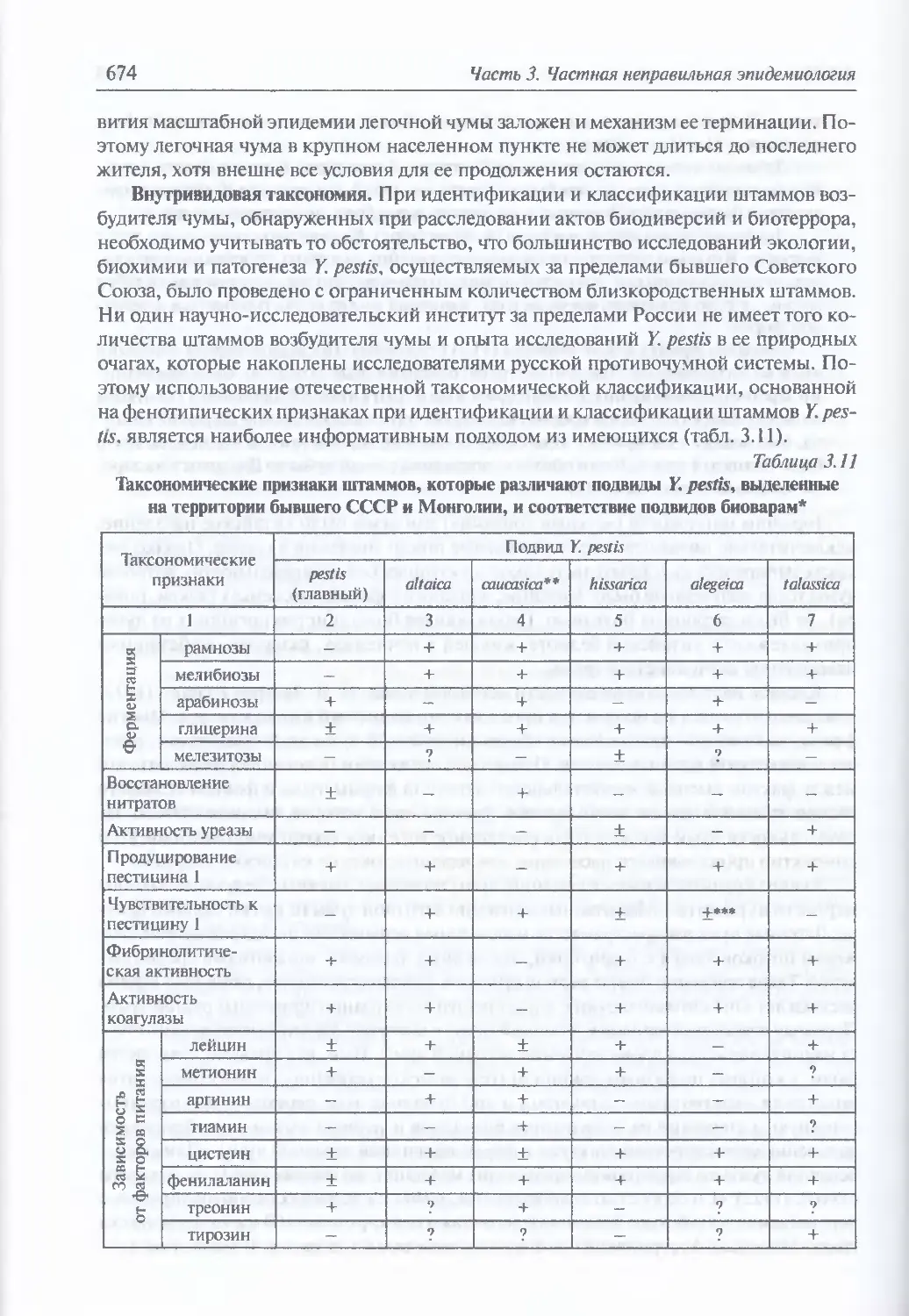
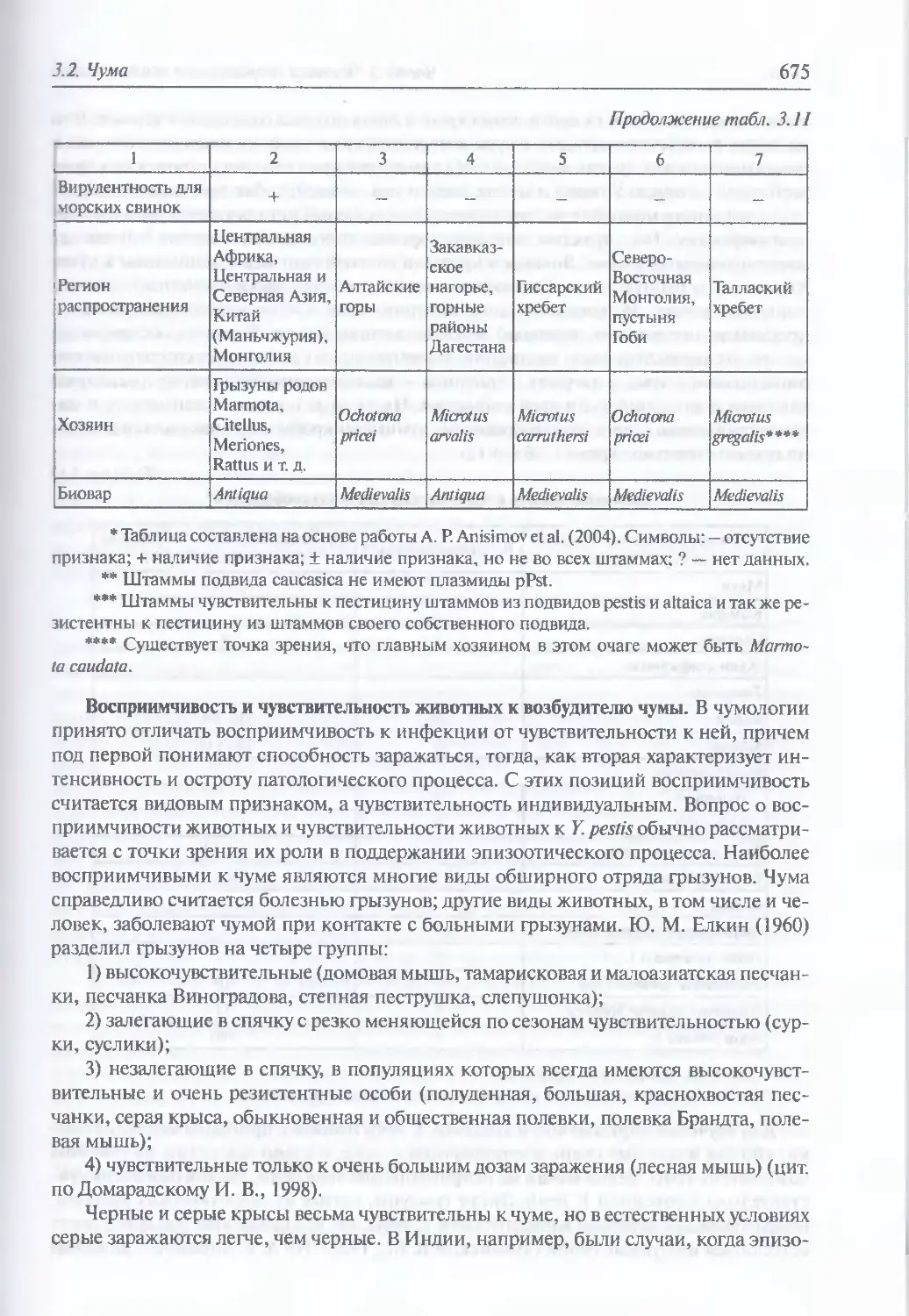
3.2. Чума..................................................................644
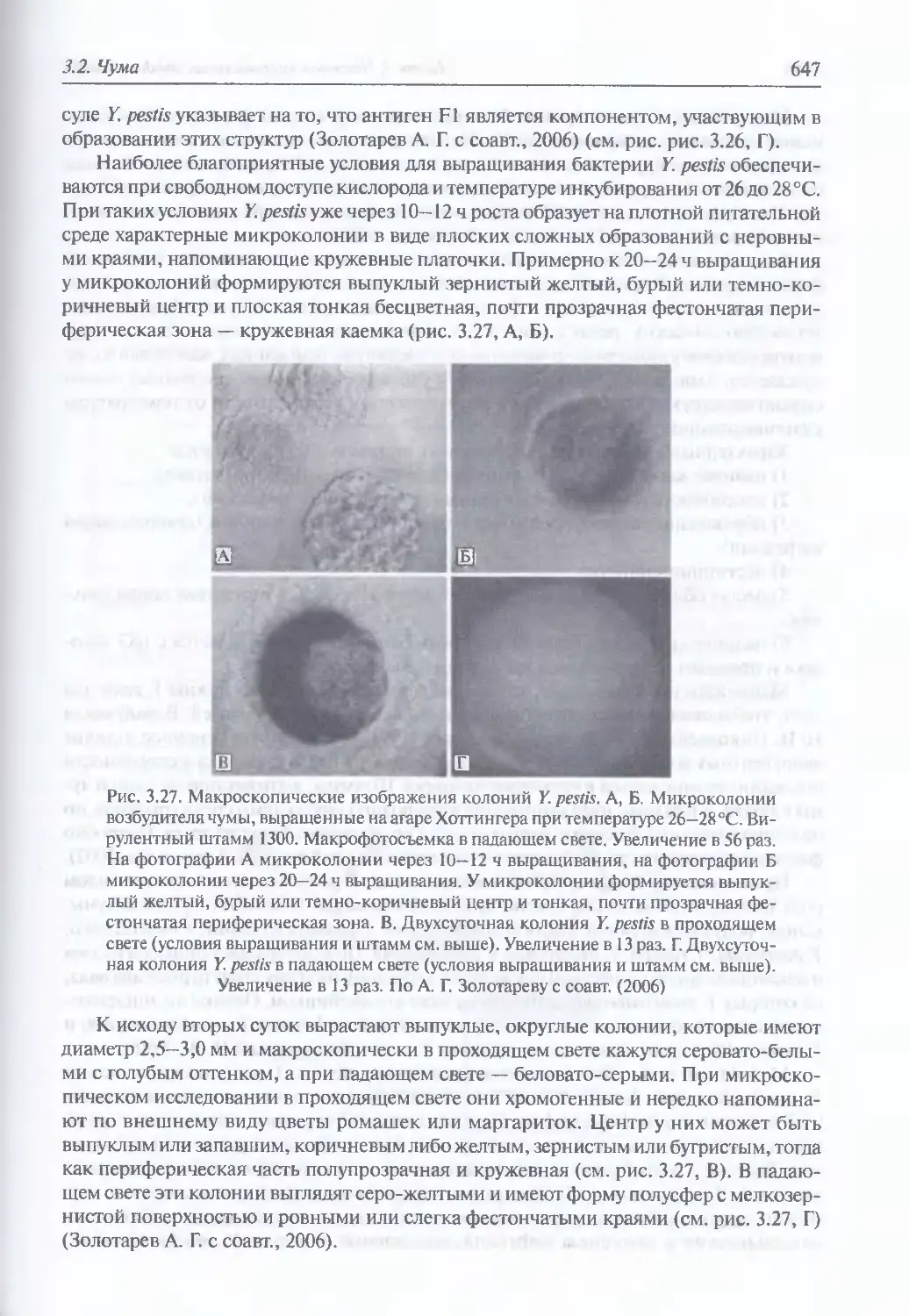
Микробиология К pestis (645). Происхождение и молекулярная
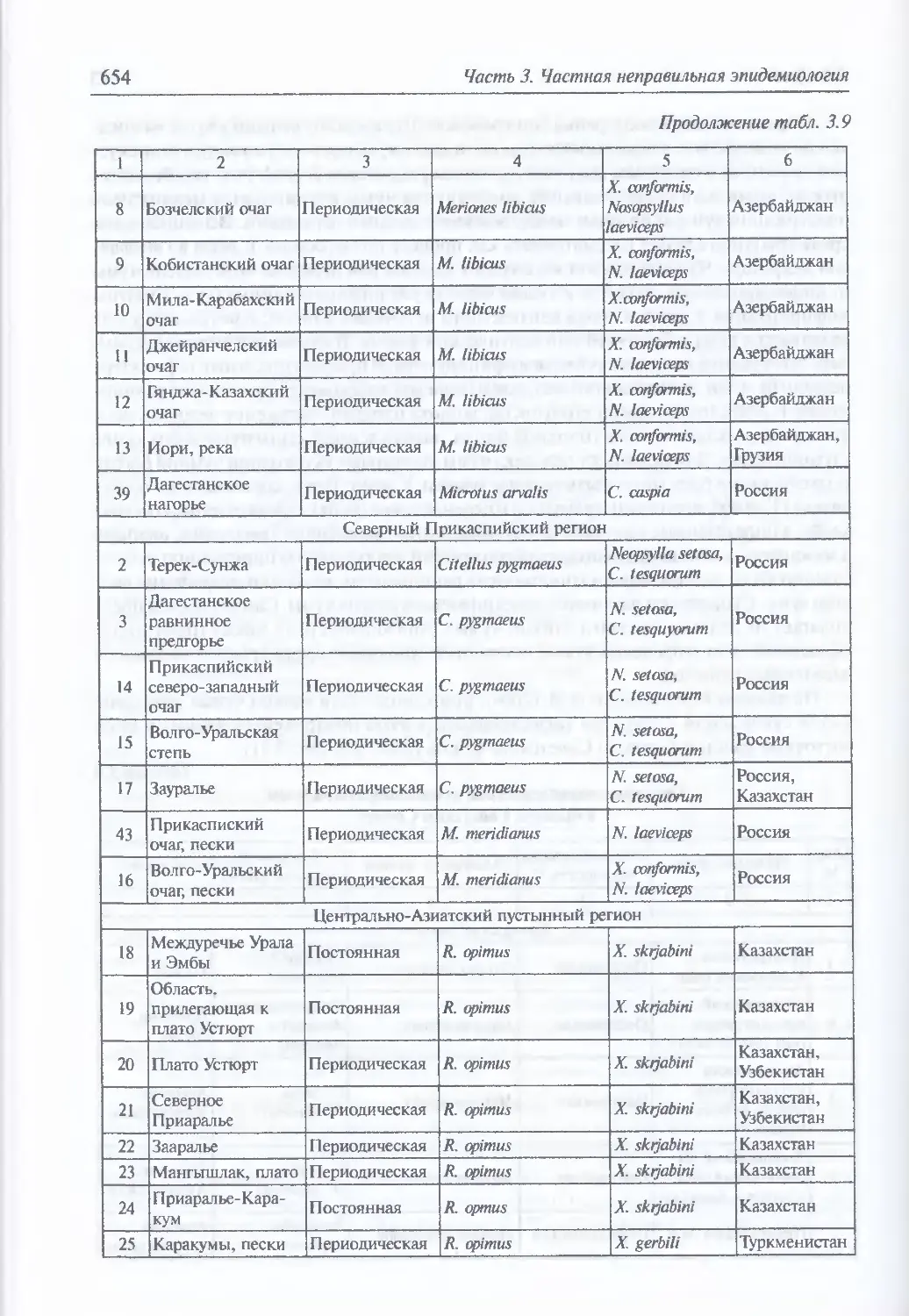
эпидемиология (648). Экология и естественная эпидемиология Y. pestis (652).
Внутривидовая таксономия (674). Восприимчивость и чувствительность
животных к возбудителю чумы (675). Инфицирующие дозы (677).
Иммунитет (677). Устойчивость Y. pestis во внешней среде и в трупах
погибших (677). Антибиотикорезистентность (678). Обнаружение генетически
измененных штаммов У. pestis (680). Патогенез (681). Клиническая картина чумы
у людей при естественном инфицировании Y. pestis (682). Моделирование
искусственного заражения У. pestis (696). Диагностика искусственного
поражения возбудителем чумы (703). Иммунопрофилактика (704). Лечение (705)
3.3. Туляремия.............................................................706
Микробиология F. tularensis (707). Факторы патогенности (709). Происхождение
и молекулярная эпидемиология возбудителя туляремии (711).
Экология и естественная эпидемиология F. tularensis (713). Восприимчивость
и чувствительность животных к возбудителю туляремии (717). Инфицирующие
дозы для животных и людей (718). Иммунитет (719). Устойчивость
F. tularensis во внешней среде и в трупах погибших животных (719).
Антибиотикорезистентность (720). Обнаружение генетически измененных
штаммов F. tularensis (720). Патогенез (722). Клиническая картина туляремии
у людей при естественном инфицировании F. tularensis (723). Моделирование
ингаляционного заражения F tularensis (728). Диагностика искусственного
поражения возбудителем туляремии (733). Иммунопрофилактика (734).
Лечение (735)
3.4. Бруцеллез.............................................................735
Микробиология бруцелл (736). Факторы патогенности (738). Таксономия,
происхождение и молекулярная эпидемиология бруцелл (741). Экология
и естественная эпидемиология бруцелл (743). Восприимчивость к бруцеллам
и инфицирующие дозы для животных и людей (745). Иммунитет (746).
Устойчивость бруцелл во внешней среде (746). Антибиотикорезистентность
бруцелл (747). Обнаружение генетически измененных штаммов бруцелл (747).
Патогенез бруцеллеза (748). Клиническая картина бруцеллеза у людей при
естественном инфицировании (752). Моделирование ингаляционного заражения
бруцеллами (760). Диагностика искусственного поражения бруцеллами (760).
Иммунопрофилактика бруцеллеза (761). Лечение бруцеллеза (761)
3.5. Сап...................................................................762
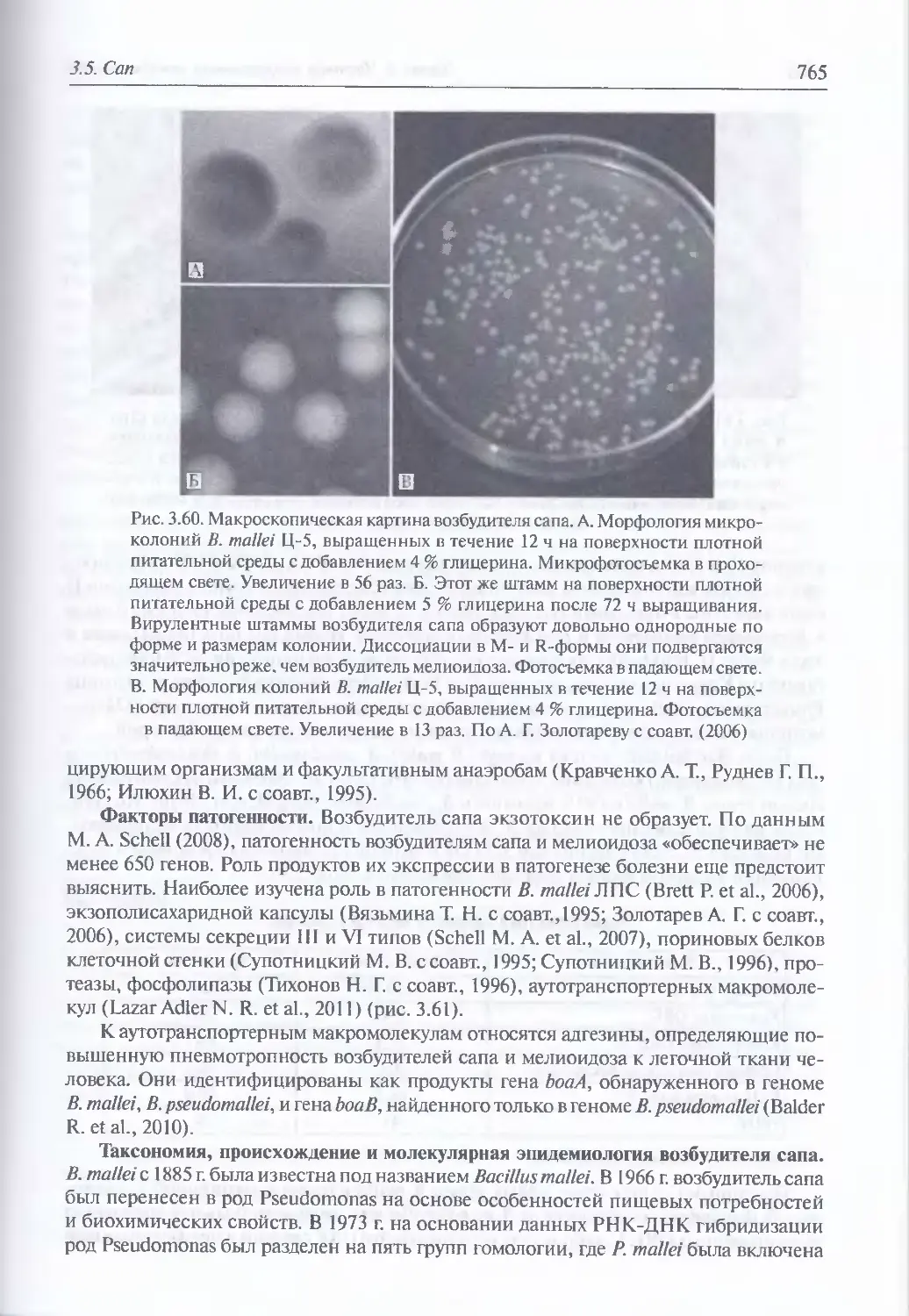
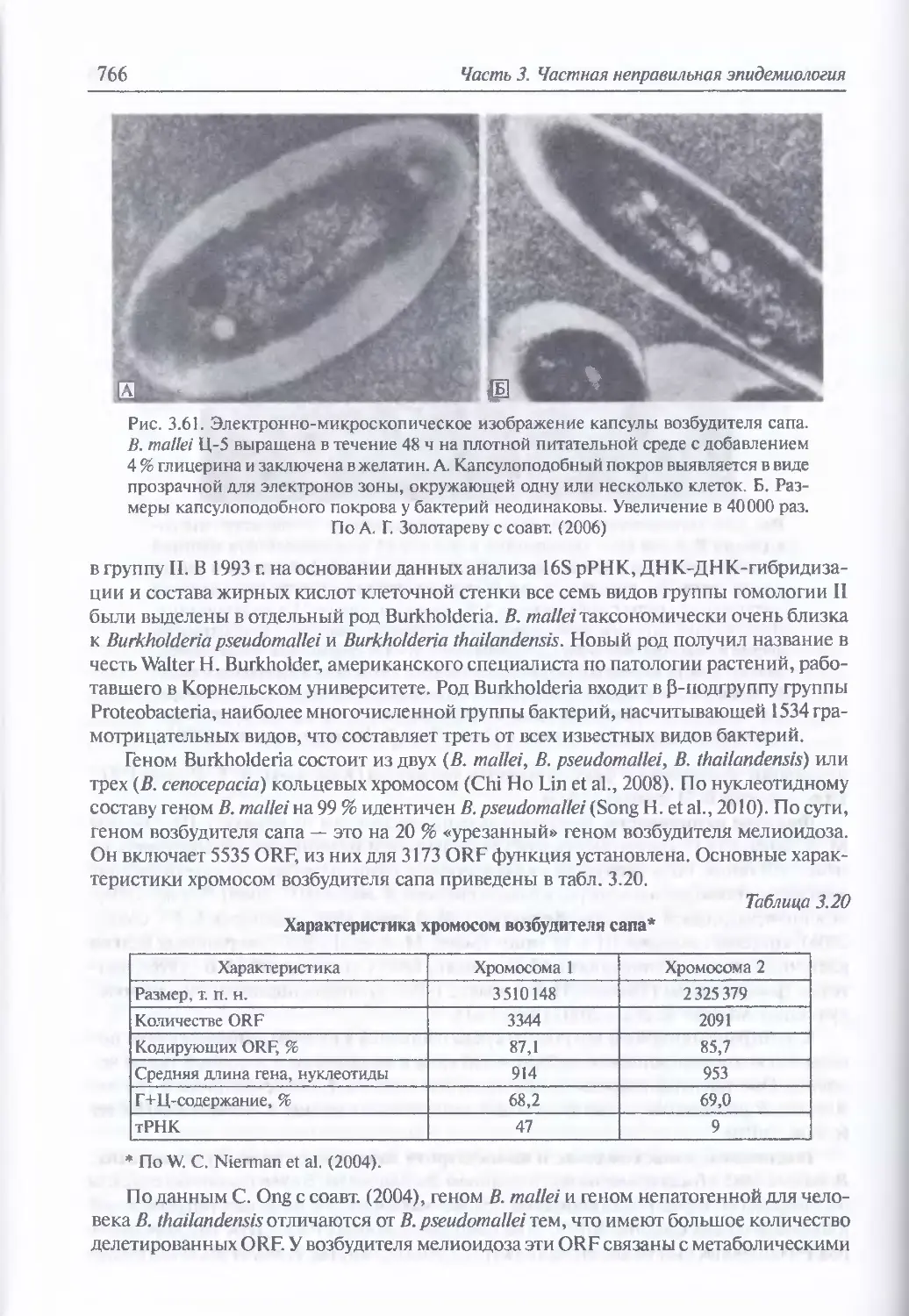
Микробиология возбудителя сапа (763). Факторы патогенности (765). Таксономия,
происхождение и молекулярная эпидемиология возбудителя сапа (765). Экология
и естественная эпидемиология возбудителя сапа (767). Сап у лошадей (769).
Восприимчивость к возбудителю сапа и инфицирующие дозы для животных
и людей (772). Иммунитет (773). Устойчивость возбудителя сапа во внешней
Содержание
9
среде (773). Антибиотикорезистентность (774) Обнаружение генетически
измененных штаммов возбудителя сапа (774) Патогенез сапа (775). Клиническая
картина сапа у людей при естественном инфицировании (776). Моделирование
ингаляционного заражения сапом (777). Клиническая картина сапа у людей при
искусственном инфицировании (778). Диагностика искусственного поражения
сапом (782). Иммунопрофилактика сапа (783). Лечение сапа (783)
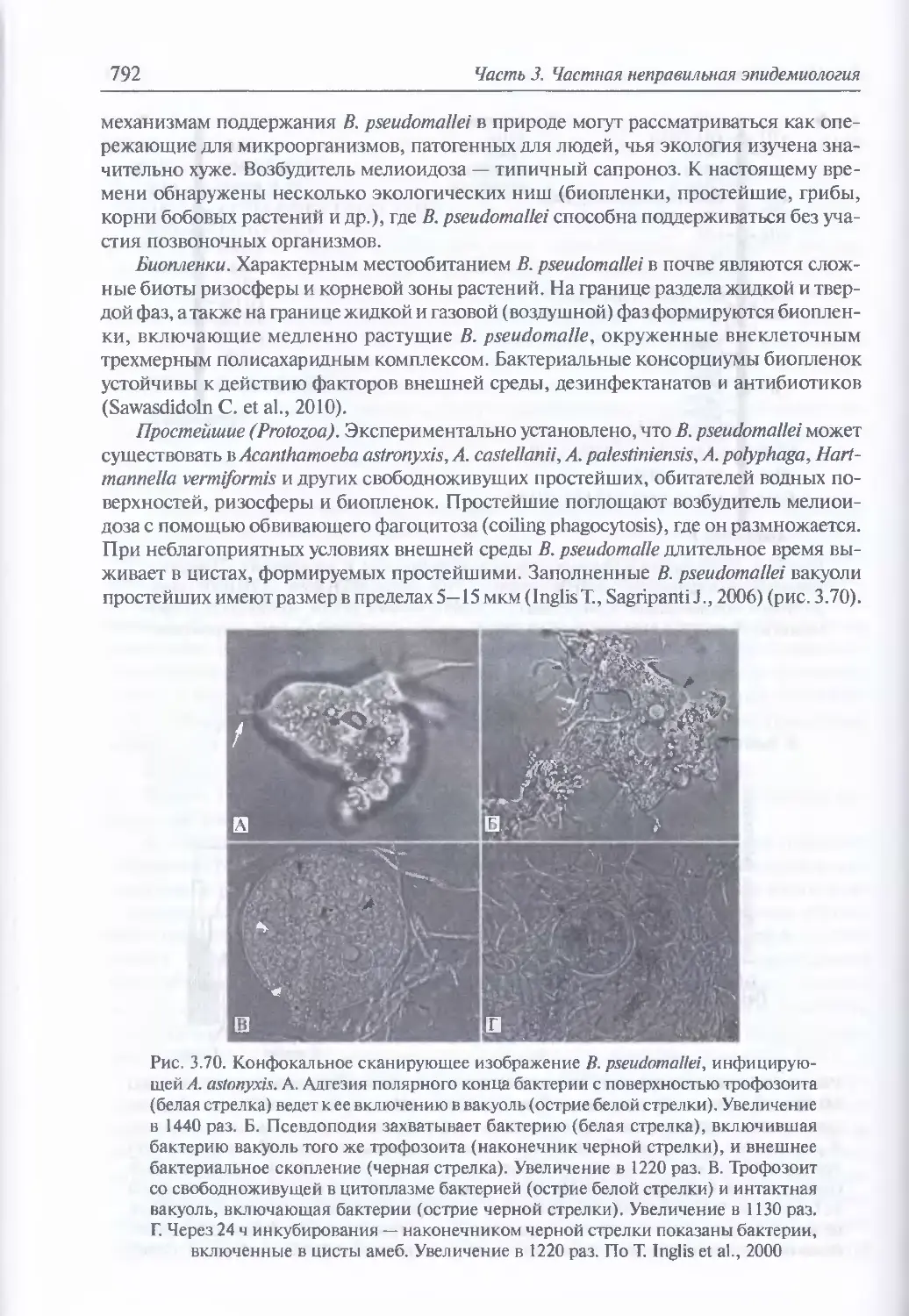
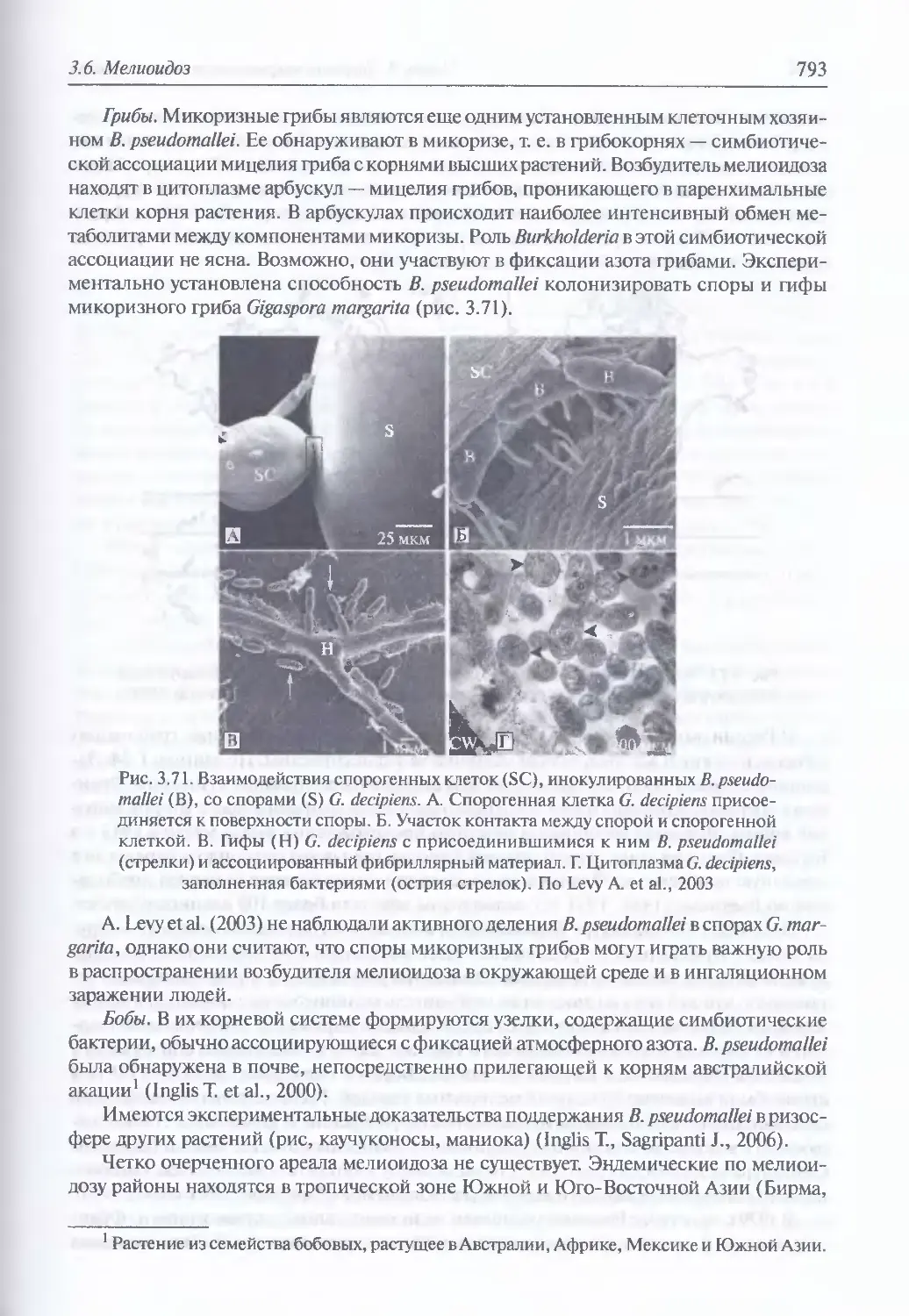
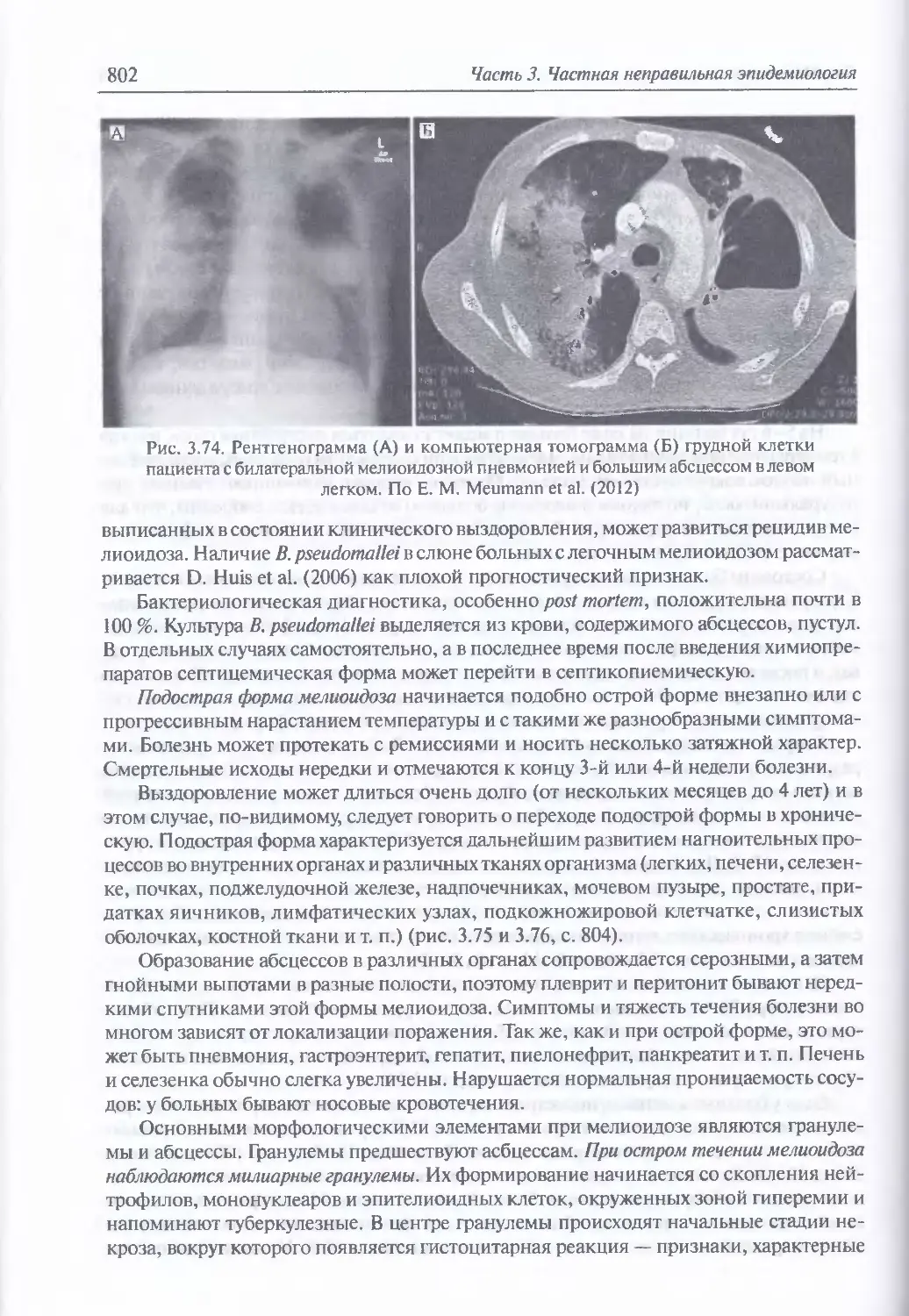
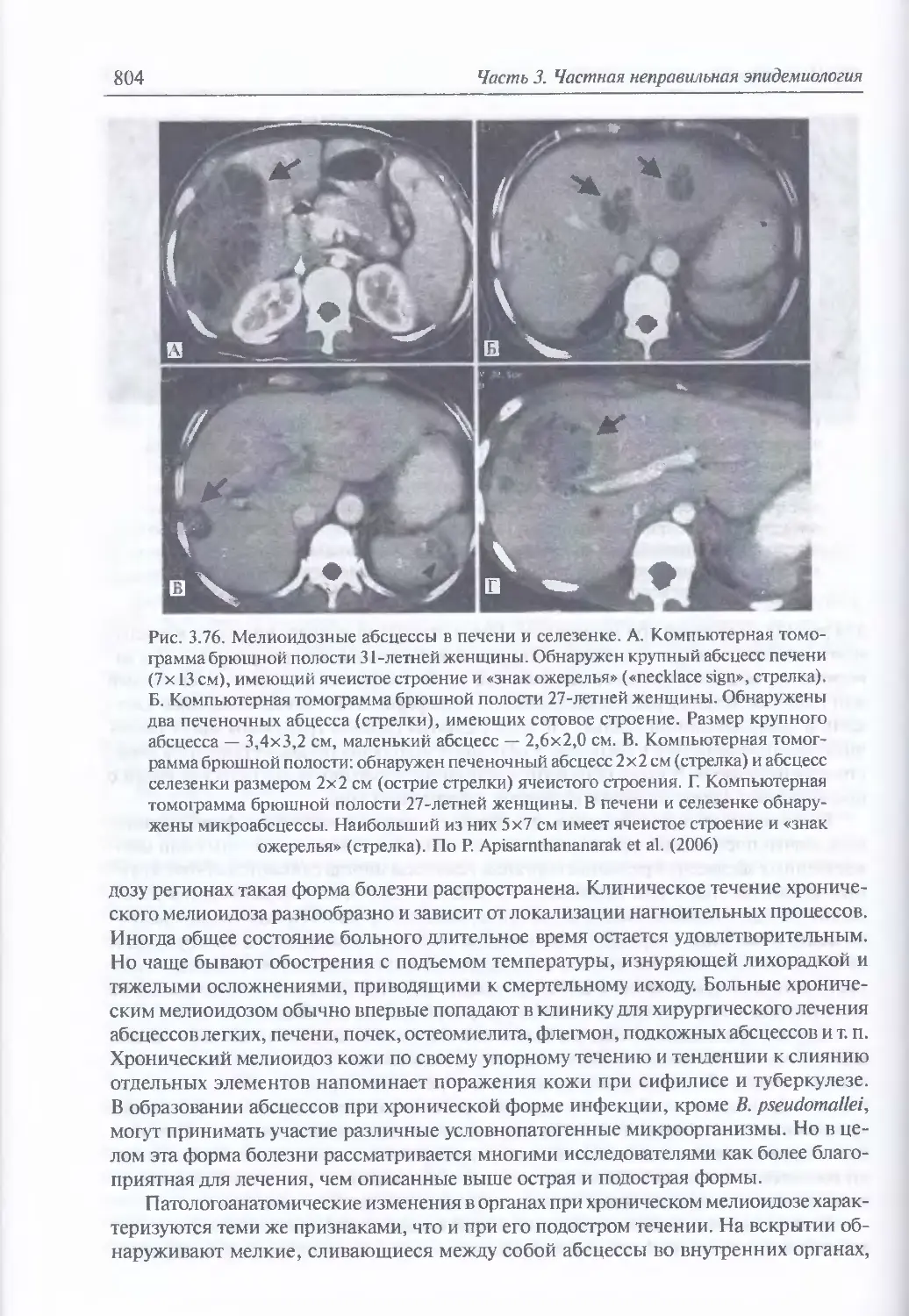
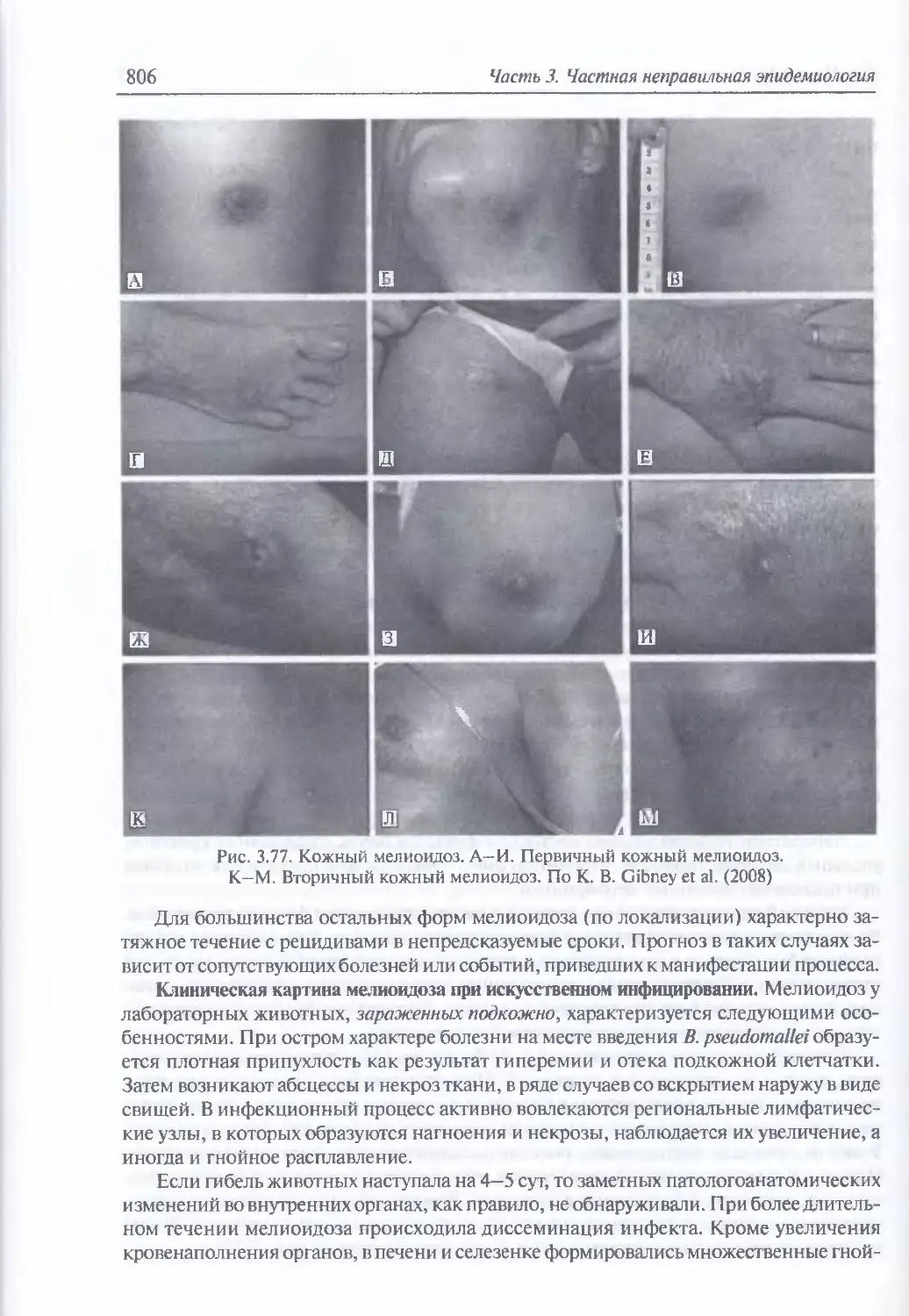
3.6. Мелиоидоз..............................................................784
Микробиология возбудителя мелиоидоза (785). Факторы патогенности (788).
Таксономия, происхождение и молекулярная эпидемиология возбудителя
мелиоидоза (790). Экология и естественная эпидемиология возбудителя мелио-
идоза (790). Восприимчивость к возбудителю мелиоидоза и инфицирующие
дозы для животных (795). Иммунитет (797). Устойчивость возбудителя мелио-
идоза во внешней среде (797). Антибиотикорезистентность (798). Обнаружение
генетически измененных штаммов возбудителя мелиоидоза (798). Патогенез
мелиоидоза (799). Клиническая и патологоанатомическая картина мелиоидоза
у человека при естественном инфицировании (799). Клиническая картина
мелиоидоза при искусственном инфицировании (806). Диагностика
искусственного поражения возбудителем мелиоидоза (807). Иммунопрофи-
лактика мелиоидоза (808). Лечение мелиоидоза (808)
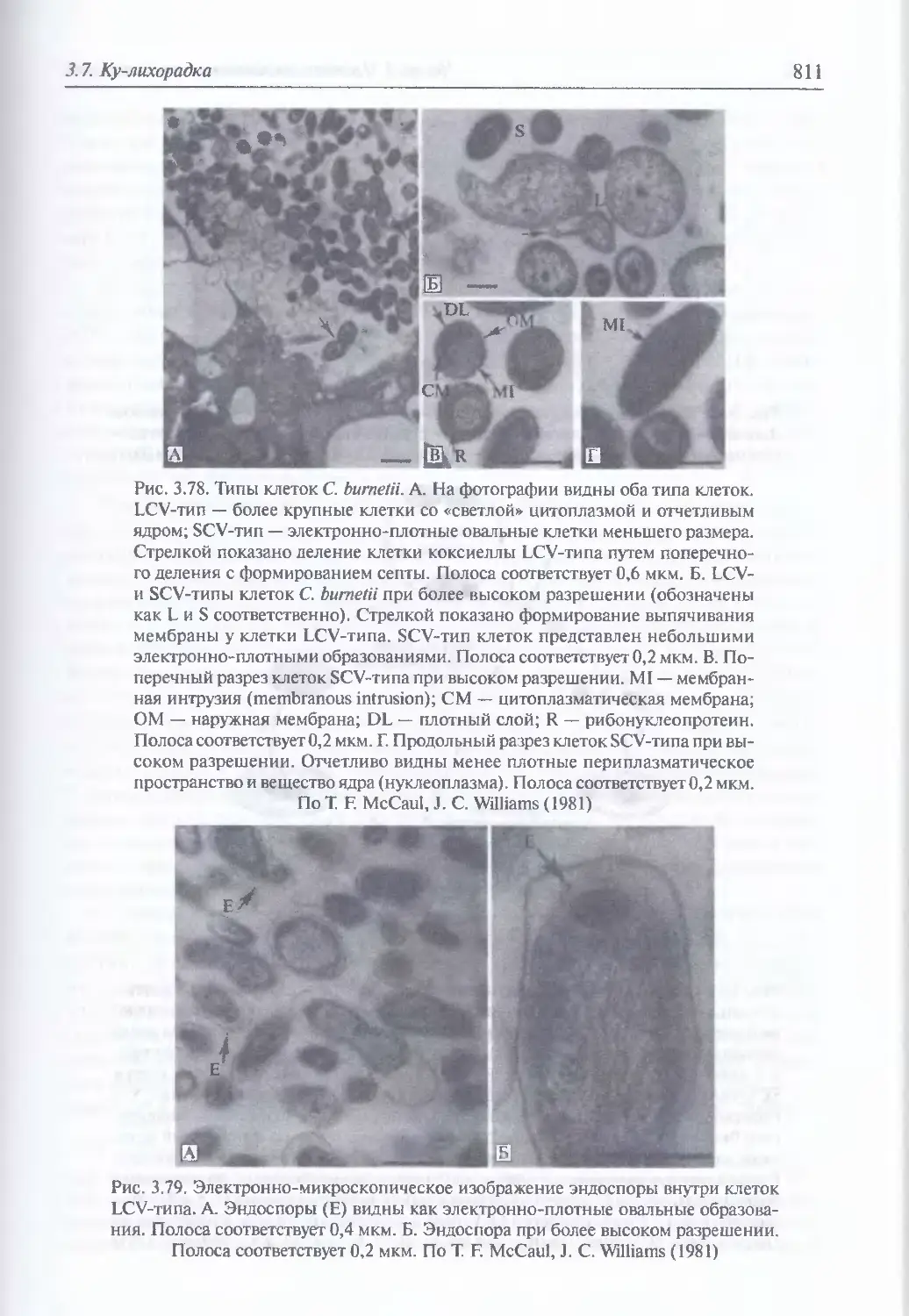
3.7. Ку-лихорадка...........................................................809
Микробиология возбудителя Ку-лихорадки (809). Факторы патогенности (813).
Таксономия, происхождение и молекулярная эпидемиология возбудителя
Ку-лихорадки (814). Экология и естественная эпидемиология возбудителя
Ку-лихорадки (816). Восприимчивость к возбудителю Ку-лихорадки
и инфицирующие дозы для животных и людей (818). Иммунитет (819).
Устойчивость возбудителя Ку-лихоралки во внешней среде (820).
Антибиотикорезистентность (821) Обнаружение генетически измененных
штаммов возбудителя Ку-лихорадки (821). Патогенез Ку-лихорадки (822).
Клиническая и патологоанатомическая картина Ку-лихорадки у человека
при естественном инфицировании (823). Клиническая картина Ку-лихорадки
у животных при искусственном инфицировании (829) Клиническая
картина Ку-лихорадки у людей при искусственном инфицировании (832).
Диагностика искусственного поражения возбудителем Ку-лихорадки (833).
Иммунопрофилактика Ку-лихорадки (833) Лечение Ку-лихорадки (833)
3.8. Натуральная оспа, оспа обезьян.........................................834
Микробиология возбудителя натуральной оспы (836). Таксономия
и происхождение возбудителя натуральной оспы (839). Экология
и естественная эпидемиология возбудителя натуральной оспы (844).
Экология и естественная эпидемиология возбудителя оспы обезьян (853).
Оспа коров (856). Восприимчивость к возбудителю натуральной оспы людей
и животных (857). Восприимчивость к возбудителю оспы обезьян
у животных (860). Иммунитет к поксвирусам (861). Устойчивость возбудителя
натуральной оспы во внешней среде (863). Антивирусные препараты (863).
Обнаружение генетически измененных штаммов ортопоксвирусов (865).
Патогенез ортопоксвирусных инфекций (866). Клиническая
и патологоанатомическая картина натуральной оспы у человека при
естественном инфицировании (868). Клиническая и патологоанатомическая
картина оспы обезьян у человека при естественном инфицировании (877).
Клиническая картина и патоморфология натуральной оспы и оспы обезьян
у животных при искусственном инфицировании (878). Клиническая картина
натуральной оспы у людей при искусственном инфицировании (883).
Диагностика искусственного поражения возбудителем натуральной
оспы (883). Иммунопрофи тактика натуральной оспы (884). Профилактика
после экспонирования к вирусу натуральной оспы и вирусу оспы обезьян
и лечение развившейся болезни (887)
10
Содержание
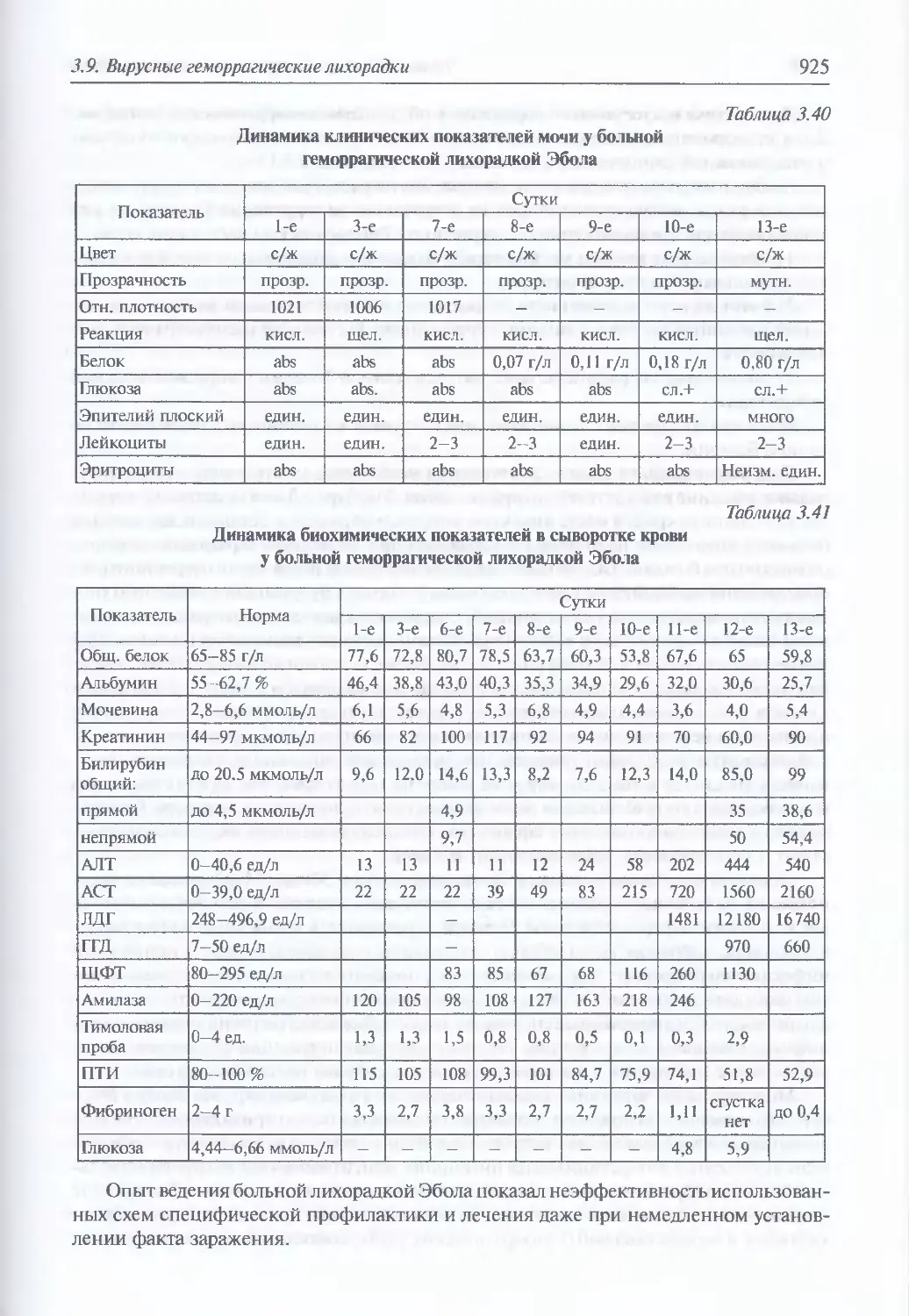
3.9. Вирусные геморрагические лихорадки....................................887
Микробиология возбудителей геморрагических лихорадок (891).
Экология и естественная эпидемиология возбудителей геморрагических
лихорадок (891). Восприимчивость людей и животных к возбудителям
геморрагических лихорадок (903). Иммунитет к возбудителям вирусных
геморрагических лихорадок (904). Устойчивость возбудителей геморрагических
лихорадок во внешней среде (907). Антивирусные препараты (907),
Обнаружение генетически измененных штаммов возбудителей геморрагических
лихорадок (908). Патогенез геморрагических лихорадок (909). Клиническая
и патологоанатомическая картина вирусных геморрагических лихорадок
у человека при естественном инфицировании (911). Клиническая картина
и патоморфология вирусных геморрагических лихорадок у животных
при искусственном инфицировании (915). Клиническая картина вирусных
геморрагических лихорадок у людей при искусственном инфицировании (920).
Диагностика искусственного поражения возбудителями геморрагических
лихорадок (926). Иммунопрофилактика вирусных геморрагических
лихорадок (927). Лечение вирусных геморрагических лихорадок (927)
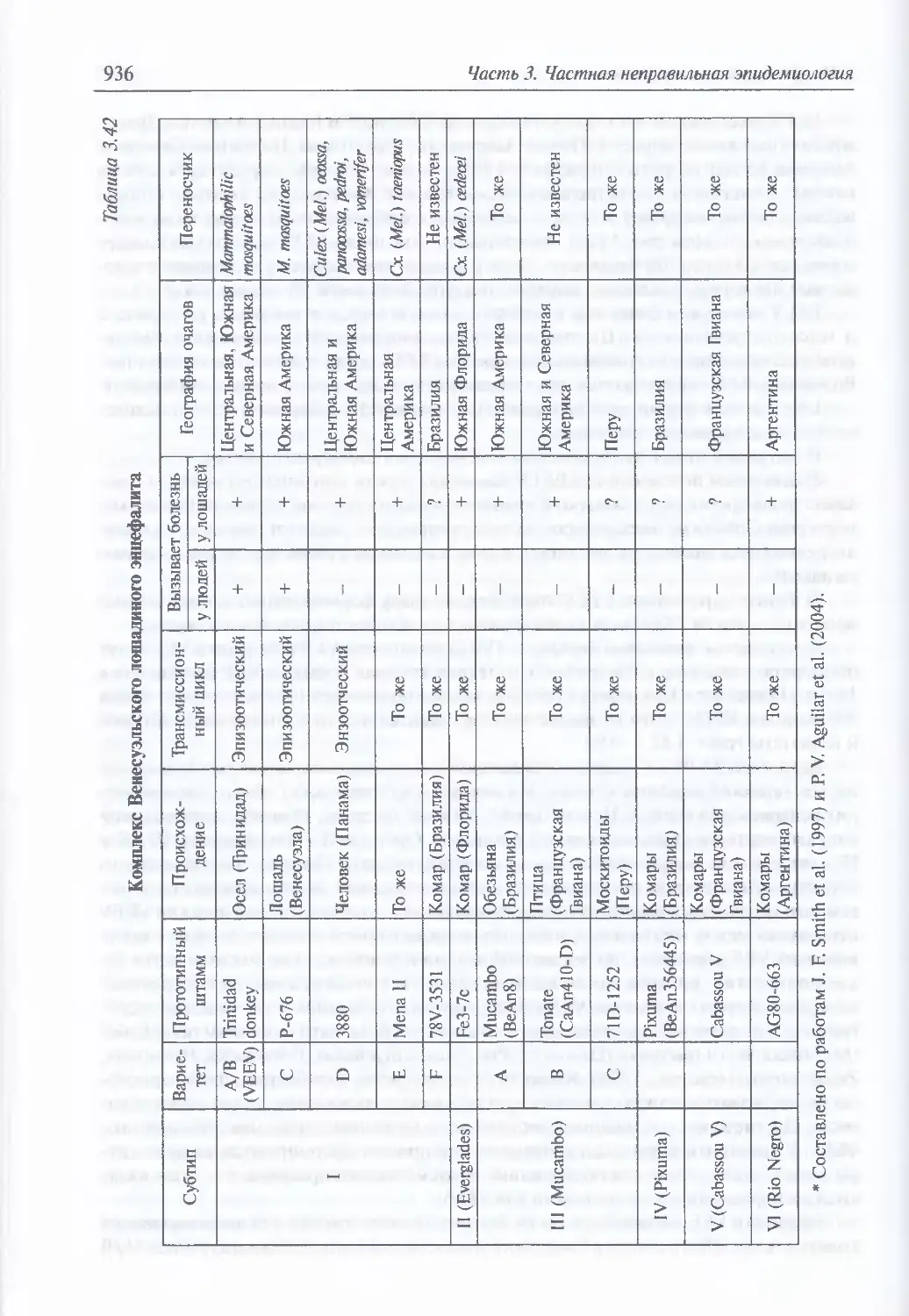
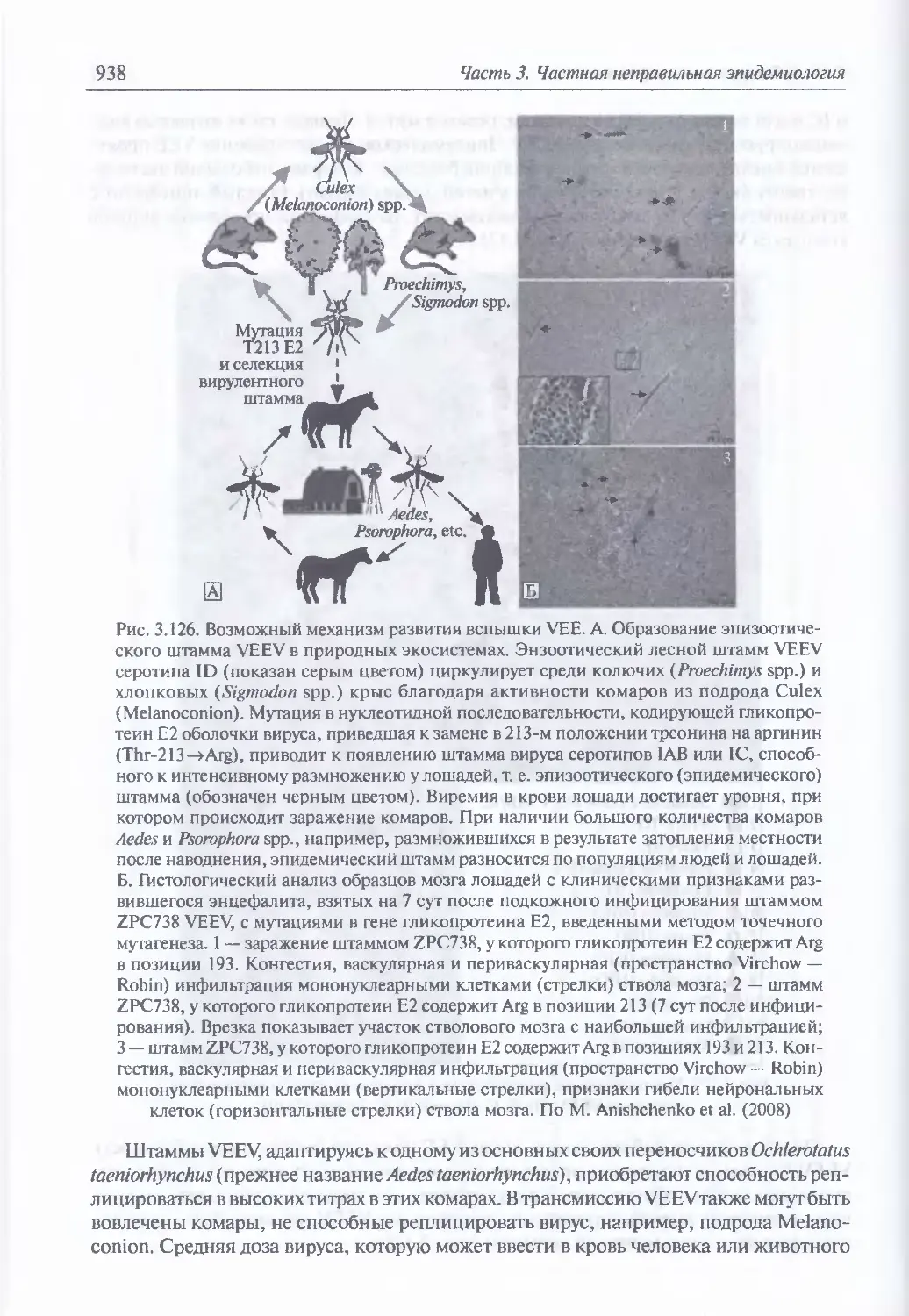
3.10. Арбовирусные энцефалиты ........................................... 928
Микробиология вирусов, вызывающих арбовирусные энцефалиты (929).
Экология и естественная эпидемиология возбудителей лошадиных
энцефалитов (933). Восприимчивость животных, клиническая картина
и патоморфология арбовирусных энцефалитов у животных при
искусственном инфицировании (939). Иммунитет к возбудителям
арбовирусных энцефалитов (942). Устойчивость возбудителей
арбовирусных энцефалитов во внешней среде (942). Обнаружение
генетически измененных штаммов возбудителей арбовирусных
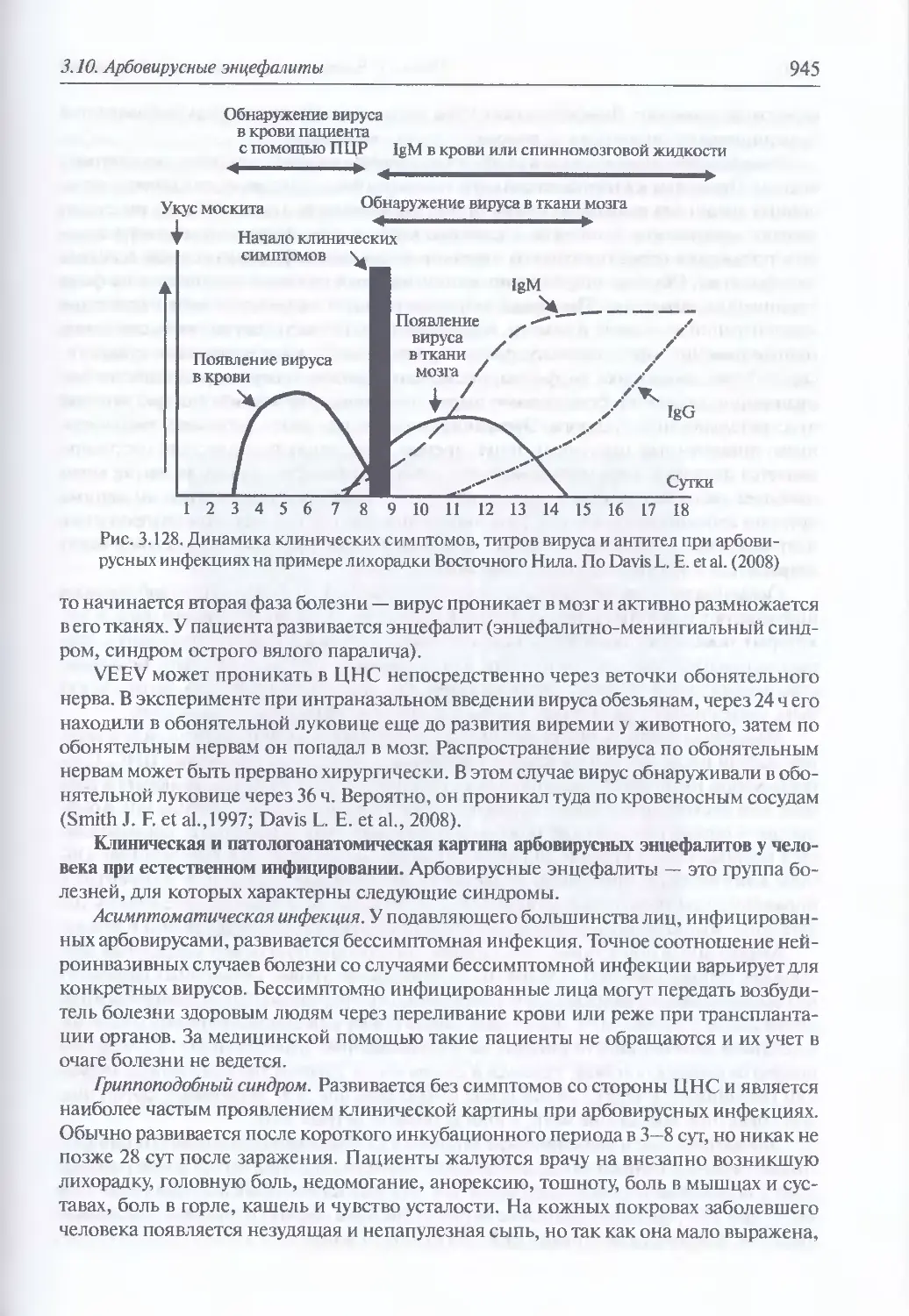
энцефалитов (943). Патогенез арбовирусных энцефалитог (944). Клиническая
и патологоанатомическая картина арбовирусных энцефалитов у человека
при естественном инфицировании (945). Диагностика искусственного
поражения возбудителями лошадиных энцефалитов (948).
Иммунопрофилактика арбовирусных энцефалитов (949).
Лечение арбовирусных энцефалитов (949)
3.11. Рицин................................................................949
Виды рицина (949). Молекулярная структура и механизм действия (950).
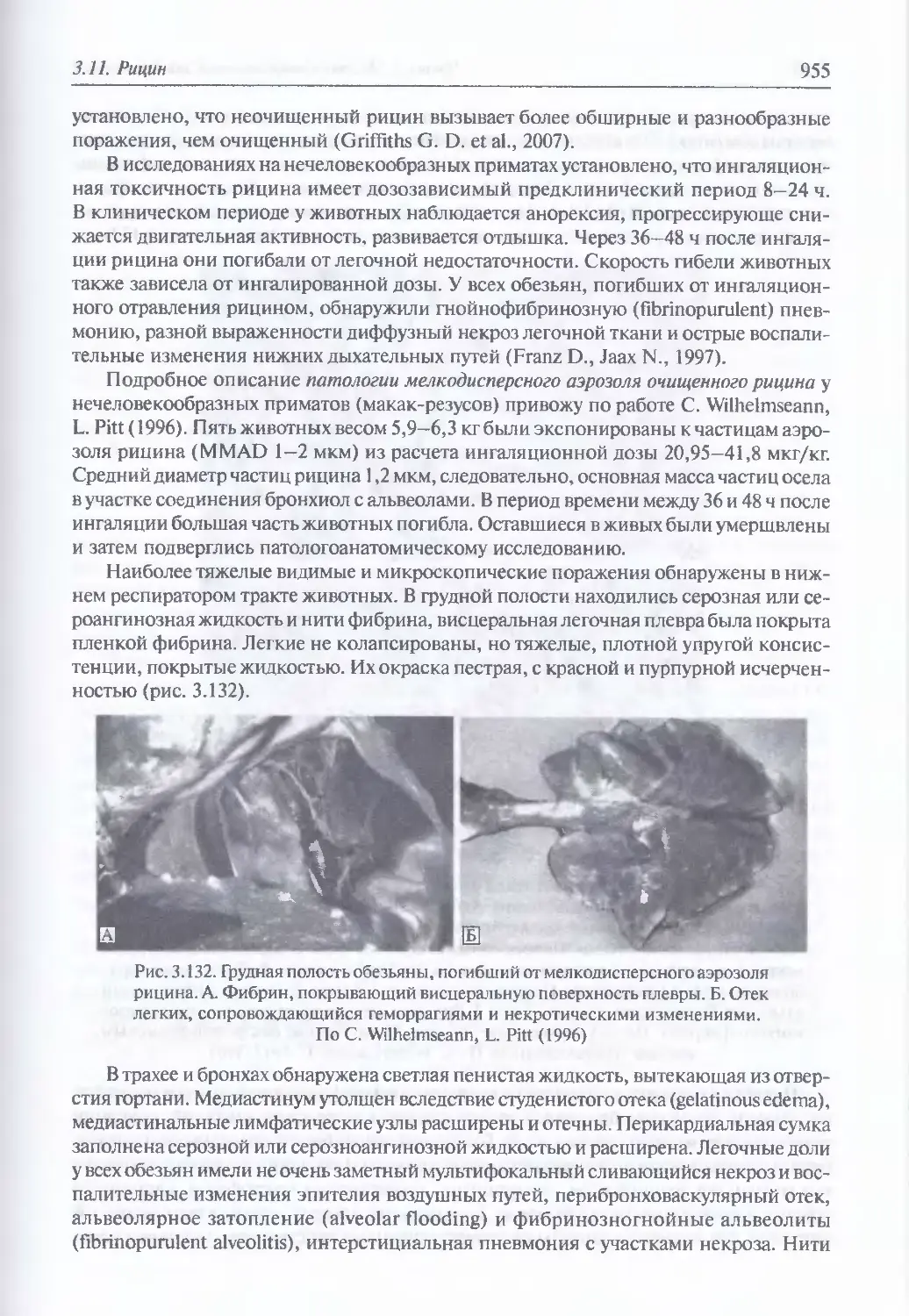
Токсические свойства (951). Поражающие свойства (953). Клиника
энтерального поражения (954). Клиника ингашционного поражения (954).
Клиника парентерального поражения (957). Диагностика поражения
рицином (958). Иммунопрофилактика (960). Лечение (960)
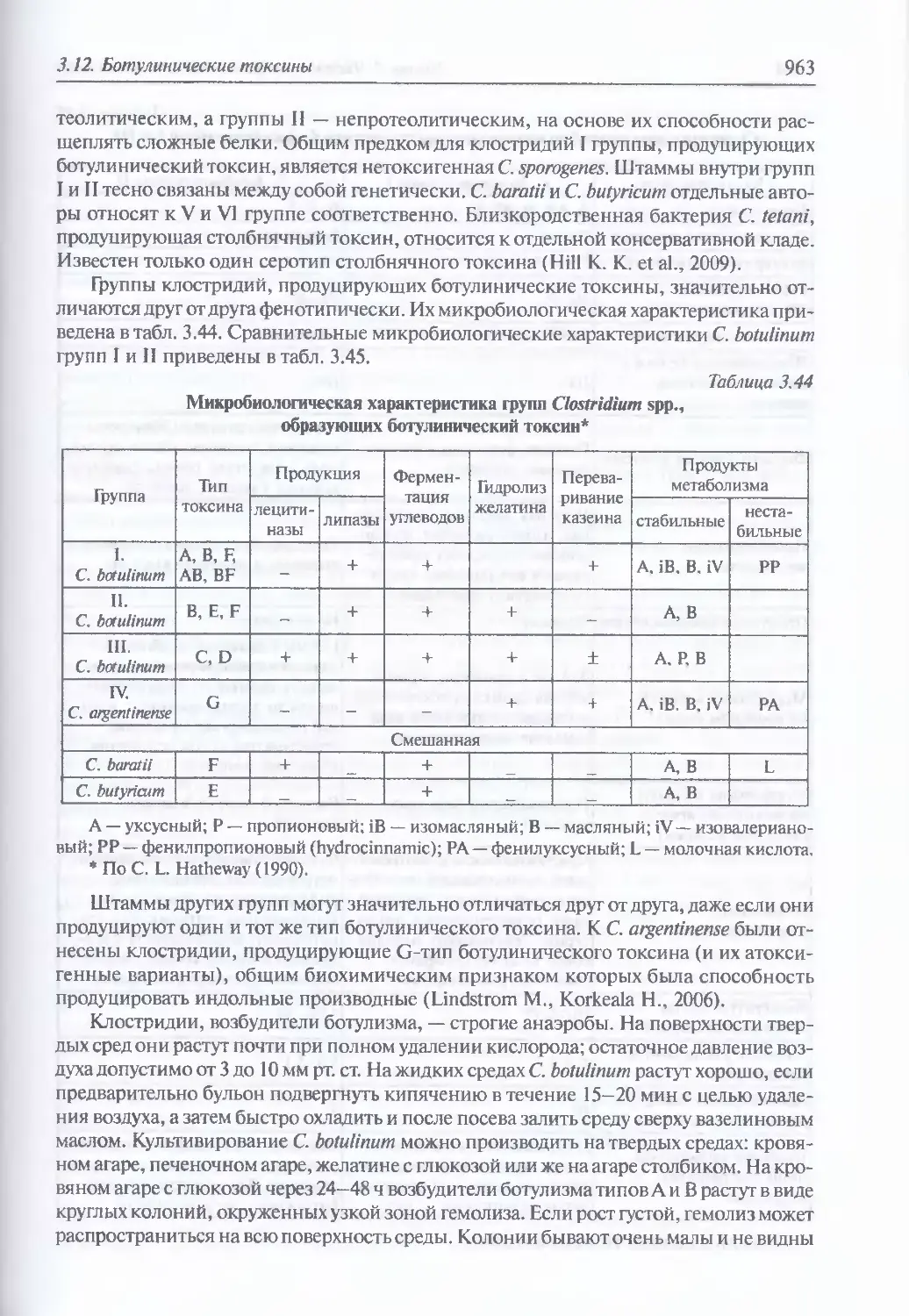
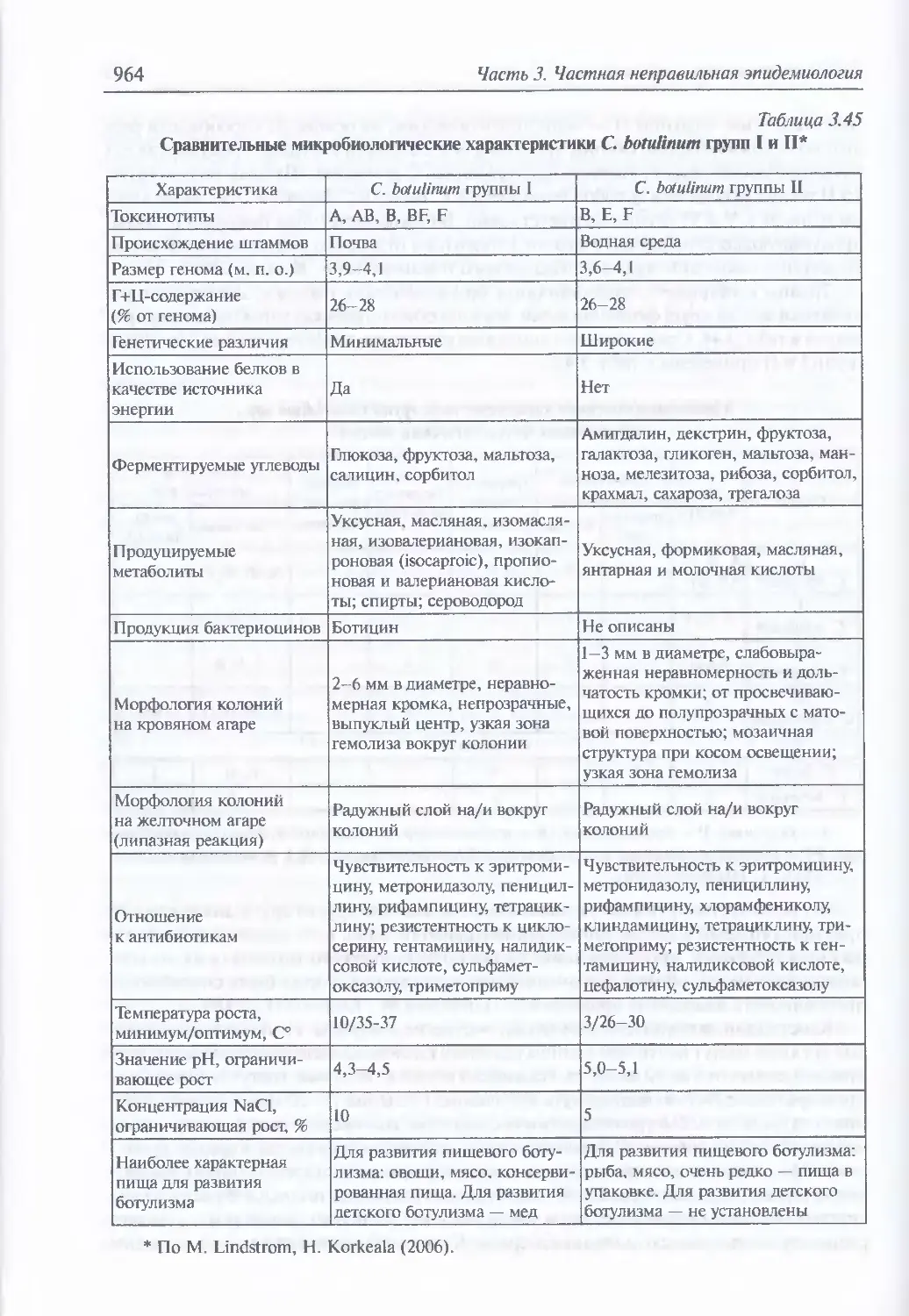
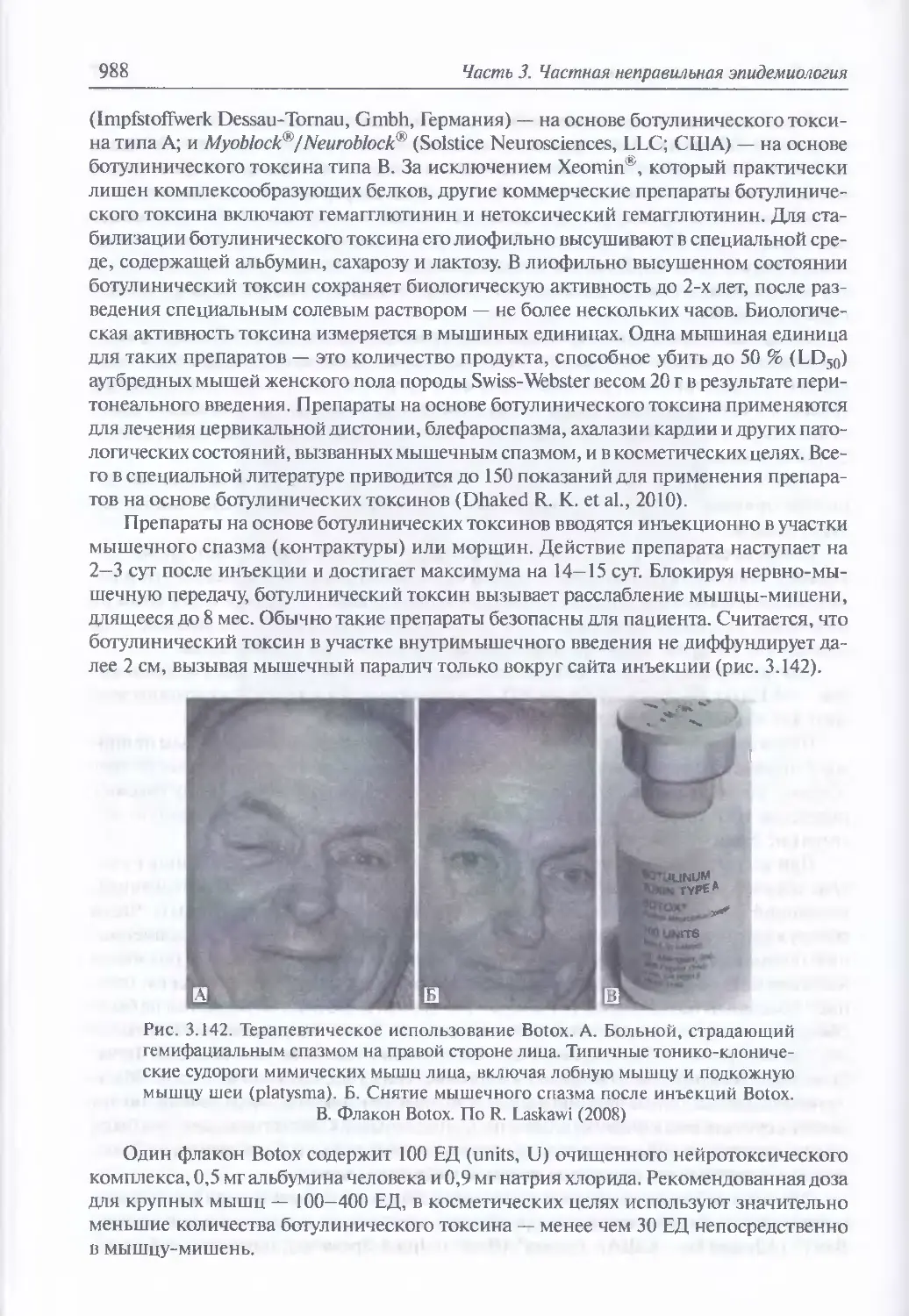
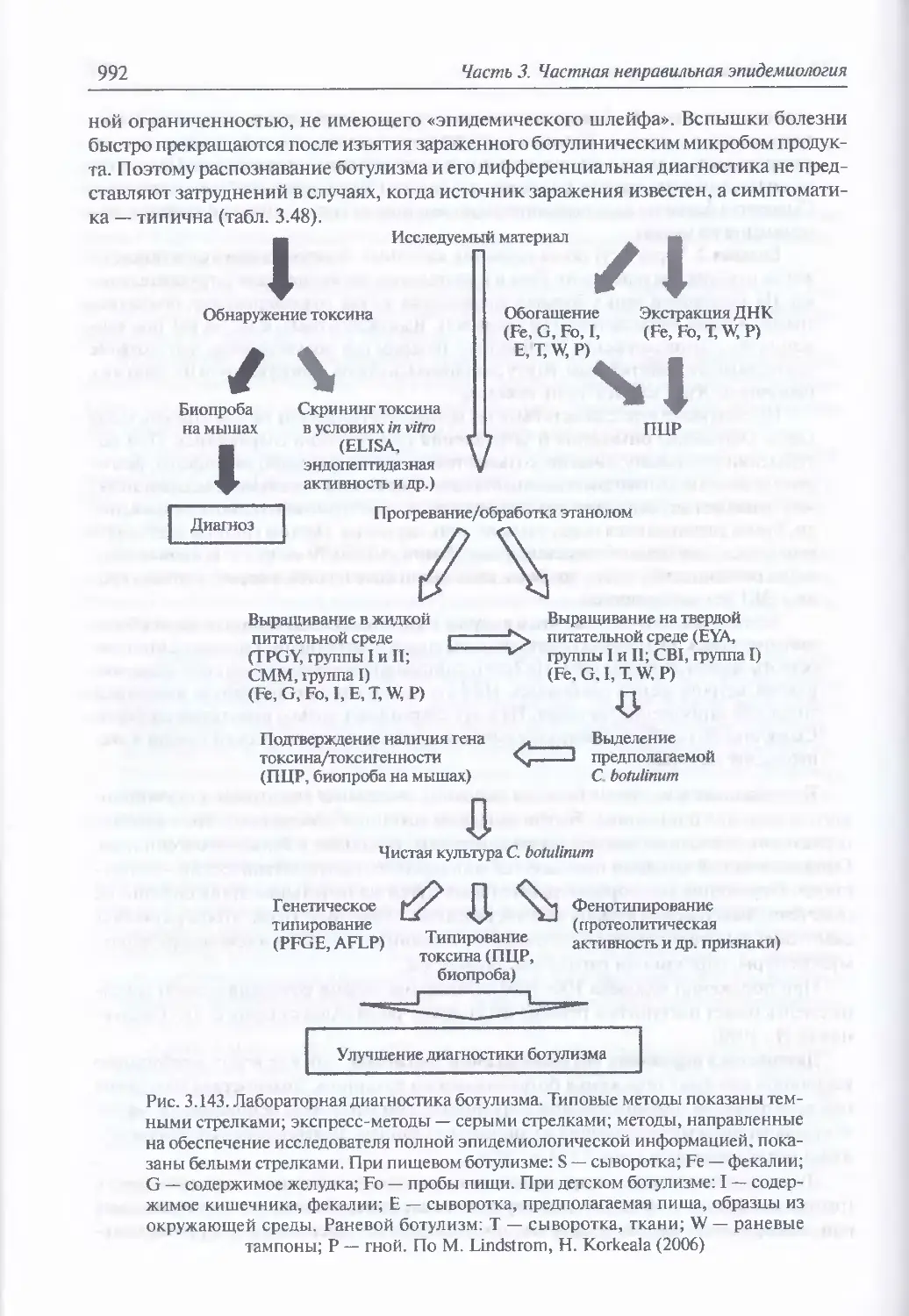
3.12. Ботулинические токсины...............................................961
Микробиология ботулинического микроба (962). Эпидемиология
ботулизма (966). Молекулярная структура и механизм действия
ботулинических токсинов (969). Генетика токсинообразования (971).
Токсические свойства ботулинических токсинов (974). Патогенез
ботулизма (974). Устойчивость ботулинических токсинов во внешней
среде (977). Поражающие свойства ботулинических токсинов (978).
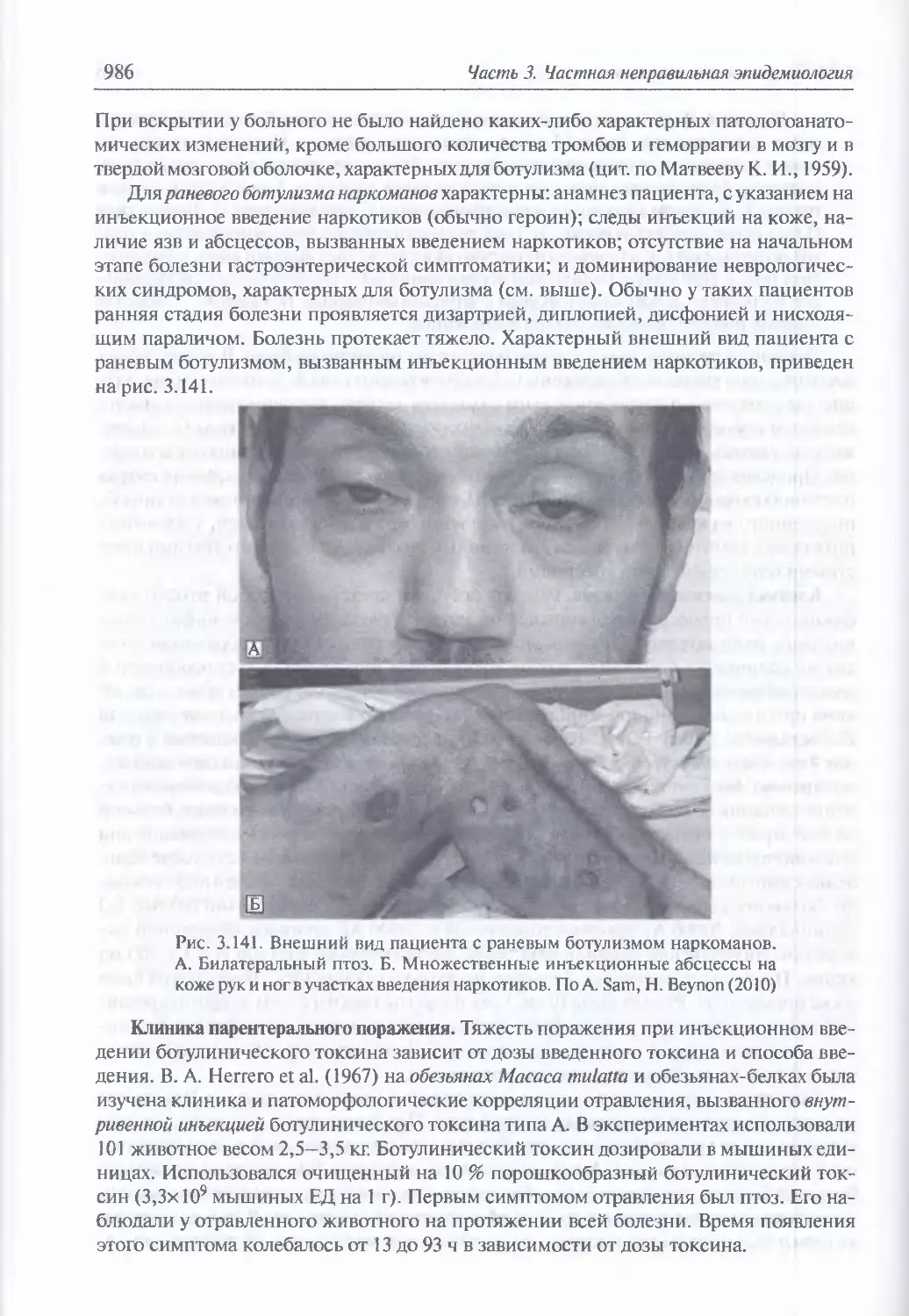
Клиника энтерального поражения (978) Клиника раневого ботулизма (985).
Клиника парентерального поражения (986). Клиника ингаляционного
поражения (989). Диагностика поражения ботулиническими
токсинами (991). Иммунопрофилактика (996). Лечение (996)
3.13. Стафилококковый энтеротоксин В.......................................998
Токсины-суперантигены (999) Молекулярная структура и механизм
действия токсинов-суперантигенов стафилококков и стрептококков (999).
Токсические свойства SEB (1000). Поражающие свойства SEB (1000).
Клиника энтерального поражения SEB (1001). Клиника конъюнктивального
поражения SEB (1001). Клиника и патанатомия ингаляционного
Содержание
11
поражения SEB (10(4). Клиника парентерального поражения SEB (1005).
Диагностика поражения SEB (1005). Иммунопрофилактика (1008).
Лечение (1008)
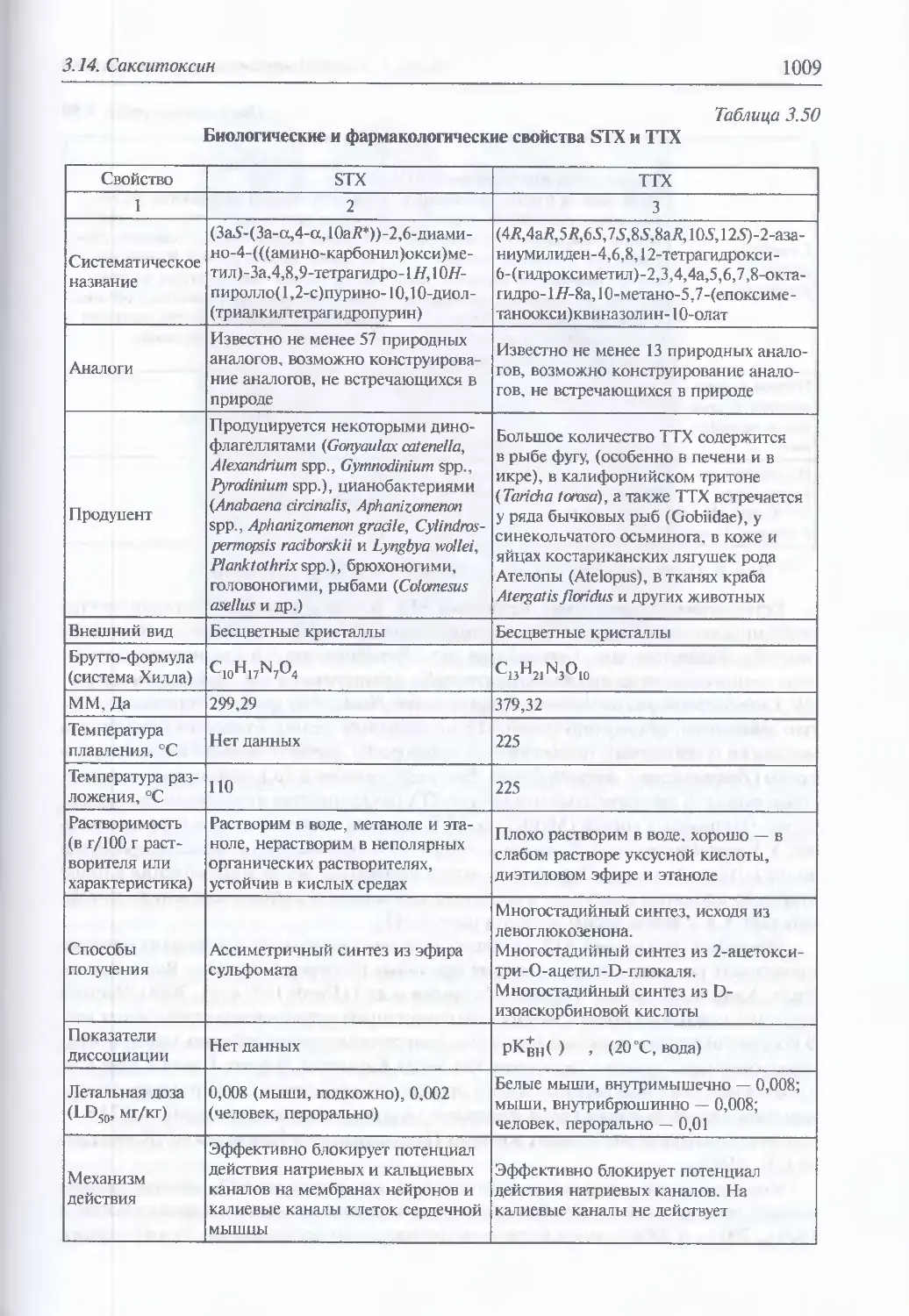
3.14. Сакситоксин........................................................1008
Естественная эпидемиология отравления сакситоксином (1010).
Молекулярная структура и механизм действия (1011). Генетика
токсиноосразования (1015). Токсические и поражающие свойства (1016).
Клиника энтерального поражения (1017). Диагностика поражения
сакситоксином (1018). Иммунопрофила дика (1019). Лечение (1019)
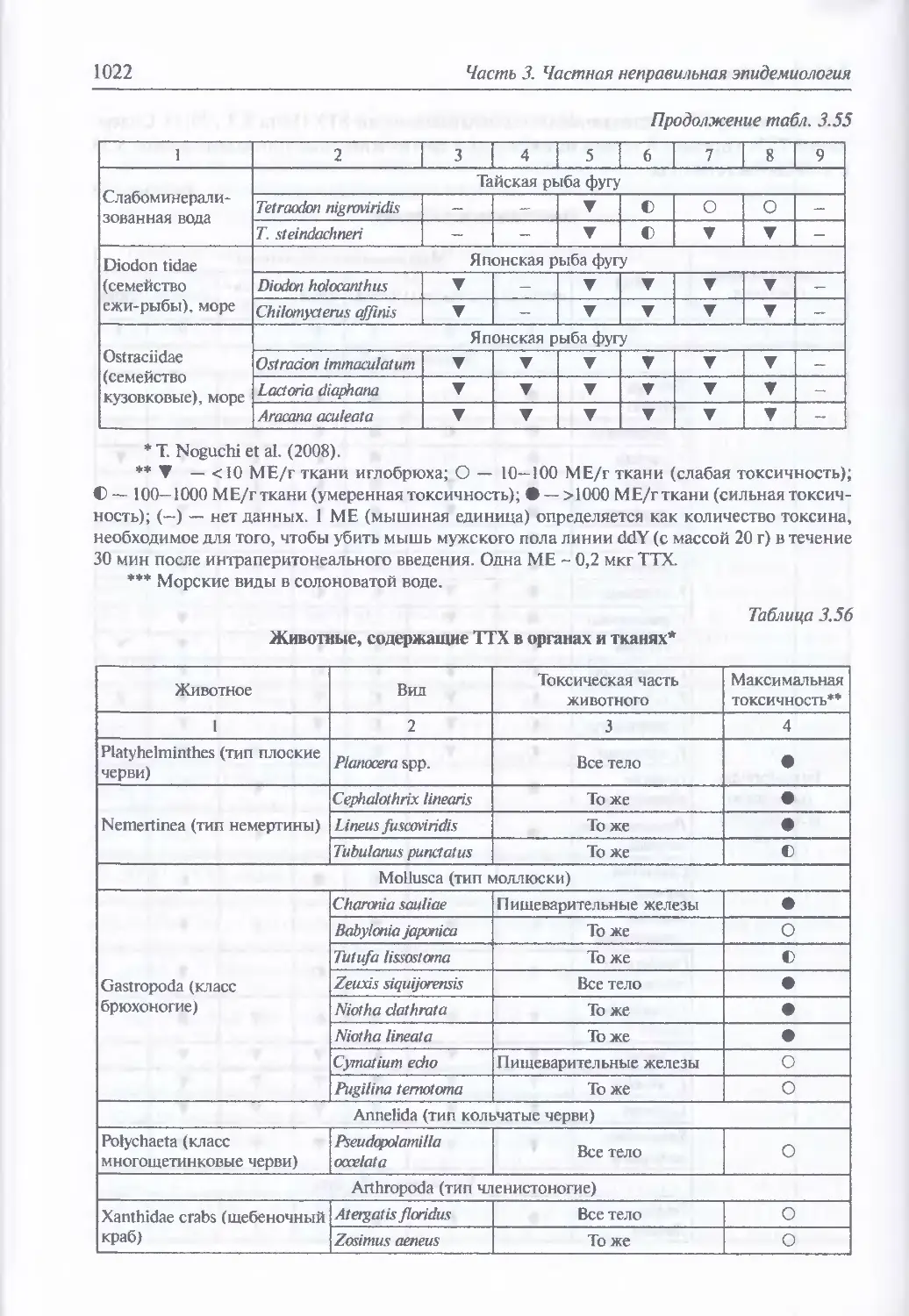
3.15. Тетродотоксин......................................................1020
Естественная эпидемиология отравления тетродотоксином (1020).
Молекулярная структура и механизм действия (1028). Токсические
и поражающие свойства (1051). Клиника энтерального поражения (1032).
Диагностика поражения тетродотоксином (1033).
Иммунопрофилактика (1034). Лечение (1034)
3.16. Палитоксин....................................................... 1034
Естественная эпидемиология отравления палитоксином (1035).
Молекулярная структура и механизм действия (1037). Токсические
и поражающие свойства палитоксина (1040). Клиника пищевого
отравления (1043). Клиника ингаляционного поражения (1044).
Клиника контактного поражения (1045). Диагностика поражения
палитоксином (1045). Иммунопрофилактика (1047). Лечение (1047)
СЛОВАРЬ ТЕРМИНОВ.......................................................... 1049
СПИСОК ЛИТЕРАТУРЫ..........................................................1067
ПРИЛОЖЕНИЕ А. Бактериологическая война. Доклад специального консультанта
по вопросам бактериологической войны Д. У. Мерка военному министру
по состоянию на 03.01.1946 г...........................................1103
ПРИЛОЖЕНИЕ Б. Показания пленных американских летчиков,
применявших бактериологическое оружие в 1952 г. во время войны
на Корейском полуострове...............................................1110
Пока )ания американского военнопленного летчика, старшего лейтенанта
Инока, об его участии в бактериологической войне, ведущейся
американскими войсками в Корее (SIA/14)............................... 1110
Показания американского военнопленного летчика, старшего лейтенанта
Квинна, об его участии в бактериологической войне, ведущейся
американскими войсками в Корее (SIa/15)................................1115
Показания американского военнопленного летчика, старшего лейтенанта
О’Нила, об его участии в бактериологической войне, ведущейся
американскими войсками в Корее (1SGK/4)................................1119
Показания американского военнопленного летчика, старшего лейтенанта
Кнчсса, об его участии в бактериологической войне, ведущейся
американскими войсками в Корее (ISCK/5)................................1130
Светлой памяти академика РАМН
П. Н. Бургасова посвящается эта книга
Злокачественная агрессия свойственна исклю-
чительно человеку, и она не порождается животными
инстинктами. Она не нужна для физиологического
выживания человека и в то же время представляет
собой важную составную часть его психики.
Эрих Фромм
Анатомия человеческой деструктивности, 1973
ВВЕДЕНИЕ
Применение биологических средств для поражения людей — трудно раскры-
ваемый вид преступлений. Осуществляться они могут в военных (биологическая вой-
на, биодиверсия), политических (биологический теракт), криминальных целях (био-
криминал), а также в рамках стратегии непрямых действий, цель и организаторов
которых очень трудно установить. Однако в криминалистической литературе отсут-
ствуют типовые схемы их расследования. Не в лучшем положении находятся и те, кто
первым столкнется с последствиями таких преступлений — медицинские работники
низового звена. Именно от их внимательности и способности выявлять необычные
звенья в эпидемической цепочке известной инфекции зависит оперативность в об-
наружении искусственного характера вспышки инфекционной болезни или массового
отравления. Но и медицинские работники ограничены в своих действиях рядом лож-
ных или неконкретных представлений о поражающей способности биологических
агентов и о вызываемых ими инфекционных и эпидемических процессах, позаимст-
вованных в основном из брошюр и статей декларативного характера. Эти обстоятель-
ства ставят в неравные условия российских врачей и работников правоохранительных
органов по отношению к их западным коллегам, да и к самим террористам, имеющим
более полные и объективные источники информации по данной проблеме.
Особенно «разрыв» в качестве и полноте информации бросается в глаза при сопо-
ставлении руководства «Medical Aspects of Chemical and Biological Warfare», вышедшем
в США в 1997 г. под редакцией Е R. Sidel et al., монографии G. Zubay et al. (2005) и,
например, с руководством «Противодействие биологическому терроризму», опубли-
кованном в 2003 г. в России под ред. Г. Г. Онищенко.
Отдельным отечественным руководствам по эпидемиологии в тех их разделах, в
которых речь идет о последствиях совершенных биотеррористических актов или при-
менения биологического оружия, характерны своеобразные «родовые травмы». На-
пример, эпидемиология искусственно вызванной вспышки инфекционной болезни
отождествляется с эпидемиологией инфекционной болезни, распространившейся в
военное время. Даже известные российские ученые-эпидемиологи микробную куль-
туру называют оружием, а культивирование бактерий в колбе с питательным бульоном,
его производством (например, Б. Л. Черкасский, 2000). Табличные перечни биоло-
гических агентов, «наиболее вероятных для применения террористами», составляются
по учебникам для младших курсов медицинских вузов и, как правило, на основе та-
Введение
13
кого лабораторного показателя, как «летальная доза», без учета возможностей их мас-
сового производства, достижений рецептуростроения, аэробиологических и других
характеристик. При этом никого из авторов таких работ не смущает то обстоятельство,
что, уже ставшие известными, биологические диверсии и теракты проводились с ис-
пользованием других биологических агентов и/или имели совершенно не следующие
из этих таблиц последствия. Нормой литературы подобного рода стали безответствен-
ные ссылки типа «некоторые исследователи считают». А за первый «достоверный
факт » применения биологического оружия принято описание нотариуса генуэзских
работорговцев де Мюсси причин чумы в Кафе в 1347—1348 гг., сделанное им на осно-
ве учения о миазмах (см. Б. Л. Черкасский, 2000; Г. Г. Онищенко, 2003). Из этого и
подобных исторических мифов произошел другой, современный, последствия кото-
рого мы видим на примере Ирака — миф о доступности биологического оружия ма-
лоразвитым странам. За пределами научных интересов авторов большинства работ по
ликвидации последствий биотеррористических актов находятся поражения биологи-
ческими токсинами (например, этим страдает работа Г. Г. Онищенко с соавт., 2003).
И уже совсем недопустимым мне представляется использование для планирова-
ния мероприятий по ликвидации биотеррористических актов и диверсий такого «про-
дукта холодной войны», как миф о «выбросе спор сибирской язвы из предприятия по
производству БО в Свердловске в 1979 г.». К сожалению, он нашел свое мутное отра-
жение в работах И. В. Цомарадского (1996); Б. Л. Черкасского (2000, 2002, 2003);
и даже Г. Г. Онищенко с соавт. (2003).
Кроме перечисленных выше субъективных препятствий на пути расследования
биотеррористических актов и биодиверсий, имеются еще и объективные, обусловлен-
ные принципиальными различиями между эпидемиологией искусственных и есте-
ственных эпидемических процессов. Современное деление эпидемиологии на общий
и частный разделы сложилось более чем 100 лет назад, когда была установлена роль
микроорганизмов в развитии эпидемических процессов, и еше не существовало уг-
розы искусственных эпидемий, вызванных применением биологического оружия или
актами биотеррора. Однако за прошедшие годы эпидемические угрозы качественно
изменились. Во-первых, начавшаяся в 1930-х гг. гонка биологических вооружений дала
возможность ряду развитых оран накопить опыт по созданию и применению биоло-
гического оружия Во-вторых, так называемая «почтовая эпидемия сибирской язвы»,
вспыхнувшая осенью 2001 г в США, показала, что в мире имеются силы, способные
в рамках стратегий непрямых действий профессионально осу ществлять биотеррори-
стические акты, оставаясь при этом неустановленными1. В-третъих. в настоящее вре-
1 Биотеррористический акт, совершенный в 2001 г. в США, не был раскрыт. Как и в случае
с убийством президента Джона Кеннеди, ФБР вело следствие «вширь», хотя использованный
террористами иммунитет, преодолевающий штамм возбудителя сибирской язвы («vaccine-
resistant Bacillus anihracis») Ames, имелся только в нескольких лабораториях, работавших по
программам Министерства обороны США. С 2002 г. в роли подозреваемого проходил микро-
биолог Стивен Хатфилл (Steven Jay Hatfill), но доказать его причастность к теракту в суде не
удалось. В 2003 г. он подал иски против ФБР и Министерства юстиции и отсудил 5,8 млн дол-
ларов компенсации («... violating his constitutional rights and for violating the Privacy Act»). В 2008г.
в качестве подозреваемого оказался микробиолог Брюс Айвинс (Bruce Edwards Ivins), но и его
вина не была доказана в суде. В июне Брюса нашли в коме в ванной комнате своего дома, от-
равленного туленолом. Через два дня, не приходя в сознание, он умер от почечной и печеноч-
ной недостаточности. После чего федеральный обвинитель, прокурор Джеффри Тэйлор
(Geoffrey Taylor), официально сделал утверждение, что Брюс совершил самоубийство, и он был
14
Введение
мя способы и средства биологического поражения людей по своим возможностям,
вышли (алеко за пределы эпидемических процессов, которые являются объектами
изучения общей и частной эпидемиологии. Поэтому цель данной работы состоит в
обосновании нового раздела эпидемиологии — эпидемиологии искусственных эпи-
демических процессов и других, умышленно вызванных биологических поражений
(неправильной эпидемиологии) , способного разрабатывать подходы к их распозна-
нию и устранению последствий.
Монография состоит из трех взаимосвязанных частей. Первая посвящена истории
применения биологических средств для поражения людей. Ее задача — показать эво-
люцию таких преступлений, приведшую к необходимости выделения неправильной
эпидемиологии из общей и частной эпидемиологии. Приведены примеры, показы-
вающие принципиальные различия в течении инфекционных процессов и развитии
вспышек инфекционных болезней, вызванных естественным и искусственным рас-
пространением их возбудил елей.
Вторая часть монографии собственно и является обоснованием неправильной
эпидемиологии. В ней рассматриваются объект, цель и >адачи неправильной эпиде-
миологии как третьего раздела эпидемиологии. Для их понимания сопоставлены этио-
логические, клинические и эпидемические аспекты искусственных и естественных
эпидемий (вспышек) инфекционных болезней. Обоснован термин «неправильная
эпидемиология». Показана ее связь с другими науками. Приведена классификация
биологических агентов, отражающая современные достижения в генетике микроор-
ганизмов и в развитии нанобиотехнологий. Дано краткое описание биологических
агентов как вызывающих инфекционный процесс (микроорганизмы), так и тех, ко-
торые проявляют свое действие специфическим биологическим поражением без раз-
вития инфекционного процесса (токсины, искусственные генетические конструкции,
наночастицы). Далее разбираются основы эпидемиологического расследования ис-
кусственных эпидемических процессов и поражений нереплицируюшимися агента-
ми. Рассматриваются характерные особенности эпидемиологии искусственных эпи-
демических процессов и поражений нереплицируюшимися агентами и возможные
подходы к их эпидемиологическому расследованию. Последний раздел второй части
монографии посвящен обнаружению биотеррористической организации.
Третья часть монографии посвящена особенностям неправильной эпидемиоло-
гии вспышек инфекционных болезней, вызванных возбудителями инфекционных
болезней (сибирская язва, чума, туляремия, бруцеллез, сап, мелиоидоз, Ку-лихорад-
ка, вирусные геморрагические лихорадки, арбовирусные энцефалиты), и поражений
токсинами (рицин, ботулинические токсины, стафилококковый энтеротоксин В,
сакситоксин, палитоксин, тетродотоксин), считающихся специалистами наиболее
опасными потенциальными агентами биологического оружия. Поэтому она названа
частной неправильной эпидемиологией.
Описание потенциальных агентов биологического оружия максимально, насколь-
ко это позволяют открытые источники, унифицировано. Оно включает краткую
единственным преступником, организовавшим биотеррористический акт в 2001 г. Газет а «New
York Times» по этому поводу задала вопрос: «А что если бы доктор Хэтфилл совершил само-
убийство в 2002 г? Достаточно ли это было бы для правосудия, чтобы объявить о смерти пре-
ступника?» (более подробно см. в разд. 1,12.4).
1 Термины «эпидемиология искусственных эпидемических процессов и других, умышлен-
но вызванных биологических поражений» и «неправильная эпидемиология» используются как
синонимы.
Введение
15
характеристику микроорганизма (токсина) как биологического поражающего аген-
та. Далее приводятся сведения о его микробиологии, факторах патогенности, таксо-
номии, происхождении, молекулярной эпидемиологии, экологии, естественной эпи-
демиологии, инфицирующих (поражающих) дозах для животных и людей, патогенезе,
устойчивости во внешней среде, резистентности к антибиотикам или антивирусным
препаратам. Диагностика искусственного поражения биологическим агентом осно-
вывается на сопоставлении эпидемиологии, клиники и патоморфологии, характер-
ных для случаев болезни, являющихся результатом естественного инфицирования
(отравления), с теми, которые могут наблюдаться в результате различных способов
преднамеренного введения (ингаляционного, энтерального и парентерального) мик-
роорганизма или токсина в организм человека. Рассматриваются подходы к обнару-
жению генетически измененных штаммов возбудителей инфекционных болезней —
потенциальных агентов биологического оружия. Приводятся истории болезни случаев
искусственного и естественного заражения (отравления) и результаты моделирующих
экспериментов на животных.
Книга предназначена широкому кругу ученых, врачей, организаторов здравоох-
ранения, работников правоохранительных органов и МЧС, но в основном она ори-
ентирована на специалистов их низового звена, обычно первыми сталкивающимися
с последствиями всевозможных диверсий и террористических актов
У меня нет «конфликта интересов» с другими учеными, данная работа не оплачи-
валась никакими фондами или грантами, я никогда не работал в организациях, имею-
щих коммерческие интересы в какой-либо сфере медицинского сервиса. Книга не
содержит информации, ставшей мне известной в рамках прошлой или нынешней
профессиональной деятельности. К ее подготовке не привлекались закрытые источ-
ники, все изложенные в ней сведения могут быть легко проверены читателем по цити-
руемой литературе. Критические замечания и пожелания можно направлять по элект-
ронной почте непосредственно автору (msupotnitskii@mail.ru).
СПИСОК СОКРАЩЕНИЙ
РУССКИЕ
ACT — аспартатаминотрансфераза
АМР — аденозинмонофосфат
АД — артериальное давление
АЛТ — аланинаминотрансфераза
АТф — аденозинтрифосфат
БО — биологическое оружие
БПА — биологические поражающие агенты
В/б — внутрибрюшинное введение
В/в — внутривенное введение
ВИЧ — вирус иммунодефицита человека
ВНО — вирус натуральной оспы
ВНС — вегетативная нервная система
ВПСП — возбуждающий постсинаптический потенциал
ВЭЖХ — высокоэффективная жидкостная хроматография
ВЭЛ — вирус энцефалита лошадей
ГАБК — у-аминобутировая кислота (у-аминомасляная кислота)
ГТФ — гуанозинтрифосфат
ГЭБ — гематоэнцефалический барьер
ДВС-синдром — синдром диссеминированного внутрисосудистого свертывания крови
ДДТ — дихлордифенилтрихлорметилметан
днДНК — двунитевая ДНК
ДНК — дезоксирибонуклеиновая кислота
Ж — женщина
ЖКТ — желудочно-кишечный тракт
И/н — интраназальное введение
И/т — интратрахеальное введение
И ВЛ — искусственная вентиляция легких
ИЛП — иммунобиологический лекраственный препарат
КБТО — см. Конвенция 1972 г.
КВЖД — Китайско-Восточная железная дорога
КОЕ — колонии образующая единица
Конвенция 1972 г. — Конвенции о запрещении разработки, производства и накопления запасов
бактериологического (биологического) и токсинного оружия и об их уничтожении
КРС — крупнорогатый скот
ЛПС — липополисахарид
М — мужчина
м. п. о. — миллион пар оснований
МАП — мембрано-активный пептид
мАт — моноклональное антитело
МИК — минимальная ингибирующая концентрация
мкм — микрон, микрометр
ММ — молекулярная масса
ММГ — N-моно-метил-гидразин
МПА — мясопептонный агар
МПБ — мясопептонный бульон
МП КП — миниатюрный потенциал концевой пластинки
МСД-пептиды — пептиды, дегранулирующие тучные клетки
МФГ — N-метил-т-формилгидразин
Список сокращений
17
НИУ — научно-исследовательское учреждение
нм — нанометр
нМ — наномоль
ОВ — отравляющее вещество
П/к — подкожно
ПДК — предельно допустимая концентрация
ПЭГ — полиэтиленгликоль
РГАСПИ — Российский государственный архив социально-политической истории (Москва)
РНК — рибонуклеиновая кислота
РСК — реакция связывания комплемента
РЭС — ретикулоэндотелиальная система
сАМФ — циклический аденозинмонофосфат
СОЭ — скорость оседания эритроцитов
ССС — сердечно-сосудистая система
т. п. о. — тысяча пар оснований
ТМП — триметоприм
ФОС — фосфорорганические соединения
ХАО — хорион-аллантоисная оболочка куриных эмбрионов
ХБО — химико-биологическое оружие
ХО — химическое оружие
цАМФ — циклический аденозинмонофосфат
ЦВД — центральное венозное давление
ЦНС — центральная нервная система
ЦТЛ — цитолитические Т-лимфоциты
ЧД — частота дыхания
ЧСС — частота сердечных сокращений
ЭПР — эндоплазматический ретикулум
ЭПС — эндоплазматическая сеть
АНГЛИЙСКИЕ
aacA, aadA, aph (genes encoding resistance to aminoglycosides) — гены, кодирующие резистентность
к аминогликозидам
АСЕ (angiotensin-converting enzyme) — ангеотензин-конвертирующий фермент
AChR (nicotinic acetylcholine receptors) — никотиновый ацетилхолиновый рецептор
ADE (antibody-dependent enhancement) — феномен антителозависимого усиления инфекции
AFLP (amplified fragment length polymorphisms) — полиморфизм длин амплифинированных фраг-
ментов
AIDS (acquired immunodeficiency syndrome) — синдром приобретенного иммунодефицита
ALP (alkaline phosphatase) — щелочная фосфатаза
alpha-LTX, a-LTX (alpha-latrotoxin) — а-латротоксин (нейротоксин каракурта «Черная вдова»)
ALT (alanine aminotransferase) — аланинаминотрансфераза
AMI (American Media Inc.) — Американская медийная компания
AML (acute myelogenous leukemia) — острая миелоидная лейкемия
ANP (acyclic nucleoside phosphonate) — ациклический нуклеозидфосфонат
Ap (ampicillin) — ампициллин
APC (antigen presenting cell) — антигенпрезентирующая клетка
AST (aspartate aminotransferase) — аспартатаминотрансфераза
ATCC (American Type Culture Collection) — Американская коллекция типовых культур
attL, attR — левый и правый присоединяющие сайты
AVA (Anthrax Vaccine Adsorbed) — адсорбированная сибиреязвенная вакцина
BAL (bronchoalveolar lavage) — бронхоальвеолярный лаваж
ВВВ (blood-brain barrier) — гематоэнцефалический барьер
BCDPC (Bureau of Chronic Disease Prevention and Control) — Бюро по профилактике и контролю
хронических болезней (США)
beta-BTX, р-ВТХ (P-bungarotoxin) — р-бунгаротоксин
bGHpA (bovine growth hormone polyadenylation sequence) — полиаденилапионная последователь-
ность бычьего гормона роста
18
Список сокращений
B1V (bovine immunodeficiency virus) — бычий вирус иммунодефицита
Bl (bleomycin) — блеомицин
BL2 (biosafety level 2) — второй уровень биобезопасности
WaVIM (gene encoding a VIM metallo-p-lactamase) — ген, кодирующий VIM-металло-Р-лактамазу
BUN (blood urea nitrogen) — азот мочевины крови
CA (capsid protein) — капсидный белок
CAEV (caprine arthritis encephalitis virus) — вирус артрита и энцефалита коз
CAR (coxsackievirus-adenovirus receptor) — рецептор коксакивирусов и аденовирусов
cat (gene chloramphenicol acetyltransferase) — ген, кодирующий хлорамфениколацетилтрансферазу
CAV (canine adenovirus) — собачий аденовирус
CBI (С. botulinum isolation medium) — среда для изоляции ботулинического микроба
CBS (Columbia Broadcasting System; в настоящее время CBS Broadcasting Inc.) — американская
телерадиосеть
CDC (Center for Disease Control and Prevention) — Центр контроля и предотвращения инфекцион-
ных болезней, Атланта, штат Джорджия, США
CFU/ml (colony forming units per ml) — количество формирующих колонии единиц в мл
CHDF (continuous hemodiafiltration) — непрерывная гемодиафильтрапия
cHS4 (chicken cHS4 P-globin insulator) — инсулятор куриного Р-глобина
СК (creatine phosphokinase) — креатинфосфокиназа
CLDT (cytolethal distending toxin) — цитолетальный растягивающий токсин
Cm (chloramphenicol) — хлорамфеникол (левомицетин)
CMM (cooked-meat medium) — среда на основе перевара мяса
CMV(cytomegalovirus) — цитомегаловирус
CNF (cytotoxic necrotizing factor) — цитотоксический некротизирующий фактор
CNS (central nervous system) — центральная нервная система
CNT (carbon nanotube) — углеродная нанотрубка
СРК (creatine phosphokinase) — креатинфосфокиназа
cPPT (central polypurine tract) — центральный полипуриновый тракт
CPXV (Cowpox virus) — вирус коровьей оспы
Cr (serum creatinine) — сывороточный креатинин
CRE (creatinine) — креатинин
CRP (C-reactive protein) — С-реактивный белок
CS (constant sequences) — константные последовательности
СТ (Connecticut) — Коннектикут
СТЕ (constitutive transport element) — конститутивный транспортный элемент
CTS (central termination sequence) — центральная терминирующая последовательность
CAT (chloramphenicol acetyltransferase) — хлорамфениколацетилтрансфераза
DAG (diacylglycerol) — диацилглицерол
DBD (DNAbinding domain) — ДНК-связывающий домен
DBP (DNA-binding protein) — ДНК-связывающий белок
DBP (dynein-binding peptide) — дениин-связывающий белок
DC (dendritic cell) — дендрическая клетка
DC-Chol (dimethylaminoethane-carbamoyl-cholesterol) — диметиламиноэтанкарбамоилхолестерол
deltaU3, AU3 (self-inactivating deletion in U3 region of 3' LTR) — самостоятельно инактивирующаяся
делеция в U3 регионе 3' LTR
DIS (dimerization initiation site) — сайт инициации димеризации
DL50 и LD50 — см. ЛДМ в Словаре терминов
Dox (doxycycline) — доксициклин
DR (direct repeat) — прямой повтор
DSE (distal sequence element) — элемент периферической последовательности
dsRNA (double-stranded RNA) — двуцепочечная РНК
DT (diphtheria toxin) — дифтерийный токсин
EAST (enteroaggregative E. coll heat-stable toxin) — энтероаггрегативный температуростабильный
токсин E. coli
EDX, EDRS или EDS (energy-dispersive X-ray spectroscopy) — метод энергодисперсионной
рентгеновской спектроскопии
Список сокращений
19
EF (edema factor) — отечный фактор сибиреязвенного токсина
EFla (human elongation factor 1-a promoter) — промотор фактора элонгации 1-a человека
EF2 (elongation factor 2) — фактор элонгации 2
EGFP (enhanced green fluorescent protein) — белок с усиленной зеленой флюоресценцией
EGFR (epidermal growth factor receptor) — рецептор эпидермального фактора роста
EIAV (infectious anemia virus) — вирус инфекционной анемии лошадей
ELISA (enzyme-linked immunosorbent assay) — твердофазный иммуноферментный анализ
Env (envelope glycoprotein) — оболочечный гликопротеин
ER (endoplasmic reticulum) — эндоплазматический ретикулум
Ery (erythromycin) — эритромицин
ESI-MS (electro spray ionization mass spectrometry) — ионизация распылением в электрическом поле
FACS (fluorescence activated cell sorting) — флуоресцентный сортинг, сортировка клеток с
активированной флуоресценцией
FCV (F. ft'/wenws-contaimng vacuole) — вакуоля, содержащая F. lularensis
FDA (Food and Drug Administration) — Управление по санитарному надзору за качеством
пищевых продуктов и медикаментов (США)
fevR (Francisella effector of virulence regulation) — эффектор регуляции вирулентности Francisella
FIV (feline immunodeficiency virus) — вирус иммунодефицита кошачьих
FL (Florida) — Флорида
FPI (Francisella pathogenicity island) — остров патогенности Francisella
GALV (gibbon ape leukemia virus) — вирус лейкемии гиббонов
GAS (interferon-gamma activated sequence) — у-интерферон-активированная последовательность
GCS (The Glasgow Coma Scale) — шкала комы Глазго
GFAP(glial fibrillary acidic protein) — глиальный фибриллярный кислый белок
GFP (green fluorescent protein) — зеленый флюоресцирующий белок
Gm (gentamicin) — гентамицин
GM-CSF (granulocyte-macrophage colony stimulating factor) — i ранулоцит-макрофаг колониести-
мулирущий фактор
gRNA (viral genomic RNA) — вирусная геномная РНК
HBS (HEPES-buffered saline) — HEPES-забуференный солевой раствор
НС (hemorrhagic colitis) -- геморрагические колиты
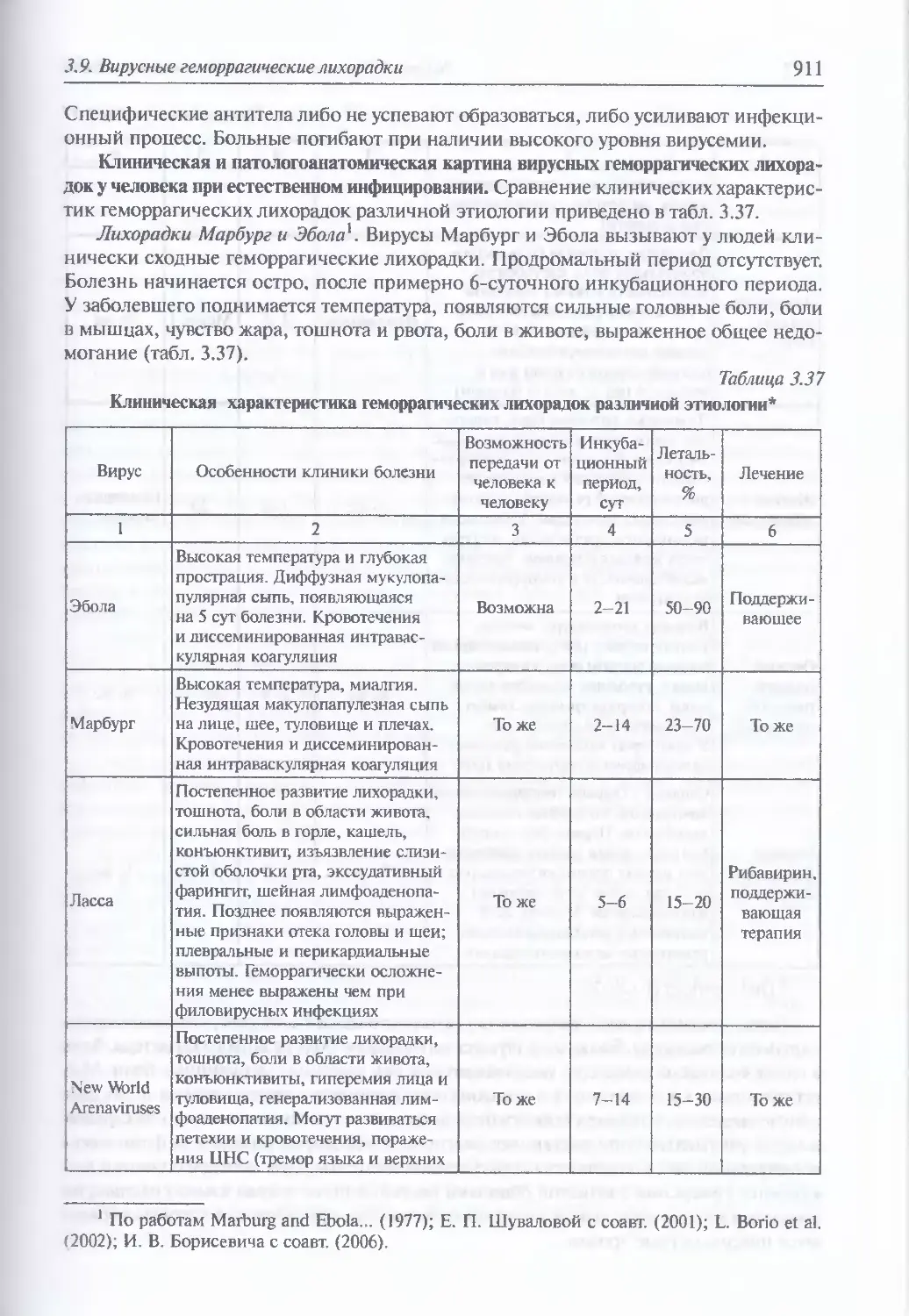
HD-Ads (helper-dependent adenovirus vectors) — хэлперзависимый аденовирусный вектор
HDF (hemodiafiltration) — гемодиафильтрация
H EK 293T (human embryonic kidney 293T cells) — эмбриональные клетки почки человеь а линии 293Т
Hfr (high frequency of recombination) — высокая частота рекомбинации
Hg (mercuric ions) — ионы ртути
HIV (human immunodeficiency virus) — вирус иммунодефицита человека
HOS (human osteosarcoma cells) — клетки остеосаркомы человека
hp (hybrid promoter) — гибридный промотор
HPLC (high performance liquid chromatography) — жидкостная хроматография с высоким разреше-
нием
HSPC (hematopoietic stem/progenitor cell) — гематопоэтические стволовые/прародительские клетки
HSPGs (heparan sulfate proteoglycans) — гепаринсульфат протеогликаны
HUS (hemolytic uremic syndrome) — гемолитический уремический синдром
HUVEC (human umbilical vein endothelial cells) — эн ютелиальные клетки пупочной вены
человека
ICAM-1 (intercellulai adhesion molecule 1) — молекулы межклеточной адгезии-1
ICEs (integrative conjugative elements, conjugation transposons) — интег ративные коныогативные
элементы, конъюгативные транспозоны
IDLV (integration-deficient lentiviral vector) — интеграционно-дефицитнй лентивирусный вектор
IL2RG (interleukin-2 receptor subunit y-gene) — ген у-субъединицы рецептора интерлейкина-2
IL- 4 (interleukin 4) — интерлейкин 4
IN (viral integrase) — вирусная интегра ia
IN(n) (integrase multimer) — интеграза-мультимер
lnt\ (class 1 integrase gene) — ген интегразы первого типа
iPS (induced pluripotent stem cell) — индуцируемые плюрипотентные стволовые клетки
20
Список сокращений
IR (inverted repeat) — инвертированный повтор
IRES (internal ribosome entry sequence) — внутренний сайт связывания рибосомы
ISG (interferon stimulated genes) — ген, стимулируемый интерфероном
ISRE (interferon stimulated response element) — стимулируемый интерфероном промоторный
элемент
JDV (Jembrana disease virus) — вирус болезни Джембрана (лентивирус. вызывающий иммунодефицит
у крупного рогатого скота)
JSRV (Jaagsiekte sheep retrovirus) — ретровирус овей Jaagsiekte
К (kalium) — калий крови
Km (kanamycin) — канамицин
KRAB (Kriippel-associated box) — белковый домен KRAB (Kruppel)
LC-MS (liquid chromatography-mass spectrometry) — жидкостная хроматография-масс-спектромет-
рия
LCMV (lymphocytic choriomeningitis virus) — вирус лимфоцитарного хорименингита
LDH (lactate dehydrogenase) — лактатдегидрогеназа
LEDGF/p75 (lens epithelium-derived growth factor) — фактор роста эпителия хрусталика глаза
LF (lethal factor) — летальный фактор сибиреязвенного токсина
LLC (Limited liability company) — в русском языке соответствует понятию «Общество с ограничен-
ной ответственностью» (ООО)
LOX-1 (lectin-like oxidized LDL receptor) — лектинподобный окисленный LDL-рецептор
LPH1 (latrophilin 1) — лактофилин 1
LT (heat-labile toxin) — температуро-лабильный токсин
LTR (viral long terminal repeat) — вирусный длинный терминальный повтор
LVS (live vaccine strain) — живой вакцинный штамм — так в США называют советскую туляремий-
ную живую вакцину на основе штамма 15 НИИЭГ, переданную им в 1950-х гг.
МА (matrix protein) — матриксный белок
МАРКК (mitogen-activated protein kinase kinase) — митогенактивированная протеинкиназакиназа
МСР (membrane cofactor protein) — белок мембранного кофактора (см. CD46)
MD (Maryland) — Мериленд
МЕЕ (multiple-locus enzyme electrophoresis) — многолокусный ферментативный электрофорез
mEGF (murine epidermal growth factor) — мышиный эпидермальный фактор роста
MglAB (macrophage growth locus) — локус роста в макрофагах (для Francisella)
МНС II (major histocompatibility complex class II) — главный комплекс гистосовместимости
второго класса
migR (macrophage intracellular growth regulator) — макрофаговый внутриклеточный регулятор
роста (для Francisella)
MLD (median lethal dose) — средняя летальная доза (то же что и LD50)
MLEE (multilocus enzyme electrophoresis) — мультилокусный ферментативный электрофорез
MLST (multiple-locus sequence typing) — многолокусное типирование последовательностей
MLV (Moloney Murine leukemia virus) — вирус мышиной лейкемии Молони
MLVA (multiple-locus variable-number tandem repeat analysis) — анализ многолокусных вариабель-
ных тандемных повторов
MMAD (mass median aerodynamic diameter) — основной средний аэродинамический диаметр
MSCs (mesenchymal stem cells) — мезенхимальные стволовые клетки
MTD (maximum tolerated dose) — максимальная переносимая доза
MW (Maedi-Visna virus) — вирус Висна-Маэди
MWCNT (multiwall carbon nanotube) — мультистеночная углеродная нанотрубка
nAChRs (nicotinic acetylcholine receptors) — никотиновые ацетилхолиновые рецепторы
NBC (National Broadcasting Company) — Национальная широковещательная компания
NC (nucleocapsid protein) — нуклеокапсидный белок
NCLDVs (nucleocytoplasmic large DNAviruses) — нуклеоцитоплазматические большие ДНК-вирусы.
В эту группу входят следующие семейства: Ascoviridae, Asfarviridae, Iridoviridae, Mimiviridae,
Phycodnaviridae, Poxviridae, Marseillevirus
NGF (nerve growth factor) — фактора роста нервов
NJ (New Jersey) — Нью-Джерси
NK (natural killer cells) — естественные киллеры
Список сокращений
21
NLS (nuclear localization sequence) — локализованная в ядре последовательность
NPC (nuclear pore complex) — поровый комплекс ядра клетки
Nrampl (natural resistance-associated macrophage protein gene 1) — первый ген макрофагального
белка, асссциированного с естественной резистентностью
nsP (nonstructural protein) — Неструктурный белок
NY (New York City) — Нью- Йорк
OAS (phenomenon of original antigenic sin) — феномен первичного антигенного греха. Другое
название — феномен антигенного импринтинга
ORF (open reading frame) — открытая рамка считывания
ORMOSIL (amino-terminated organically modified silica) — кремний, модифицированный путем
добавления аминогрупп
ОТС (ornithine transcarbamylase) — орнитинтранскарбамилаза
Р (internal promoter for transgene expression) — внутренний промотор для трансгенной экспрессии
РА (Pennsylvania) — Пенсильвания
рА (polyadenylation signal) - сигнал полиаденилации
РА (protective antigen) — протективный антиген сибиреязвенного токсина
PBS (primer binding site) — праймерсвязывающий сайт
PC (antibioticassociated pseudomembranous colitis) — антибиотикоассоциированные псевдомембра-
нозные колиты
РСЕ (post-translational control element) — посттрансляционный контролирующий элемент
РЕ (oseudomonas exotoxin) — экзотоксин псевдомонад
РЕСАМ-1 (platelet/endothelial cell adhesion molecule-1) — молекула-1 клеточной адгезии тромбоци-
тов/эндотелия
PEI (polyethylene imine) — полиэтиленимин
Per os — через рот (о приеме лекарства)
PFGE (pulsed-field gel electrophoresls) — пульс-электрофорез
PIC (viral preintegration complex) — вирусный преинтеграционный комплекс
PILs (PEGylated immunoliposomes) — ПЭГилированные иммунолипосомы
PKC (protein kinase С) — протеинкиназа C
PLC (phospholipase С) — фосфолипаза C
PLL [poly(L-lysine)] — поли-Е-лизин
PM (plasma membrane) — плазматическая мембрана
PMQR (plasmid-mediated quinolone resistance) — связанная с плазмидой резистентность к хинолонам
polyA (polyadenylation signal) — полиаденилационный сигнал
PPT (polypurine tract) — полипуриновый тракт РНК
PR (viral protease) — вирусная протеаза
PRE (post-transcriptional regulatory element) — посттранскрипционный регуляторный элемент
PSE (proximal sequence element) — проксимальный элемент
PSTs (paralytic shellfish toxins) — паралитические токсины моллюсков
PTSAgs (pyrogenic toxin superantigens) — пирогенные токсические суперантигены
qae (gene encoding resistance to quaternary ammonium compounds) — ген, кодирующий резистент-
ность к четвертичным аммониевым соединениям
qacEDX (truncated version of qacE) — усеченная версия гена qacE
qRT-PCR (quantitative reverse transcriptase polymerase cham reaction) — количественная обратнотранс-
криптазная полимеразная цепная реакция
RAPD (random amplified polymorphic DNA) — случайно а модифицированная
полиморфная ДНК
RCL (replication competent lentivirus) — репликационно компетентный лентивирус
RCM (rolling circle mode) — репликация «по типу катящегося кольца»
RD114 — эндогенный ретровирус кошек, не связанный с болезнью
RFLP (restriction fragment length polymorphism) — исследование полиморфизма длин рестрикцион-
ных фрагментов
RIPs (ribosome inactivatingproteins) — белки, инактивирующие рибосомы (обычно растительные яды
с димерной структурой: рицин, абрин модецин, вискумин, волькенсин)
RNAP II (RNA polymerase II) — РНК-полимераза II
RNAP III (RNA polymerase III) — РНК-полимераза III
22
Список сокращений
RRE (Rev response element) — Rev-отвечающий элемент
RRV (Ross River virus) — вирус Росс Ривер
RT (reverse transcriptase) — обратная транскриптаза
RTC (viral reverse transcription complex) — вирусный обратнотранскрипционный комплекс
rPHK (ribosomal ribonucleic acid) — рибосомная нуклеиновая кислота
SA (splice acceptor site) — сплайсингакцепторный сайт
SCID-X1 (X-linked severe combined immunodeficiency) — тяжелый комбинированный иммуноде-
фицит
SD (splice donor site) — сплайсинг-донорный сайт
SEB (staphylococcal enterotoxin В) — стафилококковый энтеротоксин В (третья буква обозначает
серотип токсина: SEA, SED и др.)
SeV (Sendai virus) — Сендай вирус
SF (scarlet fever) — скарлатина
SFV (Semliki Forest virus) — вирус леса Семлики
shRNA (small hairpin RNA) — маленькая шпилечная РНК
SIN (self-inactivating) — самоинакгивирующийся (обычно имеется в виду вектор)
siPHK (small/short interfering РНК) — короткие интерферирующие РНК
SIPRI (Stockholm International Peace Research Institute) — Стокгольмский институт исследования
проблем мира
siRNA (small interfering RNA) — малая интерферирующая РНК
SIV (simian immunodeficiency virus) — вирус иммунодефицита обезьян
Sm (streptomycin) — стрептомицин
SNAP-25 (synaptosomalassociated protein) — синаптосомал-ассоциированный белок
SNARE (sensitive factor attachment protein receptor) — чувствительный фактор присоединения
белкового рецептора
SNPs (single-nucleotide polymorphisms) — отдельные нуклеотидные полиморфизмы
SPG — сиаловые протеогликаны
SPICE (smallpox inhibdoi of complement enzymes) — ингибитор ферментов комплемента BHO
SRT (польск. Samodzielny Referat Techniczny) — Отдельное техническое управление
SSS (scalded skin syndrome) — синдром шелушения кожи
ST (heat-stable toxin) — температурно-стабильный токсин
ST (sequence type) — сиквиенс-тип
STAf (signal transducers and activators oftranscription) — сигнальные трансдуцеры и активаторы транс-
крипции
STX (saxitoxin) — сакситоксин
Su (sulphonamide) — сульфонамид
sull (gene conferring resistance to sulphonamides) — ген резистентности к сульфонами гам
SVMPs (snake venom metalloproteinases) — металопротеаз) i змеиных ядов
SWCNT (single-wall carbon nanotube) — одностеночная углеродная нанотрубка
TAR (transactivation response element) — элемент, отвечающий на трансактивационный сигна t
Tb (tobramycin) — тобрамицин
TBLB (transbronchial lung biopsy) — трахобронхиальная легочная биопсия
Tc (tetracycline) — тетрациклин
TCR (T-cell receptor) — Т-клеточный рецептор
tetR (Tet repressor) — репрессор гена резистентности к тетрациклинj
TER (Toll-like receptor) — Toll-подобный рецептор
ТМ (thrombomodulin) — тромбомодулин
Tm (trimethoprim) — триметоприм
TMP-SMX (trimetoprim-sulfametoxazol) — триметоприм-сульфаметоксазол
TPGY (tryptone-peptone-glucose-yeast extract medium) — среда на основе
триптоно-пептон-глюкозо-дрожжевого экстракта
TSS (toxic-shock syndrome) — синдром токсического шока
TSST (toxic shock syndrome toxin) — токсин синдрома токсического шока
tTA (Tet-controlled transactivator) — контролируемый тетрациклином транскрипционный транс-
активатор
Список сокращений
23
tTS (Tet-controlled transcriptional repressor) — контролируемый тетрациклином транскрипционный
репрессор
ТТХ (tetrodotoxin) — тетродотоксин
TU/ml (transducing units per ml) — количество транскрипционных единиц в мл
U3 (LTR У unique element) — LTR З'-уникальный элемент
U5 (LTR 5' unique element) — LTR 5'-уникальный элемент
USAMRI1D (U.S. Army Medical Research Institute of Infectious Diseases) — Медицинский исследо-
вательский институт инфекционных болезней армии США
USDA (U.S. Department of Agriculture’s) — Министерство сельского хозяйства США
USPS (United States Postal Service) — Почтовая служба США
UTI (urinary tract infection) — инфекция уринарного тракта
UTR (untranslated region) — нетранслируемый регион
VA (Virginia) — Виргиния
VACV (Vaccinia virus) — вирус вакцины
VAMP (vesicle-associated membrane protein) — везикуло-ассоциированный мембранный белок
VARV (Vanola virus) - вирус натуральной оспы
VATS (video-assisted thoracic surgery) — оперативное вмешательство на грудной клетке под видео-
наблюдением
VEGF-F (Vascular endothelial growth factor) — васкулярный эндотелиальный фактор роста
VGSCs (voltaged-gated sodium channels) — потенциалзависимые натриевые каналы
VLS (vascular leak syndrome, Clarkson syndrome) — синдром пропускания сосудов
VNTR (variable-number tandem repeat) — варьирующие по числу тандемные повторы
VSV-G (Vesicular stomatitis virus GP) — гликопротеин вируса везикулярного стоматита
W (Visna virus) — вирус висны
w/v (per cent of weight in volume) — процент веса вещества в объеме раствора
WGST (whole genome sequencing and typing) — полногеномное секвенирование
и типирование
WHV (woodchuck hepatitis virus) — вирус гепатита североамериканского лесного сурка
WPRE (woodchuck hepatitis virus post-transcriptional regulatory element) — постранскрипционный
регуляторный элемент вируса гепатита североамериканского лесного сурка
X-SC1D (X-linked severe combined immunodeficiency) — тяжелый комбинированный
иммунодефицит, связанный с Х-хромосомой
у (viral RNA packaging signal) — сигнал для упаковки РНК-вируса
ЧАСТЬ 1
КРАТКАЯ ИСТОРИЯ ПРИМЕНЕНИЯ БИОЛОГИЧЕСКИХ
СРЕДСТВ ДЛЯ ПОРАЖЕНИЯ ЛЮДЕЙ
Убийства людей с применением средств, существующих в живой природе, нача-
лись задолго до открытия микроорганизмов и получения высокоочищенных токсинов.
Понимание их истории дает нам возможность осознать причины, побудившие разви-
тие БО и способов ведения биологической войны на протяжении почти всего прошло-
го столетия и, возможно, уловить их основные тенденции в ближайшем будущем.
1.1. РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ О ПРИРОДЕ БИОЛОГИЧЕСКИХ ЯДОВ
Античные мифы. Ядовитые грибы. Ведение войны. Змеи.
Умопомешательства. Средние века. Поиски ядов.
Случайное использование в пищу ядовитых растений и грибов, укусы ядовитых
животных и насекомых, с незапамятных времен очертили круг токсических биологи-
ческих субстанций.
Античные мифы. При их анализе обращает на себя внимание следующее. Почти во
всех мифах речь идет об использовании для убийств людей и богов реально существую-
щих зоотоксинов. В зависимости от имеющихся возможностей для их введения жертве
(в мифах всегда излагаются реально осуществимые способы), античные боги исполь-
зуют токсины, активные при парэнтеральном введении, либо обладающие кожно-
резорбтивным действием. Источником яда в большинстве мифов обычно называется
убитая Гераклом Лернейская гидра (второй подвиг героя) — гигантская змея, обитав-
шая в болоте и пожиравшая окрестные стада.
Стрелой, отравленной ядом Лернейской гидры, Геракл убил стоглавого дракона
Ладона, охранявшего золотые яблоки в саду гесперид — дочерей Атланта, живших в
сказочном саду, где росла яблоня, приносившая золотые плоды. Бессмертный кентавр
Хирон, случайно раненный отравленной стрелой Геракла, страдая от мук, жаждал
смерти и отказался от своего бессмертия в пользу Прометея. Гостеприимно приняв-
ший Геракла у себя в пещере кентавр Фолос погиб по недоразумению. Он уронил себе
на ногу стрелу Геракла, пропитанную ядом Лернейской гидры, и умер.
В конце концов, и сам Геракл погиб от этого яда. А дело было так. Кентавр Несс,
уцелевший после победы Геракла над кентаврами, нашел себе спокойную работу ло-
дочника и переправлял за плату путников через реку Эвен. Переправляя жену Герак-
ла Деяниру, он не удержался в дозволенных лодочнику рамках и посягнул на ее честь.
Геракл с большого расстояния ранил его стрелой, отравленной ядом Лернейской гид-
ры. Перед смертью коварный Несс, бывший, благодаря характеру своей деятельно-
сти, в курсе всех слухов и сплетен, ходивших по Греции, посоветовал Деянире собрать
его кровь, которая якобы поможет ей чудодейственно сохранить любовь Геракла. Дея-
нира так и поступила. Боясь, что Геракл покинет ее ради своей любовницы Иолы —
1.1. Развитие представлений о природе биологических ядов
25
дочери убитого Гераклом эхалийского царя Эврита, она прислала ему хитон (нижняя
одежда из льняной ткани, имевшая вид сорочки), пропитанный отравленной кровью
Несса. Надетый хитон тут же прирос к телу Геракла. Яд стал проникать сквозь кожу,
причиняя ему нестерпимую боль. После чего Геракл разжег костер на горе Этна и взо-
шел на него, пламя вознесло его на Олимп.
Однако что же могла представлять собой Лернейская гидпа? В мифах она описы-
вается как змея, но на древних изображениях схватки Геракла с Лернейской гидрой
она мало чем отличается от известных нам пресноводных гидр, представителей ки-
шечнополостных животных. Их характерной особенностью является наличие стре-
кальных клеток (нематоцист), вырабатывающих ядовитый секрет и служащих для
умерщвления добычи и защиты от врагов. Эти секреты вызывают жгучую боль в мес-
те введения и обладают летальной, гемолитической и дермонекротической актив-
ностью. Стрекальные клетки легко можно очистить вместе с токсинами. По данным
Б. Н. Орлова и Д. Б. Гелашвили (1985), в результате аутолиза 3,8 л щупалец физалии
можно получить до килограмма конечного продукта (примерно 55 млн заряженных
нематоцист) (рис. 1.1).
Рис. 1.1. Геракл побеждает Лернейскую гидру. А. Древнегреческое изображение
противоборства Геракла с гидрой. Б. Современное изображение гидр 1 — почкую-
щаяся; 2 — с яйцами. Для получения яда греки использовали стрекательные
клетки кишечнополостных организмов
Тем же способом, что Несс отравил Геракла, Медея (персонаж одного из древней-
ших греческих сказаний — об аргонавтах, жена Ясона) умертвила Главку, дочь корин-
фского царя Креонта, когда Ясон (фессалийский герой, стоявший во главе экспеди-
ции аргонавтов) захотел на ней жениться. Медея подарила сопернице пропитанный
ядом свадебный пеплос (длинная достигавшая земли одежда, надевавшаяся прямо на
тело и оставлявшая один бок открытым), который ее «испепелил», т. е. вызвал обшир-
ные поражения кожи, от которых Главка умерла.
Яд, использованный Медеей, не имеет отношения к Лернейской гидре. Его про-
исхождение уходит в легенду о Цербере, трехголовом псе, охранявшем царство мерт-
вых. Из ядовитой пены лающего Цербера появился цветок аконит (ядовитое расте-
ние, содержащее в клубнях и корнях высокотоксичный алкалоид аконитин}, который
Медея заварила в свое колдовское зелье, убившее Главку. Аконитин действительно
обладает кожно-резорбтивным действием Соприкосновение с корнями аконита вы-
зывает мучительные ожоги на коже. При приеме с пищей аконитин еще опасней —
26
Часть 1. Краткая история применения биологических средств...
его смертельная доза для человека не более 2-5 мг/кг. При инъекционном введении
(например, смазанный ядом наконечник стрелы) смертельная доза токсина раз в
10 меньше, чем при приеме с пищей. Причиной смерти при отравлении аконитином
обычно бывает остановка сердца. Среди отравителей древности его популярность не
знала границ. Из корней аконита на Древнем Востоке получали яд бик. В Индии его
считали самым сильным ядом из имевшихся и использовали для смазывания нако-
нечников стрел.
В легенде о Цербере также нашли свое отражение как древняя практика зараже-
ния людей бешенством с помощью слюны бешеной собаки, так и видимые послед-
ствия применения еще каких-то сложных ядов, обладающих кожно-резорбтивным
действием и способных вызывать тяжелые поражения кожи, например, содержащих
кантаридины и высокотоксичные алкалоиды.
Вот любопытное описание приготовления яда для наконечников стрел, подготов-
ленное греческим писателем Элианом, жившем в Риме при императорах Траяне и
Адриане (около 98—138 гг.): «Скифы носят с собой самок гадюк, иногда совсем малень-
ких, и каждые несколько дней умерщвляют часть из них. Когда они достаточно пере-
гниют, скифы наливают человеческую кровь в небольшой котелок и, накрыв его
крышкой, помещают в навоз. Когда кровь также перегнивает, выделившуюся на по-
верхность жидкость смешивают с полученным гноищем гадюк — и таким образом
получается смертельный яд».
Скифы эмпирически пришли к токсическим композициям, обладающим комби-
нированным действием. Они обмазывали наконечники стрел сгнившей кровью, со-
держащей трупные яды птомаины и большое количество спор возбудителей столбняка
и газовой гангрены, обычно прису гствуюших в больших количествах в лошадином
навозе (A. G. Robertson, L. D. Robertson, 1995).
Ядовитые грибы. Использование ядовитых грибов как для индивидуальных
убийств, так и для убийств массовых практикуется «заинтересованными лицами» с
древнейших времен. С их помощью было отравлено немало претендентов на престол
и самих носителей верховной власти. По дошедшим до нас документам, во времена
Римской империи в I в. н. э. была известна некая Локуста (правильнее Лукуста —
Lucusta), которая готовила смертоносный порошок из ядовитых грибов и изучала его
действие в опытах на животных (козлятах, свиньях) и в экспериментах на рабах.
К услугам Локусты неоднократно прибегала принцесса Агриппина (Випсания) — дочь
Марка Випсания Агриппы и Цецилии Аттики. Будучи женщиной властолюбивой,
вероломной и развратной, и еще жаждущей верховной власти, она задумала женить
на себе овдовевшего императора К лавдия. Для этого Агриппина избавилась от соб-
ственного мужа Пассивна Крисия, отравив его грибным ядом. Выйдя замуж за импе-
ратора, Агриппина принудила Клавдия усыновить своего сына Нерона и лишить права
наследования престола сына император? Клавдия — Британикса. Вскоре Агриппина
решилась на убийство и самого императора Клавдия Используя свое положение,
Агриппина и Локуста вместе с врачом императора Ксенофонтом в блюда со съедоб-
ными грибами добавили соус, приготовленный из ядовитых грибов. Клавдий «ел до
рвоты с тем, чтобы вновь приняться за пищу до новой рвоты и т. д.», таковы были нраъы
той эпохи. Однако даже после неоднократного приема ядовитого соуса во время пир-
шества спустя несколько часов Клавдий почувствовал лишь недомогание. Тогда по
настоянию врача Ксенофонта, якобы для улучшения самочувствия Клавдия, при по-
мощи раздражения корня языка гусиным пером был вызван рвотный рефлекс. Улу-
чив удобный момент, Ксенофонт ввел в опустевший желудок императора новую пор-
1.1. Развитие представлений о природе биологических ядов
27
цию ада, и Клавдий умер. С помощью грибного ада Агриппина устраняла окружаю-
щих Нерона лиц, с ее точки зрения, потенциально опасных и претендовавших на трон.
Гак были отравлены дети Клавдия — пятнадцатилетний Британикс и его сестра Ок-
тавия (жена Нерона); внуки императора Августа — Юниус и Маркус Силанусы. Счи-
тая потенциально опасным для Нерона капитана императорской охраны из знатного
рода Серенуса, Агриппина решается на его убийство. Серенус не только не любил
грибы, но и не употреблял их в пищу ни в каком виде. Но ничто не могло остановить
Агриппину в ее коварных замыслах. Зная, что капитан питается из общего котла со
своим отрядом, грибным адом была отравлена вся пища, предназначавшаяся для его
центурии: Серенус вместе со 100 воинами погиб мучительной смертью. Расплатой за
содеянное зло было убийство Агриппины ее собственным сыном Нероном, который
уже не доверял своей матери. «Специалист» по микотоксинам Локуста провела пос-
ледние годы жизни в заключении, охраняемая специально отобранными воинами-
микофобами. После смерти Нерона по приказу императора Гальба в 69 г. н. э. Локуста
была обезглавлена. По дошедшим до нас историческим документам, это была самая
большая в истории серия убийств, когда в качестве орудия преступления использо-
валс я ад бледной поганки, включающий алкалоиды аманитотоксины. Летальная доза
аманитотоксиновлля взрослого человека с массой тела 70 кг при пероральном введе-
нии составляет 7 мг (С. Г Мусселиус, А. А. Рык, 2002).
Ведение лойны. Применение адов на войне уже с античных времен у некоторых
цивилизаций имело правовую регламентацию. Законом запрещалось отравление
пищи и использование яда в качестве оружия. Греки и римляне осуждали примене-
ния ядов в войне как нарушение «ius gentium» — закона наций. Яды и другие виды
оружия были запрещены как негуманные в Индии по закону «Ману» еще в 5и0 г. н. э.,
позднее — у сарацинов. К идее запрещения применения ядов в войне вернулся изве-
стный государственный деятель Голландии Гуго Гротис в своем труде «Закон о войне
и мире» (1625). В 1675 г. в Страсбурге было подписано соглашение между Францией и
Германией о запрещении использования отравленных пуль при ведении боевых дей-
ствий. Историк Джон Элис Мун утверждает, что негативное отношение к яду, как
оружию, было ослаблено лишь в XVIII в. в связи с замешением моральных принци-
пов в государственной политике военной необходимостью. Государства все более скло-
нялись к использованию любых возможных способов для достижения целей войны
(L. A. Cole, 1996).
Впрочем, не следует идеализировать и древних сторонников «правильной войны».
Когда умелое использование фактора яда полководцем приводило его к победе, то
сочувствующие ему историки спешили поведать миру о необычных составляющих
его военного таланта. Например, древнегреческий писатель Павсаний (Pausanias;
11 в. н. э., автор труда «Описания Эллады» в 10 книгах) привел пример удачного мас-
штабного применения в военных целях ада, содержащегося в чемерице. Чемерица —
это растение из семейства Liliaceae; его корни содержат алкалоид веатрин, обладаю-
щий слабящим и рвотным действием. В 600 г. до н. э. войска дельфийско-пилейской
амфиктионии (греч. amphiktionia, от amphiktiones—жители окружающих областей, ре-
лигиозно-политический союз племен и городов в Древней Греции), возглавляемые
Соленом (знаменитый афинский реформатор и законодатель, 640—559 гг. до н. э.), на-
чали войну против сиргарийцев. Воды реки 11л ист протекали по каналу в город Кирру
(древний город в Фокиде — оиласть в Средней Греции). Солон дал указание отвести
поток воды в сторону от города. Сиргарийцы продолжительное время выдерживали
осаду, употребляя дождевую воду и пользуясь имевшимися в городе колодцами. В это
28
Часть 1. Краткая история применения биологических средств...
время солдаты Солона собрали большое количество корней чемерицы и бросили их в
созданное ими водохранилище. После того, как ядовитые вещества, содержащиеся в
растении, растворились в воде, Солон распорядился направить поток отравленной
воды по прежнему руслу. Сиргарийцы, долгое время испытывавшие жажду, наброси-
лись на отравленную воду. В результате у большинства воинов, защищавших город,
возникло острое желудочное расстройство. Вследствие непрекращающегося поноса,
они побросали охраняемые ими посты, и амфиктионцы легко овладели городом.
Греки не были оригинальны, используя растительные яды в военных целях. Бла-
годаря работам британского исследователя Джозефа Нидхэма (Needham J., 1956),
исследовавшего древние китайские хроники и летописи, стало известно, что в Китае
уже в начале IV до н. э. использовались в военных целях ядовитые дымы, полученные
из растений. Например, дым из горчичных и других семян, содержащих вещества,
обладающие раздражающим действием, закачивался китайцами с помощью мехов во
вражеские рвы, окружающие города. Во II в. китайцы начали использовать слезото-
чивый газ из порошковой извести. Открытый в XI в. порох китайцы научились исполь-
зовать для диспергирования различных ядов. Для этого они добавляли в пороховой
заряд токсические вещества, извлеченные из растений и тканей животных и соеди-
нения мышьяка. И после помещения такой смеси в бамбуковую трубку получалась
отравляющая бомба. Тогда же китайцы научились создавать «паралитическо-удушаю-
щие газы» и использовали их на полях сражений при помощи бумажных змеев и бам-
буковых трубок.
Более подробную информацию о древних отравляющих бомбах можно найти в
работе П. Джеймса и Н. Торпа (1997), ссылающихся на «Военную энциклопедию»,
написанную в Китае в 1044 г. полководцем Сэн Кун Ляном. В своей книге они приво-
дят описание бомбы, которую сегодня можно было бы назвать «биолого-химической».
Бомба забрасывалась на позиции противника катапультой. С помощью подрыва по-
рохового заряда предполагалось диспергирование следующего состава:
человеческие испражнения, сухие, измельченные в порошок и просеянные —
15 фунтов;
горец крючковатый — 8 унций;
аконит — 8 унций;
кротоновое масло — 8 унций;
стручки мыльного дерева (как дымообразующее средство) — 8 унций;
окись мышьяка — 8 унций;
сульфид мышьяка — 8 унций;
порошок из шпанских мушек — 8 унций;
зола — 8 унций;
тунговое масло — 8 унций.
Такие бомбы широко применялись при ведении боевых действий, поэтому их
поражающее действие было хорошо известно китайским полководцам. Сэн Кун Лян,
рекомендовавший своим ученикам использовать их при штурме городов, отмечал
способность отравляющего состава проникать сквозь щели в латах и вызывать силь-
ное раздражение кожи, а при непосредственном попадании на нее — волдыри, что
соответствует токсическим свойствам описанных им компонентов. Эта композиция
обладала еще и общеядовитым действием.
Европейцы столкнулись с массированным применением отравляющих веществ в
ходе боевых действий не под Ипром в 1915 г., как это обычно принято считать, а го-
раздо ранее. В 1241 г. татары, направляясь от Киева в Венгрию, опустошили Малую
1.1. Развитие представлений о природе биологических ядов
29
Польшу, взяли Сандомир, разбили польское рыцарство под Хмельником, разграбили
Краков, Вроцлав, а 9 апреля под Лигницем разгромили войска Генриха Благочести-
вого. Профессор Юзеф Влодарский, историк из Гданьского университета, обратил
внимание на фрагмент хроники историка Яна Длугоша (Dlugosz, 1415-1480; автор
фундаментального исследования «Histoiia Polonica»), имеющего отношение к этому
сражению: «И была там в их войске среди других хоругвей одна очень большой вели-
чины. На вершине ее древка висела глава зело уродливая и чудовищная с бородой, и
когда татары единой стаей показали тыл и собрались отступать, знаменосец начал
главой этой махать изо всех сил, и в тот же миг с нее повалил густой дым, причем та-
кой смрадный, что когда среди войска разошлась эта убийственная вонь, поляки со-
млели и еле живые стояли и не способными стали для битвы». Влодарский считает,
что ивкусство применения отравляющих веществ татарами было позаимствовано от
китайцев, а сама химическая атака под Лигницем была произведена захваченными в
Китае специалистами по боевым отравляющим веществам того времени.
Змеи. Убийства с помощью насылания ядовитых змей практиковала еще Артеми-
да — богиня плодородия, покровительница охоты и растительности, защитница це-
ломудрия и, разумеется, «ветеран» Троянской войны (со стороны троянцев). Первая,
ставшая известной, попытка применить ядовитых змей в реальных боевых действиях
была предпринята в 334 г. до н. э. греками во время войны Александра Македонского
с Персией. Осадив город Галикарнасе, греки стреляли из катапульты бочонками с
ядовитыми змеями, которые разбивались при падении на территории города, и рас-
ползающиеся пресмыкающиеся деморализовали его защитников. Одновременно
осаждающие предприняли попытки вызвать в городе эпидемию (см. ниже).
В 190 г. до н. э., в ходе подготовки морского боя против флота правителя TlepiaM-
ского царства Эвмена, Ганнибал (247—182 гг. до н. э.) приказал собрать в глиняные
горшки ядовитых змей. В самый разгар битвы с кораблями этого римского союзни-
ка, воины Ганибалла забросали ими вражеские корабли. Пергамцы сначала не обра-
тили на них никакого внимания, однако, когда на кораблях появились ядовитые змеи,
они не выдержали и сдались.
Умопомешательства. Массовое применение ядов и умышленного заражения для
уничтожения людей имело место во время масштабных коллективных умопомеша-
тельств. Историк Э. Литтре (Paul-Maximihen-Emile Littre, 1801—1881) упоминает о
странном коллективном безумии, сведения о котором обнаружены им в работе Эпи-
томия Ксифилина. Этот ученый составил по поручению византийского императора
Михаила Дуки (1071—1078) извлечение из римской истории Диона Кассия. Ксифи-
лин ссылался на утраченную книгу Кассия, где тот писал следующее: «В правление
римского императора Коммода (180—192) появилась самая сильная изо всех мне изве-
стных болезней, и часто умирали в Риме по две тысячи людей в день. К тому же мно-
гие, не только в столице, но и во всей империи, пали жертвами преступных деяний;
ибо злоумышленники натирали иголки ядом и таким путем распространяли заразу,
делая это за деньги, что случалось также в правление Домициана (81—96). Некоторые
люди стали колоть ядовитыми иголками кого попало, и много умирало от этих уко-
лов, даже не чувствуя их; но, с другой стороны, многие из виновных были уличены и
наказаны. Это преступление совершалось не в одном Риме, а, можно сказать, во всей
вселенной» (Литтре Э., 1873).
Средние века. В эту эпоху отравители в общественном сознании прочно ассоции
ровались с колдунами. Считалось, что ведьмы и колдуны, пройдя школу у самого дья
вола или же руководствуясь собственными сверхъестественными талантами (каково
30
Часть 1. Краткая история применения биологических средств...
бы ни было их происхождение), умели смешивать разнообразные отравы или же обла-
дали способностями наделять вполне безобидные вещества такими свойствами, ко-
торые позволяли уничтожать не только отдельных людей, но и целые общины. Иногда
они выливали свое зелье: в колодцы, иногда рассеивали по воздуху, а то и просто маза-
ли им стены и дверные ручки. Уже тот факт, что слово «отравительница», означало то
же самое, что и «ведьма», говорит сам за себя (Киттеридж Дж. Л., 2005) (рис. 1.2).
Дж. Л. Киттеридж (2005), подробнейшим образом исследовавший материалы су-
дебных процессов по знаменитым колдунам-отравителям, поишел к выводу, что они
меньше всего напоминали «невинные жертвы инквизиции». Многие из людей, кото-
рые были казнены за ведовство, действительно имели преступные намерения. Прак-
тически каждый из тех, кто был казнен по этой причине, верил в существование по-
добного преступления — вне зависимости от того, считал он себя виновным или нет.
Поиски ядов Английский король Иоанн Безземельный (John Lackland, 1167—1216)
был отравлен жабьим ядом (его действие сходно с таковым у сердечных гликозидов —
см. разд. 2.2.3.5) монахом, который подмешал яд в заздравную чашу. Если отвлечься
от чисто технической стороны преступления, а поискать в нем мистическую состав-
ляющую, хорошо понятную современникам 25-го английского короля, чье правление
считается одним из самых катастрофических за всю историю Англии, то это убийство
Рис 1.2. Ведьмы за колдовским занятием
(из книги Ульриха Молитора «De Lamiis»,
1489). Древнее искусство приготовления
биологических ядов всегда представляло
собой тайное знание посвященных
выглядит как совершенное с «особой жес-
токостью». Жаба, согласно предрассудкам,
распространенным у средневековых евро-
пейцев, символически и телесно представ-
ляла самого дьявола. Убить с помошью ее
яда короля, означало еше и сознательно
передать его душу дьяволу.
Однако случаи использования одного
какого-то яда в практике отравлений сред-
невековой Европы были редки. Раститель-
ные и животные яды, как правило, отра-
вители смешивали между собой и/или с
минеральными ядами (соли мышьяка и
ртути, мышьяковистая кислота) по пропи-
сям, известным немногим посвященным.
В конце XVII в. на юге Италии в арсенале
отравителей появились весьма ядовитые
токсические композиции, не имевшие за-
паха и цвета, собирательно называемые
аква тофана (или тоффана; также акветга
ди Наполи; ди Перуджия или делде Тнф-
фа). Изобретате гьницей этого ядовитого
напитка считается некая сицилианка То-
щана, жившая прежде в Палермо, но затем,
когда обратила на себя внимание властей,
бежавшая в Неаполь. Она продавала его
женщинам, желавшим ускорить смерть
своих мужей. Яд содержался в особых бу-
тылочках с портретом святого Николая.
Описывается современниками как жид-
1.1. Развитие представлений о природе биологических ядов
31
кость, имеющая вид воды, без запаха, вкуса и цвета. Относительно его химического
состава известно только то, что одна из композиций включала разбавленную в дис-
тиллированной воде мышьяковистую кислоту (смертельная доза 0,1—0,3 г) и канта-
ридины (см. разд. 2.2.3.5); другую получали из экстрактов ядовитых растений, воз-
можно, с добавлением все той же мышьяковистой кислоты. Горелли, лейб-медик
Карла VI (1711—1740), утверждал, что он слышал из уст самого императора, которому
были предложены на рассмотрение бумаги, касающиеся процесса лзоиретательни-
цы тофаны, что этот яд представляет собой не что иное, как водный раствор мышья-
ковой кислоты с прибавлением растения Herba cymbalariae (?). Действие «аква тофа-
ны» мало напоминало острое отравление мышьяком. Оно проявлялось, прежде всего,
поражением легочной ткани. У человека, отравленного большим количеством яда,
развивалось острое воспаление легких, и его смерть выглядела как смерть больного
пневмонией. Последнее обстоятельство говорит в пользу того, что в составе «аква
тофаны» находились соли таллия. Хотя этот металл выделен в чистом виде Вильямом
Круксом (Crookes, 1832—1919) только в 1861 г., его соли могли быть известны соста-
вителям ядов как ядовитые компоненты минералов, например, различных колчеда-
нов или некоторых слюд. Редко встречающийся колчедан крукезит, так названный в
честь Крукса (crookesite, Crookes), содержит до 17 % таллия. Мышьяк вте годы также
получали из колчеданов. Содержание мышьяка в мышьяковистом колчедане не бо-
лее 1 %, но отравителям его хватало.
Хронические отравления «аквой тофаной» установить было невозможно. При
ежедневном добавлении 6—7 капель яда в пищу, у жертвы возникали сначала общая
слабость и потеря аппетита, затем человек делался вялым, впадал в депрессию и втече-
ние нескольких месяцев умирал на фоне общей и необъяснимой слабости. Практико-
валось отравление и сверхмалыми дозами. Известный писатель Стендаль (1793— 1842)
утверждал, что капля «аква тофаны» в неделю приводила к смерти через два года («Про-
гулки по Риму», 1828). Как пример отравления этим ядом, им приводится смерть Папы
Климента XIV (Antonio Ganganelli, 1769—1774). Хотя этот яд оказался весьма востре-
бованным для решения многих жизненно важных проблем (старые или жадные му-
жья, наследство, политические противники и др.), к концу XVIII в. секрет его приго-
товления был утрачен. Сама Тофана в 1709 г. была поймана, подвержена пытке и
задушена в тюрьме.
Особое место на этой кухне убийц в продолжение нескольких тысяч лет занима-
ли трупные яды — птомаины (от греч. ptoma — труп). Они представляют собой азоти-
стые продукты распада тканей трупов, образовавшиеся под влиянием действия на них
различных микроорганизмов. В XX в. было выяснено, что представление о птомаи-
нах, как о действующих началах трупного яда, ошибочно, так как токсичность боль-
шинства из них невелика. Ядовитость же продуктов гниения белков обусловлена на-
личием в них, помимо птомаинов, сильных бактериальных токсинов. Наиболее
опасными из птомаинов считаются кадаверин и путресцин; в месте введения оба со-
единения вызывают сильное воспаление и омертвение тканей.
Для получения трупных ядов отдельные органы животных оставляли гнить, пос-
ле чего они приобретали ядовитые свойства. Приготовление птомаинов имело свою
мистическую составляющую. Популярностью пользовались трупные яды, получен-
ные из легкого лягушки, крови (особенно человеческой и бычьей) или частей ядо-
витой змеи. По мнению Р. Вильнева (1998), Чезаре Борджия (1474-1506) имел рецепт
сложного яда средневековых отравителей, включающий птомаины и мышьяковис-
тую кислоту. Борджия несколько «усовершенствовал» его приготовление. Он заме-
32
Часть I. Краткая история применения биологических средств...
нил легкое жабы на внутренности свиньи, перемешанные с мышьяковистой кисло-
той. Для получения трупного яда свинью подвешивали за задние ноги и забивали
палками до смерти. Перегнившая масса в высушенном или жидком состоянии со-
ставляла основной элемент его смертоносной кантареллы. Отравителями очень це-
нился «трупный яд», получаемый из отравленных мышьяком животных.
Древние описания отравлений, даже тогда, когда в источниках указывается про-
исхождение яда, содержат в себе немало клинических деталей, которые нельзя объяс-
нить на основании современных представлений о клинике отравления этим же ядом.
Древнегреческий философ Сократ (469—399 до н. э.), обвиненный афинянами в
безбожии и введении «новых демонов», должен был выпить чашу яда, полученного
из растения Cicuta virosa. Платон (430—347 до н. э.) так описал последние мгновения
из жизни своего учителя: «...сначала Сократ мог ходить, но потом, сказав, что ноги его
отяжелели, он лег на спину — так приказал человек, давший ему яд. Палач вниматель-
но осмотрел его руки и ноги, затем, сильно сжав ступню, спросил, чувствует ли он что-
нибудь. Сократ ответил, что не чувствует. Потом палач стал сжимать его ноги, подни-
маясь все выше, таким образом, показал нам, что тело остывает и деревенеет. ...Через
некоторое время тело его содрогнулось в конвульсиях, палач полностью раскрыл его.
Его взгляд был неподвижным. Увидев это, Критий закрыл Сократу рог и глаза» (Фе-
дон, LXV и LXVI).
В современных описаниях случайных отравлений людей корнями этого же рас-
тения, первые симптомы проявляются рвотой, головокружением, галлюцинациями,
сильными клоническими судорогами, затем наст> пает потеря сознания, остановка
дыхания и смерть. Сократ же не испытывал никаких физических страданий, в кли-
нике отравления отсутствовал и весьма характерный для Cicuta virosa судорожный
синдром, что позволяет утверждать о наличии в данном ему яде еще каких-то сильно
действующих наркотических веществ и миорелаксантов.
До сегодняшнего лня поддерживаются древние традиции использования дурма-
на в тайных культах и для совершения преступлений. В средневековой Европе так на-
зываемые ведьмы изготовляли свои «волшебные мази», добавляя в них сок и толченые
части белладонны и дурмана. Втирая эти мази в свое тело, колдуньи подвергались нар-
котическому воздействию, во время действия которого некоторые из них ощущали
чувство полета, а другие — любовного экстаза с самим Сатаной во время шабашей.
Профессиональные отравители в Индии обычно используют семена дурмана Datura
alba. Их всегда много в созревших плодах растения. Так как эти семена горькие, то их
добавляют в острые блюда или блюда с богатым вкусом.
* * *
С глубокой древности у различных групп населения накапливались тайные зна-
ния в области токсикологии, передаваемые от поколения к поколению «посвящен-
ных», сегодня маскируемые обрядами нетрадиционной медицины, колдовства и
магии. Колдунами, целителями и отравителями для убийства людей использовались
(и используются) в разных сочетаниях следующие «природные средства»: ядовитые ра-
стения (дурман, белена, белладонна, мандрагора, болиголов, волчий корень или ако-
нит, купальница, опиум, маковый сок, ядовитые грибы, наперстянка, безвременник,
чемерица, ломонос, молочай, паслен, ветреница, рута); яды животного происхожде-
ния (выделения златки, порошок шпанской мушки, слизь старой жабы, трупные яды,
слюна бешеной собаки; яды змей, кишечнополостных животных, саламандры, три-
тона, электрического ската); яды минерального происхождения (различные соединения
1.2. Развитие представлений о природе эпидемий
33
мышьяка, ртути, таллия и их сочетания между собой и с органическими и биологи-
ческими ядами). Одновременно с накоплением знаний по токсическим свойствам
различных веществ природного происхождения формировались традиции подбора
композиций ядов, имеющих определенное предназначение (использование в военных
целях, массовые убийства, ритуальная казнь, ликвидация под видом естественной
смерти и др.).
1.2. РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ О ПРИРОДЕ ЭПИДЕМИЙ
Античные мифы. Появление миазматического учения. Колдуны, ведьмы,
демоны и наказание Божье. Антисемитские предрассудки. Появление
контагионистического учения. Локалисты. «Овеществленный» контагий.
Все бедствия, сопровождающиеся массовой гибелью людей, на уровне бытового
сознания у самых разных народов с глубокой древности считались искусственными.
«Мор», «язву» и «чуму» они считали либо карой, ниспосылаемой разгневанными бо-
гами за провинности правителей, народа и несоблюдение обрядов, либо проявлени-
ем действия злых сил, результатом колдовства или чьих-то волшебных чар.
Античные мифы. В античной мифологии способностью вызывать «чуму» облада-
ют главные олимпийские боги. Как правило, их «биотеррористическая деятельность»
ограничивалась отдельными местностями, видимо, бывшими в то время эндемичны-
ми по чуме. Например, в первой главе «Илиады» (ХП—VII в. до н. э.) читаем о собы-
тиях, произошедших вблизи Трои1 следующее: «Сын громовержца и Леты — Феб2,
царем прогневленный, язву на воинство злую навел. ...Пусть нам поведают, чем раз-
дражен Аполлон небожитель? Он за обет несвершенный, за жертву стотельчую гне-
вен? Или от агнцев и избранных коз благовонного тука требует бог, чтоб ахеян изба-
вить от пагубной язвы?»
Суть дела состояла в том, что афинянин Агамемнон похитил дочь одного священ-
ника и не хотел ее возвращать отцу. Девушка пожаловалась одному из богов, Аполло-
ну, и тот спустился с Олимпа и 9 дней метал в афинян зараженные чумой стрелы, пока
те поняли, что от них требуется.
Богиня мудрости и справедливой войны, Афина, прибегала к «биотеррору» уже не-
посредственно в самой Греции. Когда ее жрица Ауга, дочь аркадского царя Алея, роди-
ла от Геракла сына Телефа, она была вынуждена спрятать ребенка в храме Афины из-
за страха перед отцом, ожидавшего неприятности от рождения внука. Богиня, соб-
ственноручно содравшая во время войны с гигантами кожу с поверженного Палланта,
а затем одевшая ее на себя (отсюда и ее прозвище — «Паллада»), разгневалась на Алея
и без колебаний наказала всю Грецию чумой. Тогда царь, чтобы спасти страну от мора,
велел выбросить младенца на дорогу, где его подобрала лань Артемиды.
Бог плодоносящих сил и виноградарства Дионис покарал расположенную рядом
с Афинами Аттику чумой за убийство пьяными пастухами афинянина Икария, кото-
рому он покровительствовал.
Бог морей Посейдон, не удовлетворившись жертвой, принесенной ему критским
царем и врагом Трои, Идоменеем, наслал на Крит моровую язву. Используя приня-
1 Древний город на северо-западе Малой Азии.
2 В древнегреческой мифологи одно из имен Аполлона.
34
Часть 1. Краткая история применения биологических средств...
тую у специалистов датировку античных мифов, можно предположить, что все при-
писываемые античным богам эпидемии чумы вспыхнули либо во время Троянской
войны (около середины XIII в. до н. э.), либо сразу после ее окончания (рис. 1.3).
Рис. 1.3. Античные боги, прибегавшие к «биотеррору». А. Апполон (помогал троянцам).
Б. Афина (была на стороне греков). В. Дионис (во время войны страдал безумием,
но по ее окончании выздоровел). Г. Посейдон (помогал троянцам)
Появление миазматического учения. Попытки понять «естественное происхожде-
ние» эпидемий можно обнаружить в трудах древнегреческих ученых более позднего
времени, например, у Гиппократа (460-377 до н. э., см. его работу «Эпидемии»). Обыч-
но они сводились к выявлению «неблагоприятных времен» и «зловредных испарений»,
принесенных ветрами из нездоровых местностей. Эпидемия, эпидемическое проис-
хождение и распространение болезни понимались античными учеными как клиниче-
ски сходные заболевания людей на определенной территории, вызванные воздухом,
содержащим миазмы, принесенные с зараженных мест, либо испарениями, подни-
мающимися из-под земли. А так как одним и тем же воздухом дышат одновременно
много людей, то они поражаются одинаковой болезнью — возникает эпидемия.
Разумеется, современники оценили «военное значение» миазмов. В 334 г. до н. э.,
во время войны Александра Македонского (356-323 до н. э.) с Персией и осады им
города Галикарнасса (современный Бодром в Турции), греки, в полном соответствии
с взглядами Гиппократа на эпидемический процесс, пытались вызвать мор в осажден-
ном городе. Они стреляли по городу из катапульт трупами собак и полуразложивши-
мися частями других павших животных. Распространение зловония в восприятии
защитников крепости послужило причиной вспыхнувших среди них болезней.
Не все ученые придерживались взглядов Гиппократа на природу лихорадок. Арет-
ций Каппадокийский (Aretaeus, 30—90 до н. э.) различал передачу болезни через не-
посредственное соприкосновение и на расстоянии (Сталлибрасс К„ 1936). Однако в
те годы были и более авторитетные ученые, предлагавшие простые и понятные объяс-
нения их причин. Римлянин Гален (130-201) развил древнее учение о «пневые» — тон-
чайшем и легчайшем веществе, своеобразной переходной ступени от духовного к
материальному. Это вещество, проникающее в организм при дыхании, дифференци-
1.2. Развитие представлений о природе эпидемий
35
руется в нем на три различных субстрата: «жизненную пневму» с локализацией в сердце;
«психическую пневму», находящуюся в мозгу; и «физическую пневму», локализующую-
ся в печени. Но под «эпидемическими» или «лоимосородными» болезнями им понима-
лось появление в одно время многих одинаковых случаев болезни и смерти (к учению
Галена об эпидемиях мы еще вернемся).
Первая пандемия чумы (чума Юстиниана, 531-588) также объяснялась на основе
учения «миазматиков» (Гезер Г., 1867). Эти представления древних врачей о причинах
эпидемий оказались очень уст ойчивыми. Через почти полтора тысячелетия после Гип-
пократа и спустя четыре столетия после Юстиниановой чумы, Авиценна (980—1037)
придерживался той же точки зрения на эпидемический процесс, что Гиппократ и Га-
лен. Он считал, что причиной загнивания воздуха являются «примеси дурных, зем-
листых тел, которые смешиваются с нею и создают во всем ее составе дурное качество».
Нередко причиной этого бывает ветер, приносящий в здоровые местности дурной дым
из отдаленных областей, где имеются стоячие болота или лежат разлагающиеся тела
людей, погибших в сражениях или от убийственного мора, не закопанные и не сож-
женные, а порою причина находится близко от данного места или существует в нем
самом. «Иногда же гнилостности возникают под землей по причинам, частности ко-
торых для нас незаметны, и переходят на воду и в воздух».
Колдуны, ведьмы, демоны и наказание Божье. В средневековой Европе материали-
стические представления не только об эпидемических процессах, но и об окружаю-
щем мире приживались с величайшим трудом. По наблюдению Дж. Л. Кигтериджа
(2005), вера в колдовство является всеобщим наследием человечества и позиции тех
людей, кто в Средние века верил в его возможность, были логически и теологически
лучше подкреплены, чем позиции тех, кто его отрицал. Христианские мыслители того
времени, не отвергая естественное происхождение моровых болезней, учитывали
возможность участия в их появлении дьявола, злых и добрых ангелов и действующих
посредством дьявола колдунов. Например, профессора-инквизиторы Я. Шпренгер и
Г. Иститорис (1486) описали четыре вида проступков, связанных с эпидемиями, где
участвуют все выше перечисленные силы:
1) служащие кому-либо на пользу;
2) вредящие;
3) колдовские;
4) естественные.
Первый вид проступков — это такие, которые совершаются с помощью добрых
ангелов. Второй вид производится с помощью злых ангелов. Моисей поразил егип-
тян десятью казнями (в их числе эпидемии и эпизоотии), опираясь на силу добрых
ангелов, а египетские Mai и сопеоничали с ним с помощью злых духов. Трехдневная
чума из-за греха Давида, по случаю переписи народа, была делом рук ангелов Господ-
них, почитающих Создателя и знающих его. Колдовскими проступками называются та-
кие, которые совершаются дьяволом через посредство ведьм и колдунов. Естествен-
ные же проступки зависят от влияния светил небесных, и именно от менее значитель-
ных из них. Эти явления выражаются в смертности, не люжайности полей, градобитии
и тому подобном.
Далее они, заботясь исключительно об эффективности работы судей-инквизито-
ров, учили их приемам дифференцирования таких проступков. Например, дух-вре-
дитель излил на Иова несчастия, среди которых была и «проказа», но пост упок демона
нельзя назвать колдовским, а только вредительским. Почему? Да потому что Иов по-
стратал исключительно от дьявола без посредничества колдуна или ведьмы (рис. 1.4).
36
Часть 1. Краткая история применения биологических средств...
Кроме поиска, изобличения и сжигания колдунов, оба инквизитора не чурались
и непосредственного участия в противоэпидемических мероприятиях. Например,
один из них был вызван в некий город, опустошенный чумой. В народе бытовало убеж-
дение, что мор носит искусственный, а вернее, колдовской характер. Горожане утвер-
ждали, что одна недавно похороненная женщина заглатывает свой саван уже в могиле,
и что мор не кончится, пока она его совсем не проглотит. По совету инквизитора бур-
гомистр велел разрыть могилу, и оказалось (тому было большое количество свидете-
лей!), что действительно покойница заглотала почти половину савана. При виде это-
го безобразия бургомистр выхватил меч, снес голову трупу и вышвырнул ее далеко из
могильной ямы. Мор, конечно, сразу прекратился.
Рис. 1.4. Демон чумы. Цветная гравюра на дереве Германия, XVI в.
(The Granger Collection, New-York)
Даже в 1679 г. эпидемию чумы, поразившую Вену, местные власти объясняли на
основе энциклопедических познаний в демонологии — оказывается, «таковые повет-
рия причиняют злые духи, евреи, гробокопатели и ведьмы» (Трахтенберг Дж., 1998).
Как наказание от Господа за грехи воспринимались эпидемии на Руси. Псковский
летописец в 1341 г. записал: «Грехов наших, бяше мор зол на людех во Пскове и в Из-
борске: мряху бо старые и молодые люди, и чернцы и черницы, мужи и жены и
малыя детки... и где место воскопают или мужу или жены, и ту с ним положат, малых
деток, семеро или осмеро голов в един гроб» (Псков. I; пит. по В. М. Рихтеру, 1814).
Распространение чумы также приписывали колдовству татар. В 1386 г. другой русский
летописец сделал следующую запись: «Иземце (татары) бо сердце человеческое мо-
чаху во яду аспидном и полагах в водах, и от сего воды вся в яд обратишася и аще кто
от них пияще, абие умираше, и от сею великий мор бысть по всей Русской земле»
(Воскр. лет.; цит. по В. М. Рихтеру, 1814).
Антисемитские предрассудки. Вера в способность евреев вызывать мор была наи-
более прочным антисемитским предрассудком у средневековых европейцев. Соглас-
но популярному представлению масс того времени, за болезни были ответственны
1.2. Развитие представлений о природе эпидемий
37
демоны и колдовство. В силу этого медицина считалась законной вотчиной колдунов
и чародеев. Поскольку основную массу врачей того времени составляли евреи, то их
успехи влечении больных обычно закрепляли за ними репутацию опытных магов, со
всеми вытекающими из этого суеверия последствиями. Если же именитый больной
не выздоравливал^ то это означало, что врач-еврей убил его «в соответствии с требо-
ваниями своего закона». А так как для средневекового сознания понятия «яд» и «ле-
карство» были синонимами, то на бытовом уровне вера в то, что все доктора-евреи —
отравители, планирующие истребить чумой как можно больше христиан и христиан-
ский мир, была всеобщей (Трахтенберг Дж., 1998).
Появление контагионистического учения. До «черной смерти» (пандемия чумы
1346—1351 гг.) понимание инфекционных болезней как контагиозных, т. е. способ-
ных передаваться от заболевшего человека к здоровому как непосредственно, так и
через предметы, находившиеся в общем пользовании, в Европе не было распростра-
ненным. Мы находим свидетельства близкого к современному нам пониманию кон-
тагиозности отдельных полезней только тогда, когда речь идет о каких-то глубоких и
длительных кожных поражениях. Но даже в этом случае больных относили к «нечис-
тым», а не к «заразным». Соответственно люди, с ними соприкасавшиеся, станови-
лись «нечистыми». В этом смысле «контагиозной» всегда считалась «проказа», либо
та болезнь, которую так тогда называли. О понимании контагиозности «проказы»
говорят принимаемые против нее меры, подробно описанные в Левит, третьей книге
Моисея (надзор левитов за заболевшими людьми, их изоляция, самообъявление о
болезни и т. п.).
В раннее средневековье широко практиковалось отправление прокаженных в спе-
циальные больницы—лепрозории (первый лепрозорий в Европе был открыт во Фран-
ции в 570 г.), где их содержали за счет церкви и государства, переосвидетельствуя раз
в 5 лет для выявления симулянтов. Однако ни чума, ни натуральная оспа по разным
причинам контагиозными не считались.
В мае 1347 г., накануне появления «черной смерти» во Франции, Парижский ме-
дицинский факультет по повелению короля Филиппа обнародовал свое мнение о чуме
«для публичного поучения народа, как следует держать себя относительно болезни».
Документ опубликован в 1860 г. французским богословом Ж. Мишоном (1806—1881)
по оригиналу, хранящемуся в Парижской императорской библиотеке, и весьма любо-
пытен для выяснения представлении того времени о причинах эпидемий. Члены Па-
рижского медицинского факультета уведомили короля Филиппа о том, что: «...в Ин-
дии и в странах великого моря небесные светила, которые борются с лучами солнца и
с жаром небесных огней, оказывают специально их влияние на это море и сильно
борются с его водами. От того рождаются испарения, которые помрачают солнце и
изменяют его свет в тьму. Эти испарения возобновляют свое поднятие и свое падение
в течение 28 дней непрерывно; но, наконец, солнце и огонь действуют так сильно на
море, что они вытягивают из него большую часть вод и превращают эти воды в испа-
рения, которые поднимаются в воздух, и если это происходит в странах, где воды ис-
порчены мертвыми рыбами, го такая гнилая вода не может быть ни поглощена теп-
лотою солнца, ни превратиться в здоровую воду, град, снег или иней; эти испарения,
разлитые в воздухе, покрывают туманом многие страны. Подобное обстоятельство
случилось в Аравии, в части Индии, в равнинах и долинах Македонии, в Албании,
Венгрии, Сицилии и Сардинии, где ни одного человека не осталось в живых; то же
самое будет во всех землях, на которые будет дуть воздух, зачумленный Индийским
морем, пока солнце будет находиться в знаке льва» ^Архангельский Г. Ф., 1879).
38
Часть 1. Краткая история применения биологических средств...
Из этого описания следует, что в 1347 г. ученые все еще придерживались взглядов
Гиппократа на причины моровых болезней. Впрочем, как мы покажем ниже, даже
такое «научное объяснение» причин чумы 1346-1351 гг. оказалось непосильным для
менталитета народных масс той эпохи.
Натуральной оспой тогда, хоть раз в жизни, но болели все, и поэтому ее относи-
ли, как это сейчас принято говорить, к соматическим болезням. В трудах Гиппократа
нет ясных указаний на его знакомство с натуральной оспой. Авиценна (980—1037),
живший в период ее очередного «возвращения», объяснял патогенез болезни следую-
щим образом: «В крови человека иногда возникает брожение вследствие загнивания,
сходное с брожениями, которые происходят в выжатых соках плодов и приводят к тому,
что их частицы отделяются одна от другой. Причиной такого брожения являются
остатки питательного вещества месячных женщины, образовавшегося при беремен-
ности или зародившегося после нее от мутных, нехороших яств, которые разжижают
состав крови и волнуют ее, так что ее вещество становится гуще прежнего и явствен-
ней Оно волнует кровь и сильно смешивает с ней соки, а после этого возникает ки-
пение и бурление». Оспенные высыпания на коже, по его мнению, яв.тяются следст-
вием выделения излишков образующихся при гниении и брожении крови газов.
Нетрудно заметить, что это объяснение сделано под влиянием галеновского учения
о «пневме».
Загадочное природное явление средины XIV в., тогда же названное «черной смер-
тью», на время обесценило миазматическое учение. Это сегодня понятно, что в сред-
невековых городах чума распространялась блохами из вторичных крысиных очагов
и протекала по типу домовых эпидемий. Но тогда люди, находящиеся друг с другом в
контакте, обнаруживали, что болезнь, появившаяся у одного, неминуемо появляется
и у других. К тому же у некоторых горожан бубонная чума еще осложнялась пораже-
ниями легких и затем распространялась уже как первичная легочная чума. Многократ-
но сделанные наблюдения «перехода» отдельных заболеваний чумой в масштабные
эпидемии привели к рас пространению среди врачей представлений о «прилипчивом
заражении», т. е. эстафетной передаче какого-то «болезненного начала» от одного че-
ловека к другому, находящемуся с ним в контакте. Причем, по убеждению некоторых
врачей того времени, хватало только одного взгляда больного в сторону здорового,
чтобы тому передалась его болезнь. Эта теория, даже не предполагавшая знания под-
линных механизмов распространения чумы, а построенная лишь на аберрации тво-
римого ею ужаса, составила важный этап в развитии эпидемиологии в целом и в по-
явлении подходов к ведению бактериологической войны, в частности. Людей,
заболевших во время эпидемий Ч] мы, стали считать зара >ными и отделять от здоро
вых, а в охваченных чумой городах появились первые биотеррористы, или, как их тогда
называли, «сеятели чумы», или «демоны эпидемий».
Но многие врачи, наблюдая развитие эпидемий бубонной чумы, не обнаружили
доказательств контагиозности этой болезни. Поэтому они придерживались прежне-
го учения о происхождении заразного вещества вне организма и объясняли развитие
эпидемии испорченными гнилостными веществами, образующимися в почве и по-
падающими в организм из воздуха. Передачу заразы посредством зачумленных ве-
щей, а также через животных, они вполне допускали, но все-таки объясняли ее тем,
что чумной яд поступает в организм только вместе с воздухом, т. е. через легкие, а не
через кожу.
Так к традиционным мистическим представлениям европейцев о причинах эпиде-
мий, добавились материалистические. Придерживающиеся их ученые разделились
1.2. Развитие представлении о природе эпидемий
39
на два антагонистических лагеря — контагионис тов и миазматиков. Эти представле-
ния, нередко в весьма причудливой смеси друг с другом, до сих пор присутствуют в
умах тех, кто ищет «дешевое, но мощное оружие бедных».
Однако перейдем к спорам ученых XVI в. Крайние контагионисты полностью
отрицали участие и значение воздуха в передаче заразы (чумного яда) и признавали
только одну возможность заразиться — при непосредственном прикосновении к боль-
ному или к предметам, к которым он прикасался. В 1546 г. Джиорламо Фракасторо
(1483-1553) вкниге «О контагии, контагиозных болезнях и лечении» заявил, что един-
ственная причина чумы — «специфический яд (контагий), который выходит вроде
испарины из тела больных, распространяется в воздухе только на самых малых рас-
стояниях; но взамен того, прилипает ко всем вещам и предметам; что некоторые из
них могут сохранять в себе чумной яд до тридцати и больше лет и, следовательно, та-
кие вещи и предметы могут быть переносимы на безграничные пространства, и по-
служить к заражению целых городов».
В объяснение причин распространения чумы вмешалась католическая церковь.
Учение о «контагии и прилипчивом заражении» потребовалось Папе Римскому Пав-
лу ill (1534-1549) в качестве инструмента для политического шантажа. Папе нужно
было найти предлог для перевода Вселенского собора из протестанского Тридента
Южный Тироль) в католическую Болонью. Запуганные Павлом Ш члены Собора
поспешили оставить город и собрались в 1547 г. в относительно благополучной по чуме
Болонье, и, главное, подальше от настойчиво требовавшего серьезных уступок про-
тестантам германского императора Фердинанда I (1503—1564). Папе Павлу 111 удалось
сохранить систему безусловного повиновения паствы Святому престолу, одновремен-
но он санкционировал своею непогрешимостью учение о прилипчивости чумы. Ин-
квизиция, в свою очередь, поддержала это учение кострами и страхом, и, таким обра-
зом, оно было принято и введено в Европе как основание для предохранительных мер
и репрессий (Архангельский Г. Ф., 1879).
Взгляды Фракасторо на эпидемиологию натуральной оспы мало отличались от
взглядов Авиценны. И тот и другой считали оспу эндогенной болезнью, ставшей след-
ствием поражения, воспринятого в утробе матери из менструальной крови. Оно
(т. е. «поражение») выделяется путем такого вскипания и гниения, благодаря чему
кровь очищается, как бы путем кризиса, произведенного природой. По этой причи -
не почти все люди переносят это страдание, ибо все они выносят это менструальное
поражение из утробы матери.
Здесь возникла любопытная коллизия ( столкновение) эпидемиологических пред-
ставлений у ученых. Бубонная чума, неспособная передаваться от человека к человеку
при непосредственном контакте, после пандемии «черной смерти» считается ими
весьма контагиозной, так как эпидемия довольно быстро охватывает многих лиц,
находящихся на определенной территории или в отдельных домах, затем исчезает на
много лет. Поражающие свойства вещей больных чумой и их выделений в представ-
лениях «сеятелей чумы» и властей гипертрофируются; меры, направленные на борь-
бу с распространением контагия чумы, становятся просто чудовищными. А весьма
контагиозная натуральная оспа для них эпидемична, но не контагиозна, так как ею
болеют, как правило, все люди и преимущественно до года, и она постоянно присут-
ствует среди них.
Со средины XVI в. «яд» (или «специфический контагий») стал вытеснять «загнив-
шую пневму» чз представлений ученых о причинах повальных болезней. Убедитель-
ные дока $ательствй, приводимые контагионистами в пользу «прилипчивого зараже-
40
Часть 1. Краткая история применения биологических средств...
ния» после каждой «большой чумы», неблагоприятное общественное мнение и сло-
жившаяся противоэпидемическая практика превращали миазматиков в объекты на-
смешек коллег, и даже, в жертвы судебных преследований властей. И опять мы видим
коллизию эпидемиологических представлений. В конце XVI в. по каким-то неизвес-
тным сегодня причинам контагиозность циркулирующего среди людей вируса нату-
ральной оспы уменьшилась. Но по мере стихания оспенной пандемии в XVII в., т. е.
появления детей и взрослых, не перенесших в младенчестве натуральную оспу, все
яснее стала видна контагиозность ее возбудителя. Сначала бельгийский врач и теософ-
мистик Ван Гельмонт (Joh. Bapt. van Helmont, 1578-1644) пришел к выводу, что «оспа
происходит от яда и несет с собою заразу, которой пропитывает кровь и заражает лиц,
а особенно детей, окружающих больного». Затем Герман Бургав (Boerhave, 1668—1738)
подошел вплотную к современному пониманию заразности оспы. Накопившиеся в
XVIII в. «сухие дрова эпидемии», т. е. взрослые люди, восприимчивые к инфицирова-
нию вирусом натуральной оспы, вновь сделали возможным ее искусственное распро-
странение с помощью вещей больных людей. Контагионистические взгляды достигли
пика своей популярности. Единственное, что тогда сдерживало «сеятелей чумы», так
это отсутствие у них в руках самого контагия в виде какого-то вещества, а именно так
они себе его представляли.
Локалисты. В средине XIX в. у контагионистического учения вновь появился
серьезный конкурент, а если быть точнее в формулировках — он «вернулся». Благо-
даря накоплению эпидемиологической статистики и работам Макса Петтенкофера
(1818—1901) по эпидемиологии холеры, стало развиваться локалистическое (почвенное)
учение, имевшее в своей основе взгляды миазматиков на передачу болезнетворного
начала. Петтенкофер считал, что почва содержит некий фактор (Y), благодаря кото-
рому «холерный зародыш» (X) превращается в «холерный яд» (Z). Он утверждал, что
«холерный яд» выходит из почвы вместе с парами, он даже указал на связь холерных
эпидемий с понижением почвенных вод (М. Петтенкофер, 1878, 1885).
Тем самым Петтенкофер указал на участие в поддержании и распространении
возбудителей инфекционных болезней людей еще каких-то неизвестных факторов,
содержащихся в почве. Это его наблюдение нашло подтверждение в трудах других
известных ученых того времени (например, Р. Вирхова — в отношении заболеваемо-
сти брюшным тифом в Берлине; Н. К. Щепотьева — в отношении заболеваемостью
чумой в низовьях Волги).
«Овеществленный» контагий. Контагионистическое и локалистическое (миазма-
тическое) учения конкурировали между собой почти до конца XIX в., т. е до откры-
тия возбудителей холеры, чумы, брюшного тифа, сибирской язвы, туберкулеза и дру-
гих инфекций. Причем перевес в научных дискуссиях в 1860— 1870-х гг. был на стороне
локалистов. Открытие же патогенных бактерий в конце XIX в. изменило ситуацию в
пользу контагионистов. Бактерии прекрасно «состыковывались» со средневековым
учением о контагии. Психологически они воспринимались учеными как тот же кон-
тагий, но теперь стало ясно, что это живой организм (contagium vivum), а не «яд», и
что его можно получать в большом количестве и изучать в лабораторных условиях. Эти
взгляды оказались не менее прочными, чем в свое время взгляды миазматиков. На-
пример, возбудитель сибирской язвы даже в средине 1930-х гг. называли «сибиреяз-
венным контагием» (см. Н. Г. Олсуфьев, П. П. Лелеп, 1935).
Позиции локалистов были сильно поколеблены, но само локалистическое учение
не было разгромлено окончательно. И дело тут не в том, что локалисты не соглаша-
лись с доказательствами, приводимыми их противниками в пользу отсутствия в при-
1.2. Развитие представлений о природе эпидемий
41
роде какой-то пахучей миазмы, приводящей к болезни. На их стороне, как правило,
оказывалась медицинская статистика. Результаты анализа статистических данных,
характеризующих динамику, географию, напряженность эпидемических процессов,
их пространственно-временные особенности, свидетельствовали в пользу наличия в
природе каких-то еще других (кроме «прилипчивого заражения») очень сложных ме-
ханизмов поддержания и распространения возбудителей инфекционных болезней
(см., например, работу Эрисмана Ф. Ф., 1893; и комментарии к книге А А. Генрици,
2002). Но по мере развития медицинской бактериологии локалистические представ-
ления стали считаться учеными умозрительными, так как их сторонники не облада-
ли методическими возможностями для экспериментальной проверки основных сво-
их положений. «Contagium vivum», т. е. выращенный на искусственной питательной
среде микроорганизм, стал рассматриваться ими как единственная причина сокруши-
тельных эпидемий.
В начале XX в. о локалистах просто забыли, не потрудившись ответить на вопро-
сы, поставленные теми на основе изучения детальных статистических отчетов об эпи-
демиях. У исследователей возникли новые интересы, новая положительная мотива-
ция. У них появилась возможность разрабатывать вакцины и сыворотки против
возбудителей опасных инфекций и совершенствовать методы их диагностики. Все, а
вернее, почти все в эпидемиологии чумы, холеры, туберкулеза, сибирской язвы и дру-
гих, известных в те годы опасных инфекций, оказалось понятным.
Возвратом к раннесредневековой легенде о распространении чумы кораблями
стало учение, появившееся в начале XX в., предписывающее разнос чумы корабель-
ным крысам (подробнее о его первоначальном варианте, объясняющем появление
чумы в Марселе в 588 г., см. у Гезера Г., 1867) (рис. 1.5).
Рис. 1 5. «Купец» («The Merchant»)». Цветная гравюра на дереве Ганса Гольбей-
на Старшего (1460—1524) из цикла «Пляска смерти» («The Dance of Death», 1538).
Картина показывает купца, схваченного смертью в благодарность за доставлен-
ный товар. Выбор в качестве фона морского порта обусловлен тем, что в те годы
именно купеческим кораблям приписывалась основная роль в распространении
чумы (The Granger Collection, New-York)
42
Часть J. Краткая история применения биологических средств...
Такие мифы и мистификации подталкивали и стимулировали развитие новой и
не афишируемой области научного знания — военной микробиологии. В конце 1920-х гг.
во многих странах мира началась подготовка к масштабной бактериологической вой-
не. С позиций учения о «контагии» она казалась технически весьма простой.
* * *
К началу XX в. существовало как минимум три «подхода» к осуществлению био-
террора и биодиверсий, а соответственно, и три периода практического применения
накопленных в этой области знаний. Мистический — когда отдельные люди, знающи-
еся с некой высшей силой (боги, жрецы, пророки, колдуны, знахари, шаманы, евреи-
врачи и др.), путем выполнения тайных обрядов, произнесения заклинаний и совер-
шения актов колдовства или магии участвуют в распространении моровых болезней.
Такое мировоззрение нашло свое отражение в древних поэтических сказаниях (напри-
мер, «Илиада»), назидательных священных книгах (Ветхий Завет), в ритуальных пред-
метах и обрядах («обереги», молитвы, статуи богов-защитников и др.), а также в дея-
тельности инквизиции. Попытки объяснить развитие эпидемий естественными
причинами привели к появлению представлений о «миазме», как о неком материаль-
ном элементе, враждебным природе людей и способным вызывать у них болезнь.
Обычно миазматическими считали гнилостные запахи и все предметы, их издающие.
Отсюда техника первых попыток умышленного распространения инфекционных
болезней — забрасывание в стан врага гниющих трупов людей и животных, в том числе
«отравление» таким же способом колодцев и небольших водоемов — это миазмати-
ческий период в истории биотеррора и биодиверсий. Контагионистический период на-
чался во времена «черной смерти» (чумы 1346—1351 гг.). Такое восприятие биотерро-
ра и биодиверсий подразумевает поражение людей путем распространения между
ними специфических контагиев, роль которых с конца XIX в. стали играть микроор-
ганизмы — возбудители опасных болезней. И эти подходы хорошо прослеживаются
в действиях отдельных «сеятелей чумы».
1.3. «БИОТЕРРОР» МИАЗ МАТИКО В И КОЛДУНОВ
Пандемия «черной смерти» (1346-1351 гг.). флагеллянты. Удобное
объяснение причин распространения чумы в Европе.
Фридрих 1 Барбаросса (1152—1190) в период битвы при Тортоне в 1155 г. приказал
забрасывать разлагающиеся трупы людей в стан противника. В1339 г., в самом начале
Столетней войны, французы забрасывали разлагающиеся тела людей и животных с
помощью метательных машин в замок Тин на реке Шельда во время его осады. В1346 г.
татарский хан Джанибек поступил таким же образом при осаде генуэзского города
Кафы (Феодосия) на побережье Крымского полуострова. Война совпала с началом
второй пандемии чумы, и поэтому его действия до сих пор отдельные «ведущие» уче-
ные рассматривают как «первое достоверное свидетельство применения бактериоло-
гического оружия» (более подробно ниже)1. Аналогичный эпизод имел место в Европе
1 Эта чудовищная эпидемическая катастрофа до сих пор считается искусственно вызван-
ной ханом Джанибеком (см.: Черкасский Б. Л., 2000; Ефременко В. И. с соавт., 2000; Kortepe-
ter М. et. al., 2001; Wheelis M., 2002; Онишенко Г. Г. с соавт., 2003а, 20036; Нетесов С. В., 2004
и др.). Среди пропагандистов мифа цвет российской эпидемиологии.
1.3. «Биотеррор» миазматиков и колдунов
43
в 1426 г, при осаде Каролстейна. Поданным A. G. Robertson и L. D. Robertson (1995),
город забросали телами сод чат, убитых во время осады, и навозом, привезенным дву-
мя сотнями телег (рис. 1.6).
Рис. 1.6. Забрасывание в осажденный город мертвой лошади с помощью
метательной машины (требюше). По сути — это биологический террор на
основе миазматических представлений о распространении эпидемических
болезней. С рисунка Леонардо да Винчи (1445—1520)
Свой «вклад» в ухудшение эпидемической ситуации в Европе стремились внести
и колдуны. Так, в подозрении в коррупции и в колдовстве, с целью умерщвления от-
дельных должностных лиц, был арестован и осужден в 1290 г. Адам де Стратон, канц-
лер английского казначейства. Важной уликой против него был обнаруженный во
время обыска шелковый мешочек, содержащий обрезки ногтей, человеческие воло-
сы, жабьи и кротовые лапки и другие «дьявольские штучки». По заключению «экс-
пертов», весь этот набор предна шачался для «распространения моровых болезней»
(Киттередж Дж. Л., 2005). Однако «биотеррористическая деятельность» канцлера ан-
глийского казначейства оказалась сущей забавой, в сравнении с той, с которой еще
предстояло столкнуться европейцам в XIV в.
Пандемия «черной смерти» (1346—1351 гг.). В основе противоэпидемических дей-
ствий современников пандемии лежали антисемитские предрассудки, миазматиче-
ское учение и непоколебимая вера европейцев в колдовство. В основе же эпистоляр-
ной активности наших современников нет ни мистики, ни антисемитизма и тем более
научных подходов, а только здоровая любовь к бюджетному финансированию при
полном отсутствии интереса к истинным причинам возникновения пандемии чумы
«черной смерти».
Рост антисемитских предрассудков в начале XIV в. Еврейские noi ромы периоди-
чески вспыхивали в Европе задолго до эпидемии чумы 1346—1351 гг. Уже вХП в. воо-
душевление крестоносцев биться за Гроб Господень с сарацинами проявлялось мас-
совыми еврейскими погромами по пути следования их отрядов. В 1161 г. в Богемии
было сожжено 68 евреев, обвиненных в попытке извести христианское население
путем распространения яда. В 1267 г. на Соборах в Бреслау и в Вене недвусмысленно
было заявлено, что продукты питания, покупаемые христианами у евреев, могут быть
44 Часть 1. Краткая история применения биологических средств...
отравлены. В простом народе было распространенно суеверие, что если евреи даже
и не помещали яд в мясо, продаваемое христианам, то они все равно отравляли его
своими проклятиями, чтобы христиане болели и умирали. Обвинения евреев в отрав-
лении колодцев воды предъявлялись еврейским общинам в 1308 г. в Во (кантон в
Швейцарии), в 1316 г. в районе Эйленбурга (город в прусской провинции Саксонии)
и в 1319 г. во Франконии (область по Рейну, Неккару и Майну) (Гезер Г., 1866; Трах-
тенберг Дж., 1998.
В этом же столетии был популярен рассказ о том, как евреи Прованса, желая ото-
мстить христианам за эдикт, принятый на втором Соборе в Сан-Руфе в 1337 г. (эдикт
запрещал смешанные браки между евреями и христианами, а также обращение к ус-
лугам врачей и аптекарей-евреев), специально завезли чуму из Индии, которая унич-
тожила несколько деревень в Провансе, после чего распространилась по Европе (Трах-
тенберг Дж., 1998)1.
«Переквалифицирование» европейским населением евреев из «отравителей» еще
и в «сеятелей чумы» (т. е. в распространителей каких-то смертоносных болезней) обя-
зано необычайно упорной пандемии проказы, или той болезни, которую тогда счи-
тали проказой. Проказа достигла своего максимума в Европе примерно через 200 лет
после начала Крестовых походов — в XIII в. К моменту смерти Людовика VIII (1229)
во Франции, занимавшей тогда территорию вдвое меньшую, чем теперь, насчитыва-
лось уже до 2-х тыс. лепрозориев. Все началось с выявления «заговора» прокаженных.
«Заговор» просаженных 1321 г. Само существование «заговора» прокаженных (1321)
засвидетельствовано многими авторами того времени. Версия «заговора» подтверж-
далась многочисленными и убедительными для современников «доказательствами».
Например, один из свидетелей утверждал следующее: «Мы сами своими глазами ви-
дели такую ладанку в одном из местечек нашего вассальства. Одна прокаженная,
проходившая мимо, боясь, что ее схватят, бросила за собою завязанную гряпку, кото-
рую тотчас понесли в суд и в ней нашли голову ящерицы, лапы жабы и что-то вроде
женских волос, намазанных черной, вонючей жидкостью, так что страшно было раз-
глядывать и нюхать это. Когда сверток бросили в большой огонь, он не мог гореть:
ясное доказательство того, что это был сильный яд».
Вокруг «сотрудничества» прокаженных и евреев среди европейцев всегда ходило
много слухов, один ужаснее другого. Поэтому об участии в «заговоре» прокаженных,
еще и евреев, властям Франции стало известно довольно скоро. Сир де Партенэ пи-
сал королю, что один «важный прокаженный», схваченный в своем поместье, признал-
ся, что какой-то богатый еврей дал ему денег и некоторые снадобья. Они состояли из
человеческой крови и мочи с примесью тела Христова. Эту смесь сушили и измельча-
ли в порошок, зашивали в ладанки (сумочки с ладаном или какой-либо святыней,
носимые вместе с крестом на шее) с тяжестью и бросали в источники и колодцы.
Хронист из Сан-Дени сообщал, что «знаменитый среди своих соплеменников и
богатый еврей» подкупил прокаженных, чтобы те осуществили на месте еврейский
замысел. Он дал им рецепт яда, в состав которого входили следующие ингредиенты:
«человеческая кровь и моча; три вида трав, названия которых виновный не помнил
или не желал раскрыть», а также «тело Христово». Эти сведения выдавались инкви-
зиторами за признания прокаженных, пользовавшихся величайшим уважением и
почетом. Согласно другому документу, в состав яда, кроме вышеназванныхкомпонен-
1 Данное событие свидетельствует о том, что активизация европейских природных очагов
чумы началась еще в начале XIV в.
1.3. «Биотеррор» шиазматиков и колдунов
45
тов, входили также «гадючьи головы, жабьи лапы и женские волосы», значительно
усиливавшие его смертоносное действие (Трахтенберг Дж., 1998).
То, что описанные современниками зловонные снадобья и типовые наборы пред-
метов, используемых в ритуалах черной магии, не могли вызвать эпидемии чумы, не
имело никакого значения на фоне слухов, суеверий и «доказательств» того времени.
Это сегодня мы знаем, что они не имеют отношения к чуме, пока еще знаем.
Э. Дрюмон (1895), известный специалист по французскому антисемитизму, утвер-
ждал, что основным слухом в начале XIV в., которому верили европейцы, был следую-
щий. Король гренадских мавров, с горестью видя, что его так часто побеждают хри-
стиане, задумал отомстить за себя, сговорившись с евреями погубить христиан. Но
евреи, будучи сами слишком подозрительны, обратились к прокаженным и при по-
мощи дьявола убедили их уничтожит ь христиан. Предводители прокаженных собра-
ли последовательно четыре совета, и дьявол через евреев дал им понять, что так как
прокаженные считаются самыми презренными и ничтожными существами, то хоро-
шо бы было устроить так, чтобы все христиане умерли или стали бы прокаженными.
Замысел всем понравился; каждый, в свою очередь, поделился им с другим... Многие
из прокаженных, подкупленные евреями ложными обещаниями царства, графств и
других благ земных, говорили и твердо верили, что так и случится.
Также было перехвачено письмо, адресованное королю сарацинов, владыке Вос-
тока и Палестины. В нем некие лица ходатайствуют о заключении дружеского союза
между евреями и сарацинами и, в надежде, что когда-нибудь эти два народа сольются
в одной религии, просят короля возвратить евреям землю их предков.
«Когда мы навсегда поработим христианский народ, вы нам возвратите наш ве-
ликий град Иерусатим, Иерихон и Ай (иначе Гай — город в древней Палестине, упо-
минаемый еще во время Авраама и завоеванный Иисусом Навином, см. [Быт. 12:8]),
где хранится священный ковчег. Амы возвысим ваш престол над царством и великим
городом Парижем, если вы нам поможете достигнуть этой цели. А пока, как вы може-
те убедиться через вашего заместителя, короля Гренады, мы действовали в этих видах,
ловко подсыпая в их питье отравленные вещества, порошки, составленные из горьких
и зловредных трав, бросая ядовитых пресмыкающихся в воды, колодцы, цистерны,
источники и ручьи для того, чтобы все христиане погибли преждевременно от действия
губительных паров, выходящих из этих ядов. Нам удалось привести в исполнение эти на-
мерения, главным образом, благодаря тому, что мы роздали значительные суммы не-
которым бедным людям их вероисповедания, называемым прокаженными. Но эти
негодяи вдруг обратились против нас и, видя, что другие христиане их разгадали, они
обвинили нас и разоблачили все дело. Тем не менее мы торжествуем, ибо эти христи-
ане отравили своих братьев; это верный признак их раздоров и несогласий».
В 1321 г. еврейские общины в Европе отделались только испугом. По мнению
Дж. Трахтенберга (1998), кровавого исхода не случилось потому, что основным объек-
том ненависти европейцев были прокаженные. Тем не менее чума или любой другой
мор уже не только ожидались в Европе в течение нескольки < предшествующих «чер-
ной смерти» десятилетий, но были даже известны, как сегодня говорят, их «заказчи-
ки» и «исполнители».
«Черная смерть». С началом эпидемии «черной смерти» в 1346 г. появились новые
подробности «заговора» — по всей Европе заходила молва, что евреи были подстре-
каемы к этому преступлению посланными им письмами от таинственных старшин из
Толедо в Испании. Кроме того, знающие люди уверяли, что найдено письмо, напи-
санное во время распятия Спасителя, и посланное иерусалимскими евреями к своим
46
Часть 1. Краткая история применения биологических средств...
братьям, например, в Ульян (Uljan — один из далматинских островов). Содержание
письма было таковым, что вызвало ярость и жажду мщения христианского населения
Европы (Гезер Г., 1866).
В мае 1348 г. в трех городах Франции начались еврейские погромы, однако тогда
они еще не носили всеобщий характер. Ситуация изменилась осенью, когда в сентябре
в Шильоне (городок у Женевского озера) еврейский врач Балавигнус во время крова-
вых истязаний признался инквизиторам в том, что он и несколько членов еврейской
общины — Якоб Паскатэ из Толедо, Пеэрет из Шамберли и некий Абогет — пригото-
вили в одном из городков на юге Франции ядовитое зелье. В его состав входили весь-
ма зловещие ингредиенты: сердца христиан, пауки, ящерицы, человеческое мясо и
освященные гостии (облатки из пресного пшеничного теста, большей частью с изоб-
ражением агнца и креста как символов распятого Спасителя; употребляются при при-
частии в католической и лютеранской церквах вместо хлеба). Злоумышленники для
распространения чумы подсыпали полученный зловонный порошок в реки и ручьи,
из которых христиане брали воду. В некоторых местах Европы распространение чумы
приписывали не только ядам, приготовленным евреями, но и их колдовству.
Весть о «еврейском заговоре» в Шильоне быстро распространилась по всей Европе.
Многих евреев обвинили в организации массового отравления колодцев, пытками за-
ставили сознаться в подготовке преступления, судили, и на основании «суда и закона»
осужденных вместе с не подвергавшимися суду единоверцамг: сожгли на кострах.
Затем власти отбросили формальности суда и действовали с помощью толпы. На
фоне ужасов чумы страх перед евреями и помешательство стали всеобщими. В Базеле
было специально построено деревянное здание, куда собрали всех евреев и сожгли.
То же было во Фрейбурге и во многих городах Эльзаса. В резиденциях короля Карла IV
все еврейское население было перебито, имущество же их было продано магистрата-
ми «по закону и справедливости» Массовые сожжения евреев имели мес го в Аугсбурге,
Констанце, Берне, Галле, Мюнхене, Зальцбурге, Тюрингене, Эрфурте и др} гих герман-
ских городах. В Париже чернью было перебито огромное количество евреев, не похо-
роненные трупы которых долго служили пищей волкам в окрестных лесах. В Катало-
нии погромы разразились в разгар чумы 1348 г. в городах Барселоне, Тарреге, Лериде
и др. В Тарреге с криками «смерть изменникам!» толпа казнила более 300 евре гв.
Папа Климент VI (1342—1352) пытался остановить еврейские погромы. В своей
булле от 26 июля 1348 г. он приводил такие аргументы: если это обвинение справед-
ливо, то почему израильтяне становятся жертвами чумы, а эпидемия свирепствует в
равной степени там, где живут только евреи? Однако, по свидетельству де Венета (де
\fenette — французский хронист и поэт; 1307—1369), общественное мнение Европы все
более склонялось к тому, чтобы обвинять евреев в «черной смерти». Так, к ужасам
эпидемии были присоединены ужасы многотысячных сожжений и избиений еврей-
ского населения. Хронист Диссенгофен засвидетельствовал: «В течение года были
сожжены все евреи от Кельна до Австрии» (цит. по Лозинскому С. Г., 1986) Всего же
в Европе в те годы было уничтожено 50 крупных и 150 мелких еврейских общин и
устроено 350 погромов (Гезер Г., 1866). В тех городах Германии, где не было евреев —
в Магдебурге, Лейпциге — обвинение в отравлении колодцев было возведено на мо-
гильщиков. Но маховик паники и массового сумасшествия раскручивался все сильнее.
Флагеллянты. Не все христиане Европы обвиняли в случившемся бедствии евре-
ев, прокаженных, мавров и могильщиков. Церковный раскол порождал «еретиков»,
объявлявших уже самого Папу главным виновником Божьего гнева и кары, разразив-
шегося над миром. Люди, чье сознание помутилось от ужасов чумы, для смягчения
1.3. «Биотеррор» миазматиков и колдунов
47
Божьего гнева стали сами налагать на себя наказания. Огромные толпы «бичующих-
ся» (флагелланты) перемещались по европейским городам, распевая псалмы, стегая
себя ремнями с железными остриями, повсюду грабя и убивая евреев. Но папство, на
словах осудившее еврейские погромы, не могло себе позволить, чтобы паства вела
оголтелую пропаганду против Святого Престола. Папа Климент VI в 1349 г. в особой
булле осудил флагеллантов, называя их опасными еретиками, подлежащими суду
инквизиции. В течение года с ними было покончено ме годами, естественными и за-
конными для того времени.
Последующие «возвращения» чумы объясняли также просто. Когда в 1357 г. чума
вновь стала свирепствовать во Франконии (область Германии по Рейну, Неккару и
Майну), в ее распространении обвинили местных евреев, отравивших воду с помо-
щью яда. В 1401 г. евреи Фрайбурга были обвинены в распространении чумы посред-
ством отравления воздуха. Подобные обвинения в адрес европейских евреев выдви-
гались почти до концаXVII в. (см. в разд. 1.2 «Колдуны, ведьмы, демоны и наказание
Божье»), Но в XIX в. даже вспоминать об этих событиях в «просвещенной Европе»
стало неприлично, и судьба подбросила европейцам и евреям «палочку-выручалочку».
5 тобное ооъяснение причин распространения чумы в Европе. В 1842 г. А. Геншелем
(1790—1856), профессором медицинского факультета университета в Ьреславле, ев-
реем, принявшим христианство ради профессорской кафедры (более подробно о нем
см. в «Еврейской энциклопедии», 1913), было опубликовано свидетельство нотариу-
са Габриэля де-Мюссе, пережившего начало чумы «черной смерти» в городе генуэз-
ских работорговцев — Кафе. Рукопись была им обнаружена в Гедигеровской библио-
теке (Бреславль), и она ранее не только не была известна специалистам, но даже не
была датирована ее автором. Нотариус написал в полном соответствии с научными
представлениями своего времени, что причиной чумы в крепости стало зловоние,
исходящее от заброшенных татарами трупов больных чумой, т. е. миазмы: «Татары,
измученные чумой, ошеломленные и потрясенные смертью товарищей, гибнущих без
всякой надежды на выздоровление, приказывали заряжать трупы в метательные ма-
шины и забрасывать им город Кафу, чтобы эти непереносимые снаряды положили
конец защитникам города. Город забросали горами мертвецов, и христианам некуда
было убежать, и некуда было спрятаться от такого несчастья... Они предавались мер-
твым волнам. Вскоре весь воздух был заражен, отрав генная и испорченная вода ста-
ла загнивать. Усилилось нестерпимое зловоние».
Судя по клинической картине, подробно описанной де Мюссе, среди осаждавших
город татар и его защитников, преобладала бубонная форма чумы, распространив-
шаяся из вторичных крысиных очагов посредством инфицированных блох. На ее фоне
развивались вторичные легочные осложнения, но они не были доминирующими для
этой эпидемии. Однако современные толкователи этого текста обращают свое вни-
мание, прежде всего, на следующее замечание де Мюссе: «Родные, друзья и соседи
поспешили к нам, но мы принесли с собой убийственные стрелы, при каждом слове рас-
пространяли мы своим дыханием смертельный яд». Отсюда делается вывод о распрост-
ранении легочной чумы по Европе здоровыми носителями, прибывшими из Кафы. и
упорно замалчиваются свидетельства о бубонных формах болезни (хотя бы того же
Боккаччо, 1351), имевших место во всех европейских городах.
Для де Мюссе «убийственная атмосфера» — это не мелкодисперсный аэрозоль воз-
будителя чумы, что было бы характерным для применения БО, и не воздушно-капель-
ная инфекция, встречающаяся в природных очагах чумы, а «загнившая пневма», так
как пользовался он пневматическими представлениями о распространении даже
48
Часть 1. Краткая история применения биологических средств...
не контагия чумы, а еще миазмов в понимании Авиценны, Галена и Гиппократа. Иного
объяснения тех событий ни он, ни кто другой в те годы просто не воспринял бы, — до
книги Фракасторо «О контагии, контагиозных болезнях и лечении» еще должно было
пройти 200 лет. В настоящее время версия де Мюссе, неизвестная современникам «чер-
ной смерти» и такая же лживая и безграмотная (с точки зрения современных знаний
об экологии возбудителя чумы), как и версия о распространении «черной смерти»
посредством отравления рек и воздуха евреями, активно популяризируется в престиж-
ных научных изданиях российскими «эпидемиологическими светилами» (см., напри-
мер, работы Черкасского Б. Л., 2000; и Онищенко Г. Г. с соавт., 2003а, 20036). Что де-
лает в принципе не нужными любые исследования, направленные на выяснение
истинных причин масштабных эпидемий чумы на европейском континенте. Она так-
же используется западными политиками как доказательство доступности БО странам
третьего мира и, соответственно, как инструмент политического давления на эти стра-
ны путем выдвижения в их адрес обвинений в создании БО. Наглядный при-
мер — обвинения в создании БО, предъявляемые в течение 15 лет Саддаму Хусейну,
вызвавшие две войны НАТО с Ираком. Это уже установившийся стандарт профана-
ции проблемы биотерроризма. Автору пришлось самому убедиться в том, что ника-
кие другие объяснения причин «черной смерти» в Европе в «солидные» российские
научные журналы не пропускаются по соображениям «самоцензуры» редакторов и
рецензентов.
1.4. КОНТАГИОНИСТЫ «СЕЮТ ЧУМУ»
«Сеятели чумы». «Чисто биологическое убийство» Петра II? Биологические
диверсии конца добактериологической эпохи.
Наблюдения за заболеваемостью в период «черной смерти» позволили многим уче-
ным прийти к мысли о возможности «прилипчивого заражения». Попытки вызвать
эпидемии («поветрие») посредством «дурного запаха» еще имеют место, но с XVв. для
достижения этой цели уже активно используется «контагий». Поданным A. G. Robert-
son и L. D. Robertson (1995), Цезальпино1 сообщил о неудачной попытке вызвать мор
среди французских войск, имевшей место во время Неаполитанской кампании 1435 г.
Тогда испанские солдаты раздали французам вино, в которое была подмешана кровь
людей, больных проказой. Польский воевода Сименович предложил в 1540 г. запол-
нять полые ядра слюной бешеных собак и другими веществами, «способными отра-
вить атмосферу и вызвать эпидемии». Однако внимание властей теперь перехватыва-
ют «сеятели чумы».
«Сеятели чумы». Э. Литтре (1875) считал, что «посев чумы», наравне с колдовством,
принадлежит к разряду мнимых преступлений, значение которых невозможно стро-
го определить. Под влиянием настроенного известным образом воображения, страх
заразы или зла, проистекающего из сношения со злыми духами, мог разрастаться до
бесконечности, равно и требование более и более жестоких кар для преступников.
Однако и он задавался страшными вопросами. Если нельзя сеять чуму, существовали
ли сеятели ее, подобно тому, как были колдуны, хотя и не было колдовства? Правда
1 Andrea Cesalpino (1519—1603) — известный врач, естествоиспытатель и философ; еще до
Гарвея он открыл систему кровообращения.
1.4. Контагионисты «сеют чуму»
49
ли, что были злоумышленники, делавшие мнимые, конечно, но тем не менее факти-
ческие попытки для распространения заразы? Не было ли среди «сеятелей чумы» боль-
ных, обреченных на смерть и мстящих таким образом здоровым людям?
Теоретическая простота контагионистического учения на практике обернулась
террором. Э. Литтре привел выдержку из «Истории Женевы» Бонниварда*. Вот что тот
писал о событиях, которые наблюдал лично.
«В этом году (1530) чума свирепствовала в Женеве и, людям, по-видимому, недо-
статочно было наказания, посланного Господом за их грехи; злоба человеческая, не
удовлетворяясь страданиями, ниспосланными свыше на человеческий род, старалась
еще усугубить их; и факт этот показался мне столь достойным сохранения в памяти,
что я решился занести его в эту летопись.
Необходимо упомянуть, что в Женеве существовала и еще существует больница для
зачумленных во время эпидемии; в этом госпитале есть надзиратель, который вместе
с тем и хирург, чтобы перевязывать больных; духовное лицо, чтобы исповедовать и
утешать больных; и прислужники, получающие хорошее вознаграждение за опасность,
которой они подвергаются, в том числе и женщины для ухода за больными и для со-
держания помещений в чистоте. Этих женщин называют сиделками (sureresses), но не
потому чтобы они оказывали помощь из милосердия, напротив того, они получают
хорошее содержание и еще пользуются незаконными барышами, которыми делятся с
надзирателем и священником.
Однако, по милости божьей, чума стала ослабевать, что пришлось не по сердцу
этим лицам, ибо люди, извлекающие пользу из зла, не могут желать добра и всегда
предпочтут поддержать первое. Тут они вспомнили об одном юноше из хорошей фа-
милии, который занимался всякого рода плутнями. Он даже хвастался ими и гордил-
ся прозвищем злого, если его в то же время считали умным; но он еще не совершил
такого поступка, за который подвергался бы телесному наказанию. Имя его было
Михаил Каддо. Наконец, он до того довел себя своим плутовством, что очутился без
пристанища, и никто из родных и знакомых не хотел пускать его к себе в дом; тогда
ему пришлось прибегнуть к отчаянной штуке, чтобы выпутаться из затруднения, он
притворился заболевшим чумой, чтобы найти себе убежище и пропитание Его немед-
ленно отправили в больницу, с приказанием иметь за ним хороший уход, что и было
исполнено, и его лечили даже больше вином, чем микстурами. Тут он сообразил, что
эта жизнь дарового угощения не может продлиться более сорока дней, по истечении
которых его спровадят из больницы, и он придумал средство, продлить ее С этою це-
лью он сталь убеждать надзирателя де Фосижи, поддерживать чуму, которая, вредя
другим, была им столь выгодна.
Во-первых, они решили отравлять, или иным образом ускорять смерть привози-
мых в госпиталь пациентов, в случае если бы они выказывали расположение к выздо-
ровлению.
Потом они стали вырезать нарывы с тел покойников, превращали их в порошок
и, смешав его с другими составами, давали принимать больным под видом лекарства.
Этого мало, они посыпали таким порошком вышитые носовые платки, красивые под-
вязки и тому подобные вещи, а Михаил Каддо разносил и разбрасывал их ночью по
городу, выбирая преимущественно дома, где предвиделась богатая нажива, и даже на-
тирал порошком замки дверей. Утром, когда слуга или служанка выходили из дома,
им бросались в глаза эти красивые платки и подвязки; они радовались своей находке,
1 Bonnivard (1496—1570); им написаны книги «Les Chroniques de Geneve» и «De I’ancienne
et nouvelle police de Geneve».
50
Часть 1. Краткая история применения биологических средств...
а вечером запирали на замок натертые двери дома или лавки и не редко прикасались к
своим господам. Так попадали они в сети, болезнь их приносила выгоды Каддо, над-
зирателям, священникам, фельдшерам и сиделкам.
Это оставалось скрытым некоторое время, но дьявол более радеет об уве личении
числа грехов, нежели о сокрытии их. Когда Каддо достаточно поработал ночью, то он
не утерпел, чтобы не продолжать свое дело днем, и однажды, в постный день следую-
щего года, кинул сверток с порошком посреди Констанцкой улицы, воображая, что
никто его не заметит.
Однако нашелся человек, который это увидел, и, не помышляя, что m может быть
опасность, а предполагая скорее шутку, сказал: «Этот кот, Михаил Каддо, что-то бро-
сил сюда, чтобы подтрунить над народом», и хотел поднять сверток. Но другой более
рассудительный человек сказал: «Не годится в нынешнее время дотрагиваться до не-
известной вещи, подними ее чем-нибудь, но не руками, и посмотрим, что это такое.
Они достали щепки и с их помощью подняли и открыли сверток, из которого немедля
распространилась страшная вонь. Все были в изумлении и не могли постичь, в чем дело,
за исключением одной бедной женщины, которая не задолго перед тем выписалась из
больницы, и она сказала: «Наверно, господа, это сделано из чумного нарыва». И все
крайне изумились, и пошли известить о том синдиков, которыми в то время были
Иоганн Базлард, Иоганн Ами, Ботемер, Перрин Вильмет и Иоганн Лерье; и они со-
звали совет, обсудить дело и дали приказание немедленно схватить Михаила Каддо.
Сольтье, полицейский агент, поймал его в ту самую минуту, когда он думал укрыться в
доме Рива.
Он был схвачен и заключен в тюрьму, где синдики (греч. syndikos или synegoros —
в то время должностное лицо, ведущее судебные дела какого-нибудь учреждения) вме-
сте с другими делегатами совета, учинили ему допрос и требовали его сознания. И на
первых порах, он представлял из себя шута, говоря синдикам, что следует приготовить-
ся к исповеди (здесь игра слов, Confesser un crime сознаться в преступлении и испове-
дываться): «Подождите до Пасхи, и я все расскажу вам». Синдики отвечали ему. «Вы
должны, прежде всего, сознаться нам». Видя, что он виляет, поднесли к нему веревку.
Тогда он стал объяснять, что в брошенном свертке была материя из раны, бывшей у
него на ноге. Koi да же у него спросили, с какою целью он это сделал, то он ответил:
«Нац моею раною насмехались, и я хотел наказать насмешников». Синдики, не удов-
летворись этим ответом, подвергли его пытке, и тогда у него развязался язык; он об-
личил надзирателя, сиделок и фельдшеров и открыл, с помощью какого предохрани-
тельного средства они могли прикасаться к чуме, не подвергая себя опасности; об этом
средстве уже было опубликовано, так что не стоит упоминать о нем здесь.
Немедленно правительство распорядилось арестовать его сообщников, которым
делали допросы, очные ставки и которых подвергали пытке. Они все говорили на один
лат, за исключением одного прислужника, по имени Лентиль, которому удалось спа-
стись; ему, впрочем, не придавали особого значения и не давали себе труда отыскивать
его. Заключенные в тюрьме дожили до Пасхи, по прошествии которой их казнили, но
не всех за раз и в один день. Их возили на телеге по всему городу, привязанных к стол-
бу и обнаженных до пояса. И палач держал на телеге готовый огонь, в котором он ка-
лил свои щипцы, и когда они накалялись, то на каждом перекрестке вырывал у них
кусок мяса. После того их привезли на площадь Моляр, где им отрубили головы на
эшафоте; тела же их четвертовали, и части эти разнесли так, чтобы выставить в раз-
личных местах, за исключением сына надзирателя, которому, во внимание к его мо-
лодости, только отрубили голову. Он признался, что умеет составлять микстуру отца,
и его лишили жизни не ради мести, но чтобы помешать распространению зла» (Les
Chroniques de Geneve, т. II, с. 395-401).
1.4. Контагионисты «сеют чуму»
51
Случай, приведенный Литтре, отличается от попыток распространения чумы, при-
писываемых евреям и прокаженным во время пандемии чумы «черной смерти». Два
века контагионистических предс гавлений о распространении чумы не прошли даром.
Заговорщики в Женеве действуют теперь более осмысленно. Они подбрасывают лю-
дям не колдовские ладанки с головами ящериц и лапками жаб, а вещи, пропитанные
гноем, извлеченным из чумных бубонов, т. е. они пытаются привести их к контакту с
«чумным контагием».
Чума воспринималась современниками как террор «сеятелей чумы». Власти их
целенаправленно ловили и уничтожали. В 1536 г. в Италии ни у кого не было сомне-
ния в том. что чуму возобновили сорок ведьм, наносившие специальные мази на двер-
ные ручки и косяки, от чего люди вымирали целыми семьями (Киттеридж Дж. Л.,
2005). Подобные процессы пов горились в Женеве в 1545 г. уже с участием колдунов.
Один человек под пыткой признайся, что намазал ступню повешенного волшебной
мазью, после чего натер ею запоры дверей, в результате по городу распространилась
чума. Кальвинисты стали искать заговор и, разумеется, нашли и заговор и заговор-
щиков. В ход пошли суровые наказания: у осужденных мужчин щипцами срывали с
костей п. ють. осужденным женщинам отрубали перед сожжением правую руку. С осо-
бым пристрастием пытали и допрашивали бедняков «дурной репутации», дабы выяс-
нить, не содействовали они распространению чумы. Тех «негодяев», которые от-
казывались признать свою вину, приковывали к стене и оставляли умирать. Не менее
43 человек тогда были обвинены в распространении заразы и 39 из них казнены. В мар-
те 1545 г. женевский палач Jean Grranjat принужден был казнить свою мазь, обвинен-
ную в распространении чумы. Скромный муниципальный сл\жащий все сделал так,
как это требовал от него закон: сначала он отрубил ей правую руку, а потом сжег за-
живо. Однако вспышки чумы повторялись, что свидетельствовало о плохой работе
инквизиции. Жан Кальвин (1531—1564) сетовал по этому поводу: «Тем не менее, кон-
спираторы не прекращают покрывать дверные замки своей мазью. Смотрите же, ка-
кие опасности нас окружают» (цит. по Китгериджу Дж. Л.. 2005).
В 1567— 1568 гг. в Женеве казнили еще 13 «разносчиков чумы», в 1571 г. — 36. В том
же году городской врач Жан-Антуан Саразен издал трактат о чуме, в котором он под-
тверждал, что эпидемия являлась делом рук «разносчиков чумы». В Шамбери (город
во Французском департаменте Савойя) в 1572 г. патрули получили приказ стрелять в
разносчиков заразы. В Фисиньи в 1571 г. по этому обвинению 5 женщин были сожже-
ны, 6 отлучены от церкви, а 25 преданы суду (Делюмо Ж., 1994).
В 1580 г. на юге Франции, в Эксе (Прованс), был «разоблачен» очередной «еврей-
ский заговор». Власт и «выяснили», что причиной эпидемии чумы стал яд, которым
евреи натирали дверные молотки. Этот яд был получен очень сложным способом, по-
этому он оказался таким эффективным. Оказывается, летом того же года, в один из
самых знойных дней, евреи заманили в свое жилище англичан ина, имевшего рыжие
волосы, привязали его к кресту, вставили ему в рот деревянную распорку, чтобы тот
не имел возможности его закрыть, и натравили на него несколько гадюк, которые жа-
лили его в голую спин}' Несчастный англичанин скоро скончался, и злоумышленни-
ки собрали слюну, вытекавшую у него изо рта, и приготовили из нее болезнетворную
мазь (Трахтенберг Дж., 1998).
Милан пережил страшное испытание «злонамеренно вызванной» чумой в 1630 г.
Люди были уверены, что на стены и двери общественных зданий и частных домов было
нанесено ядовитое вещество. В городе упорно поддерживались слухи, что этот яд из-
готовлен из змей и жаб, слюны и гноя больных чумой. Конечно, такую отраву могли
52
Часть 1. Краткая история применения биологических средств...
приготовить только по внушению дьявола те, кто вступил с ним в сговор. Но бдитель-
ные граждане все же обнаруживали «союзников дьявола». Пожилой мужчина молил-
ся на коленях в церкви. Захотев сесть, он вытер скамью подолом плаща. Увидев это,
женщины завопили, что он нанес на скамью отраву. Собралась толпа, мужчину изби-
ли, потащили в тюрьму и подвергли пытке. Достоверна трагическая кончина милан-
ского комиссара здравоохранения Пьяцца и цирюльника Мора, обвиненных в нане-
сении на стены и двери домов города подозрительного желтого жирного вещества,
вызывающего чуму. В Милане в 1630 г. была установлена монументальная колонна с
надписью на латинском языке: «Здесь, на этом месте некогда стояла лавка цирюль-
ника Джанджакомо Мора, вступившего в сговор с комиссаром здравоохранения Гу-
льямо Пьяцца и другими во время страшной чумы и посредством смертоносной мази,
которую они повсюду наносили, истребили множество народу. Посему они были
объявлены Сенатом врагами родины. Их пытали каленым железом, переломали кос-
ти и отрубили правые руки. Затем четвертовали, а через шесть часов умертвили и со-
жгли. Чтобы не осталось никакого следа от этих преступников, их имущество было
продано с торгов, а прах брошен в реку. И чтобы люди помнили об этом событии, Сенат
повелел снести дом, в котором замышлялось преступление, и на его месте воздвиг-
нуть колонну «Позора». «Сторонись, сторонись, честный гражданин, из страха всту-
пить на эту опозоренную землю. Август 1630 г.». Колонна простояла до 1778 г., напо-
миная, что люди, замышляющие против родины, заслуживают самого сурового
наказания (Делюмо Ж., 1994).
Каноник, записавший историю чумы 1630 г. в Бусто-Арсицио (Ломбардия), рас-
сказывал следующее: «Сначала французы одержали победу в Мантуе, но император-
ские войска их остановили. Тогда враги (автор рукописи был сторонником дома Габ-
сбургов) замыслили погубить жителей зараженным околдованным хлебом». Сначала
автор не верил в подобное злодейство, но это было настолько очевидно, «потому что
во многих местах находили этот хлеб, и сам я стал очевидцем этого» (Делюмо Ж., 1994).
В одном из своих трактатов основатель немецкого протестантизма Мартин Лю-
тер (1483—1546) писал о еще одном явлении, которое, по его мнению, имеет место во
время эпидемий чумы: «Существуют еще худшие преступники. Некоторые люди, чув-
ствуя, что заболевают, ничего об этом не говорят и общаются со своими собратьями в
надежде передать им пожирающую их заразу. Проникшись этой мыслью, они бродят
по улицам, входят в дома, даже пытаются поцеловать прислугу или детей, надеясь
таким способом спастись. Хочется верить, что эти люди действовали по наущению
дьявола, что только его следует обвинять. Однако я слышал, что зависть и отчаяние
толкает этих горемык на подобное преступление, потому что они не хотят болеть одни.
Право, не знаю, верно ли это? Но если это действительно так, то, кто же мы, немцы,
люди или демоны?» (цит. по Делюмо Ж., 1994).
По утверждению Д. Дефо (1722), среди врачей, наблюдавших Великую лондонскую
чуму 1665 г., шли споры о причинах появления людей, старавшихся передать свою
болезнь другим. Некоторые из них утверждали, что причина такого поведения людей
кроется в самой сущности болезни. Каждый больной одержим злобой и ненавистью
кдругим людям. Вредоносность болезни проявляется не только в физических призна-
ках, но и искажает саму натуру человека, подобно ворожбе или дурному глазу, или
подобно тому, как ведет себя взбесившаяся собака, которая до болезни была добрей-
шим животным, а теперь кидается и кусает любого, кто попадается ей на пути, вклю-
чая тех, к кому раньше была очень привязана. Другие врачи относили это явление на
счет испорченности человеческой природы.
1.4. Контагионисты «сеют чуму»
53
Доктор Ходжес, оставивший детальное описание Великой лондонской чумы, на-
стоятельно рекомендовал исключить любую возможность намеренного заражения
чумой здоровых горожан больными: «При чумной заоа $е, что может быть более безот-
лагательным, чем отделить здоровых от больных? И особенно при заболевании, ко-
торое проникает не только в тело, но и отравляет дыхание; ведь в таком случае дыха-
ние больного губит здоровых людей, и даже на пороге смерти заболевшие норовят
передать другим тот яд, что сразил их самих. Этим бредовым стремлением и объясня-
ются всяческие проделки с подсовыванием здоровым заразы из болячек зачумленных;
не говоря уже о той женщине, которая заключила в объятья своего несчастного мужа
и заставила его окончить жизнь вместе с ней» (пит. по Атаровой К. Н., 1997).
«Чисто биологическое убийство» Петра II? Убийство под видом естественной смер-
ти всегда было козырной картой в политических интригах. «Горе»-убийц в таких слу-
чаях «безмерно», и они не навлекают на себя немедленной мести и преследования со
стороны других политических групп. Фаворит Петра I и Екатерины I, светлейший
князь А. Д. Меншиков (1673—1729), понимал, как много в его жизни и жизни членов
его семьи зависит от царствования преемника Петра Великого — Петра 11 (сын царе-
вича Алексея и принцессы Софьи-Шарлотты Бланкенбургской — единственный ле-
гитимный русский царь после Петра I в ХУШ в.).
Меншиков усиленно хлопотал о передаче престола царевичу и и его браке со сво-
ей дочерью, надеясь таким образом сохранить влияние при дворе. Среди прочих сво-
их действий он принимал меры по ограждению юного государя от «моровых напастей».
В январе 1727 г. Меншиковым было подготовлено распоряжение, чтобы никого на
Васильевский остров (тогдашнюю резиденцию императора) из тех домов, где свиреп-
ствует оспа, не пропускали. Пытаясь примириться со старыми родами, он приблизил
к Петру II князей Долгоруких, те воспользовались ситуацией и восстановили импе-
ратора против Меншикова. Вскоре тот был арестован и с женой, сыном и дочерьми
сослан в Восточную Сибирь, в поселок Березов, где и умер 12 ноября 1729 г. Сам Петр II
перебрался в Лефортовскую слободу в Москве. Здесь он попал в брачную западню,
поставленную на него князем Алексеем Григорьевичем Долгоруким. Дальше проис-
ходит следующее. По сведениям историка медицины В. М. Рихтера (1767—1822), князь
Сергей Петрович Долгорукий (1697—1761), не обращая внимания на то, что дети его
болели оспой, продолжал упорно являться ко двору в Лефортовский дворец, что было
запрещено всем, кто находится в подобных обстоятельствах. Так и осталось неизвест-
ным, заразился ли юный император оспой от Долгорукого или из другого источника,
но эта болезнь оказалась для него роковой. В «Манифесте от Верховного Тайного Со-
вета» от 4 февраля 1730 г. говорится: «Понеже по Воле Всемогущего Бога, Всепре-
светлейший Державный Великий Государь, Петр Второй Император и Самодержец
Всероссийский, болезнуя оспою Генваря от 7 дня, от врямянного в вечное блаженство
того же Генваря 18 числа, в первом часу по полуночи отыде».
Историки игнорируют свидетельство В М. Рихтера. Мотивация действий князя
Сергея Петровича не очень понятна в свете распространенных интерпретаций этих
событий. Однако роль других князей Долгоруких в этих событиях также была длитель-
ное время неизвестной. Князья Сергей и Иван Григорьевичи и Василий Лукич Дол-
горукие сочинили подложное духовное завещание Петра II, но были случайно разоб-
лачены только в 1738 г. Иваном Алексеевичем Долгоруким (сын Алексея Григорьевича;
1708-1739) во время пыток, осуществлявшихся по отношению к нему при дознании
по другому делу. Видимо, участникам заговора, в котором участвовал Сергей Петро-
вич, удалось сохранигь его в тайне. О нем самом, по историческим источникам, уда-
54
Часть 1. Краткая история применения биологических средств...
ется собрать очень немногое. Сергей Петрович принадлежал к иной ветви Долгору-
ких, чем Алексей Григорьевич, и был женат на княгине Ирине Петровне Голицыной
(1700—1751). Ее родственник, Дмитрий Михайлович Голицын (1667-1737), сыграл ос-
новную роль в приглашении герцогини курляндской Анны на российский престол на
«ограничительных условиях». Сергей Петрович сделал успешную карьеру при импе-
ратрице Анне Иоанновне и умер своей смертью в своей постели. А вот Дмитрий Ми-
хайлович умер в Петропавловской крепости, куда его заключили по надуманному
предлогу, «лишив всех имений». В России же после смерти Петра II династия Рома-
новых по мужской линии оборвалась, и, по сути, произошел государственный пере-
ворот, приведший к власти Брауншвейгскую династию в лице Эрнест- Иоанна Бирона
(1690—1772). Так, изящным и, возможно, «чисто биологическим убийством» закон-
чилась эпоха Петра Великого.
Биологические диверсии конца добактериологической эпохи. Твердая уверенность
в прилипчивости контагия привела к тому, что, с целью заражения людей, злоумыш-
ленники активно стали использовать одежду больных опасными инфекционными бо-
лезнями либо людей, бывших с ним в контакте. Образовавшаяся в XVII в. неиммун-
ная к натуральной оспе прослойка среди населения способствовала результативности
таких преступлений, проводимых посредством вещей, находившихся в пользовании
оспенных больных. В июне 1763 г. сэр Джефри Амкерой послал полковнику Генри
Буквиту, командующему войсками в Пенсильвании, письменное предложение об
«изобретении способа распространения оспы среди недружественных индейских
племен», угрожавших Форту Питту. В своем ответе от первого июля Буквит сообщил,
что он предпринял попытку заразить одеяла, которые могли попасть в руки индей-
цев. Из этого ответа можно понять, что Буквит уже знал о том, что произошло. Капи-
тан Симеон уведомил Буквита о вспышке натуральной оспы в Форте Питте в начале
июня. Далее он записал в своем дневнике от 24 июня, что два одеяла и платок из
госпиталя оспенных больных были подброшены индейцам. На следующую весну оспа
широко распространилась среди индейцев, и были случаи смерти в племенах минго,
делавер и шони.
Однако распространить другие опасные инфекции с помощью одежды больных
не удавалось по непонятным для современников причинам. Селон А. Пруст сообщил,
что тунисские племена, захватившие равнинную часть Туниса, были поражены в 1785 г.
чумой. Они сделали несколько безнадежных попыток заразить этой болезнью хри-
стиан в Ла Калле, забрасывая туда через стену одежду умерших от чумы. В 1863 г. в
Вашингтоне контрразведкой северян был задержан доктор Блекберн (Luke Blackburn),
врач конфедератов и в будущем губернатор штата Кентукки. Он попытался вызвать
вспышки натуральной оспы и желтой лихорадки в войсках северян, продавая воен-
нослужащим одежду людей, больных этими болезнями. Был доказан, по крайней мере,
один случай гибели офицера северян от натуральной оспы, носившего одежду, куп-
ленную у Блекберна Вызвать же таким способом вспышку желтой лихорадки докто-
ру не удалось.
Начавшаяся во второй половине XIX в. массовая вакцинация и ревакцинация
войск и населения против натуральной оспы, обесценила этот способ биодиверсий
окончательно. В 1870 г. во время осады Парижа пруссаками французский врач пред-
ложил командованию оставить зараженную оспой одежду в месте, оставляемым фран-
цузской армией, чтобы пришедшие туда прусские войска могли заразиться. Тогда это
предложение не имело действия (Mollaret Н., 1995). Однако через год оспа уже без
всякого усилия со стороны человека, распространилась по Европе, России и северо-
!.5. Появление военной микробиологии
55
американскому континенту, не делая различий между городами население которых
было дважды вакцинировано, и теми, где вакцинация не проводилась, придержива-
ясь каких-то своих, неизвестных нам правил (Бразоль Л. Е., 1875).
Тем не менее угроза искусственного распространения эпидемий осознавалась ев-
ропейцами. По инициативе русского царя Александра П (1855—1881) ими даже под-
писывались различные соглашения, ограничивающие применения ядов и заразного
материала в будущих войнах. Среди них Петербургская декларация (1868), запретив-
шая употребление в европейских армиях разрывных пуль, Брюссельская декларация
о законах и обычаях сухопутной войны (1874) и так называемые Оксфордские реше-
ния (1880). Они включают среди других пунктов «отказ от применения в качес гве ору-
кия войны некоторых методов (подразумеваются яды), которые бесполезно увеличи-
вают страдания людей или неизбежно вызывают их смерть, равно как и тех, которые
могут распространяться на гражданские элементы».
* * *
Попытки биологических диверсий, предпринимавшиеся в добактериологический
период, как правило, оказывались неудачными, за исключением тех редких случаев,
когда в высоковосприимчивых популяциях людей распространялась одежда больных
натуральной оспой. Тем не менее массовое поражение людей искусственно вызванны-
ми эпидемиями считалось возможным как среди контагионистов, так и миазматиков
из-за большого количества мистификаций, неправильного понимания механизмов
происходящих на их глазах эпидемических катастроф и веры в сверхъестественные
явления.
1.5. ПОЯВЛЕНИЕ ВОЕННОЙ МИКРОБИОЛОГИИ
Аэробиологические эксперименты на форту «Александр I». Бактериологические
уоииства. Способы и устройства для массового производства патогенных микро-
организмов. Технологии и устройства для длительного хранения микроорганизмов
в жизнеспособном состоянии. Взгчяды на ведение бактериологической войны.
Сродства защиты от бактериального нападения и ликвидации его последствий.
Чума. Сибирская язва. Сап. Холера.
Открытие во второй половине XIX в. микроорганизмов — возбудителей опасных
инфекционных болезней, перевело средневековый контагий из абстрактного поня-
тия в материальный объект, бактериальную культуру. Ее свойства можно было изучать
в паборатории и применять для достижения различных целей, включая ведение био-
логической войны. Но «материали юция» контагия еще не означала появления в руках
военных нового оружия. Они далеко не сразу даже поняли, какие эксперименты ста-
ли начальной точкой в развитии, так и не пожелавшей до сих пор легализоваться об-
ласти научного познания, военной микробиологии.
Аэробиологические эксперименты на фор гу «Александр I». Толчком к взрывному раз-
литию медицинской бактериологии послужили работы немецкого ученого Роберта
Коха (1843—1910), разработавшего к началу 1890-х гг. способ получения чистых куль-
тур бактерий и большинство основных приемов работы в бактериологических лабо-
раториях. Они используются и поныне.
В рамках любой технологии или методологии всегда существуют так называемые
«опережающие объекты», вбирающие в себя все новинки научной и технической
56
Часть 1. Краткая история применения биологических средств...
мысли своего времени. «Опережающим объев гом» исследования в медицинской бак-
териологии на начало 1900-х гг. был возбудитель чумы (Yersinia pestis), открытый в
1894 г. в Гонконге А. Йерсеном (J. Е. Yersin, 1863—1943).
Такой интерес со стороны ученых именно к этому микроорганизму вызвало неожи-
данное возвращение чумы, с конца 1850-х гг. считавшейся «вымершей болезнью».
Тысячи трупов людей, умерших от чумы, собранных с улиц Кантона, Гонконга, Амоя
и других южно-китаиских портовых городов, позволили исследователям единодуш-
но утверждать о начавшейся в конце XIX в. «третьей пандемии чумы». Мир жил ожи-
данием чумной катастрофы, развивающейся по типу «черной смерти» 1346—1351 гг.,
когда основную роль в массовой гибели людей сыграли легочные формы болезни.
Однако с самой легочной чумой по-прежнему было много неясного. Вспышка ле-
гочной чумы в станице Ветлянская (1878) прекратила споры между научными авто-
ритетами о самой ее возможности. Выяснилась абсолютная смертельность этой фор-
мы болезни, и в то же время осталось непонятным ее происхождение. Еще во времена
«черной смерти» контагионист Ибнулкатиб утверждал, что многие люди, несмотря на
контакты с больными с «кровохарканием» (один из симптомов легочной чумы), ос-
тавались здоровыми, и, наоборот, «получали чуму вообще безо всяких контактов» (цит.
по Гезеру Г., 1867). Наблюдения эпидемиологии, клиники, и даже патологической
анатомии легочной чумы, сделанные разными исследователями в конце XIX и в на-
чале XX вв., не совпадали между собой. Легочная чума могла оказаться очень зараз-
ной, что и наблюдалось во время вспышки чумы среди киргизов в 1900 г. в Таловском
округе Внутренней Киргизской Орды (Страхович В. и Гос В. И.,1907). Но другие уче-
ные приходили к выводу о ее незаразности для европейцев, как это обнаружено в ин-
дийских городах Пали в 1836 г. (Гирш А., 1853) или в Бомбее в 1900—1901 гг. (Берест-
невН. М., 1907). Имелись наблюдения о незаразности легочной чумы как в отношении
туземцев, так и европейцев. Например, во время вспышки чумы в горном таджикском
кишлаке Анзоб в 1899 гг. случаев передачи легочной чумы между жителями или от них
к врачам, не наблюдалось, хотя никаких предохранительных мер не предпринималось
(Финкельштейн Я. М„ 1906). В селе Колобовка, где в 1899 г. вспыхнула легочная чума
среди русского населения, не было доказано ее распространение «по цепочке» (Роза-
нов П. Г., 1900). Анализ эпидемических цепочек, прослеженных во время вспышки
легочной чумы в станице Ветлянская (1878), не выявил какого-либо влияния на рас-
пространение болезни соседских или внутридомовых отношений между заболевши-
ми людьми (Минх Г. С1., 1898). Для выяснения причин появления таких эпидемий
требовалось воспроизведение легочной чумы на лабораторных животных.
Впервые в эксперименте удалось воспроизвести легочную чуму русским исследо-
вателям В. К. Высоковичу и Д. К. Заболотному в Бомбее в 1897 г. (Wyssokowitsch,
Zabolotny, 1897). Для этого они через зонд вводили возбудитель чумы в трахею усып-
ленных хлороформом обезьян. Животные погибали через 2—4 сут., обнаруживая все
явления первичной чумной пневмонии. Через два года после этих экспериментов, Ба-
заров (Batzaroff, 1899), сотрудник Института Пастера (Париж), вызвал пневмонию у
морских свинок, нанося им на слизистую оболочку носа сухую чумную пыль стеклян-
ной палочкой с ватной обмоткой. Животные, как он полагал, погибали от первичной
чумной пневмонии, но при этом наблюдались подчелюстные или шейные бубоны вто-
ричного происхождения. Только в отдельных случаях они имели вид первичных, лег-
кие же были мало поражены.
Сопоставление результатов патологоанатомического исследования лабораторных
животных, погибших от экспериментальной легочной чумы, и людей, умерших от
1.5. Появление военной микробиологии
57
ле1 очной чумы во время чумных эпидемий, по-прежнему не проясняло для ученых
основного вопроса, — каким же образом возникает легочная чума в эпидемических
очагах? Childe (1898) описал исключительно легочные поражения, Durck (1901) при-
вел доказательства развития чумной пневмонии как следствие бубонной, a Albrecht и
Gohn (1900) считали, что развитие такой пневмонии начинается с поражения верх-
них дыхательных путей, т. е., по сути, она является вторичным процессом. Методо-
логия же экспериментальных исследований Высоковича, Заболотного и Базарова во-
обще мало соответствовала условиям заражения чумой в эпидемических очагах.
Экспериментальные работы с возбудителем чумы в России были начаты в 1896 г.
в Санкт-Петербурге, в ветеринарной лаборатории Императорского института экспе-
риментальной медицины (ИИЭМ). Через два года эти исследования по настоянию
влиятельного родственника царя, председателя Высочайше утвержденной комиссии
о мерах предупреждения и борьбы с чумною заразой (сокращенное название «Комо-
чум», утверждена 11 января 1897 г. по старому стилю), принца А. П. Ольденбургского
(1844—1932), перенесли в специально созданную лабораторию, с более высоким уров-
нем биобезопасности. Она была расположена вблизи Кронштадта, на территории
форта «Александр I», выведенного за штат военного ведомства (рис. 1.7).
Рис. 1.7. Форт «Александр I». Верхняя фотография сделана Н М. Бсрестеневым
в 1907 г. Нижняя — форт в 2003 г., взята с сайта http://www.nortfort.ru
Форт основательно переоборудовали для работы с возбудителями особоопасных
инфекций (кроме возбудителя чумы, там работали с возбудителями сапа и холеры).
Достаточно сказать, что система обработки стоков из лабораторий форта, мало чем
отличается от аналогичных, имеющихся в современных лабораториях того же назна-
чения. По условиям работы и мерам охраны, новая лаборатория (полное название
58
Часть 1. Краткая история применения биологических средств...
«Особая лаборатория Императорского института экспериментальной медицины по
заготовлению противобубонночумных препаратов в форте “Александр I”») представ-
ляла собой типичное режимное учреждение, в котором часть сотрудников носили
военную форму (рис. 1.8).
Рис. 1.8. Бактериологические лаборатории второго этажа форта «Александр I». На
правой фотографии видна система вентиляции. Однако неизвестно была ли она
приточно-вытяжной. По Н. М Берестеневу (1907)
Одним из основных направлений работ лаборатории было экспериментальное
моделирование механизмов заражения людей во время вспышеклегочной чумы. Про-
водя эти эксперименты, русские ученые случайно открыли «сосуд Пандоры», за что
некоторым из них пришлось заплатить своими жизнями. Дело в том, что вызвать забо-
левание чумой у животного, распыляя культуры возбудителя чумы, оказалось не такой
уж простой задачей, как первоначально думали. Животные либо не заболевали легоч-
ной чумой, либо их болезнь напоминала сепсис и не затрагивала дыхательные пути.
Но механика создаваемых для заражения животных аэрозолей была тогда неизвестна
ученым, и, соответственно, меры специальной техники безопасности, необходимые
при работе с такими аэрозолями, не были внедрены в микробиологическую практи-
ку. Трагедии не заставили себя долго ждать.
В1903 г. легочной чумой заболел заведующий лабораторией, Владислав Иванович
Турчинович-Выжникевич (1865—1904). Расследование обстоятельств его гибели спе-
циальной комиссией показало, что он «занимался с 28 по 31 декабря 1903 г. опытами
по заражению животных распыленными культурами и участвовал в приготовлении
чумного токсина путем растирания тел чумных микробов, замороженных жидким
воздухом». Оба эти опыта могли с лужить моментом, способствующим заражению
ученого, но их методология и результаты не опубликованы. Судя по патологоанато-
мической картине, заражение Турчинович-Выжникевича произошло в результате
проникновения в его верхние дыхательные пути крупнодисперсного аэрозоля возбу-
дителя чумы.
1.5. Появление военной микробиологии
59
Агробиологические эксперименты на форту «Александр I» были продолжены
В. И, Госом в 1905 г. Сначала он попытался осуществить аэрозольное инфицирова-
ние экспериментальных животных «сухой чумной пылью». Гос отказался от методоло-
гии Базарова, полагая, что манипуляция внесения «пыли» в носовые отверстия мор-
ской свинки стеклянной палочкой с ватной обмоткой влечет за собой повреждение
слизистой оболочки носа. Поэтому он попытался инфицировать морских свинок
путем вдыхания ими значительного количества «пыли» в особом аппарате. И сразу же
получил принципиально важные результаты: в носовых отверстиях свинки были вид-
ны скопления «чумной пыли», однако чумой они не заболевали. Но когда перед опы-
том животному наносилась травма слизистой оболочки носа (царапины), то живот-
ное погибало от чумной бубонной инфекции, причем пневмония (вторичная) встре-
чалась не чаще, чем при подкожном заражении малыми дозами возбудителя чумы.
Таким образом, Гос опроверг результаты Базарова и установил, что когда «чумная
пыль» попадает на неповрежденную слизистую оболочку носа, заражение не насту-
пает Следовательно, для заражения чумой через легкие возбудитель должен попасть
в более глубокие его отделы. Нов какие ? Чтобы установить через какие отделы легко-
го происходит заражение чумой, он пошел по пути уменьшения размера частиц ин-
фицирующего аэрозоля.
Контролировать и задавать размер частиц «сухой чумной пыли» Гос не умел. По-
этому им был построен прибор для аэрогенного инфицирования экспериментальных
животных возбудителем чумы, в котором «животное могло бы вдыхать мельчайшую
водяную пыль, состоявшую из брызг чумной бульонной культуры или разжиженного
сока чумного легкого при том условии, чтобы само животное не покрывалось бы зара-
зой». Дисперсность частиц аэрозоля он уменьшал, повышая давление воздуха, пода-
ваемого на сопло распылителя. Конструкция прибора показана на рис. 1.9.
Рис. 1.9. Прибор В. И. Госа для инфицирования лабораторных животных
мелкодисперсным аэрозолем возбудителя чумы
Прибор состоял из двух частей:
1) из рас пылителя «Paroicm», употреблявшегося в те годы врачами для ингаляции
лекарственных растворов; Госу этот аппарат служил для распыления чумных культур;
2) из особой приемной камеры, приспособленной им к аппарату «РагоНп», в ко-
торую поступали аэрозольные частицы и фиксировалась голова животного.
60
Часть 1. Краткая история применения биологических средств...
Приемная камера состояла из стеклянного цилиндра (или простой колбочки Эр-
ленмейера без дна) (I), на одном конце которого надет резиновый колпачок с отвер-
стием, плотно обхватывавшим трубку аппарата «Parolein» (11), из которой поступал в
цилиндр аэрозоль («влажная пыль»). Другой конец цилиндра плотно закрыт резино-
вой пробкой, через которую проходили две стеклянные трубки: одна из них (1), изог-
нутая, с расширением, в котором помещалась гигроскопическая вата, служила пре-
дохранительным клапаном для выхода избытка воздуха, нагнетаемого баллоном (2),
другая (3) отводила мельчайшую «влажную пыль» в воронку (4), соединенную с труб-
кой (3) короткой резиновой трубкой (5). Широкая часть воронки закрывалась рези-
новым колпачком, плотно обхватывавшим воронку. В середине колпачка сделано от-
верстие (6), через которое просовывался «конец морды» животного, причем резина
плотно обхватывала морду. Голова животного крепко фиксировалась в станке и, кро-
ме того, воронка прикреплялась к голове шнурками (7—7). Чтобы лучше обеспечить
изоляцию между воронкой и наружным воздухом, голова животного обкладывалась
слоем ваты. Аппарат (1) фиксировался в штативе и вместе с животным обволакивал-
ся мокрой марлей, сложенной в несколько слоев. Распыление продолжалось в тече-
ние 5 мин. После этого аппарат оставлялся в покое в течение 5 мин, чтобы все капельки
успели осесть на стенки аппарата, затем воронка осторожно освобождалась от голо-
вы животного, и аппарат поступал в стерилизацию. Голова животного обмывалась
раствором сулемы, и животное помешалось в клетку.
Картина поражения тканей легких после инфицирования мелкодисперсным аэро-
золем возбудителя чумы, установленная Госом, значительно отличалась от той, кото-
рую описывали в те годы под названием «чумная бронхопневмония» у людей, забо-
левших легочной чумой во время эпидемий этой болезни. При типичных бронхопнев-
мониях воспалительный процесс в бронхах и в альвеолярной ткани был выражен
одинаково сильно, причем часто воспалительный процесс с бронхов переходил на ок-
ружающие альвеолы. При чумной пневмонии, развивающейся вследствие проникно-
вения мелкодисперсного аэрозоля возбудителя чумы непосредственно в альвеолы, на
первое место выступает воспалительный процесс в альвеолах, изменения же бронхов
незначительны. К патологоанатомическим и патоморфологическим отличиям различ-
ных форм чумной пневмонии мы еще вернемся в разд. 3.2, здесь же заметим, что спо-
соб инфицирования экспериментальных животных возбудителем чумы, выбранный
Госом (также как и способы, использованные Высоковичем и Базаровым), невозмо-
жен в ее природных очагах. Для дальнейшего изложения материала (см. разд. 1.9) нам
важны два его следующих обобщения:
1. «Главный путь, по которому зараза в наших опытах проникала в организм, —
это бронхиальная трубка, по которой воздух доносил зародышей до глубоких частей
легкого, до альвеол. Если бы палочки чумы не проникали до альвеол, а оседали бы
только на стенках бронхов, то, конечно, наиболее резкие изменения были бы в брон-
хах, а паренхима легкого участвовала бы в процессе вторично, т. е. была бы типичная
бронхопневмония. Однако этого не наблюдалось.
2. Меньшая подверженность слизистых бронхов действию возбудителя чумы, не-
смотря на то, что на нее он должен попадать в больших количествах, чем в альвеолы,
объясняется бактерицидными свойствами секрета слизистых оболочек верхних ды-
хательных путей. Строма альвеол в сравнении с бронхами во много раз богаче сосу-
дами; поэтому воспалительные явления, выражающиеся при чуме, главным образом,
сосудистой реакцией с ее последствиями: появлением экссудата, эмиграцией лейко-
цитов и пр., несравненно резче выражены в альвеолах».
1.5. Появление военной микробиологии
61
Через три года после опубликования Госом своих результатов, эпидемия первич-
ной легочной чумы в Маньчжурии расставила все на свои места. По данным вскрытий
70 человек, умерших от чумы в Харбине, Г. С. Кулеша(1912,1924) (сотрудникД. К. За-
болотного), нашел, что для первичной легочной чумы в эпидемических очагах харак-
терны следующие патологоанатомические явления. Возбудитель чумы, находясь в
воздухе в виде капельной инфекции, оседает при ингаляции не в самой легочной тка-
ни, а на слизистых оболочках рта, глотки, трахеи и крупных бронхов, что неизбежно
ведет к легочной чуме, но не прямо, как в опытах Госа, а через посредство первичного
поражения миндалин, слизистой трахеи и бронхов. Причем чумная инфекция транс-
портируется в легкие уже не через воздух, а при помоши крови, т. е. гематогенно. При
своем эпидемическом распространении легочная чума вызывает преимущественно
долевую пневмонию, правильнее, плевропневмонию, ввиду постоянного участия
плевры. Понятно, что заболевания такого происхождения возможны лишь при нали-
чии очень тесных контактов с больными легочной чумой, заражающими своим каш-
лем окружающий их воздух и обусловливающих нахождение в нем так называемой ка-
пельной инфекции с миллиардами клеток возбудителя чумы в каждой капле. Так как
результаты Госа не соответствовали столь очевидной и многократно подтвержденной
другими исследователями патологоанатомической картине, о них забыли.
Бактериологические убийства. Начались практически сразу после обнаружения
бактерий и их токсинов. В 1910 г. в России был арестован Патрик О’Брайн, польский
аристократ ирландского происхождения, намеревавшийся убить всю семью своей же-
ны для получения наследства. Он также подкупил польского врача для того, чтобы тот
убил его двоюродного брата с помощью инъекции дифтерийного токсина. В 1913 г. в
Германии был осужден за серию убийств Карл Хопф, пытавшийся убить свою третью
по счету жену с помощью возбудителей холеры и брюшного тифа (Atlas R. М., 1999).
Во Франции в октябре 1921 г. разбиралось дело Жирара. Вместе с ним были аресто-
ваны его жена, любовница, водитель его автомобиля и виноторговец. В 1910 г у 5 че-
ловек и в 1913 г. еще у нескольких человек этим преступникам удалось вызвать тяже-
лое заболевание путем дачи им культуры бацилл брюшного тифа и ядовитых грибов
(Дробинский И. Р., 1940).
В 1932 г. в Индии были осуждены Бенойендра Чандра и доктор Таран Бхачариа за
убийство сводного брата Бенойендра путем инъекционного введения ему культуры
возбудителя чумы в руку во время ссоры из-за наследства отца (Atlas R. М., 1999).
Разумеется, эти бесхитростные бактериологические убийства отдельных людей,
к тому же в обыденном сознании являющиеся не более чем криминалом, не могли по
своему исполнению дать какие-то технические подходы к убийствам массовым и тем
самым принести славу своим исполнителям. Ниже мы рассмотрим технические воз-
можности для ведения бактериологической войны, существовавшие перед Первой
мировой войной.
Способы и устройства для массового производства патогенных микроорганизмов.
Основным способом массового полу чения возбудителей инфекционных болезней пе-
ред Первой мировой войной было их выращивание на поверхности плотной пита-
тельной среды в пробирках, чашках Петри и в бактериологических матрацах. Выра-
щивание в жидкой питательной среде осуществлялось в пробирках или колбах и не
давало высокого выхода биомассы бактерий. Например, Gosio получал чумные вак-
цины из трехсуточных бульонных культур, выращенных в плос кодонных колбах. Сред-
ний выход влажной бактериальной массы на литр бульона у него не превышал 1,05 г
(цит. по Габричевскому Г. Н., 1907). Gosio даже не пытался определить, как такой
62
Часть 1. Краткая история применения биологических средств...
Рис. 1 10 Защитный колпак.
По С. И. Златогорову (1916)
способ культивирования влияет на вирулент-
ность и жизнеспособность чумных бацилл.
Ни в одном из каталогов бактериологическо-
го оборудования того времени не упомина-
лись устройства для искусственной аэрации
растущих бактериальных культур, ли^о дру-
гие устройства для масштабирования процес-
са культивирования микроорганизмов до пре-
делов, превышавших лабораторные нужды.
Выращивание микроорганизмов произ-
водили при строго определенной температуре
в специальных термостатах, которых к этому
времени существовало уже большое количе-
ство модификаций. Они были оснашены тер-
морегуляторами, подогревались газом, керо-
сином или электричеством. Главное внимание
в их конструкции обращалось на то, чтобы
температура в различных частях прибора была
одинаковая, и чтобы прибор хорошо сохранял
температуру. Термостаты делались различной
величины от небольших ящиков, вмещающих
литровую колбу, до больших термостатов-
комнат. Работа с опасными микроорганизма-
ми осуществлялась в простых боксах, называ
емых защитными колпаками (рис. 1.10).
По крайней мере, до эпидемии легочной чумы в Маньчжурии в 1910—1911 гг., на
лицах врачей, контактирующих с людьми и животными, больными чумой, невозмож-
но увидеть защитные маски, а на пуках специальные перчатки. На одной из фото! ра-
фий, сделанных в форте «Александр 1» в 1907 г., показана группа врачей без каких-либо
средств индивидуальной защиты кожи, глаз или органов дыхания, вскрывающих по-
гибшего от чумы верблюда (рис. 1.11).
Концентрирование суспензий микроорганизмов проводилось с помощью неболь-
ших центрифуг различных типов.
Технологии и устройства для длительного хранения микроорганизмов в жизнеспособ-
ном состоянии. Первые попытки высушивания микроорганизмов были предприняты
еще в средине 1870-х гг., т. е. тогда, когда представления о них самих были еще весьма
смутными. Объектом таких экспериментов стали различные неклассифицированные
ортопоксвирусы, применяемые для противооспенной вакцинации населения. Разу-
меется, производители таких вакцин не знали ни о существовании вирусов, ни о ка-
ких-либо их видах.
Эксперименты носили исключительно эмпирический характер. Оспенный детрит
(«вакцинная лимфа», «вакцинная материя»), снятый с кожи теленка, помещали в не-
большую чашку, а ту, в свою очередь, ставили в специальный эксикатор, в котором
влата поглощалась хлористым кальцием, серной кислотой, фосфорным ангидридом
и другими химическими поглотителями. В таких эксикаторах вакцинная лимфа «вы-
сыхала» через сугки-двое. Ее «прививная сила» сохранялась гораздо дольше, чем при
хранении по «прежнему способу» — между стеклянными пластинками или в трубоч-
ках (Миллер Н. Ф., 1887).
1.5. Появление военной микробиологии
63
Рис. 1.11. Вскрытие сотрудниками форта «Александр I» верблюда,
погибшего от чумы. По Н. М Бересте не ву (1907)
С получением в начале 1890-х гг. первых лечебных сывороток, появилось осоз-
нание необходимости разработки технологий их длительного хранения. Прежний
способ практиковался, но уже считался малопроизводительным и дорогим. В 1894 г.
М. Чарльс, профессор физиологии в Сиднее (Австралия), описал простой и быстрый
способ высушивания иммунных сывороток и сохранения их в стерильном виде. По-
средством небольшого стеклянного аппарата и насоса он высушивал до литра сыво-
ротки в течение 24 ч при температуре 40 °C.
Компания Маррель-Соуль (Сиракузы, США) в 1906 г. зарегистрировала патент-
ную заявку (№ 30062) с описанием аппарата для «выделения твердых веществ из жид-
кости посредством высушивания». Суть изобретения состояла в следующем. Бакте-
рии, фиксирующие азот, культивируются в стерилизованном снятом молоке, причем
полученная жидкость распылялась и высушивалась, смешиваясь е воздухом, нагре-
тым до температуры 65 °C. При этом бактерии оставались живыми. Однако изобрете-
ние не оказалось востребованным. Жидкость распылялась неравномерно, и высуши-
вание потоком воздуха не могло в принципе дать однородную сухую бактериальную
массу, особенно при работе с ее разными партиями. К тому же сухие бактерии тогда
не имели коммерческой ценности, основным спросом на рынке биопрепаратов
пользовались сухие сыворотки. Последнее обстоятельство и предопределило приори-
теты развития сушильной аппаратуры не только на все предвоенные годы, но и прак-
тически до начата 1930-х гг.
Сначала производители иммунопрепаратов стремились к выпуску концентриро-
ванных (сгущенных) сывороток, используя для этой цели «слабые» сыворотки, т. е. с
низким титром протективных антител. При этом они стремились к тому, чтобы вво-
дить человеку меньше белков для уменьшения последствий тяжелой сывороточной
болезни — анафилаксии. В масштабированном варианте высушивание сывороток
(налитых в кюветы и чашки Петри) и других биопрепаратов производилось теплым
воздухом (при температуре от 30 до 40 °C).
64
Часть 1. Краткая история применения биологических средств...
Наиболее популярным перед Первой мировой войной был сушильный термостат
А. Форнета, позволяющий контролировать эффективность процесса высушивания.
На боковой стенке он имел рычажные весы высокой чувствительности, сообщавши-
еся с термостатом, где к ним была подвешена чашка с навеской. По мере высыхания
биопрепарата чашка с навеской облегчалась, и стрелка на циферблате скользила вниз,
указывая количество удаленной влаги. Для более глубокого высушивания сывороток
были разработаны вакуум-термостаты. Они отличались от обычных сушильных шка-
фов тем, что имели для откачки водяных паров вакуум-масляные насосы и сушка
происходит при пониженном давлении и при температуре, близкой к 100°C. Однако
получаемые в таких аппаратах сухие препараты сывороток имели много недостатков.
Они представляли собой плохо растворимые в физиологическом растворе сухие плен-
ки или комочки, которые еще надо было размалывать и просеивать. Как правило, в
процессе сушки они обсеменялись посторонней микрофлорой. Попытки высушива-
ния таким способом микроорганизмов редко оказывались удачными. Уменьшение
содержания влаги в плазме бактериальной клетки приводило к увеличению концен-
трации солей, а это способствовало конгломерации белковых молекул и их необра-
тимой агрегации. Методические заделы к накоплению больших количеств микроор-
ганизмов, способных длительно храниться в жизнеспособном состоянии, перед
Первой мировой войной сделаны не были.
Взгляды на ведение бактериологической войны. Об их существовании можно судить
по ставшей известной практике распространения опасных для человека и животных
бактерий. По-прежнему наиболее распространенным способом бактериологической
войны оставалось сбрасывание трупов в общие источники воды. Правда, с открыти-
ем возбудителей инфекционных болезней загрязнять источники воды стали более
«осмысленно». Поданным Н. Блюменталя (1932), во время Англо-бурской войны
1899—1900 гг. обе воююшие стороны поступали следующим образом: при отступлении
воинских частей в колодцы и реки преднамеренно забрасывались трупы людей, по-
гибших от холеры. Такой же факт имел место в 3-й болгарской армии во время Бал-
канской войны (1912). И действительно, по крайней мере, во время Англо-бурской
кампании холера получила достаточно широкое распространение на театце военных
действий. Однако какую роль в этом сыграла «бактериологическая война», судить
трудно, так как эпидемиология холеры по сей день имеет много неясного.
Определенный отпечаток на взгляды военных стратегов того времени наложил
«просвещенный XIX век» с его идеями «правильной войны». Хотя договор о соблю-
дении Брюссельской декларации (1874) так и не был заключен, тем не менее она ста-
ла прологом более важного события — Гаагской мирной конференции 1899 г.
Инициатива по созыву Гаагской конференции и на этот раз принадлежала русско-
му царю. В августе 1898 г. министр иностранных дел России граф М. Н. Муравьев
(1845—1900) передал приглашение всем правительствам, аккредитованным при рус-
ском дворе в Санкт-Петербурге, принять участие в конференции с «целью предэтвра -
щения событий, создающих угрозу всему миру». По завершении конференции все
представленные на ней страны, за исключением США, подписали Конвенции «О за-
конах и обычаях сухопутной войны», к которой было добавлено «Приложение о законах
и обычаях сухопутной войны», состоящее из четырех отделов. В отделе II приложения,
имеющего подзаголовок «О военных действиях», в главе I «О средствах нанесения вреда
неприятелю, об осадах и бомбардировках», в ст. 22 записано: «Воюющие не пользуются
неограниченным правом в выборе средств нанесения вреда неприятелю». А ст. 23 по-
ясняла это положение: «Кроме ограничений, установленных особыми соглашениями,
1.5 Появление военной микробиологии
65
запрещается также: а) употреблять яд или отравленное оружие...; е) употреблять ору-
жие, снаряды и вещества, способные причинять излишние страдания...».
Через восемь лет, 5 (18) ноября 1907 г., в Гааге 46 странами были подписаны 13 кон-
венций и 1 декларация, касающиеся правил ведения войны. Конвенция «О законах и
обычаях сухопутной войны» имела «Приложение о законах и обычаях сухопутной войны».
В нем «слово в слово» были повторены вышеприведенные положения. Этим дипло-
матическим документом также было запрещено и бросание снарядов или взрывчатых
веществ с аэропланов или каким-нибудь другим подобным способом. Страна, нару-
шившая Конвенцию, даже обязывалась к выплате компенсаций.
Средства защиты от бактериального нападения и ликвидации его последствий. На-
кануне Первой мировой войны медицинская наука располагала почти всеми сегодня
принятыми подходами к распознанию инфекционных болезней и к противодействию
их распространения. В эпидемических очагах уже осуществлялась массовая вакци-
нация населения, были предложены вполне эффективные м< тоды дезинфекции, де-
зинсекции и дератизации. Кроме бактериоло! ических методов, в рутинной практике
диагностических лабораторий применялись и серологические исследования (см., на-
пример, работы Златогорова С. И., 1916; Габричевского Г. Н., 1907). В ряде стран,
и особенно в Российской империи, был накоплен уникальный организационный опыт
ликвидации крупных эпидемических катастроф.
Апофеозом противоэпидемических усилий России стала ликвидация маньчжур-
ской эпидемии легочной чумы 1910—1911 гг. По отношению к отдельным возбудите-
лям инфекционных болезней, эти знания были распределены неравномерно. Но в
отличие от ситуации сегодняшнего дня, в России того времени, по традиции сложив-
шейся со времени Екатерины II, все противоэпидемические мероприятия, предпри-
нимаемые во время крупных эпидемий, тщательно обобщались, собранные материа-
лы открыто публиковались. Бла1 одаря этому обстоятельству в распоряжении русских
врачей имелась великолепная эпидемиологическая литература, в которой приводи-
лись описания реальных, а не вымышленных эпидемических процессов (см., напри-
мер, труды Архангельского Г. Ф., 1874; Минха Г. Н. 1898; Кашкадамова В. П.,1902;
Богуцкого В. М., 1911; Касторского В. Н., 1911 и др.). Состояние средств специфи-
ческой профилактики и лечения инфекционных болезней, возбудители которых могли
быть использованы во время Первой мировой войны для поражения людей и живот-
ных, можно охарактеризовать следующим образом.
Чума. Возбудитель болезни носил тогда название Bacilluspestis. Различали три глав-
ные клинические формы инфекции: чуму бубонную, пневмоническую и кожную.
Природным резервуаром возбудителя болезни считались грызуны, а его переносчи-
ками инфицированные от больных грызунов блохи. Основную роль в распростране-
нии чумы отводили крысам, причем не только утверждалось, но и не подвергалось
никакому сомнению в принципе, что крысы, случайно проникшие на корабль, могут
разносить чуму на тысячи километров и вызывать эпидемии в крупных городах. Дру-
гие механизмы развития эпидемии чумы были изучены значительно хуже, но счита-
лось, что больной легочной чумой может заразить окружающих при кашле. О том, как
начинаются и почему угасают эпидемии легочной чумы, было известно мало. Резуль-
таты, полученные В. И. Госом (1907) при инфицировании животных мелкодиспер-
сным аэрозолем возбудителя чумы, не сыграли никакой роли в объяснении причин
распространения легочной чумы в Маньчжурии в 1910—1911 гг. и уже были забыты.
Первая лечебная противочумная сыворотка получена Иерсином в 1896 г. от лоша-
ди. Животных сначала иммунизировали эмульсиями из агаровых культур, убитых при
66 Часть I. Краткая история применения биологических средств...
температуре 58 °C, затем живыми бактериями, вводимыми прямо в шейную вену. Су-
ществовали противочумные сыворотки, полученные в результате иммунизации жи-
вотных отдельными очищенными антигенами (например, сыворотки Люстига и Гал-
леоти). Однако попытки их применения в очагах болезни показали отсутствия лечеб-
ного эффекта у больных с легочной чумой и очень сомнительный лечебный эффект у
больных с бубонной формой болезни. В основном противочумные сыворотки приме-
няли для экстренной специфической профилактики лиц, контактировавших с боль-
ными чумой, и для пассивной иммунизации медицинского персонала, длительное
время работающего в очагах чумы.
Более широкое распространение получила вакцинация населения убитой вакци-
ной Хавкина. В те годы были предложены и другие убитые вакцины (Безредки, Gosio,
Pfeiffer, Lustig и Galeotti и др. ученых). Считалось, что хавкинская вакцина создает
активный иммунитет, соответствующий перенесенной инфекции. Его продолжитель-
ность оценивалась в 3—6 и более месяцев. Первый сигнал о неэффективности убитых
вакцин в эпидемических очагах был получен во время маньчжурской чумной катаст-
рофы 1910— 1911 г., когда легочной чумой заболевали люди, дважды вакцинированные
вакциной Хавкина. Разочарование в противоэпидемической эффективности убитых
вакцин наступило только в 1930—1940 гг., после неудач в борьбе с эпидемиями чумы
на острове Ява и в Маньчжурии.
В те же годы велись работы по созданию живых противочумных вакцин. Однако
их считали непригодными для иммунизации людей из-за возможности случайного
восстановления вирулентности вакцинным штаммом.
Сибирская язва. Было установлено, что заразиться сибирской язвой можно при
контакте со шкурами больных животных и при употреблении зараженного мяса.
Ингаляционный путь заражения тоже был вполне очевиден для эпидемиологов в связи
со случаями болезни среди лиц, занимавшихся обработкой шерсти. Все существовав-
шие к началу Первой мировой войны средства специфической профилактики и ле-
чения предназначались только для животных. Для их иммунизации использовались
живые вакцины Л. Пастера (1822—1895) и Л. С. Ценковского (1822—1887). Противо-
сибиреязвенные сыворотки того времени предназначались только для снижения ко-
личества погибших животных после прививок живыми вакцинами. На практике это
достигалось комбинированным введением рогатому скоту сыворотки и вакцины.
Сап. Считалось, что естественное заражение возбудителем сапа (Bacillus mallei)
наблюдается почти исключительно у домашних животных из семейства лошадиных,
и только у этого семейства болезнь постоянно поддерживается и проходит все стадии
своего развития. Точное распознавание болезни достигалось комбинацией трех спо-
собов: получением чистой культуры микроба; путем прививок другим здоровым жи-
вотным, восприимчивым к сапу (жеребятам, ослам, собакам, кошкам); и путем вве-
дения маллеина. Сап в большинстве случаев встречался у людей, по роду своих занятий
приходящих в частое соприкосновение с лошадьми. Точно было установлено, что их
заражение происходило от содержимого нарывов и сапных изъязвлений, встречаю-
щихся у лошадей. Болезнь легко распространялась при тесном и большом скоплении
лошадей через корм, подстилку, сбрую, попоны, щетки, скребницы, губки и пр. Ле-
чение сапа у лошадей было запрещено санитарным законодательством всех стран, «так
как применение средств с гадательными результатами только может способствовать
распространению сапа». Применялась радикальная мера — уничтожение всех боль-
ных животных. Острые формы сапа у людей почти всегда заканчивались смертью.
Вакцин и сывороток нет и сегодня.
1.6. От бактериальных средств поражения к бактериологическому оружию...
67
Холера. Возбудитель болезни, называемый тогда Bacillus cholerae asiaticae, открыт
Робертом Кохом в 1883 г. Его относили к сапрофитам, размножающимся в воде, осо-
бенно если она богата органическими веществами. Было установлено, что заражение
холерным вибрионом человека может произойти только через желудочно-кишечный
тракт, так как Bacillus cholerae не выдерживает высушивания и потому не может нахо-
диться в живом состоянии в воздухе и вызывать заражение организма, например, че-
рез дыхательные пути. Серотерапия считалась бесполезным методом лечения холеры
ввиду быстротечности болезни. Определенные надежды возлагались на серопрофи-
лактику. Эффективным способом снижения заболеваемости в очагах холеры, счита-
ли противохолерную вакцинацию убитой вакциной.
Наиболее приемлемыми средствами в профилактике холеры считались: изоляция
больных, тщательная дезинфекция испражнений и всех зараженных вещей (главным
образом, платья и белья), и меры охранения питьевой воды от заражения и обеззара-
живание ее посредством различных физических и химических средств.
♦ * *
Сотрудник «Особой лаборатории» бактериолог В. И. Гос (1907) установил, что для
искусственного заражения чумой человека необходимо использовать мелкодисперс-
ные аэрозоли возбудителя болезни, способные достигать альвеол легких. Но понима-
ние «военно-прикладного назначения» биологических аэрозолей такой дисперснос-
ти придет только в конце 1940-х гг. Своими экспериментами Гос, по сути, показал крах
эпидемической значимости контагия, действовавшего по принципу «прикоснулся,
заболел». «Контагий чумы» забивал ноздри экспериментальных животных, но не вы-
зывал у них болезнь. Последующий анализ работ, предполагавших ингаляционное
инфицирование экспериментальных животных возбудителями опасных инфекций,
показал, что публикация Госа была проигнорирована учеными, занимавшимися сход-
ной тематикой. И это не удивительно. Понимать передачу возбудителей инфекцион-
ных болезней с позиций контагионистического учения Средневековья не только про-
ще, но, в части, касающейся вопросов создания БО, находясь на этой позиции, можно
с полной убежденностью верить в мифы о его дешевизне, доступности и эффектив-
ности. Однако именно эксперименты российского ученого Госа по ингаляционному
инфицированию животных, осуществленные на форту «Александр I», можно считать
исходным пунктом в развитии военной микробиологии.
1.6. ОТ БАКТЕРИАЛЬНЫХ СРЕДСТВ ПОРАЖЕНИЯ К БАКТЕРИОЛОГИЧЕСКОМУ
ОРУЖИЮ - БАКТЕРИОЛОГИЧЕСКИЕ ДИВЕРСИИ
ВРЕМЕН ПЕРВОЙ МИРОВОЙ ВОЙНЫ
США. Россия. Румыния. Испания. Аргентина. Великобритания.
Италия. Франция. Появление БО?
Развитие бактериальных средств поражения в годы Первой мировой войны про-
исходило на фоне триумфа химического оружия. К 1918 г. газобаллонные атаки были
почти полностью вытеснены массированными обстрелами химическими снарядами.
С появлением ОБ кожно-нарывного действия типа иприта, химическое оружие ста-
ло использоваться не только с тактическими, но и с оперативными целями. Его стали
применять в маневренной войне, как средство обороны, предназначенное для за-
ражения различных участков местности в полосе наступления противника. Всего
68
Часть 1. Краткая история применения биологических средств...
на полях сражений противоборствующими сторонами было испытано 44 боевых ОВ.
Ни одна из сторон, противодействующих Германии, не достигла столь впечатляющих
успехов в разработке боевых ОВ и в эффективности их применения. Одновременно
германскими военными были сделаны первые шаги в оценке поражающей эффек-
тивности другого потенциального средства массового поражения людей, ВО. И хотя
многие разоблачения того времени случайны и противоречивы, в большинстве сво-
ем они не выходят за пределы реальных возможностей военной бактериологии того
времени. Ниже мы рассмотрим их по странам, соблюдая хронологический порядок
предполагаемых бактериологических диверсий.
США. Объектом германских диверсий стали лошади и крупный рогатый скот.
Среди диверсантов, раскрытых американской контрразведкой на территории США,
оказался экстравагантный капитан ВМФ Германии Стейнметц (Erich von Steinmetz),
любивший выдавать себя за женщину. Он многократно пытался заразить сапом ло-
шадей, отправляемых на Западный фронт из портов США, но безрезультатно. Тогда у
него возникли сомнения в вирулентности используемых им культур, и, представля-
ясь исследователем, он отдал их в одну из лабораторий, чтобы определить, действи-
тельно ли они способны вызывать болезнь и смерть животных. Культуры оказались в
полном порядке. О дальнейшей судьбе Стейнметца мне не известно.
Другой бактериологический диверсант, Дилгер (Anton Dilger), получил медицин-
ское образование в США и даже специализировался на лечении ран в клинике при
Университете Джона Гопкинса (Мэриленд). После начала войны он отправился слу-
жить в германскую армию, но вскоре из-за нервного расстройства был отправлен
назад, в Виргинию, к родителям. Так как США тогда соблюдали нейтралитет в войне,
то проблем с перемещениями между странами у него не было. За свое кратковремен-
ное пребывание в германской армии он попал в поле зрения германской разведки и
был привлечен к программе бактериологических диверсий на территории США. Из
Германии Дилгер привез культуры возбудителей сибирской язвы и сапа. С помощью
своего брата Карла устроился в лабораторию частного колледжа в Chevy Chase (Ме-
риленд), где, не привлекая к себе внимания, занялся наработкой этих бактерий.
Забирал из «лаборатории Тони» («Tony’s lab») культуры бактерий сибирской язвы
и сапа некий капитан Хинш (Frederick Hinsch). Он жил в Балтиморе (штат Мериленд)
на пересечении улиц Charles и Redwood. Хинш инфицировал ими лошадей, которых
собирали в Балтиморе перед отправкой в Европу. Дилгер попытался создать вторую
лабораторию по производству биоагентов в St. Louis (Миссури), но отказался от этой
идеи после того, как из-за холодной зимы все его бактериальные культуры замерзли
и погибли. О бактериологических лабораториях и диверсиях немцев в США стало из-
вестно только после войны (Side! Е R. et al., 1997).
Россия. Дориан Блер, во время войны выполнявший в России задания разведок
Антанты, в своих «Записках шпиона и диверсанта» (1937) утверждал, что в 1915 г. в
Россию неким Грегерсеном, немецким агентом, прибывшим из США в Архангельск,
были доставлены культуры возбудителя чумы. В дальнейшем предполагалось в лабора-
тории Самарского университета ими заразить крыс и выпустить их в Петрограде. Блер.
являвшийся участником этого заговора, одновременно работал и на русскую контрраз-
ведку, поэтому проект заражения чумой населения Петрограда не был осуществлен.
В декабре 1916 г. норвежская полиция арестовала на границе с Россией бывшего
офицера шведской лейб-гвардии барона Розена и еще нескольких лиц. При них бы-
ли обнаружены 10 пачек взрывчатого вещества в виде плотничьих карандашей и тру-
бочки с возбудителем сапа (Звонарев К. К., 2005).
1.6. От бактериальных средств поражения к бактериологическому оружию...
69
Румыния. По данным Марценака (1935), в августе 1916 г„ накануне объявления
войны Румынией Австрии, немецкий консул австрийского, пограничного с Румыни-
ей, городка Кронштадта (ныне румынский Брассов), через специального курьера дип-
ломатической почты отправил посылку из нескольких ящиков и одной коробки в ад-
рес болгарского посольства в Бухаресте под официальными печатями своего
консульства. На багаже имелась надпись на немецком языке: «Совершенно секрет-
но. Сверхосторожно. Господину М. Костову для военного атташе при императорской
эолгарской миссии в Бухаресте, господину полковнику А. М. Самаржиеву». Посыл-
ку сопровождало запечатанное письмо с такой же надписью: «Совершенно секретно»,
адресованное атташе при германской миссии полковнику А. М. фон Гаммерштейну.
Таким образом, окольными путями, через болгарское посольство, ящики пришли
германскому военному ат гаше. Они не привлекли внимания румынских властей к гер-
манским агентам и были спрятаны в подвальном помещении германского посольства.
Но неожиданно, на 8 дней раньше, чем это предполагали, началась война между
Румынией и Германией. Спешный отъезд персонала немецкого посольства не позво-
лил использовать "аинственную посылку или передать ее в надежные руки д ля соот-
ветствующего применения. Защита германских интересов в Румынии, помещения и
архивы посольства были переданы представителю САСШ (США) в Бухаресте, а об-
ременительные и компрометирующие ящики наспех зарыты в саду посольства.
Получив туманные сведения об этих действиях от оставшегося неизвестным ис-
точника, префект полиции в Бухаресте Корбеско после длительных переговоров по-
учил 5 октября 1916 г. разрешение на производство обыска в помещении германско-
го посольства. Обыск был произведен немедленно в присутствии первого секретаря
американского посольства М. Виллиама Эндрюса.
Агенты М. Маркус и А. Марфтей, оставленные для охраны германских материаль-
ных ценностей, выкопали в присутствии представителей властей спрятанные пред-
меты, которые тут же были отправлены в компетентные лаборатории для анализа.
В ящиках оказалось взрывчатое вещест во — тринитротолуол (50 шашек по кило-
грамму каждая и шнуры Бикфорда); назначение его для диверсионных целей вполне
очевидно. В коробке же под слоем ваты находились продолговатые деревянные фут-
лярчики, и в каждом из них была стеклянная ампула, наполненная желтоватой жид-
костью. В сопроводительной бумаге на немецком языке разъяснялось следующее:
«Здесь прилагается одна склянка для лошадей и четыре — для рогатого скота. Приме-
нять— как согласовано. Каждая ампула рассчитана на 200 животных. По возможности
прививать прямо в рот; если это невыполнимо — примешивать к корму. Просьба —
кратко известить о результате. Приезд г-на М. К. (Костова) на один день желателен».
Вторая находка также не представляла никаких двусмысленностей для полиции
и контрразведки Румынии. Такие указания, как вирулентные свойства для разных
животных, дозировка, способ употребления, позволили им сделать вывод, что здесь
идет речь не о лабораторных опытах, а о подготовке бактериологической диверсии.
На это же указывали способ пересылки, секретная сопроводительная и соседство с
взрывчатыми веществами. Требуемый отчет о достигнутых результатах позволил сде-
лать предположение, что лица, которые должны были использовать данные материа-
лы, hi ради лишь роль апробаторов нового метода войны. Попытки же бактериологиче-
ских диверсий должны были расширяться, и в какой-то мере масштаб этого расшире-
ния зависел от полученных результатов.
Исследование содержимого ампул было поручено румынским правительством
доктору Бабешу (V. Babes, 1854—1926) — известному ученому и директору Института
70
Часть 1. Краткая история применения биологических средств...
патологии и бактериологии в Бухаресте. В своем докладе Бабеш дал следующее зак-
лючение: «Присланные для исследования образцы содержат, один - кульгуру сиби-
реязвенной палочки, другой — культуру бацилл сапа». По мнению Марненака, эти
находки представляют собой первые, ставшие известными и точно установленные
попытки ведения бактериологической войны. В свое время о них было сообщено
Министерством иностранных дел Румынии союзным правительствам и нейтральным
государствам, и поэтому же поводу был заявлен энергичный протест германским вла-
стям, одиозно злоупотребившим дипломатической неприкосновенностью.
Румынские власти не смогли раскрыть всю диверсионно-бактериологическую
сеть в стране. Уже после войны стало ясно, что не только в румынской, но и в грече-
ской кавалерии среди конюхов было много германских агентов. «Конюхи» регулярно
снабжались культурами сапа которые они подмешивали в корм лошадям. Поэтому
огромное количество конского поголовья обеих армий погибло от сапа (Дробин-
ский И. Р., 1940). Если принять во внимание то обстоятельство, что бульонные куль-
туры возбудителя сапа н< хранятся юлго, особенно вне холодильных камер, то можно
предположить, что на территории Румынии и Греции действовали тайные бактерио-
логические лаборатории, которые и поставляли инфицирующий материал герман-
ским диверсантам.
По свидетельству Liepmann (1937), над румынскими городами германские летчи-
ки сбрасывали шоколад и другие продукты питания, а население питалось этими
продуктами, предполагая, по-видимому, что о нем заботится какая-гобла!О1Воритель-
нал организация. В дальнейшем оказалось, что шоколад был отравлен ботулиничес-
ким токсином и вызвал ряд тяжелых отравлений среди жителей.
Испания. В книге Л. Ривьера (1937), посвященной деятельности германской раз-
ведки в Мадриде за годы Первой мировой войны, приводится ряд фактов, свидетель-
ствующих о широком использовании с 1916 г. бактериальных препаратов для зараже-
ния лошадей и мулов, закупаемых для французский армии. Причем сами препараты
(видимо, возбудители сапа и сибирской язвы) получали на месте, и, как сообщал гер-
манский военный атташе в Берлин, они оказались «удачными».
Аргентина. По данным Л. Ривьера (1937). агентом, известным ему как «Арнольд»,
полученные в Испании бактериальные препараты были вывезены в 1916 г. в Арген-
тину, где они широко использовались для заражения скота, предназначенного для
экспорта в страны Антанты. «Арнольду» помогал в этом деле германский бактерио-
лог, доктор Герман Фишер, которого он представил к награде. В сентябре 1917 г. «Ар-
нольд» отправил в Месопотамию четыре судна, имевшие на борту 4,5 тыс. мулов, за-
раженных сапом. В феврале 1918 г. «Арнольд» с полным правом мог сказать в своем
донесении, что, благодаря его деятельности, экспорт лошадей из Аргентины во Фр ш-
цию полностью прекратился.
Великобритания. Поданным F R. Sidel с соавт. (1997), наиболее «удачной» из осу-
ществленных в годы войны бактериологических диверсий в огношении британски::
войск, оказалась германская диверсия в Месопотамии в 1917 г., когда немцам удалое:
вызвать вспышку сапа среди 4500 мулов. Видимо, Sidel et ai. и Ривьер описали одну и
ту же диверсию.
Италия. V. Ferret! (1931) привел доказательства попыток немцев вызвать сап сре-
ди конского поголовья итальянской армии. Е R. Sidel et al. (1997) упоминают о вспыш-
ках холеры в расположении итальянских частей, вызванных германскими агентами.
Франция. В марте 1917 г. попытки заражения армейских лошадей сапом были рас-
крыты уже на Западном фронте. У немецкого агента, задержанного в зоне военных
1.6. От бактериальных средств поражения к бактериолоё ическому оружию...
71
действий, был найден набор для осуществления бактериологических диверсий. Он
ключал: металлическую трубу со стеклянной Oj гылкой продолговатой формы, напол-
ненную микробной культурой; и кисточку, укрепленную на железном прутике, изог-
нутом на противоположном конце в виде ручки. В инструкции, найденной у агента,
। называлось, что «бульонную культуру употреблять, или, вы гивая ее на фураж, пред-
зазначенный для немедленного скармливания лошадям, или способом носового сма-
зывания посредством кисточки, смоченной жидкостью; по возможности делать же-
лезным прутиком царапину в ноздре лошади» (Звонарев К. К., 2005).
Попытки заражения лошадей сапом продолжались. Уже 6 июня 1917 г, т. е. менее
чем через 3 месяца после первого официального извещения, циркуляр Главной квар-
тиры вновь подтвердил вышеизложенные факты, констатируя «дальнейшие неприя-
тельские попытки рассеивания контагия в различных точках нашей территории».
Sartoiy A., Sartoiy R. (1935) утверждали, что на французском фронте было предпри-
нято несколько подобных попыток (один из них, в силу своих военно-служебных
обязанностей, имел в руках эти предметы, которые были ему даны в целях анализа и
установления их предназначения). В некоторых районах Франции, очищенных от
германцев в октябре 1918 г., французскими контрразведчиками были найдены мате-
риалы, предназначенные для бактериологических диверсий. Циркуляр Главной квар-
тиры по поводу этих находок сообщал, что обнаружены деревянные ящики с надпи-
сью «осторожно, заразные материалы». Трубочки, упакованные в этих ящиках,
заключали в себе культуры вирулентных микробов, что можно заключить из текста
наклеек на них. «Открывать опасно. Безотлагательно передавать в нетронутом виде в
армейские лаборатории» (Марценак, 1935).
На французской территории в 1917 г. также была осуществлена первая успешная
попытка заражения скота вирусной инфекцией - ящуром (Liepmann. 1937). Таким
образом, в начале 1917 г. бактериологическая война была вынесена за пределы зоны
военных действий и перенесена в глубь страны, деморализуя мирное население и
нарушая снабжение французской армии.
Организация и осуществление биодиверсий против войск Антанты, видимо, сто-
или жизни многим германским микробиологам и диверсантам из-за крайне высокой
опасности работы с сапом. Вакцин, защищающих от заражения возбудителем сапа,
нет и сегодня; лечение сапа после случайного заражения возможно только антибио-
тиками, но в ге годы их не существовало. Летальность среди заразившихся людей, без
лечения антибиотиками, достигает 10и %. В конце 1918 г., уже после окончания вой-
ны, все работы с возбудителем сапа в Германии были запрещены из-за невозможно-
сти обеспечения безопасности персонала микробиологических лабораторий.
Появление БО? В 1929 г. видный английский хирург, лорд Бэрклей Майниган
(Berkeley George Andrew Moynihan, 1865-1936), опубл,ковал в одном из медицинских
журналов письмо, в котором бросил упрек германской армии, действовавшей во вре-
мя войны в 1918 г. на Западном фронте, в том, что германская авиация сбрасывала в
расположение английской армии специальные бомбы, «начиненные микробами,
сходными с чумными палочками». В ответ на это сообщение Майнш ана появилось
опровержение английского бактериолога Комманса (Commins), сообщившего, что он
считает этот факг совершенно невероятным, принимая во внимание то обстоите п>
ство, что самим немцам угрожа га бы серьезная опасность, если бы инфицированные
крысы, перебежали бы из английских окопов в немецкие. Кимманс та^же счи гал, что
бакгерии, выброшенные при взрыве снаряда или бомбы, погибнут от воздействия
высокой температуры, которую развивает взрывчатка. Немцы не вступили в дискус-
72
Часть J. Краткая история применения биологических средств...
сию английских ученых. Если такой факт имел место, то он мог быть частью экспе-
риментальных исследований возможности поражения людей аэрозолем, образую-
щимся при взрывном диспергировании специальным боеприпасом бактериальной
суспензии. По мнению А. Н. Де-Лазари (1935), конструкция германских химических
снарядов не предполагала интенсивного воздействия пламени взрыва на ОВ. Оно
находилось в свинцовой специальной емкости, вкладываемой в цилиндрическую
часть снаряда. Так как после войны и почти до конца 1930-х гг. весьма активно обсуж-
далась возможность использования химических снарядов для применения бактери-
альных агентов, ниже приведены их схематические изображения (рис. 1.12).
Рис. 1.12. Германские химические снаряды конца Первой мировой войны.
А. Схема химического снаряда «синий крест» (1917—1918 гг.): 1 — ОВ (дифе-
нилхлорарсин); 2 — футляр для ОВ; 3 — разрывной заряд; 4 — корпус снаряда.
Б. Схема снаряда «двойной желтый крест» (1918 г.): 1 — ОВ (иприт); 2 — диа-
фрагма; 3 — разрывной заряд 4 — корпус снаряда (Военно-хим. дело, 1942)
Приведенные выше факты не отражают всех установленных случаев использова-
ния бактериальных средств в Первую мировую войну. В послевоенной литературе
упоминается о таких фактах, как заражение в 1915 г. в Вашингтоне фуража, направ-
ляемого в Европу; о «известном цюрихском процессе о бульонных культурах, выбро-
шенных в реку»; и ряде других подобных эпизодов. Однако не приводится никаких
деталей таких акций либо делаются ссылки на недоступные сегодня источники.
Кроме прямого диверсионного применения бактериальных агентов, бактериоло-
гическая воина велась непрямыми, т. е. организационными методами. Известно, что в
ту войну кавалерийские части противника преднамеренно оттеснялись в районы,
наиболее пораженные сапом; пехота — в самые завшивленные села, пораженные
сыпным тифом и т. д. Этими приемами пользовались все противоборствующие сто-
роны (Дробинский И. Р., 1940).
* *
В годы Первой мировой войны бактериальные агенты использовались для осуще-
ствления небольших диверсий, проводимых диверсантами-одиночками с помощью
бульонных или агаровых культур, приготовленных за несколько суток до их диверси-
онного применения, либо посредством распространения зараженных животных.
О результативности таких диверсий немецкий ученый Н. Klotz (1937) писал следую-
щее: «Мы признаем, что тогдашние (в 1918 г.) бактериологические эксперименты
предприняты с недостаточными средствами и были направлены не непосредственно
1.7. Становление бактериологического оружия между мировыми войнами
73
против людей, а лишь против лошадей и рогатого скота противника. Но это ограни-
чение (к сожалению) обусловливалось не моральными соображениями или гуманны-
ми побуждениями, а представляло лишь нежелательное явление, связанное с недоста-
точным в то время развитием бактериологической науки». Тем не менее обращает на
себя внимание умелая организация немцами биодиверсионного подполья на терри-
ториях стран, противостоящих Германии. Своей многочисленностью оно компенси-
ровало ограниченные возможности диверсионного способа применения бактериаль-
ных агентов для заражения скота и фуража. Ввиду невозможности длительного
хранения жидких бактериальных культур, их готовили в тщательно замаскированных
лабораториях, размещенных на территории противника или нейтральных погранич-
ных стран. Грамотный выбор бактериальных агентов и способов их применения сре-
ди реально возможных в то время, а также искусное планирование таких операций и
их масштабность свидетельствуют о привлечении к подготовке бактериологических
диверсий лучших германских бактериологов. Эти события Первой мировой войны еще
ждут своих исследователей.
Сегодня не так уж и важно, было ли применение описанных Б. Майниганом бомб
в действительности или нет. В ту войну людей убито более 10 миллионов и без приме-
нения БО. Главное в его заявлении то, что в 1918 г. на Западном фронте произошло
качественное изменение в представлениях о средствах и способах биологического
поражения людей. На смену бактериальным средствам поражения в виде жидких куль-
тур возбудителей опасных инфекций, распространяемых диверсантами путем зараже-
ния продуктов питания, скота, фуража и водоисточников, пришло понимание воз-
можности создания БО, т е. специальных боеприпасов и боевых приборов со
средствами и системами доставки, способных на обширных территориях вызывать по-
ражение людей биологическими агентами. В то же время понимание механизмов зара-
жения людей и распространения эпидемий после применения БО оставалось на уров-
не контагионистических заблуждений прошлого, что создавало иллюзию быстрого
успеха в создании недорогого оружия массового поражения. Гонка биологического
оружия началась.
1.7. СТАНОВЛЕНИЕ БАКТЕРИОЛОГИЧЕСКОГО ОРУЖИЯ
МЕЖДУ МИРОВЫМИ ВОЙНАМИ
Подведение итогов - Женевский протокол 1925 г. Послевоенные взгляды на
ведение бактериологической войны. Микроорганизмы - потенциальные агенты
БО. Способы и устройства 1930-хгг., предназначенные для ведения бактерио-
логической войны. Критические взгляды на БО. Мистификация Стида. Ошибки
профессора Триллья (A. Trillat). Повышение вирулентности бактерий. Техноло-
гии и устройства для производства и длительного хранения микроорганизмов
в жизнеспособном состоянии. Бактериологические диверсии в СССР. Военно-
биологическая программа Польши Возобновление бактериологических дивер-
сий в ходе военных действий. А как же немцы?
После окончания Первой мировой войны стала очевидной принципиальная воз-
можность создания нового вида оружия — бактериологического. Возбудитель инфек-
ционной болезни виделся военным теоретикам в качестве «контагия», способного
самостоятельно распространяться по войскам противника и проникать в глубь его
74
Часть 1. Краткая история применения биологических средств...
территории, вызывая сокрушительные эпидемии среди населения. Сама же бактерио-
логическая война представлялась им весьма эффективной и не требующей особых
затрат, особенно если начать ее первыми.
Подведение итогов - Женевский протокол 1925 г. После окончания Первой миро-
вой войны Германию принудили к подписанию в июле 1919 г. Версальского договора.
Статья 171 этого договора содержала следующее положение’ «Учитывая, что приме-
нение удушающих, ядовитых или других газов, или же аналогичных жидкостей, ве-
ществ или средств запрещено, Германии строго запрещается изготавливать или им-
портировать их». Аналогичные формулировки были включены державами-победи-
тельницами в мирные договоры, заключенные со всеми союзниками Германии.
В феврале 1922 г. в Вашингтоне была проведена международная конференция,
первоначально имевшая своей целью ограничение численности ВМС. В США ей пред-
шествовали обширные исследования в области химического оружия, по результатам
которых американское руководство пришло к выводу, что оно должно был запрещено.
Поэтому США предложили Вашингтонской конференции резолюцию, запрещавшую
химическое оружие, по своей сути повторявшую аналогичные положения Версаль-
ского договора. Большинство представителей стран — участниц конференции, резо-
люцию утвердили, но отметили, что она не имеет силы договора, и, следовательно, ее
ожидает участь Гаагской конвенции. Сенат же США немедленно утвердил резолюцию,
не приняв во внимание разногласия между странами.
Мысль о возможности применения патогенных бактерий в военных целях была
публично высказана экспертами сразу после войны, когда стали обобщаться сведе-
ния о бактериологических диверсиях германских агентов. Но только в 1923 г. смешан-
ная комиссия по разоружению при Лиге Наций заинтересовалась опасностью приме-
нения БО. Ею были привлечены к работе по уяснению технически возможных форм
ведения бактериологической войны крупные бактериологи того времени: Мадсен,
Бордэ, Пфейффер, Каннон (Madsen, Bordet, Pfeiffer, Cannon). В целом эксперты от-
неслись отрицательно к целесообразности создания БО, но не к самой возможности
его создания. Свою позицию они обосновывали тем. что действие БО невозможно
измерить и локализовать, а также постоянной опасностью переброски инфекцион-
ных болезней через линию фронта от неприятеля к своим войскам. У экспертов выз-
вало серьезное опасение возможность продолжения эпидемии и по завершению во-
енных действий. Эксперты высказали некоторые частные мнения:
Каннон признал совершенно осуществимым мероприятием заражение микроба-
ми почвы и уничтожение урожая.
Бордэ указал на микроб мальтийской лихорадки (возбудителя бруцеллеза), как на
идеальный агент для бактериологической войны.
Пфейффер считал возможным распространение патогенных бактерий гранатами
и шрапнелью.
Главные же выводы комиссии экспертов, представленные в докладе Лиги Наций,
состояли в следующем:
1) действие БО не может быть ни изменено, ни локализовано;
2) с применением культур тифа и холеры для заражения водных источников мож-
но бороться путем фильтрации воды;
3) распространение чумы посредством мышеи опасно также для нападающего, как
и для подвергающегося нападению;
4) опасность распространения сыпного тифа посредством вшей значительно пре-
увеличена;
7.7. Становление бактериологического оружия между мировыми воинами
75
5) бактериологическое оружие не допускает возможности получения решительных
результатов, так как при современном состоянии гигиены и микробиологии можно
прекратить распространение эпидемии.
Из этих выводов следует, что эксперты не исключали возможности бактериоло-
гической войны, хотя и старались смягчить ее значение. Однако любопы гным пред-
ставляется то обстоятельство, что уже в 1923 г. БО, еще не применявшееся даже в так-
тических операциях, рассматривалось как оружие массового поражения. Например,
в докладе этой же комиссии было стмечено. что «химические и бактериологические
методы придают будущей войне особенно бесчеловечный характер и доводят опа-
сность войны до крайнего предела, вплоть до угрозы существованию человечества и
цивилизации». Собрание Лиги Наций, заслушав этот доклад, ограничилось тем, что
поручило совету: 1) опубликовать доклад комиссии и 2) озаботиться самой широкой
популяризацией этого вопроса (Блюменталь Н., 1932).
В результате прений в Женеве был выработан следующий протокол.
«Нижеподписавшиеся полномочные представители от имени соответствующих
государств:
считая, что применение на войне удушливых, отравляющих или подобных газов,
так же всяких аналогичных жидкостей или веществ, по справедливости было осужде-
но общим мнением цивилизованного мира;
считая, что запрещение этого применения сформулировано в договорах, к кото-
рым присоединилась большая часть государств мира:
в целях заставить признать во всем мире это запрещение включением как неотде-
лимую час гь в международное право, которое одинаково обя сательно для сознания и
для практики государства, — заявляют:
что Договаривающиеся Высокие Стороны, поскольку они еще не присоединились
к этим договорам, признают это запрещение, принимают распространение этого запре-
щения на ере детва бактериологической войны и согласны рассматривать себя, как свя-
занных между собой, согласно этой декларации.
Договаривающиеся Высокие Стороны приложат все усилия, чтобы побудить дру-
гие страны примкнуть к настоящему протоколу. Это присоединение будет сообщено
Президенту Французской Республики и через него всем государствам, подписавшим-
ся и примкнувшим к этому протоколу. Это присоединение будет действительным со
дня оглашения, сделанного правительством Французской Республики.
Настоящий протокол составлен в Женеве 17 июня 1925 г.»
Протокол подписали: Германия, США, Австрия. Бельгия, Британская империя,
Канада, Ирландия, Франция, Индия, Болгария, Чили, Китай, Колумбия, Дания,
Египет, Испания, Эстония, Абиссиния, Финляндия, Греция, Венгрия, Италия, Лит-
ва, Латвия, Люксембург, Норвегия, Панама, Голландия, Персия, Польша, Португалия,
Румыния, Сальвадор, Сиам, Швеция, Швейцария, Сербия, Чехословакия, Турция,
Парагвай, Венесуэла. Однако в 1920-х гг. ратифицировати этот протокол только Фран-
ция, Венесуэла, Австрия, Египет и Италия. США не ратифицировали протокол, так
как при его обсуждении в сенате возникла политическая оппозиция, этот вопрос был
снят с повестки дня и не обсуждался до 1969 г. СССР ратифицировал его 5 апреля 1928 г.
Единственной крупной страной, не ратифицировавшей протокол, была Япония.
Протокол не предусматривал процедур контроля или рассмотрения жалоб, а так-
же санкций за его нарушение. Он запрещал лишь применение, но не запрещал изго-
тавливать и хранить в неограниченных количествах химические и бактериологические
агенты и оружие.
76
Часть 1. Краткая история применения биологических средств...
Для понимания существовавших после Первой мировой войны представлений о
БО, обратимся к докладам экспертов специальному комитету по химическому и бак-
териологическому оружию, сделанным на конференции по разоружению, проведен-
ной в 1924 г. Лигой Наций.
Профессор ди-Нола (di-Nola), работая совместно с профессорами Майером
(Мауег) и Зильбершмидтом (Silberschmidt), рассмотрели вопросы, касающиеся под-
готовки к бактериологической войне, и пришли к вполне определенным выводам.
«Эта подготовка, — утверждали они, — должна быть строго запрещена». Затем они
утверждали, что в действительности невозможно воспрепятствовать какому-либо го-
сударству проводить такую подготовку.
Далее эксперты поставили вопрос о том, каков будет характер бактериологиче-
ской войны. Ниже по работе Sartory А. и Sartory R. (1935) воспроизводится их мнение
по этому вопросу.
«Проблема бактериальной войны представляется совершенно иной, чем пробле-
ма войны химической. В области химической войны у нас есть уже пережитый нами
опыт. Напротив, бактериальная война представляется для нас еще загадочной. Мы
можем только представлять себе, какова она может быть, каким образом она может
быть подготовлена, и какими способами следует предохранять себя от нее.
Мы знаем, что именно представляет собой внезапно вспыхнувшая эпидемия. Мы
знаем, что она может быть вызвана, например, заражением вод вирулентными бацил-
лами. Таковы эпидемии, вызываемые бациллой Эберта (возбудитель брюшного тифа)
или холерным вибрионом. Некоторые эпидемии вызываются распространением па-
разитов, носителями которых являются или очень распространенные во всем мире
животные, как крыса (чума), или насекомые, имеющие жало, как комар-анофелес
(малярия) и т. д. Таким образом, о средствах распространения спонтанной (внезапной)
эпидемии у нас уже есть представление, но о том, что такое экспериментальная эпи-
демия, поражающая людей, мы совсем не знаем. Следовательно, мы не располагаем
достаточными данными, чтобы судить о размерах и важности возможной бактериоло-
гической войны.
О действии бактериальных возбудителей и вообще о заразных болезнях мы знаем
лишь то, что эти болезни различаются между собою длительностью инкубационного
периода и часто весьма типичными признаками, а также то, что отдельные индивиду-
умы чувствительны к заразным болезням в неодинаковой степени. Следовательно,
действие бактериального нападения не может быть заранее вполне реально предусмот-
рено, и опасность будет, конечно, колебаться в зависимости от положения страны (тро-
пики, Европа), от того или иного времени года, от того или другого индивидуума».
Обращаясь затем к опасности бактериологической войны, эксперты заявили:
«Принимая во внимание наши современные познания в области вакцинации, мы
не думаем, что возможный агрессор сможет вакцинировать свое население против
заразной болезни и пользоваться затем этой болезнью для совершения агрессии про-
тив другой страны. В отношении всех эпидемий, кроме оспы, — этой, очевидно, един-
ственной болезни, против которой можно эффективно вакцинировать людей, — аг-
рессор сам будет подвергаться большому риску заражения. Нужно прибавить, что, если
он вызовет эпидемию на территории противника, а затем сам займет эту территорию,
он подвергнется большому риску. Его собственные войска могут сделаться носителями
возбудителей и передать заболевание в тыл и внутрь с граны. Немногим меньше будет
опасность и для обороняющейся страны, если она станет применять бактериальное
оружие хотя бы лишь в случае отступления, потому что эпидемия распространится ведь
1.7. Становление бактериологического оружия между мировыми войнами
77
не только по фронту, но и в глубь страны. Теоретически возможно предположить пе-
редачу возбудителя на расстоянии с целью избежать опасности для самой страны, но
практически будет невозможно ограничить вызванную эпидемию одной только при-
фронтовой полосой.
Эффективная борьба против бактериальной войны может быть с наибольшим ус-
пехом достигнута в государстве, где придается огромное значение вопросам гигиены.
Хорошо поставленная в мирное время санитарная служба может быть лучшим оружием
против бактериального заражения. Чем больше будет развита санитария, тем меньше
будет опасность от применения средств бактериальной войны. Но мы не можем утвер-
ждать, что даже в совершенстве налаженная санитарная служба сможет наверняка
справиться с эпидемией, вызванной искусственным путем (epidemic experimentale)».
Дальше профессора ди-Нола, Майер и Зильбершмидт поставили следующий во-
прос: «При помощи каких средств может быть осуществлена бактериальная война?»
По их мнению, возможно, с одной стороны, использовать бактериальные культуры,
вирулентность которых сохранена или повышена в лабораториях. С другой стороны,
агрессор может взять патологический вирулентный материал, добытый от людей или
животных; с этой целью он использует, например, фекальные массы, мочу, гной, тру-
пы и т. д. Нужно также иметь в виду возможность передачи возбудителей через таких
носителей, как насекомые и грызуны; вошь может быть использована для передачи
сыпного тифа, комар-анофелес—для передачи малярии, крыса — для передачи чумы,
инфекционной желтухи и т. д. «Имеются ли средства осуществить контроль над этим
патогенным материалом?» — спрашивают далее эксперты.
«Практически — это невозможно в настоящее время во всех случаях и во всех стра-
нах. Можно получить и сохранить вирулентные культуры в каждом госпитале и вооб-
ще везде, где имеются инфекционные заболевания. Лаборатории, готовящие вакцины,
обязаны сохранять культуры заразных болезней и даже повышать их вирулентное гь.
Само изучение бактериологии требует культур и свободного обмена этих культур
между лабораториями различных стран. Нельзя же мешать развитию бактериологии,
ограничивая опыты, приводящие в действие вирулентные культуры, так как эти ис-
следования ведут, прежде всего, к гуманной цели. Нельзя запретить обмениваться
определенным опытом, имея в виду борьбу против какой-либо предполагаемой опас-
ности. Поэтому контроль над подготовкой к бактериологической войне нам кажется
трудно осуществимым.
Каждое государство должно сознавать моральную ответственность, вытекающую
из факта производства на его территории бактериолог ических исследований. Оно дол-
жно проводить на своей собственной территории особый надзор над общественными
и частными институтами, занимающимися бактериологическими исследованиями,
таким порядком, чтобы быть в состоянии, в случае необходимости, дать ответ, если бы
возник вопрос об его ответственности».
Таким образом, протоколом от 17 июня 1925 г. большая часть государств мира
единодушно осудила бактериологическую войну. Одновременно эксперты признали
невозможность со стороны наднациональных структур контролировать создание БО.
Дтя них таким оружием являлись не специальные боеприпасы и боевые приборы со
средствами и системами доставки, а бактериальные культуры, инфицированные жи-
вотные и их эктопаразиты. Сам же возбудитель инфекционной болезни виделся ими
в качестве «контагия», отсюда у них эта убежденность в способности искусственно
вызванных эпидемий «перебрасываться» через линию фронта, «поражать свои вой-
78
Часть 1. Краткая история применения биологических средств...
ска», «распространяться в глубь территории» и т. п. Крысиная теория поддержания
возбудителя чумы в природе превращала его в глазах экспертов в самый опасный
микроорганизм из тех, которые можно использовать для ведения бактериологичес-
кой войны. Сама же бактериологическая война представлялась им как масштабное
распространение чумных крыс.
Послевоенные взгляды на ве, [ение бактериологической войны. Интерес к бактерио-
логической войне достиг своего апогея в начале 1930-х гг., когда стало ясно, что мно-
гие страны обзавелись своими наступательными программами по БО. По мнению ряда
западных исследователей того времени, БО имело следующие преимущества перед
химическим оружием и обычными видами вооружений.
Ниже они приведены в обобщении И. Р. Дробинского (1940):
возможность соблюдения секретности при подготовке к биологической войне: круп-
ные военные металлургические заводы и химические производства трудно скрыть,
микробиологические лаборатории, особенно частные, могут существовать длитель-
ное время, никак не обнар] живая себя;
быстрота под. отовки к бактериологической войне: считалось возможным получить
в течение относительно короткого промежутка времени большое количество патоген-
ных бактерий;
относительная дешевизна изготовления БО: стоимость изготовления БО рассчиты-
вали из цены питательно бульона для наработки бактериальной массы;
трудности индикации и быстрого обнаружения патогенных микробов в воздухе, воде,
в пище и пр.: гарантия того, что противник не сможет вовпемя обнаружить примене-
ние БО;
возможность распространения возникших заболеваний самими заболевшими: приме-
нение БО создает эпидемические очаги, из которых от человека к человеку распрос-
траняется возбудитель инфекционной болезни;
сильное моральное воздействие на противника: бактериальная война рассматрива-
лась как один из факторов дезорганизации противника;
большое экономическое значение последствий применения БО: в этом аспекте пробле-
мы у европейцев уже был опыт — заражение германскими диверсантами лошадей в
США, Греции, Румынии и Италии сапом, во Франции ящуром и сапом нанесло боль-
шой экономический ущерб этим странам. Теперь же предполагалось заражать не толь-
ко животных, но и сельскохозяйственные растения;
наличие технических возможностей для массированного применения БО: авиация
может пепеорасывать бактериологические боеприпасы на большие расстояния, что
было еще невыполнимым в Первую мировую войну. Тем самым открылась возмож-
ность применять БО вне полосы непосредственного соприкосновения со своими вой-
сками;
простая самозащита от собственного БО: сторона, применяющая БО, может иметь
заранее разработанные вакцины и сыворотки и другие средства защиты по отноше-
нию к возбудителю инфекционной болезни — агенту БО
Касаясь цели бактериологической войны, британский специалист М. \felu (1935)
писал: «Микробная война, к сожалению, возможна. Ничто не воспрепятствуез при-
менению этого боевого метода. Это оружие, конечно, не заменит других, по оно по-
может разрежению личного состава войск, дезорганизации транспорта и, особенно,
деморализации армии и тыла. Оно относится к тем фактам, которые могут внести в
наиболее методичные планы панику и точнейшие расчеты превратить в катастрофу»
(цит. по Дробинскому И Р., 1940).
1.7. Становление бактериологического оружия между мировыми войнами
79
Объектами нападения с применением БО могут стать люди, животные и растения,
различные населенные пункты и места сосредоточения войск. Можно ожидать зара-
жения водоисточников, молочных, консервированных и других пищевых продуктов,
помещений, почвы, фуража и воздуха (Саркисов И. 3., 1940).
D. Klein (1935) подчеркивал важное значение массированного применения БО как
наиболее эффективного способа ведения бактериологической войны. Подполковник
медицинской службы польской армии J. Karyszkowski (1935) утверждал, что ареной
бактериологической войны станет вся страна противника, его гражданское население
и войска. Этот апологет польского БО, видимо по прошлому опыту германского «стар-
шего брата», считал развертывание массовых бактериологических диверсий основным
способом ведения бактериологической войны.
В 1930-х гг. наиболее эффективным способом применения БО большинством ученых
считалось распыление бактериальных агентов в воздухе. Особое значение придавалось
комбинированному применению бактерий и отравляющих веществ, а также бактерий
и дымов. Предполагалось, что ипритные поражения кожи и дыхательных путей облег-
чают инфицирование людей. В некоторых случаях комбинированное применение
бактерий с отравляющими веществами может способствовать распространению ин-
фекционных заболеваний среди населения и войск противника (Саркисов И. 3., 1940).
Процесс подготовки к бактериологической войне виделся перед Второй мировой
войной довольно в упрощенном свете. Многими прожектерами того времени (как,
впрочем, и нынешними) считалось, что поскольку наработка возбудителей инфекци-
онных болезней, относящихся, как тогда выражались, к группе «боевого назначения»
(возбудители чумы, холеры, дизентерии, сыпного и брюшного тифов, сапа, мелиои-
доза), не требует оборудованных бактериологических лабораторий, то достаточно
иметь простенькую лабораторию, которую можно замаскировать другим предназна-
чением или законспирировать (Саркисов И. 3., 1940?. А еще лучше переместить ее на
1 ерриторию противника для обеспечения деятельности диверсионных групп (Karysz-
kowski J., 1935).
Все тот же J. Karyszkowski превзошел по гнусности всех других европейских апо-
логетов биологической войны. Польский подполковник предложил использовать
лагеря военнопленных «для экспериментального изучения путей распространения
возбудителей инфекционных болезней и обоснования необходимых для бактериоло-
гической воины данных» (цит. по Кроткову Ф. Г., 1939; и Зам П., Линкину Г., 1936).
Видимо, безнаказанное массовое истребление десятков тысяч русских пленных, за-
хваченных поляками во время Советско-польской войны 1920 г., придало некоторым
польским «стратегам» уверенность в том, что такие преступления можно совершать,
исходя из любой практической необходимости, и не бояться возмездия. Karyszkowski
цинично пояснил свою позицию: «Много чудовищных вещей творится на свете».
Невозможность использования БО в связи с риском, сопряженным для применяю-
щей стороны, полностью отрицал Н. Klotz (1937). По этому поводу он писал следую-
щее: «Вопрос о том, может ли это, в принципе пригодное, оружие быть использовано
в отдельном случае, или же особые обстоятельства делают его применение опасным
для собственных войск и собственного населения, является, в сущности, только вопро-
сом тактики. В этом отношении БО ничем не отличается от многих других боевых
средств. Существуют на войне положения, в которых приходится отказываться от
использования того или иного средства, однако это ни в коей мере не означает обще-
го отказа от данного оружия, являясь только следствием тактических соображений в
определенных условиях. Так, например, в условиях позиционной войны, когда про-
80 Часть 1. Краткая история применения биологических средств...
тивники отделены расстоянием всего в несколько метров, приходится отказываться
от обстрела окопов неприятеля тяжелой артиллерией, чтобы не поразить собственных
войск; далее, в позиционной войне приходится также отказываться от обстрела хи-
мическими снарядами в тех случаях, когда их приходится направлять против ветра.
Однако эти тактические мероприятия для зашиты собственных войск ни в какой мере
не обрекают на неудачу действия тяжелой артиллерии или химических снарядов.
Между тем в других условиях тяжелая артиллерия, химический снаряд оказались при-
годным и часто единственно действительным боевым средством. На войне встреча-
ются такие обстоятельства, в которых использование какого-либо определенного
оружия представляется столь же неправильным, как в других обстоятельствах оно
является правильным и необходимым. Мы повторяем: вопросотом, имеются ли в кон-
кретном случае предпосылки для применения какого-либо определенного оружия,
как, например, бактериологического оружия? Ответ на него дает тактика на основа-
нии особых условий данного положения, как направление и сила ветра, метеороло-
гические условия, характер почвы и местности, близость собственных войск и насе-
ления. Ответ в одном случае может быть, безусловно, утвердительным, в другом —
безусловно, отрицательным, а в третьем — условным. Но этот вопрос ни в какой мере
не затрагивает проблемы принципиальной возможности ведения бактериологической
войны, т. е. ее научной, технической и военной возможности... В суровой военной дей-
ствительности, о которой мы говорим, дело будет только в том, чтобы тщательно взве-
сить, достаточно ли оправдана вероятность поражения собственного населения при
несомненном поражении противника».
Flick (1927) полагал, что БО в будущей войне может применяться на фронте, в по-
лосе непосредственного соприкосновения с противником. Однако местом его преиму-
щественного использования будет глубокий тыл противника, где опасность переноса
эпидемии на свои войска значительно меньше. Причем свои войска и население долж-
ны быть предварительно проиммунизированы в отношении используемого агента БО.
По мнению И. 3. Саркисова (1940), возможны два крайних варианта войны, при
которых целесообразно применение БО.
Вариант 1-й — война длительная, на истощение.
Вариант 2-й — блицкриг, война молниеносного удара, для уничтожения главных
сил и быстрого приведения в расстройство его тыла.
И. 3. Саркисов (1940), видимо, по результативности немецких химических атак
1915 г., считал, что БО целесообразнее применять при втором варианте ведения вой-
ны, так как неожиданность его действия даст наибольшую эффективность поражения
противника. Применять его второй раз будет малополезным занятием для нападаю-
щей стороны из-за того, что все необходимые меры противником уже будут приняты.
Микроорганизмы — потенциальные агенты БО. Обсуждение в открытой научной
литературе биологических свойств патогенных микроорганизмов на предмет их при-
годности для ведения бактериологической войны стало активно проводиться с конца
1920-х. гг. Исследователям уже тогда была ясна «военно-биологическая ущербность»
наиболее смертельных из них. Однако даже наиболее авторитетными учеными того
времени считалось, что недостатки того или иного агента БО можно в будущем (при-
чем в ближайшем) «подкорректировать», а пока целесообразно найти им место в пред-
стоящей войне «по способностям».
Итальянский бактериолог Феррети (Ferrati V., 1931) к микроорганизмам, пригод-
ным для целей бактериологической войны, относил возбудителей кишечных инфек-
ций (вирулентные штаммы кишечной палочки, брюшного тифа, паратифов, дизенте-
1.7. Становление бактериологического оружия между мировыми войнами
81
рии, холеры), чумы, сибирской язвы, сапа, столбняка. По его мнению, из них наиболее
удовлетворяют требованиям бактериологической войны возбудители холеры, брюш-
ного тифа, чумы; для поражения животных — возбудители сибирской язвы и сапа.
Первым детальную разработку вопроса о потенциальных агентах БО сделал Люс-
тиг (Lustig, 1931; нит. по Блюменталю Н., 1932). К числу возбудителей инфекций,
которые могут быть использованы в бактериологической войне с наибольшей эффек-
тивностью, он относит возбудителей сапа и сибирской язвы. Он считал, что военное
значение возбудителя сапа обусловлено его значительной контагиозностью для лоша-
дей и высокой восприимчивостью к нему людей. Отсутствие коммерческих специфи-
ческих профилактических и лечебных сывороток и вакцин создает огромное преиму-
щество в бактериологической войне неприятелю, создавшему эти средства первым.
Люстиг знал о низкой выживаемости возбудителя сапа в окружающей среде, но он
считал, что его высоковирулентной культурой легко «произвести затравку пастбищ,
питьевой воды и фуражных запасов на площадях значительных размеров».
Что касается возбудителя сибирской язвы, то к его основным достоинствам, как
агента БО, Люстиг относил устойчивость во внешней среде и способность противо-
стоять действию дезинфицирующих средств, жара, водяного пара и солнечного све-
та. Однако он видел у сибиреязвенного микроба и серьезный недостаток, ограничи-
вающий его применение в бактериологической войне в ближайшее время — низкую
вирулентность для людей и сельскохозяйственных животных. «Если бы удатось как-
нибудь повысить вирулентность возбудителей сибирской язвы так, чтобы имеющие-
ся в настоящее время специфические средства предупреждения и борьоы оказались
бы недействительными или слабо действительными для противника, то эта инфек-
ция могла бы стать для него чрезвы аайно тяжелой», — писал он. Поэтому основное
военное применение В. anthracis — заражение пастбищ, фуража, питьевой воды и
пищевых продуктов.
Возбудители желудочно-кишечных инфекций (брюшной тиф, холера, дизенте-
рия), по его мнению, имеют ограниченные возможности для применения в качестве
агентов БО из-за наличия вакцин. Однако и они заслуживают внимания, поскольку
вакцинация при массированном заражении не предупреждает заболеваемость и не
снижает смертность. Особенно опасным агентом БО мог бы быть возбудитель сып-
ного гифа, «если бы удалось найти возбудителя инфекции и получить его культуру».
Возбудитель чумы считался им слишком опасным из-за возможности непрогно-
зируемого распространения на свои войска. Поэтому Люстиг предполагал целесооб-
разным его использование только после создания надежных средств специфической
профилактики и лечения чумы. Средствами же распространения инфекции могли
быть культуры возбудителя чумы, а также зараженные им крысы. Последний способ,
т. е. использование промежуточных хозяев для распространения инфекций среди
людей при ведении бактериологической войны, Люстиг считал наиболее универсаль-
ным. По его мнению, лошади могли разносить сап, собаки — бешенство, вши — сып-
ной тиф и т. д.
Майор медицинской службы американской армии Фокс (Fox, 1933; цит. по Сар-
кисову И. 3., 1940) считал пригодными для использования в военных целей возбу-
дителей газовой гангрены и бактериальные токсины, в частности ботулинический.
Использование бактерий кишечной группы (возбудители брюшного тифа, холеры и
дизентерии), по его мнению, могло оказаться малоэффективным, так как против за-
ражения источников водоснабжения и колодцев можно бороться с помощью фильт-
рации и обеззараживания, но он допускал диверсионные действия в отношении
82
Часть 1. Краткая история применения биологических средств...
пищевых продуктов. Наибольшее военное значение Фокс придавал инфекциям, пе-
редающимся насекомыми (сыпной тиф, желтая лихорадк„, болезнь Денге) и возбу-
дителю сибирской язвы. Фокс допускал возможность заражения почвы на месте сра-
жения возбудителями столбняка, газовой гангрены и сибирской язвы, «поскольку они
в почве хорошо и долго сохраняются».
Фокс также, как и Люстиг, и, видимо, независимо от него, считал распростране-
ние чумы среди войск противника опасным экспериментом для нападающего. Легче
и проще, по его мнению, вызвать чуму среди гражданского населения тыла путем
сбрасывания с самолетов зараженных крыс. Такой же точки зрения на возбудитель
чумы придерживался Марценак (1935), видимо, вообще мало что знавший об эпиде-
миологии чумы. Поэтому считавший, что при пря1 юм заражении человека от чело-
века или через блох возбудитель чумы распространяется с чрезвычайной легкостью,
его способность к рассеиванию так же велика, как велик процент смертности и мас-
совость эпидемических вспышек.
Генерал-майор медицинской службы французской армии Ромье (Romieu, 1934;
цит. по Саркисову И. 3., 19ч0) классифицировал возбудители инфекционных заболе-
ваний с военной точки зрения на три группы.
К первой он относил возбудителей чумы, холеры, дизентерии, брюшного тифа, па-
ратифов, сибирской язвы, мальтийской шхорадки (бруцеллез), считая эту rpvnny, как
бы «группой боевого назначения» или «действующей группой».
Вторая, вспомогательная группа, включала возбудителей дифтерии, эпидемиче-
ского цереороспинального мениш ита и желтой лихорадки. Эта группа, по его мнению,
может сыграть роль в благоприятных случаях.
К третьей группе, «неизвестных микроорганизмов», он относил возбудителей опа-
сных инфекций, еще не выделенных в чистых культурах (натурал! ной оспы, гриппа,
скарлатины, свинки и сыпного тифа). Их военное значение он считал минимальным.
A. Sartory и R. Sartory (1935) разделили возбудителей инфекционных болезней на
две категории:
1 Микробы, патогенные для человека, способные вызывать эпидемии среди войск;
2 . Бактерии, поражающие животных и вызывающие эпизоотии.
В отличие от других исследователей, они решительно отвергали любую возмож-
ность использования для ведения бактериологической войны микроорганизмов, ма-
лопатогенных для людей и живот ных. По их мнению противник будет пытаться вы-
звать настоящие эпидемии, «носяшие характер внезапных вспышек». Такие эпидемии
могут в решительный момент не только остановить, но и гадержать на более или ме-
нее длительный срок те войсковые части, которые предназначены для от правки на оп-
ределенные пункты фронта, где положение может стать критическим; такие эпиде-
мии могут также уничтожить базы снабжения или же сильно их расстроить.
J. KaAszkowski (1935) классифицировал потенциальные агенты БО, исходя из их
«диверсионных свойств». К первой группе он относил возбудителей инфекций, пере-
даваемые через воду и продукты питания; ко второй — возбудителей «эпизотехниче-
ских заболеваний» (к ним он относил сап и бруцеллез); и к третьей группе — микро-
организмы, передающиеся через укусы насекомых. Наиболее пригодными для
диверсионного применения он считал возбудителей сапа и бруцеллеза, наименее —
микроорганизмы третьей группы, как «требующих громадной работы, которую труд-
но осуществлять в неприятельской стране» (цит. по Зам П. и Линкину Г., 1936).
Ф. Кротков (1939) отмечал, что количество видов патогенных микроорганизмов,
пригодных для использования с военными целями, весьма ограничено. К возмож-
1.7. Становление бактериологического оружия между мировыми войнами
83
ности открытия возбудителей новых инфекционных заболеваний, имеющих военное
значение, он относился весьма скептически. Важное военное значение, по его пред-
ставлениям, имеют эндемические очаги, которые могут быть использованы в условиях
военного времени для развертывания эпидемий в стране противника.
Наиболее благоприятными для бактериологической войны, с точки зрения со-
хранения вирулентности в лабораторной культуре, по мнению Кроткова, является
«микрококк мальтийской лихорадки» (т. е. возбудитель бруце-щеза). Военное значе-
ние последнего усиливается в связи с возможностью заражения бруцеллезом через
кожу, слизистые оболочки и дыхательные пути. Хотя бруцеллез не является смертель-
ной болезнью, однако продолжительное и тяжелое течение делает его возбудитель
весьма заманчивым средством войны, и борьба с ним требует больших уси лий и ма-
лоэффективна.
По принятой в советской виенно-научной литературе традиции, Кротков, ссыла-
ясь исключительно на мнения зарубежных исследователей, осоио останавливается на
поражающих свойствах возбудителя сибирской язвы. По его мнению, он отвечает
большинству требований, предъявляемых к потенциальному агенту БО:
поражает человека и животных;
оказывает быстрое действие;
обладает высокой вирулентностью;
устойчив в отношении воздействий внешней среды;
способен проникать в организм человека и животного различными путями: через
дыхательные пути, ЖКТ и поврежденные кожные покровы.
Серьезное военное значение, утверждал Кротков, имеет и то обстоятельство, что
споровые формы антракса переносят кипячение и небрежно выполненную стерили-
зацию. Наконец, сибирская язва удовлетворяет еще одному важному требованию: воз-
будитель инфекции очень неприхотлив в отношении культивирования и легко может
быть получен в любой лаборатории1.
Далее он приводит мнение Н. Klotz (1937), называвшего споры сибирской язвы
«излюбленным средством немецкой военной бактериологии», и А. и R. Sartory (1935),
указавших на важное значение передачи сибирской язвы через фураж, особенно если
«подмешать к зараженной пище животного колющие или режущие средства (остат-
ки репейника, ячменные колосья и т. п.), вызывающие царапины пищеваритель-
ного тракта».
В 1940 г. И. Р. Дробинский резюмировал, «по данным иностранной литературы»,
требования, предъявляемые к возбудителям инфекций, пригодным для использова-
ния в качестве средств ведения бактериальной войны. Обра з потенциального агента
для ведения бактериологической войны (кстати, никогда так и не созданного) к на-
чалу Второй мировой войны выыыдел следующим образом:
он должен быть стойким в окружающей среде;
выдерживать воздействие различных физических и химически факторов, в част-
ности — дезинфицирующих растворов;
длительное время оставаться высоко и стабильно вирулентным для человека или
животного. Ошибочно считалось, что именно вирулентность (способность вызывать
болезнь в небольших дозах) обеспечивает контагиозность (т. е. передачу от человека
к человеку) возбудителя инфекционной болезни.
1 Это в вегетативной форме. Процесс спорообразования сибиреязвенного микроба требует
дорогих специальных сред и времени, а соответ< гвенно никак не «не прихотлив».
84
Часть 1. Краткая история применения биологических средств...
Вызываемая потенциальным агентом БО, болезнь должна характеризоваться сле-
дующими чертами:
легко и постоянно передаваться от человека к человеку, от животного животному,
и от животного к человеку;
иметь короткий инкубационный период (время, проходящее с момента зараже-
ния до появления первых клинических признаков болезни);
тяжело протекать, сопровождаться высокой летальностью заболевших либо при-
водить их к длительной уграте боеспособности;
вызываться трудно обнаруживаемыми во внешней среде и малоизученными воз-
будителями, с которыми противник незнаком или которым свойственна повышен-
ная вирулентность;
эпидемии должны предотвращаться нападающей стороной посредством вакцина-
ции своих войск против применяемого возбудителя или другими надежными средства-
ми защиты, причем эти профилактические и лечебные меры борьбы с инфекцией
должны быть не известны противнику.
На основании выше приведенных точек зрения на свойства потенциальных аген-
тов БО, по предпочтениям военных бактериологов 1930-х гг., их можно проранжиро-
вать следующим образом:
возбудитель чумы — рассматривался как «суперагент» БО, применение которого в
принципе невозможно из-за риска разноса чумы по Европе инфицированными кры-
сами и мышами. Последнее ожидание было не более чем ложным постулатом, осно-
ванном на страхах Средневековья перед чумой и учении о контагии Дж. Фракасторо.
В России еще в начале XX в., благодаря работам Д. К. Заболотного и его последовате-
лей, такие представления об эпидемиологии чумы считались ложными;
возбудитель сибирской язвы — идеализировался из-за своей якобы устойчивости в
окружающей среде. Но некоторых ученых уже тогда смущала его низкая вирулентность
для человека и животных;
возбудитель сапа—доказал свою опасность для лошадей и контактирующих с ними
людей еще до Первой мировой войны. После войны его значение как диверсионного
агента постоянно обесценивалось моторизацией армий и сокращением армейского
конского поголовья. Кроме того, исследователи отмечали его плохую сохраняемость
в окружающей среде и высокий риск заражения лабораторного персонала;
возбудители бруцеллеза — идеализировались как агенты БО благодаря хорошей
сохраняемости вирулентных свойств при поддержании в лабораторных условиях
Однако их было трудно получать в больших количествах, а вызываемая ими болезнь
давала невысокую летальность;
возбудитель мелиоидоза (тропический сап) — рассматривался как весьма опасный
малоизученный микроорганизм, способный вызывать смертельное поражение чело-
века, проникая в легкие.
Эти же «предпочтения» свидетельствуют и о том, что «достоинства» одного аген-
та БО не становились «достоинствами» другого, а были лишь его частным свойством
при остальных недостатках. Тем не менее отождествление потенциального агента БО
(попросту говоря, штамма микроорганизма) с самим БО (техническим устройством)
сыграло плохую службу эпидемиологии в целом. Информация об эпидемиях и о ве-
дущихся разработках вакцин стала сначала засекречиваться, а потом забываться на
полках опечатанных железных шкафов. Три поколения российских эпидемиологов
учились не на примерах реальных эпидемий, а на их упрощенных схемах, что сказа-
лось при осмыслении причин, механизмов и тенденций ВИЧ/СПИД-пандемии.
1.7. Становление бактериологического оружия между мировыми войнами
85
Способы и устройства 1930-х гг., предназначенные для ведения бактериологической
войны. Сегодня любой англоязычный источник, где хотя бы кратко описывается ис-
тория БО, относит появление первых масштабных программ по его созданию на дату,
более позднюю, чем 1932 г. (например, Dando М., 1994; и Sidel Е R. с соавт., 1997).
Обычно авторы таки:, публикаций ссылаются на то, что первыми стали разрабатывать
БО Япония и Германия. В Японии, действительно, военно-биологическая програм-
ма начата в 1932 г. Однако союзникам по антигитлеровской коалиции (за исключе-
нием СССР) о японских работах в этой области стало известно только в 1942 г.
В Германии же, лаже после ее оккупации союзническими армиями, так и не уда-
ось найти каких-либо материальных свидетельств проведения работ по разработке,
производству и хранению БО. Историки отряда № 731 (например, Моримура Сэйти,
1983) фиксируют тот факт, что и самой идеей создания БО Исии Сиро загорелся имен-
но после возвращения из Европы в 1928—1930 гг. Знакомясь с европейской научной
литературой, посещая бактериологические лаборатории и беседуя с ведущими бакте-
риологами. он пришел к твердому убеждению, что в Европе идет гонка исследований
по созданию БО. И на то у него были серьезные причины.
Например, итальянский военный врач Casarini (1928), ссылаясь на мнение ряда
ученых, утверждал, что «Италия никогда не будет инициатором бактериологической
войны, но нельзя отставать от воинственных народов». Далее он отмечал выступле-
ние английского премьер-министра в феврале 1927 г., заявившего в парламенте, что
пока державы не дали взаимной гарантии воспрещения применения в качестве бое-
вого оружия ядов и болезнетворных бактерий, английское правительство вынуждено
применять все возможные меры и методы нападения и защиты (цит. по Лихачеву, 1928).
Правда сегодня в Соединенном Королевстве дату начала наступательной биологиче-
ской программы переносят на 1940 г. (см. Dando М., 1994).
Об организации в Бухаресте в 1927 г. специальных лабораторий и полигонов для
изучения научных и технических вопросов, относящихся к химической и бактериоло-
гической войне, много писали уже в начале 1930-х гг (см. работу Блюменталя Н., 1932).
Н. Liepmann (1937) указывал на существование в Европе специальных центров,
занимающихся вопросами подготовки бактериальной войны и соответствующих кон-
цернов, изготовляющих в массовом масштабе лечебные препараты против таких ин-
фекций, как холера и др., но которые практически в настоящих условиях не ветре
чаются. Об опытах с БО в лабораторных и полигонных условиях в Европе писал
J. Karyszkowski (1935).
Ниже по работам В. Феррети (1931), Н. Блюменталя (1932), Марценака (1935),
А. Сартори и Р Сартори (1936), Г. Клотца (1937), Ф. Г. Кроткова (1939), И. 3. Сарки-
сова (1940) и И. Р. Дробинского (1940) обобщены основные способы и технические
устройства, которые в те годы считались европейскими военными бактериологами
наиболее приемлемыми для использования в бактериологической войне.
Стрельба артиллерийскими бактериальными снарядами. Такой способ ведения бак-
териологической войны пришел в головы военным первым и, несомненно, по анало-
гии с химическими снарядами. Считалось, что высокая температура и давление, свя-
занные с силой разрыва снаряда или бомбы, действуют слишком кратковременно, и
поэтому они не могут привести к гибели микроорганизмов, находящихся внутри обо-
лочки снаряда. В подтверждение этих взглядов обычно приводились результаты, ка-
ких-то экспериментов, например, показывающих выживаемость бактерий «чудесной
палочки» (В. prodigiosus) на пуле револьвера Браунинга после выстрела. Большие на-
дежды у военных бактериологов вызывал тот факт, что устойчивость бацилл чумы
86
Часть 1. Краткая история применения биологических средств...
выше, а устойчивость спор сибиреязвенных бацилл и анаэробов — возбудителей га-
зовых инфекций — еще более велика, чем у В. prodigiosus. Правда, Клотц (1937), не
отрицая возможности применения бактериальных средств войны с помощью обстре-
ла, считал этот метод нападения малоэффективным.
Сбрасывание авиационных бактериальных бомб. В научной литературе того време-
ни описываются две принципиально различающиеся конструкции таких бомб —
итальянская (Феррети В., 1931) и германская (Клотц Г., 1937). Первая представляла
собой обычную химическую бомбу, внутри которой был помещен сосуд с питатель-
ной средой, в котором размножались микробы. В головной части бомбы помещался
маленький аппарат с кислородом, непосредственно соединенный с сосудом. Этот
кислород непрерывно подавался в питательную среду и поддерживал жизнь микро-
организмов. Бомба сохраняла жизнеспособные микроорганизмы в течение 36 ч с
момента ее снаряжения. По замыслу разработчиков, подобная бомба, брошенная с
самолета, должна распространить при взрыве «миллионы болезнетворных микробов,
рассеивающихся на большое расстояние и, особенно на сырой почве и сыром возду-
хе, сохраняющих на долгое время свою способность массового заражения».
Бактериальная бомба германского типа состояла «из наполненного культурами
бактерий плотного резервуара, сбрасываемого на парашюте с почти неограниченной
высоты. Парашют раскрывается автоматически, на любой установленной высоте
(обычно 10, 20 или 50 м над землей), резервуар автоматически раскрывается, и нахо-
дящееся под легким давлением содержимое медленно выпускается». Феррети и Клотц
писали об этих бомбах, как об уже существующих.
Метание с самолета стеклянных ампул и «беби-бомб». Предполагалось использо-
вать тонкостенные стеклянные ампулы весом от 5 до 20 г и специальные стеклянные
шары весом до 250 г («беби-бомбы»), наполненные культурами возбудителей опасных
инфекционных болезней. Стеклянные ампулы предназначались для сбрасывания с
самолета с высоты 5—6 км. Разбиваясь при падении, они освобождали содержимое в
воздух, почву или в воду. «Беби-бомбы» должны были иметь взрыватель дистанцион-
ного действия и взрываться в воздухе. «Беби-бомбы» в химическом снаряжении впер-
вые появились в Великобритании в начале 1920-х гг. (Де Лазари А. Н., 1935, 2008); в
СССР в 1934—1934 гг. пол названием АК-1 и АК-2, но без дистанционного взрывате-
ля (Широкорад А. Б., 2004).
Комбинированное сбрасывание осколочных бомб и бактериальных ампул или в сочета-
нии с распылением микробного тумана. Одновременное осколочными бомбами счита-
лось возможным сбрасывание с самолетов разбивающихся бактериальных ампул или
распыление оседающего тумана из бацилл газовой гангрены, столбнячных палочек и
других анаэробов. Ожидалось, что произойдет массивное заражение ран с последую-
щим развитием в ряде случаев раневой анаэробной инфекции.
Распыление микробной взвеси, создание бактериального тумана в воздухе, выпуск так
называемых «микробных облаков» и «микробных капелек». Технически это можно было
выполнить самолетами при помощи специальных выливающих приборов (ВАПов), рабо-
тающих без давления или распыляющих струю жидкости под давлением. Способ ка-
зался очевидным, тем более что для применения ОВ он был уже хорошо отработан
(рис. 1.13.).
Феррети утверждал, что разбрызгивание над военными объектами бактериальных
культур и ОВ является логическим следствием стремления к обеспечению максималь-
ного эффекта поражения. С военной точки зрения метод нападения посредством
«микробных капелек», по его мнению, имеет значительные преимущества, поскольку
1.7. Становление бактериологического оружия между мировыми войнами
87
Рис. 1.13. Применение иприта с помощью ВАПов. А. Само юты, оборудованные
ВАПами, выливают иприт. Б Опорожненный ВАЛ, сброшенный итальянским
самолетом в Абиссинии (1936 г). ВАПы, существовавшие в конце 1930-х гг., пред-
ставляли резервуар обтекаемой формы В хвостовой части ВАПа имелось большое
отверстие, которое закрывалось крышкой. В передней части он имел большое
наливное отверстие Такие приборы подвешива шсь под плоскостями или фюзе-
ляжем самолета. Крышка хвостового отверстия соединялась тросами с открываю-
щими приспособлениями, нах одящимися в кабине летчика-наблюдателя. Когда
самолет подходи i к цели, летчик-наблюдатель открывал прибор, и жидкое ОВ вы-
ливалось из прибора самотеком. Прибор опорожнялся за несколько секунд. Быстро
вылившееся большой массой жидкое ОВ дробилось на капли различной величины
потоками встречного воздуха и в виде дождя и тумана оседало на землю. Средний
размер капли иприта составлял 1,6 мм Фотография 1930-х гг. По Я. Жикур (1936)
избегается влияние на микробы высоких давлений и температур, неизбежных при
использовании артиллерийских снарядов и авиационных бомб. Немалую роль игра-
ет также и снижение «мертвого веса», т. е. веса оболочки бомбы или артиллерийского
снаряда. Ориентировочный подсчет, проведенный Феррети, показал, что при разбрыз-
гивании бактериальных культур на 9 весовых частей полезного груза приходится
только одна весовая часть оболочки. В авиационных бомбах «мертвый вес» пример-
но в 2 раза превышает «полезный вес», а в артиллерийских химических снарядах со-
отношение весовых частей заряда и оболочки равняется 1:8.
В те годы считалось, что «метод разбрызгивания» для ОВ и бактерий может быть
использован с высоты 4000 м и даже еще с большей. Например, Клотц, ссылаясь на
экспериментальные исследования немецких ученых, привел следующие данные.
«Максимальная скорость падения водяной капли достигается при постоянном радиусе
последней в 2,5 мм. При этом радиусе возможность расщепления капли исключена.
Для других жидкостей с другими соотношениями плотности следует избирать другие
радиусы. Так, например, для л юизита, удельный вес которого равен 1,12, наивыгодней-
ший радиус капли будет 2,3 мм: для технически чистого иприта, удельный вес кото-
рого при температуре 20°С равен 1,19, наивыгоднейший радиус — 2,1 мм. Наконец,
для зараженной бактериями воды, удельный вес которой равен 1,1, наиболее благо-
приятных: радиусом будет 2,35 мм. При таких радиусах максимальная скорость паде-
ния капли равна, в среднем, 4,8 м/с (капля, сброшенная с высоты 4000 м, достигает
земли через 8 мин)». Клотц утверждал, что возможность сноса этих капель ветром,
более значительная, нежели для авиационных бомб, должна и может быть учтена.
Потеря вещества испарением, судя по доступным ему экспериментальным данным,
88
Часть 1. Краткая история применения биологических средств...
не выходила за пределы известных границ и не имела решающего значения. Для мас-
сированного заражения объекта, утверждал Клотц, требуется одномоментный выпуск
бактериальной жидкости из значительного числа сопел при заранее рассчитанном
давлении. При выпуске необходимо следить за тем, чтобы отдельные капли не
приходили в соприкосновение и не сливались друг с другом. По его мнению, метод
разбрызгивания особенно пригоден для заражения объектов нападения спорами си-
бирской язвы в сочетании с ОВ.
Для заражения воздуха микроорганизмы могут быть введены в атмосферу не только
в виде мельчайших капель тумана, но и в виде пыли из высушенных бактерий. Но уже
тогда было известно, что ряд патогенных микроорганизмов (возбудители холеры, чумы
и гриппа) плохо переносит лиофильное высушивание, а поэтому их распространение
посредством летучей пыли невозможно. Считалось также, что для большинства воз-
будителей инфекционных болезней дыхательных путей — способ распыления их
в сухом виде применим в малой степени, в силу быстрой гибели этих микробов при
высушивании или «неспособности их распыляться» (Дробинский И. Р., 1940).
Комбинированное применение отравляющих веществ и возбудителей инфекционных
болезней. Одним из свойств фосгена, иприта и люизита является ослабление местно-
го иммунитета пораженных тканей. Поэтому всякий отравленный ими или выздоро-
вевший от подобного отравления, неизбежно погибает в случае заболевания гриппом
и другими инфекциями. Острое отравление фосгеном, ипритом, люизитом часто со-
провождается бронхитами, пневмонией, гангреной легких, придающими особую тя-
жесть клиническому течению и исходу отравления. Опытным путем было выяснено
также и то, что большинство ОВ, в частности иприт и фосген, обладают весьма слабым
бактерицидным действием, и в атмосфере этих ОВ патогенные бактерии хорошо вы-
живают в течение довольно длительного времени.
Сбрасывание на парашютах зараженных животных. Пфейффер, выступив в 1923 г.
в Лиге Наций в качестве эксперта по бактериальной войне, на вопрос комиссии о са-
мом опасном средстве бактериологической войны заявил следующее: «Наиболее вер-
ным средством создания искусственной эпидемии является выпуск зачумленных крыс
в страну противника, что очень нетрудно сделать при помощи самолетов». Практи-
чески во всех выше перечисленных трудах утверждается, что эпидемию бубонной чумы
очень легко вызвать в окопах противника, «где крысы так и кишат; внутри страны тоже
нетрудно создать ряд очагов эпидемии». Предполагалась возможность сбрасывания
с самолетов в автоматически открывающихся парашютах корзин с крысами, заражен-
ными возбудителем чумы. Корзина при ударе о землю ломается или автоматически
открывается, а животные разбегаются и могут в дальнейшем заражать местных крыс.
В том, что вслед за эпизоотией чумы среди крыс следует и вспышка бубонной чумы у
человека, почти никто тогда не сомневался («почти» — за исключением Conrich, 1931;
и Fox L., 1933). Кроме того, считалось возможным распространять инфекционные
болезни на территории противника путем сбрасывания с самолетов животных, зара-
женных спирохетой Sodoki, возбудителями туляремии, бруцеллеза, бешенства, мели-
оидоза, а также выпуском птиц, зараженных пситтакозом (Karyszkowski J., 1935).
Выпуск с самолетов зараженных насекомых. Ряд опытных проверок, проведенных
на аэродромах в местах, эндемичных по желтой лихорадке и малярии, показал, что их
переносчики-насекомые прекрасно приспосабливаются к измененным условиям сре-
ды; они легко переносят длительные и высотные полеты в аэропланах.
Заражение водоисточников и пищевых продуктов на оставляемой территории. При
отступлении противник не будет оставлять в благополучном санитарном состоянии
1.7. Становление бактериологического оружия между мировыми воинами
89
территории, источники водоснабжения, пищевые продукты, жилую площадь и пр.
Наоборот, следует ожидать, что он всячески будет стремиться загрязнять оставляемую
территорию, заражать почву и пищевые продукты и т. д.
Наземный выпуск зараженных животных: грызунов, зараженных туляремией, бруцел-
лезом и собак, зараженных бешенством. Возможно также оставление неприятелем круп-
ного и мелкого рогатого скота, лошадей, свиней и пр., зараженных и болеющих раз-
личными эпизоотическими заболеваниями i бруцеллез, сап, мыт, энцефалит, эпи-
зоотический лимфангоит, инфекционная анемия лошадей, ящур, сибирская язва,
чума свиней и др.), из которых многие могут передаваться человеку.
Диверсионные способы применения БО с целью уничтожения сельскохозяйственных
животных. Должны осуществляться посредс твом прививок недоброкачественных
вакцин и заражения фуража, а также уничтожения различных технических культур
путем контаминации или распространения переносчиков заразных болезней расте-
ний; и, наконец, заражения агентами противника в тылу пищевых продуктов, воды,
тканей, медикаментов, средств транспорта и др. Сюда же относили оставление инфек-
ционных больных на покидаемой территории.
Оценивая перечисляемые иностранными авторами способы бактериального на-
падения с точки зрения их технической реальности, И. Р. Дробинский (1940) конста-
тировал, что эти способы столь же реальны, как и вообще способы авианападения, и
что «технически нет препятствий к тому, чтобы противник сбрасывал бактериальные
бомбы, рассеивал микробный туман или спускал зараженных животных».
Критические взгляды на БО. Важное шествие апологетов бактериологической вой-
ны было омрачено двумя немецкими учеными, набравшихся смелости противопо-
ставить свое «антинаучное» мнение, мнению «ученого большинства». Дискуссия на-
чалась после статьи профессора Люстига (Lustig, 1931), преподнесшего новое направ-
ление в создание средств массового поражения людей, как уже почти решенное дело.
Против Л юстига в том же журнале через несколько месяцев выступили сначала Нейс-
сер (Neisser М., 1931), потом Конрих (Conrich, 1931). Нейссер (1869-1938) привел
конкретные факты, показывающие невозможность использования чистых культур
микроорт анизмов для массового заражения ими людей. Он обратил внимание иссле-
дователей на многочисленные неудачи, сопровождавшие попытки бороться с грызу-
нами, заражая какую-то их часть бактериями. Он также привел результаты опытов,
показывающих невозможность контаминации возбудителем болезни вызвать рожко-
вую эпидемию у растений. Нейссер увидел и другую отрицательную сторону профана-
ции БО. «Ведь может дойти даже до того, что потребовался бы контрольнад производ-
ством на сывороточных фабриках и лабораториях, а это было бы действительно невы-
носимо» (цит. по Блюменталю Н„ 1932). Он также утверждал, что гигантские эпидемии
не могут решить исхода войны.
Но еще более любопытно мнение профессора Конриха, референта по гигиене при
германском военном министерстве. Конрих совершенно конкретен. Он высказал три
возражения против БО:
1) невозможно в лабораторных условиях удержать в течение долгого времени воз-
будитель инфекционной болезни в вирулентном состоянии;
2) наличие вирулентности у микроорганизмов недостаточно, чтобы вызвать эпи-
демию;
3) при рассеивании микробов значительное количество их погибает от несоответ-
ствующих условий. Первые два возражения еще можно было в те годы обосновать
ссылками на открытые источники, но третье, несомненно, свидетельствует в пользу
90
Часть 1. Краткая история применения биологических средств...
того, что Конрих знал о гибели микроорганизмов при переводе их в мелкодисперс-
ный аэрозоль. Он был в курсе каких-то агробиологических экспериментов, показав-
ших, что заразить экспериментальное животное бактериальным аэрозолем вдали от
его источника весьма проблематично.
Кроме технической возможности применения БО, Конрих подверг критике и его
военное значение. Он, в частности, утверждал, что различная и иногда неопределен-
ная продолжительность инкубационного периода, колеблющаяся в широких преде-
лах при одной и той же болезни, не позволяет точно определить окончание срока
действия БО, что делает его даже тактически непригодным. Сомневался он и в воз-
можности сохранения подготовки к бактериологической войне втайне, «если придет-
ся производить предохранительные прививки войскам и населению по поводу 10 раз-
личных инфекций». По адресу работы Люстига он отметил, что «каждый специалист
будет считать ее не чем иным, как мнением профана». А Конрих, несомненно, был
специалистом.
Позже появились и другие скептики, отрицавшие возможность бактериологиче-
ской войны. Фокс (Fox, 1933) указывал на то, что во время Первой мировой войны
было изготовлено 50 биллионов пуль; этого количества было достаточно для того,
чтобы 50 раз убить все человеческое население земного шара; однако многие из жив-
ших тогда дожили до сих пор. Легко рассчитать военную продукцию, количество от-
литых пуль или количество инфекционных доз, которые может взять самолет, но это
еще не означает, что количество убитых будет равным количеству отлитых пуль, а
количество зараженных — взятому самолетом количеству инфекционных доз. Мож-
но распылить в виде тумана бактериальную культуру, но далеко не все вдохнувшие этот
туман обязаны заболеть (цит. по Дробинскому, 1940). Итальянский профессор Рей-
тано (Reitano U., 1937) утверждал, что одним из главных элементов подготовки бак-
териологической войны является быстрое получение необходимого количества пато-
генных культур. Последние должны сохранять вирулентность после посевов на
искусственных питательных средах. Трудности решения этой задачи являются глав-
ным препятствием для практического использования микробов в военных целях. По
его мнению, «переход от теоретической концепции к практическому осуществлению
связан с многочисленными и сложными препятствиями» (цит. по Кроткову Ф., 1939).
Однако даже наличие технических способов применения БО еще не решает пол-
ностью вопроса об его эффективности. Рейтано, разбирая условия, необходимые для
развития отдельных инфекционных заболеваний и перехода их в эпидемию, отмечал,
что недостаточно иметь запасы патогенных и вирулентных микробов и разработан-
ные средства их распространения; считаться приходится с многочисленными факто-
рами, при отсутствии которых невозможно развитие эпидемических болезней. Среди
них он называл географические, климатические, социальные факторы; индивидуаль-
ный и коллективный иммунитет и др. Например, холера или сыпной тиф могут рас-
пространяться только при определенном комплексе социальных, климатических и
других условий; натуральная оспа вообще не может быть распространена ни в виде
отдельных заболеваний, ни эпидемий среди населения, которому сделаны прививки
ит. д. (цит. по Дробинскому, 1940).
Были и другие сомнения в осуществимости искусственных эпидемических про-
цессов. Так, R. Rodriguez (1937) считал необходимым для возникновения и искусст-
венного распространения эпидемий соблюдение, по меньшей мере, пяти условий:
1) наличие микроба с фиксированной вирулентностью;
2) наличие восприимчивых организмов;
7.7. Становление бактериологического оружия между мировыми войнами
91
3) наличие среды, благоприятствующей передаче возбудителя инфекционной
болезни;
4) наличие условий, позволяющих распространяться инфекции;
5) отсутствие профи тактических мероприятий.
Но даже первое из перечисленных им условий, не выполнимо в принципе.
A. Rochaix (1935) указывал на то, что «искусственно вызываемые эпизоотии сре-
ди животных, против которых человек может применить все способы для достижения
успехов, тогда как грызуны не располагают средствами защиты, — далеко не всегда
успешны и зачастую отрицателгны». Тем более сложной задачей является успешное
искусственное распространение эпидемии среди людей, что должно зависеть от еще
большего количества и комплекса важнейших условий и случайностей, чем искусст-
венная эпизоотия среди грызунов и других животных.
Подверглись сомнению взгляды на возбудитель чумы, как на абсолютный пора-
жающий агент БО. Конрих считал, что здесь наиболее наво. 1яшим ужас является толь-
ко само слово «чума». Он сомневался в том, что заражение людей чумой посредством
крыс может произойти таким же именно путем и с такими же результатами, как это
происходит в природных условиях. Фокс тоже указывал на преувеличение угрозы
распространения чумы. Строгое соблюдение мер профилактики в сочетании с проти-
воэпидемическими мероприятиями надежно гарантирует население и войска от за-
болевания чумой. В качестве примера он привел данные о чуме в Пенджабе. За 1924 г.
среди европейского населения провинции было зарегистрировано только 10 случаев
чумы, в то гремя как среди туземцев заболело 500 тыс., что состав!' ло 1/40 населения.
Касаясь возможности военного использования биологических токсинов, Фокс
отметил, что, действительно, токсин ботулинического микроба наиболее смертелен
для человека в ничтожных дозах и вы »ывает отравление при любом способе его вве-
дения. И именно его имеют в виду отде. шные исследователи, когда утверждают, что
«самолет в состоянии донести достаточные количества токсина для отравления целого
города». Но если бы введение токсина людям было таким простым делом, как и его
перевозка, этот токсин представлял бы одно из наиболее эффективных средств вой-
ны. В действительности же, легко установить только летальную дозу токсина, нетруд-
но определить, какое его количество дойдет до намеченной цели. Сбрасывание ток-
сина с воз гуха может вызвать и человеческие жертвы, пишет автор, но больше всего и
в первую очередь пострадают птицы, домашние животные и грызуны. Следователь-
но, боевой эффект такого нападения будет незначительным.
Взгляды Нейссера, Конриха, Реигано, Фокса и других скептиков бактериологи-
ческой войны не сыграли никакой роли в развитии представлений о БО в 1930-х гг.
Авторитетным большинством ученых предвоенного десятилетия они были признаны
как «чрезвычайно поверхностные, старающиеся умалить возможность и значение бак-
териологической войны». Нейссера и Конриха обвинили еще и в стремлении «скрыть
подготовку Герман ии к бактериологической войне». И, как часто бывает в науке, тол-
чком к дальнейшему развитию военной бактериологии послужили мистификации и
экспериментальные ошибки.
Мистификация Стада. В1934 г. в июльском номере журнала «The Nineteenth century
and after» («Девятнадцатое столетие и после») была опуоликована статья бывшего глав-
ного редактора газеты «Times», известного английского журналиста Уильяма Стида
(Steed Wickham). В ней он поьедал миру о германских опытах, предназначенных для
подготовки к бактериальной войне. Его «разоблачение» тогда наделало много шуму,
да и сегодня оно цитируется как каноническое. Эти два обстоятельства заставили меня
92
Часть 1. Краткая история применения биологических средств...
найти оригинал статьи Стида и самостоятельно вникнуть в суть им написанного.
Признаюсь, я был удивлен прочитанному.
Если заставить себя абстрагироваться от приведенных Стидом деталей, то в са-
мом общем виде, и особенно в других газетных пересказах, «разоблачение» выглядит
весьма убедительно. Стид категорически заявил, что в его распоряжении имеются
подлинные секретные германские документы, доказывающие, что с приходом к вла-
сти канцлера Брюнинга, Германия стала заниматься во многих странах эксперимен-
тами по оценке возможности применения химических и бактериологических средств
поражения в людных местах. Хотя подлинники секретных документов не имеют при-
вычки валяться на подоконниках, тем не менее Стид утверждал, что они исходили от
секретного отдела германского военного министерства (Luft-Gas-Angriff — отдел воз-
душно-химического нападения) и были адресованы нескольким заводам химичес-
кой промышленности. Первый из этих документов датирован июлем 1932 г. По «дан-
ным» Стида, в 1931 г. несколько секрета ых германских агентов занимались изучением
силы втягивания воздуха при входах в метрополитен Парижа и направлениями дви-
жения воздуха в различных коридорах. Исследование предпринято немцами с целью
установления наиболее благоприятных условий для распространения патогенных
микроорганизмов и ОВ.
С теми же намерениями немецкие экспериментаторы приступили к исследовани-
ям скорости падения жидкостей. Результат работ показал, что выгоднее всего было бы
применять, возможно, более крупные капли, быстрота падения которых была бы наи-
большей, а улетучивание — наименьшим. Далее Стид приводит якобы секретные экс-
периментальные данные о скоростях падения капель жидкого иприта разного разме-
ра с большой высоты. Их нет смысла приводить здесь, так какони уже приведены нами
(даже подробнее, чем у Стида) в разделе «Способы и устройства 1930-х гг., предназ-
наченные для ведения бактериологической войны» со ссылкой на работу Г. Клотца
(1937), который сам ссылается на опубликованную раньше работу немецких ученых.
Получается, во-первых, что эти куцые данные Стидом взяты не из секретных докумен-
тов, а из общедоступных источников. Во-вторых, такого заимствования тогда никто
почему-то не заметил. С этим приемом мистификаций мы еще встретимся при рас-
смотрении так называемой «свердловской язвы» 1979 г. и грандиозной компании при-
митивной лжи, осуществленной для обоснования двух войн против Ирака.
Теперь, когда вроде бы способ воздушного нападения германцам стал ясен, надо
показать и объекты их нападения. У Стида оказался под рукой другой «секретный
документ», содержащий список из 20 французских городов, над которыми отдел воз-
душно-химического нападения германского военного министерства предлагает иссле-
довать движение воздушных потоков. Среди них следующие: Тионвиль, Мец, Страс-
бург, Бельфор, Нанси, Бриэ, Верден, Туль, Эпиналь, Париж, Тулон, Марсель, Лион,
Гавр, Руан, Канн, Нант, Крезо и др.
А далее начинается самое интересное, что никак не следует из разнообразных
пересказов публикации Стида. Все его интерпретаторы дружно твердили в 1930-е гг.
и твердят сегодня о том, что Стид смог расшифровать секретные немецкие докумен-
ты. Правда, тогда получается, что вместе с зашифрованными документами ему пере-
дали и коды для их дешифровки. Уже одно это обстоятельство должно было поставить
в тупик любого любителя строить версии. Ведь разработка секретного документа и его
шифровка выполняются разными лицами, работающими в разных подразделениях и
даже службах. Кроме того, Стид, имея такой сверхважный для обороны Соединенно-
го Королевства разведывательный инструмент, как коды для дешифровки корре-
1.7. Становление бактериологического оружия между мировыми войнами
93
спонденции врага, сообщил о нем этомуже врагу. Британцы всю Вторую мировую вой-
ну тщате. [ьно скрывали, что обладают германской шифровальной машиной «Эниг-
ма», и даже позволили немецкой авиации разбомбить Ковентри, лишь бы противник
не заподозрил, что его секретные переговоры, осуществляемые по каналам радиосвя-
зи, каким -то образом расшифровываются.
Все оказалось проще, стоило только заглянуть в статью самого Стида. Он ничего
не расшифровывал, а только интерпретировал. Объектом его воображения было со-
держание «двойного листка почти нормального размера» с карандашными записями
«без даты и подписи». В интерпретации Стида эти карандашные пометки преврати-
лись в зловещие результаты германских экспериментов по изучению возможности
заражения Парижа возбудителями опасных инфекций. Здесь нам уже нужны детали.
Вот очна из его интерпретаций «германского эксперимента»: «Площадь Согласия
является исходной точкой для тестов и координации всех измерений, причем воздви-
гнутый на ней обелиск — центр точки координации. Было сделано несколько кругов
по площади Согласия и распылено примерно 210 единиц, содержащих по миллиарду
микроорганизмов Micrococcus prodigiosus каждая. Операция проведена с помощью
помощников, а ровно через 6 ч определены результаты. Само распыление происхо-
дило 18 августа 1933 г. в 2 ч 47 мин пополудни». Далее он описывает метеоусловия
«эксперимента». Под ними он понимает всею лишь напра! ление и скорость ветра. По-
следняя величина указывается равной 8,75 м/с, по показанию анемографа, что край-
не неблагоприятно для применения химического и биологического оружия В подтвер-
ждение достигнутого результата Стид приводит подсчеты колоний на чашках Петри,
расставленных в 3,5 км от площади Согласия через 6 ч после «распыления» бактерий.
Устройство, использованное «немцами» для распыления бактериальных культур,
Стидом не описано (возможно, в его представлении это было нечто вроде пульвери-
затора, который он видел в парикмахерской), зато количество бактерий, наверное, его
самого впечатлили. Для гуманитария же, газетчика, действительно, миллиард звучит
завораживающее, и Стид не мог знать, много это или мало. Но что такое «миллиард
микробов» для бактериолога? Это 1 см3 слегка мутной жидкости, 1 ЕД по оптическому
стандарту. С одной пробирки скошенного агара можно смыть в 10—100 раз больше.
Следовательно, «германские агенты» использовали в столь масштабных экспери-
ментах бактерии, смытые с двух, как бактериологи их называют, «косячков».
Затем Стид утверждал, что результаты «немцами» были оценены положительно,
так как на контрольных чашках (на которых можно было зарегистрировать присут-
ствие колоний бактерий) «были сосчитаны 4231 микробная колония». Как могут вы-
плеснутые на мостовую бактериальные культуры дать какое-то количество колоний
на плотной питательной среде, разлитой в чашках Петри, да еще на столь большом
расстоянии от места их пролива, трудно себе представить (другие факторы внешней
среды, препятствующие распространению бактериальных аэрозолей, будут рассмот-
рены в разд. 1.11). Дело еще осложняется тем, что в воздухе всегда полно всевозмож-
ных бактерий, которые дают цветные колонии на плотной питательной среде (жел-
тые, синие, розовые), плюс плесень, покрывающая поверхность питательной среды
причудливыми узорами. Но вот когда этих колоний больше сотни на чашку, то их и
без плесени весьма сложно подсчитать (любой бактериолог не раз с этим сталкивался).
Стид «насчитал» их столько, что немецким агентам нужно было разложить на мосто-
вой, по крайней мере, от 20 до 40 чашек Петри, снять с них крышки и сидеть рядом,
отгоняя зевак, жандармов и контрразведчиков, способных ненароком на них насту-
пить. Но это еще не все!
94
Часть 1. Краткая история применения биологических средств...
В других «наинтерпретированных» Стидом «секретных экспериментах» счет ко-
лониям идет уже на сотни тысяч! Вот только сами немцы в лице профессора Конриха
знали, что бактерии (бактериальные культуры), диспергированные с тем усилием,
которое переводит их в мелкодисперсный аэрозоль, способный проникать в глубокие
отделы легкого человека, в основной своей массе гибнут уже в момент самого распы-
ления и по 6 ч «летать» вдоль улиц Парижа в жизнеспособном состоянии не могут.
Немцы пытались убедить в этом и других, но им никто не поверил (см. «Критические
взгляды на БО»).
Любопытно время суток, выбранное «немецкими агентами» для проведения аэро-
биологических экспериментов. По мысли Стида, распылять смертоносные бактерии
и отравляющие вещества надо тогда, когда людей на улицах города больше. Разуме-
ется, это логично и разумно для человека, получившего гуманитарное образование.
Поэтому у него все германские эксперименты «осуществляются» днем или в «часы
пик», лишь бы направление ветра соответствовало замыслу диверсии. Но вот о вер-
тикальной устойчивости приземного слоя воздуха, определяемого в первую очередь
температурным градиентом, Стид даже и не слышал. В разгар дня поверхность земли
(асфальт или мостовая в условиях города) разогревается солнечными лучами, поэто-
му нагревшийся воздух начинает подниматься вверх, и аэрозоль быстро рассеивает-
ся. Такое состояние вертикальной устойчивости атмосферы называется конвекцией.
Как и следовало ожидать, «разоблачения» Стида вызвали бурную реакцию в Ев-
ропе. Германские послы в Лондоне и Париже растерянно хранили молчание. В мае
1940 г. Стид был внесен Вальтером Шелленбергом, начальником отдела АМТ IV-Е
РСХА (отвечавшим за контршпионаж), в «Специальный розыскной список по Вели-
кобритании». В списке указаны фамилии еще 2819 человек как англичан, так и бежен-
цев из Европы, которых следовало арестовать или взять под надзор в случае успеха
германского вторжения в Великобританию (рис. 1.14).
Американским специалистам в области биологических вооружений, Т. Розбери и
Э. Кабату, также был понятен сомнительный характер «разоблачений» Стида. В 1942 г.
в секретном докладе Национальному исследовательскому совету они называют их
«неубедительными по своим подробностям» (доклад был рассекречен и опубликован
в американской печати в 1947 г., переведен на русский язык и опубликован в СССР
отдельной книгой в 1955 г.). Сотрудник Института Пастера (Франция) Е Frischknecht
(2003) признал, что вся эта история, выдуманная Стидом, нужна была британскому
правительству для расширения собственных военно-биологических исследований.
Что касается автора приведенных «разоблачений», то он спокойно отнесся к кри-
тике в свой адрес в журнале «Correspondance diplomatique et politique», видимо, считая
журналистику «второй древнейшей профессией». При интервьюировании его круп-
ными парижскими газетами Стид сделал следующее заявление: «Германскому прави-
тельству никогда не удастся опровергнуть факт, что его агенты производили и продол-
жают производить многочисленные опыты, касающиеся средств истребления непри-
ятельского населения, давая в руки своим авиаторам столь ужасное оружие, как
микробы и яды». Информационный повод для интенсификации исследований в об-
ласти бактериологических вооружений был получен. Теперь у «демократий» Европы
был враг, готовившийся переступить через Женевский протокол 1925 г., следовательно,
и самим уже не было необходимости его соблюдать. Именно в эти годы идея созда-
ния БО нашла своего яркого приверженца в Японии в лице генерала Сиро Исии.
А нам осталось найти тот мутный источник, из которого сам Стид наполнился
верой в способность бактерий летать на дальние расстояния, подобно птицам.
1.7. Становление бактериологического оружия между мировыми войнами
95
1. SfBngler, Josef, 9.8.9В Elbogon, nilfeorbcitor, Barry UUn, 17 Cartel
Street, RSHA IV A1, TV A1 b.
S. Stahl, Johannes, 25.6.00 HciligcrMtMdt, Decknemo: Prana, vcnw't,.
England (Am-Apparat), "tSIIA IV A 2.
8. Stampfer, Friedrich, 8.9.74 Вrlinn, Schriftstollcr, Emigrant, vermutl.
England, RSrfA ПВ5.
4 itanczyk, Mitgl. d. poln. Regierung, vcrnzutl. Eaglfcad (poln. Regie-
rung). RSHA IV D 8.
8. StanlslawskL Jan, 16.5.98 Woyek, poln. Agent, vennutl. England.
RSHA IV ЕГ>. itapo КваКп.
& Stanton. Edmond, 21.14.81 Cotit in Iriaod, Kaufmann, Lotrion W.l.
Cavendish Court, RSHA IVE 4.
7. von 'ttarhemberg, ROdiger Ernst, 10.5.99 Efending, Leiter d. Csterr.
Hcinrwchr, vennutl. England (to. Legitimist), RSHA IV A8.
a Starbemberg, Ftlrat, brit. FliegeroIfWor, Emigrani ’SH. И В 5.
9. Starzynski, Adam, 10.12.93 Jutrosch n, nhem. poln. Beomter. rmull.
E jgland, RSHA IV E 5, Stapp Dortmund.
10. Stassen, J. W, zulctzt: Holland, vermutl. England (Tlterkzeiz:
irijneo). RSHA IV E 4.
*11. Stedman, P. S, MiB, vennutl England, RSHA VIG i.
1'12. Steed, Wikbam, 1030.71, Jp .nalirt, London, RSHA IVE4. IV A5.
IVB4, HI В16, VIG1. ’
13. Steel, С E, 1. Botacnaf sckroMr. England, RSHA TVE<
14. Ste— rhtig Campbell, Алла, 10.Й.61 Sorol/Canada, Rentier, Eng-
* d, RSHA Tv E 4, stannk . Berlin.
15. Steenbergbe, Maximilian Paul Leon, 8Л.99 Leiden, dhem. hoUtnd.
Wirtechatuminister, guletat: Dem Haag, vennutl. England.
Ю4ПД1.1В.
16. Steer, G. L, vermutl. England, RSHA VIG1.
17. Steen, S.S. J, uuletzt: Rcv-il/Estland, vennutl. England (brit. ND.
L Hand). RSHA VIC2.
1Л Stegemann, B, xote’et: Winterewyk/HoUand, vermutl England
Sritetkreis; Bnir-i,, RFnA IV E.
teln, Anna, geb. Ublir, Э1ВЛ2 Wien, BOroangeateUte, vennutl. Eng-
land, RSHA 1VA1, IV A lb.
80. Stein, H. K, SekrcUlr, London E.C.8, St Helen's Court, Shell Hex
end B.P.Ltd., RSHA IVEC
Ы. Stein, Kurt, 20.2.00 London, L lano, RSHA IV E 4.
88. Stein, Morita, 4.3.15 Leipzig, poln. N.-Agent, vennutl. England, RSHA
IV E 5, Stapp Danzig.
28. Stein, Oskar, poln. Agent, ISHA VE6.
84. Steiner, JoseT, 23.12X8 Altenmark, BrotsustrSger, London, RSHA
IV A.1 Stapr S
85. Steinfets, Hilda, 17.8.03 Btrkenfeld, vermutl. England. RSHA IV A 1.
86. StempeL Baron, NachrichtenagentJ vornrutl. England, RSHA IV E 4.
87. Stepanek, Paul, Fikneebauspieler, Emigrant, vermutl England, RSHA
IIВ Ci
88. Stephen, Campbell, 1881 Politiker, vennutl E14 RSHA VI Gl.
89. Stephens, David, brit Beamier, England, R&HA IV E 4, Stapo
I™ Лгш* .
80. Stephens, Philipp Pembroeke, 23.9.03 Little, Nii. mdan/Eng ш ,
Journalist, England, RSHA IV E 4.
81. Stenka, Pierre, zulctzt: Brabant, тепппй. England, RSHA IV E 4.
IS. Stem, Karl, Dr, 1906, London, RSHA ША1.
Рис. 1.14. «Специальный розыскной список по Великобритании», составленный
в 1940 г. Вальтером Шелленбергом (Шелленберг В., 2005). Стид под № 12 (список
алфавитный и разделен на несколько разделов, поэтому положение в списке не
означает важность человека для РСХА). Против каждой фамилии стоит номер
отдела РСХА, в документах которого этот человек числился. Стид «проходил» по
пяти отделам сразу, его судьбу в Третьем Рейхе не трудно представить
Ошибки профессора Триллья (A. Trillat). В начале 1930-х гг. среди эпидемиологов и
клиницистов господствовала концепция контактной инфекции. На ее основе были
разработаны рациональные способы борьбы с инфекционными болезнями и асеп-
тическая хирургия. Возможность переноса инфекции по воздуху ассоциировалась со
средневековыми миазмами и окуриваниями. Становление военной аэробиологии нача-
лось даже раньше, чем начала развиваться эпидемиология аэрогенных инфекций.
В ноябре 1931 г. профессор А. Триллья представил в Медицинскую академию
Французской Республики записку, в которой обобщил результаты своих исследова-
ний «заразительности вирулентной эмульсии 24-часовой культуры куриной холеры».
96
Часть 1. Краткая история применения биологических средств...
Триллья сообщил, что в насыщенном влажностью воздухе, смешанном с небольшим
количеством выдыхаемых газов или таких питательных веществ, как бульон, смерт-
ность кур поднимается до 100 % при вдыхании продолжительностью менее двух ми-
нут. Следовательно, этот воздух способен вызвать смерть животных путем простого
вдыхания его настолько же безотказно, как если бы применялась техника прививки
вирулентной культуры путем непосредственной инъекции. Для объяснения таких
явлений Триллья допустил несколько предположений, которые почти полтора деся-
тилетия принимались многими разработчиками БО за истину:
микроскопические капли способны мгновенно проникать в легочные альвеолы;
бактерии повышают свою вирулентность в результате действия самого воздуха;
воздух способен содействовать активному размножению микробов, т. е. «появле-
нию молодых и очень активных микробов».
С 1920 по 1939 г. Триллья с сотрудниками (Mallein, Kaneko и др.) опубликовали
десять работ, посвященных экспериментальному изучению так называемого «микроб-
ного аэрозоля» или «микробных облаков». Суть этих работ может быть резюмирова-
на в следующих положениях:
бактериологическая война возможна только посредством заражения людей через
легкие;
возбудители болезней, не способные вызвать поражения людей путем передачи
через воздух, могут использоваться исключительно для проведения мелких диверсий;
бактериологическая война может произвести эффект, почти аналогичный эпиде-
мии; например, легочная чума может распространиться у применившего бактерии
агрессора с такой же силой, как и у его противника (здесь он почему-то сослался на
пандемию «испанского гриппа», распространившегося по всей Европе, игнорируя
границы между государствами).
В 1936 г. итальянский ученый de Alessi, подтвердил результаты, полученные груп-
пой Триллья. В частности, он привел данные, указывающие на проходимость возду-
хоносных путей для возбудителей таких инфекций, как брюшной тиф, паратифы,
бруцеллез и др. Попутно им было установлено, что предварительное вдыхание раздра-
жающих, прижигающих и наркотизирующих веществ значительно увеличивает опас-
ность заражения возбудителями инфекционных болезней. Здоровые легкие и воз-
духоносные пути с ненарушенными покровами обладают надежными элементами
локальной защиты против инфекции.
В те же годы разными авторами для возбудителей многих инфекций в лаборатор-
ных условиях была установлена радующая разработчиков БО закономерность — ин-
фицирующие дозы при ингаляционном введении в организм экспериментального
животного были значительно меньшими, чем при введении того же микроорганизма
подкожно или энтерально (иногда на несколько порядков). Причем, даже очень ко-
роткое пребывание в «микробном облаке» оказывалось достаточным для инфициро-
вания животных. Теоретические расчеты, данные практических наблюдений показы-
вали, что разбрызганные или распыленные бактерии могут находиться в воздухе до
нескольких часов. Перенос же микробов по воздуху зависел от степени дисперсности
микробных капелек.
Опыты Триллья, его учеников и последователей были выполнены в статических,
строго контролируемых условиях небольших аэрозольных камер и не только не мо-
делировали условия применения бактериального агента БО по реальной цели на теат-
ре военных действия, но даже по технике выполнения значительно уступали опытам
русского ученого Госа (1907). Неудивительно, что уже в средине 1930-х гг. нашлись
7.7. Становление бактериологического оружия между мировыми войнами
97
скептики, которые усомнились в возможности столь широкой интерпретации резуль-
татов опытов Триллья. Например, Koenigeru и Hutshison с удивлением обнаружили,
что разбрызганные возбудители большей частью в течение первого получаса оседа-
ют на пол. Muller утверждал, что еще меньшие шансы на длительную возможность
инфицирования имеются у «бактериального тумана» на открытом воздухе. Flugge
указал, что мелкие инфицированные капельки, а также зараженные бактериями пы-
линки, «вследствие быстрой смены воздуха, сильно разрежаются, и вдыхание возбу-
дителей или соприкосновение с ними представляют собой редкое исключение, ли-
шенное значения». Даже сам Триллья заметил, что «количество колоний, выросших
из сухого воздуха, было близко к нулю» (по Дробинскому И. Р., 1940).
Уже результаты этих опытов должны были вызвать у апологетов ЬО осторожное
отношение к возможности его боевого применения. Однако все вышло «с точностью
до наоборот». На протяжении 1930-х гг. они использовались «заинтересованными
сторонами» в качестве противовеса «немецкой аргументации». Например, мне встре-
чалось такое утверждение: «Заявления германских ученых о неосуществимости пред-
намеренного заражения вирулентной культурой лабораторного производства можно
считать сильно скомпрометированными в свете работ, опубликованных проф. Trillat.
Последний, как известно, добивался заражения и смерти подопытных животных
в результате ингаляции 24-часовой культуры куриной холеры» (цит. по Дробинско-
му И. Р„ 1940).
Для большей убедительности возможности создания БО, «притягивались» резуль-
таты одних эпидемиологических наблюдений, «упускались» результаты других. В ка-
честве доказательства возможности распыленных во влажном воздухе чумных бакте-
рий вызывать масштабную чуму, приводился пример появления вспышек легочной
чумы в Маньчжурии (1910—1911) как явления, возможного именно в холодные меся-
цы года. Однако игнорировалось то обстоятельство, что, под держивавшаяся среди ки-
тайского населения Владивостока (1921) на протяжении всех летних месяцев, легоч-
ная чума пришла к своему завершению с началом осени.
В обоснование возможности бактериологической войны приводились примеры
аэрогенных оаражений персонала бактериологических лабораторий. Отсюда делался
однозначный вывод: «Если заражение происходит даже в лабораторных условиях, где
принимаются специальные меры защиты, то в естественных же условиях заражение
может произойти еще легче, ибо о нем не всегда могут вовремя знать, и соответствен-
ные защитные меры могут не быть своевременно приняты» (Дробинский И. R, 1940).
И. Р. Дробинский (1940), заканчивая свою работу, приводит мнение английского
ученого A. Rochaix (1935): «Следует считать, что в определенных случаях при извест-
ных условиях, и особенно при условиях, могущих встретиться в военное время, —
вдыхание бактериального тумана может вызвать появление опаснейших инфекцион-
ных заболеваний: чумы, бруцеллеза, туляремии, пневмонии, гриппа, сапа, сибирской
язвы, цереброспинального эпидемического менингита, дифтерии. Кроме этого, для
заражения воздуха могут также служить малоизученные возбудители, например, ба-
циллы Wbitmore’a — возбудителя мелиоидоза, или тропического сапа. Итак, распыле-
ние патогенных культур в воздухе не встречает особых трудностей, а заражение челове-
ка может, к сожалению, в ряде случаев при этом произойти». Зарождающаяся военная
аэробиология сразу же сулила новому оружию сат те заманчивые перспективы.
В эти годы появились работы по эпидемиологии аэрогенной инфекции. В 1934 г.
W. F Wells выдвинул концепцию капельного ядра, поколебавшую доминирующее
положение концепции контактной инфекции. Он показал, что благодаря капельным
98
Часть 1. Краткая история применения биологических средств...
ядрам, содержащим микроорганизмы, происходит распространение кори, гриппа,
дифтерии и некоторых других инфекционных болезней. Он начал проверять свои идеи
путем установки источников УФ-излучения в школах Филадельфии, но вскоре, пос-
ле нескольких неудачных экспериментов, интерес к его работам снизился. Возрож-
дение интереса к эпидемиологии аэрогенной инфекции произошло уже после Второй
мировой войны (см. разд. 1.9).
Повышение вирулентности бактерий. Повышенная вирулентность бактерий воен-
ными бактериологами начала 1930-х гг. увязывалась с потенциальной возможностью
контактной передачи вызываемых ими болезней и способностью преодолевать спе-
цифический иммунитет. Flick (1927) утверждал следующее: «Задачей государств, го-
товящих этот способ войны, является усиление вирулентности культур, применение
симбионтов с резко усиленной вирулентностью и разработка способов иммунизации
войск и населения против всех этих бактерий».
Британский ученый К. Сталлибрасс (1936) подчеркивал, что усиление вирулент-
ности микроба для данного животного можно добиться, пассируя его через это жи-
вотное, но его вирулентность не увеличивается для других животных. Он объяснял это
явление необходимостью приспособления микроба к росту в особых условиях, пре-
доставляемых модельным животным. Сам процесс пассирования бактерий на живот-
ных Сталлибрасс рассматривал как «обратный в отношении длительного культиви-
рования на искусственных питательных средах, но он происходит быстрее, вероятно,
в результате большей гибели бактерий в теле животного, чем in vitro, и отсюда боль-
шей строгости отбора».
В те годы усиление вирулентности отдельных бактерий добивались и при их вы-
ращивании в целлулоидных мешочках, помещенных в брюшную полость животного
или при культивировании микроба на среде, содержащей сыворотку или кровь того
животного, в отношении которого желательно повышение его вирулентности.
Вирулентность микроорганизмов, пригодных для использования в целях бакте-
риологической войны, подвергалась очень внимательному изучению. Например,
Комбиеско (Gombiesco, 1928) нашел, что исходно вирулентные палочки возбудителя
сибирской язвы, которые давно не «проводились» через животное, лишены капсул,
не вирулентны и легко поглощаются фагоцитами. Однако если те же микробы поме-
стить в стеклянные капилляры (закупоренные проницаемой пробкой) и ввести пос-
ледние в брюшную полость морской свинки, то бациллы становятся резистентными —
«анимализируются» и образуют капсулы. Эти «капсулированные» микробы не погло-
щаются фагоцитами восприимчивых животных и вызывают септический процесс у
экспериментального животного в течение нескольких часов.
Таким образом, к началу 1930-х гг. разработчикам БО были известны различные
способы повышения вирулентности бактерий. Причем некоторые из них не могли
быть осуществлены без экспериментов на приматах.
Технологии и устройства для производства и длительного хранения микроорганизмов
в жизнеспособном состоянии. В вопросе производства бактерий у апологетов бактерио-
логической войны 1930-х гг. вообще не возникало никаких опасений. Бактерии рас-
сматривались ими в качестве контагия и, следовательно, их для войны не требовалось
много. Так, Romieu (1934) отмечал: «Приготовление (разведение) бактерий в большом
количестве во многих случаях не является трудной задачей и требует весьма немного
времени. Некоторые виды бактерий могут быть приготовлены водной единственной
лаборатории в кратчайший срок в количестве до 100 млрд единиц. Их массовое при-
готовление не требует ни сложных установок, ни крупных заводов».
1.7. Становление бактериологического оружия между мировыми войнами
99
Легкомысленные рассуждения об «одной единственной лаборатории», способной
создать БО, можно прочитать и сегодня, особенно когда речь идет о БО Саддама Хусей-
на. Действительно, если разделить эти миллиарды бактерий на одну инфицирующую
дозу, например, для возбудителя чумы для человека она составляет от 100 до 3000 кле-
ток (см. разд. 3.2), то теоретически уже этим количеством бактерий можно убить мил-
лионы человек. Но реально 100 млрд любых бактерий (если кто хочет подержать их в
руках) — это то их количество, которое вмещается в объеме одной таблетки брюшно-
тифозной. холерной, дизентерийной вакцин того времени, предназначенных для
энтеральной иммунизации.
В 1930-х гг. технологии глубинного культивирования бактерий в условиях аэрации,
способные обеспечить наработку микроорганизмов в количествах, достаточных для
ведения масштабной биологической войны, находились в «зачаточном состоянии».
Производственная деятельность бактериологических институтов заключалась в полу-
чении агаровой микробной эмульсии в сравнительно небольшой емкости плоских
склянок (бактериологических матрацев), бьющихся и лопающихся в автоклаве. Един-
ственно возможный путь, по которому шли в те годы разработчики способов нара-
ботки бактериальной массы, заключался в увеличении общей площади агара, одно-
моментно используемой для культивирования микроорганизмов. В Крымском
санитарно-бактериологическом институте (Севастополь) для получения бактерийных
препаратов с начала 1930-х гг. применялся оригинальный аппарат Н. Г. Щербиной,
представлявший собою серию посевных плоскостей из алюминия, уложенных в осо-
бый футляр (рис. 1.15).
Рис 1.15. Аппаратдля получения бактериальной биомассы. А Наружныйвид.
Б. Разрез. Аппарат мог иметь различные размеры. Общая посевная поверх-
ность типового аппарата около 9000 см2; при толщине слоя агара в 0,5 см в нем
использовалось около 4,5 л среды на каждый посев. По Н. Г. Щербиной (1932)
К этим конструкциям культиваторов бактерий мы еше вернемся при рассмотре-
нии технологий, используемых для создания БО в отряде 731 (см. разд. 1.9) Важно
посмотреть и на то, каким образом могли в те годы длительно храниться в >кизнеспо-
собном состоянии запасы бактерий.
Технология высушивания в вакууме сред, содержащих бактерии (см. разд. 1.6),
применявшаяся еще до Первой мировой войны, к началу Второй мировой войны была
значительно усовершенствована. В практику были введены высокопроизводительные
вальцовые (вакуумные) сушилки (рис. 1.16).
Появились и новые технологии. В1921 г. Свифтом был описан метод консервации
бактериального препарата, заключающийся в быстром предварительном заморажи-
вании жидкой культуры и в последующем выдерживании застывшей массы в глубоком
100
Часть 1. Краткая история применения биологических средств...
Рис. 1.16. Вальцевая сушилка. По Н. Н. Титову (1945)
вакууме до полного высушивания. Выживаемость микроорганизмов в конечном пре-
парате резко возросла. Причиной такого качественного скачка в технологии высуши-
вания стало удаление влаги из замороженного материала путем превращения льда
непосредственно в пар, минуя жидкую фазу. С понижением атмосферного давления
интенсивность испарения воды увеличивалась, скорость высушивания резко возра-
стала. Поэтому внутриклеточные белки не подвергались воздействию высоких кон-
центраций солей и не денатурировали. Высушенные бактерии представляли собой губ-
чатую массу, объем которой приблизительно соответствовал объему первоначально
замороженной бактериальной культуры или концентрированной суспензии. Так как
колоидно-химическая структура препарата не нарушалась, при добавлении разво-
дящей жидкости (обычно это изотонический буферный раствор) высушенный мате-
риал легко растворялся — отсюда произошло название технологии — лиофилизация
В 1930-е гг. разработано большое количество модификаций способов лиофилизации
и сушильных аппаратов, работающих на этом принципе. Даже наметились два направ-
ления их технического эволюционирования (рис. 1.17).
Вначале 1930-хгг. была значительно усовершенствована технология распылитель-
ной сушки. Высушивание биологических материалов производилось в закрытой ка-
мере, в которой жидкость, распределенная под действием центробежной силы или
давления тонким сплошным слоем, высушивается посредством потока пара, прохо-
дящего над жидкостью или под ней (рис. 1.18).
Однако исходным предназначением всех перечисленных технологий было кон-
сервирование лечебных сывороток, т. е. биопрепаратов, не содержащих живые микро-
организмы. При их применении к микроорганизмам, и в особенности к вирулент-
ным, исследователя поджидали неприятные неожиданности. Например, по данным
Н. Н. Титова (1945), в его собственных экспериментах по высушиванию культур саль-
монелл Данича, выполненных в 1936 г. на распылительном аппарате, основная масса
1.7. Становление бактериологического оружия между мировыми войнами
101
Рис, 1.17. Аппараты для лиофильного высушивания биопрепаратов конца 1930-х гг.
А, .Аппарат камерного типа конструкции Гревеса и Эдера: 1 — компрессорная уста-
новка; 2 — сушильник; 3 — конденсатор (змеевик), 4 — вакуум-масляный насос.
Б. Коллекторный аппарат Флосдорфа и Мэца: 1 — коллектор с ампулами; 2 — ван-
на для сухого льда, 3 — конденсатор; 4 — вакуум-масляный насос; 5 — манометр
Мак-Леода. По Н. Н. Титову (1945)
бактерий погибала, а вирулентность оставшихся в живых по отношению к крысам
была не более 50 % от исходной. Такие же результаты были получены в 1940 г. в лабо-
ратории по борьбе с грызунами, с амбарными и другими вредителями Института мик-
робиологии Всесоюзной академии сельскохозяйственных наук (Ленинград), с этими
же бациллами, но высушенными лиофильно.
Рис 1.18. Распылительная центрифуга. Посредством диска Ь, имеющего большое
число оборотов, жидкость, притекающая по трубке а, мелко распыляется горизон-
тальным слоем. Ниже этого слоя по трубе с подается из распределителя пар, высуши-
вающий распыленную жидкость, твердые частицы которой улавливаются на наклон-
ных стенках кожуха е и собираются в приемник g. Выделившиеся пары осаждаются
на крышках i и h, вокруг которых имеется желобок для конденсационной воды.
Из желобков вода удаляется по трубе К. По Н. Н. Титову (1945)
102
Часть 1. Краткая история применения биологических средств...
Выяснилось, что некоторые микроорганизмы не выносят быстрого охлаждения,
что большое значение для сохранения их в жизнеспособном состоянии имеют среда
высушивания, возраст культуры, концентрация бактерий в высушиваемых суспензиях,
режим высушивания, содержание остаточной влаги в препарате, условия последую-
щего хранения и еще многие другие факторы.
Оказалось также, что способ тяжело масштабируется из-за проблем с равномер-
ным замораживанием субстрата. Коллекторные сушилки в принципе не могут быть
высокопроизводительными. Камерные сушилки при манипуляциях с сухим микроб-
ным материалом создают значительные количества бактериального аэрозоля, пред-
ставляющего опасность для персонала. Их разгрузка не может произойти без наруше-
ния вакуума, что приводит к повышению влажности высушенного материала и к
интенсивному отмиранию микроорганизмов. К концу 1930-хгг. ситуацию с производ-
ством «контагия» для «оружия бедных» можно охарактеризовать поговоркой: «Оста-
лось только начать и кончить». Титов писал в 1945 г. о неудачах технологий высуши-
вания бактерий, что ситуация «может быть изменена подбором сред, на которых
изготовляется культура перед высушиванием». Остальные проблемы еще только пред-
стояло дифференцировать друг от друга. В 1945 г. американские специалисты счита-
ли необходимым иметь для нормального выполнения ооевых задач мощности по про-
изводству биоагентов в количестве не менее чем 5 з/сутки (Dando М.. 1994).
Бактериологические диверсии в СССР. В качестве основного критерия для «отфил ь-
трования» вымышленных бактериологических диверсий 1930-х гг от реально осу-
ществленных, как и при описании бактериологических диверсий Первой мировой
войны, мы будем использовать соответствие замысла преступления реальным возмож-
ностям как бактериологии того времени, так и лиц, уличенных в этом преступлении.
Например, обвинения в заражении ботулиническим токсином консервов, постав-
.гяемых в Красную армию, предъявленные в 1937 г. И. М. Великанову (1898—1938;
начальник Биотехнического инсл итута РККА), мог ут быть только вымытом, так как,
во-первых, в технологии их производства предусмотри но автоклавирование, во время
которого токсин разрушается высокой температурой; во-вторых, сам ботулинический
токсин очень нестабилен при хранении. Но в литературе предвоенных лет (например,
в работе Саркисова И. 3., 1940) есть упоминание о завозе в СССР из США породи-
стых сельскохозяйственных животных, зараженных бруцеллезом, вызвавших вспышку
бруцеллеза среди людей и сельскохозяйственных животных. В те же годы считалось,
что паразит виноградников — филлоксера, был намерено завезен в 1929 г. в СССР ли-
цами, враждебными советской влас™.
Особое место среди такой литературы занимают материалы судебного процесса
над «бухаринско-троцкистскими бандами» (март, 1938), содержащие упоминания об
организации масштабных биологических диверсий в предвоенном СССР. В течение
70 последних лет обвинения, предъявленные этим заговорщикам в желании развалить
СССР и получить власть в обмен на часть его территории, воспринимались как «высо-
санные из пальца» А. Я. Вышинским (1935-1939 гг. — прокурор СССР), как «вздорные
и нелепые измышления сталинско-молотовской пропаганды». Дьявол всегда делает
вид, что не существует. Сейчас, когда именно это и произошло, те обвинения уже не
представляются столь необоснованными. Впрочем, судите сами вот выдержка из по-
казаний на процессе бывшего наркома земледелия СССР М. А. Чернова (1891—1938):
«...В части, касающейся семян, мы вкзючиливсвою программу—запутать семен-
ное дело, смешать сортовые семена и гем самым понизить урожайность в стране.
В части, связанной с севооборотом, путем неправильного планирования посевных пло-
1.7. Становление бактериологического оружия между мировыми войнами
103
щадей поставить колхозное крестьянство в такое положение, при котором колхозники
фактически не могли бы осуществлять правильный севооборот и были бы вынужде-
ны занимать под посевы луга и пастбища Это должно было привести к уменьшению
урожая в стране и вместе с тем вызывало бы озлобление крестьян, которые никак
не могут понять, почему их заставляют распахивать луга и пастбища, когда колхозы хо-
тят развивать животноводство, и нужна кормовая база.
...В части животноводства были поставлены задачи — вырезать племенных произ-
водителей, добиваться большего падежа скота, не давать развиваться кормовой базе,
особенно использовать для падежа скота искусственное заражение скота различного
рода бактериями.
Вышинский. Расскажите более подробно о падеже скота, откуда получали бактерии,
какие и так далее.
Чернов.. Для тоги чтобы добиться падежа скота в Восточной Сибири, я предло-
жил начальнику Ветеринарного управления Гйнзбургу, участнику организации правых,
а через него начальнику Ветеринарного снабжения, также участнику организации
правых, не завозить противоязвенные биопрепараты в Восточную Сибирь, зная о том,
что в Восточной Сибири очень опасно по части сибирской язвы. Препараты эти туда
завезены не были. Подготовка эта велась в 1935 году, и когда весной 1936 года там
вспыхнула сибирская язва, то оказалось, что действительно препараты туда завезены
не были, и тем самым было погублено — я точно не могу сказать, во всяком случае,
больше 25 тыс. лошадей.
Второе. Я поручил Гинзбургу и заведующему бактериологическим отделом Бояр-
шинову произвести искусственное заражение в Ленинградской обласл и свиней рожей,
а в Воронежской и Азово-Черноморском крае — чумой. Выбрал я эти две бактерии
потому, что прививка эта производится свиньям не убитыми микробами, а живыми,
только ослабленными. Поэтому технически организовать искусственное заражение
было довольно просто.
Во-вторых, потому что по этим двум болезням существует правильный порядок,
при котором в случае заболевания чумой в той или иной местности подлежит поголов-
ной прививке все находящееся в этой местности поголовье свиней. Это давало возмож-
ность сразу придать массовый характер этому заболеванию. С этой целью были выде-
лены, по моему предложению, три фабрики: Кашинцевская, Орловская и Ставрополь-
ская Там были изготовлены биопрепараты с неослабленными бактериями под
особыми номерами серий. Номера этих серий были сообщены Бояршинову, который
сообщил их на местах тем начальникам ветуправлений, на которых можно было по-
ложиться в этом случае, а они, в свою очередь, передали их в районы тем ветврачам,
которые были антисоветски настроены, и в случае большого падежа скота, не стали
бы поднимать большого шума. Таким образом, эти биопрепараты были завезены, и ис-
кусственная прививка была произведена в этих трех областях. Трудно оценить резуль-
таты, но, во всяком случае, нужно считать, что благодаря этому диверсионному акту
было погублено несколько десятков тысяч свиней*'.
Военно-биологическая программа Польши1. Приведенное выше циничное заяв-
ление польского подполковника J. Karyszkowski о целесообразности оценки эффек-
тивности БО в экспериментах на военнопленных, не было ни бравадой, ни заявлени-
ем о намерении, оно отражало уже сложившуюся практику таких исследований во
Второй Речи Посполитой.
1 Раздел в основном написан по работе Анджея Краевского (2009), использовавшего рас-
секреченные послевоенные документы польского МИДа, Управления безопасности и
польской прокуратуры.
104
Часть 1. Краткая история применения биологических средств...
Польская разведка еще в 1925 г. забила тревогу по поводу угрозы Польше со сто-
роны БО русских. Была получена информация о том, что Советы экспериментируют
с опасными микроорганизмами в военных целях. С этого момента польские дипло-
маты под руководством Александра Юзефа Скшиньского (Aleksander Jyzef Skrzycski,
1882—1931) инициировали в Лиге Наций вопрос о включении запрета на применение
БО в переговоры о разоружении, которые шли в Женеве. Польские разведывательные
спецслужбы с гали внимательно следить за всеми случаями отравления в воинских
частях. Было отмечено, что в гарнизонах ежегодно происходили массовые случаи саль-
монеллеза. Появились подозрения, что это не случайности, а действия немецкой или
советской разведки, испытывающей возможности ослабления армии Республики
Польша путем бактериологических диверсий. По инициативе II Отдела Генерально-
го штаба в начале 1930-х гг. в Варшаве в Военном институте защиты от газов (Wojskowy
Instytut Przeciwgazowy, ул. Людная, 11) была организована секретная лаборатория
(«двойка»), которая занималась изучением поражающего действия опасных бактерий
и бактериальных токсинов. Лабораторию возглавил врач-биолог Альфонс Островский.
В 1933 г. руководителем «двойки» стал доктор Ян Гольба. Штат был небольшим, в ла-
боратории работали три бактериолога, одна лаборантка и одна уборщица.
Первоначально польскими военными бактериологами исследовались поражаю-
щие свойства возбудителей чумы, холеры, дизентерии и сапа и ботулинического ток-
сина (они называли его «колбасный яд»). После войны в ходе допросов в варшавском
Управлении безопасности (УБ) Островский показал, что летом 1933 г., по приказу
курирующего работу лаборатории капитана Игнапия Харского, он взял с собой 0,2 г
ботулинического токсина и отправился в поселок Луней, где находился гарнизон
Корпуса пограничной стражи (КПС). «В Луньце на посту КПС мне показали челове-
ка около 40 лет, русской национальности, среднего роста, брюнета, интеллигентного
вида. Этому человеку дали ботулинический токсин, угостив бутербродом с ливерным
паштетом», — показал Островский. Советский шпион, захваченный при попытке не-
легального пересечения границы, умер через два дня. По словам Островского, мерт-
вое тело перевезли к пристани на реке Припять и на лодке доставили к другому бере-
гу, где уже начиналась территория СССР. Там тело бросили в воду и толкнули на
советскую сторону.
Аналогичный «эксперимент» в то же лето Островский провел на посту КПС в Глем-
боке. «Там, по приказу начальства, тоже приготовил для лица в возрасте от 30 до 35 лет,
со следами побоев, колбасный яд в бутерброде с ливерным паштетом», — показал он.
При этом Островский подчеркнул: «В тот момент, когда его кормили токсином, меня
не было рядом, но способ подачи ему ботулотоксина был со мной согласован. Потом
я увидел его дважды: через 12 ч и через 24 ч. Я видел признаки отравления: расшире-
ние зрачков, нарушение зрения, головокружение, сухость в горле. Но смерть не на-
ступила, дальнейшая судьба этого человека мне не известна. После возвращения в
Варшаву, я подал рапорт капитану Хаскому, который был недоволен ходом опыта.
Колбасный яд оказался отравой, которая смогла причинять огромные страдания но
была мало эффективна».
В работе «двойки» появились первые успехи. Доктор Генбарска-Межвиньская
разработала метод хранения культур микробов при помощи лиофильного высушива-
ния. Столь же новаторским достижением оказалось получение ботулинического ток-
сина в виде порошка. Удалось также масштабировать способ размножения бактерий,
вызывающих брюшной тиф.
1.7. Становление бактериологического оружия между мировыми войнами
105
Подозрение о том, что соседи Польши разрабатывают новые виды оружия массо-
вого поражения, привело к тому, что 11 Отдел Главного штаба решил увеличить фи-
нансирование научных исследований. В 1935 г. в Варшаве было организовано От-
дельное техническое управление (Samodzielny Referat Techniczny, SRT). Его первым
начальником стал капитан Игнаций Харский, уже зарекомендовавший себя поло-
жительно убийствами русских людей с помощью ботулинического токсина. На осна-
щение этого тайного исследовательского центра выдали немалую д ля тех времен сум-
му — около полумиллиона крон. В 1937 г. в управлении работало семь офицеров и
около шестидесяти научных и технических специалистов. Параллельно велись ис-
следования в области боевых ОВ и других токсических веществ. В составленном про-
куратурой СССР обвинительном акте можно прочитать, что там работали над: «Уве-
личением вирулентности болезнетворных бактерий группы сальмонелла, в том числе:
тифа, паратифа А, пара-В, пара-С, Гертнера, группы дизентерии, таких как Shiga-
Kruze, Fletnera, Stronga и разработкой методов заражения этими бактериями людей,
животных, пиши и воды». Сам Гольба объяснял попытки увеличения вирулентности
бактерий необходимостью опенки их возможного поражающего действия и разра-
ботки способов защиты.
В то время Гольбе пришла в голову мысль новаторского эксперимента. По просьбе
Гольбы руководитель механических мастерских SRT Ян Кобус установил на автомо-
биле компрессор с распылителем. С его помощью распылили безопасные для людей
бактерии в нескольких точках Варшавы, в том числе и в железнодорожном туннеле
под Варшавой. Замысел этого незамысловатого эксперимента Гольба «позаимствовал»
из вранья британского журналиста Стида (см. «Мистификация Стида»). Он расста-
вил на площади Любельской Унии в Варшаве своих людей с чашками Петри, сам же,
ведя автомобиль вокруг площади, распылял бактерий при помощи аппаратуры, встро-
енной в автомобиль Кобусом. «Целью этих опытов было установление количества
бактерий в воздухе и на чашках, с целью определения, сколько времени будет зара-
жен воздух в данном месте», — отмечено в следственном протоколе.
Польская разведка сотрудничала с японской разведкой с 1925 г. Обе службы об-
менивались информацией о СССР, включая данные по БО. Новаторские идеи Голь-
бы вызвали живой интерес японцев. В 1936 г. на территории «двойки» в Варшаве со-
стоялась тайная конференция, на которую прибыла японская делегация научных
работников из Главной базы Управления по водоснабжению и профилактике частей
Квантунской армии в Харбине1. Во время встречи Гольба прочитал доклад о возмож-
ности заражения людей во время военных действий возбудителями брюшного тифа,
сыпного тифа, дизентерии, сибирской язвы и сапа. У объединенных общим русским
врагом «товарищей по оружию» не было никаких проблем с пониманием друг друга,
потому что на совещании все бегло говорили... по-русски.
Гольба находился под нарастающим давлением военного руководства Польши,
требующего создания чудесного оружия. Чтобы ускорить исследования, он предло-
жил построить в Брестской крепости более крупную лабораторию, оснащенную гер-
метичной камерой, в которой можно было бы вести аэробиологические эксперимен-
1 А. Краевский (2009) пишет о Центре исследований в области биологического оружия,
находящемся в Пинфане, но это ошибка. Объект в Пинфане в 1936 г. японцы только начали
строить. Исследования по БО в 1936 г. они проводили в «отряде Камо», дислоцировавшемся в
Бэйиньхэ (район на юго-востоке Харбина) (см. разд. 1.8).
106
Часть 1. Краткая история применения биологических средств...
ты на животных. Пожелание его исполнилось, и в 1937 г. в Бресткой крепости было
построено «помещение с каменными стенами, внутри окрашенное масляной краской,
площадью приблизительно 9 кв. м». Дверь была усилена железом. В одной из стен
находилось окошко и отверстие для установки распыляющего прибора.
После первых успешных экспериментов на животных начальник «двойки» пол-
ковник Тадеуш Пелчиньский потребовал проведения исследований также на людях.
Между польской и японской разведками постоянно шел обмен информацией. Навер-
ное, полковник Пелчиньский решил, что Польша не может позволить себе отстать в
этой гонке от союзника. Важной уликой, указывающей на то, что в Бресте на самом
деле проводили эксперименты на людях, является письмо, которое в мае 1955 г. док-
тор Гольба послал Генеральному прокурору ПНР.
«Я действительно проводил на опытной станции в Бресте на Буге опыты с болез-
нетворными микробами над индивидуумами. Это факт, которого я не отрицаю», —
писал Гольба. Потом объяснял: «Задача на выполнение этих исследований давалась
мне моими начальниками в виде военного приказа. Перед совершением опытов мои
начальники утверждали, что лица, над которыми данные опыты будут проводиться,
приговорены к смертной казни, и их дела апелляции не подлежат». Далее он объяс-
нял, что был убежден, что так может наилучшим образом служить родине, которой
угрожают внешние враги. По протоколам, составленным «сталинскими следователя-
ми»1, семь неопознанных лиц, ставших подопытными в экспериментах, доставлял в
Брест начальник III секции «двойки» подполковник Юзеф Скшидлевский.
После смерти заключенного тело растворяли в специальной керамической ванне,
наполненной смесью соляной кислоты с добавками сероуглерода и концентриро-
ванной азотной кислоты. Краевский приводит еше один фрагмент записи допроса2:
«После того как нас, т. е. меня и доктора Гольбу с разрешения руководителя S.R.I.
пропустили в место заключения—доктор Гольба надел противогаз и отправился к ван-
не, в которой находились человеческие останки, подвергшиеся действию соляной кис-
лоты. Спустя короткое время Гольба вернулся от ванны, заявляя, что не наступило пол-
ное разложение останков, подвергшихся действию кислоты, так как в ванне сохрани-
лись еще человеческие кости. Гольба рассказывал мне, что человек, останки, а точнее,
кости которого находились в ванне, умер от полученного бутерброда с мясом, который
перед этим был насыщен колбасным ядом, т. е. ботулином. Я фамилию умерщвленно-
го таким образом человека не знаю Я знаю, однако, что этот человек был уничтожен
II Отделом, а растворен в соляной кислоте для того, чтобы скрыть следы убийства. Во
время моего пребывания в указанном месте заключения я видел там одного арестован-
ного, находящегося за решеткой. Затем мы покинули месго заключения и вернулись в
Варшаву».
Несмотря на интенсификацию исследований и эксперименты на людях, к биоло-
гическому оружию польские военные не приблизились. В сентябре 1939 г. в Брест
вошли части Красной армии, польским бактериологам удалось бежать. Гольба и Ост-
ровский через Румынию добрались до Франции. Генбарская-Межвиньская осталась
в оккупированной Польше. После поражения Франции в 1940 г Островский оказался
1 Странная логика у пана А. Краевского. С одной стороны, присутствуют некие «сталин-
ские следователи», расследующие массовые убийства — это как бы плохо. С другой — серий-
ные убийцы, использовавшие для убийств русских людей биологические агенты, но ведь они
жертвы «сталинских следователей».
2 Видимо, Островского.
1.7. Становление бактериологического оружия между мировыми войнами
107
в немецком плену, зато Гольбе удалось перебраться в Соединенное Королевство. Там
бактериолог по поручению подполковника Станислава Гано, тогдашнего начальника
«двойки», написал доклад, в котором описал, чем занималась его лаборатория. Плод
его творчества был передан в Портон Даун (см. разд. 1.9 «Начало программы по со-
зданию БО в Соединенном Королевстве»), в британский тайный центр исследований
в области биологического оружия. В июле 1941 г., англичане попросили Гольбу деталь-
но описать технологию высушивания микробов и их хранения в виде порошка. Воз-
можно, достижения польских бактериологов были использованы британскими уче-
ными в работе над получением сухих рецептур возбудителя сибирской язвы.
Гольба два раза отказывал англичанам, предлагающим ему работу в своих лаборато-
риях, потому что для него это означило бы необхо, [имость службы в британской армии
и, может быть, потерю навсегда возможности возвращения в Польшу. Поэтому он
предпочел остаться обычным военным врачом в элитной Отдельной парашютной бри-
гаде им. Станислава Сосабовского. Вместе с другими британскими солдатами в 1944 г.
Гольба приземлился на парашюте под Арнеме. Там прославился управлением под об-
стрелом врага полевым госпиталем, за что получил орден «Крест Доблести» и звание
подполковника.
В Польшу Гольба вернулся только в 1947 г. и занял пост руководителя бактерио-
химической лаборатории в Повятовой клинической больнице. Вернулся и Остро-
вский. Он открыл врачебный кабинет в Плоньске. Оба почему-то надеялись на то, что
коммунисты дадут им жить спокойно.
УБ на след бывших работников бактериологической лаборатории SRT, по мнению
A. Kpat вского, вывел бывший подполковник Скшидлевский. После разгрома немцами
в 1939 г. польской армии ему удалось бежать во Францию, но в 1940 г. он попал в не-
мецкий плен и в офлаг (Oflag II D Gross-Bom — лагерь для пленных офицеров союз-
нических армий в Гродеке). Опз да в 1945 г. его освободила Красная армия. Свобода
не продолжалась долго, потому ч го этот бывший представитель «двойки» быстро попал
в руки УБ. Из его показаний в УБ очень много узнали о работе SRT. В ноябре 1951 г.
одновременно арестовали: доктора Гольбу в Щецине, Островского в Плоньске, Ген-
барску-Межвиньскую в Воломине и Кобуса в Прушкуве. Никго из этих лиц не попал
в тюрьму, но они были изолированы от мира и друг от друга где-то недалеко от Вар-
шавы в отдельных коз геджах. Через двадцать месяцев власти подготовили процесс,
который должен был быть самым большим показательным процессом в истории ста-
линской Польши. Лично заботился об этом и Якуб Берман, управлявший страной
совместно с Болеславом Берутом. Что они хотели достигнуть, показывает приказ,
который послал прокурорам в начале 1952 г. вице-шеф Министерства госбезопасно-
сти Роман Ромковский. «Процесс группы работников SRT должен доказать обществу
суть польской разновидности фашизма — пилсудчины, ее методы действий в облас-
ти внутренней и внешней политики». В Российском государственном архиве социаль-
но-политической истории (РГАСПИ, Москва) сохранилась записка, датированная
сентябрем 1952 г, подписанная министром иностранных дел Андреем Вышинским и
Генеральным прокурором СССР Григорием Сафоновым и направленная непосред-
ственно Иосифу Сталину. Из нее можно понять, что, согласно решению ЦК ВКП(б),
принятому 10 июня 1952 г., в Варшаву послали представителя Прокуратуры СССР
П. А. Кульчицкого с целью «ознакомления с материалами дела против бывших работ-
ников довоенного польского Главного штаба, который являлся организатором под-
готовки к бактериологической войне против Советского Союза, а также в целях уста-
новления целесообразности проведения такого процесса».
108
Часть 1. Краткая история применения биологических средств...
Этот представитель во время пребывания в Варшаве сошелся с Яковом Берманом
и Франнишком Мазуром. Они сообщили ему, что нужен образцовый судебный про-
цесс, дискредитирующий довоенные власти Польши. Берман просил также, чтобы
прислали на выручку советских специалистов в области бактериологии, токсиколо-
гии и химии. «Они констатировали, что не располагают специалистами, которыми
можно бы было бесконтрольно доверить расследование этих дел», — доложил по воз-
вращении Кульчицкий.
После этого рапорта в сентябре 1952 г. Политбюро ВКП(б) приняло постановле-
ние о крайней желательности процесса. Кроме того, оно рекомендовало: «доверить
Генеральному прокурору СССР тов. Г. Сафонову откомандировать в Польшу началь-
ника Следственного отдела для специальных дел Главной военной прокуратуры Со-
ветской армии полковника тов. Кульчицкого в целях оказания дальнейшей помощи
в подготовлении и организации упомянутого выше судебного процесса».
На скамью подсудимых должен был сесть довольно широкий круг людей, потому
что как, в свою очередь, сообщал в Москву полковник Кульчицкий: «Заочно под след-
ствие попали: бывший начальник II Отдела Главного штаба полковник Пелчиньский
(живет в Лондоне); бывший начальник Отдельного Технического Управления II От-
дела Главного штаба капитан Харский (живет в Эдинбурге в Англии); бывший руко-
водитель токсикологической лаборатории того же управления (живет в Нью-Йорке)».
Правительство Народной Польши обращалось в 1952 г. к правительствам США и Ве-
ликобритании об экстрадиции вышеупомянутых людей, но не получило по этому
требованию никакого ответа.
Но эти усилия оказались излишними. Скшидлевский умер в мокотовской тюрь-
ме. А, когда подготовка к процессу завершилась, в марте 1953 г. внезапно умер Иосиф
Сталин. Двумя месяцами позже письмо, визированное Советом министров СССР,
неожиданно велело Болеславу Беруту прекратить образцовый процесс. В конце сен-
тября 1953 г. перед воеводским судом Варшавы начался процесс только над четырьмя
работниками SRT.
Заседания суда были закрытыми. Адвокатов часто удаляли из зала суда. Тройку
бактериологов и руководителя механических мастерских SRT Яна Кобуса обвинили
в «проведении экспериментов на людях, в виде заражения их бактериями тифа, пере-
даваемыми с пищей и распыляемыми в специальной камере на территории Брест-
ской крепости». Доктора Гольбу обвинили в убийстве пяти деятелей компартии, Ост-
ровского — в двух убийствах. Янину Генбарскую-Межвиньскую и Яна Кобуса судили
за соучастие.
Сломленные в процессе следствия научные работники признали свою вину. Уже
19 октября 1952 г. возглавляющий заседание судья Мариан Стемпчиньский во время
оглашения приговора заявил, что «подсудимые приняли участие в одном из самых
величайших преступлений. Это преступление против своего собственного народа и
против всего человечества». Камеру в Бресте сравнили с «камерами Освенцима, Май-
дане ка, Треблинки». Но суд проявил снисходительность и осудил Гольбу и Островского
на 13 лет тюрьмы, Генбарсхую-Межвиньскую на 7 лет, Кобуса на 4 года заключения.
Затем, ссылаясь на закон об амнистии от 22 февраля 1947 г., судья сократил пригово-
ры наполовину, а период предварительного заключения засчитал в срок наказания.
В течение года осужденных, кроме Яна Гольбы, освободили. Гольба в мае 1955 г.
направил Генеральному прокурору ПНР прошение о пересмотре дела. Предложение
рассмотрели положительно. Вскоре также и доктор Гольба вышел на свободу. До кон-
ца жизни он жил в Щецине, работая на должности заместителя директора воеводской
1.7. Становление бактериологического оружия между мировыми войнами
109
санитарно-эпидемиологической станции, больше его не трогали. Судьба оставшей-
ся тройки неизвестна.
Возобновление бактериологических диверсий в ходе военных действий. Первое со-
общение о начавшейся на Дальнем Востоке бактериологической войне было опубли-
ковано в 1938 г. в ноябрьском номере немецкого журнала «Gasschutz und Luftschutz».
Оно основывалось на заявлении научного руководителя Токийского эпидемиологи-
ческого института, профессора Микава, вернувшегося из Китая. Микава обвинил
китайскую армию в преднамеренном заражении возбудителем холеры водоисточни-
ков, используемых японскими вооруженными ситами. В подтверждение своего заяв-
ления он привел следующие факты:
1. До отступления китайцев из Киукианга и занятия последнего японцами, в го-
роде и прилегающих к нему районах не было холерных заболеваний.
2. Все случаи холеры среди японских военнослужащих связаны с употреблением
воды из отдельных колодцев.
3. Холерные заболевания среди японцев появились одномоментно, что явно про-
тиворечит естественному развитию эпидемии.
4. В одном из колодцев была обнаружена стеклянная ампула, предназначенная для
хранения холерной культуры.
В специальном документе, переданном японским МИДом иностранным дипло-
матам в Токио 25.09.1938 г., приводятся следующие факты:
1. Китайские пленные военнослужащие 27.07.1938 г. признались, что каждому
китайскому полку придается особая бактериальная часть, которая через местных
жителей заражает одежду и пищу японцев патогенными бактериями.
2. После занятия Лаошана японцы установили, что перед своим отступлением
китайцы произвели заражение колодцев холерными вибрионами, в результате чего ряд
японских солдат и большое число городских жителей заболели холерой.
3. Холерная вспышка 1938 г. в Кайфыне была вызвана китайскими войсками при
помощи холерного вибриона.
4. При обследовании 75 городских колодцев в Тунг-Ли-Тцун, 14 из них оказались
зараженными возбудителем холеры.
В материалах SI PR1 (The Problem of Chemical and Biological Warfare, 1970) co ссыл-
кой на американский документ 1945 г. эта информация подтверждается. Утверждает-
ся, что американцы располагают переводом документа разведывательного отдела
Квантунской армии, в котором сказано, что китайцы в период с сентября 1937 г. по
август 1939 г. скрытно и весьма искусно применяли химические и бактериологичес-
кие материалы против личного состава японской армии, а также для заражения жи-
вотных, источников водоснабжения и продуктов питания. В этом документе указа-
но, в частности, на установление 8 случаев применения возбудителя холеры, и двух
случаев применения возбудителя сибирской язвы. Во время эпидемии холеры кото-
рую связывали с заражением колодцев, было зарегистрировано 650 случаев болезни
(500 — местное население, 150 — японские военнослужащие).
А как же немцы? О наступательной германской военно-биологической програм-
ме писать сложно, так как официальных германских документов о том, что она суще-
ствовала, нет. А официальные заявления германских ученых о невозможности созда-
ния БО, воспринимались «общественным мнением» в 1930-е гг. как попытки скрыть
такую программу.
После прихода Гитлера к власти бактериологический институт имени Роберта Коха
(Берлин) был переподчинен военному министерству. Данные разведки союзников,
но
Часть 1. Краткая история применения биологических средств...
озвученные, разумеется, уже после войны, показали, что Гитлер в 1939 г. запретил
исследования по наступательным аспектам БО, и этот запрет неоднократно подтвер-
ждался им в ходе войны. На Нюренбергском процессе к германским руководителям
не выдвигалось обвинений в разработке и применении БО (Robertson A. G., Robert-
son L. J., 1995; Bernstein В. J., 1988).
С 1943 г. Германия имела свою военно-биологическую программу, предназначен-
ную для противодействия биологическим диверсиям. Разработке программы предше-
ствовали какие-то события 1942 г., вызвавшие у германского командования подозре-
ния в применении противником биологических агентов в Польше и Чехии. Известно
об обнаружении сотрудниками гестапо в Варшаве арсенала средств польских парти-
зан, предназначенных для осуществления биологических диверсий (Mobley J. А.,
1995). Учитывая эпопею SRT (см. выше «Военно-биологическая программа Польши»),
в обнаружении такого арсенала в Варшаве нет ничего необычного.
В некоторых американских источниках утверждается, что партизаны в Польше и
России применяли против немецких войск такие биологические агенты, как ботули-
нический токсин, возбудители брюшного тифа, дизентерии, сапа, холеры, сибирской
язвы и паратифа (The Problem..., 1970). Эта тема ждет своих исследователей в воен-
ных архивах Германии, Польши и России.
В отличие от других европейских держав, в Германии посчитали идею создания
дешевого «оружия массового поражения» нереальной и слишком дорогой для осу-
ществления при имеющемся уровне знаний в области военной бактериологии. Зато
в области других вооружений германским руководством были выбраны весьма вер-
ные приоритеты. После войны еще не менее двух десятилетий в НАТО и странах Вар-
шавского блока использовались германские разработки в области ракетостроения,
авиации, подводного флота, бронетанковой техники, противотанковых средств, ПВО,
радиолокации, химического оружия и др.
♦ **
Если взять за точку отсчета в развитии бактериологии как науки, разработанный
Робертом Кохом метод получения чистых культур на плотных питательных средах
(1888), то к началу работ по созданию БО прошло не более 40 лет. Еще были живы и
работали в лабораториях ученые, помнившие выращивание бактерий на кусочках
мяса, ломтиках моркови и картофеля или пластинках застывшего крахмального клей-
стера. Поэтому успехи бактериологии первой половины XX в. казались ученым про-
сто головокружительными. У них появилось ощущение того, что в мире микроорга-
низмов все уже идентифицировано и расставлено по своим местам. Одновременно
произошло упрощение эпидемиологических представлений. В отличие от ученых
добактериологической эпохи, видевших эпидемический процесс как сложное взаи-
модействие факторов внешней среды (климатических, атмосферных, почвенных, гео-
логических и др.) и человека, эпидемический процесс после открытия бактерий был
сведен к простым и, по сути, все тем же контагионистическим схемам. Правда, теперь
вместо «контагия» по цепочке с посчитанными и пронумерованными звеньями пе-
ремещается микроорганизм — возбудитель инфекционной болезни. Осталось только
заменить одно звено на искусственное, либо поменять их местами, и распростране-
ние «контагиев» среди войск и населения противника не станет представлять проблем.
И тогда можно устроить им что-то вроде «черной смерти» (1346-1351), или маньчжур-
ской чумы (1910-1911). Этим и занялся японский генерал Исии, искренне поверив-
ший в «полноту знаний» своих европейских учителей.
1.8. Крал итрлда №731
111
1.8. КРАХ ОТРЯДА № 731
Начало японской военно-биологической программы. Отряд № 731. Отряд № 100.
Генерал-лейтенант медицинской службы Исии Сиро. Работа с «бревнами». Разработ-
ка средств бактериологического нападения. Выбор возбудителей инфекционных бо-
лезней в качестве агентов БО. Повышение вирулентности бактерий. Производствен-
ные мощности. Разработка специальных боеприпасов и устройств для диспергиро-
вания блох и бактерий. Лабораторные и полигонные заражения персонала. Поиск
диверсионных отравляющих веществ. Возможные масштабы попыток применения
БО. Бактериологические диверсии против СССР. Бактериологическая война против
Китая. Чума от дьявола в Чандэ. Действия советской разведки. Конец «кухни дьяво-
ла». Судебный процесс по делу японских военнослужащих, обвиняемых в подготовке
и применении БО. Дальнейшая судьба плененных сотрудников отряда Ns 731.
Не надо иметь особого творческого воображения, чтобы понять, какую информа-
цию извлек майор медицинской службы Сиро Исии из научных журналов, газет и
многозначительных намеков, полученных из источников в «бактериологических кру-
гах» во время посещения им Европы и США в 1928-1930 гг. Массовое поражение
людей с помощью бактерий казалось ему таким же простым делом, как и средневеко-
вому колдуну распространение чумы посредством «обрезков ногтей, человеческих
волос, жабьих и кротовых лапок».
Начало японской военно-биологической программы. В своем докладе начальнику
армейской секции Бюро военных дел Министерства армии генерал-майору Нагате
Тэцудзанэ (1884—1935), сделанном по окончании командировки, Исии сообщил, что
в Европе, и особенно в Германии (до 1934 г. Япония считала себя союзником Англии
и США), активно разрабатывается БО. Он особенно подчеркнул, что «если Япония
срочно не начнет фундаментальные исследования в этом направлении, она может
опоздать на поезд». Исии также утверждал, что поскольку Япония — страна бедная
природными ресурсами, то чтобы уравнять шансы в войне с индустриально развиты-
ми странами, ей необходимо «дешевое, но мошное оружие». В августе 1932 г. Исии
возглавил организованную им при Военно-медицинской академии лабораторию про-
филактики эпидемических болезней в японской армии, которая располагалась в То-
кио, в квартале Вакамацу района Синдзюку.
В апреле 1933 г. началось строительство лаборатории, на которое правительством
ассигновано 200 тыс иен. Закончилось оно в октябре того же года. Было построено
железобетонное двухэтажное здание площадью 1795 м2 с подсобными помещения-
ми. В этом же году по указу японского императора Хирохито в Бэйиньхэ (район на
юго-востоке Харбина) начинается строительство Главной базы Управления по водо-
снабжению и профилактике частей Квантунской армии. В районе строительства был
установлен режим военной зоны. Новую организацию назвали «отрядом Камо» (на-
чальник отряда Бусикава). Майору Исии было поручено возглавить отряд «Эй» № 8604
в Нанкине (Центральный Китай), которым он руководил до 1936 г.
С 1935 г. идею подготовки Японии к биологической войне активно поддерживает
начальник 1-го стратегического управления Генерального штаба японской армии
полковник Сузуки Еримичи. В июне 1936 г. по секретному указ! японского импера-
тора Хирохито в районе близ поселка Пинфань провинции Биньцзян, удаленного от
центра Харбина к югу приблизительно на 20 километров, начато строительство круп-
ного военного объекта, вошедшего в историю Второй мировой войны как «Маньчжур-
112
Часть 1. Краткая история применения биологических средств...
ский отряд 731» (далее — отряд № 731). Отряд подчинялся непосредственно коман-
дующему Квантунской армии Ямаде. Исследовательской работой в области подготов-
ки бактериальных средств войны руководили 1-й Оперативный отдел Генерального
штаба Японии и Военно-медицинское управление1.
Отряд № 731. Располагался в 8 км от станции Пинфань в центре между деревня-
ми Саньтунь. Сытунь и Утунь, в обширной зоне, имевшей форму квадрата со сторо-
ной, приблизительно равной 6 километрам. На строительство военно-бактериоло-
гического комплекса ушло более года. Это было громадное военное сооружение,
окруженное рвом и забором с колючей проволокой, по которой был пропущен ток вы-
сокого напряжения. Вот как описывает его Хироси Акаяма (1958), бывший вольно-
наемный служащий японской армии, попавший в 731-й отряд в марте 1945 г.:
«В самом центре однообразной плоской равнины стояли высокие современные
здания, чего я никак не ожидал. В центре над всеми другими строениями возвышалось
огромное четырехугольное, облицованное белыми плитками здание. Таких больших
зданий я не видел ни в Осака, ни в Синьцзине, ни в Харбине, через которые мы проез-
жали по пути сюда. Озаренное лучами солнца, оно казалось ослепительно белым и
высоко вздымалось, заслоняя небо. Здание было обнесено кирпичной стеной, поверх
которой в несколько рядов была натянута колючая проволока. Оглянувшись, я уви-
дел, что несколько дальше, позади нас, возвышался высокий земляной вал с колючей
проволокой, и понял, что весь этот городок изолирован от внешнего мира. Этот вал,
как я потом узнал, тянулся на пять километров, а центральное здание по объему было
в три раза больше здания Марубиру в Токио. ...К востоку от здания, находившегося в
центре, возвышалась огромная труба. Из нее клубами валил черный дым. Вдали за
трубой виднелся аэродром. К западу от здания выстроились в ряд какие-то белые дома,
похожие на больничные корпуса, склады и жилые постройки европейского типа».
«Отряд Камо» перебазировался сюда в 1939 г. Здесь его на некоторое время пере-
именовали в «отряд Того» (Того Хэйхатиро — японский адмирал, который руководил
Цусимским морским сражением с русской эскадрой в 1905 г. и был кумиром Сиро
Исии). Свое зашифрованное название, «Маньчжурский отряд 731», он получил в ав-
густе 1941 г.
Общее представление о Главной базе Управления по водоснабжению и профилак-
тике частей Квантунской армии дают: фотография, сделанная с самолета (рис. 1.19),
и краткая схема, впервые опубликованая в книге Моримура Сэйти (рис. 1.20).
Ниже приведено пояснение к схеме на рис. 1.20.
«Блок ро» в центре схемы (окруженное забором прямоугольное трехэтажное зда-
ние и несколько других строений). На его территории находились: хозяйственное
управление, 1-й отдел, 4-й отдел, лечебный отдел.
«Блок ро»: 1-й этаж. 4-й отдел: группа Ариты, группа Карасавы, группа Асахины
(производство бактерий и блох).
2-й этаж. 1-й отдел: здесь располагались лаборатории группы Иосимуры, а также
группа Минато (исследование холеры), группа Окамото и группа Исикавы (обе зани-
мались проблемами патогенеза инфекционных болезней). Далее шли группа Эдзимы
(исследование дизентерии), группа Ооты (исследование сибирской язвы) и группа
Утими (исследование сыворотки крови). Отсюда был вход в секционную. Это поме-
щение было тем местом, где вскрывали живых людей.
1 Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от
23.04.1949 г. Сов. секретно. АП РФ.Ф. 3. Оп. 66. Д. 1065. Л. 11-21 (Кошкин А. А., 2011).
1.8. Крах отряда № 731
113
Рис. 1.19. Общий вид Главной базы управления по водоснабжению
и профилактике частей Квантунской армии у станции П инфань. В центре
«блок ро». На переднем плане котельная и электростанция. Снимок
сделан со стороны аэродрома. По Р. Wiliams, D. Wallace (1989)
3-й этаж. 1-й отдел: группа Танабэ, группа Футаки, группа Кусами.
1. 1-й этаж. Лечебный отдел: лечебница.
Хозяйственное управление: группа печати исследовательского отдела, отделение
жандармерии, канцелярия исследовательского отдела, кабинет начальника исследо-
вательского отдела, съемочная группа исследовательского отдела, отдел контроля,
отдел кадров, группа Акисады (бывшая группа Эдзимы).
2-й этаж. «Выставочная комната». Конференц-зал. Хозяйственное управление:
бухгалтерия, канцелярия, «комната усопших», плановый отдел, адъютантская, каби-
нет начальника отряда.
2. 1-й этаж. Почта. Телеграф.
Хозяйственное управление: военно-топографическая группа исследовательского
отдела, группа изысканий исследовательского отдела, библиотека исследовательского
отдела.
2-й этаж. Учебный отдел: классы для проведения практических занятий.
3. Книгохранилище.
4. Группа Танаки.
114
Часть 1. Краткая история применения биологических средств...
Железная,
Общежития для
по китайских
. fcDi5°4
О 0«
(Сяяфан
33 п
33
25
,,
ПОДРОБНАЯ СХЕМА
ГЛАВНОЙ БАЗЫ УПРАВЛЕНИЯ
ПО ВОДОСНАБЖ1 НИЮ
И ПРОФИ ТАКТИКЕ ЧАСТЕЙ
КВАНТУНСКОЙ АРМИИ
Сельскохозяйственные
угодья
I отдел
21
1 отдел
03301
[1
26
Аэродром
' 3 отдел
73
20 0
!
°о
о
Манеж
ш
Железная icpora яа Пинфань
и к авиачасти 8382
Отдел
материального
снабжения
Схему выполнил бывший сотрудник
во нно-топотафичсской группы
исследовательского отдела
хозяйственного управления
«отряда 731» Тадзио Ёсида
4 отдел
1 отдел
Хоз. управление
Лечебный корпус'
J59 «Деревня Того»
Спортплощадка
I?
67
70
51c£j50 rj
учебтмотдел
5^,56535?,
Схема восстановлена на основании
аэрофотосъемки, п] отведенной
в августе 1940 г.
58
33
59
Рис. 1.20. Подробная схема Главной базы управления по водоснабжению
и профилактике частей Квантунской армии. По Моримуре Сэйти (1983)
м
5. Группа Иосимуры. Холодильная камера.
6. Котельная. Электростанция. Водонапорная станция. Мастерские.
7 «Бревна», группа Ариты.
8. «Бревна».
9. Группа Касахары.
10. Группа Такахаси.
11. Секционный зал.
12. Печь для сжигания трупов.
13. Виварий спецгруппы.
14. Комната взятия крови.
15. Конюшни,
16. Группа Потуги
17. Группа инженерных работ Ямагутч.
18. Газораспределительная камера.
19. Газгольдер.
1.8. Крах отряда № 731
115
2-й отдел, 3-й отдел.
20. Караульное помещение.
21. 2-й отдел; метеогруппа.
22. Авиа1 руппа.
23. Ангары.
24. Радиогруппа.
25. Группа Ягисавы.
26. Взлетно-посадочная полоса.
27. Гаражи.
28. 3-й отдел: транспортная группа.
Отдел материального снабжения.
30. Склады «блока ро» (помещение для грузовых машин, хранилище вакцин, сте-
рильные холодильные камеры и термокамеры, термокамера высокой температуры,
канцелярия «блока ро»). *
31. Канцелярия отдела материального снабжения.
32. Арсенал
33. Складские помещения.
36. Стеклодувная фабрика
37. Склад угля.
38. Пруд для хранения живой рыбы, предназначенной в пишу (объекты под но-
мерами 39—49 на плане не обозначены, так как они находились в Харбине, в районе
Биньцзянского вокзала).
Учебный отдел.
50. Помещение штаба.
51. Складское помещения (общежитие для подростков-стажеров).
52. Классы для учебных занятий.
53. Кухня. Баня.
54. Свинарник. Гауптвахта.
55. Склад циновок.
56. Казармы.
57. Учебный корпус, где велась подготовка санитаров.
58. Учебный корпус.
59. Общежитие подростков-стажеров.
«Деревня Того», включая помещения, назначение которых не выяснено.
60. Синтоистский храм Того.
61. Лечебница для членов семей сотрудников отряда.
62. 1-й этаж: кухня.
2-й этаж: столовая для старших чинов и чиновников второго класса.
63. 1-й этаж: большой лекционный зал, столовая для младших чинов, рабочая
группа сцены, магазин отряда; 2-й этаж: кинозал.
64, Баня.
65. Квартиры сотрудников отряда.
66. Квартиры холостых сотрудников отряда.
67. Закусочная.
68. Прачечная и котельная.
69. Квартиры Л1я старших чинов.
70. Пульт подачи электроэнергии.
71. Начальная школа «деревни Того».
116
Часть 1. Краткая история применения биологических средств...
72. Газовая камера.
73. Теннисный корт.
74. Водонапорная башня.
75. Ограждение из колючей проволоки.
76. Земляной вал. Ров без воды. Ток высокого напряжения.
77. Земляной вал. Ров без воды.
Сооружения отряда делились на 6 блоков. Первый блок — главное здание, где разме-
щались 1-й (2 и 3 этажи) и 4-й (первый этаж) отделы, — назывался «блоком ро», по-
скольку внешне он напоминал знак японской азбуки «ро», имеющий форму квадрата.
К «блоку ро» примыкали два здания: в одном из них (1-й корпус) помещались
хозяйственное управление и лечебный отдел, а в другом — отдел материального снаб-
жения. В центре «блока ро» находилось двухэтажное бетонное сооружение (7-й и
8-й корпуса) — тюрьма для 300—400 «бревен».
Тюрьма имела прочные стены толщиной 40 см и железные двери, защищенные
решетками окна. Внутри каждого корпуса по периметру шел просторный коридор. На
первом этаже имелись общие камеры, куда «бревен» помещали на время. Второй этаж
представлял собой систему 12 одиночных камер, используемых в экспериментальных
целях. В двери каждой одиночной камеры было сделано окошко, открывавшееся сна-
ружи, через которое велось наблюдение за подопытными. Во все камеры протянули
специальный воздуховод для подачи, в случае необходимости, отравляющих веществ.
Эта система была использована по назначению в марте 1945 г. при подавлении бунта
русских заключенных.
Территория, обнесенная забором с колючей проволокой, по которой был пропу-
щен ток высокого напряжения (на схеме она очерчена жирной черной линией), на-
зывалась штабом.
Жилой комплекс для семейных служащих отряда носил название «деревня Того».
Здесь же находились общежитие для холостяков и построенный отрядом синтоист-
ский храм Того.
В ведении 2-го отдела отряда № 731 были метеогруппа и авиагруппа. Последняя
имела в своем распоряжении аэродром.
Взлетно-посадочная полоса аэродрома примыкала к взлетно-посадочной полосе
авиаотряда 8372. В углу аэродрома, удаленного приблизительно на один километр от
«блока ро», находилось два ангара, в которых стояло одиннадцать самолетов. В авиа-
группе состояло около 60 человек, включая сотрудников метеослужбы, связистов и
авиатехников. Среди самолетов преобладали бомбардировщики. Они предназнача-
лись для проведения экспериментов и практического применения БО.
Самолеты авиагруппы были следующих типов: бомбардировщик «Донрю» («Дра-
кон»), тяжелый бомбардировщик 97, тяжелый бомбардировщик 97 модель 2, легкий
двухмоторный бомбардировщик 99, легкий одномоторный бомбардировщик 99, учеб-
ный бомбардировщик, истребитель, учебный транспортный самолет, санитарный
самолет, пассажирский самолет АТ и неисправный самолет «Айкоку» («Патриот»).
В районе Биньцзянского вокзала города Харбина располагался шестой блок.
К нему относилось здание госпиталя (так называемый 7-й отряд Квантунской ар-
мии) с двумя примыкающими зданиями. Одно, похожее на склад, обычно называли
Южным корпусом, другое принадлежало 3-му отделу. По воспоминаниям бывших
сотрудников отряда, Южный корпус имел просторный полуподвал. Здесь с 1933 г. и
до ввода в строй комплекса в Пинфане размещалась специальная тюрьма для «бре-
вен». Так что эксперименты над людьми начались еще со времен «небольшой лабора-
1.8. Крах отряда Ns 731
117
тории». Эти жертвы не вошли в число тех трех тысяч человек, которые погибли во
внутренней тюрьме «блока ро» с 1942 по 1945 гг.
Начальником отряда в 1936-1942 гг. и с марта 1945 г. до конца войны был гене-
рал-лейтенант Сиро Исии; в период с 1942 по февраль 1945 г. — генерал-майор Ма-
садзи Китано. Структурно отряд был разбит на несколько подразделений (отделов).
Хозяйственное управление (общий отдел). По показаниям возглавлявшего его в
1941 г. Кавасимы Киоси, этот отдел, кроме вопросов распределения кадров, финан-
сов, планирования работ отряда, занимался организацией обеспечения «бревнами»
экспериментальной деятельности отряда. Начальник — майор Накатомэ (затем его
сменил полковник Оота).
1-й отдел. Бактериологические исследования. Начальник отдела -генерал-майор
Кикути. Специально занимался выращиванием для целей бактериологической вой-
ны возбудителей холеры, газовой гангрены, сибирской язвы, брюшного тифа, пара-
тифа и других. Сотрудниками отдела проводились опыты не только над животными,
но и над людьми.
2-й отдел. Экспериментальный. Начальник отдела — полковник Акира Оота (по
совместительству)1. Осуществлял проверку эффективности БО в условиях полигона
и в боевой обстановке. Ему подчинялась специальная авиационная часть с самолета-
ми, оборудованными диспергирующей аппаратурой; полигон на станции Аньда; и
отделение по культивированию и размножению насекомых, предназначенных для рас-
пространения возбудителя чумы. Отдел разрабатывал специальные виды вооружений
для распространения бактерий. Среди них распылители в виде автоматических ручек,
тросточек, фарфоровые авиационные бомбы и т. п.
3-й отдел. Изготовление фильтров для воды. Начальник отдела — подполковник
Эгуги. Это был единственный отдел, который занимался вопросами водоснабжения.
Однако в нем были организованы производственные мастерские, вырабатывавшие
корпуса для «авиабомб системы Исии».
4-й отдел. Производство бактерий для целей бактериологической войны. Началь-
ник отдела — генерал-майор Киоси Кавасима (предстал перед военным трибуналом
Приморского военного округа в 1949 г.). Об этом отделе ниже.
Учебный отдел. Обучение служащих отряда. Готовил для боевых подразделений
японской армии и диверсионных групп специальные кадры, умеющие обращаться с
БО. Начальник отдела — полковник Сонода (затем его сменил подполковник Тоси-
хидэ Ниси).
Отдел материального снабжения. Начальник отдела — генерал-майор Ооя.
Лечебный отдел. Отрядная лечебница. Начальник — полковник Нагаяма.
Помимо этого, отряд № 731 имел четыре филиала, расположенных вдоль советско-
маньчжурской границы, и полигоны для испытаний БО. Филиалы дислоцировались
в Хайларе, Линькоу, Суньу и Муданьцзяне, а испытательный полигон-аэродром на-
ходился на станции Аньда.
В Дальнем (Далянь) располагался Научно-исследовательский центр санитарной
службы Южно-Маньчжурской железной дороги, которым руководил японский ге-
нерал-лейтенант Андо. Этот центр подчинялся непосредственно командующему
Квантунской армии и работал в тесном контакте с отрядом №731, изготовляя чумную
1 С августа 1936 г. (после назначения Исии начальником Управления по водоснабжению
и профилактике частей Квантунской армии) по февраль 1943 г. Оота возглавлял отряд «Эй»
№ 1644 в Нанкине.
118
Часть 1. Краткая история применения биологических средств...
вакцину и проводя эксперименты по массовой иммунизации китайского населения
различными типами чумных вакцин. Его специалисты не бежали в Японию в августе
1945 г., а вполне легально находились в Маньчжурии после войны и даже наблюдали
в 1947 г. за действиями советских военных бактериологов из НИИЭГ (г. Киров), иссле-
довавших эффективность стрептомицина при лечении легочной чумы.
После завершения в 1939 г. строительства первой очереди сооружений в отряде
№ 731 научно-исследовательской работой по подготовке и ведению бактериологиче-
ской войны было занято более 2600 человек. Значительную часть их составляли науч-
но-исследовательские работники и ученые, присланные с медицинских факультетов
высших учебных заведений, из медицинских институтов и гражданских научно-ис-
следовательских учреждений Японии. Их официальный статус — «вольнонаемные
японской армии» и «ученые-специалисты».
Оперативные исследовательские группы. В отряде их существовало более 20. Мо-
римура Сэйти впервые опубликовал перечень тех групп, о которых ему удалось что-
либо выяснить (табл. 1.1).
Таблица 1.1
Задачи оперативных исследовательских групп отряда № 731
Исследовательская группа Задачи
1-й отдел
Специальная группа Занималась «бревнами»
Группа Касахары Исследование вирусов
Группа Танаки Исследование насекомых
Группа Иосимуры Исследование обморожения
Группа Такахаси Исследование возбудителя чумы
Группа Эдзимы (позже группа Акисады) Исследование возбудителя дизентерии
Группа Ооты Исследование возбудителя сибирской язвы
Группа Минато Исследование возбудителя холеры
Группа Окамото Исследование патогенеза
Группа Исикавы Исследование патогенеза
Группа Утими Исследование сыворотки крови
Группа Танабэ Исследование тифа
Группа Фу таки Исследование туберкулеза
Группа Кусами Фармакологические исследования
Группа Ногути Исследование риккетсий
Группа Ариты Рентгеновская съемка
Группа Уты Неизвестно
2-й отдел
Группа Ягисавы Исследование растений
Группа Якэнари Производство керамических бомб
4-й отдел
Группы Карасавы Производство бактерий
Группа Асахины Исследование возбудителя сыпного тифа и производство сыпнотифозной вакцины
1.8. Крах отряда № 731
119
В то время, когда отряд именовался еше «отрядом Того», все эти исследователь-
ские группы имели соответствующие структуре названия: «отделение бактерий 1-го
отдела», «отделение патогенеза 1 отдела» и т. д. Однако вслед за переименованием
«отряда Того» в отряд № 731 официальные названия отделений в целях сохранения
военной тайны были засекречены, а группы стали называться по фамилиям их ру-
ководителей. Эти названия были своего рода кодом, употреблявшимся внутри отря-
да. Официально же группы, например, 1-го отдела назывались так: группа Танабэ —
1 -е отделение, группа Минато — 2-е отделение, группа Эдзимы — 4-е отделение, груп-
па Такахаси — 5-е отделение, группа Исикавы — 7-е отделение, группа Иосимуры —
8-е отделение, группа Футаки — 1-е отделение.
Первой была названа специальная группа, занимавшаяся «бревнами». Опасаясь,
что тайна специальной тюрьмы может просочиться наружу, Исии укомплектовал спец-
группу, в ведении которой находилась тюрьма, своими родственниками и свойствен-
никами. Ею руководил вольнонаемный Такэо — старший брат Сиро Исии.
У Сиро Исии было три брата: самый старший — Торао, затем Такэо и Мицуо. Все
братья Исии, кроме уже умершего к тому времени Торао, занимали ответственные
посты в отряде № 731.
Моримура Сэйти, путем расспросов бывших служащих отряда, установил, что
спецгруппа насчитывала около 50 человек, включая охрану, надзирателей, кухню и
канцелярию. Большинство сотрудников спецгруппы были земляками Исии — выход-
цами из поселка Сибаяма уезда Самбу (префектура Тиба). Все они были безземель-
ными вторыми и третьими сыновьями из крестьянских семей (в японских крестьян-
ских семьях по традиции весь земельный участок, принадлежащий семье, наследовал
первый, то есть старший сын). В отряд их привез с собой Сиро Исии. Члены спецгруп-
пы получали 70 %-ную надбавку к своему месячному окладу.
Пансионат «Береза». Располагался в центральном районе Харбина, в Новом горо-
де, в большом трехэтажном здании из красного кирпича. Это был секретный пункт
связи отряда с внешним миром. Здание было построено в виде буквы П и имело внут-
ренний двор с воротами, через которые въезжали и выезжали военные грузовики и
автобусы отряда.
Прежде чем отправиться в Харбин, служащие отряда на автобусах или грузовиках
приезжали в «пансионат». Затем, переодевшись в гражданское платье, они выходили
из ворот внутреннего двора и смешивались с толпой на городских улицах. По возвра-
щении в отряд все это проделывалось в обратном порядке. Официально здание при-
надлежало правительству Маньчжоу-Го, а об истинном его назначении даже среди
японцев знали лишь единицы.
В Харбине в то время существовало много учебных заведений, в том числе япон-
ские начальные школы «Ханадзоно» и «Момодзоно», Харбинская средняя школа,
Харбинская женская школа. Учившихся в них детей сотрудников отряда на военных
автобусах также доставляли сначала во внутренний двор «пансионата».
Полигон у станции Аньда. От места расположения отряда № 731 до станции Аньда
около 120 км. На полигоне были врыты в землю железные столбы на расстоянии 5-10 м
один от другого. К ним привязывали людей, тела которых в зависимости от условий
эксперимента защищались одеялами и щитами.
Для совершенствования бактериологических боеприпасов различных типов их
взрывали под разными углами к земле и на заданной высоте. Это давало возможность
получить точные данные о зависимости между точкой взрыва бомбы и районом бакте-
120
Часть 7. Краткая история применения биологических средств...
риального заражения. На предельно близком расстоянии от подопытных взрывали
бомбы со шрапнелью, контаминированной возбудителями газовой гангрены. Людей
экспонировали к аэрозолям возбудителей инфекционных болезней, создаваемых
пролетающими на разных высотах самолетами.
Бывший служащий отряда рассказал Моримуре Сэйти следующее: «Эксперимен-
ты по заражению газовой гангреной проводились многократно. И не только эти....
Проводились также эксперименты с применением бактериологического пистолета в
форме авторучки. Ставились и более простые опыты. Например, людям обнажали
бедра, вблизи взрывали ручные гранаты и потом изучали, каким образом осколки
входят в тело; стреляли в голову под разными углами из винтовки, после чего выни-
мали и препарировали мозг; иногда людей просто убивали ударом дубины, а затем
исследовали поврежденную ткань...».
Бывший начальник филиала № 673 подполковник медицинской службы Ниси
Тосихидэ во время предварительного следствия на допросе в 1-й хабаровской тюрьме
6 декабря 1949 г. показал:
«Кроме тою, в январе 1945 года при моем участии был произведен опыт по зара-
жению десяти военнопленных китайцев газовой гангреной. Целью опыта было выяс-
нить возможность заражения людей газовой гангреной в условиях мороза в 20 °C. Тех-
ника проведения этого опыта была следующая: 10 китайцев-военнопленных были
привязаны к столбам на расстоянии 10—20 метров от бомбы шрапнельного действия,
зараженной газовой гангреной.
Чтобы люди сразу не были убиты, их головы и спины защищались специальными
металлическими щитами и толстыми ватными одеялами, а ноги и ягодицы оставля-
лись незащищенными. После включения тока бомба разорвалась, засыпав площадку,
где размещались подопытные, шрапнелью с бактериями газовой гангрены. В резуль-
тате все подопытные были ранены в ноги или ягодицы и по истечении 7 дней умерли
в тяжелых мучениях».
Как проводились эксперименты по заражению «бревен» чумой, рассказал 25 де-
кабря 1949 г. в Хабаровске трибуналу Приморского военного округа подсудимый
Кавасима Киоси (бывший начальник 4-го отдела отряда № 731 генерал-майор меди-
цинской службы):
«Вопрос: Криме опытов в лабораторных условиях производились ли в отряде дру-
гие опыты над живыми людьми?
Ответ: Да, производились в полевых условиях.
Вопрос: Где производились эти опыты?
Ответ: На специальном полигоне, имевшемся на станции Аньда.
Вопрос: Расскажите все, что вам известно об этих опытах.
Ответ: Это было после моего назначения для работы в 731-й отряд, т. е. летом 1941 г.,
когда на станции Аньда производились эксперименты по применению фарфоровых
бомб “системы Исии”, начиненных чумными блохами.
Вопрос: Продолжайте свои показания.
Ответ: Место, использовавшееся для опытов, тщательно охранялось, и проход че-
рез него запрещался Вокру! него стояли специальные посты, которые охраняли это
место, чтобы никто посторонний не мог пройти туда. Подопытные люди использо-
вавшиеся д ля этих экспериментов, в количестве 15 человек были доставлены из внут-
ренней тюрьмы отряда и привязаны на территории, где производился опыт, к специ-
ально врытым в землю столбам. Для того чтобы самолеты могли легче ориентироваться,
легче заметить полигон, на полигоне были вывешены флажки и пущен дым. Со стан-
ции Пинфань прилетел специальный самолет. Он пролетел над расположением поли-
1.8. Крах отряда № 731
121
гона и, когда находился над полигоном, сбросил около двух десятков бомб, которые,
не долетев до земли от 100 до 200 метров, разорвались, и из них выпали чумные блохи,
которыми были начинены бомбы. Эти чумные блохи распространились по всей тер-
ритории.
После того как бомбометание было произведено, выждали значительный проме-
жуток времени, для того чтобы блохи могли распространиться и заразить подопытных
людей. Потом этих людей дезинфицировали и на самолете привезли на станцию Пин-
фань во внутреннюю тюрьму, где над ними было установлено наблюдение, чтобы вы-
яснить, заражены ли эти люди чумой.
О результатах экспериментов должен сказать следующее: мне известно, со слов
ответственного руководителя опытов полковника Оота, что эксперимент не имел хоро-
ших результатов, что было вызвано высокой температурой — большой жарой, отчего
активность блох была очень слабой. Вот все, что я могу сказать об этом эксперименте.
Вопрос: Кто составлял приказ об этом эксперименте?
Ответ: Приказ об этом составил начальник 2-го отдела1. Я как начальник общего
отдела, т. е. секретариата отряда, ознакомился с этим приказом и представил его на-
чальнику отряда на утверждение. Начальник отряда утвердил приказ.
Вопрос: Какие бактерии испытывались наиболее часто в условиях полигона9
Ответ: Бактерии чумы».
Однако этот полигон не был единственным. На допросе во время предваритель-
ного следствия 23 октября 1949 г. Кадзицука Рюди показал, что наиболее опасные
эксперименты Исии проводил «на какой-то необитаемой территории», место распо-
ложения которой он ему не сказал.
В 2005 г. китайскими учеными в автономном районе Внутренняя Монголия был
обнаружен ранее неизвестный крупный японский полигон, предназначенный дтя
испытания химического и бактериологического оружия. Анализ почвы и другие спе-
циальные исследования дали ученым возможность определить границы полигона. Его
площадь составила около 110 км2. Возможно, это тот полигон, о котором в своих по-
казаниях упомянул Кадзицука Рюди, а возможно, еще один.
«Ящик смерти» Наличие этого объекта установил в ходе бесед с бывшими служа-
щими отряда № 731 Моримура Сэйти. Этот объект располагался приблизительно в
4 км к северо-востоку от сооружений отряда № 731. В ложбине находился участок, ого-
роженный где кирпичной стеной, где изгородью из болотной травы амперы. Кругом
расстилалось ровное поле. Участок издали напоминал заброшенный склад, однако на
самом деле здесь проводились эксперименты с боевыми отравляющими веществами.
Бывший сотрудник отряда № 516 (был еще и такой, но о нем почти ничего не из-
вестно) рассказал Моримуре следующее1 «Опыты с ядовитыми газами проводила груп-
па Иосимуры в тесном сотрудничестве с отрядом № 516. Для них использовали пре-
имущественно заключенных, уже подвергавшихся экспериментам по получению
сыворотки крови или экспериментам по обморожению. Большинство подопытных
людей были либо с ампутированными конечностями, либо крайне изможденные.
Привозили их на специальной машине».
Эксперименты проводились в небольших, особым образом сконструированных
камерах. Их было две — малая и большая, — соединенные в одну систему.
Большая камера представляла собой квадрат со стороной 3,6 м. Ее стены были
сделаны из листового железа толщиной 5 мм. К камере был непосредственно присо-
единен генератор, позволяющий накачивать в нее иприт, цианистый водород и окись
1 Полковник Акира Оота.
122
Часть 7. Краткая история применения биологических средств...
углерода. В самой камере регулировалась только концентрация отравляющего веще-
ства. Для этого на потолке камеры находился большой воздушный винт — смеситель
воздуха. По потолку и полу шли две трубы диаметром 50 см. Они соединяли большую
камеру с малой. По ним циркулировал воздух с определенной концентрацией газа.
Чтобы направить его из большой камеры в малую, достаточно было лишь повернуть
выключатель.
Малая камера, за исключением задней стенки и потолка, была сделана из особо-
го, пуленепробиваемого стекла. Длина стенки составляла полтора метра (чуть боль-
ше телефонной будки). Находясь снаружи, сквозь прозрачные стены камеры можно
было наблюдать за тем, что происходит внутри, а также снимать это на пленку. Каме-
ра имела дверцу, к которой были подведены рельсы. Подопытных людей помещали в
небольшую вагонетку и привязывали к стойкам. Вагонетка передвигалась по рельсам.
Когда дверца камеры открывалась, наружные рельсы состыковывались с рельсами
внутри, вагонетка въезжала в камеру, дверца закрывалась.
Усыпальница героев. Находилась перед входом в общий отдел. Перед уничтожени-
ем отряда в усыпальнице хранились габлички с именами более трехсот человек, по-
гибших на фронте или в результате заражения в лабораториях. На возвышении, по-
хожем на большую эстраду, площадью 120-150 м2, тесными рядами стояли большие
фотографии в черных рамках.
Финансирование. Интендант отряда Хотта на процессе в Хабаровске показал, что
в 1945 г. отряду была определена сумма в 10 млн иен (что приблизительно составляло
тогда 4,7 млн долларов). Они распределялась следующим образом: 7 млн шло на на-
учные исследования и производство БО, 3 млн на содержание личного состава. На
содержание филиалов уходило до 300 тыс. иен.
Помимо этого отряду время от времени выделялись от 500 тыс. до 1 млн иен на
«непредвиденные секретные расходы». Секретные ассигнования положили начало
финансовым злоупотреблениям, следствием которых стало отстранение Исии от ру-
ководства отрядом в 1942 г.
Денежное довольствие сотрудников. Было очень высоким, и, несмотря на войну,
доходило до них полностью и без задержек. Например, еще вчерашний школьник
Хироси Акияма (1958), занимавший самую низшую должность в отряде, получил от
вербовщика в качестве подъемных 350 иен, что было не малой суммой для японца из
сельской местности. «Мне было известно, — писал Хироси, — что старший сын на-
ших соседей, который, окончив среднюю школу, поступил на службу в сельскую уп-
раву, получает всего тридцать пять иен в месяц, а месячное жалованье директора на-
чальной школы было что-то около ста иен». По прибытию в отряд Хироси удивился
еще больше: «Вечером за ужином нас накормили до отвала свининой, сладким пиро-
гом и другими вкусными блюдами, о которых мы в Японии могли только мечтать. Так
же сытно и вкусно кормили нас и потом». Его месячный оклад помимо основного
оклада в 100 иен, включал фронтовые надбавки и надбавку за службу на опасной тер-
ритории. Всего набиралось около 300 иен в месяц. В своей деревне Хироси перед вер-
бовкой в отряд был вынужден есть даже траву.
Кадры отряда. Хироси Акияма (1958) пишет следующее:
«У нас было три генерал-лейтенанта медицинской службы, пять или шесть гене-
рал-майоров, около шестнадцати полковников, больше двадцати подполковников и
майоров, а м тадших офицеров и кандидатов в офицеры — почти триста человек. Очень
многие служили здесь по вольному найму. Говорили, что в отряд почти со всей Япо-
нии собраны доктора медицины, работающие в области бактериологии, причем при-
1.8. Крах отряда №731
123
равненных к генералам среди них было больше десяти человек, а к полковникам —
более тридцати. Всего в отряде насчитывалось свыше двух тысяч человек. Мне прихо-
дилось слышать, что именно отряд 731 был тля военных врачей единственным путем
к карьеое. Не послужив здесь, им нечего было надеяться на продвижение по службе».
Среди профессоров Харбинского медицинского института было немало таких,
которые одновременно работали и в отряде № 731. В целях соблюдения режима сек-
ретности, военнослужащие, сотрудники отряда, не носили знаков различия, указы-
вающих на их принадлежность к медицинской службе.
Кремация сотрудников. Когда умирал кто-либо из сот рудников отряда, тело умер
шею отправляли в крематорий, находившийся в Синьцзине, где его подвергали кре-
мации, соблюдая весь церемониал. Печи, где сжигали трупы «бревен», для трупов
японцев не использовали.
Выбор места дислокации отряда. В те годы не сушест вовало надежных технологий,
позволяющих накапливать и длительно хранить агенты БО, поэтому японские воен-
ные были вынуждены приближать его производственную базу к местам предполагае-
мого применения. Расположению отряда № 731 именно в Маньчжурии способство-
вало еще одно весьма существенное обстоятельство, о котором рассказал подсудимый
Кавасима во время судебного заседания в Хабаровске 25 декабря 1949 г.:
«Вопрос: Вследствие каких причин подготовка бактериологической войны велась
в Маньчжурии, а не в Японии?
Ответ. Маньчжурия является страной, сопредельной с Советским Союзом, и в
случае начала войны оттуда легче и удобнее всего использовать бактериологические
средства. Кроме того, Маньчжурия очень удобна для экспериментов по изучению
средств бактериологической войны.
Вопрос: В чем, собственно, заключалось это “удобство” для проведения экспери-
ментов в Маньчжурии?
Ответ: Маньчжурия являлась очень удобной потому, что там было достаточно по-
допытного материала.
Вопрос: Что значит “подопытного материала”? Людей, которые доставлялись в
отряд для опытов?
Ответ: Именно так».
Отряд Xs 100. Другое название отряда — «Управление противоэпизоотической
защиты конского поголовья Квант некой армии». Организован в конце 1935 г. Всего
в отряде работало 600-800 человек. Штаб отряда с основной частью личного состава
дислоцировался в 10 км от города Чанчунь1, в районе поселка Мынцзятунь, а его
филиалы были в городах Дайрене и Хайларе, но ближе к концу войны хайларский
филиал был перемещен в город Кэйшань. Отряд № 100 не зависел от отряда № 731 и
подчинялся непосредственно командующему Квантунской армии Ямаде. Первона-
чально он имел своими задачами изготовление вакцин и сывороток главным образом
для защиты конского поголовья и изучения инфекционных болезней животных.
Оперативно-стратегический план войны против СССР был разработан японским
Генеральным штабом в 1940 г. В японских секретных документах он проходил под
сокращенным наименованием «Кан-Току-Эн». После нападения Германии на СССР
этот план начал воплощаться в жизнь. В соответствии с решением императорского
1 Вблизи станции Чанчунь у японцев еще с 1910 г. был развернут крупный, охраняемый
войсками чумной пункт, где работали с возбудителем чумы (более подробно см. в книге Су-
потницкого М. В., Супотницкой Н. С., 2006).
124
Часть 1. Краткая история применения биологических средств...
совещания от 2 июля 1941 г. Генеральный штаб армии и Военное министерство Япо-
нии разработали план мероприятий, направленных на форсирование подготовки к
проведению наступательных операций против советских войск на Дальнем Востоке
и Сибири. В сентябре 1941 г. перед отрядом № 100 были поставлены задачи подготовки
бактериологической войны и бактериологических диверсий на территории СССР, в
том числе и с использованием самолетов. При штабе каждой армии, находившейся в
Маньчжурии, дополнительно были созданы «эпизоотические отряды». Начальниками
этих отрядов назначили специалистов-бактериологов, направленных из отряда № 100.
Инициатором создания таких отрядов являлось первое оперативное управление Ге-
нерального штаба японской армии. Задачами «эпизоотических отрядов» являлись
подготовка и ведение бактериологической войны против СССР (Кошкин А. А., 2011).
Отрядом № 100 командовал генерал-майор ветеринарной службы Вадзиро Вака-
мацу. Отряд состоял из четырех отделов: общий отдел с канцелярией — ему подчинялся
изолятор, где находились подопытные люди; 1-й отдел — занимался разработкой
методов ведения бактериологической войны, 2-й отдел — вел работу по нескольким
направлениям и был головным в отряде (20 офицеров, 30 научных сотрудников и
50 человек технического персонала). Структура отдела была следующей:
1-е отделение — исследование и производство бактерий сибирской язвы.
2-е отделение — исследование и производство бактерий сапа.
3-е и 4-е отделения — исследование и производство возбудителей других эпизоо-
тических заболеваний.
5-е отделение — исследование и производство головневых грибов и вирусов
мозаики.
6-е отделение — производство бактерий, вызывающих заболевания рогатого ско-
та; разработка методов бактериологических диверсий и их осуществление в пригра-
ничных районах с СССР; исследование и производство боевых ОВ.
В отряде работали с возбудителями сибирской язвы, сапа, чумы рогатого скота,
овечьей оспы, мозаики и красной ржавчины. О производственных мощностях отря-
да подсудимый Такахаси Такаацу (бывший начальник ветеринарной службы Квантун-
ской армии, генерал-лейтенант ветеринарной службы — осуществлял непосредствен-
ное руководство деятельностью отряда № 100) сообщил военному трибуналу
следующее (27.12.1949):
«Вопрос: Докладывали ли вы Умезу о конкретном количестве бактерий, которое
может производить отряд № 100?
Ответ: Я докладывал, что отряд № 100 может производить в год: бактерий сибир-
ской язвы — 1000 килограммов, бактерий сапа — 500 килограммов, бактерий красной
ржавчины — 100 килограммов. Эти цифры я докладывал, и это было возможно при
достаточном оборудовании. Затем оборудование стало прибывать с декабря 1943 года
и стало устанавливаться в 6-м отделении 2-го отдела, но этот план выполнить не уда-
лось, и к концу марта 1944 года я докладывал главнокомандующему о том, что бакте-
рий сибирской язвы изготовлено только 200 килограммов, сапа — 100 килограммов и
красной ржавчины — 20-30 килограммов.
Вопрос: Считали ли вы, что то количество бактерий, которое вырабатывается от-
рядом № 100, обеспечивает потребности бактериологической войны?
Ответ: Нет, не считал достаточным.
Вопрос: Во всяком случае, после 1941 года по вашему настоянию были приняты
меры к увеличению массового производства бактерий для целей бактериологической
войны?
Ответ: Да».
1.8. Крах отряда №731
125
«Если в начале военных действий против СССР японской армии, в силу сложив-
шейся обстановки, необходимо будет отступить в район Большого Хингана, то на ос-
тавляемой территории все реки, водоемы, колодцы должны быть заражены бактери-
ями или сильнодействующими ядами, все посевы уничтожены, скот истреблен», —
такова была главная задача, которую поставил перед отрядом № 100 штаб Квантун-
ской армии.
Отряд № 100 финансировался по двум источникам: Военное министерство Япо-
нии выделяло средства на содержание личного состава отряда и на изготовление пре-
паратов для профилактической работы в Квантунской армии. На изыскание и про-
изводство БО средства отпускались по линии секретных фондов штаба Квантунской
армии, через 2-й разведывательный отдел.
