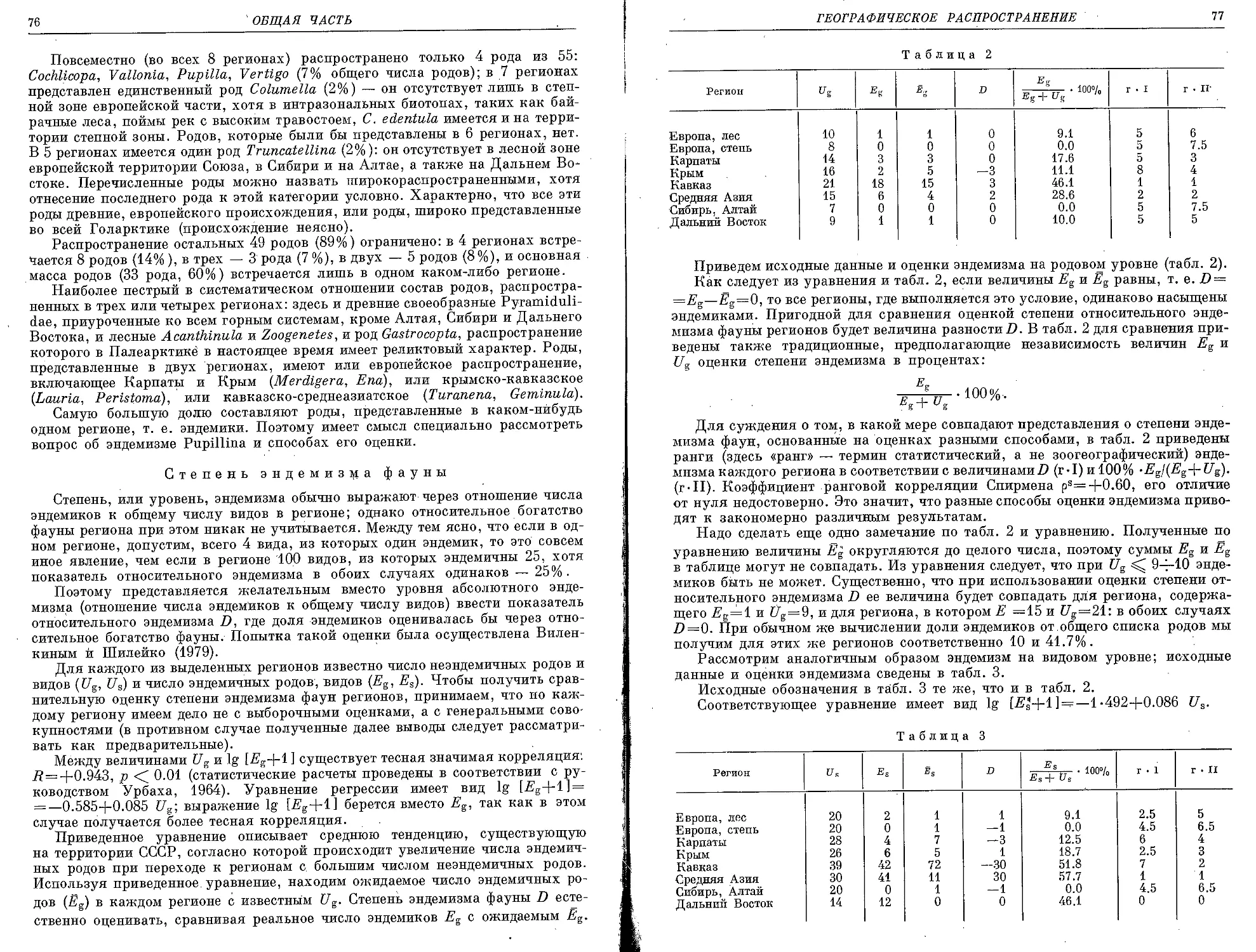
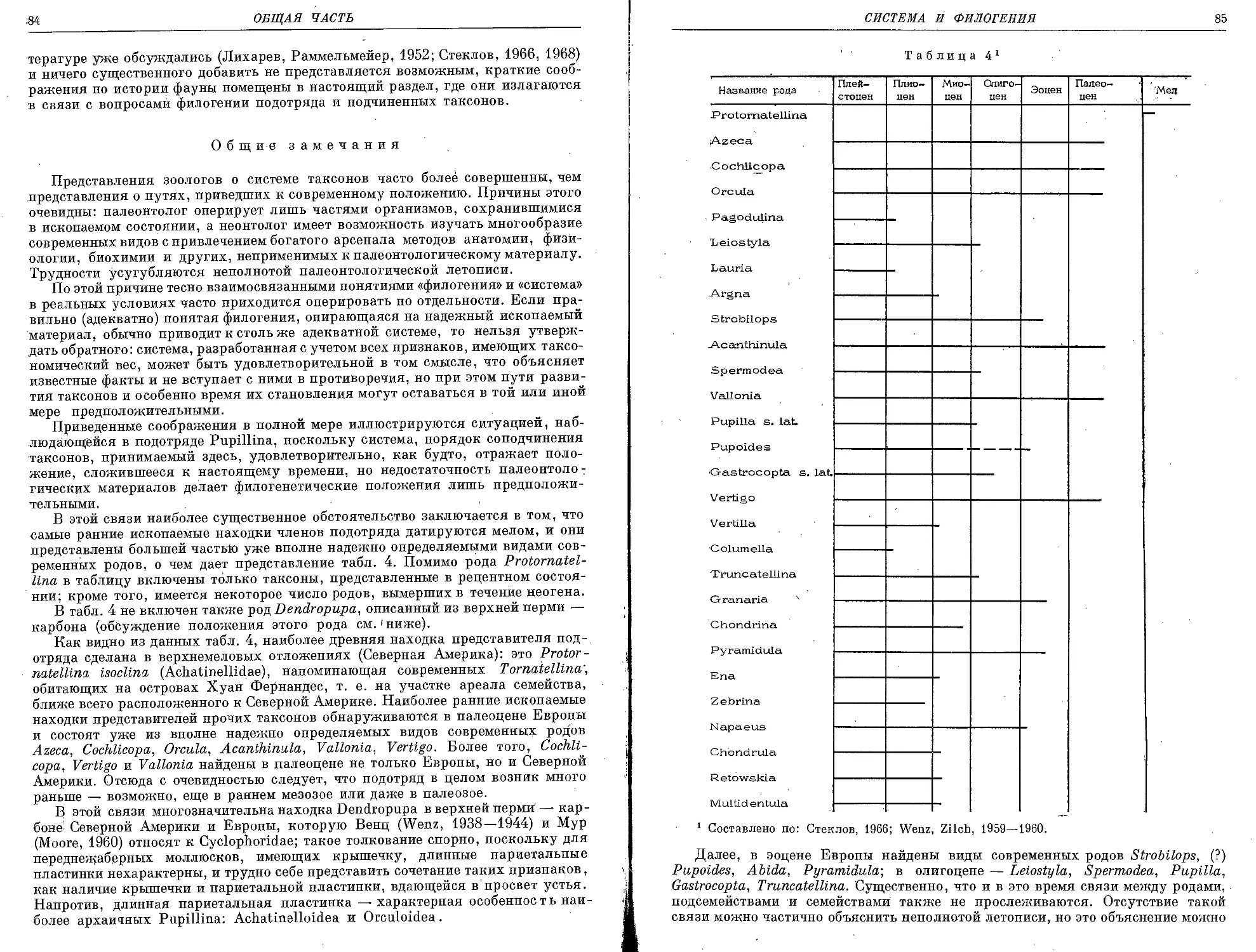
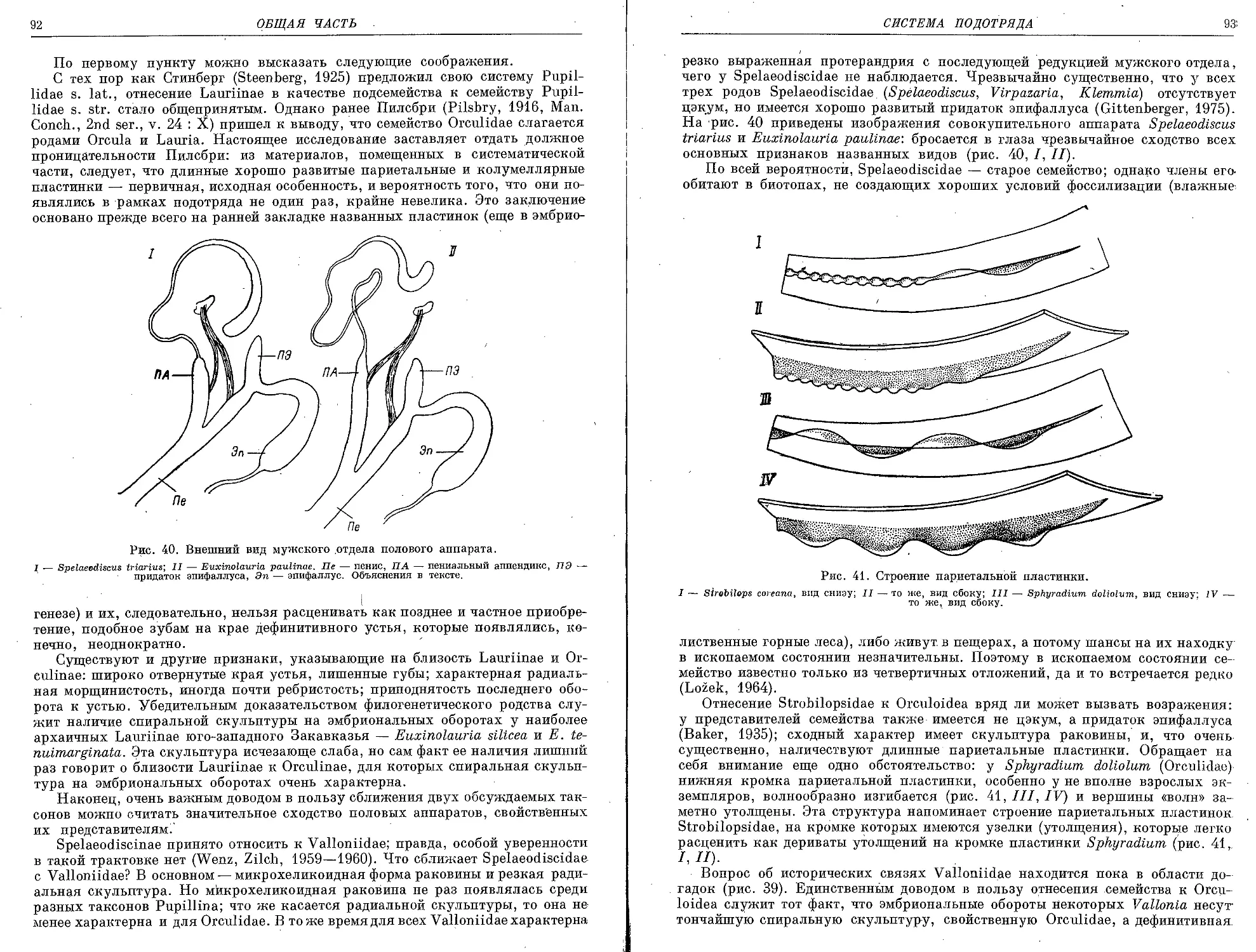
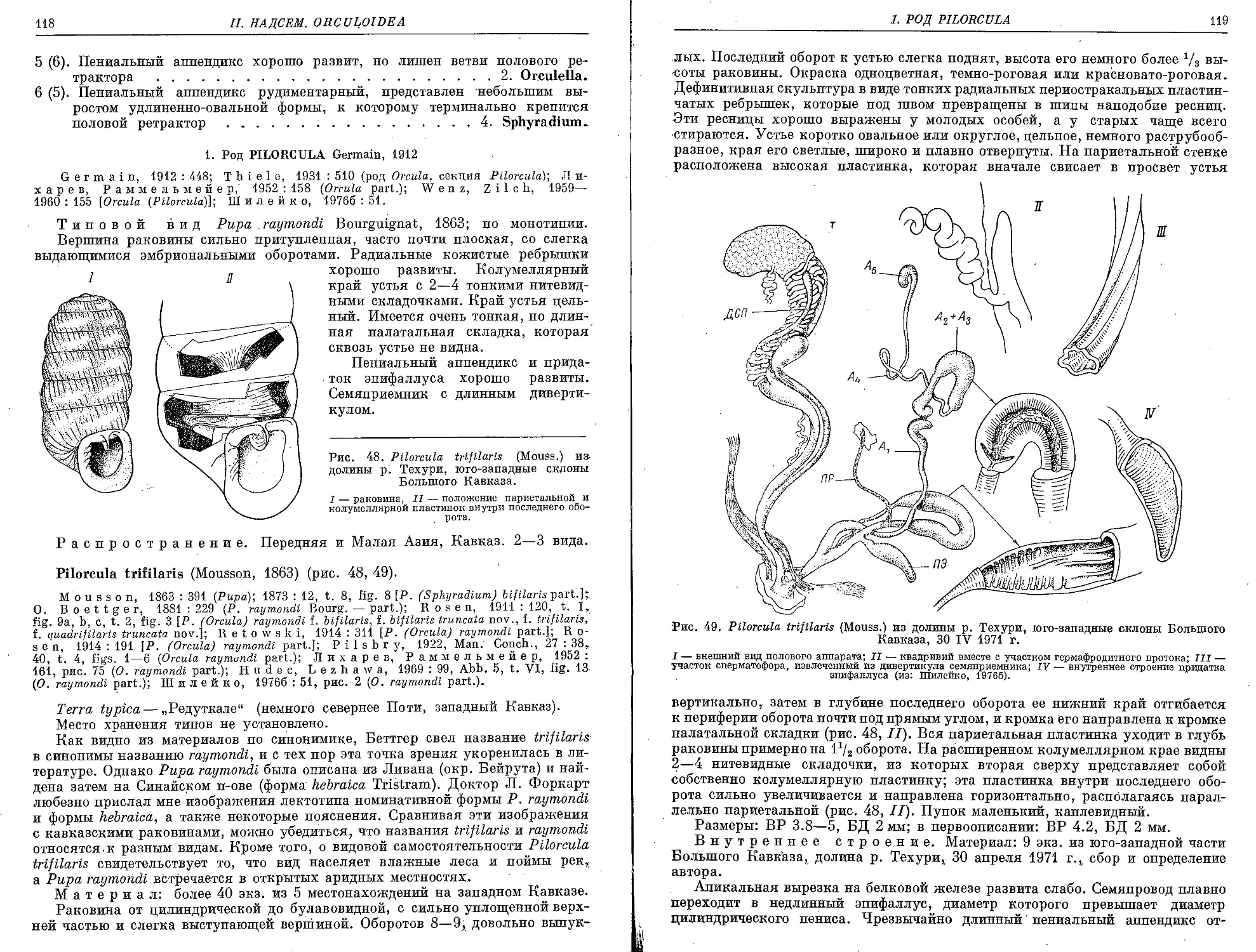
/











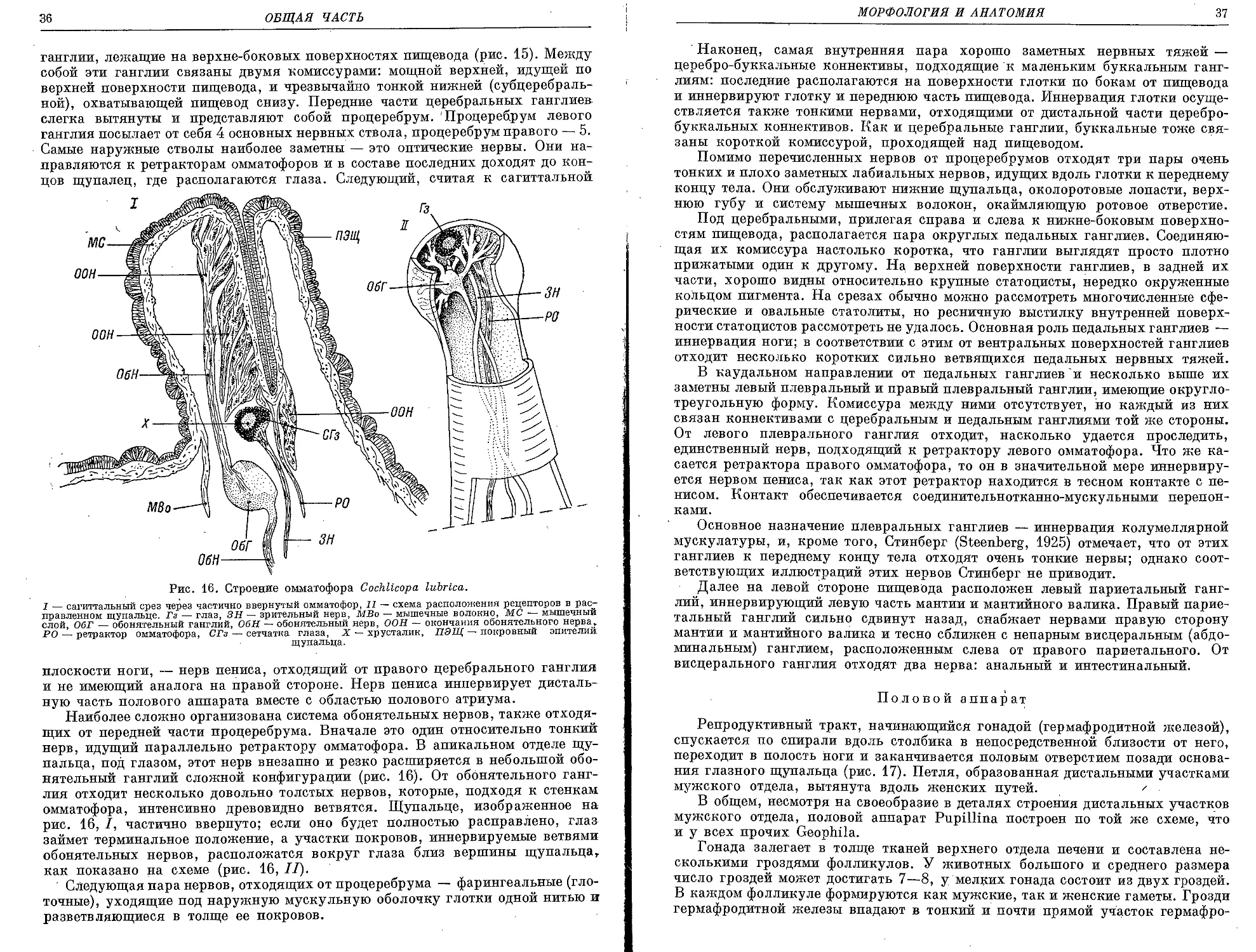
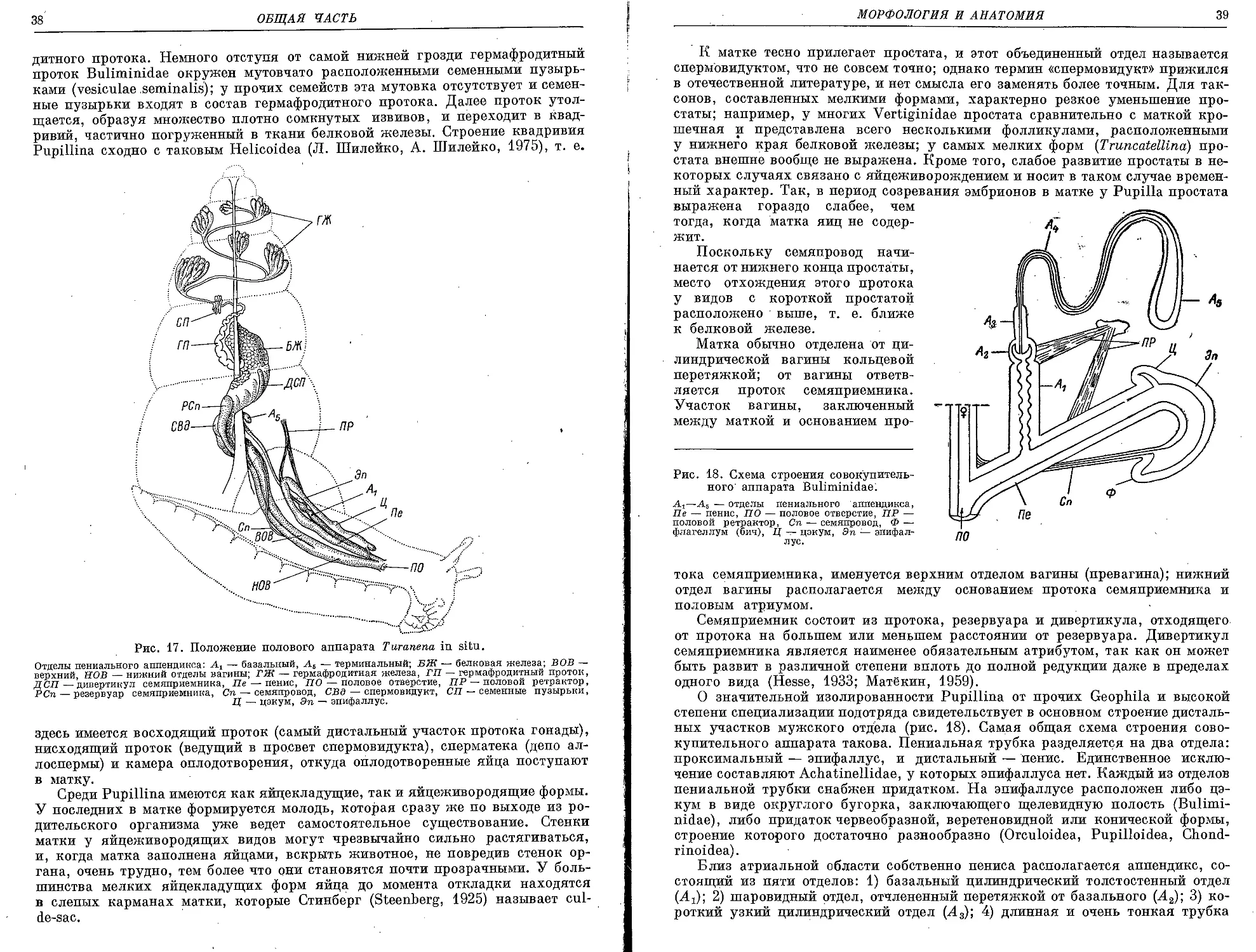














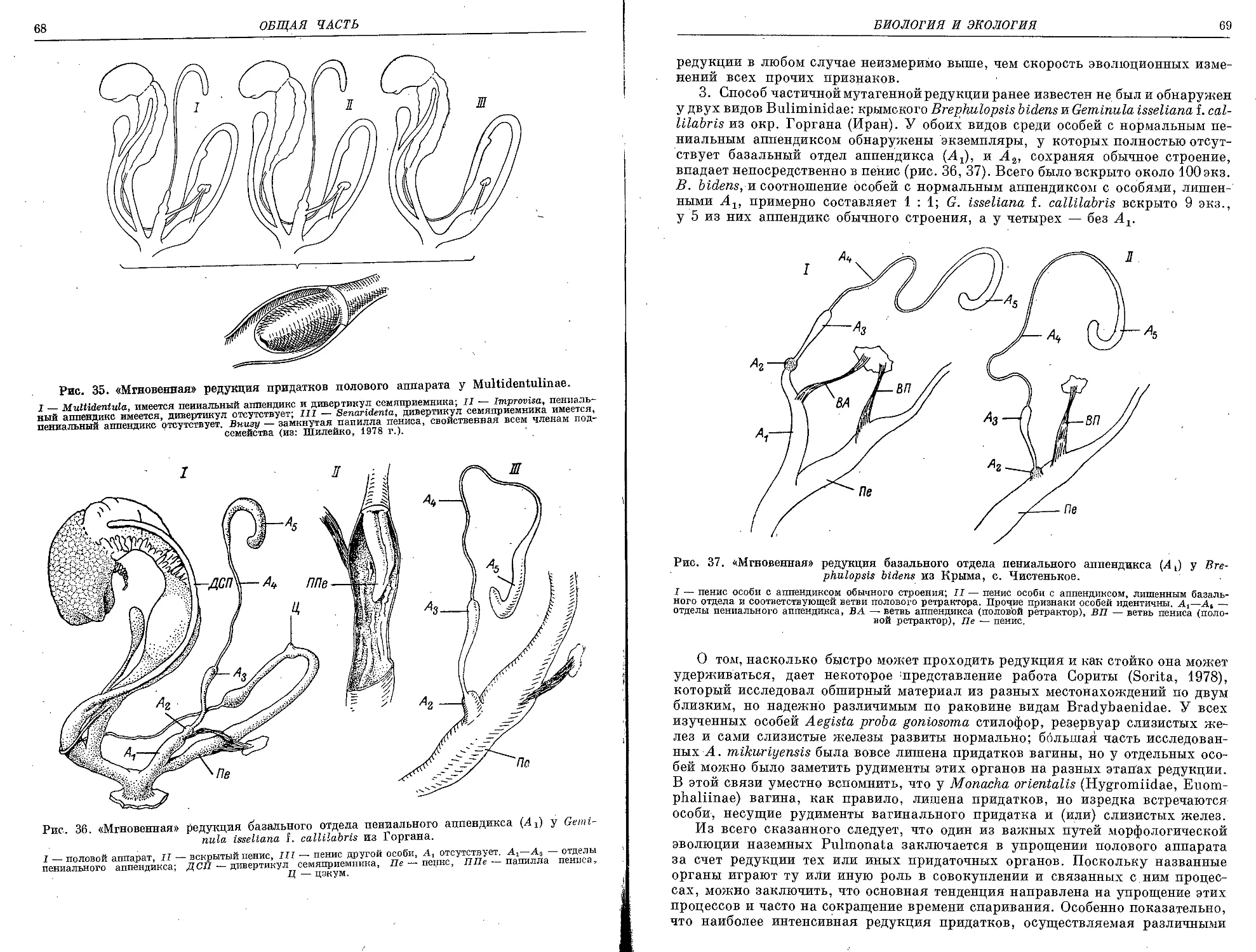


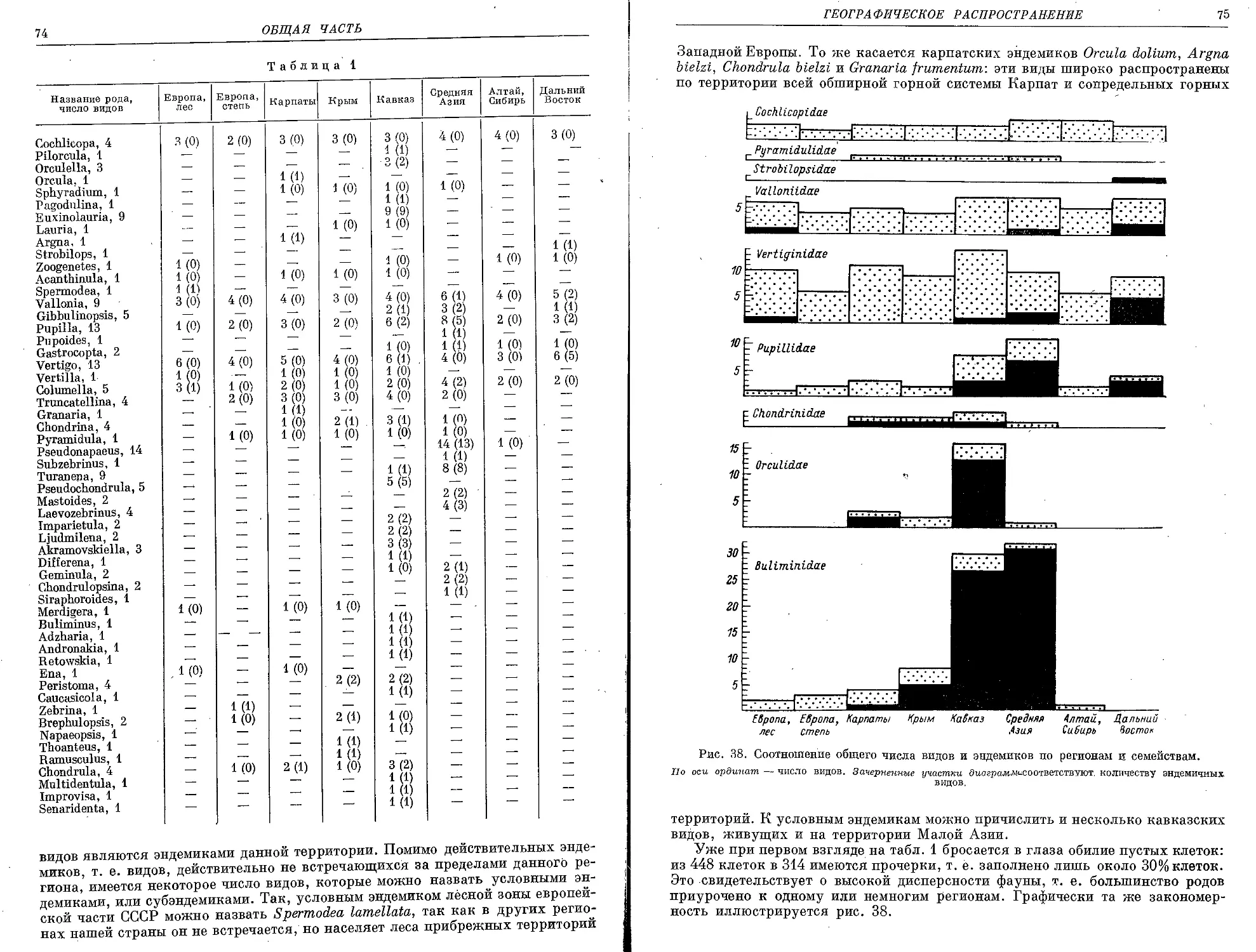






































































































Текст
ФАУНА
СССР
моллюски
ш
выпуск
з
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ, № 130
ФАУНА СССР
МОЛЛЮСКИ
том III, вып. 3
А. А. ШИЛЕЙКО
НАЗЕМНЫЕ МОЛЛЮСКИ ПОДОТРЯДА PUPILLINA
ФАУНЫ СССР (GASTROPODA, PULMONATA,
GEOPHILA)
№
ЛЕНИНГРАД
„НАУКА"
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
19 8 4
УДК 594.38 (47-{-58) (088.7)
Фауна СССР (Нов. сер., № 130). Моллюски. Т. III, вып. 3. Ши л ей к о А. А. Назем-
ные моллюски подотряда Pupillina фауны СССР (Gastropoda, Pulmonata, Geophila). —
Наука, 1984.—399 с.
Систематический обзор всех видов Pupillina, обитающих ныне на территории СССР
(170 видов и подвидов). Общий очерк организации представителей подотряда. На базе морфо-
функционального анализа сделана попытка реконструировать ход исторического развития
группы в целом и проведена таксономическая ревизия; в результате система подотряда
сильно перестроена. Приведены таблицы для определения семейств, подсемейств, родов и
видов. По каждому виду приводятся рисунки раковины, а также внутреннего и внешнего,
строения полового аппарата. Лит. — 400 назв., ил. — 279 рис., табл. — 4.
Главный редактор серии «Фауна СССР»
директор Зоологического института АН СССР
О. А. СКАРЛАТО
Редакционная коллегия:
И. М. Лихарев (отв. ред. серии), О. Л. Крыжановский (зам. отв. редактора),
С. В. Василенко, И. М. Громов, В. Ф. Зайцев, И. М. Кержнер, Л. А. Кутикова,
Г. С. Медведев, М. Е. Тер-Минасян, Н. А. Филиппова
Редактор выпуска
И. М. Лихарев
Рецензенты:
Я. И. Старобогато в, Ю. С. М и н и ч е в
2005000000-648
042(02)-84
290-84-1
© Издательство «Наука», 1984 г-
ПРЕДИСЛОВИЕ
В решениях I Всесоюзного совещания по изучению моллюсков, состоявше-
гося 20—22 ноября 1961 г. в Зоологическом институте АН СССР, среди прочих
вопросов отмечено: «В первую очередь необходимы. . . монографии по груп-
пам, имеющим важное народнохозяйственное значение, как. . . Enidae, Pupil-
lidae» (Тр. Совещания,'с. 10). Предлагаемая работа отвечает этому решению,
расширяя одновременно границы поставленной задачи: исследованием охва-
чены виды всех 9 семейств подотряда Pupillina, обитающие на территории
Советского Союза.
Значение изученных групп в народном хозяйстве неоднозначно. Многие
виды играют важную роль как переносчики нематодозов и трематодозов сельско-
хозяйственных животных; некоторые крупные виды иногда наносят ущерб
всходам полевых и огородных культур, хотя их роль в этом отношении сравни-
тельно невелика и не сравнима с вредом, наносимым, например, слизнями.
Виды, массами населяющие полупустынные и степные ландшафты, служат
немаловажным источником пищевых водных ресурсов для других животных,
особенно позвоночных. Наиболее широко представленные в ископаемом со-
стоянии группы наземных моллюсков — это.в первую очередь мелкие представи-
тели данного подотряда. Поэтому значительна их роль в геологических дисци-
плинах: стратиграфии, палеоклиматологии, палеогеографии.
Развиваемые здесь взгляды на таксономическую и филогенетическую струк-
туру семейств, в первую очередь самого большого и сложного семейства Bulimi-
nidae (Enidae), во многом противоречат представлениям, бытовавшим ранее
и более или менее прочно укоренившимся в литературе. Этот пересмотр тра-
диционных взглядов связан с тем, что для разработки системы привлечены
признаки, на которые раньше либо вообще было не принято обращать внимание,
либо отдельные признаки отмечали, но для таксономических целей не исполь-
зовали.
Так, богатейший материал был получен при исследовании организации сово-
купительного аппарата; существенно, что здесь возникла необходимость в осо-
бенно подробном рисунке, поскольку иногда детали, на первый взгляд малосу-
щественные, давали важную информацию, и эту информацию необходимо было
донести до читателя в минимально искаженном виде. Особое внимание, которое
уделялось такой технической детали, как иллюстрация, вызвано тем, что зна-
ние тонких нюансов строения иной раз чрезвычайно важно, а соответствующих
сведений в литературе удручающе мало.
В систематической части приведены описания 167 видов и 3 подвидов Pupil-
lina, характеристика которых осуществлена на основании личного знакомства
автора с материалом, за исключением Adzharia renschi, известному по двум эк-
земплярам, хранящимся в Зоологическом музее Университета им. Гумбольдта
в Берлине, и Imparietula schelkovnikovi. Кроме видов, обнаруженных на терри-
тории нашей страны, в монографию включено И видов, в Советском Союзе не
найденных. Помещая материалы по этим видам, автор исходил из того, что со
временем часть из них может быть обнаружена на нашей территории; другие
виды включены как типовые для родов, представленных в фауне СССР; наконец,
•третьи — в силу особого интереса, который они представляют для понимания
4
ПРЕДИСЛОВИЕ
систематической структуры того или иного таксона. Кроме того, признано целе-
сообразным самостоятельное описание каждой из 8 форм полиморфного средне-
азиатского вида Pseudonapaeus albiplicatus.
Спиртовым материалом по нескольким видам я не располагал; для полноты
картины в таких случаях использованы иллюстрации, взятые из литературных
источников. Однако надо иметь в виду, что насколько важны и ярки детали
внутреннего строения мужского отдела полового аппарата в одних таксонах
(Orculidae, Buliminidae), настолько они незначительны в других (Pupillidae,
Valloniidae, Vertiginidae). Поэтому внутреннее строение освещается в тех слу-
чаях, когда оно имеет дифференциальную ценность или когда есть основания
думать, что такая ценность существует.
Все иллюстрации без ссылки на источник изготовлены автором. Основу фак-
тического материала составляют оригинальные сборы; в полной мере исполь-
зованы также коллекции Зоологического института АН СССР и Зоологического
музея Московского государственного университета; за предоставление возмож-
ности работать с этими богатейшими коллекциями я очень благодарен админи-
страции названных учреждений.
Кроме того, привлечены материалы, присланные по моей просьбе д-ром
Л. Форкартом (L. Forcart, Basel) и д-ром Дж. Геллером (J. Heller, Jerusalem).
Из числа советских коллег, любезно предоставивших коллекционные мате-
риалы, с благодарностью хочу назвать С. Альмухамбетову (Павлодар), 3. Из-
затуллаева (Душанбе), Е. И. Компанцеву (Москва), А. Б. Мухитдинова (Ленин-
абад), К. К. Увалиеву (Алма-Ата), Е. В. Шикова (Калинин).
Особую признательность я выражаю Н. Н. Акрамовскому, И. М. Лихареву
и Я. И. Старобогатову, чью постоянную и разнообразную помощь трудно пере-
оценить. Научное редактирование книги, осуществленное проф. И. М. Лихаре-
вым, способствовало сильному улучшению первоначального текста.
Множество коллег помогало мне советами, дружеской критикой и просто
благожелательным отношением; несмотря на то что всех их перечислить невоз-
можно, я глубоко благодарен всем, кто так или иначе облегчал мне работу над
книгой.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
I. Надоем. COCHLICOPOIDEA Pilsbry, 1900
Сем. COCHLICOPIDAE Pilsbry, 1900
1. Род Cochlicopa Ferussac, 1821
1. С. nitens (Gallenstein, 1852).......................................... 112
2. С. pseudonitens Uvalieva, 1967 . . 1.................................. 113
3. C. lubrica (Muller, 1774).............................................. 114
4. C. lubricella (Porro, 1838)............................................ 115
II. Надоем. ORCULOIDEA Steenberg, 1925
I. Сем. ORCULIDAE Steeuberg, 1925
I. Подсем. Ore ul inae Steeuberg, 1925
1. Род Pilorcula Germain, 1912
P. trifHaris (Mousson, 1863)............................................... 118
2. Род Orculella Steeuberg, 1925
1. Подрод Meso'rculella Schileyko, 1976
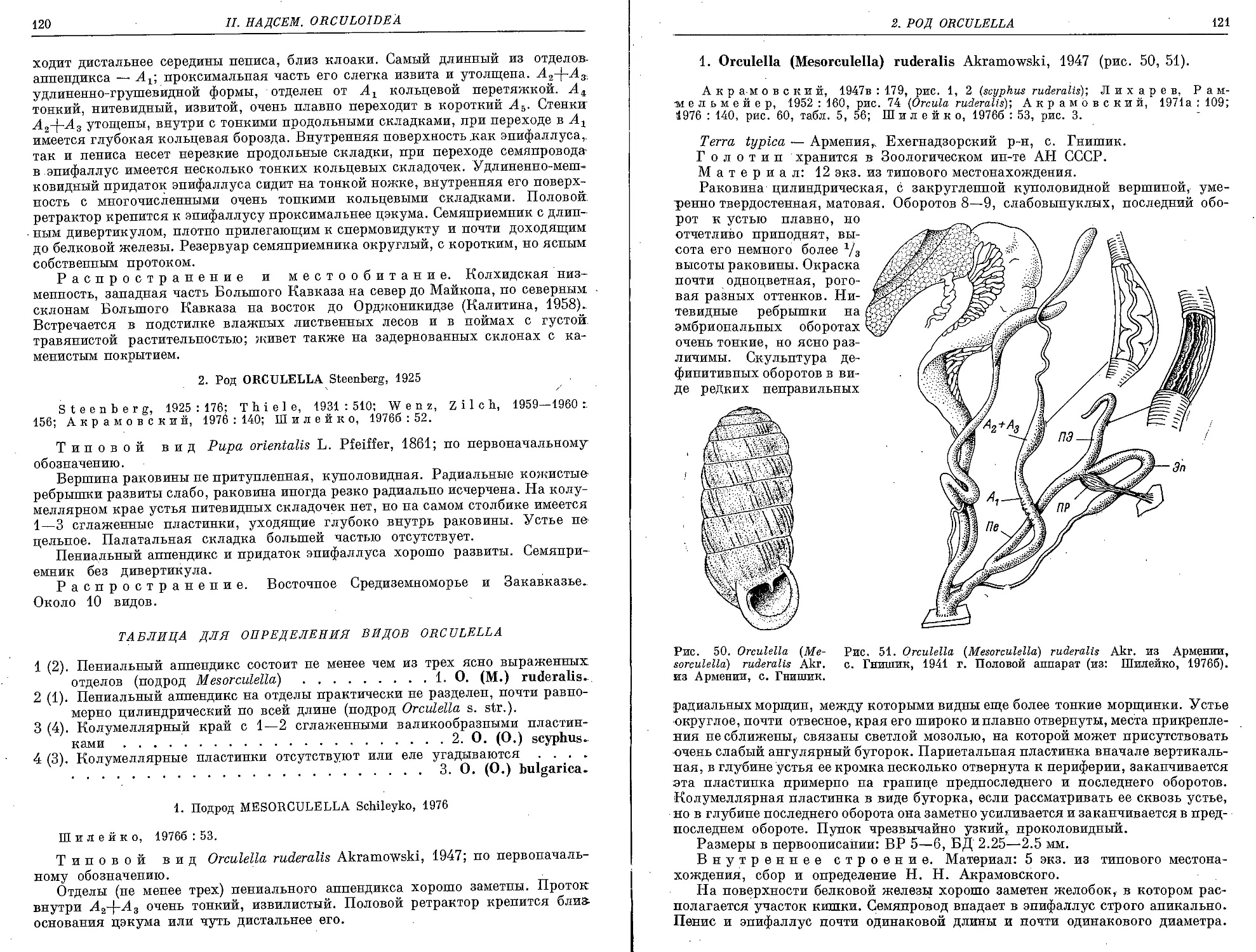
1. О. (M.) ruderalis Akramowski, 1947 ...................................... 121
2. Подрод Orculella s. str.