Обвиняемый Такахаси Такаацу во время предварительного следствия 6 декабря
1949 г. показал следующее:
«Я хорошо помню, что Военным министерством Японии на период с 1 апреля
1944 года по 1 апреля 1945 года на содержание личного состава отряда Ns 100 и на из-
готовление профилактических средств было отпущено 60 тыс. иен. На изыскание и
производство бактериологического оружия 2-м отделом штаба Квантунской армии на
указанный период было отпущено 1 млн иен. Однако эта сумма ни в коей мере нас не
лимитировала, ибо при необходимости было бы отпущено столько средств, сколько
бы потребовалось. Какие средства отпускались до 1944 года, сейчас не помню».
Отряд № 100 имел секретное скотоводческое хозяйство. Оно находилось недале-
ко от советско-маньчжурской границы, километрах в 80 северо-западнее города Хай-
лара. Стадо насчитывало 500 голов овец, 100 голов коров и лошадей. Отряд закупил
их у местного населения Северо-Хинганской провинции и выкармливал вплоть до
окончания войны. Скот предназначался исключительно для осуществления биологи-
ческих диверсий.
В апреле 1944 г. отряд № 100 направил в Северо-Хинганскую провинцию секрет-
ное подразделение, которому было приказано ознакомиться с обстановкой в провин-
ции и произвести подсчет всего имеющегося в этом районе домашнего скота. В ре-
зультате выяснилось, что в провинции его насчитывалось около полутора миллиона
голов. Цель, с которой в отряде № 100 содержали скот, была следующей: «В случае
начала войны между Японией и СССР советские войска, вторгнувшись в Северо-
Хинганскую провинцию, непременно угонят из нее в качестве трофея весь скот. Япон-
ские войска, отступая, выпустят на волю лошадей и овец, которых заразят сапом. Через
неделю или две в местах скопления скота вспыхнет эпизоотия...». По крайней мере,
так они предполагали.
Генерал-лейтенант медицинской службы Исии Сиро. Родился в 1892 г. в деревне
Тиеда-Осато (ныне город Сибаяма) префектуры Тиба. Учился сначала в местной част-
ной школе Икэда, где удивлял преподавателей своими способностями; он мог за од-
ну ночь выучить весь учебник чуть ли не наизусть. Затем поступил в общеобразова-
тельную школу в Тиба. Исии был четвертым сыном в помещичьей семье, у которой
вся округа брала в аренду землю. В молодости он увлекался идеями буддийской сек-
ты Нитирэн и разделял идею о «гармонии небесных и земных законов», изложен-
ную в «Сутре лотоса». Еще он преклонялся перед германским «железным канцле-
ром» Бисмарком.
Окончив школу в Тиба, а затем среднюю школу в городе Канадзаве префектуры
Исикава, Исии поступил на медицинский факультет Императорского университета
в Киото, по окончании которого пошел на военную службу в качестве стажера —
126
Часть 1. Краткая история применения биологических средств...
кандидата на командную должность Из армии он был направлен в целевую аспи-
рантуру Киотского императорского университета. Видя незаурядные способности
Исии к наукам и возлагая на него большие надежды, ректор университета отдал ему в
жены свою дочь (рис. 1.21).
Рис 1.21. Исии с семьей в 1938 г. Слева направо братья: Такэо,
Мицуо и Сиро. По Р. Williams, D. Wallace (1989)
После окончания аспирантуры Исии начал быстро продвигаться по службе. Ниже
мы приводим автобиографию Исии, написанную им по просьбе американского сле-
дователя в Токио 17 января 1946 г.
«Дата рождения — 25 июня 1892 г.
1920 год, декабрь — окончил медицинский факультет Императорского универси
тета в Киото.
1921 год, 20 января — 9 апреля — военная подготовка в качестве офицера-стажера
3-го пехотного полка гвардейской дивизии.
1921 год, 9 апреля — назначен военврачом в чине лейтенанта в 3-й гвардейский
пехотный полк.
1922 год, 9 апреля — назначен в 1-й армейский госпиталь в Токио.
1924 год. 20 августа — присвоено звание капитана медицинской службы.
1924— 1926 годы — аспирантура при Императорском университете в Киото по про-
блемам серологии, бактериологии, эпидемиологии и патологии.
1926 год, 1 апреля — назначен в армейский госпиталь в Киото.
1928 год, апрель — 1930 год, апрель — научно-ознакомительная поездка за грани-
цу. Посетил Сингапур, Цейлон, Египет, Грецию, Турцию, Данию, Италию, Францию,
Швецию, Германию, Австоию, Венгрию, Норвегию. Чехословакию. Бельгию, Голлан-
дию, Швейцарию, Финляндию, СССР, Эстонию, Латвию, Восточную Пруссию, Л ме-
рику, Канаду, Гавайи.
1930 год, 1 августа — присвоено звание майора медицинской службы, назначен
преподавателем Военно-медицинской академии.
1935 год, 1 августа — присвоено звание подполковника медицинской службы.
1936 год, 1 августа — назначен начальником Управления по водоснабжению и про-
филактике частей Квантунской армии.
1.8. Крах отряда № 731
127
1938 год, 1 марта — присвоено звание полковника медицинской службы.
1940 год, 1 августа — начальник Управления по водоснабжению и профилактике
частей Квант унской армии и одновременно преподаватель Военно-медицинской ака-
демии.
1941 год, 1 марта — присвоено звание генерал-майора медицинской службы.
1942 год, 1 августа — начальник 1-го отдела Военно-медицинского управления.
1943 год, 1 августа — преподаватель Военно-медицинской академии.
1945 год, 1 марта — присвоено звание генерал-лейтенанта медицинской службы,
назначен начальником Управления по водоснабжению и профилактике частей Кван-
тунской армии.
1945 год, 1 декабря — уволен в запас».
По воспоминаниям сослуживцев, собранных Моримурой, Исии был ростом бо-
лее 1 м 80 см, носил пышные усы, одет был всегда в военный мундир с орденами. Он
произносил речи громким голосом, держа микрофон в руках и щеголяя выправкой.
Воодушевляясь собственным красноречием, он расхаживал по сцене, а длинный про-
вод микрофона тянулся за ним. В такие минуты Исии был похож на огромную обезь-
яну, висящую на веревке. Для восточных людей «обезьяна» в переносном смысле оз-
начает необычайно хитрого человека: «Папаша был самой хитрой из всех обезьян,
ученой обезьяной», — говорили сотрудники отряда.
Исии был «совой», т. е. человеком ярко выраженного «ночного» типа. Днем он
крепко спал в своем кабинете, а к ночи просыпался, полный сил для работы. «Если у
кого-то есть какое-либо предложение, даже самое маленькое, касающееся работы
отряда, пусть приходит ко мне с ним даже ночью», — любил повторять Исии. Со вре-
менем у него развилась еще и «генеральская болезнь», что в сочетании с ночным об-
разом его жизни весьма раздражало других руководящих работников отряда № 731.
Глубокой ночью стоило только какой-нибудь новой мысли о подготовке бактерио-
логической войны мелькнуть в голове начальника, как он вызывал адъютанта и при-
казывал: «Собрать руководство... Проведем срочное совещание». И адъютант вы-
нужден был звонить по телефону и бегать по всей «деревне Того», собирая начальст-
вующий состав.
Первый раз Исии попал в поле зрении жандармерии в начале 1932 г., когда о «мо-
лодом кутиле, сорящем деньгами», сообщил в местную полицию владелец одного из
увеселительных заведений. Как только «кутила» узнавал, что в заведение поступила
15— 16-летняя девушка, он немедленно требовал от содержательницы прислать ее. При
этом он выкладывал сто иен, как будто это была мелочь, а между тем сто иен в то время
соответствовали трехмесячному жалованью молодого служащего. Следователями жан-
дармерии было установлено, что Исии пролоббировал «Акционерной имперской ком-
пании по производству медицинского оборудования» заказ на производство фильт-
ров очистки воды собственной конструкции, что привело к быстрому росту и обога-
щению компании как поставщика армии. Представители компании и сам Исии были
арестованы. Факт получения им взятки подтвердился, причем выяснилось, что ее
сумма была по тем временам огромной — 50 тыс. иен. Однако Исии удалось выкру-
титься, помогли связи с влиятельными людьми в Министерстве обороны, особенно
заступничество полковника Тэцудзана Нагаты (начальник управления службы войск;
впоследствии генерал-лейтенант и заместитель министра обороны Хаяси по кадрам).
Непосредственной причиной для возбуждения следствия в 1942 г. послужили зло-
употребления. имевшие место при строительстве одного из сооружений отряда —
большого лекционного зала в корпусе 63. По свидетельству осведомленных лиц, между
128
Часть 1. Краткая история применения биологических средств...
чертежами проекта Большого лекционного зала, представленными в штаб Квантун-
ской армии, и тем, что было построено в действительности, обнаружились значитель-
ные расхождения. С этого, сообщает Моримура Сэйти, и начал разматываться клу-
бок должностных преступлений Исии.
В то же время Исии не обирал своих подчиненных Показательно то, что даже
подросток Хироси Акияма за пять месяцев службы в отряде сумел скопить 1000 иен.
Исии в отношениях со своими подчиненными придерживался какого-то кодекса че-
сти, а они платили ему за это преданностью.
Работа с «бревнами». Еще в начале XX в. опыты над заключенными не рассматри-
вались европейскими учеными как нечто в принципе недопустимое по моральным
соображениям. В основе обоснования таких экспериментов лежала научная целесо-
образность. Особенно этот принцип соблюдался тогда, когда эксперименты выпол-
няли на «туземцах». Знаменитый бактериолог В. А. Хавкин (1860-1930) испытывал в
1897—1898 гг. свою убитую чумную вакцину (впоследствии ее называли «лимфой Хав-
кина») на заключенных Умеркадской и Бикульской тюрем в Бомбее и на заключен-
ных тюрьмы города Дервара. Британцу Стронгу для испытания живой чумной вакци-
ны филиппинские власти предоставили в 1906 г. в Маниле 200 человек, приговоренных
к смерти. Возможность использования военнопленных в опытах по созданию БО, без
ложной скромности, обсуждалась в Европе еще в средине 1930-х гг. Поэтому Исии
только довел до логического завершения, т. е. до массовых убийств людей «во имя
науки», то, о чем он читал в европейских научных журналах.
Обширные медицинские и военные познания, сложенные логическим мышлени-
ем в некий оптимальный алгоритм экспериментов, привели его к пониманию того,
что, по крайней мере, три вида экспериментов без подопытных людей не выполнишь
на должном уровне. Это — повышение вирулентности бактерий, оценка поражающей
эффективности отдельных образцов БО и проверка эффективности различных средств
защиты от БО (ниже на них мы остановимся более подробно).
В том, что опыты по созданию БО необходимо было проводить с использованием
людей, были убеждены и низшие чины отряда. Хироси Акияма (1958), попавший в
отряд за пять месяцев до его ликвидации, сформулировал свое отношение к таким
преступлениям следующим образом: «Я вспомнил, что в книгах по бактериологии,
которые я просматривал в лаборатории в свободные минуты, было написано, что при
одной и той же вирулентности возбудителя ход болезни у животного очень час го рез-
ко отличается от течения ее у человека». Далее этот смышленыш делает еще один
вывод: «При опытах на человеке можно гораздо быстрее получить достоверные резуль-
таты, чем при работе, например, с мышами». Ничего личного в отношении жертв эк-
спериментов - только забота о достоверности экспериментов.
Японскими властями в Маньчжурии была создана эффективная система достав-
ки «подопытного материала» в оз ряд. Для того чтобы жандармы знали, кого надо на-
правлять в отряд в качестве «бревен», для них была разработана весьма содержатель-
ная инструкция (приведена ниже). Само мероприятие называлось «особой отправкой»
(по-японски — «Токуи-Ацукай»),
Полковник жандармерии Тачибана, выступивший свидетелем на судебном про-
цессе в Хабаровске, дал 28.12.1949 г. следующие показания:
«Государственный обвинитель: Документ, который я вам предъявил, вы опознае-
те как официальный документ, регулировавший порядок “особых отправок”?
Ответ: Да, опознаю.
1.8. Крах отряда № 731
129
Вопрос: Знаете ли вы тех сотрудников жандармерии, которые принимали участие
в установлении категории лиц, подлежащих “особым отправкам”?
Ответ Да, могу назвать.
Вопрос: Пожалуйста
Ответ: Предъявленный мне документ был составлен в 1943 году. Я в то время ра-
ботал в Главном управлении Квантунской жандармерии в качестве сотрудника управ-
ления. Я в то время работал в уголовном отделе Главного управления Квантунской
жандармерии, и из штаба Квантунской армии поступило указание, предписывающее
составить подобный документ. В марте 1943 года я выезжал в командировку в гор.
Мукден для инспектирования жандармского управления. В мое отсутствие майор
Цузимото, работавший в моем отделе, редактировал документ. По возвращении я до-
кумент видел и сейчас подтверждаю его подлинность.
Вопрос: Какая существовала практика оформления отправок в порядке “Токуи-
Ацукай”? Какие документы составлялись по этому поводу жандармерией?
Ответ: Только что предъявленный мне документ в размноженном виде был разос-
лан по жандармским управлениям различных городов Маньчжурии. Людей, которые
подлежали “особой отправки”, содержали в местах заключения при жандармских уп-
равлениях. Затем выдержки из протоколов допросов и справки с ходатайством о раз-
решении подвергнуть их “особой отправке” направляли в Главное жандармское управ-
ление. Там это дело рассматривалось, решался вопрос, и тому жандармскому управ-
лению, которое сделало запрос, направлялось приказание об отправлении этих людей
в отряд № 731 под видом “особых отправок”. Когда такие документы поступали из
местного управления в Главное управление жандармерии, то через секретариат они пе-
редавались в уголовный отдел, оттуда в секцию контрразведки, начальником которой
был я. Работник моей секции Цузимото рассматривал эти документы, выносил реше-
ния, затем эти решения предоставляли мне. Я утверждал это и направлял дальше, на-
чальнику уголовного отдела. Начальник уголовного отдела после получения санкции
командующего квантунской жандармерией направлял приказание от имени командую-
щего жандармерией в то управление жандармерии, откуда этот документ поступил.
Вопрос: Санкционировали ли вы за время вашей работы в штабе квантунской
жандармерии передачу людей для “особой отправки” в отряд № 731?
Ответ: Я рассматривал эти дела, обсуждал их. Я помню, что при мне было направ-
лено свыше 100 человек.
Вопрос: Производились ли отправки людей для уничтожения в отряд № 731 жан-
дармерией с санкции командующего Квантунской армией?
Ответ: Конечно, квантунская жандармерия делала это согласно указаниям коман-
дующего Квантунской армией. Обычно жандармерия направляла дела о преступни-
ках в суд или военный трибунал, но в этих случаях особый приказ заменял закон, и
людей направляли без суда.
Вопрос: Вы подтверждаете, что в соответствии с разработанной вашим подчинен-
ным Цузимото инструкцией “особым отправкам” в отряд № 731 для уничтожения
подвергались лица, связанные с национально-освободительным движением?
Ответ: Люди, которые подвергались этим отправкам, были преступники различ-
ных категорий. Среди этих категорий были также участники национально-освободи-
тельного движения, но все они были коммунисты или националисты (в зале гул воз-
мущения).
Вопрос: Расскажите, свидетель Тачибана, как отправляли людей в отряд № 731,
какой существовал порядок отправки, конвоя, охраны, конспирации и т. д.?
Ответ: Как я уже говорил, жандармское управление посылало в Главное жандарм-
ское управление запрос о разрешении на “особую отправку”. Для этого составля-
130
Часть 1. Краткая история применения биологических средств...
лась в трех экземплярах справка, один экземпляр которой оставался в местном прав-
лении жандармерии, а два экземпляра направляли в Главное жандармское управление.
После того как официальная санкция на “особую отправку” была получена из Глав-
ного жандармского управления, оттуда возвращался один экземпляр справки, а зак-
люченного оставляли при жандармском управлении. Затем, после того как из отряда
№ 731 поступало требование о присылке подопытного материала, т. е. людей, обречен-
ных на “особую отправку”, заключенного с одним экземпляром справки направляли
в Харбин, где на Харбинском вокзале заключенного передавали сотрудникам жандар-
мерии. Людей этих конвоировала жандармерия».
Из показаний военнопленных японцев: генерал-майора медицинской службы
Кавасима, подполковника медицинской службы Ниси, майоров медицинской служ-
бы Карасава и Оноуэ, ефрейтора Сайто и других установлено, что бактериологические
опыты японцами проводились и над советскими гражданами, по различным причи-
нам оказавшимися на территории Маньчжурии. Бывшие сотрудники Харбинской
военной миссии1 и жандармских органов: майор Иидзима, капитан Кимура и сержант
Ямагучи подтвердили на допросах, что ими лично в разное время направлялись для
истребления в отряд № 731 сотни людей, в том числе и советские граждане.
Сержант Ямагучи показал:
«Для уничтожения в отряд № 731 мною лично было дост шлено из различных мест
заключения свыше 120 арестованных лиц различных национальностей, в том числе до
10 человек советских граждан, как военнослужащих, так и гражданских лиц, насиль-
но захваченных во время пограничных конфликтов или сбежавших с территории
СССР, а также граждан Советского Союза, проживавших в городе Харбине. Эти лица
направлялись в отряд № 731 потому, что не желали „авать показания, что они являют-
ся советскими агентами, а также отказывались от сотрудничества с японскими развед-
органами».
Допрошенный начальник Сахалинского жандармского отряда капитан Кимура во
время предварительного следствия показа т:
«В отряд № 731 жандармерия направляла для истребления тех лиц, которые отка-
зывались служить японским интересам и отказывались давать сведения о Советском
Союзе, что могли сделать только те, которые ранее проживали в СССР или яв лялись
советскими гражданами и патриотами своей Родины. С тем чтобы замести следы, бо-
ясь разоблачений за насильный захват советского i ражданина, за издевательства и пыт-
ки во время допроса и другие незаконные действия против советского человека, япон-
ская жандармерия производила такие “особые отправки” в данный отряд под видом
того, что отправляемый болен заразной болезнью и требуется его изоляция. В среднем
ежемесячно органами жандармерии отправлялось в отряд 150- -180 человек, из них, мне
запомнилось около 20 случаев отправки советских граждан для истребления.
Я подтверждаю, что я лично подписал 3 справки на советских граждан, китайцев
по национальности, с представлением их к “особой отправке” в отряд № 731, после
того, как их допрашивали, применяли к ним избиения и, наконец, пытались их завер-
бовать. После всего этого было неудобно передавать советским властям, чтобы не
вызвать осложнений и недоразумений, поэтому были приняты меры к тому, чтобы их
ликвидировать в отряде без всяких осложнений».
Как установлено предварительным следствием, отряд № 731 получал подопытных
людей, главным образом, из лагеря для советских граждан, оказавшихся в Маньчжу-
1 Орган японской военной разведки.
1.8. Крах отряда № 731
131
рии по ряду причин, именовавшегося лагерем «Хогоин». Выявленный среди военно-
пленных японцев, содержащихся в лагере МВД Хабаровского края, майор Иидзима,
бывший начальник лагеря «Хогоин», на допросе показал:
«Я признаю свою вину за совершенные мною преступные действия как начальник
лагеря, Я подтверждаю, что допрос советских граждан, содержавшихся в лагере “Хо-
гоин”, следователями лагеря проводился с применением к ним всевозможных пыток,
людям насильно наливали в рот и в нос воду в большом количестве, били их палками
и прочее. Я также подтверждаю, что по распоряжению бывшего начальника Харбин-
ской военной миссии генерал-майора Акикуса и его заместителя подполковника Яма-
сита Цутому мною было отправлено в отряд N° 731 около 30 советских граждан».
Аналогичные показания дали военнопленные японцы: Тани-заки, Сато, Тагучи
и Игараси, которые, будучи следователями лагеря «Хогоин», допрашивали советских
граждан с применением мер физического воздействия1.
Национальный состав «бревен». По единогласному признанию бывших служащих
отряда, национальный состав заключенных был таким: почти 70 % — китайцы, око-
ло 30 % — русские; немного корейцев, монголов, англичан, австралийцев и американ-
цев. Возраст в подавляющем большинстве — от 20 до 30 лет, максимум 40 лет.
Накопители. Так каь вместимость внутренней тюрьмы отряда была небольшой, а
«бревна» требовались ежедневно, то между Пинфанем и Харбином, в районе япон-
ского синтоистского храма, находился специальный «накопитель» — пункт для сбо-
ра «бревен». Некоторые сотрудники отряда в беседах с Моримурой вспомнили его на-
звание — Шоушидзян. В этом сборном пункте постоянно содержалось до 300 «бревен»,
и когда в материале для экспериментов начинала ощущаться нехватка, отсюда их не-
медленно переправляли в отряд. Для перевозки использовались грузовики с черным
закрытым кузовом без окон. Транспортировка производилась ночью под охраной
жандармов. На сборном пункте заключенных содержали без наручников и кандалов.
Принимались специальные меры для того, чтобы люди не поняли, что их ожидает.
Среди них были ничего не подозревавшие молодые китайцы, которых обманом зав-
лекли на этот пункт, сказав, что их приглашают для устройства на работу.
Расход «бревен». Подопытн! im людям по прибытию в отряд надевали ножные кан-
далы, присваивали номера и распределяли в качестве материала для экспериментов
между всеми группами отряда. Их фамилии не интересовали «экспериментаторов»,
они нигде не фиксировались, поэтому не сохранились. Предела в зверствах в отно-
шении этих людей сотрудники отряда № 731 не знали. Совершались любые варвар-
ские эксперименты, которые могли прийти в головы их организаторам. Нередко с на-
учной точки зрения они были полностью бессмысленными. Например, Исии
похвалялся американцам, что он лично заставлял людей, зараженных чумой, носить
тяжелые мешки. Ставились опыты, совсем не требовавшие использования людей, а
тем более живых (опыты по высушиванию живых людей горячим воздухом с целью
установить вес «твердого остатка»).
Широко практиковалось вскрытие живых людей, так как именно таким способом
можно узнать, как происходят во времени изменения в живых тканях после воздей-
ствия патогенного микроорганизма или яда. Ниже приведен один из примеров тако-
го «вскрытия», рассказанный Моримуре сотрудником отряда.
1 Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от
23.04 1949 г. Сов секретно. АП РФ.Ф. 3. Оп. 66 Д. 1065. Л. 11-21 (Кошкин А. А., 2011).
132
Часть 1. Краткая история применения биологических средств...
«Однажды в 1943 году в секционную привели китайского мальчика. По словам
сотрудников, он не был из числа “бревен”, его просто где-то похитили и привезли в
отряд, но точно ничего известно не было.
Мальчик сидел на корточках в углу секционной, как загнанный зверек, а вокруг
операционного стола стояли в белых халатах более десяти сотрудников отряда, под-
няв кверху готовые к операции руки. Один из них коротко приказал мальчику лечь на
операционный стол. Мальчик разделся, как ему было приказано, и лег на стол спиной.
Тотчас же на лицо ему наложили маску с хлороформом. С этого момента он не ведал,
что творят с его телом. Когда наркоз окончательно подействовал, все тело мальчика
протерли спиртом. Один из опытных сотрудников группы Танабэ, стоявших вокруг
стола, взял скальпель и приблизился к мальчику. Он вонзил скальпель в грудную клетку
и сделал разрез в форме латинской буквы Y. Обнажилась белая жировая прослойка.
В том месте, куда немедленно были наложены зажимы Кохера, вскипали пузырьки кро-
ви. Вскрытие заживо началось.
...Из тела мальчика сотрудники ловкими натренированными руками один за дру-
гим вынимали внутренние органы: желудок, печень, почки, поджелудочную железу,
кишечник. Их разбирали и бросали в стоявшие здесь же ведра, а из ведер тотчас же
перекладывали в наполненные формалином стеклянные сосуды, которые закрывались
крышками.
Блестел скальпель, лопались пузырьки крови. Один из вольнонаемных, искусно
владеющий инструментом, быстро опустошил нижнюю половину тела мальчика. Вы-
нутые органы в формалиновом растворе еще продолжали сокращаться.
“Смотрите! Да они еще живые!” — сказал кто-то.
После того как были вынуты внутренние органы, нетронутой осталась только
голова мальчика. Маленькая, коротко остриженная голова. Один из сотрудников груп-
пы Минато закрепил ее на операционном столе. Затем скальпелем сделал разрез от уха
к носу. Когда кожа с головы была снята, в ход пошла пила. В черепе было сделано тре-
угольное отверстие, обнажился мозг. Сотрудник отряда взял его рукой и быстрым
движением опустил в сосуд с формалином. На операционном столе осталось нечто,
напоминавшее тело мальчика, опустошенный корпус и конечности. Вскрытие закон-
чилось».
Кодзо Окамото, один из руководящих сотрудников отряда, после окончания вой-
ны на допросе в штаб-квартире американских войск в Токио показал, что только в
1945 г. вскрытию были подвергнуты не менее тысячи человек. Вот как описывает опыт
над человеком Хироси Акияма (1958), который он наблюдал сам незадолго до ликви-
дации отряда.
«Открыв первую дверь, я очутился в помещении, где находился дезинфицирую-
щий душ. Здесь я прошел обработку раствором карболовой кислоты, затем надел ре-
зиновую спецодежду, похожую на легкий водолазный костюм, и уже в ней снова про-
шел обработку дезинфицирующей жидкостью.
Эта одежда почти не отличалась от той, которой мне приходилось пользоваться при
проведении опытов на животных. Она плотно облегала все тело. За спиной находился
маленький кислородный аппарат. Открыв вторую дверь, я снова доложил, что готов
приступить к работе.
Посредине комнаты к черной, словно вымазанной дегтем, железной койке при-
вязывали человека (“бревно”), который отчаянно бился в руках ассистентов. Люди в
такойже, как и у меня, спецодежде держали жертву равнодушно, как автоматы. “Брев-
но” защищалось со страшной силой, хотя руки и ноги его уже были связаны. Это был
здоровый человек нормальной упитанности. Человек, используемый для опыта, как и
подопытное животное, должен быть здоровым, так как если он болен и ослаблен, ре-
1.8. Крах отряда Ns 731
133
зультаты опыта не будут отражать картину течения болезни в обычных условиях. По-
этому в тюрьме нашего отряда особое внимание обращалось на питание и гигиену
В этом заключалось ее главное отличие от обычных тюрем.
Я подносил, как мне было приказано, медикаменты и инструменты и наблюдал,
как военный врач делает инъекцию. Я не знал, с какой целью производится опыт, что
вводят человеку в кровь, но, судя по тому, как хронометражист внимательно следил за
течением болезни после инъекции, я понял, что проверяется скорость действия каких-
то бактерий. Несчастный корчился в страшных муках, издавая душераздирающие воп-
ли, но, наконец, затих, видимо силы его иссякли. Опыт продолжался два-три часа,
показавшиеся мне вечностью.
Вечером мне приказали вывезти его труп. Он выглядел ужасно. Вся грудь и лицо
подопытного приобрели темно-лиловый оттенок из-за подкожных кровоизлияний.
Лимфатические железы под мышками и в паху страшно вздулись, а кожа в этих местах
стала темно-красной, как у спелого персика.
...Труп окунули в крепкий раствор карболки, и я. взвалив мертвеца на тележку,
повез его. ...Высокая труба, назначения которой я до сих пор не знал, оказалась трубой
крематория. Туда был проведен электрический звонок, возвещавший об окончании
опыта, поэтому, когда я добрался до открытых дверей, меня уже ждали трое служащих».
Нельзя сказать, что «бревна» не понимали, зачем их привезли в эгу тюрьму, либо
вели себя пассивно. Сотрудники отряда рассказывали Моримуре, что им так и оста-
лось не понятным, как заключенные разных камер поддерживали между собой связь
и передавали из камеры в камеру раз 1ичные предметы. Был такой случай. По каме-
рам с целью эксперимента раздали сладкие пирожки, зараженные бактериями брюш-
ного тифа. Но те, кто знал о подобных опытах, передали по всем камерам, что пирож-
ки, возможно, заражены бактериями. Ни один заключенный к ним не притронулся.
Происходили и еще ботее удивительные случаи. Так, чтобы подкрепить больного
заключенного, ему в камеру по цепочке передавали сухие фрукты, входившие в раци-
он узников. По тюрьме ходили клочки бумаги, на которых было записано содержа-
ние экспериментов. В одиночную камеру, куда только что заключили пленного коман-
дира Народно-освободительной армии Китая, были переданы в качестве «знака
внимания» маленькие китайские туфельки, которые «бревна»-китаянки плели из
скрученных полосок бумаги.
Прореживание. В ходе экспериментов неизбежно появлялись «бревна», которые
после инфицирования бактериями чумы все-таки выживали Были «бревна», приоб-
ретавшие такого рода иммунитет и к другим микроорганизмам (о каких именно воз-
будителях болезней идет речь, Моримура не сообщает). Для исследования процесса
появления иммунитета эти подопытные представляли значительную ценность, но, с
другой стороны, оказалось, что если их оставлять в живых, то некуда будет помещать
новые «бревна», которые постоянно прибывали. И тогда прибегали к методу скорост-
ного умерщвления заключенных, который назывался «прореживанием».
Выживших подопытных заключенных сотрудники спецгруппы выводили из ка-
мер и вели в помещение рядом с ванной комнатой. Имевшие горький опыт неодно-
кратных экспериментов, люди были крайне насторожены, поэтому вывести их из
камер было нелегко, тогда «экспериментаторы» прибегали к обману: «Тебя сейчас
освободят, но для этого нужно сделать предохранительную прививку». Так их выма-
нивали из камер и вели под охраной сотрудника спецгруппы, вооруженного писто-
летом. Приведя «бревно» в специально оборудованную комнату, ему быстро вкалывали
в запястье 20 см3 хлороформа. Менее чем через секунду у подопытного начиналось
134
Часть 1. Краткая история применения биологических средств...
удушье, глаза выкатывались из орбит, тело покрывалось гусиной кожей, и наступала
смерть. Такому «прореживанию» подвергалось до 20 «бревен» в месяц.
Подсчет жертв. Осуществлен весьма приблизительно по рентгеновским снимкам
подопытных людей, поступавших в отряд. Как только новых заключенных помеща-
ли в специальную тюрьму, группа Ариты делала их рентгеновские снимки. Прежде чем
сделать снимок, на груди заключенных, как мужчин, так и женщин, сотрудники груп-
пы Ариты черной тушью писали порядковый номер арабскими цифрами. Нумерация
начиналась с трехзначной цифры, то есть со 100, и продолжалась до цифры 1500. Это
составляло один цикл. Зафиксированный на снимке номер сохранялся за узником в
течение всего пребывания в тюрьме. Когда нумерация доходила до 1500- счет опять
возобновлялся со 100.
Бывший служащий отрада рассказал Моримуре следующее: «Насколько я помню,
в мае 1942 г. нумерация заключенных выражалась цифрой порядка 700... За 1000 она
перевалила к концу 1943 г. В начале 1944 г. она быстоо достигла 1500, а затем снова
началась со 100. С этого времени циркуляция заключенных стала особенно интенсив-
ной. К концу 1944 г. порядковый номер превысил 1000, а к весне 1945 г. он выражался
примерно цифрой 1400. Поэтому только на моей памяти нумерация заключенных
прошла два цикла (2800), и говорить о том, что общее количество заключенных со-
ставило 3000, отнюдь не преувеличение».
Однако были и такие подопытные, которых направляли на эксперимент, не под-
вергнув предварительно рентгеновской съемке, некоторых умертвили сразу же по
прибьп ии в отряд, в это число не вошли люди, замученные в период 1933—194? гг. Ни
одному подопытному человеку за все время существования отряда не удалось спастись.
Из протокола допроса бывшего начальника лагеря «Хогэин», сотрудника япон-
ской контрразведки Иидзимы Есио, допрошенного 20 октября 1949 г. в качестве сви-
детеля, стала известной фамилия одного советского военнослужащего, погибшего
в отряде № 731 — это красноармеец Демченко, отказавшийся отвечать на вопросы
Иидзимы и отправленный им в отряд для уничтожения. Иидзима долго пытал этого
человека лично, поэтому он запомнил его фамилию. Только в 1945 г. в отряд было
отправлено для уничтожения 40 советских граждан. Никаких дел в суд на них не офор-
млялось (Материалы судебного процесса..., 1950).
Моримураутверждает, что большинство научных медицинских работ, написанных
сотрудниками отряда № 731 на основе экспериментов над живыми людьми, опубли-
кованы в «Журнале японского общества изучения патогенеза», без указания на то, что
в качестве экспериментальных моделей ими использовались люди.
Представление японских военных о роли БО в войне В октябре 1936 г. на совещании
с руководящим составом начальник отряда Бусикава определил место БО в пред-
стоящей войне с СССР как средства поражения стратега тактических целей (Сака-
ки, 1952).
Мацумура Томокацу, начальник оперативно-стратегического отдела штаба Кван-
тунской армии, допрошенный во время предварительного следствия 7 декабря 1949 г.
как свидетель, показал следующее:
«Вопрос: Скажите, каким образом предполагалось применение бактериологичес-
кого оружия Квантунской армией против Советского Союза?
Ответ Применение бактериологического оружия против Советского Союза дол-
жно было быть в соответствии с указаниями Генерального штаба. В оперативные пла-
ны применение бактериологического оружия против Советского Союза не включалось.
1.8. Крах отряда № 731
135
так как применение смертоносных бактерий являлось только одним из видов оружия,
предназначенного для осуществления уже имевшихся оперативных планов. Однако эти
вопросы oocj ждались в соответствующих отделах штаба. Так, лично я по оперативно-
му отделу после соответствующего изучения этого вопроса представил свои стображе -
ния о способа/, применения бактериологического оружия против СССР начальнику
штаба Касахара Юкио. Я доложил Касахара о том, что в случае возникновения с Со-
ветским Союзом военных действий бактериологическое оружие должно быть приме-
нено с помощью авиации. В частности, я доложил Касахара о том, что на случай вой-
ны с Советским Союзом бактериологическое оружие должно быть применено в районе
городов: Ворошилова (Уссурийск), Хабаровска, Благовещенска и Читы т. е. в тыло-
вых районах Советского Союза.
Заражение этих районов должно было производиться путем сбрасывания бакте-
риологических бомб и распыления бактерий с самолетов. Для этой цели должны были
быть использованы самолеты авиационных соединений Квантунской армии.
Вопрос: Однако для этой цели должны быть специальные самолеты?
Ответ: Нет, для этой цели могли быть использованы обычные самолеты. Для сбра-
сывания бактериологических бомб — обычные бомбардировщики, а для распыления
бактерий — любые самолеты, так как аппаратура, необходимая для распыления бак-
терий, была очень несложной, и ее можно было приспособить на любой самолет.
Эти свои соображения, как я уже сказал, я высказал начальнику штаба Касахара,
и он в общих чертах их одобрил. Все эти мероприятия не включались в оперативный
план, но при изучении этих вопросов я исходил из общей обстановки операций, на-
меченных оперативным планом.
Этот доклад Касахара я сделал после полученных указаний Военного министерства
об увеличении производства бактериологического оружия. Дело в гом, что после по-
лучения указаний об увеличении производства бактериологического оружия Касаха-
ра мне поручил изучить вопрос о возможностях его применения против Советского
Союза в случае необходимости. Я этот вопрос изучил и сделал соответствующий док-
лад Касахара, который, как я уже сказал, одобрил мои соображения.
Вопрос: Какие основные виды бактериологического оружия были приняты на во-
оружение Квант) некой армией?
Ответ: К 1945 г. в качестве основных видов бактериологического оружия были ут-
верждены: бактериологическая бомба, способ распыления бактерий с самолетов и на-
земный способ—диверсии. Я думаю, что эти основные способы были утверждены Ге-
неральным штабом Японии.
Вопрос: Какие виды бактериологического оружия были рассмотрены и утвержде-
ны главнокомандующим Ямада?
Ответ: В бытность главнокомандующим Ямада им были рассмотрены и затем ут-
верждены два основных способа, которые были как бы окончательно усовершенство-
ваны к этому времени. Я имею в виду бактериологическую бомбу “системы Исии ” и
способ распыления с самолетов блох, зараженных чумой.
Вопрос: Каким образом предполагалось практическое применение бактериологи-
ческого оружия?
Ответ: Мне как начальнику оперативно-стратегического отдела штаба Квантун-
ской армии известно, что практическое применение бактериологического оружия дол-
жно было осуществляться с помощью специальных кадров, имевшихся в отрядах и их
филиалах. Другими словами, в случае необходимости отряды и филиалы должны были
выделить соответствующее количество специалистов, в распоряжение которых долж-
ны были быть приданы прошедшие специальную подготовку еще в мирное время сол-
даты. Эти солдаты в мирное время набирались из различных частей японской армии
136
Часть 1. Краткая история применения биологических средств...
и направлялись на курсы в отряды № 731 и № 100, где они вместе с санитарным делом
проходили специальную подготовку и приобретали необходимые навыки по бактерио-
логии. На эти курсы отбирали наиболее преданных солдат, но техники от бора я не знаю.
После окончания курсов, в мирное время, они направлялись либо в филиалы, либо в
отряды профилактики и водоснабжения, имевшиеся при частях и соединениях япон-
ской армии. В военное время они должны были быть использованы для применения
бактериологического оружия».
Бывший начальник штаба Квантунской армии генерал-лейтенант Хата Хикоса-
Ьуро на допросах во время предварительного следствия отрицал наличие наступатель-
ного плана применения бактериальных средств, но он признал намерения Генераль-
ного штаба Японии использовать эти средства в диверсионных целях в глубоком тылу
противника1.
Бывший главнокомандующий Квантунской армии Ямада Отозоо во время пред-
варительного следствия 17 ноября 1949 г. рассказал, каким образом осуществлялось
принятие на вооружение японской армии новых средств ведения биологической вой-
ны, и кем планировалось их применение:
«Для изучения способов применения бактериологического оружия создавались
специальные комиссии, в состав которых входили: начальник штаба Квантунской
армии, начальник оперативно-стратегического отдела, соответствующий начальник
бактериологического отряда № 100 или № 731 и отдельные штаб-офицеры. Предсе-
дателем этих комиссий был начальник штаба. Командующий Квантунской армией в
работе комиссий не участвовал. Решения комиссий по применению бактериологичес-
ких средств войны представлялись командующему Квантунской армией и после его
утверждения докладывались в Генеральный штаб Японии. О принятии того или иного
способа на вооружение Генеральный штаб ставил в известность штаб Квантунской
армии, в свою очередь штаб Квантунской армии отдавал соответствующее распоря-
жение отрядам № 731 и № 100 о массовом производстве необходимых эпидемических
бактерий. Обеспечение бактериологических отрядов № 731 и № 100 всем необходи-
мым оборудованием, сырьем и материалами осущест елялось Военным министерством
Японии в соответствии с требованиями Генерального штаба о производстве того или
иного вида бактериологического оружия. Применение бактериологического оружия
и формирование необходимых частей, предназначенных для практического исполь-
зования бактериологического оружия, планировались Генеральным штабом Японии
на основании соответствующей информации штаба Квантунской армии».
На судебном заседании 26 декабря он признал, что БО в случае возникновения
военных действий должно быть применено против Советского Союза с помощью
авиации для заражения тыловых районов и путем проведения диверсионных меро-
приятий по линии отряда № 100. Ямада также заявил, что если бы военные действия
с Советским Союзом не возникли, то БО могло бы быть применено против США и
других западных стран (Материалы судебного процесса..., 1950).
Любопытно то, что представления пленных японских высокопоставленных воен-
ных о роли БО в войне исходили из уверенности в его реальном существовании в япон-
ской армии. Исии откровенно преувеличил военную значимость своего отряда в гла-
зах командования Квантунской армии. Например, бывший начальник отдела кадров
Квантунской армии, полковник Тамура, прибывший в мае 1945 г. с инспекционной
проверкой, показал на суде в Хабаровске 28 декабря следующее:
1 Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от
23.04.1949 г. Сов. секретно. АП РФ.Ф. 3. Оп. 66. Д. 1065. Л. 11—21 (Кошкин А. А., 2011).
1.8. Крах отряда № 731
137
«Вопрос: Я прошу вас, свидетель, рассказать о том, что доложил Исии вам при
инспектировании вами отряда № 731 о целях отряда, с одной стороны, и о готовности
отряда выполнять боевые задания — с другой.
Ответ: Исии рассказал мне, что эффективность бактерий проверена в опытах над
живыми людьми, как в лабораторных условиях, так и в полевых, и что бактериологи-
ческое оружие является наиболее мощным оружием в руках Квантунской армии. Он
информировал меня о том, что отряд находится в полной боевой готовности, и в слу-
чае необходимости, когда начнется война, отряд в состоянии обрушить непосредствен-
но на войска противника громадные массы смертоносных бактерий, что отряд может
также при помощи авиации провести операции по бактериологической войне в тылу
противника над его городами».
Причем в понимании японских военных БО обладало какими-то постоянно со-
храняющимися поражающими свойствами, которые позволяли «осуществление опе-
ративных планов» путем «заражения этих районов». Попросту говоря, прикоснулся к
чему-либо в «зараженном районе», то сразу же заболел — возбудитель инфекционной
болезни (инфицированная им блоха) рассматривается ими также как и европейцами
в 1930-х гг. — как «контагий».
Разработка средств бактериологического нападения. Эти работы в отрядах № 731 и
№ 100 включали:
1) выбор среди возбудителей инфекционных болезней тех, кого можно использо-
вать в качестве потенциальных агентов БО;
2) получение штаммов возбудителей инфекционных болезней с повышенной ви-
рулентностью;
3) масштабирование процесса культивирования микроорганизмов до получения
их количеств, необходимых для ведения бактериологической войны;
4) стабилизацию свойств наработанной бактериальной массы и ее длительное
хранение;
5) разработку специальных боеприпасов и технических устройств для дисперги-
рования (распространения) бактерий, способов их применения.
Для использования в диверсионных целях японцами исследовались поражающие
свойства малоизвестных в те годы биологических токсинов, разрабатывались тех-
нологии их очистки и способы скрытого применения (группы Кусами и Сэкитори).
Оценивалась возможность поражения злаковых растений в СССР путем рассеи-
вания с воздуха головневых грибов (группа Ягисавы).
Выбор возбудителей инфекционных болезней в качестве агентов БО. Не зря сотруд-
ники Исии считали своего шефа «самой хитрой из всех обезьян, ученой обезьяной».
Так оно и было! Узнав во время своей поездки по Европе о том, что в сознании пра-
вящих кругов европейских стран из поколения в поколение, как осложнение после
тяжелой болезни, передается ужас, связанный с эпидемиями чумы Средневековья,
Исии решил, что в этом ужасе заключается его шанс. Формально-логически выбор им
возбудителя чумы в качестве агента БО безукоризнен. Летальность при легочной фор-
ме чумы тогда достигала 100 % от числа заболевших людей. История человечества на
тот период времени не знала инфекционной болезни, способной распространяться
на столь огромные территории и вызывать такое количество жертв, как чума. Эпиде-
мия чумы «черной смерти» 1346—1351 гг. в Европе, истребившей до четверти ее насе-
ления, до сих пор считается вызванной забрасыванием трупов больных чумой людей
в генуэзскую крепость Кафу (см. разд. 1.3). Эпидемии легочной чумы в Маньчжурии
1910—1911 гг. и 1921 г., проредившие ее население более чем на 100 тыс. человек, про-
изошли на глазах того поколения, к которому принадлежал сам Исии.
138
Часть 1. Краткая история применения биологических средств...
Большой оптимизм ему внушали результаты исследований эпидемиологии чумы,
полученные после открытия возбудителя болезни в 1894 г. Авторитетными учеными
того времени было «убедительно показано», что в природе чума поддерживается сре-
ди грызунов, и в особенности среди крыс; а передается человеку посредством инфи-
цированных возбудителем чумы блох. Этими же авторитетами заявлялось, что пере-
мещение больных чумой крыс с кораблями стало причиной возникновения эпидемий
чумы в портовых городах в конце XIX в. и в начале XX в. — так называемой портовой
или третьей пандемии чумы (см. рис. 1.5). В те годы в любом поргу можно было уви-
деть суда, оснащенные специальными ловушками, которые должны были ограничить
распространение чумных крыс по миру (рис. 1.22).
Рис. 1.22. Специальные приспособления на судах, устанавливаемые в 1920— 1930-хгг.для
превращения распространения чумы посредством крыс. По U. A. Wu Lien Ten et al. (1936)
Далее «самая хитрая из всех обезьян» пришла к следующему бесхитростному вы-
воду. Раз страх перед чумой заставил в 1930-х гг. европейские страны с большой осто-
рожностью относиться к бактериям чумы, как к агенту БО, то европейцы в принципе
не могут иметь надежные средства защиты от бактериологического нападения с ис-
пользованием этого агента. Таким образом, именно с помощью возбудителя чумы
Исии собирался сделать БО способным решать оперативные задачи и уравнять шан-
сы Японии в войне с индустриально развитыми странами.
Из других возбудителей опасных инфекционных болезней Исии остановился на
следующих:
возбудители кишечных инфекций: брюшного тифа, паратифа, дизентерии, холеры —
диверсионное заражение продуктов питания и водоисточников;
возбудители сибирской язвы и сапа — применение диверсионными методами и
диспергирование специальными боеприпасами и устройствами с целью массового
уничтожения людей и животных;
возбудители анаэробных инфекций (газовой гангрены и столбняка) — контамини-
рование ими осколков снарядов и шрапнели с целью вызвать анаэробные инфекции
у раненых;
возбудители чумы рогатого скота и овечьей оспы (отряд № 100) — применение ди-
версионными способами с целью поражения сельскохозяйственных животных про-
тивника.
В обоих отрядах исследовали на предмет их военного применения еще и другие
бактерии и вирусы — возбудители инфекционных болезней человека (туберкулез.
1.8. Крах отряда № 731
139
дифтерия, сыпной тиф, гнойные инфекции), но работы с ними не велись так интен-
сивно, как с возбудителями инфекций, указанных выше. В частности, в письме ми-
нистра внутренных дел С. Н. Круглова Иосифу Сталину упоминается о проведении
работ по оонаружению возбудителя новой оолезни, именуемой «Сонго»1 * * * * *.
Теперь, когда в умах японских военных определилось место БО в войне, и ими
были выбраны направления его создания, осталось только создать это бесхитростное
«оружие бедных». И для борьбы с законами природы экспериментаторам никак не
препятствовали законы человеческие.
Повышение вирулентности бактерий. Идея бактериологической войны в начале
1930-х гг. была столь прив лекательна для военных, что на ее ослепительном фоне сами
бактерии им казались какими-то «недоделанными» природой маленькими существа-
ми. Еще в октябре 1936 г. начальник отряда Бусикава (предшественник Исии) на од-
ном из служебных совещаний подчеркивал, что для создания оружия на основе бак-
терий, надо значительно поднять их вирулентность и сократить инкубационный пе-
риод болезни (Сакаки, 1952). Обычно историки отряда (Моримура Сэйти, 1983;
Williams Р., Wallace D., 1989 и др.), кто с гордостью за приоткрывшуюся им тайну, кто
с ужасом от возможностей науки, покорившей человеку невидимый микроскопиче-
ский мир, пишут, что Исии сумел поднять вирулентность возбудителя чумы (т. е. сни-
зить его инфицирующую дозу для человека) в 60 раз. Невольно рисуется мрачная кар-
тина несостоявшейся бактериологической войны, где люди Исии мизерными коли-
чествами возбудителя чумы поражают тысячи людей. Однако если внимательно читать
воспоминания «смышленыша» Хироси Акиямы (1958), то ситуация с вирулентнос-
тью возбудителя чумы представляется совсем в ином свете:
«Вирулентность возбудителей чумы самой страшной из всех инфекционных болез-
ней, выращенных искусственным путем, оказывалась низкой, а их хранение делом
сложным и хлопотливым. Для ведения бактериологической войны необходимо было
найти надежный способ консервации, чтобы в течение долгого времени бактерии не
теряли своей вирулентности, и выра :тить такие бактерии, которые сохраняли бы жиз-
неспособность при высушивании. С этой целью искусственно выращенные бациллы
чумы вво 1или в организм человека, используя селезенку и кровь подопытного как
питательную среду. Затем эти микробы вновь вводили здоровым людям. Другими сло-
вами , путем многократного пассажа через живой организм выращивали жизнеспособ-
ные, вирулентные микробы. Для определения вирулентности и токсичности чумных
бацилл, а также для приготовления сыворотки использовали мышей и морских сви-
нок. В этих случаях опыты с учетом индивидуальных особенностей животных прово-
дились по несколько раз. После этого результаты их проверялись на людях и только
тогда считались окончательными».
В понимании Исии БО тогда будет способно конкурировать с обычными видами
оружия, когда вызываемые им эпидемии будут охватывать огромные территории
1 Речь шла об острой вирусной природно-очаговой болезни, характеризующейся лихорад-
кой, общей интоксикацией, поражением почек и развитием громбогеморрагического синд-
рома, известной сегодня как геморрагическая лихорадка с почечным синдромом ^синонимы
геморрагический эпидемический нефрозонефрит, эпидемический нефрозонефрит, болезнь
Чурилова, дальневосточная геморрагическая лихорадка, корейская геморрагическая лихорад-
ка, лихорадка Сонго, маньчжурская эпидемическая геморрагическая лихорадка). Вирусная
природа геморрагической лихорадки с почечным синдромом была доказана в 1944 г. А. А. Смо-
родинцевым, однако лишьв 1976 г. южно-корейскому ученому Н. W. Lee (1976) удалось выде-
лить из легких грызуна Apodemus agrarius согеае вирус Hantaan (по названию реки Хантаан).
140
Часть 1. Краткая история применения биологических средств. .
и убивать сотни тысяч человек. Исторические источники и мнения знаменитых бак-
териологов подсказывали ему только одну возможность — попытаться вызвать эпи-
демии легочной чумы типа маньчжурской эпидемии 1910-1911 гг. Но бубонная чума,
вызываемая при передаче возбудителя болезни блохами (выбор такого способа веде-
ния бактериологической войны мы рассмотрим ниже), неконтагиозна. Значит нуж-
но было добиться того, чтобы бубонная чума переходила во вторично-легочную, а
потом уже распространялась как воздушно-капельная инфекция, т. е. давала самосто-
ятельные вспышки первично-легочной чумы. Даже сегодня иные «маститые ученые»,
важно рассуждающие на темы биологической войны, любят «покрасоваться» прогно-
зами типа: «Успехи науки позволяют получать штаммы бактерий, способные поражать
по цвету кожи, разрезу глаз» и по т. п. критериям. Но у Исии задача была гораздо про-
ще — научиться воспроизводить легочную чуму, вспышки которой постоянно то там,
то здесь вспыхивали в Маньчжурии без его участия.
В конце 1930-х гг. не было четких представлений о механизмах развития легоч-
ной чумы, нет их и сегодня. Одни авторы настаивали на пневмотропности штаммов
чумного микроба. И даже получали такие штаммы путем многократного пассирова-
ния чумного микроба через экспериментальных животных. Например, профессором
А. М. Скородумовым (1888—1939) при изучении штамма, выделенного в 1927 г. во
время эпидемии легочной чумы в Монголии, получены «свойства пульмонтропнос-
ти» у возбудителя чумы, выраженные в организме морской свинки. Такой штамм
вызывал вторичную пневмонию у этих животных при подкожном или внутрибрюш-
ном заражении (Шунаев В. В., 1934). Несомненно, Исии знал эти работы и пытался
их воспроизвести на людях по схеме, описанной Хироси Акиямой.
Другие ученые считали, что главным фактором в воспроизведении той или иной
клинической формы чумы являются входные ворота инфекции, и приводили убеди-
тельные патологоанатомические данные в пользу этой точки зрения (Кулеша Г. С.,
1912, 1924).
Третьи видели причину перехода бубонной чумы в легочную в способности штамма
возбудителя болезни быстро вызывать септицемию. Поданным сотрудников Особой
лаборатории на форту «Александр 1» С. И. Златогорова и Л. В. Падлевского (1915),
чумной микроб, выделенный от больных легочной чумой, отличается некоторыми
особенностями: он реже мутит бульон, образует компактные хлопья, позже дает коль-
цо по краю пробирки. При заражении животных обладает значительной вирулент-
ностью и вследствие этого быстрее вызывает септицемии, чем имевшиеся для срав-
нения штаммы, полученные от больных в очагах бубонной чумы в Бомбее, Астрахани
и Одессе.
Проверка каждой такой версии требовала огромное количество «бревен» и экспе-
риментальных животных. Один из служащих отряда рассказывал Моримуре, что было
зарегистрировано заражение легочной чумой «бревен», находившихся в одной камере
с больным легочной чумой, однако при масштабировании этих экспериментов, т. е. в
условиях реального применения БО по китайскому городу, вспышек легочной чумы
вызвать не удалось (см. «Чума от дьявола в Чандэ»). Распространения через контакт
других инфекционных болезней от искусственно зараженных людей Исии не удава-
лось добиться даже при тесном содержании заключенных в камерах тюрьмы отряда.
Однако «смелая мысль» ученых отряда № 731 попытками распространения легоч-
ной чумы не ограничивалась. Им нужно было еще «научить» возбудитель чумы пре-
одолевать защитное действие противочумной вакцины, т. е. выражаясь современным
языком, получить иммунитет-преодолевающий штамм Y. pestis.
1.8. Крах отряда № 731
141
Такие исследования на людях проводила группа Такахаси. По сведениям, собран-
ным Моримурой в ходе бесед с работниками отряда, эксперименты по повышению
вирулентности возбудителя чумы и его способности преодолевать искусственный
противочумный иммунитет проводились следующим образом. Несколько «бревен»
инфицировали возбудителем чумы, в результате большинство из них погибали. Сы-
воротку выживших людей японцы считали «сильной». Поэтому сыворотку крови
выжившего подопытного А. вводили подопытному Б. Потом заражали Б. сывороткой,
содержащей возбудитель чумы, полученной от погибшего в предыдущем эксперименте
«бревна». В результате «борьбы бактерий чумы, уже имеющих повышенную вирулен-
тность, с антителами в организме подопытного Б.», как полагали сотрудники 1 -го от-
дела, появлялась еще более «сильная» и способная преодолевать специфический им-
мунитет культура бактерий, которую таким же образом вводили третьему подопытному
В. Разумеется, этим примером такие исследования вирулентности бактерий в отряде
№731 не исчерпывались.
Сакаки (1952) и Моримура (1983) утверждают, что путем повторных пассажей
бактерий через организм экспериментального животного сотрудникам отряда № 731
удалось сократить инкубационный период инфекции у зараженных животных до 48 ч,
тогда как у исходных штаммов он растягивался до 4-х сут. Но на этом яркая сторона
этого дела сменяется освещенной лишь всполохами отдельных намеков.
Первым обнаружил феноменальную способность возбудителя чумы быстро терять
свою вирулентность при пересевах на искусственных питательных средах тот, кто его
и открыл — швейцарский бактериолог А. Иерсен (1894). Это же явление, но приме-
нительно ко всем микроорганизмам, потенциальным агентам БО, имел в виду рефе-
рент по гигиене при Германском военном министерстве, профессор Конрих (1932) в
своем первом возражении против возможности создания БО.
Получив «высоковирулентные штаммы» в какой-то серии экспериментов, сотруд-
никам Исии приходилось решать другую, еще более сложную задачу — сохранять их
высоковирулентными до момента проникновения в организм жертвы. И вот с реше-
ния этой задачи в технологической цепочке создания запасов БО «дешевое и мощное
оружие» стало стремительно дорожать, а его «мощность» вызывать сомнения у «низ-
ших чинов» отряда. Возбудитель чумы, как следует из мемуаров Хироси Акияма(1958),
утрачивал вирулентность в процессе хранения, при многократном посеве в культива-
торах, при масштабировании процесса производства бактерий, и даже, при лиофиль-
ном высушивании и применении в качестве агента БО.
Сегодня можно предложить объяснение этому феномену. Гены «факторов пато-
генности» семейства Yersinia находятся на хромосоме бактерии в виде отдельных кла-
стеров из функционально связанных групп генов, отличающихся от большей части
генома по нуклеотидному составу. Такие дискретные структуры называют «острова-
ми патогенности». У возбудителя чумы — это нестабильный участок протяженностью
102 т. п. о., включающий несколько самостоятельных регионов. Делеция может захва-
тить как весь участок, так и один из генов участка (Mecsas J., Strauss Е. J., 1996). Этот
«остров патогенности» играет важную роль при паразитировании возбудителя чумы
в своих первичных хозяевах — простейших обитателях почвы, тем и обеспечивается
его стабильность в геноме микроорганизма. При культивировании на искусственных
питательных средах положительное селективное давление на «остров патогенности»
со стороны хозяина паразита отсутствует, и он быстро делегируется либо целиком,
либо постепенно, отдельными генами, тогда «вирулентность» возбудителя чумы ут-
рачивается постепенно («ступенчато»).
142
Часть 1. Краткая история применения биологических средств...
Нестабильность «острова патогенности» усиливается и в организме млекопитаю-
щего, но уже по другой причине. Возбудитель чумы, проникая в клетки белой крови,
использует те же механизмы инвазии, что и для проникновения в почвенные простей-
шие. Однако фагоцитирующие клетки (макрофаги и нейтрофилы), в отличие от своих
эволюционных предшественников, не являются свобод ножи вущими организмами, а
представляют собой часть иммунной системы более сложного организма и функцио-
нируют под ее управлением. Иммунная система распознает фагоциты, используемые
возбудителем чумы для своего паразитирования, и уничтожает их через систему Т-кле-
точных ответов. Тем самым оказывается сильное отрицательное давление на «остро-
ва патогенности» в популяции возбудителя чумы, вызвавшего эпидемию болезни.
Постепенно маловирулентные штаммы начинают преобладать в эпидемических це-
почках. В конце каждой крупной эпидемии чумы от заболевших людей, как правило,
высеваются маловирулентные штаммы возбудителя этой болезни. Сама же болезнь в
конце эпидемии протекает легко и поддается лечению средствами, оказывающими-
ся бесполезными на пике эпидемии. Возможно, этот механизм снижения вирулент-
ности возбудителя чумы способствовал тому, что эпидемия легочной чумы в Маньч-
журии 1910-1911 гг. внезапно оборвалась «сама» в феврале 1911 г., как раз тогда, когда
уже всему медицинскому персоналу было ясно, что противоэпидемические мероприя-
тия потерпели крах. Исии, конечно, об «островах патогенности» не знал, но Конрих
интуитивно понимал, что существуют какие-то неизвестные ограничители вирулен-
тности у возбудителей опасных инфекционных болезней, попавших за пределы сре-
ды своего естественного обитания. Следовательно, и второе его возражение против
возможности создания БО («наличие вирулентности у микроорганизмов недостаточ-
но, чтобы вызвать эпидемию» — см. «Критические взгляды на БО» в разд. 1.7), можно
признать справедливым.
Исииже, исходя из эмпирических наблюдений и стереотипов представлений того
времени об эпидемическом процессе, был вынужден постоянно повышать вирулент-
ность штаммов возбудителя чумы пассажами на «бревнах» и экспериментальных
животных. Возможно, ему удавалось получать и более вирулентные штаммы, чем тот,
о котором пишут историки отряда, но и проблем у него становилось больше.
П ытаясь селекционировать возбудитель чумы по одному признаку — вирулентно-
сти для людей, он неизбежно попадал в еще одну ловуш ку, кстати, к тому времени уже
хорошо известную селекционерам животных и растений. Русским ученым-эволюци-
онистом И. И. Шмальгаузеном (1938) она была названа «платой за селекцию». Суть
этого явления в следующем. Специализация в процессе направленного отбора одних
признаков и свойств неизбежно, по причине существования отрицательных корреля-
ций в системе целостного онтогенеза, сопряжена с ослаблением и ухудшением дру-
гих. Ю. П. Алтухов (2003) иллюстрирует этот феномен на примере узкоспециализи-
рованных сортов пшеницы. Отобранные по признаку урожайности они оказались
неустойчивыми к воздействиям внешней среды и потребовали массированных доз ми-
неральных удобрений, а также ядохимикатов для борьбы с болезнями и вредителями.
Сам того не желая, Исии добивался не только повышения вирулентности возбудите-
ля чумы, но и потери им внутрипопуляционного генного разнообразия. Нетрудно
представить, с какими проблемами, кроме внезапной и необъяснимой утери вирулен-
тности штаммами возбудителей, подготовленных для бактериологической войны, он
сталкивался в ходе своих экспериментов.
Попробуем экстраполировать хотя бы некоторые из них, опираясь на работу
Ю. П. Алтухова (2003). Несомненно, новые «высоковирулентные штаммы» потребо-
1.8. Крах отряда № 731
143
вали от японских бактериологов коррекции состава питательных сред в сторону уве-
личения количества входяпц х в их состав компонентов при постоянно снижающей-
ся «урожайности» «производственного штамма». Резко снизилась жизнеспособность
бактерий, увеличилась скорость их отмирания во время лиофильной сушки, при хра-
нении и в аэрозоле.« Бичом» процесса наработки биомассы высоковирулентных мик-
роорганизмов стала посторонняя непатогенная микрофлора, хорошо размножающа-
яся на «богатых» питательных средах
Эти и другие проблемы заставляли японских военных бактериологов искать но-
вые штаммы взамен «заселекционированных», посылать за ними специальные отря-
ды в очаги чумы. Количество работы в отряде № 731 по поддержанию вирулентных
штаммов микроорганизмов постоянно росло. Требовалось все больше «бревен», экс-
периментальных животных, научных сотрудников, лаборантов, низших специалис-
тов, переводчиков, жандармов, дорогих питательных сред, электроэнергии, пара,
кормов для животных, помещений для содержания животных, складов для кормов,
работников для складов, продовольствия для работников и т. д. и т. п.
Вся эта «борьба за вирулентность» возбудителя чумь продолжалась до ликвида-
ции отряда. Хироси Акияма (1958) так описывает знакомство со своим рабочим мес-
том весной 1945 г.:
«Я работал в здании, где содержались подопытные животные. Это здание по фор-
ме и размерам напоминало школу. В нем держали лошадей, коров, овец, свиней, кур и
кроликов, но больше всего там было мышей, крыс и морских свинок. Мне один раз
пришлось увидеть все своими глазами, и, хотя пословица говорит, что лучше один раз
увидеть, чем сто раз услышать, я не мог представить себе, как много их там было.
“Сколько всего их здесь?” — спросил я как-то Сагава, и он, молча покачав голо-
вой, все же ответил: “Сколько, говоришь? Да одних мышей, пожалуй, будет около ста
тысяч”. ...На корм животным шло огромное количество зерна, проса и сои. Железо-
бетонный склад, в котором хранился корм, достигал высоты пятиэтажного дома».
Немного поработав в первом отделе, смышленый Хироси Акияма стал понимать,
для чего это все нужно:
«Чтобы активность чумных бацилл не уменьшалась, необходимо было, по край-
ней мере, раз в месяц вводить их в живой организм. Поэтому одно хранение бактерий
было ужасно обременительным делом. Я слышал, что только для культивирования и
хранения бактерий требовалось более тысячи человек».
Получается, что почти половина отряда занималась поддержанием вирулентнос-
ти штаммов микроорганизмов, предназначенных для «дешевого, но мощного оружия
бедных». Природа посмеялась над людоедской вседозволенностью Сиро Исии и его
«высоковирулентными штаммами».
Производственные мощности. На уже упомянутом выше совещании сотрудников
отряда, состоявшемся в октябре 1936 г., его начальник Бусикава высказался в том
смысле, что в такой бедной стране, как Япония, где ощущается всесторонний недо-
статок материальных ресурсов, БО является оптимальным видом оружия. Далее он
предположил, что, «имея крошечную лабораторию в несколько квадратных метров и
пробирки, легко можно наработать боевое оружие, способное уничтожить десятки
тысяч жизней». Бусикава был большим контагионистом, чем Фракосторо с Папой
Павлом III, вместе взятые (см. разд. 1.2. «Появление контагионистическогоучения»),
С приходом Исии к руководству, в отряде возобладали более реалистичные взгля-
ды на те количества бактерий, которые необходимо накопить для ведения бактерио-
логической войны.
144
Часть 1. Краткая история применения биологических средств...
Технологии глубинного культивирования бактерий находились еще в «зачаточном
состоянии» и для масштабного производства агентов БО японцами не использовались.
Основным технологическим приемом стало выращивание бактерий на поверхности
плотной питательной среды в специальных культиваторах «системы Исии». Они пред-
ставляли собой помещенные в кожух закрепленные друг над другом формочки, в ко-
торые заливался питательный агар. После того как агар застывал, на него производил-
ся посев бактерий. По своей сути этот культиватор ничем не отличается от описанного
в 1932 г. аппарата оставшейся безвестной Н. Г. Щербиной (см. рис. 1.15), и его конст-
рукция была просто заимствована Исией из русского журнала.
Производство бактерий и блох. Осуществлялось сотрудниками 4-го отдела. О про-
изводственных возможностях отдела сообщил военному трибуналу в Хабаровске его
бывший начальник, подсудимый Кавасима (26 декабря 1949 г.):
«Вопрос: Опишите имевшееся в 4-м отделе производственное оборудование.
Ответ: В 4-м отделе имелись две системы оборудования, которые имели одинако-
вую мощность.
Вопрос: Какая была емкость котлов для варки питательной среды первой системы?
Ответ: 8 котлов по одной тонне каждый.
Вопрос: Следовательно, это будет 8 тонн питательной среды, которую 4-й отдел в
одной системе мог давать одновременно?
Ответ: Да, правильно.
Вопрос: А емкость котлов второй системы выражалась в каком общем объеме?
Ответ: Такой же емкости.
Вопрос: Сколько камер культивирования имелось в первой системе и сколько ка-
мер культивирования имелось во второй системе?
Ответ: В первой — 5, во второй — 4.
Вопрос: Какое общее количество культиваторов системы Исии имелось в 4-м
отделе?
Ответ: Точного количества не помню, но их было достаточно для массового про-
изводства бактерий.
Вопрос: Когда вы говорили относительно возможной выработки смертоносных
бактерий, из чего вы исходили?
Ответ: Из емкости котлов, из мощности прочего оборудования, а также из коли-
чества культиваторов.
Вопрос: Какое время было необходимо для выращивания бактерий определенно-
го вида: бактерий тифа, холеры, сибирской язвы и чумы?
Ответ: Для чумных бактерий и бактерий сибирской язвы требовалось 48 ч, а для
бактерий холеры, тифа и пр. — 24 ч.
Вопрос: Какое количество бактерий сибирской язвы снималось с одного культи-
ватора?
Ответ: От 50 до 60 граммов.
Вопрос: Какое количество бактерий тифа снималось с одного культиватора?
Ответ: От 40 до 45 граммов.
Вопрос: Чумы?
Ответ: 30 граммов.
Вопрос: Холеры?
Ответ: Приблизительно 50 граммов.
Вопрос: Как хранились изготовленные 4-м отделом бактерии?
Ответ: При кратковременном хранении они хранились в холодильниках.
Вопрос: Когда вы отправляли бактерии для бактериологических атак против ки-
тайского населения, как тогда упаковывали эти бактерии?
1.8. Крах отряда № 731
145
Ответ: Бактериями наполнялись специальные бутылочки, в каждую бутылочку
входило по 50 граммов. Затем эти бутылочки укладывались в металлические футляры,
несколько этих футляров упаковывалось в специальные большие ящики, которые
внутри обкладывались льдом.
Вопрос: Опишите методы и специальное оборудование, применявшееся в отряде
731 для выращивания в большом количестве блох.
Ответ: Для массового размножения блох 2-й отдел имел четыре специальных по-
мещения. В них поддерживалась определенная температура, +30 °C. Для размножения
блох использовались металлические банки в 30 сантиметров высотой и 50 сантимет-
ров шириной. Для содержания блох в эти банки засыпалась рисовая шелуха. Когда
была закончена такая подготовка, в банку сначала помещалось несколько блох, а так-
же для их питания помещалась белая крыса, которая закреплялась так, чтобы она не
могла повредить блохам. В банке поддерживалась постоянная температура +30 °C.
Вопрос: Какое количество блох получалось за один производственный цикл из
одного культиватора?
Ответ: Точно не помню, но припоминаю, что от 10 до 15 граммов.
Вопрос: Сколько времени длится такой производственный цикл?
Ответ: Два или три месяца.
Вопрос: Какое количество культиваторов имелось в специальном отделении по
выращиванию паразитов?
Ответ: Точной цифры я не помню, но могу сказать, что от 4000 до 4500.
Вопрос: Следовательно, при имевшемся оборудовании за один производственный
цикл отряд мог дать 45 килограммов блох?
Ответ: Да, правда.
Вопрос: Что имелось в виду делать с этими блохами в случае бактериологической
войны?
Ответ: Они должны были заражаться чумой.
Вопрос: И применяться в качестве бактериологического оружия?
Ответ. Да, правильно.
Вопрос: Каким путем хотели применять чумных блох как бактериологическое ору-
жие?
Ответ: При мне самым эффективным способом считался способ сбрасывания блох
с самолета.
Вопрос: Во время экспедиции в Китай блох тоже сбрасывали с самолета?
Ответ: Да, это было так.
Вопрос: Это были блохи, зараженные чумой?
Ответ: Правильно. Бактериологическая атака с помощью чумных блох в Китае
должна была вызвать эпидемию чумы».
Процесс получения бактерий держался в тайне. Кроме сотрудников группы майора
Карасавы, ни один служащий отряда без особых причин не мог проникнуть на «фаб-
рику». 26 декабря 1949 г. подсудимый Карасава Томио показал следующее:
«Вопрос: Я прошу вас описать производственное оборудование отряда, его мощ-
ность и способы производства бактерий.
Ответ: Я работал в качестве начальника производственного отделения 4-го отдела
и поэтому расскажу сначала о производственном оборудовании этого отделения Обо-
рудование для массового производства бактерий в этом отделении состояло из двух
систем. Сначала я расскажу о первой системе. Это, прежде всего, котлы для изготовле-
ния питательной среды для выращивания бактерий. Таких котлов было четыре, каждый
из них был емкостью примерно около одной тонны. Питательная среда помещалась в
специальные культиваторы “системы Исии”, которые помещались в специальные
146
Часть 1. Краткая история применения биологических средств...
автоклавы, таких автоклавов было 14. каждый из автоклавов вмешал около 30 куль-
тиваторов. Таким образом, было возможно при полной загрузке поместить одновре-
менно в автоклавы 420 культиваторов. Для охлаждения культиваторов существовало
два холодильника. Затем, после того как питательная сре га остывала, производили
посадку оакгерий. Эти бактерии размножались, затем производили снятие их. Для
этого имелись две специальные комнаты.
Вопрос: Какое количество бактерий производилось за один месяц?
Ответ При максимальном использовании производственной мощности 4-го от-
дела и наилучших условиях возможно было теоретически производить чумных бакте-
рий в течение месяца до 300 килограммов, но практически для этого использовались
всего только 500 культиваторов, которые давали возможность производить за один
цикл 10 килограммов чумных бацилл, каждый культиватор давал по 20 граммов.
Вопрос: Сколько бактерий брюшного тифа можно было производить при макси-
мальной загрузке?
Ответ: За месяц от 800 до 900 килограммов.
Вопрос: Сибирской язвы?
Ответ: Около 600 килограммов.
Вопрос: Холеры?
Ответ: Около одной тонны.
Вопрос: Паратифа?
Ответ: Так же. как и тифа.
Вопрос. Дизентерии?
Ответ: Так же.
Вопрос: Я правильно вас понял, что во всех котлах, которые имелись в отделе № 4.
можно было изготовить 8 тонн питательной среды?
Ответ: Да, правильно. В первой системе 4 тонны, во второй системе также можно
было изготовить 4 тонны».
На утреннем заседании трибунала 29 декабря 1949 г. подсудимый Карасава уточ-
нил свои показания:
«Я так же хочу внести в заключение экспертной комиссии поправку относительно
производственной мощности отряда 731. Есть место, где сказано, что отряд произво-
дил обычно в месяц до 300 килограммов бактерий. Но это не было обычным. Это было
возможно только при оптимальных условиях использования всей производственной
мощности оборудования отряда. Обычно же практически производилась только одна
треть из этого количества, а 300 килограммов бактерий не производились».
Здесь нужно пояснить, что сотрудники отряда 731 понимали под «килограммами
бактерий» — речь идет о весе густой сметанообразной бактериальной массы, непо-
средственно смытой с поверхности питательной среды (Материалы судебного про-
цесса..., 1950). Следовательно, не менее чем на 90—95 % это была вода (т. е. какая-то
разводящая изотоническая жидкость). Для того чтобы определить реальную «произ-
водственную мощность» отряда, необходимо понимать то обстоятельство, что бакте-
рии начинали отмирать на поверхности питательной среды уже по достижении ими
фазы стационарного роста. С момента их «смыва», скорость их отмирания возраста-
ла. Чтобы остановить этот процесс, бактерии лиофилизировали (с 1944 г.), но в резуль-
тате такой операции до 50 % бактерий погибало, а вирулентность оравшихся была
сомнительной (см. ниже). Следовательно, реальную месячную «производственную
мощность» отряда № 731 при сушке бактериальной массы «до постоянного веса»,
можно оценивать как 15—30 кг. Здесь надо учитывать еще и то, что большую часть
лиофилизата занимают компоненты питательной среды. Для мысленного распро-
1.8. Крах отряда № 731
147
странения «контагия» это количество бактерий может быть даже избыточным, но
для ведения бактериологической войны оно вряд ли достаточно. По приведенным в
разд. 1.7. оценкам американских военных, для того чтобы при существующих в те годы
способах применения бак гериальных агентов вести бактериологическую войну, тре-
оовалось производить не менее 5 тонн бактерий в сутки.
Стерильная камера в которой, как выразился Карасава, «производили посадку
бактерий», представляла собой комнату со стеклянными степами площадью прибли-
зительно 45 м , Входившие в нее сотрудники должны были предварительно пройти
стерилизационную камеру, представлявшую собой квадрат со стороной 7 м. С потолка
этой камеры разбрызгивалась дезинфекционная жидкость, которая обез зараживала
все тело сотрудника. Это была мера предосторожности, предпринимаемая для того,
чтобы на агар-агар не попали другие бактерии, кроме культивируемых.
Для посева бактерий на агар-агар (плотную питательную среду) использовался так
называемый ватный помазок — дюралюминиевый прут толщиной в карандаш и дли-
ной 50 см. На конце его была намотана вата. Обильно пропитав эту вату бактериями,
нужно было быстро, в один прием, перенести их и равномерно рассеять на поверхно-
сти агар-агара. Работа требовала навыка.
На первом этаже «блока ро» находились облицованные плиткой ванная комната
и комната для переодевания. Сотрудники группы Карасавы сначала переодевались,
затем шли в ванную комнату и только после этого выходили на свое рабочее место.
В комнате для переодевания они раздевались догола, затем надевали белые хала-
ты, маску из 7—8 слоев марли на лицо, белую шапочку, резиновый фартук от шеи до
щиколоток и резиновые сапоги до колен. Последними одевали резиновые перчатки
и специальные очки. В такой одежде они входили в ванную комнату. Неглубокий ре-
зервуар был заполнен раствором карболовой кислоты. Они входили в этот раствор и
шли по колено в нем. Пройдя через всю ванную комнату, они полностью стерилизо-
вались от колена и ниже. Во время работы с микроорганизмами сотрудникам запре-
щалось разговаривать, объяснялись только жестами.
После завершения посева бактерий культиваторы перемещалась в культивацион-
ную камеру. Это оыло обширное помещение, обитое листовой медью. Камера была
темной, высоко на потолке горело только две лампочки. Температура в ней легко ре-
гулировалась в пределах от 20 до 80 °C с помощью прибора, находившегося у входа.
Такая регулировка была необходима потому, что для культивирования каждого вида
бактерий требуется своя температура. Во время культивирования открывать дверь
камеры категорически запрещалось.
По истечении определенного времени сотрудники группы Карасавы начинали
сбор бактерий. Его производили скребком шириной 5—7 см, укрепленным на конце
дюралюминиевого прута длиной 50 см. Скопления бактерий соскребали в специаль-
ные химические стаканы диаметром 10 см и высотой стенок 30 см. Питательную среду
после снятия с нее бактерий снов i загружали в автоклавы и после полного обеззара-
живания жидкий агар-агар выбрасывали. Таков был цикл производства бактерий. При
желании повторно стерилизованный агар-агар можно было вторично использовать
как питательную среду, но на третий раз он обычно терял свои питательные свойства.
В технологии подготовки плотной питательной среды и ее последующей утили-
зации была какая-то недоработка. Питательная среда, помещенная в автоклав, изда-
вала тошнотворный запах, чего не должно быть в принципе. Зловоние распространя-
лось по bcpmv «блоку ро», а иногда, в зависимости от направления ветра, достигало
большого лекционного зала.
148
Часть 7. Краткая история применения биологических средств...
Блохи. Отряд со всеми филиалами производил (или мог производить) в течение
3-х месяцев 45 кг блох — много это или мало? Установлено, что один килограмм блох
это приблизительно 3 млн особей. Одна фарфоровая бомба вмещала 30 тыс. блох.
Вроде бы много. Я знаю человека, который заболел чумой через 8 ч после укуса одной
блохи. Но здесь нужно понимать то обстоятельство, что для целей бактериологичес-
кой войны Исии требовались не просто блохи, а блохи, инфицированные возбудите-
лем чумы и способные нападать на людей. В практике бактериологической войны ин-
фицированная блоха, это блоха «блокированная». Насосавшись крови на чумном
грызуне или больном чумой человеке, блоха не сразу приобретает способность зара-
жать людей. Необходимо, чтобы бактерии размножились в пищеварительном тракте
блохи. В процессе размножения они могут склеиваться и образовывать глыбки, со-
стоящие из чумных бацилл, видимые под микроскопом в виде черных пятен. Эти глыб-
ки, склеиваясь друг с другом, могут закупорить просвет желудка и преджелудка бло-
хи. Возникает так называемый «чумной блок». «Блокированная» блоха продолжает
испытывать голод и пытается сосать кровь хозяина. Однако заглатываемая кровь не в
состоянии протолкнуть «блок» в кишечник блохи. Поэтому блоха должна отрыгнуть
его в кровь хозяина, что несчастное животное и делает, если у него хватает на это сил
после длительного обезвоживания. Вместе с кровью в ранку вымывается и какое-то
количество бактерий чумы (рис. 1.23).
Рис. 1.23. Изображение «блокированной» блохи. 1 — комки бактерий
чумы («чумной блок»); 2 — желудок; 3 — преджелудок; 4 — пищевод,
запробкованный бактериями чумы. По L. Е Hirst (1953)
Но уже при инфицировании блох появляется проблема, связанная с тем, что она
лишь случайно вовлекается в цепочку «грызун — блоха — человек» во время эпиде-
мий чумы и ее роль в поддержании в природе Y. pestis сильно преувеличена чумолога-
ми начала XX в. Для заражения блохи чумой от грызуна, необходимо, чтобы болезнь
у него протекала в септической форме, а это происходит далеко не всегда. Грызун
может погибнуть от инфекционно-токсического шока раньше, чем чума перейдет в
септическую форму, либо болезнь у него ограничится лимфатическим узлом, бли-
жайшим к месту проникновения Y. pestis. Тогда образовавшийся бубон претерпит об-
ратное развитие и заместится соединительной тканью. Сам процесс «блокирования»
блох не имеет четких временных рамок, а зависит от температуры окружающей сре-
ды, влажности воздуха и частоты повторного питания. Поэтому он образуется в раз-
ные сроки: от 3 до 200 суток. Возможен также размыв (рассасывание) блока, возоб-
новление у блох способности питаться. В случае успешного «блокирования» блох,
возникает другая проблема — продолжительность их жизни резко сокращается, так
1.8 Крах отряда Ns 731
149
как насекомое не может пить, пока не отрыгнет «чумной блок». Поэтому Исии весь
производственный цикл по производству 60 надо запускать заново, предварительно
проверив возбудитель чумы на вирулентность, а в случае ее утраты, снова ее восста-
новить (см. «Повышение вирулентности бактерий»).
«Блокированная» блоха должна нападать на человека, а не на собаку или грызуна.
А вот с выбором таких блох (а их тогда энтомологи насчитывали 6 родов, включаю-
щих 25 видов), гак же обстояло не просто. На людей нападают так называемые чело-
веческие блохи (Pulex irritans), на крыс — крысиные (Xenopsylla cheopis). А это означа-
ет, что если использовать для распространения Y. pestis первых, то очень трудно вызвать
крысиную эпизоотию чумы. Риску инфицирования подвергнутся только отдельные
люди и то в течение нескольких дней, пока блохи сохраняют активность. Если вто-
рых, то нет никакой гарантии того, что в случае развития чумной эпизоотии среди
крыс, она «перекинется» на людей. В конечном итоге своих экспериментов, видимо
добавив к экспериментальным данным еще и формально-логические допущения,
японцы остановились на блохах Р. irritans. Они давали им шанс заразить чумой отдель-
ных людей, а вот шансов вызвать чумные эпидемии в городах у разработчиков БО по-
прежнему не оставалось. Существует много препятствий биологического характера,
ограничивающих возможность даже единичных случаев передачи возбудителя чумы
между людьми посредством блох Р. irritans. И основное из них опять же «блокирова-
ние» блох — чтобы блоха заразилась чумой от человека, он должен страдать септиче-
ской формой чумы, а с людьми во время болезни происходит то же самое, что и с кры-
сами (см. выше). Впрочем, ничего этого японской армии и не понадобилось. Когда
9 августа 1945 г., т. е. после начала войны с СССР, Исии потребовал от Оноуэ Масано
(начальник муданьцзянского филиала отряда № 731) доставить в штаб отряда всех
имеющихся блох, тот смог отправить ему только 25 г столь необходимого для ведения
бактериологической войны биотехнологического продукта. А их еще Исии надо было
«блокировать» вирулентным для человека штаммом возбудителя чумы и т. д. и т. п.
Следовательно, «дешевыми» могут быть только штучные и здоровые блохи на бро-
дячей собаке (вид Pulex canis). Блохи же, зараженные чумой, — «удовольствие» весьма
дорогое даже для состоятельных соискателей «оружия бедных». Цена «удовольствия»
еще более возросла, когда японцы узнали, что с таким трудом размноженные, зара-
женные и ^оставленные до цели чумные блохи при падении с высоты в основной своей
массе ломают лапки и не могут атаковать потенциальную жертву (см. «Разработка
специальных боеприпасов...»).
Стабилизация свойств уже наработанной бактериальной массы. Имеющиеся дан-
ные не позволяют считать, что японцам удалось добиться больших успехов в решении
этой сложной технической задачи. Моримура утверждает, что к концу войны (1944) в
отряде были освоены простые технологии лиофильного высушивания бактерий; япон-
цы могли их хранить в сухом виде и разводить перед применением специальной жид-
костью. Лиофильная сушка микроорганизмов доставила много проблем сотрудникам
отряда. Поразительно наблюдательный и обладающий хорошей памятью, Хироси
Акияма (1958) рассказывал о виденном им учебном фильме, посвященном этой техно-
логии, и снятом в отряде.
«В частности, в нем рассказывалось о способности некоторых видов бактерий пе-
реносить замораживание и высушивание. Микробы особенно устойчивы к действию
низких температур. Бациллы чумы, холеры, различные гноеродные кокки и ряд других
бактерий при замораживании сохраняют жизнеспособность и после оттаивания вновь
начинают размножаться. Сопротивляемость же бактерий к высушиванию очень низка.
150
Часть 1. Краткая история применения биологических средств...
Некоторые из них погибают уже через несколько часов после высушивания. Особен-
но нестойки к высушиванию возбудители гриппа, чумы и холеры, сравнительно стойки
кокки или палочки тифа и дифтерии. Палочки Коха и стафилококки в высушенном
виде не погибают довольно долго от нескольких десятков дней до нескольких месяцев».
Объяснение этому феномену было получено только в 1970-х гг. Оказалось, что
действие лиофилизации на Д Н К бактериальной клетке сходно с действием рентгенов-
ского излучения — в цепочках ДНК лиофильно высушенных бактерий образуются
однонитевые разрывы. При последующем их восстановлении репарационной систе-
мой клетки, возникают мутации, серьезно нарушающие их жизнеспособность. Конеч-
но, изменением технологий культивирования, подбором сред и режимов заморажи-
вания/высушивания, оптимизацией способов последующего восстановления можно
было повысить жизнеспособность возбудителя чумы, но все эти эксперименты тре-
бовали времени и средств. К тому же природа умеет страховать себя от таких энергич-
ных ее преобразователей, каким был Сиро Исии. Возбудитель чумы после лиофиль-
ного высушивания снижал не только жизнеспособность, но вирулентность. В отличие
от других патогенных, но малоопасных микроорганизмов, например сальмонелл,
снижение вирулентности у него было не в разы, а на порядки (Heckly R. J. et al., 1958).
А во что выходит восстановление утраченной вирулентности у бактерий, мы уже рас-
смотрели выше (см. «Повышение вирулентности бактерий»). Но потом, после восста-
новления вирулентности, их снова надо сушить.
Японцы не подошли и к тому, что В. В. Мясников с соавт. (1984) называют «био-
логической рецептурой». Для поражения людей они использовали бактериальные
агенты в виде бульонных культур, которые подращивали перед применением (напри-
мер, уже в бомбе перед ее сбрасыванием); суспензий культур, смытых с поверхности
плотной питательной среды изотонической жидкостью; в конце войны — суспенди-
рованные в изотонической жидкости бактерии, хранившиеся в виде лиофилизата.
Однако даже приготовленные с таким трудом бактерии и чумные блохи было невоз-
можно использовать для биологической войны.
Разработка специальных боеприпасов и устройств для диспергирования блох и бак-
терий. Исии относился к тем ученым, которые «сидят на литературе» и строго обосно-
вывают все свои действия перед начальством не собственными «домыслами», а имею-
щимися «литературными данными». А «литература» начала 1930-х гг. говорила ему то.
что бактерии в аэрозоле «размножаются и набираются вирулентности», попасть в их
«облако» — значит погибнуть («знаменитые опыты» профессора Триллья). В печати
сообщали о германской бактериологической бомбе так, как будто ее все уже видели
лежащей на газоне у Британского музея.
Журналист Стид «расшифровал» секретные немецкие документы об испытаниях
бактериальных аэрозолей в Париже. Они не только ни у кого не вызвали ни малейшего
сомнения, но и еще сообщали важный «результат» этих экспериментов — оказывается,
бактерии способны после распыления пульверизатором «пролететь» 3,5 км в призем-
ном слое нагретого от поверхности земли поднимающегося вверх воздуха парижских
улиц и оставаться живыми 6 ч под воздействием солнечных лучей. Как с «вирулент-
ностью», «культивированием» и «лиофильной сушкой», Исии снова попал в ловушку
несоответствия «общепринятых научных представлений» реальному знанию, получа-
емому лишь в строгих экспериментах.
Сначала работа по созданию БО пошла бойко, так как использовались наработ-
ки, полученные при создании химического оружия. Первая созданная отрядом бомба
представляла собой простую модификацию уже стоявшего на вооружении химиче-
1.8. Крах отряда № 731
151
ского снаряда. Позже появились бомбы, разработанные по собстъенным проектам.
К 1940 г. в отряде было разработано и испытано в полевых условиях девять типов
авиабомб, предназна11енных для рассеивания «контагия». В их число входили бомбы
для заражения поверхности земли; бомбы для распыления крупнодисперсного аэро-
золя; бомбы осколочного действия, предназначенные для заражения человека спо-
рооьразующими бактериями через открытые раны: бомба с распылителем для ОВ
(видимо, того типа, который тогда называли «германским») и еше ряд «мертворож-
денных» конструкций (о них ниже).
Результаты же потогонных испытаний больше свидете шствовали «о принципи-
альной возможности создания БО», чем позволяли продемонстрировать командова-
нию его эффективные образцы. О некоторых трудностях того периода создания БО
рассказал Хироси Акияма (1958):
«...опыты с обычными бомбами, по-видимому, не давали значительного эффекта,
так как из-за высокой температуры и давления возникающих при взрыве, большая
часть бактерий погибала, а оставшиеся быстро утрачивали активность Даже бактерии
газовой гангрены и сибирской язвы, сравнительно более устойчивые ктеплу, почти на
70 % погибали, а из бактерий, особенно плохо переносящих сухую среду, таких, как бак-
терии чумы, сохранялось не более 10-20 %».
Уже к 1941 г. Исии испытывал явное разочарование и по поводу возможности ис-
кусственного распространения эпидемий. На допросе во время предварительного
следствия 23 октября 1949 г. генерал-лейтенант медицинской службы Кадзицука Рюди
и обвиняемый на этом процессе (бывший начальник санитарного управления Кван-
тунской армии — непосредственно руководил работой отряда № 731) сообщил о
разговоре с Исией, состоявшемся в феврале 1941 г.:
«...Исии сказал мне, что после всех проведенных под его руководством исследо-
ваний он пришел к выводу, что умышленное распространение эпидемии — не такая
легкая вещь, как кажется некоторым людям или как ранее он считал сак:. В природе
естественное распространение эпидемии происходит очень легко, но искусственное
распространение эпидемии встречает целый ряд препятствий, которые приходится
преодолевать ино1 да с большим трудом. Успех подобных предприятий зависит, по его
мнению, от индивидуальной подверженности людей различным инфекционным бо-
лезням, и он решил заняться изучением этой проблемы».
Кризис аэробиологических упрощений произошел в отряде также где-то в начале
1941 г. Большое недоумение у Исии вызвали неудачи опытов по инфицированию лю
дей распыленными с самолетов бактериями. Эпидемии легочной чумы в Маньчжу-
рии 1910—1911 гг. и 1921 г. распространялись на огромные территории «сами собой».
Попытки вызвать их искусственно с помощью аэрогенного инфицирования людей
показали обманчивость многих, существовавших тогда представлений как о патоге-
незе и эпидемиологии легочной чумы, так и о поражающих свойствах самого БО.
Атакам с воздуха с применением аэрозолей возбудителей чумы и сибирской язвы
командование Квантунской армии предполагало подвергать передовые позиции про-
тивника и его основные коммуникации. В этом их видении места LO на поле боя
прослеживается аналогия с применением авиацией химического оружия. Но чтобы
эффективно применить бактериальные аэрозоли для поражения людей, Сиро Исии
необходимо было основательно «переделать» выливные авиационные приборы
<ВАПы), используемые для «поливки» местности ипритом или ипритно-люизитны-
ми смесями, в совершенно иные устройства.
152
Часть 7. Краткая история применения биологических средств...
ВАПы, существовавшие в конце 1930 х гг., представляли резервуар обтекаемой
формы. В хвостовой его части имелось большое отверстие, которое закрывалось крыш-
кой. В передней части он имел большое наливное отверстие. Такие приборы подве-
шивались под плоскостями или фюзеляжем самолета. Крышка хвостового отверстия
соединялась тросами с открывающими приспособлениями, находящимися в кабине
летчика-наблюдателя. Когда самолет подходил к цели, летчик-наблюдатель открывал
прибор, и жидкое ОВ выливалось из прибора самотеком. Прибор опорожнялся за
несколько секунд. Быстро вылившееся большой массой жидкое ОВ дробилось на кап-
ли различной величины потоками встречного воздуха и в виде дождя и тумана оседа-
ло на землю. Средний размер капли иприта составлял 1,6 мм.
Иприт и люизит проникают в организм человека через кожные покровы, а при
заражении ими местности поражение людей достигается еще и за счет паров ОВ.
Поэтому размер капель ОВ, создаваемых ВАПами, никак не находится в связи с не-
обходимостью проникать в глубокие отделы легких человека. Само же заражение
местности ОВ осуществляли в те годы с высоты до 100 м. Увеличение высоты увели-
чивало площадь заражения, но снижало плотность заражения местности.
Помимо конструирования принципиально иных приборов, переводящих биоло-
гическую рецептуру (или то, что Исии вместо нее получал) в аэрозоль определенного
фракционно-дисперсного состава, ему необходимо было их еще и оптимально разме-
стить на самолете. Аэрозольное облако должно равномерно накрывать цель. Но какую
выбрать компоновочную схему самолета, чтобы учесть влияние на бактериальный
аэрозоль воздушного потока, создаваемого винтом самолета или вихревых шнуров,
зависящих от характеристик его крыла? Мощность разработанных диспергирующих
устройств должна быть достаточной для того, чтобы при данной компоновочной схе-
ме самолета, его скорости, высоте полета, атмосферной турбулентности, влиянии
экрана земли, влажности и температуре окружающего воздуха, и еще десятков различ-
ных факторов, облако бактериального аэрозоля было достаточно «плотным» для ин-
фицирования людей, попавших в него на очень непродолжительное время.
Исии еще предстояло выбрать тип диспергирующего устройства, исходя из раз-
мера капель, которые ему нужно получить для эффективного поражения людей; за-
тем изучить влияние самих диспергирующих устройств на жизнеспособность бакте-
рий в аэрозоле. Для формулирования требований к диспергирующим устройствам
необходимо знать механизмы проникновения бактериального аэрозоля в глубокие
отделы легких человека, а они стали известными только в 1950—1960 гг. Такие устрой-
ства надо было калибровать, но надежных методов определения размера диспергиру-
емых капель не было лаже в 1960-х п К тому же нужны были сами бактерии в количе-
ствах, достаточных для аэрозольного способа применения БО, а их невозможно
получить с помощью «культиваторов Исии».
Время шло, неудачи следовали одна задругой, убивать людей сотнями с помощью
БО не получалось. По свидетельству бывшего сотрудника отряда, приведенному
Моримурой, даже в конце войны они не понимали того, что происходило с бактери-
альным аэрозолем, создаваемым диспергирующим устройством, установленном на
самолете. На полигоне близ станции Аньда японские исследователи расстилали бе-
лые полотнища, затем распыляли с самолета водную суспензию из двадцати тысяч
яичных желтков, чтобы выяснить, как будет распределяться по поверхности земли
распыляемый материал. Хотя им и удалось подобрать условия диспергирования, при
которых капельки такой жидкости обнаруживали на белых полотнищах, но от созда-
ния реальных подходов к массовому поражению людей бактериальными аэрозолями
1.8. Крах отряда № 731
153
они отдалились еще дальше. Получалось то, о чем предупреждал профессор Конрих
еще в 1932 г. — заражения людей во время их контакта с бактериальным аэрозолем не
происходило (см. его третье возражение в разд. 1.7).
Гибель бактерий чумы в процессе хранения и диспергирования, а также невозмож-
ность применения аэрозольного способа распространения бактерий в условиях про-
тиводействия ПВО противника, вынудили Исию искать какую-то оболочку либо но-
ситель, способные доставлять возбудитель чумы в организм человека живым и не
потерявшим вирулентность. В конечном итоге своих поисков он был вынужден оста-
новиться на блохах.
Кадзицука Рюдзи на допросе от 23 октября 1949 г. показал следующее:
«В феврале 1941 г. начальник отряда № 731 полковник медицинской службы Исии
в городе Чанчунь в моем служебном кабинете во время доклада мне о работе отряда с
разрешения командующего Квантунской армией Умедзу также рассказал о том, что
было проделано отрядом по подготовке к бактериологической войне .. Исии расска-
зал мне, что с большой высоты сбрасывать бактерии нельзя, так как они гибнут, но при
малой высоте площадь распыления бактерий получается слишком незначительной.
От Исии я узнал, что сбрасывание бактерий холеры на территории противника
малоэффективно и трудноосуществимо, во-первых, потому, что сбрасывать нужно с
малой высоты, что дает возможность противнику свободно обстреливать самолеты, а
во-вторых, потому, что для этого потребовалось бы слишком большое количество бак-
терий. Далее Исии сказал мне, что гораздо более эффективно сбрасывание бактерий
не в “голом” виде, а вместе с посредниками — насекомыми и, в частности, с блохами.
Блохи, являющиеся наиболее живучими насекомыми, заражались чумой и сбрасыва-
лись с самолетов, и бактерии чумы, оставаясь в блохах, благополучно опускались вме-
сте с ними на землю. Это обстоятельство уменьшало уязвимость самолета со стороны
зенитной артиллерии противника.
При этом Исии заявил мне, что исследования по этому вопросу еще не законче-
ны, оставался, например, невыясненным вопрос, на какую площадь распространяются
блохи, сброшенные с большой высоты».
Какие представления об экологии и эпидемиологии чумы существовали в отря-
де, наглядно показывает учебный фильм, предназначенный для подготовки бактери-
ологических диверсантов, кратко изложенный в книге Хироси Акиямы (1958).
«Действие в фильме развертывается так. Переодетые диверсанты под покровом
ночной темноты с корзинами, похожими на те, в которых обычно переносят голубей,
проникают в деревню, находящуюся в “глубоком тылу противника”. В корзинах зара-
женные чумой крысы с паразитирующими на них блохами. Укрываясь в тени постро-
ек и деревьев, диверсанты быстро открывают крышки корзин. Из корзины выпрыгива-
ет одна крыса, за ней вторая, третья... Внутри корзина разделена на несколько отделе-
ний с таким расчетом, чтобы при каждом открывании крышки оттуда мог выскочить
только один зверек. Зараженные чумой крысы очень худые, а глаза у них сверкают, как
стеклянные бусинки. Выпушенные крысы прячутся в кладовых, за домашней утварью,
в щелях стен и смешиваются с местными крысами. Блохи, паразитирующие на крысах,
зараженных чумой, перескакивают на здоровых. Вскоре среди крыс этой деревни вспы-
хивает чума. Многочисленные блохи, насосавшись крови больных крыс, а также мухи
разносят инфекцию. Через два-три дня в деревне появляются первые больные чумой.
На экране видны мухи, бесчисленные тучи мух... Они садятся на мокроту, слюну, вы-
деления из носа и экскременты больных чумой, а затем перелетают на продукты и
переносят на них бациллы чумы, которые содержатся в выделениях больных Человек,
употребляя эти продукты, заболевает легочной формой чумы».
154
Часть 1. Краткая история применения биологических средств...
Рис. 1.24. Керамические бомбы
«системы Удзи». Обнаружены в
руинах отряда № 731. Фотография
из «Доклада международной науч-
ной комиссии по расследованию
фактов бактериологической вой-
ны в Корее и Китае», Пекин, 1952
Сделав ставку на зараженных чумой блох, Исии активно стал разрабатывать кон-
кретные приемы и технические средства для их применения. Однако и здесь он на-
толкнулся на еще нерешенные технические задачи. Вот только две из них. Идея сбра-
сывания бомб, начиненных чумными блохами, как это обычно у Исии было, формаль-
но-логически выглядела безукоризненной, но практически и ее было трудно воплотить
в жизнь. Чтобы самолет с бомбами не смогла сбить зенитная артиллерия противни-
ка, необходимо поднять потолок действия бомбардировщиков, но тогда блохи поги-
бали из-за недостатка кислорода. Погибали они и от высокой температуры, развива-
ющейся при взрыве бомбы.
Для решения первой технической задачи Исии решил подкачивать кислород в бомбу
во время полета самолета; для решения второй — заменить металлические корпуса
бомб на керамические. Металл в осколках ктому же
оставлял доказательства применении БО, а керами-
ка рассыпалась при небольшом заряде взрывчато-
го вещества, не оставляя следов. Насекомые после
взрыва оставались, как он полагал, невредимыми.
По сведениям, полученным Моримурой от сотруд-
ников отряда, мысль о керамической бомбе пришла
Исии в голову поздней ночью. Он был настолько
возбужден, что немедленно приказал собрать руко-
водство отряда.
Керамическая бомба «системы Удзи», взрываю-
щаяся с помощью запального шнура, стала основ-
ной в арсенале средств бактериологического пора-
жения людей, разрабатываемых в отряде № 731.
Бомбы этого типа имели два ра змера: на 2 и 4 кг. Не-
большие количества взрывчатого вещества разме-
щали снаружи бомбы в специальных желобках в
оболочке. Оболочки рассыпались на осколки менее
1 см2 с широкой площадью распространения насе
комых (рис. 1.24).
Эта бомба стала родоначальником целого се-
мейств японских бомб с легко разрушаемыми не-
металлическими корпусами, используемыми не
только для распространения блох, но и для дис-
пергирования суспензий бактерйй и боевых отрав-
ляющих веществ. Во время войны в Корее такие
бомбы не применялись, однако они послужили
прототипом для американских контейнеров аналогичного назначения типа «яичная
скорлупа». О них более подробно рассказано в разд. 1.11.
Производство керамических бактериологических бомб было налажено в 3-м отделе
озряда и в армейском арсенале в Мукдене (Шэньяне). О технологии производства
такой бомбы свидетель Сегоси показал на Хабаровском процессе 29 декабря 1949 г.
следующее:
«Гос5 царственный обвинитель. Скажите, свидетель Сегоси, вы работали в качестве
лаборанта 4-го отделения материального отдела отряда № 731?
Свидетель Сегоси: Гак точно, я работал в качестве лаборанта 4-го отделения мате-
риального отдела.
1.8. Крах отряда Ns 731
155
Вопрос: Чем занималось это отделение?
Ответ: 4-е отделение занималось производством фильтров, керамических корпу-
сов для бактериологических бомб и фарфоровых колбочек.
Вопрос: Вы видели, как изготовляются керамические корпуса бактериологичес-
ких бомб?
Ответ- Да, приходилось. Я занимался исследованием состава глины, применявшей-
ся для изготовления этих корпусов.
Вопрос: Дайте описание Зомбы и техники ее изготовления.
Ответ: Для изготовления бомбы брали глину, размельчали ее в порошок, разбав-
ляли водой, затем приготовляли состав необходимой густоты. Этот состав вливался в
специальную гипсовую форму. Эта форма имела рид снаряда. Ввиду того, что гипс
впитывает в себя влагу, поверхностный покров этого состава засыхал. Затем гипсовую
форму снимали, вылйваЛи при этом оставшуюся внутри жидкую массу и по-.гучали
керамические изделия в виде корпусов снарядов. Изготовленные корпуса бомб высы-
хали в специальных печах. Эти бомбы были от 70 до 80 см длиной и 20 см диаметром,
внизу было нарезное отверстие Внутренность корпуса была полая. В нарезное отвер-
ст ие вставлялась дистанционная трубка. На поверхности стенок этих корпусов дела-
лись зигзагообразные каналы. В верхней части бомбы были приспособления для при-
крепления стабилизаторов. В каналы заключали взрывчатое вещество, с помощью
которого бомбы взрывались. Эти бомбы, сбрасываемые с самолета, должны были взры-
ваться над землей.
Вопрос: Чем начинялись бомбы?
Ответ: Со слов начальника отделения майора Судзуки мне известно, что в эти кор-
пуса помещались колбочки, начиненные чумными блохами.
Вопрос: Каким образом конструкция бомбы обеспечивала сохранение чумных блох
при разрыве и предохраняла их от действия высокой температуры?
Ответ: Майор Судзуки мне рассказал, что поскольку корпус этих бомб керамичес-
кий и гонкий, то взрывчатого вещества требовалось очень мало, поэтому взрыв полу-
чается небольшой силы, и это спасало блох от уничтожения, так как на них практи-
чески не действовали ни сила взрыва, ни сопротивление воздуха, ни температура».
С 1939 г. и до лета 1945 г., то есть почти до самого окончания войны, на полигоне
близ станции Аньда проводились многочисленные эксперименты по применению
керамических бомб, начиненных чумными блохами Pulex irritans. Один из бывших
служащих отряда рассказал Моримуре следующую подробность функционирования
«кухни дьявола»:
«Не всегда рассеивали зараженных блох. Это было опасно для находившихся на
станции Аньда служащих отряда. Поэтому некоторые опыты проводились с условно
зараженными блохами... Для того чтобы насекомые добрались до “бревен” и начали
сосать их кровь, требовалось четыре-пять часов. “Бревна”, видя, как несметное коли-
чество блох сначала впивается им в ноги, а затем распространяется по всему телу, и,
думая, что блохи заражены чумой, отчаянно бились и кричали, но, поскольку их руки
и ноги были привязаны к столбам, они ничего сделать не могли. В каждой бомбе на-
ходилось около 30 тысяч блох».
В материалы Хабарсвского процесса не попали сведения о бактериологических
бомбах более совершенных конструкций. Об их устройстве и технических характери-
стиках Исии сообщил американским следователям («Джи-2») в 1946 г. В 1980-х гг.
протоколы этих допросов исследовал Моримура.
Бомба «системы Удзи, тип 50» состояла из трех основных конструктивных эле-
ментов:
156
Часть 1. Краткая история применения биологических средств...
1) дистанционного взрывателя, помещенного в хвостовую часть, с зарядом три-
нитротолуола;
2) керамического резервуара, содержавшего бактериальную жидкость;
3) целлулоидных стабилизаторов.
Общий вес бомбы — около 25 кг, емкость — 10 л. Бомбу с бактериальной жид-
костью сбрасывали с самолета, выдернув предохранитель. Помещенный в хвостовой
части взрыватель срабатывал на высоте 200—300 м. Поскольку имевшиеся в то время
в японской армии дистанционные взрыватели были низкого качества, Исии продуб-
лировал их, установив в головной части бомбы еще и взрыватель ударного действия.
Вдоль керамического корпуса шли канавки, в которых помещался детонирую-
щий шнур общей длиной 4 м. Он крепился к корпусу бомбы цементом. Шнур сам по
себе являлся взрывчаткой, и, если дистанционный взрыватель срабатывал, бомба еще
в воздухе разлеталась на мельчайшие осколки, а содержавшаяся в ней бактериальная
жидкость капельками рассеивалась в воздухе. Длина корпуса бомбы «системы Удзи,
тип 50» была около 700 мм, диаметр 180 мм, толщина стенки резервуара 8 мм. После
заполнения десятилитрового резервуара культуральной жидкостью общий вес бом-
бы составлял 35 кг. По показаниям Исии, в результате испытаний установлено, что
при взрыве бомбы на высоте 200-300 м и при скорости ветра 5 м в секунду, макси-
мальна объем зараженного пространства составлял 40x60x800 м.
Если при той же скорости ветра взрывали бомбу, установленную неподвижно на
высоте 15 м, то объем рассеивания бактерий составлял 20x30x500—600 м. О результа-
тах взрыва бомбы, заполненной жидкостью с бактериями сибирской язвы на высоте
15 м, Исии сообщил своим будущим американским коллегам следующее: «Погибли
70 % лошадей и 90 % овец, которые паслись, в течение одного-двух часов с подвет-
ренной стороны». На допросе Исии высказал уверенность в том, что «при усовер-
шенствовании бомбы «системы Удзи, тип 50» она могла быть превращена в чрезвы-
чайно эффективное бактериологическое оружие». Но это был очередной его блеф.
Исии надо было показать американцам свою значимость как специалиста и тем са-
мым избежать участи военного преступника. Трудно себе представить, чтобы советс-
кие солдаты после применения японской армией такого БО, заражались посредством
поедания инфицированной им травы.
Исии дал показания и о трудностях, с которыми столкнулись создатели бактерио-
логической бомбы. Прежде всего, ее керамический корпус не обладал достаточной
ударостойкостью, и поэтому с бомбой нельзя было обращаться так, как обычно обра-
щаются с боекомплектами при транспортировке. Увеличивать же толщину стенок
керамического резервуара нельзя было из опасения, что бомба может оставить оскол-
ки, то есть доказательства ее применения. Кроме того, потребовалось бы увеличение
массы взрывчатки, что повело бы к повышению температуры при взрыве, а следова-
тельно, к гибели органического материала.
Были и другие трудности. Диатомит, служивший материалом для корпуса бомбы,
обжигался неравномерно, и бактериальная жидкость часто протекала в месте сочле-
нения металлической горловины, через которую она заливалась в корпус. Материал
не позволял делать корпуса строго одинаковыми по весу, что сказывалось на траекто-
рии полета бомбы и снижало точность попадания в цель. Чтобы при скачке темпера-
туры увеличивавшаяся в объеме бактериальная жидкость не выливалась, емкость за-
полнялась на 70 %. Воздушный мешок (30 % объема) служил причиной произвольного
вращения бомбы в полете, что также снижало точность попадания в цель. Использо-
1.8. Крах отряда № 731
157
вание керамических стабилизаторов создало новые проблемы. При обжиге они коро-
бились, а на холоде становились хрупкими и в полете нередко отваливались от кор-
пуса. Несовершенство же взрывателей не давало возможности точно задать высоту
взрыва. Но Исии гарантировал американцам, что «если исключить несовершенство
дистанционного взрывателя, основные конструктивные недостатки можно было бы
считать преодоленными».
Последнее его утверждение являлось уже откровенным бахвальством. В бакте-
риологической бомбе «системы Удзи, тип 50» использовались жидкости с возбудите-
лями красной плесени, холеры, чумы, дизентерии, сыпного тифа, сибирской язвы.
В протоколе допроса указывалось: «Эксперименты, проведенные Исией в Пинфане,
показали, что при взрыве бомбы на высоте 200—300 м минимальный объем рассеива-
ния бактерий составлял 20—30x60x700 м».
Увеличенной модификацией бомбы «системы Удзи, тип 50» была бомба «системы
Удзи, тип 100», вместимостью около 25 л. Всего изготовлено около 300 таких бомб,
которые с 1940 по 1942 г. были широко испы ганы теми же методами, что и «тип 50»
Недостатком бомбы были ее большие размеры, которые повышали опасность повреж-
дения корпуса при обращении с ней. Считалось, что она ниже по практической цен-
ности, чем «тип 50». Кроме бомб «системы Удзи», были еще бомбы типа ГА. В них
вместо керамического корпуса, использовался стеклянный.
Величина капель жидкости, распылявшихся при взрыве бомб обоих типов, бы-
ла разной: от крупных и мелких дождевых капель до микроскопических, диаметром
50 мкм1. По оценке лиц, допрашивавших Исию, баллистические качества бомбы «си-
стемы Удзи» были приемлемыми, но полностью удовлетвориться ими было нельзя.
Один из недостатков бомбы состоял в том, что невозможно было точно рассчитать
время взрыва из-за неодинакового рельефа местности в районах бомбардировок. На-
дежного высотомера для бомбометания в японской армии не было, вследствие чего
результаты всякий раз получались разными. Именно поэтому японские военные при-
ступили к разработке устройства, названного «мать и дочь».
Бомба типа «мать и дочь» — это бомба нового типа, специально предназначав-
шаяся для бактериологической войны. Она сконструирована в 1944 г., но не в отряде
№ 731, а в 9-м армейском научно-исследовательском институте поручиком Гондо
(о деятельности этого института никакой информации в открытой печати нет). Уст-
ройство представляло собой комбинацию двух бомб: крупногабаритной (материн-
ской) и малогабаритной (дочерней). Первая связывалась со второй системой, подаю-
щей радиосигналы. Когда первая достигала земли, вторая еще продолжала падение.
При взрыве материнской бомбы радиосигналы прекращались, и срабатывал меха-
низм, осуществлявший надземный взрыв дочерней бомбы. Сочетание наземного и
надземного взрывов позволяло получить устойчивое бактериальное заражение мест-
ности и приземного слоя воздуха, независимо от рельефа.
Комплект бомб «мать и дочь» был испытан, но их не успели применить, поскольку
война закончилась. Любопытна судьба идеи поручика Гондо. Предложенная им двух-
компонентная система боеприпасов типа «мать и дочь» оказалась востребованной в
1990-х гг. при создании субэлементов кассетных боеприпасов, способных детониро-
вать на требуемом расстоянии от поверхности земли в условиях сложного рельефа
местности (см. например, европейский патент № 91107842 с приоритетом от 1991 г.)
1 Частицы такого размера не проникают в глубокие отделы легких (см. разд. 1.9).
158
Часть 1. Краткая история применения биологических средств...
Шрапнельная бомба. Предназначалась для заражения людей и животных палочка-
ми столбняка и газовой гангрены или спорами сибирской язвы через раны и для зара-
жения почвы возбудителем сибирской язвы. В материалах SI PRI (The Problem of Che-
mical and Biological Warfare, 1970) она описана более детально, чем у Моримуры. Общий
вес бомбы 41 кг, корпус выполнялся из стали. Ее снаряжали 5 кг шрапнели и 0,5 л
бактериальной культуры. Зона разбрасывания осколков и шрапнелей составляла около
40 м. Бомба предназначалась для применения на передовых позициях. Это направле-
ние создания бактериологических бомб японцы разрабатывали до ликвидации отряда.
Устройство для распространения зараженных чумой крыс. Было разработано к 1936 г.
Оно представляло собой специальный метательный снаряд, представляющий собой
бумажную трубку, раскрывающуюся горизонтально. Внутри полость трубки разделя-
лась на 2 или 3 отделения. К верхней части трубки прикреплялся бумажный или из
искусственного щелка парашют. К нижней части привешивалось грузило с пистоном,
который через фитиль был соединен с пороховницей в нижней части трубки. В трубку
помещали 12 домашних и 6 диких мышей, зараженных возбудителем чумы. Туда также
помещали блох, сосавших кровь зараженных чумой мышей (их завертывали в бумагу).
Такие устройства сбрасывали в тылу противника над его базами, воинскими час-
тями и большими городами. При соприкосновении с землей трубка раскрывалась на
две части, мелкие животные и блохи разбегались. Пистон поджигал фитиль, порох
вспыхивал и бумажная трубка с парашютом сжигались дотла, не оставляя никаких
следов. Японцы были твердыми сторонниками «крысиной теории» распространения
чумы, и поэтому они надеялись, что в результате такой диверсии обязательно возник-
нет сначала чумная эпизоотия среди местных грызунов, а потом эпидемия бубонной
чумы среди местного населения, которая, в свою очередь, даст вспышку легочной чумы
(см. сюжет учебного фильма выше). Однако ни того, ни другого, ни третьего не про-
исходило при практическом применении таких бомб.
Плавучая бутылка. Являлась сосудом с длинным горлышком, вместимостью око-
ло 5 л, предназначенным для наполнения культуральной суспензией бактерий. На
конце горлышка был установлен дистанционный взрыватель с небольшим количе-
ством пороха. Находясь в воде, такая бутылка, почти вся погружалась в воду, оставляла
только кончик горлышка на поверхности воды. Эти сосуды предназначались для атаки
речных линий, если определенно известно, что враг пользуется этой речной водой для
питья или купания. В материалах SIPRI (The Problem of Chemical and Biological Warfare,
1970), co ссылкой на Scientific and Technical... (1945), указывается на то, что японцами
в конце войны была создана и экспериментальная кассетная бактериологическая бомба,
но более подробной информации о ней в других источниках нет.
«Японский метод». Так тогда называли наиболее часто используемый японцами
прием распространения чумных блох среди китайского населения. Его суть в следую-
щем. Рано утром самолеты сбрасывают зараженных блох, затем в течение целого дня
производят бомбардировку и обстрелы населенного пункта, чтобы заставить людей
длительное время находиться в бомбоубежищах. Вечером, когда они возвращаются
домой из убежищ, масса блох уже успевает рассеяться, и люди не замечают ничего по-
дозрительного.
Но природа защищается от людоедов. После каждого шага в направлении созда-
ния БО оно отдалялось от своих разработчиков минимум на два. Так, генерал-майор
медицинской службы Китано Масадзо, сменивший Исию в 1942 г., просто не мог по-
верить в то, что метод распространения чумных блох, вознесенный Исией на верши-
ну бактериологической войны, в его, Китано, руках не работает. На предварительном
1.8. Крах отряда №731
159
следствии 23 октября 1949 г. подсудимый Кадзицука Рюдзи сообщил о разговоре с Ки-
тано, состоявшемся в марте 1944 г.:
«...Китано в ответ на мой вопрос о работе отряда рассказал мне, что во время его
пребывания на посту начальника отряда № 731 были достигнуты некоторые успехи,
В частности, он рассказал о том, что группа работников отряда № 731 выезжала на
китайский фронт в районе южнее Шанхая, где с большой высоты сбрасывала на тер-
риторию с китайским населением массы блох, зараженных чумой. Эти блохи остава-
лись живы, в том месте, где они упали, возникла эпидемия чумы. Китано пояснил,
большой эпидемии не возникло, но этот способ бактериологической войны нужно счи-
тать эффективным».
Оптимизм Китано исходил из формально-логической экстраполяции естествен-
но развившихся эпидемий на искусственные. Из истории эпидемий он знал, что бу-
бонная чума, возникающая после укуса человека чумными блохами, истребила гораздо
больше людей, чем легочная чума времен «черной смерти». Но почему этого не про-
исходит теперь, когда героическая японская армия увязла в боях с многомиллионным
противником? Эксперименты по диссеминации блох повторяли, меняя их условия
почти до самого конца войны, и ясности, что с ними происходит после подрыва бом-
бы, по-прежнему не было. В этой связи любопытны показания подсудимого Куроси-
мы Юдзи, служившего в отряде № 731 с октября по ноябрь 1944 г., данные им трибу-
налу 28 декабря 1949 г.:
«Вопрос: Что вы делали в отряде 731 после окончания kvdcob?
Ответ: Я был охранником во время маневров.
Вопрос: Расскажите об этих маневрах.
Ответ: Эти маневры проводились путем сбрасывания с самолета бомб, начинен-
ных блохами. Маневры заключались только в этом.
Вопрос: В каком месте проводились эти маневры?
Ответ: Точно определит! расстояние я не могу, но приблизительно в трех километ-
рах от отряда 731, в местечке Утунь.
Вопрос: Проверялось во время маневров рассеивание блох на месте маневров?
Ответ: Да, проверялось.
Вопрос: Расскажите, при помощи каких специальных средств это делалось?
Ответ: На месте маневров были расположены ящики примерно в интервалах
два метра друг от друга, точное число ящиков я не помню, но приблизительно было
около тысячи штук.
Вопрос: На какой примерно площади?
Ответ: Точно я не могу сказать, но предполагаю, что на площади около двух квад-
ратных километров. Внутри этих ящиков находилась клейкая бумага, и рассеиваемые
при взрыве блохи попадали в эти ящики на бумагу, после чего их подсчитывали.
Вопрос: Вы сами подсчигывати блох на дне этих ящиков?
Ответ: Да, подсчитывал.
Вопрос: Вам известно, с какой целью производились эти испытания?
Ответ: Так точно, известно.
Вопрос: Расскажите об этом.
Ответ: Хотя эти блохи и не были заражены бактериями, но я знал, что это делается
в целях подготовки к бактериологической войне, в случае которой эти блохи, заражен-
ные чумой, будут сбрасываться с самолетов.
Вопрос: На каком примерно расстоянии от земли происходили разрывы бактерио-
логической бомбы, начиненной блохами?
Ответ: Приблизительно на высоте сто метров».
160
Часть I. Краткая история применения биологических средств...
Эксперименты по аэрозольному заражению людей «контагием» все больше захо-
дили в тупик. «Бревна» не заболевали чумой даже тогда, когда их помещали в облако
из капелек чумной культуры. На судебном заседании 26 декабря 1949 г. подсудимый
Карасава Томио рассказал об опыте, в котором он сам участвовал весной 1944 г.:
«... В 10 метрах от этих подопытных поместили баллон, в котором находиласьжид-
костьс чумными бактериями. Этот баллон взрывался. Но, как мне известно, после этих
опытов выяснилось, что заражение не удалось, что инфекция через дыхательные орга-
ны не проникла».
Воспроизвести на людях эффектные опыты профессора Триллья не удалось. Прав-
да, Ити Такэмура, редактор книги Хироси Акиямы (1958), писал в своих примечани-
ях к ней, что продлись война еще два-три года, эти опыты, несомненно, были бы до-
ведены до конца; впрочем, такое ощущение времени преследует всех соискателей
«дешевого оружия массового поражения» (см. разд. 1.10).
Лабораторные и полигонные заражения персонала. В то время как крупных эпиде-
мий среди китайского населения сотрудникам отряда № 731 вызвать не удавалось,
среди них самих было до 30 случаев лабораторного заражения в год. Причина послед-
него лежит в конструктивной непродуманности «блока ро», исключающего надежную
физическую защиту сотрудников при работе с возбудителями опасных инфекцион-
ных болезней и инфицированными ими насекомыми. Использованные японцами сы-
воротки, вакцины, схемы лечения и профилактики были неэффективными. На фо-
тографии «блока ро» нет признаков наличия систем приточно-вытяжной вентиляции,
не упоминают о ней в своих воспоминаниях и сотрудники отряда. Внутреннее про-
странство блока сообщалось с уличным воздухом через окна, затянутые сетками для
защиты от мух (!).
Опыты, которые описывает Хироси Акияма, проводились на обычных лаборатор-
ных столах (а не в вытяжных шкафах или боксах). Безопасность сотрудников зависе-
ла от их умения работать с бактериальными культурами, зараженными животными и
насекомыми. Даже за те 5 месяцев, что Акияма пробыл в отряде, от чумы погиб один
подросток, работавший с блохами, инфицированными возбудителем чумы. Устано-
вить, как он заразился, не удалось. Не помогли ему и лечебные мероприятия, приня-
тые в отряде, основывавшиеся на введении противочумной сыворотки и массивных
доз сульфаниламидных препаратов.
О заражении персонала во время полигонных экспериментов сообщают материа-
лы SIPRI (The Problem of Chemical and Biological Warfare, 1970). В частности, в них
утверждается, что два летальных случая сапа среди сотрудников отряда № 731 в 1937 г.
привели к прекращению полевых опытов с возбудителем этой инфекции, так как спо-
собов терапии сапа, пригодных для людей, тогда не существовало. Больных же сапом
лошадей никогда не лечили, а пристреливали. Во время полевых испытаний 1944 г.
среди сотрудников было два летальных случая чумы, однако такие опыты не прекра-
щались, так как профилактическое введение противочумной сыворотки давало надеж-
ду на некоторую защиту от случайного заражения возбудителем этой болезни.
Поиск диверсионных отравляющих веществ. Целью таких исследований было по-
лучение стойкого, удобного в обращении отравляющего вещества, пригодного для
диверсионной работы. Объектом контаминации рассматривались продукты питания.
В ходе исследований сотрудники отряда № 731 обратили внимание на устойчивый к
высокой температуре яд рыбы фугу — тетродотоксин (открыт в 1909 г. японским ис-
следователем Тахарой; более подробно об этом токсине см. в разд. 3.15).
1.8. Крах отряда № 731
161
Отрад № 731 и Военно-медицинская академия совместно работали над получе-
нием концентратов этого яда. Было обнаружено, что тетродотоксин эффективно дей-
ствует на человека и может быть использован для диверсий. Отработка технологии
производства концентрата тетродотоксина осталось незаконченной. В ноябре 1944 г.
эксперименты были прерваны налетом американских бомбардировщиков В-29 на
Токио, а в апреле 1945 г. Военно-медицинская академия была разрушена пожаром, и
работы по изучению диверсионных свойств токсинов здесь прекратились из-за otcjt-
ствия лабораторий.
Для исследования диверсионной пригодности биологических токсинов в отраде
№731 была создана оперативная исследовательская группа Кусами (фармакологиче-
ские исследования). Исследования по синтезированию химических ОВ велись имен-
но в его группе, для чего в отраде находилось 18 дипломированных фармацевтов.
По свидетельству бывших сотрудников, в отраде № 731 была еще одна родствен-
ная подразделению Кусами оперативная исследовательская группа — Сэкитори. В ней
занимались разработкой техники покушений на высший командный состав против-
ника и испытанием новых орудий убийств. Группа Сэкитори работала в условиях стро-
жайшей секретности даже внутри отрада. Сотрудники этой группы, как и их шеф,
после войны предпочли не попадаться на глаза ни американцам, ни русским. Сведе-
ний о них нет.
Изучением действия различных ядов занимались и в отраде № 100. Воз что пока-
зал подсудимый Митомо (старший унтер-офицер этого отрада) 27 декабря 1949 г. на
судебном заседании трибунала:
«Вопрос: Где содержались подопытные люди в отряде № 100?
Ответ: Эти люди содержались в изоляторе при караульном помещении отряда.
Вопрос: Кому персонально был подчинен этот изолятор?
Ответ: Этот изолятор подчинялся начальнику канцелярии общего отдела отряда.
Вопрос: Расскажите все, что вам известно относительно опытов над живыми людь-
ми, которые производились в отряде № 100.
Ответ: Эксперименты над живыми людьми проводились в августе-сентябре меся-
це 1944 года. Содержанием этих экспериментов было незаметно от подопытных лиц
давать им снотворные средства и яды. Подопытных людей было семь-восемь человек
русских и китайцев В числе медикаментов, использованных в опыта*, были: корей-
ский вьюнок, героин и зерна касторника1. Эти яды примешивались к пище За две не-
дели каждому подопытному такая пища с я том давалась пять или шесть раз. В суп
примешивался, главным образом, корейский вьюнок, в кашу, кажется, героин, в та-
бак примешивался героин и бактал. Подопытные, которым подавался суп с корейским
вьюнком, через 30 минут или через час засыпали на пять часов. Все подопытные через
две недели ослабевали после проводимых над ними опытов, и их больше использовать
нельзя было.
Вопрос: Что тогда делали с ними?
Ответ: С целью конспирации все эти подопытные умерщвлялись.
Вопрос: Каким путем?
Ответ: Один подопытный русский по приказу научного сотрудника Мацуи был
умерщвлен путем введения ему одной десятой грамма цианистого калия.
Вопрос: Кто его умертвил?
Другое название растения — клещевина (Ricinus communis L.), растение из семейства мо-
точайных (Euphorbiaceae). Его зерна содержат токсин рицин (см. разд. 3.11).
162
Часть I. Краткая история применения биологических средств...
Ответ: Я ввел ему цианистый калий.
Вопрос: Что вы сделали с трупом этого русского, которого вы умертвили?
Ответ: Я анатомировал труп на скотомогильнике в отряде.
Вопрос: Что вы сделали потом с этим трупом?
Ответ: Зарыл этот труп.
Вопрос: Где была вырыта яма?
Ответ: На скотомогильнике на задах отряда.
Вопрос: Там же, где хоронились туши скота?
Ответ: Место одно и то же, но ямы другие. (В зале движение, гул возмущения.)
Вопрос: Расскажите, каким способом был убит вами этот человек, как вы осуще-
ствили это убийство?
Ответ: Для введения этому подопытному человеку цианистого калия у него, по
указанию Мацуи, был вызван понос, это и послужило предлогом для введения циани-
стого калия.
Вопрос: Это значит, что вы обманули этого человека? Говоря ему, что вы сдела-
ете ему укол с целью лечения, вы в действительности ввели цианистый калий, это бы-
ло так?
Ответ: Правильно.
Вопрос: Это был единственный человек, убитый вами, или вы убивали и других
людей?
Ответ: Один подопытный китаец съел кашус ядом, который мной был примешан,
после чего он умер, пробыв несколько часов в невменяемом состоянии.
Вопрос: Какой яд был подмешан к каше?
Ответ: Один грамм героина.
Вопрос: Вы знали, что подмешиваете в кашу смертельную дозу яда?
Ответ: Знал.
Вопрос: Значит, это было убийство, совершенное вами вполне сознательно?
Ответ: Правильно.
Вопрос: Где был похоронен труп этого человека, убитого вами?
Ответ: Там же был зарыт, где и русский.
Вопрос: То есть на скотомогильнике?
Ответ: Да.
Вопрос: Вам известны другие случаи убийства подопытных людей?
Ответ: Два русских подопытных и один китаец жандармерией были расстреляны
на том же месте.
Вопрос: То есть эти люди были расстреляны прямо на скотомогильнике?
Ответ Да.
Вопрос: За что они были убиты жандармами?
Ответ: Я думаю, для того чтобы сохранить тайну».
Возможные масштабы попыток применения БО. Один из бывших сотрудников от-
ряда, касаясь показаний Исии о количестве проведенных экспериментов с бактерио-
логическими бомбами, заявил Моримуре следующее:
«Испытания бактериологических бомб проводились в течение десяти лет: снача-
ла просто осуществлялись наземные взрывы, затем стали проводить эксперименты на
полигоне Аньда с использованием “бревен” и, наконец, бомбы начали применять в
ходе боев в Пекине и в районах Северного Китая. Самолеты отряда № 731, несущие
на борту бомбы с бактериями сибирской язвы, делали по несколько вылетов в неде-
лю. Самолет-разведчик, тип 94, для одного вылета брал на борт 4 бактериологические
бомбы, а самолет-бомбардировщик — 12 бомб. Общее число экспериментов никак не
две и не три тысячи, а десятки тысяч...».
1.8. Крах отряда Ns 731
163
Учитывая огромную протяженность фронтов и нерешенность многих технических
проблем по производству и накоплению биологических агентов, японцы были вынуж-
дены создавать многочисленные организации, занимающиеся их наработкой в непос-
редственной близости от места предполагаемого применения БО.
Бывший начальник штаба Квантунской армии генерал-лейтенант Хата Хикоса-
буро на допросах во время предварительного следствия показал, что отряды, анало-
гичные отряду № 731, были созданы в японской экспедиционной армии в Китае и в
армии, действовавшей в районах южных морей1.
Американскими следователями в 1946 г. было установлено, что Управление по
водоснабжению и профилактике японской армии имело в своем составе обычные
(штатные) и оперативные части в самой Японии и на местах боевых действий за пре-
делами страны. К июлю 1938 г. вне Японии они были размещены в пяти городах:
а) Управление по водоснабжению и профилактике частей Квантунской армии —
отряд № 731 (Харбин);
б) Управление по водоснабжению и профилактике частей армии в Северном Ки-
тае (Пекин);
в) Управление по водоснабжению и профилактике частей армии в Центральном
Китае (Пекин);
г) Управление по водоснабжению и профилактике частей армии в Центральном
Китае, другое название — отряд «Эй» № 8604 (Нанкин), где Исии начинал свою ка-
рьеру военного бактериолога;
д) Управление по водоснабжению и профилактике частей армии в Южном Китае,
другое название — отряд «Нами» № 8604 (Кантон);
е) Управление по водоснабжению и профилактике частей армии в южных стра-
нах (Сингапур).
Такие части, как Управления по водоснабжению и профилактике, были приданы
армии каждого направления. Они подчинялись командующему армией по месту дис-
локации. Исии подчинялся одному из командующих армий, о деятельности других
«Исий» нет никаких сведений. Отделы профилактики в самой Японии, имевшие опе-
ративный характер, включали дивизионные отделы по водоснабжению и профилак-
тике и отделы военных округов; вне Японии это были полевые и дивизионные отделы
по водоснабжению и профилактике. Так же как и постоянно дислоцированные орга-
ны управления, его оперативные части были приданы соответствующим войсковым
соединениям и непосредственно подчинялись командующим этими соединениями.
Масштабы деятельности таких отрядов лишь немного уступали деятельности
отряда № 731. Например, отряд «Эй» № 8604 (до 1936 г. его начальником был Исии)
имел еще 12 филиалов, общая численность его сотрудников достигала полутора ты-
сяч. В нем проводились такие же эксперименты над людьми, как и в отряде № 731.
О «производственной деятельности» таких отрядов можно судить по показаниям
обвиняемого Сато Сюндзи (бывший начальник санитарной службы 5-й армии гене-
рал-майор медицинской службы), данных им во время предварительного следствия
6 декабря 1949 г.:
«Действительно, под моим руководством имевшийся в нанкинском отряде “Эй”
№ 1644 учебный отдел готовил кадры бактериологов ежегодно до 300 человек с целью
использования их в бактериологической войне. Я, с марта 1944 года, будучи началь-
1 Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от
23.04.1949 г., Сов. секретно. АП РФ.Ф. 3. Оп. 66. Д. 1065. Л. 11—21 (Кошкин А. А., 2011).
164
Часть 1. Краткая история применения биологических средств...
ником санитарного управления 5-й армии, входившей в состав японской Квантунской
армии, оказывал активную помощь и поддержку филиалу № 643 отряда № 731 в уве-
личении производства бактериологического материала. С этой целью в мае 1945 года
мною был издан специальный приказ частям 5-й армии о вылавливании грызунов, не-
обходимых для производства бактериологического оружия и направлении их в фили-
ал № 643 отряда № 731.
Вопрос: Уточните свои показания относительно производственной мощности
нанкинского отряда “Эй” № 1644 и о наличии в нем технического оборудования.
Ответ: Производственная мощность нанкинского отряда “Эй” № 1644 по изготов-
лению смертоносных бактерий была до 10 кг за цикл. Для изготовления этого количе-
ства бактерий в отряде “Эй” имелось техническое оборудование: культиваторов Исии
около 200, одна инкубатор-комната размером 5x5x3 метра, два автоклава цилиндри-
ческой формы размером в диаметре 1,5 метра, длиной 2,5 метра, инкубаторов, пример-
но 40—50, паровых стерилизаторов 40—50, котлов Коха, примерно 40—50, и для варки
питательной среды в отряде имелись бол ьшие стеклянные колбы. Но сколько их было,
я сейчас не помню».
К этим «управлениям» надо добавить еще «походные лаборатории», которые про-
изводили бактерии буквально на месте диверсии (см. ниже показания Хиразакура
Дзенсаку). Так, на запрос по отряду № 731 министра внутренних дел С. Н. Круглова,
начальник штаба Забайкало-Амурского военного округа генерал-лейтенант Е. Г. Тро-
ценко 14 февраля 1947 г. сообщил следующее:
«По данным санитарной разведки, после занятия войсками фронта города Ванъ-
емяо в августе 1945 г. на юго-западной окраине города вблизи японского военного го-
родка было обнаружено полусгоревшее каменное здание (это было единственное со-
жженное здание в Ванхъемяо).
С восточной стороны здания на расстоянии 15 м размещалось деревянное строе-
ние — склад химикалий и лабораторное оборудование. В складе хранились большие
запасы лабораторной посуды (пробирки, чашки Петри, пастеровские пипетки и пр.)
питательных сред (агар-агар, пептон). Там же в транспортной установке стояла мощ-
ная дезкамера и рентгеноустановка. С западной стороны в 15 м от здания размещался
аптечный склад. Между аптечным складом и складом химикалий размещалось два
примитивных строения, по-видимому, для мелкого скота (коз, овец).
С этой же стороны, в двух-трех метрах от стены в траве, на площади 5 на 30 м ва-
лялись груды набитой лабораторной посуды (пробирки, чашки Петри), причем боль-
шинство из них с питательными средами и ясно выраженными бактериальными куль-
турами. Судя по форме колоний, культуры были брюшнотифозные и паратифозные.
Основное здание двухэтажное: аптека, клиническая лаборатория, пять комнат типа
госпитальных палат, уборная, душевая, мощная телефонная станция. Мощная опера-
ционная и предоперационная, склад лабораторного и научного оборудования, стери-
лизационная с вмонтированным в стену большим автоклавом и больших размеров
моечная комната для лабораторной посуды.
На втором этаже, сгоревшем во время пожара, по остаткам обуглившейся лабора-
торной посуды, структуре комнат, моечных корыт для посуды и проч, можно сделать
вывод, что здесь размещалась крупная бактериологическая лаборатория. В оставшихся
шкафах и сейфах в здании никаких документов обнаружено не было.
В 1945 г. в Ванъемяо была вспышка чумы. По заключению китайских и японских
врачей, Ванъемяо в течение последних 10 лет был вне зоны заболевания чумой. Вспыш-
ка чумы могла явиться следствием того, что:
а) были выпущены из клеток подопытные крысы, зараженные чумой;
1.8. Крах отряда № 731
165
б) многочисленная лабораторная посуда с неубитыми бактериями была выброшена
возле здания, китайцы могли подбирать эту посуду, занести ее домой, чем вызвать за-
болевания. Следует отмстить, что первые заболевания чумой среди населения появи-
лись в пунктах, близлежащих к зданию бывшей санитарно-бактериологической лабо-
ратории».
Японская армия имела не только отряды № 731 и 100, но и подобные им форми-
рования в Северном, Центральном и Южном Китае и в странах южных морей. По-
этому полная картина попыток применения японцами во Второй мировой войне БО,
а в особенности тупых зверств, совершаемых для проверки его поражающих свойств,
вряд ли может быть восстановлена.
Бактериологические диверсии против СССР. Начались с операций специальных
отрядов смертников еще во время первых столкновений японских и советских войск
под Халхин-Голом в 1939 г. Ниже приведена выдержка из показаний подсудимого Ниси
Тосихидэ, данных им трибуналу 26 декабря 1949 г.:
«Вопрос: Что вам и >вестно о применении бактериологического оружия?
Ответ: Мне известно о применении отрядом Исии бактериологического оружия
во время инцидента у Халхин Гола В июле 1944 года я из филиала Суньу был переве-
ден на должность начальника учебного отдела 731-го отряда на ст. Пинфань. Работу я
принимал от своего предшественника полковника Санода. В тот же день полковник
Санода выехал в Японию. Я вскрыл его сейф и нашел документы, говорившие о при-
менении бактериологического оружия во время Номанганского инцидента, т е. у реки
Халхин-Гол. Тут имелись негативы фотографий того времени, список смертников, при-
нимавших участие в этой операции, и приказ майора Икари. Я помню сейчас, что в
отряд смертников входили два офицера, около 20 унтер-офицеров и рядовых. Под этим
списком шли подписи, сделанные кровью.
Вопрос: Чья подпись была первой?
Ответ: Начальника отряда Икари. Далее следовал целый ряд детализирующих при-
казов Икари, а именно, как рассаживаться на автомашины, как использовать банки
из-под керосина, затем несколько указаний о том, как возвращаться. Из этих двух
документов мне стало понятным, что отряд смертников из 20-30 человек заразил бак-
териями реку Ха ixa».
Акияма знает об этих событиях не из материалов Хабаровского процесса, а от «ста-
рожила» отряда, техника- лаборанта Сагавы (имя им изменено), поэтому в его изло-
жении история выглядит иначе:
«Однажды Управлению приказали заразить воду в верховьях реки Халхин-Гол —
источника воды для всего прилегающего к ней района, бактериями тифа, холеры и
чумы с целью вынудить противника к отступлению. Это было смертельно опасное за-
дание При его выполнении погибло более тридцати армейских и вольнонаемных вра-
чей. Возможно, некоторые из них зара шлись сами, когда пускали смертоносные бак-
терии в воду, но большинство их погибло от неприятельского огня во время совершения
этой операции. ...Слушая его с затаенным дыханием, мы узнали, что в бассейне реки
Халхин-Гол от Номонхана до озера Буир-Нур сразу же вспыхнула эпидемия, и япон-
ская армия оказалась в очень выгодном положении. ...После событий на Халхин-Голе,
где Квантунской армии пришлось вести самые тяжелые в ее истории бои, командова-
ние армии стало придавать бактериологической войне очень серьезное значение».
Обе истории совпадают, по крайней мере, в двух моментах:
1) бактериологическая диверсия во время боев у реки Халхин-Гол была подготов-
лена и проведена;
166
Часть I. Краткая история применения биологических средств...
2) количества «смертников» (по Тосихиде) и погибших во время операции «вра-
чей» (по Акиме), сходятся.
А ответ на вопрос, была ли она настолько результативной, как об этом пишет
Акияма, дадут исследования архивных документов, если, конечно, кого-то из совре-
менных военных историков эти события заинтересуют. Не случайно в своих воспо-
минаниях бывший начальник Главного военно-санитарного управления Красной
армии Е. И. Смирнов (1979) писал, что, учитывая особенности театра военных дей-
ствий, время года, а также неразборчивость агрессора в средствах борьбы, Военно-сани-
тарное управление командировало в Монголию дополнительно бригады специалистов
эпидемиологов-бактериологов и токсикологов. Смирнов имел информацию о воз-
можности применения противником биологических и химических средств пораже-
ния и тщательно готовил войска к такому развитию событий (см. ниже «Действия
советской разведки»).
В соответствии с планом «Кан-Току-Эн» (план развертывания Квантунской ар-
мии для нападения на СССР, принятый летом 1941 г.), отрядами № 731 и 100 была
развернута подготовка офицеров и унтер-офицеров по освоению и применению БО.
К этому времени были испытаны первые модели бактериологических бомб системы
Удзи, разработана и освоена технология получения микроорганизмов в количествах,
достаточных для осуществления небольших бактериологических диверсий.
С 1942 г. японцами проводилась тщательная разведка пограничных районов СССР
именно в целях подготовки к масштабным бактериологическим диверсиям. К 1945 г.
в отряде № 731 уже имелся огромный запас географических карт, собранных для ве-
дения бактериологической войны с СССР. На них подробно обозначались тактиче-
ские объекты заражения бактериями: источники питьевой воды, реки, колодцы. Та-
кие карты составляла группа описания военных объектов, входившая в хозяйственное
управление. Обвиняемый, бывший сотрудник отряда № 100 поручикХиразакура Дзен-
саку, показал на судебном заседании 6 декабря 1949 г. следующее:
«В июле — августе 1942 года я принял участие в экспедиции в район Трехречья, на-
зывавшейся “летними маневрами”. Эта экспедиция имела своей целью исследовать
возможности применения бактерий сибирской язвы и сапа в естественных условиях,
приближенных к району вероятных военных действий — у границ Советского Союза.
Во время этой экспедиции проводились опыты по заражению сапом реки Дербул и
водоемов, а также опыты по заражению сибирской язвой почвы и травяного покрова.
Микробы для этой цели вырабатывались в походной лаборатории и испытывались на
лошадях, овцах и морских свинках.
С июня 1944 года я в группе научных сотрудников отряда № 100 находился в Севе-
ро-Хинганской провинции и по приказанию командования Квантунской армии зани-
мался сбором сведений разведывательного характера, а именно: выявлял наличие и
количество скота у населения в пограничных с Советским Союзом и Монгольской
Народной Республикой районах, устанавливал состояние этого скота, наличие летних
и зимних пастбищ, участков сенокошения, состояние дорог и водоемов. Эти сведения
необходимы были японскому командованию для того, чтобы в случае войны с Совет-
ским Союзом произвести массовое заражение скота с целью бактериологических ди-
версий.
Как мне было известно от начальника отряда № 100 генерал-майора Вакаману
авиация на основании собранных мною сведений будет производить распыление бак-
терий сапа, сибирской язвы и вируса чумы крупного рогатого скота для заражения
скота в пограничных с Советским Союзом и Монголией районах в случае возникно-
вения войны».
1.8. Крах отряда № 731
167
Бывший служащий о гряда № 100 о событиях лета 1942 г. рассказал Моримуре
следующее:
«Официально эта операция носила название “Летние маневры 6-го отделения от-
ряда 100”. Сотрудники отряда взяли тогда с собой 12 килограммов бактерий сапа Судя
по тому, что группа по возвращении доложила: “Эксперимент проведен, все 12 кило-
граммов были посеяны в реку”. О том, что вследствие этого произошло в нижнем те-
чении реки, рядовым служащим отряда известно не было. Подобного рода экспери-
менты — и более крупные, и более мелкие по масштабу — проводились “отрядом 100”
постоянно».
Отрядом № 100 по указанию штаба Квантунской армии систематически направ-
лялись на границы СССР бактериологические отряды, производившие на протяже-
нии ряда лет в диверсионных целях заражение пограничных водоемов в частности рек
в районе Трехречья. Диверсии планировалось осуществлять и в зимнее время. Тот же
Хиразакура Дзенсаку 6 декабря 1949 г. на допросе во время предварительного следствия
показал следующее:
«Признаю себя виновным также и в том, что во время нахождения в Северо-Хин-
ганской провинции я по приказанию генерал -майора Вакамацу закупил скот (10 те-
лят) для использования его в опытах, проводимых ранней весной 1945 года в районе
реки Южный Хангол. От участника этих опытов майора Ямагучи мне известно, что во
время опытов, именуемых «зимними маневрами», проверялось действие вируса чумы
рогатого скота и овечьей оспы в зимних условиях путем их распыления по снегу и по
разбросанному по нему корму.
Опыты проводились в условиях, в которых должны были осуществляться бакте-
риологические диверсии против Монгольской Наро дной Республики, ибо известно,
что скот в Монголии в зимнее время питается подножным кормом».
Фактически это была необъявленная бактериологическая война против СССР.
Бактериологическая война против Китая. Осуществлялась как в ферме попыток
масштабного применения БО, так и диверсионными способами. Документально ус-
тановлено, что масштабное применение БО японской армией в боевых действиях в
Китае осуществлялось уже в 1940 г.
Летом 1940 г. специальная бактериологическая экспедиция, возглавляемая началь-
ником отряда № 731, генералом Исией, была направлена в район бо< вых действий в
Центральный Китай. В районе Нимбо самолеты отряда № 731 произвели заражение
территории противника чумой с помощью чумных блох, в результате чего там вспых-
нула эпидемия чумы. Обвиняемый Карасава Томио, допрошенный 6 декабря, во вре
мя предварительного следствия в Хабаровске показал:
«...Во второй половине 1940 года от моего непосредственного начальника майора
Судзуки я получил приказание изготовить 70 кг бактерий брюшного тифа и 50 кг бак-
терий холеры. При этом майор Судзуки мне пояснил, что распоряжение об изготов-
лении бактерий он получил от начальника отряда генерала Исии, который готовился
для осуществления специальной экспедиции отряда по применению бактерий против
китайской армии... Мною это распоряжение было выполнено. Одновременно, со слов
сотрудников 2-го отдела, мне было известно, что для экспедиции генерала Исии 2-м
отделом было выращено 5 кг блох, зараженных чумой, как распространителей этой
инфекции В сентябре месяце 1940 года генерал Исии с группой других офицеров от-
ряда выехали в гор. Ханькоу и возвратились обратно в декабре 1940 года. Офицеры,
выезжавшие с генералом Исии, по возвращении в отряд рассказывали, что примене-
ние блох, зараженных чумой, дало положительные результаты. В результате распро-
168
Часть 1. Краткая история применения биологических средств...
странения блох вспыхнула эпидемия чумы. Один из участников этой экспедиции, май-
ор Нодзаки, в качестве доказательства показал мне китайскую газету, где была поме-
щена статья, в которой указывалось, что в районе города Нимбо вспыхнула эпидемия
чумы. Далее автор статьи делал правильный вывод о том, что виновниками этой
вспышки чумы являются японцы, так как очевидцы видели японский самолет, проле-
тавший в этом районе и что-то сбрасывавший на небольшой высоте. Указанную ста-
тью я читал лично сам».
Обвиняемый подполковник медицинской службы Ниси Тосихидэ показал 6 де-
кабря во время предварительного следствия, что он лично видел в отряде № 731 сек-
ретный документальный кинофильм, отображавший боевые действия японского бак-
териологического отряда против китайских войск:
«...На экране были показаны следующие кадры: к самолетам прикрепляют специ-
альные сосуды, причем в пояснении говорилось, что в этих сосудах помещены блохи,
зараженные чумой... Самолет снят в воздухе над расположением противника. На зем-
ле видно какое-то движение китайских войск и населенный пункт.
Самолет возвращается на аэродром. Следует надпись "операция закончена”. Из
самолета выходят Исии и Икари. Следует надпись “результаты”. На экране показана
китайская газета и японский перевод. В китайской газете написано, что в районе Ним-
бо внезапно вспыхнула сильная эпидемия чумы».
Всего в Нимбо тогда чумой заболело 99 человек, из них 98 умерли. Жители города
видели среди белого дня японские самолеты, которые низко кружились над городом
и сбрасывали какие-то предметы. После налета в одной семье даже обнаружили блох
во дворе на поверхности воды в аквариуме.
Помимо приведенных выше показаний обвиняемых Карасава и Ниси, факт пе-
реброски специальной экспедиции в район боевых действий в Центральном Китае
подтверждается обнаруженными в архивах японской Квантунской армии документа-
ми. В частности, был обнаружен приказ бывшего командующего японской Квантун-
ской армией генерала Умедзу от 25 июля 1940 г. № 659-Хэй, которым начальнику по-
левой железной дороги Квантунской армии предписывалось произвести переброску
в Центральный Китай группы сотрудников отряда № 731 и специального особо сек-
ретного груза. Советскими следователями был найден, изданный во исполнение это-
го приказа, приказ начальника полевой железной дороги Квантунской армии гене-
рал-лейтенанта Кусаба от 26 июля 1940 г. за № 178. В нем также была подчеркнута
особая секретность груза, ввиду чего предлагалось не вписывать в график его наиме-
нования, и был указан маршрут станция Пинфань — Харбин — Мукден — Шаньхай-
гуань и Тяньцзин.
В 1941 г. состоялась экспедиция отряда № 731 в Центральный Китай в район города
Чандэ. Во время этой экспедиции японские самолеты произвели заражение местно-
сти чумными блохами (см. ниже «Чума от дьявола в Чандэ»).
В 1942 г., в момент отступления японских войск, на одном из участков боевых дей-
ствий в Центральном Китае отрядом № 731 была организована еще одна экспеди-
ция. О ее подготовке обвиняемый Карасава во время предварительного следствия
показал следующее:
«...Экспедиция под руководством генерала Исии против китайских войск состоя-
лась в середине 1942 года. ...Перед этой экспедицией, для ее осуществления, под моим
руководством, по распоряжению того же майора Судзуки было изготовлено 130 кг
бактерий паратифа и сибирской язвы. Насколько мне известно, в этой экспедиции
использовались и блохи как распространители эпидемий... Для проведения экспеди-
1.8. Крах отряда № 731
169
ции генерал Исии с группой выезжал в Центральный Китай, где в это время японские
войска отступали. Используя отступление войск, участники экспедиции распростра-
няли бактерии на оставляемой территории д ля поражения эпидемиями наступающих
китайских войск».
Показания обвиняемого Карасава во время предварительного следствия подтвер-
дил другой обвиняемый, Кавасима Киоси:
«...В июле 1942 года, после предварительной подготовки, экспедиция нескольки-
ми партиями направилась в Центральный Китай... Способ применения бактериоло-
гического оружия в этом случае являлся наземным, и заражение территории произво-
дилось по принципу диверсионных действий...
...Наступающие китайские войска вступали в зараженную зону и подвергались дей-
ствию бактериологического оружия».
В этой операции, как подтвердил во время предварительного следствия Мисина
Такаюки, бывший начальник информационно-разведывательного отдела штаба 13-й
японской армии, участвовали сотрудники бактериологического отряда «Эй».
А вот что рассказал 28 декабря 1949 г. трибуналу о технике бактериологических
диверсий в этой операции свидетель Фуручи, санитар филиала № 643:
«Вопрос: Чем занималась экспедиция в Центральном Китае?
Ответ: Работа экспедиции, в которой я участвовал, заключалась в следующем: это
была бактериологическая атака путем заражения водоемов, колодцев, построек бак-
териями тифа и паратифа. Отряд 731 направил для этой экспедиции указанные бакте-
рии, производившиеся в массовом количестве в 4-м отделе этого отряда. Бактерии
были помешены в бутылки из-под пептона. Эти склянки помещались в ящики, на ко-
торых было написано “Водоснабжение”. Эти ящики на самолете были направлены в
Нанкин.
Вопрос: Что же вы делали с содержимым этих ящиков?
Ответ: По прибытии в нанкинский отряд часть бактерий, находящихся в склян-
ках, мы помещали в металлические фляги, какие обычно применяются для питьевой
воды, а остальную часть оставляли в склянках. Все эти фляги вместе со склянками
складывали в ящики, затем самолетами направляли их на то место, где предполагалось
делать атаки. Атаки производились следующим образом: фляги и склянки бросали в
колодцы, болота, в дома жителей поселков. Часть пептоновых склянок была исполь-
зована для размножения бактерий на специальном бульоне. Состава этого бульона я
не помню.
Вопрос: Чем лично вы занимались во время этих диверсионных актов"5
Ответ: Я участвовал в разбрасывании фляг, наполненных бактериями, в колодцы,
болота, дома мирных жителей.
В то время в этом месте были два лагеря китайских военнопленных общей числен-
ностью около трех тысяч человек. Были приготовлены три тысячи специальных було-
чек; в приготовлении булочек участвовали члены экспедиции. Через некоторое время
эти булочки с помощью шприца заражались бактериями.
Вопрос: Кто заразил эти булочки при помощи шприца?
Ответ: Это делали сотрудники 1-го отделения 1-го отдела, участвовавшие в экспе-
диции.
Вопрос: В частности, вы сами принимали в этом участие?
Ответ: Да, принимал участие.
Вопрос: Какими бактериями были заражены эти три тысячи булочек?
Ответ: Были применены бактерии тифа и паратифа.
Вопрос: Чю дальше сделали с этими тремя тысячами человек?
170
Часть 1. Краткая история применения биологических средств...
Ответ: После того как булочки были заражены бактериями, их направляли в лаге-
ря, где военнопленным китайцам эти булочки раздал переводчик Касуга, владевший
китайским языком. Отом, что эти булочки были переданы китайцам, мне известно хотя
бы из того, что я видел фотокарточку, где был изображен китаец с булочкой в руках.
Вопрос: Значит, когда китайских военнопленных кормили булочками, зараженны-
ми паратифом, это одновременно фотографировали как акт благодеяния?
Ответ: Так точно.
Вопрос: Как потом поступили с китайскими военнопленными, после того как их
покормили булочками, зараженными бактериями?
Ответ: Их всех распустили из лагерей с тем, чтобы вызвать эпидемию тифа и пара-
тифа».
Бактериологические нападения продолжались и в дальнейшем, о чем, в частно-
сти, свидетельствуют показания подсудимого Кадзицука Рюдзи, данные им на пред-
варительном следствии 23 октября 1949 г. (см. выше). Их осуществляли практически
везде, где японские войска вступали в соприкосновение с противником.
Военнослужащий фармацевтической службы японской армии Тояма сообщил
Моримуре обстоятельства, при которых в разгар военной операции при Сянгуе в 1944 г.
на юге Китая среди личного состава японской армии вспыхнула эпидемия холеры. От
своего друга, подпоручика Сугиты, Тояма узнал, что по приказу генерального штаба
в верховьях Янцзыцзян было сброшено большое количество бомб с бактериями хо-
леры. Трудно установить, какой урон нанесен в результате этого китайской армии, что
же касается японцев, то лишь в госпитале на берегу реки Янцзыцзян, где работал То-
яма, ежедневно умирало более десяти человек.
Способы бактериологических диверсий Хироси Акияма (1958) описывал следую-
щим образом:
«Применялись и другие способы заражения населения. Чумные бациллы помеща-
ли в пробирки, которые затем бросали в колодцы и водоемы. На обочинах дорог “те-
ряли” автоматические ручки, в колпачки которых были помещены возбудители чумы.
Переодевшись в платье китайских крестьян, члены диверсионных групп ходили по
деревням и раздавали детям сладкие пирожки с “начинкой”, то есть зараженные чу-
мой. Особенно страшным является заражение колодцев. В Маньчжурии колодцы яв-
ляются основным источником снабжения водой. В некоторых ее районах лишить на-
селение колодцев — все равно, что вынести ему смертный приговор. Все деревни, где
вспыхивала эпидемия чумы, обычно сжигались дотла. В некоторых случаях вирулент-
ность чумных бацилл была слабой, и болезнь принимала затяжной характер: опухоль
лимфатических узлов опадала, и наступало как бы полное выздоровление, но через
двадцать с небольшим дней болезнь вспыхивала с новой силой.
...Все диверсии планировались с таким расчетом, чтобы вспышки чумы выгляде-
ли естественно, и чтобы ни у кого не возникло и мысли о возможности диверсии. Этим
же, вероятно, можно объяснить и то, что операция в Нунане у жителей не вызвала
подозрений».
Хироси Акияма (1958) подробно описал бактериологическую диверсию, в кото-
рой участвовал сам 1 июля 1945 г.:
«...Пришли в деревню около полудня. В ней насчитывалось не больше двадцати
дворов. ...Все жители этой деревни имеют специальное разрешение штаба армии на
право проживания здесь. Ведь эта деревня находится в каких-то 8-10 километрах от
нашего отряда, — объяснил Сагава тем, кто стоял неподалеку от него.
Старики, месившие под навесом глину, босоногие ребятишки, выглядывавшие из
домов женщины — все с любопытством рассматривали нас. Когда мы расположились
1.8. Крах отряда № 731
171
в тени и развернули свои завтраки, ребятишки, до сих пор застенчиво державшиеся
поодаль, подошли ближе. Некоторые из нас пытались завязать беседу на их родном
языке и писали китайские слова катаканой. Сагава, как и следовало ожидать, поко-
рил всех ребят знанием их родного языка. Достав из вещевого мешка сладкие пирож-
ки с бобовой начинкой, он оделил ими каждого. Со стороны это выглядело бы довольно
з рогательно, если бы не странное выражение лица Сагава: оно выражало жестокость
и какую-то напряженность. Но тогда я не придал этому почти никакого значения, хотя
и должен был знать, что угощение ребят — один из приемов работы диверсионных
групп, как это показывали нам в одном учебном кинофильме.
...Прошло несколько дней. Однажды утром мы, придя на службу, заметили необыч-
ное оживление в секции. По коридору взад и вперед сновали озабоченные вольнона-
емные. По отрывочным словам, доносившимся из лаборатории, я понял, что в дереь-
не, где мы были, вспыхнула чума. ...Работники лаборатории потом по очереди должны
были ездить в эту деревню, чтобы наблюдать за тем, как протекает эпидемия, и на месте
испытывать эффективность лечебных препаратов. На этот раз в качестве ассистента
послали и меня. Одетые с целью самозащиты в специальные резиновые костюмы, мы
под палящими лучами яркого солнца чувствовали себя словно в бане На улице не было
ни одного жителя. Деревня строго охранялась солдатами нашего отряда. Дома, где были
больные чумой, с целью предупреждения распространения инфекции мухами были
обнесены мелкой белой металлической сеткой, похожей на сетку от москитов.
...Все дети, которых Сагава оделил сладкими пирожками, заболели легочной фор-
мой чумы, и некоторые уже умерли. Один за другим в деревне появлялись новые боль-
ные. Значит, тучи мух и полчиша блох уже разнесли чумные бациллы по всей деревне.
Когда я вместе с военным врачом Цудзицука вошел в один из домов, обнесенный ме-
таллической сеткой, перед нами открылась такая страшная картина, что даже я, ви-
давший уже немало на многочисленных опытах и фотоснимках, от ужаса то и дело
закрывал глаза. Когда мне казалось, что среди трупов я узнаю какого-нибудь мальчу-
гана из тех, которые в тот день так беззаботно играли среди нас, я готов был громко
разрыдаться.
...Моя обязанность заключалась в том, чтобы помогать при вскрытии трупов с
целью извлечения легких, селезенки и других органов для изготовления препаратов,
необходимых при проведении исследований. У меня не было никакого желания вни-
кать в порученную мне работу, и я выполнял ее механически, по привычке. Сквозь
стены до нас доносились жалобные стоны умирающих, а за проволочной сеткой слы-
шалось громкое жужжание мух, привлеченных сюда запахом человеческих внутрен-
ностей.
В зачумленной деревне мы провели не более часа, но у меня от нервного напряже-
ния кружилась голова, и подкашивались ноги. Нервы мои были напряжены до преде-
ла. Я чувствовал себя совершенно разбитым, когда вернулся в расположение отряда.
Примерно через неделю деревня была сожжена. Возможно, не все жители умерли от
чумы, но так как опыт следовало держать в строгой тайне, то говорили, что все остав-
шиеся в живых были расстреляны. Судя по кинофильму, о котором я уже упоминал,
подобные жуткие опыты на людях в широких масштабах проводились несколько раз
до и после этого случая».
Хироси Акияма и здесь точен. Еще во время чумы в Маньчжурии 1910—1911 гг.
было установлено, что первичная легочная чума («чумная бронхопневмония») у чело-
века возникает посредством первичного поражения Y. pestis миндалин и слизистой
глотки. Причем чумная инфекция транспортируется в легкие не через воздух, а при
помощи крови, т. е. гематогенно (Кулеша Г. С., 1912, 1924). Кишечная форма чумы,
казалось бы, вполне естественная при таком пути проникновения возбудителя чумы
172
Часть 1. Краткая история применения биологических средств...
в организм человека, большинством авторов в качестве самостоятельной клинической
формы болезни не признается (см. в кн. Шуваловой Е. П. с соавт., 2002).
Навыки, приобретенные в отряде, пригодились некоторым его бывшим служащим
уже после войны. 26 января 1948 г. Садатоси Хирасава, бывший сотрудник отряда
№731, выдав себя за представителя некой организации по борьбе с эпидемиями, ко-
торой необходимо было провести вакцинацию служащих Имперского банка в Токио,
обманом принудил 16 банковских служащих принять цианистый калий. Он сам им
сделал инъекции яда, предварительно протерев кожу жертвы ватой со спиртом. После
того как 12 из них умерли сразу, бандит спокойно забрал деньги (164 тыс. иен налич-
ными и чеки на сумму 17 тыс. иен) и скрылся. Все сотрудники отряда, жившие в то
время в Японии, попали под подозрение. Во время допроса одного их них следова-
тель, взглянув на него и на фоторобот, воскликнул: «Ого! Как похож на преступника
из Имперского банка! Просто одно лицо!».
Чума от дьявола в Чандэ. Эта операция с применением БО спланирована на осно-
ве представлений, отводящих крысам и блохам основную роль в поддержании эпиде-
мии чумы в городах. Непосредственно руководил операцией полковник Акира Оота.
Цель операции — нарушить коммуникации китайских войск, важным пунктом кото-
рых являлся Чандэ. По замыслу Исии сброшенные с самолетов зараженные чумой бло-
хи должны были вызвать сначала чумную эпизоотию среди городских грызунов, и от
них чума должна массово, посредством уже заразившихся блох, живущих на местных
грызунах, распространиться среди населения города в бубонной форме. У части ки-
тайцев болезнь проявит себя во вторично-легочной форме, которая, в свою очередь,
даст случаи первичной легочной чумы, образующие, как правило, далеко тянущиеся
самостоятельные цепочки этой клинической формы болезни. Все вместе они приве-
дут к чумной бойне, наподобие той, которая произошла в 1910-1911 гг. в Маньчжу-
рии. Замысел операции был основан на надежной научной литературе. Для надеж-
ности самого эксперимента Исии и Оота выбрали в качестве целей для БО густона-
селенные бедные кварталы этого города. Даже время нападения — поздняя осень,
выбрано ими потому, что именно тогда домашние грызуны заканчивают свое переме-
щение с полей в дома людей на зимовку и скапливаются там в больших количествах.
Около 5 ч туманным утром 4 ноября 1941 г. одиночный японский самолет, проле-
тавший на небольшой высоте над китайским городом Чандэ, сбросил вместо бомб
зерна пшеницы и риса, кусочки бумаги, ватную набивку и некоторые не установлен-
ные мелкие объекты. Эти материалы упали главным образом в Цзиясяне на улице
Гуанмяо (район «А» по карте) и вокруг района восточных ворот (район «Б» по карте)
города. После отбоя воздушной тревоги (в 17 ч), образцы рисовых зерен были собра-
ны и посланы полицией в госпиталь Гуандэ для микробиологического исследования
(рис. 1.25).
До 11 ноября ничего не случалось. Но через 7 дней после «воздушного инциден-
та» первый случай чумы привлек внимание медицинской службы города. Заболевшей
была девочка 11 лет, проживающая на улице Гуанмяо (район «А»), которая стала жа-
ловаться на жар (тепература 39,1 °C) утром этого дня. Она поступила в госпиталь Гу-
андэ. За исключением присутствия в крови микробов, морфологически похожих на
Pasteurella pestis (так тогда называли К pestis), других положительных клинических
данных не было обнаружено. Девочка умерла 13 ноября, и только вскрытие дало очень
веские подозрения на наличие в городе чумы. В 3-х мазках из внутренних органов, так
же как и в мазке крови, были обнаружены биполярные палочки, морфологически
похожие на Р. pestis.
1.8. Крах отряда Ns 731
173
КАРТА УЕЗДНОГО ГОРОДА ЧАНДЭ
Рис. 1.25. Схема китайского города Чандэ с обозначением районов, подвергшихся
бактериологическому нападению с воздуха 4 ноября 1941 г. (из «Доклада Междуна-
родной научной комиссии по расследованию фактов бактериологической войны
в Корее и Китае», Пекин, 1952)
12 ноября внимание врачей привлекли два других случая. Оба больные были с
высокой температурой и с увеличением лимфатических узлов в паху, похожих на чум-
ные бубоны. При исследовании мазков из пунктатов бубонов, в обоих случаях уста-
новлено присутствие Р. pestis. Один больной умер 13-го, другой — 14 ноября. Они
проживали в районе восточных ворот (район «Б»),
13 ноября неожиданно умер другой больной. При опросе родственников устано-
вили, что у него с 11 ноября держалась высокая температура. Была произведена пунк-
ция печени. Исследование мазка обнаружило присутствие микроорганизмов, похо-
жих на чумные бациллы. Больной проживал по улице Чанцин, в районе восточных
ворот (район «Б»).
Следующий больной поступил в госпиталь 19 ноября с высокой температурой,
бредом, бубонами, последние замечены были 18 ноября. Больной умер в госпитале в
день поступления. Вскрытие не позволило обнаружить явные патологические изме-
нения, характерные для чумы.
Последний больной, мужчина 28 лет, проживавший по улице Гуанмяо (район «А»),
23 ноября слег в постель с высокой температурой, слабостью и наличием бубонов, на
следующий день он умер. Он был обследован доктором Чэнь Вэнь Гуем (специалист
по чуме, получивший подготовку в Индии), заведующим экспертной группой школы
по подготовке военных медработников, который приехал в район Гуйяна с обследо-
вательским отрядом. Чэнь Вэнь Гуй установил при посмертном вскрытии, что болезнь
является бубонной чумой. Этот диагноз был подтвержден заражением животных и
выделением культуры Р. pestis (сведения о больных приведены в табл. 1.2).
Так как Чандэ ранее никогда не был поражен чумой, то одновременное возник-
новение таких заболеваний в районах города, куда были сброшены какие-то предме-
ты с японского самолета, вызвало у китайских властей подозрение о применении
японской армией БО.
Развитие искусственной эпидемии чумы в городе Чандэ, подвергшегося бактериологическому нападению*
Таблица 1.2
Больной (возраст и пол) Место возникновения Дата заболевания (смерти) Признаки болезни Диагноз Осмотрены
Цай Тао Эр (11 лет, жен.) Улица Гуанмяо 11 (13) ноября 1941 г. Высокая температура: мазок крови (окраска по Райту), морфологически положителен на Р. pestis. Вскрытие: селезенка и печень увеличены; мазок (окраска по Райту) морфологически положителен на Р. pestis ? чума Д-ром Тань Сюэ Хуа (госпиталь в г. Чандэ). Прозектор Цян Бао Кан
Цай Юйчжен (27 лет, жен.) Район Восточных ворот 11(13) Высокая температура, осмотрена мертвой. Мазок из пунктата крови (окраска по Райту) морфологически положителен на Р. pestis ? чума Д-ром Кентом
Не Шу Шэн (58 лет, муж.) Район Восточных ворот 12(13) Высокая температура. Увеличение паховых желез, мазок (окраска по Райту) морфологически положителен на Р. pestis ? чума Цян Бао Кан
Сюй Лао Сан (25 лет, муж.) Район Восточных ворот 12(14) Высокая температура. Увеличение паховых желез, мазок (окраска по Райту) морфологически положителен на Р. pestis ? чума Д-ром Фан Дэ Чжэном, д-ром Тань Сюэ Хуа
Ху Чжун Фа (? лет, муж.) Утица Гуанмяо 18(19) Высокая температура, бред; увеличенные паховые железы. Данные вскрытия: мазок печени (окраска по Граму — отрицателен; посев — отрицателен) ?? чума Д-ром Ли Цин Цзэ. Прозекторы: Лю Пэй, Сюэ Цингой
Гун Цао Шэн (28 лет, муж.) Район А 23 (24) Высокая температура, слабость, правые паховые железы увеличены и болезненны. Вскрытие: селезенка увеличена, на поверхности геморрагии, геморрагия печени и кишок. Выпот в плевре и в перикарде. Мазки крови сердца, правой ингвинальной железы, печени. Окраска по Граму и карболотиониновой синькой положительны на Р. pestis, что подтверждено посевом и заражением морской свинки чума бубонная Бактериологический посев и введение культуры Р. pestis морской свинке проводились Чэн Вэнь Гуй
♦Доклад Международной научной комиссии по расследованию фактов бактериологической войны в Корее и Китае, Пекин, 1952.
Часть 1. Краткая история применения биологических средств...
1.8. Крах отряда № 731
175
Эпидемиологическое расследование показало следующее. Вспышка чумы в Чан-
дэ не могла возникнуть вследствие заноса инфекции из соседних эпизоотических
районов. Ближайшим к нему зараженным чумой городом тогда являлся Чжусянь, рас-
положенный в южной провинции Чжэцзян. Он находится примерно на расстоянии
2 тыс. км по суше и по речным коммуникациям. Кроме того, все заболевшие в Чандэ
являлись местными жителями и не покидали город и его окрестности.
Блох, зараженных возбудителем чумы, не нашли. Этому способствовало несколько
обстоятельств. Во-первых, до появления чумы в городе их и не искали. Во-вторых,
после налета на город прошло еще почти 12 ч, прежде чем была отменена воздушная
тревога. Блохи могли за это время покинуть тряпье и зерна и скрыться в близлежащих
домах с более подходящей для них температурой и влажностью. Это расселение блох
могло закончиться задолго до того, как зерна и тряпье были подметены и сожжены
после окончания воздушной тревоги. Потому местные жители, уже давно привычные
к насекомым, не обратили внимания на наличие насекомых среди сброшенных с са-
молета предметов. Для того чтобы привлечь крыс, вместе с блохами японцы сброси-
ли зерна риса и пшеницы. Тем самым они надеялись вызвать среди крыс чумную эпи-
зоотию и увеличить количество инфицированных возбудителем чумы блох среди
местного населения. Но этого не случилось, так как все заболевшие чумой жители
были обнаружены в течение 15 суток после «воздушного инцидента». Обычно забо-
левания чумой среди людей в Китае начинаются, по крайней мере, двумя неделями
позднее крысиной эпизоотии, которая также требует некоторого времени для разви-
тия (приблизительно около 2 недель).
Если зараженные блохи были сброшены с самолета, то, что же помешало им выз-
вать эпизоотию среди местных крыс? Во-первых, в естественных условиях крысы ин-
фицируются из неизвестных сегодня источников возбудителя чумы, не имеющих от-
ношения к их эктопаразитам. Поэтому наличие блох, зараженных чумой, еше не
определяет возможность развития чумной эпизоотии. Во-вторых, для инфицирова-
ния людей возбудителем чумы японцы вынуждены были использовать человеческую
блоху (Pulex irritans), а они нападают на грызунов лишь в условиях эксперимента.
Наиболее вероятно, что зараженные блохи, сброшенные с зерном и другими пред-
метами, непосредственно нападали на людей и только таким образом они вызвали
небольшую вспышку чумы. У большинства больных описываемой вспышки инкуба-
ционный период продолжался от 7 до 8 сут. Больные подверглись укусам зараженных
чумой блох, вероятно, 11 и 12 ноября, т. е. спустя семь и восемь дней после «воздуш-
ного инцидента».
Первые два случая заболевания людей чумой возникли 11 -го числа, т. е. через 7 сут.
после налета. Третий и четвертый — 12-го числа, т. е. через 8 сут после налета. Пятый
погибший, диагноз болезни у которого вызывал сомнение, заболел 18 ноября. Шес-
той больной, у которого была вне всяких мнений бубонная чума, работал в близлежа-
щей деревне. Он пришел в город и в одном из зараженных районов прожил 5 дней: с
19 ноября по 23 ноября. Заболел он через 15 сут после «воздушного инцидента». Эти
сроки укладываются в продолжительность жизни блох, инфицированных возбудите-
лем чумы. Однако конечный результат всей операции вряд ли мог удовлетворить ам-
биции Исии.
Официальные коммунистические китайские историки, заинтересованные в «сгу-
щении красок», указывали на то, что бактериологическому нападению со стороны
японцев было подвергнуто 11 уездных городов: 4 в провинции Чжэцзян, по 2 в про-
винциях Хэбэй и Хэнань и по одному в провинциях Шаньси, Хунань и Шаньдун.
176
Часть 1. Краткая история применения биологических средств...
В 1952 г. китайцы исчисляли количество жертв от искусственно вызванной чумы с
1940 по 1944 г. приблизительно в 700 человек (Доклад Международной научной ко-
миссии по расследованию фактов бактериологической войны в Корее и Китае. —
Пекин, 1952; сейчас, конечно, журналистами называются другие цифры, какие угод-
но, и 200 тыс.., и более, по обстоятельствам). Оно оказалось даже меньше количества
загубленных «бревен», и только в два раза превышает количество самих японцев, по-
гибших в экспериментах по созданию БО (около 300 человек).
Действия советской разведки. Харбин рассматривался руководителями Советской
России как крупнейший центр белой эмиграции, набитый контрреволюционными
организациями, намеренными пересмотреть результаты Гражданской войны Здесь
же находилась Харбинская военная миссия Японии — крупнейший на материке центр
японской разведки. Активно действовали спецслужбы западных государств, рас-
сматривавших разные варианты реализации известного заявления Черчилля, что
«с большевиками надо что-то делать». Поэтому совершенно естественно, что уже в
1920-х гг Манчьжурия была покрыта густой агентурной сетью обеих советских разве-
док — военной (Разведупр) и политической (иностранный отдел ОГПУ).
Со средины 1920-х гг. советская разведка располагала японскими и китайскими
шифрами и через свою агентуру контролировала линии почтовой связи, используе-
мые японской секретной почтой. После начала японской агрессии в Маньчжурии в
1931 г. к уже имевшейся в этом регионе агентуре обеих разведок добавляются новые
резидентуры и агентурные сети. В 1932 г. к ним присоединилась «третья сила» — пар-
тийная разведка Коминтерна (отдел международных связей), до этого успешно дей-
ствовавшая в Европе и Китае. Идет не только наблюдение за усилением и перемеще-
нием частей Квантунской армии, но и проникновение в структуру японских военных
органов на азиатском континенте. О численности и вооружении японских войск в
Маньчжурии Кремль знал все. Через собственные источники в японских военных
кругах (псевдонимы «Кротов», «Сук», Простак», «Пауль», «Тур» и др. агентов, остав-
шихся не разоблаченными японской контрразведкой) в СССР поступала секретная
информация стратегического значения, например, мобилизационные планы япон-
ской армии и др. (Горбунов Е. А., 2002). Однако о том, какой информацией о деятель-
ности отряда № 731 располагали в Москве и с какого времени, пока можно судить
только по косвенным сведениям.
По свидетельству Г. Г. Пермякова, бывшего сотрудника Хабаровского краево-
го центра МВД и переводчика на Хабаровском процессе 1949 г. (см. ниже), отряд
№ 731 попал в поле зрения советской военной разведки с самого начала его строи-
тельства, после того как японцы проложили новую дорогу от Харбина до захудало-
го поселка Пинфань. Есть основания считать, что гибель во время репрессий конца
1930-х гг. И. М. Великанова (1898—1938) — начальника Биотехнического института
РККА (Перхушково, Московская область), связана с попыткой советской военной
разведки выйти в 1934 г. на программу создания японского БО (6Oj tee подробно в книге
Супотницкого М. В., Супотнипкой Н. С., 2006).
Начальник Главного военно-санитарного управления Красной армии Е. И. Смир-
нов, был информирован о возможности применения японцами бактериологическо-
го и химического оружия под Халхин-Голом, а сами японцы понесли большие поте-
ри при осуществлении этой акции.
В весьма достоверных мемуарах Хироси Акиямы (1958) упоминается о похище-
нии в июне 1945 г. служащего отряда «иностранной разведкой». Страх перед «иност-
ранной разведкой» часто проглядывает со страниц его книги (в ней есть даже глава «За
1.8. Крах отряда Ns 731 177
нами охотятся»). Любопытно и то, что в средине июля 1945 г. по отряду ходил слух о
требовании СССР расследовать его деятельность, что связывали с какими-то матери-
алами отряда, захваченными «Советами» в побежденной Германии. Это еше одна ин-
тересная тема для исследований военных историков.
Конец «Кухни дьявола». На 1 января 1945 г. в отряде № 731 было 3559 солдат, офи-
церов и гражданских лиц. По сведениям Моримуры, вновь занявший в марте 1945 г.
пост начальника отряда Сиро Исии изменил шифр отряда с № 731 на № 25202, а в мае
на совещании руководства отряда он отдал распоряжение «об увеличении производ-
ства», в котором говорилось: «Война между Японией и СССР неизбежна. Теперь от-
ряд должен мобилизовать все силы и в короткий срок увеличить производство бакте-
рий, блох и крыс». Иными словами, стадия экспериментов на людях закончилась,
теперь начинается реальная бактериологическая война и нужно наращивать произ-
водство БО в ожидании «дня X».
Хироси Акияма (1958) писал, что к войне с СССР в отряде начали готовиться уже
в июле. Рядом с аэродромом готовили капониры для самолетов. Для этого было мо-
билизовано около тысячи китайских кули, да и сами сотрудники иногда целыми днями
были заняты рытьем окопов и других укрытий.
Бывший служащий отряда рассказал Моримуре следующее:
«Приказ значительно увеличить в течение ближайших двух месяцев производство,
прежде всего бактерий чумы и тифа для заражения колодцев и водоемов, холеры и
сибирской язвы для заражения рек и пастбищ, был отдан 10 мая. В результате увеличе-
ния числа сотрудников группы Карасавы. работавшей на “фабрике по производству
бактерий”, и перехода на 24-часовой производственный цикл, одних только бактерий
чумы было произведено не менее 20 килограммов, а если учесть и ранее произведен-
ные, и сухие бактерии, то, я думаю, общая масса составляла 100 килограммов».
Хиразакура Дзенсаку 6 декабря 1949 г. на судебном заседании показал следующее:
«Одновременно с указанной работой я по приказанию генерал-майора Вакамацу
летом 1945 года закупил у населения Северо-Хинганской провинции 500 овец, 100 го-
лов рогатого скота и 90 лошадей на отпущенные для этой цели 80 тыс. иен От гене-
рал-майора Вакамацу мне было известно, что в случае войны с Советским Союзом этот
скот будет заражен сибирской язвой, сапом, чумой рогатого скота и овечьей оспой и с
диверсионной целью оставлен в тылу советских войск, чтобы вызвать вспышку ост-
роинфекционных заболеваний. Мне было известно, что для этой цели в места нахож-
дения закупленного мною скота будет на самолетах доставлено необходимое количе-
ство вышеперечисленных бактерий, и скот будет заражен созданными диверсионными
группами».
В отряде, во всех его помещениях, даже в жилых, были установлены ограждения
высотой около метра, внутри которых круглосуточно велось размножение грызунов.
Отряду была поставлена задача: произвести 300 кг чумных блох, то есть около милли-
арда особей. В группе Танаки имелось 4,5 тыс. контейнеров для размножения блох,
которые могли обеспечить получение 100 млн особей в течение всего лишь несколь-
ких дней. В результате напряженной работы, к лету 1945 г. в отряде № 731 имелся зна-
чительный запас бактерий, включавший, помимо 100 кг бактерий чумы, большое ко-
личество бактерий тифа, холеры, дизентерии, сибирской язвы.
Хироси Акияма (1958) со своей должности так видел эти события:
«В середине июля часть личного состава отряда, примерно одна десятая каждого
отдела, во главе со вторым отделом переехала в Тунхуа. Ходили слухи, что и весь наш
отряд вскоре передислоцируется туда же. К части отряда, переведенной в Тунхуа, были
178
Часть 1. Краткая история применения биологических средств...
прикомандированы вольнонаемные Асабу из секции Такаги и Сато из учебного отде-
ла. Причину передислокации нам объяснили тем, что скоро могут начаться военные
действия, но тогда никто из нас еще не представлял, какая опасность нависла над
Японией. Работы стало значительно больше, и все чаще приходилось работать в лабо-
раториях всю ночь напролет. Из-за недосыпания и переутомления участились несча-
стные случаи при работе со смертоносными бактериями, и число инфекционных за-
болеваний среди личного состава возросло».
По свидетельствам, собранным Моримурой, руководство отряда имитировало
усиленную подготовку к бактериологической войне на глазах у советской разведки.
В русском квартале Харбина отряды японцев демонстративно ловили мышей и крыс,
подтверждая тем самым и без того уже давно ходившие здесь слухи, что на станции
Пинфань у них есть завод по производству БО. По замыслу японского командования
для Советского Союза эти сведения должны были быть предостережением, что Кван-
тунская армия усиленно готовится к бактериологической войне. Это, как они пола-
гали, должно вызвать настороженность у командования советских войск, концентри-
рующихся на границе с Маньчжурией, и тем самым отсрочить начало военных дей-
ствий. Но они ошиблись.
После воздушных налетов на железнодорожные узлы, военные объекты и аэродро-
мы в ночь с 9 на 10 августа советские войска неожиданно для японцев перешли в
наступление силами трех фронтов. На некоторых участках фронта за первый день
наступления они продвинулись на 50 километров. Командующий Квантунской арми-
ей Ямада потерял контроль над обстановкой на фронтах и ни о каком массированном
применении БО не могло быть и речи. Отряду № 731 было предложено действовать
«по собственному усмотрению», а проще говоря, «замести следы» и бежать.
В день начала войны, 9 августа 1945 г., небо над отрядом было затянуто плотными
облаками, почти весь день шел дождь. Ближе к полудню Хироси Акияма услышал
далекий гул мотора советского самолета-разведчика. Из-за сплошной облачности
самолета не было видно. Работы по эвакуации отряда начались в ночь с 10 на 11 авгу-
ста. В памяти бывших сотрудников отряда они запечатлелись по-разному. Их воспо-
минания, рассказанные Моримуре, сходны только в одном: «Это были ужасные дни,
похожие на кошмарный сон».
Моримура установил, что на совещании, собранном 10 августа, по вопросам, свя-
занным с эвакуацией отряда, резко столкнулись мнения Исии и генерал-майора Ки-
кути — начальника 1-го отдела.
План эвакуации, на котором настаивал Исии, включал следующие пункты:
1) главная задача — сохранение тайны отряда № 731;
2) для этой цели в филиалы отряда в Хайдаре, Линь-коу, Суньу, Муданьцзяне, на-
ходящиеся на пути наступления советских войск, уже посланы связные во главе с
подполковником Ниси (начальником учебного отряда) с приказом уничтожить все
материалы и другие вещественные доказательства, а всему личному составу покончить
жизнь самоубийством;
3) следует также приказать покончить жизнь самоубийством и всем членам семей
служащих отряда, проживающим в «деревне Того»;
4) находящихся в настоящее время в заключении подопытных всех до одного унич-
тожить, здания «блока ро» сровнять с землей, остальные сооружения отряда взорвать,
прибегнув к помощи саперов;
5) после этого личному составу, включая подростков-стажеров, организованно от-
ступить на юг, в Тунхуа.
1.8. крах отряда № 731
179
Рис. 1.26. Колба с цианистым
водородом. Химические гра-
наты такого типа применялись
всеми воюющими сторонами в
годы Первой мировой войны.
По R. Sidell et al. (1997)
Причиной резкого спора между Исией и Кикути были второй и третий пункты
плана. Генерал-майор Кикути настаивал на том, что, «поскольку в филиалах отряда
№ 731 много видны* исследователей, нужно принять меры к их спасению, а не за-
ставлять людей покончить жизнь самоубийством». Он в резкой форме потребовал
«принять меры для эвакуации членов семей сотруд-
ников отряда в Японию, которая должна проводит ь-
ся под личным руководством начальника отряда».
В итоге Исии уступил Кикути, однако судьба
пленников уже была решена ими без всяких колеба-
ний. К 9 августа заключенных в отряде было около
40 человек1. Моримуре удалось выяснить обстои-
те ъства их гибели. Ликвидацию «бревен» поручили
сотрудникам отряда № 516. Орудием убийства по-
допытных людей стал цианистый водород, т. е. си-
нильная кислота. Сотрудники отряда № 516 пере-
ходили от одной камеры к другой и бросали через
смотровые окошки «бьющиеся» гранаты с синиль-
ной кислотой. Практически одновременно с тем, как
сосуды разбивались о пол, бурно испарявшийся ци-
анистый водород наполнял камеры (рис. 1.26).
«Забрасывание колб в камеры продолжалось ми-
нут пятнадцать. Мы знали, что уничтожение подо-
пытных людей — первый шаг на пути подготовки
отряда № 731 к эвакуации. Всего мы, помнится, за-
бросили 9 колб, потому что были ведь и пустые каме-
ры, а кое-где одной колбой уничтожались 3—4 человека. Некоторые не умирали сразу,
они кричали и стучали в стальные двери камер, издавали страшное рычание, раздирали
себе грудь. Похоже, было, что перед нами обезумевшие гориллы в клетке», — расска-
зал Моримуре бывший служащий отряда, который своими глазами видел истребле-
ние подопытных. Сотрудники спецгруппы подходили и хладнокровно расстреливали
в упор из маузеров агонизирующих людей. Вместе с заключенными были расстреля-
ны и все переводчики-китайцы. О том, что происходило дальше, сообщил Моримуре
другой бывший служащий отряда:
«Убитых заключенных за ноги волокли в большую яму, вырытую около корпуса 7.
Когда трупы заполнили ее, их облили бензином и мазутом и подожгли... Помнится, это
было 11 числа после полудня. В печи, где обычно сжигали трупы, теперь сжигали пре-
параты, агар-агар, огромное количество документов и приборов. В яме трупы горели
плохо. Но поскольку была дорога каждая минута, обгоревшие тела кое-как забросали
землей. Из земли то здесь, то там виднелись руки и ноги, поэтому задачу сокрытия
преступления никак нельзя было считать выполненной. В связи с этим руководство
отряда отдало приказ “снова выкопать трупы и сжечь их полностью”. Служащие от-
ряда, выполняя эту работу, старались не смотреть на обезображенные трупы заключен-
ных и с трудом сдерживали приступы тошноты».
1 Один из исполнителей этого массового убийства старший ефрейтор Каваминами, на
допросе утверждал, что подопытных заключенных на тот момент было не 40 человек, а 500. См.
Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от 23.04,1949 г.
Сов. секретно. АП РФ.Ф. 3. Оп. 66. Д. 1065. Л. 11—21 (Кошкин А. А., 2011).
180
Часть 1. Краткая история применения биологических средств...
Затем поступила команда: «Кости немедленно собрать в мешки, вывезти и вы-
бросить». Пепел и кости были очень горячими, к ним нельзя было прикоснуться ру-
кой, сбор костей затянулся до самого вечера. Кости были сложены в десятки мешков,
вывезены на грузовых машинах и сброшены в реку Сунгари в разных местах. Вместе
с костями на дно реки пошли сотни наручников и канда лов, тысячи стеклянных со-
судов с препаратами частей человеческого тела.
Акияма сообщает другую дату завершения ликвидации «бревен» — ночь с 9 на
10 августа, детали расправы над подопытными людьми в его книге не отличаются от
приведенных Моримурой. В эту же ночь советские самолеты сбрасывали осветитель-
ные бомбы в районе дислокации отряда, но сам отряд не бомбили. По данным Акия-
мы, «блок ро» был взорван японцами в девять часов утра 10 августа 1945 г. (рис 1.27).
Рис. 1.27. Развалины отряда № 731. А. Руины котельной. Б. Бактериологические бомбы
Исии (реконструкция). В. Руины «блока ро». Г. Фильтры Исии. В настоящее время на
месте отряда находится музейный комплекс жертв преступлений японских военных.
Фотографии предоставлены А. Э. Пильниковым (Забайкальский край, Краснокаменск)
L 8. Крах отряда Ns 731
181
В ночь с 1G на 11 августа в окрестностях Харбина в пределах видимости с террито-
рии отряда № 731 был сброшен советский парашютный десант. В трудах советских
историков я не нашел упоминания об этом десанте, но Акияма (весьма достоверный,
когда пишет о деталях японской бактериологической программы) так описывает эти
события:
vOt осветительных бомб и ракет стало так светло, что мы ясно видели высокий
памятник павшим воинам, который находился от нас более чем в двадцати километ-
рах. Несмотря на моросивший дождь, были отчетливо видны спускавшиеся парашю-
ты, напоминавшие падающие хлопья снега. Воздушный бой становился все более
ожесточенным. Голубоватый свет осветительных бомб и ракет придавал нашим лицам
мертвенно-бледный оттенок. ...Через несколько часов поступило сообщение, что все
парашютисты противника уничтожены танкистами, оборонявшими Харбин. Услышав
об этом, все запрыгали от радости».
Бактериологические диверсии осуществлялись везде, где было это возможным в
сложившейся обстановке. Вот что показал 29 декабря на процессе в Хабаровске воль-
нонаемный отряда № 100, свидетель Кувабара:
«Государственный обвинитель: Где вы служили?
Кувабара: В филиале № 2630 отряда № 100.
Вопрос: Расскажите суду, что вам известно о факте заражения лошадей сапом.
Ответ: Это было 20 августа 1945 года. Я направился тогда в конюшню нашего фи-
лиала и там увилел шесть работников нашего филиала Это были сотрудники отряда
Кубота, Икеда, Яда, Кимура, Исии и Хасегава. В этих конюшнях имелось 60 лошадей,
которые содерж, лись при отряде. Я не успел подойти к этой группе, как меня предуп-
редили, что они произво 1ят заражение этих лошадей сапом путем введения бактерий
сапа в овес. Тогда я ушел обратно в лабораторию филиала. Вернувшись в лабораторию,
я увидел там пустые пробирки из-под культур сапа; затем я спросил у научного сотруд-
ника Кимура, производили ли они заражение лошадей именно этими бактериями. Он
подтвердил это и сказал, что лошадей заразили сапом.
Вопрос: Как потом поступили с лошадьми?
Ответ: Группа, производившая заражение, сломала изгороди и распустила лоша-
дей в разные стороны. Все лошади разбежались по ближайшим селениям и по разным
дорогам.
Вопрос: Не должны ли были зараженные сапом лошади служить источником воз-
никновения эпидемии сапа?
Ответ: Да.
Вопрос: Этот факт имел место 20 августа 1945 года?
Ответ: Да.
Вопрос: То есть после приказа о капитуляции японской армии?
Ответ Да.
Вопрос: Где были выращены бактерии сапа, которыми были заражены животные?
О1 вет: Для этого использовались бактерии сапа, выращенные в бактериологичес-
ком отделении отряда № 2630»
Прежде чем покинуть Дальний, японцы выдернули из журналов, хранящихся в
университетских и институтских библиотеках, все листы со статьями, которые могли
иметь какое-то отношение к бактериологической войне.
После отправления эшелона с личным составом, вольнонаемными и членами их
семей в направлении Харбина, Исии проследил за его движением с самолета и на этом
же самолете несколько раньше эшелона прибыл в Пусан. Из Пусана Исии отплыл в
Японию на специально зарезервированном для этого эсминце японских ВМС.
182
Часть 1. Краткая история применения биологических средств...
В Японии он руководил операцией по уничтожению препаратов, находившихся в Ла-
боратории профилактики эпидемических заболеваний японской армии в квартале
Вакамацу в Токио, в Императорском университете в Киото и в медицинском инсти-
туте города Канадзавы. Одновременно Исии организовал временную базу отряда
№ 731 на территории медицинского института в Канадзаве.
Попав в плен к американцам, Сиро Исии, Масадзи Китано и Вадзиро Вакамацу
не стали отрицать свою причастность к разработке БО. Наоборот, они поделились с
новыми хозяевами всем тем опытом, который накопили в результате своих экспе-
риментов на людях и тем самым сохранили свои жизни. Дело шло к схватке между
сверхдержавами, и на фоне надвигающихся событий двумя-тремя повешенными
японцами больше или меньше, уже не имело значения для американцев. Для большей
искренности с их стороны, американцы никогда не давали им гарантий, что не вы-
дадут русским. Победил циничный американский прагматизм. В феврале 1947 г. к
представителю СССР в Союзном Совете в Токио, генерал-лейтенанту К. Н. Деревян-
ко, обратились из штаба генерала Дугласа Макартура с просьбой о передаче им двух
японских генералов, находящихся в плену в СССР, Китадзава Тэйдзиро (командир
123-й пехотной дивизии) и Икэтани Хандзиро (начальник штаба 3-й армии). Амери-
канцы обещали предать их суду как известных им военных преступников. По согла-
сованию с МИДом Советское правительство дало согласие передать генералов аме-
риканцам, при условии получения от штаба Макартура бывшего начальника отряда
№ 731 генерал-лейтенанта медицинской службы Исии Сиро и полковника Оота,
бывшего начальника 2-го отдела этого же отряда (место их нахождения советской
стороне было известно)1. Однако ответ на это предложение от американцев последо-
вал отрицательный. Советской стороне было передано заключение, что «место пре-
бывания руководства отряда № 731, в том числе и Исии, неизвестно и обвинять от-
ряд в военных преступлениях нет оснований».
Еще раньше американцы «надавили» и на своих китайских союзников. Когда осе-
нью 1945 г. американский судья прибыл в город Чунцин (столица гоминьдановского
Китая) для сбора материалов о преступлениях японцев на территории Китая, ему
бактериологом Чэнь Вэнь Гуем был передан доклад о причинах чумы в Чандэ. Но го-
миньдановское правительство не выдвинуло обвинений, поэтому Международный
трибунал в Токио не рассматривал вопросов применения Японией БО.
После войны официальные источники США и влиятельные газеты представляли
материалы судебного процесса в Хабаровске как фальшивку и «дымовую завесу»,
пущенную «экстенсивной русской пропагандой». Даже в 1979 г. М. Rothenberg, науч-
ный руководитель испытательного полигона БО армии США в Дагуэе (штат Юта),
называл материалы Хабаровского процесса «вздором» и «совершенной ерундой».
Такая позиция была общей для официальных лиц и СМИ США до начала 1980-х гг.,
пока J. W. Powell на основания Закона о свободе информации не добился рассекречи-
вания почти 8000 страниц совершенно секретных документов, a Tsuneishi частично их
опубликовал. Окончательно информационная блокада на сведения о бактериологи-
ческой войне Японии против СССР и Китая и о преступлениях, совершенных сотруд-
никами отряда № 731 в ходе подготовки этой войны, была прорвана только 1985 г.
показом сначала в Великобритании, а затем в других западных странах передачи «От-
ряд 731 — знал ли император?» (Гайслер Э., 1986).
1 Письмо министра внутренних дел С. Н. Круглова Иосифу Сталину № 1717/к от
23.04.1949 г. Сов. секретно. АП РФ.Ф. 3. Оп. 66. Д. 1065. Л. 11—21 (Кошкин А. А., 2011).
1.8. Крах отряда Ns 731
183
Среднее и низшее звенья личного состава от-
ряда № 731, т. е. те люди, которых Исии хотел в
конце войны убить, в течение долгого времени
выполняли три его последних приказа:
1) по возвращении в Японию скрывать свою
службу в отряде №731;
2) не занимать официальных постов;
3) сотрудникам отряда связей между собой не
поддерживать всю свою жизнь.
После смерти Исии в 1958 г. его могила ста-
ла для них объектом поклонения. На централь-
ном кладбище Токио установлен памятник со-
трудникам отряда № 731, погибшим при работе
с опасными микроорганизмами (рис, 1,28).
Японское правительство не раскрыло доку-
ментов по японским работам в области БО, за
исключением документа в три страницы, выпу-
щенного в 1982 г., где была указана численность
персонала отряда на январь 1945 г. и приведены
общие фразы о «исключительности военного
времени». Среди японских политиков до сих пор
не нашлось желающих каяться за преступления,
совершенные сотрудниками отряда. Женевский
протокол 1925 г. о неприменении БО первыми Япония подписала только в 1970 г.
Судебный процесс по делу японских военнослужащих, обвиняемых в подготовке и
применении БО. После разгрома Квантунской армии в плен попало почти 600 тыс.
японских военнослужащих. Советскими органами госбезопасности была проведена
огромная работа по «фильтрации» всей этой массы пленных и выявлению среди них
лиц, имевших отношение к японским исследованиям в области БО. Как следует из
материалов судебных заседаний и письма министра внутренних дел С. Н. Круглова
Иосифу Сталину (№ 1717/к от 23.04.1949 г.), были выявлены даже жандармы, зани-
мавшиеся «спецотправками» и расстрелами подопытных людей. С ними особенно не
церемонились. Вот как об этих событиях вспоминает Георгий Георгиевич Пермяков:
Рис. 1.28. Памятник сотрудникам
отряда № 731, погибшим при работе
с опасными микроорганизмами
«В 1945 г. я приехал в Хабаровск, меня назначили старшим переводчиком УВД. При
управлении был особый лагерь № 2045 с улучшенными условиями, там содержались
особо важные военные преступники, высшие чины. Я с ними работал. В 1946 г. из
Москвы пришла шифровка — просили Хабаровский краевой центр МВД собирать
материал о бактериологическом оружии, то есть допрашивать военнопленных и брать
письменные показания. И тут мы “раскопали” 731 отряд. И установили, что в нашем
лагере для военнопленных находятся три генерала, которые руководили этой работой.
Они стали давать показания. Но не сразу.
Всего мы беседовали с 1000 военнопленных. От показаний рядовых солдат мы шли
к допросам старших чинов и, в конце концов, с помощью очных ставок „раскололи”
этих трех генералов. Мы выезжали в Харбин, опрашивали китайцев. Мы собрали ог-
ромный материал, которым гордились. Мы узнали, что принцип отряда — “Корни
лотоса”. Посадив одно семечко лотоса, можно увидеть, как этот цветок заполонит все
озеро. Отделений отряда № 731 было много. Весь этот материал мы готовили для То-
кийского процесса — восточного “Нюрнберга”. Но там он не был использован.
184
Часть 1. Краткая история применения биологических средств...
20 октября 1949 г. я получил приказ явиться в приемную генерал-лейтенанта Дол-
гих. Когда я пришел, там уже было десять известных мне переводчиков. Нас вызвали
в кабинет, и Долгих сказал нам, что в Хабаровске будет проведен суд над японскими
преступниками-бактериологами. И началась работа.
Японцы содержались в 1-й хабаровской тюрьме, нам там выделили помещение.
Приехали большие следователи из Москвы. Я был старшим переводчиком, японцы все
рассказывали без давления, допросы шли с 9 утра до 12 ночи. Выматывались все — и
следователи, и переводчики, и заключенные.
...Недели за две до начала процесса в хабаровской тюрьме, где мы вели допросы,
появилась группа хорошо одетых людей, державшихся особняком. Меня попросили
быть переводчиком между этими людьми и японцами, ожидавшими суда. Я понял, что
эта группа специалистов академика Н. Н. Жукова-Вережникова, и они великолепно
разбираются в проблемах бактериологического оружия. Но я был переводчиком только
на начальном этапе их знакомства с японскими генералами. Официально эта группа
считалась экспертами на процессе».
Судебные процессы над лицами, причастными к преступлениям, совершенным
в отряде № 731, началисьеще в 1948 г. Из письма министра внутренних дел С. Н. Круг-
лова Иосифу Сталину следует, что причастные к «специальным отправкам» жандар-
мы Иидзима, Тани-заки, Сато, Тагучи и Игараси были осуждены военным трибуна-
лом войск МВД Хабаровского округа на сроки от 15 до 20 лет исправительно-трудовых
лагерей еще 14.04.1948 г. Круглов отметил, что МВД СССР считает целесообразным
провести над руководящими работниками противоэпидемического отряда открытий
судебный процесс. Дела рядовых работников отряда, по согласованию с органами
прокуратуры, рассматривать на закрытых судебных заседаниях военных трибуналов.
С 25 по 30 декабря в Хабаровске, в окружном Доме офицеров, прошел судебный
процесс по делу 12 военнослужащих, обвиняемых в подготовке и применении БО.
Дело рассматривалось в открытых судебных заседаниях военным трибуналом При-
морского военного округа в составе председательствующего генерал-майора юстиции
Д. Д. Черткова и членов: полковника юстиции М. Л. Ильницкого и подполковника
юстиции И. Г. Воробьева. Государственное обвинение поддерживал государственный
советник юстиции 3 класса Л. Н. Смирнов. У подсудимых были адвокаты. Судебно-
медицинская экспертиза на процессе осуществлялась группой экспертов во главе с
действительным членом АМН СССР Н. Н. Жуковым-Вережниковым. Однако само
определение вопросов экспертам было сделано трибуналом только 28 декабря 1949 г.
Судебный процесс проходил очень интенсивно. Г. Г. Пермяков вспоминал, что
руководил процессом начальник МВД полковник Карлин. Последнее заседание суда,
когда был оглашен приговор, было «до упора», на завтра его не переносили. А когда
первого января 1950 г. вышел Указ о введении в СССР смертной казни, Пермякову
стало понятно, что бактериологических преступников просто спасли от высшей меры,
которую они заслуживали. Значит, они были нужны живыми.
Хироси Акияма (1958) объясняет бросающуюся в глаза откровенность обвиняе-
мых и свидетелей тем, что им обещали сохранить жизнь при условии сотрудничества
с трибуналом. При написании своих воспоминаний он в качестве справочного мате-
риала пользовался официальными советскими материалами процесса, изданными в
виде отдельной книги на японском языке. Акияма не отрицал ни одного факта, из
приведенных в ней, он только их дополнял.
Военный трибунал округа признал виновность всех подсудимых в совершении
преступлений, предусмотренных ст. 1 Указа Президиума Верховного Совета СССР
1.8. Крах отряда №731
185
от 19 апреля 1943 г., доказанной и, руководствуясь статьями 319 и 320 Уголовно-про-
шссуального кодекса РСФСР, с учетом степени виновности каждого подсудимого,
приговорил:
Ямада Отозоо на основании ст. 1 Указа Президиума Верховного Совета СССР от
19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 25 лет.
Кадзицука Рюдзи на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 25 лет.
Такахаси Такаацу на основании ст 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 25 лет.
Кавасима Киоси на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 25 лет.
Ниси Тосихидэ на основании ст. 1 Указа Президиума Верховного Совета СССР от
19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 18 лет
Карасава Томио на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 20 лет.
Оноуэ Macao на основании ст. 1 Указа Президиума Верховного Совета СССР от
19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 12 лет.
Сато Сюндзи на основании ст. 1 Указа Президиума Верховного Совета СССР от
19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 20 лет.
Хиразакура Дзенсаку на основании ст. 1 Указа Президиума Верховного Совета
СССР от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на
10 лет.
Митомо Кадзуо на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 15 лет.
Кикучи Норимицу на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 2 года.
Курусима Юдзи на основании ст. 1 Указа Президиума Верховного Совета СССР
от 19 апреля 1943 г. заключить в исправительно-трудовой лагерь сроком на 3 года.
Официальные материалы процесса опубликованы в 1950 г. в СССР тиражом в
50 тыс. экземпляров и переведены на китайский, японский и английский языки.
Дальнейшая судьба плененных сотрудников отряда № 731. По данным Г. Г. Пермя-
кова, рядовых сотрудников отряда № 731, не осужденных трибуналом, вскоре пере-
дали китайским властям. Он рассказал о таком факте. Второго июня 1950 г. ему при-
казали явиться на станцию Хабаровск-2. На путях стоял длинный состав вагонов,
окрашенных в красный цвет. Ему сказали, что это японские военнопленные, имею-
щие отношение к отряду № 731 и к другим бактериологическим подразделениям быв-
шей японской армии. СССР передает их Китаю, а ему поручено сопровождать их,
поскольку он владеет и японским, и китайским языками. Причем, как выяснилось,
сами японцы не знали, что их везут в Китай. В эшелоне было 1002 человека.
Лица, осужденные трибуналом на непродолжительные сроки, отбыли их полно-
стью, и были отправлены на родину Карусиму Юдзи перед отъездом еше и повозили
по Москве, показав ему достопримечательности советской столицы Осужденные на
длительные сроки отбыли в заключении в тюрьме в Иваново только 7 лет, причем в
достаточно комфортабельных условиях. Перед отправкой на родину в 1956 г. их одели
по последней моде, в Хабаровске в их честь был устроен пышный банкет. Вернувшись
в Японию ни один из японских генералов, причастных к разработке БО, не написал
мемуаров о «сталинских застенках», хотя им за это предлагались большие деньги.
186
Часть 1. Краткая история применения биологических средств...
* * *
Япония не была ни первым разработчиком БО, ни даже вторым. В 1920—1930-х гг.
не только Бугсикава считал, что, «имея крошечную лабораторию в несколько квад-
ратных метров и пробирки, легко можно наработать оружие, способное уничтожить
десятки тысяч жизней». Абстрактное восприятие возбудителей инфекционных болез-
ней как «контагиев» создавало иллюзию легкого успеха в создании «оружия массово-
го поражения бедных», которой поддались военные круги Великобритании, Франции,
Румынии, Италии, Польши и некоторых других государств. Научные журналы и СМИ
усиливали эти ожидания. Инициатор японской программы по созданию БО — Исии
Сиро, лишь стремился «не опоздать на поезд», добросовестно доведя дело до реаль-
ного испытания БО на людях. Так как работал он с «контагием», то разрабатываемые
им боеприпасы и способы ведения бактериологической войны до начала 1940-х гг.
копировали применяемые в химической войне. Основной упор им делался на зара-
жение местности путем диссеминации бактерий (бактериальный туман, дождь и т. п.),
что методически мало чем отличалось от смазывания дверных ручек содержимым
чумных бубонов средневековыми «сеятелями чумы». Отличием было го, что для рас-
пространения «контагия» японскими военными использовались модифицированные
из химических боеприпасов снаряды, бомбы и ВАПы. Исии, как и его западные учи-
теля начала 1930-х гг., считал, что достаточно человеку войти в соприкосновение с
«контагием» или вдохнуть его, чтобы началось развитие инфекции (см. разд. 1.7).
Когда ему стало ясно, что такой подход показывает лиш^ «принципиальную возмож-
ность создания БО», он попытался разработать средства ведения бактериологической
войны, исходя из необходимости проникновения возбудителей инфекционных болез-
ней непосредственно в кровь потенциальных жертв. Полностью провалив аэробио-
логическое направление своих исследований, Исии выбрал еще более тупиковое — он
попытался решить проблему массового заражения людей через использование крово-
сосущих насекомых. И чем дальше японцы продвигались в направлении создания
образцов БО, пригодных для использования в военных целях, тем больше перед ни-
ми вставало неразрешимых технических противоречий. Контагионистический пери-
од в развитии БО закончился в Маньчжурии в дождливую ночь с 10 на 11 августа
1945 г. в яме с останками людей и несгоревшей нефти
1.9. БИОЛОГИЧЕСКОЕ ОРУЖИЕ ПЕРЕД КОРЕЙСКОЙ ВОЙНОЙ
Взгляды на применение БО, существовавшие в годы Второй мировой войны.
Начало программы по созданию БО в США. Начало программы по созданию БО
в Соединенном Королевстве. Военно-биологическая программа Франции. Био-
диверсии в Европе в годы Второй мировой войны. Несостоявшиеся химические
и биологические войны. Производство биоагентов. Разработка технических
средств для индивидуальных убийств. Разработка технических средств цля мас-
сового биологического поражения людей. Аэрозоли. Моделирование масштаб-
ного применения БО.
Когда читаешь военные журналы начала 1930-х гг., складывается впечатление, что
работы над созданием БО идут полным ходом в большинстве западных стран, однако
современные источники пытаются нас убедить в том, что эти программы начались
только в ответ на аналогичные, предпринятые фашистской Германией и милита-
1.9. Биологическое орхжие перед Корейской войной
187
растской Японией в годы Второй мировой войны. С лицемерием крокодила, перева-
ривающего 50 млн жертв ковровых бомбежек, массовых расстрелов и голода концент-
’'апионных гагерей, «цивилизованное сообщество» до сих пор стесняется своих раз-
работок так и не созданного оружия.
Взгляды на применение БО, существовавшие в годы Второй мировой войны. В 1947 г.
был рассекречен и опубликован в «The Journal of Immunology» доклад о перспективах
создания БО, подготовленный сотрудниками Колумбийского университета Теодором
’озбери (Theodor Rosebury) и Эльвином А. Кабатом (Elvin Kabat) и представленный
ими 8 июня 19421 Национальному исследовательскому совету США На русском язы-
ке он был опубликован в 1955 г. отдельной книгой. Это емкий и обстоятельный доку-
мент, являющийся выражением американской точки зрения начала 194и-х гг. на био-
логическую войну Розбери и Кабат были активными участниками американской
военно-биологической программы. Поэтому интересны те их представления о био-
югической войне, которые стали основой для разработки образцов БО в годы Вто-
рой мировой войны.
По мнению Розбери и Кабата, бактериологическая и химическая война похожи
одна на другую тем, что они используют средства, которые могут быть распростране-
ны на большой территооии и таким образом оказать одновременное действие на зна-
чительное число людей Эти средства невидимы и неосязаемы, поэтому действие обоих
видов оружия коварно и оказывает на личный состав вооруженных сил большой де-
морализующий эффект. Основное преимущество БО над химическим они видели в
«эпидемичности» возбудителей инфекционных болезней, т. е. в их способности пе-
г гдаваться от одного человека к другому. Следовательно, одна эффективная бактери-
ологическая атака, даже сильно ограниченная как по территории, так и по количеству
использованных бактериологических средств, может вызвать эпидемию, распростра-
няющуюся на значительное пространство. На основе такого представления о БО они
рассматривали шесть целей, для достижения которых оно может быть использовано,
1. Для подавления изолированных крепостей (укрепленных пунктов, баз). Для подав-
ления островных, морских, воздушных баз и других районов, в отношении которых
ставится цель сделать их непригодными для противника, но которые нет необходи-
мости захватывать или занимать, бактериологические средства представляют значи-
гельные преимущества и почти не имеют тактических недостатков. Может быть ис-
пользован любой возбудитель, подходящий по климатическим и прочим условиям
местности, включая возбудителей с большой потенциальной ретроактивностью, на-
пример, чумы.
2. Для подавления осажденных городов и при других формах позиционной войны. По-
давление осаждающей армией города-крепости, когда ставится задача скорее пройти
мимо этого города, не захватывая его. Но в упомянутых условиях обе стороны могут
эффективно использовать бактериологические средства поражения. Войска, находя-
щиеся в городе, чтобы сломить осаду, могут принимать подобные же меры с анало-
гичными предосторожностями. Здесь, опять- гаки при наличии благоприятных кли-
матических условий, могут быть использованы некоторые из менее ретроактивных
средств, как, iтапример, ботулинический токсин, возбудители бруцеллеза, туляремии,
лептоспироза и болезней, передаваемых клещами. В умеренной зоне в конце лета или
осенью Розбери и Кабат предлагали использовать возбудитель лихорадки денге, рас-
тространяемый комарами.
3. Для дезорганизации промышленных районов в тылу противника. Подобного рода
задача также, по их мнению, может быть решена средствами бактериологической вой-
188
Часть 1. Краткая история применения биологических средств...
ны. Особые условия, возможно, не допустят использования при этом средств, обла-
дающих сильным ретроактивным действием, таких, например, как возбудитель чумы.
Однако другие возбудители, передаваемые по воздуху, вследствие большой скученно-
сти людей в закрытых помещениях и вследствие того, что эти районы обычно нахо-
дятся в умеренном или холодном климате, могут быть весьма успешно применены с
этой целью. Ботулиническим токсином могут быть отравлены водохранилища, питаю-
щие водой эти районы.
4. Для поражения армейских лагерей и иентров войсковой подготовки. Эта задача
подобна предыдущим, за исключением того, что плотность населения, по-видимому,
здесь меньше, пребывание в закрытом помещении не так обязательно, а климатиче-
ские условия более разнообразны. Использованные при таких обстоятельствах воз-
будители, обладающие большой заразительностью и передаваемые по воздуху, будут
направлены, в частности, на бараки и применены, по-видимому, в ночное время. Если
имеются бассейны для купанья или пруды, то Розбери и Кабат рекомендовали для
заражения воды использование лептоспир и ботулинические токсины.
5. Для применения политики «выжженной земли» при стратегических отступлениях.
Для этих целей, по ихмнению, требуется возбудитель, обладающий слабой тенденцией
к распространению, а также не обладающий большой ретроактивностью, что являет-
ся весьма важным условием благополучия войск в том случае, если противник, прой-
дя через зараженный район, войдет в соприкосновение с сопротивляющимися ему
силами. Возбудители сибирской язвы, бруцеллеза и туляремии, а также возбудители
инфекционных болезней, которые передаются клещами, и, возможно, возбудители,
заражающие воду, при соответствующих условиях могут быть с успехом использова-
ны для решения этих задач.
6. Для истребления сельскохозяйственных животных и растений. Бактериологиче-
ские атаки, направленные на животных и растения, употребляемые человеком в пищу,
а также на технические культуры, могут вызвать голод среди населения, и противни-
ку придется капитулировать. В дополнение к возбудителям, считающимся специфи-
ческими с точки зрения поражения только животных, некоторые патогенные микро-
организмы, предназначаемые для поражения человека, могут воздействовать и на
животных (возбудители бруцеллеза, сибирской язвы, сапа).
В целом же представления Розбери и Кабата о ведении бактериологической вой-
ны (как, впрочем, и других ученых того времени: см. разд. 1.7 и 1.8) основывались на
распространении «контагия». Человек контактирует с микроорганизмом, искусствен-
но введенным в окружающую его среду. Микроорганизм вызывает болезнь, и заболев-
ший становится источником инфекции для другого человека, так формируется искус-
ственная эпидемическая цепочка, позволяющая решать боевую задачу, возможно, за
более длительный период времени, чем при применении обычного оружия или даже
химического, но без особых затрат и усилий. Интересны и их рекомендации по отбо-
ру более вирулентных и устойчивых по отношению к лекарствам и дезинфицирующим
средствам штаммов возбудителей инфекционных болезней. При столь оптимистичес-
ких взглядах на возможность биологической войны, дело все же осталось за малым,
необходимо было создать само БО. Но микроорганизмов, опасных для человека, до-
машних животных и сельскохозяйственных растений, сотни. Способы же их распро-
странения в среде, окружающей человека, столь разнообразны, что надо было выб-
рать то оптимальное, на чем можно было бы и остановиться, а для этого исследования
надо было ввести в какие-то рамки — так стали появляться различные программы по
созданию БО.
1.9. Биологическое оружие перед Корейской войной
189
Начало программы по созданию БО в США. В открытой литературе еще не публи-
ковалось детального обзора работ, выполненных в США в годы Второй мировой вой-
ны по программе создания БО. Однако благодаря разумной политике в области засек-
речивания информации, ключевые моменты выполнения этой программы в самом
общем виде попали на страницы научных журналов и в открытые официальные до-
кументы (см. Report from the United States of America..., 1992).
F. Sidell et al. (1997) относят начало работ в США по созданию БО на 1918 г., когда
стало ясно, что Германия имеет свою военно-биологическую программу (см. разд. 1.7).
В качестве ответной меры на применение ОВ Германией предполагалось применение
рицина — растительного токсина, получаемого из касторовых бобов (см. разд. 3.11).
детальная доза неочищенного рицина, введенного животному парэнтерально, на
несколько порядков ниже, чем у синильной кислоты или иприта при том же способе
введения. На первый взгляд это давало рицину преимущество перед германским ОВ
того времени, однако американской армии применить рицин во время Первой миро-
вой войны не удалось. Химической службой военного ведомства было проведено ис-
следование поражающих свойств рицина, примененного двумя способами:
1) шрапнельными пулями артиллерийских снарядов;
2) распылением в порошкообразном состоянии.
При первом способе применения было установлено, что, во-первых, препараты
рицина, способные приклеиваться к шрапнельным пулям, могут быть приготовлены
без особых затруднений; во-вторых, рицин не утрачивает своей активности в резуль-
тате выстрела, не происходит быстрой его инактивации и при хранении таких боепри-
пасов. Однако исследователями не было получено доказательств того, что шрапнель-
ные пули, покрытые рицином, наносят более тяжелые ранения, чем обычные. И без
рицина, ранения, причиненные шрапнелью, обычно становились смертельными для
людей (Hunt R., 1918).
При втором способе применения рицина исследователи столкнулись с тем, что
распыленный в порошкообразном состоянии рицин не проникал в глубокие отделы
респираторного тракта экспериментальных животных и не вызывал у них смертель-
ных поражений (Hunt R., 1918). Почему это происходило, они разобрались только
через 30 лет. К тому же рицин было трудно получить в количествах, достаточных для
проведения масштабных экспериментов. Исследования поражающих свойств рици-
на в 1918 г. зашли в тупик, но они не прекращались после войны 1914—1918 гг., что
позволило СШ А в годы Второй мировой войны изготовить 1,7 тонны этого токсина.
В начале 1920-х гг. от некоторых исследователей поступали предложения, чтобы
химическая служба занялась исследованием поражающей способности патогенных
бактерий. Однако возглавлявший тогда химическую службу генерал Амос Фрайс (Amos
Fries) вполне обоснованно отклонял эти предложения, указывая на то, что БО боль-
ше является темой для разговоров в кабинетах, а не реальным боевым средством, так
как нет эффективных способов применения бактерий на поле боя (Sidell Е et al., 1997).
Эта точка зрения господствовала среди американских военных и политиков в 1930-е гг.
В докладе, направленном в 1942 г. Т. Розбери и Э. Кабатом (1955), содержится много
обзорного материала по опасности бактериологической войны, свидетельствующего
об опасении авторов, что их «не поймут». Тем не менее анализ темпов внедрения неко-
торых технологий уже во время Второй мировой войны указывает на то. что отдель-
ные направления, смежные с химическим оружием, все же развивались в США и до
войны. Среди них те, которые имеют отношение к растительным и бактериальным
токсинам.
190 Часть 1. Краткая история применения биологических средств...
По официальным данным, исследовательская работа по созданию БО началась в
США в августе 1941 г. под эгидой химической службы Министерства ооороны США,
когда в арсенале Эджвуд по предложению военного министра Генри Л. Стимсона
(Henri L. Stimson) было создано подразделение, известное как «Отдел медицинских
исследований» или «Специальный отдел». В ноябре 1941 г., так же по инициативе
Стимсона Национальная академия наук создала комитет по оценке опасности био-
логической войны (комитет WBC). В комитет WBC вошли представители Националь-
ной академии наук (среди них такие известные биологи, как Эдвин Б. Фред из Вис-
консинского университе га и Стэнхоуп Бэйн-Джоунз из Йельского университета),
Министерства сельского хозяйства, Федерального ведомства здравоохранения, Управ-
ления начальника военно-медицинской службы, Медицинского управления ВМС,
Артиллерийско-технической службы и разведки армии США (Берстайн Б. Дж., 1987).
В феврале 1942 г. комитет WBC пришел к заключению, что атака противника с
применением БО может привести к массовой гибели людей, посевов и домашнего
скога. В отчете комитета говорилось в основном об оборонительных мерах и работе
по созданию вакцин и средств защиты систем водоснабжения, однако комиссия так-
же рекомендовала, хотя и не столь явно, чтобы в США начали проводиться исследо-
вания наступательного потенциала БО (рис. 1.29).
The vwlae of < lolAclw.l vwrftre will bn • debatable .nuatlon imtil
it bne been clourV proTra. or by w orle^ce. Suci «ncnirlenc» «ay
Th» “ le* l»i - rt! tm 1» thr.t nay wcthod which ippcare to
ff«r «Jraata e* to * natlo i at war will b* wlgorrwlj < iployed by that
в ТЫте i« but onr 1О(Дсл1 осcuts to p trfee» ruua.1. to ьГ.'Ду the
s- ilULJLo» or , ieb w-iI4re n jrwrjr < ril»( жй rstry sreoaratioa for
lie effective-.»'.« and t.. r iLy . wctaoe tlJ 11-uliBooc of ita um.
ta or&rr to plan euch. -r^-xiration. It Is. dmuntapeoua to tri.» the point of
Tier of the _r»i»ir and to felww o*r»‘Ml ultentica to the chKraetorlstlea
»M*h a biologic off-nnlv a.ght ham.
» to. nr. iu О r ’
Рис. 1 29 Выдержка из рекомендации, содержавшихся в докладе Национальной
академии наук США от 1942 г., в которой обосновывается проведение исследований
по бактериологическому оружию Доклад, вероятно, убедил президента Рузвельта
создать военную исследовательскую службу. Штамп слева — «РАССЕКРЕЧЕНО».
Перевод текста: «Вопрос о значении бактериологического оружия будет оставаться
спорным до тех пор, пока это оружие не проверят в действии. Возможно, что вскоре
это будет сделано. Разумно допустить, что любой метод, дающий преимущества
воюющей стране, будет ею незамедлительно использован. Существует только один
логический путь, всесторонне исследовать возможности бактериологического ору-
жия, сделать все необходимые приготовления для уменьшения его эффективно-
сти и тем самым снизить вероятность его применения (противником). В планиро-
вании этих действий целесообразно учитывать точку зрения агрессора и уделить
особое внимание характеристикам, которыми могло бы обладать наступательное
бактериологическое оружие». По Б. Дж. Берстайну (1987)
Ссылаясь на предупреждения ученых, Стимсон стремился добиться одобрения
президента Рузвельта на осуществление программы разработки БО, в которой коор-
динирование и руководство всей исследовательской работой по заданию правитель-
ства выполняла бы небольшая консультативная группа из г ражданских лиц. «Мы
должны быть готовыми, — писал Стимсон Рузвельту в своем меморандуме в апреле
1942 г., — все работы проводить в строгой секретности и как можно быстрее». Стим-
/ 9. Биологическое оружие перед Корейской войной
191
сон хорошо знал общественное мнение в отношении БО и стремился отвести от себя
будущие обвинения в том, что его министерство стремилось использовать БО в этой
войне. Поэтому в меморандуме президенту Стимсон нигде не упомянул, что хими-
ческая служба военного министерства уже приступила к таким исследованиям; сам
же президент либо ничего не знал о них, либо не хотел, чтобы знали, что знал. Хими-
ческой службе были выделены миллионы долларов из бюджета военного министер-
ства, и она стала играть большую роль в осуществлении программы разработки БО,
чем небольшая консультативная группа гражданских лиц, формально руководившая
исследован иями.
Стимсон предложил включить (и тем самым «спрятать») консультативную груп-
пу. занимающуюся разработкой «микробного оружия», в состав вновь созданного
учреждения, называемого Федеральным агентством социального обеспечения, кото-
рое контролировало деятельность системы здравоохранения и социальных служб. Он
хотел, чтобы программой руководил какой-нибудь известный ученый, знакомый с
университетской системой научных исследований и имевший опыт административ-
ной работы. Рузвельт уклонялся оставлять какие-то свои «следы» на любых планах по
созданию БО, ограничиваясь лишь устными указаниями. После совещания с минист-
рами 15 мая 1942 г. он сообщил им, что еще не ознакомился с планом военного мини-
стра, однако сказал, что Стимсону следует продолжать осуществлять свой замысел.
Неделю спустя Стимсон обсудил свои идеи с министром сельского хозяйства
К. Р. Уикардом и добился от него согласия участвовать в исследовательской работе,
координируемой консультативной группой. П. В. Макнатт, возглавлявший Федераль-
ное агентство социального обеспечения, также дал свое согласие на участие агент-
ства в работах по созданию БО.
К середине лета три кандидата (экономист У. У. Стюарт, председатель Рокфелле-
ровского фонда; географ И. Бауман, ректор Университета Джонса Гопкинса; эконо-
мист Э. Э. Дэй, ректор Корнельского университета) отказались от предложения воз-
главить новую группу. Наконец в августе Дж. У. Мерк (George W. Merrck), президент
фармацевтической фирмы (Merck & Со., Inc.), согласился принять его (рис. 1.30)1.
До 1942 г. масштаб исследований в области БО был незначителен. В середине
1942 г. консультативная группа, получившая скромное название «Военная иссле-
довательская служба» (ВИС; в некоторых источниках приведены другие названия:
«Отдел военных исследований», «Служба военных исследований») и ассигнования
на сумму в 200 тыс. долл., начала свою деятельность. Широкие контакты между извест-
ными биологами и врачами позволили правлению, состоявшему из 8 человек, органи-
зовать проведение секретных исследований примерно в 28 американских универси-
тетах, включая Гарвардский, Колумбийский, Корнельский, Чикагский, Северо-За-
падный, Висконсинский, Стэнфордский и Калифорнийский, а также Университет
шт. Огайо и Университет Нотр-Дам (Берстайн Б. Дж., 1987).
Первая задача ВИС состояла в разработке мероприятий по обороне на случай
возможного нападения с применением БО. Ее главным достижением явилась реали-
зация программы научно-исследовательских и опытно-конструкторских работ, имев-
шей целью расширить рамки существующих знаний о средствах ведения биологичес-
кой войны. Был сделан вывод о том, что достижение этой цели невозможно без
увеличения масштаба исследований.
1 В приложении А приведен перевод доклада Джорджа У. Мерка министру обороны США
по состоянию исследований по БО на 3 января 1946 г.
192
Часть 1. Краткая история применения биологических средств...
Рис. 1.30. Советники по бактериологическому оружию, собравшиеся в Форт-Детрике
на совещание с работавшими там учеными. В центре группы — Дж Мерк, президент
фирмы Merck & Со., Inc. Слева направо — директор по научной части И. Л. Болду-
ин, капитан Н. С. Прайм, бригадный генерал У А. Борден, контр-адмирал Дж. Зю-
рер, капитан 3-го ранга У. Б. Сарлз, полковник Вулперт и подполковник Н. Пайл
По Б. Дж. Берстайну (1987)
В ноябре 1942 г. ВИС предложила химической службе армии взять на себя обя-
занности по расширению программы проводимых исследований, включая проекти-
рование и эксплуатацию лабораторий и пилотных установок. До этого армия прини-
мала участие лишь в координации деятельности ВИС. Местом для строительства таких
лабораторий был избран Кемп Детрик, г. Фредерик в штате Мериленд. ВИС передала
химической службе армии руководство всеми работами, но продолжала осуществ-
лять координацию работ по всей программе создания БО. К январю 1943 г. ВИС за-
ключила договор с У. А. Хаганом из Корнеллского университета на проведение ис-
следований по использованию ботулинических токсинов как агентов БО, а также с
Дж. X. Мюллером из Гарвардской медицинской школы на изучение пригодности воз
будителя сибирской язвы для целей биологической войны.
В 1942—1943 гг. химической службе были выделены миллионы долларов на строи-
тельство исследовательских лабораторий, для работы в которых были приглашены
многие известные ученые. Лаборатории начали строить в апреле 1943 г., и уже через
7 месяцев они были готовы для провеления исследовательских работ в области созда-
ния БО. Некоторые современные авторы неправильно отождествляют Фот-Детрик
только с US Army Medical Research Institute of Infectious Diseases (USAMRIID). Во время
Второй мировой войны этот институт представлял собой скромное учреждение, за-
нимающее несколько деревянных зданий.
ВИС совместно с химической службой армии США установила контакты с ана-
логичными службами Канады и Соединенного Королевства. Хотя Макнатт основное
внимание уделял работе социальных служб, из его поля зрения не ускользала и дея-
тельность ВИС, «спрятанной» в его агентстве. В феврале 1943 г. он сообщил Рузвель-
ту, что сумма в 200 тыс. долл., выделенная ВИС, почти израсходована. По его словам
требовалось согласие президента на «расширение исследований по двум или трем...
уже осуществляемым проектам». В конце марта 1943 г. Макнатт запросил, с одобре-
ния Стимсона, еще 25 тыс. долл, на 1943 г. и всего 350 тыс. долл, на 1944 г. Через два
дня от Рузвельта пришел лаконичный ответ: «О’кей. Ф. Д. Р». Спустя несколько ме-
9. Биологическое оружие перед Корейской войной
193
сяцев Рузвельт дал согласие на увеличение бюджета ВИС на 1944 г., который в резуль-
тате достиг 460 тыс. долл. (Берстайн Б. Дж., 1987).
Макнатт соблюдал строгую секретность в отношении исследовательской програм-
мы по БО и «деликатность» в отношении президента. Нигде, даже в корреспонден-
ции Рузвельту, он не писал о проектах по созданию БО или их деталях. В собственном
архиве Рузвельта исследователями обнаружено не более десяти писем и записок
относительно БО. В тех из них, которые датированы 194? и 1943 гг., речь идет в основ-
ном о небольших ассигнованиях и организационных вопросах, связанных с деятель-
ностью ВИС. Возможно, что Макнатт и Стимсон, а также начальник штаба сухо-
путных войск генерал Дж. К. Маршалл во время встреч с Рузвельтом устно информи-
ровали его о дополнительных суммах в миллионы долларов, выделенных ВИС на
1зработку БО. Ни один из известных документов не содержит сведений о том, что
Рузвельт получал и визировал такие отчеты (Берстайн Б. Дж., 1987).
В декабре 1943 г. Стратегическое управление сообщило в своем донесении Объ-
единенному комитету начальников штабов об имеющихся признаках подготовки гер-
манского командования к применению БО. В начале 1944 г. эксперты разведслужб
ггран антигитлеровской коалиции высказывали опасения, что Германия, возможно,
собирается использовать свои новые мощные самолеты-снаряды V-1 для бомбарди-
ровки Великобритании, и не исключено, что эти снаряды будут нести бактериологи-
ческий заряд. Поэтому в январе 1944 г. переоценка деятельности вероятного против-
ника в области БО привела к значительному расширению усилий США, и разработка
всей программы разработки БО была полностью передана химической службе. В ее
рамках было создано специальное управление по исследованиям в области БО. В рас-
поряжении управления находилось 3900 сотрудников, из которых примерно 2800 со-
стояли на службе в армии, примерно 1000 — в военно-морском флоте и примерно
100 человек были гражданскими лицами (US Army activity..., 1977).
В мае 1944 г. Стимсон и Макнатт предложили Рузвельту упразднт ть ВИС и наз-
начить Мерка консультантом Стимсона. Президент с готовностью одобрил это прет-
ожение, и последовавшие затем изменения еще больше отдалили его от секретной
программы по разработке БО. Благодаря усилиям Стимсона, химическая служба по-
следовательно расширяла свои технические возможности в разработке, испытании и
производстве БО. Кроме лабораторий в Форт-Детрике и относящейся к нему терри-
тории площадью в 220 га, к январю 1944 г. была введена в действие испытательная
станция на острове Хорн в Паскагуле (шт. Миссисипи), где был построен полигон
площадью 882 га для полевых испытаний. Вблизи испытательного полигона Дагуэй
(шт. Юта) химической службе была выделена территория площадью 70,6 тыс. га для
бомбардировочных испытаний. Эта станция, Грэнит Пик Инстелейшн, была введе-
на в действие в июне 1944 г. и после этого использовалась для проведения всех основ-
ных полевых испытаний патогенных микроорганизмов. В мае 1944 г. завод в Терр От,
штат Индиана, был переоборудован для производства биологических агентов и агент-
ных имитаторов.
В июне 1944 г. решено форсировать проводившиеся исследования по программе
БО. Соыасно решению президента, ответственность за выполнение программы БО
была возложена на военное министерство. По распоряжению военного министра на
шмическую службу была возложена ответственность за исследования агентов биоло-
гической войны, за сбор разведывательных данных по БО и за противобиологическую
защиту. Начальник медицинской службы армии получил приказ координировать свою
деятельность по вопросам защиты от БО с химической службой. Работы по программе
194
Часть 1. Краткая история применения биологических средств...
продолжались совместно со специалистами ВМФ и представителями других федераль-
ных министерств. Осуществление программы Н ИОКР по БО значительно ускорилось
в связи с введением в строй научно-исследовательских центров для полигонных испы-
таний и завода для производства БО. Когда военное министерство взяло на себя от-
ветственность за осуществление всей программы БО, военный министр Стимсон
назначил Мерка специальным консультантом по вопросам БО. В октябре 1944 г. был
создан консультативный орган — Комитет США по вопросам биологической войны
во главе с Мерком, в него вошли представители и других военных служб. Комитет кон-
сультировал военного министра и председателя Комитета начальников штабов в об-
ласти БО, а также поддерживал связи с Межведомственным подкомитетом по БО
Соединенного Королевства. Комитет США по вопросам биологической войны суще-
ствовал до октября 1945 г., когда его функции были переданы Управлению исследо-
ваний и разработок военного ведомства (US Army activity..., 1977).
Параллельно совершенствованию структуры организаций, занимающихся про-
блемами биологической войны, совершенствовалась технология производства био-
логических агентов. При техническом содействии Великобритании химическая служ-
ба вооруженных сил США получила значительную возможность самой изготавливать
бактериологические бомбы и в конце 1943 г. начала работы по созданию кассетных
бактериологических бомб массой около 225 кг, с субэлементами, снаряженными воз-
будителями сибирской язвы (в более поздних интерпретациях — бруцеллеза). Каж-
дая из этих бомб содержала 108 субэлементов массой около 2 кг. Б. Берстайн (1987)
утверждал, что эти бомбы не испытывались, поскольку и без того было ясно, что на-
чиненные возбудителями сибирской язвы, они несут неизбежную смерть, однако это
было явным преувеличением возможностей нового оружия. К бактериологическим
бомбам мы еще вернемся (см. ниже «Разработка технических средств для массового
биологического поражения людей»). Химической службе быстро удалось разработать
полупромышленную технологию получения ботулинического токсина (рис. 1.31).
Хотя снимок на рис. 1.31 назван Б. Берстайном (1987) «Группа ученых...», но для
1943 г., на который он его относит, оборудование, расположенное за спиной этих уче-
ных, выглядит не только впечатляющим, но и мало соответствующим официальной
хронологии разработки БО в США. Показанный культиватор не мог быть «заимство-
ван» ни из производственной линии, созданной для получения ботулинического ана-
токсина, так как для этого он слишком большой; ни из технологической линии по
производству антибиотиков (технология их получения требует аэрации питательной
среды во время культивирования). За один цикл работы такого аппарата (5-7 сут)
американские исследователи могли получать до 3 кг пасты ботулинического токси-
на. Запечатленная на снимке технологическая обвязка свидетельствует о существова-
нии каскада таких культиваторов. В расстановке оборудования видна продуманность,
даже металлические трубы соединены сварными швами с плавными линиями изгиба
для удобства пропаривания. Кроме этого цеха (культивирования) на предприятии
должны быть как минимум еще два — цех приготовления стерильных питательных
сред и цех по концентрированию ботулинического токсина. Если программа по созда-
нию БО начата Министерством обороны США только в 1942 г., то полупромышлен-
ная технология производства пасты ботулинического токсина была разработана дру-
гими организациями раньше.
В начале 1944 г. химическая служба, окрыленная успехом в полупромышленном
получении ботулинического токсина и возбудителя сибирской язвы, начала настаи-
вать на том, чтобы ей дополнительно было выделено 2,5 млн долл, на производство
г 9. Биологическое оружие перед Корейской войной
195
Рис. 1.31. Группе ученых осматривает оборудование в лаборатории в Форт-Детрике.
. первом ряду на снимке (слева направо) — Н. Р. Хаусон из Университета шт. Огайо,
Г. Б. Рид из Куинского университета, Ч. А. Митчелл из Министерства сельского хо-
зяйства США; Э. Г. Д. Мюррей из Университета Макгипла и полковник О. К. Вул-
перт. Во втором ряду — Дж. Крэги (справа) из Торонтского университета и полков-
ник А. Т Томпсон. Посмотрим внимательно на попавшее в кадр оборудование:
1 — культиватор на 1,5 м3для анаэробного выращивания бактерий, имеющий ниж-
ний слив. 1акой аппарат может использоваться только для культивирования анаэро-
ба — ботулинического микроба и накопления ботулинического токсина; 2 — смот-
ровое стекло для наблюдения за поверхностью жидкости в культиваторе; 3 — паровая
труба; 4 — паровой вентиль; 5 — уровнемер; 6 - труба для удаления газообразных
продуктов метаболизма, 7 — труба для отбора проб и введения посевного материала;
3 — труба для теплоносителя По Б. Дж. Берстайну (1987)
бомб с возбудителями сибирской язвы и ботулизма. По словам ее представителей,
такая сумма позволила бы ежемесячно производить либо 275 тыс. бомб с ботулини-
ческим токсином, либо 1 млн бомб с возбудителями сибирской язвы. Проблема за-
лючалась, однако, в том, чго на строительство заводов требовалось время, поэтому
оружие могло быть произведено в достаточном количестве лишь к 1945 г. По прогно-
зам военных стратегов, война к тому времени должна была вестись только с Япони-
ей. Тем не менее, по причинам указанным ниже, химическая служба получила запро-
шенную ею денежную сумму.
На момент создания Управление по исследованиям в области БО курировалось
Комитетом обороны. Впоследствии оно функционировало под руководством Коми-
тета США по вопросам биологической войны (в некоторых источниках упоминается
как Комитет по биологическому оружию).
О паоотах в Форт-Детрике Рузвельта «не информировали». В мае 1944 г. Стимсон
и Макнатт представили Рузвельту краткий отчет об исследованиях, содержавший всего
пять строк о научных результатах. В этот отчет не попало многое. Например, то, что
завод по производству бомб, снаряженных возбудителем сибирской язвы, получил
заказ на производство миллиона бомб, а в лабораториях Форт-Детрика успешно разра-
батывапись средства рассеивания на небольшой площади ботулинического токсина.
196
Часть 1. Краткая история применения биологических средств...
Тем не менее к началу операции «Оверлорд» (06.06.1944) у США имелось всего
несколько бомб с возбудителями сибирской язвы, и то предназначенные для полигон-
ных испытаний. Бактериологических бомб, которые могли бы использоваться для
удара по противнику, у них определенно не было. Однако военное командование сде-
лало «хорошую мину при плохой игре», организовав проведение прививок нелицен-
зированным ботулиническим анатоксином 100 тыс. солдат, надеясь тем самым убе-
дить Германию, что союзные войска готовятся нанести ответный удар биологическим
оружием, в случае если на его применение решится Германия. Но если бы Германия
действительно нанесла удар биологическим оружием, то англо-американские силы,
вероятно, ответили бы на него устаревшим химическим оружием времен Первой ми-
ровой войны. Далее можно продолжить строить сценарии войны, в которых учиты-
вается фактор применения Германией химического оружия на основе ФОБ, но, к сча-
стью для всех воюющих сторон, Германия не разрабатывала и не применяла БО, а
союзники не совершили смертельной для них ошибки с применением иприта (см.
ниже «Нссостоявшиеся химические и биологические войны»).
В ноябре 1944 г. Мерк направил Стимсону и Маршаллу (но не Рузвельту) отчет
всего в 12 строк, где в скрытой форме говорилось о работе по получению еще четырех
«средств против человека». Судя по другим источникам, ими, вероятно, были возбу-
дители бруцеллеза (мальтийской лихорадки), пситтакоза (попугайной болезни), ту-
ляремии и сапа. По словам Мерка, химическая служба занималась также разработкой
«по крайней мере, пяти средств, пагубно воздействующих на растения». Эти средства
на самом деле представляли собой химические вещества, но вто время они были вклю-
чены в программу разработки БО, поскольку способны уничтожать посевы. Шестое
средство, тиоцианид аммония, предназначалось для уничтожения «японских садов».
Отчет Мерка был обнаружен в рассекреченных документах военного министра в Ва-
шингтоне. Свидетельства того, что сам Стимсон или президент интересовались на-
учной стороной программы, разумеется, отсутствуют (Берстайн Б. Дж., 1987).
В июне 1945 г. арсеналу Эджвуд были выделены дополнительные денежные сред-
ства на строительство холодильных установок. Расходы США на создание учрежде-
ний, занимающихся разработкой БО, во время Второй мировой войны составили
45-50 млн долларов. В послевоенные годы программа США по БО была резко сокра-
щена. В 1946— 1947 гг. ассигнования на научные исследования по ХБО в целом состави-
ли 6 млн долларов; в 1948-1949 гг. ассигнования лабораторий Форт-Детрика не превы-
шали 0,86 млн долларов. Тем не менее, в те годы в Форт-Детрике проводились иссле-
дования с возбудителями сибирской язвы, сапа, бруцеллеза, туляремии, мелиоидоза,
чумы, пситтакоза, кокцидиоидомикоза, с рядом патогенных микроорганизмов рас-
тений (Piricularia oryzae — болезнь риса; Helminthosporium oryzae—красная пятнистость
риса; Phytophthora iirfestans — фитофтороз; Puccinia graminis — линейная ржавчина зла-
ковых) и с возбудителями болезней животных и птиц (вирус чумы рогатого скота, вирус
ньюкаслской болезни, вирус классической чумы птиц) (Rosebury Т., 1949). Катим
возбудителям, а этот список, бесспорно, не является исчерпывающим, следует доба-
вить токсины С. botulinum, вирус менингопневмонии, который изучали, главным обра-
зом, как модель вирусных агентов, подобно В. globigii и .S’, marcescens, которые исполь-
зовали как модели бактериальных агентов (Rosebury Т, 1947).
Помимо Военного министерства разработкой вопросов, связанных с БО, занима-
лось Военно-морское министерство США, организовав собственные центры. Одним
из них стал Военно-морской медицинский центр № 1, функционировавший при фа-
культете бактериологии Калифорнийского университета. Основным объектом его
9. Биологическое оружие перед Корейской войной
197
^следований был возбудитель чумы. Одновременно работы проводились с некото-
лми вирусами. В конце войны в лаборатории работало 75 специалистов. В январе
*46 г. она составила ядро группы, представленной большей частью специалистами
калифорнийского университета, которой была поручена разработка программы БО
ВМФ США. В 1950 I. группа была переведена в центр снабжения ВМФ в Окленде и
:е основе была создана биологическая лаборатория ВМФ. На момент прекраще-
на наступательной программы по БО в 1968 г. она оставалась основным центром
ВМФ США в области исследований по БО (The Problem..., 1970).
После войны отдельные ученые сочли своим долгом объяснить общественности
с сое участие в столь «недостойном деле», как создание БО. Розбери писал: «Мы боро-
ть с огнем (державами «оси Берлин — Рим») и неизбежно рисковали — либо быть
запачканными, либо сожженными». В 1942 г. он был назначен руководителем иссле-
ований в лаборатории химической службы в Форт-Детрике и внес очень большой
-клад в развитие американской программы создания БО. Среди персонала лабора-
торий Форт-Детрика за период 1943—1946 гг. было зарегистрировано 25 случаев забо-
йния сибирской язвой, 17 — бруцеллезом, 7 — туляремией, 6 — сапом и 1 — псит-
~1козом; но обошлось без летальных исходов. Полученные научные данные не были
«умерщвлены» в архивах под предлогом секретности. Всего за период с октября 1945 г.
о январь 1947 г. американскими учеными опубликовано 156 научных статей, посвя-
щенных проблемам защиты от БО. В целом работы американских ученых, участво-
_лвших в программе создания БО, серьезно обогатили мировую науку.
Начало программы по созданию БО в Соединенном Королевстве. Соединенное
Королевство признало, что оно имело программу разработки БО с 1940 г. по конец
950-х гг. (Report from the United Kingdom..., 1992). Есгественно, британский интерес
и озабоченность по поводу БО связаны с подозрением о проведении Германией ис-
следований в этой области. Даже в работах 1990-х гг. необходимость создания БО до-
казывалась «расследованиями» Стида (см. разд. 1.7). По официальным данным, еще в
*36 г. при Комитете обороны Соединенного Королевства было создано специаль-
ное учреждение, в задачи которого входило изучение вопросов, связанных с био-
логической войной.
Когда в 1939 г. началась война с Германией, канадский нобелевский лауреат Фре-
дерик Бантинг (Frederick Banting, 1891—1941), один из откр! шателей инсулина, убеж-
денный в том, что лица, контролировавшие германскую армию, безжалостны и кро-
вожадны, а уже по этим причинам они будут обязательно использовать БО против
западных демократий, и что Британия не готова выдержать атаки с применением БО,
направил британскому правительству аргументированный план по исследованиям в
•бласти создания биологических вооружений и использованию БО. Бантинг нарисо-
вал очень впечатляющую картину последствий применения немцами БО по городам
Соединенного Королевства с аэропланов и, что интересно, распространением порош-
ка из сухих бактерий (в 2001 г. в США нечто подобное называли «белым порошком»)
через почтовые конверты.
В 1940 г., в разгар германских воздушных атак, лорд Хенкеу (Maurice Hankey),
бывший секретарь британского кабинета министров, весьма серьезно воспринял идеи
Бантинга и убедил кабинет начать программу по разработке БО (Guillemin J., 2006).
Работы по созданию британского БО, по официальным данным, начаты в 1940 г.
на базе Королевской инженерной испытательной станции (Royal Engineers Experi-
mental Station) в Нортон Дауне (графство Уилтшир). Станция была создана еще в 1916 г.
для проведения исследований по созданию химического оружия. Разработку БО воз-
198
Часть 1. Краткая история применения биологических средств...
главил сэр Пауль Филдс (Sir Paul Gordon Fildes, 1882—1971), известный к тому време-
ни своими работами в области сульфаниламидов, гемофилии и сифилиса, член Ко-
ролевского научного общества. Персонал станции, занимавшийся БО, насчитывал
45 человек, включая 15 ученых. Основные усилия в области химического оружия были
сосредоточены на изучении поражающих свойств азотистого иприта. Задача создания
БО не казалась особенно сложной. Для руководства исследованиями в области созда-
ния БО британское правительство создало специальную организацию — Департамент
микробиологических исследований, входящий в Департамент химических оборонных
исследований.
Основываясь на представлениях о химической войне, британскими специалиста-
ми было решено, что наиболее эффективным способом применения БО должно быть:
«...диссеминирование аэрозоля с размером частиц, обеспечивающим их задержку в
легких, из жидкой суспензии бактерий за счет взрыва боеприпаса, такого как авиабом-
ба, с таким расчетом, чтобы на площади цели любой, находящийся там, мог вдохнуть
эффективную концентрацию...». Британский подход к созданию биологических бое-
припасов мало чем отличался от японского. Но в качестве наиболее перспективного
агента БО ими рассматривался не возбудитель чумы, а возбудитель сибирской язвы;
в меньшей степени их интересовал ботулинический токсин. Так, по крайней мере,
пишется в официальной истории центра (см. Carter G. В., 1993). Британцы гораздо
тщательнее прятали полученные результаты, чем американцы. По официальным дан-
ным, в результате работ в Портоне за годы войны всего-то и были определены инфи-
цирующие дозы возбудителя сибирской язвы, достаточные для гибели нескольких
видов животных — работа для двух сотрудников максимум на месяц (один держит
животное, другой вводит этому животному возбудитель инфекции). Еще они обычно
сообщают, что в ходе испытаний на полигонах Нортона с использованием безобид-
ного микроорганизма Bacillus subtilis была разработана методика применения бомб для
эффективного рассеяния спор возбудителя сибирской язвы. На основании этих дан-
ных ими рассмотрена необходимость проведения испытаний с реальным агентом в
безопасном месте острова Груинард в Шотландии.
Далее британцы рассказывают о тупиковых направлениях своих исследований,
выдавая их за очень успешные. Как, например, то, что в этих опытах использовалась
модифицированная 30-фунтовая химическая авиабомба с высокоактивным взрывча-
тым веществом, снаряженная примерно тремя литрами суспензии сибиреязвенных
спор, рассеиваемых возле земли, а если точнее, то разбрызгиваемых по поверхности
почвы и по шерсти экспериментальных животных. Испытания 1942 г. продемонстри-
ровали якобы возможность поражения овец с помощью такого оружия, что если
и происходило, то только в результате поедания этими овцами травы на полигоне
У исследователей из Портон Дауна сложилось убеждение в том, что оно более мощ-
ное, чем химическое оружие или обычный боеприпас такого же размера, по крайней
мере, так они представляли дело перед чиновниками Департамента химических обо-
ронных исследований. Поэтому в 1943 г. Филдс получил финансирование для иссле-
дования 4-фунтовых бомб, которые могли бы «быть еще более эффективными, если
бы применялись в виде кассет». Затем работы над кассетной бомбой, субэлементы
которой снаряжены культурой возбудителя сибирской язвы (N-бомба), были перене-
сены в Соединенные Штаты и Канаду. Так началось длительное сотрудничество между
этими странами в разработке БО.
Фактически до 1943 г. группа Филдса была лидером в разработке БО, обладая все-
ми методологическими данными, собранными и направленными в Соединенные
1.9. Биологическое оружие перед Корейской войной
199
Штаты, где они послужили «основой для последующей программы США в области
БО» (Carter G. В., 1991). Правда, за все время Второй мировой войны требования Во-
енного кабинета о создании в Портоне БО возмездия реализовались лишь в производ-
стве пяти миллионов кормовых брикетов, контаминированных спорами возбудителя
сибирской язвы. Их предполагалось сбрасывать на пастбища Германии с самолетов,
используя корпуса осветительных бомб. Брикеты в упакованном виде, обеспечиваю-
щем герметичность, хранились на секретном британском исследоват ельском объек-
те вблизи Портона и после войны были уничтожены. По замечанию G. В. Carter (1991),
о возможных последствиях применения этого оружия для мясомолочной промышлен-
ности Германии остается только предполагать, но оно имеет историческое значение
как, возможно, первое достоверно созданное БО.
Для исследований поражающих свойств БО, во время войны в Портоне было по-
строено кирпичное здание, имеющее в законченном виде площадь 19,5 тыс. м2, кото-
рое в течение десятилетий считалось самым лучшим по конструкции и оснащению
микробиологическим институтом в мире. В период существования мер строжайшей
экономии после войны на этот единичный элемент программы по БО было выделено
2,25 млн английских фунтов стерлингов.
Наверное, исключительно из-за реальной боевой эффективности нового оружия,
более мощного, чем химическое», служба вооружений после окончания войны была
особенно озабочена «необходимостью достижения лучшего понимания факторов,
влияющих на выживаемость патогенных микроорганизмов в аэрозоле».
Счит алось, что добиться «лучшего понимания» наиболее безопасно можно в море,
и в ноябре 1948 г. были проведены испытания в районе Багамских островов, в кото-
рых для проверки эффективности боеприпасов использовались пробоотборные уст
ройства и животные, находящиеся в резиновых лодках с подветренной стороны от
устройства. С помощью такого способа устанавливали корреляцию между фактора-
ми, влияющими на физический распад, жизнеспособность и вирулентность патоген-
ных микроорганизмов в аэрозоле как в реальных условиях, так и в условиях лабора-
торных исследований.
Аналогичные испытания проводились в 1952—1953 гг. недалеко от побережья
Шотландии и в 1954—1955 гг. вновь вблизи Багамских островов. По утверждению
М. Dando (1994), такой масштаб ресурсов, расходуемых на программу БО в то время,
отражал официальную озабоченность, выразившуюся в стремлении получить пра-
вильное представление о его потенциальной опасности.
Военно-биологическая программа Франции. Крупномасштабная программа иссле-
дований в области БО существовала в стране с начала 1930-х гг. до ее разгрома в июне
1940 г. В основном она была направлена на изучение вььживаемости бактерий и виру-
сов при хранении и взрывном диспергировании. Когда немцы захватили в 1940 г.
французскую агробиологическую лабораторию в Ля Буше, они осознали, что далеко
отстали в научных исследованиях по БО (Hugh-Jones М., 1992). Однако франщ зам БО
(или то, что они за него тогда принимали) не помогло в войне с Германией. После
войны они не комментировали германские обвинения военного времени. Имеются
неясные сведения об испытании французского БО в Алжире в начале 1960-х гг.1, но в
целом послевоенные наступательные программы Французской Республики в области
БО были плотно прикрыты контрразведкой и никогда не сопровождались «утечками
информации» или какими-то скандалами.
1 Например, в книге Е. Я. Сатановского (2012).
200
Часть 1. Краткая история применения биологических средств...
Биодиверсии в Европе в годы Второй мировой войны. Ни один из ставших извест-
ным эпизодов не противоречит реальным возможностям военной микробиологии того
времени. Наиболее очевидной биодиверсией многие специалисты считают ликвида-
цию 27.02.1942 г. английскими спецслужбами Рейнгарда Гейдриха — заместителя
имперского протектора Богемии и Моравии, слишком энергично решавшего «еврей-
скую проблему» на территории, вверенной ему фюрером.
Исследования ботулинического токсина составляли часть программы по созданию
БО Соединенного Королевства. В октябре 1941г. британская секретная служба (Secret
Intelligence Service, SIS) попросила Пауля Филдса помочь в проведении операции
«Антропоид», т. е. в ликвидации Гейдриха. Группа Филдса изготовила специально
модифицированную ручную противотанковую гранату № 73. Верхняя треть гранаты
была обернута клейкой лентой, пропитанной веществом «ВТХ», т. е. ботулиническим
токсином. Операция «Антропоид» в целом была неудачной. Чешские диверсанты
действовали неумело, выданное им британцами оружие давало осечки. Ранения Гей-
дриха, полученные в результате взрыва гранаты, оказались незначительными (оскол-
ки в области груди и селезенки), и он даже был в состоянии преследовать нападавших.
Хирургическая операция Гейдриху была проведена хорошо, даже по современным
стандартам. В послеоперационном периоде его состояние было удовлетворительным,
в ране был установлен дренаж, ухудшения его состояния ничего не предвещало, хотя
и была небольшая температура. Его состояние стаю ухудшаться на седьмой послеопе-
рационный день и на следующий день, рано утром, он умер (Sidel Е et al., 1997).
Вскрытие показало отсутствие каких-либо патоморфологических причин смерти.
Не было ухудшения состояния раны, признаков перитонита, абсцессов, инфициро-
вания раны или не извлеченных инородных предметов. Сердце и легкие не имели
видимой патологии. Главный германский патологоанатом, присутствовавший на
вскрытии Гейдриха, записал в протокол вскрытия: «...смерть произошла вследствие
занесения бактерий или яда осколком бомбы».
Хотя и нет официальных документов, но известно свидетельство самого Пауля
Филдса, как-то заметившего коллегам, что он имел отношение к смерти Гейдриха.
И его смерть стала «первой зарубкой» на его пистолете (Davis R. А., 1971; Mobley J. А.,
1995). Сколько было еще таких «зарубок» у британских производителей ботулиниче-
ского токсина?
Свой собственный вариант биологической войны осуществляли партизаны
Польши и Чехии. В декабре 1942 г. гестапо обнаружило в Варшаве целый арсенал,
предназначенный для биологическихдиверсий против германских войск (Mobley J. А.,
1995). Германия неоднократно заявляла о том, что русские используют биологические
агенты в диверсионных целях (Hugh-Jones М., 1992).
В период военных действий на Восточном фронте германское командование осу-
ществляло биологические диверсии, по своему исполнению более характерные для
Первой мировой войны, но весьма эффективные и не нарушающие явным образом
Женевский протокол 1925 г. Одна такая диверсия была раскрыта зимой 1942 г., когда
из лагеря для военнопленных был организован побег красноармейцев, среди которых
были сильно завшивленные и больные сыпным тифом. Этих людей немцы свободно
пропустили через линию фронта на территорию, занятую нашими войсками. Они
были обнаружены, на пути их следования возникло несколько случаев заболеваний
сыпным тифом среди контактировавших с ними лиц (Болдырев Т. Е., 1955).
Другая раскрытая биологическая диверсия планировалась немецким командова-
нием в еще больших масштабах. В 1944 г. наступающие части одного из Белорусских
1.9. Биологическое оружие перед Корейской войной
201
фронтов обнаружили в районе местечка Озаричи Полесской области, недалеко от
переднего края немецкой обороны, три концентрационных лагеря, в которых в ужа-
сающих условиях содержалось около 33 тыс. человек. Среди военнопленных свиреп-
ствовал сыпной тиф. Расследованием Чрезвычайной государственной комиссии был
установлен искусственный характер всех трех вспышек. Для их инициации из различ-
ных районов Полесской, Гомельской, Минской и других оккупированных областей
БССР в лагеря свозили гражданских лиц, больных сыпным тифом, и размещали их
среди здоровых военнопленных. Расчет был сделан на то, что переброска больных
сыпным тифом из лагерей военнопленных втыловые районы страны, вызоветтам эпи-
демии сыпного тифа, а непосредственное соприкосновение с ними личного состава
наступающих частей Красной армии повлечет распространение сыпного тифа на
фронте (Сборник сообщений..., 1946).
В конце 1941 г. в концентрационном лагере Бухенвальд было создано экспери-
ментальное учреждение для установления эффективности различных живых сыпно-
тифозных вакцин. Эта учреждение размещалось в блоке 46. Курировал работу Ин-
ститут гигиены войск СС в Берлине, возглавляемый Иоахимом Мруговским. Однако
при попытке поддерживать культуры риккетсий Провачека вирулентными и нараба-
тывать их в количествах, хотя бы достаточных для осуществления экспериментов,
немцев преследовали те же неудачи, что и японцев (см. разд. 1.8). Недостатка в чело-
веческом экспериментальном материале у них не было, с осени 1942 г. до лета 1943 г.
для этих опытов было использовано около 500 заключенных лагеря Бухенвальд. А вот
заражать заключенных было нечем. Уже в апреле 1943 г. штурмбанфюрер СС д-р Динг
сообщил Мруговскому, что вызвать заболевание у заключенных введением им виру-
лентного еще год назад штамма «Мательска» не удается. Нарабатываемые на кури-
ных эмбрионах риккетсии Провачека утратили способность вызывать сыпной тиф у
людей. До конца войны немцы не смогли решить эту проблему. Для заражения людей
сыпным тифом они использовали внутривенные инъекции 2 мл только что взятой
крови больного сыпным тифом. Также им не удалось создать вакцину, эффективно
защищающую от сыпного тифа (СС в действии..., 1969). Использование немцами
советских военнопленных в опытах с возбудителями различных болезней, сопровож-
давшихся их гибелью, на Нюренбергском процессе не было отнесено к применению
БО (The Problem..., 1970).
Несостоявшиеся химические и биологические войны. Вторую мировую войну нача-
ли страны, располагавшие колоссальными запасами химического оружия и амбици-
ями в области БО. За 7 лет мировой бойни ее участники неоднократно стояли перед
порогом, за которым война могла выйти за рамки Женевского протокола 1925 г., мно-
гократно увеличив число своих жертв и, возможно, поменяв местами победителей и
побежденных.
По итогам Первой мировой войны безусловным «лидером» среди ОВ стал иприт.
Поэтому работы стран-победительниц велись в направлении совершенствования ОВ
кожно-нарывного действия и средств их применения. Эти работы ознаменовались
получением в 1918 г. люизита, а в 1935—1936 гг. «азотистых ипритов» (N-Lost) и «кис-
лородного иприта» (O-Lost). Но они не выходили за рамки общего уровня знаний того
времени. До самого конца войны союзники СССР по антигитлеровской коалиции не
смогли установить реальных разработок немцев в области фосфорорганических ОВ
(табун, 1936; зарин, 1939; зоман, 1944).
В СССР, возможно, знали о многих программах по германскому химическому ору-
жию благодаря Вильгельму Леману (1884—1942), сотруднику гестапо, работавшему
202
Часть 1. Краткая история применения биологических средств...
на иностранный отдел ОГПУ под оператигным псевдонимом Брайтенбах. Леман с
1936 г. руководил контрразведывательным обеспечением ряда направлений военной
промышленности Германии. Информация, поступавшая советской разведке от Лема-
на, касалась германской ракетостроительной программы, программы строительства
подводных лодок, засекреченного завода по производству боевых ОВ, новых проти-
вогазов и др. Леман передал разведчику-нелегалу В. М. Зарубину (1894—1972) копию
инструкции, в которой перечислялись 14 видов новейшего вооружения Германии,
находившегося на стадии изготовления или проектирования (Владимиров С., 2010).
По мнению Н. С. Антонова (1994). благодаря новому поколению боевых ОВ, Гер-
мания получила явное преимущество перед своими противниками в области хими-
ческих вооружений. В случае развя идвания химической войны противниками Герма-
нии, применение немецкой армией зарина, зомана и табуна поставило бы перед
союзниками неразрешимые до конца войны проблемы зашиты войск и населения от
этих ОВ. Ответное применение иприта, фосгена и люизита, составлявших основу
химического арсенала США, Великобритании и СССР, не обеспечило бы адекватно-
го эффекта. У союзников отсутствовали соответствующие антидоты, газосигнализа-
торы, дегазирующие растворы, импрегнированное обмундирование. Война могла
пойти по этому сценарию уже в конце 1940 г., если бы Гитлер решился на операцию
«Морской лев». После войны стало известно, что на высшем уровне британского руко-
водства было принято решение использовать боевые ОВ в такой ситуации в качестве
последнего средства, «если все другие обычные способы обороны окажутся не состо-
ятельными». Британцами планировалось с помощью авиации применить иприт по
плацдармам, захваченным германскими войсками (см. вкн. Ширера У., 1991; с. 171).
Малая химическая война против Красной армии велась немцами с первых дней
Великой Отечественной войны. В средине 1930-х гг. германской фирмой Rhemmetall-
Borsig был разработан 7,92-мм патрон 318 с остроконечной бронебойно-химическо-
трассирующей пулей SmKH-Rs-L’spur. Пуля массой 14,55 г содержала бронепроби-
вающий сердечник из особо твердого сплава с большим удельным весом (15 г/см3),
состоящий из 92 % вольфрама, 4 % никеля и 4 % углерода. В донной части пули в зап-
рессованном состоянии находилось 0,3 г хлорацетофенона (ОВ раздражающего дей-
ствия) и трассер. Для стрельбы такими патронами использовалось противотанковое
ружье PzB39. Начальная скорость пули составляла 1115 м/с. С расстояния 50 м она
пробивала 30-мм броню, установленную под углом 30 градусов Попадание такой пули
в область размещения боекомплекта, бензобака или двигателя приводило к пожару или
подрыву боевой машины. Если повреждения механизмов не происходило, поражение
экипажа происходило в результате выделения хлорацетофенона. Внутренний объем
легкого и среднего советского танка составлял 14 тыс. л и 22 тыс. л. В этих объемах
создавалась концентрация хлорацетофенона 0,021 мг/л и 0,014 мг/л соответственно.
Этого было достаточно, чтобы экипаж покинул практически исправную боевую ма-
шину. Химическая составляющая патрона 318 была установлена только к началу ян-
варя 1942 г. после проведения исследований в Ленинградском химико-технологичес-
ком институте и Государственном институте прикладной химии. К этому времени
СССР уже потерял большую часть бронетехники, на которую в предвоенный период
возлагались большие надежды (Понамарев Ю., 2011).
Большая химическая война могла начаться осенью 1941 г. на советско-германском
фронте. Известен факт обстрела в 1941 г. под Керчью советских позиций из реактив-
ных минометов Nevelwerfer-41 химическими снарядами в ответ на применение совет-
i.9. Биологическое оружие перед Корейской войной
203
скими войсками ракетно-зажигательных снарядов PC3-132. Снаряд PC3-132 снаря-
жался 36 зажигательными элементами термита марки «6» массой 4,2 кг каждый и пред-
назначался для стрельбы из установки залпового огня БМ-13-16, известной еще как
«Катюша» (конструктор установки В. Н. Галковский). В одном залпе «Катюша» выс-
треливала 1500 зажигательных элементов. Для взаимного перекрытия секторов раз-
лета элементов устанавливали различное время срабатывания неконтактных взрыва-
телей. При воздушном подрыве PC3-132 на позициях противника создавалось
множество очагов пожаров, потушить которые было невозможно. Температура горе-
ния термитной смеси достигала 2700°С. Попадая в снег, горящий термит разлагал воду
на кислород и водород, образуя «гремучую смесь» газов, увеличивая и без того силь-
ное горение. При попадании термита на броню танков и стволы орудий легирован-
ная сталь меняла свои свойства, и боевая техника уже не могла быть использована.
Именно такие снаряды были использованы во время впечатляющего «дебюта» «Ка-
тюши» под станцией Орша 14 июля 1941 г. Осуществив обстрел химическими снаря-
дами позиций советских войск под Керчью, немцы продемонстрировали советскому
командованию, что готовы пойти на нарушение Женевского протокола 1925 г., если
применение снаряда PC3-132 будет продолжено. До конца войны этот снаряд совет-
ской армией больше не применялся (Ардашев А. Н., 2009).
С. Херш (1970), ссылаясь на мемуары Д. Лилиенталя (1964), бывшего председате-
ля комиссии по атомной энергии, писал, ч го начальником Генерального штаба армии
США генералом Дж. Маршаллом в конце войны были подготовлены рекомендации
по применению химического оружия против Японии. Господство в воздухе ВВС США
делало план весьма заманчивым и относительно безопасным на Тихоокеанском теат-
ре военных действий. Однако реализовать его не пришлось из-за жесткого противо-
действия У. Черчилля. Британский премьер хорошо усвоил уроки химической войны,
преподнесенные германскими военными химиками войскам Антанты в 1915-1918 гг.,
и серьезно опасался, что Германия вновь применит боевые ОВ против Соединенного
Королевства, Поэтому американским командованием было принято решение исполь-
зовать химическое оружие только в качестве ответной меры при нарушении Японией
Женевского протокола. Таким образом, от развязывания химической войны, по край-
ней мере, одну из воюющих сторон, удерживал не Женевский протокол 1925 г., а эле-
ментарный страх возмездия.
Но порог, за которым война из обычной превращается в химическую и бактерио-
логическую, едва не был пройден в августе 1945 г. уже после разгрома Германии. Спу-
стя несколько лет после войны, Трумэн в письме одному из своих помощников наме-
кал на то, что если бы война в Тихом океане затянулась до середины августа 1945 г., то
он одобрил бы решение о применении как бактериологических средств, так и отрав-
ляющих веществ. Санкционированная им атомная бомбардировка имела гораздо
более губительные последствия, применение других видов оружия массового пора-
жения не потребовалось. Но еще 3 августа 1945 г., т. е. за три дня до бомбардировки
Хиросимы, генерал-лейтенант И. К. Икер (заместитель командующего ВВС США
генерала X. X. Арнольда) потребовал от химической службы представить ему доклад
о возможности уничтожения посевов риса на японских островах с воздуха. Доклад
должен был включать сведения о самых сильнодействующих химических веществах
и доступных средствах и способах их доставки к цели. Доклад он получил 10 августа
1945 г., на следующий день после бомбардировки Нагасаки. Через 4 дня война в Ти-
хом океане закончилась (Берстайн Дж., 1987).
204
Часть 1. Краткая история применения биологических средств...
Производство биоагентов. Расширение исследований по БО в 1940-х гг. потребо-
вало увеличения мошностей по производству биоагентов. Первоначально наращива-
ние объемов производства бактериальной массы шло за счет увеличения посевной
поверхности аппаратов тоги же типа, что применялся сначала Н. Г Щербиной, а за-
тем и Исии (рис. 1.32).
Рис. 1.32 Сотрудники Порт Дауна получают бактериальную массу. На заднем плане
аппарат, устроенный по принципу аппарата Н. Г. Щербиной (1932), представляющий со-
бою серию посевных плоскостей, уложенных в особый футляр. Снимок начала 1950-х гг
Такие объемы производства могут удовлетворить лишь производителей вакцин.
Только для ударов по Берлину химическая служба США должна была подготовить к
1945 г. 5 тыс. кассетных боеприпасов, снаряженных спорами возбудителя сибирской
язвы. Для снаряжения сотен тысяч биологических боеприпасов, которых могла по-
требовать биологическая война с упорным и защищенным противником, каким был
тогда СССР, технология выращивания микроорганизмов на поверхности плотных сред
не могла быть масштабирована в принципе. Тем более что разработчики БО должны
были учитывать интересы тех, кто его применяет. Военные стремятся или будут стре-
миться к тому, чтобы рассеивать большие количества биологических агентов в расчете
на достижение предусмотренных результатов прямым путем через вдыхание биоло-
гического агента или контакт с ним, а не косвенно через эпидемии. Они будут при-
менять БО массированно, создавая избыточные дозы агента в аэрозолях из-за неуве-
ренности в достижении необходимого эффекта.
Поэтому интерес исследователей еще в годы войны стал смещаться в сторону тех-
нологий глубинного культивирования микроорганизмов, уже достаточно отработан-
ных для получения пенициллина в промышленных масштабах. Однако путем заим-
ствования технологий, разработанных для производства антибиотиков, получить
биологический компонент для «мощного оружия бедных» вновь не удалось. В ходе
экспериментов выяснилось, что потребность бактерий в аэрации гораздо выше, чем
у плесневых грибков, что связано с их более интенсивным метаболизмом. Изучение
этого вопроса показало непригодность методов, используемых для аэрации грибко-
вых культур, для культивирования бактерий. «Выходы» бактерий получались низки-
1.9. Биологическое оружие перед Корейской войной
205
ми. Патогенные микроорганизмы, будучи типичными гетеротрофами, способны асси-
милировать азот только в виде аминного соединения из высокомолекулярных азоти-
стых веществ, являющихся продуктами белкового распада. К ним принадлежат пепто-
ны, полипептиды и аминокислоты. Белковые вещества легче усваиваются бактериями
после предварительного расщепления их до аминокислот. Процесс расщепления бел-
ковых молекул происходит под действием протеолитических ферментов, выделяемых
микробными клетками. Однако протеолитическая активность бактерий чрезвыч im-
ho разнообразна. Например, она резко выражена у возбудителей анаэробной инфек-
ции и очень слабо — у возбудителя чумы. Военным биотехнологам необходимо было
заново изучать максимальные потребности бактерий в кислороде и углекислом газе,
азотистых веществах и других компонентах среды, создавать принципиально новые
конструкции реакторов, масштабировать и контролировать процессы, происходящие
во время культивирования (рис. 1.33).
Приемная
бутыль
Вакуумный
насос
Флакон с запасом Флакон для непрерывного
питательной среды культивирования
Нис. 1.33. Схематическое изображение первого лабораторного культиватора для непре-
рывного глубинного выращивания бруцелл. Разработан в Кемп-Детрике в 1945 г. Фил-
липом Герхардом: 1 — фильтр; 2 — зажим; 3 — разбрызгиватель; 4 — стандартное отвер-
стие; 5 — капиллярная трубка Культиватор работал следующим образом. Запасы
стерильной питательной среды для выращивания бруцелл находятся в специальном
флаконе. Ее пополнение происходит в асептических условиях с помощью трубки для
отбора проб. Среда (400 мл) поступает в сосуд для культивирования через капиллярную
трубку. Перетекания среды между сосудами вследствие изменения гидростатического
давления (hydrostatic pressure) не происходит из-за компенсирующего эффекта возду-
ха, поступающего через впускающий капилляр, достигающего дна флакона с питатель-
ной средой. Процесс культивирования бруцелл регулировался скоростями поступления
питательной среды и отбора продукта. Воздух добавлялся с постоянной скоростью —
400 мл/мин. Пенообразование контролировалось добавлением пеногасителей. Систе-
ма достигала равновесия в течение 24 ч. Продукт собирался каждые 8 ч в большой при-
емный флакон, после чего в сосуд для культивирования подавалась питательная среда,
и цикл культивирования повторялся. Аппарат мог работать в таком режиме неделями,
количество бруцелл достигало 40х109 клеток/мл. По F. Gerhard (1946)
Констпукция реакторов для производства агентов БО принципиально отличалась
от используемых при производстве антибиотиков. В 1990-х гг. и в первые годы этого
столетия, когда администрации США необходимо было обосновать причины напа-
дения на Ирак, в СМИ «запускалась» информация о том, что нет ничего проще, чем
206
Часть 1. Краткая история применения биологических средств...
перепрофилировть заводы по производству антибиотиков, пальмового масла и пиво-
варенные производства на производство биологических агентов. Однако в самих США
в 1940—1950 гг. по такому пути не пошли, так как перевод этот связан с непреодолимы-
ми трудностями. Ферментационные процессы при производстве антибиотиков про-
водятся под давлением, чтобы не допустить загрязнения кулгтуры плесневых грибов
посторонними микроорганизмами. Это представляется вполне возможным, посколь-
ку небольшая утечка плесневых грибов наружу не грозит неприятностями персоналу.
При производстве агентов БО соображения безопасности вынуждают осуществлять
процесс культивирования при отрицательном давлении в культиваторах, что приводи!
к быстрому загрязнению микробных культур посторонней микрофлорой. При пере-
ходе к крупномасштабному производству агентов БО, помимо отработки биотехноло-
гических процессов, военным биотехнологам необходимо было обеспечить гермети-
зацию зданий, оборудования и коммуникаций, эффективную стерилизацию стоков
и отработанного воздуха, сжигание погибших животных, утилизацию использован-
ных питательных сред и т. п.
Для глубинного культивирования микроорганизмов в промышленных масштабах,
военным биотехнологам потребовались сотни тонн жидких стерильных питательных
сред. Для приготовления и хранения сред им необходимо было сконструировать со-
ответствующие емкости, оборудование для стерилизации и транспортировки и еще
очень многое другое. Вот только несколько примеров трудностей, возникающих при
масштабирования процессов получения агентов БО.
Прежде всего, произошло снижение воспроизводимости получаемых результатов.
Замена дорогих питательных сред дешевыми привела к трудностям контроля качества
входящих в их состав компонентов. Увеличился риск кон гаминации посторонней
микрофлорой готовой питательной среды. Так, один жизнеспособный микроорганизм
в 1000 л среды эквивалентен 100 жизнеспособным микроорганизмам в 100 тыс. лит-
рах среды. Но при лабораторном производстве из ста 10-литровых ферментеров дол-
жен контаминироваться один, в то время как 100000-литровая партия контаминиру-
ется вся, если в обоих случаях используется одинаковое количество среды и тепла,
расходуемого на стерилизацию (Buckland В. С., 1984).
Во время стерилизации питательной среды могут происходить важные изменения
в ее составе, а некоторые питательные вещества вообще способны разрушаться. Ско-
рости разрушения компонентов среды и контаминирующих ее спор, различны. Энер-
гия активации, необходимая для деструкции факторов роста среды, находится в
диапазоне 10—30 ккал/моль. Для гидролиза казеина она составляет 20,6 ккал/моль.
В то же время энергия активации, необходимая для тепловой деструкции спор, дос-
тигает 100 ккал/моль (Buckland В. С., 1984).
К этому «букету» из трудно разрешимых технических противоречий, возникаю-
щих в процессе культивирования бактерий, следует добавить необходимость разра-
ботки нового специального оборудования для концентрирования бактериадьных кле-
ток и спор, сублимационной и распылительной сушки, измельчения, приготовления
соответствующих рецептур, снаряжения боеприпасов или боевых частей, захолажг -
вания, стерилизации и т. п. Каждое устройство, систему, технологию, режим воен-
ным биотехнологам пришлось разрабатывать самостоятельно, учитывая специфику
работы с каждым микроорганизмом и опираясь лишь на фундаментальные науки.
Например, получение возбудителя сибирской язвы для снаряжения субэлементов
кассетных боеприпасов требовало от них не просто получения тонн культуральной
жидкости, содержащей В. anthracis, а проведение процесса в таких условиях, когда
.9. Биологическое оружие перед Корейской войной
207
бациллы будут превращаться в споры. Потом надо было оптимизировать эти условия
таким образом, чтобы выход спор был наибольшим. Далее предстояло разработать
критерии для оценки «качества» получаемого «продукта», например, изучить способ-
ность спор вызывать инфекционный процесс при разных способах заражения. Изу-
чить их способность переносить лиофильное высушивание, измельчение лиофилиза-
та, перевод в аэрозольное состояние и т. п. Затем надо было воспроизвести достигнутое
таким трудом со следующей партией питательной среды или при масштабировании
процесса. Все эти процессы, технологии, передаточные шлюзы, гидрозатворы, силь-
фонные вентили, сальниковые уплотнения, системы тепловой обработки сточных вод,
проходные автоклавы, специальная техника безопасности потребовали от стран, раз-
рабатывавших БО, незаурядных людей и немалые деньги.
Разработка технических средств для индивидуальных убийств. Очен ь немногие био-
логические агенты способны вызвать быструю смерть человека. Среди них нет ни
одного, который бы оказался способен проникнуть через неповрежденную кожу.
Поэтому в годы Второй мировой войны спецслужбами воюющих стран было разра-
ботано специальное оружие и стреловидные боеприпасы к ним, имеющие либо на
наружной поверхности специальные желобки для нанесения химических или биоло-
гических агентов, либо они имели специальную полость для заполнения таким и аген-
тами (рис. 1.34).
Специалистами Управления специальных операций (Special Operation Executive,
SOE) Соединенного Королевства среди прочих неджентльменских инструментов ве-
дения войны была создана специальная авторучка, выбрасывающая струю ядовитого
вещества (Кукридж Э., 2003).
Из биологических токсинов в те годы наиболее доступными для спецслужб были
ботулинические. Их тогда было известно несколько серотипов (А, В, С, D, Е). Не
существовало методов обнаружения ботулинического токсина в теле погибшего чело-
века, если он вводился парэнтерально. Программы исследований по повышению вы-
хода токсинов А- и В-типов из культур С. botu-
linum осуществлялись в США, Соединенном
Королевстве и Канаде (см. рис. 1.34). Они со-
провождались исследованиями по распыли-
тельной и сублимационной сушке токсинов,
измельчению сублимированных препаратов и
превращению их в тонкий порошок (The Prob-
lem..., 1970).
Тогда же были предприняты первые попыт-
ки выделить очищенную токсическую субстан-
цию сакситоксина (см. разд. 3.14), что оказа-
лось трудоемким делом. В некоторых случаях
для получения одного грамма чистого токсина
приходилось перерабатывать до 8 т моллюсков.
Тем не менее сакситоксин был получен в коли-
чествах, достаточных для осуществления инди-
видуальных убийств. Этот низкомолекулярный
яд хорошо растворяется в воде, по токсичности
он не уступает рицину, но в отличие от него вы-
держивает кипячение и даже автоклавирова-
ние при 120°C. Его жертвы историкам следует
Рис. 1.34. Стреловидные элементы аме-
риканских спецслужб, предназначен-
ные для индивидуальных убийств с по-
мощью химических или биологических
агентов. Такое оружие было запрещено
«Конвенцией о законах и обычаях су-
хопутной войны», подписанной США
в 1907 г. в Гааге. По R. Sidell et al. (1997)
208
Часть 7. Краткая история применения биологических средств...
поискать среди политических и военных деятелей Третьего рейха, скоропостижно
скончавшихся на фоне расстройства мышечной координации, нарастания слабости,
головокружения и сонливости.
Управлению специальных операций Соединенного Королевства удалось добыть
у индейцев Венесуэлы яд кураре, которыми те смазывают наконечники своих стрел
Яд был доставлен в Лондон в специальной бамбуковой упаковке. В каких операциях
он применялся, разумеется, не сообщается (Кукридж Э., 2003).
После войны разработка токсинов и технических средств для индивидуальных
убийств продолжилась. В феврале 1975 г. в хранилище отдела технического обслужи-
вания ЦРУ было обнаружено 11 г сакситоксина (этого количества достаточно для того,
чтобы убить, по меньшей мере, 14 тыс. человек) и 8 г яда кобры. При расследовании
этого факта незаконного хранения токсичных материалов выяснилось, что в 1948 г.
по соглашению между ЦРУ и руководством Министерства обороны США в бывших
биологических лабораториях армии США в Форт-Детрике был создан особо секрет-
ный отдел специальных операций, который работал в интересах ЦРУ.
Задачи отдела:
создание и поддержание запаса летальных и выводящих из строя биологических
агентов в готовности для специального оперативного применения;
разработка специальных средств для тайного применения биологических агентов,
изготовление их и содержание в готовности для оперативного применения;
разработка микробиоинокуляторов для скрытого введения биологических аген-
тов человеку;
выполнение исследований по тематике, представляющей интерес для ЦРУ, ока-
зание технической и консультативной помощи.
Отдел просуществовал до 1971 г. (Hearings before the select committee..., 1975).
Разработка технических средств для массового биологического поражения людей.
Попытки еще в 1918 г. адаптировать химические боеприпасы для применения рици-
на не удались. Но причины, по которым распыленный рицин не вызывал поражения
людей, в средине 1940-х гг. понимались смутно. Ясно было то, что его токсичность в
аэрозолированном состоянии среди прочих причин как-то зависит от степени диспер-
сности аэрозоля, однако не было методической возможности эту зависимость уста-
новить. Но война шла без перспектив на быструю победу. Противник преподносил
новые сюрпризы в виде баллистических ракет, быстроходных океанских подводных
лодок, реактивных самолетов. И на все это необходим был быстрый ассиметричный
ответ, оружие на тот случай, когда «другие обычные способы обороны окажутся не
состоятельными». Поэтому для создания БО использовалось то, чем уже располагала
химическая служба — кассеты зажигательных боеприпасов и ВАПы.
Биологические боеприпасы. Японские специалисты считали основной причиной
своих неудач в применении БО — гибель бактерий во время взрывного диспергиро-
вания. Они пытались свести к минимуму гибель биологического агента путем исполь-
зования хрупкого контейнера и небольшого заряда сухой или жидкой рецептуры, от-
носительно удаленного от взрывчатого вещества. Причем керамическая оболочка
контейнера по замыслу Исии должна была экранировать биологический агент от тер-
мического воздействия взрыва (см. разд. 1.8). Специалисты Великобритании, Кана-
ды и Соединенных Штатов эмпирически подошли к такому же техническому реше-
нию проблемы сохранения биоагента при взрыве. Их работа в годы Второй мировой
войны над проблемой распыления частиц взрывом концентрировалась главным об-
разом на оптимизации количества снаряжаемой рецептуры, конфигурации разрыв-
. 9. Биологическое оружие перед Корейской войной
209
ч’о заряда, характеристик корпуса и типа взрыва. Были проведены испытания ряда
: «хпериментальных боеприпасов. К концу войны представлялось вероятным, что
- зиболее соответствует целям биологической войны небольшая бомба размера руч-
- ой гранаты весом около 1 кг, выполненная из легкого пластического материала. Она
казалась наиболее эффективной при снаряжении жидкими рецептурами. Успешно
тошл и испытания две 4-фунтовые бомбы с металлической оболочкой, разработан-
тые английскими и канадскими специалистами. По официальным материалам, эти
бомбы оказались эффективными с точки зрения образования аэрозолей и контроля
' 13 лера частиц, сведения о скорости гибели биологических агентов в аэрозоле отсут-
. -вуют (The Problem..., 1970).
О создании в США в период Второй мировой войны боевой техники для примене-
на БО почти ничего неизвестно. Но, как мы увидим далее, засекречивание имело
чисто статусный характер и не имело никакого отношения к эффективности этого вида
“хжия. Если судить по открытым данным, то в США много опытов было проведено
; 4-фунтовой бомбой, по своей форме пригодной для снаряжения 500- и ЮОП-фун-
?вых кассет, разработанных для рассеивания 4-фунтовых зажигательных бомб М-69
и М-74. Бомба под названием Ml 14, стала первым бактериологическим боеприпасом,
принятым на вооружение армии США. В 500-фунтовую кассету МЗЗ кластировалось
«8 суббоеприпасов Ml 14 (рис. 1.35 и 1.36).
Рис. 1.35. Бактериологический суббоеприпас Ml 14. Это была небольшая трубчатая
бомба длиной 21 дюйм и диаметром 1,62 дюйма. По ее оси располагался цилиндр со
сжиженным газом (как правило, с углекислым газом; испытаны были и некоторые
взрывчатые вещества), а вокруг цилиндра 300—400 г сухой рецептуры с определен-
ным размером частиц (The Problem..., 1970) или 320 мл жидкой рецептуры Brucella
suis. Носовое основание имело гексагональную форму для облегчения формиро) ания
кластеров таких бомб. Оно включало высокочувствительный всепогодный взрыва-
тель, срабатывающий при ударе о поверхность и вызывающий нарушение герметич-
ности цилиндра со сжиженной углекислотой. На верхнем изображении показан бое-
припас с раскрытым стабилизатором. При ударе такого боеприпаса о землю газ осво-
бождался и выбрасывал заряд рецептуры через отверстие в хвостовой части бомбы,
которое обычно закрыто срезывающимся диском. По R. Sidell et al. (1997)
Испытания боеприпасов Ml 14 с разными агентами продолжались на Дагуэйском
полигоне еще в 1950-х гг. По имеющимся данным, результаты полевых испытаний этой
системы показали, что до 10 % агента диспергировалось в виде инфекционного аэро-
золя. При сбрасывании 4 тонн таких бомб в условиях полигона создавалась 50 %-по-
ражающая доза на площади в 1 кв. милю (The Problem..., 1970).
На основе этой бомбы была создана 500-фунтовая, так называемая перьевая бом-
ба (feather bomb) Ml 15, оставившая свой сибиреязвенный «след» на Корейском по-
уостровев 1952 г. (см. разд. 1.10). Поданным R. Sidell et al. (1997), бомба заполнялась
1Гхими рецептурами, дезагрегированными легким и сухим носителем — птичьими
210
Часть 1. Краткая история применения биологических средств...
Рис. 1.36. Кассетная 500-фунтовая биологическая бомба МЗЗ.
Включала 108 суббоеприпасов Ml 14. По R. Sidell et al. (1997)
перьями. На вооружение американской армии она была принята уже после испыта-
ния в Корее, в 1953 г. Сам боеприпас М114 дал целое «семейство» биологических бое-
припасов, взрывающихся при ударе о поверхность (см. разд. 1.11).
После Второй мировой войны были разработаны и другие системы боеприпасов.
Например, для увеличения площади поражения и повышения однородности создава-
емого облака биологического аэрозоля американскими военными была создана по-
луфунтовая бомба Е61R4, заполняемая только 35 мл биологического агента, но 4таких
бомбочки (bomblets) по площади поражения дважды перекрывали площадь поражения
одной М114. Их в количестве 544 штук кластировали в 750-фунтовую кассетную бом-
бу E133R3 (Sidell R. et al., 1997).
Скопировав боеприпас, используемый японцами в годы Второй мировой войны,
американцы создали 80-фунтовую баллонную бомбу, предназначенную для пораже-
ния растений. Бомба представляла собой цилиндр диаметром 32 дюйма и 24 дюйма
высотой, который выполнял роль корзины. Его внутренне пространство представляло
собой 5 изолированных отсеков, каждый заполнялся перьями и сухими рецептурами
биологических агентов, поражающих растения. Агенты в контейнерах были сгруппи-
рованы вокруг нагревателя химического типа. Контейнер открывался благодаря ба-
рометрическому механизму на определенной высоте над уровнем моря или механиче-
скому таймеру через заданное время, биологическая рецептура рассеивалась по полям,
контейнер поджигался химическим нагревателем (Sidell R. etal., 1997).
Разрабатывались другие типы биологических боеприпасов, например, предназна-
ченные для распространения инфицированных насекомых. Они испытывались в ре-
альных боевых условиях во время войны на Корейском полуострове и будут описаны
в разд. 1.10. В работе R. Sidell et al. (1997) приведены только те, что были приняты на
вооружение американской армии.
ВАПы. В период Второй мировой войны работы по созданию распыляющих уст-
ройств проводились в биологических и химических лабораториях Великобритании,
Канады и Соединенных Штатов. Инициатива в данном вопросе принадлежала специ-
алистам по химическому оружию. Основная проблема, с точки зрения разработчиков
БО того времени, состояла в создании устройств, в которых распыляемая биологи-
ческая рецептура подвергалась бы минимальным воздействиям. В опытах использо-
19. Биологическое оружие перед Корейской войной
211
вались суспензии бактериальных агентов и сухие рецептуры с определенным разме-
эм частиц. В качестве имитатора токсинов применялись растворы альбумина курн-
ого яйца. Но результаты экспериментов союзников мало отличались от получаемых
сией. Когда удавалось добиться хороших результатов по выживаемое ги биоагентов
эрозоле (по результатам подсчета колоний на чашках), не происходило инфици-
эвания экспериментальных животных. А если оно все же и происходило, то дости-
галось таким количеством биоагента, которое в принципе невозможно доставить на
воле боя ни ВАПами, ни кассетными боеприпасами, а сами поражения носили харак-
-ео септицемии, патоморфология поражения не соответствовала поражениям, воз-
можным при проникновении возбудителя болезни через воздухоносные пути.
К началу 1950-х гг. стало ясно, что инфицирующими свойствами обладает аэро-
золь с размером частиц, не превышающих 5 мкм (см. ниже «Аэрозоли»). Однако тип
используемого тогда аэрозольного генератора с соплом, работающим под давлением
те. гидравлического типа; см. разд. 1.11), в принципе не позволял добиться эффек-
’ивного поражения людей биологическим аэрозолем в боевой обстановке. Существо-
вало множество модификаций аэрозольных генераторов данного типа. Но если для
'юбого из них распределение размеров частиц аэрозоля на выходе из сопла генератора
редставить в виде графика, то получалась гаусовская кривая, где количество частиц
тиаметром менее 5 мкм составляло «в лучшем случае» не более 5 %. Для одноканаль-
аого сопла распылителя, чтобы добиться эффективности на уровне даже 5 %, требо-
валось создать давление воздушной струи не менее 300 фунтов/дюйм2 (21 кг/см2).
Данные же о проценте жизнеспособных частиц (т. е. способных к инфицированию
экспериментальных животных) еще предстояло получить в условиях, соответствую-
щих применению БО по реальным целям (The Problem..., 1970; Patrie W III., 2001).
В случае сухой рецептуры регулирование размера частиц можно было осуществить
угем регулирования дисперсности порошка. Еще Исии считал диспергирующие
авиационные приборы, снаряженные сухой рецептурой, потенциально наиболее
эффективными для ведения бактериологической войны. В этом случае:
1) можно снарядить средство доставки большим количеством материала;
2) можно эффективнее контролировать размер частиц, в отношении некоторых
патогенных микроорганизмов;
3) можно избежать губительного воздействия некоторых элементов атмосферных
условий. ВАПы для сухих и жидких рецептур, с помощью которых можно дисперги-
ровать биологические аэрозоли, 90 % частиц которых имеют диаметр менее 5 мкм, дей-
'вительно были созданы в США, но только в начале 1960-х гг. (см разд. 1.11). А в конце
940-х гг. ситуация была критической. О победных реляциях времен войны, в кото-
вых БО представлялось как существующее, старались не вспоминать — самые опас-
ные микроорганизмы, оказавшись в аэрозоле, утрачивали свою смертоносность.
Финансирование программы по БО было значительно уменьшено.
Аэрозоли. Во второй половине 1940-х гг. экспериментальным путем удалось уста-
новить, что создаваемый имевшимися боеприпасами и ВАПами биологический аэро-
золь не проникает в глубокие отделы легких экспериментальных животных. Напри-
мер, сотрудник лаборатории в Форт-Детрике L С. Ferguson (1945; неопубликованная
эзбота, цитируется по работе Barnes J. М., 1947) заражал ингаляционно мышей спо-
рами сибирской язвы и забивал их за несколько часов до наступления септицемии.
При тцатсльном исследовании многочисленных срезов тканей легких им обнаружены
’олько два небольших очага воспаления в оболочке альвеолы. Сотруднику Портон
Дауна J. М. Barnes (1947) удалось заразить ингаляционно три вида животных (кролик.
212
Часть 1. Краткая история применения биологических средств...
морская свинка, мышь) так называемым «движущимся аэрозольным облаком» в ап-
парате Гендерсона (Henderson D. W., 1943; неопубликованная работа, цитируется по
работе Barnes J. М., 1947), но у них не оказалось никаких специфических легочных
поражений. Животные погибали через 2—4сут. после инфицирования, при вскрытии
обнаружена картина, характерная для острой сибиреязвенной септицемии, наблюдае-
мой при подкожном введении спор. Отличалась лишь инфицирующая доза — при
«ингаляционном заражении» она на 3—5 порядков превышала ту, что вызывала гибель
животных при подкожном заражении. Но был получен и неожиданный результат. Bar-
nes установил, как сам же и выразился, «поразительный факт» — вирулентность вдыха-
емых спор возрастала в 16 раз, если распыление производилось в 0,5 %-растворе ком-
мерческого детергента торгитола (Torgitol). Сам детергент на легкие животного ника-
кого действия не оказывал, в этом он убедился путем многократных экспериментов.
Но при воздействии на животное аэрозолем спор возбудителя сибирской язвы в тер-
гитоле, менялась патоморфология болезни — наблюдались признаки поражения, ха-
рактерные для ингаляционной сибирской язвы. Размножение вегетативных форм
сибиреязвенного микроба происходило в лимфатических узлах трахей, бронхов еще
до размножения в самих легких (см. разд. 3.1). Barnes не понял, каким образом терги-
тол увеличивает вирулентность аэрозолированных спор. Однако он убедился в том.
что и другие поверхностно-активные агенты, и полимер «Солвар», обладают таким же
действием. Несомненно, опубликованные результаты его экспериментов—«верхушка
айсберга» исследований, проводимых в Нортон Дауне по разработке жидких сибире-
язвенных рецептур.
Barnes повторил работы В. И. Госа (1907), выполненные на форту «Александр I»,
но так же, как и русский исследователь, он не понял, что добился инфекции пугем
создания мелкодисперсного аэрозоля, способного достигать глубокие отделы легких.
Оба исследователя достигли этой пели разными путями — Гос за счет увеличения дав-
ления воздуха на небольшие объемы жидкости; Barnes — за счет снижения поверхно-
стного натяжения жидкости, образующей каплю, что при той же затраченной на рас-
пыление энергии, позволило ему раздробить жидкость на капли с меньшим радиусом.
Но ситуация в 1947 г. была уже не та, что 40 лет назад. Исследованиями бактериаль-
ных аэрозолей занимались десятки коллективов, они буквально «наступали друг дру-
гу на пятки», и механизм проникновения аэрозоля в глубокие отделы легких челове-
ка вскоре стал понятен (рис. 1.37).
Пока военные взрывали свои боеприпасы на полигонах и в специальных камерах,
подсчитывая на чашках Петри количество колоний микроорганизмов, выживших
после взрывного диспергирования различных рецептур, инициатива в агробиологи-
ческих исследованиях перешла к гражданским ученым. W. F. Wells et al. (1948) первыми
заявили о решающем значении для развития туберкулезной инфекции не количества
вдыхаемых бактерий, а величины частиц аэрозоля, в которых содержатся эти бактерии
Они показали, что только мелкие частицы бактериального аэрозоля, по существу от-
дельные бактериальные клетки, могут инфицировать легкие кролика, чего нельзя
сказать о крупных частицах, содержащих большие количества живых микроорганиз-
мов. М. D. Lurie et al. (1950) расширили их работу, показав, что каждая инфекционная
частица, которая вдыхается и задерживается в альвеолах, дает отдельный туберку-
лезный бугорок. Тогда вспомнили о довоенной работе немецкого ученого W. Fin-
deisen’a (1935), выполнившего первое серьезное изучение возможности проникнове-
ния частиц разных размеров в дыхательные пути. На его работу военные биологи ра-
нее не обращали внимания, так как она выполнена с использованием инертных
9. Биологическое оружие перед Корейской войной
213
Рис. 1.37. Так называемый «8-й шар» («8-Ьа11»). Это стальной шар о&ь.мом
один млн литров, построенный в 1949 г. в Форт-Детрике. Использовался аме-
риканскими военными для изучения поражающих свойств биологических
аэрозолей и эффективности создаваемых образцов БО. В настоящее время
считается памятником истории США По J. van Aken, Е. Hammond (2003)
частиц. Findeisen предложил принцип математического моделирования процессов
проникновения и оседания аэрозольных частиц в дыхательных путях с учетом анато-
мического строения, броуновского движения, седиментации и инерционности час-
тиц при осаждении. Но его расчеты не «вписывались» в существовавшие тогда пред-
ставления о контактной передаче возбудителей инфекционных болезней. Сам он не
имел отношения к эпидемиологии. Findeisen всего лишь рассчитал процент осажде-
ния частиц различных размеров во всех отделах дыхательного тракта для модели, со-
стоящей из разветвляющихся трубок и заканчивающихся сферическими альвеолами
рахея, главные бронхи, бронхи 1-, 2- и 3-го порядков, терминальные бронхиолы, ды-
хательные бронхиолы, альвеолярные ходы, альвеолярные мешочки). Рассчитанные им
на основе выведенных формул вероятности осаждения частиц в каждом отделе лег-
ких за счет диффузии, седиментации и импакции, показаны в табл. 1.3.
Таблица 1.3
За хержка частиц попавших в трахею, % *
Участок легкого Радиус частиц, мкм
0,03 0,01 0,3 1 3 10 30
Проводящая часть 8,4 4,2 2,7 7,9 44,9 98 100
Дыхательная часть 57,6 30,9 30,5 88,5 55,1 0 0
Общая задержка 66 35,1 33,2 96,4 100 98 100
* По W. Findeisen (1935).
Findeisen не учитывал объем альвеолярных ходов и мешочков, это повлияло на
точность его расчетов для частиц в субмикронном диапазоне. Однако в диапазоне
0,3-30 мкм его расчеты оказались удивительно верными. Данные W. F. Wells et al. (1948)
хорошо объяснялись расчетами W. Findeisen в той их части, где речь шла о частицах в
иапазоне размеров 1—3 мкм. Теперь можно было рассчитывать задержку частиц
214
Часть I. Краткая история применения биологических средств...
биологического аэрозоля в дыхательных путях и экспериментально подбирать условия
для наиболее эффективного инфицирования поражаемых объектов, исходя не из био-
логических критериев (количество живых клеток возбудителя инфекции, оставших-
ся после взрывного диспергирования), а физико-химических, т. е. размера частиц,
образующих аэрозоль. Осталось только уточнить отдельные детали процесса проник-
новения биологического аэрозоля в глубокие отделы легких. «Детали» стали прояс-
няться, как только изменились экспериментальные подходы. Н. A. Druett et al. (1953)
в лабораторных условиях экспонировали морских свинок и обезьян к монодисперс-
ным аэрозолями спор возбудителя сибирской язвы. Ими было установлено, что наи-
большей инфекционностью обладал аэрозоль с размером дисперсной фазы, прибли-
жающейся к размеру споры (примерно 1,2 мкм). Когда частички аэрозоля имели
размер более 5 мкм, его инфекционность быстро снижалась. В опытах на морских
свинках инфицирующая доза аэрозоля из частичек с размером примерно 1 мкм, была
в 17 раз меньшей, чем у аэрозоля с размером частичек 12 мкм. Для обезьян это соот-
ношение держалось как 1:14. Опубликованные результаты Н. A. Druett et al. (1953).
так же, как и работы J. М. Barnes (1947), можно рассматривать как слабое отражение
масштабных экспериментов по изучению инфицирующих свойств мелкодисперсных
аэрозолей микроорганизмов, проведенных в военных биологических центрах за пе-
риод с 1948 по 1953 г. Их результаты объясняли предыдущие неудачи и открывали
перед военными новые перспективы в разработке БО.
С обнаружением значения дисперсности вдыхаемых частиц для проявления по-
ражающего действия биологических агентов, военные биологи вошли в область не-
познанных природных явлений, для которых аналогии с химическим оружием уже не
подходили. Дело тутвтом, что аэрозоли биологических агентов и капли ОВ ввоздуш-
ной среде, образующиеся в результате взрывного диспергирования или при распыле-
нии ОВ с помощью ВАПов, представляют собой принципиально различные системы.
Капли ОВ движутся через воздух, а аэрозольные частицы биологических агентов дви-
жутся вместе с воздухом. Последнее обстоятел ьство в сочетании с необходимостью соз-
дания аэрозоля биологического агента с очень узким диапазоном дисперсности меня-
ло всю методическую базу таких исследований, вынуждало исследователей изменить
подходы к создаваемым биологическим боеприпасам, средствам и способам их при-
менения.
Моделирование масштабного применения БО. Война, начавшаяся на Корейском
полуострове 25 июля 1950 г., побудила американское военное руководство вновь на-
править свои усилия на поиски средств эффективного ведения бактериологической
войны, разумеется, в рамках нанесения «ответного удара». Военным надо было про-
демонстрировать законодателям, утверждающим бюджеты министерств и ведомств,
реальные угрозы, исходящие от нового оружия массового поражения. И тогда ими
были проведены эксперименты по оценке возможности вероятного противника (т. е.
СССР) осуществить масштабное применение БО по городам США, результаты ко-
торых по сей день не критично переписываются новым поколением мистификаторов
биологической войны.
В сентябре 1950 г. была оценена уязвимость города Сан-Франциско при биологи-
ческом нападении на морской порт. В качестве агентных имитаторов использовали
жидкие рецептуры спорообразующего микроорганизма Bacillusglobigii (BG), обычно
моделирующие аэробиологические характеристики возбудителя сибирской язвы
Имитация нападения осуществлялась путем диссеминирования аэрозоля BG из ли-
нейного источника. Для этого небольшой военный корабль на заходе солнца распылял
1.9. Биологическое оружие перед Корейской войной
215
жидкую рецептуру BG вдоль линии протяженностью 2 мили и на расстоянии 2-х ми-
ги от берега. Распространение аэрозоля по Сан-Франциско контролировалось с по-
мощью сотен пробоотборников-импинжеров (рис. 1.38).
Рис. 1.38. Имитация биологического нападения на Сан-Франциско в сентяб-
ре 1950 г. 1 — зона размещения пробоотборников; 2 — прибрежный район
Сан-Франциско; 3 — зона условного заражения. По W. Patrie III (2001)
В первом опыте отмечалась сильная инверсия при слабом ветре со скоростью около
10 миль/ч. Оказался условно зараженным центр Сан-Франциско; пробоотборники
указывали, что в литре воздуха присутствовало более 10 тыс. спор BG. Организаторы
эксперимента впоследствии поясняли всем, кого это интересовало, что такая кон-
центрация спор могла привести к инфицированию свыше 60 % населения. Также кон-
таминированным спорами BG, хотя и в значительно меньшей степени, оказался район
Беркли. Испытатели пришли к выводу, что этот тест был чрезвычайно успешным,
т. е. он обосновывал их существование. В то же время им стало ясно, что при распы-
лении аэрозоля BG, даже в самых наилучших атмосферных условиях, трудно заранее
предсказать, где пройдет его облако. Способность «дошедшего» до цели аэрозоля
вызывать инфекционный процесс, они не исследовали в силу понятных причин.
Следующий тест проводился в условиях неустойчивой воздушной массы. И снова
рецептура BG распылялась вдоль линии в 2 мили. Количество BG, использованного
в опыте, было тем же. Высокая концентрация спор отмечалась только в двух кварта-
лах города. Нестабильная воздушная масса препятствовала достижению намеченного
числа «пораженных», указывая на то, что при ударе по открытой цели метеорологиче-
ские условия также важны для успеха нападения, как и сам агент, конструкция боепри-
паса и система диссеминирования. В качестве агентного имитатора неспорообразу-
ющих агентов БО (возбудители чумы, сапа, мелиоидоза, бруцеллеза, туляремии и др.),
был использован небольшой вегетативный микроорганизм, бактерия Serratia marces-
cens. Линия распыления и общие условия испытания были похожими на те, которые
были в опытах с BG. Пуск аэрозоля проводился при хороших метеорологических ус-
ловиях, включая умеренную инверсию и скорость ветра 12 миль/ч. Однако импинжеры
показывали, что в первых кварталах города присутствовало всего около 25 клеток/л
воздуха, свидетельствуя о том, что «нападение» не было успешным. В последующих
опытах испытатели установили тот факт, что даже при заходе солнца интенсивность
216
Часть I. Краткая история применения биологических средств...
ультрафиолета все еше достаточна, чтобы убивать вегетативные клетки. Тесты прово-
дились с участием микробиологов, аэробиологов, метеорологов и разработчиков бое-
припасов, все они сделали определенные выводы. Но «широкой общественности»
сообщили только о первом эксперименте, наглядно свидетельствующем о том, что
«Америка в опасности» (Patrie W. III., 2001).
Уже в октябре 1950 г. военному командованию была дана санкция на строитель-
ство центра по производству БО в арсенале Пайн Блафф (штат Арканзас). Проекти-
рование этого объекта было ускорено, строительство начато в феврале 1951 г., общая
сумма затрат на строительство завода составила 69 млн долларов. В ноябре 1951 г.
министр обороны США произвел переоценку состояния готовности к химической,
биологической и радиологической войне и сделал вывод о том, что разработка бое-
припасов, снаряжаемых химическими и биологическими агентами, требует более вы-
сокой степени готовности и привлечения значительно больших ресурсов. 21 декабря
1951 г. Министерством обороны США была издана директива о повышении готовно-
сти к химико-биологической и радиологической войне, касающаяся всех родов
войск. В 1952 г. объем НИР по военно-биологическим программам возрос в два раза
(U. S. Army activity..., March 8 and May 23, 1977).
В апреле 1952 г. было проведено более обширное, чем 1950 г., испытание с целью
выяснения поведения крупных аэрозольных облаков в различных метеорологических
условиях. В одном из опытов при распылении 200 кг флуоресцирующих частиц с ко-
рабля, шедшего вдоль берега на расстоянии 156 миль на территории в 13900 миль2,
были созданы дозы порядка 1000—10000 частиц/л воздуха (The Problem..., 1970).
По сообщению S. Hersh (1968), результаты этих полевых испытаний были пред-
ставлены комиссии Конгресса, когда химическая служба пыталась получить повы-
шенные ассигнования на 1952—1953 гг. Дополнительным аргументом химической
службы были успехи в области БО, якобы достигнутые Советским Союзом. Приве-
денные доводы показались кошрессменам убедительными и на 1953 г. были предус-
мотрены значительные ассигнования на создание крупных полигонов и лаборатор яй
по производству биологических агентов.
* * *
Если заимствовать терминологию из эволюционной биологии, то развитие БО
перед войной на Корейском полуострове можно охарактеризовать как адаптивную
радиацию (см. «Словарь терминов») Теперь ситуация с разработкой БО значительно
отличалась от той, что была в годы Первой мировой войны и даже перед Второй ми-
ровой войной, когда масштабы применения биологических агентов ограничивались
мизерными возможностями их лабораторного производства и отсутствием способов
сохранения наработанных агентов. К концу 1940-х гг. у военных биотехнологов при
тех же контагионистических взглядах на БО появилась возможность получать биоло-
гические агенты в промышленных масштабах, а авиация могла применять их по кон-
кретным целям за тысячи километров от линии фронта. Тайное прозябание БО закон-
чилось. Оно вошло в полупустую нишу оружия, предназначенного для массового и
неконтактного поражения людей, с сохранением всех созданных ими материальных
ценностей. Начались исследования возможностей нового вида оружия для решения
различных боевых задач. Появилось множество его примитивных разновидностей
Конструктивно новые образцы оружия напоминали своих предшественников среди
химических боеприпасов, не обладая при этом их эффективностью. Первые биоло-
гические боеприпасы походили на химические примерно так же, как первые млеко-
1.10. Корейская бактериологическая война
217
питающие на ящеров. Надежды разработчиков БО на Западе (так же, как и в Японии;
см. разд. 1.8) в 1940 гг. в основном возлагались на «контагий». Кассетные биоло-
гические боеприпасы, переделанные из зажигательных, разбрасывают субэлементы
с биологической начинкой так же, как и с термитной. Но если загоревшийся термит
может поджечь поверхность, на которой он оказался, то крупнодисперсный биоло-
гический аэрозоль оседает на нее, не принося вреда противнику. Биологические ре-
цептуры, распыленные ВАПами, в те годы имеют дисперсность, необходимую для
успешного применения иприта, но не агентов БО, и не проникают в глубокие отделы
легких человека. Насекомых и грызунов, которых тогда считали основными резервуа-
рами и переносчиками возбудителей наиболее опасных инфекций людей, заражают
микроорганизмами, опасными, прежде всего, для жизни их самих. Затем уже больны-
ми в надежде, что само их пребывание в среде, окружающей человека, вызовет эпиде-
мический процесс, распространяют с помощью штатных средств типа бомб для лис-
товок или просто коробок и ящиков. И в этой чаще запутанных заимствований и
аналогий в начале 1950-х гг. «проросла» главная в XX в. эволюционная ветвь нового
вида оружия массового поражения — аэробиологическая.
1.10. КОРЕЙСКАЯ БАКТЕРИОЛОГИЧЕСКАЯ ВОЙНА
Боевые действия в Корее в начале 1952 г. От ядерного оружия к химическому и
бактериологическому. «Дьявол всегда делает вид, что он не существует». Поче-
му оказалось невозможным скрыть факты применения БО в войне на Корейском
полуострове? Бактериологические боеприпасы. Попытки вызвать эпидемии чумы.
Попытка вызвать вспышку холеры. Попытки вызвать кишечные инфекции. Как это
все делалось. Использование аэрозолей.
Эволюционная радиация средств ведения биологической войны обещала военным
большой выбор образцов БО и биологических агентов, пригодных для решения раз-
ных боевых задач. Но их реальные возможности надо было установить в конкретной
боевой обстановке. Поэтому применение БО во время войны на Корейском полуост-
рове носило характер импровизаций, некоторые эпизоды бактериологической войны
становились известными, другим в ходе боев северокорейцы не придали значение, а
потом их забыли, третьи удавалось скрыть. Полную картину корейской бактерио-
логической войны восстановить сегодня невозможно. Мы попытаемся «собрать по
крупицам» хотя бы то, что еще осталось в опубликованных официальных документах
Боевые действия в Корее в начале 1952 г. К началу 1952 г., т. е. за полтора года не-
прерывных боев на Корейском полуострове, вооруженные силы ООН (70 % американ-
цы, 25 % южные корейцы, 5 % союзники США) так и не смогли добиться перелома в
войне. Военный гений генерала Дугласа Макартура предотвратил катастрофический
разгром войск ООН в августе 1950 г. Но победа, одержанная им после высадки десан-
та в Инчхоне, оказалась хуже поражения. Благодаря ей участие континентального
Китая в войне стало неизбежным. Уже 25 октября 1950 г. силами китайских доброволь-
цев серьезное поражение было нанесено трем южнокорейским дивизиям. Через ме-
сяц, неожиданно для американцев, началась Пхеньян-Ханнамская операция, в резуль-
тате которой северокорейские и китайские части вернули Пхеньян и вышли на всем
протяжении фронта к 38-й параллели. В этих боях общие потери войск ООН соста-
вили до 36 тыс. человек. Предчувствие военной катастрофы среди американского
218
Часть 1. Краткая история применения биологических средств...
командования было тогда столь велико, что генерал Макартур потребовал от Вашин-
гтона нанести ядерные удары по крупным китайским городам. Вечером 31 декабря
началась знаменитая Сеульская операция, приведшая к паническому бегству амери-
канских войск и их союзников на юг полуострова и к освобождению Сеула.
В январе 1951 г. американцы организовать серию мощных контрударов своих сил
на западном и восточном участках фронта. Но в феврале им вновь пришлось испы-
тать горечь отступления. Л ишь в средине марта после тяжелых боев войска ООН вновь
захватали Сеул. В начале апреля президент США Гарри Трумэн сместил генерала Ма-
картура с должности командующего американскими войсками на Дальнем Востоке и
назначил на нее генерала Мэтью Риджуэя. Успех не сопутствовал новому командую-
щему. В мае китайские и северокорейские войска предприняли мощное наступление
на трех направлениях и вновь вышли к Сеулу, но потом потерпели поражение и отошли
на рубежи 38-й параллели.
Осознание невозможности выиграть эту войну заставило воюющие стороны идти
на переговоры в надежде, что дипломатическим путем удастся выторговать лучшие
условия мира, чем кровопролитием. С 10 июля 1951 г. в городе Кэсон на территории
КНДР (38-я параллель) начались переговоры воюющих сторон. В августе они были
прерваны, затем последовало масштабное наступление войск ООН с целью давления
на китайско-северокорейскую сторону. Однако как это, так и аналогичное осеннее
наступление ооновцев, были отбиты. В конце октября переговоры возобновились в
деревушке Паньмыньчжон. Уже в ноябре сторонами достигнуто соглашение о демар-
кационной линии несколько севернее 38-й пара пели (в основном по линии сопри-
косновения войск), общее же соотношение сил сторон, несмотря на потери, поне-
сенные в августе — сентябре, почти не изменилось. Оказавшись неспособными
одолеть противника на поле боя, американцы применили свой излюбленный прием,
они стали уничтожать корейцев и китайцев с воздуха. С января 1952 г. начался пери-
од войны, называемый в китайских источниках «войной на удушение» (Орлов А. С.,
Гаврилов В. А„ 2003).
От ящерного оружия к х.гчическому и бак~₽рио тогическому, По мере того как война
для США становилась все менее успешной, возможность применения оружия массо-
вого поражения все чаще обсуждалась в Пентагоне и в Белом доме. Памятная запис-
ка министра обороны CI ИА «Военная эффективность применения атомного оружия
в тактических целях в Корее» от 14.08.1951 г. свидетельствует о рассмотрении амери-
канским политическим руководством возможности применения атомной бомбы в
случае, «если американские войска будут стоять перед лицом катастрофы». В октябре
1951 г. американская авиация даже провела операцию под кодовым названием «Хад-
сон Харбор», в ходе которой осуществлялись учебные атомные удары в Корее. Види-
мо понимая, что такие действия вызовут более чем негативную реакцию СССР, США
готовились нанести превентивный удар и по нему. Об этом, в частности, свидетель-
ствует обнаруженный недавно в Великобритании секретный рапорт директора мор-
ской разведки вице-адмирала Эрика Лонгли-Кука, составленный в средине 1951 г.
В нем сообщается, что американцы готовятся к началу военных действий против
СССР «где-то в средине или конце 1952 г.», независимо от согласия или противодей-
ствия других стран НАТО. Но уже в октябре 1951 г. холодным душем, пролившимся
на разгоряченные головы американских стратегов, стали результаты исследования
изотопного состава радиоактивных осадков, выпавших в Арктике после третьего ис-
пытания советской атомной бомбы на полигоне под Семипалатинском. Оказалось,
что Советы создали комбинированный ядерный заряд на основе плутония и ура-
1.10. Корейская бактериологическая война
219
на-235 с мощностью около 50 килотонн в троти ювом эквиваленте. Взрыв произво-
дился в воздухе — бомбу сбросили со стратегического бомбардировщика Ту-4 По
оценкам ЦРУ, кстати, как потом выяснилось, оказавшихся ошибочными, к началу
1952 г. СССР должен был построить около тысячи таких бомбардировщиков, а к се-
редине 1952 г. русские могли иметь не менее 100 совершенных атомных бомб. Данные
радиоперехватов свидетельствовали об интенсивном строительстве в Сибири раз-
ветвленной сети аэродромов, предназначенных для стратегической авиации. Военный
иитимизм сменился разочарованием Разговоры о нападении на СССР прекратились,
а боязнь разозлить русского медведя стала столь сильной, что американские полити-
ческие круги скрыли от американского общества даже сведения о роли русской ис-
ребительной авиации и ПВО в Корейской войне (Пиблз К., 2002).
По ряду причин применение американской армией химического оружия в период
войны на удушение» не носило массированный характер. Не использовались и со-
временные боевые ОВ. Американским генералам приходилось учитывать то обстоя-
тельство, что применение химического оружия могло вызвать аналогичный ответ со
стороны КН Р. К такой возможности американцы относились очень серьезно. По дан-
ным С. С. Лотоцкого с соавт. (2000), в составе американских войск в Корее действо-
вал 401-й технический отряд химической разведки, который собирал материалы по
противохимической защите и химическому вооружению противника. В постоянной
готовности к проведению дегазационных работ находились химические дегазацион-
ные роты.
Снижению интереса со стороны американского командования к применению ОВ
способствовала умелая тактика, выбранная китайскими и северокоречскими войска-
ми для противодействия налетам авиации. Особенно эффективным оказалось исполь-
зование подземных галерей. Большое количество изолирующих противогазов, срочно
поставленных им Москвой, делали боевые О В бесполезным средством ведения войны,
но зато они превращались в сильный раздражающий фактор в международных делах.
В большинстве случаев применение ОВ американскими вооруженными силами
проводи, гось во время артобстрелов и бомбежек одиночными химическими снаряда-
ми или сбрасывалось небольшое количество химических бомб. По неполным данным,
приведенным С. С. Лотоцким с соавт. (2000), с 27 февраля 1952 г. и до конца июня
1953 г. отмечено свыше 100 случаев применения химических и снарядов и бомб толь
ко по китайским добровольцам, ь результате чего 1095 человек получили отравление,
из них 145 погибли. Более 40 случаев применения отравляющих веществ было отме-
чено и по военнопленным. Симптомы поражения (удушье, слезотечение, опухание
глаз, рвота, быстрая потеря сознания) свидетельствуют о том, что в химических бое-
припасах применялись окислы азота в смеси с синильной кислотой, дифенилцианар-
сином или дифенилхлорарсином. Непреодолимого психологического барьера в те
годы не было лишь в отношении одного вида оружия массового поражения — бакте-
риологического, разумеется, при условии его тайного применения.
«Дьявол всегда делает вид, что ин не сушдствует». О самой войне на Корейском полу-
острове 1951—1953 гг. известно мало, а уж о ее «бактериологической составляющей»,
еще меньше. Официальные круги США и западных стран, а также их «свободная прес-
са» в освещении этих преступлений вели себя точно также, как и в отношении япон-
ских преступлений, связанных с испытанием на китайских и русских пленных БО в
1933—1945 гг. в Маньчжурии (см. разд. 1.8.). Они либо замалчивали сам факт приме-
нения БО в этой войне, либо подвергали осмеянию любые материалы, имеющие отно-
шение к расследованию фактов его применения. То же самое мы видели при осве-
220
Часть 1. Краткая история применения биологических средств...
щении западными СМИ массовых убийств сербов в Косово (1990), русских в Чечне
(1991 — 1994), осетин и российских миротворцев в Южной Осетии (2008).
Для тех «писак», кто злоупотреблял «свободой слова» при освещении событий
войны на Корейском полуострове, существовала так называемая комиссия сенатора
Дженнера, последовательного маккартиста, «занявшегося» газетами, радиовещани-
ем, учебными заведениями, кино и театром. Жертвой комиссии Дженнера стал жур-
налист Пауэлл — известный в 1950 гг. специалист по Китаю. Сенатору попалась на
глаза его статья «Бактериологическая война, которую американская армия вела в
Корее, уходит корнями в отряд 731». Комиссия вынесла определение, что Пауэлл
«нанес вред интересам США, сфабриковав статью, не имеющую под собой никаких
оснований», и приняла меры для его изгнания из органов массовой информации
(Моримура Сэйти, 1983).
В качестве примера независимого журналистского расследования, «не нанесше-
го вреда интересам США», рассмотрим труд Терри Вольтона (2000). Его суть сводит-
ся к следующему.
Кремль и китайские коммунисты через коммунистическую прессу распространи-
ли дезинформацию о том, что якобы войска ООН применяют БО против войск и на-
селения Северной Кореи и КНР. Международная ассоциация юристов-демократов
(созданная Советами в 1946 г.) поспешила подтвердить это ужасное сообщение, а
Всемирный Совет Мира по инициативе КН Р назначил «Международную научную ко-
миссию по расследованию фактов, касающихся бактериологической войны в Китае
и Корее» для «объективного изучения проблемы» (далее — Комиссия). В Маньчжу-
рию и Северную Корею отправилась Комиссия, составленная из пяти известных уче-
ных, занимавших прокоммунистические позиции. Далее Вольтон дурашливо пишет,
что руководил делегацией председатель англо-китайской ассоциации общества друж-
бы Джозеф Нидхем (ему это имя, естественно, ничего не говорит). И в сентябре 1952 г.
Комиссия представила свой отчет на 700 страницах, довольно неприятный для аме-
риканской армии документ. Соединенные Штаты обвинялись в том, что они забро-
сили на территорию Северной Кореи насекомых и крыс, зараженных вирусом оспы.
Далее, видимо думая, что никто никогда не найдет этот документ, а если и найдет, то
ничего не поймет, не утруждая себя доказательствами, Терри всего на двух страничках
объявляет все написанное на 700 страницах ложью. Но следуя принципу добираться
до первоисточников и не полагаться на вполне убедительные их интерпретации (вроде
сделанных журналистом Стидом; см. разд. 1.7 «Мистификация Стида»), я нашел и этот
документ. Он называется «Доклад Международной научной комиссии по расследова-
нию фактов бактериологической войны в Корее и Китае» (Пекин, 1952). Результаты
его анализа представлены ниже. А сейчас приглядимся к председателю, якобы «по-
досланной Кремлем» Комиссии, и к методам ее работы.
Комиссия Нидхема. Джозеф Нидхем (Joseph Needham, 1900-1995) — британский
ученый с мировым именем, биохимик. Он был одним из самых молодых членов Ко-
ролевского научного общества и к 37 годам, когда увлекся Китаем, успел опублико-
вать несколько книг, в том числе историю эмбриологии, которую преподавал в Кемб-
ридже. В СССР «История эмбриологии» была опубликована в 1947 г. Нидхем свободно
владел классическим китайским языком и с 1942 г. работал советником по науке при
посольстве Великобритании в Чунцине (столица гоминьдановского Китая; столица
коммунистического Китая находилась тогда в Яньане). В 1950-х гг. в Кембридже вы-
шло несколько десятков томов, основанной им серии книг «Science and Civilization in
China» («Наука и цивилизация в Китае»). После завершения работы в Комиссии Нид-
7.10. Корейская бактериологическая война
221
кем занял пост директора Отдела естественных наук в ЮНЕСКО, где никто никогда
его научную репутацию не подвергал сомнению и о связях с Кремлем не намекал.
О заслугах Нидхема перед мировой наукой говорил в своей нобелевской речи лауреат
Нобелевской премии 1977 г. по химии Илья Пригожин. В Кембридже и сегодня дей-
ствует институт его имени. Возглавляемая Нидхемом Комиссия работала с 23 июня
по 6 августа 1952 г. Члены Комиссии изучили большое количество официальных до-
кументов, опросили многочисленных свидетелей, в их числе были пленные летчики
и диверсанты; проводили совешания с корейскими и китайскими учеными. Беседы
и опросы протоколировались и записывались на магнитный носитель, свидетели
фотографировались, устанавливалась их личность, Нидхем посетил лаборатории в
Пекине, Шэньяне и Пхеньяне, где проводились исследования подобранных образцов
БО и инфицированных материалов, а также деревни в Китае, в районе которых бы-
и сброшены инфицированные чумой грызуны. Китайская сторона предоставила
Комиссии архивные материалы по применению БО японской армией в 1940—1945 гг.
и сохранившиеся образцы этого оружия, изготовленные в отряде № 731.
Комиссия учитывала «труднодоказуемость» фактов применения БО. Поэтому
членами Комиссии был разработан алгоритм анализа информации, с помощью ко-
торого можно было бы достоверно установить факт применения БО (рис. 1.39).
Положительное
бактереологическое
— исследование —
Самолет—Контейнер — непьяных —
Замечен Слышен
_ Другие
вопросы
Случаи
заболеваний —
и эпидемий
животных или
предметов
Замечен Найден
Маршрут
полета
Сезонная
аномальность
Зоологическая
и географическая
аномальность
Сбит
С нетронутым Признание
грузом летного
состава
Прочие
необычные
элементы
Аномальность
сезонная
Аномальность
типов иболеваний
Другие
аномальности
Рис. 1.39. Алгоритм обобщения информации при изучении
каждого отдельного случая применения БО. Доклад... (1952)
Приведенная схема в основном применялась для изучения фактов применения
БО, когда в качестве переносчиков возбудителей инфекций, использовались насеко-
мые. В то же время Комиссия считала, что этот алгоритм обобщения информации мало
применим для установления фактов использования опасных микроорганизмов аэро-
золями. Во-первых, такой способ поражения людей не требует контейнеров и бомб,
фрагменты которых можно идентифицировать и представить общественности в ка-
честве доказательства применения БО; во-вторых, для осуществления аэрозольного
способа нападения не требуются насекомые или другие переносчики, также являю-
222
Часть 1. Краткая история применения биологических средств...
щиеся вещественными доказательствами таких преступлений; в-третьих, аэрозоль не-
видим, и трудно найти свидетелей, чьи бы показания могли зафиксировать время и
место применения биологических агентов. Поэтому Комиссия использовала разные
способы анализа причин вспышек опасных инфекционных болезней, каким-либо
образом связанных с действиями американской авиации. Среди них данные о харак-
тере болезни, о типе возникшего очага, показания пленных летчиков и др. Однако,
как и в случаях применения насекомых в качестве переносчиков возбудителей инфек-
ций, основное внимание уделялось выявлению признаков неправильной эпидемиоло-
гии болезни (см. разд. 2.3).
Комиссия зафиксировала попытки вызвать вспышки чумы, холеры, дизентерии
и сибирской язвы. Возможно, использованных агентов БО было больше, но доказа-
тельств не было собрано. Опубликованные материалы предварительно обсуждались
и согласовывались всеми членами Комиссии. Тексты на разных языках подвергались
лингвистическому разбору, а их переводы согласовывались между специалистами.
В доклад включены только те факты применения БО, которые признаны всеми чле-
нами Комиссии.
В западных источниках работа Комиссии Нидхема освещается с позиции «двой-
ных стандартов». Она осуждается как «позорная», огромный материал, представлен-
ный Комиссией, игнорируется. В то же время несколько страничек текста журнали-
ста Стида с «интерпретациями», противоречащими даже законам физики, цитируются
по сей день. За свое участие в работе Комиссии по расследованию применения аме-
риканской армией БО во время войны на Корейском полуострове, Нидхем подвергся
преследованиям со стороны американского правительства. Въезд в США был для него
невозможен даже в 1970-х гг. Биографы Нидхема объясняют этот «позорный эпизод»
в его биографии приверженностью к христианскому социализму в молодости, одна-
ко они признают то, что к 1930-м гг. он утратил интерес к этому учению. Сам же Нид-
хем до конца своей жизни последовательно придерживался точки зрения, высказан-
ной им в «Докладе...», покаяния и отречения от него не дождались даже в обмен на
всевозможные научные почести, Нидхем дорожил своей научной репутацией.
Кстати, он не писал о «насекомых и крысах, зараженных вирусом оспы», как это
пишет Терри Вольтон (2000). В «Докладе...» таких материалов нет, Нидхем отлично раз-
бирался в эпидемиологии и иммунологии натуральной оспы, что следует уже из его
трудов по истории оспопрививания и вакцинации в Китае.
Почему оказалось невозможным скрыть факты применения БО в войне на Корейском
полуострове? Ответ на этот вопрос, помимо простого установления истины, крайне
важен сегодня еще и в связи с необходимостью разработки критериев для как можно
более раннего распознания биологических диверсий и терактов и привлечения их
организаторов к уголовной ответственности.
Теоретически, БО может применяться скрытно. В местностях, где проживают люди
или действуют войска, могут существовать природные очаги возбудителей инфекци-
онных болезней и их переносчики (насекомые). Поэтому анализ эпидемической об-
становки за предыдущие годы, проведенный по публикациям в научных журналах и
по результатам многолетних наблюдений эпидемиологов, способен подсказать про-
тивнику возбудитель, распространение которого не вызовет особых подозрений у
стороны, подвергшейся бактериологическому нападению. Однако когда дело доходит
до практического применения БО в конкретной войне и на конкретной местности в
данный период времени, то, судя по опыту, накопленному Исией, возникают следу-
ющие препятствия:
1.10. Корейская бактериологическая война
223
1) природные очаги уже неактивны;
2) поддерживающиеся в этих очагах возбудители инфекционных болезней не мо-
гут быть применены теми средствами, которыми располагает нападающий;
3) поражающий потенциал этих возбудителей не соответствуют замыслу плани-
руемой операции.
Следовательно, расследование даже единичного факта опасной инфекционной
болезни в местности, где ее раньше не было, уже может вызвать у противной стороны
подозрение об искусственном характере этого случая. Для достижения основной цели
биологической войны — массового поражения войск и населения противника, «еди-
ничные случаи» не нужны. Нужно такое количество больных и умерших, которое
могло бы реально сказаться на решении хотя бы оперативно-тактических задач. По-
этому бактериологическая война неизбежно масштабируется — требуются в больших
количествах «средства доставки» и «технические средства применения» БО, т. е. не
виртуальный контагий или миазма, а различные технические устройства, которые,
являясь материальными объектами, неизбежно попадают в поле зрения противника.
Также нужны военнослужащие, использующие эти средства, попадающие не только
в «поле зрения» противника, но и в плен. Любое применение каких-либо даже хоро-
шо задуманных и эффективных «технических средств», сопряжено с ошибками и не-
удачами, когда ими пользуются призванные на войну люди. Диверсанты могут быть
перехвачены контрразведкой, средства доставки БО могут быть сбиты ПВО или зах-
вачены во время наземных наступательных операций, отдельные бактериологические
боеприпасы могут попасть в руки противника неразорвавшимися из-за ошибок в их
применении, неудачности конструкции и т. д.
Например, инфицированных одними военными грызунов необходимо доставить
к цели в тот же день, так как они быстро погибают от развившейся болезни. Но до-
ставку производят уже другие военные, у которых собственные начальники и инст-
рукции. Вылет самолета может быть задержан по метеоусловиям. Пилоты ночью мо-
гут сбиться с курса и сбросить секретный груз не туда, куда им было приказано или
не с той высоты. Меняется направление ветра. Ошибочные показания высотомеров
приводят либо к преждевременному открытию контейнеров, либо они не успевают от-
крываться.
Так, утром 5 апреля 1952 г. жители четырех компактно расположенных деревень
Ганьнанского уезда китайской провинции Хэйлуньцзян обнаружили никогда не встре-
чавшихся им ранее полумертвых и мертвых полевок на крышах своих домов и во
дворах. Всего в деревнях и их окрестностях в полосе 5 на 15 км было собрано около
715 погибших животных. Бактериологическое исследование показало, что полевки
погибли от чумы. Подозрение вызвало прежде всего неправильная эпидемиология этой
болезни. Обычно полевки вида Microtusgregalis встречались намного западнее. В этом
районе Китая чума не была известна, а сами полевки данного вида не были известны
как переносчики чумы в тех районах Северо-Восточного Китая, где чума эндемична.
В тех же районах, где чума эндемична, чум ные эпизоотии среди грызунов фиксируются
в июне — июле (см. работы Николаева Н. И., 1949; и Wu Lien-Ten U. A. et al., 1936).
Ночью китайской службой воздушного наблюдения был зафиксирован пролет ист-
ребителя типа F-82 (рис. 1.40).
Операция по распространению чумы в районе города Ганьнань была спланирована
на основе ошибочного представления ученых того времени о ключевой роли грызу-
нов в поддержании чумы в природе. Ее целью было вызвать чумные эпизоотии среди
весеннего молодняка грызунов и попытаться сделать регион эндемичным по чуме.
224
Часть 1. Краткая история применения биологических средств...
Рис. 1.40. Карта маршрута американ-
ского самолета, вторгшегося в воздуш-
ное пространство г. Ганьнань 4 апреля
1952 г. Черным кружком обозначен
пункт, где были сброшены зараженные
полевки. Доклад... (1952)
Однако животных после заражения возбудите-
лем чумы, по каким-то причинам передержа-
ли в контейнерах и сбросили уже больными и
не в том месте, где планировалось; разбежать-
ся по полям и вызвать чумную эпизоотию сре-
ди других грызунов они уже не могли.
Другим примером неправильной эпидемио-
логии были вспышки чумы, когда в эпидемиче-
ской цепи, где принимает участие человече-
ская блоха Pulex irritans, не хватало несколько
звеньев. Обычно сначала возникает эпизоотия
чумы среди грызунов, затем следуют вспышки
чумы среди людей, после которых Р. irritans ин-
фицируется возбудителем чумы. Только тогда
человеческая блоха приобретает способность
заражать чумой людей. В материалах Доклада
содержатся документы, свидетельствующие о
появлении крупных масс инфицированных
возбудителем чумы Р. irritans без эпизоотий
среди грызунов и случаев чумы среди людей.
Как правило, такие «находки» имели место
после пролета американских самолетов (см.
«Попытки вызвать чуму»).
Некоторые виды насекомых, по меньшей
мере, никогда ранее не были обнаружены в тех
районах боевых действий, где их находили в
больших количествах в январе — марте 1952 г.
Но Комиссия даже не пыталась доказать факт
их искусственного распространения, если ра-
нее не были определены фаунистические обла-
сти их распространения в Китае и Корее. Од-
нако Комиссия выявила факты аномального
появления масс насекомых в стадии развития,
не соответствующей этому времени года и температуре, атакже случаи ненормальной
скученности в одном месте насекомых с разной экологией.
Неоднократно скопления насекомых обнаруживали после пролета американских
самолетов в необычных для насекомых местах. Блохи (Р. irritans) должны находиться
в жилищах людей и поблизости от них. Как правило, даже в домах, где существуют
антисанитарные условия для жизни людей, их находят не более 50 штук за раз; выплод
человеческих блох обычно немногочисленный. Но после появления американских
самолетов над городами Фушунем, Куаньданем и другими населенными пунктами, их
находили на снегу в полях десятками тысяч. Регулярно американские пилоты сбра-
сывали на снег и других зараженных насекомых. На снегу они замерзали, покрывая
овальную площадь различного размера с характерной зоной наибольшей кучности в
одном из фокусов эллипса. Комиссия установила, что сообщения об обнаруженных
скоплениях насекомых обычно исходят из районов линий коммуникаций, используе-
мых китайскими войсками, в основном вдоль северного побережья реки Ялу Ляодунь-
ской провинции (рис. 1.41).
7.10. Корейская бактериологическая война
225
Рис. 1.41. Ликвидация последствий бактериологических нападений в Северной
Корее в 1952 г. А. Организованный женский отряд бонификаторов собирает и
сжигает насекомых, сброшенных американскими самолетами Б. Дезинфекция
в городе после биологического нападения. Доклад... (1952)
Первое сообщение о начавшейся на Корейском полуострове бактериологической
войне, поступило в СМИ 22.02.1952 г. от официального органа КНР — телеграфного
агентства Синьхуа. О том, что китайцам и северокорейцам быстро стало известно о
применении БО американской стороной, можно судить и по показаниям пленных.
Американское командование очень интересовала результативность бактериоло-
гической войны. Поэтому на территорию противника засылались агенты с целью
сбора и передачи информации, касающейся якобы эпидемической обстановки. Аген-
тов вакцинировали (около 20 инъекций разных вакцин), давали с собой таблетки
антибактериальных препаратов и инструктировали относительно тех инфекционных
болезней, сведения о которых интересовали американское командование. Подлинную
цель задания от них скрывали. Перейдя границу, агенты вскоре понимали, зачем их
послали. Например, арестованный 25 мая 1952 г. северокорейской контрразведкой
южнокорейский агент Ли Бин Ним показал Комиссии, что когда он 31 марта приехал
226
Часть I. Краткая история применения биологических средств...
в город Ончин с целью найти определенного человека, оказалось, что тот был на «про-
тивоэпидемической работе». Повсюду в городе были расклеены листовки, в которых
население предупреждалось о ведении противником бактериологической войны.
Любопытные показания об осведомленности американского командования по
поводу действий противника по ликвидации последствий применения БОдал сбитый
летчик, старший лейтенант Пауль Р. Книсс. Во время инструктажа о правильном при-
менении нового оружия, проведенном 21 марта на базе К-46 (база для F-51, пример-
но в 5 милях севернее от Вонжу, Южная Корея), инструктор, капитан Маклаффин,
сказал пилотам следующее:
«Такие бомбы будут всегда сбрасываться с четырех самолетов одновременно. Вы
спикируете с высоты 10000 футов до 6000 футов и сбросьте свои бомбы над целью. Бом-
ба взорвется, примерно, в 100 футах от земли и бактерии рассеются, примерно в окруж-
ности 100 ярдов. Если бомба не взорвется в воздухе, а взорвется на земле, то бактерии
будут убиты взрывом. Если бомба взорвется в воздухе, то бактерии будут рассеяны си-
лой взрыва. Такие бомбы будут сброшены недалеко от города, но не в самом городе, так
как северокорейцы широко применяют дезинфицирующие вещества в своих городах, ко-
торые убивают бактерии. Мы сбрасываем наши бомбы вблизи больших городов с та-
ким расчетом, чтобы люди и животные могли занести болезни в города, где бактерии
должны распространяться, но эти бактерии должны попасть на животных или людей в
течение трех часов, иначе они погибнут».
Таким образом, по крайней мере, с конца февраля 1952 г. бактериологическая
война на Корейском полуострове и в Северо-Восточном Китае велась совершенно
открыто. Обе стороны учитывали действия друг друга, а о применении БО знали, и
даже, непосредственно с ним сталкивались, миллионы людей. Бактериологическому
нападению подвергались отдельные позиции китайских и северокорейских войск,
частично объекты войскового тыла, линии коммуникаций, узлы железных и грунто-
вых дорог, отдельные промышленные и сельскохозяйственные районы, порты, водо-
источники. И, если судить по показаниям Книсса, к марту месяцу американская армия
уже осознала, что бактериологическая война, это не такое простое дело, как понача-
лу казалось.
Использование насекомых. Комиссия пришла к выводу, что бактериологическая
война на Корейском полуострове велась, в основном, путем распространения опас-
ных микроорганизмов посредством переносчиков (различных видов насекомых и
грызунов) и в меньшей степени через сброшенные предметы (перья, листья), назна-
чение некоторых из них осталось неизвестным. Самих насекомых либо инфициро-
вали, либо пытались использовать в качестве механических переносчиков возбуди-
телей болезней. Одна из причин выбора такого способа ведения войны изложена выше
(разд. 1.8 «Разработка специальных боеприпасов и устройств для диспергирования
блох и бактерий»), другая же кроется в изменении характера военных действий.
С начала 1952 г. война перестала быть маневренной и все больше уходила под зем-
лю. Американцы пытались выжигать напалмом кислород у входов в туннели, в кото-
рых скрывались северокорейские и китайские войска, и достигали некоторого успе-
ха. Но с появлением у тех русских изолирующих противогазов, эффективность таких
ударов резко снизилась. Инфицированные грызуны и их паразиты казались амери-
канским военным тем смертоносным фактором, который может проявить себя в ус-
ловиях невероятной скученности людей, подземных туннелях. Кроме того, для амери-
канцев весьма важным представлялось уничтожение нелояльного населения на этом
театре военных действий.
10. Корейская бактериологическая война
227
По данным Комиссии, американская авиация стала распространять насекомых в
Северной Корее в начале февраля. На территории Китая и Кореи выявлено 18 видов
зсекомых и пауков, сбрасываемых с американских самолетов. Бактериологические
сследования доказали, что, по крайней мере, 9 из них были заражены патогенными
икроорганизмами. В отношении других у Комиссии не осталось уверенности в том,
~э они не были заражены, так как всегда трудно выделить патогенные микроорга-
низмы из неизвестного материала, особенно когда не знаешь, что на то искать
Районы распространения насекомых американскими самолетами были обширны.
В районах Северо-Востока Китая, на севере — до реки Нахэ и горы Кэшань, на юге —
до реки Чжуанхэ и уезда Фусянь, на западе — до уездов Фусинь и Цзиньчжоу, на вос-
токе — до уездов Чанбай и Аньдун. В этих районах, лишь после вторжения американ-
<их самолетов, неожиданно появлялись скопления насекомых. Кроме этого, с 6 марта
гериканские самолеты неоднократно сбрасывали насекомых на город Циндао. По
мнению Чэнь Вэнь-гуя, председателя юго-западного филиала Китайского медицин-
ского общества, изучавшего последствия применения БО японской армией, масштабы
спользования в 1952 г инфицированных возбудителем чумы блох были значитель-
но большими, чем это имело место в Китае в годы Второй мировой войны.
Комиссия, анализируя видовой состав сброшенных американской авиацией на-
секомых, пришла к выводу, что в этой войне американской стороной могли приме-
няться насекомые, малоизвестные и совсем неизвестные в качестве переносчиков
возбудителей инфекционных болезней. Причем приоритет оставался за теми видами,
для которых не былч отработаны методы их истребления.
Многие эпизоды бактериологической войны остались неизвестными из-за такти-
< г бомбовых ударов, используемых ооновской авиацией. Обычно налеты на корей-
ские города происходили по одному сценарию. Сначала появлялось несколько мелких
рупп истребителей F-51, которые с небольшой высоты обстреливали дома и улицы,
стараясь убить как можно больше жителей. Такой расстрел длился обычно 2- 3 часа.
Затем в течение 3—4 часов нац городом последовательно проходили несколько групп
по 5-8 самолетов) бомбардировщиков В-29, которые сбрасывали фугасные и зажи-
"ательные бомбы, баки с горючей смесью, стараясь охватить пожарами весь город.
Затем снова появлялись F-51 и ран треливали спасательные команды и людей, пы-
^авшихся выбраться из этого ада. В общей сложности город мог находиться под не-
прерывным огнем авиации до 12 часов. Но иногда, перед рассветом, над городом на
малой высоте появлялись самолеты, диссеминировавшие зараженных насекомых.
Затем начинались многочасовые налеты. Когда гавался отбой воздушной тревоги, как
~равило, сброшенные насекомые уже расползались. Такой способ применения БО
называли тогда «японским».
Бактериологические боеприпасы, Комиссия занималась и установлением типов
контейнеров, бомб и других технических средств, применявшихся при ведении бак-
ериологической войны. Комиссия признала, что результаты ее работы не могут счи-
таться полными, так как многие из использованных контейнеров представляли собой
саморазрушающиеся конструкции, они либо разваливались на мелкие кусочки при
• паре об землю, либо самосяЛидлись после освобождения от груза. Даже тогда, когда
падение такого контейнера происходило на глазах местных жителей, им не всезда
'авалось его обнаружить, так как они не знали, что искать Не обладали полными
знаниями по этому вопросу и пленные летчики, им такую информацию не доводили
как ненужную для выполнения задания. Однако Комиссии все же удалось охаракте-
избвать массово использовавшиеся образцы такого вооружения
228
Часть 1. Краткая история применения биологических средств...
Взрывающаяся в воздухе бомба для листовок с дистанционным взрывателем — при-
близительно тех же размеров и форм, как обыкновенная 250 кг бомба (размер 1,4 м:
диаметр 0,4 м; обшивка стальная), но ее вес составлял около 75 кг. Головка бомбы
конической формы, на конце которой находился дистанционный взрыватель. В голов-
ке имелся небольшой пустой отсек, за ним распола! алея цилиндрической формы кор-
пус бомбы, имевший 4 отсека обшей емкостью 72 л. Корпус разделялся продольно. На
корпусах бомб были заводские надписи «ЕВРТУ», «ВОМВ-LEAFLET», «500LB».
По описанию пленных летчиков, створкам бомбы полагалось открываться на высоте
30 м, распространяя содержимое на площади около 100 м в диаметре. Один из очевид-
цев видел 26 марта, как американский самолет кружился над Егвоном и сбросил две
такие бомбы при пикировании. Бомбы раскололись после взрыва в воздухе на две
части и стали причиной появления большого количества насекомых на пространстве
размером около 200х 100 м. В Корее применение таких бомб зафиксировано в провин-
циях Пьеганнам, Кангвон, Хванхе. В Китае вЧанбае, провинции Ляодун, в конце мар-
та крестьяне нашли на своих полях три таких неразорвавшихся бомбы, окруженных
расползающимися насекомыми (Комиссия опросила этих крестьян).
Когда фото! рафии собранных на китайских полях бактериологических бомб по-
явились в печати, американцы поспешили заявить, что их применяют исключитель-
но для распространения листовок. Но все испортил председатель подкомитета по ас-
сигнованиям конгресса Сайкс. Заслушав секретный (!) доклад Буллена, командующего
химическим корпусом США, Сайкс заявил прессе следующее: «Он (Буллен) говорил,
что средства распространения бактерий на вражеской земле просты. Такое снаряже-
ние имеется у нас в армии сейчас в большом количестве..., как, например, контейне-
ры, используемые для сбрасывания листовок» (Ассошиэйтед Пресс, 05.04.1952 г.).
Контейнер «яичная скорлупа». 21 марта за чертой города Куаньдяня в провинции
Ляодун местными жителями было найдено свыше 200 обломков контейнера, сделан-
ного из какого-то известкового вещества, вместе с чашеобразной металлической
пластинкой и металлическим стержнем, прикрепленным к центру ее вогнутой поверх-
ности. Вблизи контейнера было обнаружено большое количество черных мух (Ну1е~
myia spp.), пауков (Tarentula spp.) и перьев. Бактериологическое исследование пока-
зало, что все они заражены возбудтелем сибирской язвы. Эти насекомые по своей
экологии не должны были встречаться в данной местности в это время. Изучение
обстоятельств обнаружения контейнера позволило Комиссии утверждать, что он был
сброшен с американского самолета 12 марта. Металлические части и известковые ку-
сочки были предметом тщательного анализа, проведенного в Институте современной
физики и в Институте прикладной физики Академии наук Китая. Анализ выполнял-
ся с целью установления первоначальной формы этой конструкции (рис. 1.42).
Китайские ученые пришли к выводу, что неповрежденный контейнер имел цилин-
дрическую форму и, по крайней мере, один из его концов имел куполообразную фор-
му. Общая длина контейнера составляла более 40 см, стержня — 28 см. Радиус кри-
визны стальной чашеобразной пластинки — 12,5 см; ее диаметр — 17,4 см. Радиус
известкового тела контейнера — 14 см. Толщина известковых стенок — 2 мм. Весь
контейнер снаружи был покрыт алюминиевой краской. Исследования рентгеном
показали, что материал стенок состоял преимущественно из углекислого калия. При
спектрографическом анализе обнаружено наличие некоторого количества магнезии,
химический анализ показал наличие неизвестного органического вещества, возможно,
служащего для придания материалу пластических качеств. Присутствие маленьких пор
на гладкой внутренней поверхности меловых осколков и несколько желобков на них,
10. Корейская бактериологическая война
229
Рис. 1.42. Карта маршрута американских самолетов, вторгшихся в воздушное
пространство уезда Куаньдянь 12 марта 1952 г. Черным кружком обозначен на-
селенный пункт, в котором обнаружены зараженные насекомые. Доклад. (1952)
л также обнаружение следов спайки на краю полушарообразной стальной пластины,
озволяло предположить, что внутри мелового слоя имелась хрупкая рама или какие-
то оболочки. Но те, кто занимались сбором осколков или других предметов на месте,
-е нашли остатков этих частей бомбы (рис. 1.43).
Рис. 1.43. Контейнер «яичная скорлупа». А. Фрагменты контейнера.
Б. Схематическое изображение контейнера, составленное китайскими
учеными по найденным фрагментам. Доклад... (1952)
230
Часть 1. Краткая история применения биологических средств...
Несомненно, что конструкция контейнера и выбранные для его изготовления
материалы, предполагали уничтожение всех следов применения этого устройства для
распространения спор сибирской язвы — разлетевшиеся кусочки оболочки бомбы
должны были разрушиться под действием воды, а согнутый ржавый стержень ника-
кой информационный ценности сам по себе не имеет. Конструктивно бомба «яичная
скорлупа» представляет изощренное развитие японских керамических бомб «систе-
мы Удзи» с легко разрушающимися корпусами, предназначенных для рассеивания
зараженных насекомых (см. рис. 1.24).
Кроме Куаньдяня, применение таких бомб зафиксировано вблизи Пектона (Се-
верная Корея) и Бэйцзинцзы (Китай). Комиссия пришла к выводу, что такие бомбы
применялись на обоих берегах реки Ялу в марте и июне месяце 1952 г. Возможно, это и
есть перьевая бомба (feather bomb) Ml 15, о существовании которой вконце 1940-хг.,
по официальным американским документам, стало известно только в 1997 г. Она при-
менялась американской армией для распространения зараженных насекомых и сухой
сибиреязвенной рецептуры (см. «Использование аэрозолей»). Найти ее изображения
в открытых источниках информации мне не удалось.
Картонный цилиндр с шелковым парашютом — это единственный тип контейнера
с парашютом, который видела Комиссия. Он представлял собой картонный цилиндр
со стенками толщиной около 1 см, около 36 см длины и 13 см в диаметре. Образцы
были помечены «USC5/l-l-11952-Lotl00F6». Прикрепленный к ним шелковый па-
рашют имел в диаметре лишь 70 см. Видимо, он предназначался только для того, чтобы
затормозить падение и ослабить удар об землю. На картоне не было следов огня. Кон-
тейнер данного типа предназначался для распространения комаров Orthocladius, слиш-
ком хрупких, чтобы ими снаряжать контейнеры двух вышеприведенных типов.
Из разных источников Комиссии стало известно о существовании еще несколь-
ких типов бактериологических бомб (контейнеров), которые могли применяться в
этой войне, но они не были обнаружены.
Пленные летчики показали, что на лекциях по применению БО им рассказывали
о возможности распространения зараженных насекомых следующими устройствами:
1) взрывающейся в воздухе бомбе для листовок, снабженной пропеллером, вызываю-
щим взрыв после определенного числа оборотов;
2) бомбы для листовок с дверцами, открывающихся пропеллером;
3) бомбы, распадающиеся на две половины после удара о землю.
Кроме того, в статье майора Лиохи Сакаки в «Сэндей Майничи» в январе 1952 г.
был описан саморазрушающийся бумажный контейнер с бумажным парашютом. Соглас-
но этому описанию, контейнер был сделан из прочной бумаги, имел несколько отде-
лений, загружался балластом и обладал взрывателем, приспособленным для поджи-
гания в надлежащий момент как самого контейнера, так и его парашюта, сделанного
из бумаги или импрегнированного шелка. Предназначался контейнер для распрост-
ранения зараженных чумой грызунов. Животные выпускались после раскрытия кон-
тейнера, происходящего при его соприкосновении с землей, затем, после определен-
ного периода времени, происходило воспламенение, и никаких следов от контейнера
не оставалось. Устройство можно было приспособить таким образом, чтобы освобо-
дить груз контейнера на высоте 10 м над землей, вслед за чем он относился ветром в
сторону, воспламенялся и сгорал. По мнению Комиссии, именно этого типа контей-
неры использовались для распространения зараженных чумой полевок в китайской
провинции Хэйлуньцзян.
10. Корейская бактериологическая война
231
У Комиссии имелись доказательства применения других типов контейнеров.
3 частности, для транспортировки зараженных грызунов применялись цилиндриче-
екие садки из проволочной сетки и деревянные ящики, правда, не всегда было понят-
- о. как их доставляли к цели.
Диссеминирование зараженных насекомых с низколетящих самолетов — было наи-
более технически доступным и поэтому наиболее распространенным способом веде-
-ия бактериологической войны в Корее. Так как налеты осуществлялись чаще всего
-очью, то о рассеивании зараженных насекомых можно было вообще ничего не уз-
ать, если бы пилоты не ошибались и не сбрасывали насе комых на снег. Имели место
случаи, когда такие налеты осуществлялись днем на глазах многочисленных свидете-
ей. Например, 11 февраля китайскими добровольцами был замечен американский
-амолет, рассеивающий насекомых над Чорвоном с высоты 300 м. Насекомых снесло
-етром от города и раскидало на полях по снегу, на участке продолговатой формы
размером 10 км на 5 км. Население их сжигало и закапывало.
Помимо зараженных животных и насекомых, американская авиация сбрасывала
специальные бомбы, предназначенные для взрывного диспергирования бактерий над це-
мо. Они поставлялись на авиабазы уже снаряженные 100 фунтами «бактериального
желе» и оснащались дистанционными взрывателями. «Желе» перед загрузкой бомбы
самолет разводили специальной жидкостью. О захвате таких бомб неразорвавши-
мися, китайцы и северокорейцы не сообщали. Об использовании таких бомб в этой
войне известно по рассказам людей, наблюдавших их разрывы над землей и по пока-
заниям пленных летчиков.
Особой тайны в «желе» нет. Американские пилоты ничего не напутали, а просто
^правильно поняли Дело в том, что в те годы желатин в фосфат ном буфере считал-
ся наиболее удачной средой для приготовления жидких бактериальных рецептур.
Например, еще до войны на Корейском полуострове, сотрудниками Форта Детрик на
модельном неспорообразующем микроорганизме В. prodigiosum было показано, что
щи распылении его культуры, суспендированной в дистиллированной воде, выжи-
вало в среднем 0,9 % бактерий. В то время как при распылении культуры, суспенди-
~ эванной в растворе желатина, этот показатель возрос до 14,1 % (Rosebury Т. Е., 19ч 7).
желатин в фосфатном буфере в те годы использовался не только для приготовления
эецептур, диспергируемых взрывом, но и рецептур, диспергируемых выливными при-
6 эрами, использующими форсунки с соплами гидравлического типа. Образующий-
ся в том и другом случае биологический аэрозоль имел весьма широкий диапазон
частиц. Но почти 15 % материала после испарения воды переходило в частицы с диа-
мезром 5 мкм; желатин защищал бактерию от неблагоприятных факторов внешней
среды (см. у Zentner R. J., 1961).
Попытки вызвать эпидемии чумы. В те годы распространение с самолетов блох
Pulex irritans), инфицированных возбуди гелем чумы, считалось в своем роде «клас-
сикой» ведения бактериологической войны (примерно так же, как и забрасывание
-излагающихся трупов в крепости во время господства миазматических воззрений на
эпидемии). Убежденность в возможности такого способа распространения чумы было
основано, как тогда казалось, на надежном экспериментальном фундаменте, заложен-
ном еще сотрудниками отряда № 731.
Прежде всего, внимание Комиссии обратила на себя неправильная эпидемиология
чумы в Северной Корее. За последние пять столетий в Корее не было вспышек чумы.
Самыми близкими центрами эндемического распространения чумы являлись Севе-
232
Часть 1. Краткая история применения биологических средств...
ро-Восток Китая, находящийся в 450 км от места происшествия; и провинция Фуд-
цзянь в южной части Китая, отдаленная на 1500 км от того же места. Комиссия отме-
тила, что появление чумы в феврале месяце означает, что она появилась на три меся-
ца раньше, чем это обычно бывает в странах, обладающих таким же климатом, как
климат Северной Кореи. Блохи, обнаруживаемые в естественных условиях после на-
летов американской авиации, не являлись крысиными, а относились к виду Р. irrnans,
т. е. к человеческим блохам.
Комиссии стало известно о семи попытках вызвать чуму среди населения Север-
ной Кореи. Первая вспышка чумы, находящаяся в причинно-следственной связи с
внезапным появлением больших масс блох и предшествующими налетами американ-
ской авиации, зафиксирована 11 февраля. Бактериологически подтвердить инфици-
рованность блох возбудителем чумы тогда не удалось, так как санитарно-эпидемио-
логическая служба Северной Кореи еще не была готова к оактериологическим атакам.
Во всех остальных случаях были получены бактериологические подтверждения инфи-
цирования возбудителем чумы человеческих блох Р. irritans, собранных после проле-
тов американских самолетов.
Комиссия занималась изучением трех попыток вызвать чуму среди населения
Китая и Кореи. Вспышку в Ганьнане (Северо-Восточный Китай), мы уже рассмотре-
ли выше. Далее мы рассмотрим инциденты в деревне Кансукун и в районе Хойяна (Се-
верная Корея). Реальная бактериологическая война поражает своей обыденностью.
Инцидент в деревне Кансукун. Комиссии был представлен доклад начальника Цент-
рального противоэпидемического отряда министру здравоохранения КНДР от
17 апреля 1952 г. о его деятельности по ликвидации вспышки чумы в Кансукуне, атак-
же пять документов, упомянутых в докладе. Суть представленной Комиссии инфор-
мации заключается в следующем.
Второго апреля 1952 г. Пак Ион Хо, крестьянин верхнего села Кансукун (всего
936 жителей) заболел с симптомами головной боли и озноба. Больного изолировали
в отдельное помещение 3 апреля. Все дома в селе были подвергнуты дезинфекции и
дератизации. Больной умер 4 апреля, вскрытие произведено 6 апреля. Внутренние
органы трупа были отправлены в лабораторию для гистологического исследования
Мазки были приготовлены из внутренних органов трупа, и проведен посев посмерт-
ных материалов на скошенный агар. Мазки и скошенный агар отправили в лаборато-
рию для бактериологического исследования. Его результат однозначно свидетельство-
вал, что Пак умер от чумы. Окончательный диа! ноз — септицемия на почве бубонной
чумы правых лимфатических узлов.
При исследовании обстоятельств заражения чумой этого крестьянина, было ус-
тановлено, что утром 25 марта на поверхности воды в чане у колодца недалеко от сво-
его дома, куда Пак обычно ходил умываться, он обнаружил десятки блох «как будто
мертвых». Так как около 4 ч утра Пак был разбужен шумом самолета, кружившим на
небольшой высоте над селом, он решил, что блохи сброшены этим самолетом и не-
медленно доложил о случившемся председателю Сельского народного комитета. Че-
рез несколько дней он заболел чумой. Признаки неправильной эпидемиологии болез-
ни были установлены довольно быстро. Из найденных в чане блох 20 отобрали на
бактериологическое и энтомологическое исследование. По данным экспертизы, эти
блохи принадлежали к человеческому виду Р. irritans и были инфицированы возбуди-
телем чумы. Сотрудниками противоэпидемического отряда чумной эпизоотии среди
гры )унов установлено не было. Население не видело мертвых крыс и мышей. В ходе
дератизационных работ было поймано 143 крысы, ни одна из них не болела чумой.
10. Корейская бактериологическая война
233
Профессором Чень Вэнь-гуем из органов умершего крестьянина была выделена
^оковирулентная культура возбудителя чумы, 10—20 клеток которой (в разведении)
назывались смерте шными для морской свинки. Чень отметил, что и другие больные
•врали так же быстро, как эта жертва. Некоторые люди (Чень не указывает кто, где
когда) умирали в течение 24 ч, еще до развития бубонов.
На основании эпидемиолот ических и бактериологических исследований корей-
-ми и китайскими исследователями был сделан вывод, что случай появления чумы
Кансукуне вызван блохами, сброшенными с американского самолета.
Комиссия изучила данный доклад и другие приложенные документы и провела
G *бственное расследование инцидента. Членами Комиссии были опрошены односель-
чане Пака и научные эксперты, которые подтвердили и пояснили представленную
формацию Комиссия согласилась с выводом доклада начальника Центрального
ротивоэпидемического отряда.
Инцидент в районе Хойяна. Комиссии был представлен специально подготовлен-
ный для нее Государственным чрезвычайным противоэпидемическим комитетом
кНДР «Доклад о распространении американским самолетом человеческих блох, за-
тейных чумными палочками» от 11 июня 1952 г., а также 6 документов, упомяну-
дх в докладе. Суть представленной информации заключалась в следующем.
Около 10 ч утра 23 апреля 1952 г., примерно через 6 ч после появления над этой
местностью американского самолета, на открытых полях на склоне горы недалеко от
Согдонга в уезде Хойян, китайскими военными топографами случайно обнаружены
сятки тысяч блох. Площадь распространения составляла приблизительно 30x10 м.
На этой площади было место с большой кучностью размером в 3—4 м2, на котором
обнаружено такое количество блох, что от них почернела поверхность земли». Мес-
о, где обнаружены блохи, находилось в 100 м от ближайших жилищ и в 100 м от бли-
жайшей тропинки. В результате проведенной экспертизы было установлено, что эти
'лохи являются человеческими и инфицированы возбудителем чумы. Блохи были
эбтиты бензином и спалены. Все люди, принявшие участие в уничтожении насеко-
мых, заранее были привиты против чумы. После уничтожения насекомых они под-
верглись карантину и в течение 3-х дней принимали сульфатиазол в качестве средства
экстренной профилактики. Людей, заболевших чумой, ни среди них, ни среди насе-
ления не было установлено На основании эпидемиологических и бактериологических
•сследований был сделан вывод, что чумные блохи в Хойяне сброшены с американ-
. кого самолета.
Как и при рассмотрении инцидента в деревне Кансукун, Комиссия изучила данный
доклад и другие приложенные документы и провела собственное расследование это-
го инцидента. Были опрошены свидетели — китайские добровольцы, обнаружившие
скопление чумных блох, и заслушаны мнения научных экспертов. Комиссия согла-
силась с выводом доклада Государственного чрезвычайного противоэпидемического
комитета КНДР.
Приведенные случаи выглядят как очень незначительные в плане масштабов
применения БО, и особенно его последствий. Дело тут в том (и в Докладе это подчер-
кивается особо), что китайские и корейские коммунисты преследовали цель показать
мировой общественности только сам факт применения БО американской армией, а
не его возможности по уничтожению людей. Поэтому они предоставляли Комиссии
материалы по тем инцидентам, где были либо единичные пораженные чумой, либо
инфицированные насекомые с надежно подтвержденным в лабораторных условиях
видом искусственно распространяемого среди людей возбудителя инфекционной
234
Часть 7. Краткая история применения биологических средств...
болезни. Были местности, где после применения БО поражения носили массовый ха-
рактер, например, в районе Бальнамли, где 25 февраля вспыхнула чума и из 600 чело-
век этой деревни, 50 заболело бубонной чумой, а 36 из них погибло. Но в такие очаги
Комиссию не пускали и собственных материалов расследования ей не предоставля-
ли, так как еще шла война, и коммунисты не считали нужным информировать аме-
риканскую сторону об оптимальных с точки зрения эффективности способах ве-
дения бактериологической войны. По этой же причине Комиссия не публиковала ни
конкретных цифр относительно общего числа китайского и корейского гражданско-
го населения, погибшего от БО, ни цифр об общей заболеваемости, ни процента смер-
тности среди лиц, инфицированных в результате применения БО.
Попытка вызвать вспышку холеры. Один из инцидентов, которому Комиссия уде-
лила большое внимание, касался случаев заболевания холерой, имевших место в сель-
ской местности в мае 1952 г. в районе Тэтона.
Ранним утром, после ночи, в течение которых американский самолет долго кру-
жил над этой местностью, одна из молодых крестьянок, собирая траву на холме, об-
наружила соломенный пакет со съедобными морскими моллюсками. Она принесла
моллюсков домой и вместе с мужем съела их в сыром виде. Вечером того же дня оба
серьезно заболели и к вечеру следующего дня умерли.
Медицинское обследование показало, что супруги умерли от холеры. Позднее про-
тивоэпидемическая служба обнаружила и другие такие пакеты с моллюсками. Бакте-
риологическое исследование, проведенное китайскими и корейскими специалистами,
показало, что все моллюски были инфицированы возбудителем холеры.
Если отбросить в сторону бросающуюся в глаза ненормальность инцидента, ко-
торая состоит в том, что зараженные возбудителем холеры моллюски были найдены
на склонах холма, то остаются другие важные эпидемиологические аномалии. В дан-
ной местности холеры никогда не было, в основном ее случаи в Корее регистрирова-
ли в портовых городах и прибрежных районах. В течение XX в. был только один слу-
чай холеры в мае, очень редко она появлялась ранее августа.
Внимание Комиссии было обращено и на аномалии, касающиеся самих моллюс-
ков Meretrix meretrix.
1. С начала войны эти моллюски не появлялись на рынке.
2. Моллюски были найдены на холме на месяц раньше их возможного появления
на рынке.
3. В Корее при продаже эти моллюски никогда не завертываются в солому.
4. Створки некоторых моллюсков были разбиты как от сильного удара.
При выяснении обстоятельств обнаружения пакетов с моллюсками Комиссией
было установлено, что они найдены на расстоянии одного километра от ряда водных
резервуаров и меньше чем в 400 м от резервуара насосной станции, которая снабжает
питьевой водой многие прибрежные поселки и порт. В ночь, накануне той, в которую
были найдены моллюски, очистительная станция по соседству с насосной станцией
была аккуратно разрушена американскими самолетами, применившими бомбы не-
большой мощности, в результате чего насосы станции оказались неповрежденными.
При опросе местных жителей, выяснилось, что ночь во время второго рейда аме-
риканской авиации была темной и ветреной. Именно этой причиной можно объ-
яснить то, что самолет долго искал зеркало воды и, вероятно, не нашел его. Сама же
организация биологической диверсии обращает на себя внимание изощренностью за-
мысла, и даже наличием «японского следа». Холерные вибрионы устойчивы к повы-
шенной концентрации соли. По данным японской литературы, морские моллюски
1.10. Корейская бактериологическая война
235
ж,огут быть использованы как хорошая среда для роста холерных вибрионов. Эти мол-
1Юски при соприкосновении с пресной водой закрывают створки раковин и тем са-
мым отдаляют свою смерть. В течение этого времени внутри них размножается холер-
ный вибрион. После смерти моллюска его тело заражает воду возбудителем холеры
примерно в течение месяца.
В связи и этим Комиссия пришла к следующим выводам:
1. ВВС США разрушили водную очистительную станцию в Тэтоне, не повредив
при этом насосной станции (в ночь с 14 на 15 мая).
2. ВВС США пытались вы шать эпидемию холеры путем заражения распредели-
тельных резервуаров (в ночь с 15 на 16 мая).
Попытки вызвать кишечные инфекции До сведения Комиссии северокорейской
стороной была доведена информация о неоднократном обнаружении на склонах хол-
мов вблизи источников питьевой воды мертвых рыб, инфицированных возбудителя-
ми сальмонеллеза {Salmonella) и шигеллеза {Shigella). В этих случаях проглядывался
тот же замысел, что и в Тэтоне при попытках вызвать холеру.
Особый интерес Комиссии вызвал факт появления «лиофилизированного белко-
вообразного материала» после налета американских самолетов. Это вещество, обнару-
женное отд гльными массами у водоемов с питьевой водой, было клейким и гигроско-
пичным. Химический анализ показал, что оно состоит из продуктов распада белков.
Бактериологи выделили из него возбудитель дизентерии, ферментирующий маннит.
Во время работы Комиссии в Корее инциденты такого рода не повторялись, поэтому
Комиссии пришлось опираться на сообщения корейских ученых. Однако Комиссия
разделила мнение министра здравоохранения КНДР, сводящееся к тому, что этот ма-
териал являлся замороженной и высушенной культуральной массой бактерий Shigella.
Как это все делалось. По показаниям пленны < летчиков1, можно восстановить сле-
дующую картину работы механизма Корейской бактериологической войны.
Подютовка летчиков к бактериологической войне началась в конце августа 1951 г.
в авиационной школе на базе 3-го бомбардировочного полка (самолеты В-26) в Ива-
куни (Япония) в рамках обычных занятий по подготовке боевых кадров. Лекции были
секретными, читали их инструктора в штатском. После перевода 3-го бомбардировоч-
ного полка на базу Кунъсань (Южная Корея), чтение таких лекций было продолжено.
В основном они были посвящены поражающим свойствам существующих образцов
БО и методам ведения бактериологической войны. О применении БО летчикам го-
ворилось отвлеченно от данного театра военных действий и только в таком аспекте,
что это может сделать противник, но параллельно с чтением лекций им делались при-
вивки. Затем их просто поставили перед фактом начатой США бактериологической
войны. В конце декабря 1951 г. на базе появились незнакомые им офицеры, которые
участвовали в составлении заданий на полеты и принимали рапорта об их выполне-
нии в оперативном отделе штаба эскадрильи.
Бактериологические бомбы, разрывающиеся в воздухе, т. е., пред на шаченные для
взрывного рассеивания бактерий и насекомых, сбрасывались с самолетов типа В-26,
В-29, F-51, F-84, F-86. Бомбы с парашютом, механически рассеивавшие зараженных
насекомых, обычно сбрасывали с самолетов В-26 и В-29. Летчики не проверяли под-
веску таких бомб, часовые их к ним не допускали. Бактериологические бомбы подвеши-
вали на крылья бс.чбардировщиков люди в респираторах и перчатках. Обычные бомбы
помещали в бомбовые отсеки. Неразрывающиеся бомбы предписывалось сбрасывать
1 См. приложение Б
236
Часть 1. Краткая история применения биологических средств...
с высоты 200—500 футов и на скорости 200 миль/ч. Бомбы, взрывающиеся в воздухе,
необходимо было сбрасывать с высоты не менее 5000 футов от земли для того, чтобы
дистанционный взрыватель VT «Variable Time fuze» взорвал ее на высоте 50-100 фу-
тов от земли. Взрыватель включался из кабины пилота. Пилотам сообщали, что эти
бомбы бактериологические, но без указания того, какие бактерии в них находились.
Предупреждали их и об особой секретности задания. После выполнения задания о
таких бомбах требовалось докладывать и писать в рапортах либо как о «неразорвав-
шихся», либо как о «дистанционныхбомбах, взрывающихся в воздухе», не употребляя
слово «бактериологическая», и тщательно фиксировать на картах места их падения.
Диспергирующие устройства устанавливали на истребителях (типы F-51, F-80,
F-84, F-86), так как они имели большую маневренность на малой высоте. Распыление
рецептур бактерий производилось с высоты 500-1000 футов при скорости самолета
350 миль/ч. При распространении насекомых скорость снижалась до 200 миль/ч.
Имеющиеся на базе К-46 бактериологические бомбы хранились в подземных бе-
тонированных складах, которые находились в районах, где производилась зарядка
бомб. Склады были обнесены колючей проволокой. Бактериологические бомбы по-
гружались в самолет специальным отделом вооружения. В случае неисправности дви-
гателя самолета, несущего бактериологические бомбы, пилотам, если они летели над
«своей территорией», предписывалось сбросить такие бомбы над безлюдным районом,
затем связаться с радарной станцией, чтобы она отметила точно координаты самоле-
та в месте сбрасывания бомб, и сообщить их командному пункту. При сбрасывании
бомб, взрывающихся в воздухе, взрыватель требовалось не включать. При их вынуж-
денном сбрасывании над территорией противника, требовалось включить взрыватель.
Применение бактериологических бомб осуществлялось с целью убийства как
можно большего количества людей. От пилотов требовали бомбометание проводить
максимально точно, так как «бомбы стоят очень дорого», и по местам максимального
скопления людей (я особенно подчеркиваю это обстоятельство в связи с мифотвор-
чеством 1990-х гг. о дешевизне и доступности БО странам третьего мира).
Для выполнения заданий по распылению бактериальных рецептур или насекомых
обычно использовалось два самолета-истребителя, которые летели параллельно, на-
ходясь друг от друга на расстоянии около 200 ярдов. После того как самолет с диспер-
гирующим устройством возвращался с задания, за ним следовал специальный авто-
мобиль, который проводил его дезинфекцию «на ходу» путем распыления дезинфек-
танта, одновременно дезинфицировалась взлетно-посадочная полоса. Летчик отводил
самолет на особую стоянку; где его дезинфицировали уже более основательно силами
специального отряда. Затем летчик покидал самолет, менял одежду и принимал душ,
его одежду дезинфицировали. Персоналу базы, участвующему в подготовке таких
полетов и в самих полетах, проводили термометрию и брали кровь на анализ.
18-я бомбардировочная авиагруппа на февраль 1952 г. имела 4 самолета F-51, ос-
нащенных устройствами для распыления бактерий или распространения насекомых.
Самолеты стояли отдельно от остальных, на стоянке на северном конце боковой до-
рожки. За распылительными приспособлениями самолетов следил специальный пер-
сонал. Специальный персонал отдела вооружения отвечал за доставку и погрузку бак-
териологических бомб и контейнеров. Такие бомбы и контейнеры доставляли на базу
К-46 из Японии самолетами С-46, С-47, С-54, примерно, раз в две недели.
Некоторым бактериологическим ударам предшествовало разрушение санитарной
инфраструктуры городов. В Тетоне (Северная Корея), перед тем как сбросить в резер-
вуары насосной станции моллюсков, зараженных возбудителем холеры, американской
1.10. Корейская бактериологическая война
237
шиацией была аккуратно разбомблена очистительная станция, находившаяся рядом,
насосы же остались неповрежденными (см. «Попытка вызвать холеру»).
Перечисленные выше образцы БО не выдержали испытания войной, и о них проч-
но забыли. Но вот одно средство той войны остается востребованным и по сей день.
«Белый порошок». Печально известный по событиям октября 2001 г. в США «бе-
лнй порошок», это ни что иное, как биологическая рецептура — композиция, состоя-
шая из высушенных бактерий и специального наполнителя, облегчающего их рассе-
ивание в воздухе. Хотя технология лиофильного высушивания биологических сред
была известна еще с 1920-х гг., Исии не удалось полностью использовать ее потенци-
XI для разработки средств ведения бактериологической войны. Только в 1944 г. спе-
циалисты отряда № 731 освоили эту технологию, но к созданию эффективных био-
логических рецептур они не подошли, так как столкнулись с массой новых техничес-
ких проблем, причины возникновения которых станут известны только в 1970-х гг.
(см. разд. 1.8 «Производственные мощности»). Полученную лиофильным высуши-
ванием пористую массу бактерий японцы перец боевым применением просто разво-
дили специальной жидкостью. Создание «белого порошка» стало серьезным успехом
американской военной бактериологии конца 1940-х гг., и, как показывают материа-
лы Комиссии, Корейская война дала ему возможность пройти полевые испытания
под тривиальным названием «зараженная бактериями пыль». Термин «пыль» обыч-
но используется для описания дисперсной системы, состоящей из твердых различ-
ных по величине частичек, находящихся в воздухе во взвешенном состоянии. В данном
случае имеется в виду дисперсная система, состоящая из высушенных бактерий и
специального наполнителя, формирующих из них частички размером менее 5 мкм,
т. е. способные проникать в глубокие отделы легкого. Обнаружить такую композицию
в окружающей среде, как это удавалось в отношении зараженных насекомых, разу-
меется, невозможно. Однако ее «уши» все же «высовывались» из общей картины пре-
ступлений войны на Корейском полуострове.
Впервые о «зараженной бактериями пыли» упоминается в лекции некоего Виль-
ямса, читавшего 26 августа 1951 г. лекции по бактериологической войне пилотам и
штурманам бомбардировщиков В-26. По показаниям, данным Комиссии пленным
американским летчиком, старшим лейтенантом Кеннетом Л. Иноком (когда сбит, не
.казано; показания дал 7 апреля), среди прочих способов применения БО, Вильямс
/казал следующие: 1) сбрасывание с самолетов бомб, начиненных зараженной бакте-
риями пылью, которые раскрываются в воздухе, и эта пыль разносится ветром; 2) рас-
пространение зараженной бактериями пыли непосредственно самим самолетом с
помощью специальных распыляющих аппаратов. Касаясь диверсионных способов
применения БО, Вильямс упомянул бумажные конверты, зараженные бактериями.
Другой пилот сбитого В-26, Джон Квин, старший лейтенант ВВС США, расска-
зал Комиссии следующее. На одной из лекций, прочитанных летному составу 8-й
эскадрильи в средине декабря 1951 г. по методам ведения бактериологической войны,
гражданский специалист, г-н Ашфорк, показал фотографию реактивного самолета
F-84, рассеивающего «пыль, насыщенную бактериями» из баков, находящихся по
бортам самолета.
Сбитый 4 марта пилот истребителя F-51, Флойд Бреланд О’Нил, рассказал Комис-
сии, что на базе К-46, где он проходил службу с 19 января, имелось 4 самолета, при-
способленных для распыления бактерий. Он же воспроизвел комиссии схему рас-
пылительного аппарата, которую ему показали на одной из лекций по способам при-
менения БО.
238
Часть 1. Краткая история применения биологических средств...
Видимо, первый опыт применения «пыли» не был удачным, и диспергирующие
устройства нуждались в модернизации. Лейтенанта Книсса (когда попал в плен, не
указано; показания дал 20 июля), прибывшего 21 марта на базу К-46, инструктируют
особенностям применения бактериологических бомб. Инструктор, капитан Маклаф-
лин, сообщил ему, что 30 апреля четыре самолета авиагруппы будут отправлены в Та-
чикава (Япония) для оснащения аппаратами для распыления бактерий. Эти само-
леты предполагалось получить обратно 15 июня. Надо заметить, что в феврале 1952 г.,
благодаря действиям русских истребителей, американская авиация понесла в Корее
очень большие потери. О четырех самолетах с диспергирующими устройствами, о
которых сообщил Комиссии пилот истребителя F-51 О’Нил, капитан Маклафлин уже
не упоминает. Да и сам О’Нил уже к этому времени был сбит. Для американских лет-
чиков получить задание на применение БО не предвещало ничего хорошего. Сбрасы-
вание бактериологических бомб осуществлялось с малой высоты и на малой скорос-
ти, а применение «зараженной бактериями пыли» могло быть эсЬфе ктивным только в
том случае, если велось с предельно малой высоты. Однако тогда самолет становился
уязвимым для огня зенитной артиллерии и стрелкового оружия.
Использование аэрозолей. Масштабных поражений бактериальными аэрозолями
во время бактериологической войны в Корее не выявлено. Сообщается о нескольких
случаях инфицирования людей, которые можно считать ингаляционными. Так, в мар-
те в Ляодуне и Ляоси (Северо-Восточный Китай) выявлено 5 смертельных случаев
заболевания легочной сибирской язвой у лиц, по своей профессиональной деятель-
ности не имеющих к ней никакого отношения (железнодорожник, велорикша, школь-
ная учительница, домохозяйка, крестьянин). У всех из них болезнь протекала одина-
ково быстро, сопровождалась резкой слабостью, геморрагическим менингитом,
смерть наступала ранее 48 ч от начала болезни. До этого случая в Китае такая клини-
ческая форма сибирской язвы не встречалась. Про погибших известно, что четверо
из них участвовали в работах по ликвидации последствий применения американски-
ми самолетами БО, в частности, они занимались сбором насекомых и птичьих перь-
ев, сброшенных с американских самолетов, и работали без перчаток, маски и пинцета.
У всех из них, а также с собранныхх ими перьев, были выделены бациллы сибирской
язвы, проявлявших себя одинаково в реакциях на ферментацию углеводов.
Интересны наблюдения о типах БО, применяемых американской авиацией в этих
районах. В одном случае несколько человек видели, как было сброшено «нечто вроде
большого красного термоса, взорвавшегося на высоте 10 м над землей с появлением
дыма и неприятного запаха горящего белка». Однако наиболее часто находили мед-
ленно рассеивавшиеся ветром птичьи перья, покрывавшие землю в форме треуголь-
ника, но без обнаружения контейнера. Эти перья играли в конструкциях необнару-
женных боеприпасов две роли: они дезагрегировали «белый порошок» во время
хранения боеприпаса; и являлись его макроносителями на небольшие расстояния, тем
самым, способствуя большему заражению местности и людей спорами возбудителя
сибирской язвы. Члены Комиссии Нидхема не могли знать, о каком типе биологиче-
ского боеприпаса идет речь в случаях, когда носителями спор возбудителя сибирской
язвы стали птичьи перья. Впервые о существовании в то время в американской армии
перьевых бактериологических бомб стало известно из работы R. Sidell et al. (1997).
Как промежуточную форму применения биологического агента между жидкой
рецептурой и «белым порошком», можно охарактеризовать «лиофилизированный бел-
ковообразный материал», многократно находимый вблизи от водоисточников после
налетов американских самолетов, предназначенный для заражения открытых водо-
1.11. Биологическое оружие перед подписанием Конвенции 1972г.
239
емов. Смысл изготовления биологического агента в виде лиофилизированной массы
заключался в ее легкой растворимости. Лиофилизат предназначался для заражения
крупных водоисточников, где бы он растворялся «без следа», не вызывая у североко-
рейцев подозрений об искусственном характере вспышки кишечной инфекции.
И только случайно, благодаря ошибкам американских пилотов, стало известно об от-
дельных случаях его применения.
* * *
Результаты применения большого количества образцов БО и биологических аген-
тов во время войны на Корейском полуострове оказались провальными. Об этом пи-
шут в своей работе, рассекреченной и изданной почти через 50 лет после ее написа-
ния, С. С. Лотоцкий с соавт. (2000). Такой же точки зрения придерживались и в
военном ведомстве США В июне 1953 г., за месяц до окончания войны, министром
обороны был сделан вывод о том, что «возможности БО ограничены вследствие раз-
ных причин, но главным образом из-за пробелов знания в области биологических
наук» (U.S. Army activity..., March 8 and May 23,1977). Однако у военных вновь появи-
-ась иллюзия близкого успеха. Теперь поисковые исследования концентрировались
до двум направлениям: первое — разработка боеприпасов для применения жидких и
сухих рецептур в виде аэрозолей, способных проникать непосредственно в глубокие
отделы легких поте нциальной жертвы, т. е. с дисперсностью жидкой или твердой фазы,
не превышающей 5 мкм; второе — изучения возможностей использования насекомых
как переносчиков опасных инфекционных болезней, способных вводить их непосред-
ственно в кровь потенциальной жертвы.
Разработчики других видов оружия массового поражения подавали разработчи-
кам БО пример для подражания. Физики-ядерщики, начав свой путь к атомной бом-
бе на 10 лет позже разработчиков БО, уже испытывали термоядерные заряды. Воен-
иые химики в начале 1950-х гг. подошли к пределу развития химического оружия,
синтезировав боевые ОВ на основе фосфорорганических веществ. Успехи биотех-
нологии и начавшаяся «холодная война» создали идеальные условия для разработчи-
ков БО. По окончанию войны на Корейском полуострове наступил их «золотой век».
1.11. БИОЛОГИЧЕСКОЕ ОРУЖИЕ ПЕРВД ПОДПИСАНИЕМ
КОНВЕНЦИИ 1972 г.
Масштабирование прои звидства биологических агентов и испытаний БО. Изучение
поражающего действия мелкодисперсных аэрозолей агентов БО. Судьба проникших
в дыхательные пути человека частиц биологического аэрозоля. Технические сред-
ства применения аэрозолей агентов БО. Аэробиология мелкодисперсного аэрозо-
ля. Отдел специальных операций в Форт-Детрике. Насекомые как переносчики воз-
будителей опасных инфекционных болезней. Поражение растений. Взгляды на
ведение биологической войны. Конвенция 1972 г. Уничтожение БО. Форт-Детрик
после уничтожения БО.
Период разработки БО, начавшийся после завершения войны на Корейском по-
уострове, можно назвать аэробиологическим. Развитие биотехнологии сулило разра-
ботчикам БО тонны бактерий и биологических рецептур. Им теперь стал понятен
физический механизм инфицирования людей биологическими аэрозолями. В свою
очередь, знание размера частиц аэрозоля, способного проникать в глубокие отделы
240
Часть 1. Краткая история применения биологических средств...
легких человека, оказало революционизирующие влияние на разработку БО. Подход
к разработке биологических боеприпасов и авиационных диспергирующих приборов
принципиально поменялся. Теперь такие устройства должны были создавать аэрозоль
с размером частиц дисперсной фазы до 5 мкм; боеприпасы, позволявшие лишь при-
вести человека в контакт с как можно большим количеством не потерявших жизне-
способности микроорганизмов, ушли в прошлое. Но самих разработчиков БО по-пре-
жнему не оставляло ощущение, что они ведут еще пока какие-то предварительные
эксперименты.
Масштабирование производства биологических агентов и испытаний БО. В средине
1950-х гг. технологии глубинного культивирования бактерий окончательно вытесни-
ли простые технологии выращивания на плотных питательных средах, использовав-
шиеся «первопроходцами» БО в 1930—1940-хгг. Поданным СИПРИ, для успешного
ведения биологической войны военный потенциал развитой страны средних разме-
ров должен обеспечивать возможность производства, по меньшей мере, 100 партий
соответствующих биологических материалов, каждой из которых должно быть доста-
точно для осуществления бактериологического нападения в течение года или за бо-
лее короткий отрезок времени. При меньших производственных мощностях сущест-
вовала опасность очень медленного пополнения израсходованных биологических
агентов. Кроме того, должна обеспечиваться надежность в отношении выживаемости
или сохранности биологических агентов при хранении. Это значит, что страна, решив-
шая иметь в своем арсенале «мошное оружие бедных», должна была строить заводы
по производству опасных микроорганизмов весьма значительных размеров и обеспе-
чивать более или менее непрерывную работу большого числа ферментеров (The
Problem..., 1970).
Некоторое представление об организации разработки и производства агентов БО
в стране, реально готовившейся к биологической войне, дают опубликованные в от-
крытой печати данные об арсенале Пайн Блафф, находящемся в штате Арканзас
(США). Американской армией здесь было организовано производство отравляющих
веществ, дымовых смесей, зажигательных составов, биологических агентов и соответ-
ствующих боеприпасов в лабораторных, полу производственных и ограниченных про-
изводственных масштабах, а также хранение некоторых готовых боеприпасов. Коли-
чество сотрудников — около 1800 человек. В 1966 г. площадь территории арсенала
составляла около 6 тыс. га, на этой территории располагалось 945 зданий и соору-
жений, стоимость которых вместе с оборудованием оценивалась в те годы в 136 млн
долларов. Среднее суточное потребление воды было эквивалентно потреблению го-
рода с населением 33 тыс. человек, а суточный расход — расходу города с населением
13 тыс. человек (The Problem..., 1970).
Испытания БО — неотъемлемая часть исследований по его созданию. Они были
направлены, главным образом, на получение данных для оценки и подтверждения или
отрицания постулатов и теорий, разрабатываемых в условиях лаборатории, расчетных
параметров оборудования и математических моделей. Испытания БО, подобно всем
элементам программы БО, были уникальной областью исследований. Проведение
исследований по изучению поведения искусственно распыленного в атмосфере ма-
териала создаваемых биологических рецептур, в начале 1950-х гг. не представляло
хорошо изученной и слаженной научной дисциплины. Данных о факторах биологи-
ческого отмирания и (или) физической гибели микроорганизмов при обычных коле-
баниях погоды, о количестве микроорганизмов, необходимых, для развития инфек-
ции, о способах или оборудовании для их распространения было мало, либо они
.11 Биологическое оружие перед подписанием Конвенции 1972г.
241
зобще отсутствовали. В связи с этим военным ученым необходимо было провести
большой объем исследований, имеющих цель — сбор необходимых научных и техни-
еских данных для подтверждения теоретических выводов и заполнения пробелов в
ганиях, а также для определения степени уязвимости от БО личного состава воору-
женных сил, боевой техники, средств индивидуальной и коллективной защиты (US
Vmy activity..., 1977).
Испытания БО в те годы подразделялись на лабораторные (небольшого масшта-
ба). камерные (среднего масштаба) и полевые (большого масштаба). Каждую из этих
•лтегорий, в свою очередь, делили на испытания с использованием агентных имита-
Чэов и патогенных микроорганизмов. Дальнейшая классификация производилась по
лъекту испытания, т. е. испытание механических устройств, таких как детекторы или
; обоотборники, или испытания на живых объектах (люди, животные или растения),
зенка эффективности вакцин, анатоксинов и т. п. (рис. 1.44).
Испытания биологического оружия
Камерные
Полевые (на открытым воздухе при пярпг,ятппн. ТР
контроле метеорологических условий/1 р
11митатор Биоагент—
Имитатор Биоагент Имитатор Биоагент
Иммунологические
исследования ।-------------
Люди/ за пределами
Детекторы/ континентальной
Животные/
части США
Континентальная
часть США
Механически"
устройства
Иммунологические
исследования
Люди/
Детекторы /
Животные/
Механические Животные
устройства I
Защищенные |
Незащищенные
етекторы
робоотборники
Защитное оборудование
Оборудование для
испытания боеприпасов
Поражаемость
I осударственные Военные
территории объекты
Пробоотборники
Защитное оборудование
Оборудование для
испытания боеприпасов
Растения
Люди
.Животные
\ \ Детекторы
[робоотборники ^Защитное оборудование
Рис. 1.44. Схема испытаний БО и средств противобиологической зашиты,
практиковавшихся с начала 1950-х гг. в США. По US Army activity... (1977)
242
Часть 1. Краткая история применения биологических средств...
В США полевые испытания химического и биологическое оружия производились,
главным образом, на полигоне Дагуэй (шт. Юта/. 1ерритория полигона имела площадь
около 1500 кв. миль, периметр около 210 км. Число сотрудников: 1000 гражданских.
600 военных (Hersh S., 1968). В связи с недостаточными размерами полигона Дагуэй.
американские военные в те годы проводили испытания БО на различных островах
Тихого океана, в нейтральных водах в юго-западной части Тихого океана и в Форте
Грили на Аляске. Сообщалось об испытании Соединенными Штатами БО в откры-
тых водах юго-западной части Тихого океана, у атолла Эниветок (Eniwetok) на Мар-
шалловых островах и об опытах с насекомыми, переносчиками биологических аген-
тов, проведенными на острове Бейкер Айленд в средней части Тихого океана в 1965 г.
В Канаде полигон для испытания химического и биологического оружия существ^ -
вал при Канадском научно-исследовательском центре защиты в Саффиде (шт. Аль-
берта). Площадь полигона составляла 1000 кв. миль.
В Великобритании общая площадь научно-исследовательского центра в Портоне
вместе с испытательными отделами не превышала 10 кв. миль (The Problem..., 1970).
Более масштабные опыты с агентными имитаторами в период с 1961 по 1971 г. прово-
дились вблизи побережья Дорсета и Восточного Девона (Восточная Англия). Диспер-
гирование бактерий осуществлялось с самолетов и аэрозольными генераторами с ко-
раблей. Большинство имитаций массированного применения вероятным противни-
ком БО против Соединенного Королевства осуществлялось в зоне Лайм-Бея или
Уэйтмут-бея с прохождением бактериального облака через Дэрсетское побережье в
коридоре между Бритпортом и Портлендским мысом. В ряде экспериментов бактерии
рассеивались с корабля в направлении Девонского побережья между городом Торки
и границей Дорсета. Имитация биологических атак авиацией происходила в районе
аэропорта Тарант-Раштон (вблизи Блендсфорд-Форума). В качестве наполнителя в
бактериальной рецептуре использовались микронити (Spratt В., 1999).
Изучение поражающего действия мелкодисперсных аэрозолей агентов БО. С конца
1940-х гг. эти исследования стали развиваться по двум направлениям:
первое — изучение поражающей способности аэрозолей микроорганизмов;
второе — изучение поражающего действия аэрозолей биологических токсинов.
Исследования последовательно масштабировались — от изучения принципиальной
возможности инфицирования (поражения) людей и экспериментальных животных
мелкодисперсными аэрозолями агентов БО в лабораторных условиях, до воспроиз-
ведения этого инфицирования (поражения) у экспериментальных животных на по-
лигоне с применением специальных боеприпасов и в условиях, максимально прибли-
женных к действиям «вероятного противника».
В 1950 гг. не существовало ни одного распылительного устройства, дававшего го-
могенное облако агентов БО и которое можно было применить в масштабах полигон-
ных испытаний. При инфицировании небольших животных в лабораторных усло-
виях с 1951 г. применяли маломощные распылительные устройства, использующие
принцип вибрации. Обычно ими были капилляры, вибрирующие под действием элект-
ромагнита. Жидкость подавалась в капилляр под давлением и «срывалась» с его виб-
рирующего конца в виде струек капелек одного размера. Если скорость потока жид-
кости и частота вибрации капилляра постоянны, то все образующиеся капли имеют
одинаковый размер с небольшим геометрическим стандартным отклонением. При
правильном подборе параметров работы устройства оно давало относительно гомо-
генный аэрозоль с частицами в пределах 5 мкм (Davis J. М., 1951; Dimmock N. А..
1951). С конца 1950-х гг. для таких экспериментов стали использовать ультразвуке-
111. Биологическое оружие перед подписанием Конвенции 1972 г.
243
>ые распылители, обеспечивающие высокую
концентрацию аэрозолей с диаметром частиц
5—10 мкм. Но и они имели лишь лабораторное
применение. Инфицирование животных аген-
тами БО производили только что образовав-
шимся аэрозолем (рис. 1.45).
Для более масштабных экспериментов было
принято допущение, что если при распылении
бразуются «сопутствующие» мелкие частицы в
количестве, достаточном для того, чтобы выз-
вать легочную инфекцию у экспериментально-
го животного, то количество крупных частиц,
задержавшихся в дыхательных путях, не будет
иметь существенного значения.
Влияние дисперсности бактериального аэро-
золя на развитие биологического поражения.
Прежде всего, военными исследователями было
установлено, что с уменьшением размера частиц
аэрозоля уменьшается инфицирующая доза
кэзбудителя болезни и меняется патоморфоло-
чя инфекционного процесса.
Сотрудники Форт-Детрика Н. A. Druett et al.
(1956) продолжили исследования инфицирую-
щей способности мелкодисперсных бактери-
альных аэрозолей. Они установили, что доза
возбудителя чумы, требуемая для 50 %-ной ле-
Рис 1.45. Смерть в аэрозоле. На фо-
тографии показана чумная палочка в
частичке аэрозоля. Предметное стек-
ло было предварительно окрашено
основным фуксином, после чего со-
трудники Форт-Детрика дали воз-
можность частицам аэрозоля чумы
осесть на стекло и исследовали их с
помощью фазово-контрастного мик-
роскопа. Длина У. pestis 1—3 мкм. По
R. J. Goodlow и F. A. Leonard (1961)
дальности инфицированныхживотных, повышается в 2,5 раза, если размер частиц уве-
личивается с 1 до 12 мкм (Druett Н. A. et al., 1956а); а доза Brucella suis (один из возбу-
дителей бруцеллеза) увеличивается в 600 раз (Druett Н. A. et al., 1956b).
Кроме того, эти же ученые установили, что болезнь, возникшая при вдыхании
□розоля спор сибирской язвы с размером частиц приблизительно в 1 мкм (размер
споры), является системной с включением в патологический процесс лимфоузлов
средостения. Вдыхание же 12-мкм частиц дает локализованную инфекцию с выра-
женным отеком лица и головы. Вдыхание частиц с Y. pestis, приближающихся по сво-
ему размеру к бактерии, ведет к первичной легочной чуме, а 12-мкм частицы дают
септицемию с геморрагическим инфарктом в легких. По данным сотрудников Форт-
Детрика R. J. Goodlow и F. A. Leonard (1961), инфицирование аэрозолем возбудителя
туляремии с размерами частиц 1 и 8 мкм вызывало у обезьян поражения, локали-
зовавшиеся в концевых бронхиолах. Однако в серии экспериментов, выполненных
с гомогенным аэрозолем F. tularensis, состоящим из частиц диаметром 18 мкм, подо-
пытные обезьяны погибли без признаков поражения легких. Патологические изме-
нения свидетельствовали о распространении поражений, характерных для таких
случаев, когда воротами инфекции является носоглотка, откуда инфекция распрост-
оаняется в регионарные лимфатические узлы, далее следует сепсис, метастазирова-
ние инфекции и гибель экспериментального животного.
В классическом эксперименте, проведенном в начале 1960-х гг. в Форт-Детрике,
данные, полученные на обезьянах и морских свинках, сравнивались с данными в
экспериментах на людях. Добровольцами были члены церкви «Адвентисты Седьмо-
244
Часть 1. Краткая история применения биологических средств...
го Дня». Шла «холодная война», СССР и Красный Китай были врагами США, а мо-
лодые люди из этой церкви не желали носить оружие. Вместо службы в армии им
сделали предложение — поучаствовать в экспериментах по экспонированию к аэро-
золям микроорганизмов. Первые испытания на людях проводились с аэрозолем Coxiel-
la burnettii, возбудителем Ку-лихорадки (см. разд. 3.7). Затем стали использовать F. tula-
rensis (см. разд. 3.3), а позднее — стафилококковый энтеротоксин В (см. разд. 3.13).
Эти исследования были очень важны, поскольку вместе с добровольцами к аэро-
золям, содержащим микроорганизмы, потенциальные агенты БО, экспонировались
также обезьяны макаки-резусы и морские свинки. Таким образом, военными иссле-
дователями устанавливалось соотношение между человеческими и животными моде-
лями, которые могли затем применяться к другим болезням, при испытании которых
нельзя было использовать людей по этическим соображениям.
В табл. 1.4 в первой колонке приведены размеры аэрозольных частиц; во второй —
количества клеток возбудителя туляремии, требуемых для гибели 50 % морских сви-
нок, ингаляционная LD50, и в третьей колонке — то же для обезьян. В четвертой ко-
лонке даны количества туляремийных клеток, необходимые для инфицирования, но
не гибели, человека (ID^).
Таблица 1.4
Влияние диаметра частиц на инфекционность аэрозоля возбудителя туляремии*
Диаметр аэрозольной частицы (мкм) Морская свинка, LD5n Обезьяна, LD50 Человек, Ю50
1 2,5 14 10-52
6,5 4700 178 14-162
11,5 23000 672 —
18 125000 3447 —
22 230000 >8500 -
* По W. HI. Patrie (2001).
При аэрозоле, состояшем из 1 -мкм частиц, требуется всего 2,5 клетки, чтобы убить
морскую свинку, 14 — обезьяну и от 10 до 52 клеток, чтобы заразить человека. Если
аэрозоль состоит из 6,5-мкм частиц, для инфицирования респираторным путем тре-
буется уже больше клеток, а в случае размера частиц от 18 до 22 мкм количество кле-
ток возбудителя туляремии становится слишком большим, чтобы инфицировать че-
ловека. Таким образом, этот эксперимент показал, что аэрозоль биологического
поражающего агента обязательно должен быть не только мелкодисперсным, но еще
и находиться в очень узком диапазоне дисперсности (Patrie W. III., 2001).
Такие эксперименты проводились в начале 1960-х гг. очень интенсивно. К аэро-
золям микроорганизмов, агентов БО, экспонировались люди и животные, получен-
ные результаты сопоставлялись. Они позволили военным подсчитать инфекционные
дозы агентов БО для человека и, следовательно, определить количество конкретных
образцов БО, необходимых для применения по конкретным целям. Но оставалась
неуверенность в поражающем действии биологических аэрозолей, связанная с незна-
нием особенностей патогенеза при попадании патогенного микроба в организм не-
свойственным ему путем. Например, течение желтой лихорадки можно предсказать
довольно успешно в том случае, когда заболевание связано с укусом инфицирован-
ного комара, но желтая лихорадка, обусловленная вдыханием возбудителя, может
представлять собой совершенно иную болезнь.
1.11. Биологическое оружие перед подписанием Конвенции 1972 г. 245
Влияние дисперсности аэрозоля биологического токсина на развитие биологического
поражения. В природе не происходит контакта человека с аэрозолями очищенных био-
логических токсинов. В конце 1950-х гг. военные США имели смутные представления
о механизмах ингаляционного поражения такими токсинами. И это при том, что в
годы Второй мировой войны в США было изготовлено только рицина 1,7 т.
После опытов W. Е Wells et al. (1948), показавших решающее значение для разви-
тия инфекции не количества вдыхаемых жизнеспособных бактерий, а величины час-
тиц аэрозоля, прошло почти 10 лет, пока А. Корвин (А. Н. Corwin), сотрудник Уни-
верситета Дж. Гопкинса (Johns Hopkins University), в опытах с тонкодисперсными
порошками рицина показал такую же зависимость между размерами частиц распы-
ленного токсина и тяжестью поражения экспериментального животного. Он обна-
ружил, что аэрозоль, содержащий частицы токсина с размером, не превышающим
2.1 мкм, в 2,75 раз более токсичен, чем аэрозоль, содержащий частицы токсина с раз-
мером 4,2 мкм. Уменьшение размеров частиц аэрозоля, по его мнению, представляет
собой самый надежный путь к повышению его поражающей способности (цит. по
Lamanna G., 1961). В те же годы был обнаружен еше ряд эффектов, значительно по-
высивших интерес разработчиков БО к ингаляционному применению биологи-
ческих токсинов.
1. Оказалось, что верхние дыхательные пути являются проницаемыми для круп-
номолекулярных токсинов. Так, G. Lamanna (1961) в опытах на мышах, выполнен-
ных со столбнячным токсином, обнаружил, что закапывание в нос токсина примерно
в 10 тыс. раз более эффективно, чем закапывание в рот. Для ботулинического токсина
ими обнаружена та же закономерность, хотя естественным путем его проникновения
в организм является алиментарный. Высокая активность этих токсинов при закапы-
вании в нос указывала на их высокую способность к адсорбции из верхних дыхатель-
ных путей и носоглоточной области. Однако механизм данного явления G. Lamanna
не понял. Нетоксичные крупномолекулярные белки такой способностью не облада-
ли, а в доступной ему литературе он никаких объяснений своим данным не нашел.
2. Сотрудниками Форт-Детрика М. A. Cardella и J. V. Jemski показано, что при ин-
галяционном введении ботулинического токсина резко снижаются различия в чув-
ствительности животных к разным его серотипам, наблюдающиеся при их паренте-
ральном введении (цит. по Lamanna G., 1961).
3. При ингаляционном введении ботулинического токсина значительно увели-
чивалось количество антитоксина, необходимого для нейтрализации его действия.
Например, А. М. Яковлевым (1956) было обнаружено, что при ингаляционном вве-
дении ботулинического токсина одна единица антитоксина предохраняет лишь про-
тив одной летальной дозы, при подкожном введении — против 20 летальных доз, при
пероральном — против 50 летальных доз. Хотя при всех этих способах введения ток-
сина в организм животного гибель животного наступала в результате поражения ды-
хательного аппарата. Сходные данные получены G. Lamanna (1961) в опытах со столб-
нячным токсином. Им было установлено, что введение фиксированного количества
столбнячного антитоксина мышам увеличивает LD50 столбнячного токсина при внут-
рибрюшинном введении в 25 раз. Однако при введении того же количества токсина
через нос, LD50 увеличивается только в 5 раз.
Результаты этих экспериментов тогда не нашли общепризнанного объяснения, но
они открывали заманчивые перспективы перед разработчиками БО. Тем более что уже
не оставалось никаких неясностей в отношении судьбы проникших в дыхательные
пути частиц биологического аэрозоля.
246
Часть 1. Краткая история применения биологических средств...
Судьба проникших в дыхательные пути человека частиц биологического аэрозоля
В основном была установлена в конце 1940-х и вначале 1950-х гг. Исследования меха-
низмов ингаляционного инфицирования агентами БО и разработка самого БО шли
параллельными направлениями, оказывая постоянное влияние друг на друга. Они
значительно обогатили наши представления об анатомии, физиологии и механике
дыхательных путей. Более подробно о функционировании дыхательных путей чело-
века можно прочитать в работах М. Н. Ситникова (1968); L. Reid (1973); В. И. Огар-
кова и К. Г. Гапочко (1975); Р. Е. Morrow (1980).
Судьба проникших в дыхательные пути человека частиц биологического аэрозо-
ля определяется их физико-химическими свойствами (дисперсность, гигроскопич-
ность, электрический заряд и др.). Задержка частиц биологического аэрозоля обуслов-
ливается силой тяжести, силой инерции и броуновским движением. Эти факторы
связаны с массой частиц, т. е. практически с их дисперсностью. В верхних дыхатель-
ных путях, где воздух движется с достаточно большой скоростью, основным механиз-
мом осаждения крупных частиц являются инерция и седиментация, влияние которых
прямо пропорционально плотности и квадрату диаметра частиц. С уменьшением ве-
личины частиц влияние указанных факторов ослабевает и поэтому мелкие частицы в
верхних участках легких задерживаются меньше и проникают в более глубокие отде-
лы легких, где основным механизмом их оседания будет броуновское движение.
На степень задержки частиц в дыхательных путях влияют: концентрация вдыхае-
мого аэрозоля, глубина и частота дыхания, носовой или ротовой тип дыхания, состо-
яние органов дыхания. Глубина проникновения частиц биологического аэрозоля и их
первичное распределение в различных отделах дыхательной системы определяется
дисперсностью частиц. Только частицы величиной 1-3 мкм и меньше могут достиг-
нуть альвеол (рис. 1.46).
Оседание
аэрозольных
частиц, мкм
60 20 6 2 0,6
Трахея
Бронхи первого порядка
Бронхи второго порядка
Терминальные бронхи
Респираторные бронхи
♦ !
t
-*--Наличие оседания
-*---Отсу гствие оседания
Альвеолярный ход
Альвеолы
Рис. 1.46. Схематическое строение легочных путей и их
отношение к оседанию частиц. По М. Н. Ситникову (1968)
Степень задержки частиц определяется их размерами. Частицы размером 10 мкм
и более полностью задерживаются в дыхательных путях, в то время как частицы раз-
мером 3,2 мкм — всего на 61-80 %, а 0,4 мкм — 17-33 %.
II. Биологическое оружие перед подписанием Конвенции 1972 г.
247
Было установлено, что через слизистую оболочку концевых бронхиол или альве-
гярный эпителий микроорганизмы, используемые в качестве агентов БО, проника-
ют в основном благодаря фагоцитозу альвеолярных макрофагов и лейкоцитов. Затем
а ни вместе с макрофагам и и лейкоцитами попадают или непосредственно в кровенос-
-ые капилляры легкого и по ним разносятся по всему организму, либо поступают в
лимфатические сосуды, оттуда в регионарные лимфатические узлы и далее, при их
прорыве, в грудной проток и венозную кровь (Drinker, Hardenber, 1947).
Технические средства применения аэрозолей агентов БО. В наставлениях армии и
ВВС США по защите от БО (ТМ 3-216 и AFM 355-6, 1964) перечислены три основ-
-ых метода образования аэрозолей биологических агентов: с помощью генераторов,
: .пыление и взрыв. Основная проблема при создании устройств, образующих аэро-
золи, состояла в необходимости разрешения следующего технического противоре-
-1Ия — распыляемый материал должен иметь дисперсность, позволяющую осуществ-
лять ингаляционное инфицирование живой силы противника, но при этом он не
□лжен подвергаться физическим воздействиям, способным вызвать гибель микро-
организмов, либо инактивацию токсинов. В опытах американских военных ученых
'пользовались суспензии бактериальных и вирусных агентов, коллоидные раство-
ры токсинов и сухие рецептуры с определенным размером частиц. В качестве агент-
того имитатора токсинов применялись препараты альбумина куриного яйца.
Аэрозольные генераторы и ВАПы. В указанных выше американских наставлениях
о защите от БО объясняется, что аэрозоль может быть получен в результате пропус-
кания жидкой рецептуры в виде гомогенной суспензии через сопло распылителя при
регулируемом давлении. Такие распылительные устройства называются гидравличе-
заши. Размер создаваемых ими частиц определяется величиной давления, размером
гверстия, содержанием агента и относительной влажностью атмосферного воздуха.
Хотя конструкции сопел были весьма разнообразными, ни одно из них не давало
томогенного аэрозоля нужной для военных дисперсности. Большинство частиц по-
лучались крупными, и аэрозоль быстро оседал, не проникая в глубокие отделы лег-
ок предполагаемых жертв биологической войны (Patrie W. III., 2001).
Типичным для распыления жидких рецептур такого типа в 1950 гг. было сопло
РТ-12. Жидкость распылялась под давлением 1000 фунтов/кв. дюйм, проходя через
отверстие с острыми краями, и ударялась в установленный перед отверстием стержень.
У результате образовывалась струя в виде конуса, которая в последующем распадалась
-: капельки. В подобном устройстве лабораторного типа удавалось достигать сравни-
*ельно высокой скорости подачи жидкости, около 300 мл/мин. В качестве жидкой
среды обычно использовали желатин в фосфатном буфере. Примерно 15 % материа-
ла. распыленного таким образом, после испарения воды переходило в частицы с диа-
метром 5 мкм (Zentner Е, 1961). Несмотря на высокую производительность, диапазон
сэспыленных частиц был таков, что распылители гидравлического типа мало подко-
сили для целей биологической войны. Они не «показали» себя во времена войны на
корейском полуострове и уже к концу 1950-х гг. считались устаревшими.
Другой тип распылителя, исследовавшегося тогда на предмет пригодности для
рименения агентов БО с помощью авиации, был воздухоструйный. Принцип его дей-
ствия заключался в том, что жидкость, вытекающая из сопла, деформировалась под
,-йствием потока воздуха, имеющего высокую скорость. Диапазон размеров образую-
щихся частиц был не столь велик, как при работе с гидравлическими распылителями.
Благодаря импакции на экраны, удавалось удалять крупные частицы из аэрозоля.
Однако воздухоструйные распылители малопроизводительны. К тому же более 95 %
248
Часть 1. Краткая история применения биологических средств...
частиц имеют диаметр, выше того, который необходим даже для лабораторных экс-
периментов по оценке опасности инфицирующих аэрозолей (Mercer Т. Т et al., 1968).
В конце 1950-х гг. наиболее перспективными считались распылители, работаю-
щие на центробежном принципе. Они уже два десятка лет использовались для распы-
лительной сушки биологических материалов (см. рис. 1.18), и поэтому имелось мно-
го их различных модификаций. В таких устройствах жидкость подается на быстро
вращающуюся поверхность, установленную под прямым углом к оси вращения. Под
действием центробежной силы образуется тонкая пленка жидкости. Поток идет в ра-
диальном направлении, так что образующаяся пленка имеет не одинаковую толщи-
ну. Деление пленки на капельки происходит при той толщине, которую эта пленка
имеет на краях поверхности. От этой толщины зависит преобладающий размер капе-
лек. Диаметр диска в таких распылителях обычно составляет 5 см, скорость его вра-
щения примерно 70 тыс. об/мин. Вращение осуществляется с помощью электромо-
тора постоянного тока, либо пневматического двигателя или воздушной турбины,
поэтому они издают характерный свистящий звук, за который американские иссле-
дователи прозвали их «турбинками». Обычно такие распылители используют для гене-
рирования частиц в диапазоне 20—100 мкм, но варьируя скоростью вращения диска
и составом распыляемой жидкости, исследователям удавалось получать относитель-
но однородный аэрозоль с дисперсностью в 5 мкм. Центробежные распылители дают
монодисперсные а »розоли с геометрическим стандартным отклонением 1,1. Их не-
достатками являются ненадежность, низкая производительность и сложность эксп-
луатации (Marple V. A., Rubow К. L., 1980).
Разрабатывались комбинации воздухоструйных и центробежных распылителей.
R. Fraser et al. (1963), используя вращающуюся поверхность диска в комбинации с
воздушной струей, направленной по нормали к свободной поверхности жидкости,
установили возможность контролируемой дезинтеграции жидкости. Фракционно-
дисперсный состав образующихся капель зависел от толщины пленки жидкости, чем
она тоньше, тем тоньше распыление данного устройства.
Регулирование размеров диспергируемых частиц для сухих рецептур поражающих
агентов БО можно осуществлять путем регулирования дисперсности рецептуры, по
крайней мере, рассуждая формально-логически. В своих показаниях, данных амери-
канским следователям, Исии утверждал, что диспергирующие авиационные прибо-
ры, снаряженные сухой рецептурой, должны быть потенциально более эффективны-
ми при боевом применении, так как в этом случае можно снарядить средство доставки
большим количеством материала, можно эффективнее контролировать размер час-
тиц, а в отношении некоторых патогенных микроорганизмов можно избежать губи-
тельного воздействия некоторых элементов атмосферных условий. Сам он «довес-
ти до ума» такие устройства не успел, ими занялись его новые хозяева (см. FM 3-10;
NWIP 36-2; AFM 355-4; FMFM 11-3, 1966). Но оказалось, что и в этом вопросе Исии
оказался пустым теоретиком и выдавил желаемое за действительность.
Тонкодисперсный порошок получить очень трудно, еще труднее его сохранить.
Такой порошок легко слеживается и образует агломераты частиц значительно боль-
шего размера, чем это нужно для инфицирования личного состава вооруженных сил
противника К тому же создание авиационных распыляющих устройств таких порош-
ков упирается вряд сложных технических проблем. В момент распыления воздушным
потоком, по мере того как частицы поднимаются вверх из порошка, последний уп-
лотняется воздушным потоком и принимает округлую обтекаемую форму. Для того
чтобы возооновить удаление частиц из слоя порошка, необходимы уже более высо-
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
249
кие скорости воздушного потока. Исследования процессов диспергирования тонко-
дисперсных порошков уже после окончания Второй мировой войны потребовали
почти три десятилетия экспериментов и теоретических проработок, некоторые пер-
вые обобщения можно прочитать в работе J. М. Hidy (1984).
В самых простых устройствах для распыления тонкодисперсных порошков поток
воздуха пропускается через перемешиваемый слой порошка, и частицы уносятся воз-
душным потоком в аэрозольную камеру. При использовании таких устройств трудно
контролировать концентрацию частиц и их распределение по размеру. С более слож-
ными устройствами читатель может ознакомиться в специальной литературе. Их ос-
новное отличие от простых устройств заключается в использовании различных под-
ходов к дозированию распыляемого порошка и к его предварительному дроблению
перед распылением (см. работы Fontanges R., Founier J., 1971; Marple V. A., Rubow К. L.,
1980; Hidy J. M., 1984). К 1960-м гг. конструирование различных систем для примене-
ния сухих рецептур возбудителей опасных инфекционных болезней стало приоритет-
ным направлением военно-биологической программы США.
По данным С. Херша (1970), значительная часть важнейших исследований в об-
ласти средств применения биологических рецептур авиацией была осуществлена от-
делом прикладных наук компании «Лит гон индастриз инк» (Litton Industries Inc.).
В одном из отчетов о разработке этой компанией на протяжении четырех лет ВАПа
А/В 454-4 для распыления сухой рецептуры говорится, что «целью провидимых ра-
бот было проектирование, изготовление и испытание распылителя — автоматически
действующего агрегата, представляющего собой контейнер, предназначенный для на-
ружной подвески на самолетах F-100, F-105 и F-4C. Испытания показали, что этот
распылите п>, как и проектировалось, по механическим качествам, а также в отноше-
нии электропитания и конструктивных решений удовлетворительно увязывается с
самолетом-носителем; судя по результатам, эффективность распыления рецептуры
также удовлетворительна». Упомянутый отчет датирован февралем 1966 г. Примерно
в это же время компания «Литтон индастриз инк» работала над проектом «Распыле-
ния сухих биологических рецептур при сверхзвуковых скоростях».
В одном из докладов компании Министерству обороны за январь 1967 г. излага-
лись результаты испытания, проведенного на самолете F-4C, имеющем максималь-
ную скорость 2600 км/ч. В отчете, в частности, говорилось, что «были проведены
ограниченные летные испытания, в ходе которых летчик-испытатель не выявил от-
рицательных характеристик... Прибор хорошо распылял рецептуру биологического
агента над водной поверхностью при скорости самолета 654 км/ч (350 миль/ч) с
наполненным баком, а также при скоростях самолета 561, 795 и 935 км/ч (300, 425 и
500 миль/ч), когда содержимое бака было в основном израсходовано». В отчете отме-
чалось также, что «распылитель был бы полезен в случае, если США примут решение
осуществить скрытное внезапное биологическое нападение», не оставляя улик. При-
бор «обязательно разрушался при ударе о воду, и на воде не удавалось обнаружить
плавающих его частей или обломков». Американские разработчики БО придержива-
лись принципа генерала Исии — применение БО не должно оставлять материальных
«следов» в виде фрагментов боеприпасов (см. керамическую бомбу Исии в разд. 1.9 и
«меловую бомбу» в разд. 1.10). Указанная компанией «Литтон индастриз инк» скорость
самолета, применяющего сухие рецептуры агентов БО диспергированием из подвес-
ных авиационных приборов, в точности соответствует той, о которой дали показания
Комиссии Нидхема пилоты американских самолетов, сбитых в 1952 г. русскими ис-
требителями (см. разд. 1.10 и приложение Б).
250
Часть I. Краткая история применения биологических средств...
В 1960 гг. значительное развитие получили работы, связанные с изучением техни-
ческих средств применения жидких биологических рецептур. Продолжались иссле-
дования по разработке ВАПов для применения жидкой рецептуры агента БО. Среди
прочих перед разработчиками ставилась цель уменьшить заражение самолета, с ко-
торого распыляется такая рецептура, что, судя по показаниям американских летчи-
ков, данных Комиссии Нидхема, было проблемой во время войны на Корейском по-
луострове.
Учитывая потери самолетов, применявших БО во время войны на Корейском
полуострове, и последующие откровенные показания их пилотов в плену, Министерст-
ва обороны США в 1960-м г. дало заказ одному из основных американских предприя-
тий по производству авиационных и космических средств «Аэроджет дженерал аэрон-
тикал дивижн» в Дауни (штат Калифорния) на разработку для армии нового бес-
пилотного самолета, предназначенного для доставки к цели и распыления химических
и биологических средств. Первоначально спроектированные для использования в
целях разведки поля боя, эти беспилотные самолеты в дальнейшем были приспособ-
лены для снаряжения рецептурой биологического агента в количестве 90 кг и достав-
ки ее на расстояние свыше 180 км. К подобным средствам доставки относится, напри-
мер, многоцелевой телеуправляемый самолет MQM58A, созданный в конце 1950-х гг.
для химической службы армии США. Этот радиоуправляемый аппарат весом 0,5 т
имел радиус действия 185 км. Вес его боевой части составлял около 100 кг.
В марте 1967 г. ученые в Форт-Детрике завершили программу по разработке голов-
ной части тактической ракеты «Сержант» (дальность полета до 160 км), снаряжаемой
биологическими средствами. В уставе армии США FM 3-10 «Применение химического
и биологического оружия», изданном в марте 1966 г., отмечается, что типичная био-
логическая ракетная система включает головную часть, которая может быть раскры-
та на заранее определенной высоте. При этом из головной части освобождаются био-
логические бомбочки (bomblets), имеющие на своей поверхности лопасти, которые
придают им вращательное движение во время падения. Благодаря такому приспособ-
лению обеспечивается их рассеивание по территории противника (см. «Боеприпасы»).
В раннем издании устава FM 3-10, относящемся к февралю 1962 г., не содержится
данных о возможности применения биологических средств с помощью ракет.
Боеприпасы. Специалисты армии США продолжали работу над авиационными
кассетными бомбами, предназначенными для применения агентов БО, а также совер-
шенствовали субэлементы для таких бомб и головных частей ракет. Создавались бое-
припасы для боевого применения жидких и сухих рецептур микроорганизмов и ток-
синов на больших площадях. Чтобы в самых общих чертах понять их конструктивные
особенности, необходимые экспертам, участвующим в расследовании преступлений
с применением БО, рассмотрим такие боеприпасы с той обстоятельностью, которую
позволяют описания к патентам.
К 1955 г. была создан и запатентован боеприпас для взрывного диссеминирова-
ния рицина и ДДТ (US3207071). По форме и размерам он соответствовал Ml 14 — суб-
элементу кассетного боеприпаса МЗЗ (см. рис. 1.38 и 1.39). Но отличался от него тем,
что диспергирование биологической (химической — для ДДТ) рецептуры производи-
лось не расширением сжатого газа, а взрывом заряда, расположенного по оси бое-
припаса. Степень дисперсности агента регулировалась силой взрыва. Емкость вокруг
заряда заполнялась жидкостью или суспензией частиц нужной дисперсности в спе-
циальной жидкости. Жидкость должна была быть более летучей, чем агент, и не раз-
рушать контейнер. Обычно для твердых токсических агентов (рицин) в качестве та-
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
251
кой жидкости использовали четыреххлористый углерод. Взрывное распыление такой
суспензии давало значительно лучшие результаты, чем распыление сухого порошка
рицина или его водного раствора (рис. 1.47).
Рис. 1.47. Боеприпас для взрывного диссеминирования рицина или ДДТ
А. Общий вид боеприпаса. Б. Продольное сечение боеприпаса по линии 2—2
(см. А). В. Поперечное сечение боеприпаса по линии 3—3 (см. Б). Г. Попе-
речное сечение боеприпаса по линии 4-4 (см. Б). По патенту US3207071
Е
Сам цилиндр (1) был выполнен из фенолформальдегидной пластмассы, проклеен-
ной крафт-бумагой. Длина цилиндра 215/6 дюйма, наружный диаметр 1 11/16дюйма.
Пропорции подобраны экспериментально, как обеспечивающие наилучшую диссе-
минацию агента. Нижний конец боеприпаса закрыт нижней пробкой (3), снабжен-
ной заполняющим отверстием (5). Верхняя пробка (7) определяет верхнюю границу
полости бомбы. Пробки (3) и (7) сделаны из слоистой резины и зацементированы в
оболочку (1). Пространство между пробками занимает взрывная труба (13), закрытая
в ее нижнем конце алюминиевым диском (11). Труба выполнена из латуни и содер-
жит гранулы взрывчатого вещества тетрила (15).
Верхний конец взрывной трубы имеет запирающее устройство (17) и войлочную
пробку (19). Детонатор (21) вставляется в верхний конец взрывной трубы. Когда для
каких-то целей требуется меньше тетрила, то освободившееся во взрывной трубе про-
странство заполняют фетровыми прокладками.
Основная полость цилиндра (23) заполняется суспензией рицина или ДДТ в че-
тыреххлористом углероде. Верхний конец бомбы включает высокочувствительный
всепогодный взрыватель, описанный ниже. Оболочка взрывателя (25) вкручивается
в верхнюю пробку (7) и содержит колпачок (27). В пределах оболочки имеется цилиндр
капсюля (29), который содержит капсюль (31), удерживаемый на поверхности кап-
252
Часть 1. Краткая история применения биологических средств...
сюльной прокладки (33). В пределах цилиндра имеются поджигающий ударник (35),
несущий поджигающий боек (37), который удерживается на безопасном расстоянии
от капсюля пружиной ударника (39). Ударник (35) включает в себя поперечный ка-
нал (41), в который вмонтированы шары безопасности (43). В не боевом состоянии,
показанном на рисунке, шары удерживаются снаружи посредством чеки (47), предот-
вращающей движение ударника в направлении капсюля. Чека-зашелка (51) прижи-
мается пружиной, расположенной вокруг нее. Герметизирующий колпак (53) закры-
вает верхний конец корпуса цилиндра (1), и он снабжен герметизирующей прокладкой
(53). Боевая пружина (63) удерживается в сжатом состоянии колпаком (53). Кассет-
ная бомба собирается через кластеризацию таких цилиндров-субэлементов с исполь-
зованием зажимов (59), расположенных на боковой стороне цилиндра. Когда клас-
тер цилиндров фрагментируется, зажимы (59) высвобождаются, позволяя пружине
(63) отжать вверх колпак (53). Чека извлекается, позволяя шарам (43) перекатиться во
внутрь. Воспламеняющий ударник (35) высвобождается и движется в направлении
капсюля (31) и наоборот. Под воздействием удара о поверхность, чека будет выбивать-
ся, вследствие инерции движения цилиндра (29) или ударника (35). Воспламенение
капсюля (31) вызывает подрыв детонатора (21) и затем тетрила (15).
Подрыв такого боеприпаса, снаряженного суспензией рицина, приводил к обра-
зованию частиц рицина со средним размером в пределах 7 мкм и к их распростране-
нию до 50 ярдов от места взрыва.
Боеприпас, запатентованный в мае 1958 г. в США (LS3188954), представлял со-
бой другое направление эволюции субэлемента Ml 14. В нем для диспергирования
сухой биологической рецептуры использовалась не энергия взрыва тетрила, а энер-
гия, образующаяся при переходе сжиженного углекислого газа в газообразованное
состояние (как у Ml 14), но переход жидкой углекислоты в газообразное состояние
ускорялся благодаря инициированию в баллоне с углекислотой специального нагре-
вающего химического элемента. Боеприпас устроен следующим образом (рис. 1.48).
Рис. 1.48. Боеприпас для взрывного диссеминирования сухих биоло-
гических рецептур. А. Общий вид боеприпаса. Б. Поперечное сечение
боеприпаса по линии 2—2 (см. А). По патенту US3188954
Он включает оболочку (1), которая состоит из цилиндрической части (2), присо-
единенной (3) к носовому узлу (4), и хвостовую секцию (5). В пределах оболочки (1)
вставлена армированная втулка (6). Бомба имеет носовую часть (7) и хвостовую часть
с распылителем-стабилизатором (9). Носовая часть (7) включает носовую внешнюю
чашу (11) и носовую внутреннюю чашу (13). Носовой узел (4) присоединен к основа-
нию (7) с помощью винтов (16). Он (4) поддерживает газовый цилиндр и взрыватель,
описанные ниже.
Носовое основание (7) и стабилизатор, расположенный в хвостовой части бомбы
(15), имеют гексагональную форму для облегчения формирования кластеров таких
бомб в кассете. Хвостовой стабилизатор бомбы (15) установлен на хвостовой кони-
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
253
ческой части бомбы (5). Крепежная заклепка (19) используется для закрепления хво-
стового стабилизатора (15) к конической части бомбы (5). Там же расположены муф-
та распылителя (22), втулка (24) хвостового стабилизатора (15) и распылитель (9).
Взрыватель включает чеку (штырь) (23) и пружину (25), которая используется для
эжекции штыря (23). Армированная проволока (21) удаляется, когда бомба будет кла-
стированна вместе с другими для использования. Штырь (23) после этого удержива-
ется на месте за счет зажима (26). Стопорные шары (27) сжимают чеку (23) и тем са-
мым блокируют инициирование взрывателя (29). Втулка (31) и поджигающий капсюль
(33) непосредственно прилегают к удерживающим шарам (27). Удерживающая пру-
жина (35) расположена между втулкой (31) и капсюлем (37). Капсюль (37) располо-
жен в пределах досягаемости ударника взрывателя (39).
На рис. 1.48 (А) поршень (40) отделен от ударника взрывателя (39) небольшим
пространством. Основание поджигающего стержня (41) запирается поршнем (40).
Поршневая пружина (42) расположена между поршнем (40) и цилиндром (44), кото-
рый содержит жидкую углекислоту. Пружина поршня (42) не позволяет поджигающе-
му стержню (41) нанести случайный удар и преждевременно разорвать первый среза-
ющийся диск (47).
Нагревающий детонатор (49) присоединен к первому срезывающемуся диску (47)
и установлен в первом концевом участке нагревающей трубы (51). Держатель нагре-
вателя (53) также смонтирован в первом концевом участке нагревательной трубы (51)
и присоединен к нагревающему детонатору (49). Внешний держатель нагревателя
имеет пазы (55). Крышка нагревателя (59) расположена во втором концевом участке
нагревающей трубы (51). Жидкий углекислый газ (61) содержится в цилиндре (44),
расположенном вокруг нагревающей трубы (51). Гофрированный держатель нагрева-
теля (53) проходит через цилиндрическое отверстие в концевом участке цилиндра (44)
и вокруг нагревающей трубы (51). Жидкий углекислый газ (61) содержится в цилин-
дре (44) и расположен вокруг нагревающей трубы (51). Гофрированный держатель
нагревателя (53) проходит через отверстие в конце цилиндра (44). Его пазы формиру-
ют продольные каналы, которые сообщаются с внутренним пространством цилинд-
ра (44). Наружные выходы этих каналов закрыты первым срезающимся диском (47).
У этой бомбы имелись еще две частные модификации, не представляющие осо-
бого интереса с точки зрения эффективности при боевом применении. С ними мож-
но ознакомиться по описанию к патенту U S3188954.
Во время полета все три хвостовых стабилизатора раскрываются потоком возду-
ха. Когда кассетная бомба раскрывается, зажим (26) выдергивается из гнезда, и чека
(23) выталкивается из бомбы посредством разжимающего действия армированной
пружины (25). Это позволяет стопорным шарам (27) свободно перемещаться. При
ударе бомбы о грунт или о другое препятствие, шары движутся вниз, высвобождая
ударник взрывателя (39), который вследствие инерционного движения преодолевает
сдавливающее действие пружины (35) и ударяет по капсюлю (33), вызывая его поджог.
Взрыв детонатора (37) приводит в движение поджигающий стержень (41), который
проходит через первый срезающийся диск (47) и детонирует зажигающий капсюль
(49). Нагревательная труба (51) содержит смесь калия перхлората (КС1О4), древесно-
го угля и масла. Эта смесь инициализируется посредством детонатора (49) и быстро
загорается, выделяя огромное количество тепла, вызывающего взрывное испарение
жидкой углекислоты (61) в цилиндре (44). Давление углекислоты разрушает срезаю-
щийся диск (47), и газ прорывается по желобкам (55), выходит через отверстия (65),
разрывая герметизирующую ленту (69), и проникает в пространство боеприпаса (71)
254
Часть 1. Краткая история применения биологических средств...
между цилиндром с углекислотой (44) и оболочкой (1). в котором содержится сухая
рецептура (70). Разрушается второй срезающийся диск (73) и потоком углекислого газа
сухую рецептуру выбрасывает в коническое отверстие в хвостовой части боеприпаса
(5), при этом скорость выброса смеси углекислоты и сухой рецептуры увеличивается,
через хвостовое отверстие (75) рецептура попадает в атмосферу.
Распылительный стабилизатор (9) устроен таким образом, чго для большего по-
ражающего эффекта он делит мелкодисперсный аэрозоль, выброшенный через отвер-
стие (75), на три облака.
В начале 1960-х гг. интерес разработчиков БО к биологическим боеприпасам, взры-
вающимся при ударе о поверхность, снизился. Их внимание стала занимать задача
равномерного диссеминирования биологического аэрозоля в приземном слое возду-
ха. Ее решение позволило бы исключить потери дорогостоящего биологического ма-
териала, которые при наземном подрыве боеприпаса были значительными, к тому же
аэрозоль, образовавшийся непосредственно у поверхности земли, не имел возможно-
сти распространиться на большие расстояния. В годы Второй мировой войны подоб-
ные попытки предпринимались. Например, известно о создании японскими военны-
ми из 9-го армейского научно-исследовательского института, биологического бое-
припаса типа «мать и дочь», решающего ту же задачу на уровне технологий 1940-х гг.
(см. разд. 1.8). Его можно считать относящимся к первому поколению таких боепри-
пасов. Второе поколение получило название барометрических. В качестве примера
такого боеприпаса рассмотрим боеприпас., запатентованный в мае 1961 г. в США
(US3170393). Боеприпас предназначен для диспергирования химических, радиоактив-
ных и биологических агентов на разных высотах без применения взрыва или нагре-
вания (рис. 1.49).
Боеприпас состоит из корпуса (1), на поверхности которого имеются пазы (7),
обеспечивающие вращение боеприпаса вокруг горизонтальной оси во время полета.
Канал (9) расположен в корпусе (1) таким образом, что обеспечивает поступление
во щуха внутрь барометрического боеприпаса. Корпус (1) и стакан (35) обра >уют ка-
меру (101), в которую снаряжается диспергируемый материал. Этими материалами
могут быть сухая или жидкая биологические рецептуры, отравляющие и радиоактив-
ные вещества.
Две полусферы, составляющие корпус боеприпаса, удерживаются вместе замком,
несмотря на действие пружины (87). Этот замок состоит из чеки (67), стопорных
шариков (69), стопорной втулки (61), втулки чеки (59), шариков (57) и планок шари-
кового стопора (55). Замок удерживает стакан (35), соединенный с первой полусфе-
рой корпуса. В подготовленном к действию боеприпасе, т. е. при соединенных полу-
сферах, пружина (87) сжата и удерживается в таком состоянии замком
Узел сцепления может быть установлен на любую необходимую высоту путем со-
здания соответствующего давления воздуха на барометрический спусковой механизм
через канал (9). Под действием этого установочного давления буртик (выемка) (27)
хвостовой втулки устанавливается на определенном расстоянии от буртика втулки
сцепления (29). После этого давление снимается и узел сильфона можно считать ус-
тановленным для срабатывания при определенном атмосферном давлении (но не
более низком, поскольку под действием более низкою давления не произойдет сжа-
тие сил! фона, коль скоро пружина сильфона (45) через узел сцепления установлена
на более высокое давление).
Поскольку установка производится при помощи давления воздуха, то можно од-
новременно устанавливать большое число боеприпасов на подрыв на заданной высоте.
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
255
Рис. 1.49. Барометрический боеприпас для диссеминирования биологических, хи-
мических и радиоактивных агентов на разных высотах. А. Продольный разрез баро-
метрического боеприпаса. Б. Поперечный разрез боеприпаса по линии 2—2 (см. А).
В. Частичный разрез боеприпаса по линии 3—3 (см. А). Г. Частичный разрез боеп-
рипаса по линии 4—4 (см. А). Д. Поперечный разрез по линии 5 (см. Б). Е. Фронталь-
ный вид удерживающего крепления (arming weight mount). По патенту US3170398
Л тя этого их помещают в контейнер, создают в нем необходимое давление, а затем
стравливают воздух. В качестве контейнера можно использовать кассету для таких
Зомб или боеголовку ракеты.
Устройство срабатывает следующим образом. Атмосферное давление сжимает
сильфон (43), преодолевая действие пружины сильфона (45). Через канал (9) давле-
ние в стакане уравновешивается с атмосферным давлением. При сжатии сильфона (43)
втулка (47) перемещается вправо. В результате перемещаются вправо крышка (51),
которая соединена со втулкой, и пальцы (53). которые удерживают звездочку (73), и
она может свободно поворачиваться. После сбрасывания барометрического боеприпа-
са пазы (7) вызывают его вращение вокруг оси. По мере снижения боеприпаса, давле-
ние воздуха через канал (9) на сильфон возрастает. В обычном положении звездочка
под воздействием своей пружины (86) находится в зацепленном состоянии и связана
с пальцами (53). Однако перед снижением до предусмотренной высоты центробеж-
ные плашки (77) под действием центробежных сил, возникающих вследствие враще-
ния боеприпаса, поворачиваются на штифтах (81). Этот поворот плашек (77) вызы-
вает поворот звездочки (73), так как штифты звездочки (75) входят в пазы плашек (77).
Звездочка поворачивается и отходит от пальцев (53), и последние получают возмож-
ность перемещаться вправо при поджатии сильфона (43), не касаясь звездочки (73).
256
Часть 1. Краткая история применения биологических средств...
Когда боеприпас снижается до предусмотренной высоты, атмосферное давление
сжимает сильфон (43), поджимая при этом пружину сильфона (45), в результате втулка
(47) сильфона перемещается вправо вместе с крышкой (51) и пальцами (53). При дви-
жении пальцев вправо высвобождаются и отпадают стопорные планки (55), в резуль-
тате чего высвобождаются стопорные шарики (57). Шарики освобождают внутреннюю
стопорную втулку (61), которая перемещается вправо под воздействием пружины (63).
Когда левый конец внутренней стопорной втулки (61) освобождает шарики (69), пос-
ледние выпадают и высвобождают втулку (59). В результате втулка (59) может свободно
перемещаться по чеке (67), которая соединена с пластиной (89). Теперь пружина (87)
ничем не удерживается. Упираясь с одной стороны в тарелку (89), а с другой стороны
в стакан (35), она разъединяет полусферы барометрического боеприпаса в стыке (93).
поскольку тарелка прикреплена к пластине (91), которая соединена с левой полусфе-
рой, а стакан (35) — корпусу сцепления, который соединен с правой полусферой.
После разъединения полусфер освобождается и диспергируется материал, которым
была снаряжена камера (101).
Заявленная патентом US3170398 конструкция субэлемента кассетного боеприпаса
стала основой для развития семейства бомб малого калибра с автоматическим рассеи-
ванием, срабатывающих на заданной высоте. К семейству относится биологическая
бомба Е-120, представляющая собой устройство сферической формы диаметром 10см.
снаряжаемое 50—100 г порошкообразного биологического материала и предназначен-
ное для снаряжения кассетных бомб или реактивных снарядов (рис. 1.50).
Рис. 1.50. Биологическая бомба Е-120. Разработана в США в начале 1970 гг., т. е.
перед окончанием программы по созданию БО А. Внешний вид бомбы. Ее фор-
ма такова, что при падении она раскручивается вокруг своей оси с нарастаю-
щей скоростью, как детская игрушка юла. Благодаря смещенному центру тя-
жести, она не кувыркается в полете, что позволяет агенту распыляться из
верхней части бомбы. Б. Поперечный разрез бомбы. По R. Sidell et al. (1997)
В 1960-х гг. специалисты Форт-Детрика разработали устройство «ротор Флеттне-
ра» («Flettner rotor»), в котором сочетается принцип эжекции газа и принцип устройств
Олсонана времен Второй мировой войны. Последние представляли собой небольшие
трубки, содержащие жидкий химический агент и углекислый газ под высоким давле-
нием. При срабатывании открывались отверстия на обоих концах трубок, освобож-
дая жидкость и газ. Когда трубки падали на землю, они начинали вращаться под дейст-
вием выходящих струй (рис. 1.51).
11. Биологическое оружие перед подписанием Конвенции 1972 г
257
В роторе Флеттнера сухая рецептура биоло-
дческого материала выбрасывается в атмосфе-
ру углекислым газом, находящимся в неболь-
_'оч баллоне, в результате в процессе снижения
гтройства по спирали создается аэрозольное
облако. Роторы Флеттнера были приспособ-
ены для использования в кассетах Гладейе и
Садейе. По-видимому, их можно было исполь-
зовать и для снаряжения боевых частей реактив-
ных снарядов, которые в данном случае могли
срабатывать по типу кассеты Гладейе. т. е. пос-
гдовательно через установленные интервалы
освобождать блоки трубок («роторов Флеттне-
ра»), каждый из которых разрывался бы на за-
j энной высоте (The Problem..., 1970).
Перечисленными конструкциями боепри-
1Сов арсенал биологического оружия, разрабо-
•анного в период увлечения аэрозолями биоло-
гических агентов, разумеется, не исчерпывает-
ся. В табл. 1.5 приведена сводка технических
средств применения агентов БО, созданных в
США до подписания Конвенции 1972 г.
Общими характеристиками боеприпасов
бомб, генераторов, реактивных снарядов), со-
Рис. 1.51. Биологический боеприпас,
созданный с использованием принци-
па движения ротора Флеттнера. Пред-
ставляет летающую по спирали 6om6j
длиной 7 дюймов, снаряженную сухой
или жидкой рецептурой биологиче-
ского агента. Разрабатывался специ-
ально для кластеризации в кассетных
бомбах. Возможности боеприпаса ис-
следовались в 1960-х гг., но на воору-
жение армии США он не принимался.
По R. Sidell et al. (1997)
таено американским данным, являются, во-
первых, сила (мощность) источника аэрозоля и, во-вторых, коэффициент гибели ин-
фекционного агента (в процентах) при его хранении в боеприпасе (контейнере). Для
эчечного источника аэрозоля (бомбы, неподвижный генератор) сила источника
[ражается в средних инфицирующих дозах (IDS0), образующихся в точке распыле-
ия, а для линейного источника — в инфицирующих дозах на погонный метр выпус-
ка (полета) и др. (Ситников М. Н., 1968).
Таблица 1.5
Технические средства применения агентов БО, созданные в США до 1972 г.*
Тип Механизм действия Примечание
Генератор аэрозоля E44R2 Нет данных В стадии разработки с 1965 г.
Боеголовка наводимой ракеты М210 Кассетные элементы в боеголовке (М143) В стадии разработки с 1967 г.
Выливной прибор для к.ыкого агента A/B45Y-1 Распыление Для тактических истребителей, в 1956 г. находился на стадии разработки
Распиливающая емкость ня сухого агента A/B45Y-4 Диспергирование Испытывался для применения энтеро- токсина стафилококков
Кассетная бомба Е133 Суббоеприпас кас- сетных бомб (E61R4) В 1958 г. находился в стадии разработки
С .бооеприпас E61R4 Для Е133 То же
Схббоеприпас Ml43 Для М210 Принят на вооружение в середине 1960-х гг.
По материалам SIPRI (1973).
258
Часть 1. Краткая история применения биологических средств...
Разработчики БО обычно используют выгодные им частные критерии, например,
такой как диссеминирующая эффективность боеприпаса. Для боеприпаса, снаряжен-
ного рецептурой возбудителя сибирской язвы (шифр возбудителя — TR), диссемини-
рующая эффективность определяется как «отношение числа жизнеспособных спор,
переведенных в аэрозольные частицы диаметром менее 5 мкм, к числу жизнеспособ-
ных спор в снаряженном боеприпасе». В подборке документов «Joint СВ Technical Data
Source Book» отмечается: «...ЭффективностьаэрозолированияТЯ израспыливаюших
приборов AB45Y-1 и А/В45Н-4 получена на основании экспериментов с биологичес-
ким имитатором и составляет 17,2 и 49,1 % соответственно». Аналогичным образом,
в том же источнике, но относительно агента PG (стафилококкового энтеротоксина),
сообщается, что эффективность его диссеминирования с помощью распиливающе-
го прибора A/B45Y-4 составляет: «... 63 %, если учитываются размеры всех аэрозоли-
рованных частиц, и 26 %, если в расчет принимаются только частицы размером ме-
нее 5 мкм».
Определенное представление о возможных концентрациях микроорганизмов в
воздухе дают испытания на американских самолетах специальных распылителей. При
скорости 500 км/ч при подвешивании одного генератора создавалось 3, IxlO9 ID50 на
1 м полета, а при двух генераторах — в два раза большая концентрация. В экспери-
ментах, а также по расчетным данным, концентрации для туляремийного микроба
в сформировавшемся аэрозольном облаке могут достигать 1х10б микробных клеток в
1 л воздуха (Ситников М. Н., 1968).
Но, как мы увидим из материалов, обобщенных ниже («Аэробиология мелкодис-
персного аэрозоля»), ни этот критерий, ни многие другие, характеризующие биоло-
гический агент до распыления и после распыления (которые в данной работе не рас-
сматриваются), не гарантируют успеха в применении БО.
Аэробиология мелкодисперсного аэрозоля. В начале 1960-х гг. разработчики ядер-
ного оружия эффектно взрывали в космосе ядерные устройства мощностью в десят-
ки мегатонн. Но разработчикам «мощного оружия бедных» еще только предстояло
масштабировать свои эксперименты с мелкодисперсными аэрозолями на ровных и
размеченных площадках Дагуэйского полигона. Как оказалось, найти там место для
«диссеминирующей эффективности» и прочих формальных критериев лабораторной
практики весьма проблематично. Невозможным оказалось и применение опыта во-
енных химиков для изучения поведения аэрозоля опасных микроорганизмов. Его по-
ведение как системы определялось уже не только законами физики, но и еще пока
неизвестными биологическими закономерностями.
Высокопроизводительных устройств, создающих аэрозоль нужной дисперсности
на территориях, сопоставимых с площадью поражения тактическим ядерным бое-
припасом, не было. Один из простейших способов диссеминирования биологических
поражающих агентов — распыление рецептуры биологического агента при помощи
одноканального сопла, использующего энергию газа. Однако для такого сопла, что-
бы добиться эффективности на уровне 5 %, требуется давление минимум 300 фунтов
на кв. дюйм (21 кг/см2) (Patrie W. III., 2001). Но уже при давлении в десять раз мень-
шем количество выживших после диспергирования бактерий ничтожно (Rosebury Т.,
1947). Увеличение давления в распылителе приводит к возрастанию эффективности
распыления; в то же время клетки бактерий подвергаются воздействию срезывающе-
го усилия. По мере дальнейшего увеличения давления возрастает количество погибаю-
щих клеток под действием срезывающего усилия. Гибель бактерий происходит быст-
рее, чем увеличение эффективности распыления (Hatch М. Т, Wolochow Н., 1971).
II Биологическое оружие перед подписанием Конвенции 1972 г
259
Такая же закономерность обнаружена при имитировании применения сухих ре-
-Мтур поражающих агентов БО. В табл. 1.6 показана взаимосвязь между жизнеспо-
постью агента и размерами частицы на примере сухих рецептур S. marcescens (SM).
Таблица 1.6
Взаимосвязь между размером частиц, количеством жизнеспособных клеток
на частицу и количеством жизнеспособных клеток на 1000 частиц аэрозоля*
Размер аэрозольных частиц Кол-во SM на частицу аэрозоля Кол-во жизнеспособных SM на частицу аэрозоля Частота присутствия жизнеспособных клеток SM на 1000 частиц аэрозоля
0,8 1,8 0,001 0,5
1,3 4,2 0,01 2,6
3,0 18,0 0,2 15,6
6,5 73,0 2.5 38,0
11,5 195,0 7,7 14,0
16,0 350.0 11,0 60,0
*ПпУУ III. Patrie (2001).
Если аэрозоль содержит частицы размером 0,8 мкм, то на каждую из них прихо-
кгея в среднем 1,8 клетки SM, но их выживаемость составляет 0,001 %. При увели-
чении размеров частиц увеличивается и выживаемость клеток в частицах аэрозоля,
ако они теряют способность проникать в глубокие отделы легких и вызывать ин-
щионный процесс у людей и животных (Patrie W. III., 2001).
Оказалось, что практически невозможно контролировать размер частиц, образую-
щихся при любом способе диспергирования и любом типе используемых рецептур. Части-
зы аэрозоля сразу же после диспергирования изменяют свои размеры. Частицы жид-
- го аэрозоля из-за потери воды могут уменьшаться в размерах почти в 2,5 раза, а
сухие, наоборот, в результате регидратации могут увеличиваться в 4 раза. При этом
держащиеся в них микооорганизмы подвергаются быстрым температурным воздей-
а виям — при дегидратации они охлаждаются, при гидратации нагреваются (Ситни-
DB М. Н., 1968).
Казалось бы, можно посчитать примерный размер частиц диспергируемой жидкой
?едептуры, с учс гом их возможного «высыхания». Степень обезвоживания частицы
ззана с соотношением давления паров воды на поверхности частицы и в воздухе и
- [ходится в линейной зависимости. При 40 % относительной влажности равновесие
гэстигается к моменту, когда частица утрачивает 85 % исходного веса сорбированной
_мы. При 97 % относительной влажности потеря составляла не более 10 %.
Но обезвоживание частиц аэрозоля приводит не только к уменьшению их разме-
ра, но и к изменениям в структуре протеинов бактериальной клетки, в первую оче-
едь ферментов. Не исключается и другой механизм губительного действия обезро-
гивания, связанный с повышением концентрации токсических веществ в клетке.
Следовательно, чем ниже относительная влажность, тем выше скорость гибели мик-
робной клетки. Частицы аэрозоля при низкой относительной влажности воздуха при-
обретают нужный размер 3-5 мкм, однако это сопровождается гибелью части мик-
роорганизмов, в них содержащихся (HaykawaJ., Poon С.,1965). Чем крупнее частица
Аэрозоля, тем больше воздействие происходящих в ней физических процессов на
биологический агент. Просчитать такие закономерности можно при подготовке опы-
тов в полигонных условиях. В условиях боевого применения БО трудно ожидать, что
260
Часть 1 Краткая история применения биологических средств...
относительная влажность воздушной среды на территории, по которой оно применя-
ется, будет соответствовать заранее рассчитанной
В 1959 г. S. Webb установил, что в течение первой секунды после распыления гиб-
нет подавляющее число микроорганизмов. В последующие девять секунд скорость их
инактивации значительно уменьшается. В дальнейшем, в течение ближайшего часа
и позже, процесс отмирания микробов идет очень медленно (цит. по Огаркову В. И..
Гапочко К. Г., 1975).
Причины, по которым происходит гибель микроорганизмов в аэрозоле, интенсив-
но выяснялись в 1950-1960 гг. Разными исследователями было установлено, что ги-
бель микроорганизмов во время распыления и сразу после него зависит от таких фак-
торов, как вид микроорганизма и свойства конкретного штамма; условия культиви-
рования и сушки; физиологический возраст культуры; условия и продолжительность
их хранения перед диспергированием; наличие примесей в среде диспергирования;
условия самого процесса диспергирования; физико-химические свойства частиц
атмосферные условия (относительная влажность, температура, освещенность, инсо-
ляция, состав атмосферы и наличие в ней загрязняющих веществ).
Все эти годы, до принятия решения о свертывании наступательных программ по
БО, разработчиками такого оружия пелись поиски способов и средств, которые све-
ли бы к минимуму повреждающее действие выше приведенных факторов на аэрозс
лированные микроорганизмы. С этой целью ими подбирались более устойчивые к
аэрозолированию штаммы агентов БО, оптимизировались условия их культивирова-
ния и сушки, испытывались в качестве добавок к бактериальным и другим суспензи-
ям различные химические вещества (глицерин, желатин, глюкоза). Но попытки ста-
билизировать аэрозоль давали только новые научные открытия, но не уверенность в
успешном применении БО при ведении военных действий. В конце 1950-х гг. было
обнаружено, что даже кратковременное пребывание в составе аэрозоля сказывается
на поражающих свойствах биологических агентов Такие многообещающие в качестве
агентов БО микроорганизмы, как возбудители чумы и туляремии, при нахождении в
воздухе в темное время суток, наиболее благоприятное для сохранения их жизнеспо-
собности, теряли вирулентность (см. обзоо Goodlow R. J , Leonard Е А., 1961). Обна-
ружился удручающий для разработчиков БО феномен — по мере старения бактери-
ального аэрозоля процесс отмирания клеток бактерий происходит медленнее, чем
утрата ими инфекционности и вирулентност и. «Старый» аэрозоль, несмотря на сохра-
нение жизнеспособных клеток (т. е. прорастающих на питательной среде), утрачива-
ет способность вызывать инфекционный процесс у экспериментальных животных.
W. D. Sawyer et al. (1966) изучили это явление на обезьянах и людях-добровольцах
ингаляционно инфицированных штаммом возбудителя туляремии, и установили, что
величина инфицирующей дозы для обезьян по мере старения аэрозоля увеличивалась
через 180 мин в 10 раз. Сходные результаты были получены другими авторами на раз-
ных экспериментальных животных (см. Огарков В. И , Гапочко К. Г., 1975). Здесь я
напомню читателю, что снижение вирулентности микроорганизмов, рассматривае-
мых военными в качестве агентов БО, происходит не только при старении аэрозоля.
Оно начинается сразу после их вытеления из природного источника в качестве чис-
тых культур при последующем поддержании на искусственных питательных средах,
затем в процессе культивирования и высушивания (см. «Критические взгляды на БО»
в разд. 1.7 и «Повышение вирулентности бактерий» в разд. 1.8).
Труднопрогнозируемым оказалось поведение мелкодисперсного аэрозоля как
физической системы. Генерал Исии Сиро, используя ВАПы, переделанные из тех, кото-
11. Биологическое оружие перед подписанием Конвенции 1972г. 261
- .>е используют военные химики для поливки противника ипритом, хотя бы имел
можность обнаруживать на белых простынях крупные капли бактериальных куль-
>р, осевшие после распыления с самолетов. Но поведение аэрозоля с нужной для
»зработчиков БО дисперсностью определялось уже не скоростью прохождения ка-
ть через газовую фазу, а кинетикой самой газовой фазы. Рассмотрим это на приме-
диспергирования аэрозоля из линейного источника.
Аэрозоль агентов БО представляет интерес разработчикам БО только в том слу-
«е, если он распространяется в приземном слое воздуха на высоте роста человека. Под
инейным источником аэрозоля понимается генератор аэрозоля, движущийся пер-
;ндикулярно направлению ветра, сила которого достаточна для того, чтобы увлечь
бразующийся аэрозоль с собой. Это наиболее эффективный способ доставки био-
тического агента к цели. В неподвижном воздухе статических камер скорость Осе-
тия сферических частиц зависит от их плотности и диаметра. Если допустить, что
ифекционные частицы имеют приблизительно сферическую форму, то скорость
.едания частиц в неподвижном воздухе имеет следующие значения (табл. 1.7).
Таблица 1.7
Скорость оседания частиц в неподвижном воздухе*
Диаметр частиц, мкм Скорость оседания, см/мин Время падения на 1 м высоты
100 153 3 с
10 15 6 мин
5 5 22 мин
1 0,21 8ч
0,1 0,02 (мм/мин) 14 сут.
♦ По М. Н. Ситникову (1968).
В ходе полигонных экспериментов установлено, что физическую устойчивость
аэрозольного облака, содержащего инфекционные частицы, определяют не только
дисперсность и агрегатное состояние аэрозоля, но и метеорологические условия (тем-
ературный градиент, скорость ветра и атмосферные осадки) и даже характер мест-
ости (рис. 1.52).
В период времени, следующий сразу за диспергированием (либо с самолета, либо
t использованием любого другого распыляющего устройства), аэрозоль приходит в
’авновесие с атмосферными условиями. Крупные аэрозольные частицы выпадают из
чего, оседают на поверхность и образуют с ней прочные связи в результате адгезии.
Такие частицы весьма трудно поддаются реаэрозолированию с образованием вторич-
ного аэрозоля, и они не представляют опасности для человека. В приземном слое
воздуха остается первичный аэрозоль, состоящий из частиц в магическом диапазоне
размеров от 1 до 5 мкм, которые ведут себя как газ и движутся вместе с газовой фазой.
Находясь во взвешенном состоянии, они должны вызывать инфекцию у человека, но
нет никакой гарантии, что они не пройдут над целью даже при самых благоприятных
метеоусловиях и ровной местности (Patrie W. III., 2001).
В 1960 гг. влияние атмосферных факторов на биологические аэрозоли исследова-
тось очень интенсивно. Для имитирования полевых условий были созданы разнооб-
разные сложные экспериментальные камеры (статические и динамические), которые
позволяли обеспечивать точное регулирование температурного градиента, скорости
ветра, влажности и воздействия солнечного света. На основе данных, полученных
262
Часть 1. Краткая история применения биологических средств.
Период уравновешивания аэрозоля
Крупные частицы
аэрозоля оседают
Первичный аэрозоль
Частицы размером 1—5 мкм движутся
вместе с воздушным потоком
Рис. 1.52. Поведение аэрозоля, созданного линейным источником. Средство
доставки рассеивает аэрозоль по линии, перпендикулярной направлению вет-
ра, цель находится в одном или нескольких километрах с подветренной сторо-
ны. Ветер проносит инфекционные частицы над целью. По W. III. Patrie (2001)
в полевых условиях и в экспериментальных камерах, специалисты Форт-Детрик в
начале 1960-х гг. пришли к ясным представлениям об оптимальных условиях для при-
менения БО, которые они рекомендовали военным. Эти условия отражены в настав-
лении FM 3-10 (1962). Наилучшими считались: скорость ветра не более 4 м/с при
изотермических и инверсионных метеорологических условиях атмосферы, чаще все-
го наблюдающихся в ночное время при безоблачном небе или зимой при сплошной
облачности, атакже в ранние и вечерние часы в летнее время. Высота «выпуска» аэро-
золя с летальных аппаратов по-прежнему, как и во время войны на Корейском полу-
острове, рекомендовалась не более 30 м, что позволяло эффективно применять БО
только против крупных масс войск противника, сконцентрированных на узких участ-
ках фронта.
Но, как оказалось, факторы, контролирующие биологическое отмирание аэрозо-
лей, были еще не полностью понята. Расширение экспериментов по изучению пове-
дения биологического аэрозоля в указанном в FM 3-10 диапазоне метеоусловий, по-
зволило открыть дополнительные факторы, ускоряющие биологическое отмирание
аэрозолей, например, «фактор открытого воздуха». Он обнаружен британскими во-
енными исследователями из Нортон Дауна и описан в литературе в 1968 г., т. е. уже на
«закате» наступательной программы (Druett Н. А., Мау К. R., 1968).
Фактор обусловливает непредвиденно высокую скорость отмирания вегетативных
форм бактерий и вирусов на открытом воздухе в ночное время. Им оказалось метаста-
бильное химическое вещество, наподобие того, которое может генерироваться при
реакции озона с углеводородами, выделяющимися при работе автомобильных двига-
телей. Вещество полностью исчезает из наружного воздуха, подаваемого в лаборато-
рию. Обнаружение такого фактора означало, что скорость отмирания микроорганиз-
мов в экспериментальных камерах не соответствует скорости их отмирания на
открытом воздухе. Особенно высокую чувствительность к «фактору открытого возду-
ха» показали имеющие низкие инфицирующие дозы агенты БО — F. tularensis, В. suis,
и некоторые арбовирусы. Чем меньше был размер частицы аэрозоля, содержащего ви-
русы, тем быстрее происходила их инактивация. Споровый микроорганизм — возбу-
дитель сибирской язвы, оказался нечувствительным к «фактору открытого воздуха»,
.11. Биологическое оружие перед подписанием Конвенции 1972 г.
263
- о его инфицирующая доза в самых оптимальных метеоусловиях в тысячи раз выше,
-<м у возбудителей бруцеллеза и туляремии (The Problem1970).
Трудности возникли у разработчиков БО и при попытках диспергирования до
зрозольного состояния биологических токсинов. В экспериментальных условиях при
ингаляционном введении рицин в два раза токсичней вещества «VX» — самого ток-
сичного из боевых ОВ. Но его сухие рецептуры оказалось невозможным диспергиро-
1йто с помощью разрывных зарядов из-за потери активности. Дробление взрывом
" дстворов и суспензий рицина так же встретило свои проблемы. Рицин, будучи гли-
копротеином, растворим только в водных системах, но сам разлагается водой, и сна-
ряженные боеприпасы быстро утрачивают эффективность. К тому же водные растворы
"Ч'цина замерзают, что создает дополнительные проблемы для их боевого примене-
ния. Суспензии рицина в четыреххлористом углероде испытывались в качестве мо-
дельных систем при изыскании методов перевода в аэрозольное состояние других
энологических агентов. Но любые суспензии сами по себе являются неустойчивыми,
расслаиваются и меняют баллистические характеристики боеприпасов. Попытки
применить рицин в виде тонкодисперсного порошка с размером частиц 5 мкм и менее,
казались сопряженными с проблемами, создаваемыми его слеживанием и комкова-
нием. К тому же рицин в виде распыленного порошка или раствора оказался подвер-
жен дезактивирующему действию ультрафиолетового излучения — получасовая экс-
позиция рицина к ультрафиолету приводит к снижению его активности в 1000 раз
Антонов Н. С., 1994).
Влияние биотехнологических факторов, условий диспергирования и нахождения
атмосфере на выживаемость и инфицирующую (поражающую) способность биоло-
гических агентов обобщено в ряде обзоров (Ситников М. Н., 1968; Антонов Н. С.,
994), в данной книге нет смысла их рассматривать детально. К отдельным аспектам
-роблемы мы вернемся в третьей части книги (см. разд. 3.11).
Отдел специальных операций в Форт-Детрике. Основан в 1948 г. (см. разд. 1.9 «Раз-
работка технических средств для индивидуальных убийств»). На финансирование
этого отдела только в период с 1953 по 1973 г. ЦРУ израсходовало около 3 млн долла-
ров. В интересах ЦРУ в отделе изучались такие проблемы, как устойчивость биологи-
ческих агентов, возможность приготовления сухих рецептур биологических агентов,
возможность приготовления их в виде порошка для загрязнения одежды, подушек,
’петельного белья и т. д. Отделом ра щаботан11 и изготовлены устройства для рассеи-
вания биологических рецептур в виде чемоданчиков, зажигалок, авторучек, стартеров
"юминесцентных ламп. Подобные устройства изготовлялись не только для ЦРУ, но и
для диверсантов других ведомств. Специально для ЦРУ разраоотан и изготовлен мик-
робиоинокулятор, не оставлявший следов, которые обнаруживались бы при осмотре
и вскрытии трупов, а также различные уг-’-ройства для диверсионных актов с целью
"орчи продуктов питания, загрязнения нефтехранилищ и т. д.
Длительное время в отделе велись поиски альтернатив стандартным ампулам с
цианистым калием. Было изготовлено 80 штук изделий в виде булавок с токсином
моллюсков (палитоксин) для применения агентами ЦРУ в чрезвычайных обстоятель-
ствах, а также пилотами разведывательных самолетов «U-2». Такой отравленной бу-
авкой был снабжен, в частности, летчик Пауэрс, у которого она была спрятана в се-
ребряном долларе. Сотрудники КГБ СССР обнаружили эту булавку и таким образом
положили конец данной программе. Официальные лица, в частности, директор ЦРУ
Колби, утверждали тогда, что это был единственный случай использования подобного
устройства в операции (разумеется!).
264
Часть I. Краткая история применения биологических средств..
В конце 1960-х гг. Отдел специальных операций Форт-Детрика поддерживал для
ЦРУ запас 15—20 биологических агентов и токсинов в количествах от 100 мг до 100 г.
В январе 1970 г. в записке, адресованной директору ЦРУ, заместителю директора
ЦРУ по планированию Карамессинес указал следующие материалы:
Bacillus anthracis — 100 г;
Pasteurella tularensis — 20 г;
вирус венесуэльского энцефаломиелита лошадей — 20 г;
Coccidioides immitis — 20 г;
Brucella suis — 2—3 г;
Brucella melitensis — 2—3 г;
Mycobacterium tuberculosis — 5 г;
Salmonella typhimurium — 10 r;
вирус натуральной оспы — 50 г;
стафилококковый энтеротоксин — 10 г;
ботулинический токсин, тип А — 5 г;
паралитический токсин моллюсков — 5,193 г;
яд змеи Bungarus candidis — 2 г;
токсин Microcystis aeruginosa — 25 мг;
токсиферин — 100 мг.
Поданным Министерства обороны, все перечисленные материалы были уничто-
жены вместе с запасами биологических лабораторий в Форт-Детрике за исключени-
ем токсина моллюсков, который в нарушение распоряжения президента США был
передан ЦРУ. Больше того, к 5,193 г палитоксина сотрудники ЦРУ добавили налич-
ный запас этого токсина из другого подразделения Форт-Детрика (Hearings before the
select committee..., 1975).
Насекомые как переносчики возбудителей опасных инфекционных болезней. Еще с
1930-х гг. чумная блоха рассматривается военными как в своем роде «классика БО»
Гипнотизирующее действие на разработчиков БО оказывали исторические хроники,
сообщающие о гибели крупных воинских контингентов и даже целых армий от бубон-
ной чумы, малярии, желтой лихорадки, сыпного и возвратного тифов, основную роль
в распространении которых играли насекомые. С точки зрения формальной логики
создание энтомологического оружия не должно было вызвать особых сложностей, и
его эффективность представлялась разработчикам БО очень высокой. Известно свы-
ше 100 инфекционных болезней, которые передаются более чем 2000 видами члени-
стоногих. Большинство насекомых сохраняет способность заражать человека и жи-
вотных в течение от нескольких недель до 2—3 мес. Клещи живут несколько лет и
способны передавать инфекцию новому поколению. Сторонники такого способа рас-
пространения инфекционных болезней утверждали, что зашита от биологических
аэрозолей сравнительно проста, поскольку противодымные фильтры современных
противогазов полностью задерживают частицы аэрозоля микронного диапазона. А вот
насекомые способны обойти эту защиту и доставить биологические агенты «по назна-
чению» через неповрежденную кожу. Поскольку насекомые остаются жизнеспособ-
ными в течение определенного времени, весь этот период район цели сохраняет опас-
ность для пребывающего в нем личного состава вооруженных сил. Кроме того, за счет
активного перемещения (миграции) насекомых и грызунов размеры первоначальных
очагов заражения могут увеличиваться (Ситников М. Н., 1968; Dando М., 1994).
Результаты применения энтомологического оружия во время войны на Корейском
полуострове были признаны американским командованием положительными. Хотя
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
265
сброшенные американскими самолетами инфицированные насекомые не вызвали
массовых эпидемий среди войск и населения противника, но все же они показали
«принципиальную возможность» такого поражения. Поэтому после окончания вой-
ны в Форт-Детрике осуществлены несколько крупных программ по разработке средств
энтомологической войны. Сводка полевых испытаний, во время которых изучались
возможности насекомых в качестве переносчиков агентов БО, приведена в табл. 1.8.
Таблица 1.8
Полевые испытания насекомых в качестве переносчиков агентов БО,
проведенные американскими военными в 1950—1960-х гг.*
Условное обозначение испытаний Дата Описание
«Big Itch» («Сильный зуд») 1954 Испытание боеприпаса Е-14 ыя рассеивания блох
«Big Buzz» («Громкое жужжание») 1955 Демонстрация возможности массового производства комаров и их рассеивания с самолета. Оценка выживаемости комаров и их способности вызывать укусы у человека
«Мау Day» («Майский день») 1956 Рассеяние комаров из наземного источника за пределами города Саванна, шт. Джорджия
«Bellwether I» («Вожак I») 1959 Определение влияния факторов окружающей среды на час- тоту укусов голодными неоплодотворенными самками кома- ров A. aegypti личного состава войск, расположенных на открытом пространстве
«Bellwether И» («Вожак 11») 1960 Серия опытов по определению влияния расстояния до жертвы, движения и т. д. на частоту укусов комарами (с участием до сотни военнослужащих в каждом опыте)
«Bellwether III» («Вожак III») 1962 Сравнение штаммов A. aegypti по склонности к укусам, рассеянию и проникновению в жилища
* Таблица составлена М. Dando (1994) на основании публикации W. Н. Rose (1981).
По данным, собранным М. Dando (1994), начатая в 1953 г. в Форт-Детрике про-
грамма по изучению возможности применения членистоногих для распространения
биологических агентов, предполагала изучение способности комара Aedes aegypti,
переносчика вируса желтой лихорадки, вызывать вспышки желтой лихорадки среди
населения Средней Азии СССР. Предполагалось, что успех программы обеспечен
высокой восприимчивостью населения СССР к вирусу желтой лихорадки и тем, что
в условиях СССР может оказаться невозможным быстро осуществить программу мас-
совой иммунизации населения.
Возможность использования комаров для заражения людей была оценена путем
выпуска неинфицированных самок в населенную зону в районе города Саванна, штат
Джорджия, и наблюдения за т ем, сколько их попадет в дома и укусит людей (испыта-
ние «Мау Day»). Кроме того, в 1956 г. 600 тыс. неинфицированных комаров было вы-
пущено с самолета на полигоне для бомбометания, и в течение дня насекомые пре-
одолели расстояние в одну-две мили и перекусали много народа. Дальнейшие испы-
тания показали, что насекомые могут рассеиваться с вертолетов, из устройств,
сбрасываемых с самолетов, или с земли. Сообщено, что в лабораториях Форт-Детри-
ка в те годы могло выращиваться до полумиллиона особей в месяц. Использованный
для экспериментов по инфицированию насекомых вирус желтой лихорадки был по-
лучен от человека, заболевшего в Тринидаде в 1954 г., и размножен путем инфициро-
вания обезьян. Комары, в свою очередь, инфицировались вирусом путем погружения
их личинок в сыворотку зараженных обезьян макак-резус. Вирулентность инфици-
266
Часть 1. Краткая история применения биологических средств...
рованных комаров проверялась путем заражения ими мышей. Технический комитет
Химического корпуса США классифицировал систему «вирус желтой лихорадки —
комары A. aegypty» в качестве стандартизованного агента БО в июне 1959 г. Предпола-
галось использовать его по способу, отработанному во время войны на Корейском по-
луострове — снаряжая кассетные авиабомбы, сбрасываемые с самолета.
Однако в 1960-х гг. интерес военных к энтомологическому оружию ослаб. Настав-
ление FM 30-10 (1962) наиболее эффективным способом распространения агентов БО
считает применение аэрозоля. Частично сложность использования блох, инфициро-
ванных возбудителем чумы, для искусственного распространения чумы среди людей
мы рассмотрели в разд. 1.8 («Производственные мощности»).
М. Н. Ситников (1968) объяснял утрату интереса к разработке энтомологиче-
ского оружия следующим образом. Главный недостаток использования насекомых-
переносчиков состоит в том, что они не могут быть доставлены к цели без их массо-
вой гибели. Поведение доставленных к цели насекомых-переносчиков не только
нельзя проконтролировать, но и как-то предвидеть. Применение переносчиков ог-
раничено определенными метеорологическими и природными условиями, непред-
виденных моментов при их изменении может оказаться даже больше, чем при напа-
дении с применением аэрозоля. К тому же массивные дозы биологического агента,
достаточные для преодоления у человека искусственного иммунитета, можно создать
только при использовании мелкодисперсных аэрозолей. Следовательно, разработ-
чики БО в 1960-х гг. не видели у насекомых никаких преимуществ перед аэрозолями
биологических агентов.
Поражение растений. В период с 1951 по 1969 г. в США производились три био-
агента-возбудителя болезней сельскохозяйственных растений. Это — возбудители
стеблевой ржавчины пшеницы (рецептура ТХ), ржи и возбудитель пирикуляриоза риса
(рецептура LX). В течение 1951-1957 гг. споры возбудителя стеблевой ржавчины пше-
ницы и ржи получали путем заражения посевов на участках государственных опыт-
ных станций. Собранный материал отправляли в арсенал Эджвуд (Мэриленд), для
классификации, сушки и хранения. Работа была завершена в 1959 г. при участии спе-
циалистов ВВС. Споры возбудителя стеблевой ржавчины пшеницы выращивали на
государственных опытных плантациях в период с 1962 по 1969 г. Необработанный ма-
териал доставлялся в арсенал Роки Маунтин, где его очищали, классифицировали и
хранили в замороженном виде. Все споры возбудителя ржавчины пшеницы были
уничтожены к февралю 1978 г.
Возбудителя пирикуляриоза риса получали методом глубинного культивирования
по контракту. Контракт на производство был заключен в марте 1965 г. Производство
возбудителя пирикуляриоза было прекращено в июне 1966 г. Готовый биоагент упа-
ковывали и хранили в Форт-Детрике. Весь запас возбудителя пирикуляриоза риса был
уничтожен в период между 17 января и 18 мая 1972 г.
Взгляды на ведение биологической войны. В «Наставлении по боевому применению
химического и биологического оружия» (FM 30-10, 1962) БО рассматривается как
реально существующее, его поражаюшие свойства оцениваются выше, чем химичес-
кого оружия. Согласно FM 30-10, БО должно применяться для поражения противни-
ка на территории в несколько тысяч квадратных километров, на которой цели точно
не выявлены. Возможность распространения биологических агентов с помощью
средств, срабатывающих на значительном расстоянии от района цели и доставка аген-
та к цели благодаря перемещению воздуха, обеспечивает скрытность и внезапность
применения БО. Наличие инкубационного периода между моментом проникновения
1.11. Биологическое оружие перед подписанием Конвенции 1972 г.
267
биологического агента в организм человека и началом болезни обеспечивает замед-
ленное действие БО. Возможность проникновения биологического аэрозоля в различ-
ные инженерные сооружения позволяет поражать личный состав вооруженных сил
противника, не разрушая этих сооружений, что не может быть достигнуто с помощью
фугасных бомб и снарядов и ядерных боеприпасов малого калибра.
Для исключения возможности гибели биологического агента во время хранения,
FM 30-10 рекомендует условия регулируемого охлаждения, т. е. то, что в разд. 2.4.2
будет описано как «холодовая цепь». Для избежания гибели микроорганизмов в мо-
мент распыления наставлением рекомендуется температура воздуха в районе приме-
нения БО около 10°С, а относительная влажность ниже 85 % (для сухих рецептур). При
этом применение БО рекомендуется проводить в темное время суток или в дни с силь-
ной облачностью.
Аэрозоли могут быть образованы при помощи биологических боеприпасов взрыв-
ного действия, механических генераторов и распылительных устройств.
Отличительным тактическим свойством БО составители FM 30-10 считали «воз-
можность без значительных затрат поражать живую силу противника на больших
площадях». Ее они делили на: личный состав войск противника; гражданское насе-
ление на территории противника, направляющее свои усилия на повышение боеспо-
собности войск противника; и население на территории занятой противником, вы-
нужденное поддерживать противника, т. е. применение БО считалось целесообразным
как против гражданского населения стран, ведущих войну с США, так и против соб-
ственного населения (вернее, населения европейских партнеров по НАТО) на окку-
пированных Советами территориях. Стратегическое и тактическое применение БО
должно было планироваться таким образом, чтобы его применение в одном районе
не отражалось на эффективности применения в другом районе. Само БО должно при-
меняться только массированно.
Во время тактических наступательных операций БО должно использоваться для
поражения личного состава:
а) резервов и поддерживающих подразделений и частей;
б) тыловых подразделений;
в) подразделений, обслуживающих линии связи и коммуникации;
г) транспортных подразделений;
д) учебных подразделений;
е) частей и подразделений, совершающих марш или находящихся на отдыхе;
ж) партизанских отрядов;
з) воздушных и морских десантов.
В обороне БО предполагалось применять против всех подходящих целей во втором
эшелоне противника, чтобы ослабить эффективность его наступательных действий.
Стратегическое применение БО предполагало поражение живой силы противника:
а) в городах и поселках городского типа;
б) в промышленных центрах;
в) в районах отдыха, перегруппировки или обучения войск;
г) персонала ПВО;
д) в важнейших портах;
е) на железнодорожных узлах и станциях водоснабжения;
з) в научно-исследовательских центрах;
ж) на нефтепромыслах;
и) в местах пуска ракет.
268
Часть I. Краткая история применения биологических средств...
Применение более одного агента БО считалось более эффективным, так как уси-
ливало поражающее действие БО и затрудняло идентификацию агентов и лечение
пораженных.
Конвенция 1972 г. Женевский протокол 1925 г. был, по сути, соглашением между
подписавшими его государствами о неприменении БО и химического оружия первы-
ми. Что касается распространения этих видов оружия, то Женевский протокол не пре-
дусматривает соглашения по их контролю или разоружению. Он не содержит ограни-
чений по разработке и производству БО и не включает каких-либо предложений по
режиму проверки. Таким образом, протокол не был правовым барьером, например,
для разработки Англией и США БО в 1940-х гг. Кроме того, Соединенные Штаты стали
участниками этого протокола лишь в 1975 г. и применение ими БО во время войны на
Корейском полуострове в 1952 г. не было его нарушением.
Действующий в настоящее время режим по ограничению БО, помимо Женевско-
го протокола 1925 г., включает «Конвенцию о запрещении разработки, производства
и накопления запасов бактериологического (биологического) и токсинного оружия
и их уничтожении» от 1972 г. Конвенция стала результатом интенсивной дипломати-
ческой деятельности в конце 1960-х и в начале 1970-х гг. Подробно об истории разра-
ботки и подписания Конвенции государствами, имеющими программы в области со-
здания БО, можно прочитать в работе М. Dando (1994).
Уничтожение БО. В соответствии с решением президента США Ричарда Никсона
от 25 ноября 1969 г. и рекомендациями Управления вооружений армии США специ-
алисты бывших биологических лабораторий армии США в Форт-Детрике в 1970 г.
разработали планы уничтожения запасов биологических агентов и снаряженных ими
боеприпасов в следующих 4 учреждениях Министерства обороны США:
в арсенале Пайн-Блафф (шт. Арканзас);
в арсенале Роки-Маунтин (шт. Колорадо);
на базе ВВС США Бил (шт. Калифорния);
в биологических лабораториях армии США в Форт-Детрике (шт. Мэриленд).
Часть материалов была оставлена для осуществления научно-исследовательских
работ в области биологического оружия, причем в количествах, вполне достаточных
для выполнения намеченных программ.
Уничтожению подлежали:
1) запасы биологических агентов для поражения живой силы противника и сна-
ряженные ими боеприпасы на складах отдела биологических операций арсенала
Пайн-Блафф;
2) запасы биологических средств поражения сельскохозяйственных растений и
снаряженные ими боеприпасы в арсенале Роки-Маунтин, на базе ВВС Бил и в био-
логических лабораториях армии США в Форт-Детрике.
Кроме того, подлежали уничтожению небольшие запасы биологических агентов
для поражения живой силы, биологических средств поражения растений и снаряжен-
ных ими боеприпасов, которые были подготовлены для испытаний в Форт-Детрике
и на полигоне Дагуэй (шт. Юта).
Работы по уничтожению запасов БО в указанных учреждениях были осуществле-
ны в период с 10 мая 1971 г. по 1 мая 1972 г. Эти работы, включая последующую обра-
ботку производственных и складских помещений, лабораторий и оборудования, обо-
шлись в 10,8 млн долл. Несчастных случаев и аварий не было. Методы уничтожения
варьировали в зависимости от видов биологических агентов и токсинов. Например,
изделия и емкости с рецептурой, содержащей ботулинический токсин или сакситок-
1.11 Биологическое оружие перед подписанием Конвенции 1972 г.
269
син. плавили в печах с температурой около 1100°C. Рецептуры с высушенными спо-
рами В. anthracis извлекали из боеприпасов, смешивали с 2 %-ным раствором каусти-
ка и в течение 3 ч прогревали при температуре около 140 °C. Дета ш расснаряженных
боеприпасов и освобожденные контейнеры плавили в печах при 1100 °C. Материа-
лы. полученные после упомянутых операций, нейтрализовали, засевали непатоген-
иыми культурами бактерий из почвы, речной воды или стоков и оставляли на биоде-
стэукцию. После биодеструкции растворы еще раз в течение 3 ч стерилизовали при
температуре около 140 °C. далее производили проверку на стерильность, пастеризо-
али при 100— 120°С и сливали в приемники для вторичной биодеструкции. Обрабо-
танные таким образом материалы перегружали на выпарные устройства для сушки.
Сухие остатки закапывали в землю при помощи дисковых культиваторов.
Разнообразные контейнеры, отдельные детали боеприпасов и тару резали или
ломали различными способами, лом прокаливали в печах и плавили при 1100 °C.
Металлические остатки боеприпасов собирали, стерилизовали и, убедившись в сте-
рильности, складировали в специально отведенных местах.
После уничтожения запасов биологических агентов и снаряженных ими боепри-
пасов все производственные и складские помещения, лаборатории и оборудование
очищали и дезинфицировали при помощи методов, которые гарантировали последую-
щее безопасное пребывание в них и работу неиммунизированного персонала. Возду-
ховоды и дренажные галереи минимум 16 ч обрабатывали газообразным формальде-
гидом. Технологические системы производственных помещений стерилизовали паром
с температурой около 120 °C.
Форт-Детрик после уничтожения БО. Он был выведен из подчинения командова-
нию материально-технического обеспечения армии США и переподчинен вновь
сформированному медицинскому командованию армии США в Форт-Сэме (Хьюс-
тон, штат Техас). В 1973 г. на территории Форт-Детрик находилось шесть НИУ, пред-
назначенных для выполнения несекретных исследований: Научно-исследовательский
центр по изучению рака, (JSAMRIID, Научно-исследовательская и опытно-конструк-
торская лаборатория медицинской биотехнологии, Лаборатория ВМФ, Научно-
исследовательская лаборатория фитопатологии, Отдел борьбы с вредителями расте-
ний — подразделение химической лаборатории арсенала Эджвуд. И еще 11 учрежде-
ний, не имеющих отношения к научным исследованиям и выполняющих различные
задачи, не связанные с секретностью. Среди них: Архив медико-санитарной службы
США, Статистико-информационный отдел медико-санитарной службы США, Управ-
.ение начальника работ по демилитаризации химического имущества командования
материально-технического снабжения армии США и разтичные отделы Министер-
ства жилищного строительства и развития городов и Министерства внутренних дел
(Fort Detrick converting..., 1973) Так как США при ежегодных обменах информации в
области биологической защиты в соответствии с Конвенцией 1972 г. сообщают толь-
ко о программах Министерства обороны, то секретные или пользующиеся плохой ре-
путацией программы обычно передаются в ЦРУ, Министерству энергетики или в дру-
гие ведомства и подрядные организации (Rosenberg В. Н., 2002).
* * *
«Золотой век» разработчиков БО, начавшийся после окончания войны на Корей-
ском полуострове, закончился в самый разгар «холодной войны» подписанием в 1972 г.
«Конвенции о запрещении разработки, производства и накопления запасов бактерио-
логического (биологического) и токсинного оружия и их уничтожении». Хотя тогда
270
Часть 1. Краткая история применения биологических средств...
политиками было сделано много трогательных заявлений о приверженности к мир)
в котором режим по ограничению БО должен сыграть чуть ли не спасительную роль,
но я рискну высказать предположение, что подписание Конвенции было вызвано
другими причинами.
Прежде всего, «мощное оружие бедных» стало не по средствам богатым странам.
Затраты на его разработку, производство и хранение не оправдывались его эффектив-
ностью при боевом применении. Основным препятствием в применении БО тради-
ционного типа стало именно наличие в нем биологического компонента. В том диапа-
зоне размеров частиц аэро юля, в котором микроорганизмы и биологические токсины
могут вызвать поражение людей, не только сами частицы как физические системы
ведут себя непредсказуемо, но еше менее предсказуемо ведут они себя как биологиче-
ские системы. БО не стало в 1960-х гг. оружием массового поражения, сравнимым по
эффективности с ядерным. Поглотив миллиарды долларов на исследования, оно ос-
талось оружием шантажа диверсий и террористических актов. А предназначавшую
ся ему нишу оружия, способного уничтожать миллионы людей, не затрагивая созда
ваемые ими материальные ценности, стали занимать хорошо проверенные виды ору-
жия. Среди них нейтронные ядерные боеприпасы и высокоточное оружие. Изменился
и сам характер боевых действий. Столь удобное для применения БО концентрирова-
ние больших масс войск на узких участках фронта, которое имело место в годы Вто-
рой мировой войны и во время войны на Корейском полуострове, ушло в прошлое.
Уже на завершающем этапе вьетнамской войны боевые действия приобрели характер
воздушно-наземных операций. При проведении таких операций четко обозначенные
линии соприкосновения войск стирон отсутствуют, имеются большие промежутки и
разрывы в их оперативном построении. Личный состав, как правило, рассредоточен
на большую глубину, укрыт в боевых и специатьных машинах и обеспечен совершен-
ными средствами противохимической и противобиологической защиты. Для истреб-
ления населения появились новые средства поражения, которые к оружию, в привыч-
ном его понимании, отнести сложно — неконтролируемая пандемия ВИЧ/ СПИДа,
информационные и сетевые технологии, наркотизация населения и другие.
1.12. БИОДИВЕРСИИ И БИОТЕРРОР ВРЕМЕН ПРОФАНАЦИИ
БИОЛОГИЧЕСКОГО ОРУЖИЯ
После подписания в 1972 г. «Конвенции о запрещении разработки, производства
и накопления запасов бактериологического (биологического) и токсинного оружия
и их уничтожении», это оружие было изъято из арсеналов создававших его стран и
уничтожено. Во всяком случае, с такими заявлениями выступали представители стран,
подписавших Конвенцию. Механизмы контроля выполнения Конвенции не были
оговорены, но нет и оснований утверждать, что прежние арсеналы малоэффективно-
го БО поддерживались на том же уровне, что до ее подписания. Постепенно из воен-
ной биологии ушли люди, создававшие эту науку в 1940—1960-х гг. В отсутствие их
опыта и знаний и при существующих конвенциальных ограничениях БО стало при-
обретать виртуальный характер, когда математические модели заняли место реальных
экспериментов, осущесэ вленных в полигонных условиях. Представления о БО силь-
но упростились. Политиками был востребован миф о его доступности и дешевизне, и
он вновь мутными потоками пролился на страницы научных и популярных изданий.
На этом фоне профанации БО активно преследовали свои интересы спецслужбы раз-
12. Биодиверсии и биотеррор времен профанации биологического оружия
271
ных стран Одновременно появились организации и даже страны, руководители
* спорых приняли за «чистую монету» политические игры вокруг распределения бюд-
жетных денег и геополитических интересов сверхдержав. По обыкновению «смутно-
времени», к ним присоединились странные личности, типа тех, что в Средние века
.иали дверные ручки гноем бубонов больных чумой. Обстоятельства совершенных
ли планируемых ими преступлений обобщены в табл. 1.9.
1.12.1. Искусственная вспышка сальмонеллезного
гастроэнтерита в Далласе в 1984 г.
Мотив преступления. «Правильная эпидемиология» сальмонеллеза. Появление подозрений
в искусственном происхождении вспышки сальмонеллеза в Далласе. Определение «случая
инфицирования». Изучение вспышки. Лабораторные методы. Исследование окружающей
среды. Эпидемиологическое определение места совершения теракта. Исследование выде-
ленных штаммов сальмонелл. Уголовное расследование. Наказание.
Материалы расследования этой вспышки опубликованы Т. J. Torok et al. (1997),
е ^рудниками CDC (США).
Мотив преступления. В 1981 г. ученики Бхагван Шри Раджниша приобрели боль-
31 ое ранчо в округе Уаскодля строительства новой международной штаб-квартиры для
индийских гуру. Создание общины было спорным с самого начала; основными во-
росами конфликта были культурные ценности и проблемы землепользования. Части
1ччо они хотели присвоить статус города Раджнишпучам, но грамота о присвоении
t' .туса города оспаривалась в судах, что очень мешало новому строительству. Члены
тещины считали, что исход выборов в городке Даллас (население 10,5 тыс. человек)
представителей в округ Уаско (население округа 21 тыс. человек) 6 ноября 1984 г. ока-
жет влияние на дальнейшее решение вопроса о землепользовании. И тогда они решили
_орвать выборы, вызвав среди избирателей вспышку сальмонеллеза.
«Правильная эпидемиология» сальмонеллеза. Человек инфицируется сальмонелла-
**и, употребляя в пищу инфицированные продукты животного происхождения (мяс-
ныеи молочные продукты, яйца). Меньшее эпидемиологическое значение имеют
>'ба и рыбные продукты, а также продукты растительного происхождения. Вода пре-
“ушественно принимает опосредованное участие в передаче сальмонелл. Природ-
ным резервуаром сальмонелл являются сельскохозяйственные животные — крупный
эогатыи скот, свиньи, реже овцы. Весьма значительна роль птиц (особенно уток и кур)
как носителей возбудителя сальмонеллезной инфекции. Обсеменение мяса сальмо-
-еллами происходит после убоя при нарушении правил убоя, разделки и хранения
мчса. Однако инфицирование исходных продуктов животного происхождения и по-
фабрикатов из них обычно исключается проведением ветеринарно-санитарной эк-
спертизы на мясоперерабатывающих предприятиях, бойнях, в местах первичного
£ 5ора молока и на пищевых контрольных станциях. Заражение продуктов питания
»ожет происходить при неправильной их кулинарной обработке, изготовлении пищи
рязных столах, в грязной посуде. Сальмонеллез распространен повсеместно. Ре-
истрируются спорадические случаи и эпидемические вспышки (Таршис М. Г., Чер-
касский Б. Л., 1997).
Появление подозрений в искусственном происхождении вспышки сальмонеллеза в
Дугласе. В течение месяца сатьмонеллезом заболел 751 человек. При эпидемиологи-
сском обследовании были идентифицированы средства передачи возбудителя в виде
Таблица 1.9
Обстоятельства совершенных или планируемых преступлений с использованием биологических агентов*
Организатор Мотивация Идеология (цель) Объект (ы) Агент (ы) Как осуществлено или планировалось осуществить Результат
1 2 3 4 5 6 7
Weather Under- ground (1970) «Гроза под землей» Временная инкап- ситация жителей американских го- родов для демон- страции «неспособ- ности федеральных властей» Революционное движение, оппозиционное американскому империализму и войне во Вьетнаме Жители городов США Биологические агенты пытались получить в USAMR1ID путем шантажа военно- служащего гомо- сексуалиста Планировалось подбросить инкапси- тирующие биологи- ческие и химические агенты в воду город- ского водопровода Возможно, это недостоверный случай
R.I.S.E (1972) Террористиче- ская органи- зация «Райс» Уничтожить боль- шинство людей для предотвращения разрушения приро- ды и создать расу сверхлюдей Преступники были студентами коллед- жа, подвергшимися влиянию идеологии экотерроризма и наркокультуры 1960-х гг. Теракт планирова- лось совершить в отношении «миро- вой популяции», затем сузили круг жертв до жителей 5 штатов вокруг Чикаго Восемь микроб- ных патогенов, включающих аген- ты, вызывающие сыпной тиф, диф- терию, дизенте- рию и менингиты Планировалась аэрозольная атака с самолета и конта- минация запасов воды, поступающей в городской водо- провод Атака сорвалась, когда были обнару- жены микробные культуры; два глав- ных подозреваемых бежали на Кубу
Болгарские спецслужбы (1978). Покушение на болгарских политических эмигрантов Внешне это выгля- дело как расправа коммунистическо- го режима Болга- рии над своими «политическими противниками» Дискредитация болгарских и советских спецслужб Болгарские имми- гранты В. Костов (Париж) и Г. Мар- ков (Лондон) Рицин Пуля с рицином, выпущенная из неустановленного оружия Марков погиб, Костов случайно остался жив (см. разд. 3.11)
Red Army Fraction (1980) «Фракция Красной армии» Планировалась био- логическая атака против ведущих за- падногерманских политиков и бизне- сменов Марксистская революционная идеология Конкретные цели не известны Преступники культивировали ботулинический токсин на явоч- ной квартире в Париже Неизвестно Сведения, впослед- ствии опровергнутые правительством ФРГ
Часть 1. Краткая история применения биологических средств...
///iiiJh i m i nut iihib i I V
1 2 3 4 5 6 7
Rajneeshe Cult (1984) Религи- озная секта Раджнишистов Планировалось инкапситацией избирателей выиг- рать местные выборы Индийский религи- озный культ, возгла- вляемый харизмати- ческим гуру Постоянные жители городков Dalles и Wasco County, штат Орегон (США) 5. typhimurium И нфицировалась пища салатных баров ресторанов. Инфи- цирован 751 человек, никто не погиб Полиции о заговоре стало известно от информатора. Эпидемиологи пред- положили теракт на основе собственного расследования
Minnesota Patriots Council (1991) «Совет патриотов Минесоты» Нанесение вреда федеральным властям, личная месть Анти правительствен - ный протест; правое крыло движения «патриотов» Должностные лица Рицин, извлеченный из касторовых бобов, приобретенных по почте Нанесением на кожу рицина, растворен- ного в диметилсуль- фоксиде, в препарате алоэ или в составе сухого аэрозоля В группу был внедрен информатор ФБР; четыре члена группы были арестованы
«Аум Синоике» (1990-1995) Реализация проро- ческого предсказа- ния, устранение врагов, соперников, колеблющихся сектантов Новая жизнь после Страшного суда, культ становления теократического государства в Япо- нии с харизмати- ческим лидером Социальные группы, отдельные оппоненты культа Биологические агенты (В. anthra- cis, ботулиниче- ский токсин, ви- рус Эбола) и хи- мические агенты (зарин, VX. HCN) По крайней мере, 10 попыток аэрозоль- ного применения биологических аген- тов. Все оказались неудачными Теракты с примене- нием химических агентов привели к гибели, по крайней мере, 20 человек: пострадали более 1000 человек
Диана Томпсон (1996) Личная неприязнь к коллегам Нет Сотрудники медицинского центра в Техасе Возбудитель ди- зентерии редкого серотипа был добавлен в оладьи Дизентерией заболело 12 человек, никто не погиб Преступница установлена сотрудниками ФБР
Ларри Харрис (1998) Обосновать угрозу для американцев со стороны биологи- ческого оружия Ирака Связь с группой «Арийская нация» Неопределенные угрозы в адрес американских властей Пытался получить возбудители чумы и сибирской язвы (вакцинный штамм) и другие бактерии Обсуждал диссеми- нацию биологиче- ских агентов с по- мощью самолетов- опылителей и дру- гими способами Арестован после то- го, как публично зая- вил о желании совер- шить биологический теракт в отношении официальных лиц США
? Биодиверсии и биотеррор времен профанации биологического оружия
Продолжение табл. 1.9
1 2 3 4 5 6 7
Сибирская язва в США в 7001 г. Брюс Айвинс — бывший сотрудник USAMRIID Напоминает круп- номасштабную провокацию, имеющую цель втятуть США в войну с Ираком и Афганистаном Неизвестна СМИ, известные политики Рецептура спор возбудителя сибирской язвы, пригодная для применения в военных целях Рассылка по почте конвертов, содержа- щих рецептуру сибир- ской язвой Заболело 22 челове- ка, 5 из них погибло
Рициновые письма «Падшего ангела» (2003) Требование к пра- вительству США отменить измене- ния, внесенные в правила грузопе- ревозок автотран- спортными компа- ниями Her «Белый дом» Рицин Пор< шок рицина был помещен в два метал- лических флакончика и отослан с помощью обычных конвертов. На флакончиках была наклейка, предупреж- дающая о наличии в нем рицина Пораженных рици- ном не было. За по- мощь в поимке пре- ступника назначено вознаграждение в 120 тыс. долларов, но обвинение никому предъявлено не было
Саад Хусейн, ВИЧ-инфицир- ованный беже- нец из Эфио- пии, житель Чикаго, 2008 г. Религиозная Нет. Организатор и исполнитель пре- ступления оказался душевнобольным Кандидат в прези- денты США Барак Обама и несколько его помощников ВИЧ. Бумага с каплями крови самого терро- риста Президенту Обаме и нескольким его помощникам были отправлены письма с приглашением на собрание сторонников Обамы в ночь выборов Арестован полицией
Неизвестны, декабрь 2009 г. Неизвестна Неизвестна Потребители героина в Европе Возбудитель сибирской язвы В героин были добав- лены споры возбуди- теля В. anthrads турецкого происхож- дения В Шотландии зара- жено 47 человек, из них погибло 13, еще один наркоман погиб в Германии при сход- ных обстоятельствах
Неизвестны, апрель 2013 г. Судя по тексту письма, — выходка душевнобольного Нет Президент США и члены Конгресса Рицин Три конверта с порошком рицина Перехвачены спецслужбами
* За основу взята таблица из работы J. Tucker (1 °99).
Часть 1. Краткая история применения биологических средств...
12. Биодиверсии и биотеррор времен профанации биологического оружия
275
г эдуктов в многочисленных заку< очных самообслуживания с овощными блюдами
я возможные периоды контаминации. Общие механизмы, посредством которых могло
роизойти инфицирование людей в закусочных, были исключены. После закрытия
всех закусочных с овощными блюдами произошло резкое снижение заболеваемости
городке, и, возможно, именно это вмешательство положили конец вспышке. Одна-
ко эти факты трудно было увязать с тем, что в санитарно-гигиенической практике
-р“частных к вспышке ресторанов не было выявлено грубых нарушений, что участ-
ники частных банкетов, пользовавшиеся закусочными с овощными блюдами, не по-
"адали, и что большинство ресторанов, связанных со вспышкой, не имели общих
сточников поставок пищевых продуктов. Эпидемиологической службой штата Оре-
гон были рассмотрены и др) гие возможные способы передачи возбудите.тя инфекции,
-рели них искусственное распространение сальмонелл. Одновременно ФБР получи-
ло информацию от осведомителя о проведенном теракте.
Определение «случая инфицирования». Случай инфицирования определялся как
заболевание с диареей и, по меньшей мере, с тремя из следующих перечисленных
симптомов: тихоратка. озноб, головная боль, тошнота, рвота, боль в животе или кро-
__вый понос или по культуре 5. typhimurium, полученной из фекалий. Больного рас-
сматривали как случай, ассоциированный со вспышкой, если начало симптомов или
сбор положительных на 5. typhimurium проб фекалий имели место в период с 9 сентября
но 10 октября 1984 г., и больной жил или посетил Даллас в этот период. Случай у че-
1 эвека, который питался в ресторане в Далласе в течение 7 сут. до начала >аболевания
ли который работал в ресторане в Далласе, рассматривали как случай, ассоцииро-
ванный с рестораном. Единичное экспонирование в ресторане означает, что только
одно экспонирование в ресторане имело место в течение 7 сут. до начала симптомов.
Отучай считался вторичным, если он имел место у лица, который не питался и не
работал в ресторане в Дал пасе в течение 7 сут. до начала симптомов, но имел контакт
с первичным больным в течение этого интервала времени.
Тридцать восемь ресторанов в Далласе были разделены на три группы на основа-
нии количества посетителей с единичным экспонированием и с культурально подтвер-
жденным диагнозом. Рестораны первой группы были определенно связаны с зараже-
нием, и в них были зарегистрированы, по меньшей мере, 3 посетителя с единичным
экспонированием и 3 — с культурально подтвержденным диагнозом. Рестораны груп-
пы 2 были, возможно, связаны с заражением, и в нчх был отмечен, по меньшей мере,
один посетитель с единичным экспонированием, но менее 3 посетителей с культураль-
но подтвержденным диагнозом. Рестораны группы 3 не были связаны с заражением,
и в них не было зарегистрировано посетителей с единичным экспонированием.
Изучение вспышки. Случаи инфицирования были идентифицированы путем пас-
сивного контроля. Пресс-релизы между больными и медицинскими работниками
g зособствовали сбору материалов о заболеваниях. Эпидемиологи беседовали с потен-
циальными больными относительно симптомов и факторов риска и получили исчер-
пывающие данные о еде в ресторане в течение 3 дней до начала симптомов. Посети-
телей с единичным экспонированием опрашивали с целью выявления всех других лиц,
с которыми они ели в ресторане. Данные были получены и от лиц, которые не были
больны и не сообщали о другом посещении ресторана, и были выбраны в качестве
контроля для связанных с едой исследований по типу случай-контроль. Потенциаль-
но экспонированные группы людей, такие как участники банкетов и постоянные
чэсетители, берущие обеды на дом, были выявлены по документации ресторанов, и
были предприняты попытки провести опрос этих людей.
ПЬ Часть! Краткая история применения биологических средств..
Работников ресторанов первой группы опрашивали дважды. Во время вспышки
врачи опрашивали работников, когда впервые возникло подозрение о ее связи с рес-
торанами. В октябре 1984 г. сразу же после вспышки всех работников попросили от-
ветить на вопросы анкеты, заполняемой опрашиваемым лицом. Календарные планы
работы были получены на основании анализа карточек табельного учета, бесед с ру-
ководителями ресторанов и рассмотрения заявлений о выплате страхового возмеше-
ния по нетрудоспособности.
Лабораторные методы. Образцы фекалий были отправлены в местные и регионалг -
ные лаборатории дтя культивирования на наличие кишечных патогенов. От работни-
ков ресторанов первой группы требовали принести кал на анализ для культивирова-
ния или требовали их отстранения от работы. У больных работников с одной отрица-
тельной пробой стула до возвращения на работу требовали сдать вторую пробу стула
для подтверждения результатов анализа. Санитарно-гигиенические лаборатории
штатов Орегон и Вашингтон серотипировали изоляты сальмонелл и определяли ан-
тибиотикочувствительность проб изолятов. Репрезентативную пробу чзолятов дан
ной вспышки, основанную на эпидемиологических критериях, отправили в CDC для
дополнительного биохимического исследования и анализа плазмидного профиля
путем переваривания рестрикционной эндонуклеазой Hindlll. Санитарно-гигиени-
ческая лаборатория штата Орегон отправила в С DC также изоляты 5. typhitnurium.
полученные при других вспышках и в спорадически появляющихся в 1984 г. и в тече-
ние всего времени случаях, не связанных с данной вспышкой, для сравнения со штам-
мом настоящей вспышки путем плазмидного анализа. Департамент сельского хозяй-
ства и санитарно-гигиеническая лаборатория штата Орегон культивировали образцы
взятые из подозреваемых пищевых продуктов.
Штамм, выделенный при Далласской вспышке, сравнивали с изолятами, получен-
ными от людей, которые анализировали в ходе двух национальных обследований на
сальмонеллы в 1979—1980 гг. и в 1984-1985 гг. Для идентификации возможного резер-
вуара среди животных CDC охарактеризовал все имеющиеся ветеринарные изоляты
5. typhimurium, идентифицированные в период с 1 октября 1984 п по 30 сентября 1985 г.
лабораторией Национальной ветеринарной службы Министерства сельского хозяй-
ства США в Эймсе, штат Айова.
Исследование окружающей среды. Санитарные врачи местного отдела здравоохра-
нения и представители FDA выяви ш оптовых торговцев и подлинных поставщиков
пищевых продуктов в рестораны, в которых происходило инфицирование людей. Все
эти рестораны были инспектированы санитарными врачами. Были просмотрены за-
писи работы системы городского водоснабжения за сентябрь 1984 г. Во время вспышки
сальмонеллеза для анализа были взяты пробы водопроводной воды в ресторанах. Был
проведен анализ температур охлаждения в закусочных. Эти исследования позволили
исключить из числа возможных причин вспышки проникновения возбу (ителя инфек-
ции из внешней среды или с оптовыми поставками продуктов. Стало ясно, что ин-
фицирование происходило непосредственно в ресторанах.
Эпидемиологическое определение места совершения теракта. Эпидемиологи рабо-
тали независимо от агентов ФБР, имевших информацию от осведомителя Однако
благодаря кропотливости проведенного исследования, полученные данные совпали
с теми, которые получило ФБР оперативным путем (см. «Уголовное расследование»).
Эпидемиологи выявили F51 больного, которые соответствовали определению слу-
чая инфицирования (см. выше); 441 (59 %) из них составляли женщины и 310 (41 %) —
мужчины. Возрастной диапазон больных был широким: от новорожденных до 87 лет
.12. Биодиверсии и биотеррор времен профанации биологического оружия
277
в среднем 33 года). По меньшей мере, 45 человек (6 %) были госпитализированы;
этальных исходов не зарегистрировано. Эпидемическая кривая была двуфазной. Пик
вервой волны заболевания (9—18 сентября) приходился на 15 сентября, а пик второй
юлны (19 сентября — 10 октября) наблюдался 24 сентября. Из 674 больных (90 %) с
(вестной датой появления симптомов 88 (13 %) заболели во время первой волны и
5®6 (87 %) — во время второй волны. Было отмечено 692 случая, ассоциированных с
zторанами (92 %), 11 вторичных случаев (1 %) и 48 случаев (6 %) с неполными дан-
тыми об экспонировании в ресторанах. Среди лиц со случаями, ассоциированными
. ресторанами, 101 человек (15 %) были работниками и 591 (85 %) — посетителями,
“ятьсот девятнадцать посетителей имели единичное экспонирование в ресторане и
"2 шсетителя — несколько (рис. 1.53).
Сентябрь Октябрь
Рис. 1.53. Развитие вспышки гастроэнтерита в Далласе. Вспышка имела две волны
заболеваемости: с 9 по 18 сентября и с 19 сентября по 10 октября. Разрыва между вол-
нами не было. Первая волна была связана с «испытаниями» агента террористами,
вторая предназначалась для срыва выборов. Как правило, террористы применяли
агент в каждом ресторане по нескольку раз. По Т. J. Torok et al. (1997)
Десять из 38 ресторанов в Далласе были определенно подвергнуты заражению
группа 1) (табл. 1.10).
В двух ресторанах группы 1 были зарегистрированы постоянные посетители с
el [иничным экспонированием с культурально подтвержденным диагнозом в период
первой волны (рестораны А и В), но все 10 ресторанов были связаны с заражением в
период второй волны. Эти 10 ресторанов ассоциировались с 494 посетителями с еди-
ничным экспонированием (95 %), с 69 посетителями со множественными экспони-
рованиями в ресторане (96 %) и с 91 работником (90 %) (рис. 1.54).
Двенадцать ресторанов были, возможно, подвергнуты заражению (группа 2) и
несли ответственность за 25 посетителей с единичным экспонированием (5 %). Три
посетителя с несколькими экспонированиями в ресторане (4 %) указывали на то, что
они питались в ресторане группы 2, но не в ресторане группы 1. Шестнадцать ресто-
ранов не были подвергнуты заражению (группа 3). Группирование ресторанов по рай-
онам не проводили, но даты экспонирования для случаев с культурально подтверж-
денным диагнозом разбили на группы (см. рис. 1.54). Связь ресторанов со вспышкой
Часть 1. Краткая история применения биологических средств
Таблица 1.10
Ассоциированные со вспышкой в Далласе стучаи сальмонеллезного
гастроэнтерита, распределенные по группам и месту Jдепонирования
Группа и место экспонирования Случаи с культурально подтвержденным диагнозом (% от общего количества) Количество клинических случаев (% от общего количества) Общее количество случаев (% от общего количества)
Работники
Рестораны группы 1 (п = 10) 74 (9,9) 17 (2,3) 91 (12,1)
Рестораны группы 2 (п = 12) 4 (0,5) 5 (0,7) 9 (1,2)
Рестораны группы 3 (п = 16) 1 (0,1) 0 (0,0) 1 (0,1)
Постоянные посетители
Рестораны группы 1 (п = 10) 227 (30,2) 267 (35,6) 494 (65,8)
Рестораны группы 2 (п = 12) 6 (0,8) 19 (2,5) 25 (3,3)
Рестораны группы 3 (п = 16) 0 (0,0) 0 (0,0) 0 (0,0)
М ножественное экспонирование в ресторанах* 32 (4,3) 40 (5,3) 72 (9,6)
Вторичные случаи 4 (0,5) 7 (0,9) Н (1,5)
Случаи с неполными данными 40 (5,3) 8(1,1) 48 (6,4)
Всего 388 (51,7) 363 (48,3) 751 (100,0)
* Всего 60 (95,8 %) из 72 посетителей с экспонированиями в нескольких ресторанах со-
общили о посещении одного или более ресторанов группы 1,
Случаи
с единичным
Ресторан экспонированием
Дни, указанные посетителями
А В 87 63 о • • • • • • • о о о • о • • • • •
о • • •
С 26 • • • • •
D 47 о о • о •
Е 7 • о о
F 45 о • • • • •
С 149 о о о • • • •
Н 26 о оее о о
I 22 о • • о •
J 22 о о о • • •
• о
1617 18 19 20 2122 23 24 25 26 27 28 29 30
91011 12 13 14 15
Сентябрь
о
о
о
о
3 4 5 6
1
2
Октябрь
Рис. 1.54. Даты экспонирования в ресторанах первой группы для 494 посетите-
лей с единичными экспонированиями в ресторане. Черные кружки означают
дни, когда у одного или более посетителей отмечали культурально подтвержден-
ную инфекцию, обусловленную 5. typhimunum. По Т. J. Torok er al. (1997)
ассоциировалась с наличием закусочной, где подавали овощные блюда. Восемь (80 %)
из 10 ресторанов группы 1, по сравнению с 3 (11 %) из 28 других ресторанов в Далласе,
имели закусочные с овощными блюдами (RR, 7,5 %; 95 % CI, 2,4 -7; Р < 0,001). Таким
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
279
образом, было установлено, что наибольшее количество заболевших сальмонеллезом
связано с ресторанами, в которых имелись закусочные с овощными блюдами.
Аналогичным образом получены доказательства и того, что два ресторана без за-
кусочных были ответственны за самое небольшое число заболевших. Эти наблюдения
подтвердили предположение о том, что инфицирование людей осуществлено имен-
но в закусочных.
В дв1,х ресторанах из числа тех, с которыми связывают наибольшее число инфи-
цированных, были банкетные залы. Но ни одного случая среди посетителей 20 бан-
кетов в период вспышки выявлено не было. Продукты для банкетных закусочных с
овощными блюдами готовились в той же кухне, что и продукты для общих закусоч-
ных, но банкетные закусочные имели более ограниченный набор продуктов и рабо
тали только полтора ч/день Источники продуктов, поставляемых в общие закусоч-
ные с овощными блюдами, были те же, что и для банкетов. Эти наблюдения
исключили возможность инокулирования сальмонелл в пищу непосредственно в про-
цессе ее приготовления.
Затем эпидемиологами были получены доказательства инфицирования пищи
посетителями ресторанов. Во время второй волны вспышки из сливочной добавки к
салату с голубым сыром одного из ресторанов была выделена S. typhimurium, но ее не
обнаружили в сухой смеси, использованной для приготовления приправы к салату.
S typhimurium также не была выделена из культур салата латука, который был полу-
чен из той же партии, использованной во время вспышки, но ее обнаружили в салате,
изъятом из салат-бара Стал понятен и способ, которым террористы инфицировали
продукты. Работники обычно выставляли ь закусочной полные контейнеры с каким-
нибудь свежеприготовленным пищевым продуктом и не контролирова ш их в даль-
нейшем. Этим пользовались террористы.
Исследование выделенных штаммов сальмонелл. 5. typhimurium была выделена из
проб фекалий 388 больных (52 %). Выделенный во время вспышки штамм не фер-
ментировал дульцит — необычное биохимическое свойство, обнаруженное только
примерно у 2 % небрюшнотифозных сальмонелл. Этот штамм был чувствителен к
ампицил зину, цефалотин натрию, хлорамфениколу, гентамицинсульфату, канамицин-
сульфату, налидиксовой кислоте, сульфидоксазолу и триметоприм сульфаметоксазо-
лу. Была отмечена умеренная чувствительность к ' етрациклину и сульфату стрепто-
мицина. Были определены плазмидные профили для 52 изолятов, связанных со
вспышкой, включая изолят, по крайней мере, от одного заболевшего работника и
одного заболевшего посетителя, полученный в каждом ресторане первой группы. Все
полученные во время вспышки изоляты имели одинаковый плазмидный профиль,
при этом масса одной плазмиды составляла приблизительно 60 мДа.
Ни один из шести изолятов S. typhimurium. собранных в штате Орегон во время
спорадически возникаюших случаев заболевания в период с июля по декабрь 1984 г.,
не был похож на штамм, выделенный во время вспышки в Далласе. Два изолята
5. typhimurium, полученные во время двух других вспышек, которые произошли в штате
Орегон в то время, были похожи на далласский штамм по метаболизму дульцита, ан-
тибиотикограмме, плазмидному профилю и длине рестриктазных фрагментов плаз-
ми дной ДНК.
Одна из двух вышеупомянутых вспышек болезни произошла в августе 1984 г. и
пора шла 26 человек, которые заболели после посещения больничного кафетерия в
центре Уитламетт-Валли, штат Орегон. Болезнь ассоциировалась с употреблением
приправы к мясным блюдам в закусочной с овощными блюдами. Другая вспышка,
280
Часть 1. Краткая история применения биологических средств...
обусловленная 5. typhimurium, произошла после банкета в гостинице Портленда, штат
Орегон, в декабре 1984 г. и поразила, по крайней мере, 36 человек; болезнь ассоции-
ровалась с употреблением в пищу не прожаренного ростбифа. Ранее, в ноябре 1984 г,
вспышка, вызванная S. typhimurium, произошла после банкета в той же портлендской
гостинице и поразила, по крайней мере, 73 человека; болезнь также была связана с
употреблением в пищу непрожаренного ростбифа. Штамм, полученный во время этой
вспышки, имел одну плазмиду, сходную с плазмидой далласского штамма, но даль-
нейший сравнительный анализ данных лабораторного исследования проведен не был.
и связи между этими вспышками и вспышкой в Далласе не установлено.
Национальный отчет об исследовании случаев сальмонеллеза в 1979—1980 гг. вклю-
чал описание 233 штаммов S. typhimurium (за исключением варианта Копенгаген); ни
один из них не имел антибиотикограмм, аналогичных далласскому штамму. Нацио-
нальный отчет об исследовании случаев сальмонеллеза за период 1984—1985 гг. вклю-
чал описание 175 штаммов S. typhimurium (за исключением варианта Копенгаген);
35 штаммов имели антибиотикограммы, аналогичные далласскому штамму, и 6 из них
не ферментировали дульцит. Один из 6 штаммов был эпидемически связан со вспыш-
кой в Далласе, но остальные 5 изолятов не имели достоверной связи со вспышкой
в Далласе.
Из 34 изолятов £ typhimurium от животных из лаборатории Национальной вете-
ринарной службы, собранных в период с июня 1984 г. по ноябрь 1984 г., один изолят
S. typhimurium, не ферментировавший дульцит, был получен от индейки в июне 1984 г.
и соответствовал далласскому штамму по результатам рестрикционного анализа плаз-
мид, но он не имел эпидемической связи с далласской вспышкой.
Источник штамма £ typhimurium, вызвавшего вспышку, был, наконец, идентифи-
цирован в октябре 1985 г. Во время полицейского расследования сотрудник Орегон-
ской санитарно-гигиенической лаборатории обнаружил в клинической лаборатории,
находившейся в ведении религиозной общины, открытую пробирку с чистой культу-
рой £ typhimurium. Согласно имеющейся в клинике документации, эта пробирка была
приобретена до вспышки, и лабораторные исследования, проведенные в последую-
щие месяцы, продемонстрировали, что изолят соответствовал штамму, вызвавшему
вспышку. Этот штамм £ typhimurium по антибиотикограмме, биохимическим марке-
рам, плазмидным профилям и рестриктам плазмидной ДН К был неотличим от штам-
ма, выделенного во время вспышки.
Таким образом, характерная чувствительность к антибиотикам, результаты био-
химического анализа и плазмидный профиль убедительно свидетельствовали о том,
что причиной вспышки был один единственный штамм, обнаруженный в лаборато-
рии террористов, и исключали возможность одновременного возникновения незави-
симых вспышек.
Уголовное расследование. Администрации ресторанов, в которых произошло ин-
фицирование людей, агентами ФБР были заданы вопросы по поводу необычных ин-
цидентов или недовольных работников. О подозрительных событиях сообщали в по-
лицию штата Орегон и шерифу округа Уаско. ФБР контролировало действие местной
полиции по расследованию. По завершении эпидемиологического исследования и
после распада общины учеников Раджниша ФБР, при техническом содействии сани-
тарно-гигиенической лаборатории штата Орегон, провело расследование в клиничес-
ких и лабораторных отделениях в Раджнишпураме. Следователи конфисковали откры-
тую пробирку, содержащую стандартный штамм £ typhimurium (АТСС 14028, Роквилл,
штат Мэриленд) и хранившуюся в клинической лаборатории в Раджнишпураме.
7.12. Биодиверсии и биотеррор времен профанации биологического оружия
281
Согласно имеющейся в клинике документации, лаборатория получила эту пробирку
от коммерческого поставщика биологических продуктов еще до вспышки.
Свидетельские показания осведомителя предоставили дополнительную информа-
цию о мотивах преступного сговора и о ходе осуществления террористического акта.
Согласно свидетельским показаниям, планировалось провести несколько преступных
акций в определенных ресторанах. В некоторых ресторанах были также контамини-
рованы жидкие сливки для кофе, продукты в бакалейно-гастрономическом магази-
не, и планировалось контаминировать городской водопровод.
Источник инфекции для работников, которые заболели до того, как было доку-
ментально подтверждено экспонирование посетителей, остается неизвестным. Эти
шучаи болезни могли быть результатом первой неудавшейся попытки контаминации,
которую не удалось доказать. По словам осведомителя, террористы были разочаро-
заны, когда их первоначальные попытки не вызвали широкого распространения бо-
лезни, и в более поздних акциях они увеличили количество инокулята.
Не было установлено наличие обиженных работников, которые могли иметь же-
1ание отомстить своим хозяевам. Уголовное расследование подтвердило, что работ-
ники ресторанов не участвовали в попытках контаминации.
Наказание. В марте 1986 г. два члена общины были обвинены по обвинительному
акту во вступлении в преступный сговор с целью порчи пищевых продуктов путем их
заражения в нарушение федерального закона против фальсификации продуктов.
В апреле 1986 г. обвиняемые признали себя виновными и в июле 1986 г. они были
приговорены к 4,5 годам тюремного заключения. По отбытии наказания террористы
пехали в Европу.
1.12.2. Искусственная вспышка дизентерии Шмидца среди сотрудников
лаборатории медицинского центра в Техасе в 1996 г.
«Правильная эпидемиология» Shigella dysenteriae 2 типа. Мотив преступления. Появление
подозрений в искусственном происхождении вспышки дизентерии. Эпидемиологическое
расследование. Исследование выделенных штаммов возбудителя дизентерии Шмидца.
Для анализа хода эпидемиологического расследования искусственной вспышки
тизентерии Шмидца использованы материалы, подготовленные специалистами CDC
и BCDPC (Kovalic S. A. et al., 1997).
«Правильная эпидемиология» Shigella dysenteriae 2 типа. Возбудитель известен так-
же как бацилла Шмидца (Schmitz). Это относительно редкий микроорганизм, способ-
ный вызвать бациллярную дизентерию. О возбудителе дизентерии этого серотипа в
медицинской литературе информации очень мало. Впервые он описан Schmitz в 1917 г.
после зимней эпидемии диареи в Восточной Европе, в лагере для военнопленных.
Вспышки дизентерии, вызванные возбудителем данного серотипа, никогда не при-
обретали массового характера ни в США, ни в Европе. В США первая ставшая извест-
ной локальная вспышка дизентерии Шмидца произошла в 1938 г. в городе Wales сре-
ди пациентов психиатрической клиники.
Спорадические случаи и вспышки инфекции, вызванной 5. dysenteriae второго
типа, не характерны для развитых стран. В конце 1980-х гг. небольшая вспышка ин-
фекции имела место среди членов одной семьи в Сицилии, где S. dysenteriae типа 2 не
обнаруживали с 1953 г. Данные наблюдений CDC, выполненные за период с 1985 по
1995 г., показывали, что в Соединенных Штатах ежегодно регистрируются не более
282
Часть 1. Краткая история применения биологических средств...
20 индивидуальных случаев дизентерии, вызванной S. dysenteriae типа 2. Последняя
известная вспышка произошла в США в 1983 г. среди работников кафетерия Меди-
цинского центра в Мэриленде.
Болезнь, вызываемая S. dysenteriae типа 2, не имеет детального описания. Хотя она
может заканчиваться смертельными исходами, S. dysenteriae типа 2, по сравнению с
5. dysenteriae типа 1, вызывает значительно менее опасное заболевание. В отличие от
S. dysenteriae типа 1, которая ассоциируется с гемолитическим уремическим синдро-
мом и токсическим мегаколлоном, S. dysenteriae типа 2, не продуцирует шигатоксин.
Мотив преступления. Основным мотивом была личная неприязнь лаборантки
Дианы Томпсон к сотрудникам лаборатории, с которыми она вместе работала не-
сколько лет.
Появление подозрений в искусственном происхождении вспышки дизентерии. В пе-
риод между 29 октября по 1 ноября 1996 г. у 12 сотрудников лаборатории большого
медицинского центра в Техасе развилось тяжелое желудочно-кишечное заболевание,
которое они связали с употреблением выпечки, оставленной без присмотра в комна-
те отдыха персонала с ночи до утра следующего рабочего дня 29 октября. В фекалиях
больных была обнаружена 5. г/ухеШеттаетипа 2, обычно не встречающаяся в США при
вспышках дизентерии. Обследование хранилища-холодильника в лаборатории позво-
лило предположить, что нарушена сохранность культуры S. dysenteriae типа 2.
Эпидемиологическое расследование. К заболевшим дизентерией Шмидпа были
отнесены сотрудники лаборатории с симптоматикой диареи, появившейся 29 октяб-
ря 1996 г. или после этой даты, у которых из стула была выделена S. dysenteriae или с
оральной температурой более 37,8 °C. К возможно заболевшим отнесены сотрудники
лаборатории с диарейным синдромом, с оральной температурой 38 °C или меньше; или
с отрицательными результатами исследования стула. Был опрошен персонал лабора-
тории, работавший утром 29 октября или вечером прошлой смены, оценены демог-
рафические данные, прослежено происхождение выпечки, определены признаки и
симптомы болезни.
Образцы стула заболевших сотрудников культивировали на агаровой среде, содер-
жащей эозинметилен голубой и ксилозолизиндезоксихолат. Выросшие культуры на-
правляли вместе с образцами пищи в Bureau of Laboratories of the Texas Department of
Health для дальнейшего анализа и серотипирования. Штоковая культура медицин-
ского центра также была направлена в Texas Department of Health. Пища исследова-
лась по общепринятой в США процедуре.
В дополнение был выполнен пульс-электрофорез переваренной рестриктазами
Xbal и Notl хромосомной ДНК, выделенной из изолятов 5. dysenteriae типа 2, полу-
ченных из стула, продуктов питания, а также штоковой культуры, имевшейся в гос-
питале. Все эпидемиологические данные были собраны в стандартизованные формы,
затем их ввели в компьютер и анализировали, используя программу Epi Info Version 6.
При исследовании обстоятельств инфицирования людей было установлено, что
комната отдыха, в которой была оставлена на ночь купленная сдоба, отделена от лабо-
ратории, и в нее нельзя попасть, не зная секретного кода. Две коробки сдобы содер-
жали черничные оладьи и набор пончиков.
У 12 сотрудников, которые ели пончики и оладьи, появилась диарея с лихорадкой,
головная боль или рвота. Из них 11 признаны как больные, а один рассматривался как
сомнительный случай. Средний возраст заболевших сотрудников составил 41 год (воз-
растной интервал 33—52 года), 9 из них (75 %) — женщины. Все заболевшие сообщили,
что ели мучные кондитерские изделия между 7.15 ч утра и 1.30 ч пополудни 29 октяб-
7.72 Биодиверсии и оиотеррир времен профанации биологического оружия 283
ря. Один сотрудник не ел выпечку на работе, но взял ее домой и съел совместно с семь-
ей около 19 ч. Диарейный синдром у сотрудников лаборатории проявился за период с
21 ч до 4 ч утра 1 ноября (рис. 1.55).
Рис. 1.55. Случаи шигеллеза в 1996 г среди работников Техасского медицинского
центра, начавшиеся с диарейного синдрома. Динамика появления больных характер-
на для эпидемиологии «точечного источника», предполагающей контакт с возбуди-
телем в одно и то же время. Кружками обозначены случаи болезни. Подтверженные
бактериологически окрашены в черный цвет. Квадратом показан сомнительный
случай. Указанны даты начала болезни. По S. A. Kuvalic et al. (1997)
Средний инкубационный период составил 25 ч (интервал: 11-66 час). Ему пред-
шествовали т ошнота, ощущение дискомфорта и переполнения в эпигастральной об-
ласти. Пять сотрудников, обратившихся в отделение «Скорой помощи», вскоре были
отпущены домой, четверо других были госпитализированы (средний срок госпитали
зации составил 4 суток; интервал 2—10 суток). Восьми заболевшим сотрудникам внут-
ривенно вводились жидкости. Культуры из стула были получены у 11 больных; в 8
культурах была обнаружена S dysenteriae типа 2, 3 культуры были негативными. По-
рученные изоляты 5. dysenteriae оказались чувствительными ко всем антибиотикам,
использованным в тестах на чувствительность. Одиннадцать больных лечили ципроф-
локсацином, один принимал гомеопатические средства.
Смертельных исходов среди заболевших сотрудников не было. Однако эффектив-
ность атаки составила 100 %. Все 12 человек, которые ели оладьи, заболели. Напро-
тив, 33 человека, не употреблявшие в пищу эти оладьи, а только пончики, остались
здоровыми. Питание в кафетерии госпиталя и посещение общественных собраний в
течение недели с 25 по 31 октября также не были связаны с повышенным риском раз-
вития болезни.
Исследователи не зафиксировали случаев вторичной передачи болезни, но члены
семьи одного сотрудника все же заболели после того, как съели оладьи, которые он
принес им из комнаты отдыха персонала. Этот сотрудник сам съел только несколько
кусочков и заболел. Из его стула и стула членов его семьи получена культура Л1, dysen-
teriae типа 2.
При проверке лабораторных запасов различных референс-культур микроорганиз-
мов, включающих 5. dysente.iae типа 2, установлено, что все они хранились в низко-
температурном хранилище Центра и находились в Microbank-бутылочках. Каждая
бу гылочка имела 25 пор со специальными бусинками, которые были импрегнированы
284
Часть 1. Краткая история применения биологических средств...
микроорганизмами. Культуры 5. dysenteriae гипа 2 включались в банк на протяжении
1980-х гг. На момент расследования вспышки было обнаружено только 19 бусинок,
хотя они никогда не использовались в работе.
Инфицирующая доза не была установлена, однако исследователи предположи-
ли, что для того, чтобы вызвать шигеллезную инфекцию у людей, требуется от 10 до
100 микроорганизмов.
Штоковые культуры S’, dysenteriae типа 2 хранились в Медицинском центре много
лет, и их точное происхождение не известно. Однако они не были серийно получен-
ными культурами. Культуры могли быть выделены от больных и ранее использовать-
ся для развития навыков работы с микробными культурами у медицинского персо-
нала. Ни одного случая шигеллеза, вызванного S. dysenteriae типа 2, за последние пять
лет в лаборатории госпиталя зарегистрировано не было. Не проводились и исследо-
вания этого микроорганизма.
Следовательно, нельзя объяснить возникновение этой вспышки грубой лабора-
торной ошибкой. Контаминация оладий во время покупки также исключена. Не опи-
саны вспышки болезни, вызванные S. dysenteriae типа 2, ни в Техасе, ни в любом дру-
гом штате США.
Исследование выделенных штаммов возбудителя дизентерии Шмидца. S. dysenteriae
типа 2 выделена фактически в чистой культуре из бульона обогащенной культуры,
полученной из образцов оладий. Эта же шигелла была выделена из стула 8 больных.
Обе культуры 5. dysenteriae типа 2 по культурально-морфологическим и серологичес-
ким свойствам соответствовали референс-культуре Медицинского центра.
С помощью пульс-электрофореза обнаружено, что штоковые культуры по рестрик-
там ДНК неотличимы от культур, полученных из стула больных людей и из оладий,
но отличались от двух изолятов б1, dysenteriae того же типа, выделенных в других окру-
гах Техаса в 1995 и 1996 гг. и не причастных к данной вспышке. От тех штаммов, кото-
рые вызвали вспышку болезни среди персонала Центра, они отличались более чем
по пяти полосам (рис. 1.56).
Рис. 1.56. Результаты пульс-электрофореза рестрикционных переваров ДНК
из изолятов S. dysenteriae. Xbal-рестрикты ДНК-изолятов 5. dysenteriae типа 3:
дорожки 1—5 (панель А); ДНК-изолятов 5. dysenteriae типа 2: i золяты от за-
болевших сотрудников, дорожки 6, 8—15 (панели А, Б); ДНК госпитальной
штоковой культуры, дорожка 7 (панель А); ДНК-изолятов из сдобной булки,
дорожки 17,18 (панель Б): и ДНК-изолята, не относящегося кданной вспыш-
ке, дорожка 16 (панель Б). Notl-рестрикты ДНК из изолятов 5. dysenteriae ти-
па 3: дорожки 27—31 (панель В); ДНК5. dysenteriaeтипа 2: госпитальная што-
ковая культура, дорожки 19, 23, 32, 34, 36 (панели Б, В); ДНК-изолятов из
сдобной булки, дорожки 25, 26 (панель Б), и ДНК-изолята, не относящегося
кданной вспышке, дорожка 24 (панель Б). По S. A. Kovalic et al. (1997)
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
285
Результаты пульс-электрофореза рестрикционных переваров ДНК из изолятов
S’, dysenteriae подтвердили «невозможную эпидемиологию» вспышки шигеллеза сре-
ди работников Техасского медицинского центра — источником S. dysenteriae типа 2,
послужила штоковая культура, хранившаяся в лаборатории этого же Центра Таким
образом, характерными эпидемическими особенностями вспышки шигеллеза в Техас-
ском медицинском центре, позволившими американским эпидемиологам сделать
вывод об ее искусственном происхождении, были: сжатая эпидемическая кривая с пи-
ком появления заболевших в течение 25 ч (эпидемиология «точечного источника»);
«невозможная эпидемиология» — источником микроорганизмов S. dysenteriae типа 2,
вызвавших данную вспышку, послужила штоковая культура Медицинского центра в
Техасе; локализованность эпидемического очага — все лица, у которых обнаружен ши-
геллез, работали в одной лаборатории, сообщили одинаковые сведения об обстоятель-
ствах экспозиции к возбудителю болезни.
Эпидемиологами не были установлены мотив преступления и метод контамина-
ции, однако ими была составлена характеристика человека, который мог осуществить
такой террористический акт. По их мнению, им мог быть кто-то, кто имел доступ к
хранилищу бактериальных культур, обладал навыками лабораторной работы, позво-
лившими ему получить культуру с бусинок и инокулировать ее в оладьи. Кроме того,
он еще имел доступ в закрывающуюся на кодовый замок комнату отдыха персонала.
Благодаря этой характеристике, сотрудниками ФБР был установлен непосредствен-
ный исполнитель этого преступления — сотрудница лаборатории Диана Томпсон.
Дело Томпсон не единственное в своем роде. В 1966 г. в Японии в сходном пре-
ступлении был изобличен некий доктор Сузуки, врач-бактериолог. Он в течение по-
чти года давал фрукты и кексы, зараженные возбудителями дизентерии и брюшного
тифа, пациентам и сотрудникам клиники, в которой работал, поскольку чувствовал
себя дискриминируемым ими. Всего от его действий пострадало 120 человек, четверо
из них погибли (Atlas R. М., 1999). Такой способ совершения преступлений более
распространен, чем принято думать. Но они остаются безнаказанными из-за отсут-
ствия соответствующей настороженности и понимания того, каким образом надо
действовать, если такие подозрения появились.
1.12.3. Инцидент с возбудителем сибирской язвы
в районе Kameido Токио в 1993 г.
Для анализа обстоятельств инцидента использованы материалы, подготовленные
сотрудниками National Institute of Infectious Diseases (Япония) и Northern Arizona
University (США) H. Takahashi et al. (2004).
Мотив организаторов инцидента. Члены секты «Аум-Синрикё» («Aum Shinrikyo»)
полагали, что вызвав масштабную вспышку ингаляционной сибирской язвы в Токио,
они спровоцируют мировую войну, которая приведет их лидера Секо Асахару к управ-
лению всем миром.
Обстоятельства инцидента. Утром 29 июня 1993 г. пять жителей Камедо, восточ-
ного района Токио, обратились в одно из местных отделений по охране окружающей
среды с жалобой на неприятный запах на улицах района и в их квартирах. Сотрудни-
ки отделения быстро установили, что неприятный запах распространяется с восьмо-
го этажа главного офисного здания религиозной секты «Аум-Синрикё». Эта секта уже
была заподозрена властями в похищении силой лиц, пытающихся выйти из состава
286
Часть 1. Краткая история применения биологических средств..
ее членов, но отсутствие строгих доказательств криминальной активности секты и
препятствия со стороны законодательства не позволяли ограничить ее деятельность.
Сотрудники местного отделения по охране среды не решились предпринять какие-
то действия по осмотру здания. Но уже 30 июня ими была зарегистрирована 41 жало-
ба со стороны местных жителей на тошноту, рвоту и утрату аппетита, вызванных не-
прекращающимся зловонием. Недовольные жители района потребовали осмотра
щания секты «Аум -Синрике», но члены секты отказали им в эюм. Тогда сотрудники
местного отделения по охране среды стали проводить заборы воздуха вокруг здания
чтобы выяснить, чем же занимаются внутри здания. Но ничего вредного, кроме не-
приятного запаха, тогда обнаружено не было.
Утром первого июля местные жители сообщили в местное отделение по охране
среды об облаке, с шумом вырывающемся из одной из двух башен охлаждения, рас-
положенных на крыше офисного здания секты. В этот день жители 118 квартир, рас-
положенных южнее, пожаловались на отвратительный запах (рис. 1.57).
Рис. 1.57. Обстоятельства инцидента в Камедо. А. Облако, выброшенное из башни охла-
ждения офисного здания секты «Аум-Синрике» (фотография сделана 01.ОУ1993 г. со-
трудниками Department of Environment, Koto-ward). Б. Зона риска инфицирования
людей возбудителем сибирской язвы, очерченная по жалобам жителей района на отвра-
тительный запах. Ветер в пределах 2—4 м/с. Менялся в течение утра с северного на се-
веро-восточный. Ранним утром прошел небольшой дождь. По Н. Takahashi et al. (2004)
Отдельные жители заявили, что они и раньше видели «желатиноподобный мас-
лянистый серый туман» над этой башней охлаждения, а также вытекающую из нее
жидкость. Сотрудники отделения охраны окружающей среды Камедо собрали пробы
воздуха в жидкую питательную среду для последующего тестирования, а лидер сек-
ты, Секо Асахара, заверил их в том, что прекратит использовать сооружения крыши
офисного здания для своих экспериментов, а также ее помоет и очистит. Когда 16 июля
инспекторы поднялись на крышу, то там они не нашли ничего подозрительного.
«Слепота» токийской полиции в отношении сектантов «Аум-Синрикё» не может
не удив 1ять. Этот инцидент, будучи зафиксированным большим количеством свиде-
телей, заснятый фотокамерами и даже расследуемый сотрудниками отделения охра-
ны окружающей среды Камедо, оставался незамеченным токийской полицией вплоть
до распыления сектантами зарина в марте 1995 г. в тоннелях токийского ме тро. Толь-
ко тогда полиция стала искать свидетельства причастности секты к актам биотерро
ра. Сам Асахара вывел полицию из затруднительного положения, заявив 26.05.1996 г.
о том, что члены его секты изучали, какой возможный ущерб городу может нанести
распыление суспензии спор возбудителя сибирской язвы.
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
287
Эпидемиологическое расследование. Проводилось ретроспективно. Во время само-
го инцидента связь с ним каких-либо болезней в Токио не изучалась, а бактериологи-
ческое исследование, если и проводилось, то без нацеленности на обнаружение си-
биреязвенного микроба. Не было обнаружено и случаев ингаляционной сибирской
язвы. В ноябре 1999 г. после длительных переговоров руководители местной службы
контроля окружающей среды согласились передать в Northern Arizona University
(США) оставшиеся жидкие пробы, собранные 6 лет назад. Тестовые пробирки содер-
жали по 2,6 мл мутной темной жидкости. Проведенные микроскопические исследо-
вания показали наличие в ней бактериальных спор, вегетативных бактерий и боль-
шого количества разрушенных клеток. После высева этой жидкости на чашки Петри
с плотной питательной средой, содержащей эритроциты барана, было обнаружено,
что она содержит смешанную бактериальную флору, но до 10 % составляют микроор-
ганизмы, образующие колонии, сходные с сибиреязвенными*. Из этих колоний были
получены чистые культуры, которые затем были исследованы посредством MLVA.
MLVA-ПЦР проводилась с помощью праймеров, специфичных к 8 ампликон-сайтам
(amplicon sites), уникальным для В. anthracis, два из которых являются генами сиби-
реязвенного токсина и капсулы. Анализ чистых культур, полученных из 48 колоний,
отобранных на плотной питательной среде, показал, что они относятся к возбудите-
лю сибирской язвы одного генотипа. Все культуры содержали плазмиду рХ01, несу-
щую ген сибиреязвенного токсина, но не содержали плазмиду рХ02, кодирующую
капсулу. Микроорганизм был охарактеризован как вакцинный штамм В. anthracis
Sterne 34F2, обычно используемый в Японии в качестве коммерческой вакцины. Одна-
ко действительно ли сектанты принимали бескапсульный вакцинный штамм В. anthra-
cis за вирулентный, или он появился только в образцах, переданных японцами своим
американским «друзьям» через 6 лет (!) после инцидента, остается догадываться.
По мнению Н. Takahashi et al. (2004), в основе неудачи этого теракта лежат следую-
щие факторы: I) использование ослабленного по вирулентности штамма В. anthracis;
2) низкая исходная концентрация спор в аэрозоле; 3) неэффективный способ диспер-
гирования; 4) неблагоприятные метеоусловия; 5) инактивация спор возбудителя си-
бирской язвы под воздействием солнечного света.
1.12.4. Искусственная вспышка сибирской язвы в США в 2001 г.
Предшествующие вспышке события в области контроля над БО. Кошмар из сна. Общий ход
событий. «Белый порошок». Расследование биологического террористического акта. Пове-
дение людей. Эпидемиологическое расследование вспышки сибирской язвы. Два случая
ингаляционной сибирской язвы во Флориде. Сортировочный центр в Вашингтоне. Сибир-
ская язва в Капитолии. Загадочный случай в Нью-Йорке. Загадочный случай в Оксфорде.
Исследование окружающей среды. Идентификация. Постэкспозиционная неспецифическая
профилактика. Брюс Эдвард Айвинс. Эпидемиологические особенности вспышки.
Это первый, ставший известным во множестве подробностей, умело осуществ-
ленный акт биотеррора, к тому же имеющий очень серьезную политическую подо-
плеку. Рассмотрим его с максимально возможной для данной книги обстоятельностью.
1 По моим наблюдениям, споры вакцинных штаммов В. anthracis, суспендированные в
физиологическом растворе, хранятся при температуре 2—6°С в течение нескольких месяцев,
не более. Споры В. anthracis, загрязненные посторонней микрофлорой, хранятся менее про-
должительное время.
288
Часть 1. Краткая история применения биологических средств...
Предшествующие вспышке события в области контроля над БО. После 6 лет пере-
говоров в Женеве специальная группа правительственных экспертов (в том числе и
из России) разработала план соглашения по контролю над БО, известный специа-
листам как Протокол. Он насчитывал 280 страниц. В нем устранен ряд недостатков
Конвенции 1972 г. Подписавшие Протокол стороны обязывались ставить друг друга
в известность о предприятиях по производству вакцин; о наиболее крупных объектах
биологической зашиты; об объектах, проводящих генно-инженерные исследования
и осуществляющих изучение аэрозолей биологических агентов, которые с наиболь-
шей вероятностью могут использоваться в качестве агентов БО. Был разработан ме-
ханизм принятия решения об инспекциях «подозрительных объектов» на территории
всех стран, подписавших Конвенцию 1972 г., кстати, гораздо более «мягкий», чем пре-
дусмотрен «Конвенцией о запрещении разработки, производства, накопления и при-
менения химического оружия и о его уничтожении» (1993). Участники международ-
ных переговоров в Женеве стремились завершить работу над Протоколом к ноябрю
2001 г. Для американских дипломатов и экспертов неприятной неожиданностью ста-
ла русская позиция на переговорах, предусматривавшая инспекцию биологических
объектов не только на территории России, что подразумевалось американскими экс-
пертами как «само собой разумеющееся», но и на территории самих США. Резкое не-
приятие американской стороны вызвало предложение русских экспертов договорить-
ся о «пороговых количествах» биологических рецептур, необходимых для проверки
средств зашиты от БО. Возможность обладания ими для «профилактических, защит-
ных или других мирных целей» оговорена в статье I Конвенции 1972 г., но надо было
исключить возможность двусмысленности в ее толковании. Запасы таких рецептур не
должны превышать порог, за которым возможно их боевое применение. Американс-
кая позиция по этому вопросу была пафосной и благородной — любое количество ре-
цептуры уже есть нарушение Конвенции 1972 г.
В июле 2001 г. американская сторона в одностороннем порядке отказалась от за-
вершения работы над согласованным проектом Протокола, обосновывая свое реше-
ние тем, что тогда США «окажутся открытыми для других государств, но ничего не
получат взамен», и «все эти открытые меры могут быть использованы правительства-
ми других стран для выведывания американских секретов». Так экспертам по конт-
ролю над БО недвусмысленно было указано на того, «кто в мире хозяин» и кто кого
должен контролировать1.
Кошмар из сна. К 2001 г. идея совершения биологического террористического акта
путем рассылки по почте возбудителя опасной инфекционной болезни была извест-
на специалистам не менее полувека. Еще в самом начале Второй мировой войны ка-
надский нобелевский лауреат Фредерик Бантинг (Frederick Grant Banting, 1891-1941),
один из открывателей роли инсулина в лечении сахорного диабета, обосновывал бри-
танскому правительству план исследований в области создания БО потенциальными
угрозами, среди которых не последнее место занимала возможность применения нем-
цами почтовых рассылок с порошком из сухих бактерий. Об инструктажах, во время
которых в качестве способа ведения бактериологической войны упоминались конвер-
ты с «бактериальной пылью», в 1952 г. сообщили Комиссии Нидхема американские
летчики, применявшие БО на территории Северной Кореи.
1 Мне рассказывал один из участников этих переговоров, что американский представитель
просто швырнул текст Протокола на стол и молча вышел из зала, где собрались эксперты из
других стран.
/2. Биодиверсии и биотеррор времен профанации биологического оружия
289
Идея рассылки почтовых конвертов с возбудителем сибирской язвы могла попасть
сознание организаторов теракта из художественной литературы и продуктов «мас-
совой культуры». В 1986 г. в США вышел популярный роман известного писателя
Стивена Кинга, в котором описывается эпидемия, вызванная утечкой из военной
Моратории штамма опасного микроорганизма. Болезнь распространялась так назы-
ваемыми «письмами счастья», которые надо было переписывать и рассылать другим.
В средине романа автор раскрывает название болезни — антракс (anthrax — принятое
США сленговое название сибирской язвы). На момент совершения биологическо-
го теракта в сентябре — октябре 2001 г. в США даже существовала трэш-металличес-
кая группа с весьма примечательным названием — «Антракс». В 1987 г. группа выпу-
гтша диск «Среди живых» («Among the Living»). В одноименной композиции с этого
диска есть такие слова: «Зараза, зараза, распространяется зараза с помощью капита-
-а Трипса (Captain Trips — еще одно сленговое название сибирской язвы, принятое в
США; trip — путешествие). Он поставил весь мир на колени. Сила, да, сила, он по-
зал свою силу!». Любопытно то, что в другой композиции этого диска упоминается
разбомбленный до основания мегаполис, в котором легко угадываются последствия
! 1 сентября 2001 г. Капитан Трипс упоминается и в книгах Стивена Кинга (Воронин А.,
Зимкин Н., 2001). В 1998 г., после выхода фильма «Эпидемия», в США разразилась
;рвая «почтовая эпидемия» сибирской язвы. Противники абортов принялись рас-
. тать в клиники, где их проводили, конверты с неким порошком и указанием, что
. нупри находятся «споры язвы». Если в 1997 г. известен лишь один такой случай, то в
998 г. — 150 (!) случаев. Кроме того, в том же году были отмечены еще 146 угроз при-
менения сибирской язвы со стороны различных фанатиков и психически больных
юдей. В начале 1990-х гг. таких угроз было не более десятка в год. Да еще Бен Ладен
а 1999 г. в интервью журналу «Newsweek» пообещал следующее: «Если нам удастся
заполучить ядерное, химическое или биологическое оружие, мы не будем считать это
• реступлением. Наша священная земля оккупирована израильскими и американски-
ми войсками. Мы имеем полное право защищать себя и бороться за освобождение
лашей земли всеми средствами» (ВолковА., 2001).
По заказу ЦРУ в том же 1999 г. Уильмом Патриком III (William С. Patrick III), ве-
дущим специалистом США по разработке рецептур для биологических боеприпасов,
был подготовлен закрытый доклад о последствиях для США почтовых рассылок, со-
держащих реальную биологическую рецептуру. В отчете предполагалось, что терро-
рист выберет оптимальное количество рецептуры, соответствующее 2,5 г (The New York
Times, 12 декабря 2001 г.).
Следовательно, в массовом сознании населения США какая-то катастрофа, свя-
занная с распространением сибирской язвы почтовыми отправлениями, присутство-
ала задолго до описываемых событий — как некий кошмар из сна. В ЦРУ такой тер-
• эристический акт был смоделирован. Кому-то осталось только его организовать, что
я было сделано.
Общий ход событий. Искусственная вспышка сибирской язвы началась 22 сентяб-
ря. когда 31-летняя служащая почтового офиса газеты «Нью-Йорк пост» обнаружила
у себя на левом предплечье небольшую эритематозную папулу, быстро развившуюся
везикулу. Но что произошло в действительности, стало ясно только 5 октября, когда
о Флориде (городок Ьока-Рэйтон) умер от ингаляционной сибирской язвы Роберт
Z. ивенс, 63-летний фоторедактор газеты «The Sun» («Солнце»), входящей в издатель-
ский холдинг «American Media Inc.» («АМ1»), Он был уже восьмым американцем, за-
болевшим сибирской язвой, вызванной террористическим актом, только тогда этого
290
Часть 1. Краткая история применения биологических средств...
никто не знал. Последующее расследование показало, что Стивенс вскрыл письмо,
пришедшее в редакцию по почте, но ему не предназначавшееся. Письмо было напраЕ
лено в редакцию газеты «Нейшенел Инкуайер» («Nation Enquirer» — дословно «На-
родный опрос») по ее старому адресу, но йогом было переслано в офис издательского
холдинга. Когда Стивенс вскрыл конверт, ему на руки и на клавиатуру компьютера вы-
сыпался какой-то «порошок». Как потом было установлено, этим «порошком» оказа-
лась рецептура возбудителя сибирской язвы, предназначенная для снаряжения амери-
канских биологических боеприпасов. В течение сентября и окгября было обнаруже-
но еще четыре конверта с таким «порошком», направленных в СМИ и известным
политикам (рис. 1.58).
ОЯ 11-01
ым ног Stop де.
• е n*vf тн антнщи
' o<j oiE now.
To. о?
Ре*ги Г»
Оечтн го
Анди is «ВВАТ.
3
Рис. 1.58. Письма с рецептурой спор возбудителя сибирской язвы. А. Конверт
письма на имя ведущего телекомпании Эй-Би-Си Тома Брокау (Tom Brokaw)
Б. Конверт письма на имя сенатора Томаса Дашли. В. Письмо сенатору Дашли
Письма в «Эн-Би-Си» и «Нью-Йорк пост», отправленные 18.09 2001 г., представ-
ляли собой ксерокопии. Также ксерокопиями были письма, отправленные 09.10.2001 г.
сенатору от штата Южная Дакота и лидеру сенатского демократического большинства
Тому Дашли (Thomas A. Daschle) и сенатору от штата Вермонт, демократу Патрику
Лихи (Patrick J. Leahy). Письма с рецептурой сибирской язвы были проклеены лип-
кой лентой, видимо, с целью предотвратить преждевременное высыпание их содер-
жимого на почте.
По крайней мере, трое из получателей писем с рецептурой сибирской язвы, полу-
чили еще письма-мистификации с безобидным порошком. Они были отправлены по
почте после писем с рецептурой, но еще до того, как первые сообщения о случая
ингаляционной сибирской язвы просочились в прессу. Адреса на них были написа-
ны печатными буквами, с явным стремлением исказить почерк. Тексты по тонально-
сти и ошибкам были схожи друг с другом и с текстами писем, содержаших споры си-
бирской язвы.
С самого начала этих событий преступник (или преступники?) вел себя хладнок-
ровно и так, как будто был уверен в том, чти власти до него никогда не доберутся. Он
повторил рассылку писем с рецептурой возбудителя сибирской язвы из Трентона уже
12. Биодиверсии и биотеррор времен профанации биологического оружия
291
дэсле того, как его начали искать как террориста, способного применить оружие пае-
вого поражения. А когда началась паника — напоминал о себе рассылками писем-
- истификаций. Ряд фактов говорит в пользу того, что преступник хотел «возложить»
зтветственность за биотеррористический акт на исламских террористов. Еще до по-
яснил в печати сведений о совершенном биотеррористическом акте (первые пуб-
икации 12-13 сентября), в военную полицию на базе морской пехоты США в Куан-
• . о (Quantico) пришло анонимное письмо, обвинявшее в терроризме американского
•ясного египетского происхождения д-ра Ауда Асаада (Ayaad Asaad), бывшего учено-
го из USAMR1ID. Письмо было написано на хорошем английском языке и содержа-
? описание значительных познаний Асаада в военной биологии, деталей его личной
кизни и работы в USAMRI1D. Сотрудниками ФБР письмо было показано Асааду и
его адвокату. Впоследствии ФБР сняло с египтянина все подозрения.
Отдельные письма со спорами возбудителя сибирской язвы и письма-мистифи-
гаиии содержали угрозы в адрес Израиля. Например, письмо, направленное сенато-
ру Томасу Дашли, гласило: «Нас не остановить. У тебя теперь сибирская язва. Ты те-
перь помрешь. Испугался? Смерть Америке. Смерть Израилю. Аллах велик». Министр
збороны США, Дик Чейни, появившийся на публике 12 октября впервые с начала
эомбардировок Афганистана (7 октября), сообщил, что случаи сибирской язвы в США
могут быть связаны с Усамой бен Ладеном. 15 октября некоторые представители ФБР,
расследующие это дело заявили, что следы эпидемии ведут в Ирак и что в распростра-
нении сибирской язвы виновен Саддам Хусейн. Так же предполагалось, что Бен Ла-
зай мстит американским СМИ за оскорбления в свой адрес, а для такой мести у терро-
риста № 1 были основания. Газета «Glob» опубликовала портрет Бен Ладана с под-
писью «Разыскивается живым или мертвым», а слово «живым» было зачеркнуто.
Ч газета «Нейшенел Инкуайер» («Nation Enquirer») проводила рекламную кампанию
туалетной бумаги с его изображением.
Преступник, возможно, не имел намерений убить какое-то конкретное лицо или
совершить массовое убийство людей. В письмах сообщалось о сибирской язве или о
необходимости приема антибиотиков («09-11-01...This is next... Take penacilin [sic]
now...» — из письма Тому Брокау). Однако рассылаемый в конвертах штамм сибире-
язвенного микроба был устойчив к пенициллинам и цефалоспоринам, и все амери-
канцы, пораженные ингаляционно, получавшие эти антибиотики в первые дни бо-
лезни, погибли (см. ниже «Диагностика, клиника и лечение ингаляционных случаев
сибирской язвы»). Поэтому такая приписка вводила потенциальную жертву в заблуж-
дение, но знал ли об этом преступник, осуществивший террористический акт, неиз-
вестно. Его основной целью было создание в стране обстановки страха. Выбор средств
массовой информации и сенаторов в качестве адресатов делался преступником созна-
тельно с тем, чтобы гарантировать широкую публичность всего того, что касается этого
теракта. Личности самих сенаторов в этой истории далеко не случайны. Сибиреязвен-
ный теракт произошел в тот момент американской истории, когда президент Буш
пытался «протолкнуть» через Конгресс так называемый «Патриотический акт 1»
(«Patriot Act 1»), позволяющий начать боевые действия в Афганистане. Сенаторы Даш-
ли и Лики были основным препятствием для прохождения этого документа в Конг-
рессе в том виде, в котором его хотел видеть Буш. Они оба энергично защищали граж-
данские свободы, гарантированные американцам Конституцией США, и не хотели
их ограничения под предлогом борьбы с мировым терроризмом.
Мистифицированные письма рассылались по такому же замыслу, что и с рецеп-
турой возбудителя сибирской язы — в СМИ и отдельным политикам. В «Нью-Йорк
292
Часть 1. Краткая история применения биологических средств
пост», «Нью-Йорк тайме» и «Сант-Петерсбург тайме» они могли быть отправлены по
почте 5 октября из Сант-Петерсбурга или же ранее, 20 сентября, когда такое же пись-
мо было отправлено «Эн-Би-Си». В это же время (18 сентября) из Трентона были ото-
сланы сибиреязвенные письма в «Эн-Би-Си» и «Нью-Йорк тайме». Среди адресатов
писем-мистификаций Джудит Миллер (J. Miller), журналистка из «Нью-Йорк тайме»
Она автор и соавтор многочисленных публикаций по вопросам контроля над БО, пс
замыслу «неприятных» для правительства США и крайне поверхностных по содержа-
нию (см., например, Miller J. et al., 2001). Последняя ее работа в «Нью-Йорк тайме»
(05.09.2001 г.), опубликованная перед сибиреязвенным терактом, была посвящена
обличению усилий американского правительства создать некий «суперштамм» воз-
будителя сибирской язвы для целей биологической войны. Ей его и прислали.
Прежде чем приступить к описанию расследования теракта и эпидемиологии от-
дельных вспышек болезни, рассмотрим содержимое «белого порошка». Благодаря его
составу «исламский след» в этих событиях был потерян.
«Белый порошок». Пробы «порошка» изучались в «Институте патологии Вооружен-
ных сил» («Armed Forces Institute of Pathology») в Вашингтоне ив USAMRIID. Вслед-
ствие паники октября — ноября 2001 г. и отсутствия на тот период четких указаний
со стороны властей, что публиковать, а что скрывать, в СМИ появилась несколько ис-
каженная (но достаточная для понимания специалистами) информация о содержи-
мом конвертов. «Белый порошок» состоял из следующих основных компонентов:
1) спор возбудителя сибирской язвы штамма Ames в виде частиц диаметром 1,5—3 мкм.
2) и более мелких частиц наполнителя, дезагрегирующего эти споры и по химическо-
му составу представляющего собой «какую-то форму двуокиси кремния». Именно на
личием в рецептуре частиц менее 5 мкм объясняется ее способность вызывать инга-
ляционную форму сибирской язвы у людей.
Под микроскопом споры возбудителя сибирской язвы выглядели не измельчен-
ными. Сама рецептура не содержала примесей фрагментов спор или вегетативных
клеток В. anthracis, но содержание самих спор было очень высоким. К тому же споры
имели «покрытие», о природе которого в СМИ не сообщалось. Для того чтобы части-
цы не образовывали агломераты, с них был снят электрический разряд. Уильям Пат-
рик (о нем см. выше «Кошмар из сна») признал, что сухая рецептура возбудителя си-
бирской язвы, найденная в конвертах, изготовлена по разработанной им секретной
технологии. Все конверты содержали один и тот же материал по составу компонен-
тов. Наиболее «качественным» по дисперсности он оказался в конверте, присланном
сенатору Дашли — «летучий талькоподобный порошок». Именно данное обстоятель-
ство привело к значительно большему количеству случаев ингаляционной формы
сибирской язвы среди почтовых служащих по пути следования конверта, чем это
имело в случаях болезни, связанных с рассылкой от 18 сентября (см. ниже «Эпиде-
миологическое расследование вспышки сибирской язвы»), В конвертах, отправлен-
ных в «Эн-Би-Си» и «Нью-Йорк пост», обнаружена увлажненная, образовавшая аг-
ломераты, рецептура. Она выглядела как мелкие комочки грязно-коричневого цвета
В начале 2002 г. обсуждение состава «белого порошка» было поставлено под контроль
властей США. В научных публикациях его стали описывать как «В. anthracis — positive
powder» (см., например, работу Jemigan D. В. et al., 2002). По смысловому содержа-
нию это примерно то же, что и «белый порошок» (Roseberg В. Н., 2002).
Обнаружение штамма Ames в рецептуре, распространенной злоумышленником
также вызвало у военных ученых США некоторое замешательство. Дело в том, что этот
штамм имеет ряд неафишируемых свойств, делающих его очень удобным для боевого
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
293
применения в таких странах, как Россия. Он устойчив к некоторым р-лактамным ан-
тибиотикам (например, к цефтриаксону), обычно используемым у нас для лечения
сибирской язвы (см., например, схему лечения в учебнике Шуваловой Е. П. с соавт.,
2001). И способен преодолевать защитное действие некоторых коммерческих вакцин,
т. е. он относится к так называемым «vaccine-resistant В. anthracis» (см. Ivins В. Е. et al.,
990). Эти данные свидетельствуют об огромной работе, проведенной специалиста-
ми LISAM R1ID с целью выбора штамма возбудителя сибирской язвы, пригодного для
боевого применения против стран с устаревшей системой противобиологической за-
щиты и низким уровнем развития биотехнологии.
Штамм В. anthracis, использованный для получения «белого порошка», известен
как Ames Ancestor. Поданным J. Ravel (2009), его выделили в феврале 1981 г. от погиб-
шей от сибирской язвы коровы специалисты техасской компании А&М, и уже в мае
он приобретен USAMRIID. Изолят упаковали в коробку и на ней ошибочно написа-
ти старый адрес, указав А&М, отсюда его название — Ames. Для того чтобы отличать
его от производных штаммов, ему присвоено уточняющее название Ancestor, т. е. пра-
родитель. Ancestor был передан в лабораторию в Военно-биологического центра в
Портон-Дауне (Соединенное Королевство). Там на его основе получили штамм Ames
Porton. Он не содержит обеих плазмид В. anthracis (pXOl, рХО2) и невирулентен.
Штамм, выделенный из содержимого «белого порошка», называют А2012 или Ames
Florida. Фактически это тот же Ames Ancestor. Полная последовательность генома
В. anthracis Ames Ancestor включает 5227419 п. о. хромосомной ДНК, 181677 п. о. ДНК
тлазмиды pXOl и 94830 п. о. ДНК плазмиды рХО2. Плазмида pXOl имеет три копии
на клетку, плазмида рХО2 — две копии на клетку.
Геномные последовательности депонированы в GenBank под номерам и АЕ017334,
ХЕ017336 и АЕ017335 для хромосомы и плазмид pXOl, рХО2 соответственно.
Хотя сегодня нуклеотидная последовательность штамма Ames доступна в GenBank,
сам он малораспространен среди научных центров. В основном с ним работают по
правительственным контрактам не более 20 лабораторий в США, Центр обороны
Канады и лаборатория в Портон-Дауне. Но только четыре организации в США спо-
собны получить рецептуру, найденную в конвертах.
Всего в каждом конверте было примерно по 2 г сухой рецептуры В. anthracis, т. е.
почти столько, сколько указал как оптимальное количество для совершения террори-
стического акта Уильям Патрик ill в отчете, составленном им в марте 1999 г. для ЦРУ.
Расследование биологического террористического акта. Уже 18 октября ФБР и Поч-
товая служба США назначили вознаграждение в миллион долларов за информацию,
которая поможет выявить виновных в распространении спор сибирской язвы. В конце
октября 2001 г. ФБР были получены данные по составу рецептуры. Они убедили ФБР в
ом, что сибиреязвенный террористический акт является внутренним делом. 31 октяб-
ря было сообщено, что США отклонили резолюцию ООН, предложенную Франци-
ей, с осуждением сибиреязвенного теракта на том основании, что он может быть ак-
том внутреннего терроризма. По данным из правительственных источников, ФБР
.становило, что в программах США, имеющих отношение к сибирской язве, в тече-
ние пяти предшествующих лет было задействовано почти 200 ученых. Из них 50 уче-
ных участвовали в военных программах. На всех из них ФБР получило информацию
«из заслуживающих доверия внутренних источников» (т. е. от осведомителей в науч-
ной среде) и составило списки лиц, подозреваемых в совершении террористического
акта. Но Бюро не нашло того человека, которому можно предъявить обвинение. По-
этому 9 ноября оно передало в СМ И информацию с описанием преступника как тер-
294
Часть 1. Краткая история применения биологических средств...
рориста-одиночки мужского пола, американца по происхождению, очевидно науч-
ного сотрудника, работавшего в лаборатории с высоким уровнем биозащиты и спо-
собного обращаться с опасными материалами. В. Н. Rosenberg (2002), специалист по
контролю над БО из унивеоситета штата Нью-Йорк в Пачасе (Purchase) и руководи-
тель рабочей группы по БО в Независимой федерации американских ученых, обоб-
щила их следующим образом:
участник программ США по биозашите, имеет докторскую степень в соответст-
вующей области биологии;
американец средних лет;
имеет опыт и мастерство работы с опасными патогенными микроорганизмами,
включая возбудитель сибирской язвы, способен избегать инфицирования этими па-
тогенами;
работает в организации-подрядчике ЦРУ в Вашинггоне, район округа Колумбия;
имеет действующую вакцинацию против возбудителя сибирской язвы;
имеет свободный доступ к секретной информации;
в прошлом работал в USAMR1ID или имеет свободный доступ туда сейчас;
знает Уильяма Патрика и, вероятно, узнал от него что-то, касающееся из1 отселе-
ния сухой сибиреязвенной рецептуры, неофициально;
имеет навыки и опыт в сокрытии доказательств;
мог иметь связь с UNSCOM (Специальное командование ООН);
имел спор с правительственным органом или ведомством,
имеет частный участок земли, где собирались и готовились материалы для совер-
шения теракта;
работал над письмами один или с сообщником и помощником на периферии;
подходит под описание, составленное ФБР;
имеет необходимый опыт, доступ и служебную характеристику, в которой указы-
ваются его способности и черты характера;
имел беседу с сотрудниками ФБР
То, что В. Н. Rosenberg относит потенциального террориста не к военной органи-
зации, какой является USAMRIID, а к ЦРУ, объясняется тем, что США при ежегод-
ных обменах информацией с другими странами в области биологической защиты в
соответствии с Конвенцией 1972 г. обязаны сообщать только о программах Министер-
ства обороны. Пользующиеся плохой репутацией программы обычно передаются в
ЦРУ, Министерству энергетики или в другие ведомства и подрядные организации.
В начале декабря 2001 г. ФБР заявило, что оно провело проверку 1 осударственных
и подрядных лабораторий на предмет обладания ими штамма Ames, а также лиц, ра-
ботающих в них и имеющих доступ к этому штамму В, anthracis. 16 декабря ФБР со-
общило, что оно обратило особое внимание на подрядчика, который работал с ЦРУ,
и ряд гражданских лиц, вакцинированных против сибирской язвы. 17 декабря пред-
ставитель Белого дома, Эри Флейшнер (Ari Fleischer), заявил, что «все большее рас-
пространение получает мнение, что это был внутренний источник». Но дальше ФБР
повело себя «странно», на что обратили внимание многие эксперты в США. Напри-
мер, Нью-Джерси, откуда были разосланы все письма с рецептурой сибирской язвы,
был охвачен расследованием только в средине января 2002 г. Это расследование вы-
разилось тем, что в городе были расклеены объявления, призывающие население со-
общать в Бюро любую информацию о возможном преступнике. Вместо того, чтобы
искать среди специалистов по БО в учреждениях МО США, гражданских организаци-
ях, работающих по правительственным контрактам, и в подразделениях ЦРУ по спе-
12 Биодиверсии и биотеррор времен профанации биологического оружия
295
киальным операциям, ФБР вдруг стала «раскидывать широкую сеть» и использовать
..ктику «облавы». Бюро занялось проверкой на причастность к этому преступлению
атги 40 тыс. американских микробиологов. За информацию, позволяющую поймать
- реступни ка, вознаграждение было повышено до 2,5 млн долларов, но его так нико-
му и не выдали.
В конце 2и01 г. представители США на переговорах в Женеве отказались от рабо
ты над Протоколом и от дальнейшего рассмотрения механизмов контроля над БО в
елом, несмотря на то, что их создание могло устранить наиболее важный недоста-
ток Конвенции 1972 г. — невозможность проверки ее исполнения. Постепенно све-
рил о расследовании перестали проникать в СМИ, и о нем забыли. Только в августе
2)8 г. ФБР объявило, что преступник, которого так долго искали в связи с биотерро-
.тическим актом 2001 г., найден мертвым (см. «Брюс Эдвард Айвинс»).
Поведение людей. Когда жители городка Бока-Рэйтон (шт. Флорида) узнали при-
чину скоропостижной смерти Боба Стивенса, они только насторожились. Боб был
любителем рыбалки, велосипедных поездок по окрестностям и работы в саду, в экзо-
тические страны не ездил. Поэтому диа1 ноз «сибирская язва» всем, кто его знал, ка-
зался недоразумением. Когда 8 октября стало известно, что у 72-летнего сортировщика
чты редакции «The Sun», попавшего несколько дней назад в больницу округа Майа-
ми-Дэйд с диагнозом «пневмония», также обнаружена сибирская язва, население за-
беспокоилось. Дом Стивенса находился всего в 1,5 км от учебного аэродрома, где
- рактиковался Моаммед Ата, один из угонщиков пассажирского авиалайнера 11 сен-
тября. А в 60 км от него расположился учебный центр сельскохозяйственной авиации,
де так же учились террористы, причастные к событиям 11 сентября. Среди населе-
ия поползли слухи, что они интс оесовались распы тителямидля сельскохозяйствен-
ной авиации. Когда сведения о начавшейся в стране вспышке искусственно вызванной
сибирской язвы попали в прессу, население Бока- Рэйтона бросилось в ап теку скупать
антибиотики. Местный фармацевт Джошуа Люин рассказал журналистам, что если
ш I раньше распродавал в неделю не более чем 200 таблеток антибиотиков, то за день
12 октября он распродал 2500 упаковок таблеток антибиотиков, все свои запасы.
Вечером 12 октября в Нью-Йорке был зафиксирован еще один случай заболева-
ния сибирской язвой. Кожная форма болезни с большим опозданием была диагно-
е гирована у женщины, которая занималась корреспонденцией для вечерних hi «вестей
национального телеканала NBC (заболела 25 сентября). Сообщение о том, что тер-
рористы пытались убить сибирской язвой, разосланной по почте, ведущего вечерних
н >востей Тома Брокау, вызвало у населения США страх перед любыми почтовыми
отправлениями. Информация о распространяющемся постране «антраксе» приобрела
тавинообразный характер. Обывателю уже было невозможно отличать правдивые
сообщения СМИ от ложных. В субботу 13 октября СМИ сообщили, что во Флориде
признаки сибирской язвы обнаружены еще у 5 человек, что, как потом выяснилось,
Ни соответствовало действительности. Информация была подана так, что эти люди
якобы контактировали с Робертом С гивенсом, первой жертвой бактериологической
.таки на США. Кроме того, споры возбудителя сибирской язвы были найдены у тро-
их жителей Нью-Йорка — двух биологов и полицейского, которые. как предполага-
ги, держали в руках зараженное письмо, найденное в NBC. Споры выявили у них на
j ице. Сибирская язва в восприятии не специалистов стала представляться заразной,
присутствующей везде.
Настоящая паника началась 15 октября, когда письмо с рецептурой сибирской
язвы вскрыли в офисе лидера демократов, Томаса Дашли. Тысячи людей бросились
296
Часть 1. Краткая история применения биологических средств...
в медицинские учреждения сдавать крозь «на сибирскую язву». Промышленный
индекс Dow Jones упал на 1,71 %. Индекс Nasdaq упал на 2,9 %. Ранее в течение дня на
биржевых торгах наблюдался подъем: через два часа после открытия бирж индекс
NASDAQ вырос на 4,62 %, «промышленный» индекс Dow Jones — на 1,84 %. Ин-
декс S&P, который впервые с 11 сентября превысил свою докризисную отметку в
1092,54 пункта, составив по итогам торгов 1097,43 пункта, после сообщений о сибир-
ской язве упал на 1,68 % (18,48 пункта).
К 15 октября во многих компаниях США были приняты повышенные меры безо-
пасности при сортировке почты. Сотрудникам многих издательств предписывалось
сообщать ФБР о любой подозрительной корреспонденции. ФБР также выложило на
своем сайте специальные «Рекомендации» в формате .pdf для всех, кто получил какие-
нибудь подозрительные письма. «Рекомендации» представляли собой плакат, кото-
рый можно при желании распечатать. В качестве признаков «подозрительного пись-
ма» ФБР выделяло: неправильно написанные слова; отсутствие обратного адреса или,
наоборот, присутствие оного, но указывающего на зарубежное происхождение пись-
ма; странный запах, масляные пятна, потертости, наличие кристаллов на конверте и
прочее. В том случае, если получатель счел-таки письмо подозрительным, ему следо-
вало обращаться с письмом аккуратно — не бросать и не трясти, не открывать, не
нюхать и не пробовать на язык, и позвонить по телефону 9-11.
Британский «Интернет» потрясла волна бактериологического спама. В электрон-
ных посланиях, рассылаемых по Сети в огромных количествах, неизвестный автсг
призывает никогда не открывать «голубые конверты» с письмами из некоего «Фонда
Кринглемана».
Утром, во вторник, 16 октября Америку потрясло еще одно известие —кожная фор-
ма болезни обнаружена у 6-месячного сына продюсера «АВС» (заболел 29 сентября).
18 октября у помошницы ведущего новостей американской телекомпании «NBS»
Дэна Разера была установлена кожная форма сибирской язвы (заболела 1 октября).
В ее обязанности, в частности, входила работа с почтой телезвезды.
Как и в Средние века во время эпидемий чумы, в США во время паники вызван-
ной биотеррористическим актом, появись личности, желающие усугубить эпидеми-
ческую ситуацию и обстановку страха в стране. За первые 18 дней октября власти от-
реагировали на 3300 сообщений о биотерроризме, несмотря на то, что многие из
обращений изначально были очень похожи на ложные.
Люди стали бояться вскрывать любые письма. Многие жители американских го-
родов заклеили щель во входных дверях, куда обычно опускают почту. Взамен они
выставили у входа прозрачные пластиковые ящики, содержимое которых долго изу-
чали, прежде чем их открыть. Телекомпания «CBS» объявила о закрытии отдела пи-
сем В Капитжтии с 18 октября была приостановлена доставка почты, после того как
стало известно о появлении письма с подозрительным порошком, а потом его вооб-
ще закрыли (рис. 1.59).
Почтовой службе США (United States Postal Service, USPS) был нанесен огромный
урон. Годовой оборот USPS составляет не менее 900 млрд долларов. От почты прямо
или косвенно зависит 9 млн рабочих мест. Опасаясь за свое здоровье, почтовики не-
скольких штатов начали предупредительные забастовки. В результате задержек в до-
ставке корреспонденции стали опаздывать миллионы чеков, которыми американцы
традиционно погашают счета за электроэнергию и погашают кредиты. Только у од-
ной энергетической компании «Рерсо», обеспечивающей энергией 2 млн человек в Ва-
шингтоне и его пригородах, «зависло» 30 тыс. счетов.
12 Биодиверсии и биотеррор времен профанации биологического оружия
297
Рис. 1.59. Здание Конгресса США в первых числах ноября 2001 г., закрытое
на санитарную обработку в связи с биологическим террористическим актом.
Фотография из статьи А. Воронина и Н. Зимина (2001)
С 21 октября в Вашингтоне, после лабораторного подтверждения диагноза «ин-
галяционная сибирская язва» у сортировщиков почты, был закрыт крупнейший сор-
тировочный центр Брентвуд. Через него ежесуточ но проходило не менее миллиона пи-
сем, включая всю почту законодательной власти (см. ниже «Сортировочный центр в
Вашингтоне»). Центр открыли только через три года.
Паника, вызванная случаями заражения сибирской язвой в США, вышла за пре-
делы страны. Письма, из которых сыплется подозрительный порошок, в октябре —
ноябре получают всему миру известные политики, телеведущие, бизнесмены и про-
сто граждане, которых кто-то решил напугать.
Почтовая служба США в срочном порядке ищет частные компании, которые смо-
гут взять на себя по контракту облучение всей корреспонденции, поступающей в орга-
ны федеральной власти посредством мощного потока лучей «электронной пушки» или
мощной рентгеновской установки. Одна такая компания найдена в штате Огайо и
приступила к облучению почты 27 октября.
Ощущение невидимой угрозы пронизало американские города. В магазинах во-
енных товаров население стало скупать индивидуальные средства защиты, включая
противогазы, которые валялись на складских полках еще со времен войны во Вьетна-
ме. В стране в два раза вырос спрос на антибиотики. В октябре около 32 тыс. амери-
канцев начали принимать антибиотики, опасаясь заболеть сибирской язвой. В то же
время врачами шестидесятидневный курс антибиотиков рекомендовано пройти все-
го лишь 5 тыс. человек во Флориде, Нью-Джерси, Нью-Йорке и Вашингтоне. Намного
увеличилась выписка рецептов на успокоительные средства.
Панические настроения требовали успокоительной информации. Страничка в
Интернете, посвященная сибирской язве и методам борьбы с ней, стала самой посе-
щаемой. В библиотеках население разобрало книги по эпидемическим болезням.
Корреспонденты журнала «Итоги», А. Воронин и Н. Зимин (2001), были свидетеля-
ми, как в библиотеке две старушки почтенного вида вцепились в книгу Альбера Камю
298
Часть I. Краткая история применения биологических средств...
«Чума», не желая уступать ее одна другой. Борьба продолжалась до тех пор, пока им
не объяснили, что это художественное описание эпидемии чумы, а не руководство по
ее лечению, и что чума — не сибирская язва. По наблюдению этих же корреспонден-
тов, в начале ноября в Вашингтоне царил ужас и хаос. Судьи Верховного суда США
впервые с 1935 г. должны были работать вне родных стен. До средины ноября было
закрыто на дезинфекцию несколько зданий Конгресса.
Эпидемиологическое расследование вспышки сибирской язвы. Всего с 4 октября по
20 ноября 2001 г. в США было выявлено 22 случая сибирской язвы, связанной с рас-
пространением почтовыми отправлениями спор возбудителя болезни. Хронология
случаев болезни показана на рис. 1.60.
3
в*
S
8
7
6
5
4
3
2
1
0
П Округ Колумбия
'м Ингаляционная форма болезни
Ш1 Кеннектикут
Письма в Нью-Йорк
4
Письма в округ
Колумбия
С Нью-Джерси
И Флорида
И Нью-Йорк
16 19 22 25 28 1 4 7 10 13 16 19 22 25 14
Сентябрь Октябрь Ноябрь
Рис. 1.60. Хронология случаев болезни сибирской язвой, вызванных биотеррористиче-
ским актом в США в 2001 г. За основу взята схема из работы Jernigan D. В. et al. (2002)
Из этих случаев 11 были ингаляционной сибирской язвой, 11 — кожной, 5 случа-
ев ингаляционной формы болезни закончились летальным исходом. Двадцать забо-
левших (91 %) были либо работниками почты (mail handlers), либо заболели в резуль-
тате экспонирования к спорам возбудителя сибирской язвы на рабочем месте при
обработке почтовых отправлений или при работе с письмами. Общая характеристи-
ка всех случаев заболевания сибирской язвой приведена в табл. 1.11.
В марте 2002 г. обнаружен дополнительный, 23 случай кожной формы сибир-
ской язвы у сотрудника лаборатории, работавшего с пробами, полученными из окру-
жающей среды во время ликвидации террористической вспышки сибирской язвы
осени 2001 г.
Подтвержденным случай сибирской язвы (кожная, кишечная или ингаляционная
формы) считали в том случае, если:
1) из образцов ткани пациента удавалось получить культуру возбудителя болезни—
В. anthracis',
2) наличие сибиреязвенной инфекции было подтверждено двумя другими лабо-
раторными тестами. Предполагаемым считали случай болезни, когда он клинически
соответствовал сибирской язве (и не имел альтернативных диагнозов), но В. anthracis
не была выделена, или: I) лабораторные доказательства наличия в тканях больного
В. anthracis были получены на основании одного лабораторного теста; или 2) случай
болезни имел эпидемиологическую связь с возможностью экспонирования к В. anthra-
cis, находящейся в окружающей больного среде.
Лабораторными критериями для определения случая болезни как сибирской яз-
вы, были:
1) получение культуры В. anthracis из клинических образцов тканей пациента с
подтверждением вида бактерии прямым окрашиванием флуоресцентными антитела-
ми (fluorescent-antibody staining) и лизисом у-фагами;
Таблица Lil
Краткая гюракгёдостика случаев сибирский язвы, i изваяв я бншеррористмческим актом в США в 2001 г.*
Случай п/п** Дата начала болезни Дата подтверждения диагноза лабораторным путем Штат Возраст (лет) Пол Раса Занятие Статус случая Проникновение возбудителя болезни Исход болезни
1 2 3 4 5 6 7 8 9 10 11
1 22.09 19.10 NY 31 । Ж белая служащая почто- вого офиса в Нью-Йорке предполагаемый через кожу выжила
2 25.09 12.10 NY 38 Ж белая NBC, ассистентка подтвержденный через кожу выжила
3 26.09 18.10 NJ 39 М белый USPS, механик машины предполагаемый через кожу выжил
4 28.09 15.10 FL 73 М Белый испанского происхождения AMI, работник почтового офиса подтвержденный ингаляционно выжил
5 28 09 18.10 NJ 45 Ж белая USPS, почтальон подтвержденный через кожу выжила
6 28.09 12.10 NY 23 Ж белая NBCTV, специалист предполагаемый через кожу выжила
7 29.09 15.10 NY 0,6 м белый АВС, сын продюсера подтвержденный через кожу выжил
8 30.09 04.10 FL 63 ~ м белый AMI, фоторедактор подтвержденный ингаляционно умер
9 01.10 18.10 NY 27 ж белая NBC, ассистентка подтвержденный через кожу выжила
10 14.10 19.10 РА 35 м белый USPS, рабочий сортировочной машины подтвержденный через кожу выжил
7.72. Биодиверсии и биотеррор времен профанации биологического оружия
Продолжение табл. 1.11
1 2 3 4 5 6 7 8 9 10 11
11 14.10 28.10 NJ 56 Ж черная USPS, рабочая сортировочной машины подтвержденный ингаляционно выжила
12 15.10 29.10 NJ 43 Ж азиатка USPS, рабочая сортировочной машины подтвержденный ингаляционно выжила
13 16.10 21.10 VA MD 56 М черный USPS, работник почты подтвержденный ингаляционно выжил
14 16.10 23.10 MD 55 М черный USPS, работник почты подтвержденный ингаляционно умер
15 16.10 26.10 MD 47 м черный USPS, работник почты подтвержденный ингаляционно умер
16 16.10 22.10 MD 56 м черный USPS, работник почты подтвержденный ингаляционно выжил
17 17.10 17.10 NJ 51 ж белая бухгалтер через кожу выжила
18 19.10 19.10 NY 34 м Белый испанского происхождения NY Post, разносчик почты предполагаемый через кожу выжил
19 22.10 22.10 VA 59 м белый управляющий рабочий сортиро- вочной машиной подтвержденный ингаляционно выжил
20 23.10 28.10 NY 38 м белый ПОЧТОВЫЙ служащий в Нью-Йорке подтвержденный через кожу выжил
21 25.10 30.10 NY 61 ж азиатка подсобная рабочая в больнице подтвержденный ингаляционно умерла
22 14.11 21.1] СТ 94 ж белая уединенно жила в своем доме подтвержденный ингаляционно умерла
Часть 1. Краткая история применения биологических средств...
* Таблица взята из работы D. В. Jernigan et al. (2002).
* * По данным A. R. Hoffmaster et al. (2002), все случаи сибирской язвы, имевшие отношения к биотеррористическому акту в США
в 2001 г., вызваны одним и тем же штаммом В. anthracis.
12. Биодиверсии и биотеррор времен профанации биологического оружия
301
2) другими специфичными к В. anthracis лабораторными тестами, включающими:
। . получение посредством ПЦР доказательства наличия в тканях пациента ДНК
- anthracis-, б) обнаружение В. anthracis в клинических образцах посредством имму-
•эгистохимического окрашивания (immunohistochemical staining, 1НС); или в) по по-
ложительным серологическим тестам на сывороточный иммуноглобулин G (IgG), спе-
.ифичный к протективному антигену (РА) токсина возбудителя сибирской язвы
; токсине см. в разд. 3.1).
Случаи искусственно вызванной сибирской язвы зарегистрированы у жителей
хми штатов, расположенных вдоль Восточного побережья США. В Коннектикуте —
1ин случай; во Флориде — два случая; в Мэриленде — три; в Нью-Джерси — пять; в
S w York City — восемь (включая случай в Нью-Джерси с жителем, инфицированным
New York City); в Пенсильвании — один; и в Виргинии — два. Средний возраст па-
кентов — 46 лет (возрастной интервал заболевших от 7 месяцев до 94 лет). Паци-
гты с ингаляционной формой сибирской язвы были старше, чем с кожной (56 лет и
•5 лет соответственно; р< 0,01). Мужчины составили 55 % от количества всех боль-
jx сибирской язвы (12 случаев болезни), 15 заболевших были белыми (68 %), 5 че-
-.эвек погибли — 23 % от общего числа заболевших, но среди них белых было 40 %.
Все погибшие страдали ингаляционной формой болезни. Смертность при такой кли-
- («ческой форме болезни составила 45 %. Для 6 случаев ингаляционной сибирской
1вы у работников почты оказалось возможным установить дату их первого экспони-
рования к В. anthracis, произошедшего во время работы с почтой на высокоскорост-
- ой сортировочной машине. Средняя продолжительность между экспонированием и
□явлением симптомов ингаляционной сибирской язвы составила 4,5 суток (интер-
- т 4—6 суток).
Из 11 случаев кожной формы сибирской язвы 7 (64 %) отнесены к подтверж-
-•нным. Два из них подтверждены изоляцией В. anthracis из клинических образцов
один — из крови, второй — из раны). Пять других подтверждены результатами им-
муногистохимического окрашивания или ПЦР биоптатов; ПЦР крови; и подъемом
сывороточных IgG, специфичных в РА. К предполагаемым случаям кожной формы
сибирской язвы отнесли те 4 случая, для которых инфицирование В. anthracis подтвер-
ждалось только одним тестом. Кожные поражения были локализованы на лице, ру-
IX. грудной клетке. Два случая представляли собой множественные поражения.
К работе с почтовыми отправлениями имели отношение 12 (55 %) пациентов
в с ингаляционной и 4 с кожной формами болезни). Из них 9 случаев пришлось на
служащих USPS, 1 на штат обработки правительственной почты; работники почто-
вых комнат медийных компаний дали 2 случая сибирской язвы. Шесть (27 %) паци-
ентов (один с ингаляционной формой болезни, пять с кожной) работали в медийных
компаниях на участках, где осуществлялся прием почты (AMI — 1 случай; CBS —
1 случай; NBC — 2 случая; и New York Post — два случая). Оставшиеся четыре случая
(18 %) — 2 ингаляционных и 2 кожных — были классифицированы как «другие». Они
включают семимесячного гостя American Broadcasting Company (ABC); 61-летнюю
женщину, подсобную рабочую Manhattan Hospital; 51-летнюю женщину, бухгалтера
из Нью-Джерси; и 94-летнюю жительницу Коннектикута.
Из 22 лиц, заболевших сибирской язвой, 20 (91 %) либо держали в руках конвер-
ты с сухой рецептурой сибирской язвы, либо экспонировались к спорам В. anthracis
по месту работы, где с конвертами проводились различные операции. Было установ-
лено, что не менее четырех конвертов содержали рецептуру сибирской язвы, «марш-
рут движения» каждого обнаруженного конверта был установлен (рис. 1.61).
302
Часть 1. Краткая история применения биологических среде те...
?
г “ - • Случай 5, NJ
18 сентябре , ; < случай 3 NJ
конверты । w улучай з, го
Трентон" Распределительный
—— центр в Гамильтоне —q
9 октября, 1 “ZTZZ
конверты । 91В
• ?
•“ " О Случай 17, NJ
L?- []Случай 21, NY
1 ?
ь - □ Случай 22, СТ
Почтовая с.ляля. ami НВ
г-4в Вест-Пальме, FL------АМ1 И"
Офис Лихи
ф ф О Кожная форма болезни
ВВП Ингаляционная форма болезни
Установлсню :й пул! рецептуры в конвертах
“ “ “ Предполагаемый путь рецептуры в конвертах
Намеченная цель
Г"^1 Место доставки конверта с рецептурой
? Неустановленный путь контаминированных конвертов
Рич. 161. Случаи сибирской язвы в США, вызванные почтовыми отправлениями
в сентябре — октябре 2001 г. По D. В. Jemigan et al. (2002)
Все четыре обнаруженных конверта, содержащих рецептуру спор В. anthracis, были
стандартными конвертами почтовой службы США. Все отправлены из Трентона (Нью-
Джерси). Первые два направлены в СМИ: одно — в NBC, ведущему Тому Ьрокау; дру-
гое — в New York Post. Обе организации находятся в Нью-Йорк Сити (New York City).
На конвертах этих писем стоял штамп с датой отправления — ^September 18, 2001 >.
Следующие два письма были адресованы сенаторам Томасу Дашли и Томасу Лихи в
Вашингтон (Washington, D.C.), на конвертах штамп — «October 9,2001». Каждое пись-
мо содержало текст: «09-11-01 ...You can not stop us. We have this anthrax. You die now. Are
you afraid?». Но конверты, контаминированные снаружи спорами В. anthracis, найде-
ны на других почтовых участках в Нью-Йорк Сити, во Флориде и в Коннектикуте
Сентябрьские письма были транспортированы через различное почтовое оборудова-
ние и доставлялись по узкому маршруту в Нью-Джерси и медийными компаниями в
Нью-Йорке. Они были обработаны почтовой службой Трентона (L). S. Postal Service
Trenton Mail Processing), затем в распределительном центре в Гамильтоне (Distribution
Center in Hamilton, New Jersey). После чего отосланы в I (ентральную почтовую аппа-
ратную Моргана в Нью-Йорке (Morgan Central Postal Facility), где были рассортиро-
ваны и доставлены адресатам. Оба этих пункта обрабоз ки почты и, по крайней мере,
еще пять других в Нью-Джерси, аффилированные с Гамильтоном, были контамини-
рованы спорами сибиреязвенного микр< ба Хотя установленные письма с рецепту-
рой сибирской язвы посланы только в АВС и CBS, В. anthracis обнаружена в здании
AMI. где работал Р. Стивенс, и, по крайней мере, в шести почтовых отделениях по пути
1.12. Быодиверсии и биотеррор времен профанации биологического оружия
303
доставки писем в AMI. Эти данные предполагают возможность существования пято-
го письма с рецептупой, отправленного в средине сентября 2001 г. в AMI, но оно не
было обнаружено (Jemigan D. В. et al., 2002).
Письма, отправленные 9 октября, обработаны в несколько этапов. Сначала в Га-
мильтоне (Нью-Джерси). Затем их транспортировали в почтовую службу Брентвуда
(U.S. Postal Service Brentwood Mail Processing). Оттуда они попали в сортировочный
центр в Вашингтоне (Distribution Center in Washington. D.C.). Конверты в Гамильтоне
и Брентвуде были обработаны с помощью высокоскоростных сортировочных машин,
что привело к аэрозолированию сухой рецептуры спор В. anthracis, содержащейся в
конвертах. То же произошло и в Вашингтоне. Конверты, находящие в контакте с кон-
вертами, содержащими рецептуру, были контаминированы снаружи рецептурой спор
сибиреязвенного микроба и отправлены в почтовые офисы правительственных уч-
i еждений.
Один из конвертов, содержащих сухую рецептуру спор В. anthracis, из сортировоч-
ного центра в Вашингтоне (см. ниже) был доставлен в офис сенатора Дашли в здании
Hart Senate и открыт служащим офиса 15 октября. Но так как сотрудник уже был пре-
упрежден о возможности получения таких писем, он немедленно закрыл конверт и
сообщил о нем полиции (см. ниже).
С 15 октября у 625 человек, которые потенциально могли быть экспонированы к
спорам В. anthracis в Hart Senate, брались мазки из носовых ходов. 28 октября было
обнаружено, что помещения Hart Senate и почтовой службы в Брентвуде сильно конта-
минированы спорами В. anthracis. К ним добавились еще почти 25 помещений прави-
тельственных зданий, почтовой службы, почтовых офисов, связанных с Брентвудом.
Другое письмо, отправленное 9 октября сенатору Лихи, случайно задержалось на
почте и не было вскрыто в офисе сенатора. Также не были вскрыты и другие письма,
имеющие в почтовом штемпеле дату отправления «October 9, 2001» и адресованные
сенатору Лихи. 17 октября 2001 г. они были изъяты почтовой службой и переданы ФБР,
дальнейшая их судьба неизвестна (см. «Сортировочный центр в Вашингтоне»).
D. В. Jernigan et al. (2002) распределили случаи инфицирования людей спорами
В. anthracis на два кластера. Первый включает 9 случаев сибирской язвы, начавшихся
приблизительно через 4 суток после 18 сентября и два случая во второй половине ок-
тября (случаи 1—9, 18 и 20 соответственно; табл. 1.11). Все 7 случаев сибирской язвы
в Нью-Йорке (New York City) и Нью-Джерси были представлены кожной формой
болезни; 5 из заболевших в Нью-Йорке (New York City) были сотрудниками медий-
ных компаний и посетителями этих компаний. Оба заболевших в Нью-Джерси были
с. жащими почты. Два случая во Флориде были представлены ингаляционной сибир-
ской язвой и заболевшие были сотрудниками медийной компании. Восемь из девяти
человек были заражены сибирской язвой на рабочем месте. Один человек (почтальон-
женщина в Нью-Джерси) предположительно заразился от конверта, перекрест но кон-
таминированного В. anthracis на почте (случай 5, табл. 1.11). Среднее количество дней,
прошедших от даты на штемпеле письма с рецептурой спор В. anthracis и началом
болезни лиц, бывших с ним в контакте, укладывается в 10 суток (4—13 суток). Почти
все случаи болезни первого кластера (за исключением 18-го и 20-го) развились еще до
того, как была распознана их причина — В. anthracis. Роль сибиреязвенного микроба
в их появлении была установлена только четвертого октября, когда была выявлена
легочная сибирская язва во Флориде у Боба Стивенса. D. В. Jemigan et al. (2002) счи-
тают, что последние два случая кожной сибирской язвы в Нью-Йорке (New York City)
случаи 18 и 20 в табл. 1.11) также имеют отношение к письму, отправленному
304
Часть 1. Краткая история применения биологических средств...
18 сентября в «New York Post», когда оно перемещалось по почтовым отделениям в сре-
дине октября, не будучи еще распознанным как содержащее рецептуру возбудителя
сибирской язвы.
Болезни второго кластера (случаи 10—16, и 19; табл. 1.11) начались спустя 5 суток
после отправки девятого октября писем с рецептурой сибирской язвы. Все 5 случаев,
выявленных в Вашингтоне, представлены ингаляционной формой сибирской язвы
Последнее связано с лучшим «качеством» рецептуры, помещенной в конверт для се-
натора Дашли, чем она была в конвертах, направленных в СМИ Нью-Йорка. Все за-
болевшие имели отношение к почтовому оборудованию, обрабатывавшему письма,
отправленные девятого октября.
Из трех случаев в Нью-Джерси второго кластера, два были представлены ингаля-
ционной формой сибирской язвы, третий — кожной. Они были обнаружены у работ-
ников почты. Третий заболевший (кожная форма болезни) работал бухгалтером в близ-
ко расположенном к почте офисе коммерческой фирмы.
Случаи 21-й и 22-й D. В. Jernigan et al. (2002) относят к «загадочным» (см. ниже».
Среднее количество суток, прошедших после отправки писем девятого октября,
до появления случаев сибирской язвы — 7 (интервал 5—13 суток), за исключением
пациентов, источник заражения которых не определен, атакже заразившихся от кон-
вертов, отправленных 18 сентября.
Два случая ингаляционной сибирской язвы во Флориде. Во Флориде сибирская язва
у людей встречается крайне редко. За все XX столетие выявлено 8 таких случаев, пос-
ледний случай кожной формы сибирской язвы зарегистрирован в 1974 г. Поэтому,
когда 2 октября 2001 г. Боб Стивенс был госпитализирован в тяжелом состоянии,
ингаляционная сибирская язва не была распознана сразу (случай 8, см. табл. 1.11).
Только 4 октября специалисты из Florida Department of Health (FDOH) Bureau of
Laboratories подтвердили наличие В. anthracis в культурах, полученных из цереброспи-
нальной жидкости пациента. Однако версия террористического акта тогда не возник-
ла. Специалисты Департамента здравоохранения Флориды просмотрели сотни меди-
цинских карт пациентов, поступивших в разные клиники в Pain. Beach County, но лиц
с клиникой, похожей на сибирскую язву, не обнаружили. Не поступило сообщений и
от ветеринарной службы о сибиреязвенных эпизоотиях. Не было установлено случа-
ев сибирской язвы среди людей и животных в близлежащих штатах. Только тогда был
заподозрен террористический акт, а окончательно установить факт его совершения
удалось благодаря тщательному опросу сотрудников редакции, где работал Роберт
Стивенс. Они сообщили о конверте, который был получен редакцией 19 сентября и
из которого на клавиатуру его компьютера и канцелярские принадлежности посыпал-
ся белый талькоподобный порошок. И Роберт какое-то время рассматривал этот по-
рошок, приблизив к нему лицо.
Эрнесто Бланко, работник почтового офиса редакции «The Sun», второй больной
ингаляционной сибирской язвой, обнаруженный во Флориде (случай 4; табл. 1.11),
не смог связанно вспомнить, открывал или не открывал он какие-то конверты, содер-
жащие «порошок». Механизм его ингаляционного инфицирования спорами возбуди-
теля сибирской язвы остался неустановленным.
Эрнесто ежедневно занимался тем, что носил из почтового фургона в почтовую
комнату редакции «The Sun» по 10—15 тыс. писем. По своим должностным обязанно-
стям он вскрывать письма не мог. Как потом было установлено, и почтовый фургон
где Эрнесто брал письма, и почтовая комната, где он работал, были сильно контами-
нированы спорами В. anthracis.
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
305
В. anthracis был идентифицирован в двух из 12 проб, отобранных 5 октября с по-
верхности компьютерной клавиатуры Роберта Стивенса и его почтового ящика в по-
чтовой комнате компании. Только у двух человек из 1076, отнесенных во Флориде в
группу риска, были выделены из носовых ходов культуры В. anthracis. Одним из них
был Эрнесто Бланко, носивший письма Роберту Стивенсу и заболевший на два дня
раньше шефа, но переносивший ингаляционную сибирскую язву «на ногах» в тече-
ние 6 суток (см. случаи 4 и 8; табл. 1.11). Вторым оказался сортировщик писем из это-
го же почтового офиса, не имевший ни симптомов сибирской язвы, ни антител к про-
тективному антигену токсина В. anthracis.
Почти все сыворотки, полученные от 436 работников холдинга 10 и 17 октября,
не содержали IgG-антител к протективному антигену токсина В. anthracis. Единствен-
ным исследованным, у кого оказался повышенным титр таких антител, оказался чу-
дом выживший Эрнесто.
Сортировочный центр в Вашингтоне. В течение октября четыре случая ингаляци-
онной сибирской язвы были обнаружены в крупном центре сортировки почты в Ва-
шингтоне, двое заболевших работников погибли (случаи 13—16, см. табл. 1.11). Ин-
фицирование людей спорами В. anthracis стало возможным из-за того, что через центр
прошло два письма, содержащих сухую рецептуру возбудителя сибирской язвы, ад-
ресованных американским сенаторам в Hart Senate Ofce Building. «Движение» этих
писем через 46 тыс. м2 рабочей территории центра было реконструировано Р. Dewan
et al. (2002). Схема этажа сортировочного центра, где происходила сортировка почты
и инфицирование людей, представлена на рис. 1.62, с. 306.
Письма поступили в здание центра обычной почтовой доставкой вечером 11 ок-
тября или ранним утром 12 октября через дверь загрузочного дока поблизости от офи-
са, где работал один из сотрудников центра, первым заболевший ингаляционной си-
бирской язвой. Поступившие в центр конверты были свалены на желоб большой
сортировочной машины и 12 октября между 7.05 ч и 7.30 ч пропущены через маши-
ну — сортировщик штриховых доставочных кодов (deliveiy bar code sorter, DBCS) № 17
(производительность — 30 тыс. писем/ч). DBCS-машина двигала конверты вдоль
внутренней конвейерной ленты (internal conveyor belts), бегущей по роликам со скоро-
стью 51 км/ч (с серией поворотов) и сортировала письма по мешкам. После двукратно-
го прохождения через DBCS-машину № 17, конверт с рецептурой, адресованный сена-
тору Дашли, был сброшен в мешок писем, направляемый в Капитолий. Его увезли в
правительственную секцию центра, где письма сортировались вручную между 7.30 ч
утра и полуднем, затем, через погрузочный док (см. рис. 1.62, Е) письмо с рецептурой
В. anthracis было передано в службу почтовой доставки Капитолия. В правительствен-
ной секции ингаляционной сибирской язвой заболел один работник.
Письмо, адресованное сенатору Лихи, было отсортировано DBCS-машиной не-
правильно. Машина отправила его в Департамент штатов (U.S. State Department, State
Annex 32), имевший небольшую службу почтовой доставки в Виргинии. Точный марш-
рут блужданий этого письма с 12 по 17 октября, неизвестен. Но 17 октября ФБР по
всей стране изъяло письма, адресованные в Капитолий, и поместило их в герметиче-
ские контейнеры для последующего изучения содержимого. В одном из этих контей-
неров было найдено злополучное письмо. В Виргинии заболел работник из State
Annex 32 (случай 19, см. табл. 1.11), но попадало ли туда именно это письмо, неизвест-
но. Пробы, взятые с поверхностей в помещениях State Annex 32, показали их значи-
тельную контаминацию спорами В. anthracis, сходную с таковой в сортировочном
центре в Вашингтоне.
306
Часть 1. Краткая история применения биологических средств...
Сектор высокоскоростной Сектор
сортировочной машины правительственной почты
Рис. 1.62. Схема этажа центра сортировки почты в Вашингтоне, где прово-
дилась обработка писем с рецептурой сибиреязвенного микроба. Серые
ромбы с буквами показывают участки, где могли находиться письма с рецеп-
турой. Черные кружки с цифрами — место работы инфицированных людей
на 12 октября 2001 г. Участок, где инфицирование людей было наиболее
вероятно, показан ромбом С, расположение людей с наиболее вероятным
инфицированием показано открытыми кольцами. По Р. Dewan et al. (2002)
Всего в сортировочном центре в Вашингтоне работало 20 DBCS-машин в три
смены каждый день в неделю. Каждое утро с 7.30 ч до 10.00 ч они обычно открыва-
лись и продувались воздухом от компрессора для удаления пыли. Операторы DBCS-
машины Ns 17 не были инфицированы В. anthracis. Однако у оператора DBCS-маши-
ны № 14, который находился в 27-30 м от машины № 17, и у рабочего в комнате для
срочной сортировки почты, которая имела две двери и находилась от машины № 17 в
55 м, развилась ингаляционная сибирская язва. Эти рабочие могли периодически
проходить рядом с машиной Ne 17 в туалетную комнату или по своим обязанностям
Два других, заболевших ингаляционной сибирской язвой, работали в пределах не-
скольких метров от машины № 17.
Еще до приезда ученых из CDC, 18 октября, подрядчик, нанятый U PS для иссле-
дования центра на наличие спор В. anthracis, собрал сухими тампонами образцы ма-
териала с поверхности оборудования и помещений. Вокруг мешков для отсортирован-
ной правительственной почты и с DBCS-машин № 16,17,18 и 19 им отобрано 29 проб.
Оказались положительными на В. anthracis 16 образцов: 10 из 17 в участке правитель-
ственной почты и 4 из 12 взятых с поверхности DBCS-машин. Оба образца, собран-
12. Биодиверсии и биотеррор времен профанации биологического оружия
307
г с машины № 17, оказались положительными на В. anthracis. Отбор проб с объектов
жружаюгцей среды не привел к случаям ингаляционной или кожной сибирской язвы
.-рели сотрудников, выполнявших эту работу.
Персонал из здания центра был эвакуирован 21 октября после того, как у первого
обогника почты лабораторно был подтвержден диагноз — ингаляционная сибир-
ская язва.
Из 2403 сотрудников центра 1961 работали в здании, где происходила обработка
•ггы. Остальные работали в административной и торговой зонах. У всех четырех
болевших сотрудников центра симптомы болезни появились на четвертые сутки
жкле обработки писем с сибиреязвенной рецептурой. Все они были инфицированы
зремя сортировки почты. Наибольшее расстояние от источника спор В. anthracis,
котором заражение стало возможном, — 60 м от DBCS-машины № 17, но оно опре-
?'_тено теоретически по их месту работы в соответствии с занимаемой должностью;
болевшие ингаляционной сибирской язвой могли находиться и ближе к машине,
v. да проходили через цех. Количество заболевших было небольшим (2,04 на ЮООче-
век) по двум причинам: 1) доза спор в аэрозоле оказалась недостаточной, чтобы
вать инфекционный процесс у большего количества работников; 2) всем работ-
- • ам цеха по сортировке писем втечение 10 сут. проводилась экстренная профилак-
- пса антибиотиками (Sanderson W. Т, 2004).
Серологическое наблюдение за лицами, потенциально экспонированными к аэро-
«ю спор В. anthracis, осуществлялось с целью выявить людей, инфицированных по-
роговыми дозами возбудителя болезни. Сыворотки были получены от 202 работников
хитра. Никто из них не имел через 6 недель после возможной экспозиции выявляе-
мого уровня специфических IgG к протективному антигену токсина сибиреязвенно-
микроба, включая даже трех сотрудников, которые ранее вакцинировались против
ирской язвы.
Сибирская язва в Капитолии. Преодолев многочисленные трудности транспорти-
ровки и сортировки, и, частично рассыпав свое смертельное содержимое, 15 октября
- .ьмо сенатору Дашли добралось до его офиса в Капитолии. В 09.45 ч оно было
ккрыто его почтовым помощником. Мелкая пыль вырвалась из конверта сразу, как
-ддько он был надрезан. Помощник увидел вложенное в конверт письмо и понял, что
- столкнулся с тем самым «белым порошком», о котором уже три дня сообщали СМИ.
- немедленно закрыл письмо и положил его на пол, затем вызвал полицию Капито-
Офицеры полиции прибыли на место происшествия втечение пяти минут. После
; эобшений о первых случаях заражения людей сибирской язвой в Нью-Йорке им вы-
1ЛИ индивидуальные средства защиты органов дыхания и рук. Поэтому, войдя в по-
чтовый офис сенатора Дашли, они быстро их надели. Затем сотрудниками полиции с
<ощью коммерческого теста дважды было выполнено тестирование «белого порош-
• на наличие в нем спор В. anthracis. Оба теста оказались положительными, вся про-
_едура тестирования заняла 15 мин. В последующем правильность их теста подтвер-
-_ти специалисты USAMRIID.
Приблизительно в 10.30 ч вентиляционная система Капитолия была выключена.
Медицинские специалисты из Office of the Attending Physician (OAP) начали делать
ки на наличие В. anthracis из носовых ходов у всех сотрудников сенатора Дашли и
: этрудников, работавших в прилегающих офисах, принадлежащих сенатору Расселу
Эайнголду (Russell Feingold). В качестве первой меры постэкспозиционной профи-
тктики (postexposure prophylaxis) всем этим служащим назначили трехдневный курс
-ипрофлоксацина, по 500 мг дважды в день.
308
Часть 1. Краткая история применения биологических средств.
Только сотрудник, открывший конверт с рецептурой сибирской язвы, был отправ-
лен на полную санитарную обработку, включавшую мытье с мылом и водой. Для всех
остальных ограничились мытьем рук с мылом. Затем их всех перевели в другое зда-
ние, где сделали мазки из носовых ходов и с одежды. В 15.00 ч их всех отпустили до-
мой. Сотрудники других офисов работали до конца рабочего дня. Утром 16 октября
юго-западное квадратное здание Капитолия было закрыто. Специалисты ОАР продол-
жили отбор проб на В. anthracis у всех сотрудников офиса сенатора Дашли, присут-
ствовавших на рабочих местах 15 октября. Более 2 тыс. сотрудникам Капитолия был
рекомендован трехдневный курс ципрофлоксацина, 625 сотрудникам, которых посчи-
тали находившимися в зоне экспозиции (exposure area), рекомендовано принимать
ципрофлоксацин в течение 60 суток.
Отбор проб из носовых ходов сотрудников офисов сенаторов Дашли и Файнголь-
да показал, что вентиляционная система здания сыграла небольшую роль в рас-
пространении спор В. anthracis. Позитивными на В. anthracis (т. е. микроорганизм
был высеян из носовых ходов этих людей) оказались сотрудники на 5-м и 6-м этажах
офиса сенатора Дашли, соединенных открытой лестничной клеткой. Закрытая дверь
между офисами стала причиной незначительной обсемененности офиса сенатора
Файнгольда.
В первый день инцидента был сделан 71 мазок (в течение первого часа) из носо-
вых ходов сотрудников Капитолия, находящихся в непосредственной близости от от-
крытого конверта. У других сотрудников такое исследование было проведено в тече-
ние 4 дней. Всего за период с 15 по 18 октября были взяты мазки на В. anthracis из
носовых ходов 2172 сотрудников, включая 625 человек, включенных в группу риска.
Из 71 сотрудника, находившегося вблизи вскрытого конверта между 10.30 и 19.00 ч.
у 28 из носовых ходов был высеян В. anthracis. Все они заходили либо в помещения
сенатора Дашли, либо сенатора Файнгольда, за исключением одного сотрудника, ко-
торый был в это время в коридоре между офисами сенаторов, находившимися на раз-
ных этажах.
Лиц, заболевших сибирской язвой в результате доставки письма с рецептурой
В. anthracis в Капитолий, не было. Мазки, взятые на седьмой день из носовых ходов
тех 28 сотрудников, у которых в день совершения террористического акта возбудитель
сибирской язвы был обнаружен, оказались отрицательными на В. anthracis. В сыво-
ротке крови этих людей, взятой на 7,21 и 42 сут после теракта, обнаружить IgG-анти-
тела к протективному антигену токсина В. anthracis, не удалось. V. Р. Hsu et al. (2002)
объясняют удачный для сотрудников Капитолия результат теракта, своевременно
начатой постэкспозиционной профилактикой.
Загадочный случай в Нью-Йорке. Не ясен источник инфицирования спорами
В. anthraciswewiimnA (легочная форма болезни), работавшей подсобной рабочей скла-
да госпиталя в Манхэттене (случай 21; табл. 1.11). В. Jemigan etal. (2002) считают, что
источником инфекции стало письмо, перекрестно-контаминированное рецептурой
возбудителя сибирской язвы во время обработки писем. Однако Holtz Т. et al. (2003)
отрицают такую возможность по следующим причинам:
1) нет никаких доказательств присутствия спор В. anthracis у нее дома и на рабо-
чем месте;
2) хотя потенциально перекрестно-контаминированное письмо после прохожде-
ния сортировочной машины в Трентоне было доставлено в ее дом, контаминирован-
ные таким же образом письма доставлены тысячам других адресатов в Нью-Йорке.
Вашингтоне и в других частях США.
12. Биодиверсии и биотеррор времен профанации биологического оружия
309
Известен только один ингаляционный случай, зарегистрированный во время этой
<пышки, который предположительно можно считать связанным с перекрестной
энтаминаиией почты — у 94-летней женщины в Оксфорде (см. ниже). Holtz Т. et al.
_i ЮЗ) выдвинули версию возможности еще одного маломасштабного террористичес-
• их» акта, в результате которого погибла эта женщина. Возможность перекрестно-кон-
аминированных писем вызывать заражение адресатов спорами сибирской язвы, они
..нивают как сомнительную.
Загадочный случай в Оксфорде. Случай ингаляционной сибирской язвы у 94-лет-
-ей женщины, уединенно жившей в своей квартире в Оксфорде (Коннектикут), за-
'гистрирован 14 ноября (случай 22; см. табл. 1.11). Данные молекулярно-генетичес-
• ию исследования свидетельствуют о том, что женщина была инфицирована тем же
таммом В. anthracis, что и другие жертвы этого теракта. Но источник ее экспониро-
гния к рецептуре возбудителя сибирской язвы, обнаружен не был, хотя его поиски
елись очень интенсивно как в ее доме, так и в других местах. Положительными на
- аличие спор В. anthracis оказались только центр обработки почты в Валлингфорде
• U.S. Postal Service Wallingford Mail Processing) и сортировочный центр в Валлингфорде,
Коннектикут (Distribution Center in Wallingford, Connecticut). На почтовое оборудова-
-ле в Валлингфорде попадали письма с контаминированного рецептурой спор
anthracis почтового оборудования в Гамильтоне (Hamilton, Нью-Джерси), поэтому
-го рассматривают основным источником перекрестной контаминации писем, дос-
тавленных в Оксфорд.
Регистрационные данные высокоскоростной почтовой сортировочной машины
а Гамильтоне показали, что письмо адресату в Коннектикуте прошло через нее через
15 секунд после прохождения письма с рецептурой, адресованного одному из сенато-
эов. Письмо было доставлено на адрес в четырех милях от того, по которому прожи-
гала женщина, погибшая от ингаляционной сибирской язвы. Этот конверт был най-
ден и на нем обнаружены споры В. anthracis, но сибирской язвы у получателя письма
они не вызвали.
Griffith К. S. et al. (2003) считают, что и этот случай трудно привести в причинно-
следственную связь с известными рассылками писем сенаторам Дашли и Лихи. Но они
полагают, что у проводимого ими эпидемиологического расследования есть принци-
пиальные методические ограничения, затрудняющие достоверность получаемых ре-
ультатов по контаминации спорами В. anthracis окружающей среды.
Прежде всего, неизвестен порог выявления использованными методами возбуди-
*еля сибирской язвы. Отсутствие культуры возбудителя сибирской язвы в отобранных
пробах, по их мнению, не означает отсутствия его спор в окружающей среде. Дом в
Оксфорде был сильно запылен и незначительные количества спор В. anthracis могли
остаться не выявленными. К тому же эти авторы признают, что они не смогли отсле-
дить движение всех писем, бывших в контакте с письмами сенаторам Дашли и Лихи.
Да и само исследование окружающей среды на наличие спор В. anthracis в Оксфорде
начато спустя месяц после того, как споры попали в почтовую систему через Трентон.
D. В. Jemigan et al. (2002) не нашли доказательств причастности конвертов, кон-
таминированных рецептурой сибирской язвы, для случаев 17, 21 и 22 (см. табл. 1.11).
Случай 5 (почтальон в Нью-Джерси) не связан с инфицированием от почтового
оборудования в Гамильтоне либо с любым другим рабочим местом, контаминиро-
ванным спорами В. anthracis. Их классифицировали как относящиеся к рассылкам от
18 сентября только потому, что люди заболели сибирской язвой до рассылки от девя-
того октября.
310
Часть 1. Краткая история применения биологических средств
Исс ледование окружающей среды. Результаты исследований распространения спор
В. anthracis в окружающей среде после их преднамеренного диссеминирования, мож-
но обобщить следующим образом.
Концентрация спор возрастала по мере приближения к их источнику.
Споры оставались на поверхностях механизмов машины № 17 и вокруг нее спус-
тя два месяца после того, как через нее прошли письма, холя ее очищали вакуумом и
мыли раствором натрия гипохлорита (sodium hypochlorite solution). Сибирской язвы
улиц, соприкасавшихся с поверхностями, контаминированными спорами В. anthracc
не возникало.
Мытье и очистка вакуумом поверхностей, контаминированных спорами В. ant-
hracis, хоть и не освобождали их полностью от спор, но значительно снижали их кон-
центрацию.
Ложные отпипательные результаты исследования образцов на В anthrat is могут
быть получены вследствие неаде кватных методов сбора спор с поверхностей, их под-
ращивания в питательной среде и последующего анализа.
Вакуумный сбор спор В. anthracis с поверхностей дает более достоверные резуль-
таты, чем получение их другими способами (ватные тампоны и т. п.). Для бактерио-
логического исследования должна быть взята вся проба.
Пробы воздуха в сортировочном центре в Вашингтоне не содержали спор В. anthra-
cis, но их смывали с поверхностей помещений через 12 сут., после того как контами-
нированные письма прошли через сортировочную машину, и спустя трое суток, после
того как вентиляция в сортировочном центре была выключена. Эти данные показы-
вают, что, по крайней мере, в течение нескольких дней споры исчезают из воздуха
контаминированных помещений.
Нет достоверных данных в пользу того, что инфицирование людей возбудителем
сибирской язвы может произойти в результате реа >розолирования спор, осевших на
поверхности предметов, окружавших человека в момент формирования первичного
аэрозоля.
Идентификация. При расследовании биотеррористической вспышки в США в
2001 г. были заложены основы нового направления в криминалистике — судебной
микробиологии. Случаи сибирской язвы среди людей в сентябре - октябре 2001 г.
выявлены в семи штатах, расположенных вдоль Восточного побережья США. Сибир-
ская язва встречалась в разных клинических формах. Механизм инфицирования лю-
дей, по крайней мере, для двух из них, так и остался неясным. Широко используемые
в 1990-х гг. для дифференциации штаммов микроор! анизмов такие методы, как мно-
голокусный энзимэлектрофорез (МЕЕ), многолокусное типирование последователь-
ностей (MLST) и анализ длины амплифицированных фрагментов (AFLP), оказались
недостаточно чувствительными для выявления генетических различий между штам-
мами В. anthracis.
Поэтому для субтипирования В. anthracis был использован разработанный Р. Keim
(2000) анализ многолокусных вариабельных тандемных повторов (multiple-locus
variable-number tandem repeat analysis, MLVA). В отличие от AFLP, его можно исполь-
зовать только для субтипирования штаммов сибиреязвенного микроба, для изучения
филогенетических связей между видами, представителями рода Bacillus, он не при-
годен (см. разд. 3.1).
MLVA позволяет определять количество копий вариабельно-количественных тан-
демных повторов (variable-number tandem repeats, VNTR) восьми генетических локу-
сов В. anthracis (шесть на хромосоме и по одному на каждой из плазмид). Способ
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
311
относительно простой и позволяет типировать множество штаммов в одном геле за
8 ч. Благодаря его использованию, ученым из CDS удалось установить, что все 22 слу-
чая сибирской язвы, выявленные в сентябре — октябре 2001 г., вызваны штаммом
Ames. Благодаря его обнаружению:
1) в содержимом почтовых конвертов;
2) в образцах тканей пострадавших от сибирской язвы людей;
3) на поверхности объектов окружающей среды, было доказано, что эти 22 случая
имеют террористическое происхождение (Hoffmaster A. R. et al., 2002).
Одновременно был очерчен круг подозреваемых — лица, имевшие доступ к штам-
му Ames (Rosenberg В. Н., 2002).
Постэкгпочиционная неспецифическая профилактика. Заключалась в обязательном
60-суточном приеме антибиотиков. Необходимость столь разру шительного для здо-
ровья человека мероприятия объяснялась длительным инкубационным периодом ин-
галяционной сибирской язвы. Якобы втечение этого периода споры В. anthracis «про-
растают» в легких человека. Столь продолжительный инкубационный период при
ингаляционной сибирской язве не был известен эпидемиологической науке до опуб-
ликования в 1994 г. статьи М. Meselson et al. (1994) о причинах вспышки сибирской
язвы в Свердловске в 1979 г. В истории нашей страны это было время, когда Запат и
российская «пятая колонна» совместно демонтировали военный потенциал, остав-
шийся России в наследие от СССР Статья М. Meselson et al. (1994) играла роль инфор-
мационного повода для уничтожения биотехнологических объектов на территории
России. Она подводила «научную базу» под обвинения СССР (России) в нарушении
Конвенции 1972 г., сделанные Госдепартаментом США еще весной 1980 г. Поэтому о
ее научной достоверности никто не заботился. Достаточно было и того, что статья
напечатана в престижном научном журнале «Science».
Эпидемиология сибирской язвы в Свердловске в 1979 г. меньше всего походила
на результат «выброса» с военного объекта (см. Супотницкий М. В., 2000). Фальси-
фикаторов смущала только продолжительность вспышки — 69 суток (почти 10 недель),
так как уже более 20 лет было известно, что инкубационный период при ингаляцион-
ной сибирской язве не превышает одной недели (см., например, работу Druett Н. А.
et al., 1953; обзор Albrink W. S., 1961; и официальный документ ВОЗ — «Health aspects
of chemical...», 1970). Поэтому M. Meselson et al. (1994) совершили научный подлог —
они привели в своей работе ссылку на статью добросовестных исследователей —
Р. Brachman et al. (1966), изучавших профессиональные риски работников предприя-
тий по переработке шерсти, как доказывающую возможность двухмесячного «прора-
стания» спор В. anthracis в легких человека «после выброса».
Р. Brachman et al. (1966) в числе профессиональных рисков рассматривали воз-
можность инфицирования людей аэрозолем спор сибирской язвы, образующемся при
переработке uiepci и, но методология их эксперимента не предус матривала однократ-
ное ингаляционное инфицирование животных с целью последующего определения
инкубационного периода болезни. Они изучали возможные обстоятельства инфици-
рования людей спорами возбудителя сибирской язвы на таком прои шодстве в Южной
Каролине. Исследователи помешали клетки с обезьянами в тех участках цеха фабри-
ки, где предполагали возможность образования аэрозоля спор В. anthracis. О том, что
этот микроорганизм содержится в поставляемой из Индии и Пакистана козьей шер-
сти, свидете .ьствовади 19 случаев кожной формы сибирской язвы, зарегис грирован-
ные на предприятии за предшествующие эксперименту 2,5 года. Обезьяны играли
роль живых датчиков, «срабатывающих» на опасные для здоровья людей пороговые
312
Часть 1. Краткая история применения биологических средств...
концентрации спор В. anthracis. Животные экспонировались к возбудителю сибир-
ской язвы не одномоментно, что было бы характерно для эпидемической модели «вы-
броса», а весь период времени их нахождения в цехе (за исключением выходных дней,
когда оборудование не работало). Большинство животных погибли в период 14—46 сут.
от начала экспонирования. Добавив к 46 сут. еще две недели М. Meselson et al. (1994)
«получили» как раз ту продолжительность инкубационного периода при ингаляци-
онной сибирской язве, которая позволила Госдепартаменту США обвинить СССР в
«выбросе с объекта по производству БО», а соответственно и в нарушении Конвен-
ции 1972. Фальсификаторы прекрасно отдавали себе отчет в том, что им нужен не
«период экспонирования», а «инкубационный период», так как ветер в Свердловске
на дату назначенного ими «выброса» постоянно менял направление, что они сами
подтверждают в своей работе данными местных метеостанций. Л. М. Гринберг (1995)
признает, что вся выстроенная М. Meselson et al. (1994) модель «выброса» действи-
тельна только в том случае, если допустить длительность инкубационного периода
при ингаляционной сибирской язве не мене 60 суток.
Работа М. Meselson et al. (1994) содержит еще и другие подлоги. Например, авто-
ры привели не ту схему локализации случаев сибирской язвы в городе, которую им дали
в ФСБ Екатеринбурга, а ту, которую «сочинили» сами с учетом направления ветра «на
юг» и 60-суточного инкубационного периода. Исходную схему привел в своей статье
другой участник «расследования», J. Guillemin (2002). Придуманная М. Meselson et al.
(1994) схема «выброса» могла вписаться в эпидемиологию точечного источника при
декларируемом направлении ветра «на юг» только в том случае, если бы инкубацион-
ный период при ингаляционной сибирской язве действительно составлял не менее
60 суток. Авторам не было стыдно рассуждать о перемещении по воздуху 1 г рецептуры
спор В. anthracis (столько было ее «выброшено», по их оценке) на 50 км (до деревни
Абрамово). В 1994 г. ни М. Meselson, ни его коллеги, не предполагали, что их подлоги
через 7 лет могут стать «руководством к действию» в самих США. Практически во всех
публикациях, посвященных постэкспозиционной профилактике во время террори-
стической вспышки сибирской язвы сентября — октября 2001 г., ссылки делаются на
работу М. Meselson et al. (1994). Если даже ее нет в списке цитированных источников
какой-то статьи, то на нее есть ссылка в статье, на которую «опираются» авторы работы
при выборе схемы постэкспозиционной профилактики (см., например, Dewan Р. etal.,
2002; Barakat L. A. et al., 2002).
Когда случился биотеррористический акт в самих США, все что было в их статье
написано по продолжительности инкубационного периода ингаляционной сибир-
ской язвы, никто из фальсификаторов опровергать не решился. 60-дневный курс пос-
экспозиционной профилактики ципрофлоксацином (ciprooxacin) был рекомендован
10,3 тыс. лицам, потенциально экспонированным к спорам В. anthracis. Еще почти
20 тыс. человек принимали ципрофлоксацин по собственной инициативе, из-за опа-
сения заболеть сибирской язвой (Jemigan D. В. et al., 2002). Но многие из тех, кому он
был назначен, принимали его только в первые дни после назначения. Сотням служа-
щих почт, потенциально экспонированных к спорам В. anthracis, ципрофлоксацин
назначили спустя почти месяц после биотеррористической атаки. Например, почто-
вые служащие в Трентоне экспонировались к аэрозолю спор сибиреязвенного мик-
роба 18 сентября, центр закрыли 18 октября, а посэкспозиционную профилактику
им назначили только 20 октября, но к 18 ноября среди них новых заболевших инга-
ляционной сибирской язвой не появилось. Поэтому эпидемиология ингаляционной
сибирской язвы в США не свидетельствовала об инкубационном периоде в 60 сут,
. 12. Биодиверсии и биотеррор времен профанации биологического оружия
313
что заставило отдельных ученых задуматься о целесообразности такой продолжитель-
ности схемы посэкспозиционной профилактики. Но подлог оказался настолько жи-
вучим, что только в 2009 г. сотрудникам USAMRIID N. J. Vietri et al. (2009) удалось
окончательно установить необязательность схемы 60-дневного приема антибиоти-
ков после экспонирования к аэрозолю спор возбудителя сибирской язвы.
Брюс Эдвард Айвинс (Bruce Edwards Ivins, 1946—2008/ Это сотрудник USAMRI ID,
де он проработал 18 лет. Брюс занимался различными аспектами генетики, патогенеза
и иммунологии сибирской язвы. В 1980—1990-х гг. он проводил генно-инженерные
эксперименты с возбудителем сибирской язвы и близкородственными бациллами.
Брюс не подходит под описание преступника, данного искушенной в военно-биоло-
гических интригах В. Н. Rosenberg (2002) Он был всего лишь военным ученым и не
имел отношения к ЦРУ (см. выше «Расследование биологического террористического
акта»). ФБР на основе косвенных улик определила его как организатора и единствен-
ного исполнителя биотеррористического акта в США в 2001 г.
Представитель ФБР объяснил позицию своего ведомства тем, что Бюро подозре-
вало Брюса с 2002 г., поскольку тот разрабатывал сибиреязвенную вакцину и мог быть
заи а гересован в развитии вспышки сибирской язвы для привлечения интереса к своим
разработкам. Это заявление не вяжется с тем, что у Брюса было еще несколько интерес-
ных и самодостаточных направлений исследований. В их числе:
1) подбор штаммов возбудителя сибирской язвы, способных «пробивать» защит-
ное действие коммерческих сибиреязвенных вакцин;
2) изучение защитной эффективности антибиотиков нового типа при ингаляци-
онной сибирской язве.
Кроме того, он не имел никакого отношения к реализации сибиреязвенных вак-
цин. Этим занимались коммерческие структуры, непосредственно связанные с ми-
нистром обороны США, но не с USAMRIID. Патенты на два изобретения Брюса при-
надлежали не ему лично, а Правительству США, и не использовались вследствие того,
что к производству лицензированной сибиреязвенной вакцины AVA они не имели
отношения. Возможно, Брюс действительно попал в поле зрения спецслужб в 2002 г.,
но по другим причинам. В разгар официальной истерии о причастности исламских
террористов к терактам сентября — октября 2001 г., он отказался дать заключение, что
в _остав «белого порошка» входит бентонит. Это была, с его стороны, серьезная по-
литическая ошибка. Уже несколько лет бентониту американская разведка громогласно
приписывала роль наполнителя биологических рецептур Саддама. Брюс не был ло-
мтиком. он настаивал на том, что бентонит по своим физико-химическим свойствам
не может входить в состав таких рецептур. Совершив одну политическую ошибку, он
сразу же допустил другую, еще более непростительную. Он опознал субстанцию, на-
званную в ФБР «какой-то формой двуокиси кремния» и игравшую роль наполнителя
в рецептуре, рассылаемой в почтовых конвертах. Оказалась, что она хорошо ему зна-
кома по секретным работам его коллеги, Уильяма Патрика III (William С. Patrick III,
1926—20101), патриарха и ведущего специалиста США по разработке рецептур для био-
логических боеприпасов. Брюс также сыграл важную роль в изучении реальной про-
должительности иню бационного периода при ингаляционной сибирской язве. Он
был в числе сотрудников USAMRIID, экспериментально установивших необязатель-
ность схемы посгэкспозиционной профилактики, предполагавшей 60-суточный при-
ем антибиотиков после экспозиции к аэрозолю спор возбудителя сибирской язвы.
1 William С. Patrick III в 1950-е гг. возглавлял американскую программу по созданию БО.
314
Часть 1. Краткая история применения биологических средств...
Утром 27 июля 2008 г. Брюса нашли в бессознательном состоянии на полу в ван-
ной комнате собственного дома. Через два дня он умер в госпитале от почечной и
печеночной недостаточности, вызванных передозировкой Тайленола PM (Tylenol РМ).
6 августа 2008 г. федеральный обвинитель, прокурор Джеффри Тэйлор (Taylor), офи-
циально сделал утверждение, что Брюс был единственным преступником, организо-
вавшим биотеррористический акт в 2001 г. Более подробно о его жизни и смерти см.
в статье М. В. Супотнинкого (2009).
Эпидемиологические особенности вспышки. Характерными эпидемическими осо-
бенностями вспышки сибирской язвы в США в 2001 г. было появление несвязанных
между собой очагов болезни, представленной в двух клинических формах — ингаля-
ционной и кожной. Появление случаев болезни было растянуто во времени почти на
месяц. Притом, что инкубационный период сибирской язвы после экспонирования
человека к аэрозолю спор В. anthracis не превышал 4,5 сут. По предложенному нами
ранее делению сценариев биологического теракта на те, когда биологический агент
применяется террористами однократно, и на те, когда террористы применяют его
многократно (Супотницкий М. В., 2000), этот террористический акт относится к сце-
нариям с многократным применением агента БО. Эпидемиология такой вспышки ока-
залась очень непростой для своевременного установления ее причин. Первые семь
случаев сибирской язвы не были распознаны как искусственно вызванные. Трудно-
сти эпидемиологам добавила кожная форма сибирской язвы, не только не считавша-
яся возможной при сценариях биологических террористических актов, описанных
прежде (см., например, работу Franz D. et al., 1997), но и оказавшаяся совершенно
неожиданной для городских эпидемиологов. Основную роль в раннем распознании
искусственного характера вспышки сибирской язвы сыграли письма террористов.
Однако «невозможная эпидемиология» болезни также обратила на себя внимание ме-
дицинских работников, по крайней мере, со случая 8. Ни для одного больного с кож-
ной формой сибирской язвы невозможно было установить связь с естественным для
болезни в такой клинической форме источником возбудителя (мясо и шкуры боль-
ных животных, почва), а для больных с ингаляционной формой болезни — связь с про-
изводственными процессами по переработке шерсти. Количество заболевших людей
возрастало по мере приближения к источнику аэрозоля. Наибольшее расстояние от
источника спор В. anthracis, на котором заражение стало возможном — 60 м (при ин-
тенсивном встряхивании конвертов с рецептурой). При выяснении этиологии болезни
наиболее информативным оказалось использование ПЦР. Но определение масшта-
бов биотеррористического акта было выполнено методом анализа многолокусных
вариабельных тандемных повторов штаммовой принадлежности В. anthracis, высеян-
ной из образцов, собранных в очагах болезни. Не получено достоверных данных в
пользу того, что инфицирование людей возбудителем сибирской язвы может произой-
ти в результате реаэрозолирования спор В. anthracis, осевших на поверхности предме-
тов, окружавших человека в момент формирования первичного аэрозоля.
♦ * *
Установление факта умышленного применения биологических агентов для пора-
жения людей и его расследование — чрезвычайно сложная задача, решение которой
возможно только при координации действий эпидемиологических служб, научно-
исследовательских организаций и правоохранительных органов. В то же время эпиде-
миологический метод может оказаться единственным, который позволит обнаружить
сам факт совершения такого теракта или диверсии. При расследовании инцидента
1.12. Биодиверсии и биотеррор времен профанации биологического оружия
315
в медицинском центре в Техасе основными признаками умышленного однократного
применения биологического агента стали сжатая эпидемическая кривая, «невозможная
эпидемиология» и локализованность эпидемического очага. Прм многократном приме-
нении биологического агента в городке Даллас в 1984 г. и во время «почтовой эпидемии»
2001 г. в США основное доказательное значение искусственного характера вспышки
инфекционной болезни имело установление ее «невозможной эпидемиологии».
Установление «невозможной эпидемиологии» искусственных вспышек инфекци-
онных болезней стало возможным благодаря тому, что эпидемиологи не смогли об-
наружить естественные источники их возбудителей и проследить всю цепочку, по
которой они проникали в человеческие популяции. Например, такую цепочку не уда-
ось замкнуть между продуктами питания, вызвавшими инфицирование людей в
Далласе и в медицинском центре в Техасе, и исходными продуктами или полуфабри-
катами. Не соединялись ее звенья и во время вспышки сибирской язвы в США в 2001 г.
Для больных с кожной формой болезни не установлены контакты с больными живот-
ными, их мясом, шкурами, почвой. Для больных с ингаляционной формой болезни
не установлены эпизоды, когда могло произойти их экспонирование к аэрозолю спор
возбудителя сибирской язвы, образующемуся во время производственных процессов
по переработке шерсти или лабораторных экспериментов с аэрозолями. Также при
расследовании этих случаев умышленного применения биологических агентов для
инфицирования людей эпидемиологами были получены доказательства того, что
вызвавшие вспышки штаммы микроорганизмов являются «невозможными» для дан-
ного региона или механизма инфицирования.
Для получения доказательств идентичности штаммов, обнаруженных у подозре-
ваемых, штаммов, вызвавших вспышку, и для сравнения штаммов микроорганизмов,
выделенных из разных источников, использовали культуральные, биохимические,
серологические и молекулярные методы исследования. Наиболее достоверные резуль-
таты при определении их идентичности были получены при сопоставлении плазмид-
ного состава бактерий, рестрикционным анализом обнаруженных у бактерий плаз-
мид, пульс-электрофорезом рестриктов хромосомной ДНК, ПНР и анализом много-
локусных вариабельных тандемных повторов (для В. anthracis).
Однако эпидемиологический метод ограничен по своей сути; он дает возможность
выявить опасность и установить возможный способ контаминации, но не может оп-
ределить мотив преступления, конкретных исполнителей и организаторов теракта.
ЧАСТЬ 2
ОБЩАЯ НЕПРАВИЛЬНАЯ ЭПИДЕМИОЛОГИЯ
Подобно классической эпидемиологии инфекционных болезней, неправильная
эпидемиология может быть разделена на два раздела, логически следующих из сути
рассматриваемого материала - общего и частного. Как и любая другая наука, непра-
вильная эпидемиология должна иметь объект исследования, цель и задачи.
2.1. ОБЪЕКТ, ЦЕЛЬ И ЗАДАЧИ НЕПРАВИЛЬНОЙ ЭПИДЕМИОЛОГИИ
Эпидемиология искусственных эпидемий (вспышек). Этиология искусст-
венных эпидемий и поражений токсинами. Клинические проявления ис-
кусственных эпидемий (вспышек). Ограниченность общей и частной эпи-
демиологии при описании новых средств биологического поражения
людей. Терминологическое определение неправильной эпидемиологии.
Для понимания объекта, цели и задач неправильной эпидемиологии сопоставим
этиологический, клинический и, собственно, эпидемический аспекты искусственных
и естественных эпидемий (вспышек) инфекционных болезней (табл. 2.1).
Таблица 2 1
Сопоставление естественных и искусственно вызванных эпидемий
(вспышек) по основным признакам*
Признак Естественная Искусственная
1 2 3
Эпидемиологический аспект
Масштабность и длительность эпидемии Определяют: сложность и эшелони- рованность природных резервуаров возбудителей инфекционных болез- ней; многообразие эпидемических цепочек, дублирующих друг друга; продолжительность активности при- родного очага возбудителя инфекци- онной болезни; наличие высоковос- приимчивой к возбудителю популя- ции людей; эффективность противо- эпидемических мероприятий Определяют: общий замысел преступ- ления; кратность применения биоло- гического агента; количество, качество и физико-химические свойства приме- няемой биологической рецептуры: метеоусловия; биологические свойства самого агента (вирулентность, резис- тентность к лекарственным препара- там, способность преодолевать специ- фический иммунитет и др.)
Завершение эпидемии Угасание природного очага возбуди- теля инфекционной болезни. Само- торможение эпидемического процес- са из-за формирования иммунной прослойки среди населения. Эффек- тивная работа противоэпидемиче- ской службы Достижение цели, которую ставили перед собой злоумышленники. Эффек- тивная работа противоэпидемической службы. Пресечение деятельности био- террористической (диверсионной) орга- низации правоохранительными орга- нами
2.1. Объект, цель и задачи неправильной эпидемиологии
317
Продолжение табл. 2.1
1 2 3
Информацион- ная составляю- щая эпидемии Формируется стихийно как отра- жение представлений об эпидемиях в массовом сознании населения, коррупционных и коммерческих интересов групп, пытающихся «заработать» на эпидемии Планируется заранее и осуществляется последовательно как основная состав- ляющая биологического террористиче- ского акта
Этиологический аспект
Свойства этиологиче- ского агента Этиологическим агентом является патогенный микроорганизм с характерными для него биологическими свойствами Этиологическим агентом является спе- циальная рецептура, имеющая опреде- ленную дисперсность частиц и содер- жащая патогенный микроорганизм. Биологические свойства возбудителя инфекционной болезни могут отли- чаться от встречающихся во время природных вспышек (устойчивость к отдельным лекарственным препаратам, повышенная вирулентность, способ- ность преодолевать иммунитет, вызван- ный вакцинацией и др.)
Инфициро- вание человека этиологиче- ским агентом Осуществляется способом, характерным для эпидемиологии данной инфекционной болезни Осуществляется способом, выбранным преступниками (ингаляционно, энтерально или инъекционно)
Поддержание этиологиче- ского агента Обычно поддерживается в при- родных очагах, на территории которых проживает периодически инфицируемая популяция людей; иногда имеют место «завозные случаи», но для них можно про- следить эпидемическую цепочку, начинающуюся с территорий природных очагов этой болезни Инфицирование людей осуществляется штаммами, селекционированы ми и поддерживавшимися в лабораторных условиях
Идентифи- кация этио- логического агента Проводится для установления причины эпидемии, нозологии болезни, выявления эпидемических цепочек и др. подобных целей; выявление тонких генетических отличий между штаммами, вызвав- шими вспышку, не обязательно Выявление тонких генетических отличий между штаммами, вызвавшими вспышку, обязательно, так как позво- ляет установить факт искусственного вмешательства в геном микроорга- низма, определить масштабы теракта (диверсии) и получить доказательства причастности отдельных лиц к их орга- низации
Клинический аспект
Роль возбудителя инфекционной болезни в развитии клиники болезни Характерная нозологическая фор- ма. Особенности клинического течения болезни определяет спе- цифичность возбудителя,его вирулентность, доза заражения, и особенности реакции иммунной системы индивидуума на возбудитель ба гезни На клинику болезни также будет влиять способ применения биологического агента; физические и химические свой- ства биологической рецептуры, в соста- ве которой оудет находиться биологиче- ский агент
318
Часть 2. Общая неправильная эпидемиология
Продолжение табл. 2.1
1 2 3
Роль возбудителя болезни в патогенезе болезни Возбудитель инфекционной болезни является пусковым механизмом патогенеза болезни, уже известного из наблюдений над прошлыми эпидемиями Патогенез болезни будет зависеть от места аппликации возбудителя болезни, от физических и химических свойств биологической рецептуры, в составе которой он будет находиться
Патологоанато- мическая и патоморфо- логическая диагностика Посмертная диагностика инфекци- онной болезни Установление механизма и пути инфи- цирования жертвы биологического нападения, установление технического уровня, достигнутого нападавшей сто- роной при создании БО и организации биологического нападения
* При рассмотрении признаков естественной эпидемии использована работа Б. Л. Черкас-
ского (2001).
Эпидемиология искусственных эпидемий (вспышек). Внешнее сходство между ес-
тественными и искусственными эпидемическими процессами заключается в появле-
нии на определенной территории за определенный период времени людей с одним и
тем же инфекционным заболеванием, однако единой внутренней логики у них нет.
Масштабность и длительность этих процессов зависят от различных факторов. Клас-
сическая триада эпидемического процесса предполагает наличие: источника возбу-
дителя инфекции (например, эпизоотии среди диких животных), механизма передачи
возбудителя инфекции (наличие инфицированных насекомых, способных паразити-
ровать на людях и животных; например, блохи как переносчики возбудителя чумы от
больных чумой крыс к людям) и восприимчивости населения кданной инфекции. Слож-
ная и эшелонированная организация природных резервуаров возбудителей инфек-
ционных болезней, многообразие эпидемических цепочек, дублирующих друг друга,
продолжительная активность природного очага возбудителя инфекционной болезни
могут превратить естественно возникшую эпидемию в катастрофу. Но выключение
(искоренение) любого из факторов эпидемического процесса приводит к его оста-
новке (Черкасский Б. Л., 2001). Уже по этим причинам о механизмах развития искус-
ственного эпидемического процесса, о его структуре, движущим силам, проявлени-
ям и типам, нельзя судить на основе представлений о естественном эпидемическом
процессе.
Триада при искусственных эпидемических процессах иная. Источниками возбу-
дителя инфекционной болезни при применении БО и осуществления биологиче-
ских убийств или терактов становятся специальные устройства, разработанные для
ведения биологической войны, осуществления биологических убийств и терактов
(см. рис. 1.24,1.34—1.36,1.47—1.52,1.58). Например, при воздушно-капельном инфи-
цировании возбудителем инфекционной болезни в природном очаге источником
инфекции является больной человек. Дисперсность капелек мокроты (200—50 мкм),
образующихся при кашле или чихании, не позволяет им распространяться на значи-
тельное расстояние от источника. Капли такого размера движутся через воздух (газо-
вую фазу) и быстро оседают. Как правило, заболевают люди, находящиеся в непосред-
ственной близости от больного. Масштабируют вспышки воздушно-капельных
инфекций любые факторы внешней среды и санитарного состояния территорий,
которые приводят к скученности людей. При применении биологического агента тер-
рористами его источником является техническое устройство. Это устройство форми-
11. Объект, цель и задачи неправильной эпидемиологии
319
7>ет механизм передачи возбудителя инфекционной болезни — облако частиц мелкодис-
персного биологического аэрозоля. Такие частицы движутся вместе с воздухом (газо-
на фазой), т. е. ведут себя как газ и могут распространяться на значительно большие
сстояния, чем капельки мокроты больного человека. Критическую роль в масшта-
ровании вспышки болезни играют совершенно иные факторы, чем при воздушно-
капельной инфекции. Прежде всего, это: 1) физическая и биологическая устойчивость
:эрозоля; 2) дисперсность вдыхаемого человеком аэрозоля (см. ниже); 3) масштаб-
ость и кратность биологических атак (Ситников М. Н., 1968) (см. рис. 1.52).
При распространении возбудителя инфекционной болезни или токсина с пищей
механизм передачи будет заложен преступниками в технологической цепочке приго-
-овления продукта. Масштаб и длительность искусственной вспышки инфекционной
болезни определяются не только техническими характеристиками источника возбу-
дителя инфекции, но и замыслом самого преступления (см. рис. 1.52). Следователь-
-о, основной особенностью искусственных эпидемических процессов, позволяющей
"тличать их от природных эпидемических процессов, является их неправильная эпи-
демиология, т. е. обнаружение случаев инфекционных болезней среди людей при от-
сутствии в природе условий для их появления (Супотницкий М. В., 2000).
Выявление источника возбудителя инфекции, путей и факторов его передачи
ачинается с эпидемиологического обследования эпидемического очага. Но поня-
-ие «эпидемический очаг» как место пребывания источника возбудителя инфекции
больной человек при кори; больной человек и вши — при сыпном тифе; суслики,
сурки и мышевидные грызуны, вовлекающиеся в эпизоотии чумы и блохи, передаю-
щие чумную палочку от них в популяции людей) и прилегающая территория, в пре-
делах которой возбудитель инфекции предается к людям (подробно см. Зуева Л. П.,
Яфаев Р. X., 2006), не может быть применено при эпидемиологическом обследова-
нии очагов инфекционных болезней, образовавшихся в результате биологического
нападения. В этом случае протяженность очага, его границы будут определяться не
особенностями поддержания в природе возбудителя инфекционной болезни и усло-
виями социально-бытовой обстановки в очаге, а техническими возможностями пре-
ступников по применению биологических агентов; выбранным ими объектом престу-
пления; моделью территории, на которой совершено биологическое нападение;
метеоусловиями в момент применения биологического агента и другими подобными
факторами (см. рис. 1.62).
Эпидемический очаг в общей эпидемиологии считается ликвидированным тог-
да, когда обезврежен или блокирован источник возбудителя инфекции (обсервация,
изоляция, госпитализация заболевших людей); окружающие проверены на носитель-
ство возбудителя болезни и свободны от него; в очаге проведены мероприятия по
уничтожению возбудителя болезни или переносчиков инфекции (дезинфекция, де-
зинсекция, дератизация); когда истекает максимальный срок инкубации при данной
болезни и среди лиц, общавшихся с больным, нет больше случаев болезни. Для лик-
видации же искусственного эпидемического очага этих мер недостаточно. Искусст-
венный эпидемический очаг может возникнуть вновь без всякой связи с природными
очагами возбудителя болезни. Появление таких очагов будет продолжаться либо до
ликвидации террористической (диверсионной) организации, либо до достижения
террористами поставленной цели (см. рис. 1.53). Прекращение искусственно вызван-
ной эпидемии (вспышки) или серии убийств с помощью биологического токсина за-
висит уже не от действий эпидемиологов, а от способности правоохранительных ор-
ганов обнаружить и ликвидировать террористическую (диверсионную) организацию.
320
Часть 2. Общая неправильная эпидемиология
Этиология искусственных эпидемий и поражений токсинами. Даже при одной нозо-
логической форме болезни, ее этиология при искусственных и естественных эпиде-
миях может различаться. Инфицирование людей возбудителем инфекционной болез-
ни при естественно развившихся эпидемиях происходит в их природных очагах либо
трансмиссивно — посредством переносчиков (блохи для возбудителя чумы); контак-
тно, через повреждения кожи и слизистых поверхностей — например, при разделке
туш животных, погибших от сибирской язвы; алиментарным путем — употребление
в пищу продуктов и воды, контаминированных возбудителями кишечных инфекций
воздушно-капельным путем — первичная легочная чума. При любом из перечислен-
ных способов инфицирования человека, способность микроорганизма вызывать ин-
фекционный процесс и его летальная доза зависят от его биологических свойств (ви-
рулентность, токсигенность и др.); биологии переносчика, если он присутствует в
эпидемической цепочке; и восприимчивости к возбудителю инфекции человека, зам-
кнувшего эпидемическую цепочку.
При ингаляционном применении того же микроорганизма (токсина) его способ-
ность вызывать поражение людей будет зависеть, прежде всего, от физико-химических
свойств биологической рецептуры1 и способа ее применения. Именно эти два фактора
определяют «полосу свободы» в действиях террористов. Поражение людей сибирской
язвой во время «почтовой эпидемии» 2001 г. в США стало возможным благодаря
использованию террористами биологической рецептуры, предназначенной для сна-
ряжения кассетных биологических боеприпасов американской армии. После вскры-
тия конверта рецептура распространялась воздушными потоками по помещениям.
Другим способом в условиях существовавших тогда в Конгрессе США мер безопас-
ности вызвать вспышку опасной инфекции среди конгрессменов было невозможно
При ингаляционном заражении определяют летальную дозу возбудителя инфек-
ционной болезни или токсина не только их биологические свойства, но глубина про-
никновения частиц биологического аэрозоля в дыхательные пути человека и их пер-
вичное распределение в различных отделах дыхательной системы. Только частицы с
размером 1-3 мкм и меньше могут достигнуть альвеол. В альвеолах микроорганизмы
фагоцитируются альвеолярными макрофагами и лейкоцитами, и вместе с ними по-
падают или непосредственно в кровеносные капилляры легкого и по ним разносятся
по всему организму; либо поступают в лимфатические сосуды, оттуда в регионарные
лимфатические узлы, и далее в грудной проток и венозную кровь (см. рис. 1.46).
Если размер частиц аэрозоля увеличивается с 2—3 мкм до 12 мкм, то LD502 для
возбудителя чумы повышается в 2,5 раза (Druett Н. A. et al., 1956а); для возбудителя
сибирской язвы — в 17 раз (Druett Н. A. et al., 1953), а для возбудителя бруцеллеза
Brucella suis — в 600 раз (Druett Н. A. et al., 1953b). Токсичность рицина снижается, если
увеличивается средний диаметр аэрозольных частиц. При попадании в трахею рицин
высокотоксичен. Но этот же токсин, из-за большого размера диспергированных час-
тиц застрявший в носовых ходах, показывает очень низкую токсичность для экспе-
риментальных животных (цит. по Lamanna G., 1961; см. разд. 3.11).
1 Биологическая рецептура — это многокомпонентная система, содержащая патогенные
микроорганизмы (токсины), наполнители и стабилизирующие добавки, обеспечивающие
повышение их устойчивости при хранении, применении и нахождении в аэрозольном состо-
янии. В зависимости от агрегатного состояния рецептуры могут быть сухими или жидкими.
2 LD5o (англ, lethal dose, 50 %) — средняя доза вещества, вызывающая гибель половины
членов испытуемой группы.
2 Объект, цель и задачи неправильной эпидемиологии
321
Врач может столкнуться с тем, что штамм возбудителя инфекционной болезни,
. гальзованный для биологического нападения, прошел селекцию в лабораторных
•_товиях. Благодаря этому, он оказался способен преодолевать иммунитет, создавае-
«ый коммерческими вакцинами, либо вызванная им болезнь не поддается лечению
типовым схемам1. Ситуация с этиологией болезни еще более запутывается, когда
.•пользованный для биологического нападения штамм микроорганизма генетичес-
< и изменен (см. разд. 2.2.2).
Бактериальный токсин может быть подвергнут модификации, в результате чего его
- имуногенные свойства не совпадут с иммуногенными свойствами анатоксинов,
.-пользуемых для специфической профилактики поражения или антитоксинов, вво-
гмых пациенту для специфического лечения. Либо будет изменена специфичность
j ействия этого токсина (см. разд. 2.2.3.6).
Штамм возбудителя сибирской язвы Ames, обнаруженный в составе биологиче-
. гой рецептуры, примененной для осуществления террористического акта в США в
I •»! г., оказался устойчив к отдельным антибиотикам Р-лактамного ряда и обладал
_ юсобностью «пробивать» защитное действие коммерческих сибиреязвенных вакцин.
Хотя он имеет природное происхождение, но в природных очагах сибирской язвы
таммы В. anthracis с такими свойствами встречаются исключительно редко. Отобран-
-ый по признакам антибиотикорезистентности и вирулентности, он вызвал болезнь,
-е поддававшуюся лечению антибиотикам Р-лактамного ряда (см. разд. 2.2.3.6 и 3.1).
Для врачей-инфекционистов возможности технологий селекции и генной инже-
ерии микроорганизмов и их токсинов означают, что они могут оказаться в ситуации,
< мда: 1) будет невозможно установить этиологию инфекционной болезни; 2) типо-
вые схемы профилактики и лечения поражений, вызванных микроорганизмами и
-оксинами, окажутся не эффективными.
При расследовании искусственно вызванных эпидемий (вспышек) инфекцион-
ных болезней идентификация этиологического агента проводится не только для того,
чтобы установить этиологию болезни, но и для: 1) определения масштабов теракта
диверсии); 2) для представления в суде доказательств причастности отдельных лиц к
его организации. Отсюда следуют более высокие требования к точности межштаммо-
вой дифференциации возбудителей инфекционных болезней. Для идентификации
возбудителя сибирской язвы, выделенного в очагах болезни от заболевших людей и
животных, достаточно традиционных биологических и иммунологических методов
исследования. Но установить масштабы биотеррористического акта 2001 г. в США
стало возможным благодаря субтипированию В. anthracis по его ДНК с помощью
М LVA (см. разд. 1.12.4).
Клинические проявления искусственных эпидемий (вспышек). Роль возбудителя
инфекционной болезни в развитии клиники болезни при естественных и искусствен-
ных эпидемиях (вспышках) также различна. Для эпидемий, возникающих естествен-
ным путем в природных очагах, механизм инфицирования и связанная с ним доми-
нирующая клиника болезни обычно хорошо известны врачам. Например, инфи-
цирование возбудителем чумы в природных очагах этой болезни происходит через
кожу посредством укуса блохи. Возбудитель чумы лимфогенным путем достигает ре-
гионарных лимфатических узлов, где задерживается и интенсивно размножается
В лимфатическом узле развивается воспалительный процесс, внешне это проявляется
1 Он просто не может быть иным, так как в лабораторных условиях быстро теряет виру-
лентность при пересевах на искусственных питательных средах.
322
Часть 2. Общая неправильная эпидемиология
первичным бубоном. Большинство случаев чумы в ее природном очаге развиваются
как бубонные (Шувалова Е. П. с соавт., 2001).
При эпидемиях, вызванных искусственно, клиника болезни зависит от способа
введения возбудителя инфекции в организм человека, выбранного террористами.
В этом случае врач столкнется с клинической формой болезни, ранее ему неизвест-
ной, т. е. с невозможной клинической формой болезни. «Невозможная» она в том смысле,
что не встречается при естественном заражении данным возбудителем. Например,
кожная форма сибирской язвы хорошо известна инфекционистам. Она наблюдается
среди людей, контактирующих с мясом и шкурами болевших сибирской язвой жи-
вотных. Ее характерное проявление — сибиреязвенный карбункул, обычно появляю-
щийся на открытых частях тела. Однако при подкожном и внутримышечном введении
В. anthracis человеку развивается незнакомая инфекционистам локальная форма сибир-
ской язвы, протекающая без развития карбункула, но с обширными некрозами и оте-
ками в участке инъекции.
При в/в введении В. anthracis смерть человека клинически напоминает «апоплек-
сический удар». При вскрытии — никакой специфики. Выявляются только признаки
серозного или серозно-геморрагического полиаденита, полнокровие и дистрофиче-
ские изменения в паренхиматозных органах и кровоизлияния в легких. Такие случаи
сибирской язвы были зарегистрированы в 2009 г. в Шотландии у наркоманов, купив-
ших афганский героин одной партии, к которому были подмешаны споры возбуди-
теля сибирской язвы турецкого происхождения (Супотнинкий М. В., 201 Г, Hicks К. W
etal., 2012; Price Е. Е. etal., 2012).
Частным случаем невозможных клинических форм болезни является «патология
мелкодисперсного аэрозоля», т. е. патология, развивающаяся у человека при инфици-
ровании его аэрозолем возбудителя инфекционной болезни с размером дисперсной
фазы менее 5 мкм. Например, в природных очагах чумы первичная легочная чума
распространяется воздушно-капельным путем (Маньчжурия, 1910 г.). Воротами ин-
фекции являются миндалины и лимфатическая ткань глотки. Поэтому инфекцион-
ный процесс клинически проявляет себя нисходящей бронхопневмонией с пораже-
нием верхних долей легких. При поражении мелкодисперсным аэрозолем развивается
катаральная узелковая пневмония (см. разд. 2.5.1).
Поражение человека, вызванное вдыханием аэрозоля спор В. anthracis с размеров
частиц биологической рецептуры, приближающихся к размеру споры (1,2 мкм), кли-
нически проявляется системной болезнью с вовлечением в инфекционный процесс
паренхимы легких и лимфатических узлов средостенья. Вдыхание такого же аэрозо-
ля, но с размером частиц 12 мкм и более, приводит к развитию локализованной ин-
фекции с выраженным отеком лица (Jemigan J. A. et al., 2001; Lindler L. E. et al., 2005)
При инфицировании обезьян Macacca mulatto аэрозолем с жидкой фазой возбу-
дителя туляремии штамма Shu-S-4 с дисперсностью частиц 1—8 мкм, первичный па-
тологический процесс и размножение агента начинаются в области респираторных
бронхиол, где развивается специфический бронхиолит, распространяющийся в при-
лежащую легочную ткань и захватывающий альвеолярные пространства, вызывая
развитие мелкоочаговой пневмонии. Экспонирование животных к монодисперсно-
му туляремийному аэрозолю с размером дисперсной фазы 18 мкм ни у одной из обе-
зьян не вызвало поражения со стороны легких. Воротами инфекции служили носог-
лотка с поражением регионарных лимфоузлов, откуда возбудитель попадал в кровоток,
вызывая гематогенную диссеминацию и гибель животного (White J. D. et al., 1969)
Ангинозно-бубонная форма туляремии возникает при ингаляционном инфицирова-
Объект, цель и задачи неправильной эпидемиологии 323
аэрозолем с размером частичек более 5—10 мкм, задерживающихся в носоглотке,
• инфицирующих миндалины. Глазо-бубонная форма туляремии развивается при
«ипакции на конъюнктиву относительно крупных частиц аэрозоля (более 25 мкм)
Згарков В. И., Гапочко К. Г., 1975).
Клиника ингаляционной формы сибирской язвы, вызванной мелкодисперсны-
аэрозолями разного происхождения, но одинаковой дисперсности, так же может
-е совпадать из-за фактора рецептуры, компоненты которой сами могут обладать ток-
. -еским действием. За много лет наблюдений над вспышками ингаляционной си-
i - ?ской язвы естественного происхождения ни разу не было обнаружено патологи-
- .ких процессов в паренхиме легких. До биотеррористического акта 2001 г. в США
_ «галось, что первично-легочной сибирской язвы не существует. На рентгенограмме
— дной клетки пациента может быть обнаружено только выраженное расширение ме-
стинальных узлов при отсутствии признаков первичной пневмонии (Бурга-
П. Н., Рожков Г. И., 1984; Braude A. I. et al., 1986). Но изучение в США в 2001 г.
«следствий ингаляционного поражения людей сухой рецептурой сибирской язвы по-
казало, что в их легких образуются легочные инфильтраты (отмечены у семи пациен-
- из 12). При этом клиническое течение болезни было очень тяжелым (см. разд. 3.1).
К свойствам биологической рецептуры, определяющим полиморфизм клиниче-
_• й картины в очаге искусственно вызванной болезни, может быть отнесено ее ка-
хтво в момент применения. Во время искусственной вспышки сибирской язвы в
ША в 2001 г. все пострадавшие контактировали с одним и тем же «белым порошком».
Но случаи сибирской язвы в Нью-Йорке и Нью-Джерси были представлены кожной
:ормой болезни; а случаи, выявленные в Вашингтоне, представлены ее ингаляцион-
- * формой. Последнее связано с лучшим качеством рецептуры, помещенной в кон-
верт. предназначавшийся для сенатора Томаса Дашли (Thomas A. Daschle), чем рецеп-
туры, содержавшейся в конвертах, направленных в редакции СМИ Нью-Йорка.
: конвертах, полученных редакциями «Эн-Би-Си» и «Нью-Йорк пост», следователя-
ФБР обнаружена увлажненная, образовавшая агломераты, рецептура. Она оказа-
. .ь неспособной проникать в глубокие отделы легких человека и вызывала только
с эжную форму болезни у лиц, имевших небольшие повреждения кожных покровов
Roseberg В. Н., 2002).
Место аппликации возбудителя болезни меняет не только ее патогенез, но соот-
ветственно патоанатомию. Поэтому во время искусственно вызванных эпидемий
вспышек) патологоанатомические и патоморфологические исследования исполь-
тются не только для посмертной диагностики инфекционной болезни, но и для
«следования обстоятельств и масштабов террористического акта; выявления тех-
-ического уровня, достигнутого террористами в разработке средств биологического
зражения. Например, патология «мелкодисперстного аэрозоля» показывает высо-
кий уровень подготовки биологического нападения. Также они могут позволить от-
личить непосредственных жертв биологической атаки от людей, заразившихся от них
«по цепочке». При искусственно вызванных вспышках легочной чумы, для первых
5удут характерны признаки поражения мелкодисперсным аэрозолем (катаральная
•зелковая пневмония), для вторых — проявления воздушно-капельной инфекции
шумная бронхопневмония).
Ограниченность общей и частной эпидемиологии при описании новых средств био-
тогического поражения людей. Если на момент подписания Конвенции 1972 г. общая
частная эпидемиология включали меры борьбы с эпидемическими процессами,
-ызываемыми микроорганизмами, рассматриваемыми специалистами как потенци-
324
Часть 2. Общая неправильная эпидемиология
альные агенты БО, то в настоящее время средства биологического поражения уже
вышли за рамки их методологии С конца 1990-х гг. развиваются принципиально иные
средства биологического воздействия на людей, не предполагающие развития инфек-
ционного процесса и при этом не являющиеся ни живыми организмами, ни биочо-
гическими токсинами. Прежде всего, это искусственные генетические конструкции
и наночастицы (см. разд. 2.2.4 и 2.25). Первые разрабатываются в рамках технологи?*
соматической генной терапии наследственных и инфекционных болезней и способ-
ны использовать те же механизмы проникновения в клетки, что и микроорганизмы,
но они вызывают патологические процессы без развития инфекционного процесса.
Вторые создаются как наноматериалы, но обладают биологической специфичностью,
обусловленной их сходством по размеру с рецепторами клеток, а их структура позван I-
ет им взаимодействовать с этими рецепторами специфическим образом. Некоторые
направления соматической генотерапии предполагают конструирование искусствен-
ных генетических конструкций, имеющих размеры менее 100 нм, т. е. наночастиг
Терминологическое определение неправильной эпидемиологии. Приведенные данны.
показывают, что этиология, клиника и эпидемиология естественных и искусственно
вызванных эпидемий (вспышек) и других массовых искусственных поражений насе-
ления, имеющих биологическую природу, имеют принципиальные отличия и moi г
рассматриваться как самостоятельные объекты эпидемиологического исследования
Таким образом, неправильная эпидемиология составляет самостоятельный раздел
эпидемиологии инфекционных болезней, для которого объектом исследования явля-
ются искусственно вызванные эпидемии (вспышки) инфекционных болезней и дру-
гие массовые поражения населения, имеющие биологическую природу.
Общая эпидемиология инфекционных болезней и общая неправильная эпиде-
миология используют метод эпидемиологического анализа, т. е. специфическую сово-
купность приемов изучения закономерностей появления множества событий (забс
леваний, смертей и др.), позволяющих установить причины возникновения и
распространения патологических состояний любой природы в популяции людей
Эпидемиологический анализ в неправильной эпидемиологии основывается на зна-
нии характерных особенностей эпидемиологии искусственных эпидемических про-
цессов и других массовых поражений населения, имеющих биологическую природу
Подобно классической эпидемиологии инфекционных болезней, неправильная
эпидемиология может быть разделена на два раздела, логически следующих из сут?
рассматриваемого материала — общего и частного. Как и любая другая наука, непра-
вильная эпидемиология должна иметь объект исследования, цель и задачи.
Цель неправильной эпидемиологии как самостоятел! лого раздела в эпидемиоло-
гии инфекционных болезней — установление искусственного характера вспышки
(эпидемии) инфекционной болезни, поражений биологическими токсинами, искус-
ственными генетическими конструкциями и наночастицами.
Основные задачи неправильной эпидемиологии сводятся к следующим:
выявление новых биологических угроз и биологических агентов, которые могут
быть использованы для преднамеренного поражения людей;
изучение общих закономерностей развития искусственных эпидемических про-
цессов, поражений биологическими токсинами, искусственными генетическими кон-
струкциями и наночастицами;
изучение потенциальной опасности и условий, реализующих потенциальную
опасность биологических агентов при их преднамеренном применении для пораже-
ния людей;
12 Биологические поражающие агенты
325
разработка подходов к эпидемиологическому расследованию искусственных эпи-
чических процессов, поражений биологическими токсинами, искусственными ге-
гическими конструкциями и наночастицами;
выявление эпидемиологических последствий преступления, совершенного с по-
эшью биологических агентов;
разработка критериев для установления деятельности биотеррористических орга-
- заций (проводится совместно с правоохранительными органами);
систематизация специальных знаний, необходимых для проведения экспертиз по
-атовным делам, возбужденным в связи с расследованием преступлений, совершен-
ых с помощью биологических агентов.
Неправильная эпидемиология, как раздел в эпидемиологии инфекционных болез-
ней, находится в тесной методической связи с клинической медициной (инфекцион-
ные и соматические болезни), микробиологией, общей и частной эпидемиологией,
'эксикологией, молекулярной биологией, аэробиологией, соматической генной те-
шией, наноматериаловедением, судебной медициной, оперативно-розыскной дея-
тельностью и криминалистикой.
* * *
Неправильная эпидемиология — это система научных знаний, обосновывающих
распознавание искусственно вызванных вспышек (эпидемий) инфекционных бо-
язней, поражений биологическими токсинами, искусственными генетическими кон-
гчрукциями и наночастицами, а также устранение (минимизацию) их последствий и
дзание содействия правоохранительным органам в выявлении организаций и пре-
-3 пников, осуществивших применение биологических агентов для поражения лю-
щ. В отличие от общей и частной эпидемиологии, в компетенцию неправильной эпи-
демиологии не входит предотвращение вспышек (эпидемий) инфекционных болезней
• поражений другими биологическими агентами. Их предотвращение является зада-
чей правоохранительных органов.
2.2. БИОЛОГИЧЕСКИЕ ПОРАЖАЮЩИЕ АГЕНТЫ
К биологическим поражающим агентам относятся специально отобранные воз-
'ители инфекционных болезней, вещества биологического происхождения и веще-
ства, действующие как молекулы биологического происхождения, способные при
мышлением применении вызывать массовые поражения людей, животных и расте-
ний. Биологические поражающие агенты — это «вещь в себе». Обычно специалисты,
тишущие на темы биологических угроз, перечисляют в табличной форме десятки
микроорганизмов и токсинов, способных вызывать поражения людей. Но оценить по
формальным признакам потенциальную опасность какого-то конкретного из них и
казать, что биологический агент, прописанный под № 5, опасней того, что пропи-
сан под № 10, будет серьезным упрощением проблемы противодействия актам умыш-
зенного биологического поражения людей. Основное преимущество преступников
черед системой здравоохранения страны и правоохранительными органами — фак-
эр внезапности. Именно преступники, сообразуясь с обстоятельствами, будут выби-
вать время и место совершения преступления. Например, секта раджинишистов,
мело используя отсутствие контроля за продуктами в салат-барах городка Даллас,
добилась инкапситации 751 человека возбудителем сальмонеллеза, не рассматривав-
326
Часть 2. Общая неправильная эпидемиологи11
мого в качестве потенциального агента БО. Возбудитель сибирской язвы вызвал па-
нику в 2001 г. в США не количеством жертв, а уже своим появлением в почтовых рас-
сылках известным в США политикам и журналистам.
2.2.1. Классификации биологических поражающих агентов
Классификация биологических поражающих агентов по степени важности для националь-
ной безопасности США. Российская классификация биологических поражающих агентов.
Классификация биологических поражающих агентов по их способна ти к репликации в орга-
низме человека. Группа реплицирующихся агентов. Группа нереплицирующихся агентов.
Классификаций биологических поражающих агентов пред тожено много. Ив зави-
симости от цели, для которой она разрабатывается, подбирается принцип делени'
(principium divisionis) биологических агентов.
Классификация биологических поражающих агентов по степени важности для нацио-
нальной безотсности США. Представлена в табл. 2.2.
Таблица 2 2
Биологические поражающие агенты, ранжированные по степени важности
для национальной безопасности США*
Категория А Категория В Ka , тория C
Variola major (натуральная оспа) Coxiella burnetii (Ку-лихорадка) Вирус Nipah
Bacillus anthracis (сибирская язва) Brucella spp. (бруцеллез) Хантавирусы
Yersinia pestis (чума) Burkholderia mallei (сап) Вирусы комариных геморрагических лихорадок (Tickbome hemorrhagic fever viruses}
Токсины Clostridium botulinum (ботулизм) Альфавирусы, включая вирусы венесуэльского энцефаломиелита лошадей, восточного и западного энцефаломиелита лошадей Вирус клещевого энцефалита
Francisella tularensis (туляремия) Рицин — токсин из растения Ri- dnus communis (касторовые бобы) Bi.pyc желтой лихорадки
Вирусы, вызывающие геморраги- ческие лихорадки: филовирусы, включая вирусы геморрагических лихорадок Эбола и Марбург Эпсилон-токсин Clostridium petfringens Полирезистентный Myco- bacterium tuberculosis
Энтеротоксин В стафилококков
Аренавирусы, включая вирусы лихорадок Ласса, Аргентинской геморрагической лихорадки и связанные с ними вирусы Salmonella spp.
Shigella dysenteriae
Escherichia coli O157:H7
Vibrio cholerae
Cryptosporidium parvum
А (наивысшая степень), В (вторая наивысшая) и С (третья наивысшая).
* По J. Guarner, S. R. Zaki (2006).
Агенты, классифицированные как «категория А», могут легко быть диссемини-
рованы злоумышленниками или обладать способностью к трансмиссии между людь-
ми; вызывают высокую летальность среди пораженных лиц, вызванные ими эпиде-
мии разрушительны для системы здравоохранения и способны вызвать панику сред».
2 2. Биологические поражающие агенты
327
вселения и социальные эксцессы. Эффективное противодействие таким эпидеми-
требует реагирования всей системы здравоохранения страны. Агенты, включенные
•категорию В», представлены опасными микроорганизмами и токсинами, поддаю-
имися диссеминированию труднее, чем агенты «категории А». Они вызывают ме-
.•е тяжелую по клинике болезнь с меньшим количеством смертельных исходов. Тре-
« категория, «С», включает недавно выявленные микроорганизмы или те из извест-
ях, к которым отмечен повышенный интерес современных исследователей. Они
. ~особиые вызвать массовую заболеваемость и смертность, нанести серьезный удар
> здравоохранению, если в будущем будут разработаны технологии их массового про-
• -водства и диссеминации. В большинстве зарубежных публикаций последних лет по
—юблемам биологической безопасности, «привязка» свойств биологических агентов
существляется применительно к агентам «категории А» этой классификации.
Российская классификация биологических поражающих агентов. Предложена
* Г. Онищенко с соавт. (2003), охватывает только поражающие агенты бактериальной,
русной и риккетсиозной природы, ранжирует их по вероятности использования для
свершения биотеррористических актов (табл. 2.3).
Г. Г. Онищенко с соавт. (2003) относят возбудители натуральной оспы, сибирской
• ’вы и чумы к наивысшей категории значимости из-за значительной летальности вызы-
лемых ими болезней. Возбудители натуральной оспы и чумы (только при легочной
:орме болезни) обладают контагиозностью. Высоко и их психологическое воздей-
.твие на население. Возбудитель чумы, по мнению этих авторов, легко может быть
одифицирован. Возбудитель сибирской язвы считается ими весьма устойчивым к
действию внешних факторов на всех этапах приготовления биологической рецепту-
ры. При ингаляционном поражении человека он вызывает крайне тяжелую болезнь,
_аже с лечением заканчивающуюся смертью в 80—45 % случаев.
К категории высокой значимости Г. Г. Онищенко с соавт. (2003) относят возбуди-
ели геморрагических лихорадок Эбола, Марбург, Ласса, оспы обезьян и холеры. Воз-
будители данной категории вызывают экзотические для России болезни с крайне тя-
. елым течением и исходом (за исключением вируса оспы обезьян).
Возбудители, отнесенные ккатегории повышенной значимости, менее опасны, чем
•збудители, отнесенные к двум предыдущим категориям, однако в странах вероят-
ного противника России они уже исследовались в качестве потенциальных поражаю-
дих агентов БО. Поэтому могут быть использованы в террористических актах, если
интересы государственных структур и отдельных террористических организаций со-
падут. Остальные возбудители, отмеченные в табл. 2.3, отнесены к категории низкой
значимости.
Классификация биологических поражающих агентов по их способности к реплика-
ции в организме человека. Обе приведенные классификации биологических пора-
жающих агентов отражают взгляды на БО, сформировавшиеся в 30-х гг. прошлого века
см. разд. 1.7), и построены на экспертных оценках специалистов, моделирующих при-
емами формальной логики некий идеальный террористический акт. Они полезны для
определения приоритетов в планировании исследований в области разработки средств
биологической защиты, где наибольшие предпочтения будут отданы самому опасно-
му агенту. Но у них есть и серьезный недостаток — выбор преступниками биологиче-
ского агента для совершения конкретного преступления зависит не от его места в «рей-
тинге», а от замысла и цели преступления, да и возможностей самих террористов.
К. тому же современные средства биологического поражения уже вышли за рамки обе-
их классификаций.
328
Часть 2. Общая неправильная эпидемиология
Таблица 2.3
Биологические поражающие агенты бактериальной ивирусной природы,
ранжированные по вероятности их использования для совершения
биотеррористического акта*
Наивысшая значимость Высокая значимость Повышенная значимость Низкая значимость
Variola major (натуральная оспа) Вирус Эбола (лихорадка Эбола) Вирус венесуэльского энцефаломиелита лоша- дей (энцефаломиелит лошадей) Legionella pneumophila (легионеллез)
Bacillus anthracis (сибирская язва) Вирус Марбург (лихорадка Марбург) Вирус лихорадки долины Рифт (лихорадка долины Рифт) Вирус Крым-Конго (Конго- Крымская лихорадка)
Yersinia pestis (чума) Вирус Ласса (лихорадка Ласса) Burkholderia mallei (сап) Вирус Хантаан (геморрагиче- ская лихорадка с почечным синдромом)
Вирус оспы обезь- ян (оспа обезьян) Burkholderia pseudomallei (мелиоидоз) Вирус желтой лихорадки (желтая лихорадка)
Vibrio cholerae (холера) Franasella tularensis (туляремия) Вирус Денге (лихорадка Денге)
Rickettsia prawazekii (эпи- демический сыпной тиф) Вирус мачупо (Боливийская геморрагическая лихорадка)
Coxiella burnetii (Ку- лихорадка) Вирус Хунин (Аргентинская ' геморрагическая лихорадка)
Brucella spp. (бруцеллез)
Salmonella typhi (брюшной тиф)
Вирус клещевого весенне- летнего энцефалита (клещевой весенне-летний энцефалит)
* По Г. Г. Онищенко с соавт. (2003)
Поэтому для рассмотрения свойств биологических поражающих агентов мы бу-
дем использовать классификацию, не учитывающую «рейтинги» двух предыдущих, но
удобную медицинским работникам и сотрудникам правоохранительных органов для
установления факта совершенного преступления (рис. 2.1).
Группа реплицирующихся агентов. Представлена живыми организмами (бактерии,
риккетсии, вирусы). Их специфическое поражающее действиех зависит от способнос-
ти агента вызывать специфический инфекционный процесс у человека. Установле-
ние факта преступления с использованием агентов данной группы начинается с вы-
явления инфицированных ими людей.
Труппа нереплицирующихся агентов. Представлена химическими соединениями и
макромолекулами разной сложности организации (токсины, искусственные генети-
ческие конструкции, нанообъекты). Их поражающее действие проявляется способ-
ностью специфическим образом воздействовать на определенные мишени в организме
1 В данном контексте специфическое поражение — поражение, которое вызвано именно
этим фактором, имеющее характерный для действия этого фактора механизм развития и кли-
нико-анатомические проявления.
12. Биологические поражающие агенты
329
чел эвека. Установление факта преступления, совершенного с использованием аген-
тов данной группы, начинается с выявления людей, специфическое поражение кото-
-ых произошло без развития инфекционного процесса. Общие свойства приведены
в разд. 2.2.3, 2.2.4 и 2.2.5. В третьей части монографии более подробно будут рассмот-
рены свойства рицина, стафилококкового токсина, ботулинических токсинов, сак-
.итоксина, тетродотоксина и палитоксина.
Рис. 2.1. Классификация биологических поражающих агентов
по их способности к репликации в организме человека
* * *
При оценке последствий преступлений, совершаемых с помощью биологических
агентов, основное значение имеют не отдельные свойства использованного биологи-
ческого агента, а оптимизированные под эти свойства замысел и цель преступления
и возможности самих террористов. Современные средства биологического пораже-
ния невозможно систематизировать в рамках привычных с 1930-х гг. классификаций
биологических агентов, предполагающих применение в качестве агентов БО мик-
роорганизмов и токсинов. Поэтому как первый шаг к созданию современной клас-
сификации биологических поражающих агентов, их целесообразно разделить по
способности к репликации в организме человека на две группы: реплицирующиеся и
череплицирующиеся агенты. Факт их применения можно предполагать на основе по-
явления у людей признаков инфекционного процесса либо специфических пораже-
ний, протекающих под «маской» соматической болезни соответственно.
2.2.2. Реплицирующиеся биологические поражающие агенты
Механизмы вмешательства в геном бактерий. Трансдукция. Трансформация. Технологии
рекомбинантной ДНК в получении штаммов бактерий с неправильной эпидемиологией. Ре-
зистентность к антибактериальным препаратам у бактерий. Механизмы развития резистен-
тности к антибактериальным препаратам у бактерий. Механизмы вмешательства в геном
вирусов. Резистентность вирусов к химиотерапевтическим препаратам. Ограничения в кон-
струировании штаммов реплицирующихся биологических поражающих агентов с изменен-
ными свойствами. Неспецифическая профилактика и лечение поражений реплицирующи-
мися биологическими поражающими агентами.
Обнаружение необычного штамма известного микроорганизма в тканях жертв
биотеррористического акта на месте преступления или в лаборатории преступников
не означает, что его можно однозначно отнести к созданным искусственно. То же са-
мое касается трудно объяснимых вспышек инфекционных болезней естественного
330
Часть 2. Общая неправильная эпидемиология
Рис. 2.2. Электронная микрофотогра-
фия двух конъюгирующих бактерий.
Между удлиненной донорной клеткой
Hfr (F+) и круглой реципиентной клет-
кой (F~) половой ворсинкой обра-
зован тонкий соединяющий мостик,
по которому осуществляется перенос
ДНК. По Г. Стент и 3. Кэлиндар (1981)
происхождения. Кратко, но в объеме, достаточном для назначения экспертиз, мы рас-
смотрим различные варианты появления таких микроорганизмов.
Механизмы вмешательства в геном бактерий. Их не менее двух: образование мута-
ций — внезапно возникающих естественных (спонтанных) или вызываемых искусст-
венно (индуцированных) стойких изменений в геноме микроорганизмов;
генетический перенос — новая генетическая информация добавляется в геном бак-
терии одним из следующих способов: конъюгацией, трансдукцией и трансформацией
Гены бактерий и риккетсий могут быть изменены как путем индуцированного му-
тагенеза, так и путем образования спонтанных мутаций. Среди таких мутантов мож-
но отобрать штаммы, устойчивые к отдельным антибиотикам. Но значительно боль-
шие возможности в конструировании новых штаммов микроорганизмов дают методы
генетического переноса.
Конъюгация. Так называется половой процесс у бактерий. Он заключается в пере-
носе генетического материала от одной бактериальной клетки к другой. При этом две
бактерии с помощью пилей (половых ворсинок, формирующихся на поверхности
донорных клеток) соединяются тонким мостиком, через который из одной клетки
(донора) в другую (реципиент) переходит по-
следовательность ДНК. Наследственные свой-
ства реципиента изменяются в соответствии с
количеством генетической информации, зак-
люченной в переданной последовательности
ДНК (рис. 2.2).
Конъюгация возможна благодаря: плазми-
дам (plasmids), внехромосомным генетическим
элементам, способным автономно размножать-
ся и существовать в цитоплазме бактериальной
клетки; и так называемым интегративным конъ-
югативным элементам (integrative conjugative
elements, ICEs; известны еще как конъюгатив-
ные транспозоны, conjugation transposons), по-
добно плазмидам перемещающимся между бак-
териями посредством конъюгации, но в отличие
от плазмид, интегрирующихся с ДНК хромосо-
мы бактерии и реплицирующихся вместе с ней
(Wozniak R. А. Е et al., 2009).
Плазмиды представляют собой молекулы
ДНК с ММ от 1 до 400 мДа, как правило, зам-
кнутые в кольцо и находящиеся в клетке в сверх-
спирализованной форме. Термин «плазмида»
введен американским генетиком Дж. Ледербер-
гом в 1952 г. для обозначения полового фактора
бактерий (F-фактора, F-плазмиды), обнару-
женного в клетках культуры кишечной палочки и ответственного за их способность
быть донорами генетического материала (молекул ДНК) при конъюгации с клетка-
ми-реципиентами (F ), не содержащими полового фактора.
В зависимости от способности или неспособности передаваться из одной бакте-
риальной клетки в другую в процессе конъюгации, различают конъюгативные (транс-
миссивные) и неконъюгативные (нетрансмиссивные) плазмиды. Конъюгационный
2 2 Биологические поражающие агенты
331
еренос плазмид обеспечивается большим набором генов, объединившихся в tra-one-
роя. содержащий не менее 35 ORF. Продукты этих генов необходимы для осуществ-
;ния двух основных этапов процесса конъюгации: 1) образования скрещивающихся
zap; 2) переносаирепликацииДНК. Неконъ-
югативные плазмиды, имеют относительно
зстую генетическую организацию. Моле-
тярная масса большинства таких плазми i
- е превышает 20 мДа. Однако и они могут
6 оггь перенесены в другую бактерию благода-
реномену мобилизации — процесса перено-
са нетрансмиссивных плазмид с помощью
рансмиссивных. Некоторые плазмиды могут
е определенной частотой включаться (интег-
рироваться) в бактериальный геном и затем
размножаться (копироваться) вместе с ним
как его составная часть. Конъюгационный
•’ еренос плазмид может осуществляться меж-
бактериями отдаленных видов, играя важ-
ную роль в их эволюции. Детально эти про-
з ессы описаны в работах А. П. Пехова(1977);
Г. Стента и 3. Кэлиндара (1981); П. Брода
1982); М. Lorenz и W. Wackemagel (1994).
По работе A. Reddy et al. (1987) рассмот-
рим простые эксперименты по получению
измененных штаммов возбудителя сибир-
ской язвы и близкородственного ему ма ге-
патогенного микроорганизма Bacillus cereus
। рис. 2.3).
Ни В. anthracis, ни использованный в дан-
ной работе штамм В. cereus не имеют конъ-
югативных плазмид. Авторы использовали
обнаруженную ими способность плазмид че-
тырех подвидов непатогенной для человека
бациллы Bacillus thuringiensis1 мобилизовать
неконъюгативные плазмиды для переносов в
близкородственные микроорганизмы. Круп-
ные плазмиды В. thuringiensis рХО13, рХО14,
рХО 15 и рХО 16 (их ММ от 50 до 120 мДа) ока
зались способными не только передаваться
конъюгацией в В. anthracis и В. cereus, но и
осуществлять мобилизацию плазмид своих
Рис 2.3. Электрофорез в агарозном геле
плазмидной ДНК из штаммов бацилл,
использованных для демонстрации моби-
лизации плазмидой рХО14 плазмид рХО1,
рХО2 и рВС16 Номера напротив светя-
щихся полос в геле соответствуют назва-
ниям плазмид: 1 — рХО1 и т. д. Широкая
полоса соответствует фрагмент ам хро-
мосомы. Дорожки 1—4 показывают плаз-
миды в штаммах, использованных для
демонстрации переноса рХО 1; дорож-
ки 5—7 показывают плазмиды в штам-
мах, демонстрирующих перенос рХО2.
По A. Reddy et al. (1987)
новых хозяев. Вирулентные штаммы возбудителя сибирской язвы дополнительно к
хромосоме содержат две крупные плазмиды, включающие гены сибиреязвенного ток-
сина и полимерного вещества капсулы, рХО1 и рХО2 соответственно. На электрофо-
реграмме результаты разделения ДН К пре тставлены следующим образом: 1 — В. thurin-
giensis subsp. toumanoffi 4059 UMl(pXO14, pBC16), донор, 2 — В. anthracis Weybndge
1 В. thuringiensis используется в сельском хозяйстве в качестве биопестицида.
332
Часть 2. Общая неправильная эпидемиология
UM44-l(pXOl), реципиент; 3 — В. anthracis Weybridge UM44-1 trl69A-4(pXOl, рХО14,
рВС16), штамм-транспипиент, производный в результате скрещивания между штам-
мами, электрофорез ДНК которых показан надорожках 1 и 2; 4 — В. anthracis Weybridge
A UM23C1-1 tr357A-16(pXOl, рХО14, рВС16), штамм-транспипиент, производный в
результате скрещивания между штаммами, электрофорез ДНК которых показан на
дорожке 3 (донор) и излеченного от плазмиды В. anthracis Weybridge A UM23C1-E
5 — В. anthracis 4229 UM12(pXO2), реципиент; 6 — В. anthracis 4229 UM 12 tr4O7A-7
(рХО2, рХО14, рВС16), штамм-трансципиент, производный в результате скрещивания
между штаммами, электрофорез ДНК которых показан на дорожках 1 и 5;
7 — В. cereus 569 UM20-1 tr412A-1 (рХО14, рХО2, рХОЗ, рВС16), штамм-трансципиент.
производный в результате скрещивания между штаммами, электрофорез ДНК кото-
рых приведен на дорожке 6 (донор) и В. cereus 569 UM20-1.
Результаты этих экспериментов весьма показательны. Возбудитель сибирской
язвы в результате конъюгации получил не только неконъюгативную плазмиду рВС16
(дорожка 3), придавшую ему устойчивость к антибиотику широкого спектра дей-
ствия — тетрациклину (Тсг), но и приобрел способность передавать эту плазмиду в
другие штаммы сибиреязвенного микроба (дорожка 4). Конъюгативная плазмида
рХО14, ставшая для рВС16 плазмидой-помощником, легко может быть элиминиро-
вана из штамма-трансципиента отработанными еще с 1960-х гг. способами (напри-
мер, серийными пассажами с новобиоцином при температуре 42 °C при селективном
давлении на ген Тсг). Сама же рВС16 представляет собой специальный многокопий-
ный вектор для клонирования генов в бациллах (более подробно о плазмиде см. в ра-
ботах Bernhard К. et al., 1978; Гловера Д. с соавт., 1988). Поэтому ген Тсг только один
из возможных генов, которые можно передать сибиреязвенному микробу с помощью
этой плазмиды. Не менее интересны результаты передачи конъюгацией рХО2 от
В. anthracis к В. cereus (дорожка 7). Получив эту плазмиду от В. anthracis, В. cereus при-
обрел способность синтезировать фактор вирулентности сибиреязвенного микроба —
капсулу. Изменилась ли вирулентность трансципиента, авторы не сообщают.
Интегративные конъюгативные элементы. Обнаружены в грамположительных и
грамотрицательных бактериях. По сравнению с плазмидами или фагами, они изуче-
ны хуже. После интеграции ICE может «вырезаться» из хромосомы и сформировать
замкнутое в кольцо молекулу, вовлекающуюся в конъюгативный перенос. Эти элемен-
ты несут гены, определяющие специфические фенотипы их хозяев (резистентность к
антибиотикам, фиксация азота и др.). Наиболее изучен конъюгативный транспозон
SXT, впервые обнаруженный в Vibrio cholerae 0139, первом холерном вибрионе не-01
серогруппы, вызвавшем эпидемии холеры на Индийском субконтиненте в 1992 г. Он
кодирует резистентность к нескольким антибиотикам, включая сульфаметоксазол и
триметоприм, ранее использовавшихся в схеме лечения холеры. Более подробно об
интегративных конъюгативных элементах см. в обзоре R. А. Е Wozniak et al. (2009).
Трансдукция. Заключается в переносе генетического материала из одной клетки в
другую с помощью вируса. У бактерий в этом процессе участвуют бактериофаги —
группа вирусов сложной морфологической и генетической структуры, способных
размножаться только в бактериях-хозяевах. Бактериофаги обладают основными при-
знаками вирусов, но называются бактериофагами потому, что могут вызывать лизис
инфицированной ими бактерии (греч. phagos — пожиратель; буквально — пожирате-
ли бактерий). Каждый тип фага инфицирует (в зависимости от его действия или спе-
цифичности) различные виды бактерий, причем фаговая специфичность связана с
антигенной структурой бактерии.
- 2 Биологические поражающие агенты
333
На электронно-микроскопических снимках фа1 овне частицы напоминают голо-
истиков. Они имеют головку, состоящую из белковых субъединиц, и сложно устро-
_ 1ный отросток, с помощью которого фаг прикрепляется к стенке бактерии. Головка
; ~а содержит плотно упакованную гигантскую молекулу ДНК, которая инъециру-
ется в клетку по каналу, находящемуся во внутреннем пространстве отростка (рис. 2.4).
Рис. 2.4. Электронные микрофотографии Е coli и ее фагов. А Клетка кишечной
палочки с прикрепленными к ней с помощью отростка частицами фага Т5. Б. Мо-
лекула ДНК Т-четного фага, освободившегося из головки фага под воздействи-
ем осмотического шока. Длина этой молекулы фаговой ДНК составляет около
50 мкм, что в 550 раз превышает длину содержащей ее головки фага. По Г. Стент
и 3. Кэлиндар (1981). В. Проникновение бактериофага Т4 в бактериальную клет-
ку: 1 — адсорбция бактериофага на поверхности клетки; 2 — инвагинация через
наружную мембрану (от) до достижения внутренней мембраны (im); 3 — слияние
с мембраной; 4 — инъекция ДНК и истечение ионов К+, как результат наруше-
ния герметичности бактериальной стенки; 5 — герметизация пространства, об-
разовавшегося между мембранами и хвостом фага. По В. Dreiseikelmann (1994)
Особые бактериальные вирусы—умеренные фаги — в процессе вегетативного раз-
множения способны случайно захватывать и переносить в другие клетки любые уча-
стки ДНК лизируемых, т. е. разрушаемых ими бактерий (общая или неспецифическая
трансдукция), но при этом не лизировать клетку своего нового хозяина. Такая бакте-
рия становится лизогенной, т. е. она содержит генетический материал бактериофага
в неинфекционной форме профага, передающего потомству способность к построе-
нию бактериофага при соответствующих физиологических условиях. Одновременно
_рофаг придает бактерии иммунитет против инфекции соответствующим гомологич-
334
Часть 2. Общая неправильная эпидемиология
ным фагом. Длина переносимого (трансдуцируемого) отрезка ДНК определяется раз-
мером белковой оболочки фаговой частицы и обычно не превышает 1—2 % бактери-
ального генома. Переносимый фагом фрагмент генома бактерии может содержать
несколько генов. Поэтому фаги играют важную роль в горизонтальном переносе ге-
нов между бактериями. Если в результате заражения друюй бактериальной клетки
умеренным фагом меняются ее биологические свойства (например, клетка приобре-
тает способность синтезировать какой-то белок), то этот феномен называют лизоген-
ной конверсией.
Внесенный фрагмент ДНК может существовать в клетке-реципиенте в виде допол-
нительного генетического элемента, эписомы, обладающего функциональной актив-
ностью. Поскольку такой фрагмент не способен воспроизводиться, при каждом кле-
точном делении он передается лишь в одну из дочерних клеток. За исключением этой
клетки свойства всего остального потомства остаются без изменений (абортивная
трансдукция). В дальнейшем фрагмент может быть либо разрушен, либо включен в
хромосому бактерии, заменив в ней гомологичный участок ДНК. В последнем случае
новые признаки, приобретенные клеткой-трансдуктантом, будут свойственны всему
потомству этой клетки (полная трансдукция).
Гены отдельных профагов, не нужные им для цикла собственного размножения,
но несущие дополнительный для клетки-реципиента груз генов и определяющие
феномен лизогенной конверсии, называют моронами (morons) или генами лизогенной
конверсии (lysogenic conversion genes). Многие мороны в профагах кодируют белки, для
которых установлена или подозревается причастность к факторам вирулентности
патогенных бактерий (табл. 2.4).
Таблица 2.4
Факторы вирулентности бактерий, наиболее часто
передающиеся бакгериофа1лми-‘
Белок Ген Фаг Бактерия-хозяин
1 2 3 4
Внеклеточные токсины
Дифтерийный токсин tax Р-фаг С. diphtheriae
Ботулинический нейротоксин Cl Фаг Cl С. botulinum
Шига-токсин stxl, stx2 H-19B Е. coli
Энтерогемолизин hly2 ФРС3208 Е. coli
Цитотоксин ctx фСТХ Р. aeruginosa
Энтеротоксин see, sei NA S. aureus
Энтеротоксин Р sep ф№15 S. aureus
Энтеротоксин А entA ф13 S. aureus
Энтеротоксин А sea фМи50А S. aureus
Эксфолиативный токсин А eta фЕТА S aureus
Токсин типа А speA Т12 S. pyogenes
Токсин типа С speC CS112 S. pyogenes
Холерный токсин ctxAB СТХф V. cholerae
Лейкоцидин pvt fPVL S. aureus
L2. Биологические поражающие агенты
335
Продолжение табл. 2.4
1 2 3 4
'уперантигены speA], speA3, speC, spel, speH, speM, speL, speK, ssa 8232.1 5. pyogenes
Цитолетальный разбухающий токсин cdt Без названия E. coli
Белки, меняющие антигенность бактерии
Мембранные белки Л/п-подобные Pnml N. meningitidis
Гтикозилаза rfb e34 S. enterica
Гликозилаза Str P22 S. entenca
О-антигенацетилаза oac Sf6 S. flexneri
Гли козилтрансфераза gtrll sni, SfV, SIX S. flexneri
Эффекторные белки, вовлеченные в инвазию
Тип 111-эффектор sppE SopE<]> S. enterica
Тип П1-эффектор ssel (gtgB) GIFSY-2 S. enterica
Тип III-эффектор sspHl GIFSY-2 S. enterica
Ферменты
Сулероксиддисмутаза sodC Sp4, 10 E. colt 0157
Супероксиддисмутаза sodC-I GIFSY-2 5. enterica
Сулероксиддисмутаза sodC-Hl Fels-1 S. enterica
Нейраминидаза nanH Fels-l S. enterica
Гиалуронидаза hylP H4489A S. pyogenes
Лейкоцидин pvl фРУГ S. aureus
] Стафилокиназа sak ф13 S. aureus
Фосфолипаза sla 315.4 S. pyogenes
Д Н Каза/стрептодорназа sdn, sda 315.6, 8232.5 S. pyogenes
Резистентность к сыворотке крови
ОМР (наружный мембранный белок) bor A. E. coli
ОМР eib Х-подобный E. coli
Другие
Митогенные факторы mf2, mf3, nf4 370.1, 370.3, 315.3 S. pyogenes
Митогенный фактор toxA Без названия P. multodda
Митогенный фактор Без названия phisc 1 S. cam's
Вирулентность gtgE GIFSY-2 S. enterica
Антивирулентность grvA GIFSY-2, Fels-1 S. ent erica
* По Н. Brussow et al. (2004).
Ниже, на примере приобретения гена шига-токсина1 нетоксигенной кишечной
палочкой, рассмотрим один из вариантов механизма лизогенной конверсии бактерий
по генам опасных токсинов. Кишечная палочка — грамотрицательная энтеробакте-
1 Шигеллы — название произошло от имени японского ученого К. Шиги (правильно —
Сига). Род микроорганизмов из семейства энтеробактерий. Включает различные виды возбу-
дителей дизентерии у обезьян и человека.
336
Часть 2. Общая неправильная эпидемиология
рия, обычный комменсал кишечника человека. Среди многочисленных штаммов
Е. coli обнаружены несколько штаммов, являющихся патогенными для человека. Ре-
зультатом лизогенной конверсии стало появление шига-токсин-продуцирующих
Е. coli (Shiga-toxin producing Е. coli, STEC).
STEC-штамм О157:Н7 впервые описан в 1982 г. (Mead Р. S., Griffin Р. М., 1998).
Е. coli О157:Н7 является патогеном слизистой кишечника и продуцирует несколько
факторов вирулентности. Среди них ген, по крайней мере, одного фактора — шига-
токсина (Stx), локализован в геноме профага (six). Ген другого фактора вирулентнос-
ти Е. со//О157:Н7 — TTSS, расположен на «островке патогенности» вблизи от профа-
га 933L. Он отвечает за синтез белка, вызывающего специфические изменения
слизистой в месте взаимодействия с ней бактерии (Рета N. Т. et al., 1998).
Шига-токсин (другое название — веротоксин) до 1982 г. был известен как токсин,
продуцируемый возбудителем дизентерии Shigella disenteriae первого серотипа. Отно-
сится к токсинам, ингибирующим синтез белка. Имеет типичную АВ структуру’. Эн-
зиматически активная А-субъединица (35 кДа) состоит из фрагментов А1 и А2, свя-
занных между собой дисульфидным мостиком. А2-фрагмент нековалентно связывает
А1 с В-пентамером (7,5 кДа). В-субъединичный пентамер связывает голотоксин с
эукариотической клеткой через специфические гликолипидные рецепторы. После
интернализации полипептид А расщепляется на энзиматическую часть (А1) и фраг-
мент А2. А1 обладает N-гликозидазной активностью и расщепляет N-гликозидную
связь у аденина в положении 4324 на 288-рибосомальной РНК. В результате проис-
ходит отщепление 400 нуклеотидов с З'-конца рРНК, что служит препятствием для
присоединения аминоацил-тРНК, синтез белка прекращается, и клетка гибнет (Вер-
тиев Ю. И., 1996).
Ключевым моментом лизогенной конверсии стало замещение профагом участка
гена rfb, кодирующего фермент, необходимый для синтеза О-стороны цепей бактери-
альных липополисахаридов (O-side chains of the bacterial lipopolysaccharides). Механизм
лизогенной конверсии E. coli по гену Stx, показан на рис. 2.5.
Шига-токсин-продуцирующие Е. coli вызывают у человека диарею, развитие ге-
моррагического колита и гемолитического уремического синдрома. Проникая через
кишечный эпителий Stx, попадает в кровь и повреждает клетки сосудов толстой киш-
ки, почек и ЦНС. В экспериментах со штаммом Е. coli, содержащим профаг Н-19В.
было показано, что вирионы фага, образующиеся в пределах кишечника мыши, спо-
собны осуществлять там же лизогенную конверсию нетоксигенных штаммов кишеч-
ной палочки по гену шига-токсина (Acheson D. W. et. al., 1998). Более подробно о био-
логии и эпидемиологии STEC можно прочитать в обзоре С. L. Gyles (2007).
Трансформация. В бактериологии к трансформации относят перенос генетической
информации в бактериальные клетки с участием плазмид или без них, но всегда — без
участия вирусов. В свою очередь делится на естественную, например, наблюдаемую
в природных экосистемах, и индуцированную, когда бактериальная клетка готовится
к переносу ДНК специальными способами.
Трансформация в природе занимает особое место среди других способов генети-
ческого переноса. Конъюгация «предъявляет» самые высокие требования к условиям
своего осуществления. Клетка-донор генетической информации должна иметь конъ-
югативный элемент (плазмиду, конъюгативный транспозон), между донором и реци-
пиентом должен установиться физический контакт, достаточно стабильный, чтобы
1 О структуре токсинов в разд. 2.2.3.6.
22 Биологические поражающие агенты
337
Рис. 2.5. Механизмы лизогенной конверсии Е. coll по гену Stx и экспрессии
Шига-токсина в кишечнике человека и животных. А. Модель регулирования
экспрессии гена six в геноме профага Н19В. pR, — промотор для позднего оперо-
на stxAB (late-gene operon). Обычно транскрипция заканчивается посредством
терминатора tR Антитерминатор Q позволяет транскрипции избегать влия-
ния tR' и экспрессировать весь оперон. Может существовать дополнительный
регулирующий промотор Fur. А- и В-субъединицы токсина включают секреци-
онные сигналы. В форме «созревшего» АВ-токсина они собираются в пери-
плазматическом пространстве. Высвобождение молекул токсина из периплазма
осуществляется путем лизиса клетки. IM — bhj грснняя мембран, ОМ — наруж-
ная мембрана. Б. Лизогенная конверсия в STEC нетоксигенных штаммов Е. coli,
присутствующих среди флоры кишечника. По Brussow Н. et al. (2004)
звонить осуществиться переносу ДНК. Должно совпасть и то, чтобы обе клетки
снор и реципиент) оказались метаболически активными. Генный перенос посред-
гщом трансдукции требует метаболической активности донорной клетки, в которой
вбираются частицы трансдуцирущих фагов. Реципиент может находиться вне тес-
-ого контакта с донором, так как генетическая информация может сохраняться в
тансдуцирующих фагах. Бактериофаг резистентен ко многим физическим и хими-
еским агентам и длительно сохраняется в окружающей среде, адсорбируясь на по-
верхности минералов и твердых частицах. Для инфицирования интенсивной метабо-
лической активности от реципиента не требуется. Но он должен быть связан с донором
зошей чувствительностью к одним и тем же бактериофагам. Перенос генов путем
рансформации не требует живых клеток-доноров, так как ДНК, высвобождаемой из
«летки во время ее гибели или лизиса, обычно оказывается достаточно, чтобы попасть
окружающую среду. Благодаря стабильности молекулы, ДНК может длительное вре-
мя находиться в окружающей среде и распространяться на некоторое расстояние от
«летки-донора. Для восприятия чужеродной ДНК клетка-реципиент должна быть
физиологически активна, однако тесной генетической связи между донором и реципи-
ентом для успешной трансформации ДНК не требуется. Таким образом, трансформа-
_ия менее других способов генетического переноса (из известных в природе) зависит
от случайных факторов. Что делает ее важным инструментом генного переноса между
бактериями, поддерживающимися в водных и почвенных экосистемах (рис. 2.6).
338
Часть 2. Общая неправильная эпидемиология
| Почва |
Рис. 2.6. Схема генного переноса посредством свободной ДНК среди бактерий
водных и почвенных экосистем. По М. G. Lorenz, W. Wckemagel (1994)
Трансформация у бактерий является физиологическим процессом. Она начинает-
ся с развития у оактерии состояния компетентности, т. е. способное! и клетки прини-
мать ДНК из окружающей среды. Трансформация как процесс, может быть разделен
на этапы или стадии, общие для всех бактерий: развитие компетентности у бактери-
альной клетки; связывания ДНК с поверхностью клетки; проникновение молекулы
ДНК в бактерию-реципиент; интеграция ДНК в хромосому через рекомбинацию (или
ее самостоятельная репликация, когда трансформация выполнена плазмидной ДН К):
экспрессия донорских генов. Более подробно о трансформации бактерий в природ-
ных экосистемах можно прочитать в обзорах М. G. Lorenz, W. Wckernagel (1994) и
В. Dreiseikelmann (1994).
Индуцированная трансформация используется для получения генетически изме-
ненных штаммов бактерий в условиях лабораторного эксперимента. Трансформация
кишечной палочки плазмидной ДНК впервые продемонстрирована S. N. Cohen et al
(1972). Наиболее распространенными способами получения искусственной компе-
тентности для Е. coh стало инкубирование бактерий вместе с ДНК при низких темпе-
ратурах в среде, содержащей двухвалентные катионы (так называемый «кальциевый
метод»). Для В. subtilis — культивирование в специальных средах и трансформация
протопластов. Способы трансформации, разработанные для Е. coli и В. subtilis перено-
сились на патогенные бактерии. В 1980 гг. их разработано большое количество для
грамположительных и грамот рицательных патогенных бактерий. Частично они обоб
щенны в руководствах Т. Маниатиса с соавт. (1984) и Д. Гловера (1988). В 1990-х гг.
в лидеры «выбился» способ электропорации (электротрансформации) бактерий
сегодня вытеснивший собой остальные. Он основан на способности высоковольт-
ного электрического разряда образовывать поры в клеточных мембранах. Через эти
поры в клетки проникает "ужеродн 1я ДНК. Способ универсален — используется дш
индуцированной трансформации прокариотических и эукариотических клеток. На
рис. 2.7. показано устройство для электротрансформации бактерий, собранное авто-
ром данной книги (Супотницкий М. В. с соавт., 1992).
U. логические поражающие агенты
339
Рис. 2.7. Схема устройства для электротрансформации бактерий
плазмидной ДНК (за основу взята схема Speyer J. Е, 1990)
В данное устройство входят блок питания постоянного тока, обычно используе-
для приборов электрофореза (1), конденсаторный блок (2), камера для электро-
"алии (3), самописец (4). Электропорационное пространство камеры образуется
- _ южением друг на друга двух отполированных до «зеркального блеска» пластинча-
-=» >, эп< :ктродов, выполненных из нержавеющей стали (электроды В и С), которые
I _ деляются прокладками из поливинилхлоридной изоляционной ленты. Электриче-
.. контакт осуществляется через неполированную стальную пластину D, на кото-
расположен электрод С, и через цилиндр, изготовленный из латуни А, прижимаю-
_ • й электрод В к прокладке. Расстояние между электродами 0,24 мм, напряженность
i г екгропорационном пространстве задается в пределах 4000-21000 В/см2. Устрой-
работает следующим образом. От блока питания на конденсаторный блок пода-
ется напряжение. В электронорационное пространство камеры вносится 10—40 мкл
сериальных клеток, суспендированных в водном растворе полимера (ПЭГ 6000,
д:чсгран и др.) и смешанных с раствором плазмидной ДНК, после чего производит-
_» разряд. Его длительность и величина фиксируются самописцем. Наиболее высо-
I частота трансформации была у клеток, взятых на стадии логарифмического рос-
-х Но способ оказался эффективным и для клеток в стационарной фазе роста, смытых
ерхности плотной питательной среды.
технологии рекомбинантной ДНК в получении штаммов бактерий с неправильной
। ’пемиологией. Генетический потенциал плазмид и фагов, распространяющихся сре-
бактерий в природных экосистемах, не предназначался природой для генетических
пуляций с бактериями в лабораториях. Он представляет собой «камень», от ко-
рого «скульптор», т е. генетик, должен «отсечь все лишнее». Целенаправленно
. лить биологические свойства бактерии можно путем доставки в нее рекомбинант-
1НК, т. е. молекул ДНК, полученных объединением в условиях in vitro разнород-
j ж, вместе нигде в природе не существующих фрагментов ДНК. Конструирование
рекомбинантной Д НК происходит с использованием специальных векторов — само-
зеплицирующихся молекул ДНК, полученные на основе бактериальных плазмид и
лользуемых для переноса генов (групп генов) от организмов-доноров к организмам-
теципиечтам. В практике генно-инженерных экспериментов с бактериями исполь-
ются векторы разного назначения: клонирующие, интегративные, челночные, экс-
“эессии и др. Их применение для получения штаммов микроорганизмов с неправиль-
>й эпидемиологией мы рассмотрим на «прикладных примерах».
340
Часть 2. Общая неправильная эпидемиология
Клонирующие векторы. Их используют для клонирования определенных нуклео-
тидных последовательностей в специальных репипиентных штаммах для последую
щего выделения этих нуклеотидных последовательностей в индивидуальном состоя-
нии. Как правило, они обладают следующими свойствами: имеют ограниченное
(предпочтительно одно) число мест расщепления рестриктазами второго типа (про-
изводят «разрезы» ДНК в специфических последовательностях); содержат генетичес-
кие маркеры (устойчивость бактерии к антибиотику и др.), которые могут быть исполь-
зованы для отбора клонов, несущих гибридные ДНК; и обеспечивают эффективную
репликацию гибридной ДН К в реципиентных штаммах. На рис. 2.8 показана рестрик-
ционная карта плазмиды pBR322 — стандарта таких векторов в начале 1980-х гг.. я
общая схема конструирования молекул рекомбинантной ДНК.
Клонирующий
вектор, обработанный
рестриктазой II типа
Фрагменты ДНК,
обработанные
рестриктазой II типа
Трансформированные дочерние клетки
Рис. 2.8. Общая схема конструирования молекул рекомбинантной ЦНК с помощью
клонирующих векторов. А. Рестрикционная карта клонирующего вектора pBR322. За
начало отсчета нуклеотидов принят центр участка узнавания рестриктазы FcoRl.
Отсчет осуществляется по часовой стрелке. Для рестриктаз — номер первой пары нук-
леотидов участка узнавания фермента. Б. Клонирование рекомбинантной ДНК. До-
норную ДНК расщепляют рестрицирующей эндонуклеазой II типа и встраивают в
клонирующий вектор. Полученную конструкцию вводят в популяцию клеток-хозяев,
идентифицируют те клетки, которые содержат рекомбинантную ДНК
Представленный на схеме А клонирующий вектор pBR322 несет гены двух мар-
керов, по которым можно вести отбор рекомбинантных клонов — устойчивость к
ампициллину (Ар1) и к тетрациклину (Тсг). В пределах каждого гена находятся еди-
ничные участки расщепления ДНК рестриктазами второго типа. На схеме Б показа-
но расщепление ДНК вектора и клонируемой ДНК по сайту рестриктазы EcoRI. Если
рассматривать схему применительно к вектору pBR322, то сайт находится в пределад
гена Тсг. Все образовавшиеся после расщепления рестриктазами фрагменты ДНК
22 Биологические поражающие агенты
341
»-гют «липкие концы», т. е. взаимно комплементарные одноцепочечные участки
IHK. выступающие по концам двуцепочечной молекулы. Следующий этап — гибри-
.шия. Температуру смеси понижают до 4-6°С. За счет образования водородных
Вя й между «липкими концами» фрагментов ДНК происходит самосборка молекул.
_jнный процесс еще называется «отжигом». Далее водородные связи, образовавши-
sc. между основаниями «липких концов», переводят в ковалентные действием фер-
мента ДНК-лигазы фага Т4. Образуется смесь разных фрагментов, сшитых между
. «бой в произвольном порядке. Среди них молекулы клонирующего вектора, кото-
- ые снова замкнулись в кольцо. Их два типа: кольцо вектора замкнулось само на себя
• донорной ДНК не содержит; и химерная (рекомбинантная) плазмида, когда коль-
. с вектора замкнулось, включив фрагмент донорной ДНК. Полученной смесью транс-
: • -миру ют клетку-реципиент и ведут селекцию по какому-то маркеру. Им может быть
маркер вектора, так и маркер клонированного фрагмента ДНК. Применительно
4. вектору pBR322 таким маркером будет устойчивость к ампициллину. Среди полу-
ных колоний Арг-трансформантов отбирают те, которые содержат рекомбинант-
- плазмиды. Принцип отбора прост. Клонирование ДНК ведут по сайту рестрик-
1 ы EcoRl, расположенном в гене Тсг. Это значит, что в тех случаях, когда произошла
ставка в вектор донорской ДНК, ген Тсг будет инактивирован. Обираются штаммы
~ансформантов, устойчивые к ампициллину, но чувствительные к тетрациклину (Tcs).
2 лее работа по изучению рекомбинантной ДНК идет уже только с ними. Чтобы
ъеньшить вероятность замыкания вектора «на себя» или включения в сайт рестрик-
;зы EcoRl другой полноразмерной молекулы вектора и получения их димеров, ва-
- руют соотношениями ДНК донора и вектора. Существуют другие методы, специ-
льно разработанные для того, чтобы еше более снизить вероятность повторного
мыкания в кольцо клонирующего вектора, или для того, чтобы отличить рекомби-
шты от нерекомбинантов с помощью генетических методов.
В качестве примера «прикладного использования» клонирующего вектора для
ллучения микроорганизмов с измененными свойствами, рассмотрим получение
- Е. Ivins1 и S. L. Welkos (1986) сенной палочки (Bacillus subtilis), обладающей способ-
чостью продуцировать протективный антиген (РА) возбудителя сибирской язвы и раз-
множаться, по крайней мере, в организме морской свинки.
Возбудитель сибирской язвы синтезирует трехсоставной токсин типа А1-В-А22.
Он состоит из В-субъединицы, называемой протективным антигеном и двух фер-
м ентативных субьединиц (А-субъединиц), одна из которых — отечный фактор (каль-
одулинзависимая аденилатциклаза); другая—летальный фактор, является металло-
ротеазой. РА используют в качестве антигенного компонента химических сибире-
язвенных вакцин типа американской вакцины AVA.
В 1983 г. М. Vodkin и S. Н. Leppla клонировали ген РА в Е. coli. используя схему
конструирования рекомбинантной ДНК, почти полностью воспроизводящую схему,
триведенную на рис. 2.8. Небольшое отличие их экспериментов заключалось в том,
что фрагмент плазмиды рХО1 В. anthracis Sterne (см. рис. 2.3) размером 6 т. п. о., коди-
рующий РА, они клонировали в векторе pBR322 по сайту для рестриктазы BamHI.
Иммунологическими методами ими были отобраны два трансформанта Е. coli (pSE24
1 Это и есть тот самый Брюс Айвинс, которого после смерти американское правосудие
эбъявило «единственным организатором и исполнителем» сибиреязвенного теракта в 2001 г.
см. разд. 1.12.4).
2 Более подробно в разд. 2.2.3.
342
Часть 2. Общая неправильная эпидемиология
и pSE36), содержащие рекомбинантные плазмиды. Оба трансформанта синтезиро-
вали биологически и серологически активный РА в количествах от 5 до 10 нг/мл, что
почти в 1000 раз меньше, чем синтезировал штамм Sterne. Причем РА обнаруживался
в лизатах клеток, в среду культивирования он не попадал. Но М. Vodkin и S. Н. Leppla
ставили перед собой задачу доказать только то, что ген РА локализован на одной из
плазмид сибиреязвенного микроба — они это доказали. Причины низкого уровня
экспрессии клонированного гена РА были им понятны: ни клонирующий вектор, ни
система экспрессии, использованная ими в кишечной палочке, не могли эффективно
использоваться для экспрессии генов белков бацилл. В. Е. Ivins и S. L. Welkos поста-
вили перед собой задачу добиться уровня экспрессии клонированного гена РА, сопо-
ставимого с тем, что наблюдали у штамма Sterne, и на основе аспорогенного штамма
В. subtilis 1S53 получить рекомбинантный штамм, пригодный для использования
качестве живой сибиреязвенной вакцины. Для этого они использовали систему экс-
прессии, разработанную для генов бацилл. Схема эксперимента приведена на рис. 2.9.
О pUBUO pSE36
Расщепление Расщепление
рестриктазой BamHI рестриктазой BamHI
Обработка фосфатазой BamHI 6(0т.п о_ BamHI
Ваты 4 5Т.П.О. BamHI
I-------------1 , 4,3о т. п. о. ,
^Лигирование х
Трансформирование
В. subtilis 1S53
Селекция штаммов,
резистентных к канамицину
Отбор колоний,
продуцирующих РА
Рис. 2.9. Схема клонирования гена РА в аспорогенном штамме В. subtilis 1S53.
А. Вектор pUBl 10 и рекомбинантная плазмида pSE36 из Е. coli фрагментированы
рестриктазой BamHI. Полученные фрагменты ДНК лигировали ДНК-лигазой фага
Т4 и ими трансформировали В. subtilis 1S53. Отобранные Ктг-трансформанты иссле-
довались иммунологическими методами на продукцию РА-антигена. Б. Рестрикци-
онная карта рекомбинанта размером 10,5 т. п. о., полученного на основе pUBl 10 и
фрагмента ДНК, кодирующего РА — антиген сибиреязвенного микроба. Условные
обозначения рестриктаз на карте: В — BamHI; Е — EcoRI; Р — Pvull; Hd — Hindlll;
Нр — Hpall. По В. Е. Ivins и S. L. Welkos (1986)
Биологические поражающие агенты 343
Бациллы, как объект генетических манипуляций, в 1980 гг. были менее изучены,
«м кишечная палочка. В Е. Ivins и S. L. Welkos не могли воспользоваться удобным
и -пирующим вектором типа pBR322. Для переклонирования гена РА из кишечной
L очки в В. subtilis они взяли более простой вектор pUBHO, главным достоинством
рого было его происхождение, — он получен на основе плазмиды, выделенной
1 грамположительного микроорганизма S. aureus. Вектор реплицировался в бациллах
ммел уникальные сайты рестрикции, позволяющие использовать его для клониро-
лния ДНК, но ни один такой сайт не приходился на маркер, инактивация которого
ставкой ДНК позволила бы исследователем легко отобрать рекомбинанты ie клоны,
i. Е. Ivins и S. L. Welkos для исключения повторного замыкания линеаризованной ДНК
нирующего вектора обработали его щелочной фос фатазой. Смысл этой операции
итоял в том, что ДНК-лигаза катализирует образование фосфодиэфирной связи
кду соседними нуклеотидами двуцепочечной ДНК лишь тогда, когда один из
- к содержит 5'-фосфатную группу, а другой — З'-гидроксильную группу. Удаление
-фосфатной группы с обоих концов линейной молекулы вектора исключает его за-
икание в кольцо под действием ДНК-лигазы. Зато если такая группа сохранена на
I мшах фрагментов клонируемой ДНК, то фрагменты ДНК сшиваются с З'-гидро-
. пьной группой линейной молекулы вектора. При этом они образуют кольцо с двумя
^цепочечными разрывами, так как ковалентная связь формируется в участках «со-
исосновения» З'-гидроксильных 1 рупп ДНК вектора и 5'-фосфатных групп клони-
емой ДН К. Эффективность трансформации кольцевой ДНК, даже содержащей од-
кпочечные разрывы, значительно выше, чем линейной плазмидной ДНК.
В. Е. Ivins и S. L. Welkos получили два клона В. subtilis IS53, синтезирующие РА,
обозначенные ими как РА1 и РА2. Они содержали рекомбинантные плазмиды: рРА101
г имером 7,8 т. п. о. (штамм РА1), и рРА102, размером 6,1 т. п о. (штамм РА2). Выход
k | жидкой среде составлял от 20,5 до 41,9 мкг/мл, что уже приближалось к уровню
тамма Sterne. Два рекомбинантных штамма В. subtilis оказались способными размно-
жаться в организме морских свинок и вызывать у них протективный иммунный от-
вет, защищающий от заражения летальными дозами возбудителя сибирской язвы1.
Интегративные векторы. Если смотреть на эксперименты с клонирующими век-
шами, проиллюстрированные работой В. Е. Ivins и S. L. Welkos (1986), с точки зрения
оз ложности получения генетически измененных патогенных микроорганизмов, то
они имеют серьезные ограничения. Реплицирующийся вектор с клонированным ге-
эм будет существовать в бактериальной клетке до тех пор, пока на него оказывается
селективное давление по маркеру антибиотика. У pUBl 10 роль такого маркера играл
;ен Кт1г. Для стабилизации рекомбинантной плазмиды в сенной палочке исследова-
тели добавляли в среду культивирования 10 мкг/мл канамицина. В конце 1980-х гг. для
того чтобы избежать необходимости постоянного селективного давления на реком-
1 Эксперимент был как бы точкой бифуркации в жизни самого Брюса. Показав «принци-
«альную возможность» синтеза РА в сенной палочке, ему на этом стоило и ограничиться, как
делали его коллеги М. Vodkin и S. Н. Leppla, открыв плазмиды у сибиреязвенного микроба.
Иммуногенность полученных рекомбинантных штаммов В. subtilis не достигла иммуногенно-
сти вакцинного штамма Sterne. Брюс пытался потом использовать их для синтеза РА, но ре-
комбинантные штаммы синтезировали изоформы РА Он потратил почти 20 лет на поиски
-казательств того, что разные изоформы РА также пригодны для иммунизации людей, как и
РА, получаемый из вакцинных штаммов В. anthracis. Этим он как раз и занимался в 2001 г. По
мнению американского правосудия, тогда никто, кроме него, не был заинтересован в реализа-
ции сибиреязвенных вакцин. И Брюс умер.
344
Часть 2. Общая неправильная эпидемиолога •
бинантный микроорганизм по маркеру вектора, стали использовать векторы, обеспе-
чивающие интеграцию клонированной ДНК в геном реиипиентной бактерии1. В ка-
честве примера «прикладного использования» интегративно! о вектора для получени.
микроорганизмов с измененными свойствами рассмотрим опыты S. М Hailing et al
(1991) по получению антибиотикорезистенного штамма возбудителя бруцеллеза, од-
новременно (и, видимо, по случайному стечению обстоятельств) ставшего менее за-
метным для иммунной системы человека.
Возбудителей бруцеллеза (В. melitensis и В. suis) L. Е. Lindler et al. (2005) рассмат-
ривают в качестве инкапситирующих биологических поражающих агентов. Бруцел-
лез — хроническое и длительное страдание, сопровождающееся изнурительной лихо-
радкой. В настоящее время не существует безопасной и эффективной вакцины,
которую можно использовать для предупреждения развития бруцеллеза у людей. Ле-
чение этой болезни антибиотиками требует длительного применения их определен-
ной комбинации. К тому же число ел) чаев возврата болезни после завершения курс I
антибиотикегерапии достигает 10 % (см. разд. 3.4).
S. М. Hailing etal. (1991) экспериментировали с аттенуированным штаммом B.abor-
tus S19, использующимся в США в качестве вакцинного для профилактики бруцел-
леза среди сельскохозяйственных животных; и вирулентным штаммом В. abortus S2308
применяемым там же для проверки эффективности бруцеллезных вакцин. Суть их
эксперимента заключалось в инактивации хромосомных генов В. abortus, кодирующих
31 кДа иммуногенный периплазматический белок BCSP31, значительно повышаю
щий иммунный ответ на возбудитель бруцеллеза в ходе инфекционного пропесси
(о белке BCSP31 см. работу Pugh G. W. et al., 1990). Для осуществления эксперимента
ими был сконструирован интеграционный вектор pBA31-R7/Kan7 (рис. 2.10).
PBA31-R7
R S рКК223-3 SRH Н HR I BCSP31 ( j
(т. п. о.) 4,6 1 1 2,2 1,3 0,73
R S pBA31-R7/Kan7 рКК223-3 SRH Н пео/кап Н R
(т. п. о.) 4,6 IH 2,2 1,4 ),73
Рис. 2.10. Диаграммная рестрикционная карта плазмид рВАЗ 1-R7 и рВАЗ 1 -R7/Kan7.
Нуклеотидная последовательность В. abortus S19 выделена штриховкой. Ген Kmr
(пео/кап) показан открытым прямоугольником Вектор рКК223-3 показан сплош-
ным прямоугольником. Условные обозначения рестриктаз на карте, R — EcoRl;
Н - Hindlll; S - Sall. По S. М Hailing et al. (1991)
Схема конструирования плазмиды pBA31-R7/Kan7 была следующей. Сначала н.
основе вектора pBR322 (см. рис. 2.8) J. Е. Mayfield et al. (1988) получена плазмид
рКК223-3 В нее по сайту EcoRl вставлен фрагмент ДНК Brucella размером 4,2 т. п. о
1 Опубликованная работа В. Е. Ivins и S. L. Welkos по клонированию РА не более чем част-
ное отражение других неопубликованных работ с другими генами и другими микроорганиз-
мами, проводимыми в те годы специалистами USAMRIID с использованием технологии ре-
комбинантной ДНК. Это видно «невооруженным глазом». Получив РА в препаративных
количествах из культуры В. subtilis 1S53, его надо было отделять еще от канамицина, что зна-
чительно усложняет и удорожает получение химических сибиреязвенных вакцин.
2 2 Биологические поражающие агенты
345
дарующий белок BCSP31. Плазмида получила название pBA31-R7*. S. М. На1-
_ et al. (1991) заменили у плазмиды pBA31-R7 HindIII-фрагмент ДНК размером
3 т. п. о., включающий ген белка BCSP31, на 1,4 т. п. о. HindIII-фрагмент ДНК с
геном устойчивости к канамицину, полученному из транспозона Тп5. Сконструиро-
нную плазмиду они назвали pBA31-R7/Kan7. Так как она была производной от
?BR322, имеющей репликон плаз лиды Col El, а поэтому очень узкий круг хозяев, то
могла реплицироваться в бруцеллах. Но от нее это и не требовалось. С помощью
х-.ек.ропорации (см. рис. 2.7) pBA31-R7/Kan7 ввели в штаммы S2308 и S19 и получи-
клоны, устойчивые к канамицину/неомипину. Их исследование показало инакти-
лию гена белка BCSP31 в хромосоме трансформированных В. abortus за счет гомо-
вгичной рекомбинации ДНК бруцелл, включенной в вектор. На место гена белка
8CSP31 встроился ген Ктг. Исследование ДНК трансформантов показало, что они
-е содержат дополнительных плазмид (как это можно наблюдать в экспериментах по
редаче В. anthracis плазмиды рВС16; см рис. 2.3). Не обнаружено ДНК плазмиды
BR322 в геноме трансформантов методами ДНК-гибридизации. Штаммы бруцелл,
~ратившие ген белка BCSP31, по способности размножаться и поддерживаться в
тиниях клеток HeLa и макрофагах J774, не отличались от исходных штаммов.
Челночные векторы — векторы, обладающие двумя разными репликонами, благо-
*ря чему они могут реплицироваться в разных микроорганизмах — например, в Е. coli
в каком-то патогенном микроорганизме. Обычно используются в тех случаях, когда
.роще клонировать гены в методически простой системе, не требующей соблюдения
жестких условий биологической безопасности, а только затем перенести их в плохо
зученные или опасные реципиенты. В качестве примера «прикладного использова-
ния» челночного вектора рассмотрим вектор, разработанный D. A. Rasko et al. (2007)
_тя изучения поведения в F. tularensis генов, полученных из различных источников.
Возбудитель туляремии, F. tularensis, рассматривается Е. Lindler et al. (2005) в ка-
,тве инкапситируюшего биологического поражающего агента. Чрезвычайно низ-
<ая инфицирующая доза аэрозолированного возбудителя туляремии для человека
иеньшает расход биоло! ической рецептуры при поражении конкретной цели. Но как
юъект генетических манипуляций, он не удобен из-за трудоемкости процессов куль-
тивирования и необходимости соблюдения строгих мер биологической безопасности.
Возможно, по этой причине он мало исследован как объект генно-инженерных экс-
.ериментов. На средину прошедшего десятилетия наиболее распространенными
греди генных инженеров, занимающихся туляремийным микробом, были векторы на
основе критической плазмиды pFNLlO1 2 близкородственного ему микроорганизма,
F. novicida (Maier Т. М. et al., 2004). Они имели узкий спектр потенциальных хозяев, и
их применение ограничивалось представителями рода Francisella. D. A. Rasko et al.
2007) сконструировали три новые плазмиды, не имеющие peruiHKOHapFNLlO: pCU 18,
oCUGl и pCUG2. Плазмида pCU18 представляет собой челночный вектор Е. coli/
F. tularensis. Он получен на основе имеющего много уникальных сайтов клонирова-
ния вектора pUC18, в основном используемого для клонирования генов в Е. coli', и
с лазмиды рС194, выделенной из 5. aureus и имеющей широкий круг хозяев (в ихчис-
ле даже В. anthracis).
1 Для лучшего понимания читателем технологий конструирования рекомбинантных плаз-
мид, я специально привожу описания экспериментов, где показаны разные варианты исполь-
зования одних и тех же векторных молекул.
2 Критической плазмида считается, если ее роль в фенотипе бактерии неизвестна.
346
Часть 2. Общая неправильная эпидемиология
Векторы семейства pCUG были созданы путем комбинации: 1) слабых промото
ров (promoterless) гена зеленого флюоресцирующего белка (green fluorescent protein.
GFP), полученного из плазмиды pFPV25; 2) репликона широкого круга хозяев:
3) кассеты хлорамфениколрезистентности (Стг) из рС194 Клонирующий сайт у век-
торных плазмид pCUG находится впереди гена GFP. Клонируемая ДНК расщепля-
ется (частично!) рестриктазой А1и\ и ее фрагменты вставляются в соответствующий
сайт типовым способом конструирования рекомоинантных ДНК (см. рис. 2.8). Но
отбор промотора происходит уже не по маркеру, показывающему рекомбинантные
клоны в условиях in vitro (например, по резистентности к антибиотику), а по маркеру
проявляющемуся во время инфекционного процесса. F. tularensis в организме чело-
века размножается в макрофагах. Чем более активно идет этот процесс, тем более ви-
рулентным является штамм туляремийного микроба. Поэтому D. A. Rasko et al. (2007)
инфицировали культуры макрофагов трансформантами F. tularensis и методом проточ-
ной цитометрии отбирали флюоресцирующие клетки. Затем макрофаги лизировали
и получали чистые культуры трансформантов туляремийного микроба, содержащие
рекомбинанты pCUG с клонированными промоторами, запускающими экспрессию
генов во время внутриклеточного размножения F. tularensis в макрофагах. Гены, ко-
торые могут быть поставлены под контроль такого промотора и затем «вставлены» в
хромосом) бактериального возбудителя инфекционной болезни по схеме, приведен-
ной на рис. 2.10, мы рассмотрим далее в этом разделе.
Векторы экспрессии. Наряду с амплификацией клонированной ДНК, эти векто-
ры обеспечивают еще правильную и эффективную экспрессию содержащихся в этой
ДН К генов в определенной фазе клеточного цикла. Частные случаи таких клонирую-
щих векторов будут рассмотрены ниже (см. «Технологии рекомбинантной ДНК в по-
лучении штаммов вирусов с неправильной эпидемиологией»).
Резистентность к антибактериальным препаратам у бактерий. При исследовании
штаммов бактерий, обнаруженных в очаге биологического нападения, необходимо
иметь в виду, что наиболее доступной модификацией их биологических свойств се-
годня является приобретение ими антибиотикорезистентности.
Диффундирующие сигнальные молекулы. Бактерии в естественных условиях образуют
коммуникативные сообщест ва, включающие представителей разных таксонов, в кото-
рых и между которыми обмен информацией осуществляется посредством так назг i-
лаемых диффундирующих сигнальных молекул (diffusible signal molecules). Это низ-
комолекулярные вещества, взаимодействующие с системой клеточных рецепторных
структур (мишеней). К их числу относятся и антибиотики — низкомолекулярные
органические молекулы (~3 гыс. Да), получившие такое название из-за их бактерг .-
цидного или бактериостатического действия в концентрациях, обычно не встречаю-
щихся в природных экосистемах. Бактерии используют диффундирующие сигналь
ные молекулы для: 1) мониторинга своей популяционной плотности (the process of quo-
rum sensing); 2) зашиты своей экологической ниши; 3) координации своего поведения
в отношении каких-то изменившихся условий внешней среды; 4) «наблюдения» за
другими коммуникативными сообществами (Ryan R. Р., Dov'J. М , 2008). Биосинтети-
ческие пути антибиотиков сформировались у микроорганизмов еще до образования
многоклеточной жизни. Например, у актиномицетов биосинтетические пути эритро-
мицина и стрептомицина существуют не менее чем 500 млн лет (Baltz R. Н , 2007)
Количество клеточных мишеней для диффундирующих сигнальных молекул
огромно. Ими являются не только рецепторные белки на поверхности бактерий, но и
макромолекулярные структуры цитоплазмы. Первичный механизм, контролирующий
12. Биологические поражающие агенты
347
псции бактериальной клетки, — регуляция транскрипции отдельных генов. Напри-
мер, любой из 50 рибосомальных белков или РНК может быть специфическим рецеп-
эром, связывающим такие молекулы, какамкногликозиды и макролидные антибио-
к <ки. Экспериментально установлено, что субингибирующие концентрации антиби-
отиков либо увеличивают, либо, наоборот, уменьшают количество транскриптов в
различных бактериальных клетках (Goh Е. В. et al., 2002; Yim G. et al., 2007). Отдель-
ie примеры взаимодействия антибиотиков с внутриклеточными рецепторами пере-
метены в табл. 2.5.
Таблица 2.5
Примеры взаимодействия антибиотиков с внутриклеточными рецепторами*
Биохимический процесс Функциональный рецептор Мишень Антибиотик
Трансляция Рибосомы ЗОБ-субъединица Стрептомицин, гентамицин
Спектиномицин
Тетрациклин
595-субъединица Хлорамфеникол
Эритромицин
Виргиниамицин
Линкомицин
Рибосомо-связываюшие факторы Фактор элонгации Ти Кирромицин (kirromycin)
Рибосомо-ассоциирован- ные факторы Изолейцил-тРН К-синтетаза Мупироцин (mupirocin)
Транскрипция РНК-полимераза Р-субъединица Рифампицин
Р- и Р'-субть единицы Стрептолидигин
р'-субъединица Микроцин J25
Репликация ДНК-гираза Субъединица А Новобиоцин
Субъединица В Микроцин Л 7
Синтез клеточной стенки Синтез пептидогликана Пенициллинсвязываюшие белки Р-лактам
МигА Фосфомицин
ЛПС Полимиксин
Аланинреацемаза и D-аланил- D-аланинлигаза D-циклосерин
♦ G. Yim et al. (2007).
Формирование резистентных к антибиотикам биопленок — основная стратегия
п] ютиводеиствия грамотрицательных бактерий стрептомицетам. Приобретение рези-
стентности к какому-то антибиотику ведет к изменению реакции бактерии на опре-
деленный сигнал. Антибиотикорезистентные мутанты бактерий так же, как и исход-
ные чувствительные штаммы, реагируют на данный антибиотик. Но набор транскрип-
тов у них иной, чем у исходных штаммов. Спонтанное приобретение резистентности
к гем или иным антибиотикам способствует образованию других бактериальных эко-
типов (Ryan R. Р.. Dow J. М., 2008).
Мобильные генетические элементы Древность генетических механизмов биосин-
теза антибиотиков предполагает наличие не менее древних генетических механизмов
348
Часть 2. Общая неправильная эпидемиология
защиты от них. Распространение генов антибиотикорезистентности между бактери-
ями происходит благодаря активности мобильных генетических элементов. Их два
типа: генетические элементы, которые могут самостоятельно перемещаться между бак-
териями (конъюгативные плазмиды и конъюгативные транспозоны); и генетические
элементы, перемещающиеся в пределах бактериальной клетки (по хромосоме, от хромо-
сомы к плазмиде и наоборот: транспозоны, генные кассеты, интегроны и др.). Плаз-
миды и конъюгативные транспозоны являются своеобразной платформой, на которой
посредством различных рекомбинационных систем бактериальной клетки происхо-
дит сбор ей и сортировка генов антибиотикорезистентности. В табл. 2.6 дана характе-
ристика мобильных генетических элементов бактерий, вовлеченных в распростране-
ние генов антибиотикорезистентности в природе.
Таблица 2.6
Характеристика мобильных генетических элементов бактерий, вовлеченных
в распространение генов антибиотикорезистентности в природе*
Элемент Характеристика Роль в распространении генов резистентности
Конъюгата вные плазмиды Кольцевые, автономно реплицирующи- еся генетические элементы, передаю- щие гены между бактериями благодаря конъюгации Перенос генов резистентности; | мобилизация переноса других генетических элементов, вклю- чающих гены резистентности
Конъюгата вные транспозоны Интегрирующиеся элементы, которые могут «вырезаться» из хромосомы в ви- де нереплицирующегося кольцевого производного, способного к конъюгации Такая же
Мобилизуемые плазмиды Носители генов, которые могут перено- ситься между бактериями благодаря конъюгационному аппарату конъюга- тивных плазмид Перенос генов резистентности
Нерепликативные единицы Bacteroides Интегрированные в геном бактероидов элементы, которые самостоятельно не способны из него «вырезаться» и пере- носиться между бактериями; но их «вы- резание» и перенос могут иницииро- ваться конъюгативными транспозонами Такая же
Транспозоны Могут перемещать фрагмент ДНК из одного участка генома бактерии в другой Перемещение генов антибиоти- корезистентности от плазмиды к хромосоме и наоборот
Генные кассеты Кольцевые нереплицирующиеся ДНК- сегменты, включающие только ORF; интегрируются в интегроны Включение генов резистент- ности
Интегрон Интегрировавшийся в хромосому бактерии фрагмент ДНК, включающий ген интегразы, промотор, и сайт инте- грации для генных кассет Формирование кластеров генов резистентности, находящихся под контролем промотора инте- грона
Инсерционные критические последовательности (rSCR-элементы) Образование комплексных интегронов первого класса Мобилизация генов, прилегаю- щих к lSCR-элементу
По A. Salyers, С. Е Abile-Cuevas (1997); Р. М. Bennett (2008).
2 2. Биологические поражающие агенты
349
Мобильные генетические элементы участвовали в переносе генов резистентности
к антибиотикам между бактериями еще до их внедрения в клиническую практику. Опи-
сан штамм кишечной палочки, выделенный до 1937 г., содержащий плазмиду (R-фак-
тор), определяющую устойчивость к тетрациклину и стрептомицину (Smith D. Н.,
1967). Плазмиды и коннотативные транспозоны (см. выше) являются своеобразной
платформой, на которой посредством различных рекомбинационных систем проис-
ходит сборка и сортировка генов антибиотикорезистентности. Но эти гены не суше-
^гвуют и не переносятся между бактериями и/или плазмидами «сами по себе».
Транспозоны (Тп) — мобильные генетические элементы, несущие структурные
гены, детерминирующие функции, не связанные с самим процессом перемещения
х:ежду бактериями. Отдельные транспозоны включают гены антибиотикорезистент-
* ости, различающиеся между собой по структуре, происхождению и механизмам
Транспозиции. Механизмы перемещения транспозонов по геному бактерии не тре-
буют гомологии между их ДНК и ДНК хромосомы бактерии в участках интеграции,
хотя для некоторых транспозонов обнаружены предпочтения по интеграции среди
таких участков. Известны транспозоны двух типов:
1) представляющие собой модульные системы. Их называют композитными транс-
позонами (composite transposons). Они включают пару IS-элементов и между ними —
неспособную к транспозиции последовательность ДНК, содержащую структурный
ген, изменяющий фенотип бактерии (рис. 2.11, с. 350);
2) представляющие собой комплексные системы, в которых транспозиционная и
нетранспозиционная функции не отражены четким модульным расположением нук-
готидных последовательностей.
Транспозоны этой группы (complex transposons) более древние, чем композитные.
Они имеют следы множественных рекомбинационных актов, включая вставки и де-
теции. Последние приходятся на участки, не имеющие отношения к транспозиции,
что делает такие транспозоны компам ными и увеличивает частоту их транспозиции
рис. 2.12, с. 351).
Транспозоны реагируют на изменения во внешней среде, окружающей бактерию.
Например, частота транспозиции транспозона Тп501 существенно увеличивается при
наличии в среде ртути, а транспозона Тп55/ — в присутствии эритромицина. Приме-
ры транспозонов, способных перемещать между бактериями гены антибиотикорези-
стентности, приведены в табл. 2.7, с. 352.
Ишпегроны и генные кассеты. Бактериальные интегроны представляют собой ген-
ные системы, которые вместо транспозиции (как это делают транспозоны) исполь-
зуют для распространения по геному бактерии сайт-специфическую рекомбинацию.
Интегроны включают специальную рекомбинационную систему, кодирующую фер-
мент интегразу (ген inf), осуществляющий сайт-специфическую рекомбинацию; и
сайт, в котором находится короткая последовательность ДНК — генная кассета (Rec-
chia G. D., Hall R. M., 1995' Такое название она получила потому, что в наибольшей
степени приспособлена только для отдельного гена, вставленного посредством интег-
разы. В процессе перемещения от одного интегрона к другому, или от одного сайта в
интегроне к другому сайту в том же интегроне, генная кассета существует как неболь-
шая автономная и неспособная к репликации двунитевая кольцевая молекула ДНК
(Bennett Р. М., 1999).
Интегроны очень древние генетические элементы. Кроме генов антибиотико-
устойчивости кассеты, захватываемые интегронами и суперинтегронами, могут содер-
жать гены, кодирующие факторы патогенности, гены метаболизма, или гены, коди-
350
Часть 2. Общая неправильная эпидемиология
1
0
2
3
4
5
т. п. о.
О 1534 п.о. I ,
|ия1 Р
IR Нефунк- э *
ционирующий
I Активн ей О
----|—iS5OR-|
IR--IR
Полицистронная
мРНК
1522 258 93
8 т. п. о.
0
2
4
6
Нефункционирующий Активный
tetR tetA te/Cted?
IS/OL [-
1329 п. о.
Внутренний
Внешний
IR
E
IR
| 1S/P правый |
' JPHK - ранспоза^ы^
•—pOUT
Антисмысловая РНК
Рис. 2.11. Композитные транспозоны. Композитные транспозоны. А, Схема транс-
позона Тп5, включающего гены резистентности к канамицину (кт), блеомицину (W)
и стрептомицину (sm). О и I — обозначения наружной и внутренней границ терми-
нального инвертированного элемента IS50(L — левый; R — правый), вмещающих
короткие терминальные инвертированные повторы (IRs), определяющие границы
IS50M генов резистентности к антибиотикам. Б. Схема транспозона Tn/О, включа-
ющего ген резистентности к тетрациклину. IS10 — инвертированный терминальный
повтор. IS/0L — левая половина IS10, представляющая собой копию IS 10, не функ-
ционирующую из-за множественных мутаций в гене транспозазы. IS/OR — право-
сторонняя копия IS 10, кодирующая активную транспозазу и антисмысловую моле-
кулу РНК, снижающую экспрессию гена транспозазы. tetA — ген, кодирующий
резистентность к тетрациклину по типу «выкачивающего насоса» («efflux pump»);
tetC mD— гены, корегулируемые с tetA; tetR — ген, кодирующий транскрипционный
репрессор, необходимый для индуцибельной экспрессии тетрациклинового «efflux
pump» tetA. Черными кружками показаны промоторы. По Р. М. Bennett (2008)
рующие рестрикционные ферменты. Различные генные кассеты содержат интеграци-
онные сайты (aitC), которые негомологичны друг другу. Интегроны бактерий различ-
ных видов, даже принадлежащих одному роду, могут содержать разные генные кассеп
(Смирнов Г. Б., 2007).
Большинство итегронов антибиотикорезистентности соответствуют мобильной
генетической структуре, известной какинтегроны класса 1 (class 1 integron). Они вклю
чают два концевых невариабельных региона, называемых константными последова-
тельностями (constant sequences, CS), и высоковариабельный центральный участок
В одном конце интегрона — 5' -CS, обычно находятся: int — ген интегразы; attl— ре-
комбинационный сайт (локус) для кассеты; и промотор, от которого экспрессируют-
ся гены кассеты. В другом конце — З'-CS, находится часть гена qacEDl, который ин-
тактным несет устойчивость четвертичным аммониевым соединениям. За ним
расположен ген sul, определяющий резистентность микроорганизма к сульфанилам’ -
дам, и два других гена с неустановленной функцией, обозначенных как orf5 и otf6
Биологические поражающие агенты
351
и 1 2 3 4 3
__________I_________1__________J__________I
39 п. о. 39 п. о.
IR IR
tnpA tnpR ЫаТЕи.
merR merTPCAD
• Промотор
Интегрон
1 класса attl
suil aadA int
tnpR tnpA
38 п. о.
IR
Е
38 n. o.
IR
pint Pc
Рис. 2.12. Комплексные транспозоны. А. Схема транспозона Tn5, включающего
гены резистентности к ампициллину и к некоторым другим антибиотикам р-лак-
тамного ряда. IR обнаружены по краям участка транспозона, необходимого для
транспозиции; /лрА — ген, кодирующий компонент транспозазы, tnpR — ген, ко-
дирующий сайт-специфическую рекомбиназу, позволяющую перемещение коин-
тегрированной структуры, образовавшейся в результате транспозиции; blaTEM_j —
ген, кодирующий ТЕМ-1 Р-лактамазу. Б. Схема транспозона Тп2/, включающего
гены резистентности к стрептомицину, спектиномицину, сульфониламидам и
ионам ртути: merTPCAD — гены, кодирующие резистентность к ионам ртути и от-
дельным органортутным соединениям; merR — ген, кодирующий транскрипци-
онный репрессор индуцибельного mer-оперона; suil — ген, кодирующий резис-
тентность к сульфониламидам; aadAl — ген, кодирующий резистентность к стреп-
томицину и спектиномицину; int — ген интегразы; attl — вставочный сайт для
генной кассеты интегрона; tnpA — ген, кодирующий транспозазу Tn27; tnpR — ген,
кодирующий сайт-специфическую рекомбиназу; pint — rm-promoter (промотор для
интегразы); Рс — промотор для кассеты интегрона и suil. Стрелки на обеих схемах
показывают направление и протяженность транскрипции. Черными кружками
показаны промоторы. По Р М. Bennett (2008)
е. открытые рамки считывания 5 и 6). CS-регионы фланкируют вариабельные ре-
*ионы и последовательность, включающую генные кассеты отдельного интегрона. Ко-
т»чество генных кассет у разных интегронов различное. Интегроны первого класса
ожно представить в виде следующей формулы:
inti attl (r59b)nqacE\l suil orfS orf6,
де r59b соответствует кассете резистентности, л показывает количество генных кас-
сет, интегрировавшихся в локус attl (Peter М., 1999).
Кассеты генов резистентности не содержат промоторы, от которых может начаться
йх экспрессия. Промотор для кассеты является частью интегрона. Поэтому вставки
кассет в интегрон должны быть обязательно ориентированы таким образом, чтобы
.-гарт гена, расположенного на кассете, начинался вблизи от int. Генные кассеты мо-
ут быть вставлены в сайт attl одна после другой, придавая бактерии резистентность
к различным антибиотикам. Приобретение интегроном кассеты, дополнительной к
.же имеющимся, создает новый интегрон (рис. 2.13, с. 353).
352
Часть 2. Общая неправильная эпидемиология
Таблица 2.
Транспозоны, способные перемещать между бактериями гены
антибиотикорезистентности*
Транспозон Размер (т. п. о.) Терминальный элемент Маркер
Композитные транспозоны
Грамотрицательные бактерии
Тп5 5,7 IS5fl(IR) KmBISSm
ТпР 2,5 IS/(DR) Cm
Тп/0 9,3 IS/d(lR) Tc
TnPft? 3,1 1S903 (IR) Km
Тп1525 4,4 IS/5(DR) Km
Тп2350 10,4 IS/(DR) Km
Грамположительные бактерии
Тп400/ 4,7 IS256 (IR) GmTbKm
Тп40й? 3,6 1S257 (DR) Tb
Комплексные транспозоны
Грамотрицательные бактерии
Тп/ 5 38/38 Ap
ТпЗ 5 38/38 Ap
Тп2/ 20 35/38 SmSuHg
Тп501 8,2 35/38 Hg
Тп1721 11,4 35/38 Tc
Тп3926 7,8 36/38 Hg
Грамположительные бактерии
Тп55/ 5,3 35 Ery
ТпР/7 5,3 38 Ery
ТМ451 6,2 12 Cm
* По Р. М, Bennett (2008).
Разнообразие генных кассет трудно оценить. Недавно были идентифицированы
генные кассеты, включающие гены, кодирующие металло-Р-лактамазы, придающие
бактерии устойчивость к таким мощным карбапенем-Р-лактамам как имипинем и
меропинем (см. рис. 2.13, В). Известны суперинтегроны, включающие десяток ген-
ных кассет, однако к деятельности человека они отношения не имеют. Среди бакте-
рий, имеющих клиническое значение, большинство интегронов содержит менее пяти
генных кассет (Rowe-Magnus D. A. et al., 2001; Nordmann Р., Poirel L., 2002).
Инсерционные криптические последовательности (ISCR-elements, IS-cryptic sequen-
ces). Это небольшие криптические последовательности, по размеру сходные со мно-
гими IS-элементами. Ноу них совершенно иной, чем у транспозонов механизм транс-
позиции, названный транспозицией катящегося кольца (rolling circle transposition). Он
объединяет RC-репликацию, используемую отдельными бактериальными плазмидами
и бактериофагами, и рекомбинацию транспозиции IS-элементов (Tavakoli et al., 2000.
Garcillian-Bracia et al., 2002).
12. Биологические поражающие агенты
353
Л GURRY (сайт рекомбинации) [Б]
5-CS
З'-CS
int \дасЕБ_suil orf5/orjb
- attC
» Конец
генной кассеты
Генная
кассета
*Интегрон 2|
Интегрон 1
• — attl о — attC/59-be
Рис. 2.13. Интегроны и генные кассеты. А Схема интегрона резистентности клас-
са 1. Б. Схема поглощения генной кассеты бактериальным интегроном. В. Приме-
ры организации генной кассеты в бактериальных интегронах класса 1. inti — ген
итегразы класса 1; qac — ген, кодирующий устойчивость к четвертичным аммо-
ниевым соединениям; attl— вставочный сайт для генной кассеты; attC — вставоч-
ная последовательность генной кассеты; qacEDl — усеченная версия гена qacE-,
sull — ген, несущий резистентность к сульфаниламидам; orj — возможно, ген
неустановленной функции, blaVIM — ген, кодирующий VIM-металло-р-лакта-
мазу; Ыа1МР— ген, кодирующий IMP-металло-р-лактамазу; aacA, aadA, aph — ге-
ны, кодирующие резистентность к аминогликозидам; cat — ген, кодирующий
резистентность к хлорамфениколу. По Р. М. Bennett (2008)
ISCR-элементы впервые обнаружены как последовательности, ассоциирующие-
ся с интегронами класса 1, но при этом от них отличающиеся. Большинство из IS-
элементов имеют четкие границы благодаря коротким инвертированным нуклеотид-
ным повторам (см. рис. 2.10 и 2.11), функционально равнозначащим и заменяемым.
Они действуют как сайты связывания и рассечения для родственной транспозазы.
Большинство, если не все из комплексных транспозонов, имеют те же характеристи-
ки. В противоположность им, ISCR-элементы утратили концевые терминальные по-
вторы, характерные для IS-элементов. Их терминальные последовательности обозна-
чены как orilS и terlS. Они служат уникальными сайтами для инициации и терминации
репликационной стадии транспозиции, соответственно. Особенностью этих систем,
имеющих отношение к распространению генов антибиотикорезистентности среди
Зактерий, является то, что их репликация не является точной. В процесс репликации
ISCR-элемента может вовлекаться Д НК бактерии, прилегающая к terlS за пределами
инсерционной криптической последовательности. Поэтому вместе с ISCR-элемен-
354
Часть 2. Общая неправильная эпидемиология
том может быть транспозирована последовательность в несколько раз большая, чем
он сам, но всегда прилегающая к terlS (Tavakolietal., 2000)1. То есть мобилизация ДНК
является характерным признаком механизма RC-транспозиции ISCR-элемента; и
мобилизована может быть любая ДНК, прилегающая к iterlS2. Этот механизм можно
считать ключевым для образования комплексных интегронов 1-го класса (рис. 2.14)
и---------i.....I I —I г I II------------1
gacEEI/sulIls ISCRI gacE£>J/sulI orf5/6
Вероятный источник: DHA-1 Morganella morganii
FOX-4 Aeromonas spp.
CMY-1 Aeromonas spp.
CMY-8 Aeromonas spp.
МОХ-1 Aeromonas spp.
rerIS-2? ferIS-1 on IS
Рис. 2.14. Комплексные бактериальные интегроны. А. Примеры комплексных бак-
териальных интегронов первого класса с копиями ISCR1 и дупликациями З'-CS
Ломаная линия показывает 5'-CS и вариабельные регионы интегронов 1 класса, став-
шие частью комплексных интегронов DHA-1, FOX-1, CMY-1,8, МОХ-1 — гены, ко-
лирующие р-лактамазы, которые придает бактерии резистентность к третьему по-
колению цефалоспоринов, вероятно рекрутированных из хромосом перечисленных
бактерий. Другие обозначения см. в подрисуночном тексте к рис. 2 13 Б Мобили-
зация интегронов первого класса посредством ISCRJ. 1 — границы ISCR1 очерче-
ны через терминальные последовательности terIS-1 и or/IS. Их наличие у интегрона
предполагает, что копии ISCR1 были транспозированы в сайт, близкий к 3'-С8-концу
интегрона первого класса; 2 — затем делецией (показана пунктирными линиями)
удалена часть З'-CS (включая orf5,6), образуя характерную З'-CS-ISCRl-nepecTaHOB-
ку в интегроне первого класса. Теперь границы 1SCR1 очерчены терминальными
последовательностями terIS-2 и onIS. По Р. М. Bennett (2008)
1 Репликация — это синтез комплементарных нитей ДНК по матричной молекуле ДНК
Когда говорят о репликации «по типу катящегося кольца» (rolling circle model, RCM), то име-
ется в виду ее вариант, наб подаемый у плазмид. Особенностью данного типа репликации г
ляется ее однонаправленность и асимметричный характер, выражающийся в разобщении син-
теза ведущей и запаздывающей нитей плазмидной ДНК по времени. В результате че?
образуется два продукта: 1) двунитевая молекула ДНК, состоящая из родительской |—] и вноз:
синтезированной [+] нити и 2) однонитевая молекула ДНК, представляющая собой вытеснен-
ную (+] родительскую нить. Затем на матрице вытесненной [+] нити синтезируется двуните-
вая молекула ДНК Фрагменты ДНК соединяются ДНК-лигазой и суперскручиваются за сче*
активности ДНК-гиразы. Более подробно о RCM см. в работе М. А, Титок (2004).
2 Транспозиция принципиально иной процесс, чем репликация Он представляет собой
дупликацию мобильного генетического элемента, который остается в прежнем сайте а ев
копия перемещается в сайт встраивания. Транспозиция мобильных генетических элементе
включает этап оепликации ДНК, ограниченный фланкирующими их участками.
2 Биологические поражающие агенты
355
Такой интегрон имеет две особенности:
1) приблизительно половину ею структуры занимает последовательность ДНК
ггегрона 1 класса, начавшаяся от 5'-С8-конца;
2) концевая последовательность З'-CS и повтор вариабельного региона «следуют»
"ез копию ISCR7. За ней наход гтся другой вариабельный регион, давший прибе-
• ише различным п нам резистентности, включая Ыасмг- и />/йС7У-варианты, qnrA- и
’4-варианты, и catAII, которые несут резистентность к длинному ряду пефалоспо-
тпнов, флюорохинолонов (таких как ципрофлоксацин), триметоприму и хлорамфе-
IIколу. Эти вариабельные регионы находятся в «повернутом состоянии» за повтором
Taleman М A. et al. 2006). Способы генетического переноса, благодаря которым гены
ггибиотикорезистентности могут распространяться среди бактерий в природе и в
•кспериментальных условиях (трансформация, конъюгация и трансдукция), мы рас-
. «трели выше.
Механизмы развития ре шстентности к антибактериальным препаратам у бактерий.
Их можно классифицировать следующим образом:
1) модификация антибиотиков (другое название процесса — детоксикация). Зак-
опается в разрушении антибиотика еше до его проникновения в цитоплазму клетку
внешняя среда, периплазматическое пространство грамотрицательных бактерий),
“ри этом мишени антибиотиков в цитоплазме клетки остаются интактными. Осуще-
ствляется бактерией с помощью специфических ферментов, расщепляющих антибио-
так до структур, не представляющих для нее опасности;
2) уменьшение проницаемости стенки бактерии для антибиотиков и/или выкачива-
де его из клетки («efflux pump») быстрее, чем антибиотик поразит свои мишени. Апа-
тичным образом действует судовая пом1 ia, выкачивающая из трюма корабля заборт-
ную воду;
3) структурные изменения с молекулах, являющихся мишенями доя антибиотиков.
Проникший в клетку антибиотик не находит свои мишени и не может блокировать
* иохимические процессы;
4) продукция бактерией альтернативных мишеней, которые резистентны к ингиби-
ощему действию антибиотика. Они связывают антибиотик и лишают его возмож-
оста поразить настоящие мишени. Обычно в качестве ложных целей выступают фер-
енты (рис. 2.15).
Рис. 2.15. Основные механизмы резистентности к антибактериальным
препаратам у бактерий. По Р. W. Hawkey (1998)
356
Часть 2. Общая неправильная эпидемиология
Устойчивость к антибиотикам, обусловленная плазмидами, преимущественно
обеспечивается ферментами, модифицирующими антибиотики. Устойчивость к суль-
фаниламидам и триметоприму1 вызвана тем, что плазмиды детерминируют дублир* ъ ь
щие ферменты биосинтеза витаминов, нечувствительные к этим лекарственным пре-
паратам. Хромосома бактерии благодаря транспозиции также может содержать гены
модифицирующих антибиотики ферментов (Брода П., 1982). Но и для других меха-
низмов резистентности показана возможность передачи посредством мобильных ге-
нетических элементов. Даже структурные изменения мишеней для антибиотиков
могут быть вызваны вставочной активностью транспозонов. В обобщенном виде ме-
ханизмы действия антибактериальных препаратов и механи >мы резистентности книг
у бактерий приведены в табл. 2.8.
Таблица 2 •
Механизмы действия антибактериальных препаратов и механизмы
пезистентности к ним у бактерий*
Группа антибактериаль- ных препаратов Пример Механизм действия Механизм резистентности
1 2 3 4
Аминогликозиды Стрептомицин, гентамицин, тобрамицин, амикацин Ингибируют синтез белка путем связывания с частью субъединицы 30S бактери- альной рибосомы Большин- ство из них обладают бакте- рицидным действием (на- пример, вызывают гибель бактериальных клеток) Модифицирующие фер- менты (ацетилирование, аденилирование, фосфо- рилирование). Понижен- ная проницаемость или зависимое от энергии по- глощение. Пониженное связывание антибиотика рибосо1.гами
Бацитрацин Ингибирует продуцирование клеточной оболочки путем блокирования этапа в про- цессе (рециклинг носителя мембранного липида), необ- ходимого для добавления новых субъединиц клеточ- ной оболочки Пониженная проницаемость
Р-лактамные антибиотики Пенициллины: ампициллин, амоксициллин, тикарциллин. Цефалоспорины: цефтриаксон, цефокситин. цефалексин, цефтазидим. Монобактамы: азтреонам Карбепенемы: имипенем, меропенем Ингибируют образование бактериальной клеточной оболочки путем блокиро- вания перекрестного свя- зывания структуры клеточ- ной оболочки в пеницил- линсвязывающих белках (пептидогликановые син- тетические ферменты) Измененная мишень (пенициллин-связываю- ший белок).Пониженная проницаемость р-лакта- маз, модификация р-лак- тамаз
1 Сульфаниламиды и триметоприм — вещества немикробного происхождения, поэтом,
относят к химиопрепаратам.
2.2 Биологические поражающие агенты
357
Продолжение табл. 2.8
1 2 3 4
|лорамфеникол Блокирует перенос амино- кислот в пептидные цепи в субъединице 50S бактери- альной рибосомы; ингиби- рует синтез белка Пониженная проница- емость. Активный отток. Инактивирующий фер- мент (ацетилирование)
чкопептиды Ванкомицин Препятствует развитию клеточной оболочки путем блокирования прикрепления новых субъединиц клеточ- ной оболочки (мурамил- пентапептиды) Измененный сайт связы- вания предшественника пептидогликана
косамиды Клиндамицин, линкомицин Блокируют перенос амино- кислот в пептидную цепь в субъединице 50S бактери- альной рибосомы ингиби- рует синтез белка Пониженное связывание рибосом (метилирование рибосомальной РНК). Пониженная проницае- мость. Модифицирующие ферменты
' кролиды Эритромицин, кларитромицин, азитромицин Ингибируют транслокацию субъединицы 50S бактери- альной рибосомы на иРНК; ингибируют белок Пониженная проницае- мость. Модифицирующие ферменты. Слабое связы- вание с рибосомами (ме тилирование рибосомаль- ной РНК)
t гронидазол Разрушает структуру нуклеиновой кислоты Изменение механизмов активации препарата
• сазолидононы Линезолид, эперезолид Ингибируют синтез белка в 23S рРНК субъединицы 50S бактериальной рибосомы Измененная мишень
хинолоны Ципрофлоксацин, левофлоксацин, офлоксацин, норфлоксацин, налидиксовая кислота, спарфлоксацин Блокируют синтез ДНК ДНК-гиразой, топоизо- меразой внутривенно Изменение мишени (ДНК-гираза, топо- изомераза внутривенно). Пониженная проница- емость. Активный отток
гфампицин Ингибирует синтез РНК ДНК-зависимой РНК-по- лимеразой и таким образом синтез белка Слабое связывание с РНК-полимеразой
ульфаниламиды Сульфаметоксазол, сульфацетамид, сульфадоксин Конкурентное ингибирова- ние синтеза дигидрофолата из р-аминобензойной кислоты в дигидроптероат- синтетазе Измененная дигидропте- роатсинтетаза. Повышен- ный уровень р-аминобен- зойной кислоты. Пони- женная проницаемость
Стрептограмины Хинупристин/ далфопристин Блокируют вытеснение вновь синтезированных пептидных цепей из субъ- единицы 50S бактериальной рибосомы Слабое связывание с рибо- сомой (метилирование рибосомальной РНК). Пониженная проница- емость. Модифицирующие ферменты
358
Часть 2. Общая неправильная эпидемиологи*
Продолжение табл. 2 •
1 2 3 4
Тетрациклины Тетрациклин, окситетрациклин, доксициклин, миноциклин Ингибируют связывание переносимой РНК на субъ- единице 30S бактериальной рибосомы; ингибируют син- тез белка Детоксикация препарата. Барьеры проницаемости. Активный отток. Изме- ненная мишень (рибо- сома)
Триметоприм Ингибирует восстановление дигидрофолата до тетрагид- рофолиевой кислоты в дигидрофолатредуктазе Измененная дигидрофо- латредуктаза. Повышен- ный уровень р-аминобен- зойной кислоты. Пони- женная проницаемость
* По J. В. Woods (2005)
Резистентность к пенициллинам и цефалоспоринам. В ее основе лежит разрушение
пенициллинов обширной группой ферментов, называемых пениииллиназами иля
^-лактамазами. Эти ферменты разрывают Р-лактамные связи в молекулах пеницил-
линов, приводя к образованию неактивных производных (рис. 2.16).
ПЛАЗМИДА ОТО ТСТВУГ1
Внешний—> Наружная—> Периплазма- —Пенициллин- ____________>. Ингибирование
Р-лактам мембрана <— тическое связывающий синтеза
пространство белок пептидогликана
ПЛАЗМИДООПОСРЕДОВАННЫЙ СИНТЕЗ Р-ЛАКТАМАЗЫ -
.ПЛОХО ПРОНИКАЮЩИЙ Р-ЛАКТАМ»
Внешний—> Поры ► Периплазма- Периплазматический
Р-лактам наружной тическое —лакгамаэяый
мембраны пространство ГИДрОЛИЗ
Суоингибиторная
перилл азматическая
Гидролизат
концентрация
Р-лактама
I
Рост
ПЛАЗМИДООПОСРЕДОВАННЫЙ СИНТЕЗ Р-ЛАКТАМАЗЫ -
«ХОРОШО ПРОНИКАЮЩИЙ р-ЛАК ГАМ»
Внешний—>Поры —► Периплазм Периплазматический Гидролизованный
Р-лактам наружной X ► р-лакгамазный —► Р-лакгам
мембраны гидролиз
внешней Высвобо> а :ние Пенипиллин Пичис кпетпк
концентрации* Р-лактамазы связывающий ►
антибиотика
I
Рост
Рис. 2 16. Инактивация пенициллинов и цефалоспоринов р-лактамазой. В обоих слу-
чаях р~лактамная связь разрушается с помощью гидролитического механизма, но если
образующаяся при этом пенициллоиновая кислота относительно стабильна, то соот-
ветствующий продукт гидролиза цефалоспоринов чрезвычайно нестабилен и распа-
дается с образованием сложной смеси соединений. По Т. Франклину и Дж. Сноу (1984)
2Д Биологические поражающие агенты
359
Наиболее распространенными среди 0-лактамаз являются р-лактамазы ТЕМ-1 и
~ЕЧ-2, что связывают с локализацией их генов на транспозонах, переносимых меж-
I • бактериями плазмидами. У грамположительных и грамотрицательных бактерий
-.зистентность к пенициллинам ф-лактамам) осуществляется преимущественно че-
рез детоксикацию антибиотика. У хорошо изученной грамположительной бактерии
ureus основная часть синтезируемой Р-лактамазы уходит в окружающую среду и
1 разрушает Р-лактамные молекулы (рис. 2.17).
ПЛАЗМИДА ОТСУТС1ВУ Ы
чаЛНЫЙ
_=тибиотик
ж клетки
Диффузия ______Пенициллин- Ичгиьироваюк
через _ связывающие ___синтеза
пептидогликан белки пептидогликана
Активация
аутолитических
ферментов
ПЛАЗМИДООПОСРЕДОВАННЫЙ СИНТЕЗ Р-ЛАКТОМАЗЫ
истамаза
'.утри
' стернальной
клетки
Цитоплазма-
тическая
мембрана
«Встреча» Его
с р-лактамом гидролиз ►
Диффузия
через
пептидогликан
Внешняя
Р-лактамаза
С} бингибато] ные
концентрации —► “ост
Р-лактама
Гидролиз
► Р-лактамов
Рис. 2 17 Механизм развития плазмид-специфическои резистентности
к р-лактамным антибиотикам у грамположительных бактерий. По L. Bryan (1980)
Фундаментальной причиной, способствующей закреплению в ходе эволюции за
^положительными бактериями устойчивости данного типа к антибиотикам, явля-
ется отсутствие в их клеточной стенке периплазматического пространства. Уровень
-г.тибиотикорезистентности грамположительных микроорганизмов зависит от сце-
ничности фермента и того его количества, которое он может позволить себе экс-
пгссировать. р-лактама )ы таких бактерий, как правило, имеют высокий аффинитет
к Р-лактамным антибиотикам.
У грамотрицательных микроорганизмов деструкция Р-лакгамных антибиотиков
, ществляется в периплазматическом пространстве. Поэтому их уровенп резистент-
усти зависит от скорости, с которой р-лактамазы проникают в периш 1азматическое
—остранство, и скорости осуществляемого ферментами гидролиза Такие бактерии
лчно синтезируют меньшее количество фермента. Кроме того, он имеет меньшую
субстратную специфичность (аффинитет), чем у грамположительных бактерий. Боль-
шинство патогенных видов бактерий, встречающихся в стационарах, чувствительно,
о меньшей мере, к одному классу Р-лактамовых антибиотиков (Fluit А. С. et al., 2000).
Менее распространенной причиной резистентности бактерий к пенициллинам и
_зфалоспоринам могут быть мутации в генах пенициллинсвязывающих белков. Они
-риводят к пониженной аффинности этих белков к р-ллктамовым антибиотикам.
°еже резистентм >сть к таким антибиотикам вызывается их пониженным поглощением
леткой из-за изменений в ее оболочке или активного «откачивания» из бактериаль-
ной клетки проникшего антибиотика (Fluit А. С. et al., 2001).
Резистентность к аминогликозидам. Плазмидная резистентность к аминоглико-
идным антибиотикам (гентамицин, стрептомицин и др.) связана либо с их энзим-
360
Часть 2. Общая неправильная эпидемиология
ной модификацией, либо с нарушением проницаемости клеточной стенки бактерии
для этих антибиотиков при отсутствии видимых признаков их энзимной модифика-
ции. Существует более 50 ферментов, способных к N-ацетилированию, О-фосфорл-
лированию или О-нуклесл идилированию различных аминшликозидных антибиоти-
ков (Fluit А. С. et al., 2001).
Детоксикационный механизм плазмидной резистентности к антибиотикам дан-
ной группы характерен как для грамположительных, так и для грамотрицательных
микроорганизмов (Bryan L., 1980). Однако у последних резистентность носит более
выраженный характер благодаря наличию полисахаридных внеклеточных слоев. Бла-
годаря тому, что полисахариды несут отрицательный заряд (особенно это касаетс
мукоидных штаммов псевдомонад), они способны связывать отдельные амино-
гликозы, препятствуя их дальнейшему проникновению в клетку. После проникнове-
ния через наружные слои клеточной стенки аминогликозиды диффундируют через
поры, распложенные в наружной мембране, и достигают пептидогликана, не пред-
ставляющего для них серьезного барьера. Проникнув через клеточную стенку в пери-
плазматическое пространство, аминогликозиды связываются с полярными группа-
ми фосфолипидов цитоплазматической мембраны. Здесь они вступают в контакт с
модифицирующими энзимами. Модифицированный аминогликозид не способен свя-
заться рибосомами и тем самым блокировать синтез белка в бактериальной клетке
(Bryan L., 1980).
Резистентность к хлорамфениколу (левомицетину). Хлорамфеникол связываете-
с 50S рибосомной субъединицей и ингибирует этап пептидилтрансферазы при син-
тезе белка. Плазмидная резистентность к хлорамфениколу ассоциируется с наличи-
ем у них генов хлорамфениколацетилтрансферазы (cat). Широкое распространение
устойчивости к этому антибиотику связано с распространением транспозона Тп4
Хлорамфениколацетилтрансферазы у грамотрицательных и грамположительных мик-
роорганизмов обладают высокой степенью сходства по аминокислотным остаткам
активного центра. Для резистентных к хлорамфениколу бактерий обеих типов харак-
терна высокая детоксикационная активность в отношении этого антибиотика во
внешней среде. Однако для некоторых грамотрицательных бактерий, например, псев-
домонад, ее не наблюдается. У грамотрицательных бактерий иногда наблюдаются
пониженная проницаемость внешней мембраны к хлорамфениколу или его активный
отток из клетки (Bryan L., 1980; Hawkey Р. W, 1998).
Резистентность к тетрациклину. Тетрациклины проникают в бактериальные клет-
ки путем пассивной диффузии. Тетрациклин действует путем связывания с 30S ри-
босомальной субъединицей, приводя к ингибированию синтеза белка. Гены, опре-
деляющие устойчивость к тетрациклину (tet) грамотрицательных бактерий, часто
обнаруживаются в транспозоне Тп 10, передаваемом между ними крупными конъюгз-
тивными плазмидами. У грамположительных микроорганизмов резистентность к тет-
рациклину обычно ассоциируется с плазмидами с молекулярной массой 2,7 и 2,8 мД:
(например, у стафилококков). Плазмидная резистентность к тетрациклину связана с
уменьшением его аккумуляции клеткой, обратным транспортом (у грамогрицатель-
ных — гены tetA-tetE, tetG и tetH', у грамположительных — гены tet Ки tetL), внутрикле-
точной инактивацией (tetX) и защитой рибосом-мишеней (ГеГЛ/ или tetQ) (Fluit А. С
et al., 2001).
Но большинство генов tet кодируют один из двух механизмов резистентности к
тетрациклинам: либо отток антибиотика из клетки, либо рибосомную защиту. Рибо-
сомная защита включает синтез белка, подобный фактору элонгации G. Он взаимо-
j 2 биологические поражающие агенты
361
: . вует с рибосомой, не препятствуя синтезу белка, но и не давая тетрациклину
фовать этот синтез. У некоторых видов бактерий обнаружено окислительное
- сушение тетрациклина. Тем не менее детоксикационная инактивация антибиотика
1роде не играет важную роль (Fluit А. С. et al., 2001).
Резистентность к фторхинолонам. Фторхинолоновые антибиотики обнаружива-
лтибактериальное действие путем ингибирования некоторых бактериальных го-
>зомераз, а именно: ДНК-гиразы (бакгериальная топоизомераза II) и топоизоме-
жзы IV. Эти важные бактериальные ферменты изменяют топологию двунитевой ДНК
ДНК) в клетке. ДНК-гираза и топоизомераза IV являются гетеротетрамерными
ли, состоящими из двух субъединиц, А и В. Гены, кодирующие субъединицы А
В называютсяgyrA и gyrB (ДНК-гираза) или рагС ирагЕ (ДНК-топоизомераза IV)
р- А и grlB у S. aureus). ДНК-гираза является единственным ферментом, который
- ет влиять на образование сверхспирали ДНК. Ингибирование этой активности
| .орхинолонами ассоциируется с быстрым убиванием бактериальной клетки. Топо-
юмлраза IV также модифицирует топологию днДНК, но в то время как ДНК-гираза
еет, по-видимому, важное значение для поддержания образования сверхспирали,
- юизомераза IV преимущественно ответственна за разделение нитей дочерней ДНК
ри делении клеток (Fluit А. С. et al., 2001).
Механизмы бактериальной резистентности к фторхинолонам делятся на две ос-
- зные категории: изменения в ферментах-мишенях для препарата; и изменения,
ггорые ограничивают проникновение препарата в мишень. Ферменты-мишени
чень часто изменяются в доменах около активных сайтов фермента, и в некоторых
учаях была продемонстрирована пониженная способность к связыванию с препа-
том. У грамотрицательных микроорганизмов ДНК-гираза является, по-видимому,
. 1овной мишенью для всех хинолонов. У грамположительных микроорганизмов
«новной мишенью является топоизомераза IV или ДНК-гираза, в зависимости от
-усматриваемого фторхинолона, т. е. структура хинолона определяет механизм его
ггибактериального действия (Fluit А. С. et al., 2001).
Плазмидную резистентность к хинолонам (plasmid-mediated quinolone resistance,
MQR) обнаружили L. Martinez et al. (1998). Наиболее хорошо исследована резис-
тнтность бактерий к хинолонам, детерминируемая группой генов qnr. Они кодиру-
< пентапептидповторяющиеся белки (pentapeptide repeat proteins), снижающие чув-
. вительность бактерии к хинолонам посредством защиты от них комплекса днДНК
ДНК-гиразы или комплекса днДНК и топоизомеразы IV. Большинство qnr-к-
зв (qnrA, qnrB и qnrS) идентифицированы на плазмидах размеров от 54 до более чем
180 т. п. о., обнаруживаемых в Е. coli, Enterobacter spp., К. pneumoniae и Salmonella spp.
Часто они ассоциированы с генами, кодирующими р-лактамазы. Менее изученными
PMQR-механизмами являются: 1) механизм, в работу которого вовлечена аминогли-
пиданетилтрансфераза с двумя аминокислотными заменами, позволяющими ей
жтивировать ципрофлоксацин через ацетилирование; 2) кодируемый генами oqxAB
qepA «насос», обеспечивающий обратный транспорт проникшего в клетку хиноло-
а (Strahilevitz J. et al., 2009).
Подробно о механизмах резистентности к антибиотикам и сульфаниламидам
у бактерий можно ознакомиться по работам L. Bryan (1980); Р. W. Hawkey (1998);
О. Skold (2001); А. С. Fluit et al. (2001); Е Depardieu et al. (2007); H. Girgis et al. (2009).
Механизмы вмешательства в геном вирусов. Эксперименты по конструированию
модифицированных вирусов ведутся уже не менее 50 лет в рамках подходов по созда-
иию новых вакцин. За два последних десятилетия интерес к конструированию моди-
362
Часть 2. Общая неправильная эпидемиология
фииированных вирусов значительно увеличился в связи с разработкой методов сома-
тической генной терапии. Технология клонирования генов в геном вирусов только
самом общем виде соответствует схеме, приведенной ранее (см. рис. 2.8). Большой
размер вирусного генома делает задачу создания рекомбинантной конструкции в ус-
ловиях in vitro, чрезвычайно трудной для решения. К тому же изолированная вирус-
ная ДНК неудобна для трансформации клеток из-за ее низкой инфекционное™, по-
скольку интактные вирусные частицы содержат не только ДНК вируса, но еше и
ферменты, необходимые для транскрипции вирусных генов. Проблему введения чу
жеродной ДНК в геном вируса удалось решить в инфицированных вирусом клетках
используя гомологичную рекомбинацию между рекомбинантными плазмидами, со-
держащими вирусные последовательное™, и геномом вируса (рис. 2.18).
Рис. 2.18. Получение рекомбинантных вирусов на примере вируса осповакцины.
Клетки, инфицированные вирусом осповакцины, трансформируют ДНК рекомби-
нантных плазмид. Эти конструкции содержат промотор вируса вакцины (на схеме
обозначен белым прямоугольником), состыкованный в соответствующей ориента-
ции с кодирующей частью чужеродного гена (волнистая линия); такой функциональ-
ный блок в составе плазмиды фланкирован последовательностями вирусной ДНК.
Между фланкирующими последовательностями и вирусным геномом происходит
гомологичная рекомбинация, вследствие чего чужеродный ген оказывается в соста-
ве вирусной ДНК. Рекомбинантные геномы упаковываются в вирусные частицы —
формируются рекомбинантные вирусы. ПоД. Гловеру (1988)
Частное™ такой технологии применительно к решению отдельных задач мы рас-
смотрим на конкретных примерах ниже. Кроме технологий генной инженерии, у ис-
следователей имеются большие возможности вмешательства в геном РНК-вирусов с
помощью методов генетического обмена, имитирующих генетические события, про-
исходящие среди вирусов в природе: реассортации и рекомбинации.
Колонирование в вирусах генов токсинов. Привлекательным объектом для военно-
биологических исследований являются гены токсинов (см. разд. 2.2.3.6). В литерату-
ре, посвященной опасное™ распространения БО, уже не раз высказывалась мысль о
том, что технологией рекомбинантной ДНК могут воспользоваться террористы для
получения бактерий, синтезирующих смертельные токсины (см. Dando М., 1994; Гай-
22 Биологические поражающие агенты 363
скр Э. с соавт., 1986). В действительности это только благие пожелания, основанные
а формальной логике и на непонимании патогенеза инфекционных болезней, вызы-
вных бактериями. Наиболее токсичные из токсинов имеют сложное субъединичное
строение, большую молекулярную массу и активны только тогда, когда их молекула
• меет определенную конформационную структуру. Последнее достигается благода-
-1 дисульфидным связям, образующимся между цистеиновыми остатками молекулы
- ?ксина. Бактерии неэффективно катализируют образование дисульфидных связей
между участками белковых молекул. Поэтому в бактериях очень сложно добиться
. интеза крупных и имеющих сложную конформационную структуру белковых мо-
- екул, сохраняющих свою биологическую активность. Ту же задачу можно решить
рулим путем, например, используя для сборки молекул токсина ткани самой жерт-
.. В качестве примера разберем работу S. Gorman et al. (1997), посвященную разра-
ботке подхода к экспрессии в клетках млекопитающих гена смертельного нейроток-
. «на змей — к-бунгаротоксина (kappa-Bgt, к-Bgt)1.
Для достижения этой цели авторы использовали векторную систему на основе
. г.новируса. Аденовирусы относятся к группе ДНК-вирусов и вызывают респиратор-
- JTO инфекцию. Генных инженеров они заинтересовали благодаря своей способности
‘нфицировать делящиеся и неделящиеся клетки, включая клетки печени, легких и
эпителия. Векторы первого поколения реплицировались как эписомные (экстрахро-
мосомные) элементы в ядре клетки хозяина. Они обеспечивали очень высокую экс-
рессию, но на непродолжительное время (5—10 сут) из-за противодействия иммун-
ной системы реципиента, реагировавшей на синтез аденовирусных белков трансфе-
дированными клетками. Поэтому было предложено второе поколение аденовирусных
эекторов, у которых были удалены гены, вовлекающие вирус в репликацию. Такие
эекторы оказались способными к более длительной экспрессии клонированных ге-
4ов, но и у них она практически не определялась через 20—40 сут. Эта идея получила
свое дальнейшее развитие созданием «выпотрошенных» (gut-less) векторов, из кото-
рых были удалены все вирусные гены и оставлены только элементы, определяющие
-ачало и конец генома аденовируса и вирусную пакующую последовательность. Гены,
'ереносимые такими векторами, экспрессировались до трех месяцев. Аденовирусные
эекторы имеют широкий спектр хозяев, реплицируются независимо от клеток хозяи-
на, обладают большой емкостью (до 30 т. п. о.), могут вводиться в организм человека
перорально (в виде покрытой оболочкой капсулы), в составе аэрозоля в легкие и внут-
римышечно. Недостатком таких векторов, ограничивающим их использование в
генотерапии, является неспособность к интеграции в хромосому рецип иента, в резуль-
тате вектор утрачивается по мере продолжения деления клеток (см. разд. 2.2.4).
к-Bgt входит в состав яда многополосого крайта (Bungarus multicinctus), весьма
опасной для людей змеи, обитающей на юге Китая, в Индии и Бангладеш (Faiz A. etal.,
2010). В яде В. multicinctus наиболее представлен а-бунгаротоксин (a-Bgt). Он с очень
высоким аффинитетом связывается с никотиновым ацетилхолиновым рецептором
(AChR) мышечного типа, блокируя активацию рецептора ацетилхолином и вызывая
мышечный паралич. к-Bgt содержится в яде полосатого крайта в значительно мень-
1 Известны четыре бунгаротоксина. P-Bgt действует пресинаптически, а- и y-Bgt действуют
постсинаптически (антагонисты ацетилхолина в постсинаптической мембране), к-Bgt — бло-
кирует нейрональные никотиновые рецепторы. LD50 для мышей к-бунгаротоксина не превы-
шает 25 мкг/кг. Для сравнения LD50 для данного вида животных цианида калия и боевого ОВ
зомана, составляют 10000 и 100 мкг/кг соответственно.
364
Часть 2. Общая неправильная эпидемиология
ших количествах, чем a-Bgt, побуждая таким образом ученых искать другие пути его
получения, включая биотехнологические. Он имеет общую гомологию на аминокис-
лотном уровне с a-Bgt, но связывается с другим классом AChR, локализованных глав-
ным образом в периферической нервной системе. Для препаративного получения
активного к-Bgt исследователям можно было пойти по «накатанному пути» и эксп-
рессировать кодирующий его ген в дрожжах рода Picchia (см. работу Fiordalisi J. J. et
al., 1996). Но S. Gorman et al. (1997) решали еще одну задачу — им надо было обеспе-
чить длительную экспрессию гена токсина в тканях млекопитающего и, в частности,
в ЦНС. Этим обстоятельством и был обусловлен выбор аденовируса в качестве век-
тора экспрессии (см. разд. 2.2.4).
к-Bgt содержит 10 цистеиновых остатков на общее количество 66 аминокислот
В просвете эндоплазматического ретикулума клетки млекопитающего имеются хоро-
шие условия для образования дисульфидных связей. Но для секреции активной кон-
формации к-Bgt в клеточную культуральную сред; токсин должен был иметь провод-
ника, выводящего его за пределы клетки. По этой причине кДНК к- Bgt была соеди-
нена с З'-концом ДНК сигнальной последовательности бычьего пролам ина (R к-Bgt).
обеспечившей перенос активного к-Bgt через мембрану клез ки в клеточную культу-
ральную среду.
Клонирование к-Bgt в ДНК аденовируса осуществлено следующим образом.
кДНК R-K-Bgt была субклонирована в рекомбинантный плазмидный вектор, pAdlox.
в сайт справа от промотора цитомегаловируса (CMV). Плазмида была линеаризована
путем гидролиза рестриктазой Sfil и вместе с ДНК рекомбинантного аденовируса
трансфецирована в клетки НЕК-293, постоянно экспрессирующие CRE-рекомбиназ.
(клетки CRE8). Последовательность <р5 (упаковочный сигнал, РАС) аденовируса была
модифицирована таким образом, чтобы содержать элементы 1охР на каждой стороне
упаковочного сигнала. В результате внутримолекулярная рекомбинация, катализиро
ванная рекомбиназой клеток НЕК-293, устраняла упаковочный сигнал аденовируса,
вследствие чего аденовирус без кДНК R-K-Bgt не упаковывался в вирусную частицу.
В противоположность этому межмолекулярная рекомбинация между вирусом и плаз-
мидным вектором (pAdlox), содержащим упаковочный сигнал и только один сайт 1охР.
приводила к стабильному включению в аденовирусный вектор упаковочного сигна-
ла, несущего с собой желаемую кДНК вместе с ее промотором (рис. 2.19).
Через 8 сут из подвергнутых трансфекции клеток CRE8 методом замораживания
размораживания был приготовлен лизат. Вирус еще раз был пассирован через клетки
CRE8, очищен методом бляшек на монослоях НЕК-293 и затем использован в экспе-
риментах. Активный рекомбинантный к-Bgt секретировался в среду инфицирован-
ных клеток при концентрации 10-100 нМ.
Использованный авторами вектор относился ко второму поколению аденовирус-
ных вею оров. Аденовирус был модифицирован путем делеции двух генов (Е1А и Е1В'
необходимых для репликации его ДНК. Таким образом, рекомбинантный аденовирус,
кодирующий к-Bgt, мог пролиферировать только в искусственных системах (in vitro) —
клетках НЕК-293 или в культурах человеческих клеток, экспрессирующих Е1А и Е1В.
Однако делеция этих генов не мешает вирусу проникать в клетки млекопитающих в
условиях in vivo, включая ЦНС, и руководить экспрессией гена белка к-Bgt, не обо-
значая себя инфекционным процессом. Однако эти данные S. Gorman et al. (1997) в
своей работе не привели.
Аналогично подходу к клонированию генов в аденовирусах, описанному выше,
технология рекомбинантной ДНК применяется для получения инсектицидов на
2 ? Биологические поражающие агенты
365
Аденовирус (Adlox) Линеаризованная плазмида (pAd-lox/NBT)
Рис. 2.19. Конструирование рекомбинантного аденовируса, кодирующего к-бунгаро-
токсин. На схеме показан Sfil-фрагмент pAd-lox к-Bgt, который содержит, слева на-
право, инвертированный терминальный повтор (ITR), упаковочный сигнал (РАС),
CMV-промотор (CMV), кДНК к-Bgt и сайт 1охР. Фрагмент был подвергнут котранс-
фекции в специально созданной линии клеток (HEK-293/CRE8) вместе с модифици-
рованным аденовирусом, содержащим два сайта 1охРна каждой стороне упаковочного
сигнала. CRE-опосредованная внутримолекулярная рекомбинация дает в результа-
те не поддающуюся упаковке вирусную ДНК. Межмолекулярная рекомбинация с
плазмидным фрагментом дает функциональный вирус. По S. Gorman et al. (1997)
основе бакуловирусов, вот только цели экспериментов формулируются более кон-
кретно. Бакуловирусы в течение многих лет используются в программах борьбы с на-
секомыми в полевых условиях. Их основным недостатком является относительно мед-
ийное умерщвление «мишеневого насекомого», что мешает быстрому прекращению
•рона, наносимого в результате питания насекомых. П. Керлей с соавт. (2000) вклю-
чили синтетический ген специфического к насекомому нейротоксина среднеазиатского
скорпиона или паука Segestria, находящегося под контролем бакуловирусного промо-
~ора, в рекомбинантный вирус ядерного полиэдроза. Далее они обнаружили, что если
последовательность, кодирующую токсин, слить с последовательностью, кодирующей
сигнальный пептид, то можно добиться секреции нейротоксина за пределы клетки, в
которой размножается вирус (то же самое с геном к-Bgt сделали S. Gorman et al., 1997;
см. выше). Когда таким вирусом инфицировали насекомых, их гибель происходила
ча 25 % быстрее, чем при использовании рекомбинантных бакуловирусов, не снаб-
женных последовательностью секреторного сигнала для нейротоксина (109 и 85 ч
соответственно). Для удобства применения рекомбинантного вируса, экспрессирую-
щего нейротоксин скорпиона на сельхозугодьях, исследователи включили его в состав
специальной композиции, «состоящей из множества мелких частиц, охватывающих
бакуловирусы» с «предпочтительным размером частиц от 5 до 25 мкм».
Повышение вирулентности вирусов на основе нарушения регуляции иммунных отве-
тов макроорганизма. Такие эксперименты проводятся под предлогом изучения тон-
ких механизмов иммунных реакций на вирусы, разработки стратегий для контроля
аутоиммунных реакций и др. Они основаны не на формальном понимании вирулент-
ности вирусов как функции так называемых «генов вирулентности», т. е. чем больше
366
Часть 2. Общая неправильная эпидемиология
таких генов у вируса, тем больше его вирулентность; и не на прямолинейном вмеша-
тельстве в геном вируса, когда его «заставляют» секретировать ген какого-то токсина
животных из инфицированной клетки (эксперименты по клонированию в вирусах
генов нейротоксинов; см. выше). В основе таких подходов лежит знание тонких ме-
ханизмов воздействии на систему иммунных ответов организма на вирус во время
инфекционного процесса. В качестве примера разберем работу R. J. Jackson et al.
(2001), исследовавших изменение вирулентности вируса мышиной оспы (вирус экт-
ромелии1) после клонирования в его геноме гена интерлейкина-4 (IL-4)2.
В экспериментах авторы использовали линии мышей двух типов; чувствительных
к мышиной оспе (например, BALB/c), у которых болезнь развивается как острая сис-
темная инфекция с высокими вирусными титрами в печени и селезенке, обусловли-
вающими некроз и высокую смертность; и резистентных к мышиной оспе (например.
C57BL/6). У последних оспа развивается в субклинической форме и характеризуется
низкими уровнями вирусной репликации во внутренних органах и появлением не
смертельно опасных поражений.
Генетическая устойчивость мышей к вирусу эктромелии характеризуется эффек-
тивной реакцией естественных киллеров (NK) и ранним появлением сильной реак-
ции цитолитических Т-лимфоцитов (ЦТЛ) CD8+. R. J. Jackson et al. (2001) клониро-
вали в геноме вируса эктромелии ген IL-4 и изучили влияние экспрессируемого
вирусом мышиного IL-4 на клеточно-опосредованную иммунную реакцию во вре-
мя инфекции. Ими было отмечено, что у мышей, генетически устойчивых к вирус}
эктромелии (исходному вирусу), экспрессирование 1L-4 рекомбинантным виру-
сом эктромелии подавляло цитолитические реакции NK и ЦТЛ и синтез последними
у-интерферона. Инфицирование генетически устойчивых мышей модифицирован-
ным вирусом эктромелии приводило к развитию у них симптомов острой оспы, со-
провождающейся высокой летальностью, подобно заболеванию, наблюдаемому у
мышей, генетически чувствительных к этому вирусу.
Реакции иммунной памяти у мышей, ранее иммунизированных против вируса
эктромелии, также ингибировались IL-4, синтезированным вирусом эктромелии. Во
внутренних органах иммунных мышей развивалась неконтролируемая репликация
модифицированного вируса и, как следствие, у них появлялись классические симп-
томы острой мышиной оспы. В противоположность этому, предсушествующий им-
мунитет к вирусу эктромелии был достаточен для ограничения реинфекции, вызыва-
емой вирулентным вирусом. Предыдущие исследования с использованием близкород-
ственного осповируса — вируса осповакцины, показали, что и у него коэкспрессия
IL-4 ассоциируется с задержкой выведения вируса из основных органов-мишеней и
повышением вирулентности для мышей (Bembridge G. Р. et al., 1998).
Реассортация и рекомбинация вирусов. Это два особых, хотя и не исключающих друг
друга способа генетического обмена, встречающихся в природных условиях у РНК-
вирусов. Первый, реассортация — построение генома дочернего вируса из фрагмен-
тов геномов разных родителей. Встречается при смешанной инфекции вирусами
1 Вирус эктромелии (семейство Poxviridae, род Orthopoxvirus) является естественным па-
тогеном лабораторных мышей, вызывает у них системную болезнь, называемую мышиной
оспой. Используют для моделирования натуральной оспы в опытах на мышах.
2IL-4 относится к лимфоцитарным цитокинам противоспалительного действия. Ингиби-
рует функции моноцитов, макрофагов и NK-лифоцитов (более подробно о его функциях см
в работах Галактионова В. Г., 2005; и Кузника Б. И., 2004).
. 1 Биологические поражающие агенты 367
. ссагментарными геномами. Заключается в обмене одной или нескольких дискрет-
ен к молекул РНК. Реассортация обнаружена у семейств с 2 (арена- и бирнавирусы),
ньявирусы) 6, 7 или 8 (ортомиксовирусы) или 10, 11 или 12 (реовирусы) геном-
сегментами. В клетках, инфицированных двумя родственными вирусами, внут-
каждого семейства может происходить обмен сегментами с образованием жизне-
•_ эбных стабильных реассортантов (Worobey М., Holmes Е. С., 1999).
Второй процесс, рекомбинация, обнаружен у сегментированных или несегментиро-
<_1ных вирусов, когда «донорная» нуклеотидная последовательность вводится в един-
чную прилегающую «акцепторную» молекулу РНК, продуцируя новую РНК, со-
кр кашую генетическую информацию более чем из одного источника. Если донорная
жкледовательность точно замещает гомологичный участок акцепторной последова-
-»тьности, оставляя ее структуру неизменной, то такой процесс называют «гомологи-
рекомбинацией», поскольку он включает не только гомологичные родительские
и: лекулы РНК, но и обмен гомологичными участками. Наблюдается образование
идных последовательностей в результате аберрантной гомологичной рекомбина-
_ • (когда похожие вирусы обмениваются последовательностями без соблюдения
рогого соответствия) и негомологичной рекомбинации (рекомбинация между не-
• лственными PH К-последовательностями) (Worobey М., Holmes Е. С., 1999).
Реассортацию вирусов с середины 1960-х гг. используют для получения аттенуи-
- -иных штаммов вируса гриппа, пригодных для использования в качестве живой
•акцины. Суть способа заключается в том, чтобы заранее полученным и охарактери-
нным аттенуированным штаммам вируса придать необходимую антигенность в
•ответствии с антигенностью актуальных полевых изолятов вируса гриппа. Для это-
от вирусов гриппа — доноров аттенуации, берут гены, кодирующие внутренние
>тки и ограничивающие репликацию, от актуальных эпидемических (эпизоотичес-
ких) штаммов вируса заимствуют гены, кодирующие поверхностные белки (гликоп-
.-отеины). Способ хорошо отработан и есть разные варианты его осуществления.
В природных условиях генетический обмен приводил к образованию новых ви-
.* нюв, способных вовлекаться в эпидемические процессы. Например, группа вирусов
стадного энцефалита лошадей является продуктом единичной рекомбинации меж-
ту вирусом восточного энцефалита лошадей и синдбис-подобным вирусом, имевшей
* «сто около 2000 лет назад (Weaver S. С. et al., 1997). Оба этих процесса (рекомбина-
ция и реассортация) являются важным источником генетической вариабельности
вирусов в рамках вида. Однако происходит ли сегодня образование таким путем но-
-ых вирусов, в каких хозяевах «работают» эти способы генетического обмена и вызы-
sajor ли эпидемические процессы появившиеся благодаря им новые вирусы, мне най-
конкретной информации не удалось (см. разд 3.10).
Резистентность вирусов к химиотерапевтическим препаратам. Штаммы вирусов,
резистентные к химиотерапевтическим препаратам, представляют серьезную проб-
сму для здравоохранения и без их целенаправленного конструирования «заинтере-
эванными лицами». Популяция любого вируса гетерогенна из-за мутаций в его ге-
номе, возникающих в процессе его репликации. Поэтому развитие у вирусов резис-
тентности к химиотерапевтическим средствам антивирусной терап ии происходит под
селективным давлением этих же средств. М. Nijhuis et al. (2009) выделяют три фазы в
“оявлении резистентных к химиопрепаратам штаммов вирусов: 1) в квазиспецифи-
еской популяции (quasispecies population) вирусов селекционируются штаммы, ме-
нее чувствительные к химиопрепаратам, чем наиболее распространенные среди
зывающих вспышку инфекции; 2) селекционированные действием антивирусных
368
Часть 2. Общая неправильная эпидемиология
препаратов варианты вирусов приобретают дополнительные мутации, делаюшие их
значительно более резистентными; 3) в популяции вируса накапливаются компенса-
торные мутации, позволяющие преодолеть снижение репликативной активности
вируса, резистентного к химиопрепарату.
Блокирование размножения вирусов невозможно без создания сходных проблем
(например, побочные действия) для хозяина. Большинство вирусов генетически про-
сты по сравнению с бактериями. Количество специфических мишеней для антиви-
русных препаратов у них ограничено. Механизмы действия антивирусных препара-
тов и механизмы резистентности к ним у вирусов, обобщены в табл. 2.9.
Таблица 2.9
Механизмы действия антивирусных препаратов и механизмы
резистентности к ним у вирусов (кроме ВИЧ)*
Антивирусный препарат Механизм действия Механизм резистентности
Ацикловир Валацикловир Фамцикловир Пенцикловир Ганцикловир Аналоги гуанозина. Ингибируют вирусспенифи- ческую ДНК-полимеразу Модифицированная тимидинкиназа. Модифицированная ДН К-полимераза
Цидофовир Аналог цитозина. Ингибирует вирусспецифиче- скую ДНК-полимеразу Модифицированная ДНК-полимераза
Идоксуридин и трифтортимидин Аналоги тимидина. Ингибируют вирусспецифи- ческую ДНК-полимеразу Модифицированная ДНК-полимераза
Ламивидин Аналог дезоксинуклеозида. Ингибирует обратную транскриптазу (например, ВИЧ), ингибирует ДНК-полимеразу вируса гепатита В Модифицированная обратная транскриптаза Модифицированная ДНК-полимераза
Рибавирин Аналог пуриннук леозида. Механизм действия не полностью понятен. Конкурентное ингибирова- ние ферментов хозяина приводит к пониженным внутриклеточным концентрациям гуанозинтри- фосфата и замедленному синтезу ДНК. Ингиби- рование комплекса вирусной РНК-полимеразы Неизвестен
Фосклрнет Пирофосфатный аналог фосфоноуксусной кислоты. Неконкурентный ингибитор вирусной ДНК-полимеразы и обратной транскриптазы Модифицированная ДНК-полимераза
Амантадин и ремантадин Высокая доза ингибирует раннюю стадию инфек- ции, включающую слияние между вирусной обо- лочкой и мембраной вторичных лизосом. Низкая доза ингибирует сборку вируса путем взаимодей- ствия с гемагглютинином Модифицированный трансмембранный домен белка М2
Оселтамивир и занамавир Аналоги сиаловой кислоты. Ингибиторы нейра- минидаз вируса гриппа А и В Модифицированная нейраминидаза
Интерфероны Гликопротеиновые цитокины. Иммуномодули- рующие, противоопухолевые и антивирусные свойства Модифицированная нейраминидаза
* По J В. Woods (2005).
Резистентность к антивирусным химиопрепаратам может быть придана вирус;
искусственно и без особых методических сложностей. Ниже рассмотрим этот процесс
на примере получения штаммов вируса вакцины, резистентных к ациклическим
_. Био логические поражающие агенты 369
гозидфосфонатам (acyclic nucleoside phosphonate, ANP), новым препаратам, по-
_ j-пим надежды на эффективное лечение поксвирусных инфекций. D. В. Gammon
al 2008) в течение года пассировали вирус вакцины (штамм Lederle) до 40 раз в
ере*е. содержащей возрастающие количества (Y)-l-[3-hydroxy-2-(phosphonometho-
r >pyl)-2,6-diaminopurine [(5)-HPMPDAP]. В результате им удалось селекциониро-
мутант вируса, примерно в 15 раз более резистентный к (Y)-HPMPDAP. чем
- одный. Анализ ДНК последовательности резистентного штамма позволил DeHa-
rt жить, что этот вирус содержит мутации в гене E9L (ДНК-полимераза), представляю-
-г .обой комбинацию двух замен оснований (A684V и S851Y) (рис. 2.20).
|- polBexo Ч |------ДНКро1В ------1
АгаС PAA Aph Aph
N-------Ч|---------- И IB | |---------С
IV II VIIII I V
Штаммы:
Lederle (дикий тип) A AS
Lederle (мутант) T V Y
Положение 314 684 851
Рис. 2. 20. Карта расположения мутаций в гене E9L штамма вируса вакцины, ре-
зистентного к HPMPDAP. На карте показан участок гена, кодирующий высоко-
консервативный мотив ДНК-полимеразы. Мутации в положениях 684 и 851, Ра-
нее картированная мутация находится в положении 314, она придает вирусу
вакцины резистентность к другому ингибитору полимеразы — цидофовиру. «ро1
В ехо» и «DNApol В» — участки гена, отвечающие за активность 3'-щ-5'-экзонук-
леазы и 5'-Го-3'-полимеразы соответственно. По D. В. Gammon et al. (2008)
Затем было изучено влияние этих мутаций (каждой по отдельности и обеих вмес-
. на резистентность вируса к различным ANP. Новые мутанты сравнили с Мутанта-
H. полученными ранее (мутации А314Т и A684V), резистентными к цидофовиру
с Jofovir, (5)-l-[3-hydroxy-2-(phosphonomethoxypropyl)cytosine|. Исследования пока-
ти возможность комплексной резистентности у ортопоксвирусов. Вирус с двумя
мутациями в гене E9L обладал значительно большей резистентностью к аналогам
. еоксиаденозина (deoxyadenosine), чем к аналогам деоксицитидиннуклеотида (deoxi-
. ktdine nucleotide analogs). Вирусы с мутацией S851Y показывали низкую резистент-
зсть к dCMP-аналогам, но высокую к 6-[3-hydroxy-2-(phosphonomethoxy)propoxy]-
2.4-diaminopyrimidine. Наиболее эффективным против этих мутантных вирусов
чазался (5)-9-[3-hydroxy-2-(phosphonomethoxy)propyl]-3-deazaadenine. Исследова-
я полученных мутантных штаммов вируса вакцины в условиях in vitro показали, что
’ации A684V и S851Y частично подавляют рост вируса, но все мутантные вирусы
^оя'щяют разную степень снижения вирулентности для мышей при их интраназаль-
:м инфицировании. Инфекция у мышей, вызванная этими лекарственно-резистен-
тными вирусами, блокировалась высокими концентрациями ANP.
Ограничения в конструировании штаммов реплицирующихся биологических пора-
жающих агентов с измененными свойствами. За внешней простотой вмешательства в ге-
ом бактерий и вирусов прячется много «ловушек», которые очень трудно обойти
террористам из импровизированных террористических организаций типа «Аум-Син-
рикё». В разд. 1.8 мы рассмотрели две такие «ловушки» — утрата микроорганизмом
ярулентности при поддержании на искусственных питательных средах, и так назы-
ie лая «плата за селекцию», которую приходится отдавать за селекцию штамма по
370
Часть 2. Общая неправильная эпидемиология
одному признаку. Количество «ловушек» значительно увеличивается при вмешатель-
стве в геном микроорганизма генно-инженерными методами.
Успех генетического переноса рекомбинантной плазмиды не является предопре-
деленным из-за систем рестрикции-модификации бактерии После проникновения
в бактерию-реципиент чужеродная ДНК подвергается атаке специальными фермен-
тами (рес грикционными эндонуклеазами), которые «признают ее как чужеродную»
и расщепляют на небольшие фрагменты. ДНК самих реципиентных клеток не под-
вергается такому расщеплению, так как она химически «модифицирована», т. е.
отдельные ее основания (аденин или цитозин) метилированы присоединением ме-
тильной группы. В природе эти системы являются своего рода ограничителями про-
никновения чужеродной ДНК в бактериальную клетку. В случаях, когда небольшой
части плазмидной ДНК удается избежать рестрикции, она может быть сегрегирован-
на из-за случайного распределения между дочерними клетками или оказаться вовле-
ченной в рекомбинационный процесс. И в том и в другом случае плазмида может быть
утеряна бактерией вместе с клонированным геном (Маниатис Т. с соавт., 1984).
В экспериментах по клонированию генов .тля научных целей используют не так
называемые «дикие» штаммы бактерий (т. е. полученные из природных источников),
а искусственные клонирующие системы, включающие специально сконструирован-
ные реципиентные ш гаммы; и плазмидные векторы, способные в них размножатьс'
Для предотвращения рекомбинации плазмиды используют штаммы с нарушенной
способностью к рекомбинации; а для предотвращения рестрикции ее ДНК, штаммы
с нарушенными системами рестрикции-модификации. Для предотвращения сегрега-
ции пла !миды на штамм оказывают селективное давление по маркерному для плаз-
миды антибиотику, тем самым не давая размножаться бактериям, ее утратившим
При удачном эксперименте по генетическому переносу рекомбинантной плазми-
ды (см. рис. 2.3) ее дальнейшая судьба может зависеть от собс гвенных плазмид реци-
пиента. Если среди его плазмид есть такая, у которой общая с рекомбинантной пла: -
мидой регуляция репликации, то в этом случае произойдет утрата либо одной из них,
либо их обеих (феномен несовместимости плазмид) (Брода П., 1982).
Попытки обойти проблемы, связанные с нестабильностью плазмид путем интег-
рации чужеродной ДНК в хромосому реципиентной бактерии, заводят эксперимен-
татора только в новые «ловушки». Прежде всего, снижается количество экспрессирую-
щихся копий клонированного гена, так как плазмидные векторы в бактериях обычно
поддерживаются деся гками копий. Стабильность интегрировавшегося сегмента ДНК
в геноме бактерии (см. рис. 2.5 и 2.10) самим фактом интеграции не гарантирует -
Стабильность определяется свойствами фланкирующих его нуклеотидных последо-
вательностей. Если фланкирующие последовательности не благоприятствуют выре-
занию фрагмента (например, отсутствуют протяженные прямые повторы), то интег-
рированный фрагмент стабилен. При этом сегмент ДНК может быть стабильным (или
нестабильным) совершенно независимо от присутствия в нем экспрессирующегося
гена, влияющего на фенотип бактерии Введение в бактериальную клетку структур,
включающих IS-элементы и прямые повторы1 (плазмиды, интегроны, транспозоны
и др.), увеличивает нестабильность генома и приводит к утрате важных биологических
свойств бактерией. Прямые повторы стимулируют делеции, дупликации и трансло-
кации, а инвертированные повторы — инверсии. В качестве прямых повторов, гене-
рирующих делеции и другие перестройки, могут выступать IS-элементы (рис. 2.21
1 Прямые повторы всегда фланкируют IS-элементы.
’ 2 Биологические поражающие агенты
371
Рис. 2.21. Геномные перестройки у бактерий, стик улируемые IS-элементами.
1 — внедрение TS-элемета в геном; 2 — распространение IS-элемента в гено-
ме за счет транспозиции. 3 — реципрокные транслокации (участок а меняется
на участок б); 4 и 5 — делеции и образование плазмид. По Г. Б. Смирнову (2007)
Сумев обеспечить стабильность и эффективную экспрессию клонированного гена
новом хозяине, исследователь решает новые проблемы. Выше приведены примеры.
- . казывающие (см. рис. 2.11—2.14), что устойчивость бактерии даже к одному анти-
ггику зависит от действия многих генов. Но возможности клетки по экспрессии
- « еродных генов ограничены возможностью развития у нее метаболической перегруз-
а — нарушения метаболизма в результате экспрессии чужеродной ДНК. К метабо-
j чческой перегрузке приводят:
увеличение числа копий и/или размера плазмид и связанное с этим у величение
количества энергии, необходимой для их репликации и сохранения;
недостаток растворенного кислорота в среде и невозможность обеспечения им всех
..таболических реакций и процесса экспрессии плазмидных генов;
гиперпродукция чужеродных белков, приводящая к истощению пула некоторых
_миноацил-тРН К;
перегрузка системы экспорта и нарушение правильной локализации жизненно
> кных белков хозяйской клетки вследствие «перепроизводства» чужеродного белка,
кспортируемого из цитоплазмы к клеточной мембране или в периплазматическое
“остранство;
наличие у организма-хозяина необычных метаболических свойств (например,
высокая дыхательная активность), что делает его более чувствительным к различным
воздействиям, чем обычные клетки;
непосредственное влияние чужеродных белков на функционирование хозяйской
клетки (Жимулев И. Ф., 2002).
При работе с вирусами подобных трудностей будет не меньше, но «полоса свобо-
ды »у вирусолога всегда уже, чем у бактериолога из-за трудностей и специфики рабо-
п с вирусами. Путем реассортации вирусов гриппа уже более 50 лет не удается полу-
щть эффективной гриппозной вакцины, хотя недостатка кадров и финансирования
эти исследования никогда не испытывали. Повышение вирулентности ВНО не ведет
к увеличению его контагиозности (см. работу Noble J., Rich J., 1969). Получение рези-
-гентного ортопоквируса в лабораторных условиях, как это описано D. В. Gammon
372
Часть 2. Общая неправильная эпидемиологи*
et al. (2008), по сути привело к появлению дефектного вируса с ослабленной способ-
ностью к репликации. Когда такие штаммы образуются естественным путем, то это
происходит в популяциях вирусов. Эпидемический процесс вызывают популяции ви-
русов, а не их отдельные штаммы. В таких популяциях накапливаются компенсатор
ные мутации, позволяющие преодолеть снижение репликативной активности виру-
са, резистентного к химиопрепарату. Но этот процесс невозможно воспроизвести в
лабораторных условиях.
Количество трудностей, которые надо преодолеть разработчикам биологическо-
го поражающего агента с измененными свойствами, не уменьшается при удачном ис-
ходе экспериментов по получению нового штамма микроорганизма, а еще больше
возрастает. Теперь им нужны большие и квалифицированные коллективы для отра-
ботки технологий его массового производства, высушивания, хранения, ренептури-
рования и боевого применения. Разумеется, обладающие специальным оборудовани-
ем и опытом.
Неспецифическая профилактика и лечение поражений реплицирующимися биолож-
ческими поражающими агентами. Ведущую роль в лечении таких поражений буду*
играть антибиотики и химиотерапевтические препараты. Краткий перечень схем ле-
чения и профилактики болезней, вызываемых биологическими поражающими аген-
тами, приведен в табл. 2.10.
До настоящего времени описан только один случай использования для соверше-
ния биотеррористического акта необычного штамма известного микроорганизма —
это штамм Ames возбудителя сибирской язвы (более подробно о нем см. в разд. 1.12 А
Штамм устойчив к антибиотикам |3-лактамного ряда и способен «пробивать» действе .
некоторых сибиреязвенных вакцин. Однако он не относится к созданным искусствен-
но. Его наличие в iосударственных лабораториях США обязано проводимой в это
стране с 1927 г. систематической и кропотливой работе по исследованию штаммов
В. anthracis, вызывающих эпизоотии сибирской язвы у домашнего скота.
Опыт ликвидации террористической вспышки сибирской язвы в США в 2001 1
показал, что наличие резистентности у штамма микроорганизма к антибиотику, ко-
торым лечат жертвы террористического акта, при ингаляционной форме поражен
приводит к смертельному исходу. Поэтому наиболее эффективными оказались схемы
лечения, использующие комбинацию антибактериальных препаратов и комплекс
патогенетических лечебных мероприятий, включающих даже хирургическое вмеша-
тельство (например, торакоцентез, дренирование плевральной полости). Более под-
робно эти схемы мы рассмотрим в третьей части книги.
* * *
Различия по морфологическим, культуральным, биохимическим, антигенным
вирулентным свойствам межту штаммами патогенного микроорганизма, обнаруже -
ного в новом эпидемическом очаге, и уже известными, не могут однозначно считать-
ся доказательствами искусственности возникновения этой вспышки. Предположи
возможность вмешательства в геном такого микроорганизма можно в том случае, если
удастся установить, что эти «новые свойства» решают какую-то новую задачу для тех.
кто организовал это преступление, например, затрудняют лечение пораженных вслед-
ствие приобретения микроорганизмом устойчивости к антибиотикам (противовирус-
ным препаратам), входящим в типовую схему лечения вызываемой им инфекцион-
ной болезни; позволяют преодолевать иммунитет, создаваемый коммерчески',
вакцинами и т. п.
hill IHUil Ill
Краткий перечень схем неснепифической ирофилак1ики и лечения болезней, вызываемых реплицирующимися
биологическими поражающими агентами*
Заболевание Лечение Профилактика Комментарии
1 2 3 4
Сибирская язва Ципрофлаксацин по 400 мг в/в каждые 12 ч или доксициклин по 200 мг в/в, затем по 100 мг в/в каждые 12 ч и два или более дополнительных антибиоти- ков. Следует избегать использования только одних пенициллинов Ципрофлоксацин по 500 мг per os 2/сут в течение по меньшей мере 60 сут или доксициклин по 100 мг перорально 2/сут в течение по меньшей мере 60 сут. Можно сократить курс до 4 нед, если вакцини- рованы (или начата вакцинация)** Обсуждается вопрос об использовании амоксициллина для профилактики у детей или беременных женщин, если штамм является чувствительным. Доксициклин противопоказан при беременности
Холера Регидратационная терапия перорально в период значительной потери жидко- сти. Тетрациклин по 500 мг каждые 6 ч в течение 3 сут или доксициклин по 300 мг одноразово или по 100 мг каж- дые 12 ч в течение 3 сут. Ципрофлок- сацин по 500 мг каждые 12 ч в течение 3 сут. Норфлоксацин по 400 мг каждые 12 ч в течение 3 сут — Вакцина не рекомендована для рутинной защиты в эндемичных районах (50 %-ная эффективность кратковременная). Альтер- нативы для химиотерапии: эритромицин, триметоприм и сульфаметоксазол и фура- золидон. Хинолоны для штаммов, резистентных для тетрациклина и доксициклина
Лихорадка Ку (острая стадия) Тетрациклин по 500 мг перорально каждые 6 ч в течение 5—7 сут с про- должением в течение минимум 2 сут после нормализации температуры или доксициклин по 100 мг per os каждые 12 ч в течение 5-7 сут с продолжением в течение минимум 2 сут при нормаль- ной температуре Тетрациклин по 500 мг per os 2/сут в тече- ние 5 сут (начало через 8-12 сут после экс- понирования) или доксициклин по 100 мг per os 2/сут в течение 5 сут (начало через 8—12 сут после экспонирования) Фторхинолоны могут быть надежной альтернативой при острой стадии заболевания
Мелиоидоз/ сап Цефтазидим плюс TMP-SMX в/в до клинически достаточно хорошего состояния для перехода на докси- циклин+TMP-SMX per as в течение 20 нед После экспонирования для профилактики можно попробовать TMP-SMX Больших исследований схем лечения на людях не проводилось из-за редкой встречаемости заболеваний в природе
1 Биологические поражающие агенты
Продолжение табл. 2.10
1 2 3 4
Туляремия Стрептомицин 7,5-10 мг/кг вД. 2/сут втечение 10-14 сут. Гентамицин 3-5 мг/кг/сут в/в в течение 10-14 сут Ципрофлоксацин по 400 мг в/в каждые 12 ч до улучшения, затем по 500 мгper os каждые 12 ч в течение 10-14 сут Ципрофлоксацин по 750 мг per as каждые 12 ч в течение 10—14 сут Доксициклин по 100 мг per os 2/сут в течение 14 сут. Тетрациклин по 500 мг per os 2/сут в течение 14 сут Ципрофлоксацин по 500 мг per os каждые 12 ч в течение 14 сут -
Вирусные энцефалиты Поддерживающая терапия: анальгетики и спазмолитики парентерально - —
Вирусные геморра- гические лихорадки Рибавирин (CCHF/JIacca) (IND) 30 мг/кг в/в (первоначальная доза); затем 16 мг/кг в/в каждые 6 ч в течение 4 сут; затем 8 мг/кг в/в каждые 8 ч в течение 6 сут Рассматривается использование рибавирина Пассивные антитела к вирусам аргентинской (AHF), боливийской (BHF) геморрагических лихорадок Ласса и Конго-Крымской (CCHF) геморраги- ческой лихорадки
Натуральная оспа Отсутствие эффективной химиотера- пии, кроме поддерживающей; цидо- фовир (эффективен in vitro)', продолжа- ются испытания на животных Иммуноглобулин против вируса вакцины 0,6 мл/кг в/м (в течение 3 сут после экспонирования, лучше всего в течение 24 ч после инфицирования) Введение вакцины в течение до семи суток после экспонирования — наилуч- ший способ профилактики (лучше всего в течение 24 ч после инфицирования)
* По М. Kortepeteret al. (2001).
* * Публикуемые в западных источниках схемы экстренной неспецифической профилактики и лечения ингаляционной сибирской язвы
запутаны попытками объяснить длительность вспышки сибирской язвы в Свердловске в 1979 г. с точки зрения Госдепартамента США, т. е.
как результат «выброса спор сибирской язвы с военного объекта русских» (см. разд. 3.1). Рекомендуемая автором продолжительность не-
специфической профилактики сибирской язвы в течение 60 сут не имеет научного обоснования. Схема экстренной неспецифической про-
филактики при ингаляционном поражении возбудителем сибирской язвы нуждается в экспериментальном уточнении.
Часть 2. Общая неправильная эпидемиол
Биологические поражающие агенты
375
Получить доказательства генно-инженерного вмешательства в геном микроор-
зма можно:
а) молекулярно-генетическими способами диагностики по отдельным нуклеотид-
ы последовательностям с известной историей (например, известный по экспери-
ментам с кишечной палочкой итегративный вектор в геноме возбудителя чумы или
гн нейротоксина скорпиона в геноме вируса оспы обезьян);
б) полным секвенированием, сопоставив с эталонной нуклеотидную последова-
-ельность всего генома микроорганизма, выделенного в очаге искусственно вызван-
~|>й эпидемии, и обнаружив нехарактерные для него кодирующие нуклеотидные пос-
е j вательности (вирулентности; антибиотикорезистентности; белков, участвующих
регуляции иммунных ответов; прионовых белков; известные векторные конструк-
и системы экспрессии генов и т. п.).
Обнаружение генетически измененного высоковирулентного агента свидетель-
ствует о том, что он получен в организации, накопившей большой опыт разработки
средств ведения биологической войны.
2.2.3. Токсины и яды
Термин «яд» известен с глубокой древности, термин «токсин» появился в научном
юороте в конце XIX в. Термины несут разную смысловую нагрузку. Токсикологи к ядам
тносят химические соединения, отличающиеся высокой токсичностью, т. е. способ-
-ые в минимальных количествах вызывать тяжелые нарушения жизнедеятельности
травление) или гибель животного организма. Токсинами после открытия Эмилем Pj
• Александром Иерсиным в 1888 г. дифтерийного токсина традиционно называли
белковые вещества, образуемые преимущественно микроорганизмами и некоторыми
животными и обладающие ядовитым действием. Одним из характерных качеств ток-
синов считалось наличие у них антигенных свойств. Значительное расширение чис-
ла токсинов, выделенных из природных объектов в XX в., среди которых оказалось
много небелковых, заставили ученых пересмотреть критерии термина «токсин».
В настоящее время термин «токсин» чаще всего относят к индивидуальному химичес-
кому веществу (независимо от его природы), полученному из микроорганизмов, гри-
бов, растений, животных или биотехнологическим путем, в низких концентрациях
’оксичного для позвоночных животных. Тем самым разграничиваются действующее
-.ачало яда и яд, являющийся, как правило, многокомпонентной смесью различных
биологически активных веществ.
2.2.3.1. Общие свойства токсинов
Классификации токсинов. Токсикометрия токсинов. Фармакокинетика токсинов.
Фармакодинамика токсинов. Токсины как поражающие агенты.
Классификации токсинов. Наиболее широкое распространение получили класси-
фикации токсинов по их происхождению, клиническим проявлениям отравления и
химической природе. В зависимости от источника происхождения все токсины под-
разделяются на четыре группы.
Микотоксины — токсины, образуемые грибами. Они представляют собой вторич-
ные метаболиты несовершенных грибов, образующиеся из первичных метаболитов
мелонат, аминокислоты, ацетат и др.) в процессе окисления, восстановления, алки-
лирования, галогенизации, конденсации, в низких концентрациях токсичные для
позвоночных животных. Микотоксины надо понятийно отличать от других продук-
376
Часть 2. Общая неправильная эпидемиология
тов метаболизма грибов. Те из них, которые токсичны главным образом для микро-
организмов, называют антибиотиками; а токсичные для растений — фитотоксинами.
У животных и человека, вследствие воздействия микотоксинов, возникают отравле -
ния — микотоксикозы. Микотоксины — природные загрязнители зерна злаковых
бобовых, семян подсолнечника, а также овощей и фруктов. Они могут образовывать-
ся при хранении во многих пищевых продуктах под действием развивающихся в них
микроскопических грибов.
В организм человека и животных микотоксины чаще всего попадают алиментар-
ным путем при непосредственном употреблении зараженных продуктов питания и по
пищевым цепочкам. Возможен ингаляционный путь проникновения микотоксинов
в организм человека. Среди наиболее опасных для людей микотоксинов С. А. Куценко
(2004) выделяет: эрготоксины, продуцируемые грибами группы Claviceps (спорынья,
маточные рожки); афлатоксины (Bb В2, Gb G2) и близкие к ним соединения, выде-
ляемые грибами группы Aspergillus; трихотеценовые микотоксины (более 40 наиме-
нований), продуцируемые несколькими родами грибов; орахотоксины (В, С); пату-
лин и др. Многие высшие грибы синтезируют опасные для человека токсические
вещества. Например, аманитин, аманин и фаллоидин, содержащиеся в бледной по-
ганке , вызывают отравления среди людей с летальностью до 10 % (подробно об отраг -
лениях грибами см. в фундаментальной работе Мусселиуса С. Г., Рыж А. А., 2002). Из
исторических источников известно, что отравления грибами в криминальных целях
практикуются с глубокой древности, количество отравленных при совершении таки'
преступлений может достигать сотен человек (см. разд. 1 1). Микотоксин Т-2 рассмат-
ривается военными специалистами в качестве потенциального агента химического
оружия (Антонов Н. С., 1994).
Зоотоксины — токсические соединения, содержащиеся в ядах животных. Высо-
кая поражающая способность большинства ядов животного происхождения связана
с присутствием в их составе токсинов, характеризующихся высокой специфичностью
действия на определенные биосубстраты. Таковы, например, нейротоксины змей,
нарушающие передачу возбуждения в нервно-мышечных синапсах; токсины амфи-
бий (батрахотоксин), рыб (тетродотоксин), простейших (сакситоксин), блокирующие
распространение нервного импульса по нервному волокну; ферменты ядов гадюк и
гремучников, воздействующие на систему свертывания крови, и др. Из-за высокой
токсичности зоотоксины иногда включают в группу ультраядов — химических ве-
ществ, токсичность которых выше цианистого калия и не превышает 1 мг/кг. «Уме-
ло» составленные композиции таких ядов могут проявить синергическое действие
Например, комбинация батрахотоксина с ядом скорпиона позволяет повысить ток-
сичность яда в 12 раз (Grimes Н., 2006).
Зоотоксины позволяют ученым воспроизводить в условиях in vivo некоторые рас-
пространенные патологические состояния. С помощью яда гремучей змеи можно
моделировать инфаркт миокарда; яды некоторых гадюк хорошо воспроизводят тром-
богеморрагический синдром; пчелиный яд моделирует спазм коронарных сосудов
сердца; яд скорпиона вызывает развитие эпилептиформных состояний и панкреати-
тов. Отсюда понятен тот интерес, который проявляют к зоотоксинам военные токси-
кологи и спецслужбы различных государств (Орлов Ь. Н., Гелашвили Д. Б., 1985).
Яды и токсические соединения животного мира предоставляют преступникам
огромные возможности для скрытой реализации своих замыслов. Для большинства
зоотоксинов нет сертифицированных способов их идентификации в тканях жертв
отравления. Клиника вызываемых ими поражений при парэнтеральном способе вве-
1-i Биологические поражающие агенты 377
может отличаться от той, которую наблюдают врачи при ведении человеку цель-
зго яда, произошедшего в результате контакта с ядовитым животным. Состав яда
е “ко модифицировать. Расследование преступлений, совершенных с использованием
<ютоксинов чрезвычайно затрудняет разнообразие их структуры и механизмов их
телсгвия. Как правило, любой зоотоксин представлен большим количеством анало-
’стречающихся даже у представителей одного вида, обитающих в разных геогра-
ьских регионах. Среди токсических веществ животного происхождения можно
бгретить алифатические и гетероциклические соединения, алкалоиды, стероиды,
с энзиматические полипептиды и ферментативные белки. Некоторые из этих соеди-
щй можно условно назвать «истинными токсинами» в том смысле, что они не встре-
тится в организме реципиента и являются для него ксенобиотиками, т. е. чужерод-
ми веществами. В этом случае, по выражению М. Барбье (1978): «Отравление
•штядит как несчастный случай, вызванный столкновением двух несовместимых
-шов метаболизма» (цит. по Орлову Б. Н. и Гелашвили Д. Б., 1985). Но и при «совме-
стимости типов метаболизма» могут наступать патологические эффекты. В составе
»в животных находятся биологически активные вещества, одновременно присут-
"ующие в организме реципиента (ацетилхолин, биогенные амины и др.). Токсиче-
- >иое действие этих веществ связано с их передозировкой. Важную роль в патогенезе
давления играют аутофармакологические реакции организма на компоненты зоо-
синов (интенсивный выброс макрофагами кининов, медиаторов циклических нук-
.отидов, эндогенных опиатов, гормонов).
Токсины высших растений {фитотоксины) — ядовитые химические соединения,
. «держащиеся в растениях. Среди нихтоксальбумины, алкалоиды, глюкозиды, эфир-
- ы!! масла (терпены, камфара), органические кислоты, ангидриды органических кис-
(синильная кислота), лактоны и другие, менее изученные токсические соедине-
ая. Среди токсальбуминов особое значение для военной токсикологии имеет рицин,
-держащийся в семенах клещевины {Ricinus communis L.). С 1918 г. рицин изучается
качестве биологического поражающего агента. Неоднократно применялся для ин-
нвидуальных убийств и совершения террористических актов. По мнению Б. Н. Ор-
' :ва с соавт. (1990), в сравнительном аспекте растения превосходят животных по раз-
образию химических соединений, выполняющих роль токсинов, и, следовательно,
^химических реакций, ведущих к их синтезу.
Бактериальные токсины — токсины, синтезируемые бактериями. Как правило, это
юокомолекулярные соединения белковой, пептидной или липополисахаридной
^ироды, обладающие антигенными свойствами. В настоящее время выделены и изу-
чены более 150 токсинов бактерий. Многие из них относятся к числу самых ядовитых
лешеств, известных ученым. Среди них ботулотоксины, столбнячный токсин, диф-
терийный токсин, стафилококковые токсины и др. Эти токсины характеризуются
высокой специфичностью действия на определенные биосубстраты. Например, бо-
гулинические токсины связываются с рецепторами на поверхности пресинаптичес-
кой мембраны двигательных нейронов периферической нервной системы и вызыва-
ют протеолиз белков в нейронах. Дифтерийный токсин и экзотоксин А псевдомонад
инактивируют фактор элонгации 2 и тем самым подавляют синтез белка в клетках.
Ботулинические токсины А-, В- и С-типов и стафилококковый энтеротоксин В ис-
следовались в разных странах мира как потенциальные агенты БО.
Основные отличия токсинов от ОВ. Токсины отличаются от классических химичес-
ких поражающих агентов источником получения, физическими и биологическими
характеристиками (табл. 2.11).
378
Часть 2. Общая неправильная эпидемиологи,
Таблица 2.11
Сравнительная характеристика химических агентов и токсинов*
Токсины Химические агенты
Естественное происхождение Создаются искусственно
Трудоемкое, мелкосерийное производство Крупная индустрия
Твердое агрегатное состояние Жидкость
Не летучие Летучие
Высоко к >ксичные Менее токсичные
Кожно-резорбтивное действие не характерно** Обладают кожно-резорбтивным действием
Легально используются в медицине Не используются в медицине
Без вкуса и запаха Имеют запах и вкус
Большое разнообразие токсических эффектов Меньшее разнообразие токсических эффектов
Большинство — иммуногенны Иммуногенность не выражена
Наиболее эффективно поражение аэрозолем 0,5—5 мкм Аэрозоль 1—5 мкм/туман/капли/дымы
Аэрозолирование взрывом и термической возгонкой неприменимо Аэрозолирование взрывом и термической возгонкой возможно
Индивидуальная зашита — аэрозольный фильтр Индивидуальная зашита — противогаз и средства защиты кожи
Индикация и идентификация в реаль- ном масштабе времени невозможна Индикация и идентификация в реальном масштабе времени
* По Н. С. Антонову (1994), R. Sidel et al. (1997) и D. Franz (1998).
** За исключением трихотеценовых микотоксинов, лунгбуатоксина (lyngbyatoxin), пали -
токсина и отдельных токсинов синезеленых водорослей.
Токсикометрия токсинов. Важнейшей характеристикой токсинов является токсич-
ность — свойство химического вещества в минимальном количестве вызывать патоло-
гические изменения, ведущие к нарушению основных процессов жизнедеятельности
организма и приводящие к его гибели. Токсичность — один из основных количествен-
ных параметров, отражающих биологическую активность токсинов. Количественно
токсичность оценивается дозой. Токсическая доза (токсодоза), вызывающая пораже
ния, равные по тяжести, зависит от свойств яда или токсина, пути их проникновения
в организм, от вида организма и условий применения токсина или яда.
Для веществ, проникающих в организм в жидком или аэрозолированном состоя-
нии, через кожу, ЖКТ или раны, поражающий эффедт для каждого конкретного вида
организма в стационарных условиях зависит только от количества яда или токсина,
которое можно выражать в любых массовых единицах. В химии ОВ токсодозы обыч-
но выражаются в мг.
Токсические свойства ядов и токсинов определяют экспериментальным путем на
различных лабораторных животных. Токсикологи чаще пользуются понятием удель-
ной токсодозы — дозы, отнесенной к единице живой массы животного и выражаемой
в мг на кг. Токсичность одного и того же яда, даже при проникновении в организм
одним путем, различна для различных видов животных, а для конкретного животного
будет отличаться в зависимости от способа поступления в организм. Поэтому обычно
_ Биологические поражающие агенты
379
_ - определенных в эксперименте доз указывается способ введения токсина (яда) и
животного. Различают смертельные, выводящие из строя, и пороговые дозы.
Смертельная (летальная) токсодоза (LD, от лат. letalis — смертельный) — это коли-
• хтво яда или токсина, вызывающее при попадании в организм смертельный исход
. определенной вероятностью. Обычно используют понятия абсолютно смертельной
• вызывающей гибель организма с вероятностью 100 % (LD100); исреднесмертель-
. дозы (медианно-смертельной), летальный эффект от введения которых экспери-
ментальным животным наступает у 50 % пораженных (LD50).
Выводящая из строя токсодоза (ID, салат. incapacitate — вывести из строя) — это
сзличество яда или токсина, вызывающее при попадании в организм выход из строя
'ределенного процента пораженных как с временно, так и со смертельным исходом.
Е обозначают как 1D1OO или ID50.
Пороговая токсодоза (PD, от лат. primary — начальный) — это количество яда или
токсина, вызывающее начальные признаки поражения организма с определенной
эероятностью. Пороговые токсодозы обозначают как PD100 и PD50.
В дозах, меньших чем LDS0, яд или токсин вызывает поражения различной степе-
тяжести: тяжелые при 0,3—0,5 LD50; средние при 0,2 LD50 и легкие при 0,1 LDS0.
При характеристике токсичности экстрактов тканей или органов ядовитых живот-
-ых часто пользуются мышиными единицами (м. е.) — количество вещества, которое
-еобходимо, чтобы убить мышь массой 20 г в течение 20 мин. Обычно активность
экстракта выражают в количестве м. е., содержащихся в I мг продукта. Например,
активность тарихотоксина из яиц калифорнийского тритона Taricha torosa составля-
ет 3000 м. е./мг. Это означает, что 1 мг вещества достаточно, чтобы убить 3000 мышей
массой 20 г (Орлов Б. Н., Гелашвили Д. Б., 1985).
Вышеприведенные подходы к количественной оценке биологической активности
"оксинов и ядов предполагают, что эти вещества проникли в организм человека (жи-
вотного) именно в том количестве, которое и вызвало поражение. Такое возможно в
том случае, если яд или токсин вводятся в организм инъекционно или энтерально в
строго контролируемых условиях, например, при сравнительной оценке их токсично-
сти для определенных видов экспериментальных животных. Однако боевое примене-
ние токсических веществ предполагает, прежде всего, использование аэрозолей. Но
тогда часть вдыхаемого распыленного токсического вещества будет поглощаться орга-
низмом, часть выдыхаться. Поражающее действие токсического вещества будет зави-
сеть только от той его части, что была поглощена организмом человека. Чтобы разре-
шить это противоречие, еще во время Первой мировой войны было сделано допуще-
ние, что ингаляционная токсодоза ОВ прямо пропорциональна его концентрации (С)
во вдыхаемом воздухе и времени дыхания (/). Кроме того, было предложено учиты-
вать интенсивность дыхания (И), зависящую от физической нагрузки и состояния че-
ловека. В спокойном состоянии человекделает примерно 16 вдохов водну мин и, сле-
довательно, в среднем поглощает 8—10 л воздуха. При средней физической нагрузке
1 марш) потребление воздуха увеличивается до 20—30 л/мин, а при тяжелой (бег, зем-
ляные нагрузки) — до 60 л/мин.
Таким образом, если человек массой G (кг) вдыхает воздух, содержащий токсичес-
кое вещество с концентрацией С (мг/л) в течение времени t (мин) при интенсивности
дыхания V(л/мин), то удельная поглощенная доза токсического вещества D (мг/кг) бу-
дет равна
D CtV/G.
380
Часть 2. Общая неправильная эпидемиология
Германский химик Ф. Габер предложил упростить это выражение. Он сделал до-
пущение, что для людей или конкретного вида животных, находящихся в оди наковых
условиях, отношение V/G постоянно (вентиляция легких пропорциональна его весу).
Разделив на него обе части уравнения, он получил выражение
Т Ct,
где Т — ингаляционная доза, измеряемая величиной мгхмин/м3 или мгхмин/л; С —
концентрация токсического вещества в воздухе (мг/м3 или мг/л); t — время экспози-
ции (мин).
По мнению В. Н. Александрова и В. И. Емельянова (1990), это произведение не
является ингаляционная дозой в строгом смысле слова, так как часть ОВ, попавшего
в организм с вдыхаемым воздухом, выдыхается обратно, а часть обезвреживается орга-
низмом. Поэтому более правильно называть его относительной токсичностью веще-
ства при ингаляции. Кстати, сам Ф. Габер называл произведение Ct коэффициентом
активности или коэффициентом смертоносного действия (см. у Эддере Ш., Истэн М.,
1939). Но определение Ct как «ингаляционная доза» прочно вошло в научный оборот,
так как позволяет сравнивать различные токсические вещества по их ингаляционной
токсичности. Например, расчетное значение летальной дозы сакситоксина для чело-
века при ингаляции равно 5 мгхмин/м3, или оно в 2 раза меньше, чем у вещества VX
(10 мгхмин/м3), и в 15 раз меньше, чем у зарина (75 мгхмин/м3).
Поскольку действие токсических веществ воспринимается каждым человеком по-
разному, то их доза, вызывающая разные степени поражения, может значительно ва-
рьировать у разных людей. Исходя из этого, при расчетах военные химики обычно
определяют среднее количество ОВ, необходимое для нанесения заданной степени
ингаляционного поражения не всему личному составу противника, расположенному
в районе цели, а только половине его (Ротшильд Д.,1966).
Средняя смертельная ингаляционная доза О В обозначается как Lct50 (первые бук-
вы слов Lethal concentration, time — летальная концентрация, время). Предполагает-
ся, что при ингаляционном воздействии на незащищенный личный состав против-
ника средней смертельной дозы, смерть наступит у 50 % пораженных людей. Соот-
ветственно обозначаются выводящие (lct50) и пороговые дозы (Pct50). Например,
ингаляционная доза ботулинического токсина, выраженная величиной Lct50, состав-
ляет 0,02—0,1 мгхмин/м3. По этому показателю ботулинический токсин не менее чем
в 750 раз токсичнее зарина и в 100 раз токсичнее вещества VX (Антонов Н. С.,1994).
Фармакокинетика токсинов. Фармакокинетика токсинов изучает закономерности
всасывания, распределения, метаболизма и выведения токсинов из организма. При
оценке токсичности любых токсинов важное значение приобретают пути их введе-
ния в организм. Существующие пути введения обычно подразделяют на энтеральные
(через пищеварительный тракт) и парентеральные (минуя пищеварительный тракт).
В естественных условиях пути введения токсинов в организм жертвы определяются
особенностями биологии организма-продуцента и химической природой токсинов.
В случае умышленного их применения для поражения людей, эти особенности не
будут соблюдаться.
Белковые токсины змей, насекомых и паукообразных вводятся с помощью воо-
руженного ядовитого аппарата парентерально, так как многие из них разрушаются
пищеварительными ферментами. Напротив, токсины небелковой природы эффектив-
ны и при энтеральном введении (токсические алкалоиды амфибий, токсины рыб,
моллюсков и др.).
2.2. Биологические поражающие агенты
381
Яды растений, микотоксины (за исключением трихотеценовых микотоксинов,
лунгбуатоксина, палитоксина и отдельных токсинов синезеленых водорослей, способ-
ных проникать через кожу) и некоторые бактериальные токсины (ботулинические)
попадают в организм человека энтеральным путем. Но для бактериальных токсинов
более характерно их образование в организме человека на фоне развившегося специ-
фического инфекционного процесса (шига-токсин при шигеллезе, дифтерийный
токсин во время дифтерийной инфекции, столбнячный токсин при столбняке).
В каждом из этих случаев специфическое действие токсинов определяет основные
симптомы развивающейся болезни.
Попавший в организм яд (токсин) распределяется весьма неравномерно. Суще-
ственное влияние на распределение оказывают биологические барьеры, к которым
относят стенки капилляров, клеточные (плазматические мембраны), гематоэнцефа-
лический и плацентарный барьеры. При парентеральном введении (укус, ужаление,
инъекция, ингаляция) в месте инокуляции образуется первичное депо яда, из кото-
рого происходит поступление токсинов в лимфатическую и кровеносную систему.
Скорость дренирования яда во многом определяет быстроту развития токсического
эффекта.
Большинство токсинов подвергается в организме плохо изученным химическим
превращениям. Биотрансформация в определенной мере обусловливает биологичес-
кую резистентность некоторых животных к зоотоксинам. В последнее время в крови
некоторых грызунов были обнаружены белковые факторы, инактивирующие гемор-
рагическое действие змеиных ядов. При выделении токсинов из организма основную
тяжесть на себя принимают почки — отсюда и широкая распространенность нефри-
тов при отравлении зоотоксинами, микотоксинами и ядами растений. Частично ток-
сины могут выводиться с молоком кормящей матери.
Фармакодинамика токсинов. Фармакодинамика токсинов изучает наиболее типич-
ные эффекты этих веществ, локализацию и механизмы действия. Действие токсинов
может носить местный и резорбтивный характер. Действие токсинов, развивающе-
еся в месте его первичного воздействия (кожа, ткань в участке введении яда, ЖКТ,
легкие), называется местным. В зависимости от химической природы ядов местное
действие может быть сильным (яды кишечно-полостных, гадюк, гремучников, жало-
носных перепончатокрылых и др.) и в месте укуса животного вызывать жгучую боль,
отек; в тяжелых случаях приводящее к некрозу (омертвению) пораженной ткани. На-
несение на кожу раствора Т-2 токсина (микотоксин) вызывает покраснение, отек и
глубокий некроз тканей.
При попадании яда, обладающего местным действием в организм с пищей, разви-
ваются острые поражения ЖКТ (сапонин- и гликозидсодержащие растения). В дру-
гих случаях превалируют общие симптомы отравления, развивающиеся после всасы-
вания яда из первичного депо (яды элапид, каракурта; ботулинические токсины; яды,
содержащиеся в белене, дурмане и др.). Особую опасность для людей, ставших мише-
нью террористов, представляет ингаляционное поражение легочной ткани ядами,
обладающими местным действием. Специалистами USAMRIID было обнаружено, что
сакситоксин при ингаляционном введении мышам в 10 раз токсичнее, чем при внут-
ривенном или внутрибрюшинном введении. Аналогичный эффект наблюдается и у
других токсинов, способных поражать легкие (например, у Т-2 токсина). Этот фено-
мен Н. С. Антонов (1994) объясняет тем, что при ингаляции действие сакситоксина
нацелено на орган (легкие), а не на организм животного в целом. Истинно местное
действие, как правило, не наблюдается, так как определенное количество токсинов
382
Часть 2. Общая неправильная эпидемиология
всасывается и оказывает общетоксический эффект. Скорость проявления резорбтив-
ного эффекта зависит от путей введения токсинов и их способности проникать через
биологические барьеры.
Важное значение в формировании интегральной картины отравления токсинами
имеет и их рефлекторное действие, связанное с влиянием на экстеро- или интероре-
цепторы и с соответствующим изменением функционального состояния нервных
центров либо эффекторных органов. Использование тонких инструментальных ме-
тодов исследования позволяет изучать места связывания и механизмы действия ток-
синов на организменном, органном, клеточном и молекулярном уровнях. При этих
исследованиях в первую очередь выявляют биологический субстрат, на который на-
правлено действие того или иного токсина. Иными словами — обнаружение рецеп-
торов токсинов в сложных биосистемах. Это позволяет не только понять патогенез
отравления, но и использовать токсины для выделения и характеристики этих рецеп-
торных структур. Примеры такого использования зоотоксинов — характеристика
холинорецептора с помощью нейротоксинов змеиных ядов и компонентов натриевых
каналов нервных мембран с использованием токсинов амфибий, рыб и скорпионов.
В случае, когда эффект зоотоксинов на рецепторы аналогичен физиологическому,
говорят о действии токсинов как агонистов. Это действие может быть стимулирую-
щим или угнетающим данную физиологическую функцию. Часто токсины выступа-
ют в роли антагонистов эндогенных физиологических регуляторов, вызывая наруше-
ние деятельности системы, органа или всего организма. Высокая специфичность
действия токсинов определяет во многих случаях конкурентный характер их антаго-
низма по отношению к эндогенным веществам (медиаторам и др.) (Орлов Б. Н., Ге-
лашвили Д. Б., 1985).
Обратимый или необратимый характер действия токсина зависит от связи лиганд-
рецептор, которая может быть прочной (ковалентной) или менее прочной (электро-
статической, гидрофобной и др.). Важной характеристикой отдельных токсинов яв-
ляется избирательность (селективность) их действия, т. е. способность повреждать
определенные клетки-мишени, не затрагивая другие, даже если оба вида клеток на-
ходятся в непосредственном контакте. В основе избирательности лежит сродство ток-
сина к рецептору. Сродство токсина к данному типу рецепторов определяется струк-
турно-пространственной организацией молекулы и ее активных функциональных
группировок, обеспечивающих узнавание рецептора и связывание с его активным
центром. В простейшем случае, когда одна молекула токсина [Т] соединяется с одним
рецептором [Р] (при избытке токсина), образование комплекса «токсин—рецептор»
[ТР] описывается уравнением
[т]+[р]?=^[тр],
*-1
где к} — константа скорости образования комплекса [ТР]; к_х — константа скорости
диссоциации этого комплекса. Отношение констант прямой и обратной реакции:
кх/к_х = Кс называют константой сродства, а обратную ей величину — константой
диссоциации Сд. Численно Сд равна концентрации вещества, дающего 50 % эффекта,
и может быть найдена по графику зависимости доза—эффект. Чем меньше значение
ЛТд, тем выше сродство токсина к данному рецептору. КЛ — аналогична константе
Михаэлиса (Кт), характеризующей диссоциацию комплекса фермент—субстрат при
описании кинетики обратимых ферментативных реакций в биохимии (Орлов Б. Н„
Гелашвили Д. Б., 1985).
2.2. Биологические поражающие агенты
383
Токсины как поражающие агенты. По мнению Н. С. Антонова (1994), токсинное
оружие занимает промежуточное положение между биологическим и химическим
оружием. Токсины исследуются в лабораториях микробиологического профиля на-
ряду с бактериями, вирусами и риккетсиями. Это роднит токсинное оружие с био-
логическим. Но токсины являются веществами, имеют строго детерминированный хи-
мический состав и строение. Некоторые из них могут быть получены химическим
путем (палитоксин, Т-2 токсин и др.). Эти свойства более характерны для ОВ. Леталь-
ные дозы для экспериментальных животных большинства токсинов при любом спо-
собе введения оказались значительно меньшими, чем у боевых ОВ (табл. 2.12).
Таблица 2.12
Сравнение летальных доз отдельных токсинов и боевых ОВ*
Агент LDS0 (мкг/кг)** ММ (Да) Источник
Ботулинический токсин 0,001 150000 Бактерия
Шига-токсин 0,002 55000 Бактерия
Столбнячный токсин 0,002 150000 Бактерия
Абрин 0,04 65000 Растение
Дифтерийный токсин 0,10 62000 Бактерия
Маитотоксин 0,10 3400 Морские динофлагелляты
Палитокин 0,15 2700 Зоантарии
Сигуатоксин 0,40 1000 Рыбы, морские динофлагелляты
Текстилотоксин 0,60 80000 Змеи из семейства элапид
Токсины Clostridium petfringens 0,1-5,0 35000-40000 Бактерия
Батрахотоксин 2,0 539 Амфибии
Рицин 3,0 62000 Растение Ridnus communis
а-конотоксин 5,0 1500 Брюхоногие моллюски
Тайпоксин 5,0 46000 Змеи из семейства элапид
Тетродотоксин 8,0 319 Рыба фугу
а-титиустоксин 9,0 8000 Скорпионы
Сакситоксин 10 (ингал. 2,0) 299 Морские динофлагелляты
VX 15,0 267 ОВ
Энтеротоксин В стафилококков (обезьяны резус/аэрозоль) 27,0 28497 Бактерия
Анатоксин A(s) 50,0 500 Цианобактерии
Микроцистин 50,0 994 Цианобактерии
Зоман (GD) 64,0 182 ОВ
Зарин (GB) 100,0 140 ОВ
Аконитин 100,0 647 Растения Aconitum
Т-2 токсин 1210,0 466 Грибы
♦По R. Sidel et al. (1997).
♦♦ LD5o оценена в экспериментах на мышах, за исключением энтеротоксина В стафило-
кокков.
384
Часть 2. Общая неправильная эпидемиологи. f
Простое арифметическое сравнение летальных доз токсинов и боевых ОВ поро-
дило у военных ученых много ожиданий в 1940—1960-х гг. Токсины рассматривались
как более эффективные агенты, чем химические вещества при поражении незащищен-
ной живой силы противника на больших площадях. Но по мере накопления экспе-
риментальных данных во время полевых испытаний, стало ясно, что эти ожидания
изначально были иллюзорными. Н. С. Ан гонов (1994) приводит следующий пример
По Lct50 ботулинический токсин не менее чем в 750 раз токсичней зарина и в
100 раз токсичней вещества VX (см. выше «Токсикометрия токсинов»). Но при поле-
вых испытаниях, проведенных США, в ходе которых ботулинический токсин диспер-
гировался над группой привязанных животных, по числу смертельных поражений он
не намного превосходил ОВ нервнопаралитического действия, примененные тем же
способом в тех же условиях. Н. С. Антонов подчеркивает, что сравнение поражающе-
го действия ботулинического токсина и ОВ проводилось в эксперименте, моделирую-
щим нападение на незащищенную живую силу противника. При активном противо-
действии нападению, например, при своевременно одетом противогазе, результат бы
был в пользу ОВ. В докладе консультантов ВОЗ приводится сравнение площадей по-
ражения ботулиническим токсином и веществом VX, диспергированным до частиц
размером 5 мкм, при расходе каждого из них по 4000 кг. Применительно к этому сце-
нарию, ботулиническим токсином поражалась площадь 12 км2, в то время как веще-
ство VX нанесло поражение на площади до 40 км* (Медико-санитарные аспекты....
1972). Но и в этом сценарии не использованы все возможности ОВ данного типа
Применение VX преследовало цель нанесения только ингаляционных поражений.
При применении VX в виде грубодисперсного аэрозоля для поражения через кож;
площадь поражения будет значительно большей.
Несоответствие между уровнями токсичности и площадями поражений ботули-
ническим токсином и веществом VX, отмечаемое в результатах условного примене-
ния этих поражающих агентов, вызвано следующими причинами. Во-первых, при
переводе в аэрозольное состояние токсин разлагается в большей степени, чем веще-
ство VX. Во-вторых, фракции аэрозоля токсина, наиболее эффективные при ингаля-
ционном воздействии и имеющие размер 5—10 мкм и менее, рассеиваются в верхних
слоях атмосферы, не достигая поверхности земли, не вызывая поражений ни при
попадании их на кожные покровы, ни при ингаляции. В-третьих, в рассматриваемом
сценарии имел место дальний перенос аэрозоля токсина атмосферными потоками в
ходе которого часть токсина подверглась детоксикации за счет факторов окружающей
среды (Антонов Н. С., 1994).
2.2.3.2. Токсины высших растений [фитотоксины]
Алкалоиды. Гликозиды. Эфирные масла. Органические кислоты. Лактоны. Токсальбуми-
ны. Закономерности ядообразования у растений. Клиническая классификация ядовитых
растений. Роль отдельных токсических веществ в формировании клинической картины
отравления растительными ядами. Патологоанатомическое исследование. Красавка.
Белена. Дурман (Datura). Болиголов пятнистый. Вех ядовитый. Аконит (Aconitum). Без-
временник. Чемерица (Veratrum). Мак (Papaver). Строфант. Чилибуха.
Токсичность ядовитых растений для человека и животных зависит от образования
и наличия в них ядовитых химических соединений. По большей части их значение в
жизнедеятельности растений остается неизвестным. Сами фитотоксины представляю г
собой вещества с различным строением и неодинаковой биологической активностьк
Рассмотрим их по работам С. А.Куценко (2004) и И. Л. Гусынина (1962).
2.2. Биологические поражающие агенты
385
2Ш1каловды. Наиболее часто встречаются в качестве действующих веществ ядови-
тых растений. Они представляют собой сложные органические гетероциклические
основания с атомами азота в цикле. Простейшие алкалоиды содержат до 10 атомов
углерода, наиболее сложные — более 50. Алкалоиды образуют кристаллические соли
с кислотами, например с серной, соляной, салициловой, щавелевой и др.; этот про-
цесс используется для их выделения и очистки. Большинство из них содержит кисло-
род; кислородсодержащие алкалоиды являются твердыми веществами. В составе не-
которых алкалоидов, например кониина (из омега пятнистого), никотина (из табака),
анабазина (из ежовника) и других, кислорода нет. Бескислородные алкалоиды яв-
тяются жидкими летучими веществами. В свободном состоянии большинство алка-
лоидов трудно растворимы в воде и легко — в органических растворителях: спирте,
эфире, хлороформе и др. Соли их, наоборот, легко растворяются в воде. В растениях
алкалоиды находятся в виде солей различных растительных кислот (яблочной, лимон-
ной, щавелевой, янтарной) и легко всасываются, попадая в желудочно-кишечный ка-
нал человека и животных. С некоторыми кислотами, такими как кремневольфрамо-
вой, фосфорномолибденовой, фосфорновольфрамовой, танином и другими вещества-
ми, алкалоиды дают труднорастворимые, а потому трудновсасываюшиеся осадки.
Этим свойством пользуются при лечении алкалоидных отравлений. Во флоре земно-
го шара алкалоиды распространены неравномерно: в одних семействах (маковые,
пасленовые, лютиковые, бобовые и т. п.) они содержатся во многих видах растений, в
других алкалоиды пока не обнаружены. В одном и том же растении обычно встреча-
ется несколько алкалоидов, близких по строению, число которых может доходить до
15—20 (опийный мак, кора хинного дерева) и меняться в зависимости от условий про-
израстания растения. Содержание алкалоидов в растениях колеблется от долей про-
цента до нескольких процентов (табл. 2.13, с. 386).
Особую группу алкалоидов составляют соланины (от лат. solanum — паслен), рас-
тительные гликозиды (гликоалкалоиды), у которых агликоном служит стероидный
алкалоид соланидин. Их углеводная часть представлена 1—3 моносахаридами (глю-
коза, галактоза, рамноза). Содержатся в растениях рода Solanum семейства паслено-
вых (особенно их много в верхушках побегов и цветках картофеля). Обладают горь-
ким вкусом.
Строгой ботанической специфичности алкалоидных соединений нет. Одни и те
же алкалоиды могут встречаться в различных ботанических группах (видах, родах и
даже семействах). Например, алкалоид анабазин найден в ежовнике безлистном и в
табаке; алкалоид лупинин — в лупине и в ежовнике; алкалоид берберин найден во
многих растениях, принадлежащих к пяти различным семействам.
Гликозиды. Это продукты соединения (посредством легко гидролизуемых глико-
зидных связей) циклических 5- и 6-членных сахаров с веществами типа спиртов или
фенолов. Гликозидные связи образуются между ацетальным гидроксилом сахара и
спиртовой группой неуглеводного компонента так называемого агликона (при отщеп-
лении молекулы воды). В зависимости от участия атомов S, О или N в образовании
гликозидной связи различают S-, О- или N-гликозиды. Две последние группы глико-
зидов наиболее распространены в тканях растений и животных, а- и р-гликозиды раз-
личаются пространственным положением агликона по отношению к плоскости коль-
ца углеводного компонента; их гидролиз протекает при участии ферментов а- и
р-гликозидаз. Классификацию гликозидов затрудняет разнообразие веществ, высту-
пающих в роли агликона, определяющего специфическое биологическое действие
гликозида. Содержащие глюкозу гликозиды называются глюкозидами. К гликозидам,
386
Часть 2. О чцая неправильная эпидемиология
содержащим дисахаридные остатки, относится красящее вещество шащрана — кро-
цин (содержащий генциобиозуд и желтое красящее вещество семян Rhamnus infects-
ria — ксанторамнин (рамнозу). Полисахариды, также образуемые гликозидными свя-
зями, часто относят к гликозидам, а ферменты, их расщепляющие (например,
амилазу), — к гликозидазам.
Тадлица. 2.13
Основные группы алкалоидов и продуцирующие их растения*
Группа алкалоидов Важнейший представитель Растение
Пиридиновые и пиперидиновые Кониин Никотин Лобелии Болиголов Табак Лобелия
Пирролидиновые и пиперидиновые Гиосциамин Скополамин Белена Скополия
Пирролизидиновые Платифиллин Сенецифи шн Крестовник То же
Хинолиновые Эхинопсин Мордовник
Бензилизохинолиновые Папаверин Мак
Фенантрснизохинолиновые Морфин Кодеин Мак
Дибен.зили юхинолиновые Даурицин Луносемянник
Бензофенантридиновые Хелидонин Сангвинарин Чистотел То же
Индольные Галантамин Вчнканин Эрготамин Подснежник Воронова Барвинок Спорынья
Имидазольные Пилокарпин Пилокарпус
Пуринпвые Кофеин Теофиллин Чай То же
Дитерпеновые Аконит Дельсимин Борец То же
Стероидные Соланин Йервин Картофель Чемерица Лобеля
Ациклические Эфедрин Эфедра
Колхициновые Котхипин Безвременник
* По Б. Н. Орлову с соавт. (1990).
Структурное своеобразие гликозидов заключается в том, что они легко распада -
ются на составляющую их углеводную (сахаристую) часть и аглюконы. Углеводной
частью гликозидов может быть глюкоза, галактоза, рамноза; чаще — первач. Аглюко-
новый же компонент может быть весьма разнообразным как по составу, так и по хи-
мическому характеру. Токсична вторая часть гликозидов. Расщепление гликозидов
может происходить при кипячении с водой, лучше при нагревании с разведенными
минеральными кислотами, щелочами или под действием определенных, сопутствую-
щих гликозидам в растениях ферментов (гликозидаз).
Гликозиды представляют собой твердые, большей частью кристаллические, реже
аморфные соединения; лишь некоторые из них бывают окрашены; в большинстве
случаев они растворимы в воде; на вкус обыкновенно горькие, что определяет горь-
кий вкус растений. По химическому составу аглюконов можно различать:
2.2. Биологические поражающие агенты
387
1) гликозиды с аглюконами, не содержащими азот. Из этой группы гликозидов
отравление могут вызывать гликозиды растений группы наперстянки (наперстянка,
ландыш, зимовник, олеандр);
2) гликозиды с аглюконами, содержащими азот (нитрилглюкозиды, цианглюко-
зиды). Эти гликозиды имеют большое токсикологическое значение, так как при их
расщеплении образуется синильная кислота;
3) гликозиды с аглюконом, содержащим азот и серу (тиогликозиды, горчичные
гликозиды), при расщеплении их образуются горчичные масла;
4) особую группу гликозидов составляют сапонины (сапонингликозиды).
Сапонины так же, как и гликозиды, распадаются на сахаристую и несахаристую
часть — сапогенин. Все сапогенины делятся на две большие группы: стероидные и
тритерпеноидные. Характерной особенностью стероидных сапогенинов являет-
ся превращение их в физиологически активные вещества типа половых гормонов.
И. Л. Гусынин (1962) считал, что на основе этой особенности сапонинов и продуктов
их расщепления — сапогенинов в растениях образуются вещества, обусловливающие
нарушения половой деятельности животных. Одним из свойств сапонинов является
то, что водные растворы их при взбалтывании дают стойкое образование пены. Са-
понины, обладающие токсическим действием, называются сапотоксинами.
Эфирные масла. В химическом отношении представляют смесь разнообразных
соединений; обычно это летучие жидкости характерного, часто приятного запаха.
В состав их входят углеводороды, главным образом терпены, которые нередко явля-
ются единственной составной частью эфирного масла. Из соединений, содержащих
кислород, составными частями эфирных масел могут быть спирты, альдегиды, кетоны
(камфары), эфиры, а также соединения, содержащие серу (например, в горчичном
эфирном масле).
Эфирные масла легко (например, при охлаждении) выделяют свои твердые состав-
ные части, а при долгом хранении, окисляясь, осмоляются и приобретают желтую или
бурую окраску. Они нерастворимы в воде, но растворяются в органических раствори-
телях (спирте, эфире, хлороформе, бензоле). Удельный вес их обычно меньше удель-
ного веса воды, и только эфирные масла с содержанием веществ, богатых кислородом,
тяжелее воды. В растениях они содержатся или в чистом виде, или в виде гликозидов
(горчичные гликозиды), при расщеплении которых выделяются в свободном состоя-
нии. Эфирные масла легко перегоняются с водяным паром; этим способом они обыч-
но получаются из растительного материала.
Органические кислоты. Особое токсикологическое значение имеют:
1) синильная кислота — продукт ферментативного распада цианогенных гликози-
дов, образующихся у некоторых дикорастущих и культивируемых растений;
2) щавелевая кислота, находящаяся в большом количестве в щавелях (Rumex асе-
tosa L., R. acetosella L.), в кислице (Oxalis acetosella L.), в листьях свеклы и др.; встреча-
ется главным образом в виде солей, реже в свободном состоянии.
Лактоны. Обычно это ангидриды у-оксикислот. Из ядовитых растений, содержа-
щих лактоны, большое токсикологическое значение имеют полыни (таврическая,
цитварная) и лютики.
Токсальбумины. Представляют собой растительные токсические вещества бел-
кового характера. Они делятся на две группы: гемитоксины, или токсина класса I.
Содержат только каталитические домены (сапорин, бриодин 1, боунганин, гелонин);
и голотоксины, или токсины класса II. Содержит обе субъединицы — связывающую
и каталитическую (рицин, абрин, лектин омелы, модецин, вискумин, волькенсин)
388
Часть 2. Общая неправильная эпидемиология
(Kreitman R. J., 2006; Griffiths G. D. et al., 2007). По своей организации сходны с би-
нарными токсинами бактерий (см. разд. 2.2.3.6). Из токсальбуминов особое значе-
ние для токсикологии биологических поражающих агентов имеет рицин, содержа-
щийся в семенах клещевины (Ricinus communis L.; см. разд. 3.11). Токсальбумины об-
ладают иммуногенными свойствами, т. е. способностью при соответствующих методах
введения их животным вызывать в организме образование антител.
Закономерности ядообразования у растений. Ядовитые растения одного и того же
вида в зависимости от природных условий и стадии своего развития могут содержать
различные количества ядовитых веществ или даже не содержать их. Эти «тонкости»
хорошо известны отравителям. Закономерности ядообразования у растений обобще-
ны И. Л. Гусыниным (1962):
1) практика культивирования алкалоидных растений свидетельствует о возмож-
ности повышения содержания алкалоидов при затенении растений. Следовательно,
растения, выросшие в затененных местах, будут более ядовитыми, чем растения, вы-
росшие на местах, открытых для солнца. Даже смена дня и ночи влияет на процессы
образования ядовитых веществ, в частности алкалоидов. У некоторых растений
(белена, дурман) эти процессы более интенсивны ночью, и растительный материал,
собранный утром, содержит больше действующих веществ, чем собранный вечером;
2) в некоторых случаях фактором, способствующим образованию и накоплению
ядовитых продуктов в растениях, может быть резкое изменение метеорологических
условий, ведущее к нарушению роста растений или к увяданию. Это наблюдается,
например, у растений, образующих синильную кислоту;
3) количество ядовитых соединений в растениях может изменяться во время силь-
ного дождя: ядовитые вещества в этом случае как бы вымываются из листьев расте-
ний и могут находиться в спадающей с растений дождевой воде. Это доказано для
глюкозидов горькой полыни (Artemisia absinthium L.);
4) содержание действующих начал в ядовитых растениях не остается постоянным
в продолжение всего периода их развития. Наиболее высоким оно бывает обычно лишь
в определенной стадии жизни растений. Например, из наземных частей чемерицы
наиболее ядовиты молодые, нераспустившиеся ростки, питающиеся еще за счет кор-
невища; у некоторых видов дурнишника наиболее ядовиты самые первые листочки.
В маке наибольшее количество алкалоидов содержится в млечном соке не совсем зре-
лых головок; по мере созревания головок количество алкалоидов уменьшается. Мо-
лодые части дурмана богаче алкалоидами, чем вызревшие. Содержание алкалоида
эфедрина в Кузьмичевой траве (хвойник) на разных фазах развития ее колеблется от
0,3 до 2,5 %;
5) наибольшее ядообразование имеет место только в определенной фазе развития
растения. С прекращением жизнедеятельности растения (естественное отмирание)
ядообразование в его наземных частях прекращается; оно может продолжаться толь-
ко в подземной части (корневище, корни двулетних, многолетних растений);
6) у двудомных растений синтез ядовитых веществ зависит от стадии развития и
пола. Например, у хвойника накопление алкалоидов до периода цветения примерно
одинаковое как у мужских, так и у женских растений; после опыления образование
алкалоидов у мужских уменьшается, а у женских к периоду созревания плодов дости-
гает максимума;
7) распределение действующих начал в различных частях развившегося растения
не равномерно. Находясь в том или ином количестве во всех органах растения, дей-
ствующие начала локализуются в определенных частях. У аконита, веха, чемерицы яд
2.2. Биологические поражающие агенты
389
вкладывается преимущественно в корневище, у наперстянки — в листьях, а у пикуль-
ников, софоры, куколя, гелиотропа — в семенах. Пример очень узкой локализации
действующих веществ можно видеть у среднеазиатского растения эминиума Леман-
на. Луковица этого растения весьма ядовита и используется местным населением для
отравления волков и лис; отходящие же от луковицы корни без вреда используются
’астухами для утоления жажды;
8) во время сушки и последующего хранения, многие соединения, находящиеся в
свежих растениях, могут изменяться или даже полностью разрушаться, ь результате
высушивания токсичность ядовитых растений может уменьшаться или исчезать; кли-
нически отравление может протекать иначе, чем вызванное зелеными растениями.
Большинство ядовитых растений, содержащих алкалоиды, сохраняют свою ядови-
тость при высушивании. Растения с летучими действующими веществами, как, на-
пример, лютиковые, при высушивании теряют их. Относительную нестойкость дей-
ствующих веществ обнаруживают глюкозидсодержашие растения. Глюкозиды могут
егко разлагаться при высушивании и хранении растительного материала, особенно
в условиях, дающих возможность развития ферментативных процессов (длительное
высушивание, повторные намачивания). Например, листья ландыша при хранении
в течение года теряют до 77 % биологической акгивности;
9) быстро высушенные растения сохраняют большее количество действующих
веществ, чем растения, подвергшиеся медленной, длительной сушке. Например, дур-
иан при медленной сушке (температура 19 °C I теряет до 22 % алкалоидов;
10) при высушивании растений химический состав некоторых действующих ве-
ществ может изменяться, в результате чего образуются новые токсические соедине-
ния. Находящиеся в высушенном материале наперстянки глюкозиды (дигитоксин,
гитоксин) представляют собой лишь продукты гидролитического расщепления более
сложного соединения, находящегося в свежем растении.
Клиническая классификация ядовитых растений. В данной работе я использую кли-
ническую классификацию И. Л. Гусынина (1962) как наиоолее удобную из имеющих-
ся классификаций (ботаническая, химическая и др.) для раннего установления при-
чины умышленного отравления и сбора улик против лиц, совершивших такое
преступление.
И. Л. Гусынин (1962) обосновывал ее следующим образом. Клиническая картина,
развивающаяся в организме в результате отравления ядовитым растением, определя-
ется, прежде всего, характером действия на организм находящихся в нем токсичес-
ких веществ. При внимательном анализе клинической картины отравлений ядовиты-
ми растениями или полученными из них ядами, в каждом отдельном случае из всего
комплекса симптомов можно выде. гить один или несколько основных, ведущих при-
знаков — признаков преимущественного действия на какой-либо орган или систему
органов. Часто такой признак, повторяясь в клинической картине отравлений не-
сколькими ядовитыми растениями, становится общим при отравлении этими расте-
ниями. Беря за основу принцип сходства клинических признаков, ядовитые растения
можно распределить на отдельные более или менее близкие группы. Созданию такой
классификации может оказать помощь сходство клинической картины, которое час-
то наблюдается при отравлениях ботанически родственными растениями или расте-
ниями с близким химическим составом токсических веществ. Например, причиной
отравлений, протекающих с клинической картиной отека легких, чаще бывают крес-
тоцветные растения; отравлений с картиной аноксемических явлений (явлений задуше-
ния) — растения семейства злаковых; отравлений с картиной поражения сердца — груп-
390
Часть 2. Общая неправильная эпидемиология
па растений, содержащих так называемые «сердечные» гликозиды. Клиническая кар-
тина отравлений растениями, содержащими в своем составе алкалоиды группы
атропина, отлична от клинической картины, наблюдаемой при отравлениях растени-
ями, содержащими алкалоиды группы морфина и др.
Схема клинической классификации ядовитых растений представлена И. Л. Гусы-
ниным (1962) в следующем виде.
I. Растения, вызывающие преимущественно симптомы поражения ЦНС:
1) растения, вызывающие возбуждение ЦНС (красавка белладонна, белена, дур-
ман, скополия, пузырница, хвойник, соляноколосник, секуринега, вех);
2) растения, вызывающие возбуждение ЦНС и одновременно действующие на
пищеварительный тракт, сердце и почки (можжевельник, пижма, туя, багульник,
полынь, лютик, рогоглавник, калужница, ветреница, прострел, ломонос, воронец,
борщевик, девясил, ясенец);
3) растения, вызывающие угнетение и паралич ЦНС (мак, чистотел, глациум,
хохлатка, ремерия, дымянка, плевел, пикульник, антирринум, железница, чистен.
посконник, латук, молокан, ленец, чина, конопля, датиска, ковыль опьяняющий);
4) растения, вызывающие угнетение и паралич ЦНС и одновременно действую-
щие на желудочно-кишечный тракт и сердечно-сосудистую систему (аконит, живость,
табак, анабазис, безвременник, чемерица, тисс, термопсис, ракитник, жарновец, пу-
зырик, софора, аммоденрон, аммотамус, пажитник, пинтант, лонтица, самшит, хаме-
дафне, подбел, могильник, козлятник, чернокорень, окопник, синяк, анхуза, нарцисс,
панераций, подснежник, блоцветник, штернбергия, унгерния, болиголов, бутень, ко-
корыш, омежник, поручейник, рододенрон, токсикодендрон, клещевина, робиния,
кирказон, хлопчатник).
II. Растения, вызывающие преимущественно симптомы поражения желудочно-
кишечного тракта и одновременно действующие на ЦНС и почки (норичник, мыт-
ник, льнянка, погремок, марьянник, молочай, пролесник, аронник, блокрыльник,
эминиум, переступень, очиток, тамус, дискорея, мыльнянка, тысячеголов, смолевка,
колючелистник, очный цвет, дряква, плющ, скабиноза, эремурус, триллиум, стальник,
золототысячник, каперцы, ворсянка, гледичия, подмаренник, патриния, володушка,
клопогон, лен слабительный, парнолистник, марь, горец, вьнок, куколь, паслен, по-
вилика).
III. Растения, вызывающие преимущественно симптомы поражения органов ды-
хания и пищеварительного тракта — растения, образующие горчичные масла (резуш-
ка, жерушник, горчица, капуста, сурепка, редька, репник, ярутка, клоповник, деску-
рения, вайда).
IV. Растения, вызывающие преимущественно симптомы поражения сердца (на-
перстянка, ландыш, горицвет, желтушник, желтофиоль, олеандр, морозник, сирения,
вороний глаз, птицемлечник, купена, лаконос, волчье лыко, кендырь, копытень, бе-
ресклет, обвойник, джут, цинанхум, ластовень, вязель, будра, тюльпан).
V. Растения, вызывающие преимущественно симптомы поражения печени (кре-
стовник, золотарник, нардосмия, люпин, гелиотроп, триходесма).
VI. Растения, вызывающие аноксемические явления (явления удушения):
1) растения, образующие (при определенных условиях) синильную кислоту (сор-
го, манник, колоняк, бор, бухарник, триостренник, лен, вика, клевер, лядвенец, мин-
даль, абрикос, черемуха);
2) растения, образующие (при определенных условиях) низшие окислы азота
(свекла, дуб, крапива).
2.2. Биологические поражающие агенты
391
VII. Растения, сенсибилизирующие (повышающие чувствительность! животных
к действию солнечного света (зверобой, гречиха, якорцы, просо, клевер, муреция;.
VIII. Растения, вызывающие признаки геморрагического диатеза (донник, смо-
лоносипа).
IX. Растения, вызывающие нарушения половой деятельности животных (псора-
лей, воробейник, клевер, паслен птичий, юкка, подсолнечник).
X. Растения, вызывающие заболевания с характером витаминной недостаточно-
сти (папоротник-ордяк, хвощ).
XI. Растения, вызывающие симптомы нарушения солевого обмена (щавель, кис-
лица, гребенщик, кумарчик, 1алагетон).
Роль отдельных токсических веществ в формировании клиническ ш картины отрав-
ления растительными ядами. Детально исследована И. Л. Гусыниным (1962). В огром-
ном большинстве случаев клиническая картина отравления представляет сложную
комбинацию видимых клинических признаков, проявляющихся одновременно или
последовательно как результат действия, возможно, не одного какого-либо вещества,
а ряда токсических веществ, заключенных в ядовитом растении.
При отравлении многими растениями выступают на первый план и преобладают
над всеми другими внешними проявлениями отравления симптомы поражения нерв-
ной системы. Характер поражения нервной системы может быть различным. В одних
случаях ядовитые вещества растений возбуждают деятельность различных отделов
нервной системы, в других, наоборот, быстро и сильно угнетают их или совершенно
парализуют (см. пример с отравлением Сократа в разд. 1.1).
В клинической картине отравления растительными ядами, вызывающими пре-
имущественно симптомы поражения ЦНС, преобладают или признаки повышенно-
го возбуждения, проявляющиеся в различном по силе и форме двигательном беспо-
койстве, маниакальном состоянии (в виде безудержного стремления вперед без
всякого учета находящихся впереди препятствий), судорогах, возбуждении дыхания,
усилении кровообращения (отравления красавкой, дурманом, беленой, полынью, ве-
хэм); или, наоборот, признаки угнетения в виде затрудненности произвольных дви-
жений, понижения обшей чувствительности, угнетения рефлекторной спосооности,
сонливости, параличей (отравления плевелом опьяняющим, пикульниками). При
сильных отравлениях возбуждение ЦНС часто является лишь первой стадией действия
яда, за которой следуют, иногда очень быстро, тяжелое угнетение и паралич ЦНС.
Равным образом, внешнее возбуждение иногда может быть не результатом возбужде-
ния ЦНС, а следствием се угнетения или паралича. Некоторые растения оказывают
преимущественное действие на спинной мозг.
Действие ядовитых растений на нервную систему редко бывает гыражено в чистом,
обособленном виде. Обычно оно осложняется расстройствами других органов и сис-
тем, наступающими вследствие непосредственного раздражающего действия яда на
эти органы (отравления ядами, полученными из растений, содержащих эфирные
масла), или в результате воздействия яда на соответствующие отделы периферической
нервной системы (отравления аконитом, табаком, чемерицей, атропинсодержашими
растениями). Местные нарушения, в свою очередь, выступают в роли факторов, вли-
яющих на ЦНС. На нервную систему действуют преимущественно алкалоидные рас-
тения (группы мака, аконита, живокости, чемерицы, табака, ежовника, безвремен-
ника, эфедры, омега и др ) и растения, содержащие эфирные масла.
Значительное количество ядовитых растений оказывает сильное раздражаю-
щее действие на слизистую оболочку пищеварительного тракта. Они вызывают опуха-
392
Часть 2. Общая неправильная эпидемиология
ние слизистой оболочки, гиперемию и ограниченные или разлитые кровоизлияния.
В одних случаях токсическое действие таких растений носит характер местного по-
ражения пищеварительного тракта и мало затрагивает другие органы и системы; в дру-
гих — влияет также резорбтивно, и тогда одновременно с поражением пищеваритель-
ного аппарата или вслед за ним возникает ряд расстройств со стороны других органов
и систем, главным образом ЦНС и почек (отравления молочаями, гликозидными,
сапонин- и соланинсодержащими растениями). Большое токсикологическое значе-
ние из этой группы могут иметь сапонинсодержашие растения.
Наиболее важные симптомы отравления растениями данной группы: слюнотече-
ние, тошнота, рвота, боли в животе, общая слабость; в тяжелых случаях (сапонино-
вые растения) — слабость сердца, затруднение дыхания, судороги, нарушение созна-
ния, паралич ЦНС; при выделении через почки сапонины сильно раздражают их.
Сильное раздражение пищеварительного тракта производят растения, содержащие
горчичные гликозиды (крестоцветные).
Клиническая картина отравления многими растениями, преимущественно крес-
тоцветными, сопровождается симптомами поражения дыхательных путей. В этом
случае учащенное, усиленное дыхание, одышка, беспокойство, кашель, цианоз выс-
тупают как основные симптомы отравления наряду с поражением ЖКТ.
Картина нарушений сердечной деятельности в виде усиления сердечных сокра-
щений, увеличения или уменьшения сердечных сокращений в единицу времени, из-
менения ритма, появления симптомов слабости (усиленный сердечный толчок при
очень слабом, малозаметном, частом пульсе, остановка сердечной деятельности) мо-
жет наступить или как первичный симптом в результате влияния растений, обладаю-
щих преимущественным действием на сердце (группа наперстянки), или как вторич-
ный признак в результате сильного токсического дейсз вия других ядовитых растений.
В результате действия ядов растений, содержащих летучие вещества, выделяющи-
еся через почки (эфирные масла, протоанемонин лютиков, горчичные масла крестоц-
ветных, госсипол хлопчатника), могут наступить тяжелые расстройства в состоянии
мочевыделительной системы — почек, мочевого пузыря. Для таких расстройств харак-
терно частое отделение мочи (иногда только небольшими порциями) или, наоборот,
замедление отделения мочи (и уменьшение ее количества), изменение окраски мочг;
и появление в ней крови.
Нарушение беременности и аборты возникают или вследствие непосредственно-
го действия токсических веществ растений, вызывающих судорожные сокращения
матки, или как вторичные явления, обусловленные нарушениями кровообращения в
матке и питания плода.
В результате действия синильной кислоты (цианогенные растения) и низших со-
единений азота нарушается газовый обмен, развивается кислородное голодание тка-
ней, приводящее при тяжелых отравлениях к смерти от задушения. Эти отравления
могут протекать весьма быстро. Такие случаи отравления описаны ветеринарами.
Животные погибают в течение нескольких часов и даже минут.
Основой клинической картины отравления некоторыми растениями является
тяжелое нарушение кроветворной деятельности костного мозга с последующим раз-
витием лейкопении и тромбоцитопении, обусловливающим возникновение явлений
геморрагического диатеза (множественные кровоизлияния в различных органах и
тканях), нарушения питания, исхудания, кахексии.
Есть большая группа растений (крестовники, гелиотропы, люпины), вызывающих
при определенных условиях глубокие поражения печени, клиническими признака-
2.2. Биологические поражающие агенты
393
ми которых являются нарушения пищеварения, желтуха, мозговые явления, расстрой-
ства мочеиспускания, сердечной деятельности, opiaHOB дыхания.
Патологоанатомическое исследование. Вскрытие отравленных растительными яда-
ми людей, чаще всего не даст основания для точного диагноза. И. Л. Гусыниным (1962)
отмечено, что изменения, характерные для общего токсикоза, не представляют в боль-
шинст ве случаев особенностей, специфических для отравления тем или иным опре-
деленным ядовитым растением. В лучшем случае патологоанатомические данные
имеют групповое значение, указывая на характер поражений, которые могут возник-
нуть от действия ядовитых соединений представителей той или иной группы ядови-
тых растений. Но и в этом смысле они играют роль лишь подтверждающего фактора,
учитываемого в сопоставлении с другими данными, используемыми для постановки
диагноза. Поражения преимущественно пи-
щеварительного тракта наблюдаются при от-
равлениях главным образом гликозидными и
сапонинсодержашими растениями. При от-
равлении алкалоидными растениями выра-
женных патологоанатомических изменений не
бывает. Поэтому результаты вскрытия могут
иметь решающее значение лишь тогда, когда
они указывают на непосредственную причину
отравления (нахождение в содержимом пи-
щеварительного тракта частей ядовитых рас-
тений). Ниже кратко описаны токсические
свойства отдельных растений1, с древности ис-
пользуемых для отравления людей.
Красавка. Другие названия: сонная одурь,
белладонна, волчья ягода. Многолетнее травя-
нистое растение семейства пасленовых. В Ев-
ропе распространены два вида: Atropa bella-
donna L. и A. caucasica kreyer. Оба вида белла-
донны встречаются только в горах в Крыму, на
Карпатах, на Кавказе, где растут или единич-
ными экземплярами под пологом буковых ле-
сов, или небольшими зарослями по лесным
полянам, опушкам, по окраинам дорог, по реч-
кам на высоте 200—1000 м. Кроме того, расте-
ния культивируются. По клинической класси-
фикации ядовитых растений И. Л. Гусынина
(1962), относится к ядовитым растениям, вы-
зывающим возбуждение ЦНС (рис. 2.22).
Атропин — его основным фармакологичес-
ким действием является блокада М-холино-
реакгивных систем как на периферии, так и в
ЦНС. Смертельная доза атропина — 0,6—0,1 г.
Рис. 2.22. Красавка обыкновенная. Кус-
тарниковое травянистое многолетнее
растение с многоглавым корневищем и
деревянистым прямым стеблем высо-
той 1—2 м, от конца которого отходят
черешки листьев. Цветет темно-фиоле-
товыми колоколообразными цветами;
ягоды блестящие, черные, размером
примерно с вишню. Содержит токсиче-
ские алкалоиды тропанового ряда: ат-
ропин, левовращающий гиосциамин
(при выделении переходит в рацемат
атропина), атропамин. скополамин и
кумариновое производное скополетин.
По А. Ф. Гаммерману, И. И. Грому (1976)
1 Использовались работы И Л. Гусынина (1962); Л. Ф. Гаммериана и И И. Грома (1976);
Д. А. Муравьевой (1983); Р. Коберта (1888); Б. Н. Орлова и Д. Б. Гелашвили (1990); В. Ф Бого-
явленского и И. Ф. Богоявленского (2002); X. Альтмана (2004).
394
Часть 2 Общая неправильная эпидемиология
Наиболее извест ными периферическими эффектами атропина являются уменьшение
секреции слюнных, бронхиальных, желудочных потовых желез, поджелудочной же-
лезы, увеличение частоты сердечных сокращений, понижение тонуса гладких мышц
полых органов, мидриаз и паралич аккомодации. Центральные эффекты атропи-
на менее выражены, чем периферические, и считаются более возбуждающими: атро-
пин повышает групповую токсичность фенамина, провоцирует делириозный синд-
ром, усиливает дыхание в высоких дозах и активирует холинергические влияния в
малых дозах за счет блока холинэстеразы. При остром отравлении смерть наступает
от остановки дыхания, при хроническом — при явлениях анорексии, олигодипсии,
олигурии, лихорадки, анемии, гиперхолестеринемии и алкалурии от угнетения ды-
хания и судорог.
Гиосциамин — природный L-гиосциамин — левовращающий изомер, в то время
как выделение его из сырья приводит к образованию рацемической смеси, называе-
мой атропином. В настоящее время атропин рассматривают как рацемическую фор-
му, состоящую из равных частей лево- и правовращающего гиосциамина. Гиосциамин
в 2 раза активнее, чем атропин действует на мышцу, суживающую зрачок; в 3 раза
активнее, — на холинорецепторы сердца; в 10 раз — на холинорецепторы кишечни-
ка. Таким образом, гиосциамин, повторяет действие атропина, но более эффективно
в эквивалентных дозах.
Скополамин — в медицине применяется как антихолинергическое средство. Бес-
цветные прозрачные кристаллы или белый кристаллический порошок. Легко раство-
рим в воде (1:3), растворим в спирте (1:17). Выдерживает кипячение. Стерилизуют при
температуре 100 °C 30 мин. Химически скополамин близок к атропину: является слож-
ным эфиром скопина и троповой кислоты. Близок к атропину по влиянию на пери-
ферические холинореактивные системы. Подобно атропину вызывает расширение
зрачков, паралич аккомодации, учащение сердечных сокращений, расслабление глад-
ких мышц, уменьшение секреции пищеварительных и потовых желез. Оказывает так-
же центральное холинолитическое действие. Обычно вызывает седативный эффект:
уменьшает двигательную активность, может оказать снотворное действие. Картина
интоксикации сходна с таковой у атропина. Характерным свойством скополамина
является вызываемая им амнезия. Скополамин очень популярен в преступном мире,
особенно в странах Латинской Америки. Прежде всего, он используется преступни-
ками для подавления воли людей. В преступном мире известны разные способы его
применения, в том числе весьма изощренные. Будучи добавленным в напиток жерт-
вы или еду, г зависимости от дозы, он вызывает либо смерть жертвы, либо ее наркоти-
ческое опьянение, позволяющее преступником добиваться от жертвы необходимой
информации, подписей на документах и др. Спецслужбы используют скополамин в
технике наркодопроса в качестве «сыворотки правды». Для этого допрашиваемом>
внутривенно и медленно вводят 0,5-1 мл 0,05 % раствора скополамина. Эффект обыч-
но проявляется в чувствах парения и уменьшения веса; субъект не контролирует себя,
становится избыточно общительным, предельно благодушным и очень откровенным,
выбалтывая то, что он в обычном состоянии скрывает (см. у Ронина Р., 1997).
Основное токсикологическое значение при отравлении красавкой имеют атропин
и гиосциамин. Все части растения ядовиты, но содержание алкалоидов в них различ-
но. Больше всего их в корнях (0,4—1,3 %), листьях (0,14-1,20 %) и плодах (до 0,7 %).
Естественные отравления встречаются у детей и взрослых летом вследствие употреб-
ления в пищу сладких ягод растения. Смертельная доза для взрослых — более 10 ягод.
Оба алкалоида сильно возбуждают ЦНС (головной и продолговатый мозг). Клиниче-
2.2. Биологические поражающие агенты
395
ски обусловливая двигательное беспокойство, буйство, повышение тем пературы тела,
ускорение дыхания, повышение АД. Парализуют окончания парасимпатических нер-
вов, вызывая расширение зрачков, нарушение зрения, сильное учащение работы серд-
ца. ограничение секреции желез (слюнных, желудка, кишок, потовых), замедление пе-
ристальтики кишок и других органов с гладкой мускулатурой (желчного пузыря, мо-
чевого пузыря, матки). Симптомы отравления появляются через 10—20 мин после
приема атропина или содержащего его растения. У отравленого обращает на себя
внимание розовая кожа, хриплый голос, гиперемированное лицо, максимально рас-
ширенные зрачки, выраженное возбуждение, беспокойство, неадекватное поведение.
Может быть коматозное состояние. Понос и рвота (иногда). Смерть наступает от па-
ралича дыхания. В случае выздоровления — ретроградная амнезия (последствия дей-
ствия следовых количеств скополамина). При вскрытиях животных, погибших от
отравления красавкой, обращали на себя вни-
мание серо мая отечность вокруг почек, сероз-
но-желатинозное пропитывание внутреннего
жира, серозно-геморрогическая отечность лим-
фатических желез. У людей, кроме сильного
отека мозга и его оболочек хровью и состояния
зрачков, не находят ничего характерного; толь-
ко в единичных случаях, где отравление было
вызвано употреблением в пищу ягод, наблюда-
лось темно-синее окрашивание слизистой же-
лудка и кишок (от сока ягод). Встречаются еще
одиночные экхимозы в сердце.
Белена. Род (Hyoscyamus) большей частью
двулетних и однолетних травянистых растений
семейства пасленовых. Около 20 видов в Евро-
пе, Азии, Африке (до центральной части) и на
Канарских островах. В России и в странах СНГ
произрастают 8 видов, из которых наибольшее
токсикологическое значение имеет белена чер-
ная (Н. niger) — двухлетнее растение, растет по-
всюду на пустырях, у дорог, иногда как сорняк
на полях. По клинической классификации ядо-
витых растений И. Л. Гусынина (1962), относит-
ся к ядовитым растениям, вызывающим воз-
буждение ЦНС (рис. 2.23).
Алкалоидами белена беднее красавки- в ли-
стьях 0,045-0,1 %, в корнях — 0,15—017 %, в се-
менах — 0,06—0,1 %. Естественные отравления
встречаются в основном у детей. Их привлека-
ют семена белены — маслянистые и сладкие на
вкус. Белена встречается около жилья по сор-
ным местам, выгонам, паровым полям почти по
всему СНГ. К северу она «захотит» до 60° с. ш.,
выше встречается в виде одиночных растений.
На востоке России встречается в Забайкалье, в
Приморском крае белена уже редкость. Белена
Рис. 2.23. Белена черная. Двухлетнее
травянистое растение со стержневым
корнем и прямым стеблем, доверху по-
рытым листьями. Достигает высоты
40—80 см. Листья, крепящиеся к стеб-
лю, имеют серо-зеленый цвет, яйце-
видную или продолговатую форму,
выемчато-пильчатые и, как стебель,
покрыты клейко-волосистыми волос-
ками. Цветы грязно-желтые, образуют
правильно-ветвистые колосовидные
соцветия. В одном плоде — двухкамер-
ной коробочке содержится до 200 се-
мян Содержит алкалоиды тропаново-
го ряда, главным образом гиосциамин
и следы других близких алкалоидов
(скополамин, атропин). Все растение
распространяет неприятный запах По
А. Ф. Гаммерману, И. И. Грому (1976)
396
Часть 2. Общая неправильная эпидемиология
Рис. 2.24. Дурман обыкновенный. Од-
нолетнее травянистое растение высот ой
30—100 см с яйцевидно-заостренными
неравномерно-выямчато-пильчатыми
листьями. Плод — яйцевидная прямо-
стоящая коробочка, усаженная много-
численными шипами, раскрывающаяся
четырьмя створками. Содержит боль-
шое количество мелких палочковидных
семян темно-коричневого цвета. Все ча-
сти растения содержат ядовитые алка-
лоиды тропанового ряда (гиосциамин,
скополамин, атропин и др.). Растение
распространяет неприятный запах. По
А. Ф. Гаммерману, И. И. Грому (1976)
издавна употребляется е различных колдовских культах и мошенничествах лицами,
выдающими себя за колдунов и ведьм для вызова сильных зрительных и слуховых
галлюцинаций (часто без критики к ним, нередко устрашающего характера), бреда,
нарушения ориентации в пространстве и времени, временного отключения памяти.
Симптомы отравления появляются быстро, в пределах 20 мин после употребления
белены. Они сходны с отравлением красавкой. Сначала приходит ощущение сильной
сухости во рту, граничащей с онемением. Это похоже на первые несколько секунд
после укола новокаина при лечении зубов. Ощущения очень быстро распространя-
ются на носоглотку, гортань, опускаются все дальше. Садится, а иногда и вообще про-
падает голос, появляется затрудненное глотание, проблемы с дыханием. Наблюдает-
ся расширение зрачков, причем очень явное. Зрачки человека, пострадавшего от ядов
белены остаются сильно расширенными даже на ярком свету. Начинается Светобоязнь
и нарушение ближнего зрения. Иногда предметы, особенно белые, кажутся окружен-
ными светящимся ореолом. Нервное возбуждение переходит в расстройство сознания.
У отравленного появляются галлюцинации, бред, нарушение ориентации в прост-
ранстве и обшей координации движений. Есть
даже выражение «белены объелся». Психоз
может продолжаться больше суток. Нередко
это состояние сопровождается судорогами,
сильным учащением сердцебиения. Дыхание
становится частым, поверхностным, а затем
прерывистым. Население использует ядови-
тые вещества белены для самолечения в виде
так называемого «беленного масла». Его полу-
чают настаиванием листьев белены на подсол-
нечном масле и используют местно как обез-
боливающее.
Дурман (Datura). Род растений семейст-
ва пасленовых. Травы, реже кустарники и де-
ревья. На территории бывшего СССР встре-
чается дурман обыкновенный или вонючий
(D. stramonium). Растет как сорняк по краям па-
шен, на мусорных местах и т. п., а также по бе-
регам рек и ручьев в европейской части Глав-
ным образом в южной половине), на Кавказе,
в Западной Сибири и Средней Азии. В Сред-
ней Азии и I /рибалтике распространен дурман
фиолетовый (£>. tatura L.). По классификации
ядовитых растений И. Л. Гусынина (1962) от-
носится к ядовитым растениям, вызывающим
возбуждение ЦНС (рис. 2.24).
Распределение ядовитых алкалоидов по
частям растения не одинаковое. В листьях и
семенах главным образом содержится гио-
сциамин (0,2-0,45 % и 0,21-0,25 % соответ-
ственно). Скополамин содержится в корнях
растения (0,21—0,25 %). Много алкалоидов в
верхушках побегов (0,6—0,7 %). Клиника
2.2. Биологические поражающие агенты 397
отравления сходная с той, что наблюдают при отравлении красавкой. Вскрытие по-
казывает большую или меньшую степень раздражения слизистой желудка, воспале-
ние или эктравазаты слизистой желудка.
Существуют древние традиции использования дурмана в тайных культах и для
совершения преступлений. В средневековой Европе так называемые ведьмы изготов-
^ыли свои «волшебные мази», добавляя в них сок и толченые части белладонны и дур-
мана. Втирая эти мази в свое тело, колдуньи подвергались наркотическому воздей-
ствию, во время действия которого некоторые из них ощущали чувство полета, а дру-
гие — любовного экстаза с самим Сатаной во время шабашей. Профессиональными
отравителями в 11ндии в преступных целях обычно используют семена дурмана D. alba.
Их всегда много в созревших плодах растения. Так как эти семена горькие, то их до-
бавляют в острые блюда или блюда с богатым вкусом. У ацтеков семена дурмана счи-
тались священными, их клали на алтари или в специальные священные коробочки,
которые подносили в качестве даров ацтекским божествам. Почти все индейские пле-
мена Центральной и Южной Америки использовали растение, юбавляя его в особые
напитки, употребляемые при ритуальных обрядах, инициации и колдовстве. Дурман
был также весьма популярным средством народной медицины. Используя его анес-
тезирующий эффект, знахари применяли питье из дурмана при примитивных хирур-
гических операциях, делая иногда даже трепанации черепа.
На островах Карибского моря дурман под названием датура используется как
магическое растение. Здесь он известен как «herbe aux sorciers» — трава колдунов и
«concombre-zombi» — зомби-огурец. Сами названия эти указывают на область при-
менения дурмана — зомбирование. Жертвами таких практик обычно становятся пре-
ступники, на которых не действовали иные меры наказания. Тогда их превращают в
зомби (см. разд. 3.15).
В России на бытовом уровне растение используют наркоманы как заменитель
наркотика. Для полноценных жестких галлюцинаций обычно наркоманы употребля-
ют одну чайную ложку семян дурмана или чуть меньше (зависит от индивидуальной
чувствительности и вида дурмана). В этом качестве дурман пропагандируется россий-
ской молодежи на специальных интернет-сайтах и в книгах Карлоса Кастанеды (у по-
следнего как растение, именуемое «травой дьявола»). Его часто использует для дости-
жения эффекта полета персонаж книг Кастанеды — колдун дон Хуан.
Болигслов пятнистый. Входит в род болиголов (Coniunt) семейства зонтичные
(Apiaceae) порядка зонтикоцветные (Apiales). Для убийств и самоубийств использует-
ся с глубокой древности. Историки Древнего Рима Плиний и Тацит свидетельствова-
ли, что именно болиголов в Греции использовали для казни преступников, и этот вид
наказания был очень распространен. Предполагают, что казнь ядовитыми растения-
ми ввели в начале правления 30 тиранов (404—403 гг. до н. э.) в период распада Афин-
ского государства. Римляне называли ядовитый напиток из сока болиголова «sorbito
cicutae». По клинической классификации ядовитых растений И. Л. Гусынина (1962),
болиголов относится к растениям, вызывающим угнетение и паралич ЦНС и одно-
временно действующие на ЖКТ и ССС (рис. 2.25).
В природе ареал вида охватывает Алжир, Марокко, Тунис и Эфиопию; всю тер
риторию Европы; умеренные районы Азии от Турции до Китая. Произрастает на лес-
ных опушках, заливных лугах, известняковых склонах, как сорное растение в посе-
вах и огородах, на залежах, около жилья, у дорог и заборов, на свалках, по склонам
оврагов, по полотну железных дорог. В плодах болиголова содержатся до 2 % алкалои-
дов: кониин (а-пропилпиперидин), метилкониин, конгидрин, псевдоконгидрин
398
Часть 2. Общая неправильная эпидемиология
Рис. 2.25. Болиголов пятнистый. Расте-
ние высотой 1—2 м образует беловатые
веретенообразные корни с вертикаль-
ными трубчатыми стеблями, покрыты-
ми узкими бороздками. Нижняя часть
стебля покрыта пурпурно-красными
пятнами. Цветки белые. Плод неболь-
шой, овальный. Легко распадается на
два сжатых с боков полуплодика. Со-
держит ядовитые алкалоиды, главным
из которых является кониин. В жаркую
погоду распространяет вокруг себя не-
приятный запах мышиной мочи. По
А. Ф. Гаммерману, И И. Грому (1976)
и жирное масло, в состав которого входят гли-
цериды петрозелиновой и петрозелидиновой
кислот. В листьях содержится до 0,1 % алка-
лоидов, до 0,08 % эфирного масла и кофей-
ная кислота. Из цветов выделены кемпферол
и кверцетин. Аналогичная смесь алкалоидов
найдена в кокооыше (другое название расте-
ния — собачья петрушка или зноиха; род рас-
тений семейства зонтичные). Основной алка-
лоид и ядовитое начало болиголова пятнисто-
го — кониин. Смертельная доза для взрослого
человека при пероральном приеме в пределах
0,13—0,20 г. ММ — 127,3 Да. Химическая фор-
мула — CgH17N. Кониин при комнатной тем-
пературе представляет собой бесцветную жид-
кость с резким запахом мышиной мочи. За-
стывает при температуре 2,5 °C, температура
кипения — 166,5 °C. Хорошо растворим в орга-
нических растворителях, слабо — в воде; окис-
ляется на воздухе, в результате этого приобре-
тает бурую окраску. Содержится но всех час-
тях растения, главным образом в его плодах
и семенах (до 1 %). Образуется в клетках рас-
тения из остатков уксусной кислоты и амино-
кислоты лизина. Кониин и другие алкалоиды
содержатся во всех частях болиголова, однако
наиболее богаты кониином плоды этого рас-
тения до их полного дозревания.
Естественные отравления встречаются из-
за слу тайного употребления в пищу корня бо-
лиголова (вместо хрена) или его листьев (вме-
сто петрушки). Отмечены случаи отравления
плодами болиголова при ошибочном применении их вместо плодов аниса. Кониин
быстро всасывается в кровь из пищеварительного тракта. Первые симптомы появля
ются очень рано, иногда через несколько секунд; так же быстро наступает в тяжелых
случаях и смерть от паралича дыхания (через несколько часов или даже минут). Ко-
ниин выз! [вает паралич окончаний чувствительных и двигательных нервов, а также
поражает ЦНС, сначала возбуждая, а затем парализуя ее. После приема кониина уси-
ливается слюнотечение может появиться тошнота, рвота, понос, головокружение,
расстройство зрения. Весьма характерен так называемый «восходящий» паралич, со-
провождающийся полной потерей чувствительности и подвижности охваченных им
частей тела. Он начинается со ступней и далее распространяется вверх на туловище,
пока не достигнет диафрагмы, вследствие паралича которой и наступает смерть от
удушья. Возможно, что соком именно этого растения, а не цикуты (т. е. веха ядови-
того), был отравлен Сократ. По клиническим проявлениям отравления этот яд от-
личается от кураре тем, что вызывает сильные клонические судороги в начальной ста-
дии отравления. Патологоанатомическая картина при отравлении кониином специ-
фичной не является.
12 Биологические поражающие агенты
399
Вех ядовитый. Другие названия — цикута (Cicuta), водяной болиголов, кошачья
ггрушка, вяха, омег, омежник, водяная бешеница, мутник, собачий дягиль, гориго-
това. Род растений семейства зонтичных. Многолетние водные или болотные травы
с дважды- и четыреждыперистыми листьями. Насчитывают до 20 видов веха. На тер-
ритории СНГ широко распространен евроазиатский вид — вех ядовитый (С. virosa).
Произрастает на болотах, по заболоченным берегам рек, озер и канав, на сырых лугах
и в ольшаниках. По клинической классификации ядовитых растений И. Л. Гусынина
< 1962), относится к растениям, вызывающим возбуждение ЦНС. Одно из наиболее
ядовитых растений флоры России. Основное токсическое вещество — цикутотоксин.
ММ 258,38 Да. Это безазотистое, смолистое вещество, растворимое в эфире, спиртах,
хлороформе, в значительном количестве также в кипящей воде. По современным дан-
ным, он не является ни алкалоидом, ни гликозидом, а производным пирона. Отно-
сится к классу ядовитых спиртов, вызывающих смерть через расстройство ЦНС. Био-
химически идентифицирован как мощный неконкурентный антагонист рецептора
сйромедиатора — у-аминобутановой (аминомасляной) кислоты (ГАМК). Наиболее
опасно корневище растения. В свежевыкопанном корне его содержится до 0,2 %, в
высушенном — от 1,5 до 3,5 %. Корневище веха особенно ядовито ранней весной и
поздней осенью. Однако и в другое время года ядовитость его сохраняется и остается
высокой. Цикутоксин и другие ядовитые вещества веха не разрушаются ни под воз-
действием высокой температуры, ни при длительном хранении.
Цикутотоксин — нейротоксин. Он повышает рефлекторную возбудимость и вы-
зывает у теплокровных животных сильные судороги в результате возбуждения судо-
рожного центра и спинного мозга. Одновременно он возбуждает другие центры про-
долговатого мозга (сосудодвигательный, блуждающего нерва), обусловливая.этим
нарушение дыхания, кровяного давления, работы сердца. Алкалоиды в этом растении
по токсическому эффекту играют вспомогательную роль. Естественные отравления
встречаются в основном у детей. Их привлекают сладкие на вкус и легко выдергиваю-
щиеся из земли корни. Отравления людей также возникали из-за того, что болиголов
принимали за сельдерей, пастернак или корень петрушки (болиголов издает запах, на-
поминающий запах петрушки). Случаи отравления животных вехом чаще всего встре-
чаются весной среди крупнорогатого скота. Смертельная доза цикутотоксина для
взрослого человека содержится в 2—3 г свежевыкопанного корня растения. В отноше-
нии лабораторных животных (кошек) смертельной дозой являются 7 мг при внутри-
венном введении и 50 мг перорально на I кг веса животного. Симптомы отравления
развиваются спустя 15-20 мин после приема растения в пищу с чувства тошноты,
жжения во рту и глотке, головной боли, болей в животе. Начинается рвота. При тя-
желом отравлении характерно чувство холода во всем теле, нарушение равновесия,
понижение кожной чувствительности. По мере развития отравления отмечаются кло-
нико-тонические судороги, обильное выделение густой слюны. Смерть может насту-
пить от остановки дыхания на фоне острой сердечно-сосудистой недостаточности.
При вскрытии трупов животных, погибших от отравления вехом, находят кровоизли-
яния в подкожной соединительной ткани, слизистой оболочке дыхательных путей, под
эпи- и эндокардом, под капсулой и в паренхиме почек, на слизистой мочевого пузы-
ря. Сильное раздражение слизистой оболочки желудка и кишок, выраженная гипе-
ремия легких и мозга, точечные кровоизлияния под плеврой.
Аконит (Aconitum). Другое название: борец. Род многолетних травянистых расте-
ний семейства лютиковых. Около 300 видов в Северном полушарии. На территории
бывшего СССР около 75 видов. Почти все ядовиты. По клинической классификации
400
Часть 2. Общая неправильная эпидемиология
Рис. 2.26. Аконит. Травянистое много-
летнее растение, у которого зимует ре-
повидный корень. Вертикальный сте-
бель достигает высоты 50—150 см и не-
сет темно-зеленые пальчатосложные,
пяти-, семидольные надсеченные лис-
тья. Цветки шлемовидной формы име-
ют цвет от темно-синего до темно-фи-
олетового, появляются с июня до сен-
тября. Корень, листья и цветки содер-
жат ядовитый алкалоид аконитин. По
А. Ф. Гаммерману, И. И. Грому (1976)
ядовитых растений И. Л. Гусынина (1962),
аконит относится к растениям, вызывающим
угнетение и паралич ЦНС и одновременно
действующим на ЖКТ и ССС. Считается са-
мым ядовитым растением Европы (рис. 2.26).
Из корней аконита на Древнем Востоке
получали яд бик (Bikh или Bish, Tubera Aconiti
indica). В Индии его считали самым сильным
ядом из имевшихся. Добывался из A. ferox,
A. palmatum, A. napellus и A. luviduum. Особен-
ной популярностью он пользовался у отдель-
ных народностей Индостана, которые смеши-
вали растертые корни аконита с соком Dillenia
speciosa (диллениевые — распространенные в
Индии вечнозеленые деревья и кустарники)
и полученным ядом смазывали наконечники
стрел. Естественные отравления встречаются
вследствие того, что растение было принято
за корни сельдерея или хрена, а также из-за
использования листьев аконита для салата.
Иногда корни аконита принимают за корни
другого горного растения—любистока (Levis-
ticum spp.), употребляемого во многих горных
местностях Европы для приготовления травя-
ного ликера.
Действующими веществами аконитов яв-
ляются алкалоиды дитерпенового ряда. Их
состав у различных видов различается. Основ-
ное токсическое действие оказывают алка-
лоиды аконитин и зонгорин. Аконитин — ли-
пидрастворимый токсин. Е1а внешний вид —
тонкие, белые кристаллы, растворимые в хлороформе, бензоле, слабо — в спирте и
эфире, очень слабо — в воде. Обладает судорожно-паралитическим действием, кото-
рое обусловлено стойким повышением натриевой проницаемости возбудимых (не-
рвных и мышечных) мембран и их деполяризации вследствие этого LD50 0,035 мг/кг
(собаки, внутривенно); 0,25—0,4 мг/кг (собаки, с пищей); 2-5 мг/кг (человек, перо-
рально); 0,25 мг/кг (крысы, внутрибрюшинно).
В эксперименте аконитин используется для моделирования аритмии сердца. Быст-
ро поглощается через слизистые, очень быстро кишечником. Действие аконитина и
близких ему алкалоидов состоит в первую очередь в кратковременном возбуждении
ЦНС, в особенности дыхательного центра, и периферических нервов — окончаний
двигательных, чувствительных, секреторных и возвратного нервов. Вслед за возбуж-
дением нервной системы наступает ее угнетение и паралич (потеря чувствительно-
сти, сухость слизистых оболочек, паралич сердца, паралич дыхания). После больших
доз причиной смерти может быть первичная остановка сердца (в диастоле). Аконит
обладает кожно-резорбтивным действием и уже при сборе растения может проник-
нуть в кожу и вызвать воспалительные явления и зуд с последующей анестезией по-
раженного участка. Количество алкалоидов в аконитах сильно колеблется в зависи-
2.2. Биологические поражающие агенты
401
мости от вида, от положения в ареале, стадии роста, от органов растения. Есть
акониты, содержащих до 4 % алкалоидов (аконит Фишера, аконит высокий). Наи-
большее накопление аконитов в аконитах носатом и восточном происходит на высо-
те 1400 м над уровнем моря. Симптомы отравления у человека развиваются спустя
10—20 мин после перорального приема: жжение и зуд во рту, в пальцах рук и ног, рас-
пространяющееся затем на кожу всего тела, связанные с потоотделением и ознобом;
позднее типичное онемение; нечувствительность к боли и ощущение ледяного холо-
да, температура тела понижае гея. Кроме того, встречаются такие симптомы как рво-
та, коликоподобный понос, боли в голове, шее, спине и области сердца, шум в ушах.
Зрачки у отравленного расширены, цветовосприятие нарушено («зеленые и желтые
пятна перед глазами»), аритмия (вплоть до фибрилляции желудочков). Отмечаются
мышечные фибрилляции и судороги конечностей, повышенное мочеотделение.
Смерть наступает при сохранившемся сознании от паралича дыхания и сердца. На
вскрытии явления острого гастроэнтерита, дегенеративные изменения мышцы серд-
ца и почек, кровоизлияния на слизистой ЖКТ и на поверхности околосердеч-
ной сумки. Сходными токсическими свой-
ствами обладает другое растение изсемейства
лютиковых — живость или шпорник (Delphi-
nium spp.). В 1940-х гг. в токсикологической
лаборатории при 12-м отделе НКВД СССР
(руководитель Г. М. Майрановский) акони-
тин использовался для заполнения специаль-
ных полых пуль, предназначенных для стре-
льбы из бесшумного оружия (см. у Колпаки-
ди А. И. и Прохорова Д. П., 2000).
Безвременник. Другие названия: зимов-
ник, колхикум (Colchicum), род многолетних
грав семейства лилейных. Считался ядови-
тым еще с античных времен. Названия «сын
раньше отца» (filius ante patrem) или «безвре-
менник» этим растениям придаются потому,
что их плоды появляются весной как бы рань-
ше цветков, распускаюшихся осенью или
даже в начале зимы, и притом без зеленых
листьев. Описано 60 видов безвременника,
главным образом в странах Средиземномо-
рья, в Иране и на Востоке — до Северной
Индии. На территории бывшего СССР обна-
ружено 10 видов, большей частью в предгорь-
ях и горах Кавказа, некоторые в Крыму, Сред-
ней Азии. По классификации ядовитых рас-
тений И. Л. Гусынина (1962), безвременник
относится к растениям, вызывающим угнете-
ние и паралич ЦНС и одновременно дейст-
вующим на ЖКТ и ССС (рис. 2.27).
В семенах и клубнелуковицах безвремен-
ника содержатся алкалоиды: колхицин, ис-
пользуемый в генетике и селекции для искус-
Рис. 2.27. Безвременник. Каждое рас-
тение ра ввивает с августа до октября
1—3 сидящих у земли цветка с беловатой
трубкой и крупными лепестками, имею-
щими цвет от нежно-розового до лило-
вого и расположенные воронкообразно.
Цветущее растение не имеет листьев,
они развиваются лишь следующей вес-
ной, имеют длину 20—30 см, широкую
ланцетную форму и параллельные жил-
ки. Одновременно с ростом листьев за-
вязь образует продолговатую яйцевид-
ную коробочку длиной до 6 см, в которой
созревают темно-коричневые семена.
По А. Ф. Гаммерману, И. И. Грому (1976)
402
Часть 2. Общая неправильная эпидемиология
ственного получения полиплоидов и в медицине для лечения подагры, а также колха-
мин (омаин), применяемый при лечении некоторых болезней крови и ряда злокаче-
ственных новообразований.
В семенах безвременника количество колхицина может достигать до 1 %, в клуб-
нях его содержание значительно меньше, в листьях еще меньше, чем в клубнях. Кол-
хицин также содержится в растениях родов Merendera и Gloriosa (семейство мелан-
тиевых, Melanthiaceae). Колхицин — белый или белый с желтоватым оттенком мел-
кокристаллический порошок, темнеющий на свету, ММ 399,44 Да. 1 г растворяется в
25 мл воды или в 220 мл эфира; легко растворим в этаноле и хлороформе. Является
сильным антимитотиком, связывающимся с белком тубулином, образующим микро-
трубочки и, вследствие этого, блокирующим деление клеток на стадии метафазы. Дей-
ствует угнетающе на лейко- и лимфопоэз. Так же он представляет собой сильный ка-
пиллярный яд. При отравлении содержащими его растениями колхицин вызывает
нарушения местного кровообращения, в частности сильную гиперемию слизистых
оболочек желудка и кишок, поражает почки, вызывая полиурию, альбуминурию, ге-
матурию. В тяжелых случаях — анурию. Нарушает нейромышечную передачу, стиму-
лирует функции ЖКТ, угнетает дыхательный центр, суживает сосуды и повышает АД,
понижает температуру тела. Практически не связывается с белками плазмы и не за-
держивается в системном кровотоке. В высоких концентрациях накапливается в поч-
ках, печени и селезенке. Экскретируется преимущественно с желчью и через почки.
При внутривенном введении побочные эффекты со стороны ЖКТ не развиваются.
Поврежденные клубнелуковицы безвременника могут вызвать ожоги на руках.
Естественные отравления встречаются в основном у детей. Часто листья безвре-
менника осеннего дети принимают за листья съедобной черемши. Отравления воз-
можны вследствие употребления в пищу молока овец и коз после того, как животные
ели безвременник. В качестве смертельной дозы различные исследователи указыва-
ют 20-40 мг колхицина или 5-10 семян безвременника (примерно 6 г). Колхамин в
10—18 раз менее токсичен. Шесть граммов семян безвременника содержат смертель-
ную дозу его алкалоидов. Симптомы отравления безвременником вследствие медлен-
ного всасывания яда наступают спустя лишь 2-6 ч. Сначала отравленный ощущает
тошноту, першение и жжение во рту, испытывает трудности при глотании. Часто раз-
виваются рвота с поносом и судорогами, в моче кровь, сильное ощущение жажды. Пос-
ле всасывания яда развиваются поднимающийся паралич, удушье, отек легких, по-
чечная недостаточность. Если отравленный выживет, то у него возможно выпадение
волос. Данные вскрытия обычно соответствуют клинической картине отравления без-
временником. Слизистая оболочка желудка и кишок сильно воспалена и имеет боль-
шое количество кровоизлияний. В тяжелых случаях эти изменения могут носить ге-
моррагический характер. Кровоизлияния имеются на слизистых и серозных оболочках
других органов. Почки и мозг сильно гиперемированы и имеют кровоизлияния. Сер-
дечная мышца изменена.
Чемерица Oferatrum). Род растений семейства лилейных. Около 25 видов в умерен-
ном, редко в холодном поясах Северного полушария. На территории бывшего СССР
обнаружено 9 видов. Чемерица Лобеля (V. lobelianum) растет в лесной и лесостепной
зонах по сырым, часто пойменным лугам, светлым лесам и кустарникам. Чемерица
черная (V nigrum) — преимущественно в степной зоне по склонам, лесным, реже пой-
менным лугам, кустарникам. В Карпатах на горных лугах встречается чемерица белая
(И album). Ядом чемерицы белой еще с античных времен пропитывали наконечники
стрел. По классификации ядовитых растений И. Л. Гусынина (1962), чемерица отно-
2.2. Биологические поражающие агенты
403
сится к растениям, вызывающим угнетение и
паралич ЦНС и одновременно действующим
на ЖКТ и ССС (рис. 2.28).
Ядовитые алкалоиды содержатся во всех
частях растения, но преимущественно в кор-
невищах и корнях (до 2 %). В наземной час-
ти растения в весеннее время наиболее ядо-
виты прикорневые листы, причем в основа-
нии листа найдено 0,9—1,5 % алкалоидов,
выше в пластинке, только 0,1-0,3 %. Летом
содержание алка юидов в листьях падает и в
фазу плодоношения они уже не обнаружива-
ются. Наиболее ядовиты (особенно в моло-
дом состоянии) чемерица Лобеля и чемери-
ца белая, которые вызывают тяжелые отрав-
ления и гибель лошадей, крупного рогатого
скота и домашней птицы. Из-за неприятно-
го вкуса листьев и корней растения естествен-
ные отравления людей чемерицей крайне
редки. Если они происходят, то большинство
из них вызвано тем, что корень чемерицы
принимают за корень горечавки, либо ис-
пользуют его для самолечения. Алкалоиды
чемерицы, преимущественно протовератрин
и йервин, первоначально возбуждают ЦНС, а
в последующем парализуют ее и окончания
парасимпатических, чувствительных и секре-
торных нервов. После попадания чемерицы
в организм людей, у них появляются дрожь,
судорожные подергивания мышц, слюноте-
чение, чувство жжения на языке, першение
и покалывание в глотке, понос, рвота, затруд-
Рис. 2.28. Чемерица Лобеля. Многолет-
нее травянистое растение с толстым ко-
ротким вертикальным корневищем, уса-
женным многочисленными длинными
корнями; имеются пучки рафид. Сте-
бель толстый, высотой 1,5 м и более. Ли-
стья очередные, крупные, широкоэлип-
тические, цельнокрайние. Соцветие —
длинная верхушечная метелка. Цветки
невзрачные, зеленоватые. Плод в виде
трехгнездной коробочки с многочислен-
ными семенами. Растение ядовито. По
А. Ф. Гаммерману, И. И. Грому (1976)
нение глотания, потеря чувствительности, чувство холода во всем теле, нарушения
ды <ания и работы сердца. Протовеатрин вызывает сильное сужение зрачка. Симпто-
матика отравления может сохраниться в течение нескольких дней.
Мак (Papaver). Род однолетних, двулетних или многолетних растений семейства
маковых. Около 100 видов, распространенных преимущест венно в Северном полуша-
рии. Для по. [учения семян, содержащих масло растительное, и опия возделывают один
вид — мак снотворный (Р. somniferum), подразделяемый на 8 подвидов. На терр] [то-
рии бывшего СССР культивируют 5 подвидов — евразийский, тянь-шаньский, джун-
гарский, тарбагатайский, китайский. Снотворное действие мака было известно древ-
ним грекам: добывание сока из мака описано Феофрастом, Диоскоридом, а за ними
и Плинием. В Индии и Китае снотворное свойство мака и его культура стали извест-
ны гораздо позже. По клинической классификации ядовитых растений И. Л. Гусынина
(1962), мак снотворный относится к ядовитым растениям, вызывающим угнетение и
паралич ЦНС. Мак снотворный — растение высотой 80—150 см, пронизано сосудами
с белым млечным соком. Из коробочек получают алкалоиды. Их не менее 25: морфин,
кодеин, папаверин, тебаин, протопин, криптопин, неопин, папаверамин, лантопин,
404
Часть 2. Общая неправильная эпидемиология
наркотин и др. Сок этого мака, известный в сухом виде под именем опиума, добыва-
ется следующим способом. Незрелые головки слегка надрезывают острым ножом, из
надрезов вытекает сок, который через сутки твердеет, принимая буроватый цвет. Та-
ких надрезов производят постепенно на каждой головке от 6 до 8; сухой сок соскре-
бывают, переминают в ступках, выделывают из него круглые лепешки и, посыпав
плодами щавеля ( Rumex orientate), обвертывают листьями того же мака. В свежем со-
стоянии опиум мягковат, но затем твердеет. Он буро-красного цвета, на изломе глян-
цевит, неприятного запаха и горек на вкус. Важнейшим веществом опиума признают
морфин, который в высоких сортах восточного опиума может доходить до 24 % от
общего веса. Он там находится в соединении с меконовой кислотой. Опий и морфий
оказывают весьма характерное влияние на перистальтику кишечника, выражающее-
ся в продолжительном запоре. Последний имеет двоякую причину, из которых одна
заключается в уменьшении отделения слизистой оболочки, а другая в прекращении
перистальтики. Подобным же параличом мышечного и анестезией слизистого слоя
объясняется также столь часто наблюдаемая задержка мочеиспускания и растяжение
мочевого пузыря. По преданию, в 1037 г. от передозировки опия погиб знаменитый
арабский врач Авиценна (Ибн-Сина). Заболев дизентерией, он велел приготовить
ученику лекарство, в которое тот по ошибке внес слишком большую дозу опия. Слу-
чаем передозировки опия историки объясняют и смерть Вольтера. Смертельная доза
морфина для взрослого человека — 0,2—0,5 г при внутривенном введении, 0,5—1,0 г
при приеме peros. К пиника отравления опиумом складывается из суммарных эффек-
тов входящих в него алкалоидов:
морфин — центральное воздействие (болеутоляющее, успокаивающее воздействие
на дыхательный и сердечный центр; снижение секреторной функции; успокоение и
эйфория); периферическое воздействие: повышает тонус гладкой мускулатуры. Из-
вестно большое количество наркотических производных морфина, например, геро-
ин (героиновая зависимость гораздо сильнее морфиновой; смертельная доза героина
для взрослого человека, ранее не принимавшего этот наркотик, — 0,12—0,5 г);
кодеин — угнетает кашлевой рефлекс;
тебаин — яд, вызывающий судороги;
папаверин — периферическое расслабляющее воздействие на гладкую мускулатуру;
наркотин — усиливает наркотическое воздействие морфина на ЦНС; возбуждаю-
ще действует на дыхание, оказывает периферическое расслабляющее действие на глад-
кую мускулатуру.
Наркоманам известны разные способы получения опия и его суррогатов. Большое
количество русскоязычных сайтов для наркоманов помогают им выбрать способы
приготовления наркотических суррогатов «по средствам». Симптомы отравления
опием развиваются в течение часа после приема яда. У отравленного появляется ощу-
щение тяжести в голове, сухость во рту, головокружение, рвота, растущее помрачне-
ние сознания и общая слабость, понижение температуры тела, задержка мочи и сту-
ла, а также появляющееся в начале, но постоянно усиливающееся угнетение дыхания
и ослабление сердечной деятельности. Характерными симптомами острого отравле-
ния опиатами являются выраженное нарушение дыхания и сужение зрачков. После
перорального приема яда летальный исход часто наступает спустя лишь много часов
в результате паралича дыхательного центра. При вскрытии находят обильные массы
крови в мозговых оболочках и в пазухах, часто также накопление Liquor cerebrospinalis
в желудочках мозга и прилив крови в легких; кровь темного цвета и жидкой консис-
тенции, а мочевой пузырь сильно растянут мочой. Иногда ясно слышится запах опия
2.2. Биологические поражающие агенты
405
при вскрытии тела отравленного; такой же
запах издает часто и кишечный канал. Зрач-
ки могут быть еще сужены или же снова рас-
ширены.
Строфант. Strophanthus — род растений
семейства кутровых. Известны 50—60 видов,
в гропической Африке, на Мадагаскаре, в
Южной и Юго-Восточной Азии. Восточной
Африке. По клинической классификации
ядовитых растений И. Л. Гусынина (1962),
строфант относится к растениям, вызываю-
щим преимущественно симптомы пораже-
ния сердца (рис. 2.29).
Все части строфанта ядовиты, но преиму-
щественно семена. При давлении семена да-
ют маслянистую жидкость. В семенах стро-
фанта Комбе (5. kombe Oliv) содержится до
2 % К-строфантозида, до 0,6 % К-строфан-
тина-0, до 0,3 % цимарина, 0,28 % цимарола.
Помимо этого, семена содержат периплоци-
марин, гельветикозид, эмицимарин, глюко-
гельветикозоид, гликоцимарол. Семена стро-
фанта щетинистого содержат К-строфанто-
зид, цимарин до 1,47 % и, кроме того, холин,
тригонеллин, сапонины, жирное масло до
30,5 %. Кора корней содержит сердечные гли-
козиды и тригонеллин. В семенах строфанта
щетинистого содержится сердечный глико-
зид субаин (G-строфантин) — до 6 %. В ме-
дицине применяют К-строфантин-0 и G-
строфантин. Строфантин в ЖКТ почти пол-
ностью разлагается, однако при большой дозе
он все же может вызвать смертельный исход.
При внутривенном введении он быстро фик-
сируется миокардом и через 5—6 мин начи-
нает оказывать специфический эффект. Мак-
Рис. 2.29. Строфант Комбе (5. kombe Oliv).
Встречается в Юго-Восточной Африке
по реке Замбези и ее притоку Хире на се-
вер до больших озер. Деревянистые лиа-
ны, реже кустарники, с супротивными
цельными листьями. Цветки в полузон-
тиках. Чашечка пятизубчатая. Венчик
трубчатый, пятилопастной; его лопасти у
многих видов длинные, шнуровидные,
часто скрученные Плод состоит из двух
листовок, в зрелом состоянии расходя-
щихся горизонтально и вместе достигаю-
щих иногда 1 м длины. Семена с волосис-
тым хохолком. 1 — ветка с цветками; 2 —
плод (сложная листовка). 3 — семя с ле-
тучкой, 4 — семя в продольном разрезе.
По Д. А. Муравьевой (1983)
симум действия наблюдается через 20—40 мин. Действие строфантина значительно
ослабляется через 4—6 ч. При введении через желудок LD5r строфантина для белых
мышей 17 мг/кг. Смертельная доза строфантина для взрослого человека при внутри-
венном введении около 1 мг.
С древности строфант используется для получения яда. На Западном побережье
Африки (современные Габон, Гвинея, Сенегал и Гамбия) яд носил название инэй —
местное название строфанта. Теплокровное животное, пораженное стрелой, пропи-
танной соком инэя, погибает через 8—10 мин, если острие остается в ране. Клиника
отравления включает затрудненное дыхание, рвоту, упадок сил и адинамию. Сердце
останавливается в систоле ранее прекращения дыхательных движений. Анатомиче-
ские изменения ничтожны или вовсе отсутствуют. Сердце обычно сокращено; поло-
сти его бывают io пусты, то полны кровью, нормальной по цвету и свертываемости.
406 Часть 2. Общая неправильная эпидемиология
В 1960 гг. в СССР проводились исследования по выявлению строфантина в отече-
ственном растительном сырье. Строфантин обнаружен в горицвете золотистом (Ado-
nis chrysocyanthus), кендыре коноплевом (Аросупит cannabinum), кендыре проломнико-
листном (Аросупит androsaemifolium). Сходные со строфантином гликозиды содержат-
ся в наперстянке (дигитоксин, дигоксин и др.), майском ландыше (конваллатоксин,
конвалламарин, конвалозид и др.), морском луке (сцилларен А и др.), горицвете ве-
сеннем (цимарин, адонитоксин), олеандре (олеандрин, нериин) и ряде других расте-
ний. На сердце они действуют хронотропно (замедляют ритм сердечных сокраще-
ний), инотропно (повышают сократимость и тонус сердечной мышцы), дромотроп-
но (угнетение проводимости), батмотропно (повышение возбудимости). Гликозиды
воздействуют и на ЦНС и центр блуждающего нерва. Принятыеperos они раздража-
ют ЖКТ. Смертельная доза листьев наперстянки 2,25-4 г. Отравление гликозидами
дает о себе знать рвотой центрального происхождения, болями в надчревной обла-
сти, отдышкой, цианозом, поносом (чаще от горицвета весеннего), нарушениями сер-
дечного ритма. Отравленный испытывает сильную слабость, сухость во рту, мучи-
тельную головную боль, головокружение, шум в ушах, сжимающие боли в сердце.
У него возникают цветовые галлюцинации в желтом и зеленом спектре, изменяется
восприятие величины предметов, мелькание «мушек» перед глазами. Характерные
признаки отравления гликозидами — аритмии (суправентрикулярная, желудочковая)
и нарушение артриовентрикулярной проводимости вплоть до полной атриовентри-
кулярной блокады. Вначале появляются брадикардия, замедление атриовентрикуляр-
ной проводимости. Позднее желудочковые экстрасистолы, бигеминия, тригеминия,
политопные желудочковые экстрасистолы, тахикардия, мерцание желудочков, оста-
новка сердца.
Чилибуха. Strychnos nux-vomica — тропическое вечнозеленое деревце высотой до
12 м из рода стрихнос семейства логаниевые (Loganiaceae). К этому роду относятся
около 65 видов, распространенных под тропиками обоих полушарий. Семена дере-
ва —рвотные орешки — являются основным источником ядовитых алкалоидов стрих-
нина и бруцина. Другие названия — кичилибуха, кучелябка, кучеля. Произрастает в
тропических лесах Южной Азии (Индокитай, Индостан, остров Шри-Ланка), на ост-
ровах Малайского архипелага и в Северной Австралии. Культивируется в африкан-
ских тропиках. Семена чилибухи содержат 2—3 % индоловых алкалоидов, из которых
немного меньше половины (47 %) приходится на стрихнин и почти столько же на
бруцин; и небольшие количества других производных индола: вомицин, псевдострих-
нин и др. Помимо алкалоидов, в семенах рвотного ореха содержится хлорогеновая
кислота, гликозид логанин, тритерпеноид циклоарсенал, стигмастерин (рис. 2.30).
Яд из растений рода Strychnos традиционно используют для охоты на животных и
для убийств людей в регионах, где оно произрастает. Одним из таких ядов является яд
кураре. Кураре — это не конкретный яд, а собирательное понятие, означающее груп-
пу ядов, используемых индейцами Амазонки и Ориноко для охоты и войны. Первые
записи о кураре были сделаны испанским священником, отцом д’Акунья, во время
его посещения бассейна Амазонки в 1693 г. Но о нем ничего не знали, кроме того, что
яд был чрезвычайно опасным для людей. Название «кураре» происходи! от индей-
ского слова, обозначающего яд. Изготовление яда кураре было прерогативой шама-
на племени, нарушение этого правила каралось немедленной смертью провинив-
шегося. Индейские племена варьировали названиями растений, служащих сырьем
для производства кураре; яд называли вурари, вурара, курари. куруру, урали, вурали
и т. п. Только в 1938 г. американскому ученому Ричарду Гиллу (Richard Gill) удалось
2.2. Биологические поражающие агенты
407
Рис. 2.30. Чилибуха. Листья супротив-
ные, кожистые, блестящие. Мелкие зе-
леновато-белые цветки собраны в зон-
тиковидные цимозные соцветия. Плод
ягодовидный, ярко-оранжево-красный,
крупный, похож на небольшой поме-
ранец, с 2—8 очень твердыми (отсюда
названием — «орех») дисковидными се-
менами с блестящими шелковистыми
волосками. 1 — цветущая ветка; 2 — рас-
крытий венчик; 3 — плод в ра грезе; 4 —
семя; 5 — семя в продольном разрезе.
По Д А. Муравьевой (1983)
идентифицировать одно из растений, являв-
шееся источником кураре — Chondodendron
tomentosum из семейства Menispermaceae. Со-
держащиеся в этом растении кураре-алкалои-
ды являются производными бисбензилхолина.
Само растение представляет собой крупную
лиану, одеревеневший стебель которой дости-
гает 10 см в диаметре. Дальнейшие исследова-
ния позволили уточнить, что индейцы исполь-
зовали кураре трех видов, разделяя их и по
симптомам вызываемой ими смерти, и по ис-
ходному сырью, и по способам хранения при-
готовленного экстракта. Наиболее быстро и
сильно действующий яд, который надлежало
хранить в специальных трубочках, изготов-
лялся из листьев и корней С. tomentosum. Такой
кураре принято называть трубочным. Клас-
сический способ его приготовления преду-
сматривает экстрагирование на медленном
огне измельченных листьев, стеблей и корней
С. tomentosum, иногда с добавлением крови
ядовитых животных и пресмыкающихся (на-
пример, ядовитых лягушек; их кожа содержит
нейротоксические алкалоиды). Кипящую мас-
су постоянно перемешивали, доводя ее до за-
густения. Легкий яд, необходимый для охоты
на небольших животных, был светлым, Самый
сильный представлял собой темно-коричне-
вую или черную массу липкой или даже почти
твердой консистенции, имеющую отчетливый
смолистый запах. Этой субстанцией смазыва-
ли длинные колючки или специально обработанные палочки, которые для поражения
цели, с силой выдували из трубочек 25 см длиной. Поскольку кураре, расфасованный
в трубках из пустотелых растений, обладал самым сильным фармакологическим дей-
ствием, его главный алкалоид был назван тубокурарином (D-tubocurarine, он же ту-
барин). Гидрохлорид тубокурарина сегодня применяется в хирургии в качестве мио-
релаксанта. Человек, раненный стрелой, пропитанной трубочным кураре, умирал в
течение 5-10 мин при следующих клинических проявлениях действия яда. В первую
очередь у него переставали работать пальцы на ногах и руках и веки, затем наруша-
лись зрение и слух, парализовались лицевые мышцы, мышцы шеи, рук и ног Смерть
наступала от удушья Во время агонии кожа приобретала характерный синюшный
оттенок. Чтобы смертельный яд начал свое действие, необходимо его проникнове-
ние в кровь. Поэтому индейцы считали, если быстро вытащить стрелу и лизнуть язы-
ком рану, то можно остаться в живых (рис. 2.31).
Растения рода Strychnos индейцами Амазонки и Ориноко использовались для
получения двух других видов кураре: пот-кураре (горшочный кураре). Его получали из
коры 5. castelniaeana и, вероятно, видов Chondrodendron и исполь ювалидля охоты на
птиц; и калабаш-кураре (тыквенный кураре). Важной част ью экстракт а являются алка-
408
Часть 2. Общая неправильная эпидемиология
Рис. 2.31. Индейские стрелы с ядом кураре. По П. Маккинису (2008)
лоиды коры сильно ядовитого растения 5. toxifera Schomb. Калабаш-кураре наиболее
ядовит и применяется для стрел и наконечников копий при охоте на крупного зверя
и людей. Его хранили в плодах мелкой посудной тыквы. В начале Второй мировой
войны британским Управлением специальных операций (Special Operation Executive,
SOE) была получена партия яда кураре, произведенная индейцами одного из племен,
обитающего на берегах Амазонки и Ориноко в Венесуэле (Кукридж Э., 2003).
Кураре-алкалоиды, получаемые из растений рода Strychnos, ареал естественного
произрастания которых Южная Америка, являются, как и стрихнин, производными
индола, но сам стрихнин в этих растениях не содержится. Они плохо изучены. Мне
не удалось найти описания отравлений такими алкалоидами, выделенными в чистом
виде. Сегодня в медицинской практике применяют азотнокислую соль стрихнина —
стрихнина нитрат (Strychnini nitras). Представляет бесцветные блестящие игольчатые
кристаллы или белый кристаллический порошок. Имеет чрезвычайно горький вкус.
Трудно растворим в воде (1:90 в холодной) и спирте, легко растворим в кипящей воде
(1:5), нерастворим в эфире. Водные растворы имеют нейтральную или слабокислую
реакцию. Растворы стерилизуют при температуре 100 °C в течение 30 мин; 0,1 % ра-
створ в ампулах подкислен 0,1 н раствором хлористоводородной кислоты и имеет
pH 3,0—3,7. Смертельная доза стрихнина для взрослого человека — 0,03 г. Действие
стрихнина связано с облегчением проведения возбуждения в межнейронных синап-
сах спинного мозга. Он действует преимущественно в области вставочных нейронов.
По современным представлениям стрихнин блокирует действие аминокислотных ней-
ромедиаторов, главным образом глицина, играющих роль тормозящих факторов в пе-
редаче возбуждения в постсинаптических нервных окончаниях в спинном мозге. Бло-
кируя торможение, стрихнин оказывает таким образом «возбуждающий» эффект.
Рефлекторные реакции становятся более генерализованными, при больших дозах
стрихнина различные раздражители вызывают появление сильных болезненных
тетанических судорог.
Бруцин — индольный алкалоид, содержащийся в некоторых видах стрихноса, где
он содержится в количестве 1-1,5 %. ММ 394,47 Да. Бесцветные кристаллы, тем-
пература плавления: 175-178°С (тетрагидрата — 105°C). Плохо растворим в воде
(1:1320), в этиловом спирте — 1:3, в хлороформе — 1:5, диэтиловый эфир — 1:187.
2.2. Биологические поражающие агенты
409
Бруцин возбуждает центральную нервную систему, вызывает судороги двигательной
мускулатуры конечностей, шеи и лица, которые усиливаются при действии звука и
света. По фармакологическому действию бруцин напоминает стрихнин, но менее
ядовит. Смертельная доза для человека при приеме внутрь составляет 0,1—0,3 г.
При введении яда чилибухи теплокровным животным, они испытывают все бо-
лее и более сильное беспокойство, делаются в высшей степени чувствительными ко
всякому стуку, прикасанию и т. д., внезапно впадают в мышечный столбняк, голова
запрокидывается назад, и туловище изгибается дугой кпереди благодаря столбняку как
затылочных, так и спинных мышц. Такие приступы судорог повторяются периодиче-
ски, перерываясь интервалами расслабления мышц. В одном из таких приступов жи-
вотные погибают от удушья, наступающего из-за неподвижного положения судорожно
сокращенной диафрагмы и грудной клетки, находящихся в фазе вдоха. Клинические
наблюдения показывают сходную картину отравления у человека, причем лицо в
момент задержки дыхания в фазе вдоха делается багровым, глазные яблоки выпячи-
ваются из орбит. Отравленные стрихнином люди испытывают чувство ползания му-
рашек, болезненные подергивания, оцепенение, высокую чувствительность к свету,
звуку, ощущение тяжести, боли, мышечного напряжения и страха как бы перед гро-
зящим задушением, сопровождающимся сильным цианозом лица. Во время присту-
па судорог глазные яблоки неподвижны, зрачки расширены, шейные вены набухают,
рот плотно сомкнут, и тело, согнутое дугою и опирающееся только на затылок и пят-
ки, от времени до времени подбрасывается при усилении судорог. В одном из таких
приступов человек погибает. В интервалы, свободные от судорог, функции головного
мозга не представляют никаких значительных расстройств ни в сфере чувствований,
мышления и воли, откуда легко заключить, что действие стрихнина направлено не на
головной, а спинной мозг. Кроме двигательных центров спинного мозга, стрихнин
повышает возбудимость центров продолговатого мозга, а именно дыхательных и со-
судодвигательных центров, что выражается учащением дыхательных движений и по-
вышением давления крови вследствие всеобщего спазма сосудов. Так как мышечные
сокращения сопровождаются развитием тепла, то стрихнинные судороги влекут за
собою резкое повышение температуры тела, доходящее у собаки до 44,8 °C.
2.2.3.3. Микотоксины
Классификация грибов. Роль отдельных токсических веществ несовершенных грибов
(микотоксинов) в формировании клинической картины отравления. Фузариотоксины
(трихотецены, зеараленон, монилиформин). Афлатоксины. Миротециотоксины (дондро-
дохины). Спородесмины. Эрготоксины. Охратоксины. Микотоксины грибов рода
Pemcilhum. Патогенез и патологической анатомия микотоксикозов.
Грибы (Fungi или Mycetes), обособленная группа (отдел) низших растений, лишен-
ных хлорофилла. Грибам присущи признаки как растений (неподвижность, неогра
ниченный верхушечный рост, способность к синтезу витаминов, наличие клеточных
стенок), так и животных (тип питания, наличие хитина в клеточных стенках, запас-
ных углеводов в форме гликогена, образование мочевины, структура цитохромов). Для
своего развития грибы нуждаются в готовых органических веществах, т. е. являются
гетеротрофными организмами. Они обычно поселяются на растениях, животных или
их остатках. В зависимости от питательного субстрата (живого или мертвого) грибы
подразделяются на паразитов и сапрофитов. Насчитывается свыше 100 тыс. видов
грибов. Наука о грибах называется микологией. Большинство грибов (за исключени-
410
Часть 2. Общая неправильная эпидемиология
ем некоторых форм внутриклеточных паразитов) имеют вегетативное тело в виде
мицелия (грибницы), т. е. оно состоит из тонких ветвящихся нитей, или гиф, расту-
щих своими концами и распространяющихся на поверхности или внутри питатель-
ного субстрата. Развивающаяся грибница часто имеет вид нежного, пушистого или
паутинистого налета, тонких пленок или ватообразных скоплений; может изменять-
ся в зависимости от условий развития.
Классификация грибов. Грибы классифицируются на четыре класса (подробную
таксономическую классификацию грибов см. в работе Петровича С. В., 1991).
I класс. Низшие грибы, или фикомицеты (Phycomycetes). Мицелий неклеточный
или с небольшим количеством перегородок, у наиболее примитивных в виде голого
комочка протоплазмы — амебоида или в виде одной клетки с ризоидами.
II класс. Сумчатые грибы, или аскомицеты (Ascomycetes). Мицелий многоклеточ-
ный, гаплоидный, на нем развиваются конидиальные спороношения. Характерно
образование сумок с аскоспорами — основных органов размножения.
III класс. Базидиомицеты (Basidiomycetes). Мицелий многоклеточный, как пра-
вило, дикариотический; конидиальные (гаплоидные) стадии редки. Характерно об-
разование базидий, несущих на стеригмах базидиоспоры. Включает подавляющее
большинство употребляемых человеком в пищу, а также ядовитых грибов и многих
паразитов злаков и др. культурных и диких растений.
IV класс. Несовершенные грибы (Deuteromycetes или Fungi imperfect! родов Fusa-
rium, Aspergillus, Myrothecium, Stachybotrys, Trichoderma, Trichothecium, Penicillium и
др.). Мицелий многоклеточный, размножение только бесполое — конидиями. Поло-
вого размножения не имеют (не образуют ни сумок, ни базидий). Один из порядков
этих грибов (Mycelia sterilia) характеризуется полным отсутствием каких-либо спор, в
том числе и конидий.
Роль отдельных токсических веществ несовершенных грибов (микотоксинов) в фор-
мировании клинической картины отравления. Микотоксины трудно классифицировать
по токсическому действию из-за огромного количества их самих, их химических струк-
тур, вариантов биосинтетического происхождения, и разнообразных биологических
эффектов, регистрируемых после их введения млекопитающим. Клиницисты клас-
сифицируют микотоксины в зависимости от клиники, преобладающей в развитии
токсикоза и связанной с наиболее пораженным органом (гепатотоксины, нефроток-
сины, нейротоксины, иммунотоксины и т. д.), подробнее о таком делении микоток-
сикозов см. ниже «Патогенез и патологическая анатомия микотоксикозов». Кле-
точные биологи делят микотоксины на общие классы в зависимости от эффекта, ока-
зываемого ими на наследственность позвоночных организмов: тератогены, мутагены,
карциногены и др. Химики-органики классифицирует микотоксины по химической
структуре (например, лактоны, кумарины и др.). Биохимики — в соответствии с био-
синтетическим происхождением (поликетиды, производные аминокислот и т. д.);
микологи по продуцентам (например, токсины Aspergillus, токсины Penicillium). Поэто-
му в данном разделе не будет приведено их непротиворечивой классификации.
Мы рассмотрим характеристику и механизм действия основных токсических
метаболитов, вызывающих микотоксикозы, только как введение в этот класс соеди-
нений, крайне опасных с точки зрения их умышленного применения для пораже-
ния людей. В качестве основных источников информации использованы работы
Б. Н. Орлова с соавт. (1990), С. В. Петровича (1991), М. Peraicaetal. (1999), J.W. Bennett
и М. Klich (2003).
2.2. Биологические поражающие агенты
411
Большинство микотексинов попадает в организм людей и животных путем вса-
сывания из ЖКТ. Затем токсичные метаболиты через воротную вену проникают в
печень, где происходит процесс их детоксикации. Продукты метаболизма микоток-
синов с желчью выводятся в экскременты или через почки в мочу, а далее — из орга-
низма. Весь сложный процесс биотрансформации микотоксинов в организме бази-
руется на двух основных этапах — метаболи (ации и конъюгации. В процессе метабо-
лизации микотоксины подвергаются гидролиз.,, окислению, восстановлению и другим
различным реакциям. В результате метабо гизации появляются новые функциональ-
ные группировки, являющиеся активными центрами конъюгации. В процессе конъ-
югации метаболиты микотоксинов соединяются с различными веществами (амино-
кислотами, серной, глюкороновой кислотами и др.), что блокирует функциональные
группы -СООН и -ОН и приводит к снижению токсичности. Однако в процессе ме-
таболизации образуются и более токсичные вторичные продукты, чем исходный ми-
котоксин. Это явление называется метаболической активацией. При значительных
количествах микотоксина, поступившего в организм, защитные силы не в состоянии
провести его детоксикацию. Токсические характеристики отдельных микотоксинов
приведены в табл. 2.14.
Таблица 2.14
Токсические характеристики отдельных микотоксинов*
Opi анизм-продуценты Микотоксин Природный субстрат Характер токсического действия
Грибы из рода Aspergillus (A. flavus, A. parasiticus) Афлатоксины Bl, B2, G1,G2 Арахис, кукуруза и др. зернобобовые (семена), семя хлопчатника, орехи, овощи, расти- тельные корма Гепатотоксическое и гепатокарциноген- ное, мутагенное и иммунодепрессивное
Грибы рода Aspergillus и Peni- cillium, способность к охрато- ксигенезу наиболее выражена у A. ochraceum и Р. viridicatum Охратоксины А, В и С Зерновые, кофе, сыры, корма Нефротоксическое, тератогенное
В основном гриб Р. expansum. Однако и другие виды мик- ромицетов из родов Penicil- lium и Aspergillus могут выделять этот метаболит Патулин Фрукты, овощи и продукты их переработки (соки, пюре, джемы и др.) Нейротоксическое, мутагенное, тератогенное
Грибы из родов Fusarium, Trichoderma, Trichothecium, Myrothecium и Stachybotrys Трихотененовые микотоксины (более 40) Зерновые, корма, сено и т. п. Нейротоксическое, геморрагическое, лейкопеническое, иммунодепрессивное, дерматотокс ическое
F. graminearum Зеараленон Кукуруза, ячмень, пшеница, сорго, корма Эстрогенное, тератогенное
Claviceps purpurea Эрготоксины Зерновые Нейротоксическое
* По Б. Н. Орлову и др. (1990).
Токсины грибов могут накапливаться в мицелии, органах спороношения и пита-
тельном субстрате. Условия, оптимальные для роста и токсинообразования, для раз-
ных грибов различны. Условия максимального синтеза ряда токсинов не совпадают с
условиями, обеспечивающими максимальный рост мицелия.
412
Часть 2. Общая неправильная эпидемиология
Фузариотоксины (трихотецены, зеараленон, монилиформин). Грибы из рода Fusarium
продуцируют более 40 трихотепеновых токсинов и зеараленон. Трихотецены также
продуцируют грибы родов Trichoderma, Trichothecium, Myrothecium и Stachybotrys.
Термин «трихотецетен» («trichothecene») является производным от трихотецина (tri-
chothecin) — первого идентифицированного члена этого семейства микотоксинов. На
начало XXI в. обнаружено более 200 трихотеценов различного происхождения, но
только некоторые из них известны как контаминанты пищи и фуража (Kimura М. et
al., 2007). По химической структуре они разделены на четыре группы (А-D) (рис. 2.32).
«скелет» имеет пять позиций, в которых могут быть добавлены различные группы
(С-3, С-4, С-15 и С-8). Трихотецены делятся на группы: А — отдельные замены в
С-8, тип В — кетогруппа в С-8, тип С — эпоксидная группа в С-7 и С-8 и тип
D — макроциклическое кольцо между С-4 и С-15. По М. Kimura et al. (2007)
2.2. Биологические поражающие агенты
413
Наиболее часто в природе встречаются Т-2 токсин, дезоксиниванол, ниваленол и
диацетоксискирпенол. Физико-химические свойства отдельных трихотеценов пред-
ставлены в табл. 2.15.
Таблица 2.15
Физико-химические свойства трихотеиенов
Токсин Молекулярная формула ММ Точка плавления,°C
Т-2 466 150 151
НТ-2 С22Н32О8 424 —
Неосоланиол ^19^26^8 382 150-151
Диацетоксиск ирпенол с, 9н26о7 366 162-163
Ниваленол ^15^20®7 312 222-223
Фузаренон-Х ^17^22^8 354 91-92
Дезоксиниваленол С)5Н20О9 296 131-153
С учетом химической формулы обширную группу трихотеценовых микотоксинов
подразделяют на две группы. Группа А включает Т-2, НТ-2, неосоланиол, Т-2 тетра-
ол, изоноосолениол, скипентриол, диацетотоксискирпенол, моноацетоксискирпенол
и др.; группа В — ниваленол, дезоксиниваленол (вомитоксин), фузаренон-Х, нива-
ленолдиацетат, моноацетоксидезоксини валенол.
Большинство штаммов грибов, продуцирующих трихотецены, вырабатывают не-
сколько токсинов. Например, гриб F. sporotrichiella вырабатывает Т-2, НТ-2, Т-1 -ток-
сины, неосоланиол, Т-2-триол и др. Как правило, продуценты трихотеценов группы А
не способны продуцировать токсины группы В (табл. 2.16).
Таблица 2.16
Продуцирование некоторых трихотеценов различными видами Fusarium
Вид гриба Трихотеценовые микотоксины
Тип A Тип В
Т-2-ток- син Неосола- ниол Диацетокси- скирпенол Ниваленол Фузаренон-Х Дезокси- ниваленол (вомитоксин)
F. tridmium + + + — — —
F. sporotrichioides + + + — — —
F. роае + + + — — —
F. acuminatum + + — — — —
F. graminearum — — — + + +
F. nivale — — — + + —
F. lateritium + + + — —
F. equiset i — + 4- + + —
F. semitedum - + + + + -
«+» — продукция токсина установлена; «—» — не установлена.
В механизме действия этой группы токсичных метаболитов грибов прослежива-
ются два основных пути. Трихотецены с выраженным токсическим действием (Т-2,
нипаленол, веррукарин А и др.) ингибируют процесс инициации трансляции, т. е.
414
Часть 2. Общая неправильная эпидемиология
включаются на этапе перед образованием комплекса между рибосомой, информаци-
онной РНК, метионил-тРНК, включая пептидилтрансферазу, необходимую для об-
разования первой пептидной связи. Другие трихотецены (триходермол, крптоцин,
крихотецин, протокол и др.) ингибируют связывание тРНК с рибосомами или не
допускают освобождения полипептидов от рибосом и комплексов и РНК-рибосомы.
Трихотеценовые микотоксины не только ингибируют синтез белка, но и активность
пептидилтрансфераз путем конкуренции за места связывания на рибосомах. Отдель-
ные трихотецены могут ингибировать синтез ДНК и оказывать повреждающее дей-
ствие на лизосомы клеток кроветворных и иммунокомпетентных органов. Общими
проявлениями алиментарного отравления трихоцетенами у людей обычно бывает
подавление иммунных ответов, тошнота и иногда рвота (табл. 2.17).
В регионах, в которых встречаются такие микотоксикозы среди людей, 5-40 %
проб зерна содержат F. sporotrichoides. В регионах, где такие микотоксикозы не встре-
чаются, F. sporotrichoides находят в 2—8 % проб зерна. Тяжелые вспышки трихотеце-
новых микотоксикозов обычно связаны с длительным употреблением загрязненного
микотоксинами зерна. Трихотиценовые микотоксины находят в пробах воздуха, ото-
бранных из вентиляционных систем частных домов и офисных сооружений, на сте-
нах домов, эксплуатирующихся в условиях высокой влажности.
Наибольшей токсичностью из т рихотеценовых микотоксинов обладает веррука-
рин J (vemicarin J). В опытах по острой токсичности на мышах его средняя летальная
доза 0,5 мг/кг при внутривенном введении. Средняя летальная доза (LD50) Т-2 ток-
сина и ниваленола для мышей 5,2 и 4,1 мг/кг (внутрибрюшинно) соответственно.
В условьях in vivo токсичность Т-2 токсина и ниваленола приблизительно в 10 раз
выше, чем у деоксиниваленола (Thompson W. L., Wannemacher R. W., 1986).
В экспериментах на животных было показано, что ингалированные трихотетеце-
ны более чем в 40 раз токсичнее, чем те же трихотецены, введенные орально (Smora-
giewicz W. et al., 1993).
Стахиботриотоксины. Группа токсических метаболитов, относится к трихотеце-
нам. Имеют сходство с сердечными ядами растительного и животного происхожде-
ния. В последнее время расшифрована природа нескольких микотоксинов, продуци-
руемых грибом Stachybotrys alternans', роридин Е, сатратоксин Н, сатратоксин D,
сатратоксин С, сатратоксин Е веррукарин А. Различные штаммы продуцируют не-
одинаковые токсины. Стахиботриотоксины устойчивы к воздействию высоких темпе-
ратур, действию света, ультрафиолетовых лучей и минеральных органических кислот.
При воздействии температуры 120°С в течение 2 ч токсичность стахиботриотоксинов
практически не снижалась. В то же время стахиботриотоксины очень чувствительны
к воздействию щелочей. Присутствие Stachybotrys в помещениях ассоциировалось с
легочными кровотечениями у детей.
Зеараленон, или F-2 токсин, является лактоном резорциловой кислоты (6-[ 10- hyd-
roxy-6-oxo-trans-l-undecenyl]-B-resorcyclic acid lactone). Многочисленные про-
изводные зеараленона (дегидрозеараленон, зеараленон, зеарален, деметилзеарален
и др.) — продукты метаболизма многих грибов из рода Fusarium. Зеараленон отлича-
ется от других микотоксинов наличием гормоноподобных свойств и отсутствием
острого токсического действия, приводящего к летальному исходу. Он и его метабо-
литы конкурентно взаимодействуют с эстрадиолсвязывающими рецепторами в клет-
ках, они также стимулируют синтез РНК, ДНК и белка в матке и молочной железе.
LD50 для морских свинок составляет 5 тыс. мг на 1 кг массы тела, для крыс — более
10 тыс. мг/кг массы тела, для цыплят более 15 тыс. мг/кг.
Таблица 2.17
Вспышки трихотеценовых микотоксикозов с оральной экспозицией*
Страна Год Количе- ство пост- радавших/ экспониро- ванных Клинические симптомы и проявления Начальные симптомы/ продолжи- тельность Источник токсина Вид грибов Токсин
СССР 1932-1947 _а «Алиментарная токсическая лейкопения»; гиперемия слизистой поверхности ротовой полос! и и фарингит, гастриты, гастроэнтериты, обильное слюиовыделение. боли в эпигастраль- ной области и по ходу пищевода Несколько ч/2-З сут Зерно Fusarium sporotricho- ides -
общее недомогание, головокружение, ощущение неприятного вкуса во рту, прогрессирующая лейкопения, гранулоцитопеьия и лимфоцитоз 3- 4 нед - - -
геморрагический диатез, ангина, петехиальная сыпь, катаральная дифтерия, гангренозные фарингиты, язвенные и гангоренозные ларингиты, афония, асфиксия Несколько сут/2 нед - - —
Япония 1965 25; 0ь «Токсикоз плохого зерна»: тошнота, рвота, сонливое состояние - Рис F. roseum, F. nivele -
Китай 1961-1985 7818/°; 0 «Токсикоз плохого зерна»: рвота, абдоминальные боли, диарея, головокружение, головная боль * Пшеница, кукуруза Fusarium spp. Диоксиниваленол, зераленон
Китай 1984-1985 463/600 0 Тошнота, рвота, боли в животе, диарея, головокружение, головные боли 5—30 мин Кукуруза, пшеница Fusarium spp. Диоксиниваленол, зераленон
Индия 1987 97/224;0 Слабые абдоминальные боли, ощущение сыто- сти, раздражение гортани, диарея, кровь в стуле 15 мин — 1 ч/1 -2 сут Пшеница Fltsarium spp., Aspergillus flavus Ниваленол, деокси- ниваленол, Т-2, аце- тилдеоксиниваленол
Китай11 97/165;0 Тошнота, рвота, озноб, абдоминальные боли, стеснение в груди, диарея До 30 мин Рис F. heterospo- rum, F. gra- minearum Т-2
Примечание. а — заболели десятки тысяч человек со смертностью до 60 %;ь — выделенные цифры означают количество погибших лю-
дей; с — количество экспонированных к токсину людей не приведено; d — в первоисточнике год вспышки не указан.
* По М. Peraica et al. (1999).
2.2. Биологические поражающие агенты 415
416
Часть 2. Общая неправильная эпидемиология
Монилиформин — токсичный метаболит, в основном продуцируемый грибами
F. moniliforme, F. oxyporum, F. acuminatum и др. LD50 монилиформина составляет для
однодневных цыплят и 5—7-дневных утят — 50 мг/кг массы тела; для куриных эмб-
рионов LD50 — 2,8 мкг на яйцо. Кроме описанных микотоксинов, грибы из рода
Fusarium продуцируют фузариоципы А и С, бутенолид.
Афлатоксины. Токсические метаболиты, образуемые грибами из рода Aspergillus
(A.flavus, A. parasiticus). Грибы образуют эндо- и экзотоксины. Афлатоксины отно-
сятся к классу вторичных метаболитов. Начало синтеза афлатоксинов в культуре гри-
ба совпадает с окончанием периода активного роста мицелия и началом образования
грибных спор. Обычно в растущей культуре афлатоксины начинают обнаруживаться
на 2—3 сутки инкубации. В процессе инкубации гриба количество афлатоксинов в
культуре достигает максимума, после чего снижается. При этом время достижения
максимального количества афлатоксинов и их общее количество в значительной сте-
пени зависят от количества спорового инокулята. Интенсивность образования ток-
синов находится в зависимости от штамма, состава питательной среды и температу-
ры. Являются безазотистыми веществами, имеют метоксисоединения, несколько кар-
бонилов и лактоновое кольцо. Это производные дегидрофурана. Известно четыре
основных представителя афлатоксинов — Bj В2, G(, G2h более десяти соединений,
производных основной группы. Четыре основных афлатоксина могут быть одновре-
менно образованы одной культурой. Большинство грибных штаммов, которые про-
дуцируют афлатоксин G1, также вырабатывают афлатоксин В1. Наиболее токсичен
В( который нарушает метаболические функции печени, что проявляется в уменьше-
нии содержания гликогена и витамина А. Афлатоксины слаборастворимы в воде, но
хорошо растворяются в хлороформе, этаноле. В водных растворах легко разрушают-
ся на свету. Вместе с тем афлатоксины устойчивы к воздействиям высоких темпера-
тур. Нагревание до температуры 160°С не приводит к снижению токсичности. Тем-
пература плавления афлатоксинов находится в пределах 237-289°С. В ультрафиоле-
товых лучах афлатоксины флюоресцируют зеленым и голубым цветом. Токсические
вещества чувствительны к воздействию кислот, щелочей, ультрафиолетовых лучей,
которые инактивируют афлатоксины.
В основе действия афлатоксинов лежит взаимодействие с ДНК, блокирование
синтеза ДНК и ДНК-зависимого синтеза РНК. Наряду с этим афлатоксины блоки-
руют процесс терминации синтеза пептидной цепи, нарушают движение рибосом
вдоль информационной РНК и процесс их освобождения. Кроме того, афлатоксины
оказывают повреждающее действие на структурные элементы клетки: мембрану, ми-
тохондрии, рибосомы, эндоплазматический ретикулум и ядра.
При развитии гриба-продуцента афлатоксины способны накапливаться в кон-
центрациях, достигающих нескольких мг на 1 кг продукта. Наиболее часто встречают-
ся в арахисе, продуктах переработки хлопка (масло, шрот), кукурузе, различных оре-
хах, реже в зерне хлебных злаков и др. растительных объектах. При концентрации
0,25— 1 мг на 1 кг продукта вызывают афлатоксикозы. Допустимые концентрации аф-
латоксинов в продуктах питания и кормах для животных в большинстве стран огра-
ничены в законодательном порядке в пределах 5—200 мкг/кг.
Острые токсические случаи, вызванные афлатоксинами, среди людей наблюда-
ются редко, но могут приобретать масштабный характер (табл. 2.18).
В 1974 г. в Индии погибло от острого токсического гепатита почти 100 человек,
питавшихся кукурузой, сильно загрязненной афлатоксинами. По ретроспективным
оценкам, отдельные взрослые индийцы потребляли в день от 2 до 6 мг афлатоксина.
Таблица 2.18
Вспышки афлатоксикозов*
Страна Коли- чество случаев Клинические симптомы и проявления Экспозиция Образец Токсин Гистопатология печени
источник продолжи- тельность токсин
Уганда 1;1я Абдоминальные боли, отеки ног, пальпируемая печень, на ЭКГ пролонгация Р-R интер- вала, блокада правой ножки пучка Маниока 5-30 сут Афлатоксин — — Центрилобулярный некроз, полиморфонуклеарная ин фильтрация и фибрин в си- нусоидах, жировые измене- ния
Индия 397; 106 Эпизодически повышение температуры до фибрильной, рвота, анорексия, желтуха, асциты, отеки ног, массивные кровотечения в ЖКТ Маис Несколько недель Афлатоксин В, (5/5)” Сыворотка Афлатоксин В1 (2/7) Пролиферация желчного про- тока с перидукальным фиб- розом многоядерными гигант- скими клетками, пенящаяся цитоплазма, стаз желчи, рас- ширение желчных канальцев
Моча Афлатоксин В, (0/7)
(7/70)с — —
Индия 994; 97 Лихорадка, рвота, отеки ног, желтуха, спленомегалия, асцит, гепатомегалия Маис - Афлатоксин Bt(13/14) — — Холангголярная пролифера- ция, перивенозный колла- геноз, ограниченная олите- рация, экстенсивный фиброз, трансформация гепатоцитов в гигантские клетки
Кения 20; 12 Эпизодическое повышение температуры, рвота, абдоми- нальный дискомфорт, ано- рексия, желтуха, отеки ног, асцит, тахикардия, болезнен- ность печени, «черный стул», кровотечения в ЖКТ Маис Несколько недель Афлатоксин В, (2/2) Афлатоксин В2(2/2) Печень (аутопсия) Афлатоксин В,(2/2) Явный ценрилобулярный некроз, невыраженная жировая инфильтрация
США 1; 0 Незудящая макулярная сыпь, тошнота, головная боль Очищенный афлатоксин В, 2 сут Афлатоксин В,, 5,5 мгй - - Без изменений
Примечание.а — выделенные цифры означают количество смертельных исходов; ° — цифры в круглых скобках показывают количество
позитивных анализов из общего числа проведенных;с — образцы кукурузы взяты у поваженных микотоксинами семейств через год после
вспышки афлатоксикоза; d — общая доза.
* По М. Peraica et al. (1999).
2.2. Биологические поражающие агенты
418
Часть 2. Общая неправильная эпидемиология
В последующем была подсчитана острая летальная доза (peros) афлатоксина для взрос-
лого человека — от 10 до 20 мг (Pitt J. L, 2000).
Экспозиция к афлатоксинам, содержащимся в употребляемой человеком пище,
является доказанным фактором риска в развитии первичной гепатоцеллюлярной кар-
циномы. Риск развития рака печени многократно увеличивается в случае гепатита,
вызванного вирусом гепатита В. В ингаляционных экспериментах на животных до-
казано, что афлатоксины являются легочными карциногенами. В специальной лите-
ратуре афлатоксины рассматриваются в качестве биотеррористических агентов (см.
Bennett J. W, Klich М., 2004).
Миротециотоксины (дондродохины) Токсические метаболиты, выделяемые гриба-
ми Myrothecium verrucaria, М. roridum и М. leucotrichum. Дифференцированы два веще-
ства, выделяемые этими грибами: веррукарин А и роредин А. Полученные в чистом
виде, они представляют собой кристаллический порошок или желтое масло. Эти ве-
щества обладают выраженным дермацидным действием. Токсичные метаболиты хо-
рошо растворяются в хлороформе, бензине, эфире, этаноле, метаноле и хуже в воде и
четыреххлористом углероде, оказывают токсическое действие на культуры клеток.
Спородесмины. Токсические метаболиты, образуемые грибом Pithomyces chartarum,
вызывающие питомицестоксикоз у овец и крупного рогатого скота. В (истом виде
выделены споро, (есмин, спородесмин В, Е, G и Н. Спородесмин генерирует суперок-
сидные радикалы, обладает выраженными цитологическими свойствами и вызывает
дегенерацию культур клеток. Сдержится в основном в спорах гриба. Из лабораторных
животных к спородесмину наиболее чувствительны кролики и морские свинки.
В условиях эксперимента введение овцам спородесмина в дозе 1 мг/кг массы гела при-
вело к гибели 90 % животных.
Эрготоксины. Токсические метаболиты, образуемые грибами Claviceps purpurea и
Cl. paspali. Представлены двумя видами клавиналкалоидов и пептидными алкалоида-
ми. В табл. 2.19 перечислены алкалоиды спорыньи.
Алкалоиды спорыньи и их производные отличаются фармакологическими, ток-
сическими и другими свойствами. Одни из них (эрготоксин, эрготамин, эргозин)
суживают зрачок и кровеносные сосуды, парализуют двигательные симпатические
нервы, другие (эргометрин) — вызывают сильное сокращение мускулатуры матки.
В фармакологическом плане прослеживаются два основных действия эрготоксинов:
сокращение мускулатуры, особенно матки у беременных животных, и специфичес-
кое блокирование симпатической нервной системы.
Полный синтез лизергиновой кислоты осуществлен Р. Вудвордом в 1954 г. Диэти-
ламид лизергиновой кислоты, известный под названием ЛСД, впервые был получен
в 1938 г. А Гофмаоном (Швейцария). ЛСД, попадая в организм, является конкурент-
ным антагонистом серотонина — одного из регуляторов центральной нервной систе-
мы. Обладает сильным галлюциногенным действием. Психомиметическое действие
проявляет при попадании его в ЖКТ. при вдыхании аэрозолей, при проникновении
в кровь через раны и при всасывании через кожу Рассматривается военными специ-
алистами в качестве диверсионного агента.
Гриб С. paspali паразитирует на злаковых растениях и имеет тот же цикл развития,
что и спорынья. Токсические вещества (треморгеновые микотоксины и алкалоиды)
содержатся в склероциях гриба (покоящаяся стадия гриба, образующаяся в неблаго-
приятных условиях) и вместе с ними они попадают в пищу животного или человека.
Отравление ими животных известно ветеринарам под названием клавицепстоксико-
за. Основные характерные симптомы отравления, отмечаемые у всех видов животных
2.2. Биологические поражающие агенты
419
Таблица 2.19
алкалоиды спорыньи
Алкалоиды лизергиновой и изолизергиновой кислот Клавиновые алкалоиды
Эргин Агроклавин
Эргинин Секаклавин ,ханоклавин-1)
Эргокорнин Хиноклавин-П
Эргокорниньн Косгаклавин
Эргокристин Циклоклавин
Эргокристинин Д игидролозергол-1
а-, р-эргокриптины Элимоклавйн
Р-эргокриптины Элимоклавин-О-Р-Д-фруктозид
Эргометрин, эргометринин Фестуклавин
Эргосекалин, эргосекалинин Фумигаклавины А и В
Эргозин, эргозинин Изоханоклавин-1
Эргостин, эргостинин Изолизергол
Эрготамин, эрготаминин Изоленниклавин, пенниклавин
Лизергиновая кис юта Изосетоклавин
Изолизергиновая кислота Лизерген, лизергин, лизергол
Лизергид Гетоклавин, норсетоклавин
и птицы: повышенная рефлекторная возбудимость, судороги и параличи. От эрготизма
клавипепстоксикоз отличается отсутствием гангренозной формы оолезни.
Охратоксины. Токсические метаболиты, продуцируемые различными грибами
Aspergillus и Penicillium, однако способность к охратоксигенезу наиболее выражена у
A. ochraceum и Р. viridicatum. Эти микотоксины являются изокумаринами, связанны-
ми амидами с L-фенилаланином. В кормах животных обычно встречаются охраток-
син А и редко охратоксин В. В лабораторных условиях выделены охратоксин С, ме-
тиловый эфир охратоксина А, метиловый и этиловый эфиры охратоксина В. При
ферментативном и кислотном гидролизе образуются охратоксин А и L-фенилаланин.
Охрзтоксины термостабильны, и их детоксикация представляет определенные труд-
ности. Охратоксины служат конкурентным ингибитором фенилаланин-тРН К-синте-
тазы и подавляют синтез полипептидной цепи на уровне трансляции. Почки являются
основным органом-мишенью для охратоксинов. В почках охратоксин А подавляет
глюконеогенез путем ингибирования активности фосфоснолпируват карбоксилазы.
Охратоксин А обладает нефротоксическим действием в отношении всех исследован-
ных видов животных и, скорее всего, таким же образом действует на почки людей.
Кроме нефротоксического действия, охратоксин Атоксичен для печени, он подавляет
иммунную систему, и обладает тератогенным и карционогенным действием (Kuiper-
Goodman Т., Scott Р. М., 1989). Охратоксины имеют отношение к болезни людей, из-
вестной как эндемическая балканская нефропатия. Обычно это прогрессирующие
хронические нефриты, встречающиеся в популяциях людей, проживающих в долине
реки Дунай, на территориях Румынии, Болгарии и бывшей Югославии.
Микотоксины грибов рода Penicillium. Гриб Р. islandicum продуцирует более 20 ток-
сических метаболитов, из которых наиболее часто встречаются лютеоскирин и цик-
лохлоротин.
420
Часть 2. Общая неправильная эпидемиология
Лютеоскирин представляет собой желтые кристаллы с ММ 574 Да и точкой плав-
ления 287 °C. Микотоксин нерастворим в воде, но хорошо растворяется во многих
органических соединениях. Подавляет синтез информационной РНК путем взаимо-
действия с ДНК, нарушает функции митохондрий клеток печени, что приводит к
накоплению в них триглицеридов. Интоксикация лютеоскирином приводит к обра-
зованию некрозов и дегенерации печени.
Циклохлоротин — хлорсодержащий пептид с М М 537 Да. Точка плавления состав-
ляет 251 °C. Циклохлоротин хорошо растворяется в воде и С-бутаноле. Токсическое
действие проявляется в поражении печени и канцерогенной активности. Характери-
зуется высокой токсичностью.
Цитреовиридин — токсичный метаболит гриба Р. citreoviride. Это карбоцикличе-
ский полиен с точкой плавления 107°С. Метаболит не растворяется в воде, но хорошо
растворим в этаноле, эфире, бензоле, ацетоне, хлороформе. Характеризуется выражен-
ным нейротоксическим действием, сопровождающимся нарушениями центральной
нервной системы, дыхательной, сердечно-сосудистой систем, развитием параличей
Цитринин продуцируется Р. citrinum, Р. purpurogenum и другими грибами из рода
Penicillium. Точка плавления 175°С. Хорошо растворяется в большинстве органиче-
ских растворителей. Характеризуется тропизмом в отношении почек, вызывая деге-
неративные изменения почечных канальцев у всех животных, в опытах на которых
исследовалась его нефротоксичность. LD50 цитрина для уток — 57 мг/кг; для цып-
лят — 95 мг/кг; для кроликов — 134 мг/кг. Цитрин действует синергистически с охра-
токсином А, подавляя синтез РНК в почках мышей.
РубратоксинА и рубратоксин В — высокотоксичные метаболиты, продуцируются
грибами Р. nibrum и Р. purpurogenum. Точка плавления для рубратоксина А составляет
210-214°С, для рубратоксина В 168-170°С. Рубратоксин В оказывает действие на
митохондрии, подавляя активность АТ-фазы и блокируя электронно-транспортную
цепь. Кроме того, рубратоксин ингибирует активность микросомальных оксидаз
Рубратоксины обладают выраженным гепатоксическим, мутагенным и тератогенным
действием. LD50 при внутрибрюшинном введении мышам составляет для рубраток-
сина А — 6,6 мг/кг, для рубратоксина В — 3,0 мг/кг.
Патулин продуцируется в основном грибом Р. expansum. Однако и некоторые лр> -
гие виды микромицетов из родов Penicillium и Aspergillus могут выделять этот метабо-
лит. Патулин хорошо растворим в воде и органических растворителях. Токсичный
метаболит обычно обнаруживается в подгнивших фруктах, ягодах и овощах. Кожа
кролика чувствительна к патулину и позволяет обнаружить его в количестве 500 мкг
Минимальная концентрация токсина, вызывающая гибель рыб гуппи, составляет
5,0 мкг/мл. При концентрации 25 мкг/мл отмечается 100 %-ная гибель рыб. LD50 для
белых мышей при однократном введении per os достигает 35,9 мг/кг живой массы.