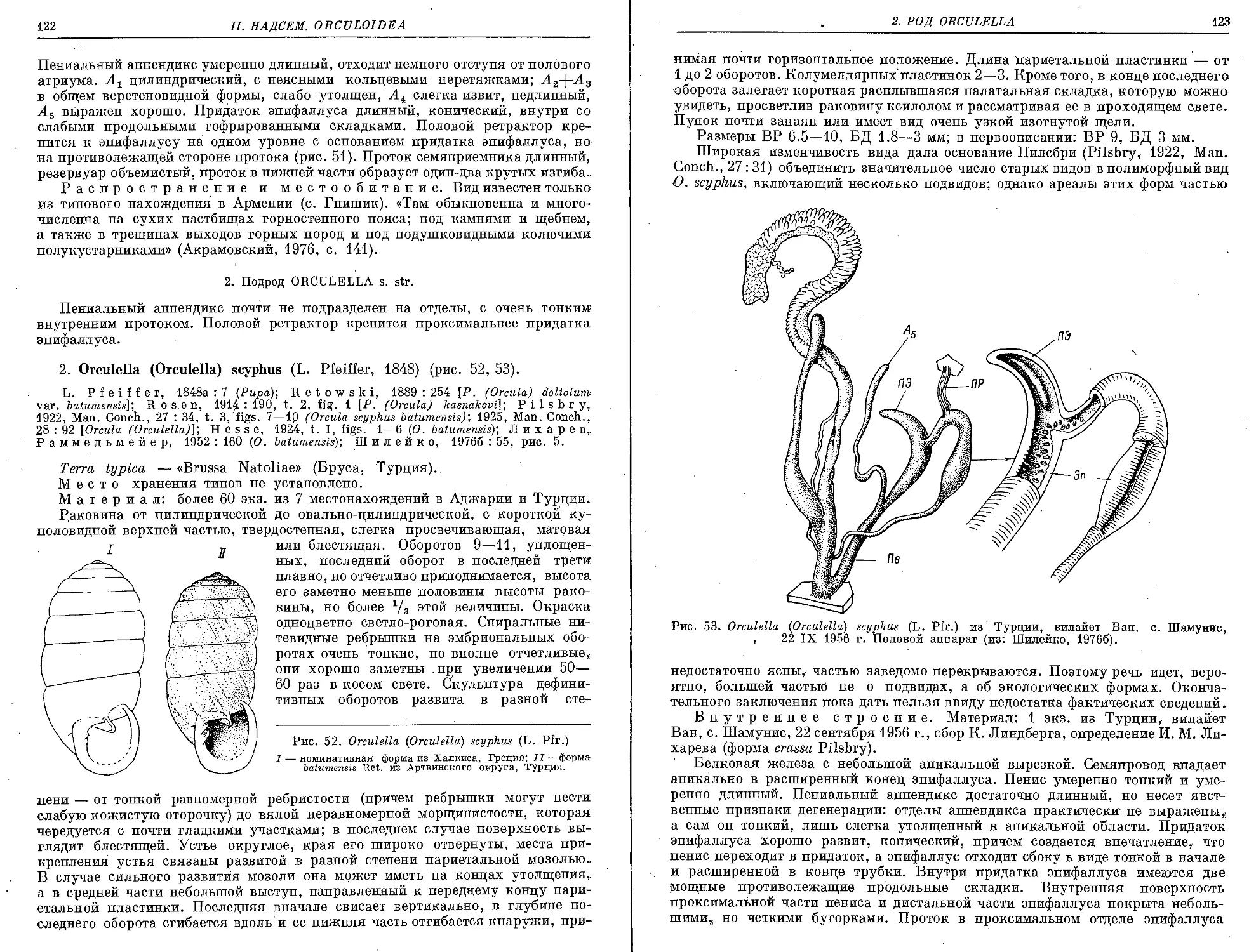
2. 0. (0.) scyphus (L. Pfeiffer, 1848)...................................... 122
3. 0. (0.) bulgarica, (Hesse, 1915)...................................... 124
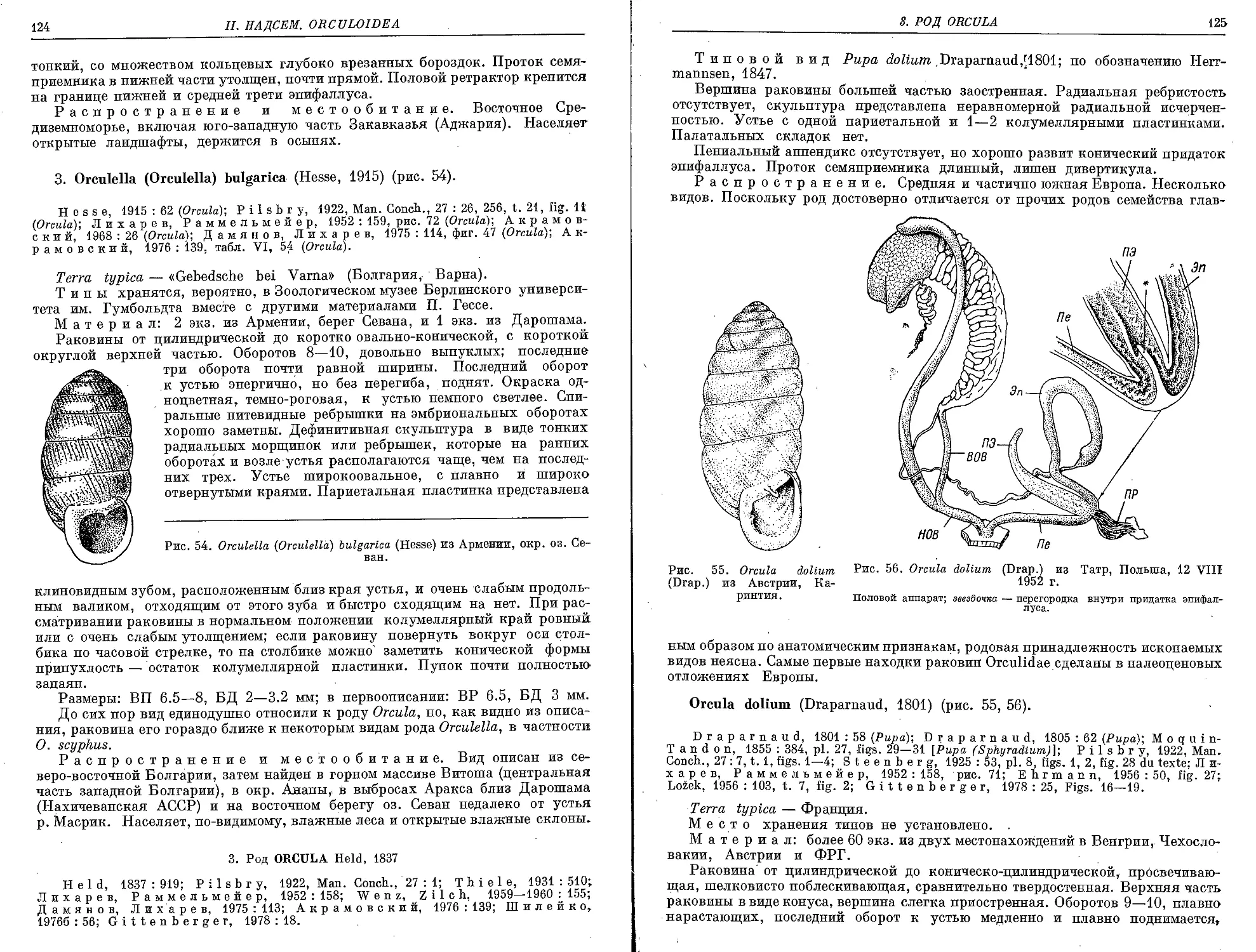
3. Род Orcula Held, 1837
0. dolium (Draparnaud, 1801)................................................ 125
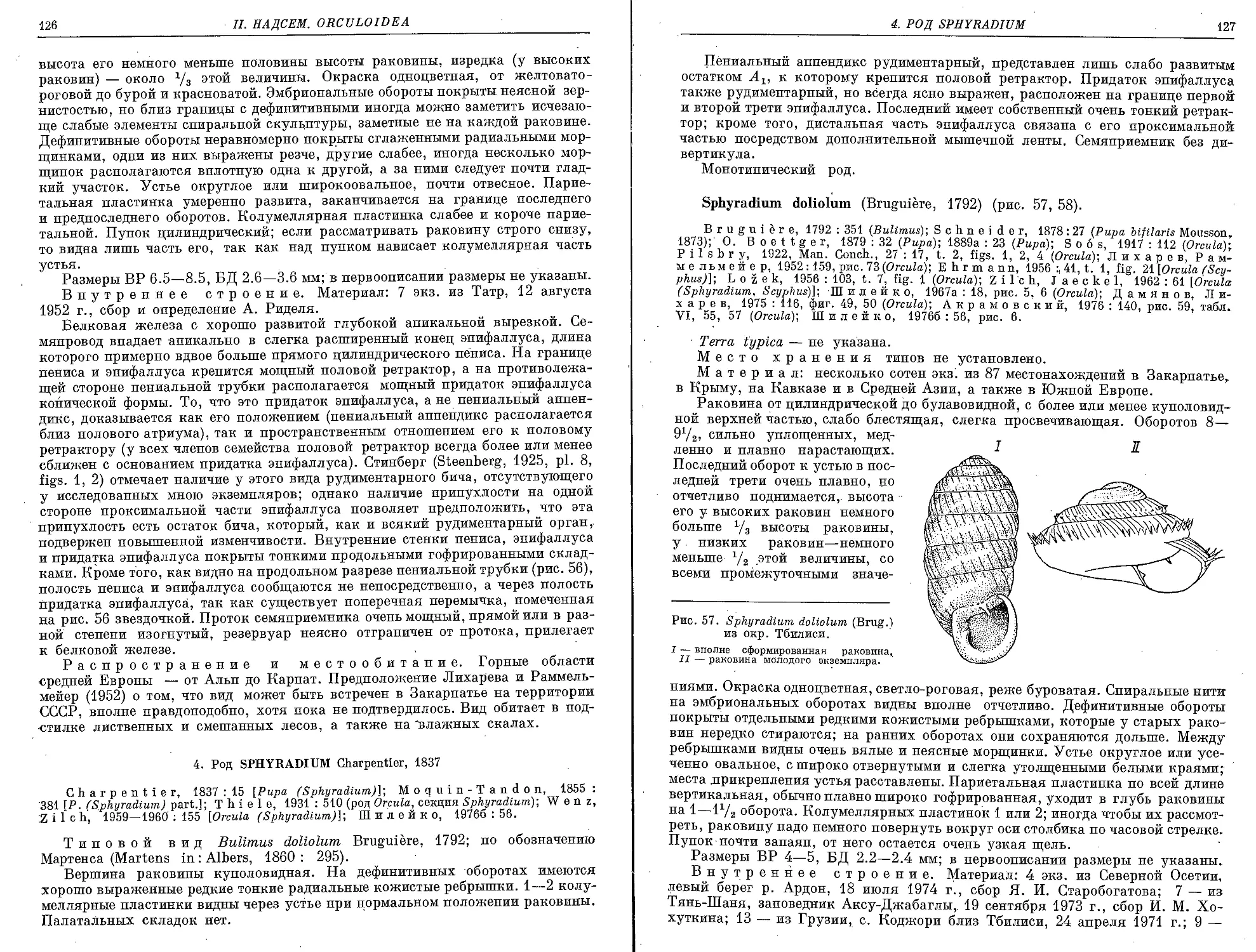
4. Род Sphyradium Charpentier, 1837
5. doliolum (Bruguiere, 1792) . .......................................... 127
II. Подсем. Pago dulin inae Pilsbry, 1924
1. Род Pagodulina Clessin, 1876
1. Пбдрод Pagodulina s. str.
1. P. (P.) pagodula (Des Moulins, 1830)..................................... 129
2. Подрод Crystallifera Schileyko, 1976
2. P. (C.) lederi (O. Boettger, 1886)...................................... 13(
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
III. Подсем. Lauriinae Steenberg, 1925
1. Род Euxinolauria Lindholm, 1924
1. Подрод Matschachelia Schileyko, 1975
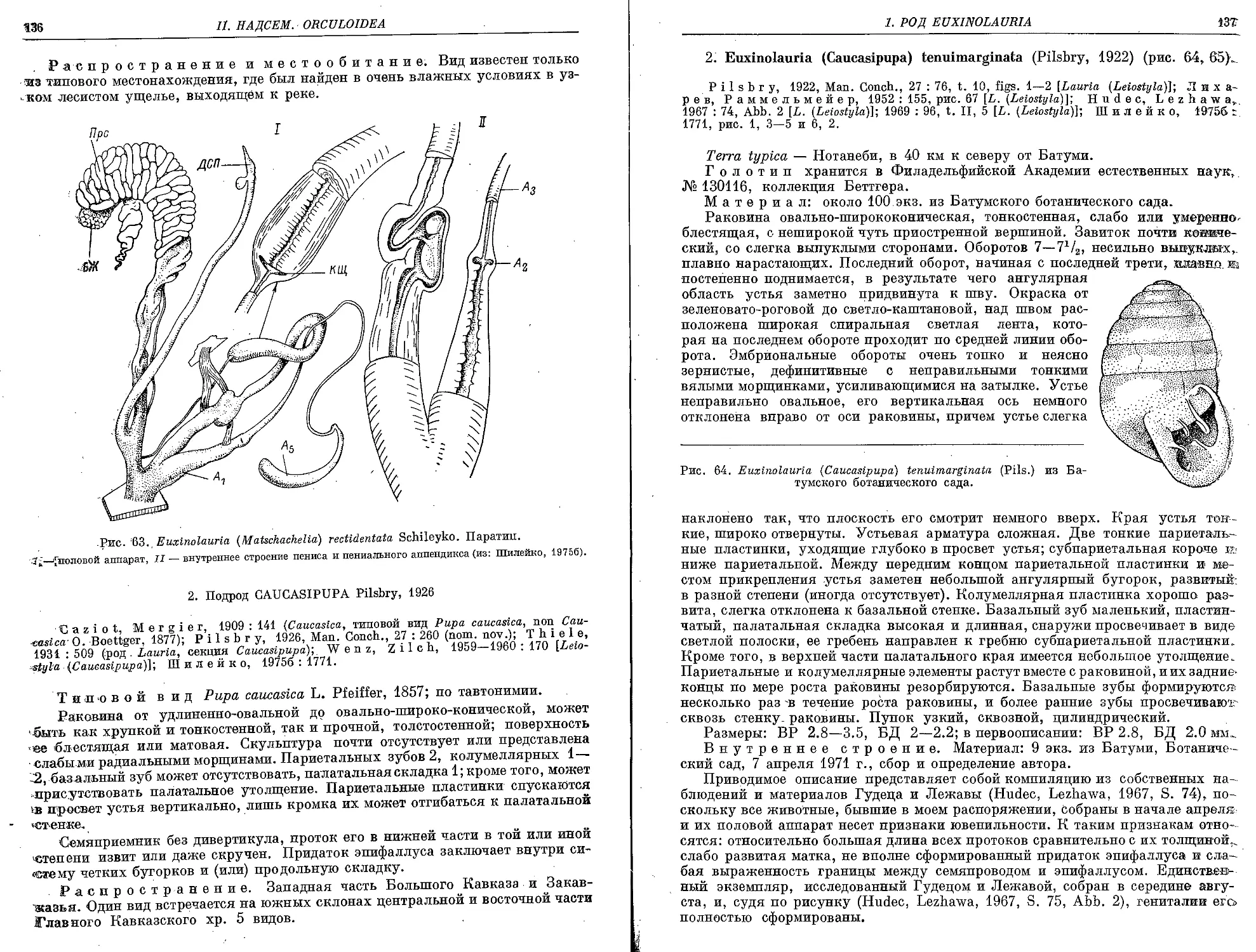
1. E. (M.) rectidentata Schileyko, 1975 ...................................... 13->
2. Подрод Caucasipupa Pilsbry, 1926
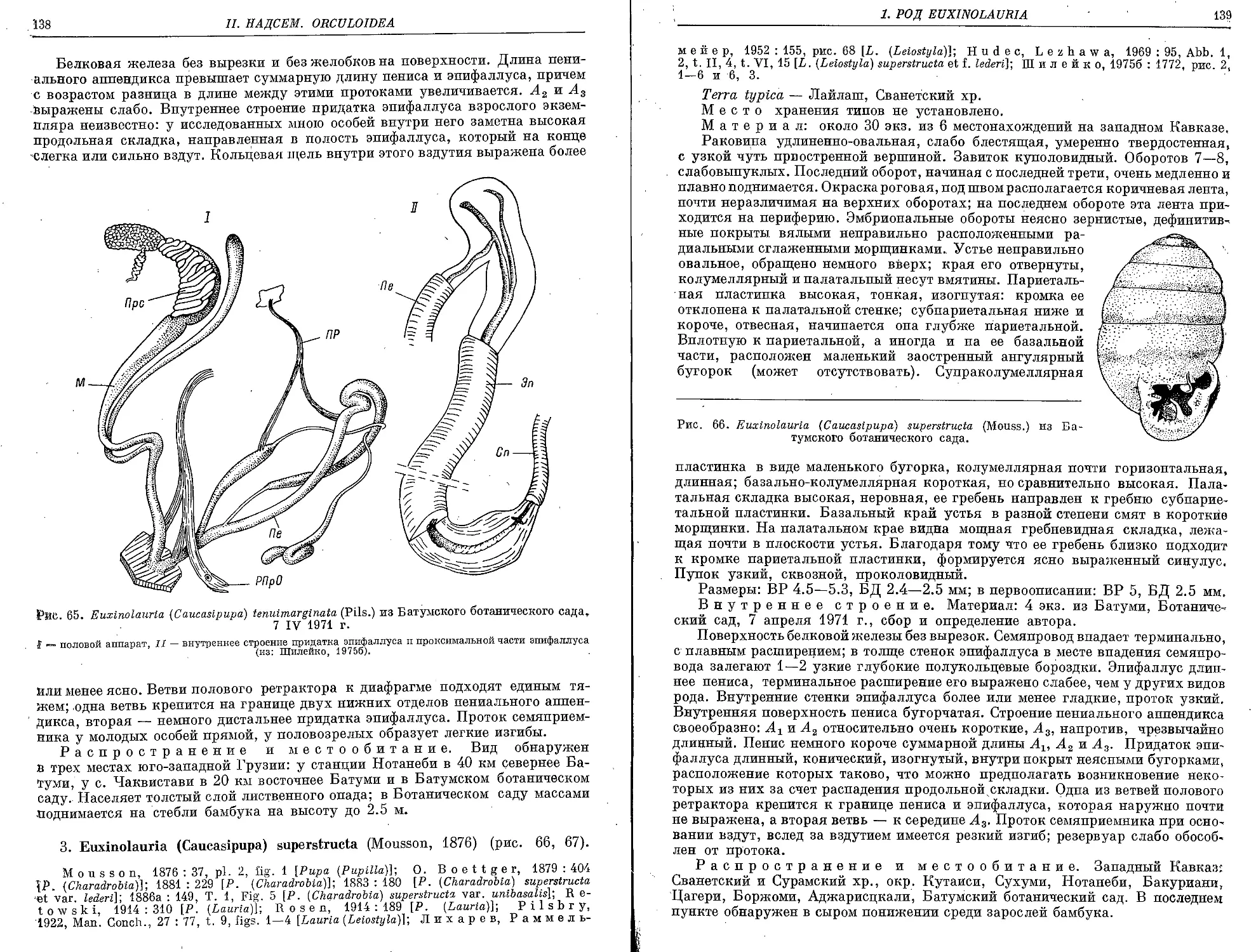
2. E. (C.) tenuimarginata (Pilsbry, 1922)...................... 137
3. E. (C.) superstructa (Mousson, 1876)......................... 138
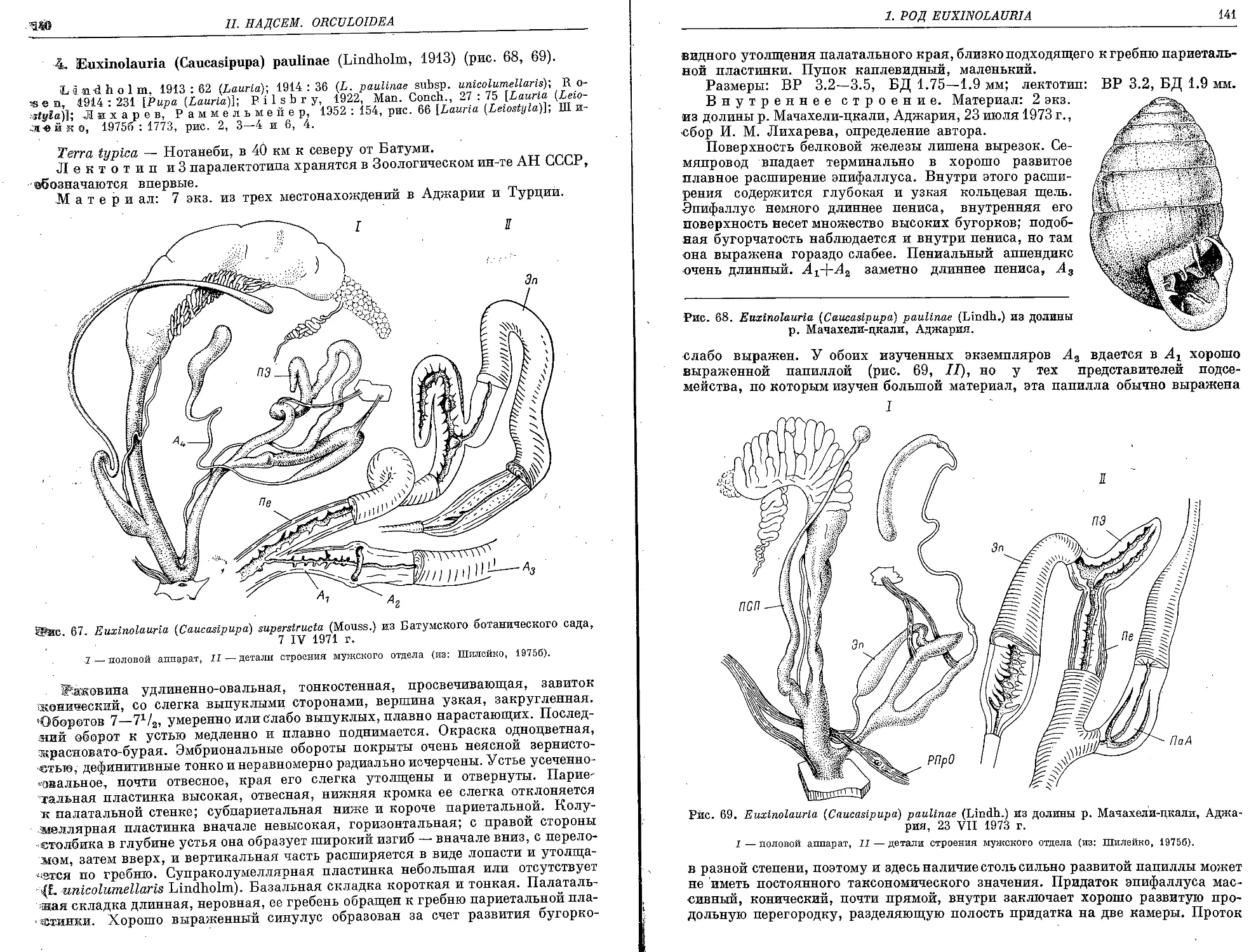
4. E. (C.) paulinae (Lindholm, 1913)............................ 140
5. E. (C.) caucasica (L. Pfeiffer, 1857)........................ 142
6. E. (C.) sinangula Schileyko, 1975 ........................... 143
3. Подрод E u x i n o 1 a u r i a s. str.
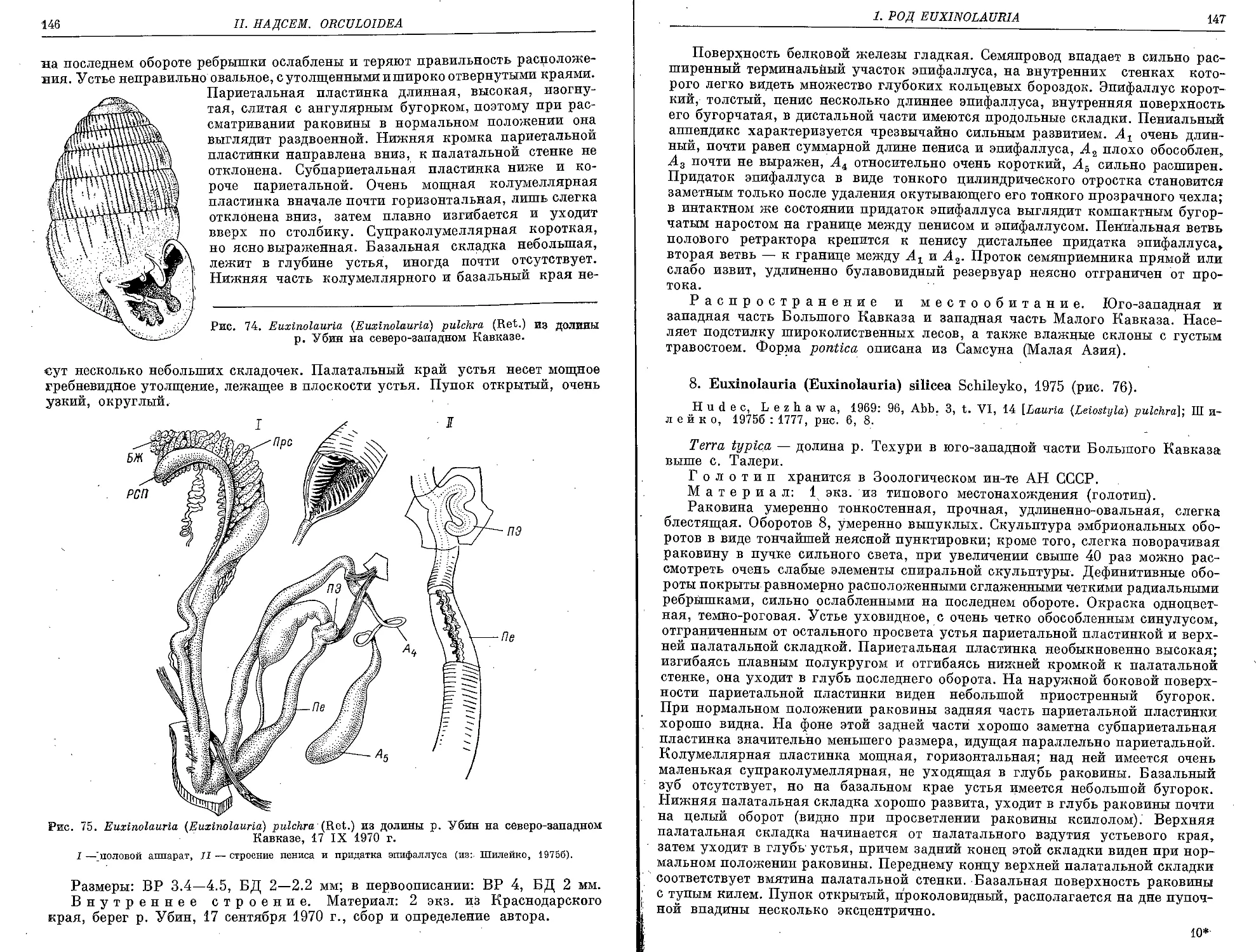
7. E. (E.) pulchra (Retowski, 1883) ........................... 145
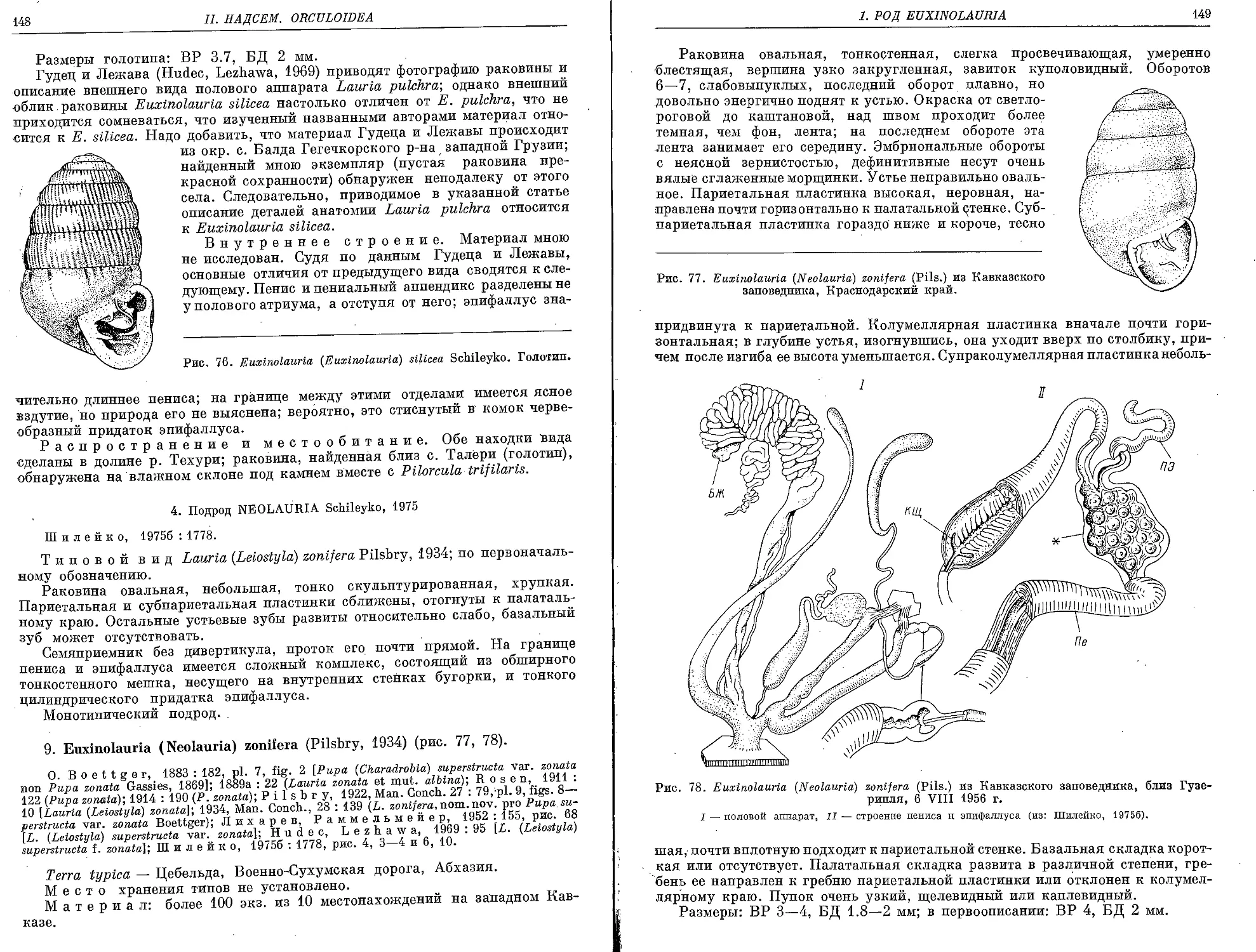
8. E. (E.) silicea Schileyko, 1975 ............................ 147
4. Подрод Neolauria Schileyko, 1975
9. E. (N.) zonifera (Pilsbry, 1934)........................................... 148
2. Род Lauria Gray, 1840
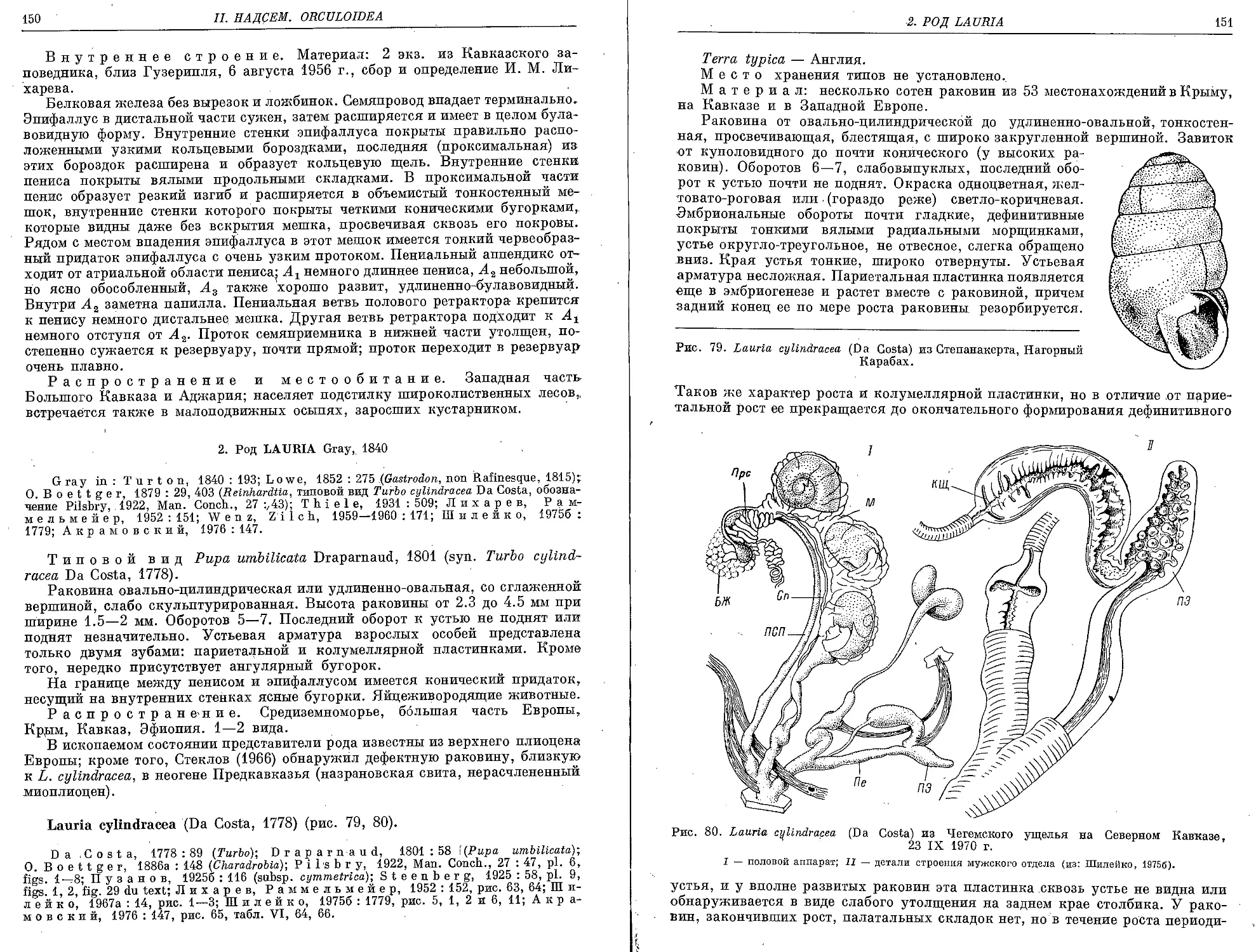
L. cylindracea (Da Costa, 1778)............................................... 150
IV. Подсем. Argninae Hudec, 1965
1. Род Argria' Cossmann, 1899
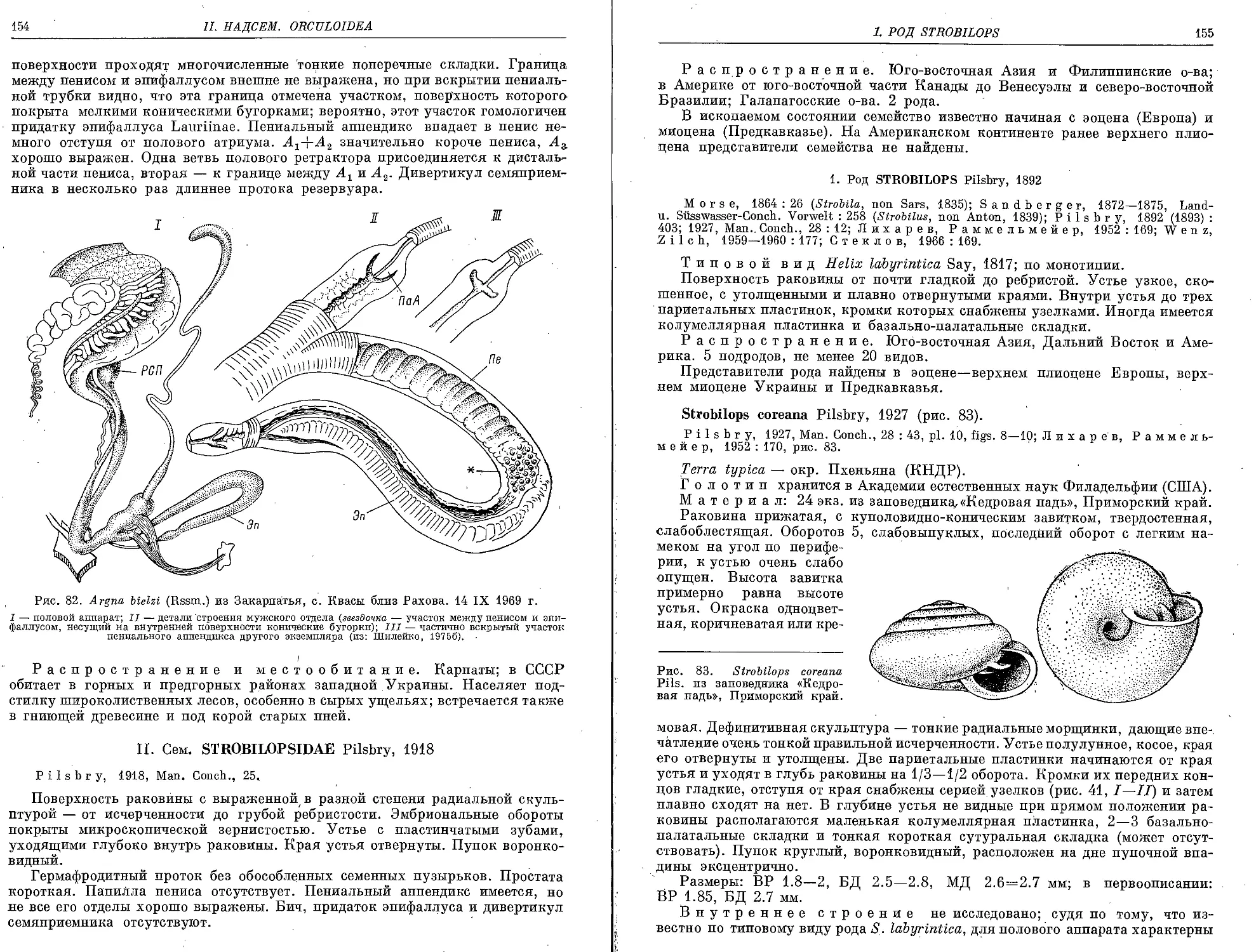
A. bielzi (Rossmaessler, 1859) ......................................’. 153
II. Сем Strohilopsidae Pilsbry, 1918
1. Род Strobilops Pilsbry, 1892 '
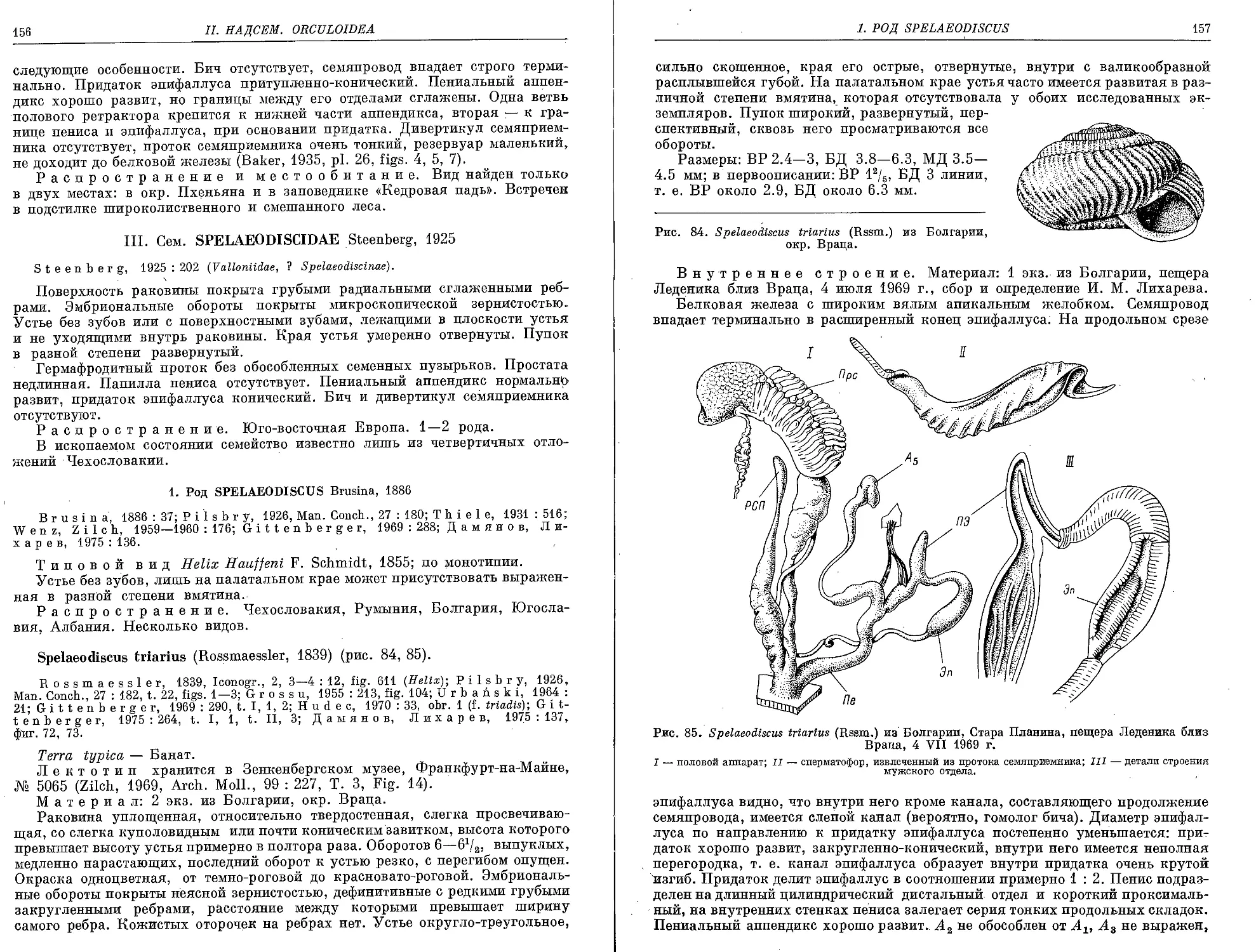
S. coreana Pilsbry, 1927 ............................................... 155
III. Сем. SPELAEODISCIDAE Steenberg, 1925
1. Род Spelaeodiscus Brusina, 1886
S. triartus (Rossmaessler, 1839)........................................ 156
IV, Сем. VALLONIIDAE Morse, 1864
I. Подсем. Acanthinulinae Steenberg, 1917
1. Род Zoogenetes Morse, 1864
Z. harpa (Say, 1864) 159
2. Род Acanthinula Beck, 1847
A. aculeata (Miiller,' 1774).............................................'.................. 160
3. Род Sperniodea Westerland, 1902
S. lamellata (Jeffreys, 1833)............................................................... 162
II. Подсем. Valloniinae Morse, 1864
1. Род Vallonia Risso, 1826
1. Подрод Vallonia s. str.