Патогенез и патологическая анатомия микотоксикозов1. Токсические эффекты ми-
котоксинов изучены в основном на примерах из ветеринарной практики. Патогенез
микотоксикозов определяется действием продуктов метаболизма грибов, динамикой
развития патологического процесса и патологоанатомическими изменениями в раз-
личных органах и тканях организма. В патогенезе заболевания важную роль играют
не только токсичные метаболиты, но и сам гриб. Гистологические исследования при
стахиботриотоксикозе часто обнаруживали элементы гриба в различных органах
1 Описание патогенеза и патанатомии микотоксикозов подготовлено по работе С. П. Пет-
ровича (1991).
2.2. Биологические поражающие агенты
421
и тканях. Токсические продукты гриба, поступая в организм энтерально, вызывают
первичные воспалительно-некротические изменения и создают благоприятные усло-
вия для проникновения элементов гриба в пораженные участки ЖКТ и далее в кровь.
В легкие элементы гриба могут попасть не только с током крови, но и ингаляционным
путем при вдыхании живот ным спор гриба.
При стахиботриотоксикозе и фузариотоксикозе, главенствующим началом в раз-
витии патологического процесса являются токсические вещества. В отношении же
других грибов нельзя высказаться так категорично. Например, развитие патологиче-
ского процесса при аспергиллотоксикозе ооусловливается токсичными веществами.
Однако грибы A.flavus и A.fumigatus могут быть не только продуцентами микотокси-
нов, но и в определенных случаях, обладая выраженными вирулентными свойства-
ми, вызывать развитие типичных микозов.
Микотоксины обладают местным и общим действием. Местное действие прояв-
ляется в воспалительно-некротических изменениях кожи губ, слизистой ротовой
полости и других отделов пищеварительного тракта. Наиболее ярко эти явления вы-
ражены при стахиботриотоксикозе и фузариотоксикозе. При афлатоксикозе местное
действие микотоксинов слабее и проявляется воспалением слизистой оболочки ды-
хательных путей и конъюнктивитами. Всасываясь в кровь и лимфу, токсичные ме-
таболиты грибов разносятся по всему организму. Резорбтивное действие токсинов
многообразно и выражается в развитии геморрагического диатеза, патологических
процессов в различных органах и тканях, расстройстве нервной системы, нарушении
деятельности пищеварительной, сердечно-сосудистой и других систем. Это приводит
к выраженному нарушению обмена веществ В результате снижается общая реактив-
ность. Естественные защитные силы организма уже не могут противостоять внедре-
нию и развитию различной микрофлоры. Резорбтивное действие микотоксинов про-
является угнетением клеточных и гуморальных факторов иммунитета. Введение в
организм животного микотоксинов приводит к снижению антителообразования, на-
рушению фагоцитоза. У животных, вакцинированных против бактериальных и вирус-
ных инфекций на фоне микотоксикозов, отмечают прорывы иммунитета.
Резорбтивное действие микотоксина сказывается на поражении различных орга-
нов и систем организма. При стахиботриотоксикозе отмечают изменения в нервной
системе, резкие нарушения в картине крови и развитие множественных некрозов,
являющихся одним из основных патологических процессов. Поверхностные некро-
тические изменения слизистой пищеварительного тракта и на коже головы связаны
с местным действием токсина. Глубокие поражения, которые бывают в более поздние
сроки, обусловлены резорбтивным действием токсина. В основе геморрагического
диатеза лежат нарушения функции капилляров, физико-химических и биологичес-
ких свойств крови. Геморрагический процесс более выражен при многократном от-
равлении микотоксином. Резкие нарушения кроветворной системы, выражающиеся
в развитии лейкопении, носят функциональный характер, после выздоровления ко-
личество лейкоцитов может вновь подниматься до исходного уровня.
При фузариотоксикозе резорбтивное действие микотоксина проявляется в первую
очередь поражением ЦНС и выраженным геморрагическим диатезом, угнетением
гемопоэза, тяжелыми поражениями печени. Выделение токсических веществ с мочой
отрицательно действует на почки.
При афлатоксикозе развитие заболевания также протекает различно в зависимо-
сти от характера поступления микотоксинов в организм. Массивное однократное
введение токсических веществ приводит к появлению острого токсикоза. При мно-
422
Часть 2. Общая неправильная эпидемиология
гократном поступлении микотоксина развивается подострое и хроническое течение
болезни. При этом на передний план выступают такие явления, как энтерогепатия.
Токсические продукты гриба могут вызывать развитие доброкачественных папилло-
матозныхи фибринозных образований. Встречается появление и злокачественных
опухолей. При афлатоксикозах отмечают кровоизлияния и дегенерацию эпителия в
канальцах почек, некрозы и тромбы в печеночных сосудах, кровоизлияния и пневмо-
нию, гиперплазию в лимфатических узлах. Выражена общая дистрофия.
Патогенез питомикотоксикоза изучен подробно. Споридесмин всасывается в ки-
шечнике и проникает в печень через портальную вену. Болезнь протекает в две ста-
дии: первая — интоксикация, вторая — поражение печени и почек. В первой стадии
под влиянием споридесмина наблюдают незначительные поражения в гепатоцитах
(нарушение синтеза протеина в митохондриях) и большие изменения в желчном пу-
зыре и желчных протоках. Под влиянием микотоксина происходит отечное воспале-
ние и набухание эпителия желчных протоков и окружающей ткани, что приводит к
закупорке желчных протоков и желчного пузыря. На стенках желчных протоков об-
разуется рубцовая ткань. В сыворотке крови появляется глютаминат щавелевой
трансаминазы. В результате неспособности печени связывать филлоэритрин из кро-
ви проявляется фотосенсибилизация. Филлоэритрин, находясь в крови, адсорбирует
энергию солнечного света и, хотя сам не участвует в реакциях повреждения кожи,
передает энергию другим соединениям, участвующим в кожных поражениях. Пора-
жения печени приводят к развитию желтухи, затем появляются цистит мочевого пу-
зыря, исхудание. В результате поражения печени в крови повышаются уровень били-
рубина, холестерола, активность щелочной фосфатазы, происходит увеличение
содержания адреналина; возрастает содержание общего протеина и у-глобулиновой
фракции. Альбуминовая фракция снижается. Количество лейкоцитов, в частности
нейтрофилов, увеличивается, а эритроцитов остается в норме.
При эрготизме токсические вещества спорыньи воздействуют на гладкую муску-
латуру, изменяют тонус сосудов, что сопровождается нарушением питания тканей
В свою очередь, это приводит к цианозу и гангрене определенных участков тела. Ха-
рактерным является сильное сокращение мускулатуры матки под воздействием ал-
калоидов. Изменения в мозговой ткани сопровождаются дегенерацией нервных кле-
ток, перерождением стенок сосудов.
Патогенез клавицепстоксикоза характеризуется тем, что токсичные вещества гриба
поражают преимущественно центральную нервную систему, у отравленных животных
появляются: тремор, судороги, парезы, параличи. Патологические изменения, отме-
чаемые во внутренних органах, не бывают следствием непосредственного действия на
них токсических веществ. Они носят вторичный характер и обусловлены нарушени-
ем деятельности центральной нервной системы.
Некоторые токсические метаболиты, продуцируемые грибами из родов Penicillium
и Aspergillus при попадании в организм животного вызывают отравление, сходное по
клинике с клавицепстоксикозом: тремор тела, сменяющийся атаксией и мышечной
ригидностью. В механизме действия треморгенов важное значение имеет их действие
на подкорковые центры. Они нарушают процесс освобождения нейротрансмиттеров
в синалтосомах, снижают концентрацию у-аминомасляной кислоты в нейронах го-
ловного мозга. По химической структуре треморгены подразделяются на четыре груп-
пы: пенитремы; фумитреморгены — веррукулоген; паспалин; триптоклавин. Поми-
мо указанных метаболитов к треморгенам относятся триптоквивалон, территремы А
и В, рокфортин, пенитрем С, паспалин, янтитремы В и С.
2.2. Биологические поражающие агенты
423
Развитие болезни при микотоксикозах может протекать различно, что связано с
количеством микотоксинов, попавших в организм. При острых токсикозах патоло-
гоанатомические изменения в органах и тканях обычно бывают выражены не очень
отчетливо. При отравлении сублетальными дозами токсина в течение длительного
времени, болезнь приобретает затянутый, подострый характер. В таких случаях пато-
тогоанатомические изменения более выражены.
В развитии микотоксикоза обычно можно выделить четыре условные стадии: пер-
вую стадию, связанную непосредственно с воздействием микотоксина; вторую ста-
дию, когда развитие патологического процесса связано не только со специфическим
действием токсина, но и с изменениями реактивности организма, а также поступле-
нием вторичных токсичных метаболитов, связанных с некрозами; третья стадия ха-
рактеризуется усугублением течения заболевания вторичной бактериальной инфек-
цией; четвертая стадия — выздоровления, если в организм попали несмертельные дозы
микотоксинов и проведена рациональная терапия.
Токсичные вещества несовешенных грибов содержатся только в определенных
морфологических структурах. Например, при эрготизме токсичными являются толь-
ко склероции, а другие стадии развития гриба (сумчатая и конидиальная) безвред-
ны. У грибов из рода Aspergillus токсические вещества содержатся в мицелии и кони-
диях. В большинстве случаев микотоксины в природных источниках встречаются не
по отдельности, а в комбинации, и обнаружение даже одного микотоксина на уровне
ПДК — достаточно веское основание полагать о присутствии других микотоксинов в
опасных концентрациях. Комбинации микотоксинов вызывают смешанные симп-
томы микотоксикозов у животных и людей, а это значительно усложняет диагности-
ку болезни. Комбинации микотоксины опасные еще тем, что они способны к синер-
гическому взаимодействию друг с другом (взаимному усилению действия). Наиболь-
ший эффект токсического синергизма обнаружен у фузариевых микотоксинов.
Фузариевая кислота, редко анализируемая в кормах для животных из-за низкой ток-
сичности, усиливает токсичность дезоксиниваленола. Синергический эффект уста-
новлен у Т-2 токсина и афлатоксина В, у зеараленона и дезоксиниваленола. Известно
также, что микотоксины способны накапливаться в организме по мере их постоян-
ного поступления. Постоянное поступление нескольких микотоксинов в количествах
в 2—3 раза ниже ПДК оказывает более негативный эффект, чем непродолжительное
поступление одного микотоксина в высокой дозе.
2.2.3.4. Яды грибов
Роль отдельных токсических веществ грибов-макромицетов в формировании
клинической картины отравления. Фаллоидиновый синдром. Гиромитрино-
вый синдром. Паксилюсный синдром. Орелланиновый синдром. Прокси-
мановый синдром. Судориновый синдром. Психомиметические синдромы.
Под грибами население понимает шляпочные грибы (макромицеты). Таксоно-
мически такие грибы относят к двум классам (классификацию грибов см. выше):
II класс — сумчатые грибы или аскомицеты; III класс — базидиомицеты. Они состо-
ят из мицелия (грибницы) и плодового тела — собственно гриба (это хорошо всем зна-
комые шляпка и ножка гриба). Мицелий (грибница) — вегетативное тело гриба, со-
стоит из тонких (1,5—10 мкм в поперечнике) разветвленных нитей (гиф). Находится
в верхних слоях почвы на глубине 6-10 см от ее поверхности. Мицелий может быть
неклеточным (у фикомицетов) и многоклеточным (у сумчатых, базидиальных и несо-
424
Часть 2. Общая неправильная эпидемиология
вершенных грибов) Мицелий выполняет роль питающих корней в образовании пло-
дового тела гриба. Шляпку и ножку гриба образуют переплетенные гифы. Шляпка
содержит органы, которые производят споры — носители наследственности гриба.
Основной из них гименофор — особая структура плодового тела гриба, расположен-
ная с нижней стороны шляпки. Верхняя ее часть примыкает к мякоти; нижняя — сво-
бодная. В зависимости от строения гименофора грибы-макромицеты делят на труб-
чатые и пластинчатые. У трубчатых грибов гименофор состоит из трубочек и ячеек, у
пластинчатых — из пластинок. Гименофор у базидиомицетов покрыт слоем микро-
скопических мелких клеток (базидий), несущих на поверхности споры. У аскомице-
тов споры образуются в особых мешочках — асках. Ножка гриба предназначена для
поднятия над поверхностью почвы гименофор со спороносным слоем. Плодовое тело
некоторых грибов в начале своего развития заключено в покрывало. В процессе рос-
та гриба покрывало разрывается. Верхняя его часть остается на шляпке в виде чешуек
и хлопьев. Нижняя часть образует своеобразный чехол или мешковидное образова-
ние (вольву), представляющую собой клубневидное вздутие. Таким образом, природ-
ное предназначение плодового тела (гриба) состоит в образовании спор. Спора явля-
ется источником роста мицелия, а в последующем и плодового тела на новом участке
почвы (Мусселиус С. Г., Рык А. А., 2002).
Роль отдельных токсических веществ грибов-макромицетов i формировании клини-
ческой картины отравления. Токсическое воздействие ядовитых грибов на внутренние
органы обусловлено либо прямым повреждающим эффектом, либо опосредованно,
через цикл метаболических превращений. Химическая структура токсинов и ядов
отдельных видов грибов изучена с разной глубиной. Так, ядовитые вещества, содер-
жащиеся в бледной поганке и строчках, уже не представляют загадки для токсиколо-
гов. Но у свинушки тонкой и многих других грибов их еще предстоит установить.
Различие химической структуры токсических соединений грибов определяет их спе-
цифический тропизм к отдельным органам и системам организма и, соответственно,
патологические синдромы, наблюдаемые врачами у лиц, отравившихся грибами.
В данной работе рассмотрены наиболее распростране иные синдромы таких отрав-
лений: фаллоидиновый, гиромитриновый, паксилюсный, орелланиновый, судорино-
вый и психомиметические (микоатропиновый, псилоцибиновый и др.). В качестве
основных источников информации использованы работы С. Г. Мусселиуса и А. А. Рык
(2002); и Б. Н. Орлова с соавт. (1990).
Фаллоидиновый синдром. Развивается при употреблении грибов, содержащих на-
бор токсических веществ, основными из которых являются аманитотоксины. В зави-
симости от количества токсических веществ, имеющихся в I г свежих грибов, все гри-
бы, содержащие аманитотоксины, делят на две группы: I группа — высокотоксичные
грибы, летальная доза для взрослого человека составляет 0,5—1,5 г свежих грибов на
1 кг массы тела (грибы рода мухоморов: бледная поганка, Amanita phalloides', мухомор
весенний, A. verna', мухомор зловонный, A. virosa', мухомор двуспоровый, A. bisporigera)',
II группа — токсичные грибы, летальная доза — 5—8 г/кг (грибы рода лепиота: лепи-
ота гельвсола, Lepiota helveola\ лепиота брюннеолилацина, L. brunneolilacina', лепиота
гребенчатая, L. cristata’, лепиота субинкарната, L. subincamata; галерина осенняя, Gale-
rina autumnaiis', галерина маргината. G. marginata' Коносиб филарис, Conocybe filans).
К аманитотоксинам относятся следующие токсические вещества.
Аманитины — циклопептиды, состоящие из 8 аминокислот (октапептиды). Трип-
тофан и цистеин в такой молекуле соединены серосодержащим мостиком, разделяю-
щим молекулу циклопептида как бы на две части. Общим для всех аманитотоксинов
2.2. Биологические поражающие агенты
425
является наличие в положении 1 изолейцина, в положениях 3 и 5 — глицина, в поло-
жении 4 — изолейцина, в положении 7 — радикала аспарагина, в положении 8 — трех
соединений гидроксипролина, а также наличие окиси серы, связывающей триптофан
и цистеин. ММ аманитинов колеблется в пределах 10U0 Да. В бледной поганке содер-
» ится 8 гидрофильных аманитинов, в зловонной поганке — 9. Из 8 аманитинов блед-
ной поганки 5 обладают высокотоксическими свойствами, отчего получили назва-
ние — аманитотоксины. Аманитотоксины теряют свою токсичность при нарушении
бициклической структуры строения — утрате окси-серного мостика или при пере-
стройке молекулы в свои изомеры. В строении трех нетоксичных аманитинов — ама-
нуллина, амануллиновой кислоты и проамануллина — вместо изолейцина в состав
молекулы входит гидроксиизолейпин. Химическая структура аманитотоксинов пред-
ставлена на рис. 2.33.
т
СН3^ ^СН— сн2—R2
СН
I
CH— СО — NH—ch—СО —NH —СН2 —СО
NH—
I
СО
н^"Дн
Rs— ch I
Н2с-^ |
СО — СН —NH— СО — CH —NH — СО — СН2— NH
I
СН—со —R3
Рис 2.33. Химическая структура аманитотоксинов.
По С. Г. Мусселиусу и А. А. Рык (2002)
Распределение аманитинов в отдельных частях гриба неодинаково. В 1 г сухой
массы бледной поганки в пластинках содержится 2,6 мг аманитотоксина, в шляпке —
1,9 мг, в ножке — 1,7 мг, в вольве — 0,6 мг. Суммарное количество аманитотоксинов в
бледной поганке непостоянно и определяется местом ее произрастания, погодными
и экологическими условиями. Содержание аманитотоксинов в различных видах по-
ганок отличается в значительной степени, что и определяет клиническое течение от-
равления. Суммарное количество аманитотоксинов в 1 г сушеной бледной поганки
составляет около 3 мг, тогда как в таком же количестве мухомора весеннего их содер-
жание в 2 раза меньше (0,5—1,5 мг). Летальная доза аманитотоксинов для взрослого
человека с массой тела 70 кг составляет 7 мг. Такое количество токсинов содержится в
30—50 г свежей бледной поганки и в 150—200 г мухомора весеннего.
кинетика аманитотоксинов. Резорбцированные в кишечнике аманитотоксины
свободно циркулируют в крови, не связываются с альбуминами. Распределение в
организме — преимущественно внеклеточное пространство. Аманитины обладают
высокой афинностью к ферменту РНК-полимеразе II. РНК-полимераза I нечувстви-
тельна к нему, а PHК-полимераза III чувствительна слабо. Попадая в клетку, аманитин
связывает этот фермент, блокируя его работу, что, в свою очередь, приводит к прекра-
426
Часть 2. Общая неправильная эпидемиология
щению синтеза белков и к разрушению клетки (цитолизу). В человеческом организ-
ме при отравлении больше всего страдают энтероциты, клетки печени и почек.
Значительная часть резорбцированных в кишечнике аманитотоксинов, пройдя в
печени внутриклеточный цикл, с желчью вновь попадает в кишечник с последующим
повторным токсическим воздействием на гепатоциты. При высокой концентрации
аманитотоксинов в кишечнике отмечается длительная циркуляция токсинов по сле-
дующему пути: кишечник — гепатоциты - желчь — кишечник. Энтерогепатическая
циркуляция аманитотоксинов обусловливает неравномерность цитолиза гепатоцитов,
а пролонгированность токсического действия — всевозрастающую с течением времени
патологию в печени.
Поражение слизистой пищеварительного тракта — следствие как минимум двух
причин:
1) всосавшиеся аманитины нарушают внутриклеточные обменные процессы, раз-
рушают энтероциты, что проявляется распространенными некрозами слизистой ки-
шечника;
2) оказывая выраженное губительное воздействие на сапрофитную флору кишеч-
ника, аманитины создают условия для интенсивного роста патогенной микрофлоры,
которая, проникая в глубокие слои слизистой, разрушает ее.
Оба фактора участвуют в патогенезе развития синдрома острого гастроэнтерита.
Поврежденная слизистая кишечника утрачивает барьерную функцию в отношении
выделяемого бактериями токсина. Возникшая бактериальная токсемия в бассейне
портального кровотока оказывает разрушительное влияние на функциональное со-
стояние печени, уже подвергшейся разрушительному действию грибного токсина.
Прямое токсическое действие аманитотоксинов на другие органы весьма незна-
чительно. Изменения в почках носят функциональный характер и возникают на ран-
нем этапе вследствие водно-электролитных и гемодинамических расстройств, а в более
позднем периоде как результат тяжелых изменений в печени и развиваются по типу
гепаторенального синдрома. В литературе имеются сообщения, не исключающие
прямого токсического воздействия аманитина на проксимальные отделы канальцев.
Однако, как показывает клинический опыт С. Г. Мусселиуса и А. А. Рык (2002), если
удаются эффективная коррекция водно-электролитных нарушений, кислотно-щелоч-
ного состава и восстановление функции печени, то почечная недостаточность не раз-
вивается. Изменения в сердце и поджелудочной железе являются вторичными, их
развитие обусловлено нарушениями гомеостаза, агрегатного состояния крови, дис-
семинированным внутрисосудистым свертыванием крови (ДВС-синдром) и эндоген-
ной интоксикацией. Такие проявления поражения ЦНС, как психомоторное возбуж-
дение, делирий, галлюцинации, эйфория и обнабуляция, не имеют четких доказа-
тельств прямого токсического воздействия грибного токсина на клетки головного
мозга. Развитие астении и сосудистый коллапс, по всей видимости, напрямую связа-
ны с повреждением аманитинами коры надпочечников. В зависимости от степени
тяжести и сроков отравления функциональная активность надпочечников может ко-
лебаться в значительном диапазоне.
Фа иоидины — циклические олигопептиды, состоящие из 7 аминокислот (гепта-
пептиды). Общим для всех видов фаллоидинов является наличие внутреннего серо-
содержащего мостика, свя 1ующего молекулы цистеина и триптофана, а также распо-
ложенные в позициях 4, 5, 7 молекулы лейцина, пролина и аланина. ММ в пределах
730 Да. Различия в 5 радикалах определяют кинетику каждого вещества. В группу фал-
лоидинов входят: фаллоидин, фаллоин, профаллоин, фаллизин, фаллацин, фаллаци-
2.2. Биологические поражающие агенты
427
дин, фаллизацин. Все эти соединения теряют свою токсичность при разрушении хотя
бы одного из некоторых пептидных колец или серного мостика. Фаллоидин связыва-
ется с актином клетки в его полимерной форме и препятствует его деполимеризации,
блокируя такие функции, как сокращение и миграция. Все фаллоидины разрушают-
ся в кислой среде. На основании этого считается, что при пероральном употреблении
бледной поганки и мухомора весеннего все фаллоидины, обладающие токсическими
свойствами в эксперименте, подвергаются разрушению в кислой среде желудка, и
поэтому они не являются токсичными для человека. Однако смерть человека может
наступить при внутривенном введение токсина. В опытах на экспериментальных
животных показано, что под действием фаллоидинов отмечается сморщивание мем-
браны гепатоцитов и повреждение лизосом. Изменения в эндоплазматическом рети-
куломе проявлялись вакуолизацией отдельных фрагментов. Нарушения электролит-
ного состава характеризовались утратой внутриклеточного калия и кальция.
Фаллин (фаллолизин) — малоизученное вещество белковой природы, ММ около
30 тыс. Да. Теряет свою токсичность при нагревании до температуры свыше 60°С, а
также разрушается в кислой среде при значении pH 5. В экспериментальных иссле-
дованиях при внутривенном введении фаллина смерть животных наступала от острой
печеночно-почечной недостаточности и острого гемолиза, приводящего к гиперка-
лиемии и нарушению сердечного ритма. Другие органы оставались интактными.
Виротоксины — содержатся только в мухоморе зловонном и представляют собой
циклические гептапептиды. Выделены следующие разновидности этого соединения:
вироидин, дезоксивироидин, вироизин и дезоксивироизин. По химической структу-
ре и токсическому воздействию виротоксины близки к фаллоидинам.
Клиника отравления при фаллоидиновом синдроме. Характеризуется фазностью раз-
вития патологических симптомов. С. Г. Мусселиусом и А. А. Рык (2002) выделены
следующие пять периодов развития болезни: латентный; период острого гастроэнте-
рита; период мнимого благополучия; период острой печеночной, печеночно-почеч-
ной недостаточности; период выздоровления.
Продолжительность латентного периода составляла 6—9 ч, в редких случаях
10-15 ч и крайне редко 16—36 ч. Сроки до появления первых симптомов отравления
определялись не только количеством поступившего в организм токсина, но и его кон-
центрацией в одновременно принятой пищевой массе, а также ее составляющими
компонентами. Сопутствующие отравлению гастрит, энтероколит, холецистит, пан-
креатит существенно изменяли клинику отравления. У этой категории пострадавших
ранняя реакция (до 6 ч) никак не была связана с грибным токсином, а являлась след-
ствием неспецифического воздействия грибной клетчатки на нездоровые органы
пищеварения, что в значительной степени затрудняет своевременную диагностику
отравления. Латентный период завершался неспецифическими, общеклиническими
проявлениями — общей слабостью, недомоганием, разбитостью, что расценивалось
как предвестник дальнейшего развития клиники отравления.
В период острого гастроэнтерита в клинике отравления преобладали симптомы
поражения желудочно-кишечного тракта. Это проявлялось внезапной обильной,
порой «фонтанирующей» рвотой съеденной пищей. Рвота приобретала характер не-
укротимой, была мучительной доя отравленного. В отсутствие пищевых масс содер-
жимым становились желчь, секрет слизистой желудка и 12-перстной кишки. Посто-
янная тошнота. При нарастающих спастических болях в животе развивалась диарея,
в тяжелых случаях отравления носящая холероподобный характер. Частота стула мо-
жет достигать 20—25 раз в сутки. Испражнения водянистые, со слизью и примесью
428
Часть 2. Общая неправильная эпидемиология
крови. В организме происходили значительные нарушения водного баланса. Потеря
жидкости и солей вызывала сухость во рту, жажду. Отравившиеся отмечали слабость,
головную боль, головокружение. Гематокрит превышал 50 %. Прием жидкости толь-
ко усиливал рвоту. Электролитные нарушения проявлялись гипонатриемией, гипо-
калиемией, гипохлоремией В крови отмечался метаболический ацидоз. Пациенты
становились адинамичными, отмечали нарастание мышечной слабости. Развивалась
олигурия. Функциональная почечная недостаточность проявлялась умеренной азо-
темией. Гипотония и тахикардия. В случае тяжелого отравления бледной поганкой
поражение желудочно-кишечного тракта приводило к глубоким расстройствам гоме-
остаза и жизнеугрожающему нарушению сердечно-сосудистой деятельности. Эти
осложнения наиболее тяжело переносили дети и люди преклонного возраста, компен-
саторные возможности которых в значительной степени ограничены. Продолжитель-
ность периода составляла 2—6 сут.
В период мнимого благополучия прекращаются мучительные боли в животе, те-
незмы, тошнота и рвота. «Улучшение состояния» порой расценивалось врачами как
тенденция к выздоровлению, но спустя некоторое время (от нескольких часов до
1—2 сут) состояние отравившегося ядом бледной поганкой вновь ухудшается. В тя-
желых случаях отравления «улучшения состояния» не происходит, и болезнь непос-
редственно переходит в следующую стадию.
Период острой печеночной, печеночно-почечной недостаточности. Степень на-
рушения функции печени и почек носит различный характер: от отсутствия каких-
либо клинических проявлений до развития жизнеугрожающих симптомов. Токсиче-
ская гепатопатия развивается уже в первые сутки отравления, что подтверждалось
лабораторным путем. Клинические проявления в виде гепатомегалии, желтухи, ДВС-
синдрома и др. отмечались спустя трое суток после появления первых клинических
симптомов отравления.
Патологическая анатомия при фаллоидиновом синдроме. По данным, собранным
С. Г. Мусселиусом и А. А. Рык (2002), на аутопсии у людей, умерших от отравления
бледной поганкой, отмечаются желтушность кожных покровов и слизистых, множест-
венные петехиальные, мелкоточечные и очаговые кровоизлияния в кожу, склеры, со-
единительные оболочки глаз, кровоизлияния в слизистые. При вскрытии у всех умер-
ших выявляются грубые изменения в печени, почках и тонком кишечнике — органах,
в наибольшей степени подвергшихся воздействию аманитинов.
В головном мозге — дистрофия нейронов и клеток глии. В тяжелых случаях острой
печеночно-почечной недостаточности отмечаются отек и набухание вещества голов-
ного мозга, мелкоточечные кровоизлияния.
В сердце—дистрофия миокарда, кровоизлияния субэндокардиальные и в миокар-
де, жировая дистрофия мышечных волокон.
В легких — субплевратьные кровоизлияния, дистрофические изменения, распро-
страненные микротромбы.
В печени — макроскопическая карт ина соответствовует остро™ желтой атрофии.
Капсула морщинистая, на разрезе ткань дряблой консистенции, желтоватого цвета,
иногда с сальным блеском и диффу >ным рисунком мускатного ореха ярко-красного
цвета. Если смерть наступила в конце 1-й недели, наблюдалось некоторое западение
поверхности зон красного цвета. При гистологическом исследовании обнаруживают
грубые поражения печени — массивные центролобулярные некрозы. Во всех случаях
некротические изменения носят диффузный характер; реже массивные некрозы сли-
ваются, оставляя интактными небольшие очажки отдельных гепатоцитов и небольших
2.2. Биологические поражающие агенты
429
фрагментов долек в перипортальной зоне. Особенностью некрозов при отравлении
аманитинами было отсутствие какой-либо клеточной реакции (ареактивность). В по-
граничных с некрозами зонах печени отмечались выраженные дистрофические и дест-
руктивные изменения трабекул вплоть до их дискомплексации, разрушение синусои-
дов, некробиоз гепатоцитов. Большая часть гепатоцитов в пограничной зоне находится
в состоянии дистрофии (белковой, гидропической или жировой). Часто выявляются
признаки холестаза, желчные тромбы. Гепатоциты перипортальных трактов наиболее
сохранны. Они содержат гликоген, который распределяется неравномерно. В других
зонах гликоген отсутствует. К концу 1-й недели в портальных трактах отмечается ин-
фильтрация, преимущественно из лимфоцитов, которая вдальнейшем распространя-
ется и на другие зоны. К концу 2-й недели наблюдаются выраженная пролиферация
холангиол портальных трактов, регенерация гепатоцитов—появление крупных много-
ядерных гепатоцитов. Структурной основой острой печеночной недостаточности яв-
ляются массивные диффузные центролобулярные некрозы.
В почках— при макроскопическом исследовании отмечается увеличение их массы
до 400—520 г. Ткань набухшая, на разрезе кора расширена, выбухает за капсулу, конт-
растирует с полнокровными пирамидками. Вся паренхима почек имеет, как правило,
диффузное желтушное прокрашивание. При этом в пирамидках иногда выявляются
радиальная темно-бурая или зеленоватая исчерченность, в слизистой лоханок — мел-
когочечные или пятнистые кровоизлияния. При гистологическом исследовании об-
наруживают диффузную гидропическую дистрофию нефроцитов как проксимальных,
так и дистальных канальцев с признаками тубулярного отека и канальцевой метаплазии
эпителия капсул клубочков. Эпителий извитых канальцев набухший, в состоянии бел-
ковой (зернистой) и жировой дистрофии, в клубочках скопление белковых масс. Мор-
фологические изменения почек в основном соответствуют холемическому нефрозу.
В просвете дистальных канальцев и в собирательных трубочках белково-пигментные
шлаки, в составе которых преобладают желчные пигменты. Слабая бензидинположи-
тельная реакция на гемоглобин отдельных цилиндров обусловлена, по-видимому, вы-
раженным геморрагическим синдромом у этих больных. Количество белково-пигмет-
ных шлаков в просвете нефронов зависит от диуреза. При олигоанурии количество
цилиндров значительно больше, чем при восстановительном диурезе. При полиурии
просветы нефронов в основном свободны. Однако при этом отмечается выраженный
интерстициальный отек и признаки осмотического нефроза. Структурной основой
острой почечной недостаточности является холемический нефроз.
В кишечнике — на аутопсии обнаруживают энтерит. Изменения носят как диффуз-
ный, так иногда и очаговый характер. При гистологическом исследовании обнаружи-
вают тромбы в слизистой и подслизистой оболочке толстых кишок, что сопровождается
язвенно-некротическим процессом, в редких случаях происходит распространение
воспалительного процесса на мышечный слой. В слизистой определяются признаки
аутолиза, на фоне очаговых кровоизлияний и резко выраженного отека отмечаются
диффузная инфильтрация кишечной стенки лимфоцитами и плазматическими клет-
ками. Из-за отека толщина кишечной стенки местами в 2—4 раза превосходит нормаль-
ные размеры. В гладких мышечных клетках выявляются грубые дистрофические из-
менения. Мышечная пластинка слизистой оболочки местами как бы разрыхлена.
В этих участках наблюдается большое скопление колоний микроорганизмов. Они рас-
полагаются в сосудах, диффузно инфильтрируя прилежащие отделы слизистой, под-
слизистую и мышечную пластинку слизистой оболочки в зоне ее разрыхления. Одна-
ко мышечная пластинка слизистой оболочки редко подвергается деструкции на
большом протяжении. Бактериальная инвазия сочетается с воспалительными и дест-
руктивными процессами и является фактором, усугубляющим эндотоксикоз. Струк-
430
Часть 2. Общая неправильная эпидемиология
турная основа энтерита — выраженные расстройства кровообращения, васкулиты,
массивная воспалительная инфильтрация преимущественно в подслизистой и мышеч-
ной оболочках тонкой кишки.
В надпочечниках — макроскопические изменения выражаются в атрофии и кро-
воизлияниях в корковом и мозговом веществе. Микроскопически обнаруживают об-
ширные жировые включения, расстройство кровообращения, атрофию и деструкцию
секреторных клеток. В участках кровоизлияний структура ткани надпочечника не оп-
ределяется. Атрофические изменения выражаются в резком уменьшении или исчез-
новении липидов из клеток клубочковой и пучковой зон, клетки имеют сильно вакуо-
лизированный вид. Спонгиоциты располагаются совершенно беспорядочно, а часть
их находится в пренекротическом состоянии.
В поджелудочной железе — гипертрофия инсулинпродуцируюших 0-клеток, дегра-
нуляция и уменьшение а-клеток островков Лангерганса.
Гиромитриновый синдром. Вызывают грибы класса аскомицетов рода гиромитра
(Gyromitra): строчок обыкновенный (G. esculenta), строчок гигантский (G. gigas), стро-
чок осенний (G. infiila). Гиромитриновый синдром развивается также при приеме не-
которых грибов семейства гельвелловых: лопастника кудрявого (Helvetia crispas), ло-
пастника ямчатого (Н. lacunosa). Список грибов, вызывающих гиромитриновый
синдром, не окончательный, однако наиболее распространенные и тяжело проте-
кающие отравления возникают при употреблении в пищу строчков, в меньшей сте-
пени — сморчков.
Токсическим веществом, вызывающим гиромитриновый синдром является гиро-
митрин. В составе грибной массы гиромитрин находится в виде свободной фракции
и различных соединений: гиромитрин N-метил-М-формилгидразон ацетальдегид
(или этилиден-гиромитрин, C4H8N2O), 3-метилбутилиден-гиромитрин, пентилиден-
гиромитрин, гексилиден-гиромитрин и др. Наиболее токсичен этилиден-гиромитрин.
Его масса в клетчатке свежих грибов достигает 87,5 % от общего количества гиромит-
рина. В процессе жизни и роста гриба в нем неферментным путем происходят пре-
вращения этилиден-гиромитрина в токсичное соединение — N-метил-М-формил
гидразин (МФГ), которое в дальнейшем превращается в более токсичное соедине-
ние — N-моно-метил-гидразин (ММГ). В свежих строчках содержание ММГ может
варьировать в больших пределах в зависимости от сезона, состава почвы, погодных
условий и степени зрелости гриба. Концентрация ММГ может колебаться от 50 до
300 мг/кг, но в отдельных случаях достигает 1200—1600 мг/кг.
При попадании в организм человека гиромитрин образует ряд соединений, обла-
дающих высокими реакционными возможностями. В кислой среде желудка при
pH 2—3 происходит гидролиз гиромитрина. В результате отщепления уксусного аль-
дегида образуется МФГ, который под воздействием ферментных систем организма
превращается в ММГ. Действующим началом МФГ и ММГ в организме человека яв-
ляются гидразины, метаболиты гидразина и гидроксиламин. Они ингибируют в эн-
тероцитах диаминоксидазу, активно участвующую в обезвреживании биогенных ами-
нов, образующихся в кишечнике при распаде белков. В клетках печени гидразины
в результате метаболических превращений образуют нестойкие диазосоединения, в
частности нитрозоамиды. Являясь источником образования свободных радикалов,
нитрозоамиды обладают мутагенной активностью и онкогенным действием. В орга-
низме человека нитрозоамиды под действием микросомальных оксидаз клеток обра-
зуют активные метаболиты, вступающие в реакции с макромолекулами ДНК, РНК и
белков клетки. Производные нитрозоамидов, будучи активными азотсодержащими
2.2. Биологические поражающие агенты
431
соединениями, ошибочно включаются в полинуклеотидную цепь ДНК и нарушают
последовательность азотистых оснований, а также комплементарность аминокислот.
Специфическое воздействие гидразинов на гепатоциты проявляется развитием
цитолиза с последующим некрозом ткани печени.
Разными исследовательскими коллективами было установлено, что МФГ воздей-
ствует на генетический код крыс: у последующих поколений были выявлены пороки
развития. Ежедневное введение хомякам ММГ приводило к снижению активности
цитохрома Р-450, разрушало структуру ДНК, повышало проницаемость клеточных
мембран, угнетало внутриклеточные обменные процессы. Одновременно происходи-
ло нарушение синтеза триглицеридов, фосфолипидов и холестерола. На вскрытии у
животных обнаруживали злокачественные опухоли в печени, желчном пузыре, желч-
ных протоках, слепой кишке и мочевом пузыре. Производные гидразина, например
метилгидразин, обладая функцией свободного амина, активно реагируют с карбониль-
ными группами пиридоксальфосфата — соединения, включающего пиридоксин (ви-
тамин В6). Снижение активности пиридоксалевого фермента, являющегося коэнзи-
мом глютаматдекарбоксилазы, приводит к нарушению синтеза у-аминобутировой
кислоты (ГАБК), играющей важную роль в передаче нервного импульса в мозговой
ткани. В эксперименте дефицит ГАБК проявлялся развитием судорожного синдрома.
Смертельная концентрация гиромитрина для взрослого человека составляет
20—50 мг/кг, для детей 10—30 мг/кг. При пересчете на количество свежих грибов смер-
тельная концентрация достигается при приеме взрослым человеком 400—1000 г гри-
бов. Максимальное количество высокотоксичных соединений образуется через
2—2,5 ч после приема грибов в процессе метаболизма гиромитрина, протекающего
по пути «летального синтеза». При отваривании грибов, так же как и при длитель-
ном их консервировании, содержащиеся в грибах токсичные соединения переходят
в раствор. Наиболее высокая токсичность отвара достигается при кипячении в нем
сменяющих одну за другой нескольких порций грибов В этих случаях отравления
людей протекают наиболее тяжело, как правило, с развитием острой печеночной, а
иногда и полиорганной недостаточности, приводящей к смерти.
Клиника отравления при гиромитриновом синдроме. Латентный период составляет
3—25 ч. У всех отравившихся гиромитрином отмечали боли в эпигастрии, тошноту,
рвоту, частый жидкий стул (до 8 раз в сутки). У части пациентов были выявлены гемо-
лиз, гемоглобинурия. На 2 или 3 сутки присоединяются поражения печени и почек.
Отравившийся может впасть в состояние сопора. Кожные покровы и склеры у него
могут быть интенсивно желтушными с множественными кровоизлияниями. Его со-
стояние прогрессивно ухудшается, развиваются печеночная кома, гипотония.
Смерть наступает от остановки сердечной деятельности и дыхания.
На аутопсии обнаруживают выраженную дистрофию печени, почек, миокарда;
распространенные некрозы в печени; очаги кровоизлияния в слизистой желудка и
кишечника; полнокровие в легких; отек головного мозга.
Паксилюсный синдром. Вызывают грибы семейства свинушковые (Paxillaceae),
рода свинушка; виды: свинушка тонкая (Р. involutes) и свинушка толстая (Р. atrotomen-
tosus). Более выраженными токсическими свойствами среди грибов этого рода обла-
дает свинушка тонкая. Токсическое вещество грибов рода свинушки изучено плохо.
При экспериментальных исследованиях введение крысам перорально и внутрибрю-
шинно экстракта, приготовленного из свинушки тонкой, вызывало признаки отрав-
ления. Препарат вводили в сроки от 4 до 12 сут. Доза препарата и продолжительность
его введения имели прямую корреляционную зависимость с симптомами отравления.
432
Часть 2. Общая неправильная эпидемиология
Латентный период составлял 4—10 сут. В течение этого времени происходила ку-
муляция токсического вещества и сенсибилизация организма. В процессе экспери-
мента снижалась активность животных, шерсть становилась влажной, отмечалось
вздутие живота. Паралич задних лап был предвестником гибели животного. При ги-
стологическом исследовании внутренних органов отмечались атрофия и некроз сли-
зистой желудка и кишечника, к 10—14 сут выявлялись дистрофия и некрозы гепато-
цитов и нефроцитов. Изменения в клетках паренхиматозных органов при отравлении
свинушкой тонкой были иными, чем при отравлении бледной поганкой.
Токсин свинушки тонкой является дициклопентеноном. Полипептид термола-
билен. В желудочно-кишечном тракте способен преодолевать кишечный барьер. Ток-
сический эффект предположительно реализуется через цикл иммуноаллергических
реакций, приводящих к повреждению эритроцитов с развитием гемолиза, гемолити-
ческой анемии и печеночно-почечной недостаточности. Поэтому считается, что ток-
син обладает кумулятивным действием. Первичное употребление грибов не вызыва-
ет симптомов отравления, новый прием грибов спустя какое-то время приводит к
развитию тяжелой токсико-аллергической реакции. У некоторых лиц повторный
прием грибов может оказаться фатальным.
Клиника отравления при паксилюсном синдроме. Клинические проявления отрав-
ления токсином свинушки тонкой начинаются через 1—3 ч после употребления в пищу
неотваренных жареных грибов, в редких случаях через 5—6 ч. Отравившиеся жалуются
на распространенные спастические боли по всему животу, у них развивается тошнота,
рвота и частый жидкий стул. На фоне обезвоживания наблюдается общая слабость,
иногда похолодание конечностей, боли в пояснице. В течение последующих 2—6 сут
состояние либо постепенно улучшается и отравившийся выздоравливает, либо симп-
томы прогрессируют, у пациента развивается ДВС-синдром, гепатопатия и нефропа-
тия, летальный исход.
Оре лланиновыи .гивдром. Развитие синдрома у людей связано с употреблением и ми
в пищу грибов семейства паутинниковых, род паутинник (Cortinarius). В природе
насчитывается более 26 разновидностей грибов Cortinarius. Токсические вещества:
орелланин (ди-М-оксид-1, Г-тетрагидрокси-3,3',4,4'-бипиридин-2,2'). Представляет
собой сложный полипептид, устойчивый к действию высокой
температуры, изменению кислотности среды и высушиванию
(рис. 2.34).
Токсичность орелланина резко снижается при воздействии
на него ультрафиолетового и солнечного облучения. В 1 г вы-
сушенной грибной массы паутинника оранжево-красного со-
держится в среднем 15 мг орелланина, в паутиннике красивей-
шем в 2 раза меньше; кортинарин А и кортинарин В — по
химическому составу представляют собой полипептиды. Их
роль в патогенезе заболевания до конца не изучена.
В эксперименте на животных (собаках, кошках, кролика:
крысах и белых мышах), которым вводили энтерально и парен-
терально гомогенат паутинника оранжево-красного и паутинника красивейшего, че-
рез 2—4 сут развивался интерстициальный нефрит без повреждения клубочков. Об-
щеклинические симптомы отравления у животных протекали с большими различиями
и зависели не только от вида гриба и его дозы, но и от пола и возраста животного.
Основным органом-мишенью орелланина и его метаболитов являются почки. Под их
действием в клетках почечного эпителия происходят процессы образования свобод-
2 2. Биологические поражающие агенты
433
ных радикалов с повреждением клеточных мембран, угнетение щелочной фосфатазы
и синтеза белка, нарушения в структуре РНК и ДНК. LD5,| для мышей и крыс состав-
“ нет 5—90 мг/кг. Изменения в организме человека аналогичны данным, полученным
в эксперименте.
Клиника отравления при орелланиновым синдроме. Отличается от клиники отравле-
ний другими видами ядовитых грибов длительным латентным периодом, развитием
острой почечной недостаточности при относительно малых нарушениях со стороны
функции печени.
В развитии отравления С. Г. Мусселиусом и А. А. Рык (2002) выделены 4 стадии:
1 -я стадия — латентная, продолжительность от 36 ч до 17 сут. Тяжесть отравления
во многом определяется продолжительностью инкубационного периода. В легких
случаях отравления латентный период составлял 10—17 сут, при отравлении средней
гжести 6-10 сут и при тяжелых отравлениях 2—3 сут. Первые симптомы отравления
роявлялись общей слабостью, гипертермией, болями в поясничной области и по все-
му животу, жаждой. При короткой латентной стадии — в пределах 2—3 сут, как правило,
развивается острая почечная недостаточность с длительным олигоанурическим перио-
дом; отравления протекают наиболее тяжело у пожилых людей и детей.
2-я стадия — преренальная, проявляется желудочно-кишечными расстройствами,
неврологическими и общими симптомами. Клинические симптомы отравления: тош-
нота, рвота, боли в животе, головная боль, шум в ушах, озноб, сонливость, затормо-
женность, быстрая утомляемость.
3-я стадия — ренальная (почечная), протекает в виде острой почечной недоста-
точности и развивается у 30-46 % пострадавших. На фоне ухудшения общего состо-
яния отмечаются постепенное снижение диуреза, анасарка, скопление жидкости в
брюшной и плевральной полостях. Развиваются симптомы нарушения центральной
и периферической нервной системы — спутанность сознания, парестезии и боли в
конечностях, тремор, кома. В крови отмечается подъем мочевины, креатинина, оста-
эчного азота. В моче определяется белок, достигающий 2,5—9 г/л, в микроосадке —
клетки почечного эпителия, неизмененные эритроциты и лейкоциты. Даже в отдален-
ные сроки от момента отравления (на 10 сут) в крови человека, отравившегося ядом
паутин никовых еще может быть определен орештанин.
При электронно-микроскопическом исследования пунктатов почек и аутопсий-
ного материала выявляются следующие изменения: а) отечные канальцы с очагами
не кроза и десквамацией эпителия, выраженный пикноз ядер, отсутствие митоза, в
"росвете канальцев — эпителиальные цилиндры, фибрин; б) диффузный отек меж-
уточной ткани с воспалительной инфильтрацией; в) скопления эозинофилов, плаз-
моцитов, фиброцитов, склероз; г) интралобулярные артерии с фибринозным наложе-
нием; д) отек и набухание эпителиальных клеток афферентных и эфферентных
сосудов, распространенные тромбы в интратобулярных венах. При иммунофлюорес-
ценции иммунные отложения отсутствуют. В отдаленные сроки (через 1—6 мес.) от-
мечаются распространенный склероз кортикального и мозгового слоев почки, тубу-
лоинтерстициальный нефрит с выраженной деструкцией канальцевого эпителия и
“аспространенным интерстициальным склерозом.
4-я стадия — выздоровления или остаточных явлений. Выздоровление медлен-
иое. в течение нескольких месяцев. Длительно сохраняющиеся явления нефропатии
£ 30—50 % случаев завершаются развитием хронической почечной недостаточности.
Выведение с мочой орелланина незначительное, из-за реабсорбции в проксималь-
ном отделе почечных канальцев. Разрушение нефроцитов сопровождается накопле-
434
Часть 2. Общая неправильная эпидемиология
нием орелланина и его метаболитов в паренхиме почек. Орел танин и его метаболиты
могут быть обнаружены в тканях почек через 6 мес. после отравления.
Проксиминовый сив аром. Развивается при употреблении в пищу мухомора насущ-
ного (Amanitaproximo). Токсическое вещество неизвестно. Клиника отравления вклю-
чает инкубационный период 8—48 ч. Начальные симптомы отравления проявляются
острым гастроэнтеритом с последующим развитием острой почечной недостаточно-
сти и гепатопатии средней степени тяжести.
Судориновый синдром. Название происходит от sudor — потеть Развивается пос-
ле употребления в пищу грибов с высоким содержанием мускарина. Его вызывают
грибы следующих родов:
грибы рода волоконница (Inocybe): волоконница патуйяра (In.patouillardii), воло-
конница земляная (In. geophylla), волоконница разорванная (In. lacera) и волоконни-
ца волокнистая (In. fastigiata)-,
грибы рода говорушка (Clitocybe): говорушка восковатая (Cl. cerussata), говоруш-
ка беловатая (Cl. dealbata), говорушка ривулоза (Cl. rivulosa).
Токсические вещества: мускарин и его изомеры (эпимускарин, алломускарин, эпи-
алломускарин, мусказон, мусцимол) (рис. 2.35).
С Н2—N (СН3)з
Алломускарин
Рис. 2.35. Химическая структура мускарина, его изомеров и ацетилхолина
Ацетилхолин
Мускарин (2-метил-3-гидрокси-5-триметиламмоний) — алкалоид, органическое
соединение алифатического ряда, состоит из одно! о тетрагидрофильного кольца и
образования, напоминающего по структуре ацетилхолин. Мускарин и его изомеры
термостабильны, водорастворимы, обладают низкой липофильностью. При варке
грибов выходят в раствор,, сохраняя биологическую активность. Смертельная доза для
человека около 500 мг. В организме человека мускарин (а из изомеров эпиалломуска-
рин) в наибольшей степени оказывает М-холиномиметическое действие, возбуждая
холинергические рецепторы мускаринового типа1. Под действием мускарина повы-
шается проницаемость синаптической мембраны рецепторов парасимпатической и
1 Встетативная иннервация (внутренние органы, кровеносные сосуды, железы» в зависи-
мости от медиатора, выделяющегося в нейро-эффекторных синапсах, подразделяется физио-
логами на холинергическую или парасимпатическую (медиатор — ацетилхолин), и адренер-
гическую или симпатическую (медиатор — адреналин). Холинорецепторы разной локализации
обладают неодинаковой чувствительностью к фармакологическим веществам. На этом осно-
вано выделение так называемых мускариночувствительных и никотинчувствительных холи-
норецепторов (М -холинорецепторы и Н-холинореиепторы). Никотин — алкалоид из листьев
таиака. Он оказывает избирательное действие на холинорецепторы, иные чем те, на которые
2.2. Биологические поражающие агенты
435
некоторых отделов симпатической нервной системы, чувствительных к ацетилхоли-
ну. Мускарин не разрушается ферментом холинэстеразой. Переход ионов через кле-
точную мембрану приводит к ее деполяризации и возникновению потенциала дей-
ствия в виде длительной поляризации мускаринчувствительных рецепторов. Действуя
на передние ветви парасимпатических ганглий, мускарин оказывает периферический
парасимпатомиметический эффект. Из-за низкой липофильности мускарин не про-
никает через гематоэнцефалический барьер. По механизму действия антагонистом
мускарина является атропин. Терапевтической ценности мускарин не представляет.
Грибы рода волоконница содержат больше мускарина по сравнению с грибами
рода говорушка, причем наибольшее количество мускарина выявлено в грибах воло-
конница патуйяра и волоконница земляная. В грибах, вызывающих судориновый
синдром, содержится мускарина в десятки раз больше, чем в мухоморе пантерном
(A. pantherina) и мухоморе красном (Л. muscaria). В связи с этим определение «муска-
риновый синдром» в большей степени соответствует клинике отравления, вызываемой
грибами рода волоконница и рода говорушка, чем мухоморами пантерным и красным.
Клиника проксиманового синдрома. Симптомы отравления — мускариноподобные.
Обычно их принято систематизировать по органам и системам. Для симптомов по-
ражения ЖКТ характерны тошнота, рвота, диарея, кишечная колика, гипертонус
ЖКТ. Для симптомов гиперсекреции — потливость, слюнотечение, слезотечение,
ринотечение. Нарушение сердечно-сосудистой деятельности проявляется синусовой
брадикардией, периферической вазодилатацией, гипотонией. Со стороны дыхатель-
ной системы: бронхоспазм, бронхорея, нарушение дыхания по астматическому типу.
ЦНС страдает редко. Отравившиеся мускарином остаются в сознании, и изменения
со стороны ЦНС (тревога, страх и др.) — это в большинстве случаев эмоциональные
проявления, возникающие как ответ на общеклинические симптомы интоксикации.
Описаны случаи нарушения деятельное! и ЦНС с развитием не только дисфории, но
и грубой патологии — поведение отравившегося становилось агрессивным, у него
развивался делирий. На основании этого некоторые авторы относят от равления вы-
шеуказанными грибами к синдрому психодислептическому делириозному.
Отравление мускарином протекает тяжело. В период от 15 мин до 2 ч после при-
ема грибов с высоким содержанием мускарина при ясном сознании у отравившегося
отмечаются усиленная болезненная перистальтика кишечника (по типу кишечной
колики), тошнота, рвота, частый жидкий стул. Повышается функция всех секретор-
ных органов: обильное потоотделение, слезо-, рино- и слюнотечение. Гиперсекреция
желез желудка и кишечника усиливает диарею. Бронхорея и бронхоспазм приводят к
расстройству дыхания, возможны астматические проявления. Синусовая брадикар-
дия и периферическая вазодилатация в тяжелых случаях обусловливают развитие ос-
трой сердечно-сосудистой недостаточности. Возникают нервно-психические рас-
стройства: эйфория, тревога, парастезии, мышечная дрожь, в редких случаях делирий.
Зрачки сужены, внутриглазное давление снижено, конвергенция невозможна. Моче-
воздействует мускарин. М-холинорецепторы расположены в постсинаптической мембране
клеток эффекторных органов у окончаний постганглионарных холинергических (парасимпа-
тических) волокон, а также имеются в ЦНС (в коре головного мозга, ретикулярной формации).
Н-холинорецепторы находятся в постсинаптической мембране ганглионарных клеток у окон-
чаний всех преганглионарных волокон (в симпатических и парасимпатических ганглиях),
мозговом слое надпочечников, синокаротидной зоне, концевых пластинках скелетных мышц
и ЦНС (в нейрогипофизе, клетках Реншоу и др.) (Харкевич Д. А., 1987).
436
Часть 2. Общая неправильная эпидемиология
испускание учащенное В большинстве случаев исход отравления благоприятный,
через 3—5 ч все симптомы самостоятельно постепенно проходят. Летальный исход
встречается в 2—6 % случаев.
Психомиметические синдромы. Развиваются при употреблении грибов, воздейст-
вующих на нейросенсорные отделы коры головного мозга. Механизм действия содер-
жащихся в грибах веществ имеет специфическую направленность: одни виды грибов
преимущественно повышают двигательную активность человека; другие — оказыва-
ют в выраженной степени наркотическое действие; третьи — вызывают галлюцино-
генный эффект. Интоксикация может протекать по смешанному типу. В клинической
токсикологии, касающейся отравлений грибами, синдром называют по латинскому
названию гриба. В некоторых случаях, когда близкая клиническая картина развива-
ется при отравлении несколькими видами грибов, синдрому чают название соответ-
ственно основному токсическому веществу, содержащемуся в грибах. В зависимости
от этиологического фактора патогенеза развития и клинических проявлении инток-
сикации С. Г. Мусселиус и А. А. Рык (2002) описали следующие психопатологичес-
кие синдромы.
Психотонический (микоатропиновый) синдром. Развивается при употреблении му-
хомора красного (A. muscaria), мухомора пантерного (A. pantherina), мухомора желтого
(A. junquillea), мухомора пышного (A. regalis). Именно этот синдром многими автора-
ми описан под названием «мускариновый». Наиболее распространены отравления
после употребления мухомора красного, мухомора пантерного и мухомора желтого.
Наиболее тяжелая клиническая картина отмечается после употребления мухомора
пантерного. Летальность составляет 4—10 %. Токсические вещества:
иботеновая кислота (а-амино-З-гидрокси-5-изоксазолацетиловая кислота) —
воздействуя на глутаминчувствительные рецепторы головного мозга, повышает дви-
гательную активность, в больших концентрациях вызывает судорш и;
мусцимол (энолбетаин 5-аминометил-З-гидроксизоксазол) — образуется после
декарбоксилирования иботеновой кислоты. По составу близок к ГАБК, участвует в
передаче нервного импульса в головном мозге. Мусцимол как активное соединение
вытесняет из реакции ГАБК, блокируя тем самым рецепторы головного мозга. Вызы-
вает угнетение эмоционально-психической сферы деятельности мозга человека;
метилтетрагидрокарболинкарбоновая кислота — оказывает галлюциногенное
действие;
мускарин и его изомеры — содержание этих алкалоидов в мякоти гриба не более
2 % от общего химического состава. Поэтому в клинике микоатропинового синдро-
ма они значения не имеют (рис. 2.36).
В развитии интоксикации играют роль и малоизученные вещества: мусказон (по-
тенцирует эффект мусцимола), мускарифин, стизолобовая и стизолобиновая амино-
кислоты, холин, бетаин, герцинин. Мусказон, стизолобовая и стизолобиновая ами-
нокислоты обладают антихолинергическим действием (атропинопоцобный эффект).
Клинику отравления определяют иботеновая кислота и мусцимол. Инкубацион-
ный период короткий, составляет от 15 мин до 4 ч. Отравление протекает по типу
холинолитического синдрома. К ранним симптомам относят тошноту, расстройства
движения. В дальнейшем отравившийся становится беспокойным. Симпатомимети-
ческий эффект проявляется тахикардией, мидриазом, сухостью слизистых. Возника-
ют визуальные гатлюцинации. Поведение вычурное: речь бессвязная, больной кри-
чит, громко поет. Состояние такого пациента может быть принято за алкогольное
опьянение. Постепенно его настроение становится подавленным. Дисфория сменя-
2. Биологические поражающие агенты
437
Декарбоксилирование
Мусцимол
Мусказон
Рис. 2.36. Химическая структура иботеновой кислоты, мусцимола и мускарина
ется чувством страха, усиливается атаксия, развивается психомоторное возбуждение,
переходящее в делирий (параноид). Развитие желудочно-кишечных расстройств от-
«чают при тяжелых формах отравления мухомором пантерным.
В результате разнонаправленного действия токсических веществ, концентраци-
онные соотношения которых в мякоти гриба варьируют в больших пределах, в кли-
нической картине отравления могут наблюдаться также умеренная брадикардия,
-- лчвация, миоз. Продолжительность клинических признаков отравления 10—15 ч.
3 тяжелых случаях развиваются коллапс и судороги. Смерть наступает от остановки
сердечной деятельности и дыхания.
В редких случаях из-за необычайно повышенного содержания в грибах мускари-
на и его производных отмечаются симптомы, связанные с возбуждением М-холинер-
ттческих рецепторов: умеренное сужение зрачков, повышенная секреция потовых
желез, гиперсативапия. В отличие от наркотических препаратов при приеме выше-
перечисленных мухоморов у больных отсутствуют эйфория и чувство безграничного
с тастья, а ведущими клиническими признаками являются подавленность настроения
чувство страха.
Психодислептические синдромы. Проявляются развитием психических расстройств
и галлюцинаций зрительных, слуховых и зрительно-слуховых (синестических). На-
считывается более 100 видов грибов, при употреблении которых развивается пси-
<одислептический синдром. В соответствии с клинической картиной отравления
С. Г. Мусселиус и А. А. Рык (2002) выделили следующие частные синдромы:
псилоцибиновый синдром (наркотический с синестическими галлюцинациями). Хи-
мический состав грибов, отравление которыми вызывает псилоцибиновый синд-
ром, включает индолсодержащие соединения' псилоцин, псилоцибин, триптамин
рис. 2.37).
В развитии синдрома псилоцибин является одним из основных химических со-
единений. Эти вещества обнаружены в грибах семейства строфариевых, рода псило-
-ибе (Psilocybe): псилоцибе мексикана (Р. tnexicana), псилоцибе запотекорум (Р. zapo-
'есогит), псилоцибе полуланпевидная (Р. semilanceata), псилоцибе каллоза (Р. callosa),
'силоцибе пелликулоза (Р. pelliculosa), а также в грибах панеолус сфинктринус (Рапа-
eolus sphinctrinus), панеолус суббалтеатус (Р. subbalteatus), коносиб цианопус (Сопо-
суbe cyanopus). Степень выраженности наркотического действия вышеперечисленных
грибов различна. У одних видов грибов это действие постоянное, у других оно спора-
дическое.
Наибольшее количество индолсодгржаших алкалоидов обнаружено в псилоци-
бе полуланцетовидной. В российских обра щах псилоцибе полуланцетовидной содер-
жание псилоцибина высокое — 1,1—1,6 % на 1 г сухой массы. Метаболиты псило-
иибина, обладая химической структурой, схожей с нейромедиатором серотонином,
438
Часть 2. Общая неправильная эпидемиология
Рис. 2.37. Химическая структура токсических веществ,
вызывающих псилоцибиновый синдром
воздействуют на серотонинергические рецепторы головного и спинного мозга, нару-
шая тем самым естественные процессы передачи нервного импульса. Несбалансиро-
ванность активности нейрональных связей, относящихся к эмоциональной сфере
деятельности, приводит к повышению возбудимости коры головного мозга, которое
проявляется острым психозом. Вегетативные расстройства связаны с симпатомиме-
тическим действием токсических веществ гриба.
Латентный период составляет от 30 мин до 3 ч. К начальным проявлениям отно-
сятся изменения в настроении в виде эйфории, иногда взволнованности. Окружаю-
щие предметы представляются ярко окрашенными. Появляется чувство необычайно
приятного состояния, беспричинное веселье, удовлетворенность собой, высокие идеи
стимулируют созидательную активность, но она беспорядочна. Нарушаются коорди-
нация движений и равновесие, походка становится шаткой. Развивается картина пси-
хических расстройств. Обостренное зрительное и слуховое восприятие сменяется зри-
тельными галлюцинациями, возникают цветные видения, калейдоскопы. Происходит
расстройство восприятия времени и места. Нарушается ощущение собственного тела,
части которого могут восприниматься как чужие. Беспричинный смех и веселье сме-
няются приступами тревоги и ужаса. Возможны агрессивность и суицидальные по-
пытки. При обследовании выявляется мидриаз, возможна умеренная брадикардия,
снижение наполнения пульса, тенденция к гипотонии. При неосложненном течении
развивается сонливость, переходящая в глубокий сон. Максимальные проявления
психических и соматических расстройств достигаются через 2—4 ч после приема гри-
бов. Через 6-12 ч отравленный постепенно выходит из этого состояния.
При тяжелом клиническом течении отравления отмечаются случаи развития ост-
рой почечной недостаточности, инфаркта миокарда. В крайне тяжелых случаях отрав-
ления грибами-галлюциногенами развивается кома. Смерть наступает от острой сер-
дечно-сосудистой недостаточности. У детей при отравлении грибами наркотического
действия, помимо психических расстройств, отмечаются гипертермия, головокруже-
ние, боли в животе, тошнота, рвота. В тяжелых случаях развиваются судороги, кома;
2.2. Биологические поражающие агенты
439
стерилпироновый синдром (со зрительными галлюцинациями). Химический состав
грибов, вызывающих синдром, включает производные стерилпирона. Основное из
них — меконовая кислота. Входит в состав опия. Механизм действия меконовой кис-
лоты на организм человека мало изучен, тем не менее воздействие на ЦНС связыва-
ют именно с этим химическим веществом. Производные стерилпирона содержатся в
строфарии синезеленой (Stropharia aeruginosa), строфарии украшенной (5. coronilla),
чешуйчатке ворсистой (Pholiota squarrosa), гимнопиле видном (Gymnopilus spectabilis)-,
индоламиновый синдром (со слуховыми галлюцинациями). Развивается при приеме
некоторых грибов рода дождевик (Lycoperdon): дождевик кандидум (L. candidum),
дождевик круциатум (£. cruciatum), дождевик микстекорум (£. mixtecorum) и дождевик
маргинатум (L. marginatum). Последний и в настоящее время используется в Мексике
при совершении обрядов и ритуальных процессий. Токсическими веществами пред-
положительно являются производные индоламина, воздействующие на серотонинер-
гические рецепторы головного мозга.
Через 15—45 мин после употребления препарата на основе мякоти такого гриба, у
человека возникает психомоторное возбуждение, появляются галлюцинации зритель-
ные (стерилпироновый синдром), слуховые (индоламиновый синдром). Слуховые
галлюцинации становятся настолько правдоподобными, что принявший грибы всту-
пает в переговорный контакт. Подобная развернутая клиническая картина отмечена
при употреблении грибов дождевика микстекорума и дождевика маргинатума. При
приеме других грибов симптомы воздействия на ЦНС не столь выражены.
2.2.3.5. Зоотоксины
Классификация ядовитых животных. Роль отдельных токсических веществ в форми-
ровании клинической картины отравления ядами животных. Сакситоксин. Гимберо-
токсин. Суберитин. Яд тонионем. Яд физалий. Яд хиронекс. Яд хризаоры. Яд циа-
неи. Лофотоксин. Яд актиний. Эквинотоксин. Теалиатоксин. Палитоксин. Яд конусов.
Мурексин. Макулотоксин. Яд скорпионов. Яд пауков-птицеедов. Яд каракурта. Яд
тарантула. Яд пчел. Кантаридин. Яд сколопендры. Яддиамфидий. Яд морских ежей.
Яд скатов. Яд бородавчатки. Тетродотоксин. Сигуатоксин. Батрахотоксин. Буфоте-
нин. Кобротоксин. Яд морских змей. Яды гадюк (виперотоксин) и гремучников (кро-
талотоксин). Яд гремучей змеи Crotalus durissus terrificus. Мойяветоксин. Яд буме-
ланга Dispholidus typus. Яд древесной змеи Thelotornis kirtlandi. Яд подвязочной змеи
Thamnophis elegans vagranus. Яд ядозубов. Модификация ядов животных.
По уже устаревшим данным С. В. Пигулевского( 1975), ядовитых животных насчи-
тывается не менее 5 тыс. видов, в том числе простейших — 21, кишечнополостных —
93, паразитических червей — 16, кольчатых червей — 50, членистоногих — 4 тыс.,
моллюсков — 91, иглокожих — 26, рыб — 500, амфибий — 40, рептилий — 102, мле-
копитающих — один вид1. В пределах территории бывшего СССР встречается около
1,5 тыс. видов ядовитых животных.
1 В более поздней работе Б. Н. Орлова и Д. Б. Гелашвили (1985) приводятся другие данные
по численности ядовитых млекопитающих. Они представлены одним видом утконосов и ше-
стью видами ехидны, относящихся к подклассу яйцекладущих млекопитающих. Их ядовитый
аппарат представлен бедренной железой, которая протоком соединяется со шпорой, распо-
ложенной на запястье задней лапы с внутренней стороны Ядовитый секрет имеет белковую
природу и действует угнетающе на сердечно-сосудистую систему млекопитающих. Кроме того
440
Часть 2. Общая неправильная эпидемиология
Классификация ядовитых животных. Е. Н. Павловский (1950) предложил всех ядо-
витых животных разделить на две большие группы: первично-ядовитые и вторично-
ядовитые. Эта классификация используется и сегодня (рис. 2.38).
Ядовитые животные
Первично-ядовитые Вторично-ядовитые
Активно-ядовитые'^''4''4''
Пассивно-ядовитые
Вооруженные Невооруженные
Рис. 2.38. Токсинологическая классификация ядовитых животных
К первично-ядовитым относят животных, вырабатывающих ядовитый секрет в
специальных железах или имеющих ядовитые продукты метаболизма. Ядовитость
первично-ядови гых животных является видовым признаком и встречается у всех осо-
бей данного вида. К вторично-ядовитым относят животных, аккумулирующих экзо-
генные яды и проявляющих токсичность только при приеме их в пищу. Примером тому
могут служить пластинчато-жаберные моллюски, накапливающие в своем теле яд
некоторых динофлагеллят; насекомые, питающиеся на ядовитых растениях; рыбы,
вызывающие отравления типа сигуатера и др.
Первично-ядовитые животные различаются по способам выработки яда и его
применения и делятся на активно- и пассивно-ядовитых. Активно-ядовитые живот-
ные, имеющие специализированный ядовитый аппарат, снабженный ранящим уст-
ройством, называются вооруженными. В типичном случае ядовитый аппарат таких
животных имеет ядовитую железу с выводным протоком и ранящее приспособление:
зубы у рептилий, жало у насекомых, колючки и шипы у рыб. В деталях строение ядо-
витого аппарата может варьировать, однако для всех вооруженных животных харак-
терно наличие ранящего приспособления, позволяющего вводить ядовитый секрет в
тело жертвы парентерально. Такой способ введения яда наиболее эффективен для
ядообразующего организма. Другую группу активно-ядовитых животных составляй т
организмы, ядовитые аппараты которых лишены ранящего приспособления — нево-
оруженные ядовитые животные. Примерами могут служить кожные железы амфибии,
анальные железы насекомых, кювьеровы органы голотурий. Ядовитые секреты таких
желез вызывают токсический эффект при контакте с покровами тела жертвы. Чем
энергичнее идет всасывание ядов с таких покровов (особенно слизистых), гем эффек-
тивнее его действие. У пассивно-ядовитых животных ядовитые метаболиты вырабаты-
ваются в организме и накапливаются в различных органах и тканях (пищеваритель-
ных, половых), как, например, у рыб, хвостатых амфибий, моллюсков, насекомых.
Пассивно-ядовитые и вторично-ядовитые животные представляют опасность
только при попадании в пищеварительный тракт, однако существенным различием
имеются данные, что слюна подчелюстных желез отдельных видов насекомоядных (щелезу-
бы, землеройки) обладает нейротоксическим действием. Однако ядовитость не свойственна
представителям млекопитающих, что авторы объясняют высоким совершенством их нервной
системы по сравнению со всеми эволюционно предшествующими классами позвоночных. Это
позволило им выработать достаточно эффективные средства защиты и способы нападения
путем различных рефлекторных и поведенческих реакций.
2.2. Биологические поражающие агенты
441
между ними является постоянство ядовитости (видовой признак) для первых и ее
спорадический характер для вторых. Яды и токсины животных различных таксоно-
мических групп, изучены и представлены в научной литературе весьма неравномер-
но. Для понимания «полосы свободы» преступников, использующих зоотоксины, нам
важно иметь общие представления о путях эволюции ядовитости среди животных.
Эволюция ядовитости среди животных. На ранних этапах эволюции животных
(простейшие) в качестве ядов выступали только метаболиты, либо выделявшиеся в ок-
ружающую среду, либо накапливающиеся в организме. С развитием специализиро-
ванных структур или органов одновременно происходило развитие специализирован-
ных структур, продуцирующих яд. Вначале — за счет усиления защитной функции
эктодермы (немертины, полихеты, иглокожие, кишечнополостные). Затем возникно-
вение таких органов происходит на базе эндокринных или экзокринных желез. У пе-
репончатокрылых ядовитый аппарат филогенетически связан с половой системой, у
головоногих моллюсков и змей — с пищеварительной.
У многих видов (рыбы, амфибии, насекомые) сохранились черты примитивной
ядовитости. Они накапливают ядовитые метаболиты в тканях и органах (Орлов Б. Н.,
Гелашвили Д. Б., 1995).
У кишечнополостных впервые на эволюционной лестнице встречаются яды
белковой природы. Среди них имеются как токсические неэнзиматические полипеп-
тиды, так и ферменты с различной субстратной специфичностью. Среди токсинов
кишечнополостных имеются цитотоксины и нейротоксины. Последние уже харак-
теризуются высокой специфичностью взаимодействия с ионными каналами (на-
пример, у актиний). У паукообразных в составе яда обнаружены видоспецифические
нейротоксины (нейротоксины млекопитающих, насекомых и ракообразных у скор-
пионов). Между химической природой зоотоксинов, морфологическими особенно-
стями ядовитого аппарата и биологической спецификой действия того или иного яда,
естественным отбором поддерживается определенное соответствие (табл. 2.20, с. 442).
Ядовитые секреты подавляющего большинства вооруженных активно-ядовитых
животных представляют собой сложные смеси токсических полипептидов и фермен-
тов (яды кишечнополостных, змей, перепончатокрылых, пауков, скорпионов и др.).
Эти яды активны при парентеральном введении, при введении внутрь они расщеп-
ляются пищеварительными ферментами.
Ядовитые животные, обладающие невооруженным ядовитым аппаратом, обычно
имеют яды небелковой природы (амфибии, муравьи, жуки, многоножки и др.). Яд
пассивно-ядовитых животных небелковой природы. В естественных условиях он эф-
фективен при попадании в организм жертвы с пищей (тетродотоксин, суругатоксин,
токсины динофлагеллят и др.).
Б. Н. Орлов и Д. Б. Гелашвили (1985) отмечают наличие связи между биологией
животного и особенностями продуцирования им яда. Как правило, хищнический
образ жизни сопровождается развитием вооруженной формы ядовитого аппарата
(кишечнополостные, змеи, скорпионы, пауки, актинии, осы). Во многих случаях яды
таких животных имеют выраженную нейротропную активность, поскольку предназ-
начаются для обездвиживания жертвы. Это нейротропное действие сопровождается
высокой избирательностью, обусловливающей пищевую специализацию животного.
В яде скорпионов имеются видоспецифические нейротоксины, избирательно дейст-
вующие на млекопитающих, насекомых и ракообразных. У животных с невооружен-
ным ядовитым аппаратом продуцируемые ими яды выполняют в подавляющем боль-
шинстве случаев защитные функции, т. е. являются отпугивающими веществами —
442
Часть 2. Общая неправильная эпидемиология
Таблица 2.20
Химическая природа зоотоксинов*
Таксон Небелковые Белковые
изо- и гетеро- циклические алифа- тические олиго- и поли- пептиды фермен- ты
Тип саркомастигофоры (Sacomastigophora)
Класс жгутиконосцы (Mastigophora) +
Тип губки (Spongia) + + +
Тип кишечнополостные (Coelenterata)
Класс гидрозои (Hydrozoa) + + +
Класс сцифоидные медузы (Scyphozoa) + + +
Класс коралловые полипы (Anthozoa) + + + +
Тип немертины (Nemertim)
Подкласс невооруженные (A.iopla) +
Подкласс вооруженные (Enopla) +
Тип кольчатые черви (Anne'ida)
Класс многощетинковые (Poluchaeta) + + + +
Тип членистоногие (Arthropoda)
Класс ракообразные (Crustacea) +
Класс паукообразные (Arachnida) + + +
Класс многоножки (Myriapoda) + + + +
Класс насекомые (Insecta) + + + +
Тип моллюски (Mollusca;
Класс брюхоногие (Gastropoda) + + + +
Класс двустворчатые (Bivalvia) +
Класс головоногие (Cephalopoda) + + +
Тип иглокожие (Echinodermata)
Класс морские звезды (Asteroidea) +
Класс морские ежи (Echinoidea) +
Класс голотурии (Holothuroidea)
Тип хордовые (Chordata)
Класс круглоротые (Cyclostomata) +
Класс хрящевые рыбы (Chondrichthyes) +
Класс костные рыбы (Osteichthyes) + + 4-
Класс амфибии (Amphibia) + + + +
Класс рептилии (Reptilia) +
Класс млекопитающие (Mammalia) +
* По Б, Н. Орлову и Д. Б. Гелашвили (1995).
репеллентами и имеют простой состав (стероиды, органические кислоты, эфиры
и цр.). Эволюционные механизмы формирования разнообразия среди зоотоксинов
описаны в работах J. Zhang (2003), V. J. Lynch (2007), Y Moran et al. (2008), A. Koz-
minsky-Attas et al. (2008) и др.
2.2. Биологические поражающие агенты
443
Роль отдельных токсических веществ в формировании клинической картины отрав-
ления ядами животных Для ядов животных организм человека представляет «мозаич-
ную мишень». Основные их компоненты представлены следующими веществами1.
Адреналин (кожный секрет жаб) — обладает выраженным кардиотоническим, прес-
сорным, гипергликемическим и пирогенным действием, вызывает сужение сосудов
кожи, почек, расширяет коронарные сосуды, сосуды скелетных мышц, гладкой мус-
кулатуры, бронхов, желудочно-кишечного тракта и т. д.
Алкалоиды (кожа и секреты амфибий) — обладают нейротоксическим и кардио-
токсическим действием (батрахотоксин, гефиротоксин, самандарин и др.).
Ацетилхолин (яд шершня, яд элапид) — сочетание гистамина и ацетилхолина обес-
печивает сильный болевой (аллогенный) местный эффект яда. Присутствие в яде
биогенных аминов (тирамин, дофамин, норадреналин, гистамин) и ацетилхолин,!
объясняет его гипотензивное действие. При внутривенном введении такой компози-
ции у экспериментального животного падает АД и периферическое сопротивление,
увеличивается сердечный выброс, стимулируется дыхание.
Ацетллхолинэстераза (яд скорпионов, яд элапид) — усиливает действие нейро-
токсинов.
Брадикинин (яд ос, кожа лягушек) — вызывает вазодилятацию и падение АД.
Васкулярный эндотелиальный фактор роста, VEGF-F (яды змей) — фактор роста,
содержащийся в яде змей, обладает выраженным гипотензивным эффектом и повы-
шает сосудистую проницаемость у отравленного. Структуры VEGF тканевого типа
высоко консервативны, однако VEGF, содержащиеся в яде змей, высоко вариабель-
ны, особенно в регионах вокруг рецептор-связывающих петель и С-конца предпола-
гаемого кирецепторного региона (Yamazaki Y. et al., 2009).
Гиалуронидаза (яд скорпионов, тарантулов, пчел, муравьев, рыбы-бородавчатки,
гадюк, ядозуба) — гидролизует гиал} роновую кислоту. Оптимум действия по pH раз-
ный. Способствует распространению яда по тканям. Вызывает некроз и отек тканей.
Гистамин (яд дискомедуз, скорпионов, шершня, пчел, муравьев, ядовитого сек-
рета гусениц, морских дракончиков и др.) — в основном содержится в чде беспозво-
ночных. При проникновении в организм человека оказывает сильное действие на
афферентную парасимпатическую иннервацию. В результате этого возникает сокра
щение матки и усиливается перистальтика кишечника. Например, при отравлении
я том перепончатокрылых нередко наблюдаются расстройства стула. В то же время
гистамин вызывает симптомокомплекс, напоминающий анафилактическое состоя-
ние, и может вызвать аллергические явления (отравления ядом пчел и муравьев). Эти
явления выражены очень сильно и приводят к тяжелым симптомам местной и общей
интоксикации. Вызывает сильное болевое ощущение в месте введения, расширение
капи ияров и повышение их проницаемости, сужение крупных сосудов, сокращение
гладкой мускулатуры, падение АД. Резко повышает секрецию соляной кислоты в
желудке Действие гистамина непродолжительно.
Дофамин (яд шершня, пчел) — является катехоламином, предшественником но-
радреналина. Оказывает специфическое влияние на дофаминовые рецепторы, для
1 Здесь приведено описание небольшой части биологически активных компонентов, вхо-
дящих в состав ядов животных и формирующих клиническую картину отравления. Только в
яде пауков их открыто не менее сотни (см. работу Vassilevski A. A. et al.. 20091 Поэтому читате-
лям, интересующимся этой темой более глубоко, я рекомендую обращаться к специальной
литературе.
444
Часть 2. Общая неправильная эпидемиология
которых является эндогенным лигандом, однако в больших дозах он стимулирует так-
же а- и |3-адренорецепторы. Клинически это проявляется повышением АД, спазмом
периферических артерий, тахикардией, желудочковой экстрасистолией, стенокарди-
ей, диспноэ, головной болью, психомоторным возбуждением. Местное действие, ско-
рее всего, проявляется некрозами.
Ингибиторы протеаз (змеи, насекомые, анемоны) — содержатся в ядах многих
ядовитых животных. Возможно, защищают белковые токсины от деградации проте-
иназами тканей жертвы.
Кардитоксины (яд нематоцист цианеи, дискомедуз, конусов, змей) — обычно это
мембрано-активные пептиды (см. ниже).
Кинины — гипотензивные аллогенные пептиды (брадикинин, веспакинин X, вес-
пакинин М, политескинин и др.). Вызывают повышение сосудистой проницаемости
и обратимое сокращение гладкой (несосудистой) мускулатуры. Важный патогенети-
ческий фактор, ответственный за развитие общих и местных симптомов отравления
у позвоночных.
Меллитин (яд пчел) — сильный цитолитический полипептид. Взаимодействует с
мембранами многих клеток, вызывая широкий спектр физиологических реакций.
Среди них прямой гемолиз эритроцитов, высвобождение гистамина из тучных кле-
ток и др.
Мембрано-активные пептиды, МАП (яд кобр и бунгарусов) — собирательное по-
нятие, включающее пептиды ядов животных, активно воздействующие на мембраны
клеток. Их не менее трех типов: гемолизины, цитотоксины и кардиотоксины. Одним
из характерных свойств МАП является одновременное присутствие в яде фосфоли-
пазы А2, действующей синергидно с МАП. МАП «вскрывают» поверхностные струк-
туры мембран, обнажая фофолипидный субстрат для атаки фосфолипазой А2.
МСД-пептид, пептид 401 (яд ос, пчел) — белковый компонент яда пчел деграну-
лируюший тучные (мастоцитные) клетки. Вызывают высвобождение из них гистами-
на. Состоит из 22 аминокислотных остатков, ММ 2588 Да. Проявляет ярко выражен-
ные основные свойства (изоэлектрический оптимум р! 12), зависящие от наличия
9 основных аминокислот. С-конец молекулы амидирован. Большое структурное свой-
ство с апамином. И в том и другом соединении имеются две дисульфидные связи, их
молекулы близки по своей длине, не содержат ароматических аминокислот, метио-
нина, серина и треонина. Однако на этом сходство и заканчивается, так как физио-
логическая активность существенно различается. Апамин не оказывает действия на
тучные клетки, а пептид 401 не влияет на нехолинергическую и неадренергическую
нервную регуляцию гладких мышц и не дает нейротоксических эффектов при систем-
ном введении в организм. Его внутрижелудочковое введение крысам производит эф-
фекты, если доза превышает таковые для апамина в 10 раз. Различие в эффектах обе-
спечивается различием вторичной и третичной структуры пептидов. По своей актив-
ности высвобождать биогенные амины из тучных клеток МСД-пептид значительно
превосходит мелитин и фосфолипазу А (в 10—100 раз). При этом механизм такой ак-
тивности различен. Если фосфолипаза и мелитин высвобождают секрет, разрушая
клеточные и внутриклеточные мембраны, то МСД-пептид высвобождает гистамин из
гранул тучных клеток посредством слияния гранул с клеточной мембраной, приводя-
щего к экзоцитозу — содержимое гранул выбрасывается во внеклеточное простран-
ство без лизиса тучных клеток.
Нейротоксины — собирательное понятие, включающее различные токсины, спе-
цифически действующие на нервные клетки путем взаимодействия с ионными кана-
2.2. Биологические поражающие агенты
445
лами и белками электровозбудимых мембран. Б. Н. Орлов и Д. Б. Гелашвили (1985)
делят нейротоксины животных на 3 группы: аксональные, пресинаптические и пост-
синапические1.
Аксональные нейротоксины (батрахотоксин, лофотоксин, макулотоксин, пали-
токсин, сакситоксин, сигуатоксин, тетродотоксин; нейротоксины немертин, морской
анемоны, скорпионов и др.) демонстрируют самый древний механизм блокирования
прохождения нервного импульса по аксону — через блокирование ионных каналов.
Клеточные мембраны простейших электровозбудимы. У них же сформировались ион-
ные каналы (пассивные и активные). В ходе эволюции они наследовались многокле-
точными организмами всем «пакетом». Это объясняет тот факт, что синтез веществ,
взаимодействующих с ионными каналами, закреплен естественным отбором, начи-
ная еще с одноклеточных (сакситоксин динофлагеллят) и первых многоклеточных жи-
вотных (палитоксин зоонтарий). У более высокоорганизованных организмов с раз-
витой нервной системой они закупоривают натриевые каналы аксонов, в результате
чего нервные волокна теряют способность проводить импульсы. Функция органов и
тканей, которые они иннервируют, нарушается (рис. 2.39).
Рис. 2 39. Схема действия аксональных нейротоксинов. 1 — блокирование
тетродотоксином ионотранспортного участка натриевого канала; 2 —блоки-
рование такого же канала батрахотоксином; 3— взаимодействие нейроток-
сина скорпиона с компонентами «воротного механизма» натриевого канала.
По Ю. А. Овчинникову (1980)
Среди аксональных нейротоксинов нет действующих только на периферическом
уровне. Сакситоксин и тетродотоксин блокируют натриевые каналы аксонов нейро-
на кальмара при наружном применении, но у интактных животных они оказывают
прямое угнетающее действие на дыхательный и сосудодвигательный центры. Нейро-
токсины скорпионов вызывают одновременное возбуждение симпатического и пара-
симпатического отделов ВНС. Деполяризующее действие отдельных аксональных
нейротоксинов распространяется на мышечные мембраны (батрахотоксин, яд брю-
1 Классификация нейротоксинов Б. Н. Орлова и Д. Б. Гелашвили (1985) несовершенна из-
за того, что она не учитывает современные данные по рецепторному составу холинэргических
и адренергических синапсов. Но дальнейшее деление понятия «нейротоксины» по рецептор-
ной специфичности (principium divisionis) необычайно усложняет изложение материала и не
соответствует цели работы. Более подробно о механизмах похождения сигналов по нервным
клеткам можно прочитать в работе X. Люльмана с соавт. (2008).
446
Часть 2. Общая неправильная эпидемиология
хоногих моллюсков), что проявляется сильным кардиотоксическим эффектом. Из-за
многообразия ионных каналов и различной специфичности аксональных нейроток-
синов по отношению к этим каналам, выделить в клинике отравления у человека ка-
кие-то симптомы, характерные именно для нейротоксинов этого типа, очень сложно.
Предположить их участие в отравлении можно по быстро развивающимся (до 30 мин
после проникновения в организм токсина) симптомам поражения периферических
нервов: онемение губ, языка, кончиков пальцев, сопровождаемых мышечной слабо-
стью с последующим параличом дыхательной мускулатуры (сакситоксин); развитие
параличей конечностей (сигуатоксин); атаксия и мышечный паралич (лофотоксин);
покалывание и онемение языка и губ, извращение температурной чувствительности,
нарушение координации движений, параличи и парезы скелетной мускулатуры (си-
гуатоксин); онемение слизистых, мышц лица и шеи, афония, дисфагия, быстро раз-
вивающийся паралич дыхательной мускулатуры (макулотоксин); судороги, мышеч-
ный тремор, эпилептические припадки у детей (нейротоксины скорпионов) и др.
Точкой «приложения» пресинаптических нейротоксинов является пресинаптиче-
ское нервное окончание. Такие нейротоксины содержатся в ядах актиний, паука-ка-
ракурта, слюне клеща Ixodes holocyclus, гремучих и гадюковых змей, тайванской коб-
ры, индийских змей рода Bungarus (Р-бунгаротоксин из яда полосатого крайта),
австралийских змей (тайпоксин из яда тайпана, нотексин из яда тигровой змеи) и др.
Для большинства пресинаптических токсинов характерно двухфазное действие.
Первая фаза действия нейротоксина на синаптическую передачу заключается в уси-
лении выброса медиатора (нейротрансмиттера) в ответ на одиночное раздражение
нерва, приводящее к усилению амплитуды возбуждающего постсинаптического по-
тенциала (ВПСП) и усилению постсинаптической реакции. Вторая фаза связана с
развитием стойкой деполяризации мембраны и блокированием проведения возбуж-
дения. Под действием такого нейротоксина яда паука-каракурта (а-ларотоксин,
a-LTX) сначала достигается 600—1000-кратное увеличение частоты миниатюрного по-
тенциала концевой пластинки (МПКП), но через 30—50 мин оно сменяется полным
блоком нервно-мышечной передачи.
Блокирование синаптической передачи пресинаптическим токсином может осу-
ществляться по нескольким дублирующим механизмам. Рассмотрим их на примере
действия a-LTX. Молекулы a-LTX, взаимодействуя с липидной мембраной в услови-
ях in vitro, формируют пору — гомотетрамерные комплексы с каналом в центре. Но в
биологических мембранах a-LTX не формирует пор без взаимодействия со специфи-
ческими рецепторами в плазматической мембране. По данным Y. A. Ushkaryov et al.
(2008), три протеина с различной структурой способны связывать a-LTX: неурексин
la (neurexin la), латрофилин 1 (latrophilin 1)ирецепторподобныйбелоктирозинфос-
фатаза a (receptor-like protein tyrosine phosphatase a). a-LTX вызывает Са2+-зависимое
и независимое высвобождение нейротрансмиттера. Как только токсин связывается с
таким сайтом, часть его действует по Са2+-зависимому механизму вследствие форми-
рования поры и поступления Са2+. Вхождение ионов кальция внутрь пресинаптиче-
ского окончания вызывает освобождение из везикул медиатора, диффундирующего
через синаптическую щель и реагирующего с рецепторами постсинаптической мемб-
раны. Другой механизм действия a-LTX основывается на рецептор-опосредованном
сигнализировании, включающим стимуляцию фосфолипазы С, образующей 1РЗ и
диацилглицерол (diacyl glycerol), с соответствующим высвобождением запасов Са2+
и активацией протеинкиназы С. Далее происходит фосфорилирование синапсина,
приводящее к отсоединению везикул и их связыванию с пресинаптической мембра-
2.2. Биологические поражающие агенты
447
ной. Этот механизм наиболее вероятно осуществляется через латрофиллин 1 (LPH1).
Оба процесса — формирование пор и рецепторопосредованные сигналы, усиливаются
благодаря взрывному высвобождению Са2+. В отсутствие Са2 a-LTX только связыва-
ется с LPH1 и РТРс (рис. 2.40).
Сигнал
через
латрофи-
линовый
рецептор
Терминаль нерва
СС
LTXzx
Кальций отсутствует
Вставка LTX
через LPH1
или РТРо
Сигнал
через
латрофи-
линовый
рецептор
Любой
рецептор
Са2+
Na1
LTXzx
Вставка LTX
через LPH1
или РТРо
РКС?
Na
Вода?
LPH1
Экзоцитоз
Повреждение J мембраны
Любой
рецептор
Кальций присутствует
♦ Запас
• кальция «
РКС
’3+DAG
PLC
Рис. 2.40. Механизм действия пресинаптического токсина на примере a-LTX.
Правая часть рисунка — Са2+ представлен в среде; левая часть рисунка — среда
без кальция. Экзоцитоз медиатора вызывают: 1 — высокая концентрация Na+,
мимикрирующая под содержание Са2+ в среде; 2 — интернализованные домены
a-LTX, взаимодействующие с компонентами механизма, осуществляющего
экзоцитоз (Е); 3 — a-LTX, вставившийся непосредственно в везикулу. PLC —
фосфолипаза С; DAG — диацилглицерол; РКС — протеинкиназа С; SV — синап-
тическая везикула; МС — митохондрии; LTX 4х — a-LTX-тетрамеры; LTX 2х —
a-LTX-димеры; СС — Са2+-каналы; РТРо — рецепторподобный белок тирозин-
фосфатаза о. LPH1 — латрофилин 1-реиептор. По Y. A. Ushkaryov et al. (2008)
В действии пресинаптических токсинов элапид, обладающих фосфолипазной
активностью (например, нотексин, полученный из яда среднеазиатской кобры Naja
oxiana), выделяют три фазы. Начальная фаза характеризуется снижением частоты
МПКП и обусловлена связыванием токсина с пресинаптическим окончанием. Во вре-
мя второй фазы наблюдается значительное увеличение частоты МП КП (в случае фос-
фолипазы А2 из N. oxiana в сотни раз). Третья фаза действия таких токсинов характе-
ризуется прогрессивным падением частоты МПКП, вплоть до полного исчезновения.
Конечная стадия третьей фазы связана с истощением запаса медиатора в пресинапти-
ческой терминали, что в итоге приводит к развитию блока синаптической передачи.
Кротоксин, выделенный из яда южноамериканской гремучей змеи Crotalis durissus
terrificus, наряду с трехфазным действием, способен блокировать мышечное сокраще-
ние при прямом раздражении (Орлов Б. Н„ Гелашвили Д. Б., 1985).
Экспериментальные данные, полученные R. W, Dixon, J. В. Harris (1999), поз-
волили им предположить, что истощение синаптических везикул при отравлении
Р-бунгаротоксином является результатом сочетанного действия повышенного выс-
448
Часть 2. Общая неправильная эпидемиология
вобождения трансмиттера и замедления заполнения трансмиттером опустошенных
везикул. Кроме того, после экспонирования животных к Р-бунгаротоксину ими об-
наружено значительное повреждение терминалей мотонейронов и прогрессирующая
во времени аксонопатия, которая приводила к деструкции внутримышечных аксо-
нов в мышце диафрагмы. Эти наблюдения объясняют дегенеративные процессы в
мышечной ткани пациентов, перенесших отравления Р-бунгаротоксином. Если
шванновские клетки мотонейронов «переживали» отравление, то они способствова-
ли регенерации аксонов и восстановлению функции мышц за период не менее 2 нед.
М. Rigoni et al. (2008) показали другой механизм развития аксонопатии в результате
действия пресинаптических нейротоксинов змей с активностью фосфолипазы А2. Та-
кие нейротоксины способны проникать в мотонейроны спинного мозга и мозжечко-
вые гранулярные нейроны (cerebellar granule neurons) и селективно связываться с ми-
тохондриями, подвергая их глубоким изменениям (рис. 2.41).
Пресинаптические нейротоксины действуют медленнее, чем аксональные. Гибель
животных при введении смертельных доз таких токсинов происходит не в течение
нескольких часов, а суток и более. Так же как и у аксональных нейротоксинов, дей-
ствие пресинаптических токсинов на клеточные мембраны проявляется разнообраз-
ными патологическими эффектами, находящими свое отражение в клинике отравле-
ния. Пресинаптические нейротоксины актиний в наномолекулярных концентрациях
вызывают положительный инотропный эффект на изолированном сердце теплокров-
ных, в токсических концентрациях они вызывают нарушение сердечной деятельнос-
ти (аритмию, фибрилляцию, остановку сердца в систоле).
Увеличение высвобождения медиаторов из пресинаптических терминалей под
действием яда обеспечивает развитие соответствующих постсинаптических реакций
(клонические и тонические сокращения скелетной и гладкой мускулатуры, измене-
ние тонуса сосудов и деятельности сердца). Действие ос-ларотоксина каракурта про-
ходит сначала через стадию возбуждения, затем наступает атаксия, развитие бронхо-
спазма, клонических и тонических судорог, сменяющимися парезами, профузным
слюно- и слезотечением Аналогичным образом (т. е. стадию возбуждения ЦНС и ВНС
сменяет стадия их торможения) развивается картина отравления Р-бунгаротоксином
(яд полосатого крайта) и нотексином (змеи рода Notechis). Период короткого возбуж-
дения, сопровождающийся двигательной активностью отравленного, резким учаще-
нии пульса и дыхания, сменяется угнетением его двигательной активности и сонли-
востью, пульс резко ослабляется, появляется затруднение дыхания. Смерть может
наступить от паралича центра дыхания.
Постсинаптические нейротоксины специфически блокируют Н-холинорецепто-
ры субсинаптической мембраны скелетной мускулатуры и некоторых отделов ЦНС
и ВНС (яд морских змей, кобр рода Naja, бунгарусов, мамб, некоторых австралийских
змей). Благодаря их большей афинности к рецепторам ацетилхолина, чем у самого
ацелилхолина, они препятствуют его связыванию с рецепторами субсинаптической
мембраны (Ruan К. Н. et al., 1990). Постсинаптические нейротоксины змей делят на
две группы: короткие (тип I) и длинные (тип II), возникающие в результате дуплика-
ции генов коротких токсинов. Короткие нейротоксины состоят из 60-62 аминокис-
лотных остатков и стабилизированы 4 дисульфидными связями, их ММ около 7 кДа.
Длинные нейротоксины состоят из 71—74 остатков аминокислот и имеют 5 дисуль-
фидных связей. В яде кобр присутствуют нейротоксины обоих типов, тогда как в яде
бунгарусов только длинный нейротоксин — а-бунгаротоксин. Короткие нейроток-
сины быстрее связываются с холинрецепторами скелетных мышц по сравнению
2.2. Биологические поражающие агенты
449
Рис. 2.41. Электронно-микроскопические фотографии нервных окончаний мышцы диа-
фрагмы мыши, отравленной летальной дозой p-бунгаротоксина. Мышам введено в хво-
стовую вену 15 мг токсина в 0,15 мл NaCl (0,9 % w/v). Животные были умерщвлены, ког-
да их дыхание практически прекратилось. А. Контроль. В мышечной ткани четко видны
синаптические везикулы (v) и митохондрии (ш). Внутренние кристы митохондрий хоро-
шо определяются. Отростки шванновских клеток (Schwann cell processes) хорошо разли-
чимы (р) и находятся в стороне от синаптической щели. Терминали нервов из мышц
мышей, экспонированных к Р-бунгаротоксину, показаны на фотографиях Б—Д. Как пра-
вило, на них не видно везикул, хотя некоторые терминали содержат небольшое количе-
ство крупных везикул (lv), многие из которых погружены в электронно-плотный мате-
риал, возможно клатрин (clathrin, cv; см. Б, В и Д). В сложных концевых пластинках (см.
В), отдельные терминали содержат только большие покрытые везикулы (coated vesicles,
cv) и другие содержащие небольшие кластеры неизмененных везикул (v). Отростки шван-
новских клеток (р) вторгаются в синаптическую щель (В, Г и Д). Мембраны терминалей
нервов показаны профилем буквы «омега» (£2), и многие содержат поврежденные ми-
тохондрии (damaged mitochondria, dm на Д) На В показана увеличенная центральная
терминаль, показанная на Б. По R. W. Dixon, J. В. Harris (1999)
450
Часть 2. Общая неправильная эпидемиология
с длинными нейротоксинам и, однако последние имеют более прочные параметры свя-
зывания. Постсинаптические нейротоксины избирательно воздействуют только на
холинорецепторы субсинаптической мембраны, не затрагивая функционирование
пресинаптических нервных терминалей. Этот эффект проявляется на периферичес-
ком уровне и в ЦНС. Ганглионарные холинорецепторы по сравнению с нервно-
мышечным соединением более устойчивы к действию постсимпатических нейроток-
синов элапид, хотя специфическое связывание последних с симптическими ганглия-
ми имеет место (Dufton М. J., Hider R. С., 1983).
Отравление постсинаптическими нейротоксинами по быстроте развития симп-
томов и клинике напоминает отравление аксональными нейротоксинами. Характер-
ный пример — отравление ядом морских змей. Такие нейротоксины составляют ос-
новной токсический компонент их яда. Действие постсинаптического нейротоксина
проявляется параличами отдельных групп скелетной мускулатуры, развивающимися
в течение часа от момента поступления яда в организм человека. Заканчивается от-
равление параличом диафрагмы и полным прекращением дыхания у отравленного.
Почти у всех пациентов, отравленных ядом морских змей, отмечался паралич мышц
гортани и у некоторых также мышц языка. Отмечалась слабая ригидность мышц, осо-
бенно конечностей. Имели место судороги. Яд элапид (кобры, бунгарусы, мамбы)
после короткой фазы возбуждения ЦНС вызывает вялый паралич скелетной и дыха-
тельной мускулатуры, приводящий к полной остановке дыхания.
Норадреналин (яд шершня, пчел, морских ежей) — катехоламин, местное действие
проявляется некрозом. Преобладание а-адреномиметического действия приводит к
сужению периферических сосудов и к повышению системного АД.
Опиоидные пептиды, дерморфины (кожа жабы) — их анальгетическая активность не
менее чем в 11 раз больше, чем у морфина.
Протеазы змеиных ядов (гадюки, гремучники) — играют ведущую роль в развитии
местных повреждений тканей, геморрагических отеков, мионекрозов. Обладают фиб-
риногенолитическим, фибринолитическим, коагулирующим и брадикининлибера-
торным действием. Их делят на две группы: сериновые и металлопротеазы. Сериновые
протеазы — термолабильные эндопептидазы, активные по отношению к синтетиче-
ским субстратам аргининовых эфиров. Ингибируются фосфорорганическими инги-
биторами и по характеру действия близки к тромбиноподобным ферментам и кинино-
геназам. Играют важную роль в действии змеиных ядов на свертывающую систему
крови, приводя к тромбоэмболиям и геморрагиям. Металлопротеазы — термолабиль-
ные мультидоменные 2п2+-зависимые протеиназы, гидролизующие белковые субстра-
ты (казеин, гемоглобин, инсулин и др.). Их активность проявляется в присутствии
двухвалентных металлов, например, цинка, необходимых для поддержания биологи-
чески активной конформации молекулы фермента. Ингибируются комплексонами.
Общим для металлопротеаз змеиных ядов является предпочтительный гидролиз пеп-
тидных связей, содержащих аминогруппы лейцина и фенилаланина.
Серотононин (яд дискомедуз, скорпионов, стрекающих, ос, шершня, морских дра-
кончиков, ядозуба, кожа лягушек) — оказывает влияние на нервную деятельность,
вызывает сокращение гладкой мускулатуры кишечника, матки, бронхов, а также су-
жение сосудов. Вызывает сильную гипергликемию. Местное действие, скорее всего,
проявляется некрозами.
Синильная кислота (многоножки) — попадая в организм через дыхательные пути,
реже — через кожу, блокирует дыхательный фермент цитохромоксидазу и вызывает
кислородное голодание тканей. Очень ядовита.
2.2. Биологические поражающие агенты
451
Стероиды (кожа и секреты амфибий) — обладают кардиотоническим действием.
В яде жаб представлены агликонами, производными циклопентанпергидрофенанг-
рена, имеющими в качестве боковой цепи шестичленное ненасыщенное лактонное
кольцо. Действие сходное с таковым у растительного сердечного гликозида строфан-
тина. Вызывают брадикардию, повышение АД, связанное с усилением сердечных
сокращений. В больших дозах вызывает остановку сердца.
Сфингомиелинидаза D (аранеоморфные naj, ки) — вызывает дерматонекроз у поз-
воночных, лизирует эритроциты, высвобождает серотонин из тромбоцитов, в резуль-
тате чего происходит агрегация тромбоцитов и развивается внутрисосудистое свер-
тывание крови. Гемолитическая анемия, в свою очередь, приводит к почечной недо-
статочности.
Тахикинины (кожа лягушек) — фармакологически активные пептиды. Действие как
у брадикинина, но в отличие от него тахикинины не разрушаются в малом круге кро-
вообращения, с чем связано их более длительное гипотензивное действие. Они так же
вызывают быстрое сокращение внесосудистой мускулатуры (см.: физалемин, уперо-
леин, церулеин).
Уперолеин (кожа жаб Uperoleia rugosa, U. marmorata) — тахикинин.
Факторы, воздействующие на систему комплемента (яд кобры) — их не менее трех.
Вызывают стойкое падение уровня компонента комплемента СЗ в крови отравленного
ядом животного.
Фасцикулин (яд зеленой мамбы) — сильный ингибитор ацетлхолинэстеразы поли-
пепидной природы. Блокирует нейромышечное соединение.
Фибринолизин (яд фезалий) — один из факторов свертываемости крови, растворя-
ет тромбы.
Физалемин (филломедузы, кожа жаб Physalaemus fuscomaculatus, Р. centralis) — та-
хикинин, вызывает сильную вазодилятацию и длительную гипотензию, оказывает
выраженное деполяризующее действие на мотонейроны.
Фосфодиэстераза (яд скорпионов, яд скатов) — оказывает кардиотоксическое
действие.
Фвсфолипаза А (яд скорпионов, муравьев, морских змей, элапид, ядозуба) — ока-
зывает миотоксически й эффект. Вызывает структурные нарушения в диафрагме (не-
кроз миофибрилл, инфильтрация фагоцитами, набухание митохондрий). Усугубляет
тяжесть отравлений.
Фосфолипаза А2 (яд шершня, пчел, гадюк, гремучих змей) — гидролизует фосфо-
зипиды с образованием цитолитика лизолецитина, который лизирует мембраны мно-
гих клеток, в том числе эритроциты, тучные клетки, вызывая при этом соответствую-
щие патологические эффекты. Поэтому фосфолипазу А2 иногда называют непрямым
гемолизином. Так же как и некоторые другие фосфолипазы ядов животных (напри-
иер, яда кобры; см. выше), фосфолипаза А2 обладает нейротропными свойствами и
нарушает процесс высвобождения медиаторов из пресинаптических терминалей.
В составе яда пчел действует синергически с цитоли гическими компонентами (напри-
мер, с меллитином), модифицируя клеточные мембраны.
Фосфолипаза В (яд скорпионов, муравьев) — нет ясности в отношении молекуляр-
ной структуры этой фосфолипазы. Возможно, что она представляет собой смесь фер-
ментов, обладающих свойствами фосфолипаз А! и А2 (см. выше).
Цитотоксины (яд физалий, коралловых полипов, нематоцист цианеи, дискомедуз,
горгонарий, немертин) — выраженное мембранотропное действие. Такие токсины
связываются с мембраной клетки и разрушают ее в определенных участках, вызыва-
452
Часть 2. Общая неправильная эпидемиология
ют деструкцию митохондрий, лизосом, выделение гистамина тучными клетками. На
рис. 2.42 в качестве примера показано формирование поры стихолизинами — цито-
токсинами многощупальцевого кораллового полипа S. helianthus (отряд Actiniari; оби-
тает в Карибском море) (рис. 2 42).
Рис. 2.42. Схематическое изображение трансмембранного участка поры, сформи-
рованной молекулами стихолизинов. Участок представлен как цилиндр 5 нм дли-
ной (погруженный на всю толщину мембраны) и 2 нм диаметром. Вокруг канала,
образованного субъединицами токсина (4—5 а-спиралей), формируется торои-
дальная липидная пора. Липиды индуцируют положительное искривление (кони-
ческой формы «головой» в основание) перпендикулярно плоскости основания
мембраны. А. Боковая проекция. Б. «Вид сверху». По С. A. Valcarcel et al. (2001)
Церулеин (кожа жаб рода Hyla) — гипотензивный пептид, стимулирующий глад-
кую мускулатуру, усиливающий секрецию желудка и поджелудочной железы.
Холин (содержится в яде беспозвоночных) — действует на парасимпатическую
нервную систему как антагонист адреналина.
Щелочная фосфатаза А (яд скорпионов, кобр) — вызывает гемолиз отмытых эрит-
роцитов, обладает кардиотоксическим действием.
Ниже кратко и в порядке эволюционного усложнения самих ядовитых животных
(см. выше «Эволюция ядовитости среди животных») рассмотрим биологические и
медицинские свойства отдельных ядов и зоотоксинов. Особое внимание уделяется
регионам возможности естественного поражения этим токсином (ядом) и рассмот-
рению имеющейся в литературе экспериментальной клиники отравлений, как воз-
можной экстраполяции при умышленном применении яда1.
Сакситоксин. Нейротоксин, водорастворимое производное пурина. Смертельная
доза для взрослого человека — 0,3—1 мг. Первичные продуценты токсина в морской
воде — простейшие отряда Dinoflagella, в пресноводных водоемах — отдельные виды
цианобактерий. Аккумулируют токсин по пищевым цепям двустворчатые моллюски
и крабы, обитающие на коралловых рифах Регионы естественного поражения вклю-
чают побережье стран, где моллюски входят в традиционный пищевой рацион и где
происходят так называемые «красные приливы» (побережье Аляски, Вашингтона.
Чили, Канады, Испании, Тайваня, Исландии и др.) (Deeds J. R., et al., 2008). Первые
симптомы отравления через 30 мин после приема в пищу моллюсков. Онемение язы-
1 Использовались работы Пигулевского С В (1966, 1975); Б. Н. Орлова с соавт. (1990);
Б. Н. Орлова и Д. Б. Гелашвили (1995) и др. авторов.
2.2. Биологические поражающие агенты
453
ка, губ, кончиков пальцев. Прогрессивно развивается сердечно-сосудистая и дыха-
тельная недостаточность. Смертельный исход через 1—12 ч вследствие остановки ды-
хания. Клиника в условиях эксперимента: прямое угнетение дыхательного и сосудод-
вигательного центров, паралич дыхательной мускулатуры. Более подробно о
поражающем действии сакситоксина см. в разд 3.14.
Гймберотоксин. Структура неизвестна. Источник токсина — динофлагелляты
Gymnodinum breve. Регион возможности естественного поражения включает побере-
жье штата Флориды (США). Симптомы отравления включают воспаление слизистых
оболочек, чиханье, кашель, расстройство дыхания. Клиника в условиях эксперимен-
та: после парэнтерального введения токсина у мышей развиваются судороги, охваты-
вающие мускулатуру тела, затем развивается паралич.
Суберитин. Белок, ММ 28 кДа. Обладает нейротоксической активностью. Субе-
ритин гемолизирует эритроциты, способен гидролизовать АТФ. Смертельная доза для
собак 10 мг/кг. Источник токсина — пробковая губка Suberites domuncula. Регион воз-
можности естественного поражения включает побережье Японского, Охотского, Бе-
рингова морей. При контакте с губкой у человека развивается зуд и слабый отек паль-
цев. Клиника в условиях эксперимента: при в/в введении собакам и кроликам вызы-
вает рвоту, расстройство ЖКХ, нарушение координации движении и дыхания. На
вскрытии геморрагические очаги во внутренних органах.
Яд гонионем. Оказывает холиномиметическое, гистаминлибераторное и серото-
нинпотенцирующее действие. Источник яда — гидромедуза Gonionemus vertens («кре-
стовичок»). Регион возможности естественного поражения включает побережье вбли-
зи Владивостока. Поражения кожи в виде «ожогов». Наиболее часто их получают
купающиеся среди зарослей водных растений. Резкая боль, гиперемия в местах «ожо-
га», через 10—15 мин появляется сыпь. Характерный симптом — падение тонуса мышц
конечностей, распространяющееся на дыхательную мускулатуру. Поражения со сто-
роны ЦНС: помрачение сознания, психомоторное возбуждение, бред, галлюцинации
(как при отравлении ЛСД) и др.
Яд физалий. Сложный яд, летальный эффект вызывает белковая фракция яда. LDS0
цельного яда для мышей 50—70 мг/кг. Источник яда — сифонофора Physalia physalis,
другое название «Португальский кораблик». Регион возможности естественного по-
ражения включает тропические и субтропические зоны Атлантического океана. Ожоги
с сильным поражением кожи и резкой болью. В тяжелых случаях наблюдаются рас-
стройства дыхания, сердечной деятельности, судороги. Клиника в условиях экспери-
мента: сильная боль в месте инокуляции яда, атаксия с постепенным развитием вялого
паралича, охватывающего дыхательную мускулатуру. Непроизвольная дефекация и
мочеиспускание. Атриовентрикулярный блок. Перед наступлением смерти судороги
аноксической природы. При экспериментальном отравлении на вскрытии: ишемиче-
ские легкие; сердце, остановленное в систоле; очаги геморрагии в брюшной полости.
Яд хиронекс. Яд нематоцист медузы. Содержит белки, углеводы и производные
индола. Обнаружены белковые фракции, ответственные за летальное, гемолитичес-
кое и дермонекротическое действие яда. Яд щупалец обладает кардиотоксическим и
гемолитическим действием. Содержит гистамин и гистаминлибераторы, серотонин,
кининоподобные соединения. Дермонекротическими свойствами не обладает. Источ-
никяда — кубомерузаСЬ1гопех/1ескеп («морская оса»). Регион возможности естествен-
ного поражения включает побережье Австралии. Обычно у пострадавшего на коже
видны ярко-красные полосы воспаления с плотными папулами, оставленные щупаль-
цами медузы. Через некоторое время на них начинаются образовываться изъязвления
454
Часть 2. Общая неправильная эпидемиология
и зоны некроза. Жгучая боль в участках пораженной кожи. Появляются сильные спаз-
мы мышц груди, бедер и живота. Дыхание становится затруднительным. Обильное
потоотделение, сухость во рту, рвота. При тяжелом отравлении развивается острая
сердечная недостаточность. Могут появиться сильные судороги и развиться коллапс.
Клиника в условиях эксперимента: подкожное введение яда нематоцист вызывает
развитие некроза кожи. Внутривенное введение крысам яда вызывает нарушения в
проводящей системе сердца: развитие атриовентрикулярного блока с последующей
фибрилляцией желудочков. Смерть от сердечной и дыхательной недостаточности. При
экспериментальном отравлении на вскрытии обнаруживаются отечные легкие с оча-
гами геморрагий на них.
Яд хризаоры. Обнаружены летальный, кардиотоксический, дермонекротический
и гемолитический факторы белковой природы. Яд нематоцист также содержит про-
стагландины, гистамин и гистаминлибераторы, серотонин, кининоподобные соеди-
нения. Источник яда — дискомедуза Chrysaora quinquecirrha («морская крапива»). Ре-
гион возможности естественного поражения включает субтропические и тропические
воды Тихого, Индийского и Атлантического океанов. Мучительные «ожоги» с возник-
новением папулезного и эритематозного дерматитов и явлений некроза. В тяжелых
случаях развивается сердечно-сосудистая недостаточность. Клиника в условиях экс-
перимента: симптомы, связанные с выраженным нейротоксическим и кардиотокси-
ческим действием яда.
Ядцианеи. Токсическая фракция, выделенная из нематоцист, содержит смесь бел-
ков с ММ около 70 кДа. Яд оказывает необратимое гипотензивное действие, поража-
ет проводящую систему сердечной мышцы. На гладкую мускулатуру яд оказывает
необратимое спазмолитическое действие. Источник яда — дискомедуза Cyanea capil-
lata. Регион возможности естественного поражения включает холодные и теплые воды
Мирового океана. Контакт со щупальцами цианеи уже через несколько секунд при-
водит к возникновению жгучей боли. Через 10—20 мин к ней присоединяются эрите-
ма и отек. Линии ожогов длинные, иногда достигают 60 см. Через некоторое время
они приобретают синий и даже черный цвет. У пострадавших начинается озноб, дыха-
ние ослабевает, развивается острая мышечная слабость. В тяжелых случаях — паралич
скелетной мускулатуры и мышц сердца. Клиника в условиях эксперимента: яд вызы-
вает затрудненное дыхание, судороги и смерть, которая при введении дозы 0,7 мг/кг
наступает через 30 мин, а при дозе 0,3 мг/кг — через 24 ч. У крыс при внутривенном
введении яда в дозе 1,6 мг/кг происходило резкое и необратимое снижение АД. Мень-
шие дозы вызывали поражение проводящей системы сердца и развитие ишемии сер-
дечной мышцы. При вскрытии у мышей, погибших от яда цианей, наблюдались за-
стойные явления во внутренних органах и в правой стенке сердца.
Лофотоксин. Необратимо блокирует нервно-мышечную передачу. ЬВ50для мышей
8 мкг/кг. Источник яда — коралловые полипы рода Lophogorgia (£. alba, L. cuspi-
data, L. rigida, L. chilensis). Регион возможности естественного поражения включает
прибрежные воды Мексики и Калифорнийского побережья США. «Ожоги» стрекаль-
ными клетками обычно получают сборщики губок и аквалангисты. Клиника в усло-
виях эксперимента: подкожное введение мышам вызывает у них атаксию, мышечный
паралич и смерть от остановки дыхания.
Яд актиний. Включает пептиды большой молекулярной массы, обладающие ней-
ротоксической и цитотоксической активностью. Нейротоксины, ММ 5 кДа. Цито-
токсины, М М от 10 до 80 кДа. Источник яда — акитинии Anemonia sulcata, Anthopleura
xanthogrummica, Stoichactisgigantea, Actinodendron plumosum. Регион возможности естест-
2.2. Биологические поражающие агенты
455
венного поражения включает мелководье тропических и субтропических областей.
Стрекальные клетки актиний поражают хожу человека, вызывая зуд и жжение на месте
контакта, позднее распространяющееся дальше по телу. На месте «ожога» образуется
папула роговой консистенции с последующим некрозом тканей. В тяже лых случаях
развивается лихорадка, головная боль, общая слабость. Клиника в условиях экспери-
мента: в токсических концентрациях нейротоксины актиний вызывают нарушение
сердечной деятельности — аритмию, фибрилляцию и остановку сердца в систоле.
Эквинотоксин Яд получают из щупальцев актинии Actinia equina. Токсический
белок, ММ 20 кДа. LD50 для мышей 33,3 мкг/кг. Регион возможности естественного
поражения включает побережье Черного моря. Зуд и жжение кожи в участке контак-
та с актинией. На месте «ожога» развивается папула с последующим некрозом тканей.
В тяжелых случаях лихорадка, головная боль, слабость. В экспериментах на живот-
ных обнаружено две фазы действия токсина: снижение АД, брадикардия и апноэ.
Затем АД повышается, нар)таегся сердечная деятельность.
Теалиатоксин. Получают из щупалец актинии Trealia felinu. Выраженное гистами-
нолитическое, гемолитическое и кардиотоксическое действие. Белок ММ 7,8 кДа.
LD50 для мышей частично очищенного токсина 69 мг/кг. Регион возможности есте-
ственного поражения включает побережья Варенцова, Карского и Берингова морей.
В эксперименте на животных токсин вызывает повышение АД, брадикардию, брон-
хоспазм, затрудненное дыхание. Кардиотоксический эффект токсина проявляется в
развитии ино- и хронотропного действия вплоть до полной остановки сердца.
Палитоксин.Одинизсамыхсильныхдепочяризующихядов. ММ2,7 кДа. ЬО50для
мышей 0,15 мкг/кг. Источники токсина — зоонтарии Pafythoa toxica, Р. tuberculosa,
Р. caribacorum, Р. mammillosa-, рыбы Alutera scripta. Регион возможности естественного
поражения включает Гавайские острова и Таити. Издавна известен аборигенам этих
островов как сильный яд. Использовался ими для приготовления травленного ору-
жия. Места обитания этих кораллов держались аборигенами в тайне. Клиника в ус-
ловиях эксперимента: внутривенное введение токсина кошкам в дозе 0,2 мкг/кг вы-
зывает сначала резкое повышение АД, затем у животных необратимо развивается
гипотензия. Характерным действием токсина является сильный спазм коронарных со-
судов сердца. Более подробно см. в разд. 3.16.
Яд конусов. Яд С. striatus обладает выраженным нейротропным действием. ГО50для
мышей составляет 2,3 мг/кг. Из него выделяют кардиотинический компонент — гли-
копротеин с ММ 25 кДа. Активное начало яда С. geographus составляют пептиды с
ММ в пределах 1,5—2 кДа, названные конотоксинами I и II. LD50 для мышей коно-
токсина 1 12 мкг/кг. Источник яда — брюхоногие моллюски рода Conus. Регион воз-
можности естественного поражения включает тропические и субтропические зоны
Мирового океана (преимущественно в Индийском и Тихом океанах). Обычные жер-
твы яда конусов — ловцы моллюсков и неопытные коллекционеры раковин. Харак-
терные симптомы отравления ядом конусов включают сильную боль в месте укола
ядовитым зубом, сопровождающуюся чувством онемения в окружающих тканях.
Онемение быстро распространяется по телу, охватывая мышцы рта и конечностей. По
мере всасывания яда наступают общие симптомы отравления: тошнота, пвота, одыш-
ка, расстройство зрительных и слуховых ощущений, нарушение координации движе-
ний, головокружение, загрудинные боли, саливация, лакримацич В тяжелых случаях
развивается паралич дыхательной мускулатуры, приводящий к гибели. Эксперимен-
тальное отравление мышей ядом С. striatus вызывает у них парезы, диспноэ, тониче-
ские спазмы скелетной мускулатуры и прогрессирующую дыхательную недостаточ-
456
Часть 2. Общая неправильная эпидемиология
ность с последующей остановкой сердца. Конотоксины С. geographus вызывают у
мышей вялый паралич и смерть от остановки дыхания.
Мурексин. Эфир холина. Его действие подобно действию ацетилхолина. LD50 для
мышей при внутривенном введении 8,1—8,7 мг/кг. Источник яда — брюхоногие мол-
люски рода Мигех. Регион возможности естественного поражения включает побере-
жье Адриатического моря. Отравления развиваются при употреблении моллюсков в
пищу. Отравление характеризуется тяжелым гастроэнтеритом, наблюдались судоро-
ги. Экспериментальное отравление мышей мурексином вызывает падение АД, бра-
дикардию, стимулирует дыхание и моторику кишечника. Усиливается слюноотделе-
ние и т. д.
Макулотоксин. Небелковый нейротоксин, выделенный из задних слюнных желез
осьминога. По физико-химическим свойствам и физиологическому действию близок
к тетродотоксину. ММ примерно 540 Да. LD50 для мышей 50 мкг/кг. Источник яда —
головоногий моллюск (осьминог) Hapalochlena maculosa. Регион возможности есте-
ственного поражения включает побережье Австралии. Отравление возникает после
укуса осьминога Н. maculosa. Симптомы отравления включают быстро развившийся
(через 2—10 мин) паралич. Вначале отравленный ощущает онемение слизистых, мышц
лица и шеи. Развиваются афония и дисфагия. Постепенно паралич охватывает дыха-
тельную мускулатуру, что может привести к летальному исходу. В одном случае смерть
человека была зарегистрирована через 90 мин после укуса осьминога. Клиника в ус-
ловиях эксперимента: внутрибрюшинное введение макулотоксина животным вызы-
вает у них беспокойство, атаксию, затруднение дыхания и смерть, которой предше-
ствует судорожный припадок из-за асфиксии.
Яд скорпионов. LD50 дтя мышей в пределах 5,1—83,0 мг/кг. Основным компонен-
том яда скорпионов являются токсические пептиды (в основном, нейротоксины), их
активность в несколько раз превосходит токсичность целого яда. Нейротоксин М10
скорпиона Buthus eupes имеет ММ 7 кДа, LD50 для мышей 0,72 мг/кг. Источник
яда — скорпионы (Scorpiones) (рис. 2 43).
Регион возможности естественного поражения включает местности в полосе
48° с. ш. и 54° ю. ш. Место ужаления нетрудно установить по точечному проколу кожи.
Характерная черта интоксикации ядом скорпиона — быстрота возникновения симп-
томов после ужаления насекомого. Первые признаки отравления развиваются уже
через несколько минут. Ужаление скорпиона вызывают чрезвычайно сильные мест-
ные болевые ощущения (жгучая, иррадиирующая боль, отек, гиперемия тканей в месте
ужаления, реже возникновение пузырей с серозным содержимым). К общетоксичес-
ким симптомам относятся: головная боль, головокружение, слабость, нарушение со-
знания, расстройство терморегуляции, судороги, мышечный тремор, затруднение
дыхания, тахикардия. Изменение АД, профузное слезо- и слюнотечение, обильное
выделение из носа, бронхиальная гиперсекреция. Нередки панкреатиты и миокарди-
ты. У детей малые эпилептические припадки. Клиника в условиях эксперимента: вско-
ре после введении мышам в хвостовую вену прогретого яда скорпиона Buthus eupes,
их походка нарушалась, дыхание учащалось, начинались судороги (особенно задних
конечностей), и наступала смерть. Смерть может наступить в результате отека легких.
Кровоизлияний в месте инъекции прогретого яда не образуется. Сублетальные дозы
яда вызывают у крыс изменения энцефалограммы, характерные для малого эпилеп-
тического припадка. У отравленных животных на вскрытии обнаруживают застойные
явления в легких, мышца сердца резко сокращена. Крови в левом желудочке нет. Из
внутренних органов яд энергичнее всего действует на печень. При гистологическом
2.2. Биологические поражающие агенты
457
Рис. 2.43. Представители отряда скорпионов. A. Buthus eupeus. Б. Andractonus
crassicauda. Количество яда у различных видов скорпионов не одинаково. У круп-
ного африканского экземпляра В. australis удается получить 10 капель, дающих при
высыхании 3 мг сухого яда. От самых крупных европейских и среднеазиатских эк-
земпляров В. occitanas, В. eupeus. В. caucasicus можно получить до 8 мг жидкого яда.
У мексиканских видов Centruroides noxius средний объем капли, выделенной с по-
мощью электрического тока, составлял 0,485 мм3, а вес — 0,625 мг. Почти такое
же количество — 7—8 мг жидкого яда - удается получить и у южноамериканских
видов Tityus bachiensis и Т. serrulatus. От крупных тропических видов скорпионов
Африки и Индии — Pandihui imperator и Heterometrus cyaneus получали от 15—20 до
30—40 мг жидкого яда. Количество яда, выделяемое скорпионами при ужалении,
сравнительно невелико. Например, при ужалении В. eupeus, В. caucasicus и В. occi-
tanus выделяется около 0,2—0,4 мг яда. При ужалении Т. bachiensis, Т. serrulatus
и С. noxius выделяется около 0,3 мг По С В. Пигулевскму (1975)
исследовании в печени обнаруживается большое количество жироперерожденных
клеток. Оно намного превосходит подобное же поражение этого органа при отравле-
нии змеиным ядом. В почках, помимо жирового перерождения клеток извитых ка-
нальцев и некротических процессов, в отдельных участках наблюдается еще крово-
излияние в полосе баумановской капсулы (капсулы Шумлянского). В клубочках почек
геморрагии внутри самой капсулы, а также сероальбуминовые отложения. В извитых
канальцах некроз эпителия. Клетки сильно вакуолизированы, цитоплазма с гранула-
ми. В прямых канальцах клеточные изменения выражены значительно слабее. Отме-
чаются хроматолиз, кариолиз, небольшая вакуолизация цитоплазмы.
Яд ncvKOB-птицеедов. При подкожном введении цельного яда LD50 для мышей
1,5 мг/кг. Яд содержит гистамин и серотонин. Токсический нейротропный компо-
нент яда — белок с ММ 10,5 кДа. LD50 для мышей 0,1 мг/кг. Содержание токсина в
яде не превышает 1,5 % Источник яда — пауки-птицееды (Pterinochilus spp.). Регион
возможности естественного поражения включает Западную, Центральную, Восточ-
ную Африку и Мадагаскар. Укус паука вызывает сильную местную боль, однако спустя
несколько минут болевой симптом ослабевает, примерно через четверть часа начина-
ют развиваться симптомы общей интоксикации. Отравленный человек испытывает
сильную усталость, его движения становятся вялыми, некоординированными, появ-
яется сонливость. В результате резкого торможения ЦНС он производит впечатле-
ние больного летаргическим энцефалитом. В случаях тяжелой интоксикации ядом
458
Часть 2. Общая неправильная эпидемиология
паука явления торможения распространяются и на жизненно важные центры про-
долговатого мозга, вследствие чего пульсация сердца и дыхательные экскурсии лег-
ких ослабляются. Однако ни мышцы сердца, ни дыхательная мускулатура не поража-
ются ядом птицееда. Не приходя в сознание и не просыпаясь, отравленный незаметно
переходит от состояния жизни к смерти. Смертельных исходов среди людей после
укуса птицееда немного, так как среднее количество яда у паука недостаточно для
отравления человека. Яд паука Р. barrowi применяется бушменами для отравления
стрел. Клиника в условиях эксперимента: мыши, получившие смертельную дозу яда,
испытывали состояние глубокого торможения. Они ложились возле стенки террари-
ума и не проявляли никакой активности, вплоть до самой смерти. На месте инъекции
яда не было заметно отека, красноты или других признаков воспаления. Функции
организма, втом числе дыхания и сердцебиения, постепенно снижались и затем совер-
шенно прекращались. У экспериментальных животных токсин вызывает паралич зад-
них конечностей и резкое угнетение дыхания. При вскрытии у животных, погибших
от яда птицеедов, отмечались только поражения ЦНС.
Яд каракурта. Источник яда — пауки-каракурты (Latrodectus spp.). В состав яда
входят нейротоксины белковой природы, гиалуронидаза, фосфодиэстераза, холин-
эстераза, киназа и др. Всего в яде каракурта обнаружено не менее 86 уникальных бел-
ков, включая гомологи латротоксина, токсичные для насекомых и ракообразных
(DuanZ. G. et al. 2006). LD^ цельного яда для мышей 220 мкг/кг. Основное действую-
щее начало для позвоночных организмов — пресинаптический нейротоксин а-лат-
ротоксин. Его ММ 139 кДа. LD50 а-латротоксин для мышей 45 мкг/кг. У каракуртов
L. pallidus и L. dahli обнаружен fi-латротоксин, ММ 75 кДа.
Регион возможности естественного поражения включает Среднюю Азию, Аравию,
Северную и Западную Африку. В момент укуса чаще всего ощущается мгновенная
жгучая боль, уже через 15-30 мин распространяющаяся по всему телу. Укушенный
каракуртом человек обычно жалуется на невыносимые боли в области живота, пояс-
ницы, грудной клетки. Характерно резкое напряжение мышц брюшного пресса. По-
являются симптомы общего отравления: одышка, сердцебиение, учащение пульса,
головокружение, головная боль, тремор, рвота, бледность или гиперемия лица, потли-
вость, чувство тяжести в грудной и подложечной областях, экзофтальм и мидриаз. При
тяжелом отравлении тяжелая депрессия, затемнение сознания, бред. Смертность
2—4 %. Клиника в условиях эксперимента: а-латротоксин у позвоночных животных
воспроизводит все эффекты действия цельного яда, однако не действует на беспоз-
воночных. Сначала происходит возбуждение вегетативной нервной системы (мидриаз,
потоотделение, повышение АД и др.), потом развивается атаксия, клонические и то-
нические судороги, сменяющихся парезами, а также обильным слюно- и слезотече-
нием. Смерть животных наступает в результате бронхоспазма и отека легких.
Яд тарантула. В его состав входят токсические полипептиды и ферменты (гиалу-
ронидаза, протеазы, эстеразы аргининовых эфиров, киназа). Кроме того, обнаружены
спермин, спермидин, путресцин и кадаверин. LD50 цельного яда для мышей 15 мг/кг.
Вызывает сокращение гладкой мускулатуры. Источник яда — пауки-тарантулы (Lyco-
sidaespp.). Регион возможности естественного поражения включает пустынные, степ-
ные и лесостепные зоны. В момент укуса ощущается значительная болезненность.
Хорошо различимы места прокола кожи холицерами паука. В месте укуса развивают-
ся гиперемия и отек. Боль сохраняется в течение нескольких суток, но имеет местный
характер. Пострадавшие жалуются на общую тяжесть тела, апатию, сонливость. Мо-
жет наблюдаться озноб, учащение пульса, потливость. Клиника в условиях экспери-
2.2. Биологические поражающие агенты
459
мента: после введения мыши в хвостовую вену 1 мг яда место введения через 5 мин
припухало, шерсть животного поднималась дыбом, движения становились вялыми,
дыхание участилось. Через 10 мин мышь легла. У нее началось мелкое дрожание все-
го тела, появились судороги и удушье. Через 15 мин мышь погибла. При очень боль-
ших дозах наступающий коллапс обрывает жизнь раньше, чем успевают развиться
значительные анатомические изменения и особенно местные У мышей, погибших от
яда тарантула, на вскрытии в месте введения яда обнаруживали резкий капилляро-
спазм и обильное пропитывание кровью тканей подкожной клетчатки. Кровоизлия-
ния отмечались в печени, легких и частично в ЦНС. Смерть наступала от асфиксии.
Яд пчел. Его активные компоненты делят на следующие группы Первая — белки
с ферментативными свойствами (фосфолипаза А;, гиалуронидаза и кислая фосфата-
за). Вторая — токсические полипептиды (меллитин, анамин, МСД-пептид, тертиа-
пин, секапин). Третья — биогенные амины (гистамин и, в незначительных количе-
ствах, дофамин, норадреналин и др.). Состав яда меняется в зависимости от возраста
пчелы. Источник яда — медоносная пчела (Apis tnellifera L.). Распростоаььны повсе-
местно. Клиническая картина отравления пчелиным ядом зависит от количества ужа-
лений, ихлокализации и функционального состояния организма. Как правило, основ-
ными являются местные симптомы отравления: боль и отек. Строгой закономерности
в проявлении отдельных симптомов общей интоксикации не наблюдается. При общем
отравлении пчелиным ядом пульс учащается и становится аритмичным. Отмечает-
ся тахикардия. Появляется боль в области сердца, чувство стеснения грудной клетки,
спазм гортани, слабость и потеря сознания, «сердечная тоска» и страх смерти. Отрав-
ленные часто жалуются на спазматические боли в области сердца. Но яд пчел обычно
не вызывает деструктивных изменений в мышце сердца и отравленные погибают от
паралича центра дыхания. До 2 % ужаленных людей дают аллергические реакции на
пчелиный яд.
Клиника в условиях эксперимента: при введении летальных доз апамина смерть
наступает в условиях генерализованных судорог, охватывающих и дыхательную мус-
кулатуру. При вскрытии место введения яда (ужаления) обычно представляло собой
плотный инфильтрат, пропитанный серозным содержимым. Соматическая мускула-
тура никаких изменений не имела. Мышца сердца без видимых поражений. Все внут-
ренние органы резко гиперемированы, особенно легкие. Сосуды мозга расширены.
В мозговых сосудах кровянистые экссудаты. Гемолиз эритроцитов.
Кантаридин. Небелковый яд кожно-нарывного и нефротоксического действия.
LD50 для кошек и собак 1 мг/кг. Смертельная доза для человека при приеме внутрь
20-30 мг. Для убийства людей используется с глубокой древности. Источник яда -
жуки-шпанки (Lytta). Распространены повсеместно. Нанесение на кожу человека
0,1 м] кантаридина вызывает типичный дерматит с образованием пузырей, заполнен-
ных выпотной жидкостью. При повышении дозы проникает в кровь, вызывая явле-
ния общей интоксикации с резорбтивным действием на различные органы. При при-
еме внутрь интенсивно всасывается и приводит к быстро развивающемуся отравлению
часто со смертельными исходами. У отравленных людей появляется чувство жжения
во рту, затрудненное глотание, рвота. При проникновении яда в кровь развиваются
судороги, падение пульса, иногда параличи и кома. Гломерулонефриты и циститы —
типичные последствия отравления кантаридином в сублетальных дозах. Клиника в
условиях эксперимента: после приема кантаридина в дозе 0,03 мг три-четыре раза в
сутки у человека появлялся озноб, головокружение, слабость, пот, падение пульса и
усиленное мочеотделение. У людей, погибших от отравления кантаридином, на вскры-
460
Часть 2. Общая неправильная эпидемиология
тии отмечается резкая гиперемия слизистых покровов, образование язв и геморраги-
ческих очагов. Диффузные очаги поражения в печени и почках.
Яд сколопендры. В яде присутствуют ацетилхолин, гистамин, серотонин, гиалу-
ронидаза, холинэстераза, кининаза, ВАЕЕ-эстераза. LDS0 цельного яда для мышей
22,5 мг/кг. Источник яда — кольчатые сколопендры (Scolopendra cingulata). Регион
возможности естественного поражения включает юг бывшего СССР. Укусы кольчатой
сколопендры вызывают жгучую боль, развивается отек, недомогание, озноб. Среди
системных эффектов — сильная гипергликемия, вызванная серотонином яда (усилен-
ный гликогенолиз в печени и мышцах). Клиника в условиях эксперимента: у отравлен-
ных животных развивается адинамия, депрессия, саливация, сужение зрачка, фибрил-
лярное подергивание отдельных групп мышц, затруднение глотания, в итоге — смерть.
Яд диамфидий. Из куколок выделяют один из сильнейших природных токсинов —
диамфотоксин, обладающий гемолитическим действием. ММ 60 кДа. LD50 для мы-
шей 25 пг/г. Гибель кроликов через 3 ч после введения 1,5—2,8 г токсина. Источник
яда — диамфидии (Diamphidia locusta, D. nigro-omata). Естественное поражение мало-
вероятно. Бушмены северной части Калагри (юг Африки) с древности используют яд
диамфидий для смазывания наконечников стрел. Выкопанные куколки измельчают
ножом, смешивают с растертыми семенами растения Schwarzia madagaseariensis и со-
ком коры акации. Обработанный такой смесью (калгарский стрельный яд) наконеч-
ник стрелы сохраняет свою ядовитость в течение года. Одной стрелы достаточно, что-
бы убить жирафа весом 500 кг. Клиника в условиях эксперимента: при подкожном
введении калгарского стрельного яда на месте инъекции развивается прогрессирую-
щее воспаление с последующим некрозом тканей кожи и подкожной клетчатки. Кли-
ническая картина отравления начинается резко выраженной адинамией. Животные,
которым был введен яд, теряют способность к движению, их мышечный тонус пада-
ет, нарушается координация движений. Дальнейшее развитие гемолитического про-
цесса приводит к развитию нефрита и гемоглобинуриемии. Затем появляются судо-
роги и параличи. Смерть наступает от паралича сердечной мышцы.
Яд морских ежей. Яд Toxopneustes pileolus является смесью токсических белков —
урихитоксинов. Белки обладают различной биологической активностью (паралитиче-
ской, спазмогенной, кардиотоксической) и ММ от 30 до 80 кДа. Источник яда — мор-
ские ежи (Echinoidea). Регион возможности естественного поражения включает суб-
тропические и тропические зоны Тихого и Индийского океанов. После неосторожного
прикосновения к 7. pileolus у человека развиваются очень сильная боль в руке, про-
должающаяся около часа; паралич языка; век; лицевой мускулатуры; расстройство
речи и сильная слабость, особенно в ногах. В тяжелых случаях — асфиксия. Агония
длится иногда несколько часов. Характерны несчастные случаи, когда аквалангисты
или ныряльщики, получив неожиданный болезненный укол морского ежа, теряли со-
знание и тонули.
Яд скатов. Токсин ядовитого секрета ската Urolophus halleri представляет собой
весьма лабильный белок гемонейротропного действия с ММ примерно 100 кДа. Он
может быть электрофоретически разделен на 10 фракций, из которых, по крайней
мере, 5 токсичны. LD50 ядовитого секрета ската для мышей 28 мг/кг. Источник яда —
скаты-хвостоколы (Dasyatiformes). Регион возможности естественного поражения
включает побережья Атлантического, Индийского и Тихого океанов, а также и откры-
тые моря. Некоторые индейские племена, обитающие в бассейне Амазонки, исполь-
зуют зазубренные шипы пресноводных скатов (Potamotrygon motoro) для изготовления
наконечников копий и стрел. Чаще всего страдают от яда скатов рыбаки, акваланги-
2.2. Биологические поражающие агенты
461
сты и просто купающиеся. Хвостокол наносит большие рваные раны. Боль в месте
удара шипом ската-хвостокола бывает очень сильной. Вокруг раны развиваются отек,
лимфангоит, осложнения вторичной инфекцией. Отравление характеризуется паде-
нием АД и нарушением деятельности сердца, развитием слабости, иногда потерей
сознания, диареей, развитием судорог и нарушением дыхания. Укол шипом хвосто-
кола в грудь и живот может закончиться смертельным исходом. Гибель людей после
укола шипом хвостокола происходила на фоне развившегося коллапса. Клиника в
условиях эксперимента: в малых дозах яд U. halleri при внутривенном введении кош-
кам вызывает у них брадикардию, появление атриовентрикулярного блока, желудоч-
ковые экстрасистолы и узловой ритм. Смертельные дозы яда вызывают очень быстро
возникающие атриовентрикулярные блоки, блокаду ножки пучка Гиса, уменьшение
амплитуды комплекса QRS, что указывало на ишемию и повреждение миокарда. На-
блюдалось резкое паден ие АД и остановка сердца. Также имело место прямое действие
яда на дыхательный центр.
Яд бородавчатки. Содержит: гиалуронидазу; фактор, увеличивающий проницае-
мость мембраны сосудистых капилляров; летальный фактор, вызывающий также ги-
потензивный эффект; болевой фактор. Целый яд блокирует нервно-мышечную пере-
дачу и действует на капиллярную проницаемость. LD50 для мышей 200 мкг/кг. Источ-
ник яда — рыбы-бородавчатки (Synancejae). Регион возможности естественного
поражения включает Индийский и Тихий океаны. Место укола иглой бородавчатки
хорошо заметно. Ранка окружена едва заметным валиком. Сначала в месте укола разви-
вается очень сильная боль, доводящая человека до исступления. Через короткое время
сам участок ранения и место вокруг него (до 1,5 см) «немеют». Но вокруг этого участ-
ка боль очень сильна и часто она может длиться до нескольких суток. На месте пораже-
ния развивается ишемия, отек, которые сохраняются от нескольких дней до несколь-
ких недель и заканчиваются некрозом тканей. По красным тяжам на коже хорошо
заметен лимфангоит. Отравленный ощущает «жар во всем теле» и «звон в ушах». В тя-
желых случаях отравленный теряет сознание, дыхание затрудняется, кожные и сли-
зистые поверхности становятся цианотичными, иногда наблюдаются судороги. Перед
смертью, которая может наступить через 5 ч после укола иглой бородавчатки, отрав-
ленный впадает в кому. Клиника в условиях эксперимента: при введении лаборатор-
ным животным яд вызывает дерматонекроз, отек легких, геморрагии в грудной поло-
сти. Наблюдается падение АД, периферическая вазодилатация, снижение сократи-
тельной функции сердца, коронарная недостаточность. При вскрытии обнаруживают
сердце, остановленное в систоле.
Тетродотоксин (ТТХ). Небелковый нейротоксин, сходный по своему действию с
сакситоксином. Представляет собой соединение аминопергидрохиназолина с гуани-
диновой группой. ММ 379 Да. Получают из тканей рыб Fugu ocellatus ohscurum.
F. niphobles, F. poecilonotum. Блокирует проведение нервного импульса в возбудимых
тканях. LD5(i для мышей 10 мкг/кг. Аналогами ТТХ являются тарихатоксин (LD50 для
мышей 9 мкг/кг), содержащийся в яйцах калифорнийского тритона, и ряд других ток-
сических соединений. Источник яда — иглобрюхие (Tetraodontidae). Регион возмож-
ности естественного поражения включает Японию и Океанию. Первые симптомы от-
равления после приема икры, молоки или печени рыбы фугу (Fugu spp.) появляются
в интервале от нескольких минут до 3 ч. Вначале отравленный ощущает странное по-
калывание и онемение языка и губ, которое может распространятся и на тело. Затем
он начинает жаловаться на головную боль, боль в животе и руках. Походка становит-
ся шатающейся, появляется рвота, причем ее отсутствие свидетельствует о неблаго-
462
Часть 2. Общая неправильная эпидемиология
приятном прогнозе. Вскоре после рвоты развивается атаксия, отравленный человек
стремится лечь. Наблюцается ступор и афазия. Дыхание затруднено, АД снижено,
характерно понижение температуры тела, развивается цианоз слизистых и кожи От-
равленный впадает в коматозное состояние, и вскоре после потери сознания насту-
пает остановка дыхания, сердечная деятельность еще продолжается некоторое вре-
мя. Ведущими симптомами отравления ТТХ у экспериментальных животных явля-
ются быстро наступающая гипотензия, угнетение дыхания, паралич скелетной
мускулатуры. Падение АД происходит в результате паралича гладкой мускулатуры
сосудистой стенки. Нарушение функции дыхания имеет или периферическую при-
роду («восходящий» паралич дыхательной мускулатуры), или связано с угныаюшим
действием ТТХ на дыхательный центр. Более подробно см. в разд. 3.15.
Сигуатоксин. Липофильное вещество с ММ 1,5 кДа. LD50 для мышей 10 мкг/кг.
Симптомы отравления делят на 4 группы:
1) желудочно-кишечные — рвота, диарея;
2) сердечно-сосудистые — гипотензия, брадикардия;
3) неврологические — извращение чувствительности;
4) прочие — артралгия. Источник яда — сигуатеровые рыбы (Ctenot haetus stria-
tus — рыба-хирург; Lutianus bonar, L. monostiema, Glabrilutanus nematophorus — лутиа-
ны; Sphyraenu barracuda — барракуда). Токсин также продуцируют динофлагелляты
Gambierdiscus toxicus. Регион возможности естественного поражения включает тропи-
ческие островные районы Мирового океана. В зависимости от дозы признаки отрав-
ления развиваются через несколько часов, а иногда и на следующие сутки после упот-
ребления рыбы. Вначале ощущается покалывание и онемение языка и губ, металли-
ческий привкус ч сухость во рту, тошнота. Отравленный человек жалуется на сильные
боли в животе, конечностях и суставах. Иногда на коже образуются сыпь и волдыри.
Очень характерным неврологическим нарушением является извращение температур-
ной чувствительности, когда больному холодные предметы кажутся горячими и на-
оборот. При тяжелом отравлении наблюдаются: нарушении координации движений;
параличи и парезы скелетной мускулатуры; кома и смерть. Выздоровление длитель-
ное. Желудочно-кишечная форма отравления вызывается травоядными рыбами
(рыба-хирург и др.), а сердечно-сосудистая — хищными (лу гиан, барракуда). Клини-
ка в условиях эксперимента: симптомы отравления у мышей следующие — включают
снижение двиг ягельной активности, диарею, лакримацию, гиперсаливацию, диспноэ,
паралич и смерть в пределах от 40 мин до 48 ч.
Маитоксин. Выделен вместе с сигатоксином, но в отличие от него растворим в воде.
Источник яда — сигуатеровые рыбы (Ctenochaetus striatus — рыба-хирург). Содержится
и в динофлагеллятах G. toxicus. LDS0 для мышей 0,17 мкг/кг. Регион возможности ес-
тественного поражения включает тропические островные районы Мирового океана.
Баграхог“чсин (В ГХ). Стероидный алкалоид. Представляет собой эфир батрахо-
токсина А с 2,4-диметилпиррол-З-карбоновой кислотой; батрахотоксин является
дериватом стероида прегнина. ММ 540,7 Да. LD50 для мышей при подкожном введе-
нии 2,0 мкг/кг. Обладает сильным кардиотоксическим действием, вызывая экстра-
систолии и фибрилляцию желудочков сердца; свойственно также паралитическое дей-
ствие на дыхательную мускулатуру, сердечную мышцу и мышцы конечностей. Стойко
и необратимо повышает проницаемость покоящейся мембраны нервных и мышеч-
ных клеток для ионов Na+, вызывая изменение электрического потенциала клерки.
При этом блокируется аксонный транспорт, и клетка больше не может передавать
нервные импульсы. При попадании в кровь через слизистую оболочку или трещину
2.2. Биологические поражающие агенты
463
в коже яд вызывает аритмию (экстрасистолию), ведущую к остановке сердца, в ре-
зультате которой наступает летальный исход. При комбинации с ядом скорпиона ток-
сичность яда повышается в 12 раз. Источник яда — неотропические лягушки рода
Phyllobates (Р aurotaenia, Р. bicolor, Р. terribilis, Р. lugubris, Р. vittatus). Наибольшее содер-
жание яда зафиксировано у Р. terribilis, в одной особи может содержаться до 500 мкг
батрахотоксина, 300 мкг гомобатрахотоксина и 200 мкг батрахотоксина А. В 2004 г. в
Новой Гвинее был обнаружен вид жуков, которые содержат много батрахотоксинов.
По предположению ученых, древолазы вполне могут питаться встречающимися в Ко-
лумбии близкими родственниками этих жуков, таким образом, накапливая яд в своем
организме. Яд может синтезироваться самими древолазами или бактериями-симби-
онтами. Регион возможности естественного поражения включает Центральную и
Южную Америку. Секрет кожных желез колумбийской лягушки (Р. terribilis), содер-
жащий ВТХ, используется туземцами Колумбии в качестве яда для стрел. Они сма-
зывают ядом стрелы д ля духовых трубок перед охотой, проводя наконечником по спи-
нам древолазов. Смерть жертвы наступает через 4—8 мин после попадания одной стре-
лы. Естественные отравления людей жабьим ядом редки. В основном из-за случайного
попадания яда на слизистые поверхности, особенно глаз. Вызывает сильное раздра-
жение, боль, конъюнктивит и кератит. Клиника в условиях эксперимента: у мышей
преобладают кардиотоксические свойства ВТХ (экстрасистолы, желудочковая фиб-
рилляция и смерть).
Буфотенин. Галлюциноген. Индольный алкалоид [Г4,М-диметилсеротонин, 5-гид-
рокси-3-(2-диметиламиноэтил)индол]. ММ 204 Да. Структурно схож с главным че-
товеческим нейромедиатором — серотонином. Источник яда — жабы Bufo (В. alvarius
и В. tnarinus). Содержится в листьях Amanita тарра, семенах Piptadenia peregrina, гри-
бах Amantia muscaria. Регион возможности естественного поражения включает север-
ные и умеренные климатические зоны. У людей буфотенин вызывает психические
нарушения, близкие к тем, что вызывает ЛСД. Психозы, вызванные буфотенином
(2—16 мг/кг), могут длиться до 3 сут. Подробно о токсических алкалоидах кожи ам-
фибий можно прочитать в обзоре J. W. Daly (1998).
Кобротоксин. Яд кобры Naja naja. В основном представлен постсинаптическими
нейротоксинами и мембрано-активными полипептидами с широким спектром фар-
макологической активности (гемолитической, кардиотоксической и цитотоксичес-
кой). В состав яда входит ацетилхолинэстераза, разрушающая ацетилхолин и тем са-
мым усугубляющая действие нейротоксинов. LD50 для мышей 74,0 мкг/кг. Вследствие
того, что в состав яда элапид и морских змей входят нейротоксины, смерть отравлен-
ных животных и человека наступает, как правило, от недостаточности функции внеш-
него дыхания на фоне поражения ЦНС (рис. 2.44).
Источник яда — змеи элапиды (Elapidae). Регион возможности естественного
поражения включает Индию. Другие виды кобр распространены в Средней Азии. Укус
кобры происходит неожиданно для человека. На месте укуса остаются следы наруше-
ния целостности кожных покровов в виде точечных проколов. Через несколько ми-
нут после укуса в месте внедрения яда возникает острая боль. Благодаря спазму кро-
веносных сосудов яд блокируется и образуется первичное депо, в котором часть яда
связывается клеточными элементами крови и окружающими тканями. В следующие
минуты яд начинает дренироваться лимфатической системой, которая и сама остро
реагирует на кобротоксин. Возникает лимфангоит, а по мере продвижения яда к ре-
гионарным лимфатическим узлам, лимфаденит. Формируется вторичное депо яда.
Тогда же проявляется гемолитическое действие яда, которое приводит к падению АД,
464
Часть 2. Общая неправильная эпидемиология
Рис. 2.44. Представители рода кобр. А. Среднеазиатская кобра (Naja oxiana).
Б. Очковая змея или собственно кобра (A. naja или N. tripudians). В. Место укуса
кобры. Яд кобры добывается путем выдавливания секрета железы у живой,
либо подвергнутой наркозу, реже у мертвой, только что убитой змеи. Свежедо-
бытый секрет ядовитой железы — слегка опалесцирующая вязкая чуть желто-
ватая жидкость нейтральной реакции без запаха. Удельный вес 1046. Свежий
яд и водные его растворы очень нестойки. Уже в течение первых суток он те-
ряет до 25 % токсичности. Токсические свойства свежего яда сохраняются
несколько суток при разведении его глицерином. Наиболее радикальным ме-
тодом сохранения токсичности яда является его высушивание. Высушенный
яд буреет и превращается в тонкую пластинку, растрескивающуюся на нерав-
ные частицы, напоминающие кристаллики. По С. В. Пигулевскому (1966)
одновременно снижается температура тела. Почти одновременно с падением АД на
месте укуса возникает отек и появляются симптомы общей интоксикации и пораже-
ния ядом ЦНС. Первоначальное раздражение кобротоксином различных отделов
мозга вызывают тошноту, рвоту, непроизвольное мочеиспускание, дефекацию, нару-
шение зрительного и слухового восприятия. Изменяется поверхностная и даже так-
тильная чувствительность, нарушается координация движения, появляются головные
боли и головокружение. Но потом отравленные становятся вялыми. Они перестают
реагировать как на внешние раздражители, так и на проявления тяжелого поражения
жизненно важных органов. В таком состоянии наступает смерть от остановки дыха-
ния. Сердце еще несколько минут продолжает сокращаться. Чем острее интоксика-
ция, тем меньше клинических симптомов. При попадании яда в крупную артерию,
например, при укусе кобры в шею, смерть может наступить мгновенно на фоне кол-
лапса. Клиника в условиях эксперимента: внутривенное введение кобротоксина со-
бакам вызвало у них резкое падение АД, внутрисосудистое — тромбообразование. Про-
гретый кобротоксин сохраняет нейротоксическую активность, но утрачивает гемоли-
тическую активность. На животных он действует наподобие яда кураре, поражая
нервную и двигательную системы. Смерть наступает от остановки дыхания. У погиб-
2.2. Биологические поражающие агенты
465
ших от кобротоксина людей при вскрытии обнаруживают тяжелое поражение орга-
нов и тканей. Большинство органов и тканей, в том числе и печень, претерпевают
жировое перерождение. В печени и почках находят участки некроза. Со стороны лег-
ких отмечается заполнение целых участков кровью, благодаря чему они становятся
более твердыми и не содержат воздуха. Сосуды легких расширены и заполнены кро-
вью, альвеолы забиты экссудатом. Сердце останавливается, как правило, в систоле.
Яд морских змей. По химическому составу напоминает яд элапид (см. выше). LD50
для мышей: яд Alpysurus laevis — 0,13 мкг/г; яд Hydrophis belcheri — 0,24 мкг/г; яд
Н. elegans — 0,12 мкг/г; яд Н. omatus — 0,16 мкг/г. В составе яда находятся нейроток-
сины постсинаптического действия. Источник яда — морские змеи (Hydrophidae).
Регион возможности естественного поражения включает побережье Индийского оке-
ана и тропическую зону Тихого океана. Укусы морскими змеями происходят при ку-
пании, при вылове их сетями и при других обстоятельствах. Они не более болезнен-
ны, чем укол булавкой. На месте укуса — точечные следы проколов. Сразу после укуса
морской змеи местные явления, как правило, отсутствуют. Небольшое покраснение
и припухлость. Иногда небольшой лимфангоит. Первые признаки отравления появ-
ляются спустя 30 мин после укуса змеи. Появляется боль в месте укуса и распростра-
няется на мышечную систему всей конечности и даже половины тела. На месте укуса
развивается отек, который может захватить большую часть тела. Кожа в месте укуса и
нередко на большой площади принимает пятнистую темно-красную с синюшностью
окраску. У отравленного появлялись резкая сухость во рту и горле, жажда, спазмы, зат-
рудняющие дыхание. Характерными симптомами отравления являются двигательные
расстройства, затруднение речи, дыхания, тонические спазмы жевательной мускула-
туры, птоз, увеличение гематокрита, миоглобинурия (особенно при укусах Е. schistosa).
Смерть наступает от периферического паралича дыхательной мускулатуры. Клиника
в условиях эксперимента: у кроликов яд вызывает паралич дыхательной мускулатуры.
При вскрытии обширный некроз скелетной мускулатуры и поражение почек.
Яды гадюк (виперотоксин) и гремучников (кроталотоксин). Характеризуются высо-
ким содержанием протеолитических ферментов различной субстратной специфично-
сти и наличием нейротоксических и кардиотоксических компонентов, обладающих
высокой активностью. Минимальная смертельная доза виперотксина для человека
колеблется от 0,4 до 0,8 мг/кг в зависимости от вида гадюки. Минимальная смертель-
ная доза кроталотоксина для человека — 20 мг. Источник яда — гадюки (Vipperidae) и
ямкоголовые (гремучие) змеи (Crotalidae). Регионы возможности естественного по-
ражения: гадюки распространены на территории всей Африки (без Мадагаскара) и в
Евразии; ямкоголовые змеи распространены на юге и востоке Азии, в Северной и
Южной Америке. Отравления характеризуются геморрагическим некрозом и отеком
в зоне инокуляции яда. Наряду с местными симптомами (отек, местная боль, лимфа-
денит) при большом количестве введенного яда на первый план выступают признаки
системного отравления (общая слабость, нарушение пульса, парестезия, головокру-
жение, обмороки, тошнота, рвота, падение АД, гипокоагуляция, нарушение дыхания,
снижение гемоглобина, уменьшение числа тромбоцитов), из которых наиболее угро-
жающим жизни отравленного является шок. Среди основных факторов, способных
привести к развитию шока у отравленного человека, следующие: развитие сердечной
недостаточности; снижение венозного возврата; нарушение структуры и функций
форменных элементов крови (сгущение крови; тромбоэмболии; дисбаланс электро-
литов), аутофармакологические реакции, угнетение функции ЦНС, снижение (увели-
чение) периферического сопротивления, уменьшение объема циркулирующей крови.
466
Часть 2. Общая неправильная эпидемиология
Смерть наступает на фоне апатии и сонливости отравленного. При благоприятных
исходах отравления выздоровление может оказаться неполным из-за поражения пе-
чени и почек, и о зразования некротического участка тканей в месте укуса змеи. Кли-
ника в условиях эксперимента: внутривенное введение яда степной гадюки экспери-
ментальным животным вызывает у них паралич капилляров и резкое падение АД. При
вскрытии у отравленных людей и животных обнаруживают обильные внутренние
кровоизлияния (легкие, печень, головной мозг и др. органы), тромбоэмболию, пора-
жения почек (гломерулонефрит), жировое перерождение печени. Отравления крота-
лотоксином могут быть причиной развития мелкоочаговой пневмонии.
Яд гремучей змеи Crotalus durissus terrificus. Содержит субъединичный нейрототок-
син кротоксин. LD50 для мышей — 0,09 мкг/г; и еше два токсических компонента.
гироггоксш, (ММ 33 кДа) и кротамин (ММ 4,8 кДа). Регион возможности естествен-
ного поражения включает Южную Америку. Клиника отравления цельным ядом сход-
на с отравлениями ядом доугих гремучников (см. выше). Клиника в условиях экспе-
римента: гиротоксин вызывает у мышей поражение вестибулярного аппарата (лаби-
ринтов); кротамин вызывает у мышей клонические судороги.
Мойяветоксин. Содержится в яде гремучей змеи Crotalus scutulatus scutulatus. Нейро-
токсин пресинаптиического действия. LD50 для мышей в пределах от 0,25 до 3,3 мг/кг,
в зависимости от места отлова змеи. Регион возможности естественного поражения
включает штаты Калифорния, Юта, Невада, Аризона. Симптомы отравления ядом
С. s. scutulatus и его токсичность отличаются у змей, отловленных в разных регионах
США. Яд змей, обитающих в штатах Калифорния, Юта, Невада и на юго-западе штата
Аризона более токсичен и дает при отравлении неврологические симптомы. Отрав-
ление ядом змей, обитающих между городами Тусон и Феникс (Аризона) проявляет-
ся местными симптомами (отеки и некрозы) Яд последних не содержит мойявиток-
сина и представлен протеолитическими ферментами.
Ядбумеланга. ( ильный прокоа! улянт, вызывающий превращение проторомбина
в тромбин. Активное коагулирующее кровь начало яда — термолабильный i ликопеп-
тид, ММ 55 кДа. LD50 для мышей целого яда — 0,72 мг/кг; гликопептида —0,78 мг/кг.
Источник яда — у» еобразная змея бумеланг Dispholidus typus. Регион возможности ес-
тественного поражения включает страны Африки. Отравление напоминает картину
поражения ядом гадюк, но менее выражено. Клиника в условиях эксперимента сход-
на с наблюдаемой при отравлении ядом гадюк (см. выше).
Яд древесной змеи. Чрезвычайно сильная дефибринирующая активность. ММ
токсического белка яда — 85 кДа. LD50 для мышей цельного яда 0,25 мг/кг. Источник
яда — древесная змея Thelotomis kirtlandi. Регион возможности естественного пора
жения включает страны Африки. Отравление напоминает картину поражения ядом
гадюк, но менее выражено по клинике. Клиника в условиях эксперимента: уже через
10 мин при введении экспериментальному животному цельного яда древесной змеи в
дозе 10 мкг/кг содержание фибриногена в плазме у него падает почти до нуля.
Яд подвязочной змеи. Обладает сильным геморрагическим действием. LDS0 для мы-
шей цельного яда — 13,9 мг/кг. В яде отсутствует фосфолипаза А и оксидаза L-амино-
кислот, обычно присутствующие в яде других змей. Источникяда — подвязочная змея
Thamnophis elegans vagranus. Регион возможности естественного поражения включает
страны Африки. Отравление напоминает картину поражения ядом гадюк, но менее
выражено по клинике. У животных, погибших от этого яда, отмечаются локальные
очаги геморрагии в месте инъекции яда, а также диффузные геморрагии в легких, ки-
шечнике, диафрагме.
2 2. Биологические поражающие агенты 467
Яд ядозубов. Содержит серотонин, оксидазу L-аминокислот, гиалуронидазу, фос-
фолипазу А, аргининэстеразу. Обладает нейроторопным и брадикининлибераторным
действием. Смертельная доза цельного яда для мыши 0,5-1 мг/кг. Для человека она
оценивается в 5—8 мг/кг, но может быть и ниже. Источник яда — ядовитые ящерицы-
ядозубы Heloderma suspectum и Н. horridum. Регион возможности естественного пора-
жения включает сухие каменистые предгорья и полупустыни Северной Америки от
юго-запада Мексики и Гватемалы. Укусы ядозуба редки. В месте укуса развивается
зтек, укушенный испытывает сильную боль, которая может длиться до 8 ч. Отравлен-
чые ядом ядозуба жалуются на слабость, тошноту, головокружение. Слизистые си-
юшные, дыхание учащено, АД низкое, тромбоцитопения. Очень часто место укуса
длительно кровоточит. Явления интоксикации держатся до трех суток. При большой
дозе яда смерть наступает от паралича диафрагмы, после чего через некоторое время
~рекрашает работу и сердце. Если доза несмертельная, то отравленный помимо боли,
сопровождающей воспаление, испытывает своеобразное сопорозное состояние, ко-
торое приводит его ко сну. После длительного сна последствий интоксикации не ос-
*ается. Клиника в условиях эксперимента: яд вызывает диспноэ, спонтанные судо-
роги, гипотензию. Смерть наступает от острой дыхательной недостаточности. При
вскрытии погибших от яда животных у них находят геморрагические очаги в ЖКТ и
лазах, застойные явления в легких, отек легких.
Модификация ядов животных. Ниже приведено несколько примеров предельно
лростой модификации ядов животных, приведенных в работе С. В. Пигулевского
(1966,1975). Яды животных можно фракционировать и очищать по физико-химичес-
ким свойствам (изоэлектрическая точка, размер молекулы, ее масса, гидрофобность
или гидрофильность и др.). Но для таких экспериментов необходимы современные
лаборатории, оснащенные дорогостоящим оборудованием. Уже не менее 100 лет из-
вестна и другая альтернатива. Эмпирическим путем обнаружено, что компоненты ядов
животных отличаются друг от друга способностью выдерживать прогревание при тем-
чературах, превышающих физиологическую. Яд скорпиона не может быть введен в
организм животного инъекционно «без оставления следов» из-за действия на сокра-
тимость мышц стенки сосудов. Гладкая мускулатура сначала испытывает резкое воз-
буждение, которое затем сменяется сильным торможением. В результате происходит
парез, а в тяжелых случаях интоксикации — паралич гладкой мускулатуры сосудис-
тых стенок. Сквозь стенки резко расширенных сосудов в окружающую ткань выхо-
лит плазма и частично форменные элементы крови. На гистологических срезах хоро-
шо видны сильно расширенные сосуды возле места введения яда. Прогревание яда
скорпиона при температуре 80°С в водяной бане позволяет устранить из него компо-
ненты, влияющие на кровь и кровеносные сосуды. Прогретый яд уже не дает ни ге-
молиза, ни спазма и последующего расширения сосудов, однако он сохраняет нейро-
токсическую активность. Гибель животного после введения такого яда происходит при
картине отравления, свидетельствующей о нейротропном характере отравления (па-
резы, параличи, судороги, особенно задних конечностей). На вскрытии застойные
явления в легких, мышца сердца резко сокращена. Гемолитических явлений, крови в
тевом желудочке и кровоизлияний в месте инокуляции яда, нет.
Сходные результаты были получены с пчелиным ядом и ядом кобры. При нагре-
вании раствора пчелиного яда до температуры 100°С в течение 15 мин разрушаются
те его компоненты, которые вызывают местное (кожное) действие, но сохраняются
компоненты, вызывающие у экспериментальных животных развитие судорог и пара-
личей. При прогревании яда при температуре в 100°С в течение 30 мин разрушаются
468
Часть 2. Общая неправильная эпидемиология
и те компоненты, которые вызывают судороги, в результате чего подопытные живот-
ные погибают от паралича. И только нагревание яда до температуры 150 °C в течение
15 мин полностью инактивирует яд. Кратковременное, не более 20 мин, нагревание
яда кобры в кислой среде до температуры 72 °C способствует разрушению геморраги-
нов и гемолизинов, но сохраняет нейротоксины1.
2.2.3.6. Бактериальные токсины
Древность бактериальных токсинов. Организация и общий механизм действия молекул
бактериальных токсинов. Классификация бактериальных токсинов. Порообразующие
токсины. Токсины, ингибирующие синтез белка. Токсины, генерирующие образование
вторичных мессенджеров (посредников). Протеолитические токсины. Активаторы им-
мунного ответа. Гибридные и модифицированные токсины. Повышение поражающего
действия токсинов Иммунотоксины. Максимально возможная токсичность.
Токсины бактерий определяют основные симптомы некоторых инфекционных
болезней у людей и животных. Это дифтерия, коклюш, холера, сибирская язва, боту-
лизм, столбняк, гемолитический уремический синдром и др. К настоящему времени
накоплены данные, показывающие возможность выполнения бактериальными ток-
синами функций, не имеющих отношения к инфекционным процессам. Среди них:
использование бактериями токсинов как средства антагонизма в микробных сообще-
ствах (холерный токсин оказывает ингибирующее действие на ряд бактерий); участие
токсинов в авторегуляторных процессах в бактериальных популяциях (энтеротоксин
С. perfringens) и др. (Бухарин О. В., Лигвин В. Ю., 1997). Но их рассмотрение не вхо-
дит в задачу данной работы.
Древность бактериальных токсинов. Любопытны сроки появления наиболее изу-
ченных токсинов. Анализ молчащих замещений нуклеотидных последовательностей
генов холерного, LT- и ST-токсинов кишечной палочки показал, что эти гены дивер-
гировали не менее чем 130 млн лет назад — в эпоху расцвета пресмыкающихся (Вер-
тиев Ю. В., 1996). И уже в знакомом нам «виде» они пережили не только их вымирание,
но и расцвет млекопитающих в миоцене, и их массовое вымирание в конце плиоце-
на. Если это так, тогда появление самого прототипного холерного токсина теряется
где-то в глубине геологического времени. А ведь до сих пор возбудитель холеры счи-
тается врачами антропонозом (см. учебник Шуваловой Е. П. с соавт., 2001). О древ-
ности известных бактериальных токсинов говорит удивительно точная «пригнан-
ность» их структур к выполняемой ими функции. Даже одиночные замены в S1-субъ-
единице коклюшного токе ина снижают ее ферментативную активность и токсичность
в 1000 раз, множественные — в 106 раз. Одновременно резко снижается иммуноген-
ность (Loosmore S. et al., 1990), нарушается пространственная структура и способность
взаимодействовать с олигомером (Lobet Y. et al., 1989).
Ю. В. Вертиев (1996) обратил внимание ученых на то, что бактериальные токси-
ны, интерфероны, бактериоцины и гормоны обнаруживают сходство в отношении
целого ряда важных свойств: I) синтезируются одним типом клеток, в то время как
воздействуют на другие типы клеток; 2) действуют на клетки в чрезвычайно низкой
концентрации (10-11—10~,4М); 3) обладают сходной молекулярной организацией: т. е.
состоят как минимум из двух функционально и структурно различных белков (доме-
нов) — энзиматического и рецепторного (см. ниже); 4) имеют сходные звенья моле-
1 Модификация токсинов генно-инженерными способами описана в разд. 2.2.3.6.
2.2. Биологические поражающие агенты
469
кулярного механизма действия (связывание с рецепторами, активация, транслокация
внутрь клетки и модификация клеточных мишеней); 5) обнаруживают сходную кине-
тику биологического эффекта — одноударный эффект; 6) все эти вещества токсичны.
Организация и общий механизм действия молекул бактериальных токсинов. Токси-
ны бактерий представляют собой либо отдельные белки, либо олигомерные белковые
комплексы, организованные в А-В-структуры. Такая структура молекулы токсина
предполагает наличие двух компонентов: А- и В-субъединиц, поэтому их еще назы-
вают бинарными. А-субъединица проявляет энзиматическую (токсическую) актив-
ность в клетке хозяина. В-субъединица доставляет А-субъединищ в клетку-мишень.
В-субъединица состоит из двух функциональных доменов: рецептор-связываюшего
домена, определяющего тропизм молекулы токсина к определенным клеткам; и транс-
локационного домена, доставляющего А-субъединицу через липидный бислой, либо
на плазматическую мембрану или в эндосому клетки-мишени. Структура В-доменов
тесно связана со структурой рецепторов-мишеней, с которыми взаимодействует ток-
син. А-субъединицы более консервативны, чем В-субъединицы, особенно в участках,
критических для проявления их ферментативной активности (Henkel J. S., 2010; Fin-
lay В., Falkow S.,1997) (рис. 2.45).
Дифтерийный токсин
Связывающий
домен
Транс юка- АДФ-рибозилирующий
ционный домен фермент
Экзотоксин псевдомонад
Рис. 2 45. Схематическая структура бактериальных токсинов. Токсин Pseudomonas
представляет собой отдельную белковую цепь из 613 аминокислот, включающей три
фунциональных домена. Домен 1а (аминокислоты 1—252) представляют собой связы-
вающий домен; домен II (аминокислоты 253—364) — транслоцирующий домен, домен
III (аминокислоты 400—613) включает АДФ-рибозилирующий энзим (ADPribosylating
enzyme), вызывающий гибель клетки путем инактивации в цитозоле фактора элон-
гации 2 (EF2). Домен 1b отделен от доменов II и III и содержит аминокислотные
последовательности 365- 399. Дифтерийный токсин представляет собой полипептид-
ную цепь из 535 аминокислот и состоит из энзиматического домена А (аминокисло-
ты 1—193) и связывающего домена В (аминокислоты 482—535). Транслокационный или
трансмембранный домен Т расположен между ними. По R. J. Kre.tman (2009)
Кроме взаимодействия с рецептором клетки-мишени, по мнению Ю. В. Вертие-
ва (199ь), В-субъединица имеет еще одну важную функцию — предохранителя, пре-
_ отвращающего «случайный выстрел». Она экранирует ферментативную субъедини-
цу, исключая случайное ее взаимодействие с субстратом в собственной клетке и за
пределами клетки-мишени. Активация токсической субъединицы происходит после
протеолитического расщепления В-субъединицы (В-прекурсорак наступающего пос-
ле ее взаимодействия с клеткой-мишенью (рис. 2.46).
470
Часть 2. Общая неправильная эпидемиология
Рис. 2.46. Основные механизмы клеточной интоксикации бинарными токсинами Clos-
tridium и Bacillus. Связыванию с клеткой В-прекурсоров предшествует их активация
через протеолитическое расщепление непосредственно на поверхности клетки. Расщеп-
ление фурином протективного антигена (РА83) В. anthracis в межклеточной жидкости
является своего рода уникальным случаем, гак другие В-прекурсоры активируются
только после связывания с поверхностью клетки Акз ивированные В-субъединицы
токсинов взаимодействуют со специфическим рецептором на поверхности клетки либо
уже сформировавшимся кольцевым гептамером, либо мономерами, формирующими
гептамеры в последующем Ферментативная А-субъединица погружается в клетку вме-
сте с гептамером, и комплекс «рецептор-голотоксин» захватывается ранними эндосома-
ми через рецептор-опосредованный эндоцитоз. Внутренняя среда этих вакуолей затем
закисляется АТФазами, что индуцирует конформационные изменения В-гептамера
и последующую его вставку в эндосомальную мембрану с формированием канала.
Через него А-субъединица транспортируется в цитозоль. По Н. Barth et al. (2004)
Частным случаем А-В-структур являются трехкомпонентные токсины со струк-
турой типа А1-В-А2, где В — это субъединица, участвующая в связывании токсина с
рецептором; А1 и А2 — субъединицы, проявляющие различную энзиматическую (ток-
сическую) активность в клетке хозяина. Трехкомпонентные токсины используют об-
щую В-субъединицу, обеспечивающую ферментативным субъединицам единый ме-
ханизм проникновения в цитозоль. Такая организация молекулы необходима для
проявления синергидного эффекта токсического действия ферментативных субъеди-
ниц (Супотницкий М. В., 2000). Наиболее изученным токсином данного типа явля-
ется сибиреязвенный (о нем более подробно см. в разд. 3.1). Большинство из токси-
нов, рассматриваемых специалистами по защите от БО, с точки зрения их возможного
применения для поражения людей, представляют собой A-В структуры.
Наличие А и В субъединиц (доменов) в структура:, молекул подавляющего боль-
шинства белковых токсинов бактерий свидетельствует о том, что они, как правило,
являются крупными функциональными белковыми агрегатами. Эволюционно обра-
зование таких агрегатов стало возможным путем объединения двух или более белков
в результате как нековалентных взаимодействий (сибиреязвенный, коклюшный и
другие токсины), так и путем образования ковалентной связи между ними (боту-
линический и столбнячный токсины). В свою очередь, отдельные субъединицы ток-
22. Биологические поражающие агенты
471
синов так же состоят из некогда различных белков, объединенных в пептид, прояв-
ляющий несколько активностей сразу. Например, тяжелые цепи ботулинического и
столбнячного токсина содержат два домена — регион, необходимый для транслока-
ции токсина и регион, необходимый для связывания с клеткой. Белки такого типа об-
разуются слиянием соответствующих генов в один ген, кодирующий большую поли-
пе птидную цепь.
Для бактериальных токсинов характерно сходство их субъединиц на молекулярном
и макромолекулярном уровнях. Оказалось, что хотя холерный токсин и относящий-
ся к его семейству температуролабильный энтеротоксин кишечной палочки (LT-ток-
син) имеют по пять идентичных В-субъединиц, а коклюшный токсин имеет четыре
различных В-субъединицы, но две из В-субъединиц коклюшного токсина свертыва-
ются аналогично В-субъединичным пентамерам семейств холерного токсина и шига-
токсина (Finlay В., Falkow S., 1997).
Сходство обнаружено как между ферментативными субъединицами токсинов, так
и ферментами эукариотов. По крайней мере пять АДФ-рибозилирующих токсинов
'коклюшный, холерный и дифтерийный токсины, температуролабильный токсин
кишечной палочки и экзотоксин А псевдомонад) имеют общий НАД-связывающий
сайт (Finlay В., Falkow S., 1997). Участок протяженностью в 100 аминокислот фермен-
тативного домена CNF1 (цитотоксический некротизирующий фактор первого типа),
гомологичен участку ферментативного домена дермонекротического токсина борде-
телл. Оба одинаковых участка выполняют одинаковую функцию — активируют Rho
(субсемейство маленьких ГТФ-связывающих белков, участвующих в модификации
регуляторов актина цитоскелета) и являются активными сайтами токсинов (Lemichez
Е. et al., 1997). Отечный фактор сибиреязвенного микроба представляет собой адени-
татциклазу, которая не только по кинетическим, но и по антигенным свойствам име-
ет сходство с действующей на ту же мишень кальмодулинзависимой аденилатцикла-
зой эукариотических клеток (Petosa С. et al., 1997; Trescos Y, Tournier J.-N., 2012).
Сходство между А- и В-субъединицами токсинов различных бактерий, между ними и
различными белками вирусов или эукариотических организмов, выполняющих сход-
ную функцию, скорее является проявлением конвергентной гомологии молекул под-
вергавшихся однонаправленному действию естественного отбора, чем свидетельством
их общего происхождения (Супотницкий М. В., 2000).
Двухкомпонентный состав и одноударность действия бактериальных токсинов
можно объяснить, сделав предположение, что способность синтезировать токсины
позволяла бактерии реализовывать какую-то сигнальную функцию в их экосистемах.
Преимущество такой структуры для передачи сигналов в том, что при ее распростра-
нении из центра, сигнал не ослабляется на большом расстоянии. Если бы передача
сигнала осуществлялась структурами, не способными к лиганд-специфическому вза-
имодействию, то сигнал ослабевал бы по мере диффузии сигнальных молекул. Отсю-
да, как следствие, способность воздействовать на другие типы клеток в чрезвычайно
низких концентрациях.
Классификация бактериальных токсинов. Бактериальные токсины некоторыми
исследователями подразделяются на группы по секреторным механизмам (1 —VII),
посредством которых токсины выходят за пределы си нтезируюших их бактериальных
клеток (например, Henkel J. S., 2010). В данной работе используется классификация
С. Schmitt et al. (1999), подразделяющая все токсины на 5 групп по механизму действия
и упрощающая понимание их поражающего действия в качестве потенциальных аген-
тов БО (табл. 2.21, с. 472 и рис. 2.47, с. 474).
Таблица 2.21
Характеристика бактериальных токсинов*
М икроорганизм/Токси н Механизм действия Мишень Болезнь Участие токсина в болезни LD50 на кг
1 2 3 4 5 6
Повреждающие мембраны
A. hydrophila/Аэролизин Пороформирующий Гликопротеин Диарея (Да) -7 мкг (м)
С. petfringens/ Перфринголизин О То же Холестерин Газовая гангрена ?
Е. а>///Гемолизин6 То же Плазматическая мембрана Инфекция уринарного тракта (Да) -
L. monocytogenes 1 Листериолизин О То же Холестерин Системные пищевые инфекции, менингиты (Да) 3 -12 мкг (м)
5. aureus/а-токсин То же Плазматическая мембрана Абсцессы3 (Да) 40-60 нг (м)
5. рпеитод/ае/Пневмолизин То же Холестерин Пневмония3 (Да) -1,5 мкг (к)
5. дус^едет/Стрептолизин О То же Холестерин Стрептококковая ангина, скарлатина3 ? -8 мкг(м) 1-2 мкг (к)
Ингибиторы белкового синтеза
С. diphtheriae/Дифтерийный токсин АОФ-рибозилтрансфереза Фактор элонгации-2 Дифтерия Да -1,6 мкг (м) ЛОО нг (ч)
Е. coli/S. dysenteriae/ Шига- токсин N-гликозидаза 28SpPHK Геморрагический колит, гемо- литический уремический синдром Да -
Р. aeruginoSii/Экзотоксин А АОФ-рибозилтрансфераза Фактор элонгации-2 Пневмония3 (Да) -3 мкг (м)
Активация путей вторичных мессенджеров
Е. coli: Цитотоксический никротизирующий фактор Деамидаза Rho G-белки Инфекция уринарного тракта ? -
Термостабильный токсин6 Стимуляция гуанилатциклазы Гуан илатцикл азный рецептор То же Да
Часть 2. Общая неправильная эпидемиология
Продолжение табл. 2.21
1 2 3 4 5 6
Термолабильный токсин АДФ-рибозилтрансфераза G-белки Диарея Да —
FAST ST-подобный0 Неизвестна То же 9 —
В. anthracis/Отечнът фактор Аденилатциклаза АТФ Сибирская язва Да —
В. ре/Тимк/Коклюшный токсин АДФ-рибозилтрансфераза G-белки Коклюш Да 21 мкг (м)
Дермонекротический токсин Деамидаза Rho G-белки Риниты (Да) —
С. botulinum/CI-roKCHH АДФ-рибозилтрансфераза Мономерный G-актин Ботулизм ? —
С. botulinim/СЗ-токсин То же RhoG-белок Ботулизм 9 —
С diffidle/Токсин А Гликозилтрансфераза ИЪоО-белок(и) Диарея/РС (Да) —
Токсин В То же То же То же ? —
V. сйо/егае/Холерный токсин АДФ-рибозилтрансфераза О-белок(и) Холера Да -250 мкг (м)
Активаторы иммунного ответа
5. aureus/ Энтеротоксины Суперантиген ТСЯиМНСП Пищевое отравление Да 20—50 мкг (о)
Эксфолиативный токсин Суперантиген (и сериновая протеаза?) То же Синдром шелушения кожи Да -
S. pyogenes 1 Пирогенный экзотоксин То же То же Скарлатина/синдром токсического шокаа Да 3—6 мг (м)
Токсин синдрома токсического шока Суперантиген То же Синдром токсического шокаа Да -
Протеазы
В. олгйгаол/Летальный фактор Металлопротеаза МАРКК1/МАРКК2 Сибирская язва Да <114 мкг (м)
С. 2юг«//7ш/п/Нейротоксины А-С Цинк - м еталлопротеазы VAM Р/синаптобревин, SNAP-25, синтаксин Ботулизм Да 0,5-1,2 нг (м)
С. ге/ат'/Столбнячный токсин То же VAMP/синаптобревин Столбняк Да -1 нг(м)
2.2. Биологические поражающие агенты
Да — строго доказанная связь между токсином и болезнью; (Да) — роль в патогенезе была показана на животных моделях или клеточ-
ных культурах; ? — неизвестно; а — другие болезни также ассоциированы с этим организмом; 6 — токсин продуцируется и другими семей-
ствами бактерии; (м) — мышь; (к) — кролик; (ч) — человек; (о) — обезьяны.
* По С. Schmitt et al. (1999).
474
Часть 2. Общая неправильная эпидемиология
Рис. 2.47 Сопоставление механизмадействиябактериальныхтоксинов разных групп.
А. Повреждение клеточных мембран а-токсином 5. aureus. После связывания и оли-
гомеризации, ножка похожего на шампиньон гептамера а-токсина вставляется в
клетку-мишень и вызывает приток или, наоборот, отток из клетки ионов (обозначе-
ны как темные и светлые кружки, соответственно). Б. Ингибирование белкового
синтеза клетки шига-токсином (Stx). Голотоксин, который состоит из энзиматиче-
ски активной субъединицы (А) и пяти связывающих субъединиц (В), входит в клетк)
через глоботриазилцерамидный рецептор (Gb3). Затем А-субъединица, обладающая
N-гликозидной активностью, отсекает аденозиновый остаток с 28S рибосомальной
РНК, что останавливает белковый синтез. В. Примеры бактериальных токсинов,
активирующих пути вторичных мессенджеров. Связывание температурно-стабиль-
ного энтеротоксина (ST) с рецептором гуанилатциклазы приводит к увеличению ко-
личества цГМФ, который обращает в обратна ю сторону ток электролитов. Посредст-
вом АДФ-рибозилирования или гликозилирования (соответственно), экзоэнзим СЗ
С. botulinum и токсины A (CdA) и В (CdB) С. difficile инактивируют небольшие Rho
ГТФ-связывающие белки. Цитотоксический некротизирующий фактор (CNF) £, coli
и дермонекротический токсин (DNT) рода Bordetella активируют Rho через дезами-
нирование. Для всех токсинов характерна «мультидоменная» структу ра Это означает,
что разные их части состоят из некогда отдельных глобулярных белков, объединенных
естественным отбором в ходе эволюции. По С. Schmitt et al. (1999)
Порообразующие токсины. К ним относят бактериальные токсины, функциони-
рующие посредством вставки в плазматическую мембрану хозяина и формирующие
в ней трансмембранные поры (каналы), приводящие клетку к лизису. Такие токсины
еще называют RTX-семейством (repeats in toxin) из-за наличия в их молекулах боль-
шого количества повторов (Finlay В., Falkow S , 1997). Механизм их действия хорошо
прослеживается на примере а-токсина S. aureus, рассматриваемого как прототип оли-
гомеризующегося пороформирующего цитотоксина (Schmitt С. et al., 1999). а-токсин
синтезируется как прекурсорная молекула из 319 аминокислот, содержащая N -терми-
нальную последовательность из 26 аминокислот. Секретируемый бактерией, «зрелый»
токсин (протомер) является гидрофильной молекулой с массой 33 кДа, утратившей
цистеиновые остатки (Bhakdi S., Tranum-Jensen J., 1991). Протомер «узнает» клетку-
2.2. Биологические поражающие агенты
475
мишень по высокоаффинным рецепторам или неспецифически сорбируется в участ-
ках плазматической мембраны, содержащих фосфатидилхолин или холестерин. На
мембране семь протомерных токсинов собираются в пору, формируя грибоподобный
гептамер (232 кДа), включающий три различных домена (Song L. et al., 1996). Шляп-
ка и ободочная область гептамера а-токсина располагаются на поверхности плазма-
тической мембраны, в то время как ножка служит трансмембранным каналом. Обра-
зовавшаяся пора позволяет маленьким молекулам и ионам двухстороннее движение,
что, в конечном итоге, приводит клетку к вздутию и гибели от осмотического шока
(Schmitt С. et al., 1999). а-токсин действует цитолитически в отношении различных
типов клеток (рис. 2.48).
Рис. 2.48. Действие а-токсина 5. aureus на эритроциты кролика и искусственные
липидные мембраны. А. Негативно окрашенный фрагмент эритроцита кролика,
лизированный а-токсином 5. aureus. На поверхности мембраны наблюдаются
многочисленные кольцеобразные струк пры размером 10 нм (показаны стрелка-
ми). Б. И юлированные гексамеры токсина в растворе детергента. В. Лектиновые
липосомы, включающие реинкорпорированные гексамеры токсина (последние
выглядят как штырьки вдоль краев липосомной мембраны — показаны стрелка-
ми). Звездочкой показана липосома, избежавшая включения молекул а-токсина.
Черные полосы соответствуют 100 нм. По S. Bhakdll, J. Tranum-Jensen (1991)
а-Токсин S. aureus способен лизировать лимфоциты, эритроциты, тромбоциты,
моноциты и эндотелиальные клетки человека (Bhakdi S. et al., 1996), но его роль в раз-
витии стафилококковых инфекций неизвестна.
К другим членам RTX-семейства относят гемолизин £. coli (Н1уА), аденилатпик-
лазу В. pertussis, лейкотоксин Pasterella haemoLtica. Это семейство токсинов является
также частью консервативного механизма секреции I типа, отвечающего за их транс-
порт из бактериальной клетки (Finlay В., Falkow S., 1997). Образование поры включа-
ет целый каскад вторичных реакций, приводящих к другим патологическим послед-
ствиям. Среди них активация эндонуклеаз, высвобождение цитокинов и медиаторов
воспаления, синтез эйкозаноидов и др. (.Bhakdi S. et al., 1996).
Способность формировать белковые каналы в клеточной мембране не является
уникальным свойством оактериальных токсинов. Посредством поры осуществляет-
ся транспорт специфических ионов через мембраны эукариотических и прокариоти-
ческих организмов. К образованию пор способны белки слияния вирусов (Филдс Б.,
Найп Д., 1989). Аналогичным образом «действуют» цитолитические белки ядов жи-
вотных ( рис. 2.42).
476
Часть 2. Общая неправильная эпидемиология
Токсины, ингибирующие синтез белка. Субстратами для этих токсинов служат фак-
торы элонгации и рибосомальная РНК. Дифтерийный токсин и экзотоксин А псев-
домонад являются дифтамид-специфическими АДФ-рибозилтрансферазами, кото-
рые рибозилируют фактор элонгации-2 и, таким образом, инактивируя его, подавляют
синтез белка в клетках. Шига-токсин (Stx-токсин) продуцируется 5. disenteriae пер-
вого серотипа и Stx-продуцирующими штаммами Е. coli (STEC). Подробно механизм
действия таких токсинов объяснен в разд. 2.1.2.2.2 (см. «Трансдукция»), На рис. 2.49
показаны механизмы интоксикации клеток-мишеней дифтерийным токсином и экзо-
токсином псевдомонад.
Рис. 2.49. Механизмы интоксикации клеток-мишеней дифтерийным токсином
(DT) и экзотоксином псевдомонад (РЕ). После интернализации РЕ и DT подвер-
гаются протеолизу и редукции дисульфидных групп, благодаря чему из связываю-
щих доменов (домен 1а для РЕ, В-цепь для DT) обнажаются каталитические (до-
мен HI у РЕ, A-цепь у DT). РЕ подвергается обоим процессам — редуцированию
карбоксильной группы у терминальной молекулы лизина (К) и процессингу между
остатками 279 и 280, в результате чего образуется 37 кДа фрагмент токсина с кар-
боксильной группой (carboxy terminal toxin fragment), заканчивающийся остатком
REDL (Arg-Glu-Asp-Leu). Этот фрагмент транспортируется внутрь клетки через
KDEL(Lys-Asp-Glu-Leu)-peuenTop от комплекса Гольджи (Golgi) до эндоплазма-
тического ретикулума (endoplasmic reticulum, ER), где он транслоцируется в ци-
тозоль. DT подвергается процессингу между остатками 193 и 194. Каталитическая
A-цепь (аминокислоты 1—193) затем транслоцируется в цитозоль через эндосому
с помощью транслокационного домена Т (остатки 326—347 и 358—376), которые
формируют ионный канал. Клеточная гибель, вызванная токсином, осуществля-
ется по механизму апоптоза. EF2 — фактор элонгации 2. По R. J. Kreitman (2009)
Токсины, генерирующие образование вторичных мессенджеров (посредников). Бак-
териальные токсины могут влиять на функцию; отдельных белков эукариотической
клетки, не приводя ее к гибели. Для этого они активируют так называемых вторич-
2 2. Биологические поражающие агенты
477
ных посредников, способных в большой степени усиливать и искажать клеточную
реакцию на внеклеточные сигналы (Schmitt С. et al., 1999). Рассмотрим механизм дей-
ствия таких токсинов на примере цитотоксического некротического фактора (CNF).
CNF первого и второго типов (CNF1/2) относятся к группе бактериальных ток-
синов, модифицирующих Rho (Aktories К., 1997). Ген CNF1 у Е. coli локализован на
хромосоме и располагается на «острове патогенности» (Blum G. et al., 1995). Токсин
синтезируется как гидрофильный полипептид (115 кДа). Он остается цитоплазма-
тическим из-за отсутствия сигнальной последовательности и имеет связывающий
(N-терминальная половина CNF1) и ферментативный (С-терминальная половина
CN F1) домены (Lemichez Е. et al., 1997). В клетки хозяина он попадает с помощью сек-
реторного механизма 111 типа. Было показано, что CNF1 деаминирует глутаминовый
остаток Rho в положении 64. Такая модификация приводит к преобладанию актив-
ности Rho, неспособного гидролизовать связанный с ним ГТФ. Эукариотические
клетки, подвергнутые воздействию CNF1, приобретают характерный вид. У них на-
блюдается «рифление» мембраны, формируется локальное сжатие актиновых нитей.
Репликация ДНК, при отсутствии клеточного деления, приводит к образованию мно-
гоядерных клеток. Внутрикожное введение CNF1 вызывает длительное воспаление и
образование некротического очага (Schmitt С. et al., 1999).
Установлена критическая роль небольших ГТФ-связывающих белков не только в
регуляции цитоскелета (семейство белков Rho), но и в везикулярном транспорте (се-
мейство Rab), и в регуляции роста и дифференциации клеток (семейство Ras). Не
исключено, что существуют пока еще неизвестные нам токсины, способные воздей-
ствовать на основные клеточные процессы через эти белки (Finlay В., Falkow S., 1997).
Протеолитические токсины. Ботулинический и столбнячный токсины (оба цинк-
металлоэндопротеазы), в опытах на животных обнаруживают наименьшую из изве-
стных LD50. Удивительно, насколько различную клиническую картину дают пора-
жения этими токсинами, имеющими столь значительное сходство в структуре, энзи-
матической активности и мишенях среди клеток нервной системы, но различающиеся
путями проникновения в макроорганизм. Например, ботулинический токсин прони-
кает в организм энтерально и вызывает вялые параличи периферических нервов. Стол-
бнячный же токсин, образуясь на поверхностях ран, колонизированных С. tetani, при-
водит к спастическим параличам через поражение ЦНС (Finlay В., Falkow St, 1997).
Но эти наблюдения отражают естественное возникновение и течение отравления
ботулиническим и столбнячным токсинами. Их диапазон действия гораздо шире.
Например, при введении столбнячного токсина в область четверохолмий головного
мозга или в блуждающий нерв на шее животные погибают в короткое время при яв-
лениях острого отека легких без типичных признаков столбняка. При инъекции стол-
бнячного токсина в область теменной зоны коры головного мозга у животных, поми-
мо своеобразных двигательных расстройств, развиваются кровоизлияния в области
конечностей (Плецитный Д. Ф., 1958).
Эти самые смертельные токсины имеют и наиболее сложные молекулы среди
известных. Оба синтезируются в виде неактивных полипептидов массой 150 кДа и
высвобождаются из лизировавшихся клеток. Затем они активируются посредством
протеолитического расщепления открытой петли в структуре своей молекулы. Каж-
дая активная молекула нейротоксина включает тяжелую (100 кДа) и легкую (50 кДа)
цепи, связанные посредством межцепочечной дисульфидной связи. Тяжелые цепи
обоих токсинов содержат два домена — регион, необходимый для транслокации ток-
сина (N-терминальная последовательность), и регион, необходимый для связывания
478
Часть 2. Общая неправильная эпидемиология
с клеткой (С-терминальная последовательность). Легкие цепи обоих токсинов со-
держат цинксвязывающий мотив, необходимый для цинкзависимой протеазы, акти-
вирующей молекулу (Schiavo G., Montecucco С, 1997).
Ботулинические токсины связываются с рецепторами на поверхности пресинап-
тической мембраны двигательных нейронов периферической нервной системы и
вызывают протеолиз белков в нейронах. Это приводит к ингибированию высвобож-
дения ацетилхолина и к предотвращению мышечных сокращений — возникает вя-
лый паралич (Halpern J., Neale Е., 1995). Столбнячный токсин сначала связывается с
рецепторами на пресинаптической мембране моторных нейронов, но затем, с помо-
щью ретроградного везикулярного транспорта, он перемещается в нейроны спинно-
го мозга. Спастический паралич возникает из-за того, что рассечение везикуло-
ассоциированных белков и синаптобревина в нейронах нарушает высвобождение
глицина и у-аминобитуриковой кислоты, прекращающих мышечное сокращение
(Amon S., 1997). Более подробно о механизме действия и поражающих свойствах бо-
тулинического токсина см. в разд. 3.12.
Активаторы иммунного ответа. Отдельные бактериальные токсины могут действо-
вать непосредственно на Т-клетки и антигенпрезентирующие клетки иммунной сис-
темы. Самое большое семейство токсинов данного типа называют пирогенными ток-
синами-суперантигенами (PTSAg). Это стабильные, секретируемые токсины с ММ в
пределах от 22 кДадо 30 кДа. К ним относятся стафилококковые энтеротоксины серо-
типов А-Е, G и Н; пирогенные экзотоксины стрептококков группы А серотипов А-С
и F; стафилококковый TSST-1. Как правило, иммуностимулирующий потенциал та-
ких токсинов является следствием их способности связывать различные участки бел-
ков главного комплекса гистосовместимости II типа, экспрессированных на поверх-
ности антигенпрезентирующих клеток и VP-элементы на Т-клеточном рецепторе
(Schmitt С. et al., 1999). В частности, В-домен стафилококкового TSST-1 связывает
a-цепь антигена DR1 человеческого лейкоцита. Одновременно его А-домен специ-
фически связывается с Vp-элементами Т-клеточного рецептора (Schlievert Р., 1997).
Связывание TSST-1 с VP2 приводит к массивной пролиферации более 20 % перифе-
рических Т-клеток. Следствием Т-клеточной экспансии является массивное высво-
бождение интерлейкинов (1, 2 и 6 типов), у-интерферона, факторов некроза опухо-
лей (а и Р) и др. (Schlievert Р., 1997). Совместно они вызывают гипотензию, гипертер-
мию и диффузные эритематозные высыпания (Schmitt С. et al., 1999). Токсины данного
типа характерны для случайных и факультативных паразитов позвоночных.
Гибридные и модифицированные токсины. К ним относятся токсины, состоящие из
субъединиц разных токсинов или включающие модифицированные субъединицы.
При описании экспериментов по созданию гибридных и модифицированных токси-
нов исследователи единодушно проявляют скромность в отношении публикации дан-
ных по их поражающему действию, когда оно оценивается на лабораторных живот-
ных. В то же время этими данными они обстоятельно делятся в тех случаях, когда такие
токсины создаются для более эффективного истребления насекомых. Инсектицидные
токсины отличаются от токсинов, поражающих млекопитающих, только иной специ-
фичностью действия.
Повышение поражающего действия токсинов. Опережающим объектом среди ин-
сектицидных токсинов, позволяющим заглянуть в непубликуемые исследования по
рекомбинантным токсинам, опасным для людей и животных, являются поробразую-
щие токсины семейства Cry (Bt-токсины, 8-эндотоксины). Они продуцируются Bacil-
lus thuringiensis (Bt) и используются в сельском хозяйстве как безопасные для окружа-
2.2. Биологические поражающие агенты
419
ющей среды средства контроля над численностью насекомых. По механизму дейст-
вия на клетки такие токсины схожи с поробразующими токсинами стафилококков
(рис. 2.48).
Cry-токсины представляют собой семейство глобулярных белков, включающее три
домена, объединенных через простой линкер. Их организация типична для бинарной
А-В-структуры бактериальных токсинов. Роль токсической субъединицы (А) играет
порообразующий домен I; роль связывающей субъединицы (В) — домены II и III.
Домен I представляет собой разветвляющуюся структуру, состоящую из 7 «-спира-
лей, из которых центральная спираль (а5-спираль) окружена внешними спиралями.
а5-спираль высококонсервативна в пределах всего семейства Cry-токсинов. Этот
домен вовлечен в формирование ионной поры в клеточной мембране эпителия кишеч-
ника насекомого. Домен II состоит из трех антипараллельных P-слоев, упакованных
вокруг гидрофобного центра, формирующего p-структурную призму. Домен II опреде-
ляет специфичность токсина и является наиболее вариабельным из его доменов.
Аминокислотные остатки, вовлеченные в контакт между доменами I и II (находятся
в а7-спирали и pi-слое), высококонсервативны для всего семейства Сгу-токсинов.
Домен III сформирован двумя антипараллельными P-слоями. Этот домен участвует в
специфическом узнавании клеток-мишеней токсином. Высококонсервативны белко-
вые последовательности, создающие контакт между доменами II и III (соответствует
pi 1-слою и р12-слою) и образующие внутреннее пространство домена III (соответству-
ет Р17-слою и Р23-слою) (de Maagd R. A. et al., 2001).
Прототоксин CrylA, синтезируемый Bt, представляет собой параспоральный (ло-
кализованный рядом со спорой) восьмигранный кристалл. Заглоченный гусеницей
чешуекрылого насекомого, он растворяется в содержимом его среднего кишечника и
рассекается протеазами, активируя 60 кДа мономерный токсин. Активированный ток-
син связы вается со специфическим рецептором на поверхности клеток, выстилающих
средний отдел кишечника гусеницы (de Maagd R. A. et al., 2001). Рецептор индуциру-
ет второе протеолитическое расщепление токсина, удаляющее al-цепь. После чего
молекула токсина становится полностью активна и, ассоциируясь с другими такими
молекулами, формирует олигомерную структуру с ММ 250 кДа — препору (prepare).
Аффинитет олигомера к вторичному рецептору возрастает более чем в 200 раз по срав-
нению с аффинитетом отдельной субъединицы токсина. Вставка препоры в микро-
домены клеточной мембраны ведет к формированию полноценной литической поры
и к гибели клетки (Pardo L. et al., 2006).
Первичными рецепторами для CrylA-токсина являются белки, сходные с кадхе-
ринами (cadherms, CADR) — трансмембранными кальцийзависимыми гликопротеи-
нами, осуществляющими адгезивные межклеточные контакты. Большинство кадхе-
ринов представляют собой однократно пересекающие плазматическую мембрану
гликопротеины (700—750 аминокислот). Большая внеклеточная часть полипептидной
цепи обычно свернута в пять доменов (около 100 аминокислот каждый), часть из ко-
торых гомологична и содержит кальцийсвязывающие сайты. Внеклеточный домен,
наиболее удаленный от плазматической мембраны, опосредует межклеточную адге-
зию. Взаимодействие Cry-токсинов с кадхерином осуществляется через комплекс
связывающих сайтов: CADR7, CADR11 и CADR12 (Pardo-Lypez L. et al., 2009).
Вторичными рецепторами для Cry 1 А-токсина являются два протеина, «заякорен-
ных» через мостикв мембране клетки-мишени: гликозилфосфатидилинозитол (glyco-
sylphosphatidyl-inositol, GP1) и либо аминопептидаза N (aminopeptidase N, APN), либо
щелочная фосфатаза (alkaline phosphatase, FAL) (Bravo A. et al., 2004).
480
Часть 2. Общая неправильная эпидемиология
Отдельные насекомые имеют видовую устойчивость к Cry-токсинам либо приоб-
ретают ее в результате систематического применения таких токсинов для регуляции
их численности. В этом случае в популяциях насекомых накапливаются особи с мути-
ровавшими кадхериновыми рецепторами, не взаимодействующими с токсином. Раз-
работчики инсектицидных токсинов вынуждены решать две технические задачи: рас-
ширять спектр поражающего действия Сгу-токсинов и увеличивать их токсичность в
отношении конкретных видов насекомых. К настоящему времени разработано не-
сколько экспериментальных подходов, позволяющих решать эти задачи (рис. 2.50).
Ингибиторы сериновых протеаз
Хитиназа
Рецептор для кадхерина
Кристалл
Олигомер токсина
J Cyt IAa-токсин
| Мономер токсина
GPI-заякоренный
рецептор q CR12-MPED
Введение дополнительных
сайтов связывания _
( । Протоксин
Введение новых
сайтов рассечения
Деления
спирали
а-1
Рис. 2.50. Механизм действия Сгу-токсинов и экспериментальные подходы к повы-
шению их инсектицидной активности. 1 — кристалл токсина растворяется в среднем
кишечнике насекомого; 2 — происходит активация прототоксина и его транслокация
через перитрофную мембрану (peritrophic membrane); 3 — токсин связывается с ос-
новным рецептором и протеаза рассекает спираль а-1 домена I; 4 — происходит оли-
гоимеризация токсина и связывание олигомера (препоры) с вторичными рецептора-
ми; 5 — токсин (препора) вставляется в мембрану и формирует пору. Для повышения
эффективности Сгу-токсина в отношении насекомых используются следующие прие-
мы: А. К токсину добавляется хитиназа, облегчающая сборку токсина на эпите-
лиальной мембране. Б. К токсину добавляются ингибиторы сериновых протеаз, сни-
жающие возможность деградации токсина и рецепторов, с которыми токсин взаимо-
действует. В. В молекулу токсина вводится сайт расщепления, улучшающий его
связывание с рецептором. Г. К токсину добавляются добавочные связывающие сай-
ты, такие как СК12-МРЕ-пептид из кадхеринового рецептора или CytlA-токсин, улуч-
шающие его контакт с рецепторами. Д. Из домена 1 делегируется участок молекулы,
формирующий спираль а-1, что вызывает олигомеризацию токсина и способствует
его взаимодействию с кадхерином. По L. Pardo-Lopez et al. (2009)
Методические подходы, использованные для вмешательства в структуру молекул
токсинов семейства Cry, включают сайт-направленный мутагенез отдельных доменов
токсина, конструирование гибридных токсинов, внедрение в молекулу токсина сай-
тов для расщепления протеазами, делеции его отдельных регионов и др. (рис. 2.51).
. 2. Биологические поражающие агенты
481
Рис. 2.51. 3D-модели токсинов семейства Cry с модифицируемыми участками.
А. Токсин Cryl А. Стрелками показаны точечные мутапии в доменах I и П. Деле-
гированный N-терминальный регион обозначен черным цветом (CrylAMod-
токсин). Мутации N372A или N372G, локализованные в петле 2 домена II, ведут
к восьмикратному увеличению токсичности по отношению к Lymantria dispar
Тройной мутант — N372A, A282G и L283S — показывает 36-кратное увеличе-
ние токсичности. Б. Токсин СгуЗА. Стрелками показаны точечные мутации в
доме II, улучшающие его инсектицидную активность. Созданный протеолити-
ческий сайт, включенный в домен I, показан черным цветом. В. Токсин Сгу2А
Черным цветом показан N-терминальный регион токсина, делегированный для
улучшения его инсектицидной активности. По L. Pardo-Lopez et al. (2009)
Сайт-направленный мутагенез используется для изучения структуры Cry -токси-
нов более 20 лет. D. Wu и A Aronson (1992) описали мутацию (H168R) в спирали а-5
_эмена I CrylAa-токсина, вызывающую трехкратное возрастание его активности
-ротив личинок каролинского бр‘г!жнила Manduca sexta (Wirth М С. et al., 1997). Вдоме-
чах II и III токсина обнаружены мутации, увеличивающие токсичность CrylAb в
отношении личинок отдельных видов насекомых. Так, мутации N372A или N372G,
локализованные во второй петле домена II, ведут к 8-кратному увеличению токсич-
ности CrylAb по отношению к непарному шелкопряду Lymantria dispar (Schnepf Е.
et al., 1998). Тройной мутант — N372A, A282G и L283S показывает 36-кратное уве-
тичение токсичности по отношению к этим же насекомым (Rajamohan Е et al., 1996)
см. рис. 2.51, А). У СгуЗА-токсина важную роль в связывании токсина с рецептором
lграет петля 1. Получены два мутанта этой петли, названные А1 и А2. Каждый мутант
содержит несколько мутаций: Al — R345A, Y350F, Y351F; А2 — R 345А и делецию
.1Y350Y35. Они обнаружили 3- и 11-кратное увеличение токсичности против жест-
кокрылого насекомого — большого мучного жука (Tenebrio molitor). Мутации в пет-
ле 3 СгуЗА-токсина проявили себя умеренным увеличением поражающего действия
токсина (в 2,4 раза) по отношению к личинкам Т. molitor (Wu S J. et al., 1992; LiuX. S.
et al., 2006) (см. рис. 2.51, Б).
Конструирование гибридных токсинов направлено на создание новых токсинов
с широким спектром целей или повышенной токсичностью. В отношении Сгу-ток
синов оно осуществляется посредством замены домена 111 на такой же домен из ток-
сина, обладающего нужной исследователям специфичностью (рис 2.51, А). Отделы
ные Cryl-токсины с низкой специфичностью по отношению к Spodoptera exigu,
включая CrylAb, Cryl Ac, Cryl Ba и Cryl Ea, становятся для них токсичными, если до-
482
Часть 2. Общая неправильная эпидемиология
мен III будет заменен аналогичным доменом из CrylCa. Например, CrylAb-токсин не
токсичен ддя S. exigua, но замена его III-домена на I П-домен Cry 1С приводит почти к
десятикратному росту его токсичности для S. exigua (deMaagd R. А., 1996). Замена
участка молекулы CrylAa-токсина в пределах аминокислотных остатков 450—612 (до-
мен III) на такие же из Cry 1 Ас-токсина, ведет к 300-кратному увеличению токсично-
сти CrylAa в отношении табачной огневки Heliothis virescens (Сагашоп Т. et al., 1991;
Pardo-Lopez L. et al., 2009).
Активация токсинов в новом для него хозяине может быть достигнута внедрени-
ем в его молекулу сайта для расщепления протеазами. Вставка в петлю СгуЗА-токси-
на между спиралью а-3 и спиралью а-4 сайта для хемотрипсин/катепсина (chymo-
trypsin/cathepsin G-сайта) позволила в 3 раза увеличить его токсичность для личинок
D. virgifera (Walters Е S. et al., 2008) (см. рис. 2.51, Б).
Деления в аминотерминальном регионе, включав >щая спираль ос-1 CrylA-токси-
на, делает возможной олигомеризацию токсина в отсутствие кадхериновиго рецеп-
тора. Такие модифицированные токсины (CrylAMod) убивают насекомых, у кото-
рых развилась резистентность к CrylA-токсинам, вызванная мутациями в гене кад-
херина. Олигомерные структуры, формируемые CrylAMod-токсинами, обладают
переформирующей активностью, сходной с таковой f природного («дикого») токси-
на (см. рис. 2.51, А).
Токсичность Сгу2-токсина была повышена для египетской хлопковой совки {Spo-
doptera liitoralis) и совки ипсилон (Agrotis ipsilon) не менее чем в четыре раза делецией
42-х аминокислот из аминотерминального региона молекулы, предшествующих ами-
нокислотным последовательностям, формирующим а-1-спираль. Этот участок моле-
кулы прототоксина в кишечнике насекомого отщепляется трипсином, после чего «об-
нажается» прежде закрытый регион токсина, соответствующий связывающему реги-
ону Сгу2-токсина (Mandal С. С. et al., 2007) (см. рис. 2.51, В1.
Иммунотоксины. Как частный случай гибридного токсина, могул рассматривать-
ся иммунотоксины — химерные белки, включающие два домена, один из которых об-
ладает свойствами антитела, а другой свойствами ферментативной субъединицы ток-
сина Первый домен обеспечивает связывание химерного белка со специфической
молекулой или клеткой, второй инактивирует молекулу-мишень или убивает клетку
Появление иммунотоксинов вызвано трудностями прицельного терапевтического
воздействия на злокачественно переродившиеся клетки крови. Первым таким сред-
ством были немеченые мАт, индуцировавшие апоптоз клеток-мишеней после взаи-
модействия с их рецепторами или вызывавших их гибель по другим механизмам
(rituximab и alemtuzumab). Однако выяснилось, что более чем у половины пациентов
раковые клетки устойчивы к действию мАт. Тогда их стали конъюгировать с радио-
нуклидами. Эти агенты оказались эффективными для лечения пациенте» со злока-
чественными новообразованиями, резистентными к мАт, но не могли использовать-
ся широко из-за их дозозависимой токсичности для клеток костного мозга. Третий тип
препаратов для прицельного терапевтического воздействия на злокачественные клет-
ки представлял собой мАт с конъюгированными химиопрепаратами. Они оказались
более эффекгивными и вызывали меньшее количество неспецифических ос тожнений
при лечении острой миелоидной лейкемии (acute myelogenous leukemia, AML), чем
мАт, конъюгированные с радионуклеотидамл. Но и к ним были обнаружены резис-
тентные опухолевые клетки, что потреоовало дальнейшего совершенствования пре
паратов, действующих через специфическое узнавание рецепторов на поверхности
опухолевых клеток (Kreitman R. J., 2006).
-2 Биологические поражающие агенты
483
Способность иммунотоксинов к прицельному уничтожению раковых клеток
инается с одной молекулы ферментативной субъединицы токсина, доставленной
к поверхности клетки-мишени специфическим антителом (см., например, работу
Yimaizumi М. et. al., 1978). Поэтому иммунотоксины представляют собой некий пре-
лел в развития таких средств терапии злокачественных опухолей крови. В качестве
? ерментативной молекулы при конструировании иммунотоксинов используют фер-
. ггативные субъединицы (домены) токсинов, блокирующие белковый синтез в зло-
качественных клетках. Среди растительных токсинов наиболее часто используются
рицин и абрин; среди бактериальных — одноцепочечные токсины: дифтерийный и
jotokchh псевдомонад1.
Разработчики иммунотоксинов учитывают то обстоятельство, что обе субъедини-
_uJ токсина содержат большое количество карбоксильных групп, связывающих их с
гизмененными тканями, и, в частности, с тканями печени и почек. Поэтому прежде
чем конъюгировать их с антителом, их дегликозилируют различными способами (по-
ручая рекомбинантные формы токсина в кишечной палочке, химическим путем и др.).
Оптимальным объектом для лечения иммунотоксинами являются опухолевые забо-
< мания системы крови. Злокачественные клетки расположены внутри сосудов и до-
ступны для препарата при внутривенном введении. К тому же иммунная система та-
<нх пациентов часто утрачивает способность к эффективному антительному ответу на
там иммунотоксин Конструирование иммунотоксинов, используемых в медицинских
талях, предполагает повышение их специфичности именно к опухолевым клеткам и
1к можно большую их безопасность для клеток нормальных тканей. В клинике ток-
тачность таких препаратов оценивают по так называемой максимальной переноси-
мой дозе (maximum tolerated dose, MTD). Признаки ее превышения неспецифичес-
кие: астения, подъем уровня печеночных трансаминаз в крови, гипотензия. При
“оявлении таких симптомов дозу препарата снижают. При более высокой дозе имму-
отоксина у пациента может развиться синдром пропускания сосудов (vascular leak
syndrome, VLS; Clarkson syndrome), вызванный высвобождением цитокинов после
г [бели в коже пеоиваскулярных Т-клетск. Обычно VLS встречается без отека легких
Количества иммунотоксинов, вызывающих осложнения, находятся в пределах десят-
ков мкг на 1 кг веса пациента (Kreitman R. J., 2006).
Мьксимально возможная токсичность Завершая раздел по токсинам, нам нужно
'пределиться с их максимально достижимой токсичностью. Выше мы уже приводи-
ли примеры повышения поражающего действия токсинов, но где пределы их «усовер-
шенствования»?
Пытаясь понять причину остановки роста токсичности новых боевых ОВ во вто-
рой половине XX столетия, Н. С. Антонов (1994) исследовал имеющиеся в научной
-итературе данные по токсичности большого количества структурно-родственных ве-
ществ со сходным характером токсического действия. Он установил, что каждая такая
группа веществ имеет границу минимальных летальных доз (ГМЛД), которая влогариф-
чических координатах «LD50 (моль/кг) — молекулярная масса» представляет собой
параболовидную кривую, причем наиболее токсичным веществам множества соответ-
ствуют строго определенные значения молекулярной массы. Используя приемы мате-
матической статистики, Антонов установил еще одну закономерность. Если на евкли-
1 Механизмы токсического действия ферментативных субъединиц экзотоксина А псевдо-
монад и дифтерийного токсина описаны выше, механизм токсического действия рицина при-
веден в разд. 3.11.
484
Часть 2. Общая неправильная эпидемиология
довой поверхности в логарифмических координатах «LD50 — молекулярная масса*
отобразить весь массив веществ, для которых летальные дозы экспериментально ус-
тановлены, то плоскость графика оказывается разделенной на две части: на одной из
них сосредоточены отображения всех взятых веществ, в то время как другая часть
плоскости графика остается свободной, ибо в природе не существует веществ, кото-
рые по величине летальной дозы и ММ соответствовали бы этой части плоскости гра-
фика (рис. 2.52).
Логарифм молекулярной массы (Да)
Рис. 2.52. График токсоиды по Н. С. Антонову. Максимальная токсичность «суперток-
синов» достигается за счет предельного увеличения размеров и сложности их молекул
Между указанными частями плоскости графика отчетливо прослеживается гра-
ница, формируемая отображениями наиболее токсичных веществ в соответствующих
интервалах изменения величины молекулярной массы. Сохранены отображения толь-
ко наиболее токсичных веществ и проведена кривая, огибающая эти отображения со
стороны минимальных летальных доз. Эта кривая названа им «токсоидой». Наиболее
токсичному веществу — ботулиническому токсину, соответствует и наибольшая мо-
лекулярная масса. Эффект возрастания биологической активности по мере увеличе-
ния молекулярной массы отмечался ранее Н. И. Кобозевым у алкалоидов, гликози-
дов, гормонов, витаминов и синтетических лекарственных веществ. Им было по-
казано, что путем вариации состава и строения молекул можно добиться некоторого
увеличения активности веществ. Но если требуется добиться увеличения активности
в десятки и более раз, одних структурных изменений молекул уже недостаточно, тре-
буется переход к соединениям с большей величиной молекулярной массы (Анто-
нов Н. С., 1994), т. е. более сложно организованных.
У ботулинического токсина ММ составляет 150 кДа. ММ типичной белковой цепи
достигает 50 кДа, размер лишь немногих пептидов превышает эту среднюю величину
(Альберте Б. и др., 1994). ММ ботулинических токсинов приближается к верхнему
12. Биологические поражающие агенты 4S5
тределу возможных ММ белков. Из закономерностей, установленных Н. С. Антоно-
вым и Н. И. Кобозевым следует, что ММ токсинов с LD50, меньшей на один порядок,
ем у ботулинического, должна превышать 1,5 мДа (описано несколько белков с та-
чои ММ), — на два порядка — 15 мДа (такие белки не описаны).
* * *
Огромное разнообразие биологических ядов и токсинов и трудности их иденти-
фикации ставят на первое место в распознании умышленного отравления исследова-
ние личности и анамнеза пациента, чье состояние здоровья внезапно ухудшилось без
признаков инфекционного процесса или обострения ранее имевшейся соматической
хтезни. При этом не следует недооценивать активности «народных отравителей»,
имеющих знания, накопленные поколениями лиц, занимавшихся целительством,
магией, оккультизмом, участниками тоталитарных сект и последователями бионега-
"ивных учений.
Модификация токсинов возможна в направлении изменения спектра их целей и
ммуногенности. LD50 гибридных и модифицированных токсинов, даже при повы-
1 ении их токсичности для отдельных экспериментальных животных, будет находить-
л в пределах, характерных для токсических веществ данного диапазона молекуляр-
ных масс. Токсичность ботулинического токсина является предельной не только дтя
бактериальных токсинов, но и для природных токсических веществ.
2.2.4. Искусственные генетические конструкции
Искусственные генетические конструкции. Ретровирусные векторные системы.
Векторные системы на основе аденовирусов. Адено-ассоциированные вирусные
векторы. Векторные системы на основе ортопоксвирусов. Векторные системы на
основе РНК-вирусов, не принадлежащих семейству ретровирусов. Искусствен-
ные векторные системы. Введение генов в ЦНС. Введение генов в легочную ткань
в составе аэрозоля. Введение генов в клетки кишечного эпителия.
Этот раздел монографии меня побудила написать статья J. L. Black (2003), пред-
ставленная от Клиники и фонда Мэйо (Mayo Clinic and Foundation, Rochester, США)1.
По его мнению, проект «Геном человека», будучи объединенным с бурно развиваю-
щимися технологиями генной терапии, позволяет разрабатывать совершенно иные
виды биологического оружия, чем запрещенные Конвенцией 1972 г. J. L. Black назы-
вает его «геномным оружием» и поясняет, что данный термин не идентичен понятию
«генное оружие», упоминаемое в СМИ и в некоторых научных статьях, поскольку
слово «ген» соответствует лишь части ДНК в ядре клетки, ответственной за выработ-
ку белков как основы этого оружия. Он считает такое определение слишком узким,
не учитывающим возможности использования для поражения людей геномных после-
довательностей, способных вызывать различные болезни, не имеющие ничего обше-
ю с инфекционным процессами или поражениями биологическими токсинами.
В качестве средств доставки в геном человека генов, способных вызвать патоло-
гические процессы, J. L. Black рассматривает искусственные генетические конструк-
ции, уже разработанные для лечения наследственных и инфекционных болезней.
1 Сайт Клиники Мэйо — http://www.mayoclinic.org/rochester/; русский перевод статьи
J. L. Black доступен в глобальной сети — http://www.cbio.ru/v5/modules/news/article.php2sto-
ryid= 1666.
486
Часть 2. Общая неправильная эпидемиология
В то же время он не приводит исходной информации, на основе которой делает свои
выводы, что просто обязывает нас «потянуть за концы» его рассуждений.
Искусственные генетические конструкции. Генная терапия наследственных и ин-
фекционных болезней предполагает доставку генов в клетки-мишени. Но ни ДН К, ни
РНК не могут быть использованы в «голом виде» для достижения этой цели. Сахаро-
фосфатный остов обоих типов молекул нуклеиновых кислот располагается по их пе-
риферии полярными группами наружу и придает им анионные свойства. При физио-
логических значениях pH, нуклеиновая кислота несет отрицательный заряд и не
способна непосредственно взаимодействовать с отрицательно заряженными наруж-
ными поверхностями клеточных мембран. Из-за высокой гидрофильности нуклеино-
вой кислоты все гидрофобные основания «повернуты» вовнутрь ее молекулы, поэто-
му она не может проникнуть через гидрофобный барьер клетки-мишени. В сыворотк
крови нуклеиновая кислота быстро деградирует под воздействием нуклеаз. Период
полужизни немодифицированной интерферирующей ГНК в сыворотке крови укла-
дывается в 5—60 мин, для ДНК — он составляет не более 10 мин. Кроме того, нук-
леиновые кислоты не способны специфически узнавать клетки-мишени (Seow Y.,
Wood М. J., 2009). Поэтому для доставки генов в эукариотические клетки, с начала
1980-х гг. разрабатываются искусственные генетические конструкции. В настоящее
время сформировалось два альтернативных направления их создания — на основе
вирусов и на основе искусственных векторных систем (таб i. 2.22).
Ретровирусные векторные системы. Ретровирусы являются группой вирусов, РНК-
геном которых в инфицированных клетках конвертируется в ДНК. Геном ретровиру-
сов включает три структурных гена, обозначенные как gag, pol и env, фланкированых
элементами, названными длинными терминальными повторами (LTR). В LTR содер-
жатся регуляторные элементы, выполняющие важные функции в жизненном цикле
ретровируса. Эти повторы необходимы для интеграции ДНК-копии генома вируса в
геном хозяина. Они определяют, где начало и где конец вирусного генома. LTR также
служат энхансер-промоторными последовательностями, т е. они контролируют экс-
прессию генов вируса. Относительно большой геном ретровирусов облегчает генети-
ческие манипуляции.
После инфицирования клетки-мишени копия ретровирусной ДНК стабильно
интегрируется с ее геномом строго определенным образом. Практически все инфи-
цированные клетки способны экспрессировать гены, привнесенные вирусом. Мощ-
ные транскрипционные энхансеры существенно повышают уровень экспрессии ге-
нов, клонированных в клетках различных типов. С их помощью можно переместить
до 8 т. п. о., что в большинстве случаев более чем достаточно для синтеза крупномоле-
кулярных белков. Удобным для исследователя является то обстоятельство, что ретро-
вирусные векторы можно размножать, достигая их высокой концентрации в неболь-
шом объеме — более 109 вирусных частиц/см3. В опытах по инфицированию ретро-
вирусами мозга, печени, мышц, глаз или клеток панкреатических островков грызунов
показана устойчивая экспрессия трансз енов в течение более 6 мес. (Inder М. V., Ni-
kunj S., 1997). Ранние этапы жизненного цикла ретровирусов и векторов на их основе
показаны на рис. 2.53.
Векторы на основе ретровирусов с самого начала их разработки предназначались
для введения через неповрежденные клетки за счет механизмов слияния, обеспечи-
ваемых поверхностными белками оболочки вируса. Чувствительность дыхательного
эпителия к ретровирусным инфекциям подразумевает возможность ингаляционною
пути введения в организм человека таких конструкций.
lati uiini 2.22
Сравнение свойсп наиболее распространенных генотерапевтических векторных систем*
Характеристика у-ретровирус- ные (MLV, FLV и др.) Лентивирусные (ВИЧ-1 и ВИЧ-2, FIV, CAEV, E1AV, JDV, MWhap.) Аденовирус- ные (Ad) Вирус герпеса первого типа (HSV-1) Аденоассоцииро- ванный вирус (AAV) Плазмидная ДНК (в искусственной векторной системе)
Размеры частицы, нм 80-100 80-100 70-90 120-300 20 15-5000
Максимальный размер вставки, т. п. о. 7-7,5 7-7,5 До 30 До 30 (на основе ви- руса), до 150 т. п. о. на основе ампликона 3,5-4,0 Неограничен
Концентрация (коли- чество вирусных час- тиц в мл) >108 >108 >10" До 108 (на основе ампликона) >1012 Неограничена
Схема доставки генов Ex vivo Ex/in vivo Ex/in vivo In vivo Ex/in vivo Ex/in vivo
Интеграция с ДНК реципиента Возможна Возможна Невозможна Невозможна для ампликонов Возможна/ Невозможна Маловероятна
Экспрессии in vivo Короткая Продолжительная Короткая Konoi >ия Продолжительная Корот! ая
Стабильность Хорошая Не проверялась Хорошая Нестабилен Хорошая Очень хорошая
Легкость приготовления Пилотные установки размером 20-50 л Неизвестно Легко масш- табируется Неизвестно Трудно очищать от клеточных фрагмен- тов и масштабиро- вать производство Легко получать в больших количествах
Иммунологические проблемы Незначи- тельны Незначительны Серьезны Незначительны для ампликонов Неизвестны Нет
Наличие иммунитета у реципиента Малове- роятно Маловероятно, за исключением больных СПИДом Возможно Возможно Возможно Невозможно
Проблемы безопасности Инсер- ционный му гагенез? Инсерционный мутагенез9 Воспалитель- ный ответ, токсичность Незначительны для ампликонов Воспалительный ответ, токсичность Преодолимы в рамках подбора нетоксичных компонентов системы
’Таблицасоставлена по работам М. V Inder, S. Nikunj (1997); М. Sena-Esteves (2000); J. Nemunaitis, J. Edelman (2002); S. Boeckle, E. Wagner
(2006); K. Pluta, M. M. Kacprzak (2009).
22 Биологические поражающие агенты 487
488
Часть 2. Общая неправильная эпидемиология
РНК
Оболочка
вируса
Вирусная
частица
Капсид
Проникновение вируса в клетку
Цитоплазма
Клеточная
мембрана
ДНК
Обратная g^~1N/nA
транскрипция „ „ ' ’
Ядерный транспорт
I
п Интеграция
Нуклеус j I
х 7 Хромосомная ДНК
Провирусная ДНК
Рис. 2.53. Ранние этапы жизненного цикла ретровирусов и векторов на их основе.
Проникновение ретровируса в клетку сопровождается освобождением капсида
от его оболочки и началом синтеза двуцепочечного ДНК-генома (провируса) на
матрице вирусной РНК с помощью входящей в состав вирусного нуклеокапсида
РНК-зависимой ДНК-полимеразы (обратной транскриптазы). Затравкой (прай-
мером) этой многоступенчатой реакции служит клеточная тРНК (для большин-
ства MuLV — пролиновая т PH К), комплементарная PBS (primer binding site) уча-
стку, расположенному в 5'-концевой части вирусной РНК. Конечным продуктом
полимеразной реакции является двуцепочечный ДНК-провирус, содержащий все
вирусные гены и фланкированный 3'- и 5'-LTR. Вирусная ДНК, интеграза (IN)
и отдельные вирусные и клеточные белки образуют вирусный преинтеграци-
онный комплекс, который импортируется во внутрь нуклеуса. IN катализирует
интеграцию вирусной ДНК с ДНК генома клетки. По D. Rene, J. A. Smith (2008)
Критическим ограничением использования у-ретровирусных векторов (MLV, FLV
и др.) в генотерапевтических процедурах является их неспособность инфицировать
неделящиеся клетки, т. е. ткани мышц, мозга, легких и печени. Вирусный преинтег-
рационный комплекс этих векторов, ответственный за интеграцию вирусной кДНК
в геном клетки, имеет слишком большие размеры, чтобы проникать через поры в
оболочке клеточного ядра. Такая возможность ему предоставляется только при деза-
грегации оболочки клеточного ядра во время митоза. В качестве других недостатков
специалисты отмечают генетическую нестабильность у-ретровирусных векторов, про-
являющуюся делециями или точковыми мутациями. Способностью проникать через
поры мембраны ядра без митоза обладают представители другого семейства ретрови-
русов — лентивирусы (Nienhuis A. W, 2008).
Лентивирусы (ВИЧ-1 и ВИЧ-2, FIV, CAEV, EIAV, JDV, MW и др.) инфицируют как
делящиеся, так и не делящиеся клетки. Наиболее изученным лентивирусом является
ВИЧ. Он рассматривается как потенциальный вектор для переноса генов в условиях
in vivo. Подобно простым ретровирусам, геном ВИЧ включает гены gag, pol и env, он
так же содержит гены еще шести регуляторных белков, названных tat, rev, vpr, vpu, nef
и vif. Гены отдельных белков могут быть убраны без снижения способности вируса к
размножению и инфицированию клеток. Концепция построения современных лен-
тивирусных векторных систем доставки генов в организм млекопитающих, представ-
лена на рис. 2.54.
2.2 Биологические поражающие агенты
489
Плазмида, несущая пакующую кассету
|CMV H
[CMV 11 8-igpoI
|CMV~H gagpol
D1S,
pBS SD 4> cPPT/CTS
SA
Оболочечный вектор
Векторная ллазмида
для переноса генов
Non SIN
рртдиз
5-^gag
P tew
Рис. 2.54. Лентивирусная векторная система для доставки генов в организм млекопи-
тающих. А. Лентивирусный вектор в сравнении с ВИЧ 1 — схематическое изображе-
ние провируса ВИЧ-1, 2 — лентивирусный вектор. Типичные для ВИЧ ^-активирую-
щиеся элементы маркированы выше схематического изображения вектора Ниже
приведены примеры дополнительных модификаций вектора, позволяющих повысить
титр вируса и/или экспрессию его отдельных генов. Б. Пакующая система. 1 — плаз-
мида, несущая пакующую кассету (packaging vector plasmids); 2 — векторная плазмида
для переноса генов (transfer vector plasmids); 3 — оболочечный вектор (envelope vector);
4 — схема векторной РНК; 5 — интегрировавшийся в геном клетки-мишени провирус
с трансген-экспрессирующей кассетой. По К. Pluta и М. М. Kacprzak (2009)
pfaT Pe- W |vPr |Vpu |Nef pa | 1-я генерация
2-я генерация
Жм~] jEFiahRev'y 1?A~1 3-я генерация
DIS,
PBS SD, У cPPT/CTS
RE ___
и рА
SIN
диз
jttiRtRS Схема векторной РНК
Ннтегриоовавшчгся в геном клетки-мишени провирус
с трансген-экспрессирующей;
кассетой
диз
Сборка (упаковка) лентивирусного вектора происходит в так называемых упако-
вывающих клетках. Они представляют собой перевиваемые клетки, в которых осуще-
ствляется синтез вирусспецифических белков Пакующая система таких клеток вклю-
490
Часть 2. Общая неправильная эпидемиология
чает как минимум три типа экспрессирующих кассет (пакующая, векторная и оболо-
чечная), ни одна из которых без участия других кассет не может «собрать» вирусную
частицу, способную проникать в клетку-мишень. В то же время, их одновременная экс-
прессия не приводит к образованию ретровирусных частиц, вызывающих инфекци-
онный процесс у человека.
Пакующая система работает следующим образом. Введенная в пакующие клетки
в составе плазмиды (packaging vector plasmid) пакующая кассета под контролем про-
мотора цитомегаловируса (CMV-промотора) экспрессирует гены ВИЧ, необходимые
для формирования инфицирующей вирусной частицы. Но из кассеты исключен ген
env, кодирующий белки-предшественники оболочки ВИЧ (Env), определяющие его
способность выходить за пределы клетки. Первые ленти вирусные векторные систе-
мы, разработанные в 1996 г., содержали в пакующих кассетах гены регуляторных бел-
ков ВИЧ (Vpu, Vpr, Vif, Nef, Rev, Tat). При дальнейшем усовершенствовании вектор-
ной системы из пакующих кассет были удалены гены регуляторных белков ВИЧ, LTR,
а также последовательности, кодирующие пакующий сигнал (у) и праймерсвязы-
вающий сайт (PBS). Это позволяло избежать случайную упаковку полной мРНК ВИЧ
в вирусную частицу. Векторы второй генерации включали только гены Tat- и Rev-бел-
ков. Третья (современная) генерация пакующих систем включает обычно две плазми-
ды: одна кодирует Gag- и Gag-Pol-белки (Gag формируют сердцевину ВИЧ, Pol — его
ферментную систему соответственно); вторая — Rev-белок (избирательно активиру-
ет синтез структурных белков вируса и обеспечивает экспорт из ядра длинных моле-
кул вирусной РНК).
Векторная плазмида (transfer vector plasmids) содержит кассету, экспрессирующую
мРНК, включающую все crs-активирующиеся элементы и пакующий сигнал. Эти
плазмиды содержат трансген-экспрессирующую кассету с геном, предназначенным
для экспрессии в новом хозяине и находящимся под контролем внутреннего промо-
тора, обычно позиционированного между З'-Tat/Rev SA-сайтом и З'-LTR.
Для формирования оболочки векторной вирусной частицы в систему включен
оболочечный вектор (envelope vector). Он содержит кассету, определяющую синтез
гликопротеинов оболочки вируса.
Образующиеся в таких системах вирусные частицы являются своего рода инфек-
ционным ретровирусом одноразового действия. При заражении ими клеток-мишеней,
не содержащих экспрессирующийся ретровирусный провирус, способный выполнять
транс-функции, осуществляется тот же порядок событий, что и при заражении обыч-
ным вирусом вплоть до образования интегрированного провируса. Однако, так как
вектор не содержит структурных вирусных генов gag,pol и env, образования вирусных
частиц не происходит. Другими словами, такие клетки-мишени, содержащие экспрес-
сирующийся интегрированный вектор, не могут передавать гены, приобретенные в
составе вектора, незаряженным клеткам по горизонтали. В то же время интегрирован-
ный вектор передается дочерним клеткам по наследству, т. е. по вертикали. Посколь-
ку процесс переноса генов с помощью рекомбинантных ретровирусных частиц закан-
чивается на клетках-мишенях, а не распространяется дальше, подобно тому, как это
происходит в случае репликационно-компетентных вирусов, то принято говорить не
об инфекционном переносе, а о трансдукции генов в составе вектора (Прасолов В. С.,
Иванов Д. С., 2000). Схема переноса генов с использованием ретровирусных векто-
ров показана на рис. 2.55.
Ретровирусные (лентивирусные) векторы можно перенацеливать на разные типы
клеток и разные участки генома человека. Первая задача решается благодаря псевдо-
22 Биологические поражающие агенты
491
Ретровирусная часть вектора
LTR Ц LTR
(1)
^тонированный ген/гены
Е. coli
ДНК ДО Трансфею (ия
------l...... >—И-ЛЛ/
ДО Транскрипция
РНК ,___' .
Упаковывающая
клетка
(3)
Упаковываний' вирусных
частиц и выход их в среду
Культуральная
среда
ДО Трансдукция
РНК __ ~ V __ ,____.
(4) \
Обратная транскрипция
Клетки-мишени
днк Ц Ичтлрация
WV«B--------CZ=ZJ----------ИВчАЛ/ I
Рис 2.55. Схема переноса генов с использованием ретровирусных чектороЕ;. 1. Кло-
нирование гена в ДНК ретровирусного вектора (transfer vector plasmids — см. рис. 2.54)
с помощью стандартных генно-инженерных методов. На данном этапе векгор ведет
себя как обычный плазмидный вектор. Амплификация ДНК вектора и селекция
осуществляются в клетках Е. coli. 2, Перенос высокоочищенной ДНК ретровирусного
вектора в упаковывающие клетки. 3. Формирование ретровирусных частиц и их вы-
ход в культуральную среду. Сбор культуральной среды упаковывающих клеток, кон-
центрирование ретровирусов. 4 Трансдукция гена в составе ретровирусного вектора
в клетки-мишени. По В. С. Прасолову и Д. С.Иванову (2000)
типированию вируса. Смысл этого методического приема в следующем. Так как из
лентивирусной векторной системы удален ген env ВИЧ, то оболочечный вектор
(см. выше) может содержать ген, направляющий синтез гетерологичного гликопро-
теина, включающегося в частицы вируса, продуцируемые пакующей системой. Такие
вирусы называют псевдотипированными (pseudotyping) или вирусами-обманщиками.
Они имеют ряд преимуществ перед ВИЧ, если рассматривать их с точки зрения ис-
пользования в качестве векторных систем:
1) увеличивают безопасность вектора вследствие удаления последовательностей,
гомологичных с вирусом дикого типа;
2) расширяют или изменяют тропизм векторного вируса в отношении клеток-
мишеней;
3) улучшают стабильность синтезированных вирусных частиц на этапах концент-
рирования, хранения и применения.
492
Часть 2. Общая неправильная эпидемиология
В зависимости от используемого в пакующей системе гликопротеина, формиру-
ются различные псевдотипы вируса (Pluta К., Kacprzak М. М., 2009).
Например, химерный гликопротеин RD114, полученный благодаря слиянию
трансмембранного и внеклеточного доменов гликопротеина эндогенного ретровируса
кошачьих (feline endogenous virus RD 114) с цитоплазматическим доменом гликопро-
теина амфотропного вируса MLV 4070а, позволяет псевдотипированному вектору
эффективно трансдуцировать мезенхимальные стволовые клетки (mesenchymal stem
cells, MSCs) с очень низкой цитотоксичностью (Zhang X. Y. et al., 2004).
Лентивирусные векторы, псевдотипированные с гликопротеинами оболочки ви-
руса бешенства (PV-штамм), приобретают тропность к нейрональной ткани и спо-
собность к ретроградному транспорту в условиях in vivo. Введенный в периферери-
ческом участке нервной системы псевдотипированный лентивирусный вектор по
нейрональным аксонам доставил трансгены в ЦНС (Federici Т. et al., 2009). Подробно
стратегия использования таких векторов для целей генотерапии описана в работах
Е. Verhoeyen, Е L. Cosset (2004); J. Cronin et al. (2005); C. Frecha et al. (2008); K. Pluta,
M. M. Kacprzak (2009). В табл. 2.23 обобщены сведения по мишеням псевдотипиро-
ванных лентивирусов.
Нацеливание вектора на клетку еще не означает транскрипции доставленного в
эту клетку гена. Для того чтобы транскрипция трансгена в клетке-мишени была эф-
фективной, генотерапевтами разработаны методы транскрипционального таргетинга
(transcriptional targeting), предполагающие введение в такие векторы специфических
для данных тканей промоторных последовательностей. Примеры таких промоторов,
используемых в тканеспецифических лентивирусных векторах, приведены в табл. 2.24.
Перенацеливание ретровирусных векторов с одних участков геном человека на дру-
гие, стало возможным благодаря выяснению механизмов интеграции провируса с ге-
номом человека. Интеграция ретровирусной ДНК с геномом клетки-мишени катали-
зируется вирусным ферментом — интегразой (см. рис. 2.53) Однако этот фермент не
специфичен и интеграция провируса не носит специфический характер. Тем не ме-
нее у различных ретровирусов обнаружены «предпочтения» в сайтах для интеграции
провирусной ДНК. По таким предпочтениям ретровирусы можно разделить на три
группы К первой относятся ВИЧ-1, ВИЧ-2, SIV и FIV. До 70 % ДНК их провирусов
интегрируются с функционирующими генами (transcription units). Вторая группа
включает MLV и FV. Они «предпочитают» 5'-концы транскрипционных единиц и
CpG-островки. Третья группа представлена ретровирусами ASV, HTLV-1 и MMTV —
у них не обнаружено никаких «предпочтений» (Rene D., Smith J. А., 2008).
В первом десятилетии текущего века наиболее распространенным подходом к на-
целиванию ретровирусного вектора на конкретные участки в геноме человека было
использование в составе вирусной частицы слитых белков, образующих вирусный пре-
интеграционный комплекс, импортирующийся вовнутрь нуклеуса. Такие белки обыч-
но включают lN-белок, полученный из ВИЧ-1 или ASV; и ДНК-связывающие по-
следовательности клеточных и бактериальных белков, слитых с N- или С-концом 1N.
Чтобы вирус не утратил инфекционное™, слитые белки вводят в вирион ВИЧ-1 вме-
сте с исходным IN (wild-type IN). Одна из используемых моделей нацеливания ретро-
вирусных векторов на конкретные участки ДНК в геноме человека (таргетинговой
интеграции) показана на рис. 2.56.
Для увеличения специфичности геномного таргетинга IN был слит с полидактил-
цинк-пальчиковым белком Е2С (polydactyl zinc finger protein E2C), связывающимся
2.2. Биологические поражающие агенты
493
Таблица 2.23
Клетки и органы — мишени для псевдотипированных леитивирусов*
Мишень Гликопротеин Примечание
Печень VSV-G Снижена токсичность вектора
LCMV Снижена токсичность вектора, уменьшена воспалительная реакция на его введение
RRV Приобретен тропизм к клеткам печени, не являю- щимся гепатоцитами — Купферовым клеткам
SeVF
Легкие Вирус Эбола Трансдукция апикальной поверхности эпителия дыхательных путей
Вирус Марбурга
SeV F и HN
JSRV
Островки Лангер- ганса поджелудоч- ной железы LCMV Снижена токсичность вектора
пне VSV-G Нацеливание вектора преимущественно на нейроны
LCMV Нацеливание вектора на астроциты
Вирус бешенства Нацеливание вектора на предшественники/ стволовые клетки нейрональной ткани
Вирус Mokola (один из членов семейства Lyssavirus, вызываю- щих бешенство) Ретроградный транспорт вектора из перифери- ческой нервной системы в ЦНС
Ретроградный (от синапса к телу нейрону) и антероградный (от тела нейрона к синапсу) транспорт вектора в пределах ЦНС
Сетчатка глаза VSV-G Трансдукция фоторецепторов и ретинального пигментного эпителия
Вирус Mokola Специфичность ограничена ретинальным пигментным эпителием
Миониты/мышцы Вирус Mokola Трансдукция кардиомиоцитов плода (in utero)
Вирус Эбола То же
Гематопоэтиче- ская система RD 114 Более эффективен и менее токсичен, чем VSV-G
GALV Увеличение стабильности вектора в сыворотке
Раковые клетки GALV Веретенообразный (fusogenic) гликопротеин
LCMV Избирательность для клеток глиомы
* По J. Cronin et al (2005).
с уникальной 18 т. п. о. последовательностью в геноме человека. Вирус, содержаший
Е2С, слитый с С-конпом, IN в 7 раз чаше интегрировался с хромосомой вблизи Е2С-
связывающего сайта, чем исходный. Этот же белок, слитый с N-концом IN, показал
более чем десятикратное предпочтение в интеграции с Е2С-связывающим сайтом, чем
исходный IN. И хотя эффективность таргетинговой интеграции пока ниже, чем у
вирусов с диким типом IN, эти эксперименты свидетельствуют о сформировавшемся
направлении таких исследований, способном серьезно повлиять на технологии сома-
тической генотерапии (Rene D., Smith J. А , 2008).
494
Часть 2. Общая неправильная эпидемиология
Таблица 2.24
Селективные таканеспецифические промоторы, используемые
в лентивирусных векторах*
Ткань- мишень Промотор Источник промотора Примечание
1 2 3 4
Ретиналь- ная ткань Мышиный CD44 Ген, кодирующий трансмембран- ный гликопротеин и клеточный поверхностный рецептор для гиалуроновой кислоты Активен в глиальных Миллеровских клетках
Человеческий и мышиный GFAP Ген, кодирующий человеческий и мышиный глиальный фибрилляр- ный кислый белок То же
Мышиный VIM Ген, кодирующий виментин (vimen- tin) — главный субъединичный белок промежуточных нитей соеди- нительных тканей и других тканей мезодермального происхождения То же
1RPB1783 Ген, кодирующий интерЛотореиеп- тор ретиноидсвязывающий белок Активен в колбочке сетчатки глаза
GCAP292 Ген, кодирующий гупнилциклозо- активирующий белок 1 То же
ШОР500 Ген родопсина Активен в палочке сетчатки глаза
Нервная ткань СашКП Ген, кодирующий кальций/кальмо- дулин-зависимую протеинкиназу II Активен в нейронах пе- реднего мозга взрослого человека
SYN Ген синапсин 1-фосфопротеина Активен в отдельных регионах гиппокампа
NSEp 1ен, кодирующий нейронспецифическую энолазу Активен в стриатуме и гиппокампе
GfaABCID Ген глиального фибриллярного кислого белка Активен в глии ЦНС
[3-клетки Инсулин Ген инсулина человека Активен в р-клетках и линиях клеток мышиной инсулиномы
Печень ALB Ген альбумина Активен в гепатоцитах
ET Синтетический промотор Более активен, чем ALB; минимальная активность в селезенке ;
ApoA-II Ген человеческого аполипопротеина А-П Индуцибелен
al-AT Ген человеческого al-антитрипсина Активен в гепатоцитах
Система крове- творения WASp Ген, кодирующий белок Wiskott- Aldrich-синдрома Активен в Т-клетках, В-клетках дендритных клеток
Tie2 Ген, кодирующий расположенный на поверхности клетки рецептор ангиопоэтина Активен в эндотелиальных клетках и моноцитах
22. Биологические поражающие агенты
495
Продолжение табл. 2.24
1 2 3 4
1 промоторы: анкирин-1, а-спектрин, р-глобин. ^-глобин; энхансеры: GATA-I, Р-глобин LCR, интрон 18, а-глобин HS40, у-глобин интрон Эритроид-специфические гены Активен в эритроидных линиях
HLA-DRa Ген, кодирующий а-субъединицу человеческого лейкоцитарного антигена DR Активен в антигенпрезен- тирующих клетках
proximal lek Ген, кодирующий специфический Т-клеточный протоонкоген, lek Активен в Т-клетках
Сердце ANF Ген, кодирующий человеческий атриальный натрийуретический фактор Активен в кардиомиоцитах
MLC2v Ген, кодирующий легкую цепь чело- веческого вентрикулярного миозина То же
Легкие SP-C Ген сурфанктантного белка С второ- го типа альвеолярных эпителиаль- ных клеток (АТ-2 клетки) Активен в АТ-2 клетках
Простата PSAp Ген, кодирующий специфический антиген простаты Не известно
* По К. Pluta, М. М. Kacprzak (2009).
DBP Г~~~Г
DBD
J IN
TS
DBD
1DBD-IN
] IN-DBD
Ретровирусная ДНК
IN-DBD(wihDBD-IN)
Хромосомная ДНК
С
Рис. 2.56. Модель таргетинговой интеграции ретровирусных векторов. Интеграция
осуществляется посредством использования модифицированной интегразы слитой
гетерологичными ДНК-связывающими доменами. DBD — ДНК-связывающий до-
мен (DNA-binding domain); IN — интеграза. TS — белок, образовавшийся в резуль-
тате слияния DBD и IN. Преинтеграционный комплекс интегрируется с участком
хромосомной ДНК (target site, TS), узнаваемым DBD. По D. Rene, J. A. Smith (2008)
496
Часть 2. Общая неправильная эпидемиология
В последние годы в практике генной терапии стали использовать лентивирусные
векторы, дефицитные по интегразе (integration-deficient lentiviral vectors, IDLVs). Их
геном содержит мутантный ген IN, продукт которого не способен обеспечить инте-
грацию лентивисуса с геномом человека. Вектор поддерживается в трансдуциро-
ванной клетке в виде кольцевой эписомы, не имеющей репликационного сигнала.
В процессе деления клетки эписома утрачивается, однако стабильно сохраняется в
неделяшихся клетках. Такие векторы считаются более безопасными, чем интегрирую-
щиеся (рис. 2.57).
Интегрированный
провирус
Рис. 2.57. Образование эписомы из лентивирусного вектора. В результате обратной
транскрипции РНК вектора в клетке формируется линейная двуцепочечная ДНК
(linear double-stranded DNA, dsDNA) c LTR на обоих концах, у самоинактивирую-
щихся векторов, содержащих делеции в иЗ-регионе (на схеме показана как dLTR).
Эта ДНК импортируется в ядро как часть вирусного пре интеграционного комплек-
са. А. Традиционный лентивирусный вектор, включающий функционирующую IN,
интегрируется с геномом как провирус. Однако в клетке он может поддерживаться
и в кольцевой форме. Для этого у линейной двуцепочечной ДНК провируса есть,
по крайней мере, две возможности: через негомологичное сшивание LTR-концов,
образуя 2-LTR-3nncoMy; или через лигирование «ников» (молекул с однонитевыми
разрывами — показаны стрелками с кружками) — образуя l-LTR-эписому. Б. Когда
провирусная интеграция блокирована через мутантный IN, в ядре клетки возрастает
количество векторных эписом. att — сайт присоединения интегразы в концевых
участках провирусной ДНК. По К. Wanisch, R. Ynez-Mucoz (2009)
Функциони-
рующая IN
Наиболее тяжелые генетические последствия наблюдали при терапевтическом
использовании ретровирусных векторов для лечения пациентов с тяжелым комбини-
рованным иммунодефицитом, связанным с Х-хромосомой (X-linked severe combined
immunodeficiency, X-SCID). Иммунодефицитное состояние у таких пациентов возни-
кает из-за мутации в гене у-субъединицы рецептора интерлейкина 2 (interleukin-2
2.2 Биологические поражающие агенты 497
vceptor subunit gamma gene, IL2RG), общего для рецепторов интерлейкинов IL-2, IL-4,
IL-7. IL-9, IL-15 и IL-21. Эти интерлейкины и их рецепторы вовлечены в процессы
„вития Т- и В-лимфоцитов. Без функционирования Т-клеток невозможна активи-
шия В-клеток. Ген IL2RG находится на Х-хромосоме. Мутация в гене IL2RG приво-
дит к инактивации общей у-цепи рецепторов интерлейкинов, и, как следствие, нару-
1ются процессы сигнализации, в которых они участвуют. Происходит почти полный
саз иммунной системы с отсутствием или очень малым количеством Т-лимфоци-
- 'в, NK-клеток и нефункциональными В-лимфоцитами. Дети с X-SCID погибают от
(фекнии в течение года (Strauss В. Е., Costanzi-Strauss Е., 2007).
Гак как X-SCID вызван мутацией в одном хорошо охарактеризованном гене, его
ечение путем введения пациенту полноценного гена в конце 1990-х гг. представля-
юсь исследователям одним из самых перспективных направлений в генотерапии.
эуппой французских исследователей из костного мозга 9 пациентов в возрасте от 1 до
- мес. были получены аутологичные гемопоэтические стволовые клетки CD34+. Эти
<летки культивировали в условиях ex vivo и трансдуцировали ретровирусным векто-
~ом на основе вируса мышиной лейкемии Молони, в который включили ген lL2Rec
еловека. Затем клетки вводили обратно пациентам. Функция Т-клеток постепенно
восстановилась. В течение нескольких месяцев количество Т-клеток и натуральных
«иллероз в крови у 8 пациентов достигло нормы. Был установлен минимальный по-
эог количества трансдуцированных клеток, которое необходимо ввести пациенту для
остижения терапевтического эффекта — lx 106—3 х 106х СО34+-клеток/кг массы тела.
Однако почти 11 -летнее наблюдение дало обескураживающие результаты: у четырех
пациентов в течение 31—68 мес. развился острый Т-клеточный лейкоз, и один из них
умер (Hacein-Bey-Abina S. et al., 2002; 2010). Причиной развития острого лейкоза ста-
з интеграция ретровирусного вектора вблизи нескольких протоонкогенов и пере-
стройка генома клеток-мишеней, приведшие к многокаскадной активации протоон-
когенов и, как следствие, неконтролируемой пролиферации Т-клеток (рис. 2.58).
Наиболее часто вовлекаемый в онкогенез ген LMO2 (LIM domain—only 2) лока-
лизован у человека на 11-й хромосоме. Он экспрессируется в ядре клетки и действует
к ак транскрипционный активатор факторов, необходимых для дифференциации
эритроцитов в костном мозге. В норме ген LMO2 экспрессируется в различных тка-
нях, таких как почки, печень, легкие, селезенка и ЦНС, участвует в ангиогенезе. Одна-
ко его активизация в Т-клетках в результате встраивания ретровируса или хромосом-
ной транслокации приводит их к неконтролируемому размножению и развитию
острого Т-клеточного лейкоза. Химиотерапия оказалась успешной у тоех пациентов
с развившимся лейкозом. Она позволила добиться восстановления поликлональных
трансдуцированных Т-клеточных популяций (McCormack М. Р., Babbitts Т. Н., 2004;
Strauss В. Е., Costanzi-Strauss Е., 2007; Hacein-Bey-Abina S. et al., 2008). Потенциаль-
ные механизмы вставочного мутагенеза, вызванного интеграцией ретровирусных век-
торов с геномом человека, приведены на рис. 2.59.
Цели подобных исследований неоднозначны. Помимо изощренности в «наце-
ивании» лентивирусных векторов на клетки и органы-мишени и даже, на участки
генома человека, критические для его здоровья, обращает на себя внимание исполь-
зование для получения псевдотипированныхлентивирусов, гликопротеинов оболочки
вирусов, считающихся потенциальными агентами БО (вирусы геморрагических ли-
кэрадок Эбола, Марбурга, Росс-Ривер и др.), вызывающих вспышки с высокой смерт-
ностью. Последнее свидетельствует о том, что работа с лентивирусными вектора-
ми, предназначенными для лечения редких наследственных болезней у обывателей-
498
Часть 2. Общая неправильная эпидемиология
Клонированный ген
S Транс-
дукция
Цитокиновый
рецептор
Рис. 2.58. Развитие острого лейкоза у четырех пациентов после интеграции в геном
их ретровирусного вектора. А. Терапевтический эффект. Б. Развитие острого лейкоза.
У двух пациентов (№ 4, 5) геном бластных клеток1 включает активный вектор вбли-
зи протоонкогена LMO2. У пациента № 10 бластные клетки содержат интегрировав-
ший вектор вблизи протоонкогена LMO2m второй интегрировавший вектор вблизи
протоонкогена ВМП. У пациента № 7 бластные клетки содержат интегрировавший
вектор вблизи протоонкогена CCND2. Хромосомные транслокации, вызванные
интеграцией вектора, у некоторых пациентов привели к активизации протоонкогена
NOTCH 1, делении гена CDKN2, супрессирующего развитие опухолей, и к пере-
стройке гена SIL-TALI, приведшие к неконтролируемой пролиферации Т-клеток.
У пациентов № 4 (умер) и 7 обнаружены обширные делеции хромосомы 6ql6-16.
По Р. Noguchi (2003) и S. Hacein-Bey-Abina et al. (2008)
инвалидов, ведется в лабораториях, по уровню биологической безопасности пригод-
ных для исследования традиционных агентов БО. Одновременно проводятся экспе-
рименты по изучению возможности массового применения таких конструкций, на-
пример, путем использования аэрозолей (см. опыты по трансдукции апикальной
поверхности эпителия дыхательных путей; табл. 2.23).
К июлю 2013 г. 19,4 % протоколов экспериментов по генной терапии в мире (из
1902) предполагали использование векторов на основе ретровирусов. Их по попу-
лярности среди генных терапевтов «перегнали» только векторы на основе аденовиру-
сов — 23.2 % (www.wiley.co.uk/genmed/clinical/).
Векторные системы на основе аденовирусов. Аденовирусы — ДН К-вирусы, способ-
ные инфицировать оба типа клеток (делящиеся и неделяшиеся), включая клетки пе-
чени, легких и эпителия. Они реплицируются в ядре инфицированной клетки как
1 Blast cells — незрелая клетка-предшественник.
2.2. Биологические поражающие агенты
499
Экзон
Направление транскрипции вектора
♦
ntiiiiii«««а•'
Аберрантные транскрипты • »
*к
Экспрессия
эндогенного
промотора через
энхансерную
активность LTR
Pro Tumor
5 LTR— Трансген жЗТТКн
Супрессор
Нарушение экспрессии
эндогенного гена
В
в
Рис. 2.59. Потенциальные механизмы вставочного мутагенеза, вызванного интег-
рацией ретровирусных векторов с геномом человека A. LTR ретровирусного век-
тора, вставившегося перед хромосомным геном, выполняет функцию промотора
(Pro) и запускает транскрипцию этого гена, что приводит к получению химерных
транскриптов, включающих всю или часть кодирующей последовательности гена,
расположенного в направлении 3' от LTR (процесс синтеза РНК протекает в направ-
лении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движет-
ся в направлении 3'->5). Такая последовательность «чтения» вызвана тем, что по-
лиаденилационный сайт в пределах R региона LFRs является относительно слабым
и неэффективным. Если геном вектора содержит активный донорный сайт сплай-
синга (active splice donor, SD), аберрантный сплайсинг может приводить к образо-
ванию усеченных транскриптов. Клетка вместо физиологических транскриптов
(белков) накапливает большое количество усеченных транскриптов (белков), чье
действие может привести к развитию патологии Б. Активация промотора через
ЕТК-^нхансер. Провирусный геном интегрируется в хромосомный ген в любой ори-
ентации. При правильной ориентации LTR-энхансера ( т. е. перед промотором хро-
мосомного гена) он может увеличить уровень его экспрессии. Этот механизм ле-
жит в основе увеличения экспрессии протоонкогена LMO2, наблюдаемой у
пациентов № 4 и 5 с X-SCID (см. рис. 2.58). В. Уменьшение или прекращение экс-
прессии гена через внутригенную вставку генома ретровирусного вектора, приво-
дящую к разрушению гена, либо через индукцию преждевременной полиаденила-
ции в сайте в пределах генома вектора или через запуск аберрантного сплайсинга,
который изменяет или элиминирует продукт гена. По A. W Nienhuis (20%)
500
Часть 2. Общая неправильная эпидемиология
эписомные (экстрахромосомные) элементы. По связыванию со специфическими сы-
воротками аденовирусы разделены на 51 серотип. На основе их способности агглю-
тинировать эритроциты у людей, кроликов и мышей; и по онкогенности для грызу-
нов их подразделяют еще на 6 подтипов или субгрупп (от А до F). Исследование
филогении аденовирусов дает дополнительные основания для их деления внутри суб-
групп. Субгруппа В дополнительно разделена на субгруппы В1 и В2 (Segerman А.
et al., 2003). Аденовирусы разных подгрупп поражают различные органы и ткани че-
ловека. Например, вирусы подгрупп В1, С и Е главным образом вызывают респира-
торные болезни; вирусы подгрупп В, D и Е способны поражать ткани глаза; субгруп-
па вирусов F вызывает гастроэнтериты; В2-вирус инфицирует почки и уринарный
тракт (Russell W. С., 2009) В табл. 2.25 обобщены сведения по тропизму аденовирусов
различных субгрупп.
Таблица 2.25
Тропизм аденовирусов различных субгрупп*
Субгруппа Серотип П реобладающий тропизм Известные рецепторы узнавания
А 12, 18, 31 ЖКТ CAR
В1 3, 7, 16, 21, 50 Респираторная система CD46, CD80/86, рецептор X, HSPG
В2 11, 14, 34, 35 Почка CD46, CD80/86, рецептор X, HSPG
С 1, 2, 5, 6 Респираторная система CAR, HSPG, MHC-I, VCAMI, интегрины
D 8, 9, 10, 13, 15, 17, 19,20, 22-31, 33, 36-39, 42-49, 51 Ткани глаза CAR, сиаловые кислоты, CD46
Е 4 Респираторная система, ткани глаза CAR
F 40, 41 ЖКТ CAR
* По работам F. Sakural (2008) и A. Sharma et al. (2009).
Исследования по созданию на основе аденовирусов векторов, предназначенных
для использования в целях генотерапии, активизировались с начала 1990-х гг. Этому
способствовали следующие обстоятельства:
1) ДНК-вирусы представлялись генотерапевтам как более простые объекты для
генно-инженерных манипуляций, чем вирусы с РНК-геномом;
2) они реплицируются в неделящихся клетках, например, клетках мышц.
Конструирование векторов на основе аденовирусов во многом повторяло путь,
пройденный создателями лентивирусных векторов. Дефектные по репликации аде-
новирусы получали посредством замены гена Е1, который необходим для репликации,
наген, представляющий интерес для исследователя и энхансер-промоторный элемент.
Такие рекомбинантные векторы способны эффективно размножаться в клетках, экс-
прессирующих продукт гена Е1 более 1011—1012 аденовирусных частиц/см3 (InderM. V.
Nikunj S., 1997).
Неспособные к репликации аденовирусные векторы используют для введения
трансгенов в условиях in vivo. Они обеспечивают очень высокую экспрессию, но на
непродолжительное время (5—10 сут) из-за противодействия иммунной системы ре-
ципиента. Возможно так же и то, что клетки-мишени содержат факторы, которые
2.2. Биологические поражающие агенты
501
осиливают синтез аденовирусных белков, приводящих к иммунному ответу. Для того
ч гобы обойти эту проблему, было предложено второе поколение аденовирусных век-
'оров, у которых дополнительно удалили гены, вовлекающие вирус в репликацию.
Такие векторы способны к более длительной экспрессии, но и у них она практически
не определялась через 20—40 сут (Inder М. V., Nikunj S., 1997). Эта идея получила свое
дальнейшее развитие созданием третьего поколения аденовирусных векторов («third
generation») — «выпотрошенных (gut-less)» векторов, из которых были делегированы
все вирусные гены и оставлены только элементы, определяющие начало и конец ге-
нома и вирусную пакующую последовательность. Их емкость для клонируемой ДН К —
до 35 т. п. о. (Sharma A. et al., 2009).
Трансгены, переносимые такими векторами, экспрессировались в течение сут
Inder М. V., Nikunj S., 1997). При экспрессии аденовирусными векторами чужерод-
ных генов могут использоваться эукариотные промоторы, такие как аденовирусный
Ela-промотор, прямой ранний промотор цитомегаловируса, или LTR-промогор ви-
-чса саркомы Рауса (Russell W. С., 2009).
Основные усилия разработчиков таких векторов направлены на:
1) преодоление иммунитета к аденовирусам, имеющегося практически у каждого
зрослого человека;
2) изменение тропизма вектора и более точное его нацеливание на клетки-мишени.
При разработке аденовирусных векторов, способных преодолевать иммунитет к
аденовирусам, обычно исследователями учитывается то, что у людей наиболее часто
стречаются антитела к аденовирусу пятого серотипа (AdS-антитела). Векторы, про-
изводные от аденовирусов других серотипов, способны избегать нейтрализацию эти-
ми антителами (см. табл. 2.25). Реже всего у людей обнаруживают антитела к адено-
ирусу 35-го серотипа. Гены, трансдуцированные экспериментальным животным
сгорами на основе вируса 35-го серотипа, способны длительно экспрессироваться
-есмотря на присутствие антител к вирусу 5-го серотипа (Zaiss А. К. et al., 2009).
В связи с тем, что вектор в любом случае должен формировать вирусную частицу,
способную узнавать специфический рецептор на клетке-мишени, бесконечно «упро-
ть» вирус и пакующую систему невозможно. Поэтому в последние годы для сниже-
ния заметности вируса для иммунной системы, стали использовать более тонкие под-
ходы. Например, введение точечной мутации в петлю гексонового белка AdC-вектора,
отвечающую за эпитоп доминирующих нейтрализующих антител. Такая мутация зна-
ятельно снижала способность вектора связываться с нейтрализующими антителами
.словиях in vitro, но в условиях in vivo он блокировался специфическими антитела-
ми (Pichla-Gollon S. L et al., 2009). Замена всей гексоновой петли вируса AdHu5 на
-дкую же петлю из вируса AdHi i48, предотвращает связывание вектора AdHuS-ней-
I рализующими антителами и позволяет эффективно использовать этот вектор для
-ереноса генов в геном животных, в крови которых уже имеются антитела к AdHu5
Roberts D. М. et al., 2006).
Для изменения тропизма аденовирусных векторов исследователями используют-
ся две стратегии: физический таргетинг (physical targeting) и генетическая модифика-
ция вируса. Кратко они обобщены в табл. 2.26, с. 502.
Физический таргетинг заключается в том, что вирусная частица покрывается спе-
1 иальной оболочкой, изменяющей ее природный тропизм и делающей ее неузнавае-
мой для иммунной системы. В качестве такой оболочки используют полимеры (поли-
этиленгликоль или poly-[N-(2-hydroxypropyl)methacryIamide, рНРМА]) или вирус-
ню частицу включают в биодеградируемую альгинатную микрочастицу. Природный
502
Часть 2. Общая неправильная эпидемиология
Стратеги.! изменения тропизма аденовирусных векшрии
Таблица 2.26
Вектор Модификация вектора Причина изменения тропизма вируса Тип клеток, использованных в экспериментах Результат 1
Ad5-pk7 Введение полили- зинового мотива (рК7) в «набал- дашник» капсида HAd5 Повышенное содер- жание SPG на по- верхности раковых клеток способствует трансдукции Клетки глиомы человека, переса- женные мышам Повышенные трансдукция и экспрессия маркера
Ad5/ 3-RGD Введение в набал- дашник капсида HAd5 последова- тельности из «набалдашника» HAd3, содержащей RGD-домен Повышение трансду- цируюшей способно- сти вектора в отно- шении раковых кле- ток с высоким уров- нем экспрессии транс- дуцированного гена То же 1000-кратное увеличение инфекти внести вектора
CAV-2 (Ad собак) Изменение тропизма вектора Использование альтернативных рецепторов In vivo-- респира- торный тракт мы- шей, ex vivo — пульмснарный эпителий человека Эффективная трансдукция пульмонарного эпителия, «ускользание* от антител к HAd5, незначительная воспалительная реакция в месте введения вектора
HAd5 mEGF полимер- ная оболочка Селективная трансдукция раковых клеток, богатых EGFR Перитонеальная модель ксено- транспл антантов овариальных раковых клеток человека у мышей Ограниченные векторный тропизм и токсичность. Повышенная антираковая эффективность
HAd5 Биспецифические антитела (АЬ), нацеливающие «наба .дашник» капсида Had на эндолгин человека Биспецифические антитела соединяют Ad с васкулярным эндол- гином в ангиогенных зонах опухолей П реимущественно эндотелиальные клетки и HUVEC- клеточные линии Повышенная селективность и CAR-независимая трансдукция HUVEC
* По A. Sharma et al. (?000)
тропизм вектора заменяется селективным таргетингом (selective targeting), т. е. в эти
полимеры вводятся таргетирующие лиганды (пептиды, белки или антитела).
Другой эффективной стратегией для физического таргетинга аденовирусного век-
тора стало использование биспецифических адапторных молекул (биспецифическис
антитела или белки слияния), состоящих из двух компонентов — один из них с высо-
кой специфичностью связывается волокнистым набалдашником (fiber knob) капсид?
аденовируса, другой с высокой специфичностью связывает рецептор клетки-мишени.
Однако обе стратегии таргетинга дают плохо воспроизводимые результаты. Поэтом
физическому таргетингу исследователи предпочитают генетические модификации
аденовируса, т. е. генетический таргетинг (genetic targeting) (Sharma A. et al., 2009).
2 2 Биологические поражающие агенты 503
В экспериментах по генетическому таргетингу объектом модификации генно-
^верными способами обычно является волокнистый «набалдашник» капсида,
рающий основную роль в специфическом узнавании Ad клеток-мишеней и, в час-
-мости, расположенные на нем два участка: С-конец и HI-петля. Вставка в эти учас-
тки RGD или полилизина ведет к повышению инфективносзи аденовирусного век-
ра в отношении широкого круга клеток-мишеней, экспрессирующих на своей
рхности интегрины или HSPG. Кроме модификации волокнистого «набалдаш-
- *а> капсида, для изменения тропизма вектора модифицируются другие оболочеч-
с белки аденовирусов: гексон, пентон, р1Х или рШ.
Другой удачной схемой изменения тропизма аденовирусных векторов, производ-
-ых от HAd5. стала замена волокнистого «набалдашника» капсида на аналогичную
структуру, взятую из вирусов других серотипов, использующих для интернализации
гые рецепторы, чем CAR. Этот прием изменения антигенной специфичности вируса
тмзок к псевдотипированию, описанному выше на примере лентивирусных векто-
(Sharma A. et al., 2009).
Недостатком аденовирусных векторов, ограничивающим их использование в
ютерапии, является неспособность к интеграции с хромосомой реципиента, в ре-
ьтате вектор утрачивается по мере продолжения деления клеток. Этот недостаток
этично устраняется использованием векторов на основе AAV
S. Е. Raper et al. (2003) описали случай летального исхода при лечении пациентов
. врожденным дефицитом орнитинтранскарбамилазы (ornithine transcarbamylase,
DTC). Мужчине 18 лет, участвовавшем в пилотном проекте по оценке безопасности
юго метода генной терапии, был введен вектор на основе человеческого аденови-
са 5 типа с делегированными генами Е1 (необходим для репликации вируса) и Е4
•аствует в регуляции транскрипции вирусной ДНК), включающий клонирован-
- ую кДНК гена ОТС человека. Пациенту было введено через правую печеночную арте-
- по 6x10” векторных частиц/кг. Приблизительно через 18 чбыли замечены изменения
его психике и развитие желтухи, чего не наблюдалось у первых 17 пациентов, уча-
_ ювавших в этом же исследовании. В дальнейшем у пациента развились диссеми-
- *рованная внутрисосудистая коагуляция (disseminated intravascular coagulation), син-
дром системного воспалительного ответа (systemic inflammatory response syndrome) и
множественная органная недостаточность, приведшие его к смерти через 98 ч после
зедения аденовирусного вектора с клонированным геном ОТС человека. При посмер
“ом исследовании вектор с клонированным геном обнаружен почти во всех тканях
“эгибшего пациента. Причины его смерти авторами не объяснены. Ими только сде-
1н вывод, что такие способы лечения нуждаются в значительно большем количестве
•ытов на животных, чем было проведено, и изучения возможных иммунных oust
пзв у пациента.
‘деноассоциированные вирусные векторы (AAV). Относительно недавно использу -
. мый для генотерапии AAVявляется простым не патогенным одноцепочечным ДНК-
вдрусом. Его два гена (cap и гер) расположены между двумя терминальными инверти-
ванными повторами, которые определяют начало и конец вируса и содержат
“акующую последовательность. Ген cap кодирует оболочечные белки вирусного кап-
смда, а продукт гена гер вовлекается в вирусную реплш аиию и интеграцию. AAV нуж-
дается в дополнительных генах для своей репликации в пакуюших клетках. Они обес-
печиваются хэлперными вирусами (обычно это аденовирусы или вирус простого
ерпеса) (Inder М. V., Nikunj S , 1997) (рис. 2.60, с. 504).
504
Часть 2. Общая неправильная эпидемиология
Рис. 2.60. Жизненный цикл AAV. AAV размножается в клетках, коинфици-
рованных аденовирусом. В отсутствие аденовируса AAV поддерживается в
латентном состоянии, интегрировавшись с хромосомой 19 (AAVS1). Латент-
ный AAV может снова стать репликационно- активным при условии су-
перинфицирования клетки аденовирусом. По S. Daya, К. 1. Berns (2008)
Трансдукция вектора в клетки-мишени происходит различными путями, вклю-
чающими интеграцию с геномом хозяина, экспрессию трансдуцированных генов с ли-
нейных и кольцевых форм вируса (Nakai Н. et al., 2000), гомологичную (Russell D. W.,
Hirata R. K., 1998) и негомологичную рекомбинацию ДНК вектора с хромосомной
ДНК (Miller D. G. et al., 2005). Для изменения тропизма AAV используются те же при-
емы, что и для изменения тропизма аденовирусных векторов (Daya S., Berns К. I., 2008).
Исследования «предпочитаемых» сайтов интеграции AAV с геномом человека, пока-
зали, что в основном они располагаются в пределах CpG-островков и в начале генов,
а его рекомбинация с хромосомной ДНК человека в основном носит негомологичный
характер. В отличие от ретровирусного вектора (см. выше), вектор на основе AAV не
содержит интегразу, и он может интегрироваться с хромосомой только при наличии в
ее участках одно- и двунитевых разрывов. CpG-островки проявляют себя измененной
структурой хроматина и имеют повышенную чувствительность к нуклеазам. В репли-
кации ДНК они участвуют в качестве участков начала репликации с высокой вероят-
ностью появления разрывов в цепи ДНК. Этот феномен объясняет интеграционное
предпочтение AAV к CpG-островкам (Miller D. G. et al., 2005).
AAV может инфицировать различные типы клеток. В присутствии продукта гена
гер вирусная ДНК способна интегрироваться преимущественно с хромосомой 19 че-
ловека. В векторах, создаваемых на основе AAV, гер и cap гены замещаются на транс-
гены Выход вирусных частиц составляет 1011—1012/см3. Емкость AAV-вскторов не
превышает 3,5—4,0 т. п. о. чужеродной ДНК, что исключает их использование для
клонирования крупных генов. AAV-вектор, содержащий ДНК человеческого факто-
ра IX, был использован для трансфецирования печени и мышц иммунокомпетентных
мышей. Трансфепированные клетки продуцировали в кровь мышей терапевтические
количества белка фактора IX более 6 мес. (Inder М V, Nikunj S., 1997).
2 Биологические поражающие агенты
505
Векторные системы на основе вируса герпеса простого. Клонирующие векторы на
.пове вируса герпеса простого (herpes simplex virus type 1, HSV-1) имеют простую
-трукпию и разработаны хуже, чем векторы на основе аденовирусов. Вирус вклю-
эолее чем 80 генов, один из которых (IE3) может быть замещен с целью создания
тора. Делегированы могут быть и некоторые другие гены, что позволяет исполь-
• вать вектор для клонирования нескольких трансгенов. Выход вирусных частиц
ч. гавляет приблизительно (108—109)/см3. Развитие таких векторов затормозили та-
е препятствия, как: 1) непродолжительная экспрессия клонированных генов (до не-
.кольких недель); 2) их способность вызывать цитопатические эффекты и ответы со
гороны иммунной системы.
Поэтому развитие получили ампликоны, многократно повторяющиеся последо-
3.x ельности HSV, включающие мономерные последовательности, организованные по
* шу «голова к хвосту» (конкатемеры). Мономеры включают, по крайней мере, один
.ток начала репликации вирусной ДНК (oriS или oriL) и последовательность для
наковки ДНК в вирусную частицу (рас). Клонирование этих двух последовательно-
csefl в бактериальную плазмиду позволяет создать вектор, в пакующих системах упа-
• Бывающий ДНК в HSV-1 вирион. Такие векторные системы способны упаковывать
Ж) 150 т. п. н. чужеродной ДНК, что позволяет одним вектором доставлять в клетку-
[ишень десятки транскрипционных единиц, и в их числе гены других вирусов, не вы-
звал иммунных ответов и цитопатических эффектов на сам вектор. Продолжитель-
-ость экспрессии клонированных генов достигает года. HSV-1 рассматривается
. зециалистами как основной при конструировании вирусных векторов, предназна-
-енныхдля генотерапии мозга (Sena-Esteves М. et al., 2000).
Векторные системы на основе ортопоксвирусов. Ортопоксвирусы — крупные виру-
сы. содержащие двунитевую ДНК. Отдельные виды могут иметь либо очень широкий,
тибо очень ограниченный круг хозяев. Емкость поксвирусного вектора позволяет
источить 25 т. п. н. чужеродной ДНК без делений генома самого вируса. Векторные
с штемы на их основе не имеют широкого применения из-за того, что эукариотиче-
-кие промоторы неэффективно распознаются транскрипционными механизмами
оксвирусов, и для эффективной экспрессии рекомбинантных генов в клетке-рени-
-ненте должны быть использованы поксвирусные промоторы. Кроме того, поксви-
ч}сные транскрипты не подвергаются сплайсингу, поэтому генетический материал,
зонированный в поксвирусах, должен быть в форме кДНК. Вследствие большого
'.сзмера и неинфекционной природы поксвирусной ДНК, чужеродные гены клони-
?. ются в поксвирусах путем рекомбинации в условиях in vivo (Bienkowska-Szewczyk К.,
SzevczykB., 1999).
Неожиданным для вирусологов стало открытие у ортопоксвирусов природного
гропизма к опухолевой ткани. Поэтому в настоящее время их рассматривают в каче-
стве векторов для доставки генов в опухолевую ткань. И в этом аспекте вирус вакци-
ны (vaccinia virus) имеет значительные преимущества перед другими вирусами, на
эснове которых конструируют векторы для генотерапии:
1) у него эффективный жизненный цикл; уже через 6 ч в инфицированной клетке
формируются зрелые вирионы;
2) в условиях in vivo он быстро распространяется от клетки к клетке;
3) несет свои строгие промоторы (stringent promoter), способные обеспечивать вы-
сокий уровень экспрессии трансгенов;
4) может инфицировать различные ткани человека, не вызывая патологических
процессов (Zeh Н. J., Bartlett D. L., 2002).
506
Часть 2. Общая неправильная эпидемиология
Векторные системы на основе РНК-вирусов, не принадлежащих к семейству ретро-
вирусов. Разрабатываются со второй половины 1990-х гг. в рамках создания новых
технологий иммунизации. Для конструирования вакцин используются векторы на
основе вирусов гриппа, бешенства, везикулярного стоматита, кори, респираторно-
синтициальной болезни, лихорадок Синдбис, Сендай, ВЭЛ идр. Основными преиму-
ществами таких векторных систем являются следующие:
1) в условиях in vitro возможно использовать легко селектируемые нецитопатичес-
кие РНК-векторы;
2) векторы данного типа, содержащие репликон, могут длительное время экспрес-
сировать высокие уровни гетерологичных белков, и поэтому они могут рассматривать-
ся как эффективная вакцинная векторная система;
3) векторы данного типа не содержат полного набора вирусных генов и благодаря
этому не могут образовывать вирусные частицы, способные вызвать генерализован-
ную инфекцию в других тканях; т. е. эти векторы могут удовлетворять строгим требо-
ваниям безопасности;
4) так как компоненты вектора, являющиеся производными вируса, имеют неболь-
шой размер и не экспрессируют структурные белки, то ответ иммунной системы на
вектор, вероятно, будет ограниченным. Следовательно, продолжительная экспрессия
чужеродных белков, не будет приводить к цитолизу клеток;
5) благодаря тому, что РНК-вирусы утратили ДНК-фазу, они не могут интегриро-
вать чужеродные гены в хромосомную ДНК. Это делает невозможной трансформацию
клеток векторами на основе РНК-вирусов (Palese R, 1995).
Искусственные векторные системы. Разрабатываются по трем направлениям:
1) комплексы, образованные нуклеиновыми кислотами и катионными полимера-
ми (подиндексы);
2) комплексы, образованные нуклеиновыми кислотами и липидами (липоплексы);
3) комплексы на основе наночастиц двуокиси кремния.
На рис. 2.61 показаны механизмы формирования первых двух комплексов и их
проникновения в клетку. Основные ограничения для доставки генов в геном челове-
ка с помощью искусственных векторных систем и способы их преодоления обобще-
ны в табл. 2.27.
Полиплексные композиции (polyplexes) получают путем конденсации нуклеиновых
кислот полимерами. Полимеры, используемые для конструирования подиндексов,
делятся на две группы: деградируемые и недеградируемые. Из недеградируемых по-
лимеров разработчики искусственных векторных систем наиболее часто используют
полиэтиленимин (polyethylenimine; PEI, 22 кДа). Он растворим в воде и состоит из
множества этиленаминовых блоков (ethylenamine units), несущих положительный
заряд. Высокая плотность положительного заряда такого комплекса создается ами-
ногруппами этиленамина. Р. Ghana et al. (2004) описали использование PEI в качестве
носителя гена субъединицы А дифтерийного токсина (DTA), эффективно экспресси-
рующего его под контролем промотора гена Н19 (распространенный онкофетальный
ген) в раковых клетках мочевого пузыря двух пациентов.
Деградируемые полимеры менее токсичны, чем недеградируемые. К ним относят-
ся поли-Ь-лизин [poly(L-lysine), PLL] и его аналоги. Архитектура этих полимеров
включает группы, чувствительные к гидролизу (сложные эфиры, ацетали или гидра-
зоны), разрушающиеся внутри клетки под воздействием ферментов и продуктов ме-
таболизма (Halama A., Kulinski М. et al., 2009).
12. Биологические поражающие агенты
507
Рис. 2.61. Механизмы формирования искусственных векторных систем и их проник-
новения в клетку. А. Формирование комплекса. Имеющие отрицательный заряд нук-
леиновые кислоты взаимодействуют с катионными липидами или поликатионами
через электростатические взаимодействия. В результате формируются положитель-
но заряженные липоплексы или подиндексы. Б. Положительно заряженные неболь-
шие частицы связываются со специфическими рецепторами или непосредственно
с поверхностью клетки. Комплексы проникают в клетку по механизму эндоцитоза.
После высвобождения из эндосомы и разборки комплекса, нуклеиновая кислота
проникает через поры в ядро клетки. По A. Halama, М. Kulinski et al. (2009)
В конце 1990-х гг. разработаны технологии, позволяющие получение нанораз-
черных форм полиплексов (~15 нм, т. е. размера глобулярных белков, см. работу
Thurmond К. В. et al., 1999). Нанополиплексы размером меньше 25 нм эффективно
трансфецируют неделящиеся клетки, так как они меньше, чем размер пор в мембране
ядра. Нанополиплексы стабильны в солевых растворах. Конденсированный пептид,
содержащий в своей структуре тридцатимерный полилизин с цистеином на N-конце,
к которому «пришит» 10 кДа ПЭГ, при температуре 4°С стабилен в солевом раство-
ре более трех лет; 9 мес. он сохраняет основные свойства при комнатной температуре
и один месяц при температуре 37°С (Davis Р. В., Cooper М., 2007). В табл. 2.28 приве-
дено описание полимеров, наиболее часто используемых для создания искусствен-
ных векторных систем.
После специфического взаимодействия полиплексов с рецепторами поверхно-
сти кпетки, они захватываются эндосомами. Но в отличие от вирусов, подиндексы
508
Часть 2. Общая неправильная эпидемиология
Таблица 2.27
Основные ограничения для доставки искусственными векторными
системами генов в геном человека и способы их преодоления*
Ограничение Способ преодоления
Доставка генов к клетке и клеточный таргетинг
Деградация ДНК нуклеазами сыворотки крови Конденсация ДНК или ее инкапсуляция катионными полимерами или липидами
Неспецифическое взаимодействие с компонентами крови Экранирование поверхности ДНК оболочкой из гидрофобных полимеров
Неспецифическое связывание тканями векторов, включающих трансген Введение специфических лигандов в поверхность обо- лочки трансгенной конструкции, использование особен- ностей васкуляризации отдельных тканей (например, ра- ковых опухолей)
Доставка генов вовнутрь клетки
Неэффективное поглощение ДНК клеткой Введение в поверхность оболочки трансгенной конструкции специфических лигандов, усиливающих рецептор-опосредованный эндоцитоз
Невыход трансгена из эндосомы Включение мембрано-активных пептидов в оболочку трансгена (например, аналогов меллитина). Поликатионные «протоновые губки» (например, РЕ1)
Затрудненный транспорт трансгена из цитозоля в ядро Включение трансгена в наночастицы
Подавление экспрессии гена Использование миникольцевой ДНК тишенной бактериальных последовательностей
Утрата трансгена Минихромосомы с эписомальными элементами для поддержания. Экспрессионная кассета для трансгена, интегрирующегося с хромосомой человека
Токсичность конструкции
Острая токсичность Экранирование поверхности для избежания агрегации в крови. Удаление контаминирующих поликатионов, бактериальных ДПС
Цитотоксичность и длительное токсическое действие Использование в качестве носителей генов биодеградируемых катионов
Клеточный иммунный ответ Временная иммуносупрессия
* По S. Boeckle, Е. Wagner (2006).
обладают плохой способностью освобождаться из эндосом и подвергаются деграда-
ции в лизосомальных компартментах (lysosomal compartments) клетки (Suk J. S. et al.,
2007). Поэтому в их состав вводят непротонированные амины (например, PEI). Они
создают так называемый протон-поглощающий эффект (proton sponge effect), препят-
ствующий закислению среды в эндосомах (Akinc A. et al., 2005).
Протон-поглощающий эффект вызывает приток в эндосому ионов хлора и уси
ление осмотического давления внутри эндосомы. Эндосома разбухает, ее мембрана
разрывается, векторная система проникает в цитоплазму клетки-мишени. Подробно
о принципах конструирования подиндексов, реагирующих на температуру, pH и ос-
мотическое давление можно прочитать в работе F. Heath et al. (2007).
Конструирование подиндексов имеет технические ограничения. Производные PEI
с ММ, большей чем 25 кДа, формируют подиндексы, эффективно трансфецирующие
клетки-мишени, но токсичные для клеток. Полимеры с меньшей ММ (5—25 кДа)
2 2 Биологические поражающие агенты
509
Таблица 2.28
Полимеры, наиболее часто используемые для создания
искусственных векторных систем*
Полимер Аббревиатура Основное свойство, принимаемое во внимание разработчиками
j Poly(ethylenelglycol PEG Инертность
Pjlyethylenimine PEI Катионные свойства
। Dithiobis(succ inimidylpropionate) DSP Биодеградируемость PEI
. )imethyl-3,3'-dithiobispropionimidate DTBP То же
Poly(ethyleneimine) biscarbamate PEIC Биодеградируе мость PEI
Poly(L-lysine) PLL Катионные свойства
Гистидинмодифицированный PLL Биодеградируемость
"’olyCN -vinylpyrrolidone) PVP Нейтральный заряд
Poly(propylenimine) PPI Дендромерность
Poly(amidoamine) РАМАМ То же
Poly(amido ethylenimine) SS-PAEI Биодеградируемость
Triethylenetetramine TETA Катионные свойства
Puly(P-aminoester) — Биодеградируемость
Poly(4-hydroxy-L-proline ester) PHP То же
Poly(allylamine) — Катионные свойства
Poly(a-[4-aminobutyl]-L-glycolic acid) PAGA Биодеградируемость
Poly(D,L-lactic-coglycolic acid) PLGA То же
Poiy(N-ethyl-4-vinylpyridiniuin bromide) Катионные свойства
Poly(phosphazene)s PPZ Биодеградируемость
Poly(phosphoester)s PPE То же
Poly(phosphoramidate)s PPA То же
Poly(N-2-hydroxypropylmethacrylamide) pHPMA Катионные свойства
Poly (2-(dimethylamino)ethyl methacrylate) Катионные свойства
P01y(2-aminoethyl propylene phosphate) pDMAEMA Биодеградируемость
Хитозан PPE-EA Нс токсичность, биодеградируе- мость и биосовместимость
jalactosylated chitosan То же
N-Dodecylated chitosan То же
Dextran-spermine D-SPM Уменьшение заряда на поверхности ли поп лекса
* По М. S. Al-Dosari, X. Gao (2009).
Армируют менее токсичные искусственные векторные системы, однако их трансфе-
дирующая активность не высока. Высокомолекулярные PEI, производные от низко-
молекулярных PEI, менее токсичны, и искусственные векторное системы на их ос-
• ове более эффективно трансфецируют клетки-мишени, чем на основе РЕ1 той же
ММ. Полиплексы, полученные путем комплексации ДЕ1К с линейными PEI, форми-
руют векторные системы, очень активные при внутривенном введении. Ветвящиеся
’роизводные РЕ1 имеют большую токсичность и меньшую трансфецирующую актив-
510
Часть 2. Общая неправильная эпидемиология
ность, чем линейные, однако на их основе можно формировать стабильные наночас-
тицы, пригодные для ингаляционной доставки генов в организм человека (см. ниже
«Введение генов в легочную ткань в составе аэрозоля»).
Улучшить таргетирующую способность искусственных векторных систем можно
конъюгированием PEI с инертными полимерами, такими как ПЭГ (5 кДа) и декст-
ран, уменьшающими заряд на их поверхности и, соответственно, неспецифическое
взаимодействие подиндекса с тканями организма человека (Al-Dosari М. S., Gao X.,
2009; Wang Yu-Qiang et al., 2012).
Эффективность трансфекции таких систем увеличивают путем повышения их
специфичности. Для этого в поверхность частиц вводят лиганд, специфически взаи-
модействующий с рецепторами на поверхности клеток-мишеней. Y. Hu et al. (2012) для
конструирования безопасного, но высокоэффективного и специфичного к рецепто-
рам фактора роста фибробластов вектора, использовали низкомолекулярный PEI
(РЕ1600, ММ 600 кДа), перекрестно сшитый с [3-циклодекстрином (P-CyD)1. В каче-
стве лиганда в векторную конструкцию они включали пептид CY11, который соеди-
няли с PEI-P-CyD, используя в качестве линкера N-succinimidyl-3-(2-pyridyldithio)
propionate. Электронно-микроскопическое исследование показало, что CY11-РЕ1-Р-
CyD способен эффективно конденсировать плазмидную ДНК (трансген) во внутрен-
нее пространство наночастицы размером около 200 нм.
Как альтернатива PEI, рассматривается природный катионный полисахарид хито-
зан, получаемый щелочной N-деацилацией хитина. Хитозан обладает высокой био-
совместимостью и низкой цитотоксичностью, однако при физиологических значени-
ях pH он утрачивает заряд и плохо растворяется, что не позволяет использовать его
для конструирования полиплексов. С. Liu Ch. et al. (2012) получили амфифильный
хитозан (N-octyl-N-quatemary chitosan, OTMCS) с небольшой ММ, затем перекрестно
его сшили с низкомолекулярным PEL В результате они создали биоразлагаемое амфи-
фильное производное хитозан-PEI, позволяющее создавать векторные конструкции,
обладающие высокой трансфецирующей активностью и низкой цитотоксичностью.
Липоплексные композиции (lipoplexes) первоначально получали ковалентным со-
единением липида и нуклеотидных последовательностей. Проникновение данного
соединения в клетку осуществлялось в результате неспецифического слияния гидро-
фобного (липидного) «хвоста» и липидов мембраны. Другим вариантом формирова-
ния липоплексных композиций стало включение плазмидной ДН К в катионные ли-
посомы. Липоплексы менее стабильны, чем подиндексы (Feigner Р. L. et al., 1987).
Липоплексы имеют общую структуру: положительно заряженная гидрофильная
«голова» и отрицательно заряженный гидрофобный «хвост» соединяются через лин-
керную структуру. Наиболее часто в таких соединениях в качестве гидрофильной «го-
ловы» используются вторичные и четвертичные амины, или четвертичные аммоние-
вые соли; реже — гуанидиновые, имидазольные, фосфорные, пиримидиновые и
мышьяковистые группы. Положительно заряженная «голова» необходима для связы-
вания с отрицательно заряженными фосфатными группами нуклеиновых кислот. Для
уменьшения агрегации липосом исследователи избегают увеличения их концентра-
ции в коллоидном растворе более 5 мг/мл (по содержанию ДНК) и добавления к ним
хелатирующих агентов и солей. Такие растворы должны обладать нулевой ионной
силой и содержать декстран (Al-Dosari М. S., GaoX., 2009).
1 Декстрин — продукт неполного гидролиза декстрана. Характеризуется хорошей усво-
яемостью тканями.
2 2. Биологические поражающие агенты
511
Гидрофобные «хвосты» можно разделить на две группы в зависимости от их гид-
рофобных групп: алифатические цепи и холестериновые (холестерин и другие типы
стероидных колец). Между гидрофильными и гидрофобными группами через эфир-
ные, сложноэфирные, карбаматные и аминные группы формируются биодеградиру-
емые связи. В табл. 2.29 приведены катионные липиды, наиболее часто используемые
для формирования искуссз венных векторных систем.
Таблица 2.29
Катионные липиды, наиболее часто используемые для создания
искусственных векторных систем*
Липид Аббревиатура Основное свойство, при- нимаемое во внимание разработчиками
1 2 3
l,2-Dioleoyl-sn-glycero-3-phosphatidy!choline DOPC Помощник
l,2-Dioleoyl-sn-glycero-3-pfibSphatMylethanolamine DOPE Тоже
Cholesterol (холестерин) — Тоже
N-[1-(2,3- Dioleyloxy)propyl]N,N,N-trimethylammonium chloride DOTMA Катионные свойства
1,2-Dioleoyloxy-3-trimethylammonium-propane DOTAP То же
Dioctadecylamidoglycylspermine DOGS Тоже
N-(3-Aminopropyl)-N,N-dimethyl-2,3-bis(dodecyloxy)- 1-propanaminium bromide GAP-DLRIE То же
Cetyltrimethylammonium bromide СТАВ То же
6-Lauroxyhexyl omithinate LHON То же
l-(2,3-Dioleoyloxypropyl)-2,4,6-trimethylpyridinium 20c Тоже
2,3-Dioleyloxy-N-[2(spermniecarboxamido)-ethyl]-N,N- dimethyl-1 -propanaminium trifluoroacetate DOSPA То же
l,2-Dioleyl-3-trimethylammonium-propane DOPA Тоже
N-(2-Hydroxyethyl)-N,N-dimethyl-2,3-bis(tetradecyl- эху)-1-propanaminium bromide MDRIE То же
Dimyristooxypropyl dimethyl hydroxyethyl ammonium bromide DMRI То же
3p-[N-(N',N'-Dimethylaminoethane)-carbamoyl]chole- sterol DC-Chol То же
Bis-guanidium-tren-cholesterol BGTC То же
1.3- Diol coxy- 2-(6-carboxy-spermyl) -propylamide DOSPER То же
Dunethyloctadecylammoniur.i bromide DDAB То же
Jioctadecylamidoglicylspermidin DSL То же
rac-[(2,3-Dioctadecyloxypropyl)(2-hydroxyethyl)]-dimet- hylammonium chloride CLIP-1 То же
rac-[2(2,3-Dihexadecyloxypropyl-oxymethyloxy)ethyl]tri- inethylammonium bromide CLIP-6 То же
Ethyldimyristoylphosphatidylcholine EPMPC То же
l,2-Distearyloxy-N,N-dimethyl-3-aminopropane DSDMA Тоже
1,2- Dimyristoyl-trimethylammoniumpropane DMTAP Тоже
O,O'-Dimyristyl-N-lysyl aspartate DMKE Тоже
512
Часть 2. Общая неправильная эпидемиология
Продолжение табл. 2.29
1 2 3
N-Palmitoyl D-erythro-sphingosyl carbamoyl-spermine CCS To же
N-t-Butyl-NO-tetradecyl-3-tetradecylammopropionamidine diC14-amidine To же
Octadecenolyoxy[ethyl-2-heptadecenyl-3 hydroxyethyl] imidazolinium chloride DOTIM To же
Nl-Cholesteryloxycarbonyl-3,7 -diazanonane-1,9-diamine CDAN To же
2-(3-[Bis-(3-ammo-propyl)-amino]propylamino)-N- ditetradecylcarbamoylme-ethyl-acetamide RPR209120 To же
* По М. S. Al-Dosari, X. Gao (2009).
Распространенными в генотерапевтическом конструировании колипидами явля-
ются холестерол и DOPE. Вклад этих липидов-помощников (helper lipids) в катионо-
посредованный эндоцитоз зависит от структуры катионных липидов. Некоторые ли-
пиды в формировании структур, трансфецирующих клетки-мишени, зависимы от
DOPE. В то же время, другие катионные липиды, имеющие два остатка жирной кис-
лоты (double fatty chains), способны самостоятельно формировать двухслойные или
мицеллярные структуры, обладающие высокой трансфекнионной активностью.
Катионные липосомы, содержащие холестерин, разрабатывались для применения
в условиях in vivo. Присутствие в мембране липосомы холестерина стабилизирует ее
структуру и снижает деструктивную активность компонентов крови. Высокой транс-
фекционной способностью обладают липосомы с выраженной мембранной текуче-
стью. Использование DOPE в качестве колипида увеличивает трансфецирующую
способность липосом и облегчает освобождение липоплекса из эндосомы через дес-
табилизацию ее мембраны. Далее происходит декомплексация липосомы с высвобож-
дением из нее ДНК трансгена. Вследствие избыточного поверхностного заряда, пе-
риод полужизни катионных липосом при их циркуляции в крови непродолжител :н
(Al-Dosari М. S., GaoX., 2009).
Липоплексы накапливаются в кровеносных сосудах легочной ткани, преимуще-
ственно трансфецируя сосудистый эндотелий и эпителиальные клетки дыхательных
путей. Для увеличения длительности циркуляции липоплексов в крови человека и
увеличения количества поражаемых клеток-мишеней, находящихся вне легочной
ткани, разработчики липоплексов понижают заряд их поверхности путем добавления
гидрофильных и нейтрально заряженных полимеров, таких как ПЭГ. Введение в со-
став липосомы галактолипидов повышает их тропность к паренхиматозным тканям.
Липосомы, содержащие dimethylaminoethane-carbamoyl-cholesterol (DC-Chol), обла-
дают повышенной тропностью к клеткам гиппокампа головного мозга1 (Harvie Р.
et al., 2000). Другой путь таргетироваия липоплексов — введение в их состав специ-
фических антител. Такие комплексы называются ПЭГилированными иммуносомамг
(PEGylated immunoliposomes, PILs). Они сохраняют коллоидную стабильность при
внутривенном введении и эффективно доставляют гены в ЦНС (Pardridge W. М., 2002,
Zhang Y. el al., 2003). На рис. 2.62 приведено схематическое изображение методиче-
ских подходов, используемых для включения нуклеиновой кислоты (на примере
siPHK) в ПЭГилированные липоплексы.
1 Гиппокамп — центральная часть лимбической системы — комплекса структур среднего,
промежуточного и конечного мозга, участвующих в организации висцеральных, мотивацион-
ных и эмоциональных реакций организма.
2.2. Биологические поражающие агенты
513
Растворение сублимационно-
высушенного матрикса
Спонтанное формирование
везикул
siPHK
Липиды + ПЭГ в водном
в 90% этаноле растворе
Контролируемое
смешивание
siPHK в липидных частицах
Дальнейшее разбавление
для стабилизации частиц
Концентрирование частиц
центрифугированием
Диализ
Самосборка
Липиды
Растворение —
многослойные
липосомы
Вытеснение —
формирование
липидных частиц
микронного размера
siPHK
+
протамин
После добавления
ПЭГ инкубирование
при Г 50 °C 10 мин
siPHK
сахароза
Липиды +
ПЭГ
в t-бутаноле
Липиды/
siPHK
Сублимационное П
высушивание
Лиофилизо-
ванный
матрикс
Перед использованием
растворяют в воде
Рис. 2.62. Методические подходы, используемые для включения нуклеиновой
кислоты в липидные частицы, содержащие ПЭГ. По S. Y. Wu, N. A. J. McMillan (2009)
Присутствие большого количества групп ПЭГ на поверхности липоплекса предот-
вращает неспецифическое взаимодействие между ним и клеточной мембраной и при
наличии на его поверхностности специфических лигандов, увеличивает способность
векторной конструкции клиганд-рецепторному взаимодействию с клеткой-мишенью.
В практику конструирования введено большое количество химических групп, позво-
ляющих деэкранировать липоплекс в кислой среде эндосомы. Основное, что мешает
практическому использованию липоплекс-опосредованной трансфекции в генной
терапии, это высокая токсичность таких комплексов и непродолжительная экспрес-
сия трансгенов (Al-Dosari М. S., Gao X., 2009).
Третьим направлением получения искусственных векторных систем, развитие ко-
торого прослеживается только с 2005 г., стало использование наночастиц двуокиси крем-
ния в качестве носителя терапевтических генов. Их основными преимуществами перед
липо- и подиндексами разработчики считают низкую токсичность, контролируемость
размеров и простоту приготовления векторной системы (Bharali D. J. et al., 2005;
Cheang Tuck-yun et al., 2012).
Наряду с разработкой типовых структур искусственных векторных систем для на-
правленной доставки трансгенов в целевые клетки (в печень, в мозг и надпочечники,
в медуллярные двигательные нейроны, в костные ткани, в гладкую мускулатуру кро-
веносных сосудов, в опухоли эндотелия и др.), в 1990 гг. была оптимизирована струк-
тура трансгена. Для того чтобы трансген мог автономно функционировать в данной
ткани, в него включают регионы транскрипции и инициации мРНК, регуляции тер-
514
Часть 2. Оищая неправильная эпидемиология
минации транскрипции. Кратко рассмотрим искусственные специфические вектор-
ные системы на примере систем, доставляющих трансгены в мозг, легочную ткань и
эпителий кишечника.
Введение генов в ЦНС. Технология управления деятельностью мозга человека пу-
тем введения в различные отделы ЦНС генов биологически активных веществ, раз-
вивается в рамках создания новых подходов к лечению нейродегенеративных болез-
ней. Однако далеко не все ставшие известными эксперименты по доставке генов в
ЦНС, однозначно можно объяснить заботой о здоровье человека. Еще в 1990-х гг. в
Калифорнийском университете якобы с чисто научными целями разработаны спосо-
бы введения в глубокие отделы мозга генов нейротоксинов в составе векторов, полу-
ченных на основе нейротропных вирусов (см., например, рис. 2.19).
Искусственные векторные системы, доставляющие гены в ЦНС, по сравнению с
векторами на основе нейротропных вирусов, обладают значительно меньшей транс-
фецирующей активностью, но их специфичностью можно тонко управлять, меняя
химические соединения, входящие в состав оболочки векторной конструкции. Кро-
ме того, они безопаснее векторов на основе нейротропных вирусов, так как примене-
ние последних предполагает риск развития воспалительных реакций и демиелиниза-
ции нервных волокон. Для того чтобы ген не только проник в заданный генным тера-
певтом отдел мозга и эффективно там экспрессировался, он должен преодолеть три
барьера: ГЭЬ, клеточно-мембранный барьер (КМБ) и барьер мембраны ядра клетки
(БМЯ) (Boado R. J., Pardridge W. М., 2011).
Разработаны два пути доставки нуклеиновых кислот в нейроны ЦНС, минуя ГЭБ:
прямой и периферический. Первый предполагает введение генов непосредственно в
анатомический регион мозга, где развивается патологический процесс. Он имеет два
варианта выполнения: интракраниальный, когда ген в составе векторной конструк-
ции вводится в участок мозга, расположенный внутри черепной коробки; и интрат ;-
кальный — векторная конструкция вводится в интратекальное пространство вокруг
спинального хорда. Второй путь предполагает периферическое введение векторных
систем. Благодаря ретроградному транспорту, гены, введенные внутримышечно, про-
никают в ЦНС, минуя IЭБ (рис. 2.63).
J. М. Bergen et al. (2008) считают ретроградный транспорт невирусных векторов,
основным способом, позволяющим прицельно доставлять гены в глубокие отделы
мозга. S. Wang et al. (2001) продемонстрировали способность комплекса РЕ1/ДНК,
введенного внутримышечно, интернализироваться через аксонные терминали подь
язычного нерва ретроградно в нейрональную сому в стволовой части мозга. Экспрес-
сия трансгена в стволе головного мозга обнаружена уже через 18 ч после инъекции в
мышцу языка и продолжалась не менее 2 нед. Однако движение векторной конструк-
ции, наблюдаемое в большинстве таких систем, является следствием ее транспорта в
пределах эндосомы, имеющей кислую среду, вызывающую деградацию вектора. Прак-
тически невозможно создать искусственную векторную конструкцию, способную
избежать деградации в эц юсомах аксонной терминали, если размер ее частицы пре-
вышает 30 нм. Прямое взаимодействие с моторными белками-динеинами1 позволяет
1 Динеины — группа моторных белков, способных перемещаться по поверхности микро
трубочек цитоскелета и транформирующих химическую энергию, содержащуюся в АТФ, в
механическую энергию движения, перенося грузы (cargo) — везикулы, митохондрии и др.
Динеины движутся по микротрубочкам от плюс-концов к минус-концам, которые, как пра-
вило, закреплены в районе клеточного центра.
2.2. Биологические поражающие агенты
515
Рис. 2.63. Основные пути доставки генов в ЦНС в «обход» IЭБ. А. Интракрани-
альное введение. Б. Интрагекальное введение. В Интрамускулярное введение.
По J. М. Bergen et al (2008)
частицам меньшего размера покидать еще только сформировавшиеся эндосомы и
проникать в цитоплазму аксона, где они подвергаются активному ретроградному
транспорту в глубокие отделы мозга (рис. 2.64).
Рис. 2.64. Механизм ретроградного транспорта в ЦНС частиц искусственных вектор-
ных систем А. Вектор, проникший в дистальным участок нерва, подвергается актив-
ному ретроградному транспорту по двум механизмам: а — в составе эндосом; б — через
прямое взаимодействие с моторными белками-динеинами, формирующими микро-
трубочки. Б Искусственная векторная система, включающая дениин-связывающий
белок (DBP — dynein-binding peptide). По J М. Bergen et al. (2008)
Для преодоления искусственными векторными системами ГЭБ, разрабатывают-
ся иммунолипоссмы (ПЭГилированные иммуносомы), способные к рецептор-опо-
средованному трансцитозу через эндотелиальные клетки сосудов мозга. Один из ва-
риантов конструирования таких липосом предполагает использование следующих
516
Часть 2. Общая неправильная эпидемиология
компонентов: 93 % 1-palmitoyI-2-oleoyl-sn-glycerol-3-phosphocholine (РОРС), 3 %
didodecyldimethylammonium bromide (DDAB), 3 % distearoylphosphatidylethanolamine
(DSPE)-PEG2000 и 1 % DSPE-PEG2000-maleimide. Малеимидные функциональные
группы позволяют через тиоэфирную связь (thioether linkage) ковалентно сшивать с
PEG2000 тиолированные моноклональные антитела (мАТ), играющие роль таргети-
рующего компонента иммунолипосомы. Обычно это мАТ, специфичные к рецепто-
рам трансферрина (поверхность эндотелиальных клеток сосудов мозга) или инсули-
на (глия) (Bergen J. М. et al., 2008; Boado R. J., Pardridge W. M., 2011). На рис. 2.65
показаны подходы к конструированию иммунолипосом, способных проникать в ядра
нейронов после внутривенного введения.
Рис. 2.65. Схема конструирования иммунолипосом, способных после внутривен-
ного введения проникать в ядра нейронов. А. Суперскрученная плазмидная ДНК,
включенная в иммунолипосому. Кодирующая трансген последовательность (coding
sequence, cds) экспрессируется под контролем промотора (promoter, Pro) вируса SV40
и полиаденилирующей последовательности (polyadenylation sequence, рА). Прибли-
зительно 1—2 % цепей PEG сшиты с мАТ, специфичными к таргетируемым рецеп-
торам (R), запускающим транспорт иммунолипосомы через биологические барье-
ры в условиях in vivo. R1 — рецептор трансферрина, опосредующий транспитоз че-
рез сосудистый эндотелий; R2 — рецептор инсулина, опосредующий эндоцитоз
иммунолипосомы через мембраны клеток интракраниальной нейроэпителиальной
опухоли (глиомы). Б. Трансмиссионная электронная микроскопия иммунолипо-
сомы. Мышиные IgG, сшитые с концами цепей PEG2000 на поверхности имму-
нолипосомы и частицами золота диаметром 10 нм. Полоса соответствует 20 нм.
В. Трехбарьерная модель генной терапии мозга. noR.J. Boado, W. М. Pardridge (2011)
Наибольшую эффективность при доставке генов в ЦНС показали подиндексы на
основе PEI (см. табл. 2.28). Их трансфецирующая активность превосходит таковую у
векторов, полученных на основе ВИЧ-1, сопоставима с трансдуцирующей активно-
стью аденовирусов, но уступает векторам на основе HSV (Berry В. et al., 2001).
Тонкое управление специфичностью искусственных векторных систем стало воз-
можным благодаря обнаружению на поверхности нейронов нескольких классов ре-
цепторов, с которыми взаимодействуют нейропептиды, нейротрофины и нейроток-
сины. Для перенацеливания искусственных векторных систем с одного участка мозга
на другой в их состав включают таргетинговые группы, соответствующие отдельному
классу рецепторов (табл. 2.30).
2.2. Биологические поражающие агенты
517
Таблица 2.30
Нейрон-таргетирующие лиганды, используемые при конструировании искусственных
векторных систем, предназначенных для доставки генов в ЦНС*
Класс лиганда Примеры лигандов Таргетируемые популяции нейронов
Нейропептиды Нейротенсин Нигростинальная и мезолюмбальная системы
{ейротрофины Фактор роста нерва Дорсальный корневой ганглий, симпатические сенсор- ные нейроны, нейроны базального отдела переднего мозга
। Нейропептиды Столбнячный токсин Все периферические нейроны, относящиеся к вегетатив- ной нервной системе, моторные и сенсорные
* По J. М. Bergen et al. (2008).
Эффективная нейрон-специфическая трансфекция искусственных векторных
систем достигается введением в их оболочку химерного белкового комплекса, вклю-
чающего четвертую петлю фактора роста нервов (nerve growth factor, NGF) или шпи-
ечный мотив, содержащий петли 1 и 2 NGF, слитый с Д11К-связывающим доменом
из 10 лизиновых последовательностей (Zeng J., Wang S., 2005). Трансфецируюшая
активность в отношении нейронов, сравнимая с трансдуцирующей активностью HSV,
в условиях in vivo показана д ля наночастиц (~30 нм), созданных на основе кремния,
модифицированного путем добавления аминогрупп (amino-terminated organically
modified silica, ORMOS1L) и комплексированных с плазмидной ДНК (Bharali D. J. et
al., 2005). Селективные таканеспецифические промоторы, осуществляющие экспрес-
сию генов в разных отделах мозга человека, ранее приведены в табл. 2.24.
Введение генов в легочную ткань в составе аэрозоля. Ингаляционный способ вве-
дения генов в организм человека в контексте данной работы можно рассматривать как
аналогию ингаляционному поражению традиционными агентами БО. Предпосылки
к созданию различных систем доставки генов в организм человека в составе аэрозо-
ля, были заложены еще во второй половине 1980-х гг. Тогда же стали разрабатываться
высокоэффективные способы введения плазмид в клетки тканевых культур и аэрозо
лируемые лекарственные препараты. Носителями как тех, таки других, являлись скон-
струированные по одному принципу катионные липосомы.
Идея неинвазивной терапии наследственных болезней аэрозолями генов была
очень привлекательной для генотерапевтов. Для ее реализации требовались: наслед-
ственное легочное заболевание, патогенез которого связан с мутацией в отдельном
гене; и ген, достаточно охарактеризованный, чтобы можно было ввести его в пора-
женную ткань для компенсации утраченной функции дефектного гена. Такое заболе-
вание уже было известно — это цистический фиброз (муковисцидоз). В основе его па-
тогенеза лежит недостаточный транспорт солей в легочном эпителии, связанный с
ут ратой CF1 R-гена. Этот ген был клонирован и охарактеризован, а цистический фиб-
роз стал основной моделью для проверки эффективности разрабатываемых способов
генотерапии легочных наследственных болезней. Тогда же были разработаны сухие
препараты (рецептуры) искусственных векторных систем, предназначенные для инга-
ляционного применения. По утверждению их разработчиков (см. Leaf Н., 1996), ис-
пользование сухих препаратов повысило более чем в 50 раз эффективность введения
трансгенов в трахеальные и легочные ткани экспериментальных животных.
К концу 1990-х гг. сформировалось два альтернативных направления ингаляци-
ОННО1 о введения генов в организм человека. Первое предполагает традиционный под-
518
Часть 2. Общая неправильная эпидемиология
ход — введение трансгена в глубокие отделы легких человека в составе мелкодис-
персного аэрозоля, частицы которого имеют аэродинамический размер в пределах
5,5—1,4 мкм. Второе, наоборот, исключает применение мелкодисперсного аэрозоля.
Введение генов осуществляется через обонятельные луковицы, расположенные в по-
верхностном эпителии носовых ходов, далее по аксонам обонятельного нерва они
«поднимаются» в отделы переднего мозга, минуя ГЭВ. Наибольшее развитие полу-
чило первое направление. Рассмотрим его более подробно.
Введение трансгенных конструкций в глубокие отделы легких преследует три дек-
ларируемые генными терапевтами цели: первая — лечение местных патологических
процессов, например, муковисцидоза или рака; вторая — проникновение через легоч-
ную ткань в кровь и тканевой таргетинг за пределами легкого; третья — экспрессия
биологически активных веществ непосредственно эндотелием кровеносных сосудов
легких. Последнее направление связано с тем, что поверхность эндотелия сосудов
легочной ткани занимает не менее 130 м2, это 30 % всей эндотелиальной ткани чело-
веческого организма (Simionescu М., 1991).
Эндотелий легочных сосудов выполняет роль нетромбогенного полупроницае-
мого барьера д ля газового обмена. Он покрыт гликокаликсным слоем (glycocalyx layer),
содержащим большое количество сульфатных групп, создающих отрицательный заряд
на его поверхности, позволяющий катионным поли- илипоплексами неспецифически
взаимодействовать с эндотелием. Эндотелий содержит рецепторы, специфические
для отдельных макромолекул. Среди них основные: тромбомодулин (thrombomodulin,
ТМ; другое название — CD141 или BDCA-3); молекула-1 клеточной адгезии тром-
боцитов/эндотелия (platelet/endothelial cell adhesion molecule-1, РЕСАМ-1); ангеотен-
зин-конвертирующий фермент (angiotensin-converting enzyme, АСЕ); молекулы меж-
клеточной адгезии-1 (intercellular adhesion molecule 1, 1САМ-1); лектинподобный
окисленный LDL-рецептор (lectin-like oxidized LDL receptor, LOX-1) и Р-селектин.
В качестве лигандов к этим рецепторам в поверхность искусственных векторных кон-
струкций вводят специфические антитела (Kuruba R. et al., 2009). Данная технология
уже используется для разработки способов защиты от ХО. Например, для создания
защиты от фосфорорганических соединений (ФОС) необходима длительная цирку-
ляция бутирилхолинэстеразы (butyrylcholinesterase) в легочной ткани, являющейся
первичным барьером при экспонировании к ФОС. Введение в составе липоплексов
гена бутирилхолинэстеразы в эпителий легких морских свинок в течение 1—2 недель
обеспечивало им интенсивную защиту от всех известных холинэстеразных ингиби-
торов (Alien N. et al., 2001).
После ажиотажа 1990-х гг., связанного с тем, что катионные липосомы обладают
выраженной тропностью к эндотелию сосудов паренхимы легких, разработчиками
искусственных векторных систем была установлена их непригодность для использо-
вания в терапевтических целях у людей. Липосомы вызывают сильный воспалитель-
ный цитокиновый ответ в участке введения вектора, сопровождающийся прекраще-
нием экспрессии трансгена. В основе развития воспалительного процесса лежит
избирательное поглощение таких комплексов макрофагами и их реакция на немети-
лированные CpG-участки ДНК. Этот недостаток значительно менее выражен у кати-
онных полимеров. Поэтому подиндексы рассматриваются как наиболее эффективные
средства доставки генов в эндотелий сосудов легочной ткани (Li S. et al., 2000).
Трансген, введенный в составе полиплекса на основе PEI (LPEI), эффективно
трансфецирует эндотелиальные и эпителиальные клетки легкого. Наиболее серьезным
ограничением применения LPEI для создания полиплексов является его небиодегра-
2.2. Биологические поражающие агенты
519
дируемость, проявляющаяся токсическим действием на печень. Значительно меньшее
токсическое действие на печень оказывают новые биодеградируемые полимеры на
основе полигидроксилалкиленаминов (polyhydroxylalkyleneamines), полученные пу-
тем связывания с полиаминовым каркасом (polyamine ЬаскЬопе, РНА) гидроксильных
(hydroxyl) и а ткилгидроксильных (alkylhydroxyl) групп. РНА-полимеры по структуре
остова сходны с LPE1, но имеют «регулируемое пространство» между аминогруппа-
ми. Плотность заряда у них меньше, чем у LPEI, благодаря наличию большего коли-
чества гидроксильных и алкил гидроксильных групп. Биодетрадабельность РНА обус-
ловлена наличием в остове молекулы дисульфидных связей Kuruba R. et al., 2009).
Кассеты трансгенов, предназначенные для ингаляционного введения, имеют свои
особенности. Для генотерапии наследственных легочных болезней разработчики еще
с 1990-х гг. включают в экспрессионные кассеты в качестве премоторных элементов
ранний промотор вируса обезьян 40 (S V40), промотор вируса Рауса, основной поздний
промотор аденовируса и ранний промотор 1 цитомегаловируса (Lee Е. et al., 1998).
В тех случаях, когда необходимо контролировать запуск транскрипции трансгена, в
экспрессионную кассету включают индуцибе льные промоторы (это промоторы генов
Р-интерферона, теплового шока, металлотионеина или генов, запускаемых стероид-
ными гормонами). Оба типа промоторов (индуцибельный и конститутивный) могут
использоваться в одной экспрессионной кассете, в зависимости од замысла ее разра-
ботчика (Debs R. J. et al., 1997; Kuruba R. et al., 2009).
При необходимости ограничить эффективную экспрессию трансгена только оп-
ределенными легочными тканями, например, стволовыми (для гена одного из фак-
торов пролиферации ствотовых клеток — фактор эпидермального роста, фактор ан-
гиогенеза и др.)т разработчики таких методов генотерапии используют сочетания
высокоспецифических премоторных или энхансерных элементов с неспецифически-
ми. Так, для легочного эпителия высокий уровень специфичности и экспрессии од-
новременно достигается «слиянием» тканеспецифического промотора из гена регу-
лятора трансмембранной проводимости легочного эпителия и гетерологичного
энхансерного элемента SV40 (Lee Е. et al., 1998).
В тех случаях, когда требуется длительная экспрессия трансгена (что наиболее
вероятно), его либо интегрируют с геномом клеток-мишеней с помощью ретровирус-
ного вектора, либо экспрессионная кассета длительно поддерживается в эпителии
мелких бронхов в составе стабильной эписомы (см. рис. 2.57). Интеграция с геномом
значительно облегчается, если экспрессионная кассета находится в линейной форме
и фланкирована 5'- и З'-последовательностями, гомологичными ДНК клетки- мише-
ни. Для более эффективной интеграции кассеты с геномом клетки-мишени, в состав
трансгенной конструкции может быть включена ретровирусная интеграза. Для дос-
тижения длительной экспрессии трансген могут вводить в клетки-мишени в составе
конструкции, содержащей нетрансформирующие последовательности вируса Эпш-
тейна — Барра (опРм EBNA-V). Их достаточно для поддержания длительной эписо-
мальной репликации трансгенной конструкции. Введенная в клетки легочного эпи-
телия экспрессионная кассета может предназначаться как для действия за пределами
этих тканей — на другие органы и ткани (гены нейротоксинов, биорегуляторов и др.),
так и на сами клетки ткани-мишени (гены индукторов апоптоза и др.). В первом слу-
чае в экспрессионную кассету обязательно включают ген, кодирующий лидерную пос-
ледовательность секретируемого белка (Kuruba R. et al., 2009).
Подиндексы, предназначенные для ингаляционного введения в организм челове-
ка, конструируются таким образом, чтобы они были устойчивыми к срезывающему
520
Часть 2. Общая неправильная эпидемиология
воздействию форсунки, возникающему во время их диспергирования. Частицы аэро-
золя имеют размер в пределах 0,5—5 мкм. Стабилизация полиплекса достигается фор-
мированием глютаральдегидом или карбодиимидами поперечных сшивок между об-
разовавшими полиплекс катионными макромолекулами (обычно это ветвящиеся
производные PEI). Чем больше таких сшивок, тем прочнее носитель, тем менее он
подвержен деградации в растворе и тем прочнее в него «упакована» экспрессирующая
кассета. Весовое содержание нуклеиновой кислоты в таких носителях достигает 30 %
(W/W). Важным критерием является сохранение биологической активности аэрозо-
ля трансгенов. Устойчивость к действию факторов диспергирования и стабильность
в аэрозоле конструкции, собранной за счет электростатического взаимодействия,
видимо, будут зависеть от многих факторов, в том числе еще не изученных. Среди них
величины противоположных зарядов нуклеиновой кислоты и носителя, определяе-
мые, прежде всего, их химическим составом; размер и дзета-потенциал частиц; их
агрегатное состояние. Уменьшение агрегации полиплексов в растворе достигают пу-
тем увеличения количества поперечных сшивок между катионными макромолекула-
ми, либо их конъюгированием с ПЭГ 5000. Лиофилизованные сшитые подиндексы в
водном растворе обычно сами дезагрегирутдо исходного размера частиц (Roy К. et al.,
1999). По данным Е Thierry (1999), субмикронные липосомы выдерживают перевод в
аэрозоль. Их устойчивость в таком состоянии достаточна, по крайней мере, для тера-
певтических целей. Более тонко аэродинамический размер частиц образуемого аэро-
золя полиплексов с жидкой дисперсной фазой регулируется с помощью диспергирую-
щего устройства (Manunta М. D. et al., 2011).
Получить сухие порошкообразные препараты искусственных векторных систем,
предназначенные для применения в виде сухого аэрозоля, гораздо сложнее, чем лио-
филизованные препараты, предназначенные для растворения перед диспергировани-
ем. Потребность создания порошковых лекарственных форм для целей генотерапии
вызвана значительно большей стабильностью их свойств, чем у растворенных лиофи-
лизатов. Аэродинамический диаметр частиц векторных систем, образующих такие
порошки, должен быть в пределах 1—5 мкм, но такие порошки слеживаются из-за
адгезионного взаимодействия между частицами, а если их удается распылить, то ди-
сахариды, обычно используемые в качестве среды высушивания, быстро вбирают в
себя влагу и увеличивают размер частиц за пределы 5 мкм. Поэтому для получения
порошковых форм липоплексов в последние годы используют распылительно-суб-
лимационное высушивание, а в качестве липопротектанта — бычий сывороточный
альбумин (Tsukamoto М. et al., 2012). С развитием технологий получения векторных
систем на основе наночастиц двуокиси кремния получение их порошковых лекар-
ственных форм значительно упростится (Cheang Tuck-yun et al., 2011). Каких-то срав-
нительных количественных характеристик устойчивости в аэрозоле, например липоп-
лексов, полиплексов и вирусов, в доступной литературе, не обнаружено.
Введение генов в клетки кишечного эпителия. Привлекательность кишечного эпи-
телия для генотерапевтов обусловлена:
1) большим количеством клеток, которые могут быть использованы как мишени
для искусственных генетических конструкций;
2) легкостью доступа к ним через кишечные протоки.
Поверхность протоков граничит с внешней средой (просветом кишечника), а их
базолатеральная поверхность с внутренним окружением. Такое расположение эпи-
телия идеально для прямого введения нуклеиновых кислот и белков, а также для се-
креции белков, синтезируемых с помощью введенной трансгенной конструкции.
2.2. Биологические поражающие агенты
521
Последние способны секретироваться непосредственно в кровеносную систему с ба-
золатеральной поверхности клеток. Площадь поверхности кишечного эпителия зна-
чительно возрастает из-за наличия длинных, язычковоподобных структур, известных
как ворсинки. Клетки эпителиальных ворсинок постоянно обновляются из клеток,
появляющихся из крипт (ямкоподобных структур), которые окаймляют основание
ворсинки. Каждая ворсинка получает питание от десяти и более крипт. Пролифери-
рующие клетки занимают нижние две трети крипты. Предшественниками как клеток
крипт, так и клеток ворсинок являются стволовые клетки, расположенные в основа-
нии крипты. Благодаря этому у генотерапевтов появляются две новые возможности:
1) добиться длительной экспрессии генов путем их введении в стволовые клетки;
2) получить кратковременную экспрессию генов их введением в другие клетки
крипт или ворсинок (Wirtz S., Neurath М. Е, 2003; Henning S. J., Ledley Е D., 1998).
Таким образом, технологии соматической генотерапии болезней кишечника с
самого начала их развития основываются на введение чужеродных генов непосред-
ственно в стволовые клетки эпителия кишечника, тем самым создаются условия как
для размножения трансгена, так и для пожизненной экспрессии введенных в организм
человека генов.
Возможности технологии генотерапии для энтерального введения генов хорошо
иллюстрирует патент США № 5821235, полученный Baylor College of Medicine. Спе-
циалистами этой организации еще в первой половине 1990-х гг. разработан способ ге-
нотерапии путем введения генов в стволовые клетки кишечного эпителия. Этот спо-
соб предполагает использование специального «векторногораствора», включающего
ДНК-вектор, способный эффективно экспрессировать чужеродную нуклеотидную
последовательность в стволовых клетках и компонент, облегчающий проникновение
вектора в стволовые клетки в условиях in vivo — фактор пролиферации стволовых
клеток. В качестве ДНК-вектора ими заявлена конструкция, включающая следующие
элементы: промотор; 5'мРНК-лидерная последовательность; сайт инициации транс-
крипции; экспрессирующая кассета с рестрикционным сайтом, З'-нетранслируемый
регион и сигнал полиаденилирования. Расположение этих элементов зависит от гена,
который должен быть экспрессирован в стволовых клетках кишечника. В качестве
альтернативы ДНК-векторам, сотрудники Baylor College of Medicine заявили векто-
ры на основе вирусов различных групп. Среди них векторы на основе кишечных виру-
сов, таких как парвовирусы, ротавирусы, норвалк-вирусы и другие, способные вызы-
вать кишечные инфекции, а также традиционные векторы на основе аденовирусов,
аденоассоциированных вирусов и ретровирусов. Наиболее предпочтительными для
введения генов в кишечник специалисты этой организации считают использование
ретровирусов. Заявленный Baylor College of Medicine ретровирусный вектор является
дефектным вирусом, способным к интеграции в геном стволовых клеток-мишеней,
но не реплицирующийся в них. Он включает регуляторные элементы, обеспечиваю-
щие экспрессию чужеродного гена в клетках. Кассета экспрессирует CAT и представ-
ляет собой плазмиду — производное вируса лейкемии мышей Moloney, в котором осу-
ществляется транскрипция CAT-вставки. Такие плазмиды размножаются в специаль-
ных пакующих линиях клеток (Henning S. J., Ledley F. D., 1998).
Обе конструкции в качестве промоторов могут включать LTR-промотор цитоме-
галовирусов, малатдегидрогеназный промотор, дигидрофолатредуктазный промотор
и аденозиндезаминазный промотор. По мнению разработчиков, наиболее пред-
почтительными промоторами для экспрессии генов в стволовых клетках кишечника
являются те, которые для них специфичны. Например, промотор белка, связываю-
522
Часть 2. Общая неправильная эпидемиология
щего кишечные жирные кислоты; промоторы дисахаридаз (сахарозо-изомальтазный
и мальтаза-гликоизомеразный тактаза-флоризин-г.щролазный промоторы); аполи-
попротеиновый промотор (аполипропротеин-A-I, аполипропротеин-А-П и аполип-
ропротеин-А IV промоторы); промотор обогащенного цистином кишечного белка.
В качестве факторов пролиферации стволовых клеток рассматриваются факторы, уси-
ливающие или способствующие пролиферации стволовых клеток (фактор эпидер-
мального роста, фактор ангиогенеза, инсулинподобные факторы роста 1 и 2, транс-
формирующий ростовой фактор и гастрин) (Henning S. J., Ledley Е D., 1998).
Энтеральное введение трансгена позволяет осуществлять доставку генов за пре-
делы кишечного эпителия. Вектор, сконструированный на основе коронаровируса и
введенный энтерально свиньям, был обнаружен в паренхиме легких, экспрессирущим
трансген (Alonso S. et al., 2002).
* * *
Теперь, после того как мы убедились, что опасения, высказанные J. L. Black (2003)
о возможности создания принципиально нового вида биологических агентов, осно-
ваны на реальных технологиях и достижениях соматической генной терапии, снова
вернемся к его работе и рассмотрим его представления о том, как может вестись вой-
на с помощью таких агентов.
J. L. Black считает, что сценарии боевого применения геномного оружия опреде-
ляются военными задачами, состоянием окружающей среды и целями, против кото-
рых это оружие используется. Возможны два типа нападений с применением таких
поражающих агентов, а именно: нападение с кратковременным или продолжитель-
ным латентным периодом. В первом случае гены могут доставляться при помоши век-
тора, который инфицирует хозяина и вызывает быструю экспрессию трансгена, ока-
зывающего вредное для здоровья действие (векторы на основе аденовирусов или AAV,
искусственные векторные системы). Этот вид нападения похож на использование
традиционных агентов БО. Наиболее вероятно, что он будет осуществлен с помощью
мелкодисперсных аэрозолей. Продолжительность латентного периода может зависеть
от того, насколько скоро начнет экспрессироваться ген, вызывающий патологичес-
кий процесс, а сама экспрессия определяется типом используемого вектора и исполь-
зуемой дозой.
Еще больший интерес представляет сценарий J. L. Black, предполагающий дли-
тельный латентный период после нападения, поскольку он означает возможность
ведения скрытой масштабной биологической войны, осуществляемой диверсионны-
ми способами. При реализации такого сценария население подвергается нападению
посредством искусственных генетических конструкций через продукты (энтерально),
лекарственные и иммунобиологические препараты (парэнтерально, например, под
предлогом массовой вакцинации). Наиболее вероятно применение векторов на основе
лентивируса. Его осуществление приведет к инкорпорированию трансгена в геном
подвергшихся такому нападению людей и, как следствие, к появлению обширных
популяций людей с психическими изменениями, онкологическими и неврологически-
ми болезнями, генетическими поражениями и даже наследственно передаваемыми
дефектами генов.
Это реальный сценарий диверсионно-террористической войны против России,
когда невозможно не только установить с территории и в интересах какого государ-
ства эта война ведется, кто противник, но и не иметь ясности при ответе на вопрос:
«Война это или не война»?
2.2. Биологические поражающие агенты
523
2.2.5. Нанообъекты
Границы наномира. Первое боевое применение частиц нано- и субмикронных
размеров. Особенности наночастиц, обусловливающие их поражающее действие.
Пуги проникновения наночастиц в организм человека. Случай группового пора-
жения людей наночастицами двуокиси кремния.
Нанообъекты по характеру поражающего действия не имеют аналогов ни среди
боевых О В, ни среди традиционных агентов БО или искусственных генетических кон-
струкций. Они обладают биологической специфичностью, обусловленной их сход-
ством по размеру с рецепторами клеток, а их структура позволяет им взаимодейство-
вать с этими рецепторами специфическим образом. D. М. Goncalves et al. (2011) пишут
о необходимости формирования новой ветви токсикологии — нанотоксикологии, т. е.
науки, изучающей потенциальную опасность наноструктур и наномеханизмов, но к
какому виду оружия (биологическому или химическому) эксперты в будущем отне-
сут специальные устройства, использующие поражающее действие наночастиц, сей-
час трудно предугадать. Свою задачу в рамках этой книги я вижу только в обозначе-
нии контуров проблемы их поражающего действия1.
Гранины наномира. Наномир представлен структурами нанометрового диапазона
размеров (1 нм = 10~9 м = 10~6 мм = 10-3 мкм), но это не значит, что если объект изме-
рять нанометрами, то мы проникнем в наномир. Размер атомов и атомных молекул
около 0,1 нм, но наноструктурой отдельный атом не является. В химии принято счи-
тать, что наноструктурой становится результат самоконденсации атомов и молекул в
малые атомные агрегации (кластеры), являющиеся промежуточным звеном между
изолированными атомами и молекулами (с одной стороны), и массивным (объемным)
твердым телом, с другой. Отличительной чертой кластеров атомов (молекул) от исход-
ных атомов (молекул) является «немонотонная зависимость свойств от количества
атомов в кластере». Переход к твердому телу осуществляется через укрупнение класте-
ров — минимальное число атомов в кластере равнодвум, верхняя граница соответству-
ет числу атомов, когда добавление еще одного атома не меняет свойства кластера, так
как переход количественных изменений в качественные уже закончился. Обычно та-
кая структура соответствует 1-2 тыс. атомов и она является границей между класте-
ром и изолированной наночастицей (рис. 2.66).
Следовательно, нижняя граница наномира начинается в диапазоне размеров от 1 до
4 нм, т. е. с образованием твердого тела — наночастицы. Верхнюю границу наномира
определяют свойства приповерхностного слоя наночастицы Дело тут в том, что доля
атомов (а), находящихся в тонком приповерхностном слое (-1 нм) растет с уменьше-
нием размеров частички вещества R, поскольку а » S/V~ R2/R3 = \/R (здесь 5 —
поверхность частички, V— ее объем, R — радиус). Атомы, дислоцирующиеся на по-
верхности частички, обладают свойствами, отличающимися от «объемных», посколь-
ку они связаны с окружающими их атомами по-иному, нежели в объеме. В результате
ненасыщенности связей на поверхности наночастицы может произойти атомная ре-
конструкция и появится новый порядок расположения атомов. На свободных поверх-
ностях могут находиться атомы и молекулы, адсорбированные из внешней среды; до-
полнительные особенности появляются в окрестностях атомов, находящихся на
1 Частично материал этого раздела в августе 2009 г. был представлен на Всероссийской
научно-практической конференции, посвященной 60-летию филиала ФГУ «48 ЦНИИ Мино-
бороны России — ЦВ7П БЗ» (Супотницкий М. В., Поклонский Д. Л., Паныгина С. А., 2009).
524
Часть 2. Общая неправильная эпидемиология
10 мкм
1 мкм
Крупнозернистые материалы
8 9
-2 10 -10
100 нм
=• Субмикрокристаллические материалы
6 7
-10 -10
10 нм
] нм
Нанокристаллические материалы
I----------------------------------- 500-1500
- Икосаэдрические металлические кластеры
-------------------------------------- 20-70
: Молекулярные кластеры (фуллерены и др.)
• з___4
Атомы и (2—3)-атомные молекулы .
100 пм
Рис. 2.66. Классификация веществ и материалов по размеру (D) частиц.
По А. И. Гусеву (2007)
краях моноатомных террас, уступов и впадин. Взаимодействие электронов со сво-
бодной поверхностью тоже приводит к возникновению специфических приповерхно-
стных явлений. Возникают и другие эффекты, более подробно о них можно узнать в
специальной литературе (например, из монографий Головина Ю. И., 2007; Суздале-
ва И. П., 2009). Все это вместе взятое позволяет химикам рассматривать приповерх-
ностный слой частиц, у которых соотношение числа атомов (молекул), лежащих на
поверхности, больше или равно ооъемным, как некое новое состояние вещества.
Внешне оно проявляется резким увеличением химической и каталитической актив-
ности поверхности, увеличением ее сорбционной емкости и другими эффектами.
Поэтому в химии под наночастицами понимают те, у которых отношение числа по-
верхностных атомов (молекул) к объемным > 1. При таком определении наночасти-
цами низкомолекулярных веществ считаются объекты с размером до 10 нм, для высо-
комолекулярных до 100 нм (в их число входят искусственные генетические конструк-
ции; см. разд. 2.4.4)1.
Первое боевое применение частиц нано- и субмикронных размеров. После появле-
ния в 1916 г. на фронтах мировой войны у воюющих армий угольных противогазов,
многие военные специалисты считали, что теперь «противогаз победил газ». Однако
в июле 1917 г. германской армией во Фландрии впервые были применены новые от-
равляющие вещества раздражающего действия — арсины (щфенилхлорарсин, адам-
сит, дифенилцианарсин), от которых угольный противогаз не защищал. Объясняется
это следующим. В отличие от использовавшихся ранее фосгена, хлора, синильной
кислоты, в тех же условиях арсины представляют собой твердые вещества. Темпе-
ратура кипения дифинилхлорарсина 333°C; адамсита — 277,5°C; дифенилцианар-
1 Частицы размером более 100 нм относят к субмикронным частицам.
2.2. Биологические поражающие агенты
525
сина — 346 °C. Их плавили и заливали в стеклянные емкости, похожие по форме на
советские кефирные бутылки, затем эти емкости помещали в корпуса снарядов и за-
ливали тротилом (см. рис. 1.12). Новые химические снаряды маркировались синим
крестом. При взрыве снаряда арсины превращались в пар и конденсировались с об-
разованием частиц размером менее 100 нм (предел разрешения методов измерения
размеров частиц в то время). Такой способ образования наночастиц называется сегод-
ня самоконденсацией, правда, для получения наночастиц заданного размера и струк-
туры он выполняется в специальных камерах и атмосфере инертного газа низкого
давления с последующей конденсацией пара в охлаждающей среде или на охлаждаю-
щих устройствах. Но в 1917 г. размер и структура получаемых частиц арсинов герман-
ское командование не интересовали.
Целью применения таких снарядов было заставить солдат противника сбросить
противогазы и вдохнуть пары фосгена и дифосгена, образовавшиеся в результате об-
стрела химическими снарядами другого типа, маркировавшихся зеленым крестом. Ча-
стицы арсинов свободно преодолевали угольные фильтры противогазов и вызывали
у солдат сильный рвотный рефлекс. Борьба с такими «дымами» оказалась более слож-
ной, чем с ОВ, применявшимися в газообразном состоянии. До конца войны не было
создано удовлетворительных противодымных фильтров. Артиллерийская подготов-
ка, проводимая одновременно химическими снарядами синего и зеленого крестов, по-
лучила у германских военных красивое название — «стрельба разноцветным крестом»
(Де Лазари А. Н., 1935, 2007).
Особенности наночастиц, обусловливающие их поражающее действие. Прежде всего,
это химическая и каталитическая активность поверхности наночастиц, отсутствую-
щая у этого же вещества при более крупной дисперсности частиц. Второй особенно-
стью наночастиц, проявляющейся их токсичностью, является их высокая концент-
рация в воздухе при незначительном количестве самого распыленного вещества.
Например, 10 мкг/м3 вещества образует более чем 1 х 106 частиц/см3 при их размере в
20 нм. Третья особенность наночастиц, проявляющаяся их токсическими свойства-
ми, это их способность к ингаляционному, грансдермальному, транснейральному и
энтеральному проникновению в любые органы и ткани человека, включая ЦНС. Чет-
вертая особенность наночастиц, проявляющаяся генотоксическим действием, — их
способность проникать в ядра неделяшихся клеток, если диаметр частиц менее 40 нм,
т. е. размера внутренней полости поры ядра клетки (Brown D. М. et al., 2001; Гусев А. И.,
2007). Физические свойства наночастиц, определяющие их распространение в орга-
низме человека и поражающее действие, приведены в табл. 2.31.
Наночастицы по размеру сходны с рецепторами клеток и молекулами, осущест-
вляющими сигнальную функцию внутри клетки. Исследования, проведенные в усло-
виях in vitro с использованием различных клеточных систем, показали развитие у
клеток, экспонированных к наночастицам, провоспалительных и связанных с окис-
лительным стрессом реакций (Brown D. et al., 2001; Donaldson К., Tran T, 2002; Li N.
et al., 2003; Goncalves D. M. et al., 2011) (рис. 2.67).
Токсическое действие наночастиц на культуры клеток зависит от типа клетки-
мишени, условий среды, в которой произошло взаимодействие наночастицы с клет-
кой, химического состава наночастицы, ее размера, формы, химии поверхности и дозы
(Smith Р. et al., 2012).
В условиях in vivo наночастицы вызывают различные патологические реакции.
В зависимости от способа проникновения в организм человека и физических свойств
(см. табл. 2.31), наночастицы могут оказывать разрушительное действие на легочную
526
Часть 2. Общая неправильная эпидемиология
Таблица 2.31
Физические свойства частиц наноразмеров, определяющие их ра< пространение
в организме человека и поражающее действие
Физическое свойство Распространение наночастиц в организме человека Поражающее дей- ствие наночастиц
Размер Определяет ее биодоступность и продолжительность циркуляции в кровеносном русле. Частицы размером <5—10 нм удаляются из организма путем почечного клиренса; >200 нм депонируются (sequestered) в селе- зенке. Частицы размером до 70 нм могут пенетри- ровать капилляры. Нанокомплексы размером в пре- делах 35—120 нм собираются в лимфатических узлах. Частицы менее 40 нм способны проникать в ядро клетки. Размер частицы определяет механизм ее интернализации в клетку (Лагоцитоз, эндоцитоз, пикноцитоз) и внутриклеточную локализацию, а также оудет ли она опсонизирована белками сыво- ротки плазмы, фагоцитирована макрофагами или удалена клетками РЭС Варьирование разме- ром наночастицы позволяет более точно нацелить на клетки, содержащие специфические сайты для наночастицы, тем самым уменьшить поражающую дозу
Мультифунк- циональность Размер частицы определяет ее биодоступность и создание более высоких концентраций в органах- мишенях, структура той же частицы (например, нано- трубка) позволяет использовать ее в качестве носи- теля токсического соединения в клетку-мишень, пов< рхность частицы определяет ее сродство к опре- деленному рецептору и др Повышение специ- фичности поражаю- щего действия нано- "астиц
Большая площадь поверхности Плохо растворимые в воде токсические вещества могут быть сорбированы на поверхности частиц Расширение круга поражающих агентов
Высокий по- верхностный заряд Поверхность клетки имеет отрицательный заряд, при- дание частице положительного заряда повышает ее проникающий потенциал. Позволяет повысить эф- фективность доставки в клетки химических и биоло- гических соединений, имеющих отрицательный заряд Снижение поражаю- щей дозы и расши- рение круга пора- жающих агентов
Формирование стабильных структур, спо- собных инкап- сулировать химические и биологиче- ские соедине- ния (нанокап- сулы и нано- трубки) Инкапсуляция позволяет защитить биологическое или химическое соединение от действия факторов внешней среды при хранении, переводе в аэрозоль, нахождении в аэрозолированном состоянии и от воздействия факторов организма (кислая среда желу- дочного сока, нуклеазы сыворотки крови, иммунные ответы и др.), а также доставлять в клетки плохо рас- творимые в воде соединения. Но только частицы раз- мером менее 100 нм могут попасть вовнутрь везикулы в ходе эндоцитоза, что определяет наиболее опти- мальный размер наночастиц, включающих биоло- гические активные соединения (от 10 до 100 нм) То же
Возможность биофункциона- лизации Достигается путем присоединения наночастиц к био- активным молекулам. Например, присоединение к ПЭГ (пэгилирование) продляет длительность цирку- ляции наночастицы в крови, уменьшает их взаимо- действие с опсонизирующими белками; конъюгиро- вание с белками-лигандами повышает эффективность проникновения наночастиц в клетки-мишени Снижение поражающей дозы
2.2. Биологические поражающие агенты
527
Рис. 2.67. Гипотетический механизм клеточного взаимодействия с частицами нано-
размера. Проникшие в клетку наночастицы металла вызывают окислительный стресс,
тем самым увеличивая внутриклеточное содержание кальция и активируя отдельные
гены. EGFR (epidermal growth factor receptor) — рецептор эпидермального фактора
роста. NF-кВ1 (ядерный фактор «каппа-би») — основной транскрипционный
активатор воспалительных цитокинов. По К. Donaldson, Т. Tran (2002)
ткань, проявлять нейро-, дермо- и генотоксичность. Без экспериментального иссле-
дования на лабораторных животных наночастиц конкретного вещества, структуры,
определенного диапазона дисперсности и степени агрегации частиц, их патологичес-
кое действие можно только предполагать. Однако, даже если оно установлено в
эксперименте или эпидемиологическим исследованием, необходимо понимать, что
это действие воспроизводимо только в тех условиях, в которых оно обнаружено. На
другие условия эксперимента мы можем его только экстраполировать, как это пока-
зано на рис. 2.68 для наночастиц, оказывающих нейротоксическое действие.
Например, при исследовании в условиях in vitro ответов микроглии на частицы
кварца размером 200 нм, обнаружено, что они не обладают токсическим эффектом
даже при высоких концентрациях (292 мкг/мл). Частицы того же вещества размером
50 нм уменьшают количество живых нейронов дозозависимым образом (Ariano Р. et
al., 2011). Наночастицы меди размером 40 нм обладают максимальным токсическим
эффектом на соматосенсорные нейроны в сравнении с таким же эффектом, оказыва-
емым наночастицами меди размером 60 и 80 нм (Prabhu Е М. et al., 2010).
В патологических эффектах наночастиц, проявившихся в условиях in vivo при од-
ном способе введения, прослеживается определенная специфичность, обусловленная
их структурой и химическим строением. Например, агрегированные одностеночные
1 NF-кВ — универсальный фактор транскрипции. Нарушение его регуляции вызывает
воспаление, аутоиммунные заболевания, развитие вирусных инфекций и рака.
528
Часть 2. Общая неправильная эпидемиология
Активация микроглии
и астроцитов „
а Окислительный
Токсичность
Индукция медиаторов воспаления,
нейротрансмиттеров, генов апоптоза
Нейровоспаление
Нейтральный Анион Катион
Нейродегенерация
Рис. 2.68. Нейротоксическое действие наночастиц. А. Предполагаемый меха-
низм нейротоксического действия наночастиц. По Tin-Tin Win-Shwe, Н. Fuji-
maki (2011) Б. Общие закономерности проявления нейротоксичности при
экспонировании к наночастицам. Нейротоксичность выше при меньшем раз-
мере частиц, большей концентрации, большей величине положительного
заряда и длительности экспонирования По D. Н. Joetal (2011)
углеродные нанотрубки в легочной ткани мышей индуцируют образование гранулем,
главным образом связанных с гипертрофией эпителиальных клеток. Эти же трубки,
но в диспергированном состоянии, вызывают развитие диффузного интерстициналь-
ного фиброза с утолщением стенок альвеол (Shvedova A. et al., 2005).
Внутренний объем нанотрубок может быть заполнен токсическими соединения-
ми биологической и химической природы. Стенки нанотрубок исключат взаимодей-
ствие биологических компонентов с клетками иммунной системы и последующие
иммунные ответы. Благодаря достижениям в конструировании таких структур стала
возможной доставка биологически активных веществ и генов в клетки, очень трудные
для трансфекции, включая В-клетки и первичные нейроны (Mataraza С. et al., 2005;
Goldberg М. et al., 2007).
Пути проникновения нанообъектов в организм человека. Их не менее шести — че-
рез легкие, обонятельный эпителий, кожу, желудочно-кишечный тракт, барьеры гла-
за и парэнтерально. Наиболее доступны для наночастиц легкие. Они состоят из двух
различных частей — воздушных путей, транспортирующих воздух в легочную ткань
(трахеобронхиальный и назофарингеальный регионы); и составляющих легочную
ткань альвеол, где происходит газообмен. Легкие человека содержат около 2300 км
воздушных путей и 300 млн альвеол. Общая поверхность легких взрослого человека
140 м2, что больше теннисного корта. Воздушные пути защищены от проникновения
крупных частиц благодаря активному эпителию и вязкому слизистому слою на его
поверхности. Но в газобменной области альвеол барьер между альвеолярной стенкой
и капиллярами тонок, всего 500 нм и легко проницаем для наночастиц (рис. 2.69).
2.2. Биологические поражающие агенты
529
Рис 2.69. Схематическое изображение строения альвеолы По Р. Hoet et al. (2004)
Распространение наночастиц по дыхательным путям может показаться весьма
неожиданным для специалистов в области защиты от БО, привыкших моделировать
эти процессы в диапазоне 1—10 мкм (рис. 2.70).
Носовые
воздушные пути
Респираторные
Зронхи
Альвеолярные
трубочки
Альвеолярно-
капиллярное
ложе
Альвеолярные
мешочки
Легочные
артерии
Рис. 2.70. Предпочтительное фракционное отложение ингалированных частиц
в назофарингеальном (1), трахеобронхиальном (2) и альвеолярном (3) регионах
респираторного тракта при носовом дыхании. По G. Oberdorster et al. (2005)
Трахея
Бронхи
6.0001 0,001 0,01 0.1 1 10 100
Диаметр (мкм)
Например, до 9(1 % ингалированных 1 -нм частиц оседает в назофарингеальном
тракте. Частицы размером 5 нм распределяются относительно равномерно по назо-
фарингеальному тракту, трахеобронхиальному тракту и альвеолам (~ по 30 %). Час-
тицы диаметром в 20 нм наиболее эффективно оседают в альвеолах (~50 %). В то же
время, в трахеобронхиальном и назофарингеальном регионах задерживается ~ по 15 %
от их общего количества. Такое дифференцированное распределение наночастиц раз-
ных размеров в различных разделах легких неизбежно сказывается на их экстрапуль-
монарном распределении (рис. 2.71).
530
Часть 2. Общая неправильная эпидемиология
Альвеолярные мешочки
Респираторные
бронхиолы
Глотка
Ганглий блуждающего нерва
Легочные вены
Альвеолярно-
капиллярное
ложе
Рис. 2.71. Экстрапульмонарное распределение наночастиц разных раз-
меров. 1 — проникновение в сенсорные нейроны через обонятельный эпи-
телий; 2 — в желудочно-кишечный тракт; 3 — интерстиций легкого, 4 —
лимфатические узлы; 5 — кровеносное русло По G Oberddrster et al. (2005)
Носовые воздушные пути
Лимфатические
Гортань узлы
Трахея
Бронхи
Нереспираторные бронхиолы
Легочные артерии
Ингалированные наночастицы проникают в кровеносное русло. Скорость этого
процесса может варьировать для наночастиц разных размеров и химического соста-
ва. Но в некоторых случаях этот процесс может осуществляться очень быстро.
A. Nemmar et al. (2002) установили, что ингалированные углеродные частицы разме-
ром менее 100 нм уже через одну минуту после экспозиции можно обнаружить в кро-
ви экспериментального животного.
Поданным Choi Н. S. et al. (2010), некатионные наночастицы размером менее чем
-34 нм способны перемещаться из легких в лимфатические узлы средостения. Такие же
наночастицы размером <6 нм проникают из легких в кровь, а затем выводятся из орга-
низма почками. Наночастицы, покрытые сурфактантом, выстилающим стенки аль-
веолы, более эффективно проникают через альвеолярно-капиллярную мембрану, чем
частицы, не имеющие такого покрытия (Banerjee R., 2012).
Наибольшим токсическим потенциалом для легочной ткани обладают углеродные
нанотрубки (carbon nanotubes, CNT). Их диаметр измеряется в нанометрах, длина
может достигать сотен нанометров. Такие структуры имеют сходство с нитями асбес-
та. Экспериментальные результаты, полученные в условиях in vitro Е A. Murphy et al.
(2012), показывают, что прямое экспонирование к CNT ведет к значительному цито-
киновому выбросу макрофагами, но не клетками мезотелия. Выраженность этих про-
воспалительных ответов макрофагов зависит от длины нитей CNT, но она меньше, чем
ожидалось при проведении аналогии с нитями асбеста. К тому же действие CNT на
клетки мезотелия носит непрямой характер Мезотелиальные клетки реагируют на
цитокины, выброшенные макрофагами во время незавершенного фагоцитоза CNT.
Поэтому авторы выдвинули гипотезу, что этот механизм лежит в основе воспаления,
развивающегося в плевральной полости человека в ответ на проникновение туда на-
нонитей. Данные по острой легочной токсичности CNT приведены в табл. 2.32.
«Новинкой» для токсикологов, именно для частиц нанодиапазона, является воз-
можность их проникновения в ЦНС по нервным волокнам, идущим от обонятельно-
го эпителия и сетчатки глаза. Более 60 лет назад было установлено, что 30-нм полио-
вирус, введенный экспериментальному животному интраназально, может через лу-
ковицу обонятельного нерва проникать непосредствс нно в ЦНС (Bodian D., Howe Н.,
1941). Наночастицы способны проникать в ЦНС этим же путем (рис. 2.72).
2.2. Биологические поражающие агенты
531
Таблица 2.32
Результаты исследования остро» легочной токсичности CNT* **
Животное Нано- частица Путь введения/ продолжительность исследования Доза Патологический эффект/повреждение
Мышь MWCNT** И/т; через 1, 2, 3, 7 и 30 сут 1; 2,5 и 5 мг/кг Зависимые от дозы летальность и вос- паление: зависимые от дозы и времени экспонирования фиброз и гпанулемы
Морская свинка MWCNT И/т; 90 сут 15 мг/ животное Неспецифическая десквамативная интерстициальная пневмония
Мышь SWCNT То же 0,1 и 0.5 мг/ животное Гибель животных в группах с наиболь- шей дозой, прогрессирующие и зави- сящие от дозы мультифокальные эпи- телиальные гранулемы
Крыса MWCNT И/т; 1 и 2 мес. 0,5; 2,0 и 5 мг/ животное Дозозависимое развитие воспаления и фиброза
Мышь SWCNT Фарингеальная аспирация. 1. 3. 7. 28 и 60 сут 40 мкг/ животное Транзиентные воспалительные ответы, зависящие от дозы эпителиоидные гранулемы, интерстициальный фиброз и утрата легочной функции
Крыса SWCNT 1 И/т; 24 ч, 1 нед, 1 и 3 мес. 1 и 5 мг/кг Смерть при наиболее высокой дозе, вызванная механической асфиксией из-за обструкции верхних дыхательных путей, вызванной агрегацией наночас- тиц; транзиентные воспалительные от- веты и неспецифические повреждения; непрогрессирующие и не зависящие от дозы мультифокальные гранулемы
* По S. Stem, S. McNeil (2008).
** MWCNT (multiwall carbon nanotube) — мультистеночные углеродные нанотрубки;
SWCNT (single-wall carbon nanotube) — одностеночные углеродные нанотрубки.
клетки
Рис. 2.72. Проникновение наночастиц в мозг через обонятельный
эпителий носовых ходог По G. Oberddrster et al. (2005)
Скорость транспорта полиовирусов и наночастиц по обонятельному нерву при-
мерно одинакова — 2,4—2,5 мм/ч Ide Lorenzo, 1970), что говорит об одном и том же
532
Часть 2. Общая неправильная эпидемиология
механизме, задействованном для их транспорта (см. рис. 2.64). Помимо транснейраль-
ного проникновения в ЦНС, наночастицы легко преодолевают гематоэнцефаличе-
ский барьер (Borm Р. et al., 2006).
D. Н. Jo et al. (2011) показали проникновение в мозг через сетчатку глаза наноча-
стиц золота размером 20 нм, введенных внутривенно.
Существуют три пути проникновения наночастиц через кожу, в промежутки между
клетками, через клетки и через волосяные фолликулы. Например, липоплексы с раз-
мерами в пределах от 20 до 200 нм легко «проходят» между клетками. Проникновение
в организм человека через кожные покровы для наночастиц облегчается тонкостью
верхнего слоя кожи, эпидермиса. Лежащий же под ним слой — дерма, богат макро-
фагами крови и тканей, лимфатическими узлами, дендритными клетками и в него
«выходят» окончания сенсорных нервов пяти различных типов; все эти «обитатели»
дермального слоя способны поглощать и распространять нанообъекты за пределы их
первоначальной аппликации (рис. 2.73).
Клетки Лангерганса
и дендритные клетки
Волос
Волосяная папилла
Нервное волокно
Кровеносные
и лимфатичес
сосуды
Сальная железа
Волосяной фоликул
Дерма
Вена
Эпидермис
Рис. 2.73. Проникновение наночастиц через кожу По G. Oberddrster et al. (2005)
Роговой слой
Тигментный'''
слой
Базальный
слой
Потовая пора
Дермальная папилла
Окончание сенсорных нервов
Макрофаги
Подкожная жировая
ткань (гиподермис)
Наночастица
В том, что любые незначительные механические повреждения кожи делают ее
«проницаемой» для наночастиц, нет ничего неожиданного. Но S. S. Tinkle et al. (2003)
продемонстрировали способность неповрежденной кожи в местах сгиба, например,
в области запястья, становиться проницаемой для наночастиц. L. Mortensen et al. (2008)
опубликовали экспериментальные данные, показывающие значительное увеличение
проницаемости кожи для ноночастиц после ультрафиолетового облучения.
При внутривенном введении способность нанообъектов распространятся в организ-
ме животного и человека зависит от их размеров. Например, частицы золота разме-
ром 10 нм при внутривенном введении крысе были обнаружены в крови, печени, селе-
зенке, почках, яичках, тимусе, сердце и мозге. Частицы этого же вещества, но более
крупного размера (50, 100, 250 нм), обнаружены только в крови, печени и селезенке
экспериментального животного (Krug H.F., Wick Р., 2011). D. Н. Jo et al. (2011) пока-
зали способность введенных внутривенно частиц золота размером 20 нм проникать
2.2. Биологические поражающие агенты
533
во все слои сетчатки глаза, включая нейрональный, эндотелиальный и периэндоте-
иальные глиальные клетки, однако частицы золота размером 100 нм не проникали в
ткани сетчатки. Из циркулирующих в крови наночастиц ГЭБ способны преодолевать
катионные наночастицы и в высоких концентрациях (!) анионные наночастицы
(Lockman Р. R. et al., 2004).
Кишечник более проницаем для наночастиц, чем для частиц микронного разме-
ра, благодаря тому; что поверхность его слизистой оболочки имеет поры диаметром
до 100 нм. Положительно заряженные наночастицы проникают в кровеносное русло
через кишечник значительно быстрее, чем отрицательно заряженные. Чем меньше
размер частицы, тем меньшее влияние на скорость ее проникновения оказывает ее за-
ряд (Frohlich Е., Roblegg Е., 2012)
Подробно с токсическими свойствами нанообъектов, производимых промышлен-
ным путем, можно ознакомится по работам S. Stern, S. McNeil (2008), J. S.Bonner
(2010), Y. Morimoto et al. (2010), M. Skocaj et al. (2011). В табл. 2.33, с. 534 приведены
примеры острой системной токсичности нанообъектов в условиях эксперимента.
Случай группового поражения людей наночастицами двуокиси кремния1. С января
2007 г. по апрель 2008 г. в Beijing Chaoyang Hospital в Пекине поступили 7 женщин в
возрасте от 19 до 47 лет (средний возраст 29 лет), работавших на одном предприятии.
Все они жаловались на отдышку и загрудненное дыхание. Объективно при поступле-
нии: поверхностное дыхание; гипоксемия, но без задержки углекислоты; на лице,
кистях рук и предплечьях интенсивная зудящая сыпь. Ультразвуковое исследование
позволило обнаружить у всех пациенток плевральные выпоты, у пяти из них перикар-
диальный выпот глубиной 4—8 мм. Других патологических изменений со стороны
сердца, почек и печени, не выявлено.
Ранее они проходили лечение в местных больницах, которое включало многократ-
ный торакоцентез, выполняемый для удаления жидкости из плевральной полости;
антибиотикотерапию; антитуберкулезное лечение; преднизолон и метилпреднизолон.
Все пациентки работали в одном плохо вентилируемом помещении площадью 70 м2,
с одной дверью и без окон. В помещении установлена машина для воздушного рас-
пыления полиакрилатов на листы оргстекла и последующего нагревания и высуши-
вания полученных поверхностей. Поликрилатные покрытия обычно содержат связую-
щее вещество, пигменты, растворители, наполнители и добавки. Наполнители и
добавки представляют собой композиционные материалы, содержащие диоксид крем-
ния, сульфат бария, карбонат кальция, силикаты (тальк, слюда или каолин)
Даже спустя 20 мес. после поступления в Beijing Chaoyang Hospital у пациенток
обнаруживали ослабленное дыхание, продолжающееся истечение жидкости в плевру
и медленно прогрессирующий легочный фиброз. У двух пациенток (случаи 1 и 5) плев-
ральный выпот образовывался чрезвычайно быстро, что потребовало установить не-
прерывный дренаж плевральной полости. В сутки врачи откачивали 300—800 мл плев-
ральной жидкости. Наиболее тяжелой 19-летней пациентке (случай 5) через 18 мес.
после начала болезни было выполнено оперативное вмешательство на грудной клет-
ке под видеонаблюдением (VATS). Процедура включала фенестрацию (прокол) пери-
карда и откачивание почти 170 мл жидкости из перикардиального пространства, де-
кортикацию плевральных фиброзных пластинок (decortication of pleural fibrous lamina),
клиновидную резекцию плевры, легочную биопсию и дренаж плевральной жидкости.
Через 24 ч после VATS у пациентки развился тяжелый пневмоторакс, на 10 сут — острая
1 По работам Y. Song et al. (2009, 2011).
534
Часть 2. Общая неправильная эпидемиология
перикардиальная тампонада, которые не удалось купировать. Пациентка умерла на
16 сут после хирургического вмешательства на фоне легочной недостаточности. Тече-
ние болезни у 29-летней пациентки (случай 5) было сходным с вышеописанным. Она
умерла от легочной недостаточности в местном госпитале через 21 мес. после появле-
ния первых симптомов болезни.
Таблица 2.33
Острая системная токсичность нанообъектов в условиях эксперимента*
Животное Нанообъект Путь введения LDS0 (г/кг) Патологический эффект
Мышь Фуллерен (С6„) в/б >2,5” Смертельных исходов не наблюл (лось, фуллерены обнаружены в селезенке и печени, гипертрофия перисинусои- дальных клеток печени
Мышь G7, катионный полиамидоамин- ный дендример в/б >0,045 Погибла 1/5 часть животных, вакуолизация печени***
Mi гшь G3, катионный меламиновым дендример в/б 0,040-0,160 Некроз печени
Крыса Полисульфо- нированный фуллерен (С60) в/б 0,6 Лизосомальная перегрузка почек, нефроз
Мышь Полигидрокси- лированный фуллерен (С60) в/б * 1,2 Отмечено зависимое от дозы увеличение веса печени
Мышь Zn, 58 нм peros >5 Погибли 2 из 20 экспериментальных животных вследствие непроходимости кишечника. Расширение канал! цев почек, отечная дегенерация печени (hydropic degeneration)
Мышь Си, 25 нм peros 0,4 Проксимальный тубулярный некроз по- чек, жировая дегенерация печени, атро- фия селезенки
Мышь Т1О2, 25, 80 и 155 нм peros >5” Печень — отечная деген ;рация, очаговый некроз; почки — расширение клубочков
* По S Stem, S. McNeil (200?).
** Указана максимальная доза.
*** Жирным шрифтом выделен орган-мишень.
Общие анализы мочи у пациенток при поступлении в стационар были в пределах
нормы. Общий анализ крови обнаружил моноцитоз с увеличением абсолютного коли-
чества моноцитов и нейтропению, с уменьшением абсолютного количества нейтро-
филов, повышенную СОЭ, гипопротеинемию, повышенные уровни ALT и AST. Спи-
рометрия показала снижение легочного объема. Иммунологические тесты были в
пределах нормы. Рентгенография и компьютерная томография грудной клетки пока-
зали наличие у всех пациенток плеврального выпота, у 5 из них обнаружен перикар-
диальный выпот. У 4 пациенток обнаружены диффузные затемнения в легких, у 6 —
узелки в легочном интерстиции, у трех — аденопатия лимфатических узлов (lymph node
adenopathies). Не обнаружено плевральных бляшек (pleural plaques), утолщений плевры
(pleural thickening) и узлов (pleural nodules). У всех пациенток выявлено воспаление
2.2. Биологические поражающие агенты
535
легочного интерстиция и легочный фиброз. После 7 мес. наблюдения у двух пациен-
ток (случаи 1 и 5) замечено быстрое прогрессирование легочного интерстициального
фиброза и у одной — образование кальцификатов в плевральной полости (случай 1).
Всем пациенткам была выполнена бронхоскопия, у 4 обнаружено увеличение и
гиперемия tunica mucosa bronchiorum1. В жидкости бронхоальвеолярного лаважа ко-
личество макрофагов снижено, лимфоцитоз, нейтрофилез, количество эозинофилов
соответствовало норме. Трахеобронхиальная легочная биопсия показала скопления
фагоцитов, воспалительных клеток и белковые выпоты в альвеолярных пространствах,
набухшие и расширенные альвеолярные септы с нейтрофильными лейкоцитами и
участки легочного фиброза. Tunica mucosa bronchiorum инфильтрирована воспали-
тельными клетками. Торакоскопия, проведенная у 3-х пациенток, позволила обнару-
жить у двух из них ровные уплотнения на поверхности плевры, не имеющие отноше-
ния к злокачественным образованиям. Биопсия плевры и жидкости показала наличие
гранулем, вызванных наличием инородного тела (foreign-body granulomas), геморра-
гии, воспалительных клеток.
Изучение грудной полости с помощью VATS позволило обнаружить отсутствие
слипания плевральных листков. На биопсию были взяты участки плевры. С легочной
поверхности плевры, где рассеяны маленькие туберкулы, взяты гистологические об-
разцы из участков, соответствующих верхней, средней и нижней долям легкого. Све-
товая микроскопия гистологических препаратов не дала ничего специфического:
фиброзные уплотнения на поверхности плеврального листка; эмфиземо-подобные
изменения легочных альвеол; расширение альвеолярных септ; отложение коллагено-
вых нитей; капилляры расширены, заполнены лимфоцитами; альвеолярные про-
странства инфильтрированы фагоцитами; в альвеолах встречаются многоядерные
гигантские клетки; пролиферация альвеолярных эпителиальных клеток II типа.
Трансмиссионная электронная микроскопия дала более интересные результаты. Ле-
гочная ткань была повреждена. Обнаружены легочный интерстициальный отек; при-
знаки гиперплазии и дегенерации пневмоцитов второго типа, почти утративших мик-
роворсинки; уплотнение базальной мембраны альвеолярных эпителиальных клеток
первого типа. Клетки сосудистого эндотелия имели признаки повреждения: они были
увеличены, наблюдалась вакуолизация клеточных органелл и уплотнение базальной
мембраны. В газобменной области альвеол барьер между альвеолярной стенкой и
капиллярами был нечетким и значительно более толстым, чем в норме (-500 нм).
В поздней стадии болезни повреждения легких, описанные выше, были более
выраженными. Барьер между альвеолярной стенкой и капиллярами в газобменной
области альвеол был практически неразличим. В альвеолярном пространстве находили
эритроциты, что указывало на наличие альвеолярной геморрагии и отека. Обнаружен
легочный фиброз с пролиферацией фибробластов и фибринозный некроз. Плевраль-
ные мембраны отечные, рыхлые, с признаками фиброза и неясной клеточной струк-
турой. Большое количество макрофагов образовывали агрегаты, что показывает их
участие в развитии патологического процесса.
У пациентов на ранней стадии болезни (три месяца после появления первых сим-
птомов) в цитоплазме, ядре и органеллах макрофагов, микрососудах легких, эндоте-
лиальных клетках легочных сосудов и в микролимфатических сосудах в жидкости
плеврального выпота были обнаружены наночастицы неизвестной природы диамет-
ром -2-20 нм. В альвеолярных эпителиальных клетках и интерстициальных тканях
1 Tunica mucosa bronchiorum — слизистая оболочка бронхов.
536
Часть 2. Общая неправильная эпидемиология
их тоже находили, но в значительно меньших количества. Размер наночастиц в жид-
кости плеврального выпота был меньшим, чем в тканях легких, что говорит о разме-
розависимой транслокации наночастиц по тканям легких. Для того чтобы установить
химическую природу наночастиц, использовался метод энергодисперсионной рент-
геновской спектроскопии (EDX). Он показал, что большинство наночастиц состоит
из двуокиси кремния (рис. 2.74).
Рис. 2.74. Обнаружение в легочной ткани человека наночастиц двуокиси кремния
и вызванные ими гистологические изменения. А. Трансмиссионные электронно-мик-
роскопические изображения наночастиц в легочной ткани (случай 5). Наночастицы
имеют размер 20—21 нм в диаметре, рассеяны по цитоплазме макрофага (размер
маркерной полосы 200 нм). Б. Артефакт, который можно принять за наночастицы.
Поэтому с помощью EDX устанавливается химический состав наноструктур (размер
маркерной полосы 150 нм). В. Ранняя стадия болезни Агрегация макрофагов в аль-
веолярном пространстве, набухание альвеолярной септы и легочный фиброз (окра-
шивание гемотокслином и эозином, увеличение в 100 раз). Г. Поздняя стадия болез-
ни. Легочные альвеолы частично эмфизематозны с агрегированными макрофагами
и пролиферированными альвеолярными эпителиальными клетками второго типа.
Альвеолярные септы и кровеносные сосуды расширены и застойны (окрашивание
гемотокслином и эозином, увеличение в 100 раз). По Y. Song et al. (2011)
В поздней стадии болезни (18 мес. от начала появления симптомов) небольшое
количество наночастиц двуокиси кремния было обнаружено в пульмонарных клетках
(pulmonary cells), ткани интерстиция (interstitial tissue) и в макрофагах. Однако в яд-
рах и цитоплазме альвеолярных эпителиальных клеток были обнаружены отдельные
нитчатые наноструктурные включения (длиной ~70 нм), сформировавшие сложные
агломераты или пересекающие мембраны ядер альвеолярных эпителиальных клеток.
Используя EDX, исследователи установили, что нитчатые наноструктуры содержат Fe,
Са, Mg, но не Si. Эти структуры не были обнаружены в легочной ткани пациентов,
что говорит о том, что они не имеют отношения к легочной патологии.
2.3. Основы эпидемиологического расследования искусственных... 537
* * *
Изучение патологических процессов, вызванных проникновением в организм
человека объектов наноразмера, еще только начинается. Однако уже накопленно до-
статочно данных, позволяющих предположить, что нанотехнологии обладают боль-
шим потенциалом для создания средств массового поражения, серьезно отличающих-
ся по механизмам поражения, путям проникновения в организм человека, способам
применения и целям ведения войны, от тех, что разрабатывались в XX в. примени-
тельно к микроорганизмам, токсинам и ОВ. Нанообъекты могут вызывать массовые
смертельные поражения людей, проявляющиеся клиникой болезни, неизвестной
врачам, либо она может маскироваться под уже известные описания, однако иметь
совершенно иную этиологию, чем та, что традиционно рассматривается врачами при
выборе способов лечения пациентов. Наибольшую опасность в настоящее время пред-
ставляют нанообъекты, имеющие размер менее 50 нм; формирующие инкапсулирую-
щие структуры и подвергнутые биофункционализации. Их применение возможно как
непосредственно по цели, путем распыления специальными устройствами, так и
скрытно, в рамках непрямой стратегии действий, через лекарственные препараты, вак-
цины, продукты питания и различные материалы, контактирующие с людьми.
2.3. Основы эпидемиологического расследования искусственных
эпидемических процессов и поражений биологическими токсинами
Накопленный опыт эпидемиологических расследований искусственных эпидеми-
ческих процессов свидетельствует о наличии характерных для них особенностей.
2.3.1. Характерные особенности эпидемиологии искусственных
эпидемических процессов и поражений биологическими токсинами
Эпидемиология искусственного эпидемического процесса, вызванного однократным
применением биологического агента. Эпидемиология искусственного эпидемического
процесса, вызванного многократным применением биологического агента. Особенно-
сти искусственного эпидемического процесса, характерные для обоих сценариев био-
логического нападения. Малораспространенные способы осуществления преступлений
с использованием биологических агентов. Объекты преступления при биологическом
нападении. Модели территорий, на которых наиболее вероятно применение биологи-
ческих агентов для поражения людей. Психосоциальные реакции у людей, вызванные
акцией с применением биологических агентов.
Данные по динамике террористических вспышек инфекционных болезней, приве-
денные вразд. 1.12, и опубликованные модели террористических актов (Kaufmann A. F.
etal., 1997; Franz D. R. etal., 1997; PavlinJ. A., 1999; DembekZ., 2005) позволяют выде-
лить два возможных сценария биологического теракта или диверсии — когда биоло-
гический агент применяется преступниками однократно и когда его применяют мно-
гократно. Соответственно, будет различаться и эпидемиология искусственного
эпидемического процесса.
Эпидемиология искусственного эпидемического процесса, вызванного однократным
применением биологического агента. Характерной особенностью эпидемиологии такого
террористического акта или диверсии можно считать сжатую эпидемическую кривую
с пиком заболеваемости, «укладывающимся» в среднее значение инкубационного периода
538
Часть 2. Общая неправильная эпидемиология
болезни (эпидемиология «точечного источника»). При большинстве естественно возни-
кающих эпидемий наблюдается постепенный рост числа заболевших (предэпидеми-
ческая стадия)1, так как люди постепенно вступают в контакт с возрастающим чис-
лом больных, насекомых — переносчиков инфекции или объектов в окружающей
среде, являющихся передатчиками патогенного микроорганизма (рис. 2.75).
Me «э пидеми-
ческая стадия
Количество
больных
Предэпидеми-
ческая стадия
Стадия макси-
мального подъ-
ема эпидемии
Стадия
развития
эпидемии
Стадия
угасания
эпидемии
Г
Пост'яГ’теми-
ческая стадия
Спорадические
случаи. Обычный
уровень заболе-
ваемости для дан-
ной местности
и данных условий
Нарастание
числа
случаев
болезни
Широкое распрост-
ранение болезни,
массовость поражения
Уменьшение
числа
случаев
болезни
Спорадические
случаи. Заболе-
ваемость надает
до обычного
уровня для дан-
ной местности
и данных условий
или ниже такового
Преобладают
микробоно-
сительство,
бессимптом-
ная инфекция
Проявление
типичных
форм
болезни
Типичные
формы
болезни,
острое
и сверх-
острое
течение
Типичные
формы
болезни,
острое и
подострое
течение
Появление
атипичных
форм болезни,
подострое,
хроническое,
абортивное
течение
Бессимптомная
инфекция и
микробоносн-
тельство
Иммунитет еще
сохраняется, но
началось нара-
стание числа
восприимчивых
лкшей
Много вос-
приимчивых
людей
Нарастание числа
иммунных людей
Максимальное число людей
с высокой степенью специ-
фического иммунитета, рост
численности населения
Рис. 2.75. Схематическое изображение типовых периодов циклического эпи-
демического процесса2 (за основу взята схема Бакулова И. А. с соавт., 1997)
В противоположность естественно возникающим эпидемиям, все те, кто подвергся
нападению с применением БО, должны вступить в контакт с биологическим агентом
примерно в одно и то же время. Сжатая эпидемическая кривая будет наблюдаться в
1 При отсутствии эпидемической настороженности эту стадию врачи упускают, и эпиде-
мия кажется им возникшей внезапно, но ретроспективное расследование позволяет устано-
вить, что эпидемия нераспознанной развивалась уже несколько месяцев. Например, первый
диагноз «бубонная чума» в Трапезунде был поставлен 7 ноября 1917 г, но потом было установ-
лено, что уже с конца лета в госпиталях русской армии от 4j мы умирали пациенты с диагно-
зом «идеопатический бубон».
2 Подробно описание циклических и нециклических инфекционных и эпидемических
процессов приведено в моей монографии (Супотницкий М. В., 2009). К циклическим отно-
сятся инфекционные процессы, лимитируемые клеточной и гуморальной иммунной системой
(чума, туляремия, оспенные инфекции, сап, мелиоидоз и др.).
2.3. Основы эпидемиологического расследования искусственных...
539
большинстве случаев экспонирования к точечному источнику инфекции (см. ниже),
включая и умышленно вызванные вспышки пищевого отравления (Franz D. R. et al.,
1997). Такая кривая индивидуальна и зависит от инкубационного периода и характе-
оа течения болезни.
В качестве примера приведу модель, разработанную А. Е Kaufmann et al. (1997),
специалистами CDC (США). Ее авторы провели теоретическую оценку последствий
биотеррористического нападения на пригород крупного города. Они исходили из того,
что нападение производится путем генерирования мелкодисперсного аэрозоля био-
логического агента вдоль линии и поперек направлению ветра (линейный источник—
см ниже). Аэрозольное облако проходит над площадью цели за период времени в пре-
делах 2 ч. Предполагалось также, что ингалируемая доза составила не менее одной ID50.
Человек, который экспонировался к облаку спор В. anthracis в любой точке в период
2 ч его прохождения, вдыхал одну Ю50, а человек, который экспонировался к В. те-
lilensis или F. tularensis вдыхал от одной до 10 IDSO, в зависимости от его близости к
источнику аэрозоля.
Эпидемическая кривая для ингаляционного поражения спорами возбудителя си-
бирской язвы, рассчитанная по суткам после экспонирования, давала 0 % случаев на
период менее одних суток. 5 % — через сутки, 20 % — через трое суток, 35 % — через
четверо суток, 20 % — через четверо суток, 10 % — через пять суток, 5 % — через шесть
суток и 5 % — через семь и более суток (рис. 2.76).
Рис. 2.76. Сжатая эпидемическая кривая после однократного применения
биологического агента (эпидемиология «точечного источника»). Динамика
появления больных сибирской язвой после однократного ингаляционного
инфицирования населения спорами В. anthracis. По модели биотеррористи-
ческого нападения на пригород большого города А. Е Kaufmann et al. (1997)
Летальность также варьировала в зависимости от суток, в которые были замече-
ны первые симптомы. Для пациентов с симптомами, появившимися в первые сутки,
она считалась равной 85 %, д,гя пациентов с симптомами, появившимися на вторые
сутки, — 80 %, при появлении симптомов на третьи сутки — 70 %, на четвертые, пя-
тые и шестые сутки — 50 %, на седьмые и последующие сутки — 70 %. Возросшая
летальность среди заболевших при инкубационном периоде семь и более суток рас-
считывалась, исходя из допущения запаздывания диагностики, и, как следствие, на-
чала лечения.
540
Часть 2. Общая неправильная эпидемиология
Эпидемическая кривая для ингаляционного поражения возбудителем бруцеллеза
по суткам после экспонирования характеризовалась 4 % случаев в интервале 0 суток,
6 % случаев в интервале восемь суток, 14 % — в интервале от 15 до 28 суток, 40 % — от
29 до 56 суток, 26 % — от 57 до 112 суток, 10 % —от 113 суток и далее. Летальность оце-
нивалась на уровне 0,5 %.
Эпидемическая кривая для ингаляционного поражения возбудителем туляремии
по суткам после экспонирования предполагалась следующей: в период менее одних
суток — 0 % случаев, одни сутки — 1 %, двое суток — 15 %, трое суток — 45 %, четверо
суток — 25 %, пять суток — 10 %, шесть суток — 3 %, семь и более суток — 1 %. Оцени-
ваемая летальность составляла 7,5 %.
Наивысший пик поступления в лечебные учреждения людей, больных сибирской
язвой и туляремией, будет иметь место на 3 сутки после однократного применения тер-
рористами сибиреязвенной или туляремийной рецептуры Пик поступления больных
оруцеллезом можно ожидать в интервале от 29 до 56 суток. Если биологический агент
вызывает такую клиническую форму болезни, когда больной способен образовать
эпидемическую цепочку (легочные случаи чумы, натуральная оспа), то после первого
пика может появиться второй вследствие того, что первоначально экспонированные
люди могут инфицировать тех, кто не экспонировался до этого (Pavlin J. А., 1999). При
использовании террористами биологических токсинов продолжительность времени,
в течение которого появится максимальное количество пораженных, будет зависеть
от скорости развития токсического действия. Для палитоксина, сакситоксина и тетро-
дотоксина — менее часа; для рицина и SEB — от нескольких часов до суток; для боту-
линического токсина — сутки и более (Franz D. et al., 1997).
Локализованность эпидемического очага. Это вторая характерная особенность ис-
кусственного эпидемического процесса, вызванного однократным ингаляционным
применением биологического агента. Вспышка болезни возникает с подветренной
стороны от возможной точки атаки аэрозолем. Если появится подозрение, что вспыш-
ка вызвана ингаляционным инфицированием людей (например, после выявления
«патологии мелкодисперсного аэрозоля»; см. ниже), то среди заболевших можно оп-
ределить круг лиц, находившихся в зоне прохождения аэрозольного облака. Зная
инкубационный период болезни, можно с точностью до одних суток установить дату
биологического нападения. Полоса поражения будет зависеть от способа рассеивания
биологических агентов и высоты источника аэрозоля над местностью. Известны три
способа рассеивания биологических агентов (см. FM 30-10).
1. Рассеивание точечным источником — предполагает рассеивание аэрозоля ста-
ционарным генератором из одной точки.
2. Рассеивание линейным источником — аэрозоль возбудителя инфекционной
болезни рассеивается по одной линии, либо по множеству линий, «прочерченных»
поперек направлению ветра. Например, такими линейными источниками будут уста-
новленные или сброшенные с одного или нескольких самолетов (других транспорт-
ных средств) источники аэрозоля (бомбы малого калибра или другие диспергирующие
устройства). В последнем случае формируется линия небольших точечных источни-
ков, дающих широкую полосу аэрозоля по поверхности земли. Распределение людей,
получивших ингаляционное поражение, будет зависеть от высоты линейного источ-
ника аэрозоля (рис. 2.77).
3. Рассеивание многоточечным источником — источники аэрозоля расположены
на местности (или над местностью) в произвольном порядке. Каждый из них пред-
ставляет собой небольшой точечный источник, зона поражения одного источника
2.3 Основы эпидемиологического расследования искусственных...
541
Рис. 2.77. Полоса поражения и уровни поражения аэрозолем, выпущенным из ли-
нейного источника. А. Линия выпуска находится на поверхности земли или на
малой высоте. Б. Линия выпуска удалена от поверхности земли. По FM 30-10 (1966)
Линия
выпуска
Линия 1% поражения
Линия 60% поражения
Линия 30% по^яжения
Линия 20% поражения
Линия 1% поражения
Линия 30% поражения
Линия 60% поражения
перекрывается другим. Наиболее вероятно использование такого способа для при-
менения биологических агентов с низкой вирулентностью, например спор сибирской
язвы. Их инфицирующая доза для человека при ингаляционном поражении нахо-
дится в пределах 8—50 тыс. спор, что значительно больше, чем у других потенциаль-
ных агентов БО. Локализованность эпидемического очага может иметь место и при
умышленном распространении возбудителя инфекции продуктами питания и водой
(см. в разд. 1.12.2).
Эпидемиология искусственного эпидемического процесса, вызванного многократным
применением биологического агента. Уже накопленный опыт эпидемиологического рас-
следования биологических террористических актов показывает, что характерной осо-
бенностью психологии преступников, решившихся на такое преступление, является
их способность к многократным попыткам его осуществления на протяжении меся-
цев и даже лет. Будучи не удовлетворенными полученными результатами, они повто-
ряли попытки инфицирования людей до тех пор, пока не получали нужный результат
или убеждались в невозможности его получения. Кроме того, одновременно мотуг дей-
ствовать несколько групп биотеррористов. Многократным контаминированием пре-
ступниками пищи в салат-барах была вызвана масштабная вспышка сальмонеллез-
ного гастроэнтерита в Да гласе в 1984 г., длившаяся почти полтора месяца. Почтовая
рассылка не менее четырех конвертов с сухой рецептурой спор возбудителя сибирской
язвы стала причиной вспышек ингаляционной и кожной форм сибирской язвы в США
в сентябре-октябре 2001 г. Секта «Аум-Синрикё» в начале 1990-х гг. в течение несколь-
ких лет пыталась вызвать поражение населения Японии ботулиничес ким токсином
или спорами В. anthracis. И только после многочисленных неудач секта приступила к
подготовке терактов с использованием зарина. Поэтому распространенное среди спе-
циалистов представление о сжатой эпидемической кривой биологического террори-
стического акта верно не для всех возможных сценариев. При многократном исполь-
зовании террористами биологических агентов возникнет несколько первичных очагов
542
Часть 2. Общая неправильная эпидемиология
инфекционных болезней, появление которых будет растянуто во времени, значитель-
но превышающим средний инкубационный период болезни (см. рис. 1.53). Так как
террористы способны подвергнуть биологической атаке несколько объектов, находя-
щихся на значительном удалении друг от друга, то эпидемиологам не удастся очертить
границы одного очага поражения, привязанного к прохождению аэрозольного облака.
Для такого террористического акта (диверсии) характерной особенностью будет рас-
тянутая эпидемическая кривая, отражающая многократное экспонирование людей к
биологическому агенту, и обширная территория, на которой их придется выявлять (см.
рис. 1.60). При осуществлении биологической диверсии многократное применение
биологического агента может иметь своей целью имитацию природной вспышки ин-
фекционной болезни. В этом случае основным критерием для установления искусст-
венного эпидемического процесса станет его «невозможнаяэпидемиология» (см. ниже).
Впоследствии, когда картина вспышки прояснится, можно будет проследить сжатые
эпидемические кривые в ее отдельных локализованных очагах, как это сделали аме-
риканские эпидемиологи, расследуя вспышку сибирской язвы 2001 г. (см. разд. 1.12.4).
Характерным для такого террористического акта или диверсии будет одновременное по-
явление сразу нескольких инфекционных болезней с «невозможной эпидемиологией».
Особенности искусственного эпидемического процесса, характерные для обоих сцена-
риев биологического нападения. К наиболее важным из них можно отнести следующие.
Невозможная эпидемиология. Эта особенность искусственных эпидемических про-
цессов, свидетельствующая о том, что на момент вспышки инфекционной болезни в
природе не существовало условий для ее развития. Например, эпидемиология лоша-
диных энцефалитов у людей тесно связана с экологией вирусов — возбудителей бо-
лезни в естественно возникающих эндемических очагах. Сведения о широком рас-
пространении случаев инфицирования вирусом ВЭЛ среди людей за пределами
известных эндемических районов, при отсутствии условий для природной трансмис-
сии — комаров-переносчиков и наличия этой болезни у лошадей, следует рассматри-
вать как свидетельствующие о неестественном попадании вируса в кровь людей (см.
разд. 3.10). К «невозможной» («неправильной») эпидемиологии также нужно относить
все случаи появления болезни, которая является необычной для данного географи-
ческого района, особенно если таких болезней появляется сразу несколько (Franz D.
et al., 1997). К ним целесообразно отнести и те вспышки, когда болезнь возникает вне
рамок обычного сезона, благоприятного для распространения ее возбудителя или в
отсутствие возможности естественного переноса обычно выполняющими эту функ-
цию переносчиками (Pavlin J. А., 1999).
Кожная форма сибирской язвы в США, вспыхнувшей в разных штатах осенью
2001 г., вызвала подозрение об искусственном происхождении после того, как эпиде-
миологами не был найден источник возбудителя инфекции, обычно выявляемый в
случаях естественного инфицирования людей (контакт с мясом или кожей погибших
от сибирской язвы животных). Эти подозрения усилились при обнаружении ингаля-
ционных случаев сибирской язвы у людей, не связанных с производственными усло-
виями, при которых возможно образование аэрозоля спор В. anthracis (см. разд. 1.12.4).
Особенно сложный характер приобретает установление «невозможной эпидемио-
логии» вспышки болезни в тех случаях, когда злоумышленники целенаправленно
пытаются замаскировать свою акцию под естественный случай, например, модели-
рованием естественного механизма передачи возбудителя или вызвать вспышку бо-
лезни в эндемическом поданному возбудителю районе. Действующие в природе меха-
низмы поддержания возбудителей инфекционных болезней и развития самих эпи-
2.3. Основы эпидемиологического расследования искусственных...
543
демий настолько сложны, что их невозможно подделать. Поэтому при появлении груп-
повых заболеваний среди людей, необходимо проследить всю цепочку, по которой воз-
будитель инфекционной болезни проник в человеческую популяцию, но уже в обрат-
ном направлении — от заболевших лиц к природному резервуару. Эпидемическая це-
почка всегда состоит из нескольких звеньев. В случае искусственного происхождения
эпидемии, отдельные звенья в цепочке отсутствуют. Кроме того, конкретный природ-
ный очаг инфекции всегда содержит определенные возбудители с характерными для
них свойствами. Природные популяции возбудителей инфекций, как правило, гетеро-
генны. Но террорист не может просто взять и заменить в существующем резервуаре
природный штамм на тот, который ему захочется, либо исключить его из действую-
щей эпидемической цепочки. И если в данном регионе ведется постоянный монито-
ринг свойств штаммов микроорганизмов (серологические, культуральные, морфоло-
гические и биохимические свойства, плазмидный состав и др.), циркулирующих в
эпидемических и эпизоотических цепочках, то отличить «своего» от «чужого» при от-
сутствующих звеньях эпидемической цепи, вполне возможно. Примеры расследова-
ния преступлений, совершенных с использованием биологического агента, когда пре-
ступники пытались замаскировать их под естественные вспышки энтероинфекции,
приведены в разд. 1.12.1 и 1.12.2. Вобоихслучаях именно несоответствие штамма мик-
роорганизма, примененного злоумышленниками для поражения людей, тому, кото-
рый мог вызвать вспышку болезни в данном регионе, послужило основанием для эпи-
демиологов предположить ее искусственное происхождение.
Обнаружение биологических агентов, измененных с помощью методов генной инжене-
рии или тех штаммов, про которые известно, что они исследуются как потенциальные
агенты БО. Наглядный пример — обнаружение преодолевающего иммунитет («vac-
cine-resistant») и устойчивого к пенициллинам третьего поколения штамма Ames во
время почтовой вспышки сибирской язвы в США осенью 2001 г. (см. разд. 1.12.4).
Однако обнаружение в очаге подозрительной вспышки резистентных к антибиотикам
штаммов возбудителей инфекционных болезней, потенциальных агентов БО, свиде-
тельствует о ее искусственном характере только в совокупности с другими эпидеми-
ческими признаками искусственного эпидемического процесса («эпидемиология
точечного источника» и/или «невозможная эпидемиология» и др.). В ином случае
такая находка может оказаться просто расширением наших знаний о живой природе.
Например, в Индии был выделен природный штамм В. anthracis, устойчивый к пени-
циллинам, обычно используемый для лечения сибирской язвы (Lalitha М., Thomas М.,
1997), а на Мадагаскаре обнаружен штамм Y. pestis, резистентный к стрептомицину
(Guiyoule A. et al., 2001).
Невозможная клиническая форма болезни. «Невозможная» в том смысле, что она ни-
когда не встречается при естественном заражении данным возбудителем. Например,
кожная форма сибирской язвы хорошо известна инфекционистам, и она наиболее
часто встречается среди людей, контактирующих с мясом и шкурами болевших сибир-
ской язвой животных. Ее характерное проявление — сибиреязвенный карбункул.
Однако при парентеральном введении В. anthracis развивается незнакомая инфекцио-
нистам локальная форма сибирской язвы без развития карбункула, но с обширными
некрозами и отеками в участке инъекции (см. в разд. 3.1). Частным случаем невозмож-
ных клинических форм болезни является «патология мелкодисперсного аэрозоля».
Патология мелкодисперсного аэрозоля. Переведение биологических поражающих
агентов в аэрозоль с дисперсной фазой 1—5 мкм — наиболее вероятный и эффектив-
ный способ их применения, будь-то: токсин, бактерия или вирус. Легочные формы
544
Часть 2. Общая неправильная эпидемиология
инфекционных болезней, возбудителей которых специалисты относят к поражаю-
щим агентам БО, при естественном пути заражения встречаются очень редко как
казуистика (легочные формы сибирской язвы, чумы, туляремии и др.) и имеют
специфическую патологию. К патологии мелкодисперсного аэрозоля я предлагаю от-
нести поражения, вызванные проникновением в глубокие отделы легких аэро-
золя биологических поражающих агентов с размером частиц менее 5 мкм. Этот раз-
мер частиц позволяет биологическому агенту длительное время находиться в воздухе
во взвешенном состоянии и после вдыхания проникать в альвеолы человека (см.
табл. 1.3—1 4 и рис. 1 46, 2.67). Инфицирующие частицы такого размера обычно не
встречаются в природных очагах инфекционной болезни. Их невозможно создать без
специального оборудования (жидкий аэрозоль) или знания засекреченных техно-
логических процессов, позволяющих получать, хранить и применять сухие биологи-
ческие рецептуры такой дисперсности. Обнаружение патологии мелкодисперсного
аэрозоля у жертв биологического нападения говорит о высоком уровне его подготов-
ки и крайней опасности для общества организации, его осуществившей (табл. 2.34).
Крупнодисперсные аэрозоли биологических поражающих агентов тоже способны
вызывать поражения у людей, но при значительно больших инфицирующих дозах.
Комментируя описанные отдельными авторами закономерности патогенеза и
клиники инфекционной болезни, вызванной аэрозолями поражающего агента с раз-
личным размером частиц, В. И. Огарков, К. Г. Гапочко (1975), основываясь на своем
опыте аналогичных исследований, отметили, что эти закономерности не являются
непреложными для каждого конкретного случая. При очень высокой концентрации
мелкодисперсного аэрозоля инфицирование человека может происходить не через
легкие, а через верхние дыхательные пути или одновременно через эти два отдела
дыхательного тракта. Вместе с тем относительно крупные частицы аэрозоля при высо-
ких концентрациях и длительной экспозиции, способны проникать в глубокие отде-
лы легких и инициировать развитие патологического процесса. К тому же такие час-
тицы способны дезинтегрироваться в органах дыхания и проникать в глубокие отделы
дыхательного тракта. В ряде случаев (энцефалиты лошадей, лихорадка скалистых гор,
желтая лихорадка, ботулинический токсин) инфицирование (поражение) аэрозолями
различного фракционно-дисперсного состава не оказывало существенного влияния
на клиническую картину болезни. Более подробно патология мелкодисперсного аэро-
золя применительно к основным потенциальным агентам БО будет рассмотрена в
третьей части данной работы.
Распространенность легочных поражений. По мнению ряда специалистов, такие
поражения свидетельствуют о террористическом акте (Wiener S. L., 1987; Pavlin J. А.,
1999). Однако легочных поражений может вообще не быть в следующих случаях:
1) возбудитель инфекционной болезни не вызывает легочных поражений при
любом способе инфицирования (например, бруцеллы, вирусы лошадиных энцефали-
тов и ботулинические токсины; см. разд. 3.4, 3.10 и 3.12 соответственно);
2) биологический агент попал в организм человека с пищей или водой (возбуди-
тели энтероинфекций), или был введен ему в кровь (токсины);
3) частицы примененной преступниками рецептуры не обладают дисперсностью,
позволяющей возбудителю инфекционной болезни проникать в глубокие отделы
легких человека.
Последний случай продемонстрировали результаты расследования биотеррори-
стического акта в США в 2001 г. Рецептура спор сибирской язвы, содержащаяся
Патология мелкодисперсного аэрозоля при поражении биологическими агентами*
Таблица 2.34
Агент Симптомы и течение болезни Рентгенология Патанатомия и патоморфология поражения
1 2 3 4
В. anthracis Средний инкубационный период 3-8 сут Недомогание, слабость, лихорадка, миалгия и сухой ка- шель (иногда с белой мокротой), тошнота, рвота. Прогрессирующее развитие респираторной недоста- точности (в течение 1-2 сут), циа- ноза, «профузный пот». Не соблю- дается двуфазность клинического течения болезни Медиастинальное рас- ширение, паратрахе- альные утолщения, хиларные утолщения, плевральные выпоты и перикардиальные вы- поты, паренхимальные инфильтраты В легких наиболее постоянными изменениями являются отек, кровоизлияние и полнокровие. В одних случаях явления отека обнаруживали только при микроскопическом исследовании; в других — при сдавливании легочной ткани с поверхности разреза стекала кровянистая пенистая жидкость. В трахее и крупных бронхах только незначительное полнокровие слизистой оболочки. Отек клетчатки средостенья. Перикардиальный выпот. Выражен- ный некроз и геморрагии лимфатических узлов, интраальвеоляр- ный и интерстициальный отек с фокальными геморрагиями и фибриновыми отложениями в легких, некроз селезенки
Y pestis Средний инкубационный период 1—2 cjt. Недомогание, слабость, озноб, сильная головная боль, лихорадка, рвота, тахикардия, боли в груди и сильная одышка, кашель с кровянистой мокротой Множественные затем- нения в паренхиме легкого, преимуще- ственно в его нижних отделах, при больших дозах возбудителя обнаруживают плев- ральные выпоты Мультифокальная двусторонняя гнойная пневмония, без значи- тельного поражения бронхов, спленомегалия. При больших дозах возбудителя вовлекаются трахобронхиальные лимфатические узлы, формируются фибринозные плевральные отеки. В паренхиме легкого альвеолярные уплотнения, вызванные миграцией поли- морфноядерных нейтрофилов и небольшого количества одноядер- ных клеток; обширная инфильтрация нейтрофилами перегородок альвеол и междольчатой, перибронхиальной и периваскулярной соединительной ткани. Очаги воспаления и некроза в эпителии бронхиол. В центрах сливных участков пневмонии геморрагии и очаговый некроз альвеолярных перегородок
В. melitensis, В. suis Инкубационный период при бруцеллезе длится от 7 до 30 сут. Симптомы бруцеллеза у людей, инфицированных бруцеллами посредством аэрозоля или любым другим способом, одинаковы Малоинформати вно** Специфических поражений легочной ткани нет. В патологический процесс вовлечены мезентеральные, трахеобронхиальные и подче- люстные лимфатические узлы. При больших дозах мультифо- кальная дегенерация и некроз эпителия бронхов с умеренным лимфогистиоцитическим и нейтрофиличеекчм бронхиолитом и перибронхиолитом, альвеолярный отек
F. tuiarensis Инкубационный период 3—7 сут. Проявляется бронхопневмониче- ским вариантом течения болезни. В эксперименте животные Тяжелая лобулярная инфильтрация. Со вре- менем затемнение легких увеличивается, Легкие гиперемчрованы и покрыты большими участками уплот- нений с множеством некротических очагов на поверхности Брон- хиальные лимфатические узлы имеют желтый цвет, плотные. Селе- зенка и печень включают много небольших очагов, окруженных
2.3. Основы эпидемиологического расследования искусственных...
Продолжение табл. 2.34
1 2 3 4
погибли через 4—8 сут после экс- понирования к аэрозолю образуются некрозы и абсцессы неровными участками некроза. Некротические фокусы в легких, трахее, бронхах и медиастинальных лимфатических узлах, мышцах сердца, диафрагме, брыжейке, эпителиальной поверхности уринар- ного тракта. Микроскопически эти участки повреждения представ- ляют собой хорошо отграниченные области паренхимальной деге- нерации и некроза с утратой архитектуры ткани. Содержат боль- шие количества макрофагов
В. pseudomallei Инкубационный период 3-4 сут. Резкий подъем температуры, кашель, боли в груди, отдышка, кровохарканье, понос, рвота, прострация, быстрая генерали- зация инфекционного процесса и прогрессирование болезни Малоинформативно. В течение первых 3-5 сут после экспони- рования возможны затемнения в нижних отделах легких. Моду- лярные инфильтраты, каверны, экссудатив- ный плеврит Специфических поражений легочной ткани нет. Генерализованные септические или септико-пиемические поражения, абсцессы в различных органах, гнойные выпоты в полостях тела
В. mallei Болезнь первые две недели проте- кает в острой форме с явлениями озноба, бронхита и пневмонии, из носа выделяются сукровичная жидкость и гной Нет данных. Возможно сходство с аналогичным поражением при ингаляционном мелиоидозе То же
С. burnetii Инкубационный период 3-19 сут. Лихорадка, недомогание, потли- вость, отсутствие аппетита, понос, головная боль, кашель и боль в груди Немногочисленные дискретные легочные поражения Вовлечение тканей легких в инфекционный процесс без пораже- ния верхних дыхательных путей. Диффузная интерстициальная пневмония с сильным альвеолярным отеком; гепатит (острый или подострый)
Вирус натуральной оспы Инкубационный период 5—14 сут. Течение болезни нетипичное для натуральной оспы. Болезнь начи- нается не с поражения кожи и слизистых, а с поражения легких. В течение 3—5 сут от начала пер- вых симптомов у пациента в ниж- них отделах легких развивается На начальной стадии болезни затемнения в нижних отделах легких, характерные для мелко- очаговой пневмонии Фибринонекротическая бронхопневмония, пролиферативный и некротизирующий дерматит, лимфоидный некроз. Энантемы и экзантемы с некротическими изменениями в лимфоидных орга- нах, коже, слизистых, репродуктивных органах. Пустулы в альве- олах, на альвеолах, бронхиолах и эпителии основных бронхов. Пустулы представляют собой широкую центральную зону некроза, внутри которой обнаруживаются некротизированные элементы; их окружает внешняя зона, образованная нейтрофилами и макрофа-
Часть 2. Общая неправильная эпидемиология
продо. мнение табл. 2 i4
1 2 3 4
очаговая пневмония, переходящая в фибронекротическую бронхо- пневмонию и системные поражения ra.M i. Соседние альвеолы гиперемированы и заполнены отечной жи костью, нейтрофилами и свободными альвеолярными фагоцитами Ос-рый артериит бронхиол и мелких бронхов с пролиферацией клеток эндотелия, макроскопическим отеком мышечной оболочки и некрозом и нейтрофильной инфильтрацией наружного слоя стенки кровеносных сосудов
Вирус Эбола и вирус Марбург Инкубационный период 3—4 сут. Острое начало лихорадки, анорек- сия, летаргия, кожная сыпь Множественные затем- нения в паренхиме лег- кого, преимущественно в нижних его отделах Поверхность легких гиперемирована и отечна, перикардиальный выпет Интерстициальная пневмония, совершенно отчетливо ори- ентированная вокруг терминальных бронхиол. Внутриальвеоляр- ные геморрагии, диффузный лейкоцитоз, васкулит, мульфокаль- ный септальный некроз, тромбозы, отек и увеличенное количество альвеолярных макрофагов. Поражения трахеобронхиальных лим- фатических узлов Иммуногистохимическое исследование легких обнаруживает наибольшую концентрацию антигеного материала в участках герминальных бронхиол
Венесуэль- ский энцефа- лит лошадей, восточный и западный энцефалиты лошадей Инкубационный период 1-6 сут Клинические признаки энцефа- лита Патологических изме- нений в легких не находят На вскрытии находят обширное и неоднородное поражение раз- личных участков и структур головного мозга. Факт ингаляцион- ного заражения мелкодисперсным аэрозолем можно установить ретроспективно, используя методы иммуно-гистохимического окрашивания эпителия мелких бронхов, бронхиол, либо альвеол. У людей, ингаляционно инфицированных ВЭЛ, возможно обна- ружение патологических изменений в медиастинальных лимфати- ческих узлах
Рицин Инкубационный период не пре- вышает 24 ч. Прогрессирующая легочная патология, проявляю- щаяся отеком легких, отдышкой, кровохарканием. Геморрагические явления в сетчатке глаз Признаки диффузной некротизирующей пневмонии с интерстициальным и альвеолярным воспалением и отеком Верхний респираторный тракт воспален незначительно. В грудной полости серозная или сероангинозная жидкость и нити фибрина, висцеральная зегочная плевра покрыта пленкой фибрина. Легкие не колапсированы, но тяжелые, плотной упругой консистенции, покрытые жидкостью. Их окраска пестрая, с красной и пурпурной исчерченностью. Медиастинальные лимфатические узлы расшире- ны и отечны. Легочные доли имеют признаки мультифокального сливающегося некроза. При микроскопическом исследовании обнаруживают перибронховаскулярный отек, альвеолярное зато- пление, фибринозногнойные альвеолиты, интерстициальную пнев- монию с участками некроза. Бронхиолы почти полностью утрачи-
3. Основы эпидемиологического расследования искусственных...
Продолжение табл. 2.34
1 2 3 4
вают эпителий, сохранив только гладкие мышцы. Терминальные бронхиолы некротизированы и «раздуты» миграцией эозинофилов, эритроцитов, альвеолярных макрофагов, клеточного дебриса и отложением нитей фибрина. Большинство альвеол без подкладоч- ного эпителия
Ботулиниче- ский токсин Инкубационный период 1-3 сут. Анорексия, летаргия, атаксия, за- трудненное дыхание, дисфагия, птоз, регургитация пиши через нос, парезы, кашель, ротовое ды- хание, нарастающая дыхательная недостаточность. Офтальмический синдром минимален, гастроэнте- рический отсутствует Патологических изме- нений в легких не находят Поражений со стороны органов и тканей, которые можно связать с прямым действием ботулинических токсинов, не бывает. У по- гибших экспериментальных животных поражения легких были незначительными и соответствовали аспирационной пневмонии, вызванной параличами/парезами мышц грудной клетки
Стафилокок- ковый энте- ротоксин В (SEB) Инкубационный период 3-12 ч. Основные симптомы поражения: лихорадка, головная боль, сухой кашель, озноб, боли в груди, ми- алгии, тошнота, рвота, анорексия, диарея. Температура у отдельных пациентов достигала 41 °C. При смертельной дозе токсина болезнь носит двуфазный характер. При- близительно через 48 ч после экс- понирования к SEB, внешне уже выздоровевшие животные впадают в летаргическое состояние и поги- бают в течение 4 ч на фоне про- грессирующей летаргии и дыха- тельной недостаточности Признаки наличия избытка жидкости или накопления слизи в небольших участках дыхательных путей (перибронхиальные затемнения или перибронхиальные утолшения) Легкие диффузно отечны, влажны, их поверхность покрыта муль- тифокальными петехиальными геморрагиями. Имеются участки ателектазов. Кишечник покрыт петехиальными геморрагиями, сли- зистая эрозирована. Лимфатические узлы слегка опухшие, на по- верхности разреза обычно влажные и выпуклые. При микроскопи- ческом исследовании легочной ткани признаки поражения легких в виде отека (мультифокального или сливного), охватывающего не- сколько долей легкого. Мультифокальная и ассиметричная эрозия эпителия бронхов с почти полной потерей эпителия бронхиол. Некоторые бронхиолы можно узнать только по их гладкомышеч- ной мускулатуре. Альвеолярные пространства мультифокально заполнены отечной жидкостью, фибрином, умеренным количест- вом больших пенистых альвеолярных макрофагов и эритроцитами. Маловыраженная острая интерстициальная пневмония, характе- ризующаяся умеренными нейтрофильными инфильтратами с не- большим количеством эозинофилов в пределах альвеолярных септ и альвеолярных пространств
Часть 2. Общая неправильная эпидемиология
* Подробно патология при разных способах введения поражающего агента описана в разд. 3.
* * Малоинформативно — имеется в виду применение данного способа диагностики для установления патологии мелкодисперсного аэро-
золя.
2.3. Основы эпидемиологического расследования искусственных...
549
в отдельных почтовых конвертах «скомковалась», и поэтому она вызвала только кож-
ные формы болезни у людей, подвергшихся экспонированию (см. разд. 1.12.4).
Как и в случае с антибиотико-резистентными штаммами возбудителей инфекци-
онных болезней, обнаружение больных с поражениями легких свидетельствует о ее
искусственном характере только в совокупности с другими признаками искусствен-
ного эпидемического процесса («эпидемиология точечного источника» и/или «невоз-
можная эпидемиология», «патология мелкодисперсного аэрозоля» и др.).
Более высокие показатели заболеваемости и смертности, нехарактерные для данной
болезни. Бол Ы1 <е имеют отношение к клинике инфекции и чувствительности к ней по-
пуляции, чем к ее эпидемиологии. Они зависят от выоранного преступниками возбу-
штеля инфекционной болезни, условий осуществления и цели террористического
акта или диверсии. Например, в результате преднамеренного заражения людей в Дал-
ласе в 1984 г и в лаборатопии медицинского центра в Техасе в 1996 г., погибших не было
(см. разд. 1.12.1 и 1.12.2 соответственно). Клинические симптомы возникшей болез-
ни могут быть забыты из-за ее длительного отсутствия в регионе (как это произошло
с Конго-Крымской лихорадкой, вспыхнувшей в 1999 г. в станице Обливской Ростов-
ской области). По этой причине никто из специалистов при появлении первых боль-
ных не сможет сказать, что характерно для нее, а ч го нет. Сегодня среди российских
специалистов очень мало тех, кто видел больных с натуральной оспой, лихорадками
Эбола и Марбург и даже с кожной формой сибирской язвы. Кроме того, многие, если
не большинство инфекционных болезней, вызываемых биологическими агентами, не
имеют специфических признаков и симптомов в первые часы их проявления.
Более низкие показатели заболеваемости у персонала, защищенного от экспонирова-
ния (например, у тех, кто находился внутри здания). Эти показатели можно уверенно
сопоставить только после окончания эпидемии, и когда станет известно направление
движения аэрозольного облака. Теракт может быть осуществлен и непосредственно в
здании (см. разд. 1.12.4).
Малораспространенные способы осуществления преступлений с использованием
биологических агентов. Как правило, это либо использование агентов, не рассматри-
вающихся в качестве потенциальных агентов БО, либо малораспространенные спо-
собы инфицирования людей. В январе 1999 г. некий В. Stewart был осужден в США за
умышленное инфицирование своего 11-месячного ребенка кровью больного СПИ-
Дом. Как он сам объяснил на суде — это преступление он совершил, чтобы не пла-
тить за содержание ребенка. Канадский аспирант-паразитолог Е. Kranz в 1970 г пы-
тался убить своих соседей по арендуемой квартире, заразив их пищу паразитом Ascaris
lumbris со ides (Atlas R. M., 1999). В Шотландии в декабре 2009 г. при введении инъек-
ции афганского героина заразились сибирской язвой 47 человек, из них погибло 13
(Powell A. et al., 2011; Ramsay С. N. et al., 2011). Технологическая цепочка получения
героина из опиума содержит режимы, исключающие выживание в конечном продукте
спор возбудителя сибирской язвы, попавших в исходное сырье. По данным Е. Е. Price
et al. (2012), штамм возбудителя сибирской язвы, вызвавший вспышку среди нарко-
манов — потребителей афганского героина, был т урецкого происхождения.
Объекты преступления при биологическом нападении. По аналогии с объектами
преступления химических террористов, классифицированных М. М. Авхименко с
соавт. (1999), выделим следующие:
1) случайный (неограниченный) набор лиц, т. е. толпа, поток людей или любой
случайный набор людей (попытки сектантов из «Аум-Синрикё» вызвать вспышки си-
бирской язвы в Токио в 1993 г.);
550
Часть 2. Общая неправильная эпидемиология
2) конкретная группа лиц, т. е. группа, по отношению к каждому члену которой
одновременно должно быть произведено летальное или инкапситирующее действие
биологического агента с одинаковым конечным эффектом (вспышка дизентерии
Шмидца среди сотрудников лаборатории медицинского центра в lexace в 1996 г.;
вспышка сальмонеллезного гастроэнтерита в Далласе в 1984 г.; попытки сектантов из
«Аум-Синрикё» вызвать массовое поражение людей ботулиническим токсином на
свадьбе японского кронпринца);
3) убийство конкретной персоны (убийство Р. Гейдриха ботулиническим токси-
ном; направление конвертов с рецептурой возбудителя сибирской язвы сенаторам
Thomas A. Daschle и Patrick J. Leahy).
Поданным J. Tuckera (1999), именно первые два объекта стали предпочтительной
мишенью террористов в конце 1990-х гг.
Модели территорий, на которых наиболее вероятно применение биологических аген-
тов для поражения людей. По аналогии с моделями территорий, наиболее опасных с
точки зрения осуществления химического теракта в условиях города, классифициро-
ванных М. М. Авхименко с соавт. (1999), выделим следующие:
1. «Комната» — замкнутое, плохо вентилируемое помещение объемом до 400 м3.
2. «Зал» — помещение, имеющее объем более 400 м3.
3. «Здание» — конструкция, отличающаяся от модели «зал» наличием вертикаль-
ных воздушных потоков и возможностью герметизации отдельных помещений.
4. «Туннель».
5. «Метро» — как сложная совокупность «залов» и «туннелей».
6. «Ландшафтный желоб» — пространство, протяженное в одном направлении
и ограниченное по краям таким образом, что воздухообмен через эти границы зат-
руднен.
7. Единичное транспортное средство.
Данные территории дают возможность моделировать мероприятия, которые не-
обходимо осуществлять при:
1) действии спасательных служб;
2) прогнозировании санитарных потерь;
3) разработке схем вероятных зон поражения конкретных мегаполисов;
4) планировании мероприятий по специфической и неспецифической профилак-
тике поражений и лечению лиц, подвергшихся воздействию биологического агента.
Психосоциальные реакции у людей, вызванные акцией с применением биологических
агентов. Терроризм — это форма психологической войны. В сознании людей с древ-
ности присутствует страх эпидемий. Уже сама мысль о возможности инфицирования
невидимыми и неосязаемыми возбудителями страшных инфекционных болезней
является пугающей. Люди не могут знать, прекратилось ли действие на них биологи-
ческих агентов, инфицированы они или нет, и будут ли они живы через несколько
часов. Поэтому информационная составляющая биологического террористического
акта планируется его организаторами заранее и осуществляется последовательно
«Конвертная атака» 2001 г. в США тому подтверждение. В качестве мишеней органи-
заторами теракта были выбраны редакции известных СМИ, политики и скандальные
журналисты, вроде Джудит Миллер.
Психосоциальные последствия вспышки инфекционной болезни на большую
популяцию людей были продемонстрированы в 1994 г. во время вспышки легочной
чумы в Сурате (Индия). Эта вспышка стала причиной бегства из Сурата 600 тыс. че-
ловек и последующей массовой паники в Дели, находящемся примерно в 600 км от
2.3. Основы эпидемиологического расследования искусственных...
551
него. Все больницы вДели оказались переполненными, «везде царил хаос, и балом
правили слухи». Одним из важных следствий этого страха и неопределенности стали
скупка и накопление запасов тетрациклина. Аналогичная ситуация сложилась и в
США в результате рассылки по почте сибиреязвенных спор в 2001 г., когда значительно
возросло использование населением ципрофлоксацина и в некоторых штатах стал
наблюдаться его дефицит (Dembek Z., 2005).
Этот же страх заставляет людей искать во всех доступных источниках информа-
цию о примененных биологических агентах и способах защиты от них. Свойства са-
мих агентов мистифицируются, «вес» приобретают мнения людей с паническими
реакциями. Появляются слухи о якобы большом количестве жертв, которые власти
скрывают. Находятся десятки очевидцев обработки города «специальной токсической
сывороткой» с вертолетов и т. п. SMS-сообщениями и блоггерами распространяется
ложная информация о заражении инфраструктуры города, например, водопровода.
Начинается бегство населения из «зараженных мест», паническая скупка воды в бу-
тылях и консервированных продуктов и т. п. (рис. 2.78)
Рис. 2.78. Бегство лондонцев из города при первых вспышках чумы 1665 г. Худож-
ник XVII в. воссоздает атмосферу ужаса лондонцев перед лицом стольких смертей,
невообразимого смешения живых и мертвых. Смерть продолжает преследовать бе-
гущих людей и распространяется вместе с ними (рисунок из книги Дефо Д., 1732).
Страх и массовая истерия привели к бегству сотен тысяч людей из индийского го-
рода Сурата в 1994 г., после того, как там вспыхнула эпидемия легочной чумы. Для
бегства населения из Саратова в декабре 2009 г. факт присутствия чумы в городе
вообще не потребовался. Хватило слухов, запущенных одним блоггером (!), о якобы
начавшейся в городе эпидемии легочной чумы
Когда биотеррористический акт станет достоянием СМИ, и о нем сообщат в но-
востях, отдельные лица как подвергавшиеся риску, так и не подвергавшиеся, могут
испытывать резкое возбуждение. Признаки и симптомы мускульного напряжения,
тахикардия, быстрое дыхание (возможна гипервентиляция), потение, тремор и дур-
ные предчувствия, вероятно, вынудят любого проявить заботу о своем здоровье. Эти
признаки и симптомы могут быть ошибочно связаны врачами с инфекцией или ин-
токсикацией. Лица, находящиеся в стрессовом состоянии и с такой симптоматикой,
создадут дополнительные трудности при сортировке пациентов во время начальной
стадии кризиса. По оценке Z. Dembek (2005), к каждому человеку, обращающемуся
за медицинской помощью в госпиталь, вследствие ранения или экспонирования к
биологическим или химическим агентам, будет добавляться, по меньшей мере, еще
552
Часть 2. Общая неправильная эпидемиология
6—10 лиц с выраженными психическими травмами. В период с 18 января по 28 фев-
раля 1991 г. 39 иракских ракет «Scud» было выпущено по Израилю. Большинство из них
цель не поразили или не сработали, некоторые достшли Тель-Авива и его окрестно-
стей. От самих ракет не погиб ни один человек. И все же пострадало около 1000 че-
ловек, среди которых двое погибли от сердечных приступов. Среди пострадавших
544 обратились за медицинской помощью с жалобами на симптомы, характерные для
поражения отравляющими веществами, хотя их применение не было зафиксирова-
но. Считая себя пораженными ФОВ, люди делали себе инъекции антидота (атропин)
по принципу «чем больше, тем лучше». Были госпитализированы 230 человек с сим-
птомами, характерными для передозировки атропина (Напу С. et al., 1997).
Существует возможность того, что медицинскую помощь в очаге эпидемии ока-
зывать буде с некому из-за бегства медицинских работников, опасающихся за свою
жизнь или жизнь своих близких. Поведение врачей на реальной эпидемии сильно
отличается от «киношного». Обычно героев среди них оказывается мало. Бывший
главный санитарный врач СССР П. Н. Бургасов (1915—2006), возглавлявший комис-
сию Минздрава СССР по установлению причин вспышки сибирской язвы в Сверд-
ловске в 1979 г., так описывал поведение местных врачей: «Сейчас многие врачи из ме-
дицинского персонала, работавшие в то время, выглядят такими смелыми, серьезны-
ми людьми, они говорят, что работали, когда мы приехали. Фактически большинство
врачей от страха и паники, а паника была страшная, потому что на их глазах умирали
люди без проявления какой-либо клиники, не выходили на работу. Их можно было
понять. Врачей в больнице было мало, потом они стали появляться. Всю работу по
лечению выполнял Владимир Николаевич Никифоров1 с моим сыном, а также рабо-
тал средний медперсонал, который был в этой больнице» (Бургасов П Н., 2001).
Имеющийся опыт проведения спасательных операций показывает, что наиболее
сильное травмирующее действие на психику медицинского персонала оказывают
случаи, связанные со смертью детей и сотрудников. Чрезвычайное происшествие
большого масштаба, вызванное применением биологических агентов, будет приводить
к множеству жертв именно среди этой категории населения. Определенные эмоцио-
нальные и психические проблемы у работников спасательных служб может вызвать
обращение с трупами. Во время ликвидации последствий биологического нападе-
ния они будуг сталкиваться с ситуацией, когда бо льшое количество жертв, с которы-
ми им приходится иметь дело, не связано с тем, что для них достаточно обычно, т. е. с
разрушениями и взрывами зданий, пожарами, авиационными катастрофами и т. п.
(Simon J. D.,1997; Напу С. et al., 1997).
Другой психологический аспект последствий масштабного биологического напа-
дения вызван тем, что ощущение неизбежной в ближайшем будущем смерти резко
снижает законопослушность граждан. По замечанию историка Фукидида, пережив-
шего чуму в Афинах в 430 г. до н. э.: «...ни страх перед богами, ни закон человеческий
не могли больше удержать людей от преступлений, так как они видели, что все поги-
бают одинаково и поэтому безразлично, почитать богов или нет С другой стороны,
никто не был уверен, что доживет до той поры, когда за преступления понесет нака-
зание по закону»
Наблюдения более поздних авторов ничем не отличаются от сделанных Фукиди-
дом. Внезапно развившаяся эпидемия с большим количеством смертельных исходов
среди населения всегда приводила к одним и тем же последствиям: трусость одних
1 В то время главный инфекционист РСФСР.
2.3. Основы эпидемиологического расследования искусственных...
553
людей сочеталась с цинизмом других — настоящих стервятников, уверенных из-за
отсутствия репрессивного аппарата в своей безнаказанности. Джованни Боккаччо,
очевидец эпидемии чумы 1348 г., пишет: «Позволено делать все, что заблагорассудит-
ся». По его наблюдениям, многие злодеяния совершались в Милане людьми специ-
альной службы, которые забирали трупы из лазаретов и домов или подбирали их на
улице и затем отвозили их месту захоронения. Они же сопровождали больных в лаза-
рет, сжигали вещи умерших или зараженных. Эти люди действовали без какого-либо
контро.гя, могли, например, за определенную мзду, оставить больного дома, и безна-
казанно грабили дома Во время чумы в Марселе в 1720 г. работу по уборке трупов
выполняли каторжники, на чей счет ходило много зловещих слухов. В домах они та-
щили все, что видели, чтобы не возвращаться по нескольку раз в дом, 1де были боль-
ные, они бросали в повозку и мертвых и умирающих. В городе появились ложные «чер-
ные вороны», они ездили по домам и в отсутствие хозяев грабили их (Делюмо Ж., 1994).
Такие же наблюдения есть и у Д. Дефо (1732), описавшего чуму 1665 г. в Лондоне. По
этим причинам трудно ожидать, что все наши граждане станут сотрудничать с влас-
тями после акции биотеррористического устрашения.
2.3.2. Эпидемиологическое расследование искусственного
эпидемического процесса
Подходы к эпидемиологическому расследованию. Текущее эпидемиологическое
расследование в очаге искусственного эпидемического процесса. Ретроспективное
эпидемиологическое обследование очага искусственного эпидемического процес-
са. Распознание поражения людей нанообъектами.
Эпидемиологическое расследование складывается из последовательного выпол-
нения трех этапов: сбор эпидемиологических данных; описательный этап и аналити-
ческий этап. Эпидемиологическое расследование искусственного эпидемического
процесса проводится с целью выявления всех эпидемических обстоятельств и послед-
ствий преступления, совершенного с помощью биологических агентов.
Подходы к эпидемиологическому расследованию. На основании накопленного опы-
та расследования преступлений, совершенных с использованием биологических аген-
тов (см. разд. 1.12), можно выделить два подхода к эпидемиологическому расследова-
нию искусственных эпидемических процессов:
текущее — начинается после выявления признаков искусственного эпидемиче-
ского процесса (эпидемиология «точечного источника», «неправильная эпидемио-
логия» и др.; см. разд. 2.3.2) на любом этапе эпидемиологического расследования
вспышки инфекционной болезни, считавшейся возникшей естественным путем. Эпи
демиологическое расследование проводится заново;
ретроспективное — проводится в отношении завершившейся эпидемии после того
как правоохранительным органам станет известна причастность к ней какой-либо
преступной организации.
Текущее эпидемиологическое расследование в очаге искусственного эпидемического
процесса. Опыт, наколенный в ходе расследования искусственных вспышек сальмо-
неллеза (1984), дизентерии Шмидца (1996) и сибирской язвы (2001), показывает, что
распознание искусственного характера вспышки возможно еще на этапе сбора эпи-
демиологических данных (см. разд. 1.12) После установления в эпидемическом процессе
особенностей искусственного эпидемического процесса, прежде всего, меняется по-
нимание «эпидемического очага». В контексте развития искусственных эпидемических
554
Часть 2. Общая неправильная эпидемиология
процессов (поражений) — это территория, на которой произошло применение (пред-
намеренное распространение) биологического агента, способного вызывать инфек-
ционный процесс или другие поражения у людей. Протяженность такого очага, его
границы определяются замыслом преступления; техническими возможностями пре-
ступников; выбранным ими объектом преступления; моделью территории, на кото-
рой совершено биологическое нападение; метеоусловиями в момент применения
биологического агента и др. подобными факторами.
Далее вносятся коррективы в уже проведенный сбор эпидемиологических данных.
Учитывая то обстоятельство, что деятельность преступников еще не пресечена, необ-
ходимо выделить случаи искусственного эпидемического процесса. То есть в стандарт-
ное клиническое определение случая болезни внести дополнения, отражающие все ва-
рианты течения инфекционного процесса (поражения), зависящие от особенностей
применения данного биологического агента, включая еще не обнаруженные клини-
ческие случаи. Например, при расследовании биотеррористического акта в США в
2001 г. вспышки кожной и ингаляционной форм сибирской язвы с неправильной
эпидемиологией, наблюдаемые ь разных штатах, были объединены в одно эпидеми-
ологическое расследование (см. табл. 1.11). Впоследствии установлено, что они вызва-
ны одной и той же рецептурой, но имеющей разный агрегатный состав (более подроб-
но об определении клинических случаев для нозологических форм см. в разд. 1.12.4).
Для сбора данных об обстоятельствах, приведших к поражению биологическими
агентами, проводится анализ научной литературы на предмет описания сходных слу-
чаев и построения аналогий: анализируются истории болезни в стационарах и поли-
клиниках, поднимаются протоколы патологоанатомических вскрытий. По ним вы-
являются больные (умершие), которые могли иметь клинику болезни, подходящую
под описание случая искусственного эпидемического процесса; проводятся опросы
(анкетирование) пораженных людей; выполняются медицинские осмотры лиц, на-
ходившихся на территории эпидемического очага и пока не имеющих клинических
признаков болезни.
Вопросы анкеты формулируются в зависимости от особенностей вспышки (см.
разд. 1.12.1). При опросе заболевших (либо родственников погибших), надо выяснить,
где заболевшие (погибшие) были в период времени, соответствующий среднему ин-
кубационному периоду болезни, особенно выявляя те места, где состояние атмосфе-
ры или окружающей обстановки могли быть благоприятными для поражения аэро-
золем. Составить почасовую раскладку территориальных перемещений пациента в
течение нескольких дней за период не менее 2/3 среднего инкубационного периода
болезни (объем проводимой работы будет зависеть от количества пациентов и ресур-
сов эпидемической службы). Нанести эту информацию на карту по улицам и здани-
ям по каждому заболевшему (умершему) в результате применения биологического
агента. При выявлении территориальных совпадений в перемещении больных и по-
гибших, исследовать участки их пересечения с максимально возможной точностью.
Выполняются лабораторные исследования материала от пораженных людей, жи-
вотных и объектов внешней среды и др. мест предполагаемого применения биологи-
ческого агента. При сборе информации об искусственном эпидемическом процессе
надо иметь в виду, что в отсутствие эпидемической настороженности врачами могут
быть поставлены другие диагнозы, не противоречащие наблюдаемой ими клинике
болезни. Например, больные с бубонной формой чумы или туляремии могут регист-
рироваться как больные с неспецифическими лимфаденитами и лечиться в хирурги-
ческих стационарах. Больные с кожной сибирской язвой пройти под диагнозом «дер-
2.3. Основы эпидемиолигическогорасследования искусственных...
555
матит», больные с кишечной сибирской язвой остаться нераспознанными под диаг-
нозом острая токсикоинфекция. Больные с легочной формой туляремии — как ОРЗ;
очаговая, сегментарная или долевая пневмония.
На описательном wane эпидемиологического расследования искусственного эпиде-
мического процесса выполняется исследование интенсивности, динамики, простран-
ственной характеристики и структуры заболеваемости в очагах. Сопоставляются дан-
ные морфологического, иммунологического, биохимического и генетического иссле-
дования штаммов микроорганизмов, вы целенных в очагах вспышек (от больных и
погибших; из окружающей среды в местах экспонирования людей к биологическому
агенту; находящихся на остатках устройств, с помощью которых они были примене-
ны; обнаруженных в лаборатории преступников). Данные по интенсивности эпиде-
мического процесса на определенных территориях (в коллективах) дают возможность
эпидемиологам установить масштаб вспышки, их количество и возможные объекты
преступления. Данные по динамике эпидемического процесса (распреде гение абсо-
лютных чисел или частотных показателей во времени) позволяют установить крат-
ность применения биологического агента (см. рис. 1.53). Пространственная характе-
ристика эпидемического процесса позволяет получить представление о модели тер-
ритории, на которой произошло применение биологического агента; и о способе
рассеивания биологического агента. Структура заболеваемости дает представление
об объекте преступления (если распределение частотных показателей проводится в
отношении каких-то групп населения); о качестве использованной биологической
рецептуры (распределение частотных показателей проводится в отношении клини-
ческих форм болезни — см. случаи ингаляционной и кожной форм сибирской язвы в
США в 2001 г.); о расстоянии жертв биологического нападения от источника аэрозо-
ля (распределение частотных показателей проводится в отношении клинических форм
болезни, вызванных крупнодисперсными и мелкодисперсными аэрозолями). Уста-
новление причастности одних и тех же штаммов во >будителя инфекционной болез-
ни к разным вспышкам и обнаружение их в лаборатории преступников позволяет
получить полную картину путей распространения возбудителя инфекционной болез-
ни в искусственном эпидемическом процессе.
На аналитическом этапе эпидемиологического расследования искусственного эпи-
демического процесса анали шруются технические аспекты биологического нападе-
ния. По результатам этого анализа выдвигается предположение о техническом устрой-
стве или способе, использованном для осуществления биологического нападения. По
уровню осуществления биологического нападения, клинике, патанатомии, эпидемио-
логии и др. данным, полученным при эпидемиологическом расследовании в очаге
искусственного эпидемического процесса, составляется описание причастной к пре-
ступлению организации с указанием уровня образования ее членов, их профессио-
нальных навыков и потенциальных возможностей; строится модель биотерр^ристи-
ческого акта (диверсии).
Ретроспективное эпидемиологическое обследование очага искусственного эпидеми-
ческого процесса. Типичным примером такого расследования можно считать рассле-
дование инцидента с возбудителем сибирской язвы в районе Kameido Токио в 1993 г.
(см. разд. 1.12.3). Проведение такого расследования предполагает те же этапы, что и
при текущем расследовании искусственного эпидемического процесса. Однако у эпи-
демиологов может не оказаться образцов, взятых с объектов окружающей среды, чи-
стой культуры возбудителя инфекционной болезни и большого количес гва информа-
ции об эпидемии от непосредственно вовлеченных в нее лиц.
556
Часть 2. Общая неправильная эпидемиология
Распознание поражения людей нанообъектами. Грань между наночастицами и ис-
кусственными генетическими конструкциями (см. разд. 2.2.4) становится все более
условной. Игнорировать угрозу умышленного применения и тех, и других с целью
депопуляции населения России и подрыва генофонда населяющих ее народов, не
создавая долговременной и прочной научно-методической оазы для противодействия,
означает создание оптимальных условий для деятельности престу пников. Самым труд-
ным в распознании таких поражений будет определение случая поражения, т. е. форми-
рование стандартного клинического, паталогоанаюмического и патоморфологичес-
кого описания случая болезни, вызванного определенным типом нанообъекта.
Сделать предположение о поражении нанообъектами (среди других причин) отдель-
ных лиц или групп населения можно на основе сопоставления их анамнеза и клини-
ческих проявлений поражения. Например, развитие однотипных онкологических
процессов у объединенных общей идеей (врагом) политических деятелей, или разви-
тие медленно прогрессирующей легочной патологии у рабочих, занятых на одном
производстве. Однако для установления фактора нанообъекта в развитии патологии
пациента неооходимо: 1) тля наночастиц известных веществ, полученных дезагреги-
рованием или конденсацией — обнаружение этих наночастиц в пораженных тканях;
2) для доставленных в клетки-мишени векторных конструкции — обнаружение век-
торных молекул вместе с трансгенами и с соответствующими регионами транскрип-
ции и инициации мРНК.
Первый подход хорошо иллюстрируется приведенным в разд. 2.2.5 описанием
группового поражения людей наночастицами двуокиси кремни, где в установление
связи между патоморфологией и наличием наночастиц основную роль сыграла транс-
миссионная электронная микроскопия, а в установлении химической природы нано-
частиц — метод EDX. Второй подход иллюстрирован примерами с поражениями лю-
дей векторными конструкциями на основе ретровируса мышиной лейкемии Молони
и человеческого аденовируса 5 типа (см. разд. 2.2.4). И в том и в другом случае в тка-
нях-мишенях жертв этих экспериментов были обнаружены векторные конструкции
с клонированными в них трансгенами и регуляторными элементами. Случай с разви-
тием острого лейкоза у детей показателен для разработки подходов к выявлению таких
поражений. Для гена LM02ранее было показано участие в развитии острого лейкоза,
когда в основе механизма его активизации в Т-клетках лежат спонтанные перестрой-
ки (транслокации) хромосомы 11. Однако в случае искусственно вызванной у детей
лейкемии было обнаружено, что этот ген в Т-клетках находится под контролем LTR1
вируса мышиной лейкемии Молони, в геноме человека никогда не встречавшегося.
Для сбора данных об обстоятельствах, приведших к поражению нанообьектами и
масштабах поражения, проводится анализ научной литературы на предмет описания
сходных случаев и выстраиваются аналогии. Также анализируются истории болезни
в стационарах и поликлиниках, поднимаются протоколы патологоанатомических
вскрытий, собираются и исследуются соответствующими методами образцы тканей
пациентов, подпадающих под описание данного случая поражения нанообьектами.
Проводятся опросы (анкетирование) установленных пораженных и их лечащих вра-
чей; выполняются медицинские осмотры лиц, находившихся на территории, где про-
изошло поражение, или взаимодействующих с факторами, которые могут привести к
такому поражению, но пока не имеющих клинических признаков болезни.
1 Область U3 каждого LTR несет промотор, запускающий экспрессию, находящегося вбли-
зи гена.
2.4. Обнаружение биотеррористической организации
557
* * *
Воспроизвести естественный эпидемический процесс невозможно. Диагностика
искусственного поражения основывается на сопоставлении эпидемиологии, клини-
ки и патоморфологии, характерных для случаев болезни, являющихся результатом
естественного инфицирования (отравления), с теми, которые могут наблюдаться в
результате преднамеренного введения (ингаляционно, энтерально и парентерально)
в организм человека микроорганизма или токсина. Поэтому в основе эпидемиологи-
ческого расследования искусственных эпидемических процессов и поражений био-
логическими токсинами должны находиться знания о характерных особенностях их
эпидемиологии и о клинических и патологоанатомических особенностях искусствен-
ных биологических поражений, определяющих случаи искусственного эпидемического
процесса. Ключевым для обнаружения искусственного эпидемического процесса мож-
но считать выявление в общей эпидемической картине признаков «невозможной
эпидемиологии» болезни. В зависимости от кратности применения преступниками
биологического агента, в пользу версии об искусственном происхождении эпидемии
будут свидетельствовать сжатая эпидемическая кривая с пиком заболеваемости, «ук-
ладывающимся» в средние значения инкубационного периода болезни (эпидемиоло-
гия «точечного источника») и обнаружение эпидемического очага, локализованного
с подветренной стороны от возможной точки атаки. Многократное экспонирование
к агенту проявляет себя растянутой эпидемической кривой без четкой локализации
очагов. О высоком уровне подготовки и исполнения преступления свидетельствуют:
обнаружение у жертв патологии мелкодисперсного аэрозоля обнаружение биологи-
ческих агентов, измененных с помощью методов генной инженерии или тех их штам-
мов, про которые известно, что они исследуются как потенциальные агенты БО; и
установление факта применения нанообъектов. Р аспознание искусственного проис-
хождения эпидемии возможно еще на этапе сбора эпидемиологических данных.
2.4. ОБНАРУЖЕНИЕ БИОТЕРРОРИСТИЧЕСКОЙ ОРГАНИЗАЦИИ
Подготовка биотеррористического акта начинается с создания преступниками
особой криминальной структуры — биотеррористической организации.
2.4.1. Биотеррористическая организация (сообщество)
Убеждения и мотивы биотеррористов. Типовая структура биотеррористической орга-
низации. Характерные признаки биотеррористической организации. Характеристика
членов биотеррористической организации.
Типовой для таких организаций на сегодняшний день можно считать «Аум-Син-
рикё» («Учение истины Аум») — террористическую организацию, добившуюся успе-
хов в производстве биологических и химических поражающих агентов и распростра-
нившую свою деятельность на многие страны мира (Австралия, США, Украина,
Германия, Югославия), в том числе и на Россию.
Убеждения и мотивы биотеррористов. Судя по приведенным в табл. 1.9 данным, у
лиц, использующих биологические агенты для поражения людей, могут иметься раз-
ные убеждения — от религиозных до политических; от левых до правых, и даже зеле-
ных; от мести всему миру до сведения счетов с отдельными лицами. Во второй поло-
вине 1990-х гг. наметилась тенденция к переориентации их совершения от леворади-
558
Часть 2. Общая неправильная эпидемиология
кальных к сепаратистско-националистическим группам или отдельным личностям,
а также к специальным группам, ь основе действий которых лежит месть. Кроме того,
заметен рост числа инцидентов с участием сект и культов, верящих в апокалиптичес-
кие предсказания. Однако анализ, сделанный J. Tucker, охватывает деятельность орга-
низаций и отдельных лиц, занимавшихся биотеррористическими акциями на «люби-
тельском уровне». Первый биотеррористический акт «профессионального исполне-
ния» (см. разд. 1.12.4) больше похож на крупномасштабную провокацию, имеющую
своей целью втягивание США в войну с Ираком и Афганистаном, чем на реализацию
каких-то бредовых замыслов идейных террористов.
Когда преступление реализуется в форме биологического террористического акта,
мотивы террористов, предопределяют ряд характерных особенностей таких акций:
масштабность и нередко бредовость замысла преступления («демонстрация неспо-
собности властей», «истребить большинство людей и создать расу господ», «реализа-
ция пророческого предсказания» и т. п.);
о таких терактах предварительно не объявляют и за них не берут на себя ответст-
венность;
беспредельная жестокость замысла преступления, стремление уничтожить как
можно больше людей, направленность исключительно на гражданское население;
выбор в качестве объекта преступления известных политиков, журналистов, ре-
дакций СМИ, федеральных органов власти;
тенденции к эскалации террористических действий во времени и к использова-
нию новых поражающих агентов или видов оружия и тактических приемов
Масштабное резонансное преступление, например, когда его объектом предпо-
лагаются известные политики, либо случайный набор людей по принципу «чем боль-
ше, тем лучше», могут готовить преступники, чьи мотивы носят иррациональный
характер. Это, с одной стороны, затрудняет обнаружение таких лиц еще до соверше-
ния преступления, с другой — делает их удобным объектом для манипуляций «третьей
силой», решившей в рамках непрямой стратегии осуществить биологическую дивер-
сию или террористический акт «чужими руками».
Типовая структура биотеррористической организации. Секта «Аум-Синрикё» осно-
вана Сёко Асахарой в 1987 г. IShoko Asahara; его настоящее имя — Тидзуо Мацумото,
родился 2 марта 1955 г.), бывшим иглоукалывателем, хозяином нескольких рестора-
нов дешевой китайской кухни в Японии. В русском переводе «Асахара» означает «Си-
яющий свет в долине конопли». После женитьбы Асахара работал в китайской аптеке
города Фунабаши префектуры Чиба. За мошенничество при выписке счетов на сум-
му 6,7 млн иен привлекался к судебной ответственности. В 1981 г. открыл свою апте-
ку в этом же городе и организовал продажу фальшивых лекарств, предлагая лечение
страдающим ревматизмом людям старше 60 лет. На продаже фальшивых лекарств
«заработал» около 40 млн иен. Был арестован и по суду оштрафован на 20 тыс. иен.
Есть много неясного в том, как почти слепой Асахара, судимый за мошенничество
и не имевший почти никакого образования, кроме школы для слабовидящих детей,
смог создать организацию, легко инфильтрировавшуюся во атастные круги многих
стран, включая и Россию. Скорее всего, реальных кукловодов во всей этой истории с
сектой мы не узнаем. В России секта объявилась сразу после расчленения СССР на
прозападные государства-лимитрофы. Российский филиал «Аум-Синрикё» был заре-
гистрирован и начал работу в начале 1992 г. Он не имел проблем с деньгами. Просле-
дить источники финансирования таких организаций, как «Аум-Синрикё», очень
сложно. Нити финансирования обычно обрываются в другой подобной организации.
2.4. Обнаружение биотеррористической организации
559
Про «Аум-Синрикё» известно, что ее финансировала влиятельная на Корейском по-
луострове и опасная секта «Тоицу Кекай» (название в переводе звучит, как «Общество
за еди нство»), уже действовавшая на территории России, и особенно активно — на Са-
халине. Более половины недвижимости «Аум-Синрикё» в японской столице (здания
и земельные участки) перешли к этой организации на неясных условиях от «Тоицу
Кекай». В 1993—1994 гг. «Аум-Синрикё» была в России самой широко рекламируемой
религиозной сектой. В первые несколько месяцев своей деятельности секта заплати-
ла около миллиона долларов на рекламу, обеспечив себе ежедневную часовую програм-
му на радиостанции «Маяк» и получасовую еженедельную передачу на канале «2x2».
Громадные средства были потрачены сектой на рекламу в печати, миллионными ти-
ражами издавались цветные листовки и плакаты, рекламные объявления транслиро-
вались по всей радиосистеме Московского метрополитена. До 1995 г. только в Моск-
ве секта имела 6 центров. Постоянно действовали киоски «Аум-Синрикё» внутри
станции метро «Библиотека им. Ленина», «Тургеневская», «Октябрьская», «Китай-го-
род». Много влиятельных российских политиков того времени «засветились» своим
избирательным покровительством этой секте, хотя, возможно, все было наоборот —
это секта им покровительствовала.
Секта была построена по строго централизованному принципу. В ее структуре
реализуется абсолютное подчинение нижестоящих адептов вышестоящим. Практи-
ковались глубокая конспирация, режим секретности внутри культа и тотальная слежка
среди адептов. Все происходящее с адептами культа учитывалось и фиксировалось.
Каждый новый адепт культа заносился в компьютерный каталог, ему присваивался
личный номер и, после инициации, новое имя. Анкета поступающего в секту росси-
янина напоминала анкету поступающего в подпольную диверсионную группу. В ней
требовалось указать наличие водительских прав, область интересов (перечень из двад-
цати четырех подпунктов, включая мореплавание, сельское хозяйство и металлове-
дение), группу крови, а также куда можно звонить открыто, куда можно звонить, не
называя себя, и куда можно звонить в случае острой необходимости. Управление сек-
той в Москве было хорошо организовано. Москву сектанты поделили на зоны, те, в
свою очередь, на районы. Во главе каждого подразделения стояло ответственное лицо.
Руководящие посты занимали в основном японцы, имелось несколько русских, но
больше азербайджанцев.
Адепты секты «Аум-Синрикё» во многих документах именовали себя «буддийс-
кой общиной». Асахара часто путешествовал за границу и знакомился с разными из-
вестными учителями йоги и буддийскими религиозными представителями, такими
как Далай-лама XIV, Калу Ринпоче (лама тибетской школы Кагью) и Кхамтрул Рин-
поче (бывший генеральный секретарь Совета по культурным и религиозным делам
правительства Тибета в изгнании).
С Далай-ламой XIV Асахару связывали очень близкие дружественные отношения.
До событий в токийском метро, они, начиная с 1987 г., встречались 5 раз и находи-
лись в постоянной переписке. Во время первой встречи Далай-лама обратился к Сё ко
Асахаре со следующими словами: «Дорогой друг, буддизм в Японии пришел в упадок,
если так будет и впредь, то буддизм исчезнет с земли Японии. Ты обязан распростра-
нять настоящий буддизм на твоей родине, ты — самый подходящий человек, посколь-
ку ты понимаешь мысль Будды. Я рад, что именно ты несешь ответственность за эту
работу, так как тем самым ты помогаешь мне».
В 1989 г. Асахара подарил Далай-ламе XIV 100 тыс. долларов, получив взамен от
него диплом и рекомендательные письма японскому правительству. Благодаря чему
560
Часть 2. Общая неправильная эпидемиология
«Аум-Синрикё» была признана в Японии. Деятельность последователей «Аум», наце-
ленная на популяризацию буддийских текстов была также отмечена правительства-
ми Шри-Ланки, Бутана и Тибетского правительства в изгнании, расположенного в ин-
дийском городе Джарамсал.
Однако традиционный российских буддизм в лице геше Джампа Тинлея, духов-
ного представителя Далай-ламы в России, Калмыкии, Бурятии и Монголии, не одоб-
рил деятельности секты на территории России и не считал ее «буддийской». В опре-
делении Архиерейского собора русской православной церкви «О псевдохристианских
сектах, неоязычестве и оккультизме» (декабрь 1994 г.) «Аум-Синрикё» обозначена как
псевдорелигиозная организация.
Асахара не мелочился в планах на свое политическое будущее. Он ставил перед
собой задачу захватить власть сначала в Японии, а затем во всем мире. Поэтому струк-
тура организации включала «министерства» и «секретариат», которые должны были
стать основой будущего государства. В 1989 г. японские власти официально зарегист-
рировали секту, в 1990 г. она приняла участие в местных выборах, затем в парламент-
ских. В секту привлекались состоятельные люди, жертвовавшие значительные сред-
ства, но сами они, как правило, оставались на периферии организации. Ядро «Аум»
составляли молодые ученые, организаторы и специалисты, которых привлек Асаха-
ра. Вот краткие биографии сектантов, участвовавших в зариновой атаке на токийское
метро 20 марта 1995 г.:
Икуо Хаяси (1947 г. р.). X. Мураками (2006) охарактеризовал его как безупречно-
го выходца из элиты японского общества. До вступления в «Аум-Синрикё» Хаяси
был доктором в японском Министерстве науки и техники. Хаяси учился в Универси-
тете Кейо и позже работал в госпитале Кейо. В 1990 г. он ушел с работы, чтобы присо-
единиться к секте «Аум-Синрикё» и стать «министром исцеления» Асахары. Был от-
ветственен за некоторые «лечения» членов секты, которые подозревались в нелояль-
ности. «Лечения» проводились электрошоком и пентоталом натрия^ заканчивались
смертью «пациентов». Распылял зарин на станция Син-Отяно-Мидзу. В результате его
действий погибло два человека, 231 человек получил отравления. Приговорен к по-
жизненному заключению.
Кэнъити Хиросэ (1966 г. р.). Окончил университет Васэда, специализировался на
прикладной физике. В организации был причислен к Министерству науки и техни-
ки, считался главным специалистом в области химии. Несмотря на сильные колеба-
ния, 20 марта проткнул оба пакета с зарином на станции Отяно-Мидзу. В результате
погиб один человек, 358 получили отравления различной степени тяжести. Пригово-
рен к смертной казни через повешение.
Тору Тоёда (1968 г. р.). Учился в Токийском университете, специализировался по
прикладной физике, добился выдающихся успехов. Защитил кандидатскую диссер-
тацию в элитной лаборатории. В секте принадлежал к Министерству науки и техни-
ки и действовал в бригаде химиков. Отвечал за производство зарина. Несмотря на то,
что он полностью представлял последствия зариновой атаки, не мог себе позволить
1 Пентотал натрия (тиопентал натрия) — средство для неингаляционной общей анестезии
ультракороткого действия. Представляет собой смесь натриевой соли (К8)-5-(1-метилбутил)-
5-этил-2-тиобарбитуровой кислоты с безводным натрия карбонатом. В США передозировка
тиопентала натрия в некоторых штатах используется для казни через смертельную инъекцию.
Спецслужбы и преступные организации применяют его как «сыворотку правды», т. е. для по-
лучения скрываемых человеком сведений.
2.4. Обнаружение биотеррористической организации
561
пойти вопреки приказу Асахары. Проткнул пакеты между станциями Нака-Мэгуро и
Эбису. Погиб один человек. 532 получили отравления. Приговорен к смертной казни
через повешение.
Масато Ёкояма (1965 г. р.). Окончил отделение прикладной физики политехни-
ческого факультета университета Токай. Один из заместителей начальника Министер-
ства науки секты. Проткнул пакеты на подъезде к станции Ёцуя. Около 200 человек
получили отравления. Приговорен к смертной казни через повешение.
Ясуо Хаяси (1957 г. р.). Хаяси родился в Токио, окончив токийскую вечернюю
школу, поступил в институт Когакуин, где изучал искусственный интеллект. После
выпуска на постоянную работу не устроился, много путешествовал. В Индии увлекся
религией. Вскоре после начала занятий йогой повстречался с вербовщиком «Аум-
Синрикё», вступил в секту, ушел из дома. Занимал третье место по рангу в Министер-
стве науки и техники. На подъезде к станции Акихабара многократно проткнул все
три взятых пакета. Скончались 8 человек, отравления получили 2475 человек. Ясуо
Хаяси за большое количество жертв в японских СМИ прозвали «машиной убийств».
Приговорен к смертной казни через повешение.
Для легализации «научной» деятельности «Аум-Синрикё» в Японии был создан
научный центр, специалисты которого якобы проводили многочисленные исследо-
вания и на них обосновывали уникальные способности, появляющиеся у людей в
результате занятий духовной практикой. Асахара прикрывал свою деятельность по
разработке оружия массового уничтожения утверждениями, что его ученики облада-
ют сверхъестественными способностями, такими как возможность видеть чакры и
энергетические состояния людей, читать мысли и др.
Сначала деятельность секты, как, впрочем, и ее создателя, носила обычный для
тоталитарных сект криминальный характер (целительство, присвоение имущества
сектантов, вымогательство и т. п.). С конца 1980-х гг. секта обозначила себя более
тяжкими преступлениями, организовав убийство члена группы, пытавшегося из нее
уйти. Тогда же (в октябре 1989 г.) были провалены переговоры секты с Цуцуо Сакамо-
то — юристом, борющимся с японскими сектами и грозившим судебным процессом
против «Аум-Синрикё», который мог обанкротить секту. В октябре 1989 г. Сакамото
создал «Ассоциацию жертв «Аум-Синрикё». В том же месяце было записано интер-
вью с Сакамото в телевизионном шоу на японской телестанции «TBS». Но секта по-
лучила запись этого интервью раньше, чем оно было показано по телевидению, и
оказала давление на «TBS», чтобы отменить показ интервью Сакамото. Он, его жена
и ребенок были похищены сектантами из их дома в Иокогаме. И хотя некоторые из
коллег Сакамото публично высказывали свои подозрения о причастности «Аум-Син-
рикё» к этому преступлению, полиция проявляла странную беспомощность в его рас-
следовании. Только в 1995 г. стало известно, что Сакамото и его семья были убиты, а
их тела уничтожены членами секты. Секта считается причастной к убийствам еще не-
скольких лиц, критиковавших секту. Среди них главы других буддийских сект Сока
Гаккай и Института исследования человеческого счастья (Кофуку Кагаку). Секта при-
частна к попытке убийства художника комиксов Ёсинори Кобаяси в 1993 г. и еще к
ряду подобных преступлений.
Антигосударственный характер деятельность секта приобрела после 1990 г., ког-
да Асахара и 24 других ее члена безуспешно пытались баллотироваться в палату пред-
ставителей парламента Японии. В 1992 г. «министр толкования» «Аум-Синрикё» Кио-
хиде Хаякава опубликовал трактат «Законы гражданской утопии», где объявил войну
японской конституции и гражданским институтам. В то же самое время Хаякава начал
562
Часть 2. Общая неправильная эпидемиология
осуществлять «дружественные визиты» в разваливаемую извне и изнутри Россию,
чтобы в обстановке нарастающих деструктивных процессов обзавестись различными
изделиями военного назначения, включая автоматы АК-47 и военно-транспортные
вертолеты Ми-17. Так же он пытался приобрести компоненты для ядерной бомбы.
Будучи введенными в заблуждение информационной кампанией западных СМИ
против Ирака и России, обвиняющей эти страны в нарушении Конвенции 1972 г., и
тиражируемым западными политиками мифом о доступности БО для стран Третьего
мира и террористов, сектанты пытались создать его «по книжкам». БО стало первым
приоритетом в их программе разработки оружия массового поражения. Ботулиниче-
ский токсин привлек внимание Асахары и его «молодых гениальных ученых» таблич-
ными данными, свидетельствующими о его самой высокой токсичности среди изве-
стных токсических веществ (см. табл. 2.12).
В 1995 г., уже после атаки секты на токийское метро, полиция обнаружила в цент-
ре «Аум-Синрикё» почти 500 томов книг по биохимии и биотехнологии, в том числе
книги по культивированию бактерий. Найдены были также культиваторы, запасы пи-
тательной среды для выращивания ботулинического микроба и многое другое из того,
что требовалось для получения ботулинического токсина. В апреле 1990 г. сектанты
распыли ди ботулинический токсин (или то, что ими считалось ботулиническим ток-
сином) вокруг здания японского парламента. Для осуществления террористического
акта секта оборудовала автомобиль системой для рассеивания ботулинического ток-
сина. В июне 1993 г. сектанты пытались расстроить свадьбу японского кронпринца
путем диспергирования ботулинического токсина с помощью той же автомобильной
системы (Atlas R. М., 1999).
Однако результаты этой и последующих подобных попыток разочаровали сектан-
тов. Тогда они стали искать возможности для поражения людей с помощью других
биологических агентов. В октябре 1992 г. Асахара с сорока своими учениками ездил в
Заир с официально заявленной целью «исцелять людей, пораженных вирусом лихо-
радки Эбола». На самом деле им нужен был штамм этого опасного вируса. «Досдали»
они его или нет, не известно. «Клюнув» на миф о выбросе возбудителя сибирской язвы
в Свердловске в 1979 г., Асахара пытался организовать такой же «выброс» в восточном
районе Токио в июне 1993 г., небезуспешно (см. разд. 1.12.3). А еще раньше, 15марта,
члены секты последний раз попытались использовать ботулинический токсин для
массового убийства лютей. Они оставили в метро в Токио три портфеля с целью рас-
пространения там ботулинического токсина. «Результата», в их понимании, не полу-
чилось (Atlas R. М., 1999). После этих привалов деятельность «Аум-Синрикё» повер-
нулась в сторону боевых ОВ. Осталось только найти технологии для их производства.
И они быстро «нашлись», на распродажу' тогда была выставлена одна шестая часть
суши. Американское агентство «Ассошиэйтед пресс» сообщило, что во время суда над
Асахарой и исполнителями террористического акта в токийском метро, член «Аум-
Синрикё» Йосихиро Иноуэ заявил в суде, что чертежи установки по производству
зарина были куплены сектой у одного бывшего вице-премьера правительства России
за 79 тыс. долларов. Чертежи доставлены в Японию в 1993 г. Иноуэ подчеркнул, что
без помощи из России «Аум-Синрикё» не смогла бы получить зарин. Так это или нет,
официальных подтверждений не поступало. Достоверно известно только, что с кон-
ца 1993 г. секта начала тайно производить ОВ непвнопаралитического действия зарин
и затем вещество «VX».
«Ученые» из «Аум-Синрикё» тестировали зарин на овцах в отдаленных пастушес-
ких владениях в Западной Австралии, убив 29 овец; и на собственных адептах. На их
2.4. Обнаружение биотеррористической организации
563
химическом мини-заводе, расположенном в 100 км от Токио, зарин производился ку-
старно, с нарушениями технологического процесса и мер безопасности, что не раз
приводило к человеческим жертвам. По свидетельству ряда приверженцев Асахары,
на территории «поместья» по ночам происходили тайные захоронения, без ведома
властей. Возможно, это были жертвы производства или испытания зарина. Недостатка
в «материале» для проведения испытаний на людях не было: в этом центре «Аум-Син-
рикё» проживало иногда до 900 человек.
И зарин, и вещество «VX» были использованы сектантами в нескольких убийст-
вах и попытках убийств в 1994-1995 гг. Наиболее заметное произошло ночью 27 июня
1994 г. в городе Мацумото, когда секта применила ОВ внутри офисного здания. От-
равления зарином получили около 200 человек, 4 из них погибли. Расследование те-
ракта пошло по «ложному следу» (Olson К. В., 1999).
Зариновая атака в токийском метро 20 марта 1995 г. осуществлена сектантами
следующим образом. Непосредственно в атаке участвовали пять пар сектантов, состо-
ящих из исполнителя и водителя. Основной обязанностью исполнителя было войти
в вагон метро, проехать определенное количество станций, опустить пластиковые
пакеты с зарином на пол вагона и перед выходом проткнуть их специально заточен-
ным концом зонта. Пакеты были завернуты в газету. Водители ждали на указанной
станции и должны были увезти исполнителей в безопасное место. Каждый исполни-
тель теракта имел при себе два пакета с зарином емкостью 1 л, за исключением Ясуо
Хаяси, у которого было 3 пакета. Никто из исполнителей теракта не отклонился от на-
меченного плана. Детали теракта и судьбы 60 выживших его жертв описаны X. Мура-
ками (2006).
В марте 1995 г. против «Аум-Синрикё» по решению Генеральной прокуратуры Рос-
сийской Федерации было возбуждено уголовное дело, и 15 марта 1995 г. Останкинский
межмуниципальный суд Москвы вынес решение наложить арест на имущество этой
организации. С 18 апреля 1995 г. российские последователи Асахары перешли на не-
легальное положение. При этом подавляющее большинство саманов (монахов)
остались верны секте и не вернулись домой, к семьям, по-прежнему проживая наявоч-
ных квартирах, ревностно выполняя аумовские ритуалы и практически бесплатно
работая на свое руководство. Уже после запрета «Аум-Синрикё», по сообщению Ко-
митета по спасению молодежи от псевдорелигий, среди ее адептов прокатилась вол-
на непонятных смертей, когда скоропостижно умирали мужчины 28-36 лет, и стран-
ных самоубийств. Например, с балкона выбросился 12-летний мальчик, вовлеченный
в «Аум-Синрикё» своей матерью.
Японская полиция арестовала более 200 сектантов, но секта не уходила просто так
и из Японии. 22 марта 1995 г. в одном из зданий «Аум-Синрикё» полиция обнаружила
пятьдесят человек, находившихся в состоянии комы. В конце марта совершено по-
кушение на начальника Главного полицейского управления Кодзи Кунимацу, возглав-
лявшего комиссию по расследованию террористического акта в токийском метро.
Неизвестный выпустил в него несколько пуль и скрылся. Спланировавший химиче-
скую атаку в метро глава научно-технического бюро секты Мураи был убит в апреле
1995 г. ультраправым экстремистом за несколько минут до ареста. В декабре 1995 г. по
решению суда был наложен арест на все банковские счета и недвижимость этой сек-
ты, а весной 1996 г. «Аум-Синрикё» объявлена банкротом. Штаб-квартира «Аум-Син-
рикё» в Токио была закрыта, имущество подлежало распродаже, а вырученные сред-
ства должны были пойти на выплату компенсаций жертвам террористического акта
в метро. На момент его совершения в марте 1995 г. оно оценивалось не менее, чем
564
Часть 2. Общая неправильная эпидемиология
в 2,5 млрд иен, или 29 млн долларов. Асахара (или те, кто стоял за его спиной) оказал-
ся обладателем земельных участков общей площадью 136 тыс. м2, 16 зданиями в То-
кио и десяти в префектурах страны.
В марте 1997 г. японские власти приступили к демонтажу комплекса зданий в
бывших владениях «Аум-Синрикё» у подножия национальной святыни — горы Фуд-
зияма. В общей сложности там планировалось снести 32 здания. По решению властей
предполагалось временно сохранить только три здания, в которых сектанты произ-
водили зарин и некоторые другие химические вещества, примененные «Аум-Синрикё»
в нескольких химических атаках на жителей Японии.
Сёко Асахара арестован в мае 1995 г. Ему было предъявлено 13 обвинений, вклю-
чая организацию убийств и похищений 26 человек. В 2003 г. государственный обви-
нитель потребовал смертной казни. В 2004 г. после восьми лет процесса Токийский
окружной суд признал Асахару виновным и приговорил его к смертной казни через
повешение. В ходе судебного процесса израсходовано более 3 млн долларов. Судом
заслушано более 170 свидетелей. Сам Асахара с ноября 1999 г. отказался от показаний
и не признал себя виновным по большинству пунктов обвинения. Верховный суд
Японии 15.10.2006 г. отклонил последнюю апелляцию и окончательно утвердил смерт-
ный приговор Асахаре. В июне 2012 г. его исполнение было отложено в связи с даль-
нейшим арестам членов «Аум-Синрикё».
В январе 2000 г. «Аум-Синрикё» изменила название на «Азеф» — по первой букве
семитских алфавитов. Также был изменен и логотип. Убраны и наиболее одиозные
части доктрины, в то время как базовые, главные аспекты остались нетронутыми.
Организация, мимикрировав, продолжает существовать.
Характерные признаки биотеррористической организации. Используя аналогию с
«Аум-Синрикё», можно утверждать, что такими признаками являются следующие:
высокая степень организованности, включающая наличие руководящего ядра
(лидера, гуру, старца, учителя и т. п.), дифференциация функций исполнителей внут-
ри организации;
жесткая дисциплина, соблюдение конспирации и режима секретности, неукосни-
тельное подчинение четко регламентированным правилам иерархических отношений
внутри организации:
наличие связей (или стремление иметь их) с коррумпированными представителя-
ми администраций различного уровня (в том числе и за пределами страны), с госпред-
приятиями, коммерческими структурами, научно-исследовательскими организаци-
ями, используемыми для прикрытия преступной деятельности;
включение в биотеррористическуюорганизацию представителей нетрадиционных
для преступной среды слоев общества — ученых, инженеров, врачей;
высокий интеллектуальный уровень руководителей биотеррористической органи-
зации и наиболее активных ее членов среднего звена; наличие мощной духовной,
религиозной, националистической и иной мотивации в их деятельности;
тщательно скрываемая связь руководителей организации с криминальной и эк-
стремистской средой внутри страны и за ее пределами;
явное и неявное использование руководителей таких террористических органи-
заций спецслужбами, политическими, религиозными, националистическими груп-
пировками различных стран в своих целях;
тщательное планирование терактов и длительная подготовка к их совершению;
стремление к использованию других видов ОМП, считающихся «дешевыми», на-
пример, химического оружия;
2.4. Обнаружение биотеррористической организации
565
осуществление разведывательных мероприятий и контрмер, препятствующих
внедрению в террористическую организацию представителей правоохранительных
органов;
устранение лиц, способных раскрыть сведения о преступлениях террористов и
отступников.
Как и всякое организованное преступное сообщество, биотеррористическая орга-
низация имеет среди своих членов юристов, банкиров, экспертов, вербовщиков, бо-
евиков и др., каждый из которых осведомлен о ее деятельности только в рамках вы-
полняемой им функции (о преступных сообществах см. работу А. Ф. Волынского с
соавт.,1999).
«Полезные услуги» такой организации, не подозревая об ее истинном назначении,
могут оказывать различные коррумпированные и некорумпированные работники
властных и управленческих органов (в том числе и правоохранительных). И что ха-
рактерно проявило себя за время деятельности «Аум-Синрикё» в России, в число «по-
лезных» для биотеррористической организации людей могут входить отдельные вли-
ятельные политики, депутаты, представители общественных организаций, юристы,
журналисты, деятели литературы и искусства. По мнению К. Н. Салимова (1999),
иногда они могут даже не осознавать неправомерность своего поведения, действуя в
защиту террористического сообщества, создавая условия, препятствующие раскры-
тию его преступной деятельности, компрометируя отдельных представителей власти
и управления. Кроме того, они нередко принимают меры к освобождению задержан-
ных членов террористической организации от следствия и суда. Это особенно харак-
терно для лиц, сочувствующих националистическому терроризму.
Характеристика членов биотеррористической организации. Специфика работы с
биологическими агентами предполагает существование организации следующих лиц.
Организаторы. Организаторы, лидеры — это вершина пирамиды биотеррористи-
ческой организации. Для биотеррористических организаций, вследствие сложности
подготовки таких террористических актов, вероятна их связь с «кукловодами» из за-
рубежных спецслужб непосредственно либо через лидеров других террористических
организаций. Верхушка террористической организации занимается организационны-
ми вопросами, выработкой общей стратегии и тактики террористической деятельно-
сти. Как и подлинная цель биотеррористического акта, их деятельность может иметь
неявный характер и осуществляться через многочисленных посредников. В «поле
зрения» правоохранительных органов они могут попасть в том случае, если удастся
определить, в чьих интересах осуществлено преступление. Слава биологического тер-
рориста — это не та слава, к которой стремятся политические лидеры, «вожди», «учи-
теля» и другие подобные деятели. Судя по результатам расследования биотеррорис-
тического акта в США в 2001 г., полный разгром «верхушки» биотеррористической
организации осуществить очень сложно.
Производители биологических поражающих средств. Это специалисты среднего
звена организации, которых организаторы теракта привлекли к разработке, производ-
ству и накоплению биологического агента. Как правило, они будут вне поля зрения
правоохранительных органов, так как не относятся к традиционной криминальной
среде. Сама специфика работы с опасными патогенными микроорганизмами требу-
ет привлечения в террористическую организацию квалифицированных специалистов.
Для их деятельности будет характерна корыстная и/или идейная мотивация.
Исполнители биологических терактов. К ним можно отнести тех, кто непосредст-
венно будет применять биологический агент с помощью технических устройств. Сое-
566
Часть 2. Общая неправильная эпидемиология
тав исполнителей зависит от замысла теракта. При сложном и многоэтапном осуще-
ствлении теракта с использованием сложных технических устройств его исполните-
лями могут быть специалисты среднего звена организации. В иных случаях ими могут
оказаться и религиозные фанатики, совершающие «акт возмездия» и преступные эле-
менты, нанятые за определенную сумму. Для таких людей существует очень высокий
риск поражения биологическим агентом в момент его применения. Это «расходный
материал» биотеррористической организации.
2.4.2. Признаки деятельности биотеррористической организации
Признаки функционирования подпольной биотеррористической лаборатории. Ха-
рактеристика следов, связанных с деятельностью биотеррористической организа-
ции. Распознание примененного биологического агента по косвенным признаком.
Поиск запасов произведенных биологических агентов. Техника специальной безо-
пасности при осуществлении следственных и иннх действий в лабораториях био-
террористов. Назначение экспертиз и выбор экспертов.
Биотеррористические организации не являются абсолютно непрозрачными и
неуязвимыми. Они оставляют следы еще до применения ими биологических агентов.
Признаки функционирования подпольной биотеррорш тической лаборатории В от-
личие от лабораторий, занимающихся производством наркотиков серьезную микро-
биологическую лабораторию трудно скрыть.
Внешними признаками функционирования подпольной микробиологической
лаборатории могут быть следующие:
работающее приточно-вытяжное вентиляционное оборудование, особенно в хо-
лодную погоду или ночью;
наличие в вентиляционных системах предприятия, не имеющего отношения к
микробиологическому производству, специальных фильтров тонкой очистки возду-
ха, например, типа ФЭТО-750 и др. (обычно монтируются в специальных вентиля-
ционных камерах);
специфический запах (питательных сред, хлорсодержаших соединений, переки-
си водорода, фенола и др.);
наличие крупногабаритных и особенно двусторонних автоклавов (обнаруживают
себя паром, сбрасываемым на улицу или в систему вентиляции) (рис. 2.79);
разреженность воздуха в отдельных помещениях (некоторые негерметичные эле-
менты конструкции стен издают звук, характерный для всасывания воздуха внутрь
здания);
наличие бактерицидных ультрафиолетовых светильников (характерный свет из
окна);
наличие помещений для экспериментальных животных (запах животных и про-
дуктов их жизнедеятельности);
наличие в мусоре, выбрасываемом из лаборатории, большого количества мерт вых
лабораторных животных и продуктов их жизнедеятельности, осколков микробиоло-
гической посуды, остатков питательных сред, обрывков листов бумаги с черновыми
записями экспериментов, инструкций-вкладышей к коммерческим иммунопрофи-
лактическим препаратам, диагностикумам, антибиотикам и др.;
закупки больших партий компонентов питательных сред и экспериментальных
животных (особенно, приматов), как правило, разделенные между разными постав-
щиками;
2.4. Обнаружение биоггеррористической организации
567
Рис. 2.79. Схема двустороннего автоклава (стерилизатора), а — подача пара в полость
автоклава; б — подача пара в рубашку автоклава; в — система вентилирования аппара-
та (. воздушна»); г — слив конденсата из полоски автоклава; д — слив конденсата из
рубашки; е — подвод охлаждающей воды в рубашку автоклава; ж — отвод охлаждающей
воды из рубашки автоклава; и — слив воды из рубашки автоклава; к — вакуум; л — под-
вод сжатого воздуха для уплотнения крышек; 1 — корпус автоклава; 2 — рубашка кор-
пуса автоклава; 3 — механизм пневмоуплотнения крышки автоклава, 4 — конденсато-
отводчик; 5 — смотровой фонарь; 6 — строительная перегородка между помещениями;
7 — металлический фартук в перегородке. По С. Г. Дроздов) с соавт. (1987)
попытки нелегального (через знакомых в различных НИИ и другими способами)
приоОретения штаммов опасных микроорганизмов, а также штаммов микроорганиз-
мов, предназначенных для осуществления генно-инженерных экспериментов, векто-
ров и плазмид с генами токсигенностп и антибиотикорезистентности;
закупки дорогостоящего оборудования и реактивов, предна шаченных для моле-
кулярной диагностики инфекционных болезней, особенно когда они осуществляют-
ся за наличные деньги;
заказы в птицеводческих хозяйствах больших количеств оплодотворенных яиц
(куриные эмбрионы) определенного возраста (сотни и тысячи яиц, не предназначен-
ных для перепродажи);
закупки специального биотехнологического оборудования (см. раздел «Характе-
ристика следов...»), особенно с высоким уровнем защиты окружающей среды и био-
логической безопасности;
закупки легких микронизированных порошков на основе двуокиси кремния с
размером частиц 7—40 нм;
568
Часть 2. Общая неправильная эпидемиология
инфекционные болезни у персонала лаборатории, неэндемичные для данного
региона или не поддающиеся диагностике;
библиотечные заказы на медицинскую, микробиологическую и биотехнологичес-
кую литературу, научные журналы, книги и другие источники информации соответ-
ствующей направленности, включая адреса сайтов глобальной сети «Интернет»;
большое потребление газа, мазута, воды и электроэнергии (работа автоклавов,
вентиляционных систем, рефрижераторов, крематория для сжигания животных и др.
оборудования) их пользователями.
План расположения оборудования в типовой лаборатории для изучения возбуди-
телей инфекционных болезней приведен на рис. 2.80.
Рис. 2.80. План расположения оборудования в типовой лаборатории для изучения воз-
будителей инфекционных болезней. 1 — дверь с уплотнением; 2 — виварий; 3 — авто-
клав; 4 — дезинфицирующий душ, 5 — помещение для замены вентилируемого костю-
ма, 6 — передаточная ванна с гидрозатвором; 7 — аэрозоль; 8 — морозильный аппарат;
9 — рефрижератор 10 — печь для прокаливания или сжигания; 11 — воздушный шлюз
с УФ-лампами; 12 — емкость для грязного белья с УФ-лампами; 13 — камера для пере-
дачи материалов с УФ-лампами; 14 — замена загрязненной одежды; 15 — душевая
(вода); 16 — замена загрязненной одежды; 17 — вход-выход; 18 — шкафчики для одеж-
ды персонала: 19 — вспомогательная лаборатория По С Г. Дроздову с соавт. (1987)
Однако не всякая обнаруженная микробиологическая лаборатория, работающая
с патогенными микроорганизмами, нелегальна. О легальности ее деятельности мож-
но судить по следующим документам.
Официальным документом, разрешающим осуществлять любую деятельность,
связанную с возбудителями инфекционных болезней человека в России, является ли-
цензия, порядок выдачи которой определен «Положением о лицензировании деятельно-
сти, связанной с возбудителями инфекционных заболеваний человека». Эта лицензия вы-
дается на определенный срок, не превышающий 5 лет. Органами, уполномоченными
на ведение лицензионной деятельности, связанной с возбудителями инфекционных
заболеваний человека, являются Государственный комитет санитарно-эпидемиоло-
гического надзора Российской Федерации и центры государственного санитарно-
эпидемиологического надзора в субъектах Российской Федерации.
2.4. Обнаружение биотеррористической организации
569
В лицензии указываются: наименование органа лицензирования, выдавшего ли-
цензию; наименование и юридический адрес предприятия, организации, учреждения,
получившего лицензию; вид деятельности, на осуществление которой выдается ли-
цензия; срок действия лицензии; условия осуществления данного вида деятельности;
регистрационный номер лицензии и дата выдачи.
Более детально работы с патогенными микроорганизмами регламентируются «Са-
нитарными правилами по безопасности работ с микроорганизмами» Они определяют
порядок выдачи разрешения на работу с микроорганизмами I-IV групп патогенности
и рекомбинантными молекулами ДНК при проведении экспериментальных и диагно-
стических исследований, производстве иммунологических препаратов и продуктов
микробиологического синтеза. Этот порядок распространяется на все государствен-
ные, кооперативные и иные учреждения (предприятия), независимо от их ведомствен-
ной принадлежности и форм собственности.
Любые экспериментальные, диагностические, производственные работы с возбу-
дителями I—IV групп патогенности могут выполняться только при наличии у учреж-
дения (предприятия) разрешения на право работы с ними, выданного органом или
учреждением Государственной санитарно-эпидемиологической службы России (этот
документ не отменяет необходимость получения лицензии, разрешающей осуществ-
лять деятельность, связанную с возбудителями инфекционных болезней человека!).
Разрешение выдается на каждый вид микроорганизма, используемого в работе, и явля-
ется официальным подтверждением того, что в данном учреждении (предприятии)
созданы необходимые санитарно-гигиенические условия, обеспечивающие соблюде-
ние требований биологической безопасности, защиту населения и охрану окружаю-
щей среды.
Санитарными правилами установлен следующий порядок выдачи разрешений:
Главный государственный санитарный врач Российской Федерации выдает раз-
решение учреждениям, независимо от ведомственной принадлежности, на проведе-
ние работ:
• с микроорганизмами I группы патогенности противочумным и другим специа-
лизированным учреждениям;
• с микроорганизмами II группы патогенности — экспериментальные исследова-
ния;
• с микроорганизмами II группы патогенности — диагностические (полевые) ис-
следования в лабораториях противочумных станций, отделов особо опасных ин-
фекций и вирусологических лабораториях центров Госсанэпиднадзора;
• с рекомбинантными молекулами ДНК, согласно санитарно-противоэпидеми-
ческим правилам «Безопасность работы с рекомбинантными молекулами ДНК»,
утвержденным главным государственным санитарным врачом СССР 18 января
1989 года № 5-6/2х, и Федеральному закону «О государственном регулировании в
области генно-инженерной деятельности» от 5 июля 199о г., №86-ФЗ.
Главные государственные санитарные врачи субъектов Российской Федерации
выдают разрешения всем учреждениям, расположенным на обслуживаемой террито-
рии, на проведение:
• диагностических исследований с микроорганизмами II группы патогенности,
за исключением противочумных станций, лабораторий отделов особо опасных
инфекций и вирусологических лабораторий центров Госсанэпиднадзора;
• всех видов работ с микроорганизмами III—IV групп патогенности.
570
Часть 2. Общая неправильная эпидемиология
1лавные государственные санитарные врачи субъектов России могут делегировать
права выдачи разрешений на работу с микроорганизмами III—IV групп патогенности
главным государственным санитарным врачам районов и городов. Противочумные
станции выдают разрешение на право проведения диагностических работ с микроор-
ганизмами I—II групп патогенности лабораториям эпидемиологических отрядов.
Поэтому первыми шагами работников правоохранительных органов при провер-
ке нелегально существующей микробиологической лаборатории на предмет принад-
лежности ее к биотеррористической организации должны быть изъятие и проверка
подлинности документов, разрешающих проведение экспериментов с патогенными
микроорганизмами. Если эти документы окажутся подлинными, то с привлечением
специалистов-экспертов должно быть проверено соответствие осуществляемой лабо-
раторией деятельности заявленной.
Лаборатория, осуществляющая либо нелегальную, либо несоответствующую за-
явленной работу с патогенными микроорганизмами, должна проверяться правоохра
нительными органами на предмет ее участия в деятельности биотеррористической
организации. Для этого необходимо выявить следы такой деятельности.
Характеристика следов, связанных с деятельностью биотеррористической организа-
ции. Их можно объединить в две большие группы.
1. Следы, указывающие на подготовку биологического террористического акта.
Прежде всего, это следы, указывающие на разработку биологических поражающих
агентов, их производство, хранение и подготовку к применению. О проводимых в
нелегальной лаборатории разработках биологических поражающих агентов можно
судить по комплексу признаков:
высокий уровень защиты лаборатории: лаборатории изолирующего типа с уров-
нем изоляции Р 3 или Р 4 (Руководство по лабораторной биозащите, Женева, 1983) с
соответствующим оборудованием, либо уровень защиты, превышающий необходи-
мый для заявленной деятельности;
наличие в лаборатории микроорганизмов, способных вызывать инфекционную
болезнь у человека и признаки работы с ними;
наличие в лаборатории средств индивидуальной защиты персонала (автономно
вентилируемые защитные комбинезоны или костюмы, специальные респираторы,
защитные маски и др.);
наличие в лаборатории биологических защитных боксов (особенно III класса; или
изоляторов (изоляторы из гибкой пленки, сушильные боксы, анаэробные камеры и
перчаточные боксы), дающие возможность работать в них с патогенными микроор-
ганизмами, обеспечивая защиту персонала и окружающей среды (рис. 2.81);
поражения персонала лаборатории возбудителями опасных инфекционных болез-
ней, на работ*' с которыми не было получено разрешение главного государственного
санитарного врача Российской Федерации;
клетки, помещения и боксы для содержания экспериментальных животных;
литература, черновые записи, расчеты и прочая документация, содержащая обо-
снование экспериментов, описание их хода и полученных результатов;
реактивы, питательные среды и их компоненты, позволяющие осуществление
экспериментов с патогенными микроорганизмами;
антибиотики, химиопрепараты и иммунобиологические препараты, позволяющие
осуществлять жстренную профилактику поражений персонала опасными микроор-
ганизмами;
2.4. Обнаружение биотеррористической организации
571
Рис. 2.81. Микробиологический защитный бокс III класса. А. Вид спереди. Б Вид
сбоку 1 — воздуховод приточной вентиляции; 2 - высокоэффективный воздуш-
ный фильтр; 3 — смотровые стеклянные панели; 4 — отверстия с перчатками для
рук работающего; 5 — панели подводок и регулирующих вентилей для воды, возду-
ха, вакуума, химических растворов; 6 — панель контрольно-измерительных и элект-
рических приборов, средств сигнализации, 7 — распределитель ламинарного потока
воздуха; 8 — воздуховод вытяжной вентиляции; 9 — передаточный шлюз со смот-
ровым окном и герметическим люком. Направление движения воздуха показано
стрелками. По С. Г. Дроздову с соавт. (1987)
вакцинные препараты против возбу штелей опасных и особоопасных инфекци-
онных болезней.
Важно понимать возможности обнаруженного оборудования для подготовки и
осуществления террористических актов и диверсий. Например, обнаружение авто-
номного вентилируемого защитного костюма в нелегальной лаборатории свидетель-
ствует о проведения в ней работ с возбудителями особо опасных инфекционных бо-
лезней (геморрагических лихорадок, натуральной и обезьяньей оспы, чумы, сапа,
мелиоидоза и некоторых других), сопровождающихся риском образования аэрозоля.
Проверка со стороны правоохранительных органов должна проводиться, даже если
только фиксируются факты приобретения подобного снаряжения какими-то органи-
зациями, не имеющими разрешения на работу с микроорганизмами I и II групп. Это
объясняется тем, что для работы в таком костюме требуется весьма серьезная инфра-
структура, которую трудно скрыть, но только при ее наличии есть смысл его приоб-
ретать. Это компрессоры для нагнетания воздуха в подкостюмное пространство, спе-
циальные защитные боксы, системы тонкой очистки лабораторного воздуха и др.
Микробиологические боксы III класса (изолирующий) допускают работу с мик-
роорганизмами любого уровня опасности. Они могут быть объединены в комплексы
и оснащены изолированными от окружающей среды и друг от друга ферментерами,
инкубаторами, рефрижераторами, замораживателями, центрифугами, измельчителя-
ми и другим оборудованием, которое может быть использовано для получения неболь-
ших партий биологических поражающих агентов.
572
Часть 2. Общая неправильная эпидемиология
На производство в нелегальной лаборатории биологических поражающих агентов
и биологических рецептур на их основе указывают:
приобретения бокса III класса (изолирующий) организацией, не имеющей разре-
шения на работу с микроорганизмами I и II групп патогенности (уже сам этот факт
предполагает наличие у организации соответствующей инфраструктуры и каких-то
серьезных намерений, для реализации которых нужно достижение самого высокого
уровня безопасности);
ферментеры, биореакторы, хемостаты и системы непрерывного действия, установ-
ленные в лаборатории таким образом, чтобы они могли работать без риска образова-
ния аэрозоля, особенно, если их конструкция позволяет стерилизацию паром без пред-
варительной разборки;
центробежные сепараторы или декантеры, обеспечивающие непрерывное сепа-
рирование без риска образования аэрозоля, особенно, если их конструкция позволя-
ет стерилизацию паром без предварительной разборки;
оборудование и системы фильтрации в поперечном потоке, предназначенные для
непрерывного концентрирования микробов, вирусов или токсинов, функционирую-
щие без риска образования аэрозоля и имеющие возможность стерилизации паром без
предварительной разборки;
оборудование для сублимационного высушивания и измельчения высушенных
продуктов, особенно, если их конструкция либо монтаж в специальном боксе исклю-
чают возможность образования а >розоля;
оборудование для смешивания измельченных продуктов микробного синтеза,
особенно, если их конструкция либо монтаж в специальном боксе исключают возмож-
ность образования аэрозоля;
наличие в лаборатории запасов легких микронизированных порошков на основе
двуокиси кремния с размером частиц 7—40 нм;
необычно большое количество питательных сред, пригодных для культивирова-
ния патогенных микроорганизмов;
линии культур клеток, пригодные для крупномасштабного выращивания патоген-
ных вирусов и риккетсий;
записи, фиксирующие работу приборов;
камеры для аэрозольного экспонирования животных к микроорганизмам или
токсинам.
На хранение запасов биологических агентов в нелегальной лаборатории и подго-
товку к осуществлению теракта указывают:
крупногабаритные холодильные установки;
хранящиеся в этих установках тщательно закрытые контейнеры;
оборудование, пригодное для рассеяния биологических агентов в окружающей
среде в виде аэрозоля с размером частиц 1—5 мкм, либо контаминирования ими про-
дуктов питания и воды.
2. Следы, указывающие на осуществление биотеррористического акта:
пораженные биологическим агентом люди (или их трупы), эпидемиология болез-
ни i.oiopbix характерна для искусственного эпидемического процесса;
люди, перенесшие инфекционную болезнь, эпидемиология которой характерна
для искусственного эпидемического процесса;
террористы, пораженные биологическим агентом во время его применения.
оборудование, пригодное для рассеяния биологических агентов в окружающей
среде в виде аэрозоля с размером частиц 1-5 мкм, либо для контаминации пищевых
2.4. Обнаружение биотеррористической организации
573
продуктов и воды, обнаруженное в очагах инфекционных болезней (поражений токси-
нами) с эпидемиологией, характерной для искусственного эпидемического процесса.
Распознание примененного биологического агента по косвенным признакам. Между
применением биологического агента и появлением первых пораженных проходит ка-
кое-то время — от нескольких часов до недель. Появление первых пораженных далеко
не сразу может быть воспринято как результат биотеррористического акта или дивер-
сии. Еще большее время может потребовать идентификация примененного биологи-
ческого агента. За это время террористы могут совершить другие террористические
акты. Своевременное установление использованного в качестве поражающего агента
возбудителя инфекционной болезни позволяет выбрать оптимальную стратегию борь-
бы с эпидемией и подобрать эффективную схему лечения пораженных людей. Поэтому
важно попытаться как можно быстрее распознать примененный биологический агент
по тем косвенным и разрозненным сведениям, которые могут оказаться в распоряже-
нии правоохранительных органов.
При личном обыске подозреваемого в совершении биотеррористическою акта, а
также при обыске у него дома важно обратить внимание на наличие запасов антибак-
териальных и антивирусных средств. Террористы очень рискую!, применяя биологи-
ческий агент, например, с помощью диспергирующего устройства. Поэтому они мо-
гут подстраховывать себя приемом антибиотиков или химиопрепаратов. Обнаружение
их у террориста может позволить специалистам сделать вывод о виде примененного
возбудителя еще до того, как он будет выделен от пораженного человека. Например,
обнаружение антибиотика уже говорит о том, что террористы пытались защитить себя
от агентов бактериальной природы; рибавирин свидетельствует об их опасении быть
инфицированными возбудителем лихорадки Ласса; цидофовир или метисазон — о
применении ортопоксвирусов. Эти находки надо интерпретировать осторожно и в
совокупности с другими. Кроме того, сама одежда террориста может содержать сле-
ды биологического агента. Это означает, что изъятие таких препаратов и одежды у
подозреваемого должно проводиться с соблюдением мер специальной техники безо-
пасности, в присутствии понятых и с составлением протокола.
У подозреваемых лиц, если того требует обстановка (например, известно, что со-
вершен террористический акт, но биологический агет не установлен, а задержанные
либо отказываются назвать поражающий агент, либо не могут этого сделать), целесо-
образно немедленно взять кровь из локтевой вены и передать ее в специализирован
ную организацию для серологического исследования. Люди, которые работают с воз-
будителями инфекционных болезней, обычно вакцинируются против них. Поэтому
серологический анализ крови подозреваемого лица способен в течение нескольких
часов дать много полезной информации, позволяющей установить не только исполь-
зованный в данном теракте агент, но и те агенты, с которыми такие теракты осуще-
ствлялись ранее, либо их еще предполагается осуществить. Взятие крови должно быть
осуществлено в присутствии понятых и на основании постановления следователя о
получении образцов.
Среди вопросов, которые следственным органам целесообразно поставить экспер-
там, могут быть следующие:
1) содержит ли сыворотка крови антитела к возбудителям сибирской язвы, чумы,
туляремии, Ку-лихорадки, лихорадки Эбола, натуральной оспы, бруцеллеза, желтой
лихорадки, ботулиническим токсинам типов А, В и Е (этот перечень обусловлен дос-
тупностью соответствующих иммунопрофилактических препаратов)?
2) содер» ит ли сыворотка антибиотики и химиопрепараты, и какие?
574
Часть 2. Общая неправильная эпидемиология
Информативен немедленный осмотр лаборатории квалифицированными микро-
биологами. Уже по имеющемуся оборудованию, использованным при напаботке аген-
та питательным средам, практикуемым мерам специальной техники безопасности, по
обнаруженной защитной одежде, дезинфектантам, экспериментальным животным,
учебным руководствам, оттискам статей из научных журналов и даже запахам, они
смогут сделать вывод о природе биологического агента, о его наработанном количе-
стве, и предполагаемом способе применения.
Поиск запасов произведенных биологических агентов. Биологические агенты могут
нараба гываться в регионе страны, не контролируемом эффективно Федеральным
центром, затем перевозиться в другой и применяться там. Их мотуг завозить дгя со-
вершения террористических актов из других стран. Следственным органам важно
понимать, где и по каким признакам, используя оперативные методы, необходимо
искать уже произведенные биологические агенты либо предназначенные для их про-
изводства штоковые культуры микроорганизмов. В отличие от наркотиков, подозре-
ваемые не будут прятать их в носках или в желудке. Микроорганизмы являются жи-
выми существами и для их хранения нужны низкие температуры. Такие же условия
требуются для хранения биологических токсинов, например, ботулинических. Чувст-
вительность к температуре является «родовой травмой» большинства биологических
агентов и делает необходимой поддержание террористами при их транспортировке так
называемой «холодовой цепи» — системы хранения и транспортировки биологического
агента от места изготовления до места его применения при пониженной температуре,
позволяющей сохранить его поражающие свойства.
Холодовая цепь предполагает наличие отдельных звеньев — стационарных или
передвижных рефрижераторов (вагоны, автомобили, контейнеры-холодильники
и др.). Обнаружение такой цепи, которая во всех ее звеньях будет контролироваться
людьми, принадлежащими к террористической группировке, видимо, и есть тот ос-
новной признак, по которому можно судить о начавшейся подготовке к осуществле-
нию биотеррористического акта в каком-то регионе страны. Для оперативного ра-
ботника это означает еще и то. что спрятанные или транспортируемые к месту пред-
полагаемого совершения преступления биологические агенты нужно попытаться
обнаружить в контролируемых людьми из биотеррористической группировки холо-
дильных камерах, рефрижераторных вагонах и автомобилях, хранилищах других ти-
пов, в которых постоянно поддерживается низкая температура. Наиболее вероятно,
что для хранения наработанных запасов биологических агентов в низкотемператур-
ных хранилищах террористами будут использоваться прочные герметичные сталь-
ные контейнеры.
Штоковые культуры микроорганизмов, наиболее вероятно, будут храниться в
стеклянных ампулах в виде пористой сухой массы. Для таких ампул будут характерны
признаки кустарной запайки (концы ампул имеют более темный вид, вытянутую
форму, шероховатости), хотя этикетки или надписи на них могут иметь фабричный
вид. Сами ампулы будут упакованы с большими предосторожностями в прочные кон-
тейнеры и передаваться по «холодовой цепи».
Техника специальной безопасности при осуществлении следственных и иных действий
в лабораториях биотеррориетоз. Первыми, кто «натолкнется» на такую лабораторию,
мотуг оказаться работники правоохранительных органов, специализирующиеся на
раскрытии более распространенных преступлений — незаконный оборот наркотиков
(предположение, что в лаборатории занимаются синтезом наркотиков), заказные
2.4. Обнаружение биотеррористической организации
575
убийства или бандитизм (причастность к таким преступлениям охранников лабо-
ратории), контрабанда (нелегальный ввоз в страну биотехнологического оборудова-
ния) — или действующие в чрезвычайных обстоятельствах, например, в «горячих точ-
ках» и «вышедшие» на биотеррористов случайно. Совершенно неожиданно для себя
они столкнутся с новой опасностью — возможностью биологического поражения.
Такое поражение происходит вне привычной для них обстановки и может не прояв-
ляться несколько дней, но в итоге оказаться смертельным. Поэтому им необходимо
понимать, откуда исходит такая опасность, и знать, как отличить лабораторию, в кото-
рой возможно поражение биологическим агентом.
Опасность биологического поражения исходит:
от инфицрованного возбудителем контагиозной инфекционной болезни челове-
ка (контакты с больными натуральной и обезьяньей оспой, легочной формой чумы,
лихорадками Марбург, Эбола, Ласса);
от переносчиков возбудителей инфекционных болезней и инфицированных ла-
бораторных животных (укусы инфицированных насекомых и животных);
от тканей, секретов/экскретов инфицированных людей и животных (контакты без
средств защиты, особенно при наличии травм и порезов на коже);
от аэрозоля инфекционного агента, созданного террористами с помощью специ-
ального аэрозольного генератора или образовавшегося при работе оборудования,
используемого террористами для получения биологических агентов;
от хранящихся в лаборатории или в специальном хранилище запасов биологичес-
ких агентов (неосторожное открытие содержащих их контейнеров);
от текучих порошкообразных субстанций;
от хранящихся в холодильниках рабочих и штоковых культур микроорганизмов,
находящихся в ампулах, пробирках, колбах (суспензии, лиофильно высушенные суб-
станции), а также в специальных матрасах и в чашках Петри на поверхности плотных
питательных сред (бактерии) и в культурах тканей (вирусы, риккетсии) — инфици-
рование возможно аэрозолем микроорганизмов, образовавшимся при их неосторож-
ном открытии либо разрушении;
от воздушных потоков, выходящих (!) в лабораторию из разгерметизированных
боксов — это обратный воздушный поток, который вбрасывает в лабораторию мик-
роорганизмы, копившиеся на фильтре тонкой очистки воздуха все то время, когда
движение воздушного потока было направлено в воздуховод вытяжной вентилляции;
от стеклянной посуды (пробирки, колбы, матрасы, шпатели, пипетки и др.),
средств индивидуальной защиты, приборов и оборудования, с помощью которых осу-
ществлялась работа с патогенными микроорганизмами, и которые не были просте-
рилизованы или обеззаражены — инфицирование возможно при контакте и неосто-
рожном обращении с ними;
от трупов лабораторных животных, погибших в результате экспериментов, и про-
дуктов их жизнедеятельности — инфицирование возможно через поврежденную кожу
или в результате вдыхания пыли, образовавшейся из высохших фекалий.
Минимально необходимым средством индивидуальной защиты является обычный
противогаз, который необходимо надеть даже по принципу «лучше поздно, чем ни-
когда» — тем самым уменьшить полученную поражающую дозу биологического агента.
Другой мерой, позволяющей спасти жизнь сотруднику, является немедленный док-
лад «по команде» о любых случаях повреждения им кожных покровов в такой лабора-
тории (укусы животных, порезы стеклом и др.).
576
Часть 2. Общая неправильная эпидемиология
Как отличить лабораторию, в которой возможно поражение биологическим агентом ?
Прежде всего, по поведению работающего в ней персонала. Если сотрудники лабо-
ратории не носят средств индивидуальной зашиты при осуществлении каких-то опе-
раций, значит риск поражения биологическим агентом для них в данный момент от-
сутствует. Поэтому при проведении каких-либо действий в такой лаборатории необ-
ходимо исключить возможность преднамеренного создания ее персоналом аварийной
ситуации, способной привести к поражению биологическими агентами находящих-
ся в лаборатории лиц. Для этого необходимо немедленно вывести весь персонал в
любое подсобное помещение и тщательно проследить за тем, чтобы при этом никто
из персонала не успел сломать или разбить оборудование, изменить режим вентиля-
ции в лаборатории, разгерметизировать боксы или емкости, разлить какие-то жидко-
сти или рассыпать порошки.
Однако обнаружение в лаборатории помещения с персоналом, работающим без
средств индивидуальной защиты, не означает, что и в других помещениях такая же
обстановка. Обычно лаборатории для работы с опасными микроорганизмами вклю-
чают помещения с разным уровнем биологической безопасности — так называемые
заразные зоны. Дальнейший осмотр лаборатории может привести к обнаружению
помещений, в которых люди выполняют какие-то операции в автономных вентили-
руемых или иных защитных костюмах, либо безлюдные помещения, оснащенные
оборудованием, используемым в микробиологических производствах. Для последних
дополнительными признаками опасности могут служить: оснащение их специальны-
ми тамбурами с запасами средств индивидуальной защиты, шлюзами, передаточны-
ми устройствами, гермодверями, боксами; работающее оборудование; включенные
бактерицидные светильники (характерный синий свет и запах озона); усиленный
вентиляционный режим и запах дезинфицирующих средств. В такие помещения
нельзя входить без средств индивидуальной защиты и специалистов, способных ра-
зобраться в том, какие технологические процессы в них осуществляются, или выклю-
чать работающую там вентиляцию.
Самый трудный случай — это неожиданное обнаружение в лаборатории работни-
ками правоохранительных органов, не оснащенными средствами индивидуальной
защиты, помещения, в котором персонал работает в таких средствах. Ситуация во
многом будет зависеть от того, как эти люди себя поведут, и от конкретной планиров-
ки лаборатории. Однако если это случилось, то необходимо, не заходя в такую лабо-
раторию дальше дверного проема (гермодвери), немедленно приказать всем в ней
работающим лечь на пол таким образом, чтобы они были полностью видны, затем с
помощью других сотрудников правоохранительных органов постараться как можно
быстрее найти в подсобных помещениях и надеть хотя бы самые элементарные сред-
ства защиты органовдыхания (респиратор, фильтрующий противогаз и др.), не вы-
пуская из поля зрения подозреваемых. До приезда специалистов, способных оценить
ситуацию, подозреваемые должны находиться в таком положении и не покидать ла-
бораторию, так как даже снятие изолирующих средств защиты всегда предполагает
определенный порядок их обработки дезинфектантами, обеспечивающий безопас-
ность персонала и окружающей среды. Должны быть исключены и любые действия
подозреваемых, направленные на порчу оборудования или изменение режима венти-
ляции в лаборатории. Возможна и такая ситуация, когда террористам удастся скрыть-
ся, бросив лабораторию. В этом случае в помещениях возможно обнаружение мерт-
вых людей и животных, обломков микробиологического оборудования и посуды,
открытых боксов, брошенных средств индивидуальной защиты, запасов питательных
2.4. Обнаружение биотеррористической организации
577
сред, дезинфектантов, холодильного оборудования и др. В брошенных лабораториях
необходимо соблюдать те же меры безопасности, что и в функционирующих, пока
специалисты не установят, какова их реальная опасность. Необходимо иметь в виду
возможность установки в таких лабораториях мин-ловушек.
Конкретные решения о проведении санитарной обработки, изоляции и экстрен-
ной профилактики поражений биологическими агентами у работников правоохрани-
тельных органов, обнаруживших лабораторию и участвовавших в задержании лиц,
подозреваемых в причастности к биотерроризму, будут принимать медицинские спе-
циалисты, исходя из анализа всех возможных рисков. Для ускорения консультаций со
специалистами, находящимися в других городах России, им целесообразно немедлен-
но по доступным каналам связи направить видеозаписи и фотоснимки, сделанные в
лаборатории биотеррористов.
Назначение экспертиз и выбор экспертов. Расследование биотеррористических
актов требует применения специальных познаний, в том числе и в форме экспертиз.
Характерные для биотеррористического акта вещества и материалы, их следы как
источники информации о способе и обстоятельствах совершения преступления, не
являются традиционными объектами криминалистического исследования. Ниже мы
рассмотрим назначение таких экспертиз применительно к гипотетической следствен-
ной ситуации, когда были выявлены: биотеррористический акт с применением в ка-
честве агента БО возбудителя сибирской язвы; обнаружена лаборатория биотеррори-
стов, в которой, предположительно, этот агент готовился к применению.
В данной ситуации целесообразно назначение, как минимум, следующих судеб-
ных экспертиз:
судебно-медицинской экспертизы пораженных — ее задачей должно стать установ-
ление судебно-медицинскими экспертами причин смерти (поражения, причинения
вреда здоровью) людей, как в очаге сибирской язвы, так и среди сотрудников данной
лаборатории. Среди последних также могут быть люди, получившие специфические
поражения во время работы с возбудителем сибирской язвы. В качестве примера мож-
но привести секту «Аум-Синрикё» — у них в 1994 г. погиб микробиолог, нарушивший
технику безопасности.
комплексной инженерно -технологической экспертизы обнаруженной лаборатории —
в состав экспертов, осуществляющих данную экспертизу, должны быть включены
биотехнологи, микробиологи-специалисты по возбудителям особо опасных инфек-
ционных болезней, аэробиологи, специалисты по технике безопасности в микробио-
логических производствах и другие. Их подбор будет зависеть от оонаруженного в
лаборатории оборудования и инженерных систем. Основной задачей данной экспер-
тизы должно стать обнаружение следов, указывающих на подготовку биотеррористи-
ческих актов (см. разд. «Характеристика следов, связанных с деятельностью биотер-
рористической организации»);
комплексной молекулярно-биологической экспертизы штаммов микроорганизмов —
в состав экспертов, осуществляющих данную экспертизу, должны быть включены
микробиологи-специалисты по возбудителям особо опасных инфекций, специалис-
ты по молекулярной генетике микроорганизмов, инфекционисты и эпидемиологи.
Основной задачей экспертизы должно стать установление причинно-следственной
связи между биологическими агентами, обнаруженными в лаборатории (штоковые
культуры микроорганизмов; микроорганизмами, обнаруженными на рабочих поверх-
ностях оборудования, использованного для их культивирования, концентрирования,
высушивания, диспергирования и других операций), и поражениями людей в очаге
578
Часть 2. Общая неправильная эпидемиология
инфекционной болезни с «невозможной эпидемиологией» — т. е. обнаружение сле-
дов, указывающих на осуществление биотеррористических актов.
Привлечение к расследованию специалистов. Спецификой дел данной категории
является то, что с самого начала расследования следователю необходимо плотно вза-
имодействовать со специалистами в области противобиологической защиты, биотех-
нологии, молекулярной биологии, эпидемиологии и другими, перечень которых
невозможно определить на начальной стадии расследования. Специфика биотерро-
ристических преступлений сегодня делает необходимым участие опытных специа-
листов-микробиологов и биотехнологов практически во всех следственных действи-
ях. Поиск таких специалистов следует проводить в научно-исследовательских
организациях РАН и РАМН, а также Министерства обороны. В случаях, получивших
международный резонанс, а также при расследовании деятельности международных
биотеррористических групп, целесообразно привлечение известных зарубежных спе-
циалистов в оьлас ги биологической защиты с последующим открытым опуб шкова-
нием совместных отчетов, монографий или статей, обобщающих опыт данного рас-
следования и исключающих какие-либо антироссийские инсинуации в будущем.
При расследовании данного вида преступлений такт ика обыска, допроса подозре-
ваемых и иные следственные действия также могут иметь свою специфику.
* * *
Биотеррористическая организация (сообществе) представляет собой систему
объединенных в группы лиц, осознанно участвующих своими материальными, интел-
лектуальными, служебно-должностными, организаторскими и исполнительскими
возможностями в реализации единого плана коллективной биотеррористической
активности или какой-либо ее важной части. Биотеррористическая организация, как
преступная организация, имеет ряд характерных признаков, основным из которых
является высокая степень организованности, включающая наличие руководящего
ядра и дифференциацию функций исполнителей внутри организации. Руководящее
ядро и специалисты среднего звена организации рекрутируются из нетрадиционных
для преступной среды слоев общества Мученых, инженеров, врачей), ее деятельность
может маскироваться созданием легальных научных центров. Скрыть деятельность
лаборатопии биотеррористической организации значительно труднее, чем лабора-
тории, занимающейся производством наркотиков. Такая лаборатория имеет ряд де-
маскирующих признаков, хорошо заметных специалистам в области биологической
защиты, и неизбежно оставляет следы в ходе подготовки к осуществлению биологиче-
ских террористических актов. И если бы японская полиция не проявляла странной
слепоты в отношении сектантов «Аум-Синрикё», то можно было бы избежать челове-
ческихж°ртв, вызванных дальнейшей эскалацией е° террористической деятельности
Для успешного противодействия биологическому терроризму, к нему необходимо
относиться как к новому виду преступлений, требующему разработки самостоятель-
ной методики расследования, формирования специализированных следственных
бригад и совершенствования законодательства.
ЧАСТЬ 3
ЧАСТНАЯ НЕПРАВИЛЬНАЯ ЭПИДЕМИОЛОГИЯ
Ниже в сравнительном аспекте будет рассмотрена «правильная» и «неправильная»
эпидемиология вспышек инфекционных болезней, вызываемых микроорганизмами
и токсинами, потенциальными агентами БО.
3.1. СИБИРСКАЯ ЯЗВА
Микробиология В. anthracis. Восприимчивость животных. Инфицирующие
дозы для животных и людей. Иммунитет. Происхождение и молекулярная
эпидемиология возбудителя сибирской язвы. Экология и естественная эпи-
демиология В. anthracis. Устойчивость во внешней среде. Антибиотикорезис-
тентность. Генетически измененные штаммы и их обнаружение. Патогенез.
Клиническая картина сибирской язвы у людей при естественном инфици-
ровании В. anthracis. Моделирование искусственного заражения В. anthracis.
Клинические примеры искусственно вызванной сибирской язвы. Обобщен-
ная клиника ингаляционной сибирской язвы. Диагностика искусственного по-
ражения возбудителем сибирской язвы. Иммунопрофилактика. Лечение.
Сибирская язва (Anthrax) — опасная сапрозоонозная инфекционная болезнь лю-
дей и животных, вызываемая спорообразующей бактерией Bacillus anthracis. Характе-
ризуется острым началом, тяжелой интоксикацией, лихорадкой, септицемией, воз-
никновением отеков и карбункулов. Протекает в кожной, легочной и кишечной фор-
мах. Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят
возбудитель сибирской язвы к II группе патогенности. По степени важности для на-
циональной безопасности США Б. anthracis относится к биологическим поражающим
агентам категории А (см. табл. 2.2). С Первой мировой войны В. anthracis неоднократно
применялась для осуществления биологических террористических актов и диверсий.
В США и в Соединенном Королевстве со средины 1940-х гг. возбудитель сибирской
язвы изучался как летальный агент БО. Для его боевого применения разрабатывались
специальные рецептуры и боеприпасы. В первой половине 1952 г. их применяли для
поражения войск и населения Северной Кореи и Северо-Восточного Китая. Специ-
альные рецептуры на основе В. anthracis и снаряженные ими боеприпасы находились
на вооружении американской армии до 1968 г. под шифром «N». Одна из сибиреяз-
венных рецептур, созданная в USAMR1ID, использована для осуществления биотер-
рористического акта в США в сентябре — октябре 2001 г. (см. разд. 1.12.4). Наиболее
вероятно применение посредством использования многоточечных источников и ди-
версионными (террористическими) методами.
Микробиология В. anthracis. Возбудитель сибирской язвы — анаэробная неподвиж-
ная грамположительная бактерия, относящаяся к роду Bacillus семейства Bacillaceae.
Существует в трех формах: капсульной (в организме больного животного или челове-
580
Часть 3. Частная неправильная эпидемиология
ка; или на питательных средах, содержащих кровь или сыворотку, и при выращива-
нии в атмосфере СО2); вегетативной бескапсульной (на обычных питательных средах),
споровой (во внешней среде при свободном доступе кислорода и при длительном куль-
тивировании на питательных средах). Растет при температуре 36—38°C на обычных
питательных средах: в МПБ образуются хлопья на дне пробирки; на МПА — харак-
терные колонии R-формы. Разжижает желатин, при выращивании на средах, содер-
жащих кровь, гемолиза эритроцитов не вызывает. На средах с пенициллином сиби-
реязвенные бациллы приобретают форму шаров и формируют своеобразные колонии,
называемые жемчужным ожерельем. На агаре Мак-Конки и других селективных пи-
тательных средах, содержащих соли желчных кислот, В. anthracis не растет. На поверх-
ности агара Хеттингера уже через 9 ч роста в аэробных условиях при температуре
36—38°С образуются характерные микроколонии, состоящие из завитых в один-два
слоя нитевидных клеток с отходящими в сторону цепочками. Через 18—24 ч инкуби-
рования при тех же условиях культуры вирулентных штаммов сибиреязвенного мик-
роба формируют матовые серо-белые и серебристо-серые, непрозрачные в отражен-
ном свете зернистые колонии с бахромча гыми краями и диаметром 3—5 мм (рис. 3.1).
Рис. 3.1. Макроскопические изображения колоний В. anthracis. А. Микроколония сиби-
реязвенного микроба (штамм 71/72), выращенного в течение 9 ч на агаре Хоттингера.
Микроколонии образованы нитевидными клетками, завитыми в несколько слоев. Фазо-
вый контраст, увеличение в 65 раз. Б. Морфологические особенности колоний В. anthra-
cis (СТИ-1), выращенных на агаре Хоттингера в течение 24 ч. Макрофотосъемка в падаю-
щем свете Колонии матовые серовато-белые. В. Те же колонии в проходящем свете.
Увеличение в 13 раз. Отчетливо видна шероховатая поверхность колоний, центр бугри-
стый коричневого цвета, края бахромчатые в виде плоской зернистой каймы. Г Коло-
ния капсульного штамма В. anthracis (вакцина Ценковского II), выращенного в бикар-
бонатном агаре в атмосфере угпекислого газа. Визуально колонии в падающем свете
напоминают жемчуг. При увеличении: центр колония выпуклый, периферия уплотнен-
ная, края ровные и округлые, поверхность гладкая. По А. Г. Золотареву с соавт. (2006)
3.1. Сибирская язва
581
Клетки В. anthracis, выросшие на искусственных питательных средах, имеют вид
прямых или слегка изогнутых палочек, длиной 6—10 мкм и шириной 1-2 мкм, часто
соединяющихся в нити или цепочки различной длины. Полюса у них внутри цепочки
прямые, словно обрубленные или выщербленные. Свободные полюса слегка закруг-
лены. Прокрашиваются анилиновыми красителями. В мазках хранившихся культур
и патологического материала встречаются лизированные клетки, окрашенные по Гра-
му отрицательно. В мазках крови, экссудатов, а также в отпечатках пораженных орга-
нов, В. anthracis обычно располагается поодиночке и попарно. При фазово-контраст-
ной микроскопии прижизненно окрашенных бацилл, приготовленных влажным ту-
шевым способом, можно увидеть светлые ореолы вокруг темных тел бацилл — это
капсулы, состоящие из поли-у-В-глютаминовой кислоты (poly-y-D-glutamic acid).
Споры располагаются субтерминально или центрально, имеют овальную форму, не
превышают диаметр тела клетки и не деформируют ее. Длина спор колеблется от 1,2
до 1,8 мкм, диаметр — в пределах 0,7—1,0 мкм. Окрашиваются методами Циля —
Нильсона, Ожешко и Пешкова. При фазово-контрастной микроскопии имеют вид
блестящих зерен овальной формы с резко очерченными контурами. Поврежденные
споры или споры в стадии прорастания выглядят более округлыми и диффузно по-
темневшими. Лизированные споры обычно темные, набухшие, деформированные, в
центре расположен участок просветления (рис. 3.2).
Восприимчивость животных. Наиболее широко болезнь распространена среди ко-
пытных (22 вида). Из сельскохозяйственных животных сибирской язвой болеют круп-
ный рогатый скот, овцы, козы, лошади, ослы, мулы, буйволы, верблюды, северные
олени и свиньи. Имеются сообшения о случаях болезни у птиц.
Среди диких животных сибирскую язву регистрировали у лосей, косуль, ланей,
бизонов, муфлонов, антилоп куду, зебр, жирафов, диких кабанов и бородавочников,
слонов, памирских архаров, обезьян. Наблюдали болезнь у собак, волков, белых мед-
ведей, львов, тигров, пантер, гепардов, пум, лисиц, норок, соболей, куниц, песцов,
скунсов, нутрий, енотов. Кошки болеют только в молодом возрасте. Зарегистрирова-
на сибирская язва и среди грызунов; зайцев, хорьков, серых крыс, малого, желтого и
песчаного сусликов, больших краснохвостых песчанок, красных и длиннохвостых
сурков, лесных мышей. Пресмыкающиеся, земноводные, рыбы и беспозвоночные
невосприимчивы к сибирской язве, но мотуг быть носителями возбудителя. Лягуш-
ки при искусственном заражении, если их держать в воде при температуре 35 °C, мо-
гут заболеть.
Из лабораторных животных к болезни чувствительны белые мыши, морские свин-
ки и кролики. При их искусственном инфицировании болезнь протекает в виде сеп-
тицемии и через 24- 72 ч наступает смерть. Обычно через 24 ч погибают белые мыши,
а морские свинки и кролики — через 2—4 сут, иногда на 5—7 сут.
Белые крысы невосприимчивы к сибирской язве, но инбредные крысы линии
Фишер-344 и Вистар чувствительны к токсину В. anthracis. К парентеральному введе-
нию В. anthracis чувствительны сирийские золотистые хомячки, суслики, хорьки.
Инфицирующие юзы для животных и людей. Обобщены в работе Н. В. Литусова
с соавт. (2002). Величина ингаляционно-поражающей дозы (Lct50) для человека
составляет около 20х103 спор, a Lct50 при объеме дыхания человека 10 л/мин —
20х103спорхминхл~1. Ингаляционные LD50 для морских свинок — 7,75х104, кроли-
ков — 14х10ч, обезьян — 2,24х104, овец — 1,68х104, собак — 2,21 хЮ7, лошадей —
8х 106 спор. Величина LD50 для споровой культуры обычно на порядок больше для
вегетативных клеток. Восприимчивость человека к ингаляционному поражению
582
Часть 3. Частная неправильная эпидемиология
Рис. 3,2. Микроскопическое изображение В. anthracis. А. Микроскопическая картина
клеток В. anthracis, штамм 14/41 в окрашенных мазках культур, выращенных на агаре
Хоттингера в течение 24 ч и хранившихся при температуре О—4°С 96 ч. Увеличение:
1150 раз. Целые клетки окрашены грамположительно (фиолетовый или черно-фио-
летовый цвет), лизированные клетки выглядят как грамотрицазельные (розового цве-
та). Б. Прижизненная микроскопическая картина клеток возбудители сибирской язвы
(штамм 4-7), выращенных на агаре Хоттингера в течение 24 ч. Иммобилизация кле-
ток агаровым гелем. Фазовый контраст. Увеличение: 1350 раз. Споры светлые и ова 1ь-
ные Бациллы темные, имеют форму удлиненных цилиндров, расположены поодиноч-
ке, попарно или соединены в цепочки. Внутри клеток мелкие светящиеся гранулы
липидов и крупные, ярко блестящие споры. Лизирующиеся бациллы отличаются по-
ниженной оптической плотностью цитоплазмы, В Капсула живых сибиреязвенных
бацилл штамма 4-7. Бациллы выращены на бикарбонатном агаре в атмосфере угле-
кислого газа в течение 48 ч. Влажный тушевой метод. Фазовый контраст. Увеличение:
1350 раз. Капсула препятствует проникновению туши к клеткам и по этой причине
выглядит в виде ореола, окружающего бациллы. Г. Зрелая спора В. anthracis, штамм
14/41 при электронной микроскопии. У зрелой споры кортекс электронно-прозрач-
ный. Компоненты спор: СП — спороплазма; КО — кортекс; СМ — споровая мембрана;
СО — споровая оболочка; ЭК — экзоспориум. По А. Г. Золотареву с гоавт (2006)
сибирской язвой ближе к восприимчивости кроликов, обезьян и овец, которые явля-
ются наиболее чувствительными к ингаляционному и подкожному заражению возбу-
дителем сибирской язвы.
При алиментарном заражении LD50 В. anthracis для животных, установленные
опытным путем, составляли: дтя морских свинок — 1,62х 108 (1,49х108— 1,76х108),
кроликов — 3,93х108(3.5х108--4,4х108), обезьян — 2,05x10' (1,6х107~2 6х107) и
овец — 5х107 спор. Расчетная LD50 для человека — 1,25х107 спор В. anthracis.
Иммунитез. Лица, перенесшие сибирскую язву, приобретают стойкий, но не аб-
солютный иммунитет. Известны случаи повторного заболевания кожной формой
сибирской язвы через 2 года (Никифоров В. Н., 1973).
3.1. Сибирская язва
583
Происхождение и молекулярная эпидемиология возбудителя сибирской язвы. Внут-
ри рода Bacillus, возбудитель сибирской язвы относится к группе В. cereus, включаю-
щей шесть видов бацилл: В. cereus, В. thuringiensis, В. anthracis, В. weihenstephanensis,
В. mycoides и В. pseudomycoides. Патологические процессы у людей вызывают В. cereus
и В. anthracis (Klee S. К. et al., 2010). До работы A. R. Hoffmaster et al. (2004) считалось,
что В. cereus причастен только к пищевой токсикоинфекции и в затруднительных слу-
чаях его можно отличить от сибиреязвенного микроба по наличию у последнего плаз-
мид рХО1 и рХО2. Однако эти авторы описали изолят Оациллы, не являющейся В
anthracis, но обладающей генами сибиреязвенного токсина и способной вызывать
ингаляционное поражение у A/J-мышей, патоморфологически очень похожее на
..нгаляционную сибирскую язву. Фенотипически и по ре 1ультатам 16S rRNA-анати-
за микроорганизм идентифицирован как цереус (В. cereus G9241), но у него обнару-
жена плазмида, pBCXOl, чья нуклеотидная последовательность на 99,6 % оказалась
идентичной кодирующей токсин плазмиде рХО1 В. anthracis И хотя другая плазмида
сибиреязвенного микроба, рХО2, кодирующая гены капсулы, обнаружена не была,
генный кластер, кодирующий гаку ю же капсулу, был обнаружен в ранее неизвестной
плазмиде цереуса, названной рВС218. Эволюционное древо, показывающее место
В. anthracis среди близкородственных бацилл, приведено на пис. 3.3.
ТП В. cereus
АТСС14579
В. cereus
°241
В. anthracis
В. thuringiensis
thuringiensis
В. thuringiensis
kurstaki
В. thuringiensis
totbotihii в
В. thuringiensis
israeliensis
В. cereus
АТСС 10987
Рис. 3.3. Эволюционное и микробиологическое сходство В. cereus и В. anthracis.
А. Эволюционное древо группы В. cereus. Составлено по результатам мультило-
кусного типирования последовательностей В. cereus G9241 и других представите-
лей рода Bacillus. Б. Капсула В. cereus G9241, выросшей в естественной атмосфере.
В. Капсула В. anthracis, выросшей в атмосфере, содержащей 5 % СО2 (полоска
соответствует 10 мкм). По A R. Hoffmaster et al (2004)
В. cereus
S366
Дальнейшие исследования показали наличие среди В. cereus штаммов, по плазмид-
ному составу идентичных сибиреязвенному микробу S. К. Klee et al. (2010) описали
полную последователь генома бациллы, вызвавшей в 2001—2002 гг. смертельные эпи-
зоотии среди шимпанзе, обитавших во влажных джунглях Национального парка Тай
(Tai National Park, Республика Кот-д’Ивуар). По клиническим признакам, патанато-
584
Часть 3. Частная неправильная эпидемиология
мии и гистологии пораженных тканей, болезнь однозначно приняли за сибирскую
язву, протекающую с симптомами поражения кишечника и легких. ПЦР в реальном
времени подтвердили наличие В. ол/Ллтсм-маркеров в ДНК, выделенных из различ-
ных образцов органов и тканей погибших животных. У микроорганизма были обна-
ружены две плазмиды, идентичные плазмидам вирулентности сибиреязвенного мик-
роба, рХО1 и рХО2, и еще одна небольшая плазмида, функции которой не были
установлены. В 2004 г. похожие штаммы были получены от трех шимпанзе и одной
гориллы, погибших в заповеднике Джа (Dja Reserve, Камерун), расположенном поч-
ти в тысяче миль к востоку от парка Тай. Однако все изоляты отличались от В. anthracis
рядом микробиологических особенностей:
а) они обладали подвижностью;
б) были устойчивыми к у-фагу;
в) обладали устойчивостью к пенициллину G.
Сначала их отнесли к новой группе В. anthracis (Leendertz Е Н. et al., 2006). Позже,
на основе классической 16S рДНК филогении, их переклассифицировали в В. cereus.
Эти данные предполагают наличие в природных резервуарах штаммов В. cereus, имею-
щих происхождение, независимое от классических штаммов В. anthracis. S. К. Klee et al.
(2010) предложили называть их «В. cereus variety (var.) anthracis».
В настоящее время в специальной литературе принято считать, что В. anthracis
является клональным производным древнего вида из группы В. cereus, утратившего
часть генов в ходе приспособительной эволюции (см. ниже), но который получил
патогенный потенциал, главным образом, через приобретение двух плазмид вирулен-
тности, рХО1 и рХО2 (Keim Р. et al., 2009).
Хромосома В. anthracis включает 5,23 т. п. о. (штамм Ames), средний показатель
Г+Ц составляет 35,4 %. По предварительным данным, хромосома содержит примерно
5750 генов со средней длиной около 710 нуклеотидов. Особенностью генома В. anthracis
является наличие генов белков, для которых не были определены соответствующие
фенотипы. Их функции включают гемолиз, подвижность и устойчивость к антибио-
тикам: пенициллину и тетрациклину. Основные гены вирулентности сибиреязвен-
ного микроба локализуются на двух больших плазмидах: рХО1 (181 т. п. о.) и рХО2
(96 т. п. о.). Экзотоксин, состоящий из трех субъединиц — летального фактора, про-
тективного антигена и фактора отека, обнаруживается в островке патогенности, рас-
положенном на рХО1. Кластер генов, необходимый для биосинтеза другого фактора
вирулентности — капсулы, расположен на рХО2 (Seshadri R. et al., 2005).
Возбудитель сибирской язвы относится к генетически гомогенным патогенным
микроорганизмам (genetically homogeneous pathogens). Р. Keim et al. (2009) считают, что
это вызвано короткой эволюционной историей микроорганизма, исключающей на-
копление мутационного полиморфизма и наличием споровой фазы в его жизненном
цикле. Большинство мутаций в генах любого вида образуются во время циклов реп-
ликации хромосомы. Спора же позволяет существовать В. anthracis годами, не вовле-
каясь в процесс деления, необходимый для поддержания вегетативных клеток.
Для изучения молекулярной эпидемиоло! ии возбудителя сибирской язвы исполь-
зуют два подхода: многолокусный анализ варьирующих по числу тандемных повторов (mul-
tiple locus VNTR analysis, MLVA) (Keim P. et al., 2060); и анализ полногеномного поли-
морфизма отдельных нуклеотидов (single nucleotide polymorphism, SNP, снипсы) (Van
Ert M. N. et al., 2007).
MLVA-анализ штаммов В. anthracis. С конца 1990-х rr. способ популярен среди
исследователей, занимающихся выявлением межштаммовых различий В. anthracis Он
3.1. Сибирская язва
585
основан на межштаммовом статистическом анализе VNTR. Подробно способ описан
в работах Р. Keim et al. (2000) и К. L. Smith et al. (2002). VN1 R представляют собой
короткие нуклеотидные последовательности (12 п. о.), образующие тандемы, ориен-
тированные в одном направлении. VNTR встречаются в хромосомной и плазмидной
ДНК В. anthracis и варьируют по длине у разных штаммов сибиреязвенного микроба
(табл. 3.1).
Таблица 3.1
Характеристики маркерных локусов VNTR В. anthracis*
Локуса Размер повтора в нуклеотидах6 Размер по количеству повторов Количество аллелей Индекс разнообразия (D)
наименьший наибольший
vrrA 12 2 6 5 0,50
vrrB) 9 15 23 5 0,32
vrrB2 9 11 15 3 0,34
vrrC} 36 4 12 6 0,55
vrrC2 18 17 19 3 0,50
CG3 5 1 2 2 0,35
pXOl-aat 3 4 11 8 0,81
pXO2-at 2 6 15 9 0,79
Avg 5,1 0,52
а — курсивом показаны маркеры, обнаруженные в ORF; 6 — vrrZl-повторы не являются
идентичными. Отдельные из них содержат множественные нуклеотидные различия; vrrC-мар-
керы содержат 9 вырожденных суЗповтогяющихся структур; D — индекс разнообразия под-
считывается как 1— 1 (аллельная частота)2.
* По Р. Keim et al. (2000).
У В. anthracis от двух до шести копий VNTR расположены в пределах открытой
рамки считывания гена, обозначенного как vrrA (variable region with repetitive sequence).
Ген vrrA кодирует белок с ММ 30 кДа, гомологичный микрофилярному белку SHP2
оболочки паразитного червя Litomosoides carinii (Andersen G. L. etal., 1996). To, что ген
эволюционно законсервирован среди бактерий, свидетельствует о функциональной
активности кодируемого им белка, но эта функция не установлена (Jackson Р. J. et al.,
1997). Ген vrrC кодирует полипептид ДНК-транслоказу (DNA translocase) и обычно
содержит триплетный VNTR. Обнаруженный в хромосоме бактерии триплет VNTR
указывает на локализацию в данном участке струк гурного гена (Keim Р. et al., 2009).
Такие локусы амлифицируются в ПЦР с использованием праймеров, комплемен-
тарных их фланкирующим последовательностям. Аллельные варианты VNTR-локу-
сов хорошо разделяются в агарозном геле. Размер артелей может быть установлен
путем сопоставления со стандартными маркерами ММ. Их количественный анализ
позволяет классифицировать различные штаммы В. anthracis в зависимости от числа
повторов и аллельных вариаций повторов. Сам факт присутствия VNTR-локусов в
геноме разных видов бактерий, возможно, связан с тем, что ДНК-полимераза ката-
лизирует синтез нуклеиновой кислоты через короткий тандем повторяющихся пос-
ледовательностей. Они и являются тем, что мы называем «VNTR» (Keim Р. etal., 2009).
Поданным MLVA-анализа, штаммы В. anthracis делят натри линии (А, В, С), сре-
ди которых A-клада (линия, группа, кластер) является наиболее значимой (до 90 % всех
586
Часть 3. Частная неправильная эпидемиология
штаммов) и имеет глобальное распространение. К линии В относятся ь разных иссле-
дованиях обычно менее 12 % изолятов. Встречается только в некоторых географичес-
ких регионах. Ее подразделяют еще на две группы, В1 и В2. На Юге Африки 93 % изо-
лятов В. anthracis представлены группой В1, где линия сосуществует со штаммами
A-клады. Штаммы В. anthracis группы В2 встречаются в Швейцарии, странах Южной
и Восточной Европы. Штаммы линии А подразделяют еще на 4 группы (кластера).
Изоляты кластера А1 доминируют на западе Северной Америки и в Канаде (геноти-
пы 1 и 3), встречаются в Южном Техасе (генотип 6). А2-ветвч встречается среди от-
дельных изолятов, привезенных из Пакистана. АЗ-кластер — самый распространен-
ный из всех известных (до 58 % изолятов). Вызывает вспышки сибирской язвы на юге
Африки (генотипы 30, 40, 67); в Австралии (штат Виктория, генотип 66), в Турции
(генотип 35). Этой ветви соответствуют известные вакцинные штаммы V770-NPR
(генотипы 45,46 и 49) и Стерн (генотипы 59 и 61). Ames, любимый штамм американ-
ских военных биологов (генотип 62), находится в АЗ.Ь-кластере. Кластер А4 недоста-
точно изучен. С-клада встречается редко и недостаточно изучена. Два независимых
изолята были найдены в Соединенных Штатах (Keim Р. et al., 2002, 2009). Кластерная
принадлежность штаммов, вызвавших на одной территории сибирскую язву у людей
и животных, может сильно отличаться. Например, в Швейцарии в начале 1980-х гг.
вспышки сибирской язвы среди животных вызывали штаммы В. anthracis, принадле-
жащие к кластеру В2, среди людей — штаммы кластера А4. Это было свя »ано с разным
происхождением источника их заражения сибирской язвой: животные инфицирова-
лись из местных очагов, а люди — во время переработки кожаного сырья, завезенно-
го из Индии (Pilo Р. et al., 2008).
Показана территориальная и временная зависимость выделения из образцов
штаммов В. anthracis различных линий. 30-летнее исследование молекулярных отли-
чий штаммов сибиреязвенного микроба, выделенных в Национальном парке Крюге-
ра (Kruger National Park, ЮАР), позволило установить ряд любопытных закономер-
ностей (рис. 3.4).
Национальный парк Крюгера расположен на востоке Трансвааля между реками
Лимпопо и Крокодиловая. На востоке парк граничит с Мозамбиком. Общая протя-
женность парка с севера на юг составляет 340 км. Три основные части парка (северная,
центральная и южная) сформированы реками Олифанте и Саби. Климат на террито-
рии парка субтропический, сезон дождей обычно с октября по март (включительно)
Сибирская язва поражает преимушественно копытных травоядных обитателей пар-
ка (Smith К. L. et al., 2000).
На территории ЮАР выявлено самое большое разнообразие среди штаммов си-
биреязвенного микроба, что дало основание некоторым ученым считать ЮАР «роди-
ной сибирской язвы». Характерной особенность сибиреязвенных очагов на юге
Африки является преобладание в них A-линии В. anthracis (Keim Р. et al., 1997). Гене-
тическое расстояние среди штаммов А- и В- линий, выделенных в сибиреязвенных
очагах Парка Крюгера, наибольшее среди известных коллекций ппаммов, собранных
по территориальному принципу. На территории парка Крюгера обнаружена четкая
территориальная локализация вспышек сибирской язвы, вызываемых В. anthracis раз-
личных линий: линия А преобладает в центральной части парка, линия В на севере
парка. Причем по распределению во времени количество изолятов той или иной ли-
нии может различаться. Например, линия В активно вовлекалась в эпизоотии только
в 1970 гг. на севере парка. Вспыхнувшие тогда же эпизоотии сибирской язвы в его
центральном районе, были вызваны штаммами A-линии. Эпизоотии начала 1990-х гг.
3.1. Сибирская язва
587
Рис. 3.4 Временные и территориальные зависимости выделения образцов В. anthracis
в национальном парке Крюгера по данным MLVA-анализа. А. Локализация изоля-
тов А- и В-линий: а — карта отбора индивидуальных изолятов; б — карта распреде-
ления изолятов по линиям. Б Распределение линий изолятов по годам В скобках
количество изолятов в абсолютных цифрах. В. Связь линий изолятов с pH почвы (I)
и наличием в ней солей кальция (И). По К. L. Smith et al. (2000)
вспыхнули в центральном оегионе парка, и их вызвали штаммы А-линии. Однако ког-
да эпизоотии охватили северный участок парка, в образцах также преобладали штам-
мы A-линии, что К. L. Smith et al. (2000) связали с наличием в природе неизученного
механизма, позволяющего сибиреязвенному микробу мигрировать с одной террито-
рии (экологической ниши) на другую территорию (экологическую нишу). В частности,
они указывают на проливные дожди в северной части парка, поошедшие в начале года,
и предполагают, что дожди «вымыли» экологические ниши, вмешавшие В. anthracis
В-линии.
Не менее любопытная находка исследователей — обнаружение статистически
достоверной связи между pH почвы и содержанием в ней кальция, и наличием в поч-
ве штаммов В. anthracis различных линий. Штаммы сибиреязвенного микроба А-ли-
нии вовлекаются в эпизоотии на территориях с нейтральным значением pH и умерен-
ным содержанием кальция, штаммы В-линии проникают в популяции животных с
территорий, pH почва которых близка к щелочной и содержит высокие концентра-
ции кальция. Таким образом, пространственное и временное распределение различ-
ных генотипов сибиреязвенного микроба в парке Крюгера, установленное MLVA-
анализом, показывает, что природные очаги сибирской язвы являются независимыми
друг от друга. В. anthracis различных генотипов могут перемещаться по территории,
коррелируя с определенными экологическими факторами; и/или сосуществовать на
одной территории в различных экосистемах.
Анализ полногеномного полиморфизма отдельных нуклеотидов. Под таким анализом
понимают выявление отличий последовательности ДНК размером в один нуклеотид
(А, Т, G или С) в геноме (или в другой сравниваемой последовательности) представи-
телей одного вида или между гомологичными участками гомологичных хромосом
индивида. Две последовательности ДНК — AAGCCTA и AAGCTTA — отличаются на
один нуклеотид. В таком случае говорят о существовании двух аллелей: С и Т. Снипсы
возникают в результате точечных мутаций.
588
Часть 3. Частная неправильная эпидемиология
Анализ индивидуальных снипсов, по сравнению с MLVA-анализом, менее инфор-
мативен. Однако если исследователь использует филогенетический подход к иденти-
фикации снипсов, он может эффективно дифференцировать штаммы бактерий в ге-
нетических группах с хорошо охарактеризованной популяционной структурой (Van
Ert М. N. et al., 2007). Полногеномный сиквиенс пяти штаммов В. anthracis позволил
выявить у них не менее 3,5 тыс. снипсов. Т. Pearson et al. (2004) картировали перекре-
стно почти 1000 таких снипсов в 27 различных (diverse) изолятах и составили точную
филогенетическую модель для В. anthracis, позволяющую установить филогенетичес-
кую позицию любого изолята В. anthracis. На основе их данных, М. N. Van Ert et al.
(2007) сформулировали рабочую гипотезу, в соответствии с которой разделение штам-
мов сибиреязвенного ми кроба по подгруппам, соответствующим каноническим снип-
сам (canSNPs), т. е. снипсам, локализованным в ключевых филогенетических узлах
эволюционного древа В. anthracis, способно заменить трудоемкий полногеномный
SNP-анализ и обеспечить более «высокое разрешение», чем MLVA-анализ, при диф-
ференциации штаммов в пределах линий А, В и С.
М. N. Van Ert et al. (2007) разделили изоляты линий А, В и С на 12 консервативных
групп или линий. Семь соответствуют полногеномным последовательностям штам-
мов сибиреязвенного микроба, охарактеризованных как «конечные точки» ветвей в
пределах древа канонических снипсов (С.USA.Al055, KrugerB, CNEVA.9066, Ames,
Australia 94, Vollum, Western North America). В дополнение к этим семи линиям древа
канонических снипсов (canSNP tree), canSNP-анализом они идентифицировали еще
пять субгрупп, позиции которых расположены вдоль ветвей древа. Для отдельных
линий и субгрупп характерна своя географическая локализация (рис. 3.5 и 3.6).
Древо
В-группа
С-группа
” W С.Вг.А1О55
*A/B.Br.001
В.Вг.004
-—^B.Br.CNEVA
А.Вг.009
Рис. 3.5. Взаимосвязь между canSNP, сублиниями и/или субгруппами В. anthracis.
Звездочки в дендрограмме соответствуют одной из семи последовательностей
генома В. anthracis, на основе которых построены семь его специфических линий
(«родословных»). Кружки соответствуют точкам ветвления, расположенным вдоль
таких линий, и отражают наличие в ее пределах специфических субгрупп изоля-
тов (показаны буквенно-цифровыми обозначениями). По М. N. Van Ert et al. (2007)
3.1. Сибирская язва
589
Количество Канонические группы Географическая локализация
•19
О
изолятов
• 2
ВсА1055
США
61
45,3
3
30
23
34
17
1,2
72
B.BrCNfcVA
В ВсЫ)ЗДЫ
122
В.Вг.001/002
B.BrKrugerB
B.BrAust94
А. Вг 001/002
А,ВгАтев
2
23
22
7
|а.Вг.005/006~|
Европа
Юг Африки, редко в Европе и США
Апгевтина, Чили
Юг Африки, Австралия, Турция,
Балканы, Индия, Китай
Северо-восток США, Китай,
редко в Европе
Китай, Европа, США
Британия, юг и восток Африки,
Корейский полуостров, Индонезия
Восток Африки, Европа, Китай,
Южная Америк х, Малая Азия,
Бразилия, Аргентина
Северная и Центральная Америка
Рис. 3.6. Генетическое родство и географическая локализация 1033 изолятов
возбудителя сибирской язвы. Указано количество изолятов, связанных с каждой
canSNP-группой. Черная полоса соответствует генетической дистанции между
штаммами. По М. N. Van Ert et al. (2007)
Экология и естественная эпидемиология В. anthracis. Резервуаром В. anthracis яв-
ляется почва, а вот каким образом и в каких хозяевах поддерживается этот паразити-
ческий микроорганизм, неясно. Ранее мной было высказано предположение, что воз-
будитель сибирской язвы поддерживается в почве не только в состоянии споры, но и
среди почвенных простейших. В отдельных их видах он существует в состоянии, ко-
торое называется «некультивируемой формой бактерий»; в других — размножается как
паразитический организм и, попадая в среду, богатую кислородом, спорулирует (Су-
потницкий М. В., 2000, 2009).
Для человека при вспышках естественно возникающей сибирской язвы источни-
ком возбудителя являются больные домашние животные: крупный рогатый скот, ло-
шади, ослы, овцы, козы, олени, верблюды, свиньи, у которых болезнь протекает в
генерализованной форме. Наиболее вероятен контактный путь инфицирования, реже
встречаются алиментарный, воздушно-пылевой и трансмиссивный. Кроме непосред-
ственного контакта с больными животными, заражение человека может произойти
при участии большого числа факторов передачи. К ним относятся выделения и шкуры
больных животных, их внутренние органы, мясные и другие пищевые продукты, поч-
ва, вода, воздух, предметы окружающей среды, обсемененные сибиреязвенными спо-
590
Часть 3. Частная неправильная эпидемиология
рами. В механической инокулятивной передаче возбудителя значение имеют крово-
сосущие насекомые (слепни, муха «жигалка») (Шувалова Е. П. с соавт., 2001). Одна-
ко для понимания причины вспышки сибирской язвы среди людей эпидемиологу
необходимо иметь представление о том, где и при каких обстоятельствах возможны ее
эпизоотии.
Поданным, приведенным Н. Г. Ипатенко с соавт. (1996), для нормального цикла
развития возбудителя сибирской язвы в почве необходимы: наличие органических
веществ (1—14 %), влажность не выше 50 % от обшей влагоемкости; нейтральная или
слабощелочная среда, температура не ниже 15 °C и не выше 35 °C. Почвы с большим
процентом содержания гумуса при прочих равных условиях имеют более благопри-
ятные условия для размножения В. anthracis. Подзолистые почвы, особенно с силь-
нокислой реакцией, неблагоприятны для размножения возбудителя сибирской язвы,
и наоборот, чернозем и все серолесные земли, имеющие нейтральную и близкую к
нейтральной реакции, с обязательным содержанием азота, калия, кальция, фосфора,
являются благоприятной средой для вегетации В. anthracis.
Основной путь заражения животных сибирской язвой — алиментарный. Возбу-
дитель сибирской язвы сохраняется не только в почве скотомогильника, как это при-
нято считать. Сибирскую язву часто наблюдали в стадах животных, которых пасли в
местах проведения земляных работ, в поймах рек с ра омытыми берегами, а зимой при
скармливании сена, собранного на территории, где осуществлялись земляные рабо-
ты. В >асушливые годы споры возбудителя сибирской язвы могут разноситься на боль-
шие расстояния пыльными бурями.
Корма как фактор передачи возбудителя сибирской язвы приобретают значение
в зимнее время, когда влияние других факторов (насекомых, пастбища и пр.) или ог-
раничено, или полностью исключено. Все корма на контаминацию спорами возбу-
дителя сибирской язвы исследовать невозможно. Необходимо сузить круг поиска,
пользуясь литературными данными. Н. Г. Ипатенко с соавт. (1996) считают, что наи-
менее опасными с точки зрения естественной контаминации В. anthracis являются
зернофураж и сено. Житняк, картофель, хрен, редис, турнепс способствуют размно-
жению сибиреязвенных микробов в почве и проникновению их в нижележащий слой.
С этими растениями не исключена возможность передачи возбудителя сибирской
язвы животным. Наиболее вероятным фактором его передачи в зимний период явля-
ются концентрированные корма, в которые добавлены корма животного происхож-
дения и, в частности, мясокостная мука.
Самым опасным источником В. anthracis является кровянистая жидкость, выте-
кающая из всех естественных отверстий трупа животного. Вместе с жидкостью сиби-
реязвенные бациллы проникают в почву и на открытом воздухе спорулируют. Тем са-
мым создается свежий почвенный очаг, продолжительность сохранения которого
зависит от почвенно-климатических условий данной местности. Труп животного пред-
ставляет меньшую опасность. В первые часы после гибели животного В. anthracis
быстро размножается, повышая свою вирулентность. В последующем в неповрежден-
ном трупе наступает полный анаэробиоз и бациллы не могут переходить в споровое
состояние. Кроме того, летом при высокой температуре в трупе размножаются в ос-
новном анаэробы, являющиеся антагонистами возбудителя сибирской язвы. В резуль-
тате гнилостного разложения В. anthracis погибает в течение 3—12 сут.
В России эпизоотии сибирской язвы обычно совпадают с засухой, неурожаем трав
и хлебов. В северных широтах (за 60°) эпизоотии отмечали в основном с марта по ав-
густ, с усилением в июле. В зимние месяцы болезнь не регистрировали. В районах,
3.1. Сибирская язва
591
расположенных между 60° и 55° с. ш., единичные случаи сибирской язвы наблюдали
в течение всей зимы. Количество случаев сибирской язвы увеличивалось с апреля,
максимума достигало в июле, в августе снижалось, а в декабре отмечали единичные
вспышки болезни. Для широт 50—55° с. ш. максимальное число случаев этой болезни
зарегистрировано в июле, с августа по октябрь оно снижалось. Для широт 50-45° с. ш.
характерно нарастание эпизоотии в мае, удерживалась она почти на одном уровне с
июля по август, в сентябре наступал спад. В пределах широт 30—45° с. ш. сибирская
язва в зимние месяцы появляется чаще, чем на территории указанных выше широт.
Н. Г. Ипатенко с соавт. (1996) выделяют три зоны проявления сибирской язвы.
Первая — зона спорадического проявления сибирской язвы. Территория севернее
58° с. ш. и восточные районы Забайкалья, полярный и северная часть бореального по-
яса. Почвенно-климатические факторы здесь неблагоприятны для жизнедеятельности
возбудителя во внешней среде. Температуры, благоприятные для вегетации бацилл,
удерживаются лишь 2 месяца — июнь и июль. Сезонности болезни не отмечается.
Вторая зона — зона периодического проявления сибирской язвы. Территория меж-
ду 58° и 54° с. ш. и районы западнее озера Байкал (бореальный пояс). Характеризуется
большим разнообразием почвенно-климатических факторов — от мало благоприят-
ных до благоприятных для жизнедеятельности возбудителя в почве. Температуры,
благоприятные для вегетации В. anthracis, наблюдаются 5 месяцев в году: май — сен-
тябрь. Заражение животных происходит от кормов и почвы. Отмечается сезонность
проявления болезни. Заболевания начинаются в мае, максимальное количество слу-
чаев наблюдается в июле — августе. Заболеваемость быстро снижается в сентябре.
Третья зона — зона устойчивого проявления сибирской язвы. Территория от
55-53° с. ш. до южных границ бывшего СССР, в Сибири — Алтайский край (суб-
бореальный и субтропический пояса). Почвенно-климатические условия наиболее
благоприятны для В. anthracis. Температуры, благоприятные для вегетации бацилл, ре-
гистрируются 6 месяцев в году: апрель — сентябрь. Инфицирование животных про-
исходит чаще от почвы. Отмечается четкая сезонность проявления болезни. Небла-
гополучие хозяйств по сибирской язве бывает относительно высоким во все месяцы,
но с конца мая оно начинает возрастать, достигая максимума в июле — августе, а затем,
в сентябре — октябре, медленно снижается до минимума.
В развитии эпизоотического процесса Н. Г. И патенко с соавт. (1996) различают два
периода: пассивный и активный. Пассивный период наблюдается при температуре
воздуханиже 12°С, а почвы —ниже 15°С. Почва при этих температурах обычно осво-
бождается от снега. Животных начинают выгонять на пастбище, и они могут попа-
дать на участки, контаминированные в прошлом возбудителем сибирской язвы (ско-
томогильник и пр.). Однако в это время они слабо контаминированы возбудителем
болезни. Количество неблагополучных пунктов и заболевших животных бывает ми-
нимальным. Активный период наблюдается при температурах воздуха от 12 до 30°С,
а почвы — от 15 до 35 °C. Н. Г. Ипатенко с соавт. (1996) выделяют три стадии эпизооти-
ческого процесса: подъем, максимальное развитие и угасание. Осенью, при температу-
рах воздуханиже 10°С, а почвы — ниже 13 °C, вновь наступает пассивный период эпи-
зоотического процесса. Неблагополучие по сибирской язве становится минимальным.
Естественным источником инфицирования людей и животных В. anthracis могут
быть животные со скрытым течением болезни. У свиней чаще, чем у других живот-
ных сибирская язва проявляется развитием местного патологического процесса и
значительно реже в виде септицемии. Степень опасности бациллоносительства во
многом зависит от формы болезни. При местной форме сибирской язвы возбудитель
592
Часть 3. Частная неправильная эпидемиология
болезни купируется в местах проникновения в организм животного (обычно в лим-
фатических узлах) и во внешнюю среду не выделяется. Обычно такую патологию на-
блюдают у свиней, однако при стрессовых состояниях (резкое ухудшение условий
содержания, переохлаждение, перегревание, физическое напряжение, голод и т. д.),
когда резистентность организма снижается, у животного может возникнуть инфек-
ционный процесс с развитием остро протекающей сибирской язвы. Иногда такое
явление наблюдают и при поражении кишечника. При этом воспаленные очаги мо-
гут в нем вскрываться, и возбудитель болезни будет выделяться во внешнюю сред}
Фермеры и мелкие владельцы скота могут скрыть факт эпизоотии сибирской язвы,
либо не распознать ее при молниеносном, подостром, хроническом или атипичном
течении болезни. Поэтому при выявлении звеньев эпидемической цепочки, по коте -
рым распространилась В. anthracis, необходимо путем опроса самих фермеров (хозя
ев), их работников и соседей по внешним признакам течения болезни установить
хозяйства, где были возможны эпизоотии сибирской язвы (рис. 3.7).
Рис. 3.7. Внешние признаки сибирской язвы у сельскохозяйственных животных. А. Сиби-
реязвенные поражения подчелюстных лимфоузлов с большим отеком в области шеи у
свиньи. Б. Отечность головы у коровы. Язык свисает изо рта, ноздри расширены. В. Отек
на тазовой конечности овцы. Состояние животного в момент возбуждения. Г Сибире-
язвенный отек в области живота у осла при подостром течении сибирской язвы. Д. Парез
грудных и тазовых конечностей у овцы за несколько часов до гибели от сибирской язвы
Е. Овца, павшая от сибирской язвы. По Н. Г. Ипатенко с соавт. (1996)
3.1. Сибирская язва
593
Наиболее обоснованное подозрение на сибирскую язву у фермера бывает лишь в
том случае, когда она протекает у животного в карбункулезной форме или при нали-
чии отеков в подчелюстной области. Карбункулы могут возникать в различных частях
тела и достигать иногда больших размеров. Вначале появляются плотные, горячие и
болезненные припухлости. Очень скоро они становятся холодными и безболезненны-
ми. В центре карбункула ткани некротизируются, отпадают, в результате образуется
язва. Температура тела повышается незначительно. Она может сопутствовать септи-
цемии, острому и подострому течению болезни.
При развитии у животного припухлостей и отеков говорят об ангинозной форме
сибирской язвы. Их появление — результат действия сибиреязвенного токсина. Тем-
пература тела повышается незначительно. Обычно поражаются подчелюстные, заг-
лоточные и шейные лимфатические узлы. Болезнь начинается в виде ангины. В об-
ласти шеи возникает припухлость. У одних животных она небольшая, у других,
начиная от гортани, тянется по трахее и нередко переходит на грудь и предплечье.
Кожа в области шеи приобретает синевато-красный оттенок. Обнаруживают отеки
языка и твердого неба. Из-за сильного опухания глотки животное держит голову на-
пряженно, сопит, глотательные движения нарушены. Отмечают кашель, хрипоту,
иногда рвоту. Видимые слизистые оболочки цианотичные. Состояние животного
подавленное, оно стремится зарыться в подстилку (свиньи) и подолгу лежит.
При внезапной гибели животного может вообще не быть каких-либо клиничес-
ких признаков. Но если болезнь несколько затягивается, то опрос позволит устано-
вить хозяйства, в которых гибели животных предшествовала стадия, когда животные
были возбуждены и напуганы. Так у овец и коз возникает возбуждение, они скреже-
щут зубами, при попытке идти делают резкие скачки и падают или совершают манеж-
ные движения и погибают в течение нескольких минут после появления первых сим-
птомов. Крупный рогатый скот и лошади тоже вначале возбуждены; взгляд испуганный,
температура тела повышена до 40—42°С, пульс учащенный, дыхание глубокое, пре-
рывистое, видимые слизистые оболочки цианотичные. Отмечается сокращение от-
дельных мышц в различных частях тела, а затем и групп мышц. Животные громко
мычат, некоторые ревут, бьют грудными и тазовыми конечностями о землю, упира-
ются головой в стенку, имеют испуганный вид, «стеклянные глаза», метеоризм руб-
ца, запоры или поносы, моча содержит примесь крови, появляются одышка, судоро-
ги мышц, конвульсии. Животное падает, издает глухое мычание, запрокидывает голову
на спину, иногда кладет ее на туловище или прижимает к груди и гибнет. Изо рта и носа
выделяется кровянистая пенистая жидкость. Возбуждение сменяется быстро разви-
вающимся угнетением. Животное погибает внезапно или в течение нескольких часов.
При подостром течении симптомы болезни нарастают медленно и временами
могут ослабевать. В этот период животные кажутся здоровыми: они поедают корм,
пережевывают жвачку. Однако через несколько часов состояние их резко ухудшается.
Приступы болезни могут повториться 2—3 раза, на последней стадии болезни у жи-
вотных вид испуганный, пучеглазие, застывший взгляд, зажатый корм в ротовой по-
лости. У овец и коз иногда появляются отеки в области вымени, живота и половых ор-
ганов и гиперемия кожи на внутренней стороне тазовых конечностей. Отеки бывают
и у крупного рогатого скота. Они могут быть различной формы и величины, тестооб-
разные, холодные, безболезненные. Иногда наблюдают карбункулы. Нередко при аго-
нальном состоянии у больных животных резко снижается температура тела вплоть до
субтерминальной, наблюдаются резко выраженные нарушения функций ЦНС. Бо-
лезнь длится не более 8 суток.
594
Часть 3. Частная неправильная эпидемиология
У свиней при остром и подостром течении сибирской язвы в первую очередь от-
мечают сонливость. Они отказываюгся от корма, подолгу лежат, если их сгоняют с
места, передвигаются вяло. В жаркую погоду свиньи не испытывают жажды. Более
того, они заметно избегают воды. Если же они пытаются пить, то засасывают воду
тонкой струей, которая сразу же выливается обратно. Аналогичную картину отмеча-
ют и при приеме корма. У некоторых свиней наблюдают рвоту. Как правило, бывает
понос или запор, температура тела повышена до 42 °C, дыхание учащенное. У неко-
торых свиней возникают отеки в области шеи и подгрудка. Вначале они красные, за-
тем становятся синюшными. У супоросных свиноматок возможны аборты или рож-
дение мертвых поросят. При таком течении болезни животное погибает на 2—3 сут или
неск< шько позже.
Хроническое течение сибирской язвы длится более 2 мес. и характеризуется у жвач-
ных и лошадей обычно слабым подъемом температуры тела, прогрессирующим ис-
тощением и профузным поносом. У свиней могут поражаться лимфатические узлы в
области шеи, здесь же иногда появляется небольшой отек, затем эти симптомы исче-
зают и животное кажется здоровым. Однако очаги поражения в организме сохраня-
ются и выявляют их лишь при послеубойном осмотре.
Большую трудность для своевременного распознания естественной вспышки си-
бирской язвы у людей создает атипичное течение сибирской язвы у животных. У забо-
левших животных возможно незначительное повышение температуры тела, иногда
появление небольших отеков в различных частях тела или усиление саливации, возни-
кновение симптомов острой тимпании рубца или вздутия др'/гих преджелу ikob, инва-
гинация 1 кишечника, слизисто-гнойное воспаление диафрагмы, травматическое по-
врежтение селезенки, острый геморрагический перикардит, атония желудка, гнойный
гепатит, ретчкулит, геморрагический гастрит, признаки отравления ядохимикатами.
Кишечная форма сибирской язвы у животных сопровождается расстройством функ-
ции органов пищеварения. Вначале у животного отмечают запор, сменяющийся ди-
ареей. Температура тела высокая и удерживается до 10 сут. У ослов болезнь проте-
кает остро и подостро. Клинические признаки примерно такие же, как у крупного
рогатого скота, но имеются отличия. У большинства ослов наблюдают карбункулез-
ную форму болезни. Поражения локализуются в области живота, вымени, мошонки,
подгрудка, анального отверстия. При остром течении болезни отмечают лихорадку,
кровоизлияния на слизистых оболочках, набухание век. При подостром течении жи-
вотные отказываются от корма, пульс учащен, слабого наполнения, зрачки расшире-
ны, дыхание глубокое, частое. Температура тела повышается медленно и за сутки до
гибели достигает 41-42 °C. Слизистые оболочки цианотичные. Волосы взъерошены.
Возбужденное состояние сменяется угнетением. Животное погибает от асфиксии.
Хроническое течение болезни нередко продолжается 30 сут и более. Клинические при-
знаки выражены слабо. В подкожной клетчатке появляются большие отеки: диффуз-
ные, тестообразные, разлитые, флюктуирующие. Отек вначале резко очерчен, твер-
дый, оезбо гезненный. Затем он медленно распространяется по всей брюшной полости
и переходит на подгрудок и морду. Температура тела повышается незначительно.
Появляются судороги. Из естественных отверстий вытекает кровянистая жидкость.
На фоне этих симптомов животное погибает (Ипатенко Н. Г. с соавт., 1996).
Устойчивость во внешней среде. В невскрытом трупе вегетативные клетки В. anthra-
cis разрушаются в течение 2—3 сут, в зарытых трупах они сохраняются до 4 сут. В замо-
роженном мясе при температуре минус 15 °C В. anthracis жизнеспособна 15 сут, в засо-
ленном мясе — до 1,5 мес. В навозной жиже, смешанной с сибиреязвенной кровью,
3.1. Сибирская язва
595
вегетативные клетки В. anthracis погибают через 2—3 ч, споры же остаются в ней виру-
лентными в течение нескольких месяцев. В запаянных ампулах с чистыми бульонными
культурами споры остаются жизнеспособными и вирулентными до 63 лет, в почве —
более 50 лет. Спирт, эфир, 2 %-ный формалин, 5 %-ный фенол, 5-10 %-ный хлора-
мин, свежий 5 %-ный раствор хлорной извести, 5 %-ная перекись водорода разруша-
ют вегетативные клетки сибиреязвенного микроба в течение 5 мин. Этиловый спирт
25—100 %-ный убивает споры В. anthracis в течение 50 сут и более, 5 %-ный фенол,
5-10 %-ный раствор хлорамина — от нескольких часов до нескольких суток, 2 %-ный
раствор формалина — через 10—15 мин, 3 %-ный раствор перекиси водорода — че-
рез 1 ч, 4 %-ный раствор перманганата калия — через 15 мин, 10 %-ный раствор гид-
роокиси натрия — через 2 ч. Вегетативные клетки В. anthracis при нагревании до тем-
пературы 50—5 5 °C гибнут в течение 1 ч,при температуре 60 °C погибают через 15мин,
при 75°С — через 1 мин, при кипячении — мгновенно. При медленном высушива-
нии наступает спорообразование и В. anthracis не гибнет. При температуре - 10°С бак-
терии сохраняются 24 сут, при минус 24°С — 12 сут. Воздействие прямого солнечного
света обезвреживает возбудитель сибирской язвы через несколько часов. Сухой жар
при температуре 120—140 °C убивает споры В. anthracis через 2—3 ч, при температу-
ре 150°С — через 1 ч, текущий пар при 100°С — через 12—15 мин, автоклавирование
при 110°С — за 5-10 мин, кипячение — через 1 ч (Mock М., Fouet А., 2001).
Антибиотикорезистентность. Для природных изолятов возбудителя сибирской язвы
антибиотикорезистентность не характерна. Поданным М. Е. Jones et al. (2003), иссле-
довавших 12 изолятов В. anthracis из различных районов Европы и Северной Амери-
ки (в том числе штаммы Ва7700, 7702, Sterne; RA3R и 7611R — выделенью во время
вспышки во Франции в 1999 г.), все они оказались чувствительными к ципрофлокса-
цину, цефтриаксону, эритромицину, азитромицину, доксициклину, пенициллину и
амоксициллину. В то же время в Южной Африке и Зимбабве, где выявлено самое боль-
шое разнообразие штаммов сибиреязвенного микроба, попадаются штаммы с различ-
ными уровнями антибиотикорезистентности. Среди 26 исследованных штаммов пять
конститутивно экспрессировали Р-лактамазу, но МПК по цефалоспорину (1—16 мг/л)
для этих штаммов была такой же, как и у Р-лактамазонегативных штаммов. В отдель-
ных коллекциях обнаруживают штаммы В. anthracis, резистентные к стрептомицину,
рифампицину, пенициллину, тетрациклину, хлорамфениколу (Маринин Л. И. ссоавт.,
1999). Среди них штамм Ames, устойчивый к цефтриаксону — цефалоспориновому
антибиотику III поколения широкого спектра действия (Jernigan J. A. et al., 2001).
Чувствительность подавляющего большинства изолятов В. anthracis к распространен-
ным антибиотикам говорит о том, что возбудитель сибирской язвы поддерживается в
природе вне контактов с антибиотиками. Наличие отдельных штаммов сибиреяз-
венного микроба, резистентных к антибиотикам, можно объяснить не только генети-
ческим переносом, но и клональным происхождением вида от более древнего В. cereus
по механизму дегенеративной эволюции. В пользу этой гипотезы говорит обнаруже-
ние в геноме В. anthracis генов, для которых не были определены соответствующие
фенотипы, т. е. «выключенных генов» (см. в работе Seshadri R. et al., 2005). В их числе
и гены антибиотикорезистентности. Например, Y. Chen et al. (2003) обнаружили мол-
чащие гены Р-лактамазы (Ыа! — пенициллиназа, Ыа2— цефалоспориназа) у чувстви-
тельного к пенициллинам вакцинного штамма Sterne. Вид либо неравномерно утра-
чивал ненужные ему для паразитирования в новом хозяине гены, и некоторые из них
остались интактными у части вида; либо имела место обратная мутация, восстановив-
шая экспрессию гена.
596
Часть 3. Частная неправильная эпидемиология
Генетически измененные штаммы и их обнаружение. При оценке микробиологиче-
скими методами свойств культуры В. anthracis, выделенной в очаге сибирской язвы,
ученый вынужден учитывать ее происхождение, проведение пассажей через питател!
ные среды и лабораторных животных. Различия по морфологическим, культуральным,
биохимическим, антигенным и вирулентным свойствам между штаммами В. anthracis.
выявленные в ходе таких исследований, могут быть вызваны их естественной диссо-
циацией. К постепенной утрате вирулентности приводит длительное пребывание
В. anthracis в почве.
Наоборот, вирулентность восстанавливается при 5—8-кратном пассировании ма-
ловирулентного В. anthracis через чувствительных животных. Наиболее часто изменен-
ные, ослабленные по вирулентности формы возбудителя сибирской язвы, выделяю
из организма инфицированных свиней. В природе широко циркулируют атипичные
штаммы В. anthracis, но тем не менее способные вызывать болезнь у сельскохозяй-
ственных животных.
Лиц, стоящих за биотеррористическими преступлениями, разумеется, меньше
всего интересует получение колоний В. anthracis «без завитков» и при выращивании
на жидкой питательной среде, в пробирках, формирующих аморфный осадок. Их уси-
лия по получению генетически измененных вариантов сибиреязвенного микроба
будут направлены на получение высоковирулентных штаммов, резистентных к анти-
биотикам и способных преодолевать имму нитет, создаваемый коммерческими вакци-
нами. Для конструирования генетически измененных штаммов сибиреязвенного
микроба используют способы генетического переноса и векторы, созданные для не-
патогенных бацилл. Принципиальных ограничений методического характера для их
применения на В. anthracis не существует. Препятствия «экспериментаторам» в попьг-
ках повысить поражающее действие биологических агентов создает только сам мик-
роорганизм (см. разд. 2.2.2 «Ограничения в конструировании штаммов реплици-
рующихся биологических поражающих агентов с измененными свойствами»),
С1980-х гг. для В. anthracis разрабатываются способы генетического переноса плаз-
мид. основанные на конъюгации, фаговой трансдукции и различных приемах транс-
формации1 (табл. 3.2).
Зарубежные исследователи обычно ограничиваются сообшениями о переносе
сибиреязвенный микроб тех или иных плазмид с генами антибиотикорезистентно-
сти и не касаются вопроса влияния приобретенной антибиотикорезистентности на
эффективность антибиотикотерапии сибиреязвенной инфекции. Поэтому Л. И. Ма-
рининым с соавт. (1999) были получены аналогичные трансформанты В. anthracis, и
в экспериментах по лечению на животных оценена их способность вызывать инфек-
ционный процесс. Методом серийных разведений они определили для трансформан-
тов МПК антибиотиков тетрациклиновой группы при равных условиях культивиро-
вания. Их данные показывают, что введение в составе пла >миды рВС16 (см. рис. 2.3
гена Тсг в возбудитель сибирской язвы может в 50— 1U0 раз повысить его устойчивость
к отдельным антибиотикам из группы тетрациклицина (табл. 3.3).
В экспериментах по лечению было установлено, что среднетерапевтические дозы
тетрациклина или доксициклина не предотвращают гибели золотистых хомяков, ин-
фицированных подкожно спорами В. anthracis Ч-7(рВС16), в то время как животные,
инфицированные спорами В. anthracis 4-7, выживали (Маринин Л. И. с соавт., 1999
1 Подробнее о способах генетического переноса см. в разд. 2.2.2 «Механизмы вмешатсз >-
ства в геном бактерий».
3.1. Сибирская язва
597
Таблица 3.2
Конструирование штаммов возбудителя сибирской язвы,
резистентных к антибактериальным препаратам
Резистентность к антибиотику Механизм приобретения резистентности Организация Источник информации
К тетрациклину (рВС16) и хлорамфениколу (рС194) Трансдукция USAMR11D, Fort Detrick (США) R. E. Rufel et al. (1984)
К тетрациклину (рВС16) и эритромицину (маркер плазмиды pLS20) Конъюгацией от В. subtilis совместно с крупной плаз- мидой pLS20 То же T. M Koehler et al. (1987)
К тетрациклину путем мута- генеза стрептококковым транспозоном Тп916 От Streptococcus faecalis DS16C1 к В. anthraas VNR-1 посред- ством конъюгации То же В E. Ivins et al., (1988)
К рифампицину Отбор спонтанных мутантов Институт при- кладной микро- биологии. г. Обо- ленск (Россия) А. П. Поме- ранцев с соавт. (1993)
К пенициллину, римфили- цину, тетрациклину, хлор- амфениколу, макролидам и лин комицину Трансформация рекомби- нантной плазмидой рТЕС Тоже A. V. Stepanov et al. (1996)
К хлорамфениколу Трансформация интегрирую- щейся в хромосому плазмидой рСЕТ То же А. П. Поме- ранцев с соавт. (1996)
К канамицину Плазмидная трансформация Centre for Applied Microbiology and Research, Porton Down, UK J. E. Bowen et al. (1999)
К хинолонам Отбор спонтанных мутантов, но возможна и плазмидная трансформация Armed Forces Radiobiol. Res. Inst., Maryland, США T A. Davies et al. (2001)
К рифампицину Индукция мутантов УФ-све- том + получение спонтанных мутантов Northern Arizona Univ., США A E Vogler et al. (2002)
Таблица 3.3
Величины МПК антибиотиков тетрациклиновой группы для генетически
измененного штамма В. anthracis 4-7
Штамм Антибиотик Величина МПК, мкг/см3 при разных условиях культивирования
без капсулообразования при синтезе капсулы
Тетрациклин 0,25 0.25
4-7 Доксициклин 0,025 0,025
Миноциклин* 0,025 0,05
Тетрациклин 128 128
4-7(pBC16) Доксициклин 16 16
Миноциклин* 2 8
* Миноциклин — полусинтетическое производное тетрациклина. У препарата отсутствует
перекрестная устойчивость с тетрациклинами.
598
Часть 3. Частная неправильная эпидемиология
Предположить, что выделенные от жертв биотеррористического акта или дивер-
сии штаммы В. anthracis являются генетически измененными, можно, сравнивая их
фенотипы с фенотипами известных штаммов по маркерам, характерным для генно-
инженерного конструирования возбудителей инфекционных болезней с усиленным
поражающим действием (антибиотикорезистентность, факторы патогенности, ток-
сины и т. п.). Например, штамм Ч-7(рВС16), полученный Л. И. Марининым с соавт.
(1999), отличается от наиболее часто встречающихся в природе штаммов В. anthracis
наличием резистентности к тетрациклину, а штамм Ч-7(рОВ12) — еще и способно-
стью вызывать гемолиз эритроцитов (см. ниже). Получить доказательства генно-
инженерного вмешательства в геном бактерии можно только молекулярно-генетиче-
скими способами — по наличию в бактериальной клетке нехарактерных для нее
генетических структур или генов. Методический уровень использованных для этого
способов исследования будет зависеть от технологий, с помощью которых новая гене-
тическая информация была введена реципиенту. Бациллярные клонирующие векто-
ры первого поколения, использованные в 1980-х гг. для конструирования генетически
измененных штаммов сибиреязвенного микроба, прелс гавляли собой многокопий-
ные автономно реплицирующиеся молекулы ДНК, не способные интегрироваться с
геномом бактерии-реципиента. Независимо от способа генетического переноса об-
наружить «лишнюю» плазмиду среди собственных плазмид реципиента можно по
плазмидному профилю (рис. 3.8).
Рис. 3.8. Переданная посредством фаговой трансдукции чужеродная плазмида у В. anthra-
cis и в других бациллах. Широкая диффузная полоса соответствует фрагментам хромо-
сомной ДНК. А. В. cereus GP7. В, В. anthracis Weybridge A UM18. С. В. anthracis Weybridge
A UM18(pBC16). D В. thuringiensis 40428 UM45. Е. В. thuringiensis 40428 UM45(pBC16).
F. В. thuringiensis 4060С LM473. G В. thuringiensis 4060C UM473(pBC16). H. B. ihurin-
giensisNGA. UM1.1. B. thuringiensis NG A LiMl(pBC16). J. B. cereus 5b9 UM47. К. B. cereus
569 UM47(pBC16). Перенос плазмиды pBC16 (2,8 мДа, Тсг) осуществлен бактерио-
фагом СР-51. Фенотипически наличие у реципиентного штамма плазмиды рВС16
проявляется устойчивостью к тетрациклину. По R. Е. Ruhtel et al. (1984)
На электрофореграммах ДНК разделенной в агарозном геле (см. рис. 2.3 и 3.8),
«лишняя» плазмида обнаруживается в виде фракции, дополнительной к собственным
плазмидам (для рВС16 — фракция, по подвижности соответствующая ММ 2,8 мДа).
Далее «лишнюю» плазмиду можно выделить из агарозного геля (вырезав соответствую-
щий кусочек из геля), перетрансформировать в более удобную для генетических иссле-
3. 1. Сибирская язва
599
дований бациллу, чем В. anthracis^, получить в препаративных количествах и устано-
вить ее происхождение путем молекулярного сравнения с известными плазмидами,
имеющими то же предназначение.
Появившиеся в начале 1990-х гг. интегративные векторы позволяют вставить чу-
жеродный ген непосредственно в хромосому бактерии (см. разд. 2.2.2 «Технологии ре-
комбинантной ДНК в получении штаммов бактерий с неправильной эпидемиологи-
ей»), Пример получения штаммов сибиреязвенного микроба с интегрировавшимися
в их хромосому генами сфингомиелиназы и фосфолипазы приводят Л. И. Маринин
с соавт. (1999). По их данным, В. anthracis Ч-7(рОВ12) вызывал сибиреязвенную ин-
фекцию у лабораторных животных, иммунизированных вакциной на основе штамма
СТИ-1. Однако ни методы электрофоретического разделения ДНК, ни хорошо заре-
комендовавшие себя для обнаружения инфекционных агентов диагностические тес-
ты, основанные на амплификации ДНК (ПЦР в любых вариантах исполнения), не
могут быть использованы для обнаружения генетически модифицированных патоге-
нов, если новая генетическая информация «добавлена» в их хромосому. Основной не-
достаток таких способов состоит в том, что они требуют предварительного и доско-
нального знания специфических генетических вариантов микроорганизма, изучаемо-
го на предмет генетической модификации. В последние годы эту задачу пытаются
решить на принципиально более высоком уровне — используя новые платформы сек-
венирования и, в частности, пиросеквенирование. Способ позволяет установить раз-
нообразные типы генетических изменений в геноме — инсерции и делении, единич-
ные нуклеотидные полиморфизмы, единичные тандемные повторы и вариабельные
участки генов. Chen Р. Е. et al. (2010) исследовали возможности способа для выявле-
ния генетически измененных штаммов В. anthracis Ames Ancestor1 2 и Sterne. В частно-
сти, по результатам пиросеквенирования ими в геноме штаммов сибиреязвенного
микроба выявлены вставки генной кассеты с геном резистентности к эритромицину;
мутации, предопределяющие низкоуровневую и высокоуровневую резистентность
В. anthracis к ципрофлоксацину; идентифицированы мутации, приводящие к спон-
танной резистентности к фагу и др. изменения. Эти исследования поставили некую
планку, ниже которой в ближайшем десятилетии исследования по идентификации ге-
нетически измененных агентов БО опускаться не будут.
Патогенез. В клинике сибирской язвы преобладают две группы симптомов:
1) обусловленные способностью В. anthracis к генерализации в клетках лимфоид-
ной системы с последующим развитием септической формы болезни.
1 В основном эти неудобства были связаны с гомологичной рекомбинацией и действием
рестриктирующих ферментов бацилл, приводящих к нестабильности чужеродных плазмид.
Поэтому такие эксперименты тогда старались выполнять, используя в качестве реципиента
мутантные В. subtilis с отключенными системами рекомбинации и рестрикции (штаммы МП 12,
МП 19, МП20идр.).
2 В экспериментах используется штамм Ames с искусственной устойчивостью к ципро-
флоксацину. Его природный вариант обладает резистентностью к пенициллинам третьего по-
коления и способностью преодолевать иммунитет, создаваемый коммерческими сибиреязвен-
ными вакцинами. Это его «террористы» применили в США для совершения так называемой
«почтовой атаки» осени 2001 г., сыгравшей главную роль в нагнетании в мире обстановки страха
перед якобы возрастающей угрозой биотерроризма со стороны Саддама Хусейна (см. разд.
1.12.4). Тогда ципрофлоксацин помог избежать массовой гибели американских граждан от
сибирской язвы (см. далее «Клинические примеры искусственно вызванной сибирской язвы»).
600
Часть 3. Частная неправильная эпидемиология
2) проявления местного патологического процесса, обусловленного действием эк-
зотоксина В. anthracis (повышение проницаемости капилляров, нарушения микроцир-
куляции, отек тканей и коагуляционный некроз) в первичном фокусе инфекции;
На месте внедрения возбудителя в кожу развивается сибиреязвенный карбункул
(реже — эдематозная, буллезная и эризипелоидная формы поражения кожи) в виде
очага серозно-геморрагического воспаления с некрозом, отеком прилегающих тка-
ней, регионарным лимфаденитом. Развитие лимфаденита обусловлено заносом
возбудителя подвижными макрофагами из места внедрения в ближайшие регионар-
ные лимфатические узлы. Дальнейшая генерализация В. anthracis с прорывом ее в
кровь и развитием септической формы болезни происходит при кожной форме че-
резвычайно редко. Сибиреязвенный сепсис обычно развивается при внедрении воз-
будителя в организм человека через слизистые поверхности дыхательных путей или
желудочно-кишечного тракта. Инфицировав макрофаги, споры быстро достигают эн-
досомальных компартментов, где должно происходить их разрушение. Однако неко-
торые споры остаются жизнеспособными и после прорастания превращаются в бацил-
лы, которые реплицируются внутри макрофагов. В этих случаях нарушение барьерной
функции трахеобронхиальных (бронхопульмональных) или мезентериальных лим-
фатических узлов приводит к генерализации процесса. Бактериемия и токсинемия
вызывают развитие инфекционно-токсического шока. В случаях развития легочной
формы болезни, перибронхиальные геморрагические лимфадениты блокируют
легочный лимфатический дренаж, вызывая отек легких (Dixon D. et al., 1999; Шува-
лова Е. П. с соавт., 2001).
Размножению В. anthracis в макрофагах способствует летальный фактор сибире-
язвенного токсина. Подвергнутые его воздействию макрофаги не утрачивают способ-
ности поглощать споры возбудителя сибирской язвы, однако в значительной мере
теряют способность освобождаться от них (Ribot W. et al., 2006) Патогенез сибиреяз-
венной инфекции у млекопитающих показан на рис. 3.9.
Сибиреязвенный токсин является прямой причиной гибели человека и животных
при развившейся сибиреязвенной инфекции. Введение белым крысам в хвостовую
вену сибиреязвенного токсина приводит их к гибели через 1—3 ч. Гибель животных
наступает от легочно-сердечной недостаточности, развившейся на фоне резкого по-
вышения проницаемости кровеносных капилляров, особенно в легких. Микроскопи-
ческие исследования показали, что изменения в легких были представлены отеком,
полнокровием и кровоизлияниями. Полнокровие и дистрофические изменения на-
блюдали в печени, почках, селезенке, сердце. В сердечной мышце найдены ранние
некробиотические изменения, представленные очаговой фуксинофильной дегенера-
цией. Повышенная проницаемость кровеносных капилляров и расстройства гемоди-
намики способствовали статистически достоверному по сравнению с контролем уве-
личению массы легких, почек и сердца (Литусов Н. В. с соавт., 2002).
Сибиреязвенный токсин состоит из В-субъединицы, называемой протективным
антигеном (РА; ее используют для иммунизации против В. anthracis), и двух фермен-
тативных субъединиц (А-субъединиц), одна из которых — отечный фактор (EF); дру-
гая — летальный фактор (LF). EF является кальмодулинзависимой аденилатциклазой
(calmodulin-dependent adenylate cyclase), увеличивающей внутриклеточные уровни
циклического АМФ (цАМФ) при проникновении в большинство типов клеток. В ме-
сте введения токсина в ткани экспериментальных животных нарушается водный го-
меостаз и развивается массивный отек. LF представляет собой цинкметаллопротеазу
(zinc metalloprotease). Вызывает гипервоспалительное состояние макрофагов, акти-
3.1. Сибирская язва
601
Спора
Бацилла
Бактериальные факторы вирулентности^*
Капсула
Экзотоксины
Другие факторы
Регионалгныг лимфатический узел
Макрофаг
Токсемия п „
„ - Летальный
Отечный'.оксин
lUKCMrl
О-+
+ IL-ip
+ др. цитокины
Шок
Региональный геморрагический лимфаденит
Отек легких
V
Смерть
Смерть
лимфатически т
блокада
Рис. 3.9 Патогенез сибиреязвенной инфекции у млекопитающих 1 — низкоуров-
невое прорастание и рост в участке инфицирования ведут к локальному отеку и не-
кротическому поражению кожи (образование карбункула); 2 — низкоуровневое про-
растание и рост в участке инфицирования веду к массивному выпоту, отеку
слизистой и некротическому поражению кишечника; 3 — лимфагенное и гемато-
генное распространение В. anthracis Смерть наступает от септицемии, токсемии или
легочной недостаточности через 1—7 сут после заражения. По Т. D Dixon et al. (1 °99)
вируя пути окислительного взрыва и высвобождая активированные кислородом ин-
термедиаты, а также продукцию провоспалительных цитокинов, таких как фактор не-
кроза опухолей а (TNF-а) и интерлейкин-10, вызывающие шок и смерть больного
(Dixon Т. D. et al., 1999).
Введение чувствительным животным любой из этих субъединиц по отдельности,
не приводило к патологическим последствиям. Внутрикожное введение комбинации
В-субъединицы и отечного фактора, вызывало развитие кожных отеков у морских
свинок и кроликов Внутривенное введение мышам и крысам смеси В-субъединицы
и летального фактора оказывалось для них смертельным, но при ее внутрикожном
введении образования отеков не происходило. При введении смеси трех компонен-
тов токсина, они действовали синергически в тестах проверки летальности на мышах.
В более поздних исследованиях показано, что отечный фактор вызывает только крат-
ковременное повышение внутриклеточного уровня цАМФ, поскольку он быстро раз-
рушается клеточными протеазами. Развитие сибиреязвенной интоксикации пред-
602
Часть 3. Частная неправильная эпидемиология
полагает обязательное участие всех трех компонентов сибиреязвенного токсина (Мои-
rez М. et al., 2002; Бургасов П. Н., Рожков Г. И., 1984).
В. anthracis не синтезирует активный токсин. В его формировании из субъединиц
основную роль играет сама клетка-мишень. С. Petosa et al. (1997) описали тонкие ме-
ханизмы этого процесса. РА сибиреязвенного токсина представляет собой длинную
плоскую молекулу, размером 100x50x30 А, состоящую из 4 доменов. Связывание РА
с рецептором клетки-мишени начинается с домена 4. Этот домен (остатки 596—735)
имеет первичную шпильку и спираль, за которыми следует р-сендвич с иммуногло-
булиновой складкой. Домены 1, 2 и 3 тесно связаны между собой, но домен 4 имеет с
ними ограниченный контакт. Внутри иммуноглобулиновой складки домена 4 содер-
жится доступная петля из 19 аминокислотных остатков, аналогичная антигенсвязы-
вающей CDRS-петле антител и рецепторсвязывающей петле дифтерийного токсина.
Протеолитическая активация происходит на клеточной поверхности. Поотеаза фурин
(она же используется для протеолитической активации дифтерийного токсина, экзо-
токсина А псевдомонад и некоторых вирусов) расщепляет поверхностную петлю внут-
ри домена 1. В результате высвобождается N-терминальный 20-кДа фрагмент (РА20).
Этот фрагмент не играет какой-либо дополнительной роли в интоксикации. Однако
его удаление приводит к образованию большой гидрофобной поверхности на остав-
шемся фрагменте РА (РА^). Остальная часть домена 1, называемого теперь домен 1',
образует N-окончание активного РА^. После утраты РА20, РА^ формирует гептамср,
который вставляется в мембраны при кислых значениях pH, формируя катионселек-
тивные каналы. Новая гидрофобная поверхность домена Г полностью обнажается,
формируя часть большого, плоского гидрофобного пятна на «вершине» гептамера. Эта
поверхность обеспечивает открытый сайт для связывания фактора отека и летально-
го фактора, которые теперь связывают РА63 с высоким аффинитетом. Собравшаяся
из трех субъединиц структура является тем, что понимают под названием «сибиреяз-
венный токсин». Он пенетрирует клеточную мембрану и захватывается эндосомой.
Внутренняя среда эндосомы закисляется и благодаря изменившейся конформацион-
ной структуре токсин формирует в стенке эндосомы канал и через него ферментатив-
ные субъединицы проникают в цитоплазму (рис. 3.10).
В патогенезе сибиреязвенной инфекции большую роль играют протеолитические
ферменты, коллагеназы, лейкоцидин, липаза и продукты распада самого микроба.
Клиническая картина сибирской язвы у людей при естественном инфицировании
В. anthracis. Патогенетическая классификация сибирской язвы у людей предложена
Н. В. Литусовым с соавт. (2002) (рис. 3.11, с. 604). Она используется при дальнейшем
изложении материала. Естественное инфицирование человека В. anthracis предпола-
гает его контакт с мясом или шкурами болевших сибирской язвой животных или с за-
раженной спорами сибиреязвенного микроба почвой. Входными воро гами возбуди-
теля болезни в этих случаях обычно является поврежденная кожа. В редких случаях
сибиреязвенная палочка внедряется в организм человека через эпителий дыхательных
путей и желудочно-кишечного тракта.
Кожная форма болезни составляет до 99 % всех случаев сибирской язвы, выявлен-
ных среди людей, кишечная и легочная формы сибирской язвы, встречаются как ка-
зуистические случаи (Никифоров В. Н., 1973). Заболевание может протекать в лока-
лизованной (кожной) или генерализованной (септической) формах. Ниже, в основном
по работам Е. П. Шуваловой с соавт. (2001) и Н. В. Литусова с соавт. (2002), приведе-
но краткое описание клинических форм сибирской язвы, встречающейся улюдей при
естественном инфицировании В. anthracis.
3.1. Сибирская язва
603
I
Огек
Смерть
Рис. 3.10. Модель действия сибиреязвенного токсина. 1 — протективный анти-
ген (РА) специфически связывается с рецепторами на поверхности клетки
(anthrax toxin receptor, ATR); 2 — PA рассекается мембранной фуриноподобной
протеазой, высвобождая 20-кДа субъединицу (РА20) во внеклеточное простран-
ство; 3 — оставшаяся 63-кДа субъединица (РА^) олигомеризуется и формирует
гептамер; 4 — отечный фактор (EF) и/или летальный фактор (LF) связываются
с гептамером; 5 — собравшийся из трех субъединиц токсин проникает в цитоп-
лазматическое пространство клетки в составе эндосомы; 6 — низкое значение pH
эндосомального пространства запускает конформационные изменения в геп-
тамере, вызывая формирование катион-селективного канала и транслокацию в
цитоплазму ферментативных субъединиц EF и LF. По М. Mourez et al. (2002)
При кожной форме сибирской язвы на карбункулезную разновидность приходится
99,1 % случаев, на злокачественный отек (эдематозная форма болезни) — 0,4 %, на
буллезную — 0,4 % и эрзипелоидную — 0,1 % (Никифоров В. Н., 1973). Поражаются
преимущественно открытые части тела. Инкубационный период при кожной форме
сибирской язвы независимо от локализации входных ворот инфекции длится от не-
скольких часов до 19 сут от момента заражения. Особенно тяжело протекает болезнь
при локализации карбункулов на голове, шее, слизистых оболочках рта и носа.
Обычно карбункул бывает один, но иногда количество их доходит до 10-20 и бо-
лее. На месте входных ворот инфекции последовательно развиваются пятно, папула,
везикула, язва. Пятно диаметром 1—3 мм красновато-синеватого цвета, безболезнен-
но, имеет сходство со следами от укуса насекомого. Через несколько часов пятно пе-
реходит в папулу медно-красного цвета. Нарастают местный зуд и ощущение жжения.
Через 12—24 ч папула превращается в пузырек диаметром 2-3 мм, заполненный се-
розной жидкостью, которая темнеет, становится кровянистой. При расчесывании или
самопроизвольно пузырек лопается, стенки его спадаются, образуется язва с темно-
коричневым дном, приподнятыми краями и серозно-геморрагическим отделяемым.
По краям язвы возникают вторичные («дочерние») везикулы. Эти элементы претер-
певают те же стадии развития, что и первичная везикула, и, сливаясь вместе, увели-
чивают размеры поражения кожи.
Через сутки язва достигает 8—15 мм в диаметре. Новые, «дочерние», везикулы,
возникающие по краям язвы, обусловливают ее эксцентрический рост. Вследствие
некроза центральная часть язвы через 1—2 нед превращается в черный безболезнен-
ный плотный струп. Вокруг струпа выражен воспалительный валик красного цвета.
Рис. 3.11. Патогенетическая классификация сибирской язвы у людей. Сплошные линии — наиболее распространенные
входные ворота возбудителя инфекции; пунктирные линии — редкие места внедрения В. anthracis в организм; в скобках
приведены аналоги клинико-анатомической классификации сибирской язвы или названия форм, встречающиеся
в научной литературе в случае несовпадения терминологии. По Н. В. Литусову с соавт. (2002)
Часть 3. Частная неправильная эпидемиология
3.1. Сибирская язва
605
По внешнему виду струп напоминает уголек на красном фоне, что послужило пово-
дом для названия этой болезни (от греч. anthrax — уголь). В целом поражение получи-
ло название карбункула. Поперечник карбункулов колеблется от нескольких милли-
метров до 10 см. Отек тканей, возникающий по периферии карбункула, захватывает
иногда большие участки с рыхлой подкожной клетчаткой, например на лице. Опасна
локализация карбункула на лице (нос, губы, щеки), так как отек может распростра-
ниться на верхние дыхательные пути и привести к асфиксии и смерти (рис. 3.12).
Рис. 3.12. Кожные поражения при сибирской язве. А. Сибиреязвенный карбункул
и отек в латеральной локтевой области пациента. Б. Сибиреязвенный карбункул
и отек в левой подчелюстной области пациента. По А. Н. Knight et al. (1969)
Параллельно с увеличением размеров язвочки и отека происходит увеличение
лимфатических узлов, расположенных ближе к карбункулу. Увеличение их не всегда
сопровождается болью, имеются лишь припухлоегь и изредка чувствительность; ста-
новясь плотными, лимфатические узлы сохраняю! подвижность. В подавляющем
большинстве случаев увеличение размеров язвочки, отека и лимфаденита наблюда-
ется в течение 5—8 сут, после чего начинается процесс формирования струпа на месте
карбункула: прекращается появление новых пузырьков и отделение жидкости; в цен-
тральных некротизированных участках начинается отложение фибрина (совпадает по
времени с моментом снижения температуры тела), который при подсыхании приоб-
ретает темный цвет; затем происходит уменьшение отека с периферии в направлении
к карбункулу. К этому времени общее состояние больного улучшается, температура
тела снижается до нормы. Сибиреязвенный карбункул в зоне некроза безболезнен
даже при уколах иглой, что служит важным дифференциально-диагностическим при-
знаком. Развивающиеся при кожной форме сибирской язвы лимфадениты, как пра-
вило, безболезненны и не имеют тенденции к нагноению.
Эдематозная разновидность кожной формы сибирской язвы характеризуется раз-
витием отека без наличия видимого карбункула. В более поздние сроки болезни про-
исходит некроз и формируется карбункул большого размера.
При буллезной разновидности на месте входных ворот инфекции образуются пу-
зыри с геморрагической жидкостью. После вскрытия пузырей или некротизации уча-
стка поражения образуются обширные язвенные поверхности, принимающие вид
карбункула (рис. 3.13).
606
Часть 3. Частная неправильная эпидемиология
Рис. 3.13. Буллезная разновидность кожной сибирской язвы. А. Пузыри с гемор-
рагической жидкостью перед формированием струпа. Б. Диссеминания кожных
поражений после хирургического вскрытия пузырей. По N. Bajkam et al. (2009)
Особенность эризипелоидной разновидности кожной формы сибирской язвы со-
стоит в развитии большого количества пузырей с прозрачной жидкостью. После их
вскрытия остаются язвы, подвергающиеся трансформации в струп.
Кожная форма сибирской язвы примерно у 80 % больных протекает в легкой и
средней тяжести, у 20 % — в тяжелой форме.
При легком течении болезни синдром интоксикации выражен умеренно. Темпе-
ратура тела нормальная или субфебрильная. К концу 2-3 нед струп отторгается с об-
разованием (или без него) гранулирующей язвы. После ее заживления остается плот-
ный рубец. Легкое течение болезни завершается выздоровлением.
При средней тяжести и тяжелом течении болезни отмечаются недомогание, раз-
битость, головная боты К концу вторых суток температура тела может повыситься до
39—40°С, нарушается деятельность сердечно-сосудистой системы. При благоприят-
ном исходе болезни через 5—6 сут температура критически снижается, происходит об-
ратное развитие общих и местных симптомов, отечность постепенно уменьшается,
лимфаденит исчезает, струп отпадает к концу 2- 4 нед, гранулирующая язва заживает
с образованием рубца. Тяжелое течение кожной формы может осложняться развити-
ем сибиреязвенного сепсиса и иметь неблагоприятный исход. Тяжесть течения забо-
левания не предопределяется величиной карбункула и зависит обычно от его локали-
зации и величины отека. В некоторых случаях кожная форма болезни протекает без
повышения температуры тела (см. рис. 3.12).
Легочная форма сибирской язвы при аспирационном заражении спорами В. anthracis
протекает, как правило, чрезвычайно остро. Вскоре после внедрения возбудителя —
в среднем через 2—3 сут, а иногда уже через несколько часов, начинается первая кратко-
временная фаза поражения (длится от нескольких часов до 1—2 сут), проятяющаяся
«гриппозными» симптомами (первая фаза болезни)', общим недомоганием, головной
болью, мышечными болями, слезотечением, насморком, охриплостью голоса и каш-
лем. Во второй фазе происходит бурное нарастание интоксикации, в результате чего
наступает резкое ухудшение состояния больного: беспокоят выраженная слабость,
Л1. Сибирская язва
607
разбитость, боли и стеснение в груди; усиливаются насморк и кашель; температура
тела повышается до 39—41 °C, и возникает сильный озноб. При болезненном, а порой
мучительном кашле отделяется вначале пенистая серозная мокрота, затем в мокроте
появляется все больше слизи и прожилок крови. Количество мокроты увеличивается,
она быстро густеет на воздухе. В случае примеси большого количества крови мокрота
становится похожей на «малиновое желе».
Данные медицинского осмотра обычно являются неспецифическими. Рентгено-
грамма грудной клетки обычно без инфильтратов, но можно наблюдать расширение
средостения с выпотом в плевральную полость, который может быть геморрагиче-
ским (рис. 3.14).
Рис 3.14. Рентгенограмма при легочной сибирской язве. На рентгенограмме
выраженное расширение медиастинальных узлов при отсутствии признаков
первичной пневмонии. По A. I. Braude et al. (1986)
В легких могут определяться очаги притупления, прослушивается обилие разно-
образных влажных хрипов (особенно в прикорневой зоне и нижних отделах). Нередко
в плевральных полостях обнаруживается жидкость. Может развиться гиперкардиум.
Накопление жидкости в плевральных полостях и полости перикарда бывает обуслов-
лено блокированием лимфооттока через пораженные лимфатические узлы средосте-
ния. Накапливаясь в большом количестве, жидкость создает механическое препят-
ствие дыханию и сердечной деятельности.
В конце второй фазы болезни, которая в среднем длится 1—2 сут, выявляются
жалобы больных на боли в груди, кашель, одышку, сильную слабость и тревогу. Кожа
и видимые слизистые оболочки бледны, отмечается тахикардия, температура тела
держится на уровне 38-40°С и выше.
Третья фаза болезни характеризуется, кроме описанных выше признаков, быст-
рым нарастанием сердечной и сосудистой недостаточности, появлением более выра-
женных, чем во второй фазе, симптомов отека легких и олигурией, что связано с усиле-
нием общей интоксикации. В предагональном и агональном периодах — выраженный
цианоз, одышка, отечность, сознание не нарушается. При явлениях легочно-сердеч-
ной недостаточности больные, как правило, умирают. Плохим прогностическим
признаком является снижение незадолго перед смертью температуры тела больного
до нормальной или субнормальной, сопровождающееся улучшением самочувствия,
608
Часть 3. Частная неправильная эпидемиология
не адекватным действительному состоянию больного. Длительность третьей фазы —
не более 12 ч, а всех трех фаз — от 2 до 5 сут. Эти сроки зависят от интенсивности те-
рапевтических воздействий на организм больного и его сопротивляемости.
При типичном течении развитие первичной желудочно-кишечной (кишечной) раз-
новидности сибирской язвы может быть подразделено на 3 фазы, хотя их не всегда
можно выделить из-за быстрого и тяжелого течения болезни.
Для первой кратковременной фазы (длится от нескольких часов до 1,5 сут) характер-
ны головная боль, головокружение, чувство сильного жжения в горле или боль при
глотании, недомогание, озноб и повышение температуры тела до 38-39 °C и выше.
Вторая фаза болезни характеризуется присоединением абдоминальных признаков:
боли в животе, тошнота, рвота с кровью и желчью, кровавый понос. Больных беспо-
коит выраженная слабость. Острые режущие боли в животе могут быть постоянными
или схваткообразными. Локализованы они преимущественно внизу живота, реже —
в области желчного пузыря. Живот часто вздут и при пальпации умеренно болезнен,
в особенности в области пупка. Напряжения мышц брюшной стенки не бывает.
В брюшной полости может определяться жидкость, порой в значительном количестве.
При распространении процесса на лимфатические узлы средостения жидкость появ-
ляется в плевральных полостях и в полости перикарда. Одновременно с этим присо-
единяются признаки поражения органов грудной клетки: кашель, одышка, отделение
мокроты и другие характерные для легочной разновидности симптомы болезни. Ярко
выражены явления общей интоксикации. Температура тела повышена, пульс ослаб-
лен, артериальное давление понижено. Беспокоят сильные головные боли, резчайшая
слабость, головокружение при стоянии и ходьбе. У некоторых больных определяется
увеличение надключичных и шейных лимфатических узлов, а также пастозность или
отечность подкожной жировой клетчатки шеи и передней поверхности грудной клет-
ки. Продолжительность второй фазы — от нескольких часов до 2—3 сут.
В третьей фазе болезни, кроме указанных выше признаков, отмечаются выражен-
ная бледность и синюшность кожи. На лице кожа розово-синюшная либо багровая,
склеры глаз инъецированы. Катастрофически нарастает декомпенсация сердечной
деятельности. Артериальное давление падает, пульс становится нитевидным. Дыха-
ние учащается. Больные испытывают тревогу, страх, беспокойство. Бывают бред и
эйфория. В предагональном периоде на коже могут появляться вторичные пустулез-
ные и геморрагические высыпания. Рвотные массы и кал могут содержать примесь
крови. Инкубационный период при кишечной разновидности сибирской язвы состав-
ляет от 1—2 до 3—5 сут, а продолжительность заболевания от момента появления пер-
вых клинических симптомов до смерти — от 1—2 до 4—6 сут (в среднем 3—4 сут) Обыч-
но диагноз первичной кишечной сибирской язвы устанавливается посмертно при
вскрытии трупов, а больные госпитализируются с весьма разнообразными диагноза-
ми: грипп, ангина, дифтерия, отравление пищевыми продуктами, токсическая маля-
рия, острый аппендицит, кишечная непроходимость, гастроэнтерит, холера, уремия,
перитонит, пневмония, менингит и т. д.
При употреблении в пищу сырого мяса или мясопродуктов иногда у людей наблю-
дается поражение верхних отделов пищеварительного тракта — орально-фарингеаль-
ная (ротоглоточная) разновидность сибирской язвы у человека. При этой разновид-
ности сибирской язвы инкубационный период составляет в среднем 42 ч (диапазон
от 2 до 144 ч). Средний период от момента появления клинических симптомов до гос-
питализации — 3,5 сут (диапазон от 1 до 6 дней) и среднее время пребывания в боль-
нице — 7 дней (диапазон от 7 до 10 сут) (рис. 3.15).
3.1. Сибирская язва
609
Рис. 3.15. Сибирская язва при энтеральном заражении В. anthracis. А. Орально-фарин-
геальная разновидность сибирской язвы у человека. 1 — 27-летний мужчина на 5 сут
после появления симптомов орально-фарингеальной сибирской язвы. Хорошо заметны
отек и гиперемия правой миндалины, распространяющиеся на окружающие ткани.
Белый участок показывает центр начавшегося поражения глотки; 2 — тот же пациент
на 9 сут после начала симптомов орально-фарингеальной сибирской язвы. Белые участ-
ки — развившиеся псевдомембранные образования, покрывающие язву. По Т. Sirisan-
thana, А. Е. Brown (2002). Б. Результаты компьютерной томографии (в/в введения конт-
растного материала) брюшной полости и малого таза у пациента с кишечной сибирской
язвой. 1 — выраженный асцит и концентрическое уплотнение стенки длинного сегмента
дистального отдела тонкого кишечника (показано стрелкой). Хорошо заметно увеличе-
ние большого количества лимфатических узлов корня брыжейки тонкого кишечника
(кончик стрелки); 2 — забрюшинное пространство. По М. S. Klempner et al. (2010)
Основные симптомы болезни при орально-фарингеальной разновидности сибир-
ской язвы: лихорадочное состояние (повышение температуры тела до 39°С и выше)
при поражении слизистой оболочки ротовой полости или глотки (опухание шеи из-
за увеличения лимфатических узлов и отека мягких тканей). На начальном этапе бо-
лезни в местах поражения (миндалины, небо, язычок, передние и задние дужки ро-
тоглотки) отмечаются отек и гиперемия; к концу первой недели в результате некроза
и изъязвления образуется беловатая бляшка (при лечении), которая в начале второй
недели болезни превращается в псевдомембрану, покрывающую язву. Летальность при
этой разновидности сибирской язвы (при лечении) составляет около 12 %.
610
Часть 3. Частная неправильная эпидемиология
Для иллюстрации клиники и патанатомии кишечной сибирской язвы привожу
краткие выписки из историй болезни, сделанные И. Л. Богдановым (1935).
1. И., 28 лет, поступила с 2,5-месячной беременностью в абортарий для производ-
ства аборта. На 3-й день после абразии была выписана в хорошем состоянии. На сле-
дующий день по выписке появилась головная боль, общая слабость, иногда обмороки
при вставании, тошнота. Больная слегла, а через два дня была доставлена снова в ги-
некологическую клинику уже в крайне тяжелом состоянии с жалобами на сильную
головную боль, частое наступление обморочного состояния и временами рвоту.
Больная хорошего сложения и питания. Пульс 120/мин, плохого наполнения, тя-
желое общее состояние. Живот мягкий, мало болезненный, нерезкий, асцит. Матка
немного увеличена (до 5-й нед беременности), наружный зев закрыт, выделений нет;
своды свободны, придатки не прощупываются. К 10 ч вечера внезапная потеря пуль-
са, рвота, сопорозное состояние. Ввиду подозрения на внутреннее кровоизлияние сде-
лана пробная лапаротомия. В брюшной полости большое количество прозрачной
выпотной жидкости. Цекальный отдел подвздошной кишки, слепая и особенно бры-
жейки кишок резко отечны, студенисты. В брыжейке тонких кишок имеется темно-
ватая опухоль величиной с кулак, брыжеечные железы значительно увеличены. В це-
кальном отделе подвздошной кишки имеются геморрагии. После операции пульс
продолжал падать, сознание не возвратилось, и через 2 ч больная скончалась, проле-
жав в клинике 6 ч. При вскрытии обнаружено: геморрагический летпоменингит, кар-
бункул в по, (взодшной кишке, геморрагический лимфаденит забрюшинных и брыже-
ечных желез. Посев крови из сердца дал чистую культуру сибиреязвенной палочки,
подтвердившую окончательный диагноз кишечной формы сибирской язвы. Провер-
ка культуры биологической пробой подтвердила бактериологический диагноз.
2. И., 38 лет, заболел 9 июля — жар, общая слабость, незначительная боль в живо-
те. Поступил на 5 сут болезни в крайне тяжелом состоянии с жалобами на головную
боль и боли в животе. В 1930 г. оперировался по поводу язвы желудка. При осмотре:
затемненное сознание, состояние тяжелейшей интоксикации. Пульс 124/мин, мягкий,
плохого наполнения, ритмичный. Тоны сердца глухие. Цианоз. Живот умеренно вздут,
болезненный, но без выраженных перитонеальных симптомов. Стул полуоформлен-
ный, с примесью небольшого количества крови. Рвота один раз. Селезенка увеличена
перкуторно. Менингеальные симптомы отсутствуют, зрачки сужены, не реагируют на
свет Быстро прогрессирующий упадок сердечной деятельности. За несколько часов до
смерти появилось чейнстоксовское дыхание. Температура 39 °C Смерть на 6 сут болез-
ни. Пролежал в отделении 11 ч.
На вскрытии: множественные карбункулы и очаговые геморрагические инфильт-
раты в тонких кишках и в верхнем отделе толстой кишки; явления серозного перито-
нита; геморрагический менингоэнцефалит; геморрагический лимфаденит забрюшин-
ных и брыжеечных лимфатических узлов.
Бактериоскопически в мазках из селезенки обнаружены капсульные палочки си-
бирской язвы, бактериологически также выделена культура этого микроба.
3. Щ. 51 год, поступила в клинику на 4 сут болезни в бессознательном состоянии.
Развитие заболевания быстрое, общая слабость, чувство жара. Больная производи,
впечатление отравленной: пульс очень частый, еле прощупывается, глухие тоны серд-
ца, дыхание учащенное. Живот не вздут, поноса и рвоты не было Полная прострация;
менингеальные симптомы отсутствуют, зрачки не реагируют на свет. Энергичное при-
менение сердечных средств не оказывало никакого эффекта. Смерть через 8 ч по по-
ступлении в отделение.
На секции: сибиреязвенный карбункул в верхнем отделе толстой кишки; серозный
перитонит; геморрагический лимфаденит забрюшинных и брыжеечных узлов; селе-
3.1. Сибирская язва
611
зенка увеличена, почти черного цвета. Из лимфатического узла и из селезенки через
посев на сахарный бульон выделена культура сибиреязвенной палочки.
4. Г., 31 года, поступил 19 июля. С 12 июля появились боли в животе, на 3-й день
слег в постель: температура поднялась до 39,9 °C, усилилась боль в животе, прогресси-
ровала общая слабость. Поноса не было; рвота была лишь один раз в день смерти.
Сознание потерял за 8-10 ч до смерти. Доставлен в клинику в бессознательном состо-
янии. Зрачки не расширены, не реагируют на свет. Пульс нитевидный, не поддающийся
счету. Тоны сердца еле прослушиваются. Цианоз. Перед смертью перемежающиеся
судороги, больной метался в постели. Пролежал в отделении 3,5 ч. Был сделан посев
крови на сахарный бульон, из которого на следующий день была выделена чистая куль-
тура сибиреязвенной палочки.
На секции: резко выраженный геморрагический менингоэнцефалит; карбункул в
толстой кишке с серозной инфильтрацией в окружающей ткани, на слизистой тонкой
кишки свежие и зарубцевавшиеся язвы туберкулезного характера, туберкулезный очаг
в легком; явления серозного перитонита. Из крови сердца, а также из пульпы селезен-
ки посевом на бульон выделена чистая культура палочки сибирской язвы (по эпиде-
миологическим показаниям труп был вскрыт через 3 ч после смерти).
5. К., 21 год, поступил на 4 сутболезни. На правом предплечье сибиреязвенная пу-
стула величиной с серебряный полтинник, с серозно-геморрагическим венчиком вок-
руг пустулы. Мало болезненный отек окружающей подкожной клетчатки и кожи, при-
пухлость и болезненность подмышечных желез. Общее состояние удовлетворительное.
Температура 38,0 °C.
При поступлении впрыснуто 200 см1 внутримышечно лечебной противосибире-
язвенной сыворотки. К вечеру того же дня введено добавочно еще 150 см3 сыворотки.
Через 2 дня температура упала до нормы, наступило резкое улучшение общего состоя-
ния и местных явлений, отек предплечья, припухлость и болезненность лимфатиче-
ских желез значительно уменьшились. Но на 4 сут снова подъем температуры до 37°C.
головная боль, общее недомогание На следующее утро общее состояние значительно
ухудшилось: пульс 120/мин, глухие тоны сердца, сильная головная боль; живот не-
сколько вздут, мягкий, болезненный при глубокой пальпации, стул жидкий, нечастый,
без крови. Последующий день принес еще большее ухудшение: нарастающий упадок
сердечной деятельности, цианоз; за несколько часов до смерти больной потерял со-
знание. На 3 сут от начала общего заражения — смерть.
На аутопсии: геморрагический лептоменингит, единичный карбункул в нижнем от-
резке тонкой кишки, геморрагический лимфаденит брыжеечных и забрюшинных уз-
лов. В мазках из брыжеечной железы и пульпы селезенки бактериоскопически найдены
бациллы сибирской язвы.
И. Л. Богданов (1938) отметил некоторые общие черты болезни, дающие повод
заподозрить кишечную форму антракса. Первое, на что, по его мнению, следует обра-
тить внимание врачу, это резкое несоответствие между крайне тяжелым состоянием
больного и отсутствием к тому локальных данных. При виде таких больных в разгаре
болезни в первую очередь возникает мысль о тяжелом обшем отравлении, идущем с
быстро прогрессирующим упадком сердечной деятельности. Второй признак: все боль-
ные отмечали в той или иной степени выраженные нарушения со стороны кишечни-
ка: тупые боли в животе, рвота, непостоянный жидкий стул. В последней стадии болез-
ни при тщательном исследовании больных удается обнаружить присутствие экссудата
в брюшной полости (действие токсина В. anthracis). Наконец третий признак: посто-
янные жалобы больных в разгаре болезни на жесточайшую головную боль, дающую
право думать об остром менингите, но протекающем, как правило, при отсутствии
объективных менингеальных симптомов.
612
Часть 3. Частная неправильная эпидемиология
На секции обращает на себя внимание то, что почти у всех умерших от кишечной
сибирской язвы находят картину резко выраженного геморрагического менинго-
энцефалита.
Септическая форма сибирской язвы встречается редко. Во всех случаях характери-
зуется чрезвычайной стремительностью нарастания интоксикации, обилием кожных
и внутренних кровоизлияний различной интенсивности и частым вовлечением в про-
цесс мозговых оболочек, что явилось основанием для выделения некоторыми авто-
рами менингеальной разновидности болезни. Болезнь начинается остро с потрясаю-
щего озноба и повышения температуры до 39—40°С. Уже в начальном периоде
наблюдаются выраженная тахикардия, тахипноэ, одышка. Нередко у больных имеют
место боли и чувство стеснения в груди, кашель с выделением пенистой кровянистой
мокроты. Физикально и рентгенологически определяют признаки пневмонии и вы-
потного плеврита (серозно-геморрагического). Часто, особенно при развитии инфек-
ционно-токсического шока, возникает геморрагический отек легких. Выделяемая
сольными мокрота свертывается в виде вишневого желе. В крови и мокроте обнару-
живают большое количество сибиреязвенных бактерий. У части больных появляют-
ся острые режушие боли в животе. К ним присоединяются тошнота, кровавая рвота
жидкий кровянистый стул. В последующем развивается парез кишечника, возможен
перитонит.
При развитии менин! оэнцефалита сознание больных становится спутанным
появляется менингеальная и очаговая симптоматика. Инфекционно-токсический
шок, отек и набухание головного мозга, желудочно-кишечное кровотечение и пери-
тонит могут явиться причиной летального исхода уже в первые дни болезни.
Прогноз при кожной форме сибирской язвы обычно благоприятный, при легоч-
ной, кишечной и септической формах — во всех случаях серьезный даже при своев-
ременно начатом лечении.
Диагностика сибирской язвы осуществляется на основе клинико-эпидемиологи-
ческих и лабораторных данных. Сибирскую язву дифференцируют с сапом, фурунку-
лами и карбункулами, чумой, туляремией, рожей, сепсисом иной этиологии.
Моделирование искусственного заражения В. anthracis. Наиболее обстоятельно ис-
следования по данному направлению обобщены в монографии Н. В. Литусова с со-
авт. (2002). Авторы изучали клинику, патогенез, патанатомию сибирской язвы при
парентеральном, ингаляционном и энтеральном введении спор В. anthracis различным
животным, включая приматов.
Подкожное инфицирование. В месте внедрения возбудителя сибирской язвы обезья-
нам (павианы гамадрилы) обычно обнаруживали серозный отек подкожной жиро-
вой клетчатки. В отдельных случаях происходило распространение отека из участка
заражения на другие области тела, расположенные на стороне внедрения возбудите-
ля. У почти половины животных, зараженных инъекцией в верхнюю конечность,
выявлен отек подмышечной клетчатки, переходивший на грудную клетку и набрюш-
ную стенку. Постоянным признаком сибирской язвы у обезьян и других эксперимен-
тальных животных являлся серозный или серозно-геморрагический регионарный
лимфаденит. У павианов гамадрилов, погибших после подкожного заражения в верх-
нюю конечность, выявлены поражения различных групп лимфатических узлов:
подмышечных — у 42 (95 %), трахеобронхиальных — у 15 (34 %), околоаортальных —
у 13 (29 %), брыжеечных — у 13 (27 %) и трахеальных — у 5 (11 %) животных.
Переход воспалительных изменений с лимфатических узлов на прилегающую к
ним клетчатку, как правило, обнаруживали при регионарном лимфадените. К посте-
3.1. Сибирская язва
613
янным патоморфологическим изменениям подкожного инфицирования В. anthracis
исследователи относят полнокровие печени, почек и селезенки, отмечаемое ими прак-
тически у всех животных. Среди остальных морфологических изменений с большой
частотой наблюдали полнокровие, отеки набухание головного мозга, кровоизлияния
во внутренние органы, медиастиниты и отек легких. У отдельных обезьян наблюдали
от двух до зрех геморрагических инфильтратов восходящей ободочной кишки, сероз-
ные плевриты и перикардиты. Формирования сибиреязвенного карбункула, харак-
терного для кожной формы заболевания человека, при подкожном способе зараже-
ния не происходи? .
Внутривенное инфицирование. Клинически сибирская язва протекает в форме пер-
вичного сепсиса. Рентгенологически могут быть обнаружены признаки медиастини-
та, наличие жидкости в плевральных полостях, расширение тени сердца за счет ги-
перкардиума и инфильтративных изменений различной интенсивности в легких.
Патологоанатомическое исследование 20 трупов павианов гамадрилов, погибших
после внутривенного заражения возбудителем в дозах от 10 до 1250 спор (средние сроки
гибели от 3,25 до 5 сут), показало, что у обезьян наиболее часто поражались лимфа-
тические узлы и селезенка. Как правило, серозное или серозно-геморрагическое вос-
паление захватывало от 3 до 5 групп лимфатических узлов. При внутривенном спосо-
бе заражения у павианов гамадрилов наиболее часто поражались околоаортальные и
рахеобронхиальные лимфатические узлы, несколько реже — брыжеечные и трахеаль-
ные, а подмышечные и поднижнечелюстные — лишь у отдельных животных.
Вовлечение в процесс клетчатки, окружающей лимфатические узлы, имелось толь-
ко у 5 (25 %) инфицированных В. anthracis павианов гамадрилов. У 4-х из них наблю-
дался отек клетчатки, прилегающей непосредственно к лимфатическим узлам груд-
ной части аорты, а у одного — клетчатки шеи возле пораженного подчелюстного
лимфатического узла. Селезенка у всех обезьян выглядела полнокровной и, как пра-
вило, увеличенной. Микроскопически в лимфатических узлах при серозно-геморра-
гическом лимфадените и в селезенке отмечали резкое обе знение тканей лимфоидны-
ми элементами, а также кровоизлияния и скопления бацилл.
В легких наиболее часто находили мелкие немногочисленные кровоизлия-
ния, расположенные диффузно в легочной паренхиме и под висцеральной плеврой.
Отек легких обнаружен у 20 % животных, погибших после внутривенного введения
им 1250 спор В. anthracis. У одной обезьяны наблюдали серозно-геморрагическую
пневмонию. При микрос копическом исследовании пораженных участков обнаружили
резкое утолщение межальвеолярных перегородок и уменьшение размеров альвеол,
полости которых на большом протяжении оказались заполненными эритроцитами.
Сибиреязвенные бациллы располагались в капиллярах межальвеолярных перегоро-
док. Изменения в сердце крайне незначительны и ограничивались мелкими крово-
излияниями под эпикардом. В печени и почках постоянно находили полнокровие и
дистрофические изменения. При микроскопии в почках обнаруживали явления не-
кротического нефроза. Изменения в ЖКТ найдены у двух (10 %) павианов гамадри-
лов. У одной из них в желудке и тонкой кишке имелись единичные мелкие кровоиз-
лияния, а у другой — слизистая оболочка толстой кишки была утолщена, отечна и
пропитана кровью. Повреждения поверхности толстой кишки не наблюдали.
Приведенные данные свидетельствуют о том, при заражении внутривенным вве-
дением спор возбудителя сибирской язвы на первый план выступают симптомы на-
растающего инфекционно-токсического шока с обилием кожных и внутренних кро-
воизлияний различной интенсивности и вовлечением в процесс мозговых оболочек.
614
Часть 3. Частная неправильная эпидемиология
Наиболее характерными патологоанатомическими находками являются серозно-ге-
моррагический полиаденит, полнокровие и дистрофические изменения в паренхима-
тозных органах и кровоизлияния в легкие.
Ингаляционное инфицирование. В эксперименте по ингаляционному заражению па-
вианов гамадрилов мелкодисперсным аэрозолем возбудителя сибирской язвы в дозах
5x102—ЗхЮ5 спор (штамм 4-7) гибель животных отмечалась на протяжении 20 сут
Основная часть животных (90 %) погибли в течение 8 сут (средний срок гибели
5,3 сут). Среди изменений, выявленных при наружном осмотре трупов погибших жи-
вотных, довольно часто отмечали цианоз видимых слизистых оболочек. В ряде слу-
чаев у отверстий рта и носа находили светлые пенистые, изредка окрашенные кро-
вью выделения.
В трахее и крупных бронхах наблюдалось только незначительное полнокровие
слизистой оболочки. В легких наиболее постоянными изменениями яв_1яются отек,
кровоизлияние и полнокровие. В одних случаях явления отека обнаруживали только
при микроскопическом исследовании; в других — при сдавливании легочной ткани с
поверхности разреза стекала кровянистая пенистая житкость. И наконец, при силь-
ном отеке подобную жидкость можно было обнаружить в просвете трахеи и бронхов,
а в ряде случаев — и у отверстий рта и носа. Кровоизлияния обычно были мелкими,
единичными или множественными, местами сливались в крупные очаги и распола-
гались диффузно в легочной ткани. Сибиреязвенная пневмония обнаружена у 4 %
павианов гамадрилов. При этом в половине наблюдаемых случаев поражение имело
очаговый характер. Количество очагов не превышало трех, а их диаметр достигал 3 см.
Пораженная легочная ткань была плотной, нара зрезе — безвоздушной и темно-крас-
ной. В гистологических срезах легких альвеолы на большем протяжении были запол-
нены эритроцитами с примесью нейтрофильных гранулоцитов. Просветы альвеол на
отдельных участках содержали скопления нейтрофильных гранулоцитов, местами —
отечную жидкость или глыбкообразное бесструктурное эозинофильное содержимое.
Межальвеолярные перегородки были утолщены, а их капилляры — полнокровны и
содержали множество бацилл, которые крайне редко можно было обнаружить в про-
светах альвеол.
Наблюдаемые пневмонии имели вторичное происхождение и развивались нафинс
предсушествуюшей патологии легких. Пораженные участки не превышали 0,5 см в
диаметре, были округлыми плотными темно-красного цвета. В центре узелков на раз-
резе ткань имела желтовато-белыь цвет. В околобронхиолярной ткани были видны
кровоизлияния, некрозы, инфильтрация нейтрофильными гранулоцитами и большое
количество сибиреязвенных бацилл.
Серозное или геморрагическое воспаление лимфатических узлов, расположенных
в грудной клетке, относится к наиболее постоянным изменениям при ингаляцион-
ной сибирской язве. При микроскопическом исследовании пораженных лимфати зес*
ких узлов к наиболее ранним изменениям относят некроз центров лимфатических
фолликулов, к которому по мере усиления инфекционного процесса присоединяют-
ся отек и кровоизлияния. Наиболее часто (78 %) вовлечены в процесс трахеобронхи
альные лимфатические узлы.
При переходе воспалительного процесса с лимфатических узлов на окружающую
их соединительнотканную клетчатку в ней появлялись студневидные бледно-желтые
изредка окрашенные кровью отеки. При микроскопическом исследовании поражен-
ных участков наблюдали набухание и разволокнение соединительнотканных волокон,
между которыми имелась отечная жидкость с незначительным количест вон нейтро-
3.7. Сибирская язва
615
фильных гранулоцитов, а также сибиреязвенные бациллы, локализовавшиеся как
среди измененных тканей, так и в кровеносных сосудах. При геморра! ическом вос-
палении описанные изменения дополнялись обширными кровоизлияниями. В Гболь-
шинстве случаев была затронута клетчатка, непосредственно прилегающая к лимфа-
тическим узлам (периаденит). Более распространенные поражения встречали реже.
Среди них можно назвать: отек клетчатки средостения (медиастинит), который сопро-
вождал поражения лимфатических узлов средостения, отек забрюшинной клетчатки,
связанный с изменениями лимфатических узлов в области брюшной части аорты, а
также отеки подкожной жировой клетчатки передней поверхности шеи, подмышеч-
ных впадин, передней грудной и брюшной стенок. Отеки шеи наблюла ги при пора-
жении поднижнечелюстных лиь фат ических узлов и иногда в случаях с резко выражен-
ным медиастинитом. У отдельных обезьян отек с шеи распространялся на переднюю
поверхность грядной клетки и даже на верхнюю треть передней брюшной стенки.
Отеки в области подмышечных впадин сопровожда ш воспалительный процесс в под-
мышечных лимфатических узлах и переходили на переднюю поверхность грудной
клетки, а в отдельных случаях — на брюшную стенку.
К обычным находкам у животных, погибших после ингаляционного заражения,
Н. В. Литясов с соавт. (2002) относят выраженные в различной степени экссудатив-
ные плевриты серозного или серозно-геморрагического характера. Резко выраженный
плеврит, при котором пристеночная плевра была желеобразно отечной, а в каждой
плевральной полости содержалось около 500 см3 мутноватой жидкости. Плевриты и
перикардиты в основном сочетались с трахеобронхиальным периаденитом и медиас-
тинитом. Пораженные листки плевры и перикарда были утолщены, желеобразно отеч
ны. В плевральных и перикардиальных полостях содержалась чаще серозная, реже —
геморрагическая жидкость. Иногда плевриты были серозно-фибринозными. У боль-
шинства животных процесс был двусторонним При микроскопическом исследова-
нии пораженные участки плевры и перикарда были утолщены, пропитаны отечной
жидкостью и незначительно инфильтрированы нейтрофильными гранулоцитами.
Изменения в сердце при ингаляционной сибирской язве непостоянны и ограни-
чиваются кровоизлияниями под эпикардом и в сердечной мышце.
Воспалительные изменения небных миндалин при ингаляционной сибирской
язве наблюдали только изредка у павианов гамадрилов.
Среди других внутренних органов полнокровие селезенки у ингаляционно зара-
женных животных встречалось наиболее часто. Обычно капсула органа была гладкой
и напряженной, а ткань плотной и темно-красной. Гистологические изменения в се-
лезенке идентичны поражениям лимфатических узлов, но выражены более резко.
Среди патологии головного мозга обычны полнокровие, кровоизлияния, отек и
набухание мозгового вещества. При гистологическом исследовании мягкие мозговые
ооолочки были отечны, в их тканях имелись кровоизлияния и незначительная ин-
фильтрация нейтрофильными гранулоцитами. Бациллы локализовались среди изме-
ненных тканей и в просветах кровеносных сосудов вещества мозга.
Поражения органов эндокринной системы встречаются крайне редко.
Описания поражений ЖКТ при ингаляционной сибирской язве имеются лишь в
единичных работах. Авторы в единичных случаях наблюдали поражение желудка.
Тонкая кишка у всех животных была интактной. Поражения толстой кишки сопро-
вождались геморрагическим брыжеечным лимфаденитом, а в одном случае — гемор-
рагическим лимфангитом. Воспалительные изменения в брыжеечных лимфатических
узлах встречались значительно чаще и в ряде случаев не связаны с поражениями ки-
616
Часть 3. Частная неправильная эпидемиология
шечника. Развитие воспалительных изменений в таких узлах было вызвано гемато-
генным путем, поскольку бациллы располагались в кровеносных сосудах.
Н. В. Литусовым с соавт. (2002) сделано еще одно любопытное наблюдение.
Используя большую выборку экспериментальных животных в опытах по ингаляци-
онному заражению (более 100 павианов гамадрилов), они обнаружили различия в па-
тологических процессах, вызванных споровыми и вегетативными формами сибире-
язвенного микроба. У обезьян, погибших после заражения спорами, воспалительный
процесс в лимфатических узлах в основном был i еморрагическим (97 % по отноше-
нию ко всем пораженным группам лимфатических узлов). У этих животных чаще
встречали кровоизлияния во внутренних органах, дистрофические изменения в пе-
чени и почках, соскоб пульпы селезенки, плевриты, перикардиты и отек забрюшин-
ной клетчатки.
У павианов гамадрилов, погибших после заражения вегетативными клетками
возбудителя, вследствие более быстрого течения инфекционного процесса, воспале-
ние лимфатических узлов чаще имело серозный характер (52 % по отношению ко всем
пораженным группам лимфатических узлов) и реже встречались остальные из указан-
ных изменений. Увеличение числа случаев с поражениями различных групп лимфа-
тических узлов, а также тканей, непосредственно прилегающих к таким лимфатиче-
ских узлам (периаденит, медиастинит), свидетельствовало о более быстром прижив-
лении и распространении возбудителя в организме таких животных, вследствие чего
количество животных с отеком легких в опытной группе было больше, чем в конт-
рольной группе обезьян. Обнаруженные отличия авторы объясняют более длительным
течением болезни при инфицировании спорами В. anthracis, благодаря чему эти сим-
птомы успевали развиться. Вегетативные же клетки, попадая в легкие, выделяют си-
биреязвенный токсин, который угнетает фагоцитоз, что способствует быстрому про-
никновению возбудителя в организм и его размножению.
Таким образом, в патологоанатомической картине ингаляционной сибирской
язвы наиболее постоянно наблюдаются: поражения лимфатических узлов, регионар-
ных по отношению к легким, периатенит и полнокровие паренхиматозных органов.
С меньшей частотой встречаются отек и набухание головного мозга, отек легких, ме-
диастинит, перикардит, кровоизлияния, огек подкожной жировой и забрюшинной
клетчатки, а также плеврит. В единичных случаях обнаруживаются менингит, тонзил-
лит, язвенный колит и пневмония. Первичные изменения в органах дыхания в виде
пневмоний и эрозий трахеобронхиального дерева у животных при ингаляционном
заражении не развиваются. Поражения этих органов, наблюдаемые в основном у па-
вианов гамадрилов, имели вторичное происхождение
Энтеральное инфицирование. Заражение павианов гамадрилов осуществлялось
внутрижелудочно, через зонд. На вскрытии наиболее часто изменения наблюдали в
тонкой кишке, слизистая оболочка которой постоянно была гиперемированной и
набухшей, а в просвете нередко имелась кровянистая слизь. При осмотре кишки об-
наруживали очаговые утолщения, на наружной поверхности которых были видны
расширенные и полнокровные кровеносные сосуды, а на bhj гренней — набухшие
пейеровы бляшки. Утолщения локализовались преимущественно в области подвздош-
ной кишки и несколько реже — в тощей кишке. В ряде случаев у павианов гамадри-
лов в этих местах сквозь серозную оболочку просвечивали пятна темно-красного цвета
величиной от 0,3 до 2 см, округлой или овальной формы, соответственно которым на
слизистой оболочке находили эрозии или язвы. Поверхность эрозий имела зернис-
тый вид и обычно была покрыта сероватым налетом.
3.1. Сибирская язва
617
Язвы имели округлую форму, неровные или гладкие края, возвышающиеся над
поверхностью слизистой оболочки, и темно-красное дно, которым служила подсли-
зистая основа или мышечная оболочка кишки. При микроскопическом исследовании
эрозивно-язвенных поражений наблюдали некроз слизистой оболочки, захватываю-
щий иногда подслизистую основу. Помимо эрозий и язв, в ряде случаев в тонкой кишке
(сгречали множественные мелкие кровоизлияния, располагавшиеся на участках, ог-
раниченных размерами 1—2 см.
Изменения, наблюдаемые в желудке, были менее выражены и в основном огра-
ничивались полнокровием кровеносных сосудов и небольшим набуханием слизистой
оболочки.
Поражения толстой кишки встречались лишь у отдельных животных и, вероятнее
всего, имели вторичное происхождение, поскольку они примерно с такой же часто-
той наблюдаются и при других способах заражения.
Среди органов лимфатической системы наиболее постоянно наблюдали серозное
или геморрагическое воспаление лимфатических узлов, расположенных у корня бры-
жейки тонкой кишки, и поражение селезенки, которое выражалось в полнокровии ее
тканей и некрозе лимфоидных элементов, сопровождаемых увеличением органа.
Среди лимфатических узлов других областей тела в процесс наиболее часто вовлека-
лись лимфатические узлы, расположенные в средостении (трахеобронхиальные, око-
югортальные). С меньшей частотой наблюдали поражение шейных, подмышечных
и поднижнечелюстных лимфатических узлов.
Помимо перечисленных изменений, Н. В. Литусов с соавт. (2002) отмечают пол-
нокровие печени и почек, а также плеврит, перикардит, отек легких, отек подкожной
жировой клетчатки шеи, асцит и единичные мелкие кровоизлияния под висцераль-
ной плеврой, эпикардом и в подкожной жировой клетчатке.
Клинические примеры искусственно вызванной сибирской язвы Ниже я приведу
примеры течения сибирской язвы, возникшей у людей в результате парентерального
введения спор В. anthracis и преднамеренного распространения сухой рецептуры
В. anthracis в окружающей человека среде (ингаляционная и кожная формы).
Парэнтералъная форма сибирской язвы. Вспышка парентеральной сибирской язвы
описана И. Дмитриевым (1928). Дежурным врачом губнарбольницы (Курск) 14 янва-
ря 1927 г., во время утреннего обхода Пастеровского общежития, среди пациентов
констатировано массовое заболевание, похожее на резкую анафилаксию с темпера-
турой до 39,6JC и с отеками, захватывающими брюшную полость и мошонку. Дня
расследования причин массового заболевания пастеровских больных была создана
комиссия во главе с директором Санитарно-бактериологического института В. И. Ва-
сильяновским. Комиссия осмотрела 15 больных Пастеровской станции, из которых
4 находились уже в хирургическом отделении, а 11 в здании общежития. Из бесед с
больными выяснилось, что заболели они с 10 января 1927 г. после антирабических
прививок; на месте укола появилась боль, отек, темпера ура до 40°С, а у некоторых и
рвота. У одного из больных, находившихся в хирургическом отделении, из инфильт-
рата путем прокола был взят жидкий прозрачный секрет, оказавшийся при бактери-
ологическом исследовании стерильным, почему комиссия предположила наличие
какой-то токсемии.
15 января в 6 ч 30 мин утра больная Анна Б. умерла при явлениях кол тапса и рез-
кого упадка сердечной деятельности. Кроме этого, срочно созванным консилиумом
врачей семь больных были найдены в безнадежном сос гоянии — без пульса и один
чувствовал себя плохо. В этот же день умер второй больной Б., вслед за ним Анна С.,
618
Часть 3. Частная неправильная эпидемиология
Илья П., Павел С., Николай Г. После получения сообщения о пяти смертельных слу-
чаях, вечером в тот же день состоялось совещание из представителей разных ведомств
и лечащих врачей. На данном совещании выяснилось, что смерть наступает быстро,
при резком упадке сердечной деятельности, в полном сознании, при отсутствии моз-
говых явлений, причем у некоторых отмечалась рвота и температура до 40 °C. На ме-
сте укола появлялся глубокий и болезненный отек, захватывавший половину живота
и мошонку или большую срамную губу. Решено было сконцентрировать всех бальных
в мужском и женском терапевтических отделениях и произвести детальное судебно-
медицинское вскрытие умерших с взятием для исследования крови, эксудата отечных
тканей, мозга, мочи, печени и других органов. Вскрытие трупа и последующие бакте-
риологические исследования установили, что причиной смерти послужило заражение
возбудителем сибирской язвы. Всего выявлено 27 заболевших сибирской язвой, из них
18 умерли. Ниже привожу краткую выписку из типичной истории болезни.
Больной Николай Г., 18 лет. Диагноз — отравление после укола от бешенства (ан-
тракс). Поступил 12 января. Получил 23 прививки. 3 дня назад после последнего уко-
ла появились боли в левой половине передней брюшной стенки вверху и в левой под-
вздошной области S. praes. В левом верхнем квадранте передней брюшной стенки
припухлость. В левой подвздошной области стенки живота инфильтрат. 13 января са-
мочувствие удовлетворительное, пульс 84, сердце без изменений, легкие — норма.
Пробной пункцией получена прозрачная жидкость, посланная для бактериолотичес-
кого исследования. 14 января отек меньше, вечером при нормальной температуре по-
явился пот, явления упадка сил, пульс 92/мин, правильный. В ночь с 14 на 15 января
жалобы на общую слабость, недомогание, головные боли, пульс 110/мин, мягкий,
тошнота, пот на лице, консилиумом назначено было физиологическое вливание с ад-
реналином, камфора, грелки. 16 января больной слаб, возбужден, пульс еле прощупы-
вается. В 5 ч 30 мин вечера больной умер.
В обобщении И. Дмитриева (1928) клиника инъекционной сибирской язвы, сле-
дующая:
1) наличие у больных болезненного отека, быстро распространявшегося от места
инъекции на более или менее значительное прос гранствс. В одном случае отек зани-
мал всю левую половину живота до левого подреберья, в другом спускался в область
наружных гениталий и передней поверхности бедра. Форма отеков в большинстве
овальная, в одних случаях не резко выдавалась над уровнем нормальной кожи, в дру-
гих — наблюдалось чрезвычайно резкое набухание всей кожи живота, что на первый
взгляд создавало впечатление сильно растянутого водянкой живота. На ощупь полу-
чалось впечатление тестоватости, местами — некоторой упругости, а то и деревянис-
тости При надавливании пальцем ямка тотчас же сглаживалась. Отек достигал в те-
чение 4 сут максиму ча и рассасывался чрезвычайно медленно; отечная кожа в течение
первых двух суток всегда была бледной, без всякого разнообразия оттенков. Потом
принимала розовый оттенок, дальше ярко-красный, а затем медленно приобретала
нормальную окраску. Отек мошонки наблюдался в тех впучаях*, когда В. anthracis была
случайно введена в нижнюю часть живота; в одном случае отек сопровождался боль-
шим пузырем, давшим карбункул;
2) высокая температура до 39—40 °C, обложенный язык. Увеличение селезенки не
констатировано, последнее потому, что в трех наиболее тяжелых случаях отек распо-
лагался слева и препятствовал пальпации;
3) жалобы на тошноту, головокружение и общую слабость, у двух наиболее тяже-
лых больных отмечалась упорная рвота;
3.1 Сибирская язва
619
4) быстрый упадок сосудистого тонуса и сердечной деятельности, причем обыч-
ные возбуждающие (камфора) никакого эффекта не давали; быстрое течение болез-
ни со см< ртельным исходом через 48—60 ч после начала заболевания; миокардиты в
Г' случаях;
5) цианоз, одышка, возбужденное состояние в последние часы жизни;
6) со стороны нервной системы ничего патологического не наблюдалось, кроме
головных болей и иногда 1аллюцинаиий;
7) короткий период инкубации: больные, пострадавшие от прививки, сделанной
10 января, обнаруживали первые признаки болезни 13 января.
На секции сердечные мышцы и ткань печени оказались дряблыми, селезенка уве-
личенной и в одном случае ткань ее плотна, в крови найдены сибиреязвенные бацил-
1ы. Чего-либо характерного для сибирской язвы не отмечалось.
Каким образом серия антирабической вакцины была контаминирована В. anthra-
cis, не установлено. Доктора В. И. Вас ильяновского следственные власти привлекли
к уголовной ответственности по обвинению в халатном отношении к исполнению
возложенных на него по службе обязанностей, но суд его оправдал.
Кожная форма сибирской язвы. Ниже по работе A. Freedman et al. (2002) рассмот-
рим случай кожной сибирской язвы, вызванный террористическим применением
рецептуры спор В. anthracis в сентябре — октябре 2001 г. в США (см. разд. 1.12 4), ока-
завшийся наиболее трудным для диагностики и лечения.
Случай 7*. 7-месячный, ранее здоровый мальчик, белый, был госпитализирован
1 октября 2001 г. За два дня до госпитализации на проксимально-медиальной поверх-
ности левой верхней конечности у него было обнаружено безболезненное красное
пятно с припухлостью. В последующие 24 ч рука стала быстро отекать, пятно превра-
тилось в папулу, и появились незначительные серозные выделения. Однако у больно-
го не было жара и явной боли или каких-либо других общих симптомов. До госпита-
лизации, на основании предположительного диагноза «целлюлит», ребенка лечили
амоксициллином/клавуланатом калия, но после третьей дозы, в связи с увеличением
припухлости и отделяемого из места поражения и затруднением перорального введе-
ния препаратов, ребенка пришлось госпитализировать.
В истории болезни ребенка было записано, что незадолго до происшедшего, с ним
гуляли в нью-йоркском парке и что накануне появления у него первых симптомов за-
болевания он пробыл в течение часа со своей матерью на ее рабочем месте, в помеще-
нии национальной телевизионной службы новостей.
При поступлении ребенок был бодрым, лихорадки и явных признаков недомога-
ния не наблюдалось В результате лабораторных исследований обнаружены значитель-
ный лейкоцитоз и гипонатриемия. Кровь не была отправлена для культивирования,
однако ребенку был назначен ампициллин/сульбактам внутривенно. После хирурги-
ческого разреза и дренажа, осуществленных под местной анестезией, не было выяв-
лено скрытого абсцесса, но из поражения выделялась темно-красная жидкость. Бак-
терии не культивировали.
На второй день пребывания в стационаре на левой руке мальчика был обнаружен
плотный, не вдав [ивающийся, безболезненный отек с темно-красным пятном, при-
мерно 2—3 см в диаметре. Из поражения обильно выделялась серозная жидкость жел-
того цвета, и более бледная эритема распространялась по передней поверхности груди
до грудины. Пальпация не выявила подмышечной аденопатии. В связи с гипонатрие-
1 Здесь и далее порядковая нумерация случаев сибирской язвы биотеррористического акта
сентября — октября 2001 г. соответствует их нумерации в табл. 1.11.
620
Часть 3. Частная неправильная эпидемиология
мией был ограничен прием жидкости, и в дополнение к антибиотикотерапии назна-
чен клидамицин. Была получена консультация у врача-инфекциониста. Окраска по
Граму отделяемого из поражения не выявила ни лейкоцитов, ни микроорганизмов. При
дифференциальной диагностике рассматривали инфекцию кости, мягкой ткани или
того и другого; укус ядовитого паука и закупоривающее новообразование. В результате
ультразвукового исследования левой верхней конечности были обнаружены диффуз-
ное воспаление без абсцесса и минимальная подмышечная лимфаденопатия. Доппле-
ровские исследования исключили возможность тромбоза глубокой вены или какого-
либо другого поражения сосудов конечности. Позднее в этот же день у больного повы-
силась температура (39,2 °C) и развилась значительная тромбоцитопения
В последующие два дня отек руки немного уменьшился, в центре поражения был
обнаружен 3-мм некротизированный участок, и в левой передневерхней части грудной
клетки и в подмышечной ямке появились петехии. Гематокритное число уменьшилось
до 23,3 %, при этом сохранялись слабая лихорадка, гипонатриемия, тромбоцитопения
и лейкоцитоз, но уже со значительным количеством палочкоядерных нейтрофильных
лейкоцитов. Из-за невозможности внутривенного введения цефалексин и клиндами-
цин стали вводить перорально.
К пятому дню температура снизилась, и отек руки заметно уменьшился. Пораже-
ние представляло собой очерченную эритематозную бляшку (4,5—5 см) с расположен-
ным в центре струпом менее 1 см. Магнитно-резонансное изображение верхней конеч-
ности продемонстрировало обширное воспаление мягкой ткани, распространяющееся
от левой боковой стенки грудной клетки до кисти руки, однако повреждения кости,
скопления газов или жидкости в мягкой ткани или новообразования не наблюдалось.
Гематокритное число уменьшилось до 18,7 %. Уровень фермента глюкоза-6-фосфат-
дегидрогеназы был нормальным, но мазок периферической крови обнаружил наличие
шизоцитов и фрагментарных эритроцитов. Уровень сывороточной лактатдегидроге-
назы, превышающий 5000 ЕД/л, наряду с незначительным увеличением уровней азо-
та и креатинина в мочевине сыворотки, свидетельствовал о микроангиопатической
гемолитической анемии.
К этому моменту был поставлен диагноз — «кожный и системный локсосцелизм»
(отравление ядом пауков из рода Loxosceles), поскольку течение болезни и развитие
кожного поражения в большей степени соответствовали отравлению животным ядом,
а не инфекционному процессу. Консультация врача-дерматолога подтвердила правиль-
ность этого диагноза. Было начато лечение преднизолоном (перорально), а антибио-
тики были отменены.
Хотя в последующие несколько суток отек руки спал, и температура оставалась нор-
мальной, картина крови ухудшилась. В связи с уменьшением гематокритного числа
до 14,3 % и сопутствующей тахикардией, потребовалисьдва переливания 15мл/кг эрит-
роцитарной массы. Была обнаружена коагулопатия при сохраняющейся тромбоци-
топении и наличии повышенных уровней D-димера и продуктов распада фибрина.
Персистирующая гипофибриногенемия потребовала переливания 4 ЕД криопреци-
питата. Обнаружены почечная недостаточность с повышенным уровнем азота в мо-
чевине сыворотки, гематурия, протеинурия, непродолжительная олигурия и гиперто-
ния (систолическое кровяное давление — 130 мм рт. ст. и диастолическое кровяное
давление — 85 мм рт. ст.). На 12 сут результаты лабораторных исследований нормали-
зовались, и состояние больного было клинически стабильным. Однако в центре кож-
ного поражения оставался 2-см черный струп
В тот день (12.10.2001) в Нью-Йорке был зарегистрирован первый случай кожной
формы сибирской язвы (2 и 6; см. табл. 1.11), и отдел здравоохранения Нью-Йорка был
предупрежден о возможном заражении этого ребенка сибирской язвой. На следующий
3.1. Сибирская язва
621
день была произведена биопсия пораженного участка, и два образца кожи, а также
кровь, полученная на 2 сут после госпитализации, были отправлены в Центры по борь-
бе и предупреждению болезней для диагностики посредством ПЦР и иммунопатоло-
гического исследования соответственно. Через два дня проведенные тесты дали поло-
жительные результаты в отношении В. anthracis. в пробе сыворотки было обнаружено
значительное количество сибиреязвенной ДНК, а иммуногистохимический анализ
выявил наличие в образцах ткани фрагментарных сибиреязвенных бацилл. При иссле-
довании методом вестерн-блота проб сыворотки, полученных на 13 сут болезни, была
обнаружена как реакция IgM на протективный антиген сибиреязвенного токсина, так
и реакция IgG.
На 20-й день болезни реакция IgM ослабла, но реакция IgG усилилась и распрост-
ранилась на фактор отека и летальный фактор сибиреязвенного токсина. На 17-й день
больной был выписан, ему был назначен ципрофлоксацин перорально. Количество
тромбоцитов и уровень фибриногена были в норме, но слабый гемолиз сохранялся.
После 2-недельного приема ципрофлоксацина, в качестве антибиотика стали исполь-
зовать амоксициллин. В течение последующих двух недель, через 30 дней после госпи-
тализации, гематурия, анемия и повышенные уровни D-димера постепенно исчезли.
Уровни азота и креатинина в мочевине сыворотки нормализовались через 12 дней после
госпитализации, а кожное поражение зажило, оставив к 60 дню слабо заметный рубец.
Клиническое течение кожной формы сибирской язвы у детей такое же, как и у
взрослых. Однако в рассмотренном случае болезнь осложнилась серьезными гемато-
логическими нарушениями, персистирующей гипонатриемией, признаками тяжелого
системного поражения. Признаки системного заболевания проявились через 36—48 ч
после начала антибиотикотерапии. Обнаружение В. anthracis в периферической кро-
ви свидетельствовало о том, что у больного уже имелся высокий уровень бактериемии,
хотя у него не было повышенной температуры, и результаты лабораторного исследо-
вания были в основном в норме. Роль хирургической обработки пораженной области
в распространении бактерий или токсина неясна.
Многие из признаков и симптомов болезни, включая отек, лихорадку, лейкоци-
тоз, тромбоцитопению, гемолиз, почечную недостаточность и диссеминированное
внутрисосудистое свертывание, которые редко ассоциируются с кожной формой си-
бирской язвы, врачи связывали с отравлением ядом Loxosceles reclusa. Хотя случаи
обнаружения этого паука в Нью-Йорке являются редкими, но кожная форма сибир-
ской язвы до октября 2001 г. ни разу не была диагностирована у младенца.
Ингаляционная форма сибирской язвы. Ниже по работам J. A. Jemigan et al. (2001) и
L. A. Barakat et al. (2002) рассмотрим все 11 случаев ингаляционной сибирской язвы,
вызванной применением рецептуры спор В. anthracis в сентябре — октябре 2001 г. в
США (см. разд. 1.12.4). Их номера приведены в соответствие с табл. 1.11.
Случай 4. Вечером 24 сентября 2001 г. 73-летний мужчина испанского происхож-
дения, почтовый клерк газеты, который доставлял почту пациенту, болезнь которого
описана как «случай 8», почувствмвал усталость На следующий день у него начался
сухой кашель, появилась перемежающая лихорадка, ринорея и конъюнктивит обоих
глаз. С 28 сентября по 1 октября постепенно усиливался кашель, общая слабость пе-
решла в летаргическое состояние, появились выраженные одышка, лихорадка, испа-
рина. В эпигастральной области появилась боль, сопровождающаяся рвотой, что выз-
вало беспокойство у членов его семьи. У него не было хронических болезней, за
исктючением быстро прошедшего приступа ишемической недостаточности в августе
2001 г. Он не курил. Больного госпитализировали 1 октября. Температура при поступ-
622
Часть 3. Частная неправильная эпидемиология
лении 38,5 °C, частота сердцебиения 109/мин, частота дыхания 20/мин, артериальное
давление 108/61 мм рт. ст. Он имел двухстороннюю конъюнктивальную инфекцию и
билатеральные пульмонарные хрипы. Исследования, включающие оценку функций
нервной системы, не дали ничего существенного. Поражений кожи у больного не было.
Лабораторные исследования не выявили патологических изменений в количестве бе-
лых и красных клеток крови и тромбоцитов, а так же среди химических компонентов
сыворотки за исключением сниженного (по сравнению с нормой) количества альбу-
минов (гипоальбуминемия), и повышенного уровня печеночных трансаминаз, близ-
кой к границе нормы гипонатриемии и увеличения содержания креатина. Анализ га-
зового состава артериальной крови свидетельствовал о гипоксии. Кровяные культуры,
которые попытались получить в госпитале на 2 сут после начала лечения антибиотика-
ми, не дали роста. Рентгенологическое исследование грудной клетки показало наличие
в верхней и нижней долях левого легкого инфильтратов, сходных с пневмоническими,
и небольшой плевральный выпот (рис. 3.16).
Рис. 3.16. Случай 4. Рентгенологическое исследование грудной клетки. Пока-
зывает диффузное затемнение, указывающее на пневмонию всего левого лег-
кого. Признаков медиастинального расширения нет. По!. A. Jemigan etal. (2001)
Медиастинального расширения не наблюдали. Бальному первоначально вводили
внутривенно азитромицин (azithromycin); в последующем были добавлены цефотак-
сим и ципрофлоксацин. Из первых смывов, полученных 5 октября, удалось вырастить
В. anthracis. Компьютерная томография (СТ) грудной клетки показала билатеральные
выпоты (effusions) и мультилобарную легочную консолидацию (multilobar pulmonary
consolidation), но без значительной медиастинальной лимфоаденопатии (рис. 3.17).
В образцах ткани, полученных трахеобронхиальной биопсией, с помощью имму-
ногистохимического окрашивания обнаружены капсула и антиген клеточной стенки
В. anthracis. Такие же тесты, выполненные в отношении возбудителей легионеллезов
(Legionella spp.), кислотоустойчивых бактерий, Pneumocystis carinii. Chlamydia spp.,
Leptospira, Hantavirus и отдельных вирусных патогенов были отрицательными.
Левосторонним торакоцентезом была отобрана серознокровянистая жидкость,
положительная в ПЦР на ДНК В. anthracis. Бронхоскопия показала кровянистые сек-
реты в правой нижней доле и левом легком с отдельными участками мукозальной ги-
перемии, «крапчатости» и воспаления. Из бронхиальных смывов и жидкости плевраль-
ного выпота получить культуру В. anthracis не удалось.
3.1. Сибирская язва
623
Рис. 3.17. Случай 4. Компьютерная томография грудной клетки. Показы-
вает билатеральную пульмонарную консотидацию и плевральные выпоты.
По J. A. Jemigan et al. (2001)
Во время госпитального лечения у больного периодически возникала суправент-
рикулярная тахикардия с гипотензией, максимальное количество лейкоцитов (WBC)
составило 26800/мм3, поьгорно сформировался левый плевральный выпот, который
удален торакоцентезом, и в этот участок была вставлена трубочка для дренажа.
Плевральная жидкость, отобранная при повторном торакоцентезе, была положи-
тельна на Д НК В. anthracis в П ЦР. В цитологических препаратах, полученных из плев-
ральной жидкости и плевральной биопсией, методом иммуногистохимического окра-
шивания показано наличие капсульного антигена и антигена клеточной стенки
В. anthracis. При исследовании образцов сыворотки с помощью энзимсвязанного им-
муносорбентного анализа (ELISA) было обнаружено более чем четырехкратное уве-
личение уровня сывороточных антител (IgG) к протективному антигену сибиреязвен-
ного токсина. Постепенно состояние больного улучшилось и 23 октября 2001 г. он был
выписан из госпиталя с назначением орального приема ципрофлоксацина.
Случай 8. Второго октября 2001 г. 63-летний фоторедактор, белый мужчина, рабо-
тавший в газете штата Флорида, проснулся с ощущением тошноты, перешедшей в рво-
ту, и машиной «скорой помощи» был доставлен в больницу для наблюдения. Его бо-
лезнь началась 27 сентября с недомогания, чувства усталости, лихорадки, озноба, ано-
рексии и потливости. В его медицинской книжке не упомянуты такие симптомы, как
кашель, ощущение боли в грудной клетке и животе, миалгия, удушье, диарея, кожные
поражения. В последней медицинской истории отмечены гипертония, кардиоваску-
лярное заболевание, подагра. Он не курил. Больной был встревожен и интерактивен,
но говорил нелепости. Температура была 39,2°С, частота сердцебиения — 109/мин.
Артериальное давление и частота дыхания были в норме. При первом исследовании
легкие, сердце и брюшная полость не имели отклонений от нормы. Больной не ориен-
тировался в пространстве и во времени, не понимал обращений к нему персонала кли-
ники. Сделанные при поступлении в клинику стандартные лабораторные исследо-
вания не дали никакой информации, позволяющей предположить диагноз болезни, так
все показатели были в пределах нормы, в том числе и количество лейкоцитов. Однако
количество тромбоцитов было низким, из результатов биохимических исследований
обпатили на себя внимания гипонатриемия (почти у границ нормы) и повышенный
624
Часть 3. Частная неправильная эпидемиология
уровень общего билирубина. Имелся незначительный метаболический ацидоз. Рент-
генографическое исследование грудной клетки показало верхнее расширение средос-
тенья и, возможно, небольшой плевральный выпот слева (рис. 3.18).
Рис. 3.18. Случай 8. Первая рентгенограмма. Показывает верхнее расши-
рение средостенья и, возможно, небольшой плевральный выпот слева.
По J. A. Jemigan et al. (2001)
В цереброспинальной жидкости лейкоцитов 4750/мл (81 % нейтрофилов), крас-
ных клеток 1375/мл, глюкозы 57 мг/децилитр (сывороточной глюкозы 174 мг/деци-
литр) и белков 666 мг/децилитр. Микроскопическое исследование цереброспиналь-
ной жидкости (ликвор) показало наличие большого количества грамположительных
палочек (рис. 3.19).
Рис. 3.19. Случай 8. Окрашивание по Граму цереброспинальной жидко-
сти. Показывает наличие цепочек грамположительных бацилл, похожих
на В. anthracis. По J. A. Jemigan et al (2001)
В. anthracis был выделен из ликвора после 7 ч инкубирования и в кровяной культуре
после 24 ч инкубирования. Больной был помешен в госпиталь с диагнозом менингит.
После одной дозы цефотаксима (cefotaxime) начато лечение комплексом антибиоти-
ков, включающих цефтазидим (ceftazidime), гентамицин (gentamicin), метронидазол
(metronidazole) и триметоприм-сульфаметоксазол (trimethoprim-sulfamethoxazole).
Вскоре после поступления больного в стационар он был интубирован для сохранения
проходимости дыхательных путей. На второй день госпитализации ему назначены пе-
нициллин G (penicillin G), левофлоксацин (levofloxacin) и клиндамицин (clindamycin).
3.1. Сибирская язва
625
Ампициллин, цефтазидим и триметоприм-сульфаметоксазол были отменены. Однако
температура у больного сохранялась, и он не реагировал ни на какие раздражители. Его
состояние продолжало ухудшаться, артериальное давление падало, почечная недоста-
точность нарастала. Больной умер 5 октября 2001 г. Аутопсические находки включают
геморрагические медиастинальные лимфадениты. Иммунохимическое окрашивание
показало диссеминацию В. anthracis по многим органам.
Случай 11. 14 октября почувствовала себя больной 56-летняя женщина афроаме-
риканского происхождения, работавшая сортировщицей почты в Гамильтоне (Нью-
Джерси). Болезнь началась с ощущения повышенной температуры тела (не измеряла)
и озноба, однако прием аспирина облегчения ей не принес. На следующий день у боль-
ной появились обильный пот и диарея, но вскоре они прошли. Через два дня у нее
поднялась температура до 38,4°С. Ее подъем сопровождался ознобом, развилась голов-
ная боль, больная почувствовала упадок сил. В этот же день у нее появился сухой ка-
шель, она не могла глубоко дышать из-за болей в грудной клетке на вдохе. Болей в горле
или ринореи у нее не было. 19 октября из-за упорной лихорадки и усиления болей в
грудной клетке она была вынуждена обратиться в местный кабинет неотложной по-
мощи. На момент обращения женщина выглядела очень больной и испытывала труд-
ности при дыхании, температура 38,4°C, частота сердечных ударов 120/мин. артери-
альное давление 159/95 мм рт. ст., частота дыхания 18/мин.
Дыхательные звуки на обоих основаниях были ослабленными, и она имела ранее
леченные струпы на шее 0,5—1,0 см. Первый подсчет лейкоцитарной формулы выя-
вил норму, за исключением некоторого увеличения количества палочковых форм. Ге-
матокрит, количество тромбоцитов, электролиты и креатинин сыворотки, величина
коагуляции были в пределах нормы. У больной были выявлены гипонатриемия и по-
вышенный уровень печеночных ферментов. Имелись признаки гипоксии.
В медицинской карточке из последних перенесенных болезней у нее отмечена тран-
зиентная ишемическая атака. Больная не курила. Рентгенологическое исследование
показало бибазилярные инфильтраты и небольшой правосторонний плевральный
выпот, но без расширения средостенья. Первоначальный дифференциальный диагноз
давал большие надежды в пользу атипичной пневмонии, а не ингаляционной сибир-
ской язвы. Поэтому больной был назначен левофлоксацин (levofloxacin); рифампицин
был назначен ей наследующий день. Но через два днялихорадка продолжала держать-
ся, нарастала одышка, развился большой плевральный выпот. Назначенные ранее
антибиотики были заменены на другие — ципрофлоксацин, рифампицин и ванкоми-
цин. Больной 21 и 22 октября был выполнен торакоцентез справа и откачано по 750 и
620 мл геморрагической жидкости соответственно. 29 октября ей была установлена
дренажная трубка справа. Компьютерная томография грудной клетки, выполненная
22 октября, показала медиастинальную и цервикальную лимфоаденопатию, бибази-
лярные инфильтраты и большой плевральный выпот слева. 25 октября увеличение
плеврального выпота слева потребовало установить еще одну дренажную трубку. 26 ок-
тября температура и уровень трансаминаз снизились до нормы. 30 октября и 1 ноября
были удалены дренажные трубки. 5 ноября больная была выписана из госпиталя. Кровь
была ПЦР-положительна на ДНК сибиреязвенного микроба. Иммунохимическое
исследование показало наличие капсульного антигена и антигена клеточной стенки
В. anthracis в цитологических препаратах, полученных из плевральной жидкости.
Случай 12. 15 октября 43-летняя женщина, выходец из Юго-Восточной Азии, ра-
ботавшая сортировщицей почты в Гамильтоне, но на другой машине, чем женщина,
история болезни которой описана как «случай 11», почувствовала себя больной. У нее
появилась интермиттирующая температура, ощущение озноба, сухой кашель, сопро-
вождающийся чувством дискомфорта в грудной клетке, затруднения при дыхании,
626
Часть 3. Частная неправильная эпидемиология
миалгии и слаоость. У нее была тошнота и рвота, но болей в животе и, (иареи не было.
Она жаловалась на ощущение «пустоты в голове» и сильную головную боль. 16 октяб-
ря, когда она посетила своего врача, температура у нее была 38 "С, и ей был назначен
левофлою ацин, так как врач подозревал бронхит. В ее медицинской книжке не было
ничего примечательного, она не курила. 18 октября больная обратилась в местный
кабинет скорой помощи, так как ей стало хуже. При осмотре температура 38,4°С, ча-
стота сердечных ударов 120/мин, частота дыхания 16/мин, АЦ 141/85 мм рт. ст. На-
сыщение крови кислородом 92 % при комнатном воздухе и 97 % после вдыхания 4 л
кислорода, поданных больной через носовую канюлю. Она выглядела больной. Под-
счет лейкоцитов показал увеличение количества палочкоядерных нейтрофилов. Гема-
токрит и количество тромбоцитов были в норме. У больной выявлена гипонатриемия,
но электролиты, почечная функция и коагуляционные показатели были в предела.;
нормы Повышенными были уровни печеночных ферментов. ПЦР на ДНК В. anthra-
cis, выполненная с пробой крови, взятой на второй день лечения антибиотиками, ска-
талась отрицательной. Рентгенография показала справа хиларные затемнения с уплот-
нениями (hilar opacity consistent with consolidation or mass), умеренный плевральный
выпот справа и небольшой слева. Антибиотик был заменен на ципрофлоксацин и ази-
тромицин. Ципрофлоксацин отменен спустя 24 ч. Проведенная 19 октября компью-
терная томография грудной клетки показала увеличение мягкой ткани в средостенье,
вызванное аденопатией, rigcht hilar consolidation, и большой плевральный выпот. Боль-
ной был проведен торакоцентез и удалено 500 мл серознокровянистой жидкости
В схему лечения дооавлены к шндамицин (clindamycin) и цефтриаксон. В цитологиче-
ских препаратах, полученных из плевральной жидкости, иммунохимическим окраши-
ванием обнаружены антигены клеточной стенки и капсульный антиген В. anthracis.
21 октября был проведен повторный торакоцентез и откачано 800 мл жидкости. 22 ок-
тября бронхоскопией обнаружена отечность и гиперемия слизистой бронхов. В тка-
нях, взятых, благодаря трахеобронхиальной биопсии, иммунохимическим окраши-
ванием пока ано наличие капсульного антигена и антигена клеточной стенки В. anthra-
cis. Культуру В. anthracis из назальных смывов, тканей и жидкостей больной, получить
не у талось. Лечение комплексом антибиотиков заменено на прием только одного док-
сициклина. Больная выписана из госпиталя 26 октября.
Случай 13. 16 октября 2001 г. 56-летнии афроамериканец, сортировщик службы
доставки почты, заметил у себя небольшую температуру, почувствовал озноб, боли в
горле, головную боль и недомогание. Затем к этим симптомам присоединились слабый
сухой кашель, ошу щение тяжести в груди, чувство н< хватки воздуха, ночные поты, тош-
нота и рвота. 19 октября, когда он был доставлен в местный госпиталь, температуры у
него не было, и артериальное давление было в норме. Частота ударов сердца составляла
110/.ИИН, но тахипное не было. Он не был в тяжелом состоянии, но дыхательные зву -
ки у него были ослабленными и хрипы в левом основании (base). Кожных поражений
у больного не наблюдалось. Его последняя медицинская карточка была без записей
Он не курил. Общее количество лейкоцитов было ь пределах нормы, но имелся неболь-
шой сдвиг влево в их дифференциации. Уровни печеночных трансаминаз и билирубина
были повышенными, но функция почек и данные биохимического исследования, сы-
вороточная химия были в норме. Величины показателей кислорода и углекислого газа
артериальной крови показывали адекватную оксигенацию. Рентгенография грудной
клетки показала расширение средостенья, особенно правой паратрахеальной облает
билатеральные хиларные (hilar) массы, билатеральные плевральные выпоты и неболь-
шое нижнедолевое затемнение воздушного пространства справа. Компьютерная то-
мография грудной клетки показала диффузный медиастинальный отек, усиленные па-
ратрахеальные узлы, непарное вдавление пищевода за счет аденопатии (azygo-esopha-
3.1. Сибирская язва
627
geal recess adenopathy) (наибольший измеренный узел имел в диаметре 4,2 см), и била-
теральные умеренные плевральные выпоты. В пределах 11 ч в кровяной культуре был
обнаружен рост В. anthracis. Лечение начато ципрофлоксацином, рифампицином и
клиндамицином. 21 октября у пациента развилась дыхательная недостаточность, ко-
торая была купирована введением диуретиков, системных кортикостероидов и тера-
певтическим торакоцентезом. В период госпитализации плевральные выпоты форми-
ровались повторно, поэтому потребовались еще две операции торакоцентеза. Плев-
ральная жидкость во всех трех отобранных пробах содержала кровь. Больному не
потребовалось введение дренажной трубки. Отдельные поверхностные язвочки пище-
вода были обнаружены верхней эндоскопией. 28 октября у пациента появились при-
знаки гемолитической анемии и тромбоцитопении, которые были купированы плаз-
мофорезом. Гематологические показатели в последующем улучшились. Пациент ос-
тавался в госпитале до стабилизации состояния.
Случай 14. 16 октября 2001 г. 55-летний мужчина-афроамериканец, работавший
на почтовой доставке округа Колумбия, почувствовал себя больным. У него появи-
лась температура; периодически возникающая сильная потливость; миалгия; кашель
с зеленой мокротой, но без уменьшения объема дыхания; ощущение дискомфорта в
груди и симптомов со стороны желудочно-кишечного тракта. Когда 18 октября он
обратился за медицинской помощью в первичное попечительство, температура у него
была 38.9°С, размеры сердца и величина артериального давления были в норме, час-
тота дыхания составила 24/мин. Обращающих на себя лабораторных показателей не
было выявлено. Только подсчет лейкоцитов показал, что их количество выше нормы.
В его истории болезни упоминались сахарный диабет и саркоидоз. Он не курил. Рент-
генограмма грудной клетки сделана не была. Он был отправлен домой с диагнозом
«вирусный синдром», антибиотики назначены не были. 21 октября он попал в отделе-
ние неотложной помощи в тяжелом состоянии. Больной жаловался на чувство стес-
нения в груди, утомление, озноб, миалгию, тошноту, рвоту и укороченное дыхание.
Температура больного была 38,9°С, пульс от 93 до 150/мин, нерегулярный. Частота
дыхания 20/мин, артериальное давление 119/73 мм рт. ст. Наблюдались признаки ды-
хательной недостаточности. У больного были обнаружены хрипы в правом основа-
нии (base) с диффузными хрипами и тахикардия. Количество лейкоцитов составило
18800/мм3, из них 73 % сегменто-ядерные нейтрофилы, 6 % палочки, 11 % лимфоци-
ты. 8 % моноциты. Гематокрит — 55 %. Тромбоциты — 141 тыс./мм3. Содержание на-
трия 130 ммоль/децилитр, глюкозы 425 мг/децилитр. Рентгенографическое исследо-
вание грудной клетки показало rigcht hilar и перитрахеальное переполнение мягких
тканей с инфильтрацией правой средней и нижней долей, совпадающих с пневмонией
и правым плевральным выпотом. ЭКГ обнаружила у больного фибрилляцию предсер-
дия. Пациенту была сделана интубация и искусственная вентиляция легких, назначе-
ны левофлоксацин, дилтиазем (diltiazem) и инсулин. В последний день жизни гемоди-
намическая нестабильность больного нарастала, у него остановилось сердце, и он умер.
В кровяной культуре обнаружен рост В. anthracis. Аутопсические находки включают
геморрагические медиастинальные лимфадениты. Иммунохимическим окрашиванием
подтверждена диссеминация В. anthracis.
Случай 15. 16 октября у 47-летнего мужчины афроамериканца, работника той же
службы почтовой доставки округа Колумбия, где произошло заражение людей возбу-
дителем сибирской язвы, описанные как «случаи 13. 14, 16», развился сухой кашель
средней интенсивности. Одновременно он почувствовал тошноту, перешедшую в рвоту,
спазмы в желудке. 20 октября у больного развился обморок во время службы в церк-
ви, но он не обратился за медицинской помощью. Рано утром 21 октября он сам при-
шел в отделение скорой помощи с жалобами на рвоту и профузный пот. Последние
628
Часть 3. Частная неправильная эпидемиология
записи в его медицинской книжке свитетельс гвовали о наличии у него астмы и моче-
каменной болезни. В ближайшее время лечение астмы кортикостероидами не прово-
дилось. Температуры у больного не было, отмечена ортостатическая гипотензия. Ко-
личество лейкоцитов оказалось несколько увеличенным, но коагуляционные тесты и
химические показатели сыворотки были в норме. Несколько увеличенным был уро-
вень сывороточной глютаматпируваттрансаминазы (glutamic pyruvic transaminase.
SGPT). Первый рентгеновский снимок сначала был прочитан как нормальный, но на
латеральном обзоре была замечена неясная область повышенной плотности, показы-
вающая наличие инфильтрата или масс в правой смпрахилярной (suprahilar) области.
Больной был отпущен домой после проведения вну гривенной гидратационной тера-
пии. Но утром 22 октября он пришел в отделение скорой помощи снова. В его исто-
рии было записано: миалгиаз, кашель, одышка, продолжительная рвота и случай обмо-
рока. Его температура была 35,6°C, артериальное давление 76/48 мм рт. ст., частота
сердцебиения 152/мин, частота дыхания 32/мин. Он имел вид больного человека с
испещренной кожей, которая при прикосновении казалась холодной. У больного были
признаки дыхательной недостаточности При аускультации двухсторонние хрипы,
тахикардия, живот слегка вздут, кишечные звуки не прослушиваются. При исследо-
вании крови обнаружено повышение содержания лейкоцитов — 31200/мм3, из ни>
78 % сегментоядерные нейтрофилы, 2 % палочки, 14 % лимфоциты и 3 % моноциты
Содержание натрия — 148 ммоль/л, бикарбоната — 18 ммоль/л, анионный промежу
ток (anion gap) — 21, креатинин — 2,8 мг/децилитр. Сывороточная глу гаматоксалане-
таттрансаминаза (glutamic oxalacetic transaminasa, SGOT) — 47 МЕ/л, SGPT — 33 МЕ/л.
щелочная фосфатаза — 197 МЕ/л. Протромбиновое время 13,3 с и время частичного
тромбопластина составило 40 с. Больному были назначены пенициллин, цефтриак-
сон, рифампицин и левофлоксацин. Однако респираторная недостаточность прогрес-
сировала, поэтому больному была сделана эндотрахеальная интубация и начата меха-
ническая вентиляция легких. Вскоре у него появились признаки перитонита. Значение
pH артериальной крови было 7,13; РаСО2 — 37 мм рт ст., РаО2 — 106 мм рт. ст., кисло-
родное насыщение — 95 %, после интубации — 100 % F’O2. Рентгенологическое ис-
следование показало билатеральные легочные инфильтраты, концентрирующиеся в
пределах перихиларных (perihilar) и инфрахиларных (infrahiLr) регионов без плевраль-
ных выпотог Компьютерная томография грудной клетки и брюшной полости, вы-
полненная с внутривенным контрастированием, показала большой билатеральный
плевральный выпот, перихиларные (perihilar) и супрахиларные (suprahilar) инфильт-
раты, отек средостенья и небольшие скопления интрамурального воздуха, вовлечен-
ные в отдельные части тонкой кишки (jejunum). Компьютерная томография головы
не выявила патологии. Больной умер через 6 ч после поступления в госпиталь. Рост
кровяных культур В. anthracis в пределах 18 ч. Посмертные находки включают выра-
женный геморрагический медиастинальный лимфаденит и доказательства системной
сибиреязвенной инфекции, полученные с помощью гистопатологических и иммуно-
химических методов.
Случай 16. 16 октября 56-летний мужчина афроамериканец, работник почтовой
доставки, почувствовал головную боль средней интенсивности, которая вскоре стала
постоянной, появилась ригидность шеи. В последующие три дня головные боли усили-
лись, появились субфебрильная температура, озноб, боли в горле, миалт ии, тошнота
и рвота, обильный пот, фотофобия, ухудшилось зрение. Появились плевральные боли,
средней интенсивности сухой кашель, остановка дыхания при напряжении (dyspnea
on exertion). Когда 20 октября он был доставлен в госпиталь, то температуры у него не
было, давление было в норме, пульс 127/мин, частота дыхания 20/мин. У него не было
острой недостаточности дыхания, и физические исследования ничего не обнаружили
3.1. Сибирская язва
629
за исключением ослабления везикулярного дыхания на основаниях легких. Предше-
ствующие этому заболеванию записи в его истории болезни были малоинформатив-
ны, он не курил. Результаты пабораторных исследований, в основном, оказались в
пределах нормы, за исключением повышенных уровней билирубина и печеночных
ферментов, низкого содержания альбумина и наличия признаков гипоксии.
Неконтрастированная компьютерная томография головы также не позволила
выявить патологические изменения. При окрашивании спинномозговой жидкости по
Граму микроорганизмы не найдены. Не было микробного роста и при подращивании
спинномозговой жидкости на питательной среде. Антеропостериальная рентгеногра-
фия грудной клетки показала расширения средостенья, билатеральные хиларные мас-
сы, правосторонний плевральный выпот и билатеральные перихиларные воздушные
пространства (perihilar air space disease). Неконтрастное сканирование грудной клет-
ки при помощи компьютерной томографии показало диффузный отек средостенья,
двухсторонние плевральные выпоты и заметную паратрахеальную, субкардиальную,
хиларную аденопатию и непарное вдавление пищевода за счет аденопатии. Кровяные
культуры дали рост В. anthracis в течение 15 ч. Лечение начато с ципрофлоксацина,
рифампицина и клиндамицина. 22 октября были обнаружены признаки нарастания
дыхательной недостаточности, 23 октября больной подвергся терапевтическому тора-
коцентезу, который позволил откачать кровянистую плевральную жидкость, после чего
его состояние улучшилось. Больному также были назначены системные кортикосте-
роиды для предотвращения бронхоспазма. Ему потребовалась повторная операция
торакоцентеза. Больной выписан из госпиталя 9 ноября 2001 г.
Случай 19. Вечером 22 октября почувствовал себя больным 59-летний мужчина, бе-
лый, контрактный служащий департамента сортировки почты, который получал поч-
ту из округа Колумбия (сл^-чаи 13—16). Первым симптомом у него был обильный пот.
Последующие два дня больной чувствовал слабость, выраженные мышечные боли,
повышенную температуру, озноб, головную боль, тошноту, боли в области живота и за
грудиной. У него начались приступы рвоты и кашель с белой мокротой. Недостаточ-
ности дыхания или диареи у него не было. После его помещения 24 октября в отдель-
ную палату для наблюдения температура была 38,2°С, частота сердечных сокращений
116/мин, частота дыхания и артериальное давление в пределах нормы. Количество лей-
коцитов было в пределах нормы, исследование сывороточных электролитов показало
гипонатриемию и гипокалиемию. Последние записи в его медицинской карте не пред-
ставляли ничего интересного, больной не курил. Первое рентгенографическое иссле-
дование грудной клетки показало норму. Больному был поставлен диагноз «вирусный
синдром», и он был выписан, однако у него была взята кровь на бактериологический
посев и назначен прием ципрофлоксацина. Он принял только одну дозу препарата ве-
чером, но потливость, чувство усталости, головные боли усилились. Он жаловался на
приходящие нарушения в левой визуальной области. Его супруга сообщила, что он стал
плохо ориентироваться в происходящем. Бактериологический посев дал рост В. ant-
hracis через 17 ч инкубирования; 25 октября пациента поместили в госпиталь. Темпе-
ратура при поступлении 38,2 °C, количество ударов сердца 108/мин, частота дыхания
20/мин, артериальное давление 121/60 мм рт ст., насыщение кислородом 94 % из ком-
натного воздуха. Он выглядел больным. При аускультации удалось прослушать глухое
дыхание на right base. Остальные исследования оказались малоинформативными.
Лабораторные показатели при поступлении в госпиталь включали общий анализ
крови. Количество лейкоцитов составило 9500/мм3, из них 81 % сегментированных
нейтрофилов. 9 % лимфоцитов, 9 % моноциты; тромбоциты 196 тыс./мм3, гематокрит
48,1 %, электролиты и креатинин в норме, SGOT 85 МЕ/л, SGPT 64 МЕ/л, щелочная
фосф и аза 141 МЕ/л, билирубин 1,6 мг/децилитр, альбумин 3,0 мг/децилитр.
630
Часть 3. Частная неправильная эпидемиология
На первом обзорном рентгеновском снимке обнаружено медиастинальное расши-
рение (рис. 3.20). Сделанная на следующий день компьютерная томография показала
медиастинальную аденопатию с присутствием геморрагий, нормальную легочную па-
ренхиму, небольшие билатеральные плевральные выпоты и, предположительно, не-
большое перикардиальное излияние (pericardial effusion) (рис 3.21).
Рис. 3.20. Случай 19. Рентгенограмма грудной клетки. Показывает
медиастина тьную аденопатию и небольшой левый п гевратьный выпот.
По J. A. Jernigan er al. (2001)
Рис. 3.21. Случай 19 Компьютерная томограмма грудной клетки. Пока-
зывает медиастинальную аденопатию и небольшие билатеральные
плевральные выпоты. По J. A. Jernigan et al. (2001)
К назначенному ранее ципрофлоксацину было добавлено внутривенное введение
пенициллина и рифампицина. Температура больного поднялась до 39 °C. В последу-
ющем пенициллин был отменен. Вместо него назначен ванкомицин. 26 октября у боль-
ного было обнаружено гастроинтестинальное кровотечение, которое потребовало
трансфузий крови, введения эндоскопа и прижигания язв желудка и двенадцатиперст-
ной кишки. 27 октября у больного была обнаружена фибрилляция предсердий с раз-
3.1. Сибирская язва
631
личными вентрикулярными ответами. 28 октября лихора 1ка достигла максимума —
39,4°C, затем снизилась до 38 °C. 30 октября достигла макси чума лейкоцитарная ре-
акция, количество лейкоцитов достигло 31 ЗОО/мм3. 31 октяоря отмечено расширение
правого плеврального выпота. Больному был выполнен торакоцентез и откачано 900 мл
серознокровянистой жидкост и. Больной выписан из госпиталя 9 ноября.
Случай 21.25 октября заболела 61-летняя женщина-вьетнамка, работавшая в цент-
ральном складском помещении больницы в Манхэттене (Нью-Йорк). Болезнь нача-
лась с недомогания и болей в мышцах. В последующие два дня она стала ощущать сла-
бость, появился кашель, боли в грудной клетке, стала прогрессировать дыхательная
недостаточность, кашель из сухого стал продуктивным. Больная стала отхаркивать
мокроту, слегка окрашенную кровью. Она так же обнаружила у себя повыше иную тем-
пературу. 28 октября, когда она обратилась за медицинской помощью в местный госпи-
таль, температуры у нее уже не было, но частота сердечных ударов составляла 110/мин,
частота дыхания 38/мин, насыщение крови кислородом при дыхании в комнате 92 %.
Она осознавала свое состояние, понимала, что находится в госпитале. Больная стра-
дала гипертонией. Она не курила. Абдоминальные и кардиоваскулярные исследования
показали норму, за исключением тахикардии. Подсчет лейкоцитов показал незначи-
тельное увеличение их количества, гематокрит, число тромбоцитов и коагуляционные
тесты были в норме. Почти нормальным оказался и химический состав сыворотки, за
исключением гипонатриемии, увеличенного уровня печеночных ферментов и гипоаль-
буминемии. Уровень сывороточной лактатдегидрогеназы 1,370 МЕ/л. Для поддержа-
ния адекватной оксигенации крови больной был дан кислород через маску. Рентгено-
грамма грудной клетки первоначально была интерпретирована как закупорка легоч-
ной вены (pulmonary venous congestion) с билатеральными плевральными выпотами.
Первоначально лечение больной было направлено на борьбу с сердечной недостаточ-
ностью, но эхокардиограмма, сделанная в отделении скорой помощи, не показала при-
знаков ослабления деятельности сердца. Тогда больной был поставлен диагноз «ати-
пичная пневмония» и назначен левофлоксацин. Однако дыхательная недостаточность
усилилась, и больная была интубирована. Компьютерная томография грудной клетки
показала медиастинальную лимфаденопатию, массивное медиастинальное кро-
вотечение, загущение бронхиальной слизи и билатеральные плевральные выпоты.
Больной назначены рифампицин, гентамицин и в последующем нафпиллин (nafcillin).
Утром 29 октября ей с двух сторон ввели грудные дренажные трубки и откачали 2,5 л
серознокровянистой жидкости с левой стороны и один литр с правой. В тот же день
бронхоскопия показала наличие кровянистой слизи в трахеобронхиальном дереве;
рыхлые, спадающиеся воздушные пути и гнойные секреты во многих сегментах с обе-
их сторон. 30 октября схема антибактериальной терапии была изменена. Больной на-
значены ципрофлоксацин, рифампицин, клиндамицин и цефтазидим. Последующая
эхокардиограмма показала увеличение перикардиального выпота, ведущего к тампо-
наде сердца. Состояние больной быстро ухудшалось, и 31 октября она умерла после
попытки перикардиоцетоза. Культивирование крови и плевральной жидкости дало
рост В. anthracis через 20 ч. Аутопсические находки включают геморрагический меди-
астинит. Иммунохимическое окрашивание обнаружило присутствие В. anthracis во
многих органах и тканях.
Случай 22.16 ноября 200 1 г. в лечебное учреждение города Оксфорд (шт. Коннек-
1 икут) обратилась 94-летняя женщина с жалобами на повышенную температуру, чувст-
во усталости, сухой кашель, затрудненное неглубокое дыхание, боли в мышцах. Жен-
щина болела уже три дня. В течение предыдущих двух месяцев она вела уединенный
образ жизни, страдала депрессивным синдромом, имела сниженный аппетит и, в свя-
зи с возрастом, физически была слаба. Она страдала хроническим легочным обструк-
632
Часть 3. Частная неправильная эпидемиология
тивным заболеванием, гипертонией, хронической почечной недостаточностью. Поль-
зовалась ингаляторами с сальметерол + флутиказоном (salmeterol xinafoate/fluticasone
propionate) и спреем для носа, содержащим азеластин (azelastine). Изредка употребляла
алкоголь. Конвертов, содержащих «порошок», она не открывала, и уже д лительное вре-
мя никуда не выезжала. Домашних животных в доме не было. За день до обращения к
врачу, члены ее семьи заметили, что ее сознание спутано.
При поступлении в клинику у пациентки температура 39,1 °C, артериальное давле-
ние 106/50 мм рт. ст. без ортостатических изменений, пульс 119/мин, частота дыхания
18/мин. Насыщение кислородом составило 93 % при комнатном воздухе. Она была
встревожена и понимала, где находится. Менингеальных симптомов у нее не было,
кожные покровы без патологии.
Лабораторные находки включали: лейкоциты 8100/мм3 (78 % нейтрофилы, 7 % мо-
ноциты и 15 % лимфоциты), гематокрит и тромбоциты в норме. Сывопо-очный азот
мочевины (serum urea nitrogen) составил 39 мг/децилитр (13,9 ммоль/л), сывороточ-
ный креатинин 1,3 мг/децилитр (115 мкмоль/л), сывороточные электролиты в преде-
лах нормы, за исключением натрия, составившего 134 миллиэквивалент/л. Ферменты
сыворотки в норме, за исключением аспаратаминстрансферазы — 45 ЕД/л. Анализ
мочи показал 3—5 лейкоцитов в поле зрения и умеренное количество бактерий. Задне-
передняя и латеральная рентгенограммы грудной клетки сначала были интерпретиро-
ваны как свидетельствующие о расширении медиастинума или о плевральных выпо-
тах, признаков инфильтратов в ткани легкого на них не было обнаружено (рис. 3.22).
Рис. 3.22. Случай 22. Первые инструментальные и лабораторные находки. А. Задне-
передняя рентгенограмма грудной клетки, сделанная при поступлении больной
в клинику Снимок показывает левое хиларное расширение (белая стрелка) и затем-
нение в области левого диафрагмально-реберного угла (черная стрелка), возможно,
образовавшегося в результате небольшого левого плеврального выпота. Б. Окраши-
вание по Граму кровяной культуры показывает наличие грамположительных палочек,
расположенных цепочками (увеличение в 100 раз). По L. A Barakat et al. (2002)
Однако позже, при сравнении с пленками 3-летней давности предположено рас-
ширение левого хилума (left hilum) и возможность небольшого левого плеврального
выпота. Компьютерная томография не проводилась. Кровь и моча были взяты на бак-
териологическое исследование, и больная госпитализирована с диагнозами «вирусный
синдром» и «дегидратация». Первоначальное лечение включало внутривенное введе-
ние жидкостей и наблюдение. Лечение антибиотиками в день поступления больной не
проводилось.
3.I. Сибирская язва
633
Утром 17 ноября ее состояние было стабильным. Но уже после 14 ч в высевах из
крови обнаружены грамположительные бациллы. Посевы мочи дали рост кишечной
палочки, чувствительной к ампициллину. Приблизительно через 24 ч после поступле-
ния в стационар больная получила однократно 2 г цефтазидима и ванкомицин (1 г/доза
внутривенно). Затем в схему лечения были добавлены: орально ципрофлоксацин 500 мг
каждые 12 ч; ампициллин/сульбактам (ampicillin/sulbactam) по 3 г в/в каждые 6 ч.
Кровяные культуры, взятые через 3 ч после начала лечения антибиотиками, оказались
стерильными.
18 ноября у пациентки появились признаки дыхательной недостаточности, лей-
коцитоз 13300/мм3 (80 % нейтрофилы, 11 % лимфоциты и 9 % моноциты). Насыще-
ние крови кислородом снизилось до 90 % при подаче кислорода в объеме 2 л/мин че-
рез носовую канюлю. Рентгенограмма грудной клетки отчетливо показала наличие
плеврального выпота слева (рис 3.23).
Рис. 3.23. Случай 22. А. Рентгеновский снимок, показывающий билатеральное
нижнее затемнение в легких (слева больше, чем справа), и небольшое нарушение
прозрачности слева. Б. Обширное затемнение в нижней части обоих легких, и би-
латеральное нарушение прозрачности легочной ткани. По L. A. Barakat et al. (2002)
Утром 19 ноября внутривенное введение ванкомицина и прием перорально цип-
рофлоксацина были продолжены, а введение ампициллин/сульбактама прекращено.
Внутривенно назначен эритромицин (500 мг каждые 6 ч). В этот день на основе бак-
териологического исследования у больной была заподозрена сибирская язва. У паци-
ентки нарастала гипоксия, появились признаки почечной недостаточности. Азот
мочевины и креатинина сыворотки крови составили 43 мг/децилитр (15,4 ммоль/л) и
2,6 мг/децилитр (230 мкмоль/л) соответственно. Лейкоцитоз достиг 25000/мм3 (83 %
сегментоядерные нейтрофилы, 8 % палочковые формы и 8 % лимфоциты). Рентгено-
грамма показала прогрессивное развитие левого плеврального выпота. С помощью
левого торакоцентеза выведено 800 мл кровянистой жидкости (pH 7,12, 4224 эритро-
цитов; 1463 лейкоцитов; уровень лактатдегидрогеназы 611 ЕД/л; глюкозы 14,4 ммоль/л;
белков 3,4 r/децилитр). Какие-либо микроорганизмы из плевральной жидкости вы-
сеять не удалось.
Но последующее исследование плевральной жидкости в Centers Disease Control and
Prevention показало, что ДНК В. anthracis обнаруживается ПЦР. Пациентке была сде-
лана эндотрахеальная интубация и начата механическая вентиляция легких. В допол-
нение к ванкомицину назначен клиндамицин (900 мг внутривенно, каждые 6 ч), вве-
дение эритромицина отменено, ципрофлоксацин стали вводить внутривенно. Начато
введение метилпреднизолона по 40 мг внутривенно, каждые 8 ч.
634
Часть 3. Частная неправильная эпидемиология
20 ноября получены результаты бактериологических исследований. Между 19 и
21 ноября температура у пациентки достигла 38,6°С. У нее развилась гипотензия, по-
требовавшая введение вазопрессоров. Рентгенограмма показала npoi рессирование
плеврального выпота слева и появление плеврального выпота справа (см. рис. 3.23, Б).
В левую плевральную полость была помещена дренажная трубка. Лейкоцитоз достиг
43600/мм3 (83 % сегментоядерные нейтрофилы, 12 % лимфоциты и 5 % моноциты).
Уровень креатинина в сыворотке достиг 3,7 мг/децилитр (327 мкмоль/л). Гематокрит,
количество тромбоцитов, уровень печеночных ферментов и коагуляционный профиль
остались в пределах нормы, за исключением аспартатаминотрансферазы (61 ЕД/л).
Состояние пациентки продолжало ухудшаться, и 21 ноября она умерла.
Аутопсия была выполнена спустя 8 ч после смерти. Более одного литра сероанги-
нозной жидкости было откачано из правой плевральной полости. Правое легкое име-
ло области уплотнения. Кожных поражений обнаружено не было. Медиастинальные
лимфатические узлы были увеличены и имели геморрагические изменения (геморра-
гический медиастинит). ЦНС без изменений. Органы брюшной полости, за исключе-
нием почек, в норме. Микроскопическое изучение показало выраженный некроз и
геморрагии лимфатических узлов, интраальвеолярный и интерстициальный отек с
фокальными геморрагиями и фибриновыми отложениями в легких, некроз селезенки.
Рост В. anthracis обнаружен только при исследовании содержимого медиастинальных
лимфатических узлов. Образцы сыворотки, нзятые 16,17,18 и 19 ноября, были тести-
рованы на IgG-антитела к протективному антигену сибиреязвенного токсина, но ни в
одной пробе таких антител обнаружено не было. Посмертный диагноз — ингаляцион-
ная сибирская язва.
Обобщенная клиника ингаляционной сибирской язвы. Средний инкубационный пе-
риод от момента заражения до начала симптомов у людей, пораженных сухой рецеп-
турой спор возбудителя сибирской язвы, составил 4 сут (интервал 4—6 сут). Два па-
циента (случаи 11 и 12), предположительно, были инфицированы спорами В. anthracis
9 октября, когда конверт, который, как позже было установлено, контаминированный
рецептурой В. anthracis, прошел через почтовое оборудование, на котором они рабо-
тали. Тот же конверт прошел через другое почтовое оборудование 12 октября — веро-
ятный день экспонирования других четырех человек, пораженных посредством ин-
галяции возбудителем сибирской язвы (случаи 13, 16, 14 и 15).
Больные обратились за медицинской помощью в среднем в течение 3,5 сут после
появления симптомов болезни (интервал 1—7 сут). Восемь из десяти пациентов были
на начальной стадии болезни, когда они первый раз обратились за медицинской по-
мощью. Из них 6 получили антибиотики, активные против В. anthracis, в день обра-
щения, и все шестеро выжили. У 4 пациентов, включая одного с менингитом, разви-
лись скоротечные симптомы болезни, и хотя, они все получили антибиотики, эффек-
тивные против сибиреязвенного микроба, они погибли. В табл. 3.4 суммированы
симптомы болезни первых 10 пациентов (без случая 22), ставших жертвами сухой ре-
цептуры В. anthracis. В табл. 3.5 приведен перечень основных клинических, лабора-
торных и рентгеновских находок у этих людей на начальной стадии болезни. В табл. 3.6
приведены основные патологические находки у 5 пациентов, погибших от ингаляци-
онной сибирской язвы. Продолжительность инкубационного периода и болезни у
11 больных с ингаляционной сибирской язвой, начиная от времени заражения (котда
оно известно), показаны на рис. 3.24.
Количество лейкоцитов при обращении пациента к врачу соответствовало норме
или было немного повышенным. Среднее начальное количество лейкоцитов 9800/мм3
(интервал от 7500 до 13 ЗОО/мм3). Однако возрастало соотношение молодых форм
3.1 Сибирская язва
635
Таблица 3.4
Основные симптомы у первых десяти пациентов
с ингаляционной сибирской язвой*
Симптомы п= 10 Симптомы п= 10
Лихорадка, озноб 10 Мышечные боли (миалгии) 6
Слабость, недомогание, летаргия 10 Головные боли 5
Кашель (слабый или непродуктивный) 9 Спутанность сознания 4
Тошнота или рвота 9 Абдоминальные боли 3
Одышка или чувство удушья 8 Боли в горле 2
Пот, ооытно очень интенсивный 7 Ринорея 1
Дискомфорт в груди, плевральные боли 7
* По J. A. Jemigan et al. (2001).
Таблица 3.5
Первые клинические находки у десяти пациентов
с ингаляционной сибирской я той*
Клинические находки Частота встречаемости
Объективно
лихорадка (оолее 37,8 °C) 7 из 10
тахикардия (более 100/мин) 8 из 10
гипотензия (систолическое давление менее НО мм рт. ст.) 1 из 10
Результаты лабораторных исследований сыворотки крови
лейкоциты (средняя величина) 9,8x103/мм’
дифференцированные нейтрофилы (более 70 %) 7 из 10
нейтрофилы палочковых форм (более 5 %) 4 из 5
увеличение активности трансаминаз8 (SHOT или SGPT более 59 %) 9 из 10
Гипоксемия
альвеолярно-артериальный кислородный градиент более 30 мм рт. ст. при О2 комнатного воздуха, насыщение менее 94 % 6 из 10
метаболический ацидоз 2 из 10
увеличение содержания креатинина (более 1,5 мг/децилитр) 1 из 10
Данные рентгенологического исследования
любые отклонения от нормы 10 из 10
медиастинальное расширение 7 из 10
инфильграты/уплотнения 7 из 10
плевральный выпот 8 из 10
Данные компьютерной томографии грудной клетки
любые отклонения от нормы 8 из 8
медиастинальная лимфоаденопатия, расширение 7 из 8
плевральный выпот 8 из 10
инфильтраты, уплотнения 6 из 8
® SGOT — сывороточная глутаматоксалацетаттрансаминаза; SGPT — сывороточная глу-
таматпируваттрансаминаза.
* По J A. Jemigan et al. (2001).
636
Часть 3. Частная неправильная эпидемиология
Таблица 3.6
Основные патологические находки у пяти пациентов, погибших
от ингаляционной сибирской язвы*
Патология Пациент 8 Пациент 14 Пациент 15 Пациент 21 Пациент 22
Продолжитель- ность лечения антибиоти- ками, ч 82 11 3 55 95
Кожные поражения Нет Кожная гематома Нет Нет Нет
Плевральная жидкость, мл** R > 1000 L > 1000 R 1300 L700 R250 L 500 R 2500 L1000 R 1000 L800
Медиастиналь- ные лимфати- ческие узлы Геморрагичес- кий некроз, сглаживание Геморрагиче- ский некроз, иммунобласты, нейтрофилы Некроз, гемор- рагии, имму- нобласты, нейтрофилы Геморрагии Некроз, гемор- рагии, имму- нобласты, нейтрофилы
Плевральные и интерхиларные перегородки Геморрагии Геморрагии, воспаление Геморрагии Воспаление
Легочная паренхима Фокальные гиалиновые мембраны Без видимых изменений Выступающие внутриальвео- лярные макрофаги Выступающие внутриальвео- лярные макрофаги Перибронхи- альное воспа- ление, внутри - альвеолярный отек
Селезенка Некроз Скопление иммуно- бластов Скопление иммуно- бластов и нейтрофилов Скопление иммунобластов
Кишечник Фокальные геморрагии Ьез видимых изменений Фокальные геморрагии и воспаление Без видимых изменений
Грамположи- тельные бациллы Нет Обнаружены во всех тканях Обнаружены во всех тканях Обнаружены в лимфатиче- ских узлах Нет
Локализация бацилл при окрашивании по Steiner Торакальные и абдоминаль- ные ткани Торакальные и абдоминаль- ные ткани Торакальные и абдоминаль- ные ткани Медиастиналь- ные лимфати- ческие узлы Медиастиналь- ные ткани, легкие
* По J. Guamer et al. (2003).
* * Количество плевральной жидкости в правой (R) и левой (L) плевральной полости.
нейтрофилов, и это явление можно считать ранним диагностическим критерием ин-
галяционной сибирской язвы. В течение болезни количество лейкоцитов увеличива-
лось у всех пациентов, интервал от 11300 до 46600/мм3.
Рентгенологическое исследование выявило патологию у всех пациентов, но только
8 из 11 имели изменения медиастинума. Уже на первых рентгенограммах можно было
заметить медиастинальное расширение, паратрахеальные утолщения, хиларные утол-
щения, плевральные выпоты и паренхимальные инфильтраты. В двух случаях на рент-
генограммах первыми были обнаружены плевральные выпоты и легочные инфильтра-
ты без каких-либо признаков расширения медиастинума. Компьютерная томография
3.1. Сибирская язва
637
Случай
8 ?
4 ?
13
16
14
15
19 ?
11
12 I
21 ?
22 ?
Инкубацис ниши
период
Болезнь
Появление
5 Ю 15 20 2з зо
Дни болезни
симптомов
Время от экспозиции
до начала симптомов
Время от начала симптомов
до первого контакта с врачом
Время от первого контакта
с врачом до назначения
антибиотиков
Продолжительность
npi’eva антибиотиков
Инкубационный
период неизвестен
X Смерть
Рис. 3.24 Продолжительность инкубационного периода и болезни у 11 пациентов
с ингаляционной сибирской язвой. По J. A. Jernigan et al. (2001) и L. A. Barakat et al. (2002)
грудной клетки оказалась более надежным инструментом для обнаружения патоло-
гических изменений в грудной клетке пораженного, чем рентгенографическое ис-
следование.
11леврал ьн ые выпоты обнаружены у всех 11 пациентов. Они были наиболее замет-
ной составляющей клинических особенностей ингаляционной сибирской язвы, что
свидетельствует об относительно быстром (по сравнению даже с медиастинальными
лимфатическими узлами) инфицировании плевры после ингаляции спор В. anthracis1.
Небольшими плевральные выпоты были у умерших больных, но у выживших они
имели тенденцию к прогрессивному увеличению и распространению. Дтя семи па-
циентов потребовалась операция по дренированию плевральной жидкости, четырем
из них были установлены дренажные трубки. Геморрагическая плевральная жидкость
обычно содержала относительно небольшое количество лейкоцитов. Легочные ин-
фильтраты, которые наблюдались у 7 пациентов, включали правую и левую нижние
доли (один пациент), левую верхнюю и нижнюю доли (один пациент), левую верхнюю
и нижнюю доли (один пациент) и билатеральные перихилярные (один пациент). Вось-
ми пациентам сделана компьютерная томография грудной клетки, медиастинальная
лимфоаденопатия выявлена у 7 пациентов.
У трех больных развилась аритмия предсердия (суправентрикулярная тахикардия
в «случае 4» и фибрилляция предсердий в «случаях 14 и 19»). У трех больных компью-
терной томографией обнаружен перикардиальный выпот. У одной пациентки пред-
полагалась перикардиальная тампонада. Перикардиальные выпоты могут быть след-
ствием токсин-обусловленного локального отека или воспаления, геморрагического
некроза или инфаркта, распространившегося в перикардиум из медиастинума, или
гематогенного разноса В. anthracis
Кровяные культуры В. anthracis получены у 8 пациентов еще до того, как им нача-
ли проводить антибактериальную терапию. Рост кровяных культур наблюдался в сред-
1 Аналогичную клинику болезни наблюдали у приматов Н. В. Литусов с соавт. (2002) (см.
выше «Моделирование на экспериментальных животных ситуации искусственного заражения
В. anthracis»).
638
Часть 3. Частная неправильная эпидемиология
нем за 18 ч (интервал от 12 до 24 ч). У трех пациентов кровь для получения кровяных
культур взяли после начала антибиотикотерапии. Во всех трех случаях кровяную куль-
туру В. anthracis получить не удалось. У одного заболевшего, у которого получили кро-
вяную культуру до начала лечения, получить ее через 24 ч после начала антибиотико-
терапии не удалось.
Множественные фокальные геморрагии в тонкой кишке были найдены у 8 паци-
ентов из 15, но каких-то значительных изъязвлений или мезентериальной лимфоаде-
нопатии и поражения органов ЖКТ обнаружено не было. Такая патологоанатомиче-
ская картина ингаляционной сибирской язвы не соответствует той, которую за нее
выдал екатеринбургский патологоанатом Л. М. Гринберг (1995), выполнивший вскры-
тие 42 тел жертв сибирской язвы, вспыхнувшей в Свердловске весной 1979 г. По-
ражения ЖКТ им отмечены почти у всех погибших от сибирской язвы (39 из 42).
Л. М. Гринберг также обнаружил, что такие поражения носят распространенный ха-
рактер. Тошая и подвздошная кишка были поражены у 36 умерших, желудок — у 30,
слепая и восходящая кишки — у 12, глотка и корень языка — у 3. Им было выделено
три типа изменений ЖКТ: в 21 случае процесс носил очаговый характер; в 10 — диф-
фузный; в 8 — диффузно-очаговый. При очаговой (карбункулезной) форме пораже-
ния в ЖКТ определялось множество карбункулов диаметром от 0,3 до 3.0 см, темно-
красного цвета. Крупные очаги занимали все слои стенки кишки и не были связаны
с лимфоидными образованиями кишечника. Ничего подобного исследователи не на-
блюдали у людей, умерших от ингаляционной сибирской язвы в США в 2001 г. Не
обнаружено такой патологии и у приматов, погибших в результате эксперименталь-
ного ингаляционного заражения спорами В. anthracis, выполненного Н. В. Литусо-
вым с соавт. (2002). Наоборот, эти авторы наблюдали подобную патологоанатоми-
ческую картину только при энтеральном заражении павианов гамадрилов (см. выше
«Моделирование на экспериментальных животных ситуации искусственного зара-
жения В. anthracis»).
Несовпадения в патологической анатомии ингаляционной сибирской язвы в США
в 2001 г. и сибирской язвы в Свердловске в 1979 г. свидетельствуют о том, что пути
инфицирования и патогенез сибирской язвы у людей, вовлеченных в эти вспышки,
были различными’. До вспышки сибирской язвы в США в 2001 г. считалось, что ге-
моррагический менингит сопровождает ингаляционную сибирскую язву в 50 % слу-
чаев (Бургасов П. Н., Рожков Г. И., 1984). Но менингеальные симптомы наблюдались
только у двух пораженных (менее 20 %). У них была отобрана цереброспинальная
жидкость, менингит подтвержден у одного пациента. Трое других пораженных имели
inermittent confusion. Менингеальных симптомов у них не наблюдалось, и поэтому люм-
барную пункцию им не делали. Следовательно, «шапочка кардинала» не является
специфическим признаком ингаляционной сибирской язвы.
' Эти расхождения объяснимы. В США имело место ингаляционное заражение людей, в
Свердловске — энтеральное. Патологоанатом Л. М. Гринберг (1995), приняв за абсолютную
истину политизированное объяснение М. Meselson et al. (1994) причины вспышки сибирской
язвы в Свердловске, как следствие «выброса с военного объекта русских», был вынужден «при-
нять» кишечную форму сибирской язвы за ингаляционную. Никакого секрета кишечная форма
сибирской язвы для патологоанатома не представляет еще с начала XIX в. Довольно подробно
она описана в работе Ф. Гейрота (1807), ее патанатомия имеется в старом учебнике Г. А. Ива-
шенцова (1948). Патанатомию кишечной сибирской язвы, описанную в работе И. Л. Богда-
нова (1935), я привел выше. Видимо, желание «вписаться» в ценности «Свободного мира» ока-
залось для Льва Моисеевича гораздо сильнее его представлений о научной репутации.
3.1. Сибирская язва
639
Результаты гистологического исследования органов и тканей людей, пораженных
мелкодисперсным аэрозолем возбудителя сибирской язвы, показаны на рис. 3.25
Иммунохимические тесты, специфичные к В. anthracis, выполненные на цитологи-
ческих препаратах, полученных из плевральной жидкости и тканей, отобранных
трансбронхиальной биопсией, были положительными всегда (рис. 3.25, А). Аутопсия
четырех умерших больных показала наличие у них медиастинального лимфаденита и
диссеминацию В. anthracis по органам и тканям (рис. 3.25, В - Г).
Рис 3 25. Результаты гистологического исследования органов и тканей людей, пора-
женных мелкодисперсным аэрозолем возбудителя сибирской язвы. А. На блоке, сде-
ланном из плевральной жидкости (несмертельный случай), в большом количестве
показан гранулярный антиген В. anthracis, окрашенный изнутри мононуклеарных
воспалительных клеток (иммунохимический тест выполнен с использованием моно-
нуклеарных антител к капсуле В. anthracis и проявленный щелочной фосфатазой и
нафтоловым крепким красным в собственной модификации авторов статьи). Увели-
чение в 158 раз. Б. Медиастинальный лимфатический узел, извлеченный из тела умер-
шего больного Отчетливо видны капсулярное и синусоидальное кровоизлияния (окра-
шивание гемотоксилином и эозином). Увеличение в 25 раз, В. Лимфатический узел,
извлеченный из тела того же больного. Видно болшое количество гранулярного анти-
гена В. anthracis, окрашенного изнутри мононуклеарной воспалительной клетки. Там
же бациллы (указаны стрелочкой), находящиеся в субкапсулярной геморрагической
области (иммунохимический тест выполнен с использованием мышиных монокло-
нальных антител к клеточной стенке В. anthracis и проявлен, как описано выше). Уве-
личение в 100 раз. Г. Легочная ткань, полученная от умершего больного. Верхней стрел-
кой показан гранулярный антиген В anthracis, окрашенный изнутри периферических
макрофагов. Нижней стрелкой показана бацилла, расположенная внутриклеточно.
Увеличение в 100 раз По J. A. Jemigan et al. (2001)
Клинические проявления болезни различались между собой и часто имели сход-
ство с вирусными респираторными болезнями, что создает риск диагностических
ошибок в условиях отсутствия террористической настороженности. J. A. Jemigan et al.
(2001), подводя итоги изучения клиники случаев ингаляционной сибирской язвы,
вызванной террористическим актом, обращают внимание на то, что такая форма си-
бирской язвы ранее (т. е. возникшая в результате инфицирования во время работ с
переработкой шерсти животных, больных сибирской язвой) описывалась как болезнь
с двухфазным клиническим течением. Ее начальная фаза длилась 1—4 сут и проявля-
лась следующими симптомами: недомогание, слабость, лихорадка, миалгии и сухой
кашель. За ней следовала молниеносная фаза, для которой было характерно быстрое
развитие респираторной недостаточности (в течение 1—2 сут), цианоза и обильное
640
Часть 3. Частная неправильная эпидемиология
потоотделение (Dixon Т. et al., 1999; Brachman В, Kaufmann А., 1998). Симптомы на-
чальной фазы 11 террористических случаев болезни в основном сходны с ранее опи-
санными у 18 пациентов с нетеррористическими случаями ингаляционной сибирской
язвы (Brachman В, 1980). Но есть и отличия:
1) выраженный, часто «просачивающийся пот», характерен для поражения сухой
сибиреязвенной рецептурой. Его не отмечали авторы, описавшие случаи инфициро-
вания спорами сибирской язвы в цехах по промышленной переработке шерсти;
2) во время «естественных вспышек» состояние больного между начальной и мол-
ниеносной фазами болезни может на короткое время улучшаться. Этот клинический
феномен не наблюдался у больных, пораженных сухой рецептурой В. anthracis',
3) до террористического акта 2001 г. считалось, что первично-легочной сибирской
язвы не существует и на рентгенограмме может быть только выраженное расширение
медиастинальных узлов при отсутствии признаков первичной пневмонии (Бурга-
сов П. Н., Рожков Г. И., 1984; Braude А. I. с соавт., 1986).
Однако практика биотеррора показала, что поражение сухой рецептурой сибир-
ской язвы может проявляться легочными инфильтратами (отмечены у семи пациен-
тов, наиболее показательны случаи 4,11,13,14), а являются ли они первичными, либо
представляют результат гематогенного заноса, остается предметом дискуссии среди
специалистов*. Три отличия в клинической картине поражения легких от ранее опи-
санных, возможно, связаны с физико-химическими свойствами использованных тер-
рористом (или террористами) отдельных компонентов сухой рецептуры В. anthracis.
Диагностика искусственного поражения возбудителем сибирской язвы. Все кли-
нические формы сибирской язвы, обобщенные в патогенетической классификации
Н. В. Литусовассоавт. (2002), включая даже кожную форму болезни, могут быть вызва-
ны у людей диверсионным или террористическим способами применения В. anthracis
(см. разд. 1.12.4). К ним необходимо добавить еще и «невозможные клинические фор-
мы болезни» (см. разд. 2.3.1): сибирскую язву, вызванную парентеральным введени-
ем В. anthracis', и ингаляционную сибирскую язву, вызванную мелкодисперсным аэро-
золем (патология мелкодисперсного аэрозоля).
Основным критерием для раннего распознания искусственного происхождения
вспышки сибирской язвы может быть невозможная эпидемиология болезни. Дтя ее
выявления важно при сборе анамнеза больного сибирской язвой обратить внимание
на обстоятельства, которые на первый взгляд не имеют отношения к эпидемиологии
сибирской язвы. Естественное происхождение кожной формы болезни предполагает
указание в анамнезе на контакт с инфицированным В. anthracis мясом, шкурами или
почвой. Отсутствие такого контакта при наличии сибиреязвенного карбункула на коже
пациента дает основание для выявления иных источников возбудителя болезни. На-
пример, ими могут оказаться вскрытые конверты с неизвестным порошком, подарки
или предметы, полученные им от других людей (организаций). А так как количество
сухой сибиреязвенной рецептуры, способной вызвать кожную форму сибирской язвы
обычно незначительно, и заболевший может ее не заметить, то необходимо:
1 Аналогичные результаты получили Н. В. Литусов с соавт. (2002), осуществив ингаляци-
онное заражение приматов мелкодисперсным аэрозолем спор В. anthracis (см. выше «Моде-
лирование на экспериментальных животных ситуации искусственного заражения В. anthracis»).
Сибиреязвенная пневмония обнаружена ими у 4 % павианов гамадрилов. Эти авторы счита-
ют сибиреязвенную пневмонию вторичной, т. е. возникшей в результате гематогенного зано-
са спор В. anthracis в паренхиму легких.
? 1. Сибирская язва
641
1) поинтересоваться у него наличием подобных поражений на коже у лиц из его
элижайшего окружения (родственники, коллеги по работе и др.);
2) провести сбор эпидемиологических данных среди этого «окружения» пациен-
та (разд. 2.3.2; «Текущее эпидемиологическое расследование в очаге искусственного
эпидемического процесса»),
В случае обнаружения больных с кожной формой сибирской язвы, необходимо
расспросить их по поводу возможных контактов с источниками инфицирования
В. anthracis, включая контакты с неизвестным «порошком».
Кишечная форма сибирской язвы может быть предположена при обнаружении рез-
кого несоответствия между крайне тяжелым состоянием больного и отсутствием к тому
окальных данных в сочетании с прогрессирующим упадком сердечной деятельности;
выраженными нарушениями со стороны кишечника (тупые боли в животе, рвота, не-
юстоянныйжидкий стул); обнаружением присутствия экссудата в брюшной полости
(действие токсина В. anthracis)', постоянными жалобами больных в разгаре болезни на
жесточайшую головную боль при отсутствии объективных менингеальных симптомов.
Подозрение об искусственном происхождении кишечной сибирской язвы обоснова-
ны, если выявлен «разрыв» в цепочке движения инфицированного продукта от есте-
ственного источника заражения. Например, споры В. anthracis обнаружены в котле-
тах, но отсутствуют в мясе, из которого сделан фарш для котлет. Даже если они попали
в продукт, вызвавший вспышку кишечной сибирской язвы, с какого-то оборудования
(разделочные столы, котлы, мясорубки и т. п.), то для исключения искусственной кон-
таминации должен быть выявлен природный источник возбудителя болезни, конта-
минировавший это оборудование ранее.
Любой случай ингаляционной сибирской язвы, обнаруженный среди жителей горо-
дов, не имеющих отношения к переработке шерсти и кожи животных, должен рас-
сматриваться с точки зрения его искусственного происхождения. Подозрение об ис-
кусственном характере вспышки, уже после установления диагноза «ингаляционная
сибирская язва», может быть основано на следующем сочетании данных рентгеноло-
гического исследования, клинических проявлений болезни и результатов вскрытия:
1) на рентгенограмме — наличие патологических процессов в паренхиме легких.
Им будут сопутствовать характерные для ингаляционной сибирской язвы патологи-
ческие процессы в медиастинуме (медиастинальное расширение, паратрахеальные
утолщения, хиларные утолщения и др.) и в плевре (экссудативный плеврит);
2) среди клинических проявлений болезни — в числе ранних симптомов — наличие
профузного пота; а при анализе клинической картины болезни в целом — отсутствие
короткого периода улучшения состояния больного между начальной и молниеносной
фазами болезни;
3) среди патологоанатомических находок — отсутствие признаков первичного
вовлечения в инфекционный процесс ЖКТ (карбункулы в пищеводе, желудке, тошей
и подвздошной кишке).
Парентеральное введение В. anthracis можно предположить у лиц, получавших ка-
кие-либо медицинские назначения, сопровождавшиеся инъекциями, и у инъекцион-
ных наркоманов. При подкожном инфицировании человека возбудителем сибирской
язвой возможны следующие группы симптомов: повышенная температура тела, об-
ширные воспаления и отеки подкожной клетчатки, прилегающей кучасткам, исполь-
зуемым для инъекционного введения, и признаки регионарного лимфаденита. Внут-
ривенное инфицирование В. anthracis можно предположить в случае внезапной
(«апоплексической») смерти человека, при вскрытии кот< iporo выявляются призна-
642
Часть 3. Частная неправильная эпидемиология
ки серозного или серозно-геморрагического полиаленита, полнокровие и дистрофи-
ческие изменения в паренхиматозных органах, и кровоизлияния в легких.
При установлении этиологии болезни наиболее целесообразно использование
ПЦР. Нельзя считать надежными тесты на получение гемокультур В. anthracis и на IgG-
антитела к протекгивному антигену сибиреязвенного токсина. В случае быстрой смер-
ти пациента, lgG-ан гитела к протекгивному антигену сибиреязвенного токсина мо-
гут не успеть образоваться. Рост возбудителя сибирской язвы на питательных средах
в случаях, когда кровь у пациента была отобрана на первой стадии болезни, имел место
для всех тех больных, которым не проводилась антибиотикотерапия. Но кровь стано-
вится стерильной после начала лечения пациента антибиотиками.
Обнаружение в очагах эпизоотий штаммов В. anthracis, устойчивых к от гельным
антибиотикам, даже если они вызвали вспышки кожной сибирской язвы у людей, еще
не говорит в пользу того, что штаммы были генетически изменены. Однако такие
штаммы надо рассматривать как генетически измененные и передавать специалистам
для плазмидного- и сиквиенс-анализа в случаях, если такой штамм обнаружен:
1) при оостоятельствах, которые относятся к неправильной эпидемиологии (см.
разд. 2.3.1);
2) у него обнаружена резистентность сразу к нескольким антибиотикам, исполь-
зуемым в распространенных схемах экстренной специфической профилактики и ле-
чения сибирской язвы (например, пенициллины и ципрофлоксацин, пенициллины
и тетрациклин);
3) он оказался способным пробить иммунитет, созданный у человека менее года
назад полноценной вакцинацией живыми или комбинированными сибиреязвенны-
ми вакцинами.
Выявление причин эпизоотий сибирской язвы среди сельскохозяйственных жи-
вотных не менее важно, чем вспышек среди людей. Во-первых, эпизоотии формируют
первые звенья эпидемической цепи, ведущей В. anthracis в человеческие популяции.
Их прослеживание поз! оляет сделать рабочие предположения об искусственном или,
наоборот, естественном происхождении вспышки сибирской язвы среди людей. Во-
вторых, сами эпизоотии могут быть вызваны искусственно при осуществлении сабо-
тажа (см. выше разд. 1.7; «Бактериологические диверсии в СССР»), и в рамках стра-
тегий непрямых ударов. Основную роль в таких ударах сыграет зараженное Я anthra
cis мясо. 11оследующая массовая доставка зараженного мяса потребителю способна
вызвать крупную вспышку сибирской язвы. Поэтому надо обязательно убедиться в
том, что в регионе, где выявлена сибирская язва среди животных, существуют при-
родные условия для ее возникновения (см. выше «Экология и естественная эпидеми-
ология В. anthracis»), а инфицирование животных произошло из природного резер-
вуара В anthracis благодаря наличию этих условий (нейтральная или слабощелочная
почва; температура не ниже 15 °C и не выше 35 °C; большой процент содержания гу-
муса в почве; правильная сезонность проявления болезни; концентрированные кор-
ма, в которые добавлены корма животного происхождения; животные со скрытым те-
чением болезни идр.). Далее, используя VNTR-анализ и анализ полногеномного поли-
морфизма отдельных нуклеотидов, необходимо убедиться в том, что штамм В. anthracis.
вызвавший эпизоотию, характерен для данного региона, и что именно он вызвал
вспышку сибирской язвы среди людей.
Так как нет достоверных данных в пользу того, что инфицирование людей воз-
будителем сибирской язвы может произойти в результате реаэрозолирования спор
В. anthracis, осевших на поверхности предметов, окружавших человека в момент фор-
3.1. Сибирская язва
643
мирования первичного аэрозоля (см. разд. 1.12.4), то контаминацию ими поверхнос-
тей в местах совершения биотеррористического акта не следует считать представляю-
щей опасность.
Иммунопрофилактика. В России для специфической профилактики сибирской
язвы разрешено применение двух вакцин:
вакцина сибиреязвенная живая, представляющая собой лиофилизат живых спор
вакцинного штамма В. anthracis СТИ-1. Используется для подкожного введения и
накожного скарификационного нанесения. После двукратного применения с интер-
валом в 20—30 сут вызывает формирование специфического иммунитета продолжи-
тельностью до года. Ревакцинацию проводят однократно ежегодно, подкожно;
вакцина сибиреязвенная комбинированная, представляющая собой лиофилизат сме-
си живых спор вакцинного штамма В. anthracis СТИ-1 и очищенного концентри-
рованного протективного сибиреязвенного антигена, адсорбированного на геле гид-
роокиси алюминия. Вакцинацию проводят однократно подкожно. Обеспечивает
формирование специфического иммунитета через 7 сут после прививки продолжи-
тельностью до года. Ревакцинацию проводят однократно. Первые три ревакцинации
осуществляют ежегодно. Все последующие ревакцинации проводят один раз едва года.
Вакцины производят ФГУ «48 Центральный научно-исследовательский институт
Министерства обороны Российской Федерации» (г. Киров) и его филиал в г. Екатерин-
бурге — «Центр военно-технических проблем биологической защиты».
Обе вакцины эффективно защищают от ингаляционного заражения спорами воз-
будителя сибирской язвы.
Лечение. Почти все случаи ингаляционной сибирской язвы, когда лечение начи-
налось после того, как у больных уже проявлялась значительная симптоматика, за-
канчивались смертельным исходом, независимо от способов лечения. С 1950-х гг. в
качестве основного варианта лечения рассматривался пенициллин, применяемый
внутривенно через каждые 2 ч в дозе 2 млн единиц. Большинство штаммов есте-
ственного происхождения чувствительны к эритромицину, хлорамфениколу, гента-
мицину и ципрофлоксацину. Однако в природе поддерживаются и пенициллин-ре-
зистентные штаммы, один из них, Ames, использовался американскими военными
для получения биологических рецептур, предназначенных для снаряжения биологи-
ческих кассетных боеприпасов и осуществления биологических диверсий. При лече-
нии в США в 2001 г. больных с кожной и ингаляционной формами сибирской язвы,
вызванными этим штаммом, наиболее эффективной оказалась комбинированная те-
рапия, сочетающая применение ципрофлоксацина и других антибиотиков широкого
спектра действия. Однако к настоящему времени уже разработаны подходы к полу-
чению штаммов В. anthracis, резистентных к ципрофлоксацину. Поэтому при отсут-
ствии данных об антибиотикочувствительности штаммов, вызвавших сибирскую язву,
уже при самых ранних признаках болезни должно быть назначено эмпирическое ле-
чение комбинацией антибиотиков.
Первоначально для лечения ингаляционной формы сибирской язвы рекоменду-
ется использовать либо ципрофлоксацин, либо доксициклин в сочетании с одним или
более препаратом, к которым обычно чувствителен возбудитель. После проверки чув-
ствительности и улучшения состояния больного схема лечения может быть измене-
на. Предпочтительными препаратами для лечения кожной формы сибирской язвы
являются также ципрофлоксацин или доксициклин. Пенициллин, такой как амокси-
циллин (или его сочетание с клавулановой кислотой), может быть использован для
завершения курса лечения в случае положительных результатов проверки чувствитель-
644
Часть 3. Частная неправильная эпидемиология
ности (Bell D. М. et al., 2002). К улучшению состояния больного ингаляционной си-
бирской язвой приводит дренирование плевральных выпотов посредством плевраль-
ной дренажной трубки. Поскольку эти выпоты имеют тенденцию быстро скапливаться
вновь, целесообразно вставлять одну или несколько плевральных дренажных трубок.
Клиндамицин может оказывать антитоксическое действие. Стероиды используются
для устранения отека при кожной форме заболевания и рекомендуются для лечения
менингита или обширного медиастинального отека. В качестве антитоксинов в усло-
виях in vitro исследовали протективное действие ингибиторов ангиотензин-конвер-
тирующих ферментов, блокаторов кальциевых каналов и ингибиторов фактора не-
кроза опухоли. Вспомогательными средствами лечения могут служить и антисибире-
язвенные IgG (Bell D. М. et al., 2002).
3.2. ЧУМА
Микробиология Y. pestis. Происхождение и молекулярная эпидемиология. Эколо-
гия и естественная эпидемиология Y. pestis. Внутривидовая таксономия. Воспри-
имчивость и чувствительность животных к возбудителю чумы. Инфицирующие
дозы. Иммунитет. Устойчивость Y. pestis во внешней среде и в трупах погибших.
Антибиотикорезистентность. Обнаружение генетически измененных штаммов
Y. pestis. Патогенез. Клиническая картина чумы у людей при естественном инфици-
ровании Y. pestis. Моделирование искусственного заражения Y. pestis. Диагностика
искусственного поражения возбудителем чумы. Иммунопрофилактика. Лечение.
Чума (pestis, plague, black death) — опасная сапрозоонозная инфекционная болезнь
людей и животных, вызываемая неспорулирующейся бактерий Yersinia pestis. Харак-
теризуется острым началом, тяжелой интоксикацией, лихорадкой, септицемией, по-
мрачением сознания, возникновением бубонов и карбункулов. Протекает в кожной,
легочной и септицемической формах. Карантинная инфекция. Российские санитар-
но-эпидемиологические правила СП 1.3.2322-08 относят возбудитель чумы к I груп-
пе патогенности. По степени важности для национальной безопасности США Y. pestis
относится к биологическим поражающим агентам категории А (см. табл. 2.2). Особую
опасность возбудителю чумы придает его способность при легочный форме болезни
передаваться между людьми.
Попытки диверсионного применения культур Y. pestis и зараженных чумой гры-
зунов замечены еще в годы Первой мировой войны. С начала 1930-х гг. и до оконча-
ния Второй мировой войны возбудитель чумы активно исследовался японскими во-
енными как потенциальный поражающий агент БО. Он неоднократно применялся
ими в варварских экспериментах на людях и для осуществления биологических ди-
версий против китайских войск и мирного населения. В США и в Соединенном Ко-
ролевстве со средины 1940-х гг. возбудитель чумы изучался как летальный агент БО.
Для его боевого применения разрабатывались специальные рецептуры и боеприпа-
сы. В первой половине 1952 г. Y. pestis использовалась армией США для поражения
войск и населения Северной Кореи и Северо-Восточного Китая. До 1968 г. рецепту-
ры на основе возбудителя чумы находились на вооружении американской армии под
шифром «N». В 1995 г. обнаружена заинтересованность радикальных организаций в
использовании чумы в террористических целях. В США за попытку приобретения
культуры Y. pestis по почте был арестован некий Ларри Уэйн Харрис, связанный с труп-
3.2. Чума
645
пой «Арийская нация». Харрис не скрывал своего намерения осуществить биотерро-
ристический акт против федеральных структур.
Наиболее вероятно применение Y. pestis посредством использования линейных и
многоточечных источников и диверсионными (террористическими) методами.
Микробиология Y. pestis. Y. pestis — относится к роду Yersinia семейства Enterobac-
teriaceae. Факультативный анаэроб. Растет на обычных жидких и плотных питатель-
ных средах при температуре 25—30 °C и pH 6,9—7,2. На бульоне культура растет в виде
поверхностной пленки со спускающимися вниз нитями, напоминающими сталакти-
ты, и хлопьевидного осадка на дне пробирки. Из «природных источников» возбуди-
тель чумы выделяется в своей типичной R-форме (шероховатой). При росте бактерий
на питательных средах может происходить диссоциация их колоний от R-форм, к
авирулентным S-формам, образующим на плотном агаре гладкие, ахромогенные ко-
лонии, а в бульоне равномерное помутнение. Но и R-формы Y. pestis могут необрати-
мо утрачивать вирулентность. Среди таких штаммов обычно ищут штаммы, пригод-
ные для применения в качестве вакцинных (Николаев Н. И., 1968).
Бактерии Y. pestis эндоспор не образуют, не имеют жгутиков и лишены активной
подвижности. В организме больных чумой, а также на искусственных питательных
средах при определенных условиях развития могут образовывать капсулу. Специфи-
ческая окраска для чумных микробов не разработана. Они легко окрашиваются кар-
боловыми и щелочными растворами анилиновых красок, при окраске по Граму обес-
цвечиваются. На их форму, размеры и окраску существенное влияние оказывают
условия развития микробной популяции. В мазках из агаровых культур, выращенных
при температуре 26—28 °C, Y. pestis имеет форму прямых или слегка изогнутых пало-
чек с закругленными полюсами длиной 0,8—2,0 мкм и диаметром 0,5—0,7 мкм. На
поверхности препарата они располагаются без определенной закономерности и ок-
рашены преимущественно равномерно. Возможно образование небольшого количе-
ства нитей и цепочек различной длины. В мазках из агаровых и бульонных культур,
полученных в результате выращивания при температуре 37—39°C, а также в мазках
крови и мокроты людей и животных, погибших от чумы, или в мазках — отпечатках
их органов, микробы Y. pestis имеют вид небольших палочек, овоидов или бочкооб-
разных клеток с закругленными полюсами, многие из которых окрашены биполяр-
но, что проявляется в преимущественном окрашивании полюсов и появлении нео-
крашенной крупной вакуоли в центре клеточного тела. Биполярными чаще всего
выглядят микробы, развившиеся на искусственных питательных средах при темпера-
туре 37—39°С или в организме больных чумой при остром течении инфекционного
процесса (Золотарев А. Г. с соавт., 2006) (рис. 3.26, А).
В случаях затянувшейся чумы (нагноившиеся, вскрывшиеся бубоны и язвы) либо
при неблагоприятных условиях роста (воздействие бактериофага, суббактериостати-
ческие дозы антибиотиков и солей, неоптимальное осмотическое давление) чумные
микробы утрачивают биполярность и превращаются в гетероформные структуры в
виде шаров, нитей или теней клеток (Золотарев А. Г. с соавт., 2006).
Капсулу микробы У. pestis образуют в организме восприимчивых животных или
после выращивания при температуре 37—39°С на питательных средах определенного
состава. У клеток культур, полученных при температуре 26—28 °C, капсула отсутствует.
Микроскопическая картина и размеры капсулы зависят, с одной стороны, от штам-
мовых особенностей культур, а с другой — от методики приготовления препарата.
Наиболее оптимальные условия для выявления капсулы у чумных * икробоь обеспе-
чивает прижизненная фазово-контрастная микроскопия препаратов, приготовленных
646
Часть 3. Частная неправильная эпидемиология
Рис. 3.26. Микроскопическое изображение Y.pestis. А. Биполярные палочки Y. pestis
в крови пациента с септической формой чумы. Окрашивание по Wright-Giemsa.
Увеличение в 1000 раз. По Т. Inglesby et al. (2000). Б. Капсула живых Y. pestis, выра-
щенных в течение 24 ч при температуре 37—39 °C в переваре Хоттингера с добавле-
нием витаминов и ионов кальция. Вирулентный штамм 1300. Увеличение в 1350 раз.
Фазовый контраст. В. Электронно-микроскопическое изображение клеток Y. pestis,
выращенных в бульоне Хоттингера при температуре 37—39 °C. Штамм 1300. Снару-
жи от клетки имеются остатки капсулы (К). Контрастировано напылением хромом,
увеличение в 15 тыс. раз. Г. Электронно-микроскопическое изображение ультра-
тонкого среза клетки Y. pestis, обработанной специфическими анти-Fl-иммуногло-
булинами, меченными ферритином. Показано равномерное расположение ферро-
иммуноглобулинов. Увеличение в 100 тыс. раз. По А. Г. Золотареву с соавт. (2006)
по влажному тушевому методу. В таких препаратах капсула наблюдается в виде про-
зрачной светлой зоны между клетками и темным фоном, создаваемым частицами
туши. Толщина капсулы у живых Y. pestis вирулентных штаммов возбудителя чумы ко-
леблется примерно от 1,2 до 0,3 мкм (Золотарев А. Г. с соавт., 2006) (см. рис. 3.26, Б).
Под электронным м икроскопом в препаратах, оттененных хромом, у чумных бак-
терий определяются цилиндрическая форма, сферические полюса, гладкая, без рез-
ких возвышений и углублений поверхность и равномерная электронно-оптическая
плотность цитоплазмы. По периферии клеток, особенно на полюсах, протопласт не-
сколько отходит от клеточной стенки с образованием полупрозрачной каймы, в ко-
торой видны ровные внешние контуры клеточной стенки и цитоплазматической мем-
браны. Снаружи клеточной стенки находится капсула, отличающаяся низкой
плотностью по отношению к электронам. По степени выраженности капсулы Y. pestis
гетерогенны (Золотарев А. Г. с соавт., 2006) (см. рис. 3.26, В).
В капсуле чумных бактерий располагается оболочечный или капсульный антиген
F1. Его детерминанты взаимодействуют с меченными ферритином специфическими
иммуноглобулинами. На ультратонких срезах клеток, предварительно обработанных
такими меченными антителами, гранулы комплекса ферритин — иммуноглобулин
обнаруживаются как на поверхности клеточной стенки, так и на материале капсулы.
Наличие меченых специфических иммуноглобулинов на клеточной стенке и в кап-
3.2. Чума
647
суле Y. pestis указывает на то, что антиген F1 является компонентом, участвующим в
образовании этих структур (Золотарев А. Г. с соавт., 2006) (см. рис. рис. 3.26, Г).
Наиболее благоприятные условия для выращивания бактерии К pestis обеспечи-
ваются при свободном доступе кислорода и температуре инкубирования от 26 до 28 °C.
При таких условиях У. pestis уже через 10—12 ч роста образует на плотной питательной
среде характерные микроколонии в виде плоских сложных образований с неровны-
ми краями, напоминающие кружевные платочки. Примерно к 20—24 ч выращивания
у микроколоний формируются выпуклый зернистый желтый, бурый или темно-ко-
ричневый центр и плоская тонкая бесцветная, почти прозрачная фестончатая пери-
ферическая зона — кружевная каемка (рис. 3.27, А, Б).
Рис. 3.27. Макроскопические изображения колоний Y. pestis. А, Б. Микроколонии
возбудителя чумы, выращенные на агаре Хоттингера при температуре 26-28°С. Ви-
рулентный штамм 1300. Макрофотосъемка в падающем свете. Увеличение в 56 раз.
На фотографии А микроколонии через 10—12 ч выращивания, на фотографии Б
микроколонии через 20—24 ч выращивания. У микроколонии формируется выпук-
лый желтый, бурый или темно-коричневый центр и тонкая, почти прозрачная фе-
стончатая периферическая зона. В. Двухсуточная колония У. pestis в проходящем
свете (условия выращивания и штамм см. выше). Увеличение в 13 раз. Г. Двухсуточ-
ная колония Y pestis в падающем свете (условия выращивания и штамм см. выше).
Увеличение в 13 раз. По А. Г. Золотареву с соавт. (2006)
К исходу вторых суток вырастают выпуклые, округлые колонии, которые имеют
диаметр 2,5—3,0 мм и макроскопически в проходящем свете кажутся серовато-белы-
ми с голубым оттенком, а при падающем свете — беловато-серыми. При микроско-
пическом исследовании в проходящем свете они хромогенные и нередко напомина-
ют по внешнему виду цветы ромашек или маргариток. Центр у них может быть
выпуклым или запавшим, коричневым либо желтым, зернистым или бугристым, тогда
как периферическая часть полупрозрачная и кружевная (см. рис. 3.27, В). В падаю-
щем свете эти колонии выглядят серо-желтыми и имеют форму полусфер с мелкозер-
нистой поверхностью и ровными или слегка фестончатыми краями (см. рис. 3.27, Г)
(Золотарев А. Г. с соавт., 2006).
648
Часть 3. Частная неправильная эпидемиология
Морфология колоний чумных бактерий зависит от многих факторов. При удли-
нении сроков культивирования до 72—96 ч центр колонии уплощается или западает, а
периферическая кружевная зона становится плотной либо узкой, а может и вовсе
отсутствовать Поверхность колоний зернистая, а иногда исчерченная (феномен Твор-
та). На морфологию колоний К pestis влияют также температура выращивания и со-
став питательной среды (Золотарев А. Г. с соавт., 2006).
Биохимически возбудитель чумы активен: восстанавливает нитраты в нитриты,
ферментирует с образованием кислоты без газа глюкозу, левулезу, мальтозу, галакто-
зу, ксилозу и манит, продуцирует ферменты типа дегидрогеназ и уреаз. Хорошо рас-
тет на синтетических средах с аминокислотами (в качестве источников азота) и фер-
ментируемыми углеводами. В витаминах и азотистых основаниях, как правило, не
нуждается. Аминокислотные потребности у штаммов из разных природных очагов
сильно варьируют. Кроме того, они могут меняться в зависимости от температуры
культивирования.
Характерными признаками вирулентных штаммов У. pestis являются:
1) наличие капсулы и V- и W-антигенов (антифагоцитарное действие);
2) способность синтезировать пурины (генерализация инфекции);
3) образование пигментированных колоний на среде с гемином (генерализация
инфекции);
4) пестициногенность;
5) неспособность расти на средах при температуре 37 °C в отсутствие ионов каль-
ция;
6) наличие «рН-6-антигена» (играет роль Fc-рецептора, связывается с IgG чело-
века и приводит к образованию псевдоиммунных комплексов).
Менее ясен ответ на вопрос, какие «факторы вирулентности» нужны К pestis для
того, чтобы она оказалась способной вызвать вспышку чумы у людей. В этой связи
Н. И. Николаев (1968) и И. В. Домарадский (1998) отмечают, что основное отличие
авирулентных штаммов чумного микроба от вирулентных заключается в способности
последних размножаться в организме человека. Штаммы, вызывающие вспышки чу-
мы у людей, и штаммы, циркулирующие в природных очагах чумы среди грызунов, по
наличию известных факторов вирулентности не отличаются друг от друга. Подробно
факторы вирулентности возбудителя чумы описаны в работе А. П. Анисимова (2002)
Происхождение и молекулярная эпидемиология. К pestis является типовым видом
рода Yersinia, относящимся к семейству Enterobacteriaceae. Помимо возбудителя чумы,
к этому роду относятся еще шесть видов бактерий: К pseudotuberculosis, Y. enterocolitica,
У. intermedia, Y. ruckeri, Y. frederiksenii и Y. kristensenii. По культурально-биохимическим
и некоторым другим свойствам ближе всего к чумному микробу стоят первые два вида,
из которых У. pseudotuberculosis называют даже его двойником. Однако по эпидемио-
логическим особенностях и патогенезу вызываемой инфекции К pseudotuberculosis и
У. enterocolitica существенно отличаются от У. pestis (Домарадский И. В., 1998).
М. AvAtman et al. (1999) высказали предположение, что У. pestis является клоном
К pseudotuberculosis. Аллели У. pestis были идентичны или почти идентичны таковым
у 12 штаммов У. pseudotuberculosis. Согласно таксономическим стандартам, У. pestis
можно рассматривать как Y. pseudotuberculosis, но из-за больших различий в проявле-
ниях болезни и роли Y. pestis в истории эпидемий, этой классификации не придержи-
ваются. А. Р. Anisimov et al. (2004) считают, что различия в плазмидном составе и, воз-
можно, другие небольшие генетические различия, ответственны за особенности
эпидемиологии и патогенеза инфекций, вызываемых У. pestis и У. pseudotuberculosis.
3.2. Чума
649
Филогенетическое древо К pestis, построенное М. Achtman et al. (2004) на основе
сопоставительного анализа нуклеотидных последовательностей, включает три ветви:
0, 1 и 2. Большинство изолятов биовара Orientalis соответствуют монофилетической
группе, обозначенной как LORI. Большинство штаммов биовара Medievalis соответ-
ствуют второй монофилетической группе — 2.MED. Изоляты биовара Antiqua нахо-
дят среди штаммов обеих ветвей —1 (1.ANT) и 2 (2.ANT). Штаммы Y. pestis биовара
Pestoides кластированы вдоль ветви 0 (0.РЕ1-0.РЕ4). Биохимические различия между
биоварами и регионы их выделения показаны в табл. 3.7; филогенетическое древо
приведено на рис. 3.28.
Таблица 3.7
Биовары У. pestis, идентифицированные на основе ферментации глицерина,
восстановления нитратов и окисления аммония*
Биовар Различие Регион, в котором был выделен штамм
Ферментация глицерина Восстановление нитратов Окисление аммония Активность уреазы Ферментация мелибиозы
Antiqua + + + - — Центральная Африка, Центральная и Северная Азия, Китай (Маньчжурия), Монголия
Medievalis + - - - ± Бывший Советский Союз (регион Каспийского моря)
Orientalis - + ± ± - Бирма, Южный Китай, Индия, Южная Африка, Южная Америка, Калифорния
* За основу взята таблица из работы А. Р. Anisimov et al (2004).
Рис. 3.28. Схематическое изображение филогенетического древа Y. pestis.
Вдоль каждой ветви указаны снипсы, использованные для его построения.
По М. Achtman et al. (2004)
650
Часть 3. Частная неправильная эпидемиология
S. Haenschetal. (2010), используя ДНК, выделенную из останков людей, умерших
от чумы во время второй пандемии (двое — из Англии, 6 — из Голландии, один — из
Франции), показали, что генотипы У pestis, индентифицированной в археологичес-
ких образцахXIV в., отличаются от современных Orientalis (1 .ORI), Medievalis (2.MED),
Antique (1 .ANT, 2.ANT) (см. рис. 3.28). Можно предположить, что в работе S. Haensch
et al. (2010) описаны штаммы Y. pestis, поддерживающиеся в неизвестных первичных
резервуарах очагов чумы европейского континента, угасших в начале XIX в.
Молекулярная эпидемиология У. pestis более точно устанавливается методами
MLVA-анализа. Полученные этим способом данные представлены на рис. 2.29 и 3.30.
caucasica, ulegeica, altaica,
hissarica, xilingolensis,
qinghaiensis
- Antigua foci H, KP, 38
<-| Antigua foci G, Тибет
Medievalis foci
- I, J, KI, K2
* в Medievalis foci 0,16,
18,21, 27,43, Курдистан
- Antigua foci А, В, C
'f-j Antigua Африка
- Antigua foci C, D
Antigua focus E
< включающий
DFR13 + штаммы
Orientalis, Yunnan,
Fujian, Vietnam, Myanmar
«Третья пандемия»
Microtus
Y. pestis
pestis
Рис. 3.29. Дендрограмма Y. pestis, основанная на 25 локусах VNTR. Псевдотуберкулезно-
му микробу соответствует черный круг в левом верхнем углу дендрограммы. В правой
части дендрограммы указаны новые биовары Y. pestis. Цифровые и буквенные обозначе-
ния очагов чумы соответствуют приведенным в табл. 3.9 и на рис. 3.30. По Li Y. et al. (2009)
У У. pestis под разными названиями описаны три типовые плазмиды: 9,5 т. п. о.
(pPst, pPla, pPCPl или pYP: кодирует пестициногенность, фибринолизин и плазмо-
3.2. Чума
651
Рис. 3.30. Географическое распределение штаммов возбудителя чумы различных биоваров.
Подписаны очаги, включающие штаммы биовара Microtus. Цифровые и буквенные обозна-
чения очагов чумы соответствуют приведенным в табл. 3 9 и на рис. 3.30. По Li Y. et al. (2009)
коагулазу); 70—75 т. п. о. (pCDl, pCad, pVW, pYV или pLcr: определяет зависимость
штамма от кальция, синтез VWa и других поверхностных белков); и 100-110 т. п. о.
(pFra/Tox, pFra, рТох, pMTl или pYT: кодирует Fl, «мышиный» токсин). Плазмидные
профили у штаммов, выделенных в различных природных очагах чумы, можно срав-
нить между собой по электрофоретической подвижности в агарозном геле, а выделив
отдельные плазмиды, сравнить их по результатам рестрикционного анализа.
Для штаммов, полученных в России и Азии, характерна значительная измен-
чивость плазмидного состава. До 18 % изолятов из Волго-Уральского песчаного очага
(очаг 16)* и 43 % изолятов из Таласского очага (очаг 40) содержали критические плаз-
миды, обычно с ММ 20 мДа. У некоторых штаммов из Таласского, Центрально-Кав-
казского и Прииалхашского очагов (очаги 40, 1 и 30 соответственно), а из Сенегала,
Шри-Ланки и Индонезии обнаружены критические плазмиды с ММ 1,6—31 мДа.
У отдельных штаммов отмечалось отсутствие одной или двух типовых плазмид
(Anisimov А. Р. et al., 2004).
Кроме того, размер плазмид может изменяться, что затрудняет получение ответа
на вопрос о том, является ли плаз лида, идентифицированная в клиническом изоля-
те, природным вариантом или же генно-инженерным вариантом с возможными до-
полнительными факторами вирулентности. Понимание причины изменений разме-
1ерриториальные границы очагов чумы, расположенных на территории бывшего СССР,
приведены ниже.
652
Часть 3. Частная неправильная эпидемиология
ра плазмид, обнаруженных у различных изолятов Y. pestis, важно для того, чтобы от-
личить естественное изменение от генно-инженерного, потенциально обусловливаю-
щего повышенную вирулентность или полиантибиотикорезистентность у штаммов,
содержащих такие теоретические конструкции. ММ классических плазмид Y. pesils,
характерных для штаммов различных подвидов, приведены в табл. 3.8.
Таблица 3.8
Молекулярные массы типовых плазмид Y. pestis, характерных
для штаммов различных подвидов*
Подвид Y pestis Биовар Название/номер очага Основной хозяин ММ (мДа)
pPst** pCad pFra
Y. pestis подвид apestis Antiqua Большинство очагов, кроме Тувы и Алтая Грызуны различных видов 6,4 47 67
Medievalis Все очаги, кроме очагов песчаночного типа Грызуны, за исключением песчанок 6,4 47 67
Orientalis Большинство очагов Грызуны различных видов 6,4 47 64
Antiqua Тува/37, 41 Citellus undulatus 6,4 46 67
Алтай/35 Marmota caudata 6,4 48 67
Medievalis Все очаги песчаночного типа Песчанки различных видов 6,4 48 67
Y. pestis подвида altaica Medievalis Горный Алтай/36 Ochotona pricei 6,4/- 48 72
Medievalis Южно-Кангайский, Монголия Microtus brandti 6,8 48 72
Y. pestis подвида ulegeica Medievalis Северо-Восточная Монголия, пустыня Гоби Ochotona pricei 6,4 47 67
Y. pestis подвида hissarica Medievalis Гиссарский/34 Microtus carruthersi 6/4 47 70
Y pestis подвида talassica Medievalis Талас/40 Microtus gregalis*** 6,4 46 70
Y. pestis подвида caucasica Antiqua Ленинакан/4; Присеванский/5; Занзегур-Карабах/6; Дагестанское нагорье/39 Microtus arvalis 50 82
* Таблица составлена А. Р Anisimov et al. (2004) по работам разных авторов.
** Количества pPst-изолятов Y. pestis подвида Altaica изменяются и в отдельные годы они
составляли до 10 % от общего количества изолятов.
*** Основным хозяином в этом очаге может быть Marmota caudata.
Экология и естественна* эпидемиология Y. pestis. О том, что экология и эпидемио-
логия возбудителя чумы далеко не так просты, как это обычно излагается в учебни-
ках для студентов, свидетельствует большое количество разных точек зрения на эко-
логию и эпидемиологию Y. pestis. существующую у чумологов, и высказываемых
обычно в специализированной научной литературе (см., например, монографии Сун-
цова В. В. и Сунцовой Н. И , 2006; Козлова М. П. и Султанова Г. В., 2003; Домарад-
ского И. В., 1998).
32. Чума
653
С практической точки зрения эпидемиологу, изучающему вспышку чумы на пред-
мет возможности ее искусственного происхождения, следует иметь в виду, что дискус-
сии чумологов в основном касаются первичных резервуаров Y. pestis (т. е. тех, без кото-
рых не возможно существование возбудителя чумы в природе) и механизмов
поддержания чумной палочки между эпизоотическими периодами. Вспышки чумы
среди грызунов следует рассматривать как процесс поддержания У. pestis во вторич-
ном резервуаре. Чумные эпизоотии создают условия для передачи возбудителя чумы
от инфицированных грызунов к людям через укусы инфицированных блох. Грызуны
инфицируются Y. pestis из пока неизвестного источника в почве; в результате у них
развивается чума в бубонной или септической форме. В случае септической чумы,
паразитирующие на грызунах блохи инфицируются. В преджелудке блохи (сфинктер-
подобный орган, разделяющий желудок и пищевод насекомого) происходит размно-
жение У. pestis, что вызывает его блокаду, и блоха голодает. Заражение человека про-
исходит в результате укуса голодной блохи, вынужденной «срыгнуть» в его кровь
«чумной блок». Для развития у человека чумы достаточно укуса одной чумной блохи.
Более 80 видов блох могут быть переносчиками У. pestis. Цепь, состоящую из возбу-
дителя (У. pestis), носителей (грызунов) и переносчиков (блох), принято называть «три-
адой», а определенные географические ландшафты, заселенные грызунами, видовые
и межвидовые отношения которых обеспечивают непрерывную циркуляцию чумно-
го микроба от донора через членистоногих реципиентам, называют природными оча-
гами чумы. Существуют различные классификации очагов чумы. Самая простая пред-
полагает их деление на очаги «дикой чумы» (эпизоотии среди диких грызунов) и
«крысиной» или «городской чумы» (эпизоотии возникают среди крыс, обитающих в
населенных пунктах).
По данным А. Р. Anisimov et al. (2004), природные очаги «дикой чумы» занимают
6—7 % суши Земли. Сорок три таких природных очага обнаружены в южных и юго-
восточных районах бывшего Советского Союза (табл. 3.9, рис. 3.31).
Таблица 3.9
Основные характеристики природных очагов чумы
в бывшем Советском Союзе*
Очаг № Название очага Эпизоотическая активность ** Основной хозяин Основной переносчик(и) Страна
1 2 3 4 5 6
Кавказский регион
1 Центральный Кавказский очаг Постоянная Citellus musicus Citellophillus tesquorom Россия
4 Закавказский высокогорный очаг (Ленинакан) Постоянная Microtus arvalis Callopsylla caspia, Nosopsyllus consimilis Армения, Грузия
5 Закавказский высокогорный очаг (в районе Севана) Постоянная Microtus arvalis C. caspia, N. consimilis Армения, Азербайджан
6 Закавказский вы- сокогорный очаг (Занзегур-Карабах) Постоянная Microtus arvalis C. caspia, N. consimilis Армения Азербайджан
7 Приаракский очаг Периодическая Meriones vinogradov Xenopsylla conformis Армения, Азербайджан
654
Часть 3. Частная неправильная эпидемиология
Продолжение табл. 3.9
1 2 3 4 5 6
8 Бозчелский очаг Периодическая Meriones libicus X. conformis, Nosopsyllus laeviceps Азербайджан
9 Кобистанский очаг Периодическая М. libicus X. conformis, N. laeviceps Азербайджан
10 Мила-Карабахский очаг Периодическая М lioicus X. conformis, N. laeviceps Азербайджан
11 Джейранчелский очаг Периодическая М. libicus X conformis, N. laeviceps Азербайджан
12 Гянджа - Казахский очаг Периодическая М, libicus X. conformis, N. laeviceps Азербайджан
13 Иори, река Периодическая М. libicus X. conformis, N. laeviceps Азербайджан, Грузия
39 Дагестанское нагорье Периодическая Microtus arvalis C. caspia Россия
Северный Прикаспийский регион
2 Терек-Сунжа Периодическая Citellus pygmaeus Neopsylla setosa, C. tesquoium Россия
3 Дагестанское равнинное предгорье Периодическая C. pygmaeus N. setosa, C. tesquyorum Россия
14 Прикаспийский северо-западный очаг Периодическая C. pygmaeus N. setosa, C. tesquorum Россия
15 Волго-Уральская степь Периодическая C. pygmaeus N. setosa, C tesquorum Россия
17 Зауралье Периодическая C aygrriaeus N. setosa, C. tesquorum Россия, Казахстан
43 Прикаспиский очаг, пески Периодическая M. meridianus N. laeviceps Россия
16 Волго-Уральский очаг, пески Периодическая M. meridianus X. conformis, A laeviceps Россия
Центрально-Азиатский пустынный регион
18 Междуречье Урала и Эмбы Постоянная R. opimus X. skijabini Казахстан
19 Область, прилегающая к плато Устюрт Постоянная R. opimus X. skijabini Казахе ган
20 Плато Устюрт Периодическая R, cpimus X. skijabini Казахстан, Узбекистан
21 Северное Приаралье Периодическая R opimus X. skijabini Казахстан, Узбекистан
22 Зааралье Периодическая R. opimus X. skijabini Казахстан
23 Мангышлак, плато Периодическая R. cpimus X skijabini Казахстан
24 Приаралье-Кара- кум Постоянная R. opmu s X. skijabini Казахстан
25 Каракумы, пески Периодическая R. opimus X gerbili Туркменистан
3.2. Чума
655
Продолжение табл. 3.9
1 2 3 4 5 6
26 Копет-Даг, хребет Периодическая R. opimus X. gerbilli Туркменистан
27 Кызылкум, пески Периодическая R. opimus X. gerbilli Узбекистан, Казахстан
28 Муюнкум, пески Периодическая R. opimus X. gerbilli Казахстан
29 Таукум, пески Периодическая R. opimus X. gerbilli Казахстан
30 Прибалхашье Периодическая R. opimus X. gerbilli Казахстан
42 Бет-Пак-Дала Периодическая R. opimus X. gerbilh Казахстан
Центрально-Азиатский горный регион
31 Сарыджас, река Периодическая Marmota baibadna Oropsylla silantiewi Киргизия
32 Верхний-Нарын, река Периодическая M. baibadna O. silantiewi Киргизия
33 Аксай, река Периодическая M. baibadna O. silantiewi Киргизия
34 Гиссарский очаг Периодическая Microtus carruthersi C. caspia, Amphipsylla phaiomydis, Frontopsylla elata, N. pleskei Таджикистан, Узбекистан
35 Алтай, нагорье Периодическая Mannota caudata O. silantiewi Киргизия
40 Талас, река Периодическая Microtus gregahs, Marmota caudata C. caspia Киргизия
Сибирский регион
36 Алтай, горы Периодическая Odiotona pricei fttradaxopsyllus scorodumovi Россия
37 Тува (Монгун- Тайга) Периодическая Citellus undulatus C. tesquorum Россия
41 Тува (Саглы) Периодическая C. undulatus C. tesquorum Россия
38 Забайкалье Периодическая Citellus daudats C. tesquorum Россия
* Таблица составлена А. Р. Anisimov et al. (2004) по работам разных авторов. Границы оча-
гов чумы очерчены авторами по границам чумных эпизоотий среди грызунов, зафиксирован-
ных в XX в.
* * Постоянная активность указывает на наблюдение за активностью в течение всего пе-
риода эпизоотии. Периодическая активность означает, что эпизоотии прерывались
2—10-летними периодами между вспышками.
Очаги данного типа чумы охватывают более 216 х 106 га или 8,6 % территории быв-
шего СССР. Разделение и последующую классификацию этих природных очагов чу-
мы А. Р. Anisimov et al. (2004) проводили на основе их географического распределения-
с учетом основного инфицированного хозяина и основного переносчика, обнару-
женных в каждом очаге’. Основные хозяева из различных очагов данного типа имеют
несколько общих экологических характеристик, таких как большое и устойчивое
1 Ответ на вопрос, кто «главный или энзоотический хозяин, ответственный за поддержа-
ние Г. pestis в природных очагах», и кто, соответственно, «неглавный или эпизоотический хо-
зяин», до сих пор не получил ответа, удовлетворяющего всех чумологов. А. Р. Anisimov et al.
(2004), чьей работой я здесь пользуюсь, придерживаются взглядов на процесс поддержания
656
Часть 3. Частная неправильная эпидемиология
Рис. 3.31. Географическое расположение очагов чумы на территории бывшего СССР.
Каждый природный очаг чумы отличается от другого видовым составом носителей и пе-
реносчиков К pestis, ландшафтно-географическими особенностями. Цифровые обозна-
чения соответствуют названиям очагов, приведенных в табл. 3.9. Светло-серым выде-
лены очаги чумы с основным подвидом Y. pestis, темно-серым — очаги с неосновными
подвидами Y. pestis. По А. Р. Anisimov et al. (2004)
количество хозяев-грызунов, инфекционная бактериемия как одна из стадий болез-
ни, блохи-паразиты, которые являются активными переносчиками чумы, и блохи, ко-
торые могут легко выживать в норах грызунов. А. Р. Anisimov et al. (2004) предполага-
ют, что адаптация к непрерывной передаче возбудителя чумы у различных видов
грызунов в значительной степени способствует появлению разных подвидов Y pestis,
различаемых по ферментативной активности, потребностям в питательных веществах
и по способности вызывать бактериемию и гибель различных видов животных.
Крупным эпидемиям чумы обычно предшествуют вспышки-предшественники,
отражающие процесс активизации природных очагов чумы. Например, эпидемиям ле-
гочной чумы в Маньчжурии 1910 г. и 1921 г., погубивших жизни десятков тысяч чело-
век, предшествовали локальные вспышки бубонной чумы, фиксируемые с 1860-х гг.
в Забайкалье, Маньчжурии и Монголии. Они ограничивались десятками заболевших
и были связаны с добыванием тарбаганов и работами местных жителей на покосах.
Со средины 1890-х гг. в этих же регионах врачами выявляются локальные вспышки
легочной чумы. К тому же чума стала «задерживаться» в отдельных населенных пунк-
чумы как на непрерывную эпизоотию среди «энзоотических хозяев». Другие ученые, напри-
мер, И. В. Домарадский (1998), обращают внимание на то, что десятилетиями в отдельных
очагах чумы могут не выявляться отдельные эпизоотии, но потом они внезапно становятся
массовыми. Это говорит в пользу существования совершенно иных очагов чумы, чем поддер-
живающиеся среди грызунов (их границы см. в табл. 3.9). Поэтому при расследовании преступ-
ления, вызывавшего вспышку чумы, необходимо понимать, что приглашенные эксперты-чу-
мологи могут принадлежать к антагонистическим школам.
3.2. Чума
657
тах. В 1898 г. в районе Вейчан (Южная Манчьжурия) чума в течение трех лет «проре-
живала» жителей пяти компактно расположенных деревень, где из 550 жителей от
чумы умерли не менее 400.
Активизация очагов чумы имеет пульсирующий характер: вспышки чумы сменя-
ются периодами относительного благополу тия Южно-маньчжурский очаг чумы стал
«разогреваться» с отельных вспышек бубонной чумы, начавшихся в 1924 г. к северу
и к западу от города Тунляо. В 1930-х гг. чума стала эндемичной болезнью на десятках
тысяч километрах территории Внутренней Монголии и Южной Маньчжурии. Вспыш-
ки чумы 1933 г. и 1945—1947 гг., погубившие десятки тысяч жизней, зафиксировали
максимальные пульсации очага, но вначале 1950-х гг. он угас.
В настоящее время мы имеем дело с «дремлющей чумой». Границы ее современных
природных очагов очерчены по границам чумных эпизоотий среди грызунов, зафик-
сированных в XX в. По территориальным масштабам они значительно меньше тех,
которые можно очертить по историческим хроникам XIV-XIX вв. Их возможные ipa-
ницы приведены в нашей работе (см. Супотницкий М. В., Супотницкая Н. С., 2006).
Заболевания людей чумой начинаются вслед за эпизоотиями среди крыс с разрывом
в 10—14 сут. Но до сих пор нет однозначного ответа на вопрос, как быстро развивают-
ся вспышки чумы среди людей вслед за эпизоотиями среди диких грызунов. Так же
неизвестен механизм «вброса» К pestis в популяции грызунов, обитающих на терри-
ториях очагов чумы. Периодически он приводится в действие неизвестными факто-
рами, что проявляется крупными вспышками чумы среди людей, уже забывшими о
чумных катастрофах прошлого. Вследствие этого не всякое появление чумы там, где
ее «никогда не было», можно считать результатом диверсии. Эпидемиологическое
расследование должно начинаться с выявление естественной эпидемиологии чумы.
Естественная эпидемиология бубонной чумы предполагает наличие не менее двух
звеньев эпидемического процесса. Прежде всего, должен быть обнаружен природный
источник возбудителя чумы для людей — зараженные чумой грызуны (первое звено —
источник возбудителя чумы). Люди заболевают бубонной чумой на эпизоотическом
фоне, т. е. с эпизоотии чумы среди грызунов («дикий очаг» чумы) или крыс («город
ской очаг» чумы). Наблюдается падеж грызунов и накопление инфицированных пере-
носчиков Y. pestis — блох (второе звено — механизм передачи возбудителя чумы)1.
Лишившиеся своих хозяев блохи нападают на животных и людей. Для сохранения
активности блох требуются определенные температура и влажность. Блохи X. cheopis
не могут размножаться в холодную или жаркую погоду. Благоприятная температура
для развития X. cheopis около 24°С при относительной влажности 70 %. Понижение
относительной влажности ниже 40 % вызывает гибель личинок до их окукливания.
В природных очагах чумы заражение человека чумой через укусы блох сопровож-
дается развитием бубонной и септической форм болезни. В случаях укусов блох на-
блюдается характерная локализация бубонов: наиболее распространены паховые и
бедренные бубоны — половина всех случаев; до четверти составляют подмышечные
бубоны, и до 10 % — шейные (см. Николаев Н. И., 1949). В тех очагах чумы, где дикие
грызуны являются предметом охоты (сурки, суслики, зайцы и др.), инфицирование
людей может происходить путем их непосредственного контакта с источником инфек-
ции, когда охотники снимают шкурки или разделывают тушки убитых ими больных
чумой животных. Люди могут заразиться чумой при снятии шкуры и разделки мяса
1В разд. 1.8 приведен пример вспышки чумы в китайском городе Чанде без чумной эпизоо-
тии среди крыс. Вспышка была вызвана распространением японцами зараженных чумой блох.
658
Часть 3. Частная неправильная эпидемиология
больною чумой верблюда. В таких случаях инфицирование Y. pesti. происходит через
повреждения кожи рук Первичные бубоны развиваются в группе лимфатических
узлов, ближайшей к участку поврежденной кожи, через который произошло инфи-
цирование возбудителем чумы. Характерными для заражения вызванного разделкой
больного чумой животного, являются подмышечные бубоны и вторичная чумная
пневмония (Лобанов В. Н., 1968). Вторым звеном эпиде лического процесса при та-
ком пуз и заражения будут кровь и ткани больных чумой грызунов, с которыми человек
соприкасается благодаря своей деятельности в очаге чумы. При любом из обнаружен-
ных механизмов передачи возбу штеля (через укусы блох или при контакте с кровью
и шкурами инфицированных грызунов), в случае естественного происхождения
вспышки бубонной чумы, должна быть выявлена чумная эпизоотия среди грызунов.
Эпидемия бубонной чумы в природном очаге чумы чосит выраженный сезонный
характер, и характеризуется постепенным подъемом заболеваемости. В начале чум-
ной эпизоотии в очаге чумы имеется небольшое количество блох, способных инфи-
цировать человека. Нграстание заболеваемости среди людей идет параллельно возра-
станию гибели крыс и повышению инфекционного потенциала очага, т. е. увеличению
количества блох, зараженных возбудителем чумы.
Начальная стадия чумной вспышки проходит незамеченной по разным причинам.
Основная из них — психологическая. Чума — это слово-табу. Как правило, ее никто
не ж чет, и о ней никто не помнит. При появлении первых типичных случаев болезни,
слово «чума» очень непросто произнести чиновнику или врачу, учитывая дальнейшие
последе гвия для них в случае ошибочного диагноза.
Кроме того бубонная чума трудна для диагностики. Первые случаи могут по не-
скольку месяцев приниматься за неспецифические лимфадениты. Больные попа-
дают не в инфекционные больницы, а в хирургические стационары общего профиля.
И только необычно высокая смертность среди таких больных дает основание врачам
заподозрить, что они имеют цело с какой-то более серьезной болезнью и предпринять
бактериологическое исследование.
Развитие эпидемии чумы достигает пика, затем эпидемия постепенно угасает из-
за гибели грызмнов. Погибают и лишившиеся своих хозяев блохи. Б городах, в кото-
рых чума вызвана завезенными чумными крысами, вспышка носит кратковременный
характер. Когда в городе возникает энзоотия среди местных крыс, то заболевания
людей могут иметь место в течение нескольких месяцев. В тех случаях, когда чума
переходит в септическую форму с летальным исходом, и когда в жилише ооитает мно-
го блох, вспышка может принять антропонозный хаоак гер и продолжаться длитель-
ное время из-за продолжительного периода блокообразования у блох (в среднем
13—14 сут). Если не проводить дератизацию и дезинсекцию, больные бубонной чумой
появляются в одних и тех же домах в течение 2—3 мес. На продолжительность и ин-
тенсивность вспышки будет оказывать влияние величина населенного пункта, коли-
чество проживающего в нем населения, его культурный уровень, санитарное состоя-
ние жилищ и, видимо, те неизвестные факторы, которые приводят к активизации
очагов чумы. В Москве эпидемия чумы, начавшаяся осенью 1770 г., закончилась в фев-
рале 1772 г., в Одессе чума, начавшись в августе 1812 г., прекратилась через полгода.
Ecmet твенная эпидемиология легочной чумы нуждается в дальнейшем исследовании.
Из исторических источников известно, что легочная чума способна «пропадать» на
несколько столетий, при том, что бубонная чума за этот период времени губит жизни
миллионов людей (см. у Гирша А., 1853). В настоящее время легочная чума чаще все-
го развивается на фоне вспышек бубонной чу'мы. Выделяют две клинических формы
3.2. Чума
659
легочной чумы: первичную и вторичную. Под вторичной легочной чумой понимают
чумную пневмонию, развившуюся у больных с бубонной чумой. Такие больные мо-
гут оказаться источниками воздушно-капельной инфекции для других людей. Ини-
циация вспышки первичной легочной чумы становится возможной тогда, когда у
больного вторичной легочной чумой поражение легких сопровождается приступами
кашля с обильным выделением кровянистой мокроты, содержащей К pestis. Возмо-
жен контактно-бытовой путь передачи — возбудитель чумы со слюной больного чу-
мой попадает в ротовую полость здорового человека (см. ниже «И. Легочная чума
взрывного развития»), В обоих случаях роль ворот для инфекции играет лимфатиче-
ская система глотки, гортани, крупных бронхов. Источниками заражения для людей
могут быть больные чумой кошки и собаки (Kool J. L., 2005). Легочная чума развива-
ется по типу нисходящей бронхопневмонии. Случаи легочной чумы, вызванные воз-
душно-капельной и контактно-бытовой передачей возбудителя болезни от больных
легочной чумой, называют первичной легочной чумой.
Эпидемиология легочной чумы существенно отличается от эпидемиологии бу-
бонной чумы:
1) если при бубонной чуме роль больного чумой человека как источника инфек-
ции незначительна, то при легочной чуме больной человек становится основным ис-
точником заражения Y. pestis',
2) бубонная чума не выходит за границы природных очагов чумы, легочная чума
способна распространяться вместе с потоками людей далеко за их пределы;
3) бубонная чума относительно проще диагностируется, чем легочная. Ошибки в
диагнозе бубонной чумы не приводят к столь серьезным эпидемическим последстви-
ям, к которым приводит не своевременное распознание легочной чумы у больного;
4) легочная чума без лечения антибиотиками в 100 % случаев приводит боль-
ного к смерти, тогда как при бубонной чуме летальность среди заболевших 50—60 %.
Больные на ранней стадии легочной чумы (приблизительно в течение первых
20—24 ч) не представляют большой опасности из-за небольшого количества бактерий
в выделениях из дыхательных путей и, как правило, отсутствия кашля. Наибольшую
опасность они представляют на поздних стадиях болезни, когда при кашле выделя-
ется обильная мокрота с видимой кровью и/или гноем. Благодаря антибиотикотера-
пии, мокрота больных быстро очищается от К pestis. Уже через несколько часов после
начала лечения эффективными антибиотиками пациент обычно становится неинфек-
ционным (Kool J. L., 2005).
Современные вспышки легочной чумы носят локальный характер и ограничива-
ются единичными случаями болезни. Проследить источник возбудителя чумы очень
сложно. Чаще всего врач становится перед фактом вспышки среди людей, не зная всех
звеньев эпидемической цепочки распространения Y. pestis. Приведу два примера та-
кого появления легочной чумы среди людей, одновременно показывающих, насколько
различными могут быть показатели заболеваемости при одной и той же болезни.
Случай легочной чумы в Лос-Анджелесе 1924г. (по работе Dickie W. М., 1926). В ок-
тябре 1924 г. в Лос-Анджелесе после 4-дневной болезни умерла мексиканка. Ее суп-
руг и одна из медицинских сестер тоже заболели и умерли. Причиной смерти мужа
врачи считали «двустороннюю лобулярную пневмонию». В течение двух суток имели
место еще 32 смерти от легочной чумы. В число погибших вошли лица, жившие в этом
доме, а также друзья и родные, посещавшие дом первого заболевшего, у которого, по-
видимому, была вторичная легочная чума. Среди погибших от чумы было четверо детей
в возрасте 10 лет и младше, одна женщина 50 лет, священник, медицинская сестра
660
Часть 3. Частная неправильная эпидемиология
и водитель санитарной машины, помогавший перевозить больных с легочной чумой.
У нескольких жителей этого района выявлена чума в бубонной форме. Возбудитель
чумы обнаружили у городских крыс, которые, вероятно, заразились от беличьих блох.
Установить, при каких обстоятельствах заразился чумой первый больной, не удалось.
Случай легочной чумы, вспыхнувшей в декабре 2004г. в Конго Субкаунти, округ Небби,
Уганда (по работе Begier Е.М. et al., 2004). 26 декабря 2004 г. угандийский полицейский
сообщил по телефону местному врачу Gershim Asiki о ряде случаев заболевания с быст-
рым летальным исходом, зарегистрированных в районе Западного Нила. В результате
эпидемиологического расследования были определены две пары первичных случаев
легочной чумы. Между ними не удалось установить каких-либо социальных связей.
Первой первичной больной (А1) была 22-летняя женщина, помощь ей оказывала
ее 40-летняя мать (А2). По словам членов этой семьи, болезнь началась с головной
боли, жара и озноба, которые сохранялись в течение нескольких дней. Увеличение
лимфатических узлов впервые было обнаружено на 3 сут болезни. На 5 сут болезни у
больной появился кашель, сутками позже он стал влажным, а на 7 сут уже сопровож-
дался выделением кровянистой мокроты. На 6 сут больная женщина была осмотрена
владелицей аптеки (дипломированной медицинской сестрой), которая в течение 3 сут
лечила ее хлорохином от малярии. На 9 сут она стала кашлять кровью и ночью умер-
ла. Больная А2, оказывавшая первичную помощь больной А1, заболела через 5 сут
после смерти дочери. В первый день заболевания у больной А2 появились головная
боль, жар, озноб, слабость, больвгруди и кашель с мокротой. Ее осмотрела та же вла-
делица частной аптеки, которая диагностировала язвенный фарингит. Больной вво-
дили внутримышечно пенициллин (6 раз в течение 36 ч) для лечения предположитель-
ной острой пневмонии. На следующие сутки у нее появилась обильная кровянистая
мокрота, и на 3 сут болезни она умерла.
Вторым первичным больным В1 был 25-летний мужчина, которому первичную
помощь оказывала его сестра, больная В2. Болезнь первичного больного В1 началась
с головной боли, жара и озноба. Его семья обращалась за помощью в частную аптеку
и возила его на консультацию в три государственных медицинских центра. Лимфаде-
нопатия не была выявлена, лечившие его врачи специально не осматривали его с этой
целью. Ему назначили противомалярийные средства и вводили внутримышечно
пенициллин для лечения предположительной острой пневмонии. На 5 сут болезни
кашель больного В1 стал сопровождаться выделением кровянистой мокроты, а на
6 сут он умер.
Ухаживавшая за больным В1 сестра (больная В2), заболела через 5 сут после смер-
ти В1. У нее была сильная одышка, повышенная температура и частота дыхания
(39,3 °C и 56 дыханий/мин соответственно). Она не могла передвигаться самостоятель-
но. Лимфатические узлы не прощупывались. При исследовании легких выявлены втя-
жение межреберных промежутков и двусторонние жесткие крепитирующие хрипы.
Первую врачебную помощь она получила через 29 ч после начала болезни в местном
медицинском центре, где ей были назначены хлорамфеникол (левомицетин), 2 г внут-
ривенно, и доксициклин, 100 мг перорально, затем ее отправили в специализирован-
ное лечебное учреждение.
Серия из трех фронтальных рентгенограмм грудной клетки, сделанных на 2, 3 и
18 сут болезни, наглядно подтвердила многодолевую пневмонию с плевральными
экссудатами. Наличие К pestis в мокроте выжившей больной было подтверждено
положительными результатами ПЦР-анализа маркерных генов на всех трех плазми-
дах Y. pestis.
3.2 Чума
661
Больной продолжили лечение хлорамфениколом: 1 г внутривенно каждые 6 ч
в течение 48 ч, а затем 750 мг каждые 6 ч (в течение 10 сут внутривенно). Пациентка
была выписана на 10 сут и принимала хлорамфеникол уже перорально, 750 мг каж-
дые 6 ч еще 8 сут.
Лицам, имевшим тесный контакт с первичными больными А1 и В1, не было на-
значено профилактическое лечение антимикробными препаратами, потому что, когда
было сообщено о вспышке, после смерти первичных больных прошло уже больше
недели. Двадцать пять человек имели прямой контакт с одним из первичных больных
(т. е. соприкасались) после появления у них кашля с выделением кровянистой мок-
роты и перед смертью, но заболели только два человека, оказывавшие им первичную
помощь (показатель заболеваемости 2/25, 8 %).
Во время обследования в конце декабря и начале января в нескольких деревнях
этого района поступили сообщения от жителей о гибели крыс, и семьи двух первич-
ных больных также сообщили об обнаружении мертвых крыс вблизи своих жилищ
накануне заболевания своих родственников. Это все, что удалось выяснить про ис-
точники инфицирования людей специачистам CDC, расследовавшим эту вспышку.
Источник инфицирования людей возбудителем чумы в Канго Субкаунти и в Лос
Анджелесе остался не установленным. Однако обе вспышки легочной чумы развились
на фоне эпизоотий чумы среди грызунов, а в Лос-Анджелесе были установлены слу-
чаи бубонной чумы среди людей. Важным критерием природного характера вспыш-
ки в условиях неопределенности с источником инфекции будет отсутствие патологии
мелкодисперсного аэрозоля.
Не всегда вспышка легочной чумы ограничивается единичными случаями болез-
ни. Известны относительно недавно произошедшие вспышки легочной чумы, напом-
нившие современникам ужасы «черной смерти» 1346—1351 гг. Анализ научной лите-
ратуры позволяет выделить три типовых сценария естественного развития крупных
вспышек легочной чумы.
I. Легочная чума на фоне бубонной. Обратимся к исследованию Г. Н. Минха (1898),
описавшего вспышку чумы в станице Ветлянская осенью 1878 г. Станица Ветлянская
расположена на Волге, в местах повсеместных поселений малого суслика, одного из
основных вторичных резервуаров К pestis. Какое-то заболевание, сопровождающее-
ся развитием паховых и подмышечных бубонов, появилось в Астраханской губернии
в августе 1877 г. небольшими вспышками (до 40 заболевших), не сопровождавшими-
ся летальными исходами, что вызвало много споров среди врачей по поводу того,
чума это или не чума. Так как бактериологические методы диагностики отсутствова-
ли, эти дискуссии ясности о природе болезни не дали.
Эпидемию чумы в станице Ветлянская Минх разделил на два периода. Первый
период начался в станице в конце сентября 1878 г. с единичных случаев бубонной чумы.
До 17 сентября заболело 6 казаков и казачек, из них трое погибли (летальность обыч-
ная для бубонной чумы — 50 %), потом, по выражению Минха, «болезнь смягчи-
лась» — из семи заболевших умер только один (Хритонов Макар, 27 октября). Чума
«взяла паузу». Вернулась она уже в септической форме: 8 ноября скоротечно и без
бубонов умерла молодая казачка, за ней при сходных обстоятельствах — молодой ка-
зак (12 ноября), на следующий день с аналогичными симптомами скончалась еще одна
женщина. Со второй половины ноября, злокачественность болезни возросла, в это же
время стали появляться случаи заболеваний, осложненные поражением легких. В по-
следней декаде ноября были еше два случая затянувшейся бубонной чумы с кровохар-
каньем, оба больных погибли. Однако распространения легочной чумы по станице не
662
Часть 3. Частная неправильная эпидемиология
произошло, возможно, ее вообще бы там не возникло, если бы возбудитель болезни
не попал в многочисленное семейство богатого ка ика Осипа Белова.
Его сын Федор был женат на Марье Хритоновой. Минх, писавший свой труд еще
до открытия возбудителя чумы1, не знал о существовании вторичных крысиных оча-
гов чумы и о возможности развития домовых эпидемий чумы. Его представления об
эпидемиологии чумы строились на возможности переноса «чумного контагия». Он
считал, что именно от семейства Хритоновых произошел занос «контагия» чумы в
семейство Беловых. Всего у Хритоновых в первом периоде эпидемии было поражено
бубонной чумой 9 человек, из которых четверо умерли, но ни один из них не страдал
легочной формой болезни. В домах Беловых эпидемия проявляет себя сразу массой
почти внезапных смертельных заболеваний (первыми погибли Федор и Марья) с почти
полным отсутствием бубонных осложнений. Минх подчеркивал, что это обстоятель-
ство могло бы «навести лиц, мало или совсем незнакомых с первым периодом ветлян-
ской чумы, на мысль о какой-то другой, начавшейся только с Беловых, болезни».
Сегодня же можно утверждать, что, так как в семье Хритоновых легочной чумы
не было (как, впрочем, и в других станичных семействах в это время), то занос возбу-
дителя болезни в семейство Беловых мог быть осуществлен только из крысиных оча-
гов чумы блохами. Эти очаги могли находиться как под домами Беловых, так и Хри-
тоновых, которых «по-родственному» те посещали во время болезни. Тогда странным
выглядит не типичная для данного способа заражения возбудителем чумы клиника
поражения семей Беловых. Обратимся опять к работе Мииха.
Исследуя отношение эпидемии по месту жительства пострадавших от болезни
семейств, Минх обратил внимание на то, что близкое соседство, даже с наиболее по-
страдавшими от чумы семействами, не имело никакого влияния на распространение
болезни по станице. Он отметил отсу гствие заболевших во многих домах, расположен-
ных вблизи тех, семейства которых поголовно вымерли. Просматривая метрические
записи, он увидел в списке умерших все одни и те же фамилии1 целые десятки Бело-
вых, много Лобановых, Астаховых, Писаревых, Назаровых и т. д.
Вывод, который Минх сделал из этих наблюдений, состоял в том, что «только эти
родословные могут досказать нам то, чего не могли открыть свидетели, что только
они могли дать нам ясное представление о списке умерших, представляющемся нам,
как мы сказали, без знания родословных, простым перечнем имен фамилий». Следо-
вательно, можно предположить, что второй период ветлянской чумы — легочный,
начался со случайного вовлечения в эпидемический процесс «критической массы»
человеческих генотипов, у которых инфекционный процесс проявился вторично-ле-
гочной формой болезни. Относительная генетическая однородность многочислен-
ных родов Беловых, и особенно членов семейства Белова Ивана, способствовала край-
не упорному распространению вторично-легочной чумы по станице и отбору высо-
ковирулентных штаммов ее возбудителя. После достижения «критической массы» из
вторично-легочных случаев болезни, чума сформировала самостоятельные эпидеми-
ческие цепочки (без участия эктопаразитов), состоящие уже из первично-легочных
случаев («первичные заносы» — по определению Минха), это и привело к эпи-
демической катастрофе (рис. 3.32).
На схеме показаны три разные эпидемии чумы, протекающие одновременно, но
вызванные одним возбудителем. Первая — бубонная чума, развивавшаяся среди жите-
лей станицы единичными случаями с конца сентября в результате их укусов блохами,
1 В начале 1880-х гг.
3.2. Чума
663
Рис. 3.32. Распространение чумы в станице Ветлянская. I — динамика забо-
леваний чумой в станице; 2 — движение чумы в домах Беловых; 3 — движе-
ние заносов (первичных) из домов Беловых; 4 — движение заносов чумы
из ст. Ветлянской в другие селения. По Г Н Минху (1898)
паразитировавшими на больных чумой грызунах. Вторая — «переход» бубонной чумы
во вторично-легочную в семье Беловых и образование эпидемических цепочек меж-
ду членами этой семьи и другими жителями станицы. Третья — формирование само-
стоятельных эпидемических цепочек первично-легочной чумы среди жителей стани-
цы и других селений, начавшихся от вторично-легочных случаев в семье Беловых.
Пик эпидемии пришелся на 14 декабря — в этот день погибли 36 человек. Затем
число заболеваний и смертей стало уменьшаться, 15 декабря погибли 19 человек, а
12 января 1879 г. эпидемия прекратилась1. В последнем периоде эпидемии (январь)
преобладали случаи воспаления лимфатических узлов при лихорадочных проявлени-
ях. Эпидемия закончилась, как и началась — бубонами. Всего в Ветлянке болело чумой
446 человек (25 % всего населения), из них выздоровели 82 человека, погибли 364 че-
ловека (82 % из числа заболевших).
В конце марта в Ветлянке наблюдалось еще несколько случаев boci гален ия лимфа-
тических узлов, причем иногда температура поднималась до 39 °C и выше, в других слу-
чаях она оставалась нормальной, «опухоль желез постепенно рассасывалась». Только
в одном случае нагноения шейных лимфатических узлов врачам пришлось вскрыть
абсцесс. Так как во всех случаях следовало выздоровление, то их к чумным не относили.
Сходным образом развивалась чума в изолированных популяциях таджиков в
кишлаке Анзоб (современный Таджикистан) в 1897 г. Сначала бубонные формы бо-
лезни, затем легочные, потом снова бубонные. Основными отличиями чумы в Анзобе
от Вет шнекой чумы, были: отсутствие случаев заражения от больных легочной чумой
и необычная клиника легочной чумы — заболевшие люди не выглядели тяжелоболь-
ными. Ни буйного бреда, ни прострации у них не наблюдалось. Последние часы жиз-
ни они впадали в тихий бред (Супотницкий М В , Супотницкая Н. С., 2006).
На рис. 3.32 показана динамика заболеваний всеми формами чумы в ст Ветлянская (мак-
симум 8 декабря). Пик гибели людей смещен на несколько дней (максимум 14 декабря)
664
Часть 3. Частная неправильная эпидемиология
II. Легочная чума взрывного развития. Взрывному развитию легочной чумы в Мань-
чжурии предшествовало почти 50 лет небольших вспышек бубонной и легочной чумы
(«предвестников») в Маньчжурии, Монголии и Забайкальской области. Географиче-
ская изоляция, естественным образом препятствующая распространению легочной
чумы между населенными пунк.ами, была устранена проведением Китайской-Вос-
точной железной дороги (КВЖД). Первый раз чума (в бубонной форме) появилась на
полосе отчуждения КВЖД в 1905 г. на Джаланройских каменноугольных копях, рас-
положенных в 27 км от станции Маньчжурия. Вспышку удалось локализовать еще до
тою, как чума перешла на китайское население.
Точных данных, когда появились первые больные легочной чумой в Маньчжурии,
нет. Но известно, что в конце лета 1910 г. одновременно в разных местах Маньчжурии
появились вспышки легочной чумы. Первый бактериологически подтвержденный
случай чумы констатирован врачами в поселке при станции Маньчжурия 2 октября,
но начало эпидемии не совпадает с этой датой. В местности по реке Ган в Монголии
против Старого Цурухайтуя весной охотники находили трупы своих товарищей и
около них шкурки убитых ими сурков. Затем в конце июля верстах в двухстах от стан-
ции Маньчжурия казаки замечали, что среди сурков был мор, и они гибли в большом
количестве. Видимо, и среди монголов в то время эпидемия уже свирепствовала, так
как казаки обнаружили табуны скота, бродившие без присмотра.
Поданным Э. П. Хмара-Борщевского (1912), начало эпидемии чумы на ст. Маньч-
журия относится к 12 октября 1910 г., когда председатель поселкового совета
А, Н. Никитин сообщил по телефону заведующему железнодорожной больницей док-
тору Писемскому о том, что по Александровскому проспекту, на участке № 3083, сре-
ди китайцев появились заболевания со смертельным исходом и что за последние
5 сут там умерло 9 человек. Явившись в указанный дом, доктор Писемский увидел
там такую картину: барак был оставлен жильцами, на нарах валялись в беспорядке
брошенные ими вещи и посуда, а в углу дожал прикрытый лохмотьями больной кита-
ец с высокой температурой. Перкуссией и аускультацией у него было установлено
воспаление правого легкого. Больной тотчас же был отправлен в железнодорожную
больницу, барак заколочен и сдан под охрану полиции.
По рассказам соседей, в этом доме жили китайцы-тарбаганщики, в одной поло-
вине 25 человек, в другой — 15. Среди жильцов первой группы умерло 9, а второй —
5 человек. Перед прибытием врача все жильцы дома разбежались, бросив свое иму-
щество. Доставленный в больницу охотник за тарбаганами был помешен в изолятор,
где он и умер. Вскрытием трупа, а также бактериологическим исследованием был ус-
тановлен диагноз легочной чумы.
По мнению японского профессора Абэ (1942), распространение чумы в 1910 г.
началось при следующих обстоятельствах. В Даурии существовала артель плотников-
китайцев, среди которых примерно в середине сентября умерло 7 человек с явления -
ми кровохарканья. Оставшиеся из этой артели в живых два плотника-китайца пере -
шли на работу в другую артель на ст. Маньчжурия и здесь скончались 12 октября. Затем
в этой ьторой артели среди 20 плотников к 25 октября умерли один за другим 9 чело-
век. Отсюда легочная чума распространилась по всему поселку Маньчжурия (цит. по
Николаеву Н. И., 1949).
Возможно, обе версии отражают части целого — реально произошедшие события.
С 12 октября легочная чума начала быстро прогрессировать в поселке Маньчжурия и
отсюда распространяться по железной дороге в Забайкальскую область и по всей ли-
нии КВЖД (табл. 3.10).
3.2. Чума
665
Таблица 3.10
Развитие эпидемии легочной чумы в полосе отчуждения КВЖД в 1910 г.*
Период Заболело Умерло Период Заболело Умерло
С 13 по 17 октября 36 36 С 8 по 14 ноября 48 51
С 18 по 24 октября 51 44 С 15 по 21 ноября 37 38
С 25 по 31 октября 85 73 С 22 по 28 ноября 12 13
С 1 по 7 ноября 86 98 С 29 по 3 декабря 2 3
* По М. Л. Блюменфельду (1911).
К этим 356 погибшим надо добавить еще не менее 100 человек, ставших жертва-
ми чумы с июля по октябрь (Касторский Е. С., 1911). Но и тогда приведенные сведе-
ния оказываются неточными. И. Л. Мартиневский и Г. Г. Молляре (1971) указывают
на то, что в апреле 1911г., после таяния снега, в окрестностях поселка был обнаружен
191 труп, причем 131 труп найден на полосе отчуждения и 60 трупов вне ее. Итог эпи-
демии: 647 погибших.
20 октября появились первые случаи заболевания легочной чумой среди китай-
цев на станции Даурия, 21 — в Джалайнорских копях, 26 — в Тарбагатайских копях,
27 — в Хайларе, Джалантуне, Харбине, Петровском заводе в Верхнеудинске, 31 —
в Бухеду. Из этих пунктов, в особенности из Харбина, эпидемия распространилась по
линиям КВЖД и Южно-Маньчжурской железной дороге.
Чума добралась до Харбина через 15 сут после ее появления на ст. Манчьжурия.
Первое заболевание чумой в Харбине констатировано 27 октября у прибывшего со
ст. Маньчжурия в Новый город зажиточного китайца. О больном узнал случайно бой
доктора Можаева, а последний довел об этом до сведения главного врача города. Ко-
мандированный для принятия соответственных мер прозектор городской больницы
нашел этого китайца уже мертвым, вывел лиц, бывших с ним в контакте на обсерва-
цию и, произведя частичное вскрытие трупа, бактериоскопически нашел биполярно
окрашенные палочки возбудителя чумы. Но потом почти полмесяца чума в Харбине
не показывалась.
Массовые заболевания людей легочной чумой начались в начале ноября среди
населения смежного с Харбином китайского города Фуцзядяна. Кто занес туда легоч-
ную чуму, неизвестно. Первым больным, поступи вшим 11 ноября в чумный барак Хар-
бина, был русский фельдшер, работавший в фуцзядянской чумной больницы. В тот
же день у ворот дома 10 на Пристани по Страховой улице поднят труп китайца,
умершего от легочной чумы. 13 ноября в чумной барак доставлен китаец, накануне
заболевания проведший целый день в Фуцзядяне.
С начала декабря стали систематически находить больных и трупы в кварталах
торговой части города, Пристани. С 5 по 15 декабря больные и трупы были обнару-
жены по Японской, Русской, Корейской, Торговой и Страховой улицах. Число смер-
тельных случаев от чумы в Харбине стало заметно увеличиваться. Контроль над ле-
гочной чумой был утрачен (рис. 3.33, с 666)
С 19 декабря в Фуцзядяне начинается интенсивный рост заболеваемости чумой.
Эпидемия достигла пика в середине января, когда в течение почти двух недель ежед-
невная смертность в городе составляла более сотни человек. За эти две страшные неде-
ли умерло 2180 жителей, что составило 40 % от всего количества людей, умерших в эту
эпидемию в городе. Но с 16 февраля, когда смертность достигла максимума, 173 че-
ловека в день, она начинает стремительно снижаться. Со 2 по 20 февраля (окончание
666
Часть 3. Частная неправильная эпидемиология
249
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 недели
Условные обозначения: Умершие в чумной больнице:
Трупы, поднятые за чертой города I I Китайцы
Трупы, поднятые в городе Европейцы
Рис. 3.33. Диаграмма смертности от легочной чумы по неделя".;
в Харбине и окрестностях. По В. М. Богуцкому (1911)
эпидемии) умерли 273 человека. Всего же эпидемия чумы в Фуцзядяне унесла жизни
не менее чем 5355 человек.
В Харбине эпидемия достигает кульминационной точки 12 января, в этот день от
чумы гибнет 51 человек. В кварталах, между улицами Водопроводная, Мостовая,
Китайская и Биржевая, сформировался обширный чумной очаг самополцерживаю
щихся домовых эпидемий, от 5 до 20 повторных заболеваний в одной квартире. Во
вторую половину января смертность начинает снижаться. Всего же в течение января
в Харбине от чумы погибло около 800 человек. Но потом эпидемия обрывается: 12 фев-
раля было только два смертельных случая. Всего умерло на чумном пункте, разверну-
том в Харбине, 987 человек, т. е. все те, кто туда был доставлен; поднято трупов в черте
города 217; поднято трупов за чертой города в полосе отчуждения 265; всего 1469 по-
гибших от чумы (рис. 3.34).
Во время эпидемии чумы в Маньчжурии в 1910—1911 гг. роль крыс и их блох в пе-
редаче инфекции не установлена. Все усилия русских, японских и китайских врачей
найти в местностях, лежащих вдоль железнодорожного пути, зараженных чумой крыс,
ни к чему не привели. В лаборатории профессора Сиабуро Китазато в Мукдене было
исследовано 30 тыс. крыс, из которых ни одной заболевшей чумой не обнаружено.
Максимальное количество лиц, инфицированных при непосредственном кон-
такте с больным легочной чумой, составило 40 % (из 203 человек, находившихся в со-
прикосновении с больными, заболел легочной чумой 81 человек). Однако необходи-
мо понимать, что заражение этих людей стало возможным исключительно в крайне
негигиенично устроенных квартирах: почти совершенно темных, лишенных венти-
ляции, содержащихся жильцами в высшей степени грязно, со скученным до неверо-
ятности населением. В 80 % всех записей в регистрационных карточках больных ука-
зывалось на то, что циновка, постель, пол, одежда и руки были запачканы мокротой
больного, в нескольких случаях мокротой были запачканы не только постель и руки,
но и лицо; в 11 %, прилегающие к нарам стены, были покрыты кровавой мокротой.
3.2. Чума
667
Рис. 3.34. Апофеоз чумы в Харбине в январе 1911г. А. Навес для хранения трупов
на Московском чумном пункте. Б. Русские врачи осматривают овраг, куда китайцы
тайно свозили трупы людей, умерших от чумы. По В. М. Богуцкому (1911)
Мокрота, выделенная больными чумой на предметы общего пользования, в опре-
деленных обстоятельствах приводила к вспышкам легочной чумы. Интересное на-
блюдение сделано доктором Р. А. Будбергом в деревне Сан-Диадзе (рис. 3.35).
Сначала легочной чумой заболел опиекурилыцик Юй Цзян, после смерти Юй
Цзяна его трубкой воспользовался опиекурилыцик Дан. На третий день он заболел и
умер от легочной чумы, трубка перешла к его жене, а после ее смерти к служителю
опиекурильни, тоже заразившемуся чумой и умершему. Из всего имущества покой-
ных только одна эта трубка перешла в собственность жителя той же деревни Дан Цзи-
на, выкравшего ее из «выморочной» фанзы-опиекурильни. Дан заболел чумой и умер;
из трубки курила его жена, также вследствие этого заразившаяся чумой и умершая;
от матери заразились чумой и умерли двое ее детей; после смерти жены Дана трубка
перешла к ее сестре и мужу сестры; оба умерли от чумы. Наконец трубка попала к ки-
тайцу Л и (гем же путем, что и к Дан Цзину), заболевшему чумой и умершему, потом к
его жене, которая, после смерти Ли, была отдана во вторые жены к бездетному Дан
Эр Ху и переда ia ему трубку своего первого мужа; заболели чумой и умерли все трое:
Дан Эр Ху и обе его жены.
668
Часть 3. Частная неправильная эпидемиология
Рис. 3.35. Схема развития заболеваний чумой
в деревне Сан-Диадзе. По В. М. Богуцкому (1911)
III. Легочная чума, развивающаяся по типу медленного горения. Рассмотрим ее на
примере легочной чумы, вспыхнувшей в 1921 г. во Владивостоке. Как и чума в Харби-
не в 1910— 1911 гг., чума во Владивостоке не появилась ниоткуда и внезапно. Ей пред-
шествовали очень сложные эпидемические и социальные процессы, при ретроспек-
тивном анализе наглядно свидетельствующие, что для крупной вспышки легочной
чумы нужны очень серьезные основания.
В апреле 1920 г. жители поселка Харанор известили Читинскую противочумную
станцию о падеже среди тарбаганов. Вспышка бубонной чумы началась в конце августа
1920 г. почти одновременно в четырех пунктах: в районе станции Даурия, в поселке
Абагайтуй, в поселке Каластуй и его окрестностях, в поселке Калцагайтуй и в ближай-
ших к нему местностях. Все указанные населенные пункты расположены на правом
берегу нижнего течения реки Аргунь, на расстоянии нескольких десятков верст один
от другого. В районе станции Даурия в сентябре было зарегистрировано три заболе-
вания бубонной чумой среди солдат Даурского гарнизона. В то же время обнаружено
одно заболевание бубонной чумой на разъезде № 83 (между ст. Даурия и ст. Харанор),
в семье путевого сторожа. Чума была установлена бактериологически. Одновремен-
но с этим в поселке Абагайтуй зарегистрировано четыре случая также бубонной фор-
мы, в поселке Калайстуй — два случая, в поселке Калцагайтуй — семь заболеваний.
Перечисленные 17 случаев наблюдались в конце августа, сентябре и начале октября
1920 г. Из 17 больных, двое выздоровели, остальные умерли. Этим закончился первый
период чумной вспышки в долине реки Аргунь.
В Маньчжурии дело обстояло следующим образом. С 6 октября 1920 г. вспыхнула
эпидемия бубонной чумы в Старом Хайларе (6 заболевших). Китайские власти пыта-
лись скрыть эпидемию, но она приобрела неуправляемый характер, с 15 октября по
28 октября там умерло от чумы еще 63 человека. Старый Хайлар — большой китай-
ский город, имеющий железнодорожное сообщение на линии ст. Маньчжурия — Хар-
бин — В. гадивосзок. Помимо этого пути, Хайлар является конечным пунктом боль-
шого старинного китайского тракта, соединяющего этот город с целой областью посе-
лений на запад и север от него.
В декабре, Старом Хайларе чума развивалась уже в легочной форме. С января она
стала затихать, а в дальнейшем наблюдались лишь единичные привозные случаи бо-
3.2. Чума
669
лезни. Из Хайлара чума была занесена 1 января 1921 г. в Чжалайнор (около ст. Маньч-
журия), где получила среди китайских рабочих местных угольных копей значитель-
ное распространение. Болезнь протекала исключительно в легочной форме с быстрым
смертельным исходом, поражала китайских рабочих, которых на копях было до 6 тыс.
человек; среди европейского населении (2 тыс. человек) заболеваний не было.
25 ноября 1920 г. зарегистрирован случай чумы на ст. Маньчжурия. Эпидемии в
этих двух населенных пунктах были значительными. Так, за время с 1 января по 1 марта
1921 г на станции и в поселке Маньчжурия было 507 случаев смерти от чумы, а за та-
кой же период времени на Чжалайнорских копях 622 случая.
По японским данным, ежедневная заболеваемость в поселке Маньчжурия в фев-
рале 1921 г. достигла 20—50 случаев. В марте эпидемия, видимо, стала затихать. Всего
на станции и в поселке Маньчжурия, с момента возникновения (12 января) по 1 мар-
та было зарегистрировано: больных китайцев 217, европейцев 127; из них умерло:
китайцев 216 и европейцев 121. Кроме того, обнаружено чумных трупов 163 (140 ки-
тайцев и 23 европейца). Здесь же в поселковой больнице умерли от чумы из медицин-
ского персонала два фельдшера и 5 санитаров. Е. С. Касторский (1978) указывал, что в
эту эпидемию от чумы умерло не менее 50 врачей.
В Цицикар легочная чума была занесена кули, ухаживавшим 18 января за чумны-
ми больными в Хайдаре. По возвращении к себе домой в Цицикар этот кули и вся его
семья умерли. Эпидемия охватила почти все население Цицикарской провинции.
В конце января чума была уже в Фуцзядяне. В начале февраля 1921 г. легочная чума
продвинулась из Хайлара по линии КВЖД далее на север и на запад и проникла на
территорию Дальневосточной республики.
Е. С. Касторский (1978), наблюдавший чуму 1910-1911 г., указывал на наиболее
резкие особенности в клинической картине легочной чумы в маньчжурскую эпидемию
1920—1921 гг. Это быстрота течения и отсутствие ясно выраженной локализации про-
цесса. Больные умирали в большинстве случаев в первый день. При исследовании же
и выслушивании больного каких-либо изменений в легких констатировать не уда-
валось, хотя в мокроте появлялась ржавчина или кровь. Очень часто в начале болезни
появлялась ржавая мокрота, а затем кровавая. Обильное отхаркиванье пенистой
серознокровянистой мокроты, которое считается характерным признаком чумного
воспаления легких, наблюдалось далеко не всегда. Нахождение бактерий в мокроте
(бактериологическое исследование) не всегда имело исчерпывающее диагностическое
значение. Были случаи, когда в мокроте находили чумные бактерии, а чума у больно-
го не пазвивалась, и, наоборот, бактерий не находили, а больной умирал от чумы в
тот же день.
С эпидемиологической точки зрения обращала на себя внимание необыкновенная
заразительность больных во время чумы 1920—1921 гг. при условии их совместного
пребывания со здоровыми людьми в тесном и грязном помещении. Благодаря этому
эпидемия принимала чрезвычайно опасный очаговый характер. Были случаи заболе-
вания в один день до 40 человек в одном помещении (рабочие казармы). Касторский
считал, что такая заразительность свидетельствует «об усилившейся вирулентности
чумных бацилл в эпидемию 1920-1921 гг.».
Северное направление. Уже 3 февраля был отмечен первый случай чумы на ст. Борзя
и два—в поселке Брусиловка; 18 февраля на ст. Мациевской (в 20 верстах от ст. Маньч-
журия на линии железной дороги, по направлению к Чите) зарегистрировано первое
заболевание, а 24 февраля — второе. В первых числах марта еще два случая на разъез-
де «1420 верста». О движении чумы на север, вверх течения реки Аргунь, имеются
670
Часть 3. Частная неправильная эпидемиология
сведения, относящиеся только к февралю 1921 г. Судя по ним, заболевания чумой бы-
ли отмечены в четырех пунктах Забайкалья: Калайстуи, в числе 25 случаев болезни за
время с 1 по 20 февраля, и заимках Кыра (2 случая) и Клички (6 случаев), на Нерчин-
ском заводе (1 случай).
Западное направление. На ст. Бухеду (расположена между Хайларом и Цицикаром)
эпидемия вспыхнула с 3 февраля 1921 г. В самом начале февраля были обнаружены
случаи смерти от чумы в пригородах Хароина. В самом городе Харбине чума офици-
ально началась с 13 Февраля и до 18 февраля зала около 30 заболеваний; за все время
эпидемии в Харбине было зарегистрировано около 1 тыс. заболеваний. Отсутствие
регулярных и более точных сведений не позволяет установить действительную кар-
тину движения эпидемии в рассматриваемом направлении (рис. 3.36).
Рис. 3.36. Больной легочной чумой китаец на улице Старого
Харбина, 1921 г. По U A. Wu Lien Ten et al. (1936)
Развитие и характер эпидемии легочной чумы во Владивостоке. Потребовалось зна-
чительное время, с августа месяца 1920 г. по 10 апреля 1921 г., т. е. восемь месяцев, чтобы
легочная чума от места своего «зарождения» могла дойти до В ла тивостока. Еженедель
ные сводки по смертельным случаям свидетельствуют, что эпидемия легочной чумы
во Владивостоке развивалась не взрывом как в Харбине (1910), а типом «спокойного
горения». Начавшись с десятка случаев на первой неделе, чума далее разгоралась по-
степенно увеличивающимися в течение пяти недель темпами, проявив наибольшую
силу в течение шестой недели. Затем три недели чума «догорала». Но и под пеплом
своих жертв она «тлела» еще тринадцать недель (рис. 3.37).
Первая неделя эпидемии (10- 16 апреля 1921 г.). Первым выявленным больным ле-
гочной чумой во Владивостоке оказался Лиу Чунг Шень. 10 апреля китайцы прине-
сли его на носилках в Морской госпиталь (эта дата считается началом эпидемии). Отту-
да его направили в чумной госпиталь (Первый крепостной госпиталь), но по дороге
он умер. При обследовании летучим отрядом двора дома № 193, где жил Тиу, выясни-
лось, что в этом же помещении 5 дней тому назад умер от неизвестной болезни другой
3.2. Чума
671
Количество человек
42 41
Рис. 3.37. Смертность от легочной чумы во Владивостоке
в 1921 г. по неделям. По В. П. Захарову с соавт. (1922)
китаец, которого похоронили на кладбище в районе 2-й речки. А10 апреля умерли еще
два китайца, проживавшие совместно в доме № 112 по Светланской улице. 13 апреля
летучим отрядом был обнаружен труп китайца, спрятанный в дровах на дворе дома
№ 98 по Светланской улице. Исследование трупа показало, что смерть наступила от
чумы. В тот же день был доставлен в чумное отделение китаец Лан Гун, проживавший
по Светланской улице, дом № 175, так как про неги стало известно, что он часто посе-
щал квартиру Лиу Чунг Шеня и ввиду наличия у него повышенной температуры. В ночь
на 14 апреля он был убит часовым при попытке бегства из чумного госпиталя. На
вскрытии трупа обнаружено прижизненное заболевание легочной чумой. Положитель-
ный результат на чуму дало и бактериологическое исследование.
14 апреля был доставлен труп китайца Лю Бен Вяна, проживавшего в доме № 42
по Корейской улице. Труп оказался чумным. При обследовании китайских квартир
этого же дома был обнаружен больной китаец в тяжелом состоянии, по фамилии Тун
Шу Хо, 17 лет, который проживал совместно с Лю Бен Вяном. Тун Шу-хо умер в тот
же день, при явлениях легочной чумы. На следующий день в чумной госпиталь были
доставлены 2 чумных трупа, выброшенных из дома № 42 по Корейской улице. Еще
через день (16 апреля) в чумной госпиталь были доставлены уже 5 трупов китайцев,
умерших от чумы. Всего же за первую неделю эпидемии, с 10 апреля по 16 апреля вклю-
чительно, во Владивостоке было зарегистрировано два больных легочной чумой и
обнаружено 10 трупов китайцев, умерших от чумы.
Вторая неделя эпидемии (17—23 апреля). 15 человек умерло в чумном госпитале, на
улицах подобрано 12 чумных трупов. Болезнь развивалась быстро — четыре китайца.
Переведенные из изоляционного отделения в чумное, умерли в тот же день. В эту не-
делю зарегистрирован первый случай заражения чумой медицинского персонала —
погибла медсестра Даниленко.
К концу недели стало ясно, что эпидемия вышла из-под контроля противочумной
организации. Чума «пошла» в окрестности города. 17 апреля на станции Океанская, в
38 км от города, был поднят китаец в тяжелом состоянии с явными признаками легоч-
ной чумы. На полу помещения, где он лежал, была масса характерных кровянистых
плевков мокроты. 19 апреля он умер
672
Часть 3. Частная неправильная эпидемиология
Третья неделя эпидемии (24—30 апреля). Новые случаи заболеваний легочной чу-
мой среди медицинского персонала: умерли два санитара (Годянкий и Мальцев). Боль-
ные чумой стали поступать из окрестностей города: с каменноугольных копей Скидель-
ского (30 верст от города), с 5 версты Сучанской ветки и со станции Океанская. Чума
приобрела тенденцию к распространению на побережье Приморской области.
Всего за неделю на улицах, площадях и с квартир подобрано 42 трупа, из них чум-
ными оказались 22, и 13 человек умерли в чумном госпитале. Большинство трупов было
подобрано в тех же кварталах, что и в предыдущие две недели (район скрещения улиц
Семеновской и Суйфунской — 6 трупов). Стало ясно, что там находятся чумные очаги.
Однако обследование 27 апреля китайского общежития в доме № 20 по Суйфунской
улице, ничего не дало, трупы из него были уже выброшены. Когда через 3 дня проверку
внезапно повторили, то было выявлено 8 больных чумой, 4 китайцев отправили в изо-
ляцию, 41 — в обсервацию. Из соседних помещений китайцы успели разбежаться.
Четвертая неделя эпидемии (1—7 мая). Чума «пошла» по более широкому кругу,
захватив Шлюпочную площадь бухты Золотой Рог с шаландами и шлюпками (5 тру-
пов), Первую Речку (7 трупов), северную часть Корейской улицы (7 трупов). Прочие
китайские кварталы северо-западной части города по-прежнему дали основное число
трупов. По дням число трупов стало распределяться более равномерно. В чумном гос-
питале умерло 27 человек, с улиц и площадей города поднято 50 чумных трупов. По-
гиб санитар Воробьев.
Пятая неделя эпидемии (8—14 мая). Небольшое повышение заболеваемости и смер-
тности по сравнению с четвертой. Во всех отделениях чумного госпиталя от чумы умер-
ло 26 человек, во Владивостоке поднято 59 чумных трупов, в окрестностях — 4. Погиб
фельдшер Доброжанский.
Шестая неделя эпидемии (15-21 мая). Пик развития эпидемии. Во всех отделени-
ях чумного госпиталя за неделю умерло от чумы 42 человека, по улицам и окрестно-
стям города поднято 47 чумных трупов. В основном больные поступали с тех же улиц,
что и в предыдущие недели. Погиб фельдшер Козлов. На хуторе Сехюче 15 мая умер-
ло четыре русских жителя. Все они имели контакты с китайцами, приезжавшими из
Владивостока.
Седьмая неделя эпидемии (22—28 мая). Количество больных чумой и подобранных
чумных трупов уменьшилось. Умер от чумы 41 человек, поднято 32 чумных трупа. По-
гиб санитар Хазов.
Восьмая и девятая недели эпидемии (29 мая-11 июня). Во Владивостоке эпидемия
неуклонно шла на убыль. От чумы умерло 32 человека, подобрано 28 чумных трупов.
Погиб санитар Короткевич.
Наступил период, резко отличающийся по своему эпидемическому течению и по
существу заболеваний от предшествовавшего. Его П. В. Захаров с соавт. (1922) назва-
ли «хвостом эпидемии легочной чумы». Как оказалось, в большом городе очень трудно
уловить момент прекращения эпидемии легочной чумы.
Десятая неделя эпидемии (12-18 июня). Эпидемия резко оборвалась: за неделю
было зарегистрировано только три чумных трупа. В предшествовавшую девятую не-
делю было 20 случаев, а в восьмую — 40. Возник естественный вопрос, не конец ли это
эпидемии легочной чумы, или, быть может, это только передышка? Некоторые врачи
определенно утверждали, что эпидемии окончилась, и нужно готовиться, после десяти-
суточного срока со времени обнаружения последнего чумного трупа, объявить город
угрожаемым по чуме, а затем и благополучным. Дальнейшее развитие событий показа-
ло, что такой 10-суточный срок наступил не скоро.
Одиннадцая—двадцатая недели эпидемии (19 июня-27 августа). В разных местах
города выявляются единичные заболевшие чумой и поднимаются единичные чумные
3.2. Чума
673
трупы. Дважды разрывы между зарегистрированными случаями чумы составляли 6 су-
ток (между 11 и 13 неделями).
Двадцать первая неделя эпидемии (28 августа-3 сентября). Случаев смерти в чум-
ном госпитале не было. Трупов было поднято два. Оба были вскрыты 30 августа в при-
сутствии бактериолога Левашова и японского врача Гото, подтвердивших чуму.
Двадцать вторая неделя эпидемии (4—9 сентября). Количество случаев чумы вдруг
возросло. В чумном госпитале от нее умерли 2 китайца, два трупа, подобранные на ули-
цах, оказались чумными. У властей возникло опасение, что дотянувшаяся до октября
месяца, т. е. до холодного времени года, эпидемия может вновь разгореться в легоч-
ной форме.
Двадцать третья неделя эпидемии (11—17 сентября). Последняя неделя эпидемии
чумы во Владивостоке. Два чумных трупа, обнаруженные за неделю, были последни-
ми зарегистрированными случаями чумы. Один труп китайца был поднят 11 сентября
на берегу Амурского залива проти в Новокорейской слободки, другой труп тоже китай-
ца, был поднят 15 сентября с угла Суйфунской и Нагорной улиц. В эту неделю всего
было поднято 4 трупа. Таким образом, последний случай чумы во Владивостоке заре-
гистрирован 15 сентября 1921 г.
Горючим материалом («сухими дровами») для чумы было китайское население,
исключительно питавшее и поддерживавшее пожар эпидемии в городе. Однако ни-
каких эмпирических доказательств какой-то «этнической избирательности» легочной
чумы тогда получено не было Китайцы, жившие в европейских семьях (лакеи, пова-
ра), не были затронуты болезнью. Подавляющее большинство погибших от чумы
принадлежало к китайской бедноте, жившей в ночлежках, казармах, в обстановке
невероятной скученности и грязи.
Касаясь вопроса заразительности легочной чумы, П. В. Захаров с соавт. (1922)
отмечают, что ответ на него не так прост, как это на первый взгляд кажется. Многие
факты, выявленные при изучении эпидемии легочной чумы во Владивостоке, гово-
рят о ее высокой заразительности. Однако они также дают основание утверждать, что
между фактом высокой заразительности легочной формы чумы и фактом ее малого
распространения лежит нечто третье, проходя через которое, высокая степень за-
разительности чумы все же давала «медленное горение» такого многочисленного и
компактно проживающего населения, как владивостокские китайские кварталы.
Анализ крупных вспышек легочной чумы позволяет выявить следующие законо-
мерности их развития. Масштабные эпидемии легочной чумы не имеют единого цент-
ра. Легочная чума распространяется небольшими вспышками по направлениям дви-
жения потоков людей с территорий, охваченных чумными эпизоотиями среди гры-
зунов. Такие эпидемии имеют период вспышек-предшественников, длящийся иногда
десятки лет. Они сигнализируют о нарастающей активизации природных очагов чумы.
Переходу локальных вспышек легочной чумы в масштабную эпидемию предшеству-
ет накопление «сухих дров» эпидемии легочной чумы. Ими, как правило, становятся
люди, у которых по разным причинам (генетические особенности, массивные дозы
заражении вирулентными штаммами и др.) бубонная чума переходит во вторично-
легочную, а стереотип их социального поведения и условия жизни способствуют ее
дальнейшему распространению уже в форме первичной легочной чумы. «Движение»
легочной чумы по территориям происходит медленно, по выражению П. В. Захарова
с соавт. (1922): «Со скоростью пешехода». Так, в 1921 г. в условиях неконтролируемого
перемещения китайского населения легочная чума «прошла» 100 км от Уссурийска
(тогда Никольск-Уссурийский) до Владивостока только за месяц. В механизме раз-
674
Часть 3. Частная неправильная эпидемиология
вития масштабной эпидемии легочной чумы заложен и механизм ее терминации. По-
этому ле) очная чума в крупном населенном пункте не может длиться до последнего
жителя, хотя внешне все условия для ее продолжения остаются.
Внутривидовая таксономия. При идентификации и классификации штаммов воз-
будителя чумы, обнаруженных при расследовании актов биодиверсий и биотеррора,
необходимо учитывать то обстоятельство, что большинство исследований экологии,
биохимии и патогенеза Y. pestis, осуществляемых за пределами бывшего Советского
Союза, было проведено с ограниченным количеством близкородственных штаммов.
Ни один научно-исследовательский институт за пределами России не имеет того ко-
личества штаммов возбудителя чумы и опыта исследований Y. pestis в ее природа ix
очагах, которые накоплены исследователями русской противоч) мной системы. По-
этому использование отечественной таксономической классификации, оснозанной
на фенотипических признаках при идентификации и классификации штаммов У. pes-
tis. является наиболее информативным подходом из имеющихся (табл. 3.11).
Таблица 3.11
Таксономические признаки штаммов, которые различают подвиды Y. pestis, выделенные
на территории бывшего СССР и Монголии, и соответствие подвидов биоварам*
Такс ономичес кие признаки Подвид Y. pestis
pestis (главный) altaica caucasica** hissarica alegeica talacsica
1 2 3 4 5 6 7
Ферментация рамнозы — + + + +
мелибиозы — + + + + +
арабинозы + — + — + —
глицерина ± + + + + +
мелезитозы — 9 — ± 9 —
Восстановление нитратов ± - + - —
Активность уреазы — — — ± — +
Продуцирование пестицина 1 + + - + " + +
Чувствительность к пестицину 1 - + ± -
Фибринолитиче- ская активность + — + + +
Активность коагулазы + + - + + +
Зависимость от факторов питания лейцин ± + ± — +
метионин + — + + — 9
аргинин — + + — — +
тиамин — — + — — 9
цистеин ± + ± + + +
фенилаланин + + + + + +
треонин + 9 + 9 9
тирозин — 9 9 — 9 +
3.2. Чума
675
Продолжение табл. 3.11
1 2 3 4 5 6 7
Вирулентность для морских свинок + — - - -
«Регион рас пространения Центральная Хфрика, Центральная и Северная Азия, Китай (Маньчжурия), Монголия Алтайские горы Закавказ- ское нагорье, горные районы Дагестана Гиссарский хребет Северо- Восточная Монголия, пустыня Гоби Таллаский хребет
Хозяин Грызуны родов Marmota, Citellus. Meriones, Rattus и т. д. Ochotona pricei Microtus arvalis Microtus carruthersi Ochotona pricei Microtus gregalis****
Биовар Antiqua Medievalis Autiqua Medievalis Medievalis Medievalis
* Таблица составлена на основе работы А. Р. Anisimov et al. (2004). Символы: — отсутствие
признака; + наличие признака; ± наличие признака, но не во всех штаммах. ? — нет данных.
** Штаммы подвида caucasica не имеют плазмиды pPst
*** Штаммы чувствительны к пестицину штаммов из подвидов pestis и altaica и так же ре-
зистентны к пестицину из штаммов своего собственного подвида.
**** Существует точка зрения, что главным хозяином в этом очаге может быть Магто-
ta caudata.
Восприимчивость и чувствительность животных к возбудителю чумы. В чумологии
принято отличать восприимчивость к инфекции от чувствительности к ней, причем
под первой понимают способность заражаться, тогда, как вторая характеризует ин-
тенсивность и остроту патологического процесса. С этих позиций восприимчивость
считается видовым признаком, а чувствительность индивидуальным. Вопрос о вос-
приимчивости животных и чувствительности животных к Y. pestis обычно рассматри-
вается с точки зрения их роли в поддержании эпизоотического процесса. Наиболее
восприимчивыми к чуме являются многие виды обширного отряда грызунов. Чума
справедливо считается болезнью грызунов; другие виды животных, в том числе и че-
ловек. заболевают чумой при кон акте с больными грызунами. Ю. М. Елкин (1960)
разделил грызунов на четыре t руппы:
1) высокочувствительные (домовая мышь, тамарисковая и малоазиатская песчан-
ки, песчанка Виноградова, степная пеструшка, слепушонка);
2) залегающие в спячку с резко меняющейся по сезонам чувствительностью (сур-
ки, суслики);
3) незалегающие в спячку, в популяциях которых всегда имеются высокочувст-
вительные и очень резистентные особи (полуденная, большая, краснохвостая пес-
чанки, серая крыса, обыкновенная и общественная полевки, полевка Брандта, поле-
вая мышь);
4) чувствительные только к очень большим дозам заражения (лесная мышь) (цит.
поДомарадскому И. В., 1998).
Черные и серые крысы весьма чувствительны к чуме, но в естественных условиях
серые заражаются легче, чем черные. В Индии, например, были случаи, когда эпизо-
676
Часть 3. Частная неправильная эпидемиология
отии сначала начинались среди серых крыс и лишь позднее охватывали черных. В то
же время по чувствительности к чуме популяции крыс столь же неоднородны, как и
популяции многих других животных. Из домашних животных восприимчивы к чуме
верблюды и кошки. У ослов и мулов, овец и коз, свиней, собак при введении боль-
ших доз чумного микроба в экспериментальных условиях иногда удается вызвать чум-
ную инфекцию. Но с практической точки зрения этих животных можно считать не-
восприимчивыми к чуме. Лошади и крупный рогатый скот невосприимчивы к чуме.
Хищники, за небольшим исключением (хорьки, лисицы, ласки, солонгои), невосп-
риимчивы к чуме. Из насекомоядных восприимчивы к чуме землеройки. Птицы и
пресмыкающиеся (змеи, черепахи) невосприимчивы к чуме. Ящерицы восприимчи-
вы при экспериментальном заражении. Из земноводных жаб следует считать невосп-
риимчивыми к чуме, а лягушек и тритонов — маловосприимчивыми, не имеющими
значения в эпидемиологии этой инфекции. Насекомые и паукообразные хотя и ма-
ловосприимчивы к чуме, но в их организме чумные микробы могут находиться дово ть-
но продолжительное время (табл. 3.12).
Таблица 3.12
Восприимчивость к чуме насекомых и паукообразных*
Наименование насекомого или паукообразного Восприимчивость** Продолжительность сохранения чумного микроба (сут)
Мухи ± До 18
Комары —
Саранча + 4
Жуки стафилины —
Тараканы —
Блохи ± До 396
Клопы + До 147
Вши человека:
головные +
платяные +
Вши земляного зайца + До 14
Вши тарбагана + Цо 13
Клещи:
Rhipicephalus Schultze Olen +
Ixodes autumnalis L. +
Demacentor silvarum Olen + 10
Hyalomma volgense Schultze + 11
Argos persicus F. + 100
* По В. М. Туманскому (1958).
** Условные обозначения: + восприимчивы, — невосприимчивы.
Для изучения вирулентности штаммов Y. pestis наиболее пригодны морские свин-
ки, ибо эти животные очень восприимчивы к чуме, и слабо реагируют на токсины
возбудителя чумы. Белые мыши мало пригодны для этой цели, так как они очень чув-
ствительны к токсинам Y. pestis. Дикие грызуны, взятые из энзоотических районов,
непригодны для изучения вирулентности Y. pestis, так как среди этих грызунов могут
встречаться иммунные особи (Туманский В. М., 1958). Но А. Р. Anisimov et al. (2004)
3.2. Чума
677
отмечают, что в тестах на вирулентность у морских свинок большинство штаммов
Y. pestis подвида pestis являются летальными, тогда как штаммы подвидов altaica,
caucasica, hissarica, ulegeica и talassica, как правило, обнаруживают резкое снижение
вирулентности или даже полное ее отсутствие для этих животных, тем не менее они
ci юсобны вызывать вспышки чумы среди людей, сопровождающиеся развитием пнев-
монии. Эти авторы считают, что еще невозможно реально определить, какой именно
набор генетических и фенотипических признаков указывает на вирулентность штам-
мов Y. pestis для лабораторных животных или людей.
Инфицирующие дозы. Вирулентность штаммов Y. pestis колеблется в широких пре-
телах, даже если они выделены в условиях одной эпидемии или эпизоотии. Ингаля-
ционная поражающая доза (LD100) возбудителя чумы для человека обычно находится
в тределах от 100 до 3000 клеток и зависит от очень многих факторов помимо вирулент-
ности самого штамма и дисперсности аэрозоля (Ротшильд Д., 1966; Ситников М. Н.,
1968; Van Andel R. et al., 2008). К увеличению инфицирующей дозы приводят частые
пересевы, хранение штаммов возбудителя чумы при высокой температуре, выращи-
вание на средах с антисептиками и антибиотиками, глубинное культивирование, пас-
саж через маловосприимчивых или иммунных животных, увеличение размера частиц
аэрозоля и др. Снижение инфицирующей дозы Y. pestis достигают путем пассажа че-
рез организм морской свинки или путем отбора наиболее вирулентных клеток из
микробной популяции (Николаев Н. И., 1968)1.
Иммунитет. Случаи повторно! о заболевания чумой после перенесенного заболева-
ния редки, но возможны. Подробно протективные антигены возбудителя чумы опи-
саны в монографии А. А. Бывалова (2011). Какому из них принадлежит решающая роль
в иммуногенезе чумы, ясности нет. К тому же исследователи не всегда могут быть уве-
рены в том, что данные, полученные на животных, безоговорочно приложимы к объяс-
нению процессов, происходящих в организме человека. Антитела появляются на 7 сут
от начала болезни. Особое значение имеет выявление IgM как показателя недавнего
контакта с источником чумы. Достоверно известно, что в иммунитете к чуме основ-
ная роль принадлежит клеточным факторам. Из всех вакцинных штаммов лучше всего
зарекомендовали себя TJW и EV. В отечественной практике предпочтение отдается
живой вакцине на основе штамма EV. В практике научно-исследовательских организа-
ций Министерства ооороны Ро< сии используется эффективный и безопасный спо-
соб массовой ингаляционной иммунизации живой чумной вакциной на основе штам-
ма EV. Создаваемый таким способом иммунитет к возбудителю чумы, исключает воз-
можность ингаляционного заражения человека при тех концентрациях возбудителя
чумы в аэрозоле, которые можно создать при применении БО.
Устойчивость Y. pestis во внешней среде и в трупах погибших. В выделениях больных
людей и животных Y. pestis может сохраняться продолжительное время, что зависит
от температуры и наличия других бактерий, обладающих антагонистическим дейст-
вием. В замороженной мокроте чумной микроб сохраняет свою жизнеспособность от
6 до 14 сут; в мокроте, находящейся при комнатной температуре, до 36 сут; в высушен-
ной мокроте он погибает через 4—7 сут. В гною из бубонов вне организма чумной
микроб может сохраняться до 20—30 сут. В крови, взятой у животных больных чумой,
возбудитель чумы может сохраняться длительное время при защите от высыхания до
260 сут (при температуреЮ— 12°С) (Туманский В. М., 1958).
1 Попытки японских военных ученых повысить вирулентность возбудителя чумы описа-
ны в разд. 1.8.
678
Часть 3. Частная неправильная эпидемиология
В замороженных человеческих трупах Y. pestis может сохраняться 4-5 мес. В лет-
нее время уже через 4-5 сут после смерти человека от чумы из его лимфатических
узлов, крови и внутренних органов не удается выделить культуры даже при заражении
восприимчивого животного способом втирания. Дольше всего Y. pestis сохраняется в
костном мозгу, так как он не подвергается быстрому загниванию. В шкурках грызу-
нов, погибших от чумы, микроб сохраняется в зависимости от условий обработки
хранения этих шкурок: при комнатной температуре — до 23 сут, а при температуре
14—28°С — до 17—18 сут. В жаркую летнюю погоду на солнце Y. pestis сохраняется в
шкурках до 6 ч, а в тени — до 3 сут. На круто сваренном яичном белке Y. pestis со-
храняется до 138 сут, в молоке —до 81 сут, в соленом мясе—до 130 сут, на черном хле-
бе — до 4 сут, на овощах и фруктах — 6—11 сут, на зерновых продуктах — 12—54 cyi, в
воде—до 75 сут. Но вспышек чумы, связанных с употреблением в пищу молочных про-
дуктов, хлеба, овошей, фруктов и воды, не описано (Туманский В. М., 1958).
Мясо верблюда, больного чумой, в момент его разделки неоднократно было при-
чиной инфекции, вспышек заболевания среди людей. Зерно во время больших чум-
ных эпизоотий может быть загрязнено кровью и трупами мышей, погибших от чумы.
Теоретически если это зерно в необработанном виде будет перевезено в здоровую
местность, го оно может заразить грызунов и создать среди них эпизоотию. Именно
такой способ пытались использовать американские военные для распростр щения
чумы среди жителей китайских городов (см. разд. 1.10).
В почве Y pestis сохраняется непродолжительное время, при этом сохраняемость
ее зависит от активности других микроорганизмов и от температуры. При низкой тем-
пературе чумной микроб может оставаться жизнеспособным сравнительно долгое
время—до 28 сут, при более высокой температуре он быстро погибает. Солнечные лучи
оказывают энергичное убивающее действие на Y. pestis. Она чувствительна и к высо-
кой температуре. При кипячении возбудитель чумы в бульонной и агаровой культуре
и даже в крови погибает через несколько секунд, при этом большинство микробов
гибнет раньше, чем начинает закипать жидкость (Туманский В М., 1958).
А эрозоли Y. pestis демонстрировали постоянную скорост ь распада, когда они аэро-
золировались при относительной влажности 50 % из бульона с настоем сердечной
мышцы. Происходила внезапная утрата живых микроорганизмов при повышении
отнисител! ной влажности до величин, превышающих 50 %, особенно до 85 % (Sin-
clair R. et al., 2008).
В дентине >убов методами молекулярной диагностики можно обнаружить нуклео-
тидные последовательности Y. pestis спустя несколько столетий после гибели челове-
ка от чумы (Haensch S. et al., 2010).
Антибиотикорезистентность. Обычно изоляты Y. pestis в равной степени чувстви-
тельны ко всем антимикробным препаратам, эффективным против грамотрицатель-
ных бактерий. В 1995 г. у клинического изолята Y pestis обнаружена высокая резис-
тентность к ан гимикробным средствам, включая все лекарственные препараты,
рекомендованные для профилактики и лечения чумы. Штамм (17/95) выделен в рай-
оне Амболавау в Мадагаскаре у 16-летнего юноши с симптомами иубонной чумы.
Вт< рой штамм (16/95) с высокой резистентностью к стрептомицину выделен в том же
году в Мадагаскаре в районе Ампитана от 14-летнего больного бубонной чумой. Оба
больных выздоровели после антибиотикотерапии, включающей внутримышечные
инъекции стрептомицина (2 г/сут в течение 4 сут) и пероральное применение триме-
топрим-сульфаметоксазола (2 г/сут в течение 10 сут). Более подробно свойства этих
штаммов рассмотрим по обобщающей работе М Galimand et al. (2006).
3.2. Чума 679
Штамм 17/95обнаружил высокую резистентность к восьми антимикробным пре-
паратам, вк ъочая не только рекомендованные для лечения (стрептомицин, хлорам-
феникол и тетрациклин) и профилактики (сульфонамид и тетрациклин) чумы, но
также и возможные альтернативные средства, такие как ампициллин, канамицин и
спек гиномицин.
Резистентность к ампициллину, хлорамфениколу, канамицину, сульфонамиду и
стрептомицин-спектиномицину носила детоксикационный характер1 *. Она была обу-
словлена продуцированием Y. pestis пенициллиназы ТЕМ-1, хлорамфеникол-ацетил-
трансфсразытипа!, аминогликозия-фосфот эансферазы типа I, резистентной к лекар-
ственным препаратам дигидроптероат-синта ия и аминогликозид-аденилтрансферазы
соответственно. Резистентность к тетрациклину у штамма 17/95 связана со способ-
ностью бактерии выкачивать тетрациклин из клетки быстрее, чем антибиотик пора-
зит свои мишени. Изолят сохранил чуве гвительность к цефалоспоринам, другим ами-
ногликозидам, хинолонам и триметоприму. Скорее всего, именно активность три-
метоприма, несмотря на отсутствие синергизма с сульфонамидом, и обеспечила
выздоровление больного.
Все гены резистентности штамма 17/95 содержались в конъюгативной плазмиде,
обозначенной р!Р1202 (примерно 150 т. п. о.). В эксперименте она могла быть пере-
несена с высокой частотой путем конъюгации от Y. pestis 17/95 к другим штаммам
Y.pestisn E.colv, и в обратном направлении. Плазмида р!Р 1202 принадлежит к группе
несовместимости 1пс6-С и содержит структуру, аналогичную гаковой интегронов
Тп21-подобных элементов. Содержавшиеся в ней гены резистентности близкород-
ственны по структуре переносимым плазмидами детерминантам, обычно обнаружи-
ваемым у энтеробактерий.
Штамм 16/95 обладает устойчивостью детоксикационного типа к стрептомици-
ну, но сохраняет чувствительность к спекгиномицину и другим антимикробным сред-
ствам, включая рекомендованные для лечения и профилактики чумы. Гены резис-
тентности к стрептомицину переносились между бактериями плазмидой р!Р1203
(примерно 40 т. п. о.), способной конъюгировать с другими изолятами Y. pestis. Они
являются частью кластера tnpR-strA-strB-IR, характерного для Тп5393-группы транс-
позонов. Перенос резистентности к стрептомицину от Y. pestis 16/Q5 к Y. pestis и
У. pseudotuberculosis происходил с высокой час готои, к Е. coli с более низкой частотой,
а перенос к Y. enterocolitica вообще не удалось обнаружить. Повторный перенос pl Р1203
от трансконъюгантов Y. pestis и Y. pseudotuberculosis к Y. pestis и Y. pseudotuberculosis
также происходил с высокой частотой.
Антибиотико-резистентные штаммы Y. pestis имеют несколько отличий. Они были
выделены в двух разных районах, расположенных на расстоянии 80 миль друг от дру-
га. Штамм 17/95 относится к типичному риботигу В, тогда как штамм 16/93 относит-
ся к недавно описанному, Мадагаскар-специфическому риботипу Q. Плазмида
р!Р1202 несет гены множественной резистентности к антимикробным препаратам,
принадлежит к группе 1пс6-С, тогда как р!Р1203 несет только детерминанты резис-
тентности к стрептомицину, принадлежит к группе IncP, Наконец, резистентность к
стрептомицину у штамма 17/95 обусловлена аденилированием препарата, а у штам-
ма 16/95 — фосфорилированием. Эти данные свидетельствуют о том, что антибиоти-
корезистентные штаммы Y pestis возникли независимо друг от друга.
1 См. в разд. 2.2.2 «Механизмы развития резистентности к антибактериальным препара-
там у бактерий».
680
Часть 3. Частная неправильная эпидемиология
Оонаружение генетически измененных штаммов К pestis. Подходы к обнаружению
генетически измененных штаммов возбудителя чумы те же, что и для обнаружения
генетически-измененных штаммов В. anthracis (см. разд. 3.1). Однако необходимо
учитывать, что генетика К pestis изучена не хуже, чем кишечной палочки. Методы
генетического переноса для возбудителя чумы стали отрабатываться сразу же после
обнаружения у него собственных плазмид. Для К pestis могут быть использованы ме-
тоды генетической модификации и векторы клонирования, разработанные для Е. coli
и многих других грамотрицательных микроорганизмов. В К pestis в условиях in vitro
легко переносятся плазмиды вирулентности близкородственных микроорганизмов,
и наоборот (см., например, работу Wolf-Watz Н. et al., 1985). Поэтому генетическая
модификация чумного микроба, ставшего причиной намеренного заражения людей,
может оказаться значительно более изощренной, чем у возбудителя сибирской язвы.
В очагах искусственно вызванных вспышек чумы с наибольшей вероятностью
можно встретить штаммы возбудителя чумы со следующими модификациями (и их
сочетаниями):
резистентные к антибиотикам, используемым в распространенных схемах лече-
ния чумы (введение генов, придающих бактерии резистентность к антибиотикам
преимущественно детоксикационного типа);
обладающие более высокой вирулентностью, чем исходные природные штаммы
(введение в Y. pestis генов токсинов и иммунорегуляторных белков);
с измененной антигенной специфичностью, пробивающих иммунитет, создавае-
мый коммерческими вакцинами (получение мутантов Y. pestis с V-антигеном некото-
рых сероваров Y. enterocolitica или с Л ПС с высокоацетилированным липидом А).
Дифференцировать генетически измененные Y. pestis от их природных вариантов
по фенотипическим признакам сложно из-за поддержания в природных резервуарах
большого количества атипичных штаммов Y. pestis. Генотипическое и фенотипическое
разнообразие наблюдают даже среди чумных изолятов из одного и того же природно-
го очага. Поданным А. Р. Anisimov etal. (2004), в полевых условиях появление атипич-
ных штаммов Y. pestis может коррелировать с различными фазами эпизоотического
цикла. Так называемые атипичные штаммы отличаются по некоторым из основных
признаков, обнаруженных в доминирующем варианте Y. pestis, выделенном в опреде-
ленном очаге чумы. Экологические и другие изменения, происходящие во время эпи-
зоотического цикла, способствуют появлению недоминируюших штаммов Y. pestis.
Анализ частоты появления разных форм Y. pestis, выделенных из разнообразных
природных очагов чумы, показал, что уровень изменчивости в очагах различен и ко-
леблется от 1,6 % (Волго-Уральский степной очаг), до 6,7 % (Гиссарский очаг). Наи-
более часто атипичные штаммы выделяют на пике эпизоотии среди грызунов. Было
установлено, чз о 24,4 % атипичных штаммов обнаруживают нехарактерные биохими-
ческие активности; 17,6 % были лизогенизированы бактериофагами; 13,5 % обнару-
жили измененные потребности в факторах роста; 14,27 % имели мутации в локусе hms
(для «системы хранения гемина»); 16,75 % показали пониженную вирулентность для
лабораторных животных; 9,93 % были F1-; 2,36 % оказались дефицитны по пестицину;
и 0,4 % были резистентны к литическому действию диагностических фагов (Anisi-
mov А. Р. et al., 2004).
Ответ на вопрос о том, является ли штамм Y. pestis, выделенный от жертв биотер-
рористического акта, атипичным или генетически модифицированным, затруднен
еще и отсутствием согласованной бактериальной систематики, что не позволяет оп-
ределить основную единицу биологического разнообразия — вид, не говоря уже
3.2. Чума
681
о подвиде. Нет ясности и по факторам вирулентности К pestis. Скорее всего, для от-
вета на него необходимо:
11 установить прототипный (наиболее близкий) природный штамм Y. pestis',
2) затем провести сравнение его генома с геномом штамма Y. pestis, использован-
ного для преступления;
3) пользуясь знаниями с существующих в природных экосистемах генах патоген-
ности (антибиотикорезистентности) и механизмах их передачи, провести экспертную
оценку возможности или, наоборот, невозможности искусственного происхождения
данного штамма. Например, наличие у штамма Y. pestis резистентности к стрептоми-
цину, вызванной приобретением им конъюгативной плазмиды с кластером tnpR-strA-
strB-IR, объяснимо на основе знаний о генетическом обмене среди бактерий семей-
ства Enterobacteriaceae.Однако обнаружение этого же кластера в геноме Y. pestis в
составе интегративного вектора с известной историей конструирования (см. разд. 2.2.2
«Технологии рекомбинантной ДНК в получении штаммов бактерий с неправильной
эпидемиологией») требует уже поиска лаборатории, где этот штамм был получен.
Патогенез. Отдельного «фактора патогенности», определяющего способность воз-
будителя чумы вызывать инфекционный процесс у человека, не описано (Аниси-
мов А. П., 2002). По мнению Е. П. Шуваловой с соавт. (2001), воснове патогенеза чумы
.ежат два связанных между собой процесса — септический и токсический. Септиче-
ский процесс определяется способностью Y. pestis размножаться в фагоцитирующих
клетках. После укуса блохи возбудитель чумы попадает в региональные лимфатиче-
ские узлы, где происходит его бурное размножение (1—3 сут). В этот процесс вовлека-
ется все большее число нейтрофилов, возбудитель находится в первичном очаге (если
он формируется) и региональных лимфатических узлах. Затем происходит стремитель-
ное вовлечение в процесс все возрастающего числа нейтрофилов, выход в кровь их
костномозгового резерва. Болезнь начинается остро. У больного высокая температу-
ра, симптомы выраженной интоксикации и нейтрофильный лейкоцитоз.
Основу токсических проявлений инфекции, вы тываемой возбудителем чумы, со-
ставляет действие эндотоксина — Л ПС бактериальной стенки. Клетки Y. pestis частич-
но разрушаются, и освобождается Л ПС, действующий непосредственно на клетки
крови, тканей, эндотелии сосудов и опосредованно, активируя систему комплемента
и нейтрофилы. Последние выделяют биологически активные вещества, изменяющие
микроциркуляцию и нарушающие баланс систем свертывания и противосвертывания.
Вначале это реакция местная и соответствует инкубационному периоду. Но с дости-
жением определенного количественного уровня ЛПС переводит инфекционный про-
цесс в иное качество — начинаются клинические проявления чумы1.
Бурное и интенсивное накопление ЛПС и соответственно продуктов дегрануля-
ции нейтрофилов приводит к развитию синдрома внутрисосудистого свертывания
крови (ДВС-синдром) и инфекционно-токсического шока, т. е. процесса, вызванно-
го лавинообразной экспрессией провоспалительных цитокинов, основным из кото-
рых является TNF-a. Шок проявляется тахикардией, заторможенностью, гипотони-
ей, реже рвотой. С нарастанием тяжести течения чумы появляются такие симптомы,
как бред, снижение температуры тела, петехии, понос, глухость тонов сердца, олигу-
рия. У некоторых больных развиваются одышка, судороги, акроцианоз, боли в животе,
явления отека легких.
1 Подробно о роли ЛПС возбудителя чумы в патогенезе болезни см. в диссертационной
работе Р. 3. Шахутдиновой (2008).
682
Часть 3. Частная неправильная эпидемиология
Клиническая картина чумы у людей при естественном инфицировании Y. pestis1.
В клинике чумы выделяют общие явления, характерные для всех форм болезни, и ча-
стные, на основании доминирования которых у отдельных больных, создаются кли-
нические классификации чумы.
Инкубационный период Y pestis составляет 1—6 сут. В большинстве случаев бо-
лезнь начинается остро, с сильного озноба и быстрого повышения температуры тела
до 39—40 °C. Уже на первый день болезни характерны озноб, чувство жара, миалгии,
мучительная головная боль, головокружение. Хар пстерен внешний вид больного: лицо
и конъюнктивы гиперемированы, тубы сухие, язык отечный, сухой, дрожащий, об-
ложен густым белым налетом (как бы натерт мелом), увеличен. Речь больного смазан-
ная, неразборчивая, он заторможен или, наоборот, возбужден, бредит, стремится убе-
жать, у него нарушена координация движения (иногда такие больные производят
впечатление пьяных), на лице появляется выражение страха и ужаса («facies pestica»).
На лбу выступают крупные капли пота («роса чумы»).
За счет поражения ССС, границы сердца расширены, тоны сердца глухие, прогрес-
сивно нарастает тахикардия (до 120-160 уцаров/мин), появляются цианоз, аритмия
пульса, значительно снижается АД. На ЭКГ чаще всего бывает синусовая тахикардия,
иногда с изменением интервала S-Т(уменьшение зубца 7). У некоторых больных об-
ращают на себя внимание преждевременные сокращения предсердий, а также высо-
кий зубец Р и отклонение оси вправо. Эти изменения ЭКГ трактуются как указание
на cor pulmonale, хотя изменений в легких еще может не быть (Домарадский И. В., 1998)
При тяжелом течении чумы отмечается кровавая или цвета кофейной гущи рво-
та, жидкий стул со слизью и кровью. В моче находят примеси крови и белок, развива-
ется олигурия. Печень и селезенка увеличены. В крови нейтрофильный лейкоцитоз
со сдвигом формулы влево, СОЭ повышена. Количество эритроцитов и гемоглобина
изменяется незначительно.
Смерть больного чумой наступает при прогрессирующей циркуляторной недоста-
точности, часто с отеком легких.
В соответствии с классификацией Г. П. Руднева (1970) различают следующие кли-
нические формы чумы:
1. Преимущественно локальные формы: кожная, бубонная, кожно-бубонная.
2. Внутреннедиссеминированные, или генерализованные, формы: первично-сеп-
тическая, вторично-септическая.
3. Внешнедиссеминированные (центральные, чаще с обильной внешней диссеми-
нацией): первично-легочная, вторично-легочная, кишечная. Кишечная форма как
самостоятельная большинством авторов не признается.
Описаны стертые, легкие, субклинические формы чумы.
Бубонная чума. Важнейшим признаком бубонной формы чумы является бубон —
резко болезненное увеличение лимфатических узлов. Бубон появляется на первый
день болезни. Уже в первые часы развития бубона в месте его образования больной
чувствует сильную боль, которая затрудняет движение соответствующего органа. Боль-
ной вынужден принимать неестественные позы. В первые дни на месте развивающе-
гося бубона можно прощупать отдельные лимфатические узлы, в дальнейшем они
1 В работе (Супотнипкий М. В., Супотнипкая Н. С., 2006) приведены десятки описаний
клиники и патанатомии различных форм чумы, сделанных авторами, непосредственно уча-
ствовавшими в ликвидации эпидемий чумы. В данном разделе приводятся обобщенные опи-
сания, взяв за основу работы Е. П. Шуваловой с соавт. (2001) и В. М. Туманского (1958).
3.2. Чума
683
спаиваются с окружающей клетчаткой. Маленькие бубоны обычно более болезнен-
ны, чем большие. Бубоны, появившиеся в начале болезни, называются первичными
бубонами первого порядка. Они обусловливаются внедрением чумного микроба в
лимфатический узел по лимфатическим путям извне через кожу (обычно при укусе
зараженных блох). Первичные бубоны первого порядка в большинстве случаев еди-
ничные, в редких случаях их бывает несколько; многочисленные бубоны развивают-
ся при одновременном внедрении чумного микроба в разных частях тела.
На месте внедрения инфекции при бубонной форме чумы обычно не наблюдает-
ся никаких поражений, и только в редких случаях на коже отмечаются ссадины или
маленькие покраснения, напоминающие укусы блох. Не наблюдается также воспале-
ния лимфатических сосудов (лимфангоитов), по которым чумной микроб проникает
в регионарный лимфатический узел (лимфангоиты встречаются только в тех случа-
ях, когда вместе с возбудителем чумы заносятся и другие микробы, но такие случаи
встречаются очень редко).
Помимо первичных бубонов первого порядка, различают еще первичные бубо-
ны второго порядка; последние развиваются в результате внедрения в железу чумно-
го микроба по лимфатическим путям из первичного бубона первого порядка. Эти бу-
боны появляются в разное время: почти одновременно с первичными бубонами
первого порядка, в разгар болезни и даже в конце ее. Чаще они появ [яются в первую
неделю болезни. Вторичные бубоны, обусловливаемые заносом чумного микроба по
кровеносным путям, появляются вдали от места внедрения инфекции обычно в конце
болезни. При септической и легочной форме чумы они могут появиться в первые дни
болезни.
Частота первичного поражения тех или иных лимфатических узлов зависит глав-
ным образом от площади, с которой собирается лимфа в лимфатический узел. Вместе
с тем играют роль и другие факторы: профессия больного, бытовые условия, возраст.
Чаще всего наблюдаются паховые и бедренные бубоны (55—68 %), затем подмышеч-
ные (16—22 %), шейные (5—6 %); множественные бубоны не часты (5—13 %). Подко-
ленные и локтевые бубоны встречаются очень редко. Шейные бубоны чаще всего
отмечаются у детей (рис. 3.38).
В начале образования бубона кожа над ним не изменина. По мере увеличения бу-
бона кожа натягивается и краснеет. Над большим бубоном она болезненна, натянута
и блестяща, цианотична, иногда имеет багрово-синюю окраску. При осмотре бубона
на месте нахождения пораженного узла в большинстве случаев наблюдается выпук-
лость. Иногда же, ввиду глубокого расположения пораженного узла, этой выпукло-
сти не удается отметить не только в первые, но и в последние дни болезни.
При прощупывании бубона в первый день болезни обнаруживается лишь небсль
шое, очень болезненное уплотнение. В зависимости от развития болезни, часто уже к
концу первого дня ее, бубон хорошо заметен: при наружном осмотре он имеет вид
опухоли. Кожа над ним гиперемирована и цианотична, в особенности в центральной
части. Окружающая клетчатка отечна. Бубон при прощупывании оказывается очень
плотным, имеющим консистенцию хряща. С течением времени плотность его может
уменьшиться, и на участке нагноения прощупывается размягчение, а затем опреде-
ляется Флюктуация. Ввиду вовлечения в процесс окружающей узел ткани (периаде-
нит), контуры бубона очень неясны, что еще более усугубляется за счет отечности
окружающей клетчатки. По окончании стадии формирования бубона наступает фаза
его разрешения, протекающая в одной из трех форм: рассасывание, вскрытие и скле-
розирование.
684
Часть 3. Частная неправильная эпидемиология
Рис 3.38. Бубоны у больных чумой. А. Правый шейный бубон. По В П. Кашкадамову
(1902). Б. Больной чумой с бедренными бубонами. В Скульптурное изображение свя-
того Рока — защитника европейцев от чумы (XV в.). По U. A. Wu Lien-Ten et al. (1936)
Величина бубонов различна: от горошины до размеров гусиного яйца и даже боль-
ше. Она зависит главным образом от степени развития периаденита: чем больше про-
цесс распространяется на окр; жающую клетчатку, тем больше бубон; чем более вы-
ражен периаденит, тем сильнее бубон спаян с окружающими тканями и кожей и менее
подвижен.
Развитие бубона происходит по-разному: в некоторых случаях он увеличива-
ется очень быстро и в 2—4 ч достигает величины куриного яйца, чаще же это проис-
ходит медленнее. Наибольшей величины бубон достигает к 8—10 сут болезни, реже —
к 4—5 сут.
В случае нагнаивания бубона к чумному микробу присоединяется вторичная ин-
фекция (стафилококк, стрептококк). В нагноившемся бубоне кожа над очагом нагное-
ния истончается и некротизируется, иногда на большом участке. Бубон прорывает-
ся, и из него вытекает гнойная масса с примесью крови. Заживление образовавшейся
полости (нередко довольно большой и глубокой) идет очень медленно, сплошь и ря-
дом отмечается то улучшение, то ухудшение процесса заживления. Как правило, на-
гноение начинается с первичных бубонов первого порядка. Вторичные бубоны наг-
наиваются очень редко. На месте заживления образуются рубцы (рис. 3.39).
3.2. Чума
685
Рис 3.39. Характерные изъязвления кожи больных
бубонной чумой По В. П. Кашкадамову (1902)
Рассасывание бубона идет медленно; только в редких случаях наблюдается быст-
рое рассасывание, что расценивалось в начале XX в. как неблагоприятный прогно-
стический признак. Иногда рассасывающиеся бубоны увеличиваются в размерах, при-
чем это может наблюдаться неоднократно. Иногда в период увеличения рассасываю-
щегося бубона температура повышается до 37,3—37,5°C; может также усиливаться
болезненность бубона и гиперемия кожных покровов. Иногда же увеличение расса-
сывающегося бубона не сопровождается этими явлениями. Рассасывание бубона не-
редко происходит довольно хорошо, и уже через 2—3 недели обратного развития бу-
бона на месте его остается еле прощупываемое уплотнение. Иногда даже при хорошем
самочувствии больного и нормальной температуре бубон сохраняет значительные
размеры (до куриного яйца) в продолжение долгого времени. Объясняется это скле-
ротическими изменениями в лимфатическом узле.
В зависимости от клинического течения бубонной чумы, многие авторы различают
три степени ее:
1) тяжелую;
2) среднюю;
3) легкую.
В легких случаях заболевания температура повышается незначительно; иногда
болезнь протекает даже при субнормальной температуре (амбулаторная форма).
Обычно же температура сразу повышается до 38—39 °C. Наблюдаются суточные
колебания температуры в 1—2 °C и более, при этом повышение ее бывает в вечерние,
а понижение — в утренние часы (ремиттирующий тип лихорадки). В связи с ухудше-
нием болезни, наблюдается несоответствие частоты пульса с температурой, что счита-
ется неблагоприятным прогностическим признаком.
В легких случаях течения болезни учащение пульса выражено слабее, и то только
в начале заболевания. В случаях средней тяжести пульс равен 90—110/мин при тем-
пературе 38—39°С.
686
Часть 3. Частная неправильная эпидемиология
Пульс является тонким показателем состояния ССС больного. Помимо учашения
пульса, отмечается также его слабое наполнение, часто аритмия, иногда пульс быва-
ет дикротичным. В последние часы жизни больного он становится нитевидным, мало
ощутимым.
Со стороны органов дыхания при бубонной чуме наблюдается увеличение числа
дыханий. Перед наступлением смерти в легких — застойные явления (влажные хри-
пы, притупления).
Со стороны нервной системы отмечается сильная головная боль, особенно выра-
женная в начальный период болезни. Этот симптом занимает первое место в жалобах
больного. Наблюдается также головокружение, довольно часто — рвота (мозгового
происхождения). Нередко больные жалуются на бессонницу. В тяжелых случаях на-
блюдается помрачение сознания и бред, иногда подергивания и судороги.
На коже изредка появляется геморрагическая сыпь (грудь, живот, спина, реже
конечности) темно-красного, темно-бурого, иногда даже черного цвета («черная
смерть») величиной от просяного зерна до двадцатикопеечной монеты. Иногда у бол ь-
ных отмечается желтушность кожи, которая появляется в конце острого периода бо-
лезни и держится несколько дней.
Бубонная форма чумы в некоторых случаях осложняется вторичной чумной пнев-
монией, иногда чумным менингитом; при генерализации процесса — вторичной сеп-
тицемией. Продолжительность болезни — от нескольких дней до месяца и более. Ле-
тальность без лечения — от 30 до 90 %. При рано начатом антибактериальном и
патогенетическом лечении смертельный исход наступает редко.
Ниже, по работе И. В. Страховичаи В. И. Госа (1907), приводится история болезни
и протокол вскрытия крестьянина Григория Тетерятникова, умершего от бубонной
чумы 17.12.1901 г. через 7 сут после начала болезни. Случай интересен еще тем, что у
Григория имел место патологический процесс в легких, однако клинически он не про-
являлся, кашля и кровохаркания у него не было.
Григорий Тетерятников. 62 года. Заболел 10 декабря в доме Луки, куда он перешел
после смерти Александры Тетерятниковой.
В первый день болезни; характерных явлений не замечается. Рвота, озноб, голов-
ная боль, температура 38,9°С, пульс 86. Язык бледно-розового цвета совершенно чи-
стый. Аппетит хороший. Самочувствие удовлетворительное. Сразу осунулся. Апатич-
ное выражение лица. Железы в правом паху до лесного ореха. Область селезенки при
пальпации несколько болезненна; вечером температура 39,8°С, пульс 106.
11 декабря. Общее состояние хуже, язык покрыт серым густым налетом. Сильная
жажда. Петехий нет. Железы в правом паху резко увеличены, доходя до голубиного яйца
и образуя весьма болезненный продолговатый пакет, но кожа на нем не изменена. Моча
выделяется свободно. Температура 39,5°C, пульс 102, вечером температура 39.0°С,
пульс 112. Сонливое состояние весь день.
12 декабря явления те же. Больной на ногах. Утром температура 39,0°С, пульс 112;
вечером температура 39,2°С, пульс 96.
13 декабря самочувствие хорошее, аппетит тоже, жалуется на болезненность в об-
ласти бубона.
14декабря температура 39,5 °C, пульс 94. Самочувствие удовлетворительное. Язык
обложен густым серым налетом, вечером температура 39,0°С, пульс 100.
15 декабря утром температура 39,5 °C, пульс 108. Язык чище. Самочувствие хуже.
Отсутствие аппетита. Упадок сил увеличивается, пульс 122
16 декабря температура 39,2°C, пульс частый. Аппетит лучше. Инфильтрат на
верхней трети правого бедра величиною в две ладони, красного цвета, крайне болез-
3.2. Чума 687
нен. На животе и бедрах многочисленные мелкие петехии. Конъюнктивы немного ик-
теричны, но не инъецированы. Полное сознание, аппетит хороший. Язык растрескав-
шийся, но чистый. Пульс еле ощутим и не сосчитывается.
17декабря умер в 8 ч утра в полном сознании. Перед смертью вышел из избы, вер-
нулся, напился чаю, потребовал варенья и умер. Вскрытие произведено 19 декабря,
через 52 ч после смерти. Труп сохранялся при температуре ниже 0°С.
Протокол вскрытия.
Труп ниже среднего роста, правильного телосложения, плохого питания. Кожа
бледно-серого цвета. В правом паху находится напряженная припухлость, подымаю-
щаяся выше пупартовой связки и вниз спускающаяся до границы верхней и нижней
трети бедра. На разрезе оказывается, что припухлость состоит из целого пакета увели-
ченных от боба до крупной сливы величиною лимфатических желез, спаянных отеч-
ной геморрагической клетчаткой. Железы сильно гиперемированы. При разрезе па-
кета желез из ткани вытекает серознокровянистая, слегка мутноватая, жидкость, и края
разреза расходятся. Увеличенные железы, спаянные инфильтрованною отечною тка-
нью, находятся так же и выше в правом забрюшинном пространстве. Положение орга-
нов грудной полости правильное. Сердечная сумка содержит небольшое (около 60 см3)
количество серозной жидкости. Висцеральный и периэтальный листки сердечной со-
рочки блестящи, гладки. Экхимозов нет. Длина сердца 10,0 см, ширина — 11,5 см.
Полость сердца содержит кровяные сгустки темно-красного и желтого цвета. Клапа-
ны сердца без изменений. Эндокард блестящий, гладкий. Мышца сердца буро-крас-
ного цвета с ясным желтым оттенком, дрябла. Внутренняя поверхность аорты без из-
менений. Правое и левое легкое плотно приращены в нижних своих долях к реберной
плевре. Бронхиальные железы увеличены справа, черного цвета. В плевральных поло-
стях по столовой ложке серозной жидкости. Левое легкое гиперемировано, отечно, при
разрезе вытекает кровянистая, пенистая жидкость. Никаких фокусов уплотнения не
замечается. Правое легкое в верхней и средней доле повторяет картину левого легко-
го. Вся нижняя доля правого легкого уплотнена, на разрезе серо-желтого, а в верхней
части более красного цвета. Кусочки тонут в воде. Положение органов грудной поло-
сти правильное. Брюшинный покров блестящий, гладкий.
В полости брюшины жидкости нет. Брыжеечные железы не увеличены. Селезенка
имеет в длину 12,5, в ширину 3,5 и толщину 3,0 см Капсула слегка утолщена, морщи-
ниста. Паренхима дрябла, темно-красного цвета, легко соскабливается. Трабекулы не
утолщены. Мальпигиевы тельца не ясно выступают. Поверхность печени ровная, глад-
кая; в одном месте находится узелок с булавочную головку белого цвета. Общая ши-
рина 27 см, длина правой доли 19 см, левой 17,5 см, толщина правой 6 см, левой 4 см.
Паренхима дрябла, тестовата, желто-бурого цвета, малокровна. В паренхиме такой же
узелок, как и на поверхности; оба на разрезе хрустят, плотны, по-видимому, содержат
известь. Левая почка: длина 12,5, ширина 6,5, толщина 4,0 см. Капсула снимается легко.
Корковой слой слегка утолщен 10,8 см. Бертиньевы столбы припухли, пограничная
линия сглажена. Пирамида, как и корковый слой, розового цвета с ясным серым от-
тенком. Правая почка: длина 12,0, ширина 6,0, толщина 4,0 см. Капсула местами сни-
мается с паренхимой, на поверхности небольшая кисточка с серозным содержимым.
В остальном походит на левую почку. Желудок растянут, складки сглажены, станки
атрофичны. Кишечный канал особых изменений не представляет. Половые органы без
изменений. Мочевой пузырь содержит прозрачную мочу, слизистую, бледного цвета,
остальные органы вскрыты не были.
Первично-септическая форма. Развивается бурно после короткой инкубации, со-
ставляющей от нескольких часов до 1—2 сут. Больной ощущает озноб, резко повыша-
ется температура, появляются сильная головная боль, возбуждение, бред. Возможны
688
Часть 3. Частная неправильная эпидемиология
признаки менингоэнцефалита. Развивается картина инфекционно-токсического
шока, быстро наступает кома. Продолжительность заболевания — от нескольких ча-
сов до трех суток. Случаи выздоровления крайне редки. Больные погибают при явле-
ниях тяжелейшей интоксикации и выраженного геморрагического синдрома, нара-
стающей сердечно-сосудистой и почечной недостаточности.
Иногда болезнь заканчивается смертью через несколько часов (молниеносная
форма — pestis siderans). В этих редких случаях температура у заболевшего чумой че-
ловека почти не повышается. Болезнь протекает так быстро и бурно, что его организм
не успевает или оказывается не в состоянии реагировать на инфекцию. Умерших
людей часто находят в свободных позах, свидетельствующих об их внезапной смерти
(например, с чашкой чая в руке).
Патологоанатомические изменения скудны и малоспецифичны. Наиболее вы-
раженное изменение отмечалось в селезенке: резкое увеличение, иногда с очажками
некроза. Наблюдались точечные кровоизлияния в слизистой кишечника и плевры.
Диагноз септической чумы устанавливается бактериологическими исследованиями.
По работе D. A. Margolis et al. (2008) привожу описание атипичного случая пер-
вично-септической чумы, имевший место в Восточной Калифорнии в 2008 г. В отли-
чие от типичных случаев септической чумы, когда больной в течение 1—3 сут поги-
бает от инфекционно-токсического шока, в данном случае, благодаря интенсивной
антибиотикотерапии и патогенетической терапии, жизнь пациентке удалось продлить
до 10 сут. Однако исход болезни был предопределен еще на ее начальной стадии.
Пациентке 79 лет, проживала в сельской местности. В местную больницу поступила
с температурой 38,2°С, с пульсом 110/мин, АД 121/60 мм рт. ст. и насыщением крови
кислородом 80 % в условиях воздуха помещения. Пациентка была возбуждена, ори-
ентировалась только в отношения себя самой. Аускультация легких обнаружила хри-
пы на левой верхушке (left base), живот мягкий. Л имфоаденопатии не было. Невроло-
гическое исследование показало отсутствие очаговых поражений со стороны ЦНС.
Лейкоцитоз (46 тыс. клеток/л, 19 % из них—палочковые формы), ренальная дисфунк-
ция, высокий уровень трансаминаз и сывороточного креатина. Признаков поражения
лимфатических узлов не было. Рентгенограмма грудной клетки интерпретирована ле-
чащим врачом как свидетельствующая о левом нижнем ателектазе легкого (left lower
king atelectasis). Пациентке поставлен диагноз «пневмония» (community-acquired pneu-
monia, САР) и назначено лечение антибиотиками (цефтриаксолин, ампициллин/суль-
бактам, левофлоксацин).
Через 48 ч после назначения антибиотиков, стала нарастать почечная недостаточ-
ность (повышение уровня креатинина в сыворотке крови), появились ацидоз и тром-
боцитопения (тромбоциты, 54 тыс. клеток/л). На 3 сут болезни больная была отправ-
лена в академический центр госпиталя для дальнейшего наблюдения. Там ей потребо-
валась интубация и гемодинамическая поддержка. В этот же день из другого госпиталя
поступил отчет о выделенной из ее крови культуры какой-то грамотрицательной па-
лочки. Ее интерпретировали как контаминант. Уровень лактатде гидрогеназы был по-
вышен. Сочетание клинических и лабораторных данных дало основание поставить
диагноз — тромботическая тромбоцитопеническая пурпура (thrombotic thrombocy-
topenic purpura). Начат плазмафорез и плазмаобмен. На груди и верхних конечностях
пациентки обнаружены участки некроза кожи с эритемой по краям (рис. 3.40).
Только на седьмые сутки болезни с помощью ПЦР были получены доказательства,
что грамотринательные палочки являются ни чем иным, как возбудителем чумы. Но к
этому времени на отдельных пальцах у пациентки уже началась гангрена. Прогресси-
ровала почечная недостаточность. Компьютерная томография брюшной полости
3.2. Чума
689
Рис. 3.40. Кожные и внутриорганные повреждения при первично-септической
чуме. А. Участок некротической кожи (чумной карбункул) с зоной гиперемии
вокруг. Б. Гангрена пальцев. В, Г и Д Дермальный некроз, острое воспаление и
тромбоз небольших сосудов при отсутствии видимых микроорганизмов. Субдер-
мальное расслоение кожи в участке некроза (увеличение в 200 раз). Е. Компью-
терная томограмма брюшной полости, показывающая билатеральный ренальный
кортикальный некроз. По D A. Margolis et al. (2008)
показала билатеральный ренальный кортикальный некроз. Пациентка умерла от уре-
мии на 10 сут пребывания в больнице.
После того, как диагноз «чума» был подтвержден ПЦР, на участке, прилегающем
кдому умершей от чумы пациентки, проведено эпизоотическое исследование Отлов-
лены калифорнийские и антилоповые суслики (Spermophilus beecheyi, Ammospermophilus
leucurus), с которых сняты блохи, инфицированные Y. pestis (поданным ПЦР). Среди
калифорнийских сусликов обнаружены особи с высокими титрами антител к Y. pestis.
Антитела к возбудителю чумы обнаружены и у собаки пациентки.
Вторично-септическая форма. Является осложнением других клинических форм
чумной инфекции. Характеризуется исключительно тяжелым течением, наличием
вторичных очагов, бубонов, выраженных проявлений геморрагического синдрома.
690
Часть 3. Частная неправильная эпидемиология
Развитие вторичной септицемии возможно при любой форме чумы. Прижизненная
диагностика этой формы затруднена. Безошибочно поставить диагноз можно после
обнаружения Y. pestis в тканях и жидкостях организма.
Клиника кожной формы чумы. Несмотря на то, что кожа является наиболее частым
местом проникновения возбудителя чумы в организм человека, кожная форма чумы
наблюдается редко. Обычно кожа на месте внедрения чумного микроба не дает вос-
палительной реакции1.
При развитии кожной формы чумы наблюдаются следующие явления: на коже в
месте поражения появляется красное пятно, очень болезненное, диаметром около
1 см, напоминающее след от укуса блохи Вскоре это пятно приобретает багровую ок-
раску, а через несколько часов приподнимается, вследствие накопления под ним жид-
кости. Жидкость эта почти сразу становится мутной от примеси гноя и крови, пятно
превращается в пустулу. Вокруг пустулы имеется красная воспалительная зона (ран-
няя фликтена — phlyctaenae praematura). Величина пустулы колеблется от размеров
горошины до вишни. Жидкость пустулы содержит значительное количество вирулен-
тной К pestis. С течением времени пустула и окружающая ее зона увеличиваются в
размерах и превращаются в чумной карбункул, очень болезненный, в особенности при
надавливании (рис. 3.41).
На месте вскрывшейся пустулы образуется язва с инфильтрированным плотным
дном желтоватого цвета. Язва покрывается темным струпом, который вместе с окру-
Рис. 3.41, Кожная форма чумы. А. Язва. Б. Пустула. По Г. П. Рудневу (1938)
1 Возможны отдельные вспышки чумы с преобладанием кожной формы болезни. Приме-
ром может служить чума 1920 г. в Батуми, где кожная форма составила до 80 % всех случаев бо-
лезни (Руднев Г. П., 1938)
3.2. Чума
691
жающей его темно-багровой зоной дает картину, весьма напоминающую сибирскую
язву — pestis anthraeicus. Таким образом, основываясь лишь на одном наружном виде
чумной пустулы, без лабораторного исследования пунктата, можно довольно легко
впасть в диагностическую ошибку Размеры язв при кожной форме чумы бывают раз-
личными — от монетки до ладони взрослого человека.
Первичных пустул и первичных карбункулов может быть несколько, причем они
появляются одновременно в различных местах внедрения инфекции, т. е. в местах
укуса зараженных блох. Помимо первичных, встречаются вторичные пустулы и кар-
бункулы. Они могут наблюдаться при разных формах чумы и обычно появляются в
разгар болезни или в конце ее.
Общие клинические явления при чисто кожной форме чумы такие же, как и при
легких или средних по тяжести случаях бубонной чумы. Очень часто кожная форма
чумы осложняется воспалительным процессом в регионарном лимфатическом узле.
Нередко это осложнение наступает очень быстро, так что у больного, поступившего в
госпиталь, наблюдается и первичное поражение на коже, и бубон. Последний беспо-
коит больше, чем первичное кожное поражение: обычно больной больше жалуется на
бубон, чем на кожное поражение, а нередко и совсем не упоминает о нем. Последнее
обстоятельство заставляет врача при наличии бубона тщательно искать на коже вход-
ные ворота И1 фекции. Осложненная бубоном кожная форма чумы носит название
кожно-бубонной формы.
Кишечная форма чумы. Главным признаком кишечной формы чумы является час-
тый понос (до 40 раз в сутки), нередко с обильной примесью слизи и крови. Дефека-
ция сопровождается болезненными тенезмами. Тошнота, нередко многократная рвота.
Все эти симптомы сопровождаются высокой температурой и прочими признаками,
свойственными друз им формам чумы.
Существование первичной кишечной формы чумы в настоящее время оспаривает-
ся рядом авторов. Однако при обнаружении в энзоотических районах или вблизи них
таких больных, у которых наблюдаются кишечные яатения невыясненного происхож-
дения, необходимо помнить о чуме и провести тщате пьное клиническое обследование
больного, а также бактериологическое исследование испражнений и рвотных масс.
Легочная форма чумы. Чаще всего первичная легочная чума развивается в резуль-
тате капельной инфекции. Входными вооотами при легочной форме чумы moi ут быть
миндалины, слизистая трахеи и бронхов, откуда возбудитель чумы гематогенным
путем попадает в перибронхиальные лимфатические узлы.
Первичная чумная пневмония начинается внезапно: у заболевшего появляется
озноб, сильная головная боль, температура быстро повышается. Часто наблюдается
неоднократная рвота. Затем появляются тахикардия, боли в груди и сильная одышка.
Кашель в первые дни может отсутствовать, но иногда он появляется и в самом начале
болезни. Кашель сначала репкий и сухой, затем с мокротой. Мокроты обычно бывает
очень много, иногда же она совсем отсутствует. В начале болезни мокрота не харак-
терна для чумной пневмонии; затем к ней примешивается кровь, и она становится
розовой или красной. Мокрота содержит обычно большое количество чумных бакте-
рий. Иногда она имеет ржавый вид, а изредка наблюдается даже обильное кровохар-
канье. Эти симптомы даюз повод к смешению легочной чумы с крупозной пневмо-
нией и туберкулезом легких. При объективном исследовании легких в начале болезни
характерных признаков чумы обнаружить не удается. Позже дыхание учащается до
24—32 и более раз в минуту, а при выслушивании отмечаются разбросанные сухие,
иногда влажные хрипы, число которых в процессе болезни увеличивается.
692
Часть 3. Частная неправильная эпидемиология
При перкуссии места притупления или разбросаны, или как бы совпадают с гра-
ницами доли легкого. По мере развития болезни плошадь притупления увеличивает-
ся, так как процесс захватывает соседние доли.
Данные объективного клинического и рентгенологического исследования легких
не дают возможности дифференцировать первичную чумную пневмонию от пневмо-
ний, вызванных другой инфекцией (пневмококк, стрептококк и пр.) (рис. 3.42).
Рис. 3.42. Рентгенологическое исследование грудной клетки больной с первичной
легочной чумой. Серия фронтальных рентгеновских снимков, сделанных на 2,3 и 18 сут
болезни. Показывает двустороннюю мультилобулярную пневмонию, сопровождаю-
щуюся двухсторонним плевритом (двустороннее затемнение легочной ткани преиму-
щественно в нижних отделах легких с двусторонними плевральными выпотами, без
признаков хиларной или медиастинальной лимфаденопатии). Больная получала внут-
ривенно хлорамфеникол и перорально доксициклин. По Е. М. Begier et al. (2004)
Иногда в конце болезни, реже в середине ее, на коже появляется геморрагическая
сыпь темно-красного или даже черного цвета. Продолжительность болезни в среднем
2—4 сут, иногда до 6-7 сут и дольше. В прошлом все заболевшие легочной формой чумы
погибали. В настоящее время в связи с применением для лечения чумы стрептоми-
цина почти все заболевшие первичной легочной формой чумы выздоравливают.
Первичный пневмонический очаг при легочной чуме при правильном лечении
антибиотиками может рассасываться бесследно. Если лечение начато по истечении
12 ч и более от начала температурной реакции, то развитие первичного пневмоничес-
кого очага происходит более активно. Поражение легких при этом более обширно,
иногда процесс успевает распространиться на другое легкое. Тем не менее и в этом
случае больного еще можно спасти.
Температурная кривая у больных с легочной чумой не имеет четкой картины трех
волн, как это наблюдается при раннем начале лечения. Температура во время ремис-
сий вообще может не достигать нормы. Начиная с 5—10 сут, она изобилует многочис-
ленными подъемами на фоне общего суточного снижения температуры. Специфичес-
кий процесс иногда затягивается на месяц и более. В процессе выздоровления часто
наблюдаются абсцессы в легких и бронхоэктазии. Своевременное применение анти-
биотиков и в этих случаях ведет к благополучному исходу указанных осложнений.
Клиническая картина первичной легочной чумы у людей, леченных стрептоми-
цином, имеет свои особенности, которые затрудняют диагностику болезни. Если же
принять во внимание, что уже через несколько часов после начала лечения количе-
ство микробов в мокроте резко падает и выделить их штруднительно, то в некоторых
случаях бывает очень трудно поставить правильный диагноз болезни, в особенности
у контактировавших и подвергшихся лечению (с цепью профилактики) до проявле-
3.2. Чума
693
ния болезненных симптомов. Определить появление новых очагов в корневых лим-
фатических узлах и образование абсцессов и бронхоэктазов гоже затруднительно. Все
это устанавливается главным образом на основании рентгенологического исследова-
ния и клинических симптомов.
Чума у вакцинированных больных. Характеризуется удлинением инкубационного
периодадо 10 сут и замедлением развития инфекционного процесса. Втечение 1—2сут
заболезания лихорадка субфебрильная, общая интоксикация выражена нерезко, со-
стояние больных удовлетворительное. Бубон небольших размеров, без выраженных
проявлений периаденита. Однако симптом резкой болезненности бубона всегда со-
храняется. Если эти больные в течение 3—4 сут не получают лечение антибиотиками,
то дальнейшее развитие болезни ничем не будет отличаться от клинических симпто-
мов у невакцинированных больных.
Патоморфологические признаки легочной чумы при инфицировании в эпидемических
цепочках. По данным вскрытий 70 человек, умерших от чумы в Харбине в 1910— 1911 гг.,
Г. С. Кулеша (1912) нашел, что визуально на слизистых зева, дыхательного горла и
бронхов удается обнаружить лишь к зтаральные поражения. Только в одном случае в
1 ыхательном горле наблюдались фибринозные пленки, содержащие огромное коли-
itCTBO У. pestis. Однако при микроскопическом исследовании он обнаружил в тканях
миндалин массовые скопления чумных палочек, особенно под самым эпителием
Поражения эпителия в таких местах иногда напоминали кожные пустулы, причем в
многослойном эпителии образовывались ровной величины полости, содержащие
белые кровяные тельца и чумные палочки, а подлежащая ткань миндалины густо ин-
фильтрировалась чумными палочками и круглыми клетками. Кулеша находил массо-
вые скопления чумных бактерий и в лимфоидной ткани миндалины в глубине орга-
на, а также в просветах кровеносных сосудов, где нередко наблюдалось образование
тромбов, содержащих чумные палочки. В некоторых случаях ему удавалось заметить,
что миндалины также служат местом проникновения осложняющей течение чумы по-
сторонней микрофлоры: ему встречались участки тканей, наполненные чумными па-
очками и цепочками стрептококков.
Слизистая оболочка дыхательного горла и бронхов также оказывалась значительно
измененной. Эпителий и подслизистая ткань были «пропитаны» чумными палочка-
ми, которые в цилиндрических клетках эпителия образовывали яйцевидные скопле-
ния, а в подслизистых поверхностях отлагались в больших количествах и простира-
лись вглубь, почти до самых хрящей, окружая кровеносные сосуды и прорастая в их
стенки. Эти скопления чумных палочек располагались также вокруг глубоко лежащих
лимфатических желез. Им сопутствовали кровоизлияния и мелкоклеточная инфиль-
рация в пораженной ткани, причем воспалительная инфильтрация редко была вы-
ражена особенно резко. Поражения дыхательного горла и бронхов не имели разлито-
го распространения, встречались очагами и в отдельных случаях были выражены
различно, иногда их не было вообще. Поражения бронхиальных лимфатических узлов
носили характер первичных бубонов, но разрастание Y. pestis в них не всегда прости-
ралось за пределы сумки пораженного лимфатического узла, периаденитов практи-
чески не было. В некоторых случаях разрастания чумных палочек в ткани лимфати-
ческих узлов приобретали своеобразный вид, и при рассмотрении в бинокулярную
лупу они напоминали по рисунку кожные лишаи.
В легких Кулеша всегда наолюдал долевую (лобулярную) форму воспаления, кото-
рая, по его мнению, свойственна эпидемической легочной чуме. Пораженными оказы-
вались то одна, то несколько долей легких. Одна доля была поражена 3 5 раз, несколь-
694
Часть 3. Частная неправильная эпидемиология
ко долей — 29 раз, без поражения легких было 4 случая. Верхняя правая доля была
поражена J0 раз, верхняя левая доля — 22 раза, нижняя правая — 18 раз, нижняя ле-
вая — 20 раз, средняя доля — 10 раз. Таким образом, верхние доли были поражены
52 раза, а нижние — 38 раз (на 70 исследованных случаев). Пораженной всегда оказы-
валась плевра, на которой обычно наблюдался нежный фибринозный нал“т. В 2-х слу-
чаях поражение легких сопровождалось сывороточно-фибринозным выпотом в со-
ответствующую область плевры. Пораженные доли легких обычно были менее
объемисты, чем при фибринозной пневмонии, и на разрезе не обнаруживали свой-
ственные последней зернистости. Разрез был более или менее гла 1кий, что зависело
от отсутствия фибрина в выпоте альвеол. Так называемое опеченение в большинстве
случаев было отчетливым, и вырезанные кусочки тканей тонули в воде. По давности
опеченения, в нем можно было увидеть серую и красную стадии. В светлых воспален-
ных участках, соответствующих красному опеченению, выпот в альвеолах состоял из
сывороточной, не содержащей нитей фибрина, жидкости, в которой в огромных ко-
личествах лежали У. pestis и эритроциты. Другие же клеточные элементы (клетки эпи-
телия альвеол и лейкоциты) встречались обычно в ограниченном количестве. Иног-
да скопления У. pestis достигали огромных количеств, включенные в эти массы клетки
тесно не смешивались с ними, а располагались в особых просветах, как бы в сумках;
фагоцитоза не наблюдалось.
В стадии серого опеченения состав выпота изменялся. В полость альвеолы про-
исходила энергичная инфильтрация белых кровяных телец (лейкоцитов), причем
количество К petfis в альвеолах резко уменьшалось, они постепенно исчезали, и альве-
олы, в конце концов, оказывались заполненными почти одними лейкоцитами. В этой
стадии воспаления, по-видимому, его обычным исходом было токсическое омертве-
ние ткани, что наблюдалось под микроскопом почти в каждом исследованном случае.
Омертвение сопровождалось часто обширными кровоизлияниями. Кулеша обратил
внимание на то, что У. pestis не равномерно распределялись в легочной ткани и что по
большей части они скапливались в ближайшей окружности кровеносных сосудов, и
последние (чаще всего вены) на препаратах, окрашенные синькой, представлялись
окруженными мощными синими кольцами. Подобные же скопления У. pestis замеча-
лись в перибронхиальной ткани и вокруг перибронхиа тьных кровеносных сосудов.
Такие же скопления наблюдались под плеврой: как «синие реки», стекались лимфа-
тические полосы, заполненные К pestis. Их в больших количествах находили и в дру-
гих кровеносных и лимфатических сосудах.
Кулеша в 1912 г. не пришел к окончательному выводу, считать ли найденные им
изменения слизистых поверхностей первичными или вторичными, посчитав, что
окончательно характер этих явлений поможет установить только эксперимент. Однако
он предположил, что для первичной легочной чумы в эпидемических очагах 'характер-
ны следующие патологоанатомические явления:
1) чумные палочки могут в нескольких случаях проникать в организм уже из поло-
сти рта, вызывая специфическое поражение миндалин, или со слизистой оболочки
трахеи и бронхов путем массового прорастания их сквозь неповрежденный эпителий.
Легкие поражаются в таких случаях вторично через кровь либо вследствие развития
нисходящего бронхита;
2) при своем эпидемическом распространении легочная чума вызывает преиму-
щественно долевую пневмонию, правильнее плевропневмонию ввиду постоянного
участия плевры. Эта чумная пневмония представляет большое сходство с фибриноз-
ной, отличаясь от нее главным образом отсутствием фибрина в выпоте.
3.2. Чума
695
В 1913 г., ьо время чумной вспышки в Уральской области, Кулеша (1924) произ-
вел вскрытия 7 человек, умерших от легочной чумы, из которых четверо было взрос-
лых и трое детей в возрасте от 3 до 10 лет, и получил те же результаты, что и1 года назад
в Харбине.
На слизистых оболочках при легочной чуме невооруженным глазом никаких из-
менений выявлено не было, кроме явлений острого катара на слизистых оболочках
гортани, трахеи и бронхов, зева, полости носа и конъюнктив. Тщательная микроско-
пия слизистых оболочек носа и конъюнктив не дали положительных результатов в
смысле нахождения свойственных только чуме поражений. В миндалинах, в слизис-
той трахеи и крупных бронхов при изучении их под микроскопом Кулеше вновь уда-
лось обнаружить изменения, аналогичные тем, которые описаны им во время Мань-
1журекой чумы.
В эпителиальном покрове трахеи и крупных бронхов входными воротами для чум
ной инфекций служили главным образом бокаловидные клетки, в которых появля-
лись овальные или яйцевидные тела, состоящие из множества чумных палочек.
Кроме того, им замечались непрерывные тяжи У. pestis, расположенные между
эпителиальными клетками. В дальнейшем эпителиальный покров омертвевал и от-
слаивался, а на его месте оставался только базальный слой, по большей части также в
состоянии некроза. В то же время в подслизистой ткани возникало колоссальное скоп-
ление Y. pestis, образующих сплошную подстилку эпителия и совершенно замещаю-
щих сооой подлежащую ткань. Хорошо видимыми оставались только кровеносные
сосуды с их кровью, включенные в эти сплошные бациллярные массы.
Из семи исследованных случаев, в одном случае найдено было голько поражение
миндалин и в четырех случаях поражение миндалин, трахеи и крупных бронхов.
Кулеша отметил две отличительные черты, свойственные описываемым пораже-
ниям. Первая состоит в том, что в большинстве случаев огромные отложения чумных
бацилл в тканях не сопровождаются резкой реакцией со стороны последних, где вид-
ны. бывают только слабовыраженная круглоклеточная инфильтрация, незначитель-
ные кровоизлияния и более или менее явственный некроз. Вторая сказывается тем,
что почти во всех мелких кровеносных сосудах, располагающихся в очагах скопления
К pestis, замечается их обильное проникновение в эти сосуды (капилляры и мелкие
вены) из окружности. Примесь У. pestis к крови наблюдается главным образом только
в сосудах, проходящих через бациллярные очаги, тогда как в подобных же сосудах,
лежащих вне названных очагов, обнаружи гь присутствие бацилл среди элементов
крови удается во много раз реже и труднее.
Последняя находка послужила ему фактическим подтверждением предположения
о гематогенном происхождении чумной пневмонии, которая не только может возни-
кать как воздушно-капельная инфекция, но появляется и в таких случаях, где об ин-
галяции не может быть и речи. К случаям воздушно-капельного происхождения ле-
гочной чумы Г. С. Кулеша относил лабораторную чуму, поражающую научных
работников во время экспериментов с Y. pestis, а также заболевания, проявляющиеся:
1) в начале эпидемии, как ее исходные случаи, например, у охотников за тарбага-
нами, у лиц, соприкасающихся с мясом зараженных чумой верблюдов;
2) возникающие уже в период разгара эпидемии (например, случай, описанный
Р. А. Будбергом, см. выше рис. 3.35).
К случаям же воздушно-капельного заражения человека Г. С. Кулеша отнес те,
когда возбудитель чумы, находясь в воздухе в виде капельной инфекции, оседает при
вдохе не в самой легочной ткани, а на слизистых оболочках рта, глотки, грахеи и круп-
696
Часть 3. Частная неправильная эпидемиология
ных бронхов. Такого рода оседание Y. pestis неизбежно ведет к легочной чуме, но не
прямо, а через посредство первичного поражения миндалин, слизистой трахеи и брон-
хов, причем чумная инфекция транспортируется в легкие уже не через воздух, а при
помощи крови, т. е. гематогенно. Понятно, что заболевания такого происхождения
возможны лишь при наличии больных легочной чумой, заражающих своим кашлем
окружающий их воздух и обусловливающих нахождение в нем капельной инфекции.
Мнение, что поражение чумой миндалин ведет не к легочной чуме, а обязательно к
появлению чумных бубонов на шее, Кулеша считал неправильным, так как во всех
случаях, где он установил путем микроскопического исследования наличие описан-
ных выше чумных поражений миндалин, чумные бубоны на шее отсутствовали.
Моделирование искусственного заражения Е pestis. Ситуация искусственного зара-
жения К pestis многократно моделировалась в результате несчастных случаев, произо-
шедших при работе с возбудителем чумы и в опытах на животных.
Подкожное инфицирование. Возможно в случае умышленного заражения инъекци-
онным путем. По данным М Finegold et al. (1968), в участке введения возбудителя чумы
у экспериментального животного наблюдался некроз подкожной клетчатки. Клини-
ка болезни при подкожном инфицировании возбудителем чумы мало отличается от
клиники бубонной чумы. Разница в том, что если при бубонной чуме чаще всего ос-
новным признаком болезни будет бубон в паховой области (блохи обычно пьют кровь
у человека в области нижних конечностей), то при подкожном инфицировании он
развивается в группе лимфатических узлов, ближайшей к участку инфицирования.
Энтеральное инфицирование. Возможно, при намеренном подмешивании в пищу
возбудителя чумы. Клиника чумы при таком пути проникновения Y. pestis в организм
человека напоминает нисходящую бронхопневмонию, описанную Г. С. Кулешей.
В качестве примера рассмотрим случай, имевший место в феврале 1907 г. в Кронш-
тадте на форту «Александр I» (Берестнев Н М., 1907).
12 февраля 1907 г. доктор М. Ф. Шрейбер собирал эмульсии Y. pestis с агаровых
разводок и, случайно набрав много эмульсии чумных бацилл в пипетку, замочил вату;
при этом какое-то количество клеток Y. pestis попало ему в рот. Шрейбер промыл рото-
вую полость сулемой, но никому о происшедшем не рассказал и сыворотки себе не ввел.
14 февраля вечером Шрейбер почувствовал общее недомогание, озноб, ломоту в
ногах, небольшой насморк, отсутствие аппетита, в 9 ч вечера температура была 39,4 °C.
Ночью появилось колотье в правом боку и небольшой кашель без мокроты. На утро
температура была 39,1 °C, общее самочувствие удовлетворительное, колотье в боку
больной относил к мышечным болям. В 11 ч утра он в присутствии заведующего чум-
ной лабораторией на форте, Н. М Берестнева, легко выделил в стерилизованную чашку
Петри плевок слизисто-гнойной кровавой мокроты. При перкуссии грудной клетки-
справа сзади в области лопатки было отмечено небольшое притупление, при выслуши-
вании — легкое ослабление дыхания и единичные сухие хрипы. После исследования
мокроты, в которой оказалось громадное количество овальных микробов с полюсной
окраской, не красившихся по Граму, в чумном характере заболевания не было сомне-
ния. Ему было введено 80 мг сыворотки. Температура вечером 39,8 °C. пульс 140/мин;
дыхание утром и днем 28/мин; вечером 34/мин. Кашель днем усилился, обильная кро-
вянисто-гнойная мокрота отделялась довольно легко. К вечеру, в связи с высокой тем-
пературой, одышкой и слабостью сердца, общее состояние и самочувствие ухудшились.
Объективные явления со стороны легких за день без особых перемен; притупление в
области лопатки и спереди в области грудной мышцы более выражено. К вечеру спра-
ва спереди появились влажные средне- и мелкопузырчатые хрипы.
3.2. Чума
697
16 февраля днем сильный кашель, обильная мокрота с меньшим содержанием
крови в первую половину дня. Больной очень ослабел, пульс 120-130/мин; появились
клокочущие хрипы, мокрота стала жиже и пенистой, количество мокроты значитель-
но увеличилось; кофеин и камф< >ра под кожу, но явления слабости сердца и отек лег-
ких тем не менее усилились. При выслушивании в правом легком в верхней доле больше
влажных хрипов, появились хрипы в нижней доле левого легкого. Дыхание в верхней
доле правого легкого с бронхиальным оттенком, притупление звука нейольшое. Со-
знание полное.
Ночь на 17 февраля Шрейбер провел без сна, был возбужден, ослабел настолько,
что был не в состоянии отплевывать мокроту в ведро с сулемой. Появилась синюха на
концах пальцев. Днем температура снова поднялась до 39.9°С, кашель и одышка уси-
лились, масса жидкой кровянистой мокроты. Днем больной не отпускал своих коллег
от себя, много говорил и был в полном сознании; в 11,5 часов ночи наступила смерть
при явлениях отека легких и пара 1ича сердца.
Чумная инфекция у Шрейбера сопровождалась поражением почек, мочи за сутки
было около 500—300 см3 с большим количеством белка. Белок появился в моче 15 фев-
раля, причем количество его с каждым днем увеличивалось. Жидкие, довольно обиль-
ные испражнения, были 16-го утром и 17-го днем.
На вскрытии найдено следующее: оба легких плотно сращены с ребрами и диаф-
рагмой, плевра на обоих легких фиброзно утолщена особенно на местах отпечатков
ребер. Явлений старого фиброзного плеврита не заметно лишь на передних краях лег-
ких, которые были резко эмфизематозны. На плевре переднего края верхней доли
правого легкого несколько точечных кровоизлияний и по местам нежные фибриноз-
ные отложения.
Правое легкое', верхняя его доля, за исключением переднего края, занята сплош-
ным пневмоническим фокусом красно-серого цвета. Поверхность разреза имеет мра-
морный вид, серый цвет чередуется с красным, в некоторых дольках особенно резко
выражены кровоизлияния. Кровоизлияния в легком в его периферических частях по
местам заметны через утолщенную плевру. Передняя часть верхней правой доли отеч-
на, гиперемирована, пронизана кровоизлияниями, в ней находится несколько брон-
хо-пневмонических фокусов красновато-серого цвета величиною от горошины до
лесного ореха.
С поверхности разреза верхней доли соскабливается много довольно жидкого сли-
зистого красновато-серого сока. Средняя и нижняя доля этого легкого сильно отечны
и гиперемированы.
Левое легкое отечно, в его нижней доле, пронизанной кровоизлияниями, несколь-
ко мелких бронхопневмонических фокусов, таких же, какие найдены в передней час-
ти верхней доли правого легкого. В крупных бронхах, особенно в правом легком, кро-
вянистая слизисто-гнойная пенистая мокрота. Бронхиальные железы с крупный
лесной орех величиной пронизаны кровоизлияниями.
Сердце увеличено раза в полтора, перикардий без изменений, в его полости столо-
вая ложка прозрачной серозной жидкости. Мускулатура сердца буро-красного цвета с
сальным блеском, в папиллярных мышцах тонкие продольные полосы желтого цвета
жирно-перерожденных волокон. Клапаны аорты слегка склерозированы, в дуге аор-
ты рассеяны мелкие склеротические бляшки.
Печень увеличена раза в полтора, довольно плотная, резко застойная, желтоватого
цвета. Правая доля печени в виде купола сильно вдается в грудную полость, на ее по-
верхности замечается некротический участок неправильной четырехугольной формы.
Селезенка буро-красного цвета, увеличена раза в два, довольно плотная, пульпа
соскабливается в небольшом количестве.
698
Чисть 3. Частная неправильная эпидемиология
Почки увеличены в размере, беловато-желтого цвета. Корковый слой утолщен,
мелкие вены переполнены кровью. Капсула снимается легко; в лоханках кровоизлия-
ний нет. Мочевой пузырь содержит небольшое количество мочи, на поверхности сли-
шстои оболочки пузыря несколько точечных кровоизлияний. Предстательная железа
без изменений.
Обе миндалины сине-красного цвета, несколько ’'величены. Лимфатические желе-
зы под челюстью и на шее увеличены очень незначительно. В желудке небольшое ко-
личество жидкого содержимого и кровянистой мокроты. Слизистая оболочка его слег-
ка набухла. В тонких кишках немного жидкого содержимого желтоватого цвета.
Брыжеечные железы в норме.
Полость черепа: черепная крышка толще раза в 1,5 нормальной. Тонкие мозговые
оболочки фиброзно утолщены, особенно по ходу сосудов, под ними значительное
количество прозрачной отечной жидкости. Сосуды основания мозга без изменений.
В боковых желудках около 2 столовых ложек прозрачной, серозной жидкости в каж-
дом. Мозг застоен, отечен.
На срезах, окрашенных полихромной синькой и водным эозином, было найдено
следующее: в легких — правое легкое, верхняя доля (главный первичный пневмонический
фокус): типичная картина чумной пневмонии. Многие альвеолы почти сплошь выпол-
нены массами чумного микроба, таких альвеол особенно много было под плеврой, в
других преобладал клеточный экссудат, и в них находились чумные палочки, разбро-
санные между клетками экссудата; по местам альвеолы были выполнены кровью; в не-
которых атььеолах наблюдались одновременно кучки чумных микробов, клеточный
экссудат и кровоизлияния. Сосуды в альвеолярных стенках резко гиперемированы;
некоторые альвеолы были разорваны излившейся кровью. На срезах, окрашенных по
Граму и по Weigerte’y с дополнительной окраской пикрокармином, не было констати-
ровано ни микробов, красящихся по Граму, ни нитей фибрина. Такая же микроскопи-
ческая картина, в общем, наблюдалась и в небольших бронхопневмонических фоку-
сах нижней доли левого легкого Бронхиальные железы резко гиперемированы,
пронизаны кровоизлияниями и содержали массы микробов, которые в виде причуд-
ливых сплошных тяжей пронизывали железу во всех направлениях, особенно много
микробов было в корковом слое желез. Миндалевидные железы сильно гиперемиро-
ваны, но не имели вида первичного чумного бубона, в фолликулярном аппарате их
находились лишь кое-где отдельные короткие бациллы, не окрашивающиеся по Гра-
му, зато в складках слизистой оболочки встречались лимфатические сосуды, сплошь
набитые палочковидными микробами, не окрашивавшимися по Граму, по всей веро-
ятности, чумными. На основании этой микроскопической картины можно заключить,
чю миндалины не представляли первичного места внедрения чумного бацилла и что
они лишь реагировали на заразное начало, выделяемое из легких с мокротою.
В печени на срезах возбудитель чумы не удалось обнаружить. Печеночные клетки
были жирно перерождены. Вены печени были переполнены кровью. В почках найде-
на картина резко выраженного паренхиматозного нефрита, нигде в сосудах (в клубоч-
ках) микроорганизмов не было найдено В сердечной мышце на срезах видно перерож-
дение мышечных волоконец и отложение бурого пигмента. В папилярных мышцах
полному жировому перерождению были подвергнуты тяжи волоконец, что резко бро-
салось в глаза при наружном осмотре сердца.
Вскрытие подтвердило прижизненную диагностику первичной чумной пневмо-
нии, осложненной паренхиматозным нефритом и паренхиматозным и жировым пе-
рерождением печени и сердца.
Инфицирование мелкодисперсным аэрозолем. Возможно в случае применения тер-
рористами специальных рецептуру возбудителя чумы. Поражения мелкодисперсны-
3.2. 4vMa
699
ми аэро юлями смоделированы в опытах на животных. Приорите! в таких исследова-
ниях принадлежит русскому ученому В. И. Госу (см. разд. 1.5). Впоследствии его дан-
ные, полученные на мышах, были многократно воспроизведены западными исследо-
вателями с использованием других животных, включая приматов. Ниже я приведу их
в сравнении с более поздними и детализирующими данными М. J. Finegola (1968),
полученными на макаках-резус (Масаса mulatto).
По данным В. И. Госа, воспалительные изменения в альвеолах и бронхах на на-
чальной стадии развития инфекции у мышей (до 12 ч) были незначительны. Реакция
со стороны бронхов выражалась в основном в усилении секреции слизистой оболоч-
ки и некотором перибронхиальном скоплении лейкоцитов. Альвеолярная ткань толь-
ко кое-где давала более или менее отчетливую картину воспаления (присутствие в аль-
веолах более значительного числа клеток, состоящих из слущенного эпителия и белых
кровяных телец).
М. J. Finegola обнаружил, что на этой стадии инфекции у макак каких-либо кли-
нических признаков инфекции не наблюдалось. Животные казались внешне здоро-
выми. В легких обезьян при макро- и микроскопическом исследовании не выявлено
патологических изменений. У части животных отмечали повышение температуры до
40,0—40,5°С ме:кду 60 и 72 ч и незначительное тахипноэ (60—70 дыханий/мин). В дру-
гих отношениях они были клинически здоровы без каких-либо признаков апатии или
потери аппетита. Но при аутопсии соотношение веса легкого и веса тела одного из
животных, превышало среднюю нормальную величину, что свидетельствовало о на-
чавшемся отеке альвеол. В срезах тканей периферических бронхов, бронхиол, альве-
ол, венул, вен и лимфатических сосудов ни микроорганизмов, ни структурных измене-
ний в этот период обнаружено не было даже с использованием электронно-микроско-
пическою исследования.
В дальнейших ста тиях воспаления (24,48 и 72 ч после инфицирования животного)
В. И. Гос обнаружил, что воспалительные явления нарастали не одинаково. Рас-
ширение кровеносных сосудов достигало значительной степени. Благодаря расш ирен-
ным капиллярам, границы альвеол резко выступали (особенно при дополнительной
окраске эозином). Через 24 ч был заметен очаговый характер воспаления. В экссудате
альвеол кое-где наблюдалась значительная примесь красных кровяных телец. В от-
дельных участках легочной ткани образовывались мельчайшие геморрагии. Скопле-
ния лейкоцитов вокруг сосудов продолжали увеличиваться, тогда как лимфатические
узелки паренхимы легкого оставались в том же виде, как через 12 ч после заражения.
В бронхах же воспалительные явления прогрессировали очень незначительно (24 ч).
Тяжесть течения болезни не соответствовала тяжести поражения легких.
У каждой из 4 обезьян, забитых через 72 и 96 ч, М. J. Finegola так же отмечал пнев-
монию, причем у одной из, них, забитой через 72 ч, наиболее обширная пневмония
представляла собой сливные очаги серого опеченения правой нижней доли с централь-
ным разжиженным некрозом этого участка; лежащая над ним плевра была тусклой и
непрозрачной с геморрагическими очагами. В левой верхней доле легкого этой обе-
зьяны обнархжили участок с повышенной плотностью и избыточным содержанием
жидкости. Аналогичные очаги, сходные с ранними признаками долевой пневмонии,
отмечали еще у 3 животных.
Поданным В. И. Госа, с наибольшей интенсивностью при поражении аэрозолем
чумы процесс воспаления шел в альвеолах. Группы альвеол образовывали воспалитель-
ные узелки, которые постепенно распространялись на все большие участки легкого,
сливаясь межчу собой и давая пеструю картину воспаления (48 ч). Рядом с группой
700
Часть 3. Частная неправильная эпидемиология
альвеол, теряющей уже альвеолярную структуру, попадались альвеолы с начальными
явлениями воспаления. В местах их наибольшего воспаления особенно ясно высту-
пала разница в степени воспаления альвеолярной ткани и бронхов: среди резко из-
мененных альвеол, переполненных экссудатом, выступали просветы бронхов и брон-
хиол с едва измененным эпителием и незначительным количеством экссудата (48 ч).
Через 72 ч явления воспаления становились еще резче. Очаги воспаления увели-
чивались, попадались узелки, распространяющиеся на протяжении 10—20 альвеол в
диаметре, причем форма их разнообразная, то округлая, то продолговатая или непра-
вильно угловатая. Но встречались и явления, не наблюдаемые ранее. Экссудат, зак-
люченный в альвеолах, кое-где принимал диффузную бледную окраску гематокси-
лином, содержал мало форменных элементов крови. Местами тянулись полосы не-
правильной формы таких диффузно-окрашенных масс, т. е. наблюдалось явление
нарастающего отека. Во многих местах легкого были заметны капиллярные геморра-
гии, в экссудате альвеол примеси крови. Соединительная ткань, окружающая брон-
хи и сосуды, не принимала активного участия в процессе воспаления. Ее клетки всю-
ду сохраняли свой вид. Среди воспалительных фокусов встречались бронхи до альве-
олярных ходов включительно, в которых воспалительные явления ничтожны и
которые совершенно не содержали экссудата.
М. J. Finegola, так же как и В. И. Гос, обнаружил обширные альвеолярные отеки,
содержащие огромное количество ми кробных клеток и вариабельной лейкоцитарной
инфильтрациеи перегородок и альвеол (рис. 3.43, А и Б).
В некоторых участках альвеолярных перегородок капилляры содержали очень
незначительное количество полиморфноядерных нейтрофилов, но на других участ-
ках, отдельные альвеолы были заполнены этими клетками. Первоначально в стенках
альвеол никаких изменений не было обнаружено, но после исследования электрон-
ных микрофотографий появилась возможность с помощью световой микроскопии
установить наличие подэпителиальных пузырьков на участках отека (рис. 3.43, В).
Подэпителиальные пузырьки представляли собой круглые возвышения стенки
альвеол диаметром 7—12 мкм, содержащие эозинофильную жидкость. Их природу
можно было установить только с помощью электронного микроскопа. Периваскуляр-
ные и перибронхиальные лимфатические сосуды вблизи ранних поражений оказались
растянутыми в результате заполнения жидкостью. Кроме подэпителиальных пузырь-
ков альвеол характерными признаками, прогрессирующей чумной долевой пневмо-
нии, вызванной мелкодисперсным аэрозолем Y. pestis, на гистологии являются: аль-
веолярные уплотнения (по В. И. Госу, «воспалительные узелки»), вызванные мигра-
цией полиморфноядерных нейтрофилов и небольшого количества одноядерных
клеток; обширная инфильтрация перегородок и междольчатой, перибронхиальной и
периваскулярной соединительной ткани этими же клетками; значительный рост чум-
ных палочек в альвеолах и лимфатических сосудах; очаги воспаления и некроза эпи-
телия бронхиол. В центрах сливных участков пневмонии отмечали значительные ге-
моррагии и очаговый некроз альвеолярных перегородок (рис. 3.43, Г).
Срезы участков альвеолярного отека, обнаруженного при световой, микроскопии,
выявили обилие чумных палочек внутри альвеол, однако непосредственно в тканях
ни одного чумного микроорганизма отмечено не было. При микроскопии выяатялось
много участков без каких-либо морфологических изменений, несмотря на наличие
микроорганизмов (рис. 3.44, А).
По мере развития пневмонии интерстициальный отек становился белее обшир-
ным и распространялся от околостволовой зоны на тонкие участки альвеолярных
3.2. Чума
701
Рис. 3.43. Патоморфологические изменения в легком обезьяны, пораженной мелко-
дисперсным аэрозолем возбудителя чумы, различаемые с помощью оптического мик-
роскопа. А. Легочный отек Адвентиция вены, перивискулярные лимфатические со-
суды и многие альвеолы растянуты эозинофильной жидкостью. Заметна лейкоцитарная
инфильтрация и очаговое кровоизлияние (эритроциты) Окраска гематоксилин-эози-
ном. Увеличение в 68 раз. Б. Ранняя лобулярная пневмония. Альвеолы и альвеолярные
ходы заполнены эозинофильной жидкостью, содержащей большое количество У. pestis,
Лейкоциты обнаружены в умеренных количествах в перегородках и альвеолах. Пере-
городки интактны. Окраска гематоксилин-эозином. Увеличение в 210 раз. В. Эпите-
лиальные пузырьки. Два заполненных жидкостью пузырька выступают внутрь альве-
ол из перегородки Интерстициальная локализация жидкости заметна при электронной
микроскопии. Окраска гематоксилин-эозином Увеличение в 500 раз. Г. Некротизи-
рующая пневмония. Структура альвеол стерта. Имеет место огромное количество ней-
трофилов, в периваскулярных лимфатических сосудах заметны скопления У pestis.
Окраска гематоксилин-эозином. Увеличение в 210 раз. По М. J. Finegola (1968)
Рис 3.44 Патоморфологические изменения в легком обезьяны, пораженной мелко-
дисперсным аэрозолем возбудителя чумы, различаемые с помощью электронного мик-
роскопа. А. Несколько Кpestis находится внутри альвеолы около перегородки, которая
выглядит интактной. Увеличение в 9000 раз. Б Миграция лейкоцитов. Пространство
(стрелки) в капилляре альвеолярной перегородки занято моноцитом, часть которого
остается в просвете капилляра, тогда как другая его часть находится в интерстициаль-
ном пространстве. Частицы угля, введенные внутривенно, сопровождают лейкоциты
вследствие повреждения эндотелия Увеличение в 16100 раз. По М. J. Finegola (1968)
702
Часть 3. Частная неправильная эпидемиология
перегородок. В эндотелии капилляров обнаружили незначительные очаговые цитоп-
лазматические изменения на участках отека. Прогрессирующее поражение эндотелия
наблюдали на участках некроза, где были вовлечены все элементы ткани. Эти изме-
нения представляли собой набухание клеток, помутнение материала цитоплазмы,
образование вакуолей митохондрий и деструкцию клеточных оболочек.
С помощью светового и электронного микроскопов на некоторых участках отме-
чали экстравазацию эритроцитов (выход из сосудов в ткани). Миграцию лейкоцитов
можно было проследить от капилляров и более часто от венул в промежуточное про-
странство и отсюда в альвеолы (см. рис. 3.44, Б).
Бронхиальные лимфатические узлы были увеличены, с участками жидкого некроза
(liquefactive necrosis) и геморрагиями. Повреждения печени минимальны. На поверх-
ности почек были заметны геморрагические фокусы, на срезах обнаружен тромбоз
гломерулярных сосудов. Так же были тромбированы отдельные афферентные артери-
олы и интертубулярные и медуллярные капилляры. При выраженном тромбозе наблю-
дался кортикальный некроз.
Картина воспаления альвеолярной ткани, вызванная инфицированием мелкодис-
персным аэрозолем возбудителя чумы, установленная В. И. Госом и М. J. Finegola,
отличается от той, которую описывают в природных очагах чумы при вспышках пер-
вично-легочной чумы и относится к «невозможной клинической форме болезни» (см.
разд. 2.3.1). При типичных чумных бронхопневмониях, характерных для воздушно-
капельной передачи чумных бацилл, воспалительный процесс в бронхах и в альвео-
лярной ткани выражен одинаково сильно, причем часто воспалительный процесс с
бронхов переходил на окружающие альвеолы. При чумной пневмонии, развивающей-
ся вследствие проникновения возбудителя непосредственно в альвеолы, на первое
место выступает воспалительный процесс в альвеолах, развивается пневмония. Из-
менения же бронхов незначительны. Характер пневмонии десквамативный, причем
интерстициальная ткань не принимает активного участия в процессе.
Обобщая результаты своих экспериментов, Гос сделал вывод, что чумная пневмо-
ния (под таковой он понимал пневмонию, полученную в результате инфицирования
мелкодисперсным аэрозолем возбудителя), имеет характер катаральной узелковой
пневмонии в чистом виде, т. е. без значительного поражения бронхов и без образова-
ния фибрина. Характер экссудата — десквамативный с примесью лейкоцитов и эрит-
роцитов. Последние появляются в альвеолах из расширенных ad maximum капилля-
ров. Этим объясняется кровавый характер мокроты при чумной пневмонии. Быстроту
развития процесса он объяснил отсутствием интерстициальных явлений.
Поражение только легочной ткани при инфицировании человека мелкодисперс-
ным аэрозолем возбудителя чумы, показанное В. И. Госом и некоторыми другими
исследователями, нельзя понимать буквально как единственный возможный исход
такого нападения. В экспериментах, выполненных на Macacafascicularis, R. Wn Andel
et al. (2008) показали дозозависимый эффект ингаляционных поражений возбудите-
лем чумы при среднем размере частиц аэрозоля 2—5 мкм. ED50 (т. е. доза, при которой
у экспериментального животного обнаруживается лихорадка и другие клинические
признаки) для М. fascicularis составила около 66 колониеобразующих единиц (КОЕ).
Если ингалирование экспериментальным животным 479 КОЕ возбудителя чумы
(7 ED50) вызывало у них мультифокальную двустороннюю гнойную пневмонию и сред-
невыраженную спленомегалию, то при дозе 759 КОЕ (11 ED50) в инфекционный про-
цесс вовлекались трахобронхиальные лимфатические узлы, а при дозе 3050 КОЕ у
обезьян развивались фибринозные плевральные отеки. Это важное наблюдение
3.2. Чума
703
R. Van Andel et al. (2008), показывающее зависимость патологоанатомической струк-
туры первичных поражений мелкодисперстным аэрозолем возбудителя чумы от ин-
галированной дозы, при массовых жертвах биотеррористического нападения позво-
ляет определять относительную близость людей к источнику аэрозоля и направление
движения аэрозоля.
Теперь сопоставим патологоанатомическую картину чумы, наблюдаемой при ес-
тественном инфицировании в очагах (воздушно-капельная инфекция) и искусственно
созданным мелкодисперсным аэрозолем (табл. 3.13).
Таблица 3.13
Сопоставление патологоанатомической картины легочной чумы при естественном и
искусственном инфицировании*
Поражение Естественное заражение в очагах чумы Искусственно созданным аэрозолем
Обший характер поражения Вторичное (гематогенное) развитие нисходящего бронхита, клиниче- ская картина соответствует тяжести поражения легких Первичное поражение легких, клини- ческая картина не соответствует тяже- сти поражения легких
Слизистая верхних дыхательных путей Катаральные изменения, эпителий и подслизистая ткань «пропитаны» чумными палочкам Не вовлечена
Лимфоидная ткань миндалин Густая инфильтрация возбудителем чумы, воспалена Не вовлечена
Характер пневмонии Долевая форма пневмонии с пре- имущественным поражением верх- них долей В основном катаральная узелковая пневмония. Преимущественно пора- жаются нижние доли легкого. При гистологическом исследовании в альвеолах могут быть обнаружены подэпителиальные пузырьки и др. изменения, характерные для пора- жения мелкодисперсным аэрозолем (см. выше)
Плевра Обычно поражена (фибринозный налет) Вовлекается только при очень больших дозах заражения
Пери бронхиальная ткань Скопления чумных палочек, осо- бенно вокруг перибронхиальных кровеносных сосудов; перибронхи- альные узлы могут быть значитель- но увеличены, спаяны в пакет, отечны, геморрагичны, окружают трахею, на разрезе мозговидной консистенции. Отдельные лимфа- тические узлы величиной в сливу, розового цвета, с кровоизлияниями Воспалительные процессы мало выра- жены. Их выраженность зависит от доз заражения
* Составлена по работам В. Госа (1907), Г С. Кулеши (1912, 1924), М J. Finegola (1968) и
R. Van Andel et al. (2008)
Диагностика искусственного поражения возбудителем чумы. Принципы диагностики
искусственного поражения возбудителем чумы те же, что описаны в подразделе 3.1 для
поражения возбудителем сибирской язвы. В их основе лежит выявление неправиль-
ной эпидемиологии и невозможной клиники болезни. Но есть и некоторые особен-
ности, характерные для чумы. Естественной вспышке чумы среди людей (в бубонной,
704
Часть 3. Частная неправильная эпидемиология
септической и легочной формах), даже ограничившейся единичными случаями бо-
лезни, должны предшествовать чумные эпизоотии среди грызунов. Должна просле-
живаться и эпидемическая цепочка от местностей, где чумные эпизоотии замечены,
к людям, инфицированным возбудителем чумы. Появление больных чумой грызунов
и инфицированных Y. pestis блох на территории, длительное время «свободной от
чумы», может свидетельствовать об активизации древнего и забытого очага чумы.
Поэтому, прежде чем делать предположение о возможности ее искусственного про-
исхождения, необходимо по историческим источникам убедиться в том, что вспыш-
ки чумы на данной территории имели место в прошлом. Крупным естественным
вспышкам легочной чумы предшествуют локальные вспышки бубонной и легочной
чумы на территориях активизировавшихся природных очагов чумы. «Движение» ле-
гочной чумы в таких случаях прослеживается по больным легочной чумой, выявлен-
ным на транспортных коммуникациях. Обычно крупные естественные вспышки ле-
гочной чумы не имеют единого центра развития и представляют собой растянутый во
времени процесс распространения бронхолегочной чумы. Крупные эпидемии бубон-
ной и легочной чумы имеют длительный период (до нескольких десятилетий) неболь-
ших вспышек-предшественников, свидетельствующих о постепенном «разогреве»
природного очага чумы.
Подозрения об искусственном характере вспышки бубонной чумы обоснованы
при обнаружении в общественных местах блох, инфицированных Y. pestis, связь ко-
торых с больными чумой грызунами установить не удается. Такие вспышки ограни-
чиваются единичными заболевшими.
В природных очагах чумы вторично-легочная чума развивается на фоне вспышек
бубонной чумы. Первично-легочная чума, возникшая у человека в результате воздуш-
но-капельной инфекции, в основном развивается как нисходящая бронхопневмо-
ния. В инфекционный процесс вовлекаются слизистая верхних дыхательных путей,
лимфоидная ткань миндалин, плевра, перибронхиальные лимфатические узлы. На
рентгенограммах выявляются затемнения долей легких и признаки экссудативного
плеврита. В отличие от нее, случаи первично-легочной чумы, развившиеся в резуль-
тате непосредственного экспонирования человека к мелкодисперсному аэрозолю воз-
будителя чумы, преимущественно будут представлены «невозможными клинически-
ми формами болезни» (см. разд. 2.3.1) — очаговыми поражениями в нижних долях лег-
ких (очаговые затемнения легочной ткани на рентгенограмме) без вовлечения в
инфекционной процесс слизистой верхних дыхательных путей, лимфоиднай ткани
миндалин, листков плевры, перибронхиальных лимфатических узлов. Тяжесть таких
поражений у отдельного человека (при прочих равных условиях) будет зависеть от ин-
галированной им дозы возбудителя чумы, вирулентности штамма и дисперсности
аэрозоля. Вторичные случаи первично-легочной чумы в очаге поражения мелкодис-
персным аэрозолем, т. е. возникшие в результате воздушно-капельной передачи воз-
будителя болезни от людей, заболевших легочной чумой в результате экспонирования
к аэрозолю, будут протекать как типичная чумная бронхопневмония.
Наличие двух типов течения первично-легочной чумы (как очаговой пневмонии
и как бронхопневмонии) позволит выявить лиц, ставших непосредственными жерт-
вами биологического нападения, и лиц, заразившихся чумой воздушно-капельным
путем уже от них.
Иммунопрофилактика. Для специфической профилактики чумы в России приме-
няется вакцина чумная живая лиофилизат и ваю ,ина чумная живая табле тированная.
Вакцина чумная живая лиофилизат выпускается двумя предприятиями: ФГУЗ Став-
3.2. Чума
705
НИПЧИ Роспотребнадзора и ФГУ «48 ЦНИИ Минобороны России». Таблетирован-
ную вакцину выпускает ФГУ «48 ЦНИИ Минобороны России». Вакцинация лиофи-
"изованной вакциной осуществляется накожным, подкожным и ингаляционным
способами Ревакцинация обеими вакцинами осуществляют через год. При неблаго-
приятной эпидемиологической обстановке — через 6 мес. Дозы живой вакцины при
иммунизации взрослых пациентов составляют ЗООхЮ6 живых микроорганизмов в
0,5 см3 и 3 х 109 живых микроорганизмов в 0,15 см3 для подкожного и накожного спо-
собов применения соответственно. При ингаляционной иммунизации доза должна
доставлять (5±3)х 106 живых микроорганизмов штамма EV. Последний способ счита-
ется патогенетически более корректным для предупреждения первично-легочной
чумы. Российские вакцины защищают от ингаляционного заражения возбудителем
чумы. Американская инактивированная чумная вакцина USP не эффективна в отно-
шении воздушно-капельного и ингаляционного заражения У. pestis. В США выпуск
этой вакцины был прекращен в 1998 г. По мнению многих специалистов, эра убитых
вакцин заканчивается (Вывалов А. А., 2011).
Лечение. Лечение чумы антимикробными средствами было начато в 1938 г. с суль-
фонамида (сульфамид), а в августе 1947 г. русскими врачами Н. И. Николаевым,
Д. А. Федориновым и В. И. Гороховым для лечения легочной чумы впервые исполь-
зован стрептомицин. Применение стрептомицина привело к резкому снижению ле-
тальности у людей На сегодняшний день показатель летальности при чуме в миро-
вом масштабе снизился до 8 %. Тремя основными антимикробными препаратами,
рекомендованными для лечения чумы, являются стрептомицин, тетрациклин и хло-
рамфеникол (левомицетин). Предпочтительным по-прежнему остается стрептоми-
цин, тогда как хлорамфеникол чаше всего применяется для лечения менингита. Из-
за ограниченного запаса стрептомицина в некоторых странах в качестве заменителя
используется гентамицин, и имеются данные, подтверждающие, что гентамицин или
его комбинация с тетрациклином могут служить эффективными средствами лечения
чумы у людей. Фторхинолоны рассматриваются в качестве альтернативного средства
дтя лечения чумы на основании результатов исследований in vitro и экспериментов на
мышах, в которых их эффективность оказалась аналогичной таковой аминогликози-
дов. Однако за исключением одного единственного случая чумы, которая была успеш-
но вылечена при помощи ципрофлоксацина, нет сведений об эффективности этого
препарата. Назначение одного, двух или трех препаратов зависит от тяжести симпто-
мов. Шансы больного на выздоровление непосредственно связаны со своевременно-
стью антибиотикотерапии, особенно при более тяжелых формах чумы, легочной и сеп-
тицемической.
Экстренная неспецифическая профилактика предусматривает применение суль-
фонамида, триметоприм-сульфаметоксазола или тетрациклина. Назначается лицам,
тесно контактирующим с больными легочной чумой, или родственникам больных
бубонной чумой, живущим с ними вместе или по соседству.
В случае биологического нападения с использованием аэрозолированного возбу-
дителя чумы рабочая группа по противобиологической защите гражданского населе-
ния США рекомендовала при лечении пострадавших в специально отведенном для
этого месте (т. е. при небольших количествах больных, нуждающихся в лечении) в
первую очередь применять стрептомицин или гентамицин, а уже в качестве альтерна-
тивных средств — доксициклин, ципрофлоксацин или хлорамфеникол. При массо-
вом поражении рекомендовано пероральное применение доксициклина, тетрацикли-
на или ципрофлоксацина. Предотвратить передачу инфекции от человека к человеку
706
Часть 3. Частная неправильная эпидемиология
больными легочной чумой можно при помощи стандартных мер изоляции, по край-
ней мере, в течение четырех дней после начала антибиотикотерапии.
Хотя цефалоспорины иногда относили к числу неэффективных лечебных средств
против чумы у людей, но без документального подтверждения. Кроме того, в иссле-
дованиях в условиях in vitro и в экспериментах на мышах эффективность этого анти-
микробного препарата оказалась аналогичной таковой фторхинолонов и амино-
гликозидов.
3.3. ТУЛЯРЕМИЯ
Микробиология F. tularensis. Факторы патогенности. Происхождение и моле-
кулярная эпидемиология возбудителя туляремии. Экология и естественная
эпидемиология F. tularensis. Восприимчивость и чувствительность живот-
ных к возбудителю туляремии. Инфицирующие дозы для животных и людей.
Иммунитет. Устойчивость F. tularensis во внешней среде и в трупах погиб-
ших животных. Антибиотикорезистентность. Обнаружение генетически из-
мененных штаммов F. tularensis. Патогенез. Клиническая картина туляремии
у людей при естественном инфицировании F. tularensis. Моделирование
ингаляционного заражения F. tularensis. Диагностика искусственного пора-
жения возбудителем туляремии. Иммунопрофилактика. Лечение.
Туляремия (чумоподобная болезнь, малая чума, заячья лихорадка, кроличья ли-
хорадка, лихорадка леммингов, лихорадка оленьей мухи), встречающаяся в основном
в Северном полушарии, опасная сапрозоонозная инфекционная болезнь людей и
животных, вызываемая неспорообразующей бактерией Francisella tularensis. Проте-
кает с наличием специфического лимфаденита и поражениями различных органов
(глаза, легкие, ЖКТ и др.). Возбудитель туляремии впервые обнаружен в 1911 г. у гры-
зунов в округе Туляре (Калифорния) американскими исследователями Ч. Мак Коем
(G. W. McCoy) и Ч. Чепиным (С. W. Chapin). В природных условиях туляремией зара-
жаются в результате контакта кожных или слизистых покровов человека с тканями
или жидкостями тела инфицированных животных или при укусах инфицированных
кровососущих членистоногих насекомых. Реже болезнь развивается вследствие вды-
хания контаминированной F. tularensis пыли, потребления контаминированной пищи
или воды. Заражение аэрозолем вызывает тифоидную или легочную форму туляре-
мии. Больной туляремией не представляет опасности для окружающих. Российские
санитарно-эпидемиологические правила СП 1.3.2322-08 относят возбудитель туля-
ремии ко II группе патогенности. По степени важности для национальной безопасно-
сти США F. tularensis относится к биологическим поражающим агентам категории А
(см. табл. 2.2). Туляремия не является заболеванием, подлежащим регистрации ВОЗ,
поэтому мировая заболеваемость не известна.
С конца 1920-х гг. возбудитель туляремии из-за небольшой инфицирующей дозы
для человека — примерно 10 КОЕ, считается перспективным агентом БО. В СССР
возбудитель туляремии выделен в 1926 г. сотрудниками Астраханской противочумной
станции С. В. Суворовым, А. А. Вольферцом и М. А. Воронковой. В Германию возбу-
дительтуляремии попал в 1928 г. из музея культур института «Микроб» (Саратов), бла-
годаря усилиям германского подданного Ганса Цейсса (Heinz Zeiss), заведующего
музеем живых культур Института экспериментальной терапии и контроля сывороток
3.3. Туляремия
707
и вакцин (Москва), возглавляемого Л. А. Тарасевичем (1868—1927)1. Цейсс находил-
ся в дружеских отношениях с наркомом здравоохранения РСФСР Н. А. Семашко
(1874-1949). Из 91-й опубликованной научной работы Института Тарасевича за пери-
од с января 1925 г. по январь 1930 г., Цейсс был автором или соавтором 15 работ, 8 из
них были опубликованы в германских научных журналах. Однако альтруистом этот
последователь идей пангерманизма, а с начала 1930-х гг. — активный нацист, не был.
Дружба с Цейссом стала причиной смешения в 1930 г. с должности Семашко. В 1945 г.
в восточной оккупационной зоне Германии Цейсс был задержан сотрудниками НКВД.
Его осудили за шпионаж в пользу Германии на 25 лет. В 1949 г. Цейсс умер во Влади-
мирской тюрьме (Чельный А. А., 2010).
В лабраториях Форт-Детрика поражающие свойства F. tularensis изучались с 1944 г.
В 1950—1960-х гг. на вооружении армии США состояли специальные рецептуры на
основе возбудителя туляремии и снаряженные ими боеприпасы.
Наиболее вероятно применение F. tularensis посредством использования линейных
и многоточечных источников и диверсионными (террористическими) методами.
Микробиология F. tularensis. По морфологическим свойствам между микробами
различных подвидов (рас) принципиальные различия отсутствуют. F. tularensis имеет
вид очень мелкого кокка или короткой палочки, неподвижна. Наиболее часто встре-
чающимися размерами можно считать 0,3-0,5 мкм, однако обнаруживаются и мель-
чайшие формы диаметром до 0,1 мкм. Строгий аэроб. Растет только на специальных
средах (желточная среда Мак Конки, агар с цистином и кровью и др.) и в куриных
эмбрионах (Олсуфьев Н. Г. с соавт., 1960).
При культивировании на искусственных питательных средах происходит аттенуа-
ция туляремийных бактерий и превращение их из вирулентной S-формы, в авирулент-
ную и неиммуногенную R-форму. Колонии становятся менее выпуклыми и более
прозрачными. Клетки утрачивают Vi-антигенный комплекс, становятся несколько
крупнее. Вакцинный штаммы представляют собой промежуточную форму изменчи-
вости от вирулентной S-формы к авирулентной R-форме и могут быть обозначены как
SR-вариант. Обладают остаточной вирулентностью для чувствительных к возбудите-
лю туляремии животных (первая группа; см. далее). По строению колоний ближе к
S-форме (Олсуфьев Н. Г., Руднев Г. П., 1966).
F. tularensis легко окрашивается всеми красками, обычно применяемыми в лабо-
раторной бактериологической практике, но заметно бледнее, чем бактерии многих
других видов. Специфической окраски не имеют. По Граму, окрашиваются негатив-
но. Мазки — отпечатки органов погибших животных целесообразно окрашивать по
методу Романовского — Гимза. В таких препаратах туляремийные микробы отлича-
ются от посторонней микрофлоры более нежной фиолетовой окраской. Степень про-
явления изменчивости морфологических свойств у туляремийных микробов в окра-
шенных мазках зависит от условий развития микробной популяции и их штаммовых
особенностей. Выраженный полиморфизм они проявляют чаще в культурах, свеже-
выделенных из организма животных, чем при длительных пересевах на искусствен-
ных питательных средах. Преобладающей формой в культурах, сформировавшихся на
плотных питательных средах, являются кокки и овоиды. После выращивания в жид-
ких питательных средах с аэрацией и перемешиванием большинство микробов при-
обретает палочковидную форму (Золотарев А. Г. с соавт., 2006).
1 Первый случай заболевания человека туляремий в Германии имел место в 1928 г., но его
этиология установлена была только в 1931 г. (Олсуфьев Н. Г., Дунаева 1. Н., 1970).
708
Часть 3. Частная неправильная эпидемиология
Возбудитель туляремии отличается повышенными потребностями к составу пита-
тельных сред. Колонии клеток F. tularensis можно получить на агаре, содержащем ци-
стеин или обогащенном естественными субстратами — яичным белком, кровью и т. п.
(рис. 3.45).
Рис. 3.45. Макроскопические изображения колоний F. tularensis. А. Колонии ви-
рулентного штамма Schu, выращенного в течение 48 ч на FT-arape (увеличение
в 56 раз). Б. Колонии вирулентного штамма 170, выращенного в течение 96 ч
на FT-arape. В. Колонии штамма Schu в падающем свете. Г. Колонии штамма
Schu в проходящем свете. Увеличение в 13 раз. По А. Г. Золотареву с соавт. (2006)
На таких средах хорошо оформленные микроколонии образуются после 40-48 ч
роста при температуре 36—38°С. По форме они напоминают полусферу или уплощен-
ный конус с ровными краями и гладкой поверхностью. На вершине у них могут рас-
полагаться округлые зоны центрального роста (рис. 3.45, А). Через 72—96 ч инкуби-
рования микробы F. tularensis формируют колонии (S-форма) беловатого цвета с
голубоватым оттенком, круглые выпуклые с ровными четкими краями и гладкой по-
верхностью. Диаметр изолированно расположенных колоний может достигать 2—3 мм
(рис. 3.45, Б). При микроскопическом исследовании в падающем свете они белые и
сферические (рис. 3.45, В), в проходящем и скошенном свете — желто-зеленые или
коричневато-зеленые. По внешнему виду они могут быть двух разных видов: одни из
них отличаются наличием куполообразного центра и темным цветом (рис. 3.45, Г),
другие выглядят более светлыми и плоскими без выступающей центральной части.
Края у обоих вариантов колоний ровные, поверхность гладкая, нередко покрыта не-
растворимыми компонентами питательной среды (Золотарев А. Г. с соавт., 2006).
Способность сбраживать углеводы и спирты у микроба туляремии ограничена и
может быть достоверно выявлена лишь при применении специальных плотных сред
с ограниченным содержанием белка и точно определенным pH. Среды Гисса для этой
цели непригодны, так как в них туляремийные бактерии не размножаются. Туляре-
мийные бактерии ферментируют до кислоты глюкозу, мальтозу, в ряде случаев леву-
лезу и маннозу, а американские штаммы — дополнительно еще глицерин. Лактозу,
сахарозу, рамнозу, маннит и ряд других веществ туляремийные бактерии не фермен-
тируют (Олсуфьев Н. Г, Руднев Г. П., 1966).
3.3. Туляремия
709
F. tularensis образует сероводород, не образует индол и обладает способностью
редуцировать некоторые красители, например, тионин, метиленовую голубую, мала-
хитовую зеленую и др. У туляремийных бактерий обнаружены ферменты: каталаза,
глутаминаза, аспарагиназа, дезаминаза и трансамидаза, а у вирулентных американ-
ских штаммов — еще цитруллинуреидаза (Олсуфьев Н. Г., Руднев Г. П., 1966).
Факторы патогенности. У микробиологов не вызывает разногласий предположе-
ние о том, что патогенность F. tularensis зависит от ее способности выживать в макро-
фагах. Но детали этого процесса изучены плохо. Из факторов патогенности F. tularen-
sis, играющих роль в патогенезе туляремии, трудно выделить такие, которые бы под-
ходили под их традиционное понимание как факторов, участвующих в колонизации
или инвазии тканей. У возбудителя туляремии обнаружены фибринолизины и ней-
раминидазы, найти другие секретируемые молекулы, наличие которых способно
объяснить патогенез туляремии, пока не удалось (Ellis J. et al., 2002).
Среди факторов патогенности возбудителя туляремии R. Pechous et al. (2009) вы-
деляют поверхностные структуры (капсула, ЛПС, пили), остров патогенности, транс-
крипционные регуляторы генов и дополнительные факторы вирулентности.
Капсула защищает F. tularensis от факторов комплемента, поскольку некапсули-
рованные штаммы чувствительны к лизису клеток неиммунной сывороткой (Sands-
trom G. et al., 1988).
ЛПС F. tularensis не обнаруживает свойств классического эндотоксина. Он обла-
дает низкой иммунологической активностью, не может индуцировать интерлейкин-1
из мононуклеарных клеток и слабо индуцирует продуцирование фактора некроза опу-
холи и окиси азота (NO) из макрофагов. ЛПС F. tularensis не взаимодействует с рецеп-
торами ЛПС на макрофагах (Ancuta Р. et al., 1996). Показано, что вирулентность воз-
будителя туляремии зависит от фазовых вариаций ЛПС (Pechous R. et al., 2009).
Пили у возбудителя туляремии обнаружены с помощью электронной микроско-
пии, их роль в вирулентности не изучена (McLendon М. et al., 2006).
Остров патогенности возбудителя туляремии носит название Francisella patho-
genicity island (FP1) и имеет размер приблизительно 30 т. п. о. Он дуплицирован в ви-
рулентных субтипах F. tularensis (tularensis и holarctica). Хромосома маловирулентной
F. novicida содержит только одну его копию. FPI включает приблизительно 17 ORF,
регулирующих различные аспекты вирулентности F. tularensis, в частности генов, ко-
дирующих компоненты системы секреции, сходной с системой секреции VI типа,
описанной ранее у других микроорганизмов (de Bruin О. М. et al., 2007).
Гены, локализованные в пределах FPI, организованны в две основные транскрип-
ционные единицы и индуцируются во время внутриклеточного роста F. tularensis. Они
могут экспрессироваться и вследствие внешнего воздействия, например, ионов железа
или перекиси водорода, или посредством отдельных факторов регуляции, включаю-
щих MglAB, SspA, PmrA, FevR и MigR. Первая транскрипционная единица кодирует
локус генов, обеспечивающий внутриклеточный рост (intracellular growth locus,
iglABCD) F. tularensis. Мутация в пределах любого из этих генов замедляет размноже-
ние Francisella в макрофагах, клетках насекомых и/или в организме мыши. Гены, коди-
рующие белки IglA и IglB, вовлечены в эффекторную секрецию; iglC кодирует белок
с ММ 23 кДа. Хотя точная функция этого белка не была установлена, ijg/C-мутанты
F. tularensis (tularensis, holarctica) и F. novicida почти полностью утрачивают способ-
ность к росту в условиях in vitro и in vivo. В частности, IglC необходим для ингибиро-
вания созревания фагосомы (phagosomal maturation); IglD необходим для внутрикле-
точной репликации Francisella в человеческих макрофагах (Pechous R. et al., 2009).
710
Часть 3. Частная неправильная эпидемиология
Гены pdpA и pdpB соответствуют первым двум генам второй основной транскрип-
ционной единицы (second major transcriptional unit), локализованной в пределах FPI.
Они кодируют гипотетические белки, которые необходимы для проявления вирулент-
ных свойств F. novicida в клетках насекомых, куриных эмбрионах и/или организме мы-
шей. Продукт гена dpA необходим возбудителю туляремии для нарушения процесса
созревания фагосомы, но детально их роль в вируленности Francisella не изучена
(Pechous R. et al., 2009).
Транскрипционные регуляторы участвуют в регуляции как структурных, так и дру-
гих регуляторных генов F. tularensis. Их инактивация проявляется утратой способно-
сти F. tularensis размножаться в фагоцитирующих клетках, но механизмы данного яв-
ления для каждого транскрипционного регулятора различны.
Белок MglAB (macrophage growth locus) супрессирует экспрессию кислой фосфа-
тазы, необходимую F. tularensis для проникновения в макрофаг без респираторного
взрыва. Гены mglA и mglB организованы в оперон и кодируют белки, гомологичные
SspA и SspB Е. coli и Haemophilus spp. SspA представляет собой РНК-полимеразиассо-
циированный белок (RN A polymerase-associated protein) и регулирует экспрессию раз-
личных генов Г. tularensis в ответ на стрессовое воздействие. Мутации в mglA или mglB
замедляют размножение F. tularensis вмакрофагах, амебах (Baron G. S., Nano Е Е., 1у98:
Lauriano С. М. et al., 2004), клетках дрозофил и в организме мышей. Всего же MglA ре-
гулирует экспрессию более чем 100 генов, включая те, которые включены и которые
окружают FPI (Brotcke A. et al., 2006). Разрушение mglA ослабляет способность Fran-
cisella ингибировать созревание фагосомы и проникать в цитозоль клетки, вероятно,
вследствие регуляции генов, имеющих отношение к этому процессу, включая iglABCD
(Pechous R. et al, 2009).
Белок SspA обладает высокой гомологией с MglA. У F. tularensis LVS, оба белка, SspA
и MglA, ассоциированы с РНК-полимеразой. Кроме toi о, SspA и MglAнепосредствен-
но взаимодействуют друг с другом. Ассоциация MglA с РНК-полимеразой требует
участия SspA, т. е. эти белки являются транскрипционными корегуляторами одних и
тех же генов в геноме F. tularensis (Charity J. С. et al., 2007).
ГенpmrA кодируеторфанотвечающий регулятор (orphaned response regulator), гомо-
логичный регулятору ответов (response regulator) PmrA Salmonella enterica серовара
typhimurium. Он регулирует экспрессию большой группы генов, включая гены FPI,
однако не способен регулировать MglA и наоборот (Mohapatra N. Р. et al., 2007).
Ген fevR (Francisella effector of virulence regulation) кодирует гипотетический белок
с неясным механизмом действия, но необходимый для экспрессии MglA/SspA (Brotcke
A., Monack D. М., 2008).
Ген migR (macrophage intracellular growth regulator) — недавно открытый регулятор
экспрессии генов вирулентности F. tularensis 1 VS Кодирует гипотетический белок,
способный регулировать экспрессию igl-локуса у LVS и являющийся позитивным ре-
гулятором транскрипции фактора fevR (Buchan В. W. et al., 2009).
Дополнительные факторы вирулентности возбудителя туляремии включают белок
Аср, функционирующий как кислая фосфатаза. Благодаря Аср F. tularensis проника-
ет в макрофаг без инициирования губительного для него респираторного взрыва (Reilly
Т. J. et al., 1996). АсрА блокирует взрыв респираторной активности эффективнее, чем
ранее описанные кислые фосфатазы других внутриклеточных бактерий (лейшмании,
легионеллы). Его транскрипция регулируется белком MglAB.
Другим локусом, необходимым туляремийному микрооу для выживания в макро-
фаге, является minD. MinD — это 29-кДа белок, чья роль в этом процессе предполага-
3.3. Туляремия
711
ется на основе его гомологии с белками, формирующими насос для удаления токсич-
ных ионов из бактериальной клетки (Anthony L. S. D. et al., 1994).
Происхождение и молекулярная эпидемиология возбудителя туляремии. Таксономи-
ческое положение возбудителя туляремии является сложным и при накоплении но-
вых данных может измениться. Род Francisella (семейство Franciseilaceae) относится
к классу Gammaproteobacteria. Род Francisella эволюционно связан с такими патоген-
ными для людей видами микроорганизмов, как Coxiella burnetii и Legionella pneumophila.
Семейство включает Wblbachia persica, паразита членистоногих и червей, симбионта
Dermacentor andersoni, который был первоначально выделен у лесных клещей Скали-
стых гор, и симбионта мягкого клеща Omithodorous moubata. Большинство этих эндо-
симбионтов членистоногих плохо охарактеризованы. Известно, что W. persica сущест-
вует в мальпигиевых сосудах мягкого клеща Argas arboreus и не считается патогенным
для млекопитающих. F. philomiragia (прежнее название Yersinia philomiragia) отнесена
к роду Francisella на основании сходства последовательностей их 16S рДНК и по ряду
биологических свойств. Считается вирулентной только для лиц с ослабленным им-
мунитетом (рис. 3.46).
- Coxiella burnetii
— Legionella pneumophila
Pseudomonas aeruginosa
----------Escherichia coli
----------Proteus vulgaris
100
Francisella philomiragia
100
—Francisella tularensis
Wblbachia persica
Agrobacterium tumefaciens
Рис. 3.46. Эволюционное древо Franciseilaceae, основанное на анализе последователь-
ностей 16S рДНК. Показывает связь F. tularensis с предполагаемыми членами Francisel-
laceae и с другими близкородственными членами у-подкласса Proteobacteria. Agrobacteria
tumefaciens включена как неродственная группа. По J. Ellis et al. (2002)
Основываясь на генетических маркерах, способности образовывать кислоту из
глицерина, цитруллинуреидазной активности и вирулентности F. tularensis обычно
разделяют на три подвида (subspecies): F. tularensis подвид tularensis, F. tularensis под-
вид holarctica и F. tularensis подвид mediasiatica (табл. 3.14).
Francisella novicida ранее рассматривали как подвид F. tularensis (Олсуфьев H. Г.,
Дунаева Т. Н., 1970); однако проведенный полногеномный анализ однонуклеотидно-
го полиморфизма (wholegenome single-nucleotide polymorphism analysis) показал, что
этот микроорганизм, более вероятно, является самостоятельным видом (Keim Р. et al.,
2007). С тех пор как F. novicida была выделена впервые, сообщалось лишь о шести боль-
ных с подозрением на эту инфекцию, последний случай имел место в Таиланде в 2007 г.
(Leeporn A. et al., 2008).
F. tularensis, подвид holarctica (генотип В), обнаруживается повсюду в Северном
полушарии и в основном вызывает туляремию в Европе. Его делят натрибиовара: био-
вар I (чувствительный к эритромицину); биовар П (устойчивый к эритромицину); и
биовар Japonica (Ellis J. et al., 2002).
712
Часть 3. Частная неправильная эпидемиология
Таксономия и характеристики подвидов F. tularensis*
Таблица 3.14
Современное название Предыдущее название Биовар 16S rPHK- генотип Географическое местон ахождение Активность : и итрулд и н уре идазы Ферментация глицерина Ферментация глюкозы Чувствительность к эритромицину
F. tularensis подвид tularensis F. tularen- sis A; F. tularensis подвид nearartica Непри- меним А Преимущест- венно Северная Америка + + + +
F. tularensis подвид holarctica F. tularen- sis B; F. tularensis подвид palaearctica т В Преимущест- венно Европа, Сибирь, Дальний Восток, Казахстан и Северная Америка — - + +
II В Преимущест- венно Евразия — - +
Japonica А Япония - + + +
F. tularensis подвид mediaasiatica - Непри- меним А Центральная Азия и неко- торые терри- тории бывше- го СССР + + - +
F. novicida F. tularensis подвид novicida Непри- меним Не описан Преимущест- венно Северная Америка Не описана Не описана + Не описана
* По J. Ellis et al. (2002).
F. tularensis подвид tularensis, no 16S rPHK относящийся к генотипу А, может в
будущем разделен на две клады (A.I и А.П), различающиеся по тяжести вызываемой
болезни, географической локализации очагов и способности к трансмиссии (Pechous
R. et al., 2009). Штаммы А.1 (известные так же как A-восточные) обычно обнару-
живаются на востоке Скалистых гор. Штаммы A.I1 (A-западные) встречаются в меж-
горном районе в западной части Соединенных Штатов, ассоциируются с более низ-
кими показателями летальность у людей и являются в настоящее время единственны-
ми штаммами, перенос которых от больных туляремией животных к людям связан с
оленьими слепнями (Chrysops spp.) (Petersen J. M. et al., 2008). До недавнего времени
считали, что F. tularensis подвида tularensis встречается только в Северной Америке,
но обнаружение их в Европе показывает, что подвид распространен значительно шире,
чем предполагалось. Распределение штаммов генотипа А и генотипа В в Соединен-
ных Штатах является в основном перекрывающимся, с некоторыми географическими
3.3. Туляремия
713
различиями. Что касается экологии, то эти два подвида сохраняются в разных, но не
вполне определенных циклах, причем штаммы типа А часто ассоциируются с зайце-
образными, а штаммы типа В обычно ассоциируются с грызунами и водной средой
(Petersen J. М. et al., 2008).
Третий подвид — Mediaasiatica, обнаруживают преимущественно в среднеазиат-
ских республиках бывшего СССР. Эти штаммы обладают активностью цитруллину-
реидазы, способны ферментировать глицерин и менее вирулентны, чем штаммы под-
вида Tularensis (Ellis J. et al., 2002). К этому же подвиду относится живой вакцинный
штамм 15 НИИЭГ, используемый для производства вакцины в России.
Штаммы туляремийного микроба одной и той же географической расы (подвида),
выделяемые в природных очагах от диких позвоночных и беспозвоночных животных,
а также из объектов внешней среды, обладают в высокой степени сходными призна-
ками, включая вирулентность для лабораторных животных (Олсуфьев Н. Г., Дуна-
ева Т. Н„ 1970).
MLVA-анализ и анализ полногеномного полиморфизма отдельных нуклеотидов
штаммов F. tularensis хорошо разработаны и могут быть использованы для тонкой
дифференциации штаммов, обнаруженных в очагах биопреступлений, и определения
мест их географического происхождения1 (Johansson A. et al., 2004; Vogler A. et al., 2009).
Экология и естественная эпидемиология F. tularensis2. Экология возбудителя туля-
ремии исследуется со средины 1920-х гг. Накоплено много сведений о вспышках
туляремии среди различных видов животных, проживающих в естественных услови-
ях, но вопрос о первичном природном резервуаре F. tularensis остается открытым до
настоящего времени. В фундаментальной монографии Н. Г. Олсуфьева, Т. Н. Дунае-
вой (1970) обращается внимание на невозможность поддержания в природе возбуди-
теля туляремии путем массовых эпизоотий среди высокочувствительных к нему
животных (водяные крысы, обыкновенные полевки, зайцы, хомяки, отчасти домовые
мыши и некоторые другие виды), обычно приводящих к вспышкам туляремии среди
людей. Также ими подвергнута сомнению роль в этом процессе насекомых (клещи,
комары, слепни и др.; всего на территории бывшего СССР выявлено не менее 74 ви-
дов членистоногих, переносчиков возбудителя туляремии), участвующих в передаче
возбудителя туляремии от больных туляремией животных к людям. Похожей точки
зрения придерживаются A. Tamvik, L. Berglund (2003), утверждая, что грызуны не могут
поддерживать в природе возбудитель туляремии между эпизоотиями. Почти два де-
сятилетия назад норвежскими учеными обнаружено инфицирование возбудителем
туляремии простейших, обитающих в водоемах (Berdal В. Р., 1996). Тот факт, что сво-
бодноживущие амебы могут быть буквально заполнены F. tularensis, причем в небла-
гоприятных условиях среды возбудитель туляремии может сохраняться в цистах
A. castellanii (Baron G. S., Nano E E., 1998; Abd H. et al., 2003; Lauriano С. M. et al.,
2004), свидетельствует в пользу того, что в первичных природных резервуарах он под-
держивается в простейших (Супотницкий М. В., 2009).
В России основными естественными источниками F. tularensis для людей служат
грызуны, в особенности обыкновенные полевки {Microtus arvalis), водяные крысы
(Arvicola terrestris), домовые мыши {Mus. musculus), отчасти ондатры (Ondatra zibethica)
1 Примеры использования этих методов для тонкой дифференциации штаммов и опреде-
ления мест их географического происхождения, приведены в разд. 3.1 и 3.2.
2 Естественная эпидемиология F. tularensis в основном приведена по работам Н. Г. Олсуфье-
ва, Г. П. Руднева (1966); Н. Г. Олсуфьева, Т. Н. Дунаевой (1970).
714
Часть 3. Частная неправильная эпидемиология
и хомяки (Cricetus cricetus и другие виды), а также зайцы, в особенности русак (Lepus
europaeus), беляк (L. timidus) и толай (L. total). Заражение людей возбудителем туля-
ремии происходит либо в результате соприкосновения с больными или павшими гры-
зунами и зайцами, либо вследствие укусов инфицированных кровососущих члени-
стоногих (комаров, слепней, клещей), либо через воду, пищевые продукты, солому и
другие субстраты, загрязненные выделениями больных туляремией зверьков. Изве-
стны случаи заражения людей туляремией при забое на мясокомбинатах овец и kpjti-
ного рогатого скота, на которых в большом количестве находились инфицированные
бактериями туляремии клещи.
Заболевания людей туляремией возникают почти исключительно в сельской ме-
стности. В больших городах условия для заражения людей F. tularensis отсутствуют. Но
известны заболевания на окраинах российских городов, расположенных вблизи пой-
менно-болотных очагов, а также случаи заражения горожан при поездках за город. Из
горожан чаще заболевают охотники и рыбаки, случайно попавшие в природные оча-
ги туляремии.
Выявленные на территории бывшего СССР туляремийные очаги не имеют сплош-
ного распространения. Н. Г. Олсуфьев и Т. Н. Дунаева (1970) выделили две обширные
области диффузного распространения очагов туляремии. Первая расположена в пре-
делах европейской части бывшего СССР, она охватывает северо-запад, юг, центр и
юго-восток. Вторая находится в пределах Западной Сибири и охватывает Западно-
Сибирскую низменность, предгорья Алтая и Кузнецкого Алатау (рис. 3.47).
Природные очаги туляремии характеризуются исключительной стойкостью. Оп-
тимальными для формирования природных очагов туляремии являются лесостепь,
степь и отчасти широколиственные леса, а также поймы и дельты рек, побережья озер
Рис. 3.47. Распространение природных очагов туляремии на территории бывшего
СССР. 1 — территории, занятые природными очагами туляремии; 2 — границы
провинций очаговости. По Н. Г. Олсуфьеву и Т. Н. Дунаевой (1970)
3.3. Туляремия
715
и оолота. В годы пониженной или низкой численности грызунов F. tularensis обычно
не удается обнаружить в их популяциях даже при исследовании нескольких тысяч
зверьков (Олсуфьев Н. Г., Дунаева TH., 1970).
Локализованная вспышка туляремии может быть вызвана разными подвидами и
кладами F. tularensis. В июле 2007 г. в штате КУ. а (США) произошла вспышка туляре-
мии, связанная с укусами оленьих слепней. Инфекция у людей была вызвана 2 кла-
дами (А1 и А2) туляремийно! о микроба подвида tularensis. Как показало исследование
трупов животных (зайцеобразных), обнаруженных в этом регионе, они были зараже-
ны А1 и А2, а также подвидом holarctica (Petersen J. М. et al., 2008).
Заболеваемость людей в природных очагах туляремии возникает на фоне предше-
ствовавших или одновременно протекающих эпизоотий туляремии среди мышевид-
ных грызунов в условиях возросшей их численности. Разлитые эпизоотии туляремии
отмечаются в популяции грызунов и зайцев, лишь при высокой численности зверьков,
и чем она выше, тем они бывают интенсивнее. В популяции водяных крыс эпизоо-
тии туляремии обычно наблюдаются летом, но могут начаться еще весной. Число боль-
ных туляремией зверьков в разгар эпизоотии составляет в среднем 1—3 %, но на отдель-
ных участках может достигать 30 %. Среди обыкновенных полевок интенсивные
эпизоотии туляремии обычно наблюдаются в зимнее время, а среди домовых мышей —
поздней осенью и в начале зимы. В скирдах и ометах в разгар эпизоотии число обна-
руживаемых павших от туляремии полевок может составлять 30—50 % и более к числу
живых зверьков. Среди ондатр интенсивные эпизоотии туляремии отмечались глав
ным ооразом осенью и зимой.
Естественная восприимчивость людей очень высокая. Заболевания туляремией
могут наблюдаться в виде разрозненных случаев или эпидемических вспышек той или
иной интенсивности. Частота заболеваний людей зависит от колебаний численнос-
ти грызунов и зайцев — источников инфекции, а также от количества неиммунных
лиц и их трудовой деятельности. Заражаются F. tularensis любые возрасты, включая
детей и стариков, при этом отмечается почти 100 %-ная восприимчивость к инфек-
ции. По роду занятий заболевшие могут относиться к самым разнообразным группам
населения — сельскохозяйственным рабочим, рабочим боен, охотникам, рыбакам,
домашним хозяйкам, школьникам и т. д. Эпидемиологические типы естественных ту-
ляремийных вспышек приведены в табл. 3.15.
Заболевания трансмиссивного типа возникают в результате передач F. tularensis
комарами и слепнями, реже иксодовыми клещами и отмечаются исключительно в теп-
лый период года. Форма заболеваний чаще язвенно-бубонная. Промысловые заболе-
вания возникают среди лиц, занятых охотой на водяных крыс, ондатру и других гры-
зунов, являющихся объектами промысла. Сезон заболеваний различен, но чаще это
весна и осень, когда промысел максимально развит. У больных отмечается язвенно-
бубонная или чисто бубонная форма.
Заболевания водного типа связаны с употреблением для питья или умывания воды,
взятой из ручья, неблагоустроенного колодца и т. д. после попадания в них больных
туляремией грызунов. Заражение людей F. tularensis через воду колодцев чаще наблю-
дается зимой, через воду ручьев — чаще в летнее время. Форма заболевания ангиноз-
но-бубонная, реже абдоминальная и еще реже глазно-бубонная.
Заболевания сельскохозяйственного типа могут возникнуть при переборке соло-
мы, зерна и овощей, транспортировке сена, даче корма скоту и т. д., если перечислен-
ные объекты окажутся загрязненными выделениями больных туляремией грызунов.
Заболевания возникают преимущественно поздней осенью, зимой и весной, и, так как
Таблица 3.15
Эпидемиологические типы естественных туляремийных вспышек*
Эпидемиологический тип заболеваемости (по условиям заражения и факторам передачи) Вариант Основной источник инфекции Преобладающий механизм заражения Преобладаю- щая клиническая форма болезни Тип природного очага, с которым наиболее часто связан данный тип заболеваемости
Трансмиссивный а) заражение через комаров и слепней; б) заражение через иксодо- вых клещей а) водяная крыса, зайцы; б) обыкновенная полевка, хомяки и др. грызуны, зайцы Инокулятивный Язвенно-бубонная а) пойменно-болотный; б) степной, луго-поле- вой лесной
Промысловый Водяная крыса, ондатра, хомяки Контактный Бубонная, язвенно-бубонная Пой мен но-болотный, степной, тугайный
Охотничье-пи щевой Зайцы Контактный, алиментарный Ангинозно- бубонная, абдоминальная Степной, луго-полевой, тугайный, лесной
Водный а) заражение через воду ручьев и др. открытых водоисточников; б) заражение через воду колодцев а) водяная крыса; б) домовая мышь, обыкновенная полевка Алиментарный Ангинозно- бубонная, абдоминальная а) предгорно-ручьевой, пойменно-болотный; б) луго-полевой, степной
Сельскохозя й ственный Обыкновенная полевка Аспирационный Торакальная Луго-полевой, степной
Бытовой (домовый) Домовая мышь, обыкновенная полевка Аспирационный Торакальная Степной, луго-полевой, тугайный
Продуктовый Домовая мышь Алиментарный Хнгинозно- бубонная, абдоминальная Степной, луго-полевой
Производственный а) заражение при перера- ботке сельскохозяйствен- ных продуктов; б) заражение при забое животных и разделке мяса а) обыкновенная полевка и домовая мышь; б) овцы и клеши на них. зайцы а) аспирационный; б) контактный а) торакальная; б) бубонная Заболевания возникают вне природного очага («завоз»)
Траншейный (окопный) Обыкновенная полевка, домовая мышь Аспирационный Торакальная Степной, луго-полевой
* По И Н. Майскому и Н. Г. Олсуфьеву; взято из работы Н. Г. Олсуфьева и Г. П. Руднева (1966).
Часть 3. Частная неправильная эпидемиология
3.3. Туляремия
717
заражение происходит главным образом аспирационным путем, у заболевших людей
преобладает торакальная (легочная) форма туляремии.
Заболевания людей домового (бытового) или продуктового типа чаще всего быва-
ют связаны с домовыми мышами, проникающими в жилье человека или в продоволь-
ственные магазины, склады и т. д. и загрязняющими своими выделениями предметы
домашнего обихода, пищевые продукты и т. д. Эти типы заболеваемости приходятся
на позднюю осень и зиму и наблюдаются одновременно с сельскохозяйственными
вспышками. При бытовом заражении преобладает аспирационный путь передачи,
поэтому наиболее частой является торакальная форма болезни, тогда как при зара-
жении от продуктов преобладает алиментарный путь передачи и чаще встречается
ангинозно-бубонная форма туляремии.
Заболевания охотничье-пищевого типа связаны с охотой на зайцев, разделкой их
тушек и употреблением их мяса в пищу в недостаточно термически обработанном виде.
Они наблюдаются осенью, зимой и весной. Наиболее частые формы болезни при за-
ражении от зайцев — язвенно-бубонная, ангинозно-бубонная и абдоминальная.
Заболевания производственного типа возникают при первичной переработке на
предприятиях сельскохозяйственного сырья (зерно, сахарная свекла и т. д.), загряз-
ненного выделениями больных туляремией грызунов, или же при забое на мясоком-
бинатах скота, пораженного инфицированными клешами. Сюда же относятся случаи
заражения людей на пищевых предприятиях при переработке заячьих или кроличьих
тушек. Преобладает аспирационный или контактный способ заражения.
Восприимчивость и чувствительность животных к возбудителю туляремии. По мне-
нию Н. Г. Олсуфьева и Т. Н. Дунаевой (1970), различия в степени восприимчивости и
инфекционной чувствительности определяют роль млекопитающих как источников
инфекции для людей. Установлено, что инфекционность (заразительность) животных
зависит от интенсивности накопления F. tularensis в организме больных зверьков и
количества бактерий, попадающих в кровь, или выделения.
Наибольшую восприимчивость и инфекционную чувствительность к туляремии
(первая группа) проявляют полевки, водяные крысы, зайцы, хомяки, песчанки, до-
мовые мыши, землеройки и некоторые другие грызуны и насекомоядные. Для них доза
в 1 микробную клетку (по оптическому стандарту мутности ГИСК им. Л. А. Тарасе-
вича) при подкожном введении является, безусловно, смертельной. У этих животных
заболевание протекает по типу острой септицемии, и они погибают на 6—10 сут, редко
позднее. На вскрытии выявляется необычайно интенсивное обсеменение бактерия-
ми внутренних органов и крови.
Значительно менее чувствительны к туляремии (вторая группа) суслики, серые
крысы, ежи и др., для которых наименьшая полная смертельная доза обычно равняется
1 млрд микробных клеток. Но восприимчивость животных второй группы значитель-
на — их удается заразить дозой всего в 1 микробную клетку. П ри этом у них развивается
доброкачественная инфекция с умеренным обсеменением бактериями паренхиматоз-
ных органов и крови; через 6—8 сут начинается очищение внутренних органов от воз-
будителя, животные выздоравливают, в это время у них обнаруживается высокона-
пряженный иммунитет к реинфекции.
Третью группу составляют хищные животные, такие как кошки, лисицы, хорьки
и др., даже при заражении массивными дозами заболевание может протекать без ви-
димых клинических явлений, тогда как небольшими дозами их вообще не удается
заразить. Для копытных животных, а также для птиц и холоднокровных туляремий-
ный микроб в большинстве случаев малопатогенен.
718
Часть 3. Частная неправильная эпидемиология
Высокую чувствительность к туляремии проявляют белые мыши и морские свинки
(первая группа), они погибают при явлениях септицемии спустя 3—10 сут (реже по-
зднее) после подкожного заражения. Этих животных широко используют для биоло-
гической пробы с целью лабораторной диагностики туляремии. Белые крысы и до-
машние кролики малочувствительны к туляремии (вторая группа) и погибают от нее
только при введении массивных доз возбудителя. К американским штаммам домаш-
ние кролики так же высокочувствительны, как белые мыши и морские свинки, при
подкожном введении, безусловно, смертельная доза для них составляет всего 1 мик-
робную клетку. Лабораторных животных используют также для определения вирулен-
тности туляремийных культур. Путем подкожного сражения разными дозами опре-
деляют полные или неполные смертельные дозы (например, LD50). Наиболее точные
результаты получают при одновременном испытании культуры на высокочувствитель-
ных и малочувствительных к туляремии животных белых мышах, морских свинках и
белых крысах (или кроликах).
Инфицирующие дозы для животных и людей. Штаммы F. tularensis подвида tularensis
характеризуются высокой вирулентностью для кроликов, также они считаются наи-
более вирулентными для людей. Их инфицирующая доза для человека обычно менее
10 КОЕ Смертность при нелечечных случаях кожной формы болезни (чрескожное
заражение) достигает 5 %. При ингаляционном заражении 10 КОЕ возбудителя туля-
ремии подвида tularensis смертность без лечения составляет уже от 30 до 60 % заболев-
ших (Saslaw S. et al., 1961).
Штаммы подвида holarctica (генотип В) маловирулентны .зля кроликов (при под-
кожном введении смертельная доза может достигать 1 млрд клеток), смертность при
нелеченной кожной форме болезни у людей менее 0,5 %. Их инфицирующая лоза для
людей при кожном пути заражения — до 1000 КОЕ (Олсуфьев Н. Г., ДунаеваТ. Н., 1970;
Pechous R. et al., 2009). Сравнительная вирулентность подвидов F. tularensis для чело-
века и различных видов экспериментальных животных, приведена в табл. 3.16.
Таблица 3.16
Сравнительная вирулентность подвидов F. tularensis*
Подвид LD5o, KOI.**
Люди Мыши Морские свинки Кролики
Е tularensis подвид tularensis <10 <10 <10 1-10
F. tularensis подвид holarctica <103 <1 <10 >106
F. tularensis подвид meaiaasiatica HOJ** HO НО >106
F rovidda >103 <103 <103 >10‘
* По J. Ellis et al. (2002), обобщивших данные разных авторов.
** Все дозы вводились подкожно.
*** НО — не описано.
При ингаляционном заражении макак-резусов (Масаса mulatto) жидким аэрозолем
F. tularensis с дисперсностью частиц в пределах 1—5 мкм наблюдаются четкие разли-
чия по LD50 и патогенезу болезни между шзаммами генотипов А и В. LD50 для F. tula-
rensis SCHU-S4 (подвид tularensis, генотип А; номер доступа в GenBank AJ749949)
составляла менее 50 КОЕ. У инфицированных обезьян быстро ра шивались множе-
ственные инфильтраты в легких и бронхах, а также многодолевые уплотнения (брон-
хопневмония) и общая инфекция с зонами обширного некроза в легких, селезенке,
3.3. Туляремия
719
лимфатических узлах и костном мозге. Животные погибали до развития гранулем.
LD50 для F. tularensis 425 (подвид holarctica, генотип В) не была достигнута даже при
ингаляции 106 КОЕ. Поражения легких носили характер самоограничивающихся гра-
нулем. Клинические и анатомические проявления инфекции и летальный исход при
инфицировании аэрозолем F. tularensis 425 имели место в том случае, если бактери-
альная нагрузка достигала определенного уровня (Schricker R. L. et al., 1972).
Иммунитет. Перенесенное заболевание или экспонирование к сублетальной дозе
Francisella вызывает у людей и экспериментальных животных иммунитет против по-
вторного инфицирования. Протективная роль антител против F. tularensis, обнаружи-
ваемых в сыворотке крови реконвалесцента, незначительна. Введение специфических
антител экспериментальным животным показало, что они могут препятствовать раз-
витию туляремии, но только если животное заражается маловирулентными штамма-
ми туляремийного микроба. Наиболее вероятно, что иммунитет к Francisella связан с
клеточным иммунным ответом, эффективно контролирующим развитие инфекцион-
ного процесса через активизацию у макрофагов внутриклеточных киллерных механиз-
мов. В основном он зависит от Т-клеточного иммунного ответа, проявляющегося
активизацией CD4+ и CD8+ Т-клеток (Elkins К. L. et al., 1996; Kroca М. et al., 2000).
Стойкость иммунитета против возбудителя туляремии относительна. Неоднократ-
но регистрировались случаи повторного заболевания туляремией. R. Н. Taylor (1950)
описал двух больных, заразившихся в естественных условиях. Соответственно через
1 и 10 мес. после выздоровления эти лица заболели еще раз. В обоих случаях туляре-
мия при инфекции и реинфекции протекала в язвенно-бубонной форме. Оба боль-
ных, начиная с первой недели болезни, получали антибиотики. Болезнь в обоих слу-
чаях протекала легко. При первом заражении F. tularensis больные лечения не полу-
чали. Т. W. Green, N. Т. Eigelsbach (1950) описали 6 случаев повторного лабораторного
заражения туляремией (язвенно-бубонная форма болезни). Е. Е. Overholt et al. (1961),
сами описавшие повторное ингаляционное инфицирование у одного пациента, заме-
тили, что наблюдения такого рода позволяют предполагать гораздо большее распро-
странение в эндемических очагах, чем принято думать, случаев реинфекции F. tularen-
sis, протекающих в бубонной или легочной формах болезни. И мы просто не знаем,
как часто лица, охотящиеся на диких кроликов под видом «простуды» или долго за-
живающих порезов переносят одну из форм туляремии. В случае повторного попада-
ния F. tularensis в организм человека вероятность развития болезни определяется со-
отношением таких факторов, как уровень иммунитета, количество микроорганизмов
и их вирулентность.
Устойчивость F. tularensis во внешней среде и в трупах погибших животных. По дан-
ным, обобщенным Н. Г Олсуфьевым и Г. П. Рудневым (1966), возбудитель туляремии
сравнительно долго сохраняется (но не размножается) в воде, а также в других объек-
тах внешней среды при условии низкой температуры. Так, при температуре 4°С туля-
ремийные бактерии (подвид holarctica) могут сохраняться в воде или влажной почве
свыше 4 мес. без снижения вирулентности, а при температуре 1 °C сохраняются в воде
до 9 мес. При повышении температуры сроки хранения уменьшаются. Например, при
температуре 20—25°С микробы отмирают в воде в течение 30—60 сут; F. tularensis ма-
лоустойчива к высоким температурам (при 60°С гибнет через 5—10 мин, при 100°С —
в течение 1—2 мин). В зерне и соломе F. tularensis сохраняется при температуре ниже
0°С до 6 мес., при 8—12°С — до 56 сут, а при 20—30°С — не более 20 сут. В молоке и
сливках при 8—10°С возбудитель сохраняется не менее недели, а при замораживании
этих продуктов — до 3 мес. В замороженных трупах животных, павших от туляремии,
720
Часть 3. Частная неправильная эпидемиология
возбудитель может сохраняться до 6 мес., при более высокой температуре сроки хра-
нения уменьшаются. В шкурках животных, павших от туляремии, возбудитель сохра-
няется при температуре 8—12°C более 1 мес., а при 32—33 °C—лишь втечение недели.
Туляремийные бактерии сравнительно долго противостоят высушиванию. Так, на
хлопчатобумажной ткани, пропитанной кровью и тканевой жидкостью мышей, по-
гибших от туляремии, и высушенной при температуре 13—21°С (при влажности
39-68 %), микробы оставались жизнеспособными до 35 сут (срок наблюдения). При
отсутствии защитных субстанций туляремийные микробы нестойки к высушиванию
и погибают через 1—3 сут. Под действием прямых солнечных лучей туляремийные бак-
терии погибают через 20—30 мин, на рассеянном же свету жизнеспособность их со-
храняется до 3 сут. Ультрафиолетовые лучи почти мгновенно убивают F. tularensis.
F. tularensis быстро разрушается под действием ультразвука; и нестойка к обычным
дезинфицирующим веществам — лизолу, фенолу, хлору, сулеме и т. д. Обычное хло-
рирование питьевой воды полностью обезвреживает возбудителя туляремии. Особен-
но чувствительны туляремийные бактерии к этиловому спирту, при его воздействии
они погибают менее чем за 1 мин.
Антибиотикорезистентность. Выделенные из природных источников изоляты воз-
будителя туляремии в условиях in vitro обнаруживают чувствительность к большин-
ству антибиотиков, применяемых в клинической практике. Природная устойчивость
к эритромицину рассматривается как таксономический признак. Ею обладают штам-
мы генотипа В подвида holarctica (биотип II) (Ellis J. et al., 2002). S. К. Urich, J. M. Peter-
sen (2008) исследовали 169 изолятов F. tularensis (92 изолята типа А и 77 изолятов типа
В) из Северной Америки на чувствительность к семи антибиотикам (стрептомицин,
гентамицин, тетрациклин, доксициклин, ципрофлоксацин, левофлоксацин, хлорам-
феникол), среди них не было ни одного изолята, резистентного к этим антибиотикам.
I. Ikaheimoetal. (2000) исследовали наантибиотикорезистентность 38 штаммов/’, tula-
rens подвида holarctica, выделенных в Финляндии от больных туляремией людей
и грызунов. Штаммы оказались чувствительными к стрептомицину, тетрациклину,
хлорамфениколу, аминогликозидам (тобрамицин, гентамицин), хинолонам (ципро-
флоксацин, левофлоксацин, грепафлоксацин, тровафлоксацин), но резистентентны
к Р-лактамным антибиотикам и азитромицину. Е. Е. Overholt et al. (1961) не получи-
ли доказательств возможности появления резистентных к тетрациклину штаммов
F. tularensis в процессе лечения этим антибиотиком 42 пациентов с туляремией.
Обнаружение генетически измененных штаммов F. tularensis. Подходы к обнаруже-
нию генетически измененных штаммов возбудителя туляремии в основном те же, что
и для обнаружения генетически измененных штаммов возбудителей сибирской язвы
и чумы (см. разд. 3.1 и 3.2). В отличие от возбудителя чумы, возбудитель туляремии
более однороден по фенотипическим признакам, но его генетика изучена хуже и ме-
нее подробно описана, чем у Y. pestis. Уникальное таксономическое положение рода
Francisella делает невозможным использование для генетического переноса векторов
на основе репликонов плазмид кишечной палочки. Видимо, для получения генети-
чески измененных штаммов возбудителя туляремии не пригодны и отработанные на
традиционных модельных микроорганизмах (кишечная палочка) методы генетиче-
ского переноса и векторы.
Последнее обстоятельство сулит много неожиданностей при расследовании био-
преступлений, совершенных с использованием генетически измененных штаммов ту-
ляремийного микроба, так как лишает экспертов возможности обнаружить в геноме
F. tularensis генетические конструкции с уже известным происхождением и историей.
3.3. Туляремия
721
Можно предположить, что конструирование векторных систем для переноса генов в
микроорганизмы рода Francisella будет вестись по двум направлениям:
1) на основе собственных плазмид бактерий рода Francisella (pFNLlO, 3,9 т. п. о.;
и, возможно, других) — реплицирующиеся векторные системы (пример такого век-
тора см. в работе LoVullo Е. D. et al., 20091);
2) на основе челночных конструкций, переназначенных для интеграции клони-
руемых генов в геном бактерий рода Francisella (пример такого вектора см. разд. 2.2.2
«Технологии рекомбинантной ДНК в получении штаммов бактерий с неправильной
эпидемиологией»).
В очагах искусственно вызванных вспышек туляремии с наибольшей вероятно-
стью можно встретить штаммы возбудителя туляремии со следующими модификация-
ми (и их сочетаниями):
резистентные к антибиотикам, используемым в распространенных схемах лече-
ния туляремии. Получением таких штаммов отдельные лаборатории в USAMRIID
занимаются уже почти 60 лет и даже исследуют их поражающую эффективность в
опытах по ингаляционному инфицированию экспериментальных животных. Напри-
мер, в 1960-х гг. ими в таких экспериментах использован резистентный к стрептоми-
цину, но не утративший вирулентность штамм F. tularensis SCHU-S5 (Sawyer W. D. et
al., 1966)2. Российскими исследователями на основе плазмиды pFNLlO сконструиро-
вана векторная челночная плазмида pFNLlOO, способная экспрессировать гены ре-
зистентности к тетрациклину и хлорамфениколу в вакцинном туляремийном штам-
ме (LVS, номер доступа в GenBank АМ233362) и в кишечной палочке (Pavlov V. М. et
al., 1996). La Scola В. et al. (2008) использовали в экспериментах по выявлению пиро-
секвенированием высоковирулентный штамм F. tularensis, устойчивый к хинолонам.
В геноме генетически модифицированных штаммов F. tularensis секвенирование
позволит выявить вставки генных кассет с генами резистентности к антибиотикам
1 резистентность детоксикационного типа); либо мутации в генах, имеющих отноше-
ние к проницаемости стенки бактерии для данных антибиотиков, и/или к структур-
ным изменениям в молекулах, являющихся для них мишенями;
с измененной антигенной специфичностью, «пробивающие» иммунитет, создавае-
мый живой туляремийной вакциной в течение первого года после вакцинации. Такие
штаммы F. tularensis, скорее всего, будут получены путем целенаправленного поиска
среди штаммов, выделенных от людей и животных, заболевших туляремией в ее при-
родных очагах. Их геном не будет иметь признаков, характерных для какого-то явного
генно-инженерного вмешательства. Его секвенирование позволит обнаружить только
инсерции и делеции, единичные нуклеотидные полиморфизмы, единичные тандем-
ные повторы и вариабельные участки генов, характерные для процессов микромути-
рования и пластичности генома. И нтерес экспертов должны привлечь гены, имеющие
отношение к фазовым вариациям ЛПС туляремийного микроба (например, IpxF).
1 С публикацией стоит ознакомиться. Разработанный Е. D LoVullo et al. (2009) вектор
стабилен в популяции туляремийного микроба без селективного давления антибиотиками.
2 Высоковирулентный штамм F. tularensis SCHU-S5 относится к подвиду tularensis. Выде-
лен в 1941 г. в США L. Foshay из содержимого туляремийной язвы у человека (Eigelsbach Н. Т.
et al., 1951). Его исходный вариант не является резистентным к стрептомицину. Для американ-
ских военных микробиологов он имеет то же значение, что и штамм Ames возбудителя сибир-
ской язвы. Использованный в данной работе стрептомииинрезистентный штамм F. tularensis
SCHU-S5, скорее всего, был получен путем отбора спонтанных мутантов на плотной питатель-
ной среде, содержащей стрептомицин.
722
Часть 3. Частная неправильная эпидемиология
Патогенез1. Возбудитель туляремии проникает в организм человека через кожу
(даже внешне не поврежденную), слизистые оболочки глаз, дыхательные пути и же-
лудочно-кишечный тракт. Способ проникновения возбудителя, его патогенность, доза
и иммунобиологические реакции макроорганизма определяют развитие соответс i -
вующей клинической формы болезни. При типичной язвенно-бубонной форме туля-
ремии у человека через 3—5 сут после заражения появляется кожное поражение в ме-
сте инфицирования.
Патогенность F. tularensis для млекопитающих обусловлена ее способностью ин-
фицировать макрофаги, нейтрофилы, дендрические клетки, гепатоциты, клетки ле-
гочного эпителия и др. Возбудитель туляремии проникает в фагоцитирующие клетки
благодаря необычному механизму фагоцитоза, включающему формирование обшир-
ных ассиметричных псевдоподийных петель (spacious asymmetric pseudopod loops).
Этот процесс называется петлевым фагоцитозом (looping phagi >су tosis) (Clemens D. L.
et al., 2005). Он включает перестановку нитей актина через фосфатидилинозитол-
З-киназную передачу сигналов (phosphatidylinositol 3-kinase signaling), зависящую от
наличия фактора комплемента СЗ и рецептора к комплементу CR3. Francisella может
проникать в клетку через маннозный рецептор, scavenge-рецепторы класса А типов
I и П, Fc-рецгптор. При пос ледующем продвижении в клетке фагосомы F. tularensis
предотвращает респираторный взрыв, избегает экспонирования к супероксидам. По-
сле созревания фагосомы она лишь кратковременно взаимодействует с компонентами
эндопла зматической транспортной сети. Вакуоль, содержащий F. tularensis (F. tula-
mra’s-containing vacuole, FCV), не сливается с лизосомой, F. tularensis разрушает ее пи-
пидный бислой по неизвестному механизму и попадает непосредственно в цитозоль
клетки, где она интенсивно размножается. По времени эти стадии различаются у раз-
личных видов Francisella и в различных инфицированных клетках хозяина. Те мутан-
ты, которые не способны предотвратить слияние фагосомы с лизосомой и/или не
способны проникнуть из фагосомы в цитозоль, маловирулентны в условиях in vitro и
in vivo. Приблизительно через 12 ч после инфицирования Francisella начинает репли-
цировать в цитозоле клетки. Процессы разрушения фагосомы и репликация F. tula-
rensis в цитозоле клетки зависят от генов, расположенных на FP1. Разрушение клетки
занимает примерно 16—24 ч и заканчивается аутофагией (рис. 3.48).
F. tularensis индуцирует гибель макрофага путем апоптоза. Для этого требуется
скопление значительно больших количеств клеток туляремийного микроба и больше
времени от инфицирования до индуцирования апоптоза, чем имеет место при анало-
гичном процессе, вызванном сальмонеллами, шигеллами, лигионелами и Yersinia.
Начинается лимфогенное распространение F. tularensis г Lai X. et al., 2001).
Занос F. tularensis в регионарные лимфатические узлы и ее размножение вызыва-
ют воспалительные явления — лимфаденит. Гибель бактерий сопровождается высво-
бождением эндотоксина, который усиливает развитие местного патологического про-
цесса, а при поступлении в кровь вызывает интоксикацию организма. Если барьерная
функция лимфатического аппарата оказывается нарушенной, F. tularensis проникает
в кровь (бактериемия) и распространяется по всему организму, инфицируя ретикуло-
эндотелиальные ткани. В течение нескольких дней в ретикулоэндотелиальных тканях
экспрессируется большое количество цитокинов. Однако они не способны предуп-
редить инфекцию, вызванную вирулентным штаммом туляремийного микроба.
1 Патогенез туляремии в основном описан по работам Н. Г. Олсуфьева и Г. П. Руднева
(1966); Е. П Шуваловой с соавт. (2001); J. Ellis et al. (2002); R. Pechous et al. (2009).
3.3. Туляремия
723
Рис 3 48. Размножение Гrancisella в макрофагах. Мутации в FPI-генах и ключевых
факторах вирулентности уменьшают способность Francisella покидать фагосому.
По R Pechous et al. (2009)
Генерализация инфекции сопровождается специфическими поражениями парен-
химатозных органов (селезенка, печень, легкие) и аллергизацией организма. Благо-
даря особенностям внутриклеточного размножения F. tularensis, пролиферативные
изменения при туляремии носят характер специфических гранулем, имеющих много
общего с туоеркулезными бугорками. Гранулемы образуются в местах размножения
F. tularensis и состоят из эпителиоидных и лимфоидных клеток с примесью полиморф-
ной дерных лейкоцитов и гистиоцитов. У чувствительных к туляремии животных, об-
разовавшиеся в тканях гранулемы некротизируются, при далеко зашедшем процессе
в очаге воспаления образуются обширные некрозы. Наиболее демонстративно грану-
лематозный процесс выражс н в регионарных лимфатических узлах, где развивается
первичный лимфаденит (бубон). При нагноении и вскрытии бубона на коже образу-
ется длительно не заживающая язва.
Во вторичных бубонах, возникающих при генерализации, гранулематозные и
некротические изменения не сопровождаются нагноением.
I шничесдая картина туляремии у людей при естественном инфицировании F. tula-
rensis1. Инкуоационныи период чаще колеблется в пределах от 3 до 7 сут. Однако име-
ются указания, что этот срок может растягиваться до 3 нед и сокращаться до несколь-
ких часов. Клинические формы туляремии зависят от пути проникновения в организм
человека возбудителя туляремии (табл. 3.17).
При туляремии, как и при других инфекциях, различают легкие, средней тяжести
и тяжелые, случаи болезни. К категории легких относятся атипичные, стертые, афеб-
рильные и абортивные случаи. По длительности течения рационально деление на
острые и затяжные формы (до б мес. и дольше). Схематически все течение болезнен-
ного процесса при туляремии делится на три периода:
1) начальный;
2) период разгара болезни;
3) период выздоровления.