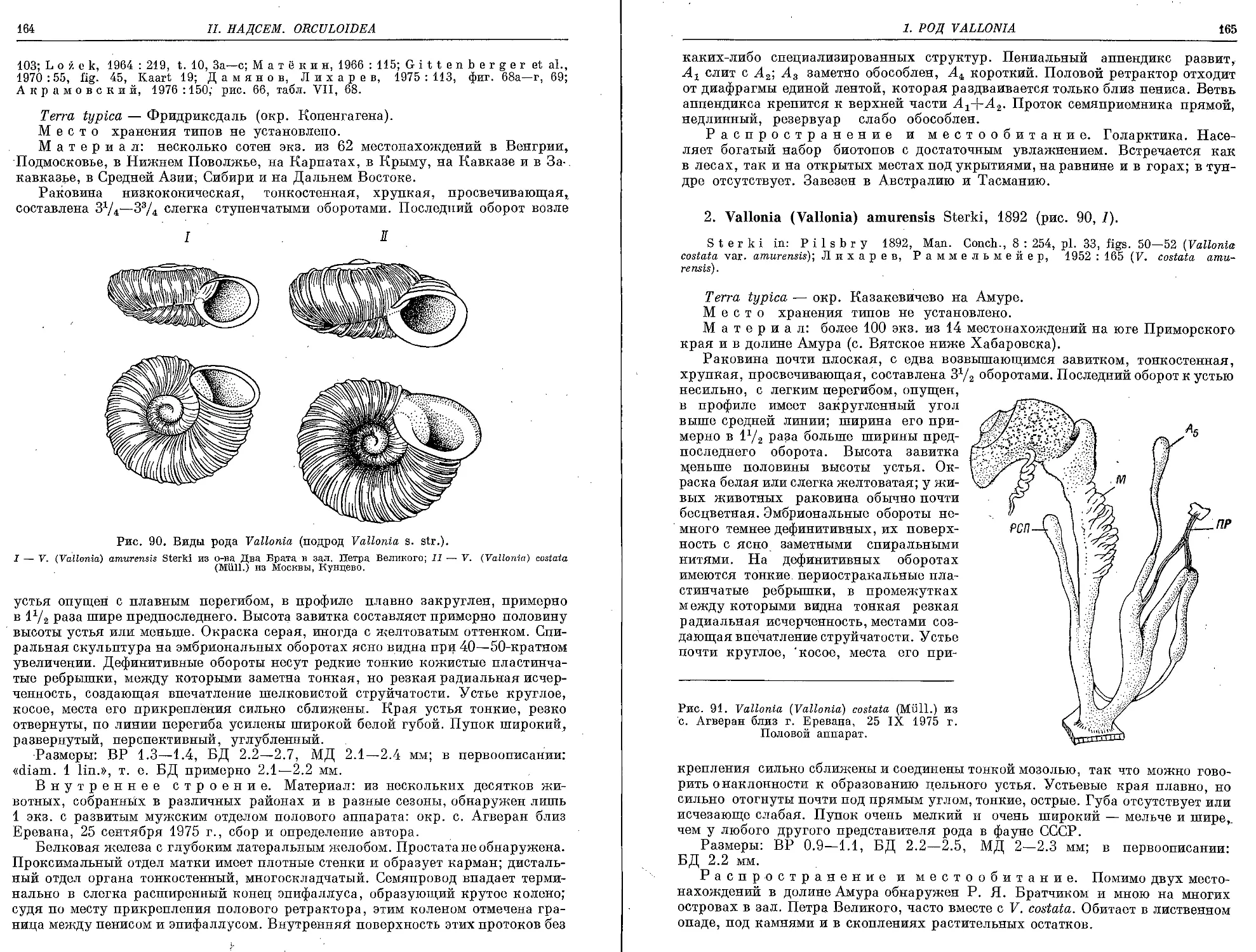
1. V. (V.) costata (Miiller, 1774)............................• • • .............., . . . 163
2. V. (V.) amurensis Sterki, 1892 ........................................................ 165
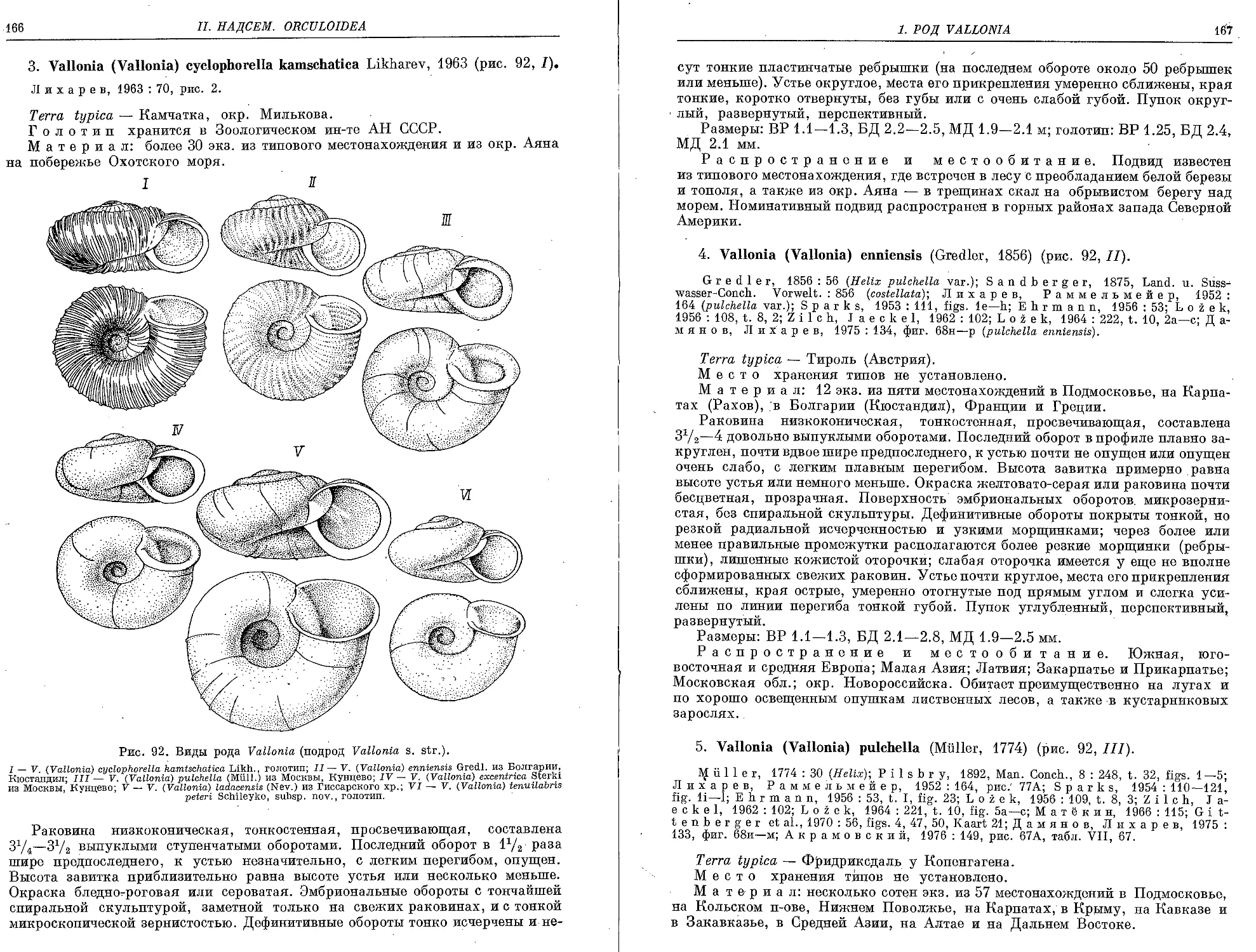
3. V. (V.) cyclo pho rella kamschatica Likharev. 1963 . . .......................... 166
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
7
4. V. (V.) enniensis (Gredler, 1856)................................... 167
5. V. (V.) pulchella (Muller, 1774)..................................... 167
6. V. (У.) excentrica Sterki, 1892 .................................... 168
7. V. (У.) tenuilabris tenuilabris (Al. Braun, 1842)..................... 169
7a. V. (V.) tenuilabris peteri Schileyko, subsp. nov..................... 169
8. У. (V.) ladacensis (Nevill, 1878)..................................... 169
2. Подрод Planivallonia Schileyko, subgen. nov.
9. V. (P.) zaru Almuhambetova, 1979 ..................................... 170
III. Надсем. PUPILLOIDEA Turton, 1831
Сем. PUPILLIDAE Turton, 1831
1. Род Gibbulinopsis Germain, 1919
1., Подрод Gibbulinopsis s. str.
1. G. (G.) eryptodon (Heude, 1880)...................................... 172
2. G. (G.) interrupta (Reinhardt, 1876)................................. 173
2. Подрод Primipupi.Ila Pilsbry, 1921
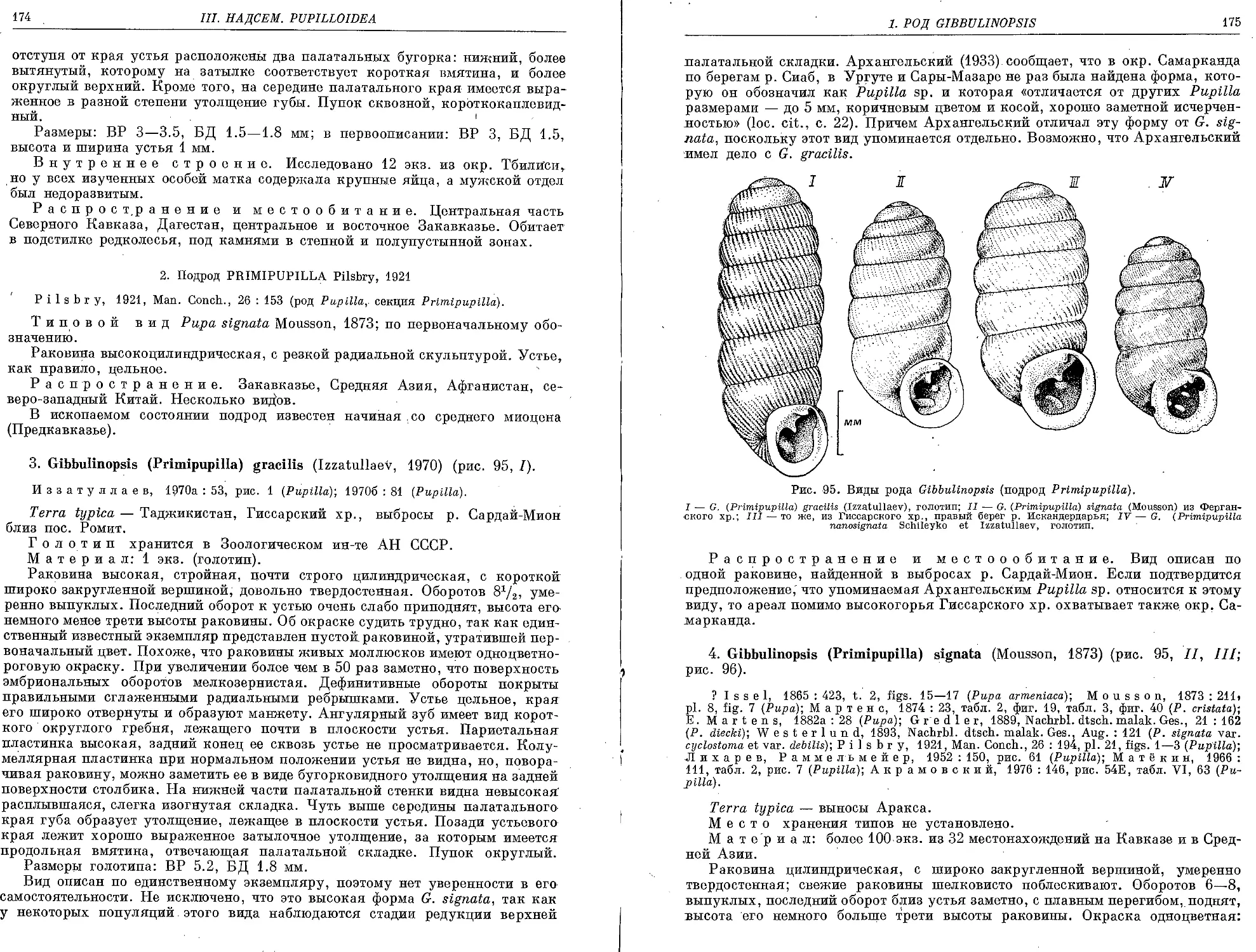
3. G. (P.) gracilis (Izzatullaev, 1970).................................. 174
4. G. (P.) signata (Mousson, 1873) . . . ............................... 175
5. G. (P.) nanosignata Schileyko et Izzatullaev. 1980 ................... 177
2. Род Pupilia Turton, 1831
1. Подрод Pupilia s. str.
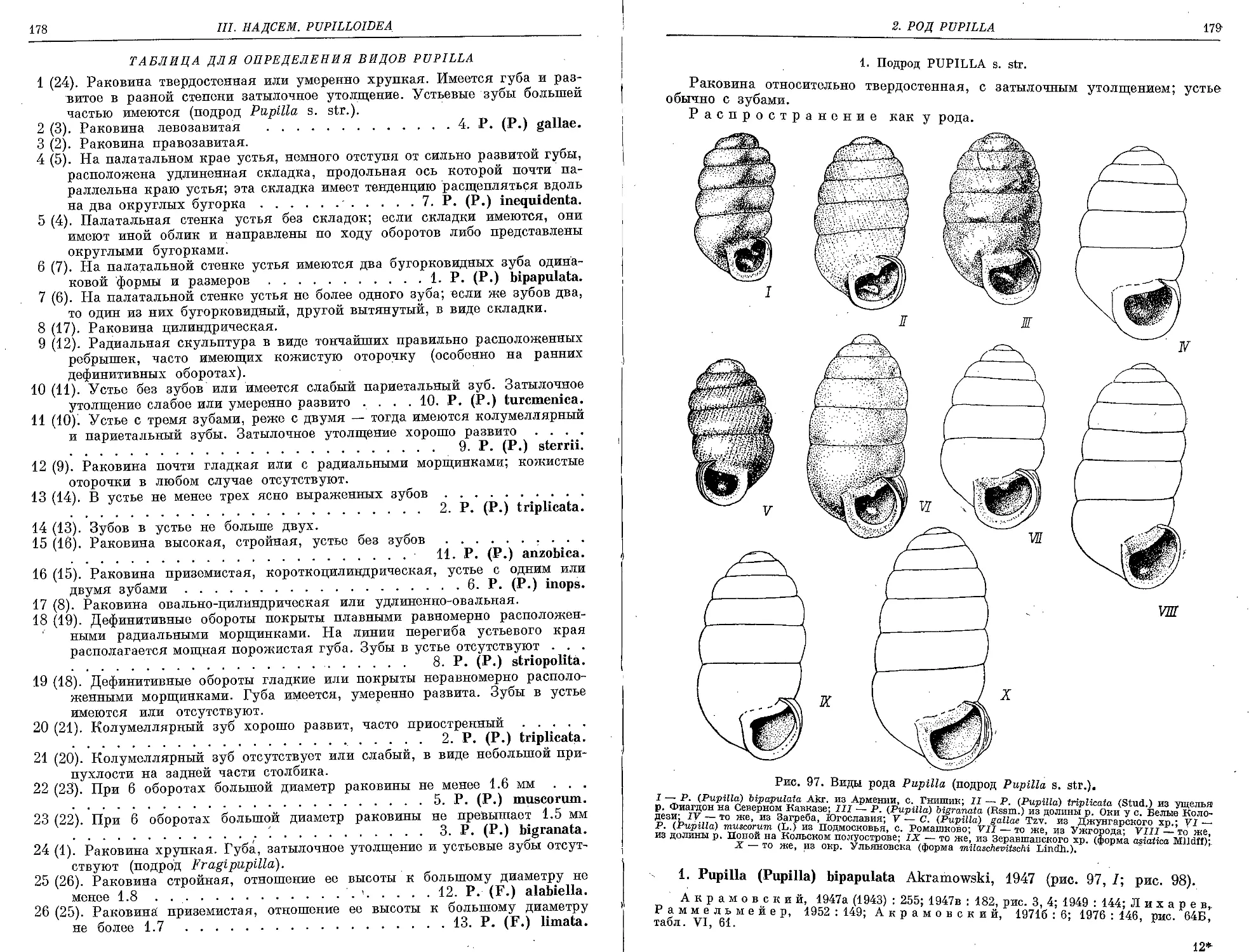
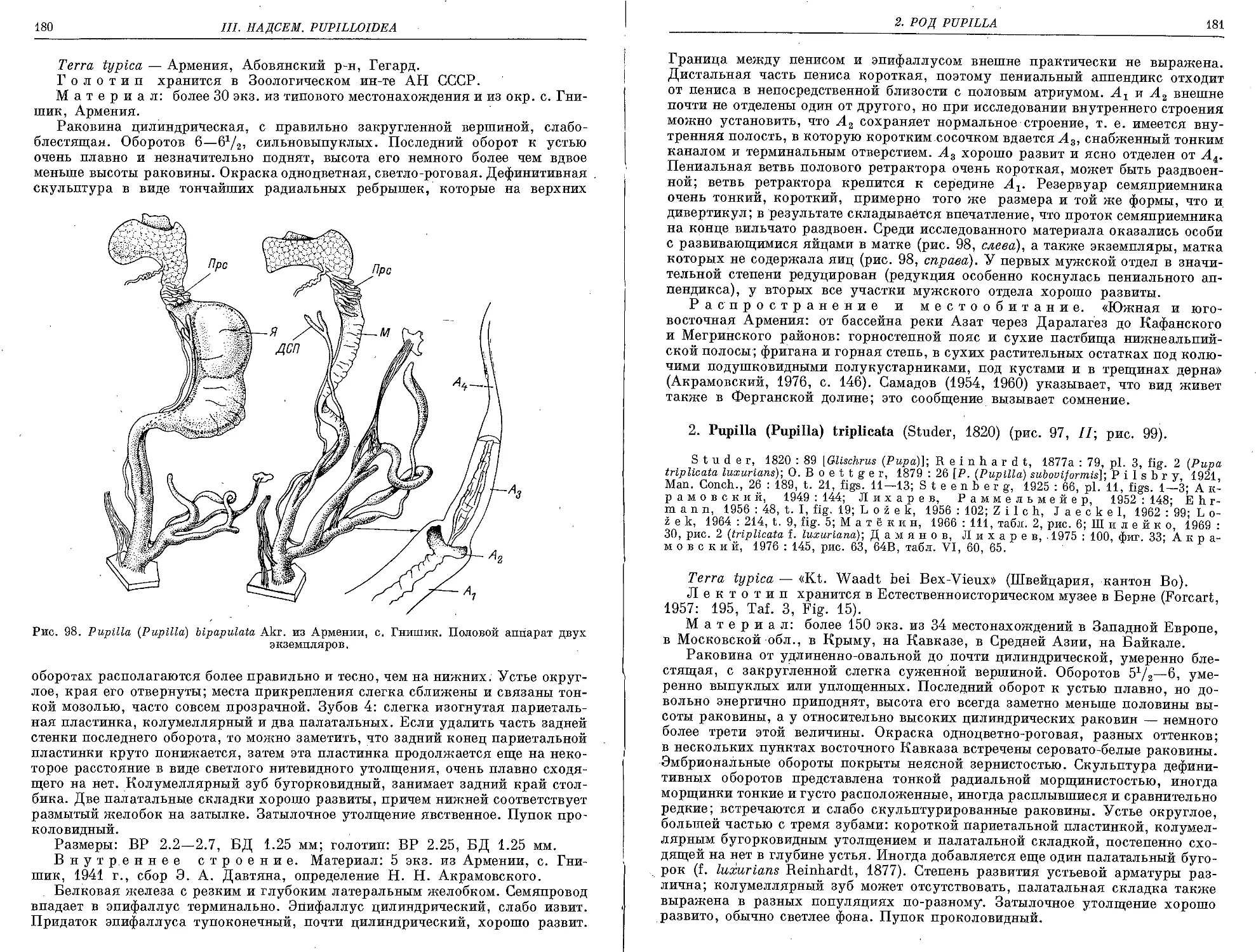
1. P. (P.) bipapulaia Akramowski, 1947 ................................. 179
2. P. (P.) triplicata (Studer, 1820).................................... 181
3. P. (P.) bigranata (Rossmaessler, 1839)............................... 182
4. P. (P.) gallae Tzvetkov, 1940 . . . 183
5. P. (P.) muscorum (Linnaeus, 1758).................................. 185
6. P. (P.) inops (Reinhardt, 1877) ..................................... 186
7. P. (P.) inequidenta Schileyko et Almuhambetova, 1979 ................ 187
8. P. (P.) striopolita Schileyko, sp. nov............................... 188
9. P. (P.) sterrii (Voith, 1840)........................................ 189
10. P. (P.) turcmenica (O. Boettger, 1889)............................... 190
11. P. (P.) anzobica Izzatullaev, 1970 ................................... 191
2. Подрод Fragipupilla Schileyko, subgen. nov.
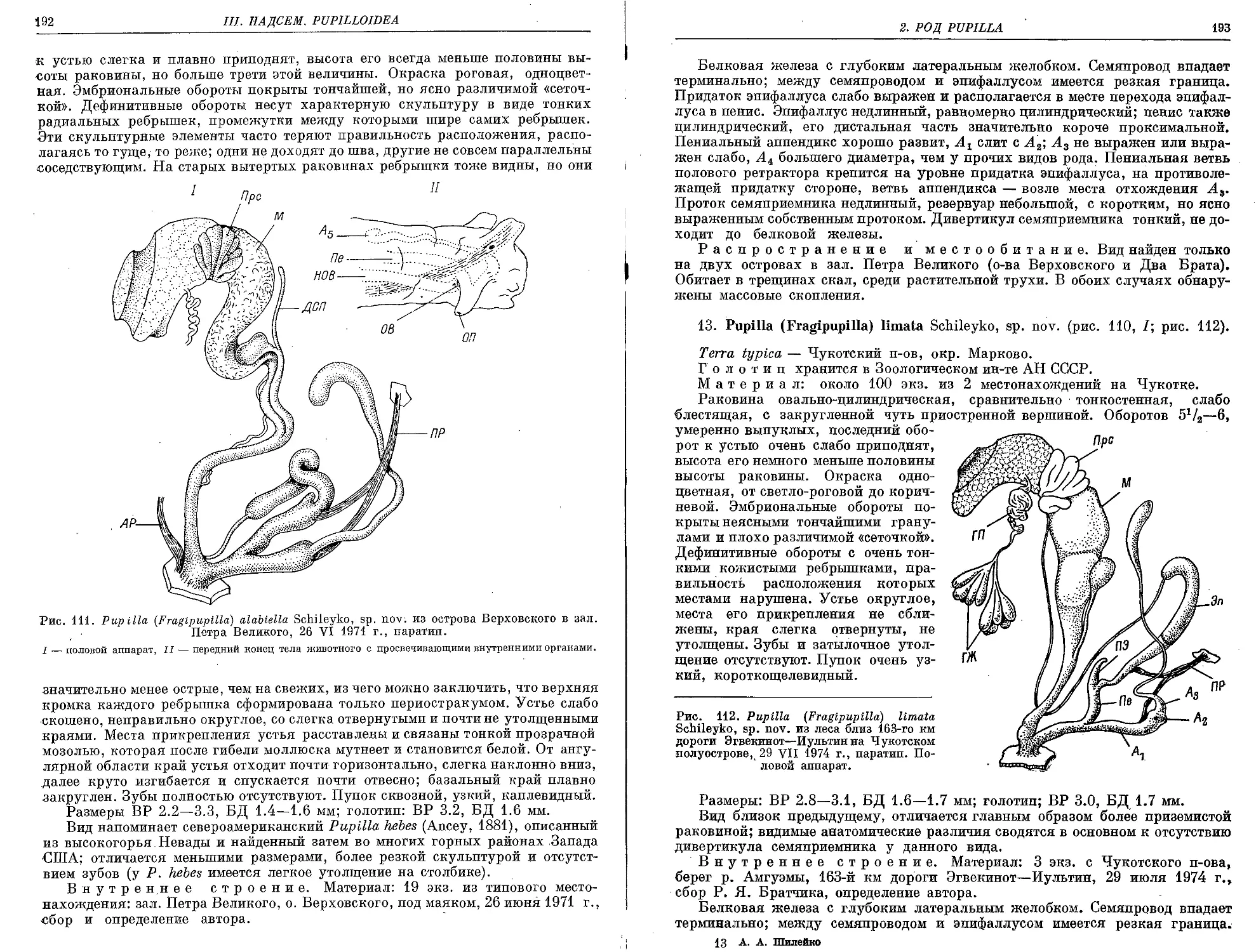
12. P. (F.) alabiella Schileyko, sp. nov.................................. 191
.13. P. (F.) limata Schileyko, sp. nov................................... 193
3. Род Pupoides L. Pfeiffer, 1854
P. coenopictus (Hutton, 1834)............................................ 194
IV. Надсем. VERTIGINOIDEA Pilsbry, 1918
Сем. VERTIGINIDAE Pilsbry, 1918
I. Подсем. Gastrocoptinae Pilsbry, 1918
1. Род Gastrocopta Wollaston, 1878
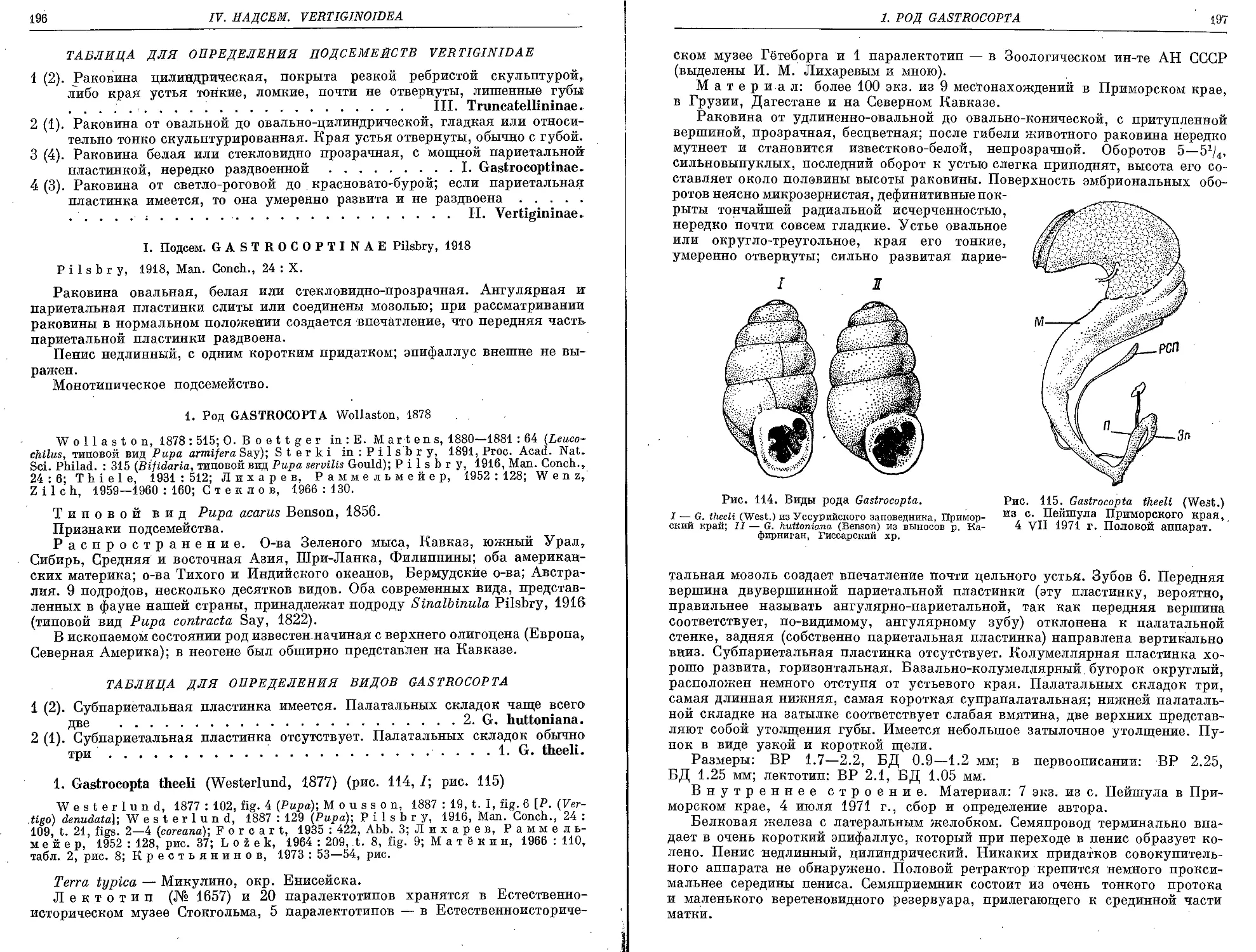
1. G. theeli (Westerlund, 1877).......................................... 196
2. G. huttoniana (Benson, 1849)......................................... 198
II. Подсем. Vertigininae Pilsbry, 1918
1. Род Vertigo Miiller, 1774
1. Подрод Vert jgo s. str.
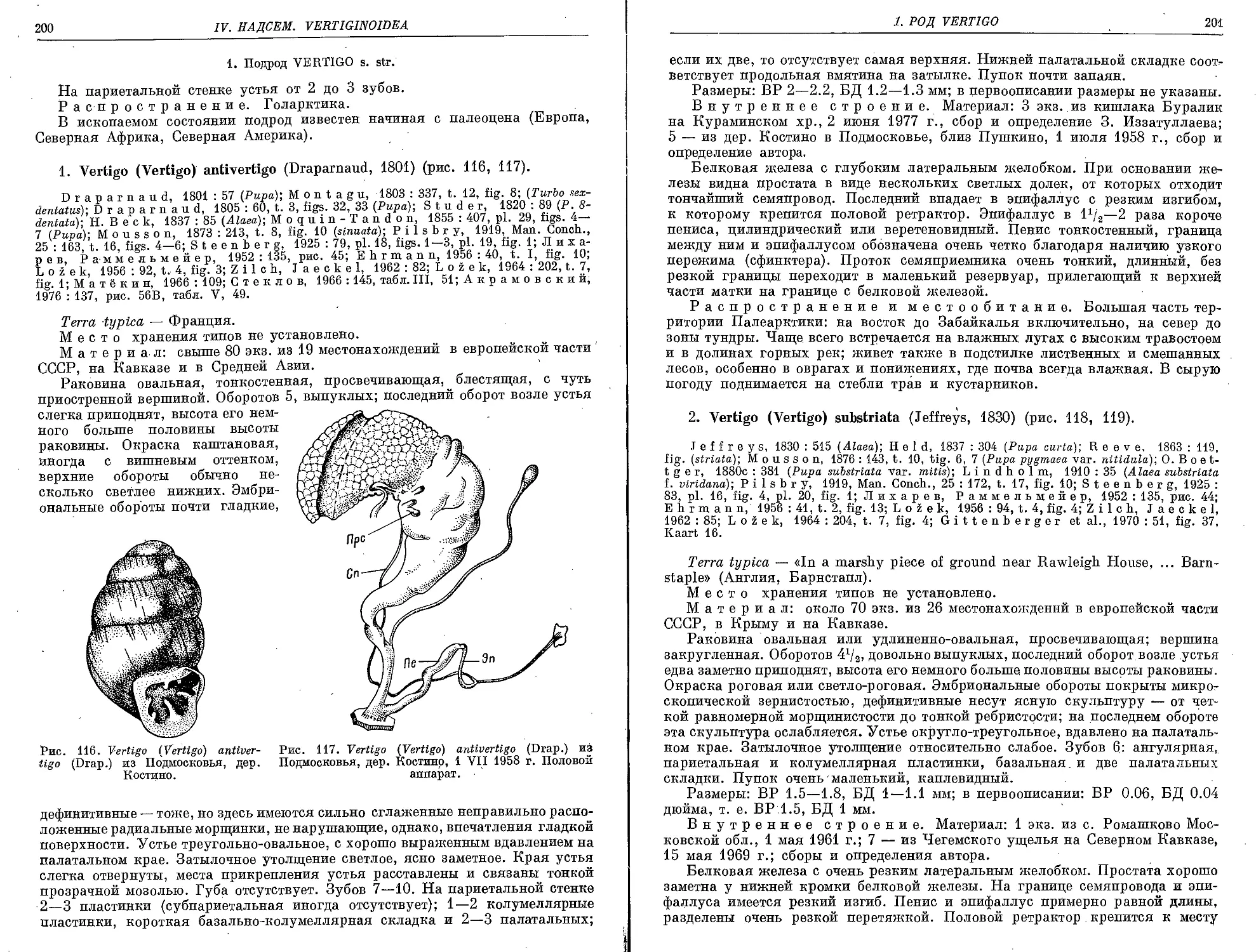
1. V. (V.) antivertigo (Draparnaud, 1801)................................. 200
2. V. (V.) substriata (Jeffreys, 1830).......................... 201
8
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
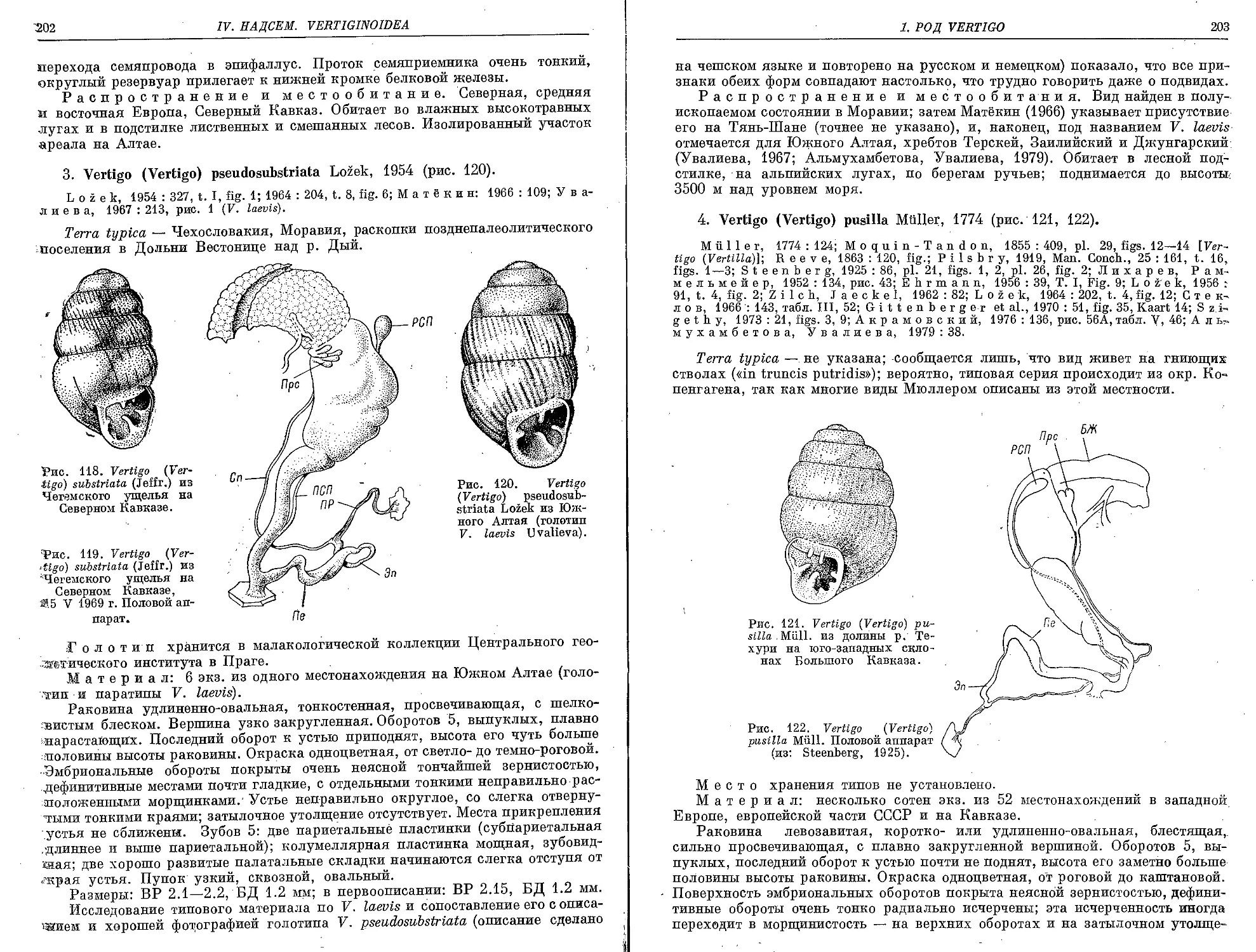
3. V. (V.) pseudosubstriata Lozek, 1954 ....................................... 202
4. V. (V.) pusilia. Muller, 1774 .............................................. 203
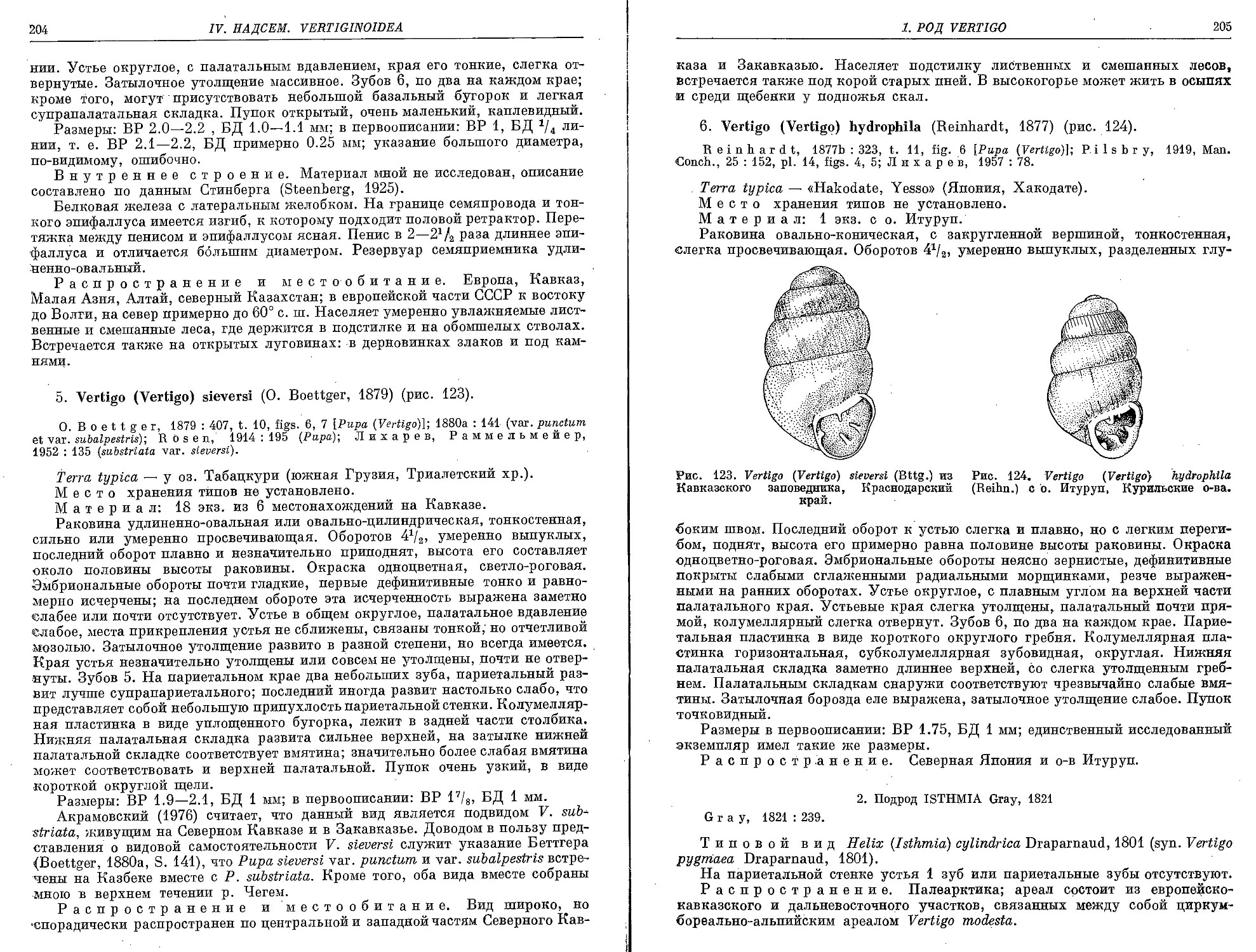
5. V. (V.) sieversi (0. Boettger, 1879)........................................ 204
6. V. (V.) hydrophila (Reinhardt, 1877)....................................... 205
2. Подрод I s thm i a Gray, 1821
7. V. (I.) moulinsiana (Dupuy, 1849) ....................................... 206
8. V. (I.) pygmaea (Draparnaud, 1801)........................................ 207
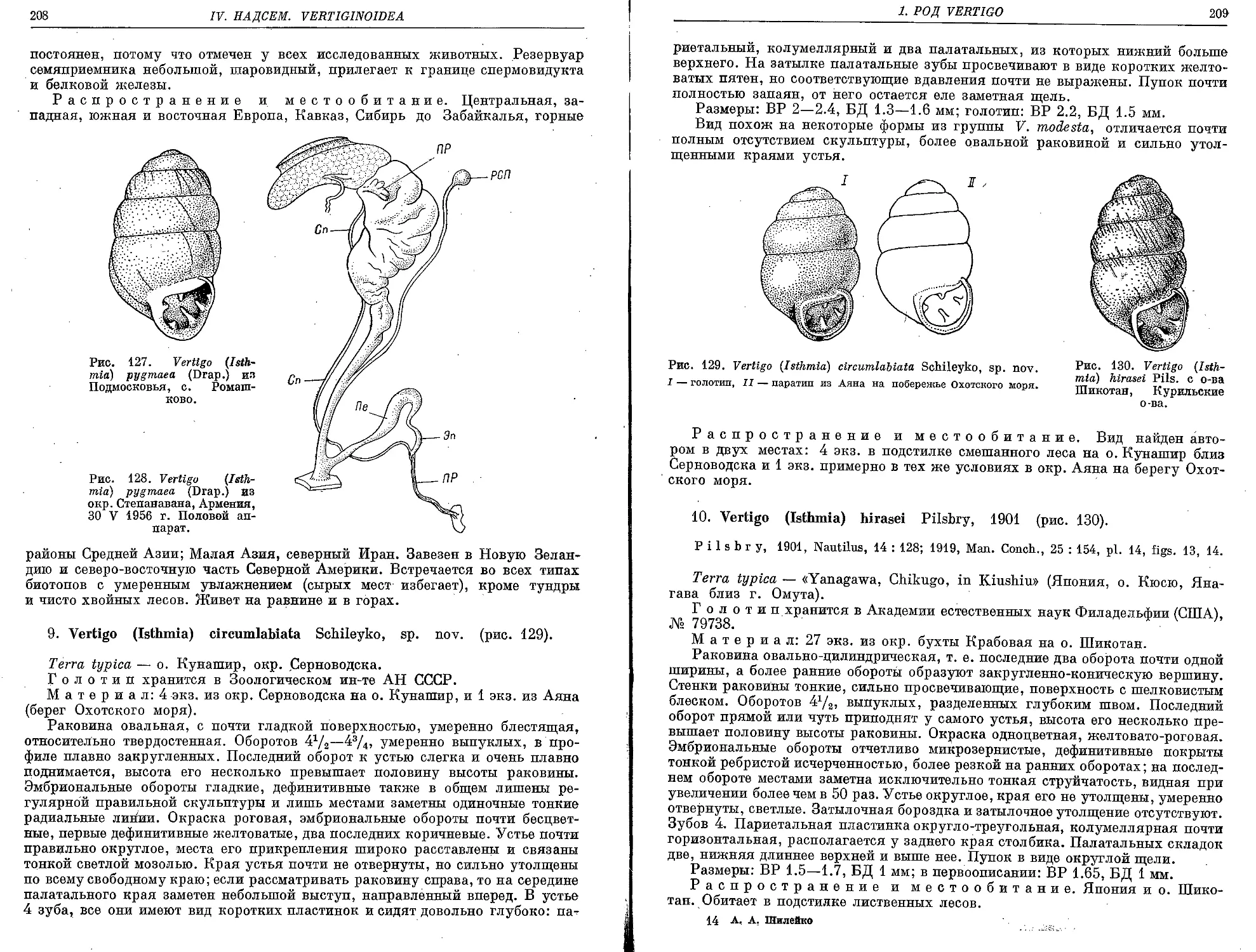
9. V. (I.) circumlabiata Schileyko, sp. nov.................................. 208
10. V. (I.) hirasei Pilsbry, 1901 ............................................ 209
11. V. (I.), japonica Pilsbry et Hirase; 1904 ............................... 210
12. V. (I.) microsphaera Schileyko, sp. nov................................... 210
13. V. (I.) modesta modesta (Say, 1824)....................................... 211
13a. V. (I.) modesta alpestris Alder, 1839 .................................... 213
2. Род Vertilla Moquin-Tandon, 1855
V. angustior (Jeffreys, 1830).................................................. 213
III. Подсей. Truncatellininae Steenberg, 1925
1. Род Columella Westerlund, 1878
1. C. columella (G. Martens, 1830)............................................. 216
2. C. edentula (Draparnaud, 1805).............................................. 217
3. C. acicularis Almuhambetova, 1979 .......................................... 217
4. C. aspera Walden, 1966 ................................................... 218
5. C. intermedia Schileyko et Almuhambetova, sp. nov........................... 219
2. Род Truncatellina Lowe, 1852
1. T. calllcratis (Scacchi, 1833).............................................. 220
2. T. costulata (Nilsson, 1822)................................................ 221
3. T. claustralis (Gredler, 1856).............................................. 222
4. T. cylindriea (Ferussac, 1807).............................................. 222
V. Надсем. CHONDRINOIDEA Steenberg, 1925
I. Сем. CHONDRINIDAE Steenberg, 1925
1. Род Granaria Held, 1837
G. frumentum (Draparnaud, 1801)................................................. 225
2. Род Abida Turton, 1831
A. secale (Draparnaud, 1801) 227
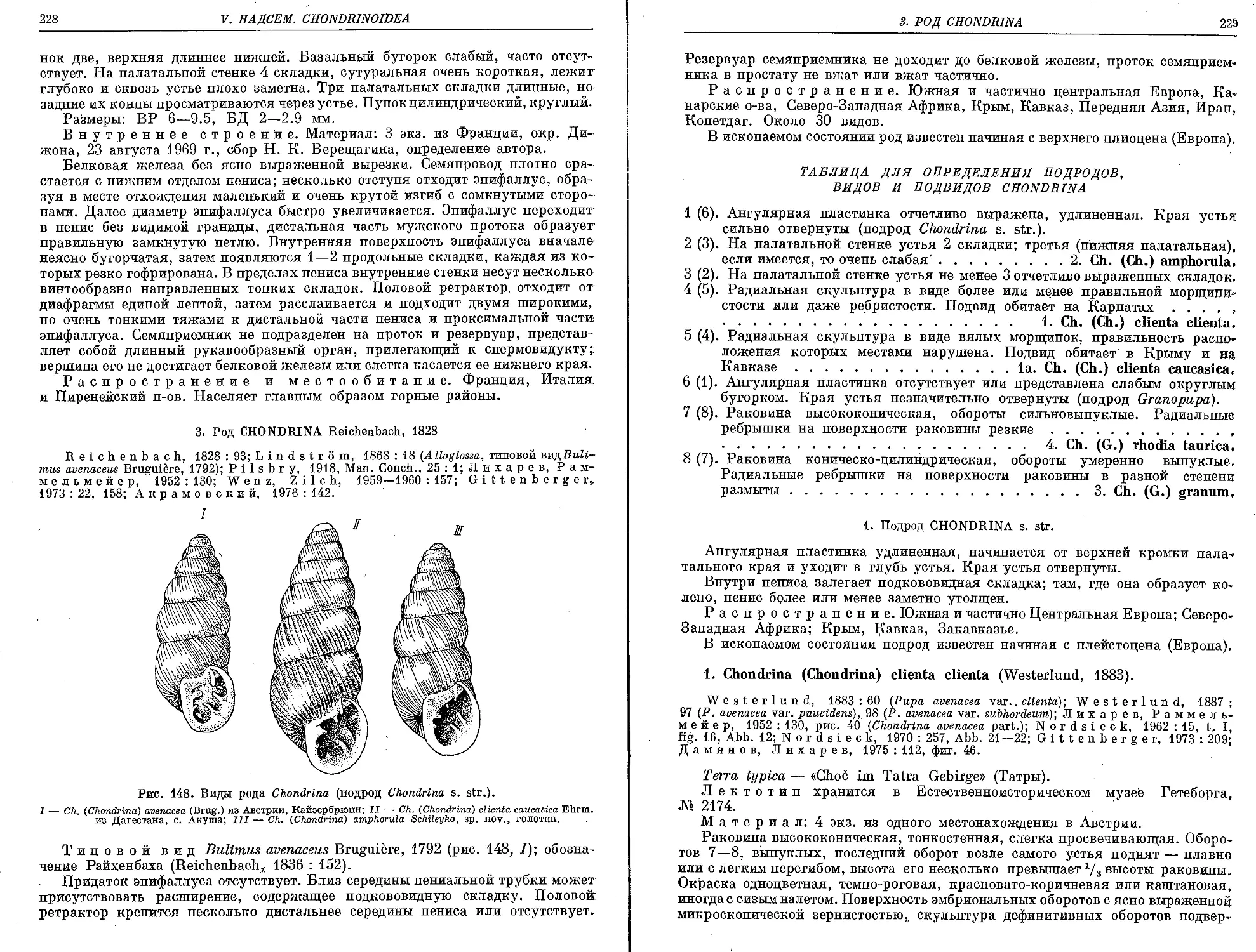
3. Род Chondrina Reichenbach, 1828
1. Подрод Chondrina s. str.
1. Ck. (Ch.) clienta clienta (Westerlund, 1883).............................. 229
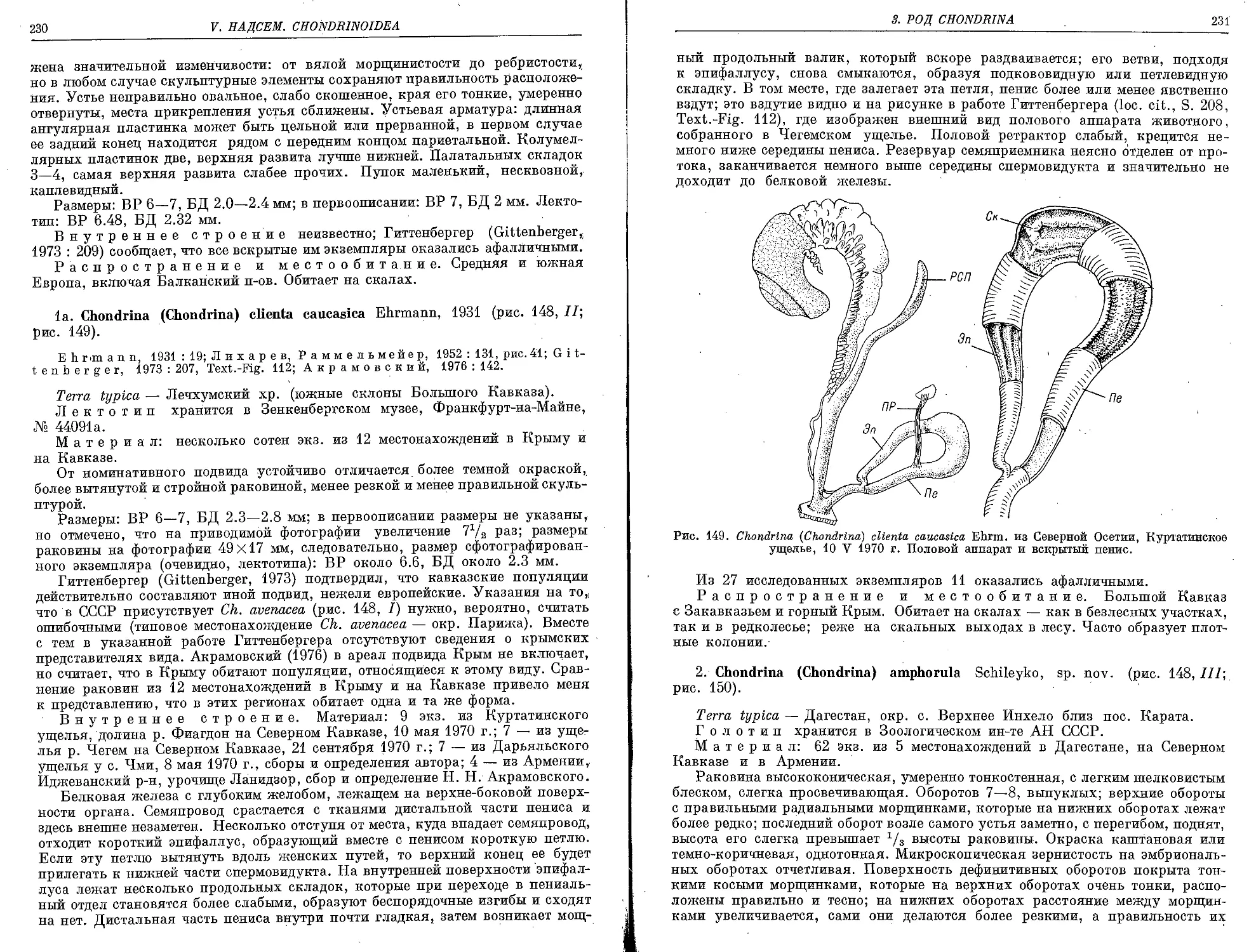
la. Ch. (Ch.) clienta caucasica Ehrmann, 1931 ................................. 230
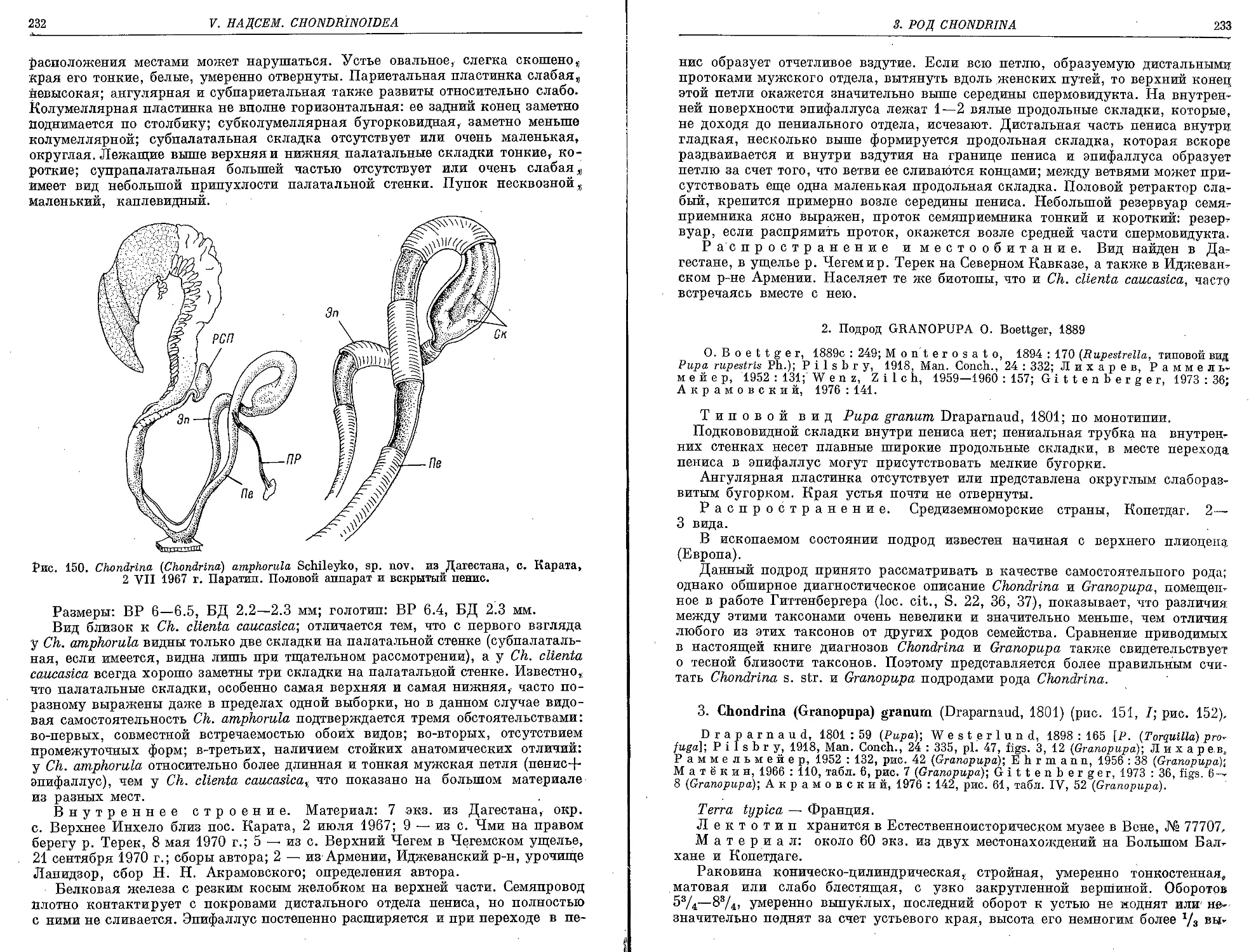
2. Ch. (Ch.) amphorula Schileyko, sp. nov..................................... 231
2. Подрод Granopupa O. Boettger, 1889
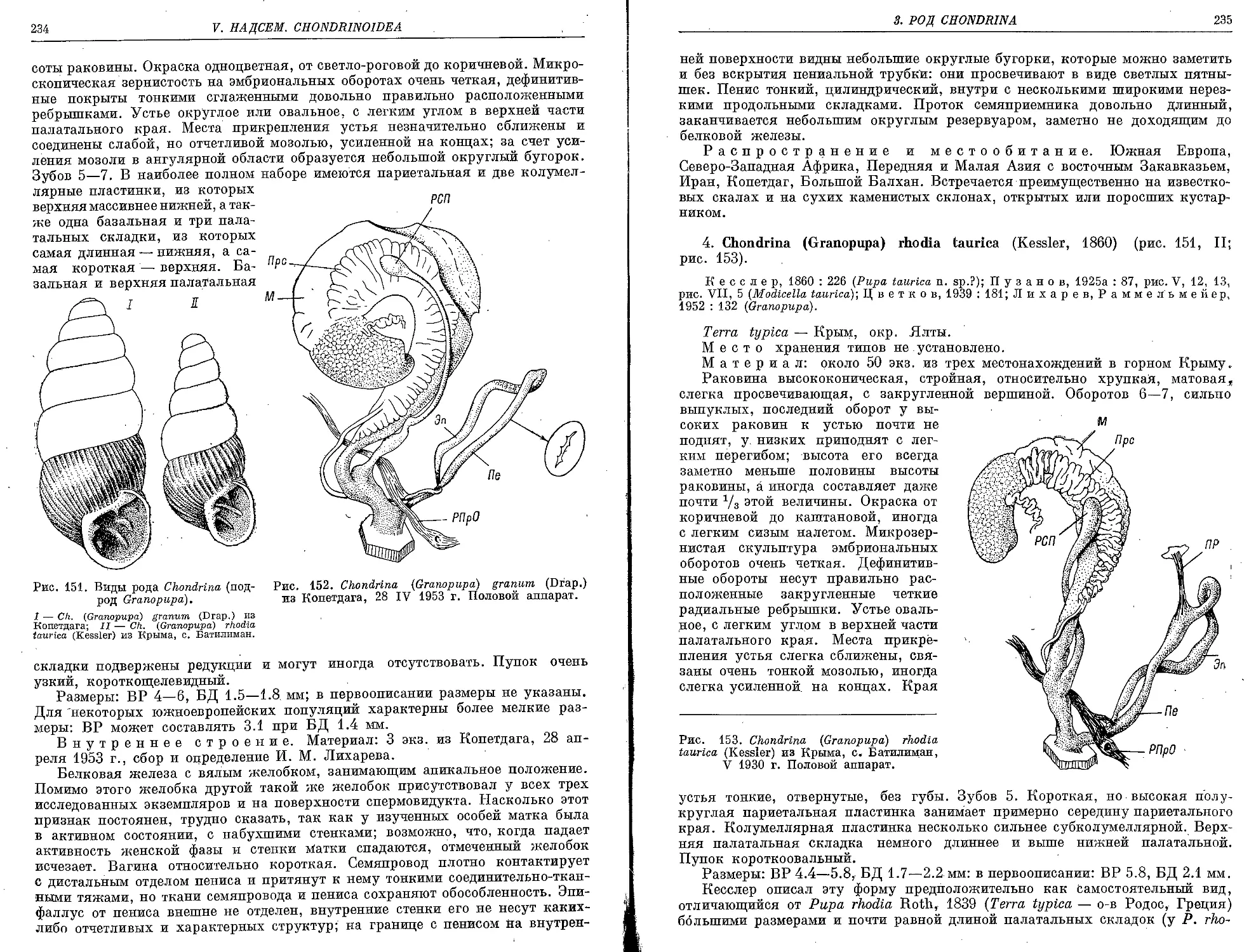
3. Ch. (G.) granum (Draparnaud, 1801)........................................... 233
4. Ch. (G.) rhodia taurica (Kessler, 1860)...................................... 235
?2. Сем. PYRAMIDULIDAE Kennard et Woodward, 1914
1. Род Pyramidula Fitzinger, 1833
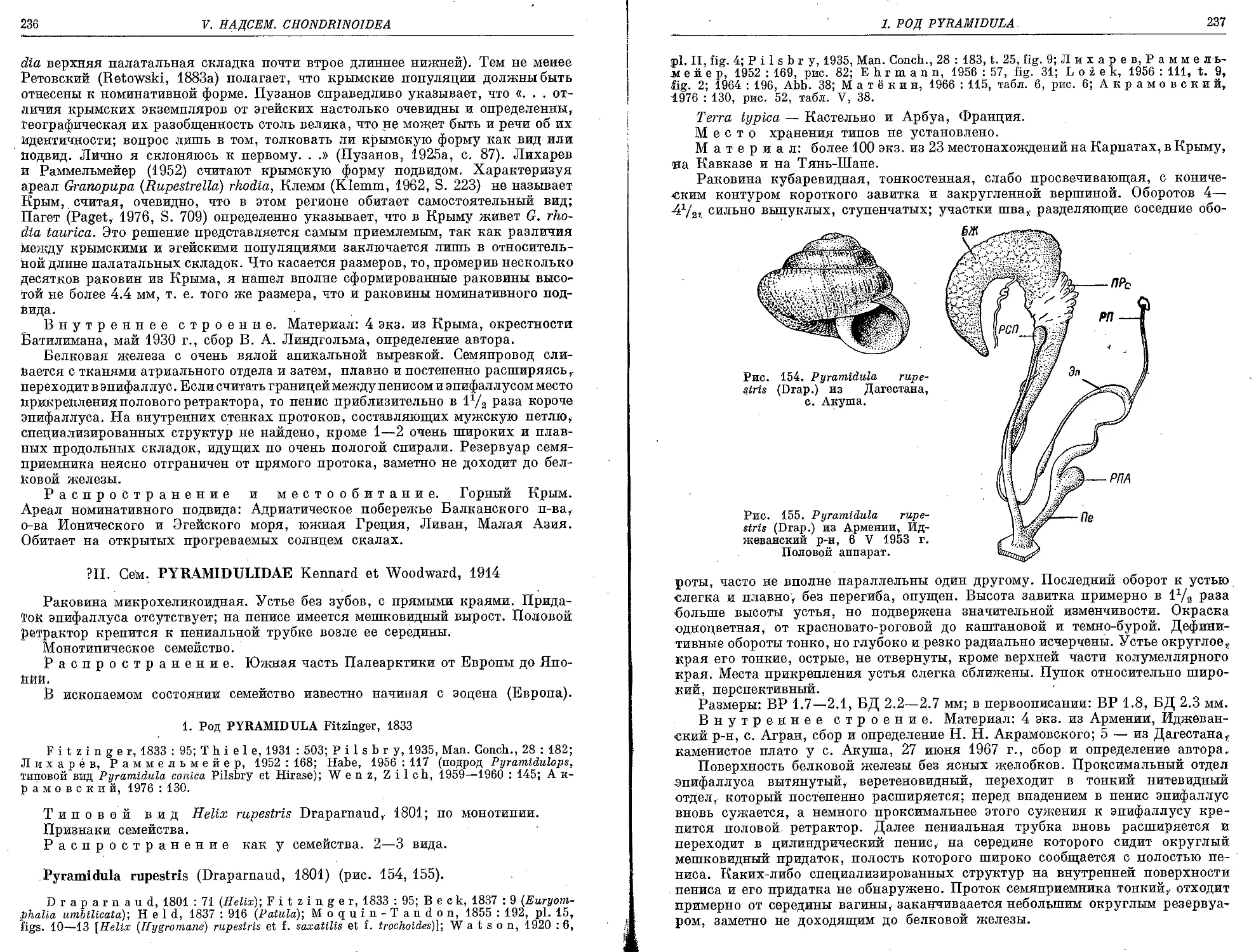
P. rupestris (Draparnaud, 1801)................................................. 236
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
VI. Надоем. BU1IMIN0IDEA Kobelt, 1880
Сем. BULIMINIDAE Kobelt, 1880
I. Подсем. Pseudonapaeinae Schileyko, 1978
1. Род Pseudonapaeus Westerlund, 1887
1. Подрод Pseudonapaeus s. str.
1. P. (P.) regelianus (Ancey, 1886).............................................. 246
2. P. (P.) albiplicatus (E. Martens, 1874) ................. . .................. 247
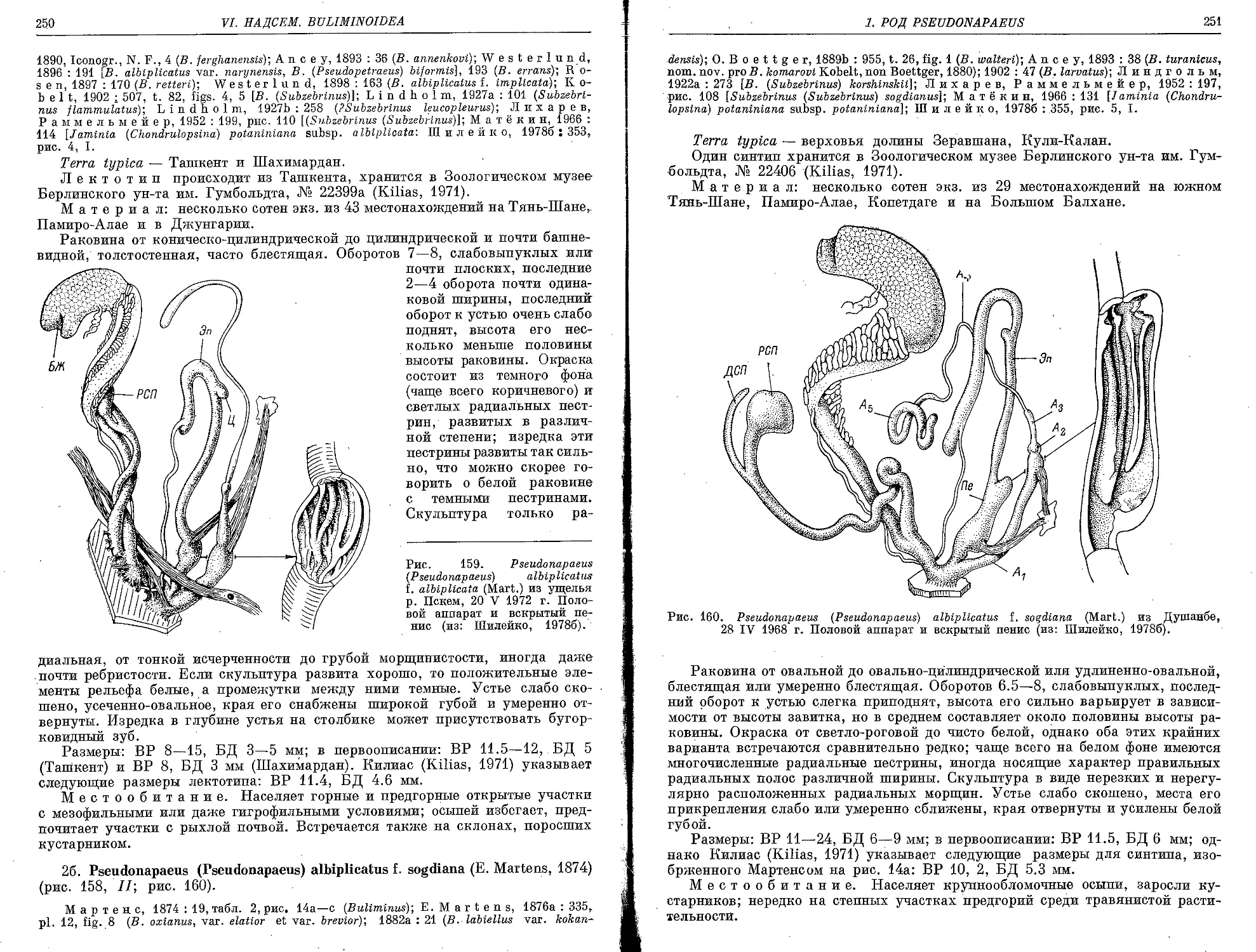
2a. P. (P.) albiplicatus f. albiplicata (E. Martens, 1874)....................... 249
26. P. (P.) albiplicatus f. sogdiana (E. Martens, 1874).......................... 250
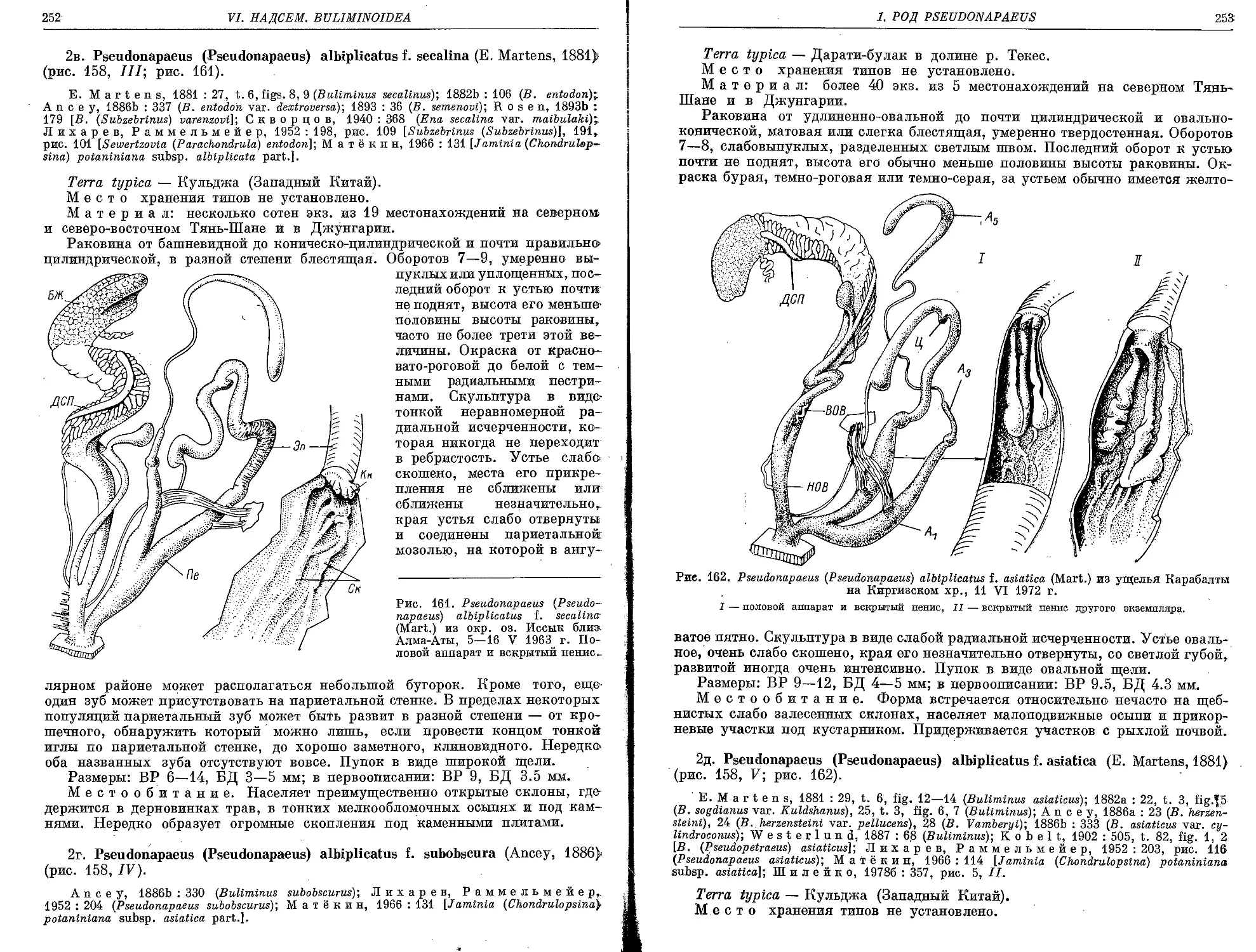
2в. P. (P.) albiplicatus f. secalina (E. Martens, 1881) . . '.................... 252
2г. P. (P.) albiplicatus f. subobscura (Ancey, 1886)............................. 252
2д. P. (P.) albiplicatus f. asiatica (E. Martens, 1881).......................... 253
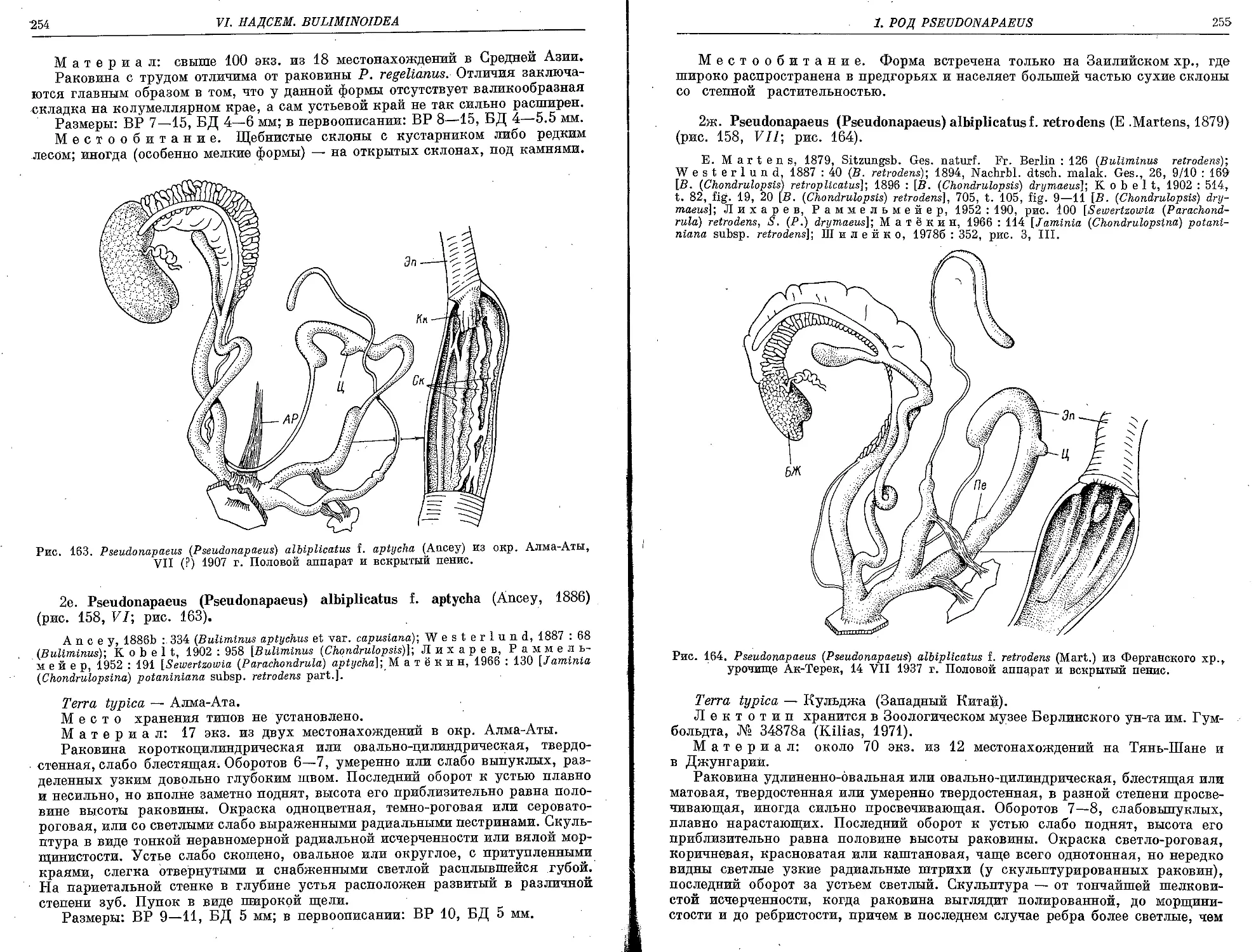
2e. P. (P.) albiplicatus f. aptycha (Ancey, 1886)................................ 254
2ж. P. (P.) albiplicatus f. retrodens (E. Martens, 1879).......................... 255
2з. P. (P.) albiplicatus f. dissimilis (E. Martens, 1882)......................... 256
3. P. (P.) miser (E. Martens, 1874)............................................. 256
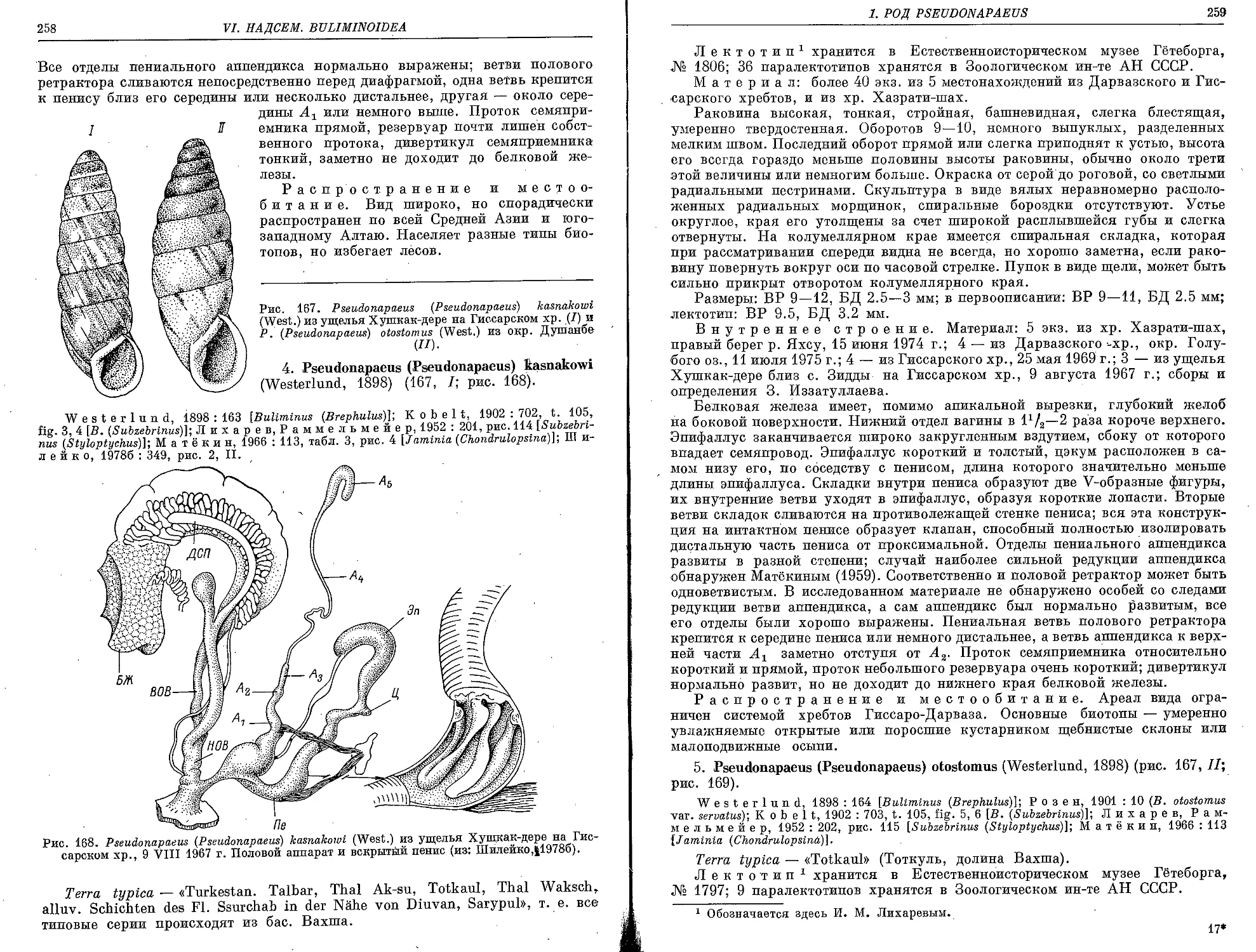
4. P. (P.) kasnakowt (Westerlund, 1898)........................................ 258
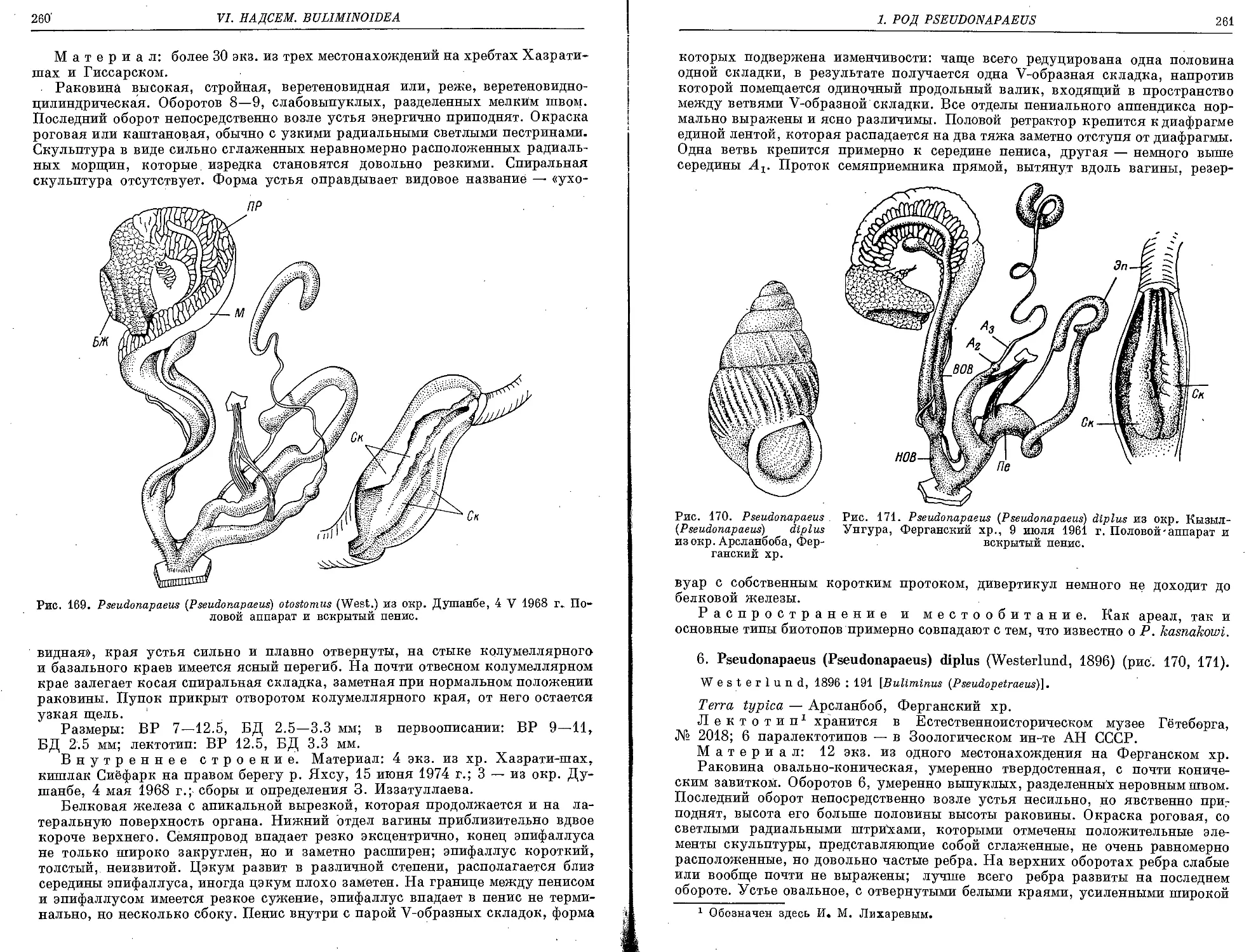
5. P. (P.) otostomus (Westerlund, 1898) ....................................... 259
6. P. (P.) diplus (Westerlund, 1896)........................................... 261
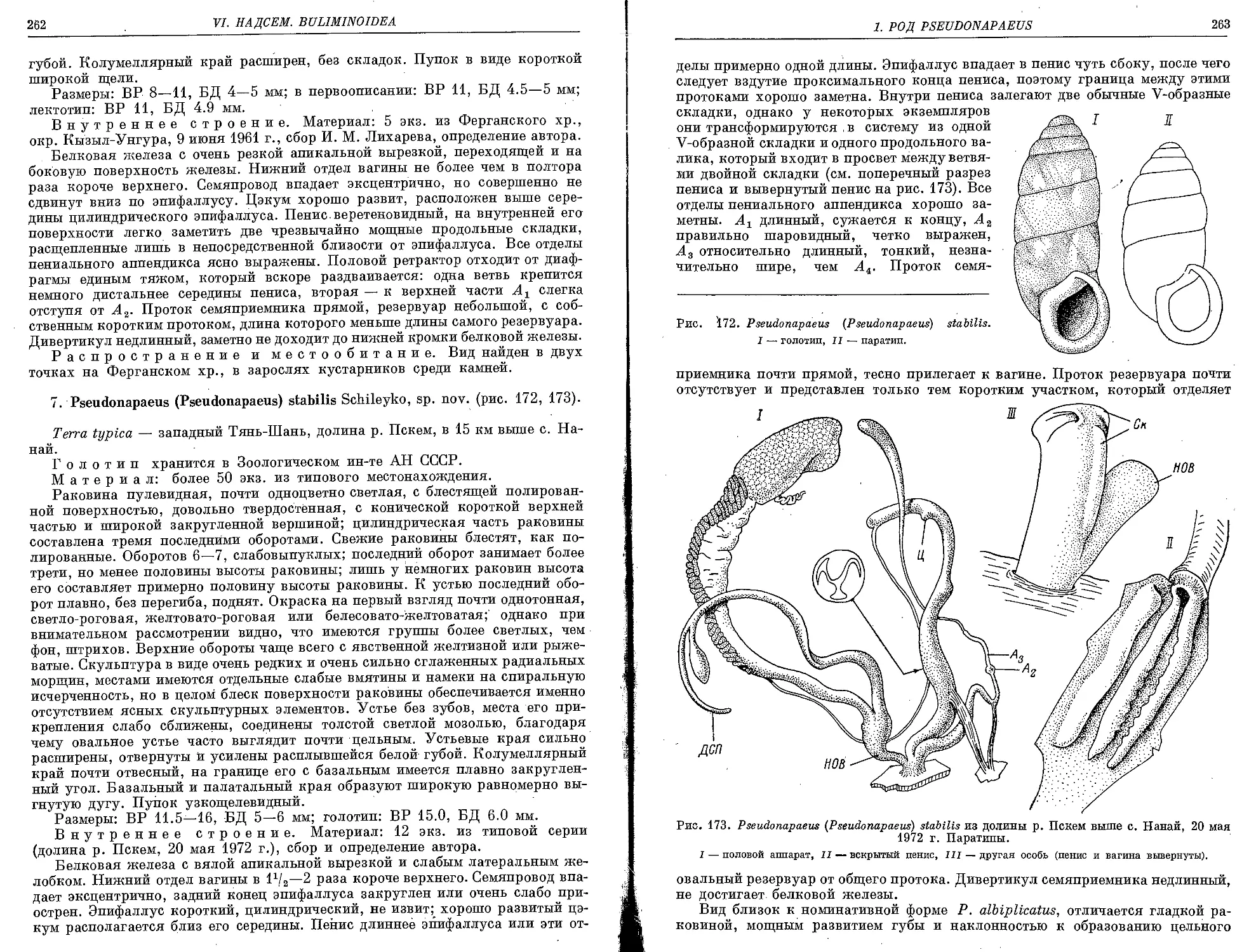
7. P. (P.) stabilis Schileyko, sp. nov......................................... 262
8. P. (P.) schnitnikovi (Lindholm, 1922)........................................ 264
9. P. (P.) galinae (Tzvetkov, 1940)........................................... 265
10. P. (P.) trigonochilus (Ancey, 1886)......................................... 265
11. P. (P.) goldfussi (Kobelt, 1893)............................................ 267
12. P. (P.) chodschendicus (Mukhitdinov, 1976).................................. 268
2. Подрод Siraphorus Lindholm, 1925
13. P. (S.) entoptyx (Lindholm, 1925)............................................ 269
3. Подрод A r i d e n u s Schileyko, subgen. nov.