1 Приведена по работам Н. Г. Олсуфьева и Г. П. Руднева (1966); Е. П. Шуваловой с со-
авт. (2001).
724
Часть 3. Частная неправильная эпидемиология
Таблица 3.17
Клинические формы туляремии в зависимости от пути заражения*
Клиническая форма Путь заражения
Бубонная При заражении через кожу и слизистые глаза
Язвенно-бубонная
Глазо-бубонная
Ангинозно-бубонная При заражении через рот
Абдоминальная (кишечная)
Легочная (бронхитический и пневмонический варианты) При заражении через дыхательные пути
Генерализованная или первично септическая форма (встречается при любом пути
заражения, особенно у ослабленных лиц)
* По Н. Г. Олсуфьеву и Г. П. Рудневу (1966).
Начальный период по своим клиническим симптомам является в основном общим
для всех форм туляремии. Он особо важен для целей ранней диагностики. При лю-
бой клинической форме туляремия начинается, как правило, остро, внезапно, без
продромального периода. Заболевание проявляется многократным познабливанием,
реже ознобом. Температура быстро поднимается до 38—39°С и выше. Больные жа-
луются на головную боль, общую разбитость, мышечные боли, чаще в области икр и
поясницы, головокружение и отсутствие аппетита. В более тяжелых случаях болезни
отмечается рвота и кровотечение из носа, позже — помрачение сознания и бред. Не-
редки тягостные или болевые ощущения за грудиной. Наиболее упорны и длительны
головные боли. Объективно отмечается гиперемия липа, иногда зева, часто конъюн-
ктивит. Кровяное давление несколько снижается. Катаральные явления обычно от-
сутствуют. В картине крови нормальный или слегка повышенный лейкоцитоз с уме-
ренным сдвигом нейтрофилов. В моче наличие белка и цилиндров. Через несколько
дней у значительного числа больных появляется сухой кашель, отмечаются скудные
сухие хрипы (при легочной форме их прослушивается значительно больше).
Поскольку начальный период заболевания туляремией не имеет патогномонич-
ных симптомов, большое диагностическое значение придается эпидемиологическо-
му анамнезу, который должен отличаться четкостью, достоверностью и исчерпываю-
щей полнотой. Общая продолжительность начального периода туляремии 2—3 сут.
Период разгара болезни. Из общих симптомов особого вни иания заслуживает тем-
пературная реакция и реакция со стороны нервной и сердечно-сосудистой систем.
Характерно относительное учащение и резкая лабильность пульса, которая длитель-
но держится даже в периоде выздоровления. Электрокардиографически установлено
наличие хотя и неглубоких, но довольно длительно отмечаемых изменений в мышце
сердца. Температура в начале болезни обычно круто повышается до сравнительно вы-
сокого уровня. В дальнейшем по характеру температурной кривой можно различать
ремитирующий, неправильно интермитирующий, постоянный и ундулирующий типы
(здесь возможны не только две, а три и иногда четыре волны, правда, более короткие).
Наиболее часты ремитирующий и неправильно интермитирующий типы лихорадки.
Падение температуры, как правило, происходит литически. Присоединившиеся ос-
ложнения или наличие патологических процессов иной природы изменяют ход туля-
ремийной температурной кривой. Встречаются случаи и афебрильной туляремии.
Длительность лихорадки при туляремии может колебаться от 5—7 до 30 и более сут,
3.3. Туляремия
725
но чаще ограничивается 2—3 сут. В периоде выздоровления типичен длительный и
беспорядочный субфебрилитет. Более чем в 70 % случаев заболевание туляремией дли-
лось от 16 до 30 сут, при этом в 40 % длительность заболевания колебалась от 3 нед
до 1 мес. При заболеваниях с преобладанием, например, бубонных форм этот срок уд-
инялся до 2—3 и более месяцев (без лечения стрептомицином).
Гематологически при туляремии отмечается лейкопения, умеренный сдвиг влево
ейкоцитарной формулы, лимфоцитоз и моноцитоз, ускорение РОЭ. Наслоение ос-
ложнений и развитие нагноительных процессов соответствующим образом изменя-
ют данные гемограммы (лейкоцитоз, нейтрофилез, усиление сдвига влево и т. д.).
Бубонная форма туляремии характеризуется образованием воспалительного про-
цесса в регионарном лимфатическом узле. Здесь бубон (лимфаденит) является не-
пременным и кардинальным признаком болезни. Патогенетически бубоны следует
рассматривать как воспалительный процесс в месте концентрации возбудителя. По
аналогии с чумой различают бубоны первичные и вторичные. Первичные бубоны воз-
никают лимфогенно и связаны с меслом внедрения возбудителя, причем возможны
случаи наличия многих бубонов. Вторичные бубоны такой территориальной связи с
' окализацией входных ворот не имеют и возникают, как правило, гематогенно, что
связано с септицемическим характером туляремийного процесса. Частота и локали-
зация бубонов при разных вспышках неодинаковы, что зависит от путей заражения.
При промысловых вспышках пре обладают подмышечные бубоны (у охотников), при
водных и алиментарных вспышках — шейные и подчелюстные. Бубоны бывают оди-
ночные и множественные, односторонние и двусторонние, значительно варьируя по
величине — от мелкого ореха до куриного яйца и больше. Как правило, в процесс
вовлекается не отдельный регионарный узел, а целая группа лимфатических узлов
данной области и отчасти прилежащие ткани.
Исходы туляремийных бубонов бывают разные: рассасывание, нагноение, изъязв-
ление с последующим рубцеванием при заживлении и, наконец, склеротизация —
стойкое уплотнение вследствие замены ткани узла соединительной тканью с после-
дующим ее сморщиванием. Рассасывание бубонов идет медленно и волнообразно, со
сменой периодов улучшения и обострения. Это отражается на общей продолжительно-
сти туляремии, которая при бубонной форме может затягиваться до 3—4 мес. и более.
У некоторых больных отмечаются воспалительные изменения кожи на мест е вход-
ных ворот инфекции. Эти изменения первичного характера обосновывают выделение
сожно-бубонной или язвенно-железистой формы туляремии, так как в этих случаях,
как правило, присоединяется регионарный лимфаденит. На коже можно последова-
тельно отметить пятно, папулу, везикулу, пустулу, язву. Язвы заживают медленно и
обычно путем рубцевания.
Кроме описанных кожных изменений первичного порядка, при туляремии может
наблюдаться вторичная сыпь, чаше аллергического характера (эритемы, папулы, ве-
зикулы, петехии и т. д. с разнообразной локализацией) нередко симметричной, чаще
во втором периоде болезни. При исчезновении высыпаний обычно наблюдается ше-
лушение, иногда огрубевидное, а иногда крупнопластинчатое. Такая вторичная сыпь
особенно часто наблюдается при генерализованной форме туляремии.
Если возбудитель проникает через слизистые оболочки глаза, возникает бубонная
форма туляремии с первичными измененими со стороны органа зрения (так называе-
мая железисто-глазная или глазо-бубонная форма). Помимо регионарного лимфаденита
(поражение околоушных, переднешейных, подчелюстных и других узлов), характерен
резкий конъюнктивит, иногда наличие папул и язвочек на оболочках глаза.
726
Часть 3. Частная неправильная эпидемиология
При ангинозно-бубонной форме имеют место местные изменения со стороны мин-
далин: гиперемия, гиперплазия, наличие налетов серовато-белого цвета, чаще на од-
ной стороне, боли и затруднения при глотании. Дужки и язычок нередко отечны и
гиперемированы. Налеты могут сначала располагаться островками, позже сливаться,
напоминая картину дифтерии зева. В дальнейшем часто развивается некротический
процесс с образованием глубоких, медленно заживающих язв. Миндалины в этих
случаях, очевидно, и являются местом первичного внедрения возбудителя. Эти слу-
чаи чаще встречаются при пищевых и особенно водных вспышках туляремии. У та-
ких больных позже появляются шейные, околоушные, подчелюстные и другие бубо-
ны с их обычным течением.
При абдоминальной форме туляремии типичны сильные боли в животе в связи с
лимфаденитом, главным образом мезентериальных лимфатических узлов. Этот вари-
ант не следует отождествлять с генерализованной формой, так как при нем явно пре-
обладает поражение лимфатических узлов определенной (абдоминальной) области.
У некоторых больных боли в области живота могут быть настолько сильными, что
весьма напоминают картину «острого живота» и могут повлечь хирургическое вмеша-
тельство из-за ложного диагноза («аппендицит», «холецистит» и т. п.).
Для генерализованной (тифоидной или септической) формы туляремии типично
развитие общих симптомов болезни без предшествующих локальных изменений, ко-
торые в дальнейшем, как правило, отсутствуют. Может развиваться в результате ин-
галяционного поражения. Это единственная клиническая форма туляремии без пер-
вичной местной и регионарной реакции. При ней характерны упорные и тяжкие
головные боли, резко выраженные адинамия и мышечные боли, стойкое снижение
аппетита. Лихорадка длится до 3 нед и более, а температурная кривая часто волнооб-
разная. При этой форме гораздо чаще, чем при других, наблюдается сыпь типа экссу-
дативной полиморфной эритемы. Сыпь появляется обычно во второй половине за-
болевания, нередко симметричными высыпаниями на нижних и верхних конечностях,
а у отдельных больных — на лице, шее, груди (в, виде «перчаток», «гетр», «носков»,
«воротника», «маски»). Сыпь имеет чаще розово-красный цвет, затем принимает баг-
рово-медный оттенок, под конец с синеватым отливом и держится 8—12 сут. Высы-
пание может сопровождаться припуханием суставов (например, кисти, стопы), их
болезненностью, отечностью пальцев, что затрудняет движение.
Легочная (торакальная) форма (поражение дыхательных путей) патогенетически
возникает при ингаляционном заражении, однако она не всегда характеризуется раз-
витием первичного воспалительного процесса в паренхиме легких или поражением
верхних дыхательных путей (рис. 3.49).
От первичной легочной формы следует отличать вторичную, которая может как
осложнение присоединиться к любой другой клинической форме туляремии, возникая
метастатически в более поздние дни и ухудшая течение болезни. Последняя, в свою
очередь, имеет два патогенетических варианта:
а) собственно альвеолярно-метастатический при метастазах (гематогенных) в пар-
енхиму легкого;
б) бронхоаденитический при лимфогенных заносах в бронхопульмональные узлы
с последующим продвижением процесса от пораженных лимфатических узлов на
паренхиму.
Альвеолярно-метастатический вариант вторичных туляремийных пневмоний (ге-
матогенно возникающий) отличается большим полиморфизмом в смысле топики
процесса, наличия многоочаговости в легких и присоединения осложнений. Бронхо-
3.3. Туляремия
727
Рис. 3 49. Рентгенологические варианты течения первичной легочной туляремии.
А. Хиларное расширение (показано стрелкой) в грудной клетке 24-летнего ферме-
ра. Из симптомов отмечалась только лихорадка. Симптомов со стироны респира-
торного тракта не было. Верхний снимок сделан через 13 сут после начала болезни,
нижний — через 10 нед после начала болезни. Б. Правостороннее затемнение в лег-
ких (показано стрелками) у 16-летнего мужчины с лихорадкой и продуктивным
кашлем. Верхний снимок сделан через 11 сут от начала болезни, нижний — ч< рез
2 мес. от начала болезни. По A. Tamvik, L. Berglund (2003)
аденитический вариант связан с так называемой «лимфотропностью» возбудителя.
Патогенетически же здесь определяется большая частота бронхоаденитов, которые
иногда являются отправными очагами для вторичных туляремийных пневмоний.
При типичной первичной легочной форме туляремии воспалительный процесс с
начала болезни развивается в легких. При этом возможны два основных варианта те-
чения заболевания: в первом случае процесс захватывает лишь крупные верхние ды-
хательные пу ги (бронхитпический, или так называемый гриппоподобный, вариант), во
зтором — более глубокие разделы легких — легочную паренхиму {бронхопневмониче-
ский вариант).
Клиническая картина разнообразна в зависимости от локализации, интенсивно-
сти и сочетания воспалительных изменений (туляремийные бронхиты, ларингиты,
трахеиты, их сочетание и т. д., что чаще диагностируется как грипп). В отличие от грип-
па здесь нет капельной передачи инфекции, обычно отсутствует начальный период
острых катаральных симптомов, отмечается скудость объективных данных, и наблю-
дается более затяжное течение.
Типичная туляремийная пневмония (пневмонический вариант течения болезни)
отличается вялым и изнуряющим течением, длится по 2 мес. и более. Патологоанато-
мически — это лобулярная, очажковая бронхопневмония. Характерна наклонность
728
Часть 3. Частная неправильная эпидемиология
к слиянию отдельных очажков, что может создать сходство с долевой пневмонией.
Объективно, помимо сухих хрипов (что наблюдается всегда при бронхитическом ва-
рианте), ясно обнаруживаются влажные, чаще крепитирующие и мелкопузырчатые
хрипы. У некоторых больных они прослушиваются даже в период выздоровления. Бла-
годаря глубокому расположению легочных очагов, клинически распознать туляремий-
ную пневмонию обычно нелегко. По литературным данным, совпадение клиничес-
ких и анатомических диагнозов в этих случаях наблюдалось в 36 % случаев.
Пневмония протекает тяжело, не имеет циклического течения, обнаруживает
наклонность к рецидивам и образованию различных специфических и иных ослож-
нений (бронхоэктазии, гангрена легких, абсцессы, плевриты сухие и экссудативные,
иногда геморрагические и т. д.). Некротизация в пораженных участках легкого может
вести к образованию полостей различной величины или туляремийных каверн. Ос-
ложнения резко затягивают течение болезни и могут вести к смерти.
Так называемый гриппоподобный, или, правильнее, бронхитический, вариант
легочной формы туляремии характеризуется поражением крупных воздухоносных
путей (трахея, крупные и средние бронхи). Объективно это разлитой бронхит, редко
большой интенсивности.
Из специфических осложнений, кроме упомянутых вторичных пневмоний, наи-
более тяжелыми, но редкими являются осложнения со стороны ЦНС (туляремийный
вторичный менингит, менинго-энцефалит и т. п.). В период выздоровления нарушения
со стороны нервной и особенно ССС продолжаются длительное время (вегетативные
неврозы, миодегенерация сердца и т. п.). Остаточные явления в легких после туляре-
мийных пневмоний удается подтвердить рентгенологически через 3—4 мес. и позже.
Наслоение обострений и рецидивы, в свою очередь, отражаются на общей длитель-
ности процесса. При этом, кроме обострений и ближних рецидивов (через 3—5 нед),
возможны поздние рецидивы (спустя ряд месяцев и даже свыше года), т. е. затяжные
или так называемые хронические формы туляремии.
Моделирование ингаляционного заражения F. tularensis. Эксперименты с аэрозоля ми
F. tularensis, однородными по дисперсности частиц, выполнены W. С. Day и R. Е Ве-
rendt (1972) на 96 обезьянах (Macaco mulatto) массой 4-5 кг. Ими исследовались осо-
бенности патологического процесса, вызванного жидкими аэрозолями F. tularensis
SCHU-S5 со средним диаметром частиц 2,1; 7,5; 12,5 или 24,0 мкм (рис. 3.50).
Животные, экспонированные к аэрозолю с частицами со средним диаметром
2,1 или 7,5 мкм, оказались инфицированы все. Начало клинических симптомов отме-
Рис. 3.50. Микрофотографии аэрозольных частиц разных размеров, содержащих воз-
будитель туляремии. А. Частицы со средним диаметром 12,5—6,5 мкм. Б. Частицы
со средним диаметром 8,0—5,0 мкм. В. Частицы со средним диаметром 1,0—2,5 мкм.
Увеличение в 2000 раз, фазовый контраст. По W. С. Day и R. F.Berendt (1972)
3.3. Туляремия
729
чали через 2—3 сут после экспонирования к аэрозолю, и первым симптомом было по-
вышение ректальной температуры до 40,0—41,1 °C. После первоначального повыше-
ния температуры, которое обычно продолжалось от 2 до 4 сут, ректальная температу-
ра снижалась до 35,0—36,7°С. Обезьяны, проявляющие такую реакцию, обычно
погибали. Постепенно происходило повышение СОЭ и количества клеток белой кро-
ви. Обнаружено интенсивное заражение нижних дыхательных путей. Рентгенологи-
ческое исследование показало тяжелую лобулярную инфильтрацию у большинства
животных. Со временем затемнение легких увеличивалось, позволяя предполагать об-
разование некрозов и абсцессов. На вскрытии легкие животных были гиперемирова-
ны и покрыты большими участками уплотнений с множеством некротических оча-
гов на поверхности. Бронхиальные лимфатические узлы имели желтый цвет и были
плотными так же, как и селезенка и печень, в которых исследователи выявили много
небольших очагов, окруженных неровными участками некроза. Животные погибли
через 4—8 сут после экспонирования к аэрозолю.
У обезьян, экспонированных к аэрозолю F. tularensis SCHU-S5 с частицами раз-
мера 12,5 или 24,0 мкм, инкубационный период был продолжительным (6—10 сут),
симптомы болезни менее выраженными, чем у обезьян, экспонированных к части-
цам меньшего размера. Первым проявлением болезни было незначительное повыше-
ние ректальной температуры, невыраженный лейкоцитоз. По мере развития болезни
у животных отмечали потерю аппетита, сопровождаемую хронической диареей. За-
тем обезьяны впадали в прострацию и погибали через 7—10 сутпосле экспонирования.
У 7 животных (из 45 животных этой группы), экспонированных к частицам аэрозоля
12,5 мкм и 24,0 мкм, развился тяжелый конъюнктивит, сопровождаемый гнойными
выделениями из носа и глаз, повышением ректальной температуры и увеличением ко-
личества бактерий в крови. Обезьяны, у которых развивался конъюнктивит, погиба-
ли между 10—21 сут после экспонирования. На вскрытии у обезьян, экспонирован-
ных к частицам 12,5 мкм и 24,0 мкм, выявлено только поражение верхних дыхатель-
ных путей с гиперемией слизистой оболочки в области носоглотки и увеличением
шейных и челюстных лимфатических узлов. У нескольких обезьян небольшие пора-
жения были обнаружены в нижних дыхательных путях, однако они не были такими
обширными как те, которые наблюдали у животных, экспонированных к аэрозолю
возбудителя туляремии со средним диаметром частиц 2,1 или 7,5 мкм. Эти пораже-
ния могли быть вызваны гематогенным распространением F. tularensis.
Показатели респираторных LD50 и ID50 повышались по мере увеличения средне-
го диаметра частиц аэрозоля туляремийного микроба от 2,1 до 24,0 (табл. 3.18).
Более поздние исследования лишь дополняли на различных экспериментальных
моделях данные W. С. Day и R. Е Berendt (1972) по частным аспектам патогенеза и
патоморфологии ингаляционного поражения обезьян аэрозолем F. tularensis. Как
правило, исследователи используют штамм SCHU-S5, средний диаметр ингалируемых
частиц в пределах 1—5 мкм (африканские зеленые обезьяны — Twenhal N. A. et al., 2009;
обезьяны мармозетки — Nelson М. et al., 2010).
N. A. Twenhal et al. (2009) характерной патоморфологической особенностью по-
ражения мелкодисперсным аэрозолем возбудителя туляремии (средняя доза 729 КОЕ)
считают наличие некротических фокусов преимущественно в легких, селезенке, тра-
хее, бронхах и медиастинальных лимфатических узлах. Сходные, но менее выражен-
ные поражения были обнаружены ими в мышцах сердца, диафрагме, брыжейке, эпи-
телиальной поверхности уринарного тракта. Гистологически эти участки повреждения
представляют собой хорошо отграниченные области паренхимальной дегенерации
730
Часть 3. Частная неправильная эпидемиология
Таблица 3.18
Респираторные LD^ и ID50 аэрозолей F. tularensis SCHU-S5 для
обезьян Масаса mulatto при различных распределениях частиц по размеру*
Средний диаметр частицы (мкм) ЬОда КОЕ 95 % доверительный интервал, КОЕ ю50, кое
2,1 14 7-27 <5
7,5 378 50-630 <240
12,5 872 250-1500 <556
24,0 4447 1600-8500 340
* По W. С. Day и R. Е Berendt (1972).
и некроза с утратой архитектуры ткани. Они содержат большие количества макрофа-
гов и жизнеспособных и деградировавших Нейтрофилов (рис. 3.51 и 3.52).
Легочная паренхима между участками некроза подвергается сжатию, и альвеоляр-
ное пространство часто оказывается затопленным в результате отека, геморрагий и
«наплыва» макрофагов с участками обильно пенящейся цитоплазмы (abundant foamy
Рис. 3.51. Патоморфо югические особенности поражения мелкодисперсным аэрозолем
возбудителя туляремии органов и тканей обезьян. А. Легкое. В пределах каждой легоч-
ной доли имеется множество сливающихся белых очагов с поперечным размером от
0,1 до 1 см, слегка выступающих над поверхностью легкого. Доли не коллапсированы с
закругленными краями. Паренхима легкого между участками поражений имеет темно-
красную окраску, что свидетельствует о застойной гиперемии и отеке. Черная полоса
соответствует I см. Б. Разрезанное легкое. Поражения в пределах паренхимы. В норме
1 ладкая плевральная поверхность вы глядит сухой и гранулированной в участках развив-
шегося туляремийного плеврита. Черная полоса соответствует 0,5 см. В. Трахеоброн-
хиальный лимфатический узел. В пределах лимфатического узла отче гливо видны мно-
гочисленные неправильной формы выпуклые поражения. Черная полоса соответствует
0,3 см Г. Селезенка. Множество участков поражения белого цвета размером от 0,1 до
0,2 см в диаметре. Черная полоса соответствует 1 см. По N. A. Twenhal et al. (2009)
3.3. Туляремия
731
Рис. 3 52. Гистологические особенности поражения мелкодисперсным аэрозолем воз-
будителя туляремии органов и тканей обезьян. А. Легкое. Множественные некротиче-
ские участки замещают паренхимальную ткань. Участок плевры отделен от участка по-
ражения паренхимы легкого. Вокруг участка поражения паренхима отечна, имеются
геморрагии. Б. Легкое. Участок поражения выходит в воздушный путь среднего размера
(звездочка). Вокруг зоны центрального некроза расположено большое количество мак-
рофагов с микровакуолизованной цитоплазмой (вставка). Воз 1ушный путь искривлен
под воздействием расширяющегося участка повреждения паренхимы. В. Кровеносный
сосуд легкого. Некротизированный васкулит кровеносного сосуда в участке повреж-
дения легочной ткани. Разрушена tunica intima. Tunica media и tunica adventitia расши-
рены клеточным дебрисом, дегенерированными нейтрофилами, микровакуолизиро-
ванными макрофагами, фибрином, отеком и геморрагиями. Г. Трахея. Клеточный
дебрис расширяет субэпителий, формируя куполообразное повреждение (звездочка).
Эго приводит кдегенерации и утрате респираторного эпителия (стрелки). Субэпителий
расширен за счет давления, создаваемого отеком. По N A. Twenhal et al. (2009)
cytoplasm). Для плеврита характерна некротизация плевры, пиогранулематозное вос-
паление, отек и геморрагии. Архитектура селезенки нарушается. Белая пульпа, крас-
ная пульпа, капсула разрушаются и заменяются участками некроза, макрофагами и
деградировавшими нейтрофилами. Медиастинальные лимфатические узлы содержат
множественные некротические участки, вплоть до полного некроза узла.
Е. L. Overholt et al. (1961)1 описали 42 случая ингаляционного заражения F. tularensis
сотрудников бактериологической лаборатории. Ниже приведена одна из наиболее
показательных историй болезни.
Случай 15. Негр, 43 года, поступил в госпиталь 16 февраля 1958 г. От первой вак-
цинации до заболевания прошло 7 лет2. Три последние ревакцинации были сделаны
за 4 года до заболевания. Титр специфических агглютининов в октябре 1957 г. состав-
лял 110. Достоверных данных о времени и обстоятельствах заражения нет
1 Ведомственная принадлежность лаборатории, где заразились эти люди, не указана, сами
авторы из Армейского медицинского центра Уолтера Рида (The United States Army Medical Unit,
Walter Reed Army Medical Center, Fort Detrick, Frederick, Maryland).
2 Вакцинировался он инактивированной туляремийной вакциной.
732
Часть 3. Частная неправильная эпидемиология
За 5 сут до госпитализации у него внезапно появился озноб, боли по всему телу,
заложенность носа и боль за грудиной. При обследовании в поликлинике через 3 ч
после начала заболевания никакой патологии обнаружено не было. Правда, температу-
ра в это время повысилась до 37,2 °C (измерение в полости рта), отмечался лейкоци-
тоз (14900) с нейтрофилезом, СОЭ — 19 мм/ч. Реакция агглютинации не выявила по-
вышения титра специфических антител. Больному назначили симптоматическое
лечение и посоветовали повторно обратиться к врачу, если самочувствие не изменит-
ся или ухудшится. В следующие 5 сут у него появилась сильная потливость, лихорад-
ка, боль в горле, кашель с мокротой, тошнота, рвота, вялость, слабость. Больной был
вынужден лечь в постель. В это время не отмечалось ни диспноэ, ни кровохарканья,
ни поноса. Температура колебалась от 40 до 41,1 °C (измерение в полости рта).
На 6 сут болезни он вновь был осмотрен врачом. Больной задыхался, казался испу-
ганным, производил впечатление внезапно тяжело заболевшего человека. Ректальная
температура — 40,6 °C, АД — 130/70 мм рт. ст., пульс — 130/мин, ритмичный, хорошего
наполнения. Слизистая носа была умеренно отечна, зев диффузно гиперемирован.
В верхних двух третях левого легкого определялся при перкуссии тупой звук, а при аус-
культации — крепитируюшие хрипы и шум трения плевры. Справа никакой патоло-
гии выявлено не было. Живот мягкий, перистальтика обычная. В день госпитализа-
ции количество лейкоцитов — 8800, имел место нейтрофилез (87 %), юных форм
насчитывалось до 70 %, СОЭ — 32 мм/ч, а реакция на С-реактивный белок дала 4+.
В моче определялся белок (2+). На прямой рентгенограмме был виден диффузный,
негомогенный инфильтрат в верхней доле левого легкого, расширение корня этого же
легкого и сетчатый инфильтрат в правом кардиодиафрагмальном углу. На рентгено-
грамме, снятой в боковой проекции, обнаружен воспалительный очаг, занимавший
весь верхнезадний сегмент верхней доли левого легкого.
Выраженность расстройства дыхания, ненормальное количество лейкоцитов, ин-
фильтрат в легком и увеличение лимфоузлов ворот легкого позволили поставить пред-
положительный диагноз туляремии. Через час после госпитализации больной получил
«ударную» дозу тетрациклина (1 гperos), а затем принимал этот антибиотик по 0,5 г
каждые 6 ч. В результате через 24 ч температура упала с 41,1 до 38,3°С. Однако по-преж-
нему сохранялись одышка (60/мин), цианоз и шумное дыхание. Во время кашля вы-
делялось немного мокроты. При аускультации по всей поверхности обоих легких
прослушивались влажные хрипы. В связи с развитием тяжелого поноса пришлось от-
менить тетрациклин и назначить больному внутривенные инъекции окситетрациклина
по 1,5 г/сут. Инъекции продолжались 5 сут. Назначена поддерживающая терапия.
Несмотря на то что температура снизилась, и не было падения деятельности ССС,
больной находился в критическом состоянии. Из-за развившегося бреда ему был на-
значен паральдегид. В это время частота пульса до 150/мин. Больного по-прежнему
мучил кашель; цианоз не уменьшался; отсосать слизь из бронхов не удавалось.
На 4 сут госпитализации больному была произведена трахеотомия. После трахео-
томии частота дыхания снизилась до 40/мин, дыхание стало тихим; при аускультации
в обоих легких прослушивалось меньше хрипов. На 6 сут госпитализации стало воз-
можным отменить внутривенные инъекции жидкостей и вновь было начато введение
тетрациклина peros по 3 г/сут. Лечение тетрациклином продолжалось в общей слож-
ности 21 сут. Рентгенограмма, снятая на 7 сут болезни, выявила в обоих легких мно-
жественные воспалительные очаги диаметром до 0,5 см. На 13 сут госпитализации тем-
пература тела и частота дыхания нормализовались; сохранялся небольшой кашель с
мокротой. На 14 сут больного беспокоила головная боль и боль в пояснице; ректаль-
ная температура 38,4 °C. Так как в это время в госпитале была вспышка гриппа, у боль-
ного взяли для бактериологического исследования смыв из зева. Удалось выделить
3.3. Туляремия
733
вирус японского гриппа типа А-57. В смывах из зева, взятых ранее, этот вирус обнару-
жен не был. Еще. через 3 сут все симптомы исчезли.
На 3 сут терапии удалось выделить штамм F tularensis, устойчивый к стрептоми-
цину. Этот микроорганизм обнаружен в крови больного, взятой на 2 сут госпи-
тализации, а также в промывных водах желудка и смывах из зева. Бактерии F. tularensis
содержались и в материале, взятом из трахеи на 6 сут лечения. Бактериологическое
исследование мокроты на 2 нед лечения дало отрицательные результаты.
Кожная проба была в 14 сут заболевания отрицательной, положительной она стала
на 28 сут. На2 нед болезни титр агглютининов поднялся с 1:160 до 1:1280, анаЗ и 4 нед
достиг 1:2560.
Месяц спустя, больной чувствовал себя хорошо, прибавил в весе 25 фунтов, одыш-
ка при нагрузке исчезла. Реакция на С-реактивный белок стала отрицательной уже на
25 сут болезни, но СОЭ оставалась ускоренной еще в течение месяца. Пневмониче-
7ские инфильтраты в легких полностью рассосались
Этот случай иллюстрирует крайнее проявление ингаляционной туляремии. Боль-
ному повезло. Врачи знали не только то, что его болезнь вызвана F. tularensis, но и об
ее устойчивости к стрептомицину. Поэтому для лечения ими сразу же был применен
тетрациклин. Можно предположить, что инфицирование сотрудника лаборатории
произошло в результате случайного экспонирования к полидисперсному аэрозолю
возбудителя туляремии, часть частиц которого проникла в альвеолы (1—5 мкм), бо-
лее крупные (> 5 мкм) задержались в верхних дыхательных путях.
Диагностика искусственногс поражения возбудителем туляремии. Принципы диаг-
ностики искусственного поражения возбудителем туляремии те же, что описаны в
разд. 3.1 для поражения возбудителем сибирской язвы. Ключевым моментом в диаг-
ностике туляремии, являющейся следствием нападения с применением F. tularensis в
качестве поражающего агента БО, может быть большое поступление больных за ко-
роткий промежуток времени со сходным системным заболеванием и непродуктивной
пневмонией. Клинические проявления туляремии могут быть тяжелыми, но неспе-
цифическими. Основным критерием искусственного происхождения вспышки туля-
ремии является ее неправильная эпидемиология. Невозможность отнести эпидемио-
логию вспышки к одному из эпидемиологических типов естественных туляремийных
вспышек (см. табл. 3.15), должно вызвать подозрение у эпидемиологов о преднаме-
ренном распространении возбудителя туляремии. Способ распространения F. tula-
rensis, выбранный преступниками, можно предположить по клинической форме
болезни (ингаляционно, алиментарным путем, парэнтерально и др.; см. табл. 3.17).
К неправильной эпидемиологии болезни нужно отнести выделение от людей возбу-
дителя туляремии не встречающегося в России подвида tularensis, если его распрост-
ранение не обнаружено в природных очагах, которые посещали заболевшие туляре-
мией. При выявлении среди людей вспышек туляремии, вызванных характерным для
России подвидом holarctica, должна быть установлена их связь с местностями, где он
циркулирует среди грызунов.
Ориентироваться при расследовании биопреступления на «невозможные клини-
ческие формы болезни» вне их связи с ее неправильной эпидемиологией невозмож-
но из-за особенностей факторов вирулентности F. tularensis и патогенеза болезни, а
также низкой инфицирующей дозы возбудителя туляремии для людей. Особенности
факторов вирулентности F. tularensis и патогенеза болезни делают маловероятными
какие-то «клинические зацепки» для выявления «невозможной клинической формы
болезни» при парентеральном введении возбудителя туляремии. Местных проявле-
734 Часть 3. Частная неправильная эпидемиология
ний, например, таких, которые дает сибиреязвенный токсин при парентеральном
введении В. anthracis, при инъекционном введении F. tularensis не будет. Болезнь ра-
зовьется в бубонной, язвенно-бубонной или первично-септической форме.
Аэрозоль с дисперсностью частиц 1 —5 мкм образуется лри разделке тушек боль-
ных туляремией животных в ничтожных количествах, однако его иногда оказывается
достаточно, чтобы вызвать легочную форму болезни с поражением глубоких отделов
легких у людей, находящихся даже на расстоянии от источника инфекции. Например,
А. М. Hauri et al. (2010) описали групповой случай ингаляционного заражения людей
туляремией, находящихся на расстоянии 5 м от места промывки струей воды гушки
больного туляремией зайца. Один из заболевших погиб с диагнозом «двухсторонняя
пневмония», несмотря на лечение моксифлоксацином (moxifloxacin). Ретроспектив-
но у него была установлена легочная форма туляремии. Поэтому клинически и рент-
генологически трудно установить различия между бронхопневмоническим вариантом
течения легочной формы туляремии у людей, вызванных заражением в ее природных
очагах, и таким же вариантом течения болезни, вызванным ингаляцией F. tularensis в
составе преднамеренно созданного аэрозоля. Кроме того, клинические и рентгено-
логические признаки легочной формы туляремии довольно изменчивы, что затруд-
няет ее диагностику независимо от того, произошло инфицирование в природных ус-
ловиях, либо оно стало результатом биопреступ^ения.
В то же время, если установлено, что вспышка легочной туляремии вызвана рас-
пылением аэрозоля, го тщательный анализ клинических особенностей болезни мо-
жет многое рассказать специалистам о типе тиспергирующего устройства, характери-
стиках примененной рецептуры, инфицирующей дозе и даже о расстоянии, на кото-
ром находились жертвы преступления от источника аэрозоля. Поражение глубоких
отделов легких — бронхопневмонический вариант туляремии вызовет аэрозоль в диа-
пазоне частиц 1-5 мкм. Бронхитический, или так называемый гриппоподобный, ва-
риант туляремии будет вызван более крупной фракцией распыленного аэрозоля. Но
этот аэрозоль имеет и более высокую скорость оседания частиц, чем аэрозоль со сред-
ним размером частиц в диапазоне 1 -5 мкм, его распространение ограничено значи-
тельно меньшим расстоянием от источника, чем аэрозоля 1—5 мкм. Существует прямая
зависимость между величиной ингаляционной дозы возбудителя туляремии и коли-
чеством первичных легочных поражений. Есть и другие тонкости агробиологии туля
ремийного микроба, известные специалистам.
При обнаружении культур возбудителя туляремии в лаборатории преступников
или изъятия у них рецептуры туляремийного микроба приготовленной в неустанов-
ленной лаборатории, необходимо провести их сопоставительный MLVA-анализ и ана-
лиз полногеномного полиморфизма со штаммами F. tularensis, выделенными от жертв
биопрсступлен ия.
Иммунопрофилактика. В России иммунопрофилактика туляремии осуществляет-
ся с помощью вакцины туляремийной живой сухой. Получена на основе F. tularensis
подвидаholarctica (тип В). Содержит лиофильновысушенную культу руживого вакцин-
ного штамма 15 НИИЭГ и стабилизаторы — сахарозу, натрий глутаминовокислый,
тиомочевину, желатин. Имеет вид пористой массы желтовато-белого цвета. Через 20—
30 сут после вакцинации обеспечивает развитие иммунитета к возбудителю туляре-
мии продолжительностью до 5 лет. Вакцинация проводится накожным и внутрикож-
ным способом. Производится ФГУП «НПО «Микроген» М3 РФ (г. Омск).
В СССР вакцина использовалась для массовой иммунизации людей с 1946 г.
В середине 1950-х гт. ее передали в США где она известна как вакцина LVS. Амери-
3.4. Бруцеллез
735
канскими исследователями показана ее высокая эффективность при ингаляцион-
ном заражении возбудителем туляремии. Добровольцев иммунизировали LVS при-
близительно за 1 год до заражения. Заражение проводили ингаляционно штаммом
SCHL S4. Доза для заражения варьировала от 200 до 20 тыс. КОЕ. Результаты экспе-
римента показали, что большинство вакцинированных добровольцев, зараженных
дозой до 2 тыс. КОЕ, не заболевали туляремией. У иммунизированных доброволь-
цев, зараженных 20 тыс. КОЕ, обнаружились более легкие симптомы заболевания по
сравнению с неиммунизированными добровольцами, инфицированными аналогич-
ной лозой SCHU S4(EigelsbachH. Т. etal., 1967). На сегодняшний день вакцина LVS —
единственная в мире эффективная вакцина против туляремии.
Лечение. Аминоглигазиды стрептомицин и гентамицин обладают бактерицидным
действием против F. tularensis и в настоящее время используются для лечения туляре-
мии. Полевое наставление по лечению пораженных боевыми биологическими аген-
тами армии США (Army FM 8-284, 2000) для антибиотикотерапии туляремии реко-
мендует одну из следующих схем:
стрептомицин, 7,5—10 мг/кг внутримышечно каждые 12 ч в течение 10—14 дней;
гентамицин, 3—5 мг/кг внутривенно ежедневно в течение 10—14 дней;
ципрофлоксацин, 400 мг внутривенно каждые 12 ч с переключением на перораль-
ное введение ципрофлоксацина (500 мг каждые 12 ч) после достижения клинического
улучшения; продолжать до завершения 10-14-дневного курса лечения;
ципрофлоксацин, 750 мг перорально каждые 12 ч в течение 10—14 дней.
Экстренная профилактика туляремии сразу после нападения включает назначе-
ние доксициклина в дозе 100 мг перорально каждые 12 ч в течение 2 недель; или тет-
рациклина в дозе 500 мг перорально каждые 6 ч в течение 2 недель; или ципрофлок-
сацина в дозе 500 мг перорально каждые 12 ч в течение 2 недель.
3.4. БРУЦЕЛЛЕЗ
Микробиология бруцелл. Факторы патогенности. Таксономия, происхожде-
ние и молекулярная эпидемиология бруцелл. Экология и естественная эпи-
демиологе бруцелл. Восприимчивость к оруцеллам и инфицирующие дозы
для животных и людей. Иммунитет. Устойчивости бруцелл во внешней среде.
Антибиотикорезистентность бруцелл. Обнаружение генетически измененных
штаммов бруцелл. Патогенез бруцеллеза. Клиническая картина бруцеллеза
у людей при естественном инфицировании. Моделирование ингаляционного
заражения бруцеллами. Диагностика искусственного поражения бруцеллами.
Иммунопрс филагтика бруцеллеза. Лечение бруцеллеза.
Бруцеллез (мальтийская, средиземноморская, неаполитанская, кипрская, ундули-
рующая, козья лихорадка; септицемия Брюса; болезнь Банга; болезнь Траума) — опас-
ная сапрозооно >ная инфекционная болезнь людей и животных с высоким потенциа-
лом к хронизации. Протекает как лимфогематогенный хрониосепсис, системный
ретикулоэндотелиоз и артериоваскупит септико-аллсргического характера. Приводит
к поражению опорно-двигательного аппарата, нервной, сердечно-сосудистой и других
систем человека. В настоя: цее время известны 6 основных видов возбудителей бруцел-
леза: Brucella melitensis, В. abortus bovis, В. abortus suis, В. neotomae, В. canis, В. ovis.
736
Часть 3. Частная неправильная эпидемиология
Бруцеллез у людей впервые описан военным хирургом Дж. Марстоном (J. A. Mars-
ton) в 1859 г., наблюдавшим его на острове Мальта во время Восточной войны. Тог-
да же эта болезнь получила название «мальтийская лихорадка». В 1886 г. Д. Брюс
(D. Bruce) обнаружил в пунктатах селезенки умершего человека мелкие бактерии
овоидной формы и дал им название Micrococcuc melitensis. В 1897 г А. Райт (А. Е. Wright)
и Д. Семпл (D. Sample) установили, что сыворотка больных «мальтийской лихорад-
кой» дает реакцию агглютинации с М. melitensis, с тех пор эта реакция под названием
реакции Райта приобрела большое значение в серодиагностике бруцеллеза. Датские
исследователи Б. Банг (В. Bang) и В. Стриболд (В. Stribolt) в том же году из около-
плодной жидкости абортировавшей коровы выделили микроорганизм, названный
В. abortus bovis. В 1914 г. из плода свиньи Дж. Траум (J. Traum) выделил возбудителя
инфекционного аборта свиней — В. abortus suis. Вследствие биологического и мор-
фологического сходства в 1920 г. возбудители аборта коров, коз, свиней и овец были
объединены в группу бруцелл (название дано в честь Д. Брюса), а само заболевание
получило название «бруцеллез». В 1957 г. в США от пустырниковых крыс были выде-
лены микроорганизмы, близкие к бруцеллам, отнесенные к новому виду—В. neotomae.
В 1970 г. к бруцеллам причислены новые виды микроорганизмов:
1) вызывающие орхоэпидидимит у баранов (В. ovis);
2) культуры, полученные от собак (В. canis). На территории дореволюционной
России, в Туркмении бруцеллез был известен под названием козьей лихорадки (Шу-
валова Е. П. с соавт., 2001; П. П. Очкур, 1951).
В природных условиях источниками инфекции для людей являются мелкий и
крупный рогатый скот, свиньи, а в некоторых местностях России и северные олени.
Наиболее важное эпидемиологическое значение принадлежит мелкому рогатому скоту
вследствие облигатной патогенности для человека В. melitensis. Заражение человека
бруцеллами обычно происходит алиментарным или контактным путями. Однако воз-
можен и ингаляционный механизм заражения, обусловленный проникновением мик-
робов через верхние дыхательные пути с пылевыми частицами шерсти, навоза, под-
стилки и земли. В интенсивных очагах, особенно овечьего и свиного бруцеллеза, ос-
новным является контактный путь заражения человека. При этом микробы проникают
через кожные покровы лиц, обслуживающих больных животных или обрабатываю-
щих сырье животного происхождения.
Больной бруцеллезом не представляет опасности для окружающих. Российские
санитарно-эпидемиологические правила СП 1.3.2322-08 относят возбудители бруцел-
леза ко 11 группе патогенности. Постепени важности для национальной безопасности
США возбудители бруцеллеза относятся к биологическим поражающим агентам ка-
тегории В (см. табл. 2.2). С начала 1920-х гг. возбудители бруцеллеза считаются иде-
альными агентами для ведения биологической войны. Исследования по изучению по-
ражающих свойств бруцелл велись в Форт-Детрике (США) с 1944 г.
Наиболее вероятно применение возбудителей бруцеллеза посредством использо-
вания линейных и многоточечных источников и диверсионными (террористически-
ми) методами.
Микробиология бруцелл. Виды бруцелл по морфологическим признакам трудно
различимы. Они не образуют эндоспор, не имеют жгутиков и при определенных ус-
ловиях (воздействие специфического бактериофага, выращивание на средах с добав-
лением 10 % иммунной сыворотки) обладают способностью продуцировать внекле-
точную субстанцию, напоминающую капсулу. Бруцеллы грамотрицательны, обладают
одинаковыми тинкториальными свойствами, легко прокрашиваются анилиновыми
3.4. Бруцеллез
737
спасителями и в отличие от других видов бактерий способны сохранять красную ок-
раску сафранином при последующем докрашивании холодным раствором малахито-
зого зеленого, что послужило основанием для разработки Е. А. Козловским специфи-
ческого для данных микроорганизмов метода окраски. При окраске по Граму бруцеллы
обесцвечиваются, а после докрашивания фуксином Пфейффера становятся красны-
ми, темно- и бледно-розовыми. Большинство из них окрашивается равномерно и не
содержит включений и вакуолей. Однако отдельные палочковидные клетки могут
отличаться интенсивной окраской одного или обоих полюсов (Вершилова П. А., 1972;
Золотарев А. Г. с соавт., 2006).
В окрашенных мазках бруцеллы имеют вид кокков или овоидов, коротких палочек
и располагаются обычно беспорядочно, а иногда — в цепочках, агрегатах либо попар-
но. Микробы В. melitensis чаще всего представлены кокками и овоидами диаметром
0.4—0,8 мкм и длиной от 0,5 до 2,2 мкм. Клетки В. abortus и В. suis несколько крупнее,
и для большинства из них характерна палочковидная форма с закругленными и эллип-
совидными полюсами. Диаметр бруцелл коровьего типа колеблется от 0,4 до 0,6 мкм,
а длина — от 0,5 до 3,0 мкм. У бруцелл свиного типа диаметр клетки находится в пре-
делах 0,6—0,8 мкм, а длина — 0,6—3,0 мкм (Золотарев А. Г. с соавт., 2006).
Нити и цепочки в культурах бруцелл обнаруживаются редко. Под влиянием аби-
отических факторов микробы этих видов могут образовывать нестабильные и стабиль-
ные L-формы, состоящие из кокков различных размеров, зерен, аморфных масс и
гетероморфных форм. Характерной особенностью микробов рода Brucella является
медленный рост на плотных питательных средах. Морфологически оформленные
микроколонии они образуют через 36-48 ч выращивания на FT-arape при темпера-
туре 36—38°C. У бруцелл в S-форме микроколонии по внешнему виду напоминают
голусферу, расположенную на круглом основании. В проходящем свете они плотные,
желтоватые, с гладкой поверхностью и ровными округлыми краями (рис. 3.53, А).
Через 72—96 ч культивирования у изученных видов бруцелл образуются одина-
ковые по морфологии колонии в S-форме. Визуально в проходящем свете они бес-
цветные, прозрачные или с перламутровым оттенком, гомогенные либо с нежной зер-
нистостью в центре, а в падающем свете молочно-белые и не имеют по форме, цвету
и структуре видовых отличий (см. рис. 3.53, Б). С возрастом колонии могут мутнеть и
становиться буропигментированными. При микроскопическом исследовании с ана-
логичным освещением колонии белые, сферической формы, с ровными, четко кон-
турированными краями и гладкой блестящей поверхностью, отражающей лучи света
(см. рис. 3.53, В). Аналогичные по морфологии колонии в S-форме образуют вакцин-
ные штаммы В. melitensis Rev и В. abortus 19 ВА. Размеры колоний бруцелл отличаются
вариабельностью. Они зависят от видовых, штаммовых и функциональных особен-
ностей микробной популяции, состава питательных сред и продолжительности куль-
тивирования. В среднем диаметр крупных колоний бруцелл достигает 4 мм, а у мелких,
точечных — колеблется в пределах от 0,05 до 0,1 мм. Вакцинные штаммы В. melitensis
Rev-1 и В. abortus 19 ВА отличаются значительно меньшей скоростью роста по срав-
нению с вирулентными штаммами, и их колонии достигают диаметра 2,5—3,0 мм лишь
к 5 сут инкубирования (Золотарев А. Г. с соавт., 2006).
Бруцеллы характеризуются выраженной склонностью к диссоциации в случае
использования питательных сред неадекватного состава, причем изменениям морфо-
логии колоний предшествуют изменения их внутренней структуры и компонентного
состава. Из числа способов, предложенных для выявления отмеченных особенностей
колоний бруцелл в процессе диссоциации, широкое практическое применение
738
Часть 3. Частная неправильная эпидемиология
Рис. 3.53. Макроскопические изображения колоний бруцелл. А. Морфология мик-
роколонии бруцелл, выращенных в течение 36 ч на FT-arape (В. suis 1300). Микро-
фотосъемка в проходящем свете. Увеличение в 56 раз. Б. Внешний вид пластинок
агара с колониями бруцелл, выращенных в течение 72 ч на FT-arape (В. suis 1300). Фо-
тосъемка в падающем свете. В. Морфология колоний вирулентных штаммов бруцелл
(В. abortus 544). Микрофотосъемка в падающем свете. Увеличение в 13 раз. Г. Морфо-
логические особенности S- и R-форм колоний бруцелл, кристаллическим фиоле-
товым окрашенных по методу Уайта — Уилсона, полное прокрашивание. Гладкие
колонии (S-формы бактерий) остаются белыми, R-формы прокрашиваются. Макро-
фотосъемка в падающем свете. Увеличение в 3 раза. По А. Г. Золотареву с соавт. (2006)
получил метод Уайта — Уилсона, основанный на различной способности S- и R-ko-
лоний окрашиваться генциановым фиолетовым. Под действием этого красителя дис-
социирующие колонии становятся темно-фиолетовыми или розовато-синими, при-
чем в зависимости от степени диссоциации они прокрашиваются полностью или
частично. Колонии в S-форме остаются бесцветными (см. рис. 3.53, Г) (Золотарев А. Г.
с соавт., 2006).
Бруцеллы биохимически активны. Имеются сведения о значительном наборе раз-
нообразных ферментов, выделяемых бруцеллами разных видов. К ним относятся ка-
талаза, уреаза, гиалуронидаза, пероксидаза, амилаза, липаза, фосфатаза, цитохромок-
сидаза, аспаргиназы и некоторые другие ферменты аминокислотного и углеводного
обмена. Эта активность выражена неодинаково у представителей разных видов бру-
целл (Вершилова Н. П., 1972).
Факторы патогенности. Бруцеллы обладают высокой инвазивностью и проника-
ют в организм человека даже через неповрежденные слизистые покровы. Вирулентные
Brucella инфицируют как нефагоцитирующие, так и фагоцитирующие клетки. В орга-
низме млекопитающих они обитают преимущественно внутри макрофагов и плацен-
тарных трофобластов. Различные виды бруцелл, и даже различные штаммы одного и
того же вида, обладают разной вирулентностью. Вирулентность бруцелл определяет-
ся той минимальной дозой, которая способна вызвать заражение всех взятых в опыт
чувствительных животных (морские свинки и белые мыши) при подкожном введении
3.4 Бруцеллез
739
(ID — инфицирующая доза). К вирулентным культурам относятся такие, которые
инфицируют указанных животных при введении им 5-10 КОЕ (Вершилова П. А.,
1972). Основные типы клеток, являющихся местом обитания В. abortus у КРС и лю-
дей, приведены на рис. 3.54,
Рис. 3.54. Основные типы клеток, являющихся местом обитания В. abortus у КРС
и людей. Внутри нефа!оптирующих клеток бруцеллы имеют тенденцию локали-
зоваться в гранулярном эндоплазматическом ретикулуме. В полиморфноядерных
или одноядерных фагоцитирующих клетках они используют ряд механизмов, позво-
ляющих избежать или подавить бактерицидные реакции. По R. М. Roop et al. (2004)
В результате генетических исследований с использованием метода транспозонного
мутагенеза у различных видов Brucella идентифицированы многочисленные генетиче-
ские локусы, необходимые для их выживания и репликации в культивированных мак-
рофагах и индуцирования и поддержания хронической инфекции у модельных мы-
шей. Однако у бруцелл идентифицировано мало генов, кодирующих традиционные
факторы вирулентности». Механизм инвазии бруцеллами нефагоцитирующих кле-
ток не совсем ясен. Роль капсулы в вирулентности бруцелл не установлена, да и само
ее существование оспаривается. Клеточные компоненты, специфически способствую-
щие клеточной адгезии и инвазии, у бруцелл не определены, а попытки обнаружить
гены инвазина, гомологичные таковым энтеробактерий, оказались безуспешными.
Большинство идентифицированных генов необходимы, по-видимому, для физиоло-
гической адаптации к условиям окружающей бруцеллы среды в репликативной фа-
юсоме. Этс явление хорошо объяснено S. Kohler с соавт. (2003), которые считают, что
характер этих iенов отражает те условия, в которых оказываются виды Brucella в этой
нише. Репликативная фагосома бедна питательными веществами и микроаэробна.
Следовательно, для длительного существования и размножения в этом компартмен-
те бруцеллам необходима метаоолическая многосторонность. Краткую сводку факто-
ров и кодирующих их генов, имеющих отношение к вирулентности бруцелл, приво-
жу ниже по работам R. М. Roop et al. (2004), J. Ko, G. Splitter (2003) и M. J. Corbel (1997).
Липополисахарид. Подобно большинству молекул липополисахарида с длинными
боковыми О-цепями, гладкий ЛПС (S-ЛПС) видов Brucella защищает эти бактерии
от комплемент-опосредованного уничтожения. Взаимодействие S-ЛПС с липидны-
ми плотами на поверхности макрофагов хозяина позволяет бруцеллам проникать в эти
740
Часть 3. Частная неправильная эпидемиология
фагоциты посредством механизма, который избегает слияния содержащей Brucella
фагосомы с лизосомами. S-ЛПС также играет важную роль во внутриклеточном вы-
живании бруцелл, так как их S-формы выживают в макрофагах гораздо лучше, чем
R-формы. S-ЛПС обладает иммуномодулирующей активностью, которая потенциаль-
но помогает бактерии в обеспечении длительного существования в макрофагах хозяи-
на. В частности, эта молекула не разрушается фагоцитами, а переносится к их клеточ-
ной поверхности, где она образует макродомены с молекулами МНС класса II.
Образование комплекса «МНС класса П—ЛПС» приводит к снижению способности
инфицированного хозяина активировать специфичные к антигенам Brucella CD4+
Т-клетки. По сравнению с энтеробактериальным ЛПС S-ЛПС бруцелл имеет много
других необычных свойств: относительно низкую токсичность для чувствительных к
эндотоксину мышей, кроликов и куриных эмбрионов; низкую токсичность для мак-
рофагов; низкую пирогенность и низкую гипоферремия-индуцирующую активность.
Он также является плохим индуктором интерферона (и фактора некроза опухоли), но,
как ни парадоксально, эффективным индуктором интерлейкина 12.
Система секреции типа IV. Штаммы В. melitensis, В. suis и В. abortus обладают си-
стемой секреции типа IV, кодируемой компонентами wrB-оперона. Хотя природа
эффекторной молекулы (молекул), транспортируемой этой системой, в настоящее
время неизвестна, имеющиеся экспериментальные данные свидетельствуют о том, что
экспрессия wrfi-оперона имеет важное значение для обеспечения длительных взаимо-
действий между содержащей Brucella вакуолью и эндоплазматическим ретикулумом,
необходимых для развития репликативной фагосомы в культивированных макрофа-
гах. Штаммы Brucella дикого типа переносятся в эти внутриклеточные компартмен-
ты, где имеются благоприятные условия для выживания и репликации, в то время как
изогенные игБ-мутанты, полученные из этих штаммов, транспортируются вфаголи-
зосомы, где они разрушаются. Данная система секреции также необходима для пре-
образования Brucella-содержащих вакуолей в репликативные фагосомы в культиви-
рованных клетках HeLa, которые, подобно плацентарным трофобластам, имеют
эпителиальное происхождение.
Эффективный биосинтез пуринов, пиримидинов и аминокислот. Позволяет бру-
целлам выживать и размножаться в репликативной фагосоме, где они испытывают
острый недостаток питательных веществ во время своего пребывания в этом компарт-
менте. Мутанты В. melitensis с нарушенным биосинтезом пуринов и некоторые ами-
нокислотные ауксотрофы В. suis (геныригЕ и агоС) обнаруживают значительную ат-
тенуацию в культивированных мышиных макрофагах, и, соответственно, эти мутанты
неспособны поддерживать хроническую селезеночную инфекцию у экспериментально
инфицированных мышей.
Фактор хозяина 1 (HF-I). Представляет собой РНК-связывающий белок, который
играет важную роль в экспрессии генов в стационарной фазе роста у многих бакте-
рий. ЛД/-мутант В. abortus Hfq3 обнаруживает плейотропный, дефектный в стационар-
ной фазе фенотип (включая резкое повышение чувствительности к низкому pH и
окислительному стрессу при переходе в стационарную фазу), напоминающий фено-
тип, описанный для этих бактериальных гро5-мутантов. hfq-мутант В. abortus также
является слабо приспособляющимся к условиям существования и репликации в куль-
тивированных макрофагах и неспособен поддерживать хроническую селезеночную
инфекцию у экспериментально инфицированных мышей.
Фермент цитохром-bd-оксидаза. Имеет высокий аффинитет к кислороду. Его спо-
собность удалять О2 из клетки позволяет бру целлам адаптироваться к микроаэробным
3.4. Бруцеллез
741
условиям, характерным для репликативной фагосомы и «переживать» окислительный
.пресс во время стационарной фазы.
Оперон bvrRS. Brucella кодирует двухкомпонентную регуляторную систему, кон-
гролирующую экспрессию генов, содействующих сохранению целостности клеточной
оболочки бактерии. Гены оперона кодируют многочисленных представителей семей-
ства белков ОтрЗ, а также продукты генов, которые модифицируют состав жирных
кислот липида А, являющегося составной частью Л ПС. Измененные свойства мемб-
раны bvrRS-мутантов В. abortus делают их менее устойчивыми к кислой среде, в кото-
рой им приходится находиться в фагосомном компартменте макрофагов хозяина на
ранних стадиях инфекции. Обнаружено также, что />гг/?5-мутанты В. abortus неспо-
собны достичь репликативной фагосомы.
Белок HtrA. Участвует в индукции ранней грануломатозной реакции на В. abortus
у мышей и ассоциируется со снижением уровней инфекции на ранней стадии инфек-
ции. Однако он не препятствует последующему увеличению количества бактерий, и
Л/гЛ-дефицитные мутанты, в конечном счете, продуцируют в селезенке уровни инфек-
ции, аналогичные продуцируемым В. abortus дикого типа.
Аденин- и гуанинмонофосф лт. Оба нуклеотида ингибируют слияние фаголизосом,
дшрануляцию и активацию миелопероксидазагалоидной системы и продуцирование
фактора некроза опухоли.
2,3-дигидроксибензойная кислота (2,3-DHBA). Используется бруцеллами в качестве
ш церофора в ответ на недостаток железа. Необходима для вирулентности В. abortus
2308 дикого типа у беременных жвачных животных. Эритритол является предпочти-
тельным источником углерода для штаммов В. abortus, а плацентарные трофобласты
жвачных животных начинают продуцировать это соединение в огромных количества*
в начале последнего триместра беременности. Именно в это время беременные жвач-
ные наиболее восприимчивы к инфекции, вызываемой бруцеллами. Способность
бруцелл эффективно метаболизировать эритритол играет важную роль в проявлении
вирулентности этих бактерий в половых путях беременных жвачных животных.
У В. abortus идентифицированы несколько генов, мутации в которых снижают их
вирулентность, но наличие самих этих генов заставляет усомниться в полноте имею-
щихся знаний об экологии бруцелл:
ген cgs кодирует циклическую Р(1 -2)гликансинтетазу [cyclic в(1—2i glycan synt-
hetase], участвующую в синтезе циклического Р(1—2)гликана [cyclic Р(1—2) glycan],
осморегулирующего периплазматического полисахарида, характерного для раститель-
ных патогенов Agrobacterium tumefaciens и Rhizobium meliloti',
ген ЬасА кодирует цитоплазматический транспортный белок. Он гомологичен
такому же гену у R. meliloti, необходимому для симбиоза ризобий с растениями.
«Острова патогенности» у бруцелл пока не обнаружены. Большинство из иденти-
фицированных факторов вирулентности необходимы бруцеллам не для нанесения
«поражения» фагоцитирующей клетке, а для физиологической адаптации к услови-
ям среды в ее репликативной фагосоме. Их поиски продолжаются.
Таксономия, происхождение и молекулярная эпидемиология бруцелл. Род Brucella
относится к семейству Brucellaceae (в него входят еще два рода: Mycoplana и Ochro-
bactrum), относящегося к порядку Rhizobiales класса Alphaproteobacteria филума Рго-
leobacteria. Класс Alphaproteobacteria включает семейства организмов, патогенные для
млекопитающих и растений. Среди родов, входящих в Alphaproteobacteria, патоген-
ными для млекопитающих являются Bartonella, Rickettsia и Ehrlichia В естественных
условиях среди млекопитающих они распространяются насекомыми. Небольшой
742
Часть 3. Частная неправильная эпидемиология
размер генома этих микроорганизмов предопределяет их облигатное внутриклеточ-
ное существование. Особенностями Brucella, отличающих их от большинства других
родов Rhizobiales, являются их способность инфицировать клетки млекопитающих и
при этом схожесть их генома с геномами бактерий, вызывающих патологические про-
цессы у бактерий (Ficht Т., 2010).
По данным J. Foster с соавт. (2009), использовавших метод молекулярных часов,
большинство видов Brucella дивергировали от их общего предка В. ovis 86—296 тыс. лет
назад. Секвенирование рРНК позволило определить филогенетическое родство рода
Brucella. Ближайшим из известных «родственников» является Ochrobactrum anthropi,
бактерия, вызывающая оппортунистические инфекции у людей. Возможно, более
близкородственным является не полностью охарактеризованный Vibrio cyclosites, обна-
руживающий >90 % сходство последовательности 5S рРНК. Менее близкородственны-
ми, но относящимися к филуму Proteobacteria, являются рода Agrobacterium, Phyllo-
bacterium и Rhizobium, которые обладают многочисленными репликонами и способ-
ностью к внутриклето тому росту. Группа Bartonella обнаруживает родство с Brucella
на основе сходства рРНК, но не ДНК. У них также замечено сходство в липидном
составе клеточной мембраны и механизме внутоиклеточного роста (Corbel М. J., 1997).
Геном Brucella содержит две хромосомы, 2,1 и 1,5 м. п. о., соотвстшвенно. Сред-
няя ММ генома бруцелл составляет 2,37х 109 Да при содержании молярного Г+Ц
58—59 %. Сам род является высокогомогенным: все его члены при попарном сравне-
нии ДНК-ДНК обнаруживают >95 % гомологию, что позволяет классифицировать
Brucella как моноспецифический род. Генетическая консервативность рода Brucella
затрудняет установление связи между их классическими видами и биоварами и обна-
ружение молекулярных маркеров для отдельных групп.
Биогипирование видов, входящих в семейство Brucella, до настоящего времени
осуществляется по фенотипическим характеристикам, таким как зависимость от СО2,
утилизация субстратов, чувствительность к фагам, антибиотикам и по антигенным
свойствам. Номенклатура, предложенная J. М. Verger et al. (1985), в соответствии с ко-
торой все типы следует рассматривать как биовары В. melitensis, не была принята в
основном из практических соображений. По этой причине, несмотря на хорошо изве
стные недостатки, старая номенклатура сохранена с прежними видовыми названиями
В. abortusbovis (бычья), В. melitensis (мальтийская), В. abortus suis (свиная), В. neotomae
(грызуны), В. ovis (овечья) и В. canis (собачья), используемыми для обозначения со-
ответствующих номенклатурных видов. Внутри видов семь биоваров установлены для
В. abortus, три — для В. melitensis и пять — для В. suis. Дрхтие виды не были дифферен-
цированы на биовары, хотя варианты существуют Существующая система биотипи-
рования не охватывает все известные варианты даже основных видов. Так, например,
описаны варианты В. melitensis. Штаммы, выделенные у морских млекопитающих
(дельфины, тюлени, малый полосатик), несомненно, составляют отдельную группу
и неофициально названы В. maris. Можно различить, по крайней мере, два варианта
этого штамма, приблизительно соответствующих штаммам, выделенным у китов и
тюленей соответственно (Corbel М. J., 1997). Хотя случаев выделения штаммов бру-
целл от морских млекопитающих до сих пор относительно немного, имеются сероло-
гические доказательства того, чго такие инфекции среди морских млекопитающих
широко распространены (Dawson С. et al., 2008).
Географическая дифференциация штаммов бруцелл, выполненная мульгилокус-
ным секвенированием по 9 генетическим локусам 160 изолятов, приведена в работе
A. Whatmore с соавт. (2007).
3.4. Бруцеллез
743
Экология и естественная эпидемиология бруцелл1. Бруцеллез традиционно счита-
ется зоонозом. Вспышка бруцеллеза среди людей возникает на фоне эпизоотии сре-
ди сельскохозяйственных животных. Основными источниками инфекции для людей
иаляются мелкий и крупный рогатый скот и свиньи, иногда — лошади, верблюды,
олени, му вы, собаки, кошки и др. С этими традиционными представ юниями входят
в 1ротиворечия данные, полученные при изучении генома бруцелл. Они свидетель-
ствуют о том, что бруцеллы могут обитать в почве и быть патогенами или симбионта-
ми растений. Скорее всего, млекопитающие являются вторичным резервуаром
Brucella, а их первичный резервуар находится в почве и водоемах. Из почвы бруцеллы
подал,ают в растения и через них инфицируют наземных животных; с гидробионтами
они попадают в организм речных и морских млекопитающих, и вызывают у них ин-
фекционный процесс. Так как бруцеллы являются облшатными паразитами, и вести
сапрофитический образ жизни они не могут, то их первичный резервуар находится
„реди простейших клеточных организмов почв и водоемов.
В. melitensis обладает патогенностью для человека и вызывает эпидемические
вспышки, тогда как В. abortus и В. suis менее патогенны и вызывают спорадические
заболевания. Поэтому мелкий рогатый скот рассматривается как наиболее опасный
источник заболевания людей бруцеллезом. Однако каждый из трех видов бруцелл
может мигрировать с одних видов животных на другие.
При совместном содержании или выпасе нескольких видов больных и здоровых
животных между ними происходит обмен различными видами бруцелл. В этом про-
цессе большое эпидемиологическое значение приобретает миграция В. melitensis с
мелкого рогатого скота на крупный рогатый скот и другие виды животных.
Возбудитель бруцеллеза проникает в организм животного через слизистые пище-
варительного тракта, половых органов, дыхательных путей, конъюнктиву, а также
кожные покровы. Для большинства животных характерна высокая чувствительность
к инфекции в период беременности. Заражение в этом периоде в большинстве случа-
ев вызывает аборт. Эпизоотологическое и эпидемиологическое значение абортов бру-
целлезной этиологии определяется тем, что выкидыш, оболочки, околоплодная жид-
кость, плацента содержат возбудитель бруцеллеза в огромном количестве. Возбудитель
выделяется также с молоком, мочой и испражнениями больных животных.
Овцы и козы имеют наибольшее эпидемиологическое значение в России. Круп-
ный рогатый скот в высокой степени восприимчив к В. abortus. Заражаемость людей
в естественных очагах бруцеллеза крупного рогатого скота достигает в отдельных хо-
зяйствах до 70 %. Положительная реакция на бруцеллез по иммунологическим пока-
зателям и клинически выраженный бруцеллез отмечаются лишь у отдельных больных.
Поэтому инфекция коровьего типа определяется как спорадический бруцеллез. Эпи-
демиологические наблюдения не подтверждают передачу инфекции от больного бру-
целлезом человека здоровому. Большое эпидемиологическое значение в естественной
передаче инфекции от животных к человеку имеют пищевые продукты и сырье живот-
ного происхождения. Наибольшую опасность представляют сырые молочные продук-
ты, мясо и сырье (шерсть, смушки и кожа) от коз и овец, больных бруцеллезом.
Коровье молоко занимает преобладающее место в заражении людей, не связан-
ных профессионально с животноводством. Однако вспышек заболеваний, связанных
с заражением коровьим молоком, не наблюдалось, за исключением тех случаев, когда
1 Естественная эпидемиология бруцелл приведена по работам П. А. Вершиловой, Г П, Руд-
нева (1964), П. А. Вершиловой с соавт. (1972), М. J Corbel (1997).
744
Часть 3. Частная неправильная эпидемиология
имелась миграция В. melitensis на крупный рогатый скот. Известны вспышки бруцел-
леза, возникшие вследствие употребления сырого козьего молока, со значительным
числом заболевших и тяжелыми клиническими проявлениями бруцеллеза.
Из молочных продуктов наибольшее эпидемиологическое значение придается
овечьей брынзе. При расследовании причины вспышки бруцеллеза необходимо учи-
тывать возможность заражения бруцеллезом при использовании кумыса и продуктов,
изготовленных из верблюжьего и оленьего молока.
Установлено эпидемиологическое значение шерсти, каракулевых смушек и кож
животных, больных бруцеллезом. Обсеменение бруцеллами шерсти зараженного жи-
вотного происходит во время аборта и родов, а также с мочой и калом, также попадают
бруцеллы на шерсть при пребывании в помещениях и на пастбищах, где находится
больной скот. Особую опасность представляют необработанные каракулевые смуш-
ки. так как их нередко снимают с абортированных и мертворожденных плодов или с
ягнят 3—4-суточного возраста. Животные, больные бруцеллезом, при выделении экс-
крементов в окружающую среду загрязняют бруцеллами почву, подстилку, корм, воду,
которые, в свою очередь, становятся факторами, обусловливающими заражение че-
ловека. Сроки сохранения бруцелл в воде весьма продолжительны. В литературе опи-
саны случаи заражения через воду при питье, мытье продуктов и столовой посуды.
Бруцеллы проникают в организм человека через слизистые оболочки пищевари-
тельного тракта, дыхательных путей, конъюнкгиву, через кожные покровы. Алимен-
тарное заражение через молоко и молочные продукты установлено во многих случа-
ях, когда полностью исключается контакт с источником инфекции (козы, овцы,
коровы). При этом предполагается, что микроб проникает в организм через слизис-
тые оболочки верхних отделов пищеварительного тракта, так как кислое содержание
желудка губительно действует на бруцелл. Под действием соляной кислоты в разве-
дении 1:1000 бруцеллы погибают в течение нескольких минут. Возможен занос мик-
робов в рот с папиросой или руками при проведении работ с зараженными животны-
ми или их сырьем, что также не исключает возможности проникновения бруцелл в
организм через слизистые оболочки верхнего отдела пищевого тракта. Возможность
проникновения бруцелл через дыхательные пути подтверждена экспериментальны-
ми исследованиями ряда авторов при вакцинации или заражении аэрозолями. Инга-
ляционный путь заражения может быть обусловлен наличием бруцелл в пылевых ча-
стицах шерсти, навоза, земли. Этот путь инфицирования возможен при стрижке,
сортировке шерсти, вычесывании пуха (разборка, вязанье и пр.), а также при уборке
помещений и территорий, где содержатся животные или обрабатывается их сырье.
Ингаляционное заражение также возможно в лабораторных условиях.
Контактный путь заражения имеет наибольшую роль в очагах бруцеллеза. Особен-
но высока возможность инфицирования при оказании животным помощи во время
родов и при абортах, когда приходится производить ручное отделение плаценты. За-
ражение через кожные покровы рук возможно при всех других работах, проводимых
с бруцеллезными животными или с объектами внешней среды, загрязненными вы-
делениями зараженных животных. При убое, обработке туш животных, больных бру-
целлезом, возможно попадание бруцелл на наружные кожные покровы рук. При этом
проникновение микроба облегчается, если руки рабочих находятся постоянно во
влажном состоянии, что способствует мацерации кожи рук.
На предприятиях мясообрабатывающей промышленности наряду с контактным
возможно также заражение алиментарным и ингаляционным путем. На предприятиях,
3.4. Бруцеллез
745
обрабатывающих шерсть, кожу, могут иметь значение в равной мере и контактный,
и ингаляционный путь. Наряду с общеизвестными путями заражения человека бру-
целлезом известны случаи заражения при изготовлении брошек из кости, что может
быть обусловлено сохранением бруцелл в костном мозгу. Отмечено также заражение
при употреблении в пищу свежей крови от бруцеллезных животных.
При бруцеллезе выявляется сезонность заболеваний, которая наиболее четко вы-
ражена в животноводческих районах. Для бруцеллеза козье-овечьего типа характерен
зимне-весенний подъем заболеваний, связанный с окотным периодом у овец и коз.
На этот период в очагах козье-овечьего бруцеллеза приходится 70—80 % всех зараже-
ний. Сезонность в естественной заболеваемости людей при заражении от крупного
рогатого скота выражена значительно меньше, чем при овечьем бруцеллезе. В период
отела количество заболеваний повышается, но незначительно, а затем несколько сни-
жается и держится почти на одних и тех же показателях весь сезон лактации, т. е. до
октября — ноября. Сезонность в заболеваемости городских жителей выражается ме-
нее четко, однако закономерность ее сохраняется, особенно в городах животновод-
ческих районов. Имеются различия в динамике заболеваний отдельных континген-
тов населения и профессиональных групп, обусловленных заражением от мелкого
рогатого скота. Учет помесячной заболеваемости бруцеллезом имеет большое прак-
тическое значение для правильного планирования профилактической работы.
Восприимчивость к бруцеллам и инфицирующие дозы для животных и людей. В на-
стоящее время имеются сообщения о восприимчивости к бруцеллезу более 60 видов
позвоночных животных. Имеются сообщения о зараженности рыб, амфибий и реп-
тилий. Имеются также сообщения о выделении культуры бруцелл от кровососущих
членистоногих. Установлена восприимчивость к бруцеллам 16 видов клещей из се-
мейств Ixodidae и Argasidae.
Морские свинки очень чувствительны ко всем видам бруцелл. При подкожном
заражении они могут воспроизводить генерализованную бруцеллезную инфекцию,
начиная с 10—20 КОЕ. При пероральном, конъюнктивальном и накожном введении
бруцелл заражение морских свинок возможно при дозах в 20-100 раз больших, чем
при подкожном заражении. Мышей удается подкожно заразить 5—20 КОЕ, но у них
выражены индивидуальные колебания в чувствительности к бруцеллам. Крысы бо-
лее резистентные к бруцеллам, чем мыши и морские свинки. Заразить их подкожно
В. abortus, В. suis можно, начиная с дозы в 1000 КОЕ, В. melitensis — с дозы 100 КОЕ
(Вершилова П. А. с соавт., 1972).
Овец (баранов) удается заразить В. melitensis подкожно, через рот, конъюнктиву,
слизистые оболочки влагалища, через скарифицированную и мацерированную кожу.
Однако воспроизводимое заражение овец через слизистые удается только при нане-
сении больших количеств высоковирулентного возбудителя: не менее 1—500 млн КОЕ
(вагинальное и конъюнктивальное заражение). Для заражения животного через сли-
зистые рта путем кормления инфицированным овсом необходимо 2-5 млрд КОЕ.
Заражение через кожу путем наложении навозной пасты наблюдалось при дозе не
менее 500 млрд КОЕ вирулентной культуры В. melitensis. Подкожное введение овцам
10—50 тыс. В. melitensis воспроизводило у них регионарное воспаление лимфатиче-
ских узлов, и только со 100 тыс. КОЕ регулярно наблюдалась генерализованная фор-
ма инфекции. По аналогии с человеком овцы и козы проявляют меньшую восприим-
чивость к В. abortus по сравнению с В. melitensis. Овец можно заразить В. abortus, на-
чиная с 1—4 млрд КОЕ В. abortus (Вершилова П. А. с соавт., 1972).
746
Часть 3. Частная неправильная эпидемиология
Человек относительно устойчив к возбудителю бруцеллеза. Наибольшей пато1 ен-
ностью для человека обладает В. melitensis и В. suis, не обладают вирулентностью для
человека В. ovis и В. neotomae.
По данным Д. Ротшильда (19б6), ингаляционная инфицирующая доза В. melitensis
для человека составляет 1300 КОЕ.
Иммунитет. Характерной чертой иммунитета при бруцеллезе является его замед-
ленное формирование и относительность. У бруцелл охарактеризовано значительное
число антигенных компонентов. Однако антигеном, который играет доминирующую
роль в реакции выработки макроорганизмом антител, является Л ПС. У гладких штам-
мов (smooth phase strains, S) S-ЛПС включает липид А (содержащий два типа амино-
гликозы; aminoglycose); характерные жирные кислоты (исключая р оксимиристино-
вую кислоту); коровую область, содержащую глюкозу, маннозу и хиновозамин, и
О-цепь, заключающую в себе гомополимер, состоящий примерно из 100 остатков
4-формамидо-4,6-дидезоксиманнозы. Различие в их соединении влияет на форму эпи-
топов Л ПС. А-доминантный тип Л ПС имеет палочковидную форму, М-доминантный
тип является петлевидным ЛПС. Штаммы, которые реагируют с антисыворотками к
обоим эпитопам, А и М, продуцируют ЛПС обоих типов примерно в равных пропор-
циях. Присутствие 4-амино-4,6-дидезоксиманнозы в ЛПС также обусловливает ан-
тигенную перекрестную реактивность с ЛПС Escherichia hermanni и Escherichia coli
0:157, Salmonella 0:30, Stenotrophomonas maltophilia, Vibno cholerae 0:1 и Jersinia entero-
colitica 0:9 (Corbel M. J., 1997).
Структура ЛПС шероховатых штаммов (R-ЛПС) в основном сходна с таковой
S-ЛПС, за исключением того, что О-цепь либо отсутствует, либо уменьшена до не-
скольких остатков. Следовательно, специфичность R-ЛПС определяется главным об-
разом коровым полисахаридом. Охарактеризованы также многочисленные белковые
антигены наружной и внутренней мембраны, цитоплазмы и периплазмы. Некоторые
распознаются иммунной системой во время инфекции и являются потенциально по-
лезными в диагностических тестах. Однако тесты, основанные на таких антигенах,
имеют низкую чувствительность, поскольку у инфицированных людей обычно раз-
вивается гораздо менее стойкая реакция на индивидуальные белковые антигены, чем
на Л ПС (Corbel М. J., 19Q7).
Бруцеллез относится к тем инфекционным болезням, при которых иммунный от-
вет не обеспечивает у значительной части больных освобождения организма от бру-
целл. Бруцеллы длительно сохраняются в метастатических очагах (поданным отдель-
ных исследователей, — год и более), из них происходит повторная многократная
диссемичация возбудителя с развитием реактивно-аллергических изменений и хрони-
зацией процесса (фаза экзоочаговых обсеменений и реактивно-аллергических измене-
ний). Суперинфекция, нередко имеющая место в эндемических очагах болезни, явля-
ется дополнительным фактором, способствующим развитию обострений, рецидивов
и хронизации болезни. Хроническое течение, сопровождающееся многократной гене-
рализацией возбудителя из метастатических очагов, придает бруцеллезу характер хро-
ниосепсиса. Длительное воздействие возбудителя и продуктов его жизнедеятельности
на органы и ткани обусловливает иммуноаллергическую перестройку организма и
комплекс проявлений, характерных для подострого и хронического бруцеллеза (Шува-
лова Е. П. с соавт. 2001).
Устойчивость бруцелл во внешней среде. Бруцеллы неустойчивы к высокой темпе-
ратуре: при 60 °C они погибают в течение 30 мин; кипячение губит бактерии момен-
тально. При низких температурах они могут сохраняться длительное время. Бруцел-
3.4. Бруцеллез
747
лы быстро погибают под воздействием прямых солнечных лучей и обычных дезинфи-
цирующих веществ в рабочих концентрациях.
Замораживание и засолка не влияют на снижение сроков сохранения бруцелл в
мясе: при —23 °C В. melitensis в мясе сохраняются 47 сут, В. suis — свыше 30 сут при той
же температуре в естественно зараженном мясе свиней. Брынза, приготовленная из
зараженного овечьего молока, сохраняет свою заразительность в течение 45 сут, если
она хранится при температуре 11 -14с С, кислотности 26—50 °Т (по Тернеру) и засолен-
ности 5—11 %. Повышение кислотности способствует сокращению срока самоочище-
ния. В шерсти овец бруцеллы могут сохранять жизнеспособность в течение 1,5 и даже
3—4 мес. Факторами, определяющими возможность заражения от навоза, являются
данные о том, что в навозе внутри штабеля при температуре 50-67 °C бруцеллы сохра-
няются в течение 25 сут, а при температуре 37—4 2 °C бруцеллы остаются длительно
жизнеспособными. Сроки выживаемости бруцелл в почве зависят от факторов, харак-
теризующих ее состав, микрофлору. Бруцеллы в почве сохраняют жизнеспособность
летом в течение 3 мес , а зимой — до 4,5 мес. В поверхностных слоях почвы открытых
мест бруцеллы сохраняются до 40 сут, а на глубине 10 см — до 60 сут.
Антибиотикорезистентность бруцелл. Клинические изоляты бруцелл, выделенные в
разное время из разных источников, в условиях in vitro чувствительны к большинству
антибиотиков, применяемых в клинической практике (тетрациклин, стрептомицин,
гентамицин, эритромицин, ципрофлоксацин, норфлоксацин, левофлоксацин и др.).
Периодически клиницисты обнаруживают штаммы бруцелл, умеренно резистентные
к рифампицину и триметаприм-сульфаметоксазолу. Например, в работе A. Turkman!
et al. (2006) такие штаммы составляли 2 и 8 из 74 исследованных соответственно. Еще
чаще встречаются штаммы бруцелл, резистентные к пенициллинам. L. Robertson с
соавт. (1973) обнаружили, что из 25 штаммов В. abortus, исследованных на антибио-
тикорезистентность, 17 оказались резистентными к пенициллинам. В то же время
лечение антибиотиками требует длительного применения их определенной комбина-
ции. К тому же число случаев возврата болезни после завершения курса антибиоти-
котерапии достигает 10 %. Это вызвано тем, что чувствительность бруцелл к антиби-
отикам в условиях in vivo значительно снижается благодаря их способности длительное
время находиться в бруцеллосомах фагоцитирующих клеток и тем самым избегать
воздействия антибактериальных препаратов (Roop R. М. et al., 2004).
Обнаружение генетически измененных штаммов бруцелл. Подходы к обнаружению
генетически измененных штаммов возбудителей бруцеллеза в основном те же, что и
для обнаружения генетически измененных штаммов возбудителей сибирской язвы и
чумы (см. разд. 3.1 и 3.2). В отличие от этих возбудителей инфекционных болезней, у
Brucella собственные плазмиды не обнаружены, однако их трансформация осуществ-
ляется без особых проблем плазмидами других микроорганизмов методами конъюга-
тивного переноса или электропорацией. Показано, что В. abortus S19 способна под-
держивать плазмиды широкого круга хозяев из микроорганизмов других таксонов и
разных групп несовместимости: рТНЮ (IncP), pSa (IncW) и R751 (IncP), а также
передавать участки своей хромосомной ДНК кишечной палочке благодаря конъюга-
тивной плазмиде pl Н10. Также был показан обмен внутри рода бруцелл приобретен-
ными плазмидами. Генетически измененные варианты бруцелл приобретали резис-
тентность к тетрациклину, канамицину, стрептомицину, триметоприму, ампициллину
(Rigby С. Е., Fraser А., 1989).
Поэтому у генных инженеров имеется большой выбор плазмид для конструиро-
вания векторных челночных конструкций, предназначенных для интеграции клони-
748
Часть 3. Частная неправильная эпидемиология
руемых генов в геном бруцелл. Пример такого вектора см. в разд. 2.2.2 «Технологии
рекомбинантной ДНК в получении штаммов бактерий с неправильной эпидемиоло-
гией». В очагах искусственно вызванных вспышек бруцеллеза с наибольшей вероят-
ностью можно будет встретить штаммы возбудителя бруцеллеза с модификациями
ЛПС, нехарактерными для данного вида. Например, ЛПС В. melitensis у менее виру-
лентной для человека В. suis, но более эффективно «преодолевающей» защиту, созда-
ваемую живой бруцеллезной вакциной на основе штамма 19. Возможно обнаружение
штаммов бруцелл, резистентных к антибиотикам, обычно используемых для лечения
бруцеллеза. Так как собственные плазмиды у бруцелл не обнаружены, то обнаруже-
ние каких-либо плазмид у штамма, выделенного при обстоятельствах, характерных
для умышленного распространения бруцелл среди людей или животных, также дол-
жно вызвать настороженность в отношении его искусственного происхождения.
Патогенез бруцеллеза1. Многократные поступления бруцелл и их эндотоксина в
русло крови проявляются определенными патогенетическими закономерностями
(табл. 3.19).
Таблица 3.19
Фазовость патогенеза и клиники бруцеллеза у человека*
Специфические раздражения Фаза Название фазы патогенеза Клиника
Первичные I Фаза лимфогенного заноса и лимфо-рецепторных раздражений Инкубация
II Фаза гематогенного заноса (первичная генерализация) и гемососудисто-рецепторных раздражений Острый период
III Фаза полиочаговых локализаций (формирование метастатических очагов) Развитие сепсиса
Повторные IV Фаза экзоочаговых обсеменений (повторная генерали- зация. чаще многократная) и реактивно-аллергических изменений Рецидивы и обострения
V Фаза конечного метаморфоза: а) рассасывание; б) фиброз; в) стойкие последствия (циррозы, рубцы) Исходы
* По работам П. А. Вершиловой, Г. П. Руднева (1964), П. А. Вершиловойс соавт. (1972).
Фаза лимфогенного заноса и лимфо-рецепторных раздражении. Бруцеллы проника-
ют в организм через кожу или слизистые оболочки, захватываются макрофагами, раз-
множаются в них. На месте входных ворот никаких клинических изменений первич-
но воспалительного порядка обычно не обнаруживается. Механизм выживания
бруцелл в макрофагах следующий. Сразу после проникновения в макрофаг бруцеллы
располагаются в подкисленном компартменте, который сливается с компонентами
ранней стадии эндосомного обмена. Там они переживают окислительный взрыв.
Продвижение содержащих бруцеллы вакуолей по эндосомно-лизосомному пути ог-
раничено, и вирулентные штаммы переносятся во внутриклеточный компартмент,
обычно называемый репликативной фагосомой. Иногда его еще называют «бруцел-
лосомой». Репликативные фагосомы возникают в результате обычного клеточного
1 Патогенез бруцеллеза описан по работам П. А. Вершиловой, Г. П. Руднева (1964),
П. А. Вершиловой с соавт. (1972) и R. М. Roop et al. (2004).
3.4. Бруцеллез
749
процесса — постоянных взаимодействий между содержащей Brucella вакуолью и эн-
доплазматическим ретикулумом макрофагов хозяина. Эти репликативные фагосомы
не только не сливаются с лизосомами, но и внутри их повышается pH; условия среды
становятся более благоприятными для внутриклеточного существования бруцелл.
Само же взаимодействие между образующимися фагосомами и эндоплазматическим
ретикулумом является механизмом, необходимым для нормального формирования
фагосом (рис. 3.55).
Рис. 3 55 Биогенез репликативной фагосомы, являющейся местом обитания
Б. abortus в культивированных мышиных макрофагах. По R. М. Roop et al. (2004)
Но межтканевым щелям, а затем по лимфатическим сосудам бруцеллы заносятся
в со< тветствующие регионарные лимфатические узлы. В узлах, где происходит филь-
трация лимфы, бруцеллы задерживаются вследствие фагоцитарной активности мест-
ных тканевых и других клеточных элементов.
Фаза гематогенного заноса и гемососудисто-рецепторныхраздражений. Л имфатиче-
ские узлы становятся резервуаром бруцелл («септические очаги»), откуда, путем лим-
фогенных метастазов, происходят многократные их прорывы в кровяное русло с пос-
ледующим формированием новых комплексных очажков.
Фаза формирований метастатических очагов. При бруцеллезных поражениях сосу-
дов в процесс вовлекаются различные слои их стенок, обусловливая разные комбина-
ции, включая панваскулит. Развивается бруцеллезный миокардит с наличием инфиль-
тративных очагов (гистиоциты, плазмоциты и лимфоидные клетки) в миокарде по хо-
ду мелких капилляров. Возможно сочетание эндокардита и миокардита. Возможно
развитие специфической этиологии при бруцеллезе перикардитов, миокардитов и
язвенно-бородавчатых эндокардитов. Миокардит обычно сопровождает поражение
эндокарда и характерен наличием очагового размножения гистиоцитов или наличи-
ем в интерстициальной ткани очагов резко выраженного экссудативно-продуктивного
воспаления. Иногда сопровождается перикардитом. Артериальная сеть страдает силь-
нее и чаще, чем венозная. Распространенное поражение сосудистой системы в раз-
ных местах и органах человека, страдающего бруцеллезом (особенно в мелких сосу-
дах), во многом обусловливает затем полилокальную органопатологию при бруцеллезе,
а отсюда в известной степени и полиморфизм симптомов в клинике. При наличии
лимфогематогенных метастазов и бактериемических «атак» селезенка и печень по сво-
им анатомо-физиологическим особенностям, как правило, вовлекаются в процесс.
Патоморфологические изменения со стороны печени при бруцеллезной инфекции
выступают в форме глубокого дегенеративно-некротического поражения гистиострук-
турных элементов данного органа, интерстициального гранулирующего и серозного
750
Часть 3. Частная неправильная эпидемиология
воспаления. Нередко они заканчиваются атрофическим циррозом. Увеличение селе-
зенки в начале процесса обусловливается гиперплазией ретикуло-энцотелиальных и
лимфоидных элементов.
Изменения в легких при бруцеллезе представляют интерес в патогенетическом
отношении, поскольку рчд авторов допускает возможность заражения человека аспи-
рационно, со вдыхаемой инфицированной пылью и т. п. Однако нет указаний на об-
разование первичного аффекта в легких при естественном заражении бруцеллезом.
В легких при бруцеллезе наблюдаются неспецифические экссудативно-воспалитель
ные изменения и очаги инфекционной гранулемы, напоминающие туберкулез. Воз-
можны первичные бруцеллезные пневмонии катарально-геморрагического характе-
ра с обнаружением бруцелл в мокроте.
Поражение почек при бруцеллезе клинически устанавливается редко. Нефрит
отмечался как в подострой, так и в хронической стадии инфекции. Он имел интер-
стициальный очаговый характер.
В клинике врачи очень часто отмечают у больных изменения со стороны локомо-
торного аппарата. Со стороны нервной системы клиницисты весьма часто отмечают
у больных бруцеллезом ряд тех или иных нарушений. Поражения периферической
нервной системы встречаются клинически гораздо чаше, но протекают легче, а пора-
жения ЦНС — наоборот. Патоморфологические изменения нервной системы проте-
кают как гиперергические поражения сосудов, дегенерация нервных волокон и кле-
ток, а также пролиферация экю- и мезоглии. Бруцеллезный менингит представляет
серозно-продуктивное воспаление с лимфо-гистиоцитарной клеточной реакцией и
наличием в мягких мозговых оболочках узелков из эпителиоидных круглоядерных
к 1еток. Бруцеллезный энцефалит обычно сочетается с менингитом и развивается в
подострой и хронической стадиях бруцеллеза. В пазоморфологическом отношении
бруцеллезный энцефалит характеризуется наличием узелков гранулемы, состоящих
из микроглии. Вторую характерную черту составляем продуктивно-деструктивный
васкулит. Помимо воспалительных поражений, нередко обнаруживаются обознача-
емые как энцефалоз дистрофические изменения центральной нервной системы.
При диагностике болезни уделяется большое внимание изменениям половых ор
ганов. В острой септической стадии бруцеллеза отмечались очаги периваскулярной
круглоклеточной воспалительной инфильтрации в яичках мужчин. У женщин — оофа-
рит, сальпингит, эндометрит, нарушение менструального цикла и прерывание бере-
менности.
П гацентарные трофобласты играют решающую роль в патобиологии аоорта при
инфекциях, вызываемых Brucella. Эти клетки явиыются компонентами эпителиаль-
ных слоев плаценты и выполняют важную функцию барьера между системами кро
вообращения матери и плода. У жвачных животных и других естественных хозяев
бруцеллы могут размножаться внутри плацентарных трофобластов на поздних стадиях
беременности. Быстрый и экстенсивный ро< т бруцелл в этих клетках хозяина приво-
дит к нарушению плацентарной целостности и инфицированию развивающегося
плода, что, в свою очередь, приводит к аборту или рождению слабого и инфициро-
ванного потомства. Как показал микроскопический анализ плацентарных тканей,
полученных при вскрытии инфицированных В. abortus коз и крупного рогатого ско-
та, бруцеллы размножаются во внутриклеточных компартментах, связанных с грану-
лярным эндоплазматическим ретикулумом в плацентарных трофобластах (рис. J.56).
Эти экспериментальные данные свидетельствуют о том, что внутриклеточный
компартмент, захваченный бруцеллами в плацентарных трофобластах, аналогичен по
3.4. Бруцеллез
751
BvrRS-зависимые
модификации
клеточной оболочки
Система секреции типа IV
BvrRS-зависимые модификации
клеточной оболочки
Гладкий ЛПС
Цитохром-
bd-оксидаза
Липидный
плот
Фаго лизосома
Гладкий ЛПС
Представление пептидного
Цитохром-
bd-оксидаза
Ранняя
фагосома
(~рН 4,0)
Репликативная
фагосома
(-pH >5,0)
Нитратное
дыхание
Кислый Недостаток
питательных <
веществ /
Механизм
^биосинтеза пуринов
и аминокислот
Окислительный
взрыв
Низкое
давление
02
антигена главного комплекса
гистосовместимости (МНС) класса II
Рис. 3.56. Схематическое изображение взаимодействий В. abortus с макрофагами
и плацентарными трофобластами на поздней стадии беременности. А. Макрофаги.
Б. Плацентарные фибробласты. Белыми стрелками указаны специфические взаимо-
действия, во время которых проявляется действие клеточных компонентов Brucella,
обсуждаемых в тексте. Условия окружающей среды, которые оказывают значитель-
ное влияние на эти взаимодействия, выделены курсивом. По R. М. Roop et al. (2004)
составу таковому компартменту, внутри которого эти бактерии обитают в макрофа-
гах хозяина. Но особенности внутриклеточного передвижения вирулентных штаммов
Brucella в культивированных плацентарных трофобластах жвачных животных изуче-
ны плохо.
Фаза экзоочаговых обсеменений. Проявляется многократными поступлениями в
русло крови бруцелл и их эндотоксина, изменением реактивностью макроорганизма
по типу характерной аллергии как одного из самых основных критериев патогенеза
бруцеллеза. Болезнь развивается по типу хрониосепсиса. Инфицированные Brucella
лимфатические узлы играют основную роль в затяжном течении бруцеллеза и обус-
ловливают рецидивы и обострения болезни.
752
Часть 3. Частная неправильная эпидемиология
Фаза конечного метаморфоза. Отражает последствия перенесенного бруцеллеза,
включая стойкие, цирротического характера изменения (постбруцеллезные циррозы
и рубцы). Это так называемый резидуальный бруцеллез.
Клиническая картина бруцеллеза у людей при естественном инфицировании*. Инку-
бационный период при бруцеллезе длится от 7 до 30 сут. Клинические проявления
болезни весьма полиморфны и могут варьировать как по длительности течения, так и
по разнообразию симптомов. В очагах инфекции нередко встречаются латентный
бруцеллез (первичная длительная латенция), при котором у зараженных людей бру-
целлы находятся в виде «дремлющей» инфекции, вызывая иммуноаллергическую
перестройку организма. Однако при тщательном клиническом обследовании у части
инфицированных лиц можно обнаружить некоторые симптомы болезни: увеличение
лимфатических узлов, функциональные расстройства нервной системы, увеличение
печени и селезенки и т. д. Серологические пробы и кожная аллергическая проба Бюрне
у таких людей положительные.
Клинически выраженные формы бруцеллеза проявляются длительной лихорад-
кой, ознобами, повышенной потливостью, гепатоспленомегалией, поражением опор-
но-двигательного аппарата, нервной, сердечно-сосудистой, урогенитальной и других
систем организма. Известно несколько клинических классификаций бруцеллеза, ос-
нованных на учете особенностей проявления этой инфекции. Е. С. Белозеров (1985),
много работавший с больными бруцеллезом melitensis в Казахстане, предложил сле-
дующую классификацию болезни:
I. Субклиническая форма («положительно реагирующие» на пробу Бюрне).
II. Клинически выраженные формы: острый бруцеллез; подострый бруцеллез;
хронический бруцеллез (с указанием органопатологии): компенсированный; субком-
пенсированный; декомпенсированный; суперинфекция; реинфекция.
III. Резидуальный бруцеллез (остаточные явления).
Острая и подострая формы бруцеллеза. В начале болезни нередко имеет место
продромальный период длительностью 3—5 сут, проявляющийся недомоганием, сла-
бостью, подавленностью настроения, быстрой физической и умственной утомляе-
мостью, легкой головной болью и понижением аппетита. К концу продромального
периода признаки интоксикации нарастают. Развиваются основные клинические про-
явления болезни, среди которых основными являются гипертермия, сопровождаю-
щаяся ознобами и проливными потами, гепатоспленомегалия. Лихорадка длительная,
в разгар заболевания температура ремиттирующего типа с подъемом во второй поло-
вине дня или в вечерние часы. Может быть ундулирующий (волнообразный) и интер-
миттирующий тип температурной реакции или длительный субфебрилитет. Несмот-
ря на длительность и высоту лихорадки, состояние больных не нарушается, самочув-
ствие остается хорошим, часто выявляются словоохотливость и эйфория.
При острой форме бруцеллеза в течение одного и того же дня у больных наблюда-
ются однократные и многократные познабливания, иногда потрясающий озноб.
В начале озноба температура тела иногда падает ниже нормы, затем возникают жар,
сухость во рту, постепенно нарастает температура. Каждый раз озноб и повышение
температуры завершаются профузным потоотделением. Лимфатические узлы, особен-
но шейные и подмышечные, увеличены от 1 —2 до 1,5—3 см, безболезненны и не спа-
яны с окружающей тканью.
1 Клиническая картина при бруцеллезе приведена в основном по работам П. П. Очкура
(1951) и Е. П. Шуваловой с соавт. (2001).
3.4. Бруцеллез
753
У большинства больных обнаруживается умеренное расширение границ сердца
влево, приглушенность его тонов, систолический шум на верхушке; в тяжелых случа-
ях выявляются миокардит, эндокардит, перикардит. Артериальное и венозное давле-
ния понижены. Частота пульса соответствует температуре. У ряда больных бывает
дикротия пульса, иногда экстрасистолия. Повышается проницаемость капилляров.
При острой форме заболевания по мере развития токсико-септического процес-
са выявляются изменения со стороны органовдыхания: катаральное воспаление верх-
них дыхательных путей, бронхиты, бронхопневмонии, бронхоадениты, узелковые
поражения легких, сопровождающиеся кальцификацией узлов (рис. 3.57).
Рис. 3.57. Узелковые поражения легких при бруцеллезе. Пациент 40 лет, активный
курильщик, поступил в стационар с острой болью в грудной клетке, но лихорадки
или респираторных симптомов не было. Рентгенологически у него обнаружены би-
латеральные легочные узлы. Компьютерная томография выявила два таких узла
слева и три справа размером от 2 до 3,5 см, имеющие хорошо отграниченные края,
концентрическую линейную кальцификацию. Из биоптата узла высеяна культура
В. melitensis. По S. S. Lypez et al. (2011)
Легочные осложнения при остром бруцеллезе наблюдаются не более чем в 15 %
случаев. Осложнения могут включать хиларную и паратрахеальную аденопатию, ин-
терстициальную пневмонию, узелковые утолщения в легких, плевральные истечения
и эмпиему плевры.
Печень и селезенка увеличенные, мягкие, болезненные при пальпации. При по-
ражении печени часто отмечаются ноющие боли в правом подреберье.
О поражении ЦНС в разгар острой формы болезни свидетельствуют головные
боли, раздражительность, эмоциональная неустойчивость, чрезмерная утомляемость,
нарушения сна. В тяжелых случаях бруцеллезной инфекции наблюдаются расстрой-
ства психики, явления менингизма и менингита. Течение менингита обычно вялое,
без четкой выраженности обшемозговых и менингеальных симптомов. Лишь в отдель-
ных случаях менингит протекает с яркой симптоматикой. Бруцеллезный менингит
имеет обычно серозный характер.
Опорно-двигательная система при остром бруцеллезе поражается лишь у части
больных в виде кратковременных артралгий, которые быстро исчезают по мере умень-
шения признаков интоксикации.
На поздних сроках заболевания появляются клинические признаки иммуноаллер-
гической перестройки организма.
У значительной части больных заболевание характеризуется наклонностью к за-
тяжному течению с переходом его в подострую и хроническую формы. Этому могут
способствовать несвоевременная диагностика и поздно начатое лечение. После про-
754
Часть 3. Частная неправильная эпидемиология
веденной этиотропной терапии в организме больных даже при полном отсутствии
клинических признаков могут сохрани гься бруцеллы, локализованные внутриклеточ-
но в системе мононуклеарных фагоцитов внутренних органов.
Рецидивы могут возникать через 1-2 мес. и в более поздние сроки после угасания
симптомов. Обычно рецидивы бруцеллеза протекают с ознобом, повышением темпе-
ратуры, усилением потооз деления. С первых же дней рецидива выявляются очаговые
поражения отдельных органов и систем, при этом наиболее постоянно поражается
опорно-двигательная система.
У больных с нарушениями иммунной системы бруцеллез может сразу приобрес-
ти генерализованный характер и привести их к гибели в течение нескольких недель
Ниже приведены компьютерные томограммы больного, страдавшего диабетом и за-
разившегося В. abortus биотип 1 через мясо больной коровы (рис. 3.58).
Рис. 3.58. Фата_!ьный системный оруцеллез с мультифокальным поражением печени
и лимфатических узлов грудной клетки, по данным компьютерной томографии.
А. Грудная клетка. Видны увеличенные легочные и плевральные лимфатические узлы,
двухсторонний плевральный выпот. Б. Брюшная полость. Наблюдаются многочислен-
ные небольшие узлы в печени, гепатоспленомегалия и конгломераты увеличенных
лимфатических узлов в ретрокруциальной зоне грудного региона Орюшной полости.
Характерные для бруцеллеза кальцификаты не успели образоваться. Смерть насту-
пила на 16 сут после поступления в стационар на фоне прогрессирующей печеночной
недостаточности и желудочно-кишечного кровотечения. По К. W. Park et al. (2007)
В качеств! иллюстрации острой формы бруцеллеза, наиболее вероятной при мас-
сивном заражении бруцеллами, привожу историю болезни пациента, завершившей-
ся смертельным исходом (по работе П. П. Очкура,1951).
Больной Д., 37 лет, служащий. Поступил 16 апреля. Умер 6 мая в больнице, через
три месяца с начала заболевания. Способ заражения не выяснен. Клинический диаг-
ноз: бруцеллез, менингоэнцефалит.
История болезни. Поступил с жалобами на высокую температуру, общую слабость,
недомогание и сухой кашель. В начале заболевания fiapT месяц) появились: озноб и
повышение температуры по вечерам, головная боль и запоры, череду юшиеся с поно-
сами. В последнее время установилась постоянная высокая температура. Наследствен-
ность без патологических уклонений. Перенес воспаление легких и малярию. Питание
пониженное. Кожабледная, с иктеричным оттенком. Периферические лимфатические
узлы: шейные, подмышечные и паховые — слегка увеличены. У края реберной дуги
пальпируется нижний полюс селезенки. Сердце: границы в норме, тоны приглушены,
пульс 78/мин, ритмичен, среднего наполнения Легкие’ при перкуссии — легочный
3.4. Бруцеллез
755
звук, при аускультации выслушивается жесткое дыхание и сухие хрипы. Органы пище-
варения. Языкобложен. зев гиперемирован, слизистая его чистая. Живот слегка напря-
жен, при пальпации болезненный. Печень и селезенка увеличены, пальпируются.
Аппетит отсутствует. Стул нормальный.
Течение болезни. Состояние больного ухудшилось. Постепенно нарастала частота
пульса до 96/мин. Температура была постоянной и держалась на высоких цифрах.
Печень и селезенка увеличивались в объеме, пальпируясь у реберной дуги. С 25 апре-
ля стали нарастать легочные явления: в области нижних долей легких наблюдалось не-
большое усиление голосового дрожания и выслушивалось ослабленное дыхание, а
также сухие и влажн ые хрипы. Появился частый и болезненный кашель с вязкой мок-
ротой.
С 26 апреля сознание затемнено. Оно лишь изредка проясняется. Ночью наблюда-
ется бред. Над областью нижней доли правого легкого отмечалось постепенное нара-
стание притупления перкуторного тона и увеличение влажных хрипов. С 1 мая разви-
лись явления геморрагического диатеза: кровоточивость из десен. Шейные и подмы-
шечные лимфатические узлы увеличены. Нарастает учащение пульса до 100/мин,
наполнение его постепенно падает. Артериальное давление составляет 100/58 мм рт. ст.
В коже области предплечий и бедер появились плотные папулы величиной с гороши-
ну, синюшного цвета. Развилось гноетечение из правого среднего уха и болезненность
при надавливании на правый сосцевидный отросток. Смерть наступила 6 мая при па-
дении сердечной деятельности с развитием цианоза губ и кончиков пальцев.
Со стороны крови отмечалась вторичная анемия, лейкопения с относительным
лимфоцитозом и сдвиг влево. РОЭ в полчаса — 5 мм, в час — 10 мм. Реакция Райта по-
ложительная в разведении 1:100. Реакция Вейль — Феликса и Видаля отрицательные.
Исследование мочи: 19 апреля удельный вес — 1,020. Белка — 0,033 %. В осадке —
лейкоциты (два в поле зрения) и единичные зернистые цилиндры.
Лихорадка сопровождалась высокой температурой постоянного, частью ремити-
рующего типа.
Протокол вскрытия. Труп молодого мужчины в состоянии сильного исхудания.
Рост средний. Конституция астеническая. Кожный покров и видимые слизистые обо-
лочки слегка иктеричны. На коже туловища и верхних конечностей имеются одиноч-
ные темно-красного цвета узелки. Оба листка брюшины блестящие. Диафрагма справа
на уровне четвертого, слева — у пятого ребра. Печень слегка выступает из-под ребер-
ного края. Селезенка не выступает из-под реберной дуги. Оба легкие частично спая-
ны с костальной плеврой плотными фиброзными спайками. В полости сердечной со-
рочки имеется около 50 см3 кровянистой жидкости. Оба листка перикарда блестящие.
ЦНС. Твердая мозговая оболочка умеренно напряжена. Мозговые извилины слегка
сглажены. Мягкие мозговые оболочки ярко-розового цвета, значительно отечные, мес-
тами утолщены, белесоватого цвета. Мозг весит 1350,0 г. Вещество мозга на разрезе
белого цвета, отечное. В полостях обоих средних ушей имеется геморрагическая жид-
кость, особенно справа.
Органы кровообращения. Сердце 12x11x3 см. Оба желудочка расширены, миокард
мясо-красный, с глинистым оттенком, дряблый. Клапаны сердца и больших сосудов —
без видимых изменений. Аорта в грудном отделе 4 см шириной. Интима ее резко имби-
бирована кровяным пигментом, багрово-красного цвета.
Органы пищеварения. Миндалины небольшие, аспидно-пигментированы. На раз-
резе в их ткани имеются беловатого цвета тяжи. Кроме того, в правой миндалине вид-
ны гнойные пробки. Слизистая пищевода без видимых изменений.
Слизистая желудка имеет шагреневый вид, розовый цвет, содержит мелкие крово-
излияния и покрыта значительным слоем слизи. Слизистая тонких кишок без види-
756
Часть 3. Частная неправильная эпидемиология
мых изменений. На слизистой слепой кишки имеются очаги кровоизлияния с некро-
зом в центре. На остальном протяжении толстого кишечника видимых изменений не
отмечается.
Печень 27x20x8 см. Капсула ее блестящая, пол капсулой просвечивают многочис-
ленные круглой формы, с горошину величиной очаги серовато-желтоватого цвета,
мутного вида, частью с геморрагическим поясом. На разрезе ткань печени буровато-
глинистого вида, дряблая. Желчные пути проходимы. Слизистая желчного пузыря
бархатистая. Поджелудочная железа дольчатого строения, желтовато-серого цвета.
Органы дыхания. На слизистой оболочке гортани в области голосовых связок име-
ются небольшие язвенные дефекты, покрытые грязно-серой некротической массой.
Слизистая трахеи и крупных бронхов цианотично-красная. Левое легкое тестовато-
губчатой консистенции, ткань его на разрезе темно-красная, сжимаемая, при этом с
поверхности разреза стекает много темной крови и пенистой жидкости. Нижняя доля
правого легкого имеет очаги уплотнения, на разрезе темно-красного и серо-красного
цвета, безвоздушные. На остальном протяжении ткань этого легкого насыщенно-крас-
ная, сжимаемая.
Органы кроветворения. Костный мозг грудины бледно-розовый, сочный. Костный
мозг бедра в центре жировой, а по периферии (вблизи кости) розовый. Селезенка (раз-
мерами 14x11x4 см), плотно спаяна с диафрагмой на всем протяжении. Ткань ее на
разрезе темно-красная, с аспидным оттенком, соскоб пульпы отсутствует. Лимфати-
ческие узлы ворот легкого и илеоцекального угла умеренно гиперплазированы. Осталь-
ные лимфатические узлы не увеличены.
Мочеполовые органы. Почки 11x6x3 см каждая. Капсула снимается без паренхимы,
ткань набухшая, корковый слой дрябл ый, с сероватым оттенком. В нем видны мелкие
очаги желтовато-серого цвета, мутного вида, с распадом в центре; слизистая лоханок
серо-розовая. Слизистая мочевого пузыря резко-бледная. Предстательная железа без
видимых изменений. В ткани яичек на разрезе видны сероватые тяжи соединительной
ткани.
Эндокринные железы. Надпочечники: корковый слой истончен, бледно-желтый,
мозговой — геморрагически размягчен. Щитовидная железа не изменена.
Патологоанатомический диагноз. Острый бруцеллез. Интерстициальный гепатит
и паренхиматозно-липоидная дистрофия печени. Острая гиперплазия селезенки и
периспленит. Двусторонняя катаральная пневмония с геморрагическим оттенком.
Паренхиматозное перерождение миокарда и почек. Серозный менингит.
Гистологическое исследование. Сердце. Мышечные волокна набухшие, содержат
белковые зерна, частью — мелкие капли липоидов, местами отмечается фрагментация
миокарда.
Легкие. В нижней доле правого легкого обнаружена картина катаральной пневмо-
нии. В очагах воспаления альвеолы выполнены лейкоцитами и макрофагами с бобо-
видным ядром и широкой зоной протоплазмы. В экссудате имеется значительная при-
месь лейкоцитов. В легочных сосудах среднего и мелкого калибра встречаются
интимальные грануломы из клеток моноцитарного типа, заключенных в сети фибри-
на. Кое-где в альвеолах видны группы сидерофагов.
Печень. В центре долек нередко встречаются очаги микронекрозов. Печеночные
клетки на периферии долек имеют в протоплазме липоидные включения в виде круп-
ных капель. В области центральных вен и печеночных капилляров видны множествен-
ные узелки из мелких клеток, с круглыми и вытянутыми, интенсивно окрашенными
ядрами. Интерстициальные прослойки печени пронизаны круглоклеточным воспали-
тельным инфильтратом. Со стороны мелких кровеносных сосудов местами видна кар-
тина пролиферативно-деструктивного васкулита: эндотелий пролиферирован и слои
3.4. Бруцеллез
757
стенки пронизаны круглоядерными «детками воспалительного инфильтрата На месте
разрушенных воспалительным процессом сосудов видны розеткообразные гранулемы.
Заключение: токсические микронекрозы печени и внутридольковые грануломы в
ней. Продуктивно-деструктивный васкулит мелких сосудов.
Селезенка. В красной пульпе отмечается наличие диффузной гиперплазии клеток
ретикуло-эндотелиальной системы с образованием свободных макрофагов и накоп-
лением экссудативных клеточных элементов — гранулоцитов, лимфоцитов и плазмо-
цитов. Ретикулярная строма частью исчезла, частью набухшая, коллагенизирована.
Нередко встречаются оча1 и кровоизлияния и серозной апоплексии с исчезанием кле-
точных элементов. Кроме того, имеются микронекрозы и одиночные анемические
инфаркты В основе последних стоит острый септический тромбангит. Лимфатичес-
кие фолликулы редуцированы.
Лимфатические узлы. Краевой синус расширен, в нем имеется значительное скоп-
ление лейкоцитов и макрофагов Скопление макрофагов и экссудативных клеток вид-
но также и в синусах между мозговыми тяжами. В мозговых тяжах преобла йют круп-
ные макрофаг ыьного типа клетки, с бобовидным или круглым ядром и широкой зоной
протоплазмы. Лимфатические фолликулы неясно контурированы.
Заключение: острый катар синусов и диффузная гиперплазия элементов ретикуло-
эндотелиальной системы.
Костный мозг диафиза бедра отечный, бедный кроветворными клеточными эле-
ментами. Последние представлены в виде рассеянных островков, состоящих из миз-
лобластов и миэлоцитов. Видны участки фиброзного запустения ткани мозга.
Почки представляют картину некротического нефроза. Эпителий извитых каналь-
цев первого порядка резко набухший, протоплазма клеток мутная, содержит много бел-
ковых зерен. Встречаются большие группы канальцев, в которых ядра клеток не окра-
шены. Наряду с этим видны септические некрозы, вызванные микотическими эмбо-
лами в артериолах. Бактериальные колонии состоят из мелкил грамотрицательных
коккобацилл. Одиночные клубочки фиброзно запустели. При этом среди коллагено-
вой ткани можно видеть остатки организующейся фибриноидной субстанции, окра-
шенной пикрииофильно. Вокруг артериол изредка встречаются муфты круглоклеточ-
ного инфильтрата.
Заключение: некротический нефроз почек, очаговый эмболический нефрит.
Яички. Межканальцевые интерстициальные прослойки расширены вследствие
значительного развития волокнистой соединительной ткани, богатой вытянутой фор-
мы клетками. Вокруг сосудов в интерстициальной ткани имеются узелки грануломы,
состоящие из клеток лимфоидного и моноцитарного типа с примесью фибробластов.
Эти гранулемы имеют связь с мелкими сосудами, возникая на фоне пролиферативно-
деструктивного васкулита. Зародышевые канальцы атрофичны. В них видны сперма-
тогонии и сперматоциты, между тем как сперматозоиды отсутствуют. Сертолиевы клет-
ки встречаются редко, имеют пикнотичные ядра
Кожа. В строме кожи видны множественные грануломы. состоящие из круглоядер-
ных и гистиоцитарных клеток с примесью эпителиоидных. Мелкие сосуды представ-
ляют картину продуктивно-деструктивного васкулита: слои их стенок не различаются
вследствие круглоядерной воспалительной инфильтрации. На месте разрушенных со-
судов образуются гранулемы. Сосочковый слой сглажен. Слой эпидермиса атрофич-
ный. В базальном слое имеется множество клеток, содержащих зерна темно-бурого
пигмента меланина.
Гипофиз. Капсула гипофиза склерозирована и местами инфильтрирована кругло-
клеточным воспалительным инфильтратом. Капилляры расширены и переполнены
кровью. Отмечается умеренный склероз интерстициальных ретикулярных прослоек.
758
Часть 3. Частная неправильная эпидемиология
В задней доле имеются очаги периваскулярной круглоклеточной воспалительной ин-
фильтрации. Железистые пузырьки передней доли состоят, главным образом, из эози-
нофильных клеток. По периферии этой доли имеются группы железистых трубок,
состоящие из крупных базофильных клеток. В центре передней доли видны немного-
численные железистые фолликулы, выстланные хромофобными клетками. Часть же-
лезистых пузырьков содержит коллоид. Средняя доля i [меет много железистых поло-
стей, кистознорасширенных и переполненных кол тоидом. В задней доле, на границе
с передней, видны i руппы внедрившихся базофильных клеток.
Надпочечники ре 1ко ггрофичны. В корковом слое встречаются очаги кровоизлия-
ний и некрозов.
ЦНС. В области полушарий мозга отмечаются очаги склеротического утолщения
мягких мозговых оболочек, а также очаговая круглоклеточная воспалительная инфиль-
трация их. В оболочках имеются обширные кровоизлияния вокруг сосудов среднего
калибра, с некротически измененными стенками. Такие же кровоизлияния встреча-
ются в белом веществе мозга. В сером веществе коры имеется распространенное по-
ражение капилляров: набухание и пролиферация эндотелия, располагающегося в не-
сколько рядов, некробиоз стенки и кольцевые кровоизлияния. Артериолы также
воспалительно изменены. В них отмечаются интимальные грануломы из одноядерных
клеток и муфты в адвентиции из круглоклеточного воспалительного инфильтрата. Под
эпендимой бокового желудочка имеется много миэлиновых шаров, видна картина
разрежения вещества мозга Ганглиозные клетки значительно н<пружены пигментом
липофусцином. Часть 1 англиозных клеток находится в состоянии хронического забо-
левания и некробиоза. Отмечается периваскулярный огек. В продолговатом мозге и
мозжечке наблюдается резко выраженный периваскулярный отек.
Заключение: очаговый серозно-лимфоцитарный летоменингит с исходом в скле-
роз; эндо- и периваскулит мелких сосудов с наличием кровоизлияний в ткани мозга.
При своевременном и правильно проведенном лечении некотаря часть больных
выздоравливает спустя 1—2 года от начала болезни, однако чаще болезнь затягивает-
ся и принимает хроническое течение.
Хронический бруцеллез. Развивается чаще всего вследствие оанее перенесенных
острой и подострой форм болезни и ее рецидивов. Однако допускается возможность
развития первично-хронического бруцеллеза непосредственно после периода первич-
ной латенции. Наиболее часто при хроническом бруцеллезе наблюдается поражение
опорно-двигательной системы в виде артритов, периоститов, перихондритов. Харак -
терны полиартриты с вовлечением в патоло! ический процесс преимущественно круп-
ных суставов — коленный, тазобедренный, локтевой, пояснично-крестцовое сочле-
нение и др., редко поражаются мелкие суставы. Отмечаются боли в мышцах и суставах,
ограничение движений. Суставы опухают, может быть гиперемия кожи за счет пери-
артрита. В суставах могут развиваться воспалительные изменения, сопровождающи-
еся накоплением выпота. При повторных поражениях суставов изменению подвер-
гаются внутрисуставные поверхности, мениски, хрящи с последующим сужением
суставной щели и ее заращением. Это приводит к артрозам, спондилоартрозам, ан-
килозу. Характерно поражение крестцово-подвздошных сочленений с развитием ан-
килоза. При хроническом бруцеллезе часто поражаются мышцы. Миозиты сопровож-
даются болями — продолжительными, разной интенсивности. Помимо миозитов, у
больных бруцеллезом часто выявляются фиброзиты (цел политы). Они локализуют-
ся в подкожной клетчатке на спине, пояснице, реже — на голенях и предплечьях. Раз-
меры их колеблются от 5—10 мм до 3—4 см. Со временем они уменьшаются и могут пол-
ностью рассосаться или склерозируются и остаются длительно безболезненными.
3.4 Бруцеллез
759
Жалобы больных на общую слабость, утомляемость, повышенную раздражитель-
ность, нарушение сна свидетельствуют о функциональном расстройстве нервной си-
стемы, протекающем по типу неврастении. Поражения периферической нервной
системы при хроническом бруцеллезе проявляются радикулитами, плекситами, меж-
реберными и другими видами невралгий, расстройствами чувствительности, пареза-
ми, невритами слухового и зрительного нервов со значительным снижением слуха и
остроты зрения.
Хроническая интоксикация ЦНС, особенно коры головного мозга, в случаях дли-
тельного течения инфекции приводит к тяжелым неврозам, реактивным состояниям,
ипохондрии, психозам. Наблюдаются кратковременные психосенсорные расстрой-
ства, оптико-вестибулярные и рецепторные нарушения. Более стойкими бывают рас-
стройства психики с астеническим и ипохондрическим синдромами, ослабление па-
мяти, понижение или повышение эмоциональной возбудимости.
При вовлечении в процесс вегетативной нервной системы нарушается тонус сосу-
дов, возникают акроцианоз, обильное потоотделение, трофические нарушения кожи.
Закономерно поражается сердечно-сосудистая система и в первую очередь крове-
носные сосуды. У больных возникают эндо-, пери- и панваскулиты, повышается про-
ницаемость капилляров. Нередко отмечаются миокардиты, эндокардиты, панкар-
диты. Печень и селезенка увеличены, уплотнены, их функции нарушены.
При хроническом бруцеллезе нередко поражается урогенитальная система: у муж-
чин возникают орхит, эпидидимит, у женщин — оофарит, сальпингит, эндометрит,
нарушение менструального цикла и прерывание беременности.
в ряде случаев выявляется нарушение функции шитовитной железы, надпочеч-
ников и других эндокринных органов.
Картина крови характеризуется лейкопенией, лимфоцитозом, моноцитозом, эози-
нопенией, тромбоцитопенией.
Для хронического бруцеллеза характерно волнообра шое течение болезни с ремис-
сиями и рецидивами. Бальные надолго утрачивают работоспособность, могут стать
инвалидами.
У вакцинированных лиц заболевание протекает легче и менее прололжительно, с
невысокой температурой, нередко с самого начала отличается локальными симпто-
мами в виде первично-хронической формы болезни. У значительного числа вакци-
нированных, заболевших первично-хроническим бруцеллезом, заболевание при-
нимает стертый характер с мало выраженными проявлениями. Наиболее постоянным
и иногда единственным симптомом в таких случаях являются артралгии.
Резидуальная фаза {клиника последствий) бруцеллеза. У лиц, перенесших бруцел-
лез, нередко наблюдаются те или иные остаточные явления, в основном функциональ-
ного характера, обусловленные иммуноаллергической перестройкой организма и
расстройствами вегетативной нервной системы. У таких больных отмечаются повы-
шенная потливость, раздражительность, изменения нервно-психической сферы, не-
редко имеются артралгии. Боли в суставах чаше непостоянного характера, при этом
видимых изменений в суставах не выявляется. Эти боли усиливаются при выполне-
нии физической работы и в связи с переменой погоды. Температура тела у больных
обычно нормальная, иногда субфебрильная. У ряда больных, перенесших бруцеллез,
наблюдаются органические изменения опорно-двигательной системы с деформация-
ми суставов за счет разрастания околосуставной ткани, что выявляется при клиниче-
ском и рентгенологическом обследовании. Стойкие и необратимые явления со сторо-
ны опорно-двигательного аппарата могут требовать оперативного вмешательства.
760
Часть 3. Частная неправильная эпидемиология
Смертность при нелеченном бруцеллезе не превышает 2 %. В большинстве случа-
ев причиной гибели является эндокардит, вызванный поражением В. melitensis.
Моделирование ингаляционного заражения бруцеллами. По данным R. Sidell et al.
(1997), симптомы бруцеллеза у людей, инфицированных бруцеллами посредством
аэрозоля или любым другим способом, одинаковы. S. L. Yingst с соавт. (2009) изучи-
ли патоморфологию ингаляционного бруцеллеза, вызванного аэрозолем В. suis со
средним размером частиц 1 мкм. Даже в дозе 5,6х108 КОЕ он не вызвал у обезьян Ма-
саса mulatto каких-либо специфических поражений легочной ткани. К седьмому дню
после заражения аэрозолем в патологический процесс были в основном вовлечены ме-
зентеральные, трахеобронхиальные и подчелюстные лимфатические узлы, что про-
являлось их умеренно выраженным отеком. Наиболее увеличенными по сравнению
с нормой были трахеобронхиальные узлы (в 4 раза), наименее — подчелюстные. Но,
по мнению авторов, эти изменения носили неспецифический характер и отражали ре-
акцию иммунной системы на антигенную стимуляцию бруцеллами. На третий день
после заражения аэрозолем В. suis, у обезьян были обнаружены: мультифокальная де-
генерация и некроз эпителия бронхов с умеренным лимфогистиоцитическим (lympho-
histiocytic) и нейтрофилическим (neutrophilic) бронхиолитом и перибронхиолитом, и
альвеолярным отеком. Через 7 сут после заражения аэрозолем В. suis у двух из трех
заболевших обезьян обнаружен лимфогистолитический гепатит. Дальнейшее течение
болезни авторы не наблюдали.
Диагностика искусственного поражения бруцеллами. Общие принципы такой диаг-
ностики те же, что описаны в разд. 3.1 для поражения возбудителем сибирской язвы.
Но надо учитывать то обстоятельство, что искусственное поражения бруцеллами но-
сит более изощренный и неявный характер, чем поражения возбудителями чумы,
сибирской язвы или туляремии. Возбудители бруцеллеза — это калечащие агенты БО
и целью их применения преступниками станет не создание обстановки страха или
паники, а нанесение длительного ущерба здоровью как можно большему числу наших
граждан. Основным критерием искусственного происхождения вспышки бруцелле-
за является неправильная эпидемиология. Ее распознание затрудняют продолжитель-
ные инкубационный и продромальные периоды болезни, сочетающиеся с неяснос-
тью и полиморфностью клинических признаков. Быстро обнаруживаемые острые
формы болезни могут составить небольшую часть выявленных случаев бруцеллеза, но
при этом их далеко не сразу распознают как бруцеллез. У большей части больных
клинические проявления бруцеллеза ограничатся только признаками продрома, в
последующем бруцеллез у них будет протекать как хроническая калечащая инфекция,
распознать которую можно только по иммунологическим реакциям. «Невозможных
клинических форм болезни», которые сразу «бросятся в глаза» при ингаляционной
чуме или сибирской язве, ингаляционное поражение бруцеллами не даст. Поэтому
очень сложно определить источник аэрозоля возбудителя бруцеллеза, идентифици-
ровать его тип, установить его территориальную локализацию, оценить масштаб пре-
ступления и принять меры к ликвидации всех его последствий. К тому же может быть
упущено время, необходимое для ликвидации преступной группы.
«Зацепкой» для установления искусственного характера вспышки бруцеллеза бу-
дет обнаружение разрыва эпидемической цепи, ведущей от природного источника
возбудителя бруцеллеза к людям (для В. melitensis — козы и овцы; В. abortis — КРС;
В. suis — свиньи, северные олени). При естественной вспышке бруцеллеза, по край-
ней мере, у части заболевших людей в анамнезе должен прослеживаться либо прямой
контакт с этим животными, либо опосредованный — через полученные от них инфи-
3.4. Бруцеллез
761
нированные бруцеллами продукты (молоко, шерсть, шкуры). Обнаружение бруцелл
в конечных продуктах сельхозпроизводства, употребляемых человеком в пищу, тре-
бует установления их в исходном продукте, т. е. установление источника контамина-
ции этих продуктов. Обязательно необходимо найти сельскохозяйственное предпри-
ятие, из которого этот продукт попал в продажу, затем установить наличие вспышки
бруцеллеза среди содержащихся там животных и подтвердить идентичность штаммов,
выделенных от людей и от животных. На самом предприятии необходимо установить
все обстоятельства появления бруцеллеза среди животных, включая выявление постав-
щиков кормов и проверку кормов на контаминацию бруцеллами.
Для оценки последствий биологической диверсии следует выявить всех инфи-
цированных бруцеллами лиц. Для этого необходимо провести серологическое иссле-
дование на бруцеллез лиц, которым в предполагаемый период в местных лечебных
учреждениях были поставлены диагнозы гриппа, ОРЗ, брюшного тифа, паратифа,
пневмонии, инфекционного гепатита, туберкулезного менингита, ревматического по-
лиартрита, лимфогранулематоза, лихорадки Ку, сыпного тифа, малярии и сепсиса.
Затем установить, где заболевшие были в период времени, соответствующий среднему
инкубационному периоду бруцеллеза, и попытаться определить:
1) локализацию источника аэрозоля (см. разд. 2.3.2 «Эпидемиологическое рассле-
дование искусственного эпидемического процесса»);
2) либо установить продукты питания (торговые точки, пункты общественного
питания), которые могут быть общими для всех инфицированных бруцеллами лиц.
Иммунопрофилактика бруцеллеза. В России производятся две вакцины:
вакцина бруцеллезная живая сухая на основе вакцинного штамма В. abortus 19ВА.
Вакцина предназначена для профилактики козье-овечьего бруцеллеза у взрослых
людей. Производитель: «Омское предприятие по производству бакпрепаратов» (фи-
лиал ФГУП НПО «Микроген» Минздрава России);
вакцина бруцеллезная лечебная жидкая. Предназначена для лечения больных с
острым, подострым и хроническим течением бруцеллеза. Представляет собой смесь
инактивированных нагреванием вирулентных микробов штаммов В. melitensis № 21,
145 и В. abortus № 544, смешанных в соотношении 1:1, консервант — фенол в концент-
рации 0,25 %. Производитель: ФГУ «48 ЦНИИ Минобороны России», г. Киров.
Вакцинацию живой вакциной проводят однократно накожно за 3—4 нед до нача-
ла работ, связанных с риском заражения бруцеллами. Ревакцинацию проводят через
10—12 мес. накожно, используя половинную дозу вакцины. Согласно исследованию
П. А. Вершиловой с соавт. (1972), вакцинация живой вакциной на основе штамма 19ВА
снижает заболеваемость бруцеллезом среди лиц, обслуживающих овец в 3—11 раза, в
зависимости от их вида деятельности. Ее защитная эффективность при ингаляцион-
ном заражении человека каким-либо видом бруцелл, мне неизвестна.
Лечебная вакцина в настоящее время используется крайне редко из-за высокой
аллергизации населения и сенсибилизирующих свойств вакцины.
Лечение бруцеллеза1. Задачами лечения больных бруцеллезом являются купиро-
вание патологического процесса, ликвидация его последствий и восстановление тру-
доспособности пациента. Терапия болезни зависит от фазы, степени компенсации
процесса и характера иммунноаллергической перестройки организма.
Для лечения острых и подострых форм бруцеллеза вначале применяют антибио-
тики: левомицетин (по 0,5 г 4 раза в сут) или рифампицин (0,9 г в сут), тетрациклин
1 По работам П. А. Вершиловой с соавт. (1972) и Е. П. Шуваловой с соавт. (2001).
762
Часть 3. Частная неправильная эпидемиология
(по 0,5 г 4 раза в сут), доксициклин (0,2 г в сут), стрептомицин (по 0,5 г 2 раза в су г
внутримышечно). Продолжительность курса антибиотикотзрапии не менее 14 сут.
В последние годы в терапии бруцеллеза с хорошим эффектом применяют препараты
из группы хинолонов (офлоксацин, дифлоксацин и др.), антибиотики комбинируют
с бисептолом.
При хроническом бруцеллезе антибиотики применяют лишь в периоде обостре-
ния болезни с выраженными клинико-лабораторными проявлениями. При отсутствии
признаков активации процесса, а также в стадии субкомпенсации хронического бру-
целлеза с устойчивой нормализацией температуры назначение антибиотиков нецеле-
сообразно.
При артритах и периартритах эффективно внутри- или периартикулярное введе-
ние гидрокортизона. При подостром и хроническом бруцеллезе с преимущественным
поражением опорно-дьигательного аппарата и периферической нервной системы
назначают нестероидные противовоспалительные средства с анальгезирующим эф-
фектом: бутадион, ацетилсалициловая кислота, анальгин, реопирин, индометацин,
бруфен, ибупрофен, вольтарен, диклофенак и др.
В стадии компенсации или резидуальных проявлений бруцеллеза больным наи-
более эффективно санаторно-курортное лечение, в особенности на бальнеологиче-
ских курортах с радоновыми и серно-радоновыми водами. При поражении суставов
эффективно грязелечение.
3.5. САП
Микробиология возбудителя сапа. Факторы патотеннисти. Таксономия, про-
исхождение и молекулярная эпидемиология возбудителя сапа. Экология и есте-
ственная эпидемиология возбудителя сапа. Сап у лошадей. Восприимчивость
к возбудителю сапа и инфицирующие дозы для животных и людей. Иммунитет.
Устойчивость возбудителя сапа во внешней среде. Антибиотикорезистен'ность.
Обнаружение генетически измененных штаммов возбудителя сапа. Патогенез
сапа. Клиническая картина сапа у людей при естественном инфицировании.
Моделирование ингаляционного заражения сапом. Клиническая картина сапа
у людей при искусственном инфицировании. Диагностика искусственного пора-
жения сапом. Иммунопрофилактика сапа. Лечение сапа.
Сап [malleus (лат.)', rotz (нем.); glanders, farcy (англ.); morve (франц.); morva (итал.);
muermo (ucn.)] — опасная инфекционная болезнь животных и человека, вызываемая
бактерией Burkholderia mallei. Протекает в острой и хронической формах. Характе-
ризуется образованием специфических клеточных гранулем на слизистых оболочках
носа, гортани, трахеи, в ткани легких и других паренхиматозных органах и кожных
покровах. Болезнь известна с древности как болезнь лошадей. Заражение сапом че-
ловека от лошади впервые описал Оскандес (Oskandes) в 1783 г. Возбудитель сапа в
срезах ткани и гное больного человека впервые обнаружен румынским ученым В. Ба-
бешем (V. Babes, 1881), а в крови больного — русским исследователем Н. П Василь-
евым (1883).
В природных условиях источниками В. mallei для людей в России являются лоша-
ди. Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят воз-
будитель сапа ко II группе патогенности. По степени важности для национальной
3.5. Can
763
безопасности США возбудитель сапа относится к биологическим поражающим аген-
там категории В. С 1915 г. он рассматривался германским Генштабом как диверсион-
ный поражающий агент и успешно использовался германской агентурой для зараже-
ния лошадей и мулов, используемых в войсках Антанты. Исследования по изучению
поражающих свойств возбудителя сапа как агента БО велись с начала 1930-х гг. в япон-
ском отряде № 731, с 1944 г. в США в Форт-Детрике. Японскими военными в годы
Второй мировой войны разрабатывались боеприпасы, снаряженные возбудителем
сапа. Он неоднократно использовался диверсионными способами против советских
войск в приграничных районах Забайкалья, Дальнего Востока и во время Маньчжур-
ской наступательной операции. Диверсии продолжались даже после капитуляции
Квантунской армии. В. mallei очень опасна для работающего с ней персонала лабора-
торий. Вакцин, эффективно защищающих от заражения В. mallei, не разработано.
С целью поражения людей наиболее вероятно применение возбудителя сапа по-
средством использования линейных и многоточечных источников и диверсионными
(террористическими) методами.
Мию 1оислогия возбудителя сапа. В. mallei—тонкая слегка изогнутая и неподвиж-
ная палочка, средние ее размеры 2—3 мкм в длину, 0,5—1 мкм в ширину. Специфиче-
ская окраска для В. mallei не разработана. Бактерии возбудителя сапа слабо прокра-
шиваются обычными водными растворами анилиновых красителей. Интенсивность
их окраски возрастает при введении в состав красок фенола или едкого калия. При
окраске по Граму обесцвечиваются и нередко окрашиваются неравномерно с чередо-
ванием на проекции тела клеток светлых и темных участков. Степень проявления не-
равномерности окраски зависит от физиологических особенностей микробов и ме-
тода подготовки препарата. При выращивании на питательных средах В. mallei может
полиморфизироваться. Наблюдаются палочки меньших размеров и длинные цепоч-
ки неразделившихся клеток. Размеры бактерий зависят от штамма и питательной сре-
ды. Описаны нити от 5 до 15 мкм и даже 50—65 мкм с разветвленными и колбовидны-
ми вздутиями на концах, что послужило некоторым авторам основанием причислить
возбудигеля сапа кстрептотрихиям или актиномицетам. Молодые клетки имеют глад-
кие контуры. В старых культурах контуры клеток неровные. Концы клеток, как пра-
вило, слегка закруглены. После вырашивания в жидких питательных средах в усло-
виях недостаточной аэрации микробы укорачиваются, утолщаются и в большом
количестве накапливают гранулы различных размеров. При длительном культивиро-
вании могут появляться длинные нити с булавовидными или почкообразными утол-
щениями на концах (Кравченко А. Т., Руднев Г. П., 1966; Золотарев А. Г. с соавт., 2006)
(рис. 3.59).
Степень проявления полиморфизма клеток В. mallei зависит и от формы течения
инфекции. При хроническом сапе, особенно на фоне химиотерапии, нередко выде-
ляют культуры с гетероморфными микробами, а также в виде стабильных или неста-
бильных L-форм (Золотарев А. Г. с соавт., 2006).
Клетки возбудителя сапа растут на обычных питательных средах с добавлением
4-5 % глицерина и в дополнительных факторах роста не нуждаются. Оптимальный
рост В. mallei возможен при температуре 37 °C. При температуре ниже 20 и выше 42 °C
развитие культуры не происходит. Реакция питательной среды может быть слабокис-
лой или слабощелочной. Оптимальный рост возбудителя сапа наблюдается при pH 6,8.
Незначительный рост культуры отмечается даже при pH 7.6 и 5,6. В. mailt i — строгий
аэроб. При выращивании В. mallei в пробирке с мясо-пептонным бульоном, содер-
жащим глицерин, в начале роста среда выглядит равномерно мутной. На 2 сут роста
764
Часть 3. Частная неправильная эпидемиология
Рис. 3.59. Микроскопическая картина возбудителя сапа. А. В. mallei Ц-5 в окрашен-
ном мазке культуры, выращенной в течение 48 ч на плотной питательной среде
с добавлением 4 % глицерина. Б. В. mallei Ц 5 в циторефрак гометрическом пре-
парате. По оптической плотности живые клетки возбудителя сапа неоднородны.
Одни из них представлены лишь оптическим эквивалентом оболочки в виде свет-
лого ободка и имеют протоплазму по цвету такую же, как фон поля зрения. Другие
отличаются наличием ярко светящихся включений различной величины. Мертвые
клетки выглядят в виде светлых тел. По А. Г. Золотареву с соавт. (2006)
зуется пристеночный осадок, превращающийся в пленки. Затем на дне пробирки
появляется белесоватый слизеобразный осадок, который при вращательном движе-
нии пробирки поднимается кверху в виде тяжей. С поверхности среды от слизистой
оболочки пленки вниз спускаются нити. С течением времени осадок на дне увеличи-
вается, среда светлеет, но полностью прозрачной не бывает. На желатине с глицери-
ном В. mallei растет на поверхности в виде прозрачного сероватого налета. В верхней
части укола заметна сероватая полоска без отростков, а на поверхности среды в месте
укола появляется желтоватая окраска. Желатин не разжижается (Кравченко А. Т.,
Руднев Г. П., 1966).
На глицериновом агаре вирулентные штаммы В. mallei через 12 ч инкубирования
при температуре 36—38 °C образуют куполообразные микроколонии с ровными округ-
лыми краями и мелкозернистой поверхностью. После 48-72 ч инкубирования выра-
стают выпуклые колонии с четкими нерасплывчатыми краями. В проходящем свете
они прозрачные, а в отраженном — серовато-белые и имеют форму полушарий или
уплощенных куполов с гладкой поверхностью и ровными краями. На вершине коло-
ний могут располагаться четко отграниченные зоны центрального роста. В отличие
от В. pseudomallei бактерии возбудителя сапа редко образуют шероховатые и слизис-
тые формы колоний (рис. 3.60).
Возбудитель сапа обладает замедленной ферментативной активностью. Некото-
рые из штаммов разлагают глюкозу, маннит, левулезу, глицерин с образованием кис-
лоты, но без газа. Не образуют индола и не восстанавливают нитраты в нитриты. На
жидких средах образуют сероводород и аммиак. Обладают каталазной активностью.
При культивировании в молоке В. mallei образует небольшое количество кислоты,
молоко свертывается на 10 сут, последующей пептонизаиии молока не наблюдается.
На картофельной среде. Возбудитель сапа образует пигмент. Анаэробно В. mallei не
размножается, за исключением условий, когда в питательной среде присутствуют
нитраты. Поэтому отдельные исследователи возбудитель сапа относят к денитрофи-
3.5. Can
765
Рис. 3.60. Макроскопическая картина возбудителя сапа. А. Морфология микро-
колоний В. mallei Ц-5, выращенных в течение 12 ч на поверхности плотной
питательной среды с добавлением 4 % глицерина. Микрофотосъемка в прохо-
дящем свете. Увеличение в 56 раз. Б. Этот же штамм на поверхности плотной
питательной среды с добавлением 5 % глицерина после 72 ч выращивания.
Вирулентные штаммы возбудителя сапа образуют довольно однородные по
форме и размерам колонии. Диссоциации в М- и R-формы они подвергаются
значительно реже, чем возбудитель мелиоидоза. Фотосъемка в падающем свете.
В. Морфология колоний В. mallei Ц-5, выращенных в течение 12 ч на поверх-
ности плотной питательной среды с добавлением 4 % глицерина. Фотосъемка
в падающем свете. Увеличение в 13 раз. По А. Г. Золотареву с соавт. (2006)
цируюшим организмам и факультативным анаэробам (Кравченко А. Т, Руднев Г. П.,
1966; Илюхин В. И. с соавт., 1995).
Факторы патогенности. Возбудитель сапа экзотоксин не образует. По данным
М. A. Schell (2008), патогенность возбудителям сапа и мелиоидоза «обеспечивает» не
менее 650 генов. Роль продуктов их экспрессии в патогенезе болезни еще предстоит
выяснить. Наиболее изучена роль в патогенности В. mallei ЛПС (Brett Р. et al., 2006),
экзополисахаридной капсулы (Вязьмина Т. Н. с соавт., 1995; Золотарев А. Г. с соавт.,
2006), системы секреции III и VI типов (Schell М. A. et al., 2007), пориновых белков
клеточной стенки (Супотницкий М. В.ссоавт., 1995; Супотницкий М. В., 1996),про-
теазы, фосфолипазы (Тихонов Н. Г. с соавт, 1996), аутотранспортерных макромоле-
кул (Lazar AdlerN. R etal., 2011) (рис. 3.61).
К аутотранспортерным макромолекулам относятся адгезины, определяющие по-
вышенную пневмотропность возбудителей сапа и мелиоидоза к легочной ткани че-
ловека. Они идентифицированы как продукты гена boa А, обнаруженного в геноме
В. mallei, В. pseudomallei, и гена boa В, найденного только в геноме В. pseudomallei (Balder
R. etal., 2010).
Таксономия, происхождение и молекулярная эпидемиология возбудителя сапа.
В. mallei с 1885 г. была известна под названием Bacillus mallei. В 1966 г. возбудитель сапа
был перенесен в род Pseudomonas на основе особенностей пищевых потребностей
и биохимических свойств. В 1973 г. на основании данных РНК-ДНК гибридизации
род Pseudomonas был разделен на пять групп гомологии, где Р. mallei была включена
766
Часть 3. Частная неправильная эпидемиология
Рис. 3.61. Электронно-микроскопическое изображение капсулы возбудителя сапа.
В. mallei Ц-5 выращена в течение 48 ч на плотной питательной среде с добавлением
4 % глицерина и заключена в желатин. А. Капсулоподобный покров выявляется в виде
прозрачной для электронов зоны, окружающей одну или несколько клеток. Б. Раз-
меры капсулоподобного покрова у бактерий неодинаковы. Увеличение в 40000 раз.
По А. Г. Золотареву с соавт. (2006)
в группу II. В 1993 г. на основании данных анализа 16S рРНК. ДНК-ДНК-гибридиза-
ции и состава жирных кислот клеточной стенки все семь видов группы гомологии II
были выделены в отдельный род Burkholderia. В. mallei таксономически очень близка
к Burkholderia pseudomallei и Burkholderia thailandensis. Новый род получил название в
честь Walter Н. Burkholder, американского специалиста по патологии растений, рабо-
тавшего в Корнельском университете. Род Burkholderia входит в Р-подгруппу группы
Proteobacteria, наиболее многочисленной группы бактерий, насчитывающей 1534 гра-
мотрицательных видов, что составляет треть от всех известных видов бактерий.
Геном Burkholderia состоит из двух (В. mallei, В. pseudomallei, В. thailandensis) или
трех (В. cenocepacia) кольцевых хромосом (Chi Но Lin et а'.., 2008). По нуклеотидному
составу геном В. mallei на 99 % идентичен В. pseudomallei (SongH. etal., 2010). По сути,
геном возбудителя сапа — это на 20 % «урезанный» геном возбудителя мелиоидоза.
Он включает 5535 ORF, из них для 3173 ORF функция установлена. Основные харак-
теристики хромосом возбудителя сапа приведены в табл. 3.20.
Таблица 3.20
Характеристика хромосом возбудителя сапа*
Характеристика Хромосома 1 Хромосома 2
Размер, т. п. н. 3510148 2325379
Количестве ORF 3344 2091
Кодирующих ORF, % 87,1 85,7
Средняя длина гена, нуклеотиды 914 953
Г 1-Ц-содержание, % 68,2 69,0
тРНК 47 9
* Пи W, С. Nierman et al. (2004)
По данным С. Ong с соавт. (2004), геном В. mallei и геном непатогенной для чело-
века В. thailandensis отличаются от В. pseudomallei тем, что имеют большое количество
делегированных ORF. У возбудителя мелиоидоза эти ORF связаны с метаболическими
3.5. Can
767
процессами и другими функциями клетки, необходимыми для выживания микроор-
ганизма в окружающей среде. Например, утрата В. mallei 2-аминобензоат КоА-лига-
зы (транспортер нитратов и нитритов) и отдельных субъединиц нитратредуктазы ли-
шила ее возможности существовать в ризосфере1.
Делеции, обнаруженные в геноме В. mallei, по сравнению с делениями В. thailan-
densis имеют характер крупных кластеров, что говорит за утрату хромосомами В. mallei
крупных сегментов. Механизм этого явления объяснен в работе Н. Song et al. (2010).
По их данным, возбудитель сапа дивергировал от возбудителя мелиоидоза 3,5 млн лет
назад. Этот процесс носил ступенчатый характер и сопровождался утратой генов, не
нужных В. mallei при ее специализации в узких экологических нишах. Разрушение
генов В. mallei происходило благодаря периодической массивной экспансии IS-эле-
ментов ]SBmal, [SBma2n IS4O7A. Каждый элемент пролиферировал по своим специ-
фическим сайтам, затем \SBmal и ISВта2 были использованы IS40Z4 для своего рас-
пространения, где они играли уже роль вторичных сайтов. Экспансия IS-элементов
увеличила нестабильность генома В. mallei и вызвала обширные делеции и переста-
новки, в которых основную роль сыграл IS40Z4 (рис. 3.62).
Н. Song et al. (2010) на примере эволюции В. mallei предложили следующую мо-
дель редукционной эволюции бактерий. Массивная экспансия IS-элементов форми-
рует множество инсерционных сайтов для экспансии какого-то мажорного IS-элемен-
та, способных делетировать и переставлять крупные участки генома бактерии через
рекомбинацию гомологичных IS-копий. Этот процесс ведет к делениям участков
ДНК, не нужных бактерии в занимаемой ею экологической нише.
Результаты географической дифференциации штаммов В. mallei и В. pseudomallei,
выполненные мультилокусным секвенированием по 14 генетическим локусам 147 изо-
лятов, приведены в работе D. Godoy et al. (2003). Однако изоляты В. mallei, выделен-
ные за последний 30-летний период в Турции, Индии, Болгарии, имели идентичные
аллельные профили и кластировались в пределах изолятов В. pseudomallei. В более
поздней работе Н. Homstra et al. (2009) этим же методом, но используя картирование
по 23 локусам VNTR, удалось провести географическую дифференциацию штаммов
возбудителя сапа, выделенных в Пакистане.
Экология и естественная эпидемиология возбудителя сапа2. Сап считается зоонозом,
однако есть основания полагать, что млекопитающие составляют только вторичный
резервуар В. mallei. Отдельные случаи сапа среди людей возникают на фоне одноимен-
ной эпизоотии среди сельскохозяйственных животных. Заболевание чаще наблюда-
ется в холодное время года. Основной контингент больных при естественной заболе-
ваемости — мужчины. Заболеваемость спорадическая, но описаны семейные вспыш-
ки. Основным резервуаром возбудителя и источником заражения человека являются
больные острой формой сапа лошади, мулы, ослы, верблюды, зебры. Значительно
реже наблюдается инфицированность представителей семейства кошачьих (львы,
тигры, барсы, леопарды), которые заражаются, поедая мясо больных сапом животных.
Хронические формы болезни у животных не представляют серьезной опасности для
человека.
1 В работе С. Ong с соавт. (2004) приведено детальное сравнения геномов возбудителей сапа
и мелиоидоза.
2 Естественная эпидемиология сапа приведена по работам Н. Е. Цветкова, В. А. Черняка.
(1935), Т. Кравченко, Г. П. Руднева (1966), Г. М. Ларионова (1995), Е. П. Шуваловой с соавт.
(2001).
768
Часть 3. Частная неправильная эпидемиология
100
Рис. 3 62. Экспансия IS-элементов по геному В. mallei. А Среднее количество
копий IS-элементов, обнаруженных у В. mallei (ВМ) и В. pseudomallei (ВР). Пока-
зано количество интактных (серый цвет) и дегенеративных (т е. частично деле-
гированных, выделено белым цветом) копий IS-элементов. Б. Распределение
трех типов IS-элементов в геноме В. mallei. ISBmal, ISBma2и IS407A могут быть
разделено на три группы: серым цветом обозначены «коровые» IS-элементы —
представлены во всех штаммах (играют основную роль в модификации генома
возбудителя сапа); черным цветом — IS-элементы, представленные более чем
в двух штаммах, но не во всех исследованных; светло-серым — IS-элементы,
обнаруженные только в одном штамме В. mallei. По Н. Song et al. (2010)
Естественное инфицирование человека сапом происходит в результате контакт-
ной передачи В. mallei при попадании гноя и слизи больных животных на поврежден-
ные места кожи и слизистых оболочек. Реже заражение В. mallei может происходить
алиментарным путем через инфицированную воду, употребляемую для питья, и еще
реже — ингаляционно (в лабораторных условиях). Не исключена возможность зара-
жения человека через слизистые пищеварительного тракта, слизистую оболочку носа
и легкие. Чаше всего заражение В. mallei носит профессиональный характер у лиц,
связанных с уходом и лечением больных животных. Возбудитель сапа устойчив в воде
и при попадании в нее из пораженных слизистых оболочек носа животных может
сохранять свою жизнеспособность до 30 сут. Использование зараженной В. mallei воды
для питья представляет для человека реальную опасность заражения.
Восприимчивость людей к сапу всеобщая. Локализация первичного аффекта для
каждого случая заражения может отличаться. По данным, приведенным Г. М. Лари-
оновым (1995), в 117 случаях сапа у людей, локализация первичного аффекта при ес-
тественном инфицировании возбудителем сапа, распределена следующим образом:
в 45 случаях первичное поражение располагалось на кисти; в 7 — на предплечье или
3.5 Can
769
плече; в 22 — на нижних конечностях (ходьба босыми ногами); в 15 — на голове, шее,
’ине; в 6 — на коже других частей тела. В остальных 22 случаях В. mallei проникла в
организм человека через слизистые оболочки глаза, рта и носа.
В настоящее время в России сап не регистрируется, но эпизоотическую ситуацию
но сапу нельзя упрощать. Во-первых, возможен занос В. mallei из азиатских стран (Ки-
тай, Монголия, Афганистан и др.). Имеются сведения об отдельных случаях сапа среди
лошадей в Иране, Монголии, Турции, Индонезии, Китае и на Филиппинах (Ларио-
нов Г. М., 1995). Занос болезни в Россию возможен и из северных областей Казахста-
на, куда с начала 1990-х гг. казахские власти для «исправления» демографической
ситуации переселяют казахов-аралманов из Монголии. Во-вторых, официальной
российской статистике, отражающей лишь благополучие по сапу, доверять не стоит.
Ни один руководитель не решится признать наличие у него в хозяйстве заболевшей
сапом лошади. Обычно в таких случаях больную лошадь без излишней огласки заби-
вают, а ее труп сжигают. Иногда эта практика приводит к большим эпизоотиям сапа.
Например, такая эпизоотия вспыхну та в 1967 г. в Ханты-Мансийском национальном
округе Тюменской области и привела к большим экономическим потерям. В 1984 г. в
совхозе «Заречный» Донидинского района Бурятии, в 5 км от Монголии, также отме-
чалась эпизоотия сапа среди лошадей. Больных животных уничтожили и закопали в
землю, а условно здоровых лошадей (без клинических признаков и не реагирующих
на маллеин) сдали на санитарную бойню для немедленного убоя. Но во всех моногра-
фиях и учебниках по инфекционным болезням можно прочитать только то, что сап в
СССР ликвидирован в 1959 г. В-третьих, неизвестен первичный резервуар возбуди-
теля сапа, а следовательно, неизвестны механизмы активизации и географическая
локализация очагов сапа.
Так как фермеры и мелкие владельцы скота могут скрыть факт эпизоотии сапа,
.либо не распознать ее при молниеносном, подостром, хроническом или атипичном
течении болезни, то для выявления звеньев эпидемической цепочки, по которой рас-
пространилась В. mallei в популяции людей, необходимо путем опроса самих ферме-
ров (хозяев), их работников и соседей по внешним признакам течения болезни уста-
новить хозяйства, где эпизоотии сапа могли иметь место.
Сап у лошадей. При естественном инфицировании лошадей В. mallei клинические
признаки сапа проявляются через 4 нед, иногда через несколько месяцев1. В некото-
рых случаях сап протекает без клинических симптомов, локализируясь только во внут-
ренних органах, и распознается биологическими и молекулярно-генетическими ме-
тодами. По локализации специфического процесса сап разделяется на носовой,
легочный и кожный. Деление на эти формы имеет только клиническое значение. Эти
формы часто развиваются постепенно одна за другой и могут быть одновременно все
три у одной лошади или только некоторые из них.
Острый сап у лошадей может протекать в двух формах: очень острый — скоротеч-
ный и острый. Скоротечный сап приводит животное к смерти, иногда не распознавае-
мый ни клинически, ни биологически. В этих случаях наблюдались язвенные пора-
жения глотки и верхних дыхательных путей, нехарактерные для сапных язв. Харак-
терным признаком этих язв являлись некротические изъязвления слизистой оболочки
вследствие смешанной инфекции. Обычная острая форма сапа возникает или пер-
вично после заражения, или является рецидивом хронического сапа. В первом слу-
чае после короткого инкубационного периода в 3—5 сут поднимается температура
1 Описание клиники сапа у лошадей дано по работе Н. Е. Цветкова, В. А. Черняка (1935).
770
Часть 3. Частная неправильная эпидемиология
до 41—42 °C; наблюдается дрожь, животное впадает в прострацию: слабый пульс, ды-
хание становится ускоренным и неравномерным, пропадает аппетит. Видимые сли-
зистые оболочки гиперемированы. После этого через 1—3 сут начинают проявляться
специфические симптомы. На слизистой носа появляются экхимозы, скоро прини-
мающие желтоватый оттенок, круглой или вытянутой формы, величиной с горошину,
переходящие в пустулы. Через немного часов содержимое их становится серозно-гной-
ным, и они превращаются в язвы с неровными подрытыми краями, кратероподобным
саловидным дном. Из носа начинает вытекать часто с одной стороны зеленовато-жел
товатый, иногда с примесью крови, гнойный секрет. Секрет, покрывая носовые от-
верстия, засыхает здесь и при кашле и фыркании лошади выбрасывается в большом
количестве и инфицирует находящиеся вблизи предметы. Одновременно с этим по-
является припухание подчелюстных желез, часто с той же стороны, с которой проис-
ходит выделение секрета. Подчелюстные железы вначале чувствительны при ощупы-
вании, и припухлость не имеет точных границ. Дыхание становится затрудненным и
сопящим. Животные быстро истощаются, теряя в весе до 40 кг в сут, и умирают обыч-
но на 2-3 нед поел** заражения. Болезнь кончается смертью тем скорее, чем неблагоп-
риятнее были условия для жизни лошади: недостаток корма, сильное утомление, ос-
ложнение другими болезнями, как, например, пироплазмозом. Но и при такой ос-
трой форме сапа иногда развитие процесса приостанавливается, патологические
признаки постепенно уменьшаются, и болезнь принимает хроническое течение.
Чаше острая форма болезни является обострением хронической. В таких случаях
процесс обычно тянется дольше и к симптомам носового сапа присоединяется кож-
ный и легочный сап. Изъязвления в носовой полости развиваются медленнее, подче-
люс1 ные железы, иногда и подъязычные, бывшие вначале чувствительными и более
или менее диффузно-припухшими, теряют свою чувствительность, но остаются уве-
личенными в размере, плотными и прирастают или к коже или к подлежащим тканям
межчелюстного пространства. В паренхиме железы прощупываются уплотненные
фокусы, иногда в виде одного, величиной до куриного яйца. К носовому сапу при-
соединяется кожный.
Кожный сап у лошади имеет много вариантов течения. В различных частях тела на
коже появляются небольшие возвышения, которые вначале прощупываются только
рукою, иногда же образуются большие абсцессы. В центре появляется небольшое
отверстие, через которое сначала вытекает желтоватый клейкий секрет. Края язвы
постепенно расходятся, принимая подрытый вид. Язва то остается небольшой, то в
течение иногда 2- 3 сут достигает десятков и сотен квадратных сантиметров. Дно язвы
покрыто серовато-саловидным секретом Лимфатические сос уды вокруг язвы утол-
щаются, выел утя в виде шнуров, по ходу их образуются возвышения, которые, рас-
падаясь, образуют язвы. Если кожные поражения развиваются на конечности, то в
процесс вовл! кается соединительная ткань, последняя, разрастаясь, делает ногу тол
стой — слоноподобной (рис. 3.63 и 3.64).
Кожный сап может выражаться иногда только в отечности подкожной клетчатки,
часто располагающейся на препуции, нижней стороне брюха, груди. О^ек спускается
вниз по конечности и также вызывает слоновость. Отеки то увеличиваются, то умень-
шаются в своей величине, и обычно на них появляются сапные язвы.
Легочный сап наиболее трудно диагностируемая форма сапа. Обычно легочные
узелковые поражения настолько незначительны по величине, что ни перкуссией, ни
аускультацией они не обнаруживаются . При легочном сапе на первый план у лошади
выступают явления общего страдания — повышение температуры, исхудание, быстрая
3.5. Can
771
Рис. 3 63. Кожные проявления сапа у лошади. Узлы и шнуры
лимфатических сосудов. По Н Е. Цветкову и В. А. Черняку (1935)
Рис. 3.64. Кожные проявления сапа у лошади. А Узлы по ходу лимфатических
сосудов. Б. Сапные язвы на коже. По Н. Е. Цветкову и В. А. Черняку (1935)
утомляемость, 1лухой, слабый, сухой кашель. Только тогда, когда поражения в легких
многочисленны, и они сливаются, образуя пневмонические фокусы, а иногда и ка-
верны, или пневмония осложняется плевритом, то при перкуссии на этом месте на-
блюдается притупление, а иногда и совершенная тупость, аускультация же дает отсут-
ствие нормальных шумов дыхания, или усиленное везикулярное, иногда бронхиальное
тыхание совместно с хрипами. Тип дыхания при этих формах больше абдоминальный.
При локализации поражений в трахее прослушиваются крупнопузырчатые влажные
фипы, при локализации в гортани судорожный кашель, болезненность при надавли-
вании, глухой голос и/или свистящее или сопящее дыхание.
Хронический сап у лошадей составляет 90 % случаев выявляемого сапа. Болезнь
протекает без всяких клинических признаков — скрытый сап. Иногда клинические
признаки сапа настолько незначительны, что не обращают на себя внимания, болезнь
772
Часть 3. Частная неправильная эпидемиология
тянется много лет. Диагностировать заболевание у таких лошадей возможно только
при помощи биологических реакций, из них наиболее постоянное показание дает
маллеин. Клинические признаки иногда выражаются в подъемах температуры реми-
тирующего характера, кашле, хронической одышке и исхудании. При хорошем содер-
жании последние признаки, и в особенности исхудание, часто отсутствуют.
В том случае, когда острый клинический сап у лошади переходит в хронический,
клинические признаки постепенно уменьшаются. Язвы на слизистой носа постепенно
заполняются соединительной тканью, не покрываясь с поверхности эпителием. Со-
единительная ткань с течением времени начинает стягиваться и образует звездчатые
рубцы. Нередко можно наблюдать одновременно с рубцующимися язвами образова-
ние новых. При доброкачественном течении сапа на слизистой носа можно видеть или
единичные звездчатые рубцы, или множественные, покрывающие значительную часть
носовой перегородки, а иногда и раковины. На месте кожных язв также, образуются
рубцы. Общее состояние лошади улучшается до следующего рецидива.
На основании клинических признаков сап надежно может быть диагностирован
у лошади тогда, когда он имеет типичные проявления болезни, а именно: при носо-
вом сапе — язвы на слизистой носа с подрытыми краями, саловидным дном, медлен-
но заживающие, с заполнением дефекта эпителия рубцовой тканью, которая впослед-
ствии, сокращаясь, образует звездчатые рубцы. Развитие язв сопровождается гнойным
истечением, иногда с примесью крови, часто односторонним и из левой ноздри. Па-
раллельно с развитием процесса в носовой полости припухают подчелюстные, а иног-
да и подъязычные лимфатические узлы преимущественно соответствующей стороны.
Впоследствии они становятся плотными, бугристыми и неподвижными. Нагноение
наблюдается очень редко.
При кожном сапе типичным проявлением болезни является незначительное при-
пухание кожи в различных частях тела, в особенности на конечностях, мошонке и
вымени. Иногда эти припухания тянутся по ходу лимфатических сосудов. Далее кожа
над припухлостями истончается, и образуются язвы с подрытыми краями и саловид-
ным дном. Язвы очень медленно зарастают с образованием рубцовой ткани. Медлен-
ность заживления язв с типичной картиной строения язвы и является основным при-
знаком кожного сапа.
При легочном сапе клинические признаки малоинформативны.
Характерными для сапа патологоанатомическими изменениями являются узлы,
гноящиеся язвы и звездчатой формы рубцы на коже и в носовой полости. В легких,
печени, селезенке, лимфатических узлах находят стекловидные, просвечивающиеся,
окруженные пояском гиперемии сапные узелки и очаги с некротическим распадом
тканей при остром течении сапа или инкапсулированные, пропитанные солями фо
кусы — при хроническом.
Восприимчивость к возбудителю сапа и инфидирхюпще дозы для животных и людей.
В естественных условиях сапом чаще всего болеют непарнокопытные животные
(лошади, ослы, мулы и значительно реже верблюды). Естественно, восприимчивы к
В. mallei также некоторые виды хищников, особенно семейство кошачьих (тигры,
львы, леопарды, рыси, барсы, шакалы, степные кошки и др.), которые инфицируют-
ся при поедании мяса зараженных сапом животных. Заболевание сапом крупного ро-
гатого скота, овец и коз в естественных условиях не отмечено. Резистентные к сапу
свиньи, птицы, домашние крысы. Из лабораторных животных сапом легко заражают-
ся хомяки (LD50 для В. mallei Ц-5 — несколько КОЕ), хорьки, морские свинки (весом
до 100 г) и кошки. Кролики более резистентны к В. mallei, они могут переносить без
' 5. Can
773
лаких-либо явлений внутривенную инъекцию миллионов КОЕ В. mallei. Белые мыши,
белые крысы, если и заражаются сапом, то только в случае введения больших доз
эозбудителя. При внутримышечном введении белым крысам 1х 108 КОЕ В. mallei Ц-5,
тишь у отдельных особей удается морфологически выявить проявления инфекцион-
того процесса, которые характеризовались наличием единичных продуктивных гра-
нулем в печени и экссудативных гранулем в селезенке. Затем продуктивные грануле-
мы подвергались резорбции, а экссудативные — рубцевались (Кравченко А. Т., Руд-
нев Г. П., 1966; Вязьмина Т Н. с соавт., 1995).
Судя по той легкости, с которой происходят лабораторные заражения сапом,
человек обладает высокой чувствительностью к возбудителю сапа. Инфицирующая
доза В. mallei для человека при ингаляционном заражении находится в пределах
300-3200 КОЕ (Ротшильд Д., 1966; Ларионов Г. М., 1995).
Иммунитет. Постинфекционный иммунитет к сапу кратковременный или отсут-
ствует. Описаны единичные случаи повторных заболеваний сапом у людей. Много-
численные попытки воспроизвести пассивный иммунитет введением специфических
сывороток, полученных от лошадей, коров, собак, не дали положительных результатов.
Введение специфического антигена обостряет инфекционный процесс, сопровож-
дающийся инфекционной аллергией, и животные погибают от развившейся инфек-
ции, несмотря на высокие титры специфических антител в их сыворотке. Положитель-
ная реакция на маллеин глаза (у животных) или кожи (у животных и человека) отра-
жает состояние инфекционной аллергии организма. Хроническое течение сапа
человека или животн ых может всту п ить в фазу, когда реакция связывания комплемен-
та (РСК) бывает отрицательной, а аллергическая реакция глаза и кожи на маллеин по-
ложительная. Этот период длится неопределенное время и может закончиться пол-
ным выздоровлением. Угасание аллергических реакций на маллеин означает полное
выздоровление организма (Кравченко А. Т., Руднев Г. П., 1966).
Устойчивость возбудителя сапа во внешней среде. Возбудитель сапа относится к
микроорганизмам, мало устойчивым во внешней среде. Прямой солнечный свет уби-
вает В. mallei в чистой культуре за 24 ч, но в выделениях больных животных возбуди-
тель сапа может сохранять жизнеспособность при действии прямого солнечного све-
та несколько недель. При высокой температуре (100°С) В. mallei погибает в течение
нескольких минут, при 55—60°С — за 1—2 ч. Бульонная культура с глицерином сохра-
няет жизнеспособность возбудителя сапа втечение четырех лет. В водопроводной воде
жизнеспособность его сохраняется до 30 сут. Помещения, в которых находились боль-
ные сапом животные, опасны для заражения в течение 6 нед. Внутри высохших ко-
мочков слизи и гноя В. mallei надежно защищен от воздействия прямого солнечного
света. В органах павших от сапа животных возбудитель болезни остается жизнеспо-
собным 24 сут. Лиофильно высушенные культуры В. mallet при глубоком вакууме со-
храняют жизнеспособность в течение длительного времени, но снижают свою виру-
лентность по отношению к восприимчивым животным. Вирулентность их быстро
восстанавливается после одного или нескольких пассажей на восприимчивых живот-
ных (хомяк, морская свинка). Употребляемые в лабораториях дезинфицирующие
растворы убивают В. mallei в чистой культуре, но не дают надежной стерилизации
гнойных выделений, содержащих большое количество слизи, либо материала, остав-
шегося в пипетках после опытов. Для практических целей дезинфекции пользуются
3 %-ным серно-карболовым раствором, 5 %-ной хлорной известью, 2 %-ным раство-
ром формалина и 3-5 %-ным раствором щелочи. Раствор сулемы и лизол как дезин-
фицирующие агенты не эффективны (Кравченко А. Т., Руднев Г. П., 1966)
774
Часть 3. Частная неправильная эпидемиология
Антибиотикорезистентность. В опытах in vitro возбудитель сапа 'гувствителен к зна-
чительному числу антибиотиков. Высокая чу вс, витепьносп. В. mallei к сульфанила-
мидам продемонстрирована еще в конце 1940-х гг. Позже показано достоверное
синергидное действие на возбудитель сапа сульфанилами, iob (сульфазина, сульфаме-
токсазола, сульфомонометаксина) в сочетании с триметопримом. Стрептомицин,
пенициллин, полусинтетические пенициллины (ампициллин, оксациллин), налидик-
совая кислота, эри громицин активностью против В. mallei не обладают (Илюхин В. И.
с соавт., 1995). Поданным D. Кеппу et al. (1999), в условиях in vitro В. maliei и В pseudo-
mallei обладают высокой чувствительностью к цефтазидиму (цефалоспориновый анти-
биотик III поколения), имипинему (карбапенем), доксициклину (группатетрацикли-
нов) и ципрофлоксацину (группа фторхинолонов II поколения). Гентамицин (анти-
биотик аминогликози 1ного ряда) был активен по отношению к возбудителю сапа, но
не мелиоидоза. Е М. Thibault et al. (2004) получили сходные результаты по цефтази-
диму, имипинему, тетрациклинам (доксициклин, миноциклин), отмстив низкую чув-
ствительность обоих микроорганизмов к аминогликозидам и фторхинолонам. Одна-
ко оценивая эти данные, следует иметь в виду, что многие из препаратов, активных в
условиях in vitro, не оказывают терапевтического действия при лечении людей и экспе-
риментальных животных. Среди них имипинем, аминогликозиды, цефалоспорины,
метациклин, рифампицин (Илюхин В. И. с соавт., 1995).
Обнаружение генетически измененных штаммов возбудителя сапа. Под коды к обна-
ружению генетически измененных штаммов возбудителя сапа в основном те же, что
и для обнаружения генетически измененных штаммов возбудителей сибирской язвы
и чумы (см. разд. 3.1 и 3.2). Возбудитель сапа легко поддается генетической модифи-
кации, но работ по данному направлению в открытой литературе мало. Эксперимен-
ты по генетической модификации В. mallei ведутся с 1960-х гг., когда путем индуци-
рованного мутагенеза были получены штаммы, резистентные к Тс и Cm. С появлением
в 1970-х гг. методов генной инженерии, возможности для генетических манипуляций
с возбудителем сапа значительно увеличились. Отсутствие собственных плазмид у
В. mallei снимает проблему несоместимости плазмид. Реципиентная активность возбу-
дителя сапа обнаружена в отношении плазмид Inc P-I-, Inc Р-2-, Inc Р-3-, Inc P-4- и
Inc Р-5-iрупп. Переданные плазмиды полноценно реплицировались в реципиентном
штамме и опосредовали высокие уровни резистентности к Кт, Тс и Ар. Плазмиды
стабильно наследовались в различных условиях поддержания и хранения реципиен-
тных штаммов, передавались конъюгативным путем другим В. mallei и близкород-
ственным микроорганизмам (Меринова Л. К., Агеева Н. П., 1995; Fiionov А. Е. et al.,
1996). Соответственно на основе репликонов этих плазмид могут быть разработаны
векторы различного назначения (см. в разд. 2.2.2 «Технологии рекомбинантной ДНК
в получении штаммов бактерий с неправильной эпидемиологией»), предназначенные
для переноса генов в В. mallei от доноров из других видов, родов и даже семейств мик-
роорганизмов.
Возбудитель сапа является донором генов вирулентности для других микроорга-
низмов. Например, R. Balder et al. (2010) клонировали ген ЬоаА, обнаруженный в ге-
номе В. pseudomallei и В. mallei, и ген ЬоаВ, найденный в геноме В. pseudomallei, опре-
деляющие пневмотропность этих микроорганизмов.
Подозрение о применении генетически модифицированной В. mallei должно воз-
никнуть при:
1) обнаружении штаммов, ре шетентных к антибактериальным препаратам, обыч-
но используемых для лечения сапа (тетрациклин, триметоприм, сульфамид);
3.5. Can
775
2) обнаружении любых плазмид у штаммов В. mallei, выделенных при обстоятель-
ствах, характерных для умышленного распространения возбудителя сана среди лю-
дей или животных.
Патогенез сапа. Возбудитель сапа проникает в организм человека через повреж-
тенную кожу и слизистые оболочки, чаще носа, глаз и дыхательных путей, реже —
пищеварительного тракта. При заражении малой дозой В. mallei или штаммом со сла-
бой вирулентностью, первичная реакция организма развивается медленно. Инфек-
ционный процесс локализуется. Образуются специфические узелки, которые сохра-
няются на месте внедрения возбудителя продолжительное время и затем подвергаются
казеозному распаду.
В большинстве случаев инфекционный процесс протекает более остро. На месте
внедрения В. mallei вслед за первичным воспалением появляется отечность прилежа-
щих тканей. Затем образуются специфические узелки разных размеров. Узелки запол-
няются гноем, вскрываются и образуются язвы. Бактерии сапа с лимфой и током крови
разносятся по всему организму, обусловливая острый сап с наличием множественных
рассеянных очагов, что быстро приводит к смертельному исходу. При остром сапе
процесс принимает септикопиемический характер с наличием очагов гнойного рас-
плавления и образования метастатических абсцессов в мышцах и внутренних орга-
нах. Отмечаются гнойные остеомиелиты, артриты и пустулезные множественные сыпи
на коже и слизистых оболочках. Возможно не только гнойное, но и геморрагически-
гнойное воспаление. При сапе гной тягучий, слизистый. В печени нередки множе-
ственные мелкие абсцессы, гораздо реже их находят в почках и в яичках. При остро и
лодостро протекающем сапе в печени отмечаются многочисленные узелки с размяг-
чением в центральной части, окруженные узкой геморрагической зоной. Микроско-
пически они представляют собой абсциссоподобные очаги. В печеночных балках меж-
ду узелками наблюдаются явления диссоциации и жировой дистрофии клеток.
В месте внедрения В. mallei образуется первичный узелок, переходящий постепен-
но в язву. Гематогенное и лимфогенное распространение возбуди геля сапа приводит
к развитию в различных органах и тканях грануляционных узелков, состоящих из
эпителиоидных клеток и нейтрофильных лейкоцитов.
При остром течении болезни гранулемы претерпевают гнойное расплавление,
процесс приобретает септикопиемический характер. Образуются метастатические
абсцессы во внутренних органах и мышцах, гнойные остеомиелиты и артриты. Раз-
вивается пустулезная сыпь на коже и слизистых оболочках. Возбудитель сапа облада-
ет выраженным тропизмом клеточной ткани. Поэтому почти все] да гематогенно и
бронхогенно в процесс вовлекаются легкие. В них появляются мелкие узелки, сход-
ные с туберкулезными 6yi орками которые в дальнейшем превращаются в небольшие
абсцессы. Возможно развитие лобулярной, очаговой, сливной и даже лобарной пнев-
моний. Нередко абсцессы образуются в печени и других органах (почки, селезенка,
миокард, костный мозг, яички).
При хроническом течении сапа в узелках преобладают пролиферативные явления.
Развиваются хронический сепсис, абсцессы во внутренних органах, полиартриты.
В легких наблюдается пневмосклероз, бронхоэктазы, хронические абсцессы.
В редких случаях при сапе поражается нервная система в виде гнойного лептоме-
нингита, эпидуральных и субду ратьных абсцессов, септического тромбофлебита си-
нусов твердой и вен мягкой мозговых оболочек, абсцессов мозга.
Одной из характерных особенностей воспаления, вызванного В. mallei, является
исход его в некроз, который очень близок к творожистому некрозу. Он может петри-
776
Часть 3. Частная неправильная эпидемиология
фицироваться, подвергаться расплавлению. Расплавление некротических масс при
сапе является предпосылкой для возникновения специфических каверн и абсцессов.
Быстрая генерализация В. mallei приводит к поражению легких. Легкие поража-
ются в 98—99 % всех случаев сапа. Наиболее типична крупноузелковая форма легоч-
ного сапа. Острый милиарный сап легких принадлежит к одной из редких форм бо-
лезни. Бронхопневмонии ацинозноподобного и лобулярного характера являются
проявлением новой вспышки старого процесса под влиянием воздействия на организм
различных неблагоприятных внешних и внутренних факторов.
Сап носовой полости может быть экссудативным или продуктивным. Основной
формой экссудативного сапа носовой полости является язвенная. Вид сапных язв
довольно характерен. Края их валиковидно приподняты над уровнем слизистой обо-
лочки и мелко гранулированы. Дно язвы неровное, как бы изъеденное, и тоже мелко
гранулировано. На слизистой носовой полости одновременно имеются и сапные узел-
ки. Лучистые и звездчатые рубцы являются результатом заживления язв. Фиброзные
тяжи этих рубцов как бы сходятся в одной точке (центр язвы) и придают им лучистое
строение.
Сап кожи, как правило, являе гея вторичным процессом, возникающим гема-
тогенно за счет очагов болезни в других органах. Поражения кожи могут иметь по-
верхностную и глубокую локализацию. При сапе довольно постоянны и характерны
поражения лимфатических узлов, которые проявляются в форме специфических и не-
специфических лимфаденитов.
Сап в паренхиматозных органах (печень, почки, селезенка) проявляется как в фор-
ме узелков, гак и крупных узлов. Обычный исход этих форм — обызвествление и осум-
кование очагов. Но при крупноочаговой форме в селезенке сапные узлы нередко под-
вергаются гнойному расплавлению с образованием абсцессов.
Клиническая картина сапа у "юдей при естественном инфицировании. Инкубацион-
ный период при острой форме сапа у человека от 1 до 5 сут, реже дольше. В. И. Илю-
хин с соавт. (1996) считаю!, что у людей, так же как и у лошадей, сап протекает в ост-
рой и хронической формах. Причем это две самостоятельные формы, т. е. хроническое
течение не является фазой острого течения сапа. Для сапа характерно острое начало
болезни, с ознобом, повышением температуры до 38—39°С с чувством жара, голов-
ными и мышечными болями, позже — болями в суставах и их припухлостью. В месте
проникновения В. mallei последовательно наблюдаются красно-багровая папула, ок-
руженная рожистоподобной краснотой, затем пустула с кровянистым содержимым и,
наконец, язвы. Язва характеризуется «сальным» дном и подрытыми краями, а также
развитием регионарною лимфангоита и (реже) лимфаденита. Через 5—7 сут наблю-
даются клинические признаки генерализации инфекции — после кратковременного
снижения температуры вновь отмечаются обострения лихорадки и многократное
появление множественных вторичных узелков и папулезных высыпаний, переходя-
щих в пустулы и язвы. В мышцах, особенно икроножных, могут развиваться некро-
тические очаги. В легких может развиться ползучая плевропневмония, сопровождаю-
щаяся кашлем с выделением кровавой мокроты. Поражение слизистых оболочек носа
(пустулы, язвы) сопровождается зеленовато-гнойными, а иногда кровянистыми вы-
делениями. Сходные поражения возможны и на слизистой оболочке рта. Состояние
больных тяжелое, позже присоединяются гнойные поражения суставов, изнуритель-
ные поносы, развиваются явления сердечной слабости. При остром сапе без лечения
смертельный исход наблюдается в 100 % случаев. Типичная история болезни острого
сапа у медицинского (ветеринарного) работника выглядят следующим образом.
3.5. Can 777
Ветеринар X. поступил в госпиталь на второй неделе после ранения руки анато-
мическим ножом при вскрытии лошади. Через сутки после ранения у него появился
озноб, температура 40 °C, головные боли, общая слабость. На тыльной поверхности
большого пальца левой руки небольшая резаная рана. Состояние с каждым днем ухуд-
шалось. Присоединились отеки века и кисти. К концу второй недели пребывания в
больнице (30 сут от начала болезни) у больного появились папулы и гнойники по телу,
непроходимость носовых ходов, артриты. На фоне постоянно ухудшающегося состо-
яния больной умер. На вскрытии у него обнаружена левосторонняя пневмония, спе-
цифические для сапа узлы в легких и носовых ходах, из которых была получена чистая
культура В. mallei.
Хроническая форма сапа у людей протекает в трех формах: кожная, легочная и
юсовая. Воспалительные процессы при хронической форме сапа развиваются мед-
гнно и характеризуются сменой фаз улучшения и ухудшения. У больных с кожной
формой (самой частой) характерны пустулы, переходящие в язвы, лимфангоиты, лим-
фадениты и множественные мышечные абсцессы холодного типа (чаще всего в ик-
роножных мышцах). Язвы багрового цвета, вяло гранулирующие, дают значительное
отделяемое, при заживлении оставляют обширные рубцы. Самопроизвольное вскры-
тие мышечных абсцессов ведет к образованию свищей с выделением в гное палочек
позбудителя сапа. При инцизии же в отделяемом свищей 3. mallei не находят. Типич-
но весьма медленное заживление и наклонность к многократным рецидивам, что уве-
личивает время течения болезни до нескольких лет.
При поражении кожи лица характерно рожистоподобное воспаление с наличием
изъязвленных сапных узелков. Для язв типичны глубокие, подрытые края и масляни-
сто-желтоватое отделяемое с примесью крови. При хронической форме сапа с первич-
ным поражением легких, кроме лихорадки, отмечается ползучая плевропневмония,
к которой позже присоединяются множественные мышечные абсцессы. При пораже-
ниях слизистой оболочки носа типичны слизисто-гнойные выделения, образование
желто-зеленых корок и глубоких изъязвлений, распространяющихся до зева, горта-
ни и трахеи.
Прогноз при хронической форме сапа требует осторожности, так как типичны
многократные рецидивы и обострения. Летальность при хронической форме сапа
достигает 50 % и выше.
Моделирование ингаляционного заражения сапом. У людей ингаляционное зара-
жение сапом в основном происходит в результате аварий в бактериологической ла-
боратории. Кратко описание такого случая ингаляционного заражения приведено в
работе В. И. Илюхина с соавт. (1996).
Бактериолог Л. заразился вследствие аварии при работе с культурой В. mallei. За-
болевание протекало в виде общего недомогания, явлений бронхита. К концу второй
недели он поступил в больницу с явлениями озноба и пневмонии 11а третий день пре-
бывания в клинике (17 сут от момента вероятного заражения) развились артриты ниж-
них конечностей. На третьей неделе на теле и в носовых ходах больного появились
гнойники. Смерть наступила на 24 сут болезни.
Патология ингаляционного заражения В. mallei у экспериментальных животных
детально изучалась Т. Н. Вязьминой с соавт. (1995). К числу отличительных особен-
ностей острой формы легочного сапа, возникшего после ингаляционного заражения,
авторы относят развитие первичного патологического комплекса в месте внедрения
В. mallei — органах дыхания, скоротечность болезни (быстрая диссеминация и гене-
рализация инфекционного процесса), ее течение по типу септикопиемии, 100 %-ная
778
Часть 3. Частная неправильная эпидемиология
летальность, выделение возбудителя во внешнюю среду. Механизм развития ингаля-
ционного сапа с острым течением ими представлен в следующем виде:
1. Начальная стадия болезни: локализация возбудителя сапа в верхних и нижних
отделах дыхательной системы. Взаимодействие возбудителя с альвеолярными макро-
фагами, перенос ими В. mallei в регионарные лимфоузлы. В ряде случаев отмечается
проникновение возбудителя сапа лимфогематогенным путем во внутренние органы и,
прежде всего, в печень и селезенку. Наблюдается размножение В. mallei в легочной тка-
ни и лимфоузлах, что сопровождается формированием первичного патологического
комплекса в месте внедрения, выявляемого на клеточном уровне в первые 3—12 ч после
заражения в виде полнокровия и геморрагии. Через 24 ч отмечается образование оча-
гов воспаления с геморрагическим синдромом. В печени и селезенке выявляются более
ранние стадии воспалительных реакций. У экспериментального животного появляют-
ся признаки интоксикации. Продолжительность начального периода развития инфек-
ционного процесса составляла 24—48 ч.
2. Генерализация инфекционного процесса. Диссеминация возбудите тя сапа, бак-
териемия, развитие бактериурии, выделение В mallei во внешнюю среду. Выделение
возбудителя сапа может происходить и с мокротой. В легких завершается формиро-
вание очагов бронхопневмонии, как правило, гнойной. Наблюдается развитие мета-
статических очагов воспаления в печени и селезенке. В этих же органах у высокочув-
ствительных к В. mallei животных образуются экссудативные гранулемы, являющиеся
специфическим морфологическим субстратом острого сапного сепсиса. У относитель-
но резистентных к заражению животных отмечается образование гранулем с выражен-
ной продуктивной реакцией. В других внутренних органах развиваются дегенератив-
ные процессы.
3. Терминальная стадия болезни. Завершается процесс септикопиемии. Наблю-
дается обсемененность всех внуз ренних органов В. mallei. Отмечаются явления инток-
сикации. Развиваются некротические процессы в очагах воспаления (легкие, печень
и селезенка), вплоть до гистолиза ткани. Выявляются расстройства гемодинамики и
дистрофические изменения в паренхиматозных органах. Смерть животных наступа-
ет на 3—14 сут в зависимости от их резистентности к заражению В. mallei, дисперсно-
сти аэрозоля и величины заражающей дозы.
Клиническая картина сапа у людей при искусственном инфицировании. Все извест-
ные случаи искусственного заражения В. mallei представляют собой случаи лаборатор-
ного заражения. В этой связи огромный интерес представляют сделанное В. В. Ни-
кифоровым с соавт. (2005) описание и анализ случая, имевшего место в Московской
области весной 2004 г.
Больная Б., 59 лет, обратилась к участковому терапевту по месту жительства с жа-
лобами на боли в грудной клетке, в шейном отделе позвоночника слева и ежедневное
повышение температуры до 39°C в вечернее время. Указанная симптоматика появи-
лась около недели назад, само заболевание пациентка связывала с переохлаждением.
Участковым терапевтом диагностирована цервикал! ия, невралгия затылочного нерва;
назначены диклофенак вну гримышечно, карбамазепин в таблетках
24.04.04 г. пациентка повторно вызвала врача на дом. При осмотре: температура
39 °C, жалобы на интенсивные боли в затылочной области слева, усиливающиеся при
малейшем движении головы; невозможно было выполнить осмотр полости рта, так как
из-за сильных болей больная не могла открыть рот. Больная была госпитализирована
в инфекционное отделение городской больницы с диагнозом «вирусная инфекция с
выраженным невралгическим синдромом».
3.5. Can 779
Из анамнеза (по амбулаторной карте) известно, что пациентка наблюдалась в дан-
ной поликлинике с 1980 г. Перенесенные заболевания: ангина, ОРВИ, хронический
левосторонний мезотимпанит, липоматоз кожи, остеоартроз. С марта 2002 г. периоди-
ческий медицинский осмотр проходит по месту работы.
В приемном отделении больная предъявляла жалобы на повышение температуры
тела в течение недели до 39 °C, с колебаниями от 37,2 до 38 °C; сильные боли в заты-
лочной области, слева усиливающиеся при малейшем движении головы, боли в пра-
вой половине грудной клетки при дыхании, боли в шейном отделе позвоночника.
Диагноз при поступлении: «Пневмония на фоне вирусной инфекции, шейно-грудной
остеохондроз, межреберная невралгия». Пациентка направлена в инфекционное от-
деление.
При осмотре в инфекционном отделении: состояние средней тяжести, температура
38°С. Кожные покровы обычной окраски, сыпи нет. Слизистая полости рта чистая,
язык влажный; небные дужки и миндалины, мягкое небо, задняя стенка глотки ярко
гиперемированы. Периферические лимфатические узлы не увеличены. Менингеаль-
ной и очаговой симптоматики нет. В легких — дыхание везикулярное, хрипов нет.
Частота дыхания (ЧД) 20/мин. Тоны сердца ясные, ритмичные. Частота сердечных
сокращений (ЧСС) 96/мин. АД 140/80 мм рт. ст. Живот при пальпации мягкий, безбо-
лезненный. Печень у края реберной дуги при пальпации безболезненная. Мочеиспус-
кание неучащенное, безболезненное. Стул один раз в сут, оформленный.
В отделении назначено: антибактериальная (ампиокс внутримышечно), анальге-
тическая, дезинтоксикационная, антигистаминная терапия, нестероидные противо-
воспалительные средства. В течение 4 сут больная получала указанное лечение и про-
шла клиническое обследование.
25.04.04 г. Рентгенография органов грудной клетки—данных, свидетельствующих
о пневмонии, не выявлено.
26.04.04 г. Анализ крови: НЬ 129 r/л, лейкоциты 5,2х 109/л, палочкоядерные 25 %,
сегментоядерные 46 %, эозинофилы 1 %, лимфоциты 23 %, моноциты 5 %, СОЭ
20 мм/ч; анализ мочи: цвез соломенно-желтый, реакция кислая, следы белка, лейко-
циты 3—4 в поле зрения, слизь +, бактерии +.
Осмотрена заведующей инфекционным отделением, диагноз: «ОРВИ Шейный
остеохондроз с корешковым синдромом».
27.04.04 г. Консультация невропатолога, заключение: «реактивный периартрит
плечевого сустава слева на фоне ОРВИ».
28.04.04 г. Рентгенография шейного отдела позвоночника: отмечается выпрямле-
ние физиологического лордоза, выраженный склероз C|V-CVI с передними остеофи-
тами, признаки деформирующего артроза в их суставных отростках.
Консультация хирурга, заключение: «остеохондроз шейного отдела позвоночни-
ка, плечелопаточный периартрит слева». После консультации с неврологом и хирур-
гом у больной диагностированы: «острая респираторная вирусная инфекция, рекон-
валесцент; шейный остеохондроз, периартрит левого плечевого сустава».
29.04.04 г. больная переведена в неврологическое отделение с коррекцией терапии:
была усилена антибактериальная терапия (цефазолин + гентамицин внутримышечно),
дезинтоксикационная терапия, внутривенно дексаметазон в дозе 8 мг/сут и витами-
нотерапия.
С 24.04.04 г. по 30.04.04 г больная, находясь в стационаре и получая большие дозы
антибиотиков, продолжала лихорадить (29.04.04 г. и 30.04.04 г. сохранялся субфебри-
литет в пределах 37,5—37,6 °C).
29.04.04 г. при осмотре на фоне лечения: «состояние удовлетворительное, темпе-
ратура 37,5°С. Кожные покровы чистые, сыпи нет. В легких дыхание везикулярное,
780
Часть 3. Частная неправильная эпидемиология
хрипов нет. ЧД 20/мин. Тоны сердца ясные, ритмичные. ЧСС 76/мин. АД 130/80 мм
рт. ст. Живот при пальпации мягкий, безболезненный. Физиологические отправления
в норме». 30.04.04 г. сделана лидокаин-дексаметазоновая блокада периартикулярных
точек левого плечевого сустава. Отмечено временное уменьшение болей и увеличение
движений в левом плечевом суставе.
Резкое ухудшение состояния наступило в воскресенье 02.05.04 г в 12 ч 30 мин (вы-
зов дежурного персонала в палату). Пациентка жаловалась на одышку, слабость, го-
ловную боль. Состояние тяжелое, температура тела 37°С. Больная заторможена, в со-
знании Кожные покровы бледные с желтушным от генком, склеры иктеричны. Язык,
губы сухие, цианоз ногтевых фаланг. ЧД 24—26/мин. В легких дыхание везикулярное
ослабленное, крепитирующие хрипы. ЧСС 76 в мин. АД 120/70 мм рт. ст. Тоны сердца
ясные, ритмичные. Живот при пальпации мягкий, безболезненный. Печень выступа-
ет из-под края реберной дуги на 1 см, при пальпации безболезненная. Диурез недоста-
точный (мало мочи), моча темного насыщенного цвета. Начата профилактика дыха-
тельной недостаточности и дезинтоксикационная терапия (гемодез, физиологический
раствор, 5 %-ный раствор глюкозы, сульфакамфокаин, кокарбоксилаза, аскорбино-
вая кислота) объемом до 1 л.
По ci to выполнены клинические исследования. Анализ крови: НЬ 125 г/л, эритро-
циты 4.4х 1012/л, цветной показатель 0,86, гематокрит 37,5 %, лейкоциты 12,6 х109/л.
Анализ мочи: цвет оранжевый, удельный вес — 1010 г/л, peai ;ция кислая, следы бел-
ка, лейкоциты 4—5 в поле зрения, эритроциты 0—1 в поле зрения, единичные гиалино-
вые и зернистые цилиндры в поле зрения, слизь — много. Биохимический анализ кро-
ви: общий белок 52,3 г/л, мочевина 14,7 ммоль/л, креатинин 157 мкмоль/л, непрямой
билирубин 36,75 мкмоль/л, прямой билирубин 27,36 мкмоль/л, глюкоза 6 5 ммоль/л.
Однако состояние больной продолжало ухудшаться, нарастала дыхательная недо-
статочность, ЧД 38—42/мин, ЧСС 78/мин, АД 140/100 мм рт. ст., и в 17 ч 30 мин боль-
ная была переведена в реанимационное отделение
При поступлении в отделение реанимации: состояние крайне тяжелое. Сознание
сопорозное, но периодически возбуждена. Жалобы на одышку, чувство нехватки воз-
духа. Кожные покровы бледные, сухие, акроцианоз. Дыхание шумное, выдохудлинен,
ЧД до 32/мин. В легких — дыхание ослабленное, выслушивается масса сухих и влаж-
ных, разнокалиберных хрипов. При перкуссии — притупление перкуторного звука над
всей поверхностью легких. Тоны сердца приглушены, ритмичные. ЧСС 96/мин.
АД 120/80 мм рт. ст. Живот при пальпации мягкий, безболезненный, не вздут. Катете-
ризирован мочеьой пузырь, моча концентрированная, 150 мл.
Для обеспечения постоянного венозного доступа и с целью проведения адекват-
ной инфузионной терапии и контроля центрального венозного давления (ЦВД) вы-
полнена катетеризация подключичной вены справа. ЦВД 30 см вод. ст. Начата интен-
сивная инфузионная терапия (вводились реополиглюкин, трентал, одногруппная плаз-
ма, 5 % ный раствор глюкозы, дексаметазон, контрикал, гепарин, эуфиллин, лазикс)
объемом до 1,5 л, с целью обезболивания введен промедол 2 % — 1,0 мл. В 18 ч по дан-
ным обследования: ЧД до 32/мин, ЧСС 112/мин, АД 130/80 мм рт. ст., SaO2 — 84 %.
На рентгенограмме органов грудной клетки определялось двустороннее снижение про-
зрачности легочных полей за счет мелкоочаговой диффузной инфильтрации (рентге-
нологическая картина альвеолярного отека легких).
Несмотря на проводимую терапию, состояние больной прогрессивно ухудшалось,
нарастала одышка (ЧД 36/мин), гипоксемия (SaO2 — 74 %). Пациентка переведена на
искусственную вентиляцию легких (ИВЛ). Выполнена бронхоскопия — слизистая
бронхов отечна, гиперемирована, поступает вязкая слизь с сукровицей. В 24 ч состо-
яние больной крайне тяжелое. На фоне ИВЛ сохраняется гипоксемия SaO2 70-66 %
3.5. Can
781
при FiO2 100—80 %. Диффузный цианоз кожных покровов. В легких выслушива-
ется масса мелкокалиберных влажных хрипов с обеих сторон. АД 60/40 мм рт. ст., ЧСС
110/мин. ЦВД — 0 см вод. ст. Учитывая данные лабораторного обследования (лейко-
циты 12.6х 109/л, полиморфоядерные 57 %, лимфоциты 7 %), тяжесть состояния боль-
ной объяснялась наличием септической пневмонии, сепсиса, септицемией, полиор-
ганной недостаточностью. 03.05.04 г. в 1 ч 30 мин произошла остановка кровообраще-
ния, реанимационные мероприятия эффекта не дали. В 2 ч больная скончалась.
О смерти пациентки было сообщено в объектовую медико-санитарную часть. Было
выяснено, что умершая в течение 2 мес. работала с культурами мелиоидоза и сапа. По
требованию сотрудников объектовой медико-санитарной части патологоанатомичес-
кое вскрытие проводилось специалистами Федерального медико-биологического аген-
тства в средствах индивидуальной защиты «Кварц». В ходе вскрытия во всех отделах
обоих легких достаточно симметрично было обнаружено большое количество узлова-
тых образований серо-розового цвета с распадающимся центром, с фестончатыми
контурами, местами имеющих тенденцию к слиянию между собой, размером от 0,5 до
1,4—1,8 см, не связанных со стенками бронхов (рис. 3.65).
Рис. 3 65. Легкие больной, умершей от сапа Макропрепарат. В паренхиме легких
отчетливо видны специфические гранулемы По В. В. Никифорову с соавт. (2005)
В печени выявлен участок с несколько мутноватой капсулой площадью около
1,7х 1,2 см. В этой зоне под капсулой обнаружена группа сливающихся дрябловатых,
сероватых очагов диаметром до 0,5—0,6 см с намечающейся полостью распада в цент-
ре отдельных очагов.
Мышцы левого плеча серо-красные, несколько плотноваты. В толще их, ближе к
области левого плечевого сустава, обнаружена полость с рыхлыми, сероватыми стен-
ками, диаметром около 2,3 см, заполненная густой, серовато-желтой жидкостью. При
вскрытии капсулы левого плечевого сустава из нее выделилась аналогичного вида
жидкость. Патологоанатомический диагноз: сап, острая форма с поражением легких,
печени, межмышечный абсцесс левого плеча. Гнойный артрит левого плечевого сус-
тава. Острая гиперплазия красной пульпы селезенки. Отек легких. Дистрофия парен-
химатозных органов.
Заключение патологоанатома: непосредственной причиной смерти явилась про-
грессирующая дыхательная недостаточность, обусловленная специфическим пораже-
нием легких на фоне интоксикации.
782
Часть 3. Частная неправильная эпидемиология
В результате расследования случая летального исхода больной Б. комиссия спе-
циалистов Федерального медико-биологического агентства пришла к «включению,
что инфицирование могло произойти в период непосредственной работ ы с возбуди
телем сапа с 22 марта по 14 апреля 2004 г.
Анализ материалов расследования, данных клинического течения заболевания и
патологоанатомического исследования позволил комиссии сделать предположение,
что одной из причин инфицирования пациентки Б. могло быть перкутанное проник-
новение возбудителя через незаметный дефект кожных покровов (ссадина, заусени-
ца, потертость и др.) или укола левой руки
Подтверждением данной версии могут служить следующие факты, факт повтор-
ного использования Б. резиновых перчаток; выявленный случай неполного погруже-
ния в дезраствор использованных в работе с сапом резиновых перчаток; первоначаль-
ные жалобы на боли в области левого плеча; наличие свежего гнойного воспаления
левого плечевого сустава; наличие гнойников в мышцах левой руки; отсутствие спе-
цифических антител в сыворотке крови сотрудников, работавших в тех же условиях и
в то же время, что и больная Б. Однако авторы статьи, основываясь на отсутствии у
больной первичного аффекта, считают, что ее заражение могло произойти ингаляци-
онным путем.
Диагностика искусственного поражения сапом. Общие принципы такой диагности-
ки те же, что описаны в разд. 3.1 для поражения возбудителем сибирской язвы. Необ-
ходимо осторожно относиться к сообщениям о выделении возбудителя сапа из объ-
ектов окружающей среды, раневого отделяемого, мочи или мокроты пациента. Иден-
тификация В. mallei требует обязательного подтверждения в специализированных
научных центрах, располагающих опытными специалистами с соответствующей под-
готовкой. Естественная заболеваемость среди людей всегда спорадическая, эпидемий
сапа не бывает. Обычно заражения В. mallei носят профессиональный характер: бо-
леют работники животноводства, лица, занятые уходом за лошадьми, ослами, мула-
ми, ветеринарные специалисты.
Пока есть регионы (страны), неблагополучные по сапу, угроза проникновения этой
болезни в Россию будет оставаться, несмотря на государственную границу. Следова-
тельно, если для заболевшего сапом человека (и если болезнь не появилась в виде
вспышки!) не удается однозначно установить источник инфекции, то не обязательно
это будет означать «неправильную эпидемиологию». Более вероятно, что источник
возбудителя болезни находится в каком-то хозяйстве или бактериологической лабо-
ратории, но его тщательно скрыли. Поэтому прежде чем делать вывод об искусствен-
ном происхождении болезни у конкретного человека, необходимо попытаться уста-
новить ее профессиональный характер — контакты заболевшего с лошадьми, ослами
и мулами либо его работу с культурами возбудителя сапа в лаборатории.
О механизме инфицирования В. mallei можно судить по клинике болезни. Для
естественного инфицирования в результате травмы или пореза при контакте с инфи-
цированным животным характерно появление в области входных ворот инфекции
красно-багровой папулы, окруженной рожистоподобной краснотой, переходящей
затем в пустулу с кровянистым содержимым и в язву.
При ингаляционном инфицировании отсутствуют поражения, характерные для
порезов и травм. Болезнь первые две недели протекает в острой форме с явлениями
озноба, бронхита и пневмонии, из носа выделяются сукровичная жидкость и гной.
Обнаружение генетически измененных штаммов В. mallei у человека или живот
ных свидетельствует об искусственном происхождении вспышки болезни.
3.5. Can
783
Лабораторная диагностика сапа включает обнаружение Р. mallei в мазках, выде-
ление культуры возбудителя, ее идентификацию, а также постановку серологической
реакции. Для серологической диагностики у людей и животных применяют РСК,
реакцию агглютинации и РПГА. Аллергическую пробу с маллеином применять у лю-
тей не рекомендуется, так как это может вызвать обострение болезни (Руководство по
зоонозам, 1983).
Дифференциальную диагностику сапа следует проводить с чумой, натуральной
оспой, сибирской язвой, фурункулезом, септикопиемией, некоторыми формами си-
филиса, а также туберкулезом лимфатических узлов.
Опыт ведения больной Б. показал чрезвычайную важность сбора и оценки эпи-
демиологического анамнеза, который в данном случае позволил бы предположить
правильный диагноз, выбрать адекватную тактику лечения и, вероятнее всего, пре-
дотвратить неблагоприятный исход.
Иммунопрофилактика сапа. Эффективных вакцин и иммуноглобулинов нет.
Лечение сапа. Исходя из данных литературы и на основании собственных иссле-
дований, В. И. Илюхин с соавт. (1995) считают, что при лечении сапа эффективны
антибактериальные препараты трех групп тетрациклины, сульфаниламиды и фтерхи-
нолоны. Природные штаммы возбудителя сапа обычно высокочувствительны к ми-
ноциклину, доксициклину, метациклину и хлортетрацикли ну. При экстренной про-
филактике сапа наиболее эффективен миноциклин. Доксициклин, хлортетрациклин,
схльфазин, сульфамонометоксин, сочетания с^льфазина и сульфамонометоксина с
триметопримом, бактрим обладали практически одинаковой химиотерапевтической
активностью. При лечении сапа наиболее эффективны сочетания сульфазина с три-
метопримом, сульфаметоксазола с триметопримом (бактрим) и миноциклин, несколь-
ко менее эффективны сульфазин, а также сочетание сульфамонометоксина с три-
метопримом (сульфатон). При появлении резистентности к сульфаниламидам Илю-
шин В. И. с соавт. (1995) рекомендуют использовать препараты других групп, например,
фторхинолоны.
Army FM 8-284 (2000) приводит схемы лечения сапа в зависимости от типа и тя-
жести клинических проявлений болезни.
При локализованной форме болезни применяется одна из следующих схем продол-
нительностью от 60 до 150 сут:
амоксициллин/клавуланат, 60 мг/(кг сут), перора тьно, тремя дробными дозами;
тетрациклин, 40 мг/(кг сут), перорально, тремя дробными дозами;
триметоприм/сульфамид (ТМП, 4 мг/(кг сут); сульфамид, 20 мг/(кг сут), оба пре-
парата перорально дробными дозами).
При локализованной форме болезни с признаками умеренной или слабой токсичности
применяется следующий порядок введения антибиотиков: комбинируются две из
казанных выше схем продолжительностью 30 сут, затем следует монотерапия либо
с амоксициллином/клавуланатом, либо с ТМР/сульфамидом. продолжающаяся от
60 до 150 сут.
При внелегочной болезни с образованием гнойников антибиотикотерапия должна
продолжаться 6—12 мес. Показан хирургический дренаж абсцессов.
При тяжелой и/или септической форме болезни антибиотики применяются по
следующей схеме: цефтазидим, 120 мг/(кг сут) тремя дробными дозами в комбинации
с ТМП/сульфамидом (1 МП, 8 мг/(кг сут); сульфамид, 40 мг/(кг сут) четырьмя дроб-
ными дозами). Сначала, в течение двух недель, терапия проводится парентерально, а
затем продолжается перорально в течение 6 мес.
784
Часть 3. Частная неправильная эпидемиология
Если по клинике (острая пневмония) и анализу мокроты подозревается чума, то
дополнительно показано применение стрептомицина.
При сапе также, как и при мелиоидозе и натуральной оспе, может возникать диф-
фузная пустулярная сыпь. Поэтому показаны строгая изоляция и карантин до тех пор,
пока не будет исключена оспа. При обращении с пациентами, имеющими поражения
кожных покровов, рекомендуется следовать мерам предосторожности от заражения
при контакте. Сап, мелиоидоз и оспа могут проявляться в виде острой легочной ин-
фекции с гнойной мокротой. Поэтому, если при анализе последней обнаруживаются
грамотрицательные бациллы с биполярными «безопасными английскими булавками»,
когда проводится окрашивание по Райту или метиленовой синью, будет благоразум-
но по гьзоваться средствами изоляции для защиты органов гыхания цо тех пор, пока
из диагноза не будет исключена чума.
3.6. МЕЛИОИДОЗ
Микробиология возбудителя мелиоидоза. Факторы патогенности. Таксономия, про-
исхождение и молекулярная эпидемиология возбудителя мелиоидоза. Экология и
естественная эпидемиология возбудителя мелиоидоза. Восприимчивость к возбу-
дителю мелиоидоза и инфицирующие дозы для животных. Иммунитет. Устойчивость
возбудителя мелиоидоза во внешней среде. Антибиотикорезистентность. Обнару-
жение генетически измененных штаммов возбудителя мелиоидоза. Патогенез ме-
лиоидоза. Клиническая и патологоанатомическая картина мелиоидоза у человека при
естественном инфицировании. Клиническая картина мелиоидоза при искусственном
инфицировании. Диагностика искусственного поражения возбудителем мелиоидо-
за. Иммунопрофилактика мелиоидоза. Лечение мелиоидоза.
Мелиоидоз (melioidosis; син.: псевдосап, сап Восточной Индии, псевдохолера,
пневмоэнтерит, септицемия морфинистов, болезнь Флетчера — Стентона, сапопо-
добное заболевание из Рангуна) — опасная сапрозоонозная инфекционная болезнь
людей и животных, вызываемая бактерией Burkholderia pseudomallei. Эндемична для
Юго-Восточной Азии, Австралии и Мадагаскара. Протекает в форме острой или хро-
нической септикопиемии. Благодаря разнообразным клиническим симптомам в ост-
рой форме напоминает сап, чуму, холеру, грипп и ряд других инфекционных болезней
с явлениями поражения различных органов, с кожными и подкожными гнойничко-
выми фокусами. Впервые мелиоидоз описал Р. W. Whitmore (1911) в Бирме (Рангун).
Из трупа умершего человека ему удалось выделить патогенный микроорганизм, ко-
торому тогда было дано название Bacillus pseudomallei. Видовое название болезни
складывается из двух латинских слов «melis» — сап и «oidos» — подобный. Термин пред-
ложили в 1921 г. А. Т. Stanton и W. Fletcher.
Возбудитель мелиоидоза поддерживается в экосистемах, сформировавшихся в
отдельных типах почв и в застойных мелких, хорошо прогреваемых водоемах. Поли-
патогенен. Вызывает инфекционный процесс не менее чем у 50 видов позвоночных.
Основными факте нами передачи В. pseudomallei для человека являются почва и вода.
В экспепименте доказана возможность трансмиссивной передачи возбудителя мели-
оидоза через укус блох и комаров. В России природные очаги мелиоидоза не описаны.
Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят воз-
будитель мелиоидоза ко II группе патогенности. По степени важности дня националь-
3.6. Мелиоидоз
785
ной безопасности США возбудитель мелиоидоза относится к биологическим пора-
жающим агентам категории В. С 1930-х гг. военными специалистами он рассматри-
вается как потенциальный агент БО. Исследования по изучению его поражающих
свойств начаты в 1944 г. в США (Форт-Детрик). В. pseudomallei очень опасна для ра-
ботающего с ней персонала лабораторий. Вакцин, эффективно защищающих от за-
ражения возбудителем мелиоидоза, не разработано.
С целью поражения людей наиболее вероятно применение В. pseudomallei посред-
ством использования линейных и многоточечных источников и диверсионными (тер-
рористическими) методами.
Микробиология возбудителя мелиоидоза. В. pseudomallei — мелкая подвижная бак-
терия, спор не образует, форма и размеры клеток возбудителя мелиоидоза зависят от
состава питательных сред, температуры и продолжительности выращивания, а также
от штаммовых особенностей культур. В мазках из 2-суточных колоний, выращенных
на плотных питательных средах с добавлением глицерина, мелиоидозные микробы
обычно выглядят как овоиды или прямые, иногда — как слегка изогнутые палочки
с закругленными полюсами и имеют длину от 1,0 до 6,0 мкм, а диаметр — 0,3—0,5 мкм.
В небольшом количестве могут встречаться нити или цепочки клеток длиной до
20 мкм. После выращивания в течение 24 ч в жидких питательных средах с аэрацией
и перемешиванием клетки В. pseudomallei становятся более однородными по морфо-
логическим свойствам. Подавляющее большинство из них составляют овоиды или
прямые палочки длиной 1,0—3,0 мкм и диаметром около 0,3 мкм. Содержание иных
морфологических форм незначительно.
Полиморфизм В. pseudomallei усиливается после длительного культивирования,
-ранения, а также в результате воздействия на них антибиотиков. Он проявляется
увеличением содержания нитей, зерен, шаровидных или других неправильных форм
и сопровождается возрастанием вариабельности окраски клеток. Многие из них те-
ряют способность воспринимать красители, становятся бледноокрашенными и мо-
гут иметь метахроматические включения. В отличие от возбудителя сапа возбудитель
мелиоидоза обладает активной подвижностью, перемещаясь при помощи жгутиков,
расположенных пучком на одном конце (лофотрих). Количество жгутиков варьируе г
от 1 до 4. Жгутиковые штаммы в бульонных культурах могут сохранять свою подвиж-
ность в течение 5 мес. (рис. 3.66).
Специфическая окраска для Я pseudomonas не разработана. Д.ы усиления их про-
крашиваемое™ применяют карболовые или щелочные растворы красок. По Граму
В. pseudomonas окрашиваются негативно. Возможна неравномерная окраска отдель-
ных микробов, обусловленная наличием внутриклеточных скоплений липидов и
волютина. Включения поли-ф-оксимасляной кислоты окрашиваются Суданом чер-
ным Б и Суданом III в мазках, фиксированных нагреванием или формалином. Грану-
лы волютана (полифосфатов) отличаются кислотоустойчивостью и окрашиваются при
низких значениях pH метахроматически (в иной цвет по сравнению с остальными
частями клеточного тела) (Золотарев а. Г. с соавт., 2006).
У В. pseudomonas как и у В. mallei отсутствует капсула, которую можно было бы
выявить известными методами окраски, специально предложенными для этих целей.
Окрашенные мазки микробов этого вида отличаются лишь наличием большого коли-
чества аморфной слизи, нередко окружающей группы клеток. Не удается обнаружить
капсулу у мелиоидозных микробов высоковирулентных штаммов и при помощи при-
жизненной фазово-контрастной микроскопии препаратов, приготовленных с исполь-
зованием влажного тушевого метода (Золотарев А. Г. с соавт., 2006).
786
Часть 3. Частная неправильная эпидемиология
Рис. 3.66. Микроскопическое изображение В. pseudomallei. А. Окрашенный по Граму
мазок культуры штамма Duc-V, выращенного в течение 48 ч на плотной питательной
сцеде с добавлением 4 % глицерина. Увеличение в 1150 раз Штаммы, отличающиеся
низком вирулентностью д ля лабораторных животных, характеризуются выраженной
способностью образовывать нити и цепочки Б. Прижизненная микроскопическая
картина клеток возбуди геля мелиоидоза (штамм С-141, условия культивирования
те же). Иммобилизация клеток желатиновым гелем. Аноптральный контраст, уве-
личение в 1350 раз. Живые клетки со светлыми оболочками и протоплазмой слегка
темнее поля зрения. В. Электронно-микроскопическое изображение клеток В. pseudo-
mallei С-141. Увеличение в 15000 раз. Контрастирование уранилацетатом. Длина жгу-
тиков различна: от 2,6 до 8 мкм, в среднем 3,8 мкм. В молодых культурах палочка часто
имеет только один или два жгутика. Накануне деления клеток жгутики появляются
на обоих полюсах. По А. Г. Золотареву с соавт. (2006)
При выращивании культур возбудителя мелиоидоза на плотной питательной среде
с добавлением 4 % глицерина наблюдается более обильный и быстрый рост, чем при
культивировании близкородственного вида В. mallei. В. pseudomallei относительно
хорошо растет на обычных питательных средах (pH 6,8—7,0) при температуре 37°C,
способна ферментировать с образованием кислоты глюкозу, сахарозу, лактозу, ман-
нит, мальтозу, декстрин и дульцит, не образует индола и сероводорода, медленно свер-
тывает молоке, обладает выраженными протеолитическими свойствами. Факульта-
тивный аэроб (Матвеев К. И., Быченко Б. Д., 1966).
На жидких средах через 24-36 ч после посева В. pseudomallei в проходящем свете
отмечают помутнение, которое спустя некоторое время становится более интенсив-
ным за счет накопления клеток. Поверхность среды покрывает тонкая прозрачная
пленка, состоящая из микробов. При добавлении индикатора нейтрального красно-
го пленка окрашивается в красно-оранжевый цвет У всех 28-часовых культур такая
же реакция с индикатором метиловым красным постоянно бывает отрицательной.
Через 2—3 сут пленка заметно утолщается, собирается в складки и утрачивает прозрач-
ность. Толщина пленки через 4—5 сут может равнял «ся 5 мм. К этому времени клетки
оседают на дно в виде обильного рыхлого осадка, а среда постепенно просветляется
(Матвеев К. И., Быченко Б. Д., 1966).
Среди наиболее ярко выраженных вариабельных свойств В. pseudomallei в первую
очередь следует назвать морфологическую диссоциацию. Большая часть его культур
при выращивании на плотных питательных средах образует, по крайней мере, три типа
колоний в S-, R- и М-формах, а также их промежуточные варианты. Выявлены неко-
16. Мелиоидоз
787
*орые факторы, способные оказывать влияние на состояние диссоциативного про-
цесса у В. pseudomallei. В частности, установлено, что выращивание при повышенной
-емпературе или продолжительное пассирование на питательных средах изменяет
соотношение S- и R-форм. Отмечено также, что изменение условий культивирования
или добавление компонентов сыворотки крови повышает мукоидность клеток. По-
видимому, способность клеток к продукции мукоида может колебаться в значитель-
ных пределах. Некоторые исследователи наблюдали в органах и макрофагах живот-
ных, зараженных В. pseudomallei, образование не только слизистой псевдокапсулы, но
и более сформированной околоклеточной структуры, которую они расценивали как
истинную капсулу (Меринова Л. К. с соавт., 1995).
Колонии шероховатого R-варианта В. pseudomallei, появляясь в виде прозрачных
мелких капелек, быстро утрачивают правильность формы и прозрачность, край ста-
новится неровным, зубчатым, поверхность сморщивается либо собирается в сухие или
слизистые складки, в центре часто образуется сосочковый выступ, округленный сли-
зистым валиком. Такие колонии также имеют металлический блеск.
Хорошо оформленные гладкие колонии В. pseudomallei в S-форме вырастают уже
через 24 ч. Визуально они выпуклые, полупрозрачные, сероватого цвета или с голубо-
ватым оттенком. При микроскопическом исследовании в падающем свете — бежевые
или желтоватые, имеют округлые ровные края, гладкую блестящую поверхность и диа-
метр около 1,5 мм. К исходу 2 сут инкубирования их диаметр увеличивается до 3 мм,
а прозрачность постепенно утрачивается. Колонии тускнеют и становятся серовато-
белыми или желто-коричневыми. У недиссоциируюших колоний сохраняется глад-
кая блестящая поверхность и ровные края (рис. 3.67).
Слизистые колонии (М-формы) отличаются слизистой консистенцией, оплывшей
конфигурацией и значительной величиной (до 6-7 мм в диаметре на вторые сутки вы-
ращивания). Они бывают прозрачными и непрозрачными, овальными или неправиль-
ной формы, иногда слегка опалесцируют и могут иметь на своей поверхности дочер-
ние колонии. С увеличением срока выращивания В. pseudomallei на плотной среде,
количество слизистых колоний возрастает. Их формирование вызвано продуцирова-
нием во внешнюю среду клетками возбудителя мелиоидоза мукоида — многокомпо-
нентного соединения, состоящего из нейтральных и кислых полисахаридов, белков
и нуклеиновых кислот (Тихонов Н. Г. с соавт., 1995).
Кроме описанных трех форм колоний, существует множество переходных форм,
которые обычно относят к переходному варианту О. К нему относятся колонии, ко-
торые одновременно обладают какими-либо признаками двух или даже всех трех вари-
антов, например плоская колония с ровным круговым краем и слизистой поверхно-
стью, разделенной множественными валиками в виде концентрических колец (Мат-
веев К. И., Быченко Б. Д., 1966).
Н. Narisara et al. (2007) объясняют большое количества форм колоний В. pseudo-
mallei ее высокой фенотипической пластичностью, реализующуюся через различия в
экспрессии поверхностных структур бактериальной клетки.
В. pseudomallei неприхотлива к питанию. Может размножаться на синтетических
средах, содержащих отдельные аминокислоты или аммонийную соль некоторых орга-
нических кислот как единственный источник углерода и азота. Растет на глицерино-
вой цитратно-аспарагиновой среде, а также на дрожжевой воде. Культуры В. pseudo-
mallei издают специфический аромат, напоминающий затхлый запах плесени (Мат-
веев К. И., Быченко Б. Д., 1966),
788
Часть 3. Частная неправильная эпидемиология
Рис. 3.67. Макроскопические изображения колоний В pseudomallei. А. Через 12 ч
инкубирования В. pseudomallei при температуре 36—38 °C образуются микроколо-
нии, имеющие форму уплощенной полусферы, расположенной на тонком плос-
ком основании с ровными краями. Центральная часть микроколонии выпуклая,
с исчерченной поверхностью, и покрыта небольшими зернами. Штамм С-141.
Микрофотосъемка в проходящем свете. Увеличение в 56 раз, Б. Внешний вид
агаровой пластинки с колониями B.pseudomallei, штамм Wang, колонии В.pseudo-
mallei по морфологическим свойствам различаются более заметно, чем колонии
В. mallei. В. R-форма колонии В. pseudomallei в ультраморщинистом вариан-
те. Отличается расщепленной поверхностью, напоминающей горный рельеф.
Штамм Duc-V. Инкубирование в течение 48 ч на агаре с добавлением 4 % гли-
церина. Микрофотосъемка в проходящем свете, увеличение в 13 раз. Г. Морфо-
логия S-форм колоний В. pseudomallei С-141 В проходящем свете такие колонии
выглядят полупрозрачными, в падающем — бежевыми или светло-желтыми и
напоминают по форме уплощенную полусферу с ровными гладкими краями
и гладкой блестящей поверхностью. Увеличение в 13 раз. Макрофотосъемка
в падающем свете. По А. Г. Золотареву с соавт (2006)
Возбудитель мелиоидоза продуцирует широкий спектр ферментов (каталазы, ци-
тохромоксидазы, дегидрогеназы, липазы, нейраминазы, нитратредуктазы, фосфата-
зы, фосфогидролазы, протеазы, лецитиназы, пероксидазы, супероксиддисмутазы
и др.), обеспечивающих ему возможность существования в различных условиях внеш-
ней среды (Cheng А. С., Currie В. J., 2005).
Факторы патогенности. Поданным М. A. Schell (2008), патогенность возбудителям
сапа и мелиоидоза «обеспечивает» экспрессия не менее 650 генов, но их иерархиче-
ские связи и механизмы регуляции в различных условиях окружающей бактерию сре-
ды неизвестны. А. С. Cheng, В. J. Currie (2005) разделили гены, определяющие пато-
генность В. pseudomallei на две группы: гены, ассоциированные с выживанием в окру-
жающей среде; и гены, ассоциированные с вирулентностью (табл. 3.21).
К клеточным структурам, участвующих в проявлении вирулентности у В. pseudo-
mallei, относят капсулярный полисахирид, ЛПС, наружные мембранные белки (проте-
интирозинфосфатаза, порины), пили I и IV типов, сидерофоры (Cheng А. С., Cur-
rie В. J., 2005), аутотранспортерные макромолекулы (Edwards Т. et al., 2010).
3.6. Мелиоидоз
789
Таблица 3.21
Гены патогенности В. pseudomallei*
Гены, ассоциированные с выживанием бактерии Гены, ассоциированные с вирулентностью бактерии
Продукты экспрессии которых участвуют в биосинтетических путях вторичных метаболи- тов: антибиотиков, сурфактантов, сидерофо- ров (вк полая fur) Определяющие типы транспортной системы секреции: I, II, III (три типа bsa, Ыр и bop) и V
Лекарственной резистентности: (3-лактамазы включая аха и репА); аминогликозидаиетил- трансферазы, гены, обеспечивающие мульти- резистентность бактерии за счет функциони- рования «efflux pumps» Кодирующие компоненты поверхности бактерии: ЛПС, капсулярные полисахариды (включая waa, rfb и web)
ОПеспечиваюшие блокирование внутрикле- точного стресса: ферменты, инактизирующие перекисные соединения (супероксиды) и окись азота Кодирующие экзопротеины: фосфолипаза С (включая />/с), металлопротеаза А и другие протеазы (включая трг) и коллагеназа
Продукты экспрессии, в которых участвуют подвижности и хемотаксисе бактерии систе- ма флагеллина (включая fli и /7g), белки, ассо- циированные с хемотаксисом (включая che) Кодирующие адгезины; поверхностные белки клетки, взаимодействующие с поверхностью клеток эукариотических хозяев. Фимбрии и пили I и IV типов, г «/-типа (включая pH)
* По А. С. Cheng, В. J. Currie (2005).
Роль экзопротеинов в вирулентности В. pseudomallei далеко не так очевидна, как
это следует из формально-логического понимания их роли в патогенезе инфекцион-
ных болезней. Наличие молниеносных форм течения мелиоидоза дало основание
ученым еще в 1930-х гг. предположить продукцию В. pseudomallei летального токсина.
При экспериментальном изучении первоначально была установлена токсичность
фильтратов бульонных культур для морских свинок. Подробнее феномен токсинооб-
разования у возбудителя мелиоидоза был изучен в 1950-1960-х гг. Однако получить
высокоочищенный токсин долгое время не удавалось. В фильтратах жидких культур
В. pseudomallei содержались различные токсические агенты, которые можно было раз-
_елить на две группы: вызывающие гибель животных; и вызывающие некроз при внут-
рикожном введении. Разделение этих факторов патогенности оказалось достаточно
сложной в техническом отношении задачей из-за близости их физико-химических
свойств, в первую очередь, — молекулярной массы. В 1990-х гг. было установлено, что
дермонекротическая активность фильтратов бульонных культур В. pseudomallei выз-
вана действием тиоловых и сериновых протеаз, обладающих коллагеназной и элас-
тазной активностью. Гибель животных вызывает содержащийся в фильтратах токсин
(Тихонов Н. Г. с соавт., 1995).
Механизм действия токсина В. pseudomallei предполагали еще до получения его
высокоочищенного препарата по характеру ультраструктурных патоморфологи-
ческих изменений в тканях экспериментальных животных, вызванных парентераль-
ным введением препаратов грубой очистки. В первую очередь исследователи отмечали
.еструкцию митохондрий, редукцию эндоплазматической сети клетки и уменьшение
числа рибосом, что свидетельствовало о торможении токсином синтеза белка (Кури-
лов В. Я. с соавт., 1987).
S. Ilausesler et al. (1998) подтвердили это предположение, получив в очищен-
ном виде мелиоидозный токсин и изучив механизм его действия на белковый синтез.
Токсин В. pseudomallei представляет собой цитотоксический экзолипид, ингибирую-
790
Часть 3. Частная неправильная эпидемиология
ший клеточный белковый синтез через АДФ-рибозилирование фактора элонга-
ции 2 (EF-2), т. е. он действует на белковый синтез клетки таким же образом, как и
дифтерийный токсин, экзотоксин А псевдомонад, шига-токсин возбудителя дизен-
терии и растительный токсин рицин (см. разд. 3.11). Однако действие только токсина
возбудителя мелиоидоза не объясняет полностью патогенез болезни. Н. Г. Тихоновым
с соавт. (1995) токсинопродукция обнаружена у всех одинн щцати исследованных куль-
тур Й. pseudomallei, в том числе и у авирулентного пуринзависимого мутанта VPA.
Сходные данные получены Р. J. Brett et al. (1994) в отношении фосфолипазы С,
гемолизина, лецитиназы, липазы, секретируемых В. pseudomallei посредством систе-
мы секреции второго типа. Выключение генов секреторной системы, выполненное
путем транспозонного мутагенеза, блокировало секрецию этих молекул, но не при-
водило к снижению вирулентности возбудителя мелиоидоза.
Можно предположить, что большинство факторов патогенности возбудителя ме-
лиоидоза играют ключевую роль в процессах адгезии, проникновения в эукариоти-
ческую клетку и в механизмах физиологической адаптации бактерии к условиям ок-
ружающей их среды в фагосоме.
Таксономия, происхождение и молекулярная эпидемиологии возбудителя ме шоичоза.
Таксономия В. pseudomallei описана в разд. 3.5. Геном В. pseudomallei включает две боль-
ших хромосомы (4,07 и 3,17 м. п. о. соответственно), имеющие разные функции в
клетке (Atkins L. С. et al., 2004). Хромосома большего размера (хромосома 1) включа-
ет гены, в основном, относящиеся к клеточному метаболизму и росту; меньшая по
размеру (хромосома 2) содержит гены, необходимые для выживания бактерии и для
проявления ее вирулентности. Обнаружение у хромосомы 1 консервативной органи-
зации генов и большого количества генов-ортологов говорит в пользу ее происхож-
дения от какого-то отдаленного предка, общего дгя многих современных видов бак-
терий. Обнаружение 71 -го геномного островка (genomic islands, G1) в обеих хромосомах
возбудителя мелиоидоза свидетельствует:
1) о большой роли горизонтального переноса генов в его эволюции;
2) о возможных генетических и фенотипических вариациях среди штаммов, вы-
деляемых в клинике и из объектов окружающей среды (рис. 3.68).
В противоположность возбудителю мелиоидоза, В. mallei не содержит геномных
островков (Nierman W. С. et al., 2004). Более подробно о механизмах эволюции воз-
будителя сапа см. в разд. 3.5. Для установления генетической связи между изолятами-
В. pseudomallei используются различные молекулярные методы исследований (PFGE,
RAPD. MLST и др.), обычно демонстрирующие сходные результаты (Cheng А. С., Cur-
rie В. J., 2005). В аспекте молекулярной эпидемиологии В. pseudomallei представляет
полную противоположность возбудителю сапа. Он слишком разнообразен, и поэто-
му трудно проследить не только клональные связи между штаммами, но и установить
границы вида. В то же время геном В. mallei слишком однороден, чтобы надежно диф-
ференцировать штаммы различного географического происхождения (рис. 3.69).
Поэтому в поздней работе D. М. Engelthaler et al. (2011), сравнивших разные моле-
кулярные способы дифференциации штаммов В. mallei и В. pseudomallei, предпочте-
ние отдается полногеномному сиквиенс-типированию (whole genome sequence typing,
WGSI), хотя исследователи и признают, что еще слишком мало расшифровано гено-
мов Burkholderia, чтобы надежно проводить между ними филогенетические различия.
Экология и естественная эпидемиология возбудителя мелиоидоза. Экология возбу-
дителя мелиоидоза интенсивно исследуется еще со времен полномасштабного вмеша-
тельства США в войну во Вьетнаме (1965—1973). Поэтому накопленные знания по
3.6 Мелиоидоз
791
l-Glila
K<M»« Gil- -GI1.GI1.1
K96243 GI2------Gna
GI3----GI3.1
GI4-
GI5-
GI6-----GI6
-Gila
-GI4.1 GI4.2, GI4.3, GI4.4
-GI5a,GI5a.l,GI5a.2
-GI6a
-G16b
GI7- -GI7.1,GI7.2,GI7.3,GI7.4
GI8| jBcTaBKHGI8.1,GI8.2, GI8.3
-GI8c
-GI8d
-GI9a
-GI9b
-GI9c,GI9c.l
GI10----GI1O, GI10.1,GI10.2,GI10.3
019-
Другие? шташы
1710b дцх,
110&а Ы16а-
MSHR668 GI16b, GI16b.l--GI16b
MSHR305 YLFBfFC-------YLF
GI13.1, GI13.2, GI13.3, GI13.4 - - GI13
GI14.GI14.1- -GI14
GI14a-
GI15.1---GI15
GI15a-
GI15b-
GI15c-
GI15d-
GI15c
GI16.1, GI16.2
К96243
GI16
Хромосома 2
Gill----GI11.1.GI11.2
GI12----GI12.1,GI12.2,GI12.3,GI12.4
Хромосома 1
Рис. 3.68. Локализация геномных островков в хромосомах В. pseudomallei G1 иденти-
фицированы в хромосомной ДНК штаммов 1710b, 1106а, MSHR668, MSHR3O5 (внут-
ри линий, обозначающих хромосомы) и К96243 (снаружи линий, обозначающих хро-
мосомы) Черными стрелками обозначены участки начала репликации хромосом.
По A. Tuanyok et al. (2008)
Рис 3.69. Взаимосвязь между 147 изолят ами Burkholderia, установленная с помощью
MLST по 7 локусам. Пять изолятов В. mallei (ST40) из Индии, Болгарии, Китая и Турции
(два изолята) имели идентичные аллельные профили и кластировались среди изЬлятов
В. pseudomallei. Два изолята Burkholderia сначала были определены как В. pseudomallei, но
при проведении этих исследований обнаружено, что они тесно связаны с В. thailandensis
(показаны как В. thailandensis?). На древе также кластированы три изолята Burkholderia
из Оклахомы Первоначально их принимали за В. pseudomallei, но их алле пьные профили
не имеют сходства ни с одним из исследованных изолятов В. pseudomallei, ни с В. thailan-
densis по всем семи локусам. ST (sequence type) - сиквинс-тип. По D. Godov et al. (2003)
792
Часть 3. Частная неправильная эпидемиология
механизмам поддержания В. pseudomallei в природе могут рассматриваться как опе-
режающие для микроорганизмов, патогенных для людей, чья экология изучена зна-
чительно хуже. Возбудитель мелиоидоза — типичный сапроноз. К настоящему вре-
мени обнаружены несколько экологических ниш (биопленки, простейшие, грибы,
корни бобовых растений и др.), где В. pseudomallei способна поддерживаться без уча-
стия позвоночных организмов.
Биопленки. Характерным местообитанием В. pseudomallei в почве являются слож-
ные биоты ризосферы и корневой зоны растений. На границе раздела жидкой и твер-
дой фаз, а также на границе жидкой и газовой (воздушной) фаз формируются биоплен-
ки, включающие медленно растущие В. pseudomalle, окруженные внеклеточным
трехмерным полисахаридным комплексом. Бактериальные консорциумы биопленок
устойчивы к действию факторов внешней среды, дезинфектанатов и антибиотиков
(Sawasdidoln С. et al., 2010).
Простейшие (Protozoa). Экспериментально установлено, что В. pseudomallei может
существовать в Acanthamoeba astronyxis, A. castellanii, A. palestiniensis, A. polyphaga, Hart-
mannella vermiformis и других свободноживущих простейших, обитателей водных по-
верхностей, ризосферы и биопленок. Простейшие поглощают возбудитель мелиои-
доза с помощью обвивающего фагоцитоза (coiling phagocytosis), где он размножается.
При неблагоприятных условиях внешней среды В. pseudomalle длительное время вы-
живает в цистах, формируемых простейшими. Заполненные В. pseudomallei вакуоли
простейших имеют размер в пределах 5—15 мкм (Inglis Т., SagripantiJ., 2006) (рис. 3.70).
Рис. 3.70. Конфокальное сканирующее изображение В. pseudomallei, инфицирую-
щей A. astonyxis. А. Адгезия полярного конца бактерии с поверхностью трофозоита
(белая стрелка) ведет к ее включению в вакуоль (острие белой стрелки). Увеличение
в 1440 раз. Б. Псевдоподия захватывает бактерию (белая стрелка), включившая
бактерию вакуоль того же трофо зоита (наконечник черной стрелки), и внешнее
бактериальное скопление (черная стрелка). Увеличение в 1220 раз. В. Трофозоит
со свободноживущей в цитоплазме бактерией (острие белой стрелки) и интактная
вакуоль, включающая бактерии (острие черной стрелки). Увеличение в ИЗО раз.
Г. Через 24 ч инкубирования — наконечником черной стрелки показаны бактерии,
включенные в цисты амеб. Увеличение в 1220 раз. По Т. Inglis et al., 2000
3.6. Мелиоидоз
793
Грибы. Микоризные грибы являются еще одним установленным клеточным хозяи-
ном В. pseudomallei. Ее обнаруживают в микоризе, т. е. в грибокорнях — симбиотиче-
ской ассоциации мицелия гриба с корнями высших растений. Возбудитель мелиоидоза
находят в цитоплазме арбускул — мицелия грибов, проникающего в паренхимальные
клетки корня растения. В арбускулах происходит наиболее интенсивный обмен ме-
таболитами между компонентами микоризы. Роль Burkholderia в этой симбиотической
ассоциации не ясна. Возможно, они участвуют в фиксации азота грибами. Экспери-
ментально установлена способность В. pseudomallei колонизировать споры и гифы
микоризного гриба Gigaspora margarita (рис. 3.71).
Рис. 3.71, Взаимодействия спорогенных клеток (SC), инокулированных В. pseudo-
mallei (В), со спорами (S) G. decipiens. А.. Спорогенная клетка G. decipiens присое
диняется к поверхности споры. Б. Участок контакта между спорой и спорогенной
клеткой. В. Гифы (Н) G. decipiens с присоединившимися к ним В. pseudomallei
(стрелки) и ассоциированный фибриллярный материал. Г. Цитоплазма G. decipiens,
заполненная бактериями (острия стрелок) По Levy A. et al., 2003
A. Levy et al. (2003) не наблюдали активного деления В. pseudomallei в спорах G. mar-
garita, однако они считают, что споры микоризных грибов могут играть важную роль
в распространении возбудителя мелиоидоза в окружающей среде и в ингаляционном
заражении людей.
Бобы. В их корневой системе формируются узелки, содержащие симбиотические
бактерии, обычно ассоциирующиеся с фиксацией атмосферного азота. В. pseudomallei
была обнаружена в почве, непосредственно прилегающей к корням австралийской
акации1 (Inglis Т. et al., 2000).
Имеются экспериментальные доказательства поддержания В. pseudomallei в ризос-
фере других растений (рис, каучуконосы, маниока) (Inglis Т., Sagripanti J., 2006).
Четко очерченного ареала мелиоидоза не существует. Эндемические по мелиои-
дозу районы находятся в тропической зоне Южной и Юго-Восточной Азии (Бирма,
1 Растение из семейства бобовых, растущее в Австралии, Африке, Мексике и Южной Азии.
794
Часть 3. Час тная неправильная эпидемиология
Малайзия, Вьетнам, Камбожа, Таиланд), на островах юго-западной части Тихого оке-
ана, Новой Гвинее, в северо-восточной части Австралии. Спорадические случаи ме-
лиоидоза выявляются на Филиппинах, в Индонезии, Иране, Индии, Сингапуре, Шри-
Ланке, Западной Австралии, Южном Китае и ряде стран экваториальной Африки.
Единичные случаи мелиоидоза завозного характера зарегистрированы в США Фран-
ции, Англии, Турции, преимущественно у военносл; жащих, вернувшихся из районов
боевых действий в Юго-Восточной Азии (рис. 3.72).
Рис. 3.72. География очагов мелиоидоза. 1 — регионы спорадического обнаружения
возбудителя мелиоидоза; 2 — основные эндемичные области. По I. Магу et al. (1995)
В России очаги мелиоидоза не обнаружены, но в Европе и в странах, граничащих
с Россией на юге и востоке, случаи мелиоидоза зафиксированы. По данным Г. М. Ла-
рионова с соавт. (1995), случаи мелиоидоза были диагностированы в Англии и Фран-
ции у военных, вернувшихся из Юго-Восточной Азии во время и после Второй миро-
вой войны. Вспышка мелиоидоза неясного происхождения имела место в 1945 г. в
Берлине. Франция стала первой страной Европы, 1де завозы мелиоидоза переросли в
серьезную проблему: во Французском экспедиционном корпусе за период пребыва-
ния во Вьетнаме (1948—1954 гг.) мелоидозом заболели более 100 военнослужащих.
В ноябре 1975 г. культуру В. pseudomallei выделили в Парижском зоопарке из тру-
па лошади Пржевальского, родившейся здесь же, а затем этоз микроорганизм обна-
ружили в навозе нескольких загонов. Эпизоотия разрасталась, и ретроспективно ус-
тановили, что за 3 года до эпизоотии возбудитель мелиоидоза уже находили в навозе
зоопарка. Затем мелиоидоз появился среди лошадей парижской национальной гвар-
дии и в скаковых клубах западной части Парижа. Далее В. pseudomallei обнаружили у
лошадей в Нормандии, в районах Монпелье Бордо и Бержерака. За 1976—1981 гг. в
стране было выявлено три случая мелиоидоза у людей. Распространению инфекции
способствовало использование навоза в качестве удобрения: В. pseudomallei стали обна-
руживать в почве хозяйств по выращиванию шампиньонов. Доставка в Париж из
Сингапура экзотических рыб сопутствовала завозу возбудителя мелиоидоза, которого
неоднократно обнаруживали в воде и песке аквариумов (Ларионов Г. М. с соавт., 1995).
В1979 г. на острове Реюньон погибла от мелиоидоза лошадь, привезенная из Фран-
ции. Вскоре В. pseudomallei обнаружили в почве на территории нескольких скаковых
3.6. Мелиоидо^
795
клубов этого острова. Серологические исследования, проведенные в 1977 и 1979 гг.,
указали на недавнее появление мелиоидоза на данной заморской территории Фран-
ции. Это пока единственный, считающийся достоверным, занос мелиоидоза, повлек-
ший за собой укоренение инфекции в новой местности. Однако установить, какому
именно завозу обязана сама Франция появлению на ее территории мелиоидоза, так и
не удалось (Ларионов Г. М. с соавт., 1995).
Интересные данные по географии мелиоидоза получены в 1978-1980 гг. сотруд-
никами Волгоградского НИПЧИ. Учитывая близость к СССР иранских очагов мели-
оидоза, ими была предпринята попытка обнаружить В. pseudomallei в Закавказье.
В этом регионе было исследовано более 5600 сывороток крови людей и животных, а
также более 2600 проб почвы и воды. На юге Азербайджана и в Грузии в крови мест-
ных жителей обнаружены специфические антитела в титрах 1:40—1:160. В этих же
районах из объектов внешней среды выделено более 100 культур микроорганизмов,
по ряду признаков сходных с возбудителем мелиоидоза, в частности, агглютинирую-
щихся видовой сывороткой в титрах 1:40—1:3200. Однако выделенные культуры ока-
зались апатогенными. Эти данные подтверждают целесообразность продолжения
поиска мелиоидоза на территории России и сопреде.1ьных с ней стран СНГ, особен-
но в районах рисосеяния и в зонах субтропиков (Ларионов Г. М. с соавт., 1995).
Более поздние данные по поиску очагов мелиоидоза на территории России и стран
СНГ мне неизвестны. Учитывая сложность экологии В. pseudomallei можно пред-
положить, что «завоз» возбудителя мелиоидоза не играет особой роли в формирова-
нии его очагов.
Естественное инфицирование людей В. pseudomallei в основном происходит в эн-
демических по мелиоидозу районах в результате попадания в организм человека че-
рез поврежденные кожные покровы (открытые раны, ожоговые поверхности, микро-
травмы и т. п.) обсемененных возбудителем воды и почвы. Намного меньшее значение
для естественного инфицирования людей имеет алиментарный путь проникновения
возбудителя. Мужчины составляют до 98 % всех заболевших в эндемических районах
(Шувалова Е. П. с соавт., 1979). В отдельных эндемических очагах возможен респи-
раторный путь заражения мелиоидозом. Например, среди заболевших мелиоидозом
во Вьетнаме американских военнослужащих 1/3 составили члены экипажей вертоле-
тов, которые заражались при вдыхании зараженной пыли, поднятой лопастями ма-
шин (Руководство по зоонозам, 1983).
Обнаружение В. pseudomullei в секрете простаты и влагалища сексуальных парт-
неров больных мелиоидозом позволяет предположить возможность полового пути
заражения возбудителем мелиоидоза.
Восприимчивост ь к возбудителю мелиоидоза и инфицирующие дозы для животных.
В бактериологической прак гике для изолирования и идентификации возбудителя
мелиоидоза обычно используют морских свинок, кроликоь и мышей. Наиболее чув-
ствительны к возбудителю инфекции хомяки и хорьки, затем следуют морские свин-
ки, кролики, щенки и кроты, мыши, белые крысы и обезьяны. Инфицирующие дозы
В. pseudomallei для человека и приматов в открытых источниках мне найти не удалось.
В табл 3.22 приведены величины LDS0 для лабораторных животных при разных спо-
собах заражения животных и клинические варианты течения экспериментального
мелиоидоза.
Оптимальной моделью, наиболее адекватной естественным условиям и пригод-
ной для воспроизведения на одном виде животного нескольких клинических форм
мелиоидоза, В. В. Алексеев с соавт. (1995) считают белую крысу.
796
Часть 3. Частная неправильная эпидемиология
Таблица 3.22
Величины LD50 для лабораторных животных при подкожном и ингаляционном
способах заражения и клинические варианты течения мелиоидоза*
Вид животного Штамм ld50, КОЕ Инфицирующая доза, КОЕ Клиническое течение болезни
Подкожное заражение
Белая мышь BALB/c В. pseudomallei 100 10,0 1,0-10,0 Острое
Золотистый хомячок В. pseudomallei 100 10,0 1,0 Острое
Морская свинка В. pseudomallei 100 10,0 10,0-30,0 Острое
Белая крыса В, pseudomallei 100 103-ю5 30,0 Острое, подострое, хроническое
Белая мышь BALB/c В. pseudomallei С-141 ю2 100,0 Молниеносное
10,0-30,0 10,0-30,0 Острое, подострое
1,0-10,0 Хроническое
Золотистый хомячок В. pseudomallei С-141 10,0 1,0 Острое
Морская свинка В. pseudomallei С-141 103-105 10,0 Острое, подострое, хроническое
Белая крыса В. pseudomahei С-141 ю9 108 Низкая летальность, выздоровление животных
Ингаляционное заражение
Белая мышь BALB/c В. pseudomallei 100 4,0 32,0 Молниеносное
2,0-4,0 Острое
Золотистый хомячок В. pseudomallei 100 21,0 50,0 Молниеносное
10,0-25,0 Острое
Морская свинка В. pseudomallei 100 3,0 30,0-50,0 Острое
1,0-5,0 Подострое
Белая крыса В. pseudomallei 100-3 Р 135,0 10,0-20,0 Острое
Золотистый хомячок В. pseudomallei 97 2,5х103 10,0-20,0 Подострое
Белая крыса В. pseudomallei С-141 2,9х103 6,0 Молниеносное
0,3-2,0 Острое, подострое, хроническое
* По В. В. Алексееву с соавт (1995).
В естественных условиях мелиоидозом болеют многие виды домашних и диких
животных. К числу животных, спонтанно зараженных В. pseudomallei в эндемичных
районах, относятся лошади, К.РС, овцы, козы, свиньи, различные виды мышевидных
грызунов, морские свинки. Наиболее чувствительны к заражению возбудителем ме-
лиоидоза верблюды, лошади, овцы, КРС, козы, свиньи, кенгуру, сумчатые медведи,
альпаки (вид ламы), олени, кошки, собаки и содержащиеся в неволе морские живот-
ные. Относительно резистентны к заражению возбудителем мелиоидоза птицы, во-
дяные буйволы и крокодилы (Cheng А. С., Currie В. J., 2005).
Характерным признаком болезни у всех животных, независимо от видовой при-
надлежности, также как и у человека, является развитие единичных или множествен-
ных абсцессов в органах и тканях. В зависимости от длительности и тяжести течения
болезни, мелиоидоз у животных подразделяется на острый, подострый и хронический
(Ширяев Д. Т. с соавт, 1976).
3.6. Мелиоидоз
797
Иммунитет. Внедрение в эпидемиологическую практику методов иммунологичес-
кого тестирования позволило выявить в эндемичных по мелиоидозу регионах боль-
шое количество серопозитивных лиц, но клинически болезнь проявлялась лишь у
отдельных жителей. В Малайзии положительные серологические реакции зарегист-
рированы у 15—20 % людей, занятых сельскохозяйственным трудом. У 50 % обследо-
ванных рисоводов Ирана в сыворотке крови обнаружены антитела к возбудителю
мелиоидоза методом РИГА. Данные этих и других аналогичных исследований позво-
лили сформулировать представление о мелиоидозе как инфекции, протекающей в ла-
тентной форме у клинически здоровых людей (Тихонов Н. Г. с соавт., 1995).
Манифестация клинических проявлений мелиоидоза может наступить через мно-
го лет после пребывания человека в эндемичной зоне. Известны случаи рецидива
болезни через 60 лет после перенесения ее в легкой форме (Ngauy V. et al.. 2005). Как
травило, В. pseudomallei не удается высеять из крови человека с латентным течением
болезни. Объяснение этого феномена может следовать из сапронозного существова-
ния мелиоидозного микроба в природе. Бактерии-сапронозы сосуществуют с простей-
шими в различных формах симбиоза. Одной из них является состояние «некультиви-
руемости», 1. е. покоящееся состояние, характеризующееся сниженным уровнем ды-
хательной активности и активности ряда ферментов, делающих невозможным их рост
на искусственных питательных средах. Бактерии, поддерживающиеся в природе в про-
стейших в состоянии «некультивируемости», при инфицировании макрофагов чело-
века переходят в это же состояние1, оставаясь незамеченными иммунной системой и
не давая роста в гемокультурах. Выход бактерии из состояния «некультивируемости^
происходит в результате воздействия на макрофаг каких-то стрессовых факторов.
Возможно, по этой причине рецидив мелиоидоза обычно происходит на фоне обостре-
ния у пациента хронических болезней, алкоголизма и тяжелых травм.
Возбудитель мелиоидоза резистентен к действию бактерицидных компонентов
сыворотки и комплемента. Выживает в профессиональных фагоцитах, включая мак-
рофаги/моноциты и нейтрофильные клеточные линии. Фагоцитоз усиливается под
воздействием комплемента и антител, однако лизис бактерий остается незавершен-
ным. У больных мелиоидозом люцей и животных формируется гиперчувствительность
замедленного типа (Тихонов Н. Г. с соавт., 1995; Cheng А. С., Currie В. J., 2005).
Гуморальное звено иммунитета включается в иммуногенез с начала инфекцион-
ного процесса. В организме человека и животных начинают вырабатываться антите-
ла, уровень которых зависит от интенсивности течения инфекции. Обнаружение спе-
цифических антител в сыворотке крови человека позволяет сделать вывод о наличии
у него мелиоидоза в прошлом, а при росте титра антител за кроткий период времени
(2—3 нед) — об активно развивающейся мелиоидозной инфекции, но прогностическо-
го значения антитела не имеют. Антитела у переболевших мелиоидозом людей и жи-
вотных не обладают протективным действием и не защищают от повторного зара-
жения В. pseudomallei. К тому же они не высоко специфичны и дают перекрестные ре-
акции с возбудителями сапа и легионеллеза (Тихонов Н. Г. с соавт., 1995).
Устойчивость возбудителя мелиоидоза во внешней среде. Возбудитель мелиоидоза
не только может длительно сохраняться в окружающей среде, но если иметь в виду
вышеприведенные данные по его экологии, то можно считать, что для некоторых тер-
ригорий он и есть часть окружающей среды.
1 Предположение о том, что возбудитель мелиоидоза может сохраняться в макрофагах в
некультивируемом состоянии, ранее высказывал В. Е. Жемчугов (2004).
798
Часть 3. Частная неправильная эпидемиология
Возбудитель мелиоидоза, помещенный во влажную почву или воду, может быть
выделен из этих объектов через 44 сут, в моче при доступе кислорода В. pseudomallei
сохраняется не менее 17 сут, в испражнениях до 27 сут, в высушенном носовом секре-
те — 2 нед, в трупах — не менее 2 нед. В почве и фекалиях сохраняется 27 сут и более,
в трупах — не менее 12 сут. Устойчивость к высоким температурам у возбудителя ме-
лиоидоза мало отличается от других видов неспорообразующих бактерий. В жидкой
среде при температуре 56 °C бактерии гибнут в течение 10—12 мин, при температуре
70°С — 5 мин, при 100°С — моментально (Ширяев Д. Т. с соавт., 1976)
Антибиотикорезистентность. У В. pseudomallei обнаружены семь генов, кодирующих
Р-лактамазы (А-, В- и D-типов), efflux-система, определяющая резистентность высо-
кого уровня к эритромицину и аминогликозидам, и еще ряд плохо изученных меха-
низмов антибиотикорезистентности. До 90 % изолятов В. pseudomallei в условиях in vitro
резистентны к амикацину, амоксициллину, ампициллину, азитромицину, азтреонами-
ну, карбенициллину, цефамандолу, цефазолину, цефокситиму, цефалексину, циклосе-
рину, эритромицину, фосфомицину, гентамицину, канамицину, налидиксовой кисло-
те, неомицину, норфлоксацину, рифампицину, сульфаметоксазолу, триметоприму.
Монотерапия может приводить к селекции штаммов возбудителя мелиоидоза, рези-
стентных к использованному антибиотику (доксициклин, цефтазидим, хлорамфени-
кол, триметоприм-сульфаметоксазол) (Cheng А. С., Currie В. J., 2005).
Мелиоидоз относится к таким инфе кциям, при которых наблюдайся отсутствие
четкой корреляции между результатами исследования чувствительности возбудителя
в условиях in vitro и in vivo. Даже комбинированное лечение мелиоидоза не всегда ока-
зывалось действенным. Клиническое выздоровление (улучшение состояния) еще не
гарантирует от вероятных рецидивов болезни (Илюхин В. И., Батманов В. П., 1995).
Обнаружение генетически измененных штаммов возбудителя мелиоидоза. Подходы
к обнаружению генетически измененных штаммов возбудителя мелиоидоза в основ-
ном те же, что и для обнаружения генетически измененных штаммов возбудителей
сибирской язвы и чумы (см. разд. 3.1 и 3.2). Учитывая совокупность поражающих
свойств возбудителя мелиоидоза как агента БО и опасность работы с ним в лабора-
торных условиях, маловероятно, что кто-то возьмется за его генетическую модифи-
кацию. Если такие исследования все же будут проведены организаторами террорис-
тического акта, то они могут быть направлены на снижение инфицирующей дозы,
сокращение инкубационного периода болезни или на придание выбранному для по-
ражения людей штамму В. pseudomallei резистентности к тем антибиотикам, к кото-
рым он окажется чувствительным.
Возбудитель мелиоидоза обладает природной компетентностью к чужеродной
ДНК и легко подвергается генетической модификации. Все известные методы гене-
тического переноса (конъюгация, трансформация, фаговая зрансдукция) отработа-
ны для В. pseudomallei в начале 1990-х гг. Еще раньше обнаружены собственные плаз-
миды мелиоидозного микроба. Векторные молекулы на основе репликонов плазмид
групп несовместимости IncQ, IncW, IncN, IncP стабильно поддерживаются в клетках
В. pseudomallei и близкородственных псевдомонад (Abaev I. V. et al., 1995).
Анализ плазмидных профилей малоинформативен для выявления генетически
измененных штаммов возбудителя мелиоидоза. Исследование плазмидного состава
штаммов В. pseudomallei, выделенных в различных географических регионах из объек-
тов окружающей среды, от больных людей и животных, проведенное в Волгоградском
НИПЧИ. показало, что клетки большинства штаммов возбудителя мелиоидоза содер-
жат плазмиды различной молекулярной массы. Четкой корреляции с географическим
3.6. Мелиоидоз
799
распространением плазмид не отмечено. Более того, в отдельных штаммах одного
географического региона выявляется по два плазмидных репликона различного
размера. Явной связи между культуральными и биологическими свойствами штаммов
и наличием плазмид у В. pseudomallei, также не обнаружено. Плазмидные ДНК, выде-
ленные из различных штаммов возбудителя мелиоидоза, обладают взаимной гомоло-
гией и гомологией с геномной ДНК В. pseudomallei и близкородственных псевдомо-
над — В. mallei и В. cepacia. В зависимости от размера генома, плазмидные ДН К обо-
значены рРМ1, рСМ2, рСМЗ и рСМ4 (Меринова Л. К. с соавт., 1995).
Из-за большого генетического разнообразия изолятов возбудителя мелиоидоза
предпочтительней осуществлять полный сиквиенс их генома с использованием тех-
нологии пиросеквенирования. Первым признаком генетической модификации воз-
будителя мелиоидоза, выявленного в очаге предполагаемой биологической диверсии
и теракта, будет обнаружение в его геноме кластера генов в составе интегративного
вектора с известной историей конструирования.
Патогенез мелиоидоза. Мелиоидоз представляет собой септицемию. Патогенез
мелиоидоза, особенно его ранние стадии, определяется способом инфицирования.
Возбудитель болезни может проникнуть в организм человека несколькими путями:
а) через слизистые оболочки желудочно-кишечного тракта вместе с зараженной
пищей;
б) через легкие вместе с инфицированными материалами, находящимися в пыле-
видном состоянии;
в) через поврежденную кожу и слизистые при контакте с заразным материалом, а
также через укусы зараженных насекомых.
Эпидемиологически значимыми являются чрескожный и ингаляционный пути
проникновения В. pseudomallei. В естественных условиях чаще всего не представляется
возможным проследить способ заражения. Развитие мелиоидоза зависит от вирулент-
ности В. pseudomallei, величины заражающей дозы и чувствительности конкретного
человека к инфекции. Сочетание этих факторов определяет патогенез, клиническую
картину и, как следствие, исход болезни.
Внедрение В. pseudomallei в макроорганизм начинается с местного воспалительно-
го процесса. В. pseudomallei захватывается фагоцитами и с током лимфы или крови по-
падает в кровяное русло и разносится по организму, проникая в нефагоцитирующие
клетки и создавая новые очаги воспаления в различных органах и тканях (рис. 3.73).
При хронических формах болезни постепенное разрушение различных органов с
образованием абсцессов тянется месяцами и даже годами. В целом наблюдаемые из-
менения у животных бывают очень сходными с таковыми у больного человека.
Клиническая и патологоанатомическая картина мелиоидоза у человека при естествен-
ном инфицировании1. Клинические проявления мелиоидоза у человека и животных
столь разнообразны, что практически все зафиксированные случаи установлены не в
результате логической постановки диагноза на основе анализа симптомов болезни, а
лишь после идентификации выделенной культуры или учета серологических реакций.
Совершенно не случайно за мелиоидозом утвердились термины «болезнь с обезьянь-
им лицом», «великий имитатор». Диагностика и лечение мелоидоза осложняются тем,
что из патологических очагов культура В. pseudomallei выделяется часто в ассоциации
1 Клиническая и патологоанатомическая картина мелиоидоза приведена в основном по
работам К. И. Матвеева, Б. Д. Быченко (1966); Д. Т. Ширяева с соавт. (1976); В. И. Илюхина,
В. П. Батманова (1995); Шуваловой Е. П. с соавт. (2001); Т. Inglis, J. Sagripanti (2006).
800
Часть 3. Частная неправильная эпидемиология
Время
ВорА
BipD
BsaZ
BsaQ
BsaLi
S' '''^Секреция IFN-P, INF-y
i SOCS3] О-анти-li
JAK-STAT —JI-CIS > —
iNO|S NOD2
------------------------------------------NO V
f __ \BimA
ген
ЛПС
5
*
2
T6SS-l/Hcpl
Rpo5
BipB?
3 PurM
HisF
PabB
Клетка 2
Рис 3.73. Отдельные этапы внутриклеточного выживания В. pseudomallei в макрофа-
гах и нефаго! датирующих клетках. 1 — проникновение В. pseudomallei в эндоцитиче-
скую везикулу; 2 — разрушение мембраны везикулы и проникновение В. pseudomallei
в цитозоль; 3 — репликация в цитоплазме клетки; 4 — слияние с мембраной клетки;
5 — апоптоз и выход В. pseudomallei из клетки. По Allwood Е. A. et al. (2011)
с другими представителями микробной флоры, а сама болезнь нередко протекает на
фоне других патологических состояний, ведущих к снижению иммунного статуса
человека. Среди них истощение, алкоголизм, сахарный диабет, хронические отравле-
ния, тяжелые травмы и операции, онкологические болезни, почечная недостаточ-
ность, системная волчанка, талассемия, хронические гранулематозные болезни, ли-
хорадка Денге, употребление напитков из кавы и др.
В специальной литературе на равных правах существуют две классификации —
первая построена целиком на скорости течения болезни (молниеносная, острая, по-
дострая и хроническая формы), вторая базируется на анатомических проявлениях
(септическая, септикопиемическая, местная, включая легочную). В данной работе
используется первая классификация клинических форм мелиоидоза.
При выраженных клинических формах с определенным моментом заражения,
например, при внутрилабораторных авариях или фиксированной травме в районах с
высокой степенью обсемененности В. pseudomallei, продолжительность инкубацион-
ного периода составляет 3—4 сут.
Для молниеносной (сверхострои) формы мелиоидоза характерно бурное начало, рез-
кий подъем температуры, сильные головные и мышечные боли, одышка. У больных
всегда нарушено психическое состояние, они нередко поступают в клинику уже с по-
терей сознания, в бреду. Понос и рвота приводят к обезвоживанию и судорогам. Объ-
ективно — в легких множественные хрипы разного типа, печень и селезенка слегка
увеличены, лейкоцитоз с нейтрофильным сдвигом влево. Болезнь стремительно про-
грессирует, и при явлениях коллапса больной погибает в течение 2-4 сут, а нередко и
через несколько часов после поступления в стационар. Окончательный диагноз
в подавляющем большинстве случаев определяется после смерти на основании выде-
ления из крови и внутренних органов культуры В. pseudomallei.
Ничего общего по своей клинической картине, кроме скорости течения, не име-
ет с молниеносной формой мелиоидоза так называемый «синдром внезапной смерти».
При этой форме практически не выявляются про, фомальные симптомы, за несколько
часов до смерти могут быть жалобы на головные боли, а сама смерть наступает ночью,
3.6 Мелиоидоз
801
окружающие слышат только явления удушья: хрипы и стоны внезапно умирающего
человека. На вскрытии отмечаются лишь некоторые отклонения от нормы со сторо-
ны почек, без особой патологии — легкие и сердце. У больных отмечаются повышен-
ные титры при постановке РПГА с мелиоидозным диагностикумом.
В клинической практике встречается острая септицемическая форма мелиоидоза.
Подобные больные попадают в стационар с диагнозом сыпного или брюшного тифов,
малярии, чумы, спирохетозов, вирусных лихорадок. Болезнь представляет собой симп-
томокомплекс тяжелого, быстро развивающегося сепсиса. Начинается внезапно с
сильного озноба и резкого повышения температуры до высоких значений (40 40,5 °C).
Лихорадка в первые 4—6 сут болезни постоянная, в последующие дни с небольшими
ремиссиями. Появляются мучительные головные боли, артралгии, миалгии, тошно-
та, часто рвота общая слабость, развивается нарушение сознания с возбуждением или,
напротив, с угнетением и бредом.
На 5—6 сут болезни на коже больного может появляться пустулезная сыпь, иногда
с геморрагическим содержимым. Элементы сыпи содержат вязкий гной, воспалитель-
ный ободок вокруг пустул отсутствует. Пустулы, нередко напоминают таковые при
натуральной оспе, но ладони и подошвы больного от сыпи всегда свободны, что для
натуральной оспы нехарактерно. Высыпания сопровождаются обычно лимфаденита-
ми, чаще шейными и подмышечными.
Состояние больного быстро ухудшается в случае присоединения рвоты и поноса.
Как правило, подобная клиника во времена отсутствия эффективных химиопрепара-
тов заканчивались летальным исходом в течение первых 10 сут. У больных этой фор-
мой мелиоидоза на фоне высокой температуры отмечаются боли в мышцах и суста-
вах, а также иные явления, связанные с формированием множественных абсцессов во
внутренних органах. Очень часто на первый план выступает легочная патология, по-
этому клиницисты нередко выделяют ее в отдельную легочную форму мелиоидоза.
Острый и подострый легочный мелиоидоз может быть первичным, т. е. развиться в
результате проникновения В. pseudomallei непосредственно в легкие, либо вторичным
вызванным гематогенным заносом возбудителя болезни в легкие у пациента с другой
локализацией первичного фокуса болезни. Первичные случаи острого/подострого
легочного мелиоидоза в Австралии составляют до 83 % легочного мелиоидоза, вторич-
ные — до 17 %. Для больных с вторичным легочным мелиоидозом характерно пора
жение нижних долей легких и мультилобарное течение пневмонии. У больных с пер-
вичным легочным мелиоидозом такой четкой зависимости не установлено. Две трети
случаев хронического легочного мелиоидоза были связаны с вовлечением в процесс
верхних долей легких (Meumann Е. М. et al., 2012).
Основные симптомы острого/подострого легочного мелиоидоза — лихорадка
(температура 39°C), кашель, потеря веса, боли в груди, кровохарканье. При лабора-
торном обследовании — лейкоцитоз (15—20 тыс/мм), гематокрит превышает 40 %.
Рентгенологическое обследование позволяет обнаружить пневмонические очаги,
абцессы, каверны, экссудативный плеврит (рис. 3.74)
Если у больных с острым/подострым легочным мелиоидозом серологические ре-
акции имеют диагностические тигры более чем в 95 % случаев, го выделение В. pseudo-
mallei из мокроты и из крови явление редкое. По данным D. Huis et al. (2006), только у
348 из 508 (68 %) пациентов с рентгенологически подтвержденным легочным мелиои-
дозом удалось обнаружить наличие В. pseudomallei в слюне. Несмотря на длительное
комбинированное лечение антибиотиками и сульфаниламидами летальность среди
стационарных больных легочным мелиоидозом достигает 50 %. У многих пациентов,
802
Часть 3. Частная неправильная эпидемиология
Рис. 3.74. Рентгенограмма (А) и компьютерная томограмма (Б) грудной клетки
пациента с билатеральной мелиоидозной пневмонией и большим абсцессом в левом
легком. По Е. М. Meumann et al. (2012)
выписанных в состоянии клинического выздоровления, может развиться рецидив ме-
лиоидоза. Наличие В. pseudomallei в слюне больных с легочным мелиоидозом рассмат-
ривается D. Huis et al. (2006) как плохой прогностический признак.
Бактериологическая диагностика, особенно post mortem, положительна почти в
100 %. Культура В. pseudomallei выделяется из крови, содержимого абсцессов, пустул.
В отдельных случаях самостоятельно, а в последнее время после введения химиопре-
паратов септицемическая форма может перейти в септикопиемическую.
Подострая форма мелиоидоза начинается подобно острой форме внезапно или с
прогрессивным нарастанием температуры и с такими же разнообразными симптома-
ми. Болезнь может протекать с ремиссиями и носить несколько затяжной характер.
Смертельные исходы нередки и отмечаются к концу 3-й или 4-й недели болезни.
Выздоровление может длиться очень долго (от нескольких месяцев до 4 лет) и в
этом случае, по-видимому, следует говорить о переходе подострой формы в хрониче-
скую. Подострая форма характеризуется дальнейшим развитием нагноительных про-
цессов во внутренних органах и различных тканях организма (легких, печени, селезен-
ке, почках, поджелудочной железе, надпочечниках, мочевом пузыре, простате, при-
датках яичников, лимфатических узлах, подкожножировой клетчатке, слизистых
оболочках, костной ткани и т. п.) (рис. 3.75 и 3.76, с. 804).
Образование абсцессов в различных органах сопровождается серозными, а затем
гнойными выпотами в разные полости, поэтому плеврит и перитонит бывают неред-
кими спутниками этой формы мелиоидоза. Симптомы и тяжесть течения болезни во
многом зависят от локализации поражения. Так же, как и при острой форме, это мо-
жет быть пневмония, гастроэнтерит, гепатит, пиелонефрит, панкреатит и т. п. Печень
и селезенка обычно слегка увеличены. Нарушается нормальная проницаемость сосу-
дов: у больных бывают носовые кровотечения.
Основными морфологическими элементами при мелиоидозе являются грануле-
мы и абсцессы. Гранулемы предшествуют асбцессам. При остром течении мелиоидоза
наблюдаются милиарные гранулемы. Их формирование начинается со скопления ней-
трофилов, мононуклеаров и эпителиоидных клеток, окруженных зоной гиперемии и
напоминают туберкулезные. В центре гранулемы происходят начальные стадии не-
кроза, вокруг которого появляется гистоцитарная реакция — признаки, характерные
3.6. Мелиоидоз
803
Рис. 3.75. Мелиоидозные абсцессы в поджелудочной железе. А. Осевое томографи-
ческое изображение, показывающее мультилокулярные панкреатические абсцессы
(кружок) в теле поджелудочной железы. Б. Осевое томографическое изображение,
показывающее множественные абсцессы (гипоэхогенные поражения), расположен-
ные в теле и хвостовой части поджелудочной железы (овал), тромбоз селезеночных
вен (длинная трубчатая гипоэхогенная структура), показанные белыми стрелками;
селезеночные микроабсцессы (пунктирный круг). Последующее аналогичное иссле-
дование, выполненное в ходе лечения пациента, показало разрешение абсцессов
и реканализацию селезеночных вен. По Chong Vui Heng et al. (2010)
для ранних изменений при мелиоидозе. Воспалительный процесс приводит к казеоз-
ному или гнойному расплавлению некротических очагов в центре гранулем. При за-
тяжном течении мелиоидоза в центральной части гранулемы завершается казеозный
или гнойный некроз с расположенными по периферии очага эпителиоидными клет-
ками и грануляционной тканью. По мере развития болезни гранулемы могут увели-
чиваться, превращаться в абсцессы, в результате чего ткань органов становится ячеи-
сто-изрешеченной. В итоге образуются обширные казеозно-некротические очаги с
последующим формированием абсцессов, содержащих гной.
При вскрытии трупов больных, погибших от острых и подострых форм мелиои-
тоза, почти постоянным признаком болезни является наличие отдельных или мно-
жественных абсцессов в различных органах. Абсцессы иногда сливаются в более круп-
ные элементы. Часто они встречаются в печени, селезенке, лимфатических узлах,
легких, почках и надпочечниках, кишечнике, мочевом пузыре. Изредка находят некро-
тические изменения в сердце. Почти никогда подобные изменения не обнаруживали
нив головном, ни в спинном мозгу, что свидетельствует о неспособности В. pseudomallei
проникать непосредственно в ЦНС. Довольно часто гнойные очаги встречаются на
коже (иногда в виде пустул), в подкожножировой клетчатке, мышцах и костях.
Гной внутри абсцессов обычно белого цвета и вязкой консистенции, но в печени
имеет зеленоватую или желтоватую окраску. В подкожножировой клетчатке слияние
многих мелких абсцессов может привести к образованию флегмон. Гной иногда по-
падает в бронхиолы из абсцессов через отверстия в слизистой оболочке. В больших
бронхах и трахее наблюдают изъязвления и гнойное воспаление слизистой. Печень и
селезенка увеличены, надпочечники гипертрофированы, в кишечнике часто отмеча-
ют воспаление слизистой оболочки с сильной инфильтрацией стенки нейтрофилами,
моноцитами и лимфоцитами, а иногда и образование эрозий и даже язв.
Хронический мелиоидо* отличается от острой и подострой форм болезни продолжи-
тельное гью течения (от 4 нед до 6 лет). По-видимому, в эндемических по мелиоидозу
804
Часть 3. Частная неправильная эпидемиология
Рис. 3.76. Мелиоидозные абсцессы в печени и селезенке. А. Компьютерная томо-
грамма брюшной полости 31-летней женщины. Обнаружен крупный абсцесс печени
(7х 13 см), имеющий ячеистое строение и «знак ожерелья» («necklace sign», стрелка).
Б. Компьютерная томограмма брюшной полости 27-летней женщины. Обнаружены
два печеночных абцесса (стрелки), имеющих сотовое строение. Размер крупного
абсцесса — 3,4x3,2 см, маленький абсцесс — 2,6x2,0 см. В. Компьютерная томог-
рамма брюшной полости: обнаружен печеночный абсцесс 2x2 см (стрелка) и абсцесс
селезенки размером 2x2 см (острие стрелки) ячеистого строения. Г. Компьютерная
томограмма брюшной полости 27-летней женщины. В печени и селезенке обнару-
жены микроабсцессы. Наибольший из них 5x7 см имеет ячеистое строение и «знак
ожерелья» (стрелка). По Р. Apisarnthananarak et al. (2006)
дозу регионах такая форма болезни распространена. Клиническое течение хрониче-
ского мелиоидоза разнообразно и зависит от локализации нагноительных процессов.
Иногда общее состояние больного длительное время остается удовлетворительным.
Но чаще бывают обострения с подъемом температуры, изнуряющей лихорадкой и
тяжелыми осложнениями, приводящими к смертельному исходу. Больные хрониче-
ским мелиоидозом обычно впервые попадают в клинику для хирургического лечения
абсцессов легких, печени, почек, остеомиелита, флегмон, подкожных абсцессов и т. п.
Хронический мелиоидоз кожи по своему упорному течению и тенденции к слиянию
отдельных элементов напоминает поражения кожи при сифилисе и туберкулезе.
В образовании абсцессов при хронической форме инфекции, кроме В. pseudomallei,
могут принимать участие различные условнопатогенные микроорганизмы. Но в це-
лом эта форма болезни рассматривается многими исследователями как более благо-
приятная для лечения, чем описанные выше острая и подострая формы.
Патологоанатомические изменения в органах при хроническом мелиоидозе харак-
теризуются теми же признаками, что и при его подостром течении. На вскрытии об-
наруживают мелкие, сливающиеся между собой абсцессы во внутренних органах,
3.6. Мелиоидоз
805
костях и под кожей. Не редки остеомиелит с образованием свищей, воспаление лим-
фатических узлов с образованием милиарных узелков или геморрагий.
Латентная (инаппарантная) форма мелиоидоза является той самой «бомбой замед-
тенного действия», из-за которой в США и ряде других стран военнослужащим, при-
бывающим из Юго-Восточной Азии, делается специальная отметка в медицинской
карточке. Суть в том, что у лиц, находившихся в служебных или туристических поезд-
ках в эндемических районах, в самые различные сроки, иногда и через 60 лет, возможно
возникновение инфекции под влиянием снижения иммунного статуса. К подобного
рода бессимптомным носителям инфекции относятся практически все лица, имею-
щие диагностические титры в РПГА, а таковых насчитывается от 1 до 30 % лиц в стра-
нах Юго-Восточной Азии и отдельных территориях Австралии, Африки, Китая, Ла-
тинской Америки. Потенциально больны мелиоидозом миллионы людей, в это число
входят и сотни тысяч людей из Европы и Северной Америки, посещавшие в разное
время эти страны.
Осложнения при мелиоидозе в том случае, если заболевшие остаются жить, зави-
сят от многих факторов, включая тяжесть клинического течения инфекции, локали-
зацию и интенсивность деструктивных процессов, сроки и выбор средств лечения, а
также наблюдение и уход за больным после его выздоровления.
К числу тяжелых осложнений относятся сливные гнойнички и абсцессы во внут-
ренних органах с обширным разрушением паренхимы и стромы органа; перерожде-
ние мышцы сердца; нарушение выделительной функции почек; нарушения опорно-
го аппарата в результате разрушения суставов; остеомиелит с множественными
секвестрами; вторичная инфекция, вызванная отдельными условнопатогенными мик-
роорганизмами или ассоциациями этих микроорганизмов; кахексия.
При экстрапулъмоналъных формах болезни в инфекционный процесс вовлекаются
мочеполовая, нервная, лимфатическая, опорно-двигательная, кровеносная системы.
Соответственно меняются области поражения, варьирует и доминирующая симпто-
матика. Для всех вариантов остается общим волнообразная лихорадка, слабость, не-
рвные расстройства, потеря веса. Абсцессы почек, простатиты, циститы, эпидидими-
ты отмечаются у почти 35 % больных острыми и подострыми формами мелиоидоза.
Лабораторные исследования состояния функции почек (содержание креатина,
удельный вес мочи, баланс электролитов) имеют реальное прогностическое значение
при проведении лечебных мероприятий.
Гнойный паротит может быть отнесен к распространенным формам мелиоидоза.
Во всех случаях паротит завершался формированием абсцесса. Характерными ослож-
нениями были свищи в слуховой канал, паралич лицевого нерва. Болезнь протекает
относительно доброкачественно, гематогенная диссеминация с вовлечением в про-
цесс других органов бывает в единичных случаях. Прогноз при правильном хирурги-
ческом и химиотерапевтическом лечении благоприятен.
Кожный мелиоидоз встречается в 13-24 % случаев зарегистрированного мелиои-
доза, делится на первичный и вторичный. Подавляющее большинство пациентов с
первичным кожным мелиоидозом в Северной Австралии имели один участок пора-
жения в виде язвы размером от нескольких миллиметров до нескольких сантиметров.
У них не выявляли бактериемию, болезнь не заканчивалась смертельным исходом.
Однако в Южной Азии такая форма болезни ассоциируется с некротизирующими фас-
циитами, сепсисом, внутренними абсцессами. Вторичный кожный мелиоидоз пред-
ставлен в основном пустулезной сыпью. Развивается при септицемической форме
мелиоидоза как результат диссеминации В. pseudomallei кровью (рис. 3.77).
806
Часть 3. Частная неправильная эпидемиология
Рис. 3.77. Кожный мелиоидоз. А -И. Первичный кожный мелиоидоз.
К-М. Вторичный кожный мелиоидоз. По К. В. Gibney et al. (2008)
Для большинства остальных форм мелиоидоза (по локализации) характерно за-
тяжное течение с рецидивами в непредсказуемые сроки. Прогноз в таких случаях за-
висит от сопутствующих болезней или событий, приведших к манифестации процесса.
Клиническая картина мелиоидоза при искусственном инфицировании. Мелиоидоз у
лабораторных животных, сраженных подкожно, характеризуется следующими осо-
бенностями. При остром характере болезни на месте введения В. pseudomallei образу-
ется плотная припухлость как результат гиперемии и отека подкожной клетчатки.
Затем возникают абсцессы и некроз ткани, в ряде случаев со вскрытием наружу в виде
свищей. В инфекционный процесс активно вовлекаются региональные лимфатичес-
кие узлы, в которых образуются нагноения и некрозы, наблюдается их j величение, а
иногда и гнойное расплавление.
Если гибель животных наступала на 4—5 сут, то заметных патологоанатомических
изменений во внутренних органах, как правило, не обнаруживали. При более длитель-
ном течении мелиоидоза происходила диссеминация инфекта. Кроме увеличения
кровенаполнения органов, в печени и селезенке формировались множественные гной-
3.6. Мелиоидоз
807
но-некротические узелки, часто сливающиеся между собой с образованием абсцес-
сов. В легких отмечали наличие очагов уплотнения серого или темно-красного цвета,
с расположенными под плеврой некротическими узелками. При хроническом тече-
нии мелиоидоза имела место уже описанная выше патологоанатомическая картина,
но с более выраженными гнойно-некротическими проявлениями в виде абсцессов и
гистолиза тканей. Больные животные заметно теряли в весе.
Экспериментальный легочный мелиоидоз у животных имел признаки, характерные
для септико-пиемического процесса, за исключением белых крыс, у которых острая
инфекция протекала как септицемия. К числу отличительных особенностей легочного
мелиоидоза следует отнести активное внедрение возбудителя в легочную ткань, пер-
вичный характер и быстрое развитие пневмонии, наступление генерализации инфек-
ционного процесса в короткие сроки, избирательное поражение печени и селезенки.
Диагностика искусственного поражения возбудителем мелиоидоза. Общие принци-
пы такой диагностики те же, что описаны в разд. 3.1 для поражения возбудителем си-
бирской язвы. «Неправильная эпидемиология» мелиоидоза может иметь ра шичные
проявления. Природных очагов возбудителя мелиоидоза на территории России нет.
Любой случай мелиоидоза, выявленный срели россиян, не выезжавших в регионы, эн-
демичные по мелиоидозу (см. рис. 3.72), либо не имевших отношения к работе с воз-
будителем мелиоидоза в лаборатории, можно рассматривать как «неправильную эпи-
демиологию болезни».
В то же время надо учитывать ограниченность имеющихся знаний по экологии
В. pseudomallei. В связи с потеплением климата, границы природных очагов мелиои-
доза могут расшириться на север, возможна активизация собственных природных
эчагов мелиоидоза, ранее неизвестных. Поэтому при диагностировании даже единич-
ного заболевания мелиоидозом, необходимо провести широкие сероэпидемические
исследования среди людей, проживающих на той территории, где этот случай был за-
регистрирован. Одновременно целесообразно провести ретроспективное исследова-
ние историй болезни в хирургических и инфекционных стационарах на предмет вы-
явления случаев инфекции с подобной клиникой, уделяя особое внимание случаям
болезни, осложнившимся септицемией, вызванной Burkholderia, которую идентифи-
цировали как синегнойную палочку.
Цаже в регионах, эндемичных по мелиоидозу, болезнь проявляет себя единичны
ми случаями среди людей, занятых сельским хозяйством и, как правило, среди забо-
левших мелиоидозом преобладают люди с хроническими болезнями. Поэтому любые
групповые заболевания мелиоидозом среди здоровых людей, также надо рассматри-
вать как «неправильную эпидемиологию» болезни и исключать возможность их умыш-
ленного инфицирования.
Информации по «невозможным клиническим формам мелиоидоза» в открытой
литературе мало. Мне не удалось найти описаний экспериментов по заражению при-
матов как мелкодисперсными аэрозолями В. pseudomallei, так и парентерально. По
данным, приведенным выше (см. «Клиническая картина мелиоидоза при искусст-
венном инфицировании»), можно предполагать, что ингаляционное поражение, по
крайней мере, в течение первых 3—5 сут после экспонирования человека к мелкодис-
персному аэрозолю возбудителя мелиоидоза, будет захватывать нижние отделы легких,
что при ретроспективном исследовании можно установить по ранним рентгенограм-
мам больного. Однако из-за высокой способности к диссеминации и пневмотропно-
сти возбудителя мелиоидоза эта фаза болезни будет кратковременной. Она перейдет
в септическую или септикопиемическую с поражениями печени, селезенки и диссе-
808
Часть 3. Частная неправильная эпидел,иология
минированной формой легочного мелиоидоза. При чрескожном заражении Б. pseudo-
mallei в естественных условиях первичного фокуса, как при сапе, не формируется.
После подкожного введения В. pseudomallei в участке введения возникают абсцессы и
некроз ткани, и у пациента необходимо выяснить кто, когда и при каких обстоятель-
ствах делал ему инъекцию. Необходимо учитывать анамнез пациента, зависимость
от наркотиков, вводимых инъекционно, и его социальный статус.
Молекулярные методы исследования (например, полногеномное сиквиенс-типи-
рование) мало что дадут для установления географического происхождения выявлен-
ного из тканей жертв биотеррористического акта (диверсии) штамма В. pseudomallei.
Однако их можно использовать для сопоставления со штаммами, обнаруженными в
лаборатории преступников, в найденных диспергирующих устройствах, объектах
окружающей среды и других источниках.
Выделение генетически измененных штаммов возбудителя мелиоидоза от чело-
века или животных однозначно свидетельствует об искусственном происхождении
вспышки болезни.
Иммунопрофилактика мелиоидоза. Не разработана.
Лечение мелиоидоза. Army FM 8-284 (2000) приводит несколько схем лечения ме-
лиоидоза в зависимости ол типа и тяжести клинических проявлений болезни:
1) при тяжелой и/или септической форме болезни порядок применения анти-
биотиков следующий: больному дается цефтазидим, 120 мг/(кг сут) тремя дробными
дозами в комбинации с ТМП/сульфамидом (ТМП, 8 мг/(кг сут) плюс сульфамид,
40 мг/(кг сут) четырьмя дробными дозами). Сначала препараты вводятся парентераль-
но в течение 2 нед, а затем — перорально в течение 6 мес.;
2) при локализованной клинической форме болезни применяется одна из следу-
ющих схем с продолжительностью лечения от 60 до 150 сут:
амоксициллин/клавуланат, 60 мг/(кг сут) в трех дробных дозах, перорально;
тетрациклин, 40 мг/(кг сут) в трех дробных дозах, перорально;
триметаприм/сульфамид (ТМП, 4 мг/(кг сут) и сульфамид, 20 мг/(кг сут) в трех
дробных дозах, перорально);
2) при локализованной клинической форме болезни с признаками умеренной или
слабой токсичности применяются следующие схемы введения антибиотиков: комби-
нируются два из вышеприведенных режимов в течение 30 сут, затем следует моноте-
рапия либо с амоксипиллином/клавуланатом, либо с ТМП/сульфамидом в течение
60—150 сут. При внелегочной гнойной инфекции антибиотикотерапия продолжается
от 6 до 12 мес. Показан хирургический дренаж абсцессов;
4) если по клинике (острая пневмония) и анализу мокроты подозревается чума,
то дополнительно показано применение стрептомицина.
В Австралии и Таиланде широко распространено использование внутривенного
введения высоких доз цефтриаксона — 2 г/сут (Cheng А. С., Currie В. J., 2005)
В National Hospital Antibiotic Guidelines (Ministry of Health Brunei Darussalam) ис-
пользуются сложные схемы лечения мелиоидоза, включающие сначала внутривенное
введение 1—2 г цефтазидима или внутривенное введение 1—2 г карбапенема. Препа-
раты вводят три раза в сутки на протяжении 4—6 нед, предпочтение отдается схеме с
цефтазидимом. Затем лечение продолжают, либо назначая пациенту комбинацию
амоксициллинклавуланиковой кислоты (amoxicillinclavulanic acid) 625 мг 2 раза/сут,
плюс доксициклин 200 мг/сут, ципрофлоксацин 500 мг 2 раза/сут; либо проводят мо-
нотерапию с котримоксазолом (сульфаметоксазол/триметаприм): 960 мг 2 раза/сут на
протяжении 6 мес. (Chong Vui Heng et al., 2010).
3.7. Ку-лихорадка
809
3.7. КУ-ЛИХОРАДКА
Микробиология возбудителя Ку-лихорадки. Факторы патогенности. Таксономия,
происхождение и молекулярная эпидемиология возбудителя Ку-лихорадки.
Экология и естественная эпидемиология возбудителя Ку-лихорадки. Восприим-
чивость к возбудителю Ку-лихорадки и инфицирующие дозы для животных и
людей. Иммунитет. Устойчивость возбудителя Ку-лихорадки во внешней среде.
Антибиотикорезистентность. Обнаружение генетически измененных штаммов
возбудителя Ку-лихорадки. Патогенез Ку-лихорадки. Клиническая и патолого-
анатомическая картина Ку-лихорадки у человека при естественном инфицирова-
нии. Клиническая картина Ку-лихорадки у животных при искусственном инфи-
цировании. Клиническая картина Ку-лихорадки у людей при искусственном
инфицировании. Диагностика искусственного поражения возбудителем Ку-лихо-
радки. Иммунопрофилактика Ку-лихорадки. Лечение Ку-лихорадки.
Ку-лихорадка (Q-fever; син.: Ку-риккетсиоз, пневмориккетсиоз, австралийский
Ку-риккетсиоз, коксиеллез, лихорадка скотобоен, болезнь Деррика-Бернета, бал-
канский грипп, среднеазиатская лихорадка, квислендская лихорадка, термезская ли-
хорадка) — острая сапрозоонозная инфекционная болезнь людей и животных, вызы-
ваемая Coxiella burnetii. Характеризуется внезапным началом с различной степенью вы-
раженности симптомов, обычно включающих ремитирующую лихорадку, озноб,
потливость, головную боль, миалгии. Смертельные исходы редки. Название Q-fever
дано болезни впервые описавшим ее австралийским исследователем Дж. Дерриком
(John Derrick) в 1935 г. Буква «Q» соответствует заглавной букве английского алфави-
та в слове «Query» — неясный, неопределенный. Распространена повсеместно.
Источниками С. burnetii для людей являются инфицированные клеши, не обрабо-
танные термически молоко и мясо, окоплодные воды и плацента больных животных,
контаминированый воздух. Заражение здоровых людей от больных случается редко и
эпидемического значения не имеет. Российские санитарно-эпидемиологические пра-
вила СП 1.3.2322-08 относят возбудитель Ку-лихорадки ко II группе патогенности. По
степени важности для национальной безопасности США возбудитель Ку-лихорадки
относится к биологическим поражающим агентам категории В. Статус потенциально-
го поражающего агента БО он приобрел благодаря низкой инфицирующей дозе для
людей —1—10 клеток, устойчивости к высушиванию и при переводе в аэрозоль (Вепеп-
son A. S., TigerttW. D., 1956; Ротшильд Д., 1966). Военными специалистами С. burnetii
рассматривается как инкапситирующий агент БО. В рамках военно-биологических
программ исследуется в США с 1950-х гг. (Ротшильд Д., 1966; Sidell R. et al., 1997).
С целью поражения людей наиболее вероятно применение С. ймгие/п посредством
использования линейных и многоточечных источников, и диверсионными (террори-
стическими) методами.
Микробиология возбудителя Ку-лихорадки. С. burnetii — грамотрицательная плейо-
морфная бактерия, являющаяся облигатным внутриклеточным паразитом простей-
ших организмов и клеток макрофагального ряда многоклеточных организмов.
До последнего времени считалось, что С. burnetii не растет на искусственных пита-
тельных средах. Для ее культивирования использовались культуры клеток, в которых
она повторяла размножение в клетках организма животных (мышиные эмбриональ-
ные фибробласты, клетки почек зеленых обезьян, культуры клеток клещей, макрофа-
гоподобные опухолевые клеточные линии J774 и P388D1) или желточные мешки кури-
810
Часть 3. Частная неправильная эпидемиология
ных эмбрионов (Васа О. G., Paretsky D., 1983). В 2009 г. A. Omsland et al. предложена
цистеиновая среда, подкисленная цитратом (acidified citrate cysteine medium, ACCM)1,
позволяющая выращивать коксиеллы вне клеток. В последующем авторы улучшили
среду и использовали ее для выделения С. burnetii из инфицированных тканей и полу-
чения генетически измененных трансформантов (Omsland A., et al., 2011).
При размножении в культурах клеток мышиных эмбриональных фибробластов и
макрофагоподобных клетках отмечен «ассиметричный» тип деления клеток. Возни-
кающие дочерние клетки становятся неравноценными. Одни из них по-прежнему
остаются инфицированными коксиеллами, тогда как другие — свободны от возбуди-
теля. Такой механизм обеспечивает персистенцию коксиелл в культуре клеток, по-
скольку зараженная клеточная культура не погибает. Осуществляется ли такой тип
деления клеток в организме теплокровных — неизвестно. Предположительно такой
особенностью взаимоотношений чувствительной клетки и коксиелл можно объяснить
хронизацию вызываемой ими инфекции у человека.
Плейоморфность С. burnetii во время инфекционного процесса проявляется нали-
чием среди вида трех различных по морфологии типов клеток, обнаруженных с по-
мощью электронной микроскопии: крупные клеточные варианты (large-cell variants,
LCV), маленькие клеточные варианты (small-cell variants, SCV) и маленькие плотные
клетки (small dense cells, SDC). Эти варианты отличаются друг от друга по морфоло-
гической структуре, антигенному составу, метаболической активности и по резистент-
ности к физическим и химическим факторам (рис. 3.78).
LCV-тип С. burnetii в оптический микроскоп выглядит как кокковидная или па-
лочковидная грамотрицательная бактерия размером от 0,25x0,5 мкм до 0,25х 1,5 мкм.
Характерен для стадии активного роста и размножения коксиеллы, и, по сути, явля-
ется ее вегетативной формой. У таких клеток диффузный хроматин, четко выражен-
ные внешняя и цитоплазматическая мембраны с ЛПС на поверхности клетки. Они
обладают большей метаболической активностью, чем клетки типов SCV и SDC, имею-
щих конденсированный хроматин (McCaul Т. Е et al., 1991).
Тип SDC — эндоспоры, обнаружены внутри LCV и выделены путем их лизиса.
Также они обнаружены в свободноживущих амебах (рис. 3.79 и 3.80, с. 812).
В лабораторных условиях С. burnetii подвержена изменчивости с образованием
форм или фаз, аналогичным гладким (1 фаза — экспрессия полной Л ПС-структуры)
и шероховатым формам (II фаза — экспрессия усеченного ЛПС) бактерий, как они,
например, образуются в процессе диссоциации у Enterobacteriaceae. Инфекционный
процесс у людей и животных вызывает С. burnetii в 1 фазе. Такие коксиеллы образуют
стойкую суспензию, не дают спонтанной агглютинации и не фагоцитируются в отсут-
ствие специфических антител. Среди отдельных штаммов отмечаются значительные
колебания вирулентности. В этой фазе клетки С. burnetii имеют поверхностный анти-
ген полисахаридной природы, извлекаемый органическими растворителями или три-
хлоруксусной кислотой (Здродовский П. Ф., Голиневич Е. М., 1972; Arrican-Bouvery N.,
RodolakisA., 2005).
На основании собственных наблюдений Т. Е McCaul и J. С. Williams (1981) пред-
ложили гипотезу, объясняющую участие трех различных по морфологии типов кле-
ток в цикле развития С. burnetii в эукариотических клетках (рис. 3.81, с. 812).
В процессе последовательных пассажей через куриные эмбрионы в геноме С. bur-
netii накапливаются спонтанно образующиеся мутации, приводящие к экспрессии
1 Подробно состав среды приведен в статьях A. Omsland et al. (2009, 2011).
3.7. Ку-лихорадка
811
Рис. 3.78. Типы клеток С. buruetii. А. На фотографии видны оба типа клеток.
LCV-тип — более крупные клетки со «светлой» цитоплазмой и отчетливым
ядром; SCV-тип — электронно-плотные овальные клетки меньшего размера.
Стрелкой показано деление клетки коксиеллы LCV-типа путем поперечно-
го деления с формированием септы. Полоса соответствует 0,6 мкм. Б. LCV-
и SCV-типы клеток С. burnetii при более высоком разрешении (обозначены
как L и S соответственно). Стрелкой показано формирование выпячивания
мембраны у клетки LCV-типа. SCV-тип клеток представлен небольшими
электронно-плотными образованиями. Полоса соответствует 0,2 мкм. В. По-
перечный разрез клеток SCV-типа при высоком разрешении. MI — мембран-
ная интрузия (membranous intrusion); CM — цитоплазматическая мембрана;
ОМ — наружная мембрана; DL — плотный слой; R — рибонуклеопротеин.
Полоса соответствует 0,2 мкм. Г. Продольный разрез клеток SCV-типа при вы-
соком разрешении. Отчетливо видны менее плотные периплазматическое
пространство и вещество ядра (нуклеоплазма). Полоса соответствует0,2 мкм.
По Т. Е McCaul, J. С. Williams (1981)
Рис 3.79. Электронно-микроскопическое изображение эндоспоры вну гри клеток
LCV-типа. А. Эндоспоры (Е) видны как электронно-плотные овальные образова-
ния. Полоса соответствует 0,4 мкм. Б. Эндоспора при более высоком разрешении.
Полоса соответствует 0,2 мкм. По Г Е McCaul, J. С. Williams (1981)
812
Часть 3. Частная неправильная эпидемиология
Рис. 3.80. Электронно-микроскопическое изображение С. burnetii внутри вакуоли
A. castellanii. Клетка С. burnetii LCV-типа — стрелка 1, спора — стрелка 2, внутрен-
няя поверхность мембраны вакуоли — стрелка 3. По В. La Scola, D. Raoult (2001)
Рис. 3.81. Схематическое изображение цикла С. burnetii в эукариотической клетке:
1 — цикл начинается с поглощения споры или SCV-типа коксиеллы фагоцитирую-
щей клеткой. При проникновении клетки SCV-типа в фаголизосому кислая среда
фаголизосомного пространства активирует метаболические процессы у С. burnetii',
2 — клетки SCV-типа начинают делиться; 3,4 — альтернативный механизм: клетки
SCV-типа дифференцируются в вегетативные клетки, т. е. в клетки LCV-типа; 5 —
происходит дальнейшая дифференциация клеток LCV-типа через стадию попереч-
ного бинарного деления (transverse binary division stage); 6 — альтернативный меха-
низм: стадия деления может совпасть со стадией дифференциации клетки в спору.
В результате эукариотическая ю.етка заполняется фаголизосомами, содержащими
большие количества С. burnetii на разных стадиях дифференциации (7, 8 и 9), 10 —
высвобождение споры из клетки LCV-типа в результате ее лизиса Созревание из
споры клетки SCV-типа. Повторение цикла. По Т. F. McCaul, J. С. Williams (1981)
3.7. Ку-лихорадка
813
клеткой усеченных форм ЛПС, утративших сахара О-антигена (Hackstadt Т et al., 1985).
Так же как и для шероховатых форм бактерий семейства Enterobacteriaceae, для С. bur-
netii фазы II характерна склонность к спонтанной агглютинации и агглютинации в
нормальной сыворотке. Они хорошо фагоцитируются в отсутствие антител, их ви-
рулентность значительно ниже, чем у коксиелл в I фазе (Здродовский П. Ф., Голине-
вич Е. М., 1972). Вызвать инфекционный процесс у иммунокомпетентного хозяина
они не могут (Moos A., Hackstadt Т., 1987).
Принято считать, что фазы взаимно обратимы и под влиянием пассажей на вос-
приимчивых животных II фаза переходит в I фазу (Arrican-Bouvery N., Rodolakis А.,
2005). Однако еще в конце 1960-х гг. А. Гениг (лаборатория П. Ф. Здродовского) на ос-
новании клонально-селекционного анализа культур показано, что С. burnetii I и II фазы
в действительности являются стойкими вариантами или мутантами данного вида. При
их выращивании в куриных эмбрионах или при их размножении в организме чувстви-
тельного животного происходит не переход одной фазы в другую, а селекционный
отбор варианта, находящего оптимальные условия размножения в данной среде (Здро-
довский П. Ф., Голиневич Е. М., 1972).
Факторы патогенности. У возбудителя Ку-лихорадки факторы патогенности изу-
чены плохо. Скорее всего, их в традиционном понимании как неких молекул, синте-
зируемых микроорганизмом для повышения своей агрессивности в многоклеточном
хозяине (токсины, ферменты, адгезины, инвазины), у С. bumeui нет, что подтвержда-
ется результатами расшифровки генома этой бактерии (см. Seshadn R. et al., 2003). По
мнению J. Shannon и R. Heinzen (2009), патогенность С. burnetii проявляется прежде
всего в ее способности инфицировать монону клеарные фагоциты в условиях in vivo.
Инфицированные клетки оказываются неспособными сдержать дальнейшее размно-
жение коксиелл, что ведет к развитию инфекционного процесса. Возможно, механизм
инвазии запускается некой оптимальной комбинацией молекул-лигандов на поверх-
ности С. burnetii первой фазы, взаимодействующих с рецепторами на поверхности
клетки-мишени. С. Capo et al. (1999) показали, что молекулы-лиганды поверхности
С. burnetii второй фазы взаимодействуют с иными рецепторами на поверхности моно-
цитов и макрофагов, чем С. burnetii первой фазы, в результате не запускается механизм
поглощения коксиеллы клеткой и последующая ее репликация. С. burnetii фазы I спо-
собна инфицировать дендритные клетки и реплицироваться в них без индукции их
созревания или продуцирования воспалительных цитокинов (рис. 3.82).
С. burnetii фазы II, наоборот, индуцирует быстрое созревание дендритных клеток
и выброс ими воспалительных цитокинов. Интересно, что единственный, подробно
описанный фактор вирулентности коксиел — полноразмерный ЛПС, также требуется
им для уклонения от стимуляции дендритных клеток. Эти данные указывают на новую
роль ЛПС как фактора вирулентности С. burnetii', заключающуюся в экранировании
микроорганизма от рецепторов дендритных клеток. В результате не происходит пре-
зентации антигенов коксиеллы Т-клеткам, и у инфицированного человека наруша-
ется формирование специфического иммунитета к возбудителю Ку-лихорадки, что
приводит к развитию персистирующей инфекции (Shannon J., Heinzen R., 2009)
1 Для ЛПС, как фактора вирулентности бактерий, основной считалась его эндотоксиче-
ская активность. Но у ЛПС С. burnetii она от 100 до 1000 раз оказалась меньшей, чем у ЛПС
Enteriobaeteriaceae. Поэтому до работы J. Shannon, R. Heinzen (2009) предполагали, что основ-
ная функция ЛПС коксиелл заключается в придании им резистентности к комплемент-опо-
средованному лизирующему действию сыворотки (см. Arrican-Bouvery N., Rodolakis А., 2005).
814
Часть 3. Частная неправа чьная эпидемиология
Рис. 3.82. Сканирующее электронно-микроскопическое изображение проникно-
вения С. burnetii фазы I (С.Ь) в дендритную клетку, производную моноцита. Полоса
соответствует 0,2 мкм. По J. Shannon, R. Heinzen <2009)
Таксономия, происхождение и молекулярная эпидемиология возбудителя ^-ли-
хорадки. Хромосома С. burnetii представляет собой кольцевую молекулу размером
1999275 п. о. Большинство изолятов С. burnetii содержат от одной до четырех плаз-
мид, названных QpHl, QpRS, QpDV и QpDG размером от 32 до 42 т. п. о. Плазмиды
включают общий «коровый» регион размером 25 т. п. о. Геном отдельных изолятов
С. burnetii включает последовательность, подобную плазмиде QpRS. IS-элементы у
собственных плазмид С. burnetii не обнаружены. Плазмидные типы ассоциированы
со специфическими геномными группами, но единства мнений среди ученых можно
ли считать плазмиды коксиелл, имеющими отношение к вирулентности, нет. Скорее
всего, эти плазмиды являются критическими. Особенности генома С. burnetii изо-
лята Nine Mile фазы I и плазмиды QpHl приведены в табл. 3.23.
Род Coxiella относится к классу у-протеобактерий (Gammaproteobacteria), входя-
щего в группу протеобактерий (Proteobacteria) — самую многочисленную группу бак-
терий, включающую как симбионты эукариот, так и большое число патогенных и ус-
ловнопатогенных микроорганизмов. С. burnetii представляет пока единственный
описанный вид коксиелл. Более 350 млн лет назад от рода Coxiella дивергировало мо-
нофилетическое семейство Rickettsiella, включающее виды, патогенные для насеко-
мых и ракообразных. С. burnetii эволюционно связана с такими патогенными цля людей
видами микроорганизмов, как F. tularensis и L. pneumophila (Cordaux R. et al., 2007).
Таксономические положение возбудителя Ку-лихорадки показано на рис. 3.83.
Для установления генетической связи между изолятами С. burnetii и проведения
молекулярно-эпидемиологических исследований используют следующие методы:
метод исследования полиморфизма длин рестрикционных фрагментов (restriction frag-
ment length polymorphism, RFLP) с разделением фрагментов ДНК электрофорезом в
пульсирующем поле. Позволил дифференцировать 80 изолятов С. burnetii, выделен-
ных от людей и животных в Европе, США, Африке и Азии (Jager С. et al., 1998);
метод исследования полиморфизма отдельных нуклеотидов (SNP). На основе сопо-
ставления десяти различных SNP метод позволил установить наличие не менее пяти
генотипов среди 40 изолятов С. burnetii, выделенных из клинических образцов и про-
дуктов питания во время вспышки Ку-лихорадки в 2007—2009 гг. на юге Гол гандии (до
4 тыс. заболевших) (Huijsmans С. et al., 2011);
3.7. Ку-лихорадка
815
Таблица 3.23
Особенности генома С. burnetii изолята Nine Mile фазы I*
Особенности генома Хромосома Плазмида QpHl
Размер, и. о. 1995275 37 393
Г+Ц-содержание, % 42,6 39,3
Стабильные рРНК 3 0
Стабильные тРНК 42 0
Гены, кодирующие белки
Количество генов, сходных с известными 1022 11
Количество генов, сходных с генами с неизвестной функцией 179 5
Количество генов, кодирующих гипотетические консервативные белки 200 1
Количество генов, кодирующих гипотетические белки, не имеющие сходства с известными 693 23
Всего 2094 40
Средний размер ORF, п. о. 849 736
% генов, кодирующих белки 89,1 78,8
♦ По Seshadri R. et al. (2003).
0,05
99
76
99
1001
100
95
59
70 Г
98 87|-
66
100 ।— Bartonella bacilliformis (М65249)
30 [ I— Bartonella quintana (M73228)
I-------Brucella abortus (XI3695)
IPOj Rickettsia conorii (L36105)
L Rickettsia prowazekii (M21789)
Orientia tsutsugamushi (L36222)
1001 — Ehrlichia phagocytophila (M73220)
93, Wolbachia spp. Av (AJ223238)
99 Wolbachia spp. Hb(AJ001603)
-----------П Wolbachia spp. Pm(AJ001607)
100|—wolbachia spp. Gp (U83090)
8% Rickettsiella armadillidii Av (Am490937)
100(1 RickettsiellaarmadillidiiHb(AM490938)
—------- 1 Rickettsiella armadillidii Pm (AM490939)
*- Rickettsiella grylli (U97547)
------Coxiella burnetii (М21291)
,------Legionella israelensis (Z32640)
100 '----Legionella pneumophilla (X73402)
-------- Francisella tularensis (L26084)
--------Acinetobacter hvoffii (U10875)
--------Pseudomonas aeruginosa (X06684)
----Buchnera aphidicola (M27039)
Escherichia coli (M24996)
- Xenorhabdus nematophilus (X82251)
—Arsenophonus nasoniae (M90801)
100
97
84
---------Mycoplasma pneumoniae (M29061)
100 ।----- Chlamydia psittaci (M13769)
'— Chlamydia trachomatis (U68443)
a
Рис. 3.83. Таксономическое положение возбудителя Ку-лихорадки поданным секвени-
рования 16S рРНК. Номер справа от видового названия микроорганизма соответствует
номеру его нуклеотидной последовательности в GenBank. По R. Cordaux et al. (2007)
816
Часть 3. Частная неправильная эпидемиология
многолокусный анализ варьирующих по числу тандемных повторов (MLVA). На основе
анализа распределения ЮУМТКвДНК 125 изолятов С. burnetii, полученных во время
той же вспышки Ку-лихорадки в Голландии от животных (вагинальные мазки, око-
лоплодные воды, плацента, молоко), было установлено 13 генотипов возбудителя Ку-
лихорадки, разделенных на два кластера (Roest Н. et al., 2011).
Экология и естественная эпидемиология возбудителя Ку-лихорадки. Н. И. Федоро-
вой (1968) было отмечено, что выделение основного источника и резервуара С. burnetii
и понимание механизмов ее циркуляции в природе затруднено из-за широкой пора-
женности этим микроорганизмом млекопитающих, птиц и огромного количества раз-
личных видов клешей. Приведение выше данных R Cordaux et al. (2007) о существо-
вании рода Coxiella еще задолго до появления млекопитающих и птиц говорит, что в
природе существует стабильный резервуар коксиелл, чем позвоночные организмы,
иммунная система которых может обрывать вызванный ими инфекционный процесс.
Для возбудителя Ку-лихорадки пока описана только одна экологическая ниша, где он
может поддерживаться неограниченно долго без участия позвоночных организмов —
простейшие (La Scola В., Raoult D., 2001), поэтому их можно считать первичным резер-
вуаром С. burnetii. Эта же экологическая ниша вмешает связанные с ним эволюционно
возбудители легионеллеза и туляремии. Территории, на которых С. burnetii поддер-
живается среди простейших, представляют собой первичные очаги Ку-лихорадки.
Вторичным резервуаром С. burnetii в природе являются более 60 видов диких мелких
млекопитающих (в основном грызуны), около 50 видов птиц и более 77 видов клешей
(в основном иксодовых и в меньшей степени — аргасовых, гамазовых и краснотелко-
вых), большинство из которых обитают на территории России. Инфекция у клещей
протекает бессимптомно и длительно, причем примерно у 25 видов установлена транс-
фазовая и трансовариальная (и даже трансспермальная) передача возбудителя Ку-
лихорадки, что, естественно, способствует ее длительному сохранению во вторичном
резервуаре. Все указанные носители выделяют риккетсии во внешнюю среду с мочой
и фекалиями. Испражнения инфицированных клещей Hyalomma asiaticum и Derma-
centor nutalli содержат жизнеспособные коксиеллы до титра 1010-1011 и сохраняют
свою инфекционность до 160 сут (Лобан К. М. с соавт., 2002). В результате, кроме
трансмиссивного пути, С. burnetii может передаваться ингаляционно (от инфициро-
ванных фекалиями и плацентарным отделяемым объектов внешней среды).
Согласиться с тем, что клещи «адаптированы» к С. burnetii, как утверждают
К. М. Лобан с соавт. (2002), нельзя из-за особенностей их иммунной системы. У чле-
нистоногих хорошо развита фагоцитарная система, однако для гуморальных факто-
ров их иммунной системы характерно отсутствие высокой специфичности, прису-
щей антителам позвоночных (Купер Э., 1980). Поэтому более вероятно то, что не
адаптация клещей к С. burnetii, а неспособность их иммунной системы выявлять ин-
фицированные коксиеллами фагоцитирующие клетки, приводит к длительной пер-
ситирующей инфекции.
Природные очаги Ку-лихорадки, являющиеся естественным источником С. burnetii
для людей, — это вторичные очаги, они существуют за счет постоянного обмена воз-
будителя болезни между клещами и их теплокровными прокормителями, играющи-
ми роль вторичного резервуара С. burnetii. Циркуляция С. burnetii в таких очагах обыч-
но описывается в виде схемы: клеши -> теплокровные -> клещи -> теплокровные (см.
у Федоровой Н. И., 1968; Лобан К. М. с соавт., 2002).
В настоящее время в России различают природные очаги Ку-лихорадки, связанные
с биоценозом лесных, лесостепных, степных, полупустынных, пустынных, ни жогор-
3.7. Ку-лихорадка
817
ных ландшафтов; и природно-антропургические, где в циркуляцию возбудителя болезни
включаются домашние животные.
Эпидемиологическое значение диких млекопитающих, птиц и клещей как непо-
средственных источников С. burnetii для человека, незначительно. В литературе име-
ются лишь отдельные указания на возможность заражения людей непосредственно в
природных очагах Ку-риккетсиоза. Поэтому в данной работе они не рассматривают-
ся, подробно о них можно прочитать в работе Н. И. Федоровой (1968).
Природно-антропургические очаги Ку-лихорадки возникают чаще всего в местах
существования инфекции в природе, когда сельскохозяйственные животные заража-
ются при нападении инфицированных клешей во время пастьбы. В таких очагах ре-
зервуаром и источником С. burnetii для человека являются домашние животные, в том
числе кошки и мелкий скот, особенно козы и овцы; меньше — лошади, мулы, ослы,
яки, верблюды, свиньи, олени, буйволы, зебу и др. Заражаются животные в природ-
ных очагах от инфицированных клещей, а в сельскохозяйственных очагах — при сов-
местном содержании с больными животными. Как и у других млекопитающих, инфек-
ция у домашних животных протекает чаще всего латентно, хотя может проявляться и
в виде легких лихорадочных острых или хронических процессов по типу бронхопнев-
моний, бронхита, мастита, абортов, ринита, конъюнктивита, офтальмии, гастрита,
перикардита и т. д., которые чаще остаются нераспознанными. Острый процесс в тече-
ние 2 мес. завершается не только выздоровлением, но и освобождением организма от
инфекции. Хронические формы болезни могут затягиваться на многие месяцы, вплоть
до 2 лет, часто напоминая бруцеллез. Больные животные выделяют С. burnetii во вне-
шнюю среду с испражнениями и молоком, а во время родов — с плацентой, что при-
водит к инфицированию ухаживающего персонала и лиц, употребляющих сырое (и
даже пастеризованное!) молоко от таких животных; заражаются также зоотехники и
ветеринарные врачи, принимающие роды у скота при отеле и ягнении (Лобан К. М. с
соавт., 2002).
Вспышки Ку-лихорадки могут возникнуть среди различных групп населения, не
имеющих отношения к животноводству, переработке сельскохозяйственного сырья
(шерсть, необработанные шкуры овец, хлопок и т. п.) и не употребляющих молоко и
молочные продукты. К. М. Лобан с соавт. (2002) их объясняют следствием ингаляци-
онного инфицирования. Причем источники и механизмы формирования инфици-
рующего аэрозоля могут быть неожиданным для врачей: инфицированный пух коз
(вспышка с 300 зараженных); шкуры овец (81 зараженный из числа работников овце-
водческого комплекса); рожающая кошка (до 80 членов семьи или лиц, оказавшихся
в этот момент в доме); отары овец, прогоняемые вблизи или на значительном рассто-
янии от альпийских деревень или же через сельские населенные пункты (до 415 че-
ловек в одном эпизоде).
И. К. Каракулов и В. Д. Борисов (1966) выделяют следующие типы естественного
инфицирования людей возбудителем Ку-лихорадки:
промышленно-производственный тип (обработка продуктов животноводства: моло-
ка, мяса, шкур, шерсти, пуха, щетины). Этот тип заражения С. burnetii распространен
у работников молокозаводов, молокосливных и приемных пунктов, убойных пунктов,
мясокомбинатов, меховых, кожевенных и шерстеобрабатывающих предприятий и др.
Входные ворота: наружные покровы, пищеварительный тракт и слизистые верхних
дыхательных путей;
сельскохозяйственно-животноводческий тип (обслуживание неблагополучных по
Ку-лихорадке животноводческих хозяйств с мелким и крупным рогатым скотом). Этот
818
Часть 3. Частная неправильная эпидемиология
тип естественного инфг пирования людей С. burnetii наблюдается среди ветеринарного
персонала, зоотехников, чабанов, пастухов, сакманщиков, стригалей, свинарок, до-
ярок, телятниц, скотников, конюхов, верблюдоводов, кролиководов и птицеводов.
Входные ворота: наружные покровы — кожные и слизистые (контактный путь зара
жения), пищеварительный тракт (алиментарный путь) и слизистые дыхательных пу-
тей (воздушный путь);
лабораторно-клинический тип (работа с живыми культурами С. burnetii, воспроиз-
ведение инфекции на экспериментальных животных, исследования инфицирован-
ных выделений крови и продуктов, оказание ветеринарной и медицинской помощи).
Лабораторное заражение может наступить у работников лабораторий, изучающих
С. burnetii’, возможно оно в ветеринарно-бактериологических лабораториях, в пище-
вых лабораториях при определении качества мясомолочных продуктов бруцеллезных
животных, в клинических и поликлинических медицинских лабораториях, а также у
работников ветеринарных клиник и лечебниц, медицинских лечебных учреждений
(больницы, клиники, диспансеры, роддома). Входные ворота — наружные покровы
и слизистые, пищеварительный тракт и слизистые дыхательных путей;
бытовой тип заражения (повседневные бытовые процессы: приготовление пищи
из инфицированных проду ктов, употребление их в недостаточно термически обрабо-
танном виде, содержание инфицированных С. burnetii домашних животных, появле-
ние зараженных домовых грызунов, возможность кормления детей инфицированным
грудным молоком, пользование инфицированной водой для питья и бытовых целей
и т. д.). Такой вид заражения может наблюдаться среди самых различных групп насе-
ления, независимо от пола, возраста и профессии. Входные ворота — наружные по-
кровы, пищеварительный тракт, а также слизистые верхних дыхательных путей.
Для лихорадки Ку характерна преимущественно весенне-летне-осенняя заболе-
ваемость. Чаще оолеют мужчины. Наибольшее число заболеваний приходится на
февраль — май, когда происходит массовый отел и окот животных, заболеваемость но-
сит непрофессиональный характер как на селе, так и среди городских жителей и ре-
гистрируется преимущественно у мужчин в возрасте 20—40 лет, которые чаще и долго
работают вне жилых и производственных помещений (Лобан К. М. с соавт., 2002).
Восприимчивость к возбудителю Ку-лихорадки и инфицирующие дозы для животных
и людей. Инфицирующая доза для человека и экспериментальных животных, варьи-
рует в пределах 1—10 клеток при любом пути введения. Признаком заболевания явля-
ется обычно повышение температуры, которое тлится от 1—5 сут до 2—3 нед. Нередки
случаи, когда инфекция протекает без температурной реакции и выявляется лишь се-
рологически. Люди более чувствите. [ьны к заражению С. burnetii, чем морские свинки,
так как дыхательный обмен у них в 100 раз больше чем у морских свинок. Данные по
определению минимальной инфицирующей дозы для морских свинок и людей при
ингаляционном экспонировании к С. burnetii приведены в табл. 3.24.
W. D. Tigertt et al. (1961) приведены исследования обратной зависимости между
дозами С. burnetii, вводимыми респираторным путем морским свинкам и людям, и ин-
кубационным периодом. На рис. 3.84 показаны три совпадающие прямые, отражаю-
щие результаты этого исследования.
Американские штаммы возбудителя Ку-лихорадки более вирулентны для лабора-
торных животных, чем штаммы из других геогрфических регионов, в особенности
штамм Найн-Майл (Nme Mile). Отечественные штаммы по вирулентности для лабо-
раторных животных и по другим свойствам близки к австралийским и европейским
вариантам этого микроорганизма (Каракулов И. К , Борисов Ь. Д., 1966).
3 7. Ку-лихорадка
819
Таблица 3.24
Минимальные инфипирумицне дозы для морских свинок и людей
при ингаляционном экспонировании к С. burnetii*
1 Доза С. burnetii (инфекционные единицы)** Сероконверсия у морских свинок (позитивные/ скспонированные) Время до начала лихорадки (сут) Сероконверсия у человека (позитиьные/экспони- рованные) Время до начала лихорадки (сут)
1 0/29 12,5 0/2 17
10 1/26 Не показано 4/5 (два случая субклини- ческого течения инфекции) 16
102 4/24 11 7/8 15
103 22/27 9 4/5 13,5
104 28/28 8,5 4/4 10,5
105 24/24 7 2/2 10
*W. D. Tigertt et al. (1961).
** Использовался штамм AD, полученный в Калифорнии из молока и пассированный на
трех морских свинках и восьми куриных эмбрионах. MMAD — 1 мкм.
о —।—I—I—।—।—I—।—।—।—I
1 ю ю2 ю3 ю4 ю5 ю6 ю7 ю8 го’ го10
Инфицирующая доза
Рис 3.84 Средние инкубационные периоды при заражении различными дозами
С. burnetii (штамм AD). Инфицирующая доза в единицах. Одна единица соответ-
ствует количеству коксиелл, которое i ызывает серо югические реакции у 50 %
морских свинок. По W D. Tigertt et al. (1961)
Установлена связь отдельных изолятов С. burnetii с разными клиническими фор-
мами болезни. Например, штамм Priscilla вызывает хроническую форму Ку-лихорад-
ки, штамм Nine Mile — острую (Yeaman М. R. et al., 1987).
Иммунитет. В настоящее время установлена роль клеточного звена иммунной си-
стемы в предотвращении и контроле над инфекцией, вызванной С. burnetii. Роль же
гуморального звена в контроле над инфекцией остается недоказанной. Антитела к
антшенам 11 фазы можно обнаружить в крови человека через несколько дней после
его инфицирования С. burnetii. Сначала появляются иммуноглобулины М (IgM), за-
тем IgA и IgG. IgM к антигенам I фазы появляются в крови у пациен та уже на стадии
выздоровления. В низких концентрациях их можно обнаружить через 2 года после за-
вершения острой формы болезни. У пациентов с хронической формой Ку-лихорадкой
обычно не удается обнаружить в крови специфические IgM, но у них всегда опре-
820
Часть 3. Частная неправильная эпидемиология
деляются высокие уровни IgG- и IgA-антител к антигенам I фазы. Специфические
антитела играют важную роль в поглощении С. burnetii макрофагами и полиморфо-
ядерными лейкоцитами. Но здесь скорее наблюдается феномен антителозависимого
усиления инфекции, а не протективный эффект. В условиях in vitro показано, что
микроорганизмы I фазы разрушаются макрофагами значительно хуже, чем микроор-
ганизмы II фазы. При добавлении антител к культурам клеток, коксиеллы могут раз-
множаться в макрофагах, причем антитела к микроорганизму I фазы стимулируют их
размножение более эффективно, чем антитела к микроорганизму II фазы. Эти данные
показывают, что антитела к С. burnetii играют важную роль в развитии инфекционно-
го процесса, но каким образом они оказывают протективное действие, не ясно. Ни
для одного типа специфических антител не установлен защитный титр (Reimer L. G.,
1993; Shannon J. G., Heinzen R. A., 2009). Переболевшие лихорадкой Ку приобретают
иммунитет, как правило, прочный и длительный (Лобан К. М. с соавт., 2002).
Устойчивость возбудителя Ку-лихорадки во внешней среде. Коксиеллы весьма устой-
чивы во внешней среде, а также к различным физическим и химическим воздействиям.
Они длительно сохраняются в сухом виде и во влажных субстратах. По данным, обоб-
щенным К. М. Лобаном с соавт. (2002), облучение ультрафиолетовыми лучами с рас-
стояния 1 м от лампы не убивает их в течение 5 ч, а температуру нагревания до 90 °C они
выдерживают более 1 ч. Показана возможность выживания риккетсий Бернета в лио-
фильно высушенных культурах не менее 8— 10 лет. На сухой поверхности они сохраня-
ются в течение 15 сут. В желудочном соке с нормальной и повышенной кислотностью
выживают и сохраняют вирулентность в течение 2 ч, в высохшей моче и крови заражен-
ных животных — от нескольких недель до полугода, в сухих фекалиях клещей — 586 сут.
На хлопчатобумажной ткани (белье, спецодежда), в том числе и при высушивании ее,
жизнеспособность и вирулентность риккетсий сохраняются до 35—40 сут, на линоле-
уме, деревянных, стеклянных и железных предметах 16—17 сут. В стерильной водопро-
водной воде риккетсии выживают свыше 160 сут, причем одинаково в условиях низ-
кой и комнатной температур, в стерильном коровьем молоке при комнатной темпе-
ратуре — 125 сут, а при 4° С — 273 сут. При пастеризации молока снижается количество
содержащихся в нем риккетсий, но не обеспечивается полная < терилизация. Послед-
няя достигается лишь при кипячении не менее 10 мин. В сыворотке от творога и дру-
гих молочнокислых продуктах риккетсии остаются жизнеспособными в течение 30 сут;
примерно до этого же срока сохраняется возбудитель и в свежем мясе зараженных жи-
вотных при температуре 4—8 °C, а в замороженном виде и более длительный срок; в
соленом мясе, при сохранении его в 10 %-ном растворе поваренной соли в условиях
ледника, — 150 сут. В масле, приготовленном из инфицированного молока, при хра-
нении его в рефрижераторе возбудитель сохраняется не менее 41 сут. Все эти данные
говорят о возможности заражения человека через инфицированные пищевые продук-
ты и воду (при хлорировании воды возбудитель гибнет).
С. burnetii выживает даже при продолжительном воздействии дезинфицирующих
растворов. В течение суток — при воздействии 1 % раствора карболовой кислоты или
4 сут — 0,5 % раствора формалина. При кипячении риккетсии Бернета гибнут в тече-
ние 1 мин, при действии 3—5 % раствора фенола или 2 % раствора формальдегида, 5 %
раствора перекиси водорода и 10 % раствора едкого натрия — через 1—5 мин; 3 % ра-
створа хлорамина инактивирует С. burnetii через 5 мин, 2 % раствора хлорной извести —
через 15 мин. В 70 %-ном спирте они гибнут через 1 мин При камерной обработке па-
ровоздушной смесью при 89—90rC С. burnetii инактивируется в течение 45 мин; паро-
формалиновой смесью — в течение 3,5 ч при 58—60°С.
3.7. Ку-лихорадка
821
Жизнеспособность С. burnetii в шерсти при температуре в 4—6 ' сохраняется до
16 мес., при температуре в 15—20 ’С — до 4 -9 мес., а при температуре 34—36°C возбу-
дитель оказывался жизнеспособным в течение 1—2 мес Высокая устойчивость рик-
кетсий Бернета во внешней среде способствует заносу их при транспортировании вме-
сте с шерстью или хлопком на далекие от эпизоотических районов расстояния.
Антибиотикорезистентность. Для терапии острой формы Ку-лихорадки рекомен-
дуется использовать антибиотики, обладающие бактериостатическим действием; для
течения хронической формы болезни — бактерицидным (Raoult D., 1993). С. burnetii
на искусственных питательных средах не растет. Определение бактерицидной и бак-
териостатической активности в условиях in vitro проводится в опытах на куриных
эмбрионах и культурах клеток. По данным A. J. Spicer et al. (1981), полученных в опы-
тах на куриных эмбрионах, высокую бактериостатическую эффективность в отноше-
нии С. burnetii показали тетрациклин и его аналоги, а также котримоксазол и рифам-
пицин. Один тестируемый штамм (Cyprus) оказался резистентным к тетрациклину.
D. Raoult et al. (1991) изучили бактериостатическое действие антибиотиков в от-
ношении 13 штаммов С. burnetii, используя HEL-клетки. По их данным, в этой экспе-
риментальной модели наибольшую бактериостатическую активность показали кот-
оимоксазол, рифампицин, доксициклин, тетрациклин, миноциклин, спарфлоксацин
(sparfloxacin) и хинолоны PD 127, 391 и PD 131,628. Из 13 штаммов чувствительными
к офлоксацину были 12, к пефлоксацину — 10, к хлорамфениколу — 10, цефтриаксо-
ну — 4, фузидиковой кислоте — 6. К эритромицину 7 из 13 штаммов показали проме-
жуточную чувствительность, 6 оказались резистентными. Амоксиллин и амикацин
смазались не эффективными для всех исследуемых штаммов.
Maurin М. et al. (1992), используя культуры клеток Р388 (линия клеток макрофа-
гов мышей), продемонстрировали, что пефлоксацин, доксициклин, ципрофлоксацин
и рифампицин не обладают бактерицидным действием в отношении С. burnetii, так
как она выживает в кислой среде лизосом. Используя амантадин, хлоридин и NH4C1,
они увеличили pH среды лизосом до 5,3, 5,7 и 6,8 соответственно. Рифампицин не
проявил бактерицдный эффект ни при одной из достигнутых величин pH. Доксипик-
ин проявил бактерицидное действие при pH 5,7 и 6,8; пефлоксацин — при pH 6,8.
Авторы сделали вывод, что величина значения pH лизосом является критической для
эффективного применения антибиотиков при лечении Ку-лихорадки, и схема ее ле-
чения должна предполагать включение либо лизосомотропных алкалирующих аген-
тов, либо кислотоустойчивые антибиотики. При лечении 20 больных Ку-лихорадкой
они использовали сочетание хлоридина (900 мг/сут) и тетрациклина (200 мг/сут), и в
течение года не отметили рецидивов болезни.
Обнаружение генетически измененных штаммов возбудителя Ку-лихорадки. Подхо-
ды к обнаружению генетически измененных штаммов возбудителя Ку-лихорадки, в
основном, те же, что и для обнаружения генетически измененных штаммов возбуди-
телей сибирской язвы и чумы (см. разд. 3.1 и 3.2). Учитывая совокупность поражаю-
щих свойств С. burnetii, как агента БО, работы по его генетической модификации мо-
гут быть направлены на повышение вирулентности и на придание выбранному для
поражения людей штамму резистентности к антибиотикам тетрациклинового ряда,
фторхинолонам, рифампицину и хлорамфениколу.
Для генетических манипуляций коксиеллы являются трудным объектом. Природ-
ной компетентностью они не обладают. До появления искусственной питательной
среды, предложенной A. Omsland et al. (2009, 2011), культивировать С. burnetii можно
было только в культурах клеток. Мощная рестрикционная система коксиелл значи-
822
Часть 3. Частная неправильная эпидемиология
тельно снижает частоту трансформации. Поэтому для введения векторных плазмид в
коксиеллы используют метод электропорации с высоким напряжением импульса,
пропускаемым через концентрированные суспензии клеток (Suhan М. L. et al., 1996;
Beare Р. et al.,2009; Beare P. A. et al., 2011).
Для обнаружения генетически измененных штаммов возбудителя Ку-лихорадки
анализ плазмидного профиля малоинформативен. Нет и четкой корреляции плаз-
мидного профиля с географическим распространением С. burnetii. Характерной
особенностью искусственно сконструированных плазмид будут маркерные гены (гены
антибиотикорезистентности, флюоресцирующих белков и др.) и IS-элементы. У соб-
ственных плазмид С. burnetii IS-элементов нет.
В настоящее время для генной инженерии коксиел в основном используют ин-
теграционные векторные системы и транспозонный мутагенез (Beare Р. A. et al., 2011).
Основным признаком генетической модификации возбудителя Ку-лихорадки, выяв-
ленного в очаге предполагаемой биологической диверсии или теракта, будет обнару-
жение в его геноме кластера генов антибиотикорезистентности в составе интегратив-
ного вектора или транспозонов Тп5 и Тп7.
Патогенез Ку-лихорадки. К. М. Лобан с соавт. (2002) рассматривают патологичес-
кий процесс при лихорадке Ку в целом как острый циклический доброкачественный
процесс. От риккетсиозов лихорадка Ку отличается отсутствием первичного пораже-
ния эндотелиальных клеток мелких сосудов и других частей кардиоваскулярной сис-
темы. Изменения в сосудах скорее могут быть особенностью хронического течения
этой болезни, чем острого, и тогда они представляются в виде периваскулита, а не
панваскулита, как это имеет место при риккетсиозах. Характерными элементами вос-
паления при коксиеллезе являются вакуолизированные клетки ретикулоэндотелиаль-
ного происхождения (мононуклеарные фагоциты), в цитоплазме которых содержат-
ся микроколонии С. burnetii. Сравнительно часто встречаются тромбы в кровеносных
сосудах, обычно в легких и селезенке, что связывается с замедлением циркуляции
крови вследствие дегенеративно-токсического миокардита.
Генерализация инфекционного процесса при лихорадке Ку объясняется К. М. Ло-
бан с соавт. (2002) постоянным выходом из разрушенных гистиоцитов и макрофагов
большого (чреспорогового) количества коксиелл Бернета в межклеточное простран-
ство, где часть их гибнет, а из оставшихся одни проникают в новые клеточные элемен-
ты, другие — в кровь или лимфу, где также гибнут или же диссеминируют в новые очаги
системы мононуклеарных фагоцитов.
Эту фазу патогенеза К. М. Лобан с соавт. (2002) называют фазой вторичной, или
большой коксиеллемии. Патологический процесс может протекать остро — цикли-
чески с быстрым выздоровлением, что имеет место у большинства больных, или же
как персистирующая инфекция (ациклически).
Патогенез острого течения лихорадки Ку К. М. Лобан с соавт. (2002) разделяют на
следующие фазы:
внедрение возбудителя болезни в организм человека аспирационным (в основном)
или иным путем;
проникновение коксиелл в организм человека через слизистые оболочки и кожу,
обычно поврежденную, без каких-либо патологических местных проявлений;
лимфогенный занос возбудителя в циркуляторную систему;
первичная, малая коксиеллемия;
диссеминация коксиелл в паренхиматозные органы с помощью моноцитарно-
макрофагальной системы;
3.7. Ку-лихорадка
823
размножение и развитие коксиелл в гистиоцитах, а затем и в клетках моноцитарно-
микрофагальной системы;
вторичная, или большая, коксиеллемия и токсикоз с диссеминацией возбудите-
ля в новые очаги макрофагальной системы (начало и разгар болезни);
развитие аллергических реакций организма с формированием иммунных комп-
лексов и их «оседанием» на тканях, например, створках сердечных клапанов, при
длительной циркуляции возбудителя в организме больных;
развитие иммунитета, как правило, достаточно выраженного и стойкого; в случа-
ях медленной иммунологической перестройки организма и слабости неспецифиче-
ских защитных реакций организма инфекционный процесс может затягиваться с тен-
денцией к хроническому течению;
выздоровление.
Патогенез персистирующей инфекции приведен выше.
Клиническая и патологоанатомическая картина Ку-лихорадки у человека при есте-
ственном инфицировании1. Инкубационный период при лихорадке Ку, по наблюдени-
ям К. М. Лобана с соавт. (2002) и данным других клиницистов, колеблется в преде-
лах 3—39 сут, но наиболее часто во время эпидемических вспышек он составляет
12—19 сут. При спорадической заболеваемости время инкубации в большинстве слу-
чаев не удается установить.
Клиника лихорадки Ку характеризуется выраженным полиморфизмом, что опре-
деляется, с одной стороны, механизмом передачи возбудителя, первичной инфици-
рующей дозой коксиелл, а с другой стороны, уровнем защитных сил макроорганиз-
ма. В результате этого при данной болезни не всегда достаточно четко определяется
ведущий или типичный симптомокомплекс, позволяющий если не диагностировать
болезнь, то хотя бы заподозрить ее. До 60 % лиц, инфицированных С. burnetii, пере-
носят Ку-лихорадку в бессимптомной форме. В течении болезни в форме острого
инфекционного процесса К. М. Лобан с соавт. (2002) выделили три периода: началь-
ный — первые 3-5 сут; период разгара болезни, или период наибольшей выраженно-
сти клинической манифестации — последующие 4-8 сут; и период реконвалесцен-
ции. Болезнь может приобретать затяжное и даже хроническое течение.
При остром течении у большинства пациентов (72—95 %) болезнь начинается
с внезапного появления озноба, нередко сильного, и повышения температуры тела в
первые 1—2 сут болезни до высоких значений — 39-40°С. Острота болезни часто на-
столько выражена, что больные указывают не только день, но даже и час ее начала.
С первых дней все или почти все больные жалуются на сильную головную боль, кото-
рая чаще бывает диффузной и реже — локализованной (в области лба или затылка),
разбитость и слабость, потливость, иногда даже профузные поты, отсутствие аппети-
та, сухой кашель или иногда с мокротой (часто он болезненный), нарушение сна,
вплоть до бессонницы, миалгии, боль в пояснице и иногда в суставах, болезненность
при движении глазами, резко выраженная боль в глазницах, глазных яблоках и рет-
робульбарные боли, причем последние имеют место почти всегда и представляются
даже многим клиницистам характерными для лихорадки Ку. Некоторые больные жа-
луются на головокружение, тошноту и рвоту, иногда возможно носовое кровотечение.
Постепенное развитие болезни наблюдается гораздо реже. Болезнь начинается с
легкого недомогания и познабливания, умеренной потливости, слабости, головной
1 Раздел написан в основном по работам Н. И. Федоровой (1965, 1968); И. К. Каракулова,
В. Д. Борисова (1966); К. М. Лобана с соавт. (2002) и Р. С. Karakousis et al. (2006).
824
Часть 3. Частная неправильная эпидемиология
боли. В этих случаях повышение температуры у пациента отмечается лишь по вече-
рам, максимальная температура составляет 37,3—37,8 °C, и больные в течение 3-6 сут
остаются на ногах. Лихорадка Ку не имеет определенного типа температурной кривой.
При более же длительном течении ухудшается общее состояние больных, нарастает
слабость, появляются боли в суставах и мышцах. Клинические проявления приобре-
тают более выраженный характер, чем в начале болезни. Наиболее часто наблюдают
ремиттирующий неправильный тип лихорадки. Реже наблюдается волнообразный
тип. Характерно наличие температурных «врезов», которые обычно сопровождаются
обильным потоотделением при падении температуры и сильным ознобом при ее по-
вышении.
Снижение температуры обычно происходит ступенеобразно за 3—4 сут. Критиче-
ское падение отмечается редко. Длительность лихорадочного периода при острых фор-
мах Ку-риккетсиоза значительно варьирует, но чаще составляет 5—15 сут. При хрониче-
ском течении болезни лихорадочная реакция может наблюдаться в течение нескольких
месяцев.
При лихорадке Ку кожные экзантемы наблю тают редко. Сыпь носит розеолезный
характер и редко имеет петехиальные элементы. Появление сыпи отмечают в период
от 3 до 13 сут болезни. Их необходимо тщательно дифференцировать от медикамен-
тозной и аллергической сыпи.
Со стороны ССС при остром течении Ку-лихорадки обычно не отмечается кли-
нических явлений, характерна лишь относительная брадикардия. Наличие приглу-
шенных тонов и систолического шума, которые нередко отмечают у больных Ку-ли-
хорадкой, не имеет органических основ и обусловливается обшей интоксикацией.
В тяжелых случаях может повышаться или, наоборот, понижаться АД. Электро-
кардиографически нередко обнаруживают миокардо-коронарные изменения. В ли-
тературе описаны отдельные случаи специфического поражения сердца и сосудов,
приведших к эндокардиту, миокардиту и перикардиту.
Поражение легких при лихорадке Ку не является частым явлением. Установлено,
что больные с легочными поражениями составляют не более 5—10 % от всех клини-
чески выраженных случаев лихорадки Ку. При развитии специфической пневмонии
основными симптомами являются: лихорадка (обычно температура тела в пределах
38,4—38,7°C), анорексия, головная боль, озноб, кашель, плевральные боли. Рентге-
нологические описания пневмонии у разных авторов не совпадают между собой.
Очаги воспаления могут быть обнаружены в прикорневой области и нижних долях,
но могут быть найдены и в верхних долях. Множественная очаговость наблюдается
редко. При поверхностном расположении пневмонического фокуса в процесс неред-
ко вовлекается плевра. Специфические изменения в легких при Ку-риккетсиозе ка-
саются в основном интерстициальной ткани и физикально дают скудные данные, в
связи с чем их обнаруживают часто лишь при рентгенологическом исследовании. При
аускультации иногда выслушивают непостоянные сухие или влажные хрипы. Боль-
ные часто жалуются на боли за грудиной и кашель, который иногда сопровождается
выделением мокроты с прожилками крови. Редкие рентгенологические проявления
пневмонии при Ку-лихорадке показаны на рис. 3.85.
Поражения ЖКТ при Ку-лихорадке не носят специфического характера. Иногда
в начале болезни появляются тошнота и рвота, боли в области эпигастрия, отсутствие
аппетита. Увеличение печени и селезенки является более постоянным признаком
лихорадки Ку и наблюдается у 50—80 % больных. Увеличение этих органов происходит
с первых дней болезни и наблюдается на протяжении всего лихорадочного периода.
3.7. Ку-лихорадка
825
Рис. 3.85. Рентгенологические проявления пневмонии при Ку-лихорадке. А. Рентгено-
грамма грудной клетки ребенка, заразившегося Ку-лихорадкой от рожавшей кошки.
Затемнение в правой верхней доле легкого (показано стрелкой). Б. Правостороннее
перихилярное затемнение (perihilar opacity) у пациента с Ку-лихорадкой (показано
стрелками) В Серия ренгенограмм, сделанных в динамике у пациента с Ку-лихорад-
кой. На рентгенограммах 1 и 2 показано множество затемнений округлой формы, на
рентгенограмме 3 — их разрешение после трех недель лечения. По Т. J. Marrie (2003)
Поражение ЦНС проявляется головной болью, которая служит ведущим симпто-
мом Ку-лихорадки и локализуется чаще в лобной, реже в височных и затылочных
областях. Помимо головной боли, отмечают ретроорбитальные и мышечные боли, а
также боли типа радикулита или полиневрита. Поражение ЦНС может проявиться
также вялостью, сонливостью, заторможенностью или, наоборот, состоянием возбуж-
дения, бессонницей, бредом. Нередко отмечают менингеальные явления. В тяжелых
случаях поражения ЦНС более выражены. Могут наблюдаться стойкие параличи глот-
ки, гортани с нарушением речи. Описаны и психические расстройства на почве забо-
левания лихорадкой Ку.
Постоянны нарушения со стороны вегетативной нервной системы: брадикардия,
озноб, потливость, запоры, жалобы на боли в животе.
Изменения со стороны периферической крови выражены нерезко. В лейкоцитар-
ной формуле наблюдают нейтропению со сдвигом влево до палочковидных, лимфо-
моноцитоз, эозинопению или анэозинофилию. СОЭ ускорена. В более тяжелых слу-
чаях отмечают нейтрофильный лейкоцитоз.
Лихорадка Ку обычно протекает доброкачественно и заканчивается выздоровле-
нием. В некоторых случаях (1 —5 %) наблюдается рецидивирующее течение с одним —
тремя подьемами температуры. Рецидивы возникают после 4—16 сут апирексии, вы-
сота и длительность температурного периода обычно меньше, чем во время первич-
ного заболевания; симптоматика та же, но течение обычно легче. В качестве ослож-
нений при Ку-риккетсиозе отмечают плевриты, легочные инфаркты, тромбофлебиты,
энцефалиты, эндокардиты, артриты, гепатиты, орхиты, эпидидимиты, остеомиелиты.
Патологическая анатомия изучена у небольшого числа умерших от данной болез-
ни. Все исследователи обращали внимание на патологию легких, что вполне объяс-
826
Часть 3. Частная неправильная эпидемиология
нимо, так как почти у всех больных смерть была обусловлена диффузной пневмони-
ей одной доли или более распространенным процессом, внешне не напоминающим
пневмококковую пневмонию. При быстро пазвившемся остром гепатите у умершего
на секции обнаруживают обширные некрозы в почечных дольках. Лимфатические
узлы обычно увеличены, в них обнаруживают гранулемы, состоящие из эпителиоид-
ных клеток. В почках отмечено набухание канальцевого эпителия, иногда небольшие
фокусы эритроцитарной инфильтрации в корковом слое и ишемический фиброз в
некоторых гломерулах. В ткани мозга находили периваскулиты и точечные геморра
гии вокруг сосудов, но иериваскулярный клеточный экссудат в пораженных областях
никем не регистрировался. В капиллярах иногда обнаруживали тромбы, их набухший
эндотелий. Возможно некоторое разрастание клеток нейроглии. Описаны энцефали-
ты с экстрапирамидными расстройствами и менингиты со значительным увеличени-
ем лимфоцитов в спинномозговой жидкости.
При рецидивирующей форме лихорадки Ку с развитием риккетсиозного эндокар-
дита на секции отмечены изменения аортальных клапанов, в которых обнаружены
риккетсии Бернета. Удается обнаружить риккетсии Бернета в легких, селезенке, яич-
ках, где они находились в макрофагах (незавершенный фагоцитоз), в головном моз-
ге — в клетках нейроглии, в почках — в эпителии канальцев, свободными в просвете
их, а также в клетках межк шальцевой сое шнитетьной ткани.
Представляет интерес случай длительного течения лихорадки Ку с летальным ис-
ходом, описанный в работе И. К. Каракулова, В. Д. Борисова (1966; (наблюдение
С. Н. Поповой).
Больной Л , 57 лет, поступил в стационар на 19 день болезни. Заболевание нача-
лось постепенно: повышение температуры, озноб, ночной пот, боль в правом боку,
кашель с мокротой. При поступлении в стационар общее состояние больного удовлет-
ворительное. Температура тела 38 °C, границы сердца немного расширены влево, тоны
приглушены. В легких — сухие хрипы. Печень выступает из подреберья на 2 см, при
пальпации отмечена чувствительность. На 32 день болезни в легких справа, в задне-
нижнем отделе,— незвучные мелкопузырчатые влажные хрипы. При исследовании
крови обнаружен умеренный лимфоцитоз и моноцитоз; СОЭ — 35 мм/ч. РСК с анти-
геном лихорадки Ку на 33 день болезни дала положительный результат в разведении
1:320. В дальнейшем температура имела волнообразный характер; состояние и само-
чувствие больного стало прогрессивно ухудшаться: нарастали общая слабость, сонли-
вость, апатичность, адинамия. На рентгенограмме легких справа и слева в верхних
долях обнаружено несколько очаговых теней по типу лобулярной пневмонии Деталь-
ные обследования больного на бруцеллез и туберкулез дали отрицательные результа-
ты. На 202 день болезни больной погиб при явлениях нарастающей сердечно-сосуди-
стой слабости и отека легких. Клинический диагноз: ложнобруцеллезная форма
лихорадки Ку.
Данные патологоанатомического вскрытия больного Л Труп пожилого мужчины,
резко истощенного, среднего роста, правильного телосложения. Кожа с землистым
оттенком, склеры слегка иктеричны. Поцкожный жировой слой атрофичный. Пари-
етальная и висцеральная брюшина гладкая и блестящая. В брюшинной полости —
700 см3 прозрачной соломенно-желтого цвета жидкости. Задненижние отделы легких
сращены с костальной плеврой; соединительно-тканные спайки легко разъединяются.
Центральная нервная система. Мягкая мозговая оболочка блестящая, застойно
полнокровная и отечная. Вещество головного мозга пассивно полнокровное и отечное.
Органы кровообращения. Сердце: 11x8, 5x5 см. Эпикард блестящий, жировой слой
атрофичный, ткань отечная. Стенки коронарных артерий склеротически утолщены
3.7. Ку-лихорадка
827
и просвет зияет. Толщина стенки левого желудочка — 2,5 см, правого — 1,5 см. Мио-
кард мясокрасного цвета, дряблый и тусклый; в нем видны белесоватого цвета про-
слойки соединительной ткани. Клапаны сердца и сосудов гладкие и блестящие. Аорта
в начальной части умеренно диффузно расширена, стенки ее склеротически утолщены.
В интиме грудного и брюшного отдела аорты — множество, большей частью, изъязв-
ленных атеросклеротических бляшек.
Органы дыхания. Слизистая оболочка верхних дыхательных путей пассивно полно-
кровная Тканьлегких в задненижних отделах также пассивно полнокровная и отечная;
здесь пальпируются рассеянные очаги уплотнения серого цвета с фестончатыми кра-
ями размером 0,5—1 см в диаметре. В нижней доле правого легкого — сливной пнев-
монический очаг серого цвета, плотной консистенции размером 3x4 см. Передневерх-
ние отделы легких губчатой консистенции, бледно-розового цвета.
Органы пищеварения. Язык суховат и обложен серым налетом. Слизистая пищевода
блестящая, застойно полнокровная. Слизистая оболочка желудка и кишечника атро-
фичная, блестящая, бледно-розового цвета.
Печень из-под реберной дуги не выступает, размеры ее немного уменьшены, вы-
пуклая поверхность гладкая, капсула блестящая, консистенция плотная; на разрезе
ткань имеет вид мускатного ореха: на коричнево-желтом фоне темно-красные точки
соответственно с застойным центром долек; в ткани печени встречаются белесоватые
прослойки соединительной ткани. Желчный пузырь не изменен, содержиттемно-олив-
кового цвета желчь; желчные пути проходимы.
Органы кроветворения. Костный мозг грудины не гиперплазирован, бледно-розо-
вого цвета. Селезенка оперативно удалена шесть лет назад вследствие разрыва. Лим-
фатические узлы области шеи, средостения, ворот легких, парааортальные и мезенте-
риальные узлы гиперплазированы, плотноватой консистенции, бледно-розового цвета,
размером 0,5—1 см в диаметре, не спаяны между собою.
Мочеполовая система. Капсула почек снимается без паренхимы, поверхность глад-
кая, ткань на разрезе серо-красная; корковый слой набухший, в нем обнаруживается
темно-красный крап. Слизистая мочевыводящих путей бледно-розового цвета, блес-
тящая. Половые железы без видимых изменений.
Эндокринные железы. Щитовидная железа атрофична, красно-коричневого цвета.
Надпочечники истончены, бледно-розового цвета, корковый слой содержит мало ли-
поидов. Гипофиз без видимых изменений.
Патологоанатомический диагноз. Лихорадка Ку: мелкоочаговая пневмония обоих
легких, преимущественно нижней правой доли; хронический продуктивный лимфа-
денитшейных, гилюсных и брыжеечных лимфатических узлов; очаговый интерстици-
альный и клубочковый нефрит; хронический интерстициальный гепатит; язвенный
атеросклероз аорты; гипертрофия обоих желудочков сердца; коронаро-кардиосклероз;
асцит; пассивное полнокровие и отек головного мозга; общее резкое истощение.
Микроскопия. Центральная нервная система. Мягкая мозговая оболочка отечная,
вокруг сосудов изредка обнаруживаются очаги круглоклеточной инфильтрации. В тка-
ни головного мозга — распространенный периваскулярный отек. Стенки артериол
утолщены вследствие пропитывания плазмой крови, эндотелий нередко размножен,
иногда наблюдаются очаги периваскулярной круглоклеточной инфильтрации. В бе-
лом веществе коры больших полушарий диффузно размножена дренажная глия, наб-
людается также очаговая пролиферация клеток микроглии.
Нервные клетки коры больших полушарий мозга атрофичны и местами разреже-
ны. В ядрах продолговатого мозга нервные клетки большей частью дистрофически
изменены: ядра сморщены и расположены эксцентрично, протоплазма однородна и
лишена нисслевской зернистости вследствие хроматолиза, который начинается в цент-
828
Часть 3. Частная неправильная эпидемиология
ре клеточного тела, в то же время на периферии протоплазмы зерна хроматофильной
субстанции сливаются в грубые глыбки. Таким образом, в центральной нервной сис-
теме резко выражен отек, вследствие повышенной проницаемости сосудов, и дистро-
фия нервных клеток, наряду с пролиферацией глиальных элементов.
Сердце и сосуды. Мышечные волокна миокарда гипертрофированы, набухшие и со-
держат в протоплазме белковые зерна. В стенках желудочков и в папиллярных мышцах
обнаружены очаги серозного воспаления с лизисом мышечных волокон и круглокле-
точной воспалительной инфильтрацией из гистиоцитов и плазмацитов. В перикарде
очаги круглоклеточной инфильтрации. Коронарные сосуды склерозированы.
В андвентиции аорты очаги периваскулярной воспалительной инфильтрации из
лимфоидных, плазмацитарных и эпителиоидных элементов. Мелкие сосуды адвенти-
ции местами облитерированы вследствие воспалительного разрастания соединитель-
но-тканных элементов интимы. В среднем слое стенки аорты встречаются капилляр-
ные сосуды, окруженные зоной воспалительной инфильтрации из лимфацитов и плаз-
моцитов. В интиме аорты много частью изъязвленных атеросклеротических бляшек.
Лимфатические узлы. Лимфатические узлы ворот легких, средостения, парааор-
тальные, брыжеечные и глубокие шейные узлы гиперплазированы. Лимфаденоидная
ткань в значительной мере замещена морфологически специфической инфекционной
гранулемой из эпителиоидных, гигантских и плазмацитарных клеток. Эпителиоидные
элементы имеют круглую или овальную формы, пузырьковидное ядро и широкую зону
бледно окрашенной протоплазмы с неясными контурами; местами эпителиоидные
клетки некробиотически изменены, лишены ядер и представляются в виде клеток-
теней. В гранулеме — многочисленные гигантские клетки круглой, грушевидной или
глыбчатой форм, с несколькими крупными гиперхромными ядрамг и бледно окрашен-
ной протошазмой. Развитие гранулемы начинается с размножения ретикулярных и
эндотелиальных элементов синусов и последующего превращения в эпителиоидные
и гигантские клетки. В грануломе встречаются очаги некроза, в которых клеточные
элементы разрушены с лизисом ядер. Наряду с некрозом в новообразованной грану-
ляционной ткани наблюдается очаговый фиброзе развитием коллагеновых волокон в
межуточном веществе. В пораженных лимфатических узлах отмечается продуктивно-
деструктивный васкулит мелких сосудов с круглоклеточной воспалительной инфиль-
трациеи их стенок.
Легкие. Легочная паренхима полнокровна; межальвеолярные перегородки расши-
рены вследствие гиперемии капилляров, размножения гистиоцитоь и разрастания
коллагеновых волокон. В альвеолярной паренхиме задненижних отделов легких, пре-
имущественно справа, обнаружено много очагов катаральной пневмонии. Здесь про-
свет альвеол выполнен серозным экссудатом и множеством макрофагов круглой и
овальной форм с крупным ядром и широкой зоной протоплазмы. Макрофаги эпите-
лиоидноклеточного типа, соединяясь отростками, образуют клеточный синцитий; в
межуточном веществе образуются коллагеновые волокна, а в альвеолах организуется
экссудат. Возникают множественные очаги карнифицирующей пневмонии; в очагах
продуктивного воспаления альвеолы выполнены эпителиоидными и гистиоцитарными
элементами с примесью фибробластов. Разрастающая грануляционная ткань из эпи-
телиоидных и гистиоцитарных элементов полностью выполняет просветы альвеол и
поры между ними и приводит к образованию сливных пневмонических очагов. Гра-
нуляционная ткань в очагах пневмонии разрушает и замещает альвеолярную легочную
паренхиму и в дальнейшем склерозируется за счет разрастания фибробластов.
Наряду с внутриальвеолярными процессами карнификации идет разрастание гра-
нуляционной ткани в межальвеолярных перегородках, а также в перибронхиальной и
периваскулярной соединительной ткани легких. Таким образом, вданном случае оча-
3.7. Ку-лихорадка
829
говая пневмония характеризуется разрастанием своеобразной гранулемы из эпители-
оидных и гигантских элементов с облитерацией просвета альвеол. Продуктивной фазе
воспаления предшествует экссудативно-катаральная фаза пневмонии.
Печень. Центральные вены и впадающие в них капилляры резко полнокровны.
Перикапиллярные пространства Диссе расширены и выполнены серозным экссуда-
том. Звездчатые клетки печени гипертрофированы и размножены. Перипортальные
прослойки соединительной ткани склерозированы; в них обнаружены очаги продук-
тивного воспаления из эпителиоидных и гигантских клеток с примесью плазмацитов.
Печеночные клетки диссоциированы, многие из них погибают, подвергаясь паренхи-
матозно-липоидной дистрофии и атрофии.
Почки. Ткань почек пассивно полнокровная. Отдельные группы клубочков увели-
чены в объеме вследствие размножения эндотелия капилляров, структура которых не
различается. В просвете капсулы отдельных клубочков и извитых канальцев много
эритроцитов. Паренхима щитовидной железы атрофичная, а строма склерозирована.
Таким образом, у больного Л. наблюдался ряд типичных морфологических изме-
нений, характерных для длительного течения Ку-лихорадки: системная гиперплазия
клеток ретикуло-эндотелиальной системы лимфатических узлов, легких, печени с
образованием характерных гранулем из эпителиоидных и гигантских клеток; очаговая
макрофагальная пневмония с исходом в карнификацию; повышение проницаемости
сосудов внутренних органов, а также эндартериит.
Хроническая Ку-лихорадка может развиться у человека через много месяцев после
инфицирования С. burnetii. В 75 % случаях болезнь проявляется эндокардитом, одна-
ко культуру С. burnetii из пораженной ткани получить не удается. Хроническая Ку-
лихорадка развивается почти исключительно у людей с иммуносупрессией или уже
имевших какие-то поражения сердца (протезы аортального или митрального клапа-
нов, сосудистые нарушения, ревматические и сифилитические поражения сердечных
клапанов и др.). Кроме эндокардита, у пациентов с хронической Ку-лихорадкой вы-
являют остеоартриты, хронический гепатит, хронические легочные инфекции, син-
дром хронической усталости. Каких-то специфических симптомов при этой форме Ку-
лихорадки нет. У 20 % больных может не быть лихорадки, но имеет место сердечная
недостаточность с расширением границ сердца и застойными явлениями в перифе-
рических сосудах, дисфункции сердечных клапанов, сыпь, гепатомегали, спленоме-
гали, гломерулонефриты, эмболии. Диагноз Ку-лихорадки ставится на основании
результатов серологических исследований (фиксация комплемента, непрямое флю-
оресцентное исследование, микроиммунофлюоресценция, EL1SA, микроагглютина-
ция и др.) (Karakousis Р. С. et al., 2006). Алгоритм диагностики острой и хронической
форм Ку-лихорадки приведен на рис. 3.86.
Клиническая картина Ку-лихорадки у животных при искусственном инфицировании.
У морских свинок Ку-лихорадку можно вызвать введением С. burnetii подкожно, внут-
рибрюшинно, внутритестикулярно, интраназально, перорально, при нанесении на
поврежденную кожу и слизистые и при кормлении на них зараженных клещей — пе-
реносчиков лихорадки Ку. Инкубационный период при любом способе заражения
колеблется в зависимости от дозы введенной С. burnetii. Наименьший инкубационный
период (от 1 до 3 сут) наблюдается при внутривенном и внутрибрюшинном способе
заражения. При подкожном, накожном, интраназальном и тестикулярном заражении
инкубационный период несколько длиннее (в среднем 4—7 сут). Наибольший инку-
бационный период наблюдается при пероральном заражении свинок и кормлении на
них содержащих возбудитель клещей (до 15-20 сут) (Федорова Н. И., 1968).
830
Часть 3. Частная неправильная эпидемиология
Рис. 3.86. Алгоритм диагностики Ку-лихорадки. Для острой Ку-лихорадки характерно
обнаружение антител против ат игенов II фазы, тогда как хроническая лихорадка харак-
теризуется присутствием в сыворотке крови антител (IgG) к антигенам I фазы. Обна-
ружение у пациента антител к антигенам II фазы: IgM > 50, IgG > 200 — свидетельствует
о развитии острой Ку-лихорадки. Выявление трансторакальной эхокардиографией по-
вреждения клапанов сердца: «да» — терапия продолжительностью не меньше года для
избежания инфекционного эндокардита; «нет» — повторение иммунологического ис-
следования через 3 мес. Погторное иммунологические тестирование через6 мес., если
антител к антигенам первой фазы не обнаружено, то дальнейшие обследования на Ку-
, шхорадку прекращаются. Если IgG или IgM к антигенам первой фазы 1800, с помощью
ГЩР в сыворотке пациента выявляют возбудитель болезни. В случае обнаружения
возбудителя Ку-лихорадки начинают лечение. По J. Hartzell et al. (2008)
Высота и длительность лихорадки зависят от способа введения и величины зара-
жающей дозы С. burnetii. При одной и той же заражающей дозе наиболее высокая тем-
пературная реакция наблюдается при внутрибрюшинном заражении. Наиболее дли-
тельный температурный период отмечается при заражении через нос, подкожно и
тестикулярно. При пероральном заражении и кормлении на свинках клещей темпе-
ратурная реакция нередко отсутствует. При недостаточной дозе при любом способе
заражения температурной реакции может также не быть. В таких случаях инфекция
протекает бессимптомно и о наличии ее можно судить лишь по серологическим дан-
ным, отражающим развитие специфического иммунитета (Федорова Н И., 1968).
При любом способе заражения у морских свинок развивается генерализованная
инфекция, которая выявляется при вскрытии зараженных животных соответствую-
щими изменениями внутренних органов. В паренхиматозных органах наблюдаются
дегенеративные изменения в виде белкового перерождения и дегенеративного ожи-
рения. Фибринозный налет на селезенке отмечается лишь при заражении большими
дозами вирулентного штамма. Патоморфологические изменения выражаются в про-
3.7. Ку-лихорадка
831
лиферативных процессах в ретикулоэндотелиальной системе. Ингаляционное зара-
жение морских свинок далеко не всегда влечет за собой возникновение первичной
пневмонии. Введение С. burnetii через дыхательные пути приводит к развитию лихо-
радочной формы болезни, как с признаками воспалительного процесса в легочной
ткани, так и без них. Вместе с тем вторичная специфическая пневмония может раз-
виваться на фоне общего инфекционного процесса, возникшего в результате любого
способа введения С. burnetii морским свинкам (Федорова Н. И., 1968).
Эксперименты по ингаляционному заражению мелкодисперсным аэрозолем
(105 МИД50 для мышей) С. burnetii 10 обезьян Gynomolgus, выполненные J. С. Gondor
et al. (1979), показали следующее:
1) у всех обезьян были обнаружены потеря аппетита, депрессия, кашель, одыш-
ка, лихорадка и учащенное дыхание. Потеря аппетита и депрессия совпадали с повы-
шением ректальной температуры, начавшимся через4—7 сут после заражения (в сред-
нем через 5,3 сут) и продолжавшимся в течение 4-6 сут. С. burnetii была выделена из
периферической крови всех обезьян на 7 сут и персистировала в крови до 13 сут (сред-
няя продолжительность 7,7 сут);
2) рентгеновские снимки четырех обезьян показали на 5 сут после заражения нали-
чие гранулярной инфильтрации в области корня легких. На 9 сут в периферических
областях наблюдались пятнистые неравномерные затемнения, усилившиеся к 12 сут.
В некоторых случаях очертания диафрагмы не просматривались. Выздоровление от
пневмонии началось на 16 сут;
3) патоморфологическое исследование двух обезьян, погибших на 7 и 14 сут пос-
ле заражения, показало наличие у них диффузной интерстициальной пневмонии с
сильным альвеолярным отеком. Был обнаружен также подострый гепатит. Обезьяна,
которая погибла на 14 сут, к моменту гибели находилась в острой фазе заболевания, и
пневмония у этого животного имела тяжелую форму. При исследовании заморожен-
ных проб ткани с помощью флюоресцентных антител исследователи обнаружили на-
личие риккетсий в ткани легких, селезенки, печени, почек, сердца и яичек этой обезь-
яны. Гистологическим исследованием тканей обезьян, забитых на 8 и 15 сут, были
установлены, соответственно, тяжелая и легкая форма интерстициальной пневмонии;
4) серологическое исследование (непрямой иммуноферментный метод) инфици-
рованных обезьян показало, что антитела к С. burnetii фазы II появляются на 7 сут (титр
1:300), а к микроорганизмам в фазе I — на 14 сут (титр Г.50). У 7 обезьян, зараженных
небольшой дозой С. burnetii, несмотря на отсутствие коксиелл в периферической кро-
ви, на 21 сут были обнаружены антитела к С. burnetii в фазах I и II. Титры антител у
этих обезьян были ниже, чем в группе животных, получивших большую дозу;
5) гематологические изменения выражались в нейтрофилии и лимфопении, из-
менений в общей концентрации лейкоцитов не наблюдалось. Наблюдавшееся увели-
чение концентрации фибриногена считали значительным (т. е. обусловленным не де-
гидратацией), на основании одновременного уменьшения в плазме соотношения
белок/фибриноген. Высокие концентрации фибриногена свидетельствуют о воспа-
лении или тканевой деструкции, и наблюдались как при незначительном поражении
печени, так и при наиболее тяжелых формах инфекции;
6) повышение уровней щелочной фосфатазы, глютамин-щавелевоуксусной транс-
аминазы и общего билирубина в сыворотке крови наблюдалось при наличии подостро-
го гепатита у обезьян, которые погибли, и тех, которые были забиты для патолого-
анатомического исследования. В данном исследовании у обезьян не обнаружили воз-
растания в сыворотке уровней глюкозы или холестерина.
832
Часть 3. Частная неправильная эпидемиология
Клиническая картина Ку-лихорадки у людей при искусственном инфицировании.
J. Е. Johnson и Р J. Kadull (1966) описали 40 случаев лабораторного инфицирования
коксиеллами, включая ингаляционное и чрескожное. Ниже привожу наиболее пока-
зательные из них.
Случай IV 34-летний мужчина, оператор аэрозольной камеры, работал с микро-
организмами С burnetii в течение одного месяца. У него внезапно возникла боль в ле-
вой ноге и в бедре и затем в течение 3-суточного периода, появились лихорадка, недо-
могание, потливость, отсутствие аппетита, понос, головная боль и, наконец, кашель
и боль в груди. Температура достигала 39,4°С. При перкуссии и аускультации в легких
первоначально не отмечали никаких изменений, но на рентгенограмме грудной клет-
ки был выявлен околохилюсный инфильтрат, распространяющийся на среднюю часть
доли левого легкого После поступления в госпиталь у больного появился сухой кашель
и при обследовании обнаруживались жесткие дыхательные шумы в области ворот ле-
вого легкого и местами влажные хрипы. Зольному проведен десятидневный курс ле-
чения хлортетрациклином. В материале глоточного смыва на 5 сут были обнаружены
С. burnetii, но гемокультура была отрицательной, В течение первых четырех суток пос-
ле начала болезни температура колебалась от 37,2 до 40,0 °C, по-видимому, под влия-
нием приема аспирина, но в течение 24 ч после приема хлортетрациклина у больного
температура упала до нормальной. Выздоровление было медленным, боли в спине и
повышенная утомляемость сохранялись в течение шести недель. Легочный инфильт-
рат рассасывался медленно в течение пяти недель после начала заболевания.
Случай VIL 28-летний микробиолог, вакцинированный ассоциированной вакци-
ной против пятнистой лихорадки Скалистых гор, лихорадки Ку и сыпного тифа (все-
го 4 инъекции), случайно поранил периферический сустав пятого пальца левой руки
иглой шприпа, содержащего суспензию С. burnetii. Палец покраснел, опух, стал болез-
ненным и на следующий день после заражения у больного появились лихорадка с оз-
нобом, слабосз ь, миалгия, боль в суставах и головная боль. Гемокультуры на второй и
третий день заболевания были положительными на С. burnetrii, а титр в РСК повысил-
ся до 1:160. Больной был госпитализирован на одну неделю, в течение которой палец
оставался сильно покрасневшим. Лечили больного хлортетрациклином. Никаких ме-
стных или общих поражений лимфатических узлов не отмечалось После выписки из
госпиталя выздоровление протекало гладко без каких-либо остаточных явлений.
Приведенные J. Е. Johnson и Р. J. Kadull (1966) данные по ингаляционному зара-
жению мелкодисперсным аэрозолем С. burnetii лабораторного персонала аналогичны
данным, полученным позже J. С. Gondor et al. (1979) в экспериментах по ингаляци-
онному заражению приматов (см. выше). Они показывают вовлечение тканей легких
в инфекционный процесс без поражения верхних дыхательных путей. Чрескожное
введение С. burnetii, поданным этих же авторов, сопровождается развитием местного
и общего инфекционных процессов без явного вовлечения регионарных лимфатичес-
ких узлов. Интересно отметить, что инкубационный период в этом случае продолжался
один день (от времени инъекции до начала клинических проявлений заболевания).
При туляремии и гистоплазмозе у человека и животных существует прямая зави-
симость между величиной инга тяционной дозы микроорганизмов и количеством
первичных легочных поражений. Эта зависимость не так очевидна при ингаляцион-
ном заражении человека С. burnetii дозами, варьирующих от минимальных до кратных
нескольким тысячам. Такие эксперименты приходится ограничивать, поскольку в
большинстве случаев экспериментально вызванная у человека Ку-лихорадки преры-
вается путем применения антиоиотикотерапии в течение суток со времени начала
лихорадки. Однако в некоторых случаях экспериментальных заболеваний при отсутст-
3.7. Ку-лихорадка
833
вии лечения и в многочисленных случаях, исследованных до применения антибиоти-
котерапии, на рентгенограмме можно было видеть лишь три или четыре дискретных
легочных поражения в отличие от многочисленных поражений, часто наблюдаемых
при легочной инфекции, вызванной другими бактериями (Tigertt W. D. et al., 1961).
Диагностика искусственного поражения возбудителем Ку-лихорадки. Принципы
диагностики искусственного поражения С. burnetii те же, что описаны в разд. 3.1 для
поражения возбудителем сибирской язвы. Естественные вспышки Ку-лихорадки
могут охватывать сотни людей и животных. Основным критерием их искусственного
происхождения является «неправильная эпидемиология». Основные признаки «не-
правильной эпидемиологии» для вспышек Ку-лихорадки следующие:
1) невозможность отнести эпидемиологию вспышки к одному из приведенных
выше типов естественного инфицирования людей возбудителем Ку-лихорадки;
2) выявление только одного генотипа С. burnetii среди десятков изолятов, получен-
ных во время вспышки.
Пневмонии у людей, вызванные инфицированием мелкодисперсным аэрозолем
С. burnetii, имеют свои признаки:
1) вовлечение тканей легких в инфекционный процесс без поражения верхних
дыхательных путей;
2) рентгенологически немногочисленные дискретные легочные поражения;
3) диффузная интерстициальная пневмония с сильным альвеолярным отеком;
4) гепатит (острый или подострый), сопутствующий легочным поражениям. Од-
нако эпидемиологам необходимо иметь в виду то обстоятельство, что специфическая
пневмония при ингаляционном заражении С. burnetii возникает далеко не всегда. По-
этому диагностическую значимость обнаружения пневмонии, соответствующей опи-
санным выше признакам, необходимо рассматривать только в связи с выявлением
признаков «неправильной эпидемиологии».
Резистентность к антибиотикам тетрациклинового ряда, фторхинолонам, рифам-
пицину и хлорамфениколу у изолятов С. burnetii, выделенных во время вспышки Ку-
лихорадки среди людей из объектов окружающей среды, продуктов питания, воды и
клинических изолятов, должна вызвать подозрение у эпидемиологов о возможности ее
искусственного происхождения и наличии в очаге болезни генетически измененных
штаммов. Особое внимание должны привлечь штаммы, обладающие резистентностью
к антибиотикам нескольких групп.
Иммунопрофилактика Ку-лихорадки. В России для специфической профилактики
Ку-лихорадки у людей используют живую вакцину на основе штамма М-44 С. burnetii
(вакцина Ку-лихорадки М-44). Вакцина представляет собой лиофилизированную
взвесь живой культуры вакцинного штамма М-44 С. burnetii, выращенных в желточ-
ных мешках развивающихся куриных эмбрионов. Имеет вид аморфной массы в виде
таблетки розового цвета с сероватым, желтоватым или коричневым оттенком. При-
меняется скарификационно. Однократное введение вакцины сопровождается разви-
тием специфического иммунитета через 3—4 нед после прививки. Ревакцинация про-
водится через год после первичной вакцинации лицам, в сыворотке которых
отсутствуют специфические комплементсвязывающие антитела. Вакцина произво-
дится на предприятии в ФГУП «НПО «Микроген» М3 РФ в Перми (614089, Пермь,
ул. Братская, д. 177).
Лечение Ку-лихорадки. Больным с подозрением на острую Ку-лихорадку назначают
тетрациклин, 500 мг каждые 6 ч, или доксициклин, 100 мг каждые 12 ч. Лечение про-
должают до клинического улучшения плюс не менее двух суток после прекращения
834
Часть 3. Частная неправильная эпидемиология
лихорадки у больного. Больным, которые не могут принимать тетрациклин, назна-
чают ципрофлоксацин. Продолжительность лечения ципрофлоксацином обычно
составляет 5—7 сут и не менее двух сут после прекращения у больного лихорадки. Для
лечения детей применять хинолоны не рекомендуется (Army FM 8-284, 2000).
Схема лечение эндокардита при хроническом течении лихорадки Ку включает
использование доксициклина в комбинации с рифампицином, офлоксацином или
ТМП/сульфаниламидами втечение не менее двух лет. Во время терапии требуется каж-
дые 6 мес. контролировать титры анти.ел. По окончании терапии контроль за анти-
те тами следует продолжать каждые 3 мес. в течение первых двух лет. Часто для излече-
ния от эндокардита при лихорадке Ку требуется замена клапана; однако эта процедура
откл адывается в связи с гемодинамическими показаниями или эмболическими ослож-
нениями (Army FM 8-284, 2000).
3.8. НАТУРАЛЬНАЯ ОСПА, ОСПА ОБЕЗЬЯН
Микробиология возбудителя натуральной оспы. Таксономия и происхождение
возбудителя натуральной оспы. Экология и естественная эпидемиология возбуди-
теля натуральной оспы. Экология и естественная эпидемиология возбудителя
оспы обезьян. Оспа коров. Восприимчивость к возбудителю натуральной оспы
людей и животных. Восприимчивость к возбудителю оспы обезьян у животных.
Иммунитет к поксвирусам. Устойчивость возбудителя натуральной оспы во внеш-
ней среде. Антивирусные препараты. Обнаружение генетически измененных
штаммов ортопоксвирусов. Патогенез ортог юг свирусных инфекций. Клиническая
и патологоанатомическая картина натуоал1 ной оспы у человека при естественном
инфицировании. Клиническая и патологпанзтомическая картина оспы обезьян у
человека при естественном инфицировании. Клиническая картина и патоморфо-
логия натуральной оспы и оспы обезьян у животных при искусственном инфициро-
вании. Клиническая картина натурал, ной оспы у людей при искусственном инфи-
цировании. Диагностика искусственного поражения возбудителем натуральной
оспы. Иммунопрофилактика натуральной ослы. Профилактика после экспонирова-
ния к ВНО и ВОО и лечение развившейся болезни.
Натуральная оспа (Variola major, Variola vera, Smallpox, Die Pocken, Blattem) — ост-
рая высококонтагиозная вирусная болезнь. В типичных случаях характеризуется
общей интоксикацией, лихорадкой, высыпаниями на коже и слизистых оболочках,
последовательно проходящими стадии пятна, пузырька, пустулы, корочки и рубца.
Вызывается вирусом натуральной оспы (ВНО; Variola virus, VAR, VARY). По вирулент-
ности для людей ВНО делится на тва подвида: Variola major (летальность среди заболев-
ших людей колеблется в пределах от 5 до 40 %); и Variola minor или алястрим (леталь-
ность 0,1—2%)*. Вспышки малой оспы отмечались параллельно и независимо от тако-
вых большой оспы. Люди, выживающие после натуральной оспы, могут частично или
1 Для названий штаммов ВНО, вызывающих малую оспу, установлены терминологические
разграничения, обусловленные различиями в их биологических свойствах и происхождении:
variola minor используется для обозначения африканских штаммов, a alastrim — для штаммов,
выделенных в Южной Америке Различий в клиническом течении вызываемой ими болезни
не установлено.
3.8. Натуральная оспа, оспа обезьян
835
полностью терять зрение, и у них на коже остаются рубцы в местах бывших язв. ВНО
относится к семейству Poxviridae, подсемейства Chordopoxviridae, рода Orthopoxvirus.
Других природных хозяев, кроме человека, для ВНО не установлено. Поэтому при ес-
тественном заражении наиболее вероятным источником вируса для человека являет-
ся другой человек, больной натуральной оспой. До конца 1950-х гг. военными специ-
алистами ВНО не рассматривался в качестве агента БО. Т. Розбери, Э. Кабат (1955) в
своем докладе, представленном Национальному исследовательскому совету США в
1942 г., писали, что ВНО непригоден для ведения войны из-за всеобщей и регулярно
проводимой вакцинации войск и населения. Кроме того, в эти годы не существовало
технологий получения ВНО в количествах, достаточных для боевого применения.
Основным способом получения ВНО было выращивание на хорионаллантоисной
оболочке (ХАО) развивающихся куриных эмбрионов. Способ позволял получать чи-
стую культуру вируса и поддерживать его в лабораторных условиях, но не более. Только
после появления в 1956 г. технологий культивирования ВНО в культурах клеток, его
стали упоминать в специальной литературе в качестве агента БО, способного «про-
бить» высокой дозой иммунитет у человека, созданный вакцинацией (Ротшильд А.,
1966). С 1980 г. массовая вакцинация против натуральной оспы отменена и в настоя-
щее время восприимчивость населения к ВНО повсеместная.
Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят ВНО
к I группе патогенности. По степени важности для национальной безопасности США
возбудитель натуральной оспы относится к биологическим поражающим агентам
категории А (см. табл. 2.2). Военными специалистами он рассматривается в качестве
потенциального поражающего агента БО из-за низкой инфицирующей дозы для лю-
дей; устойчивости к высушиванию и длительному хранению, и при переводе в аэро-
золь. Уже само слово «оспа» обладает большим психологическим эффектом. Упоми-
нание в СМ И о появлении натуральной оспы в каком-то населенном пункте способно
вызвать панику среди его населения и социальные эксцессы.
В настоящее время натуральная оспа считается ликвидированной болезнью,
официальные коллекции ВНО находятся только в двух лабораториях: Centers for
Disease Control and Prevention (CDC; Атланта, США) и в Федеральном бюджетном
учреждении науки «Государственный научный центр вирусологии и биотехнологии
«Вектор»» Федеральной службы по надзору в сфере защиты прав потребителей и бла-
гополучия человека (Новосибирск, Россия)1.
Оспа обезьян — зооноз, природно-очагововая вирусная болезнь, клинически по-
чти не отличается от натуральной оспы. Характеризуется лихорадкой, общей интокси-
1 Возможно, есть и другие, предпочитающие себя не афишировать, лаборатории-храни-
лища ВНО. Показательна следующая история. В книге воспоминаний бывшего руководителя
внешней разведки КГБ СССР Л. В. Шебаршина (1992) содержится любопытный и почему-то
упорно не замечаемый научной общественностью эпизод. Речь в нем идет о визите в феврале
1982 г. в Кабул председателя КГБ СССР Юрия Андропова. «Для Юрия Владимировича, —
пишет Шебаршин, — короткий визит в Кабул имел неожиданные и неприятные последствия.
Редкому посетителю афганской столицы удавалось покинуть ее хотя бы без желудочного за-
болевания. Андропову не повезло — он заболел оспой. Видимо, врачи не сразу поняли, с каким
заболеванием они имеют дело. По рассказам, состояние больного быстро становилось безна-
дежным. Каким-то чудом жизнь Андропова была спасена, но предстояло ему жить меньше двух
лет». В 1999 г. журналист А. Г. Ваганов связался с Шебаршиным, и тот подтвердил, что болезнь
Андропова действительно была натуральной оспой, и уточнил, что случай имел место в 1980 г.
То есть через три года после официально объявленного «последнего» случая заболевания оспой
в Сомали (1977 г.) и через семь лет после ликвидации оспы в самом Афганистане (1973 г.).
836
Часть 3. Частная неправильная эпидемиология
кацией и появлением экзантемы, сходной с высыпаниями при натуральной оспе.
Вызывается вирусом оспы обезьян (BOO; Monkeypoxvirus, MPV MPXV). Известны два
геномных варианта этого вируса. ВОО является близкородственным ВНО и вирусу
осповакцины (BOB; Vaccinia virus, W, VACV), но его эволюционная история не свя-
зана ни с ВНО, ни с BOB (Douglass N., Dumbell К., 1992) Первичный резервуар ВОО
не установлен (Hughes A. L. etal., 2010).
Впервые ВОО выделен в 1958 г. в Копенгагене от азиатских обезьян, содержащихся
в зоопарке, и поначалу считался лабораторной редкостью. В 1970 г. было установле-
но, что ВОО вызывает в тропической Африке у людей болезнь, похожую на натураль-
ную оспу. На фоне побед над ВНО, эти случаи казались досадным недоразумением.
Считалось, что оспа обезьян малоконтагиозна и большой опасности не представляет.
Однако позднее, вопреки всем своим прежним заявлениям о прекращении глобаль-
ной иммунизации против оспы, ВОЗ возобновила иммунизацию населения тропичес-
ких регионов Африки и продолжала ее до середины 1990-х гг., когда из-за резко воз-
росшего числа ВИЧ-инфицированных иммунизация населения живыми вакцинами
стала невозможна в принципе (Cohen J., 1997). В 1990-х гг. в регионе активизирова-
лась оспа обезьян. В небольшой заирской деревушке в районе Котако-Комба оспой
обезьян заболели 42 жителя из 346, трое из них погибли. Установлен больной, став-
ший источником каскада передач вируса 8 жителям, что уже сопоставимо с контаги-
озностью натуральной оспы. Все заболевшие ранее не были иммунизированы оспен-
ной вакциной (Mukinda V. et al., 1997). Но и после этого случая оспа обезьян не рас-
сматривалась как потенциальная угроза за пределами Африки. Это мнение изменилось
в 2003 г., когда ВОО был завезен в США вместе с дикими африканскими грызунами и
распространился среди людей в 11 штатах, вызвав 82 случая болезни (Chastel С., 2009).
Естественное инфицирование ВОО в России возможно в результате контакта с кон-
трабандно завезенными из тропических лесов Центральной и Западной Африки ди-
коживущими белками и обезьянами.
С целью поражения людей наиболее вероятно применение ВНО и ВОО посред-
ством использования линейных и многоточечных источников аэрозоля, и диверси-
онными (террористическими) методами. Террористами ВОО может использоваться
для имитации вспышек натуральной оспы. Клинику, напоминающую натуральную
оспу, у иммунодефицитных людей может вызвать вирус оспы коров.
Микробиология возбудителя натуральной оспы. ВНО размножается в развивающих-
ся куриных эмбрионах при различных путях введения: на ХАО, в амнион, желточный
мешок и аллантоисную полость. Использование куриных эмбрионов позволило впер-
вые выделить вирус оспы в чистой культуре и поддерживать его в лабораторных усло-
виях. В результате заражения ВНО на ХАО образуются характерные белые точечные,
куполообразные, резко отграниченные поражения (рис. 3.87).
Оптимальная температура для размножения ВНО в куриных зародышах 35 ЭС. При
температуре 37°C размножение вируса замедляется. Предельная температура разви-
тия на ХАО для Variola major — 38,5 °C; для Variola minor - 37,5 JC. При этом уменьша-
ется размер и плотность оспин. Большинство штаммов ВНО не вызывает гибели ку-
риных зародышей при первичном заражении, однако при последовательных пассажах
происходит адаптация вируса, проявляющаяся в приобретении им патогенности для
зародыша и изменении характера поражений.
ВНО размножается в тканевых культурах, вызывая развитие цитопатического
эффекта. Характер цитопатического действия вируса алястрима не отличается от тако-
вого у Variola major, однако его накопление в культуре происходит медленнее. К дейст-
3.8. Натуральная оспа, оспа обезьян
837
Рис. 3.87. Внешний вид оспенных пустул (pocks) на поверхности хорионаллан-
тоисной оболочки, образованных различными видами Orthopoxvirus. Пустулы
ВОО фотографировали после инкубирования в течение трех сут при температу-
ре 35 °C; остальные — после инкубирования в течение трех сут при 36 °C. A. Variola
major virus. Б. Vaccinia virus (штамм Листера). В. Monkeypox virus (штамм Копенга-
ген). Г. Cowpox virus (штамм Brighton). Размер пустул, образованных ВНО на ХАО,
находится в пределах 0,3—06 мм. По F. Fenner et al. (1988)
вию вируса чувствительны культуры различных тканей человека и животных. При этом
существенного значения не имеют возраст и восприимчивость к действию вируса вида
животного, от которого получена ткань, характер культуры (первичная или переви-
ваемая), особенности ткани (нормальная или злокачественная) и способ ее приготов-
ления (эксплантат, однослойная или взвешенная культура). Отличается лишь харак-
тер цитопатического действия: в первичных культурах вирус вызывает с самого начала
деструктивные изменения, а в перевиваемых — поражения начинаются пролифера-
цией ткани, и только в последующем отмечается деструкция клеток.
Для большинства однослойных культур развитие цитопатического действия харак-
теризуется появлением в неизмененном монослое очагов, в которых клетки разобще-
ны, округлены и сильно преломляют свет. Границы клеток четко очерчены, часть кле-
ток сильно увеличена в размере. С течением времени размер и количество таких очагов
увеличиваются и в дегенеративный процесс вовлекается весь монослой. Процесс за-
канчивается полной деструкцией клеточного пласта и отпадением клеток от стенок
прооирки, В зараженных клетках образуются цитоплазматические включения — тель-
ца Гварниери. Тельца Гварниери формируются уже через несколько часов после зара-
жения. В дальнейшем их количество увеличивается (как по числу включений в одной
клетке, так и по количеству клеток, содержащих включения). Скорость появления
включений в различных культурах клеток варьирует. Максимальное накопление ви-
руса отмечается обычно к 72—96 ч. ВНО способен образовывать бляшки (негативные
колонии) в культуре ткани с применением техники агарового покрытия и без него
Бляшки вируса в культуре клеток без агарового покрытия образуются через 72-96 ч
после заражения и имеют значительно меньшие, чем у вируса вакцины, размеры (до
1 мм). В культуре клеток под агаровым покрытием бляшки вируса оспы выявляются
не ранее 5-х сут с момента инфицирования культуры. Размер их не превышает 2 мм.
838
Часть 3. Частная неправильная эпидемиология
В культурах клеток, зараженных Variola major и Variola minor, наблюдается фено-
мен гемадсорбции — агглютинация эритроцитов отдельных видов животных суспен-
зиями ВНО. Наиболее вирулентные для людей штаммы ВНО (летальность заболев-
ших 14—44 %) циркулировали в Азии. Летальность среди людей, заразившихся афри-
канскими штаммами ВНО составляла 5—15 %. Для штаммов ВНО, выделенных от
людей из Ближневосточного региона, характерна низкая патогенность для куриных
эмбрионов при термоустойчивости, практически равной таковой референс-штамму
Harvey. По способности накапливаться в печени куриных эмбрионов они занимают
промежуточное положение между азиатскими и африканскими штаммами. Штаммы
ВНО, циркулировавшие в Бразилии, представляли собой типичные штаммы аляст-
рима. Объяснения этим феноменам найдено не было (Маренникова С.С., Щелку-
нов С. Н., 1998).
В отличие от большинства ДН К-содержащих вирусов животных, жизненный цикл
поксвирусов проходит в цитоплазме клетки в значительной степени автономно от ее
ядра. Для этого они, как и бактерии (!), располагают своими ферментативными сис-
темами синтеза РНК и ДНК (рис. 3.88 и 3.89).
Рис. 3.88. Вирион ортопоксвируса. А Вирион в фагосоме. 1 — мембрана фагосомы;
2 — вирионы; 3 — оболочка вириона; 4 — оболочка нуклеотида; 5 — нуклеотид;
6 — боковое тело. По П. Н Бургасову, Г. П. Николаевскому (1972). Б. Структура вне-
клеточною вириона ортопоксвирусов (на примере вируса вакцины). 1 — сердце-
вина; 2 — мембрана сердцевины; 3 — боковые тела; 4 — поверхностная мембрана;
5 — липопротеидная оболочка; 6 — трубчатые структуры поверхностной мембраны.
По С С. Маренниковой и С.Н. Щелкунову (1998)
Вирионы ортопоксвирусов (см. рис. 3.89) а цсорбируются на клетке (1) и сливают-
ся с плазматической мембраной клетки, высвобождая сердцевины в цитоплазму (2).
Сердцевины синтезируют ранние мРНК, с которых транслируются различные белки,
включая факторы роста, молекулы защиты от иммунной системы, ферменты и фак-
торы для репликации ДНК и транскрипции промежуточных генов (3) Происходит
«раздевание» сердцевины (4), и вирусная ДНК реплицируется, формируя кон кате мер-
ные молекулы (5). Промежуточные гены транскрибируются на дочерних молекулах
Д Н К и с этих мРНК транслируются факторы поздней транскрипции (6). Затем транс-
крибируются поздние гены. С их мРНК транслируются вирионные структурные бел-
ки, ферменты и факторы ранней транскрипции (7). Сборка вирионов начинается с
образования дискретных мембранных структур (8) Конкатемерные промежуточные
формы вирусной ДН К разделяются на единичные геномы и упаковываются в незрелые
3.8. Натуральная оспа, оспа обезьян
839
РНК-полимераза,
факторы ранней
транскрипции,
кэпирующий
фермент,
поли(А)-полимераза
ЦИТОПЛАЗМА
«Раздевание»
вириона
Ранние
мРНК
Репликация
—
--------*- РНК-полимераза
О®
h
О м
* Е
о О
S «
it
8.s
Промежуточные
6 мРНК
Поздние факторы
। транскрипции
7
Поздние мРНК
Поздние ферменты,
факторы ранней
транскрипции,
структурные белки
Созревание
прео юления
защитных
систем клетки
и организма
Факторы ядра?
Я Сборка
вирионов
Рис. 3.89. Жизненный цикл осповирусов (на примере вируса вакцины). Вирионы
содержат двуцепочечную геномную ДНК, ферменты, факторы транскрипции.
По С. С. Маренниковой и С. Н. Щелкунову (1998)
5
вирионы (9). Созревание приводит к образованию внутриклеточных зрелых вирионов
(10). Эти вирионы покрываются модифицированными мембранами аппарата Гольд-
жи и перемещаются к периферии клетки (11). Слияние таких вирионов с плазмати-
ческой мембраной завершается высвобождением внеклеточного оболочечного виру-
са (12). Хотя размножение вируса происходит полностью в цитоплазме, факторы ядра
могут быть вовлечены в процессы транскрипции генов и сборки вирионов (Марен-
никова С. С., Щелкунов С. Н., 1998).
Инфекционные частицы поксвирусов содержат в своем составе полную фермента-
тивную систему транскрипции, способную синтезировать функциональную мРНК в
юлиаденилированной, кэпированной и метилированной форме. Это позволяет ВНО:
1) самостоятельно осуществлять эффективную защиту от ранних специфических
реакций на инфекцию и противодействовать развивающимся позже специфическим
реакциям;
2) размножаться до высокого уровня, обеспечивающего быструю передачу друго-
му хозяину.
Таксономия и происхождение возбудителя натуральной оспы. Семейство поксвиру-
сов относится к монофилетической группе крупных ДН К-вирусов, размножающихся
в цитоплазме клетки (NCLDVs — nucleocytoplasmic large DNA viruses). В эту же группу
входят семейства Ascoviridae, Asfarviridae, Iridoviridae, Mimiviridae, Phycodnavii idae и
Marseillevirus (рис. 3.90).
Семейство поксвирусов подразделяется на подсемейства энтомопоксвирусов
(EnPV) и хордопоксвирусов (ChPV) (Entomopoxvirinae и Chordopoxvirinae), инфици-
рующих насекомых и хордовых животных соответственно. ChPVдополнительно под-
разделяются на 8 родов (Avipoxvirus, Molluscipoxvirus, Orthopoxvirus, Capripoxvirus,
Suipoxvirus, Leporipoxvirus, Yatapoxvirus и Parapoxvirus), тогда как EnPV подразделяются
840
Часть 3. Частная неправильная эпидемиология
Рис. 3.90. Филогения крупных ДНК-вирусов, размножающихся в цитоплазме
клетки. W — Vaccinia virus', FPV — Fowlpox virus-, MCV — Molluscum contagiosum virus',
MSV — Melanoplus sanguinipes entomopoxvirus; AM V—Amsacta moorei entomopoxvirus',
HcDNAV— Heterocapsa circularisquama DNA virus. По M. G. Fischera et al. (2010)
на 3 рода (А, В и С). Организация генома поксвирусов, их репликация, круг хозяев и
патогенез хорошо изучены. Краткая характеристика геномных последовательностей
отдельных штаммов ортопоксвирусов приведена в табл. 3.25.
Таблица 3.25
Краткая характеристика геномных последовательностей поксвирусов*
Вид Штамм Геном (п. о.) А + Т (%) ITR, Т. П. О. № доступа в GenBank
1 2 3 4 5 6
Хордопоксвирусы, род Orthopoxvirus (OPV)
Вирус вакцины, Vaccinia virus (BOB, W) Копенгаген 191636 12,0 М35027
MVA 177923 66,6 9,8 U94848
Тиан Тан 189274 66,8 7,5 AF095689
Вирус натуральной оспы, Variola virus (ВНО, VAR) Бангладеш-1975 186102 66,3 0,7 L22579
Индия-1967 185578 67,3 Х69198
Гарсия-1966 186986 67,3 0,6 Y16780
Вирус оспы обезьян, Monkeypax virus (BOO, MPV) Заир-96-1-16 196858 68,9 6,4 AF380138
Вирус эктромелии, Edromelia virus (ЕСТ) Москва 209771 66,8 AF012825
Нейвл (Naval) 207620 66,9 7,4
Вирус оспы верблюдов, Camel pax virus (С MPV) CM-S 202185 66,9 6 AY009089
М-96 205719 66,8 7,7 AF438165
Вирус коровьей оспы, Cowpax virus (CPV) Брайтон Рид 224501 66,6 AF482758
3.8. Натуральная оспа, оспа обезьян
841
Продолжение табл. 3.25
1 2 3 4 5 6
Lepori poxvirus
Вирус миксомы, Myxoma virus (MYX) Лозанна 161774 56,4 11,5 AF170726
Вирус фибромы Шоупа, Shope fibroma virus (SFV) Каза (Kaza) 159857 60,5 12,4 AF170722
Avipoxvirus
Вирус оспы птиц, Fawlpax virus (FPV) 288539 69,0 9,5 AF198100
Capripoxv-rus
Вирус узелковой сыпи, Lumpy skin disease virus (LSDV) Нитлинг 2490 (Neethling) 150733 73,0 2,4 AF325528
Вирус оспы коз, Goatpox virus (GTPV) Пеллор (Pellor) 149935 75,0 2,3 AY077835
G20-LKV 149695 75,0 2,2 AY077836
Вирус овечьей оспы, Sheeppox virus (SPPV) TLJ-V02127 140 995 75,0 2,2 AY077832
Штамм A 150057 75,0 2,3 AY077833
NISKHI 149 662 75,0 2,1 AY077834
Suipoxvirus
Вирус оспы свиней, Swinepox virus (SWPV) 17077-99 146454 72,0 3,7 AF410153
Molluscipoxvirus
Вирус контагиозного моллюска Molluscum contagiosum virus (MCV) Подтип 1 190289 36,0 4,7 LJ60315
Yatapoxvirus
Вирус Яба-подобного заболевания. Yaba- like disease virus (YLDV) 144575 73,0 1,9 AJ293568
Энтомопоксвирусы, Entoinopoxvirus В
Melanoplus sangwntpes (EnPVm) 236120 81,7 7 AF063866
Amsacta moorei (EnPVa) 232392 81,5 9,4 AF250284
* По G. Gubseret al. (2004).
Выполненное G. Gubser et al. (2004) сравнение геномов 26 поксвирусов показало,
что организация генома вирусов, входящих в подсемейство хордопоксвирусов, кон-
сервативна. Их центральная область кодирует почти идентичные белки для син-
теза РНК и ДНК, белкового процессинга, сборки вирионов и структурных белков.
Гены, кодируемые терминальными участками генома вирусов этого же подсемейства,
более дивергентны у различных родов, у видов внутри рода и даже у штаммов одно-
го и того же вида. Многие из этих генов кодируют белки, определяющие круг хозяев
вируса, его вирулентность или взаимодействие с иммунной системой хозяина. Несмот-
ря на выявленное сходство геномов ChPV, их длина варьирует от примерно 144 т. п. о.
у вируса YLDV до 289 т. п. о. у вируса оспы птиц (FPV), а содержание А + Т варьирует
от 75 % у рода Capripoxvirus до 36 % у рода Parapoxvirus (см. табл. 3.25). Геномы энто-
мопоксвирусов, наоборот, более дивергентны и по расположению генов отличаются
не только от CRPV, но и между различными родами EnPV.
Сравнение секвенированных поксвирусов позволило идентифицировать 90 генов,
которые консервативны у всех ChPV; при включении двух EnPV это число сокращено
до 49. У ChPV все 90 консервативных генов расположены в центральной области ге-
нома, содержащей 100 т. п. о.
842
Часть 3. Частная неправильная эпидемиология
Построенное G. Gubser et al. (2004) филогенетическое древо показывает, что Ch PV
составляют четыре группы вирусов. Первые две группы относятся к Molluscipox и Avipox
соответственно. FPV (род Avipuxvirus) является наиболее дивергентным вирусом сре-
ди ChPV. За ним следует MCV (род Molluscipoxvirus). Авипоксвирусы являются един-
ственными ChPV, которые инфицируют птиц, a MCV считается патогеном человека;
в результате эволюции оба вируса приобрели уникальные иммуномодуляторные бел-
ки, позволяющие им противодействовать иммунной системе их хозяев.
Третья, самая большая, группа ChPV, включает роды Yatapoxvirus (YLDV), Capri-
poxvirus (LSDV), Suipoxvirus (SWPV; и Leporipoxvirus (SFV и MYX). Внутри этой груп-
пы SFV и М5» X. которые тесно связаны друг с другом, расположены рядом с SWPV и
LSDV, в то время как YLDV является более дивергентным. Геномы всех этих вирусов
относительно высококонсервативны по составу генов, расположению генов и иден-
тичности аминокислот. SWPV и LSDV произошли от общего предка.
Четвертая группа ChPV — это род ортопоксвирусов (OPV). У них более крупный
геном, чем у вирусов других групп ChPV. Наиболее изученными считаются вирус вак-
цины (W-СОР) и вирус натуральной оспы (VAR-BSH).
G. Gubser et al. (2004) установили близкое родство CMPV и ВНО. Три штамма ВНО
составляют одну группу, так же как и два штамма CMPV. Группы ВНО и CMPV более
тесно связаны друг с другом, чем с любым другим видом OPV. OPV отделились от об-
щего предка позже, чем лепорипоксвирусы. При сравнении различных видов OPV
установлено, что наименьшее генетическое различие существует между штаммами
CMPV и ВНО, а наибольшее — между ЕСТ и ВНО. Интересной особенностью гено-
мов OPV является наличие большого количества генов, интактных у одного вируса,
но фрагментированных у другого. Объяснением может сл} жить тот факт, что некото-
рые OPV являются относительно новыми патогенами для их хозяев и в эволюцион-
ном масштабе времени произошли от предкового вируса сравнительно недавно, при
этом их дивергенция сопровождалась специализацией паразитизма и, как следствие,
«вык печением» генов, не нужных в новом хозяине. На рис. 3.91 показано филогенсти-
ческое древо семейства поксвирусов, основанное на анализе сцепленных (concatena-
ted) аминокислотных последовательностей 29 консервативных ортологичных белков.
Объяснению происхождения ВНО мешает представление о натуральной оспе как
о строгом антропонозе. Поэтому данные, полученные путем молекулярно-биологи-
ческих исследований генома вируса, обычно исследователи пытаются увязать с архео-
логическими данными или упоминаниями в письменных источниках о болезнях, ко-
торые они считают натуральной оспой (рис. 3.92).
В результате сопоставления столь разнородных фактов возникло противоречие в
оценке частоты мутации на пару оснований/цикл репликации (mutations per base pairs
per replication). Если в первом случае (частота мутации для ДНК-вирусов оценивает-
ся в пределах от 2х 1(Г8 до 7х 10“7) исследователи предполагают, что дивергенция ВНО
и Taterapox virus (инфицирует змей и грызунов в Западной Африке) от неустановлен-
ного предка произошла не ранее чем 50 тыс. лет назад; то во втором случае (предпо-
лагается частота мутации 4х 10"6, т. е. почти как у ВИЧ) — их эволюционная история
значительно короче — 3-4 тыс. лет (Hughes A. L. et al., 2010) Но такая частота мутаций
1 Yu Li et al. (2007), на основе анализа SNP, переносят время проникновения ВНО в челове-
ческие популяции из популяций грызунов еще на более ранний срок - 68 тыс. лет до н. э.
Появление в Южной Америке вируса алястрима они рассматривают как результат диверген-
ции ВНО, но датируют VI тысячелетием до н. э., т. е. еще до открытия Америки европейцами.
3.8. Натуральная оспа, оспа обезьян
843
Рис. 3.91. Филогенетическое древо поксвирусов. Таксономически наиболее близкими
к ВНО являются не представляющие опасности для людей ортопоксвирусы, вызы-
вающие оспенные заболевания у змей и верблюдов. Эволюционная история патоген-
ного для людей ВОО не связана ни с ВНО, ни с ВОВ. По A. L. Hughes et al. (2010)
в жизненном цикле ДНК-вирусов невозможна, так как репликация их ДНК сопро-
вождается проверочным считыванием и пострепликационной репарацией. При реп-
ликации РНК-вирусов, корректировок в синтезе РНК не происходит. Молекулы ви-
русной РНК реплицируются через ассиметричную транскрипцию от одной цепи,
исключающую большинство корректирующих механизмов, характерных для репли-
кации молекул ДНК (по репликации РНК-вирусов см. работу Holland S. et al., 1982).
Разрешение данного противоречия целесообразно искать в эволюционной исто-
рии всей монофилетической группы крупных ДНК-вирусов, рассматривая историю
поксвирусов только как ее частный случай и используя аналогии с семействами, чьи
первичные хозяева установлены.
Семейство Mimiviridae и семейство Poxviridae—дивергирующие ветви общего для
них неизвестного предка (см. рис. 3.90). Мчмивирус впервые обнаружен в амебах Асап-
thamoebapolyphaga. Диаметр зрелых частиц достигает 400 нм (La Scola В. et al., 2003).
844
Часть 3. Частная неправильная эпидемиология
Рис. 3.92. Мумия фараона Рамзеса V,
умершего в 1157 г. до н. э. На коже вид-
ны следы оспенных пустул. Мумия счи-
тается достоверным археологическим
подтверждением сушествования на-
туральной оспы в Древнем Египте,
однако далее в глубь времени фанта-
зия сторонников антропонозного про-
исхождения ВНО не идет. Рисунок из
книги F Fenner et al. (1988)
Его другое название -- мимивирус Acanthamoe-
ba polyphaga (Acanthamoeba polyphaga mimivirus',
APMV). Размер вириона мимивируса сравним
с размером микобактерии. Геном APMV вме-
шает 1,2 млн нуклеотидов (т. е. почти в 6 раз
больше генома самого крупного хордопоксви-
руса — FPV) и кодирует не менее 911 белков
(Benarroch D. et al., 2006). Е. Ghigo et al. (2008)
впервые продемонстрировали, что мимиви-
рус в условиях in vitro путем классического фа-
гоцитоза (т. е. так же как и ортопоксвирусы)
инфицирует макрофаги мышей, выживает в
них и успешно размножается. ДНК-топоизо-
меразы мимивирусов (IA, IB и ПА) сходны с
аналогичными ферментами ортопоксвирусов
(Benarroch D. et al., 2006). Репликация орто-
поксвирусов и мимивирусов происходит ана-
логичным образом. Вирусы обоих семейств
используют убиквитин-протеосомную систе-
му (Teale A. et al., 2009). Другие примеры сход-
ства мимивирусов и ортопоксвирусов можно
найти в работе М. G. Fischera et al. (2010). По-
этому поиски первичных резервуаров поксви-
русов целесообразно проводить там же, где их
нашли для мимивирусов, т. е. среди простей-
ших, а эволюцию поксвирусов рассматривать
как дегенеративную, т. е сопровождающуюся
утратой генов вследствие их большей спе-
циализации к новым хозяевам. Более подроб-
но о роли утраты генов в эволюции ортопоксвирусов, см. в работе R. С. Hendrickson
et al. (2010). Поиск первичного резервуара вируса, предкового для ВНО и Taterapox
virus и, возможно, самого ВНО, целесообразно осуществить в регионах, где произошла
их дивергенция (рис. 3.93).
Молекулярная эпидемиология ВНО хорошо изучена. С помощью метода изуче-
ния RFLP могут дифференцироваться ВНО, ВОВ, ВОС, CMPV, CPV Taterapox virus и
другие ортопоксвирусы и устанавливаться их географическое происхождение (Loparev
V. et al., 2001). Определены полные геномные последовательности большинства гео-
графических изолятов ВНО. Они объединены в базу данных для поксвирусов с откры-
тым доступом; www.poxvirus.org tLeDuc J. М. et al., 2002).
Экология и естественная эпидемиология возбудителя натуральной оспы. Экология
ВНО объявлена хорошо изученной. Сегодня принято считать, что ВНО может под-
держиваться только среди людей, используя механизм воздушно-капельной передачи
(см. Fenner F. et al., 1988). В качестве примера высокой контагиозности ВНО в научной
литературе приводятся отдельные вспышки, которые охватывают десятки человек
(см., например, работы Бургасова П. Н., Николаевского Г. П., 1972; Серенко А. Ф.,
1962). Однако когда анализируются вспышки в целом и за длительный период врс ме-
ни, то выясняется, что способность ВНО передаваться от человека к человеку в имму-
нокомпетентных популяциях ограничена. По данным V. Bhatnagar et al. (2006), обоб-
3.8. Натуральная оспа, оспа обезьян
845
Рис. 3.93. География ареалов Т. kempi и Е. ocellatus в Африке
По О. Piskurek, N. Okada (2007)
щивших статистическую информацию по 51 вспышке «завозной» натуральной оспы,
зарегистрированной в Европе и Северной Америке в период после 1945 г., в среднем
каждая такая вспышка ограничивалась четырьмя случаями болезни с одним леталь-
ным исходом. Количество передач (генераций) ВНО от заболевшего человека к здо-
ровому, редко превышало 3; а 30 % заносных вспышек вообще не сопровождалось
передачами ВНО от человека к человеку. Такую незначительную способность ВНО к
быстрому распространению в популяциях людей нельзя объяснить только предвари-
тельными массовыми вакцинациями, тем более что достаточно примеров массовых
вспышек в вакцинированных популяциях людей (например, московская вспышка
натуральной оспы 1959—1960 гг.). Вспышка натуральной оспы среди невакцинирован-
ного населения Abakalikj (Нигерия) в 1967 г. ограничилась всего 31 случаем болезни
(Eichner М., Dietz К., 2003). В свете этих данных невозможно понять, каким образом
жестокая пандемия натуральной оспы 1870—1874 гг. вспыхнула практически одно-
временно на нескольких континентах (Бразоль Л. Е., 1875), если не предполагать на-
личия первичного природного резервуара для ВНО, способного синхронно активи-
зироваться под воздействием каких-то еще неизвестных факторов. В противоречие с
распространенной точкой зрения на природный резервуар ВНО входят и данные,
полученные при молекулярно-биологических исследовании генома ВНО (см. выше),
свидетельствующие о циркуляции этого вируса еще «на заре» истории человечества-
а также странные «родственные связи» самого ВНО К его «ближайшим родственни-
кам» относятся вирусы, вызывающие болезнь у змей и верблюдов, и при этом не опас-
ные для людей (см. рис. 3.90), что говорит о случайном характере дивергенции пред-
кового вируса и патогенности ВНО для людей. Поэтому в представлениях об экологии
ВНО последнее слова еще не сказано.
846
Часть 3. Частная неправильная эпидемиология
Естественная эпидемиология натуральной оспы предполагает, что наиболее веро-
ятным источником возбудителя инфекции для человека является человек. И нкубани-
онный период болезни — 7—17 сут. Больной натуральной оспой заразен в течение всего
периода заболевания, начиная с момента возникновения лихорадки, а иногда с пос-
леднего дня инкубационного периода. Наибольшая заразительность больных прихо-
дится на период вскрытия оспенных элементов на слизистой оболочке рта и зева,
соответствующий 6- 9 сут болезни. Вместе с тем, по данным Комитета экспертов ВОЗ
по оспе, многие больные наиболее заразны на 3—4 сут болезни, т. е. к моменту появ-
ления высыпаний. Заражение может произойти от больного любой формой оспы, в
том числе и оспы без сыпи. Наибольшее эпидемиологическое значение имеют тяже-
лые формы болезни с обильными высыпаниями. Но самыми опасными в эпидемио-
логическом отношении являются именно нетипичные, вариолоидные формы забо-
леваний оспой у привитых. Больные, заболевание у которых не диагностируется,
остаются неизолированными и могут служить источником инфекции для окружаю-
щих. Они представляют особенную опасность, если среди контактирующих с этими
больными оказываются не иммунные лица. В подобных случаях эпидемический про-
цесс может затянуться на продолжительное время.
При абортивных формах болезни (вариолоид. оспа без сыпи и др.) продолжитель-
ность инфекционного периода невелика и может измеряться несколькими часами.
Несмотря на то что в корках больных оспой содержатся большие количества возбу-
дителя, наибольшее значение в распространении инфекции имеет вирус из дыхатель-
ного тракта. Основной путь передачи ВНО — воздушно-капельный: рассеивание ви-
руса с капельками слизи и слюны при разговоре, кашле и чиханье. Меньшее значение
имеет воздушно-пылевой путь: рассеивание вируса с частицами пыли, встряхивание
инфицированного белья, одежды и т. д. Сохранению вируса в пылевых частицах спо-
собствует его высокая устойчивость «о внешней среде. Вследствие этой особенности
возбудителя, зараженные вирусом оспы вещи и предметы, могу г служить источником
заражения даже при их пересылке на далеки*1 расстояния. Примером подобного рода
является несколько вспышек оспы в Англии, возникших в результате заражения че-
рез инфицированный хлопок, полученный из ОАР и Индии.
Контактный пугь передачи ВНО, хотя иногда и встречается, большого практичес-
кого значения не имеет. Трупы погибших от оспы людей могу г служить источником
заражения. Описаны случаи возникновения как генерализованной оспы, так и оспы,
протекающей по типу variola inoculata, улиц, принимавших участие в аутопсии и об-
работке материалов, взятых от трупа. Распространение натуральной оспы может осу-
ществляться насекомыми, которые выполняют роль механического переносчика ВНО.
Крупные вспышки завозной натуральной оспы происходят при следующих обсто-
ятельствах: во-первых, формируется массивный первичный очаг, длительное время
остающийся нераспознанным; во-вторых, последовательно формируются несколько
вторичных эпидемических очагов, в том числе в крупных медицинских учреждени-
ях, куда больные натуральной оспой поступают под ошибочными диагнозами. ВНО
начинает распространяться как внутрибольничная инфекция, среди компактно раз-
мещенных по палатам оо зьных людей, и от них переходить на посещающих их род-
ственников и врачей. Например, вовремя московской вспышки 19^9-1960 гг. снача-
ла сформировался семейный очаг в семье художника К., вернувшегося из Индии, затем
очаг в Московской клинической больнице им. С П. Боткина среди больных, меди-
цинского и обсл) живающего персонала, контактировавших с больным художником.
Эти очаги способствовали формированию третьего, городского, очага оспы в Москве.
3.8. Натуральная оспа, оспа обезьян
847
Около 1600 сотрудников больницы в течение 3 нед контактировали с населением го-
рода; в-третьих, очаги формируются среди населения, неиммунного в отношении
ВНО. Как правило, крупные вспышки возможны в городах, где вакцинация и ревак-
цинация против натуральной оспы либо вообще не проводилась, либо проводилась
небрежно и непоследовательно.
Самая крупная вспышка завозной натуральной оспы (99 заболевших) в послевоен-
ное время имела место в 1963 г. на территории Польской Народной Республики (ПНР).
Первым заболевшим оспой в конце мая 1963 г. был гражданин, прибывший из Индии
во Вроцлав. Местные врачи поставили ему диагноз малярии. Этот диагноз позднее был
изменен на необычно протекающую ветряную оспу, поскольку как у приехавшего, так
и у контактировавших с ним лиц появились характерные высыпания, сопровождав-
шиеся тяжелым клиническим течением, закончившимся в двух случаях смертью боль-
ных. Только через 48 сут после появления первого больного, лабораторными исследо-
ваниями был установлен диагноз натуральной оспы, а 3 сут спустя начали проводиться
необходимые противоэпидемические мероприятия. За время этой эпидемической
вспышки натуральной оспы во Вроцлаве заболело 79 человек, из них 6 человек умерли.
Инфекция была завезена также в провинцию Вроцлав, где заболели 11 человек, в про-
винцию Ополе (заболели 4 человека), в провинцию Лодзь (заболели 4 человека, один
из них умер), и один больной был зарегистрирован в Гданьске. Более четверти забо-
левших в описываемую вспышку натуральной оспы составили медицинские работ-
ники и лица из обслуживающего персонала больниц. Среди заболевших было 5 вра-
чей, 8 медицинских сестер, 5 палатных нянь, студентка-медик и 7 служащих больницы.
Из 7 умерших от оспы, 5 человектакже были медицинскими работниками (один врач,
2 медицинские сестры и 2 няни). За несколько недель в районах распространения
натуральной оспы было привито более 8 млн человек. Выборочной проверкой было
установлено, что 81 % привитых положительно реагировали на введение вакцины. Это
свидетельствовало об отсутствии у основной массы населения специфического им-
мунитета, который был утрачен, по-видимому, со временем в силу нарушения систе-
мы ревакцинации против натуральной оспы.
Ниже, по работам А. Ф. Серенко (1962) и С. Б. Дубровинского (1964), как пример
естественного течения вспышки натуральной оспы в большом городе, привожу крат-
кое описание московской вспышки 1959—1960 гг.
22/XII. 1959 г. из Дели в Москву на самолете вылетел художник К1. По прибытии в
Москву 23/ХП он почувствовал недомогание и вызвал врача. 24/XII больной К. выс-
казал врачу жалобы на слабость, боли в животе, головную боль, отсутствие аппетита.
Объективно у больного температура 38—39°С, язык обложен, в зеве и легких чисто,
тоны сердца глухие, пульс 106/мин, болевые точки в области желчного пузыря.
Был установлен предположительный диагноз гриппа, назначен постельный режим.
25/XII при повторном осмотре температура 39,1 °C, язык обложен, живот не напряжен,
в зеве чисто, пальпируется печень и селезенка, пульс 96/мин. Предположительный
диагноз: риккетсиоз? аппендицит?
26/ХП приглашен хирург, который отверг диагноз аппендицита. У больного на
животе и груди крупнопапулезная сыпь, положительный симптом Кончаловского.
Высказано подозрение на сыпной тиф, но больной не госпитализирован, так как квар-
тирные условия были вполне удовлетворительными.
1 Алексей Атексеевич Кокорекин (1906—1959) — советский график. Заслуженный деятель
искусств РСФСР (1956). Лауреат двух Сталинских премий (1946, 1949).
848
Часть 3. Частная неправильная эпидемиология
27/ХП у больного температура 37,8 °C, лицо красное, отечное, кровоизлияние в
склере, отхаркивает кровь, АД 100/80 мм рт. ст, тоны сердца глухие, пульс 92/мин.
Больного осмотрел врач-инфекционист, который остановился на диагнозе токсиче-
ского гриппа с медикаментозной сыпью. Больной госпитализирован в 13-е (инфекци-
онное) отделение Московской городской больницы им. С. П. Боткина. В дальнейшем
клиническая картина заболевания приводится согласно записям в истории болезни.
Лицо одутловатое, гиперемировано, часть спины синюшняя. На верхних конеч-
ностях. груди и животе обильные пятнисто-папулезные полиморфные высыпания, в
правой и левой подмышечных впадинах имеются элементы пузырькового характера с
гнойным содержимым. Такие же высыпания и на внутренних поверхностях верхних
конечностей, на бедрах мелкие пятнисто-папулезные высыпания. Слизистая оболоч-
ка зева гиперемирована, сочная. Язык сухой, обложен. На высоте вдоха пальпируется
край печени. Сплошное кровоизлияние в склере, конъюнктивит. Предположительный
диагноз: лекарственная сыпь.
В ночь на 28/ХП больной отхаркивал слизистую мокроту с примесью алой крови,
после приема преднизолона была рвота. 28/'ХП общее состояние больного тяжелое.
Яркая, разлитая, интенсивная гиперемия кожи. Кожа лица и верхней половины туло-
вища одутловатая, при пальпации как бы отечна. В паховых областях отдельные гемор-
рагии, в нижней трети правого бедра с наружной стороны имеется кровоподтек вели-
чиной с детскую ладонь. Живот мягкий, безболезненный при исследовании. Держится
резкий отек и набухание век, из-за резкого отека конъюнктиву не видно.
Клинически впечатление острого аллергического дерматита с выраженными яв-
лениями капилляротоксикоза. На основании анамнеза (больной 22,23,24/XII прини-
мал левомицетин, стрептомицин, биомицин, синтомицин) консультантом также под-
тверждается этот диагноз.
28/ХП присоединились явления геморрагического нефрита. Поражен весь кожный
покров, слизистая оболочка рта и конъюнктива глаз: на общем эритематозном фоне
характера эритродермии множественные геморрагические экстравазаты различной
величины и оттенков до синего цвета. На слизистой оболочке языка, щек и неба точеч-
ные белые налеты — «молочница», местами сливающаяся и образующая на небе плен-
ку. Ощущение жжения в глазах и кожных покровах. Консультация профессора. Диаг-
ноз: тяжелая токсико-аллергическая реакция на антибиотики, прогноз сомнителен.
29/ХП в 12 ч консультация отоларинголога, стоматолога, терапевта, гематолога,
окулиста, в 18 ч 30 мин состояние больного крайне тяжелое: выраженная гипоксия,
повышенная кровоточивость, пульс едва прощупывается, кожные покровы синюш-
но-серые. В 23 ч 29/ХП больной К. умер. 30/XI I было произведено патологоанатоми-
ческое вскрытие По заключению патологоанатома основным заболеванием у больного
К. являлась геморрагическая алейкия, на фоне которой развился некротический ла-
рингит, фарингит, эзофагит и колибациллярный сепсис.
Смерть больного наступила от резкой общей интоксикации, обусловленной сеп-
сисом и чрезвычайно обширными множественными кровоизлияниями,
31/ХП трчп К. был кремирован, проведена дезинфекция помещений, в которых
находился больной К., личные вещи были сожжены.
На вскрытии патологоанатомы, не отрицая диагноза токсического капилляроток-
сикоза, заподозрили на основании обнаружения биополярных палочек токсическую
чуму. 31/ХП на основании дополнительных лабораторных исследований диагноз ток-
сической чумы был отменен.
Позднее было установлено, что больной К. за 2 недели до выезда в Дели «был вак-
цинирован против оспы, но вакцинальной реакции у него не было отмечено, ранее
прививался против оспы только в детстве».
3.8. Натуральная оспа, оспа обезьян
849
Во время нахождения больного К. дома в Москве с 23 по 27/XII и позднее в боль-
нице с 27 по 29/ХИ с ним контактировали члены семьи, друзья и медицинский персо-
нал больницы им. С. П. Боткина.
Первые случаи заболеваний натуральной оспой в Москве зарегистрированы с 11 по
14/1 1960 г. Таким образом, к 14/1, т. е. через 16 сут после смерти К., было установлено,
что первый случай натуральной оспы был пропущен. Ретроспективно у больного К.
был установлен дигноз purpura variolosa. Пути распространения и формирования оча-
гов натуральной оспы в Москве в 1959-1960 гг. показаны на рис. 3.94.
Больница им. С П. Боткина. Персонал больницы и внутрибольничные заражения
31 Н132 Hl 33 Hl 34 HI 35
38
Семейный очаг
39 Н140 Н141HY42 ЙГ43 it'
-----Пути формирования очагов
Предполагаемые пути формирования очагов
В — Вакцинированные Н — Невакцинированные
О Летальный исход
О Подтвержденные случаи
О Предполагаемые случаи болезни
Рис. 3.94. Пути распространения и формирования очагов натуральной оспы в Москве
в 1959—1960 гг. В результате формирования крупного очага натуральной оспы в семье
художника К. (5 человек) сформировался очаг в московском отделении Союза худож-
ников — от сотрудника Союза художников, в Химико-технологическом институте им.
Д. И. Менделеева — от дочери художника, « Проектном институте «Теплоприбор» —
от родственника художника, ь пошивочном ателье — от близких родственников ху-
дожника. Кроме того, в семье страхового агента сформировался самостоятельный очаг
натуральной оспы, а позднее 24/1 был обнаружен очаг в Сандуновских банях. Оба
очага сформировались в результате контакта с членами семьи художника. Наряду с
этими очагами самостоятельно развивался мощный очаг в Московской клинической
больнице им. С. П. Боткина. Около 1600 сотрудников больницы в течение 3-х недель
контактировали с населением города. По А. Ф. Серенко (1962)
850
Часть 3. Частная неправильная эпидемиология
Первыми выявленными заболевшими натуральной оспой в результате контакта с
больным К. были: его знакомая Н., посещавшая К. после возвращения из Дели, и врач-
отоларинголог Т., посещавший больного К. 27и29/ХП 1959 г. Больная Н., 39 лет, про-
живает в Москве, находилась в контакте с больным К. и его семьей с момента его при-
езда г Москгу. Оспенные знаки от прививок, произведенных в детстве, отчетливы,
позднее прививкам против оспы не подвергалась. Заболела 8/11960 г., когда почувство
вала головную боль, сильную слабость, боли в суставах; температура 38,6 °C. На 4 сут
болезни при снижении температуры появилась мелкоточечная сыпь на коже в облас-
ти локтевых и коленных суставов, кистях рук с зудом и болевыми ощущениями. На
голове, лице и груди пятнистая сыпь. На 5-6-й день пятна как будто приподнялись,
образовались везикулы, наполненные желтоватой жидкостью, на ладонях и подошвах
зуд, сыпи не отмечалось. На 4-й день наблюдалась припухлость локтевых суставов.
Лабораторно из содержимого пустул и везикул выделен на куриных эмбрионах
оспенный вирус.
Диагноз: вариолоид с множественными высыпаниями.
К 26/1 на коже сохранились пигментированные пятна на месте бывших папул.
Больная лечилась в инфекционном отделении больницы им. С П Боткина, выздоро-
вела, выписана 16/П 1960 г.
Больной Т, 61 года, работает врачом-отоларингологом. В больнице им. С. П. Бот-
кина посещал больного К. 27 и 29/ХП 1959 г. Последний раз прививался против оспы
в 1914 г. 7/11960 г. почувствовал недомогание, головную боль, повышение температу-
ры. 8/1 работал, но чувствовал усталость и головную боль, ушел с работы раньше вре-
мени, к вечеру температура поднялась до 38°С. 9/1 j гром принял 1 таблетку сульфазола,
затем еще 2 таблетки. К середине дня на коже лица и туловища появилась мелкото-
чечная узелковая сыпь. Температура 8 и 9/1 держалась в пределах 38,5—39°С. 10/1 сыпь
появилась на голове и конечностях. Температура 39 °C.
11/1 поступил в инфекционное отделение больницы им С П. Боткина. При по-
ступлении в больницу состояние больного тяжелое, температура 39,3 °C. На лице, во-
лосистой части головы, туловище и конечностях обильная пятнисто-папулезная сыпь,
выраженная также на разгибательных поверхностях конечностей. В зеве легкая гипере-
мия. 12/1 температура высокая, обильная папулезная полиморфная ярко-кра< ная сыпь.
13/1 зарегистрировано падение температуры до 37,6“С, сыпь значительно увели-
чилась по всей поверхности тела, приняла геморрагический характер, появились от-
дельные везикулы. Врачи заподозрили ветряную оспу.
14/1 лицо пациента стало пастозным, сыпь на лице и волосистой части головы сде-
лалась сливной, более полиморфной. Появилась сыпь на ладонях и подошвах. Наря-
ду со свежими пузырьками, наполненными мутной жидкостью и окаймленными крас-
ным ободком, имеются пузырьки, появившиеся ранее, на некоторых из них черные
корочки. Возникло подозрение на variola vetv.
15/1 температура у больного Т. снизилась до нормальной, состояние улучшилось,
локализация сыпи осталась прежней, но на верхушках высыпаний появилось много
вдавлений в виде пупочков, количество корочек увеличилось.
Материал из пустул взят на исследование и направлен в лабораторию Московской
городской санитарно-эпидемиологической станции и в Институт им. И И Мечнико-
ва. Приглашенный к больному I. консультант диагностировал variola vera discreta.
Диагноз оспы 15/1 подтвержден вирусоскопически: обнаружены элементарные
тельца Пашена серебрением по Морозову, в последующем через 3 дня методом Пауля
и через 6 дней путем реакции задержки гемагглютинации.
В последующие дни при лечении состояние больного заметно улучшилось, отме-
чалось нагноение везикул и образование корочек. С 21/1 началось обильное образова-
ние корочек и шелушение, корочки длительно задерживались на ладонях и подошвах.
3.8. Натуральная оспа, оспа обезьян
851
К 14/11 очищение от корочек закончилось, оставались пигментированные рубцы в
большом количестве на туловище и конечностях, в меньшем количестве на лице.
Лечение проводилось пенициллином и стрептомицином, а также симптоматиче-
ское по 21/1. Выписан из больницы 16/11 на 40-й день болезни.
За период с 22/XI I 1959 г., т. е. с момента прибытия больного К. из Индии в Моск-
ву, и по 16/11 19601. различными клиническими формами оспы заболело 46 человек.
При изучении клинических проявлений натуральной оспы во время вспышки
выявлен ряд особенностей, представляющих интерес для клиницистов и эпидемиоло-
гов даже сегодня. Анергическая форма была у одного больного, сливная — у 2, рассе-
янная — у 3, вариолоид со множественными высыпаниями — у 8, вариолоид с единич-
ными высыпаниями — у 29 и заболевание без сыпи — у 3 больных (табл. 3.26, 3.27).
Таблица 3.26
Клинические формы болезни и течение натуральной оспы
во время вспышки в Москве в 1959—1960 гг.*
Клиническая форма болезни Количество случаев болезни Клиническое течение Число случаев с летальным исходом
все подтверж- денные клинически подтверж- денные лабораторно тяжелое среднее легкое
Анергическая 1 1 — 1 — — 1
Сливная 2 2 2 2 — — 1
Рассеянная 3 3 3 3 — — 1
Вариолоидная с множественными высыпаниями 8 8 7 - 3 5 -
Вариолоидная с единичными высыпаниями 29 29 6 — 3 26 -
Оспа без сыпи 3 3 — — 1 2 —
Итого 46 46 18 6 7 33 3
* По А. Ф. Серенко (1962).
Тяжелая форма отмечалась у 6 человек, средней тяжести — у 7 и легкие формы —
у 33 больных. Из общего количества (46) больных у 43 заболевание закончилось пол-
ным выздоровлением, у 3 — летальным исходом. При изучении клинического течения
болезни выявлено, что у абсолютного большинства пациентов продромальные явле-
ния отмечались в пределах от 5 до 8 сут от заражения, температурная реакция появля-
лась в период от 8 до 10 сут, высыпания — от 9 до 11 сут. По возрасту всех 44 больных
можно разделить на следующие группы: детей до 13 лет — 3 человека, в возрасте от 20 до
50 лет — 37 человек. Среди больных 21 женщина.
У подавляющего большинства больных течение заболевания было легким, только
у двух больных натуральной оспой наблюдалось тяжелое течение болезни. Инкубаци-
онный период у подавляющего большинства больных (у 31 из 44 наблюдавшихся) ко-
лебался в пределах от 7 до 9 сут.
В группе больных вариолоидом с единичными высыпаниями температура быстро
повышалась и держалась на высоких значениях 2-3 сут. В день появления папул тем-
пература снижалась до нормы; второй лихорадочной волны, связанной с появлением
папул, ни в одном случае вариолоида не отмечалось. Продромальная сыпь ранее была
зарегистрирована у 2/3 больных на высоте температуры перед появлением характер-
ных для оспы высыпаний.
852
Часть 3. Частная неправильная эпидемиология
Таблица 3.27
Течение натуральной ослы во время вспышки в Москве в 1959—1960 гг.*
День госпитализации с момента заболевания Количество больных, поступивших из города Количество больных, выявленных из контактов Явления продрома Количество больных с
температурной реакцией высыпанием
1-й — 13 — — —
2-й 1 5 — — —
3-й 3 1 — — —
4-й — 2 — — —
5-й 1 — 12 —
6-й 3 2 18 — —
7-й 2 1 10 — —
8-й 1 4 15 —
9-й 2 — 2 23 6
10-й 2 — — 2 25
11-й 1 — — - 9
12-й 2 — — - 1
13-й 4 — — - 2
Итого 22 24 46 40 43
* По А. Ф. Серенко (1962).
После отторжения корок у больных вариолоидом рубцовых изменений не остава-
лось, длительно, до 10—15 сут, держалась пигментация. Кроме общетоксических
явлений (головная боль, адинамия, боль в суставах, тошнота, рвота и др.), были отме-
чены изменения со стороны слизистой оболочки мягкого неба и глотки в виде пузырь-
ковых элементов разной величины на фоне разлитой гиперемии.
До 10-го дня болезни патологических изменений со стороны крови не выявлялось.
После 20—30 сут болезни в картине крови отмечалась тенденция к лейкоцитозу и аб-
солютному нарастанию количества лимфоцитов. Только у одной больной в моче в
период лихорадки наблюдалась альбуминурия, у остальных больных патологических
отклонений в моче не отмечено.
Один случай заболевания, закончившийся летально, вызвал серьезные разногла-
сия между клиницистами и патологоанатомами.
Больной А. во время вспышки оспы в Москве находился на лечении в Московской
городской клинической больнице им. С. П. Боткина с диагнозом лимфогранулемато-
за, впоследствии присоединилась оспа. Больной погиб на 3-й день после появления
высыпаний. Клиницисты утверждали, что больной А. погиб от основного заболевания,
патологоанатомы — от оспы, протекавшей по типу вариолоцда. Спор этот был решен
в пользу натуральной оспы в результате вирусологических исследований и выделения
вируса оспы из материала от больного А
Несмотря на дискуссионный характер диагноза у больного А., учитывая тесный
контакт больного А. с больным Б., с эпидемиологической точки зрения этот случай
заболевания рассматривался как случай натуральной оспы с последующим осуществ-
лением всех противоэпидемических мероприятий, направленных на локализацию
очага. Лабораторная диагностика во время вспышки натуральной оспы в Москве яви-
лась одним из обязательных методов. Большинство (36 случаев) заболеваний протекало
3.8. Натуральная оспа, оспа обезьян
853
по типу вариолоида, т. е. с атипичной клинической картиной, что следует объяснить
строгим проведением в нашей стране оспопрививания. Это вызвало серьезные затруд-
нения при клинической диагностике оспы и требовало исключения ветряной оспы по-
чти в каждом случае. Кроме того, лабораторная диагностика была крайне необходима
еще и потому, что вспышка протекала в условиях массовой вакцинации, что, естествен-
но, затрудняло дифференциальную диагностику в случаях с различными поствакци-
нальными осложнениями.
Вирус удавалось выделять из содержимого кожных элементов до 17 сут болезни, а
из носоглоточных смывов — на 3—9 сут болезни.
Определение антигемагглютининов в сыворотках заболевших натуральной оспой
показало, что более 60 % заболевших имели титр антигемагглютининов 1:160 и выше,
в то время как почти у 90 % здоровых, недавно вакцинированных лиц, титр антигемаг-
глютининов не превышал 1:80.
В 36 случаях диагноз натуральной оспы был подтвержден вирусоскопически пу-
тем обнаружения телец Гварниери и Пашена в материале из носоглоточных смывов и
кожных элементов.
Двое больных, у которых вирус был выделен из крови, погибли.
Три случая болезни без кожных высыпаний были подтверждены выделением ви-
руса натуральной оспы из носоглоточных смывов.
Экология и естественная эпидемиология возбудителя оспы обезьян. Инкубационный
период болезни у людей — 7—19 сут. Динамика вспышек плохо изучена. Встречается
среди людей, живущих в тропических лесах Центральной и Западной Африки. Люди
заражаются ВОО при прямом контакте с инфицированными животными (преимуще-
ственно обезьяны и белки рода Funisciurus) или больными людьми, а также при упот-
реблении в пищу термически необработанных органов больных животных. Среди жи-
вотных ВОО очень эффективно передается через воздух, экскременты (рвотные массы
и испражнения) и порезы на коже. Первичный природный резервуар ВОО неизвес-
тен. На рис. 3.95. показаны географические районы естественной встречаемости оспы
обезьян на африканском континенте и регионы, где она может быть обнаружена из-
за сходства экологических условий.
Изоляты ВОО, полученные от больных оспой обезьян, обитающих в Западной
Африке, менее вирулентны для обезьян и людей, чем изоляты этого же вируса, выде-
ленные от заболевших людей в бассейне реки Конго. Генетически эти изоляты пред-
ставляют собой разные клады ВОО (см. рис. 3.91). Смертельные исходы оспы обезьян
среди людей отмечены только в бассейне реки Конго (Chen N. et al., 2005).
В основном инфицированные ВОО животные встречаются рядом с водоемами в
зонах вторичных влажных тропических лесов, густо населенных полосатыми и сол-
нечными белками. Деревья масличной ореховой пальмы (Elaeisguineensis) обеспечи-
вают белок обильной пищей и одновременно привлекают к себе население этих ре-
гионов, занимающееся сбором орехов. Такие деревья растут в широких вторичных
лесах и на плантациях по всему западу Африки, располагаясь почти непрерывной по-
лосой 50—250 км вдоль побережья Сьерра-Леоне на западе до Южного Камеруна. Этот
пояс расширяется в Габоне и Конго, а также занимает почти весь бассейн реки Заир
На рис. 3.96 показана схема передачи ВОО из вторичного природного резервуара в
человеческие популяции.
Эпизоотическая цепочка, по которой распространяется оспа обезьян, может быть
длинной и тянуться с африканского континента в любую точку мира. Например, при-
чиной вспышки оспы обезьян среди населения штата Висконсин (США) был завоз
854
Часть 3. Частная неправильная эпидемиология
Рис. 3.95. Известные и возможные ареалы оспы обезьян на африканском континенте.
А. Территории, где выявлены случаи оспы обезьян среди людей. ВОО, встречающиеся
в Центральной Африке, более вирулентные для людей и обезьян, чем вирусы, цирку-
лирующие в Западной Африке. Б. Территории, где экологические условия сходны с те-
ми, на которых поддерживается ВОи. Более плотная штриховка показывает большую
вероятность заражения человека ВОО на этой территории. 1 — западноафриканские
штаммы; 2 — центральноафриканские штаммы По R. S. Levine et al. (2007)
► Наиболее распространенный путь м а Редкий
Менее распространенный --------Очен» редкий
Рис. 3.96. Схема передачи ВОО из вторичного природного резервуара в чело-
веческие популяции. Heliosuurus — род «солнечные белки». Funisciurus —
род «полосатые белки». Первичный природный резервуар ВОО не известен.
По L. Khodakevich et al. (1988)
3.8. Натуральная оспа, оспа обезьян
855
ВОО из Ганы с гамбийскими Хомяковыми крысами (Cricetomysgambianus). Крысы зара-
зили оспой обезьян луговых собачек1 в одном из магазинов в штате Висконсин, не кон-
тактируя с ними непосредственно. Луговые собачки были помещены в клетки, в кото-
рых ранее содержались больные крысы. Затем их в инкубационном периоде болезни
развезли по другим магазинам и продали населению. Всего оспой обезьян заболело
11 человек (5 мужчин и 6 женщин) в возрасте от 3 до 45 лет. В 10 случаях заболевание
возникло в результате контакта с больным животным (укусы, попадание вируса в ран-
ки на коже). В одном случае эпидемиологи предполагают передачу ВОО от человека
к человеку. Инкубационный период болезни занимал от 4 до 24 сут, в среднем 15 сут.
Так как ВОО относился к вирусам, циркулирующим в Западной Африке, тяжелых
случаев болезни не было (Reed К. D. et al., 2004).
В 1982-1984 гг. в Заире случаи заболевания оспой обезьян, вызванные передачей
ВОО от человека к человеку, составляли до трети всех случаев (66 из 210). Временные
периоды между контактом и появлением сыпи составляли от 7 до 23 сут. Летальность
среди детей в возрасте от 7 мес. до 7 лет достигала 11 %, в возрастной группе от 5 до
14 лет — 7,7 %. Примеры передачи ВОО между людьми приведены на рис. 3.97.
Деревни, в которых были обнаружены заболевания среди жителей
(количество жителей)
Bwene
(514)
Yamakolo
(125)
lipolo
(226)
Mwela
(1024)
— возраст/пол
19 сут — период
между контактом
и появления сыпи
lyeke
(80)
23 сут
21 сут
Рис. 3.97. Примеры цепочек передачи ВОО между людьми в Заире. Все заболевшие
оспой обезьян проживали в небольших деревнях, распложенных в зоне влажных
джунглей, и находились в тесном контакте друг с другом. Предполагается, что забо-
левший оспой обезьян человек мог быть источником ВОО для другого человека в
течение одной недели. М — мужчина, F — женщина. По F. Fenner et al. (1988)
Естественное инфицирование человека ВОО в России возможно в результате кон-
такта с коптрабандно завезенными из тропических лесов Центральной и Западной
Африки дикоживущих грызунов, белок и обезьян. Другая возможность естественно-
го инфицирования — это пребывание в эндемических по вирусу оспы регионах тро-
пической Африки.
‘Луговые собачки (Cynomys) — грызуны из семейства беличьих. Внешне напоминают сур-
ков. Живут в прерии Северной Америки.
856
Часть 3. Частная неправильная эпидемиология
Оспа коров. Это название болезни условно, спектр патогенности вируса оспы ко-
ров (CPV) оказался шире, чем считалось тогда, когда болезнь получила это название.
Перечень восприимчивых к СРУживотных продолжает увеличиваться. У большинства
из них болезнь протекает тяжело и сопровождается высокой летальностью. В значи-
тельном проценте случаев оспа коров у животных протекает в типичной для поксви-
русов форме с развитием характерных поражений (высыпаний) на коже и слизистых.
У слонов отмечался, кроме того, отек головы, задних конечностей. У отдельных живот-
ных происходила генерализация процесса, сыпь приобретала сливной характер с по-
следующим некрозом кожи и слизистых. Кожная форма инфекции у хищных семей-
ства кошачьих как и у слонов, окапи (крупное парнокопытное млекопитающее из
семейства жираф) и носорогов иногда заканчивалась летальным исходом. Оба случая
заболевания гигантских муравьедов в Московском зоопарке завершились смертью.
У кошачьих (львы, гепарды, черная пантера) болезнь может протекать в легочной фор-
ме со 100 % летальностью не позже 8 сут от появления первых симптомов. Клиника
легочной формы коровьей оспы характеризуется адинамией, отказом от пищи, затруд
ненным дыханием, одышкой, приступообразным кашлем и цианозом слизистых. На
аутопсии обнаруживают серозно-фибринозную пневмонию с массивным выпотом
(до 3 л) в плевральную полость. При легочной форме у животных, как правило, не наб-
людалось оспенной сыпи (Маренникова С. С., Щелкунов С. Н., 1998).
Для людей источником CPV могут быть домашние кошки, ламы, рыжие полевки,
белые крысы, собаки и кроты (Маренникова С. С. с соавт., 1996; Pahlitzsch R. et al.,
2006; Campe H. et al., 2009; Vogel S. et al., 2011). Природные резервуары CPV имеются
в Московской области. Ниже по работе С. С. Маренникова с соавт. (1996) привожу
краткую историю случая болезни, имевшего место в Московской области.
Девочка Л. Г., 4 лет, не вакцинированная против оспы, заболела в августе 1990 г. во
время пребывания на даче под Москвой. До заболевания в течение 2 дней она играла
с найденным на участке кротом, беря его в руки и прижимая к себе. Крот был необычно
спокойным и малоподвижным. Контакта с другими животными или больными людь-
ми не было. Через 2—3 сут девочка стала вялой, капризной, чувствовала жар. Были
очевидными ее недомогание и слабость. На 5 сут после контакта с кротом у нее появи-
лись жалобы на боль в правой подмышечной области, где при осмотре оказалось по-
краснение с узелком в центре, который через сутки превратился в пузырек с пупкооб-
разным вдавлением.
Состояние девочки ухудшалось, и родители увезли ее в Москву, где она была гос-
питализирована на 9 сут болезни с температурой 39,4 °C. При осмотре в правой под-
мышечной области была обнаружена окруженная зоной гиперемии и инфильтрации
крупная (диаметром 20х 18 мм), покрытая плотной коркой подсохшая пустула; значи-
тельно увеличенные и болезненные лимфатические узлы справа и слегка увеличенные
(до размера горошины) слева. На 10 сут рядом с первичным элементом появились че-
тыре «дочерних» везикулы, три из которых затем с ним слились. Увеличение и болез-
ненность региональных лимфатических узлов, субфебрильная температура и болезнен-
нее состояние сохранялись в течение 2 нед. После отпадения корок на месте поражения
остался отчетливый большой рубец.
При электронной микроскопии в материале из кожного поражения, взятого на
12 сут болезни, были выявлены вирионы, имевшие кирпичеобразную форму и разме-
ры. Инокуляция этого же материала на ХАО куриных эмбрионов сопровождалась фор-
мированием нежных поверхностных оспин среднего размера с яркими геморрагиями.
На некоторых оболочках можно было также заметить единичные белые оспины, выс-
тупающие над поверхностью ХАО. В целом характер поражений был сходен с таковым,
3.8. Натуральная оспа, оспа обезьян
857
вызываемым референс-штаммом CPV, однако в отличие от последнего, при 72-часовой
инкубации вторичные оспины не появились, а сами оспины были мельче. Исследова-
ние сыворотки крови девочки иммунологическими способами также свидетельство-
вало об инфицировании ребенка CPV.
Когда CPV инфицируются люди с иммунодефицитами различного происхожде-
ния, оспа коров может по клинике напоминать натуральную оспу. Такой случай опи-
сан в Германии С. Р. Czerny et al. (1991) у 18-летнего пациента. Он не был вакциниро-
ван против натуральной оспы, страдал экземой и аллергическим астматическим
бронхитом, принимал иммуносупрессивные препараты. Оспа коров у пациента раз-
вилась сразу как генерализованная инфекция с поражением легких и частично слив-
ными экзантемами, распространившимися по всему телу. Клинические симптомы
болезни были сходны с «variola pustulosa haemorrhagica». Больной погиб от легочной
эмболии в палате интенсивной терапии. Сводные данные по клинической манифес-
тации оспы коров у людей, географии и источникам их заражения в России, приве-
дены в табл. 3.28, с. 858.
П ередача CPVот человека к человеку наблюдается очень редко и происходит толь-
ко при тесном контакте с больным. Внешние признаки болезни у животных и внешние
признаки болезни у людей приведены на рис. 3.98 и рис. 3.99, с. 859.
Восприимчивость к возбудителю натуральной оспы людей и животных. Среди ие-
вакцинированных людей есть как невосприимчивые к ВНО лица, так и обладающие
повышенной восприимчивостью. Во времена крупных эпидемий второй половины
XIX в. наблюдали, что на 100 взрослых 5—6 человек не заболевают, несмотря на «пре-
бывание в заразной обстановке». Обширные наблюдения врожденной невосприим-
чивости к ВНО стали накапливаться с началом кампаний массовой вакцинации. При
систематической вакцинации в Германии некоторая часть детей обнаруживала имму-
нитет. Так, по наблюдениям Р. Коха (1880), на 58913 привитых детей 780 дали отрица-
тельный результат при однократной и повторной прививке и 48 — при троекратной.
Описаны случаи 5-, 6- и 8-кратной безуспешной прививки. Процент невосприимчи-
вых к вакцине детей, у различных авторов сильно колеблется: от 0,08 до 2,44 %, что, по
мнению Н. Ф. Гамалеи (1913), отражает частоту оспенных эпидемий в данной группе
населения в прошлом. Он объяснял это явление тем, что при долгом существовании
среди населения какой-либо опасной инфекции, наиболее поражаются и вымирают
самые восприимчивые к ней индивидуумы, остаются же и оставляют потомство те,
которые обладают сравнительно прочной иммунной системой.
Сверхвосприимчивость у людей к ВНО проявлялась во время оспенных эпидемий,
по крайней мере, шестью группами событий:
1) повторными (многократными) заболеваниями у отдельных лиц и семей;
2) высокой смертностью в отдельных возрастных группах населения;
3) крайне тяжелым типичным течением болезни у отдельных лиц, при относитель-
но легком ее развитии у большинства остальных заболевших;
4) крайне тяжелым нетипичным течением оспы или вакцинного процесса у отдель-
ных лиц при типичном их развитии у остальных заболевших;
5) высокой смертностью в отдельных человеческих популяциях (этнических
группах);
6) микст-эпидемиями — в этом случае натуральная оспа либо завершает эпиде-
мическую катастрофу, вызванную другими опасными инфекциями, либо встраивается
в последовательно развивающуюся цепочку эпидемических катастроф, где они вместе
или, следуя «друг за другом», в каком-то смысле, уподобляются «бичу Божьему».
ос
<-А
ОС
Таблица 3.28
Случаи оспы коров у людей в России*
Пациент Пол Возраст в годах Вакцинация против оспы в прошлом Профессия Источник заражения Место и время зараже- ния Клинические симптомы Наличие вируса в содержимом кожных поражений Наличие антител к CPV(EIA-A)b сыворотке крови
Е. Н. Ж 34 + Сотрудница вивария Укус больной белой крысы Москва, январь. 1987 На 3 сут — недомогание, головная боль, слабость температура до 38 °C, поражение на месте укуса Выделен CPV (изолят Ель-87) Обнаружены видо- специфические антитела к CPV
С. О. ж 31 + Ветеринар- ный врач вивария Укус больной белой крысы (через резино- вую перчатку) Москва, апрель, 1987 На 3 сут — слабость, недомогание, субфебрильная температура; неболь- шое поражение на месте укуса; уве- личение регионарного лимфоузла Обнаружены вирионы поксвируса То же
Г. А. м 10 + Учащийся Точно не уста- новлен (пред- положительно мелкие полевые грызуны) Смолен- ская обл., август, 1988 На 3-4 сут — слабость, недомогание, температура до 39 °C; подчелюстной лимфаденит; боль и поражения в области правого уха; на 12-13 сут высыпания на нижних конечностях Выделен CPV (изолят Арго-88) Тоже
Г. Е. ж 4,5 - Точно неизвестно (тесный контакт с кротом) Москов- ская обл., август, 1990 На 3 сут — вялость, недомогание, температура до 40°C; на 5 сут — боль и местные поражения в правой под- мышечной области; подмышеный лимфаденит. На 10 сут вторичные высыпания вокруг первичного очага То же (изолят GRI-90) То же
Р. Е. м 12 - Учащийся Тесный контакт с белой крысой-вирусо- носителем ** Москва, апрель, 1991 На 2-3 сут — слабость, недомогание, температура до 38 °C; местные пора- жения на плече и шее справа, под- мышечный лимфаденит справа То же (изолят Rev-91) То же
Д. К. м 10 Учащийся Тесный контакт с белой крысой-вирусо- носителем ** Москва, май, 1991 На 3 сут — слабость, недомогание, температура до 38-40 °C; поражения на туловище, на 6-8 сут — на лице и руке; двусторонний лимфаденит То же (изолят DK-91) То же
* По С. С. Маренниковой и С. Н. Щелкунову (1998).
Часть 3. Частная неправильная эпидемиология
3.8. Натуральная оспа, оспа обезьян
859
Рис. 3.98. Кожные поражения при оспе коров у животных. А. Кожные пора-
жения в виде кратера вокруг ануса ламы. По С. Cardetietal. (2011). Б. Кожные
поражения на лапках крыс. По S. Vogel et al. (2011)
Рис. 3.99. Кожные поражения при оспе коров у людей. А. Некротические пора-
жения и лимфаденит у 16-летней пациентки, ранее не вакцинированной против
натуральной оспы. Б. Кожные поражения у ее матери (40 лет). Ранее была вак-
цинирована против натуральной оспы. Оба снимка сделаны через 15 сут после
приобретения в зоомагазине белой крысы, инфицированной CPV. По Н. Campe
et al. (2009). В. Некротические поражения и лимфаденит у 16-летней пациент-
ки. Г. Раннее проявление кожного поражения у 42-летнего мужчины — язва
диаметром 35 мм с некротическим дном и лимфаденит. По S. Vogel et al. (2011)
Крайне тяжелое нетипичное течение вакцинального процесса у отдельных лиц,
при типичном их развитии у остальных вакцинированных, наблюдали в конце XIX в.
в регионах Африки, население которых сегодня поражено СПИДом. Так, из 1156 де-
860
Часть 3. Частная неправильная эпидемиология
тей, привитых британскими врачами в Стране зулусов (Зулуленд), у 240 из них вакци-
нация осложнилась генерализацией процесса. Для 16 детей она окончились смертью
при явлениях бронхопневмонии. У 20 % вакцинированных детей общее высыпание
наступило через 28—44 сут после прививки, а у 75 % на 8 сут наблюдалось появление
вторичных пузырьков вокруг первичной оспины (master-pox), а затем развивалось
общее высыпание в течение 2-3 мес. Высыпания имели все характерные признаки
оспы: сначала пятна, затем папулы и везикулы с пупком (Гамалея Н. Ф., 1913).
Инфицирующая доза ВНО для человека составляет несколько вирусных частиц,
более точные данных в открытых источниках я не нашел. Инфицирующие дозы ВНО
для приматов на несколько порядков превышают инфицирующие дозы для челове-
ка. К экспериментальному заражению большими дозами ВНО восприимчивы как
человекообразные обезьяны (орангутаны), так и ряд низкоорганизованных видов этих
животных: макака-резус, яванские обезьяны и др. Вирус патогенен для обезьян при
различных путях введения. Описаны результаты успешного заражения нанесением
ВНО на скарифицированную кожу, введением внутривенно, с использованием аэро-
золя, в тестикулы, бронхи, трахею. При интрацеребральном заражении вирус не
вызывает болезни. Инокуляция вируса ВНО обезьянам может вызывать у них в зави-
симости от дозы и способа заражения либо местный процесс, либо генерализован-
ную инфекцию различной тяжести, напоминающую отдельные клинические формы
оспы у человека (от дискретной до геморрагической). Обычно генерализованная ин-
фекция протекает в относительно легкой форме и заканчивается выздоровлением.
Инфицирующие дозы для приматов в зависимости от пути введения вируса приведе-
ны в разд. «Клиническая картина натуральной оспы у животных при искусственном
инфицировании».
По данным большинства исследователей, ВНО не патогенен для взрослых белых
мышей при различных путях введения. Выявлена восприимчивость к ВНО белых
мышей-сосунков. Вирус размножается и может поддерживаться в пассажах при внут-
рибрюшинном введении. При введении ВНО в мозг новорожденным мышам у них
наблюдается генерализованная инфекция с размножением вируса в мозгу, во внутрен-
них органах и с двумя волнами виремии. Повышение восприимчивое ги к ВНО взрос-
лых белых мышей может быть достигнуто с помощью облучения (60Со). В этих усло-
виях как клиническое течение болезни, так и вирусологические показатели становятся
такими же, как у 10—12-суточных, естественно восприимчивых к вирусу мышей.
Данные о восприимчивости к ВНО белых крыс противоречивы. Наряду с указа-
ниями на отсутствие патогенности ВНО для этих животных имеются сообщения о
способности некоторых штаммов образовывать папулы при нанесении вируса на ска-
рифицированную кожу, специфического кератита при введении вируса на роговую
оболочку и интерстициальной пневмонии с цитоплазматическими включениями в
пораженных клетках при интраназальном заражении.
К ВНО невосприимчивы морские свинки, кролики, золотистые хомячки, хлоп-
ковые крысы, свиньи, козы, бараны и телята. Помимо дозы, на тяжесть и исход болез-
ни влияет возраст животного: молодые животные более чувствительны к ВНО.
Восприимчивость к возбудителю оспы обезьян у животных. ВОО весьма патогенен
для белок, кроликов, белых мышей, хлопковых крыс, многососковых крыс, обезьян,
и, возможно, антилоп. Из обезьян наиболее чувствительны к ВОО орангутаны. Обезь-
яны могут быть бессимптомными носителями ВОО. К вирусу невосприимчивы зо-
лотистые хомячки, белые крысы и морские свинки (Маренникова С. С., Щелку-
нов С. Н., 1998). Инфицирующие дозы для отдельных видов животных, в зависимости
3.8. Натуральная оспа, оспа обезьян
861
от пути введения вируса, приведены в разделе «Клиническая картина натуральной
оспы у животных при искусственном инфицировании».
Иммунитет к поксвирусам. Основной антигенной детерминантой поксвирусов и
мишенью для протективных антител является консервативный белок L1 (другое на-
звание L1R). У вируса вакцины, вируса оспы обезьян и ВНО этот белок различается
отдельными аминокислотами. Антитела к L1 способны блокировать инвазию орто-
поквирусов в клетки. Поэтому он рассматривается учеными в качестве кандидата на
включение в перспективные противооспенные вакцины (рис. 3.100).
Рис. 3.100. Структура белка L1. А. Две проекции белка на ленточной диаграмме. Фрон-
тальный вид (верхнее изображение) показывает расположение спиралей на одной сто-
роне молекулы, на другой стороне изображены параллельные р-складки, сформирован-
ные непараллельными p-цепями, соединенными через петлю и тремя дисульфидными
связями. Нижняя проекция показывает рЗ, Р4 и <х5, упакованные вблизи центра моле-
кулы. Обозначены спирали (otl—а5), N-конеп (N), и С-конец (С). Б. Вверху показан
димер L1. Взаимодействие двух L1 молекул происходит через края р-складок. Нижнее
изображение показывает L1 со стороны р-складок (темная область). L1 — крупная
консервативная белковая молекула-мономер с несколькими конформационными
эпитопами презентируется иммунным клеткам в виде димера. По Su Hua-Poo (2005)
На заключительной стадии морфогенеза вируса образуются так называемые внут-
риклеточные зрелые вирионы (intracellular mature virions, IMVs; см. рис. 3.89), кото-
рые имеют липидные мембраны и представляют собой уже способные к инфекции
частицы вируса. Большинство IMV высвобождаются из фагоцитирующей клетки пос-
ле ее лизиса. Некоторые из них могут транспортироваться через клеточную мембрану
хозяина до лизиса клетки. После чего они остаются присоединенными к ее наружной
поверхности или отделяются от нее уже как внеклеточный оболочечный вирус (extra-
cellular enveloped virus, EEV). Главную роль в передаче вируса от одного хозяина к дру-
гому играет его IMV-форма (Su Н. et al., 2005).
L1 играет основную роль в «созревании» вирионов ортопоксвирусов. Он представ-
ляет собой миристиолированный оболочечный белок (myristoylated envelope protein),
862
Часть 3. Частная неправильная эпидемиология
состоящий из 250 аминокислотных остатков, экспрессирующийся на поверхности
IMV-формы вируса. Его С-гидрофобный сегмент погружен в вирусную мембрану, но
консервативная часть L1, включающая 185 аминокислотных остатков, локализуется
в цитоплазме клетки-хозяина. Консервативность L1 как антигена предопределяет его
ключевая роль в морфогенезе вируса. В области N-конца белка L1 формируется гид-
рофобная «каверна», необходимая для сборки вириона. После лизиса клетки этот
эктодомен экспонируется Т- и В-клеткам иммунной системы и вызывает сильный
ответ с их стороны (Su Н. et al., 2005).
Мыши, дефектные по В-клеткам (В-клетки синтезируют и секретируют специ-
фические антитела), так же как и мыши, истощенные по CD4 (помогают в активации
и трансформации В-лимфоцитов и в синтезе иммуноглобулинов), неспособны ос-
вободиться от вируса и погибают. Так же они оказались неспособны переключаться
на IgG (высокоаффинный и количественно доминирующий иммуноглобулин) или на
IgM-ответы (IgM низкоаффинный иммуноглобулин, преобладающий во внутрисосу-
дистом пуле иммуноглобулинов в начале инфекционного процесса). В ответах на ос-
трую инфекцию вирусоспецифические антитела наиболее эффективно освобождают
организм от вируса. Вирус разрушается по различным механизмам, предполагающим
участие специфических антител. Во-первых, антитела могут непосредственно связы-
вать вирус, вызывая его агрегацию и препятствуя адсорбции и интернализации в клет-
ках. Во-вторых, они могут связывать вирус, вызывая его разрушение с помощью ком-
племента или опсонизации, а затем посредством фагоцитоза. Антитела к L1 могут
связывать инфицированные клетки хозяина, вызывал цитотоксические реакции со
стороны клеток-киллеров и др. (Xu R. et al., 2004).
Кроме белка L1, в иммунном ответе на ортопоксвирусы участвуют еще два анти-
гена. Один из них, так называемый ранний антиген (ES), формируется на мембранах
инфицированных клеток до начала синтеза ДНК. Антитела, вырабатывающиеся в
ответ на введение этого антигена, реагируют с ним в РСК, но не нейтрализуют вирус.
Такой же антиген выявлен у большинства штаммов вируса вакцины, а также вирусов
оспы обезьян и оспы коров. Другой антиген вируса оспы, гемагглютинин, впервые
обнаруженный у ортопоксвирусов Барнетом, считался лишенным выраженной видо-
специфичности, хотя ряд авторов отмечали, что титры в РТГА с гомологичным виру-
сом были обычно выше, чем с гетерологичным. Лишь в самое последнее время после
расшифровки структуры гена гемагг. потинина ряда ортопоксвирусов и кодируемых
им полипептидов, удалось не только установить видоспецифические различая отдель-
ных генов гемагглютинина, но и разработать на этой основе методы межвидовой диф-
ференциации ортопоксвирусов (Маренникова С. С., Щелкунов С. Н., 1998).
Иммунитет к оспе может быть создан путем прививки оспенной вакцины. Одна-
ко напряженность и продолжительность такого иммунитета меньше, чем после пере-
несенной натуральной оспы. По данным Комитета экспертов ВОЗ по оспе (1964),
иммунитет после первичной вакцинации примерно в 10 раз слабее, чем после пере-
болевания оспой.
Пассивный иммунитет к ВНО может быть создан путем введения ИЛП, содержа-
щих противооспенные антитела (сыворотки реконвалесцентов, иммуноглобулины из
сывороток гипериммунизированных животных или недавно вакцинированных лю-
дей). Имеются доказательства эффективности со Дания таким образом иммунитета
как в эксперименте, так и в эпидемиологических условиях. Пассивный иммунитет
может быть также обусловлен трансплацентарным переносом антител от матери но-
ворожденному.
3.8. Натуральная оспа, оспа обезьян
863
Натуральная оспа повторяется у одного и того же человека очень редко — по на-
блюдениям Н. Ф. Гамалеи (1913), 1:3000 переболевших, но через 50 лет Комитет экс-
пертов ВОЗ по оспе (1964) оценивал это соотношение уже как 1:1000.
Устойчивость возбудителя натуральной оспы во внешней среде. В высушенном со-
стоянии ВНО более устойчив к воздействию высокой температуры. Так, при темпе-
ратуре 100°Сон погибает через 10 мин, а при температуре 60°С —через 1 ч, в то время
как в жидком состоянии (суспензия) вирус инактивируется полностью при темпера-
туре 60°C через 30 мин. В паровоздушной камере полное обеззараживание достига-
ется при температуре 96°C при экспозиции 45 мин, а в пароформалиновой камере —
при температуре 57°С через 1 ч при введении формалина из расчета 150 мл на 1 м3.
Для инактивирования ВНО, содержащегося в корочках больного натуральной
оспой, требуется воздействие 3 %-ного раствора хлорамина в течение 1 ч, тогда как
вирус в жидком состоянии погибает в 1 %-ном растворе хлорамина в течение 30 мин.
Для обеззараживания выделений больного применяли растворы хлорамина в большей
концентрации в связи с тем, что белковые субстанции выделений создают защитный
барьер. Для обеззараживания слюны требовался 2 %-ный раствор хлорамина с экспо-
зицией 30 мин, а для обеззараживания мокроты — 3 %-ный раствор и экспозиция 1 ч,
5 %-ный раствор хлорамина инактивирует вирус в клетках ХАО в течение 2 ч. Губи-
тельно действуют на вирус 3 %-ные растворы фенола и лизола в течение 30 мин, освет-
ленный I %-ный раствор хлорной извести, 1 %-ный раствор хлор-Р-нафтола и 0,5 %
гексилрезорцина в течение 1 ч. Чувствителен ВНО к сулеме и окислителям типа мар-
ганцовокислого калия. ВНО неустойчив к действию света. На свету он погибает через
35 сут, а при хранении в темноте — через 84 сут. В течение нескольких лет вирус выжи-
вает в 50 %-ном глицерине, не теряя активности. Оптимальной зоной pH для ВНО
является 7—7,4. Изменение pH как в щелочную, так и особенно в кислую сторону, при-
водит к снижению активности вируса. При pH 3 ВНО инактивируется в течение 1 ч.
Наиболее длительно ВНО сохраняется в оспенных корках. По данным Е. Ф. Бе-
ланова с соавт. (1997), проведших исследование оспенных корочек, хранившихся
26 лет при температуре —20°С, жизнеспособный вирус был выделен практически из
всех семи изученных проб от разных больных. На основании сделанных расчетов ав-
торы пришли к выводу, что продолжительность сохранения жизнеспособности ВНО,
находящегося в подобных условиях, может быть более 100 лет и таким образом име-
ются реальные шансы обнаружить живой вирус в трупах жертв оспы, захороненных в
мерзлотных грунтах.
Находясь во взвеси ХАО, ВНО выдерживает прогревание при температуре 50°С в
течение двух часов без резкого снижения инфекционной активности. Однако даль-
нейшее повышение температуры оказывается критическим: при температуре 60 °C он
инактивируется за 10 мин, а при температуре 70°С — за 5 мин. Кипячение разрушает
вирус практически сразу. Высушивание вируссодержащих культур (из замороженно-
го состояния под вакуумом) значительно увеличивает устойчивость вируса к темпе-
ратуре (Маренникова С. С., Щелкунов С. Н., 1998).
Антивирусные препараты. В отличие от бактериальных инфекций, контролировать
с помощью химиотерапевтических препаратов вирусные инфекции очень сложно.
Бактерии, инфицировавшие клетку, для своей репликации используют собственные
метаболические пути, и на них можно влиять с помощью антибиотиков и химиопре-
паратов. Вирусы реплицируются в клетке с использованием ее собственных метабо-
лических путей. Механизмы действия антивирусных препаратов и механизмы резис-
тентности к ним у вирусов, обобщены в табл. 2.9.
864
Часть 3. Частная неправильная эпидемиология
Выбор химиопрепаратов для лечения поксвирусных инфекций основывался на
результатах изучения особенностей репликационного цикла поксвирусов и синтеза
вирусных белков, позволяющих осуществить их избирательный таргетинг, не нару-
шая метаболических процессов инфицированной клетки. Ниже, в основном, по рабо-
те К. Sliva и В. Schnierle (2007), приведено краткое описание препаратов, обладаю-
щих активностью в отношении инфекций, вызываемых поксвирусами.
Метисазон (methisazonum, 2-(1,2-дигидро-1-метил-2-оксо-ЗН-индол-3-илиден)-
гидразинкарботиоамид; синонимы: kemoviran, marboran, metisazon, viruzona, мербо-
ран) — нарушает процесс сборки вирионов, угнетает синтез вирусных структурных
белков. До 1960-х гг. рассматривался как противотуберкулезный препарат. В 1960 гг.
активно изучался как препарат для химиотерапии натуральной оспы. Клинически
было доказано, что этот препарат не эффективен при лечении развившейся болезни,
но снижает заболеваемость и смертность, при введении с профилактической целью
людям, контактировавшим с больными натуральной оспой. Наиболее оптимальным
считался прием peros 3 г препарата двумя дозами с интервалом 8 ч (McClain D. J., 1997).
Препарат используется для лечения больных с поствакцинальными осложнениями
(вакцинальная экзема, гангренозная вакцина и др.).
Рифампицин (rifampicinum, rifampin; 3-[[(4-метил-1-пиперазинил)имино]ме-
тил]) — подавляет ДНК-зависимую РНК-полимеразу микроорганизмов. Противоту-
беркулезный препарат. Размножение поксвирусов рифампицин блокирует, предот-
вращая расщепление прекурсорных белков. Клиническая эффективность показана
только при применении очень высоких доз, что исключает его широкое применение.
Рибавирин (ribavirin, copegus, rebetol, ribasphere, vilona, virazole) — синтетический
аналог нуклеозидов, антивирусный препарат с плохо изученным механизмом действия
в отношении ДНК-вирусов.
Цидофовир (CDV-Vistide, вист ид) — ациклический нуклеозид фосфонат (acyclic
nucleosidephosphonate). Селективно ингибирует вирусную ДНК-полимеразу, тем самым
блокирует вирусную репликацию и транскрипцию. Рекомендуется для лечения покс-
вирусных инфекций, в том случае, если гипериммунный противооспенный иммуно-
глобулин оказывается неэффективным. Путем присоединения липида к молекуле
цидофовира создан модифицированный препарат, который в 100 раз более эффектив-
но, чем исходный, блокирует размножение ВНО в культурах клеток. В опытах на мы-
шах показана его значительно более высокая и длительная профилактическая эффек-
тивность при ингаляционном введении (Bray М., Roy С., 2004). Механизмы резистен-
тности поксвирусов к цидофовиру показаны на рис. 2.20.
Адефовир (adefovir dipivoxil, HEPSERA, ГЕПСЕРА) — ациклический аналог адено-
зина монофосфата. В клетке фосфорилируется под действием клеточных киназ в ак-
тивный метаболит—адефовира дифосфат. Он ингибирует ДНК-полимеразу ДНК-ви-
русов (обратную транскриптазу), конкурируя с естественным субстратом дезоксиаде-
нозина трифосфатом и вызывая обрыв цепи ДНК после его инкорпорации в вирусную
ДНК. В условиях in vitro показывает значительную активность в отношении поксви-
русов. Хорошо переносится человеком при приеме per os.
Новое направление в разработке антипоксвирусных препаратов включает раз-
работку киназных ингибиторов, ранее применявшихся только для лечения онкологи-
ческих болезней. Это новый класс препаратов, не ингибирующих репликацию покс-
вирусов, но уменьшающий их способность передаваться между клетками через бло-
кирование полимеризации актина, играющего важную роль в межклеточной адгезии
и передаче вирусов. Среди таких препаратов наиболее изучен противолейкозный цито-
3.8. Натуральная оспа, оспа обезьян
865
статический препарат иматиниб (Gleevec, STI-571, imatinib mesylate или Glivec). На
рис. 3.101 приведено схематическое изображение цикла репликации поксвирусов с
указанием «точек воздействия» антипоксвирусных препаратов.
Присоединение
и слияние
Рифампицин
Противоо' гк ними
гаммаг-зэбулин
Ассоцииро-
ванный с клеткой
оболочечный
Ядро
и эндоплазматический
ретикулум ™
fv j л ST-246 Gleevec
CI-1033
Ингибиторы протеазы
Внеклеточный
оболочечный
вирус
Рис 3.101. Схематическое изображение цикла репликации поксвирусов с указанием
«точек воздействия» антивирусных препаратов. По К. Sliva, В. Schnierle (2007)
Виеклеточны!
оболочечный А
«ирус
Р»чликация ДНК вирус?
«Раздевание»
вир”са
в белков, ю оболочку
Сборка
вириона [ липив
Обнаружение генетически измененных штаммов ортопоксвирусов. Коллекции ВНО
хранятся в двух государственных лабораториях в России и США и теоретически ВНО
не доступен для террористов. К тому же он не имеет «недостатков», которые могут
вызвать у преступников желание вмешаться в его геном, а потом запутаться в раз-
нообразных рукотворных эффектах при культивировании и хранении полученного
штамма вируса. Значительно большую опасность, чем генетически измененные штам-
мы, представляют природные антигенно-атипичные штаммы ВНО, обнаруженные в
средине 1970-х гг. (см. работу Маренниковой С. С., Щелкунова С. Н., 1998). В отно-
шении таких вирусов вакцинация может оказаться малоэффективной. Более вероят-
но, что объектом генно-инженерного вмешательства преступников станет не ВНО, а
другие патогенные и даже непатогенные для человека ортопоксвирусы. В разд. 2.2.2
«Механизмы вмешательства в геном вирусов» приводится описание таких экспери-
ментов. Подозрение о применении генетически модифицированных или специаль-
но селекционированных ВНО и ВОО должно возникнуть при:
1) обнаружении у заболевших людей штаммов вирусов, резистентных к препара-
там, обычно используемым для лечения поксвирусных инфекций;
2) установлении низкой защитной эффективности вакцинации, проведенной
живыми противооспенными вакцинами.
Достоверно установить же факт вмешательства в геном ортопоксвируса, исполь-
зованного для совершения преступления, можно по результатам секвенирования,
сопоставив выявленную нуклеотидную последовательность с эталонной. Интерес у
экспертов должны вызвать нуклеотидные последовательности генов токсинов эука-
866
Часть 3. Частная неправильная эпидемиология
риотических организмов; генов других вирусов, особенно когда обнаруживаются реп-
ликоны этих вирусов; гены белков, участвующих в регуляции иммунных ответов, но
отсутствующие в геноме эталонного вируса; гены прионовых белков.
Патогенез ортопоксвирусных инфекций. Общим для всех генерализованных орто-
поксвирусных инфекций, включая натуральную оспу и оспу обезьян, является ступе-
необразный характер распространения вируса по организму, проходящий три основ-
ных этапа: инфицирование клеток, репликация вируса и его «выброс» в кровеносное
русло. Заражение человека происходит через слизистые оболочки верхних дыхатель-
ных пу гей, куда ВНО проникает воздушно-капельным или воздушно-пылевым путем.
Не исключено также заражение через поврежденную кожу. Если заражение произошло
через поврежденную кожу, то первоначально возникает локальное поражение на ме-
сте внедрения вируса с последующей генерализацией процесса. Подобный путь зара-
жения возможен, но для вспышек естественного происхождения он не имеет практи-
ческого значения и встречается чрезвычайно редко.
Первыми клетками, вступающими в контакт с ортопоксвирусами, являются поли-
морфоядерные лейкоциты (макрофаги и моноциты). За счет градиента продуцируе-
мых ими хемоаттрактантов они «притягивают» другие фагоцитирующие клетки и за-
пускают каскад самых разнообразных реакций, которые «в сумме» рассматриваются
учеными как «иммунные». В составе макрофагов и моноцитов вирус достигает дре-
нажных лимфатических узлов примерно через 8 ч. Разрушившиеся фагоцитирующие
клетки становятся источником вируса, а через 2—3 сут его обнаруживают уже в крови
(первичная виремия), костном мозге, печени и селезенке. В пробах крови больного на-
туральной оспой в этот период можно обнаружить как инфицированные, так и неин-
фицированные лейкоциты, но в основном репликация ВНО происходит в моноци-
тах/макрофагах, тогда как Т -лимфоциты остаются неинфицированными, но при этом
имеет место выраженная лимфопения. У больного наблюдается первый подъем тем-
пературы до 39,5- 40,5 °C, сопровождающийся ознобом, тошной, рвотой, мучитель-
ной головной болью.
Вирус распространяется из лимфатических узлов фагоцитирущими клетками кро-
ви по внутренним органам через выносящие (эфферентные) лимфатические сосуды.
Обладая выраженным тропизмом к тканям эктодермального происхождения, вирус
проникает в эпителий кожи и слизистых, в печень, костный мозг и другие ткани и орга-
ны ретикулоэндотелиальной системы, где начинается его репликация Количество
вируса через 4-5 сут уже превышает емкость ретикуло-эндотелиальной системы, и он
вновь проникает в кровяное русло.
Вторичная виремия обычно соответствует началу клинической манифестации бо-
лезни. Развиваются паренхиматозные изменения во внутренних органах. На коже
начинается циклическое развитие папулезной и везикуло-пустулезной сыпи (экзан-
тем), заканчивающееся образованием корок. В тканях слизистых оболочек рта, язы-
ка, зева, гортани, трахеи и др. появляются внутренние сыпи — энантемы.
Начальный элемент сыпи на коже — макула (лат. macula — пятно) — пятно блед-
но-красного цвета, слегка возвышающееся над нормальной кожей. Развивается вслед-
ствие гиперемии дермы. Далее наступают отек, воспалительная инфильтрация со-
сочкового слоя эпидермиса и пролиферация глубоких слоев, создающая утолщения,
что ведет к образованию бесполостных узелков или папул (лат. papula — узелок) на
3—7 сут болезни, более темного цвета и диаметром около 3 мм. Эпителиальные клет-
ки глубокого слоя подвергаются коагуляционному некрозу и отчасти вакуольному пе-
рерождению, при котором протоплазма клеток разжижается, отходит от ядра, обра-
3.8. Натуральная оспа, оспа обезьян
867
зуя перенуклеарные пространства, которые заполняются жидкостью. Клетки теряют
ядра, оболочки их разрываются, и на 7-8 сут болезни образуется пузырек — везикула
(лат. vesicula — пузырек), т. е. элемент, имеющий дно, покрышку и полость. Образо-
вание везикул сопровождается дискомплекцией эпидермальных клеток, расслаива-
нием эпидермиса и формированием длинных тяжей, направленных от основания пу-
зырька к его верхушке и делящих его на камеры. Пролиферация клеток эпителия по
периферии пузырька образует валик, приподнимая его края. В центре сохраняются
тяжи, которые втягивают поверхностную оболочку пузырька. Это ведет к образова-
нию оспины — везикулы с центральным (пупкообразным) западанием.
Западание в центре оспин характерно только для оспы. Оно не встречается при
пустулезных дерматитах иной этиологии. Нарастающие явления воспаления в сосоч-
ковом слое дермы и миграции лейкоцитов из капилляров в экссудат, заполняющий
пузырек, придают последнему молочно-мутный оттенок. Содержимое везикулы, как
правило, подвержено вторичному инфицированию стафилококками, стрептококка-
ми и другими микробами, поэтому на 9— 10 сут экссудат становится серозно-гнойным
и приобретает серовато-желтую окраску; центральное западание исчезает, формиру-
ется оспенная пустула (лат. pustule — гнойничок). При повышенной проницаемости
сосудов в экссудат проникает большое количество эритроцитов, пустула становится
геморрагической.
Увеличение количества экссудата ведет к истончению стенки покрова пустулы с
последующим нарушением его целостности. Через поврежденную стенку покрова
содержи мое вытекает из пустулы и, подсыхая, образует оспенную корку. Цвет корок за-
висит от состава экссудата: если в нем преобладает серозно-гнойное содержимое, то
оно, подсыхая, образует серовато-бурые, а при геморрагическом компоненте — чер-
ные корки.
Второй подъем температуры наступает на 9— 10 сут болезни. Он обусловлен нагно-
ением содержимого пустул и связанного с ним всасывания пирогенных веществ, об-
разующихся при разрушении клеток, а также вследствие присоединившейся вторич-
ной инфекции. У больных, ослабленных оспенной инфекцией, вторичная инфекция
может послужить причиной серьезных осложнений — вызвать флегмоны, абсцессы,
рожистое воспаление, бронхопневмонию и т. д.
В основе патогенеза поксвирусных инфекций лежит способность вирусов к интер-
ференции с защитными иммунными механизмами макроорганизма. Такая стратегия
поксвирусов включает синтез инфицированными фагоцитирующими клетками вирус-
ных гомологов хемокинов или хемокиновых рецепторов и других белков, связы-
вающих хемокины, синтезируемые неинфицированными фагоцитирующими клетка-
ми. Она не оригинальна; примен ительно к фагоцитирующим клеткам такая стратегия
используется большинством ДНК вирусов (рис. 3.102; табл. 3.29).
Постоянно находясь во влажной среде, слизистые оболочки верхних дыхательных
путей мацерируются, изъязвляются, и заразное содержимое, выделяясь на влажную
поверхность слизистой оболочки, инфицирует секрет и служит материалом для об-
разования капельной инфекции. Выделение вируса из зева отмечалось, начиная с
3—4 сут, т. е. практически с появления энантемы, и продолжалось в течение 7—9 и даже
13 сут в зависимости от тяжести болезни и ее типа.
Циркулирующие в крови специфические антитела выявляются уже на 7 сут после
инфицирования человека ВНО, и их максимальный уровень достигается на 14 сут.
Одновременно с нарастанием количества специфических антител, происходит сни-
жение количества вируса в тканях.
868
Часть 3. Частная неправильная эпидемиология
Плексин С1
связывающий
вирусный хемокин
Вирусный
семафорин
Хемокин (CCL3, CCL5)
Несозревшие дендритные клетки (DC;
___ I Хемокиновый рецептор
(yv)--1 I переключается на
ф активацию
-Хемокин (CCL19,
< :XCL12)
Рис. 3.102. Модуляция хемокиновой системы ортопоквирусами. А. Экспрессия
вирусных гомологов хемокинов, рецепторов хемокинов и хемокинсвязывакмцих
белков. Б. Частный случай антихемокиновой стратегии — ингибирование мигра-
ции созревающих дендритных клеток (DC). Вирусные хемокинсвязывающие белки
интерферируют с хемокинами CCL3 и CCL5. Несозревшие DC экспрессируют
воспалительные хемокиновые рецепторы (CCR1, CXCR1) и мигрируют в инфи-
цированные ткани в ответ на такие хемокины, как CCL3 и CCL5 Созревшие DC
экспрессируют CCR7 и CXCR4 и мигрируют во вторичные лимфоидные органы
в ответ на CCL19 и CXCL12, где они представляют антиген «наивным» Т-клет-
кам. 1 — вирусный хемокин (антагонист); 2 — вирусный хемокин (агонист); 3 —
хемокиновый рецептор; 4 — рецептор вирусного хемокина; 5 — сигнальная
трансдукция и биологический эффект; 6 — клеточная миграция. По A. Alcami (2007)
Клиническая и патологоанатомическая картина натуральной оспы у человека при
естественном инфицировании.1 Течение типичных форм болезни средней тяжести у не-
привитых включает три периода:
1) инкубацию;
2) болезнь;
3) выздоровление.
Инкубационный период чаще всего длится 9— 12 сут. Максимальная его продолжи-
тельность — 15 сут.
Период болезни подразделяется на:
а) стадию предвестников (продром), в среднем длящуюся 2—4 сут;
б) стадию высыпания продолжительностью 4—5 сут;
в) стадию нагноения длительностью 7—10 сут.
Период выздоровления длится 20—30 сут. В нем выделяют стадию подсыхания и
стадию отпадения корок. В целом заболевание средней тяжести продолжается около
1 Для описания клиники и патанатомии натуральной оспы в основном использовались
работы Э. А. Гальперина (1962); П. Н. Бургасова, Г. П. Николаевского (1972); С. С. Маренни-
ковой, С. Н. ГЦелкунова (1998). Классификация клинических форм натуральной оспы взята
из работы П. Н. Бургасова, Г. П Николаевского (1972), параллельно приведена классифика-
ция из работы С. С. Маренниковой и С. Н. Щелкунова (1998).
3.8. Натуральная оспа, оспа обезьян
869
Таблица 3.29
Портрет серийного убийцы: иммуномодулирующие гены ВНО*
Иммуномодулирующее действие Variola major Variola minor Комментарии
IFN-a/p-ингибитор B20R D9R He проверено
IFN -у-ингибитор B9R H9R Ингибирование человеческого IFN-y
е!Р2а-мимикрия C3L D3L He проверено
dsRNA-BP E3L C3L He пров :рено
Ингибитор фактора некроза опухолей G2R G2R He проверено
Ингибитор NF-kB P1L P1L He проверено
TLR сигнальный ингибитор A52R A56R He проверено
IL-18-связывающий белок D5L B6L Ингибитор человеческого IL-18
Хемокинсвязывающие белки G3R A46L G3R ингибирует у людей хемокины СС
Фактор роста вирусов G3R B3R Связывает и активирует человеческий EGF-R (erbBl)
Ингибитор комплемента/SPICE D12L B18L Ингибирует каскад комплемента у людей
GM-CSF/IL-2 ингибитор A46R A49L Не проверено
Семафорин A42R A45R Не проверено
Серпины B13R, B25R, C2L D14R, D2R Не проверено
* Такое название таблице дал ее составитель — G. McFadden (2004).
6 недель, но иногда принимает затяжное течение и длится до 3 месяцев и дольше. Лег-
кие формы болезни имеют меньшую продолжительность. Известны легкие заболева-
ния, которые длятся один день (эфемерные формы оспы).
Клиническая картина натуральной оспы обусловливается тяжестью развивающе-
гося инфекционного процесса. Существуют разные классификации клинических
форм натуральной оспы. В табл. 3.30 приведена классификация из работы С. С. Ма-
ренниковой и С. Н. Щелкунова (1998). Ниже приведена классификация и клиниче-
ское описание П. Н. Бургасова, Г. П. Николаевского (1972). Эти авторы различают
следующие формы натуральной оспы:
1. Тяжелые
Геморра! ическая оспа:
оспенная пурпура — purpura variolosa;
пустулезно-геморрагическая или черная оспа — Variola pustulosa haemorrhagika;
сливная оспа — variola contluens.
2. Средней тяжести
Рассеянная оспа — variola vera discreta.
3. Легкие
вариолоид — variolois;
оспа без сыпи — variola sine exanthemate;
оспа без температуры — variola afebrilis.
Геморрагическая форма оспы — тяжело протекающая болезнь с наличием гемор-
рагического диатеза. Если кровоизлияния появляются в раннем периоде предвестни-
ков, болезнь принимает крайне тяжелую форму с летальностью до 100 %, известную
870
Часть 3. Частная неправильная эпидемиология
Таблица 3.30
Классификация клинических типог э >льшой оспы (variola major)*
Тип Основные особенности
Обычная Распространенная сыпь, элементы которой проходят характерную эволю- цию (макула —> папула -> везикула -» пустула —> корки). Подтипы сливная — сливная сыпь на лице и разгибательных поверхностях конечностей, дискретная на остальных частях тела: полусливная — сливная сыпь на лице и дискретная на л еле и конечностях; дискретная — оспины рассеяны по всему телу с нормальной кожей между ними
Модифицированная (вариолоид) Подобна обычной (может быть сливной, полусливной и дискретной), но отличается ускоренным течением и отсутствием токсикоза
Оспа без сыпи Лихорадка и другие продромальные симптомы, оспенная сыпь не разви- вается; наблюдается у лиц с высокой напряженностью иммунитета, одна- ко в редких случаях встречается и у невакцинированных; диагноз под- тверждается серологически
Плоская Элементы сыпи плоские, как бы утоплены в коже и в большинстве слу- чаев, не имеющие характерного западения в центре. Пустулы наблюда- ются очень редко. Заболевание протекает на фоне высокой температуры и тяжелого токсикоза и часто сопровождается осложнениями со стороны легких. Мохут наблюдаться отслоения кожи в местах поражений. В отли- чие от геморрагических форм оспы большую часть больных плоской фор- мой оспы составляют тети Летальность очень высока, при сливном типе плоской оспы — 100 %. Подтипы сливная, полусливная, дискретная
Геморрагическая Распространенные геморрагли в кожу и/или слизистые оболочки в одной или другой стадиях болезни ранняя - развитие геморрагии на коже и слизистых, начиная с продромальной стадии, всегда смертельна позд- няя — геморрагии на коже и/или слизистых после появления сыпи, обычно смертельна
* По С. С. Маренниковои и С. Н. Щелкунову (1998).
ную в клинике как оспенная пурпура (purpura variolosa). Появление кровоизлияний
в более поздний период болезни, в стадии пустул, ведет к развитию пусгулезно-ге-
моррагической оспы (variola pustulosa haemorrhagica) с легальностью от 70 до 100 %
(рис. 3.103).
При оспенной пурпуре инкубационный период длится не более 6-9 сут. Темпе-
ратура в первый день болезни поднимается до 41—42 °C. Характерным для этой фор
мы натуральной оспы является развитие множественных кровоизлияний в кожу и
слизистые оболочки, важным симптомом являются кровоизлияния в субконъюнкти-
вальную слизистую оболочку. Возникают обильные кровотечения из носа, рта, дыха-
тельных путей, желудка, почек и других внутренних органов. Смерть обычно насту-
пает на 2-3 сут болезни еще до развития оспенных высыпаний от резкого упадка
сердечной деятельности.
Для пустулезно-геморрагической оспы также характерны укороченный инкуба-
ционный период, высокая температура, токсикоз. Геморрагии могут появиться в пер-
вые дни болезни, в период образования папул, но, как правило, геморрагический
диатез развивается на 8—10 сут и совпадает с образованием пустул. Кровоизлияния в
содержимое пустул придают им вначале темно-коричневый, а позже черный цвет, что
и послужило причиной названия черная оспа (variola nigra) (см. рис. 3.103).
В это время у больных появляется кровь в мокроте, моче, рвотных массах. Осо-
бенно тяжело протекает заболевание у беременных женщин. В результате токсикоза
3.8. Натуральная оспа, оспа обезьян
871
Рис. 3.103. Геморрагическая натуральная оспа. А и Б. Ранняя стадия геморрагической
натуральной оспы у беременной 18-летней женщины. Тяжелая токсемия, петехиальные
экзантемы, кровотечения из полостей тела. Снимок сделан за час до смерти пациентки.
В. Поздняя стадия геморрагической натуральной оспы у молодой женщины. Крово-
течения у основания пустул и общий геморрагический диатез. По F. Fenner et а). (1988)
возникают кровоизлияния в матке, что ведет к самопроизвольному аборту и профуз-
ному кровотечению. При пустулезно-геморрагической форме оспы смерть наступает
на 8-12 сут болезни.
Сливная оспа (vanola confluens). Начало болезни характеризуется высокой темпе-
ратурой (41-42 °C), сильными головными болями, тошнотой, рвотой, болями в мыш-
цах, особенно в области крестца. Сыпь, появившись, быстро распространяется по
всему телу, особенно поражая волосистую часть головы, лицо, слизистые оболочки
верхних дыхательных путей и конъюнктиву глаз (рис. 3.104).
Для сливной оспы характерно образование большого количества высыпных эле-
ментов и ускоренное образование пустул, процесс развития которых заканчивается к
5—6 сут болезни. В периоде нагноения отдельные пустулы сливаются и образуют
сплошные пузыри, заполненные гноем, иногда величиной больше ладони. Увеличи-
вающееся количество гнойного экссудата растягивает и истончает стенку пузыря;
целостность ее легко нарушается, и содержимое вытекает наружу. Подсыхая, гной
образует корки, которые, как панцирем, сковывают тело больного. С появлением
высыпных элементов температура понижается незначительно, оставаясь в пределах
40°С. Не отмечается повышения ее и в период нагноения пустул. Поэтому типичная
для заболевания оспой двухволновая температурная кривая отсутствует. У больных
отмечается сильная интоксикация организма, появляется бред, наступает коматозное
872
Часть 3. Частная неправильная эпидемиология
Рис. 3.104. Сливная форма натуральной оспы у не вакцинированной женщины.
Снимок сделан на девятые сутки от начала болезни. На лице пустулы слились, на
туловище, руках и ногах они дискретны. По F. Fenner et al. (1988)
состояние и с нарастающими явлениями упадка сердечной деятельности бальные ча-
сто погибают. Летальность при сливной форме оспы достигает 30 %. В случае выздо-
ровления на кожных покровах и, особенно на лице, остаются глубокие обезображи-
вающие рубцы.