14. P. (A.) submucronatus (Lindholm, 1927)....................................... 271
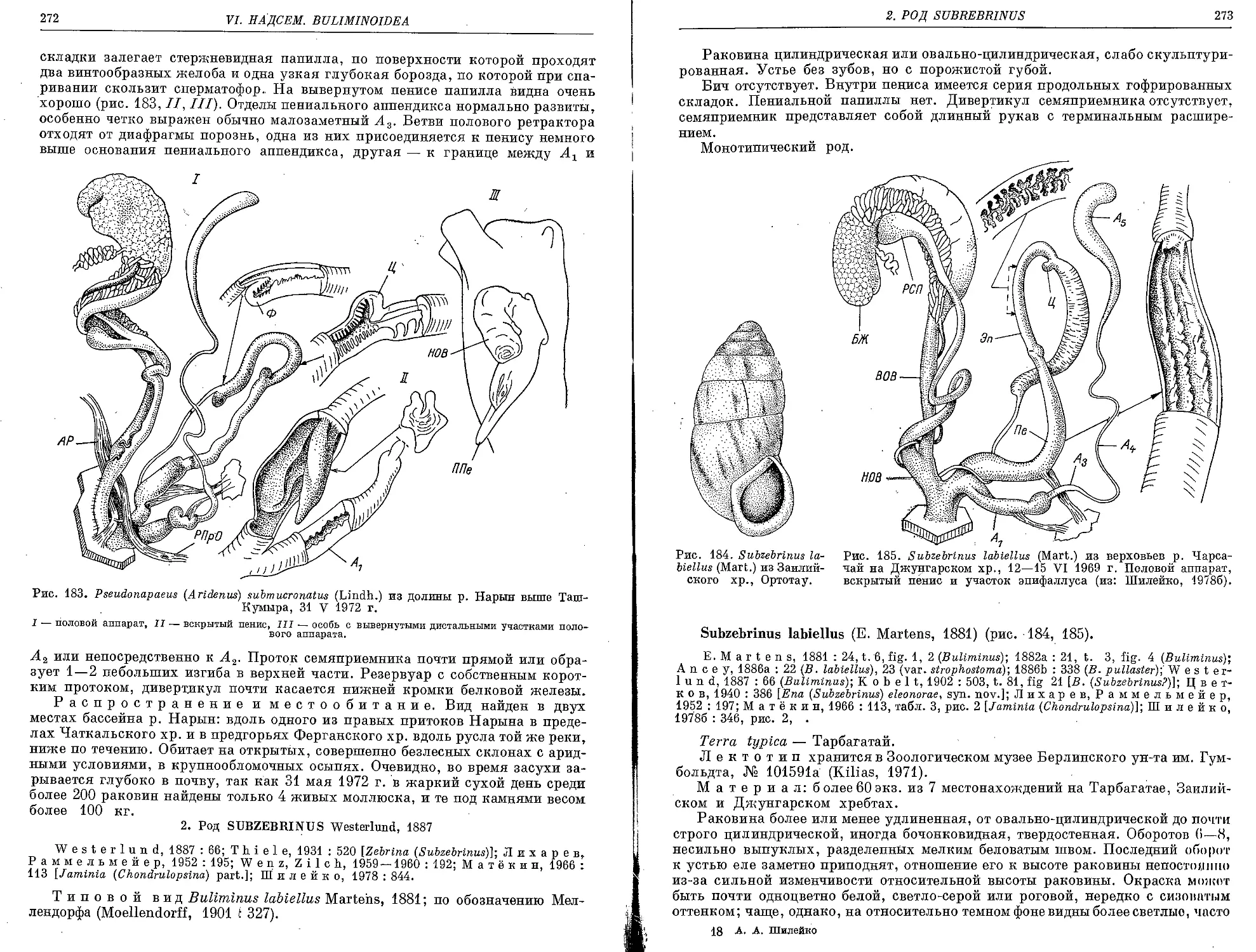
2, Род Subzebrinus Westerlund, 1887
.S’. labiellus (E. Martens, 1881)................................................. 273
3. Род Turanena Lindholm, 1922
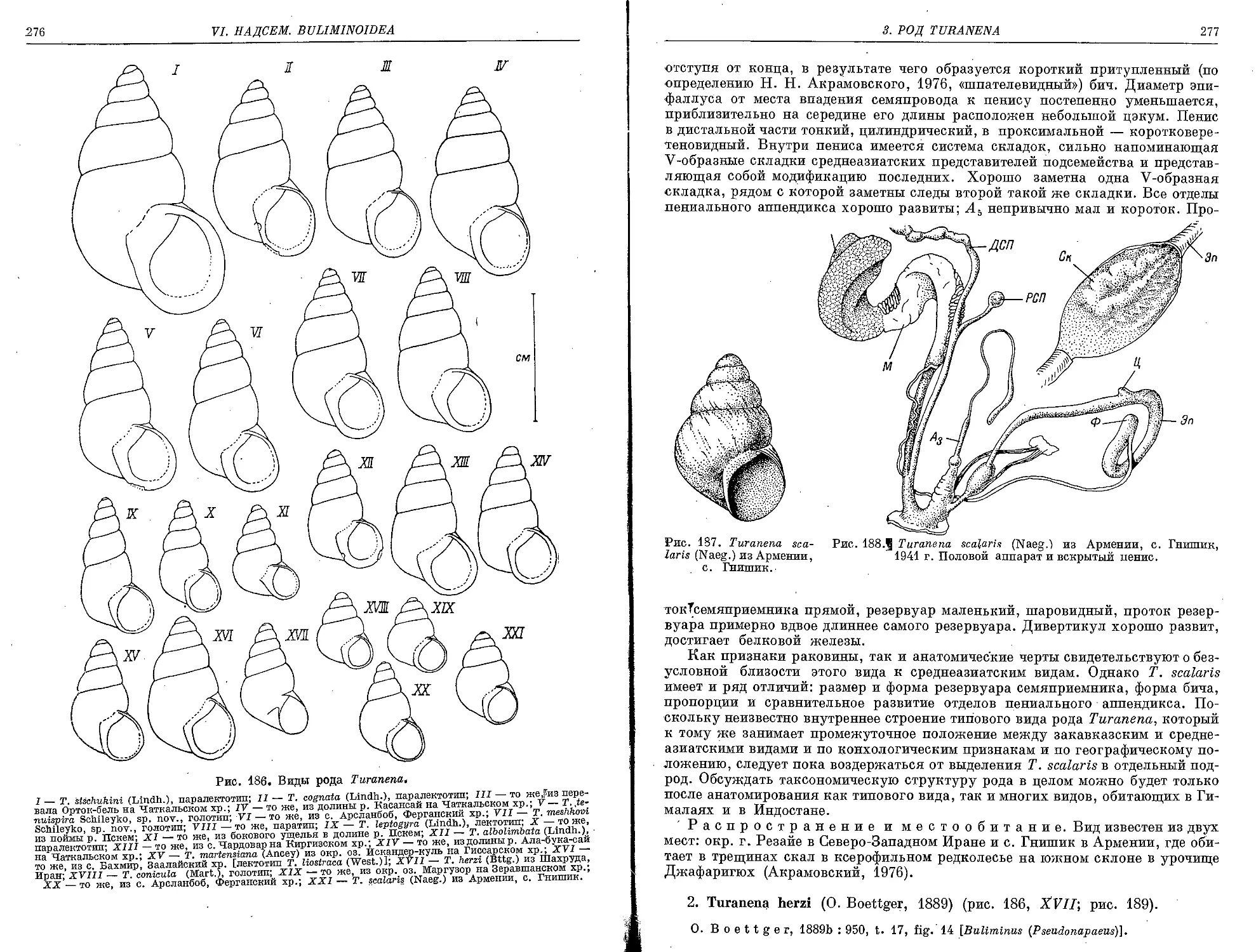
1. T. scalaris (Naegele, 1902)................................................... 275
2. T. herzi (O. Boettger, 1889).......................:.......................... 277
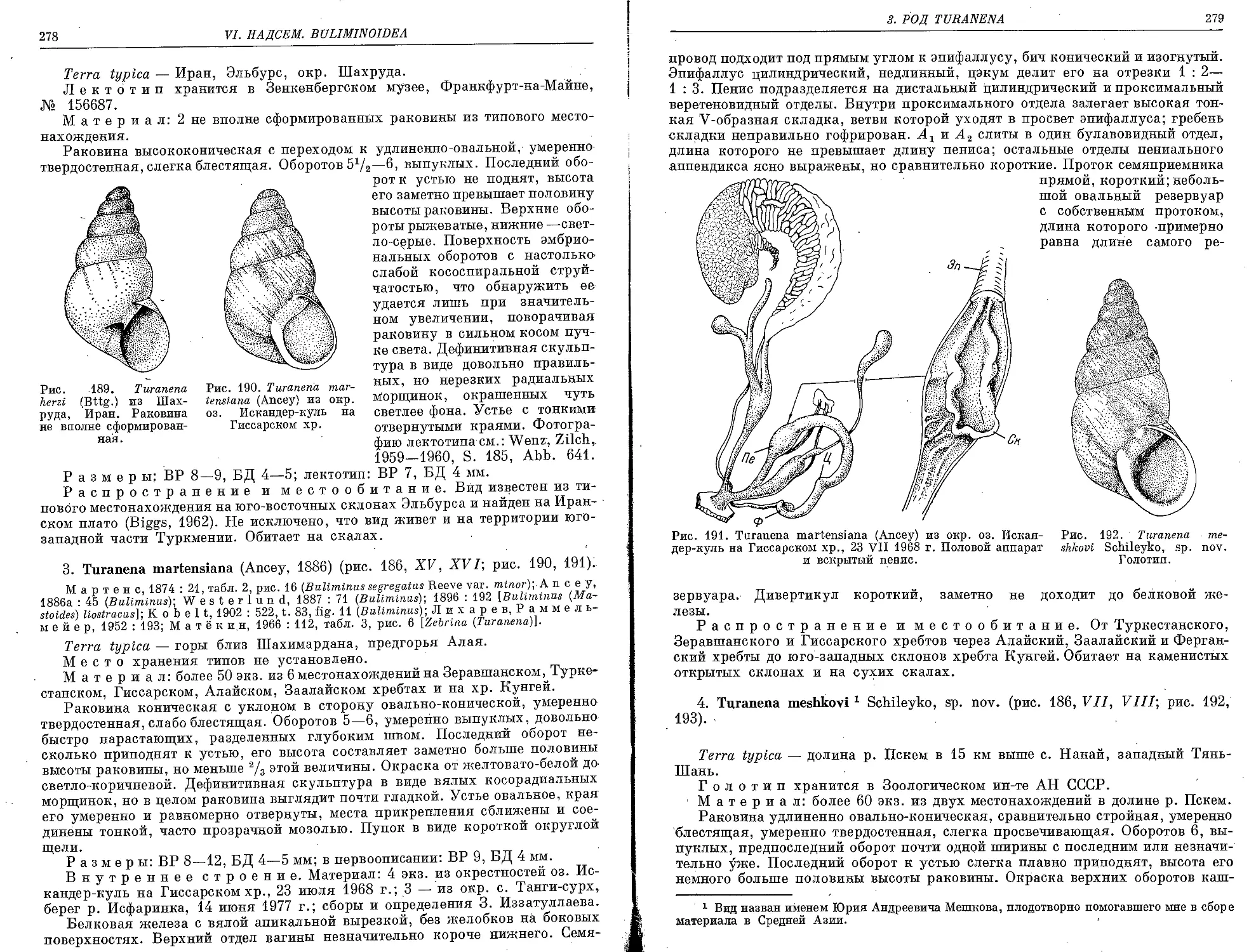
3. T. martensiana (Ancey, 1886)................................................ 278
4. T.~ meshkovi Schileyko, sp. nov............................................... 279
5. T. leptogyra (Lindholm, 1927)................................................ 281
6. T. tenuispira Schileyko, sp. nov.............................................. 282
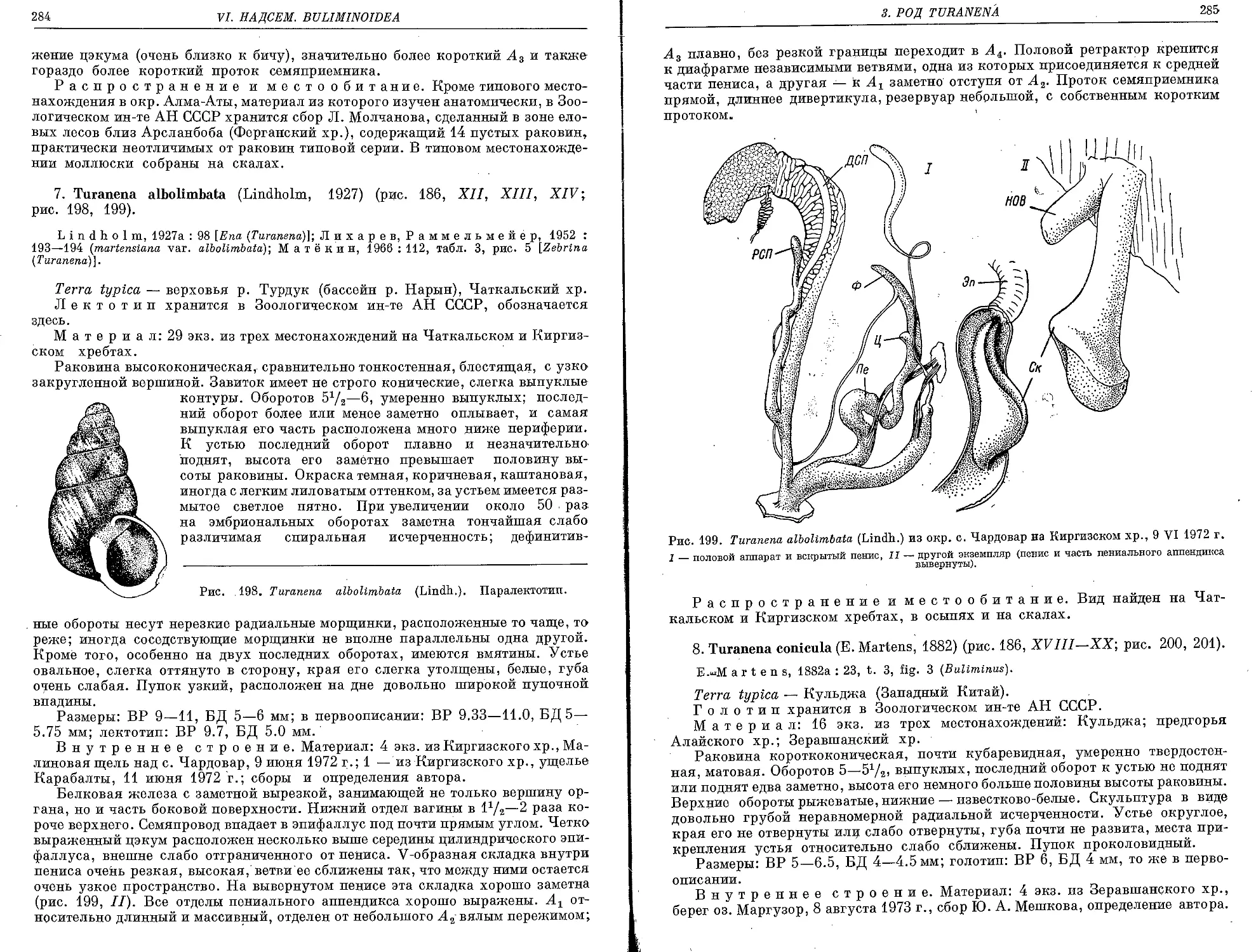
7. T. albolimbata (Lindholm, 1927)............................................... 284
8. T. conicula (E. Martens; 1882) ............................................... 285
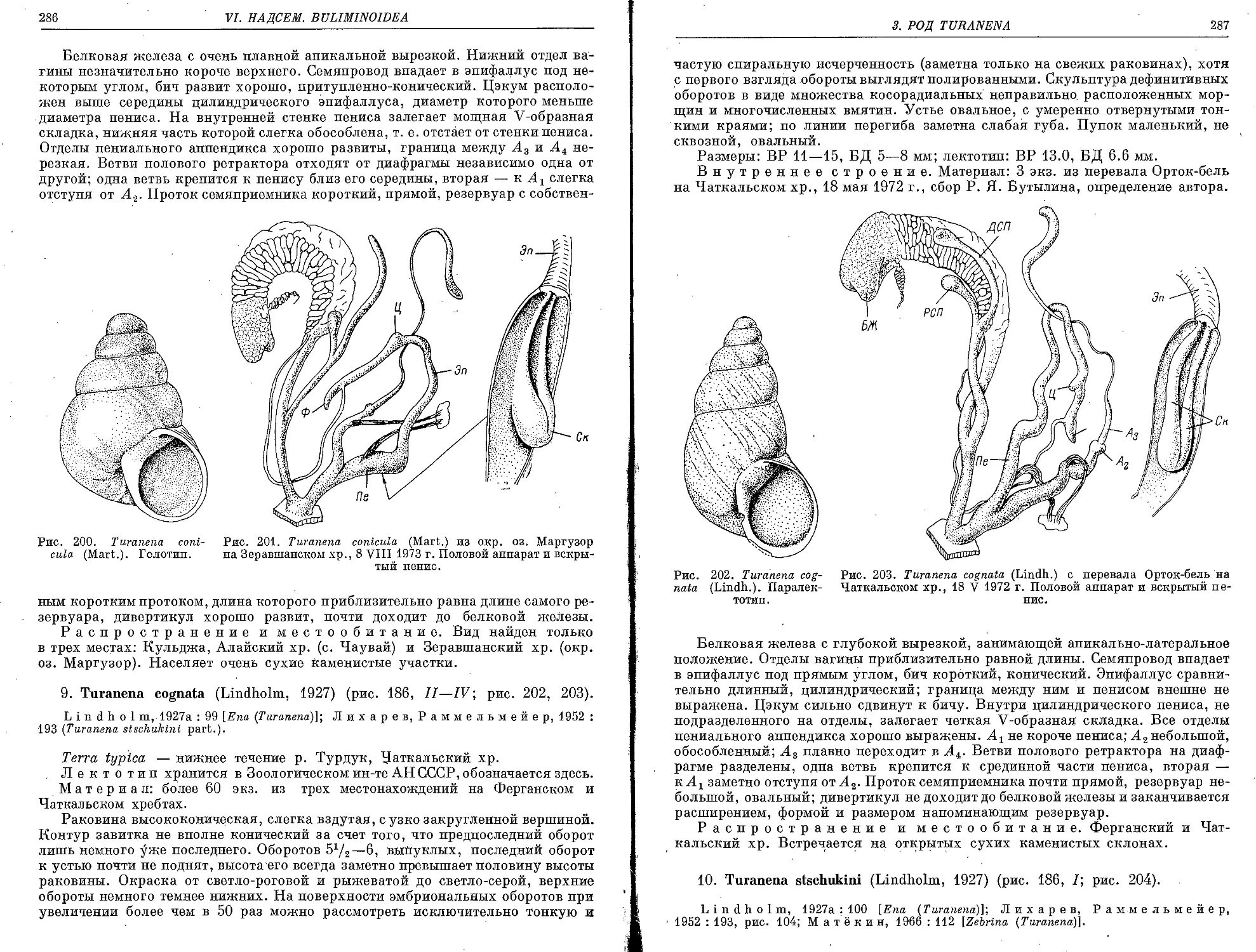
9. T. cognata (Lindholm, 1927)................................................... 286
10. T; stschukini (Lindholm, 1927).................. .• .•...................... 287
4. Род Pseudochondrula Hesse, 1933
1. P. seductilis (Rossmaessler, 1837)............................................ 289
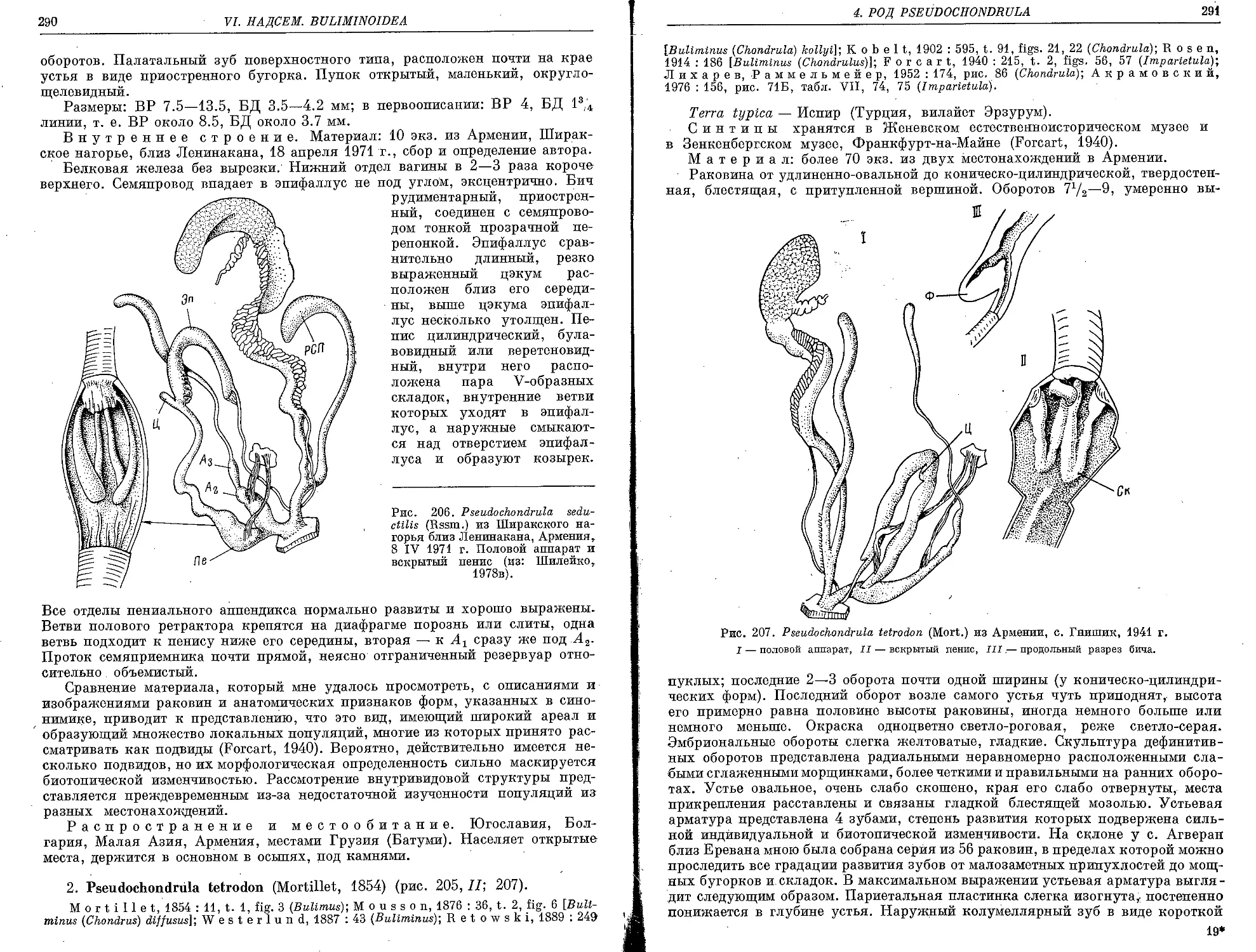
2. P. tetrodon (Mortillet, 1854)................................................. 290
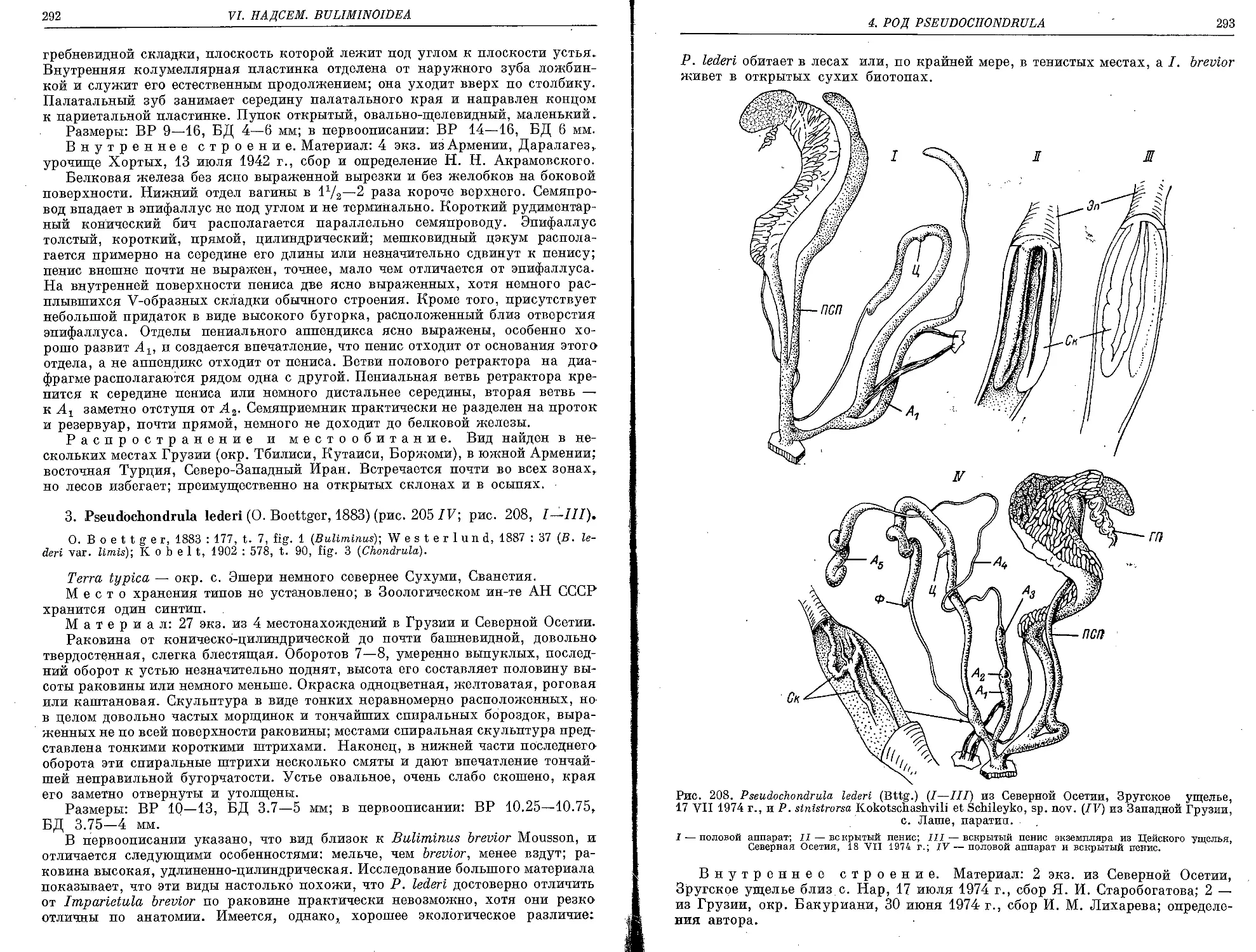
3. P. lede.ri (O. Boettger, 1883)............................................... 292
4. P. sinistrorsa Kokotschashvili et Schileyko, sp. nov.......................... 294
5. P. tuberifera (0. Boettger, 1879)............................................. 294
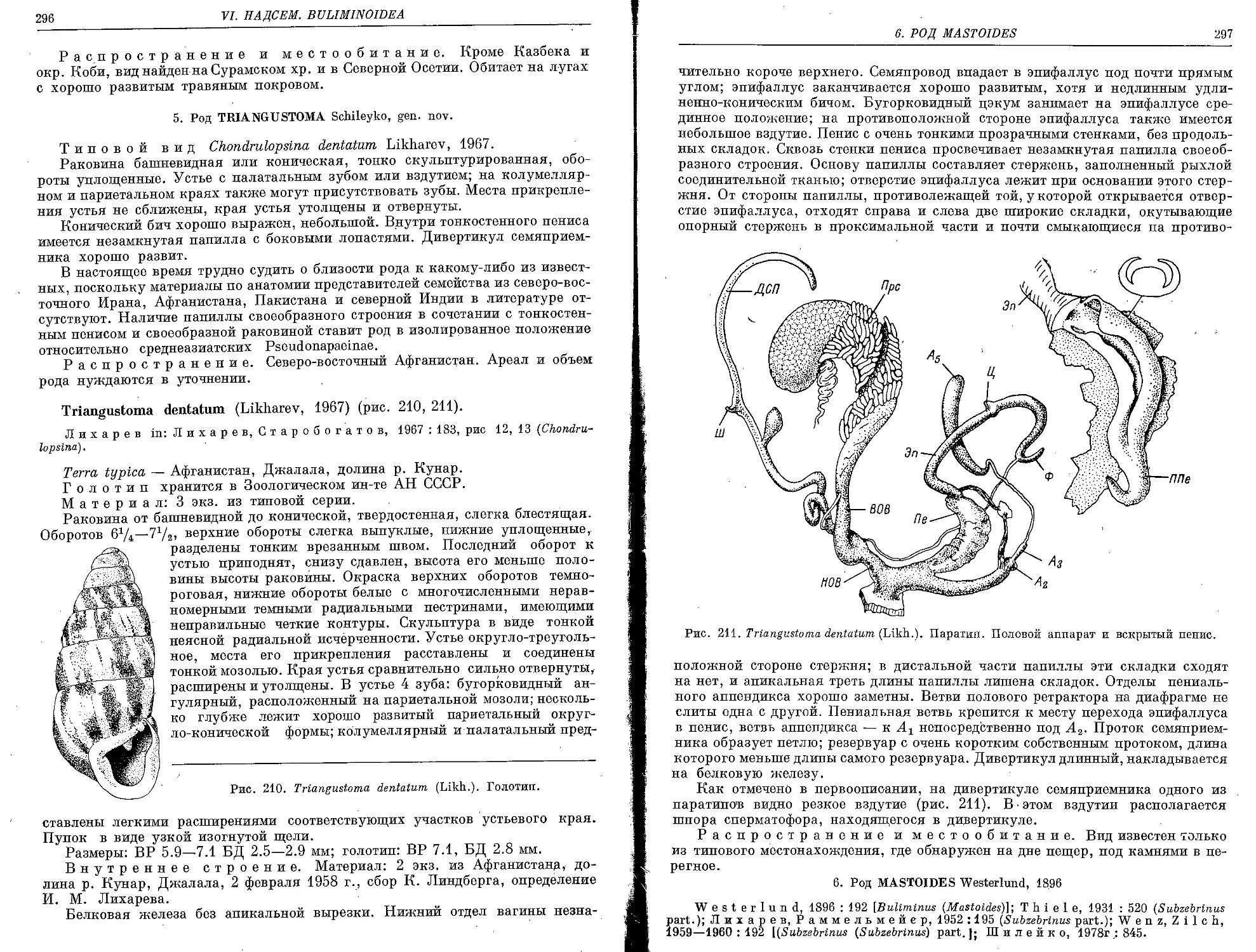
5. Род Triangustoma Schileyko, gen. nov.
T. dentatum (Likharev, 1967)...................................................... 296
6. Род Mastoides Westerlund, 1896
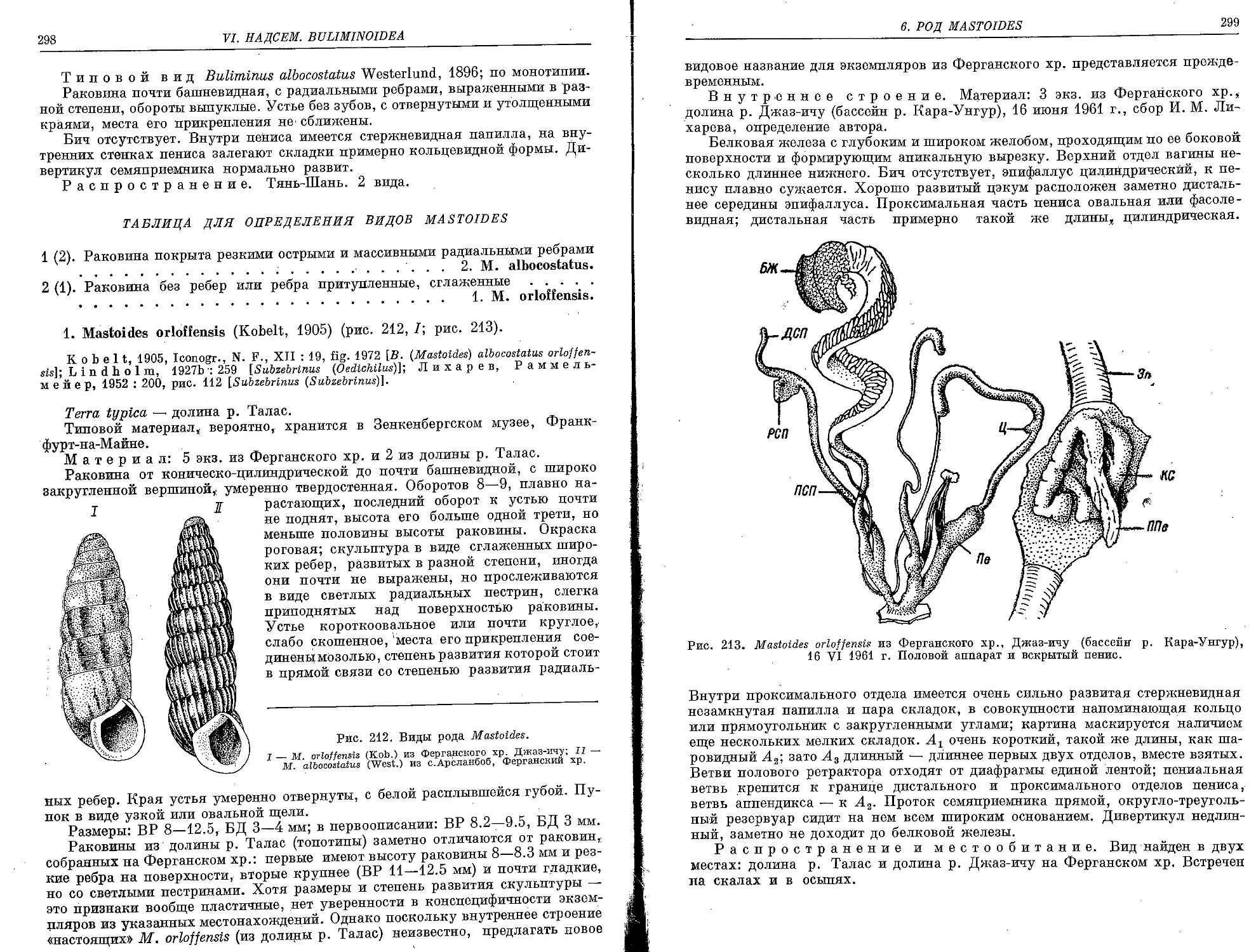
1. M. orlof fensis (Kobelt, 1905) . .......................................... . 298
2. M. albocostatus (Westerlund, 1896)......................................... 300
1(0 СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
7. Род Laevozebrinus Lindholm, 1925
1. L. eremita (Benson, 1849)............,...................................... 301
2. L. urgutensis (Kobelt, 1902)................................................ 3Q3
3. L. uifalvyanus (Ancey, 1886).......................'........................ 304
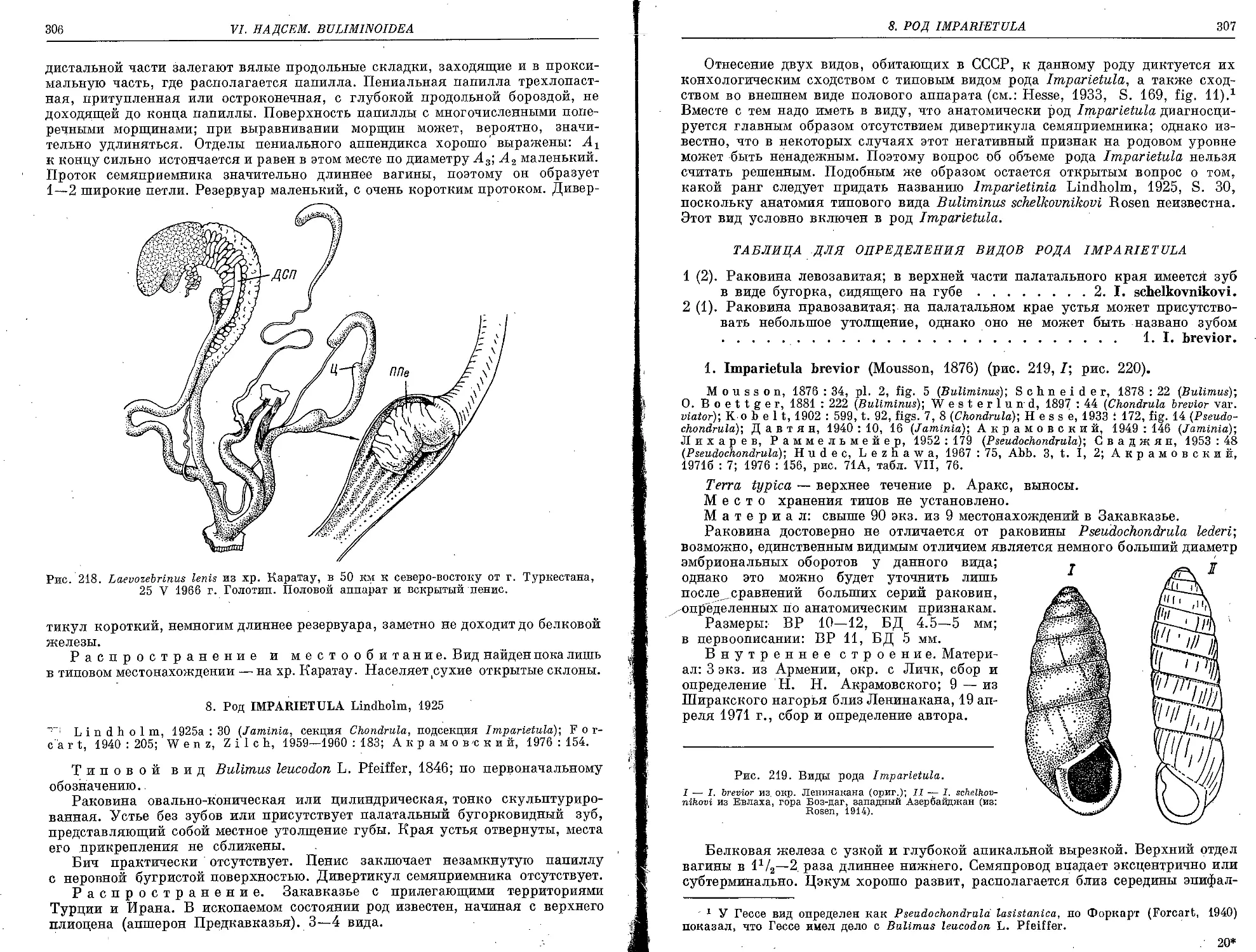
4. L. lenis Schileyko, sp. n................................................... 305
8. Род Imparietula Lindholm, 1925
1. I. brevior (Mousson, 1876).................................................. 307
2. I. schelkovnikovi (Rosen, 1914).................. . ...................... 309
9. Род Ljudmilena Schileyko, gen. nov.
1. L. sieversl (Mousson, 1873)................................................ 309
2. L. tricollis (Mousson, 1876)............................................... 312
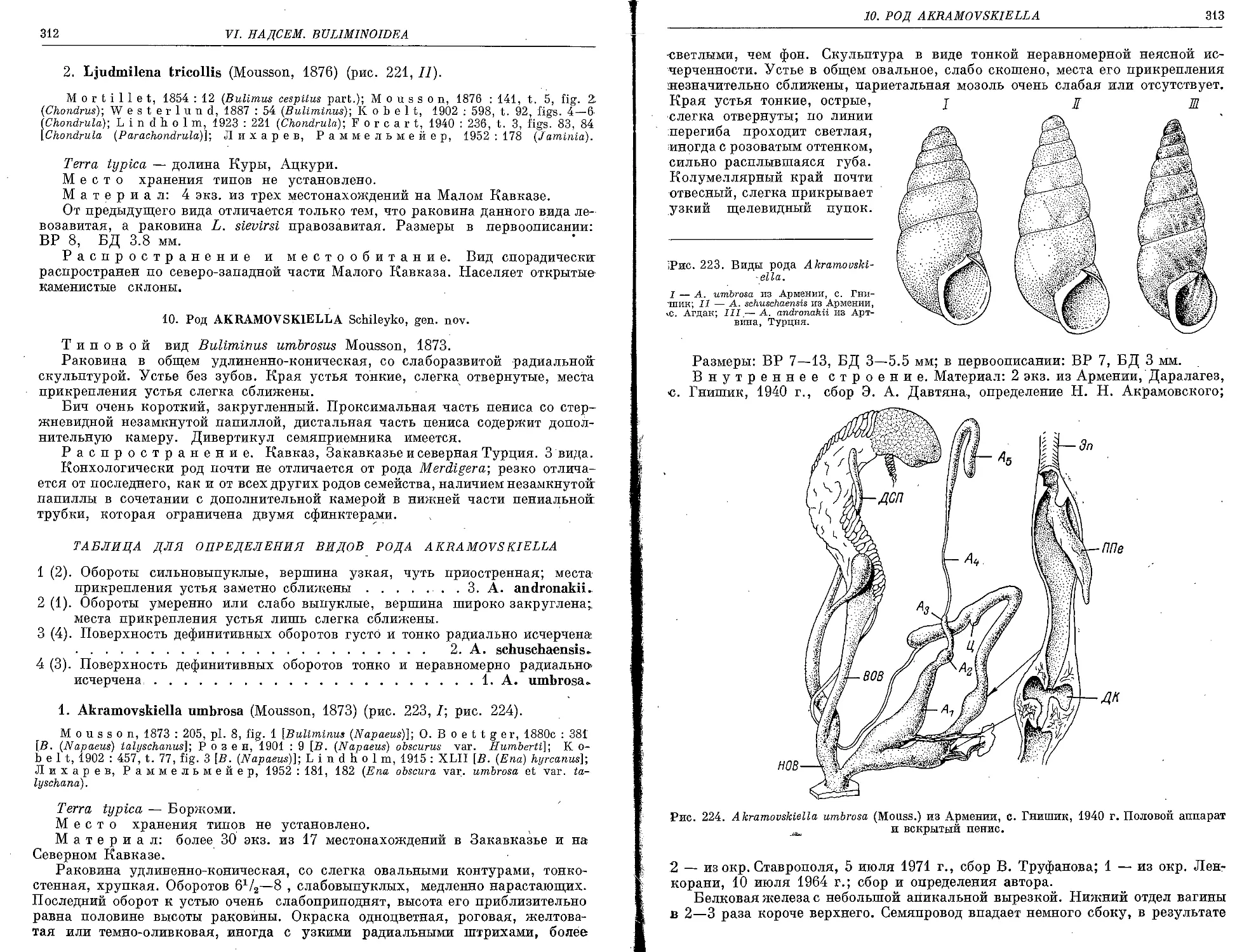
10. Род Akramovskiella Schileyko, gen. nov.
1. A. umbrosa (Mousson, 1873).................................................. 312
2. A. schuschaensis (Kobelt) 1902) . . ....................................... 314
3. A. andronakii (Lindholm, 1913).............................................. 315
11. Род Differena. Schileyko, gen. nov.
D. leucostoma Schileyko, sp. nov................................................ 316
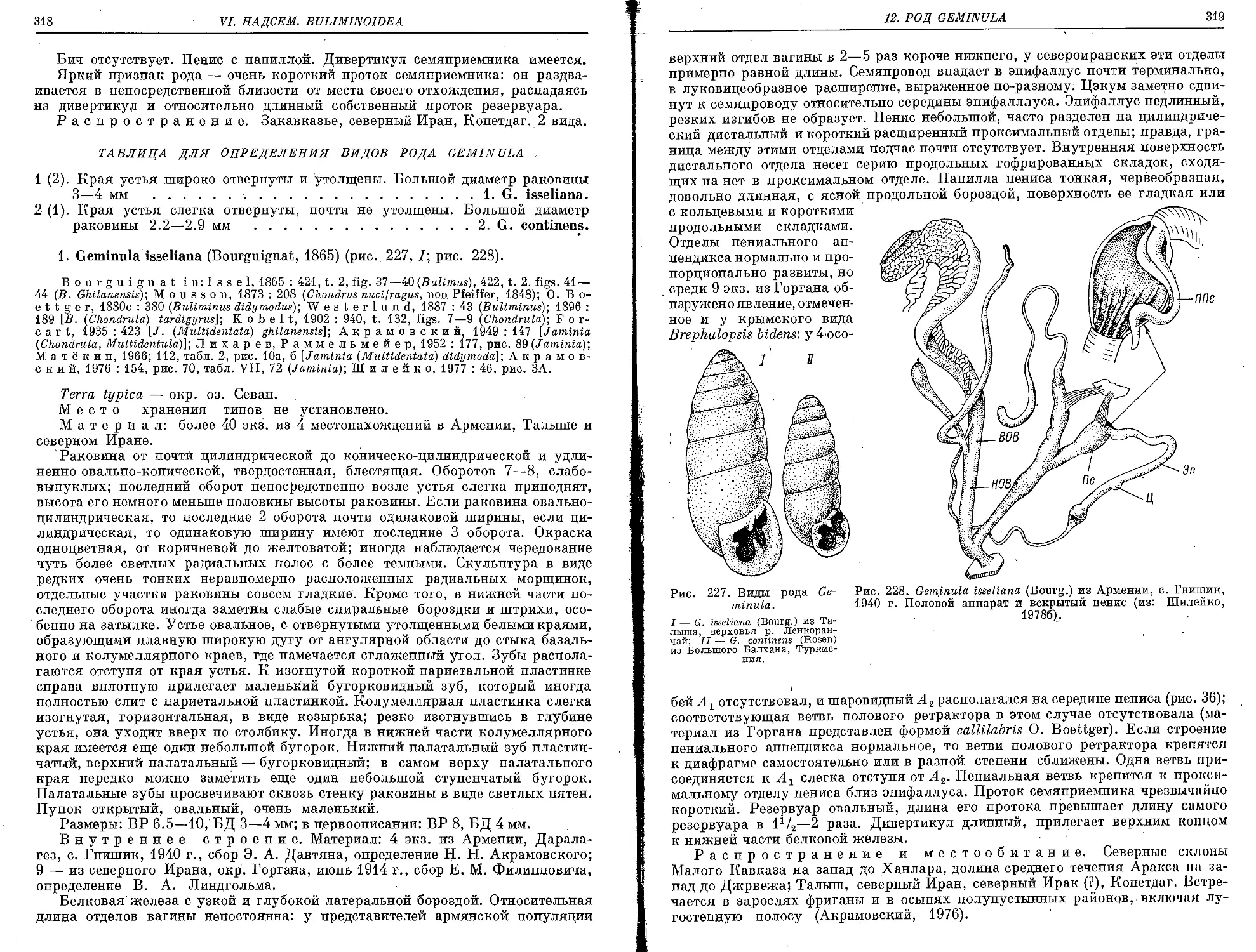
12. Род Geminula Lindholm, 1925
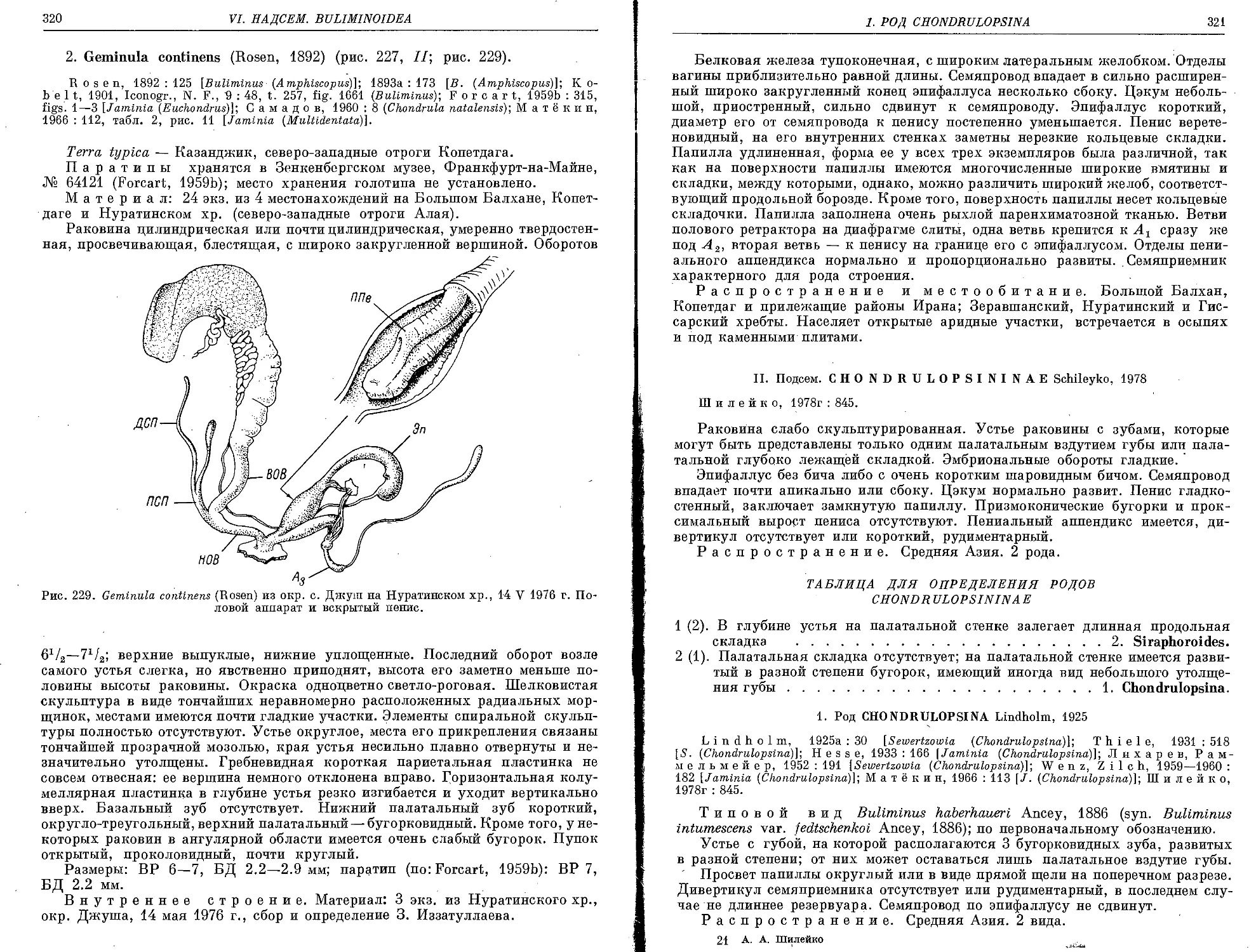
.1, G. isseliana (Bourguignat, 1865).....•...................................... 318
2. G. continens (Rosen, 1892)................................................. 320 .
I. Подсем. Chondrulop sin inae Schileyko, 1978
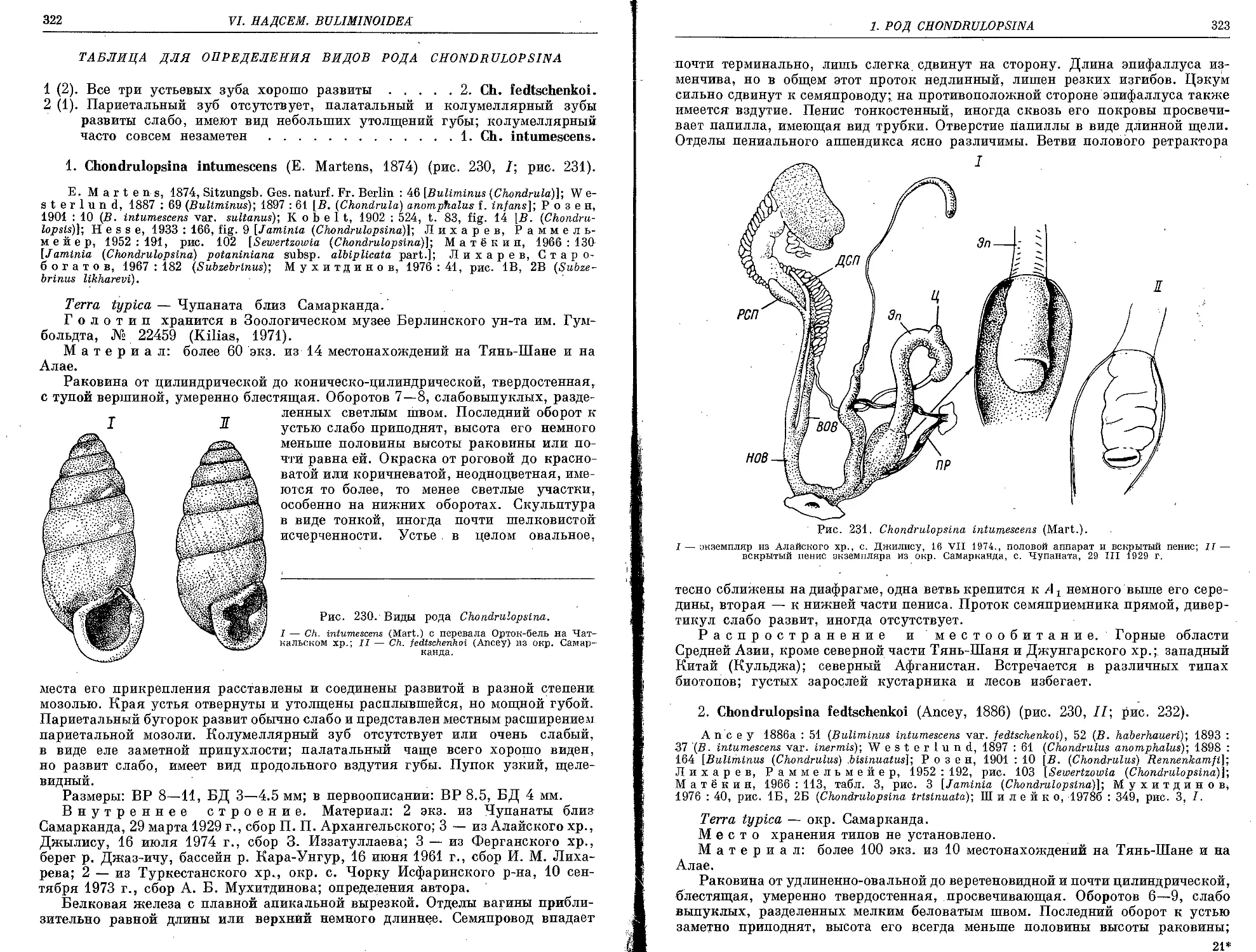
1. Род Chondrulopsina Lindholm, 1925
1. Ch. intumescens (E. Martens, 1874)........................................... 322
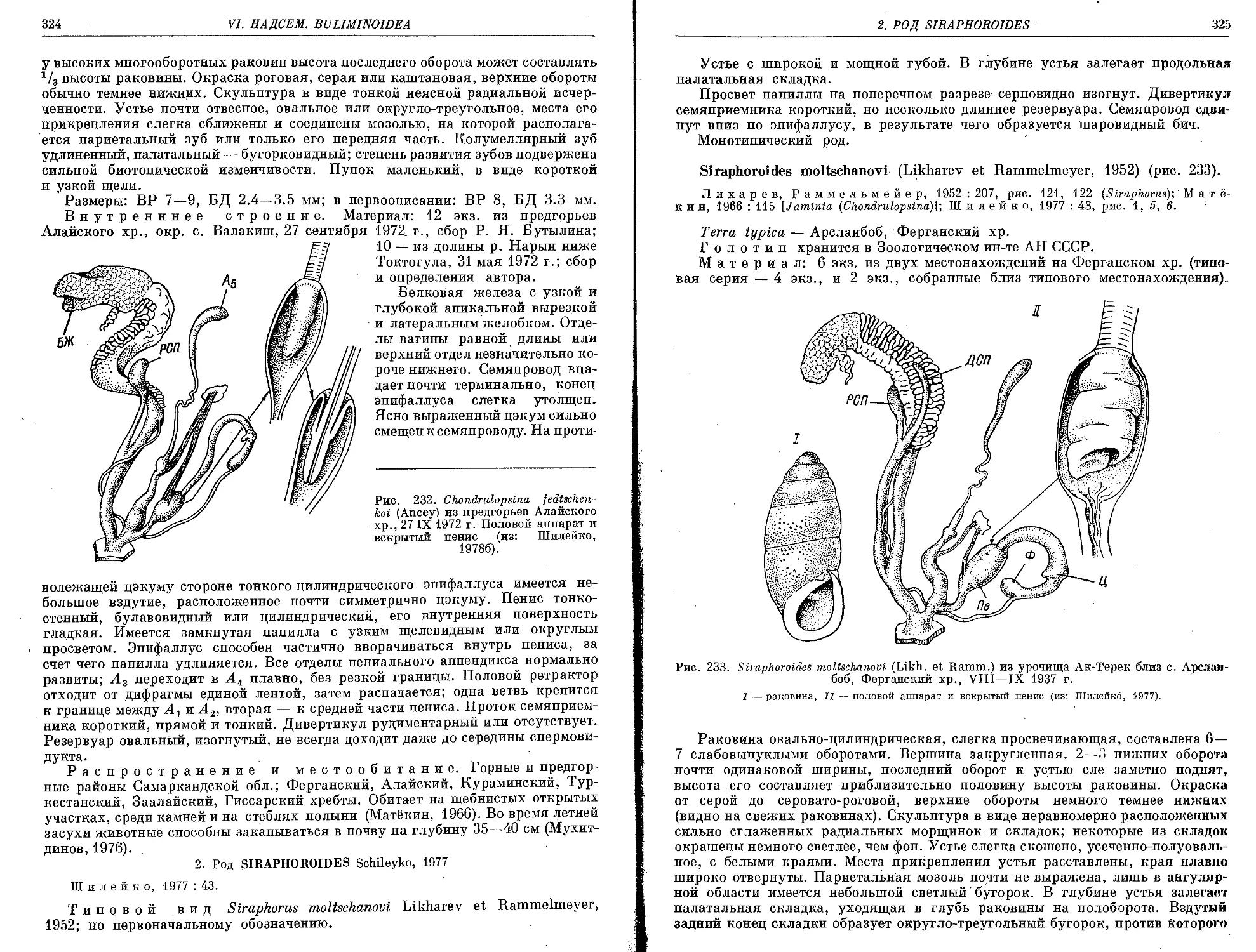
2. Ch. fedtschenkoi (Ancey, 1886) .................................. 323
2. Род Siraphoroides Schileyko, 1977
S. moltschanovl (Likharev et Rammelmeyer, 1952)................................. 325
III. Подсем. Jaminiinae Thiele, 1931
1. Род Jaminia Risso, 1826
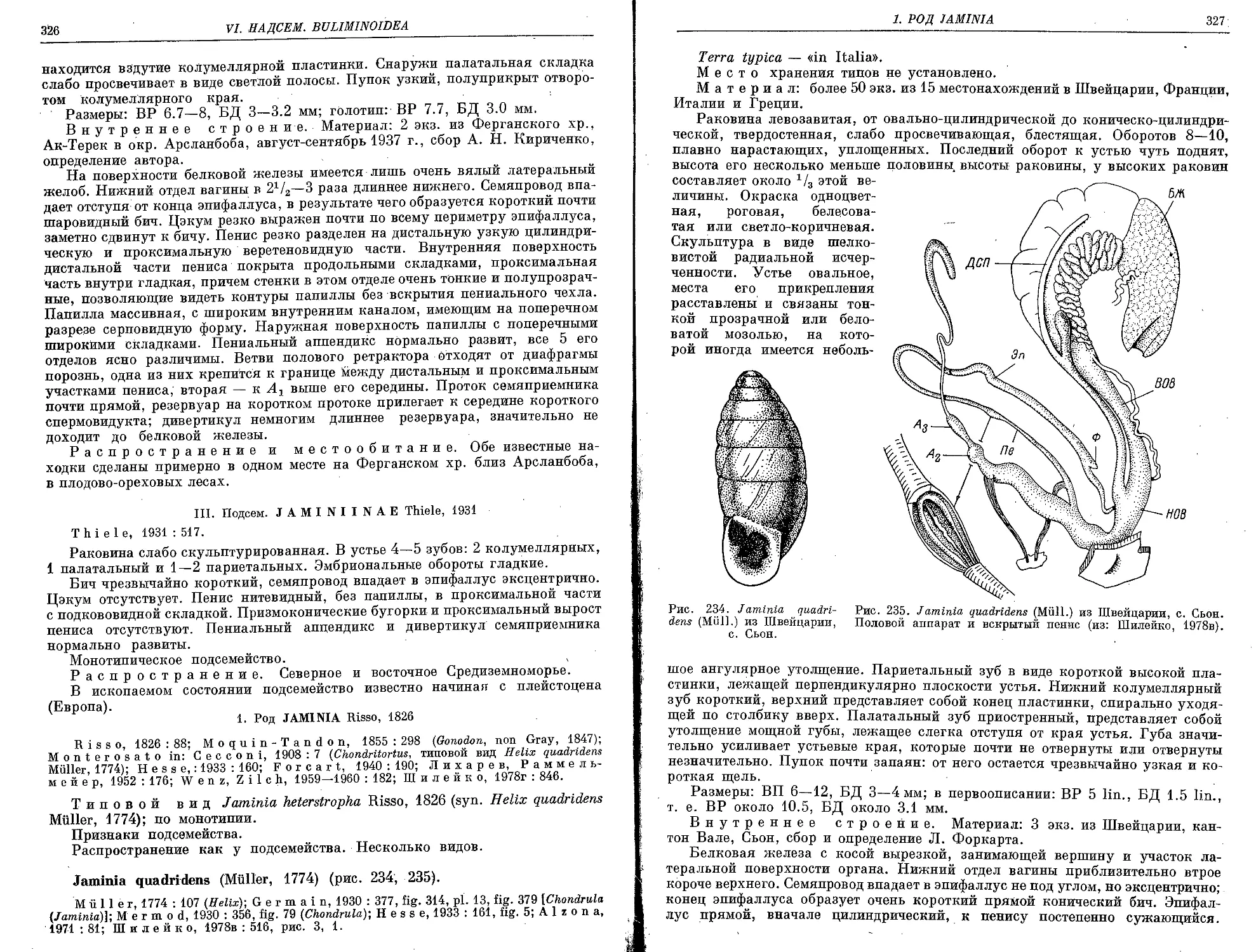
J. quadridens (Miiller, 1774).............................................. . 3 26
IV. Подсем. Merdigerinae Schileyko, subfam. nov.
1. Род Merdigera Held, 1837
M. obscura (Muller, 1774) ...................................................... 328
V. Подсем. Bulimininae Kobelt, 1880
1. Род Buliminus Beck, 1837
B. urmianus O. Boettger, 1898 .............................. 331
2. Род Adzharia Hesse, 1933
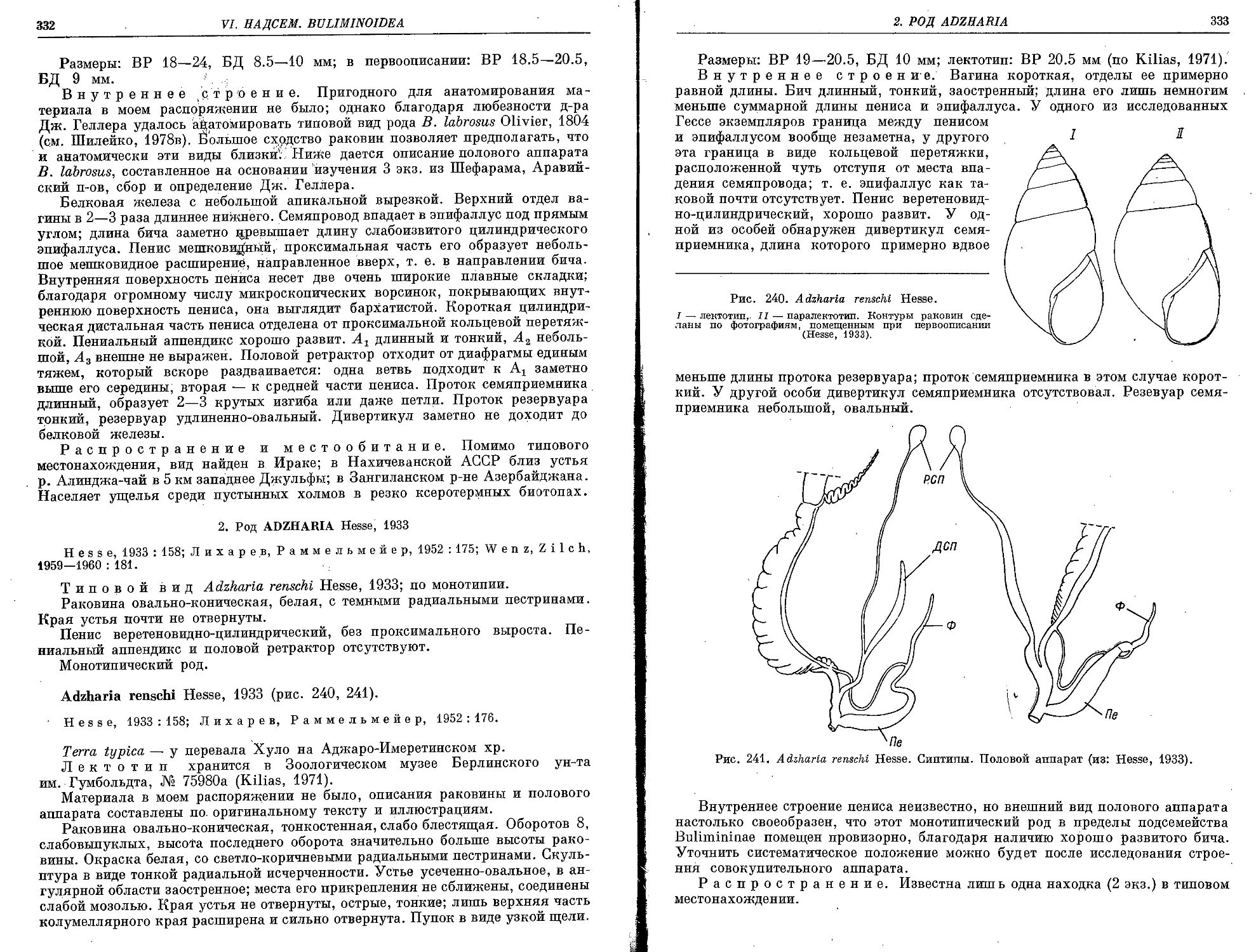
; A. renschi Hesse, 1933 , ..................................................... 332
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ Н
VI. Подсем. Retowskiinae Schileyko, 1978
1. Род Andronakia Lindholm, 1914
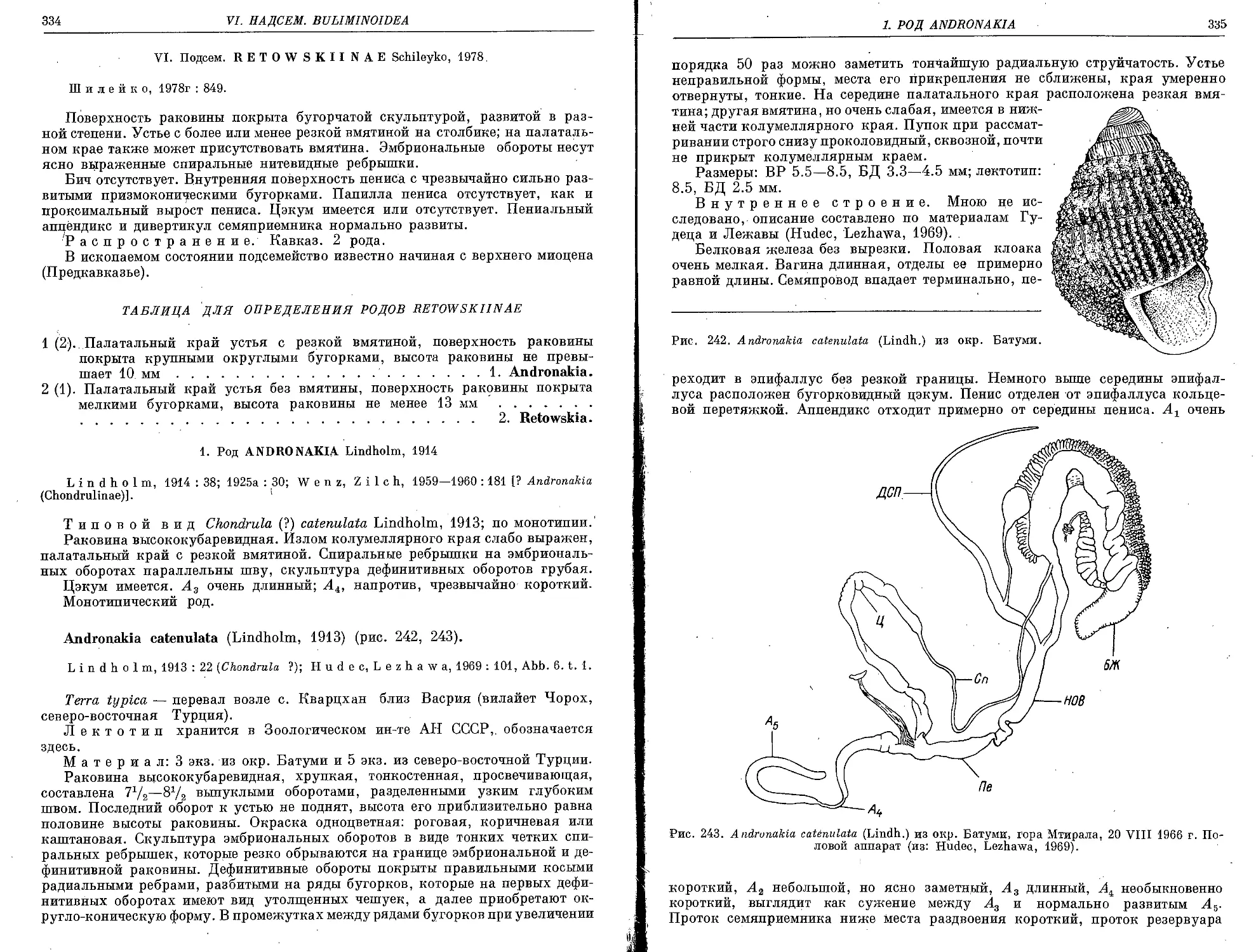
A. catenulata (Lindholm, 1913)................................................. 334
2. Род Retowskia О. Boettger,. 1881
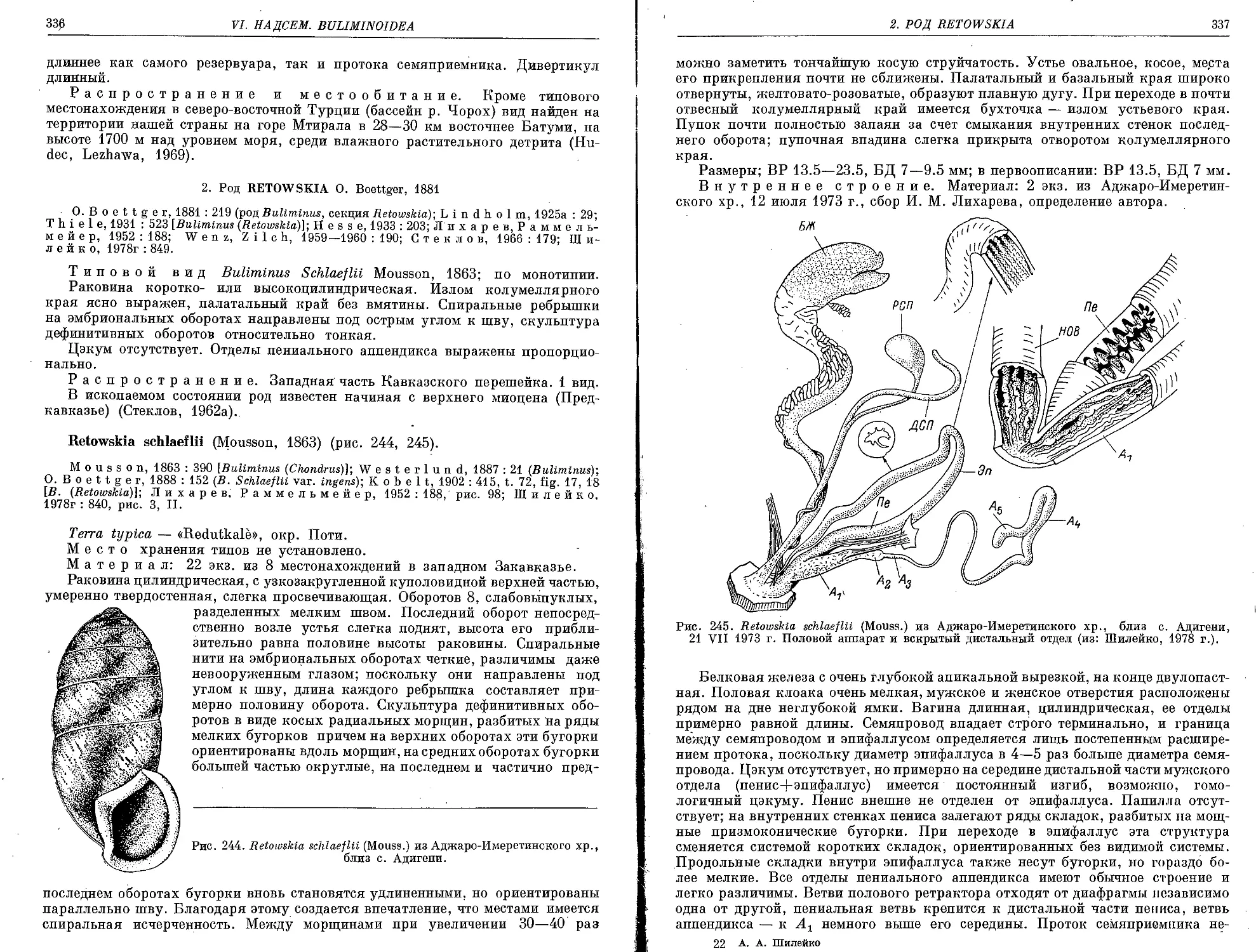
R. schlaeflii (Mousson, 1863)................................................... 336
VII. Подсем. E n i n a e Woodward, 1903
1. Род Ena Turton, 1831
E. montana (Draparnaud, 1801).................................................. 339
2. Род Peristoma Krynicki, 1833
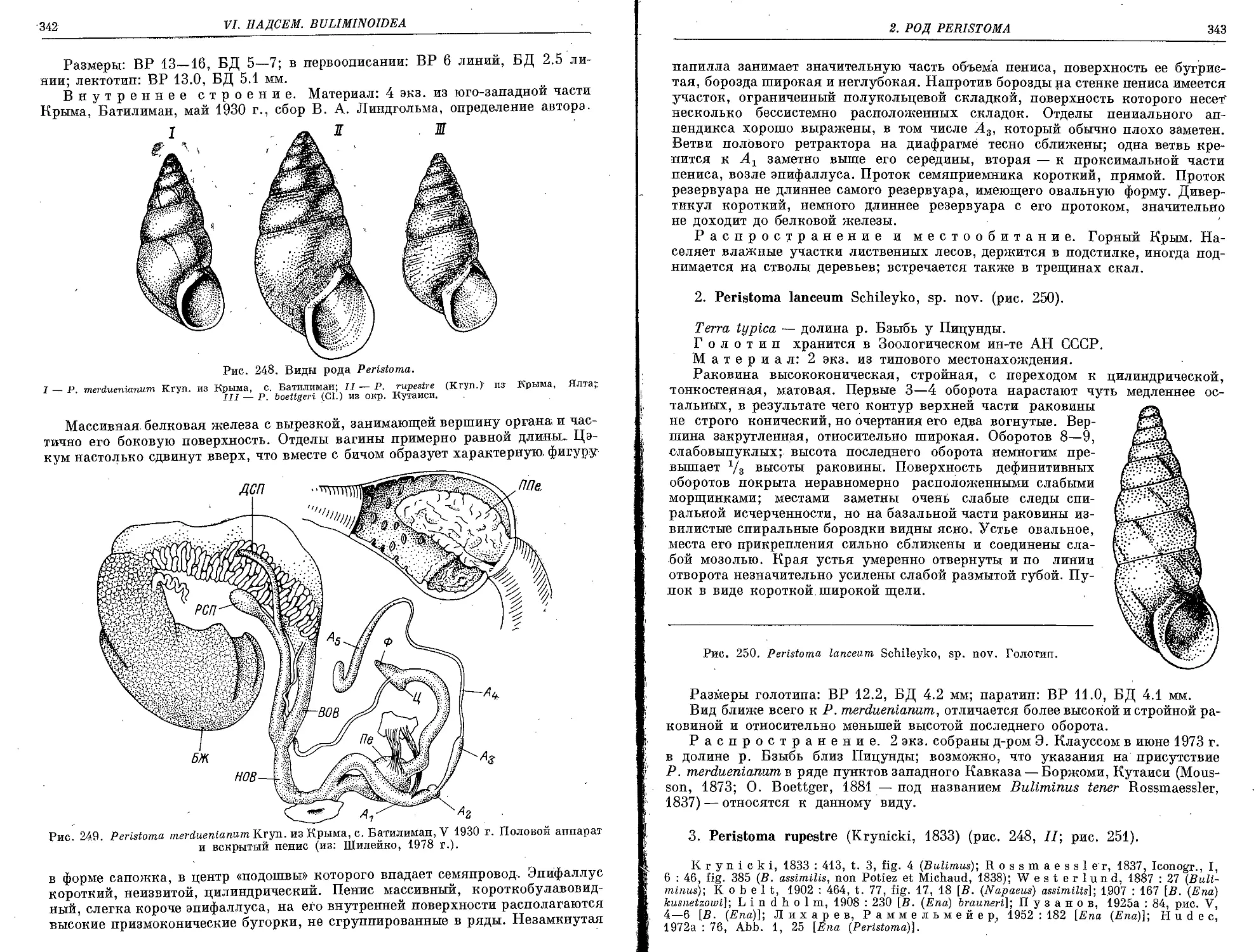
1. P. merdaeniamim Krynicki, 1833 ............................................. 341
2. P. lanceum, Schileyko, sp. nov.............................................. 343
3. P. rupestre (Krynicki, 1833)................................................ 343
4. P. boettgeri (Clessin, 1883)................................................ 345
3. Род Caucasicola Hesse, 1917
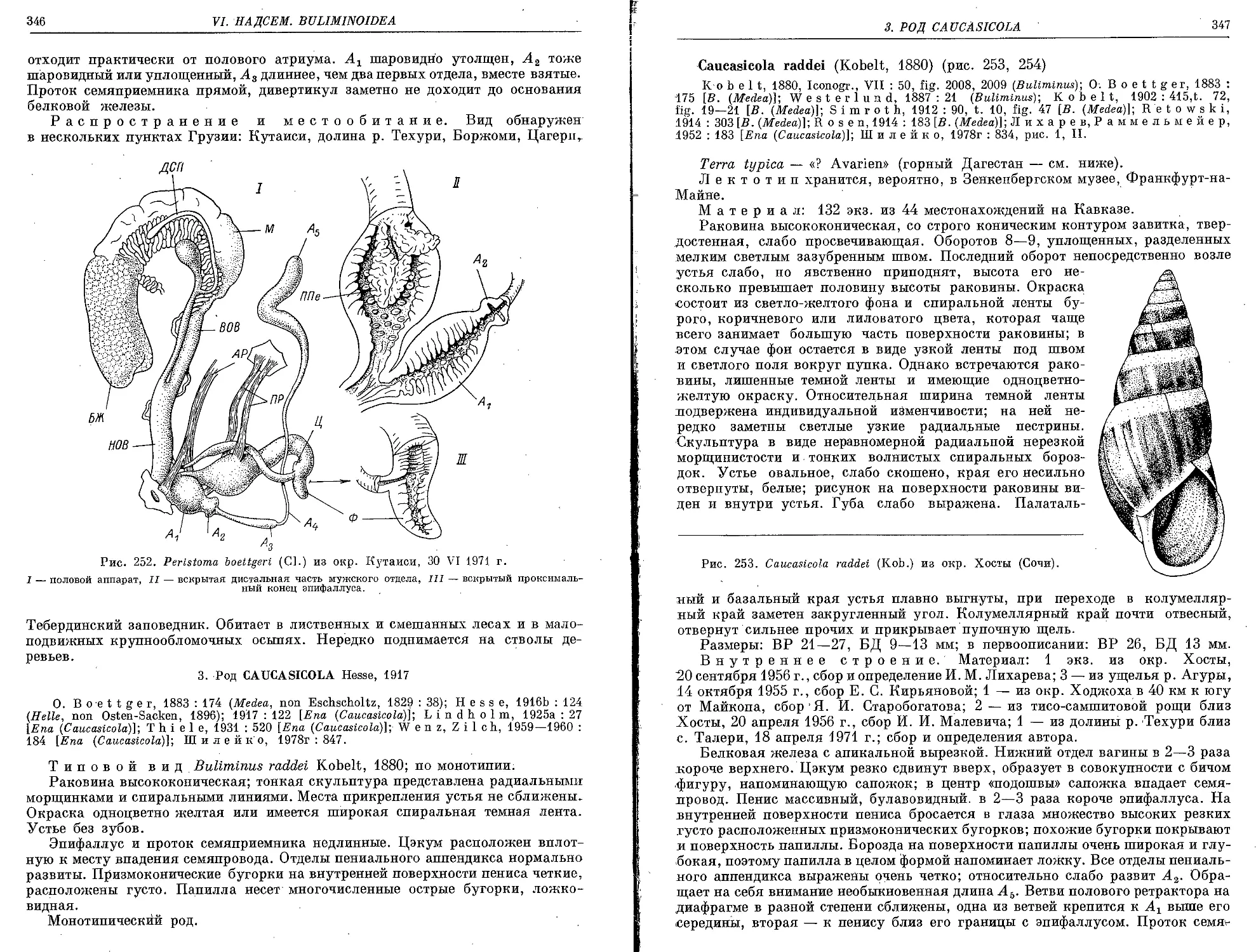
C. raddei (Kobelt, 1880)........................................................ 347
4. Род Zehrina Held, 1837
Z. deirita (Miiller, 1774)...................................................... 349
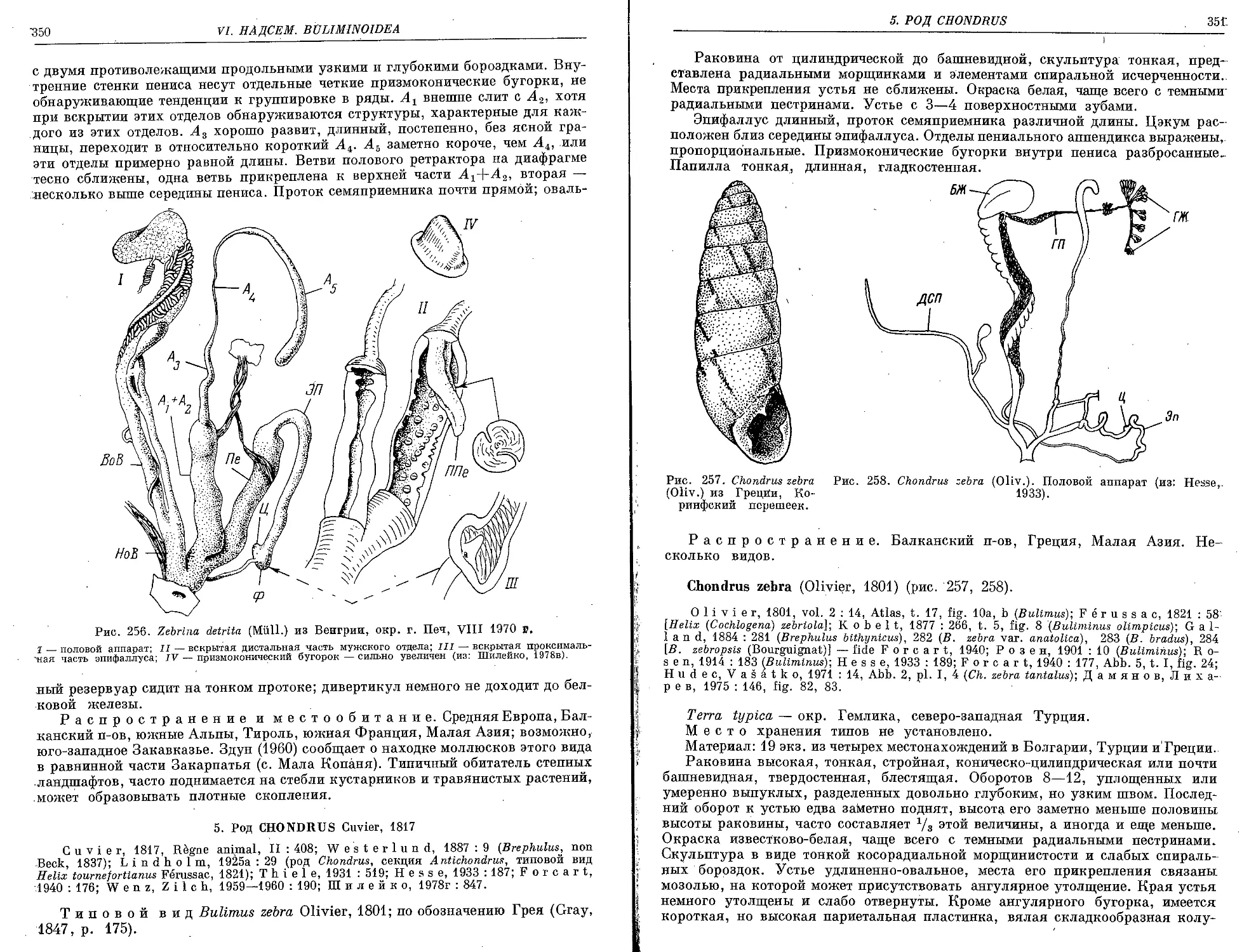
5. Род Chondrus Cuvier, 1817
Ch. zebra (Olivier, 1801)..................................................... 35J.