Оспа средней тяжести (variola vera discreta). Инкубационный период продолжи-
тельностью обычно 14 сут; иногда он может быть короче — 5-7 сут, а в редких случаях
может затянуться до 17—22 сут. В инкубационном периоде заразившийся человек чув-
ствует себя вполне удовлетворительно. Болезнь начинается внезапно потрясающим
ознобом. Это начальный, или продромальный, период заболевания, который длится
3—4 сут. У больного резко поднимается температура, достигая 39,5—40,5 °C, иногда
41 °C, учащаются пульс и дыхание, наблюдается тошнота, рвота, мучительная голов-
ная боль и характерная боль в области крестца. Слизистые оболочки мягкого неба,
носоглотки и зева резко воспалены, язык сухой, обложенный. У детей могут появиться
судороги. Печень и селезенка увеличены, диурез уменьшается, удельный вес мочи вы-
сокий. Таким образом, уже в начальном продромальном периоде ярко выступают
признаки тяжелой интоксикации организма.
На 2—3 сут болезни примерно у 40 % больных появляется продромальная, или
токсическая, сыпь, известная в клинике под названием «реш» (rose rash). Высыпания
появляются вначале на лице, затем на конечностях (как правило, на разгибательной
поверхности) и на туловище. Единичные петехии могут быть и на конъюнктивах.
Продромальная сыпь, имея вид мелких красных пятен, может по своему характеру
быть похожа на коревую или скарлатинозную — петехиальную. Характерна локали-
зация сыпи. Она располагается в области бедренного треугольника Симона: основа-
нием треугольника служит поперечная линия на уровне пупка, а вершина находится
в области коленных суставов, включая переднюю поверхность бедер, или в области
большой грудной мышцы (грудной треугольник). Особенно резко сыпь выступает на
участках кожи, где в инкубационном периоде применялись механические или хими-
ческие раздражители (наложение горчичников, повязок, смазывание йодом и други-
ми раздражающими кожу веществами) (рис. 3.105).
Обратное развитие продромальной сыпи происходит быстро, обычно в течение
12-24 ч. Сыпь бледнеет и исчезает, но иногда следы ее в виде мелких буроватых пятен
сохраняются до появления типичных оспенных высыпаний. К концу 4 сут в период
3.8. Натуральная оспа, оспа обезьян
873
Рис. 3.105. Продромальная сыпь у больного натуральной оспой. А. Локализация
продромальной сыпи. 1 — грудные треугольники; 2 — треугольник Симона.
Б. Кореподобная продромальная сыпь на латеральной стороне туловища на
четвертые сутки болезни. По F. Fenner et al. (1988)
продромальных явлений у больного резко снижается температура почти до нормы,
улучшается общее состояние, и на этом фоне появляется характерная для натураль-
ной оспы сыпь.
Такое улучшение состояния больного в период появления сыпи патогномонично
только для натуральной оспы и не встречается при других инфекциях, сопровождаю-
щихся экзантемами и энантемами. Высыпания появляются не только на кожных по-
кровах, но и на слизистых оболочках конъюнктив, щек, мягкого неба, носоглотки,
бронхов, пищевода и т. д. Раньше всего сыпь появляется на лице, волосистой части
головы, шее, затем на руках, туловище и ногах. Наиболее интенсивна сыпь на лице,
предплечьях и тыльной поверхности кисти. На ногах сыпь обильнее на голенях, чем
на бедрах. На конечностях высыпаний значительно больше на разгибательной сторо-
не, чем на сгибательной. На ладонях и подошвах также наблюдаются высыпные эле-
менты. Обнаружение их является важным диагностическим симптомом натуральной
оспы. На туловище и животе, как правило, элементы сыпи менее выражены. Подоб-
ная локализация указывает на центробежное распределение сыпи, т. е. преобладание
высыпаний на периферии (рис. 3.106).
С образованием пустул начинается период нагноения. Он сопровождается вторич-
ным подъемом температуры и резким ухудшением состояния больного. Температура
держится в течение нескольких дней вплоть до подсыхания пустул. Присоединяюща-
яся вторичная инфекция отягощает клиническую картину болезни. В этот период
наступает отек кожи, особенно на лице. Отек настолько силен, что больной не может
открыть глаза. Оспенная сыпь, располагаясь по краю века, травмирует роговицу, а
последующая вторичная инфекция вызывает воспаление роговицы и радужной обо-
лочки. Часто наблюдаются кровоизлияния в субконъюнктивальную слизистую обо-
лочку. Это ведет к развитию панофтальмита, кератита и ирита, что в дальнейшем мо-
жет привести к потере зрения. Гнойным экссудатом заполнены носовые ходы;
слизистая оболочка верхних дыхательных путей резко отечна, с многочисленными
874
Часть 3. Частная неправильная эпидемиология
Рис, 3 106 Сыпь у больного натуральной оспой. А. Характерная локализация
сыпи Б Раньше всего сыпь появляется на лице. По П. Н. Бургасову, Е П. Нико-
лаевскому (1972). В. Сыпь на ладонях и подошвах характерна для натуральной
оспы и оспы обезьян. По F. Fenner et al. (1988)
эрозиями. Обилие высыпаний создает напряжение воспаленной кожи, а сильная отеч-
ность фактически лишает больного активных движений. Резкая боль и отек настоль-
ко сильны, что оольной не может шевелить пальцами.
В этот период больные часто погибают oi бронхопневмонии, ослабления сердеч-
ной деятельности и от сепсиса, вызываемого вторичной инфекцией. С12— 14 сут гной-
ное содержимое пустул начинает подсыхать с образованием плотных корок, которые
постепенно отпадают. Процесс обратного развития оспин протекает в том же поряд-
ке, что и процесс высыпания, т. е. начинается с лица, а затем переходит на гуловище
и конечности. Дольше всего оспины остаются на подошвах и ладонях. Из-за плотно-
сти эпидермального слоя гнойный экссудат не вытекает наружу, а медленно рассасы-
вается, оставляя плотные темные пятна. Отечность, воспаление кожи и слизистых
оболочек постепенно исчезают, общее состояние больного улучшается, температура
снижается, и после полного подсыхания содержимого пустул кожа становится нор-
мальной. В этот период появляется мучительный нестерпимый зуд кожи и часто, рас-
чесывая ее до крови, сдирая корки, больной вносит инфекцию, что вновь вызывает
нагноение и подъем температуры. На месте отпавших корок метаются красноватые
пятна, а у людей со смуглой кожей — пятна депигментации, впоследствии проходя-
щие. В том случае, если нагноением были затронуты глубокие слои кожи, то после
отпадения корок ооразуются стойкие обезоораживающие рубцы, особенно заметные
3.8. Натуральная оспа, оспа обезьян
875
на лице. Развитие рубцов в глазу ведет к частичной или чаще к полной потере зрения.
В неосложненных случаях болезнь обычно продолжается 5-6 нед. Летальность при
этой форме оспы составляет до 15 %.
Легкие формы оспы. Вариолоид (variolois). Вариолоид — видоизмененная стертая
форма оспы с легким и коротким течением болезни, незначительным и абортивным
развитием сыпи. Это заболевание иногда называют оспой привитых, тем самым под-
черкивая, что оно часто встречается улиц, привитых против оспы.
Продолжительность инкубационного периода при вариолоиде, как и при нату-
ральной оспе, длится около 2 нед. Продромальный период мало отличается от такового
при типичной форме заболевания, но симптомы менее выражены, а сыпь, имеющая
пятнистый (эритематозный) характер, часто отсутствует. Заболевание начинается
внезапно с резкого подъема температуры, которая в редких случаях превышает 39°С.
У больного появляются головная боль, небольшое раздражение слизистых оболочек
глотки, зева, трахеи, боль при кашле и глотании. Большинство больных жалуются на
боли в области крестца, что характерно для оспы. Сильный токсикоз развивается край-
не редко. Сыпь появляется на 3-4 сут болезни. Она представляет собой мелкие крас-
ные пятна, быстро переходящие в папулы и пузырьки, расположенные более поверх-
ностно, чем при натуральной оспе. Пузырьки окружены узким красным венчиком.
Характерное для оспы центральное втяжение, как правило, отсутствует. На 7—8 сут
содержимое пузырька подсыхает и образуется корочка, которая быстро отпадает. На
месте отпавшей корочки остается исчезающее пигментное пятно. Рубец при варио-
лоиде формируется очень редко. С появлением сыпи на 3—4 сут болезни температура
быстро нормализуется и больше не поднимается, так как период нагноения отсутст-
вует. В редких случаях имеет место нагноительный процесс, но он всегда выражен
крайне слабо, что ведет к незначительному и кратковременному подъему температу-
ры. С установлением нормальной температуры исчезают и общие явления — голов-
ная боль, раздражение слизистых оболочек. Болезнь заканчивается на 7—8 сут, и лишь
в тяжело протекающих случаях бывают смертельные исходы. Летальность при варио-
лоиде 1,5-3 %.
Оспа без сыпи (variola sine exanthemate) — легко протекающая болезнь, при кото-
рой в начальном периоде наблюдаются характерные для натуральной оспы симпто-
мы: повышение температуры, головная боль, боль в области крестца и иногда как
результат токсикоза продромальная сыпь, однако характерная оспенная сыпь отсут-
ствует. Заболевание заканчивается через 3—4 сут.
Оспа без температуры (variola afebrilis). При этой форме оспы на теле больного
появляется скудная, быстро проходящая узелково-пузырчатая сыпь. Заболевание
проходит без повышения температуры и нарушения общего состояния заболевшего
С. С. Маренникова и С. Н. Щелкунов (1998) упомянули в своей работе об инте-
ресном наблюдении Kempe et al. (1969), которые отметили у 21 из 55 контактировав-
ших с больными натуральной оспой развитие конъюнктивита, явившегося единствен-
ным проявлением оспенной инфекции. У12 из 21 обследованного оспенная природа
конъюнктивита была подтверждена выделением из конъюнктивального экссудата
вируса оспы. В другой серии наблюдений этой же группы авторов вирус был обнару-
жен в слезах у таких лиц. При натуральной оспе описаны и так называемые субкли-
нические формы, при которых не обнаруживается никаких клинических проявлений
и диагноз может быть установлен только на основании серологического обследова-
ния. Наличие таких форм было установлено как во время вспышек variola major, так и
variola minor.
876
Часть 3. Частная неправильная эпидемиология
О существовании врожденной формы натуральной оспы было известно еще с се-
редины XVIII в. Ее неоднократно наблюдали как при вспышках variola major, так и
variola minor. Известны два варианта врожденной натуральной оспы: истинная, когда
плод рождается с явными признаками болезни; и ложная — развивающаяся уже пос-
ле родов (в пределах инкубационного периода). Описаны случаи рождения двойни,
когда один из новорожденных появлялся на свет с кожными проявлениями оспы, а
другой оставался здоровым (Маренникова С. С. и Щелкунов С. Н., 1998).
Основные патологоанатомические изменения происходят в коже. Они характеризу-
ются двумя видами нарушений, развивающихся в клетках эпидермиса: баллонирую-
щей дегенерацией и ретикулярной колликвацией. Баллонирующая дегенерация об-
наруживается в нижних слоях эпидермиса (зародышевый и частично шиловидный
слои). Она проявляется в набухании и округлении клеток эпителия, потере ими меж-
клеточных связей. В результате этого клетки напоминают скопления «баллонов» в
межэпителиальном пространстве. Ядра клеток увеличены, в цитоплазме обнаружи-
ваются тельца Гварниери. В верхних слоях эпителия развивается ретикулярная деге-
нерация с образованием небольших вакуолей в цитоплазме; протоплазма пораженных
клеток приобретает сетчатый вид. Постепенно границы клеток исчезают. За счет по-
ступления экстрацеллюлярной жидкости из окружающих тканей и дискомплексации
эпителия с образованием вертикальных тяжей формируется увеличивающаяся мно-
гокамерная полость. Эта стадия соответствует образованию папул на коже больного.
В дальнейшем поступление экссудата увеличивается, и папулы постепенно перехо-
дят в везикулы. Верхняя часть везикулы образуется из клеток зернистого и рогового
слоя эпидермиса, который не затрагивается существенно патологическим процессом.
Уже в начале формирования везикулы обнаруживается центральное западение, вы-
являющееся при макроскопическом наблюдении. Основание везикулы образуется
клетками кориума, иногда сохраняются и элементы герминативного слоя. Одновре-
менно с развитием дегенеративных изменений в оспенном элементе по периферии
пузырька происходит пролиферация. Проникновение лейкоцитов в везикулу превра-
щает ее в пустулу. Затем происходит постепенное подсыхание элемента с превраще-
нием его в корочку. Заживление может идти двояко. Если в процессе образования
оспины не был задет слой собственно кожи (сохранилась хотя бы часть клеток заро-
дышевого слоя), восстановление целостности эпидермиса происходит без образова-
ния рубца. Если же в процесс была вовлечена собственно кожа (особенно при вторич-
ной инфекции), образуется характерный соединительнотканный рубчик.
Разрушительные изменения особенно выражены на лице, где много сальных же-
лез, погибающих в результате патологического процесса. Потовые железы обычно не
повреждаются. В волосяных фолликулах может быть баллонирующая дегенерация, но
материнский, основной слой не повреждается, благодаря чему волосяные фоллику-
лы полностью восстанавливаются.
Патогистология оспенных элементов на слизистых оболочках в основных чертах
напоминает процесс их развития на коже. Различия заключаются в изъязвлении эле-
ментов оспенной энантемы, обусловленной отсутствием рогового слоя эпителия.
Изменения на слизистых оболочках обычно заканчиваются к 3-й неделе без образо-
вания рубцов. Во внутренних органах (печени, почках, сердце) наблюдаются дистро-
фические изменения, в костном мозгу — участки некроза, пролиферативные измене-
ния. Селезенка увеличена. Часто находят орхит, воспаление яичников и слизистых
оболочек труб матки. Могут иметь место очаговые пневмонии в нижних долях легких.
До применения антибиотиков у умерших от натуральной оспы нередко находили
3.8. Натуральная оспа, оспа обезьян 877
пневмонии, связанные с аспирацией отмершей ткани и вторичной инфекцией, не-
редко с исходом в абсцесс или гангрену легких.
При геморрагических формах оспы изменения носят более тяжелый характер.
В коже наблюдаются распространенные кровоизлияния, отек. В трахее и желудочно-
кишечном тракте обнаруживают субмукозные геморрагии. Кровоизлияния могут быть
под эндокардом, в почках, яичниках и тестикулах. Универсальная гиперплазия лимфа-
тических узлов. Полнокровие и отек легких. Оспенные элементы на коже не успевают
проделать полную эволюцию; они наблюдаются (если имеются) лишь до стадии папул.
Клиническая и патологоанатомическая картина оспы обезьян у человека при есте-
ственном инфицировании1. Клиника оспы обезьян у людей различается в зависимости
от того, к какой кладе принадлежит вызвавший болезнь вирус. ВОО, циркулирующие
среди обезьян, обитающих в бассейне реки Конго, вызывают опасные клинические
формы болезни, сходные с натуральной оспой; ВОО из Западной Африки вызывают
у людей легкие формы оспы обезьян.
Инкубационный период оспы обезьян укладывается в 7—19 сут. Болезнь, как пра-
вило, начинается с подъема температуры, слабости, недомогания, головной боли,
иногда появляются боли в горле, кашель. Особенностью клинического течения оспы
обезьян у человека, отличающей ее от натуральной оспы, являются лимфадениты,
которые наблюдаются у 86,4 % заболевших. Они появляются, как правило, одновре-
менно с лихорадкой, за 1-2 сут до высыпания и иногда совпадают с его началом.
В ряде случаев лимфадениты служат поводом для обращения за медицинской помо-
щью. Их локализация варьирует (подчелюстные, шейные, подмышечные, паховые),
причем, иногда они двусторонние, иногда односторонние. В 11 % случаев лимфаде-
ниты возникают в лимфоузлах разных областей (например, на шее и в подмышечной
области), чаще всего (в 63,9 %) развивается генерализованная лимфаденопатия. Че-
рез 2—4 сут от начала болезни появляется сыпь. Сыпь локализуется на всем теле с пре-
имущественным поражением лица и конечностей, включая ладони и подошвы. Как
и при оспе, высыпания отмечаются и на слизистых оболочках, а также языке. Элемен-
ты сыпи мономорфны и претерпевают те же стадии развития (папулы — везикулы —
пустулы — корки). После отпадения корок на коже остаются депигментированные
участки, на месте которых формируются отчетливые, но неглубокие рубцы, или ги-
перпигментированные пятна. Болезнь обычно длится 2—4 недели в зависимости от
клинического течения и других факторов.
В целом по характеру клинического течения оспа обезьян у человека напоминает
тип натуральной оспы, преобладавший на африканском континенте, давая картину,
близкую к обычной дискретной, полусливной и сливной ее формам (рис. 3.107).
Помимо этих основных форм, встречаются и другие. Имеется информация о ге-
моррагической форме оспы обезьян у ребенка, протекавшей аналогично оспенной
геморрагической пурпуре и закончившейся летальным исходом. Наблюдалось и очень
легкое течение оспы обезьян по типу вариолоида с единичными кожными элемента-
ми. При оспе обезьян встречается субклиническая форма болезни, описанная при
натуральной оспе. О наличии этой формы судят по появлению антител улиц, имев-
ших контакт с заболевшими оспой обезьян. Был зарегистрирован случай врожденной
инфекции у младенца, родившегося от матери, незадолго до родов перенесшей оспу
обезьян. Подобного рода наблюдения неоднократно описывались при натуральной
1 Для описания оспы обезьян у людей в основном использовалась работа С. С. Маренни-
ковой и С. Н. Щелку нова (1998).
878
Часть 3. Частная неправильная эпидемиология
Рис. 3 107. Оспа обезьян у жителей Заира. А и Б. Семилетняя девочка на 7 сут болезни.
Хорошо заметна паховая билатеральная аденопатия и увеличение подчелюстных лим-
фатических узлов на правой стороне лица. Пустулярные поражения губ. Локализация
сыпи такая же, как и при натуральной оспе В. Трехлетний мальчик с сыпью на стадии
корок. Аксиллярные лимфатические узлы все еще увеличены. По Г Fenner et al. (1988)
оспе. Среди осложнений при оспе обезьян наиболее часто встречается вторичная бак-
териальная инфекция кожи, бронхопневмония, диарея и связанная с ней дегидрата-
ция, кератиты. Последние в части случаев заканчиваются потерей зрения или его ос-
лаблением.
В 2003 г. оспу обезьян среди жителей штата Висконсин (11 заболевших) вызвал
ВОО, циркулирующий в Западной Африке, поэтому среди заболевших не было смер-
тельных случаев. Первыми признаками болезни у них были поражении кожи в участ-
ке места или царапины, нанесенных больными животными, головная боль, повышен-
ная температура тела (более 38 °C), профузный пот и сильный озноб. У 73 % заболев-
ших болезнь проявила себя сильным кашлем, у 55 % лимфоаденопатией и язвами в
гортани, у 27 % наблюдали фарингит, у 18 % диарею. Легкие, сердце, органы брюш-
ной полости, нервная система — без изменений. У пяти пациентов, преимуществен-
но с кожными поражениями, имела место узловая припухлость вокруг мест укусов и
царапин. У шести пациентов, не имевших видимых следов укусов или царапин, об-
наружены оз 1 до 50 оспин (от папул до везикулопустул) в течение 7 сут от начала бо-
лезни на лице, коже черепа, кистях руках, ногах, туловище, промежности, конъюнк-
тиве (рис. 3.108).
Клиническая картина и патоморфология натуральной оспы и оспы обезьян у жиноз -
ных при искусственном инфицировании. Натуральную оспу воспроизвести у традици-
онных лабораторных животных (грызуны, кролики) сложно, так как они маловос-
приимчивы к ВНО, и если у них удается вызвать болезнь, то она протекает в мягкой
форме и быстро обрывается. К ВОО эти животные более чувствительны, однако не на-
столько, чтобы считаться оптимальной моделью. Для моделирования клиники и па-
тогенеза натуральной оспы и оспы обезьян используют три подхода:
1) подбирают дозу и способ заражения, вызывающие у приматов болезнь, сход-
ную по патогенезу и клинике с моделируемой болезнью;
3.8. Натуральная оспа, оспа обезьян
879
Рис. 3.108. Кожные поражения при оспе обезьян вокруг участков укусов и царапин у жите-
лей штата Висконсин (США). А, Б, В Узловая припухлость вокруг участков укусов и цара-
пин. Г. Оспопоподобные высыпания. Д. Высыпания, сходные с ветряной оспой.
Ж. Кожное поражение менее чем через 24 ч после его появления. 3. Поражение через 6 сут.
Е. Поражение через 96 ч. И. Поражение более чем через 9 сут. По К. D. Reed et al. (2004)
2) модельное животное заражают не ВНО или ВОО, а другим ортопоксвирусом,
вызывающим у него инфекционный процесс, аналогичный натуральной оспе или оспе
обезьян, т. е. проводят исследование «по аналогии»;
3) используют для заражения иммунодефицитных животных, например, мышей
линии C57BL/6, утративших STAT1 — белок, участвующий в регуляции синтеза интер-
феронов.
В табл. 3.31 обобщены основные поражения у модельных животных при различ-
ных путях заражения вирусами натуральной оспы и оспы обезьян.
Вызвать у приматов болезнь, сходную по патогенезу и клинике с натуральной ос-
пой, можно внутривенным введением 109 КОЕ ВНО (штаммы Harper или India).
У макак развивается смертельная болезнь, сходная с геморрагической формой нату-
ральной оспы у людей. Уменьшение дозы вируса на порядок(10* КОЕ) позволяет по-
лучить у приматов клиническую форму болезни, соответствующую variola vera discrete
880
Часть 3. Частная неправильная эпидемиология
Таблица 3.31
Основные поражения у модельных животных при различных путях
заражения вирусами натуральной оспы и оспы обезьян*
Вирус Модельное животное Вид животных Путь заражения Доза (штамм) Основные поражения
ВНО Обезьяны Macaque cynomolgus В/в 109 КОЕ (Harper, India) Геморрагии во внутренних органах, на коже, слизистых; геморрагические выпоты, ранние энантемы/экзантемы; смерть
10s КОЕ (Harper, India) Энантемы/экзантемы
Ингаля- 11ИОННО 108-5 КОЕ (Yamada, Lee) Общая инфекция, легочные поражения, экзантемы
Yamada Общая инфекция, возможные поражения легких, редкие энантемы/экзантемы
l,8x!06—3,4xl06 (Lee); 5xl05—2xl06 (Yamada) Умеренный бронхит без некроза, лимфоидная гиперплазия, экзантемы
ВОО Обезьяны Macaque cynomolgus В/в 5xl07 KOE Везикулопустулярная сыпь, лимфо- аденопатия, спленомегалия
rhesus 2xl07 KOE Везикулопустулярная сыпь, лимфоаденопатия
cynomolgus Ингаля- ционно 10xl03- 141x10’ KOE Фибринонекротическая бронхо- пневмония, пролиферативный и некротизирующий дерматит, лимфоидный некроз
И/т 107 KOE Фибринонекротическая бронхо- пневмония, пролиферативный и некротизирующий дерматит, трахеит, некротизирующий лим- фаденит и спленит
106 KOE Умеренная некротизирующая брон- хопневмония, трахеит, лимфаденит
Земляные белки 13 линия В/б 12,6xl03 KOE Некроз центральной доли печени, интерстициальная пневмония, мультифокальный некроз селезенки
П/к 100 KOE Мультифокальный некроз печени и селезенки, легочная геморрагия, отек и острое воспаление
И/н 25,2xl03 KOE Диффузный некроз печени, интер- стициальная пневмония, мульти- фокальный некроз селезенки
Луговые собачки черно- хвостые В/б 12,6xl03 KOE Некроз селезенки, некроз цент- ральной доли печени, умеренная интерстициальная пневмония
В/в 25xl03KOE Выраженный отек легких, геморрагии и некрозы
J L. Chapman et al. (2010).
3.8. Натуральная оспа, оспа обезьян
881
у человека. У животного развивается петехиальная сыпь, затем появляются везикулы
и пустулы с таким же центробежным распределением, как и при натуральной оспе. У
погибших животных на вскрытии обнаруживают петехиальные геморрагии на
поверхности слизистых и серозных тканей, геморрагический некроз висцеральных
лимфатических узлов. Гистологически обнаруживают лимфоидное истощение и лим-
фоцитолиз лимфоидной ткани, селезенки, периферийных и висцеральных лимфати-
ческих узлов, кишечника и костного мозга, дегенерацию почечных канальцев и гепа-
тоцеллюлярную дегенерацию. аналогичную таковым, наблюдаемым при натуральной
оспе у людей. Кожа и слизистая поверхность ротовой полости включают очаги эпи-
дермальной/эпителиальной дегенерации и некроза с формированием везикул и пус-
тул, аналогичных тем, что развиваются у людей при натуральной оспе (рис. 3.109).
Рис. 3.109. Внешние проявления патологического процесса у обезьяны, вызванного
заражением ВНО. А. Диффузные геморрагии на серозной поверхности дистального
отдела кишечника и увеличение кишечных лимфатических узлов через 6 сут после
заражения. Б. Геморрагический колит, геморрагические поражения кишечных лим-
фатических узлов. В. Верхняя губа и ноздря обезьяны через 11 сут после заражения.
Синхронное развитие пустул, некоторые из них еще сохранили пупкообразное запа-
дение. Г. Ладонная поверхность левой верхней конечности обезьяны, 11 сут после
инфицирования. Дискретные и сливающиеся пустулы сохранили целостность из-за
сильно ороговевшего эпидермиса. По Р. Jahrling et al. (2004)
М. С. Lancaster et al. (1966) исследовали патологические изменения у эксперимен-
тальных животных, образующиеся после инфицирования мелкодисперсными (1 мкм)
аэрозолями вирусов группы оспы. Ими установлено, что при ингаляционном экспо-
нировании кроликов к аэрозолю вируса кроличьей оспы (Myxoma virus, MYX)1, первые
поражения в легочной ткани кролика отмечали на 4 сут. В альвеолах они представле-
ны участками геморрагий с миграцией воспалительных клеток из соседних кровенос-
ных сосудов. На следующие сутки макроскопически видимые поражения появились
на поверхности легкого, а при микроскопическом исследовании на бронхиолах мож-
но было увидеть типичные пустулы. Обнаруживались явная геморрагия и гиперемия
капилляров соседних альвеол. Поражения были окружены зоной нейтрофильной
1 Эту модель авторы считали более соответствующей течению натуральной оспы у людей
при ингаляционном заражении, чем приматов, зараженных ВНО.
882
Часть 3. Частная неправильная эпидемиология
инфильтрации и подобную инфильтрацию отмечали на поверхности наружного слоя
стенок cooi велся вуюших артерий. На 6 сут на бронхиолах появились самые крупные
пустулы, они были обнаружены также и на альвеолах; эти пустулы, хотя и были меньше
по сравнению с поражениями бронхиол, показали типичную картину центрального
некроза с окружающей инфильтрацией нейтрофилов. Наблюдались пролиферативные
изменения во внутренних оболочках бронхиальных артерий с отеком и нейтрофиль-
ной инфильтрацией наружного слоя стенки кровеносного сосуда. На 8 сут, несмотря
на то, что пустулы, прогрессировали в своем развитии, создавая большие очаги некро-
за, наиболее ярко выраженным признаком являлся широко распространенный острый
артериит с пролиферацией клеток эндотелия, макроскопическим отеком мышечной
оболочки и некрозом и нейтрофильной инфильграцией наружного слоя стенки крове-
носных сосудов. К 10 сут острая воспалительная реакция распространилась на боль-
шие участки альвеол, заполненные отечной жидкостью, нейтрофилами макрофагами
и эритроцитами. Самые ранние признаки рассасывания появились только на 20 сут.
Стенки артерий были очень отечными с воспалительными клетками в эндотелии, в
то время как вместо нейтрофилов в альвеолярных поражениях появились лимфоци-
ты. Даже на 62 сут были заметны остатки пустул; центральная некротическая зона
содержала теперь отложения кальция и все еще была окружена зоной альвеолярных
макрофагов и лимфоцитов. Отмечали повторное разрастание эпителия бронхиол, но
упорно продолжалось утолщение мышечной оболочки стенок артерий за счет ее ин-
фильтрации лимфатическими клетками из адвентициального слоя. Альвеолярные
очаги представляли собой скопление лимфоцитов и гигантских клеток.
У обезьян, инфицированных ВНО ингаляционно, через 5 сут после заражения
было хорошо заметно образование пустул в легочной ткани. Они представляли собой
типичную широкую центральную зону некроза, внутри которой обнаруживались не-
кротизированные элементы бронхиол; их окружала внешняя юна, образованная ней-
трофилами и макрофагами. Соседние альвеолы были гиперемированы и заполнены
отечной жидкостью, нейтрофилами и свободными альвеолярными фа) оцитами. Уча-
стки очагов деструкции эпителия и нейтрофильной инфильтрации также появились
и в основных бронхах Имела место пролиферация во внутренней оболочке и отек
мышечной оболочки и адвентициального слоя стенки артерий, находящихся вблизи
поражений. Очень тяжелые поражения обнаружили на 3 сут, когда пустулы представ-
ляли собой обширные очаги некроза, окруженные широкими зонами альвеол, запол-
ненных эксудатом. Просветы бронхов были заполнены нейтрофилами и продуктами
клеточного распада. Обнаружены выступающие макроскопические поражения на
плевральной поверхности, но они были, по-видимому, связаны с кровеносными со-
судами, а не с первичной инфекциеи а тьвеол. На 12 сут отмечали рассасывание пора-
жений. Вместо нейтрофилов на внешних зонах пустул появлялись лимфатические
клетки, и начиналось восстановление бронхиального эпителия.
Изменения в других тканях были обнаружены у кроликов, зараженных вирусом
кроличьей оспы; наиболее заметными оказались изменения надпочечных желез и
тестикул. Поражения представляли собой типичные пустулы, которые в надпочечной
железе были ограничены поверхностным слоем органа. При микроскопическом ис-
следовании поражения можно было обнаружить только в том случае, когда в тканях
находились большие количества вирусов.
Внутримышечное заражение обезьян Cynomolgus macaques ВОО приводит к разви-
тию системной инфекции с широким распространением везикулопустулезной сыпи
и образованием участка мышечного некроза в месте введения вируса. Внутривенное
3.8. Натуральная оспа, оспа обезьян
883
введение больших доз BOO (2х 107 КОЕ) вызывает у обезьян типичную генерализо-
ванную везикулолустулезную сыпь и выраженную лимфоаденопатию. На вскрытии:
лимфоаденопатия, спленомегалия, отек легких. Введение в легкие обезьяны ВОО в со-
ставе мелкодисперсного аэрозоля вызывает у них системную диссеминацию вируса,
сходную с таковой у людей. У погибших животных на вскрытии обнаруживают фиб-
ронекротическую бронхопневмонию, некротические изменения в лимфоидных орга-
нах, коже, слизистых, репродуктивных органах. Особенностью таких поражений яв-
ляется пролиферация эпителия в участках поражения кожи и слизистых. Остальные
поражения были идентичны таковым при в/в введении ВНО или ВОО обезьянам или
при естественном заражении человека этими вирусами. Интраназальное и ынтратра-
хеальное заражение ВОО вызывает у приматов поражения, сходные с получаемыми в
результате заражения аэрозолем BOO (Chapman J. L. et al., 2010).
Хотя в результате интратрахеального, ингаляционного и в/в введения ВОО у при-
матов возможно вовлечение легких в инфекционный процесс и развитие пневмонии,
эта патология имеет свои особенности для каждого из путей введения вируса. Аэро-
золирование и интратрахеальное введение вируса в период от 9 до 17 сут вызвало пер-
вичную фибронекротическую бронхопневмонию с системной диссеминацией возбу-
дителя болезни и смерть животного. Внутривенное введение ВОО вызывало у
животных интерстициальную пневмонию, развившуюся в результате вторичного ин-
фицирования (Goff A. J. et al. 2011).
Клиническая картина натуральной оспы у людей при искусственном инфицировании.
Искусственное инфицирование здоровых людей содержимым оспенных пустул, взя-
тых от лиц, переносящих болезнь в легкой форме, широко применялось в XVH1 в. с
целью предотвращения у заболевания оспой в тяжелой форме. Способ назывался ва-
риоляцией. Смертность среди привитых достигала 10 %, но страх перед натуральной
оспой был настолько велик, что такая «цена» не казалась чрезмерной. В 1796 г. этот
способ был заменен более безопасным — вакцинацией, т. е. прививкой содержимого
оспенных пустул на вымени коров. Патогенез заражения через кожные покровы (на-
пример, при вскрытии погибших от оспы и т. д.) принципиально не отличается от
такового при воздушно-капельном инфицировании через дыхательные пути. Одна-
ко наличие первичного поражения на коже, появляющегося на 3—4 сут, ускоряет раз-
витие инфекционного процесса. Инкубационный период болезни укорачивается до
5-7 сут (Маренникова С. С., Щелкунов С. Н., 1998).
Диагностика искусственного поражения возбудителем натуральной оспы. Появление
единственного случая натуральной оспы — чрезвычайное происшествие мирового
масштаба. Любую внезапно возникшую вспышку натуральной оспы целесообразно
рассматривать, как искусственно вызванную. Из-за отсутствия у врачей эпидемичес-
кой настороженности по натуральной оспе, вспышка может развиваться единичны-
ми случаями болезни в течение несколько месяцев, маскируясь под диагнозами «корь»,
«скарлатина», «токсический капилляротоксикоз», «ветряная оспа», «болезнь Лайма»
и др. Поэтому для установления масштабов и времени совершения преступления,
необходимо изучить истории болезни людей с такими диагнозами, обратившимися в
местные лечебные учреждения не менее чем за три месяца, предшествующих выяв-
ленному случаю натуральной оспы Особое внимание надо обратить на больных,
имевших патологию органов дыхания (бронхит, пневмония) и сыпь. У них необходи-
мо проанализировать сроки появления и характер сыпи (см. рис. 3.103—3.106), исхо-
ды болезни и данные объективного исследования в динамике (рентгенография, ком-
пьютерная томография, серология). Можно предположить, что будут выявлены два
884
Часть 3. Частная неправильная эпидемиология
типа заболевших натуральной оспой: первично инфицированные, т. е. экспонирован-
ные к мелкодисперсному аэрозолю ВНО; и вторично инфицированные, т. е. те, кто
заразился ВНО воздушно-капельным путем от первично инфицированных больных.
Соответственно будут и два пика заболеваемости: первый, наибольший, вызванный
экспонированием людей к мелкодисперсному аэрозолю; и второй, более растянутый
по времени, включающий вторично инфицированных лиц.
Для первично-инфицированных пациентов будет характерна «невозможная клини-
ка болезни» — быстрое (в течение 3—5 су г от начала первых симптомов) развитие ле-
гочного поражения, начинающееся с очаговой пневмонии в нижних отделах легкого,
переходящее в фибронекротическую бронхопневмонию и системные поражения с раз-
витием энантем и экзантем, с некротическими изменениями в лимфоидных органах,
коже, слизистых, репродуктивных органах. В случаях массированного заражения ВНО
болезнь будет развиваться без второй фазы подъема температуры. При однократном
применении ВНО для такой вспышки будет характерна сжатая эпидемическая кри-
вая с пиком заболеваемости, «укладывающимся» в средние значения инкубационно-
го периода при натуральной оспе (эпидемиология «точечного источника») и относи-
тельная локализованность эпидемического очага, т. е. выявленные пораженные
должны «пересекаться» в одно и то же время на одной территории. Необходимо отли-
чать натуральную оспу, вызванную ингаляцией мелкодисперсного аэрозоля ВНО от:
1) так называемой легочной формы оспы, когда у больных при отсутствии кож-
ных элементов развивается пневмония;
2) осложнений со стороны легких при плоской форме оспы.
Для вторично-инфицированных пациентов будет характерно типичное течение на-
туральной оспы, при которой основными являются поражения кожи и слизистых.
Поражения же легких развиваются позже как вторичные, и чаше в период нагноения.
Локализованный эпидемический очаг возможен только в организованных коллекти-
вах. Учитывая длительный инкубационный период болезни и возможность несколь-
ких генераций передачи ВНО, вторично-инфицированных больных натуральной ос-
пой, скорее всего, придется разыскивать по всей территории России.
При выявлении случаев обезьяньей оспы среди людей, ппежде всего надо искать
контакты таких больных с животными, зараженными ВОО (обезьяны, белки, грызу-
ны). Отсутствие таких контактов можно рассматривать как «невозможную эпидемио-
логию болезни». Типы больных будут те же, что и для натуральной оспы, если для пре-
ступления использован штамм ВОО, происходящий из долины реки Конго.
В месте накожного введения ВНО или ВОО быстро развиваются оспины с обшир-
ной зоной гиперемии. Под этим участком формируется плотный и отечный инфильт-
рат, который является «подножьем» для побочных оспин, одновременно развивается
лимфаденит и системная инфекция. Подкожное введение вирусов приводит к мест-
ному поражению с образованием участка некроза, отека, гиперемии, побочных оспин
и системной инфекции. Внутривенное введение вызывает типичную системную ин-
фекцию, тяжесть которой зависит от дозы введенного вируса Скорее всего, больные,
зараженные внутривенно, не будут представлять опасности в эпидемическом плане.
Обнаружение генетически измененных штаммов ВНО или ВОО однозначно сви-
детельствует об искусственном происхождении вспышки болезни.
Иммунопрофилактика натуральной оспы. Подробно о кандидатах в противооспен-
ные вакцины и об их роли в связанных с биотерроризмом стратегиях вакцинации, см.
в работе I. Wiser et al. (2007). Для вакцинации против натуральной оспы и оспы обезь-
ян в Российской Федерации используют три типа вакцин.
3.8. Натуральная оспа, оспа обезьян
885
ОспаВИР— вакцина оспенная инактивированная, лиофилизат для приготовления
раствора для подкожного применения. Представляет собой вирус вакцины, выращен-
ный на коже телят, обработанный фреоном 113 и инактивированный у-излучением
6 ’Со. Одна доза препарата содержит инактивированный антиген вируса — 50 мг и
стабилизаторы. Препарат применяется в двухэтапном методе вакцинации для созда-
ния бустериммунитета с последующим введением вакцины оспенной живой. Произ-
водится в Томске филиалом ФГУП «НПО «Микроген».
Вакцина оспенная живая сухая, лиофилизат для приготовления раствора для накож-
ного применения — препарат представляет собой вирус осповакцины, выращенный на
скарифицированной коже телят, частично освобожденный от бактериальной фло-
ры обработкой фенолом или хлоргексидина биглюканатом. Препарат содержит от
1 х 108 до 2х 109 оспообразующих единиц (ООЕ) в 1 мл. Одна прививочная доза содер-
жит не менее 1х10б ООЕ. Вакцину применяют накожно, скарификационным мето-
дом. Производится в Томске филиалом ФГУП «НПО «Микроген».
Вакцина ТЭОВак — вакцина оспенная эмбриональная живая. Представляет собой
высушенный со стабилизатором (лактоза) вирус вакцины, вырашенный в ХАО и пло-
дике куриного эмбриона и спрессованный с наполнителем в таблетки. Производится
в виде таблеток, покрытых оболочкой, делимых. Прививочная доза препарата содер-
жит 1 х 106 до 2х Ю7 ООЕ вируса вакцины. Препарат дозируют согласно инструкции,
содержащейся в упаковке. Таблетку разжевывают и держат во рту до полного раство-
рения, но не менее 2—3 мин, после чего в течение 30 мин не рекомендуется прини-
мать пищу. Препарат предназначен для ревакцинации взрослых против натуральной
оспы по эпидемическим показаниям и плановой ревакцинации лиц, работающих с
ВНО, ВОО, ВОВ и другими ортопоксвирусами, патогенными для людей. Производи-
тель Вирусологический центр НИИ микробиологии МО РФ, г. Сергиев Посад-6.
При вакцинации лиц, ранее никогда не прививавшихся против натуральной оспы
и не имеющих вакцинальных знаков, ее проводят двухэтапным методом:
первый этап вакцинации — подкожное введение инактивированной оспенной
вакцины ОспаВИР;
второй этап вакцинации — накожная прививка живой оспенной вакцины.
Экспериментально и практикой подтверждено, что, как правило, через 4—5 лет,
даже после успешной вакцинации, коллективный иммунитет резко снижается, если
не исчезает полностью. Снижение специфического иммунитета происходит еще ин-
тенсивнее, если в промежутках между вакцинациями имели место какие-либо массо-
вые инфекционные заболевания: грипп, корь и др. У отдельных лиц, успешно при-
витых за год и даже менее года до эпидемической вспышки натуральной оспы, при
очень тесном контакте с больным натуральной оспой вакцинальный иммунитет не
может предупредить возникновение и развитие заболевания оспой. В этом случае
характер клинических проявлений заболевания находится в зависимости от уровня
остаточного вакцинального иммунитета и вирулентности циркулирующего ВНО.
Частота различных клинических типов variola major и летальность среди непривитых
и привитых против натуральной оспы, приведены в табл. 3.32.
Имеются данные о низкой эффективности массовых вакцинаций против оспы
обезьян, проведенных живыми оспенными вакцинами в регионах Африки, эндемич-
ных по BOO (Катет К. L. et al., 2007).
Для лечения и профилактики натуральной оспы, для лечения поствакцинальных
осложнений после вакцинации живой оспенной вакциной, используется иммуногло-
булин человека противооспенный. Он представляет собой концентрированный раствор
886
Часть 3. Частная неправильная эпидемиология
Таблица 3.32
Частота различных клинических типов variola major и летальное гь
среди непривитых и привитых против натуральной оспы*
Клинический тип Невакцинированные Вакцинированные
Частота (% от общего количества)** Летальность (%) Частота (% от общего количества)*** Летальность (%)
Обычная 88,8 30,2 70,0 3,2
сливная 22,8 62,0 4,6 26,3
полусливная 23,9 37,0 7,0 8,4
дискретная 42,1 9,3 58,4 0.7
Модифицированная 2,1 0 25,3 0
Плоская 6,7 96,5 1,3 66,7
Геморрагическая 2,4 98,4 3,4 93,9
ранняя 0,7 100,0 14 100
поздняя 1,7 46,8 2,0 89,8
Всего 35,5 6,3
* По С. С. Маренниковои и С. Н. Щелкунову (1998).
** Общее количество больных — 3544.
*** Общее количество больных — 3398,
очищенной фракции иммуноглобулинов, выделенной методом фракционирования
этиловым спиртом при температуре ниже (J°C из плазмы крови здоровых доноров,
содержащей антитела к ортопоксвирусам в титре не менее 1.200. Препарат содержит
антитела к ортопоксвирусам в титре не менее 1:4000 и глицин в качестве стабилиза-
тора. Производится в Томске филиалом ФГУП «НПО «Микроген». Иммуноглобулин
вводят внутримышечно. Для экстренной npocj илактики однократно в дозе 3 мл, для
лечения натуральной оспы — в дозе 0,5— 1,0 мл/кг веса больного в зависимости от
тяжести клинического течения болезни один раз в сутки или дробно.
Осложнение после вакцинации:
глазная вакциния — возникает из-за случайного занесения в глаз живой противо-
оспенной вакцины. Развивается сильный конъюнктивит с везикулами и пустулами.
Могут появиться кератит и изъязвление роговицы;
генерализованная вакциния — характеризуется умеренными органическими симп-
томами и генерализованной везикулярной сыпью. Из везикул и из сыворотки боль-
ных с этим состоянием вирус вакцины методом культивирования не выделяется Па-
тогенез неизвестен, но возможно обусловлен иммунологическим механизмом. Обычно
это самоограничивающаяся реакция, и лечение не показано, но в тяжелых случаях
рекомендуется применение противооспенного иммуноглобулина;
поствакцинальный энцефалит — патогенез неизвестен, но может быть обусловлен
иммунопатологическим механизмом, так как из ткани мозга и спинномозговой жид-
кости пациентов вирус вакцины методом культивирования не выделяется. При этом
осложнении летальность равна 10—30 %, а у выживших больных наблюдаются невро-
логические последствия. Рекомендуется поддерживающая терапия;
eczema vaccinatum — тяжелый дерматит, характеризующийся рептикацией вируса
вакцины в местах экземы, псориазных, ожоговых и других хронических или сильных
кожных повреждений. Для лечения показан противооспенный иммуноглобулин.
3.9. Вирусные геморрагические лихорадки
887
vaccinia necrosum — состояние характеризуется сильной реакцией в месте вакци-
нации, прогрессирующей до локального некроза кожи. Развивается у иммуноослаб-
ленных вакцинируемых. Может привести к летальному исходу. Для лечения назнача-
ется противооспенный иммуноглобулин через каждые 2—3 сут внутримышечно в
дробных дозах в разные места на протяжении 24—36-часового периода пока не насту-
пит несомненное клиническое улучшение.
В результате введения вируса вакцины беременным женщинам может развиться
вакциния плода, прогноз плохой.
Профилактика после экспонирования к ВНО и ВОО и лечение развившейся болезни.
Army FM 8-284 (2000) рекомендует:
1) в течение 1—7 сут подвергнуть всех лиц, экспонированных или возможно экс-
понированных к ВНО, активной иммунизации живой вакциной, и пассивной имму-
низации противооспенным иммуноглобулином;
2) вакцинировать всех контактных лиц, включая медицинских работников, если
они не были вакцинированы или ревакцинированы в течение трех последних лет;
3) наложить карантин на всех лиц, непосредственно контактировавших с любым
больным натуральной оспой не менее чем на 17 сут.
Лечение натуральной оспы и оспы обезьян симптоматическое. Профилактика
заражения ВОО аналогично таковому при натуральной оспе. ВИЧ-инфицированным
и лицам с другими иммунодефицитами вводить следует только противооспенный
иммуноглобулин.
3.9. ВИРУСНЫЕ ГЕМОРРАГИЧЕСКИЕ ЛИХОРАДКИ
Микробиология возбудителей геморрагических лихорадок. Экология и естественная
эпидемиология возбудителей геморрагических лихорадок. Восприимчивость людей
и животных к возбудителям геморрагических лихорадок. Иммунитет к возбудителям
вирусных геморрагических лихорадок. Устойчивость возбудителей вирусных геморра-
гических лихорадок во внешней среде. Антивирусные препараты. Обнаружение ге-
нетически измененных штаммов возбудителей геморрагических лихорадок. Патогенез
геморрагических лихорадок. Клиническая и патологоанатомическая картина вирусных
геморрагических лихорадок у человека при естественном инфицировании. Клиничес-
кая картина и патоморфология вирусных геморрагических лихорадок у животных при
искусственном инфицировании. Клиническая картина вирусных геморрагических
лихорадок у людей при искусственном инфицировании. Диагностика искусственного
поражения возбудителями геморрагических лихорадок. Иммунопрофилактика вирус-
ных геморрагических лихорадок. Лечение вирусных геморрагических лихорадок.
Вирусные геморрагические лихорадки (viral haemorrhagic fever, VHF, ВГЛ) — группа
мало отличающихся друг от друга по клинике зоантропонозных острых инфекцион-
ных болезней, вызываемых оболочечными PH К-вирусами семейств: аренавирусы (ви-
рус Ласса, вирус Хунин, вирус Мачупо, вирус Гуанарито, вирус Сэбия); бунъявирусы
(вирус геморрагической лихорадки с почечным синдромом, вирус Конго-Крымской
лихорадки, вирус лихорадки долины Рифт); филовирусы (вирус Марбург, вирус Эбола);
флавивирусы (вирус болезни леса Киассанур, вирус омской геморрагической лихорад-
ки, вирус желтой лихорадки, вирус лихорадки Денге). Первичный резервуар этих виру-
сов не известен. Передача вирусов в природных очагах из вторичных резервуаров среди
888
Часть 3. Частная неправильная эпидемиология
млекопитающих к людям происходит либо артроподами-переносчиками (вирусы
Конго-Крымской лихорадки, желтой лихорадки, лихорадки Денге, лихорадки долины
Рифт, омской геморрагической лихорадки, болезни леса Киассанур), либо в результате
непосредственно! о контакта с больными или погибшими животными (вирусы Ласса,
Эбола, Марбург). Вирусы Ласса, Эбола и Марбург могут передаваться между людьми
воздушно-капельным путем. На территории России имеются природные очаги виру-
сов, вызывающих геморрагическую лихорадку с почечным синдромом (Башкортостан,
Удмуртия, Марий-Эл, Мордовия, Чувашия, Ульяновская и Пензенская области),
Конго-Крымскую лихорадку (Краснодарский и Ставропольский края, Астраханская,
Волгоградская и Ростовская области, республики Дагестан, Калмыкия и Карачаево-
Черкесия), омскую геморрагическую лихорадку (степные и лесостепные районы Ом-
ской, Новосибирской, Тюменской, Курганской и Оренбургской областей). Инкуба-
ционный период болезни у человека после заражения вирусами геморрагических
лихорадок составляет от 2—21 сут. Основными клиническими проявлениями ВГЛ
являются лихорадка, астения, тошнота, рвота, диарея, макулопапулезная сыпь, гемор-
рагический диатез и шок.
Общий анализ крови в большинстве случаев выявляет вначале лейкопению, за-
тем тромбоцитопению. Большинство больных умирают в состоянии ступора с про-
явлениями интоксикационного шока. Главной причиной гибели при ВГЛ является
развитие синдром диссеминированного внутрисосудистого свертывания (ДВС-синд-
ром, коагулопатия, тромбогеморрагический синдром), отека легких и головного мозга.
ДВС-синдром был описан при лихорадках долины Рифт, Марбург и Эбола, но при
оольшинстве ВГЛ этиология коагулопатии является многофакторной (например, по-
ражение печени и первичное повреждение костного мозга с нарушением процесса
мегакариоцитопоэза). Вакцин, позволяющих эффективно осуществлять специфиче-
скую профилактику в природных очагах ВГЛ, нет. Исключение составляет инактиви-
рованная вакцина против желтой лихорадки. Специфическое лечение ВГЛ не отра-
ботано (Kortepeter М. et al., 2001; Bono L. et al., 2002; Борисевич И. В. с соавт., 2006).
Количество видов и даже семейств вирусов, вызывающих геморрагические лихо-
радки, неустановленно. Они могут проявлять себя локальными смертельными вспыш-
ками, и лишь случайно распознаваться, как, например, Lujo virus (семейство Агепа-
vindae). В Йоханнесбурге этот вирус в сентябре и октябре 20и8 г. вызвал пять случаев
геморрагической лихорадки, четыре из которых закончились смертью заболевших
(Paweska J. et al., 2009). Краткая характеристика наиболее изученных вирусов, возбу-
дителей геморрагических лихорадок, приведена в табл. 3.33.
Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят
филовирусы (вирус Марбург, вирус Эбола) и аренавирусы (вирус Ласса, вирус Хунин,
вирус Мачупо, вирус Гуанарито, вирус Сэбия) к I группе патогенности. По степени
важности для национальной безопасности США к группе А относятся филовирусы,
включая вирусы геморрагических лихорадок Эбола и Марбург, и аренавирусы, вклю-
чая вирусы лихорадок Ласса, Аргентинской геморрагической лихорадки и связанные
с ними вирусы (см. табл. 2.2).
Как потенциальные агенты БО возбудители геморрагических лихорадок стали
упоминаться в научной литературе после вспышки болезни, вызванной вирусом Эбола
в Южном Судане и Заире в 1976 г. В течение нескольких недель вирус вызвал вспыш-
ки геморрагической лихорадки на территории 3 тыс. км2, погубив жизни 435 человек.
Летальность среди заболевших составила 71 %, что, естественно, произвело впечат-
ление на военных специалистов (см. 1айслер Э. с соавт., 1986). Но об исследованиях
Таблица 3.33
Вирусы, вызывающие геморрагические лихорадки*
Семейство Род Вид Вызываемая болезнь Переносчик в природе Географическое распределение очагов болезни
1 2 3 4 5 6
Filoviridae Filovirus Ebola субтипы: Zaire, Sudan, Cote d’Ivoire (Ivory Coast), Bundi- bugyo. Lloviu и Reston Лихорадка Эбола. Субтипы Lloviu и Reston для человека не опасны He известен Африка (в основном Судан, Заир, Кот-д’Ивуар, Уганда). Север Испании и Филиппи- ны — субтипы Lloviu и Reston соответственно
Marburg Лихорадка Марбург He известен Африка (в основном Ангола, Уганда, Судан, Кения, ЮАР)
Arenaviridae Are navi rus Lassa Лихорадка Ласса** Грызуны Западная и Центральная Африка
New World Arenaviridae** Геморрагические лихорадки Нового Света*** Грызуны Американский континент
Bunyaviridae Nairovirus Crimean-Congo hemorrhagic fever Конго-Крымская геморрагическая лихорадка Клещи Африка, Центральная Азия, Восточная Европа, Средний Восток
Phlebovirus Rift Valley fever Лихорадка долины Рифт Комары Африка, Саудовская Аравия, Йемен
Hantavirus Agents of hemorrhagic fever with renal syndrome Известно более 25 серологически и генетически отличающихся друг от друга хантавирусов и две клинические формы хантавирусной инфекции у людей: геморрагическая лихорадка с почечным синдромом (ГЛПС), возбу- дителем которой являются вирусы Hantaan, Seul, Puumala, и Dobrava/ Belgrade; и хантавирусный пульмональ- ный синдром, вызываемый хантавиру- сами Sin-Nombre, Black Creek, New York, Bayou, Andes, Laguna Negra Основным носителем вируса Хантаан слу- жит полевая мышь. Основным вторичным резервуаром вируса Пуумала является рыжая полевка Вирус Хантаан циркулирует в природных очагах Дальнего Востока, России, Южной Кореи, КНДР, Китая, Японии. Вирус Пуумала обнаружен в Финляндии, Швеции, в России. Франции, Бельгии Вариант Dobrava/Belgrade — циркулирует на Балканах
'.9. Вирусные геморрагические лихорадки
оо
чэ
о
Продолжение табл. 3.33
1 2 3 4 5 6
Flaviviridae Flavi virus Летние Лихорадка Денге. Денге геморрагиче- ская лихорадка, шоковый синдром при лихорадке Денге Комары Южная и Юго-Восточная Азия, Африка, Океания, Карибский бассейн
Yellow fever Желтая лихорадка Существует в двух эпидеми- ческих формах: лихорадки джунглей (передается кома- рами от зараженных обезь- ян) и лихорадки населенных пунктов (передается кома- ром Aedes aegypti от боль- ного человека здоровому) Африка, тропические зоны американского континента
Omsk hemorrhagic fever Омская геморрагическая лихорадка Резервуаром инфекции в природе являются в основ- ном водяная крыса, рыжая полевка, ондатра, а также клещи D. pictus, D. marginal us, которые могут передавать вирус потомству трансовариально Природные очаги выявлены в степных и лесостепных рай- онах Омской, Курганской, Новосибирской, Тюменской, Оренбургской областей РФ
Kyasanur Forest disease Болезнь Киясанурского леса Клещи вида Haemaphysalis spinigera Северная часть Индии
* За основу взята работа L. Borio et al. (2002).
* * Относится к вирусам геморрагических лихорадок Старого Света (Old World complex). В эту же группу входят вирус лимфоцитарного
хориоменингита (Lymphocytic choriomeningitis, LCMV), вирусы Мопея (Mopeia virus, MOPV), Мобала (Mobala virus, MOBV), Иппи (Ippy virus,
IPPV), Луджо (Lujo virus, LUJV).
* ** Группа New World Arenaviridae (Аренавирусы Нового Света) включает вирус Machupo (Мачупо), вызывающий Боливийскую гемор-
рагическую лихорадку; вирус Junin (Хунин), вызывающий Аргентинскую геморрагическую лихорадку; вирус Guanarito (Гуанарито), вызы-
вающий Венесуэльскую геморрагическую лихорадку; вирус Sabia (Сэбия), вызывающий Бразильскую геморрагическую лихорадку и др.
Часть 3. Частная неправильная эпидемиология
3.9. Вирусные геморрагические лихорадки
891
возбудителей ВГЛ как потенциальных агентов БО, включающих масштабные экспе-
рименты в полигонных условиях, не известно. С 1972 г. действует Конвенция о зап-
рещении биологического оружия, поэтому если такие исследования и проводились в
США, то они не афишировались.
Секта «Аум-Синрикё» на пике западной дезинформационной кампании о доступ-
ности странам Третьего мира «дешевого оружия массового поражения» предприняла
попытку подготовить террористический акт с использованием вируса Эбола, но пе-
реоценила умственные возможности своих «биотеррористов». После гибели их веду-
щего вирусолога, интерес к возбудителям ВГЛ у сектантов пропал. Военные специа-
листы проводят оценку потенциальной опасности возбудителей ВГЛ, исходя из общих
требований к потенциальным агентам БО. Вирусы Денге и желтой лихорадки счита-
ются ими непригодными для ведения биологической войны из-за высокой ингаляци-
онной дозы. Возбудители Конго-Крымской геморрагической лихорадки и геморра-
гической лихорадки с почечным синдромом, также не рассматриваются ими как
перспективные агенты БО из-за технических трудностей, возникающих при масшта-
бировании производства. К наиболее пригодным для использования в целях биоло-
гической воины они относят вирусы Эбола, Марбург, Ласса, Мачупо и Хунин (Borio L.
et al.. 2002). Основной способ их применения для поражения людей — создание аэро-
золя (см. Аплу FM 8-284, 2000).
Микробиология возбудителей геморрагических лихорадок. Вирусы геморрагических
лихорадок размножаются в культурах клеток. Выбор культур клеток зависит от задач
эксперимента и биологических особенностей вируса. Например, для размножения
вируса лихорадок Эбола, Марбург и Ласса, используют кле гки Vero (Halfmann Р. et al.,
2008; Alves D. A. et al., 2010; Lukashev.ch I. S. et al., 2005 соответстиенно). При имита-
ции природных резервуаров, способных поддерживать длительную персистенцию
вируса Эбола, используют мышиные линии клеток NIH 3F3 и RAW264.7 (линии кле-
ток фибробластов и макрофагов мышей соответственно) и линию клеток мексикан-
ского складчатогуба (Strong J. Е. et al., 2008). Пассируются вирусы геморрагических
лихорадок на морских свинках и в культуре перевиваемых клеток почки зеленой мар-
тышки. Их основные микробиологические свойства приведены ь табл. 3.34.
Экология и естественная эпидемиология возбудителей геморрагических лихорадок.
Из-за огромного видового и штаммового разнообразия и существования в разных гео-
графических районах в качестве эндемиков, детальное изложение таксономии,
Таблица 3.34
Микробиологические свойства вирусов геморрагических лихорадок*
Семейство Диаметр частицы, нм Морфология Геном
Размер, т. п. о. Свойства Сегментированность
Filoviridae 80 В форме бациллы 19 РНК(-), одноцепочечная Несегментированный (1 —сегмент)
Arena viridae 110-130 Сферическая 11 РНК(±), одноцепочечная Сегментированный (2 ± сегмента)
Bunyaviridae 80-120 Сферическая 11-19 РНК(-), одноцепочечная Сегментированный (3 - сегмента)
Flaviviridae 40-50 Изометрическая 10-12 РНК(+), одноцепочечная Несегментированный (1 + сегмент)
* По L. Borio et al. (2002).
892
Часть 3. Частная неправильная эпииемиология
происхождения, экологии и эпидемиологии вирусов, возбудителей ВГЛ, в данной
работе невозможно. Ниже будут приведены типовые примеры природных вспышек,
вызванных этими вирусами, необходимые д ля понимания эпидемиологических дан-
ных, собранных при расследовании вспышки на территории России.
Filoviridae:вирусы Эбола и Марбург. Семейство вирусов Filoviridae известное 1967 г.,
когда ранее неизвестный вирус вызвал вспышку геморрагической лихорадки в Мар-
бурге (ФРГ) среди работников предприятия по производству вакцин, ассоциирован-
ную с контактным заражением от приматов, привезенных из Уганды. Филовирусы
являются членами порядка Mononegavirales. Их название происходит от лат. filum
(нить), отражающего их уникальную нитчатую морфологию. Семейство Filoviridae
включает только один род, Filovirus, состоящий из двух видов — вирусы Эбола и Мар-
бург. Эти виды имеют сходную геномную организацию, но в значительной степени
отличаются друг от друга по нуклеотидной последовательности и лишены антигенной
перекрестной реактивности. Вирус Эбола имеет не менее шести субтипов (Zaire,
Sudan, Cote d’Ivoire, Bundibugyo, Lloviu и Reston). У вируса Марбург (Lake Victoria) суб-
типы не известны, но выделены от больных людей и животных, как минимум, девять
генетически отличающихся изолятов (Ci6777, Popp, Musoke, Ravn, Angola и др.). Наи-
более вирулентным для человека среди субтипов вируса Эбола является Zaire, среди
изолятов вируса Марбург — Angola. Основываясь на сопоставлении летальных исхо-
дов среди заболевших людей, D. A. Alves et al. (2010)1 считают изолят Angola вируса
Марбург более вирулентным для людей, чем субтип Zaire вируса Эбола.
Большинство случаев филовирусной инфекции распознается только в условиях
больших вспышек и при наличии эпидемической настороженности у врачей. Еще
несколько лет назад считалось, что филовирусы циркулируют в прилегающих с юга к
Сахаре странах Африки, вызывая масштабные вспышки тяжелых форм геморрагиче-
ских лихорадок; и на Филиппинах, где обнаружен не опасный для человека субтип
Reston вируса Эбола (рис. 3.110).
Однако Филиппинами и Юго-Восточной Азией географическое распространение
филовирусов не ограничивается. В трупах летучих мышей Miniopterus schreibersii (обык-
новенный длиннокрыт), собранных в одной из пещер в испанской провинции Асту-
рия, обнаружен новый филовирус, названный Lloviu (Lloviu virus, LLOV). Название
вируса происходит от названия пещеры Cueva del Lloviu. Вирус выделен из пневмо-
нических очагов в легких М. schreibersii. Хотя пещеру часто посещают люди, случаев
болезни, вызванных вирусом Lloviu, у них не выявлено. Это второй известный фило-
вирус, не опасный для людей. Филогенетический анализ указывает на существование
в прошлом обшего предка всех филовирусов, от которого Lloviu отделился, по край-
ней мере, 150 тыс. лет назад (Negredo A. et al., 2011). Возможно обнаружение других
очагов филовирусов на европейском континенте.
На электронных микрофотографиях филовирусные вирионы представляют собой
длинные нити. Длина вирусных частиц варьирует в пределах от 800 до 1400 нм, но они
имеют постоянный диаметр равный 80 нм. Филовирусы — относительно простые
вирусы, имеющие геном 19 т. п. о., кодирующий, по меньшей мере, 8 белков: нуклео-
протеид (NP), вирионные белки (VP35, VP40, VP30 и VP24), белок с полимеразной
активностью (L), трансмембранный гликопротеид (GP) и растворимый гликопротеид
(sGP). Все гены расположились в одной цепи отрицательной-смысловой РНК. Ген^р
1 Сотрудники US Army Medical Research Institute of Infectious Diseases, Fort Detrick,
Maryland, CHIA.
3.9. Вирусные геморрагические лихорадки
893
Рис. 3.110. Известная и предполагаемая география распространения филовирусов.
А. Карта известных эпидемических вспышек геморрагических лихорадок, вызванных
филовирусами с вирусологическим подтверждением диагноза. Б. Географические про-
екции экологических ниш, где могут поддерживаться филовирусы, построенные на
основе географической локализации известных вспышек филовирусных инфекций.
В и Г. Географические проекции возможных экологических ниш филовирусов, постро-
енные на основе географической локализации известных вспышек лихорадок Эбола и
Марбург соответственно. Д и Е. Географические проекции возможных экологических
ниш филовирусов на Филиппинах и в Юго-Восточной Азии, основанные на проекции
экологических ниш возбудителей лихорадки Марбург и Эбола в Африке соответственно.
Большая плотность серого цвета означает большую вероятность присутствия в регио-
не природных очагов филовирусов. Кольца и треугольники на карте африканского кон-
тинента означают географическую локализацию регионов, где встречается заболевае-
мость лихорадки Эбола и Марбург соответственно. По A. Townsend Peterson et al. (2004)
кодирует оба гликопротеида и является самым вариабельным участком генома виру-
са Эбола. Доминирующим продуктом гена является гликопротеид sGP. Синтез GP
происходит в результате корректировки РНК. Белки Np30, VP30, VP35 и L объединя-
ются с вирусной геномной РНК, образуя рибонуклеопротеидный комплекс (централь-
ный рибонуклеопротеидный кор), тогда как белки VP40, VP24 и GP ассоциируются с
вирусной мембраной (Warfield К. L. et al., 2006; Melito Р. L. et al., 2008) Белок VP35
вируса Эбола (Zaire) функционирует аналогично белку NS1 вируса гриппа, препят-
ствующего индукции интерферона типа I в инфицированных клетках. Белок VP35
может действовать путем связывания с двунитевой РНК, препятствуя таким образом
активации днРНК-ассоциированной фосфокиназы и, возможно, других антивирус-
ных белков.
Линейное расположение вирусных генов напоминает таковое у рабдо- и парамик-
совирусов. 3'- и 5'-концы генома содержат консервативные, комплементарные после-
довательности, которые являются важными цис-действующими регуляторами геном-
ной репликации, транскрипции и, возможно, упаковки вирусной частицы. Транс-
крипция отдельных генов регулируется некодирующими участками на 3'- и 5'-концах,
содержащих короткие консервативные последовательности (Bray М., Paragas J., 2002).
Цикл репликации у филовирусов похож на таковой у хорошо изученных рабдо- и
парамиксовирусов. Репликация начинается с прикрепления вириона к рецептору на
894
Часть 3. Частная неправильная эпидемиология
поверхности клетки (предположительно а-фолатный рецептор). После связывания с
рецептором вирус проникает в клетку путем рецептор-опосредованного эндоцитоза.
В результате слияния вирусной оболочки с мембраной эндосомы, центральный рибо-
нуклеопротеидный кор высвобождается в цитоплазму. Транскрипция начинается на
З'-конце генома посредством вирусной PH К-зависимой РНК-полимеразы и приво-
дит к синтезу лидерной PH К и семи полиаденилированных мРНК. Аккумуляция пер-
вых двух белков, кодируемых на З'-конце генома (NP и VP35), каким-то образом сти-
мулирует продуцирование положительно-смысловых «антигеномов» полной длины,
которые, в свою очередь служат в качестве матриц для синтеза генома. Вирион состо-
ит из центрального рибонуклеопротеидного кора, связанного с двумя матричными
белками, VP24 и VP40, покрытого двойным липидным слоем, содержащим гликопро-
теид, полученным от клетки-хозяина. Гликопротеид образует на поверхности вирио-
на тримерные шипы (Bray М., Paragas J., 2002).
В случае вируса Эбола, но не вируса Марбург, первичный продукт, кодируемый
геном gp, является усеченным белком, лишенным С-терминального гидрофобного
мембранного якоря. Этот «растворимый GP» (sGP) высвобождается из инфицирован-
ных клеток. Синтез GP вируса Эбола полной длины требует вставки дополнительного
аденозина в «редактирующем сайте» в процессе транскрипции. Во время прохожде-
ния через аппарат Гольджи, GP обильно гликозилируется и расщепляется клеточным
фуриноподобным ферментом на две единицы: внеклеточный GP] и трансмембран-
ный GP2, которые остаются связанными дисульфидной связью в виде СР)2. Сборка
новых вирусных частиц происходит на внутренней поверхности плазматической мем-
браны, где VP24 и VP40 связываются, по-видимому, не только с новым центральным
рибонуклеопротеидным кором, но и с цитоплазматическим хвостом GP2. Полный
цикл репликации длится примерно 12 ч.
Вирусы Эбола и Марбург происходят от одного предка. Несинонимические заме-
ны в гене gp вирусов Эбола и Марбург оцениваются как 3,6х 10-5 на сайт/год (per site
per year). Что в сотни раз медленнее, чем у ВИЧ или у вируса гриппа, но примерно
соответствует скорости несинонимических замен у вируса гепатита В, имеющего ДНК-
геном. Дивергенция вирусов Эбола и Марбург произошла тысячи лет назад (Suzuki Y.,
Gojubon Т, 1997). Их филогенетическое древо приведено на рис. 3.111.
Особенности этиологии, эпидемиологии и клиники геморрагических лихорадок
Эбола и Марбург приведены в табл. 3.35, с. 896.
Первичный природный резервуар филовирусов неизвестен. В течение д лительного
времени считалось, что подобно некоторым другим классическим вирусным зооно-
зам, вирусы Эбола и Марбург под держиваются у некоторых видов позвоночных в виде
хронической/персистирующей инфекции, которая не вызывает заболевание. Причем
и вид-резервуар, и вирус сохраняются в течение достаточного периода времени, чтобы
обеспечить передачу другим восприимчивым хозяевам. Предполагалось, что резервуа-
рами являются многие виды позвоночных, включая летучих мышей, мышей, земле-
роек и других, живущих на земле животных, но, несмотря на интенсивный поиск, было
обнаружено, что ни одно из них не продуцировало живые вирусы в естественных усло-
виях. Хотя вспышки лихорадок Эбола и Марбург часто ассоциировались с контакта-
ми с низшими приматами, но эти виды не являются природными резервуарами фило-
вирусов, поскольку и у них наблюдается высокая летальность при заражении. Местное
население мало чем способно помочь исследователям при установлении эпидемичес-
ких цепочек филовирусов. Традиционные поверья, распространенные в Центральной
Африке, связывают болезнь с колдовством или отравлением, но не с вирусом.
3.9. Вирусные геморрагические лихорадки
895
Марбург 94
.99/90--
1/94-•
.98
.99/58
90
i/iooi 09 DRC 1999
Ravn Kenya 1987
Ozolins Zimbabwe 1975
, 07 DRC 1999
I 05 DRC 1999
- 1379c Angola 2005
Popp Uganda/Germany 1967
• Musoke Kenya 1980
Эбола
1/1001 Zaire, Yambuku 1976
Zaire, Kikwit 1995
— Cote d’Ivoire 1994
Bundibugyo, Uganda 2007
0,5 замен/сайт
1/96
1/100
Sudan, Gulu Uganda 2000
Reston, USA 1989
1
Рис. 3.111. Филогенетическое древо вирусов Эбола и Марбург. Сравнение полных
геномов обоих вирусов разного географического происхождения выполнено с по-
мощью Bayesian analysis (байесовский анализ). Серым цветом выделен субтип Bun-
dibugy вируса Эбола, обнаруженный авторами данной статьи во время большой
вспышки геморрагической лихорадки в Западной Уганде в ноябре 2007 г. Субтипы
филовирусов выделяются в том случае, если их геномы различаются один от другого,
по крайней мере, на 30—40 %. По J. S. Towner et al. (2008)
После вспышек лихорадки Эбола в 1994 г. в республике Кот-д’Ивуар и в 1995 г. в
Демократической Республике Конго, были предприняты интенсивные попытки иден-
тифицировать природные резервуары филовирусов. Однако они не дали ответа на
вопрос, кто из животных или растений является первичным резервуаром*. Некоторые
ученые по-прежнему настаивают на том, что резервуаром, из которого происходит
инфицирование людей, являются летучие мыши. J. S. Towner et al. (2009) приводят
доказательства длительного поддержания вируса Марбург в колониях египетских
фруктовых летучих мышей (Rousettus aegyptiacus). Возможные модели трансмиссион-
ных циклов филовирусов приведены на рис. 3.112, с. 897.
До настоящего времени не идентифицированы членистоногие-переносчики, и
какие-либо другие механизмы передачи филовирусов из природного резервуара лю-
дям и низшим приматам. Неясными остаются механизмы, лежащие в основе высо-
кой чувствительности людей и низших приматов к вирусам Эбола и Марбург дикого
типа по сравнению с другими видами (т. е. мышами и, возможно, летучими мышами).
Эксперименты на приматах показали, что животные заболевают после интрана-
зального, перорального и конъюнктивального ведения вирусов Марбург и Эбола.
У людей такой путь занесения инфекции в организм возможен, но прямые доказатель-
ства отсутствуют, полученные в очагах болезни, отсутствуют. Воздушно-капельный
путь заражения скорее исключение, чем правило. В природе, судя по патологоанато-
мической картине болезни у погибших животных, вирус передается через пищу. У них
нет поражений альвеол и бронхов, в основном в патологический процесс вовлечены
кишечные лимфатические узлы, печень и селезенка (Wyers М. et al., 1999).
1 Считаю существенным упущением этих исследований то, что не были изучены почвен-
ные простейшие как возможный первичный резервуар филовирусов. Если исследователей
заинтересовала резервуарная роль растений, значит связь между ними и заражением позвоноч-
ных животных предполагалась. Филовирусы не живут в растительных клетках, но они могут
проникнуть в растения вместе с почвенной водой, содержащей инфицированные простейшие.
896
Часть 3. Частная неправильная эпидемиология
Таблица 3.35
Особенности этиологии, эпидемиологии и клиники филовирусных
геморрагических лихорадок Эбола и Марбург*
Критерий Характе- ристика Эбола Марбург
Этиология Возбудитель Частица 800 нм, полиморфен, не менее 8 белков Частица 750 нм, полиморфен, не менее 8 белков
Эпидемио- логия Эпидемии (количество заболевших) 1476—1978 гг. — Судан, Заир (602); 1985 г. — Кения (?); 1994 1995 гг. - Заир (297), Кот д’Ивуар (3); 1994-1996 гг. - Габон (90), ЮАР(>17); 2001 г. — Кения; 2003 г. - Габон (135); 2004 г. — Судан (> 20?) 1967 г. - ФРГ, СФРЮ (31); 1975—1990 гг. — Родезия (2), Кения (1); 1999 г. — Конго (Заир) (70); 2004—2005 гг. — Ангола (308, из них 284 смертельных исходов)
Сезонность Весна — лето Весна — лето?
Генерации До 5-8 2—3 (до 7—8 в Конго)
Летальность 60-100% 28-100 %
Источник Больные люди, обезьяны, летучие мыши, грызуны, растения? Больные люди, обезьяны, растения?
Путь передачи Контакты (в т. ч. половые), ингаляционно, инъекции Контакты (в т. ч. половые), ингаляционно
Контагиозность 23 % в семье, 81 % среди медицинского персонала Невысокая (Конго — высокая)
Заболеваемость 10-81 % Нет данных
Заразный период 1—27 сут болезни (до 2—3 мес. при половой передаче) 1—27 сут болезни (до 2—3 мес. при половой передаче)
Клиника Инкубацион- ный период 4—20 сут 3—20 сут
Начало Острое Острое
Первичные симптомы Лихорадка, фарингит, тошнота, боль в животе Лихорадка, боли, прострация
Фаза разгара Мелена (2—3 сут), сыпь (4—6 сут), диарея, возбуждение, кровотече- ния и кровоизлияния (3—5 сут) Фарингит, воспаление поло- вых органов, сыпь, геморра- гический диатез (8—13 сут), поражение печени и почек, миокардит
Причина гибели ДВС-синдром, отек легких и мозга, шок (8—9 сут) ДВС-синдром, отек легких и мозга, шок (8—17 сут)
Длительность заболевания 2—3 нед 3—4 нед (до 2 мес )
Длительность реконвалес- ценции 2- 3 мес. 2—3 мес.
Осложнения Анорексия, психозы Гепатит, орхит, миелит, психозы
По И. В. Борисевичу с соавт, (2006) с дополнениями из работы Р. L. Melito et al. (2008).
3.9. Вирусные геморрагические лихорадки
897
Хищничество Насекомояднь1е Контакт
——у позвоночные \———
растений
Членистоногие
Обезьяна
Человек
Человек
Обезьяна
Членистоногие
Пауки, скорпионы,
многоножки и др.
Летучие
илигрь
Контакты, укусы,
половая передача
Человек
Человек
Обезьяна
Обезьяна
Укусы ядовитых животных
Рис. 3.112. Возможные модели трансмиссионных циклов филовирусов.
А. Сдоминированием летучих мышей. Б. С доминированием артроподов
и насекомоядных. По Т. М. Monath (1999)
Больные лихорадками Эбола и Марбург очень опасны для медицинского пер-
сонала. Например, в 2000 г. во время вспышки лихорадки Эбола в Уганде (погибло
224 человека) из 22 человек медицинского персонала заболело 14 (64 %). Первичны-
ми больными с филовирусной инфекцией чаше всего были мужчины, которые рабо-
тали в лесах, котлованах или рудниках. Вирусы лихорадок Эбола и Марбург могут в
течение 101 сут и 83 сут, соответственно, выделяться с семенной жидкостью реконва-
лесцентов. Доказаны случаи полового заражения этими вирусами (Borio L. et al., 2002;
Peterson A. T. et al. 2004). Ниже рассмотрим пример молекулярной эпидемиологии
естественных вспышек лихорадки Эбола (рис. 3.113).
Циркуляция вируса Эбола среди населения Габона серологически показана еше в
1982 г. В 1994-1994 гг. были зарегистрированы три независимые друг от друга вспыш-
ки геморрагической лихорадки Эбола у людей в северо-восточной части Габона, в райо-
не лесов экваториальной провинции Огуо-Ивиндо. Каждая вспышка сопровождалась
гибелью приматов. Первая из них началась в декабре 1994 г. в сезон дождей (декабрь и
февраль) в лагере золотодобытчиков на северо-востоке Габона в районе М i nkouka меж-
ду 1'24" и 1'44" с. ш. и 12'59" в. д. около реки Нуна. Вторая — в начале февраля 1996 г.
в селении Mayibout (1'07" с. ш. и 13'06" в. д.) около реки Ивиндо (37 случаев болезни).
Наиболее близкий к ним город Makokou, расположенный в 0'33" с. ш. и 12'50" в. д.,
имел свой госпиталь, куда доставлялись больные и контактировавшие с ними жители.
Третья вспышка началась в июле 1996 г. в поселке Вооиё (0'06" ю. ш. и 11 '57" в. д.), где
выявлено большинство заболевших. Также были диагностированы отдельные случаи
898
Часть 3. Частная неправильная эпидемиология
Рис. 3.113. Географическое распределение трех эпидемий геморрагической лихорад-
ки Эоола в Габоне. Район Minkouka — первая вспышка. Mayibout — вторая вспышка.
Вооиё — третья вспышка. По М. С. Georges-Courbot et al. (J497)
g &
болезни в близлежащих городах и поселках. Всего за время третьей вспышки зарегист-
рировано не менее 43 летальных случаев геморрагической лихорадки Эбола.
Вирус Эбола легко выделялся из крови людей в культуре клеток Vtero Е6 при единст-
венном пас< аже У вирусов, выделенных от больных, определялась последовательность
гена®?. Филогенетический анализ выявил общее эволюционное происхождение виру-
сов, вызвавших вспышки в Габоне и Заире. Кроме того, полученные данные подтверди-
ли независимость друг от друга трех вспышек в Габоне. Ген gp вирусов, выделенных
время весенней вс пышки 1996 г. в Габоне, отличался от последовательности гена
вирусов вспышки конца 1994 г. четырьмя дополнительными нуклеотидами. Между
вирусными последовательностями, полученными непосредственно из клинического
материала и путем пассирования в тканевой культуре, или между последовательностя-
ми от первичных и вторичных случаев заболевания, различий не было. Наличие посто-
янных вирусных последовательностей и отсутствие генетического разнообразия меж-
ду штаммами, выделенными в пределах отдельной вспышки, было показано в 1995 г.
во время вспышки геморрагичекой лихорадки Эбола в селении Киквит (Заир) и под-
тверждено в Габоне.
Полученные М. С. Georges-Courbot М. С. et al. (1997) данные показали, что виру-
сы, вызвавшие вспышки лихорадки Эбола на разных эндемичных территориях, отли-
чаются друг от друга по нуклеотитным последовательностям гена#р, однако генети-
ческого разнообразия между штаммами, выделенными в пределах отдельной вспышки
ооычно не наблюдается. Возможны вспышки, вызванные рекомбинантами, образо-
вавшимися в пределах одного субтипа вируса Эбола (Wittmann Т. J. et al., 2007).
3.9. Вирусные геморрагические лихорадки
899
irenavu idae. Название семейству было дано по структуре вириона, который внеш-
не выглядит так, как будто посыпан песком (лат. arena — песок). Семейство состоит
из одного рода (Arenavirus), включающего не менее 23 идентифицированных вирусов,
подразделенных на основе их антигенных свойств на две группы:
1) серокомплексТакарибе (или группа Нового Света), включающий вирусы, харак-
терные для Северной и Южнпй Америки;
2) серокомплекс Ласса-лимфоцитарный хориоменингит (группа Старого Света).
Включает вирусы, характерные для Африки.
Повсеместно распространен только вирус лимфоцитарного хориоменингита.
Географическое распространение каждого аренавируса определяется диапазоном его
носителей в природе. Наиболее важными вирусными антигенами являются нуклео-
лротеид и гликопротеид, причем антигены нуклеопротеиды наиболее консерватив-
ные среди антигенов аренавирусов. Точная идентификация аренавирусов достигает
ся путем исследования в реакциях нейтрализации, которые зависят от высокоспеци-
фических эпитопов оболочечных гликопротеидов. Генетическая классификация
аренавирусов соответствует результатам серологических исследований. Семейство
состоит из 4 филогенетических линий вирусов (Chanel R. N., de Lamballerie X., 2003).
Особенности этиологии, эпидемиологии и клиники аренавирусных геморрагиче-
ских лихорадок Ласса, Боливийской и Аргентинской приведены в табл. 3.36.
Таблица 3.36
Особенности этиологии, эпидемиологии и клиники аренавирусных
геморрагических лихорадок Ласса, Боливийской и Аргентинской*
Критерий Характеристика Лихорадки Ласса Боливийская и Аргентинская лихорадки
1 2 4 4
Этиология Возбудитель Частица 50 нм, сферичен, 10—15 гранул; 13 белков Частица 70 нм, сферичен, 2—10 гранул; 13 белков
Эпидемиология Эпидемии Постоянно с 1969 г.; до 2—3 млн человек в год Постоянно; 10—30 человек в год
Завозы 1969—1988 гг. — около 20 случаев в США, Европе, Японии, Израиле Нет данных
Сезонность Отсутствует Весна — лето?
Генерации ДоЗ 1-2
Летальность 0,1-0,2 % (20-30 % среди госпитализированных) 5-30 % (10 %)
Источник Крысы Mastomys, больные люди Хомячки Calomys, больные люди
Путч передачи Контакты, алиментарно, аэрозольно Контакты, алиментарно, аэрозольно
Контагиозность Средняя Средняя
Заболеваемости Высокая Низкая
Заразный период До 70 сут До 2-3 нед
Клиника И нкубационный период 3- 21 сут (7—14 сут) 3—21 сут (14 сут)
Начало Постепенное Постепенное
900
Часть 3 Частная неправильная эпидемиология
Продолжение табл. 3.36
1 2 4 4
Первичные симптомы Лихорадка, недомогание, конъюнктивит Лихорадка, фарингит, головная боль, гингивит
Фаза разгара Интоксикация, язвенный фарингит, сыпь, геморраги- ческие проявления, лимфа- денит Полиаденит, сыпь, энце- фалит. геморрагические проявления
Причина гибели Некрозы внутренних органов (7—14 сут), острая почечная недостаточность, шок Сопутствующие осложнения (аутоиммунные) (10—14 сут), нарушение сердечно- сосудистой деятельности
Длительность болезни До 1 мес. До 1 мес.
Длительность реконвалесценции До 2—3 мес. 1—1,5 мес.
Осложнения Глухота, облысение, астения Гиперчувствительность, на- рушения статики, параличи
* По И. В. Борисевичу с соавт. (2006).
Вирион аренавируса покрыт плотной оболочкой, включающей липиды, и име-
ет крупные шипы на поверхности. Вирионы полиморфны, их диаметр в пределах
62—200 нм, в среднем, около 110 нм. Геномная (—)РНК аренавирусов двуцепочечная,
содержит два однонитевых сегмента (bi-segmented genom). Каждый сегмент кодирует
два различных белка в двух неперекрывающихся рамках считывания с противополож-
ной полярностью, разделеных некодирующим межгеномным регионом (non-coding
inteigenic region, IGR): S-сегмент — 3,5 т. п. о., кодирует, два полипептида: NP — нук-
леопротеин (63 кДа) и прекурсор поверхностного гликопротеина (surface glycoprotein
precursor, GPC; 75 кДа). GPC разрезается на два гликопрогеида GP1 и GP2;
L-сегмент — 7,3 т. п. о., кодирует два белка — РНК-полимеразу и Z-белок, содер-
жащий «Zn-связывающий палец».
Некодирующий межгеномный регион имеет стабильную шпилечную структуру.
5'- и З'-концы каждого сегмента являются некодирующими, и 19 терминальных
нуклеотидов одного конца, соответствуют обратной комплементарной последователь-
ности из 19 нуклеотидов на другом конце. Геномная (—) PH К реплицируется в (+)ан-
тигеномную и далее транслируется (-)мРНК. Жизненный цикл аренавирусов огра-
ничен цитоплазмой клетки (рис. 3.114).
В природе источником аренавирусов для людей являются инфицированные гры-
зуны. Заражение происходит алиментарным, контактным, через микротравмы кожи,
а также воздушно-пылевым (ингаляционным) путями. Благодаря высокой устойчи-
вости аренавирусов в окружающей среде и низкой инфицирующей дозе, ингаляци-
онное заражение человека может произойти на расстоянии, измеряемом десятками
метров от источника аэрозоля. При загрязнении предметов обихода или инструмен-
тария выделениями (кровавая мокрота, кровавая рвота) или непосредственно кровью
больных, содержащими вирус, возможна контактная передача возбудителя болезни,
а также заражение через медицинский инструмент. Попадание в пищу или воду зара-
женной вирусом мочи, носового и ротового отделяемого грызунов или носоглоточного
отделяемого больных людей также ведет к заражению. Хотя больной человек может
3.9. Вирусные геморрагические лихорадки
901
S РНК (3,5 т.п. о.)
GP
Геном
Репликация
L РНК (7,2 т.п. о.)
| Транскрипция
NPmPHK
3' -CZZZZZZZZa- >5'
| Трансляция
NP
Антигеном У
3'IZZZZZZZL
Z/Z/////Л 5'
Z
Транскрипция |
5'1 *ZZZZZZZZD-3'
GPmPHK
| Трансляция
TTTTTgp
L
Рис. 3.114 Организация генома и репликация аренавирусов. А. Организация генома
аренавирусов. Б. Репликация аренавирусов на примере S-сегмента. РНК-полимера-
за, ассоциированная с рибонуклеопротеидным комплексом вируса (ribonucleo-
protein, RNP), инициирует транскрипцию от геномного промотора, локализованного
в участке З'-конца (обозначен треугольником). Начальная транскрипция приводит
к синтезу мРНК NP (или мРНК L в случае L-сегмента). Затем вирусная полимераза
начинает действовать как репликаза и перемещается вдоль межгеномного региона
(IGR), образуя полную копию антигенома (antigenome, ag) S-РНК (или ag L-PHK).
Эта ag S-PHK служит матрицей для синтеза GP мРНК (ag L-PHK служит матрицей
для синтеза Z мРНК). По J. С. de la Torre (2008;
служить источником заражения для других людей, но при повторных передачах от
человека к человеку инфекция угасает, и в человеческой популяции длительной цир-
куляции аренавирусов не происходит. При расследовании внутрибольничных вспы-
шек механизм передачи вируса часто остается неизвестным. Во время первой вспышки
ихорадки Ласса в нигерийской больнице 1969 г. в населенном пункте Ласса, среди
пациентов выявлено 16 вторичных случаев болезни, однако не было доказано воздуш-
но-капельной передачи инфекции. Нет документального подтверждения передачи
аренавирусов от людей, находящихся в инкубационном периоде болезни. В то же вре-
мя вирус Ласса может выделяться спермой пациента в течение 3 мес. и мочой более
месяца от начала болезни (Шувалова Е. П. с соавт., 2001; Borio L. et al., 2002). Все аре-
навирусы, исследуемые в вирусологических лабораториях, оказывались инфекцион-
ными для человека в тех случаях, когда существовала возможность ингаляционного
заражения (Charrel R. N., de Lamballerie X., 2003).
Лихорадка Ласса встречается в Западной (Нигерия, Сьерра-Леоне, Сенегал, Мали,
Гвинея) и Центральной (Заир, Центрально-Африканская Республика, Буркина-Фасо)
Африке, где регистрируется в виде спорадических случаев и небольших вспышек на
территории природных очагов. В Западной Африке ежегодно инфицируются виру-
сом Ласса от 2 до 3 млн человек, из них от 5 до 10 тыс. погибает (Fichet-Calvet Е., Ro-
gers D. J., 2009) (рис. 3.115).
Вирус Ласса (LASV) передается людям от многососковых крыс {Mastomys spp.) при
непосредственном контакте с фекалиями инфицированных животных и/или при их
попадании на слизистую оболочку (рис. 3.116).
902
Часть 3. Частная неправильная эпидемиология
Рис. 3.115. География вспышек лихорадки Ласса в Западной Африке в 1969-1975 гг.
Лихорадка Ласса была многократно описана у заболевших людей еще в 1930-х гг под
названием «тиф саванны», но ее этиология не была установлена. Кружком обозначе-
ны преимущественно внутрибольничные вспышки болезни, квадратом — вспышки
в общинах на территориях, эндемичных по вирусу Ласса. По Т. Р. Monath (1975)
Сухой сезон
Потенциал
для развития
вспышки в не-
эндемической
области
пидемия
Дождливый сезон?
. ^Сельская
местакх гь ^Трансмиссивный
X Западной Африки цикл
(внутридомовая передача)
'^*|*>* ““‘“ПТ1*1 "ИТ** .**ГТ-~.~Т"Гчпги—'“nfllt
М. natalensi" ’
Горизонтальная (?) Контактный,
и вертикальная воздушный
передача и парентеральный
пути заражения
Внутрибольничная
инфекция
Международный
транспорт
Домашние грызуны или другие
позвоночные хозяева вируса О „ , .
у , Лабораторная инфекция
Рис. 3.116. Схематическое изображение трансмиссии вируса Ласса из природ-
ного резервуара среди грызунов в популяции людей. По Т. Р. Monath (1975)
Хотя и считается, что резервуаром вируса Ласса является Mastomys natalensis1, од-
нако в этом утверждении есть противоречие. Ареал М. natalensis включает не только
Западную Африку, но и Восточную, Центральную и Южную, но вспышки лихорадки
Ласса среди грызунов географически локализованы в отдельных регионах Западной
Африки. М. natalensis не разносит вирус Ласса по своему ареалу. Значит, есть какой-
то другой источник вируса (первичный резервуар), локализованный в отдельных ре-
гионах Западной Африки, из которого вирус проникает в популяции М. natalensis.
Филогенетический анализ показал, что вирусы Ласса состоят из четырех линий,
три из которых циркулируют среди грызунов в Нигерии, и четвертая — среди грызунов
Гвинеи, Либерии и Сьерра Леоне. Сравнительный анализ последовательностей генов
1 В московских зоомагазинах М. natalensis продают как корм для рептилий.
3.9. Вирусные геморрагические лихорадки
903
белков GP1 и GP2 позволяет предположить, что нигерийские штаммы вируса Ласса
были предками штаммов, циркулирующих в Гвинее, Либерии и Сьерра-Леоне. Гене-
тическое расстояние между штаммами Ласса коррелирует с географическим расстоя-
нием, но не со временем (Bowen М. D. et а!., 2000). Отсутствие доказательств работы
«молекулярных часов» вызвано низкой скоростью мутирования вируса. Скорость
образования мутаций у вируса Ласса в пределах от 5,6х 10~4 до 17,6х 10 4 нуклеотид-
ных замен на сайт/год (per site per year) (Ehichioya D. U. et al., 2011). Штаммы вируса
Ласса, выделенные от жертв вспышки, идентичны, если они произошли из одного ис-
точника заражения людей (Bowen М. D. et al., 2000). Возможность рекомбинации
между штаммами вируса Ласса разных линий, циркулирующих в природных резер-
вуарах, не доказана (Ehichioya D. U. et al., 2011).
Восприимчивость людей и животных к возбудителям геморрагических лихорадок. Для
испытания антивирусных препаратов и вакцин и для изучения филовирусного пато-
генеза используются три типа лабораторных животных — мыши, морские свинки и
низшие приматы. У этих разных видов животных аналогичные клетки-мишени, с
которыми специфически взаимодействую вирусы, и, соответственно, сходная симп-
томатика болезни. Низшие приматы крайне чувствительны ко всем филовирусам, в
то время как морские свинки и иммунокомпетентные мыши обладают врожденной
резистентностью к филовирусной инфекции. Это основное различие между живот-
ными моделями может стать причиной расхождений в результатах при проверке эф-
фективности лекарственных препаратов и вакцин.
Мыши. Вирусы, возбудители лихорадок Марбург и Эбола, вызывают летальное
заболевание у новорожденных мышей, но не вызывают явного заболевания у взрос-
лых иммунокомпетентных мышей. Однако в результате последовательного пассиро-
вания вируса Эбола (Zaire 76) на взрослеющих мышах-сосунках был получен вариант
вируса, который вызывает скоротечную летальную инфекцию у взрослых мышей при
интраперитонеальном заражении. Патологические признаки инфекции, вызванной
этим «адаптированным к мышам вирусом», сходны с таковыми у приматов, за исклю-
чением менее выраженной коагулопатии. У мышей отсутствует такой характерный
признак лихорадки Эбола как генерализованный тромбогеморрагический синдром,
обнаруживаемый на последней стадии в поражениях у людей и нечеловекообразных
приматов (Geisbert Т. W. et al., 2002). Мышиная модель используется для предваритель-
ной оценки вакцин и антивирусных препаратов и для изучения филовирусного пато-
генеза. У мышей с иммунодефицитом, лишенных либо врожденных, либо антиген-
специфических иммунных ответов, заражение вирусами Марбург и Эбола вызывает
быстрый летальный исход.
Морские свинки. После заражения вирусом Марбург или вирусом Эбола (Zaire) или
Эбола (Sudan) у морских свинок развивается слабо выраженная лихорадка. В результа-
те переноса вируса от животного к животному постепенно повышается его вирулен-
тность в отношении морских свинок, и после нескольких пассажей можно отобрать
штамм вируса, который постоянно вызывает инфекционный процесс, заканчиваю-
щийся гибелью животного. Основные патологические признаки летальной инфекции
у морских свинок сходны с таковыми у мышей и приматов. Морские свинки исполь-
зуются для испытания вакцин, но из-за их размера они в меньшей степени пригодны
для предварительной оценки экспериментальных препаратов, которые обычно име-
ются лишь в небольшом количестве.
Низшие приматы. Все филовирусы вызывают у низших приматов тяжелую гемор-
рагическую лихорадку. Наиболее вирулентным является вирус Эбола (Zaire), вызы-
904
Часть 3. Частная неправильная эпидемиология
вающий летальное заболевание у африканских зеленых мартышек, обезьян цино-
молгус, макак-резус и павианов. У обезьян циномолгус, являющихся широкораспро-
страненной моделью, эта инфекция характеризуется повышением температуры и сни-
жением активности на 3—4 сут после заражения; появлением с 4—5-х сут на туловище
сыпи в виде красно-фиолетовых пятен; притуплением чувствительности на 6 сут и
летальным исходом на 7—8 сут. Первоначально вирус обнаруживается в сыворотке на
3 сут, а к 5-м сут титры могут превысить 10 КОЕ/мл. Высокие концентрации вируса
обнаруживаются также в печени, селезенке и других тканях. Изменения в количествах
клеток крови и других параметрах клинических лабораторных, исследований сходны
с таковыми у людей. Обычно наб. подаются слабые кровотечения, а обильное крово-
течение является редким явлением. Вирусы Эбола (Sudan), Эбола (Reston) и Марбург
также вызывают у низших приматов тяжелую геморрагическую лихорадку, отличаю-
щуюся лишь большей продолжительностью болезни и летальностью, приближающей-
ся к '00 % (Bray М , Paragas J., 2002).
При инфицировании аэрозолем вируса Эбола (Zaire) с дисперсностью частиц
0,8—1,2 мкм, LD100 для макак-резус не превышала 400 КОЕ (Johnson Е. et al., 1995).
При ингаляционном заражении макак-крабоедов (Масаса fascicularis) разными
дозами вируса Марбург (MARV-Angola), составлявшими 2-14 КОЕ и 99-705 КОЕ,
обезьяны погибали все от любой дозы. Разница была только в продолжительности
жизни после заражения на одни сутки (Alves D. A. et al., 2010).
Парентеральное введение обезьянам 1—10 КОЕ вирусов Эбола (Zaire) или Мар-
бург (Musoki) обычно приводит их к смерти (Johnson Е. et al., 1995).
К вирусу Ласса чувствительны приматы и морские свинки. LD100 (штамм Josiah)
для обезьян М. fascicularis весом 2—3,5 кг не превышает 465 КОЕ при дисперсности
аэрозольных частиц меньше 4,5 мкм. При заражении этим же аэрозолями инбредных
морских свинок линии Hartley определить LD50 оказалось затруднительно, так как
летальность среди экспонированных животных никогда не превышала 50 %. Среднее
время до гибели (23- 37 сут) не обнаруживало значительной зависимости от концент-
рации вируса в аэрозоле. У инфицированных морских свинок находили лишь мини-
мальные симптомы болезни, такие как всклоченность шерсти и затрудненность дыха-
ния в последние 24—48 ч жизни. Однако морские свинки этой линии считаются
высокочувствительными к ингаляционному заражению вирусом Ласса, так как Ю50
для них составляет 15 КОЕ (Stephenson Е. et al., 1984).
Более точно воспроизвести в эксперименте клинику и патогенез лихорадки Лас-
са у человека, можно подкожным заражением макак-резусов (Gallis R. Т. et al., 1982).
Иммунитет к возбудителям вирусных геморрагических лихорадок. При отдельных
филовирусных, флавивирусных и буньявирусных инфекциях ответы со стороны им-
мунной системы человека на возбудитель инфекции мало похожи на те, что описаны
в учебниках по иммунологии для студентов. Основную роль в них играют феномены
антителозависимого усиления инфекции (antibody-dependent enhancement, ADE) и анти-
генного импринтинга (другое название — феномен первичного антигенного греха,
phenomenon of original antigenic sin, OAS).
Феномен ADE заключается в том, что вирусспецифические антитела усиливают
проникновение вируса в фагоцитирующие клетки посредством взаимодействия с ре-
цептором FcR и/или рецепторами комплемента на поверхности фагоцитирующих
клеток. Поэтому феномен наблюдается в двух вариантах:
а) комплемент-опосредованное антителозависимое усиление инфекции (comple-
ment-mediated ADE; C-ADE);
3.9. Вирусные геморрагические лихорадки
905
б) независящее от комплемента и связанное с Fc-рецептором усиление инфек-
ции (Fc-receptor-mediated ADE; FcR-ADE) (TakadaA., Kawaoka Y, 2003; Tirado S. M.,
Yoon K. S., 2003).
Особенности вирусов, вызывающих феномен ADE, следующие:
а) обычно такие вирусы реплицируются в макрофагах;
б) они индуцируют продукцию большого количества антител с плохой способно-
стью к нейтрализации гомологичных вирусов;
в) способны к персистентной инфекции, характеризующейся продолжительной
зиремией (Tirado S. М., Yoon К. S., 2003). Феномен ADE проявляется в ответ на обра-
зование антител изотипа IgGl (Henchai Е. A.et al., 1985).
A. Takada et al. (2001, 2002) обнаружена способность сыворотки реконвалесцен-
тов, переболевших лихорадкой Эбола (Zaire), увеличивать инфекционность вируса в
отношении клеток 293-й линии, клеток почек обезьян и эндотелиальных клеток пу-
почной вены человека. Исследователями показано, что основную роль в этом процессе
играют отдельные анти-IgM, специфичные к GP, и что выраженность феномена ADE
различна у сывороток, взятых от разных пациентов. Аналогичные данные получены
ими с сывороткой, взятой от мышей, иммунизированных ДНК-вакциной с клониро-
ваным геном gp. Феномен ADE был менее выражен для субтипа вируса Reston, чем для
вирусов субтипов Zaire и Sudan. Авторы данных работ предположили, что феномен
ADE играет важную роль в патогенезе лихорадки Эбола (рис. 3.117).
В дальнейшем A. Takada et al. (2007) показали в условиях in vitro, что ADE при
инфекционном процессе, вызванном вирусом Эбола (Zaire), развивается в результа-
те взаимодействия вирусспецифических антител с вирусом и Fc-рецептором, или
компонентом комплемента Clq и его рецептором (С-ADE вариант феномена). ADE
GP-антисыворотки связана с уровнями IgG2a и IgM, но не с уровнями IgG 1. Исполь-
зуя моноклональные антитела, авторы определили эпитопы GP вируса субтипа Zaire,
отвечающие за индукцию антител, вызывающих ADE. Они также сконструировали
химерные эпитопы, индуцирующие продукцию антител у мышей со сниженной спо-
собностью вызывать ADE, но обладающих нейтрализующей активностью в отноше-
нии вируса субтипа Zaire.
Для лихорадки Марбург феномен ADE был описан в 2011 г. Также как для вируса
Эбола, показана связь между ADE и вирулентностью штаммов вируса Марбург. Авто-
рами делается вывод, что феномен ADE лежит в основе патогенеза не только лихора-
док Марбург и Эбола, но и других филовирусных лихорадок (Nakayama Е. et al., 2011).
Среди инфекционных процессов, вызываемых флавивирусами, феномен ADE наи-
более изучен при лихорадке Денге и желтой лихорадке. Первая инфекция, вызванная
вирусом Денге, чаще всего протекает у человека бессимптомно и дает пожизненный
иммунитет к вирусу серотипа, ее вызвавшего. Однако если он сталкивается с вирусом
другого серотипа, то благодаря FcR-ADE, болезнь протекает в тяжелой форме с боль-
шой вероятностью летального исхода (до 50 %) (Boonnak К. et al., 2008). ADE рас-
сматривается как основная причина нейровирулентности вируса и тяжелого течения
желтой лихорадки. Показана связь ADE с антителами (нейтрализующими, не ней-
трализующими и протективными) к гликопротеину Е вируса желтой лихорадки (Bar-
rett A. D. Т., Gould А.,1986; Goulld Е A. et al., 1989). При лихорадке Западного Нила
ADF вызывают субклассы IgG, обладающие высокой афинностью к фактору комп-
лемента Clq (Mehlhop Е. et al., 2007).
При инфекционных процессах, вызываемых вирусами семейства Buriyaviridae, фе-
номен ADE остался не изученным. ADE смоделирован в условиях in vitro для инфек-
906
Часть 3. Частная неправильная эпидемиология
Рис. 3.117. Модель C-ADE (Clq-\DE) при лихорадке Эбола. А. Схематическое
изображение белков комплемента С1 и Clq. Молекула Clq включает глобулярный
и лигандсвязывающий домены. Глобулярный домен состоит из шести глобуляр-
ных набалдашников (globular heads), которые связываются с Fc-участком антите-
ла Лигандсвязываюший домен Clq взаимодействует с лигандом на поверхности
фагоцитирующей клетки. Аффинитет Clq к лиганду снижается при ассоциации
с С1г и Cis (сериновые протеазы). Б Механизм ADE пои лихорадке Эбола. Вирус
Эбола инициирует инфекционный процесс путем связывания со специфически-
ми рецепторами на поверхности фагоцитирующей клетки (1). Clq связывает ком-
плекс «вирус-антитело» с Clq-лигандами, расположенными на поверхности кле-
ток, вызывая взаимодействие между вирусом и рецептором (2). Диссоциация С1г
и Cis от Clq увеличивает связывающий аффинитет молекулы Clq с лигандами на
поверхности фагоцитирующей клетки (3). По A. Takada et al. (2003)
ционного процесса, вызванного вирусом Хантаан, но на этом его исследования пре-
кратились (Yao J. S. et al., 1992).
Феномен OAS состоит в том, что при повторном контакте иммунной системы с
вирусом или вакциной, иммунная система может не воспринимать различия между
вариантом эпитопа, с которым уже познакомилась ранее, и его новым вариантом.
И тогда активизируются В-клетки памяти, «запомнившие» преды тущий антиген. Вы-
работка антител происходит в отношении этого антигена, хотя реально иммунная
система с ним не контактирует. Образующиеся антитела не способны нейтрализо-
вать вирус, выработка же специфических антител тормозится из-за подавления «наив-
ных» В-клеток активизировавшимися В-клетками памяти. Феномен OAS детально
изучен при инфекционных процессах, вызванных ВИЧ, вирусом гриппа А, лимфо-
цитарного хориоменингита, лихорадки Денге и некоторых тога-, парамикс- и энте-
ровирусами. Вирус лимфоцитарного хориоменингита (LCMV) в настоящее время
3.9. Вирусные геморрагические лихорадки
907
единственный из аренавирусов, д ля которого установлена способность вызывать дан-
ный феномен. Р. KJenerman и R. М. Zinkemage (1998) показано, что последовательное
введение мышам различных штаммов LCMV, вызывало у них образование CTL-кле-
ток, специфичных только в отношении штамма LCMV, введенного первым. Среди
ВГЛ, вызванных флавивирусами, феномен OAS наблюдается при лихорадке Денге
(Halstead S. В. et al., 1983).
Исследования роли OAS и ADE в эпидемических и инфекционных процессах, и
ответах на вакцинацию малоинтересны создателям вакцин1 из-за опасений лишить-
ся финансирования, поэтому они носят выборочный характер.
Устойчивость возбудителей вирусных геморрагических лихорадок во внешней среде.
Преобладание ингаляционного пути заражения возбудителями ВГЛ среди других
механизмов заражения людей свидетельствует в пользу того, что выделяясь с экскре-
тами больных животных, они способны длительно сохраняться во внешней среде.
Основными причинами высокой резистентности к внешним воздействиям вирусов,
возбудителей геморрагических лихорадок, является наличие плотной липидной обо-
лочки и РНК-генома, который менее чувствителен к физическим факторам, чем ге-
ном ДНК-вирусов.
Стабильность и инфекционность аэрозолей возбудителей геморрагических ли-
хорадок изучены на примере вируса Ласса. Е. Stephenson et al. (1984) показали, что че-
рез 4 мин после перевода суспензии вируса в аэрозоль с дисперсностью частиц менее
4,5 мкм, при температуре окружающей среды 24°C и относительной влажности 55 %,
75,3 ±9,6 % частиц сохраняли инфекционность. При повышении температуры окру-
жающей среды до 38 °C при той же относительной влажности инфекционность аэро-
золя падала до 33,7 %. Периоды биологического полураспада аэрозоля вируса как при
температуре окружающей среды 24°C, так и при 32°С, составляли от 10,1 до 54,6 мин,
в зависимости от относительной влажности воздуха, но в любом случае инфекцион-
ность аэрозолей была достаточной для поражения людей на значительном расстоя-
нии от аэрозольного генератора.
Антивирусные препараты. Лицензированных препаратов, оказывающих эффектив-
ное профилактическое и терапевтическое действие при инфекциях, вызванных фи-
ловирусами, нет. В некоторых случаях были использованы сыворотка и цельная кровь
реконвалесцентов, но без убедительных доказательств их эффективности. При лабо-
раторных исследованиях терапевтической эффективности поли- и моноклональных
антител на обезьянах циномолгус обнадеживающих результатов получено не было.
Попытку лечения нескольких больных с использованием «-интерферона, рекомби-
нантного интерферона и экстракорпоральной крови с гемосорбентами и диализом,
можно считать успешной, но неясно, были ли только эти меры ответственны за бла-
гоприятный исход болезни. Нуклеозидный аналог рибавирин (ribavirin, Rib; 1-fl-D-
ribofuranosyl-1,2,4-triazole-3-carboxamide), эффективный при некоторых других вирус-
ных геморрагических лихорадках (см. ниже) и поксвирусных инфекциях (см. разд. 3.8),
не оказывает ингибирующего воздействия на филовирусы. Удачными оказались эк-
сперименты по терапевтическому применению антисмысловых олигонуклеотидов и
молекул короткой интерферирующей PH К (siRNA). Антисмысловые фосфородиами-
дат-морфолино-олигомеры, направленные на предполагаемые последовательности
1 Прежде всего они «не интересны» разработчикам вакцин против гриппа (феномен OAS),
ВИЧ (феномены OAS и ADE), гепатита С (феномен ADE) и энцефалитов, вызываемых фла-
вивирусами (феномен ADE).
908
Часть 3. Частная неправильная эпидемиология
в мРНК трех вирусных генов, защищали 75 % макак-резусов от вируса Эбола подтипа
Zaire, когда лечение начинали до заражения. Лечение S.RNA было успешным у мор-
ских свинок. Эффективность этих подходов может быть повышена благодаря усовер-
шенствованным системам доставки и комбинациям молекул, нацеленных на большое
количество вирусных генов. Но, несмотря на некоторые достижения в лечении фи-
ловирусных инфекций, следует признать, что лечение имело успех, если оно начина-
лось либо до, либо вскоре после заражения вирусом (Bausch D. G. et al., 2008).
Профилактическое и терапевтическое действие при инфекциях, вызванных аре-
навирусами, оказывает рибавирин. Эффективность препарата доказана эксперимен-
тально в условиях in vitro и in vivo, и он лицензирован для лечения лихорадки Ласса.
В основе механизма действия рибавирина — подавление синтеза вирусной РНК. Ри-
бовирин активен и в отношении Bunyaviridae (включая CCHF), но не эффективен по
отношению к Filoviridae или Flaviviridae. Рибавирин не проникает через гематоэнце-
фалический барьер и, следовательно, не эффективен при неврологических наруше-
ниях, вызванных арена- и буньявирусами. Внутривенное введение рибаверина в пер-
вые шесть суток лихорадочного состояния у больного с лихорадкой Ласса, с высоким
уровнем виремии, снижало летальность с 76 до 9% (Borio L. et al., 2002).
В последние два десятилетия активно идет поиск более эффективных препаратов,
способных блокировать инфекционные процессы, вызываемые аренавирусами. Среди
них: ингибиторы IMP-дегидрогеназы (IMP-dehydrogenase), S-аденозилгомоцистеин-
гидролазы (S-adenosylhomocysteine hydrolase), фенотиазидные соединения (phenotia-
zines compounds), брассиностероиды (brassinosteroids), муристиковая кислота (myris-
tic acid), отдельные антиретровирусные препараты, производные пиразина (Т-705;
6-fluoro-3-hydrody-2-pyrazinecarboxamide), бензимидазола (ST-193; benzimidazole
derivative), фосфоротиоатные олигонуклеотиды (phosphorothioate oligonucleotides) и
амфипатические ДНК-полимеры (amphipathic DNA polymers) (Lee А. М. et al., 2011).
Обнаружение генетически измененных штаммов возбудителей геморрагических ли-
хорадок. Возбудители геморрагических лихорадок Марбург, Эбола и Ласса, имеют еще
меньше «недостатков», чем ортопоксвирусы, которые бы могли вызвать у террорис-
тов желание подвергнуть себя дополнительному риску, вмешиваясь в их геном. Кроме
того, геном этих вирусов значительно меньше, чем геном ортопоксвирусов (10—19 и
144—289 т. п. о.), и представлен РНК, а не ДНК, что значительно снижает возможно-
сти генных инженеров по получению генетически измененных штаммов возбудите-
лей геморрагических лихорадок Марбург, Эбола и Ласса.
Обнаружение более вирулентных для людей рекомбинатов, образовавшихся между
вирусами Эбола одного субтипа, не следует рассматривать как проявление деятель-
ности человека, так как такие рекомбинанты встречаются в природе. Генетическое
конструирование филовирусов в основном идет в направлении получения «псевдо-
типированых вирусов», использующих механизм репликации вируса везикулярного
стоматита, вируса лейкемии мышей или ВИЧ-1, упаковывающихся в вирион вируса
Марбург или Эбола (см. табл 2.23). Такие «вирусы» не представляют опасности для
людей. С ними работают для изучения механизмов проникновения вирусов Марбург
и Эбола в клетки-мишени (Chan S. Y. et al., 2000; Bray M., Paragas J., 2002).
Значительно большую опасность, чем генетически модифицированные филови-
русы, представляют лентивирусные векторы, псевдотипированные с гликопротеинами
оболочки вирусов Эбола и Марбург, предназначенные для введения в легочную ткань
через апикальную поверхность эпителия дыхательных путей (см. табл. 2.23), но их
рассмотрение не входит в задачу данного раздела монографии.
3.9. Вирусные геморрагические лихорадки
909
Геном аренавирусов консервативен. Рекомбинанты, образовавшиеся между штам
мами вируса Ласса разных линий, циркулирующих в природных резервуарах, неиз-
вестны. Но аренавирусы могут обмениваться сегментами генома (реассортировать) в
культурах клеток, коинфицированных разными видами аренавирусов. Этот простой
методический прием, хопошо отработанный еше в 1960-х гг. при конструировании
гриппозных вакцин, в настоящее время используется учеными для конструирования
вакцин против вируса Ласса. I. S. Lukashevich et al. (2005) описали реассортантный
клон ML29 (MOPV/LASV), содержащий L-сегмент РНК из невирулентного для мор-
ских свинок вируса Мопея (МОГУ) и S-сегмент РНК из вируса Ласса (LASV). Репли-
кация ML29 в организме морских свинок и низших приматов была слабой. Все мор-
ские свинки линии 13, вакцинированные клоном ML29, выживали, по меньшей мере,
через 70 сут после заражения вирусом Ласса, не обнаруживая никаких признаков бо-
лезни или патологических гистологических изменений. Но возможны и другие ком-
бинации сегментов, например, комбинация L-сегмента из вируса Ласса и S-сегмента
из другого вирулентного для человека аренавируса. Количество видов аренавирусов
точно неизвестно и постоянно открываются новые, и весьма опасные для человека.
Например, Lujo virus, открытый в 2008 г., вызывает болезнь, приводящую в 75 % слу-
чаях к летальному исходу. Атак как S-сегмент содержит гены, определяющие анти-
генные свойства вируса, то в рамках данного методического приема открываются
широкие горизонты для фантазии преступников.
Естественная реассортация наблюдается для некоторых вирусов геморрагичес-
ких лихорадок (Bunyaviridae), таких как вирусы лихорадки долины Рифт, Конго-
Крымской геморрагической лихорадки, но для аренавирусов она не описана (Lukashe-
vich I. S. et al., 2005 ).
Подозрение о применении генетически модифицированного возбудителя лихо-
радки Ласса должно возникнуть при обнаружении у жертв вспышки болезни штам-
мов вирусов, являющихся реассортантантами с другими аренавирусами или реком-
бинантами штаммов вируса Ласса разных линий. Такое же подозрение должны вызвать
штаммы вируса Ласса, резистентные к рибовирину, если неизвестно о применении
рибовирина в данном очаге для лечения людей и при отсутствии сведений об эпизо-
отии Ласса среди грызунов.
Достоверно установить факт вмешательства в геном вирусов, вызывающих гемор-
рагические лихорадки, можно по результатам секвенирования, сопоставив выявлен-
ную нуклеотидную последовательность вируса с эталонной.
Патогенез геморрагических лихорадок. Исследования на моделях низших прима-
тов показали, что моноциты, макрофаги и дендритные клетки, «вездесущие сторожа»,
которые обычно охраняют организм от микробной инвазии, являются главными ме-
стами репликации филовирусов и играют центральную роль в патогенезе болезни.
Хотя лимфоциты не инфицируются, они разрушаются в огромных количествах в ходе
болезни в результате апоптоза, что аналогично процессу, наблюдаемому при септи-
ческом шоке. Филовирусы распространяются по организму циркулирующими моно-
цитами и макрофагами. Вирионы, высвободившиеся из этих клеток, инфицируют
соседние клетки, включая гепатоциты, клетки коры надпочечника и фибробласты
(рис. 3.118).
Виремия и высвобожденые инфицированными клетками провоспалительные
цитокины и хемокины (включая а-ФНО), вызывают лихорадку, повышают прони-
цаемость эндотелиальной выстилки кровеносных сосудов. У больного развиваются-
гипотензия, шок и системное поражение органов. Синтез клеточно-поверхностного
910
Часть 3. Частная неправильная эпидемиология
Вирус проникает через
небольшие повреждения кожи
Печеночная Пространство
Гепатоциты синусоида Дисса
Селезенка
Генерализация
инфекции через
лейкоциты
Рис. 3.118. Схематическое изображение генерализации вирусов Эбола
и Марбург в организме хозяина. По Н. Schnittler и Н. Feldmann (1998)
тканевого фактора запускает внешний механизм коагуляции. На моделях геморраги-
ческой лихорадки Эбола у низших приматов диссеминированную внутрисосудистую
коа1 уляцию наблюдали на второй день после инфицирования по появлению D-ди-
меров1 (образуются при расщеплении молекул фибрина) в плазме. Отмечали обшир-
ный некроз, особенно в печени, хотя обычно не в такой степени, чтобы быть причи-
ной смерти.
Роль специфических антител в патогенезе лихорадок Марбург и Эбола не имеет
ничего общего с их ролью при таком опасном заболевании, как натуральная оспа.
1 Выявление в плазме крови D-димера свидетельствует об активации фибринолиза. Оп-
ределение в плазме D-димера используется для исключения тромбоза и диагностики ДВС-
синдрома.
3.9. Вирусные геморрагические лихорадки
911
Специфические антитела либо не успевают образоваться, либо усиливают инфекци-
онный процесс. Больные погибают при наличии высокого уровня вирусемии.
Клиническая и патологоанатомическая картина вирусных геморрагических лихора-
док у человека при естественном инфицировании. Сравнение клинических характерис-
тик геморрагических лихорадок различной этиологии приведено в табл. 3.37.
Лихорадки Марбург и Эбола1. Вирусы Марбург и Эбола вызывают у людей кли-
нически сходные геморрагические лихорадки. Продромальный период отсутствует,
ьолезнь начинается остро, после примерно 6-суточного инкубационного периода.
У заболевшего поднимается температура, появляются сильные головные боли, боли
в мышцах, чувство жара, тошнота и рвота, боли в животе, выраженное общее недо-
могание (табл. 3.37).
Таблица 3.37
Клиническая характеристика геморрагических лихорадок различной этиологии*
Вирус Особенности клиники болезни Возможность передачи от человека к человеку Инкуба- ционный период, сут Леталь- ность, % Лечение
1 2 3 4 5 6
Эбола Высокая температура и глубокая прострация. Диффузная мукулопа- пулярная сыпь, появляющаяся на 5 сут болезни. Кровотечения и диссеминированная интравас- кулярная коагуляция Возможна 2-21 50-90 Поддержи- вающее
Марбург Высокая температура, миалгия. Незудящая макулопапулезная сыпь на лице, шее, туловище и плечах. Кровотечения и диссеминирован- ная интраваскулярная коагуляция То же 2-14 23-70 То же
Ласса Постепенное развитие лихорадки, тошнота, боли в области жигота, сильная боль в горле, кашель, конъюнктивит, изъязвление слизи- стой оболочки рта, экссудативный фарингит, шейная лимфоаденопа- тия. Позднее появляются выражен- ные признаки отека головы и шеи; плевральные и перикардиальные выпоты Геморрагически осложне- ния менее выражены чем при филовирусных инфекциях То же 5-6 15-20 Рибавирин, поддержи- вающая терапия
New World Arenaviruses Постепенное развитие лихорадки, тошнота, боли в области живота, конъюнктивиты, гиперемия лица и туловища, генерализованная лим- фоаденопатия. Могут развиваться петехии и кровотечения, пораже- ния ЦНС (тремор языка и верхних То же 7-14 15-30 То же
1 По работам Marburg and Ebola... (1977); Е. П. Шуваловой с соавт. (2001); L. Borio et al.
(2002); И. В. Борисевича с соавт. (2006).
912
Часть 3. Частная неправильная эпидемиология
Продолжение табл. 3.37
1 2 3 4 5 6
конечностей, миоклонические дви- жения, дизартрия, генерализован- ные судороги)
Лихорадки долины Рифт Лихорадка, головная боль, ретро- орбитальная боль, светобоязнь, желтушность кожных покровов, в 1 % случаев развиваются гемор- рагии или энцефалит. В 10 % случаев развивается ретинит (острый период болезни или в течение 4 нед от начала болезни) Невозможна 2-6 Менее 1 То же
Желтой лихорадки Лихорадка, головная боль, гипере- мия кожи лица, конъюнктивальные инъекции. Больные либо выздорав- ливают, либо у них наступает пе- риод короткой ремиссии, за кото- рой следует лихорадка. У больного развиваются брадикардия, желтуш- ность кожных покровов, почечная недостаточность и геморрагические осложнения То же 3-6 20 Поддержи- вающая
Омской геморра- гической лихорадки Высокая температура, кашель, конъюнктивит, папуловезикулярная сыпь на мягком небе, гиперемия лица и туловища без образования сыпи, генерализованная лимфо- аденопатия и спленомегалия. У некоторых пациентов развивает- ся пневмония и нарушения ЦНС То же 2-9 0,5-10 То же
Болезни Киасанур- ского леса Сходная с Омской геморрагической лихорадкой, но течение болезни двухфазное. Первая фаза длится 6—11 сут. за ней следует афибриль- ный период продолжительностью 9—21 сут, после чего лихорадка возобновляется. У почти 50 % пациентов с рецидивом болезни развивается менингоэнцефали г То же 2-9 3-10 То же
* По L. Borio et al. (2002).
Через несколько дней развиваются геморрагический синдром, обезвоживание,
нарушается сознание. Больные жалуются на головную боль разлитого характера, боль
в груди колющего характера, усиливающуюся при дыхании, загрудинные боли. Мо-
гут появиться кровоизлияния в конъюнктиву, фарингит и эритематозная и макуло-
папулезная сыпь, которые являются отличительными особенностями этих лихорадок
и могут учитываться при постановке диагноза. У некоторых больных на фоне маку-
лопапулезной сыпи отмечаются везикулезные элементы. При осмотре больного вы-
являются гиперемия слизистой оболочки глотки, кончик и края языка красные, на
твердом и мягком небе, языке появляются везикулы, при вскрытии которых образу-
ются поверхностные эрозии.
3.9. Вирусные геморрагические лихорадки
913
На 3—4 сут болезни на фоне болей в животе схваткообразного характера, стул ста-
новится жидким, водянистым, у половины больных отмечается примесь крови в сту-
ie или появляются признаки желудочно-кишечного кровотечения. Может также быть
овота с примесью желчи и крови в рвотных массах. Понос длится около недели; рвота
продолжается 4—5 сут. Сыпь распространяется на верхние конечности, шею, лицо.
При развитии геморрагического синдрома появляются кровоизлияния в кожу, в
конъюнктиву, слизистую оболочку полости рта, часто носовые, маточные, желудоч-
но-кишечные кровотечения. Нарастает интоксикация. На 7—9 сут заболевания инток-
сикация достигает максимального развития, появляются симптомы ди (дратации, при
тяжелом течении развивается инфекционно-токсический шок. Могут появиться су-
дороги, потеря сознания. На высоте этих проявлений больные нередко умирают.
Стандартные лабораторные исследования позволяют выявить следующие нару-
шения: раннюю лимфопению, сопровождающуюся нейтрофилией, выраженную
тромбоцитопению и пролонгацию времени свертывания, наличие циркулирующих
продуктов распада фибрина и другие признаки диссеминированной внутрисосуди-
стой коагуляции. В сыворотке крови обнаруживаются повышенные уровни связанных
с печенью ферментов, в частности аспартат-аминотрансферазы и аланинамино-
транферазы, причем первая имеет стабильно высокий уровень и может служить диаг-
ностическим признаком. У некоторых больных заметно повышено содержание сы-
вороточной амилазы, что свидетельствует о развивавшемся остром панкреатите. У ряда
больных наблюдали гипопротеинемию, которая приводила к образованию отеков
разной степени.
Геморрагический синдром проявляется в виде петехий, кровоподтеков, кровоиз-
лияний в конъюнктиву и кровотечений из мест венопункции. Обильное кровотече-
ние наблюдается редко. Сильная тошнота и рвота, крайняя слабость, тахипноэ, анурия
и понижение температуры тела свидетельствуют о клиническом ухудшении. Смерть
от шока обычно наступает через 6—9 сут после появления первых симптомов болезни.
У переживших критический период болезни пациентов постепенно клинические
симптомы затухают, период выздоровления затягивается на 3—4 нед. В это время мо-
гут периодически беспокоить боли в животе, ухудшение аппетита, длительные пси-
хические расстройства, а также облысение.
Патологоанатомические изменения свидетельствуют о пантропности вирусов
Эбола и Марбург. Они поражают все органы, но наиболее сильно печень и селезенку.
Происходит стимуляция ретикулоэндотелиальной системы и ингибирование лимфа-
тической системы, одновременно развиваются сосудистые изменения, ведущие к ге-
моррагиям и к образованию тромбов в сосудах.
Почти во всех органах, за исключением поперечнополосатой мускулатуры, легких
и костной системы, появляются очаговые некрозы, которые, как правило, не сопро-
вождаются значительными воспалительными реакциями. Самые обширные некрозы
отмечаются в печени и лимфатической системе. Во всех органах поражены в основ-
ном клетки паренхимы, в то время как мезенхимальные структуры поражены в мень-
шей степени. Патологоанатомы находят у умерших пациентов обширные участки
ЖКТ, наполненные жидкостью, напоминающей кофейную гушу и частично испраж-
нениями, похожими на деготь, при этом источник кровоизлияния, например, язвы
или эрозии, не удается обнаружить. Печень и селезенка увеличены, темного цвета. На
разрезе селезенки отмечается присутствие фолликул; пульпа мягкая и пористая. Пе-
чень рыхлая, при разрезе из нее свободно выливается кровь, и орган принимает свет-
ло-желтый оттенок.
914
Часть 3. Частная неправильная эпидемиология
При гистологическом исследовании селезенки наблюдается значительная гипе-
ремия и стаз, пролиферация ретикулоэндотелиальных элементов в красной пульпе и
образование большого количества макрофагов. Некроз красной пульпы сопповожда-
ется разрушением лимфоидных элементов. В мальпигиевых тельцах количество лим-
фоцитов заметно уменьшено.
При гистологическом исследовании печени отмечается обширная дегенерация и
некроз клеток, гиалиновые изменения. В нескольких случаях обнаружены эозино-
фильные гиалиновые некротические тельца, сходные с тельцами Кунцильмана при
желтой лихорадке. Купферовские клетки выглядят набухшими, с выпячиваниями, и
заполнены фрагментами клеток и эритроцитами. Синусы также заполнены фрагмен-
тами клеток и в сколоворотных пространствах наблюдаются скопления одноядерных
клеток. Даже в момент наибольшей активности некротического процесса в печени
находят регенерацию печеночных клеток. Мононуклеарная трансформация лимфо-
идной ткани, а также некротические поражения обнаруживаются не только в печени
и селезенке, но и в почках, поджелудочной железе, половых железах, надпочечниках,
гипофизе, щитовилной железе, почках и коже.
В легких находят мало последствий патологического процесса, за исключением
небольших геморрагий и признаков эндартериита, особенно в небольших артериолах.
Редко отмечают в различной степени выраженную бронхопневмонию
Нейропатологические изменения ограничиваются преимущественно глиальными
элементами, рассеянными по всему мозгу. В веществе головного мозга наблюдаются
множественные геморрагии. Глиальные поражения имеют либо пролиферативный
характер в виде глиальных узлов, узелков и розеток; либо дегенеративный, проявляю-
щийся в пикнозе или фрагментации ядра. Мозговой отек распространяется на все
исследуемое вещество мозга. Отчетливая гиперемия мягкой мозговой оболочки .
Выявляемые у людей, умерших от геморрагических лихорадок Эбола и Марбург,
патологические изменения ни по отдельности, ни в комбинации не характерны и не
дают возможности составить определенную картину болезни. При каждой аутопсии,
наряду с определением основной болезни, возможных осложнений и сопутствующих
заболеваний, у патологоанатомов встает вопрос о непосредственной причине смер-
ти. Но ее не всегда удается установить.
Лихорадка Ласса1. Инкубационный период болезни продолжается от 3 до 21 сут,
чаще — 7—10 сут. Болезнь начинается относительно постепенно без продромальных
явлений. Ежедневно нарастают лихорадка (температура тела через 3—5 сут достигает
39-40°С) и симптомы обшей интоксикации (астенизация, мышечная боль, расстрой-
ства сознания). С первого дня болезни пациенты отмечают общую слабость, разби-
тость, недомогание, умеренную мышечную и головную боль. Выявляется генерали-
зованная лимфаденопатия, в конце 1-й нед появляется сыпь розеолезного, папулез-
ного или макулезного характера.
Длится лихоратка 2—3 нед. Характерен общий вид больного: лицо и шея гипере-
мированы, иногда пастозны, сосуды склер инъецированы, в 80 % случаев выявляют-
ся характерное поражение зева — на дужках миндалин и мягком небе отмечаются очаги
некротически-язвенных изменений желтовато-сероватой окраски, окруженные зоной
яркой гиперемии, которые могут сливаться.
1 По работам W. С. Winn, D Н. Walker (1975,1982); R. Т. Callis et al (1982); Е. П. Шуваловой
с соам. (2001); И В Борисевича с соавт. (2006).
3.9. Вирусные геморрагические лихорадки
915
На 5—6 сут болезни нередко появляются боль в подложечной области, тошнота,
рвота, обильный жидкий, водянистый стул, иногда развивается обезвоживание.
В тяжелых случаях развиваются отеки, инфекционно-токсический шок, острая сер-
дечная недостаточность
На 2-й нед заболевания резко усиливаются симптомы интоксикации, присоеди-
няются пневмония, отеклегкого, миокардит, отек лица и шеи, геморрагический син-
дром, которые могут обусловить летальный исход. При обследовании в этот период
клинически и рентгенологически может выявляться пневмония, нередко плевраль-
ный выпот. Пальпируется увеличенная печень, возможен асцит. Повышается актив-
ность АЛТ и ACT. В крови лейкопения, тромбоцитопения, увеличивается концент-
рация азота мочевины.
Патологоанатомические находки не отражают тяжести клинической картины при
лихорадке Ласса. Постоянными, но неспецифическими находками патологоанатомов
являются следующие: гиперемия внутренних органов; отеки мягких тканей; наличие
петехий, особенно их много в органах ЖКТ; плевральные выпоты; асцит; кровь в про-
свете тонкого кишечника, но внутреннего массивного кровотечения, объясняющего
шоковое состояние больного, не обнаруживали; почки увеличены, могут иметь мес-
то геморрагии; мозг обычного веса, сосуды мозговых оболочек переполнены кровью;
отек голосовых связок. Основной причиной смерти бывает острая дыхательная недо-
статочность и, возможно, ларингоспазм.
При гистологическом исследовании в сердечной мышце находят небольшой ин-
терстициальный отек без признаков миокардита. В легких невыраженный отек без
значительных геморрагий, в некоторых случаях слабо выраженный интерстициаль-
ный пневмонит с мононуклеарными клетками и мегакариоцитоз. Иногда обнаружи-
вают фокальные интерстициальные скопления мононуклеарных клеток, но без клас-
сической интерстициальной пневмонии. В почках находят тубулярный и гломеруляр-
ный некрозы, типичный ишемический некроз не встречается. Селезенка уплотнена,
имеется атрофия белой пульпы, скопление эозинофильного материала в белой пуль-
пе, инфильтрация интимы селезеночных вен лимфоидными клетками. В лимфатиче-
ских узлах лимфоидное истощение и дегенерация, эритрофагоцитоз. На слизистой
ЖКТ обнаруживают петехии и признаки воспаления. Мозг обычно без изменений.
Наиболее распространенным и заметным гистологическими изменениями при лихо-
радке Ласса являются многоочаговый некроз печени (небольшие очаги некроза раз-
бросаны по долькам печени без зонального распределения, как это бывает при жел-
той лихорадке) и эозинофильный некроз гепатоцитов. Эти изменения сходны с теми,
что наблюдаются у людей, умерших от Боливийской и Аргентинской лихорадок.
Клиническая картина и патоморфология вирусных геморрагических лихорадок у жи-
вотных при искусственном инфицировании. Такие исследования проводятся для пони-
мания механизмов инфицирования людей возбудителями ВГЛ в природных очагах,
изучения патогенеза болезни и оценки эффективности схем специфического и неспе-
цифического лечения и профилактики.
Моделирование лихорадки Эбола. В экспериментах Е. Johnson et al. (1995) изучались
клиника, патологическая анатомия и патоморфология, болезни, вызванной у обезь-
ян-резусов (Масаса mulatto) аэрозолем вируса Эбола (подтип Zaire) с дисперсностью
частиц 0,8— 1,2 мкм. Животным вводилось не менее 400 КОЕ вируса. В опытах исполь-
зовано 6 обезьян. Обезьяны оставались активны и не проявляли никаких признаков
болезни до начала лихорадки (39,9-40,2°С) между четвертыми и шестыми сутками
916
Часть 3. Частная неправильная эпидемиология
после заражения. Лихорадка сопровождалась анорексией, летаргией, развитием кож-
ной сыпи на морде, туловище и проксимальных участках конечностей. Обезьяна,
получившая самую высокую дозу вируса (1465 КОЕ), погибла на третьи сутки от на-
чала болезни (8 сут после заражения аэрозолем вируса); остальные были умерщвле-
ны на 7, 8 и 9 сут после заражения аэрозолем вируса, когда летальный исход болезни
был очевиден. Гематогические изменения заключались в лейкоцитозе и абсолютных
нейтрофилии и лимфопении. На 7 сут после заражения аэрозо. icm вируса количество
тромбоцитов падало от 288х 103 до 124х 103/мм3, уровень лактатде! идрогеназы повы-
шался в 12—16 раз, АЛТ в 5 раз.
Патогистологическое исследование легких и легочных лимфатических узлов по-
казало отчетливые признаки патологии мелкодисперсного аэрозоля. У всех обезьян,
экспонированных к аэрозолю вируса Эбола, развилась интерстициальная пневмония,
совершенно отчетливо ориентированная вокруг терминальных оронхиол (бронхио-
лоцентрическая модель пневмонии). Обнаружены диффузный лейкоцитоз, васкулит,
мульфокальный септальный некроз (multifocal septal necrosis), тромбозы, геморрагии,
отек и увеличенное количество альвеолярных макрофагов. В циркулирующих и аль-
веолярных макрофагах найдены крупные аморфные или плеоморфные ацидофиль-
ные включения. Бронхиолоцентрическая модель пневмонии1, по мнению авторов
исследования, характерна для ингаляционного поражения приматов мелкодиспер-
сным аэрозолем, содержащим вирус Эбола. При иммуногистохимическом иссле-
довании легких этих животных распределение вирусного антигена в легочной ткани
также соответствовало бронхиолоценгрической модели пневмонии. Наибольшая кон-
центрация антигенного материала располагалась в участках терминальных бронхи-
ол Бронхиальный и бронхиолярный эпителий, альвеолярные пневмоциты (alveolar
pneumocytes) и альвеолярные макрофаги были положительны на вирус Эбола при
иммунохимическом окрашивании (рис. 3.119).
Большое количество внеклеточного антигена вируса Эбола обнаружено на слизи-
стой поверхности носовой полости, глотки, гортани и дыхательных путей. В трахеоб-
ронхиальных лимфатических узлах было обнаружено лимфоидное истощение, участки
некроза, васкулиты, трахеиты, тромбозы и геморрагии различной степени выражен-
ности. Цитоплазматические включения обнаружены в циркулирующих и фиксиро-
ванных макрофагах. Поражения трахеобронхиальных лимфатических узлов были
сопоставимы с поражениями мезентериальных узлов, что при парэнтеральном зараже-
нии не наблюдается (см. Baskerville A. et al. 1978).
Внелегочные поражения у обезьян, инфицированных аэрозолем вируса Эбола, не
отличались значительно от тех, которые описали A. Baskerville et al. (1978), заражав-
шие обезьян внутрибрюшинно. По их данным, у обезьян-резус, забитых на вторые и
четвертые сут после внутрибрюшинного инфицирования, макроскопических пора-
жений не обнаружили. У обезьян, погибшихс 5 по 12 сут после инфицирования, имела
место кожная сыпь различной интенсивности на лбу, морде и щеках, на внутренней
поверхности передних и задних конечностей и на груди. Кроме того, у этих четырех
обезьян-резус, и двух зеленых мартышек наблюдались висцеральные петехии, хотя их
распределение не было постоянным. Они чаще всего обнаруживались на деснах,
слизистой оболочке рта, на плевральной поверхности и поверхности срезов легких,
1 Термин «бронхиолоцентрическая модель пневмонии», предложенный Е. Johnson et а1
(1995), можно рассматривать как синоним используемого в этой книге термина «патология
мелкодисперсного аэрозоля».
3.9. Вирусные геморрагические лихорадки
917
Рис. 3.119. Гистология легких обезьяны, экспонированной к аэрозолю вируса Эбола.
А. Окрашивание проведено смесью, содержащей мАТ к оболочке вируса Эбола. Брон-
хоцентрическое расположение поражения, локализованное вокруг терминальных
бронхиол. Легочные макрофаги в просвете альвеол специфически окрашены. Увели-
чение в 100 раз. Б. Внеклеточные созревшие вирионы (V) в альвеолярных простран-
ствах (alveolar sac, AS) и в интерстициальном пространстве (interstitial space, IS) между
эпителиальными клетками I или II типа (Ер) и базальная пластинка (basal lamina).
Вирусные капсиды в цитоплазме эпителиальной клетки показаны стрелкой, и учас-
ток септальной клетки (двойная стрелка). Е — эритроциты; Еп — эндотелиальные
клетки. Увеличение в 15000 раз. По Е. Johnson et al. (1995)
на поверхности серозной оболочки кишечника, в слепой и толстой кишках и в обо-
лочке яичек. У животных имели место геморрагии в просвете тонкого кишечника и
толстой кишки с изъязвлением слизистой. У одного животного были обнаружены мас-
сивные геморра1ии из левого легкого в грудную полость. В желудочно-кишечном
тракте животных имелись лишь небольшие количества пищи. У трех из обезьян-ре-
зус и обеих зеленых мартышек развился перитонит с локализованными ранними спай-
ками, причем в некоторых случаях они имели место в области печени и селезенки.
Печень всех погибших обезьян оказалась темной, пятнистой и рыхлой. Брыжеечные
лимфатические узлы были увеличенными и геморрагическими.
Гистологические исследования печени позволили обнаружить многочисленные
небольшие очаги некроза, произвольно локализующиеся в долях печени и зоны коагу-
ляционного некроза диаметром до 0,8 мм и геморрагии на поверхности капсулы. В бо-
лее сильно пораженной ткани печени четырех обезьян имел место тромбоз централь-
ной и воротной вен. В капиллярах небольшого количества клубочков почки были
обнаружены тромбы фибрина, но преобладающее поражение охватывало канальцы
и более крупные кровеносные сосуды. Отмечался некроз канальцев с пикнозом ядер
и фрагментацией соединительнотканных мембран под эпителием. Это поражение
обнаруживалось в петлях Шумлянского — Генле и имело несимметричное очаговое
распределение. Телец включений наблюдать не удалось. В прямых мочевых каналь-
цах почек и протоках обычно обнаруживались амфорные отложения, цилиндры фиб-
рина и пробки из осколков клеток. Клеточная инфильтрация отсутствовала.
918
Часть 3. Частная неправильная эпидемиология
У различных животных степень поражения легких значительно варьировала. Лег-
кие обезьян, забитых на второй и четвертый день, были непораженными. В легких трех
обезьян имели место лишь небольшие отдельные очаги некроза межальвеолярных пе-
регородок и внутриальвеолярное образование отеков, но у остальных трех животных
было обнаружено сильное спадение легкого, некроз межальвеолярных перегородок,
образование отеков, отложение фибрина и геморрагии в альвеолах. В цитоплазме
некоторых эпителиальных клеток альвеол в зонах повреждения были обнаружены
тельца включения. Эпителий бронхов и бронхиол оказался неповрежденным во всех
случаях, повреждение имело место лишь на уровне альвеол и сосудов.
Некроз лимфоидной ткани мальпигиевых телец селезенки имел место до некото-
рой степени у всех обезьян, он сопровождался уменьшением количества лимфоцитов
в большинстве фолликулов и гиперемией красной пульпы. У пяти животных с наи-
более тяжелой формой болезни, мальпигиевы тельца содержали скопления эозино-
фильного аморфного материала. Кроме того, имел место некроз клеток и соединитель-
ной ткани красной пульпы.
Лимфоидная ткань имела все признаки генерализованного истощения; многие
лимфоидные фолликулы подвергались очаговому некрозу. Наиболее сильно поражен-
ные лимфатические узлы, например брыжейки, имели большие участки коагуляци-
онного некроза гкани коркового слоя и нередко также геморрагии в подкапсульных
и медуллярных синусах.
У погибших животных обнаружили гиперемию и небольшие геморрагии из кро-
веносных сосудов слизистой и подслизистой оболочек в желудке, в подвздошной и
толстой кишках. В кишечнике большинства обезьян имел место некроз с изъязвле-
нием поверхностного эпителия, ткани слизистых же. юз и lamina propria В случаях нек-
роза слизистой оболочки он нередко распространялся на лежащие под ней пейеровы
бляшки. В пораженных участках ворсинки были сморщенными и имели неправиль-
ную форму.
При микроскопическом исследовании сыпи в кожном покрове были обнаруже-
ны гиперемированные капилляры и вены и небольшие геморрагии. В некоторых уча-
стках имели место отслойка и десквамация поверхностных слоев эпидермы от базаль-
ного слоя. Клеточной воспалительной реакции не от печали.
Небольшие геморрагии были обнаружены между мышечными волокнами желу-
дочков сердца, некоторые волокна сердечной мышцы подверглись гиалиновому пе-
рерождению. Клеточные инфильтраты отсутствовали.
Моделирование лихорадки Марбург. В экспериментах D. A. Alves et al. (2010) изуча-
лись клиника, патологическая анатомия и патоморфология болезни, вызванной у
обезьян-циномолгусов (Cynomolgus macaques) аэрозолем вируса Марбург (MARV-An-
gola). Дисперсность частиц аэрозоля не указана. Животным вводилось от 2 до 705 КОЕ
вируса. Температура у животных появилась в период 2—4 сут после заражения, за ней
следовали кожные высыпания, сходные с теми, что бывают при парентеральном за-
ражении. У животных развилась выраженная макулопапулярная кожная сыпь (macu-
lopapular cutaneous rash), преимущественно вокруг глаз и на подбородке, на тулови-
ще, подмышках и паховой области. К девятым суткам после сражения все животные
погибли (рис. 3.120).
Патологические изменения были сходны с теми, что наблютали у обезьян, погиб-
ших после экспонирования к аэрозолю вируса Эбола. Поверхность легких гипере-
мирована и отечна, перикардиальный выпот 3-5 мл сероангинозной жидкости, лим-
фатические узлы увеличены, уплотнены, имеют геморрагии, печень увеличена и вы-
3.9. Вирусные геморрагические лихорадки
919
Рис. 3.120. Геморрагические высыпания у макак-резусов, вызванные внутримы-
шечным заражением вирусом Марбург (Angola). А. Сыпь на 8 сут после зараже-
ния. Б и В. Сыпь на верхней конечности обезьяны на 7 и 8 сут после заражения
соответственно. Заражающая доза — 1000 КОЕ. По Т. W. Geisbert et al. (2007)
глядит жирной, пилор и дуоденум гиперемированы либо имеют геморрагии. В легких
фибринозная интерстициальна пневмония, внутриальвеолярные геморрагии,
разрушение альвеолярных септ различной выраженности, фокальное перибронхиаль-
ное/перибронхиолярное воспаление, включающее главным образом мононуклеарные
клетки и немногочисленные нейтрофилы, смешанные с некротическим дебрисом.
В одном случае воспаление распространилось на бронхиальный субэпителий с раз-
рушением и утратой покрывающего бронхиального эпителия.
Периваскулярный отек и пульмонарную гиперемию с увеличением количества
циркулирующих нейтрофилов и мононуклеарных клеток наблюдали у всех инфици-
рованных животных. Лимфоцитолиз и лимфоидное истощение наблюдали в трахео-
бронхиальных, медиастинальных лимфатических узлах, селезенке и миндалинах. По-
ражения печени были относительно мягкими. Наблюдали умеренную гепатоцеллю-
лярную цитоплазм ическую вакуолизацию (hepatocellular cytoplasmic vacuolation),
интерпретированную как жиро/липоидная дегенерация. В почках тубулярная дегене-
рация с отложением фибрина.
Моделирование лихорадки Ласса. Описание патологии, вызванной ингаляционным
заражением вирусом Ласса, в открытой литературе найти не удалось. Поэтому ниже
будет приведено только описание патологии, вызванной подкожным заражением
макак-резусов (М. mulatto) вирусом Ласса (Josian), сделанное R. Т. Callis et al. (1982).
Обезьянам вводили по 10~ 1 КОЕ вируса Ласса, у всех животных на четвертые сут-
ки после заражения развиватись летаргия, афагия, шпор и лихорадка. На седьмые
сутки обнаруживались легкий конъюнктивит и высыпания на коже в виде многочис-
ленных красных пятен неправильной формы диаметром до 1 мм, которые особенно
были выражены на лице и веках. Максимальная температура тела 40,6- 41,1 "С отме-
чалась перед понижением до субнормального уровня, после чего следовала смерть.
Среднее время до гибели составляло 11 сут.
Макроскопические повреждения были вариабельными и, как правило, слабыми.
Они представляли собой гиперемию легких, плевральный выпот, перикардиальный
отек и гиперемии внутренних органов.
У всех животных наиболее выраженным микроскопическим поражением был
многоочаговый некроз клеток печени. Распределение некротических очагов было
произвольным, их размер варьировал. Они содержали от отдельных, до небольших
920
Часть 3. Частная неправильная эпидемиология
групп гепатоцитов, подвергающихся коагулятивному некрозу, не затрагивающему
опорную строму. Как правило, воспалительные клетки, инфильтрующие участки не-
кроза, были малочисленными, представляли собой нейтрофилы и мононуклеары,
которые обнаруживались также в окружающих синусоидах. Купферовские клетки в
очагах некроза или поблизости от них содержали продукты клеточного распада, а
иногда и эозинофильные тельца, подобные таковым Кунцильмана. У большинства
животных в воротной и печеночной венах имелись небольшие субэндотелиальные
мононуклеарные инфильтраты.
Другим поражением, обнаруженным у всех животных, была интерстициальная
пневмония различной степени тяжести. Пневмония характеризовалась утолшением
альвеолярных стенок, являющимся результатом оседания внутри перегородок эози-
нофильного белкового материала и (или) мононуклеарной инфильтрации. Кроме того,
в некоторых перегородках имелся клеточный детрит, который вызывал предположе-
ние о некрозе эндотелиальных клеток, смешанных с инфильтратом. Содержимое аль-
веол варьировало: у большинства обезьян альвеолы сказались слабо инфильтрован-
ными макрофагами и содержали варьирующие количества отечной жидкости и/или
фибрина. У некоторых обезьян были обнаружены очаговые альвеолярные геморрагии.
В легочных сосудах некоторых животных наблюдали многоочаговые периваскуляр-
ные, а иногда субэндотелиальные инфильтраты одноядерных воспалительных клеток.
У шести животных из девяти имели место поражения сердца. Они представляли
собой многоочаговые мононуклеарные инфильтраты (от минимальных до слабых) в
эпикарде и миокарде, а в некоторых случаях и в эндокарде. Эти инфильтраты состоя-
ли преимущественно из лимфоцитов с незначительным количеством плазматических
клеток и макрофагов. Кроме того, у двух животных оказались слабо инфильтрован-
ными лимфоцитами и плазмацитами перикардиальные автономные ганглии.
В тонком кишечнике всех обезьян обнаруживали эритроциты, клеточный детрит
и макрофаги в lamina propria кончиков ворсинок. У нескольких обезьян была отмечена
минимальная периваскулярная инфильтрация одноядерных клеток в подслизистую.
Селезенки всех животных содержали относительно повышенные количества од-
ноядерных клеток, которые концентрировались вокруг кровеносных сосудов трабе-
кул и в трабекулярных венах субэндотелия. У пяти обезьян в мальпигиевых тельцах в
центре или по периферии обнаруживался аморфный эозинофильный материал. По-
ражения в лимфатических узлах были незначительными и вариабельными, в основ-
ном они сводились к уменьшению количества лимфоцитов и увеличению содержания
ретикулоэндотелиальных клеток.
У всех животных обнаружены поражения почек. Они выражались в многоочаго-
вой инфильтрации вокруг кровеносных сосудов в зоне кортикомедуллярного соеди-
нения, которая распространялась на междольковые сосуды и иногда захватывала со-
седний интерстиций. У семи из девяти животных имели место поражения ЦНС. Они
характеризовались слабыми скоплениями лимфоцитов вокруг отдельных кровено-
сных сосудов головного мозга, спинного мозга и мягких мозговых оболочек.
Клиническая картина вирусных геморрагических лихорадок у людей при искусствен-
ном инфицировании. Работа с вирусом Эбола строго регламентируется санитарными
правилами, однако человеческий фактор не позволяет полностью исключить внутри-
лабораторные заражения. Одна из таких аварий произошла в Государственном науч-
ном центре вирусологии и биотехнологии «Вектор» в 2004 г1.
1 Использована работа А. А. Акинфеевой с соавт (2005).
3.9. Вирусные геморрагические лихорадки
921
Из истории болезни: больная П., 1958 г. р., была доставлена в стационар особо опас-
ных инфекций МСЧ № 163 (п. Кольцово. Новосибирская область).
Эпидемиологический анамнез: при подготовке использованных (для взятия крови у
животных, инфицированных вирусом Эбола) инъекционных игл к утилизации сотруд-
ница П. уколола ладонную поверхность левой кисти.
При поступлении (через 3 ч 40 мин после аварии): пострадавшая жалоб не предъяв-
ляет. При осмотре: состояние удовлетворительное, температура тела 36,4°С, по орга-
нам без патологии, показатели гемодинамики в пределах нормы Status localis: на ла-
донной поверхности левой кисти точечный след от укола.
С учетом данных эпидемиологического анамнеза и осмотра констатировано: со-
стояние после лабораторной аварии (при работе с культурой вируса Эбола), назначено
общеклиническое и вирусологическое обследование. Пациентка помещена в мельпе-
ровский бокс стационара особо опасных инфекций. При поступлении в приемном
покое стационара пациентке была проведена специфическая профилактика имму-
ноглобулином Эбола из сыворотки крови лошадей (суммарная доза 12 мл) и начата
комплексная терапия: противовирусный препарат (рибаверин 600 мг/сут per os),
иммунологические препараты (реаферон по 1000000 ME 2 раза в сутperos, полиокси-
доний 0,006 г/сут внутримышечно), общеукрепляющие препараты (рибоксин, аско-
рутин). тавегил. С момента поступления пациентки медицинский персонал стацио-
нара работал в защитных противочумных костюмах.
В течение 5 сут (инкубационный период) состояние пациентки оставалось стабиль-
ным, показатели термометрии и гемодинамики, данные лабораторных исследований —
в пределах нормы. На правом предплечье (в месте проведении кожной пробы на пере-
носимость лошадиного белка) сохранялась гиперемия диаметром 1.5 см На 6 сут пятно
с четкими, ровными краями на правом предплечье увеличилосьдо 2,5x3 см, не возвы-
шающееся над поверхностью, горячее на ощупь. Появились зудящие, пятнистые высы-
пания на правой ягодице (в месте введения иммуноглобулина) размерами: основное
пятно 1,0х 1,5 и 5 дочерних — до 0,5 см. Состояние расценено как сывороточная бо-
лезнь легкой степени. Пациентке скорригирована терапия: супрас .ин по 1 таб. 2/сут,
местно — гидрокортизоновая мазь, амиксин 0,125 г/сутperos, тактивин по 0,5 мл/сут
подкожно, общеукрепляющие препараты.
На 7 сут в 6 ч отмечен первый подъем температуры до 37,2 °C. При осмотре: жалоб
нет, небные дужки и миндалины умеренно гиперемированы, имеются пятнистые вы-
сыпания на правом предплечье и ягодице. Больной поставлен диагноз: геморрагичес-
кая лихорадка Эбола (?). Сывороточная болезнь легкой степени. К терапии добавлены
дексаметазон 16 мг/сут внутривенно, ципробай по 200 мг 2/cjt внутривенно (для про-
филактики бактериальных осложнений), аскорбиновая кислота 5 % — 10 мл 2/сут
внутривенно. Больной после повышения температуры до 39,1 °C (в 15 ч 30 мин) произ-
ведена катетеризация правой подключичной вены и выполнен сеанс дискретного плаз-
мафереза (взято крови 530 мл, эксфузия плазмы 320 мл). Назначена инфузионная те-
рапия: поляризующая смесь 400 мл 3/сут, раствор Рингера 400 мл + трентал 5 мл 3/сут.
реополиглюкин 400 мл/сут, гепарин по 2500 ЕД 4/сут. В результате проводимой тера-
пии состояние пациентки улучшилось, нормализовалась температура, уменьшились
(регрессировали) пятнистые высыпания, однако в 0 ч температура повысилась до 38 °C.
На второе введение ципробая отмечалась выраженная побочная реакция (тошнота,
рвота), в связи с чем он заменен на абактал 400 мг 2/сут внутривенно.
В течение 3 сут состояние больной стабильно тяжелое, сохранялась температура
до 38 °C, инъекция сосудов склер, кожные покровы чистые, бледные, небные дужки и
миндалины умеренно гиперемированы, экхимозы (сине-черного цвета) в местах
инъекций. В анализах отмечено увеличение СОЭ до 20 мм/ч, повышение уровня АЛТ
922
Часть 3. Частная неправильная эпидемиология
и ACT до 58 и 215 ед/л, снижение количества тромбоцитов до 17бх 109/л, общего бел-
ка до 53,8 г/л, протромбинового индекса (ПТИ) до 75,9 %, появились токсигенная
зернистость нейтрофилов, белок в моче 0,07 г/л. Ежедневно проводились сеансы дис-
кретного плазмафереза с увеличением объема до 0,5 объема циркулирующей плазмы
(ОЦП) в сутки.
На 9 сут были получены результаты вирусологического обследования пациентки П.
дву1 (я независимыми лабораториями: в крови больной выявлено наличие геномной
РНК вируса Эбола ПЦР-методом и наличие антигена иммуноферментным методом.
Утром на 11 сут на фоне субфебрильной температуры появились: слабость, мел-
коточечная, макулопапулезная сыпь без геморрагического компонента на лице и ту-
ловище, однократная рвота «кофейной гущей» с прожилками адой крови около 200 мл,
кровоточивость и экхимозы в местах инъекций, экхимозы на нижних конечностях в
местах соприкосновения с твердыми предметами. В связи с развернутой клиникой ге-
моррагической лихорадки Эбола пациентке изменена терапия. Отменены антикоагу-
лянты прямого действия и дезагреганты, все внутримышечные и подкожные инъекции,
начата профилактика ДВС-синдрома и печеночной недостаточности. Дискретный
плазмаферез с замещением свежезамороженной одногруппной донорской плазмой до
0,5 ОЦП ежедневно, гемостатическая терапия (дицинон 2 мл 4/сут, глюконат кальция
по 10 мл 4/сут, аминокапроновая кислота по 100 мл 3/сут), гордокс 800 тыс. ЕД/сут.
Продолжена интенсивная инфузионная терапия с увеличением объема до 3,5 л/сут
(реамберин, аминиплазмальгепа, 5 % раствор глюкозы, гидроксиэтил крахмал, соле-
вые растворы).
В 16 ч 30 мин отмечен подъем температуры до 39,7°С, однокра гчая рвота слизью с
гемолизированными сгустками крови в скудном количестве. Появились боль в горле,
сухость и боль при глотании, на коже лица и туловища обильная макулопапулезная
сыпь, местами сливная, на верхних и нижних конечностях мелкоточечная макулопапу-
лезная сыпь. При осмотре состояние больной тяжелое. Отмечаются отечность век, но-
са, губ, гиперемия лица в виде «бабочки», склеры инъецированы, иктеричны; яркая
гиперемия мягкого неба, язычка, миндалин, дужек, задней стенки глотки. В легких ды-
хание жесткое, хрипов нет; край печени выступает из-под реберной дуги на 2,5 см, при
пальпации безболезненный; из мест инъекций серозно-геморрагическое отделяемое.
На 12 сут состояние больной оставалось стабильно тяжелым, беспокоили выражен-
ная слабость, тахипноэ (ЧДД 30—32/мин), сухость и боль в горле, отсутствие аппети-
та. Пациента была заторможена, адекватна, астенична, в контакт вступала неохотно.
При осмотре: температура 39,9°С, отмечалась желтушность кожных покровов, обиль-
ная макулопапулезная сыпь распространилась на кисти и голени, экхимозы в местах
инъекций (от лилового до черного цвета) в размерах не увеличились, кровоточивости
нет; периферические лимфоузлы не увеличены; пока гатели гемодинамики в пределах
нормы. В течение суток прогрессировало увеличение размеров печени (край печени
выступал из-под реберной дуги утром на 2,5 см, вечером — на 4 см) Печень безболез-
ненная при пальпации и перкуссии, к вечеру изменился цвет мочи на темный («цвета
пива»), диурез достаточный, стул 3/сут, оформленный, обычного цвета.
По лабораторным данным: в анализе крови снизился уровень гемоглобина до
103 г/л, СОЭ увеличилось до 38 мм/ч, токсигенная зернистость +++; время свертыва-
емости крови (5 мин — 6 мин 30 с), время кровотечения увеличено (4 мин 30 с); в био-
химическом анализе крови общий билирубин 85 ммо ть/л, прямой — 35 ммоль/л, не-
прямой — 50 ммоль/л, АЛТ — 444 ед/л, ACT - 1650 ед/л, ЛДГ — 12180 ед/л, ГГТП —
970 ед/л, ЩФ — ИЗО ед/л, фибриноген-сгусток не образовался, ПТИ — 51,8 %. Учи-
тывая отсутствие геморрагического синдрома и прогрессирование печеночной недо-
статочности, врачебным консилиумом решено продолжить терапию за исключением
3.9. Вирусные геморрагические лихорадки
923
гепатотоксичных препаратов (рибавирин, абактал), увеличить число сеансов дискрет-
ного плазмафереза до 2/сут. Вечером для профилактики почечной недостаточно-
сти назначен допамин 200 мг в 400 мл физиологического раствора (1—3 мкг/кг в мин)
через дозатор.
На 13 сут состояние крайне тяжелое, резкая слабость, сохраняется стойкая гипер-
термия (39,2—39,9°C). Больная испытывала трудности при разговоре, глотании слю-
ны и воды из-за выраженной сухости и боли в горле. Пациентка была в сознании, за-
торможена, вялая, адинамичная, на вопросы отвечала скупо, чаше кивала головой.
При осмотре: лицо отечное, гиперемированное, губы цианотичные, склеры жел-
тушны, инъецированы, слева — небольшие участки кровоизлияний. Слизистая поло-
сти рта сухая, язык обложен густым бело-желтым налетом, гиперемия менее яркая.
Кожные покровы желтушные, новых высыпаний нет, ранка после взятия крови из
пальца не кровоточит, кровоизлияния регрессируют, новых нет. Тоны сердца приглу-
шены, ритмичные. АД 90/60 мм рт. ст., ЧСС 108/мин, ЦВД 120 мм вод. ст. (допамин
4,5 мкг/(кг мин)). В дыхании участвует вспомогательная мускулатура. В легких дыха-
ние жесткое, поверхностное, ослабленное в нижних отделах, хрипов нет. ЧДД в покое
32-36/мин, при вставании до 40/мин. SaO2 92-94 % на фоне ингаляции увлажненно-
го кислорода через назальный катетер. Живот вздут, безболезненный при пальпации,
перистальтика вялая. Печень выступает из-под реберной дуги на 4 см, безболезненная
при пальпации и перкуссии. Газы отходят, стул самостоятельный, жидкий, зеленова-
того цвета, без примеси крови. Мочится самостоятельно, диурез снижен (стимуляция
лазиксом 20 мг 2/сут).
По лабораторным данным: в моче увеличилось содержание белка до 0,800 г/л.
В клиническом анализе крови НЬ 93 г/л, эритроциты 2,7х 1012/л, лейкоциты 13,5х 109,
тромбоциты 180х 109/л, лейкоциты палочкоядерные 25 %, лейкоциты сегментоядерные
65 %, эозинофилы 1 %, базофилы 2 %, лимфоциты 4 %, моноциты 3 %, СОЭ 42 м/ч,
время свертывания 5—6 мин. Биохимический анализ крови: общий белок 59,8 г/л,
альбумины 25,7 г/л, мочевина 5,4 ммоль/л, креатинин 90 мкмоль/л, АЛТ — 540 ед/л,
ACT - 2160 ед/л, ЛДГ - 16740 ед/л, ГГТП - 660 ед/л, фибриноген 0,4 г/л, ПТИ -
52,9 %, билирубин 99 ммоль/л, прямой — 38,6 ммоль/л, непрямой — 54,4 ммоль/л.
Тяжесть состояния больной обусловливали интоксикационный синдром, печеноч-
но-клеточная недостаточность, паренхиматозная желтуха, дыхательная недостаточ-
ность, нарастающая почечная недостаточность, водно-электролитные нарушения.
К лечению добавлен антибактериальный препарат: максипин по 2 г 2/сут.
В 17 ч состояние больной резко ухудшилось, появились боли в животе, сильная
тошнота, позывы на рвоту. По всей поверхности тела появилась геморрагическая сыпь,
мелкоточечные кровоизлияния, на левом глазном яблоке свежие кровоизлияния в
белочную оболочку, акроцианоз. Живот умеренно вздут, при пальпации болезненный
в правом подреберье и мезогастрии, симптомов раздражения брюшины нет. Печень при
пальпации мягкая плотная, эластической консистенции, выступает из-под края ребер-
ной дуги на 4,5 см, перкуссия над поверхностью печени резко болезненна (патогно-
моничный симптом). АД 110/60 мм рт. ст., ЧСС 125/мин. Мочится самостоятельно, в
течение дня 1,1 л мочи.
В 21 ч при проведении планового вечернего сеанса дискретного плазмафереза (се-
анс начат без инфузионной поддержки ввиду высокого риска развития отека легких)
при взятии крови состояние ухудшилось, появилось головокружение, ЧДД возросла
до 40/мин (SaO2 — 88 %), АД снизилось до 60/40 мм рт. ст. (ЦВД — 120 мм вод. ст.),
ЧСС 120/мин, из места взятия крови из пальца началась сильно выраженная кровото-
чивость. В связи с этим принято решение отказаться от проведения плазмафереза. Доза
допамина была увеличена до 10 мкг/(кг мин), назначена инфузия свежезамороженной
924
Часть 3. Частная неправильная эпидемиология
донорской плазмы и эритроцитной массы, продолжена гемостатическая терапия. Боль-
ная попросила отдых и уснула. В 2 ч ночи (14 сут) была констатирована биологиче-
ская смерть. Заключительный клинический диагноз: геморрагическая лихорадка Эбо-
ла. Инфекционно-токсический шок. Прогрессирующая полиорганная недостаточ-
ность. Острая гепато-неллюлярная недостаточность. Паренхиматозная желтуха.
Респираторный дистресс-синдром. Геморрагический синдром.
Вскрытие больной не проводилось по соображениям эпидемиологической безо-
пасности. В табл. 3.38—3.41 приведены динамические показатели клинических анали-
зов крови и мочи, биохимического анализа крови, времени свертываемости и време-
ни кровотечения.
Таблица 3.38
Динамика клинических показателей крови у больной
геморрагической лихорадкой Эбола
Показатель Норма Сутки
1-е 3-е 7-е 8-е 9-е 12-е 13-е (утро) 13-е (вечер)
Гемоглобин, г/л 120-140 128 136 140 130 120 103 93 83
Эритроциты, млн 3,9 4,0 4,5 4,6 4,1 3,9 3,4 2,7 2,1
Цветной показатель 0,85-1,05 0,96 0,9 0,91 0,95 0,92 0,9 1,03 0,8
Ретикулоциты, % 2-10 — 30/00 20/00 90/00 80/00 — 70/00 —
Тромбоциты, тыс. 180-320 290,0 220,0 300,0 220,0 176,0 — 180,0 102,0
Лейкоциты, тыс.: 4-9 4,4 4,0 4,5 4,7 4,2 8,0 13,5 —
палочкоядерные, % 1-6 3 2 7 8 203 — 25 —
сегментоядерные, % 47-52 66 63 67 72 58 65 —
эозинофилы, % 0,5-5 1 2 1 1 1 — 1 —
базофилы, % 0-1 — 1 1 — 1 — 2 —
лимфоциты, % 19-37 27 28 18 17 14 — 4 —
моноциты, % 3-11 3 4 6 2 53 — 3 —
Плазматические клетки, % — — — — — 1 — —
СОЭ, мм/ч 2-15 6 6 10 20 9 38 42 —
Токсогенная зернистость - - — — — + +++ - —
Таблица 3.39
Динамика показателей свертываемости крови у больной
геморрагической лихорадкой Эбола
Сутки Показатель (норма)
Время свертывания (8-12 мин) Время кровотечения (1—3 мин)
1 2'50"—3'10" —
3 1'50"—2'30" 0'30"
7 2'30"—3'08" 2'00"
8 11'00"—12'00" 2'00"
9 11'10"—12'20" 4'00"
10 5'00"—6'30" 4'30"
13 (утро) 5'00"—6'00" —
13 (вечер) 6'30"—7'20" 24'30"
3.9. Вирусные геморрагические лихорадки
925
1аблица 3.40
Динамика клинических показателей мочи у больной
геморрагической лихорадкой Эбола
Показатель Сутки
1-е 3-е 7-е 8-е 9-е 10-е 13-е
Цвет с/ж с/ж с/ж с/ж с/ж с/ж с/ж
Прозрачность прозр. прозр. прозр. прозр. прозр. прозр. мугн.
Отн. плотность 1021 1006 1017 — — — —
Реакция кисл. шел. КИСЛ. КИСЛ. кисл. кисл. щел.
Белок abs abs abs 0,07 г/л 0,11 г/л 0,18 г/л 0,80 г/л
Глюкоза abs abs. abs abs abs СЛ.+ СЛ.+
Эпителий плоский един. един. един. един. един. един. много
Лейкоциты един. един. 2-3 2-3 един. 2-3 2-3
Эритроциты abs abs abs abo abs abs Неизм. един.
Таблица 3.41
Динамика биохимических показателей в сыворотке крови
у больной геморрагической лихорадкой Эбола
Показатель Норма Сутки
1-е 3-е 6-е 7-е 8-е 9-е 10-е 11-е 12-е 13-е
Общ. белок 65-85 г/л 77,6 72,8 80,7 78,5 63,7 60,3 53,8 67,6 65 59,8
Альбумин 55-62,7 % 46,4 38,8 43,0 40,3 35,3 34,9 29,6 32,0 30,6 25,7
Мочевина 2,8—6,6 ммоль/л 6,1 5,6 4,8 5,3 6,8 4,9 ч,4 3,6 4,0 5,4
Креатинин 44-97 мкмоль/л 66 82 100 117 92 94 91 70 60,0 90
Билирубин общий: до 20.5 мкмоль/л 9,6 12.0 14,6 13,3 8,2 7,6 12,3 14,0 85,0 99
прямой до 4,5 мкмоль/л 4,9 35 38,6
непрямой 9,7 50 54,4
АЛТ 0—40,6 ец/л 13 13 11 11 12 24 58 202 444 540
ACT 0-39,0 ед/л 22 22 22 39 49 83 215 720 1560 2160
лдг 248-496,9 ед/л — 1481 12180 16740
ггд 7-50 ед/л — 970 660
ЩФТ 80-295 ед/л 83 85 67 68 116 260 ИЗО
Амилаза 0—220 ед/л 120 105 98 108 127 163 218 246
Тимоловая проба 0- 4 ед. 1,3 1,3 1,5 0,8 0,8 0,5 0,1 0,3 2,9
ПТИ 80- 100 % 115 105 108 99,3 101 84,7 75,9 74,1 51,8 52,9
Фибриноген 2—4 г 3,3 2,7 3,8 3,3 2,7 2,7 2,2 1,Н сгустка нет до 0,4
Глюкоза 4,44—6,66 ммоль/л — — — - — - - 4,8 5,9
Опыт ведения больной лихорадкой Эбола показал неэффективность использован-
ных схем специфической профилактики и лечения даже при немедленном установ-
лении факта заражения.
926
Часть 3. Частная неправильная тчдемиология
Диагностика искусственного поражения возбудителями геморрагических лихорадок.
Должна проводиться на основе установления «невозможной эпидемиологии болезни»
и «невозможной клинической формы болезни» (см. разд. 2.3.1).
Любой иммунологически и генетически подтвержденный диагноз геморрагичес-
кой лихорадки, возбудитель которой не встречается на территории России, служит
основанием для предположения о совершенном биологическом нападении, если:
1) заболевший в течение месяца перед появлением лихорадки не выезжал в стра-
ны, эндемичные по данной болезни;
2) в этот же период времени он не находился в контакте с таким же больным, ко
торый мог контактировать с лицами, приехавшими из стран, где распространена дан-
ная болезнь;
3) заболевший не работал с возбудителем данной болезни в вирусологической
лаборатории;
4) не контактировав с животным, привезенным из местности, эндемичной по
данной болезни.
При расследовании такого преступления необходимо иметь в виду, что паренте-
ральное введение возбудителей лихорадок Эбола, Марбург и Ласса не оставляет следов,
так как очагов некроза в месте инъекции вируса не образуется. Клиническая картина
болезни в этом случае не отличается наблюдаемой у пациентов, заразившихся в при-
родных очагах болезни. Основными критериями установления преднамеренного рас-
пространения возбудителей геморрагических лихорадок будут анамнез пациента (упо-
минание им медицинских манипуляций, сопровождающихся парентеральным введе-
нием каких-то «лекарств») и доказанная «невозмож ная эпидемиологии болезни». Для
доказательства последней надо учитывать сложность экологии возбуди гелей гемор-
рагической лихорадок и возможность расширения их ареалов в связи с изменением
климата и техногенного воздействия на окружающую среду. Прежде чем выдвинуть
предположение о «невозможной эпидемиологии» обнаруженной/новой геморрагичес-
кой лихорадки, необходимо провести исчерпывающее эпидемиологическое исследо-
вание и убедиться в том, что вирус не имеет на территории, где зарегистрирована
вспышка вызванной им болезни ранее неизвестного природного резервуара. Обнару-
жение уже заде локированных в зарубежных коллекциях штаммов вирусов свидетель-
ствует о «невозможной эпидемиологии» вспышки.
Поражение мелкодисперсными аэрозолями вирусов Эбола и Марбург можно пред-
положить по наличию характерного патологического процесса в легких («невозмож-
ная клиническая форма болезни»), У людей, заразившихся этими вирусами в природ-
ных очагах, легочная ткань обычно оказывалась не вовлеченной в первичный
инфекционный процесс. Поражения легких у них были вызваны гематогенным зано-
сом возбуди.еля болезни в легочную ткань и повышением проницаемости эндотели-
альной выстилки кровеносных сосудов из-за высвобождения цитокинов и хемокинов
инфицированными макрофагами. Поэтому для таких поражений характерны мало-
выраженные геморрагии в легочной ткани и эндартериит небольших артериол.
Моделирование заражения мелкодисперсными аэрозолями вирусов Эбола и Мар-
бург, выполненное на приматах, позволяет предположить, что при поражении мелко-
дисперсными аэрозолями этих вирусов у погибших людей на вскрытии будет обнару-
жена выраженная интерстициальная пневмония, ориентированная вокруг терминаль-
ных бронхиол (бронхиолоцентрическая модель пневмонии). Возможно обнаружение
некротических изменений в эпителии бронхиол и мелких бронхов, и в трахеоброн-
хиальных и медиастинальных лимфатических узлах, сопоставимых с поражениями
3.9. Вирусные геморрагические лихорадки 927
лимфатических узлов брюшной полости. Установить по клинической и патологоана-
томической картине болезни факт ингаляционного поражения человека вирусом
Ласса сложно. Результаты моделирования болезни на приматах показывают, что лег-
кие могут вовлекаться в инфекционный процесс и при парэнтеральном заражении.
Для доказательства ингаляционного поражения человека вирусом Ласса недостаточ-
но установить только факт развития интерстициальной пневмонии. У людей, заразив-
шихся вирусом лихорадки Ласса в природном очаге болезни, пневмония начинается
на второй неделе болезни. Можно предположить, что у людей, пораженных мелкодис-
персным аэрозолем вируса Ласса, пневмония будет развиваться как первичный про-
цесс, и поэтому она может быть обнаружена рентгенологически еше в первую неделю
болезни, после появления лихорадки. У погибших пациентов, кроме очагов интерсти-
циальной пневмонии, возможно обнаружение обширных поражений трахеобронхи-
альных и медиастинальных лимфатических узлов.
Обнаружение генетически измененных штаммов возбудителей ВГЛ, однозначно
свидетельствует об искусственном происхождении вспышки болезни.
Иммунопрофилактика вирусных геморрагических лихорадок. Не разработана, и,
видимо, будет разработана еще не скоро из-за особенностей иммунных ответов на
возбудители этих болезней (см. «Иммунитет к возбудителям ВГЛ»),
Лечение вирусных геморрагических лихорадок. Лечение ВГЛ в основном сводится
к поддерживающей терапии, поддержанию водно-солевого баланса и гемодинамики.
Army FM 8-284 (2000) рекомендует обратить особое внимание на поддержание баланса
жидкостей и электролитов, противошоковые мероприятия, возмещение потери крови,
предотвращение развития почечной недостаточности у пациентов. Лечение таких
больных может потребовать интенсивный уход и специфические вмешательства, такие
как механическая вентиляция легких, диализ и неврологическая помошь. При дис-
семинированной внутрисосудистой коагуляции рекомендуется терапия гепарином.
Но ее роль считается не бесспорной, и ее рекомендуется применять только у больных
с клинически значимым кровотечением и лабораторным подтверждением диссеми-
нированной внутрисосудистой коагуляции. Аспирин и другие препараты, обладаю-
щие антиагрегантным действием, противопоказаны, как и любые внутримышечные
инъекции. Использование внутрисосудистых устройств рекомендуется рассматривать
в контексте соотношения потенциальной пользы и риска кровотечения.
Возможности специфической антивирусной терапии ограничены. Препаратов,
эффективных в отношении филовирусных инфекций, нет. Определенные надежды
возлагаются в отношении препаратов, эффективных против Arenaviridae и Bunya-
viridae, в частности, на рибавирин. Результаты исследований эффективности рибави-
рина в условиях in vitro и клинических исследований, свидетельствуют в пользу в/в
введения рибавирина при лечении лихорадки Ласса, ГЛПС (вирус Хантаан) и ККГЛ.
При лихорадке Ласса и ККГЛ назначается ударная доза рибавирина, 30 мг/кг в/в, с
последующими 16 мг/кг в/в каждые 6 ч в течение 4 сут, а затем 8 мг/кг в/в каждые 8 ч
в течение 6 сут для завершения 10-дневного курса лечения. Лечение больных с ГЛ ПС
может дать положительный результат, если лихорадка длилась не более шести дней.
Назначается ударная доза рибавирина, 33 мг/кг в/в, с последующими 16 мг/кг в/в
каждые 6 ч в течение 4 сут, а затем 8 мг/кг в/в каждые 8 ч в течение 3 сут для заверше-
ния 7-суточного курса лечения.
В качестве химиопрофилактики после экспонирования для Конго- ККГЛ и ли-
хорадки Ласса Army FM 8-284 (2000) рекомендует использовать рибавирин, 500 мг
перорально каждые 6 ч в течение 7 сут.
928
Часть 3. Частная неправильная эпидемиология
3.10. А.»БОВИРУСНЫЕ ЭНЦЕФАЛИТЫ
Микробиология вирусов, вызывающих арбовирусные энцефалиты. Экология и есте-
ственная эпидемиология возбудителей лошадиных энцефалитов. Восприимчивость
животных, клиническая картина и патоморфология арбовирусных энцефалитов у
животных при искусственном инфицировании. Иммунитет к возбудителям арбови-
русных энцефалитов. Устойчивость возбудителей арбовирусных энцефалитов во
внешней среде. Обнаружение генетически измененных штаммов возбудителей арбо-
вирусных энцефалитов. Патогенез арбовирусных энцефалитов. Клиническая и пато-
логоанатомическая картина арбовирусных энцефалитов у человека при естественном
инфицировании. Диагностика искусственного поражения возбудителями лошадиных
энцефалитов. Иммунопрофилактика арбовирусных энцефалитов. Лечение арбови-
русных энцефалитов.
Арбовирусные энцефалиты (encephalitic arboviruses) — группа мало отличающих-
ся друг от друга по клинике зоантропонозных болезней, вызванных арбовирусами, для
которых характерны гриппоподобный синдром (flu-like syndrome), энцефаломенин-
геальный синдром (encephalitis-meningitis syndrome), остро развивающийся вялый па-
ралич (acute flaccid paralysis) и, во многих случаях, бессимптомное течение инфекци-
онного процесса (asymptomatic infections) (Davis L E. et al., 2008).
Арбовирусы (от англ, arthropod-bome viruses) — это собирательный термин, обо-
значающий групп)' РНК-вирусов, переносчиками которых от животных к человеку яв-
ляются членистоногие (москиты, клещи, мухи). Арбовирусы классифицируют на
семейства и рода на основе их морфологии (размер, форма, капсидная симметрия,
наличие или отсутствие оболочки), физических (структура генома и антигенный со-
став) и биологических свойств (механизмы репликации и трансмиссии, хозяева, па-
тогенность для человека и др.). В группу арбовирусов входят четыре семейства: семей-
ство Togaviridae (род Alphavirus: вирус Восточного лошадиного энцефалита, Eastern
Equine encephalitis virus, EEEV; вирус Западного энцефалита лошадей, Western equine
encephalomyelitis virus, WEEV; вирус энцефалита Сэнт-Луис, St. Louis encephalitis virus;
вирус Венесуэльского энцефалита лошадей, Venezuelan equine encephalitis virus, VEEV;
семейство Flaviviridae (род Flavivirus: вирус Западного Нила, West Nile virus, вирус
Повассан, Powassan); семейство Bunyaviridae (род Bunyavirus: вирус Ла Кросс, La Crosse
virus, калифорнийская группа вирусов); семейство Reoviridae (род Coltivirus: вирус
колорадской комариной лихорадки, Colorado tick fever virus). Всего известно около
500 видов арбовирусов различных семейств, в основном поддерживающихся среди по-
звоночных животных тропиков. Болезнь у человека вызывает лишь несколько видов
арбовирусов, в Северной Америке их не более 10 видов (Lanciotti R. S. et al., 1999: Da-
vis L. E. et al., 2008).
Природные очаги арбовирусов имеют строгие ограничения, налагаемые на них
наличием амлифицирующих хозяев и насекомых-переносчиков, особенностей их
жизненных циклов, климатических условий и другими экологическими и биологи-
ческими факторами. Люди обычно не вовлекаются в энзоотические жизненные цик-
лы арбовирусов из-за низкого уровня виремии, развивающейся у них на пике инфек-
ционного процесса и не позволяющей насекомым осуществлять непрерывно процесс
передачи вируса от человека животным и наоборот. Исключением из этого правила
являются вирусы, вызывающие лихорадку Денге и Венесуэльский лошадиный энце-
3.10. Арбовирусные энцефалиты
929
фалит, способные создавать в организме человека высокую вирусную нагрузку. Однако
передача вирусов от человека к человеку воздушно-капельным путем или при тесных
контактах не происходит. Заражение возможно при переливании инфицированной
крови или в результате пересадки органов, взятых у инфицированного человека, а
также перинатально и трансплацентарно (Davis L. Е. et al., 2008).
На территории России в природных очагах поддерживаются арбовирусы, относя-
щиеся к семейству Flaviviridae: вирус японского энцефалита (южные районы Примо-
рья), вирус клещевого энцефалита (распространен повсеместно в средних широтах),
вирус Западного Нила (Волгоградская, Астраханская и Омская области). Природных
очагов вирусов лошадиных энцефалитов в России и странах СНГ не обнаружено (Тар-
шис М. Г., Черкасский Б. Л., 1997).
Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят ви-
русы семейств Togaviridae и Flaviviridae ко II группе патогенности. По степени важ-
ности для национальной безопасности США альфавирусы, включая вирусы VEEV и
WEEV, относятся к группе В (см. табл. 2.2)1.
Вирусы, возбудители арбовирусных энцефалитов, и особенно альфавирусы,
вызывающие лошадиные энцефалиты (WEEV, VEEV и EEEV), рассматривались в ка-
честве перспективных агентов БО еще перед Второй мировой войной (Розбери Т, Ка-
бат Э., 1955; Huxsoll D. L. et al., 1987). Причины столь раннего интереса военных к ви-
русам этого рода обобщены в работе J. Е Smith et al. (1997)2:
1) эти вирусы могут производиться в больших количествах недорогими и просты-
ми способами;
2) они относительно стабильны и сохраняют инфекционность для человека в аэро-
золях;
3) среди этой группы вирусов есть такие, которые могут быть использованы как
для инкапситации, так и смертельного поражения противника, в зависимости от се-
ротипа штамма;
4) большое количество серотипов среди вирусов VEEV и EEEV, и трудности полу-
чения иммунитета слизистых поверхностей к этим вирусам, создают проблемы раз-
работчикам вакцин.
Основной способ применения вирусов лошадиных энцефалитов для поражения
людей — создание аэрозоля. Среди них военными специалистами на основе анализа
случаев внутрилабораторных заражений персонала выделяется VEEV, как обладающий
наиболее высокой инфективностью в аэрозоле (Army FM 8-284, 2000; Zacks М. А.,
Paessler S., 2010).
Микробиология вирусов, вызывающих арбовирусные энцефалиты. Арбовирусы раз-
множаются в распространенных в лабораториях культурах клеток, чаще всего для этого
используют клетки почки детеныша хомячка (baby hamster kidney, ВНК), клетки поч-
ки зеленой обезьяны (green monkey kidney, Vero), фибробласты эмбриона утки (duck
embryo fibroblast, DEF) и клетки Aedes albopictus (C6/36).
Частицы альфавирусов, флавивирусов и буньявирусов имеют сферическую фор-
му. Их геном представлен положительно-смысловой однонитевой РНК, размером
10-12 т. п. о. Геном буньявирусов сегментирован, он состоит из трех линейных ми-
нус-цепей однонитевой РНК (ssRNA) размером -11 т. п. о. Реовирусы представляют
1 http://www.niaid.nih.gov/topics/biodefenserelated/biodefense/pages/cata.aspx.
2 Специалисты USAMRIID, Форт-Детрик, США.
930
Часть 3. Частная неправильная эпидемиология
собой безоболочечные вирусы икосаэдрической формы. Геном реовирусов состоит из
12 двунитевых линейных PHK-cei ментов, в сумме имеющих размер 19—32 гг. п. о.
Почти все арбовирусы, за исключением нескольких, принадлежащих к семейству
Reoviridae, обладают липидной оболочкой, полученной из клеток хозяина, в которой
несколько белков играют важную роль при взаимодействии с клеткой. В случае Reo-
viridae, эти белки являются частью внешней белковой оболочки вириона (Cali-
sherC. Н„ 1994).
Геном альфавирусов в пределах двух ORF кодирует четыре неструктурных белка
(nsPl-nsP4), составляющих две трети 5'-участка генома (нуклеотиды от 1 до 7514), и
четыре структурных белка (капсид, Е2 и Е1 оболочечные гликопротеины, небольшой
белок 6К), занимающие одну треть генома вируса со стороны З'-конца. Отдельные
альфавиру ы, циркулирующие в природных очагах, являются природными рекомби-
нантами. Один из таких рекомбинантов — WEEV. Нуклеотидная последовательность
капсидного белка и 80 нетранслируемых нуклеотидов З'-конца WEEV происходят от
альфавируса Нового Света — EEEV, а последовательности гликопротеинов Е2 и Е1
происходят от шьфавируса Старого Света — вируса Синдбис (Sindbis virus, S1NV).
Следовательно, WEEV образовался либо в результате рекомбинации EEEV и SINV,
либо предкового для S1NV альфавируса Нового Света, который еще не обнаружен.
WEEV сохранил энцефалопатогенность EEEV, но приобрел антигенную специфич-
ность SINV (Hahn С. S. et al., 1988). По данным S. C.Weaver et al. (1997), рекомбина-
ция между EEEV и каким-то синдбис-подобным вирусом произошла в 1300—1900 лет
назад (рис. 3.121).
Геликаза,
протеаза
Синтез
РНК
nsP2 nsP3 nsP4
Субгеномный промотор (26S)
Е2
Е1 - Ро1у[А]-3
5'сар-| ns~p7
Синтез (-)-цепи dPHK
РНК-кэппинг
РНК-зависимая I со
РНК-полимераза | ,___________|
Капсид Оболочечные
Рис. 3.121. С грукгура генома альфавирусов. А. Схематическое изображение структуры
генома альфавирусов. Показаны 5' cap 5'-нетранслируемого региона, ORF неструктур-
ного гликопротеина, субгеномный промогор, ORF структурного протеина, З'-нетранс-
лируемый регион и ро!у(А)-хвост. По М. В. Sherman, S. Weaver (2010). Б. Схематическое
изображение образования WEEV путем рекомбинации EEEV и SINV. Перекрестиями
показаны точки, по которым произошла рекомбинация. По С. S. Hahn et al. (1988)
Неструктурный белок 1 (nsPl) обладает активностью гуанин-7-метилтрансфера-
зы (guanine-7-methyltransferase) и гуанилтрансферазы (guanyltransferase). Неструктур-
ный белок 2 (nsP2) обладает геликазной активностью, необходимой для раскручива-
3.10. Арбовирусные энцефалиты
931
ния двунитевых репликативных интермедиатов РНК (dsRNA). Протеазная активность
требуется для рассечения неструктурных белков. До 50 % nsP2 обнаруживают в ядре
клетки-хозяина. Неструктурный белок 3 (nsP3) требуется для синтеза минус-цепи и
субгеномной РНК и обладает активностями АДФ-рибозы-1(")фосфат фосфатазы
(ADP-ribose l"-phosphate phosphatase) и РНК-связывающей активностью. Этот белок
имеет множество фосфоакцеп горных сайтов и легко фосфорилируется. Неструктур-
ный белок 4 (nsP4) обладает активностью РНК-зависимой РНК-полимеразы. Необ-
ходим для репликации вирусной РНК.
Капсидный белок является основным белковым компонентом нуклеокапсида.
Котранслированный капсид отсекается от структурного полипептида.
Е1-гликопротеин представлен в оболочке вируса, участвует в процессе слияния
вируса с мембраной клетки.
Е2-гликопротеин представлен в оболочке и выполняет роль лиганда, взаимодей-
ствующею с рецептором на поверхности клетки-мишени.
Созревшие Е? и ЕЗ формируются в результате расщепления протеазой фурином
прекурсорного белка рЕ2 на поздней стадии «созревания» вируса. Большинство аль-
фавирусов не сохраняют ЕЗ в вирионе.
Белок 6К может участвовать в активности протон-специфических каналов, необ-
ходимых для «бутонизации» (budding) вируса и/или его проникновения в клетку.
Альфавирусы реплицируются в цитоплазме инфицированной клетки. Проникно-
вение вируса в клетку происходит с использованием механизма рецепторного эндо-
цитоза. Поверхности оболочечных гликопротеинов Е1 и Е2 вируса инициируют эн-
доцитоз, взаимодействуя с рецептором на поверхности клетки. Вирус проникает
внутрь эндосомы кларитин-зависимым образом (clathrin-dependent manner) и вместе
с эндосомой попадает в цитоплазму клетки. Кислая среда эндосомы приводит к струк-
турным изменениям вириона и к экспонированию на его поверхности белка слияния,
входящего в гликопротеин Е1. Белок слияния вставляется в мембрану везикулы и фор-
мирует пору, через которую вирус выходит в цитоплазму, где происходит его разборка
и начинается трансляция вирусной РНК (рис. 3.122).
Первыми транслируются неструктурные белки с полноразмерной вирусной РНК.
На эндосомальных и лизосомальных мембранах формируются комплексы вирусной
репликазы. На ранней стадии инфекции преобладает синтез (-)-цепи, (+)-цепь и суб-
геномная РНК синтезируются на поздней стадии инфекции.
Консервативные элементы вирусной РНК участвуют в инициации транскрипции,
действую'! как трансляционные энхансеры и как пакующие сигналы. Структурные
белки (Е2 и Е1) транслируются с субгеномной РНК. Капсидный белок собирается в
нуклеокапсиды с полноразмерной вирусной РНК. рЕ2 (прекурсор Е2), 6К и Е1 про-
никают в mic-ретикулум (mic reticulum) и перемещаются в плазматическую мембрану
клетки. Созревание вириона начинается после того, как гетеродимеры Е2/Е1 вставите"
в плазматическую мембрану и 240 копий капсидного белка будут взаимодействовать
с одной копией геномной РНК, формируя нуклеокапсид. Этот нуклеокапсид взаимо-
действует с цитоплазматическим доменом белка Е2, расположенным в плазматиче-
ской мембране, инициируруя бутонизацию вируса. Созревший вирион «одевается» в
оболочку, состоящую из липидного бислоя, позаимствованного вирусом у клетки-
хозяина. Она покрывает 240 копий капсидного белка и 240 Е2/Е1 гетеродимеров,
выстро* иных как 80 тримерных шипов на поверхности вируса. Формируется двадца-
тигранная частица вируса диаметром 700 А, содержащая одну копию (+)-цепи РНК
размером 11,5 т. п. о.
932
Часть 3. Частная неправильная эпидемиология
Инфицирующий Вирус
Вьк вободившийся
из клетки г-ирк* н
h
Понижение
pH в эндосоме
Рецептор-
опосредованный
андошноз
Присоединение
вирионов к кчстке
Высвобождение
нуклеокапсида
из эндосомы
Нуклеокапсид
Мембранное
слияние
Нуклеокапсид
Разборка
нуклеокапсида
Цитоплазма •
Созревшие
гликопротеины
в плазматической
мембране клетки
‘ Почкующийся
вирион _
Плазматическая
_ мембрана
рЕ2Е1
+ Капсид
Геномная РНК
Глико-
протеины
транспор-
тируются
кплазма-
(тической
\мембране
Сборка
нуклеокапсида
у Геномная РНК
3',
m 43
i Транс-
ляция
Р123
Чтение
через
трансляцию
у Геномная РНК у
m?G Ар
Минус- А| Репликация
У цепь РНК '▼ у
------ 1 ..... ''Un
Транскрипция
PI234
insP2 процессинг (cis)
nsP4 &
▼ nsP4 Минус-
I,..,- .....,.z=i-nxc3 цепь
Р123 репликационный
I комплекс
nsP2 процессинг (trans)
nsPl nsP2nsP3 nsP4 цепь
Субгеномная s.
РНК m7G с
Структурный
полипротеин
репликационный
комплекс
=зхп
Трансляция
Процессинг
Капсид РЕ2 6К Е1
рЕ2Е1
Процессинг
и созревание
гликопротеинов
Аппарат
Гольджи
nsP2
Ядро
* РЕ2 3
___ ER
Гликопротеины проникают
в эндоплазматический ретикулум
Рис. 3.122. Жизненный цикл альфавируса. Жизненный цикл вируса начинается с момен-
та его прикрепления к рецептору на поверхности клетки (левая верхняя часть рисунка),
после чего запускаются механизмы рецепторного эндоцитоза, разборки вируса и высво-
бождения геномной РНК Затем транслируются белки репликации, трансляции и
процессинга (нижняя левая часть рисунка). Эти белки позволяют запустить репликацию
геномной РНК вируса и трансляцию структурных белков с субгеномной мРНК (центр
нижней части рисунка). Гликопротеины подвергаются процессингу в эндоплазматиче-
ском ретикулуме и транспортируются через аппарат Гольджи к плазматической мембра-
не (правая часть рисунка). Цитоплазматическая сборка геномной РНК вируса и капси-
да позволяет клетке собрать нуклеокапсидный кор, который ассоциируется с процес-
сированными гликопротеинами в плазматической мембране, приводя к бутонизации
вирусной частицы (верхняя часть рисунка, справа). По J. Jose et al. (2009)
3.10. Арбоеирусные энцефалиты
933
Экология и естественная эпидемиология возбудителей лошадиных энцефалитов1.
Вирусы лошадиных энцефалитов включают три серологически различающихся ком-
плекса: WEEV, EEEV и VEEV. Схематическое изображение эндемических и эпидеми-
ческих трансмиссионных циклов этих вирусов приведено на рис. 3.123, филогенети-
ческое древо на рис. 3.124.
mg.
WEEV Старые субтипы VEEV
(ID-IF, II-VI)
V
Селекция через Е2 T213R
§ I
о fa
й Я
Случайное питание инфицированно
кровью москитов семейств Ochlerotatus, ф
Culex, Aedes, Coquilletuda spp. и др. ▼ VEEV cyf ,ти, ш IAB/1C
Ochlerotatus 1
▼ gammon
* ' у.......................
at Psorophora, '
УД Ochlerotatus
jd» л " V taeniorhynchus v ')
I \ it
S
£
S
К Я
m £
Рис. 3 123. Схематическое изображение эндемических и эпидемических
трансмиссионных циклов EEEV, WEEV и VEEV. По М. Pfeffer, G R. Dobler (2010)
Middelburg
О 'nyong-nyong
Chikungunya
Ros River
Semliki Forest
Aura
Wha aroa
Kyzylagah
Sindbis
Ockelbu
Babanki
Buggy Creek
Fort Morgan
Highlands J
WEE-AR80
WEE-BR66
WEE-CA53
Рекомбинантные
потомки
EEE Северная Америка
EEE Южная Америка
WEE-IE
WEE-II
WEE-ID
WEE-IAB
WEE-IC
Новый Свез
c=:l Старый Свет
ип™ Рекомбинантные акты
Рис. 3.124. Филогенетическое древо и сценарий эволюции альфавирусов. Заштрихо-
ванные ветви показывают локализацию гипотетических предков и рекомбинацион-
ных актов между EEEV и Синдбис-подобным предком. По S. С. Weaver et al. (1997)
1 По работам С S. Hahn etal. (1988); J. F. Smith etal. (1997); P V Aguilar etal. (2004), M Pfeffer,
G. R. Dobler (2010); B. Sherman, S. Weaver (2010)
934
Часть 3. Частная неправильная эпидемиология
Западный энцефалит лошадей (WEE) вызывается комплексом вирусов, объединен-
ных под общим названием WEEV В комплекс входят шесть субтипов вирусов: WEE,
Sindbis, Y62-63, Aura, Fort Morgan и Highlands E Субтипы Sindbis и Y62-63 распрост-
ранены только в Старом Свете, на американском континенте они не обнаружены.
Остальные субтипы циркулируют в природных очагах на территории Западной Ка-
нады, США, Мехико, значительной части Южной Америки (Гвиана, Эквадор, Бра-
зилия, Уругвай, Аргентина). Генетический анализ изолятов WEEV, выделенных в оча-
гах Южной Америки (Бразилия и Северная Аргентина), пиказал, что все они представ-
ляют собой монофилетическую линию, когда их сравниваюс с изилятами из штатов
Калифорнии, Техаса и более отдаленной Монтаны.
WEEVне вызывает крупных вспышек энцефалита. В США подтверждено 639 слу-
чаев WEE за период с 1964-2005 гг. В трансмиссивный цикл WEEV в Северной Аме-
рике вовлечены домашние и дикие птицы, в которых происходит размножение виру-
са, но течение болезни бессимптомно. Основными переносчиками вируса являются
комары Culex tarsahs (см. рис. 3.123). WEEVобнаружен, по крайней мере, еще у 14 ви-
дов москитов рода Aedes и шести видов моски гов рода Culex. В Южной Америке в
трансмиссионный цикл вируса вовлекаются рисовые крысы (Oryzomys spp.), кролики
и завезенные европейские зайцы (Lepus europaeus). У вовлеченных в эпидемический
цикл людей и лошадей не развивается виремия, достаточная для инфицирования кро-
вососущих комаров. Следовательно, они не участвуют в поддержании и размножении
WEEV. Средняя величина летальных исходов при WEE улюдей в пределах 3—7 %. Со-
отношение нераспознанных случаев к распознанным меняется с возрастом заболев-
ших людей: 1:1 у детей менее одного годы; 58:1 у детей в возрасте от одного года до
четырех лет; и более 1 000: 1 у лиц старше 14 лет. От 15 до 30 % пациентов, перенесших
тяжелый энцефалит, сохраняют тяжелые неврологические расстройства, особенно
часто они встречаются у детей до года. Среди заболевших лошадей летальность дос-
тигает 50 % от числа заболевших.
В Европе существуют условия, необходимые для циркуляции WEEV. Прежде все-
го, это обилие птиц семейства воробьиных и комаров С. pipiens и Aedes vexans, однако
интродукции вируса не происходит. М. Pfeffer, G. R. Dobler (2010) объясняют это от-
сутствием миграции в Европу птиц, обитающих на американском континенте.
Восточный энцефалит лошадей (ЕЕЕ) вызывается EEEV. Крупных вспышек ЕЕЕ
не зафиксировано. В США за период с 1964-2005 гг. подтверждено 200 случаев EEEV.
Известны два антигенно различающихся варианта вируса: североамериканский и
южноамериканский. Североамериканские и карибские изоляты показали высокую
антигенную и генетическую гомогенность. Штаммы EEEV, выделенные в Централь-
ной и Южной Америке, более гетерогенны. EEEV вызывает тяжелое заболевание у
людей и лошадей. Этот вирус наиболее вирулентен среди альфавирусов, вызывающих
энцефалиты. В отдельные вспышки болезни летальность среди заболевших людей
достигает 70 %. У заболевших лошадей эта величина может юстичь 100 %. EEEV, в от-
личие от WEEV, вызывает у многих видов птиц (воробьиные, фазаны, домашние го-
луби, эму) тяжелое заболевание с развитием неврологических поражений. Кроме как
у птиц, EEEV обнаруживают у летучих мышей, однако их роль в поддержании в при-
роде вируса неизвестна, так как передачи вируса между летучими мышами не обна-
ружили. У рептилий и амфибий EEEV может поддерживаться месяцами в высоких
титрах, что говорит о возможности их вовлечения в трансмиссивные циклы вируса в
природных очагах. Эффективных и коммерчески доступных вакцин для профилак-
тики ЕЕЕ у лошадей и людей нет.
3.10. Арбовирусные энцефалиты
935
EEEV поддерживается в природных очагах Северной и Южной Америки. Транс-
миссионные циклы вируса в Южной Америке изучены плохо. На востоке Северной
Америки основную роль в циркуляции EEEV играют комары, паразитирующие на
птицах, в частности, не встречающийся в Южной Америке вид комаров Culiseta
melanura (комары жгучие), птицы из семейства воробьиных и отдельные виды водо-
плавающих птиц (см. рис. 3.123). Трансмиссионные циклы EEEV в основном поддер-
живаются в болотистой местности. Люди и лошади вовлекаются в этот процесс в пе-
риоды сильного размножения комаров, наступающие после обильных дождей.
EEEV обнаружен более чем у двадцати видов комаров, в том числе у С. pipiens и
A. vexans, встречающихся в Центральной Европе и многих других частях мира. Резуль-
таты исследований трансовариальной передачи EEEV среди комаров противоречивы.
Возможно, EEEV не передается через заражение яиц следующего поколения комаров.
Естественное формирование природных очагов EEEV в Европе и России малове-
роятно по следующим причинам:
1) миграции птиц с американского континента в Европу не происходит;
2) основным переносчиком EEEV на американском континенте является С. me-
lanura, роль других видов комаров в этом процессе минимальна. Однако в Европе ко-
мары рода Culiseta не имеют широкого распространения, сам этот род комаров мало-
численен (пять видов по всему миру), к тому же комары Culiseta spp. редко нападают
на людей;
3) птицы, зараженные EEEV, погибают, поэтому формирование искусственных
природных очагов EEEV можно обнаружить при соответствующем мониторинге.
Венесуэльский лошадиный энцефалит (VEE) вызывается VEEV. Вспышки VEE могут
приобретать характер катастрофы. Последняя крупная вспышка VEE произошла в
1995 г. в Венесуэле и Колумбии, в нее было вовлечено не менее 100 тыс. человек, из них
300 погибло. VEEV — это комплекс вирусов, разделенных на 6 антигенных субтипов
и вариететы (табл. 3.42, с. 936).
Вариететы IA/B и С обычно описываются как «эпизоотические/эпидемические
вирусы (epizootic/epidemic viruses)». Их выявляют во время масштабных эпидемичес-
ких вспышек в Северной, Центральной и Южной Америке. Они обладают высокой
вирулентностью в отношении людей и лошадей. Субтипы П—VI и вариететы ID, IE и
IF, известны как «энзоотические вирусы (enzootic viruses)». Подобно эпизоотическим
вирусам, энзоотические могут вызывать болезнь у человека, но они отличаются от них
тем, что не обладают вирулентностью в отношении лошадей. Энзоотические VEEV
передаются между небольшими млекопитающими слюной комаров. Хотя они могут
вызывать VEE у человека, но без амплификации в лошадях, их не накапливается та-
кое количество, которое могло вызвать крупную вспышку болезни при разносе
комарами. Вирусы комплекса VEEV циркулируют, в основном, в тропических и суб-
тропических регионах американского континента благодаря москитам рода Culex
(Melanoconion) и грызунам (Oryzomys, Proechimys, Sigmodon, Peromyscus, Heteromys,
Zygodontomys) (см. рис. 3.123). Кроме птиц из семейства воробьиных, цапли способ-
ны поддерживать высокую и длительную виремию, и через кровь инфицировать ко-
маров. Другие дикие и домашние животные не поддерживают трансмиссионный цикл
VEEV. У людей, вовлеченных в эпидемический процесс, формируются высокие тит-
ры вируса, достаточные для поддержания эпидемического процесса, т. е. люди явля-
ются амлифицирующими хозяевами для VEEV.
Эпидемии VEE развиваются тогда, когда создаются условия для инфицирования
комаров Aedes (Ochlerotatus) и Psorophora эпизоотическими штаммами субтипов IA/B
Таблица 3.42
Комплекс Венесуэльского лошадиного энцефалита
Субтип Варне- тет Прототипный штамм Происхож- дение Трансмиссион- ный цикл Вызывает болезнь География очагов Переносчик
у людей у лошадей
I А/В (VEEV) Trinidad donkey Осел (Тринидад) Эпизоотический + + Центральная, Южная и Северная Америка Mammalophilic mosquitoes
C P-676 Лошадь (Венесуэла) Эпизоотический + + Южная Америка М. mosquitoes
D 3880 Человек (Панама) Энзоотческий - + Центральная и Южная Америка Culex (Mel.) ocossa, panocassa, pedrvi, adamesi, vomerifer
E Mena 11 То же То же - + Центральная Америка Cx. (Mel ) taeniopus
F 78V-3531 Комар (Бразилия) То же — 9 Бразилия He известен
Il (Everglades) Fe3-7c Комар (Флорида) То же — + Южная Флорида Cx (Mel.) cedecei
HI (Mucambo) A Mucambo (BeAn8) Обезьяна (Бразилия) То же - + Южная Америка To же
В Tonate (CaAn410-D) Птица (Французская Гвиана) То же - + Южная и Северная Америка He известен
C 71D-1252 Москитоиды (Перу) То же - ? Перу To же
IV (Pixutna) Pixuma (BeAn356445) Комары (Бразилия) То же - ? Бразилия To же
V (Cabassou V) Cabassou V Комары (Французская Гвиана) То же - ? Французская Гвиана To же
V] (Rio Negro) AG80 663 Комары (Аргентина) То же - + Аргентина To же
* Составлено по работам J. Е Smith et al. (1997) и Р. V. Aguilar et al. (2004).
Часть 3. Частная неправильная эпидемиология
3.10. Арбовирусные энцефалиты
937
и IC и для передачи вирусов лошадям, ослам и мулам. Лошади также являются амп-
лифииируюшим хозяином для VEEV. Эпидемическое распространение VEE прояв-
ляется инкапситирующей Денге-подобной болезнью, которая у небольшой части за-
болевших людей, преимущественно у детей, может вызвать тяжелый энцефалит с
летальностью до 3 % от количества заболевших. География распределения вирусов
комплекса VEEV приведена на рис. 3.125.
Рис.3.125. Географическое распределение эпидемических/энзоотических
штаммов VEEV. По S. С. Weaver, W. К. Reisen (2010)
Эпидемические наблюдения в очагах VEE показали высокую вариабельность
VEEV. Отдельные нуклеотидные замены в геноме вируса могут изменять тропизм ви-
руса к отдельным переносчикам и вирулентность для лошадей, в результате чего ло-
шади становятся амлифицирующими хозяевами для VEEV. что способно «подтолк-
нуть» развитие эпидемической вспышки (рис. 3.126).
938
Часть 3. Частная неправильная эпидемиология
Рис. 3.126. Возможный механизм развития вспышки VEE А. Образование эпизоотиче-
ского штамма VEEV в природных экосистемах. Энзоотический лесной штамм VEEV
серотипа ID (показан серым цветом) циркулирует среди колючих (Proechimys spp.) и
хлопковых (Sigmodon spp.) крыс благодаря активности комаров из подрода Culex
(Melanoconion). Мутация в нуклеотидной последовательности, кодирующей гликопро-
теин Е2 оболочки вируса, приведшая к замене в 213-м положении треонина на аргинин
(Thr-213->Arg), приводит к появлению штамма вируса серотипов IAB или IC, способ-
ного к интенсивному размножению у лошадей, т. е. эпизоотического (эпидемического)
штамма (обозначен черным цветом). Виремия в крови лошади достигает уровня, при
котором происходит заражение комаров. При наличии большого количества комаров
Aedes и Рм-ophora spp. например, размножившихся в результате затопления местности
после наводнения, эпидемический штамм разносится по популяциям людей и лошадей.
Б. Гистологический анализ образцов мозга лошадей с клиническими признаками раз-
вившегося энцефалита, взятых на 7 cjt после подкожного инфицирования штаммом
ZPC738 VEEV, с мутациями в гене гликопротеина Е2, введенными методом точечного
мутагенеза. 1 — заражение штаммом ZPC738, у которого гликопротеин Е2 содержит Arg
в позиции 193. Конгестия, васкулярная и периваскулярная (пространство Virchow —
Robin) инфильтрация мононуклеарными клетками (стрелки) ствола мозга; 2 — штамм
ZPC738, у которого гликопротеин Е2 содержит Arg в позиции 213 (7 сут после инфици-
рования). Врезка показывает участок стволового мозга с наибольшей инфильтрацией;
3 — штамм ZPC738, у которого гликопротеин Е2 содержит Arg в позициях 193 и 213. Кон-
гестия, васкулярная и периваскулярная инфильтрация (пространство Virchow — Robin)
мононуклеарными клетками (вертикальные стрелки), признаки гибели нейрональных
клеток (горизонтальные стрелки) ствола мозга. По М. Anishchenko et al. (2008)
Штаммы VEEV, адапт ируясь к одному из основных своих переносчиков Ochlerotatus
taeniorhynchus (прежнее название Aedes taeniorhynchus), приобретают способность реп-
лицироваться в высоких титрах в этих комарах. В трансмиссию VEEVтакже могут быть
вовлечены комары, не способные реплицировать вирус, например, подрода Melano-
conion. Средняя доза вируса, которую может ввести в кровь человека или животного
3.10. Арбовирусные энцефалиты
939
москит, составляет примерно 2000 КОЕ. Интродукция и укоренение VEEV в Европе
теоретически возможны через инфицирование москитоидов, грызунов, птиц (цапель)
и людей. Но европейские грызуны субсемейств Murinae (мышиные) и Arvicolinae (по-
левковые) не могут служить амлифицирующими хозяевами для VEEV. Для американ-
ской разновидности комара Aedes albopictus доказана способность передавать VEEV
между людьми. Могут ли реплицировать VEEV европейские комары родов Culex и
Aedes, не известно. Ограниченная трансмиссия VEEV в Европе возможна в местно-
стях, в которых имеются большие популяции лошадей.
Восприимчивость животных, клиническая картина и патоморфология арбовирусных
энцефалитов у животных при искусственном инфицировании. Е EEV (изоляты NJ 1959 или
ArgM) использовали для ингаляционного заражения мышей Balb/c обоего пола весом
19-22 г и возраста 8—9 нед и морских свинок Hartley возраста 8—10 нед и весом около
250 г. Заражение проводили аэрозолями 1 мкм и более 6 мкм. Генерализованный вя-
лый паралич наблюдается у мышей, получивших более 1000 КОЕ вируса. У морских
свинок первые признаки инфекции после экспонирования к крупнодисперсному
аэрозолю начинались через 18—24 ч. Они включали ограниченную подвижность жи-
вотного и дорсальный тремор. Но при поражении аэрозолем с дисперсностью частиц
1 мкм, такие признаки болезни появлялись позже, через 65—72 ч. Развитие болезни
было одинаковым как при инфицировании аэрозолем 1 мкм, так и аэрозолем с дис-
персностью частиц, превышающей 6 мкм. Клиника болезни включала дрожание го-
ловы, сильное изменение походки и утрату равновесия. Болезнь быстро прогресси-
ровала. Животные совершали круговые движения, опрокидывались на бок, впадали
в кому и погибали.
Вирулентность аэрозолированного EEEV была большей для мышей, чем для мор-
ских свинок. Заражение мышей аэрозолями штаммов NJ1959 (североамериканский
штамм, New Jersey, 1959) и ArgM (южноамериканский штамм, Аргентина) давала сход-
ную летальность в группах, если размер частиц был в пределах 1 мкм. Если размер
частиц аэрозоля превышал 6 мкм, LDS0 для NJ1959 составлял 2,9х103, что было по-
чти в пять раз больше, чем LDS0 для NJ1959 при размере частиц 1 мкм (6,2х 102). LD50
для штамма ArgM при размере частиц аэрозоля более 6 мкм, не была определена, так
как летальность в таких группах животных не достигала 50 %. Для морских свинок LD50
при размере частиц аэрозоля EEEV 1 мкм, для штамма NJ1959, была большей в 17 раз,
по сравнению с LD50, определенной в тех же условиях для мышей (1,1 х 104 в сравне-
нии с 6,2 х 102 соответственно) (р=0,0098). LD50 для штамма ArgM при размере частиц
аэрозоля 1 мкм, была почти в два раза меньшей, чем для штамма NJ1959 (5,2х 103 и
1,1 х 104 соответственно) (Roy С. G. et al., 2009).
Вирусологические и иммуногистохимические исследования показывают, что ви-
рус проникает в мозг морской свинки через обонятельную систему нейронов (olfactory
system) в течение одних суток после ингаляционного заражения. Далее транснейро-
нально он распространяется в другие отделы мозга. Обонятельные биполярные ней-
роны являются основной мишенью вируса. Основные гистологические изменения,
выявленные в ткани мозга инфицированных морских свинок, включали некроз ней-
ронов, воспаление оболочек мозга, васкулиты. Минимальное количество вирусного
антигена обнаружено в тканях легких морских свинок, инфицированных NJ1959
EEEV, но вирусный антиген не был обнаружен ни у одного из животных, инфициро-
ванных штаммом ArgM. Среди морских свинок, инфицированных крупными части-
цами аэрозоля штамма NJ1959, только одно животное, подвергнутое этваназии в пер-
вые сутки после экспонирования к аэрозолю, имело незначительное количество
940
Часть 3. Частная неправильная эпидемиология
вирусного антигена в просвете небольших воздушных путей. Эти данные позволили
С. G. Roy et al. (2009) сделать вывод о высоком тропизме аэрозолированного EEEV к
обонятельным клеткам (olfactory cells) и о его низком тропизме к клеткам дыхатель-
ного эпителия.
У животных, инфицированных мелкодисперсным аэрозолем штамма NJ1959, ви-
русный антиген обнаруживали либо в эпителии воздухопроводящих путей диаметром
50—150 мкм (мелкие бронхи или бронхиолы), либо в альвеолах. В участках, где обна-
руживали вирус, одновременно находили признаки апоптоза или дегенерации клеток.
Легкие морских свинок содержали разбросанные участки скоплений макрофагов,
лимфоцитов и отдельные гетерофильные уплотнения, соответствующие участкам
воспаления, вызванных инга-тяцией мелкодисперсных твердых частиц, т. е. участки
пневмокониоза. С. G. Roy et al. (2009) обнаружили повышенную тропность EEEV не
только к обонятельным нейронам, но и к клеткам мезенхимальной линии — остеоб-
ластам однако они не наблюдали тропизма этого вируса к лимфоидной ткани ни у
мышей, ни у хомяков.
При интраназальном заражении обезьян- мармозеток (Callithrixjacchus) А. В Adams
et al. (2008) использовали североамериканский штамм EEEV FL93-393, выделенный
в 1993 г. во Флориде от комара С. melanura; и южноамериканский штамм ВеАг436087,
изолированный в 1985 г. от комара в Fortaleza (Бразилия). Животным инокулировали
по 1 х 106 КОЕ EEEVв 0,1 мл буфера. Обезьяны, инфицированные штаммом FL93-393,
как правило, погибали на пятые сутки при симптомах анорексии и поражения нервной
системы. Обезьяны, инфицированные штаммом ВеАг436087, оставались живыми,
хотя и давали пик виремии на 4 сут с титром от 2,8 до 3,1 log10 КОЕ/мл. У животных,
инфицированных североамериканским штаммом FL93-393, виремию не обнаружи-
вали, однако у погибших животных вирус находили в ткани мозга (в основном), пе-
чени и мышцах.
В ЦНС обезьян EEEV вызывал утрату нейронов (neuronal loss), нейронофагию
(neuronophagia), периваскулярные скопления клеток вокруг кровеносных сосудов,
фокальные и диффузные скопления воспалительных клеток и лептоменингит. Пато-
морфологическая картина у обезьян, ингаляционно зараженных FL93-393, напоми-
нала поражения мозга у человека при ЕЕЕ, вызванном североамериканскими штам-
мами, но не совпадала полностью.
5 людей, погибших от ЕЕЕ, сосудистые поражения более выражены, особенно в
сосудистых стенках. Часто обнаруживают сосудистые тромбы и кровоизлияния крас-
ной крови. Но у обезьян А. Р. Adams et al. (2008) не обнаружили васк] литов или фиб-
риновых тромбов (brin thrombi) в микроциркуляторной составляющей мозга (cerebral
microvasculature). Только у одной мармозетки из трех, инфицированных североамери-
канским штаммом FL93-393, они наблюдали локальные периваскулярные геморра-
гии. К областям ЦНС человека, наиболее поражаемым при ЕЕЕ, относятся кора го-
ловного мозга, базальные ганглии, таламус, гиппокамп и ствол мозга. Относительно
умеренно поражается мозжечок и спинной мозг. У мармозеток, инфицированных
штаммом FL93-393NA. наиболее серьезные поражения обнаруживали в коре голов-
ного мозга.
В мезентериальных лимфатических узлах всех трех мармозеток, интраназально
инфицированных штаммом FL93-393NA, обнаружен синусовый гистиоцитолиз (sinus
histiocytosis), в то же время только у одной обезьяны, инфицированной южноамери-
канским штаммом EEEV, мезентериальные лимфатические узлы имели признаки сла-
бо выраженного синусового гистиоцитоза. Но ни у обезьян, зараженных FL93-393NA,
3.10. Арбовирусные энцефалиты
941
ни у обезьян, зараженных ВеАг436087, не было обнаружено патологических измене-
ний в сердце, легких или скелетных мышцах (Adams А. Р. et al., 2008).
При аэрозольном инфицировании макак-циномолгусов (С. macaques) североаме-
риканским штаммом EEEV FL91-4679 (выделен от комара во Флориде, США), про-
должительность инкубационного периода болезни зависела от ингалированной дозы
(7,5 сут при ингаляции 3,65x 106 КОЕ и 3,5 сут при ингаляции 1,27х 107 КОЕ). Обезь-
яны погибали на фоне тяжелых неврологических нарушений через 36 ч после начала
лихорадки. В группе, получившей 1,27x107 КОЕ, погибли все 6 обезьян. В группе,
получившей 3,65х 106 КОЕ, погибли 4 обезьяны из 6. Виремия у заболевших животных
либо была транзиентной, либо ее вообще не могли обнаружить (Reed D. S. et al., 2007).
При исследовании вирулентности шести североамериканских изолятов WEEV
обнаружено, что при подкожном введении 103 КОЕ беспородным мышам, летальность
и среднее время гибели значительно варьируют для каждого изолята. Штаммы McMil-
lan (МсМ) вызывал 100 % летальных исходов на 5 сут после введения вируса. Штамм
Imperial 181 (IMP) не вызывал гибели животных. Такие же результаты были получе-
ны при ингаляционном, интраназальном и внутривенном введении обоих штаммов
WEEV (Logue С. Н. et al., 2009).
При ингаляционном заражении экспериментальных макак-циномолгусов разны-
ми дозами (6,3+0,5 log10 и 7,3±0,4 log]() КОЕ) WEEV СВА-87 (штамм изолирован из
мозга лошади во время эпизоотии в Аргентине) определена ингаляционная Ш50, со-
ставляющая 6,25 log10 КОЕ. WEE у обезьян развивался с преимущественным пораже-
нием ЦНС, практически без вирусемии. Лихорадка — первый и наиболее очевидный
признак начавшейся болезни после экспонирования к аэрозолю WEEV. Температура
у обезьян повышалась через 2-3 сут после заражения. Клинические признаки энцефа-
лита (тремор) появлялись на пике лихорадочного периода. Экспонирование к аэрозо-
лю WEEV проявлялось развитием энцефалита в более тяжелой форме, чем экспониро-
вание к аэрозолю VEEV. Развитие тремора у макак не всегда предсказывало фатальный
исход ингаляционного заражения WEEV (Reed D. S. et al., 2005).
Все штаммы VEEV, включая аттенуированный штамм TC-83, вирулентны для мы-
шей-сосунков и клеток эмбриона утки (duck embryo cells). Для морских свинок (внут-
рибрюшинное введение вируса) вирулентность проявляли штаммы VEEV, изолиро-
ванные от москитов во время эпизоотий VEE. Штаммы, выделенные в межэпизооти-
ческий период, оказались не вирулентными (Calisher Н. A., Maness К., 1974).
Хомяки более чувствительны к VEEV, чем мыши. При интраперитонеальном за-
ражении хомяков дозой VEEV, в десять раз меньшей, чем та, которой заражали мы-
шей (1,0х 103 КОЕ и 1,0х 104 КОЕ соответственно), мыши погибали на седьмые сут-
ки после заражения, при этом у них развивались параличи задних конечностей.
Хомяки не жили больше трех суток после заражения и погибали еше до того, как у них
развивались признаки поражения нервной системы (Jackson А. С. et al., 1991).
В случае аэрозольного экспонирования мышей к VEEV, прямое инфицирование
обонятельной области животного позволяет вирусу проникнуть в мозг, не вызывая ви-
русемии (Charles Р. С. et al., 1995).
Энзоотические штаммы VEEVменее патогенны, чем эпизоотические, по крайней
мере, при подкожном введении, которое имитировало укус комара, однако они сохра-
няют высокую инфекционность при аэрозольном заражении макак и могут вызвать у
них энцефалит с выраженными клиническими симптомами. При ингаляционном
заражении обезьян M.fasicularis энзоотическими штаммами VEEV: 68U201 (VEEV-IE;
впервые выделен в 1968 г. из мозга хомяка в Гватемале); и Мукамбо (VEEV-ША; впер-
942
Часть 3. Частная неправильная эпидемиология
вые выделен в 1957 г. у нечеловекообразной обезьяны в Бразилии), у приматов появ-
лялись симптомы активно развивающейся инфекции, включая лихорадку; вирусемию,
лимфопению. Клинические признаки энцефалита развивались в постфибрильный
период. Частности клинической картины различались в зависимости от штамма VEEV,
использованного для заражения (Reed D. S. et al., 2004).
В первый день после аэрозольного заражения VEEV-IE у макак началась лихорад-
ка, имевшая двуфазный характер. Второй, менее значительный пик, наблюдался при-
мерно через 1—2 сут после первоначального повышения температуры. Совершенно
иная картина наблюдалась после аэрозольного экспонирования к VEEV-IIIA: даже
при использовании наивысшей дозы вируса лихорадка появлялась не ранее, чем че-
рез 3 сут после экспонирования, и не была двуфазной; кроме того, пик лихорадки
наблюдался лишь через 3—4 сут после ее появления, или примерно через 7 сут после
экспонирования. У макак, экспонированных к VEEV-IE, пик вирусемии соответство-
вал по времени началу лихорадки, тогда как у макак, экспонированных к VEEV-IIIFA,
вирус уже не обнаруживался в крови к началу лихорадки. Степень лимфопении бы-
ла выше у макак, экспонированных к VEEV-IE, чем у макак, экспонированных к
VEEV-IIIA (Reed D.S. et al., 2004).
Тяжесть течения болезни зависела от вдыхаемой дозы VEEV. Но даже при макси-
мальных дозах (7,Ох Ю7 КОЕ для VEEV-IE; и 3,34х 106 КОЕ для VEEV-IIIA), ни одно
из инфицированных животных не погибло (Reed D.S. et al., 2004). VEEV, в отличие от
EEEV, имеет выраженную тропность к лимфоидной ткани и вызывает некроз лимфа-
тических узлов (Roy С. G. et al., 2009).
Иммунитет к возбудителям арбовирусных энцефалитов. В природных условиях, т. е.
тогда, когда передача альфавирусов между людьми и животными идет через укусы
комаров, развитие иммунитета к повторному заражению возможно, и основную роль
в блокировании развития инфекционного процесса играют нейтрализующие антите-
ла. Однако даже высокие титры ней грализующих поликлональных и моноклональ-
ных антител не предотвращают заражение, если оно происходит интраназально или
ингаляционно (Reed D.S. et al., 2004; Paessler S. et al., 2007; Yun N. etal. 2009) Лечение
с помощью вируснейтрализующих антисывороток не способно остановить развитие
болезни (Army FM 8-284, 2000). Исследование протекгивной активности американс-
ких вакцин ТС-83 (живая аттенуированная) и С-84 (инактивированная формалином),
проведенное на обезьянах-циномолгусах, показало, что они защищают от внутрикож-
ного заражения VEEV, но не эффективны при ингаляционном заражении (Pratt W.D.
et al., 1998). Роль Т-клеточного звена иммунитета в защите от альфавирусов так же
сомнительна. После вакцинации мышей вакциной на основе штамма ТС-83, цитоток-
сических Т-клеток (CTL) в селезенке и лимфатических узлах обнаружить не удалось.
Эти данные позволяют предположить, что CTL не играют большой роли в защите от
VEEV (Jones L. D. et al., 2003).
Устойчивость возбудителей арбовирусных энцефалитов во внешней среде. VEEV и
EEEV хорошо сохраняются в замороженном и высушенном состоянии, но термо-
лабильны: при температуре 56°C они разрушаются через 30 мин, при 60гС — через
10 мин; быстро инактивируются при замораживании и оттаивании и обработке СО2,
эфиром, дезоксихолатом.
WEEV, по сравнению с VEEV и EEEV, более устойчив к влиянию факторов среды.
При температуре 60°С он разрушается в течение 10—20 мин. При температуре 4°C ви-
рус в течение нескольких месяцев сохраняется в 50 %-ном глицерине и в суспензиях,
содержащих 2 % бычьего сывороточного альбумина или 20 % сыворотки крови кро-
3.10. Арбовирусные энцефалиты
943
ликов. В высушенном состоянии при минус 70 °C вирус сохраняет жизнеспособность
несколько лет. За 10 мин его разрушают 5 %-ные растворы хлорной извести и 2 %-ный
раствор креолина Хлороформ и 1 %-ный раствор карболовой кислоты разрушают ви-
рус в суспензии мозга за 10 сут (Таршис М. Г., Черкасский Б. Л., 1997).
Обнаружение генетически измененных штаммов возбудителей арбовирусных энцефа-
литов. Целью генетических модификаций генома вирусов, возбудителей арбовирус-
ных энцефалитов, будет повышение их вирулентности. Арбовирусы, циркулирующие
среди позвоночных, очень разнородны по вирулентности для человека и лошадей.
Вирулентность отдельных штаммов арбовирусов зависит от источника их выделения
и пассажной истории (см. вышек Поэтому к первому направлению, которое может
быть использовано преступниками для получения генетически модифицированных
вариантов арбовирусов, можно отнести выбор и селекцию среди уже существующих
в природе наиболее вирулентных штаммов. Эти штаммы, если и будут отличаться от
природных штаммов, то только отдельными нуклеотидными заменами в участках ге-
нома, некритических для проявления вирулентности вирусом Так как эта работа очень
кропотлива, требует больших материальных затрат и квалификации персонала, то,
скорее всего, такие штаммы будут похищаться из исследовательских лабораторий
крупных научных центров. Их можно распознать путем сопоставления нуклеотидных
последовательностей вирусов, выделенных в очаге с неправильной эпидемиологией,
с нуклеотидным последовательностям референс-штаммов, хранящихся в специализи-
рованных лабораториях. Второе возможное направление конструирования генетиче-
ски измененных вирусов, возбудителей арбовирусных энцефалитов, это использова-
ние точечного мутагенеза для повышения вирулентности вируса, аналогичное тому,
что показано на рис. 3.126.
Другой подход (третий) заключается в получении химерных вирусов, содержащих
либо генетический материал вирусов различных субтипов, либо вирусов разных ви-
дов. Технология конструирования вирусных химер основана на клонировании вирус-
ной кДНК в плазмидах, способных реплицироваться и совместно экспрессировать
гены вирусов в эукариотических клетках. Сборка химерных вирусов происходит по
схеме, представленной на рис. 3.122. Только считывание генетической информации
идет не с субгеномной мРНК (центр нижней части рисунка), а с кДНК вируса, кло-
нированной в этих плазмидах. Например, I. Р. Greene et al. (2005) был получен химер-
ный VEEV, у которого основная часть генома представлена генами вируса эн юотичес-
кого субтипа ID, но часть генов, кодирующих гликопротеин, «взята» из генома вирусов
эпизоотических субтипов 1С и IAB. Полученные химеры были использованы для экс-
периментального заражения лошадей. Исследователями продемонстрировано, что
такие химеры вызывают инфекционный процесс у лошадей с титром вируса в крови,
достаточным для инфицирования комаров (viremiacompetent equine phenotype).
S. Paessler et al. (2003) и N I. Haolin et al. (2007), используя эту же технологию с
целью создания безопасной вакцины против VEE, сконструировали несколько хи-
мерных вирусов Sindbis/VEEV. Их геном кодирует ферменты репликации и ««-акти-
вируемые РНК-элементы, производные от вируса Sindbis (SINV), и структурные бел-
ки VEEV (рис. 3 127).
Хотя полученные S. Paessler et al. (2003) и N. 1. Haolin et al. (2007) химерные виру-
сы имеют вирулентность меньшую, чем даже у вакцинного штамма ТС83, в природе
по такому же сценарию образовались химерные вирусы типа WEEV (см. рис. 3.121),
достаточно вирулентные для того, чтобы вызвать вспышку болезни с летальностью за-
болевших, достигающей 7 %.
944
Часть 3. Частная неправильная эпидемиология
Химерные SINV/VEEV
Рис. 3.127. Характеристики исходных и химерных SINV/VEE-вирусов. Все
участки генома, кодирующие неструктурные белки, происходят от SINV
(штамм Toto), гены структурных белков от различных штаммов VEEV (ТС83,
Trinidad donkey или TRD, ZPC738,68U201 и 78V-3531). Горизонтальные стрел-
ки показывают расположение субгеномных промоторов в геноме вируса.
Справа указан серологический субтип. По N. 1. Haolin et al. (2007)
Подозрение о применении генетически модифицированного арбовируса должно
возникнуть при обнаружении у жертв вспышки энцефалита химерных арбовирусов,
выходящих за рамки известных химер Sindbis/VEEV, либо при обнаружении в гено-
ме высоковирулентных для человек арбовирусов генов, ранее неизвестных для дан-
ного рода.
Достоверно установить факт вмешательства в геном вирусов, вызывающих вирус-
ные энцефалиты, можно путем сравнительного секвенирования их генома.
Патогенез арбовирусных энцефалитов. Арбовирусные энцефалиты имеют двуфаз-
ное течение. Первая фаза болезни начинается после того, как вирус вместе со слюной
комара попадает в кожу человека. F.EEV и VEEVхорошо реплицируются в дендритных
клетках Лангерганса, которые затем мигрируют в регионарные лимфатические узлы.
EEEV плохо реплицируется в лимфоидной ткани, но хорошо в остеобластах. Низкий
уровень вирусной нагрузки может быть обнаружен уже в первые два дня после инфи-
цирования. Он сохраняется в течение одной недели и заканчивается в связи с появ-
лением в сыворотке IgM-нейтрализующих антител (рис. 3 128 патогенез).
Если виремия быстро и эффективно устраняется иммунной системой хозяина,
болезнь имеет либо бессимптомное течение, либо у пациента развивается гриппопо-
добный синдром без энцефалита. Если иммунная система не справляется с виреми-
ей, что бывает у пожилых пациентов, либо у лиц с ослабленной иммунной системой,
3.10. Арбовирусные энцефалиты
945
Обнаружение вируса
в крови пациента
с помощью ПЦР IgM в крови или спинномозговой жидкости
Рис. 3.128. Динамика клинических симптомов, титров вируса и антител при арбови-
русных инфекциях на примере лихорадки Восточного Нила. По Davis L. Е. et al. (2008)
то начинается вторая фаза болезни — вирус проникает в мозг и активно размножается
в его тканях. У пациента развивается энцефалит (энцефалитно-менингиальный синд-
ром, синдром острого вялого паралича).
VEEV может проникать в ЦНС непосредственно через веточки обонятельного
нерва. В эксперименте при интраназальном введении вируса обезьянам, через 24 ч его
находили в обонятельной луковице еще до развития виремии у животного, затем по
обонятельным нервам он попадал в мозг. Распространение вируса по обонятельным
нервам может быть прервано хирургически. В этом случае вирус обнаруживали в обо-
нятельной луковице через 36 ч. Вероятно, он проникал туда по кровеносным сосудам
(Smith J. Е et al.,1997; Davis L. E. et al., 2008).
Клиническая и патологоанатомическая картина арбовирусных энцефалитов у чело-
века при естественном инфицировании. Арбовирусные энцефалиты — это группа бо-
лезней, для которых характерны следующие синдромы.
Асимптоматическая инфекция. У подавляющего большинства лиц, инфицирован-
ных арбовирусами, развивается бессимптомная инфекция. Точное соотношение ней-
роинвазивных случаев болезни со случаями бессимптомной инфекции варьирует для
конкретных вирусов. Бессимптомно инфицированные лица могут передать возбуди-
тель болезни здоровым людям через переливание крови или реже при транспланта-
ции органов. За медицинской помощью такие пациенты не обращаются и их учет в
очаге болезни не ведется.
Гриппоподобныи синдром. Развивается без симптомов со стороны ЦНС и является
наиболее частым проявлением клинической картины при арбовирусных инфекциях.
Обычно развивается после короткого инкубационного периода в 3—8 сут, но никак не
позже 28 сут после заражения. Пациенты жалуются врачу на внезапно возникшую
лихорадку, головную боль, недомогание, анорексию, тошноту, боль в мышцах и сус-
тавах, боль в горле, кашель и чувство усталости. На кожных покровах заболевшего
человека появляется незудящая и непапулезная сыпь, но так как она мало выражена,
946
Часть 3. Частная неправильная эпидемиология
ее часто не замечают. Лимфатические узлы увеличены. Пациенты редко обращаются
за медицинской помощью и переносят болезнь «на ногах».
Энцефалшпно-менингиальный синдром (encephalitis-meningitis syndrome, neuroinvasive
disease). Пациенты жалуются на сильную головную боль, лихорадку, ригидность заты-
лочных мышц без локальной слабости или нарушения психики. Обычно им ставят
диагноз «менингит». Пациенты с клиническими и лабораторными подтверждени-
ями поражения паренхиматозных участков мозга классифицируются как больные
энцефалитом. Обычно энцефалитно-менингиальный синдром развивается на фоне
гриппозного синдрома. Признаки энцефалита могут включать в себя нарушения
концентрации внимания и памяти, выраженную сонливость, спутанность сознания,
головокружение, бред, тошноту, рвоту, ступор, тремор, миоклонические судороги,
светобоязнь, дизартрию, дисфагию, нистагм, нарушение походки, расстройство ко-
ординации движения, болезненное выражение лица, парестезии, потерю кожной
чувствительности и судороги. Экстпапирамидные признаки включают: паркинсо-
низм, повышенный мышечный тонус, тремор. Изменение психического состояния
является отличительной чертой всех вирусных энцефалитов, однако ра житие комы
наиболее часто встречается у пациентов с ЕЕЕ и реже при энцефалитах, вызванных
другими арбовирусами. Острая фаза энцефалита длится 1—3 нед, но выздоровление
идет медленно. В зависимости от тяжести энцефалита, признаки и симптомы могут
сохраняться в течение нескольких месяцев.
Синдром острого вялого паралича (acuteflaccid paralysis, AFP). Многие из арбовирусов
инфицируют нейроны спинного мозга, в частности, нейроны переднего рога. У не-
которых тяжелых пациентов происходит гибель этих нейронов, что приводит к ост-
рым вялым параличам, похожим на те, что развиваются при полиомиелите. Большин-
ство пациентов с A FP также имеют клинические признаки энцефалита, но они могут
быть умеренными и находиться как бы в тени остро развившихся параличей.
Мышечная слабость может развиваться одновременно с энцефалитом или в тече-
ние недели после появления первых клинических признаков поражения ЦНС. Сла-
бость мышц начинается внезапно и носит ассиметричный характер, включает в себя
одну или несколько конечностей, но наиболее часто поражаются мышцы ног. Ареф-
лексия — общий клинический признак для арбовирусных энцефалитов, сопровожда-
е гея периодической утратой контроля над мочевым пузырем или кишечником. Об-
щим клиническим признаком является парестезия конечностей. У пациентов с
поражением шейного отдела спинного мозга и ствола мозга развивается слабость ды-
хательных мышц, и они нуждаются в интубации и искусственной вентиляции легких.
Анализ крови при арбовирусных инфекциях обычно показывает умеренный лей-
коцитоз, гипонатриемию (<135 ммоль/л), повышение уровня печеночных трансами-
наз (аспар гатаминотрансферазы и аланинаминотрансферазы). Содержание глюкозы
почти всегда в норме. МРТ может показывать норму или демонстрировать очаги по-
вышенной интенсивности сигнала на Т2 -взвешенных изображениях (T2-weighted
images» базальных ганглиев, таламуса и ствола мозга. Количество выявленных анома-
лий увеличивается через 2—4 нед после начала болезни. ЭЭГ показывает диффузное
или локальное замедление волн, а иногда области острых волн.
Эпизоотические и энзоотические штаммы VEEV вызывают аналогичные синдро-
мы заболевания. Однако лишь у небольшой части больных VEE (от 0,5 до 4 %) наблю-
дается поражение нервной системы. У тех, кто выжил после воспаления головного
мозга при VEE, функционирование нервной системы обычно полностью восстанав-
ливается. Клиническая картина ЕЕЕ и WEE аналогичны.
3.10. Арбовирусные энцефалиты
947
Диагностика арбовирусных энцефалитов. Диагностика почти всех арбовирусных
инфекций строится на серологическом исследовании сыворотки крови или ликвора.
Сам факт наличия антител IgG к вирусу при отсутстви и роста титра антител класса IgM
может быть показателем прошлого контакта иммунной системы с вирусом, а не ост-
рой инфекции. У человека виремия всегда имеет место в низких титрах, она заверша-
ется после появления IgM-антител. Поэтому выделить арбовирус в культуре сложно
и, как правило, диагностические лаборатории это не делают. Чувствительность ПЦР
для диагностики арбовирусов значительно ниже, чем для герпесвирусов и энтерови-
русов. ПЦР и серологические тесты должны рассматриваться как взаимодополняю-
щие, а не конкурирующие методы диагностики.
Патанатомия. Арбовирусы вызывают обширное и неоднородное поражение раз-
личных участков и структур головного мозга (нейроны коры головного мозга, базаль-
ные ганглии, ствол мозга, мозжечок, а иногда и спинной мозг). В основном это пери-
васкулярные инфильтраты и мультифокальные некрозы (рис. 3.129).
Рис. 3.129. Патологические изменения в сером веществе головного и спинного мозга
женщины, погибшей от WW. А. Область тканевого некроза в правом путаменте. Видна
инфильтрация вен (большая стрелка), макрофаги (маленькая стрелка) и мононуклеар-
ные воспалительные клетки (реакция Шифф — иодная кислота; увеличение в 125 раз).
Б. Артериола в базальном ганглии с признаками инвазии стенки мононуклеарными клет-
ками (гематоксилин эозин; увеличение в 500 раз). В. Передний рог поясничного отдела
спинного мозга. Скопления микроглиальных мононуклеарных клеток (стрелки) (реакция
Шифф — иодная кислота; увеличение в 125 раз). По В. A. Anderson (1984)
Механизм поражения нейронов не вполне ясен. Арбовирусы, как правило, не
инфицируют олигодендроциты, так что демиелинизация при арбовирусных инфек-
циях не характерна. Воспаление мозга редко бывает обширным. Тяжесть паренхима-
тозных воспалительных процессов отражает степень плейоцитоза. Лептоменинкс
(leptomeninges) внешне выглядит неизмененным, но при гистологическом исследо-
948
Часть 3. Частная неправильная эпидемиология
вании обнаруживают лимфоцитарную инфильтрацию. Мозг может выглядеть не из-
мененным либо отечным или содержать фокальные петехии в сером и белом веществе.
При летальных исходах тяжелые вирусные поражения чаще всего обнаруживают в
стволовом отделе мозга. Когда в инфекционный процесс вовлечен спинной мозг, то
патологический процесс чаще всего локализуется в передних рогах его поясничной и
шейной областей (Davis L. Е. et al., 2008).
Диагностика искусственного поражения возбудителями лошадиных энцефалитов.
Арбовирусы, вызывающие лошадиные энцефалиты (WEEV, VEEV и EEEV), не име-
ют природных очагов на территории России. Общие принципы диагностики искус-
ственно вызванных вспышек лошадиных энцефалитов те же, что и для вирусных ге-
моррагических лихорадок (см. разд. 3.9). Любой иммунологически- и генетически
подтвержденный диагноз лошадиного энцефалита, выявленного на территории Рос-
сии, служит основанием для предположения о совершенном биологическом нападе-
нии, если:
1) заболевший в течение месяца перед появлением лихорадки и/или симптомов
энцефалита не выезжал в страны, эндемичные по данной болезни;
2) в эти же сроки заболевший не работал с возбудителем данной болезни в виру-
сологической лаборатории.
Вне природных очагов вирусов лошадиных энцефалитов заражение людей WEEV,
VEEV и EEEV возможно ингаляционным и парентеральным способами. При установ-
лении способа заражения жертв биотеррористического акта необходимо учитывать,
что вирусы, возбудители лошадиных энцефалитов, не обладают тропностью к легоч-
ной ткани и не вызывают местных процессов (некрозы, карбункулы, абсцессы) в уча-
стке внутримышечной инъекции. Внутривенное введение этих вирусов дает ту же кли-
ническую картину болезни, что наблюдается в их природных очагах при трансмиссии
комарами. Поэтому основными клиническими признаками искусственно вызванных
лошадиных энцефалитов будут те же, что наблюдают врачи при естественном зара-
жении людей в природных очагах. Если принять во внимание экспериментальные
данные, полученные в опытах на модельных животных, факт ингаляционного зара-
жения мелкодисперсным аэрозолем все же можно установить ретроспективно, ис-
пользуя методы иммуно-гистохимического окрашивания эпителия мелких бронхов,
бронхиол, либо альвеол. Но кроме признаков апоптоза или дегенерации клеток в уча-
стках, положительных на вирус, трудно рассчитывать на обнаружение других гисто-
логических подтверждений ингаляционного заражения возбудителями лошадиных
энцефалитов. У жертв террористического преступления, ингаляционно инфициро-
ванных VEEV, возможно обнаружение патологических изменений в медиастинальных
лимфатических узлах.
Возбудители лошадиных энцефалитов благодаря тропности к нейронам обоня-
тельной системы, находящихся в носовых ходах, могут проникнуть в мозг, не попадая
в кровеносное русло и не вызывая вирусемии. Поэтому вблизи от источника аэрозо-
ля возможно инфицирование людей крупнодисперсным аэрозолем вируса. Клиниче-
ски такое заражения проявится быстрым, в течение 1—3 сут, развитием энцефалитно-
менингиального синдрома и синдрома острого вялого паралича без вирусемии. Мето-
ды иммуно-гистохимического окрашивания эпителия носовых ходов могут оказаться
полезными для определения участка первичной аппликации вируса.
Выделение генетически измененных штаммов WEEV, VEEV и EEEV от человека
или лошади, даже в их природных очагах, однозначно свидетельствует об искусствен-
ном происхождении вспышки болезни.
3.11. Рицин 949
Иммунопрофилактика "рбовивтсных энцефалитов. Более 8 тыс. человек были вак-
цинированы в США за четыре последних десятилетия вакциной на основе аттенуи-
рованного штамма ТС-83 VEEV. Накопленные данные однозначно показывают, что у
20—40 % вакцинированных людей развивались симптомы болезни, сходной с типич-
ным VEE. Вакцинированные чувствовали жар, недомогание и головную боль, при-
чем у половины из них самочу вствие бывало настолько плохим, что они нуждались в
1—2 сут постельного режима. Лошади, вакцинированные этой вакциной, реагирова-
ли на нее лихорадкой и развитием виремии в титрах, достаточных для инфицирова-
ния москитов и инициирования трансмиссионного цикла VEEV. Эти данные пока-
зывают наличие остаточной вирулентности для людей и лошадей у штамма ТС-83.
Перекрестный иммунитет к гетерологичным серотипам слабый или вообще отсут-
ствует (Paessler S. et al., 2003). В России вакцина на основе аттенуированного штамма
ТС-83 VEEV не разрешена к применению.
Существуют экспериментальные инактивированные формалином вакцины для
людей против VEEV, EEEV и WEEV, но они требуют многократных инъекций и явля-
ются слабоиммуногенными.
Лечение арбовирусных энцефалитов. Для альфавирусных энцефалитов не существу-
ет средств для специфической терапии, поэтому применяется симптоматическое ле-
чение. Особой проблемой, возникающей иногда у больных, инфицированных WEEV,
является крайне высокая температура, для снижения которой могут потребоваться
энергичные меры.
3 11. РИЦИН
Виды рицина. Молекулярная структура и механизм действия. Токсические свой-
ства. Поражающие свойства Клиника энтерального поражения. Клиника
ингаляционного поражения. Клиника парентерального поражения. Диагностика
поражения рицином. Иммунопрофилактика. Лечение.
Рицин (ricin toxin) — бинарный токсин, получаемый из жмыха семян касторовых
бобов Ricinus communis (клещевина обыкновенная). Его токсическое действие связа-
но с блокированием синтеза белков в рибосомах клеток-мишеней. Привлекает вни-
мание специалистов в области химического и биологического оружия с конца Пер-
вой мировой войны из-за высокой токсичности и доступности1. В годы Второй
мировой войны рицин интенсивно изучался в США и Соединенном Королевстве в
качестве средства уничтожения живой силы противника. Разработаны технологии его
выделения и очистки в кристаллическом виде. В США в годы войны было изготовлено
1,7 т рицина. Как поражающему агенту ему был присвоен шифр «W», но на вооруже-
нии ни одной из армий он не состоял. Использовался спецслужбами для индивиду-
альных убийств. В медицине широко применяется для разработки противоопухоле-
вых препаратов (Griffiths G. D. et al., 2007; Антонов Н. С., 1994).
Виды рицина. Препараты рицина различаются агрегатным состоянием, степенью
очистки и наличием изоформ (isotoxins, isoforms). Основными производителями кас-
торовых бобов являются Китай, И ндия, Бангладеш и США, где они используются для
получения касторового масла. В самом масле рицина нет, технологическая цепочка
1 См. разд. 1.9 «Начало программы по созданию БО в США».
950
Часть 3. Частная неправильная эпидемиология
его получения предполагает инактивацию токсина на1 реванием. В жмыхе семян касто-
ровых бобов содержится 0,5—5 % рицина. Получение неочищенного рицина не пред-
ставляет особых сложностей для террористических организаций. Семена касторовых
бобов гомогенизируются в солевом растворе, гомогенат закисляют до pH 4,8 и выдер-
живают ночь на шутель-аппатате при "стряхивании, после чего центрифугируют. Не-
растворимая часть осаждается, супернатант содержит грубый экстракт рицина. При
электрофоретическом исследовании методом однопространсгвенного гель-электро-
фореза в полиакриламидном геле, содержащем додецилсульфат натрия, такие препа-
раты разделяются на множество белковых фракций, среди которых рицин, обра-
зующий комплекс с агглютинином, представлен основной фракцией (Griifiths G. D.
et al., 2007; Audi J. et al., 2005).
Агглютинин представляет собой лектин гликолипидной природы. Прямым цито-
токсическим действием не обладает, но способен агглютинировать и лизировать эрит-
роциты. Клинически значимый гемолиз он вызывает только после внутривенного
введения. Для отделения агглютинина и получения чистого рицина используют хро-
матографические методы. Методы аффинной хроматографии позволяют получать до
180 мг чистого рицина из 100 г касторовых бобов. Очищенный рицин представляет
собой белый порошок без запаха, он хорошо растворим в воде и сохраняет токсичес-
кие свойства в широком диапазоне pH. В водных растворах рицин инактивируется при
температуре 80 °C в течение часа, в сухом виде он более устойчив. Получение активного
рекомбинантного голотоксина рицина в ооцитах Xenopus iaevis затруднено из-за не-
возможности экспрессии его субъединиц, сохраняющих обширные гидрофобные
зоны на своей поверхности (Bigalke Н., Rummel А., 2005; Audi J. et al., 2005).
Имеются разные сорта касторовых бобов — культивары (cultivars). Они различа-
ются по морфологии растения, бобов и изоформам рицина. В дополнение к мажорной
форме рицина (major form of ricin toxin) T. Mise et al. (1977) описали рицин E\ M. Ishi-
guro et al. (1966) — рицин D. Рицин D обычно встречается в бобах крупного размера, в
мелких бобах встречаются оба изотипа. От мажорной формы рицина они незначитель-
но отличаются по аминокислотному составу, ММ, изоэлектрической точке и сродству
к различным адсорбентам (Griffiths G. D. et al., 2007). В более поздних работах изо-
формы рицина обозначены порядковыми номерами: 1,2 и 3. Их молекулярные мас-
сы определены как 63,55, 64,03 и 62,8 кДа соответственно. В опытах на клетках Vero
показано увеличение токсичности изоформ рицина в зависимости от степени глико-
зилирования их В-субъединиц. Самой токсичной является третья изоформа рицина,
имеющая наиболее гликозилированную В-субъединицу (Senhgal Р. et al., 2011).
В касторовых бобах, кроме рицина, содержатся небольшие количества токсичес-
кого алкалоида рицинина (ricinine). Он экстрагируется совместно с рицином. Случаев
отравления очищенным рицинином людей не описано (Darby S.M. et al., 2001). В экс-
периментах на мышах показано, что рицинин вызывает конвульсии и смерть. Его ток-
сическое действие возможно связано с увеличением высвобождения глутамата в тка-
нях мозга и ингибирования субтипа А постсинаптического рецептора у-аминома-
сляной кислоты (Ferraz А. С. et al., 2002).
Молекулярная структура и механизм действия. ММ рицина — 60-65 кДа. Рицин —
типичный АВ-токсин1. На рибосомах синтезируется в виде цельной полипептидной
цепи. В вакуолях клеток растения эта цепь расщепляется в единственном сайте, об-
1 См. разд. 2.2.3.6. «Организация и общий механизм действия молекул бактериальных
токсинов».
3.11. Рицин
951
разуя две субъединицы: А (ферментативная) и В (связывающая), остающиеся соеди-
ненными через дисульфидную связь (рис. 3.130).
Рис. 3.130. Кристаллическая структура рицина. В-цепь (RTB, левый домен) рицина
представляет собой N-гликозилированный гомодимер в форме гантели (barbell),
262 аминокислоты каждого мономера формируют структуру типа р-трилистника
(P-trefoil structure). Каждый мономер содержит по одному «карману», связывающему
сахариды (saccharide binding pock). A-цепь (RTA, правый домен) рицина соединена с
В-цепью посредством дисульфидного мостика и кристаллизуется как а-р-а-сандвич.
По Н. Bigalke, A. Rummel (2005)
Субъединицы рицина сами по себе не токсичны, токсическое действие проявля-
ется только при условии кооперативного взаимодействия обеих субъединиц в составе
молекулы рицина. В-цепь представляет собой лектин, связывающийся с галактозой
или N-ацетилгалактозамином гликопротеинов и гликолипидов на поверхности клетки
мишени, вызывая эндоцитоз и проникновение молекулы рицина из ранней эндосо-
мы в транспортную сеть Гольджи (рис. 3.131). Часть рицина через сеть Гольджи прони-
кает в эндоплазматический ретикулум (endoplasmic reticulum, ER), где после разрыва
S—S-связи его молекула распадается на А- и В-субъединицы. Затем А-субъединица
(RTA) частично развертывается таким образом, чтобы пересечь мембрану ER через
8ес61р-транслокон (Sec61p translocon). А-субъединица рицина является высоко-
эффективной N-гликозидазой. Она действует через удаление специфического адени-
нового основания (А4324) вблизи З'-конца 28S рибосомальной РНК. Одна А-субъ-
единица способна в минуту инактивировать около 1500 рибосом. Аминоацилтранс-
портная РНК не соединяется с усеченной 28S рибосомальной РНК. Синтез белка
останавливается и запускается механизм апоптоза клетки (Audi J. et al., 2005; Lord M. J.
et al., 2003). Аминокислотная последовательность А-субъединицы рицина имеет об-
ширную гомологию с энзиматической субъединицей шига-токсина*. Механизмы дей-
ствия обоих ферментов одинаковы (Bigalke Н., Rummel А., 2005).
Токсические свойства. Летальные дозы рицина зависят от способа введения в орга-
низм животного, от вида животного, степени очистки рицина, условий его хранения
и наличия изоформ в препарате. При парэнтеральном введении летальные дозы ри-
цина для различных животных измеряются в широком интервале (табл. 3.43).
1 О шига-токсине см. в разд. 2.2.2. «Трансдукция».
952
Часть 3. Частная неправильная эпидемиология
Рис. 3.131. Механизм токсического действия рицина. После взаимодействия В-субъ-
единицы рицина с галактозой или N-ацетилгалактозам ином на поверхности клетки,
молекула токсина подвергается эндоцитозу (1,2). Большинство молекул рицина выво-
дится обратно на поверхность клетки благодаря механизму экзоцитоза (3). Если ве-
зикулы сливаются с лизосомами, то рицин разрушается благодаря механизму проте-
олиза (4) Часть рицинсодержащих везикул сливается с транспортной сетью Гольджи
(trans-Golgi network, TGN) (5), оттуда они также могут быть выведены на поверхность
клетки (6). Токсическое действие рицин оказывает тогда, когда его молекула распа-
дается на субъединицы и А-субъединица пенетрирует мембрану TGN и проникает в
цитозоль (7), где катализирует депуринизацию (depurination) рибосом, останавливая
синтез белка. По Н Bigalke, A. Rummel (2005)
Таблица 3.43
Летальные дозы рицина для млекопитающих*
Вид животного Способ введения LD5o, мг/кг
Морская свинка Внутрибрюшинно 0, 00005
Внутримышечно 0,0008-0,003
Подкожно 0,2
Кошка Внутримышечно 0,0002
Кролик То же 0,0005
Собака То же 0,0006
Мышь Внутривенно 0,0026
Внутрибрюшинно 0,006
Подкожно 0.0075
Крыса То же 0,02
Корова То же 0,013
Козел Го же 0,035
* По Н. С. Антонову (1994).
Lct50 аэрозоля очищенно! о рицина (MMAD 1—5 мкм) для крыс определена в пре-
делах 4,54—12,7 мгхмин/м3 (LD50 3,7—9,8 мкг/кг) (Griffiths G. D. et al., 2007). Леталь-
ные дозы рицина при ингаляционном поражении для обезьян-резусов и африканских
зеленых обезьян составили 15,0 и 5,8 мкг/кг соответственно (Wannemacher R. W.,
Anderson J. В., 2006). Минимальная ингаляционная летальнаядоза рицина длячелове-
3.11. Рицин
953
ка близка к 4,0 мкг/кг (Антонов Н. С., 1994). При энтеральном отравлении для чело-
века ее оценивают в диапазоне от 1 до 20 мг/кг (Audi J. et al., 2005).
Абсолютная летальная доза у животных находится в обратной зависимости от вре-
мени наступления смерти после ингаляции рицином (Griffiths G. D et al., 2007). Ток-
сичность рицина при оральном введении на порядки ниже, чем при введении други-
ми путями. Для мышей LD50 ингалированного рицина обычно в пределах 3—5 мкг/кг,
в то время как при оральном введении она составляет 20 мг/кг (Franz D., Jaax N., 1997).
Снижение токсичности рицина при попадании в организм через ЖКТ, вызвана его
разрушением панкреатическими ферментами. Рицин не обладает строгой селективно-
стью. Наиболее чувствительны к нему клетки ретикулоэндотелиальной системы, такие
как Купферовы клетки и макрофаги из-за большого количества на их поверхности
маннозных рецепторов. Так как все клетки являются для него мишенью, го симптомы
отравления зависят от места аппликации токсина (Bigalke Н., Rumme1 А., 2005).
Рицин обладает прямым цитотоксическим действием. Но патологические послед-
ствия сосудистых нарушений, индуцированные токсином, гораздо более серьезны,
чем прямое действие токсина на клетки. Причина смерти зависит от пути введения
рицина. Интоксикация экспериментального животного A-цепью рицина при различ-
ных путях его введения проявляется в виде синдрома проницаемости сосудов (гипо-
альбунемия, отеки), вызванного повреждением клеток эндотелия. Мыши, кролики,
приматы, ингаляционно экспонированные к высоким дозам рицина, всегда имели тя-
желые поражения легочной ткани, сопровождающиеся отеком легких и затоплением
альвеол (alveolar flooding), приводящие их к смерти вследствие гипоксемии. Появле-
ние первого клинического симптома ингаляционного отравления рицином — одыш-
ки, С. Wilhelmseann, L. Pitt (1996) связывают с начавшимся затоплениям альвеол; отек
легких — с проявлением синдрома проницаемости сосудов. Высокие дозы рицина,
введенные экспериментальным животным внутривенно, уже через 4 ч повреждали
Купферовы клетки, затем развивалось повреждение клеток эндотелия и диссемини-
рованная внутрисосудистая коагуляция. В сосудистой сети печени формировались
тромбы, приводящие к гепатоцеллюлярному некрозу. Животные погибали от гепато-
целлюлярной и ренальной недостаточности. Повреждение сосудистого эндотелия,
вызванного внутривенным введением рицина, может вызвать периваскулярный отек
легких. Заглатывание рицина вызывает некроз эпителия желудочно-кишечного трак-
та, локальное кровотечение и некроз печени, селезенки и почек. Внутримышечное
введение вызывает сильный локальный некроз мышц и региональных лимфатичес-
ких узлов с умеренным поражением висцеральных органов (Franz D., Jaax N., 1997).
Одним из первых клинических признаков отравления рицином является геморрагия
сетчатки глаз (Антонов Н. С., 1994).
Поражающие свойства. Химик Н. С. Антонов (1994) со «своей колокольни» харак-
теризует рицин как несостоявшийся поражающий агент химического оружия. При
боевом применении он оказывает поражающее действие только в аэрозольном состо-
янии, так как не обладает кожно-резорбтивным действием. Применение противни-
ком жидкого или сухого аэрозоля рицина не создает войскам проблем защиты от него,
какие имеют место при защите от жидких ОВ нервнопаралитического действия. Для
защиты от аэрозоля рицина не требуются средства защиты кожи, военнослужащему
достаточно фильтрующего противогаза. В чистом виде рицин не может диспергиро-
ваться до аэрозольного состояния с помощью разрывных зарядов боеприпасов из-за
потери активности. Дробление взрывом растворов рицина трудно осуществить прак-
тически. Будучи гликопротеином, он растворим только в водных системах, но сам
954
Часть 3. Частная неправильная эниоемиология
разлагается водой, что делает невозможным длительное хранение таких боеприпасов.
Водные растворы рицина замерзают, создавая дополнительные проблемы его боево-
му применению. Суспензии рицина в четырех: лнристом углероде оказались неустой-
чивыми, расслаивались и меняли баллистические характеристики боеприпаса. Полу-
чение эффективной сухой рецептуры рицина с дисперсностью частиц менее 5 мкм
натолкнулось на необходимость предотвращения ее слеживаемости и комкования.
Решение этих проблем сопровождалось появлением новых. Сухие мелкодисперсные
аэрозоли рицина дезактивировались ультрафиолетом солнечной радиации. Серьез-
ным недостатком рицина, ограничивающим его боевое применение, является отсут-
ствие у него быстродействия. Токсоэффект наступает не ранее 8—72 ч после экспони-
рования. По этим причинам боеприпасы с рицином реально никогда не состояли на
вооружении ни одной армии. В настоящее время отравления рицином наиболее ве-
роятны среди жертв преступлений, имеющих криминальную и политическую моти-
вацию (отдельные личности, небольшие группы людей).
Клиника энтерального поражения. Из 751 обобщенных случаев отравления касторо-
выми бобами, смертельными оказались 19 (смертность 2,5 %). При отсутствии эффек-
тивной терапии смертность достигает 6 %. Количество съеденных бобов, вызвавших
летальный исход у отравленного (или отравившегося) человека, варьирует от 10 до
20 штук. Клиника отравления однообразна. В течение нескольких часов после при-
ема в пищу касторовых бобов у отравленного появляется чувство тошноты, развива-
ются рвота и боли в брюшной полости; затем появляются диарея и кровотечение из
ануса. К ним присоединяются анурия, судороги, расширение зрачков, повышается
температура тела. Отравленного мучают чувство жажды, боли в груди, головные боли,
развивается сосудистый коллапс и шок. Смерть наступает на третий день после при-
ема касторовых бобов или позже. На аутопсии у погибших от отравления рицином об-
наруживают мультифокальное изъязвление и геморрагии на слизистой желудка и тон-
кого кишечника, которые могут быть очень выраженными; лимфоидный некроз
мезентеральных лимфоидных узлов, некроз Купферовых клеток, некротические оча-
ги в печени, диффузный нефрит и спленит (Franz D., Jaax N., 1997).
Клиника ингаляционного поражения. Ингаляционное поражение рицином людей
и экспериментальных животных возможно в том случае, если размер частиц аэро-
золя находится в пределах 1—5 мкм. Данных по ингаляционному поражению людей
рицином в досту иной литературе мне найти не удалось. Ингаляционное экспониро-
вание крыс к рицину вызывает у них диффузную некротизирующую пневмонию с
интерстициальным и альвеолярным воспалением и отеком, но не ранее 8 ч после ин-
галяции. Через 12 ч отмечаются первые признаки нарушения проницаемости воз-
душно-кровяного (аэрогематического) барьера и цитотоксического действия рици-
на: в жидкости бронхиального пространства обнаруживаются повышенное содержа-
ние общего белка и воспалительные клетки крови. Наличие воспалительного процесса
показывает и общая формула крови. Через 18 ч после ингаляции рицина уже заметен
альвеолярный отек и увеличено количество экстраваскулярной жидкости в легких.
Через 30 ч выраженная артериальная гипоксемия и ацидоз, гистопатологические до-
казательства альвеолярного отека Иммунохимические исследования показали, что
ингалированный рицин связывается с реснитчатыми клетками, альвеолярными мак-
рофагами и альвеолярными обкладочными клетками дыхательных путей (Franz D.,
Jaax N„ 1997).
При сравнительном исследовании поражений легких крыс, вызванных ингаляци-
ей рицина разной степени очистки (в сопоставимых дозах по очищенному рицину),
3.11. Рицин
955
установлено, что неочищенный рицин вызывает более обширные и разнообразные
поражения, чем очищенный (Griffiths G. D. et al., 2007).
В исследованиях на нечеловекообразных приматах установлено, что ингаляцион-
ная токсичность рицина имеет дозозависимый предклинический период 8—24 ч.
В клиническом периоде у животных наблюдается анорексия, прогрессирующе сни-
жается двигательная активность, развивается отдышка. Через 36—48 ч после ингаля-
ции рицина они погибали от легочной недостаточности. Скорость гибели животных
также зависела от ингалированной дозы. У всех обезьян, погибших от ингаляцион-
ного отравления рицином, обнаружили гнойнофибринозную (fibrinopurulent) пнев-
монию, разной выраженности диффузный некроз легочной ткани и острые воспали-
тельные изменения нижних дыхательных путей (Franz D., Jaax N., 1997).
Подробное описание патологии мелкодисперсного аэрозоля очищенного рицина у
нечеловекообразных приматов (макак-резусов) привожу по работе С. Wilhelmseann,
L. Pitt (1996). Пять животных весом 5,9—6,3 кг были экспонированы к частицам аэро-
золя рицина (MMAD 1—2 мкм) из расчета ингаляционной дозы 20,95-41,8 мкг/кг.
Средний диаметр частиц рицина 1,2 мкм, следовательно, основная масса частиц осела
в участке соединения бронхиол с альвеолами. В период времени между 36 и 48 ч после
ингаляции большая часть животных погибла. Оставшиеся в живых были умерщвлены
и затем подверглись патологоанатомическому исследованию.
Наиболее тяжелые видимые и микроскопические поражения обнаружены в ниж-
нем респиратором тракте животных. В грудной полости находились серозная или се-
роангинозная жидкость и нити фибрина, висцеральная легочная плевра была покрыта
пленкой фибрина. Легкие не колапсированы, но тяжелые, плотной упругой консис-
тенции, покрытые жидкостью. Их окраска пестрая, с красной и пурпурной исчерчен-
ностью (рис. 3.132).
Рис. 3.132. Грудная полость обезьяны, погибший от мелкодисперсного аэрозоля
рицина. А. Фибрин, покрывающий висцеральную поверхность плевры. Б. Отек
легких, сопровождающийся геморрагиями и некротическими изменениями.
По С. Wilhelmseann, L. Pitt (1996)
В трахее и бронхах обнаружена светлая пенистая жидкость, вытекающая из отвер-
стия гортани. Медиастинум утолщен вследствие студенистого отека (gelatinous edema),
медиастинальные лимфатические узлы расширены и отечны. Перикардиальная сумка
заполнена серозной или серозноангинозной жидкостью и расширена. Легочные доли
у всех обезьян имели не очень заметный мультифокальный сливающийся некроз и вос-
палительные изменения эпителия воздушных путей, перибронховаскулярный отек,
альвеолярное затопление (alveolar flooding) и фибринозногнойные альвеолиты
(fibrinopurulent alveolitis), интерстициальная пневмония с участками некроза. Нити
956
Часть 3. Частная неправильная эпидемиология
перибронховаскулярной соединительной ткани разделены эозинофильным белковым
материалом (отек). Перибронхиальные лимфатические узлы расширены фибрилляр-
ным эозинофильным материалом (отек и фибрин) с эритроцитами, нейтрофилами,
лимфоцитами и макрофагами. Лимфоциты в ассоциированных с бронхами лимфоид-
ных тканях истощенны (depleted). На отдельных бронхах эпителий остался i нтактным,
другие частично утратили подкладочный эпителий (lining epithelium) (рис. 3.133).
Рис. 3 133. Гистологические изменения в легочной ткани у обезьян, экспонированных
к мелкодисперсному аэрозолю рицина. А. Выраженный периваскулярный и периброн-
хиальный отек с лимфангиестазией (lymphangiectasia) и истощением бронхоассоцииро-
ванной лимфоидной ткани. Полоса соответствует 333 мкм. Б. Бронхоальвеолярный
некроз с альвеолярным затоплением (alveolar flooding) и гнойно-фибринозным пери-
бронхиолитом и альвеолитом. Полоса соответствует 133 мкм. В. Гнойно-фибринозный
альвеолит. Полоса соответствует 33 мкм. Г. Внутриальвеолярное отложение полимеризо-
ванного фибрина. Полоса соответствует об мкм. Окрашивание фосфовольфрамовым
кислым гематоксилином. По С. Wilhelmseann, L. Pitt (1996)
Перибронхиальные инфильтраты включают нейтрофилы, лимфоциты и макрофа-
ги. Многие маленькие бронхиолы почти полностью утратили эпителий, сохранив
только гладкие мышцы (см. рис. 3.133, Б). Терминальные бронхиолы некротизированы
(респираторные бронхиолы, альвеолярные проходы и соседние альвеолы) и «разду-
ты» миграцией эозинофилов, эритроцитов, альвеолярных макрофагов, клеточного
дебриса и отложением нитей фибрина. Большинство альвеол утратили подкладочный
эпителий. Альвеолярное септальное интерстициальное пространство утолщено эози-
3.11. Рицин
957
нофильным белковым материалом, капилляры переполнены и инфильтрированы
нейтрофилами и макрофагами (см. рис. 3.133, В). Капилляры подкладочного эндоте-
лия и большие кровеносные сосуды «раздуты». Имеет место внутриальвеолярное от-
ложение полимеризованного фибрина (см. рис. 3.133, Г).
Верхний респираторный тракт воспален незначительно. Трахея у всех обезьян и
гортань у трех обезьян имели небольшое количество мигрировавших нейтрофилов
и редкие очаги некротических эпителиальных клеток. В просвете обнаружено незна-
чительное количество воспалительного экссудата, включающего разрушенные нейт-
рофилы, макрофаги, эритроциты, слущившиеся эпителиальные клетки и муцин.
Гортань и трахея одной обезьяны имели незначительно истощенный подкладоч-
ный эпителий, утративший реснички. Подслизистая трахеи одной обезьяны была не-
значительно инфильтрирована нейтрофилами, лимфоцитами, плазматическими клет-
ками, макрофагами, что является следствием наложения острого воспалительного
процесса на уже имевшийся хронический.
Другие торакальные поражения выявлены вдоль легочного лимфатического дре-
нажного протока. У всех обезьян синусы трахеобронхиальных лимфатических узлов
утолщены отложением фибрина, гиперсегментированными нейтрофилами, макрофа-
гами, некротизированным клеточным дебрисом. В трахеобронхиальных лимфати-
ческих узлах отдельных обезьян обнаружено выраженное лимфатическое истощение
кортекса и кариолизис фолликулярных клеток. Соединительная ткань вокруг лимфа-
тических узлов и центростремительные лимфатические протоки заполнены нейтро-
филами, макрофагами, лимфоцитами, эритроцитами и фибрином. У двух обезьян об-
наружен адренокортикальный некроз.
Клиника парентерального поражения. Известна значительно лучше, чем вызванная
ингаляционным поражением, благодаря тому, что парентеральное введение рицина
использовалось для лечения онкологических больных и осуществления индивидуаль-
ных убийств. Клиника низких доз изучена в экспериментах по лечению рака. Раствор
рицина пациентам вводили внутривенно (18—20 мкг/м2). Первые симптомы отравле-
ния у них появились через 4—6 ч после инъекции. Преобладали гриппозные симптомы,
больные жаловались на тошноту, рвоту, боли в мышцах и слабость. Такое состояние
длилось у отравленных до двух суток. Смертельных исходов не было. В аналогичном
эксперименте с абрином* (растительный токсин из бобов Jequirity, сходный с рици-
ном по механизму действия), вызвавшем смертельное отравление, у пациента наблю-
дались сильные судороги и другие признаки поражения ЦНС (Fodstad О. et al., 1984).
Клиника парентерального отравления рицином задокументирована при расследо-
вании убийства в Лондоне в 1978 г. болгарского диссидента Георгия Маркова. Из не-
установленного оружия в него была выпущена пуля, содержащая примерно 500 мкг
рицина (Crompton R., Gall D., 1980). Пуля представляла собой покрытый воском пу-
стотелый шарик, состоящий на 9/10 из платины и 1/10 иридия. Воск расплавлялся при
температуре тела, высвобождая рицин (рис. 3.134 пуля с рицином).
Марков почти сразу почувствовал локальную боль в бедре, в участке, в который
попала пуля. Но он не понял, что в него стреляли. Примерно в течение 5 ч ощущение
боли прошло. В период времени 15—24 ч после выстрела у него резко повысилась
температура, появилось ощущение тошноты, началась рвота. Через 36 ч после выст-
рела Маркова доставили в госпиталь в тяжелом состоянии и с высокой температурой
1 В опытах на крысах Lct50 аэрозоля очищенного абрина (MMAD 1 -5 мкм) 4,54 мгхмин/м3
(LD50 3,3 мкг/кг) (Griffiths G. D. et al., 2007).
958
Часть 3. Частная неправильная эпидемиология
1,7 мм
Рис. 3.134. Пуля, извлеченная из тела
Г. Маркова. По В. Beckett (1982)
и тахикардией, с увеличенными и болезнен-
ными паховыми лимфоузлами. АД оставалось
нормальным. На бедре вокруг участка инъек-
ции рицина наблюдалась плотная воспаленная
зона диаметром 6 см. Спустя чуть более 2 сут,
после инъекции рицина у Маркова внезапно
упало артериальное давление (пульс 160/мин),
развились сосудистый коллапс и шок. Белые
клетки крови — 26300/мм3, анурия. Еще до за-
вершения 3 сут после инъекции рицина у него
началась рвота с кровью. ЭКГ-исследование
обнаружило полную атриовентрикулярную
блокаду сердца. Марков умер. Белые клетки
крови на момент смерти — 32 200/мм3 (Cromp-
ton R„ Gall D„ 1980).
Внутримышечное и подкожное введение
высоких доз рицина людям осуществлявшееся
с целью их убийства, обычно приводило к ло-
кальному лимфоидному некрозу, желудочно-
кишечному кровотечению, некрозу печени, диффузному нефриту и сплениту. Сходные
результаты были получены и в опытах на животных (Franz D., Jaax N., 1997).
При внутривенном введении больших доз рицина, отравление может клинически
маскироваться под сепсис (лихорадка, головная боль, головокружение, тошнота, ано-
рексия, гипотония, абдоминальные боли) (Audi J. et al., 2005).
Диагностика пзрпжения рицином. Медленное действие рицина дает возможность
прояснить многие обстоятельства преступления. Предположение об отравлении ри-
цином позволяют сделать данные анамнеза отравленного в сочетании с определенны-
ми клиническими симптомами и результатами патологоанатомического исследова-
ния (см. выше). Окончательное подтверждение диагноза остается за лабораторными
исследованиями, предполагающими обнаружение следов рицина в тканях отравлен-
ного и/или на орудиях совершения преступления. У выжизших после экспонирова-
ния к рицину подтвердить диагноз можно через две недели по обнаружению специ-
фических антител в сыворотке крови. Поэтому для измерения образования антител у
пациентов, необходимо брать парные пробы сыворотки в острый период и период
выздоровления.
Без определенной настороженности клиницистов единичное энтеральное пораже-
ние рицином, скорее всего, пройдет под видом «нераспознанного пищевого отрав-
ления». Подозрение о криминальном характере отравления должно появиться в ре-
зультате сопоставления анализа личности (социального статуса) отравленного tком-
мерсант, политический или военный деятель) и возможных мотивов к убийству у
окружающих его лиц, наличия у них легального доступа к рицину, и событий, пред-
шествовавших отравлению. В1995 г. онколог Дебора Грин из Канзас-Сити пыталась
убить своего мужа путем добавления в пищу рицина, используемого для лечения он-
кобольных в клинике, где она работала. Важны показания и самого отравленного,
например, связывание им своего болезненного состояния с приемом какой-то пищи.
На возможную причастность рицина к отравлению будет указывать сочетание симп-
томов, характерных для пищевой интоксикации (рвота и боли в брюшной полости,
диарея) и внутрисосудистой коагуляции (геморрагии, кровотечения из ануса).
3. И. Рицин
959
Парентеральное отравление рицином можно предполагать при наличии в анамне-
зе отравленного указаний на получение им каких-то царапин или уколов, вызвавших
жгучую боль в участке повреждения кожи, особенно если они были нанесены (сдела-
ны) в общественных местах или на улице неизвестными лицами, или их происхожде-
ние осталось отравленному неизвестным. На отравление рицином (кроме общих сим-
птомов отравления — тошнота, рвота, коллаптоидное состояние, признаки поражения
печени и почек и др.) будут указывать плотная воспаленная зона в участке инъекции,
воспаление лимфатических узлов, ближайших к участку инъекции, локальный лим-
фоидный некроз и симптомы внутрисосудистой коагуляции (геморрагии, желудоч-
но-кишечное кровотечение). Внутривенное введение рицина не сопровождается мест-
ными симптомами.
Предположение об отравлении рицином можно сделать на основе клинических
признаков диссеминированной внутрисосудистой коагуляции, сочетающихся с при-
знаками гепатоцеллюлярной и ренальной недостаточности.
Поражение аэрозолем рицина можно предполагать в случае:
I) появления в течение одних суток больных с геморрагическими явлениями в сет-
чатке глаз и прогрессирующей легочной патологией, проявляющейся отеком легких,
одышкой, кровохарканием, нарастающими очагами деструкции легочной ткани;
2) обнаружением на секции патологии мелкодисперсного аэрозоля (см. выше);
3) установлением эпидемиологии точечного источника и локализованности оча-
га поражения1.
Если рицин не обнаружен непосредственно в использованном для совершения
преступления орудии (например, в случае с Марковым диагностика отравления ри-
цином была облегчена обнаружением пули с рицином), то диагностировать (подтвер-
дить) лабораторными способами отравление человека рицином очень сложно. Даже в
тех случаях, когда такое отравление можно предполагать, исходя из анамнеза жертвы,
клиники и патанатомии отравления, необходимо иметь в виду то, что рицин распре-
деляется в организме неравномерно и быстро метаболизируется. Внутривенное вве-
дение крысам рицина, меченного 1251, позволило установить, что через 30 мин наи-
большее его количество находится в печени — до 45 %, в других органах и тканях его
содержание значительно меньше (в мышцах — 13 %, в селезенке — 8 %). Через 24 ч в
организме животного удается обнаружить не более 11 % от введенного количества
меченного рицина. До 70 % рицина выводится с мочой в первые сутки после отравле-
ния в виде низкомолекулярных метаболитов. Подтвердить факт рициновой ингаля-
ции, по крайней мере, через 24 ч после инцидента можно иммунологическими мето-
дами (различные варианты иммуноферментного анализа), отобрав материал из носо-
вых ходов жертвы (Franz D., JaaxN., 1997). Чувствительность ELISA порицину вполне
удовлетворительна — 1 нг/мл при определении токсина в буферном растворе, моче и
сыворотке крови (Poli М. A. et al., 1994).
Для дифференциальной диагностики необходимо исключить другие биологические
агенты, такие как стафилококковый энтеротоксин В (SEB), РТХ, возбудителей лихо-
радки Ку, туляремии, чумы, гемолитико-уремического синдрома (Е. coli О104:Н4 и
О157:Н7) и некоторые ОВ (фосген, дифосген).
Индуцируемый рицином отек легких ожидается намного позднее (1-3 сут после
экспонирования) по сравнению с таковым, индуцируемым смертельными дозами SEB
1 См. разд. 2.3.1 «Эпидемиология искусственного эпидемического процесса, вызванного
однократным применением биологического агента».
960
Часть 3. Частная неправильная эпидемиология
(около 12 ч после экспонирования) или фосгеном (около 6 ч после экспонирования).
Вероятно, интоксикация, вызванная рицином, будет прогрессировать, несмотря на
лечение антибиотиками, в отличие от инфекционного процесса со сходными симпто-
мами. При ингаляционном поражении рицином не бывает медиастинита, как в слу-
чае ингаляционной сибирской язвы. У больных, отравленных рицином, не следует
ожидать клинической стабилизации, как в случае интоксикации SEB. К дополнитель-
ным признакам, подтверждающим клиническую картину или диагноз, после аэро-
зольного экспонирования к рицину относятся следующие: двусторонние инфильтраты
на рентгенограммах грудной клетки, артериальная гипоксемия, нейтрофильный лей-
коцитоз и бронхиальный аспират, богатый плазменным белком, что характерно для
отека легких (Kortepeter М. et al., 2001).
В качестве маркера отравления рицином может использоваться алкалоид рици-
нин. Он содержится в следовых количествах практически во всех препаратах рицина,
но менее чем рицин подвержен метаболизации. Его определение в моче возможно даже
спустя двое суток после экспонирования животного к рицину (Audi J. etal., 2005). При
помощи ПЦР в большинстве рициновых препаратов можно обнаружить ДНК клеще-
вины обыкновенной (Kortepeter М. et al., 2001).
При исследовании рицинсодержащих препаратов, использованных для соверше-
ния преступления, необходимо обращать внимание не только на количественное со-
держание в них рицина, но и на их агрегатную форму (жидкость или порошок) и со-
став компонентов. В сухих препаратах целесообразно определить средний размер
частиц рицина и выявить наличие дезагрегирующего наполнителя, в жидких — ста-
билизирующие соединения. Наличие таких компонентов говорит о высоком техни-
ческом уровне лиц, подготовивших преступление.
Иммунопрофилактика. Рицин обладает выраженными иммуногенными свойства-
ми. На стадии проведения доклинических исследований находится несколько вакцин
и иммуноглобулинов, обладающих протективным действием в отношении рицина, в
том числе и при его ингаляционном применении. Подробное изложение достигнутого
уровня и истории создания рициновых вакцин и антитоксинов см. в работе G. D. Grif-
fiths et al (2007). Практического значения они пока не имеют.
Лечение. Специфического лечения отравления рицином не существует. В настоя-
щее время лечение ограничивается интенсивной поддерживающей и симптоматичес-
кой терапией. При лечении поражения, возникшего после проникновения токсина
через дыхательные пути, значительных усилий от врачей требует купирование отека
легких и поддержание нормальной легочной вентиляции. При энтеральных пораже-
ниях лучше всего проводить энергичное промывание желудка с последующим при-
менением слабительных средств, таких как цитрат магнезии. Активированный уголь
малоэффективен для сорбирования больших молекул, таких как рицин. Важное зна-
чение имеет возмещение объема по герь желудочно-кишечной жидкости. При парэнте-
ральном введении рицина в распоряжении врача остаются только поддерживающие
и реанимационные мероприятия. Гемодиализ бесполезен (Audi J. etal., 2005; Kortepe-
ter М. et al., 2001).
В экспериментах на мышах показана возможность химиотерапии при отравлениях
рицином. С этой целью могут быть использованы D-галактоза и ее производные (кон-
курирует с рицином за участки связывания на поверхности клеток-мишеней), ази-
дотимидин (AZT) и брефельдин А (блокируют транспорт рицина по сети Гольджи)
(Franz D., Jaax N., 1997).
3.12. Ботулинические токсины
961
3.12. БОТУЛИНИЧЕСКИЕ ТОКСИНЫ
Микробиология ботулинического микроба. Эпидемиология ботулизма. Молекулярная
структура и механизм действия ботулинических токсинов. Генетика токсинообразо-
вания. Токсические свойства ботулинических токсинов. Патогенез ботулизма. Устой-
чивость ботулинических токсинов во внешней среде. Поражающие свойства ботули-
нических токсинов. Клиника энтерального поражения. Клиника раневого ботулизма.
Клиника парентерального поражения. Клиника ингаляционного поражения. Диагно-
стика поражения ботулиническими токсинами. Иммунопрофилактика. Лечение.
Ботулинические токсины (botulinum toxins) — бинарные нейротоксины, образуе-
мые грамположительной анаэробной спорообразующей бактерией Clostridium botu-
linum и некоторыми ее близкородственными видами (С. butyricutn, С. baratii, С. argen-
tinense). Вызывают ботулизм — тяжелое отравление, характеризующееся поражением
нервной системы, преимущественно продолговатого и спинного мозга, протекающее
с преобладанием офтальмоплегического и бульбарного синдромов.
Первый описанный в медицинской литературе случай ботулизма имел место в Гер-
мании в 1735 г. Первая крупная вспышка ботулизма произошла там же в 1793 г. и была
вызвана потреблением кровяной колбасы. Название ботулизм — от лат. botulus, т. е.
колбаса. Впервые оно использовано в литературе XIX в. для определения заболеваний,
развивающихся после употребления колбас, в частности, кровяной колбасы. Синдром,
вызываемый ботулиническим токсином, упоминался в XIX в. как болезнь Ю. Керне-
ра, поскольку этим исследователем в 1817—1822 гг. было описано большое количество
случаев ботулизма. В России в XIX в. ботулизм неоднократно описывался под названи-
ем «ихтиизм» и связывался с употреблением соленой и копченой рыбы. В конце XIX в.
бельгийский ученый Е. Van Ermengen установил, что ботулизм вызывает водораство-
римый токсин, продуцируемый анаэробной бактерией, названной Bacillus botulinus (се-
годня С. botulinum). В начале XX в. A. Tchitchkine предположил, что растворимым фак-
тором, ответственным за развитие симптоматики ботулизма, является нейротоксин
(Pearce L. В. et al., 1997).
Различные типы ботулинического токсина были первоначально идентифициро-
ваны с использованием специфических антитоксинов. Токсины, ассоциированные с
отравлениями, различали на основании того, какие из этих специфических антител
блокировали летальность недавно выделенных экстрактов или культур. G. S. Burke
(1919) ввел номенклатуру, аналогичную используемой в настоящее время для иденти-
фикации различных серотипов токсина.
Известно семь иммунологически отличных типов (серотипов) ботулинических
токсинов (А, В, С], D, Е, F и G), четыре из них (А, В, Е и F) вызывают ботулизм у
людей, два (С! и D) — у животных и водоплавающих птиц. Ботулинический токсин
A-типа имеет пять субтипов (Al—А5). Ботулинический токсин типа С подразделяли
на субтипы С] и С2. Однако последние исследования показывают, что действие ток-
сина С2 отличается от действия других ботулинических нейротоксинов. Поэтому его
не относят к токсинам, ответственным за собственно ботулизм. Для токсинов типов
С и D, а также Е и F характерна перекрестная нейтрализация, свидетельствующая об
их антигенном родстве. Естественные отравления людей ботулиническими токсина-
ми происходят в основном через пищу (Marks J. D., 2004; Hill К. К. et al., 2009).
Токсическое действие ботулинических токсинов вызвано нарушением нейро-
мышечной передачи вследствие блокады выделения ацетилхолина в синапсах. Боту-
962
Часть 3. Частная неправильная эпидемиология
линические токсины являются наиболее сильнодействующими из известных биоло-
гических токсинов. Их средняя летальная доза при разных способах введения колеб-
лется от 5 до 50 нг/кг. Ингаляционная доза ботулинического токсина для человека, вы-
раженная величиной Lctso оценивается как 0,02—0,1 мгхмин/м3. По этому показателю
ботулинический токсин не менее чем в 750 раз токсичнее зарина и в 100 раз токсич-
нее вещества VX. Смерть от отравления ботулиническими токсинами наступает в ре-
зультате развития «бульбарного синдрома», паралича дыхательных мышц, осложнен-
ных пневмонией и токсическим миокардитом. У людей наиболее высокая летальность
наблюдается при отравлении токсинами типов А и Е (Антонов Н. С., 1994; Попелян-
ский Я. Ю. с соавт., 2000).
Российские санитарно-эпидемиологические правила СП 1.3.2322-08 относят бо-
тулинические токсины ко II группе патогенности. По степени важности для нацио-
нальной безопасности США они относятся к группе А. Ботулинические токсины при-
влекают внимание специалистов в области химического и биологического оружия с
1920-х гг. из-за высокой токсичности и кажущейся доступности получения и приме-
нения. В годы Второй мировой войны и до конца 1960-х гг. они интенсивно изучались
в США и Соединенном Королевстве в качестве возможного средства уничтожения
живой силы противника. Токсин серотипа А состоял на вооружении армии США под
шифром «X». В те же годы разработаны технологии их выделения в кристаллическом
состоянии, промышленного производства и специальные боеприпасы для примене-
ния в боевых условиях. В настоящее время ботулинические токсины не рассматрива-
ют как поражающие агенты, пригодные для ведения биологической войны из-за их
нестабильности при производстве, хранении и диспергировании. Но в качестве аген-
тов для осуществления биологических террористических актов, диверсий и действий
криминального характера, вероятность их использования оценивается специалиста-
ми как весьма высокая (Антонов Н. С., 1994; Middlebrook J. L., Franz D. R., 1997; Ar-
non S. S. et al., 2001; Dharked R. M. et al., 2010). В начале 1990-х гг. секта «Аум-Син-
рикё» пыталась осуществить в Японии серию террористических актов с использова-
нием ботулинического токсина типа А (см. разд. 2.4.1). В медицине ботулинический
токсин типов А и В используется для разработки обезболивающих и косметических
препаратов (Laskawi R., 2008).
Микробиология ботулинического микроба. Род Clostridium относится к группе
Firmicute, которая делится на анаэробные Clostridia, облигатные или факультативные
аэробы Bacilli, и Mollicutes — бактерии, не имеющие клеточной стенки вообще (среди
них самый известный род — Mycoplasma). Насчитывается не менее 83 видов Clostridia.
Хотя отдельные из них способны расти в присутствии кислорода, они отличаются от
бактерий рода Bacillus утратой ферментов каталазы и пероксидазы, оказывающих
протективный эффект от действия на бактерии кислорода, и фермента цитохромок-
сидазы, вовлеченной в аэробный механизм. Супероксиддисмутаза обнаружена толь-
ко у отдельных видов клостридий.
Термин «токсин» в отношении рода Clostridium означает биологически активный
белок, имеющий антигенную природу, и его активность может быть специфически
нейтрализована соответствующей антисывороткой (Hatheway С. L., 1990). Изучение
ДНК-гомологии клостридий, продуцирующих ботулинические токсины, позволило
разделить их на четыре генетически и фенотипически различающихся группы. Микро-
организмы в этих группах различаются настолько, что могут представлять отдельные
виды, но они классифицируются как С. botulinum из-за того, что их общей особенно-
стью является синтез ботулинического токсина. Организмы группы I относят к про-
3.12. Ботулинические токсины
963
теолитическим, а группы II — непротеолитическим, на основе их способности рас-
щеплять сложные белки. Общим предком для клостридий I группы, продуцирующих
ботулинический токсин, является нетоксигенная С. sporogenes. Штаммы внутри групп
I и II тесно связаны между собой генетически. С. baratii и С. butyricutn отдельные авто-
ры относят к V и VI группе соответственно. Близкородственная бактерия С. tetani,
продуцирующая столбнячный токсин, относится к отдельной консервативной кладе.
Известен только один серотип столбнячного токсина (Hill К. К. et al., 2009).
Группы клостридий, продуцирующих ботулинические токсины, значительно от-
личаются друг от друга фенотипически. Их микробиологическая характеристика при-
ведена в табл. 3.44. Сравнительные микробиологические характеристики С. botulinum
групп I и 11 приведены в табл. 3.45.
Таблица 3.44
Микробиологическая характеристика групп Clostridium spp.,
образующих ботулинический токсин*
Группа Тип токсина Продукция Фермен- тация углеводов Гидролиз желатина Перева- ривание казеина Продукты метаболизма
лецити- назы липазы стабильные неста- бильные
1. С. botulinum А, В, F, АВ, BF — + + + A. iB. В. iV РР
11. С. botulinum В, Е, F — + + + — А, В
III. С. botulinum С, D + + + + ± А. Р, В
IV. С. argentinense G — — — + + A. iB. В. iV РА
Смешанная
С. baratii F + + А В L
С. butyricum Е — + А, В
А — уксусный; Р — пропионовый; iB — изомасляный; В — масляный; iV — изовалериано-
вый; РР — фенилпропионовый (hydrocinnamic); РА — фенилуксусный; L — молочная кислота.
* По С. L. Hatheway (1990).
Штаммы других групп могут значительно отличаться друг от друга, даже если они
продуцируют один и тот же тип ботулинического токсина. К С. argentinense были от-
несены клостридии, продуцирующие G-тип ботулинического токсина (и их атокси-
генные варианты), общим биохимическим признаком которых была способность
продуцировать индольные производные (Lindstrom М., Korkeala Н., 2006).
Клостридии, возбудители ботулизма, — строгие анаэробы. На поверхности твер-
дых сред они растут почти при полном удалении кислорода; остаточное давление воз-
духа допустимо от 3 до 10 мм рт. ст. На жидких средах С. botulinum растут хорошо, если
предварительно бульон подвергнуть кипячению в течение 15—20 мин с целью удале-
ния воздуха, а затем быстро охладить и после посева залить среду сверху вазелиновым
маслом. Культивирование С. botulinum можно производить на твердых средах: кровя-
ном агаре, печеночном агаре, желатине с глюкозой или же на агаре столбиком. На кро-
вяном агаре с глюкозой через 24—48 ч возбудители ботулизма типов А и В растут в виде
круглых колоний, окруженных узкой зоной гемолиза. Если рост густой, гемолиз может
распространиться на всю поверхность среды. Колонии бывают очень малы и не видны
964
Часть 3. Частная неправильная эпидемиология
Таблица З.ч5
Сравнительные микробиологические характеристики С. botulinum групп I и II*
Характеристика С. botulinum группы I С. botulinum группы II
Токсинотипы А, АВ, В, BF, F В, Е, F
Происхождение штаммов Почва Водная среда
Размер генома (м. п. о.) 3,9-4,1 3,6-4,1
Г+Ц-содержание (% от генома) 26-28 26 28
Генетические различия Минимальные Широкие
Использование белков в качестве источника энергии Да Нет
Ферментируемые углеводы Глюкоза, фруктоза, мальтоза, салицин, сорбитол Амигдалин, декстрин, фруктоза, галактоза, гликоген, мальтоза, ман- ноза. мелезитоза, рибоза, сорбитол, крахмал, сахароза, трегалоза
Продуцируемые метаболиты Уксусная, масляная, изомасля- ная, изовалериановая, изокап- роновая (isocaproic), пропио- новая и валериановая кисло- ты; спирты; сероводород Уксусная, формиковая, масляная, янтарная и молочная кислоты
Продукция бактериоцинов Ботицин Не описаны
Морфология колоний на кровяном агаре 2-6 мм в диаметре, неравно- мерная кромка, непрозрачные, выпуклый центр, узкая зона гемолиза вокруг колонии 1—3 мм в диаметре, слабовыра- женная неравномерность и доль- чатость кромки; от просвечиваю- щихся до полупрозрачных с мато- вой поверхностью; мозаичная структура при косом освещении; узкая зона гемолиза
Морфология колоний на желточном агаре (липазная реакция) Радужный слой на/и вокруг колоний Радужный слой на/и вокруг колоний
Отношение к антибиотикам Чувствительность к эритроми- цину, метронидазолу, пеницил- лину, рифампицину, тетрацик- лину; резистентность к цикло- серину, гентамицину, налидик- совой кислоте, сульфамет- оксазолу, триметоприму Чувствительность к эритромицину, метронидазолу, пенициллину, рифампицину, хлорамфениколу, клиндамицину, тетрациклину, три- метоприму; резистентность к ген- тамицину, налидиксовой кислоте, цефалотину, сульфаметоксазолу
Температура роста, минимум/оптимум, С’ 10/35-37 3/26-30
Значение pH, ограничи- вающее рост 4,3-4,5 5,0-5,1
Концентрация NaCl, ограничивающая рост, % 10 5
Наиболее характерная пища для развития ботулизма Для развития пищевого боту- лизма: овоши, мясо, консерви- рованная пища. Для развития детского ботулизма — мед Для развития пищевого ботулизма: рыба, мясо, очень редко — пища в упаковке. Для развития детского ботулизма — не установлены
По М. Lindstrom, Н. Korkeala (2006).
3.12. Ботулинические токсины
965
невооруженным глазом, поэтому для обнаружения роста необходимо просматривать
поверхность чашки с помощью лупы или же стереоскопического микроскопа. Они
имеют вид круглых, в некоторых случаях блестящих образований, края бывают ров-
ные и неровные, цвет колоний слегка коричневатый или серовато-мутный, середина
вогнутая или выпуклая, выпуклость немного походит на дочернюю колонию, нахо-
дящуюся посредине основной колонии. Крупные колонии более плоские. Один и тот
же штамм может давать несколько типов колоний. Некоторые штаммы дают полный
гемолиз, другие — только обесцвечивают среду, но не растворяют эритроциты. На пе-
ченочном агаре с глюкозой ботулинический микроб дает нежный прозрачный рост или
в виде отдельных колоний, или покрывает всю поверхность среды.
Морфология палочек, имеющихся в колониях на кровяном и печеночном агаре,
немного отличается от морфологии бактерий на жидких питательных средах. Палочки
в колониях на твердых средах тоньше и короче, более ровные по сравнению с теми,
которые вырастают на жидких средах. На поверхности желатина С. botulinum дают
сероватые, блестящие колонии. Они круглые, прозрачные, иногда окрашенные слегка
в коричневый цвет и окружены узкой зоной разжижзннго желатина. При старении ко-
лонии становятся мутными и немного коричневатыми.
На агаре столбиком микроб образует один или два вида колоний. Если имеется
диссоциация на R- и S-формы, то R-форма вырастает в виде мелких чечевицеобраз-
ных колоний, а S-форма дает колонии в виде пушинок.
На жидких средах С. botulinum растет хорошо. Наиболее часто применяют для
выращивания этого микроба мясопептонный бульон с кусочками печени или мясно-
го фарша на дне. Подходящей средой является также пептонный или же мясной бу-
тьон с кусочками вареного куриного яйца. Можно использовать бульоны из различ-
ных 1 идролизатов казеина, мяса и других субстратов, содержащих белки. Оптимальная
температура для выращивания возбудителя ботулизма типа А, В, С и D — 36 °C, а для
типов Е и F — 28—30 °C. Наиболее подходящее значение pH среды от 7,3 до 7,6, если
асевают вегетативные микрооргани «мы. Если посевной материал состоит из спор, то
оптимальное колебание значений pH может быть от 6 до 7,2. Это указывает на то, что
небольшая кислотность может стимулировать прорастание спор этого микроба (Мат-
веев К. И., 1959).
Возбудители ботулизма в окрашенных препаратах и в электронном микроскопе
имеют вид палочек длиной от 4 до 8 мкм и шириной от 0,6 до 0,8 мкм с закругленны-
ми концами. Микробы образуют споры, располагающиеся субтерминально, редко
центрально. Палочки со спорами имеют вид теннисных ракеток. По морфологии раз-
личные типы не отличаются друг от друга. Подвижны. Жгутики в количестве от 3 до
20 расположены по всему телу палочки. При старении клеток жгутики могут исчез-
нуть. Палочки легко окрашиваются анилиновыми красками. Молодые клетки окра-
шиваются по Граму положительно. При старении культуры (через 4—5 суток роста)
палочки начинают окрашиваться грамотрицательно.
Ботулинические токсины синтезируются во время роста бактериальных клеток в
форме крупномолекулярных комплексов (progenitior toxin) и высвобождаются в пи-
тательную среду до или после клеточного лизиса. Крупномолекулярные комплексы
включают нейротоксин (derivative toxin), гемагглютинин и нетоксические белки (см.
ниже «Молекулярная структура и механизм действия ботулинических токсинов»).
Для изучения химических и токсических свойств ботулинических токсинов их
подвергают очистке от балластных белков среды, затем диссоциируют компоненты
крупномолекулярного комплекса, при этом токсины можно сконцентрировать и полу-
966
Часть 3. Частная неправильная эпидемиология
чить в кристаллическом виде. В зависимости от серотипа ботулинического токсина
изоэлектрическая точка лежит в пределах величин pH от 3,5 до 7,8.
Наиболее простой метод очистки крупномолекулярных комплексов ботулиниче-
ских токсинов от компонентов питательной среды — осаждение сернокислым аммо-
нием. При 50-60 % насыщении раствора они полностью осаждаются (Матвеев К. И.,
1959). Однако такой способ не гарантирует полного удаления из препаратов боту-
линического токсина компонентов питательной среды и разрушенных клеток ботули-
нического микроба (DasGupta В. R., Boroff Р. А., 1968). Поэтому их доочищают,
используя методы гель-фильтрации, ионообменной хроматографии, высокоэффек-
тивной жидкостной хроматографии и др. Для диссоциации компонентов токсиче-
ского комплекса подбирают оптимальные условия гель-фильтрации и анионообмен-
ной хроматографии (см., например, способ Грачева О. Е. с соавт., 2002). Подробно
способы исследования фармакологических свойств очищенных препаратов ботули-
нических токсинов описаны в работе L. В. Pearce et al. (1997).
Эпидемиология ботулизма*. Возбудители ботулизма обнаруживаются в почве, воде,
траве, на листьях и плодах овощей, фруктов, в кишечнике человека и животных, в
пыли домашних помещений. Наиболее обсеменены С. botulinum илистая и чернозем-
ная почвы, почва приусадебных участков, особенно в местах, удобряемых навозом.
Наиболее благоприятные условия для продуцирования токсина и его депонирования
клостридии находят в трупах животных. По данным W. Е Sonnabend et al. (1987), изу-
чавших типовой состав возбудителей ботулизма в почвах Швейцарии, С. botulinum типа
А «предпочитает» почву с травянистым покровом. Все 15 исследованных образцов
почвы из альпийских регионов содержали С. botulinum типа В, в четверти из них об-
наружили также ботулинический микроб типа G, по одному образцу почвы содержа-
ли в дополнение к С. botulinum типа G, еще и С. botulinum типов С и Е С. botulinum типа
С (а) был обнаружен вместе со штаммами серотипов В и G в одном образце почвы,
взятом вблизи от дренажной канавы. Штамм С. botulinum типа F обнаружен в образце
почвы вместе со штаммами типов А и G, взятом из луговой почвы, насыщенной водой.
С. botulinum типа G идентифицирован в 5 образцах почвы. Два из них представляли
собой садовую почву, два — почву травянистых полей, один выделен непосредствен-
но из ткани самого растения. На рис. 3.135 в качестве примера показано географиче-
ское распределение ботулинических микробов разных серотипов в почве Швейцарии
(рис. 3.135).
Поэтому естественные вспышки ботулизма, вызываемые определенным типом
токсина, могут быть связаны с определенным географическим районом. В США боль-
шинство случаев ботулизма, вызванных действием токсина типа А, обнаружены за-
паднее реки Миссисипи. Большинство же случаев, вызванных типом В — восточнее
этой реки. Наконец, вспышки, связанные с типом Е, зарегистрированы на Аляске и
в штатах, включающих район Великих озер (Sellin L. С., 1984).
По эпидемиологии случаи и вспышки ботулизма можно разделить на естествен-
ного и искусственного происхождения (табл. 3.46).
В России пищевой (алиментарный) ботулизм наиболее часто вызывается возбуди-
телями типов А и В. Наблюдается тенденция к распространению ботулизма типа Е.
Консервированные грибы, рыбные и мясные продукты, овощи и фрукты, приготов-
ленные кустарным способом с нарушением требований пищевой гигиены, являются
1 Для описания эпидемиологии ботулизма в основном использовалась работа Я. Ю. По-
пелянского с соавт. ( 2000).
3.12. Ботулинические токсины
967
Рис. 3.135. Распределение С botulinum в пробах почвы Швейцарии. — ботулини-
ческий микроб присутствует, □ — отсутствует. Заглавными латинскими буквами
указаны типы С. botulinum. По W. Е Sonnabend et al. (1987)
Таблица 3.46
Эпидемиологическая классификация ботулизма
Форма болезни Источник токсина Клинические лсооенности Преимущественно встречающийся серотип токсина
Естественного происхождения
Пищевой Консервированные грибы, рыбные и мясные продук- ты, овощи и фрукты, при- готовленные кустарным способом Симптомы поражения ЖКТ офтальмоплегический и бульбарный синдромы, паралич дыхательных мышц А, В, С, Е и F
Раневой Токсико-инфе кцион ный процесс — токсин выде- ляется в процессе роста С botulinum в инфициро- ванной ткани раны Офтальмоплегический и бульбарный синдромы, паралич дыхательных мышц Аи В
Раневой ботулизм наркоманов То же Тоже А и В
Ботулизм новорожденных Токсико-инфекционный процесс — токсин образу- ется в кишечнике детей Запор, офтальмоплегиче- ский и бульбарный синд- ромы, паралич дыхательных мышц С, F и G
Искусственный
Парентеральный Инъекции при передози- ровке препаратов Dysport или Botox (более 40 ЕД/кг) Офтальмоплегический и бульбарный синдромы, паралич дыхательных мышц А. В
Ингаляционный Экспонирование к аэро- золю ботулинического токсина Тоже Любой
968
Часть 3. Частная неправильная эпидемиология
главными источниками С. botulinum в нашей стране. Домашние консервы, отравлен-
ные токсином ботулизма, выглядят, как правило, совершенно доброкачественными,
не меняют ни внешнего вида, ни janaxa, ни вкуса. Единственным признаком раз-
множения ботулинического микроба может быть вздутая крышка на герметично за-
крытой банке.
Раневой ботулизм развивается в результате попадания в рану почвы, контамини-
рованной С. botulinum. Токсин выделяется в процессе роста С. botulinum в ткани ин-
фицированной раны. В последние годы наблюдается новая эпидемиологическая раз-
новидность болезни — раневой ботулизм наркоманов.
У детей в возрасте 2—3 мес. встречается ботулизм новорожденных. Он развивается
в результате колонизации ЖКТ спорами возбудителя ботулизма. Токсин образуется в
кишечнике детей. Запор часто является первым признаком болезни и может опере-
жать появление неврологических симптомов на три недели. К ранним симптомам
детского ботулизма относятся ослабление способности сосать и ослабления крика.
Артифицированные (т. е. искусственные) формы ботулизма стали выделять в отдель-
ную форму болезни после внедрения в клиническую практику лекарственных пре-
паратов на основе ботулинического токсина. Осложнения возникают в результате
передозировки препаратов Dysport, Botox и др. (Crowner В. et al., 2007). К ним же
целесообразно отнести и формы ботулизма, преднамеренно вызванные распростра-
нением аэрозоля ботулинического токсина (см. ниже «Клиника ингаляционного по-
ражения»),
В России пищевым ботулизмом болеют во все времена года, но максимум вспы-
шек и единичных случаев приходится на осеннее и зимнее время (70 % — октябрь,
ноябрь, декабрь). Наблюдаемая сезонность не отличается строгой закономерностью
и отражает изменения условий питания: в октябре, ноябре, декабре больше, чем в
другое время, употребляют в пищу консервированные продукты домашнего приготов-
ления. Кроме того, заболеваемость зависит от урожая грибов в том или ином регионе.
По данным Я. Ю. Попелянского с соавт. (2000), в России чаще заболевают боту-
лизмом при употреблении грибов домашнего консервирования, однако изредка от-
равление вызывают грибы, засол которых проводится традиционно открытым спосо-
бом, при котором иногда создается относительная герметичность (закупоривание
бочек при солении целлофановыми пленками). По их данным (более 300 наблюде-
ний), чаше других источниками отравления ботулиническим токсином являются
приготовленные в домашних условиях консервированные грибы (75,3 %), вяленая
рыба (5,8 %), сало (5,1 %), соленая рыба, мясо (2,5 %) и другие продукты (соленые гри-
бы, рыба холодного копчения, консервированные помидоры, колбаса, консервиро-
ванное мясо кур).
Естественные вспышки ботулизма, как правило, имеют групповой или семейный
характер. Я. Ю. Попелянский с соавт. (2000) наблюдали 67 вспышек ботулизма, в
21 случае заболевания были единичными. Каждая вспышка — это отравление 2—7 че-
ловек, только в одном случае одновременно заболело 12 человек (причина отравле-
ния — консервированные грибы). По данным этих же авторов, в России ботулизм наи-
более часто вызывается токсином типа В (56 %), реже типа А (30 %) и еще реже типа Е
(14 % случаев); ими зарегистрирован только один случай болезни, вызванный токси-
ном типа С после употребления сала домашнего приготовления. Токсины типа А и В
обнаруживаются чаще в консервированных грибах. Ботулизм, вызванный токсином
типа Е, возникает при употреблении рыбных продуктов и характеризуется тяжелым
или средней тяжести течением.
3.12. Ботулинические токсины
969
По L. С. Sellin (1984) из 766 случаев пищевого ботулизма, зарегистрированных в
Соединенных Штатах за периоде 1899 по 1977 г., 26% было вызвано типом А; 7,8 % —
типом В; 4,2 % — типом Е; 0,1 % — типом F; 0,1 % — комбинацией типов А и В; а 61,8 %
остались не идентифицированными.
Молекулярная структура и механизм действия ботулинических токе инов. Ботулини-
ческие токсины продуцируются С. botulinum в форме крупномолекулярных комплек-
сов (progenitior toxin; BoNT-комплекс), имеющих ММ около 900 кДа. Размер токси-
ческого компонента (derivative toxin, neurotoxinassociated protein) для всех серотипов
токсинов приблизительно одинаков (ММ 150 кДа). Величина нетоксического компо-
нента варьирует, поэтому молекулярные характеристики агрегатов как разных серо-
типов токсинов, так и внутри серотипа, различаются. Нетоксический компонент
включает белок, классифицированный как гемагглютинин (hemagglutinin protein, НА)
из-за его способности агглютинировать красные клетки крови; и белок, названный
нетоксическим негемагглютинином (nontoxin nonhemagglutinin protein, NTNH). Эти
белки защищают нейротоксин от воздействия окружающей среды и деградации в ЖКТ
(Marks J. D., 2004).
Токсический компонент с нетоксическим связаны в BoNT-комплексе нековалент-
но. Комплексы устойчивы при значениях pH ниже 7,2 и диссоциируют на составляю-
щие их компоненты при значениях pH выше 7,2. Молекулярная диссоциация круп-
номолекулярного комплекса на токсический и нетоксический компоненты носит
обратимый характер и может быть достигнута путем диализа смеси этих компонен-
тов против слабокислых растворов. Специфические нейротоксические свойства ком-
плекса определяются только токсическим компонентом, однако в зависимости от
способа введения токсина, нетоксический компонент играет различную роль. При-
сутствие его несущественно при парэнтеральном введении токсина и очень важно при
попадании токсина в ЖКТ. Очищенный токсин (derivative toxin) разрушается уже в
ротовой полости, но он сохраняет свою активность при парентеральном введении
(Езепчук Ю. В., 1985; Hatheway С. L., 1990).
Клостридиальные нейротоксины имеют трехдоменную структуру и активируются
посредством протеолитического расщепления открытой петли в структуре своей моле-
кулы. Каждая активная молекула нейротоксина включает тяжелую (Н-цепь, 100 кДа)
и легкую (L-цепь, 50 кДа) цепи, связанные посредством межцепочечной дисульфид-
ной связи. Н-цепи клостридальных нейротоксинов содержат два домена — регион,
необходимый для транслокации токсина (транслокационный домен) и регион, необ-
ходимый для связывания с клеткой (рецепторсвязывающий домен). L-цепи клостри-
дальных нейротоксинов содержат каталитический домен. Схематическое изображе-
ние клостридальных нейротоксинов приведено на рис. 3.136, атомная структура
ботулинического токсина серотипа А показана на рис. 3.137, механизм действия боту-
линических токсинов — на рис. 3.138.
Ботулинический токсин проникает в пресинаптический нейрон путем рецептор-
ного эндоцитоза. Рецепторсвязывающий домен С-терминального участка Н-цепи
ботулинического токсина связывается с клеточными рецепторами на поверхности
пресинаптических нейронов. Молекулярные структуры, используемые ботулиниче-
ским токсином в качестве рецепторов, неизвестны, однако установлены его два ко-
рецептора: белок и сиалоганглиозид (GDlb или GTlb). Рецепторсвязывающий домен
структурно включает N-концевой субдомен, включающий мотив типа «рулет с ва-
реньем» Jelly roll), и С-концевой субдомен, содержащий fJ-трехлистниковый мотив
(p-trefoil motif). С-терминальный домен взаимодействует со связывающим сайтом
970
Часть 3. Частная неправильная эпидемиология
Активация токсина через
рассечение дисульфидной связи
активность
транслокация
связывание
Рис. 3.13t> Трехдоменнач структура клостридиальных нейротоксинов. Каждый
иомен имеет ММ — 50 кДа. Домен, прилегающий к ГЧН2-концу, представляет
собой цинкэндопептидазу, которая неактивна, когда этот домен связан посред-
ством дисульфидного мостика с остальной молекуле токсина. Активный учас-
ток атома цинка координирован через два гистидиновых участка, молекула воды
связана с консервативным остатком глутамата и карбоксильной группой друго-
го глутамата. Домен HN отвечает за мембранную транслокацию L-цепи из вези-
кулы в цитозоль нейрона. Домен, прилегающий к СООН-концу молекулы,
состоит из двух субдоменов эквивалентного размера. Hc-N-субдомен имеет
структуру, сходную с лектинподобными белками (1). Нс-С-субдомен имеет сход-
ство с К1-каналспецифическим дендротоксином (2) По G. Schiavo (2000)
Рис. 3.137. Атомная структура ботулинического токсина серотипа А по результатам
рентгеноструктурного анализа. А. Ленточная схема. Б Пространственная модель.
1 — каталитическийдомен; 2—транслокационный домен; 3 — связывающий домен.
По J. D. Marks (2004)
ганглиозида, N-терминальный участок Н-цепи (HN) включает транслокационный
домен, состоящий из а-спирали — его основное назначение заключается в формиро-
вании поры. Низкое значение pH содержимого эндосомы вызывает конформацион-
ные изменения в домене, создающим пору, позволяющую L-пепи выйти из эндосо-
мы в цитоплазму.
L-цепь токсина является цинк-эндопептидазой. В зависимости от серотипа ток-
сина, она рассекает различные белки, входящие в семейства белков с малопонятным
без схематического изображения их функционирования названием: растворимый
N-этилмалеимид-чувствительный фактор соединения с белковым рецептором (soluble
3.12. Ботулинические токсины
971
N-ethylmaleimide—sensitive factor attachment protein receptor, SNARE). Семейство бел-
ков SNARE играет основную роль в процессе слияния синаптических везикул и высво-
бождения ацетилхолина. Токсины серотипов А и Е рассекают разные сайты в преде-
лах синаптосомально-ассоциированного белка массой 25 кДа (synaptasomal-associated
protein of 25 kd, SNAP-25), токсины серотипов В, D, F и G рассекают другие сайты в
пределах везикулоассоциированного мембранного белка (vesicle associated membrane
protein, VAMP), также известного как синаптобревин (synaptobrevin), серотип С рас-
секает белок синтаксин и SNAP-25. Эти три SNARE белка (синтаксин, SNAP-25 и
синаптобревин) взаимодействуя, формируют четырехвитковую спираль (four-helix
coiled coil) на этапе, предшествующем синаптическому слиянию. Рассечение любого
из этих белков цинк-эндопептидазой блокирует слияние и высвобождение ацетилхо-
лина. В результате слияние синаптических везикул с пресинаптической мембраной и
высвобождения ацетилхолина в синаптическую щель становятся невозможными,
развивается блокада нейромышечной проводимости, сокращения мышц прекраща-
ются, клинически это проявляется вялым параличем (рис. 3.138, с. 972).
Сравнительное изучение клинических признаков пищевого ботулизма, обуслов-
ленного токсинами типов А и В, свидетельствует о том, что тип А вызывал более ост-
рую форму ботулизма, чем тип В. Хотя количественные пропорции летальных исхо-
дов были аналогичными, больные ботулизмом типа А раньше попадали к врачу, чаще
нуждались в вентиляции и находились в больнице дольше, чем больные с отравлени-
ем ботулиническим токсином типа В. При внутрибрюшинном введении мышам эти
типы обнаруживают аналогичную токсичность (см. ниже «Токсические свойства бо-
тулинических токсинов»). L С. Sellin (1984) объясняет такие различия в выраженно-
сти клинических форм ботулизма более быстрой детоксикацией токсина серотипа В,
чем А. Он также предположил, что различные типы ботулинического токсина могут
вызывать отравления, различающиеся по клинической картине.
Генетика токсинооЬпазованих. Гены, кодирующие NTNH, в геноме клостридий
кластированы в непосредственной близости и перед генами, кодир) ющими ботули-
нический токсин (BoNT, 1,3 т. п. о.). Гены, кодирующие другие токсин-ассоциирован-
ные белки (НА-17, НА-33, НА-70, р21 и/или р47), расположены перед генами NTNH.
Кластер, включающий гены НА70, НА17, НАЗЗ, BotR, NTNH, BoNT и межгенные
последовательности, имеет протяженность 11,7 т. п. о. Установлено пять различных
генных кластеров (рис. 3.139, с. 973).
BoNT-гены обнаруживают в разных участках хромосомы или плазмид клостри-
дий, продуцирующих ботулинический токсин, однако они всегда фланкированы вста-
вочными последовательностями (insertion sequence, IS). Эти находки предполагают
существование механизма, благодаря которому в природе возможен горизонтальный
перенос BoNT-генов между клостридиями В пределах BoNT-генов и л/лй-генов воз-
можны рекомбинации генетического материала, о чем свидетельствует наличие их
мозаичных вариантов (например, bont/c/d и bont/al/aS). Так как генные кластеры,
кодирующие BoNT-комплексы, способны к горизонтальному переносу, их не исполь-
зуют для установления филогенетических связей между клостридиями, продуцирую-
щими токсины. Состав генных кластеров, кодирующих BoNT-комплексы и ассоции-
рованные белки, и их расположение в геноме С. botulinum и С. butyricum (для BoNT/
Е-продуцирующего штамма), показаны на рис. 1.140, с. 973.
Расположение йолГ-генов в геноме и плазмидах клостридий, продуцирующих бо-
тулинический токсин, показывает, что их интеграция с ДНК хозяина не происхо-
дит случайным образом. В основном участки интеграции йолГ-генов обнаруживают
072
Часть 3. Частная неправильная эпидемиология
и
Ацетилхолин
Синаптическая везикула
§
Синтаксин
Слияние синаптической везикулы
с пресинаптической мембраной
Комплекс, сформированный
белками семейства SNARE
Синаптобревин
SNAP-25
Синаптическая щель * Ацетилхолин
Рис. 3 138. Механизм действия ботулинических токсинов. А Механизм нейромышеч-
ной передачи. Синаптические везикулы, содержащие ацетилхолин, имеют на своей по-
верхности симпатобревин. Взаимодействуя с синтаксином и SNAP-25, белками из се-
мейства SNARE, он формирует четырехвитковую спираль, способствующую слиянию
синаптической везикулы с пресинаптической мембраной. Ацетилхолин высвобожда-
ется из везикулы и диффундирует через синаптическую щель и связывается с рецеп-
тором ацетилхолина, вызывая мышечное сокращение. Б. Воздействие ботулинического
токсина на нейромышечную передачу. Ботулинический токсин связывается с неизве-
стным рецептором на мембране пресинаптического нейрона, вызывая эндоцитоз. Пос-
ле проникновения в нейрон его L-цепь высвобождается в цитоплазму. В зависимости
от серотипа токсина, каталитический домен рассекает один из белков (или несколько
белков) из семейства SNARE. Слияние везикулы с ацетилхолином с пресинаптической
мембраой станоЕится невозможным. Ацетилхолин в пресинаптическую щель не про-
никает. 1 — синаптический комплекс слияния; 2 — высвобождение нейротрансмиттера;
3 — ацетилхолиновый рецептор; 4 — ботулинический токсин. По J. D. Marks (2004)
в гомологичных областях генома и там, где возможна их транспозиция. Рекомбинаци-
онные перестановки происходят в пределах генов BoNT-комплекса (л/лЛ-ген, bont-ген
и регион между этими генами), в частности множество сайтов интеграции находят в
пределах гена ntnh. Эти находки объясняют появление гена одного и того же гена bont/e
3.12. Ботулинические токсины
973
Типы А, В НА-70 НА-17 НА-33 Р-21 NTNH BoNT
1 )Г II |||| II 1
Типы С, D orf-22 НА-70 НА-17 НА 33 NTNH BoNT
Ml II II II II 1
Типы А2, F Р-21 Р-47 NTNH BoNT
г"1 г ПГ“ II ' 1
Тип G НА-70 НА-17 Р*21 NTNH BoNT ’
1 т. п. о. 1 1 1 I | 1 II HZ L
Рис. 3.139. Диаграмма пяти типов генных кластеров, кодирующих BoNT-kom-
плексы и ассоциированные белки. BoNT — ботулинический нейротоксин.
По М. D. Collins, А. К. East (1998)
Локализация
BoNT-комплекса Штамм/субтип
Хромосома CLP BL 5262 Е4 CLH Alaska E43E3 IS’orfX3o>JX2 orfXlp47 ntnh bont/e IS'
Хромосома CLO Beluga El IS' ISb oifX3 oifX2 orfXl p47 ntnh bont/e IS' •♦- lycA
Хромосома A1(B)A1 orfX3 otfX2orfXl botR p47 ntnh bont/al
Хромосома CLI Langetand F oifX3 orfX2 otfXl IS6' botRp47 ntnh bont/f lycA
Хромосома Плазмида CLM Kyoto-FA2 pBfF otfX3 orfX2orfXl IS6' botR p47 Is605' fycA orfX3orfX2 orfXllS& botR p47 ntnh bont/a2 lycA
ntnh bont/f JS3'
Плазмида pCLK Loch Maree A3 ls605'lycA otfX3 orfX2 orfXl IS6' botRp47 ntnh bont/a3 Is3'
Плазмида PCL657A4 tyAorfX3 orfX2 otfXl IM'botR p47 ntnh bont/a4
Плсзмида pCLJ657bvB hypo hypo /Ze' ha7O ha 17 ha33botR ntnh Tonf/b''* ...IS
Плазми (а pCLDOkraBl hypo hypo fia' ha70 ha!7 ha33 botR ntnh bont/b IS256'
Плазмида pB.'B hypo hypo fla’ ha70 ha 17 ha33botR ntnh bont/b IS256'
Плазмида pCLL Eklund ГЪ npB ka70 ha 17 ha33 botR ntnh bont/b IS256'
Хромосома A1(B) (B) hypo hypo fla' IS3' ha70 ha 17 ha33botR ntnh bont/b ISNCY
hypo hypo fla' IS3' ha70 ha!7 ha33borR ntnh bont/al IS3'
Рис. 3.140. BoNT-комплексы и фланкирующие их регионы у различных штаммов клос-
тридий, продуцирующих ботулинический токсин. or/T-кластер [комплекс генов orfX3-
orfX2-orfXl-(botR)-p47-ntnh-bonf\ обнаружен у BoNT/E-продуцирующих штаммов
(С. botulinum и С. butyricum), BoNT/Al у А1(В)-штамма, у серотипа F (BoNT/F и BoNT/
bvF) и BcNT/A2-A4-cy6™noB. йа-кластер (комплекс генов ha70-ha!7-ha33-botR-ntnh-
bont) обнаружен у штаммов, продуцирующих токсин В-серотипа [BoNT/bvB, BoNT/Bl,
npBoNT/B, BoNT/(B)] и BoNT/Al Фланкирующие регионы включают IS-элементы,
гены флагеллина (flagellin, Па), lycA и гипотетический белок (hypo). Прямые символы
показывают усеченные гены. BotR — регуляторный белок. Гены в пределах йа-кластера
экспрессируют белки, которые являются частью токсического комплекса, экспрессия и
функционирование orfX-белков о//Т-кластера не изучены. По К. К. Hill et al. (2009)
в геноме разных видов клостридий: С. botulinum и С. butyricum. Вставки гена произош-
ли через одинаковый у обоих микроорганизмов ten — rarA (2,7 т. п. о.). Он является
транспозон-ассоциированным геном (transposon-associated gene), обладающим ре-
974
Часть 3. Частная неправильная эпидемиология
комбиназной активностью. Другие транспозон-ассоциированные белки, резолваза
(resolvase) и сайт-специфическая рекомбиназа (site-specific recombinase), иденти-
фицированы ниже от bont/np b, в участке, где находится IS-элемент. Рекомбинации
генетического материала и вставки генетического материала в данном участке объяс-
няют большое количество серотипов и подтипов ботулинического токсина. Механизм
горизонтального переноса генов, кодирующих ботулинические комплексы остается
неясным (Hill К. К. et al., 2009).
Токсические свойства ботулинических токсинов. Данные по сравнительной токсич-
ности ботулинических токсинов представлены в табл. 3.47.
Таблица 3.47
Летальные дозы ботулинических токсинов для мыши
при внутрибрюшинном введении*
Тип токсина ММ. Да LD50, мг/кг
Домен А Домен В
А 51000 99000 4,25x10-’
В 53000 112000 4,4x10"’
С 53000 98000 2,0x10"*
D 60000 110000 6,0x10-’
Е 50000 102000 1,04 ХЮ"5
F 51000 108000 1,25x10-*
G - - 1,0x10"*
♦ По Н. С Антонову (1994).
При внутрибрюшинном введении мышам наиболее высокий уровень токсично-
сти отмечается у нейротоксина типа А. Из лабораторных животных наиболее чувстви-
тельной к действию ботулинических токсинов является морская свинка, для которой
летальные дозы при внутрибрюшинном, ингаляционном и оральном введении рав-
ны 9,4х10-7, 3,18х10~5 и 1,87х10~4мг/кг соответственно.
Зафиксированнач летальная доза ботулинического токсина для человека при
оральном введении находится в пределах 1,43х10“6-1,43х10~5 мг/кг. Используя ме-
тоды моделирования (переноса доз, полученных в экспериментах на лабораторных
животных, на человека), получены следующие величины прогнозируемых летальных
доз ботулинического токсина д.ы человека:
1) при парэнтеральном введении Зх 1СГ9 мг/кг;
2) при ингаляционном введении Зх 10~8 мг/кг;
3) при оральном введении Зх 10-6 мг/кг.
4) Lct50 0,02—0,1 мгхмин/м3.
Патогенез ботулизма*. Естественно возникающий ботулизм (пищевой, раневой,
новорожденных) обычно представляет собой токсикоинфекционный процесс. При
попадании в организм несмертельных порций токсина и ботулинического микроба
происходит размножение последнего в организме, благодаря чему вырабатываются
постоянно небольшие дозы токсина. В течение инкубационного периода, количество
токсина накапливается, что приводит к развитию типичной клиники оотулизма.
Основная масса токсина при попадании в пищеварительный тракт всасывается в же-
1 По работам К. И. Матвеева и Т. И. Булатовой (1966), L. С Sellin (1984), Я. Ю. Попелян-
ского с соавт. (2000).
3.12. Ботулинические токсины
975
лудке, в верхнем отделе тонкого кишечника. Ферменты ЖКТ активируют ботулини-
ческий токсин, превращая прототоксин в активную токсическую форму.
Токсин проникает первоначально в лимфатические пути, затем в общий крово-
ток, разносится током крови по всему организму, причем титр в лимфе всегда превы-
шает титр в плазме крови. При избыточном количестве ботулинического токсина в
крови он в первые минуты после введения сорбируется, кроме [3- и у-глобулинов, и
другими фракциями глобулинов, но при уменьшении его количества в крови обнару-
живается только во фракциях Р- и у-глобулинов.
Присутствие токсинов в крови больных ботулизмом людей и животных доказано
экспериментально. Чаще всего он обнаруживается на 2-3 сут, а иногда даже на 6—8 сут
после появления первых симптомов болезни, что указывает на образование токсина
в организме. Циркуляция токсина в крови может продолжаться от 9 до 25 сут и более.
Быстрое обсеменение организма бактериями ботулизма было установлено при зара-
жении животных через рот большими дозами спор; при этом животные погибали от
токсина, образовавшегося в организме. Введение через рот небольших доз спор живот-
ным вызывает у них бессимптомную инфекцию, сопровождающуюся иммунологиче-
скими изменениями тканей органов.
Из крови токсин проникает в органы, где поражает клетки в первую очередь нерв-
ной системы, наиболее чувствительной к ботулиническим токсинам. Резкое пораже-
ние сосудов обнаруживается при вскрытии трупов, и экспериментально установлено
на изолированных органах людей и животных. Вследствие повреждения эндотелия
капилляров и прекапилляров ЦНС токсин проникает в ткань головного мозга, вызы-
вая клинические симптомы бульбарного происхождения. При этом в ликворе людей
и животных, больных ботулизмом, токсины не обнаруживаются. У теплокровных и
холоднокровных животных токсин ботулизма типов А и В прежде всего повреждает
моторные центры передних рогов спинного мозга и продолговатого мозга, что явля-
ется истинной причиной развития ботулинического паралитического синдрома1.
Через плаценту токсин не проходит, от больных ботулизмом женщин рождаются здо-
ровые дети.
Нарушения деятельности периферических моторных иннервационных аппаратов
скелетной мускулатуры претерпевают ряд стадий. В начальные стадии ботулиничес-
кой интоксикации происходит выключение моторных центров спинного мозга. На
этой начальной стадии поражения периферические моторные проводники сохраняют
свое влияние на иннервируемые мышцы. В дальнейшем, номере развития ботулини-
ческой интоксикации, происходит постепенное выключение «тетанического» нервно-
го влияния на мышцу при полной сохранности при этом влияния «тонического».
Ботулинический токсин не блокирует проведение возбуждения по нерву или к
нервному окончанию. Несмотря на полный паралич, мышцы по-прежнему реагиру-
ют на прямую стимуляцию. Ботулинический токсин не влияет на синтез и накопле-
ние ацетилхолина, число синаптических пузырьков или ультраструктуру нервного
окончания. Токсин не препятствует также проникновению в нервное окончание каль-
ция. Хотя паралич, вызванный ботулиническим токсином, может сохраняться толь-
ко втечение нескольких недель, остаточное подавление высвобождения трансмитте-
ра может длиться месяцы.
1 Пути и даже сама возможность проникновения ботулинических токсинов в ЦНС явля-
ются предметом ожесточенных дискуссий среди специалистов. Более подробно см. в работе
Я. Ю. Попелянского с соавт. (2000).
976
Часть 3. Частная неправильная эпидемиология
Помимо подавления высвобождения трансмиттера ботулинический токсин вли-
яет на метаболические и физиологические свойства скелетных мышц, возбуждаемых
этими нервами. Непрямое постсинаптическое действие ботулинического токсина
вызывает в мышечном волокне эффекты, подобные хирургической денервации. Это
происходит в результате воздействия токсина на трофический эффект нерва, который
он обычно оказывает на мышцу. Такие воздействия ботулинического токсина могут
проявляться в виде мышечной атрофии, реинервации или изменения электрических
свойств мышечной мембраны. Эти данные свидетельствуют о том, что даже в случае
восстановления процесса высвобождения трансмиттера, ботулинический токсин
может вызывать длительные изменения в постсинаптических мышечных волокнах.
Возможны случаи отравления людей и животных одновременно несколькими
токсинами. При одновременном отравлении несмертельными дозами нескольких
токсинов происходит суммирование их патогенетического действия на организм, и
наблюдаются одинаковые клинические явления, как и при отравлении одним боту-
линическим токсином.
Нарушение функции дыхания при ботулизме вызвано поражением передних рогов
спинного мозга и соответствующих корешков и нервов. Первыми признаками дыха-
тельной недостаточности являются двигательное беспокойство, учащение дыхания,
нарушение его глубины при удовлетворительной или слегка ослабленной функции
дыхательных мышц. В легких случаях расстройства ограничиваются ощущением дав-
ления, сжатия в груди, нехватки воздуха без выраженной одышки. В тяжелых случаях
появляется одышка. Дыхание становится учащенным (до 40—44/ мин), поверхност-
ным, нарушается глубина вдоха и выдоха, вдох затруднен. Больные жалуются на не-
хватку воздуха и одышку в покое или при малейшем движении, при разговоре быстро
устают, делают паузы, чтобы отдохнуть, минутный объем дыхания уменьшается по-
чти в два раза, увеличивается амплитуда дыхания. Включаются вспомогательные ды-
хательные мышцы, больные вынуждены принимать особое положение. Нарушение
дыхания вызывается не только ослаблением межреберной мускулатуры, но и парезом,
или параличом, диафрагмы. Возможно внезапное появление патологических видов
дыхания и остановка его. Пневмонию, бронхит и ателектаз обычно вызывает аспира-
ция слюны, воды, пищи в дыхательные пути или закупорка последних слизью. Если
болезнь началась с дыхательной недостаточности, то прогноз неблагоприятный.
Нарушения функции ССС может быть связана с чрезмерной нагрузкой на миокард
в условиях гипоксемии и респираторного ацидоза и в результате прямого действия
ботулинического токсина на миокард («сердечная смерть»). Каких-либо данных, ука-
зывающих на первичное поражение сердечно-сосудистых центров ЦНС при кардио-
васкулярных расстройствах, нет.
Тахикардию, изменение уровня артериального давления, расширение границ сер-
дца отмечают даже при легких формах ботулизма. В тяжелых случаях наблюдаются
приглушенность тонов, систолический шум на верхушке сердца, акцент II тона на
легочной артерии, аритмия. Признаки поражения сердца, изменения ЭКГ при боту-
лизме возникают, как правило, на 2—3 сут болезни, держатся в течение 2—3 нед и посте-
пенно уменьшаются к концу заболевания.
Данные ЭКГ в динамике свидетельствуют о диффузном поражении сердечной
мышцы метаболического характера с развитием выраженной гипоксии миокарда (пре-
имущественно в эндокардиальных отделах); синусовой тахикардии (в период выз-
доровления); часто регистрируется отклонение электрической оси влево, снижение
вольтажа зубца Р, замедление атриовентрикулярной проводимости (удлинение интер-
3.12. Ботулинические токсины
977
вала Р — Q), снижение вольтажа зубца R, увеличение QRST. Я. Ю. Попелянский с со-
авт. (2000) обратили внимание на появление миокардиопатий различной степени че-
рез 1,5—2 года после перенесенного ботулизма.
Нарушение функции ЖКТ возникает вследствие местного воздействия токсина на
периферические вегетативные образования и как результат вовлечения в патологичес-
кий процесс стволовых отделов мозга. Симптомы поражения ЖКТ (гастроэнтерит,
метеоризм, запор) связаны с поражением гладких мышц кишечника. Возникают ино-
гда одновременно с неврологическими проявлениями, в других случаях — предшеству-
ют нервно-мышечному синдрому или появляются спустя некоторое время после него.
Нарушения функции почек и мочевыводящих путей зависят от тяжести болезни.
Ботулинические токсины не оказывают прямого повреждающего действия на почеч-
ную ткань. Нарушение функции почек при ботулизме встречается крайне редко и при
тяжелых формах заболевания. Они связаны с парезом мышц брюшного пресса, играю-
щих вспомогательную роль в акте мочеиспускания и с расстройством кровообраще-
ния, гипоксемией. При этих нарушениях удельный вес мочи, как правило, остается
нормальным и колеблется в пределах 1010—1025. Функция почек восстанавливается
при выздоровлении пациента полностью и без специальной терапии.
Устойчивость ботулинических токсинов во внешней среде. По данным К. И. Матве-
ева и Т. И. Булатовой (1966), ботулинические токсины обладают устойчивостью в
отношении физических и химических факторов. В кислой среде при значении pH
3,5—6,8 устойчивость токсинов больше, чем в щелочной среде. Ботулинические ток-
сины устойчивы к инактивирующему действию содержимого желудка и кишечника,
протеолитические ферменты желудочно-кишечного тракта не разрушают токсины
типа А, В, С, D, Е При значении pH 7,8 они значительно снижают свои токсические
свойства, в щелочной среде токсины быстро разрушаются. Токсины типа А и В явля-
ются стойкими в присутствии гнилостных микробов при воздействии света и летних
температур, когда они находятся во влажной среде.
Ботулинические токсины теплоустойчивы. Сахароза повышает устойчивость к
нагреванию токсинов типов А и В. При концентрации сахарозы от 43,5 до 87 % ток-
сины выдерживали нагревание при температуре 100°С Юмин и при 80°С—до40мин.
Значительную роль в устойчивости токсинов А и В к температуре играют растворы
солей, имеющихся в продуктах. Различные ионы или способствуют более быстрому
разрушению токсинов или, наоборот, защищают от разрушения под действием тем-
пературы. Наибольшей устойчивостью к нагреванию они обладают при значении
pH 5,0. Следовательно, невозможно предусмотреть температуру, необходимую для раз-
рушения ботулинического токсина, если не знать ионного состава, величину pH сре-
ды, в которой находится токсин. В раст ворах токсины обладаю! устойчивостью не
только в зависимости от величины pH, но также от наличия в растворе белка и нуклеи-
новых кислот.
При разведении водопроводной водой токсин серотипа А оказался менее устой-
чивым по сравнению с токсином серотипа С, который значительное время сохранял
свои токсические свойства в воде. Токсин типа А терял больше 80 % активности через
24—48 ч, типа В — за 2—3 сут, типа D — за 2-3 сут, типа Е — за 3—4 сут. Токсин типа С
сохранял 20 % активности в течение 5—7 сут. Чем больше токсин очищен, тем менее
он устойчив к действию различных химических веществ в водопроводной воде. В 0,2 %
растворе формалина токсичность ботулинического токсина типа А уменьшалась в
800 тыс. раз за 16 сут, в 0,4-0,8 % растворе формалина такое же уменьшение токсич-
ности происходило за 7 сут.
978
Часть 3. Частная неправильная эпидемиология
Токсины в жидкостях, полученных из домашних или фабричных консервирован-
ных продуктов, искусственно зараженных возбудителями ботулизма типов А и В, были
более стойки к нагреванию, чем токсины, потученныс на бульоне с глюкозой из те-
лячьего мяса. При прогревании до температуры 80 JC три образца спаржи, содержа-
щие токсины, выдерживали нагревание в течение 4—10 мин. Жидкость, полученная
из консервированной спаржи, сохраняла токсичность при прогревании до 10 мин. Два
образца кукурузного сока сохраняли токсичность при прогревании до температуры
80°С в течение 40—60 мин. Если брали различные цельные продукты вместе с жидко-
стью и кусочками овощей, то устойчивость токсина была более значительной.
Ботулинические нейротоксины быстро теряют свою активность в водных раство-
рах (см. выше), что ограничивает возможности их применения путем диспергирова-
ния жидких рецептур. В сухом состоянии измельченный ботулинический токсин мо-
жет длительное время сохраняться без потери активности и в таком виде применяться
путем распыления. Однако в аэрозолированном состоянии он быстро теряет свою
активность (Антонов Н. С., 1994).
Поражающие свойства ботулинических токсинов. По мнению Н. С. Антонова (1994),
боевое применение ботулинических токсинов ограничено несоответствием уровня их
токсичности площадям возможного поражения. Более подробно см. «Токсины как
поражающие агенты» в разд. 2.2.3.1.
Клиника энтерального поражения1 Инкубационный период при пищевом ботулиз-
ме чаще всего от 6 до 48 ч, реже от 3 до 7 сут. Описаны случаи ботулизма с очень корот-
ким инкубационным периодом, от 2 до 6 ч, и с очень длинным, до 8—10 сут. Различ-
ная длительность инкубационного периода зависит от первоначальной дозы токсина,
попавшей в организм с пищевым продуктом, и от состояния (индивидуальной чув-
ствительности) организма человека или животного. Я. Ю. Попелянский с соавт. (2000)
отмечают связь между продолжительностью инкубационного периода и тяжестью
болезни. При очень тяжелых формах он длится несколько часов, максимально — 2 сут.
При формах средней тяжести наблюдается более продолжительная инкубация — до
3 сут, при легких формах — свыше 3 сут. Однако в отдельных случаях тяжелые формы
болезни развиваются после 3—5-суточного инкубационного периода, а легкие — после
2-часового.
Болезнь начинается обычно остро. В продромальном периоде отмечается поли-
морфизм болезненных ощущений. Больных беспокоит боль в подложечной области,
тяжесть в желудке, тошнота, рвота, понос или janop, что говорит о диспепсических
явлениях. Развитие заболевания характеризуется тем, что первоначатьно возникаю-
щие неспецифические желудочно-кишечные расстройства сменяются специфичес-
кой для ботулизма неврологической симптоматикой. Основные неврологические
синдромы ботулизма: офтальмический, парез жевательной и мимической мускулату-
ры, парез бульбарной группы мышц, парез скелетных мышц, парез дыхательной мус-
кулатуры, вестибулоатактический синдром.
Начальными, наиболее ранними и частыми симптомами, являются глазодвигатель-
ные — до 93,4 %, вызванные нарушением функции внутренних (чаще) и наружных
мышц глаза (наблюдение Попелянского Я. Ю. с соавт., 2000). Такая высокая частота
поражения глазных мышц при ботулизме не случайна. Ботулинический токсин не
имеет мишеней в организме человека, кроме пресинаптических нейронов. Тонкое
1 По работам К И. Матвеева, Т. И. Булатовой (1966). W. Е Sonnabend et al. (1981), Я. Ю. По-
пелянского с соавт. (2000).
3.12. Ботулинические токсины
979
управление движением глаза требует высокого иннервационного индекса (отношение
числа нейронов, аксоны которых оканчиваются на наружной мембране мышечных
клеток, к числу мышечных клеток). Для глазной мышцы иннервационный индекс
составляет 1:3, это значит, что аксонные окончания одного мотонейрона выделяют
свой медиатор не более чем на три отдельные мышечные клетки — иннервационный
индекс бицепса всего 1:100 К числу основных глазодвигательных расстройств, харак-
терных для ботулизма, Я. Ю. Попелянский с соавт. (2000) относят нарушение акко-
модации (75 %) — ранний специфический признак ботулизма. Больной жалуется на
появление тумана и сетки перед глазами, предметы видны не четко, дрожат и др. Реже
встречаются жалобы на полную потерю зрения. Следующими по значимости в ран-
ней диагностике отравления ботулиническими токсинами авторы считают: вялую ре-
акцию зрачков на свет (70 %), ослабление реакции зрачков на конвергенцию (59 %),
птоз (58 %), мидриаз (51 %), снижение конъюнктивального рефлекса (51 %), офталь-
мопарез (49 %), диплопия (48 %), анизокория (31 %) и др.
Парез жевательной и мимической мускулатуры встречается у 96 % больных. Боль-
ной не может поднять уголки губ. В сочетании с птозом век лицо больного выглядит
сонным. При тяжелом отравлении лицо теряет выражение и становится маскообраз-
ным, застывшим. Из-за слабости напряжения мышц височной области с обеих сто-
рон, нижняя челюсть отвисает, акт жевания нарушается.
Вслед за глазными симптом зми, а нередко параллельно с ними, при прогресси-
ровании заболевания появляются бульбарные расстройства. В клинической картине
развившегося ботулизма они являются основными и часто определяют прогноз. Сте-
пень бульбарных нарушений — один из ведущих критериев тяжести болезни. У паци-
ента появляются ощущение кома и сдавления в горле, першение, иногда жжение. Ран-
ними признаками нарушения глотания бывают поперхивание и кашель во время еды,
в большинстве случаев они появляются одновременно с глазными симптомами. Далее
развиваются более грубые дисфагические расстройства: затруднение глотания сначала
вердой, затем жидкой пищи и слюны.
Наиболее доступной при первичной диагностике бульбарных расстройств явля-
ется оценка функций мягкого неба. Паралич небной занавески возможен в остром
периоде болезни при отсутствии свободного токсина ботулизма в крови больного.
Ларинго югическое обследование в этой фазе болезни выявляет функциональную
недостаточность мышц мягкого неба в виде пареза (провисание, понижение тонуса).
Паре зы и параличи мягкого неба характерны для тяжелых форм болезни, реже они
могут выявляться на 2—3 сут от начала болезни при состоянии средней тяжести: по-
являются гнусавость, поперхивание во время еды, жидкая пища вываливается через
нос, иногда аспирируется.
Одновременно с дисфагией, а нередко и предшествуя ей, возникают фоноларин-
геальные симптомы, нарушается фонация: голос становится слабым, глухим, хрип-
лым, изменяется его тембр и звучность. Поражение голосовых связок — всегда дву-
стороннее, симметричное и сочетается с другими бульбарными симптомами.
При тяжелом течении в связи с параличом мышц гортани у больных отмечается
полная афония. Дисфагия и дисфония сопровождаются нарушением речи. В резуль-
тате двустороннего поражения мышц языка движения его ограничиваются: больной
не может высунуть язык или испытывает при этом saipvflHeHHe («язык заплетается»).
Перемещение пищи в полости рта нарушается, акт жевания и глотания затрудняется
или становится совершенно невозможным. Расстройства движений губ, языка, мяг-
кою неба, наружных мышц глотки и гортани приводят к изменению речи — она стано-
980
Часть 3. Частная неправильная эпидемиология
вится гнусавой, смазанной, растянутой, без достаточной модуляции, иногда совер-
шенно непонятной для окружающих.
Температура тела в большинстве случаев нормальная, но может быть и повышен-
ной. Многие больные жалуются на головную боль, 1 оловокружение. Отмечаются зат-
рудненность движений, быстрая утомляемость, которая иногда очень быстро нарас-
тает. Слабость в некоторых случаях бывает локальной в ногах или руках. Симптомы
болезни быстро прогрессируют. Значительно ухудшается деятельность сердца. Пульс
заметно отстает от уровня температуры (брадикардия). Давление спинномозговой
жидкости, содержание в ней белка, глюкозы и клеток остаются в норме.
В заключительной фазе болезни главное место занимает нарушение дыхания. Оно
становится затрудненным, частым и поверхностным, в тяжело протекающих случаях
дыхание Чейна — Стокса. Больные принимают различные позы для усиления деятель-
ности вспомогательных дыхательных мышц. Смерть наступает от паралича дыхания
при продолжающейся сердечной деятельности. Сознание не изменяется до наступ-
ления смерти, оно помрачается иногда только в агональном состоянии. При полной
потере голоса больные стремятся выражать свои жалобы письменно или жестами.
Иногда заболевание развивается катастрофически быстро: бурно нарастают оф-
тальмические симптомы, а присоединившиеся бульбарные и дыхательные расстрой-
ства в течение суток приводят к летальному исходу; молниеносные гипертоксические
формы могут начинаться с глазо-гортанно-глоточных и дыхательных нарушений.
Патологоанатомическис изменения не представляют собой чего-либо специфиче-
ского. При вскрытии трупов людей и животных, погибших от ботулизма, обнаружи-
вается вызванная длительной гипоксией выраженная гиперемия всех внутренних
органов, сопровождающаяся множественными мелкими и крупными кровоизлияни-
ями. Оболочки головного мозга сильно гиперемированы, встречаются геморрагии.
В ткани мозга обнаруживаются тромбозы, дегенеративно-некротические, деструктив-
ного характера поражения эпителия сосудов. Стенки сосудов ткани мозга разрыхлены.
Больше всего поражаются ганглиозные клетки нервной системы. Наблюдается ваку-
олизация протоплазмы, распад ядра, форма клеток изменяется, отростки отпадают,
изменения также происходят в нейроглии.
Слизистая пищеварительного тракта гиперемирована. В слизистой желудка точеч-
ные и более крупные кровоизлияния, она набухшая, желтоватого цвета, разрыхлена,
очень легко разрывается. Сосуды кишечника резко инъецированы и придают сероз-
ной оболочке «мраморный рисунок»; слизистая кишечника набухшая, при микроско-
пии обнаруживается расширение сосудов, повреждение стенки и выход эритроцитов.
Из скелетных мышц сильнее всего поражается мускулатура грудной клетки, брюш-
ной стенки и конечностей. Мышцы имеют вареный, с сероватым оттенком вид; при
микроскопии обнаруживается расширение сосудов, стаз в капиллярах, исчезает попе-
речная полосатая исчерченность, волокна набухают, имеют вид, как при диффузном
жировом (ценкеровском) перерождении.
Ниже, на примерах историй болезни, приведенных в работах Я. Ю. Попелянского
с соавт. (2000) и W. F. Sonnabend et al. (1981), рассмотрим случаи ботулизма, вызван-
ные токсинами А-, С- и G-типов.
Случай из практики Я. Ю. Попелянского с соавт. (2000), когда в клинике отрав-
ления токсином типа А ..реобладают неврологические симптомы.
Больная М., 27 лет. Заболела остро, через 20 ч после употребления грибов домашне-
го консервирования; через 2 сут заболел муж. Появились общая слабость, нарушение
зрения, была однократная рвота. Вскоре присоединились расстройства речи и глота-
3.12. Ботулинические токсины
981
ния. Госпитализирована на 2 сут заболевания и лечилась первоначально по поводу
энцефалита. Состояние продолжало ухудшаться, нарастали расстройства зрения и
дисфония, появилось ощущение затрудненного вдоха.
При поступлении состояние тяжелое: вялость, заторможенность, адинамия. Ды-
хание поверхностное, учащенное, испытывает нехватку воздуха. Тоны сердца приглу-
шены, пульс 84/мин, АД 110/70 мм рт. ст. Язык сухой, обложен коричневым налетом,
гиперемия и сухость слизистой оболочки рта и глотки.
Анализ крови: лейкоциты 14800, СОЭ 67 мм/ч. Анализ мочи: белок 0.3 %о.
Неврологический статус: зрачки слегка расширены, реакции на свет, конверген-
цию и аккомодацию ослаблены, двусторонний симметричный птоз век, офтальмо-
плегия, дизартрия. Парез мягкого неба, дисфагия, глоточный рефлекс отсутствует.
Сила и мышечный тонус в конечностях слегка снижены. Сухожильные и периосталь-
ные рефлексы равномерно угнетены
На 2 сут заболевания из крови выделен токсин ботулизма типа А. Начато специ-
фическое, а также дезинтоксикационное лечение. Состояние ухудшалось, нарастали
дыхательные расстройства. Трахеостомия по Бьерку. На 3 сут развилась пневмония, а
вскоре из эрозированной безымянной вены возникло кровотечение. При нарастаю-
щих явлениях дыхательной недостаточности больная скончалась.
Патологоанатомический диагноз: ботулизм типа А. Кровотечение из эрозирован-
ной безымянной вены. Отек оболочек и вещества головного мозга. Двусторонняя пнев-
мония. Венозное полнокровие внутренних органов. Дистрофия паренхиматозных
органов; при микроскопическом исследовании мозга выявлено значительное домини-
рование токсических изменений, воспалительные реакции выражены меньше. Мел-
кие сосуды и капилляры из-за набухания стенок и сморщивания клеток эндотелия и
перицитов отчетливо выявлялись на фоне отечного мозгового вещества. В зонах из-
мененных сосудов видны обширные поля погибших нервных клеток, сморщивание и
пикноз глиозных элементов и множественные мелкие очажки кровоизлияний. Обра-
щал на себя внимание участок зрительного бугра с погибшими нервными клетками и
сморщенными клетками глии.
Диффузная пролиферация глии была значительно выражена в белом веществе
коры, внутренней капсулы, в белом веществе мозжечка. И хотя наблюдалась массо-
вая гибель нервных клеток (простой лизис), глиозные узелки, розетки и гранулемы
встречались редко и не в каждом препарате. Почти полная гибель нервных клеток на-
блюдалась в подкорковых образованиях, мозжечке, на фоне диффузной пролифера-
ции глии узелки при этом обнаруживались в незначительном количестве.
Нервные клетки в коре, полосатом теле, в ретикулярной формации погибали в
результате лизиса или сморщивания всего клеточного тела, при этом обнаруживались
обширные поля запустения без видимой глиозной реакции. Энцефалитические изме-
нения с воспалительной реакцией были обнаружены в стволовых отделах мозга, где
нервные клетки претерпевали уже описанное ранее «тяжелое заболевание» с гибелью
многих нервных клеток продолговатого, среднего мозга и моста мозга (варолиева мо-
ста). Воспалительная реакция наблюдалась не только в сером, но и в белом веществе,
но эта реакция носила своеобразный характер: глиозные узелки организовывались
почти исключительно из клеток олигодендроглии, занимали довольно значительные
площади в местах погибших нервных клеток или целых ядерных образований. В ство-
ловых отделах нервные клетки претерпевали изменения, сходные с описанными в пре-
дыдущем случае: нервные клетки ядра и. cuneati резко набухшие, ядра их плохо кон-
турируются и почти не выявляются, ядрышко резко увеличено, неправильных очер-
таний, субстанция Ниссля гомогенизирована, вокруг — клетки глии с полиморфной
формой ядер, последние — пикнотичны. Подобные изменения несколько слабее выяв-
982
Часть 3. Частная неправильная эпидемиология
лены в нижней оливе продолговатого мозга. Из-за токсического изменения сосудистой
стенки и нарушения ее проницаемости выявлялись наряду с мелкими геморрагиче-
скими очажками более обширные участки кровоизлияний в области ядра б пуждающего
нерва с гибелью нервных клеток и в зоне переднего рога шейного отдела спинного мозга
с распространением на клетки бокового рога. Эти очаги кровоизлияний могли явиться
непосредственной причиной смерти больной.
Патоморфологическая картина мозга обусловлена в первую очередь гипоксией
(нарушения дыхания в течение 13 сут), нарушениями сердечно-сосудистой регуляции
(грубые органические изменения сосудов), воспалительными и дистрофическими
изменениями внутренних органов. Неспецифические изменения без характерных
симптомов найдены и в путях двигательных, глубокой чувствительности, в подкор-
ковых узлах.
Случай из практики Я. Ю. Попелянского с соавт. (2006). Тяжелая форма ботулиз-
ма, вызванная токсином серотипа А. Тяжесть течения болезни определялась дыхатель-
ными нарушениями, вызванными поражением мышц, иннервируемых «бульбарной»
группой черепных и спинальных дыхательных мышц, локальные патологические оча-
ги в области ядер соответствующих нервов отсутствовали.
Больная С., 36 лет. За два дня до заболевания ела грибы домашнего консервиро-
вания. Заболела остро: появились головокружение, тошнота, изменение зрения, су-
хость ворту, голос стал осиплым, гнусавым, одновременно возникло затрудненное ды-
хание. Участковый врач не обнаружил терапевтического заболевания, посоветовал
обратиться к невропатологу, который направил больную в стационар.
Состояние при поступлении тяжелое: вялая сознание сохранено. Самостоятель-
ное дыхание затруднено. Число дыханий 26-28 /мин Пульс 66/мин, ритмичный. Тоны
сердца приглушены. АД 125/80 мм рт. ст Слизистая оболочка рта сухая. Стула нет,
живот мягкий, безболезненный. Температура нормальная. Анализы крови и мочи,
рентгенограмма грудной клетки — без особенностей.
Неврологический статус: двусторонний птоз век, мидриаз с полным отсутствием
реакции зрачков на свет, конвергенцию и аккомодацию. Движения глазных яблок рез-
ко ограничены. Гипомимия, парез мягкого неба. Толос осиплый, речь невнятная, гло-
тание невозможно, глоточный рефлекс отсутствует. Мышечная гипотония, сухожиль-
ные и периостальные рефлексы равномерно снижены. Болевая чувствительность
сохранена. Координация движений не нарушена. Диагностирована тяжелая форма бо-
тулизма. Введены поливалентная противоботулиническая сыворотка типов А, Е по
20000 ME и В — 10000 ME, дезинтоксикационные средства. В отделении реанимации
произведены трахеостомия, ИВЛ. На второй день произошла остановка сердца. После
внутрисердечной инъекции адреналина и открытого массажа сердца сердечная дея-
тельность восстановилась, однако гемодинамика оставалась нестабильной и сознание
не возвращалось. Реанимационные мероприятия были безуспешными, констатирова-
на смерть.
Анатомический диагноз: тяжелая форма ботулизма (серотип А). Выраженное ве-
нозное полнокровие внутренних органов, оболочек и отек вещества головного мозга
без локатьных очаговых поражений. Диффузные мелкоклеточные кровоизлияния в
слизистую оболочку ЖКТ. Паренхиматозная дистрофия сердца, почек, печени.
Случай из практики Я. Ю. Попелянского с соавт. (2000). Тяжела» форма ботулиз-
ма, вызванная токсином серотипа С. Нет типичного эпидемиологического анамне-
за, характерного для отравлений токсинами серотипов А и В. Заболевание ошибочно
связывают с сосудистыми процессами, стволовым энцефалитом, миастенией.
3.12. Ботулинические токсины
983
Больной Т, 57 лет, поступил с жалобами на затрудненное дыхание, нарушение гло-
тания, сухость во рту, нарушение речи. Заболел остро: появились тошнота, многократ-
ная рвота в течение дня и ночи. Со слов родственников, на второй день после неизве-
стной инъекции врачом неотложной помощи появилось затрудненное дыхание, а через
час нарушилось глотание. Болей в животе не ощущал, стул нормальный. При госпита-
лизации в одну из городских больниц отмечено изменение речи, нарушения глотания.
С диагнозом «ботулизм» переведен в больницу им. С. П. Боткина; во время транс-
портировки больной отметил преходящее двоение далеко расположенных предметов.
Эпидемиологический анамнез без особенностей: грибов, консервов не ел, род-
ственники здоровы. Общее состояние средней тяжести. Больной активен, лицо гипе-
ремировано, постоянно откашливается, вдыхает со свистом (стеноз гортани?). В лег-
ких коробочный звук, сухие и влажные хрипы. АД 130/60 мм рт. ст. Тоны сердца слегка
приглушены, пульс 76/мин, ритмичный, удовлетворительного наполнения и напря-
жения. Язык влажный, живот немного вздут, мягкий, безболезненный, стул отсутствует
два дня. Температура 36,6°C. Неврологический статус при первом осмотре невропа-
толога: сознание ясное, ориентирован, зрачки равномерные, реакция на свет живая,
речь смазанная, невнятная, мягкое небо неподвижно. Консультация отоларинголога:
парез мышц гортани и мягкого неба слева.
Лишь после этого осмотра родственница рассказала лечащему врачу, что накану-
не больной (только он один) ел сало, полученное из деревни. Невропатолог не был
информирован; с заключением «диагноз неясен, возможно, нарушение кровообраще-
ния в области ствола мозга» больной переведен в неврологическое отделение. При
повторном осмотре невропатолога глазодвигательных расстройств не отмечено. Голос
немного хриплый, несколько гнусавый. Больной не глотает, мягкое небо слегка сви-
сает, глоточный рефлекс слева снижен. Сухожильные и периостальные рефлексы на
руках чуть выше справа, коленные и ахилловы живые с обеих сторон. Чувствительность
сохранена. Координация движений не нарушена. Предположительный диагноз: атеро-
склероз, нарушение мозгового кровообращения с очагом в области ядра языкоглоточ-
ного нерва слева. Назначены дообследование, зондовое питание, сосудистая терапия.
В связи с тем что неврологическая симптоматика не была типичной для сосудис-
того поражения, больной вновь осмотрен инфекционистами.
Заключение: состояние больного тяжелое, связано с опасностью остановки дыха-
ния; глотание затруднено, резкая сухость слизистой оболочки рта, несколько сужена
левая глазная шель; небольшая слабость круговой мышцы левого глаза; легкий парез
мягкого неба слева; дисфагия; дизартрия. Парезов, расстройств чувствительности и
нарушений координации нет. Дыхание с обеих сторон ослаблено, выслушивается боль-
шое количество сухих и влажных хрипов. Тоны сердца приглушены, пульс ритмичный,
удовлетворительного наполнения и напряжения. Язык сухой, живот слегка вздут, пе-
ристальтика ослаблена, стула нет два дня. Диагноз: ботулизм с преимущественным
поражением двигательных ядер IX и X пар черепных нервов. Назначена противоботу-
линическая сыворотка типов А, Е по 20 тыс. ME и В по 10 тыс. ME. Вскоре после вве-
дения сыворотки несколько улучшилось глотание, на следующий день стала более
внятной речь. Анализ крови и мочи в норме. При дальнейшем обследовании выявле-
на двусторонняя пневмония: парез надгортанника, стоматит.
Информация ГорСЭС: из сала выделен токсин типа С. Несмотря на специфичес-
кую терапию соответствующей моновалентной сывороткой состояние ухудшалось с
присоединением пневмонии. В то же время неврологическая симптоматика медлен-
но и постепенно регрессировала. Под влиянием антибиотиков, общеукрепляющего
лечения пневмония быстро купировалась и состояние к 14-м сут после поступления в
стационар значительно улучшилось: глотание и речь восстановились, полностью исчез-
ла сухость во рту, на 28 сут больной выписался в удовлетворительном состоянии.
984
Часть 3. Частная неправильная эпидемиология
При отсутствии эпидемиологического анамнеза и характерных для ботулизма глаз-
ных симптомов следует учитывать и такие немаловажные признаки, как желудоч-
но-кишечные расстройства, сухость слизистой оболочки рта, а также интактность
пирамидных и чувствительных путей при наличии столь грубых «бульбарных» нару-
шений, что трудно объяснить ишемическим или другим стволовым очагом. В этом
случае болезнь характеризовалась преимущественно симптомами, встречающимися
при поражении мышц, иннервируемых IX, X, XII черепными нервами с расстрой-
ством дыхания.
Клиника отравления токсином серотипа G значительно отличалась от таковой при
отравлении токсинами А, В и С-типов, хотя в научной литературе утверждается, что
отравления ботулиническими токсинами дают во всех случаях сходную картину. Ниже,
по работе W. Е Sonnabend et al. (1981), приводятся описания четырех случаев скоро-
постижной смерти, вызванной ботулиническим токсином серотипа G. Все они про-
изошли в северо-восточном районе Швейцарии, на территории радиусом 50 км. Во
время исследования проб, полученных при аутопсии от лиц, доставленных в окруж-
ную больницу Сен-Галлен, Швейцария, С. botulinum типа G были идентифицирова-
ны у пяти лиц, а у трех из них в сыворотке был обнаружен токсин типа G.
Больной I. Это 24-летний мужчина, жаловавшийся на «не совсем хорошее само-
чувствие» и который «выглядел немного бледным» вечером за день до смерти, лег спать
в 22 ч. На следующее утро в 7 ч 30 мин его нашли мертвым в кровати у себя дома. При
аутопсии, произведенной через 24 ч, обнаружили сильный геморрагический отек
легких, сильный отек печени, почек и головного мозга и расширение его правого же-
лудочка. Кроме того, отмечался сильный интерстициальный отек сердечной мышцы.
В крови сердца, в легких, печени и почках были идентифицированы бактерии С. botu-
linum типа G и типа А. Реакция нейтрализации на мышах свидетельствовала о нали-
чии токсина С. botulinum типа А.
Больной 2. В середине дня 29-летний мужчина внезапно почувствовал тошноту.
Такое ощущение продолжалось весь вечер, он жаловался также на боль в верхней час-
ти живота, общую слабость и усталость. Ранним вечером он начал испытывать силь-
ную жажду и ощущать сухость во рту и горле В 9 ч вечера он лег в постель, в которой
на следующее >тро был найден мертвым. При аутопсии, произведенной через 24 ч,
обнаружили сильный геморрагический отек легких, сильную гиперемию печени, се-
лезенки, почек и головного мозга и интерстициальный отек сердечной мышцы. Из
крови сердца, из легких и тонкого и толстого кишечника были выделены С. botulinum
типа G, а в сыворотке крови был идентифицирован токсин типа G. При использова-
нии непрямого метода флюоресцирующих антител к L. pneumophila титр сывороточ-
ных антител составлял 1:256.
Больной 3. В течение трех суток до смерти 33-летний мужчина жаловался на боль
в верхней части живота, при этом он страдал от приступов головокружения, которые
его лечащий врач диагностировал как нервное расстройство. За день до смерти у него
появилась сильная жажда, которую он не мог утолить, и ощущение сухости во рту и
горле. Он лег спать дома в полночь, а в 9 ч 30 мин утра был найден мертвым При аутоп-
сии, проведенной через 24 ч, были обнаружены сильный геморрагический отек лег-
ких, интерстициальный отек сердечной мышцы и очаговые геморрагические пятна на
слизистой оболочке желудка. С. botulinum типа G были выделены из крови сердца, из
тканей легких, селезенки, сердца и печени и из содержимого тонкого и толстого ки-
шечника. Реакция нейтрализации на мышах свидетельствовала о наличии в сыворотке
токсина С. botulinum типа А. В сыворотке был обнаружен также поверхностный анти-
ген гепатита В.
3.12. Ботулинические токсины
985
Больной 4. Через 2 ч после кормления 18-недельный мальчик был обнаружен мерт-
вым в своей кроватке. Через 57 ч была произведена аутопсия. Селезенка, почки, пе-
чень и головной мозг оказались стерильными. Из сердца крови и ткани легких были
выделены Staphylococcus aureus, из легких же были выделены Е. coli и стрептококки
группы Е. В тонком кишечнике идентифицировали С. botulinum типа G. Токсин типа
G был обнаружен в сыворотке. За 3 нед до смерти ребенок был иммунизирован про-
тив полиомиелита, и из тонкого и толстого кишечника был выделен вирус полиомие-
лита типа 2. При микроскопическом исследовании было обнаружено интерстициаль-
ное воспаление легких слабой формы и внутриальвеолярные геморрагии. Слизистая
носовой полости оказалась сильно воспаленной.
Сезонного распределения случаев болезни установлено не было. В почве побли-
зости от домов умерших обнаружены С. botulinum только типа А. Больные жили в рай-
оне, где продукция и потребление сыра находятся на очень высоком уровне, в связи с
чем были проверены пробы сыра различных сортов, но С. botulinum типа G обнару-
жить не удалось. Авторам работы не удалось собрать проб пищи, принятой больны-
ми. Происхождение микроорганизмов осталось неясным. В четырех случаях смерть
наступила скоропостижно и неожиданно. После введения животным токсина типа G,
полученного из культур С. botulinum, выделенных от погибших людей, у животных
появлялись симптомы ботулизма, аналогичные таковым, вызываемым другими изве-
стными ботулиническими токсинами.
Клиника раневого ботулизма. Раневой ботулизм представляет собой токсико-ин-
фекционный процесс, аналогичный тому, который развивается после инфицирова-
ния раны возбудителями столбняка или газовой гангрены. Тяжесть поражения зави-
сит от количества попавшего в рану возбудителя ботулизма, его способности к
токсинообразованию, типа токсина, хирургической обработки раны и общего состо-
яния организма инфицированного. Клинику раневого ботулизма впервые описали
С. Thomas et al. (1951). Раневой ботулизм они наблюдали у мальчика, который в тече-
ние 4 сут после огнестрельного ранения из дробового ружья чувствовал себя удовлет-
ворительно. На 5 сут его речь стала нечеткой, в следующие дни наблюдалось затруд-
нение глотания. В дальнейшем усилилась слабость мышц, особенно шейных, больной
не мог держать правильно голову. Он жаловался на боли в горле, в следующие дни
сознание стало неясным. Смерть наступила от паралича дыхания на 4 сут после появ-
ления симптомов ботулизма. Подозрения на другие заболевания у врачей отсутствова-
ли. Больному для лечения через каждые 3 ч вводили интраперитонеально 200 тыс. ЕД
пенициллина, 20000 АЕ противостолбнячной и 20000 АЕ противодифтерийной сы-
воротки. Внутривенно вводили декстрозу, изотонический раствор NaCl и 500 мл
крови. После этого состояние больного несколько улучшилось. Температура была
ниже нормальной. Размер раны 10 см, края покрыты тонким слоем некротизирован-
ной ткани и фибрина, в глубине раны пальпировалось несколько дробинок. Клини-
ческая картина га ювой гангрены отсутствовала. Гистологически и патологоанатоми-
чески в ране было обнаружено острое воспаление.
Бактериологическое исследование подтвердило клинический диагноз. При посеве
из горла обнаружена нормальная микрофлора. При постмортальном исследовании
кусочков ткани с раневой поверхности был выделен гемолитический и негемолитиче-
ский стрептококк, а также С. botulinum типа А и С. perfringens. Видовая принадлежность
бактерий была определена на основе биологических свойств и реакции нейтрализации
на морских свинках и мышах со специфическим антитоксином. В гильзе патрона,
которым был произведен выстрел, также обнаружен возбудитель ботулизма типа А.
986
Часть 3. Частная неправильная эпидемиология
При вскрытии у больного не было найдено каких-либо характерных патологоанато-
мических изменений, кроме большого количества тромбов и геморрагии в мозгу и в
твердой мозговой оболочке, характерных для ботулизма (цит. по Матвееву К. И., 1959).
Для раневого ботулизма наркоманов характерны: анамнез пациента, с указанием на
инъекционное введение наркотиков (обычно героин); следы инъекций на коже, на-
личие язв и абсцессов, вызванных введением наркотиков; отсутствие на начальном
этапе болезни гастроэнтерической симптоматики; и доминирование неврологичес-
ких синдромов, характерных для ботулизма (см. выше). Обычно у таких пациентов
ранняя стадия болезни проявляется дизартрией, диплопией, дисфонией и нисходя-
щим параличом. Болезнь протекает тяжело. Характерный внешний вид пациента с
раневым ботулизмом, вызванным инъекционным введением наркотиков, приведен
на рис. 3.141
Рис. 3.141. Внешний вид пациента с раневым ботулизмом наркоманов.
А. Билатеральный птоз. Б. Множественные инъекционные абсцессы на
коже рук и ног в участках введения наркотиков. По A. Sam, Н. Веупоп (2010)
Клиника паренп рально! з поражения. Тяжесть поражения при инъекционном вве-
дении ботулинического токсина зависит от дозы введенного токсина и способа вве-
дения. В. A. Herrero et al. (1967) на обезьянах Масаса mulatto и обезьянах-белках была
изучена клиника и патоморфологические корреляции отравления, вызванного внут-
ривенной инъекцией ботулинического токсина типа А. В экспериментах использовали
101 животное весом 2,5—3,5 кг. Ботулинический токсин дозировали в мышиных еди-
ницах. Использовался очищенный на 10 % порошкообразный ботулинический ток-
син (3,3х 109 мышиных ЕД на 1 г). Первым симптомом отравления был птоз. Его на-
блюдали у отравленного животного на протяжении всей болезни. Время появления
этого симптома колебалось от 13 до 93 ч в зависимости от дозы токсина.
3.12. Ботулинические токсины
987
Затем появлялась мышечная слабость, о чем свидетельствовало изменение позы
животного. Обезьянам было трудно находиться в вертикальном положении. Мышцы
шеи были также поражены, и голова качалась из стороны в сторону, а также вперед и
назад. Часто голова животного опускалась на грудь. Деформация позвоночника так-
же была одним из характерных признаков. Для поддержки животные облокачивались
на стенки клеток. По мере развития болезни они поддерживали себя тем, что цепля-
лись за потолок клетки. Во время последних стадий заболевания они лежали на полу.
Глубокие сухожильные рефлексы отличались пониженной активностью на более
поздних стадиях.
Следующим признаком отравления было расстройство дыхания. По мере разви-
тия болезни отмечали часто зевоту и глубокие вдохи. Дыхательные движения стано-
вились поверхностными, абдоминальными и прерывистыми; животные дышали с
трудом и с открытым ртом. Часто наблюдали двойной вдох. Далее наблюдали выде-
ления изо рта и носа. Выделения были белого цвета, пенистыми и, чаще всего, не осо-
бенно вязкими.
Отказ от пищи и воды можно было наблюдать в более поздние стадии болезни. Ос-
новная причина — слабость жевательных и глотательных мышц. Вода выливалась
через ноздри.
Гибель представляла собой конечный результат не леченой формы ботулизма. Ли-
хорадку, запоры или поносы, рвоту, изменения в размере, форме и способности зрач-
ка приспособляться для видения далеких и близких предметов, судороги или комы не
наблюдали ни у одного животного. Обезьяны, которым вводили очень высокие дозы
токсина (20 LD 50 для обезьян), впадали в кому и погибали в течение суток.
Внутривенная LD50 для обезьян-резусов составляет 40 ЕД/кг, а для обезьян-бе-
лок — 66 ЕД/кг. Внутрижелудочная LD50 (токсин вводился в желудок животного зон-
дом) для обезьян-резусов составляет 30 тыс. ЕД/кг.
Никаких существенных отклонений клинических анализов и гемограммы от нор-
мы у отравленных животных не наблюдали. Пробы сывороток, полученные от трех
обезьян, исследовали для определения уровня антител к ботулиническому токсину,
определяя титр антител в сыворотке крови с помощью реакции нейтрализации. Ан-
титела не были обнаружены.
При вскрытии существенные морфологические изменения найдены лишь в сис-
теме органов дыхания и были обусловлены недостаточной приточной вентиляцией,
вызванной параличом дыхательных мышц (диафрагмальных и межреберных). Часто
обнаруживались очаговые участки ателектаза. Очаговую лобулярную бронхопневмо-
нию постоянно наблюдали у животных, получавших более низкие дозы, и она имела
наиболее острую форму у животных с самым продолжительным клиническим тече-
нием болезни. В большинстве случаев ранней гибели животных на вскрытии не было
обнаружено никаких морфологических изменений. Были обнаружены значительные
участки гиперемии легких, кровотечения или отеки с большим уплотнением. Тром-
бы не были обнаружены ни в одном из многочисленных исследованных срезов. Обыч-
но наблюдали застойные явления в сосудах мозга и изменения размера печени, но эти
явления считали неспецифическими и незначительными. Слизистые выделения были
скудными в трахее и бронхах и играли незначительную роль в гибели животных. У вы-
живших животных все симптомы исчезали в обратном порядке.
Отравления ботулиническими токсинами у людей могут быть вызваны внутримы-
шечным введением лекарственных препаратов, содержащих ботулинический токсин:
Botox® (Allergan Inc., США), Dysport® (Beafour-Ipsen-Speywood, Франция) и Xeomin®
988
Часть 3. Частная неправильная эпидемиология
(Impfstoffwerk Dessau-Tomau, Gmbh, Германия) — на основе ботулинического токси-
на типа А; и Myoblock®/Neuroblock® (Solstice Neurosciences, LLC; США) — на основе
ботулиническо! о токсина типа В. За исключением Xeomin'®', который практически
лишен комплексообразующих белков, другие коммерческие препараты ботулиниче-
ского токсина включают гемагглютинин и нетоксический гемагглютинин. Для ста-
билизации ботулинического токсина его лиофильно высушивают в специальной сре-
де, содержащей альбумин, сахарозу и лактозу. В лиофильно высушенном состоянии
ботулинический токсин сохраняет биологическую активность до 2-х лет, после раз-
ведения специальным солевым раствором — не более нескольких часов. Биологиче-
ская активность токсина измеряется в мышиных единицах. Одна мышиная единица
для таких препаратов — это количество продукта, способное убить до 50 % (LD50)
аутбредных мышей женского пола породы Swiss-Webster весом 20 г в результате пери-
тонеального введения. Препараты на основе ботулинического токсина применяются
для лечения цервикальной дистонии, блефароспазма, ахалазии кардии и других пато-
логических состояний, вызванных мышечным спазмом, и в косметических целях. Все-
го в специальной литературе приводится до 150 показаний для применения препара-
тов на основе ботулинических токсинов (Dhaked R. К. et al., 2010).
Препараты на основе ботулинических токсинов вводятся инъекционно в участки
мышечного спазма (контрактуры) или морщин. Действие препарата наступает на
2—3 сут после инъекции и достигает максимума на 14—15 сут. Блокируя нервно-мы-
шечную передачу, ботулинический токсин вызывает расслабление мышцы-мишени,
длящееся до 8 мес. Обычно такие препараты безопасны для пациента. Считается, что
ботулинический токсин в участке внутримышечного введения не диффундирует да-
лее 2 см, вызывая мышечный паралич только вокруг сайта инъекции (рис. 3.142).
Рис. 3.142. Терапевтическое использование Botox А. Больной, страдающий
гемифациальным спазмом на правой стороне лица. Типичные тонико-клониче-
ские судороги мимических мышц лица, включая лобную мышцу и подкожную
мышцу шеи (platysma). Б. Снятие мышечного спазма после инъекций Botox.
В Флакон Botox. По R. Laskawi (2008)
Один флакон Botox содержит 100 ЕД (units, U) очищенного нейротоксического
комплекса, 0,5 мг альбумина человека и 0.9 мг натрия хлорида. Рекомендованная доза
для крупных мышц — 100—400 ЕД, в косметических целях используют значительно
меньшие количества ботулинического токсина — менее чем 30 ЕД непосредственно
в мышцу-мишень.
3.12. Ботулинические токсины
989
Избежать осложнений от применения в клинической практике ботулинических
токсинов не удалось. Описан случай развития ботулизма после инъекций ботули-
нического токсина типа В ребенку 10 лет. У него развился птоз, фациальная дипле-
гия, слабость сгибающих и разгибающих мышц шеи, цяхательная недостаточность и
другие симптомы системного ботулизма (Partikian A., MitchellW. G., 2007). Системный
ботулизм развился при введении ребенку большой дозы ботулинического токсина
(40 ЕД/кг) (Crowner В. Е. et al., 2007). Описаны три случая развития ботулизма у па-
циентов, которым проводился повторный курс инъекций ботулинического токсина
(Dysport®) в терапевтических дозах. У пациентов через неделю после введения токсина
развивалась слабость мышц конечностей, быстрая утомляемость при ходьбе, появля-
лись трудности с глотанием пищи, регургитация пищи через нос, дисфония, у одной
пациентки — слабовыраженный левосторонний птоз. Антител к рецептору ацетилхо-
лина ни у одного из пациентов не обнаружено. Причина этих осложнений осталась
невыясненной (Bhatia К. Р. et al., 1999).
При введении ботулинического токсина типа А в дозе до 50 ЕД на процедуру, об-
разование антител к токсину не происходит. Летальная доза Botox для человека мас-
сой 70 кг составляет 2500-3000 ЕД (Dhaked R. К. et al., 2010).
Клиника ингаляционного поражения. Результаты экспериментов по ингаляционно-
му поражению обезьян R. macaques BoNT-комплексами серотипов Al и Bl, подробно
описаны в работе D. С. Sanford et al. (2010). Авторами не указана дисперсность час-
тиц ботулинических токсинов (MMAD), использованных для поражения обезьян, но
ими приведено название устройства, с помощью которого было осуществлено диспер-
гирование токсинов — распылитель Коллисона (Collison nebulizer). Его технические
характеристики предполагают не только микронный, но и субмикронный диапазон
размеров генерируемых частиц аэрозоля. В опыт брались обезьяны массой от 4 до 10 кг.
Количество токсина измерялось в мышиных ЕД/кг1 *. определенных перитонеальным
введением мышам (ЕО50для мышей; см. выше).
Для BoNT/Al ингаляционная LD50 составила 550 ЕД/кг; LD90 — 1330 ЕД/кг;
Lct50 — 3910 ЕД/кгхмин/л; Lct90 — 9680 ЕД/кгхмин/л.
ДляВоМТ/В1 ингаляционная LD50 составила 21600 ЕД/кг; LD90—148 тыс. ЕД/кг;
Lctjo — 129 тыс. ЕД/кгхмин/л; Lct90 — 1137 тыс. ЕД/кгхмин/л.
Общими клиническими признаками, наблюдаемыми у обезьян после их экспо-
нирования к аэрозолю ботулинического токсина, были анорексия, летаргия, атаксия,
затрудненное дыхание, дисфагия, птоз, регургитация пищи через нос, парезы, кашель,
ротовое дыхание, положение «лежа на боку». Для обезьян, экспонированных к низ-
ким дозам BoNT/Al или BoNT/Bl, из клинических симптомов обращала на себя вни-
мание анорексия. У выживших обезьян, экспонированных к средним дозам ботули-
нических токсинов, анорексия сопровождалась атаксией, дисфагией, кашлем,
летаргией. У погибших обезьян эти симптомы отягощались нарастающей дыхатель-
ной недостаточностью. Обезьяны, экспонированные к высоким дозам ботулиничес-
ких токсинов, погибали в начале развития симптоматики отравления. Продолжитель-
ность жизни обезьян после экспонирования к ботулиническим токсинам зависела от
ингалируемой дозы и серотипа токсина. Обезьяны, экспонированные к BoNT/Bl,
жили в среднем в два разе меньше, чем экспонированные к BoNT/Al (58,4 и 125,8 ч
соответственно).
1 Специфическая активность по BoNT/Al и BoNT/Bl, вводимого обезьянам материала,
составляла 3,2х 107 и 7,5х 107 ЕД/мг белка соответственно.
990
Часть 3. Частная неправильная эпидемиология
Для обнаружения ботулинических токсинов использовались тесты на мышах, их
минимальный порог чувствительности 2 ЕД/мл (-20—30 пкг/мл). Однако циркули-
рующий в крови животных BoNT/Al не был обнаружен даже при наибольшей ингали-
рованной дозе — 5274 ЕД/кг. BoNT/Bl не обнаруживали в крови обезьян, если ингали-
руемая доза была меньше чем 25 тыс. ЕД/кг. BoNT/B 1 обнаруживали в крови животных
через 5 ч после экспонироьания к аэрозолю токсина при дозе 25 тыс. ЕД/кг и через
2ч — при дозе 82 тыс. ЕД/кг.
Три обезьяны, получившие ингаляционно дозы токсина в интервале от 25 тыс. до
82 тыс. ЕД/кг, но у которых не был обнаружен в крови токсин, выжили. Погибли все
обезьяны, у которых BoNT/B 1 быт обнаружен в крови.
На вскрытии каких-то поражений со стороны органов и тканей, которые можно
связать с прямым действием ботулинических токсинов, найдено не было. У погибших
животных поражения легких были незначительными и соответствовали аспирацион-
ной пневмонии, вызванной параличами/парезами мышц грудной клетки.
Симптоматика ингаляционного поражения ботулиническим токсином у людей
описана Е. Holser (1962). Заболели двое мужчин и одна женщина, нарушивших в ла-
боратории специальную технику безопасности при вскрытии трупов морских свинок
и кроликов.
Накануне животные, находясь в герметических закрытых клетках, были подверг-
нуты действию аэрозоля ботулинического токсина серотипа А высокой степени чис-
тоты. Отравление возникло в результате вдыхания вторичного аэрозоля ботулиничес-
кого токсина, сохранившегося на шерсти животных.
Больной 1. В анамнезе есть указание на аллергическую реакцию в ответ на введение
лошадиной противостолбнячной сыворотки. Через 3 сут после работы в лаборатории
почувствовал «комок слизи в горле». Комок не исчезал, несмотря на откашливание и
от> зркивание. На следующий день самочувствие было таким, как в начале простудного
заболевания, однако температура тела не повышалась. Во рту было много слюны, ко-
торую больной был не в состоянии проглотить в течение всего этого дня. К вечеру
появилось затруднение при глотании: больной мог проглатывать твердые куски пищи,
только запивая их водой. Больной заподозрил у себя ботулизм.
На следующий день его состояние было подавленным, затруднение глотания воз-
росло, движения глаз стали замедленными. В этот день больного госпитализировали.
В приемном покое отметили частые глотательные движения пациента и смазанную
речь. Зрачки круглые, одинакового размера, умеренно расширенны, реагируют на свет
и на близко расположенный предмет; легкий вращательный нистагм; органы грудной
полости при рентгеноскопии в норме; пульс 92/мин; АД 135/90 мм рт. ст.; брюшная
стенка, тонус, трофика и мышечная сила в руках и ногах без заметных изменений; су-
хожильные и периостальные рефлексы, а также поверхностная и глубокая чу вс гвитель-
ность сохранены; походка несколько неуверенная.
Картина крови и ЭКГ без особенностей. В моче единичные лейкоциты и эпители-
альные клетки. Было сделано несколько пробных введений малых доз сыворотки.
Больной перенес эти инъекции хорошо. После введения преднизолона больному вве -
ли в день поступления 100 мл антитоксической сыворотки. На следующий день боль-
ной получил в несколько приемов еще 100 мл сыворотки. Помимо сыворотки, пациент у
вводили перистон (заместитель крови, производное глютаминовой кислоты) и ацетил-
холин (в первые 4 сут). На 2 сут пребывания в стационаре у больного отмечались зат-
руднения глотания, субъективные расстройства зрения и несильная головная боль, по-
давленность настроения исчезла. На 3 сут глотать стало легче, но настроение вновь
3.12. Ботулинические токсины
991
стало подавленным. На 5 сут отмечались некоторые расстройства аккомодации. В пос-
ледующие дни состояние больного постепенно улучшалось, но на 6 сут появилась ур-
тикарная сыпь, в связи с чем ему назначили антигистаминные препараты. После 9 сут
развилась типичная картина сывороточной болезни. Выздоровление было медленным.
Сыворотка больного дала положительную реакцию на ботулинический токсин в экс-
перименте на мышах.
Больная 2. Через 3 сут после вскрытия животных, вечером появилось головокру-
жение и ощущение опьянения. Речь и глотательные движения стали затруднительны-
ми. На следующий день у больной сохранилось легкое головокружение, появились
спазмы в горле, больная почти не могла есть. Вынуждена была лечь, так как при дви-
жении глаз головокружение усиливалось. Вечером она почувствовала, что глотание
опять стало затруднительным, во рту скапливалась слюна, которую почти не удавалось
проглотить. Язык казался очень тяжелым.
На следующее утро слабость была так велика, что больная не смогла отрезать кусок
хлеба. Ощущение опьянения и затруднения при глотании сохранялись. При по-
ступлении в больницу: лицо несколько отечно, зрачки умеренно расширены, реаги-
руют на свет и на близко поставленный предмет. Признаков поражения глазодвигатель-
ных мышц нет, легкий вращательный нистагм, чтение возможно только с напряжени-
ем, буквы расплываются перед глазами, речь нарушена. Органы грудной клетки при
рентгеноскопии без особенностей, пульс 72/мин, АД 130/70 мм рт. ст., брюшная стен-
ка без особенностей; тонус, трофика, сила мышц конечностей в норме. Картина кро-
ви и ЭКГ без особенностей.
Лечение аналогично тому, что и в случае 1. На второй день госпитализации у боль-
ной отмечалась преходящая подавленность, сильное затруднение глотания, снижение
остроты зрения, головная боль. На 3 сут госпитализации головная боль стала менее вы-
ражена, острота зрения повысилась. На 4 сут госпитализация пациентка жаловалась
только на затруднение глотания. На 6 сут сохранилась только небольшая слабость.
Сыворотка больной дала положительную реакцию на ботулинический токсин в экс-
перименте на мышах.
Приведенные в историях болезни описания начальных симптомов ингаляцион-
ного поражения очищенным ботулиническим токсином показывают, что в картине
отравления постоянно присутствуют симптомы, входящие в бульбарный синдром.
Офтальмический синдром проявляется минимально, гастроэнтерический — отсут-
ствует. Поражений со стороны дыхательных путей на начальном этапе болезни не
отмечено. Доза токсина в обоих случаях оказалась слишком низкой, чтобы развились
симптомы дыхательной недостаточности, связанные с ослаблением межреберной
мускулатуры, парезом или параличом диафрагмы.
При поражении человека 100-1000 летальными дозами ботулинического токси-
на смерть может наступить в течение нескольких часов (Александров В. Н., Емелья-
нов В. И., 1990).
Диагностика поражения ботулиническими токсинами. Прежде всего, необходимо
установить сам факт поражения ботулиническим токсином. Диагностика ботулизма
при естественном инфицировании ботулиническим микробом основывается на со-
вокупности эпидемиологических и клинических данных, подтвержденных лаборатор-
ными исследованиями (рис. 3.143, с. 992).
Диагноз устанавливается при выявлении симптомов отравления одновременно у
группы лиц (редко — в единичных случаях) после употребления консервированных
или недоброкачественных продуктов, проявляющегося массовостью, территориаль-
992
Часть 3. Частная неправильная эпидемиология
ной ограниченностью, не имеющего «эпидемического шлейфа». Вспышки болезни
быстро прекращаются после изъятия зараженного ботулиническим микробом продук-
та. Поэтому распознавание ботулизма и его дифференциальная диагностика не пред-
ставляют затруднений в случаях, когда источник заражения известен, а симптомати-
ка — типична (табл. 3.48).
Исследу :мый материал
Обнаружение токсина
Обогащение Экстракция ДНК
(Fe, G, Fo, I, (Fe, Fo, T, W P)
E, T,W,P)
Биопроба
на мышах
Скрининг токсина
в условиях in vitro
(ELISA,
эндопептидазная
активность и др.)
Диагноз
ПНР
Выращивание в жидкой
питательной среде
(TPGY, группы I и П;
СММ, группа I)
(Fe, G, Fo, I, Е, Т, W, Р)
Выращивании на твердой
питательной среде (EYA,
группы I и II; CBI, группа I)
(Е:, G, I,T,W,P)
Подтвержцеьие наличия гена
токсина/токсигенности
(ПЦР, биопроба на мышах)
Выделение
предполагаемой
С botulinum
Чистая культура С. botulinum
Генетическое
типирование
(PFGE, AFLP)
Типирование
токсина (ПЦР,
биопроба)
Фенотипирование
(протеолитическая
активность и др. признаки)
Рис. 3.143. Лабораторная диагностика ботулизма. Типовые .методы показаны тем-
ными стрелками; экспресс-методы — серыми стрелками; методы, направленные
на обеспечение исследователя полной эпидемиологической информацией, пока-
заны белыми стрелками. При пищевом ботулизме: S — сыворотка; Fe — фекалии;
G — содержимое желудка; Fo — пробы пищи. При детском ботулизме: 1 — содер-
жимое кишечника, фекалии; Е — сыворотка, предполагаемая пиша, образцы из
окружающей среды. Раневой ботулизм: Т — сыворотка, ткани; W — раневые
тампоны; Р — гной. По М. Lindstrom, Н. Korkeala (2006)
3.12. Ботулинические токсины
993
Таблица 3.48
Дифференциальная диагностика ботулизма*
Заболевание Основные отличия от ботулизма
1 2
Пищевые токсикоинфекции: а) сальмонеллез Эпидемиологический анамнез: контакт (прямой или непрямой) с инфици- рованными животными и следующими продуктами: мясо, сырые молоко, яйца. Клиническая картина: 1) гастроэнтерит (спастические боли в животе, понос, повышение темпе- ратуры, иногда рвота), стул обычно водянистый или полужидкий, обиль- ный; тахикардия, понижение артериального давления. 2) тифоподобная форма. Высокая температура с прострацией и сепсисом. Клиническая картина гастроэнтерита, бактериемия Протекает как брюш- ной тиф, но в более легкой форме. Посевы кала — возбудитель не выяв- ляется, гемокультура — положительная; 3) очаговые проявления: могут бьпь поражены печень, желчный пузырь, аппендикс, эндотелий сосудов, перикард, мелкие суставы, кости, мозговые оболочки, мочеполовой тракт, мягкие ткани
б) стафилококковое пищевое отравление Эпидемиологический анамнез: вспышка инфекции происходит при кон- такте инфицированной стафилококком кожи людей с приготавливаемой пищей (кондитерские изделия с кремом, молоко, мясные и рыбные продукты, сладкие соусы). Клиническая картина: сильные тошнота и рвота, спастические боли в животе, понос, иногда головная боль и лихорадка. В тяжелых случаях — гиповолемия, нарушение кислотно- основного равновесия, резкая слабость, шок
Сосудистые заболевания головного мозга В анамнезе — заболевания ССС. Клиническая картина: нарушение созна- ния, развитие несимметричных, мозаичных, очаговых неврологических симптомов; зрительные расстройства несимметричны, монокулярное зре- ние сохраняется (острота зрения каждого глаза в отдельности не страдает). Глазное дно: изменения сосудистого характера. Мышечный тонус изменен на стороне поражения (высокий или низкий). Анизорефлексия сухожиль- ных и периостальных рефлексов, пирамидные знаки, расстройства чувст- вительности, возможны менингеальные симптомы, изменения ликвора. Вазоактивные препараты улучшают состояние больного
Миастения Эпидемиологический анамнез нетипичный. Клиническая картина: начало обычно подострое, постепенное, в последующем может сопровождаться кризами. Сухожильные и периостальные рефлексы, как правило, живые, но при повторном исследовании снижаются. Характерны слабость отдель- ных групп иышц, меньше — общая слабость, миастеническая реакция на феномен Уолкера. При применении антихолинэстеразных препаратов и глюкокортикоидов наступает улучшение произвольных сокращений. Отсутствует симптом «врабатывания». ЭМГ-показатели: а) при стимуляции редкими частотами снижается амплитуда ответа вызванного потенциала; б) после введения прозерина улучшаются показатели нервно-мышечной передачи. Отсутствуют общетоксические симптомы, желудочно-кишечные и сфинктерные нарушения, сухость слизистой оболочки рта, сердечно- сосудистые расстройства, головная боль, головокружение, дисциркуля- торные расстройства, синкопальные состояния. Внутренние мышцы глаза не поражаются, мигательный рефлекс сохранен
Стволовый энцефалит Эпидемиологический анамнез нетипичный. Начинается после инфекции. Клиническая картина асимметричная стволовая симптоматика, анизореф- лексия, мышечная дистония, локальная слабость, патологические знаки, изменение чувствительности, отсутствуют желудочно-кишечные расстрой- ства и сухость слизистой оболочки рта, Внутренние мышцы глаза не стра- дают, монокулярное зрение сохраняется. Нет сфинктерных нарушений
994
Часть 3. Частная неправильная эпидемиология
Продолжение табл. 3.48
1 2
Клещевой энцефалит Эпидемиологическим анамнез: природноочаговое, сезонное (весенне-лет- нее) заболевание (Дальний Восток, Сибирь, Урал; встречается в Венгрии, Польше), резервуаром вируса являются грызуны; заболевание возникает после укуса клеща, употребления в пищу сырого козьего молока. Инкуба- ционный период 8-20 сут Клиническая картина: заболевание начинается остро с подъема температуры до 39 40°C, озноба, сильной головной боли, тошноты, рвоты. В тяжелых случаях возможна оглушенность, иногда воз- буждение, сопровождающееся бредом, слуховыми или зрительными галлю- цинациями. На 2 -4 сут появляются менингеальные симптомы, затем пара- личи периферического типа в мышцах шеи, плечевого пояса, проксималь- ных отделов рук, «свисающая» голова. На фоне ограничения движений в ногах коленные и ахилловы рефлексы повышены, нередко выявляются патологические рефлексы. Описанные симптомы при ботулизме отсутст- вуют. Как и при ботулизме, наблюдается нарушение функции стволовых отделов мозга (нарушение речи, глотания), однако в дальнейшем, когда атрофируются мышцы языка, диагноз уточняется. При клещевом энцефа- лите постоянно обнаруживаются признаки воспаления в крови и ликворе
Полиомиелит Свойственный только этой болезни эпидемиологический анамнез. Клини- ческая картина: болезнь начинается с лихорадки. Характерны избиратель- ная, несимметричная черепно-мозговая симптоматика, парезы и атрофии отдельных групп мышц, изменение цереброспинальной жидкости, измене- ния показателей ЭМГ, функции внутренних мышц глаза не изменены
Полиневропатии Нетипичный эпидемиолот ический анамнез. Клиническая картина: восхо- дящий парез или паралич скелетных мышц, боли, симптомы натяжения, нарушения чувствительности, нередко атрофия. Отсутствуют желудочно- кишечные расстройства, сухость слизистой оболочки рта, сфинктерные нарушения. Нет пареза аккомодации
Острая полирадикулонев- ропатия (синдром Гийена — Барре) Эпидемиологический анамнез нетипичный. Клиническая картина: распро- странение парезов и параличей, чаше в восходящем направлении, иногда завершается поражением черепных нервов. Бурное развитие восходящего процесса может закончиться параличом дыхательной мускулатуры с буль- барными нарушениями — восходящий паралич Ландри. Характерны сен- сорные расстройства: боли в конечностях, суставах, иногда гипестезия или гипалгезия по типу «носков» и «перчаток»; симптомы натяжения, белково- клеточная диссоциация в цереброспинальной жидкости. Устойчивая тахи- кардия. Изменения показателей ЭМГ Сфинктерные расстройства встреча- ются редко. Отсутствуют желудочно-кишечные расстройства, сухость сли- зистой оболочки рта, сфинктерные нарушения
Вариант Фишера при синдроме Гийена — Барре (сочетание остро возникшей офталь- моплегии с поли- невропатисй и атаксией) Эпидемиологический анамнез отсутствует. Клиническая картина: функция внутренних мышц глаза сохранена (в редких случаях — тонически сужен- ные зрачки, в других — синдром Горнера). Выраженная двусторонняя атаксия, постоянно исчезающие глубокие рефлексы, эффект от кортико- стероидов и анаболических гормонов. Желудочно-кишечных и сфинктер- ных расстройств нет
Дифтерия Свойственный только этому заболеванию эпидемиологический анамнез. Клиническая картина: дифтеритический налет в зеве, парез мягкого неба развивается иногда до того, как исчезнет налет (преимущественное пора- жение блуждающего и языкоглоточного нервов, реже — глазодвигатель- ного, лицевого, отводящего), гнусавая речь, нарушение глотания, афония,
3.12. Ботулинические токсины
995
Пробе гжение табл. 3.48
1 2
тахикардия Развивается невропатия нижних конечностей. Дифтеритиче- ские полиневропатии сопровождаются поражением миокарда, гипертер- мией, болезненностью тонзиллярных лимфатических узлов. В ликворе иногда обнаруживается белково-клеточная диссоциация
М иастенический синдром при тер- минальной поли- невропатии, опи- санной Б. М. Гех- том (1979) Клиническа г картина медленное (1—2 нед) начале, преобладание дисталь- ных парезов, быстро прогрессируют атрофии, парестезии, боли поражение экстрлокулярной мускулатуры, характерный тип ЭМГ, положительный эффект от кортикостероидов
Отравление рибами. бледная поганка, мухомор** строчки, сморчки При отравлении ядовитыми грибами отсутствуют доминирующие при бо- тулизме симптомы: офтальмические, бульбарные, дыхательные расстрой- ства, нервно-мышечные нарушения
Фаллоидиновый синдром (см. с. 424) Анамнез источник отравления известс н («точечный»). Клиническая картина: латентный период - 6—9 ч, в редких случаях 10—15 ч. Общая слабость и недомогание резко переходя- щие в симптомы поражения ЖКТ (фонтан, рующая неукротимая рвота, спастические боли в животе, понос с кровью), период мнимого благо- получия до 10 ч, нарастание мышечной слабости, развитие симптомов печеночно-почечной недостаточности, анурия, кома, коллапс
Гиромитиновый синдром (см. с. 430). Клиническая картина: латентный период 3—25 ч, частый жидкий стул (до 8 раз в сутки), рвота, боли в жи- воте, при пальпации печень увеличена и болезнена, острая почечная недо- статочность (гемоглобинурийный нефроз), токсический гепатит, коагуло- патия, гиперферментемия, гипопротеинемия, гиперазотемия, гемолити- ческая желтуха
♦ По Я Ю. Попелянскому с соаьг. (2000).
** Дополнено по работе С. Г. Мусселиуса и А. А. Рык (2002).
Возникновение вспышки случаев нисходящего и прогрессивного бульбарного и
скелетного паралича у безлихопадочных больных может служить основанием для пред-
положения о ботулинической интоксикации.
Для неправильной эпидемиологии ботулизма характерно сочетание хотя бы двух ука-
занных ниже условий:
1) наличие большого числа случаев острого ботулизма, не имеющего связи с кон-
сервированными продуктами (в России основной источник ботулинического мик-
роба — консервированные грибы);
2) обнаружение в пище серотипов ботулинического токсина, обычно не встречаю-
щихся в регионе, либо серотипов типа D-, F-, G- и Е-типов без связи с продуктами
водного происхождения;
3) связь вспышки с каким-то общим географическим фактором (аэропорт, кино-
театр, метро, учреждение и др.), но без указаний на прием общей пищи;
4) множество одновременно возникших вспышек без общего источника ботулини-
ческого токсина. Последние два условия характерны для ингаляционного поражения.
Парентеральное отравление ботулиническим токсином можно предполагать при
наличии в анамнезе пациента указаний на получение им каких то царапин или инъек-
ций в течение ближайших 48 ч. При умышленном внутривенном введении ботулини-
996
Часть 3. Частная неправильная эпидемиология
ческого токсина человеку в картине отравления первым специфическим симптомом
будет птоз. Симптом появляется уже через несколько часов после введения токсина.
К этому времени еще остается след от инъекции. Дальнейшее развитие симптомати-
ки представляет собой проявление развития бульбарного синдрома и дыхательной
недостаточности. Глубокие сухожильные рефлексы понижены или не определяются.
Симптомы расстройства ЖКТ отсутствуют.
Поражение аэрозолем ботулиническим токсином можно предполагать, если связь
вспышки болезни с контаминированными продуктами либо с инъекциями препара-
тов на основе ботулинического токсина, установить невозможно, в первоначальной
симптоматике преобладают симптомы бульбарных нарушений, но симптомы рас-
стройства ЖКТ отсутствуют.
Аэрозолированный токсин обычно не поддается идентификации в сыворотке или
испражнениях, хотя при пищевом ботулизме это возможно. Однако токсин может
присутствовать на слизистых оболочках носа и быть обнаруженным методом ELISA
в течение 24 ч после вдыхания (Franz D.R. et al., 1997). Краткие сводки возможностей
отдельных лабораторных способов по обнаружению ботулинических токсинов в об-
разцах продуктов и сыворотке пациентов, приведены в работах М. Lindstrom, Н. Ког-
keala (2006) и R. К Dhaked et al (2010).
Ретроспективная диагностика отравления ботулиническим токсином иммуноло-
гическими методами затруднительна. Количество токсина, способное вызвать пора-
жение человека, настолько мало, что антитела к нему не образуются.
Иммунопрофилактика. В России для иммунопрофилактики ботулизма используют
комбинированные препараты, содержащие адсорбированные на гидроксиде алюми-
ния ботулинические анатоксины типов А, В и Е, которые представляют собой обезв-
реженные формальдегидом и тепловым воздействием ботулинические токсины. Выпу-
скаются два вида комбинированных препаратов: трианатоксин и тетраанатоксин,
содержащий наряду с ботулиническими еще и столбнячный анатоксин. С профилак-
тической целью применяют также противоботулиническую сыворотку. Ее вводят
людям, употреблявшим одновременно с заболевшими продукты, вызвавшие ботулизм.
Предприятие-производитель: ФГУП «НПО «Микроген» Минздрава России.
Лечение1. Основной принцип лечения больных ботулизмом — возможно раннее
выведение токсина из ЖКТ и нейтрализация его в организме с помощью специфи-
ческих и неспецифических средств.
При выраженных бульбарных и дыхательных нарушениях желудок с оольшой
осторожностью промывается через назогастральный зонд. Для промывания кишеч-
ника применяют высокие очистительные клизмы 5 % раствором натрия гидрокарбо-
ната. Немедленно после установления диагноза начинают специфическое лечение
антитоксической противоботулинической сывороткой. Перед введением сыворотки
ставят внутрикожную пробу с разведенной сывороткой (1; 100) на чувствительность к
лошадиному белку. До установления типа токсина целесообразно начинать лечение с
поливалентной противоботулинической сыворотки (тип А, В, Е, С), а после установ-
ления типа возбудителя применять сыворотку соответствующего типа.
Лечебная доза противоботулинической сыворотки, вводимой одновременно внут-
римышечно, равна 10 тыс. ME при токсине типа А и Е и 5 тыс. ME при типе В. Если
тип токсина известен, а также после его идентификации лечение проводят (или про-
должают) только соответствующим типом противоботулинической сыворотки. В тя-
1 В основном использовался опыт, накопленный Я. Ю. Попелянским с соавт. (2000).
3.12. Ботулинические токсины
997
желых случаях заболевания сыворотку вводят каждые 6—8 ч по 10 тыс. ME типа А и Е
и по 5 тыс. ME типа В. На 2-3-и сут серотерапии кратность введения может быть
снижена до 2 раз в сутки. Курс лечения — не более 4 сут и независимо от динамики
обратного развития на весь курс лечения достаточно 80—90 тыс. ME противоботули-
нической сыворотки типа А и Е, 40—45 тыс. ME типа В.
Увеличение разовых доз, кратности введения или длительности курса сывороточ-
ного лечения, не ускоряя процесс выздоровления, повышают риск развития сыворо-
точной болезни. Данные экспериментальных исследований и клинических наблюде-
ний свидетельствуют о том, что после того, как ботулинический токсин фиксируется
тканями нервной системы, он практически не под дается нейтрализации даже в случае
введения в кровоток массивных доз сыворо-ки. Вместе с тем, учитывая возможность
длительной циркуляции токсина (а иногда и возбудителя) в крови больного, вполне
оправданным является применение сыворотки и в более поздние сроки. Во всех слу-
чаях, если неврологические расстройства не нарастают и состояние больного не ухуд-
шается, далпнейшее ее введение следует прекратить.
Данные литературы по применению антибиотиков противоречивы: одни авторы
наблюдали положительные результаты, другие такого влияния на течение заболева-
ния не отмечали. Ни один из известных химиотерапевтических препаратов (включая
антибиотики) не обладает выраженным антибактериальным и детоксицирующим
действием на возбудителя и токсин ботулизма. Аминогликозиды и клиндамицин, наз-
наченные больным с ботулизмом, могут еще более ослабить нервно-мышечную транс-
миссию и привести к ухудшению клинического состояния больного.
При лечении ботулизма широко применяются неспецифические дезинтоксика-
ционные средства (под контролем диуреза, гематокритного числа, центрального ве-
нозного давления): плазма (внутривенно), плазмозаменяющие растворы внутривенно
капельно, реополиглюкин до 1000 мл в сут. Инфузионно-дезинтоксикационная тера-
пия сочетается с назначением диуретиков (лазикс, фуросемид, бринальдикс, триам-
пур и др.), антигистаминных препаратов.
В комплекс противооогулинической терапии независимо от тяжести неврологи-
ческих и висцеральных симптомов включаются аденозинтрифосфорная кислота (спо-
собствует синтезу ацетилхолина), витамины группы В (витамин В, по 1 мл 5 % раствора
в/м), благоприятно влияющие на биоэнергетические процессы в сердечной мышце,
способствующие улучшению коронарного кровообращения и кровообращения вооб-
ще, кокарбоксилаза (по 100 мг ежедневно в течение 10-12 сут).
При острой сосудистой недостаточности рекомендуется вводить вазопрессорные
средства (ангиотензинамид), кортикостероиды, высокомолекулярные декстраны —
полиглюкин, реополиглюкин под контролем АД.
Терапевтическая коррекция нарушений гемоциркуляции и транскапиллярного
обмена требует применения вазоактивных препаратов (дифференцированно в каж-
дом случае), а также метаболических средств (церебролизин, ноотропил, энцефабол,
геровитал, гериоптил). При тяжелых формах ботулизма иногда может наблюдаться по-
вышение вязкости крови, в связи с чем некоторые авторы рекомендуют осторожное
(при абсолютных показаниях) введение антикоагулянтов.
На ранних этапах заболевания особенно эффективна гипербарооксигенотера-
пия при давлении кислорода в пределах 147,2—196,2 кПа и длительности экспози-
ции 40—60 мин. В среднем на курс лечения больным легкой формой ботулизма тре-
оуется 1—2, больным среднетяжелой формой — 3—5, тяжелой формой — 8—10 сеансов.
При тяжелых формах ботулизма главное внимание уделяется коррекции нарушений
998
Часть 3. Частная неправильная эпидемиология
дыхания и кровообращения с целью предупреждения гипоксии мозга. Продолжитель-
ность дыхательных нарушений зависит от состояния дыхательных мышц (парез), ле-
гочных осложнений и сроков начала лечения (имеется в виду своевременный пере-
вод больного на искусственное дыхание). При искусственной вентиляции легких у
больных с недостаточностью внешнего дыхания оно восстанавливается на 3—15 сут
после подключения к аппарату, однако у этой группы больных признаки легочной не-
достаточности сохраняются долго.
При нарушении глотания и отсутствии застойных явлений в желудке питание
осуществляется через назогастральный зонд, который вводится каждый раз при корм-
лении больного, так как при выраженной сухости слизистых оболочек рта, глотки и
пищевода (отсутствие саливации) возникает опасность образования пролежней. При
ослаблении моторики желудка питательные смеси вводятся парентерально.
Выписку реконвалесцента производят через 7—10 сут после исчезновения всех
клинических признаков болезни с рекомендацией щадящего режима на ближайшие
2—3 нед. Вопрос о времени возвращения к обычной трудовой деятельности после ста-
ционарного лечения решается индивидуально.
3.13 СТАФИЛОКОККОВЫЙ ЭНТЕРОТОКСИН в
Токсины-суперантигены. Молекулярная структура и механизм действия токси-
нов-суперантигенов стафилококков и стрептококков. Токсические свойства SEB.
Поражающие свойства SEB. Клиника энтерального поражения SEB. Клиника
конъюнктивального поражения SEB. Клиника и патанатомия ингаляционного по-
ражения SEB. Клиника парентерального поражения SEB. Диагностика пораже-
ния SEB. Иммунопрофилактика. Лечение.
Стафилококковый энтеротоксин В (staphylococcal enterotoxin В, SEB) — один из
семи энтеротоксинов, продуцируемых грамположительной неподвижной кокковид-
ной бактерий Staphylococcus aureus (золотистый стафилококк). В норме он присутствует
на коже и слизистых человека и животных. Инфекционистам 5. aureus известен как
основная причина пищевых отравлений, клинически проявляющихся рвотой с диа-
реей или без нее, сопровождающихся гипотензией и лихорадкой. Все энтеротоксины
5. aureus — эметики, т. е. при попадании с пищей, парентерально или ингаляционно
в организм человека они вызывают рвотный рефлекс. В российских санитарно-эпи-
демиологических правилах СП 1.3.2322-08 SEB не упоминается, но по степени важ-
ности для национальной безопасности США SEB относится к группе В.
Работы с SEB по программам военно-биологических исследований начались в
США еще в годы Второй мировой войны и продолжились в годы «холодной войны».
Токсин имел сначала кодовое обозначение PG, затем UC. SEB был одним из 7 биоло-
гических агентов, запасы которых были сделаны США для ведения боевых действий.
Предполагалось его использование в первые часы вторжения на Кубу во время Кариб-
ского ракетного кризиса в 1962 г., но потом эти планы были изменены. Согласно
Конвенции 1972 г., SEB как высокоактивный токсин отнесен к разряду запрещенных
агентов. В отличие от ботулинических токсинов, SEB относительно стабилен в аэро-
золях. Его «привлекательность» как агента БО связана со значительно более низкими
дозами, вызывающими инкапситируюший эффект при ингаляционном применении,
чем это возможно для химических агентов такого назначения. Так же он может быть
3.13. Стафилококковый энтеротоксин В
999
использован для организации диверсий с целью заражения пищевых продуктов или
небольших водных источников (Franz D. R. et al.,1997; Usamriids medical mana-
gement..., 2001).
Токсины-суперантигены. Относятся к семейству токсинов-активаторов иммунно-
го ответа (см. разд. 2.2.3.1). Как правило, это токсины стафилококков и стрептокок-
ков, составляющие три гомологичные группы:
1) энтеротоксины стафилококков серотипов SEA, SED и SEE;
2) энтеротоксины стафилококков серотипов SEB, SEC1, SEC2, SEC3 и стрепто-
кокковые пирогенные энтеротоксины А и С (SPE-A и SPE-C);
3) токсин синдрома токсического шока (toxic shock syndrome toxin, TSST-1) и
SPE-B, имеющие незначительную гомологию с токсинами двух предыдущих групп.
Каждый из токсинов-суперантигенов в условиях in vivo проявляет, как минимум,
три биологических свойства: пирогенность, суперантигенность и способность увели-
чивать летальность эндотоксина для кроликов до 100 тыс. раз. Отдельные токсины
обладают более широкой биологической активностью. Так, стафилококковые энте-
ротоксины вызывают рвоту и некроз миокарда.
Большинство белков, секретируемых S. aureus, продуцируются в питательную сре-
ду в постэкспоненциальной фазе роста. Гены токсинов-суперантигенов локализова-
ны в плазмидах, бактериофагах и гетерологичных генетических элементах, называе-
мых «островами патогенности». Их экспрессия контролируется, по крайней мере,
гремя регуляторными системами- agr (accessory gene regulator), лгг (staphylococcal acces-
sorygene regulator) и системой катаболической репрессии (catabolite repression system).
Каждый токсин транслируется в прекурсорный белок, содержащий аминотерминаль-
ную сигнальную последовательность, которая рассекается во время экспорта из клет-
ки. «Созревшие» токсины-суперантигены представляют собой негликозилированные
полипептиды с ММ в пределах от 20 до 30 кДа, ММ SEВ 28,4 кДа. Они умеренно ста-
бильны к факторам химической инактивации, протеолизу и денатурации кипячени-
ем. Данные ренгеноелру ктурного анализа показа ти, что при значительном различии
в первичной структуре белковых молекул токсинов-суперантигенов, TSST-1, SEA,
SEB и SEC имеют почти идет ичную третичную структуру (Schad Е . М. et al., 1995;
Dinges М. М. et al., 2000).
Энтеротоксины получают путем культивирования клонов-продуцентов 5. aureus
в жидкой питательной среде до стационарной фазы роста в условиях хорошей аэра-
ции при температуре 37 °C и pH 7—8. Накопившиеся в культуральной жидкости ток-
сины осаждают добавлением четырех объемов этанона, охлажденного до температу-
ры 4°С, осадок растворяют в апирогенной воде. Полученную смесь токсинов и других
компонентов питательной среды разделяют тонкослойным изоэлектрическим фоку-
сированием. Амфолиты удаляют из препарата посредством диализа. Таким способом
можно получить от 1 до 20 мг токсина из одного литра жидкой питательной среды
^Dinges М. М. et al., 2000).
Молекулярная структура и механизм действия токсинов-суперантигенов стафилокок-
ков и стрептококков. Молекула токсинов-суперантигенов стафилококков имеет эллип-
соидную форму. Она состоит из двух разных доменов (А и В), соединенных дисуль-
фидной связью. Гоксины-суперантигены связывают антигенраспознающие рецепто-
ры лимфоцитов не в участках их активных центров, а в других участках рецепторов.
SEB, TSS Г-1 и другие токсины связываются с боковыми участками a-цепи Т-клеточ-
ного рецептора (TCR) Т-лимфоцитов и одновременно с V-областыо МНС класса II.
В результате они блокируют возможное связывание с TCR специфических антигенов
1000
Часть 3. Частная неправильная эпидемиология
и вызывают активацию больших популяций Т-хелперных клеток при одновременном
обходе обычного процессинга и представления антигена. Вместо этого ими индуци-
руется выброс провоспалительных цитокинов, таких как IL-1, IL-2, IL-6, IFN-y, LT,
TNF-a, вызывающих интенсивную воспалительную реакцию в месте аппликации
токсина. Выделенные лимфоцитами цитокины являются причиной общетоксического
действия SEB, гипотензии и интерстициального отека легких (рис. 3.144).
Рис. 3.144. Механизм действия токсинов-суперантигенов. А. Суперантиген — это
бифункциональная молекула, связывающаяся одновременно с рецепторным уча-
стком МНС класса II и с рецептором Т-клетки. Б. Схематическое изображение
SEB, связанного с вариабельным Р-доменом Т-клеточного рецептора
Основная функция токсинов-суперантигенов стафилококков и стрептококков за-
ключается в блокировании защитных иммунных ответов макроорганизма.
Токсические свойства SEB. Хорошо изучены. Доза SEB, способная вызвать инкап-
ситирующий эффекту 50 % человеческой популяции1 при ингаляционном введении,
не превышает 0,0004 мг/кг. Ю50 (перорально) — 0,4 мкг/кг (вызывают у человека силь-
ную рвоту). Для сравнения ED50 для Т2 токсина — 0,1 мг/кг. По выводящей из строя
дозе Ict50 SEB в 2000 токсичнее синтетического эметика апоморфина (Ict50 апомор-
фина 1000 мгхмин/м3). Расчетная LD50 SEB для человека при ингаляционном введе-
нии — 0,02-0,25 мг/кг (Антонов Н. С., 1994; Ulrich R. G. et al., 1997). Значения Let и
LD в сотни раз выше временно выводящих из строя. Для обезьян при в/в введении SEB
эффективная доза, вызывающая рвоту и понос, равна 0,0001 мг/кг, a LD50=0,025 мг/кг,
т. е. в 250 раз выше (Александров В. Н., Емельянов В. И., 1990).
Поражающие свойства SEB. В окружающей среде SEB более стабилен, чем рицин
или ботулинические токсины. Может сохранить свою активность в кипящей воде в
течение нескольких минут. Хотя SEB не рассматривается как высоколетальный агент,
но благодаря низкой инкапситирующей дозе (ED50) и стабильности в аэрозоле, мо-
жет вывести из строя людей, находящихся на расстоянии нескольких километров с
подветренной стороны от места сбрасывания средства поражения (Usamriid’s medical
management..., 2001). Распыленные 95 кг сухой рецептуры SEB способны вызвать ин-
капситацию людей, незащищенных индивидуальными средствами защиты органов
дыхания, на площади до 2600 км2. Другое направление боевого применения SEB —
преодоление защитного действия противогазов за счет проскока через противодым-
ный фильтр, клапан выдоха и по полосе обтюрации (Антонов И. С., 1994).
1 Другое название — эффективная доза (effective dose, ED50).
3.13. Стафилококковый энтеротоксин В
1001
Большинство людей, пораженных инкапситирующими дозами SEB, могут чув-
ствовать себя удовлетворительно после первоначальной острой стадии болезни, но они
не в состоянии приступить к своим обязанностям в течение одной-двух недель.
Клиника энтерального поражения SEB. Инкубационный период чаще продолжа-
ется 2—4 ч (иногда сокращается до 30 мин и редко увеличивается до 6 ч) после приема
пищи, содержащей токсин. Наиболее постоянным признаком стафилококковых от-
равлений являются режущие схваткообразные боли в эпигастральной области. Вско-
ре появляется рвота, обычно многократная. Температура тела чаще субфебрильная
или нормальная, хотя иногда может кратковременно достигать фебрильных величин
(38-38,5°C). Поноса может и не быть, слабо выраженное расстройство стула наблю-
дается примерно у 50 % больных. Для стафилококковых отравлений характерны нара-
стающая слабость, бледность, похолодание конечностей, понижение АД. Может раз-
биться коллаптоидное состояние. Однако даже при тяжелом начале состояние больных
быстро улучшается, и к концу первых суток от начала болезни наступает выздоровле-
ние. Иногда в течение 2—3 сут сохраняется небольшая слабость. Это вызвано тем, что
благодаря быстрому эметическому действию SEB, большая его часть выводится из
организма человека вместе с рвотными массами. Летальность при стафилококковых
пищевых отравлениях не превышает 0,03 % (Holmberg S. D., Blake Р. А., 1984).
Клиника преднамеренного энтерального поражения SEB моделировалась в опы-
тах на Масаса mulatto. Двум группам животных массой 2—3 кг через зонд вводили в
желудок физиологический раствор, содержащий SEB в количестве 30 мкг (10 ED50) и
150 мкг (50 ED50). Через определенные промежутки времени животных забивали и
подвергали патоморфологическому исследованию. Первыми признаками отравления
SEB были энтерит и рвота, развившиеся через 4 и 2 ч и достигшие максимальной вы-
раженности в период от 4 до 8 ч соответственно. Состояние животных вошло в норму
в пределах 72 ч. Патоморфологические исследования показали, что воспалительные
поражения слизистой и желудка и тонкого кишечника были неспецифическими и не
имели строгой зависимости от введенной дозы токсина. По мере диффузии токсина
в слизистые поверхности, воспалительные изменения развивались в подслизистых
сосудах и мезентеральных лимфатических узлах. Изменения в подвздошной кишке
были мало выраженными (KentT. Н., 1966).
Патологии со стороны легких при энтеральном проникновении токсина в орга-
низм человека и экспериментальных животных не возникает (Rusnak J. et al., 2004).
Клиника конъюнктивального поражения SEB. Такие поражения у людей возника-
ют из-за нарушений специальной техники безопасности при работе с SEB в условиях
лаборатории. При попадании SEB на конъюнктиву в течение 8—12 ч у пациента раз-
вивается гнойный конъюнктивит, отек в области глаза и умеренные симптомы по-
ражения ЖКТ, сходные с теми, что бывают при пишевых отравлениях токсином. На
3—4 сут наступает полное выздоровление (Rusnak J. et al., 2004).
Клиника и натанктомия ингаляционного поражения SEB. R. G. Ulrich et al. (1997)
обобщили клинику 9 случаев инкапситирующего ингаляционного поражения сотруд-
ников USAMR11D, связанных с нарушениями специальной техники безопасности во
время агробиологических экспериментов с SEB. Разумеется, тип аэрозоля и диспер-
сность ингалируемых частиц, не указаны. Из девяти пациентов трое считались ими
очень тяжелыми, двое — умеренно тяжелыми, у четверых течение болезни было лег-
ким. Основные симптомы ингаляционного поражения SEB (лихорадка, головная
боль, сухой кашель, озноб, миалгии, тошнота, рвота, анорексия, диарея) начинались
в период от 3 до 12 ч после вдыхания токсина. Боли в груди и внезапно появившаяся
1002
Часть 3. Частная неправильная эпидемиология
отдышка свидетельствовали о тяжелом поражении. Лихорадка наблюдалась во всех
случаях. Подъем температуры происходил внезапно, в период времени от 8 до 20 ч
после экспонирования к аэрозолю SEB, но в среднем через 12,4±3,9 ч. Продолжитель-
ность лихорадки от 12 до 72 ч (50±22,3 ч). Температура у отдельных пациентов дости-
гала 41 °C. У 5 пациентов имела место одышка и хрипы, респираторные симптомы
появлялись через 10,4 ± 5,4 ч. Продолжительность миалгии была от 4 до 44 ч, а сред-
няя продолжительность составила 16±15 ч.
На боли в грудной клетке жаловались 7 пациентов. Ее продолжительность коле-
балась от 4 до 84 ч. Рентгенография показала интерстициальный отек легких у трех
пациентов с хрипами на вдохе и выряженной отдышкой. Рентгенологически у паци-
ентов находили перибронхиальные затемнения или перибронхиальные утопщения
(peribronchial cuffing) — рентгенологические признаки наличия избытка жидкости или
накопления слизи в небольших участках дыхательных путей. Во время выздоровле-
ния дисковые ателектазы обнаружены не были. У троих пациентов с легким течени-
ем болезни рентгенологически изменений в легких обнаружено не было.
На головные боли жаловались 8 пациентов, рвота имела место у 4 пациентов и,
обычно, была однократной. Частота пульса соответствовала температуре. Лейкоци-
тоз наблюдается у большинства пациентов в период от 12 до 24 ч после воздействия
токсина. Ни у одного пациента не был обнаружен конъюнктивит, хотя один из них
вспоминал, что его глаза были «как бы сожжены».
Данные по клинике и патологии ингаляционных поражений SEB, полученные на
приматах, более информативны, так являются результатами экспериментов, в кото-
рых варьировали количеством ингалированного токсина до летальных доз, проводили
патоморфологические и другие исследования. Рассмотрим их по работам М. Е. Matrix
et al. (1995) и R. G. Ulrich et al. (1997).
Клиника ингаляционного поражения приматов шведомо летальными дозами SEB
(5LD5O— 18LD5o) носит двуфазный характер. После экспонирования обезьян каэро-
юлю SEB, в период времени, зависящий от дозы, но не превышающий 24 ч, у них по-
являлись рвота, анорексия, диарея. Эти симптомы быстро угасали Приблизительно
через 48 ч после экспонирования к SEB, внешне уже здоровые животные впадали в
летаргическое состояние и погибали в течение 4 ч на фоне прогрессирующей летар-
гии и дыхательной недостаточности.
Посмертное макроскопическое исследование обезьян, погибших от ингаляцион-
ного поражения SEB, обнаруживало сходные поражения легких. Легкие были диф-
фузно отечными, влажными, их поверхность покрыта мультифокальными петехиаль-
ными геморрагиями (multifocal petechial hemorrhages) и имела j частки ателектазов. Из
гортани вытекала пенистая прозрачная серозная жидкостг Кишечник был покрыт
петехиальными геморрагиями, слизистая эррозирована. Лимфатические узлы слегка
опухшие, на поверхности разреза обычно влажные и выпуклые.
При микроскопическом исследовании легочной ткани находили явные призна-
ки поражения легких в виде отека (мультифокального или сливного), охватывающего
несколько долей легкого. Отдельные животные имели мультифокальную и ассимет-
ричную эрозию эпителия бронхов с почти полной потерей эпителия бронхиол. Не-
которые бронхиолы узнавали только по их гладкомышечной мускулатуре. Периброн-
хиальные, перибронхиолярные и периваскулярные пространства, заполненные со-
единительной тканью, были расширены до 5 мм за счет отека, скопления фибрина,
эритроцитов, и небольшого количества лимфоцитов, макрофагов, и нейтрофилов.
Прилегающие лимфатические сосуды заметно расширены. Альвеолярные простран-
3.13. Стафилококковый энтеротоксин В
1003
ства мультифокально заполнены отечной жидкостью, фибрином, умеренным коли-
чеством больших пенистых альвеолярных макрофагов (large foamy alveolar macro-
phages) и эритроцитами. Обнаружены подострые периваскулиты (subacute periva-
sculitis) в виде умеренных инфильтратов в адвентиции пульмонарных вен и иногда в
адвентиции артериол, включающие лимфоциты, макрофаги и нейтрофилы. Большин-
ство обезьян имели малое! 1раженную острую интерстициальную пневмонию (intersti-
tial pneumonia), характеризующуюся умеренными нейтрофильными инфильтратами
с небольшим количеством эозинофилов в пределах альвеолярных септ и альвеоляр-
ных пространств. Альвеолярные септы были растянуты из-за отека, скоплений фиб-
рина и крови. Характерной особенностью смертельного для обезьян ингаляционно-
го поражения SEB было обнаружение в пределах застойных легочных сосудов и аль-
веолярных септальных капилляров больших лимфоцитов с частыми митозами. Эти
внутрисосудистые лимфоциты имели размер 25—30 мкм в диаметре и богатую цитоп-
лазму с овальным ядром с исчерченным хроматином и от одного до двух хорошо за-
метных ядрышек. Митотическая величина составляла до одного митоза на два поля
зрения микроскопа под большим увеличением (рис. 3.145).
Эпителий дыхательных путей разных уровней оеспираторного тракта имел сход-
ные микроскопические изменения. Собственная пластинка (lamina propria — рыхлый
Рис. 3.145. Гистологические признаки поражения легких обезьяны мелкодисперсным
аэрозолем SEB. А. Перибронхиальный и периваскулярный интерстиций, заметно рас-
ширен из-за отечной жидкости, смешанной с экстравазированными эритроцитами и
неоольшим количеством воспалительных клеток. Стрелками показаны расширенные
лимфатические сосуды. Увеличено в 24 раза. Б. Адвентиций легочной вены, инфиль-
трированный лимфоцитами, макрофагами и нейтрофилами. Обратите внимание на
интраваскулярные лимфоциты. Увеличено в 200 раз. В. Растянутые отечной жидко-
стью лимфатические сосуды и большое количество лимфоцитов. Увеличено в 400 раз.
Врезка: расположенный внутри сосуда СОЗ-иммунореактивный лимфоцит. Авидин-
биотиновый метод окрашивания. Увеличено в 400 раз. По М. Е. Mattix et al. (1995)
1004
Часть 3. Частная неправильная эпидемиология
слой соединительной ткани под эпителием) трахеи и бронхов была заметно расши-
рена из-за отечной жидкости и небольшого количества инфильтрирующих лимфоци-
тов, плазмоцитов и нейтрофилов. Отек и клеточные инфильтраты распространялись
на подслизистую оболочку трахеи и средостенье. Лимфатические сосуды средостенья
выглядели застойными. Лимфатические ткани респираторного тракта имели измене-
ния, сходные с такими же в лимфоидных тканях других участков тела животного, под-
вергнутого ингаляционному воздействию SEB, с истощением В-клеточнозависимых
областей и гиперплазией Т-клеточнозависимых. У четырех обезьян из шести подсли-
зистая трахеи имела множественные дискретные лимфоидные узелки. Ассоциирован -
ные с бронхами лимфоидные ткани (bronchus-associated lymphoid tissue, BALT) трех
обезьян были редуцированы, имели признаки паракортикальной лимфоидной гипер-
плазии и инволюцию фолликулярных герминальных центров.
При улътраструктурном исследовании обнаружено небольшое количество свобод-
ных митохондрий в альвеолярных капиллярах и альвеолярном пространстве. Лимфо-
циты имели крупные ядра с неровным контуром, обильный эухроматин, окаймлен-
ный гетерохроматин и одно или два ядрышка (рис. 3.146).
Разрозненные нейтрофилы, эозинофилы и моноциты были обнаружены в капил-
лярах. Эндотелиальные клетки и пневмоциты I типа имели многочисленные, тесно
упакованные пиноцитотические везикулы (pinocytotic vesicles) и нормальное внутри-
клеточное соединение. Альвеолярные септальные интерстициальные пространства
утолщены из-за накопления отечной жидкости, иногда формирующей субэпители-
альные пузыри, которые разделяют пневмоциты первого типа от подлежащей базаль-
ной мембраны. Отечная жидкость и фибрин заполняют альвеолярные пространства.
Рис. 3.146. Ультраструктурные признаки поражения легких обезьяны мелкодис-
персным аэрозолем SEB. Крупные лимфоциты в состоянии митоза в пределах аль-
веолярного капилляра содержат цитоплазму с большим количеством полирибо-
сом и немногочисленные митохондрии. Эндоплазматический ретикулум плохо
выражен. Цитоплазма эндотелиальных пневмоцитов и пневмоцитов первого типа
содержит большое количество пиноцитотических везикул (показаны стрелками).
Электронная микроскопия, увеличение в 8000 раз. По М. Е Mattix et al. (1995)
3.13. Стафилококковый энтеротоксин В
1005
М. Е. Mattix et al. (1995) считают, что к снижению периферической сосудистой
резистентности, гипотензии и отеку легких приводят большие количества цитокинов
(IL-2, IFN-y, LT, TNF-a и др.), попавшие в кровь в результате индуцированной SEB
поликлональной активации Т-клеток и митогенеза.
Клиника парентерального поражения SEB. Внутривенное введение обезьянам SEB
вызывает развитие поражения, сходного с тем, что имеет место при их ингаляцион-
ном экспонировании к токсину: обратимая гипотензия, периферический застой крови
и летальный исход на фоне отека легких после латентного периода различной продол-
жительности. Схожи макроскопические и гистопатологические изменения легочной
ткани. Исключение составляют дегенерация и некроз эндотелиальных клеток, выс-
тилающих альвеолярные капилляры, описанные только у обезьян, которым SEB вво-
дили внутривенно (Finegold М. J., 1967).
Диагне тика поражения SEB. Энтеротоксины продуцируются коагулаза-положи-
тельными стафилококками Продукты, содержащие стафилококковый энтеротоксин,
по внешнему виду, вкусу и запаху не отличаются от доброкачественных продуктов.
Факторами, способствующими естественному образованию энтеротоксина в продук-
тах, является pH среды 6,9—7,2, наличие углеводов, белков, повышение температуры
до 25—30°С. При температуре ниже 20°С и pH 6,5 образование энтеротоксина замед-
ляется, а при температуре ниже 10°С и pH 6,0 прекращается.
Стафилококки хорошо размножаются в молоке и молочных продуктах (пирожные,
кремы). Молоко и молочные продукты могут быть контаминированы непосредствен-
но от коров, больных стафилококковыми инфекциями, реже — от больных людей.
Стафилококковые энтеротоксины образуются более интенсивно в полуанаэробных
условиях. В простокваше, кефире, твороге образования энтеротоксина не происхо-
дит вследствие повышенной кислотности этих продуктов, а в мо{ оженом — в связи с
его низкой температурой. В этих продуктах стафилококковый энтеротоксин может
быть обнаружен лишь в том случае, если эти продукты изготовлены из молока или
молочной смеси, заведомо обсемененных стафилококками и содержащих токсин.
Благоприятной средой для продуцирования энтеротоксина стафилококка является
печеночный паштет. В этом продукте стафилококковый энтеротоксин образуется при
комнатной температуре в течение 24—30 ч. Сырое и отварное мясо — менее благопри-
ятная среда для продуцирования стафилококками энтеротоксина, чем фарш.
При естественном поражении SEB должны прослеживаться:
1) энтеральный путь проникновения в организм человека продукта, контамини-
рованного токсином;
2) наличие размножившихся коагулаза-положительных стафилококков (проду-
центов токсинов) в продукте, ставшем причиной отравления;
3) резервуар стафилококка, из которого он попал в пищевой продукт.
Наличие в продукте только SEB, без размножившихся стафилококков, можно от-
нести к неправильной эпидемиологии пищевого отравления. Для подтверждения
правильности эпидемиологии в дальнейших лабораторных исследованиях целесооб-
разно проверить выделенный из продукта питания стафилококк на способность про-
дуцировать тот тип токсина, которым было вызвано поражение. В случаях стафило-
коккового пищевого отравления симптомы со стороны легких не наблюдаются.
Как и в случае ботулинических токсинов, для диагностирования интоксикации,
вызванной вдыханием мелкодисперсного аэрозоля SEB, необходимо эпидемиологичес-
кое, клиническое и патологоанатомическое исследование. При однократном приме-
нении аэрозоля SEB, у людей, пораженных токсином, клинические признаки отрав-
1006
Часть 3. Частная неправильная эпидемиология
ления дадут о себе знать в течение 3—12 ч. Поскод] ку рвота может быть первым сим-
птомом ингаляционного поражения SEB, который обратит на себя внимание врачей,
то исключить возможность массового пищевого отравления можно на основании
обнаружения связи вспышки с каким-то общим географическим фактором (аэропорт,
кинотеатр, метро, учреждение и др.), но без указаний на прием общей пищи. Послед-
нее обстоятельство говорит о неправильной эпидемиологии пищевого отравления.
Среди пациентов с симптомами пищевого отравления и с неправильной эпиде-
миологией болезни, должны обратить на себя внимание те:
1) у которых возникли симптомы ОРЗ, и, в частности кашель, выраженная отдыш-
ка с хрипами на вдохе. У таких пациентов рентгенологически можно обнаружить при-
знаки интерстициального отека легких,
2) пациенты, у которых течение болезни приобрело двуфазный характер. т. е. после
периода улучшения у них появились признаки дыхательной недостаточности и про-
грессирующей летаргии. Такое течение болезни свидетельствует об ингалировании
смертельной дозы SEB.
Поскольку симптомы SEB-интоксикации могут быть аналогичны симптомам,
вызываемым несколькими респираторными патогенами, такими как вирус гриппа,
аденовирус и микоплазмы, вначале при постановке диагноза могут возникнуть сомне-
ния. Для всех них характерны высокая температура, сухой кашель, боль в мышцах и
головная боль. В этом случае основной критерий — время. При нападении с приме-
нением SEB основное количество больных появится через период времени, не пре-
вышающий 24 ч, в то время как естественно возникающие вспышки гриппа, адено-
вирусной или микооплазменной инфекции развиваются у людей за более тигельный
промежуток времени.
Интоксикация, вызванная инкапситирующими дозами SEB, имеет тенденцию
быстро достигать довольно стабильного клиническо! о состояния, в го время как инга-
ляционная сибирская язва, туляремийная пневмония или легочная чума без лечения будут
прогрессировать. Туляремия и чума, так же как и лихорадка Ку, ассоциируются с натичи -
ем инфильтратов на рентгенограммах легких, в отличие от SEB-ингоксикации в ин-
капситирующих дозах. В летальных дозах поражение SEB проявляется клинически-
ми и рентгенологическими признаками отека легких и двуфазным течением болезни.
И нтоксикация при воздействии ОВ нервно-паралитического действия должна выз-
вать фасцикуляцию и гиперсекрецию, а иприты и ипритно-люизитные смеси должны
вызвать поражение кожи, помимо поражения легких. Одышка, связанная с ботули-
нической интоксикацией, ассоциируется с явными признаками мышечного паралича,
бульбарного паралича, отсутствием лихора. щи и сухими легочными гл тями вследствие
холинергической блокады. Затрудненное дыхание характерно для поздней стадии
ботулизма, тогда как при вдыхании SEB расстройство дыхания возникает на ранней
стадии болезни (Franz D.R. et al., 1997). Критерии дифференциальной диагностики
поражений, вызванных ОВ нервно-паралитического действия, ботулиническим ток-
сином и SEB, приведены в табл. 3.49.
Поражения легких летальными дозами мелкодисперсного аэрозоля SEB представ-
ляют собой сочетание местного суперантигенного действия токсина, приводящего к
интенсивной воспалительной реакции и нарушению проницаемости сосудов в месте
аппликации токсина, и общей реакции, вызванной индукцией токсином мощного
каскада провоспалительных цитокинов. Поэтому поражения легких мелкодисперс-
ным аэрозолем SEB в отличие от поражения, вызванного аэрозолированным рицином
(см. разд. 3.11), носят более обширный, но менее разрушительный характер, и на на-
3.13. Стафилококковый энтеротоксин В
1007
Таблица 3.49
Дифференциальная диагностика поражений нервно-паралитическим ОВ,
ботулиническим токсином и SEB*
Симптомы поражения ОВ нервно-пара- литического действия Ботулинический токсин SEB
Время после экспо- нирования, в течение которого появляются симптомы поражения Минуты Часы (24—72) Часы (1-12)
Симптомы со стороны ЦНС Конвульсии, спа- стическое сокра- щение мышц Прогрессивный паралич Головная боль, боли в мышцах
Кардиоваскулярные симптомы Брадикардия Нормальный сердечный ритм Нормальный сердечный ритм или тахикардия
Респираторные симптомы Трудности в дыха- нии, развитие непроходимости воздушных путей Сначала дыхание нормаль- ное, затем прогрессирую- щее развитие паралича дыхательной мускулатуры Сухой кашель (отдель- ные случаи), боли в грудной клетке/затруд- нения в дыхании
Желудочно-кишеч- ные симптомы Увеличение пери- стальтики кишеч- ника. боль, диарея Замедление перистальтики Тошнота, рвота и/или диарея
Окулярные симптомы Маленькие зрачки Опущение века Можно наблюдать «симптом красных глаз» (конъюн ктивальная инъецированность)
Сальвационные симптомы Профузная салива- ция, жидкая слюна В норме, но глотание затруднено Количество слюны может быть несколько увеличено
Смерть Минуты 2—3 сут Маловероятна
Реакция на введение атропина Есть Нет Введение атропина может уменьшить желудочно-кишечные симптомы
* По D.R. Franz et al. (1997).
чальной стадии болезни не ограничиваются патологией бронхиол и альвеол. При
поражении рицином на первый план выходит гнойный некроз терминальных брон-
хиол, сопровождающийся отеком, с последующим развитием мультифокального сли-
вающегося некроза долей легких, т. е. гнойно-фибринозной пневмонии. В поражении
SEB преобладают признаки нарушения проницаемости капилляров. Отек может ох-
ватывать сра sy несколько долей легкого Развивается острая интерстициальная пнев-
мония без некротических изменений в паренхиме легких. Мощный выброс противо-
воспалительных цитокинов приводит к тому, что лимфоидная ткань респираторного
тракта имеет признаки паракортикальной лимфоидной гиперплазии с истощением
В-клеточнозависимых областей и гиперплазией Т-клеточнозависимых, а в пределах
застойных легочных сосудов и альвеолярных септальных капилляров обнаруживают
крупные лимфоциты с частыми митозами.
Данные лабораторного исследования не оказывают большой помощи при диаг-
ностировании SEB-интоксикации. Неспецифический нейтрофильный лейкоцитоз
1008
Часть 3. Частная неправильная эпидемиология
и повышенная скорость оседания эритроцитов могут быть обнаружены через 12 ч по-
сле экспонирования. Токсин очень трудно обнаружить в сыворотке к моменту появ-
ления симптомов. Данные исследований на животных свидетельствуют о том, что при-
сутствие SEB в сыворотке является кратковременным; однако SEB или его антигенные
метаболиты накапливаются в моче и могут быть обнаружены через несколько часов
после экспонирования.
Поскольку у пациентов вырабатывается большое количество антител к токсину,
для ретроспективного диагноза необходимо брать пробы сывороток в острый период
и период выздоровления, так как это может помочь при установлении точного диагно-
за. Токсин может быть идентифицирован методом твердофазного иммунофермент-
ного анализа (ELISA) в носовых смывах, полученных через 24 ч после аэрозольного
экспонирования (Franz D. R. et al.,1997)1.
Иммунопрофилактика. Не разработана.
Лечение. В настоящее время лечение ограничивается поддерживающей терапией.
В тяжелых случаях с отеком легких могут потребоваться вентиляция с положительным
давлением в конце выдоха и мочегонные средства. Терапевтическое значение стерои-
дов вызывает споры среди клиницистов. Предполагается, что большинство людей,
пораженных инкапситиру ющими дозами SEB, должны выздороветь после начальной
стадии острой фазы заболевания и будут в состоянии вернуться к нормальной деятель-
ности через 1-2 недели (Franz D R. et al.,1997).
3.14. САКСИТОКСИН
Естественна эпидемиология отравления STX. Молекулярная структура и ме-
ханизм действия. Генетика токсинообразования. Токсические и поражающие
свойства. Клиника энтерального поражения. Диагностика поражения. Имму-
нопрофилактика. Лечение.
Сакситоксин (saxitoxin, STX; устричный яд, паралитический яд моллюсков) —
низкомолекулярный, близкий по химической структуре с тетродотоксином (tetro-
dotoxin, ТТХ) нейротоксин алкалоидной природы, вызывающий паралич дыхатель-
ной мускулатуры. Оба токсина в годы Второй мировой войны изучались и, возмож-
но, использовались как диверсионные яды британскими (STX) и японскими (ТТХ)
спецслужбами. Летальность среди людей, отравившихся продуктами питания, содер-
жащими STX, достигает 15%. STX рассматривается как агент для осуществления био-
логических террористических актов, диверсий и действий криминального характера.
Существует теоретическая возможность применения STX в составе аэрозоля. Анти-
дотов нет. Под условным наименованием TZ находился на вооружении армии США
(Антонов Н. С., 1994; Wikipedia). Сопоставление биологических и фармакологичес-
ких свойств STX и ТТХ приведено в табл. 3.50.
1 Главным государственным санитарным врачом России Г. Г. Онищенко утверждены Ме-
тодические указания (МУК 4 2 2429-08) «Метод определения стафилококковых энтеротокси-
нов в пищевых продуктах». Метод основан на непрямом твердофазном иммуноферментном
анализе. Согласно методическим указаниям, предел обнаружения стафилококковых энтеро-
токсинов соответствует диапазону массовых концентраций от 0,2 до 2 мкг/кг (в зависимости
от типа токсина и вида продукта).
3.14. Сакситоксин
1009
Таблица 3.50
Биологические и фармакологические свойства STX и ТТХ
Свойство STX ТТХ
1 2 3
Систематическое название (За5-(За-а,4-а,10а7?*))-2,6-диами- но-4-(((амино-карбонил)окси)ме- тил)-3а,4,8,9-тетрагидро-1Н, 10Н- пиролло( 1,2-с)пурино-10,10-диол - (триалкилтетрагидропурин) (4/?,4аЯ,57?,65,75,85,8аЛ, 105,125)-2-аза- ниумилиден-4,6,8,12-тетрагидрокси- 6-(гидроксиметил)-2,3,4,4а,5,6,7,8-окта- гидро- 1Я-8а, 10-метано-5,7-(епоксиме- таноокси)квиназолин-10-олат
Аналоги Известно не менее 57 природных аналогов, возможно конструирова- ние аналогов, не встречающихся в природе Известно не менее 13 природных анало- гов, возможно конструирование анало- гов, не встречающихся в природе
Продуцент Продуцируется некоторыми дино- флагеллятами (Gonyaulax catenella, Alexandrium spp., Gymnodinium spp., Pyrodimum spp.), цианобактериями (Anabaena dranalis, Aphanizomenon spp., Aphanizomenon gracile, Cylindros- permopsis raaborskii и Lyngbya wdlei, Planktothrixspp.), брюхоногими, головоногими, рыбами (Colomesus asellus и др.) Большое количество ТТХ содержится в рыбе фугу, (особенно в печени и в икре), в калифорнийском тритоне (Taridia torasa), а также ТТХ встречается у ряда бычковых рыб (Gobiidae), у синекольчатого осьминога, в коже и яйцах костариканских лягушек рода Ателопы (Atelopus), в тканях краба Ateigatis floridus и других животных
Внешний вид Бесцветные кристаллы Бесцветные кристаллы
Брупо-формула (система Хилла) ^10^17^7^4 C13H21N3O10
ММ, Да 299,29 379,32
Температура плавления, °C Нет данных 225
Температура раз- ложения, ’С ПО 225
Растворимость (в г/100 г раст- ворителя или характеристика) Растворим в воде, метаноле и эта- ноле, нерастворим в неполярных органических растворителях, устойчив в кислых средах Плохо растворим в воде, хорошо — в слабом растворе уксусной кислоты, диэтиловом эфире и этаноле
Способы получения Ассиметричный синтез из эфира сульфомата Многостадийный синтез, исходя из левоглюкозенона. Многостадийный синтез из 2-анетокси- три-О-ацетил-О-глюкаля. Многостадийный синтез из D- изоаскорбиновой кислоты
Показатели диссоциации Нет данных РКвн( ) > (-0 С, вода)
Летальная доза (LD50, мг/кг) 0,008 (мыши, подкожно), 0,002 (человек, перорально) Белые мыши, внутримышечно — 0,008; мыши, внутрибрюшинно — 0,008; человек, перорально — 0,01
Механизм действия Эффективно блокирует потенциал действия натриевых и кальциевых каналов на мембранах нейронов и калиевые каналы клеток сердечной мышцы Эффективно блокирует потенциал действия натриевых каналов. На калиевые каналы не действует
1010
Часть 3. Частная неправильная эпидемиология
Продолжение табл. 3.50
1 2 3
Симптомы острого отравления Первые симптомы отравления STX после приема в пищу зараженных моллюсков появляются через 30 мин и выражаются в онемении языка, губ, кончиков пальцев. Быстро развивается сердечно-сосу- дистая и дыхательная недостаточ- ность, которая может привести к смертельному исходу через 1-12 ч вследствие остановки дыхания Через 10—45 мин появляются зуд губ, языка и дру1 их частей тела, отмечаются обильное слюнотечение, тошнота, рво- та, понос, боли в животе. Возникают подергивания мышц, потеря чувстви- тельности кожи, затрудняется глотание, развивается зфония. Смерть наступает от паралича дыхательных мышц
Период полувы- ведения из орга- низма человека, мин 90 Нет данных
Нагревание до температуры 100 °C при pH 1,0 в течение 25 мин Не оказывает воздействия на токсичность Устраняет токсичность
* По J. R. Deeds et al. (2008), М. Wiese et al. <2010) и Y Kudo et al. (2012).
Естественна эпидемиология отравления STX. В морской воде STX продуцируется
свободноживущими простейшими, относящимися к отряду Dinoflagellata (Gonyaulax
catenella, Alexandrium spp., Gymnodinium spp., Pyrodinium spp.); в пресноводных водо-
емах — цианобактериями (Anabaena circinalis, Aphanizomenon spp., Apf'anizomenon gra-
cile, Cylindrospermopsis raciborskii и Lyngbya wollei, Planktothrix spp.). К вторично-ядови-
тым животным, аккумулирующим STX по пищевым цепям, относятся брюхоногие
моллюски (Gastropoda), головоногие (Cephalopoda), рыбы (Colomesus asellus и др.) и
крабы (Zosimus acneus, Atergatisfloridis, Platypodia granulose и др.), обитающие на корал-
ловых рифах. У двустворчатых моллюсков STX накапливается в пищеварительной си-
стеме. Например, у мидий (Mytilus) до 95 % токсина накапливается в гепатопанкре-
асе. У S. giganteus около 70 % токсина собирается в сифоне, где он может сохраняться
до нескольких месяцев. Отравления людей происходят из-за употребления в пищу
вторично-ядовитых животных, в основном моллюсков в периоды «цветения» диноф-
лагеллят, т. е. с конца весны до осени (рис. 3.147).
География отравлений STX включает регионы планеты, на побережьях которых
происходят так называемые «красные приливы» (побережье Аляски, Вашингтона.
Чили, Канады, Испании, Тайваня, Исландии и др.) (Deeds J. R. et al., 2008). Частота
«красных приливов» прямо связана с антропогенным загрязнением прибрежных вод.
В России токсичные «красные приливы», сопровождающиеся гибелью людей, досто-
верно отм( чены только в тихоокеанских водах Камчатки. В бухте Павла в 1945 г. от
использования в пищу вареных мидий произошло отравление со смертельным исхо-
дом двух членов экипажа судна флотилии «Алеут», в Авачинской бухте в 1973 г. —
сильное отравление нескольких жителей Петропавловска-Камчатского (Коновало-
ва Г. В., 1995).
Количество отравленных морепродуктами, содержащими STX, обычно не пре-
вышает десятка человек. Наибо tee массовые отравления зафиксированы в Китае в
1979 г., 2002 г. и 2004 г., когда количество отравленных составило 68, 50 и 55 человек
3.14. Сакситоксин
1011
Рис. 3.147. Ядовитые динофлагелляты. А. Рисунки динофлагеллят: I — Gymnodinum
breve; 2 — Gonyaulax catenella (увеличено в 1800 раз); 3 — Gonyaulax tamarensis (уве-
личено в 1500 раз); 4 — Prymnesiumparvum (увеличено в 5000 раз). По Б. Н. Орлову
и Д. Б. Гелашвили (1985). Б. Фотография G. tamarensis, сделанная сканирующим
электронным микроскопом (увеличено в 2820 раз). По J. Acres, J. Gray (1978)
соответственно. Во всех случаях причиной отравления стало употребление в пишу
морских брюхоногих моллюсков из семейства Nassariidae (Deeds J. R. et al., 2008).
Источники и концентрация STX в тканях вторично-ядовитых морских животных при-
ведены в табл. 3.51, с. 1012.
В развитых странах содержание STX в морепродуктах обязательно контролиру-
ется. В странах Евросоюза допустимые нормы содержания сакситоксина составляют
от 40 до 80 мкг/100 г пробы. В нормативном документе Госкомсанэпиднадзора РФ
(СанПиН 2.3.4.050-96 от 11 марта 1996 г. №6 «Производство и реализация рыбной про-
дукции») при производстве и реализации рыбной продукции приведены требования
к качеству продукции. В п. 6.10.4. этого документа указано: при экспорте живых дву-
створчатых моллюсков количество паралитического яда (сакситоксина), содержа-
щегося в съедобных частях моллюсков (тело целиком или любая отдельно взятая его
съедобная часть), не должно превышать 80 мкг на 100 г мяса моллюсков (Коновало-
ва Г. В.. 2006).
Молекулярная структура и механизм действия. STX входит в эмпирически соб-
ранную группу паралитических токсинов моллюсков (paralytic shellfish toxins, PSTs).
Химическая природа STX установлена в 1960—1970 гг. с помощью точных физико-
химических методов — рентгеноструктурного анализа и ЯМР-спектроскопии. Ток-
син представляет собой водорастворимое производное пурина (триалкилтетрагидро-
пурин), образующее соли с минеральными кислотами. Молекула STX состоит
1012
Часть 3. Частная неправильная эпидемиология
Таблица 3.51
Источники и концентрация SIX в тканях вторично-ядовитых морских животных*
Продуцент (Dinoflagellata) Вторично-ядовитое животное Максимальное содержание STX Географическое положение
Alexandrium acarenella Лунная улитка Льюиса 176—600 мкг STX/100 г ткани Побережье в Британ- ской Колумбии, Канада
Argobuccinum SPP- Брюхоногий моллюск 5629 мкг STX/100 г (пищеварительная система) Побережье Чили
Lunatia heros Северная лунная улитка 1450 мкг STX/100 г ткани ПобереЖоС Массачусетса, США
B. undatum Волнистый моллюск 3337 мкг STX/100 г ткани Залив Мэн, США
A. catenella Красный краб 285 мкг STX/100 г в ткани внутренних органов Побережье Вашингтона, США
Источник не известен Волосатый краб 2723 мкг STX/100 г ткани внутренних органов Побережье в префек- туре Фукусима, Япония
A. tamarense Американский лобстер 1512 мкг STX/100 г гепатопанкреаса Залив Гаспе, Канада
P bahamense Рыба-собака флорид- ская (южная разно- видность рыбы фугу) 1443 мкг STX/100 г печени; 14571 мкг STX /100 г мышечной ткани Побережье Флориды, США
A. tamarense Японская скумбрия 2800 мкг STX/100 г мышечной ткани; 500 мкг STX/100 г пече- ни; 72 мкг STX/100 г жабер Побережье Аргентины
* Составлено по работе J. R. Deeds et al. (2008).
из тетрагидропуринового кольца, соединенного с двумя оста .ками гуанидина, игра-
ющими основную роль в токсическом действии. При изменении pH установлено, что
активной формой STX является катионная, однако при повышении pH более 9 еди-
ниц, он становится неэффективным (Орлов Б. Н., Гелашвили Д. Б., 1985).
С хлористым водородом STX образует хлоршдрат. ММ STX в форме хлор гид-
рата — 354 Да, а в виде свободного основания — 299,29 Да, т. е. молекула STX только
в 2 раза больше, чем молекула зарина. Дихлоргидрат STX представляет собой белый
гигроскопичный порошок, хорошо растворимый в воде, ограниченно растворимый
в спиртах и ледяной уксусной кислоте, нерастворимый в органических растворите-
лях и жирах. В форме дихлоргидрата STX является устойчивым веществом, плавящим-
ся при температуре 200 °C с разложением. Выдерживает нагревание в водных раство-
рах в автоклавах при температуре до 120 °C без потери активности. Допускает перевод
в аэрозольное состояние диспергированием растворов и, видимо, методом термиче-
ской возгонки (Антонов Н. С., 1994).
На начало 2010 г. установлено 57 производных STX, различающихся по пяти по-
зициям в структуре молекулы. Разнообразие аналогов STX в природе вызвано присо-
единением по вариабельным позициям молекулы (variable positions) гидроксильных,
сульфатных или карбамоилатных групп. Поэтому аналоги STX структурно классифи-
цируют наследующие классы: несульфатированные (non-sulfated), моносульфатиро-
ванные (mono-sulfated), дисульфатированные (di-sulfated), декарбамоилированные
(decarbamoylated), деоксидекарбомоилированные (deoxy-decarbomoylated), L. wollei -
токсины (Z. wollei toxins), моногидроксибензоат-аналоги (mono-hydroxy-benzoate ana-
logs), дигидроксибензоат-аналоги (di-hydroxy-benzoate analogs), сульфатированные
3.14. Сакситоксин
1013
беанзоат-аналоги (sulfated benzoate analogs) и другие аналоги паралитических токси-
нов моллюсков (Wiese М. et al., 2010). Аналоги различаются между собой по токсич-
ности для животных и человека (рис. 3.148; табл. 3.52).
Рис. 3.148. Основная химическая структура STX (А) и его наиболее токсичного
аналога — зетекитоксина АВ (Б). R — вариабельные позиции. Детальный пере-
чень аналогов STX приведен в работах L. Е. Llewellyn (2006) и М. Wiese et al. (2010)
Сравнительная токсичность аналогов STX*
Таблица 3.52
Структура Токсин Относительная токсичность**
Несульфатированные 'sfx 1
NeoSTX 0,5-1,1
Моносульфатированньк GTX1 и GTX4*’* 0,39/1,09-0,48/0 76
GTX2 и GTX3*** 0,8/0,33-0,9/0,9
Декарбамоилированные dcSTX 0,43
dcNeoSIX 0,43
dcGTXl—4 0,18-0,45
Дисульфатированные Cl—4 <0,01-0,14
* По рабоп М. Wiese et al. (2010).
** Сравнительные исследования токсичности STX и его аналогов выполнены в опытах
на мышах.
*** a/p-эпимерные смеси.
Количество известных аналогов STX ”величивается по мере совершенствования
методов их идентификации и расширения географии исследования продуцентов.
Большинство из аналогов STX обнаружено у цианобактерий и динофлагеллят. У па-
намской золотой лягушки Atelopus zeteki обнаружен самый экзотический и токсичный
аналог STX, получивший название зетекитоксин АВ (zetekitoxin АВ). Зетекитоксин АВ
избирательно связывается с натриевыми каналами электровозбудимых мембран мозга,
сердца и мышц, проявляя активность в 580, 1 би и 63 раза большую, чем STX cooi вет-
ственно (Yotsu-Yamashita М. et al., 2006; Wiese М. et al., 2010).
Кроме STX, из динофлагеллят G. tamarensis (G. excavata) и Protogonyaulax catenella
были выделены токсины, очень близкие к STX по химической природе и фармаколо-
гическому действию, — гониаутоксины 1 -6 (GTX1—GTX6) и неосакситоксин (neoSTX).
Сначала они рассматривались токсикологами как самостоятельные соединения (Ор-
лов Б. Н., Гелашвили Д. Б., 1985). Но в более поздней работе М. Wiese et al. (2010) они
включены в перечень аналогов STX.
1014
Часть 3. Частная неправильная эпидемиология
STX встречается в природных продуцентах в сложной смеси со своими аналога-
ми, что делает невозможным получение специфического антидота, но позволяет пред-
положить его источник по характерному токсическому профилю (табл. 3.53).
Таблица 3.53
SIX и его аналоги в природных продуцентах*
Продуцент Токсический профиль
Dinoflagellates
Alexandrium acatenella STX
A. andersoni STX, NEO
A. angustitabulatum Неизвестная композиция токсинов
A. catenella STX, GTX1-4, NEO.B1-2, Cl-4
A. cohort iadа STX, GTX1-4
A. fundyense STX, NEO, GTX1-4, Cl-2, Bl
A. minutum (A lusitanicum) GTX1-4
A. ostenfeldii GTX2-3, B2, Cl-2
A. tamarense STX, NEO, GTX1-4, Bl, Cl, C2, C4
A. tamiyavanichi STX, GTX1-4, Bl, Cl-4
Gymnodinium cate, Mt urn STX, NEO следы GTX2-3, Bl-2, Cl-4
Pyrodimum bahamense STX, NEO, B1-B2
Cyanobacteria
Anabaena aranahs STX. GTX1-4, C1-C2, dcGTX2 3
Anabaena lemmermawtu STX
Aphanizomenon gracile STX, NEO
Aphanizomenot. issatschenkoi NEO, SfX
Cylindrospermcpsis radborskii STX, NEO, GTX2-3
Lyngbya wallet dcSTX, dcGTX2-3, ацетилированные аналоги STX
Planktothrix spp. STX
Rivularia spp. GTX2, GTX4
* По J. R. Deeds et al. (2008).
Не встречающиеся в природе аналоги STX могут быть получены химическим пу-
тем. В. М. Andresen, J. Bois (1009) описали последовательность этапов химического
синтеза, позволяющего получить карбамат-модифицированную форму STX, не отли-
чающуюся значительно по суб'тра'. рецепторному связыванию от исходного токсина.
STX можно получить путем ассиметричного химического синтеза. Синтез N,O-
ацеталя начинают из легко доступного эфира сульфомата (производное глицерина,
sulfamate ester) путем С—Н-аминирования. Полный путь синтеза STX осуществляют
на модели гетероцикла оксатиазинандиоксида ([l,2,3]-oxathiazinane-2,2-dioxide
heterocycle) для сборки полифункцианизированных амино-производных. В конечной
стадии получения токсина формируется промежуточное соединение — необычный
девятичленный циклический гуанидин. Его селективно окисляют и подвергают раз-
рушающей дегидратирующей циклизации с формированием трициклического ядра
природного STX (Fleming J. J., Du Bois J., 2006).
После употребления контаминированных моллюсков в пищу происходи ! быстрое
абсорбирование STX из желудочно-кишечного тракта. Гуанидиновый остаток по сво-
3.14. Сакситоксин
1015
им молекулярным размерам соответствует диаметру гидратированного иона Na+. Он
входит в натриевый канал электровозбудимой мембраны нервной и мышечной ткани,
застревает в нем и удерживается остальной частью молекулы, размеры которой пре-
вышают диаметр канала. В результате прерывается передача потенциалов действия в
возбудимые клетки, передача сигнала вдоль нерва прекращается. Это ведет к дисфун-
кции нервно-мышечных соединений, а в тяжелых случаях к смерти вследствие пара-
лича дыхательных мышц и остановки дыхания (рис. 3.149).
Рис. 3.149. Модель связывания (+)-STX с порой натриевого канала нейрона. На-
ружное входное отверстие канала включает фильтр для селекции ионов и рецеп-
торный сайт лля STX. В этом участке поры ею формируют пять карбоксильных
остатков (D400, Е755, Е403, Е758, D1532); их присутствие играет критическую
роль для связывания в поре 7,8,9-гуанидин-5ТХ По В. М. Andresen, J. Bois (2009)
Блокирующее действие STX развивается не только на периферическом уровне.
Эксперименты, проведенные на кошках и кроликах с различными способами введе-
ния STX (bhj гривенное и внутрижелудочковое), показали, что он может проникать
через гематоэнцефалический барьер и вызывать прямое угнетение дыхательного и
сосудодвигательного центров Существует видовая чувствительность к STX — рыбы,
моллюски и амфибии более устойчивы, чем теплокровные (например, мыши) (Bori-
son Н. L. et al., 1980; Орлов 6. Н., Гелашвили Д. Б., 1985).
I эдетика токсинообразования. Биосинтез SIX цианобактерией Cylindrospermopsis
raciborskii ТЗ определяется кластером генов (sxt) протяженностью около 35 т. п. о.,
включающим 31 ORF и гены 26 белков, выполняющие 30 каталитических функций.
Кластер включает гены О-карбамиолтрансферазы (sxtf), метилтрансферазы (sxtAl),
второй класс аминотрансфераз (sxtA4), амидотрансферазу (sxtG), дегидрогеназу (dioxy-
genases, sxtH) и др. (рис. 3.150, с. 1016).
Биосинтез STX начинается с аргинина, S-аденозилметионина (S- adenosylmethio-
nine) и ацетата посредством поликетидсинтазы (polyketide synthase) нового гипа,
котирая, как предполагается, выполняет метилирование ацетата и конденсационную
реакцию Клайсена (Claisen condensation reaction) между пропионатом и аргинином.
Следующие этапы включают катализируемые ферментами три гетероциклизации (he-
terocyclizations) и различные сшивающие реакции, которые приводят к образованию
многочисленных аналогов STX (Kellmann R. er al., 2008).
Кластер sxt-генов цианобактерий высококонсервативен. По данным S. A. Murray
et al. (2011), он возник не позже 2,1 млрд лет назад, т. е. за 1,5 млрд лет до появления
многоклеточной жизни и, разумеется, натриевых каналов нейронов. A. Stuken et al.
1016
Часть 3. Частная неправильная эпидемиология
Ч""! п.
0 4000 8и00 12000 16000 20000 24000 28000 32000 36000 38000
STX-связывание
ж Предполагаемая
ш регуляторная субъединица
Транскрипционный
регулятор
D Неизвестный ген
гл Инактивированная/частич-
HaaORF
: _/ ORF снаружи sxt-кластера
> Трансфераза
А Гидролаза
В Гидроксилаза
Оксидоредуктаза/пере-
носчик электронов
Рис. 3.150. Структурная организация .ос/-гена С. raciborskii ТЗ. 1S4 — вставочная по-
следовательность; ompR — транскрипционный регулятор белков ompR-семеиства',
hisA — двукомпонентная гистидинкиназа; orf24— ORF4. Шкала вверху показывает
протяженность генных кластеров в п. о. По R. Kellmann et al. (2008)
(2011) показали, что транскрипты sxtA динофлагелят имеют ту же доменную структу-
ру, что транскрипты sxtA цианобактерий. Но, в отличие от бактериальных гомологов,
транскрипты sxt динофлагелят моноцистронны, имеют более высокое содержание GC,
встречаются в нескольких копиях, содержащих типичные для динофлагеляты сплай-
синг-лидерные последовательности (spliced-Ieader sequences) и эукариотические
polyA-хвосты.
Сходство структурной организация sxt-кластера цианобактерий и динофлагелят,
а также присутствие генов нескольких транспозаз, связанных с 154-семейством транс-
позонов, предполагают мобильность кассет sxt-генов и возможность их горизонталь-
ного переноса между представителями этих таксонов (Kellmann R. et al., 2008).
Токсические и поражающие свойства. STX, с момента открытия, привлекал внима-
ние специалистов в области химического оружия высокой токсичностью и небольшой
для токсинов ММ. Предполагалось, что можно разработать недорогую технологию его
получения в больших количествах путем химического синтеза. Однако до настояще-
го времени такая технология не разработана. Смертельная доза STX для взрослого
человека (массой 70 кг) составляет (по разным данным) 0,3-1 мг при пероральном и
парэнтеральном введении. Смерть наступает в течение 10 мин. STX может проникать
в организм человека и через открытые раны. В этом случае летальная доза составляет
0,05 мг (Антонов Н. С., 1994; Wikipedia).
В 1989 г. специалистами USAMRIID (Форт-Детрик, штат Мэриленд) было обна-
ружено, что STX при ингаляционном введении мышам примерно в 10 раз токсичнее,
чем при внутривенном и внутрибрюшинном введении. Феномен объясняется тем, что
при ингаляции действие STX нацелено на орган (легкие), а не на организм в целом,
что происходит при внутривенном введении. Гибель животного наступает из-за на-
рушения функционипования органа-мишени. Аналогичное действие наблюдается и
у других токсинов, способных поражать легкие (например, Т-2 токсин). Наоборот, в
случаях, когда органом-мишенью являются не легкие, а другие органы, летальные дозы
токсинов при ингаляции могут быть более высокими, чем при внутривенном введе-
нии. Это наблюдается при ингаляции микроцистина, органом-мишенью которого
3.14. Сакситоксин
1017
является печень. С учетом этого наблюдения расчетное значение летальной дозы сак-
ситоксина для человека при ингаляции равно 5 мгхмин/м3, или в 2 раза меньше, чем
у вещества VX и в 15 раз меньше, чем у зарина. Но вследствие высокой скорости рас-
пада аэрозоля (-17 %/мин) и стоимости STX, наиболее вероятно его использование с
помощью специальных стреловидных элементов (Антонов Н. С., 1994; Wikipedia).
STX, как и некоторые другие низкомолекулярные токсины (ТТХ, бреветоксин,
РТХ, анатоксин и Т-2 токсин), способен вызывать поражения при накожной аппли-
кации. Летальная доза STX при действии через кожу не установлена. В случае бреве-
токсина доза, в 20 раз превышающая летальную дозу при парентеральном введении,
не вызывала смертельных исходов (Антонов Н. С., 1994). Летальные дозы STX для
различных животных приведены в табл. 3.54.
Таблица 3.54
Летальные дозы STX*
Вид животного Способ введения LD50, мг/кг
Мышь Внутривенно 0,0034
Мышь Внутрибрюшинно 0,008
Мышь Перорально 0,26
Крыса Внутривенно 0,012
Крыса Внутрибрюшинно 0,0105
Крыса Перорально 0,19
Морская свинка Перорально 0,135
Кролик Внутривенно 0,003
Кошка Внутривенно 0,0027
Кошка Перорально 0,28
Обезьяна Перорально 0,364
Собака Перорально 0,18
Цыпленок Внутривенно 0,0034
Голубь Перорально 0,091
* По Н. С. Антонову (1994).
Применение полярных растворителей, ускоряющих трансдермальный перенос
веществ, не приводило к заметному снижению величин летальных доз различных
токсинов при накожной аппликации. В литературе применение STX в виде растворов
для нанесения кожно-резорбтивных поражений не обсуж халось (Антонов Н. С., 1994).
Клиника энтерального поражения. Первые симптомы отравления STX после при-
ема в пищу зараженных моллюсков появляются в период от 5 мин и до нескольких
часов после еды, в зависимости от поглощенной дозы, но чаще всего в течение 30 мин.
Они выражаются в онемении языка, губ, кончиков пальцев. Затем немеют шея и ко-
нечности, и обнаруживается двигательная раскоординированность. Поражение череп-
ного нерва может приводить к диплопии и трудностям при речи и глотании. Другие
симптомы могут включать головокружение, ощущение «пустой головы», слабость,
спутанное сознание, потерю памяти и головную боль. Прогрессивно развивается вя-
лый паралич, сердечно-сосудистая и дыхательная недостаточность, которая может
привести к смертельному исходу через 1—12 ч от начала симптомов вследствие оста-
новки дыхания. Пострадавшие обычно сохраняют сознание и память, вплоть до на-
1018
Часть 3. Частная неправильная эпидемиология
ступления смерти. Токсин выводится из организма в течение первых суток посред-
ством почечной экскреции; однако для полного неврологического восстановления
может потребоваться от 7 до 14 суз (Usamriid’s medical management..., 2001).
По данным С. Garcia et al. (2005), описавшим четыре случая отравления STX в
Чили, первые симптомы отравления у пациентов появились через 5 мин после упот-
ребления в пищу двух сырых ребристых мидий (ribbed mussel). Они включали чувство
онемения в полости рта и кожи лица, рвоту, слабость верхних и нижних конечностей,
головную боль, абдоминальную боль. Симптомы прогрессирующей дыхательной не-
достаточности и гипотония, развились в течение 40 мин. Было установлено, что в 100 г
мяса мидий, использованных в пищу, содержалось 8066±61,37 мкг STX и его аналогов
(эпимерные пары GTX3/GTX2)1. Других паралитических токсинов в крови и моче па-
циентов обнаружено не было. Каждый из них съел в среднем 23 г мяса мидий, т. е.
отравление было вызвано 3,7 мг STX (и его аналогами).
Диагностика поражения STX. Естественные отравления STX возможны только пе-
роральным путем при употреблении в пищу морепродуктов (мясо мидий, крабов,
лобстеров, отдельных пород рыб). Такие отравления носят единичный характер, число
отравленных людей редко достигает 10. Как правило, жители прибрежных районов
знают об опасности «цветных приливов». При случайном отравлении STX они пони-
мают, что с ними произошло, и немедленно обращаются за медицинской помощью.
При умышленном отравлении STX из-за быстрого летального исхода вероя гность того,
что удастся расспросить потерпевшего об обстоятельствах отравления, значительно
меньшая, чем при поражениях рицином или ботулиническим токсином. Предполо-
жение об отравлении быстродействующим нейротоксином позволяют сделать данные
анамнеза отравленного в сочетании с определенными клиническими симптомами,
если их удастся зафиксировать. При осмотре тела надо обратить внимание на нали-
чие свежих следов уколов, инъекций или царапин. При их обнаружении целесообраз-
но провести тщательную токсикологическую экспертизу.
Эксперименты по воздействию аэрозоля STX на животных позволяют считать, что
при ингаляции развитие отравления ускоряется, и смерть наступает в пределах не-
скольких минут после экспонирования Прогноз плохой (Usamriid’s medical manage-
ment..., 2001). Более подробного описания в литературе клиники ингаляционного и
парентерального отравления STX, найти не удалось, но она должна отличаться от кли-
ники алиментарного отравления этим токсином тем, что не будут выражены симптомы
со стороны ЖКТ (рвота, абдоминальные боли) и признаки местного действия токсина
(чувство онемения в полости рта и кожи лица). При ингаляционном и парентеральном
отравлении STX основными симптомами будут симптомы поражения перифериче-
ской нервной системы (развившаяся в течение нескольких минут мышечная слабость),
признаки угнетения сосудодвигательного (глубокая гипотония) и дыхательного (оста-
новка дыхания) центров. Ингаляционные поражения STX, скорее всего, проявят себя
вспышками, когда признаки отравления появляются у группы людей в течение не-
скольких минут, и эти группы объединяет какой-то общий гешрафический фактор
(кинотеатр, аэропорт, метро, учреждение), но без указаний на прием общей пищи.
Результаты обычного лабораторного исследования клинических проб являются
неспецифичными для отравления STX. Могут развиваться нарушения сердечной дея-
тельности, однако электрокардиографические данные неспецифичны. Патолого-
1 Высокая концентрация токсина объясняется тем, что одна мидия длиной 6—7 см за час
фильтрует до 3,5 л воды.
3.14. Сакситоксин
1019
анатомические изменения у людей, вызванные отравлением STX, в доступной лите-
ратуре найти не удалось. Скорее всего, на вскрытии будут обнаружены изменения,
характерные для остро развившейся гипоксии. Диагноз подтверждается обнаружением
STX (и его аналогов) в крови и моче пациента с помощью жидкостной хроматогра-
фии высокого разрешения, ион-парной хроматографии (разновидность обращенно-
фазовой ВЭЖХ; либо методом ELISA, или посредством биоанализа на мышах. Клиш 1-
ческие пробы, которые направляют для анализа на токсин, включают содержимое
желудка, сыворотку, выделения из дыхательных путей, смывы из носоглотки (для рас-
познания ингаляционного поражения). При этом следует иметь в виду, что биологи-
ческие методы, как правило, дают более высокое содержание STX, чем химические,
что объясняется присутствием в исследуемых образцах (динофлагеллят или моллюс-
ков) наряду с STX и его аналогов.
Дифференциальная диагностика при отравлении STX включает употребление с
пищей сигуатоксина (сигуатера), ТТХ, РТХ и ботулинического токсина. Отравление
сигуатоксином происходит при потреблении крупной рифовой рыбы и, в отличие от
STX, приводит к более тяжелым симптомам со стороны ЖКТ (тошнота, рвота, диа-
рея) и к специфичному обратному восприятию температуры: горячее кажется холод-
ным, а холодное — горячим. В отличие от ботулизма, при отравлении STX. помимо
симптомов вялого паралича скелетных мышц, присутствуют симптомы поражения
периферических нервов (парестезии). Дифференцировать отравление STX от отрав-
ления ТТХ по клиническим симптомам очень сложно. Отравление ТТХ обычно про-
исходит при употреблении в пищу продуктов, полученных из рыб рода скалозубов
(Sphaeroides, Fugu, рыбы-собаки) и рода .етраодонов (Tetraodon) из семейства игло-
брюховых (Tetraodontidae) и морских брюхоногих моллюсков. Отравления STX обычно
связаны с употреблением в пишу мяса мидий, крабов, лобстеров, отдельных пород рыб
(рыбы-собаки, японская скумбрия и др.), обитающих на корситовых рифах. Отрав-
ления РТХ отличаются от отравлений, вызванных STX и ТТХ, отсутствием у пациен-
та симптомов вялого паралича скелет ных мышц и поражения периферических нервов
(парестезии). Сигуатоксин, STX и ТТХ не вызывают рабдомиолиз, характерный для
отравлений, вызванных РТХ.
Иммунопрофилактика. Не разработана.
Лечение. Критическое значение для жизни пациента имеет фактор времени, так
как токсин в течение суток выводится с мочой. Чем быстрее начата интенсивная те-
рапия, тем больше шансов на ее успех. После перорального попадания токсина в орга-
низм человека, следует применять стандартные меры для лечения пищевого отравле-
ния, включая промывание желудка и введение в ЖКТ суперактивированного угля.
Лечение отравления STX включает реанимационные, детоксикационные и поддержи-
вающие мероприятия: искусственное дыхание, аналептики, мочегонные (фуросе-
мид 40 мг в/в), вазоактивные средства (добутамин 250 мг в 25 мл изотонического ра-
створа в/в). Рекомендуется в/в вливание растворов, содержащих бикарбонат натрия,
для компенсации дыхательного ацидоза и нейтрализации самого токсина, так как STX
и его аналоги инактивируются в щелочной среде. С этой же целью Garcia С. et al. (2005)
вводили пациентам в/в ранитидин, ингибирующий продукцию соляной кислоты па-
риетальными клетками желудка. Улучшение состояния пациента наблюдается уже че-
рез 4 ч интенсивных реанимационных и детоксикационных мероприятий. Благодаря
интенсивной терапии, начатой Garcia С. et al. (2005) в течение часа после появления
первых симптомов от] явления, им удалось сохранить жизни пациентам, получившим
не менее четырех летальных доз STX (его анатогов).
1020
Часть 3. Частная неправильная эпидемиология
3.15. ТЕТРОДОТОКСИН
Естественная эпидемиология отравления ТТХ Молекулярная i труктура и механизм
действия. Токсические и поражающие свойства. Клиника энтерального поражения.
Диагностика поражения ТТХ. Иммунопрофилактика. Лечение.
Тетродотоксин (tetrodotoxin, тарихатоксин, anhydrotetrodotoxin, 4-epitetrodotoxin,
tetrodonic acid, ТТХ) — низкомолекулярный нейротоксин алкалоидной природы,
близкий по механизму действия с сакситоксином, вызывающий паралич дыхатель-
ной мускулатуры и поражение ССС. Отравления людей происходят в основном из-за
употребления в пищу рыбы фугу из семейства иглобрюхов (Tetraodontidae). ТТХ об-
наружен у калифорнийского тритона (Taricha totosa), в икре лягушки Atepolus chiri-
quensis (Коста-Рико), в коже отдельных видов саламандр, жаб, в камчатском крабе,
сине-кольцевом осьминоге и др. Сведения о ядовитости иглобрюхов в китайских
медицинских книгах восходят к хлубокой древности (-2500 лет до н. э.). Среди евро-
пейцев одно из первых описаний отравления ТТХ дал известный мореплаватель
Джеймс Кук, который вместе со своими 16 товарищами отравился иглобрюхом во
время второго кругосветного путешествия в 1774 г. Первое формальное описание
симптомов отравления ТТХ сделал С. Remy в 1883 г. Он же установил высокое содер-
жание токсина, вызывающего такие отравления, в гонадах рыбы фугу. Название «тет-
родотоксин» ввел в научный оборот в 1909 г. японский исследовате гь Tahara, получив-
ший плохо очищенный экстракт токсина из рыбы фугу (Fuhrman F. А., 1996). В 1930
г. и в первую половину 1940-х гг. экстракт ТТХ, полученный из печени фугу под на-
званием «яд фугу» изучался в отряде 731 как поражающий агент, предназначенный для
осуществления диверсий и индивидуальных убийств (Моримура Сэйти, 1983). В кри-
сталлическом виде' (ТХ был получен в 1950 г. A. Yokoo из яичников рыбы Fugu rubripes
(бурый скалозуб). Химическая структура ТТХ установлена в 1960-х гг. почти одновре-
менно четырьмя группами ученых из США и Японии (К. Tsuda, Т. Goto, R. В. Woodward
и Н S. Mosher) (Fuhrman F. А., 1996).
Летальность среди людей, отравившихся рыбой, содержащей ТТХ, в последние два
десятилетия, благодаря своевременному переводу пациентов на искусственную вен-
тиляцию легких, редко достигает 10 %. Обычно она составляет 3,3—6 % (Noguchi Т. et
al., 2008). ТТХ рассматривается как агент для осуществления биологических террори-
стических актов, диверсий и действий криминального характера. Антидотов нет (.Wiki-
pedia). На Гаити тетродотоксин в виде грубого экстракта из органов рыбы фугу мест-
ные колдуны включают в состав порошка, используемого для зомбификации (Benedek
С., Rivier L., 1989). Сопоставление биологических и фармакологических свойств STX
и ТТХ приведено в табл. 3.50.
Естественная эпидемиология отравления ТТХ. Наиболее опасными для людей ис-
точником ТТХ являются рыбы рода скалозубов (Sphaeroides, Fugu, рыбы-собаки) и
рода тетраодонов ( Tetraodon) из семейства иглобрюховых (Tetraodontidae) отряда иг-
лобрюхообразных (Tetraodontiformes). Эти рыбы не выделяют ядовитой слизи, их ко-
лючки не опасны, но их печень, икра, молоки, кишечник и кожа — содержат ТТХ.
Токсин, накапливаясь в икринках, играет роль фактора, защищающего от хищников
потомство фугу во время нереста. При нападении хищника фугу отгоняет его, увели-
чиваясь в 2—3 раза в размере и экскретируя ТТХ с поверхности кожи (Mahmud Y.
et al., 2003). Рыба может обнаружить чрезвычайно низкие концентрации ТТХ и STX в
морской среде с помощью специализированных вкусовых рецепторов. Рецепторы,
3.15. Тетродотоксин
1021
распознающие ТГХ, отличаются от тех, что распознают STX (Hara Т. J., 2011). Содер-
жание ТТХ в органах и тканях иглобрюхих и других животных приведено в табл 3.55
и 3.56 соответственно.
Таблица 3.55
Токсичность иглобрюхих*
Семейство/среда обитания Вид Максимальная токсичность**
ЯИЧНИКИ тести кулы печень кожа кишеч- ник мышцы кровь
1 2 3 4 5 6 7 8 9
Tetraodontidae (семейство иглобрюхие), море Японская рыба < >угу
Takifugu niphobles • О • € • О —
Т. poecilonotus • С • С С О —
Т. pardalr • о • С С ▼ ▼
Т. snyden • ▼ • С С О —
Т. porphyreus • ▼ • с с ▼ —
Т. chinensis • — • — — — —
Т. obscurus • ▼ С с с ▼ —
Т. exascurus • ▼ С с — ▼ —
Т. pseudommus • ▼ О о о ▼ —
Т. chrysops с ▼ с с 4 о ▼ ▼
Т. vermicularis с ▼ с с о ▼ —
Т. rubripes с ▼ с ▼ о ▼ ▼
Т. xanthoptems с ▼ с ▼ о ▼ —
Т. stictonotus с ▼ с о ▼ ▼ —
Tetraodon alboreticulatus • - о о с о -
Pleuranacanthus sceleratus • - о о с о -
Chelonodon patoca*** с с с • - с -
Arothron flrmamentum с ▼ ▼ о ▼ ▼ -
Canthigaster rivulata ▼ - о с о ▼ -
Lagocephalus lunaris ▼ ▼ ▼ с ▼ • -
L. inermis ▼ ▼ с ▼ ▼ ▼ —
L. wheeled ▼ ▼ ▼ ▼ ▼ ▼ —
L. gloved ▼ ▼ ▼ ▼ ▼ ▼ —
Sphoemides pachygaster ▼ ▼ ▼ ▼ ▼ ▼ -
Китайская рыба фугу
Takifugu flavidus • с • с с о -
1022
Часть 3. Частная неправильная эпидемиология
Продолжение табл. 3.55
1 2 3 4 5 6 7 8 9
Слабоминерали- зованная вода Тайская рыба фугу
Tetraodon nigroviridis — — ▼ C О 0
Т. steindachneri — — ▼ € ▼ ▼ —
Diodon tidae (семейство ежи-рыбы), море Японская рыба фугу
Diodon holocanthus ▼ — ▼ ▼ ▼ ▼ —
Chilomycterus affirus ▼ — ▼ ▼ ▼ ▼ —
Ostraciidae (семейство кузовковые), море Японская рыба фугу
Ostraaon immaadatum ▼ ▼ ▼ ▼ ▼ ▼ —
Lactoria diaphana ▼ ▼ ▼ ▼ ▼ ▼ —
Aracana aadeata ▼ ▼ ▼ ▼ ▼ ▼ -
* Т. Noguchi et al. (2008).
* * ▼ — <10 МЕ/г ткани иглобрюха; О — 10—100 МЕ/г ткани (слабая токсичность);
€> — 100—1000 МЕ/г ткачи (умеренная токсичность). • — >1000 МЕ/гткани (сильная токсич-
ность); (—) — нет цанных. 1 ME (мышиная единица) определяется как количество токсина,
необходимое для того, чтобы убить мышь мужского пола линии ddY (с массой 20 г) в течение
30 мин после интраперитонеального введения. Одна ME ~ 0,2 мкг ТТХ.
* ** Морские виды в солоноватой воде.
Животные, содержащие ТТХ в органах и тканях*
Таблица 3.56
Животное Вид Токсическая часть животного Максимальная токсичность**
1 2 3 4
Platyhelminthes (тип плоские черви) Planooera spp. Все тело •
Nemertinea (тип немертины) Cephalothrix linearis То же •
Lineus fuscoviridis То же •
Tubulanus punctatus То же С
Mollusca (тип моллюски)
Gastropoda (класс брюхоногие) Charonia sauliae Пищеварительные железы •
Babylonia japonica То же О
Putufa lissostoma То же с
Zeuxis siquijorensis Все тело •
Niotha dathrata То же •
Niotha lineata То же •
Cyrnarium echo Пищеварительные железы о
Pugilina temotoma То же о
Annelida (тип кольчатые черви)
Polychaeta (класс многощетинковые черви) Pseudopdamilla oaelata Все тело о
Arthropoda (тип членистоногие)
Xanthidae crabs (щебеночный краб) Atergatis floridus Все тело о
Zosimus aeneus Тоже о
3.15. Тетродотоксин
1023
Продолжение табл. 3.56
1 2 3 4
Arthropoda (тип членистоногие)
Horseshoe crab (подковообразный краб) Careinascorpius rotundicauda Яйца О
Chaetognatha (тип щетинкочелюстные)
Arrowworms (щетинкочелюстные) Parasagitta spp. Головная часть Д
Flacasagitta spp. To же Д
Echinodermata (тип иглокожие)
Starfish (класс морская звезда) Astropecten spp. Все тело С
\fertebrata подтип позвоночные)
Pisces (надкласс рыбы)
Goby (семейство бычковые) Yongeichthys criniger Кожа, внутренности, гонады С
Amphibia (класс земноводные)
Newts (группа тритоны) Tancha spp. Кожа, кровь, мышцы, яичники, икра С
Notophthalmus spp Кожа, яичники, икра О
Cynopsis spp. Кожа, кровь, мышцы, яичники, икра О
Triturus spp То же Д
Frogs (отряд лягушки) Atelopus spp. Кожа •
Colosiethus spp. То же С
Polypedates spp. Тоже с
JBrachycephaltts spp. Кожа, печень ©
Д — производное ТТХ, данных по токсичности нет.
* Т. Noguchi et al. (2008).
* । Обозначения те же, что в табл. 3.55.
Длительное время токсикологи полагали, что отравления ТТХ происходят почти
исключительно в Индо-Тихоокеанском регионе. Атлантический регион считался сво-
бодным от вторично-ядовитых животных, вызывающих отравления ТТХ до 2008 г.,
когда на побережье Португалии было установлено отравление человека мясом мор-
ского брюхоногого моллюска Charonia lampas lampas, содержащим ТТХ и его аналог
5,6,11 -trideoxy ГIX. Через несколько минут после его употребления в пищу у пациен-
та развился паралич и дыхательная недостаточность. В течение 52 ч он находился на
искусственной вентиляции легких (Rodriguez Р. et al., 2008; Fernandez-Ortega J Е et al.,
2010). Расширяется ареал рыбы фугу. В марте 2013 г. ее поймали в порту небольшого
хорватского городка Трибунь (Адриатическое море)1.
В России естественные отравления ТТХ возможны на Тихоокеанском побережье
Дальнего Востока. Очень токсичный для людей японский иглобрюх (Tetrodon vermi-
cularis) попадается рыбакам в прибрежных морских водах вблизи Владивостока.
'http://www.slobodnadalmacija.hr/%C5%A0ibenik/tabid/74/articleType/ArticleView/articleId/
205351/Default.aspx
1024
Часть 3. Частная неправильная эпидемиология
Первичными продуцентами ТТХ токсикологи считают некоторые виды морских
бактерий. Еще в 1980-е гг. было обнаружено, что Vibrio alginolyticus, выделенный изток-
сичной морской звезды Astropecten polyacanthus, штаммы Vibrio, выделенные из краба
Atergatis floadus и рыбы фугу Takifugu snyderi, продуцируют паралитический токсин в
количестве 213, 30 и 3 MU/500 мл питательной среды соответственно. Так же было
обнаружено, что морские грамотрицательные бактерии Shewanella alga и Alteromonas
tetraodonis, изолированные из красных водорослей семейства Jania, грамотрицатель-
ная бактерия Shewanellaputrefaciens, выделенная из иглобрюхих вида Takifugu niphobles,
и некоторые другие морские бактерии, изолированные от организмов, содержащих
ТТХ, самостоятельно продуцируют ТТХ (Noguchi Т. et al., 2008). Однако прогресса в
расшифровке механизмов генетической регуляции синтеза токсина этими бактерия-
ми нет. В табл. 3.57 приведен краткий перечень ТТХ-продуцирующих бактерий по
годам и источникам обнаружения. Возможный механизм накопления ТТХ в организме
морских животных приведен на рис. 3.151.
Таблица 3 57
Источники ТТХ-продуцирующих бактерий*
Год выделения ТТХ-продуцирующая бактерия Источник бактериального изолята
1987 Pseudomonas spp. Кожа Takifugu poealonotus
1987 Vibrio alginolyticus Кишечник Takifugu vermicularis
1989 Shewanella putrefaciens Кишечник Takifugu niphobles
2000 Vibrio spp.. Кишечник Takifugu vermicularis radiates
2004 Microbacterium arabimgalactanolyticum Яичник Takifugu niphobles
2004 Serratia marcescens Кожа Chelonodon patoca
2004 Vibrio alginolyticus Кишечник Takifugu alboplumbeus
2005 Actinomyces spp. Яичник Fugu rubripes
2005 Bacillus spp. Яичник, печень и кишечник Fugu rubripes
2005 Nocardiopsis dassonillei Яичник Fugu rubripes
2007 Proteobacteria, CFB группа и Spirochaetales Кожа, кишечник, яичник и печень Takifugu obscures
2010 Aeromonas spp. Яичник Takifugu obscures
2010 Bacillus spp. Яичник Fugu obscures
2010 Lysinibacillus fusiformis Печень Fugu obscures
2011 Raoultella terrigena Кишечник Takifugu niphobles
♦ По V. Chung-Hirn Yu et al. (2011)
Фугу являются вторично ядовитыми животными. Разведение таких рыб в искус-
ственных условиях позволяет устранить ТТХ из их органов и тканей. В естественных
условиях океанов ТТХ проникает в организм фугу по пищевым цепям, но сами фугу
устойчивы к токсину. При интраперитонеальном введении ТТХ токсичным (Т niphob-
les, Т. pardalis и Т. rubribes) и нетоксичным иглобрюхам (£. wheeleri, L. gloveri и Liosaccus
cutaneus) было установлено, что LD50 ТТХ для токсичных иглобрюхов от 300 до 750 раз
больше, чем для мышей. У нетоксичных иглобрюхов эта величина не превышала LD50
для мышей в 15 раз (Noguchi Т. et al., 2008). Данные по резистентности различных видов
животных к ТТХ приведены в табл. 3.58, с. 1026.
3.15. Тетродотоксин
1025
0
Рис. 3.151. Накопление ТТХ в организме морских животных. А. Бурый скалозуб
(Takifugu rubripes). Достигает длины 80 см, но обычная длина 20—40 см. Мор-
ская тепловодная рыба. Б. Возможный механизм накопления ТТХ в организме
морских животных. По Т. Noguchi et al. (2008)
Происхождение и биосинтез ТТХ земноводными не имеет однозначного объяс-
нения. Доказательства эндогенного синтеза ТТХ у этих животных основываются на
косвенных признаках. Прямых доказательств такого синтеза, полученных, например,
путем поглощения прекурсоров, имеющих радиоактивную метку, нет (Shimizu Y.,
Kobayashi М., 1983). Тритоны Taricha granulosa, содержащиеся в неволе и питающие-
ся дождевыми червями, не только поддерживали высокие уровни ТТХ в коже в тече-
ние нескольких лет, но и восстанавливали в неволе высокий уровень содержания ТТХ
(до 3 mi) за относительно короткий период времени без употребления в пищу живот-
ных, содержащих ТТХ. Аналогичные результаты были получены в сходных экспери-
ментах с жабами Atelopus oxyrhynchus, поддерживающими высокий уровень ТТХ в коже
в течение трех лет. При исследовании кожи и желез Т granulose не было обнаружено
бактерий, продуцирующих ТТХ (Hanifin С. Т, 201С)
Наиболее часто отравления ТТХ происходят в Японии. Фугу — традиционное,
очень популярное блюдо японской кухни. Чаше всего для приготовления фугу исполь-
зуется рыба вида бурый скалозуб (Takifugu rubripes)1. Рыба фугу у японцев считается
особым деликатесом, но не из-за ее вкусовых качеств, а из-за возможности рискован-
ного приключения. В небольших дозах ТТХ действует как наркотик, вызывая эйфо-
рию и приятные физические ощущения. Задачей повара является не полное удаление
тетродотоксина, а понижение его концентрации до требуемого уровня.
1 Т. rubripes встречается в водах Южного Приморья (река Туманная), залив Петра Велико-
го, залив Ольги, у Южного Сахалина.
1026
Часть 3. Частная неправильная эпидемиология
Таблица 3.58
Резистентность различных видов животных к ТТХ*
Отношение к токсину Вид LD50 (MU/20 r)**
Организмы, содержащие ТТХ
Щебеночный краб A. floridus 1000
Тропический бычок Vongeidithys criniger >300
Японский тритон Cynops pyrrhqgaster >10000
Иглобрюхи (Pufferfish)
Токсичные Takifugu niphobles 700-750
T. pcrdal's 500 550
T. rubripes (culture) 300-500
Нетоксичные или малотоксичные Lugocephalus wheeleri 15-18
L. gloveri 19-20
LtosaaMS cutaneous 13-15
Ostraaon immaculatum 0,9-1,3
Позвоночные, не содержащие ТТХ
Teleosts (костистые рыбы) Oplegnathus punctatus 0,8-0,9
O. fasaatus 0,8-1,8
Gtrella punctata 0,3-0,5
Наземные млекопитающие
Мыши Mus musailus 1,0
* По Т. Noguchi et al. (2008).
** LD50 ТТХ (MU/20 г массы тела), которая при интраперитонеальном введении убивает
100 % экспериментальных животных.
Рыба футу поставляются в японские рестораны живой и хранится в большом ре-
зервуаре, как правило, на видном для посетителей месте. Особенно японскими гур-
манами ценятся фугу, пойманные в осенне-зимний период в «дикой природе», т. е. в
море. С 1958 г. только специально лицензированные шеф-повара могут готовить и
продавать фугу для публики. Фугу-ученик проходит двух- или трехлетний курс обу-
чения, прежде чем он допускается к официальному экзамену. Экзамен состоит из пись-
менного теста, теста на идентификацию рыбы и практического теста, заключающе-
гося в том, что экзаменуемый должен приготовить самостоятельно рыбу и затем съесть
ее. Только 30 % заявителей проходят этот экзамен с первого раза. Продажа наиболее
токсичной части фугу, печени, является в Японии незаконным видом деятельности.
Профессионально приготовленное фугу содержит ТТХ в количестве, вызывающем
только ощущение легкого покалывания и онемения на языке и губах. Несмотря на
строгие правила по приготовлению фугу, иногда повара ошибаются. В 1975 г. от от-
равления ТТХ погиб Мицугора Бандо VIII, артист театра Кабуки, при жизни объяв-
ленный правительством Японии национальным сокровищем. Мицугора умер после
того, как съел четыре порции печени фугу. В 2009 г. семеро посетителей ресторана
японского города Цуруоки (севернее Токио) отравились блюдом из фугу. Фуг? — до-
рогое удовольствие. Цена одного блюда начинается от 120 долларов. Его можно отве-
дать только в лучших ресторанах Японии (рис. 3.152).
3.15. Тетродотоксин
1027
яж «г«IяяяХ
Рис. 3.152 Лицензия на приготовление блюд из фету. Надпись на вывеске
этого японского ресторана сообщает, что «город Киото ра [решает данному
ресторану подавать клиентам блюда из фугу». По Э. Р Ричиути (1979)
В некоторых отдаленных районах Японии, местными жителями используются
кустарные способы удаления ТТХ из рыбы. Они включают этапы вымачивания фу-
гу в саке и солевых растворах в течение трех лет, но точное описание способов держится
в секрете.
Жители прибрежных районов Индийского и Тихого океанов обычно знают о ядо-
витых свойствах рыбы фугу. Отравления людей происходят в отдаленных от побере-
жья районах, куда рыбу завозят недобросовестные торговцы и продают ее населению
по низким ценам. Такие отравления бывают массовыми. В материковой части Банг-
ладеш (дистрикты Narshingdi, Natore и Dhaka) в апреле и июне 2008 г. имели место три
вспышки отравлений людей рыбой фугу, проданной местному населению по «привле-
кательной цене» — всего за 50—80 центов/кг. Тогда как рыбы других сортов продава-
лись в 2—3 раза дороже. Всего было выявлено 98 отравленных ТТХ местных жителей,
из них 14 погибли (рис. 3.153).
Рис. 3.153. Рыбы-иглобрюхи на одном из базаров
Бангладеш. По N. Homaira et al. (2010)
1028
Часть 3. Частная неправильная эпидемиология
Кроме отравления фугу, в КНР и на Тайване (побережье Восточно-Китайского и
Желтого морей) регулярно фиксируются случаи отравления людей небольшими брю-
хоногими моллюсками, содержащими ТТХ. К таким моллюскам относятся Niotha
clathrata, Natica vitellus, О. miniacea, О. hirasei, Nassarius glans и Zeuxis samiplicutus. В КНР
в течение 1977—2001 гг. моллюсками, содержащими ТТХ, отравились более 300 че-
ловек, из них 16 погибли. В июне 2001 г. в провинции Чжэцзян (КНР) жертвами
отравления, вызванного употреблением в пищу небольшого брюхоногого моллюска
Z samiplicutus из семейства Nassariidae, стали 30 человек. У них развились симптомы,
характерные для отравления ТТХ (тошнота, рвота, афазия, атаксия, паралич, кома).
Токсин был идентифицирован хроматографическими методами (Sui L.M. et al., 2002).
В июле 2007 г. в Нагасаки (Япония) имело место пищевое отравление моллюском
Alectrion glan. В оставшихся не съеденными моллюсках обнаружен ТТХ в количестве
600 МЕ/г мышц и пищеварительных желез. Механизм накопления ТТХ в организме
брюхоногих моллюсков не ясен. Т. Noguchi et al. (2008) считают, что они накаплива-
ют ТТХ, питаясь внутренними органами Pufferfish, погибшими во время нереста.
Молекулярная структура и механизм действия. ТТХ представляет собой соедине-
ние аминопергидрохиназолина с гуанидиновой группой. Физические и химические
свойства ТТХ в сравнении с STX показаны втабл. 3.50. Известно большое количество
природных, полусинтетических и синтетических аналогов ТТХ. Их делят на две об-
ширные группы: гемилактальные (hemilactal) и лактоновые (lactone)1. Как правило,
аналоги менее токсичны, чем ТТХ. На рис. 3.154 показана структура ТТХ и его ана-
логов, обнаруженных у тритонов и фугу.
6-ерЛГТХ(9) 11-СН2ОН ОН
H-deoxyTTX(lO) ОН 11-СН,
ll-norTTX-6(S)-ol(ll) ОН Н
8-epr-5,6,ll-trideoxyTTX(2) Н ОН Н Н
l-hydroxy-8-epr-5,6,ll tndeoxyTTX (4) ОН ОН Н Н
5,6, H-trideoxyTTX(6) НН ОН Н
4,9-anhydro-8-epi- ОН Н
5,6,1 itndeoxyll X (3)
4,9-anhydro-5,6,11-tndeoxyTTX (7) Н ОН
Рис. 3.154. Структура ТТХ и его аналогов, обнаруженных
у тритонов (2—5, 9, 10, 12) и футу (6-8, 10, 11). По Y, Kudo et al. (2012)
1-hydroxy-4,4a-anhydro-8-ept- ОН ОН Н
5,6,11-trideoxyTTX (5)
4,4a-anhydro-5,6.11 -tndeoxyTTX (8) Н Н ОН
1 Подробно об аналогах ТТХ см. в обзоре М. Yotsu-Yamashita (2001).
3.15. Тетродотвксин
1029
ТТХ в природных источниках всегда находится в смеси со своими аналогами.
Состав таких смесей и количественное соотношение входящих в них компонентов
различаются у разных таксонов животных. Амфибии в сравнении с рыбами бедны
аналогами ТТХ. Стереоизомер 6-е/и’ТТХ является общим для класса амфибий, но ред-
ко встречается среди морских видов животных, токсичных по ТТХ (Hanifin С. Т, 2010).
Сравнительное исследование содержания ТТХ и его аналогов в тканях тритонов
(С. е. popei) и в яичниках рыбы фугу (F. poecilonotus) показало, что ТТХ и 11-deoxyTTX
присутствовали в обоих источниках Тем не менее 6-ер/ТТХ и 8-эпи-аналоги были
обнаружены только у тритона, a 5,6,11-trideoxyTTX — основной аналог ТТХ в рыбе
фугу (табл. 3.59).
Таблица 3.59
Содержание ТГХ и его аналогов (мкг/г) в тканях тритона
Cynops ensicauda popei и яичниках рыоы Fugu poecilonotus*
Токсин F. poedlonotos С. е. popei
ТТХ 134,0 49,6
6-ерПТХ <0,3 27,0
4-<?р/ТТХ 11,5 8,0
4,9-anhydroTTX 31,2 58,1
5-deoxyTTX 4.2 <0.3
11-deoxyTTX 15,9 5,7
6,11-dideoxyTTX 6 3 <0,3
5,6,11 -trideoxyTTX 108,8 Нет данных
anhydro-5,6,11 -trideoxyTTX 79,9 Нет данных
8-epz-5,6,11-trideoxyTTX Нет данных 5,8
4,9-anhydro-8-ep-5,6,11-trideoxyTTX Нет данных 7,7
1 -hydroxy-S-epi-trideoxyTTX <0,3 2.4
1 -hydroxy-4,4a-anhydro-8-ep<-5 6,11 -trideoxyTTX <0.3 8.3
ll-norTTX-6(S)-ol 9,9 <0,3
* По Y. Kudo et al (2012).
Фугу различаются по составу деоксианалогов ТТХ (5-deoxyTTX, 11 -deoxyTTX,
6,11-dideoxyTTX и 5.6,11-tndeoxyTTX). При сравнении аналогов/7, niphobles, обитаю-
щих в слабосоленых водах близи южного побережья Кореи, и Tetraodon nigm iridis и
Tetrodon biocellatus, выловленных в морях Юго-Восточной Азии, в тканях всех этих рыб
был обнаружен 5,6,11-trideoxyTTX, однако 6,11-dideoxyTTX обнаружен только у рыб
F. niphobles (Jang Jun-Но et al., 2010).
Концентрация 5,6,11 -trideoxj ТТХ в моче и сыворотке крови пациента, отравив-
шегося в 2008 г. на побережье Португалии моллюском С. larnpas larnpas, была в три раза
большей, чем ТТХ (Rodriguez Р. et al., 2008).
ТТХ в препаративных количествах может быть получен биотехнологическм —
путем культивирования морских бактерий-продуцентов ТТХ, выделением ТТХ из
органов рыбы фугу (1 г ТТХ из 100 кг яичников) и путем химического синтеза. Все эти
способы имеют хорошую патентную защиту. Подробный обзор способов химическо-
го синтеза ТТХ приведен в работе J. Chau и М. Ciufolini (2011).
1030
Часть 3. Частная неправильная эпидемиология
ТТХ обладает чрезвычайно высокой биологической активностью, его LD50 для
мышей при внутрибрюшном введении составляет всего 10 мкг/кг. Ведущими симп-
томами отравления ТТХ у животных являются быстро наступающая гипотензия, уг-
нетение дыхания, паралич скелетной мускулатуры. Падение АД происходит в резуль-
тате паралича гладкой мускулатуры сосудистой стенки. Нарушение функции дыхания
имеет или периферическую природу (паралич дыхательной мускулатуры), или связа-
но с угнетающим действием ТТХ на дыхательный центр. В основе патологического
действия ТТХ лежит его способность блокировать проведение нервного импульса в
возбудимых тканях (нервных и мышечных), чем и обусловлены разнообразные фар-
макологические эффекты токсина (Орлов Б. Н., Гелашвили Д. Б., 1985).
ТТХ, так же как и STX, в очень низких концентрациях (10-7 моль/л) блокирует
входящий натриевый ток потенциалзависимых натриевых каналов (voltaged-gated
sodium channels, VGSC) во время развития потенциала действия. Другая его особен-
ность заключается в том, что он действует только с наружной стороны мемораны аксо-
на. На основании этих данных японские ученые Као и Нишияма в 1965 г. высказали
гипотезу, что ТТХ, гуанидиновая группа которого близка по своим размерам к диа-
метру гидратированного иона натрия, входит в устье VGSC и застревает в нем, удер-
живаясь остальной частью молекулы, размеры которой превышают диаметр канала.
VGSC образованы десятью белками: девять интегральных мембранных белков с
ММ 260 кДа (а-субъединипы) и один (иногда их бывает несколько) вспомогательный
ф-субъединица). а-субъединицы формируют каналы, определяющие биофизические
и ионоселективные свойства поры, ^-субъединица способна изменять кинетику на-
триевого тока и вольтаж поры и взаимодействует с адгезионными внеклеточными
молекулами, внеклеточным матриксом и внутриклеточным скелетом (Chahine М.,
O’Leary М. Е., 2011). а-субъединицы имеют сходную структуру у разных таксонов:
четыре гомологичных домена (DI-D1V), каждый содержит шесть трансмембранных
а-спиральных сегментов (S1—S6), соединенных посредством внеклеточных и внутри-
клеточных петель. Специфические аминокислотные последовательности а-субъеди-
ниц формируют стенку поры ионного канала, датчик напряжения (voltage sensor),
ворота инактивации (inactivation gate) и белокфосфорилируемый сайт (protein phospho-
rylation sites), а-субъединицы также формируют сайт, связывающий молекулы, меня-
ющие функцию этих каналов (Ulbricht W., 2005). Схематическое изображение взаи-
модействия ТТХ с а-субъединицами мембранных белков, формирующих VGSC, по-
казано на рис. 3.155.
У млекопитающих известны девять субтипов VGSC, различающиеся по кинетике
натриевого тока и вольтажным характеристикам (voltage dependent properties), по ло-
кализации в тканях и чувствительности к ТТХ. В наномолекулярных концентрациях
ТТХ блокирует Navl,l; Navi,2; Navl,3 (гены всех трех субтипов каналов в основном
экспрессируются в ЦНС); Navi ,4 (скелетная мускулатура); Navi ,6 (нейроны ценз даль-
ней и периферической нервной системы) и Navi,7 (обнаружен в периферической
нервной системе). Такие субтипы каналов называются ТТХ-чувствительными. Для
блокирования каналов субтипов Navi,5 (их гены в основном экспрессируются в сер-
дечной мышце); Navi.8 и Navi,9 (гены обоих каналов экспрессируются в нейронах
задних корешков ганглиев) требуются микромолярные количества токсина. Такие
каналы называют ТТХ-резистентными. VGSC обнаружены не только в нейронах, но
и в других тканях в пределах нервной системы (например, в астроцитах) (Geffeney S
L. Ruben Р. С., 2006). Поэтому действие ТТХ на различные органы и ткани происхо-
дит с разной выраженностью и меняется в зависимости от его концентрации.
3.15. Тетродотоксин
1031
Рис. 3.155. Схематическое изображение а-субъединиц, формирующих VGSC и сайты
связывания IГХ. VGSC формируются а-субъедининами мембранных белков через
формирование четырех гомологичных доменов (DI—IV), каждый из которых вклю-
чает 6 а-спиральных трансмембранных сегментов (1—6). Сегмент 4 (образно) соот-
ветствует датчику напряжения. Сайты фосфорилирования протеинкиназой А (РКА;
и протеинкиназой С (РКС) представлены кругами и квадратами соответственно.
«Быстро закрывающиеся ворота» (IFM-мотив) находятся во внутриклеточной петле
межд; областями 3 и 4 и обозначены буквой h. Цифрой 7 показаны сайты, участву-
ющие в формировании рецепторов «быстро закрывающихся ворот». Р-петли распо-
ложены между спиралями 5 и 6. Они играют роль внутренней обкладки поры (ниж-
ний рисунок). Внешнее (EEDD-мотив) и внутреннее (DEKA-мотив) кольца поры
обозначены цифрами 8 и 9 соответственно. Они формируются аминокислотами, как
показано' стрелками. Молекула ТТХ взаимодействует с аминокислотными остат ками
этих двух колец в поре канала, и пора закрывается. По Nieto F. R. et al. (2012)
Механизм, обеспечивающий животным резистентность к ТТХ (см. табл. 3.58),
основывается, главным образом, на резистентности к нем)’VGSC. Ароматические ами-
нокислоты у ТТХ-чувствительных каналов расположены в пределах Р-пегли домена I,
у резистентных каналов они представлены неароматическими аминокислотами, что
резко снижает их афинность к ТТХ (Noguchi Т et al., 2008).
Токсические и поражающие свойства. Токсичность яичников и печени некоторых
видов фугу (F. ocellatus obscurutn, F. niphobles, F. poecilonotum) такова, что достаточно
2 г ткани, чтобы вызвать смертельное отравление у человека.
LD50 дл1. мышей при интраперитонеальном введении ТТХ, 11-deoxyTTX и 6,11-
dideoxyTTX, определена как 10 мкг/кг, 70 мкг/кг и 420 мкг/кг соответственно. LD99
5,6,11-trideoxyTTX для мышей составила 750 мкг/кг (Jang Jun-Но. et al., 2010).
Летальная доза ТТХ для человека массой 70 кг оценивается в пределах 1-2 мг
(Noguchi Т, Ebesu J.S.M., 2001).
1032
Часть 3. Частная неправильная эпидемиология
Клиника энтерального поражения. Первые описания симптомов отравления тетро-
дотоксином содержатся в дневнике знаменитого английского мореплавателя Джеймса
Кука (1728—1779). Восьмого сентября 1774 г. шлюп «Решительность» («Resolution»)
стоял на якоре у только что открытого острова Новая Каледония. На ужин Куку и двум
находившимся в составе экспедиции натуралистам подали свежеприготовленную
рыбу неизвестного вида. В своем дневнике Кук записал: «Процедура описания рыбы
и ее зарисовки отняли так много времени, что уже было слишком поздно; на стол
подали только печень и молоки, к которым оба Форстера и я едва притронулись. Около
трех или четырех часов утра мы почувствовали небывалую слабость во всех членах,
сопровождаемую таким ощущением, будто окоченевшие на морозе руки и ноги сразу
попали в огонь. Я уже почти ничего не чувствовал и даже утратил способность соизме-
рять тяжесть тел: кружка воды емкостью в одну кварту и перо казались в моей руке оди-
наково тяжелыми... Одна из свиней, съевшая внутренности, околела». Спустя 166 лет
после описанного события, было установлено, что продегустированная Куком и его
спутниками рыба называлась Pleuranacanthus seleratus. Она относилась к семейству
скалозубовых рыб или тетродонтид.
При энтеральном отравлении первые симптомы отравления ТТХ появляются в
интервале от нескольких минут до 3 ч после приема фугу в пищу. В острых случаях
смерть может наступить в течение первого часа, но обычно между 4 и 6 ч. Вначале
отравленный ощущает странное покалывание и онемение языка и губ, которое мо-
жет распространяться и на тело. Затем он начинает жаловаться на головную боль, боль
в животе и руках. Походка становится шатающейся, появляется рвота, причем, при
ее отсутствии прогноз неблагоприятный. Вскоре после рвоты развивается атаксия,
отравленный стремится лечь. У него наблюдается ступор, афазия. Дыхание затрудне-
но, артериальное давление обычно снижено, характерно также понижение темпера-
туры тела, развивается цианоз слизистых и кожи. Несмотря на почти полную останов-
ку всех жизненных функций, отравленный какое-то время продолжает осознавать
происходящее вокруг, но при тяжелом отравлении он впадает в коматозное состояние.
После потери сознания наступает остановка дыхания, однако сердечная деятельность
еще продолжается некоторое время. Клинические стадии отравления ТТХ представ-
лены в табл. 3.60.
Вскрытие тел двух пациентов, умерших через 2 ч после употребления в пищу брю-
хоногих моллюсков Nassaurisglans, показало отсутствие каких-либо морфологических
Таблица 3.60
Клинические стадии отравления ТГХ*
Стадия Симптомы Начало по времени
Первая Ощущение онемения кожи вокруг рта и появление желудочно- кишечных симптомов (в основном, тошнота) 5—45 мин
Вторая Онемение языка и кожи лица, параличи двигательных мышц, невнятная речь, нарушение координации движения. Рефлексы остаются нормальными 10 60 мин
Третья Генерализованный вялый паралич, афония, дыхательная не- достазочность. Зрачок фиксирован или расширен. Отравлен- ный находится в сознании От 15 мин до нескольких часов
Четвертая Тяжелая дыхательная недостаточность и гипоксия, гипотензия, брадикардия, сердечная аритмия. Возможна потеря сознания От 15 мин до 24 ч
По Leung К. et al. (2011).
3.15. Тетродотоксин
1033
изменений их органов за исключением выраженного расширения и признаков гипер-
секреции желудка. Ткани мозга, легкие и сердце имели неспецифические посмертные
изменения. Не было обнаружено ни значительной клеточной инфильтрации, ни тром-
бов, ни больших инфарктов. Yin Н. L. et al. (2005) полагают, что причиной смерти в
данных случаях был холинергический кризис, приведший пациентов к ранней смерти.
В связи с тем что VGSC Navi ,5, представленные в сердечной мышце, резистентны
к ТТХ, действие токсина на сердце человека возможно только при большой концент-
рации в сыворотке. При 80 нМ ТТХ в сыворотке крови пациента, у него не было при-
знаков нарушения деятельности сердца и падения АД. У пациента с концентрацией
ТТХ в сыворотке крови 114 нМ наблюдались брадикардия, гипотония, но без нару-
шения сердечной проводимости. Более серьезные симптомы и неблагоприятный про-
гноз при отравлении ТТХ имели место у пациентов на фоне уже развившихся у них
соматических болезней, в частности, у больных сахарным диабетом (ZimmerT, 2010).
ТТХ может имитировать полную остановку всех жизненных функций у человека.
В некоторых регионах Японии человека, отравившегося фугу, не хоронят сразу, а ждут
трое суток, чтобы убедиться, что тело его разлагается и отравленный умер. На Гаити
ткани рыбы фугу являются основным ингредиентом яда, используемым колдунами
вуду на первом этапе превращения людей в «зомби». В самом процессе зомбифика-
ции (zombication) мистики нет. Яд, содержащий ТТХ, вызывает у отравленного чело-
века «смерть как состояние». Его хоронят в присутствии родственников, и он счита-
ется ими умершим. На втором этапе зомбификации человека откапывают ночью, не
позже 12 ч после «смерти», и для инициации его «жизни после жизни» дают ему пить
другой яд, содержащий растительные галлюциногены (например, скополамин из ра-
стения Datura stramonium L.), подавляющие волю отравленного ТТХ, затем увозят в
глубинные районы Гаити, где продают в рабство. Ежедневно добавляемая доза дату-
ры поддерживает такого человека в состоянии постоянного гипноза. Зомби полностью
теряет ощущение себя и своей идентичности в окружающем мире.
С точки зрения европейской психиатрии «зомби» проявляют симптомы, сходные с
симптомами кататонической шизофрении. У них чередуются моменты ступора и дея-
тельности, выражение лица безжизненное, в голосе преобладают носовые интонации,
набор слов ограничен, речь состоит из повторения односложных выражений. Гаитян-
ское понимание «зомби» — это не человек, а его тело без характера, без воли. Уголов-
ный кодекс Гаити (ст. 246) приравнивает зомбификацию к убийству. Самих «зомби»
гаитяне не боятся. Но они испытывают панический страх перед колдунами вуду, кото-
рые могут превратить их в «зомби» за нарушение неких «кодов общества» (Davis Е. V,
1983; Craan A. G., 1988). Растения и животные, используемые колдунами вуду для
ритуала зомбификации, описаны в работе U. Р. Albuquerque et al. (2012).
Диагностика поражения ТТХ. При пероральном отравлении ТТХ диагностика и
дифференциальная диагностика в основном сходны с аналогичной диагностикой
поражения STX (см. разд. 3.13). В открытой научной литературе описания клиники
ингаляционного отравления людей («патологии мелкодисперсного аэрозоля») или экс-
периментальных животных мной не обнаружены. Естественные отравления всегда
связаны с употреблением в пищу брюхоногих моллюсков и продуктов, приготовлен-
ных из рыбы фугу. Умышленные отравления могут быть вызваны ТТХ и содержащи-
ми его продуктами. Вкус фугу трудно обнаружить среди овощей и специй. Диагноз
отравления ТТХ в основном строится на основе анализа личности и анамнеза паци-
ента, симптомов отравления и обнаружения ТТХ (его аналогов) в остатках пиши, в
моче и сыворотке крови. Количество ТТХ в моче и в крови пациента, как правило,
1034
Часть 3. Частная неправильная эпидемиология
крайне мало. Эффективным способом обнаружения ТТХ является масс-спектромет-
рия. ТТХ в основном выводится с мочой. После перорального отравления в плазме
крови он циркулирует менее 24 ч, но в моче его можно обнаружить в течение 4 суток.
Распространение культа вуду в России требует анализа личности отравленного
ТТХ. В России имеются как отдельные адепты культа вуду, так и тайные вудистские
организации, «ищущие в вуду путь к познанию мира, обретению мудрости и гармо-
нии». Отравленный ТТХ может оказаться вовлеченным в культ вуду и стать либо жер-
твой вудистского ритуала, либо преступления, совершенного с помощью яда, исполь-
зуемого в таких ритуалах.
Иммунопрофилактика. Не разработана.
Лечение. Эффективных мер медицинской помощи, кроме искусственной вентиля-
ции легких, до настоящего времени нет. Лечебные мероприятия аналогичны тем, что
проводятся при отравлении STX (см. разд. 3.13). Прогноз хороший, если пациент
поступил в реанимацию еще до остановки дыхания и если он не погиб в первые 24 ч
после появления симптомов отравления. Симптомы отравления исчезают в период от
одних до пяти суток.
3.16. ПАЛИТОКСИН
Естественная эпидемиология отравления. Молекулярная структура и механизм
действия. Токсические и поражающие свойства РТХ. Клиника пищевого отрав-
ления. Клиника ингаляционного поражения. Клиника контактного поражения.
Диагностика поражения РТХ. Иммунопрофилактика. Лечение.
Патитоксин (palytoxin, ЕА 3940, РТХ) — сильнейший геморрагический токсин
небелковой природы, поражающий сердечно-сосудистую и респираторную системы.
ЖКТ, почки. Вызывает коронароспазм. Обладае” кардиотоксическим и гемолитиче-
ским дейс гвием. Имеет сложную структуру. РТХ обнаружили в шестилучевых корал-
лах зоантариях (Polithoa); некоторых видах крабов и рыб; в красных водорослях
Chondria crispus', и в широко распространенных в Мировом океане динофлагеллятах
Ostreopsis. Продуцентами РТХ также являются морские бактерии. Впервые Pl X вы-
делен в 1971 г. из мягких кораллов Polithoa учеными Гавайского университета (Univer-
sity of Hawaii) R. E. Moore и P. J. Scheuer. Токсические свойства этих кораллов издавна
известны аборигенам Таити и Гавайских островов, которые их использовали для при-
готовления отравленного оружия. Места обитания ядовитых кораллов туземцами дер-
жались в секрете. Как поражающий агент химического оружия, РТХ детально иссле-
дован в Эджвудском арсенале армии США под шифром «вещество ЕА 3940». Хими-
ческая структура РТХ установлена в 1981 г. двумя группами исследователей.
В 1994 г. Е. М. Suh и Y. Kishi впервые синтезировати РТХ химическим путем. От-
рав юния людей в основном происходят в результате употребления в пищу морских
животных, содержащих РТХ, реже через кожу при контакте с кораллами Polithoa (Ор-
лов Б. Н., Гелашвили Д. Б., 1985; Антонов Н. С., 1994: Ramos V., Vasconcelos V. et aL,
2010). Летальность среди людей при неумышленных энтеральных отравлениях ГТХ
достигает 8 % (6 погибших из 75 отравленных в 20 инцидентах) (Taniyama S. et al., 2011).
РТХ может переводиться в аэрозольное состояние из растворов с помощью распыли-
телей или взрывом заряда взрывчатого вещества Рассматривается специалистами как
агент для осуществления биологических террористических актов, диперсий и действий
криминального характера (Антонов Н С , 1994).
3.16. Палитоксин
1035
Естественная эпидемиология отравления. РТХ и его аналоги обнаружены в зоонта-
риях Palythua spp. (Р. toxica, Р. tuberculosa, Р. carbacorum, Р. mammillosa, P.vestitus, Р. aff.
Margwitae), красных водорослям Chondrin crispus и донных динофлагеллятах Ostreop-
sis spp. (Ostreopsis siamensis, O. mascarenensis, O. ovata). Их также находят у организмов,
живущих в тесной ассоциации с колониями зоантарий. Активность РТХ обнаружена
у бактерий рода Pseudomonas (Alteromonas), ассоциированных с динофлагеллятами
Ostieopsis lenticularis (Carballeira N. M. et al., 1998); у бактерий fl. cereus и бактерий ро-
дов Brevibacterium и Acinetobacter, выделенных из зоонтарий Palythoa caribaeorum
(Seemann Р. et al., 2009).
По пищевым цепям РТХ проникает в организм животных, питающихся зоонта-
риямч, либо живущих с ними в ассоциациях животными (губки, моллюски, ракооб
разные, горгонии, многощетинковые черви, морские звезды Acanthasterplanci и др.).
Те, в свою очередь, поедаются более крупными хищниками — рыбами (сардины, ан-
чоусы, сельди и др.), морскими ежами, крупными крабами, обитающими в тропиче-
ских морях. Человек — конечное звено этой пищевой цепи (табл. 3.61).
Таблица 3.61
Случаи отравления людей морепродуктами, контаминированными РТХ*
Год (количество отравленных) Источник РТХ Симптомы/Клинические данные Лечение Исход Другие токсины
Филиппины, 1984 (1) Краб Demania reynaudii Через несколько минут после употребления в пищу мяса краба: головокружение, тош- нота, усталость, металличе- ский привкус во рту, понос, онемение конечностей, бес- покойство, рвота. После гос- питализации: чередование периодов тяжелой брадика- рдии, дыхательной недоста- точности, цианоз, почечная недостаточность, анурия Атропин, димедрол, демерол, адреналин Смерть через 15 ч после отравления ТТХ и STX не обнару- жены
Япония, 1986 (2) Рыба Scants ovifrons Боль в мышцах, одышка, судороги. Миоглобинурия, высокие сывороточные уровни КФК, ACT, ЛДГ Не описано Умер 1 Найдены небольшие количества ТТХ и его аналогов
Мадагаскар, 1994 (1) Рыба Herklotsichtys quadrimaadatus Металлический вкус мяса. Через 0—2 ч после приема внутрь: обшее недомогание, неконтролируемые рвота, диарея. Следующие 4,5 ч до смерти: чувство покалывания конечностей, брел То же Умер 1 Не описано
Япония, 2000 (11) Рыбы Epinephelus bruneus, E. fuscogattus 3—36 ч после приема внутрь: миалгия, черный цвет мочи (гемоглобинурия), боль в плечевых мышцах. Повы- шенный уровень сыворо- точной креатинфосфатазы То же Выздоров- ление Сигуаток- син не обнаружен
* По A, Tubaro et al. (2011).
1036
Часть 3. Частная неправильная эпидемиология
Риск неумышленного отравления РТХ в последние годы значительно повысился
за пределами традиционного ареала зоантарий. Установлено, что динофлагелляты
Ostreopsis, местом обитания которых ранее считались рифы тропических и субтропи-
ческих морей, распространились в Средиземном море, особенно у берегов Туниса,
Испании, Франции, Италии, Монако и Греции (Amzil Z. et al., 2012). В моря умерен-
ных широт они попадают с ба тластными водами грузовых судов и активно размно-
жаются вследствие прогрева морской воды, вызванного глобальным потеплением
климата (Shears N. Т, Ross Р. М., 2009). По оценке Z. Amzil et al. (2012), фильтрующие
двустворчатые моллюски (мидии) и травоядные иглокожие (морские ежи), обитаю-
щие в водах Средиземного моря вблизи побережья Франции, способны за летний
период накопить большие количества РТХ и его аналогов. Однако случаи отравления
РТХ у людей, употреблявших в пищу моллюски, не описаны (рис. 3.156).
Рис. 3.156. Морфология Ostreopsis spp. А и Б. Световая микроскопия. В и Г. Световая
эпифлуоресцентная микроскопия. Д и Е. Сканирующая электронная микроскопия.
В и Д. Эпитекальная область. Г Гипотекальная область. Ж. Деталь вентральной обла-
сти (вид сбоку) с вентральной порой (Vp). 3 и Е: Детали вентральной области. Поло-
са на фотографиях А—Д соответствует 10 мкм, Ж—Е — 1 мкм.По S. Sato et al. (2011)
Поданным А.В. Андрианова (2009), на морских водорослях прибрежья Владиво-
стока обнаружены Ostreopsis ovata и О. siamensis, являющиеся продуцентами РТХ и его
аналогов, а также сигуатоксина, вызывающего отравление сигуатеру. Их численность
превышает 10 тыс. клеток на один грамм сухого веса макрофита, что соответствует
максимальным значениям плотности этих видов. Ранее эти водоросли находили толь-
ко в тропических водах.
Отравления РТХ могут проходить под маской пищевого отравления, названного
клипеотоксизмом (clupeotoxism), либо массового ингаляционного отравления, изве-
стного как «синдром водорослей» («algal syndrome»). Симптомы клипеотоксизма вклю-
чаю! тошноту, рвоту, металлический вкус во рту, диарею, неострое развитие летарги-
ческого состояния. Через несколько часов у отравленного появляется ощущение
сильного жжения вокруг рта и в конечностях; мышечные спазмы, сопровождающие-
ся сильными мышечными болями; отдышка, дисфония. Возможна смерть вследствие
развития острой сердечной недостаточности. Вспышки заболевания, которому ста-
3.16. Палитоксин
1037
вят диагноз «клипеотоксизм», обычно фиксируются в бассейнах Карибского моря и
Индийского океана (Louzao М. et al., 2008). «Синдром водорослей» пока зафиксирован
в летние месяцы на итальянском побережье Средиземного моря (Тирренское море) в
периоды цветения Ostreopsis spp. Отравление проявляет себя клиникой, сходной с ОРЗ
(Tubaro A. et al., 2011).
A. S. Kerbrat et al. (2011) обнаружили РТХ и его аналог 42-гидроксипалитоксин у
мооских диазотрофных цианобактерий Trichodesmium (порядок Oscillatoriales), широ-
ко распространенных в тропиках и субтропиках, и особенно в водах Новой Каледо-
нии. По мнению авторов, случаи клипеотоксизма у местных жителей, вызванные упо-
треблением в пищу планктоноядных рифовых рыб, связаны с вовлечением в пищевую
цепь цианобактерий Trichodesmium.
РТХ и его аналоги обнаружены у коралловых полипов (Anthozoa, Hexacorallia), по-
мещенных в домашние аквариумы и вызвавшие контактные и ингаляционные отрав-
ления у людей. В отдельных образцах кораллов количество грубого токсина достига-
ло 0,5—3,5 мг/г (Deeds J. et al., 2011). В описанном К. Hoffmann et al. (2008) случае
отравления, РТХ проник в кровь пациента через поврежденную кожу пальца при кон-
такте с кораллами Parazoanthus spp., находящимися в аквариуме. Клинические симп-
томы включали отек, парестезию и онемение вокруг места повреждения, распростра-
нившиеся по руке; и признаки системного отравления, такие как головокружение,
общая слабость, боль в мышцах, нарушения на ЭКГ и рабдомиолиз. Анализ показал
чрезвычайно высокую концентрацию палитоксина (между 2 и 3 мг/г) в тканях корал-
лов, вызвавших отравление.
Молекулярная структура и механизм действия. РТХ представляет собой большую
сложную молекулу с длинным полигидроксилированным (polyhydroxylated) и частич-
но ненасыщенным алифатическим остовом, содержащим как липофильные, так и
гидрофильные участки, и 64 хиральных центра. Последняя особенность включа-
ет наличие 8 двойных групп, которые способны к с/5//гаи5-изомеризации, благодаря
чему РТХ может иметь более чем 1021 стереоизомеров (Katikou Р., 2007).
Идентифицировано, по крайней мере, 14 аналогов РТХ. Гомопалитоксин (homo-
palytoxin), бисгомопалитоксин (bishomopalytoxin), неопалитоксин (neopalytoxin), де-
опалитоксин (deopalytoxin) и 42-гидроксипализоксин (42-hydroxypalytoxin) были
обнаружены в тропических Palythoa. Остреоцин-D (ostreocin-D, 42-hydroxy-3,26-di-
demethyl-19,44-dideoxypalytoxin, OSD), оватоксин-А, -В, -C, -D и -Е (ovatoxin-A, -В,
-С, -D и -Е), маскаренотоксин-А, -В и -С (mascarenotoxin-A, -В и -С), обнаружены у
бентосных динофлагеллят Ostreopsis. Наиболее хорошо изучена химическая структу-
ра палитоксина и остреоцина-D (рис. 3.157)
R. Е. Мооге и Р. J. Scheuer (1971) экстрагировали Р^Х из необработанных сырых
кора тлев Р. toxica водным раствором 70 % этанола. Затем обращенно-фазовой хрома-
тографией на колонке с порошкообразным полиэтиленом они отделили токсин от не-
органических солей и полярных органических соединений, используя для элюции
50 %-ный водный раствор этанола. Чистый РТХ они получили путем ионообменной
гель-фильтрации элюата сначала на DEAE-Sephadex при pH 7,0, затем на CM-Sepha-
dex при pH 4,5 Выход РТХ составил 0,027 % от массы сырых кораллов. По данным
R. Е. Мооге et al. (1982), кораллы Р. toxica могут содержать до 300 мкг/г РТХ.
Очищенный палитоксин представляет собой белый аморфный гигроскопичный
порошок. В кристаллической форме токсин и его аналоги пока получены не были.
Брутто-формула (система Хилла) РТХ: Q29H223N3O54. ММ РТХ укладывается в пре-
делы от 2659 до 2680 кДа (что в 19 раз больше, чем у зарина), в зависимости от вида
1038
Часть 3. Частная неправильная эпидемиология
Рис. 3.157. С груктура палитоксина и остреонина-D. По М. Louzao et al. (2008)
Palythoa, из которого он был получен. ММ истеоцина-D — 2635 кДа. ММ маскарено-
токсина- А и маскаренотоксина-В находятся в пределах от 2500 до 2535 кДа оваток-
сина-А — 2648 кДа. Водный раствор РТХ в ультрафиолетовом спектре дает два пика
поглощения: 233 и 263 нм. РТХ термостабилен, температура разложения 300°С. Он
не инактивируется кипячением, стабилен в нейтральных водных растворах в течение
нескольких месяцев, однако быстро разрушается под воздействием кислой и щелоч-
ной среды. Растворим в диметилсульфоксиде и пиридине, но не растворим в эфире,
ацетоне и хлороформе, плохо растворим в спиртах. При растворении в воде РТХ об-
разует стойкую пену (Антонов Н. С., 1994; Katikou Р., 2007).
Механизм действия РТХ на субклеточном уровне проявляется в блокировании Na+/
К+-АТФазы, лежащей в основе работы Na+/K+-Hacoca клеток мышц сердца, эритро-
цитов и нервной системы, и поддерживающего физиологический градиент Na+ и К+
внутри и снаружи клетки. Та часть белков №+/К+-канала, которая not ружена в ци-
топлазму клетки, имеет повышенное сродство к ионам натрия. Поверхность белков,
обращенных во внеклеточное пространство, имеет повышенное сродство к калию
Внутренние сайты связывания работают тогда, когда внешние не связаны с ионами
калия. Na+/K+-Hacoc выкачивает ионы натрия из клетки, одновременно накачивая
ионы калия вну грь клетки. Он транспортирует три иона Na+ из клетки и два иона К+
в клетку, используя гидролиз АТФ в качестве источника энергии. Таким образом, обес-
печивается низкая внутриклеточная концентрация ионов натрия и высокая — калия.
Градиент концентрации ионов натрия на мембране имеет специфические функции,
связанные с передачей информации в виде электрических импульсов, а также с под-
держанием других активных транспортных механизмов и регулирования объема внут-
реннего пространства клетки.
3.16. Палитоксин
1039
Рецептором дтя РТХ является транспортный белок №+/К+-АТФаза. На внутрен-
ней поверхности мембраны она расщепляет АТФ на АДФ и фосфат. На транспорти-
ровку трех ионов натрия из клетки и одновременно двух ионов калия в клетку исполь-
зуется энергия одной молекулы АТФ, т. е. суммарно за один цикл работы насоса из
клетки удаляется один положительный заряд. Таким образом, Na /К+-насос является
электрогенным (создает электрический ток через мембрану), что приводит к увеличе-
нию электроотрицательности мембранного потенциала приблизительно на 10 мВ.
Транспортный белок выполняет эту операцию со скоростью: от 150 до 600 ионов нат-
рия в секунду. Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой,
расходуется на работу Na+/K+-Hacoca, а в некоторых наиболее активных клетках на
его работу расходуется до 70 % энергии.
Молекулы РТХ подавляют активный транспорт Na+ и К+ через клеточные мемб-
раны, связываясь с внеклеточным участком №+/К+-АТФазы. Na+/K+-Hacoc превра-
щается в неспецифический постоянно открытый ионный канал. Мембрана деполяри-
зуется и в цитозоле происходит накопление Са2+. Изменение концентрации внутри-
клеточных катионов, в частности, увеличение кальция, как правило, вызывает гибель
клеток. Внутриклеточной мишенью РТХ является цитоскелет. Показано, что сигналь-
ный каскад, индуцированный РТХ и остреоцин-D, приводит к деформации нитей
актина, формирующих цитоскелет. В условиях in vitro токсическое действие РТХ про-
является осмотическим лизисом эритроцитов с деструкцией клеточной мембраны
и высвобождением гемоглобина. Гемолиз эритроцитов можно ингибировать добавле-
нием сердечного гликозида квибаина (ouabain, g-strophanthin) — специфического ин-
гибитора мембранной №+/К+-АТФазы. Однако в условиях in vivo терапевтический
эффект от применения квибаина при экспериментальном отравлении РТХ не наблю-
дается (Munday R., 2011).
В условиях in vivo РТХ вызывает широкий спектр вторичных фармакологических
эффектов. Они включают спастическое сокращение скелетных, гладких и сердечных
мышц и другие сердечно-сосудистые эффекты, так как высвобождение нейротранс-
миттеров из терминалей нервов приводит к сокращению миоцитов. Спазм гладких
мышц сосудов вызывает инфаркт миокарда, мозга и некрозы кишечника. Прямое
действие РТХ на клетки вызывает разрушение мышечного волокна (и, как следствие,
рабдомиолиз), гемолиз эритроцитов, выброс гистамина, простагландинов, норадре-
налина, агрегацию тромбоцитов, костную резорбцию и ингибирование подвижно-
сти сперматозоидов. РТХ является промотором опухолей, но сам мутагенным дейст-
вием не обладает. РТХ вызывает необратимое увеличение цитозольной концентрации
кальция в первичных культурах клеток мозжечковых гранулярных клеток (cerebellar
granule cells, CGC), составляющих почти половину всех нейронов ЦНС (Vale-Gonza-
lez С. et al., 2007).
Смерть при отравлении РТХ наступает от разных причин в зависимости от дозы
токсина, способа его введения, проводимых лечебных и реанимационных мероприя-
тий. При остром отравлении РТХ (большая доза токсина при пищевом отравлении или
в/в введение) смерть наступает вследствие тяжелой коронарной вазоконстрикции
(спазма коронарных сосудов) и последующей сердечной недостаточности. Изменения
ЭКГ у собак и кошек после в/в введения РТХ показали выраженную ишемию мио-
карда, ассоциированную с прекращением коронарного кровообращения. Респиратор-
ная недостаточность не является при остром отравлении РТХ основной причиной
смерти. Искусственная вентиляция легких у отравленных РТХживотных не увеличи-
вала дозу токсина, необходимую для летального исхода. Расширение правого желу-
1040
Часть 3. Частная неправильная эпидемиология
дочка у мышей, отравленных РТХ, показывает выраженную легочную гипертензию,
следующую за коронарнарной вазоконстрикцией (Munday R., 2011). При раннем на-
чале реанимационных мероприятий или недостаточной для острого отравления дозы
токсина, летальный исход возможен на вторые-третьи сутки из-за кардиопульмонар-
ного шока (cardiopulmonary arrest), возникающего вследствие:
1) рабдомиолиза дыхательных мышц;
2) поражения стволовой части мозга, вызванного прямым действием РТХ на моз-
жечковые гранулярные клетки;
3) инфарктом мозга, вызванным спазмом сосудов;
4) прямым повреждением РТХ миокарда.
Начиная с третьих-четвертых суток после отравления РТХ, основную роль в ле-
тальном исходе начинает играть острая почечная недостаточность, вызванная рабдо-
миолизом (Shinzato Т. et al., 2008).
Токсические и поражающие свойства РТХ. Острая токсичность РТХ для млекопи-
тающих в основном зависит от пути его проникновения в организм. Экстремально
низкие летальные дозы отмечаются в случае внутривенного введения токсина. При
внутривенном введении РТХ примерно в 20 раз токсичнее STX и более чем в 3 тыс
раз токсичнее стрихнина. Кролики, собаки, обезьяны-резусы, крысы наиболее чув-
ствительны к РТХ при данном пути введения. Во всех случаях смерть наступает в те-
чение нескольких минут. Токсичность РТХ при трахеальном введении почти такая же,
как и при внутривенном введении. Токсичность РТХ при внутрибрюшинном введе-
нии ниже, чем при внутривенном. Даже при 5 LD50 летальный исход наблюдается не
ранее 1 ч после введения токсина. При дозе, приближающейся к LD50, мыши погибают
в течение 12—40 ч после инъекции РТХ. Летальные дозы РТХ при внутримышечных и
подкожных инъекциях сходы с теми, что получены при внутрибрюшинном введении.
Еще менее токсичен РТХ при энтеральном введении (табл. 3.62).
Таблица 3.62
Острая токсичность РТХ для животных*
Вид Штамм Способ введения РТХ Источник РТХ Параметр токсич- ности Время наблю- дения, ч Токсичность (мкг/кг)
1 2 3 4 5 6 7
Мышь ICR В/в Р. vestitus LD50 24 0,45 (0,33- 0,62)
? Р tuberculosa ld50 ? 0,53 (0,38-0,73)
? Р. toxicus ld50 ? 0,15
Крыса Wist ar- Sprague Dawley гибрид Р. toxicus LD50 24 0,089 (0,080 0.098)
Морская свинка Mongrel Р. vestitus LD50 24 0,11 (0,070-0,170)
Кролик NZ White Р. vestitus ld50 24 0,025 (0,024-0,026)
Собака Mongrel Р. vestitus LDS0 24 0,033 (0,026-0,041)
Р. tuberculosa LD50 9 0,05—0,06
Кошка ? Р tuberculosa ld50 ? Около 0,2
Макака- резус — Р. vest'tus LD30 24 0,078 (0,060-0,090)
3.16. Палитоксин
1041
Продолжение табл. 3.62
1 2 3 4 5 6 7
Мышь 1CR В/б Demania alcaiai ld50 24 1,0—1,5
1? 9 ld50 ? 0,5
? Pafythoa toxica ld50 ? 0,4
Swiss-Webster Palythoa caribaeorum LD ? 0.05-0.10
NMRI P. tuberculosa ld50 48 0,31 (0,26 -0,37)
? MLD 9 0,6
Swiss ld50 48 0,72 (0,64-0,80)
Крыса Wistar-Sprague Dawley гибрид P. vestitus ld50 9 0,63 (0,44-0,91)
Кролик Wistar-Sprague Dawley гибрид Через зонд P vestitus ld50 72 >40
Мышь Swiss P. tuberculosa ld50 24 510 (311-809)
CD-I P. tuberculosa ld50 24 767 (549-1039)
Swiss Вместе с пищей P. tuberculosa LD 24 >2500
ICR Сублингв- ально P. tuberculosa LDjo 24 >235
Крыса Wistar-Sprague Dawley гибрид В/м P. vestitus ld50 24 0,24 (0,21-0,28)
Собака Mongrel P. vestitus ld50 24 0,08 (0,05-0,14)
Крыса Wistar-Sprague Dawley гибрид П/к P. vestitus ld50 24 0,40 (0,29- 0,54)
Мышь ICR P. vestitus ld50 24 1,39 (1,07-1,80)
Крыса Wistar-Sprague Dawley гибрид Интрат- рахеально P. vestitus ld50 5 cyr 0,31 (0,17-0,54)
Крыса Wistar-Sprague Dawley гибрид P. vestitus ld50 10 cyr 0.18 (0,09-0,38)
Мышь ICR P. tuberculosa LD ? >2
Крыса Wistar P. tuberculosa LD ? 5,0-7,5
Мышь CD-I П/к P. tuberculosa LD ? 400-1700
Крыса Wistar-Sprague Dawley гибрид Интра- трахеально P. vestitus LD 24 >10
* По обзорной работе R. Munday (2011).
Клинические наблюдения, сделанные после однократного внутривенного вве-
дения летальной дозы токсина кошкам и собакам, показали следующее. Со стороны
ЭКГ — развитие брадикардии, эктопические систолы, вентрикулярная аритмия.
В последующем происходит остановка дыхания. Атаксия и пилоэрекция наблюда-
ются вскоре после внутрибрюшинного введения мышам смертельной дозы РТХ. Че-
рез 30—60 мин, в зависимости от дозы, у животных наступал паралич задних конечно-
стей. Частота дыхания постепенно снижалась, паралич распространялся на все тело.
1042
Часть 3. Частная неправильная эпидемиология
Животные не реагировали на внешние раздражители, незадолго до смерти у них на-
блюдался цианоз и экзофтальмия. В плазме крови — повышенный уровень креатин-
киназы (Munday R., 2011).
После введения РТХ через желудочный зонд (300—1697 мкг/кг) мыши линии CD-1
погибали в течение 24 ч на фоне развития дыхательной недостаточности и паралича
(LD50 767 мкг/кг). В плазме крови — повышение уровней креатинфосфокиназы и
лактатдегидрогеназы (при дозе токсина 600 мкг/кг и выше), и аспартаттрансаминаз
(848 мкг/кг и выше) (Sosa S. et al., 2009).
Самое заметное макроскопическое посмертное изменение при внутривенном вве-
дении РТХ — выраженное расширение правого желудочка. При внутрибрюшинном —
спайки брюшины, асцит и расширение тонкого кишечника: при энтеральном — гипе-
ремия нежелезистой части желудка. После нанесения РТХ на кожу у животных раз-
вивались тяжелое местное воспаление, отек и некроз. Попадание РТХ в глаз индуци-
ровало тяжелый конъюнктивит, отек, помутнение и язвы роговицы. Внутрикожное
введение приводило к выраженной бледности кожи в участке введения, что объясня-
ется сосудосуживающим действием РТХ (Munday R., 2011).
У мышей, погибших в результате интраперитонеального введения РТХ, наблюдали
признаки некроза отдельных клеток сердечной мышцы, некроз лимфоцитов в тиму-
се и селезенке. В тонком кишечнике имелись признаки кровотечения, отека и некро-
за. Электронная микроскопия обнаружила грубые изменения митохондрий и отдель-
ных органелл миоцитов, утрату микроворсинок в дистальном и проксимальном
отделах почечных канальцев и вакуолизацию ацинарных клеток поджелудочной же-
лезы. В световой микроскоп патологические изменения в клетках мышц сердца и в
скелетных мышцах (soleus) мышей, отравленных введением токсина в желудок, за ис-
ключением агрегаций митохондрий и дегенерации волокон, обнаружить не удавалось.
В нежелезистой части желудка наблюдалось воспаление слизистой поверхности, в
почках — расширение почечных канальцев, в печени — гепатоцеллюлярная вакуоли-
зация, в поджелудочной железе — истощение секретирующи> клеток (Sosa S. et al.,
2009; Munday R., 2011).
Попадание РТХ в трахею вызывало альвеолярное кровотечение (alveolar haemor-
rhage), легочный отек, эрозию слизистой желудка и кишечника и атрофию почечных
клубочков. Сходный токсический эффект был получен в результате сублингвального
введения токсина в количестве 200 мкг/кг (Ito Е., Yasumoto Т, 2009).
Острая токсичность аналогов РТХ — остреоцина D, N-ацетилпалитоксина и
42-гидроксипалитоксина при энтеральном введении несколько меньше чем у РТХ, но
разница в токсичности не существенна. LD 50 остероцина-D для мышей при внутри-
брюшинном введении составляет 0,00075 мг/кг. При энтеральном введении леталь-
ная доза остреицина-D вызывает у мышей эрозию желудка и кишечника. Нов отли-
чие от РТХ он не вызывает развития перитонита. Наиболее токсичным аналогом
является оватоксин-А (Munday R., 2011). По данным Е. Ito и Т. Yasumoto (2009), при
интратрахеальном введении мышам РТХ, смертельной была доза 2 мкг/кг, при том же
пути введения остреоцина D его смертельная доза 11 мкг/кг.
Данных по токсичности аэрозолированного РТХ в открытых источниках мной не
обнаружено. При интратрахеальном введении летальная доза РТХ лишь в 4 раза пре-
вышает таковую при внутривенном введении (см. табл. 3.62), что позволило Н. С. Ан-
тонову (1994) сделать вывод о высокой эффективности палитоксина в аэрозолиро-
ванном состоянии и его высокой поражающей способности при воздействии инга-
ляционным путем.
3.16. Палитоксин
1043
Накопленные в опытах на животных экспериментальные данные по 1 воля ют сде-
лать весьма достоверный прогноз уровня внутривенной токсичности для человека.
Со1ласно прогнозу Н. С. Антонова (1994), летальная доза палитоксина при внутри-
венном введении человек}'лежит в пределах vl,0-2,0)x JO-5 мг/кг.
РТХ может переводиться в аэрозольное состояние из растворов с помощью рас-
пылителей или взрывом заряда взрывчатого вещества. О возможности получения аэро-
золей этого токсина путем термической возгонки не сообщалось. РТХ вряд ли может
эффективно применяться в виде заблаговременно приготовленного тонкодисперсного
порошка из-за его гигроскопичности. Недостаток РТХ как потенциального пора-
жающего агента в том, что он является твердым веществом, в связи с чем существуют
проблемы перевода его в аэрозольное состояние. По той же причине уровень кожно-
резорбтивной токсичности не может быть столь высок по сравнению с ингаляционной
токсичностью. Практически при обычных температурных условиях РТХ не образует
пар в конце нграциях, при которых возможны поражения, из-за чего сковывающее
действие РТХ проявляется в значительно меньшей мере, чем у жидких отравляющих
веществ. РТХ. по крайней мере в настоящее время, не может производиться в коли-
чествах, необходимых д ля того, чтобы им снаряжали боеприпасы. Не исключено, что
в будущем РТХ получит производственную базу и будет представлять опасность при
применении в смесях с жидкими отравляющими веществами (Антонов Н. С.. 1994).
Клиника пищевого отравления. В табл. 3.61 приводится описание нескольких слу-
чаев отравления людей РТХ, содержащимся в морепродуктах. У рыб РТХ аккумули-
руется в основном в кишечнике и печени. Симптомы отравления у человека появля-
ются через несколько минут пос ле употребления в пищу отравленных морепродуктов.
Они включают общее недомогание, чувство усталости, тошноту, рвоту, понос, боли в
мышцах (в основном плеча и предплечья), мышечные спазмы, сердечную и легочную
недостаточное гь. Морепродукты на вкус кажутся пациенту гооькими или обладающи-
ми металлическим привкусом. Для отравления РТХ типично сочетание этих симпто-
мов с увеличением в сыворотке крови уровня креатинфосфокиназы (до 180 тыс. МЕ/л
на третьи сутки болезни) и других ферментов, характерных для рабдомиолиза. При по-
ступлении в стационар температура тела пациента может быть нормальной или пони-
женной (35,5°С), АД повышенно или в норме, нарушений сенсорной чувствительно-
сти и других неврологических симптомов не бывает. В течение суток АД у пациента
падает, развивается брадикардия, нарастают симптомы сердечно-сосудистой и легоч-
ной недостаточности. Лабораторные анализы показывают лейкоцитоз, в сыворотке
повышение уровня креатинфосфокиназы, аспартатаминотрансферазы (повреждение
мышечной ткани и печени), аланинаминотрансферазы (повреждение печени), лактат-
дегидрогеназы (разрушение любых тканей), миоглобина (повреждение мышечной
т кани сердца), альдолазы (повреждение скелетной мускулатуры и печени) и С-реак-
тивнего белка (повреждение гюбых тканей — обычно плохой прогностический при-
знак). В моче макрогематурия (заметное на глаз покраснение мочи), но эритроциты
в осадке не находят. В последующем появляется миоглобинурия. Появление в сыво-
ротке крови и в моче миоглобина свидетельствует о поражении сердечной мышцы.
Количество миоглобина зависит от степени повреждения миокарда. Выздоровление
пациента, отравленного РТХ, идет медленно (более месяца). Наиболее частая причина
смерти в первые несколько часов после отравления — сердечная недостаточность,
вызванная коронароспазмом и прямым повреждением миокарда РТХ. На аторые-
третьи сутки летальный исход возможен вследствие развития кардиопульмонарного
шока. В более позднем периоде опасность пациенту представляет почечная недоста-
1044
Часть 3. Частная неправильная эпидемиология
точность. У пациентов, находящихся на искусственной вентиляции легких и гемодиа-
лизе, возможна смерть мозга (Okano Н. et al., 1998; Shinzato Т. et al., 2008; Tubaro A. et
al., 2011). В описанном T Shinzato (2008) случае отравления РТХ, содержащемся в
печени скаровой рыбы (parrot fish, Scarus, рыбы-попугаи), у пациента на третьи сут-
ки после отравления развился кардиопульмонарный шок, он впал в кому, после чего
находился на механической вентиляции легких до остановки сердца. С появлением
признаков рабдомиолиза (вторые сутки госпитализации) ему проводили непрерыв-
ную гемодиафильтрацию (continuous hemodiafiltration, CHDF). Но так как гиперка-
лиемия и гиперфосфатемию (признаки почечной недостаточности) не удалось взять
под контроль, на 4 сут госпитализации CHDF заменили на гемодиафильтрацию (he-
modiafiltration, HDF) с 10 л замещающего раствора.
На 9 сут госпитализации у пациента диагностирована смерть мозга (cerebral death).
На 11 сут объем мочи стал увеличиваться и к 14 суткам достиг 1300 мл/сут, что свиде-
тельствовало о снятии состояния острой почечной недостаточности. Родственники
попросили отключить диализ, что и было сделано на 11 сут. У пациента развилось
состояние тяжелой гиперкалиемии, на 16 сут госпитализации он умер от сердечной
недостаточности, вызванной гиперкалиемией.
Посмертные патологические изменения у людей, погибших от острого отравления
РТХ, обычно включают наличие геморрагических уплотнений на слизистой тонкого
кишечника и в проксимальном участке толстой кишки. В этих же участках кишечни
ка патологоанатомы обнаруживают скопление геморрагического выпота (Tan С. Т. Т.,
Lee Е. J. D., 1988).
Клиника ингаляционного поражения. Со средины прошлого десятилетия вдоль ита-
льянской, французской и испанской береговой линии наблюдается цветение Ostreopsis
spp. Одновременно с цветением новых для этих мест динофлагеллят врачи стали от-
мечать новую массовую патологию, развивающуюся у людей, находящихся на берегу
моря во время шторма, либо участвовавших в спортивных соревнованиях, сопровож-
дающихся образованием брызг воды. Новая болезнь была клинически сходна с ОРЗ,
но развивалась летом и имела более тяжелое течение. Так как болезнь ассоциирова-
лась с цветением моря, ее назвали «синдром водорослей». Исследование воды в пе-
риоды массовых вспышек «синдрома водорослей» показало высокое содержание РТХ
и его аналогов, среди которых преобладал оватоксин-А — основной токсин, проду-
цируемый Ostreopsis ovata (Ciminiello Р. et al., 2008). В табл. 3.63 приведены данные по
двум вспышкам, вызванным вдыханием частиц морского аэрозоля, содержащего РТХ.
Большинство эпизодов отравления, вызванного ингаляцией РТХ, приходится на
средину лета — пик цветения О. ovate. Диагноз «синдром водорослей» ставят на осно-
вании анамнеза пациента и клиники болезни. В анамнезе таких пациентов имеются
указания на посещение морского курорта, и их присутствие на берегу моря не далее
90 м от воды, контаминированной цветущими водорослям Ostreopsis spp. Клинически
у пациента находят в различных сочетаниях симптомы раздражения верхних и ниж-
них дыхательных путей (иногда сопровождающиеся бронхоспазмом), конъюнктивит,
одышку, воспаление гортани, обильный насморк, лихорадку (до 38°С), слезотечение,
тошноту и рвоту, дерматит. От экспонирования к аэрозолю, содержащему РТХ, до
развития первых симптомов отравления проходит 3—4 ч. До 20 % отравленных нуж-
дались в госпитализации продолжительностью до трех суток. При поступлении в ста-
ционар у них обнаруживали умеренный лейкоцитоз и нейтрофилез. Биохимия крови
была в норме. ЭКГ и рентгенограммы грудной клетки также оставались в норме, что
позволяет классифицировать эти отравления РТХ как легкие (Tubaro A. et al., 2011).
3.16. Палитоксин
1045
Таблица 3.63
Вспышки ингаляционного отравления РТХ на морском побережье*
Год (количество отравленных) Источник токсина** Симптомы/клинические данные Лечение Последствия
Италия, Тирренское море, лето 2005 (209) О. ovata Лихорадка, воспаление горта- ни. кашель, одышка, головная боль, тошнота, слезотечение, обильный насморк, рвота, дерматиты. Умеренный лейкоцитоз, нейтрофилия Ингаляция кортико- стероидов и р-агони- стов, нестероидные противовоспалитель- ные препараты, кис- Выздоровление в течение 3 сут
То же, лето 2006 (19) О. ovata Лихорадка, воспаление гор- тани, кашель, одышка, го- ловная боль, тошнота, сле- зотечение, риноррея, рвота дородная терапия отдельным госпита- лизированным пациентам
* По A Tubaro et al. (2011).
** Обнаружение РТХ и его аналогов в микроводорослях выполнено с помощью LC-MS
Клиника контактного поражения. РТХ обладает раздражающим действием на кожу
и слизистые поверхности. Тяжесть симптомов зависит от продолжительности контакта
с кораллами, содержащими РТХ, и от наличия повреждений кожи, которые могли бы
облегчить проникновения токсина в кровь. При локальном воздействии РТХ вызывает
онемение участка кожи, контактировавшего с кораллом, эритему, отек и даже некроз
кожи. При попадании в глаз токсин вызывает лакримацию, раздражение и конъюнк-
тивит. Через сутки происходит помутнение и изъявление роговицы, пораженный глаз
закрывается. При тяжелых отравлениях добавляются общие симптомы: металличе-
ский привкус во рту, боли в груди, в брюшной полости, головокружение, слабость,
сильные мышечные боли и спазмы мышц, парестезии. Биохимия крови показывает
развитие умеренного рабдомиолиза. ЭКГ в начальной стадии отравления демонстри-
рует синусовую тахикардию без ишемии или с ишемией миокарда, в зависимости от
полученной пациентом дозы РТХ,
Диагностика поражения РТХ. Естественные отравления РТХ возможны:
1) пероральным путем при употреблении в пищу морепродуктов (крабы, сардины,
анчоусы, сельди, кузовки и др.). Как правило, такие отравления происходят в тропи-
ческих и субтропических регионах, но с потеплением климата их география может
расшириться;
2) чрескожно, при соприкосновениями с кораллами Palythoa зрр. в тропических и
субтропических регионах, а также в домах, где есть аквариумы с представителями зоан-
тарий, содержащих РТХ. Отравление обычно происходит в результате соприкоснове-
ния с зоантариями во время чистки аквариума;
3) ингаляционно в прибрежной морской полосе и не далее 90 м от воды, контами-
нированной цветущими водорослям Ostreopsis арр. Естественные чрескожные и пище-
вые отравления РТХ носят единичный характер. Естественные ингаляционные от-
равления могут быть массовыми, но, как правило, такие отравления протекают в
легкой форме и сопровождаются в основном поражением верхних дыхательных пу-
тей, так как при таком способе образовании аэрозоля не создается высоких концен-
траций частиц аэрозоля РТХ с размером дисперсной фазы 1—5 мкм. Диагностика
умышленного перорального отравления РТХ в основном сходна с диагностикой по-
ражения STX (см. разд. 3.13).
1046
Часть 3. Частная неправильная эпидемиология
Дифференциальная диагностика пищевого отравления РТХ проводится с отрав-
лениями, вызванными сигуатоксином (сигуатера), STX и ТТХ. Отравление сигуаток-
сином происходит при потреблении крупной рифовой рыбы и, в отличие от РТХ,
приводит к более тяжелым симптомам со стороны ЖК Г (тошнота рвота, циарея) и к
специфичному обратному восприятию температуры: горячее кажется холодным, а
холодное — горячим.
Дифференциальная диагностика с отравлениями, вызванными паралитическими
токсинами STX и ТТХ, основывается на отсутствии у пациента симптомов вялого
паралича скелетных мышц и поражения периферических нервов (парестезии). Для
отравления РТХ будет характерна боль в мышцах и мышечные спазмы при отсутствии
параличей и парезов на начальном этапе отравления, острое развитие сердечной не-
достаточности, биохимические признаки рабдомиолиза, длительный период выздо-
ровления. Кроме того, отравление ТТХ обычно происходит при употреблении в пищу
продуктов, полученных из рыб рода скалозубов (Sphaeroides, Fugu, рыбы-собаки) и
рода тетраодонов (Tetraodon) из семейства иглобрюховых (Tetraodontidae) и морских
брюхоногих моллюсков. Отравления STX обычно связаны с употреблением в пищу
мяса мидий, крабов, лобстеров, отдельных пород рыб (рыбы-собаки, японская скум-
брия и др.), обитающих на коралловых рифах. Сигуатоксин, £ТХ и ТТХ не вызывают
рабдомиолиза.
При искусственном ингаляционном поражении РТХ первые симптомы будут сход-
ны с теми, что наблюдаются при «синдроме водорослей» (воспаление гортани, кашель,
одышка, головная боль, гошнота, слезотечение, риноррея, рвота), однако вследствие
большей концентрации токсина в аэрозоле они будут носить более выраженный ха-
рактер и сопровождаться симптомами острой легочно-сердечной недостаточности.
Летальные исходы возможны в течение от нескольких минут до нескольких часов
после нападения.
«Патология мелкодисперсного аэрозоля», т. е характерное поражение легочной
ткани, возникающее после экспонирования к аэрозолю поражающего агента с дис-
персностью частиц 1—5 мкм, для РТХ не описано ни у людей, ни у эксперименталь-
ных животных. Но из тех лаконичных описаний экспериментов, выполненных на
животных, что есть в доступной литературе, и данных по механизму действия РТХ,
можно предположить, что у пациентов, проживших более суток от момента экспони-
рования к аэрозолю РТХ, «патология мелкодисперсного аэрозоля» будет сходна с
аналогичной патологией, вызванной поражением мелкодисперсным аэрозолем рици-
на (см. разд. 3.11). Наиболее тяжелые видимые и микроскопические поражения бу-
дут обнаружены в нижнем респиратором тракте. В легочных долях — мультифокаль-
ный сливающийся некроз и воспалительные изменения эпителия воздушных путей,
перибронховаскулярный отек, альвеолярное затопление (alveolar flooding), фибриноз-
но-гнойные альвеолиты (fibrinopurulent alveolitis), интерстициальная пневмония с уча-
стками некроза и некротизацией терминальных бронхиол (Wilhelmseann С , Pitt L.,
1996; Franz D. et al., 1997).
Отличия при ингаляционном поражении РТХ от аналогичного поражения рици-
ном будут состоять в большей выраженности симптомов острой сердечной недоста-
точности и развитии рабдомиолиза у экспонированных к РТХ лиц, не погибших в
первые сутки от острой коронарной недостаточности. Искусственные ингаляционные
поражения РТХ будут носить характер групповых вспышек, и эти группы будет объ-
единять какой-то общий географический фактор (аэропорт, кинотеатр, метро, учреж-
3.16. Палитоксин
1047
дение и др.), но без указаний на прием обшей пиши. Чем ближе расстояние до источ-
ника аэрозоля РТХ, тем больше может оказаться пациентов с острой коронарной не-
достаточностью, с поражением глаз, ринореей, дерматитами, воспалением верхних
дыхательных путей и «патологией мелкодисперсного аэрозоля».
Диагноз отравления РТХ подтверждается обнаружением РТХ (и его аналогов) в
крови и моче пациента с помощью LC-MS и биопробами. О происхождении РТХ
может многое рассказать сравнительное исследование его аналогов в образцах, полу-
ченных от отравленного и токсического профиля различных источников.
Иммунопрофилактика. Не разработана. В условиях эксперимента специфические
антитела нейтрализуют активность STX.
Лечение. Симптоматическое и патогенетическое. После перорального попадания
токсина в организм человека следует применять стандартные меры для пищевого от-
равления, включая промывание желудка и введение в ЖКТ суперактивированного
угля. В легких случаях отравления применяют ингаляционно кортикостероидные
препараты и нестероидные противовоспалительные препараты. При бронхоспазме
пациентам назначают ингаляционно Р2-агонисты (Р2-адреномиметики) вместе с
кислородной терапией. В остром периоде отравления эффективно введение вазо-
дилататоров непосредственно в сердце. В случае появления признаков легочно-сер-
дечной недостаточности необходимо перейти на контролируемую механическую
вентиляцию легких. При развитии рабдомиолиза и угрозе ра шития почечной недо-
статочности целесообразно использовать CHDF или HDF с пос гоянным контролем
активности мозга.
СЛОВАРЬ ТЕРМИНОВ
5' кэп (от англ, cap — шапочка) — это модифицированным гуанозиновый нуклеотид, который
добавляется на 5' (передний) конец «незрелой» мРНК. Эта модификация очень важна для узнава-
ния мРНК при инициации трансляции, а также для защиты от 5'нуклеаз — ферментов, разрушаю-
щих цепи нуклеиновых кислот с незащищенным 5'-концом.
CD80/CD86 — CD80 (В7-1) и CD86 (В7-2) — гликопротеины I типа и члены Ig-суперсемейства,
включают два Ig-подобных экстрацеллюлярных внеклеточных домена, связанных с трансмембран-
ным доменом, и цитоплазматический хвост. Экспрессируются на поверхности антиген-презенти-
рующих клеток, включая дендритные клетки и В-лимфоциты, и действуют как костимуляторные
сигналы для активации клеточно-зависимого иммунного ответа
CpG-островки — в GC-богатых изохорах (гомогенные участки генома) локализовано большое
количество CpG-островков — последовательностей от 500 до 2000 п. о., характеризующихся очень
высоким содержанием гуанина и цитозина (Г + Ц более 60 %). Характерным свойством CpG-ост-
ровков является их локализация вблизи структурных генов, преимущественно в их 5'-участках (регу-
ляторные последовательности, промоторы, последовательности первого экзона). Формальные
признаки CpG-островков (присутствуют в промоторных зонах примерно 60 % генов человека). Зна-
чительная часть тканеспецифических генов не имеет в своих промоторах CpG-островков: вместо
них могут присутствовать одиночные CpG-динуклеотиды. Наибольшая плотность CpG-островков
наблюдается в теломерных участках хромосом 1, 9, 15, 16, 17, 19, 20, 22. Точные молекулярные ме-
тоды регистрации CpG-островков показали, что их число в геноме человека приближается к 45 тыс.
Hfr (high frequency of recombination) — высокая частота рекомбинации. Под таким обозначе-
ние известен донорный штамм кишечной палочки (F+ Met-), который при скрещивании со стан-
дартным штаммом (F- Thr- Leu-) образовывал прототрофные рекомбинанты с частотой по край-
ней мере в 103 большей, чем исходный штамм F+ при стандартном скрещивании F+xF“.
LDS0 аэрозоля — количество клеток микроорганизма, вызывающее при ингаляционном попа-
дании в организм человека смертельный исход с вероятностью 50 %. Обычно эти величины рас-
четные, сделанные для аэрозоля с дисперсной фазой от 1 до 5 мкм. Чем меньше величина LD50, тем
больше инфицирующих доз агента содержится в определенном объеме биологической рецептуры
SNAP25 (от англ, synaptosomal-associated protein, 25-kD) — мембранный белок, компонент бел-
кового комплекса SNARE, осуществляющего стыковку синаптической везикулы с пресинаптичес-
кой мембраной нейрона и их слияние с последующим высвобождением нейромедиатора. SNAP25
является небольшим белком (около 25 кДа). Он крепится к мембране синаптической везикулы за
счет ацильных остатков пальмитиновой кислоты, находящихся в середине молекулы белка. При
образовании стыковочного SNARE комплекса с синтаксином и синаптобревином SNAP25 предо-
ставляет две из четырех а-спиралей комплекса. По структурной классификации SNAP25 относит-
ся к подгруппе Q-SNARE.
SNARE-белки (от англ, soluble NSF attachment receptor) — большая группа белков, осуществ-
ляющих слияние внутриклеточных транспортных везикул с клеточной мембраной (экзоцитоз) или
органеллой-мишенью, такой как лизосома. Насчитывается около 60 белков SNARE. Белки груп-
пы делятся на две функциональные категории: везикулярные белки (v-SNARE) и белки принимаю-
щей органеллы (t-SNARE). Новая структурная классификация подразделяет группу на R-SNARE
и Q-SNARE. Наиболее изучены белки, которые выполняют доставку синаптических везикул к пре-
синаптической мембране и их слияние. Эти белки являются мишенями опасных бактериальных
токсинов ботулизма и столбняка.
Адаптивная радиация — образование разнообразных форм организмов в пределах вида или груп-
пы родственных видов. Лежит в основе всех форм адаптациогенеза и является результатом приоб-
ретения организмами специальных приспособлений — адаптаций и проникновения в новые адап-
тивные зоны.
Адвентиция — внешняя оболочка органа, образованная в основном соединительной тканью.
В адвентиции артерий и вен проходят кровеносные сосуды, питающие стенку — «сосуды сосудов»
Словарь терминов
1049
(лат. vasa vasorum). Адвентиции вен и артериол относительно толще, чем адвентиции артерий.
Некоторые авторы называют адвентицией соединительнотканные клетки, окружающие капилляры.
Аденилатциклаза — фермент класса лиаз, катализирующий превращение АТФ в 3',5'-цАМФ
(циклическую форму АМФ) с образованием пирофосфата. В процессе передачи сигнала аденилат-
циклаза может быть активирована связанными с плазматической мембраной рецепторами, связан-
ными с G-белками (GPCR), которые передают гормональные и иные стимулы в клетку. Актива-
ция аденилатциклазы приводи! к образованию цАМФ, действующего как вторичный посредник.
цАМФ взаимодействует с протеинкиназой А, ионными каналами, свя 1анными с циклическими нук-
леотидами, и регулирует их функции. Молекулярно-биологическими методами показано существо-
вание в клетках млекопитающих, по крайней мере, шести различных аденилатциклаз. Все они
имеют ММ около 120— 130 кДа, а исследование их гидрофобной части показывает наличие 12 транс-
мембранных сегментов.
Аденопатия — заболевания лимфатических узлов.
Алкалоиды — биологически активные гетероциклические соединения с атомами азота в цикле,
преимущественно растительного происхождения, обладающие свойствами оснований. Простейшие
алкалоиды содержат до 10 атомов углерода, наиболее сложные — более 50.
Альтернативный сплайсинг — соединение экзонов данного гена в разных комбинациях с обра-
зованием различающихся зрелых молекул мРНК
Амфотропный вирус — вирус для которого характерен более широкий круг хозяев, чем свойст-
венно вирусам данного вида: это свойство связано со способностью белков оболочки такого вируса
взаимодействовать с рецепторами клеток большего числа видов — например, амфотропный MLVM
(вирус лейкоза Молони мышей) заражает клетки многих млекопитающих и птиц, а экотропный
MLVM — только клетки мышей и крыс.
Авали । полиморфизма длины амплифицированных фрагментов - вариант молекулярно-генетиче-
ского идентификационного анализа. Феномен ПДАФ проявляется в том, что в случае ПЦР-ампли-
фикации гипервариабельных мини- и микросателлитных генетических матриц (локусов), присут-
ствующих в геноме каждого вида, в ходе реакции образуются фрагменты ДНК, которые у разных
индивидуумов имеют различную длину и потому оказываются индивидуально специфичными. Эти
полиморфные по длине фрагменты, по сути представляющие разные аллельные варианты поли-
морфных локусов геномной ДНК становятся доступными для сравнительного анализа. В ряду по-
колений каждый вариант ПДАФ наследуется как простой менделевский кодоминантный признак.
Анамнез — совокупность сведений, получаемых при медицинском обследовании путем расспро-
са самого обследуемого и/или знающих его лиц. Данные анамнеза (сведения о развитии болезни,
условиях жизни, перенесенных заболеваниях, операциях, травмах, беременностях, хронической
патологии, аллергических реакциях, наследственности и др.) медицинский работник выясняет с
целью их использования для диагностики, выбора метода лечения и/или профилактики. Сбор анам-
неза является одним из основных методов медицинских исследований и универсальный метод ди-
агностики, применяемый во всех областях медицины.
Анатоксин (токсоид) — иммунопрофилактический препарат, приготовленный из бактериаль-
ного токсина, не имеющий выраженных токсических свойств, но сохранивший его антигенные и
иммуногенные свойства и способный индуцировать выработку нейтрализующих антител к исход-
ному токсину. Анатоксины используются для профилактики инфекционных болезней, в основе па-
тогенеза которых лежит интоксикация (дифтерии, столбняка, ботулизма и т. п.).
Анемограф — прибор для непрерывной автоматической записи скорости и направления (рум-
ба) ветра.
Анизокория — неравенство величины зрачков, связанное с нарушением иннервации двух глад-
ких внутренних мышц глаза (круговой мышцы — суживающей зрачок, и радиальной мышцы, рас-
ширяющей его).
Антибиотики — специфические вещества жизнедеятельности ряда микроорганизмов (бактерий,
актиномицетов, плесневых грибов), растений или животных тканей, угнетающие рост и размно-
жение микробов. Некоторые антибиотики обладают противоопухолевым действием.
Антиген онкофетальный — белок, в норме вырабатываемый только тканями плода; иногда его
присутствие в организме человека можно обнаружить в случае развития у него некоторых опухолей.
Примером такого антигена является карциноэмбриональный антиген, который служит в качестве
опухолевого маркера.
1050
Словарь терминов
Антитела — глобулины, синтезируемые в лимфоидной ткани плазматическими клетками пос-
ле введения антигена в организм. Антитела являются строго специфическими, т. е. вступают в ре-
акцию только с антигеном, индуцирующим их синтез. На специфике реакции антиген — антитело
основан один из главных современных методов диагностики инфекционных болезней (иммуно-
циггностики). Установление антител в сыворотке крови указывает на контакт данного организма с
определенными возбудителями.
'нтитоксины — разновидность антител, возникающих в сыворотке крови в ответ на попадание
в организм токсинов или анатоксинов. Способны взаимодействовать с токсинами, нейтрализуя их
активность.
Антропонозы — группа инфекционных (паразитарных) болезней, резервуаром возбудителей
которых является только организм человека.
Аптамер — небольшая молекула нуклеиновой кислоты, способная выполнять функцию высо-
коспецифичного рецептора для низкомолекулярных органических соединений.
Ареал — пространство, на котором происходит становление вида или в пределах которого он
встречается и проходит цикл своего развития.
Ауксотроф — организмы (чаще микроорганизмы), утратившие способность к самостоятельному
синтезу какого-либо метаболита (ам тнокислоты, витамина, азотистого основания и т. д.) в резуль-
тате мутации и потери способности к образованию определенного фермента. Ауксотрофы могут
расти только на средах, в которые этот метаболит добавлен Организм дикого типа, способный раз-
виваться в той же среде без добавления питательных веществ, называют прототрофом. Ауксотро-
фы широко используются в генетических и других биологических исследованиях.
Аутологичный — взятый у той же особи.
Аутотранспортеры — семейство белков секретируемых и представленных на наружной мемб-
ране бактерии, играющих важную роль в их вирулентности (инвазины, адгезины, протеазы и др.).
Аутофагия — процесс разрушения клеток лизосомами.
Ахалазия кардии — хроническая болезнь, характеризующаяся отсутствием или недостаточным
рефлекторным расслаблением нижнего пищеводного сфинктера
Аэрозоль — дисперсная система, представляющая собой газ или смесь газов, в которой взве-
шены твердые или жидкие частицы. Термин стал применяться в конце Первой мировой войны тыя
обозначения мелкодисперсных суспензий мышьякосодержаших веществ в воздухе, создаваемых
германскими химическими снарядами, т. е. он с самого начала применения в научной литературе
имел военно-прикладное значение. Аэрозоли с жидкими частицами принято называть туманами,
аэродисперсные системы с твердыми частицами, в зависимости от механизма образования, - пы-
лями или дымами. По механизму образования аэрозоли подразделяют на дисперсионные и кон-
денсационные. Первые из них образуются при диспергировании (распылении, измельчении) твер-
дых и жидких тел, вторые — в результате конденсации пересыщенных паров или газовых реакций,
следствием которых является образование нелетучих продуктов. Термин бактериальные (вирусные)
аэрозоли относится к аэродисперсным системам, жидкие или сухие частицы которых включают
микроорганизмы. Поражающее действие таких систем зависит от вида примененных микроорга-
низмов, концентрации их в воздухе, дисперсности системы, абсолютной влажности воздуха и дру-
гих факторов.
Бентонит — природный глинистый материал, относится к классу алюмосиликатов, имеет вы-
сокую дисперсность.
Биологическая безопасность — система медико-биологических, организационных и инженер-
но-технических мероприятий и средств, направленных на защиту работающего персонала, насе-
ления и окружающей среды от воздействия патогенных биологических агентов.
Биологическая война — война, в ходе ведения которой применяется биологическое оружие.
Биологическая рецептура — многокомпонентная система, содержащая патогенные микроорга-
низмы (токсины), наполнители и стабилизирующие добавки, обеспечивающие повышение их ус-
тойчивости при хранении, применении и нахождении в аэрозольном состоянии. В зависимости от
агрегатного состояния рецептуры могут быть сухими или жидкими.
Биологические поражающие агенты — патогенные микроорганизмы, токсины и их генно-инже-
нерные модификации; трансгенные организмы и тродукты из них полученные; вирусоподобные
и иные конструкции, предназначенные для переноса генов между многоклеточными организма-
ми, а также отдельные гены с системой регуляции экспрессии, способные вызывать специфиче-
ские биологические поражения у человека и животных.
Словарь терминов
1051
Биологические поражающие средства — рецептуры биологических поражающих агентов, пред-
назначенные для снаряжения биологического оружия.
Биологическое оружие - специальные боеприпасы и боевые приборы со средствами доставки,
снаряженные биологическими поражающими средствами.
Биопленки — обладающее пространственной и метаболической структурой сооошество (коло-
ния) микроорганизмоь, расположенных на поверхности раздела сред и погруженных во внеклеточ-
ный полимерный матрикс.
Боке биитогической безопасности — конструкция, используемая для физической изоляции
(удержания и контролируемого удаления из рабочей зоны) микроорганизмов с целью предотвра-
щения возможности заражения персонала и контаминации воздуха рабочей зоны и окружаю
шей среды.
Болезнь — повреждение паразитом хозяина на любом уровне: молекулярном, субклеточном,
клеточном, органном, организменном, популяционном
Бореальный пояс — природная зона умеренного географического пояса Северного полушария
с холодной зимой и теплым летом. Средняя температура воздуха в июле от 14 до 18°С. Количество
атмосферных осадков больше испаряемой влаги, поэтому господствуют хвойные леса (тайга). Бо-
реальная зона занимает широкую полосу в Евразии от Прибалтики до Охотского моря и в Г Неверной
Америке между Атлантическим и Тихим океаном. Климат резко континентальный, с сильными
длительными морозами при тонком снежном покрове, что обусловило многолетнюю мерзло-
ту грунтов.
Бульбарный синдром (бульбарный паралич) — синдром поражения черепных нервов (языкогло-
точного, блуждающего и подъязычного), ядра которых располагаются в продолговатом мозге. Буль-
барный паралич возникает при двустороннем и, в менее выраженной степени, при одностороннем
поражении ядер черепно-мозговых нервов каудальной rpj ппы (IX, X, XI и XII нервов), заложен-
ных в продолговатом мозге, а также их корешков и стволов как внутри, так и вне полости черепа.
При бульбарном синдроме отмечается дизартрия (нарушение речи, за счет поражения отвечающих
за артикуляцию нервов) и дисфагия (нарушение глотания, за счет периферического паралича или
пареза мышц языка, мягкого неба, глотки, надгортанника, гортани). Голос больных становится
слабым, глухим, гнусавым, истощающимся вплоть до полной афонии. Больные обычно поперхи-
ваются жидкой пищей, иногда не в состоянии произвести глотательное движение. В связи с этим
слюна у таких сольных нередко вытекает из углов рта. При бульбарном параличе наступает атрофия
мышц языка и выпадают г юточный и небный рефлексы (вследствие поражения ядер IX и X нервов,
которые являются частью рефлекторных дуг вышеуказанных рефлексов). У тяжелобольных с буль-
барным синдромом, как правило, раззизаются расстройства риз ма дыхания и сердечной деятель-
ности, что нередко приводит к смерти. Это объясняется близким с ядрами каудальной группы че-
репно-мозговых нервов расположением центров дыхательной и сердечно-сосудистой системы, в
связи с чем последние могут вовлекаться в патологический процесс.
Вакцин- — препарат ослабленного или инактивированного инфекционного агента (вируса, бак-
терии и т. п.) или отдельных компонентов, несущих антигенные детерминанты и способные вызы-
вать образование иммунитета (невосприимчивости) к данной инфекции у привитых объектов.
Васкулярный эндотелиальный фактор роста, VEGF — содеожашийся в яде змей гликопротеин,
связывающийся только с эндотелиальными клетками и стимулирующий их пролиферацию. Обла-
дает выраженным гипотензивным эффектом и повышает сосудистую проницаемость у отравлен-
ного. Структуры VEGF тканевого типа высоко консервативны, однако VEGF, содержащиеся в яде
змей, высоко вариабельны, особенно в регионах вокруг рецептор-связываюших петель и С-конца
предполагаемого корецеп горного регионов.
Везикула — (в цитологии) относительно маленькие внутриклеточные органоиды, мембрано-
защищенные сумки, в которых запасаются или транспортируются питательные вещества.
Вектор — молекула ДНК, способная к включению чужеродной ДНК и к автономной реплика-
ции, служащая инструментом для введения генетической информации в клетку.
Вертикальная устойчивость атмосферы — различают три степени устойчивости атмосферного
воздуха в приземном слое, высота которого принимается равной 20 м: инверсию, изотермию, кон-
векцию При инверсии температура приземных слоев воздуха постепенно повышается, а плотность
понижается по мере удаления от поверхности земли. Инверсия атмосферы наблюдается летом, после
захода солнца, или в зимние дни Она возникает вследствие более быстрого охлаждения земли по
сравнению с плохо проводящим тепло воздухом. Холодная земля охлаждает прилегающие к ней
1052
Словарь терминов
слои воздуха, которые становятся более холодными и плотными, чем вышележащие слои. Наибо-
лее сильно перепад температуры при инверсии выражен не у самой поверхности земли, а на неко-
торой высоте от нее. Это обстоятельство имеет большое значение для повеления аэрозольного об-
лака. Частички аэрозоля распространяются в слоя,- ближайших к земле, где инверсия не так велика.
Образованное при инверсии аэрозольное облако движется под влиянием ветра непосредственно у
поверхности земли. Инверсия создает наиболее благоприятные условия для сохранения и распро-
странения высоких концентраций паров ОВ, мелкодисперсных аэрозолей БПА и ядовитых дымов.
Изотермия характеризуется стабильным равновесием воздуха. Она наиболее типична дтя пасмурной
погоды, а также возникает в j тренние и вечерние часы. Изотермия, так же как и инверсия, способ-
ствует длительному застою паров ОВ, мелкодисперсных аэрозолей БПА и ядовитых дымов на ме-
стности, в лесу, в жилых кварталах городов и населенных пунктов. Конвекция — это вертикальные
перемещения воздуха с одних высот на другие. При конвекции плотность нижних прилегающих к
земле слоев воздуха меньше, а температура больше, чем плотность и температура верхних слоев.
Конвекция наблюдается в летние дни при отсутствии облачности. Она возникает вследствие более
сильного нагревания земли по сравнению с воздухом в солнечные часы. Нагретая земля отдает тепло
прилегающим к ней слоям воздуха. Эти слои, расширяясь и становясь поэтому менее плотными,
подымаются вверх, вытесняясь более холодными и плотными слоями. При конвекции восходящие
токи воздуха рассеивают пары ОВ, мелкодисперсные аэрозоли БПА и ядовитые дымы.
Виварий — специально оборудованное помещение для содержания подопытных животных.
Вирусы — облигатные внутриклеточные паразиты животных, растений, насекомых, грибов,
простейших, бактерий, микоплазм, архей и других живых существ. Представляют собой неклеточ-
ные формы жизни, обладающие собственным геномом и способные к воспроизведению лишь в
клетках хозяина. Вирусы содержат только один тип нуклеиновой кислоты (РНК или ДНК).
Галлюцинация — образ в сознании, не соответствующий внешнему раздражителю.
Гемифациальным спазм (лицевой гемиспазм, фациальный гемиспазм) — заболевание, проявляю-
щееся безболезненными непроизвольными односторонними тоническими или клоническими со-
кращениями лицевой мускулатуры, иннервируемой ипсилатеральным »т. е. расположенным на той
же стороне) лицевым нервом.
Генетические поражения — в контексте данной работы под ними понимаются патологические
процессы, вызванные преднамеренным введением в организм человека генов.
Генная инженерия — отрасль биологической науки, изучающая закономерности конструиро-
вания in vitro рекомбинантных молекул ДНК и поведение их в реципиентной клетке. Цель генной
инженерии — создание новых генетических структур рекомбинантных молекул.
Геном — совокупность генов или хромосом организма.
Гетеротрофы — организмы, существующие за счет потребления готовых органических веществ.
К гетеротрофам относятся все животные и человек, грибы, а также растения и микроорганизмы,
не обладающие способностью к фотосинтезу или хемосинтезу.
Гибридный промотор — промотор, состоящей из частей промоторов разных генов, который по-
лучен с помощью методов генной инженерии.
Гифы — микроскопические простые или разветвленные нити, из которых формируются вегета-
тивные (грибница, или мицелий) и плодовые тела грибов.
Гликозидаза - - группа ферментов класса гидролаз, катализирующих гидролитическое расщеп-
ление эфирных связей в гликозидах; участвуют в процессах превращения углеводов.
Глубинное культивирование бактерий — способ промышленного выращивания бактериальной
биомассы в специальных котлах-реакторах. Они снабжены системами поддержания температуры,
подачи в бульон различных питательных веществ, перемешивания биомассы и постоянной подачи
кислорода. Создание аэробных условий по всей толще среды способствует протеканию энергети-
ческих процессов по аэробному пути, что способствует максимальной утилизации энергетического
потенциала глюкозы и, следовательно, максимальному выходу биомассы.
Голотоксин — любой белковый токсин со структурой, собранной из нескольких субъединиц.
Гольджи комплекс — мембранная структура эукариотической клетки, органелла, в основном
предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме
Гомоплазия: 1) сходное строение органов, развивающихся у разных систематических групп
организмов в случае приспособления к одинаковым условиям существования. Возникает в резуль-
тате конвергентной эволюции; 2) однотипная изменчивость признаков и свойств у организмов раз-
личных таксономических групп при параллельных, но независимых эволюционных процессах.
Словарь терминов
1053
Гранулярные клетки — нейроны малого размера, около 10 мкм в диаметре. Гранулярные клетки
встречаются в гранулярном слое мозжечка, в четвертом слое коры головного мозга, в зубчатой изви-
лине гиппокампа, и в обонятельной луковице. Мозжечковые гранулярные клетки составляют почти
половину всех нейронов ЦНС.
ДВС-синдром (диссеминированное внутрисосудистое свертывание, коагулопатия потребленш.
вомОьгеморрагический синдром) — нарушенная свертывав иость крови по причине массивного осво-
бождения из тканей тромбопластических веществ. Может протекать бессимптомно или в виде остро
развившегося нарушения свертываемости крови (коагулопатии).
Дезинфекция — обеззараживание, уничтожение возбудителей инфекционных болезней (вирусы,
бактерии, микоплазмы, патогенные грибы и т. д.) во внешней среде путем применения физически"
и химических средств.
Деконтаминация — уничтожение патогенных микроорганизмов на поверхности тела человека
или животных и абиотических объектов окружающей среды (почва, предметы обихода, пищевые
продукты и др.).
Делирий — расстройство сознания, которое характеризуется наличием истинных, преимуще-
ственно зрительных, галлюцинаций и иллюзий, и, как следствие, — вторичным бредом; наличием
эмоционально аффективных нарушений, сенсопатиями, затрудненной ориентировкой в окружаю-
щем мире, дезориентацией во времени. При этом сохраняются осознание собственной личности и
опасностей. Эмоциональное состояние больного зависит от характера галлюцинаций. После вы-
хода из делирия — частичная конградная амнезия (амнезируются реальные события и болезненные
воспоминания). Больной может быть опасен для себя и окружающих. Может прерываться периода-
ми ясного сознания с критическим отношением к болезненным переживаниям.
Демиелинизация — термин, используемый для обозначения потери миетина, особой субстан-
ции белого вещества мозга. Приводит к нарушению функций миелиновых нервных волокон. Де-
миелинизация может быть первичной (например, при рассеянном склерозе) или развивается после
травмы черепа.
Дендример — макромолекула с симметричной древообразной с регулярными ветвлениями
структурой. Ветви дендримера имеют общую центральную группу. Дендример первого поколения
(первой генерации) имеет одну точку ветвления в каждой ветви, второго поколения — две точки
ветвления и т. д.
Дендромериость — способность образовывать разветвленные полимеры, концы которых могут
присоединять различные молекулы.
^мастер! оизомерь. - пространственные изомеры с несколькими асимметрическими центрами,
у которых конфигурация части асимметрических атомов совпадает, а части - противоположна.
Дизартрия - расстройство членораздельной речи, выражающееся затрудненным или искажен-
ным произношением отдельных слов, слогов и звуков (главным образом согласных).
Динамика эпидемического процесса — изменение интенсивности передачи возбудителя, выра-
жающееся в изменении инфекционной (паразитарной) заболеваемости (носительства) во времени
(по месяцам, годам и другим периодам); среди животных — динамика эпизоотического процесса.
Динеин — белок, ассоциированный с микротрубочками, участвующий наряду с кинезином в
их перемещении, в частности, вызывая движение аксонемы спермиев.
Диплопия — нарушение зрения, состоящее в двоении видимых предметов.
(исковидный ателектаз — в основном рентгенологическое понятие. На рентгенограмме пред-
ставляет собой линейную тень, горизонтально расположенную над диафрагмой, шириной от 0,5 до
1,5 см. Формируется в результате спадения нескольких соседних долек легкого, что часто имеет
место при диффузных поражениях бронхолегочной системы (пневмонитах, фиброзирующих аль-
веолитах, саркоидозе).
Дисфагия — расстройство акта глотания.
Дисфоиия — расстройство голосообразования при котором голос становится хриплым, слабым,
приобретает гнусавый оттенок.
Доза — количество химического вещества, поглощенного организмом.
Задача — то. что требует исполнения, разрешения.
Заразная зона — помещение или группа помещений лаборатории, где осуществляются мани-
пуляции с патогенными биологическими агентами и их хранение.
Зоонозы (зооантропонозы) — группа инфекционных (паразитарных) болезней, резервуаром
возбудителей которых являются животные, но к которым восприимчив и человек.
1054
Словарь терминов
Изотермия — см. «Вертикальная устойчивость атмосферы».
Изохоры — гомогенные протяженные участки ДНК. Когда изохоры фрагментируются во вре-
мя разрезания ДНК, выявляются четыре главных семейства фрагменте" с различным GC-содер-
жанием.
Имл-унизация — метод специфической профилактики инфекционных болезней путем создания
в организме искусственного иммунитета. Различают активную и пассивную иммунизацию.
Иммунопрофилактика - использование иммунных сывороток, анатоксинов и вакцин с целью
создания специфического иммунитета относительно конкретного инфекционного агента.
Им.лу ншрерментный анализ — лабораторный иммунологический метод качественного или коли-
чественного определения пазаичных соединений, макромолекул, вирусов и пр., в основе которого
лежит специфическая реакция антиген-антитело.
Импактор - устройство для принудительного фракционного осаждения частиц аэрозоля с це-
лью последующего определения их количества и размеров.
Импакция — инерционное осаждение аэрозольных частиц на препятствиях, вызывающих ис-
кривление линий воздушных потоков.
Импинджер — прибоодля определения запыленности воздуха, пр заставляющий собой стеклян-
ный сосуд с жидкостью, в который под давлением подается исследуемый воздух.
Инадаптация — совокупность несовершенных приспособлений, возникающая у отдельных
групп животных в ходе эволюции и обусловливающая чпос ледс гвии вымирание этих групп.
Инбредные «чистые) линий ш'юраторных животных — ограниченная совокупность наследствен-
но однородных организмов, происходящих от одного общего предка. Такие линии животных полу-
чают путем близкородственных скрещиваний в течение нескольких поколений. В результате живот-
ные, составляющие чистую линию, получаю । одинаковые копии хромосом каждой из гомологич-
ных пар.
Инверсия — см. «Вертикальная устойчивость атмосферы».
Инвертированный повтор — участок молекулы нуклеиновой кислоты, два сегмента которого
имеют одинаковую нуклеотидную последовательность, но противоположную ей ориентацию (част-
ным случаем является палиндром).
Инкубационный период — время между экспонированием к биологическому поражающему аген-
ту и появлением симптомов болезни. Этот период обусловлен многими переменными, включая
первоначальную дозу, вирулентность штамма, путь проникновения, скорость репликации и имму-
нологические факторы хозяина.
Инеерционные последовательности, IS-элементы - мобильные генетические элементы длиной
700—1800 п. н. (транспозоны, впервые описанные у прокариот). Как правило, IS-элементы не имеют
собственного выражения в фенотипе, хотя и могут содержать открытую рамку считывания для
1—2 белков (в частности, ген транспозазы), а также промотор, способный обеспечивать транскрип-
цию последующего участка донорского гена; IS-элементы ограничены короткими инвертированны-
ми концевыми повторами (10—40 п. н.) и фланкированы прямыми повторами, обнаружение таких
участков нуклеотидных последовательностей может быть свидетельством н шичия IS-элементов.
Инсуляторы — последовательности ДНК особые регуляторные элементы, которые оилалают
способностью блокировать сигналы, исходящие от окружения гена. Эта функция инсуляторов
включает две активности. Во-первых, они блокируют взаимодействие между энхансером и промо-
тором, если находится между ними. При этом инсулятор выполняет только разделительную функ-
цию и не влияет на активность энхансера и промотора. Во-вторых, инсулятор выполняет барьер-
ную функцию для распространяющегося конденсированного хроматина. Показано, что существуют
инсуляторы, как выполняющие одну из двух функций, гак и обе. Инсуляторы представляют собой
сайты связывания особых, инсуляторных, белков. Тот факт, что инсуляторы могут влиять на энхан-
сер-промоторные взаимодействия может быть рассмотрен в контексте роли этих последовательно-
стей в модулировании функций энхансера. Энхансеры не об гадают специфичностью действия,
следовательно, у эукариот должны были выработаться механизмы, обеспечивающие невозможность
активации генов в ненужном месте или в ненужное время развития энхансерами соседнего гена.
Интерстициальные заболевания легких — преимущественно хронические заболевания легочной
ткани, проявляющиеся воспалением и нарушением структуры альвеолярных стенок, эндотелия
легочных капилляров, перивазальных и перилимфатических тканей. Характерным симптомом
интерстициальных заболеваний легких является одышка, являющаяся отражением легочной не-
достаточности.
Словарь терминов
1055
Интрои — часть гена, переписывающаяся в РНК, но затем из нее удаляемая ферментативным
путем при образовании зрелой мРНК; эта часть не кодирует белок.
Инфекционный процесс — процесс взаимодействия организмов возбудителя и хозяина (челове-
ка, животного), проявляющийся клинически выраженным заболеванием или носительством.
Инфицирование — внедрение в организм (клетку) микроорганизмов, вызывающих инфекци-
онный процесс, т. е. антагонистическое взаимодействие паразита и организма хозяина, проявляю-
щееся болезнью.
Исследования диагностические — исследования объектов биотической и абиотической природы,
проводимые с целью обнаружения и идентификации возбудителя, его антигена или антител к нему.
Кадхерины — семейство трансмембранных кальцийзависимых гликопротеинов, осуществляю-
щих адгезивные межклеточные контакты. Подобно интегринам кадхерины обладают внутрикле-
точными доменами, которые через цепь взаимосвязанных цитоплазматических белков связываются
с цитоскелетными структурами контактирующих клеток. Клетки позвоночных экспрессируют один
или более типов кадхеринов, каждый из которых кодируется отдельным геном; для определенного
типа клеток характерен определенный набор экспрессируемых кадхеринов; на данный момент изв-
естно более десятка типов кадхеринов. Кадхерины — основные молекулы клеточной адгезии, удер-
живающие клетки вместе в ранних эмбриональных тканях. Кадхерины одной клетки связыва-
ются с кадхеринами того же типа соседней клетки (гомофильный механизм). Большинство кадхе-
ринов представляют собой единожды пересекающие плазматическую мембрану гликопротеины
(700—750 аминокислотных остатков). Большая внеклеточная часть полипептидной цепи обычно
свернута в пять доменов (около 100 аминокислотных остатков каждый), часть из которых гомоло-
гична и содержит кальций-связываюшие сайты. В отсутствие кальция кадхерины претерпевают
конформационную перестройку и, в результате, быстро деградируются протеазами. Внеклеточный
домен, наиболее удаленный от плазматической мембраны, опосредует межклеточную адгезию.
Карантин — система (комплекс) ограничительно-административных, противоэпизоотических,
противоэпидемических, санитарно-гигиенических и лечебно-профилактических мероприятий, на-
правленных на охрану определенной территории от заноса возбудителей инфекционных (парази-
тарных) болезней с других территорий и предупреждение их распространения за пределы эпизооти-
ческого или эпидемического очага.
Карбункул — острое гнойно-некротическое воспаление нескольких волосяных фолликулов и
сальных желез с обширным некрозом кожи и подкожной клетчатки. Карбункул, как правило, бы-
вает одиночным. Локализуется на шее, в межлопаточной области, на лице, пояснице, ягодицах, реже
на конечностях.
Клада — группа организмов, которые являются потомками единственного общего предка и всех
потомков этого предка. В русскоязычной литературе иногда можно встретить другой вариант пе-
ревода — «клад». Если клада находится верной в разных кладистистических исследованиях, кото-
рые используют разные наборы данных, она может быть принятой к таксономии и стать таксоном.
Кодоминирование — тип взаимодействия аллелей, при котором оба аллеля в полной мере прояв-
ляют свое действие. В результате, так как проявляются оба родительских признака, фенотипически
гибрид получает новый, усредненный вариант двух родительских признаков.
Компьютерная томография — метод неразрушающего послойного исследования внутренней
структуры объекта. Метод основан на измерении и сложной компьютерной обработке разности
ослабления рентгеновского излучения различными по плотности тканями.
Конвекция — см. «Вертикальная устойчивость атмосферы».
Конвенции о запрещении разработки, производства и накопления запасов бактериологического
(биологического) и токсинного оружия и об их уничтожении (КЬТО) — первый международный дого-
вор о разоружении, запрещающий производство целого класса вооружений. Ее подписание яви-
лось результатом многолетних усилий международного сообщества по созданию правовой базы,
дополняющей собой Женевский протокол (1925). КБТО была открыта для подписания 10 апреля
1972 г. и вступила в силу 26 марта 1975 г., когда 22 государства передали на хранение свои документы
о ратификации Генеральному секретарю ООН. В настоящее время ее участниками являются 163 го-
сударства, обязавшиеся не разрабатывать, не производить и не накапливать БО. Государства-участ-
ники обязуются: Статья 1: Ни при каких обстоятельствах не приобретать и не накапливать биоло-
гическое оружие. Статья II: Уничтожить или переключить на мирные цели все, что с связано с БО.
Статья III; Не передавать, не помогать никоим образом, не поощрять и не понуждать кого бы то
ни было к приобретению и накоплению БО. Статья IV: Принимать необходимые изменения в свое
1056
Словарь терминов
законодательство. Статья V Консультироваться друг с другом с целью разрешать все вопросы ка-
сающиеся выполнения положений КБТО. Статья VI: Сотрудничать в проведении любых рассле-
дований касающихся жалоб других участников КБТО, поданных в Совет Безопасности ООН. Ста-
тья VII: Оказывать помощь государствам, которые могут подвергнуться опасности в результате
нарушения Конвенции. Статья X: Выполнять все вышеперечисленное с целью мирного использо-
вания научных разработок в области бактериологии.
Контагиозность — способность инфекционной болезни передаваться от одного человека к дру-
гому при непосредственном контакте, воздушно-капельным путем или через промежуточных аген-
тов, таких как насекомые-переносчики. Это свойство предопределяет возможность распростране-
ния эпидемии после применения биологических агентов. При непосредственном контакте наиболее
опасны больные легочной чумой и натуральной оспой. Через насекомых-переносчиков легко рас-
пространяются возбудители чумы (блохи) и сыпного тифа (вши).
Короткие терминальные инвертированные повторы — короткие гомологичные нуклеотидные
последовательности ориентированные в противоположных направлениях, расположенные на
концах провирусов ретровирусов и некоторых мобильных генетических элементов. Их размер со-
ставляет 15—25, иногда 9—41 п. н., причем копии повторов могут быть не идентичными, хотя и очень
близкими по первичной структуре
Кортекс - - самый т олстый слой оболочки споры. Он состоит из пептидогликана, содержащего
мало поперечных сшивок и поэтому очень чувствительного к лизоциму. Разрушение кортекса ли-
зоцимом играет пусковую роль в процессе прорастания споры.
Крафт-бумага — высокопрочная оберточная бумага из слабопроваренной длинноволокнистой
сульфатной целлюлозы.
Культура клеток - клетки какой-либо ткани, выращиваемые в лаборатории в питательной среде.
Широко применяются при вирусологических исследованиях. Для культивирования вирусов живот-
ных можно использовать все типы эмбриональных и зрелых клеток животных и пгиц.
Купфера клетки — клетки ретикуло-эндотелиальной системы, расположенные на bhj греннеи
поверхности капилляроподоиных сосудов (синусоидов) печени.
Куриный эмбрион — оплодотворенное куриное яйцо, выдержанное в инкубаторе. Применяет-
ся для культивирования некоторых вирусов (гриппа, оспы и др.)
Лаборатория — организация или ее структурное подразделение, выполняющее эксперименталь-
ные, диагностические или производственные работы с патогенными биологическими агентами
ЛД50 (полулегальная доза, также DLS0, LD50 англ, lethal dose, 50 %) — средняя доза вещества,
вызывающая гибель половины членов испытуемой группы. Один из наиболее широко применяе-
мых показателей опасности ядовитых и умеренно-токсичных веществ. Обычно указывается в еди-
ницах веса вещества на единицу веса испытуемого субъекта. Предполагается, что исследуемый
объект находится в типичном состоянии, в нормальных условиях, без приема каких-либо антидо-
тов и других специальных мер предосторожности.
Летальность — отношение числа умерших к числу заболевших данной болезнью. Выражается
в процентах. Как правило, этот показатель применяется для характеристики заболеваний, возни-
кающих естественным путем. Летальность той или иной инфекционной болезни при различных
эпидемиях может зависеть как от вирулентности вовлеченных в нее микроорганизмов, так и раз-
ной напряженности иммунитета у людей, подвергшихся заражению. Кроме того, летальность зави-
сит от стадии эпидемического процесса (начало, разгар, завершение), достигая максимума в разгар
эпидемии и потом постепенно снижаясь. Можно предполагать, что микроорганизмы, использо-
ванные для получения биологических рецептур и введенные в окружающую среду с целью пора-
жения людей, будут вызывать однородные по клинике заболевания с заранее предвидимой степе-
нью выраженности симптомов болезни. Летальность среди людей, пораженных биологическими
агентами, будет зависеть от выбранного для применения штамма, качества рецептуры и способа ее
применения.
Летальный/инкапситирующий — эти термины предполагают статистические вероятности реак-
ций организма человека на биологический поражающий агент Например, без лечения смертность
при ингаляционным поражении возб> гителем натуральной оспы будет достигать 60 % от числа
пораженных людей, однако при таком же пути инфицирования смертность при поражении возбу-
дителем туляремии не будет превышать 8 %.
Лигазы — фермент, катализирующий соединение двух молекул с образованием новой хими-
ческой связи (лигирование). При этом обычно происходит отщепление (гидролиз) небольшой
Словарь терминов
1057
химической группы от одной из молекул. Лигазы относятся к классу ферментов ЕС 6. В молекуляр-
ной биологии лигазы подкласса 6.5 классифицируют на РНК-лигазы и ДНК-лигазы.
Макрогематурия — наличие в моче крови, видимой невооруженным глазом.
Макрофиты — крупные многоклетчатые водоросли (зеленые, бурые, красные) и морские цвет-
ковые растения (травы). Обитают на всех широтах, главным образом в прибрежной зоне, где обра-
зуют густые заросли и служат продуцентом органического вещества.
Макула — это центральная часть сетчатки глаза, которая располагается к виску от диска зри-
тельного нерва.
Маллеин — биологический препарат, аллерген, используемый для аллергического диагностиро-
вания сапа у лошадей, мулов, ослов, лошаков, верблюдов. Маллеин — стерильный фильтрат инакти-
вированной нагреванием бульонной культуры Pseudomonas mallei. Применяют для глазной и под-
кожной маллеинизации.
Малые РНК, образующие шпильки, или короткие РНК, ооразующие шпильки (англ. shRNA, small
hairpin RNA. short hairpin RNA) — молекулы коротких рибонуклеиновых кислот, ооразуюшие во
вторичной структуре плотные шпильки. ShRNA могут быть использованы для выключения эксп-
рессии генов путем РНК-интерференции Короткие РНК, образующие шпильки, вводят в клетки
при помощи вектора, использующего промотор U6, для обеспечения конститутивной экспрессии.
Такой вектор, как правило, передается дочерним клеткам и обеспечивает наследуемое выключе-
ние гена. Структура малых РНК, образующих шпильки, разрезается клеточными ферментами в
малые интерферирующие РНК (siRNA), которые далее переходят в РНК-индуцируемый комплекс
выключения гена (RISC). Этот комплекс связывается и разрезает мРНК, участки которых комп-
лементарны siRNA. Малые РНК, образующие шпильки, транскрибируются РНК полимеразой III.
Образование shRNA в клетках млекопитающих иногда вызывает интерфероновый ответ для защи-
ты клетки, как в случае вирусной атаки. Такие процессы не наблюдаются в случае miRNA (мик-
роРНК), которые транскрибируются РНК полимеразой II.
Медиастинум — см. Средостенье.
Метод эиергодиеперсионной рентгеновской спектроскопии — аналитический метод элементно-
го анализа твердого вещества, базирующийся на анализе энергии эмиссии ее рентгеновского спек-
тра. С помощью пучка электронов (в электронных микроскопах) или рентгеновских лучей (в рен-
тгеновских флуоресцентных анализаторах) атомы исследуемого образца возбуждаются, испуская
характерное для каждого химического элемента рентгеновское излучение. Исследуя его энергети-
ческий спектр, можно сделать выводы о качественном и количественном составе образца.
Механизм: 1) внутреннее устройство машины, прибора, аппарата, приво [яших их в действие;
2) система, устройство, определяющие порядок какого-то вида деятельности.
Миазмы — по старым (в «добактериальный период») представлениям, ядовитые испарения,
продукты гниения, якобы вызывающие заразные болезни.
Мидриаз — расширение зрачка.
птонофилия — происхождение данной группы организмов от общего предка; основной прин-
цип эволюции органического мира.
Нанотехнология — совокупность технических процессов, связанных с манипуляциями моле-
кулами и атомами в масштабах 1—100 нм.
Нейропептиды — разновидность пептидов, которые синтезируются в нервных клетках, напри-
мер эндорфины, энкефалины. Они выполняют в организме роль регуляторов физических функ-
ций. Используются как лекарственные средства.
Нейротрофины — общее название секретируемых белков, поддерживающих жизнеспособность
нейронов стимулирующих их развитие и активность. Эти белки, в свою очередь, входят в обшир-
ное семейство факторов роста. Их выделяют за особую роль в воздействии на нейроны, но они
экспрессируются также и в других тканях и органах, равно как и другие факторы роста, не называ-
емые нейротрофинаь.и, оказывают некоторое влияние в том числе и на нейроны.
Некультивируемое состояние бактерий — понятие подразумевает отсутствие роста бактерий на
традиционных для них питательных средах. Некультивируемое состояние обнаружено у многих
патогенных видов бактерий В связи с тем, что рутинные бактериологические методы не эффектив-
ны для их обнаружения, истинные масштабы распространения феномена в объектах окружающей
среды остаются неизвестными.
Нематоцисты — органы нападения и защиты, свойственные большинству так называемых жгу-
чих кишечнополостных (Cnidaria), а именно: всем коралловым полипам, всем полипомедузам (гид-
1058
Словарь терминов
ромедузам, сифонофорам, сцифомедузам) и некоторым ктенофорам; кроме того, нематоциты встре-
чаются у некоторых червей (а именно: у низших турбеллярий) и некоторых голых моллюсков (из
заднежаберных брюхоногих). Каждая нематоциста состоит из пузырька с двойной стенкой, продол-
жающегося в тонкую трубочку и наполненного едкой жидкостью. В покое трубочка впячена внутрь
пузырька, но под влиянием раздражения (непосредственно действующего на клеточку, заключаю-
щую нематоцисту, так называемый книдобласт, или через посредство нервной системы) трубочка
выворачивается наружу и через нее изливается едкая жидкость пузырька, парализуя или умерщв-
ляя мелких животных. Трубочки нематоцист, кроме того, обвиваются иногда вокруг добычи Бо-
лее широкая основная часть трубочки (шейка) бывает снабжена заостренными отростками, направ-
ленными в разряже ином состоянии нематоцисты к пузырьку.
Нерепликативные единицы бактероидов — интегрированные в геном бактероидов элементы, ко-
торые самостоятельно не способны из него «вырезаться» и переноситься между бактериями; но их
«вырезание» и перенос могут инициироваться конъюгатиьными транспозонами.
Нозология — наука, которая изучает болезни и их классификации.
Оонуоиляция — легкая оглушенность или помрачение сознания с затруднением концентрации
внимания и замедлением психических процессов.
Обращение- фазовая (обратно-фазовая) хроматография — вариант хроматографии, в котором
стационарная фаза является неполярной.
Объект — явление, предмет, на который направлена какая-нибудь деятельность.
Оперой — функциональная единица генома у прокариот, в состав которой входят цистроны
(гены, единицы транскрипции), кодирующие совместно или последов зтельно работающие белки
и объединенные под одним (или несколькими) промоторами.
Опухолевый промогер - соединение, которое стимулирует рост опухоли, не являясь мутагеном.
< фтологичные гены — гомологичные гены филогенетически родственных организмов, разошед-
шихся в процессе видообразования; гены которые у различных видов произошли от общего пред-
шественника и передавались вертикально в эволюции. Два гена ортологичны у двух видов, если они
появились и дале! дивергировали вместе с видами в результате видообразования. Часто (но необя-
зательно) ортологи сохраняют свои функции в процессе эволюции (см. паралогичные гены). Орто-
логи близких видов имеют одну и ту же функцию.
Осмотический шок — любое нарушение в клетке, происходящее при ее переносе в сильно гипер-
тоническую или гипотоническую среду.
Остооь патогенности — кластер 1енов чужеродного происхожденш обнапуженный в плазми-
де или хромосоме оактгрии, кодирующий группу генов вирулентности.
Открытая рамка считьп ания — последовательность нуклеотидов в составе ДНК или РНК, по-
тенциально способная кодировать белок. Основным параметром наличия ORF служит отсутствие
стоп-кодонов, в большинстве случаев — ТАД, ТСА и TAG на достаточно продолжительном участ-
ке последовательности после стартового кодона (в подавляющем случае — ATG).
Отравляющие вещества — токсичные химические соединения, предназначенные для пораже-
ния живой силы противника.
Очаг природный — наименьшая часть одного или нескольких географических ландшафтов, где
циркуляция возбудителя между животными способна осуществляться неопределенно долго без
необходимости заноса его извне
Очаг эпидемический — место нахождения источника возбудителя с окружающей его террито-
рией в тех пределах, в которых этот источник способен п< редавать возбудителя здоровым людям в
цанной конкретной обстановке; тоже очаг эпизоотический, только в отношении животных.
Панмиксия — свободное скрещивание разнополых особей с различающимися генотипами.
Папула — морфологический элемент кожной сыпи, представляющий собой бесполостное об-
разование возвышающееся над уровнем кожи.
Паразит — организм, использующий другого в качестве источника питания и (или) среды оби-
тания и обычно наносящий ему вред.
Патогенность — способность микроорганизмов вызывать инфекционный процесс у макроор-
ганизмов определенного вид? Патог шность представляет сложный комплекс свойств микроорга-
низма, сформировавшийся в процессе борьбы за существование и приспособление к паразитиро-
ванию в организмах растений, животных и человека Патогенность является видовым признаком
микроорганизмов, которые характеризуются специфичностью действия: каждый вид способен
вызывать только определенную инфекционную болезнь.
Словарь терминов
1059
Пяшеиа телыш — вирионы, обнаруживаемые в содержимом элементов сыпи у больных нату -
пальмой оспой.
Петри чашки — п суда, употребляемая в бактериологической лаборатории для выращивания
бактерий на плотных питательных средах, дтя получения изолированных колоний и определения
спектра действия микробов-антагонистов.
Пилпэрекция — сокращение мышц, поднимающих волосы; приводит к образованию «гусиной
кожи»: появлению на коже конусообразно приподнятых волосяных фолликулов. Возможна при
гиперактивации симпатической нервной системы (например, при охлаждении; или применении
симпатомиметиков.
Линоцито? j) захват клеточной поверхностью жидкости с содержащимися в ней веществами;
2) процесс поглощения и внутриклеточного разрушения макромолекул.
Пиросеквенирование — способ секвенирования ДНК, основанный на принципе «секвениро-
вание путем синтеза». Позволяет секвенировать одну цепочку нуклеотидов ДНК путем синтеза
комплементарной цепочки, при этом регистрируется присоединение каждого нуклеотида. Матрица
ДНК иммобилизована, растворы нуклеотидов А, С, G и Т добавляются и отмываются последова-
тельно после реакции При последовательном добавлении к ДН К-полимеразному' комплексу де-
зоксинуклеозидтрифосфатов, их включение в синтезируемую нить зависит от нуклеотидной пос-
ледовательности матрицы. Полимеразный синтез ДНК сопровождается выделением пирофо< фата.
Этот пирофосфат в присутствии сульфурилазы и аденозинфосфосульфата преобразуется в АТФ и
запускает окисление люциферина ферментом люциферазой, сопровождающееся биолюминесцен-
цией . Свет образуется в тот момент, когда раствор нуклеотидов соответствует первому неспаренному
основанию матрицы. Последовательность растворов, которые дают хемилюминесцентный сигнал,
позволяет определить последовательность матрицы.
Плазмида — кольцевая или линейная молекула ДНК, автономно от хромосомы реплицирую-
щаяся в клетке за счет репликационной системы клетки хозяина. Большинство плазмид кодирует
специальные белки — инициатооы репликации. Эти белки начинают процесс репликации, кото-
рый затем подхватывается и продолжается репликационной системой клетки. Плазмиды широко
используются в генной инженерии для переноса генетической информации и генетических мани-
пуляций.
Плевральный выпот — патологическое накопление жидкости в плевральной полости.
Плеоморфные микроорганизмы — микроорганизмы, способные изменять форму клетки в зави-
симости от условий внешней среды (например, бифидобактерии на разных средах)
Плеоцитоз — увеличение числа клеток в цереброспинальной жидкости, наблюдающееся при
воспалите тьном поражении вещества головного или спинного мозга или их оболочек.
Пневмонит — общее название некоторых атипичных воспалительных процессов в легких
Пневмония интерстициальная — пневмония, характеризующаяся преимущественным пораже-
нием соединительной ткани легких (межальвеолярной, перибронхиальной, периваскулярной).
Полиморфные микроорганизмы — вообще не имеющие постоянной формы, в одной и той же
культуре встречаются разные по форме клетки (например, микоплазмы, не имеющие клеточной
стенки).
Постулат — положение, которое, не будучи доказанным, принимается в силу теоретической
или практической необходимости за истинное.
Потенция т концевой пластинки , зостсиваптический потенциал) — временное изменение потен-
циала постсинаптической мембраны в ответ на сигнал, пос ^пивший с пресинаптического нейро-
на. Возникает при спонтанном выделении ацетилхолина, содержащегося в одном пресинаптичес-
ком пузырьке.
Продолжительность реабилитации — продолжительность восстановления функций организма,
нарушенных или утраченных в результате поражения биологическим агентом.
Просвечивающий (тоачсм^сьонный» электронный микроскоп - устройство, в котором изображе-
ние от ультратонкого образца (толщиной порядка 0,1 мкм) формируется в результате взаимодейст-
вия пучка электронов с веществом образца с последующим увеличением магнитными линзами (объ-
ектив) и регистрацией на флуоресцентном экране, фотопленке или сенсорном приборе с зарядовой
связью.
Протективный иммунитет — специфическая иммунная перестройка организма, выражающая-
ся в более или менее продо гжительчой невосприимчивости к повторному воздействию одного и
того же типа и вида этиолот ического агента при новом заражении.
1060
Словарь терминов
Протомер — полипептидная цепь, являющаяся исходной единицей организации структуры
белка. Протомеры гемоглобина состоят из 8 спиралей, свернутых в плотную глобулярную структу-
ру, содержащую внутреннее ги трофобное ядро и «карман» для связывания гема. Протомеры обыч-
но связаны друг с другом посредством лишь нековалентных связей
Прототрофы — микроорганизмы, не требующие для своего развития готовых витаминов, ами-
нокислот и других факторов роста, а синтезирующие их из минерального и органического сырья.
Процессинг — процесс превращения белка-предшественника в зрелый белок.
Прямые повторы — идентичные (или похожие) последовательности нуклеотидов, присутствую-
щие в двух или более копиях, одинаково ориентированные, находящиеся в одной молекуле ДНК,
но необязательно примыкающие друг к другу в частности, короткие (чаще всего 5 или 9 п. о ) пря-
мые повторы всегда фланкируют IS-элементы
Псевдотипированне — упаковка вирусного генома в «чужую» вирусную осю точку, что позволяет,
в частности, расширять круг хозяев, т. е. превращать экотропные вирусы в амфотропные.
Психомоторное возбуждение — это патологическое состояние, характеризующееся двигатель-
ным беспокойством разной степени выраженности (от суетливости до разрушительных действий),
часто сопровождающееся речевым возбуждением (многоречивость, выкрик 4 фраз, слов, отдельных
звуков). Наряду с этим характерны ярко выраженные аффективные расстройства тревога, растерян-
ность, гнев, злобность, агрессивность, веселье и так далее.
Птоз — опущение верхнего века.
Птомаины — группа азотсодержащих химических соединений, образующихся при гнилостном
разложении (с участием микроорганизмов) белков мяса, рыбы, дрожжей и пр.
Пэгилирование — ковалентное конъюгирование полиэтиленгликоля к лекарственному препара-
ту, белку или полимерному носителю, что позволяет снизить их иммуногенность и увеличить про-
должительность циркулирования в кровеносном русле.
Рабдомиолиз — синдром, представляющий собой крайнюю степень миопатии и характеризую-
щийся разрушением клеток мышечной ткани, резким повышением уровня креатинкиназы и мио-
глобина, миоглобинурией, развитием острой почечной недостаточности.
Расследовать — подвергнуть (подвергать) всестороннему рассмотрению, изучению.
Реакция непрямой гемагглютинации - метод иммунологической диагностики вирусных и бакте-
риальных инфекций. При этом эритроциты, на которых предварительно адсорбированы антиге-
ны (антитела), приобретают способность агглютинироваться в присутствии гомологических сыворо-
ток (антигенов). Эритроциты выполняют роль носителей антигенов со специфическими детерми-
нантами, агглютинация которых происходит в результате реакции антиген-антитело.
Реакция связывания комплемента — одна из традиционных серологических реакций, применя-
емая для диагностики многих вирусных и бактериальных болезней. У многих вирусов групповая
специфичность комплементсвязывающих антигенов позволяет испол! зовать РСК при массовой
диагностике.
Регургитация — быстрое движение жидкостей или газов в направлении, противоположном нор-
мальному, возникшее в полом мышечном органе в результате сокращения его стенки.
Резервуар возбудителя инфекции — совокупность биотических и абиотических объектов, являю-
щихся средой естественной жизнедеятельности паразитического вида и обеспечивающих существо-
вание его в природе.
Рестрнктазы (рестрикционные эндонуклеазы) — ферменты, расщепляющие ДНК в участках,
имеющих специфические последовательности нуклеотидов длиной обычно 4—7 п. н. Первые три
буквы в наименовании рестрнктазы указывают микроорганизм, из которого она была выделена, на-
пример Есо (Escherichia coli) четвертая буква обычно обозначает штамм, а римская цифра — конк-
ретный фермент.
Ретровирусы — РНК-содержащие вирусы, имеющие в своем составе фермент обратную транс-
криптазу, копирующий РНК в ДНК, интегрирующуюся затем с геномом организма-хозяина.
Ретротранспозоны — транспозоны, механизм транспозиции (перестановки по геному) которых
полностью совпадает с частью жизненного цикла ретровируса, однако они лишены функциональ-
ной белковой оболочки и могут перемещаться лишь внутри какой-нибудь одной клетки и быть пе-
реданы ее потомкам.
Ризосфера — слой почвы (2—3 мм), прилегающий к корню растения.
РНК-индуцируемый сайленсинг-комплекс IRNA-induced silencing complex) — мультибелковый
комплекс, в состав которого входит один из белков семейства Argonaute и малые интерферирую-
Словарь терминов
1061
щие РНК (англ. siRNa), предварительно подвергшиеся процессингу эндонуклеазой Dicer. Dicer рас-
щепляет предшественник siRNA, представляющий собой двуцепочечную молекулу РНК (dsRNA)
на одноцепочечные фрагменты. В состав RISC всегда включатся только один из них; предпочтение
о~дается тому фрагменту, 5'-конеи которого конъюгирован менее прочно. Затем RISC образует ком-
плекс с РНК-мишснью, что приводит либо к репрессии ее трансляции в случае неполной компле-
ментарное™, либо к расщеплению ее последовательности приблизительно в середине участка спа-
ривания в случае полной или почти полной комплементарности. Эндонуклеазная активность
внутри комплекса RISC опосредована РНКаза-Н-подобным доменом (piwi) в белке Argonaute. Дан-
ный процесс имеет значение как в регуляции активности генов с помощью микроРНК (microRNA,
mi RNA), так и в защите от вирусных инфекций, так как вирусы часто используют двуцепочечные
РНК как инфекционный вектор.
РНК-интерференция — процесс подавления экспрессии гена на стадии транскрипции, транс-
ляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.
Санитарная обработка — мероприятие, проводимое в эпидемическом очаге, предусматриваю-
щее дезинфекционную (дезинсекционную) обработку постельных и носильных вещей, волос, мытье
под душем.
Сегрегация: I) расхождение родительских хромосом в мейозе; 2) возникновение различных
гено- и фенотипов в потомстве гибридов, обусловленное случайным распределением гомологичных
хромосом между гаметами в мейозе, кроссинговером и случайными сочетаниями гамет при опло-
дотворении.
Секвенирование — определение последовательности нуклеотидов в молекуле ДНК.
Сепсис (гниение) — общая инфекция, состояние организма, при котором патогенные микро-
организмы, проникшие из первичного очага инфекции в кровь, размножаются в ней и заносятся
во все ткани и органы, где вызывают воспалительные и дегенеративно-некротические процессы.
Клиническая картина сепсиса не зависит от вида возбудителя инфекции.
Серологические реакции — методы иммунодиагностики, разработанные для установления в
сыворотке крови антител или антигенов. Серологические реакции являются специфическими и
основаны на взаимодействии антигена и антитела. К ним относятся реакции: агглютинации, гемаг-
глютинации, связывания комплемента, преципитации, нейтрализации, а также методы определе-
ния групп крови и преципитационные пробы в судебно-медицинской экспертизе.
Сеятели чумы ^демоны эпидемий) — так в Средние века называли людей, которые пытались
вызывать эпидемии с помощью предметов и материалов, находившихся в контакте с больными чу-
мой, либо содержащих выделения чумных бубонов. В некотором смысле они являются предшест-
венниками современных биотеррористов.
Сиаловая кислота — аминосахар, N-ацетил производное нейраминовой кислоты. Сиаловая
кислота широко распространена в тканях животных и, в меньшей степени, других организмов:
растений и грибов, дрожжей и бактерий, в основном в составе некоторых гликопротеинов и ганглио-
зиды и стенок бактериальных клетог.
Сильфон — тонкостенная металлическая трубка или камера с гофрированной (волнообразной)
боковой поверхностью. Сильфон изготавливаю? из латуни (обычно полутомпака), фосфористой и
бериллиевой бронзы и нержавеющей стали Наиболее широко сильфоны применяют в пневмо- и
гидроавтоматике в качестве чувствительных элементов, реагирующих (расширением или сжатием,
подобно пружине) на изменение давления газа или жидкости, действующего на дно сильфона (на-
пример, в датчиках температуры, давления), а также в качестве гибких соединений трубопроводов,
компенсаторов температурных удлинений, упругих разделителей сред и т. п. Стенки сильфона для
работы при больших разностях давлений и в агрессивных средах изготовляют 2-, 3- и 4-слойными.
Используются в биологических боеприпасах барометрического типа.
Симбионты — совместно существующие организмы, в определенной степени возлагающие один
на другого задачи регуляции своих отношении с внешней средой.
Синтаксин — трансмембранный белок, компонент белкового комплекса SNARE, обеспечиваю-
щего слияние синаптической везикулы с пресинаптической мембраной и высвобождение нейро-
медиатора в синаптическую щель. Существует 4 изотипа синтаксинов. Синтаксин относится к под-
группе Q-SNARE. Молекула синапсина состоит из трех доменов: N-концевой регуляторный домен
(Habc), комплексоооразуюший SNARE домен и С-концевой трансмембранный домен.
Синтения — это структурное сходство групп сцепления генов у организмов разных биологи-
ческих видов. В частности, в геномах человека и мыши известно несколько десятков синтетичных
1062
Словарь терминов
групп генов. Наличие феномена синтении позволяет суживать круг поиска места локализации ис-
следуемого гена на хромосомах, ограничивая его областью известных генов, принадлежащих к
конкретной синтеничнои группе
Синторофизм — способность двух ауксотрофных штаммов микроорганизмов расти на синте-
тической питательной среде, не содержащей необходимой аминокислоты, лишь при их совмест-
ном культивировании.
Склероций — покоящаяся стадия гриба, образующаяся в неблагоприятных условиях; окру, зое
или продолговатое тело, образуемое тесным сплетением гиф у некоторых сумчатых, базидиальных
и несовершенных грибов — спорыньи, склеротинии, бразильского трутовика, монилии и др. Разме-
ры склероция обычно от долей мм до 2 см, иногда более (так, у трутовика диаметр склероция дос-
тигает 30 см, масса 20 кг). Наружная часть склероция состоит из толстостенных и темноокрашен-
ных клеток, внутренняя — из тонкостенных бесцветных клеток. Склероций беден водой (5- 10 %)
и богат питательными м ществами. В благоприятных услог иях склероций прорастает, образуя пло-
довые тела (у сумчатых или базидиальных грибов) или грибницы с конидиями (у несовершенных
грибов). Жизнеспособность склероция может сохраняться несколько лет. Склероции спорыньи
содержат алкалоиды (эрготоксин, эргометрин, эрготамин и др.), используемые в медицине как
родовспомогательные и кровоостанавливающие (при маточных кровотечениях) средства.
Смертность — процесс убыли населения вследствие смерти. В медицинской статистике — ча-
стота случаев смерти среди определенной совокупности населения (страны, города, района), оп-
ределяемая. как правило, числом умерших за определенный период (год) на 1 тыс. населения.
Сорироика клеток с активированной флуоресценцией, флуоресцентный сортинг метод проточ-
ной цитометрии, в котором мишени (клетки, индивидуальные хоомосомы и т. п.) окрашены флуо-
ресцентным красителем, возбуждающимся под воздействием лазерного луча. Разделение осуще-
ствляется на специальном приборе-сортере, критерием для сортировки материала служат различия
в эмиссии флуоресцентного сигнала.
Спорадическая заболеваемость — характер заболеваемости, когда регистрируемые случаи еди-
ничны, разрозненны, т. е. когда преимущественно отсутствует шидемиологическая или эпизооти-
ческая связь между ними, а также с общим источником или общим фактором передачи возбудите-
ля болезни.
Споры бактерий - овальные образования, представляющие собой особую форму существова-
ния клеток некоторых видов бактерий. Служат средством сохранения вида в неблагоприятных ус-
ловиях. В почве могут сохраняться десятилетиями.
Средостение — анатомическое пространство в средних отделах грудной полости. Средостение
ограничено грудиной (спереди) и позвоночником (сзади). Органы средостения окружены жиро-
вой клетчаткой По бокам от средостения расположены плевральные полости.
Среды пит (тельные — различные искусственные среды для культивирования микробов с целью
выделения возбудителя болезни из исследуемого материала и определения его вида, для накопления
микробной массы при изготовлении биологических препаратов. Каждая питательная среда содер-
жит в достаточнсм количестве необходимые питательные вещества в стерилизованном виде.
Стерилизация — уничтожение различных микроорганизмов и их спор в различных объектах с
помощью физических и химических факторов.
Стриатум - анатомическая структура конечного мозга, относящаяся к базальным ядрам полу-
шарий головного мозга.
Сублимация - возгонка, один из методов консервирования пищевых продуктов. Заключается
в обезвоживании при низкой температуре и высоком вакууче. Продолжительно^. ь хранения субли-
мированных продуктов не более 2 лет. :Этот метод используется в производстве стабильных е дли-
тельным сроком хранения живых вакцин.
Судебная микробиология — новая область криминалистики, предназначенная для анализа дока-
зательств уголовных актов по актам биотеррора и биодиверсий, позволяющая определить ответст-
венную за них сторону и оправдать невиновных.
Таргетиш - мутирование генов в условиях in vivo.
Тахикинины - крупная группа нейропептидов. В нее входят пептиды, имеющие сходную С-кон-
цевую последовательность — Phe-X-Gly-Leu-Met-NH2, где под X обозначена алифатическая или
ароматическая аминокислота. Гены, кодирующие предшественников тахикининов, называются
препротахикинины Название группа пептидов получила из-за способности bi вывать сокращение
кишечных тканей.
Словарь терминов
1063
Тетрил (2,4,о-тринитрофени оиетилнитр иин) — бризантное взрывчатое веще< тво; скорость де-
тонации 7500 м/с при плотности 1,63 г/см3, теплота взрыва 4609 кДж/кг (1100 ккал/кг). Обычно его
используют в капсюлях-детонаторах и в качестве промеж; точных детонаторов.
кань ингергтициальная — рыхлая волокнистая соединительная ткань, образующая строму
паренхиматозных органов.
Токсинология — наука, изучающая свойства ядов животного, растительного и микробного про-
исхождения и токсический процесс, вызванный отравлением ими. Может рассматриваться как
раздел токсикологии, а также как междисциплинарная наука.
Токсины — вещества белковой природы растительного, животного, микробного или иного про-
исхождения, обладающие высокой токсичнос тью и способные при их применении оказывать пора-
жающее действие на организм человека и животных.
Токсичнс.'ть - свойство химического вещества в минимальном количестве вызывать патологи-
ческие изменения, ведущие к нарушению основных процессов жизнедеятельности организма и
приводящие его к гибели. Токсичность — один из основных количественных параметров, отражаю-
щих биологическую активность токсинов.
Торакоцеитез (плевроцентез, пункция плевральная) — прокол грудной стенки с введением иглы
или троакара в плевральную полость; проводится с диагностической или лечебной целью.
Транслокон (транс то»атор) - интегральный белковый комплекс, участвующий в транслокации
новосинтезируемой полипептидной цепи в эндоплазматический ретикулум из цитоплазмы при
экзоцитозе белков у эукариот.
Трансмиссивность — способность биологического поражающего агента передаваться кровосо-
сущими членистоногими (насекомыми и клешами).
Транспозаза - фермент, связывающий одноцепочечную ДНК и встраивающий последнюю в
геномную ДНК. Транспозоны класса 2 кодируют транспозазу, которая позволяет транспозонам быть
вырезанным из геномной ДНК и встроенным в другие места.
Транспозиция — перемещение генетического материала с одной хромосомной позиции на дру-
гую. Последоват льн ости ДНК, действительно обладающие способностью изменять свою геномную
локализацию, называются мобильными элементами или транспозиционными элементами. Суще-
ствуют два механизма транспозиции: репликативная транспозиция — увеличивает количество
транспозонов, так как исходный родительский транспозон остается на своем месте нуждается в
транспозазе (взаимодействует с концами родительского транспозона) и резольвазе (взаимодействует
с дочерней копией); нерепликативная транспозиция — транспозон перемещается физически.
Транспозон — последовательность ДНК, способная перемещаться внутри генома в результате
транспозиции.
Трансферрин — белки плазмы крови, осуществляющие транспорт ионов железа.
Трофозоиты - вегетативные формы простейших
Тучные клетки (мастоциты, лаброциты) — высокоспециализированные иммунные клетки соеди-
нительной ткани позвоночных животных, аналоги оазофилов крови. Участвуют в адап гивном им-
мунитете.
Условие — обстоятельство, от которого что-нибудь зависит.
Устойчивость — способность биологического поражающего агента сохранять жизнеспособность
под воздействием факторов окружающей среды (температура, относительная влажность, наличие
атмосферных загрязнений, солнечная радиация, кислород и др.). Количественным показателем
устойчивости является скорость гибели агента (например, «скорость распада аэрозоля»).
Фагоцитоз — процесс активного захватывания и поглощения живых и неживых частиц одно-
клеточными организмами или особыми клетками (фагоцитами) многоклеточных животных орга-
низмов.
Фактор — существенное обстоятельство в каком-то процессе.
Факторы вирулентности — факторы бактерий, способствующие проникновению бактерий в
макроорганизм, нейтрализации его механизмов зашиты, собственному размножению и развитию
болезни
Факторы соответствия - более широкое понятие, чем факторы вирулентности. Это гены, фак-
торы или аллели, найденные j бактерий, типичных представителей патогенного вида, разрушение
которых снижает возможности этих бактерий к размножению в данной среде. Их комплемента-
ционное «возвращение» в геном бактерии восстанавливает ее способность поддерживаться в дан-
ной среде.
1064
Словарь терминов
Фасцикуляция — непроизвольней сокращение отдельных пучков мышечных волокон в зоне,
иннервируемой пораженными передними корешками или передними рогами спинного мозга.
Фациальная диплегия — маскообразное выражение лица.
Фибринолизин (плазмин) — фермент класса гидролаз, присутствующий в плазме крови; ката-
лизирует расщепление фибрина, приводящее к растворению (фибринолизу) кровяного сгустка —
тромба. В крови фибрин находится главным образом в форме неактивного профермента профиб-
ринолизина (плазминогена), который под действием ферментативных активаторов превращается
в плазмин; нередко присутствует в комплексе с ингибитором — антиплазмином.
Филогенез, филогения — историческое развитие организмов. В биологии филогенез рассмат-
ривает развитие биологического вида во времени. Таксономия, классификация организмов по сход-
ству, основана на филогенезе, но методологически отличается от филогенетического представле-
ния организмов Филогенез рассматривает эволюцию в качестве процесса, в котором генетическая
линия — организмы от предка к потомкам — разветвляется во времени, и ее отдельные ветви могут
специализироваться относительно общего предка, сливаться в результате гибридизации или исче-
зать в результате вымирания.
Фуллерен (бакибол букибол) — молекулярное соединение, принадлежащее к классу аллотроп-
ных форм углерода (другие — алмаз карбин и графит) и представляющее собой выпуклые замкну-
тые многогранники, составленные из четного числа трехкоординированных атомов углерода.
Фурин — фермент, сериновая протеаза клеток животных, расположенная в аппарате Гольджи.
Фурин по структуре напоминает бактериальный протеолитический фермент субтилизин. Фурин
также известен под названием РАСЕ (англ. Paired basic amino acid cleaving enzyme — «фермент, рас-
щепляющий белок в месте спаренных основных аминокислот»). Название «фурин» произошло от
названия гена FUR (англ. FES upstream region), поскольку в 1986 г. соответствующая ему ДНК-пос-
ледовательность была обнаружена в участке выше («upstream region») кода онкогена под названием
FES. Помимо обработки клеточных белков-предшественников, фурин также участвует в активации
ряда токсинов микроорганизмов и вирусов. В частности, расщепление фурином либо фурин-
оподобными протеазами необходимо для созревания оболочечных белков таких вирусов, как ВИЧ,
вирус гриппа, папилломавирус и вирус Денге. Фурин играет важную роль в активизации экзо-
токсинов сибирской язвы и Pseudomonas. Этот фермент необходим для их расщепления лри попа-
дании в клетку, только после этого токсины становятся активными.
Химиопрепараты — химиотерапевтические средства, природные и синтетические соединения,
применяемые для лечения и профилактики инфекционных и неинфекционных болезней.
Холинергический кризис — состояние, провоцируемое гиперактивностью ацетилхолина вслед-
ствие передозировки препаратов, блокирующих фермент, который дезактивирует ацетилхолин
(препараты антихолинэстеразы). Наблюдаются подергивание мышц, паралич, потливость, повы-
шенное слюноотделение, бледность, сокращение зрачков. Таким действием обладают нервнопа-
ралитические ОВ.
Цервикальная дистония спастическ 1я кривошея) — дистоническое заболевание, вид кривошеи,
в которой неправильное положение головы вызвано патологическим напряжением мышц шеи.
Цинковый палец — тип белковой структуры, небольшой белковый мотив, стабилизированный
одним или двумя ионами цинка, связанными координационными связями с аминокислотными ос-
татками белка. Как правило, цинковый палец включает около 20 аминокислот и ион цинка связыва-
ет 2 гистидина и 2 цистеина. Цинковые пальцы являются белковыми модулями, взаимодействую-
щими с ДНК, РНК, другими белками или небольшими молекулами. Основной группой белков с
цинковыми пальцами являются ДНК-связывающие фак|Оры транскрипции.
Штамм — культура микроорганизмов одного вида с одинаковыми морфологическими и биоло-
гическими свойствами. Штаммы микроорганизмов могут отличаться друг от друга по вирулентно-
сти, чувствительности к антибиотикам, способности к токсинообразованию.
Штаммы агента с измененными или редко встречающимися свойствами — штаммы биологического
поражающего агента, имеющие свойства, затрудняющие типовое лечение вызванных ими пораже-
ний. Например, возбудитель чумы, устойчивый к стрептомицину, или возбудитель сибирской язвы,
устойчивый к антибиотикам р-лактамного ряда.
Эйфория — повышенное радостное настроение, чувство довольства, благополучия, не соот-
ветствующие объективным обстоятельствам. У лиц с эйфорией, как правило, преобладает оптими-
стическая оценка окружающего, противоположные аффекты им недоступны. Может сочетаться как
с ускоренным течением мыслей, склонностью к образному фантазированию, активной, но малопро-
Словарь терминов
1065
дуктивной деятельностью, расторможением влечений, так и с замедленностью психических процес-
сов, пассивностью, вялостью, отсутствием побуждений.
Экзон — часть гена, кодирующая участок структуры его продукта (белка, РНК). В ходе эксп-
рессии гена экзоны образуют зрелую информационную РНК.
Экзоспорий — наружная толстая оболочка споры, покрытая часто снаружи шипиками, бугор-
ками, гребнями.
Эктодерма — наружный зародышевый листок, из которого в раннем утробном периоде разви-
вается кожный эпителий или эпидермис, и нервная система.
Эндоплазматический ретикулум (эндоплазматическая сеть) — внутриклеточный органоид эука
риотической клетки, представляющий собой разветвленную систему из окруженных мембраной
уплощенных полостей, пузырьков и канальцев. При участии эндоплазматического ретикулума про-
исходит трансляция и транспорт белков, синтез и транспорт липидов и стероидов. Для эндоплаз-
матического ретикулума характерно накопление продуктов синтеза. Принимает участие в созда-
нии новой ядерной оболочки (например, после митоза).
Эндотелий — специализированные клетки животных и человека, выстилающие внутреннюю
поверхность кровеносных и лимфатических сосудов, а также полостей сердца. Представлен одно-
слойным пластом тесно прилежащих друг к друг) плоских клеток, обычно отграниченных от подле-
жащих тканей базальной мембраной.
Эндоцитоз - процесс захвата (интернализации) внешнего материала клеткой, осуществляемый
путем образования мембранных везикул. Различают фагоцитоз, пиноцитоз и рецептор-опосредо-
ванный эндоцитоз.
Эпидемиологический надзор (эпизоотологический мониторинг) — система динамического и комп-
лексного слежения за эпидемическим (эпизоотическим) процессом инфекционной болезни на
определенной территории, включающая сбор, передачу, анализ и оценку эпидемиологической
информации в целях разработки рекомендаций по профи тактике и борьбе.
Эпидемическая ситуация (эпизоотологическая обстановка) — характеристика распространенно-
сти инфекционно (паразитарной) болезни или их совокупности на конкретной территории за оп-
ределенное время.
''пидемш. - массовое поражение всего населения или отдельных его контингентов инфекцион-
ной (паразитарной) болезнью.
Эпимеры — это диастереоизомеры (см. выше), которые отличаются по конфигурации только
одного хирального центра.
Эукариоты — организмы, у которых: 1) имеется ядро, где содержатся хромосомы; 2) в цитоплаз-
ме присутствуют различные органеллы — митохондрии, хлоропласты и т. д. К эукариотам относятся
животные, растения, грибы, дрожжи, некоторые водоросли
СПИСОК ЛИТЕРАТУРЫ
Абу Бакр Мухаммад ибн Закария ар-Рази. Книга об оспе и кори // В кн. Губерта В. О. Оспа и оспопри-
вивание. СПб., 1896. С. 26—60.
Авдиев В. И. История Древнего Востока. М.. 1953.
Авиновицкий А. Я. Химическая война и оборона СССГ М., 1927.
Авиценна (Абу Али Ибн Сина). Канон врачебной науки. Книга IV Ч 1 Ташкент, 1980.
Агбунов М. Античные мифы и легенды. М., 1993.
Азарьев С. Микробное оружие // Военная мысль 1938. N° 12. С. 97—105.
Акинфеева Л. А., Аксенова О. И., Василевич И. В. с соает. Случай вирусной геморрагической лихорадки
Эбояа// Инфекционные болезни. 2005. Т. 3. № 1. С. 85—88.
Александренков Г.. Александрова Н, Бутырский М. ссоавт. Энциклопедия «Древний мир». М., 2005.
Александров В. Н., Емельянов В. И. Отравляющие вещества. М., 1990.
Алексеев В. В., Тихонов Н. Г., Вязьмина Т. Н. с соавт. Патогенез, патолщ ическая анатомия и пато-
логическая морфология мелиоидоза // Мелиоидоз. Сб. науч гр. Под ред. Н. Г. Тихонова. Волго-
град. 1995. С. 91-119.
Ачтухов Ю. П. Генетические процессы в популяциях. М., 2003.
Aibvedwib М. Ф. В магическом круге СПб., 2002
Альберте Б., БрейД., Льюис Дж. с совт. Молекулярная биология клетки. В 3 г. // Пер. с англ, под ред.
Г. П. Георгиева и Ю С. Ченцова. М., 1994.
Альтман X. Ядовитые растения. Ядовитые животные. М., 2004.
Андерсон Р., Мэи Р. Инфекционные болезни человека. Динамика и контроль. М., 2004.
Андрианов А. В. Экологическая безопасность дальневосточных морей России, Владивосток, 2009
(http://www.biosoil.ru/hydro/Xmeet/05.pdf).
Анисимов А. П. Факторы Yersiniapestis, обеспечивающие циркуляцию и сохранение возбу тителя чумы
в экосистемах природных сообществ. Сообщения I и 2 // Молекуляр. ген., микробиол вирусол.
2002. № 3. С. 3-23; № 4. С. 3-11.
Антонов Н. С. Химическое оружие на рубеже двух столетий. М., 1994.
Ардашев А. Н Зажигательное и огнеметное оружие. М , 2009.
АрожеласЛ. К. Высушивание микробов и бактерийных препаратов из замороженного состояния под
вакуумом // Труды Московского городского НИИ эпидемиологии и бактериологии. 1952. Выл. 5.
С. 298-312.
Архангельский Г. Ф. Амбулантные формы чумы (pestis ambulans) и ее значение в эпидемиологии //
Сборник сочинений по судебной медицине, судебной психиатрии, медицинской полиции, об-
щественной гигиене, эпидемиологии, медицинской географии и медицинской статистике. 1879.
Т. 1.С. 132-193.
Архангельский Г. Ф. Холерные эпидемии в Европейской России за 50-летний период 1823—1872 гг.
СПб., 1874.
Атарова К.Н. Вымысел или документ? // В кн. Дефо Д. Дневник чумного города. М., 1997.
Барбье А. Введение в химическую экологию. М., 1978.
Беланов Е. Ф., Гуськов А. А., Сокунова Е. Б. с соавт. Выживаемость вируса натуральной оспы в короч-
ках от больных И Докл. РАН. 1997. Т. 354. С. 832—834.
Берестнев Н. М. О чумных заболеваниях в лаборатории в форте Александр I в Кронштадте в февра-
ле 1907 г. //Арх. биол. наук. 1907. Т. XIII. № 3. С. 271—289.
Берестнев Н. М. Русская чумная лаборатория. М., 1907.
Беркоуз Р., Флетчер Э. Руководство по медицине. М., 1937.
Библия в гравюрах Гюстава Доре. М., 1996.
Ьилай В. И., Пидопличко Н. М. Токсинообразующие микроскопические грибы. Киев, 1970.
Блюменталь Н. Бактериологическая война (сокращенный обзор) // Военно-медицинский журнал.
1932. Т. III. Вып. 2. С.167-175.
Блюменфельд М. Л. Из официальных итогов эпидемии чумы в г. Харбине // Сибирская врачебная
газета. 1911. № 42. С. 507-509.
Список литературе
1067
Богданов И. Л. К клинике кишечной формы антракса // Клин. мед. 1935. № 8. С. 1228—1232.
Богоявленский В. Ф._ Богоявленский И. Ф. Клиническая диагностика и неотложная терапия острых
отравлении М., 2002.
Богуцкий В.М. Эпидемия чумы в Харбине и его окрестностях. Противочумное бюро. Харбин. 1911.
Болдырев Т. Е. Опыт советской медицины в Великой Отечественной войне 1941—1945 гг М., 1955.
Борисевич И. В., Воробьева М. С., Гайдерова Л А с соавт. Медицинские иммунобиологические пре-
параты М., 2010.
Борисевич И. В., Маркин В. А., Фирсова И. В с соавт. Эпидемиология, профилактика, клиника и ле-
чение геморрагических лихорадок (Марбург, Эбола. Ласса и Боливийской) // Вирусология. 2006.
№5. С. 8-16.
Борисов Л. Б. О глубинном культивировании бактерий // Важнейшие инфекционные болезни.
Л., 1959.
Бразоль Л. Е. Дженнеризм и Пастеризм, критический очерк оснований оспопрививания. Харьков,
1875.
Брода П П газмиды. М., 1982.
Брокгауз и Ефрон. Энциклопедический словарь. СПб., 1890—1907.
Бургасов П. Н. Я верил.. М , 2001.
Бургосов П Н., Николаевский Г П. Натуральная оспа. М., 1972.
Бургосов П Н, Рожков Г И. Сибирская язва. М., 1984.
Бывалое А. А Молекулярная чумная вацина. Екатеринбург, 2011.
Васильев Ц. Б, Кудрявцев С. В., Шумаков О В. Руководство по технике безопасности, методам рабо-
ты с ядовитыми змеями в зоопарковских условиях, профилактике и лечению укусов. М., 199'’
Вертиев Ю. В. Бактериальные токсины: биологическая сущность и происхождение //Журн. микро-
биол. 1996. № 3. С. 43—46.
Вершилова П.А. с соавт. Бруцеллез. М., 1972.
Вершилова П.А. с соавт. Бруцеллез. Организационно-методические материалы. М., 1968.
Вершилова П.А., Руднев Г П. Бруцеллез // Руководство по микробиологии, клинике и эпидемиологии
инфекционных болезней. М., 1964. Т. VI. С. 255—309.
Владимиров С. Наш человек в гестапо, или Кто был прообразом Штирлица // Независимое военное
обозрение. 2010. № 19. С. 13—14.
Волков А. Что там, за маревом страхов? // Знание-сила. 2001 № 12. С. 3—7.
Вопросы противобактериологической зашиты / Под ред. П. Ф. Здродовского М , 1960.
Воронин А., Зимкин Н. В ФБР и ЦРУ практически уверены: «почтовая эпидемия» сибирской язвы —
дело рук американских экстремистов// Итоги. 2001. № 44 (6 ноября). С. 13—16.
Высокович В. К., Чистович Н. Л. О природе КоЬобовскои эпидемии // Больничная газета С. П. Бот-
кина. 1900. Т. V. Вып 1. С. 131—135.
Высокович В.К. Предварительное сообщение о результатах, добытых русской экспедицией, коман-
дированной для изучения чумы в Бомбее // Русский архив патологии, клинической медицины
и бактериологии. 1897. № 4. С 387—390.
Вязьмина Т. Н., Тихонов Н. Г., Алексеева В. В. с соавт. Патогенез, патологическая анатомия и патоло-
гическая морфология сапа // Сап. Сборник научных трудов. Под ред. Н. Г. Тихонова. Волгоград.
1995 С. 50-73.
Габричевский Г. Н. Медицинская бактериология. М., 1907.
Гайслер Э. с соавт. Биологическое и токсинное оружие сегодня. SIPRI. London, 1986.
Гальперин Э. А. Клиника оспы и прививочных реакций. М., 1962.
Гамалея Н.Ф. История оспы М , 1913.
Гаммериан Л. Ф., Гром И. И. Дикорастущие лекарственные растения СССР. М., 1976.
Гезер Г История повальных болезней. СПб., 1867.
Геирот Ф. Медикопрактическое описание о болезни, так называемой Сибирскою СПб., 1807
Гирш А. Индийская чума//Военно-медицинский журнал. 1853. Часть LX1I (III). С. 29—48.
Головин Ю. И. Введение в нанотехнику. М., 2007.
Горбунов Е. А. Схватка с черным драконом. Тайная война на Дальнем Востоке. М., 2002.
Гос В. И. К экспериментальной чумной пневмонии // Архив биологических наук. 1907. Т. XIII. № 3.
С. 311-333.
Грачев О. Е., Елагин Г. Д., Дубровин М. Ю. с соавт Патент Российской Федерации 2230325( 13) С2,
МПК GO1N33/531, А61К39/08. Способ приготовления очищенного препарата ботулинического
1068
Список литературы
токсина типа А / заявитель Научно-исследовательский институт микробиологии Министерства
обороны Российской Федерации; заявл. 15,08.2002, 2002122179/13: опубл. 10.06 2004.
Гринберг Л М. Патологическая анатомия и некоторые вопросы морфо-, пато- и танатогенеза инга-
ляционной формы сибирской язвы человека. Дис.... докт. мед. наук. Екатеринбург, 1995.
Губерт В. О. Оспа и оспопрививание. СПб, 1896.
Гугерсгоф Ф Специальный каталог на бактериологические препараты и приборы (Лейпциг - Моск-
ва, Московское отделение, Рождественский бульвар, дом Маттерн) М , 1908.
Гусев А. И. Наноматериалы, наноструктуры, нанотехнологии. М., 2007.
Гусынин И.А. Токсикология ядовитых растений. М., 1962.
Де Лазари А. И. Химическое оружие на фронтах мировой войны. М., 1935.
Девис Ли. Природные катастрофы. Смоленск, 1997.
ДелюмоЖ. Ужасы на западе. М., 1994.
1ербек Ф И. История чумных эпидемий в России. СПб,, 1905.
Деревянко В. С. Влияние аэродинамических возмущений на процессы авиационного опыливания и
опрыскивания. М., 1974.
ДефоД. Дневник чумного года. М., 1997.
Джалилов К. Д. Бруцеллез. Ташкент, 1979.
Джеймс П., Торп Н. Древние изобретения. Минск, 1977.
Дженнер Э. Исследование причин и последовательного действия Variolae vaccinae, болезни, откры-
той в некоторых западных графствах Англии, преимущественно в Глочестере и известной под
именем коровьей оспы // В кн. Губерта В. О. Оспа и оспопрививание. СПб, 1896. С. 372—397.
Дмитриев И. Случай массового подкожного заражения сибирской язвой во время антирабических
прививок // Гигиена и эпидемиология. 1928. № 10. С. 64-74.
Доклад Международной научной комиссии по расследованию фактов бактериологической войны
в Корее и Китае. Пекин, 1952.
Домарадский И. В. Чума. М., 1998.
Дробинский И. Р Проблема бактериальной войны и противобактериальная защита // Труды Украин-
ского Мечниковского института. 1940. Т VI. С 87—123.
Дроздове. Г., Гарин Н С., Джиндоян Л. С. С., Тарасенко В. М Основы техники безопасности в микро-
биологических и вирусологических лабораториях. М., 1987.
Дрюмон Э. Еврейская Франция. Харьков, 1895.
Дубровинский С. Б Оспа и оспопрививание . М., 1964.
Егорышева И. В., Гончарове С. Г. Вклад принца А. П. Ольденбургского в развитие российской меди-
цины И Военно-медицинский журнал. 2004. Т. 325. № 5. С. 66—70.
Езепчук Ю В. Патогенность, как функция биомолекул. М., 1985.
Ермаков И. А., ИвановО. К., ИцковА И. и др. Очерки истории российской внешней разведки. Т. 1. М.,
1995.
Жемчугов В. Е Как мы делали вакцины. М , 2004.
ЖикурЯ. Химическое оружие в современной войне. М., 1936.
Жимулев И. Ф. Общая и молекулярная генетика. Новосибирск. 2002.
Таи И., Линкчн Г. Рецензия на статью Karyszkowski J. «Бактериальная война» // Военно-санитарное
дело 1936. Ч 12 С. 58-59.
Захаров П.В., Беляев Т.Н., Вельмовский Б.П. Эпидемиология легочной чумы в Приморской области
и в г. Владивостоке в 1921 году. Владивосток, 1922.
Звонарев К. К. Агентурная разведка. М., 2005.
Здродовский П. Ф., Голиневич Е. М. Учение о риккетсиях и риккетсиозах. М , 1972.
ЗлатогоровС. И. Учение о микроорганизмах. СПб., 1916.
Золотарев А. Г., Дармов И. В., Пименов Е. В Возбудители особо опасных инфекционных заболева-
ний бактериальной природы. Морфология и ультраструктура. М., 2006
Зуева Л П., Яфаев Р X. Эпидемиология. СПб., 2006.
Иванов Н. А., Богач В. В. Оружие вне закона: Кто готовит бактериологическую войну? Хабаровск,
1989.
Ивашенцов Г. А. Курс острых инфекционных болезней. Л., 1948.
Илюхин В. И., Батманов В. П. Клиника и лечение мелиоидоза // Мелиоидоз. Сб. науч. тр. Под ред.
Н. Г. Тихонова. Волгоград. 1995. С. 142—161.
Илюхин В И , Батманов В И., Лозовая Н. А. Клиника и лечение сапа // Сап. Сб. науч. тр. Под ред.
Н. Г. Тихонова. Волгоград, 1995. С. 85—100.
Список литературы
1069
Илюхин В. И., Замараев В. С., Каплиев В И. Микробиология Pseudomonas mallei // Мелиоидоз. Сб.
науч. тр. Под ред. Н. Г. Тихонова. Волгоград, 1995. С. 7—19.
Ипатенко И. Г., Гаврилов В. А., Зелепукин В. С с соавт Сибирская язва. М., 1996.
Капустин А. К. Бактериологическая война //Химия и оборона. 1939. № 5. С. 22.
Каракулов И К, Борисов В. Д. Эпидемиология, эпизоотология и лабораторная диагностика лихорад-
ки Ку. Алма-Ата, 1966.
Касторский Е.С. Эпидемия легочной чумы на Дальнем Востоке в 1910—1911 гг. и меры борьбы с нею.
Иркутск, 1911.
Кашкадамов В. И. Альбом снимков чумных больных СПб.. 1902
КерлейП.,ДайтЛ., ПоссиР. ссоавт. Патент2156300, RU, MF1KC12NI5/33, C12N15/86, A01N63/00.
Дата публикации: 2000-09-20.
КиттериджДж. Л. Колдовство в Средние века СПб., 2005.
Климентова А., Крук R, Ярмомчик Г. Консервирование бактериологических и биологических препа-
ратов высушиванием при высоком вакууме // Журн. микробиол. »пидемиол. иммунобиол. 1942.
№ 7. С. 14-22
Клонирование ДНК / Под ред, Д. Гловера. М., 1988.
Клотц Г. Бактериологическое оружие// Военный зарубежник. 193 7. № 12 С 19—31.
Киберт Р. Compendium практической медицины. Руководство для врачей и студентов. Дерпт, 1888
Козлов М. И., Султанов Г. В. Эпидемические проявления чумы в прошлом и настоящем Махачка-
ла, 1993.
Колпакиди А И, Прохоров Д. И. КГБ. Спецоперации советской разведки. М., 2000.
Конвенция о запрещении разработки, производства и накопления запасов оактериологического
(биологического) и токсинного оружия и об их уничтожении // Сборник документов и матери-
алов, регламентирующих обеспечение выполнения Российской Федерации международных обя-
зательств по запрещению биологического и токсинного оружия. М., 2004.
Конвенция о запрещении разработки, производства, накопления и применения химического ору-
жия и о его уничтожении. ООН, 1993.
Коновалова Г.В. «Красные приливы» у Восточной Камчатки (Атлас-справочник) Составитель
А. М. Токранов. Петропавловск-Камчатский, 1995.
Коновалова И. В., Могиленикова Т. А. Токсичный фитопланктон и содержание биотоксинов в тканях
гребешка // Фундаментальные исследования. 2006. № 9. С. 102—103.
Кошкин А. А. «Кантокуэн» — «Барбаросса» по-японски. М., 2011.
КраевскийА. Бутерброд со смертью//Focus.pl. 2009 Юиюня (http://perevodika.ru/articles/12652.htmD
Кравченко А. Т, Руднев Г.П. Сап // Руководство по микробиологии, клинике и эпидемиологии ин-
фекционных болезней. М., 1966. Т. VII. С. 327—346.
КремлевС. Россия и Япония: стравить. М., 2005.
Кротков Ф. Г. «Бактериальная война» на Дальнем Востоке // Военно-санитарное цело. 1939. № 9
С. 81-82.
Кротков Ф. Г. Угроза бактериологической войны // Военно- санитарное дело. 1939. № 4. С. 64—74.
Кузник Б. И. Физиология и патология системы крови. М., 2004.
Кукридж Э. Европа в огне. Диверсии и шпионаж британских спецслужб на оккупированных терри-
ториях 1940—1945. М., 2003.
Кулеша Г. С. К патологической анатомии легочной чумы. Поражение при легочной чуме слизистых
оболочек верхних дыхательных путей // Вестник микробиологии и эпидемиологии. 1924. Т. 3.
С. 239-243.
Кулеша Г С. О патологической анатомии легочной чумы по данным последней эпидемии в Маньч-
журии И Русский врач. 1912. Т. 11 № 15. С. 502-503.
Купер Э. Сравнительная иммунология М.. 1980.
Курилов В. Я., Фарбер С. М., Ломова Л В. с соавт. Бактериальные токсины. М., 1987. С. 81—86
Курляндский Б А., Филов В. А., Безель В. С. с соавт. Общая токсикология. М., 2002.
Куценко С. А. Основы токсикологии СПб., 2004.
Ларионов Г. М. Эпидемиология и профилактика сапа // Сап. Сб. науч. тр. Под ред Н. Г. Тихонова
Волгоград, 1995. С. 100—109.
Ларионов Г. М., Гавенский С. Д., Погасий И. И. с соавт. Эпидемиология и профилактика мелиоидоза
// Мелиоидоз Сб. науч. тр. Под ред. Н. Г. Тихонова. Волгоград 1995. С. 161—195.
Либинсон И. М. Химия в войне. М., 1926.
Литре А. Медицина и медики. СПб., 1873.
1070
Список литературы
Люпусов И.В., Васильев Н. Т„ Васильев П.Г. и др. Патоморфогенез сибирской язвы М , 2002.
Лихачев. Вопросы химической и бактериологической войны в иностранной военно-санитарной ли-
тературе И Здравоохранение. 1929 № Д. С. 100 105
Лобан К. М., Лобзин Ю. В., Лукин Е. Л. Риккетсиозы у человека. М.. 2002
Лобанов В. Н. Чума у верблюдов и ее эпидемиологическое значение. Саратов, 1968.
Лозинский С. Г История папства. М., 1986.
Лотоцкий С С. с соавт. Война в Корее 1950—1953 гг. СПб., 2000
Лоуренс Д. Р., Бенитт П. Н. Клиническая фармакология. М., 1993.
ЛюльмаХ., Мор К., Хайн Л. Наглядная фармакология М., 2008.
Макиннис П. Тихие убийцы. М., 2008.
Ммиатис Т., Фрич Э., СэмбрукДж Молекулярное клонирование. М., 1984.
Маренникова С. С. Опа. Алястрим. Ветряная оспа. Оспа животных // Руководство по микробиологии,
клинике и эпидемиологии инфекционных болезней. М., 1966. Т. VIII. С 443—502.
Маренникова С.С., Щелкунов С.Н. Патогенные для человека ортопоксвирусы. М., 1998.
Маренникова С.С., Гашников П. В., Жукова О. А. и др. Биотип и генетическая характеристика изолята
вируса оспы коров, вызвавшего инфекцию у ребенка // Журн. микробиол. эпидемиол. иммуно-
биол. 1996. № 4. С. 8-10.
Маринин Л. И., Онищенко Г. Г., Степанов А. В. и др. Микробиологическая диагностика сибирской
язвы. М., 1999.
Мартиневский И. Л., Молзяре Г. Эпидемия чумы в Маньчжурии в 1910—1911 гт. М., 1971.
Марценак. Бактериологические оружие в современной войне // Советская ветеринария. 1935. № 4.
С. 88-90-№5 С 92-95.
Матвеев К. И. Ботулизм. М., 1959.
Матвеев К И., Быченко Б Д. Мелиоидоз // Руководство по микробиологии, клинике и эпидемиоло-
гии инфекционных болезней. М , 1966. Т. VII. С. 346—375.
Матвеев К. И, Булатова Т. И. Ботулизм // Руководство по микробиологии, клинике и эпидемиологии
инфекционных болезней. М., 1966. Т VI. С. 183—210.
Медико-санитарные аспекты применения химического и бактериологического (биологического)
оружия. Доклад группы консультантов ВОЗ Женева, 1972.
Мери нова Л. К, Агеева Н П. Генетика Pseudomonas mallei // Сап. Сб. науч. тр. Под ред. Н Г. Тихонова.
Волгоград, 1995. С. 19—30.
МериноваЛ. Гришкина Т. А., Тарасова Т. Д с соавт. Генетика возбудителя мелиоидоза // Мелиоидоз.
Сб. науч тр. Под ред. Н Г. Тихонова. Волгоград, 1995. С. 27-47.
Мессарски Н. Бактериологическое оружие и защита от него. София, 1955.
Миллер Н. Ф. Оспопрививание. М., 1887.
МинхГ. Я. Чума в России (Ветлянская эпидемия 1878—1879 гг.). Киев, 1898.
МолодяковВ Э. Россия и Япония: меч на весах. М., 2004.
Молодяков В. Э. Россия и Япония: поверх барьеров. М., 2005.
МоримураСэйти. Кухня дьявола. М., 1983.
Муравьева Д. А. Тропические и субтропические пекарственные растения. М , 1983.
Мураками X. Подземка. М., 2006.
Мусселиус С. Г, Рык А. А. Отравление грибами. М., 2002.
Ненахов Ю. Ю. «Чудо-оружие» Третьего рейха. Минск, 1999.
Нетесов С. Р. Инфекции: новые угрозы в XXI веке // Мол. мед. 2004. № 4. С 41 -49.
Никифоров В. Н. Кожная форма сибирской язвы. М., 1973.
Никифоров В. В., Мельникова Л. И., Зарьков К. А. с соавт. Сап: случай из практики // Инфекционные
болезни. 2005. Т. 3. № I. С. 89—92.
Николаев Н. И. Чума в Маньчжурии. Дис. ... д-ра мед. наук. Киров, 1949.
Николаев Н.И. Чума. М.. 1968.
Носов К. С. Осадная техника Античности и Средневековья. СПб., 2003.
Огарков В. И, Гапочко Н. Г. Аэрогенная инфекция. М., 1975.
О методах и приемах иностранных разведывательных органов и их троцкисто-бухаринской агенту-
ры (сборник). М., 1937.
Овчинников Ю. А. Природные токсины в изучении основ нервной проводимости ,'/ Фундаменталь-
ные науки — медицине. М., 1980.
Олсуфьев Н Г., Руднев Г. Л., Дунаева Т. П. с соавт. Туляремия М , 1960.
Список литературы
1071
Олсуфьев Н. Г., Доброхотов В. П Туляремия // География природноочаговых болезней человека с за-
дачами их профилактики. М., 1969.
Олсуфьев Н. Г., Дунаева Т. Н. Природная очаговость, эпидемиолш ия и профилактика туляремии. М.,
1970
Олсуфьев Я. Г, Лелеп П. Я. Значение слепней в распространении сибирской язвы //Паразиты, пе-
реносчики и ядовитые животные. Л., 1935.
Олсуфьев Я. Г., Руднев Г. П Туляремия // Руководство по микробиологии, клинике и эпидемиологии
инфекционных болезней. М., 1966. Т VII. С 190—234.
ОнищенкоГ. Г. Биотерроризм как угроза безопасности. Оценка готовности медицинских учреждений
к противодействию биотерроризму // Вестник РАМН. 2003. № 4. С. 38-45.
Онищенко Г. Г., Федоров Ю. М., Тихонов Я. Г. с соавт. Противодействие биологическому терроризму
как новая проблема эпидемиологии // Эпидемиол. и инфек. болезни. 2003. № 2 С. 4—6.
Онищенко Г. Г., Шапошников А. А., Субботин В. Г. с соавт. Противодействие биологическому терро-
ризму. М., 2003.
Орлов А С, Гаврилов В. А. Тайны Корейской войны М., 2003.
Орлов Б. Я., Гелашвили Д. Б. Зоотоксинология (ядовитые животные и их яды). М., 1985.
Орлов Б.Н., Гелашвили Д.Б., Ибрагимов А К. Ядовитые животные и растения СССР. М., 1990
Очкур П. П Патологическая морфология бруцеллеза у человека. Алма-Ата, 1951.
Очкур П. Я. Патологическая морфология и патогенез Ку-риккетсиоза // В кн. Каракулов И, К., Бо-
рисов В. Д Эпидемиология, эпизоотология и лабор. диаг. лихорадки Ку. Алма-Ата, 1966.
Павлович М. Я. Химическая война и химическая промышленность. М., 1925.
Павловский Е. Я. Ядовитые животные Средней Азии. Сталинаоал, 1950.
Павсаний. Описание Эллады (перевод С. П. Кондратьева) М., 2002.
Лервушин Б. П. Сибирская язва в Омском округе в эпизоотию 1929 года //Архив Омского медицин-
ского института. 1936. Кн. 2. С. 351—377.
Петефис Ж-Х. Дело о ядах. М . 2001.
Петрович С. В. Микотоксикозы животных. М., 1991.
Петрянов-Соколов И. С., Сутугин А. Г. Аэрозоли. М , 1989.
ПеховА. П. Генетика бактерий. М , 1977.
Пиблз К. Тайные полеты. Смоленск, 2002.
Пигулев^кий С. В. Ядовитые животные. Токсикология позвоночных. Л., 1966.
Пигулевский С. В. Ядовитые кивотные. Токсикология беспозвоночных. Л., 19/5.
Покровский В. В. с соавт. ВИЧ-инфекция: клиника, диагностика и лечения. М,, 2003.
Понамарег Ю Малая химическая война Вермахта // Калашников. Оружие, боеприпасы, снаряже-
ние. 2011. № 10 С 49- 53
Попелянскии Я Ю., Фокин М. А., Пак С. Г. Поражение нервной системы при ботулизме. М., 2000.
Плецитный Д. Ф. Экспериментальное изучение патогенеза столбнячной интоксикации. М., 1958.
Прасолов В. С., Иванов Л С. Ретровирусные векторы в генной терапии // Вопр. мед. хим. 2000. № 3.
Рейтц В. К вопросу о прививании предохранительной коровьей оспы СПб., 1874.
Рейтц В. Критический взгляд на оспопрививание. СПб., 1873.
Ривер Л. Центр германской секретной службы в Мадриде в 1914—1918 гг М.. 1938.
Риверс Г. Вирусные и риккетсизные инфекции человека. М., 1955.
Рихтер В. М. Geschichte der medizin in Russland. M., 1813—1817.
Puxniep B. N. История медицины в России. М., 1814.
Ричиути Э. Р. Опасные обитатели моря. М., 1470
Розанов П Г. Отдельное мнение о Колобовской эпидемии // Больничная газета С. П. Боткина. 1900,
12 июня.
Розбери Г, Кабат Э. Бактериологическая война. М., 1955.
Ронин 3. Своя разведка: способы вербовки агентуры, методы проникновения в психику, форсиро-
ванное воздействие на личность, технические средства скрытого наблюдения и съема информа-
ции. Минск, 1997.
Ротшильд Д. Оружие завтрашнего дня. М., 1966.
Роше М. Моя разведывательная работа. М., 1938.
Руднев Г. П. Антропозоонозы. М., 1970.
Руководство к токсикологии, составленное по Рабюто «Elements de Toxicologies // Под ред. Е. Пе-
ликана. СПб., 1878.
1072
Список литературы
Руководство по вакцинопрос] илактике особо опасных инфекций / Под ред. Борисевича И В., Дар-
мова И. В. Киров. 2011.
Руководство по зоонозам / Под ред. В. М. Покровского Л., 1983.
Рыбкин В. С., Тихонов Н. Г., Жукова С. И. и др. Иммунология сапа // Сап: Сб. науч. тр. Под ред.
Н. Г. Тихонова. Волгоград, 1995. С. 73—84.
Casarini//Arch. med. Beilges. 1928. № 3.
Сакаки. Бактериологическая воина // Сэндэй Майничи 1952. № 1682 от 27 января.
Санитарно-эпидемиологические правила СП 1.3 2322-08 «Безопасность работы с микроорганизма-
ми III—IV групп патогенности (опасности) и возбудителями паразитарных болезней» (с изме-
нениями от 2 июня 2009 г., 29 июня 2011 г.) (http://base.garant.ru/12160681/).
Саркисов И. 3. Бактериальная война. Ростов-на-Дону, 1940.
Сартори А., Сартори Г Бактериологическая война, (пер. с франц. 3. Орской; под ред. и с предисло-
вием проф. И. Великанова) М., 1936.
Сатановский Е. Я. Россия и Ближний Восток. Котел с неприятностями. М., 2012.
Сборник сообщений Чрезвычайной государственной комиссии о злодеяниях немецко-фашистских
захватчиков. М., 1946.
Серенко А. Ф. Заносные вспышки натуральной оспы. М., 1962.
Ситников М. Н. Поражающее действие бактериологического оружия и некоторые медицинские воп-
росы зашиты// Журн. Всесоюз. Хим. общ. им. Д. И. Менделеева. 1968. Том XIII. № 6. С. 637-648.
Скороходов Л. Я. Материалы по медицинской микробиологии в дореволюционной России. М., 1948.
Славин С. Н. Секретное оружие Третьего рейха. М., 1999.
Смирнов Г. Б. Механизмы приобретения и потери генетической информации бактериальными гено-
мами // Усп. совр. биол. 2008. Т. 128. № 1. С. 52—76.
Смирнов Е. И. Война и военная медицина. М., 1979
Соколов Н. Химическая война. Минск, 1924.
Сперанский Н. Ведьмы и колдовство // История инквизиции. Средневековые процессы о ведьмах.
Харьков, 2001.
СС в действии. Документы о преступлениях СС. М., 1969.
Сталлибрас К. Основы эпидемиологии. М., 1936.
Страхович И. В., Гос В. И. Чумные эпидемии в Астраханской губернии в 1900—1901 гг.// Сборник ра-
бот по чуме. Вып. 2. 1907. СПб. С, 43—92.
Судебный отчет по делу антисоветского и правотроцкистского блока. М., 1938.
Суздалев И. П. Физико-химия нанокластеров, наноструктур и наноыатериалов. М., 2009.
Сунцов В. В., СунцоваН И. Чума. Происхождение и эволюция эпизоотической системы М.. 2006
Супотницкий М. В., ПолевщиковС. Н., Желнина Т. Б. с соавт. Способ трансформации микроорганиз-
мов. Патент 2052505, 25.02.92, RU, МКИ С12 N 15/87,13/00.
Супотницкий М. В., Кравец И. Д., Новикова О. В. Специфическая профилактика жспериментального
сапа порином из Pseudomonas mallei 11 Ветеринария. 1995. № 3 С. 24—26.
Супотницкий М В. Протективные свойства порообразующих белков патогенных бактерий // Вест-
ник Российской академии медицинских наук. 1996. № 8. С. 18—22.
Супотницкий М. В. Общие закономерности и типовые приемы в конструировании генно-инженер-
ных токсинов // Сборник докладов межучережденческой конференции «Средства защиты от
опасных и особо опасных инфекционных заболеваний». Киров, 1997. С. 144—162.
Супотницкий М. В. ДНК-иммунизация в профилактике инфекционных болезней сельскохозяй-
ственных животных//Ветеринария. 1998. № 5. С. 18 -24.
Супотницкий М. В. Микроорганизмы, токсины и эпидемии. М., 2000, 2005 (2-е изд.).
Супотницкий М. В., Супотницкая Н. С. Очерки истории чумы. М., 2006.
Супотницкий М. В. Эволюционная патология. К вопросу о месте ВИЧ-инфекции и ВИЧ/СПИД-
пандемии среди других инфекционных, эпидемических и пандемических процессов. М., 2009.
Супотницкий М. В., Поклонский Д. Л , Паныгина С. А. Токсичность наночастиц // Актуальные про-
блемы биологической зашиты войск и населения. Матер. Всерос. науч.-прак. конф., поев. 60-ле-
тию филиала ФГУ «48 ЦНИИ Минобороны России — ЦВТП БЗ». Екатеринбург, 2009. С. 41—44.
Супотницкий М.В. Героиновая сибирская язва в Шотландии и Германии // Биопрепараты. 2011. № 1.
С. 40-44.
Супотницкий М В Генотерапевтические векторные системы на основе вирусов // Биопрепараты.
2011. № 3 С. 15-26.
Список литературы
1073
Терри В. КГБ во Франции. М.. 2000.
Титок М. А. Плазмиды грамположительных бактерий. Минск, 2004.
Тихонов Н. Г., Фарбер С. М., Илюхин В. И. Биохимия возбудителя сапа // Сап. Сб. науч. тр. Под ред.
Н. Г. Тихонова. Волгоград, 1995. С. 45—51
Тихонов Н. Г , Илюхин В. И. История открытия и изучения возбудителя мелиоидоза // Мелиоидоз. Сб.
науч тр. Под ред. Н. Г. Тихонова. Волгоград, 1995. С. 5—8.
Тихонов Н. Г., Рыбкин В. С., Жукова С. И. с соавт. Иммунология мелиоидоза // Мелиоидоз. Сб науч,
тр. Подред Н. Г.Тихонова. Волгоград, 1995 С 119-142.
Трахтенберг Дж. Дьявол и евреи. Средневековые представления о евреях и их связь с современным
антисемитизмом. М., 1998.
Туманский В. М. Микробиология чумы М., 1958.
Устинов А. Н. К истории эпидемий Древнего мира. М., 1894.
Федорова И. И. Эпидемиология и профилактика Ку-риккетсиоза. М.. 1968.
Федорова Н. И. Риккетсиоз Q (Q-лихорадка) // Руководство по микробиологии, клинике и эпиде-
миологии инфекционных болезней. М., 1966. Т. VIII. С. 670—689.
Феррати В Uerreti И) Бактериальная война (перевод) // Военный зарубежник 1932. № 5. С. 32—42.
Филдс Б., Найп Д., Ченок Р. и др. Вирусология: В 3 т. // Пер. с англ Под ред. Н. В. Каверина,
Л. Л. Киселева. М., 1989.
Финкельштейн Я. М. Еще раз о чуме в Анзобе // Воен-мед. журнал. 1906. Т. 217. Ns 9. С 27—57.
ФордР Немецкое секретное оружие во Второй мировой войне. М., 2003.
Франклин Т., Сноу Дж. Биохимия антимикробного действия. М., 1984.
Фромм Э. Анатомия человеческой деструктивности. М , 1998
ХаркевичД. А. Фармакология. М., 1987.
Хироси Акияма Особый отряд 731. М., 1958.
Хмара-Борщевский Э.П. Чумные эпидемии на Дальнем Востоке и противочумные мероприятия
КВЖД // Воен.-мед. журнал 1913. Июль. С. 1724—1747.
Хухо Ф. Нейрохимия. Основы и принципы. М., 1990.
Цветков Н. Е., Черняк В. 3. Сап. М., 1935.
ЧелъныйА. А. Микробиолог Степан Васильевич Коршун. М., 2010.
Черкасский Б. Л. Путешествие эпидемиолога во времени и пространстве М., 2003.
Черкасский Б. Л. Руководство по общей эпидемиологии. М., 2001.
Черкасский Б. Л. Сибирская язва как биологическое оружие. М., 2000.
Черкасский Б. Л. Эпидемиология и профилактика сибирской язвы. М., 2002
Шахутдинова Р 3. Структурно-функциональная организация и генно-инженерная модификация
липополисахарида Yersinia pestis. Дис.. канл биол. наук. М., 2008.
ШебаршинЛ.В. Рука Москвы. М.. 1992.
Шелленберг В. Вторжение 1940. М., 2005.
Ширяева Д. Т, РяписЛ. А., Адамов Л. В с соавт. Мелиоидо’. М., 1976.
Шмальгаузен И. И Организм как целое в индивидуальном и историческом развитии. М., 1938.
Шпеер А. Воспоминания. Смоленск, 1997.
ШпренгерЯ., Инститорис Г. Молот ведьм // История инквизиции Средневековые процессы о ведь-
мах. Харьков. 2001.
Шувалова Е. П. с соавт. Инфекционные болезни. М., 1976.
Шувалова Е. П. с соавт. Инфекционные болезни. Ростов-на-Дону, 2001.
Шувалова Е. П с соавт. Тропические болезни. М., 1973.
Шувалова Е П с соавт. Тропические болезни. М., 1979.
Шунаев В. В. Об ингаляционном методе заряжения животных при экспериментальной чуме //
Сб. работ противочумных организаций Восточносибирского края за 1932—1933 гг (под ред.
А. М. Скородумова). Москва — Иркутск. 1934.
Шюре Э. Великие посвященные. М., 1914.
Щербина Н. Г. Аппарат для массового выращивания микробных культур //Лабораторная практика.
1934 № 1.С. 12-14.
Эддере Ш., Истэн М. Химическое оружие и его токсическое действие. М., 1939.
Эрисман Ф. Ф. Холера. Эпидемиология и профилактика. М., 1893
Яковлев А. М. Значение антитоксического иммунитета для защиты животного от респираторного про-
никновения бактериальных токсинов. I. Роль пассивного иммунитета в защите животных про-
1074
Список литературы
тив респираторного поражения, вызванного токсином Clostridium botulinum // Журн микро
биол. эпидемиол. иммунобиол. 1956. № 29. С. 904—909.
Яковлев А. М Значение антитоксического иммунитета для защиты животного от респираторного про-
никновения бактериальных токсинов. II. Тестирование токсического иммунитета у морских сви-
нок при интраназальном введении токсинов Clostridium botulinum // Журн. микробиол. эпиде-
миол. иммунобиол. 1956. № 29. С. 1068- 1072.
Abaev I. V., Astcshkin E.I., Pachkunov D M. et al Pseudomonas mallei and Pseudomonas pseudomallei:
introduction and maintenance of natural and recombinant plasmid replicons // Mol. Gen. Mikrobiol.
Virusol. 1995. № 1. 28- 36.
Abd H., Johansson T., Gotovliovl. el al. Survival and growth of Francisella tularensis in Acanthamoeba castel-
lanii // Appl. Environ. Microbiol. 2003. V 69. № 1. P. 600 -6u6.
Acheson D. W., Reidl J., ZhangX. etal. In vivo transduction with shiga toxin 1 -coding phage // Infect. Immun.
1998. V. 66. P. 4496 -4498.
Achtman M, Morelli G., Zhu P. et al. M'croevolution and history of the plague bacillus, Yersinia pestis // Proc.
Natl. Acad. Sci. U. S. A. 2004. V. 101 P. 17837-17842.
Achtman M., Zurth K., Morelli G. etal. Yersinia pestis, the cause of plague, is a recently emerged clone of Yersinia
pseudotuberculosis // Proc. Natl Acad. Sci. USA. 1999. V. 96. P. 14043-14048.
Acres J., Gray J. Paralytic shellfish poisoning//CM A J. 1978. V. 119. P. 1195—1197.
Adams A. P., Aronson J. F, TardifS. D. etal. Common Marmosets (C allithi ixjacchus) as a nonhuman primate
model to assess the virulence of Eastern equine encephalitis virus strains //J. Virol. 2008. V. 82. № 18.
P. 9035-9042.
Aguilar P. V., Greene 1. P., Coffey L. L. Endemic Venezuelan equine encephalitis in Northern Peru // Emerg.
Infect. Dis. 2004. V. 10. № 5. P. 880-888.
Aken Jan van, HammondE. Genetic engineering and biologic 11 weapons // EM BO reports. 2003. V. 4 (special
issue) P. 57—60.
AkincA., Thomas M., KlibanovA. M. etal. Exploringpolyethylenimine-mediated DNAtransfecuonandthe
proton sponge hypothesis //J. Gene. Med. 2005 V 7. P 657-663.
Akosso N. B., Yates W. E., Wilce S. E. Performance of atomizers for aircraft chemical application // Proc. 4 th
Intern. Agric. Aviation Congress. The Hague, 1971. P. 254—264.
Aktories K. Rho proteins: targets for bacterial toxins // Trends Microbiol. 1997. V.5. № 2. P. 282—288.
Albrink Ж 5. Pathogenesis of inhalation anthrax // Bact. Rev. 1961 V 25 № 3. P. 268-273.
Albuquerque U. P., Melo J. G., Medeiros M. F. et al. Natural products from ethnodirected studies: revisiting
the ethnobiology of the zombie poison // Hindawi Publishing Corporation Evidence-Based Comp-
lementary and Alternative Medicine. 2012. Article ID 202508 (doi 10 1155/2012/202508).
Al-Dosari M. S., GaoX. Nonviral gene delivery: principle, limitations and recent progress // AAPS J. 2009.
V. 11. №4. P. 671-681.
Alton N., Broomfield C. A., Doctor В. P. Induction of protection against organophosphate poisoning by liposome
mediated delivery of the human butyrylcholinesterase gene to the lungs // Proc, of Int Symp. on Appli-
cation of I nzymes in Chem. and Biol. Defense. 2001 14-18 May.
Allwood E. A., Devenish R., Prescott M etal Strategies for intracellular survival of Burkholderia pseudomallei //
Frontiers in Microbiology. Cellular and Infection Microbiology. 2011. V. 2 (doi: 10.3389/fmicb. 2011.
00170).
Alonso S., Sola L, Teifke J. P. et al. In vitro and in vivo expression of foreign genes by transmissible gast ro-
enteritis coronavirus-derived minigenomes // J. Gen. Virol 2002. V. 83. P. 567-579.
Alves D. A., Glynn A. R., Steele К. E. et al. Aerosol exposure to the Angola strain of Marburg virus causes lethal
viral hemorrhagic fever in Cynomolgus macaques // Vet. Pathol. 2010 V. 47. №5. P. 831—851
AmzilZ., Sibat M., Chomerat N. et al. Ovatoxin-a and palytoxin accumulation in seafood inrelation to
Ostreopsis cf ovata blooms on the french mediterranean coast//Mar. Drugs. 2012 V. 10. P. 477—496.
Ancuta P., Pedron T, Girard R. et al. Inability of the Francisella tularensis lipopolysaccharide to mimic or to
antagonize the induction of cell activation by endotoxins// Infect. Immun. 1996. V. 64. P. 2041—204o.
Andersen G. L., Simchock J. M., Wilson К. H. Identification of a region of genetic variability among Bacillus
anthracis strains and related species //J. Bactcriol. 1996. V. 178. P. 377—384.
Anderson B. Focal neurologic signs in Western equine encephalitis // Can. Med. Assoc. J. 1984. V. 130.
P. 1019-1021.
Andresen В. M., BoisJ. De novo synthesis of modified saxitoxins for sodium Ion channel study //J. Am. Chem.
Soc 2009. V. 131. P. 12524-12525.
Список литературы
1075
Anishchenko М., Bowen R. A., Paessler S. et al. Venezuelan encephalitis emergence mediated by a phyloge-
neticallv predicted viral mutation // Proc. Natl. Acad Sci. USA. 2008. V. 103. P. 4994-4999.
Anisimov A. P., LindlerL. E„ PierG B. Intraspecific diversity ofYersiniapestis//Clin. Microbiol Rev. 2004.
V. 17. Ne 2. P414- 464.
Anthony L. S. D., Cowley S. C., Mdluli К. E. et al. Isolation of a Francisella tularensis mutant that is sensitive
to serum and oxidative killing and is avirulent in mice: correlation with the loss of MinD homologue exp-
ression // FEMS Microbiol. Lett 1994. V. 124. P 157—166
Autigny F, Norez C., Becq F. et al. CFTR and Ca2+- signaling in cystic fibrosis,/ Front Pharmacol. 2011. V. 2
(doi: 10.3389/fphar.2011.00067).
Apisarnthananarak P. Apisamthanarak A., Mundy L. Computed tomography characterisitics of Burkholderia
pscudomallei - associated liver abscess // CID. 2006. V. 43. P. 1618—1620.
Ariano P., Zamburlin P., GilardtnoA. et al. Interaction of spherical silica nanopai deles with neuronal cells: size-
dependent toxicity and perturbation of calcium homeostasis // Small. 2011. V. 7 P. 766-774.
Army FM 8-284. Navy NAVMED P 5042 Air Force AFMAN (I) 44—156 Marine Corps MCRP4-11.1C.
2000.
Amon S. Human tetanus and human botulism I I In: Rood J. I., McClane B. A., Songer J. G., Titball R. W,
editors. The Clostridia: molecular biology and pathogenesis. San Diego: Academic Press. 1997. P. 95— 115.
AmonS. S., Schechter R., Inglesby T. V. etal. Botulinum toxin as a biological weapon: medical and public health
management //JAMA. 2001. V. 285. № 8. P. 1059-1070 (doi:10.1001/jama 285 8.1059).
African-Bouvery N., RodolakisA. Is Q fever an emerging or re-emerging zoonosis711 Vet. Res. 2005. V. 36.
P. 327-349.
Atkins L. C., Crossman T., Pitt C. et al. Genomic plasticity of the causative agent of melioidosis, Burkholderia
pseudomallei // Ptr. Natl. Acad Sci. USA. 2004. V. 101. P. 14240—14245.
Audi J., Belson M. etal. Ricin poisoning. A comprehensive review//JAMA. 2005. V. 294. № 18. P. 2343—3251.
Baca O. G., Paretsky D. Q fever and Coxiella burnetii: a model for host-parasite interactions I IM icrobiol. Rev.
1983. V. 47. P. 127-149.
Balder R, Lipski S., Lazarus J. etal. Identification of Burkholderia mallei and Burkholderia pseudomallei
adhesins for human respiratory epithelial cells // BMC Microbiology. 2010. V. 10. № 250 (http //
www.biomedcentral.com/1471-2180/10/250).
Baltz R. H. Antimicrobials from Actinomycetes back to the fu’ure // Am. Soc. Microbiol. 2007. V. 2.
P. 125-131.
Banerjee R. Nanoparticle aerosols: boon or bane for breathing? //Nanomedicine. 2012. V. 7, № 6.795—798.
Barakat L. A., Quentzel H. L., Jemigan J. A. et al. Woman fatal inhalational anthrax in a 94-year-ola Con-
necticut //JAMa 2002. V. 287. № 7. P. 863-868.
Baron G. S., Nam F. E. MglA and MglB are required for the intramacrophage growth of Francisella novicida
// Mol. Microbiol. 1998. V 29. P. 247-259.
Barrett A. D.T., Gould A. Antibody-mediated early death in vivo after infection with Yellow fever virus // J
Gen. Virol. 1986, V. 67. P. 2539-2542.
Barth H., Aktories K., Popoff M. R. et al. Binary bacterial toxins- biochemistry, biology, and applications of
common Clostridium and Bacillus proteins // Microbiol. Mol. Biol. Rev. 2004. V. 68, № 3. P. 373—402.
Baskerville A., Bowen E. T. W, PlattG. S. et al. The pathology ofexperimental Ebola virus infec.ion in monkeys
//J. Pathol. 1978. V 125. № 3. P. 131-138.
BatzaroffA. La pneumonie pesteuse experimentale //Ann. Inst. Parieur. 1899 V. 13 P 385—405.
Bausch D. G., SprecherA. G., Jeffs B. et al. Treatment of Marbutg and Ebola hemorrhagic fevers: A strategy for
testing new drugs and vaccines under outbreak conditions//Antiviral Res. 2008. V. 78 № l.P. 150 -161.
Baykam N., Ergonul O., Ulu A. et al. Characteristics of cutaneous anthrax in Turkey // J. Infect. Dev. Ctries.
2009. V. 3. № 8. P. 599-603.
Beare P. A., Samuel J. E., Howe D. et al. Genetic diversity of the Q fever agent, Coxiella burnetii, assessed by
microarray-based whole-genome comparisons//J Bacteriol. 2006. V. 188. № 7. P. 2309-2324.
Beare P. A., Sandoz К M., Omsland A. Advances in genetic manipulation of obligate intracellular bacterial
pathogens // Frontiers in Microbiology. 2011. V 2. Article 97 (doi: 10.3389/ftnicb. 20] 1.000Ч7'
Bea.e P., Howe D., Cockrell D. et al. Characterization of a Coxiella burnetii fts Z mutant generated by Himar 1
transposon mutagenesis//J. Bacteriol. 2009. V. 191. № 5. P. 1369—1381.
Beckett B. Weapons of Tomorrow. N. Y., 1982.
BegierE.M., AsikiG., AnywaineZ. etal. Pneumonic plague cluster, Uganda, 2004 // Emerg. Infect. Dis. 2006.
V. 12. № 3. P. 460 467
1076
Список литературы
Bell D. М., Kozarsky Ph. E., Stephens D. S. Clinical issues in the prophylaxis, diagnosis and treatment of anthrax
// Emeig. Infect. Dis. 2002. V. 8. № 2 P. 222-225.
Bembridge G. P., Lopez J. A., Cook R.etat Recomb nant vaccinia virus coexpressing the F protein of respiratory
syncytial virus (RSV) and interleukin-4 (IL-4) does not inhibit the development of RS V-specific memory
cytotoxic T lymphocytes whereas priming Is diminished in the presence of high levels of IL-2 or gamma
interferon // J Mroi. 1998. V. M. № 5. P. 4080-4087.
Benarroch D., Claverie J. M., Raoult D. et al. Characterization of mimivirus DNA topoisomerase IB suggests
horizontal gene transfer between eukaryal viruses and bacteria // J. Virol. 2006 V. 80. P. 314—321.
Benedek C., Rivier L. Evidence for the presence of tetrodotoxin in a powder used in Haiti for zombification
// Toxicon. 1989. V. 27. № 4. P 473—480.
BenensonA. S., Tigertt W. D. Studieson Q. fever in man //Trans. Assoc. Am. Physicians. 1956. V. 69. P. 98—104.
Bennett J. W., Klich M. Mycotoxins // Clin Microbiol Rev. 2003 V. 16, №. 3. P. 497—516.
Bennett P. M. I ntegrons and gene cassettes: agenetic construction kit for bacteria // J. Antimicrob. Chemother.
1999. V. 43. P 1-4.
Bennett P. M. Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic resistance genes in
bacteria // Brit. J. Pharmacol. 2008. V. 153. S347—S357.
BerdalB. P., MehlR., Meidell N. K. etal. Field investigations of tularemia in Norway //FEMS Immunol. Med.
Microbiol. 1996. V. 13. P. 191-195.
Bergen J. M., Park In-Kyu, Horner P. J etal. Nonviral approaches for neuronal delivery of nucleic acids //
Phar. Res. 2008. V. 25. № 5 P. 983 998
Bergen J. M., Pun S. H. Evaluation of an LC8-binding peptide for the attachment of artificial cargo to dynein
// Mol. Pharm. 200/. V. 4. P. 119-128
Bernhard K., Schrempf H., Goebel W. Bacteriocin and antibiotic resistance plasmids in Bacillus cereus and
Bacillus subtilis//J. Bacteriol. 1978. V. 133. № 2. P. 897-903.
Bernstein B. J. America’s biological warfare program in the Second World War //J. Strateg. Stud. 1988. V. 11.
№ 2. P. 292-317.
Berry B., Barrett L, Seymour L. etal Gene therapy for central nervous system repair//Curr. Opm. Mol.Ther.
2001. V 3.P. 338-349.
Bhakdi S., Bayley H. et al. Staphylococcal alpha-toxin, streptolysin-0 and Escherichia coli hemolysin: proto-
types of pore-forming bacterial cytolysins//Arch. Microbiol. 1996. V 165. № l.P. 73—79.
Bhakdll S., Tranum-Jensen J. Alpha-toxin of Staphylococcus aureus // Microbiol. Rev. 1991. V. 55. № 4.
P. 733-751.
Bharali D. J., Klejbor I., Stachowiak E. et al. Orgamcallj modified silica nanoparticles: a nonviral vector for in
vivo gene delivery and expression in the brain // Pi oc Natl. Acad. Sci. USA. 2005. V. 102. P. 11539— 11544.
Bhatia К P., Miinchau A., Thompson P D. et al. Generalised muscular weakness after botulinum toxin
injections for dystonia: a report of three cases // J. Neurol. Neurosurg. Psychiatry. 1999. V. 67 P. 90—93.
Bhatnagar V., Stoto M., Morton S. et al. Transmission patterns of smallpox: systematic review of natural
outbreaks in Europe and North America since World War II // BMC Public Health. 200b. V. 6. № 126
(http://www.biomedcentral.com/1471-2458/6/126).
Bienkowska-Szewczyk K., Szewczyk B. Expression of genes coding for viral glycoproteins in heterologous
systems // Acta Biochimica Polomca. 1999. V. 46 № 2. P. 325—339.
Bigalke H., Rummel A. Medical aspects of toxin weapons // Toxicol. 2005. V. 214. P. 210- 220
Black J. L. Genome projects and gene therapy: gateways to next generation biological weapons // Milit Med.
2003. V 108. № 11. P. 864-871.
Blum G., Falbo V., Caprioli A. etal. Gene clusters encoding the cytotoxic necrotizing factor type 1, Prs-fimbriae
and -hemolysin form the pathogenicity island II of the uropathogenic Escherichia coli strain J96 // FEMS
Microbiol. Lett 1995. V. 126. № 1. P. 189-196.
Boado R J, Pardridge IV. M. The Trojan horse liposome technology for nonviral gene transfer across the blood-
brain barrier//J. of Drug Deliv. 2011. (doi:10.1155/201 l/29t>151).
Bodian D.. Howe H. A. Experimental studies on intraneural spread of poliomyelitis virus // Bull. J. Hopkins
Hosp. 1941. V. 69. P. 248-267.
Boeckle S., Wagner E. Optimizing targeted gene delivery: chemical modification of viral vectors and synthesis
of artificial virus vector systems //AAPS J. 2006. V. 8. № 4. P. 731 -742.
Bonner J. S. Nanoparticles as a potential cause of pleural and interstitial lung disease // Pro< Am Thorac.
Soc. 2010. V. 7. P 138-141.
Borio L , Inglesby T, Peters C J. etal Hemorrhagic fever viruses as biological weapons. Medical and public
health management //JAMA. 2002. V. 287. № 18. P. 2391—2405.
Список литературы
1077
Borison Н. L., Culp W.J., Gonsalves S. F. et al Central respiratory and circulatory depression caused by intra-
vascular saxitoxin // Br. J. Pharmac. 1980. V. 68. P. 301—309.
Borm P., Robbins D., Haubold S The potential risks of nanomaterials: a review earned out for ECETOC //
Particle and Fibre Toxicology. 2006. V. 3. (http.//wwwparticleandfibretoxicology. com/content/3/1/11).
Bossi P, Tegnell A, Baka A et al Bichat guidelines for the clinical management of haemorrhagic fever viruses
and bioterrorism-related haemorrhagic fever viruses // Euro. Surveill. 2004. V. 9. № 12 (http //
www.eurosurveiltance.org/em/v09nl2/0912-235.asp).
Bowen J. E., Quinn С P. The native virulence plasmid combination affects the segregational stability of a theta-
replicating shuttle vector in Bacillus anthracis var New Hampshire//J Appl. Microbiol 1999. V. 87. № 2.
P. 270—2’8
Bowen M. D., Rollin P. E., Ksiazek T. G. etal. Genetic diversity among Lassa virus strains //J. Virol. 2000. V 74.
№ 15. P. 6992-7004.
Brachman P. S., Kaufmann A. E, DalldoreF. Industrial inhalation anthrax // Bacterio!. Rev. 1966. V. 30. № 3.
P. 646 657.
BraudeA. I., DavisC. E., FiererJ F Infectious diseases and medical microbiology. W. B. Saunders Company,
Philadelphia, 1986.
Bravo A., Gymez I., Conde J. et al. Oligomerization triggers binding of a Bacillus thuringiensis Cry 1 Ab pore-
forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains// Bio-
chim. Biophys. Acta. 2004. V. 1667. P. 38—46.
Bray M, Paragas J Experimentaltherapyoffilovirusinfections//Antiviral. Res. 2002. V.54.№ l.P. 1—17.
Brett P 1., Mah D. C, Woods D. E. Isolation and characterization of Pseudomonas pseudpmallei flagellin
proteins // Infect. Immun. 1994. V. 62. P. 1914—1919.
Brett P., Burtnick M.R, Snyder S. et al. Burkholderia mallei expresses a unique lipopolysaccharide mixture
that is a potent activator of human foll-like receptor 4 complexes human Toll-like receptor 4 complexes
I I Mol. Microbiol. 2007. V. 63. № 2. P. 379-390.
BrotckeA., Monack D. M. Identification of fevR, a novel regulator of virulence gene expression in Francisella
novicida // Infect. Immun. 2008. V. 76. P. 3473-3480.
Brotcke A., Weiss D. S., Kim С. C. et al. Identification of MglA-regulated genes reveals novel virulence factors
in Francisella tularensis // Infect. Immun. 2006. V 74. P. 6642—6655
BrownD. M., WilsonM R, Mac Nee W. etal. Size-dependent proinflammatory effects of ultrafine poly-styrene
particles: a role for surface area and oxidative stress in the enhanced activity of ultrafmes // Toxicol. Appl.
Pharmacol 2001. V. 175 P 191—199.
Brown R. F. R., White D E. Ultrastructure of rat lung following inhalation of riem aerosol//Int. J. Exp Pathol.
1997. V. 78. Ns P. 267- 276.
Brussow H., Canchava C., Wolf-Dietrich H. Phages and the evolution of bacterial pathogens: from genomic
rearrangements to lysogenic conversion // Microbiol Molecular. Biol. Rev, 2004. V. 68. № 3. P 560—602.
Bryan L. Mechanisms of plasmid mediated drug resistance // Plasmids and Transposons. N. Y., 1980. P. 51—81.
Buchan В W., McCaffrey R L., Lindemann S. R et al. Identification of migR, a regulatory element of the
Francisella tularensis live vaccine strain iglABCD virulence operon required for normal replication and
tiafficking in macrophages // Infect. Immun. 2009. V. 77. P. 2517—2529.
Buchmeier M. J., Peters C. J., de la Torre C. Arenaviridae: The viruses and their replication // Ed. Kmpe D.
M., Holey P. M / Fields Virology. 2007. P. 1792—1827.
Buckland В. C. The translation of scale in fermentation processes: the impact of computer process cont-
rol // Bio/Tecnology. 1984. V. 2. № 10. P. 875 -883.
Burke G. S. Notes on Bacillus botulinus I I J. Bacteriol. 1919. V. 4. P. 555—565.
Bums J. L., Jonas M., Chi E. Y. et al. Invasion of respiratory epithelial cells by Burkholderia (Pseudomonas)
cepacia // Infect. Immun. 1996. V. 64. P. 4054 4059.
CalisherC. H. Medically important arboviruses of the United States and Canada//Clin. Microbiol. Rev 1994.
V. 7. P. 89-116.
Calisher H. A., Maness K. Virulence of Venezuelan equine encephalomyelitis virus subtypes for various labo-
ratory hosts // Appl. Microbiol. 1974. V 28. P. 881—884.
Callis R. T, Jahrling P.B., DePaolli A. Pathology of Lassa virus infection in the rhesus monkey //Am. J. Trop.
Med. Hyg. 1982. V. 31. № 5. P 1938-1045
Campe H., Zimmermann P., Glos K. etal. Cowpox virus transmission from pet rats to humans Germany //
Emer Infect. Dis. 2009. V 15. № 5. P. 777—780.
Capo C, Lindberg F. P., Mecont S. et al. Subversion of monocyte functions by Coxiella burnetii: impairment
ofthecross-talkbetweenalphaVbeta3integrinandCR3//J.Immunol. 1999.V. !63.№ 11.P.6078 6085.
1078
Список литературы
Caramon Т, Albertini А. М., Galizzi A. I n vivo generation of hybrids between two Bacillus thuringiensis insect-
toxin-encoding genes //Gene. 1991. V. 98. P. 37—44.
CarballeiraN. M., EmilianoA., SostreA. etal. Fatty acid compos tionofbacteria associated with the toxic dino-
flagellate Ostreopsis lenticularis and with Caribbean Palythoa species // Lipids. 1998. V. 33. P 627—632.
Carded C, BrozziA. etal Cowpox virus in liama, Italy// Enter. Infect Dis. 201 l.V. 17. № 8. P. 1513-1515.
Caron E, Peyrard T., Kohler S. et al. Live Brucella spn. fail to induce tumour necrosis factor alpha excretion
upon infection of U937-derived phagocytes // Infect. Immun. 1994. V. 62. P. 5267—5274.
CarterG. B. Biological warfare and biological defence in the United Kingdom 1940-1970 // Royal United
Services Institute Journal. 1993. December, P. 67—74.
CarterG. B. The microbiological research establishment and its precursor at Porton Down: 1940—1979 //ASA
Newsletter. 1991.11 December. P. 91—96.
ChahineM., O’Leary M. E. Regulatory role of voltage-gated Na channel Subunits in sensory neurons//Front.
Pharmacol 2011. V. 2 (dorlO 3389/fphar.2011.00070).
Chan S. Y, Speck R., Ma M. et al. Distinct mechanisms of entry by envelope glycoproteins of Marburg and
Ebola (Zaire) viruses//J. Virol 2000. V. 74 No. 10. P. 4933-4937.
Chapman J. L., Nichols D. K., Martinez M. J. et al. Animal models of Orthopoxvirus infection // Vet. Pathol.
2010. V. 47. № 5. P. 852-870.
Charity J. C., Costante-Hamm M. M., Baton E. L. et al. Twin RNA polymerase-associated proteins control
virulence gene expression in Francisella tularensis // PLoS Pathog 2007.3:e84.
Charles P C., Walters E., Margolis F. etal. Mechanism of neuromvasion of Vfenezuelan equine encephalitis
virus in the mouse // Virology. 1995. V. 208. P. 662—671.
Charrel R. N., de Lamballerie X. Arenaviruses other that Lassa virus//Antiviral. Res. 2003. V. 57. P. 89—100.
ChastelC. Human monkeypox // Pathol. Biol. 2009 V. 57. № 2. P. 175—183.
Chau J , C'ufoh'ni M. A. The chemical synthesis of tetrodoxin: an ongoing quest // Mar. Drugs 2011 V. 9.
P. 2046-2074.
Cheang Tuck-yun, Tang Bing, Xv An-wu et al. Promising plasmid DNA vector based on APTESmodified silica
nanoparticles//Intern. J. of Nanomed. 2012 V. 7. P 1061—1067.
Cheers C., Janas M., Ramsay A. et al. Use of recombinant viruses to deliver cytokines influencing the course
of experimental bacterial infection // Immun. Cell Biol. 1999. V. 77. P 324—330.
Cheers C, Janas M., Ramsay A. et al. Use of recombinant viruses to deliver cytokines influencing the course
of experimental bacterial infection // Immun. Cell Biol. 1999 V. 77. P 324—330.
Chen N., Li G., Liszewski A. et al. Virulence differences between monkeypox virus isolates from West Africa
and the Congo basin // Virology. 2005. V. 340. № 1. P. 46-63.
Chen P. E., Willner К. M., Butani F. et al. Rapid identification of genetic modifications in Bacillus anthracis
using whole genome draft sequences generated by 454 pyrosequencmg // PLoS ONE. 2010. V. 5. Is. 8.
el2397 (doi:10.1371/journal.pone.0012397).
Chen Y, Sued J., Ten over F. C. etal. 0-Iactamase genes of the penicillin-susceptible Bacillus anthracis //J.
Bacteriol. 2003. V. 185. № 3. P. 823-830.
Cheng A. C., Currie B. J. Melioidosis: epidemiology, pathophysiology, and management //Clin. Microbiol.
Rev. 2005. V. 18. № 2. P. 383 416
Chi Ho Lin, Bourque G., Tan P. A comparative synteny map of Burkholderia species links large-scale genome
rearrangements to fine-scale nucleotide variation in prokaryotes // Mol Biol. EVbl. 2008. V. 25. № 3.
P. 549- 558.
Childe L. F. The pathology of plague // Brit. Med J. 1898. V. 2. P. 858-862.
Choi H. S„ Ashitate Y, Lee Rapid J H. et al. Translocation of nanoparticles from the lung airspaces to the body
// Nat Biotechnol. 2010. V 28. № 12.1300-1303.
Chong VuiHeng, Lim Kian Soon, Sharif F. Pancreatic involvement in melioidosis // Jop. J. Pancreas (Online).
2010. V. 11.№ 4. P. 365-368.
Chung-Him Yu V.,Hoi-Fu Yu P., Kin-ChungHoetal. Isolation and identification of anew tetrodotoxin-pro-
ducing bacterial species, Raoultella terrigena, from Hong Kong marine puffer fish Takifugu niphobles /
/Mar. Drugs. 201 l.V. 9 P 2384 2396.
Ciminiello P, Dell’Aversano C., Fattorusso E. et al. Putative palytoxin and its newanalogue, ovatoxin-A in
Ostreopsis ovata collected along the Ligurian coasts dui ing the 2006 toxic outbreak //J. Am. Soc. Mass
Spectrom. 2008. V 19. P. 111-120.
Clemens D. L., Lee B. Y., Horwitz M. A. Francisella tularensis enters macrophages via a novel process involving
pseudopod loops // Infect Immun. 20u5. V. 73. P. 5892—5902.
Список литературы
1079
Cohen J. Is an old virus up to new tricks // Science. 1997. V. 277. Ne 5324. P. 312—313.
Cohen S. N Chang A. C. Y., Hsu L. Non-chromosomal antibiotic resistance in bacteria: genetic transformation
of Escherichia coli by R factor DNAI I Proc. Natl. Acad. Sci. USA 1972. V. 69. P. 2110-2114.
Cole L. A. The specter of biological weapon // Sci. Amer. 1996 (Dec.). V. 275. № 6 P. 60-65.
Colebunders R., Borchert M. Ebola haemorrhagic fever — a review //J. Infect. 2000. V. 40. P. 16—20.
Coll'ns M. D„ East A. K. Phylogeny and taxonomy of the tbod-bome pathogen Clostridium botulinum and
its neurotoxins //J. Appl. Microbiol. 1998. V. 84. P. 5—17.
Corbel M. J. Brucellosis: an overview // Emerg Infect. Dis. 1997. V. 3, № 2. P 213-221.
Cordaux R., Paces-Fessy M., Raimond M. et al. Molecular characterization and evolution of arthropod-patho-
genic Rickettsiellabacteria//Appl. Envirol. Microbiol. 2007. V. 73. № 15. P. 5045—5047.
Covert I., Mathison A., Eskra L. et al. Brucella melitensis, B. neotomae and B. ovis elicit common and
distinctive macrophage defense transcriptional responses // Experiment. Biol. Med. 2009. V 234.
P. 1450-1467.
Craan A. G. Toxicologic aspects of voodoo in Haiti // Biomed. Environ. Sci. 1988. V. 1 № 4. P. 372—381.
Crompton R., GallD. Georgi Markov: death in a pellet // Med. Leg. J. 1980. V. 48. P. 51—62.
Cronin J., Zhang X. Y., Reiser J. Altering the tropism of lentiviral vectors through pseudotyping // Curr. Gene,
fhei 2005 V 5. P. 387-398.
CrovmerB. E., Brunstrom J. E., Racette B. A. Iatrogenic botulism due to therapeutic botulinum toxin a injection
in a pediatric patient // Clin. Neuropharmacol. 2007. V. 30. № 5 P. 310-313.
Czerny С. P„ Eis-Hbbinger A.M., MayrA. etal. Animal poxviruses transmitted from cat to man current event
with lethal end//Zentralbl. Vfeterrinarmed B. 1991. V. 38.№6. P. 421—431.
Daly J. W. Thirty years of discovering arthropod alkaloids in amphibian skin //J. Nat. Prod. 1998. V. 61.
P. 162-172.
Dando M Biological warfare in the 21st century. N. Y, 1994.
Darby S.M., Miller M.L., Allen R.O. Forensic determination of ricin and the alkaloid marker ricinine from
castor bean extracts //J. Forensic. Sci. 2001. V. 46. P. 1033-1042.
DasGupta B. R., Boroff P. A. Separation of toxin and hemagglutinin from cristalline toxin of Clostridium
botulinum type A by anion exchange chromatography and determination of their dimension by
gelfiltration //J. Biol. Chem. 1968. V. 243. P.1065—1070.
Davies T. A., Pankuch G. A., Dewasse В. E et al. In vitro development of resistance to five quinolones and
amoxicillin-clavulanate in Streptococcus pneumoniae // Antimicrob. Agents Chemother. 1999. V. 43.
P 1177-1180.
Davis E. V. The ethnobiology of the Haitian zombi // J. Ethnopharmacol. 1983. V. 9.№ 1. P. 85—104.
DavisJ M A vibrating apparatus for producing drops of uniform size// U S Dept. Agr., Report Et-295 U.S
Bur. Ent. and Plant Quaterly, April, 1951. P. 2.
Davis L. E., Beckham J. D., Tyler L. L. North American encephalitic arboviruses 11 Neurol Clin 2008. V. 26.
№ 3 (727—ix. doi:10.1016/j.ncl.2008.03.012).
Davis P. B., Cooper M. Vectors for airway gene delivery //AAPS J. 2007. V. 9, № 1. P Ell—E17.
Davis R. A. The assassination of Reinhard Heydrich//Surg. Gynecol. Obstet. 1971, P. 304—318.
Davletov B. A., Meunier F., Ashton A. C. et al. Vesicle exocytosis stimulated by a-latrotoxin is mediated by
latrophilin and requires both external and stored Ca2+ // EMBO Jour. 1998. V. 17. № 14. P. 3909—3920.
DawsonC., StubbeifieldEPerrettL. etal. Phenotypic and molecular characterisation ofBrucella isolates from
marine mammals // BMC Microbiology. 2008 V. 8. № 224 (doi:10.1186/1471-2180-8-224).
Day W. C., Berendt R. F. Experimental tularemia in Macaca mulatta: relationship of aerosol particle size to
the infectivity of airborne Pasteurella tularensis // Infect. Immun. 1972. V. 5, № 1. P. 77-82.
Daya S., Berns К. I. Gene therapy using adeno-associated virus vectors // Clin. Microbiol. Rev. 2008. V. 21.
№ 4. P. 583-593.
De Alessi 11 Giornale di Medicina Militare. 1936. № 2.
De Bruin O. M„ Ludu J. S., Nano F. E. The Francisella pathogenicity island protein IglA localizes to the
bacterial c> toplasm and is needed for intracellular growth // BMC Microbiol. 2007. V. 7. Ns 1.
De la Torre J. C. Reverse genetics approaches to combat pathogenic arenaviruses // Antiviral Res. 2008. V. 80,
№ 3. P. 239-250. (doi:10.1016/j.antiviral.2008.08.002).
De Lorenzo A. J. The olfactory neuron and the blood-brain barrier // In: Taste and Smell in Vertebrates
(Wolstenholme G, Knight J, eds). London, 1970. P. 151—176.
De Maagd R. A., Bravo A., Creekmore N. How Bacillus thuringiensis has evolved specific toxins to colonize
the insect world //Trends in Genet. 2001, V. 17 P. 193—199.
1080
Список литературы
De Maagd R. A., Kwa M S. G., van derKlei H. et al. Domain Ill substitution in Bacillus thuringiensis delta-
endotoxin Cryl Ab results in superior toxicity for Spodoptera exigua and altered membrane protein
recognition // Appl. Environ. Microbiol. 1996. V. 62. P. 1537—1543.
Debs R. J., Valley M., Zhu N. Патент 5641662, МПК6, Transfection of lung via aerosolized transgene delivery
I The Regents of the University of California., США; заявл. 10.03.1993, 29022; опуб. 24.06.1997.
Deeds J. R., Handy S., White K. et al. Palytoxin found in Palythoa spp. zoanthids (Anthozoa, Hexacorallia)
sold in the home aquarium trade// PLoS ONE. 2011. V. 6 (e 18235 doi:10.1371/joumal.pone.0018235).
Deeds J. R., Landsberg J H., Etheridge S M et al. Non-traditional vectors for paralytic shellfish poisoning
I I Mar. Drugs. 2008. V. 6. P. 308-348.
DembekZ. Modeling for bioterrorism incidents//Biological weapons defense. N. J., 2005.
Depardieu F., Podglajen I., Leclercq R et al. Modes and modulations of antibiotic resistance gene expression
//Clin. Microbiol. Rev. 2u07 V. 20. № 1. P. 79-114.
Dewan P., Fry A., Laserson K. et al. Inhalational anthrax outbreak among postal workers, Washington, DC,
2001 1/ Emerg. Infect. Dis. 2002. V. 8. P. 1066-1072.
Dhaked R. K., Singh M. К, Singh S. et al. Botulinum toxin: bioweapon & magic drug I I Indian. J. Med. Res.
2010. V. 132. № 5. P. 489-503.
Dickie W. M. Plague in California 1900—192511 Conf. State and Prov. Health Authorities North America.
1926. V. 21. P. 30—78.
DimmockN.A. Production of uniform droplets//Nature. 1951. V. 166. P. 686—687.
Dinges M. M., Orwin P. M., Schlievert P. M. Exotoxins of Staphylococcus aureus // Clinical. Microbiol. Rev.
2000. V. 13. №l.P. 16-34.
Dixon R. W., Harris J. В Nerve terminal damage by p-bungarotoxin its clinical significance // Am. J. Pathol.
1999. V. 154. P. 447-455.
Donaldson K., Tran T. Inflammation caused by particles and fibers // Inhal. Toxicol. 2002. V. 14. P. 5—27.
Douglass hl, Dumbell K. Independent evolution of monkeypox and Variola viruses//!. Virol. 1992. V 66. № 12
P. 7565-7567.
Dreiseikelmann B. Translocation of DNA across bacterial membranes 11 Microbiol. Rev. 1994. V. 58. № 3.
P. 293-316.
DrinkerC. K., Hardenber E. Absorption from the pulmonary alveoli//J. Exp. Med. — 1947. V. 86. P. 7—18.
DruettH 4., Henderson D W Packman L. etal. Studies on respiratory infection '.The influence of particle
size on respiratory infection with anthrax spores I I J. Hyg. Camb. 1953. V 54. P. 49—57.
DruettH A., Henderson D. W., Packman L. etal. Studies on respiratory infection: I. The influence of aerosol
particle size on infection gunea-pig with Pasturella pestis//J. Hyg. 1956a. V. 54. P 37—48.
Druett H. A., Henderson D. W., Peacock S. Studies on respiratory infection: III. Experiments with Brucella
suis//J. Hyg. Camb. 1953b. V. 54. P. 359-371.
Druett H. A., Honderson D. W., Packman L. et al. Studies on respiratory infection. I. The intluence of particle
size on respiratory infection with anthrax spores // J. Hyg. 1953. V. 51. P. 359—371.
DruettH. A., May K. R. Unstable germicidal pollutant in rural air // Nature. 1968 V. 220. № 395. P. 395—396.
Duan Z. G., Yan X J„ He X. Z. et al. Extraction and protein component analysis of venom from the dissected
venom .-lands of Latrodectus tredecimguttatus // Comp. Biochem. Physiol. B. Biochem. Mol Biol. 2006.
V 145. P. 350- 357.
Dufton M. J., Hider R. C. Conformational properties of the neurotoxins and cytotoxins isolated from Elapid
snake venom//CRC. Crit. Rev. Biochem. 1983. V. 14.P. 113—171.
Edwards T., Phan L, Abendroth J. et al. Structure of a Burkholderia pseudomallei Trimeric Autotransporter
Adhesin Head// PLoS ONE. 2010. V. 5, № 9 (el2803. doi: 10.137l/journal.pone.0012803).
Ehichioya D. U., Hass M., Becker-Ziaja Current B. et al. Molecular epidemiology of Lassa virus in Nigeria //
J. Clin. Microbiol. 2011. V. 49. № 3. P. 1157-1161.
Eichner M., Dietz K. Transmission potential of smallpox: estimates based on detailed data from an outbreak
//Amer. J. Epidemiol 2003.V 158. № 2. P. 110-117.
Eigelsbach H. T., Braun W., Herring R. Studies on the variation of Bacterium tularense I IJ. Bacterio!. 1951.
V. 61. P. 557-569.
Eigelsbach H. T., Hornick R. B., Tulis J. J. Recent studies on live tularemia vaccine 11 Med Ann Dist.
Columbia. 1967. V. 36. P. 282-286.
Elkins K.L., Rhinehart-Jones T. R., Culkin S. J. etal. Minimal requirements for murine resistance to infection
with Francisella tularensis LVS // Infect. Immun. 1996. V. 64. P. 3288—3293.
Ellis J., Oyston P., Green M. etal. Tularemia // Clin. Microbiol. Rev. 2002. V. 15, № 4. P. 631—646.
Список литературы
1081
Engelthaler D. М., Bowers J., SchuppJ.A. etal. Molecular Investigations ofa Locally Acquired Case of Melioi-
dosis in Southern AZ, USA // PLoS Negl Trop Dis. 2011. V. 5, № 10 (el347. doi:10 1371/journal.
pntd.0001347).
Fail A., Ghose A., Ahsan F etal. The greater black krait (Bungarusniger), a newly recognized cause of neuro-
myotoxic snake bite envenoming in Bangladesh//Brain. 2010. V. 133. № 11. P. 3181—3193.
Federici T, Kutner R., Zhang X. Y. et al. Comparative analysis of HIV- 1-based lentiviral vectors bearing
lyssavirus glycoproteins tor neuronal gene transfer 11 Genet. Vaccin. Ther. 2009. V 7. № 1.
FeignerP. L., Gadek T. R., Holm M. etal. Lipofection: a mghly efficient, lipid-mediated DNA-transfection
procedure // Proc Natl. Acad. Sci. USA. 1987. V 84. P. 7413—7417.
Fenner E, Henderson D. A., Arita I. et al. Smallpox and its eradication. World Health Organization, Geneva,
1988.
Fernandez-Ortega J. F., Morales-de los Santos J. M., Herrera-Gutierrez M. E. et al. Seafood intoxication by
tetrodotoxin: first case in Europe //J Emerg. Med. 2010. V. 39. № 5. P 612—617.
Ferraz A. C., Anselmo-Franci J A., Perosa S. R. et al. Amino acid and monoamine alterations in the cerebral
cortex and hippocampus of mice submitted to ricinine-induced seizures I I Pharmacol. Biochem. Behav.
2002. V. 72. P. 779-786.
Fichet-Cabet E., Rogers D. J. Risk maps of Lassa fever in West Africa I I PLoS Negl. Trop Dis 2009. V. 3. № 3
(e388. doi: 10.1371 /journal.pntd.0000388)
Ficht T. Brucella taxonomy and evolution 11 Future Microbiol. 2010. V. 5, № 6. P. 859—866.
FilonovA. E., Manzeniuk I. N., Svetoch E. A. Conjugative transfer and expression of R plasmids of the genus
Pseudomonas in the cells of Pseudomonas mallei C-5//Antibiot. Khimioter. 1996. V 41, No 3 P. 20—24.
Findetsen W. Ubei das absetzen klemer, in der luft suspendierter teilchen uder menschlichen lunge bei der
atmung I I Arch. Ges. Physiol. 1935. Bd. 236. S. 367.
Finegola M. J Pneumonic plague in monkeys an electron microscopic study 11 Amer. J. Pathol 1969. V. 54.
№ 2 P 167-185.
Finegold M. J Interstitial pulmonary edema. An electron microscopic study of the pathology of staphylococcal
enterotoxemia in Rhesus monkeys //Lab. Invest 1967. V. 16. № 6. P. 912—924.
Finegold M., PeteryJ., Berendt R. et al. Studies on the pathogenesis of plague I I Am. J. Pathol. 1968. V. 53.
№ 1. P. 99-114.
Finlay B, Falkow S. Common themes in microbial pathogenicity 11 Microbiol. Rev. 1997. V 53. № 2.
P 210-230.
Finn P. Agitation-aeration in the laboratory and in industry // Bact. Rev. 1954. V. 18. № 4. P. 254—27^
Fiordalisi J. J., James P. L., Zhang Y. P. et al. Facile production of native-like к-bungarotoxin in yeast - an
enhanced system for the production of neuronal nicotinic acetylcholine receptor probe // Toxicon. 1996.
V. 34. P. 213-224.
FirovedA. M, Miller G. E, Moayeri M. et al. Bacillus anthracis edema toxin causes extensive tissue lesions and
rapid lethality in mice //Am J. Pathol. 2005 V. 167. P. 1309—1320.
Fischera M. G., Allenb M. L., Wilson W. H. et al Giant virus v'ith a remarkable complement of genes infects
marine zooplankton // Proc Natl. Acad. Sci. USA. 2010. V. 107, № 45. P. 19508—19513.
Fleming J. J., Du Bois J. A synthesis of (+)-saxitoxin //J. Am. Chem. So. 2006. V. 128 № 12. P. 3926—3927.
Fluit A. C. Jones M. E., Schmitz F. et al. Bacteremia in European hospitals, incidence and antimicrobial
susceptibility//Clin. Infect Dis. 2000. V. 30 P. 454—460.
Fluit A. C., Visser M. R., Schmitz F. Molecular detection of antimicrobial resistance I I Clin. Microbiol. Rev.
2001 V. 14. №4. P 836-871.
Fodstab О , Kvalheim G., GodalA. et al. Phase I study of the plant protein ricin I I Cancer Res 1984. V. 44.
P. 862-865.
Fontanges R., Founier J Construction of a generator for dry aerosols // Travaux Scientifiques, Annee 1971,
Centre de Recherches du Service de Sante des Armees. P 301—302.
Fort Detrick converting to major medical research center//Arm. Res. Devel. 1973. V. 14.№4.P. 16—18.
Foster J., Beckstrom-Sternberg S., Pearson T. et al. Whole-genome-based phylogeny and divergence of the
genus Brucella//J. Bacterol. 2009. V. 191. № 8. P. 2864—2870.
Franz D. R., JahringP. B., Friedlande’ A. M. etal. Clinical recognition and management of patients exposed
to biological warfare agents I I JAMA. 1997. V. 278. № 5. P. 399—411.
Franz D., Jaax N Ricin toxin // Medical aspects of Chemical and Biological Warfare I Ed. F. R. Sidell,
E. T. Tafuqi, D R. Franz. Washington, 1997 P. 631—642.
Fraser R., Dombrowski N., Rontley J. // S. Chem. Eng Sci. 1963. V. 18. № 315. P. 323.
1082
Список литературы
Frecha С., Szecsi J, Cosset F. L et al. Strategies for targeting lentivnal vectors 11 Curr. Gene I her. 2008. V. 8.
P 449-460.
Freedman A., Afonia O., Chang M. W. et al. Cutaneous anthrax associated with microangiopathic hemolytic
anemia and coagulopathy in a 7-month-old infant // JAMA, 2002. V 287. P. 869—874.
Frohlich E., Roblega E. Models for oral uptake of nanoparticles in consumer products '/ Toxicology. 2012.
V 291 P. 10- 17.
Fuhrman F. A. Tetrodotoxin, tarichatoxin, and chiriquitoxin: historical perspectives //Ann. N. Y. Acad Sci.
1986.V.479. P. 1-14.
Galimand M., Camiel E., Courvalin P. Resistance of Yersinia pestis to antimicrobial agents 11 Antimicrob.
Agents Chemother. 2006. V 50. Ns 10. P.3233—3236.
Gallis R. T., Jahrling P. B., dePaoli A. Pathology of Lassa virus infection in the rhesus monkey 11 Am. J. Trop.
Med. Hyg. 1982. V. 31. Ne 5. P. 1038-1045.
Gammon D. B., Snoeck R , Fiten R. P. et al. Mechanism of antiviral drug resistance of vaccinia virus:
identification of residues in the viral DNA polymerase conferring differential resistance to antipoxvirus
drugs//J. Virol. 2008. V. 82. № 24 P 12520-12534.
Garcia C., Lagos M., Truan D. et al. Human intoxication with paralytic shellfish toxins: clinical parameters
and toxin analysis in plasma and urine I I Biol. Res. 2U05. V 38. P 197—205.
Garcillian-Bracia M. P., de-la Crus F. Distribution of IS91 family insertion sequences in bacter.al genomes:
evolutionary implications // FEMS Microbiol. Ecol. 2002. V. 42. № 2. P. 303—313.
Geffeney S. L. Ruben P. C. The structural basis and functional consequences of interactions between
tetrodotoxin and voltage-gated sodium channels I I Mar. Drugs. 2006. V. 4. P. 143—156.
Geisbert T. W, Daddario- DiCaprio К. M., Geisbert J. B. etal. Marburg virus Angola infection of Rhesus maca-
ques: pathogenesis and treatment with pecombinant nematode anticoagulant protein c2 // J. Infect. Dis.
2007. V. 196. P. 372-381.
Geisbert T. W., Pushko P, Anderson K. etal. Evaluation in nonhuman primates ofvaccines against Ebola virus //
Emerg. Infect. Dis. 2002. V. 8. № 5 P. 503- 507.
Georges-Courbot M. C., Sanchez A., Lu Ch. Y. et al. Isolation and phylogenetic characterization of Ebola viruses
causing different outbreaks in Gabon // Emerg. Infect. Dis. 1997. V. 3. № 1. P. 59—62.
Gerhard F. Brucella suis in aerated broth culture. III. Continuous culture studies I I J. Bacteriol. 1946. V. 52.
№ 3. P. 283-292.
Ghigo E. Kartenbeck J., Lien P. et al. Ameobal pathogen mimivirus infects macrophages through phagocytosis
11 PLoS Pathogens. 2008. V. 4. № 6. P. 1-17.
GibneyK., ChengA. C., Currie B. Cutaneous melioidosis in the tropical top end ofAustralia: aprospective study
and review of the literature // Clin. Infec. Dis. 2008. V. 47 P. 603 609.
Gitgis H„ HottesA., Tavazoie S. Genetic architecture of intrinsic antibiotic susceptibility // PI oS ONE. 2009.
V. 4, Is. 5. P. 5629.
Godoy D., Randle G., Simpson A. J et al Multilocus sequence typing and evolutionary relationships among
the causative agents of melioidosis and glanders, Burkholderia pseudomallei and Burkholderia mallei //
J. Clin. Microbiol. 2003. V. 41. P. 2068-2079.
Goff A. J, Chapman J., Foster C. et al. A novel respiratory model of infection with monkeypox virus in
Cynomolgus macaques//J. Virol. 2011. V. 85 № 10. P. 4898—4909.
Goh E B., Yim G, Tsui Ж et al Transcriptional modulation of bacterial gene expression by subinhibitory
concentrations of antibiotics 11 Proc. Natl. Acad. Sci. USA. 2002 V. 99. P. 17025-17030.
GoldbergM., Langer R , JiaX. Nanostructured material' for applications in drug delivery and tissue engineering
//J. Biomater. Sci. Polym. 2007 V. 18. Ns 3. P. 241—268.
Goncalves D. M., de Liz R-, Girard D. Activation of neutrophils by nanoparticles // The Sci. World J. 2011 V 11.
P. 1877-1885.
Gondor J. C., Kishimoto R A., Kastello M. D. et al. Gynomolgus monkey model for experimental Q fever
infection //J Infect. Dis. 1979. V. 139. № 2 P. 191—196.
Goodlow R. J., Leonard F. A. Viability and infectivity of microorganisms in experimental airborne infection
I/ Bact. Reviews. 1961. V. 25. Ne 3. P. 182-187.
GormanS, Viseshakul N., CoI.enB. et al. A recombinant adenovirus that directs secretion of biologically active
к-bungarotoxin from mammalian cells // Molecular Brain Research. 1997. V. 44, Ns 1. P. 143—146.
Gould E A, Buckley A., GroegerB. K. etal. Immune enhancement of yellow fevervirus neurovirulence formice:
studies of mechanisms involved //J Gen. Virol. 1987 V. 68. Ne 3. P. 105- 120
Green T. M., Eigelsbach H. T Immunity in tularemia: report of two cases of proved reinfection//Arch. I nt
Med. 1950. V 85. P. 777.
Список литературы
1083
Greene I. Р, Pat ssler S„ Austgen L. el al. Envelope glycoprotein mutations mediate equine amplification and
virulence of epizootic \fenezuelan equine encephalitis virus // J. Virol. 2005. V. 79. № 14. P. 9128-9133.
GreeneC. M., ReeJhuisJ., Christina Tan C. etal. Epidemiologic investigations of bioterrorism-related anthrax,
New Jersey, 2001 // Emerg. Infect. Dis 2002. V 8. № 10 P 1048—1050
Griffith K. S., Mead P, Armstrong G. L.et al. Bioterrorism-related inhalational anthrax in an elderly woman,
Connecticut, 2001 // Emerg. Infect. Dis. 2003. V. 9. № 6. P. 681—688.
Griffiths G. D., Phillips G. J., Holley J. Holley inhalation toxicology of ricin preparations: animal models,
prophylactic and therapeutic approaches to protection I I Inhal. Toxicol 2007. V 19. P. 873—887
Guamei J., Jemigan J. A., Wun-Ju Shieh etal. Pathology and pathogi nesis of bioterrorism-related inhalational
anthrax//Amer. J. Pathol. 2003. V. 163. № 2. P. 701-709.
GuamerJ., Zaki S. R. Histopathology and immunohistochemistry in the diagnosis of bioterrorism fgents //
J. Histochem Cytochem. 2006. V. 54. № 1. P. 3—11.
GubserG., HueS.. Kellam P. etal. Poxvirus genomes: a phylogenetic analysis//J. Gen. Virol. 2004. V. 85.
P. 105-117.
Guitlemin J Scientists and the history of biological weapons, a brief historical overview of the development
of biological weapons in the twentieth century // EMBO Rep 2006. V. 7. S45—S49.
Guillemin J. The 1979 anthrax epidemic in the USSR: applied science and political controversy // Proceeding
of the American Philosophical Society. 2002. V. 146. № 1. P. 19—36.
GuiyouleA., GerbaundG., BuchrieserC. etal. Transferable plasmid-mediated resistance of Yersinia pestis //
Emerg. Infect. Dis. 2001. V. 7. P. 43—48.
Gyles C. L. Shiga toxin-producing Escherichia coli: An overview // J Anim. Sc>. 2007. V 85. P. 45—62.
Hacein-Bey -Abina S., GarngujA., WangC. P. etal. Insertional oncogenesis in 4 patients after retrovirus-
mediated gene therapy of SCID-X1 //J. Clin. Invest. 2008. V. 118. P. 3132—3142.
Hacein-Bey-Abina S., Hauer J., Lim A et al Efficacy of gene therapy for X-linked severe combined
immunodeficiency // N. Engl. J. Med. — 2010. — V 363. № 4. P 355—364.
Hacein-Bey-Abina S., le Deist F, Carliei F et al. Sustained correction of X-linked severe combined immuno-
deficiency by ex vivo gene therapy // N. Engl. J. Med. 2002. V. 346. P. 1185— 1193.
Hacein-Bey-Abina S., von Kalle C., Schmidt M. et al. A serious adverse event after successful gene therapy for
X-linked severe combined immunodeficiency I IN. Engl. J. Med. 2003. V. 348. № 3. P. 255—256.
Hacein-Bey-Abina S., Von Kalle C., Schmidt M. etal. LMO2-associated clonal T-ce’J proliferation in two
patients after gene therapy for SCIDX1 [see comment] // Science. 2003. V. 302. P. 415—419.
Hackstadt T., Peacock M. G., Hitchcock P. J etal. Lipopolysaccharide variation in Coxiella burnettii: intrastrain
heterogeneity m structure and antigenicity I I Infect. Immun. 1985. V. 48. № 2. P. 359- 365.
Haensch S., Bianucci R., Signoli M. et al. Distinct clones of Yersinia pestis caused the Black Death // PLoS
Pathog. 2010. V 6. № 10. P. elOOl 134. doi 10.1371/joumal.ppat 1001134.
Hahn C S., LustigS., Strauss E. G. etal. Western equine encephalitis virus is a recombinant virus // Proc. Natl.
Acad. Sci USA. 1988. V. 85. P. 599’’ 6001.
Halama A., Kulinski M , Librowski T. et al. Polymer-based non-viral gene delivery as a concept for the
treatment of cancer // Pharm. Rep. 2009. V. 61. P. 993-999
Halfmann P., Kim Jin Hyun, Ebihara H. et al. Generation of biologically' contained Ebola viruses // Proc. N atl.
Acad. Sci USA. 2008. V. 105. №4. P. 1129-1133.
HallingS. M., DetilluxF. G., Tatum F. M. etal. Deletion ofthe BCSP31 gene ofBrucella abortus by replacement
//Infect Immun. 199LV.59.№ 11. P. 3863-3868.
Halpern J., Neale E. Neurospecific binding internalization and retrograde axonal transport // Cure Top.
Microbiol. Immunol. 1995. V. 195. Ni 1. P. 221-241.
Halstead S. B., Rojanasuphot S., Sangkawibha N. Original antigenic sin in dengue // Am. J Trop. Med. Hyg.
1983 V. 32. №1 P 154-156.
Hanifin С T. The chemical and evolutionary ecology of tetrodotoxin (TTX) toxicity in terrestrial vertebrates
I I Mar. Drugs. 2010. V. 8. P. 577-593.
Haolin N. 1., Yun N., Zacks M. A. el al. Recombinant alphaviruses are safe and useful serological diagnostic
toolls 11 Am. J. Trop. Med. Hyg. 2007. V. 76. № 4. P. 774—781.
Hara T. J. Gustatory detection of tetrodotoxin and saxitoxin, and its competitive inhibition by quinine and
strychnine in freshwater fishes // Mar. Drugs. 2011. V 9. P. 2283—2290.
Harty R No exit: targeting the budding process to inhibit filovirus replication // Antiviral. Res. 2009. V. 81.
№3. P. 189-197.
Hartzell J., Wood-Morris R., Martinesz L. et al. Q fever: epidemiology, diagnosis, and treatment I I Mayo Clin.
Proc. 2008. V. 83. № 5. P. 574-579.
1084
Список литературы
Harvie Р., Wong F. М, Bally М. В. Use of poly(ethylene glycol)—lipid conjugates to regulate the surface
attributes and transfection activity of lipid—DNA particles // J. Pharm Sci. 2000. V. 89. P. 652—663.
Hatheway C. L. Toxigenic Clostridia // Clinical. Microbiol. Rev. 199U V. 3. № 1. P. 66—98.
HauriA. M., Hofstetter I., SeiboldE. etal. Investigating an airborne tularemia outbreak, Germany// Emerg.
Infect. Dis. 2010. V. 16. № 2. P. 238 -243.
HauseslerS., Nimtz M., Domke T. et al Purification and characterisation of a cytotoxic exolipid of Burkholderia
pseudomallu// Infect. Immunol. 1998. V. 66. P 1588—1593.
Hawkey P. W. The origins and molecular basis of antibiotic resistance // BMJ. 1998. V. 317. P. 557-560.
Health aspects of chemical and biological weapons. Report of a WHO group of consultants, WHO, Geneva,
1970
Hearings before the select committee to study governmental operations with respect to intelligence activities
of the United States Senate Ninety-fourth congress, 1st session Volume 1. Unaut horized storage of toxic
agents. Sept. 16,17,18; 1975. Washington, U. S. Government Printing Office, 1975.
Hearings before the subcommittee on health and scientific research of the committee on human resources of
the United States Senate. Ninety-fifth congress 1 st session on examination of serious deficiencies in the
defense department efforts to protect the human subjects. March 8, May 23/1977. Washington, U S.
Government Printing Office, 197? P. 5—6.
Heath E, Harla P., Alexander C. Varying polimer architecture to deliver drugs // AAPS J. 2007. V. 9. № 2.
P. F235-E240.
Henchai E. A., McCown J. M., Burke D. S. et al. Epitopic analysis of antigenic determinants on the surface of
dengue 2 virions using monoclonal antibodies 11 Am. J. Trop. Med. Hyg 1985. V. 34. P. 164—169.
Hendrickson R. C., Wang C, Hatcher E. L. et al. Orthopoxvirus genome evolution: the role of gene loss 11
Viruses. 2010. V. 2. P. 1933-1967.
Henkel J. S, Baldwin M. R, Barbieri J. T. Toxins from bacteria //EXS. 2010. № 100. P. 1—29.
HenningS. J, Ledley F. D. Патент США 5821235, МПК6 A61K 31/704; A61M 5/14; C12N 15/09. Gene
therapy using the intestine / заявитель Baylor College of Medicine, США; заявл. 20.01.1995, 376473;
опубл. 13.10 1998
Herrero В. A., EcklungA. E., StreettC S. etal. Experimental botulism in monkeys — a clinical pathological
study I I Exp. Mol. Pathol. 1967. V. 6. № 1. P. 84—95.
Hersh S. Chemical and biological warfare: America’s hidden arsenal. I ondon, 1968.
Hicks K. W., Sweeney D. A., CuiX. et al. An overview of anthrax infection including the recently identified form
of disease in injection drug users // Intensive Care Med. 2012 V. 38. № 7. P. 1092—1104.
HidyJ. M. Generation of particulate clouds // In: Aerosole. An industrial and environmental science I Higy
G. M„ N. Y, 1984. Ch. 4. P 119-162.
Hill К. K., Xie C., Foley В. T. et al. Recombination and insertion events involving the botulinum neurotoxin
complex genes in Clostridium botulinum types А, В, E and F and Clostr.dium butyncum type E strains
I I BMC Biol. 2009. V. 7 (doi: 10.1186/1741-7007-7-66).
Hirst L. F. The conquest of plague. A study of the evolution of epidemiology. Oxford, 1953.
Hoel P, Bnske-Hohlfeld I., Salata O. Nanoparticles — known and unknown health risks 11 J. Nanobiotech.
2004. V. 2 (http://wwvjnanobiotechnology.com/content/2/1/12).
Hoffmann K., Hermanns-Clausen M., Buhl C. et al. A case of palytoxin poisoning due to contact with zoan-
thid corals through a skin injury// lexicon. 2008. V. 51. № 8. P. 1535—1537.
Hoffmaster A. R., Fitzgerald С. C. et al. Molecular subtyping of Bacillus anthracis and the 2001 bioterrorism-
associated anthrax outbreak, United States//Emerg Infect. Dis. 2002. V 8. № 10. P 1011-1016.
Hoffmaster A R., Ravel J., Rasko D. et al. Identification of anthrax toxin genes in a Bacillus cereus assoc^a
ted with an illness resembling inhalation anthrax 11 Proc. Natl. Acad. Sci. USA. 2004. V. 101. № 22.
P. 8449-8454.
Holland S., Spindler K., Horodlevski Г et al. Rapid evolution of RNA genomes // Science. 1982. V 215.
P. 1577-1585.
Holmberg S D., Blake P. A. Staphylococcal food poisoning in the United States //JAMA. 1984. V. 251. №4.
P. 487-489.
Holser E. Botulism durch ingalation 11 Med. Klin. 1962. V. 57. P. 1735—1738.
Holtz T. H., AckelsbergJ., Kool J. L.et «1. Isolated case of bioterroi ism-related inhalational anthrax, New York
City, 2001 /I Emerg. Infect. Dis. 2003. V. 9. № 6. P. 689-695.
Homaira N., Rahman M., Luby S. P. etal Multiple outbreaks of puffer fish intoxication in Bangladesh 2008
//Am. J. Trop. Med. Hyg. 2010. V. 83. № 2 P. 440—444.
Список литературы
10Я5
Hornstra Н., Pearson Т, Georgia S. et al. Molecular epidemiology of glanders, Pakistan // Emerg. Infect. Dis.
2009. V. 15. № 12. P. 2036-2039
Hsu К P, Lukacs S. L., Handzel T. et al. Opening a Bacillus anthracis containing envelope, Capitol Hill,
Washington, D.C.: The public health response // Emerg. Infect. Dis. 2002 V 8. № 10 P. 1039—1043.
Hu Y, TangG., Liu J etal. FGF Receptor-Mediated Gene Delivery Using Ligands Coupled to PEI-P-CyD
//J. of Biomed. and Biotech. 2012. (doi:10.1155/2012/989235).
Hughes A. L., Irausquin S., Friedman R. The evolutionary biology of poxviruses /1 Infect. Genet. Evol. 2010.
V. 10. № 1 (50. dorl0.1016/j.meegid.2009.10.001).
Hugh-Jones M. Wickhan Steed and german biological warfare research I I Intelligence and National Security.
1992 V. 7. № 4. P. 379-402.
HuijsmansC., SchellekensJ., WeverP. etal. Single-nucleotide-polymorphism genotyping of Coxiella burr.etu
during a Q fever outbreak in the Netherlands// Appl. Environ. Microbiol 2011 V.77.№6 P.2051—2057.
Huis D, Wuthiekanun V., Cheng A. et al. Short report: the role and significance of sputum cultures in the diag-
nosis of melioidosis // Am. J. Trop. Med. Hyg. 2005. V. 73. № 4. P. 657-661.
Hunt R. Ricin. Washington, DC’ American University Experiment Station; 1918 P. 107— 117.
HuxsollD. L., Patrick W.C., ParrottC. D. Veterinary services in biological disasters//J. Am. Vet. Med. Assoc.
1987. Mil, 190. № 6. P 714-722.
Ikaheimo Sytjala H., Karhukorpi J. etal. In vitro antibiotic susceptibility of Francisella tularensis isolated
from numan and animal //J. Antimicrob. Chemother. 2000. V. 46. P. 287 200
Inder M V., Nikunj S. Gene therapy promises, problems and prospects // Nature. 1997. V. 389. P. 239—242.
Inglesby T., Dennis D, Henderson D. et al. Plague as a biological weapon: medical and public health mana-
gement //JAMA. 2000. V. 283. № 17. P. 2281-2290.
Inglis T. J. J., Garrow S. C., Henderson M. et al Burkholderia pseudomallei traced to water treatment plant in
Australia // Emerg. Infect. Dis. 2000. V. 6. P. 56—59.
Inglis T, Rigby T., Robertson T. et al. Interaction between Burkholderia pseudomallei and Acanthamoeba
species results in coiling phagocytosis, endamebic bacterial survival, and escape // Infect. Lmmunity. 2000.
V. 68. № 3. P. 1681- 1686.
Inglis T, Sagripanti J. Environmental factors that affect the survival and persistence of Burkholderia pseudo-
mallei I/ Appl. Environmen. Microbiol. 2006. V. 72. № 11. P. 6865—6875.
IshiguroM., Takahashi T., FunatsuG. etal. Biochemical studies on ricin. I. Purification of ricin//J. Biochem.
(Tokyo). 1966. V. 55. P. 587-592.
Ito E., Yasumoto T. Toxicological studies on palytoxin and ostreocin- D administered to mice by three different
routes /I Toxicon. 2009. V. 54. P. 244—251.
Ivins В. E., Welkos S. L. Cloning and expression of the Bacillus anthracis protective antigen gene in Bacillus
subtilis 11 Infect. Immun. 1986, V. 54. № 2. P. 537—542.
Ivins В E., Welkos S. L., Knudson G. B. et al. Immunization against anthrax with aromatic compound-
dependent (Aro-) mutants of Bacillus anthracis and with recombinant strains of Bacillus subtilis that
produce anthrax protective antigen I I Infect. Immun. 1990. V. 58. № 2. P. 303-308.
Ivins В. E., Welkos S. L , Knudson G. B. et al Transposon Tn916 mutagenesis in Bacillus anthracis I I Infect.
Immun. 1988. V. 56. № l.P. 176-181.
lackson A. C., Sengupta S. K., Smith J. F. Pathogenesis of Venezuelan equine encephalitis virus infection in
mice and hamsters//Vet. Pathol. 1991. V 28. P. 410—418.
Jackson P. J., Ramsay A. J., Christensen C. D. et al Expression of mouse mterleukin-4 by a recomb nant
ectromeha virus suppresses cytolytic lymphocyte responses and overcomes genetic resistance to mousepox
//J. Virol. 2001. V75. № 3. P. 1205-1210.
Jackson P. J., Walthers E. A., KalifA. S. et al. Characterization of the variable-number tandem repeats in vrrA
from different Bacillus anthracis isolates//Appl. Environ. Microbiol 1997. V. 63 P. 1400 -1405.
Jager C., Willems H, Thiele D. et al. Molecular characterization of Coxiella burnetii isolates // Epidemiol.
Infect. 1998. V. 120. P. 157-164.
Jahrling P., Hensley L., Martinez M. Leduc J. W. et al. Exploring the potential of variola virus infection of
cynomolgus macaques as a model for human smallpox // Proc. Natl. Acad. Sci. USA. 2004. V. 101.
№42. P. 15196-15200.
JangJun-Ho, Lee Jong-Soo, Yotsu-Yamashita M. LC/MS Analysis of tetrodotoxin and Its deoxy analogs in
the marine puffer fish Fugu niphobles from the southern coast of Korea, and in the Brackish water puffer
fishes Tetraodon nigroviridis and Tetraodon biocellatus from Southeast Asia 11 Mar. Drugs. 2010. V. 8.
P. 1049-1058.
1086
Список литературы
Jernigaп D В., RaghunathanP L., Bell В. Р. etal. Investigation ofbioterrorism-related anthrax. United States.
2001: epidemiologic findings // Emerg. Infect. Dis. 2002. V. 8. P. 1019- 1028.
Jerntgan J. A., Stephens D. S., Ashford D. A. et al. Biotei rorism-relatod inhalational anthrax: the first 10 cases
reported in United States I I Emerg. Infect. Dis. 2001. V 7.933—942.
Job. H., Lee T, Kim J. H. Nanotechnology and nanotoxicology in retinopathy//I nt. J. Mol. Sci. 2011. V 12.
P. 8288-8301.
Johansson A., Farlow J , I arsson P. et al. Worldwide genetic relationships among Francisella tularensis isolates
determined by multiple-locus variable-number tandem repeat analysis//J. Bacteriol. 2004 V 186.№ 17.
P. 5808-5818.
Johnson E„ Jaax N., White J. et al. Lethal experimental infections of rhesus monkeys by aerosonzed Ebola
virus /1 Int. J Exp. Path. 1995. V. 76. P. 227-236.
Johnson J. E., Kadull P. J. Laboratory acquired Q fever// Am. J. Med. 1966. V. 41. № 3. P. 391—404.
Jones A. L., Beveridge T. J., Woods b. E. IntracellularsurvivalofBurkholderiapseudomallei//Infect. Immun.
1996 V. 64. P. 782-790.
Jones L. D., Bennett A. M., Moss S. R et al. Cytotoxic T -cell activity is not detectable in Venezuelan equine
encephalitis virus-infected mice II Virus Res. 2003. V 91. № 2. P. 255—259.
Jones M. E., Goguen J., Critchley I. A. et al. Antibiotic susceptibility of isolates of Bacillus anthracis, a bacterial
pathogen with a potential to be used in biowarfare // Clin. Microbiol. Infect. 2003. V. 9. P 984—986.
Jose J., Snyder J. E., Kuhn R. J. A structural and functional perspective ol alphavirus replication and assembly
11 Future Microbiol. 2009. V. 4 (837—856. doi:10.2217/fmb.09.59).
Joubert F.J. Snake venom toxins, the amino acid sequences of two toxins from Ophiophagus Hannah //
Biochem. Biophys. Acta. 1973. V. 317. P. 85—98.
Karako'tsts P. C., Trucksis M., DumlerJ. S. Chronic Q tever tn the United States // J. Clin. Microbiol. 2006.
V. 44. № 6. P. 2283-2287.
Karem К L., Reynolds M., Hughes C. et al. Monkeypox-mduced immunity and failure of childhood smallpox
vaccination to provide complete protection // Clin. Vac. Immunol. 2007. V. 14. P. 1318—1327.
Karyszkowski L. Ц Lekarz Wojskowy. 1935. № 12.
Katikou P. Chemistry of palytoxins and ostreocins 11 In Phycotoxins, Chemistry and Biochemistry; Bota-
na L.M., Ed.; Blackwell Publishing: Ames, IA, USA. 2007. P. 75—93.
Katikou P. Palytoxin and analogs ccobiology and origin, chemistry, metabolism, and chemical analysis // In
Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection, 2nd ed.; Botana, L.M., Ed.;
CRC Press/Taylor & Francis Group: Boca Raton, FL, USA. 2008c P. 631—663.
Keim P., Gruendike J. M., KlevytskaA. M. et al. The genome and variation of Bacillus anthracis //Mol. Aspects.
Med. 2009. V. 30. № 6. P. 397- -405.
Keim P, Johansson A., Wagner D M. Molecular epidemiology, evolution, and ecology of Francisella // Ann.
N. Y. Acad. Sci. 2007. V. 1105. P. 30-66
Keim P., KalifA, Schupp J. etal. Molecular evolution and diversity in Bacillus anthracis as detected by amplified
fragment length polymorphism markers I I J. Bacteriol. 1997. V. 179. P. 818—824.
Keim P., Price L. B., Klevytska A. M. et al. Multiple-locus variable-number tandem repeat analysis reveals
genetic relationships within Bacillus anthracis // J. Bacteriol. 2000. V. 182. P 2928—2936.
Kellmann R., Mihali T. K, Jeon Y. J. e; al. Biosynthetic intermediate analysis and functional homology reveal
a saxitoxin gene cluster in Cyanobacteria I I \ppl Environ. Microbiol. 2008. V. 74. № 13. P. 4044—4053.
Kenny b., Russell R., Rrogers D. etal. In vitro susceptibilities of Burkholderia mallei in comparison to
those of other pathogenic Burkholderia spp. I I Antimicrob. Agents and Chemother. 1999. V. 43. №11.
P. 2773-2775.
Kerbrat A. S., Amzil Z., PawlowiezR. etal. First evidence of palytoxin and 42-hydroxy-palytoxin in the marine
cyanobacterium Trichodesmium // Mar. Drugs. 2011. V. 9. P. 543—560.
Khodakevich L., JeeekZ., Messinger D. Monkeypox virus: ecology and public health significance // Bulletin
of the World Health Organization. 1988. V. 66. № 6. P. 747—752.
Klee S K., Brzuszkiewicz E., Nattermann H. et al. The genome of a Bacillus isolate causing anthrax in
chimpanzees combines chromosomal properties of B. cereus with B. anthracis virulence plasmids // PLoS
ONE. 2010. V. 5. el0986.
KlempnerM. S, Talbot E. A., LeeS. I. etal. Case records of the Massachusetts general hospital. Case 25-2010.
A 24-year-old woman with abdominal pain and shock// N Engl. J. Med. 2010. V. 363. P. К£—ТГ1
Klenerman P, Zinkemagel R. M. Original antigenic sin impairs cytotoxic T lymphocyte responses to viruses
bearing variant epitopes// Nature. 1998. V. 394. P. 482-485.
Список литературы
1087
Knight А. Н., Wynne-Williams J. Е., Willis А. Г Cutaneous anthrax-the non-industrial hazard//Brit. med.
JY. 1969. № 1 P. 416-418.
Ko J., Splitter G. Molecular host-pathogen interaction in Brucellosis: current understanding and future appro-
aches to vaccine development for mice and humans // Clin Microbiol. Rev 2003. V. '6, № 1. P. 65—78.
Koehler T.M., Thorne C.B. Bacillus subtilis (natto) plasmid pLS20 mediates interspecies plasmid transfer //
.1 Bacterio) 1987. V. 169. № 11. P. 5271-5278.
Kohler S., Michaux-Charachon S., Porte F. et al. What is the nature of the replicative niche of a stealthy bug
named Brucella? // Trends Microbiol 2003 — V. 11. P. 215—219.
Kool J. L Risk of person-to-person transmission of pneumonic plague // Healthcare epidenuol. 2005. V. 40.
P. 1166-1172.
Kortepeter M., Christopher G., Cieslak T. et al. USAMRIlD’s medical management ofbiological casualties
Handbook. Forth Edition Editors. US Army Medical Research Institute of Infectious Diseases
(USAMRIID), Fort Detrick, Maiyland, 2001.
Kozminsky-Atias A., Bar-Shalom A., Mishmar D. et al. Assembling an arsenal, the scorpion way 11 BMC
Evolution. Biol. 2008. V. 8: 333 (http //www biomedcentral.com/1471-2148/8/333).
Kreil T R., Eibl M. M. Pre- and postexposure protection by passive immunoglobulin but no enhancement of
infection with a flavivirus in a mouse model //J. Viror. 1997. V. 71. № 4. P. 2921 2927.
Kreitman R. J. Immunotoxins for targeted cancer therapy I I The AAPS .1 (http://www.aapsj.org). 2006. V. 8.
№ 3. E532-E55I.
Kreitman R. J. Recombinant immunotoxins containing truncated bacterial toxins for the treatment of
hematologic malignancies// BioDrugs. 2009. V. 23. № 1. P. 1-13.
KrocaM., TamvikA., SjostedtA. The proportion of circulating gammadelta T cells increases after the first week
of onset of tularaemia and remains elevated for more than a year // Clin. Exp. Immunol. 2000. V. 120.
P. 280- 284.
KrugH E, WickP. Nanotoxicology: an interdisciplinary challenge /' Angew. Chem. Int. Ed. Eng'. 2011. V. 50.
P. 1260-1278.
Kudo Y, Yasumoto T., Konoki K. et al. Isolation and structural determination of the first 8-epi-type tetrodo-
toxm analogs from the newt, Cynops ensicauda popei, and comparison of tetrodoroxin analogs profiles
of this newt and the puffer fish, Fugu poecilonotus 11 Mar. Drugs. 2012. V. 10. P. 655—667.
Kuiper-Goodman T, Scott P. M. Risk assessment ofthe mycotoxin ochratoxin A//Biomed. Environ. Sci. 1989.
V. 2. P. 179-248.
Kuruha R., Wilson F. , Gao X. et al. Targeted delivery of nucleic acid-based therapeutics to the pulmonary
circulation // AAPS J. 2009. V. 11. № 1. P. 23-30.
La Scola В , Audic S., Roberta C. etal. A giant virus in amoebae//Science. 2003. V. 299. № 5ol 5 P. 2033.
La Scola B., Elkarkouri K., Li W et al Rapid comparative genomic analysis for clinical microbiology: The
Francisella tularensis paradigm//Genome Res. 2008. V. 18. P. 742—750.
La Scola В , Raoult D. Survival of Coxiella burnetii within free-living amoeba Acanthamoeba castellanii //
Clin. Microbiol. Infect. 2001. V. 7. P. 75-79.
Lacy D. B., Lin FL, Melnyk R. A. et al. A model of anthrax toxin lethal factor bound to protective antigen //
Proc Natl. Acad Sci. USA 2005. V. 102. P. 16409-16414.
Lacy D. B., Lin FL, Melnyk R. A. et al A model of anthrax toxin lethal factor bound to protective antigen //
Proc Natl. Acad. Sci. USA. 2005. V. 102. P. 16409-16414
Lai X. H., Golovliov L, SjostedtA. Francisella tularensis induces cytopathogenicity and apoptosis in murine
macrophages via a mechanism that requires intracellular bacterial multiplication // Infect. Immun. 2001.
V.69 P. 4691-4694.
Lalitha M., Thomas M. Penicillin resistance in Bacillus anthracis // Lancet. 1997. V. 349. № 9064. P. J 522.
Lamanna G. Immunological aspects of airborne infection: some general consideration of response to inhala-
tion of toxins// Bact. Reviews. 1961. V. 25. № 3. P. 323—330.
Lamb K., Lokesh G. L., Sherman M. et al. Structure of a Venezuelan equine encephalitis virus assembly // Viro-
logy. 2010. V. 406. № 2. P. 261-269.
Lancaster M. C., Boulter E. A., Westwood J. C. N. etal. Experimental respiratory infection with poxviruses. II.
Pathological studies,/ Br J. Exp. Pathol. 1966. V. 47 № 5. P. 466—471.
Lanciotti R. S., RoehrigJ. T. Deubel V et al. Origin of the West Nile virus responsible for an outbreak of
encephalitis in the Northeastern United States // Science. 1999. V 286. P. 2333—2337
Lasaro M. L., Ertl H. New insights on adenovirus as vaccine vectors 11 Mol. Ther 2009. V. 17. № 8.
P. 1333-1339.
1088
Список литературы
Laskawi R The use of botulinum toxin in head and tace medicine An interdisciplinary field I I Head & Face
Medicine. 2008. V. 4. № 5 (doi:10.1186/1746-160X-4-5).
Lauriano С. M., Barker J. R., Yoon S. S et al. MglA regulates transcription of virulence factors necessary for
Francisella tularensis intraamoebae and intramacrophage survival // Proc Natl. Acad. Sci. USA. 2004.
V. 101. P. 4246-4249.
Lazar Adler N. R., Stevens J., Stevens M. et al. Autotransporters and their role in the virulence of Burkholderia
pseudomallei and Burkholderia mallei I 12011. V. 2. Article 151 (www.frontiersin.org.).
LeafH. Патент96/27393 (Al) ВОИС, МПК6 А61К38/Т7; A6J K48/00; A61K9/127; A61K38/17 А61К48/
00. A61K9/127. A dry powder formulation for gene therapy, заявитель Huang Lea., патентооблада-
тель Univ. Pittsburgh, США; заявл. 07.03.1995,08/400,089; опубл. 12.09.1996.
LeDucJ. M., Damon L, MeeganJ. M. etal. Smallpox research activities: U. S. Interagency collaboration//
Emerg. Infect. Dis. 2002. V. 8 № 7. P. 743-745.
Lee A. M., Pasquato A., Kunz S. Novel approaches in anti-arenaviral drug development // Virology. 2011.
V. 411. №2. P. 163-164
LeeE., Harris. D.J., Siegel J. etal. Патент США 5840710 (А), МПК A61K47/48; A61K48/00; A61K9/127;
C07J41/00; C07J43/00; C12N15/88 Cationic amphiphiles containing ester or ether-unked lipophilic
groups for intracellular delivery of therapeutic molecules I заявитель Genzyme Corp , США; заявл.
20.10.1995, 546087; опубл. 24.11.1998.
LeendertzF.H., YumluS., PauliG. etal. A new Bacillus anthracis found in wild chimpanzees and a gorilla from
West and Central Africa I I PLoS Pathogens. 2006. V. 2 № 1. P 1-4.
LeepomA., YongyodS., Yungyuen T е/о/. Emergenceof Francisellanovicidabacteremia,Thailand//Emerg.
Infect. Dis. 2008. V. 14. № 12. P. 1935-1937
Lemichez E., Flatau G., BruzzoneM. etal. Molecular localization of the Escherichia coli cytotoxic necrotizing
factor CNF1 cell-binding and catalytic domains//Mol. Microbiol. 1997. V. 24. № 7. P. 1061—1070.
Leung K, Fong B., Tsoi Yeuk-Ki Analytical challenge; determination of tetrodotoxin in human urine and
plasma by LC-MS/MS // Mar. Drugs. 2011. V. 9. P. 2291-2303.
Levine R. S., Peterson A. T., Yorita K. L. et al. Ecological niche and geographic distribution of human mon-
keypox in Africa // PLoS ONE. 2007. V 2 (№ 1: e!76. doi:10.1371/journal.pone.0000176).
Levy A., Chang B. J, Abbott L. K. et al. Invasion of spores of the arbuscular mycoriiizal fungus Gigaspora
decipiens by Burkholderia spp. // Appl. Environ. Microbiol. 2003. V. 69. P. 6250—6256
LiN., SioutasC., ChoA. etal. Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage
11 Environ. Health. Perspect. 2003 V. 111. P. 455—460.
Li S., HuangL. Nonviral gene therapy: promises and challenges I I Gene Ther. 2000. V. 7. № 1. P. 31—34.
LiS., Tan Y., Viroonchatapan E etal. Targeted gene delivery to pulmonary endothelium by anti-PECAM
antibody//Am J. Physiol. Lung. Cell. Mol. Physiol. 2000. V 278, № 3. P. 504 511.
Li Y, Cui Y., Hauck Y et al. Genotyping and phylogenetic analysis of Yersinia pesos by MLVA: insights into
the worldwide expansion of Central Asia plague foci // PLoS ONE. 2009. V. 4. № 6 (e6000. doi 10.1371/
joumal.pone.0006000).
Liepmann H. Death from the science. London, 1937.
LindlerL. E., Lebeda F J., Kerch G. W. Biological weapons defense. N. J., 2005.
Lindstrom M., Korkeala H. Laboratory diagnostics of botulism //Clin. Microbiol. Rev. 2006. V. 19. P 298-314
LiuChengchu, ZhuQing, Wu Wenhuietal. Degradable copolymer based on amphiphilic N-octyl-N-quatenary
chitosan and low-molecular weight polyethylenimine for gene delivery // Intern. J. of Nanomed .2012
V. 7. P. 5339-5350.
Liu X. S., Dean D. H. Redesigning Bacillus thuringiensis Cryl Aa toxin into a mosquito toxin // Prot. Eng.
Design. Selec. 2006. V. 19. P. 107-111.
Llewellyn L E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors // Nat. Prod.
Rep. 2006. V. 23. P. 200-222
Lobet Y., Cieplak W, Smith S. G et al. Effects of mutations on enzyme activity and immunoreactivity of the
SI subunit of pertussis toxin//Infec. Immun. 1989 V.57.№11 P. 3660 1668.
Lockman P. R., Koziara J. M , Mumper R. J. et al. Nanoparticle surface charges alter blood-brain barrier
integrity and permeability//J. Drug Target. 2004. V. 12. P. 635—641.
Logan W Mortality in the London fog incident, 1952 // Lancet. 1953. V. 1. P. 336—338.
LogueC. H., BosioC. E, WelteT. etal. Virulence variation among isolates of western equine encephalitis virus
in an outbred mouse//J Gen. Virol. 2009. V. 90 P. 1848—1858.
Loosmore S., Zealy G. R., Boux H. A. et al. Engineering of genetically detoxified pertussis toxin analogs for
development of a recombinant whooping cough vaccine // Infec. Immun. 1990. V. 58, № 5. P.3653—3662.
Список литературы
1089
Loparev И, MassungR., Esposito J. etal. Detection and differentiation of Old World orthopoxviruses: rest-
riction fragment length polymorphism of the crmB gene region // J. Clin. Microbiol. 2001. V. 39. № 1.
P. 94 -100.
Lord M. J., Jolliffe N. A., Marsden C. J. et al. Ricin. Mechanisms of cytotoxicity // Toxicol. Rev. 2003. V. 22.
P. 53 -64.
Lorenz. M, Wackernagel IK Bacterial gene transfer by natural genetic transformation in the environment I I
Microbiol. Rev. 1994 V. 58. № 3. P. 563—602.
Louzao M., Ares L, Cagide E. Marine toxins and the cytoskeleton: a new view of palytoxin toxicity // FEBS
Journal 2008. V. 275. P. 6067-6074.
Lo Vullo E D., Sherrill L. A., Pavelka M. S. Improved shuttle vectors for Francisella tularensis genetics 11 FEMS
Microbiol Lett 2009 V. 291. № I. P. 95—102.
Lukashevich I.S., Patterson J., Carrion R. et al. A live attenuated vaccine for Lassa fever made by reassortment
of Lassa and Mopeia viruses//J. Virol. 2005 V.79.Ns22.P. 13934—13942.
Ernie M. D., Hoppleston A. G., AbramsonS. etal. An evaluation of the method of quantitative air-borne infection
and its use in the study of the pathogenesis//Am. Rev. Tubore. 1950. V. 61. P. 765—797.
Lusztig11 Bacterienkrieg. zeitschrift fur desinfections und Gesundheitswesen. 1931. № 3.
Lynch V. J. Inventing an arsenal: adaptive evolution and neofunctionalization of snake venom phospholipase
A2 genes// BMC Evol. Biol. 2007. V 7 № 2 (http://www.biomedcentral.eom/1471-2148/7/2>.
Lypez S. S., Valenzuela F., Fernandez L. Nydulos puhnonares bilaterales por-brucelosis // Arch. Bronco-
neumol. 2011. V. 47. P. 320-321.
Haykawa J., Poon C. 11 Am. Ind. Hyg. Assoc. 19b5. V. 26. № 2. P. 150.
Mahmud Y., Arakawa O., Lchinose A. et al. Intracellular visualization of tetrodotoxin (TTX) in the skin of a
puffer Tetraodon nigroviridis by immunoenzymatic technique 11 Toxicon. 2003. V. 41. P. 605—611.
Maier T.M. Construction and characterization of a highly efficient Francisella shuttle plasmid // Appl.
Environ. Microbiol. 2004. V. 70. P. 7511—7519.
Mandal С. C., Gayen S., Basu A. et al. Prediction based protein engineering of domain 1 of Cry2A entomocidal
toxin of Bacillus thuringiensis for the enhancement of toxicity against lepidopteran insects // Prot. Eng.
Design. Selec. 2007. V. 20. P. 599—606.
Manunta M. D., McAnulty R J., Tagaiakis A. D. et al. Nebulisation of receptor-targeted nanocomplexes for
gene delivery to the airway epithelium 11 PLoS ONE. 2011. V. 6. № 10 (e26768. doi:10.1371/
journal pone.0026768).
Marburg and Ebola virus infections: a guide for their diagnosis, management and control. Geneva, WHO,
1977.
Margolis D. A., Bums J., Reed S. R. et al. Case report: septicemic plague in a community hospital in California
//Am. J. Trop. Med. Hyg. 2008. V. 78. № 6. P. 868-871.
Marks J. D. Medical aspects of biologic toxins //Anesthesiol. Clin. N. Am. 2004. V. 22. P. 50*3—522.
Marple V. A. , Rubow K. L. Aerosol generation concepts and parameters // In: Generation of aerosols and
facilities for exposure experiments / Ed. Willeke K., Ann Arbor Science, 1980, Ch. 1. P 3—29
Marrie T. J. Coxiella burnetii pneumonia // Eur. Respir. J. 2003. V. 21 P 713-719.
Martinez-Martinez L., Pascual A., Jacoby G. A Quinolone resistance from a transferable plasmid // Lancet.
1998.V51 P. 797- 799.
Mary I., OsterbergL., ChauP. et al. Pulmonary melioidosis //Chest. 1995. V. 108. P. 1420—1424.
Mataraza C., Qin Z. H., HuangZ. et al. Highly efficient molecular delivery into mammalian cells using carbon
nanotube spearing // Nature Methods. 2005. V. 2. P. 449-454.
Mattix M. E., Hunt R. E., Wilhelmsen C. L. etal. Aerosolized staphylococcal enterotoxin В-induced pulmonary
lesions in Rhesus monkeys // Toxicol. Pathol. 1995. V. 23. № 3. P. 262—268.
Maurin M., Benoliel A. M., Bongrand P. et al. Phagolysosomal alkalinization and the bactericidal effect of
antibiotics: the Coxiella burnetii paradigm //J Infect. Dis. 1992. V. 166. P. 1097—1102.
Mayfield J. E., Bricker B. J., Godfrey H. etal. The cloning, expression, and nucleotide sequence of a gene coding
for an immunogenic Brucella abortus protein // Gene. 1988. V. 63. P. 1—9.
McCaul T. F., Banerjee-Bhatnagar N., Williams J. C. Antigenic differences between Coxiella burnetii cells
revealed by postembedding immunoelectron microscopy and immunoblotting // Infect. Immun. 1991
V. 59. P. 3243-3253.
McCaul T. F., Williams J C Developmental cycle of Coxiella burnetii: structure and morphogenesis of
vegetative and sporogenic differentiations//J. Bacteriol. 1981. V. 147. P. 1063—1076.
McClain D J. Smallpox // Cm. Medical aspects of chemical and bio'ogical warfare / Ed. F. R Sidell, E. T. Ta-
fuqi, D. R. Franz. Washington, 1997. P. 539-559.
1090
Список литературы
McCormack М. Р., Rabbitts Т. Н. Activation of the T-cell oncogene LMO2 after gene therapy for X-linked
severe combined immunodeficiency // N. Engl. J. Med. 2004. V. 350. P. 913-922.
McLendon M., Apicella M., Allen Lee-Ann H. Francisella tularensis: taxonomy, genetics, and immuno-
pathogenesis of a potential agent of biowarfare I I Annu. Rev. Microbiol. 2006. V. 60. P. 167—185.
McMahon B. R., Tanaka K., Doyle J. E. et al. A change of heart: cardiovascular development in the shrimp
Metapenaeus ensis//Comp. Biochem. Physiol., Part A: Mol. Integr. Physiol. 2002. V. 133. P. 577-587.
Mead P. S., Griffin P. M Escherichia coli O157:H711 Lancet. 1498. V. 352. P. 1207—1212.
M-’csas J., Strauss E.J. Molecular mechanisms of bacterial virulence- type HI secretion and pathogenicity
islands//Emerg. Infect. Dis. 1996. V. 2. № 4. P. 271—290.
Medical aspects ofchemical and biological warfare/Ed. F. R. Sidell, E.T. Tafuqi, D. R. Franz .Wash., 1997.
Medina C., Santos-Martinez M. J., Radomski A. et al. Nanoparticles: pharmacological and toxicological
significance // Brit J. Pharmacol. 2007. V. 150. P. 552—558.
Melito P.L., Qiu X., Fernando L.M etal. The creation of stable cell lines expressing Ebola virus glycoproteins
and the matrix protein vp40 and generating Ebola virus-like particles utilizing an ecdysone inducible
mammalian expression system // J. Virol. Meth. 2008. V. 148. F 237-243.
MercerT. T, Tillery M. L, Chow H. К Operating characteristics ofsome compressed air nebulizers//Am. Ind
Hyg. Assoc. J 1968. V. 29. P. 68-69.
Meselson M., Gillemin J., Hugh-Jones M. etal. The Sverdlovsk anthrax outbreak of 1979 // Science. 1994.
V. 209. № 12. P. 1202—1208
Meyer K. F. Pneumonic plague /, Bact. Rev. 1961. V. 25. P. 219-230.
Middlebrook J. L., Franz D. R. Botulinum toxins // Medical aspects of chemical and biological warfare / Edit.
F. R Sidell, E. T. Tafuqi, D. R. Franz. Washington, 1997.
Miller D. G., Trobrldge G. D., Petek L. M. Large-scale analysis of adenossociated virus vector integration sites
in normal human cells //J. Virol. 2005. V 79. № 17. P. 11434—11442.
Miller J., EngelbergS., Broad W. Germs biological weapons and America’s secret war. N. Y, 2001.
Miller J., EngelbergS. Broad W. The New York Times. 2001 (05.09.2001).
Mise T., Funatsu G, Ishiguro M. et al. Isolation and characterization of ricin E from castor beans I I Agric. Biol.
Chem. 1977. V. 41. № 10. P. 2041-2046.
MockM., FouetA. Anthrax// Annu. Rev. Microbiol. 2001. V. 55 P. 647—671.
Mohapatra N. P., Soni S., Bell B. L. et al. Identification of an orphan response regulator required for the
virulence of Francisella spp. and transcription of pathogenicity island genes I I Infect. Immun. 2007. V. 75.
P. 3305-3314.
Mollaret H. Bref historique de la guerre bacteriologique // Med. et Maladies Infect. 1995. V. 7. P. 402—407
Monath T. P. Ecology of Marburg and Ebola viruses: speculations and directions for future research I I J. Infec.
Diseases. 1999. V. 179 (Sup. 1). P. 127—138.
Monath T. P. Lassa fever: review of epidemiology and epizootiology// Bull. World Health Organ. 1975. V. 52.
P. 577-592.
Moore R E., Helfrich P., Patterson G. M L. The deadlj seaweed of Hana. Oceanus. 1982. V. 25. P. 54-63.
Moore R. E., Scheuer P. J. Palytoxin a new marine toxin from a coelenterate // Science. 1971. V. 172.
P 495-498.
Moos A., Hackstadt T. Comparative virulence of intra- and interstrain lipopolysaccharide variants of Coxiella
burnetii in the guinea pig model // Infect. Immun. 1987. V. 55. № 5 P. 1144 -1150.
Moran E, Weinberger H., Sullivan J. C. et ai Concerted evolution of sea anemone neurotoxin genes is revealed
through analysis of the Nematostella vectensis genome // Mol Biol Evol. 2008 V 25. № 4. P. ТУ1—141.
Morimoto Y., Kobayashi N, Shinohara N. et al. Hazard assessments of manufactured nanomaterials // J.
Occup. Health. 2010. V. 52. P. 325-334.
Mortensen L. J., OberdorsterG., Pentland A. P. etal. In vivo skin penetration of quantum dot nanoparticles in
the murine model, the effect of UVRI I Nano Lett. 2008. Aug 8.
Mourez M., Lacy D. B., Cunningham K. et al. 2001: a year of major advances in anthrax toxin research//
TRENDS in Microbiol. 2002. V.10. № 6. P. 287-293.
Mui inda V., MwemaG., Kiluudu M. etal. Reemergence of human monkeypoxin Zaire in 1996//I ancet. 1997.
V. 349. № 9063 P. 1449-1450.
Munday R. Palytoxin toxicology: Animal studies // Toxicon. 2011. V. 57. P. 470—477.
Murphy F. A., Schinwald A., Poland C. A. et al. The mechanism of pleural inflammation by long carbon
nanotubes: interaction of long fibres with macrophages stimulates them to amplify proinflammatory
responses in mesothelial cells /7 Particle and Fibre Toxicology. 2012. V. 9, 8 (http://www.partic-
leandfibretoxicology.com/content/9/1/8).
Список литературы
1091
Murray S. А., Mihali Т.К., Nellan В. A. Extraordinary conservation, gene loss and positive selection in the
evolution of an ancient neurotoxin I I Mol. Biol. Evol. 2011 V 28 P. 1173-1182.
Nakai H., Storm T. A., Kay M. A. Recruitment of single-stranded recombinant afeno-associated virus vector
genomes and intermolecular recombination are responsible for stable transduction of liver in vivo // J.
Virol. 2000. V. 74. P. 9451-9463
Nakayama E., Tomabechi D, Matsuno K. et al. Antibody-dependent enhancement of Marburg virus infection
//J. Infect. Dis. 2011. V. 204. S. 3. P. 978-985.
Narisara H., Wuthiekanun V., Boonbumrung K. ei al Biological relevance of colony morphology and
phenotypic switching by Burkholderia pseudomallei //J. Bacteriol. 2007. V. 189. № 3 P. 807-817.
Needham J. Science and civilization in China. Cambridge University Press, 1956.
NegredoA., Palacios G., Vazquez-Moron S et al. Discovery of an ebolavirus- like filovirus in Europe // PLoS
Pathog. 2011. V. 7, № 10 (el002304. doi:10.1371/journal.ppat.l002304).
NeisserA. // Zeitschrift fur desinfections und gesundheitswesen. 1931 № 5. P. 237.
Nelson M„ Lever M. Dean R. etal. Characterization of lethal inhalational infection with Francisella tularensis
in the common marmoset (Callithrix jacchus) 11 J. Med. Microbiol. 2010. V. 59. P 1107—1113.
NemmarA., HoetP., VanquickenbomeB.etal. Passage ofinhaled particles into the blood circulation in humans
// Circulation. 2002. V. 105. P. 411—414.
Ngauy V., Lemeshev K, Sadkowski L. et al. Cutaneous melioidosis in a man who was taken as a prisoner of war
by the Japanese during World Wai II // J. Clin. Microbiol. 2005. V. 43. P. 970—972.
NienhuisA Development of gene therapy for blood disorders//Blood. 2008. V 111 P. 4431—4444.
NienhuisA. W., DunbarC E., Sorrentino В, P Genotoxicity of retroviral integration in hematopoietic cells
// Molecular Then 2006. V. 13. № 6.1031-1049
Nierman W. C., DeSharer D., Kim H et al. Structural flexibility in the Burkholderia mallei genome I I Proc.
Natl. Acad. Sci. USA. 2004. V. 101 № 39. P. 14246- 14251
Nierman W. C., DeShazer D., Kim H. S. et al. Structural flexibility in the Burkholderia mallei genome // Proc.
Natl. Acad. Sci. USA. 2004. V. 101. P. 14246- 14251.
Nieto F. R, Cobos E J., Tejada M. A. et al. Tetrodotoxin (TTX) as a therapeutic agent for pain // Mar Drugs
2012. V. 10. P. 281-305
Nijhuis M., van Maarseveen N. M., Boucher C. A. Antiviral resistance and impact on viral replication capacity:
evolution of viruses under antiviral pressure occurs in three phases // Handb. Exp. Pharmacol. 2009.
V. 189. P. 299-320.
Noble J., Rich J. Transmission of smallpox by contact and by aerosol routes in Macaca irus // Bull. Wld. Hith.
Org. 1969 V. 40. P. 279-286.
Noguchi P. Risks and benefits of gene therapy I IN. Engl. J Med. 2003. V. 348, № 3. P. 193—192.
Noguchi T, Arakawa O. Tetrodotoxin — distribution and accumulation in aquatic organisms, and cases of
human intoxication // Mai. Drugs. 2008. V. 6. P. 220—242.
Noguchi T„ EbesuJ. S. M. Puffer poisoning: epidemiology and treatment//J. Toxicol 2001. V. 20. P 1-10
Nordmann P., PoirelL. Emerging carbapenemases in gramnegative aerobes//Clin. Microb. Infect. 2002. V. 8.
P 321-331.
OberdorsterG., Oberdurster E., Oberduyster J. Nanotoxicology: an emerging discipline evolving from studies
of ultrafine particles // Environ. Health. Perspect. 2005. V. 113. P. 823—839.
Ghana P., Gofrit O., Sayesh S. et al Regulatory sequences of the H19 gene in DNA based therapy of bladder
cancer// Gene. Ther. Mol. Вю1. 2004. V. 8. P. 181—192.
Okano FL, Masuoka H., Kame1 S. et al Rhabdomyolysis and myocardial damage induced by palytoxin, a toxin
of blue humphead parrotfish// Int. Med. 1998. V. 37. P. 330—333.
Olson К. B. Aum Shinrikyo: once and future threat//Emerg. Infect. Dis. 1999. V. 5. P. 513—514.
Omslard A., Beare P. A., Hill J. et al. Isolation from animal tissue and genetic transformation of Coxiella bur-
netii are facilitated by an improved axenic growth medium // Appl Environ. Microbiol. 2011. V. 77.
P. 3720-3725.
Omsland A. etal. Host cell-free growth ofthe Q feverbacterium Coxiella burnetii//Proc. Natl. Acad. Sci. USA.
2009. V. 106. P. 4430- 4434.
Ong C., Huey Ooi C., Wang D. et al. Patterns of large-scale genomic variation in virulent and avirulent
Burkholderia species // Genome Res. 2004. V. 14. P. 2295—2307.
Overholt E.L., Tigertt W. D., Kadull P. J. et al. An analysis of forty-two cases of laboratory-acquired tularemia
// Amer. J. Med. 1961. V. 30. № 5. P. 785-806.
PaesslerS., Fayzulin R. Z., Anishchenko M. et al. Recombinant Sindbis/Venezuelan equine encephalitis virus
is highly attenuated and immunogenic Ц J. Virol. 2003. V. 77. № 17. P. 9278—9286.
1092
Список литературы
Paessler S., Yun N. E., Judy B.M. etal. Alpha-beta T cells provide protection against lethal encephalitis in the
murine model of VEEV infection//Virology. 2007. V. 367. № 2. 307-323.
Pahlitzsch R., HammarinA., WidellA. A case offacial cellulitis and necrotizing lymphadenitis due to cowpox
virus infection // Clin. Infec. Dis 2006 V. 43. P. 737—742.
Palese P. RNA virus vectors: where are we and where do we need to go? // Proc. Natl. Acad. Sci. USA. 1998.
V. 95. № 22. P. 12750-12752.
PMo-Lypez L., Gomes I., Rausell C. et al. Structural changed of of the eCry 1 Ac oligomeric pre-pore from
Bacillus thuringiensis//Biochemi 2006 V. 45. P. 10329-10336.
Pardo-Lypez L., Munoz-Garay C., Porta H. Strategies to improve the insecticidal activity of Cry toxins from
Bat illus thuringiensis // Peptides. 2009. V. 30 № 3. P. 589—595.
Pardridge W M. Drug and gene targeting to the brain with molecular Trojan horses //Nat. Rev. Drug. Discov.
2002. V. l.P. 131-139.
ParkK. W„ KimD.. ParkC.Y. etal. Fatal systemic infection with multifocal liver and lung nodules caused by
Brucella abortus // Am. J. Trop. Med. Hyg. 2007. V. 77 № 6. P. 1120— 1123.
Partikian A., Mitchell W. G Iatrogenic botulism in a child with spastic quadriparesis // J. Child. Neurol. 2007.
V. 22. № 10. P. 1235-1237.
PatckaJ., SplinoM., Mgrka V. Botulism and bioterrorism: how serious is this i roblem?//ActaMedica(IIradec
Kralove). 2005. V 48. № 1. P 23-28.
PatraG., VaissaireJ. E., Weber-Levy M. et al. Molecular characterization of Bacillus strains involved in
outbreaks of Anthrax in France in 1997 // J. Clin. Microbiol. 1998. V 36 № 11. P. 3412—3414.
Patrie W III. Biological warfare scenarios // Firepower in the Lab. Automat, in the Fight against Infect. Dis.
and Bioter. / S. P. Layne (Eds.). Washington D. C.: Joseph Henry Press, 2001. P. 215—223.
Pavlin J. A. Epidemiology of bioterrorism // Emerg. Infect. Dis. 1999. V. 5. № 4 P. 528-530.
Pavlov V. M., Mokrievich A. N. Crypbc plasmid pFNLlO from Francisella novicida-like F6168: the base of
plasmid vectors for Francisella tularensis // FEMS Immunol. Med. Microbiol. 1996. V. 13. P. 253—256.
Paweska J., SewlallN. H.‘ Ksiazek T. J. et al. Nosocomial outbreak of novel arenavirus infection: Southern
Africa//Emerg. Infect. Dis. 2009. V. 15. № 10 P. 1598-1602.
Pearce L. B., Eirst E. R., Maccallum R. D et al. Pharmacologic characterization of botulinum toxin for basic
science and medicine //Toxicon ’997. V. № 9. P. 1373—1412.
Pearson T., Busch J. D., Ravel J etal. Phylogenetic discovery bias in Bacillus anthracis using single-nucleoti-
de polymorphisms from whole-genome sequencing // Proc. Natl. Acad. Sci. US A. 2004. V. 101.
P. 13536-13541.
Pechous R., McCarthy T. R., Zahrt T C. Working toward the future: insights into Francisella tularensis
pathogenesis and vaccine development // Microbiol. Mol. Biol. Rev. 2009. V 73. № 4. P. 684—711.
Peraica M., Radic B., Lucic A. et al Toxic effects of mycotoxins in humans // Bulletin of the World Health
Organization. 1999 V. 77. № 9. P. 754—766.
Pema N. T., Mayhew G. F, Posfai G. et al. Molecular evolution of a pathogenicity island from entero-
hemorrhagic Escherichia coliO157:H7// Infect Immun 1998. V 66. P. 3810—3817
Peter M. Bennett integrons and gene cassettes: a genetic construction kit for bacteria // J. Antimicrob. Che-
mother. 1999. V. 43. P. 1—4.
Petersen J. M., Carlson J. K., Dietrich G. etal. Multiple Francisella tularensis subspecies and clades, tularemia
outbreak, Utah // Emerg. Infect. Dis. 2008. V. 14. № 12. P. 1928—1930.
Peterson A. T., Carroll D.S., Mills J.N. et al. Potential mammalian filovirus reservoirs // Emerg. Infect. Dis.
2004. V. 10. № 12. P. 2073-2081.
Petosa C., Collier P, Klimpel К R. et al. Crystal structure of the anthrax toxin protective antigen // Nature.
1997. V. 385 № 6619 P. 833- 838.
Pfeffer M., DoblerG. R. Emergence of zoonotic arboviruses by animal trade and migration // Parasites &
Vectors. 2010. V 3. № 35 (http://www.parasitesandvectors.eom/content/3/l/35).
Pichla-Gollon S. L., Lin S. W., Hensley S. E. etal. Effect of preexisting immunity on an adenovirus vaccine
vector: in vitro neutralization assays fail to predict inhibition by antiviral antibody in vivo //J. Viroi „ 2009.
V. 83. P. 5567-5573.
Pilo P., Perreten V., Frey J. Molecular epidemiology of Bacillus anthracis: determining the correct origin //
Appl. Environ. Microbiol. 2008. V. 74 № 9. P 2928—2931.
Piskuiek O., Okada N. Poxviruses as possible vectors for horizontal transfer of retroposons from reptiles to
mammals// Natl. Acad. Sci. USA. 2007. V. 104. № 29. P. 12046-12051.
Pitt J. I. Toxigenic fungi: which are important? // Med Mycol. 2000. V. 38 (Suppl. 1). P. 17—22.
Список литературы
1093
Pluta К., Kacprzak М. М. Use of НIV as a gene transfer vector//Act. Biochim. Pol. 2009. V. 56. P. 531-595.
PoliM. A., Rivera V. R., HewetsunJ F. etal. Detection ofricin by colorimetric and chemiluminescence ELISA
//lexicon. 1994. V 32. № 11. P. 1371-1377.
Pollitzer R Plague studies. 4. Pathology // Bull. World Hlth Org. 1952. V. 5. P 337—376.
Pomerantsev A.P., Staritsyn N.A. Behavior of heterologous recombinant plasmid pCET in cells of Bacillus
anthracis // Genetika. 1996. V. 32. № 4. P. 500—509.
Pomerantsev A. P, Sukovatova L.V., Marinin L.l. Characterization of a Rif- R population of Bacillus anthraco
//Antibiot. Khimioter. 1993. V. 38. № 8—9 P. 34-38.
Portnoy D., Falkow 5: Virulence-associated plasmids from Yersinia enterocolitica and Yersinia pestis // J
Bacterid. 1981. V. 148. P. 877-883.
Powell A., Crozier J., Hodgson H. et al. A case of septicaemic anthrax in an intravenous drug user // RMC
Infectious Diseases. 2011 (http://www.biomedcentral.com/1471-2334/ll/21/).
Prabhu В. M., AH S. E, Murdock R. C. etal. Copper nanoparticles exert size and concentration dependent
toxicity on somatosensory neurons of rat // Nanotoxicol. 2010. V 4. P. 150—160.
Pratt W. D., Gibbs P., Schmaljohn A. L. Use of telemetry to assess vaccine-induced protection against parenteral
and aerosol infections of Venezuelan equine encephalitis virus in non-human primates // Vaccine. 1998.
V. 16. №9-10. P. 1056-1064.
Price E. E., Seymour M.L., Sarovich D. S. et al. Molecular epidemiologic investigation of an anthrax outbreak
among heroin users, Europe // Emerg. Infect. Dis. 2012. V 18. № 8. P.1307—1313.
Promdonkoy B., Ellar D. J. Membrane pore architecture of a cytolytic toxin from Bacillus thuringiensis
Biochem J. 2000. V. 350 P. 275-282.
Pugh G. W., Tabatabat L B., Bricker B. J. etal. Immunogenicity of Brucella-extracted and recombinant protein
vaccines in CD-I and BALB/c mice // Am. J. Vet. Res. 1990. V. 51. P. 1413—1420.
Rajamohan E, Alzate O., Cotrill J. A. et al. Protein engineering of Bacillus thuringiensis delta-endotoxin:
mutahons at domain 11 of CrylAb enhance receptor affinity and toxicity toward gypsy moth larvae/1 Proc
Natl. Acad. Sci. USA. 1996 V. 93. P. 14338-14343.
Ramos V., Vasconcelos V. Palytoxin and analogs: biological and ecological effects // Mar. Drugs. 2010. V 8.
P. 2021-2037
Ramsay C. N, Stirling A., Smith J. et al. An outbreak of infection with Bacillus anthracis in injecting drug users
in Scotland // Eurosurveillance. 2010. V. 15. Is. 2(14 января).
Raoult D. Treatment of Q Fever// Antimicrob. Agents and Chemother. 1993. V. 37. № 9. P. 1733—1736.
Raoult D., Houpikian P., Tissot D. H. et al. Treatment of Q fever endocarditis: comparison of 2 regimens
con taining doxycycline and ofloxacin or hydroxychloroquine // Arch. Intern. Med. 1999. V. 159. № 2.
P. 167-173.
Raoult D., Torres H, Drancourt. M Shell-vial assay: evaluation of a new technique for determining antibiotic
susceptibility, tested in 13 isolates of Coxiella burnetii 11 Antimicrob. Agents Chemother. 1991. V. 35.
P. 2070- 2077.
Raper S. E., Chirmule N., Lee F. S. et al. Fatal systemic inflammatory response syndrome in a ornithine
transcarbamylase deficient patient following adenoviral gene transfer // Mo) Genet. Metab. 2003. V. 80
(1-2). P. 148-158.
Rasko D A., Esteban C. D., Sperandio V. Development of novel plasmid vectors and a promoter trap system
in Francisella tularensis compatible with the pFLN 10 based // Plasmid. 2007. V 58. № 2. P. 159—166.
Ravel J., Jiang L., Stanley S. T et al. The complete genome sequence of Bacillus anthracis Ames «Ancestor»
11 J. Bacteriol. 2009 V. 191. № 1. P. 445-446.
Recchia G. D., Hall R M. Gene cassettes: a new class of mobile element // Microbiol. 1995 V. 141 (Pt 12).
P. 3015-3027.
Reddy A., Battisti L., Thorne С B. Identification of self-transmissible plasmids .n four Bacillus thuringiensis
subspecies//!. Bacteriol. 1987. V. 169. № 11. P. 5263-5270.
Reed D. S., Lackemeyer M. G., Garza N. L. et al. Severe encephalitis in Cynomolgus macaques exposed to
aerosolized Eastern equine encephalitis virus // J. Infect. Dis 2007. V. 196. P 441—450.
Reed D. S., Larsen T, Sullivan L. J et al. Aerosol exposure to Western equine encephalitis virus causes fever
and encephalitis in Cynomolgus macaques//J. Infect. Dis. 2005. V. 192. P. 1173—1182.
Reed D. S., Lind С. M., Sullivan L.J.et al. Aerosol infection of cynomolgus macaques with enzootic strains
of Venezuelan equine encephalitis // J. Infect Dis. 2004. V. 189 P. 1013—1017.
Reed K. D., Melski J. W., Graham M. B. etal. The detection of monkeypox in humans in the western hemisphere
// N. Engl. J. Med. 2004. V. 350. P. 342-350.
1094
Список литературы
Reid L. The landing site// Airooms transmission and airborne infection. 1973. N. Y. Toronto, P. 165—173.
Reilly T. J., Baron G. S., Nano F. E. et al. Characterization and sequencing of a respiratory burst inhibiting
acid phosphatase from Francisella tularensis //J. Biol. Chem. 1996. V. 271. P. 10973—10983.
Reimer L G. Q Fever// Clin. Microbiol. Ret. 1993. V 6, № 3. P. 193—198.
Reitano U. Considerazioni sullaguerramicrobica//Riv. di Fanteria. 1937. V. 1. P 78.
Rene D., Smith J. A. Integrat’on site selection by retroviral vectors: molecular mechanism and clinical
consequences // Hum. Gen. Ther. 2008. V. 19. P. 557—568.
Report from the United Kingdom of Great Britain and Northern Ireland (1992). DDA/4-92/BW111
[P. 213—251], Office of Disarmament Affairs, United Nations, New York. Report from the United States
of America (1992), DDA/4-92/BW111 [P. 252—428], Office of Disarmament Affairs, UN., N. Y.
Ribor W., Panchai R., Brittingham K. et a1 Anthrax lethal toxin impairs innate immune functions of alveolar
macrophages and facilitates Вас llus anthracis survival // Infect. Immun. 2006. V. 74. №9. P 5029- 5034.
Rigby C. E., Fraser A. Plasmid transfer and plasmid mediated genetic exchange in Brucella abortus // Can. J.
\fet. Res. 1989. V. 53. P. 326- 330.
Rigoni M. Paoli M., Milanesi E. et al. Snake phospholipase A2 neurotoxins enter neurons, bind specifically
to mitochondria, and open their transition pores I I J. Biol. Chem. 2008. V. 283. № 49. P. 34013—34020.
Roberts D. M., Nanda A., Havenga M.J. et al. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-
existing anti-vector immunity//Nature "’006. V. 441. P 239—243.
Robertson A. G, Robertson L. J From asps to allegations: biological warfare in history // Militar. Med 1995.
V 160. № 8 P. 369-373.
Robertson L., Farrealalnd I. D., Hinchliffethe P. M. Sensitivity of Brucella abortus to chemotherapeutic agents
//1. Med, Microbiol. 1973. V. 6. P. 549-557.
Robriguez R. // Riq. espan, di Medic. 19.37
Rodriguez PAlfonso A., ValcC etal. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTXin the
trumpet shell Charonia lampas lampas in Europe // Anal. Chem. 2008. V. 80. P 5622-5629.
Roest H., Ruuls R., Tilbuig J. et al. Molecular epidemiology of Coxiella burnetii from ruminants in Q fever
outbreak, the Netherlands// Emerg. Infect. Dis. 2011. V 17. № 4. P. 668—675.
Rose W. H. An evaluation of entomological warfare as a potential danger to the United States and European
Nato Nations// DPG 81-41, US Army, Dugway Proving Ground, Utah. 1981.
Roseberg В. H. Analysis of the anthrax attacks //1 ederation of American Scientists. 2002, 5 february
Rosebury T. Experimental air-borne infection Baltimor, 1947.
Rosebury T., Rabat E. A. Bacterial warfare, a critical analysis of the available agents, their possible military
applications, and the means for protection against them// J. Immunol. 1947. V 1. P. 7—96.
RouxE., YersinA Contributionaretudedeladiphtheria//Annal.deITnstitutPasteur. 1888. V.2.P629 661
Rowe-Magnus D. A., GuerootA M., Pioncard P. etal The evolutionary history of chromosomal superintegrons
provides an ancestry for multiresistant integrons // Pioc. Natl. Acad. Sci. USA. 2001. V. 98. P. 652-657.
Roy C G., Reed D. S., Wilhelmsen C. L. etal. Pathogenesis of aerosolized Eastern Equine Encephalitis virus
infection in guinea pigs // Virol. J 2009. V. 6 (170 doi: 10 1186/1743-422X-6-170).
Roy K., MaoH., Truong Yu L. etal. Патент США 5972707, МПК6, C12N15/00; Genu delivery system I
заявитель The Jons Hopkins University., США; заявл. 09.07 1997,08/890599; опубл. 26.10.1999.
Ruan К. H., SurlinoJ., Quicho F. A. etal. Acetylcholine receptor- a-bungarotoxin interactions: determination
ofthe region-to-region by peptide-to -peptide interactions and molecular modeling of the receptor activity
// Proc. Natl Acad Sci. USA. 1990. V. 87. P. 6156 -6160.
Ruhfel R. E, Robillard N. J., Thorne С. B. Interspecies transduction of plasmids among Bacillus anthracis,
B. cereus. and B. thuringiensis//J. Bacteriol. 1984. V. 157. № 3. P 708-711.
Rusnak J M KortepeterM., Ulrich R. etal Laboratory exposures to staphylococcal enterotoxin В //Emerg.
Infect. Dis. 2004. V 10 No. 9.1544-1549.
Russell D. W., Hirata R. K. Human gene targeting by viral vectors // Nat. Genet. 1998. V. 18. P. 325—330.
Russell Ж. C Adenoviruses: update on structure and function // J. Gen. Virol. 2009. V. 90. P. 1—20.
Ryan R. P., Dow J. M. Diffusible signals and interspecies communication in bacteria // Microbiology 2008.
V. 154. P 1845 -1858.
Romieu I I Revue de deux mondes. 1934 T XXIII. P. 41.
Sakural F. Development and evaluation of a novel gene delivery vehicle composed of adenovirus serotype 35 //
Biol. Pharm. Bull. 2008 V. 31. № 10 F 1819-1825.
Salyers A., Abile-Cuevas C. F. Why are antibiotic resistance genes so resistant to elimination? // Antimicrob.
Agents Chemother. 1997. V. 41. № 11. P. 2321—2325.
Список литературы
1095
Sam А., Веупоп И. Wound botulism // N. Engl. J. Med. 2010. V. 363. № 25. P. 2444.
Sanderson W. T., Stoddard R. R., EchtA. S. etal. Bacillus anthracis contamination and inhalational anthrax
in a mad processing and distribution center // J. of Appl. Microbiol. 2004. V. 96. P. 1048-1056
Sandstrom G., Lofgren S., TarnvikA. A capsule-deficient mutant of Francisella tularensis LVS exhibits
enhanced sensitivity to killing by serum but diminished sensitivity to killing by polymorphonuclear
leukocytes I I Infect. Immun. 1988. V. 56. P 1194- 1202.
Sanford D. C., Bamewall R. E., Vassar M. E. et al. Inhalational botulism in Rhesus macaques exposed to
botulinum neurotoxin complex serotypes Al and Bl //Clin. andVac. Immunol. 2010 V 17. p. 1293—1304.
Sartory A., Sartor-R. La Guerre Bacteriologique. Paris, 1935.
SaslawS., Eigelsbach H. T., Prior J. A. etal. Tularemia vaccine study. II Respiratory challenge //Arch Intern.
Med. 1961. V. 107. P. 702-714.
Saslaw S., Eigelsbach H. T., Wilson H. E. et al. Tularemia vaccine study. I Intracutaneous challenge // Arch.
Intern Med. 1961. V 107. P 689-701.
SaslawS., Eigelsbach H. T., Prior J. A. etal. Tularemia vaccine study. II Respiratory challenge//Arch. Intern.
Med. 1961. V. 107. P. 702-714.
SatoS., Nishimura T., UeharaK.etal. PhylogeographyofOstreopsisalong West pacific coast, with special refe-
rence to a novel clade from Japan // PLoS ONE. 2011. V. 6. № 12 (e27983. doi:10.1371/joumal.pone.
0027983).
Sawasdidoln C., Taweechaisupapong S., Sermswan R. Wet al. Growing Burkholderia pseudomallei in biofilm
stimulating conditions significantly induces antimicrobial resistance // PLoS ONE 2010 V 5, № 2
(e9196. doi-10.1371/joumal.pone.0009196).
Sawyer W. D, Dangerfield H. G., Hugge A. L.et al Antibiotic prophylaxis and therapy of airborne tularemia
I I Bacteriol. Rev. 1966. V. 30. P. 542-548.
Sawyer W. D., Jetnski J. V., HoggeA. L et al. Effect of aerosol age on the infectivity of airborne Pasteurella
tularensis for Macaca mulatta and man//J Baci 1966. V. 91. P. 2180—2186.
Schad E. M., Zaitseva I., Zaitsev V. N et al. Crystal structure of the superantigen staphylococcal enterotoxin
type A // EMBO J. 1995. V. 14. P. 3292- 3301.
Schell M. A., Lipscomb L., DeShazerD. Comparative genomics and an insect model rapidly identify novel
virulence genes of Burkholderia mallei //J. Bacteriology 2008 V 190. №7. P. 2306—2313.
Schell M. A., Ulrich R. L., Ribot W. R. el al. Type VI secretion is a major virulence determinant in Burkholderia
mallei // Mol. Microbiol. 2007. V. 64. P. 1466-1485.
Schiavo G., Matteoli M, Montecucco C. Neurotoxins affecting neuroexocytosis // Physiol. Rev. 2000. V 80.
№ 2. P. 717-765.
Schiavo G., Montecucco C. The structure and mode of action of botulinum and tetanus toxins 11 In: Rood J.,
McClane B., Songer J., Titball R. editors. The Clostridia molecular biology and pathogenesis. San Diego:
Academic Press; 1997. P. 295-322.
Schlievert P Searching for superantigens// Immunol. Infec. 1997 V. 26. № 2. P. 283—290.
Schmitt С. K., Meysick К. C., Brien A. D. Bacterial toxins: friends or foes? // Emerg. Infect Dis 1999. V. 5.
№ 2. P. 224-234.
SchnepfE., Cnckmore N., VanRieJ. etal. Bacillus thuringiensis and its pesticidal crystal proteins // Microbiol.
Mol. Biol. Rev. 1998. V. 62. P. 705-806.
Schnittler H, Feldmann H. Marburg and Ebola hemorrhagic fevers: does the primary course of Infection
depend on the accessibility of organ-specific macrophages? // Clin. Infect. Dis. 1998. V. 27. P. 404 -406
SchrickerR. L., Eigelsbach H. T., Mitten J. et al. Pathogenesis of tularemia in monkeys aerogenically exposed
to Francisella tularensis 425 // Infect. Immun. 1972. V. 5. P. 734—744.
Scientific and Technical Advisory Section, GHO, AFPAC. Report on scientific intelligence survey in Japan.
September and October 1945. V. 5. Biological Warfare (BIOS/JAP/PR/746).
Seemann P„ GemertC., SchmittS. etal. Detection of hemolytic bacteria from Palythoa caribaeorum (Cnidaria,
Zoantharia) using a novel palytoxin-screening assay // Anton. Leeuw. Int. J. G. 2009. (doi. 10.1007/
S10482-009-9353-4).
Segerman A., Arnberg, N., Erikson At et al. There are two different species В adenovirus receptors sBAR,
common to species Bl and B2 adenoviruses, and sB2AR, exclusively used by species B2 adenoviruses I/
J. Virol 2003. V. 77. P 1157-1162.
Sellin L. C. Botulism — an update // Milit Med. 1984. V. 149. № 1. — P. 12—16
Sena Esteves M., Saeki Y, Fraefel C. et al. HSV-1 amplicon vectors — simplicity and versatility // Mol Ther
2000. V. 2. № 1. P. 9-15
1096
Список литературы
Senhgal Р., Kumar О., Katneswararao М. etal. Differential toxicity profile of ricin isotbrms correlates with their
glycosylation levels // Toxicol. 2011. V. 282. № 1—2. P. 56—67.
Seow Y., Wood M. J. Biological gene delivery vehicles: beyond viral vectors // Molecular. Therapy. 2009. V. 17.
№ 5. P. 767-777.
Seshadri R., Paulsen I. T., Eisen J. A. et al. Complete genome sequence of the Q-fever pathogen Coxiella
burnetii// Proc. Natl. Acad. Sci. USA. 2003. V. 100. № 9. P. 5455-5460.
Seshadri R., ReadT. D., Merman W. C. etal. Genomic efforts with biodefense pathogens //Biological weapons
defense. N. J„ 2005. P 417-433
Shannon J. G., Heinzen R. A. Adaptive immunity to the obligate intracellular pathogen Coxiella burnetii //
Immunol Res. 2009. V. 43. № 1—3. P. 138—148.
Shannon J., Heinzen R. Adaptive immunity to the obligate Intracellular pathogen Coxiella burnetii // Immunol
Res. 2009. V. 43. Ns 1-3. P. 138-148.
Sharma A., Lia X., Bangari D. S. et al. Adenovirus receptors and their implications in gene delivery // Virus.
Res. 2009. V. 143. Ns 2. P. 184-194.
Shears N. T., Ross P. M. Blooms of benthic dinoflagellates of the genus Ostreopsis; an increasing and
ecologically important phenomenon on temperate reefs in New Zealand and worldwide // Harmful Algae.
2009. V. 8. P. 916-925.
Sherman M. B., WeaverS. Structure of the recombinant alphavirus Western equine encephalitis virus revealed
by cryoelectron microscopy // J. Virol. 2010. V. 84. V. 19 P. 9775-9782.
Shimizu Y, Kobayashi M. Apparent lack of tetrodotoxm biosynthesis in captured Taricha torosa and Taricha
granulose // Chem. Pharm. Bull. (Tokyo). 1983. V. 31. P. 3625-3631
Shinzato T., FurusuA., Nishino T. et al. Cowfish (Umisuzume, Lactoria diaphana) poisoning with
rhabdomyolysis // Inter. Med. 2008. V. 47 P. 853-856.
Shvedova A., Kisin E., Mercer R et al. Unusual inflammatory and fibrogenic pulmonary responses to single-
walled carbon nanotubes in mice Ц Am. J. Physiol. Lung. Cell. Mol. Physiol. 2005. V. 289. L698—L708.
Sidell R. et al. — cm. Medical aspects of chemical and biological warfare / Ed. E R. S.dell, E. T. Ta-fuqi,
D. R. Franz. Washington, 1997.
Silva T,, Costa E., Paixao T. etal. Laboratory animal models for brucellosis research //J Biomed. Biotechnol.
2011, Article ID 518323. doi:10.1155/2011/518323
Simionescu M 1 ung endothelium: structure-function correlates//The Lung: Scientific Foundations, Raven,
N.Y., 1991. P. 301-331.
Simon J., Hook R. Word war I gas warfare tactics and equipment. Oxford, 2007.
Sinclair R., Boo.ieS.A., Greenberg D. etal. Persistence of category a select agents in the environment //Appl.
Environ. Microbiol. 2008. V. 74. № 3. P. 555-563.
Singer O., Verma I. M. Applications of lentiviral vectors for shRNA delivery and transgenesis // Curr. Gene
Ther. 2008. V. 8. № 6.483-488.
Sirisanthana T., Brown A. E. Anthrax ofthe gastrointestinal tract//Emerg. Infect. Dis. 2002. V 8. P. 649—651.
SkocaJ M., Filipic M, Petkovic J. et al. Titanium dioxide in our everyday life; is it safe? // Radiol. Oncol 2011.
V. 45. P. 227 -247.
SkoldO. Resistance to trimethoprim and sulfonamides//\fet. Res. 2001. V. 32. P. 261—273.
Sliva K., Schnierle B. From actually toxic to highly specific — novel drugs against poxviruses // Virol. J. 2007.
V. 4. № 8 (doi: 10.1186/1743-422X-4-8).
Smith D. H. R factor infection of Escherichia coli lyophilized in 1946 // J. Bact 1967. V. 94. P. 2071—2072.
Smith J. F., Davis K., Hart M K. et al. Viral encephalitidis // Medical aspects of Chemical and Biological
Warfare / Ed. F. R. Sidell, E. T. Tafuqi, D R. Franz. Washington, 1997. P. 561-589.
Smith K. L., Devos V., BrydenH. etal. Bacillus anthracis diversity in Kruger national park//J. Clin. Microbiol.
2000. V 38. № 10. P. 3780-3784.
Smith P., Giroud M., Wiggins H et al. Cellular entry of nanoparticles via serum sensitive clathrin-mediated
endocytosis, and plasma membrane permeabilization // Inter. J. of Nanomed. 2012. V. 7. P. 2045—2055.
Smoragiewicz W. et al Trichothecene mycotoxins in the dust of ventilation systems in office buildings //
International archives of occupational and environmental health. 1993. V. 65. P. 113—117.
Song H., Hwang J., Yi H. et al. The early stage of bacterial genome-reductive evolution in the host I I PLoS
Pathog. 2010. V. 6. № 5 (el000922 doi:10.1371/joumal.ppat.1000922)
Song L., Hobaugh M., Shustak C. et al. Structure of staphylococcal alpha-hemolysin, a heptameric trans-
membrane pore // Science. 1996. V 274. № 14. P. 1859—1866.
Song Y, Li X., Du X. Exposure to nanoparticles is related to pleural effusion, pulmonary fibrosis and granuloma
// Eur. Respir. J. 2009. V. 34. P. 559-567.
Список литературы
1097
Song Y, Li X., Wang L. et al. Nanomaterials in humans: identification, characteristics, and potential damage
//Toxicol. Pathol. 2011. V. 39. P. 841-849.
Sonnabend W. F., Sonnabend U P., Hemzle R. et al. Isolation of Clostridium botulinum type G and
identification of type G botulinal toxin in humans report of five sudden unexpected deaths //J. Infect.
Dis. 1981. V. 143. № 1. P. 22-27
Sonnabend W. E, Sonnabend U. P., Kresch T. Isolation of Clostridium botulinum type G from Swiss soil
specimens by using sequential steps in an identification scheme // Appl. Environ. Microbiol. 1987. V. 53.
№ 8. P 1880-1884.
SosaS., DelFaveroG., De Bortoli Met. al. Palytoxin toxicity after acute oral administration in mice//Toxicol.
Lett. 2009. V. 191(2-3). P. 253-259.
Solo P. J., Roessler W. G. Staphylococcal enterotoxemia: pathologic lesions in rhesus monkeys exposed by
aerosol // Technical Manuscript 226. U.S. Government Printing Office, Washington. 1965. P. 1—16.
Speyer J. F. A simple and effective electroporation apparatus // Bio Technigue. 1990. V. 8. № 1. P. 28—30.
Spicer A J., Peacock M. G., Williams J. C. Effectiveness of several antibiotics in suppressing chick embryo letha-
lity during experimental infections by Coxiella burnetii, Rickettsia typhi. and R. rickettsii // Rickettsiae
and rickettsial diseases. Ed. W. Burgdorfer и R. L. Anacker. 1981, Academic Press, N. Y. P. 375—383.
Spratt B. G. Independent review of the possible health hazards of the large-scale release of bacteria during the
Dorset defense trials. 1999 (MoD Website http: // www. mod. uk).
SteedW. Aerial warfare: secret German plans //The Nineteenth century and after. 1934. July V. 116. P. 1—15.
Stenseth N., Atshabar В. B., Began M. et al. Plague past, present, and future // PLoS Med. 2008. V. 5. № 1.
e3. doi: 10.1371/joumaLpmed.0050003.
Stepanov A. V., Marinin L. L, Pomerantsev A. P. etal. Development of novel vaccines against anthrax in man
//J. Biotechnol 1996. V. 44. P. 155-160.
Stephenson E. H., Larson E. W., Dominik J. W. Effect of environmental factors on aerosol-induced Lassa virus
infection// J. Med. Virol. 1984 V. 14. № 4. P. 295—303.
Stem S., McNeil S. Nanotechnology safety concerns revisited // Toxicol. Sci. 2008. V 101. V. 4—21.
Strahilevitz J, Jacoby G. A., Hooper D. C. et al. Plasmid-mediated quinolone resistance, a multifaceted threat
// Clin. Microbol. Rev. 2009. V. 22. № 4 P. 664-689.
Strauss В. E. et al. Combating oncogene activation associated with retrovirus-mediated gene therapy of
X-linked severe combined immunodeficiency// Brazil. J. Med. Biol Res. 2007. V. 40. P. 601 -613.
Strong J. E., WbngG., Jones S. E. et al. Stimulation of Ebola virus production from persistent infection through
activation of the Ras/MAPK pathway//PNAS. 2008. V. 105, № 46. P 17982—17987.
Stuken A., Orr R. J. S., Kellmann R.eta' Discovery of nuclear-encoded genes for the neurotoxin saxitoxin in
Dinoflagellates//PLoSONE. 2011. V. 6. № 5 (e20096. doi:10.1371/journal.pone.0020096).
Su J., Yang J., Zhao D. et al. Genome-wide identification of Francisella tularensis virulence determinants //
Infect. Immun. 2007. V 75 P. 3089- 3101.
Su K., Wang D., Ye J. et al. Site-specific integration of retroviral DN A in human cells using fusion proteins
consisting of human immunodeficiency virus type 1 integrase and the designed polydactyl zinc-finger
proteinE2C //Methods. 2009. V. 47. № 4. P. 269—276 (doi:10.1016/j.ymeth.2009 01.001.).
Suh E. M., Kishi Y. Synthesis of palytoxin from palytoxin carboxylic acid // J. Am. Chem. Soc. 1994. V. 116.
P. 11205-11206.
Suhan M. L., Chen S. Y, Thompson H. A. Transformation of Coxiella burnetii to ampicillin resistance //J.
Bacteriol. 1996. V. 178. № 9. P 2701-2708
Sui L. M., Chen K, Hwang P. A. et al. Identification of tetrodotoxin in marine gastropod implicated in food
poisoning // J. Nat. Toxins. 2002. V. 11. V. 213—220.
Suk J. S., Suh J., Lai S K. et al. Quantifying the intracellular transport of viral and nonviral gene vectors in
primary neurons// Exp. Biol. Med. (Maywood). 2007 V. 232. P. 461—469.
Sullivan N., Yang Zhi-Yong., NabelG J Fbola virus pathogenesis: implications for vaccines and therapies //
J. Virol. 2003. V. 77. № 18. P. 9733 -9737.
Suzuki Y., Gojobori T. The origin and evolution of Ebola and Marburg viruses // Mol Biol. Evol. 1997. V. 14.
№ 8. P. 800-806.
Takada A., Ebihara H., Feldmann H. et al. Epitopes required for antibocy-dependent enhancement of Ebola
virus infection//J. Infec. Dis. 2007. V. 196. P. 347—356.
Takada A., Feldmann H., Ksiazek T. G. et al. Antibody-dependent enhancement of Ebola virus infection //
J. Virol. 2003. V. 77. № 13. P. 7539-7544.
Takada A., Watanabe S. Okazak K. et al. Infectivity-Enhancing antibodies to Ebola virus glycoprotein //
J. Virology. 2001. V. 75. № 5. P. 2324-2330.
1098
Список литературы
Takahashi И., Keim Р, Kaujma.in et al. Bacillus anthracis incident, Kameido, Tokyo, 1993 // Emerg Infect
Dis. 2004. V. 10. № 1. P. 117-119.
Takahata У Neutral theory of molecular evolution // Cun. Optn. Genet. De*'. 1996. V. 6 № 6. P 767—772.
Takano T., Kawakami S., Yamada S. et al. Antibody-dependent enhancement occurs upon re-infection with
the identical serotype virus in feline infectious peritonitis virus infection//J. Vet Med Sci. 2008 V. 70.
№ 12. P. 1315-1321.
TanC. T. T.,LeeE.J. D. A fatal case ofcrabtoxin(Lophozozymus pictor) poisoning// Asia Рас. J. Pharmacol.
1988. V. 3. P. 7-9.
TaniyamaS.,ArakawaO , TeradaM. et al. Ostreopsisspa possible origin of palytoxin (PTX) in parrotfish Scares
ovifrons I I Toxicon. 2003. V. 42. P. 29—33.
TamvlkA., Berglund L. Tularaemia I I Eur. Respir. J. 2003. V 21. P. 361—373.
Tavakoli N, Comanducci A., Dodd H M. et al. IS 1294, a DNA element that transposes by RC transposition
I I Plasmid. 2000. V. 44. № 1 P. 66 84.
Taylor R. R. Report on tularemia: Its diagnosis, and treatment// J. Arkansas M. Sol. 1050. V. 47. P 47.
Teale A. et al. Orthopoxviruses require a functional ubiquitin-proteasome system for productive replication
//J. Virol. 2009. V. 83. P. 2099-2108.
The problem of chemical and biological warfare SIPRI, 1970.
Thibault F. M., Hernandez £., Vidal D. R. et al. Antibiotic susceptibility of 65 isolates of Burkholderia
pseudomallei and Burkholderia mallei to 35 antimicrobial agents //J. Antimicrob. Chemother 2004. V. 54.
P. 1134-1138.
Thierry А. Патент 5908635 (А) США, МПК6 A61K9/127B2; C12N15/8X. Method forthe liposomal delivery
of nucleic acids I заявитель .Man Thierry патентообладатель The United States of America as repre-
sented by the Dep. of Health and Human Service; заявл. 05.08.1994,08/286,730; опубл. 01.06.1999.
Thomas C., KelecherM. F., McKee A. P. Botulism a complication of Clostridium botulinum wound infection Ц
Arch. Pathol. 1951. V. 51, № 6. P. 623-628.
Thompson. IV. L., Wannemacher P. IV. Structure-function relationships of 12,-13-epoxytricothecene
mycotoxins in cell culture: comparison to whole animal lethality // Toxicon 1986. V. 24. P. 985.
Thurmond K., Remsen E. E., Kowalewski T. et. al. Packaging of DNA by shell crosslinked nanoparticles //
Nucleic. Acids. Res. 1999. V 27. № 14. P. 2966-2971.
Tigertt IV. D., BenensonA. S., Gochenour IV IS. Airborne Q fever// Bacteriol Rev. 1961. V. 25. P 285.
Tinkle S. S., Antonini J M., Rich B. A. et al. Skin as a route of exposure and sensitization re chronic beryllium
disease//Environ. Health. Perspect. 1003 V. Ill P 1202-1208.
TiradoS. M. Yoon K. S. Antibody-dependent enchancement of virus infection and disease //Viral. Immunol.
2003. V. 164. № 1. P. 69-86.
Toleman M A., Bennett P M., IValsh T. Common regionse.g orf513 and antibiotic resistance: IS91-like
elements evolving complex class 1 integrons 11 J. Antimicrob. Chemother. 2006. V. 58. P. 1—6.
Towner J. S., Amman B. R., Sealy T. К et al. Isolation of genetically diverse Marburg viruses from Egyptian
fruit bats// PLoS Pathog. 2009. V. 5. № 7 (el000536. doi: 10.1371/joumal.ppat. 1000536).
Towner J. S., Sealy T. R., Khristova M. R. etal Newly discovered Ebola vires associated with yemorrhagic fever
outbreak in Uganda// PI oS Pathog. 2008. V. 4. № 11 (e!000212 doi:10.1371/journal.ppat.i,)00212).
Townsend Peterson A., Bauer J., Mills J. N. Ecologic and geographic distribution of filovires disease // Emerg.
Infect. Dis. 2004. V. 10. № 1. P. 40-47.
Trescos Y., Tournier J-N. Cytoskeleton as an emerging target of anthrax toxins//Toxins. 2012. V. 4. P. 83—97.
TnllatA Immunization by inhalation //J. Arr... Med Ass. 1932. V. 99. № 7 P 573—574
Tsukamoto M., Okuda T. et al. Bovine serum albumin as a lyoprotectant for preparation of DNA dry powder
formulations using the spray-freeze drying method // Biol. Pharm. Bull. 2012. V. 35. № 7. P. 1178-1181.
Tuanyok A., Leadem B. R., Auerbach R. et al. G enonuc islands from five strains of Burkholderia pseudomallei
I I BMC Genomics. 2008. V. 9 (566 doi. 10.1186/1471-2164-9-566).
Tubaro A., Durando P., Del Favero G. et al. Case definitions for human poisonings postulated to palytoxins
exposure//Toxicon. 2011. V. 57. P. 478- 495.
Tucker J. Toxins: area weapons Ц Gene Wa-ch. 1985. V.2. № 2. P. 10—12.
Turkmani A., loannidisA., Christidou A. et al. In vitro susceptibilities ot Brucella melitensis isolates to eleven
antibiotics//Annals of Clin. Microbiology and Antimicrobials. 2006. V. 5 (doi: 10.1186/1476-0711-5-24).
TwenhalN. A., Alves D. A., Purcell В. K. Pa.hology of inhalahonal Francisella tularensis spp. tularensis SCHU
S4 infection in African green monkeys (Chlorocebus aethiops) // Vet Pathol. 2009. V. 46. P. 598- 706.
Ulbricht IV. Sodium channel inactivation- Molecular determinantsand modulation // Physiol. Rev. 2005. V 85.
P. 1271-1301.
Список литературы
1099
Ulrich R. G., Sidell S., Taylor T. et al. Staphylococcal enterotoxin В and related pyrogenic toxins // Medical
aspects of Chemie, and Biol. Warfare I Ed. F R. Sidell, E. T. Tafuqi, D. R. Franz. Wash., 1997. P. 621
Urich S. K., Petersen J. M. In vitro susceptibility of isolates of Francisella tularensis types A and В from North
America // Antimicrob. Agents and Chemother. 2008. V. 52. № 6. P. 2276- 2278.
US Army activity in the U.S. Biol, warfare programs V. 1 and 2. In: Hearings before the Subcommittee, un
Health and Scientific Research of the Committee on Human Resources. United States Senate. 95 Congr.
1 Session. March 8 and may 23, 1977. Washington, U.S. Government Printing Office, P 22—234.
US Army activity in the U S. Biological warfare programs. V. 1. February 24,1977 (Unclassified), P. 57—60,
202-209, 224-234.
US Departments oftne Army and Air Force, Military biology and biological agents. March 1964 (TM 3-216
andAFM 355-6)
US Departments of the Army, Navy and Air Force. Employment of chemical and biological agents March,
1966 (FM 3-10; NWIP 36-2; AFM 355-4 FMFM 11-3).
Usamriid’s medical management of biological casualties handbook (Forth Edition)/ Edit. M. Kortepeter, G.
Christopher, T Cieslak, R. Culpepper, R Darling. J. Pavlin, J. Rowe, K. McKee, E. Eitzen, USAMR1ID.
Fort Detrick, Maryland, 2U01
Ushkaryov Y. A., Rohou A., Sugita S. a-Latrotoxin and its receptors // Handb. Exp. Pharmacol. 2008. V. 184.
P. 171-206.
Valcarcel C. A., Serra M. D., Potrich C. et al Effects of lipid composition on membrane permeabilization by
sticholysin I and II, two cytolysms of the sea anemone Stichodactyla helianthus // Biophys. J. 2001. V. 80.
P. 2761-2774.
Yale-Gonzalez C., Gomez- Limia B., Vieytes Af R. et al. Effects of the marine phycotoxin palytoxin on neuronal
pH in primary cultures of cerebellar granule cells // J. Neurosci. Res 2007. V. 85. P. 90-98.
Van Andel R., Sherwood R., Gennings C. et al. Clinical and patnologic features of Cynomolgus macaques (Maca-
ca fascicularis) Infected with aerosolized Yersinia pestis // Compar Med. 2008. V. 58. № 1. P. 69—75.
Van Ert M. N., Easterday W. R., Huynh L. et al. Global genetic population structure of Bacillus anthracis //
PLoS ONE. 2007. Is. 5. e461. doi:10.1371/joumal.pone.0000461.
Van Ert M. N., Easterday W. R., Simonson T. Set al. Strain-specific single-nucleotide polymorphism assays
for the Bacillus anthracis Ames strain // J. Clin. Microbiol. 2007. V. 45. P. 47—53.
Vassilevski A. A., Kozlov S. A., Grishin E. V. Molecular diversity of spider venom // Biochem. (Moscow). 2009.
V. 74. № 13. P. 1505-1534.
Verger J. M., Grimont F„ Grimont P. et al Brucella a monospecific genus as shown by deoxyribonucleic acid
hybridization//Int. J. Syst. Bacteriol. 1985 V. 35. P. 292-295.
Verhoeyen E., Cosset F. L. Surface-engineering of lentiviral vectors // J. Gene Med. 2004. V. 6. P 83—94.
Vietri N. J., Purcell В. K., Toberry S A. et al. A short course of antibiotic treatment is effective in preventing
death from experimental mhalational anthrax after discontinuing antibiotics //J 1 nfect. Dis. '’009. V. 199.
Ne 1. P. 336-341.
Vbdkin M., Leppla S H Cloning of the protective antigen gene of Bacillus anthracis // Cell. 1983. V. 34.
P 693 -696.
Vogel S., S6rdy M., Glos K. et al. The munich outbreak of cutaneous cowpox infection: transmission by infected
pet rats//Acta Derm. Venereol. 2011. V. 91 (doi: 10.2340/00015555-1227)
Vogler A , Birdsell D., Price L. et al. Phylogeography of Francisella tularensis: global expansion of a highly fit
clone //J. Bacteriol. 2009. V. 191. № 8. P. 2474-2484.
Volger A., Busch J., Percy—Fine S. et al. Molecular analysis of rifampin resistance in Bacillus anthracis //
Antimicrob. Agents Chemother. 2002. V.46. № 2. P. 511—513.
Walker D., McCormick J, Johnson K. et al. Pathologic and Virologic Study of Fatal Lassa Fever in Man 11 Am.
J. Pathol. 1982. V. 107. P. 349-356.
Wallace M. J., Smith D. W., Broom A. K. etal. Antibody-dependent enhancement of Murray Valley encephalitis
virus virulence in mice // J. Gen. Virol. 2003. V. 84. P. 1723—1728.
Walters F. S., Stacy Ch. M., Lee M K. etal An engineered chymotrypsin/cathepsin G site in domain 1 renders
Bacillus thuringiensis Cry ЗА active against western corn rootworm larvae // Appl. Environ. Microbiol.
2008. V. 74. P. 367-374.
Wang E., Bowen R. A., Medina G. et al. Virulence and viremia characteristics of 1992 epizootic subtype Ic
\fene 'uelan equine encephalitis viruses and closely related enzootic subtype Id strains//Am J. Trop. Med.
Hyg. 2001. V. 65. № 1. P. 64-69.
Wang S., Ma N., Gao S. J. et al. Transgene expression in the brain stem effected by intramuscular injection of
polyethylenimine/DNAcomplexes// Mol. Ther 2001 V3 P.658—664.
1100
Список литературы
Wang Yu-Qiang, Su Jing, Wu Fei et al. Biscarbamate cross-linked polyethylenimine derivative with low
molecular weight, low cytotoxicity, and high efficiency for gene deliveiy// Intern. 1 of Nanomed. 2012.
V. 7. P. 693-704.
Wanisch K., Yanez-Munoz R. J Integration-deficient Lentiviral vectors: a slow coming of age // The American
Society of Gene & Cell Therapy. 2009. V. 17. № 8. P. 1316-1332.
Wanisch K., Ynez-Mucoz R. J. Integration-deficient lentiviral vectors: a slow coming of age // www.mole-
culartherapy.org. 2009. V. 17. № 8. P. 1316—1332.
Wannemacher R. W., Anderson J. B. Inhalation ncin: aerosol procedures, toxicology and therapy I I Inhalation
toxicology, 2nd ed., eds. H. Salem and S. Katz. 2006. P. 973—980.
Warfield K.L., Swenson D.L., Olinger G G., Nichols D K. et al. Gene-specific countermeasures against Ebola
virus based on antisense phosphorodiamidate morpholino oligomers // PLoS Pathogens. 2006. V. 2.
Is. l.P.5-13.
Weaver S. C., KangW.L., Shirako Y. etal. Recombinational history and molecular evolution of western equine
encepnalomvelitis complex alphaviruses//J. Virol. 1997. V. 71. P. 613—623.
Weaver S. C, Reisen W. K. Present and future arboviral threats // Antiviral Res 2010. V. 85. № 2 (328.
doi: 10.1016/j.antiviral.2009.10.008).
Webster R. Influenza: an emerging disease // Emerg. Infect. Dis. 1998. V. 4. № 3. P. 416- 419.
Wells W. F. Air-borne infection. II. Droplets and droplet nuclei//Am. J. Hyg. 1934. V. 20. P. 611-618.
Wells W. F., Rateliffe H. L., Crumb C. On the mechanism of droplet nuclei infection. II. Quantitative
experimental air borne infection in rabbits // Am. J. Hyg. 1948. V. 47. P. 11—28.
Westwood J. C. N, Boulter E. A., Bowen E. T. W. et al. Experimental respiratory infection with poxviruses. I.
Clinical virological and epidemiological studies // Br. J. Exp Pathol. 1966. V. 47. № 5. P. 453—465.
Whatmore A., Perrett L , MacMillanA. Characterisation of the genetic diversity ofBrucella by multilocus
sequencing // BMC Microbiol. 2007. V. 7:34 doi 10.1186/1471-2180-7-34.
White J. D., Rooney J. R., Prickett P. A et al. Paithogenesis of experimental respiratory tularemia in monkeys
// J. Inf Dis. 1964. V. 47. P. 277-286.
WienerS. L. Strategies ofbiowaffarae defense // Mil. Med. 1987. V. 152. № 1. P. 25—28.
Wiese M., Agostino P., Troco К. M. et al. Neurotoxic alkaloids: saxitoxin and its analogs I I Mar. Drugs. 2010.
V. 8. P. 2185-2211.
Wilhelmseann C., Pitt L Lesions of acute inhaled lethal ricin intoxication in Rhesus monkeys // \fet Pathol.
1996. V. 33. P. 296-302.
Williams D. A. RAC reviews serious adverse event associated with AAV therapy trial I I Mol. Ther. 2007. V. 15.
P. 2053-2054.
Williams P., Wallace D. Unit 731: The Jap. Army’s secret ot secrets. London, 1989.
Winn W.C., WalkerD. H. The pathology of human Lassa fever// Bull. World Health Organ. 1975 V. 52.
P. 535-545.
Win-Shwe Tin-Fin., Fujimak' H. Nanoparticles and neurotoxicity// Int. J. Mol. Sci. 2011. V. 12. P. 6267—6280.
Wirth M. C., Georghiou G. P., Federici B. A. CytA enables CryIV endotoxins of Bacillus thuringiensis to over-
come high levels of CryIV resistance in the mosquito Culexquinquefasciatus//Proc. Natl. Acad. Sci. USA.
1997. V. 94. P. 10536-10540.
Wirtz S., Neurath M. F. Gene transfer approaches for the treatment of inflammatory bowel disease // Gen. Ther.
2003. V. 10. P. 854-860.
Wiser L, Balicer R. D., Cohen D. An update on smallpox vaccine candidates and their role in bioterrorism
related vaccination strategies // Vaccine. 2007. V. 25. P.976—984.
Wittmann T J, Biek R., Hassanin A. et al Isolates of Zaire ebolavirus from wild apes reveal genetic lineage
and recombinants // Proc. Natl. Acad. Sci. USA. 2007. V. 104. № 43. P. 17123—17127.
WokerG. We wurde ein neuer Krieg aussehen? — Zunch, Leipzig, 1931.
Wolf-Watz H., Portnoy D., Bolin I. etal. Transfer of the virulence plasmid of Yersinia pestis to Yersinia
pseudotuberculosis // Infec. Immun. 1985. V. 48. N° I P. 241—243.
WoodsJ. B. Antimicrobials for biological warfare agents //Biological weapons defense. N. J., 2005. P. 285—315.
WbrobeyM., HolmesE. C. Evolutionary aspects of recombination in RNA viruses//J. Gen. Virol. 1999. V. 8.
P. 2535-2543.
Wozniak R. A. F., Fouls D. E., Spagnoletti M. et al. Comparative ICE genomics: insights into the evolution of
the SXT/R391 family of ICEs// PLoS Genet. 2009. V. 5. № 12. el000786.
Rh I)., Aronson A. I. Localized mutagenesis defines regions of the Bacillus thuringiensis deltaendotoxin
involved in toxicity and specificity // J. Biol. Chem 1992. V. 267. P. 2311—2327.
Список литературы
1101
Wu Е, Fan S., Martiniuk F. et al. Protective effects of anti-ricin А-chain antibodies delivered intracellularly
against ricin-induced cytotoxicity//World. J. Biol. Chem. 2010. V. 26. № l.P. 188—195.
Wu Lien-Fen U A., Chun J. W., Pollitzen R. etal. Plague. Shanghai, 1936.
Wu S. J., Koller C. N., Miller D.L. at al. Enhanced toxicity of Bacillus thuringiensis Cry ЗА delta-endotoxin
in coleopterans by mutagenesis in a receptor binding loop // FEBS Lett. 2000. V. 473. P. 227-232.
WuS. Y., McMillan N. A. J. Lipidic systems for in vivo siRNAdelivery//AAPSJ.2009. V. 11,№4.P.639—642.
Wyers M., Formenty P, Cherel Y. etal. Histopathological and Immunohistochemical studies of lesions
associated with Ebola virus in a naturally infected chimpanzee//J. Infec. Dis. 1999. V. 179. (Suppl 1):
S54-S59.
Wyssokowitsch et Zabolotny 11 Ann. Inst. Past. 1897 V. XL P. 665.
Yamaizumi M., Mekada £., Uchida T et al. One molecule of diphtheria toxin fragment A introduced into a
cell can kill the cell // Cell. 1978. V. 15. P. 245-250.
Yamazaki Y., Matsunaga Y., Tokunaga Y Snake venom vascular endothelial growth factors (VEGF-Fs)
exclusively vary their structures and functions among species 11 J. Biol. Chem. 2009. V 284, № 15.
P. 9885-9891.
Yao J. S., Kariwa H., Takashima I. et al. Antibody-dependent enhancement of hantavirus infection in
macrophage cell lines // Arch. Virol. 1992. V. 122. № 1—2. P. 107- 118.
Yeaman M. R., Roman E., Baca O. Antibiotic susceptibilities of two Coxiella burnetii isolates implicated in
distinct clinical syndromes//Antimicrob Agents Chemother. 1989. V. 33. № 7. P 1052—1057.
Yim G., Wang H. FL Davies J. Antibiotics as signaling molecules // Phil. Trans. R Soc. B. 2007. V. 362.
P. 1195-1200.
Yin FL L., Lin H. S., HuangC. C. et al. Tetrodotoxication with Nassauф glans: a possibility of tetrodotoxin
spreading in marine products near Pratas island 11 Am. J Trop. Med. Hyg. 2005. V. 73, № 5. P 985 990.
Yingst S. L., Huzella L. M., Chuvala L. et al. A rhesus macaque (Macaca mulatta) model of aerosol-exposure
brucellosis (Brucella suis): pathology and diagnostic implications //J. Med. Microbiol. 2010 V 59.
P 724- 730.
Yotsu-Yamashita M. Chemistry of puffer fishtoxm//J. Toxicol.: Toxin Rev. 2001. V. 20. P. 51 66.
Yotsu-Yamashita M., Kim Y. EL, Dudley S. C. etal The structure of zetekitoxin AB, a saxitoxin analog from
the golden frog Atelopus zeteki: a potent sodium channel blocker // Proc. Natl. Acad Sci. USA. 2004.
V. 101. P. 4346-4351.
Yu Li, Carroll D., Gardner S. et al. On the origin of smallpox: Correlating variola phylogenies with historical
smallpox records // Proc. Natl. Acad. Sci. USA. 2007. V. 104. P. 15787—15792.
Yun N. E, PengB. EL, Bertke A. S. etal CD4+T cells provide protection against acute lethal encephalitis caused
by Venezuelan equine encephalitis virus // Vaccine. 2009. V. 19. № 27. P 4064 4073.
ZaJcsM. A., PaesslerS. Encephalitic Alphaviruses//Ve*. Microbiol. 2010. V. 140. № 3 -4 (281. doi:10.1016/
j.vetmic.2009.08 023).
Zaiss A. K, Machadn FL, Herschman H. R. The01 influence of innate and pre-existing immunity on adenovirus
therapy//J. Cell. Biochem. 2009. V. 108. № 4 (778. doi:10.1002/jcb.22328).
ZehH. J., Bartlett D. L Development of a replication-selective, oncolytic poxvirus for the treatment of human
cancers // Cancer Gen. Ther. 2002 V. 9. P. 1001—1012.
Zeng J., Wang S. Enhanced gene delivery to PC 12 cells by a cationic polypeptide // Biomaterials. 2005. V. 26.
P. 679-686.
ZentnerR J. Techniques of aerosol formation // Bact. Rev. 1961. V. 25. № 3. P. 188—193.
Zhang J Evolution by gene duplication: an update // Trends. EcVol. 2003. V. 18. Ns 6. P. 292-298.
Zhang K. Y., La Russa V. F., Reiser J. Transduction of bone-marrow-derived mesenchymal stem cells by using
lentivirus vectors pseudotyped with moOiled RD114 envelope glycoproteins // J. Virol. 2004. V. 78.
P. 1219-1229.
Zhang Y., Colon F., Zhu C. et al. Intravenous nonviral gene therapy causes normalization of striatal tyrosine
hydroxylase and reversal of motor impairment in experimental parkinsonism // Hum. Gene. Ther. 2002.
V. 14. P. 1-12.
Zimmer T. Effects of tetrodotoxin on the mammalian cardiovascular system // Mar. Drugs. 2010. V. 8.
P 741-762.
Zinn-Justin S., Roumestand C, Gilquin B. et al. Three-dimensional solution structure of curaremimetic toxin
from Naja nigricillis venom: a proton NMR and molecular modeling study t/ Biochemistry. 1992. V. 31.
P. 11335-11347.
ZubayG., Balfour R., ChubakB. etal. Agents ofbioterrorism. Pathogens and their weaponization. N. Y., 2005.
ПРИЛОЖЕНИЕ А
БАКТЕРИОЛОГИЧЕСКАЯ ВОЙНА1
Доклад специального консультанта по вопросам бактериологической
войны Д. У. Мерка2 военному министру по состоянию на 3 янвгпя 1946 г.
Комментарий к докладу.
Разведывательные сводки, составленные военными разведывательными органами в
Японии после оккупации и полученные уже после подготовки господином Мерком доклада
военному министру, указывают на то, что в подготовке к биологической войне Япония
достигла определенных успехов. В результате проведенного расследования стало извест-
но, что с 1936 г. по 1945 г. японская армия усиленно разрабатывала биологическое оружие.
Японские военные специалисты развили интенсивную деятельность по введению БО
в практику наступательных боевых действий. Модификации различного оружия, разрабо-
танные в лабораторных условиях, испытывались на армейских полигонах, где проводились
также эксперименты по использованию бактериологических средств в диверсионных це-
лях. Несмотря на то, что Япония добилась в этой области определенного успеха, ко вре-
мени окончания войны она не достигла еще уровня, при котором эти планы наступатель-
ных действий могли бы быть реализованы.
Каких-либо данных о применении противником подобных средств ведения войны не
существует, и, конечно, неизвестно, могла ли японская армия со временем усовершенство-
вать этот вид оружия и применить его в случае продолжения войны. Однако проблема
противобиологической защиты была в этой стране предметом программы активных ис-
следований и разработок.
В настоящем докладе описываются объединенные усилия американских ученых, про-
мышленности и вооруженных сил совместно с аналогичными организациями Великобри-
тании и Канады, по разработке систем защиты от нападения противника с применением
БО. Хотя в интересах национальной безопасности сведения о военных разработках не под-
лежат разглашению, эти исследования во многом способствуют повышению уровня на-
ших знаний в отношении борьбы с болезнями, поражающими людей, животных и расте-
ния. Поэтому достигнута договоренность, по которой эти в целом ценные для человечества
данные должны стать доступными д ля научной общественности из источников, ответствен-
ных за эту работу. Это будет осуществляться средством сообщений перед научными собра-
ниями, публикаций в научных журналах и других путей, по которым информация о дос-
тижениях в науке и медицине распространяется в мирное время.
Господин министр!
Мощь вооруженных сил воюющей страны зависит не только от оружия, которое фак-
тически применяется, чтобы нанести поражение противнику, но также от основательное -
ти, с которой страна готовится к нападению любого возможного вида. США следовали этой
основной военной доктрине при ведении войны против стран оси. Вид оружия, который
мог бы быть использован во время Второ! мировой войны, — возможное средство напа-
1 US Army activity in the U.S. Biological warfare programs. V. 1 and 2. In: Hearings before the
Subcommittee on Health and Scientific Research of the Committee on Human Resources United Sta-
tes Senate. Ninety-Fifth Congress. First Session. March 8 and may 23, 1977. Washington, U.S. Go-
vernment Printing Office, 1977. P. 22—234.
2 Geoige Wilhelm Herman Emanuel Merck (1894—1957) — президент фармацевтической ком-
пании Merck & Co. c 1925 г no 1950 r.
Приложение А
1103
дения наших противников — это биологическое оружие. Биологическая война характери-
зуется использованием бактерий, грибков, вирусов, риккетсий и токсических продуктов
живых микроорганизмов (в отличие от синтетических, химических веществ, используемых
в виде газов или ядов) с целью поражения живой силы, животных или растений. Этот спо-
соб ведения войны был известен уже в период Первой мировой войны, хотя он применялся
лишь в очень ограниченном масштабе. Имеются неопровержимые гоказательства, напри-
мер, того, что в 1915 г. немецкие агенты заражали болезнетворными бактериями лошадей
и крупный рогатый скот, направляемый из портов Соединенных Штатов союзникам.
В период между Первой и Второй мировыми войнами общий интерес к возможности
развязывания биологической войны поддерживался учеными и военными специалиста-
ми во многих странах, и многие пришли к убеждению, что такой способ военных действий
возможен или даже вероятен в будущем По мере того как межвоенный период прибли-
жался к концу, мнения в Соединенных Штатах относительно возможности биологической
войны расходились, однако элементарное благоразумие диктовало лицам, ответственным
за национальную оборону, необходимость серьезного рассмотрения опасностей, которые
могут скрываться в этой области. Мнение тех, кто относился настороженно к подобным
опасностям, было официально представлено на рассмотрение военному министерству
осенью 1941 г., после чего министр Стимсон немедленно обратился в национальную ака-
демию наук с предложением создать комитет для полного и тщательного изучения суще-
ствующего положения и возможной перспективы.
После детального изучения этого вопроса комитет, известный как комитет WBC (Кон-
сультативный комитет по биологическому ору жию), пришел к выводу (в своем докладе в
феврале 1942 г.), что биологическая война определенно возможна и необходимо принятие
срочных мер для организации соответствующей обороны. В докладе, в частности, гово-
рится: «Вопрос о военной ценности биологического оружия будет спорным до тех пор, пока
его эффективность не будет определенно доказана или опровергнута на практике. Обще-
известно, что любой способ, который позволяет воюющей стране достигну ть превосход-
ства, будет широко использоваться этой страной. Следовательно, имеется лишь один ра-
зумный путь, которого следует придерживаться, а именно: изучать возможности подобного
оружия со всех точек зрения, сделать все возможное, чтобы уменьшить его эффективность
и тем самым снизить вероятность его использования».
На основании этих выводов министр Стимсон рекомендовал президенту Рузвельту
создать гражданскую организацию, которая должна возглавить все работы, связанные с
изучением проблем биологической воины. Летом 1942 г. такая организация под названием
«Служба военных исследований» была создана в составе Управления государственной
безопасности. Во главе ее был поставлен доктор Джордж У. Мерк. В интересах эффектив-
ности работы, экономии и обеспечения секретности служба военных исследований остава-
лась небольшой организацией. Она занималась главным образом координацией проводи-
мых работ, используя для этой цели научных работников, материальную базу и опыт
различных государственных и частных учреждений. Ее рекомендации облекались в форму
приказов и директив, издаваемых различными военными службами, в частности медицин-
ской службой армии и флота и химической службой армии. С различными видами воору-
женных сил, министерствами здравоохранения, сельского хозяйства и внутренних дел
поддерживались соответствующие связи. Необходимая информация доставлялась армей-
ской и военно-морской разведкой; вопросы общественной информации решались совме-
стно с бюро общественной информации при военном министерстве, управлением воен-
ной информации и управлением военной цензуры. С целью оказания консультативной
помощи службе военных исследований национальная, академия наук и Научно-исследо-
вательский комитет США создали в качестве консультативного органа по вопросам био-
логической войны специальный комитет (комитет АВС), куда вошли известные ученые.
1104
Приложение A
За несколько месяцев до этого был организован обмен информацией по вопросам БО
между США, Англией и Канадой, и, кроме того, были приняты меры к проведению в этих
странах обмена специалистами в этой области.
Первая и главная задача, которая встала перед службой военных исследований, зак-
лючалась в разработке мероприятий по зашите в случае возможного биологического на-
падения. Совместно со службами вооруженных сил были приняты меры для зашиты за-
пасов воды и продовольствия на материке, на Гавайях, на островах Карибского моря, в
частности, в зоне Панамского канала, и, наконец, на всех заокеанских базах.
Была разработана большая программа по сбору разведывательных данных по БО с
привлечением для этой цели органов разведки видов вооруженных сил, управления стра-
тегической разведки и федерального бюро расследований. Кроме того, делались приготов-
ления к засылке специально подготовленных офицеров разведки в зоны боевых действий
с целью расширения сбора данных по вопросам биологического оружия.
Однако самым значительным достижением службы военных исследований было осу-
ществление программы научных исследований и разработок, имевшей целью расширение
рамок знаний относительно использования патогенных биоагентов в качестве оружия и
средств зашиты от их возможного использования противником. Самые авторитетные уче-
ные произвели тшательное изучение и отбор всех известных патогенных биоагентов для
того, чтобы определить возможности использования их противником Отобранные болез-
нетворные биоагенты подверглись дальнейшему изучению в лабораториях ряда универ-
ситетов и частных учреждений с целью экспериментального определения их летальных
свойств, способов продуцирования и зашиты при их использовании. Однако по мере вы-
полнения программы становилось очевидно, что исчерпывающие знания о болезнетвор-
ных биоагентах и их использовании в качестве оружия, а также разработка средств заши-
ты от них невозможны без проведения крупномасштабных экспериментальных работ.
В ноябре 1942 г. служба военных исследований предложила химическому корпусу ар-
мии взять на себя ответственность за расширение программы научных исследований и
разработок, включая создание и эксплуатацию специальных лабораторий и пилотных уста-
новок. Местом, выбранным для такого объекта, был Кэмп-Детрик, Фредерик, штат Мэри-
ленд, где в апреле 1943 г. и было начато строительство лабораторий. Когда объект был
введен в действие, планы научных работ, разработанные при поддержке службы военных
исследований, были переданы химическому корпусу армии для дальнейшей их реализации
в Кэмп-Детрик. Служба военных исследований осуществляла общий контроль за всеми
работами в этой области, поддерживала фундаментальные научные исследования в уни-
верситетах и частных учреждениях и обеспечивала Кэмп-Детрик научными кадрами и обо-
рудованием.
В декабре 1943 г., стратегическое управление сообщило объединенному комитету на-
чальников штабов об имеющихся признаках подготовки германского командования к
применению биологического оружия. Хотя сведения о намерениях Германии применить
эти средства были недоказательными, имелась обширная конкретная информация, полу-
ченная в результате проводимых в США, Англии и Канаде работ, о возможности осуще-
ствления такого нападения с применением БО В связи с этим в январе 1944 г. было реше-
но форсировать все исследования в этой области, в частности, касающиеся средств защиты
войск при возможном применении противником этого вида оружия, и возложить большую
часть ответственности за выполнение программы работ по проблемам БО на военное ми-
нистерс гво. По указанию президента передача была завершена в июне 1944 г., когда на
химический корпус была возложена ответственность за реализацию разработанной в во-
енном министерстве совместно с управлением начальника военно-медицинской службы
программы исследований по некоторым важным вопросам обороны Министерство ВМС
продолжало вносить существенный вклад в выполнение этой программы, работая в тесном
Приложение А
1105
сотрудничестве с военным министерством. Осуществление программы исследований и
разработок значительно ускорилось, несмотря на запрещение производить биоагенты в
больших количествах без специального на то разрешения военного министра. В действи-
тельности больших запасов этих агентов никогда не было накоплено.
После того как военное министерство приняло на себя полную ответственность за
выполнение работ по БО, военный министр утвердил начальника службы военных иссле-
дований в должности специального консультанта по вопросам биологической войны и
учредил «Комитет США по вопросам биологической войны» во главе с Марком. Комитет
был создан в качестве консультативного органа при военном министре для рассмотрения
политических проблем и для поддержания тесной связи с английской и канадской груп-
пами, занимающимися вопросами биологической войны. В состав комитета вошли пред-
ставители химической службы и военно-медицинского управления армии; медицинско-
го управления и управления вооружения ВМС, войск армейского обслуживания, отдела
новых формирований управления специальной службы военного министерства, разведы-
вательного управления (G-2) и стратегического управления. Академия наук США и На-
учно-исследовательский совет США в качестве совещательного органа при военном ми-
нистерстве организовали также специальный комитет, так называемый ДЕФ-комитет.
Отдел специального планирования при химическом корпусе армии, который после
июня 1944 г. нес основную ответственность за выполнение программы, имел в своем рас-
поряжении около 3900 сотрудников, из которых примерно 2800 состояли на службе в ар-
мии, около 1000 — в военно-морском флоте и примерно 100 человек были гражданскими
лицами. Работы, проводимые отделом, специального планирования на всех его четырех
ооъекгах, выполнялись при тесном сотрудничестве армейского, военно-морского и граж-
данского персонала. Работы велись очень быстро и в условиях строжайшей секретности.
При этом были достигнуты весьма значительные успехи.
Первым объектом, учрежденным отделом специального планирования в апреле 1943 г.,
был основной научно-исследовательский центр в штате Мэриленд. Вторым явился центр
полигонных испытаний, созданный летом 1943 г. в штате Миссисипи. Третьим был завод
для произво 1ства биологического оружия, построенный в начале 1944 г. в штате Индиана.
Четвертый — центр полигонных испытаний был создан летом 1944 г. в штате Юта. Упомя-
нутые объекты уникальны во многих отношениях, поскольку они требовали специальных
конструктивных решений, удовлетворяющих требованиям изучения совершенно новых
проблем Необходимость в высокой степени точности и строгом соблюдении требований
техники безопасности создавала много сложных инженерных проблем. Необходимо было
разработать, сконструировать и смонтировать специальное оборудование для проведения
процессов, ранее никогда не изучаемых и не осуществляемых в таком масштабе.
Хотя в настоящее время еще невозможно раскрыть названия специфических биоаген-
тов, интенсивная работа над изучением которых наводилась в этих центрах, все же можно
рассказать об общем характере проблемы и типе информации, полученной в этой обла-
сти. Следует подчеркнуть, что хотя основной целью при всех этих попытках была разра
ботка методов зашиты от возможного применения БО, необходимо было изучить его на-
ступательные возможности, чтобы определить меры, которые можно было бы использовать
в целях обеспечения зашиты от БО. Не менее очевидно, что нельзя не принимать во вни-
мание возможность нанесения ответного удара в случае применения этого оружия против
США. Таким образом, во всех проводимых исследованиях прослеживалась тесная взаи-
мосвязь между проблемами биологического напа дения и зашиты от него. Это вытекает из
обсуждения, представленного ниже.
Рассматривался целый ряд агентов, патогенных для человека, животных и растений.
Агенты, отобранные для всестороннего исследования, делали максимально вирулентными,
выращивали в специально отобранных культуральных средах и при оптимальных условиях
1106
Приложение А
роста и испьпывали с целью определения их поражающей силы на животных или расте-
ниях. Проводили глубокое изучение многих аспектов в этой области, включая: определе-
ние продолжительности сохранения вирулентное ги различными микроорганизмами с
высокой поражающей силой и продолжительности сохранения ими жизнеспособности при
различных условиях хранения биологических, физических и химических средств защиты;
количества микроорганизмов, необходимого для поражения организма; эффективности
антибиотиков и химиотерапевтических препаратов; инкубационного периода различных
заболевании; и эффективности воздействия некоторых химических препаратов (или со-
путствующих агентов) при их использовании вместе с патогенными биоагентами или ток-
синами на поражающую силу последних. При проведении этих и других исследований
получено много новых данных, которые, после их опубликования в соответствующих жур-
налах, будут в значительной мере способе гвовать повышению уровня научных знаний.
Широкое изучение биологических и химических агентов, которые могли бы быть исполь-
зованы дтя уничтожения урожая овса, позволило сделать некоторые открытия, которые,
несомненно, окажд гея весьма ценными для сельского хозяйства.
Изучали вероятные методы и способы применения БО против США. Делалось это не
только в целях совершенствования мер защиты против диверсионных действий против-
ника, сведения о которых были представлены соответствующим гражданским и военным
властям, но и в целях выявления различных типов военного снаряжения, которое могло
бы быть использовано для распространения биоагентов. Была осуществлена разведыва-
тельная программа, i оторая оказалась весьма эффективной на всех театрах военных дейст-
вий, в результате чего были получены также основательные сведения относительно дея-
тельности Германии в этой области Таким же образом проводилось изучение деятельности
Японии. По окончании расследования можно будет в полной мере оценить работу, прове-
денную в этой области нашими противниками. На сегодняшний день все данные указы-
вают, что державы оси отставали от США, Англии и Канады в области разработки биоло-
гического оружия. Известно также, что с начала 1942 г. Германия не получала сведений
относительно деятельности США в этой области, и какой-либо серьезной утечки сведений
по данному вопросу из нашей страны не наблюдалось. В этом отношении весьма сущест-
венную роль играло разумное и искреннее сотрудничество прессы и радио с управлением
военной цензуры.
При всех работах в области БО. проводимых США, исключительное внимание уделя-
ли защите персонала от инфекции. Для предупреждения заражения было разработано мно-
жество новых методов, которые оказались весьма эффективными. Во всех центрах име-
лись больницы и амбулаторные пункты, укомплектованные специалистами армии и ВМС
и имевшие все необходимое для лечения внутрилабораторных инфекций. В результате
чрезвычайных мер было зарегистрировано только 60 доказанных случаев заражения, выз-
ванных случайным экспонированием к вирулентным возбудителям, предназначенным для
использования в качестве БО и треоовавших лечения. В 52 случаях наблюла (ось полное
выздоровление; в остальных восьми случаях процесс выздоровления проходит удовлетво-
рительно. Помимо 60 доказанных случаев, отмечено 159 случаев случайного экспониро-
вания к возоудителям в неизве стных концентрациях. Во всех случаях, кроме одного, было
начато немедленно лечение, что предотвратило развитие заболеваний В одном случае
сотрудник не сообщил об экспонировании, у него началось заболевание, однако после со-
ответствующего лечения он выздоровел.
Очевидно, ни одно из этих заболеваний не было вызвано намеренно и в этой связи не
рассматривалось как «контролируемый» эксперимент, тем не менее в результате лечения
заболевших были получены некоторые ценные сведения, особенно в отношении эффек-
тивности новых антибиотиков, химиотерапевтических препаратов и методов иммуниза-
ции, которые, если бы не случайное зара кение, могли бы в иных условиях испытываться
Лвиложение А
1107
только на животных Учитывая разнообразие обрабатываемых высокопатогенных аген-
тов, масштаб проводимых операций и сравнительно большое количество занятых при этом
людей, вопросам обеспечения безопасности в нашей программе БО поистине уделялось
значительное внимание.
Действия США в области БО, предпринимаемые по необходимости и направленные
главным образом на обеспечение безопасности своей страны и вооруженных сил в случае
возможного использования БО противником, проводились с той же слаженностью, какой
характеризовались наши действия в военное время. Объединенные общей целью вели
совместную работу армейское и военно-морское командование, гражданские ученые, уни-
верситетские и частные учреждения и некоторые министерства. Дело было весьма сроч-
ным, многие из проблем уникальны и чрезвычайно сложны. Цель была достигнута: была
разработана система защитных мер против потенциально опасных методов ведения вой-
ны, возможность неожиданного нападения с этой стороны была предупреждена. Однако,
помимо достижения целей военного значения, было получено много сведений, имеющих
неоценимое значение для блага человека. Были созданы уникальные объекты для иссле-
дования и экспериментирования с патогенными биоагентами в невиданном до сих пор
масштабе. Эти объекты будут иметь неоценимое значение для будущих биологических
исследований военного и невоенного характера. Вот в общих чертах некоторые из наибо-
лее важных выполненных пунктов программы:
1. Разработка методов и объектов для массового производства микроорганизмов и
продуктов их жизнедеятельности.
2. Разработка методов для быстрого и четкого обнаружения незначительных коли-
честв возбуди гелей инфекционных болезней.
3. Значительный вклад в знания по борьбе с возбудителями аэрогенных инфекций.
4. Производство и выделение впервые кристаллического бактериального токсина, что
дает возможность приготовить более высокоочищенный иммунизирующий анатоксин.
5. Разработка и производство, в случае необходимости, эффективного анатоксина в
достаточных количествах для защиты, в случае крупных операций.
6. Значительные успехи в отношении выработки иммунитета у людей и животных к
некоторым инфекционным заболеваниям.
7. Значительные успехи в лечении некоторых инфекционных болезней человека и
животных и в разработке эффективной защитной одежды и оборудования.
8. Разработка объектов для разведения и содержания лабораторных животных с це-
лью обеспечения огромного количества проверенных линий экспериментальных живот-
ных, необходимых для исследований.
9. Применение специальных методов фотосъемки для изучения аэрогенных микро-
организмов и обеспечения безопасности лабораторных исследований.
10. Данные о действии более 1000 различных химических агентов на живые растения.
11. Изучение происхождения и разработка мер по борьбе с некоторыми болезнями
растений.
Лицами, которые были заняты исследованиями в этой области, предпринимаются
шаги к получению разрешения на публикацию таких технических документов и донесе-
ний, которые могут быть опубликованы без нанесения ущерба безопасности страны. Эго
очень важно, поскольку значительная часть получаемой при этом информации будет иметь
большое значение для здравоохранения, сельского хозяйства, промышленности и теоре-
тических наук.
Хотя верно и то, что БО все еще относится, скорее, к области теорий, чем к дейст-
вительности, в том смысле, что оно не использовалось в боевых действиях, данные, полу-
ченные США в этой области, наряду с результатами исследовательских групп, занимав-
шихся аналогичной работой в Англии и Канаде, показывают, что такой метод ведения
1108
Приложение А
войны не может не приниматься в расчет теми, кто занимается вопросами государствен-
ной безопасности. Наши старания во время войны обеспечили получение средств для за-
щиты страны от БО с точки зрения известных в настоящее время его возможностей и по-
зволили разработать средства, нанесения ответного удара, которые могли бы быть
применены, если бы возникла такая необходимость. Хотя можно отметить значительные
достижения в изучении возможностей БО, границы такого способа ведения войны оп-
ределены далеко не полностью. Работу в этой области, обусловленную угрозой биологи-
ческой войны, нельзя игнорировать и в мирное время, она до гжна продолжаться в доста-
точном масштабе в целях обеспечения соответствующей защиты страны.
Целесообразно отметить, что, в отличие от разработки атомной бомбы и других видов
секретного оружия, разработка биологического оружия возможна во многих странах, боль-
ших и малых, без громадного расхода денежных средств или сооружения огромных объек-
тов для производства этих агентов. Очевидно, что разработка БО могла весьма успешно
продолжаться во многих странах, возможно, под видом законных медицинских или бак-
териологических исследований.
При любом обсуждении вопросов, касающихся обеспечения прочного мира во всем
мире, нельзя без риска игнорировать возможности биологического оружия.
Джордж У Мерк, консультант
ПРИЛОЖЕНИЕ Б
ПОКАЗАНИЯ ВОЕННОПЛЕННЫХ АМЕРИКАНСКИХ ЛЕТЧИКОВ,
ПРИМЕНЯВШИХ БАКТЕРИОЛОГИЧЕСКОЕ ОРУЖИЕ В 1952 г.
ВО ВРЕМЯ ВОЙНЫ НА КОРЕЙСКОМ ПОЛУОСТРОВЕ1
Показания американского военнопленного летчика, старшего
лейтенанта Инока, об его участии в бактериологической войне,
ведущейся американскими войсками в Корее (SIA/142)
В течение последних двух недель августа 1951 г. я находился в Ивакуни. В течение ав-
густа производилась переброска 3-го бомбардировочного авиаполка в Куньсань (Корея).
В последнюю очередь покинула Ивакуни авиационная школа, которая была переведена в
Куньсань в начале сентября 1951 г. Во время моего пребывания в Ивакуни школу посеща-
ло около 15 экипажей, которые только что прибыли из Соединенных Штатов. В курс лек-
ций входило то же, что и в 4400 школе по подготовке боевых кадров. Мы, штурманы, слу-
шали лекции по навигации, о самолете В-26 и о Корее с тем, чтобы лучше овладеть своей
профессией и быть лучше подготовленными для полетов в боевых условиях.
25 августа 1951 г. в 13 ч 00 мин в кабинете навигации этой школы для нас была прочи-
тана секретная лекция. На этой лекции присутствовало, насколько я помню, 10 пилотов и
15 штурманов. Из числа пилотов я припоминаю лейтенанта Броутона, лейтенанта Шмид-
та и капитана Лемака; из числа штурманов — лейтенанта Брауна, лейтенанта Харди, лей-
тенанта Дего, лейтенанта Зелинского, лейтенанта Ларсона и лейтенанта Гарвина. Я не был
знаком со всеми пилотами и штурманами и знал лишь тех, с которыми я служил вместе на
аэродроме Лангли. Нашим инструктором был Вильсон. Он был в штатском. Других инст-
рукторов на лекции не было. Вильсон сказал нам, что темой его лекции является бактерио-
логическая война. Он сказал, что наша сторона не намерена в данное время применить бак-
териологическое оружие, но тем не менее нам, может быть, придется это сделать в будущем;
поэтому лекция является секретной, и мы не должны разглашать ее содержание кому бы
то ни было или говорить о ней между собой.
Основная часть лекции Вильсона была посвящена бактериологическому оружию.
Вильсон не имел с собой никаких пособий, но рассказал нам о различных методах распро-
странения бактерий, которые в основном делятся на распространение культуры бактерий
в чистом виде и распространение бактерий путем заражения ими сбрасываемых с самоле-
тов насекомых и животных. В своей лекции Вильсон рассказал следующее.
Способы распространения культуры бактерий в чистом виде сводятся к следующему:
1) сбрасывание с самолетов бомб, начиненных зараженной бактериями пылью3, кото-
рые раскрываются в воздухе, и эта пыль разносится ветром;
2) распространение зараженной бактериями пыли непосредственно самим самолетом
с помощью специальных распыляющих аппаратов, а результате чего бактерии появляют-
ся в воздухе всюду, куда попадает пыль;
3) сбрасывание наполненных зараженной пылью контейнеров (вместилищ) — либо
бомб, которые раскрываются в воде, либо картонных коробок, которые размокают в во-
де, — в водоемы и озера, вода из которых потребляется людьми и животными и из которых
бактерии могут распространяться дальше поглотившими их насекомыми.
1 Доклад Международной научной комиссии по расследованию фактов бактериологиче-
ской войны в Корее и Китае. — Пекин, 1952.
2 Номер приложения в Докладе Международной научной комиссии... (1952).
3 Речь шла о применении сухих рецептур поражающих биологических агентов.
1110
Приложение Б
Способы распространения бактерий через насекомых следующие:
1) сбрасывание бактериологических бомб, которые похожи на обыкновенные бомбы,
но начинены зараженными бактериями насекомыми и открываются при соприкоснове-
нии с землей, освобождая содержащихся и них насекомых;
2) сбрасывание зараженных насекомых в картонных контейнерах, которые ломаются
при соприкосновении с землей, освобождая содержащихся в них насекомых;
3) распространение насекомых, находящихся на животных.
Способы распространения бактерий через живот ных сводятся к следующему:
1) сбрасывание крыс, кроликов или других мелких животных в подвешенные к пара-
шютам контейнерах, которые при соприкосновении с землей раскрываются и освобож-
дают животных. Эти животные несут на себе зараженные бактериями вшей и блох,
2) распространение таких животных с судов в территориальных водах противника1.
Существуют и другие способы распространения бактерий:
1) сбрасывание листовок, бумаги, конвертов и изделий из бумаги, зараженных бак-
териями;
2) сбрасывание зараженных бактериями мыла и одежды;
3) сбрасывание автоматических ручек с чернилами, зараженными бактериями;
4) сбрасывание зараженной пищи в районах, 1де расположены войска противника.
Можно распространять бактерии, также применяя артиллерийские (гаубичные) сна-
ряды и мины. Но поскольку такие снаряды и мины будут падать в непосредственной бли-
зости от линии фронта, это небезопасно.
Существует много видов различных бактерий, которые могут быть использованы в
качестве бактериологического оружия. Кроме множества малоизвестных и необычных
видов бактерий, могут использоваться также бактерии хорошо известных болезней, как-
то: сыпного тифа, брюшного тифа, холеры, дизентерии, чумы, оспы, малярии и желтой
лихорадки2. Существует много видов насекомых, которые могут служить переносчиками
этих бактерий. Наиболее распространенные из них — вши, блохи, мухи и комары. Вши,
блохи и мухи могут переносить бактерии тифа, холеры, оспы, чумы и дизентерии. Кома-
ры могут быть переносчиками малярии и желтой лихорадки.
Лучшей защитой против бактериологической войны является подготовленность к ней.
Всем, кому можно, необходимо сделать прививки против всех возможных болезней. Гели
будут сброшены насекомые, то следует обливать контейнеры, в которых они находятся,
керосином или нефтью и сжигать их Если насекомые уже успели покинуть контейнеры,
лучше всего опрыскивать район препаратом ДДТ, желательно с самолета. В случае если
противник распространил пыль, зараженную бактериями, — следует применять раствор
ДДТ3. Вся находившаяся в зараженном районе ничем не прикрытая пища, должна унич-
тожаться. Вся одежда и вещи должны быть вымыты горячей водой с мылом. Вся вода перед
употреблением должна быть прокипячена. Все употребляемые в пищу продукты следует
тщательно проваривать или прожаривать. Каждый, находящийся в зараженном районе,
должен каким-либо способом защищать при дыхании нос и рот и должен также, когда
приняты все остальные меры предосторожности, сменить одежду и выкупаться. Весь му-
сор в зараженном районе должен сжигаться.
Летом все окна должны быть снабжены специальными сетками для защиты от насе-
комых. Мелкие: кивотные такие как крысы—должны уничтожаться, в результате чего будет
1 Общепринятое в те годы представление о распространении пандемий чумы грызунами
(см. рис 1.22).
2 Натуральная оспа и желтая лихорадка вызываются вирусами, малярия — простейшими
рода Plasmodium, передающимися комарами рода Anopheles.
3 Он так запомнил. Вильсон рассказывал ему о сброшенных насекомых
Приложение Б
1111
уменьшаться опасность чумы, переносчиками которой являются блохи, находящиеся на
этих животных. Если противник сбросит предметы из бумаги или другие подобные вещи,
они должны немедленно сжигаться.
Все виды бактериологического оружия по своему характеру требуют, чтобы их сбра-
сывали с самолетов, летящих на возможно меньшей высоте с возможно бо iee низкой ско-
рое! ью. чтобы не повредить насекомых. Если бактериологическое оружие сбрасывается с
парашютом, самолет может лететь на значительной высоте, но не выше 1000 футов, так
парашют может быть отнесен ветром от цели.
Вильсон окончил свою лекцию вЗч(15ч00 мин/. Он напомнил нам, что о бактериоло-
гическом оружии нельзя ни с кем говорить, и затем покинул помещение школы. Эта лек-
ция была единственной в своем роде, которую нам довелось когда-либо слышать.
1 сентября 1951 г. я направился в Куньсань. В октябре 1951 г. вновь в декабре этого же
года в Куньсане майор Браунинг прочитал (продолжавшиеся по одному часу) лекции о за-
щите от бактериологического оружия. Эти лекции он читал много раз по каждому случаю,
и от каждого требовалось прослушать часовую лекцию. В декабре он прочитал ту же лек-
цию, что и в октябре. Цель, конечно, заключалась в следующем — поскольку по плану
чередования все время появлялись новые войска, также полезно было бы запомнить со-
держание его лекции. Он заявил нам, что было оы неразумным рассчитывать на то, что
противник применит против нас бактериологическое оружие. Если противник прибегнет
к этому, то будут распыляться бактерии или распространяться зараженные бактериями
насекомые, и он подчеркивал, что мы должны обновлять справки о прививках и делать
прививки. Во время лекции он указывал и на другие моменты, как я об этом уже говорил.
1 января 1952 г. офицер-инструктор оперативного отдела во время нашего очередного
инструктажа заявил нам, что должны сообщать обо всех наших «неразорвавшихся бомбах»,
а также о том, где они упадут. Это была обыкновенная процедура, и в то время это ка >алось
совершенно обычным напоминанием Об этом на инструктаже напоминал всем членам
экипажа офицер-инструктор оперативного отдела капитан Кэри. В эту ночь я из-за про-
студы не вылетел, и меня заменили дру1 им штурманом. Мой следующий вылет по распи-
санию был назначен в ночь на 6 января 1952 г. Согласно расписанию, мы должны были
лететь по зеленому 8-му маршруту (между Пхеньяном и Саривонем), и наш вылет был
намечен на 3 ч 00 мин. В составе экипажа были капитан Эймос — пшют, я — штурман и
сержант Трэйси — стрелок. Как обычно, капитан Эймос и я в 2 ч 00 мин за час до вылета,
сделали доклад инструкторскому и оперативному отделам. Там мы всегда проверяли пос-
ледние данные о погоде и по 1учали информацию о задании, которое нам предстояло вы-
полнить. В эту ночь незнакомый мне дежурный капитан заявил нам, что мы должны ле-
теть на город Хванчжу и сбросить подвешенные к крыльям самолета бомбы (которых было
две), а затем, как можно быстрее сбросить остальной груз и сразу же возвратиться в Кунь-
сань. Дежурный офицер приказал нам сбросить бомбы над Хванчжу на высоте 500 футов
и при максимальной скорости 200 миль в час. Мы обратили его внимание на небольшую
высоту, поскольку мы должны были, согласно инструкции, взять десять 500-фунтовых
бомб. Однако он заявил нам, что это является совершенно секретным, что эти бомбы бакте-
риологические и велел никому и ничего не говорить о нашем задании. Он сказал, что под
весные бомбы уже заряжены и проверены и велел не трогать их. По возвращении он велел
нам доложить о них, как о «нера юрвавшихся бомбах». Затем мы направились в оператив-
ный отдел штаба эскадрильи и встретили нашего стрелка, который не делал докладов отде-
лу и который, насколько мне известно, не знал о нашем специальном задании. Когда мы
подошли к самолету, там стоял часовой из отдела вооружения. Он сказал нам, что подвес-
ные бомбы уже проверены, о чем мы уже знали. Я проверил шесть бомб в бомбовом отсеке,
и все 6 были обычными 500-фунтовыми бомбами.
Мы вылетели в 3 ч 00 мин и канравились к Хванчжу, сбросив две бактериологиче-
ские бомбы на западной окраине города. Не было заметно никаких взрывов или чего-либо
1112
Приложение Б
необычного. Затем мы в течение двух минут продолжали полет на север и сбросили 8 обыч-
ных бомб на шоссе в 5 милях к северу от Хванчжу, а затем сразу же возвратились в Кунь-
сань. Мы вылетели в 3 ч 00 мин, наши бомбы были сброшены в 4 ч 00 мин и возвратились
в Куньсань в 5 ч 00 мин. В этот раз я впервые услышал о сбрасывании бактериологических
бомб, и мы хранили это в секрете.
Эти бактериологические бомбы выглядели точно так же, как обычные 500-фунтовые
бомбы. Возможно, что днем можно было заметить какие-либо характерные отличия, но
когда я их увидел, было темно. Я не подвешивал этих бомб и не видел, как их подвешива-
ли, однако на крыльях не было никаких специальных приспособлений. Поэтому они под-
вешивались так же, как обычные бомбы. Когда мы рапортовали в разведывательном от-
деле после выполнения задания, мы сообщили, что две бомбы 500-фунтовые (фактически
150-фунтовые) были сброшены на Хванчжу, назвав их «неразорвавшимися бомбами». За-
тем мы сообщили, где мы сбросили наши 8 разорвавшихся бомб. Бомбы назывались «не
разорвавшимися бомбами», очевидно, с целью не допустить, чтобы слишком много лю-
дей узнало о цели задания, однако высшее командование могло проверять наши рапорты
и знать, где были сброшены бактериологические бомбы.
10 января случайно или намеренно, я не знаю, меня снова назначили для выполнения
аналогичного задания с Эймосом и Трэйси. На этот раз, когда я докладывал в оператив-
ном отделе, нам сказали, что все четыре наших подвесных бомбы были бактериологичес-
кими. На этот раз нашей целью был город Чунхва, расположенный на зеленом 8-ом марш-
руте, затем должны были, как можно быстрее сбросить остальные бомбы и возвратиться
на базу. Мы все еще сохраняли наши операции в секрете и докладывали о наших бактери-
ологических бомбах как о «неразорвавшихся бомбах. Максимальной скоростью, при ко-
торой мы должны были сбрасывать бомбы, было 200 миль в час и высота — 500 футов. Сно-
ва человек из службы вооружения должен был проверить для нас бомбы, подвешенные под
крыльями самолета. Мы захватили с собой Трэйси из оперативного отдела штаба эскад-
рильи и направились к самолету. Бомбы, подвешенные под крыльями самолета, выгляде-
ли как обычные. Человек из службы вооружения сказал мне, что нам не следует беспоко-
иться за бомбы, подвешенные под крыльями самолетов, так как все они подготовлены к
сбрасыванию. Я проверил обычные бомбы в день бомбардировки. В 03 ч 00 мин мы под-
нялись с аэродрома и взяли курс прямо на Чунхва, сбросив 4 бактериологических бомбы в
04 ч 10 мин с высоты 500 футов при скорости 190 миль в час на западной окраине Чунхва.
Затем мы пролетели на юг и сбросили обычные бомбы на шоссе к севера от Хванчжу и воз-
вратились на базу в Куньсане, приземлившись в 05 ч 15 мин.
Отдавая рапорт, мы доложили, где мы сбросили 6 разорвавшихся бомб и доложили
также опять по той же причине, что и раньше, а именно, по причине секретности, — что
четыре «неразорвавшихся бомбы» сбросили в Чунхва. Ниже приводится схема типа бак-
териологических бомб, которые мы применяли.
Мне кажется, что эти бактериологические бомбы поступают из того же медицинского
центра снабжения, в котором изготовляется вакцина, употребляемая для борьбы с зараз-
ными болезнями, и я полагаю, что этот центр находится в Японии либо на острове Хон-
сю, либо на острове Кусю1.
1 Видимо, он прав. Разработка биологических боеприпасов, отбор штаммов опасных мик-
роорганизмов, создание технологий их поддержания, культивирования и хранения, осуществ-
лялось в военных НИИ на территории США. Но снаряжение биологических боеприпасов,
разведение животных и насекомых осуществлялось где-то поблизости от театра военных дей-
ствий. Сэйти Моримура (1983) пишет о воссоздании в феврале 1950 г в восточной части пре-
фектуры Саитама кооперативов по разведению животных, во время войны поставлявших их в
отряд № 731. Руководил этой работой некий Кобаяси, бывший одно время адъютантом Исии,
Приложение Б
1113
Если применить тот вид бактериологической бомбы,
которую сбрасывали мы, то она раскроется при ударе о
землю, освобождая находящихся внутри нее бактерий и
насекомых. В холодную погоду эти насекомые будут вялы-
ми, но на солнце они оживут.
Листовки сбрасываются в Северной Корее с самолетов
В-29. Эти листовки сбрасываются в коробках, которые
раскрываются в воздухе, и листовки рассеиваются на боль-
шой площади. Эти листовки могут быть использованы в
бактериологической войне.
Бактериологические бомбы сбрасывает пилот. Штур-
ман отмечает, когда, где и сколько сброшено бактериоло-
гических бомб. Эти бомбы сбрасываются путем нажатия
электрической кнопки. Когда после выполнения задания,
экипаж самолета отдает рапорт разведывательному отделу,
при этом присутствует весь экипаж, и рапорт отдают пилот
и штурман. Это неофициальный рапорт, весь экипаж сидит
у стола и докладывает представителю разведывательного
отдела, который принимает рапорт и записывает его, а за-
тем передает его своему начальнику. Вот почему о бакте-
риологических бомбах в этом рапорте говорится как о «не-
разорвавшихся бомбах», для того чтобы непосвященный в
дело персонал разведывательного отдела и экипаж самоле-
Схема бомбы, нарисованная
лейтенантом Иноком
та не узнали о секретном характере задания. Насколько мне известно, самолеты В-26 яв-
ляются единственными, с которых сбрасываются бактериологические бомбы, похожие на
обычные бомбы. Однако самолет В-26 не подходит для сбрасывания других видов оружия.
Листовки сбрасываются с самолетов В-29 и транспортных самолетов С-47 и С-46, но глав-
ным образом с В-2. Транспортные самолеты больше всего подходят для сбрасывания всех
остальных видов бактериологического оружия, как, например, картонных ящиков, сбрасы-
ваемых с парашютами контейнеров, предметов одежды, продовольствия, мыла, бумаги, ав-
томатических ручек; однако, для сбрасывания этих видов оружия могут быть использованы
также самолеты В-29.
Что касается того, когда мы впервые начали применять бактериологические бомбы,
то это было в начале года — приблизительно 1 января 1952 г., поскольку тогда всем нам
напомнили о том, что мы должны следить за «неразорвавшимися бомбами». Возможно,
что другие группы, как, например, 452-й авиаполк, начали применять бактериологическое
оружие в это же время. Решение применять бактериологические бомбы является, конечно
же, совершенно секретным. Принимая во внимание серьезный характер этого решения,
можно с уверенностью сказать, что это решение зависит от высшего командования — веро-
ятно, от штаба вооруженных сил на Дальнем Востоке, находящегося в Токио.
Кеннет Л. Инок, 7 апреля 1952 г.
и до конца войны занимавшийся материальным снабжением отряда № 731. Кооперативы раз-
водили морских свинок, крыс, кроликов, хомяков и кошек. Животных отвозили в Токио, в
отдел «Джи-2», входивший в состав подразделения американской армии, имевшего название
«Джей-2 Си-406» и в «Объединенный научно-исследовательский институт подопытных жи-
вотных», занимавшиеся якобы изучением эпидемических болезней. Кобаяси занимал долж-
ность управляющего делами и начальника финансового отдела этого института. Отряд «Джей-2
Си-406» располагался в районе Тиеда - - Маруноути, в здании корпорации «Мицубиси». В обеих
организациях работали в основном бывшие сотрудники отряда № 731. Насекомых разводили
частные японские фирмы.
1114
Приложение Б
Показания американского военнопленного летчика, старшего
лейтенанта Квинна, об его участии в бактериологической
войне, везущейся американскими войсками в Корее (SIA/15)
Я—Джон Квинн, старший лейтенант военно-воздушных сил США, личный военный
номер 17993А. Мне 29 лет. Я вступил в ряды воздушных сил 16 февраля 1948 г., когда мне
было 26 лет. Я из г. Пасадена, штата Калифорния. По окончании военно-авиационной шко-
лы 25 февраля 1949 г. я был назначен в авиационный университет. Там я прошел 6-недель-
ный курс инструктора, так называемый академический курс инструкторов. Окончив этот
курс, я был назначен в качестве сотрудника штаба в дивизию академических инструкторов.
Моя работа заключались в подготовке помощников-инструкторов, в обучении их тому, как
правильно использовать фотографии, карты, кинофильмы и проекционные фонари при
обучении других. Итак, я был инструктором, когда я получил назначение в Дальневосточ-
ные военно-воздушные силы США, чтобы летать в Корее на самолетах В-26. Мне было
приказано явиться 25 августа 1951 г. на авиационную базу Лангли для изучения пилотажа
В-26. Там я оставался восемь недель. Оттуда нас послали в лагерь Стонман, где мы ожида-
ли выезда за пределы США В лагере Стонман мне сделали прививки против сыпного тифа,
брюшного тифа, холеры и оспы. Затем мы вылетели из Соединенных Штатов и прибыли
в Японию 27 ноября 1951 г. Мы прибыли на конечную станцию авиалинии Ханеда, а отту-
да нас перевели в Фучу, район Б, где мы должны были ожидать отправки в Корею Мы
оставались в Фучу до 29 ноября, а затем были отправлены поездом на авиационную базу
Асия в Южной Японии. Наша поездка продолжалась всю ночь, и мы приехали к месту на-
значения 30 ноября. В тот же день мы были переброшены на самолете С-47 на авиацион-
ную базу Кунсан в Корее. Я был назначен в 3-е бомбардировочное звено 8-й эскадрильи
3-й авиагруппы. Третья авиагруппа состоит из трех эскадрилий: восьмой, девяностой и
тринадцатой, и была единственной авиагруппой в Кунсане.
17 декабря 1951 г. я явился в дежурную комнату 8-й эскадрильи и на доске приказов
увидел свое имя среди тех, кому нужно было идти на лекцию, которая должна была состо-
яться на следующий день в 9 часов утра. На следующий день я и штурман Ларсон, фами-
лия которого тоже была в списке, пошли на лекцию. Лекция читалась в большой комнате
здания наземной школы. Сиденья в комнате позволяли разместить 30 человек. На лекции
присутствовало 20 человек пилотов и штурманов. Кроме меня и Ларсона, там были млад-
ший лейтенант Робертс, лейтенант Шварц, старший лейтенант Роджерс, старший лейте-
нант Ватсон, капитан Лонг и капитан Даффи — все штурманы, и капитан Ховарте, капи-
тан Лэнд, старший лейтенант Шмидт, капитан Бисон, капитан Робертсон и старший
лейтенант Макаллистер — все пилоты. Я и Ларсон задержались за кофе и опоздали на не-
сколько минут, другие уже были в комнате. Капитан, который говорил с собравшимися,
кажется, был очень недоволен нашим опозданием и повторил нам, что лекция, которую
мы будем слушать, является очень важной и крайне секретной. Он сказал, что мы должны
внимательно слушать, что нам говорят, но что мы не должны обсуждать лекцию даже друг
с другом. Капитан сказал нам, что лектор прибыл из Японии и является специалистом в
своей области. Затем он познакомил нас с лектором господином Ашфорк, который не был
военным. Г-н Ашфорк был человеком средних лет, ему было 40 лет. Он был худощавым,
пяти футов десяти дюймов ростом и почти лысым.
Он начал свою лекцию с того, что объявил нам, что его темой будет биологическая
война. Он сказал, что, когда подумаешь, — это ужасная вещь, но что в эпоху атомной бом-
бы, когда наука быстро идет вперед, мы должны быть готовы ко всему. Он сказал, что мы
никогда не можем знать, как обернутся события в будущем и должны быть готовы к само-
защите, что мы должны знать, как весп; бактериологическую войну, когда это потребует-
ся. Он говорил, что он изучал бактериологическую войну в течение многих лет и что он
расскажет нам все, что он считает нужным для нас.
Приложение Б
1115
Сначала он рассказал нам, что есть много способов распространения бактерий. Бак-
терии можно распространять в любом месте и в любое время. Необходимое для распрост-
ранения оборудование также изучено и подготовлено. Он рассказал, что нельзя сбрасы-
вать одни бактерии, так как лучи солнца убивают их в течение 60 секунд, но добавил, что
бактерии могут распространяться с помощью различных видов насекомых и грызунов. Эти
насекомые и грызуны целыми поколениями разводятся искусственным образом в лабо-
раториях; их избрали потому, что они могут жить в любом месте, в любое время, даже в
самых неблагоприятных условиях. Рассказывая нам о способах распространения бактерий,
он сказал, что это можно сделать с помощью пыли, распыляя ее подобно дымовой завесе.
Распространять бактерии таким образом можно с кораблей, ведя их вблизи берега, когда
ветер дует в направлении берега1. Распространять их таким же образом могут и низко ле-
тящие реактивные самолеты, реактивные самолеты любых типов, пояснил он. Он сказал,
что бактерий могут распространять насекомые: платяные вши, блохи, мухи и москиты.
Таких насекомых можно сбрасывать многими другими способами: в коробках, которые под
воздействием тепла солнечных лучей становятся хрупкими и тем самым позволяют насе-
комым выползти, а также в бомбах. Он сказал, что поскольку мы летаем на самолетах
В-26,он оолее подробно остановится на последнем способе, то есть на распространении
инфекций с помощью бомб. Затем он нам показал фотографии реактивного самолета, ри-
сунок самолета F-84, разбрасывающего пыль, насыщенную бактериями и находящуюся в
баках по краям самолета. Он показал нам фотографию старой одежды, по которой ползли
насекомые, похожие на мух и вшей. Этим насекомым тепло в одежде, хотя добавил к это-
му, что этих насекомых можно путем отбора акклиматизировать так, чтобы насекомые
легко переносили холод и могли долго существовать без пищи.
Затем он нам показал фотографии бомб, в которых можно сбрасывать бактерии. Эти
бомбы очень похожи на наши 500-фунтовые бомбы «GPS», только у них нет взрывателей.
Он нам сказал, что размер и форма бомбы не имеют значения, важно чем она наполнена.
Он добавил, что перевоз бактерий не представляет опасности, так как бомбы крепко за-
крыты и открываются только тогда, когда их сбрасывают с самолетов. На одной из показан-
ных нам фотографий была снята 500-фунтовая бомба с очень тонкими стенками — менее
четверти дюйма (английского) в толщину. Он добавил, что над этими бомбами все еще
производятся опыты и что есть много видов таких бомб. Он нам показал фотографию од-
ной бомбы, которая раскалывается надвое при ударе о землю. Другая бомба сзади в изги-
бе бомбы, вблизи хвостового оперения, имеет дверки, которые открываются при ударе. Эти
дверки открываются маленьким электрическим мотором, соединенным с оатареей, кото-
рая приходит в действие, когда бомба ударяется о землю. Тонкий щиток из пластмассы
отделяет жидкость от металлических пластинок батареи до того, как бомба ударится о зем-
лю. Сила, с которой бомба ударяется о землю, достаточна для того, чтобы жидкость про-
рвала щиток. Жидкость затем покрывает пластинки батареи, и мотор открывает дверки.
Он нам показал фотографию бомбы, у которой отламывается хвост при ударе о землю.
Все показанные нам бомбы были сделаны так, чтобы походить на обыкновенные 500-фун-
товые бомбы, но ни у одной не было взрывателя. Он нам сказал, что есть бомбы, которые
распадаются в воздухе. Насекомые, которыми наполнены коробки, заключенные в бомбе,
могут, таким образом, до падения на землю рассеяться на большой площади. Он расска-
зал, что под воздействием солнечных лучей эти коробки становятся очень ломкими и насе-
комые (мухи, блохи и комары) выползают. Все три бомбы, которые он нам показал, были
одной и той же структуры, и все они походили на 500-фунтовыс бомбы с тонкими стенка-
ми. Первой была показана распадающаяся на две половины бомба, второй — с дверками
в задней части бомбы, вблизи хвостового оперения, а третья — с отломанным хвостом.
1 Технология считалась уже отработанной — см. рис. 1.38.
1116
Приложение Б
Схема бомбы, нарисованная старшим лейтенантом Квинном
Бомбы, которые раскрываются в воздухе, прицеплены к бомбодержателям на крыль-
ях обыкновенным стальным тросиком. В носовой части бомбы есть небольшой пропел-
лер, который из-за тросика не приходит в действие, пока бомба не сброшена. При сбра-
сывании бомбы стальной тросик остается на самолете, и пропеллер приходит в действие.
Пропеллер заводит генератор, который снабжает энергией небольшой электрический
мотор (который я описал выше). Мотор сначала открывает три дверки с вди, как и в дру-
гой бомбе, а затем открывает дверку спереди. Силы движения воздуха в бомбе достаточно
для того, чтобы выдуть насекомых, коробки, и они вылетают в разные стороны. Лектор не
показывал нам фотографии бомбы этого типа и вообще описал ее очень скудно.
Затем он рассказал нам о способах распространения бактерий. Он сказал, что почти
любое насекомое может быть использовано для распространения бактерий, но, что он нам
расскажет только о некоторых из них и что этого будет достаточно для того, чтобы защи-
тить себя от бактерий в том случае, если они распространятся и на Южную Корею. Он
сказал, что крысы разносят бубонную чуму, но, что совсем не обязательно сбрасывать крыс,
хотя это можно делать. Бактерии можно сбрасывать в любых предметах, лишь бы крысы
могли свободно проникать в них, таким образом, крысы заражаются бактериями. Сбра-
сывать насекомых совсем легко, а они переносят много видов бактерии. Мухи разносят
бактерии сыпного тифа и холеры, а блохи — бактерии чумы. Комары разносят бактерии
различных видов лихорадки, желтой лихорадки, брюшного тифа, малярии и энцефалита,
для излечения которого нет еще эффективных средств. Энцефалит также известен, как
японский энцефалит «В», который был распространен в японской армии и был впервые
занесен в Корею японской армией. О борьбе с энцефалитом пока известно очень мало, но
средства предотвращения те же, что и против малярии. Он сказал, что опишет нам. как
разносится малярия, и добавил, что другие виды лихорадки распространяются тем же спо-
собом, то есть через комаров. Он показал нам большой рисунок комара.
Эти комары безвредны, сказал он, пока они не заразятся малярией — они могут зара-
зиться, укусив больного малярией, могут быть так же заражены в лаборатории. Когда ко-
мар кусает человека, он впивается верхней и нижней губами. Когда комар сосет кровь, в
это же время слюна его вытекает в кровь человека. Если комар заражен, он передает бак-
терии в этой слюне, и укушенный человек заболевает лихорадкой. Он сказал, что плакаты
на стенах столовой (о том, как предохраняться от энцефалита) висят отнюдь не для укра-
шения. Лектор сказал, что мы должны соблюдать все правила чистоты, и в особенности в
будущие несколько месяцев. Когда нам дают лекарство атебрин1, мы должны принимать
его, а не выбрасывать. Он добавил, что мы должны своевременно сделать все необходи-
1 Синтетический лекарственный препарат применяется для лечения разных форм маля-
рии, а также как противоглистное средство и для лечения кожного лейшманиоза и красной
волчанки. В России более известен под названием акрихин.
Приложение Б
1117
мые прививки. Если мы будем исполнять все это, нам нечего бояться. Лекция началась в
9 ч и закончилась около одиннадцати. Все мы обратили внимание на то, что бактерио-
логические бомбы не будут взрываться, что это невзрывающиеся бомбы.
На инструктивном совещании 31 декабря оперативный офицер напомнил нам о том,
что по возвращении мы должны докладывать разведывательной части о всех неразорвав-
шихся бомбах. 3 января в 2 часа дня я явился в оперативный отдел авиагруппы. Кроме меня,
там было 26 пилотов, 27 штурманов, 27 радистов-бомбардиров и 27 механиков (в некоторых
случаях эти механики были одновременно пулеметчиками; в нашей эскадрильи были
3 самолета, которые могли брать пулеметчиков). В моем экипаже были старший лейтенант
Роджерс (штурман) и сержант Сейер (механик). Мы не брали с собой радиста-бомбарди-
ра, так как погода стояла хорошая Я списал с доски приказов маршрут нашего задания:
мы должны были вылетать в 2 ч 30 мин утра и бомбить район между Саривоном и Пхень-
яном Начался очередной инструктаж. На этом инструктаже оперативный офицер предуп-
редил нас, что информация, которую мы получим, является секретной и должна оставаться
в секрете, что обсуждать ее даже между собой нельзя. Затем офицер разведывательной ча-
сти рассказал нам о том, какое движение было замечено в предыдущую ночь и сколько
машин было повреждено или уничтоженс Офицер связи сухопутной армии объяснил по-
ложение на фронте, а офицер-метеоролог сообщил нам данные о погоде. Другую инфор-
мацию, относительно ветра и температуры, штурман списал с доски приказов. Во всех
отношениях казалось, что наша миссия должна была быть самой обыкновенной.
Пять минут второго я, Роджерс и Сейер встретились в оперативном отделе авиагруп-
пы, а затем я прошел в маленькую комнату, где ночью находится оперативный офицер.
Дежурил капитан Рейнолдс. Я дал ему наши имена, и он сказал, что я назначен для выпол-
нения специальной миссии. Он добавил, что, прежде всего, я должен сбросить уже подве-
шенные на крыльях бомбы, причем я должен, насколько осмелюсь, сделать это возможно
ближе к Пхеньяну. На висевшей на стене карте он показал место, в которое была воткнута
красная булавка — оно находилось в пяти милях южнее Пхеньяна и в трех милях восточ-
нее главного шоссе. Он сказал, что после этого мы должны выполнить наше обыкновен-
ное задание, как можно скорее и по возвращении доложить о всем в разведывательную
часть. Он сказал, что я должен сбросить бомбы с высоты 200 фугов или ниже, если воз-
можно, и добавил, чтобы я не волновался, если они не разорвутся, так как это неразрываю-
щиеся бомбы. Я вспомнил о лекции относительно бактериологических бомб и спросил его,
что это значит, но он ответил, что не знает и что будет лучше, если я буду делать так, как
мне сказано и меньше буду задавать вопросов о том, что и почему. Тогда я подумал, что это
все же бактериологические бомбы.
Когда мы подошли к самолету, нас встретил часовой, и я убедился, что это бактерио-
логические бомбы. Часовой просил нас не беспокоиться о бомбах на крыльях, добавив, что
они подвешены как следует. Но я посмотрел на них, когда осматривал самолет и заметил,
что штурман был прав, когда он говорил, что «бомбы на крыльях не имеют взрывателей».
Мы посмотрели друг на друга, и я сказал, что приказ есть приказ, и пусть они так остаются.
Я сказал штурману, где мы должны сбросить бомбы, и он отметил это место на своей карте.
Мы вылетели в 2 ч 25 мин и в 3 ч 30 мин были немного южнее Пхеньяна. Я повернул к
востоку от дороги, как раз южнее моста, и футов через двести Роджерс сказал, что мы ле-
тим над целью. Я быстро, одну за другой, сбросил все четыре бомбы. Они не разорвались.
Мы оба окончательно решили, что это были бактериологические бомбы.
Мы выполнили другую часть задания в 4 ч 15 мин и приземлились в Кунсане в 5 ч
10 мин. Мы покинули самолет, возвратили наше снаряжение, доложили, что самолет в пол-
ной исправности и пошли прямо в разведывательную часть при оперативном отделе авиа-
группы, чтобы доложить о выполнении задания. Мы доложили, что мы сбросили четыре
бомбы там, где нам было, приказано, с высоты 200 футов и что ни одна из них не разорва-
лась. Сержант записал это для того, чтобы утром передать в разведчасть.
1118
Приложение Б
10 января в два часа дня я был на очередном инструктаже и получил задание вылететь
в район между деревней Гуну и Канге. Р моем экипаже на этот раз были старший лейте-
нант Шварц (штурман) и сержант Сейер (механик). Нам было приказано вылететь в два
часа утра. В половине первого утра мы явились в оперативный отдел и получили обычный
инструктаж. Как обычно, я зашел сделать отметку о готовности экипажа и узнал, что у нас
вновь будет особое задание. Дежурным офицером опять оказался капитан Рейноллс. По-
мня, что я уже раз выполнял специальное задание с «неразрывающимися бомбами», он
заговорил об этом и добавил, что на этот раз задача будет такая же, как и в прошлый раз.
Только на этот раз у меня будет две «неразрывающиеся бомбы» и я должен буду сбросить
их северо-восточнее деревни Гуну. Он показал мне на висяшей на стене карте где я дол-
жен это сделать. Место находилось в трех милях севернее деревни Гуну и в пяти милях
восточнее железнодорожного пути. У самолета нас опять встретил часовой и сказал нам,
что бомбы уже подвешены. Я заметил, что бомбы на крыльях опять не имели взрывателей.
Я сказал Шварцу, где мы должны сбросить эти специальные бомбы; мы уже знали, что везем
бактериологические бомбы. Мы вылетели в 2 часа и были над деревней Гуну в 3 ч 25 мин.
Я повернул самолет в указанном мне Шварцем направлении, снизился до 200 Лугов и сбро-
сил эти две бомбы. Это были «неразрывающиеся бомбы». Мы, как можно быстрее выпол-
нили остальную часть нашего задания и в 4 ч 10 мин полетели назад в Кунсан В 5 ч 25 мин
мы приземлились в Кунсане, сдали наше снаряжение, доложили, что самолет в исправно-
сти и прошли в разведывательную часть при оперативном отделе. Там мы доложили, что
сбросили две «неразорвавшиеся бомбы» и сказали, куда их сбросили». Сержант записал
все это, чтобы передать в разведчасть.
14 декабря на доске приказов я прочитал свое имя среди тех, которые обязаны были
присутствовать на лекции, которая должна состояться в 9 ч утра на следующий день в зда-
нии наземной школы. В 9 часов я пришел в большую комнату, где собрались 25 других
офицеров и штурманов. Там были капитан Бисон, старший лейтенант Шмидт, капитан
Лонг, капитан Лэнд, капитан Ховарте, старший лейтенант Ларсон, старший лейтенант
Шварц, младший лейтенант Робертс и старший лейтенант Ватсон. Самым старшим офи-
цером в комнате был оперативный офицер базы майор Аллен, который познакомил нас с
лектором (далее нет 5 страниц текста).
Показ mi1я американского военнопленного летчика, старшего
лейтенанта О’Нила, об его участии в бактериологической войне,
ведущейся американскими войсками в Корее (ISGK/4)
Я, Флойд Бреланд О’Нил, служил младшим лейтенантом резерва военно-воздушных
сил США, личный военный номер АО 1848575. Мне 24 года. Мой домашний адрес: почто-
вый ящик 66, проспект Мойе, г. Ферфакс штата Южная Каролина. Не женат. Дома у меня
находятся мать, сестра и четыре брата. С сентября 1945 г. по июнь 1948 г учился в коллед-
же Цитадель, в г. Чарлстоне, штата Южная Каролина и по окончании колледжа я получил
ученое звание бакалавра естественных наук, и одновременно корпусом по тренировке
офицеров мне было присвоено военное звание младшего лейтенанта. В сентябре 1948 г. я
поступил в Тулэйнский университет в г. Нью-Орлеане штата Луизиана, где я занял долж-
ность ассистента кафедры химии, в мои функции входило руководить практическими за-
нятиями студентов-первокурсников по химии в лаборатории и ставить оценки их письмен-
ных работ; одновременно я готовился к защите диссертации на ученую степень магистра.
В июне 1950 г. я окончил Тулэйнский университет и получил ученую степень магис гра ес-
тественных нау к по физической химии.
В августе 1950 г. я был призван на военную службу и избрал специальность летчика.
24 августа 1950 г. я прибыл на авиабазу Рандольфа в Сан-Антонио штата Тексас, где нахо-
дился до 19 марта 1951 г. и получил основу тренировочных полетов. С 3 апреля по 15 сентяб-
Приложение Б
1119
ря 1951 г. я проходил тренировочный курс высшего пилотажа на истребителе F-51, на авиа-
базе Крейга г. Селма штата Алабама. После этого я был командирован в артиллерийскую
школу авиабазы Люк, в г. Феникс штата Аризона. Однажды, 1 декабря 1951 г., в этой шко-
ле я прослушал секретную лекцию о бактериологической войне. 15 декабря 1951 г. я окон-
чил курс артиллерийской стрельбы и 3 января 1952 г. был отправлен в лагерь Стонмэна
штата Калифорния, где ждал отправления парохода за границу. До выезда из США мне
сделали профилактические прививки против столбняка, брюшного тифа, холеры и оспы
10 января 1952 г. я уехал из лагеря Стонмэна и 12 января 1952 г. прибыл на авиабазу
Ханеда, около Токио (Япония). Я был отправлен на базу Фучу, и 16 января 1952 г. меня
отправили на авиабазы Тачикава (Япония) на базу К-10, находящуюся в Чинхае в Корее,
которая является тыловой базой 18-й бомбардировочно-истребительной группы. На базе
К-10 я был назначен в 67-ю бом Зардировочно-истребительную эскадрилью 18-й бомбар-
дировочно-истребительной группы. 19 января 1952 г я был отправлен на базу К-46 для
получения боевого задания; это была передовая база 18-й бомбардировочно-истреби-
тельной группы, которая находилась в 5 милях севернее Вонжу, откуда меня направили
в авиазвено «Айтэм» 67-й эскадрильи; здесь я слушал лекцию о порядках авиагруппы и
эскадрильи, о структуре организации, дисциплине, требованиях и порядке отпусков. На
базе К-46 я совершил два полета с целью ориентировки в обстановке. Первый полет был
одиночный для ознакомления с районом, второй раз был групповой полет с полной на-
грузкой бомб, которые были сброшены в океан, недалеко от восточного морского берега
Кореи С этого времени я был признан «подготовленным к бою».
22 января 1952 г. на базе К-46 я слушал лекцию о бактериологической войне. В то
время члены моего звена (звено «Айтэм») были в отпуску в Японии, и я должен был ждать
их возвращения. Первое задание я выполнил 28 января 1952 г. Мы вылетели на самолетах
F-51, типа «Мустанг». 15 февраля 1952 г. я выполнил первый и единственный раз задание
бактериологической войны. С 20 по 29 февраля 1952 г. я болел воспалением легких;
22 февраля 1952 г. я был отправлен в госпиталь авиабазы К-10. С 3 марта я опять начал
летать: 4 марта, во время выполнения моего тренировочного боевого задания, примерно,
в 9 ч 15 мин утра, я был сбит зенитным огнем западнее Синмака и тут же взят в плен китай-
скими добровольцами.
Я ел) шал секретную лекцию о бактериологической войне на авиабазе Люк. Секрет-
ная лекция о бактериологической войне была прочитана, всем курсантам артиллерийской
школы 1 декабря 1951 г. на авиабазе Люк в аудитории базы. На лекции присутствовали
около 70 курсантов, из них 40 курсантов были с эскадрильи самолетов F-84 и 30 KjpcaH-
тов с эскадрильи F-51. Лекция началась в 15 ч и продолжалась около получаса, закончи-
лась в 15 ч 30 мин, Среди курсантов, присутствовавших на этой лекции, насколько я при-
поминаю, были капитан Вилльям Похнер, старший лейтенант Аллен Бэттис, младший
лейтенант Джон Инглинг, младший лейтенант В. С. Санки, младший лейтенант Мэл Суза,
младший лейтенант Р. Л. Майкл, младший лейтенант Р. С. Грэйл, младший лейтенант Джек
Кук, младший лейтенант Джон Шелендер, младший лейтенант Рик Кэнэди и младший
лейтенант Джек Шепард. Все эти офицеры были отправлены на Дальний Восток; из них
младшие лейтенанты: Инглинг, Санки, Суза, Грэйл, Майкл и Шепард прибыли в Корею,
где получили назначение в 67-ю бомбардировочно-истребительную эскадрилью 18-й бом-
бардировочно-истребительной авиагруппы.
Лекция была прочитана майором военно-воздушных сил США, Бесед В. Вилльямсом.
До лекции его познакомил с нами капитан, офицер по информации и обучению на базе,
фамилию которого сейчас я не помню. Капитан сказал нам, что майор прибыл в Люк из
Вашинг гона специально для того, чтобы читать такие лекции один раз в месяц новым
курсантам артиллерийской школы, прикомандированным на базу Люк. Майор Вилльямс
служил в Генеральном штабе авиации США. Я полагаю, что он подчиняется заместителю
1120
Приложение Б
начальника оперативного отдела штаба Ростом он в 5 футов 8 дюймов, ему было около
34 лет, говорил басом, среднего телосложения, со слегка выпуклым животом.
Майор предварительно сказал нам, что тема и содержание лекции являются секрет-
ными и обо всем, что будет сказано, никому ничего не следует говорить. Он сказал нам,
что эту лекцию читает с той целью, чтобы мы имели представление о бактериологической
войне и чтобы дать нам обшие сведения о ней, с тем, чтобы в случае встречи с этим вопро-
сом в будущем для нас это не было ново.
После этого он начал свою лекцию. Он говорил, что бактериологическая война ведет-
ся двумя способами: она может вестись наземными войсками, при помощи бактериоло-
гических снарядов, или авиацией путем сбрасывания бактериологических бомб, в кото-
рых помещаются бакгерии или зараженные насекомые. Он заявил, что количество
бактерий в одной бомбе, сброшенной авиацией, гораздо больше, чем количество бакте-
рий в нескольких снарядах, используемых наземными войсками. К тому же использова-
ние таких снарядов ограничено районом фронта или близлежащими к нему районами в то
время, как авиация может сбрасывать бактериологические бомбы в отдаленные от фрон-
та районы, в глубоком тылу врага. Поэтому бактериологические снаряды используются
только против передовых частей противника, а бактериологические бомбы — против со-
средоточенных резервов в тылу врага и вражеских городов.
Майор еще сказал, что бактериологические снаряды и бомбы, во время их переноски,
не представляют большой опасности или совсем не опасны для артиллеристов и экипа-
жей самолетов, за исключением неосторожных случаев, например, если она случайно упала
и разбилась и из нее выпали бактерии или зараженные насекомые. Он сказал, что есть
особые группы, которые прошли специальную подготовку по приготовлению бактерио-
логических бомб и снарядов, они умеют обращаться с ними. Дальше он добавил, что име-
ется антитоксин для каждого вида бактерии, поэтому наши войска не будут заражаться
болезнями, вызг 1ваемыми бактериями, используемыми в бактериологической войне.
Майор нам сказал, что бактерии выращиваются на особых средах в наших специаль-
ных лабораториях. Для выращивания культуры каждого вида бактерий требуется опреде-
ленная среда и специальные условия — определенная температура и влажность. Он ска-
зал, что используемые в бактериологической войне насекомые и грызуны выбраны из
переносчиков заболеваний, известных медицинской науке, как например: мухи, блохи,
вши, клещи, мошки, комары, пауки и крысы. Эти насекомые и грызуны выращиваются в
наших специальных лабораториях в Абердине штата Мэриленд1.
Майор Вилльямс сказал, что уже подготовлены особые морозостойкие бактерии и
насекомые, и их можно использовать в бактериологический войне в холодных климатиче-
ских условиях. Бактерии были сделаны морозостойкими следующим образом: их помеща-
ют в воздушные ванны, затем постепенно периодически снижают температуру и влажность-
таким образом бактерии привыкают к все более и более низким температурным услови-
ям. После каждого охлаждения отбирают наиболее стойких бактерий, которые пригодны
для следующего этапа снижения температуры и влажности.
1 Полигон был создан в 1917 г. после вступления США в Первую мировую войну. Старей-
ший испытательный полигон Сухопутных войск США. Занимает территорию 29,3 тыс. га. Гео-
графически разделен рекой Буш на 2 зоны — северный Абердинский район и южный Эджвуд-
ский (арсенальный). Сюда подходят железнодорожные пути, есть взлетно-посадочная поло-
са. На базе постоянно работают свыше 11 тыс. человек военного и гражданского персонала
(2003). Здесь располагается крупнейшее отделение (всего в составе лаборатории пять отделе-
ний) Лаборатории сухопутных войск США ARL (Army Research Laboratory) — ведущего цент-
ра Министерства обороны США по организации и проведению фундаментальных и приклад-
ных исследований в интересах обеспечения сухопутных войск ключевыми технологиями и ана-
литическими исследованиями.
Приложение Ь
1121
Морозостойкие насекомые получаются также путем гибридизации, т. е. берут насеко-
мых-переносчиков болезней и скрещивают их с морозостойкими формами насекомых того
же вида; потомство снова скрещивают с такими же морозостойкими насекомыми и таким
образом получают морозостойкие виды, способные передавать болезни.
Майор сказал, что для этой цели в больших масштабах ведутся исследования бакте-
рий и насекомых, на что затрачиваются большие деньги, и эти исследования дали воору-
женным силам СШАжелаемое бактериологическое оружие. Он нам сказал, что эти иссле-
дования были произведены химическим корпусом и отделом вооружения сухопутных
войск, которые включены в так называемый «особый план». Отдел вооружения сухопут-
ных войск имеет большую лабораторию около испытательного полигона в Абердине шта-
га Мэриленд Большинство работ в Абердине производится людьми, которые являются
специалистами различных отраслей сухопутной армии, но здесь работают также и граж-
данские научные работники. Кроме того, видные ученые страны могут привлекаться в
качестве консультантов. Отдел вооружения совместно с авиацией конструируют бактерио-
логические бомбы. Военная авиация передает отделу вооружения требования и конкрет-
ные указания на типы бактериологических бомб, а сухопутная армия конструирует и про-
изводит бактериологические бомбы.
Майор сказал нам, что обычными видами микробов для ведения бактериологической
войны являются возбудители брюшного и сыпного тифа, холеры, бубонной чумы, маля-
рии, дизентерии, желтой лихорадки и других заболеваний. Эти возбудители не всегда вы-
зывают смерть, но они очень болезнетворны. После заражения этими микробами люди
серьезно заболевают и если вовремя не оказать медицинскую помощь, то многие умира-
ют. Если вызвать эпидемические болезни в армии и среди населения в тылу, то они причи-
няют большие потери. Армия, подвергшаяся эпидемии, не может быть использована в
войне, а гражданское население не в состоянии работать; тогда вражеские медицинские
учреждения окажутся в тяжелом состоянии и их общее положение ухудшится. Майор ука-
зал, что моральное состояние как в тылу, так и на фронте будет сильно падать, и это еще
больше ослабит силу врага. Перед майором лежали какие-то записи, куда он иногда по-
сматривал. Очевидно, эти записи были его конспектом для того, чтобы он не мог ничего
упустить, а также, чтобы содержание этих лекций было всегда одинаковым. Он сказал, что
он поедет из Люка на авиабазу Нелисса вблизи г. Лас-Вегаса, штата Невада1, где он сдела-
ет доклад в артиллерийской школе. Он еще раз обратил наше внимание на секретность
этого доклада. В 15 ч 30 мин он закончил свою лекцию. Когда мы возвращались с лекции
в наземную школу на базе Люк, мы недоумевали над тем, зачем нам нужно было слушать
такую лекцию, и все мы удивлялись содержанию этой лекции.
Лекция о бактериологической войне, которую мы слушали на базе К-46, сделал нам
капитан Маклафлин 22 января 1952 г. в маленькой комнате, находящейся за командным
пунктом отряда, где летчики докладывали о выполнении заданий. Кроме меня, присутство-
вали еще следующие летчики: младший лейтенант Пит Ниблей, младший лейтенант Джим
Хорсли и младший лейтенант Р. С. Грэйл. Когда мы возвратились в казарму, нам сказали,
что капитан Маклафлин ишет нас. Нас четверых сразу отправили в оперативный отдел
авиагруппы, где докладывают о выполнении задания и куда провел нас Маклафлин, ком-
ната эта находилась в конце здания. Это было в 14 часов. Капитан Маклафлин работал в
разведчасти, инициалов его я не знаю, мы звали его «капитан Мак». Его рост приблизи-
тельно 5 футов 10 дюймов, лет ему около 30, волосы у него черные, с проседью.
Он раньше уже читал эту лекцию всем летчикам нашей авиагруппы. Так как мы при-
были сюда недавно, то он специально прочел нам эту лекцию. Комната, где он читал нам
лекцию, была 8 футов в ширину и 10 футов в длину. В комнате были стол, несколько стульев
1 Сайт авиабазы — http://www.nellis.af.mil/.
1122
Приложение Б
и доска, которая висела на стене. Он сказал нам, чтобы мы чувствовали себя свободно, и
если захотим, то можем курить. Он ска 1ал, что если наши обычные задачи полета отно-
сятся к категории «секретно», и мы могли обсуждать их между собой и сообщать о них друг
Другу, то все, что он будет сегодня читать, не следует никому говорить и даже нельзя об-
суждать слышанное между собой.
Он сказал нам, что лекция является «абсолютно секретной», и еще раз подчеркнул, что
нам нельзя говорить о ней никому, и то, что он сейчас расскажет нам, для нас будет но-
вым, а поэтому мы должны внимательно слушать его. После этого он начал свою лекцию
о бактериологической воине.
Капитан не имел при себе никаких записей и не пользовался никакими пособиями.
Очевидно, он уже хорошо знал эту тему и прошел кур' обучения по этому вопросу. Мне
каже-.ся, что он проходил обучение в Абердине штата Мэриленд. Разведывательная шко-
ла находилась в Вашингтоне, недалеко от которого расположен Абердин. Я ни от кого не
слышал о капитане Маклафлине и никто не говорил мне, что он из себя представлял, по-
этому не знаю, где он проходил обучение.
Он сказал, что бактериологические бомбы делятся на 2 основных типа:
1-й тип — разрывающиеся в воздухе;
2-й тип — с парашютом.
Бомбы с парашютом используются для сбрасывания насекомых, зараженных бакте-
риями, а разрывающиеся в воздухе бомбы используются для сбрасывания бактерий. Можно
также распылять бактерии; можно сбрасывать насекомых, или зараженные предметы, как
например: бумагу и листья деревьев. Наша авиагруппа тогда применяла разрывающиеся в
воздухе бомбы и способы распыления.
Капитан сказал, что эти бактериологические бомбы, разрывающиеся в воздухе, по
своим размерам одинаковы с эОО-фунтовыми обычными бомбами, а на самом деле они
весят всего лишь 150—200 фунтов. Эти бомбы сделаны специально для этой цели: внутри
бомбы имеется желеобразная масса, где живут бактерии. Бомбы имеют взрывчатые веще-
ства, которые при взрыве выбрасывают желе. Однако не следует класть взрывчатого ве-
щества чрезмерно много, так как оно при разрыве может уничтожить бактерии. В эти бом-
бы также вставлены взрыватели, которые взрываются в воздухе, или дистанционные
взрыватели «VT»1, которые взрывают бомбы в воздухе на высоте 50—100 футов от земли.
Эти бомбы следуют сбрасывать с высоты не менее 5000 футов от земли с тем, чтобы дистан-
ционный взрыватель «VT» автоматически зарядился и мог взорваться на нужной высоте.
Капитан говорил, что бомбы могут быть начинены следующими видами бактерий:
возбудителями сыпного и брюшного тифов, холеры, дизентерии и чумы. В бомбы разры-
вающихся в воздухе, не помещают насекомых, так как они могут быть уничтожены при
взрыве бомбы. Бомба с дистанционным взрывателем устроена так. что если она не взор-
вется в воздухе, то должна взорваться при соприкосновении с землей. Стенки бактерио-
логической бомбы сделаны тоньше, чем стенки обычной бомбы, для того чтобы легко взор-
ваться. Бактериологические бомбы были привезены на базу и заряжены специальной
командой отдела вооружения. Имеющиеся на базе К-46, бактериологические бомбы хра-
нятся в подземных бетонированных складах, которые находятся в районах, где произво-
дится зарядка бомб. Склады обнесены колючей проволокой. Бактериологические бомбы
погружаются на самолеты специальным отрядом отдела вооружения.
Капитан сказал нам, что если по пути к цели испортится мотор самолета, и мы будем
находиться южнее линии бомбардировки или в дружественном районе, то мы должны
сбросить бактериологические бомбы над безлюдным районом. Мы должны покружиться
1 Имеется в виду радиовзрыватель VT (Variable Time fuze) — неконтактный взрыватель с
переменным временем срабатывания.
Прилоэ-сение Б
1123
Вэрь’»члтые вещества Ширина 1,5 фута
для распространения Длина 3 фута Толщина наружной оболочки
бактерий Вес около 200 фунтов 0,25 дюйма
Схема бактериологической бомбы, взрывающейся в воздухе,
нарисованная старшим лейтенантом О’Нилом
над местом, сбросить бомбы и связаться с радарной станцией, чтобы она отметила точное
место падения бомб и сообщила нам, а мы, в свою очередь, должны сообщить об этом
командному пункту авиагруппы. Тогда командный пункт сможет послать туда людей для
обезвоживания места падения бомб с тем, чтобы наши войска не очутились в опасном
положении. При сбрасывании бомб взрыватель не нужно включать. Если летчик захочет,
чтобы бомба взорвалась, то он должен нажать две электрические кнопки, установленные
в кабине управления, и тогда взрыватель включиться. Если мы будем находиться к северу
от линии фронта или во вражеском районе, то мы при сбрасывании бомбы, взрыватель
должны включить. Если место, куда сбросили бомбы, не будет являться целью, то нужно
сообщить, где находится это место.
Капитан сказал, что мы должны хорошо прицеливать я при сбрасывании бактерио-
логических бомб с тем, чтобы они попадали точно в цель, а не сбрасывать их как попало,
так как бомбы стоят очень дорого. Мы должны использовать бактериологические бомбы
в городах и местах большого скопления воинских частей, так как наибольший эффект
получается там, где имеется большое количество народа. Тогда бактерии могут заразить и
пищу и водоисточники, а также могут заразить одежду и людей. Капитан сказал, что когда
мы отправляемся для выполнения задания по бактериологической войне, то следует дей-
ствовать как обычно, как будто ничего необычного не случилось.
Затем капитан Маклафлин коротко объяснил нам устройство бактериологической
бомбы с парашютом и рассказал, как она действует. Такая бактериологическая бомба с
парашютом используется не для сбрасывания бактерий, а для сбрасывания насекомых,
зараженных бактериями. Бомба имеет приспособление, при помощи которого насекомые
выбрасываются при ударе о землю. Бактериологическая бомба с парашютом не имеет де-
тонатора и взрывчатого вещества. Если в бомбе помещается только один вид насекомых,
то процесс распространения происходит следующим образом: при ударе о землю бомба
раскалывается на 2 части; в это время парашют отделяется от бомбы, чтобы он не мешал
насекомым выползать. При ударе бомбы о землю электрический включатель, прикреплен-
ный к головке бомбы, приводит в движение маленький электрический мотор, который
выталкивает запор и бомба распадается на две половники. Электрический мотор также
может вытолкнуть шпильку, соединяющую парашют с бомбой, это позволяет парашюту
уйти в сторону.
Если бактериологическая бомба используется для распространения нескольких видов
насекомых, то внутри бомбы устраиваются перегородки. Когда бомба ударится о землю,
тверки каждой секции открываются, и через них выходят насекомые. При ударе бомбы о
землю включатель приводит электрический мотор в движение, который открывает двер-
ки для выхода насекомых: одновременно включатель выталкивает и шпильку парашюта,
и парашют свободно уходит в сторону.
Затем капитан объяснил нам приспособление для распыления бактерий или распрос. -
ранения насекомых. Аппарат распыления устроен в задней части самолета. На бензиновый
бак, который находится в фюзеляже, сзади летчика, ставятся контейнеры с бактериями или
1124
Приложение Б
Насекомые, зараженные бактериями
Электрические провода t
мотор
Длина 3 «рута
Ширина 1,5 фута
Толщина наружной оболочки
Вес 200 фунтов
0,25 дюйма
Схема бомбы с парашютом для сбрасывания насекомых, зараженных бактериями (для
помещения одного вида насекомых), нарисованная старшим лейтенантом О’Нилом
насекомыми. Так самолет приспосабливается для распыления бактерий. Существует осо-
бый отряд, который укладывает банки с бак.ериями или с насекомыми в контейнеры для
распыления.
Насекомые 'комарики)
Комары 1 Гауки
Дверки
Мухи —
Шпилька для
высвобождения
парашюта
Батарея
Маленький п а Две₽ки
электрический
мотор
Ширина 1,5 фута
Толщина наружной оболочки
0,25 дюйма
Включатель
Длина 3 фута
Вес около 200 фунтов
Схема бактериологической бомбы с парашютом для сбрасывания насекомых, зараженных
бактериями (4 камеры с дверками), нарисованная старшим лейтенантом О’Нилом
Предохранительный колпак*
Летчик
Дверка
Длина трубы около 15-20 футов,
диаметр около 3 дюймов
Длина контейнера около 3 футов,
ширина около 2 футов, может
вместиться 7000 насекомых
или 100 фунтов желе с бактериями механизмом
Рычаг, установленный в кабине
самолета, при помощи которого
летчик управляет распылительным
Приспособление для распыления на особом самолете,
нарисованное старшим лейтенантом О’Нилом
Капитан продолжал, что при распылении бактерий летчик переводит рычаг, установ-
ленный в кабине самолета, который открывает контейнер с насекомыми или бактериями,
открывает и дверки, соединяющие контейнер с трубой распыления, и открывает предох-
ранительный колпак, чтобы распылять бактерии или распространять насекомых через
трубу. При распространении насекомых, надо снять наконечник распылителя1, который
1 Форсуночный распылитель.
Приложение Б
1125
находится под предохранительным колпаком, на конце трубы, иначе насекомые не смо-
гут выйти из трубы через маленькие отверстия На наконечнике (он находится на конце
трубы) имеются маленькие отверстия для распыления бактерий. Бактерий получают в
оловянных банках, а насекомых в банках с открытыми концами с обеих сторон, но затяну-
тые сетками или материей, чтобы насекомые могли дышать. Предохранительный колпак
на конце трубы распыления прикрывается рычагом, поэтому после того, как самолет при-
землился, бакгерии или насекомые оставшиеся в трубе распыления, не могут выходить
наружу. После посадки самолета, летчик доставляет самолет в особо отведенное место.
Затем специальный отряд производит дезинфекцию самолета. Летчик сразу же меняет
одежду и моется в бане. Его одежда подвергается дезинфекции. После того как самолет воз-
вратился с выполнения задания, специальный автомобиль едет за самолетом и произво-
дит по пути следования самолета дезинфекцию путем распыления дезинфицирующего
вещества, с тем чтобы предохранить базу от опасности.
Капитан продолжал далее, что желе с бактериями можно употреблять только после
разведения водой или раствором1, для распыления бактерий или распространения насеко-
мых, зараженных бактериями, можно использовать только самолеты-истребители, так как
они имеют большую маневренность на низкой высоте. Обычно бактерий распыляют на вы-
соте 500- 1и00 футов от земли. При распылении скорое гь самолета составляет 350 миль в
час, а при распространении насекомых — 200 миль в час. Самолеты типа F-51, F-80, F-86
можно использовать для распыления бактерий или распространения насекомых, заражен-
ных бактериями.
Если испортится мотор, то летчик должен приземлиться на близком дружественном
аэродроме. Если мотор совсем испортится, то летчику разрешается спрыгнуть с парашю-
том, а самолет пусть разбивается и сгорает, во время пожара будут также уничтожены бак-
терии и насекомые. Для выполнения этих задач обычно используют два самолета, кото-
рые летают параллельно, находясь друг от друга на расстоянии около 200 ярдов, и распы-
ляют бактерий и зараженных насекомых над городами и войсками.
Капитан нам сказал, что в нашей авиагруппе имеется 4 самолета, приспособленные к
распылению бактерий или распространению зараженных насекомых. Эти самолеты ста-
вятся для стоянки на северном конце боковой дорожки и отделены от остальных самоле-
тов. За распылительными приспособлениями самолетов следят механик и особый назем-
ный обслуживающий персонал, прошедший обучение по бактериологической войне.
Кроме этих 4 человек, имеется еще специальный персонал отдела вооружения, который
отвечает за доставку и погрузку бактериологических бомб и контейнеров.
Самолеты, предназначенные для погрузки бактериологических бомб, стоят у специ-
альной боковой дорожки. Бактериологические бомбы грузятся ночью или рано утром осо-
бым персоналом. Самолеты с приспособлениями для распыления тоже доставляются сюда
для зарядки аппарата распыления. Погрузка бактериологических контейнеров на самолеты
для распыления производится при помощи подъемного крана. Прицепка бактериологи-
ческих бомб к самолету производится также обычным погрузочным приспособлением.
Капитан сказал, что бактериологические бомбы и контейнеры привозятся из Японии
самолетами, примерно, раз в две недели. Здесь не делают большого запаса. Бактериологи-
ческие бомбы привозятся из Японии транспортными самолетами С-46, С-47, С-54. Бак-
териологические бомбы, разрывающиеся в воздухе, сбрасываются самолетами типа: В-26.
В-29, F-51, F-80, F-84 и 86, а самолеты типа В-26 и В-29 очень удобны для сбрасывания
бомб с парашютом. Капитан упомянул в нескольких словах о выращивании бактерий. Он
сказал, что бактерии при выращивании требуют много хлопот и постоянного внимания.
Они растут в особых условиях, на специальных средах и требуют больших средств2.
1 Речь идет о фосфатном буфере ( см. разд. 1.0).
2 В 1950-х гг. БО дешевым оружием массового поражения не считалось.
1126
Приложение Б
Капитан еще раз предупредил нас о том, что все, что он говорит, представляет весьма
секретную информацию и нельзя об этом ни говорит ь между собою, ни тем более разгла-
шать. Когда он говорил это. у него было серьезное и строгое лицо, и мы почувствовали,
что это серьезное дело.
Лекция длилась с 14.00 до 16.00 ч, всего 2 часа, в 15 00 был сделан десятиминутный
перерыв. Из-за секретности, лекция проходила в комнате, где док 1адывали о выполнении
задания. Эта комната очень мала, поэтому лекцию слушало всего несколько человек.
В комнате для получения приказов могло вместиться 60 человек, но здесь всегда бывает
много народу, так что трудно сохранить секретность, в нашей авиагруппе только 10 ново-
прибывших, поэтому эта комната была для нас достаточна, хотя комната ограничивала
число слушателей. Чтобы объяснить содержание своей лекции, капитан Маклафлин на-
рисовал на доске мелом 7 схем бактериологических бомб и распылительного аппарата. Вс
время объяснения этих схем он показывал нам все детали. Очевидно, он уже читал эту
лекцию несколько раз, поэтому хорошо знал содержание ее и обходился без конспекюв.
Несомненно, он читает эту лекцию каждому новому летчику, прибывающему в авиагруппу.
По окончании лекции мы вышли из комнаты и вернулись в свои палатки. Никто не
сказал ни слова, каждый думал о прослушанной лекции. Я удивлялся, почему мы еще при-
меняем это ужасное оружие, в то время, когда идут мирные переговоры и военные действия
приостановлены. («The war was at a stalemate».)
Мы сели в палатке и смотрели трут на друга несколько минут, потом я предложил сыг-
рать в карты, чтобы отвлечься от содержания прослушанной лекции.
15 февраля 1952 г. 67-я эскадрилья, первая в этот день, послала авиазвено «Айтэм»
выполнить задание. Старший лейтенант Франк Харвей был назначен командиром звена,
который подобрал себе команду в составе: лейтенанта Пэджэтт, младшего лейтенанта
Р. С Грэйля и меня для выполнения этого задания. Утром около 5 ч 30 мин мы пошли по-
лучить задание. Во время дачи обычных распоряжений о бактериологический войне ни-
чего не говорили. На этот раз около 20 летчиков всей эскадрильи принимали задания Одни
должны были перерезать железную дорогу, другие — линии снабжения и т. д. Офицер опе-
ративной службы майор Кларк спросил: «Кто из «Топкик» (условный знак 67-й эскадрильи)
будет выполнять первое задание?» Когда майор услышал ответ: «Аьиазвено «Айтэм», — он
сказал: «Секретный бой»; затем он отдал другие задания и особые указания. После этого,
все мы сверили часы по точному времени и заслушали метеосводку от старшего лейтенан-
та Скотта. Затем мы заслушали данные разведки от капитана Маклафлина. Эта процедура
закончилась в 5 ч 45 мин, затем капитан Маклафлин приказал: «Авиазвену «Айтэм» взять
авиаснимки». Когда мы подошли к комнате, он провел нас в тот маленький кабинет, где
несколько дней тому назад он же нам говорил о бактериологической войне. Он дал стар-
шему лейтенанту Харвей авиационные снимки города Сибионни и сказал ему: «Вы пове-
зете с сооой бактериологические бомбы, сбросите их в западной части города и вернетесь
на базу К- 46; я буду ждать вашего рапорта; во время рапорта вам нужно будет называть их
дистанционными бомбами, взрывающимися и воздухе. Запомните, что когда будете идти
к самолетам ни в коем случае нельзя делать озабоченный или странный вид, и помните,
что это крайне секретно». О задании, которое мы будем выполнять, мы не говорили. Пос-
ле; завтрака мы вернулись в кабинет оперативного отдела эскадрильи, чтобы наметить план
нашего полета, т. е. маршрут полета, высоту, скорость и т. п. Капитан Маклафлин сказал
нам, что время нашего взлета будет 8.00, в то время как обычно первый взлет всегда начи-
нался в 7.00. Мы надели парашю.ы и пошли к самолетам. Наши 4 самолета стояли около
платформы для погрузки. Мы проверили самолеты, все было в порядке, бактериологичес-
кие бомбы уже были прикреплены под крыльями самолетов специальным персоналом из
отдела вооружения. Я только проверил шнур взрывателя бактериологической бомбы; он
был в порядке, после чего я сел в самолет.
Приложение Б
1127
Мы взлетели, набрали высоту до 9000 футов, лез ели со скоростью 250 миль в час. Ког-
да мы прилетели к месту, приблизительно на 10 миль западнее Сибионни, повернули на
восток и пошли к этому город) Долетев до города Сибионни, старший лейтенант Харвей
несколько раз набрал высоту и опустился и этим дал нам сигнал (сигнал без радио), по
которому наши самолеты выстроились в линию, затем он спикировал и сбросил бомбы.
Мы также спикировали с высоты 9000 футов до 7000 футов и сбросили бактериологичес-
кие бомбы: когда снизились до 6000 футов, снова набрали высоту до 7000 футов, и полете-
ли на свою базу со скоростью 350 миль в час. Когда мы подходили к базе К-46, — летели со
скоростью 250 миль в час. Мы взлетели в 8 часов, в 8.50 сбрэсили бактериологические
бомбы и в 9 ч 30 мин сделали посадку. Перед сбрасыванием бактериологических бомб, мы
зарядили их, затем нажатием кнопки на рычаге рулевого управлении сбросили бактерио-
логические бомбы. Каждый самолет сбросил по 2 бактериолш ических бомбы, всего на
Сибионни сбросили 8 бактериологических бомб. Мы не знали, какие бактерии были в этих
бактериологических бомбах, мы знали только, что эти бомбы были бактериологически-
ми. После посадки мы остановили самолеты на взлетно-посадочной дорожке и пошли в
кабинет оперативного отдела эскадрильи, там оставили свои парашюты и шлемы, затем
перешли через улицу и вошли в разведчасть, где нас ждал капитан Маклафлин. Он сказал:
«Я приму их!». Это значило то, что он будет принимать рапорт о выполнении задания. После
выполнения обычного задания любой офицер разведки мог слушать наш рапорт. Он про-
вел нас в маленькую комнату, где докладывали о выполнении задания, где мы утром полу-
чили авиафотоспимки и особые указания. Во время выполнения задания я заметил, что
бомбы, которые были сброшены первыми тремя самолетами, взорвались в воздухе. Коман-
дир звена видел, что бомбы, сброшенные мною, также взорвались в воздухе, поэтому мы
доложили, что 8 дистанционных бомб взорвались в воздухе над объектом бомбардировки.
Я заметил, что форма записывания докладов капитаном была обычная. Он задавал также
обычные вопросы, как например: был ли бой в воздухе с противником, ка1 ая была погода
в районе бомбардировки, какие были неисправности и трудности, встречали ли вражес-
кие самолеты, фамилия и имя летчика, номер самолета и т. д. Истинные цели нашего за-
дания не были затронуты. Мы доложили, что задание секретного боя выполнено успеш-
но. Доклад начался в 9 ч 40 мин и закончился в 9 ч 45 мин.
В тот день я вел самолет № 055 и занимал 4-е место в авиозвене, Харвэй был команди-
ром звена. Грэйл — вторым, Пэджэтт третьим После разрыва бактериологических бомб
образовался сероватый туман, дым; так нам казалось, во всяком случае, сверху вниз. Звук
взрыва бомбы в воздухе был, вероятно, не громким для тех, кто находился на земле по срав-
нению со звуком обычных бомб при разрыве. Все бактериологические бомбы были сбро
шены в западной части города Сибионни.
Я уверен, что каждый летчик 18-й авиагруппы, судя по их поведению, принимал уча-
стие в бактериологической войне. Летчики, которые прибыли недавно, вроде меня, уча-
ствовали всего несколько раз в выполнении подобных заданий, а старый состав летчиков
этой авиагруппы выполнял такие задания чаще. Лейтенанты Харвэй и Пэджэтт как будто
уже привыкли к выполнению таких заданий, а Грэйл и я нервничали и испытывали страх
при выполнении первой такой задачи Я замечал, как проявлялось сильное беспокойство
каждый раз после выполнения таких заданий. Джон В Интлин, Мэл Суза, В. С Санки,
Пит Ниблэй, Джим Хорсли и другие младшие лейтенанты прибыли со мной в одно и то же
время в авиагруппу. Я замечал их волнение, но не спрашивал их и не шутил с ними, так как
я соблюдал тайну бактериологической войны. Но по выражению и поведению старших лей-
тенантов Харвэя и Пэджэтта, я думаю, что они уже раз 5 выполняли задания бактериоло-
гической войны. Насколько мне известно, младший лейтенант Грэйл выполнил только
одно такое задание, лейтенанты Г. Б. Армстронг и М. Сатэнштэйн выполнили задание по
3 раза. Младшие лейтенанты Джим Хорсли и Пит Ниблэй выполняли такие задания 2 раза
1128
Приложение Б
Видно, что они уже привыкли сбрасывать бактериологические бомбы Среди летчиков
67-й эскадрильи я знаю только одного летчика, старшего лейтенанта С. О. Армстронга, ко-
мандира авиазвена «Н», который распространял бактерии при помощи распылителя.
Примерно, 18 февраля я заметил, что автомашина, следуя за одним самолетом по взлет-
но-посадочной площадке, проводила дезинфекцию местности. Это был единственный
случай, когда я заметил такое явление, но я не помню, было ли это в тот самый день, когда
"тарший лейтенант С. О. Армстронг распространял бактерии при помощи распылителя;
перед тем, когда я взлетал, всегда видел, как стоят 4 самолета с распылительными приспо-
соблениями на северном конце аэродрома. В состав 18-й авиагруппы входили 2-я южно-
африканская эскадрилья, 12-я, 39-я, 67-я бомбардировочно-истребительные эскадрильи.
Очевидно, в январе 1952 г. был уже разработан план сбрасывания бактериологических бомб.
Поэтому, когда я прибыл в эту авиагруппу, план мероприятий был уже разработан Насколь-
ко мне известно, я считаю, что для составления такого плана, для тренировок и подготов-
ки потребовался, по крайней мере, 1 месяц. Это доказывает, что 18-я авиагруппа, прибли-
зительно, в середине декабря 1951 г. или немного раньше уже применяла бактериологи-
ческие бомбы. Судя по всем признакам, решение о ведении бактериологической войны
американской армией в Корее было принято еще в начале осени 1951 г. Стало быть, этот
отрезок времени в несколько месяцев потребовался для составления всех мероприятий,
обеспечения необходимыми материалами, подбора служебного персонала для ведения бак-
териологической войны.
В связи с ведением бактериологической войны моральный дух нашей авиагруппы па-
дает. Я точно знаю, что никто из летчиков не желает выполнять заданий бактериологичес-
кой войны. При исполнении обычных заданий летчики, отправляясь к самолетам или
возвращаясь после полета, всегда оживленно разговаривали. Однако после выполнения
задания по бактериологической войне, они считали, что чем меньше разговаривать, тем
лучше. Летчики могут говорить только о том, куда они отправлялись для выполнения за-
дания, но сами по своей инициативе не рассказывают конкретно о выполнении своего
задания, например, о воздушном бое, количестве сброшенных бомб и т д. Во время бесе-
ды можно легко узнать тех, кто выполнял задания бактериологической войны в этот день,
так как они всегда молчат; о выполнении задания бак.ериологической воины можно ус-
лышать случайно или во время беседы близких друзей друг с другом.
Если кто-либо случайно проболтался о выполнении задания бактериологической вой-
ны. то разговор на эту тему прекращается, и сразу же переводят разговор на другую тему.
Я помню, как-то, после выполнения задания по бактериологической войне, я навес-
тил младшего лейтенанта Р. Л. Майкла, который лежал в госпитале. Он меня спросил, чем
я занимаюсь, и он по моему выражению и ответу сразу догадался, что я выполнял задание
по бактериологической войне Он сказал: «С одной стороны, я рад, что нахожусь здесь, а не
с вами и не вылетал для выполнения задания» До того, как он был ранен при аварии, он
выполнил задания 2—3 раза. Я знаю, что он человек очень религиозный, поэтому, я думаю,
что он не хочет выписываться из госпиталя, так как после этого может быть ему придется
вылетать для выполнения задания по бактериологической войне. Я сказал ему, что меня
ничто не радует. Он сказал, что вполне сочувствует мне. Летчики после выполнения задания
по бактериологической войне бывают мрачными целый день. Они часто бывают в клубе,
пьют вино и сильно переживают. Темой беседы за столом всегда являлась мысль о печаль-
ном положении американского правительства. Я помню, как вечером 15 февраля, после
выполнения задания по бактериологической войне, старшие лейтенанты Харвэй, П гджэтг
младший лейтенант Грэйл и я вчетвером разговаривали о том, как бы мы хотели увидс гь
Трумэна, который бы управля i самолетом типа «Мустанг» над Кореей. Это казалось смеш-
ным, и мы сразу дали Трумэну прозвише «Джордж Второй». Прозвище «Джордж Второй»
подходило полковнику Левинсону, командиру авиагруппы, и его называли так за спиной.
Приложение Б
1129
Однажды он, совершая полет со звеном Джорджа, занимал второе место. Когда он
долетел до цели, то сбросил бомбы как попало. В это время командир звена приказал вто-
рому самолету вести звено к базе. Однако полковник Левинсон ответил, что он уже не знает
свое точное местонахождение. Этот случай вызвал смех за его спиной, и прозвище«Джордж
Второй» сохранилось за ним навсегда.
В клубе, у стола сидели только мы вчетвером, другие, видимо, решили дать нам самим
пережить наши чувства, вызванные выполнением нашего задания. Это уже стало привыч-
кой, так как мы считаемся с чувствами тех, кто не хочет говорить о бактериологической
войне или не хочет, чтобы напоминали им об их участии в этом деле. В средних числах
февраля, однажды вечером, в офицерском клубе произошел следующий случай сидящий
рядом со столом старшего лейтенанта С. О. Армстронга, спросил его, что он сделал в этот
день Он уже выпил изрядно, поэтому Армстронг ответил громко: «Ясно, что я не распро-
странял бактерии». Командир 67-й эскадрильи, подполковник Кроу, который сидел ря-
дом с нами, услышав это, сразу встал и вывел лейтенанта Армстронга из клуба и где бесе-
довал с ним минут 15; после этого старший лейтенант Армстронг ушел совсем из клуба в
свою палатку. После этого случая все люди замолчали. Потом кто-то крикнул: «Дайте еще
пиво!» — и разговор возобновился, но каждый уже следил за своей речью.
Я не знаю, какое мнение среди солдат о бактериологической войне, так как мы встре-
чаемся с ними очень редко. Но я точно знаю, что, когда мы садимся в самолет, начальник
наземной команды бывает обычно очень разговорчивым Однако утром 15-го февраля он
мне сказал только два слова «Доброе утро», в то время как обычно при встрече с летчика-
ми начальник наземной команды приветствует словами с пожеланиями: «Доставьте, по-
жалуйста, этот самолет обратно, самолетов у нас немного» и т п.
Показания американского военной тенно а тетчика, старшего
лейтенанта Книсса, об его участии в бактериологич“'.кои войне,
ведущейся американскими войсками в Корее (ISCK/5)
Мое имя Паул Р., фамилия Книсс; я являюсь офицером резерва военно-воздушных сил
Соединенных Штатов. Мое военное звание — старший лейтенант, мой личный военный
номер АО 1909070. Я родился 29 апреля 1927 г. в Монмосе, штата Иллинойс. Адрес моей
жены: штат Техас, Сан-Антонио, юго-западная военная дорога 1103. Адрес моих родите-
лей: Иллинойс, Монмос, Южная 7-я улица 339.
В декабре 1946 г. я поступил в военно-воздушный флот, где мне дали военное звание
капрала. Потом я стал пилотом и работал в качестве инструктора при воздушной базе в
Грэйге (Graig) до 30 января 1952 г., т. е. до откомандирования меня в Корею. 21 февраля
1952 г. я прибыл в лагерь Стонман (Camp Stoneman), Калифорния, для дальнейшего на-
значения за границу с 5 другими инструкторами из Грэйга, которые также отправлялись
за границу. Их фамилии: старший лейтенант Джон Карлтон, старший лейтенант Джон
Янсен, старший лейтенант Джеймс Кэмп, старший лейтенант Роберт Маннинг и старший
лейтенант Рандолл. Из лагеря Стонман мы были посланы в Корею. Мы прибыли на К-46
(база для F-51, примерно в 5 милях севернее Вонжу) 20 марта 1952 г. Я был прикоманди-
рован к 12-й истребительно-бомбардировочной эскадрилье, 18-й истребительно-бомбар-
дировочной группы и летал на самолете типа F-51.
В июне 1951 г., когдая был еще в Грэйге, присутствовал налекции в комнате для полетов
нашей эскадрильи. Там присутствовали все пилоты 3616-й эскадрильи для тренировки.
Нашим лектором был капитан Лаури, который являлся офицером информации и воспита-
ния нашей авиачасти. Он детально разъяснил нам организацию наземной противоатомной
обороны. Он заявил, что атомная бомба наносит не больше вреда, чем обыкновенная бом-
ба, но район разрушения ею будет больше. Способами предохранения себя являются: лечь
на землю, забраться под стол или же встать за стену и тем самым предохранить себя от взры-
изо
Приложение Б
ва бомбы. По его данным, взрыв бомбы убивает человека на расстоянии 3/4 мили, если он
не спрячется за что-нибудь; радиация не нанесет вам вреда, если вы будете находиться за
полмили от бомбы. Он указал, что жгучая жара убьет каждого на расстоянии 1/4 мили от
места взрыва.
Он также сказал, что большую опасность представляет бактериологическая война,
которую намечают вести другие страны. Он заверял, что другие страны могут распростра-
нять бактерии при помощи бактериологических бомб; они могут быть также завезены вра-
жескими агентами или же будут заброшены артиллерийскими снарядами с подводных
лодок. Он выразил надежду, что в 1952 г. весь военный персонал пройдет специальный курс
бактериологической войны, им выдадут предохранительные маски и сделают специаль-
ные прививки против бактерий. Пилоты нашей эскадрильи спросили его, где он получил
такую информацию, но он не ответил и только заверял нас, что об этом мы узнаем позднее.
Через день после того, как мы прибыли в лагерь Стонман, т. е. 22 февраля, все мы, вновь
прибывшие, совместно с другими группами, которые также направлялись в Корею, были
проинструктированы в течение 15 мин капитаном Холлеманом. Старший лейтенант Чарльз
Крон, который проходил тренировку со мной на авиабазе воздушного флота в Барксвене,
штата Луизиана, также присутствовал на этой лекции; он был назначен летать на В-26 в
Корее. Всего на лекции присутствовало 50 пилотов. Капитану Холлеману около 35 лет, хо-
дит в очках, рост его около 6 футов, волосы темного цвета и слегка лысоват. Он указал, что
ходят различные слухи о том, что Америка применяет бактериологическое оружие в Ко-
рее. Он говорил, что эти слухи неправдоподобны, и мы должны отвергнуть эти слухи.
Америка имеет, как он говорил, бактериологические бомбы, и бактерии могут быть рас-
пространены путем опыления при помощи самолетов, но мы не применяем это и желаем,
чтобы все вы, военнослужащие, если услышите разговоры о бактериологической войне,
то опровергайте эти слухи. Мы имеем в Америке бактериологические бомбы «VT» (с ди-
станционными взрывателями), и мы также имеем бомбы с парашютами для ведения бакте-
риологической войны. Такие бомбы с парашютами могут быть наполнены больными жи-
вотными и насекомыми и, когда они будут выброшены из бомбы, распространяют болезни.
Мы также можем распылять бактерии прямо с самолетов. Мы можем также занести их на
вражескую территорию нашими агентами, которые могут заразить источники водоснаб-
жения всех больших и малых городов.
Капитан Холлеман дал нам всю эту информацию в лагере Стонман, в оперативной
комнате. После этой лекции я и другие пилоты нашей группы обсуждали содержание лек-
ции. Я, какидрутие военнослужащие были того мнения, что мы не применяем бактериоло-
гического оружия, а слухи же являлись пропагандой северокорейцев. Мы считали, что наше
правительство естественно желает пресечь все эти слухи.
21 марта, когда мы прибыли в Корею, на базу К-46, мы шесть пилотов: лейтенанты
Карлтон, Янсен, Кэмп, Маннинг, Рандолл и я опять прослушали одночасовую лекцию
капитана Маклафлина. Ему было около 30 лет, ростом он около 6 футов. Он был офице-
ром разведки нашей авиагруппы. Инструктаж происходил в комнате, где докладывали о
выполнении заданий; комната была закрыта на ключ. 18-я истребительно-бомбардировоч-
ная авиагруппа ведет бактериологическую войну с 1 января 1952 г. Капитан Маклафлин
заявил, что в настоящее время мы применяем два типа бомб: бактериологическую бомбу
«VT» и бомбу с парашютом для сбрасывания животных. «Мы собираемся начать распыле-
ние бактерий с самолетов в июне. 30 апреля 1952 г. мы пошлем 4 самолета нашей авиагруп-
пы в Тачикава (Япония), чтобы снабдить их аппаратами для распыления бактерий. Там,
за кабиной пилота, установят бак для помещения бактерий, откуда они будут распылять-
ся позади самолета. Самолеты будут готовы 15 июня, и тогда мы проинструктируем всех
пилотов авиагруппы, как распылять бактерии. Этот метод успешно применялся в Корее».
«В наших бомбах (VT), — как указал капитан Маклафлин, — мы будем применять три
вида болезней: брюшной тиф, малярию и бубонную чуму. Кажется, он упомянул малярию,
Приложение Б
1131
но этого я точно не помню. Эти бомбы будут доставлены из Вонжу cnet шальным грузови-
ком и будут загружаться на самолеты за 15 минут до полета (в то время, как наши самоле-
ты обычно загружаются за 2 часа до полета). Этот грузовик закрытый, и если вы увидите
его, то узнаете сразу. Самолет загружается специальной командой из отдела вооружения,
и эта команда при погрузке носит белую форму, маски и перчатки. Не бойтесь этих бомб.
На вас не будет особого снаряжения; бактерии хорошо изолированы в бомбах. Самолет
подвергнется дезинфекции после возвращения с задания».
«У нас нет, — продолжал он, — специальных самолетов для ведения бактериологичес-
кой войны; мы применяем самолеты, которые у нас имеются. По возвращении с задания
вы после рапорта сразу же примите душ, а на следующий день вам исследуют кровь с тем,
чтобы проверить все ли в порядке. Если по каким-либо причинам вы не сможете выпол-
нить задания, вы не должны сбрасывать бомбы зря, а вернитесь на базу К-46. Вы никогда
не до лжны говорить о бактериологической войне, и будете всегда, после выполнения за-
дания, докладывать так, как я вам укажу. Вы дадите подписку после этого собрания, что
вы не будете обсуждать между собой или с кем-либо этот вопрос. Содержание данной лек-
ции будет считаться «совершенно секретным». Наше правительство будет отрицать факт
ведения бактериологической войны, насколько это будет возможным. Не переживайте в
душе по поводу применения бактериологического оружия, так как все другие пилоты на-
шей авиагруппы уже применяют, его и это будет вестись в большем масштабе и в будущем».
Схема бактериологической бомбы «УТ», нарисованная капитаном
Маклафлином на черной доске (в изложении старшего лейтенанта Книеса)
Такие бомбы, — продолжал капитан, — будут всегда сбрасываться с четырех самоле-
тов одновременно. Вы спикируйте с высоты 10000 футов до 6000 футов и сбросьте ваши
бомбы над целью. Бомба взорвется, примерно, в 100 футах от земли и бактерии рассеют-
ся, примерно, в окружности 100 ярдов. Если бомба не взорвется в воздухе, а взорвется на
земле, то бактерии будут убиты взрывом. Если бомба взорвется в воздухе, то бактерии бу-
дут рассеяны силой взрыва. Такие бомбы будут сброшены недалеко от города, но не в са-
мом городе, так как северокорейцы широко применяют дезинфицирующие вещества в
своих городах, которые убивают бактерии. Мы сбрасываем наши бомбы вблизи больших
городов с таким расчетом, чтобы люди и животные могли занести болезни в города, где
бактерии будут распространяться, но эти бактерии должны попасть на животных или лю-
дей в течение трех часов, иначе они погибнут». По указанию капитана Маклафлина эти
бактерии являются паразитическими, и они не могут жить долго самостоятельно; но в
живом организме — могут жить. После успешного выполнения задания вы доложите: «За-
дание выполнено, результаты не наблюдались», а в газетах будем сообщать о «подавлении
зенитного огня». Бомба, как указал капитан Маклафлин, похожа на обыкновенную 500-
фунтовую бомбу «\Т» и не имеет каких-либо специальных пометок.
Эта бомба, как указал капитан Маклафлин, будет применяться только одним из опыт-
ных пилотов нашей авиагруппы. Он спикирует с высоты 10000 футов до 1000 футов и сбро-
сит бомбы. На задней части бомбы раскроется дверка, откуда вырывается парашют, и бомба
медленно упадет на землю При ударе бомбы о землю, бомба разъединится на две части.
1132
Приложение Б
или носекомые
Схема бомбы с парашютом, как она была нарисована капитаном
Маклафлином (в изложении старшего лейтенанта Книсса)
При раздвоении бомбы животные или насекомые выходят оттуда. В этих бомбах мы по-
мещаем крыс, вшей, мышей и блох. Такие бомбы будут сброшены также около больших
городов, но не в самом городе, так как городское население может истребить животных и
насекомых. Эту бомбу пилот, во время доклада, должен называть «неразорвавшейся». Вы
сможете узнать эту бомбу по дверке и шарнирам, которые соединяют две части бомбы
С вышеуказанными типами бомб вы не должны летать выше 12000 футов, так как живот-
ные, насекомые и бактерии могут погибнуть от холода и недостатка кислорода. Капитан
Маклафлин продолжал: «Я напоминаю вам, еще раз, что нигде не следует говорить об этой
информации, иначе вас предадут суду военного трибунала». Этими словами он закончил
свои указания. Инструктаж длился с 08.00 ч до 09.00 ч.
На этом инструктаже присутствовали следующие лица: я, старший лейтенант Джон
Карлтон, старший лейтенант Джон Янсен, старший лейтенант Роберт Маннинг, старший
лейтенант Джеймс Кэмп и старший лейтенант Рандолл. Мы все были инструкторами в
Грэйге и были направлены в эту авиагруппу одновременно.
Капитан Маклафлин после инструктажа потребовал от нас подписку о молчании. По-
сле подписания мы их ему вернули. Эти обязательства были в 10 дюймов длиной и в 2 дюйма
шириной. Насколько я помню, обязательства были следующего содержания: «Я, нижепод-
писавшийся, не буду обсуждать информацию, сообщенную на собрании 21 марта 1952 г.,
и если это случится, то я подлежу осуждению по параграфу военного устава. Подпись»
Я забыл точный номер параграфа военного устава. Офицер разведчасти нашей авиа-
группы хранит эти подписки в своем кабинете. Если кто-либо из пилотов разгласит факт
ведения бактериологической войны, то он будет предан военному трибуналу, а обязатель-
ство будет служить основанием.
Я чувствовал отвращение к себе при мысли о применении бактериологического ору-
жия, но я чувствовал, что у меня нет другого выбора и я должен делать то, что приказывают;
я также вспомнил слова капитана Холлемана, которому я верил, и на этот раз стал обду-
мывать кто прав, кто виноват в этой бактериологической войне.
27 марта 1952 г. в 5 ч 30 мин я, капитан Томас (командир нашего авиазвена), капитан
Брутон и старший лейтенант Флур и все те, которые принадлежат к 12-й истребительно-
бомбардировочной эскадрилье, получили инструкцию перед выполнением задания бак-
териологической войны. Мы были проинструктированы капитаном Маклафлином в ком-
нате, где обычно дают инструктаж. Инструктаж проходил при закрытых дверях. Капитан
Маклафлин показал нам аэрофотографию — горы с открытыми пятнами, что в 10 милях
от Саривона. Снимки были размером, примерно, в 12 дюймов длиной и в 8 дюймов ши-
риной, которые были засняты 67-й тактико-разведывательной группой. Открытые пятна
на горе были артиллерийскими позициями. Я начал рассматривать снимки с лупой, но ни
пушек, ни артиллерийских позиций не заметил. «Вашей задачей является подавление зе-
нитного огня», — сказал капитан Маклафлин. «Вы вылетите в 07.00 и будете над целью в
08.00. В пути следования к цели вы будете лететь на высоте 10000 футов. Когда вы долетите
Приложение Б
1133
до цели, вы спикируете и с 6000 футов сбросите ваши бомбы. По возвращении вы будете
докладывать мне по форме: «Задание выполнено, результаты не замечены».
Наш инструктаж д лился 15 минут. После инструктажа я прошел в рабочую комнату
нашей эскадрильи и одел свое летное снаряжение. В 06 ч 45 мин я пошел к своему самоле-
ту. Два человека в белых комбинзонах, в респираторах и в перчатках загружали мой само-
лет бомбами. Они брали бомбы с грузовика и прикрепляли их руками к бомбодержателям.
Эти бомбы грузились как обыкновенные 500-фунтовые бомбы «VT» и не имели каких-либо
особых опознавательных знаков. Когда они закончили погрузку и отъехали, я осмотрел
свои бомбы, чтобы проверить насколько прочно они держатся в бомбодержателях. Мой
самолет, как и три других самолета, имел две 500-фунтовые бомбы.
Мы стартовали в 07.00 и набрали высоту 10000 футов. В 08.00 мы прилетели к нашей
цели, которая находилась в 10 милях южнее Саривона примерно, в 100 футах от железной
дороги. Мы спикировали до 6000 футов и сбросили наши бомбы. Две бомбы взорвались
на земле, остальные шесть — в воздухе. Бомбы, которые взорвались на земле, дали серова-
тый туман дыма; дым поднялся на высоту 100 футов и через 45 секунд исчез. Мы вновь под-
нялись на высоту 10000 футов и вернулись на базу К-46. Когда мы приземлились, было
09.00; мы пошли к Маклафлину и в комнате разведчасти доложили ему: «Задание выпол-
нено, результаты не замечены». После этого экипаж принял душ. Принимая душ, я сказал
капитану Томасу: «Это может очистить мое тело, но моя совесть никогда не будет чистой
после такого преступления». 28 марта в 08.00 наш доктор сделал нам исследование крови.
О сбрасывании бактериологических бомб лично я знаю следующие факты: 29 апреля
старший лейтенант Далео сказал мне, что он и старшие лейтенанты Кюрри, Джон Янсен и
Рандолл 5 апреля сбросили 8 бактериологических бомб «VT» в 5 милях южнее Синъанжу
(все эти пилоты были из 12-й истребительно-бомбардировочной эскадрильи). Он также
сказал, что подполковник Крейн, командир нашей эскадрильи (12-й эскадрильи), 15 марта
сбросил две парашютные бомбы в 5 милях восточнее Санчон. 5 мая в 09.00 я слышал, как
подполковник Кроу, командир 67-й эскадрильи, сказал капитану Маклафлину, что он сбро-
сил две «неразорвавшиеся» бомбы недалеко от Пхеньяна. Они были бактериологическими
бомбами с парашютами. 21 мая капитан Томас сказал мне, что старший лейтенант Эд Вил-
льямс, из 67-й истребительно-бомбардировочной эскадрильи, вел звено из четырех само-
летов на позицию, что в 5 милях восточнее Кунари, для выполнения задания по подавле-
нию зенитного огня. Мы знали, что тогда они сбросили 8 бактериологических бомб «VT».
Я хочу довести до сведения читателей данного показания, что поскольку у меня имеется
чувство справедливости, и поскольку я могу отличить верное от неверного, сознание непра-
воты заставило меня рассказать об этих фактах. С того дня, как я начал совершать эти
преступления, моя совесть все время укоряла меня, и я уверен, что каждый, кто отличает
справедливость от несправедливости, будет испытывать то же самое чувство. Эта война
должна быть прекращена. Я выдвигаю эти факты перед всем миром и подтверждаю, что
вооруженные силы Соединенных Штатов применяют в Корее бесчеловечное оружие. Это
оружие применяется не только против северокорейских войск и китайских добровольцев,
но и против населения Северной Кореи. Гражданское население Северной Кореи уже
ужасно пострадало от войны, а сейчас оно подвергается еще самым бесчеловечным мето-
дам войны. Обязанность всех народов мира — обратить внимание на факты, приведенные
мною, и потребовать немедленного прекращения бактериологической войны в Северной
Корее. Народ Соединенных Штатов должен потребовать, чтобы ни одна страна не приме-
няла такого вида оружия войны. Только тогда, когда каждый человек в мире выполнит свой
долг в борьбе за запрещение войны и бесчеловечных преступлений, мы достигнем всеобще-
го мира. Все люди мира являются братьями, и до тех пор пока мы не научимся жить вме-
сте и помогать друг другу, мы не будем иметь всемирного мира, которого мы так жаждем.
Паул Р Книсс (подпись), 20 июля 1952 г.
1134
Приложение Б
Комментарии редакторов Доклада... на показания Книсса
Налицо имеется ряд противоречий между свидетельскими показаниями Книсса и
О’Нила, хотя оба пилота служили в 18-й истребительно-бомбардировочной авиагруппе,
оба летали на самолете типа F-51 и оба инструктировались офицером разведчасти авиа-
группы, капитаном Маклафлином. Основными моментами сходства в свидетельских по-
казаниях двух пилотов можно отметить:
1. Оба пилота, по прибытии на базу, были немедленно инструктированы капитаном
Маклафлином в группах по несколько человек. О’Нил заявил, что обязанностью Маклаф-
лина было инструктировать всех пилотов по ведению бактериологической войны, прибы-
вающих на базу для выполнения заданий.
2. Идентичность применяемых средств: бактериологическая «VT», бомба с парашю-
том и с насекомыми, зараженными бактериями или с мелкими животными, и распыление
бактерий непосредственно с самолетов.
3. Необходимость сохранения тайны.
4. Высота, с которой бомбы «VT» должны быть сброшены. О’Нил, показал, что он сбро-
сил с высоты 7000 футов, а Книссу было приказано сбросить бомбу с высоты 6000 футов.
5. В бактериологической войне могут быть также использованы артиллерийские сна-
ряды, но бактериологические бомбы наиболее эффективны.
6. Специальная команда занимается погрузкой бактериологических бомб.
7. Установлена особая форма доклада о результатах выполнения заданий, и что капи-
тан Маклафлин будет лично принимать их.
Расхождения в показаниях:
1. О’Нилу было сказано, что вне зависимости от взрыва бомбы «УТ» в воздухе или на
земле, бактерии останутся живыми. Книссу же было сказано, что при взрыве бомбы на
земле бактерии будут убиты силой взрыва; бактерии могут остаться живыми, если бомба
взорвется на высоте 100 футов от земли. Оба подтверждают, что бомбу «УТ» следует взор-
вать на высоте 100 футов.
2. Книсс был предупрежден не летать выше 12000 футов, так как бактерии и насеко-
мые погибнут из-за недостатка кислорода и от холода. О’Нилу про это не говорили.
3. 0’ Нилу было сказано, что бактериологические бомбы будут сброшены на города или
районы, где сконцентрировано много воинских частей. Книссу же было сказано сбрасы-
вать бактериологические бомбы около больших и маленьких городов, примерно в 5— 10 ми-
лях от города; это видно из выполненных ими заданий или же из известных ему фактов.
(Следует отметить, что Книсс был инструктирован двумя месяцами позднее О’Нила. Аме-
риканская армия к этому времени поняла, что бактериологическое оружие, применяемое
ею, по крайней мере, три месяца, не приносило желаемых результатов. Вполне понятно,
что специалисты будут искать всевозможные причины неуспеха этого дела. Ко времени,
когда инструктировали Книсса, производились попытки устранить все неблагоприятные
факторы. Американская армия пришла к выводу, что бактериологическая война не так
проста, как ей раньше казалось. Книссу было конкретно сказано не сбрасывать бактерио-
логические бомбы на города ввиду противоэпидемических мер, принятых корейским на-
родом.)
4. О’Нилу было сказано, что в случае, если ему не удается сбросить свои бактериоло-
гические бомбы в Северной Корее, то выбрать какое-нибудь безлюдное место в Южной
Корее, сбросить их там и дать знать ближайшей радарной станции, чтобы она определила
место сброса бомб, куда затем будет послана специальная команда для доставки их обрат-
но на базу. (А Книссу было сказано в таких случаях не сбрасывать бомбы и вернуться на
базу.) Подтверждением этого является сообщение всех американских телеграфных аген-
тов в начале марта о том, что в отдаленной горной деревне в Южной Корее возникла
«необъяснимая» вспышка сыпного тифа. Весьма вероятно, что это и было результатом
Приложение Б
1135
такого сброса летчиками «неразрывающихся» бомб, которые в действительности взорва-
лись во время удара бомбы о землю.
5. О’Нил говорит, что распыление бактерий было начато с середины февраля, соглас-
но инструктажу. Книсс же показал, что распыление аэрозоля начнется только с середины
июня и что самолеты будут отправлены в Японию в конце апреля для оборудования. О’Нил
и Книсс утверждают, что их показания правильны. О’Нил действительно видел самолет с
приспособлением для распыления бактерий, который находился отдельно от других само-
летов. Это объясняется двумя причинами:
а) первые пробы распыления бактерий не были успешными, и необходимо было улуч-
шить методы, поэтому их применение временно было приостановлено;
б) потери самолетов с опылительными приспособлениями были крайне большими (или
потери самолетов F-51 были настолько значительными, что самолеты с опылительными
приспособлениями были использованы для обычных боевых действий) и поэтому времен-
но прекратили осуществление этого плана. Интересно отметить, что Маклафлин сказал
Книссу, что метод распыления бактерий уже успешно применяется в Корее и что февраль
был месяцем, когда потери американских военных самолетов были наиболее высокими.
6. Книсс и другие были вынуждены дать подписку, тогда как от О’Нила и других не
потребовали этого. 22 января, когда О’ Нил получил инструктаж, корейско-китайская сто-
рона еще не указывала о бактериологической войне и действительно не было еще доказа-
но, что она уже началась. До 21 марта было много разговоров о бактериологической вой-
не, которые вызвали споры и разговоры на авиационных базах. Это видно из показаний
военнопленных, и, конечно, необходимость пресечения таких разговоров логически была
более актуальной 21 марта, чем 22 января. 22 февраля при инструктаже в лагере Стонман,
на котором присутствовал Книсс, было приказано отрицать все слухи о том, что амери-
канское правительство применяет бактериологическое оружие. Очевидно, такой инструк-
таж еще не был необходим для О’Нила во время его инструктирования.
7. Большие предосторожности были приняты для сохранения здоровья летчиков в
период, когда Книсс начал участвовать в бактериологической войне. Пилотам начали ис-
следовать кровь после выполнения заданий по бактериологической войне.
Расхождения между показаниями вышеуказанных двух пилотов показывают, что аме-
риканское правительство изменяет свои методы ведения бактериологической войны на-
ряду с усовершенствованием бактериологического оружия.
Редакторы Доклада...
Научное издание
Супотницкий Михаил Васильевич
БИОЛОГИЧЕСКАЯ ВОЙНА
Введение в эпидемиологию искусственных эпидемических процессов
и биологических поражении: монография
Главный редактор А. И. Моисеев
Научный редактор Е. В. Лебединская
Редактор А. В. Тихомиров
Оформление А. С. Щукин
Компьютерная верстка Н. С. Болтунова
Корректор Н. А. Александрова
НП ИД «Русская панорама»
109028, Москва, Серебряническая наб., 27, оф.1-03
тел. (495) 917-59-83, 917-70-94,
e-mail: in@rus-pan.ru
ООО «Издательство «Кафедра»
E-mail: info@cathedrabooks.ru
www.cathedrabooks.ru
Подписано в печать 12.08.2013.
Формат 70х100'/16. Бумага офсетная. Усл. печ. л. 91,59
Тираж 3000 экз. Первый завод 1—1000. Заказ № 3017
Отпечатано способом ролевой струйной печати
в ОАО «Первая Образцовая типография»
Филиал «Чеховский Печатный Двор»
142300, Московская область, г. Чехов, ул. Полиграфистов, д. 1
Сайт: www.chpd.ru. E-mail: sales@chpd.ru,
8(495)988-63-76, т/ф. 8(496)726-54-10
Супотницкий Михаил Васильевич — микробио-
лог, кандидат биологических наук, старший на-
учный сотрудник. С 1979-го по 2006 г. работал на
разных должностях в научно-исследовательских
учреждениях Министерства обороны СССР и
Российской Федерации. В настоящее время экс-
перт одной из федеральных организаций. Автор
книг «Микроорганизмы, токсины и эпидемии»
(2000, 2005), «Очерки истории чумы» (2006), «Словарь генетических
терминов» (2007), «Эволюционная патология» (2009).
Данное исследование представляет собой первую серьезную попыт-
ку сформировать новый раздел эпидемиологии — «Эпидемиологию
искусственных эпидемических процессов» или, как еще называет его
автор, «Неправильную эпидемиологию». Эта идея давно лежит на по-
верхности накопленных знаний по эпидемиологии, однако пока никто
кроме автора не попытался обобщить разнонаправленные материа-
лы, провести их критический анализ и синтезировать на этой базе но-
вые знания и обобщения, и таким образом превратить абстрактную
идею в самостоятельную науку, определив ее объект, цели и задачи.
Автор книги — военный ученый-микробиолог, полковник медицин-
ской службы запаса. Мне он стал известен работами, сыгравшими
важную роль в появлении принципиально нового направления в ис-
следованиях, проводимых научными организациями Войск радиаци-
онной, химической и биологической защиты Министерства обороны
Российской Федерации. Написанная им книга энциклопедична, на-
сыщена научными данными и идеями, отличается последовательным
и логичным изложением материала «от общего к частному». Даже
знакомство с ее содержанием и списком литературы показывает, ка-
кую автор выполнил огромную работу.
Желающих ознакомиться с этой книгой я предупреждаю, что в ней
не содержится простых истин по эпидемиологии и привычных мифов
об «оружии массового поражения бедных». Задача автора состоит в
том, чтобы научить читателя понимать реальные биологические угро-
зы и отличать их от придуманных. Книга держит в напряжении и вом-
хищает редким сегодня профессионализмом. И еще мне приятно,
что вопреки длящейся уже два десятилетия деградации созидатель-
ных сторон нашей российской жизни, находятся люди, способные
этой деградации противостоять, среди них и автор книги, которую вы
Генерал-полковник в отставке,
доктор технических наук С. В. Петров