6. Род Brephulopsis Lindholm, 1925
1. B. cylindrica (Menke, 1828) ................................................ 353
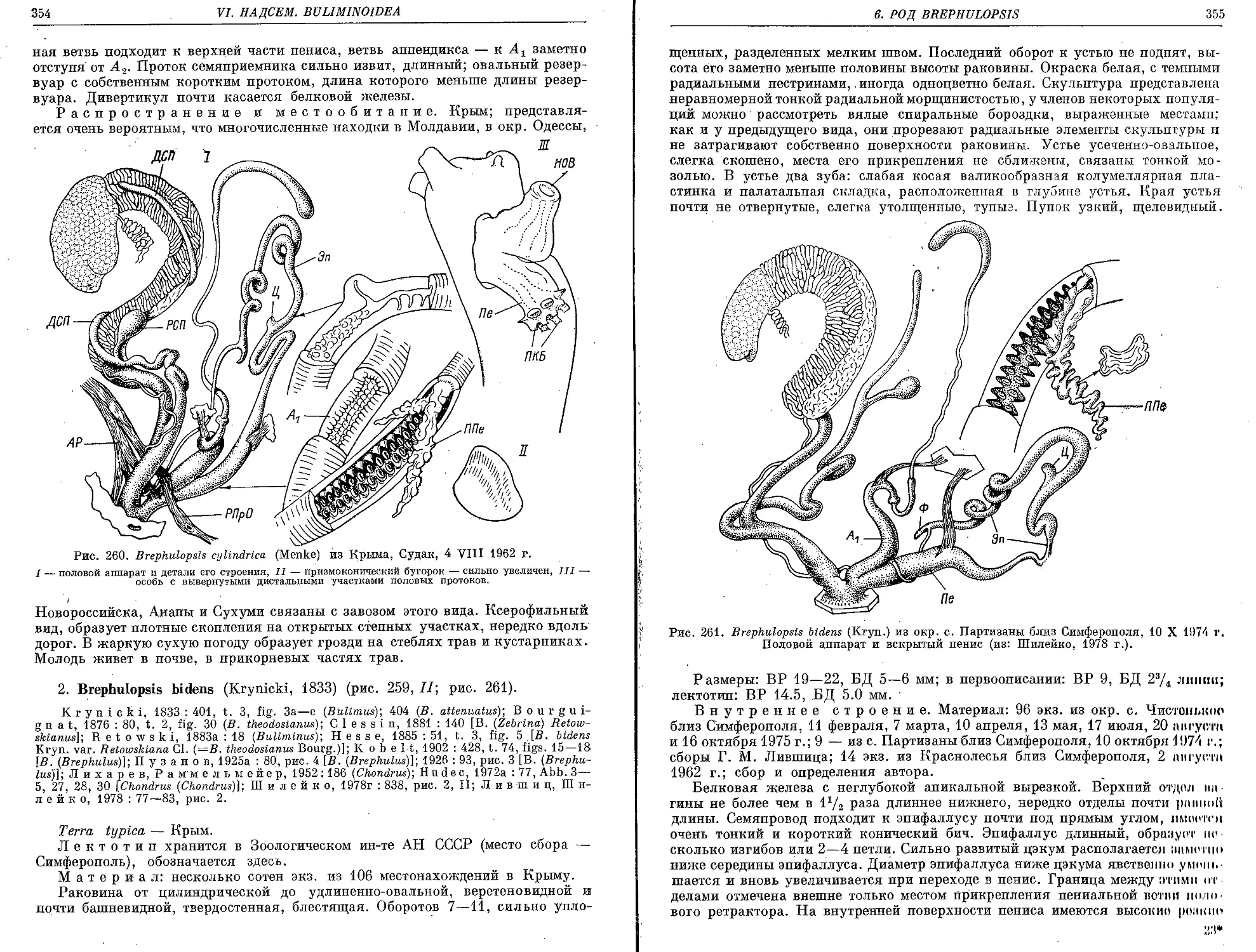
2. B. bidens (Krynicki, 1833) . •.....'. ...................................... 354
7. Род Napaeopsis Sturany et Wagner, 1914
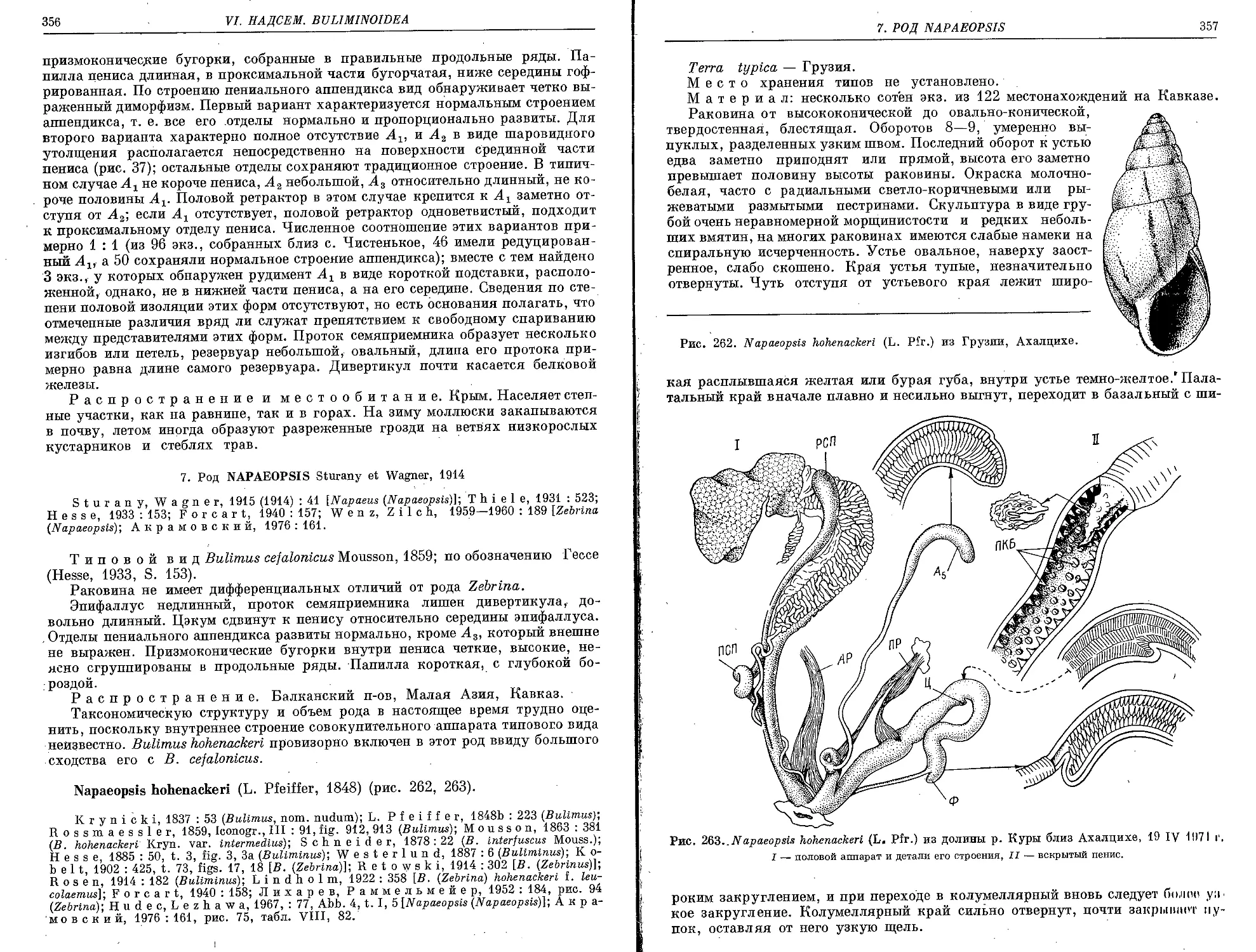
N. hohenackeri (L. Pfeiffer, 1848)............................................. 35g
8. Род Thoanteus Lindholm, 1925
Th. gibber (Krynicki, 1833).................1................................... 358
9. Род Ramusculus Lindholm, 1925
R. subulatus (Rossmaessler, 1837).............................................. 360
10. Род Chondrula Beck, 1'837
1. Подрод Chondrula s. str.
1. Ch. (Ch.) sunzhica Steklov, 1962 .......................................... 363
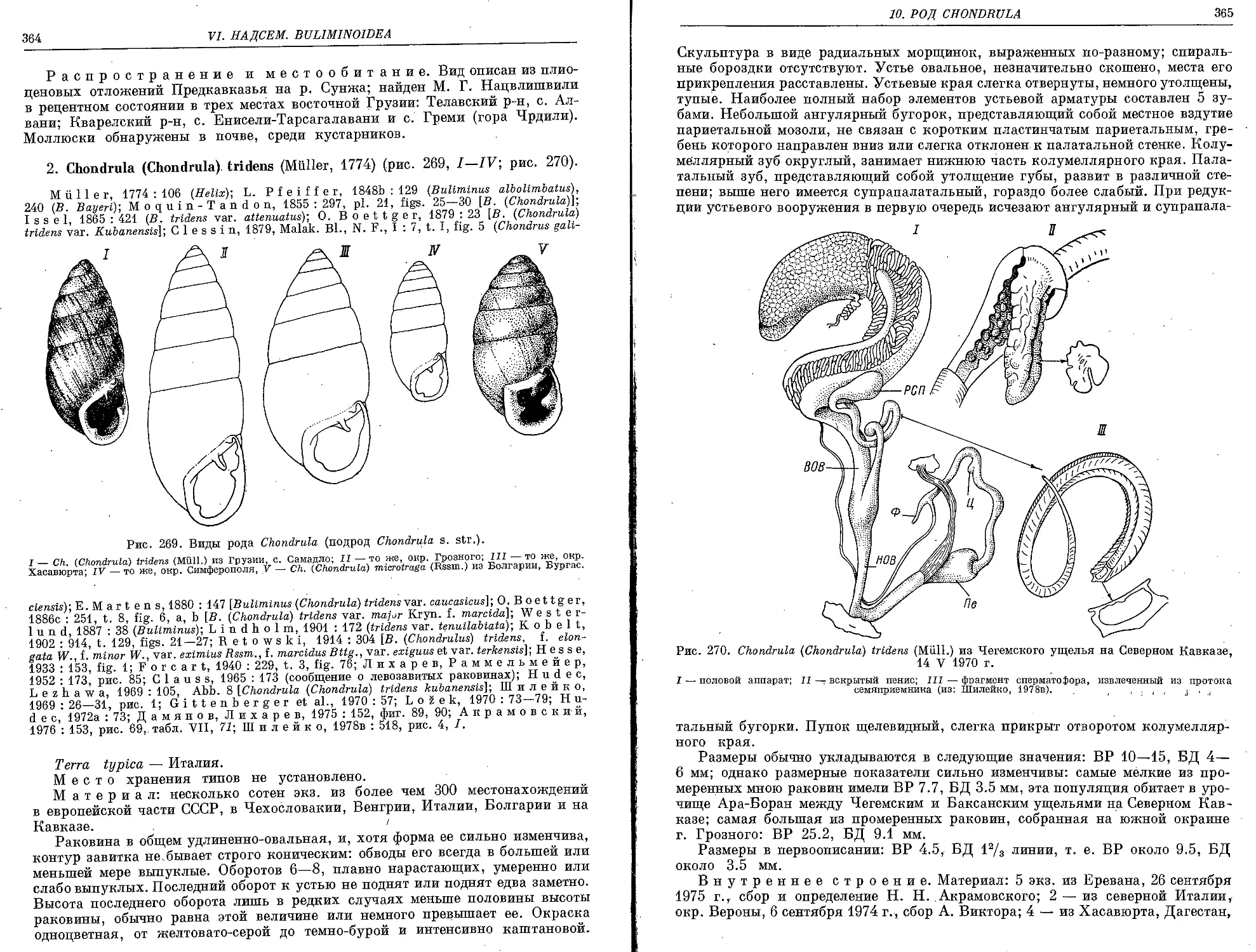
2. Ch. (Ch.) tridens (Muller, 1774) . . ....................................... 364
3. Ch. (Ch.) microtraga (Rossmaessler, 1839).................................. 366
2. Подрод Eubrephulus Wagner, 1927
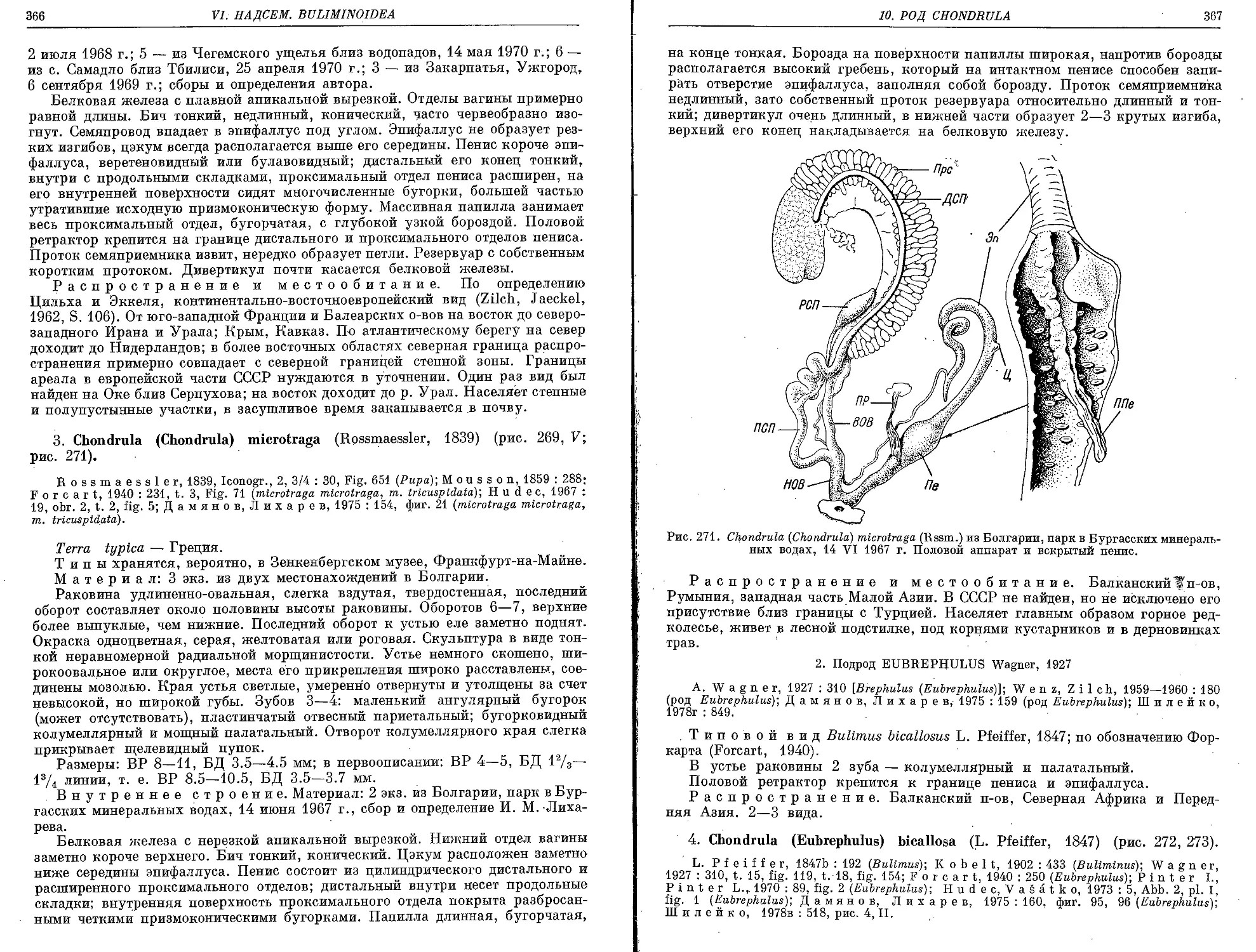
4. Ch. (E.) bicallosa (L. Pfeiffer, 1847) . . ................................ 367
3. Подрод M a s t u s Beck, 1837
5. Ch. (M.) caucasica (L. Pfeiffer, 1854).................................... -369
6. Ch. (M.) bielzt (Kimakowicz, 1890) ......... , . ............. 370
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
VIII. Подсем. Multidentulinae Schileyko, 1978
1. Род Multidentula Lindholm, 1925
М. lamellifera (Rossmaessler, 1859)...................................................... 372
2. Род Improvisa Schileyko, 1978 ""
/. pupoides (Krynicki, 1833).............................................................. 373
3. Род Senaridenta Schileyko, 1978
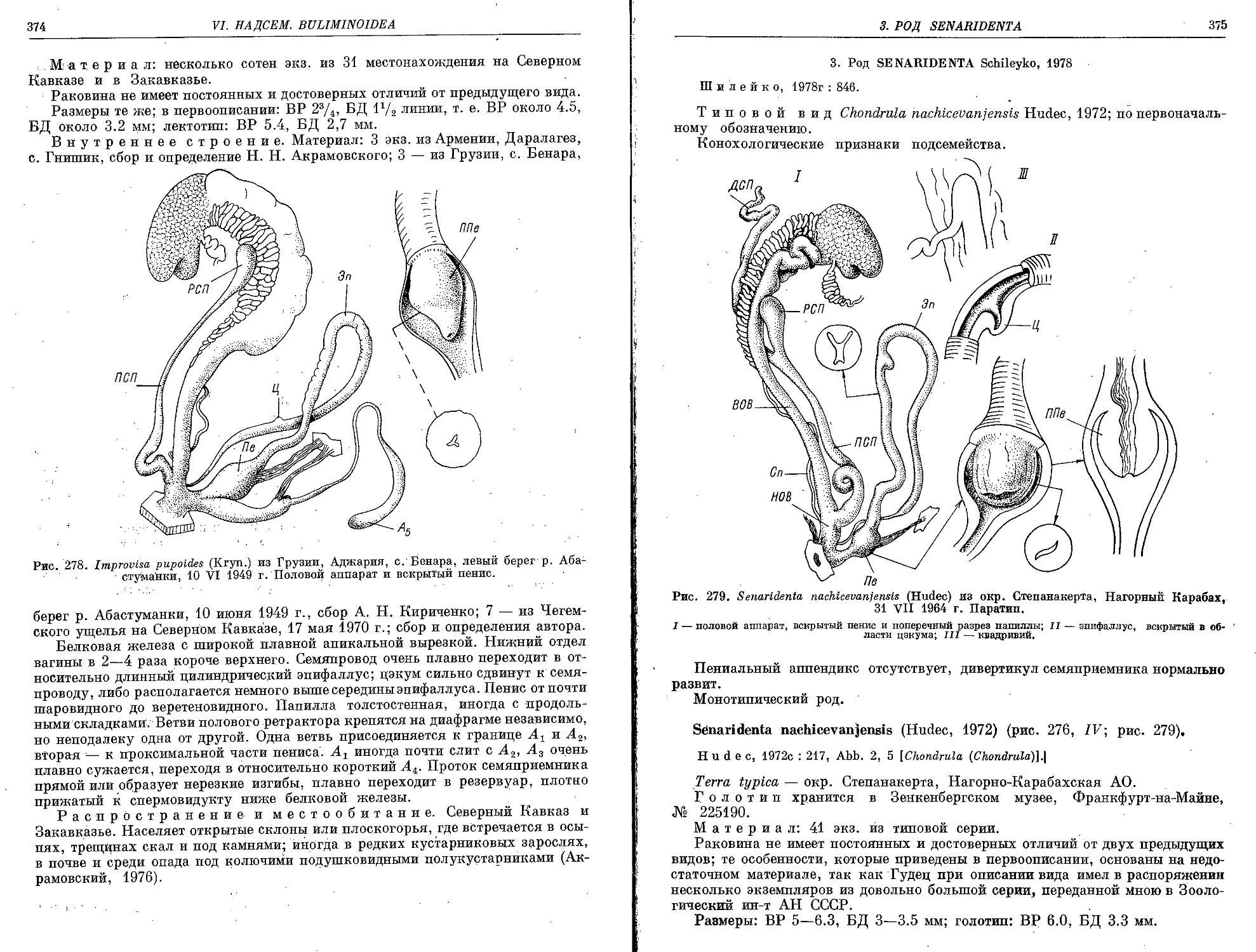
S. nachicevanfensis (Hudec, 1972)......................................................... 375
Incertae sedis
Stenogyra retteri Kobelt, 1904 ...................................>....................... 376
ОБЩАЯ ЧАСТЬ
ОБЩАЯ ХАРАКТЕРИСТИКА МОЛЛЮСКОВ ПОДОТРЯДА PUPILLINA
Животные с наружной раковиной, составленной 3—12 оборотами и никогда
не имеющей склонности к редукции. Раковина правозакрученная (декстраль-
ная, дексиотропная), значительно реже левозакрученная (синистральная, лео-
тропная). Раковина исходно и чаще всего высокая, лишь для некоторых се-
мейств характерно вторичное укорочение раковины и приобретение ею микро-
хеликоидного облика (Valloniidae, Strobilopsidae, Pyramidulidae); именно в этих
случаях число оборотов снижается до 3—4.
Последний оборот раковины в профиле чаще всего плавно и правильно за-
круглен, но у укороченных раковин может быть угловатым, у некоторых Аша-
strinae даже с резким килем. Последний оборот к устью может быть слегка или
резко опущенным (у микрохеликоидных раковин), в разной степени приподня-
тым (у высоких форм) либо прямым.
Окраска или одноцветная — от белой до темно-бурой, или светлая с тем-
ными радиальными (осевыми) полосами и пестринами. Спиральные цветовые
ленты встречаются редко (например, Caucasicola из Buliminidae, некоторые
Euxinolauria из Orculidae). Яркая окраска наблюдается только у немногих
Achatinellidae.
Эмбриональные обороты гладкие, полированные, или покрыты микроскопи-
ческой зернистостью, или, наконец, несут выраженную в разной степени спи-
ральную положительную скульптуру. На ранних дефинитивных оборотах
иногда имеются выросты периостракума наподобие треугольных шипов или
ресниц, которые иногда сохраняются и на последующих оборотах. Для дефи-
нитивной скульптуры характерно преобладание радиальных элементов — от
тонкой исчерченности или морщинистости до резкой грубой ребристости. Спи-
ральная скульптура встречается гораздо реже.
Устье обычно с выраженной в разной степени губой, нередко с зубами, число
которых может достигать 12 (если не считать мелких складочек на крае устья).
Первично имеются париетальная и колумеллярная пластинки, проходящие от
^первого до последнего оборота, причем верхние концы их лизируются по мере
роста раковины; однако у многих групп эти пластинки частично или полностью
редуцированы.
Пупок, т. е. отверстие, ведущее в полость столбика, у высоких раковин от
полностью запаянного до открытого, но узкого. У прижатых раковин пупок
нередко широкий, развернутый.
Тело моллюска состоит из трех основных отделов: головы, ноги и внутрен-
ностного мешка. На голове располагаются две пары щупалец, лишь у очень
мелких моллюсков в некоторых таксонах нижняя пара щупалец отсутствует
(Vertiginidae). Края ротового отверстия с боков окаймлены парой околоротовых
лопастей, под ротовым отверстием видна поперечная щель — отверстие педаль-
ной железы. Это отверстие особенно хорошо заметно у фиксированных особей
мелких моллюсков — представителей Valloniidae, Vertiginidae и Pupillidae.
Подошва ноги лишена продольных борозд, но по краю ее проходит кольцевая
перипедальная борозда. Хвостовая ямка отсутствует.
14
ОБЩАЯ ЧАСТЬ
Кожа тонкая, лишь у крупных ксеротермных Buliminidae достигает зна-
чительной толщины. Поверхность покровов покрыта многочисленными про-
дольными и поперечными морщинами.
В левом верхнем углу мантийного валика (у левозавитых форм — в правом
углу) расположено дыхательное отверстие — пневмостом, а также анальное
отверстие, открывающееся часто на вершине небольшого сосочка — анальной
папиллы. К анальному отверстию примыкает отверстие вторичного мочеточ-
ника, имеющего вид ресничной борозды. Мантийный валик с тремя лопастями —
верхней (анальной), правой и левой.
Почка ортуретрального типа, т. е. тело ее сильно вытянуто, занимает не
менее 2/3 длины мантийной полости. Щелевидное отверстие почки в большей
или меньшей степени смещено на боковую поверхность ее дистальной части.
От почечного отверстия отходит первичный мочеточник в виде мерцательной
борозды, примыкающей к телу почки. Дойдя до задней стенки легочной полости,
первичный мочеточник образует крутой изгиб и спускается вдоль задней кишки
до мантийного валика.
Челюсть пикногнатного типа, т. ,е. без срединного выступа и с серией попе-
речных утолщений, тенденции к редукции не имеет, хотя у мелких видов она
обычно тонка и почти прозрачна.
Осевая мускулатура составлена колумеллярным мускулом, распадающимся
на множество ретракторов. Самая внешняя ветвь колумеллярного мускула
сильно уплощена, расширена и переходит в диафрагму, которая формирует
дно легочной полости.
Центральная нервная система (ЦНС) слагается 11 ганглиями, каждый из
которых сохраняет морфологическую самостоятельность.
Гонада состоит из 2—7 гроздей фолликулов, в которых созревают как муж-
ские, так и женские гаметы. От гермафродитной железы отходит гермафродит-
ный проток, образующий в тканях белковой железы резкий изгиб. К белковой
железе тесно прилегает петля кишечника, в силу чего поверхность железы не-
сет борозду, степень развития которой зависит от функционального состояния
белковой железы. Простата в виде ленты простирается на большую часть длины
спермовидукта, но у мелких видов она редуцирована до небольшого числа
фолликулов или, редко, вообще не выражена. Дистальная часть женского от-
дела без придатков. Мужской отдел с придатками: в максимальном наборе это
бич, цэкум или придаток эпифаллуса и пениальный аппендикс, состоящий из
5 отделов. Нередко наблюдается полная или частичная редукция придатков
мужского отдела либо их видоизменение. Пенис представляет собой трубку, на
внутренних стенках которой располагаются различной формы складки; иногда
внутри пениса имеется папилла — в виде желобка или замкнутой трубки. Поло-
вой ретрактор в типичном случае двуветвистый, одна ветвь крепится к пенису,
другая — к пениальному аппендиксу. Семяприемник обычно с дивертикулом,
подверженным редукции, иногда даже в пределах одного вида.
Ареал подотряда почти всесветный, но многие семейства имеют хорошо ог-
раниченные, сравнительно узкие области распространения.
Представители Pupillina известны начиная с верхнего мела — палеоцена,
но уже тогда ряд семейств и даже отдельных родов был четко обособлен; из
перми и карбона известны Dendropupidae, которые также относятся к Pupil-
liria.
По приблизительной оценке в мировой фауне насчитывается не менее 2000
видов Pupillina, в составе фауны СССР — 159 современных видов и подвидов;
кроме того, в неогеновых ютложениях на территории нашей страны найдено
большое число представителей подотряда, частью полностью вымерших,
частью принадлежащих родам, представленным ныне за пределами террито-
рии СССР.
Подотряд Pupillina входит в состав отряда Geophila подкласса Pulmonata
класса Gastropoda. .
МОРФОЛОГИЯ И АНАТОМИЯ
15
МОРФОЛОГИЯ И АНАТОМИЯ ПРЕДСТАВИТЕЛЕЙ PUPILLINA
Внешний вид и раковина
Первое, что бросается в глаза при рассматривании расправленного мол-
люска — это расчленение его тела на два хорошо выраженных отдела: спи-
рально завитый внутренностный мешок, покрытый раковиной, и части, высту-
пающие из раковины (голова и нога, или цефалоподиум). На голове располо-
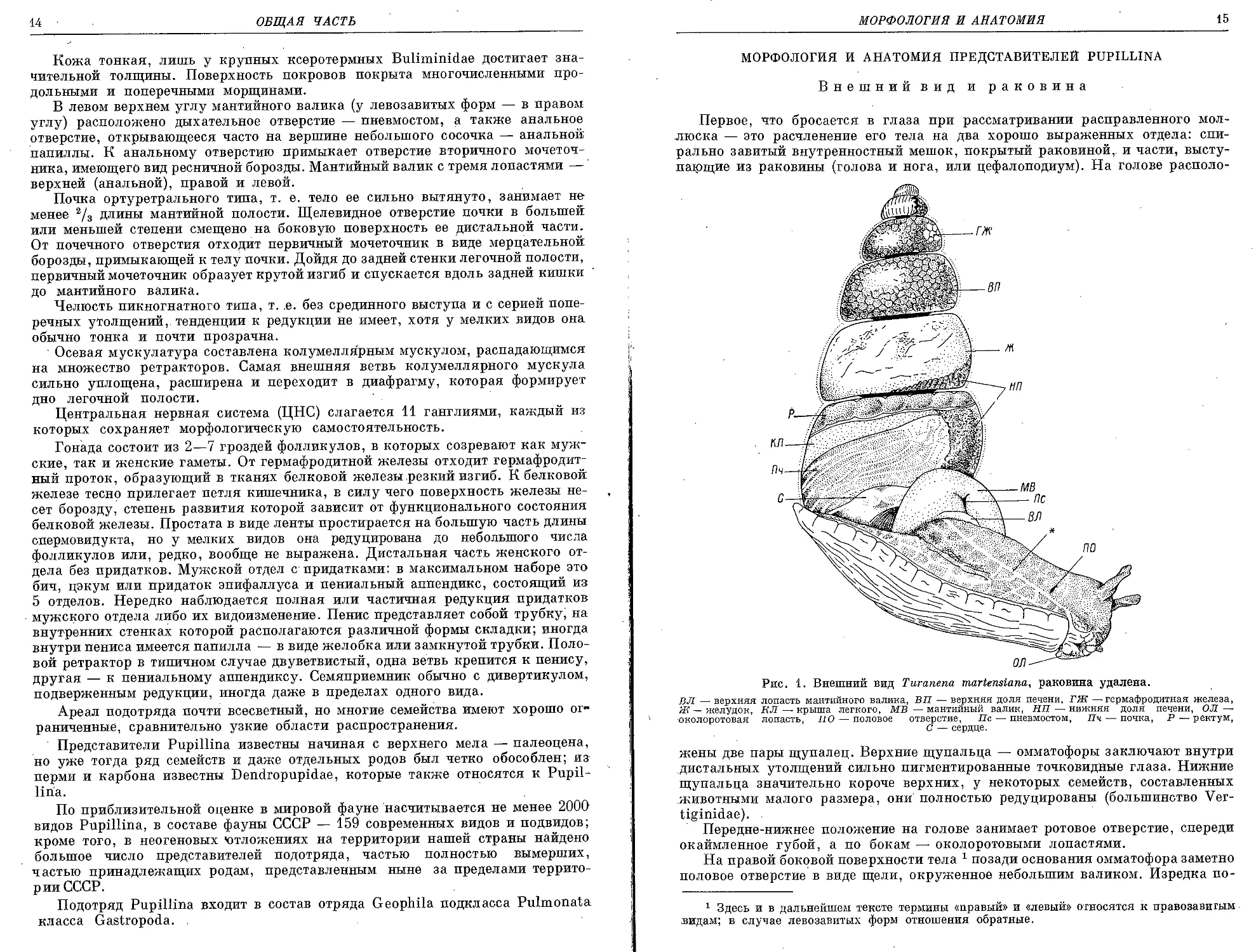
Рис. 1. Внешний вид Turanena martensiana, раковина удалена.
ВЛ — верхняя лопасть мантийного валика, ВП — верхняя доля печени, ГЖ —гермафродитная железа,
Ж—желудок, КЛ — крыша легкого, МВ—мантийный валик, НП—-нижняя доля печени, ОЛ —
•околоротовая лопасть, ПО — половое отверстие, Пс — пневмостом, Пч — почка, Р — ректум,
С — сердце.
жены две пары щупалец. Верхние щупальца — омматофоры заключают внутри
дистальных утолщений сильно пигментированные точковидные глаза. Нижние
щупальца значительно короче верхних, у некоторых семейств, составленных
животными малого размера, они полностью редуцированы (большинство Ver-
tiginidae).
Передне-нижнее положение на голове занимает ротовое отверстие, спереди
окаймленное губой, а по бокам — околоротовыми лопастями.
На правой боковой поверхности тела 1 позади основания омматофора заметно
половое отверстие в виде щели, окруженное небольшим валиком. Изредка по-
1 Здесь и в дальнейшем тексте термины «правый» и «левый» относятся к правозавигым
видам; в случае левозавитых форм отношения обратные.
16
ОБЩАЯ ЧАСТЬ
ловое отверстие заметно отодвинуто от основания омматофора и располагается
ближе к мантийному валику (Cochlicopidae).
Цефалоподиум покрыт сглаженными бугорками, разделенными бороздками
(рис. 1); эти бороздки служат для стекания слизи и увлажнения поверхности
тела. Самая постоянная борозда соединяет место контакта анальной лопасти
мантийного валика с покровами спинной стороны ноги с передне-нижней частью
головы: выходя из-под анальной лопасти, она проходит снизу от полового от-
верстия, задевая его своей верхней кромкой, и уходит под околоротовую лопасть
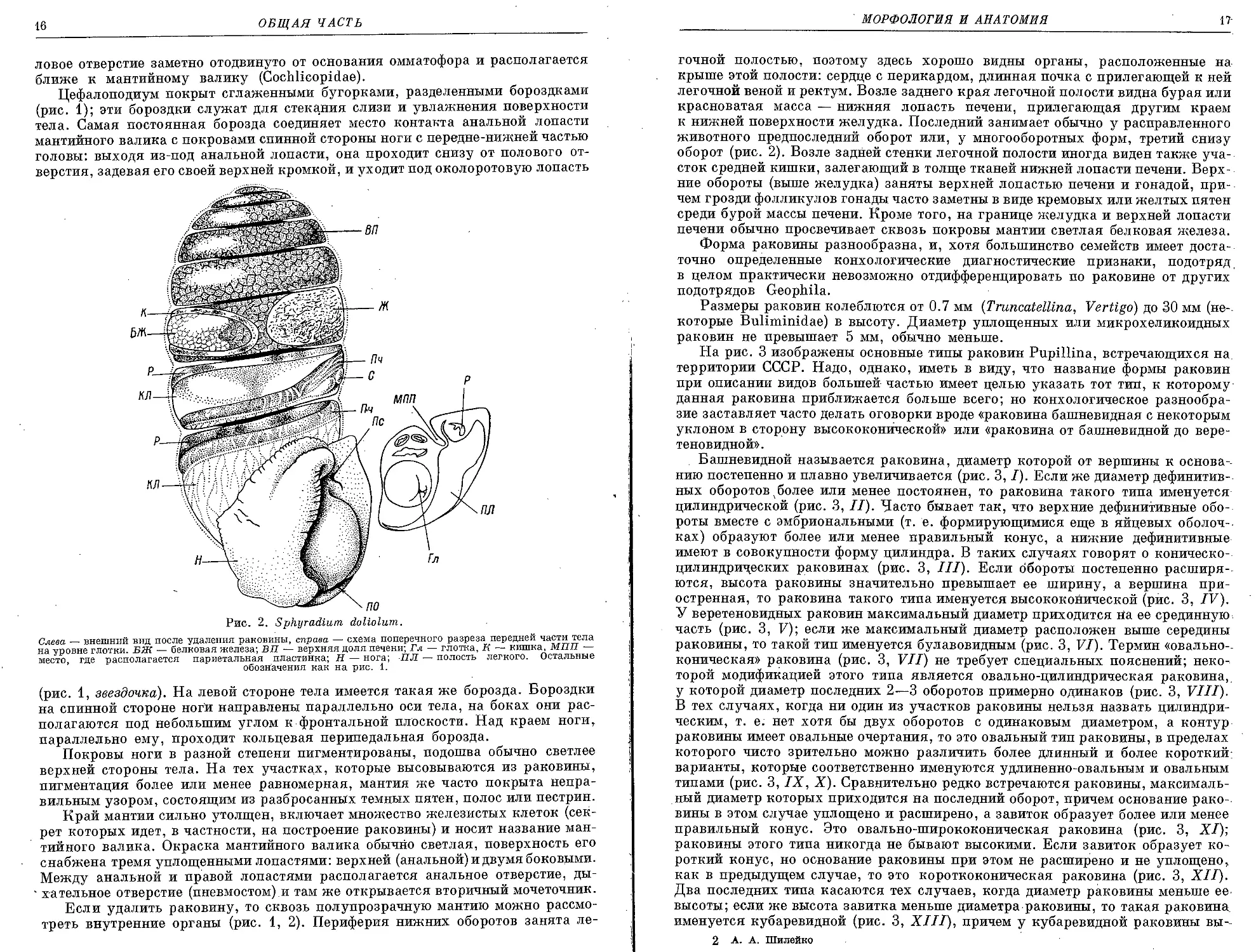
Рис. 2. Sphyradium doliolum.
Слева — внешний вид после удаления раковины, справа — схема поперечного разреза передней части тела
на уровне глотки. БЖ — белковая железа; ВП — верхняя доля печени; Гл — глотка, К — кишка, МПП —
место, где располагается париетальная пластинка; Н — нога; ПЛ — полость легкого. Остальные
обозначения как на рис. 1.
(рис. 1, звездочка). На левой стороне тела имеется такая же борозда. Бороздки
на спинной стороне ноги направлены параллельно оси тела, на боках они рас-
полагаются под небольшим углом к фронтальной плоскости. Над краем ноги,
параллельно ему, проходит кольцевая перипедальная борозда.
Покровы ноги в разной степени пигментированы, подошва обычно светлее
верхней стороны тела. На тех участках, которые высовываются из раковины,
пигментация более или менее равномерная, мантия же часто покрыта непра-
вильным узором, состоящим из разбросанных темных пятен, полос или пестрин.
Край мантии сильно утолщен, включает множество железистых клеток (сек-
рет которых идет, в частности, на построение раковины) и носит название ман-
тийного валика. Окраска мантийного валика обычно светлая, поверхность его
снабжена тремя уплощенными лопастями: верхней (анальной) и двумя боковыми.
Между анальной и правой лопастями располагается анальное отверстие, ды-
хательное отверстие (пневмостом) и там же открывается вторичный мочеточник.
Если удалить раковину, то сквозь полупрозрачную мантию можно рассмо-
треть внутренние органы (рис. 1, 2). Периферия нижних оборотов занята ле-
МОРФОЛОГИЯ И АНАТОМИЯ
17-
гочной полостью, поэтому здесь хорошо видны органы, расположенные на
крыше этой полости: сердце с перикардом, длинная почка с прилегающей к ней
легочной веной и ректум. Возле заднего края легочной полости видна бурая или
красноватая масса — нижняя лопасть печени, прилегающая другим краем
к нижней поверхности желудка. Последний занимает обычно у расправленного
животного предпоследний оборот или, у многооборотных форм, третий снизу
оборот (рис. 2). Возле задней стенки легочной полости иногда виден также уча-
сток средней кишки, залегающий в толще тканей нижней лопасти печени. Верх-
ние обороты (выше желудка) заняты верхней лопастью печени и гонадой, при-
чем грозди фолликулов гонады часто заметны в виде кремовых или желтых пятен
среди бурой массы печени. Кроме того, на границе желудка и верхней лопасти
печени обычно просвечивает сквозь покровы мантии светлая белковая железа.
Форма раковины разнообразна, и, хотя большинство семейств имеет доста-
точно определенные конхологические диагностические признаки, подотряд
в целом практически невозможно отдифференцировать по раковине от других
подотрядов Geophila.
Размеры раковин колеблются от 0.7 мм (Truncatellina, Vertigo) до 30 мм (не-,
которые Buliminidae) в высоту. Диаметр уплощенных или микрохеликоидных
раковин не превышает 5 мм, обычно меньше.
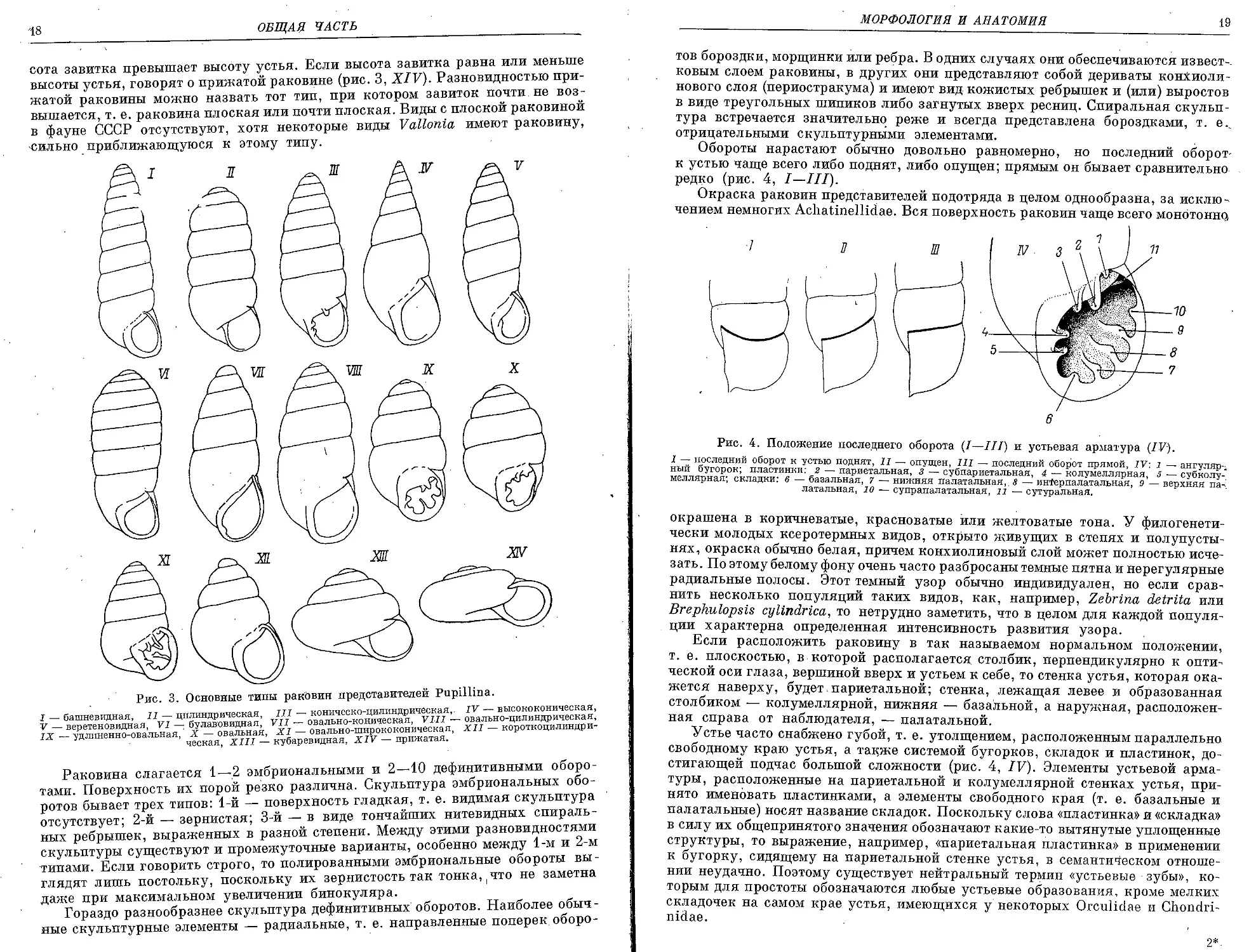
На рис. 3 изображены основные типы раковин Pupillina, встречающихся на
территории СССР. Надо, однако, иметь в виду, что название формы раковин
при описании видов большей частью имеет целью указать тот тип, к которому
данная раковина приближается больше всего; но конхологическое разнообра-
зие заставляет часто делать оговорки вроде «раковина башневидная с некоторым
уклоном в сторону высококонической» или «раковина от башневидной до вере-
теновидной».
Башневидной называется раковина, диаметр которой от вершины к основа--
нию постепенно и плавно увеличивается (рис. 3,I). Если же диаметр дефинитив-,
ных оборотов Долее или менее постоянен, то раковина такого типа именуется
цилиндрической (рис. 3, II). Часто бывает так, что верхние дефинитивные обо-
роты вместе с эмбриональными (т. е. формирующимися еще в яйцевых оболоч-
ках) образуют более или менее правильный конус, а нижние дефинитивные
имеют в совокупности форму цилиндра. В таких случаях говорят о коническо-
цилиндрицеских раковинах (рис. 3, III). Если обороты постепенно расширя-
ются, высота раковины значительно превышает ее ширину, а вершина при-
остренная, то раковина такого типа именуется высококонической (рис. 3, IV).
У веретеновидных раковин максимальный диаметр приходится На ее срединную ,
часть (рис. 3, V); если же максимальный диаметр расположен выше середины
раковины, то такой тип именуется булавовидным (рис. 3, VI). Термин «овально--
коническая» раковина (рис. 3, VII) не требует специальных пояснений; неко-
торой модификацией этого типа является овально-цилиндрическая раковина,,
у которой диаметр последних 2—3 оборотов примерно одинаков (рис. 3, VIII).
В тех случаях, когда ни один из участков раковины нельзя назвать цилиндри-
ческим, т. е. нет хотя бы двух оборотов с одинаковым диаметром, а контур
раковины имеет овальные очертания, то это овальный тип раковины, в пределах
которого чисто зрительно можно различить более длинный и более короткий:
варианты, которые соответственно именуются удлиненно-овальным и овальным
типами (рис. 3, IX, X). Сравнительно редко встречаются раковины, максималь-
ный диаметр которых приходится на последний оборот, причем основание рако-
вины в этом случае уплощено и расширено, а завиток образует более или менее
правильный конус. Это овально-ширококоническая раковина (рис. 3, XI);
раковины этого типа никогда не бывают высокими. Если завиток образует ко-
роткий конус, но основание раковины при этом не расширено и не уплощено,
как в предыдущем случае, то это короткоконическая раковина (рис. 3, XII).
Два последних типа касаются тех случаев, когда диаметр раковины меньше ее-
высоты; если же высота завитка меньше диаметра раковины, то такая раковина,
именуется кубаревидной (рис. 3, XIII), причем у кубаревидной раковины вы-
2 А. А. Шилейко
18
ОБЩАЯ ЧАСТЬ
сота завитка превышает высоту устья. Если высота завитка равна или меньше
высоты устья, говорят о прижатой раковине (рис. 3, XIV). Разновидностью при-
жатой раковины можно назвать тот тип, при котором завиток почти не воз-
вышается, т. е. раковина плоская или почти плоская. Виды с плоской раковиной
в фауне СССР отсутствуют, хотя некоторые виды Vallonia имеют раковину,
сильно приближающуюся к этому типу.
Рис. 3. Основные типы раковин представителей Pupillina.
I— башневидная, II— цилиндрическая, III— коническо-цилиндрическая,. IV— высококоническая,
V — веретеновидная, VI — булавовидная, VII — овально-коническая, VIII — овально-цилиндрическая,
IX — удлиненно-овальная,' X — овальная, XI — овально-ширококоническая, XII — короткоцилиндри-
ческая, XIII — кубаревидная, XIV — прижатая.
Раковина слагается 1—2 эмбриональными и 2—10 дефинитивными оборо-
тами. Поверхность их порой резко различна. Скульптура эмбриональных обо-
ротов бывает трех типов: 1-й — поверхность гладкая, т. е. видимая скульптура
отсутствует; 2-й — зернистая; 3-й — в виде тончайших нитевидных спираль-
ных ребрышек, выраженных в разной степени. Между этими разновидностями
скульптуры существуют и промежуточные варианты, особенно между 1-ми 2-м
типами. Если говорить строго, то полированными эмбриональные обороты вы-
глядят лишь постольку, поскольку их зернистость так тонка, что не заметна
даже при максимальном увеличении бинокуляра.
Гораздо разнообразнее скульптура дефинитивных оборотов. Наиболее обыч-
ные скульптурные элементы — радиальные, т. е. направленные поперек оборо-
МОРФОЛОГИЯ И АНАТОМИЯ
19
тов бороздки, морщинки или ребра. В одних случаях они обеспечиваются извест-.
ковым слоем раковины, в других они представляют собой дериваты конхиоли-
нового слоя (периостракума) и имеют вид кожистых ребрышек и (или) выростов
в виде треугольных шипиков либо загнутых вверх ресниц. Спиральная скульп-
тура встречается значительно реже и всегда представлена бороздками, т. е._
отрицательными скульптурными элементами.
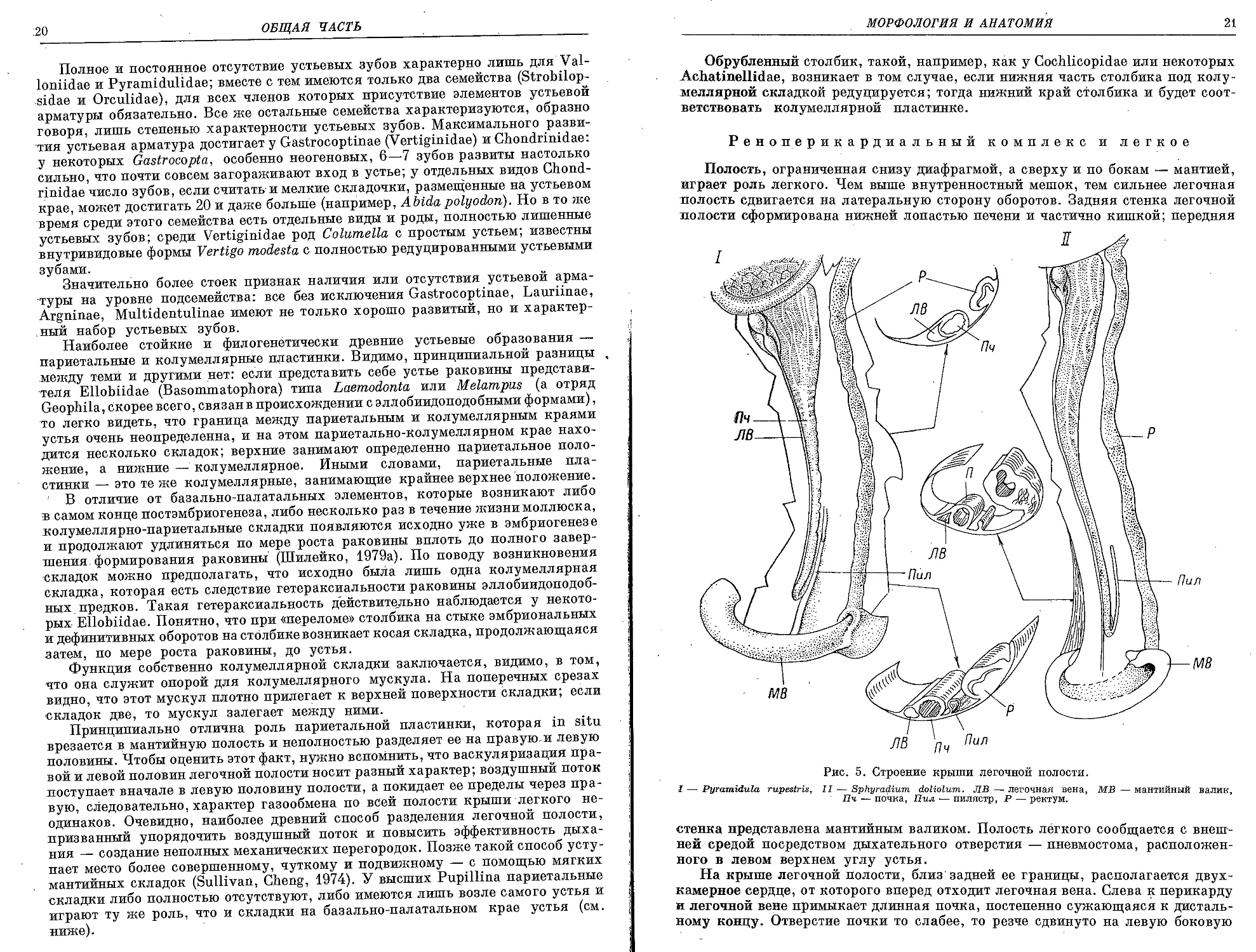
Обороты нарастают обычно довольно равномерно, но последний оборот'
к устью чаще всего либо поднят, либо опущен; прямым он бывает сравнительно
редко (рис. 4, I—III).
Окраска раковин представителей подотряда в целом однообразна, за исклю-
чением немногих Acliatinellidae. Вся поверхность раковин чаще всего монотонно
Рис. 4. Положение последнего оборота (/—777) и устьевая арматура (7.7).
1 — последний оборот к устью поднят, 11 — опущен, 111 — последний оборот прямой, IV: 1 — ангуляр-
ный бугорок; пластинки: 2 — париетальная, з — субпариетальная, 4 — колумеллярная, 5 — субколу-
меллярная; складки: в — базальная, 7 — нижняя палатальная,, л — интерпалатальная, 9 — верхняя па-
латальная, ю — супрапалатальная, н — сутуральная.
окрашена в коричневатые, красноватые или желтоватые тона. У филогенети-
чески молодых ксеротермных видов, открыто живущих в степях и полупусты-
нях, окраска обычно белая, причем конхиолиновый слой может полностью исче-
зать. По этому белому фону очень часто разбросаны темные пятна и нерегулярные
радиальные полосы. Этот темный узор обычно индивидуален, но если срав-
нить несколько популяций таких видов, как, например, Zebrina detrita или
Brephulopsis cylindrica, то нетрудно заметить, что в целом для каждой популя-
ции характерна определенная интенсивность развития узора.
Если расположить раковину в так называемом нормальном положении,
т. е. плоскостью, в которой располагается столбик, перпендикулярно к опти-
ческой оси глаза, вершиной вверх и устьем к себе, то стенка устья, которая ока-
жется наверху, будет.париетальной; стенка, лежащая левее и образованная
столбиком — колумеллярной, нижняя — базальной, а наружная, расположен-
ная справа от наблюдателя, — палатальной.
Устье часто снабжено губой, т. е. утолщением, расположенным параллельно
свободному краю устья, а также системой бугорков, складок и пластинок, до-
стигающей подчас большой сложности (рис. 4, IV). Элементы устьевой арма-
туры, расположенные на париетальной и колумеллярной стенках устья, при-
нято именовать пластинками, а элементы свободного края (т. е. базальные и
палатальные) носят название складок. Поскольку слова «пластинка» и «складка»
в силу их общепринятого значения обозначают какие-то вытянутые уплощенные
структуры, то выражение, например, «париетальная пластинка» в применении
к бугорку, сидящему на париетальной стенке устья, в семантическом отноше-
нии неудачно. Поэтому существует нейтральный термин «устьевые зубы», ко-
торым для простоты обозначаются любые устьевые образования, кроме мелких
складочек на самом крае устья, имеющихся у некоторых Orculidae и Chondri-
nidae.
2*
:20
ОБЩАЯ ЧАСТЬ
Полное и постоянное отсутствие устьевых зубов характерно лишь для Val-
loniidae и Pyramidulidae; вместе с тем имеются только два семейства (Strobilop-
sidae и Orculidae), для всех членов которых присутствие элементов устьевой
арматуры обязательно. Все же остальные семейства характеризуются, образно
говоря, лишь степенью характерности устьевых зубов. Максимального разви-
тия устьевая арматура достигает у Gastrocoptinae (Vertiginidae) и Chondrinidae:
у некоторых Gastrocopta, особенно неогеновых, 6—7 зубов развиты настолько
сильно, что почти совсем загораживают вход в устье; у отдельных видов Chond-
rinidae число зубов, если считать и мелкие складочки, размещенные на устьевом
крае, может достигать 20 и даже больше (например, Abida polyodonj. Но в то же
время среди этого семейства есть отдельные виды и роды, полностью лишенные
устьевых зубов; среди Vertiginidae род Columella с простым устьем; известны
внутривидовые формы Vertigo modesta с полностью редуцированными устьевыми
зубами.
Значительно более стоек признак наличия или отсутствия устьевой арма-
туры на уровне подсемейства: все без исключения Gastrocoptinae, Lauriinae,
Argninae, Multidentulinae имеют не только хорошо развитый, но и характер-
ный набор устьевых зубов.
Наиболее стойкие и филогенетически древние устьевые образования —
париетальные и колумеллярные пластинки. Видимо, принципиальной разницы ,
между теми и другими нет: если представить себе устье раковины представи-
теля Ellobiidae (Basommatophora) типа Laemodonta или М elam pus (а отряд
Geophila, скорее всего, связан в происхождении с эллобиидоподобными формами),
то легко видеть, что граница между париетальным и колумеллярным краями
устья очень неопределенна, и на этом париетально-колумеллярном крае нахо-
дится несколько складок; верхние занимают определенно париетальное поло-
жение, а нижние — колумеллярное. Иными словами, париетальные пла-
стинки— это те же колумеллярные, занимающие крайнее верхнее положение.
В отличие от базально-палатальных элементов, которые возникают либо
в самом конце постэмбриогенеза, либо несколько раз в течение жизни моллюск а,
колумеллярно-париетальные складки появляются исходно уже в эмбриогенезе
и продолжают удлиняться по мере роста раковины вплоть до полного завер-
шения формирования раковины (Шилейко, 1979а). По поводу возникновения
складок можно предполагать, что исходно была лишь одна колумеллярная
складка, которая есть следствие гетераксиальности раковины эллобиидоподоб-
ных предков. Такая гетераксиальность действительно наблюдается у некото-
рых Ellobiidae. Понятно, что при «переломе» столбика на стыке эмбриональных
и дефинитивных оборотов на столбике возникает косая складка, продолжающаяся
затем, по мере роста раковины, до устья.
Функция собственно колумеллярной складки заключается, видимо, в том,
что она служит опорой для колумеллярного мускула. На поперечных срезах
видно, что этот мускул плотно прилегает к верхней поверхности складки; если
складок две, то мускул залегает между ними.
Принципиально отлична роль париетальной пластинки, которая in situ
врезается в мантийную полость и неполностью разделяет ее на правую.и левую
половины. Чтобы оценить этот факт, нужно вспомнить, что васкуляризация пра-
вой и левой половин легочной полости носит разный характер; воздушный поток
поступает вначале в левую половину полости, а покидает ее пределы через пра-
вую, следовательно, характер газообмена по всей полости крыши легкого не-
одинаков. Очевидно, наиболее древний способ разделения легочной полости,
призванный упорядочить воздушный поток и повысить эффективность дыха-
ния — создание неполных механических перегородок. Позже такой способ усту-
пает место более совершенному, чуткому и подвижному — с помощью мягких
мантийных складок (Sullivan, Cheng, 1974). У высших Pupillina париетальные
складки либо полностью отсутствуют, либо имеются лишь возле самого устья и
играют ту же роль, что и складки на базально-палатальном крае устья (см.
ниже).
МОРФОЛОГИЯ И АНАТОМИЯ
21
Обрубленный столбик, такой, например, как у Gochlicopidae или некоторых
Achatinellidae, возникает в том случае, если нижняя часть столбика под колу-
меллярной складкой редуцируется; тогда нижний край столбика и будет соот-
ветствовать колумеллярной пластинке.
Рено п е рикардиальный комплекс и легкое
Полость, ограниченная снизу диафрагмой, а сверху и по бокам — мантией,
играет роль легкого. Чем выше внутренностный мешок, тем сильнее легочная
полость сдвигается на латеральную сторону оборотов. Задняя стенка легочной
полости сформирована нижней лопастью печени и частично кишкой; передняя
Рис. 5. Строение крыши легочной полости.
I— Pyramidula rupestris, II — Sphyradium doliolum. JIB—легочная вена, MB—мантийный валик,
Пч — почка, Пил — пилястр, Р — ректум.
стенка представлена мантийным валиком. Полость лёгкого сообщается с внеш-
ней средой посредством дыхательного отверстия — пневмостома, расположен-
ного в левом верхнем углу устья.
На крыше легочной полости, близ задней ее границы, располагается двух-
камерное сердце, от которого вперед отходит легочная вена. Слева к перикарду
и легочной вене примыкает длинная почка, постепенно сужающаяся к дисталь-
ному концу. Отверстие почки то слабее, то резче сдвинуто на левую боковую
22
ОБЩАЯ ЧАСТЬ
поверхность органа, но строго терминально не открывается никогда.Отверстие
почки открывается в первичный мочеточник — ресничную борозду, ограничен-
ную с одной стороны телом почки, а с другой — складкой мантии (пилястром).
Эту ресничную борозду можно обнаружить даже при прямом вскрытии, не при-
бегая к гистологическим методам. Дойдя до заднего конца легочной полости,
первичный мочеточник образует очень, крутой изгиб и направляется вперед,
тесно примыкая к задней кишке. Отдел выводных путей почки от места изгиба
до выходного отверстия именуется вторичным мочеточником (рис. 5).
ПЗП
кмк
п
Пр
РО
АВК
Рис. 6.
Жк
А
Реноперикардиальный комплекс Chondnna
clienta caucasica.
Внизу — очень сильно увеличенный срез через проток рено-
| перикардиального отверстия. А — аорта, АВИ — атриовентри-
' кулярный клапан, ВЗП — выводящая зона почки, Жк — же-
лудочек, ИМИ — конкреции мочевой кислоты, ИВ — легочная
вена, П — перикард, Пр — предсердие, ПС — почечные со-
суды, СЗП — секреторная зона почки, РО — реноперикардп-
альное’ отверстие-
взп.
РО
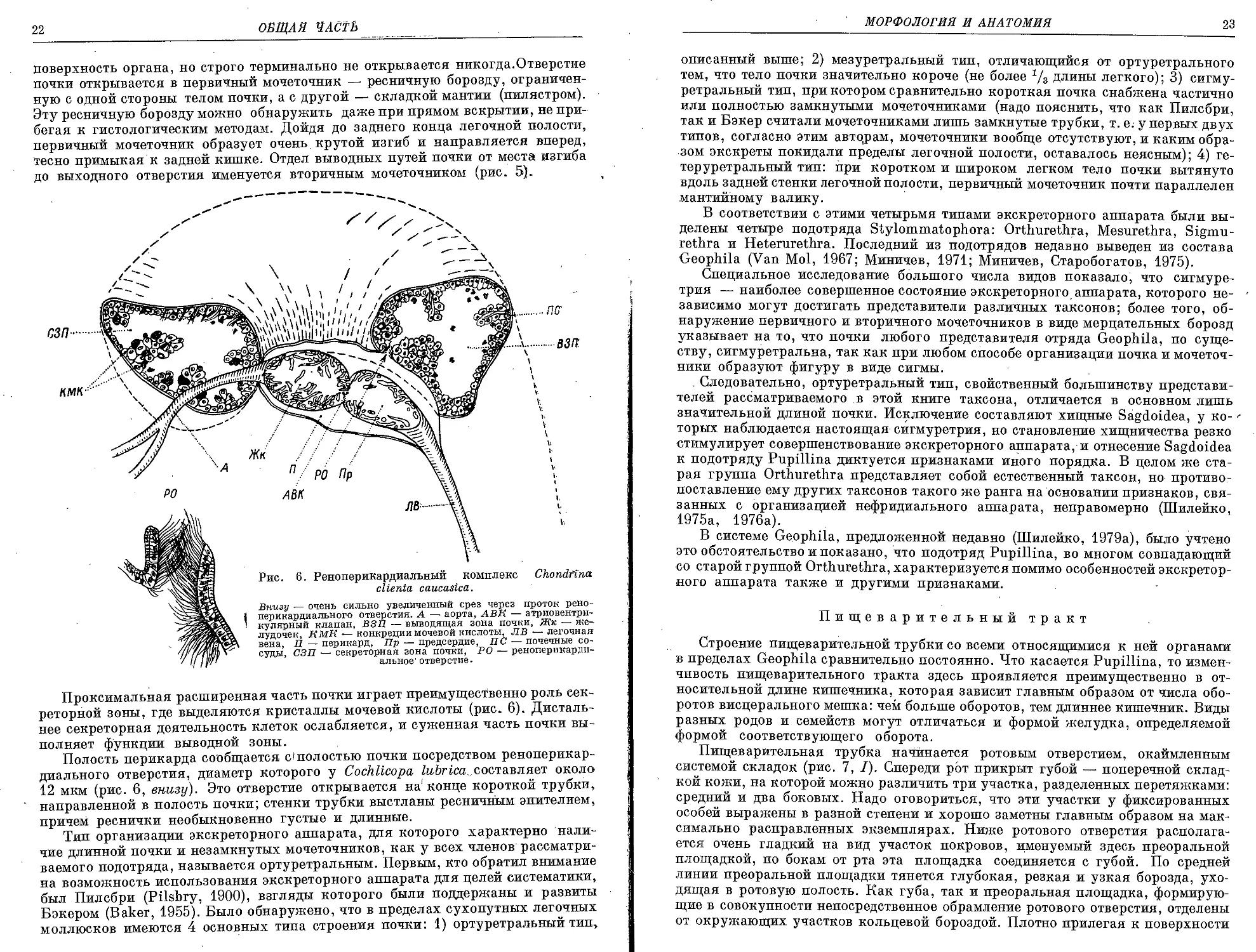
Проксимальная расширенная часть почки играет преимущественно роль сек-
реторной зоны, где выделяются кристаллы мочевой кислоты (рис. 6). Дисталь-
нее секреторная деятельность клеток ослабляется, и суженная часть почки вы-
полняет функции выводной зоны.
Полость перикарда сообщается dполостью почки посредством реноперикар-
диального отверстия, диаметр которого у Cochlicopa lubrtea составляет около,
12 мкм (рис. 6, внизу). Это отверстие открывается на'конце короткой трубки,
направленной в полость почки; стенки трубки выстланы ресничным эпителием,
причем реснички необыкновенно густые и длинные.
Тип организации экскреторного аппарата, для которого характерно нали-
чие длинной почки и незамкнутых мочеточников, как у всех членов рассматри-
ваемого подотряда, называется ортуретральным. Первым, кто обратил внимание
на возможность использования экскреторного аппарата для целей систематики,
был Пилсбри (Pilsbry, 1900), взгляды которого были поддержаны и развиты
Бэкером (Baker, 1955). Было обнаружено, что в пределах сухопутных легочных
моллюсков имеются 4 основных типа строения почки: 1) ортуретральный тип,
МОРФОЛОГИЯ И АНАТОМИЯ
23
описанный выше; 2) мезуретральный тип, отличающийся от ортуретрального
тем, что тело почки значительно короче (не более 1/3 длины легкого); 3) сигму-
ретральный тип, при котором сравнительно короткая почка снабжена частично
или полностью замкнутыми мочеточниками (надо пояснить, что как Пилсбри,
так и Бэкер считали мочеточниками лишь замкнутые трубки, т. е; у первых двух
типов, согласно этим авторам, мочеточники вообще отсутствуют, и каким обра-
зом экскреты покидали пределы легочной полости, оставалось неясным); 4) ге-
теруретральный тип: при коротком и широком легком тело почки вытянуто
вдоль задней стенки легочной полости, первичный мочеточник почти параллелен
мантийному валику.
В соответствии с этими четырьмя типами экскреторного аппарата были вы-
делены четыре подотряда Stylommatophora: Orthurethra, Mesurethra, Sigmu-
rethra и Heterurethra. Последний из подотрядов недавно выведен из состава
Geophila (Van Mol, 1967; Миничев, 1971; Миничев, Старобогатов, 1975).
Специальное исследование большого числа видов показало, что сигмуре-
трия — наиболее совершенное состояние экскреторного аппарата, которого не-
зависимо могут достигать представители различных таксонов; более того, об-
наружение первичного и вторичного мочеточников в виде мерцательных борозд
указывает на то, что почки любого представителя отряда Geophila, по суще-
ству, сигмуретральна, так как при любом способе организации почка и мочеточ-
ники образуют фигуру в виде сигмы.
. Следовательно, ортуретральный тип, свойственный большинству представи-
телей рассматриваемого в этой книге таксона, отличается в основном лишь
значительной длиной почки. Исключение составляют хищные Sagdoidea, у ко- '
торых наблюдается настоящая сигмуретрия, но становление хищничества резко
стимулирует совершенствование экскреторного аппаратами отнесение Sagdoidea
к подотряду Pupillina диктуется признаками иного порядка. В целом же ста-
рая группа Orthurethra представляет собой естественный таксон, но противо-
поставление ему других таксонов такого же ранга на основании признаков, свя-
занных с организацией нефридиального аппарата, неправомерно (Шилейко,
1975а, 1976а).
В системе Geophila, предложенной недавно (Шилейко, 1979а), было учтено
это обстоятельство и показано, что подотряд Pupillina, во многом совпадающий
со старой группой Orthurethra, характеризуется помимо особенностей экскретор-
ного аппарата также и другими признаками.
Пищеварительный тракт
Строение пищеварительной трубки со всеми относящимися к ней органами
в пределах Geophila сравнительно постоянно. Что касается Pupillina, то измен-
чивость пищеварительного тракта здесь проявляется преимущественно в от-
носительной длине кишечника, которая зависит главным образом от числа обо-
ротов висцерального мешка: чем больше оборотов, тем длиннее кишечник. Виды
разных родов и семейств могут отличаться и формой желудка, определяемой
формой соответствующего оборота.
Пищеварительная трубка начинается ротовым отверстием, окаймленным
системой складок (рис. 7, Z). Спереди рот прикрыт губой — поперечной склад-
кой кожи, на которой можно различить три участка, разделенных перетяжками:
средний и два боковых. Надо оговориться, что эти участки у фиксированных
особей выражены в разной степени и хорошо заметны главным образом на мак-
симально расправленных экземплярах. Ниже ротового отверстия располага-
ется очень гладкий на вид участок покровов, именуемый здесь преоральной
площадкой, по бокам от рта эта площадка соединяется с губой. По средней
линии преоральной площадки тянется глубокая, резкая и узкая борозда, ухо-
дящая в ротовую полость. Как губа, так и преоральная площадка, формирую-
щие в совокупности непосредственное обрамление ротового отверстия, отделены
от окружающих участков кольцевой бороздой. Плотно прилегая к поверхности
24
ОБЩАЯ ЧАСТЬ
пищевого куска, преоральная площадка и губа образуют замкнутое простран-
ство, изолируя работающую радулу от внешней среды.
По бокам от'преоральной площадки имеется пара околоротовых лопастей,
ткани которых насыщены множеством одноклеточных желез. Если поместить
моллюска на стекло и затем сквозь это стекло рассматривать ползущее животное,
легко видеть, что при поступательном движении околоротовые лопасти плотно
прилегают к субстрату. Напрашивается предположение, что роль околоротовых
лопастей (вероятно, не единственная) — контроль свойств субстрата.
Внутри ротового отверстия, на спинной его стороне, залегает полулунная
конхиолиновая челюсть пикногнатного типа, т. е. лишенная срединного выступа
Н1Ц Г ч I
Рис. 7. Cochlicopa lubrica.
I — передний отдел тела (вид снизу), II — челюсть. ВЩ — верхнее щупальце (омматофор), Г — губа,
НЩ — нижнее щупальце,- ОЛ — околоротовая лопасть, ПЖ — отверстие педальной железы, ЦП — пре-
оральная площадка, Ч — челюсть.
и с множеством тонких поперечных ребрышек (рис. 7, II). На поперечном раз-
резе видно, что рабочая пластинка челюсти имеет клиновидную форму, а ее
внутренний край, постепенно истончаясь, простирается назад довольно далеко;
в результате передняя часть нёба выстлана конхиолиновым «потолком», что
предохраняет мягкие ткани от повреждений твердыми частицами и зубами ра-
дулы. Основное назначение челюсти — служить опорой, к которой прижима-
ется пищевой кусок в то время, когда он скоблится радулой. При этом опреде-
ленная роль отводится борозде на поверхности преоральной площадки: края
борозды раздвигаются и также способствуют фиксации пищевого объекта.
Ротовое отверстие ведет в глотку, в состав которой входит радула, субраду-
лярный хрящ (одонтофор) и сложная система мышц. Глотка (рис. 8, I, II) пред-
ставляет собой компактный сильно мускулизованный орган примерно фасоле-
видной формы, обслуживаемый богатым набором мышечных пучков. Задняя
часть глотки снизу и с боков охвачена мощным глоточным ретрактором, между
правой и левой ветвями которого видно радулярное влагалище, или радуляр-
ный карман. Поверхность остальной части глотки покрыта плотным слоем
мышц с преимущественно кольцевым ходом волокон, где, однако;- имеются и
продольные и косые мышечные пучки: это компрессор (сжиматель) глотки,
Передняя часть глотки соединяется с губой посредством множества тонких пе-
редних протракторов глотки. Нижние протракторы глотки парные, начинаются
от линии прикрепления глоточного ретрактора и подходят к передней стенке
тела по бокам от ротового отверстия и несколько ниже его. Кроме того, имеются
верхние и иногда средние протракторы глотки. На вентральной поверхности
МОРФОЛОГИЯ И АНАТОМИЯ
25
глотки, между нижними протракторами, видны два ряда тонких депрессоров
(опускателей) глотки, связывающих этот орган с дном полости тела. С верхней
и верхне-боковой стенкой тела глотка связана 2—3 парами леваторов (подни-
мателей).
Наконец, к мышечному аппарату, обеспечивающему деятельность глотки,
можно отнести пару так называемых лабиальных (губных) ретракторов. На
Рис. 8. Мускульная система глотки.
I—II— внешний вид глотки Brephulopsis bidens (I — справа, II — снизу); III — фронтальный срез
глотки Gibbulinopsis signata на уровне, примерно соответствующемДптриховой линии на I', IV — схема
основной мускулатуры глотки, вид с вентральной стороны. Протракторы глотки: ВПГ — верхний, НПГ —
нижний, ППГ — передний; ретракторы одОнтофора: ВРО — верхний, НРО — нижний; ГЛ — глоточные
лакуны; протракторы одонтофора: ЗПО — задний, ППО — передний; ДГ — депрессоры, КГ — компрес-
сор, ЛГ — леваторы, РГ — ретрактор, СГ — стенка глотки, КРВ — компрессор радулярного влага-
лища, ЛР — лабиальный ретрактор, О — одонтофор, П9 — пищевод; ДО — дилятаторы, ПрО— про-
натор, СО — супинатор, ТО — тензоры одонтофора; ПРВ — протрактор радулярного влагалища, Ра —
радула, РВ — радулярное влагалище, Ч — челюсть.
самом же деле эти ретракторы крепятся не к губе, а к внутренней поверхности
преоральной площадки, под ротовым отверстием; эти тонкие мышечные ленты
есть дериваты колумеллярного мускула и отщепляются от глоточного ретрак-
тора.
Основной рабочий элемент глотки — радула, состоящая из мембраны и рас-
положенных на ней многочисленных мелких зубов. Передняя часть радулы на
поперечном разрезе напоминает дугу со слегка завернутыми внутрь краями,
которые спаяны с телом субрадулярного хряща — одонтофора. Далее в глубь
глотки радулярная мембрана поднимается по одонтофору, имеющему седло-
26
ОБЩАЯ ЧАСТЬ
видную форму, вверх, где сначала намечается, а потом становится все более за-
метным продольный перегиб мембраны. Затем радула спускается вниз, все
сильнее сгибаясь вдоль средней линии; вогнутая центральная часть мембраны
располагается между задними склонами субрадулярного хряща. Наконец,
радулярная мембрана сужается; сохраняя и усиливая продольный перегиб,
она переходит в радулярное влагалище, где происходит формирование новых
зубов взамен стершихся.
' Рис. 9. Схема строения глотки.
ЗПО — задний протрактор (пучки), ЛИО, ПКО — левое, правое крылья, ОНО — супинатор крыльев,
одонтофора. Остальные обозначения как на рис. 8.
Крылья одонтофора способны в известных пределах менять форму благодаря
наличию тензоров — мышечных пучков, пронизывающих хрящеподобную
ткань (рис. 8, III). Изменение положения субрадулярного хряща внутри глотки
и регуляция взаимного расположения крыльев одонтофора обеспечиваются тремя
группами мышц (протракторы, ретракторы, дилататоры), а также непарными
мышцами: мощным супинатором и слаборазвитым пронатором (рис. 9). Строе-
ние и механизм работы глотки в существенных чертах сходны у всех Pulmonata
(Market, 1958) и в то жевремя изучены недостаточно; в этом отношении иссле-
дованы преимущественно Helix pomatia (Trappmann, 1916), Testacella mangel
(Crampton, 1975) и некоторые пресноводные формы (Carriker, 1946; Walter,
1969; H^mbrow, 1973). Поэтому есть смысл остановиться на этих вопросах под-
робнее. Предварительно надо подчеркнуть, что изображения на рис. 8, IV и 9
представляют собой упрощенные схемы; так, основания многих мышц настолько
широки, что на рисунке, приближающемся к натуре, ониполностью заслонили бы
одонтофор и разобраться в таком рисунке было бы практически невозможно.
МОРФОЛОГИЯ И АНАТОМИЯ
27
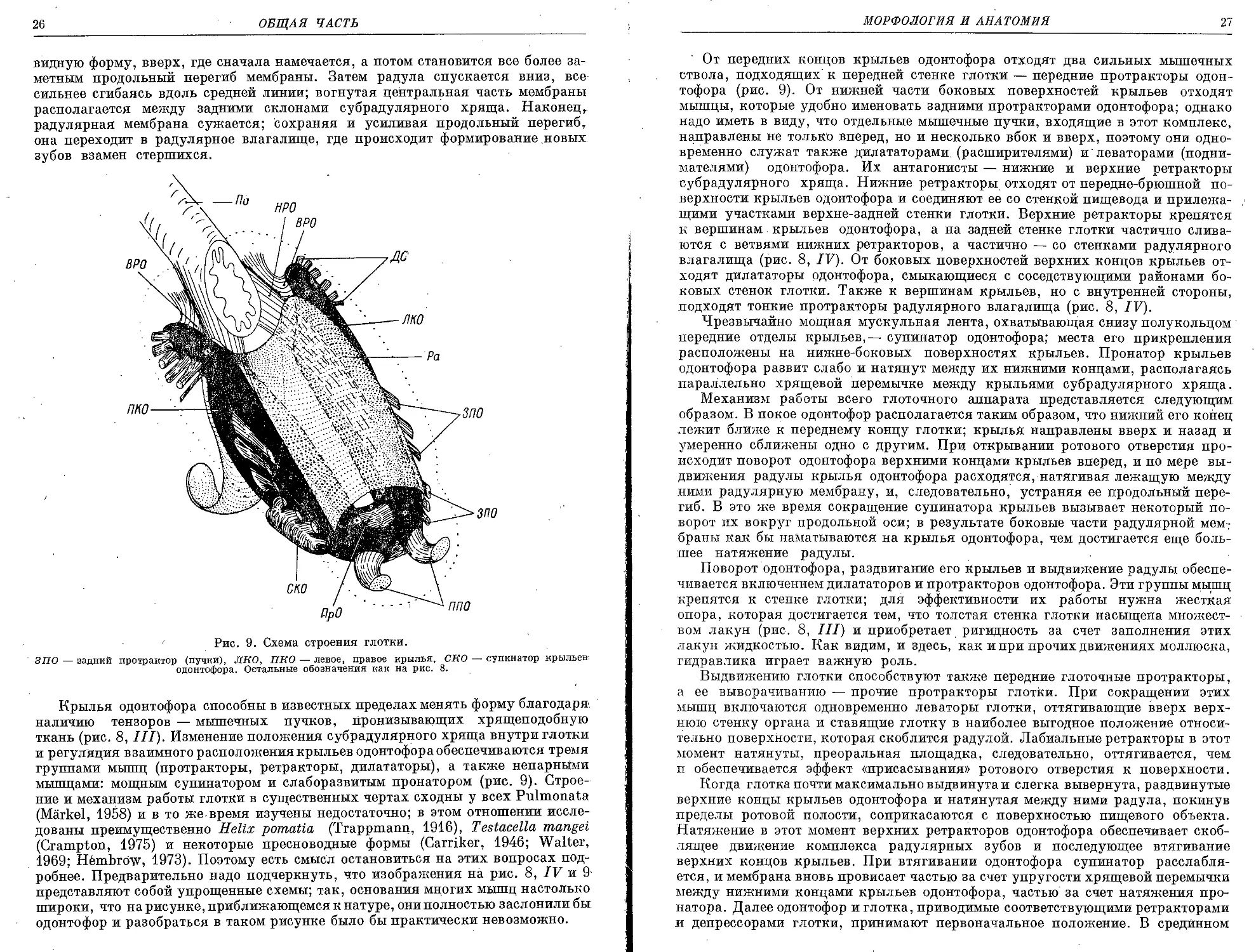
От передних концов крыльев одонтофора отходят два сильных мышечных
ствола, подходящих к передней стенке глотки — передние протракторы одон-
тофора (рис. 9). От нижней части боковых поверхностей крыльев отходят
мышцы, которые удобно именовать задними протракторами одонтофора; однако
надо иметь в виду, что отдельные мышечные пучки, входящие в этот комплекс,
направлены не только вперед, но и несколько вбок и вверх, поэтому они одно-
временно служат также дилататорами (расширителями) и'леваторами (подни-
мателями) одонтофора. Их антагонисты — нижние и верхние ретракторы
субрадулярного хряща. Нижние ретракторы отходят от передне-брюшной по-
верхности крыльев одонтофора и соединяют ее со стенкой пищевода и прилежа-
щими участками верхне-задней стенки глотки. Верхние ретракторы крепятся
к вершинам крыльев одонтофора, а на задней стенке глотки частично слива-
ются с ветвями нижних ретракторов, а частично — со стенками радулярного
влагалища (рис. 8, IV). От боковых поверхностей верхних концов крыльев от-
ходят дилятаторы одонтофора, смыкающиеся с соседствующими районами бо-
ковых стенок глотки. Также к вершинам крыльев, но с внутренней стороны,
подходят тонкие протракторы радулярного влагалища (рис. 8, IV).
Чрезвычайно мощная мускульная лента, охватывающая снизу полукольцом
передние отделы крыльев,— супинатор одонтофора; места его прикрепления
расположены на нижне-боковых поверхностях крыльев. Пронатор крыльев
одонтофора развит слабо и натянут между их нижними концами, располагаясь
параллельно хрящевой перемычке между крыльями субрадулярного хряща.
Механизм работы всего глоточного аппарата представляется следующим
образом. В покое одонтофор располагается таким образом, что нижний его конец
лежит ближе к переднему концу глотки; крылья направлены вверх и назад и
умеренно сближены одно с другим. При открывании ротового отверстия про-
исходит поворот одонтофора верхними концами крыльев вперед, и по мере вы-
движения радулы крылья одонтофора расходятся, натягивая лежащую между
ними радулярную мембрану, и, следовательно, устраняя ее продольный пере-
гиб. В это же время сокращение супинатора крыльев вызывает некоторый по-
ворот их вокруг продольной оси; в результате боковые части радулярной мем7
браны как бы наматываются на крылья одонтофора, чем достигается еще боль-
шее натяжение радулы.
Поворот одонтофора, раздвигание его крыльев и выдвижение радулы обеспе-
чивается включением дилататоров и протракторов одонтофора. Эти группы мышц
крепятся к стенке глотки; для эффективности их работы нужна жесткая
опора, которая достигается тем, что толстая стенка глотки насыщена множест-
вом лакун (рис. 8, III) и приобретает ригидность за счет заполнения этих
лакун жидкостью. Как видим, и здесь, как и при прочих движениях моллюска,
гидравлика играет важную роль.
Выдвижению глотки способствуют также передние глоточные протракторы,
а ее выворачиванию — прочие протракторы глотки. При сокращении этих
мышц включаются одновременно леваторы глотки, оттягивающие вверх верх-
нюю стенку органа и ставящие глотку в наиболее выгодное положение относи-
тельно поверхности, которая скоблится радулой. Лабиальные ретракторы в этот
момент натянуты, преоральная площадка, следовательно, оттягивается, чем
и обеспечивается эффект «присасывания» ротового отверстия к поверхности.
Когда глотка почти максимально выдвинута и слегка вывернута, раздвинутые
верхние концы крыльев одонтофора и натянутая между ними радула, покинув
пределы ротовой полости, соприкасаются с поверхностью пищевого объекта.
Натяжение в этот момент верхних ретракторов одонтофора обеспечивает скоб-
лящее движение комплекса радулярных зубов и последующее втягивание
верхних концов крыльев. При втягивании одонтофора супинатор расслабля-
ется, и мембрана вновь провисает частью за счет упругости хрящевой перемычки
между нижними концами крыльев одонтофора, частью за счет натяжения про-
натора. Далее одонтофор и глотка, приводимые соответствующими ретракторами
и депрессорами глотки, принимают первоначальное положение. В срединном
28
ОБЩАЯ ЧАСТЬ
прогибе радулы остаются соскобленные пищевые частицы; сокращением коль-
цевых волокон очень мощного компрессора глотки эти частицы прессуются и
затем в виде небольшого комка поступают в пищевод.
Совокупность всех прочих отделов пищеварительного тракта составляет ки-
шечную трубку, длина которой в большой степени зависит от числа оборотов вис-
церального мешка: чем больше оборотов, тем длиннее кишечная трубка (ср.
рис. 10, 11).
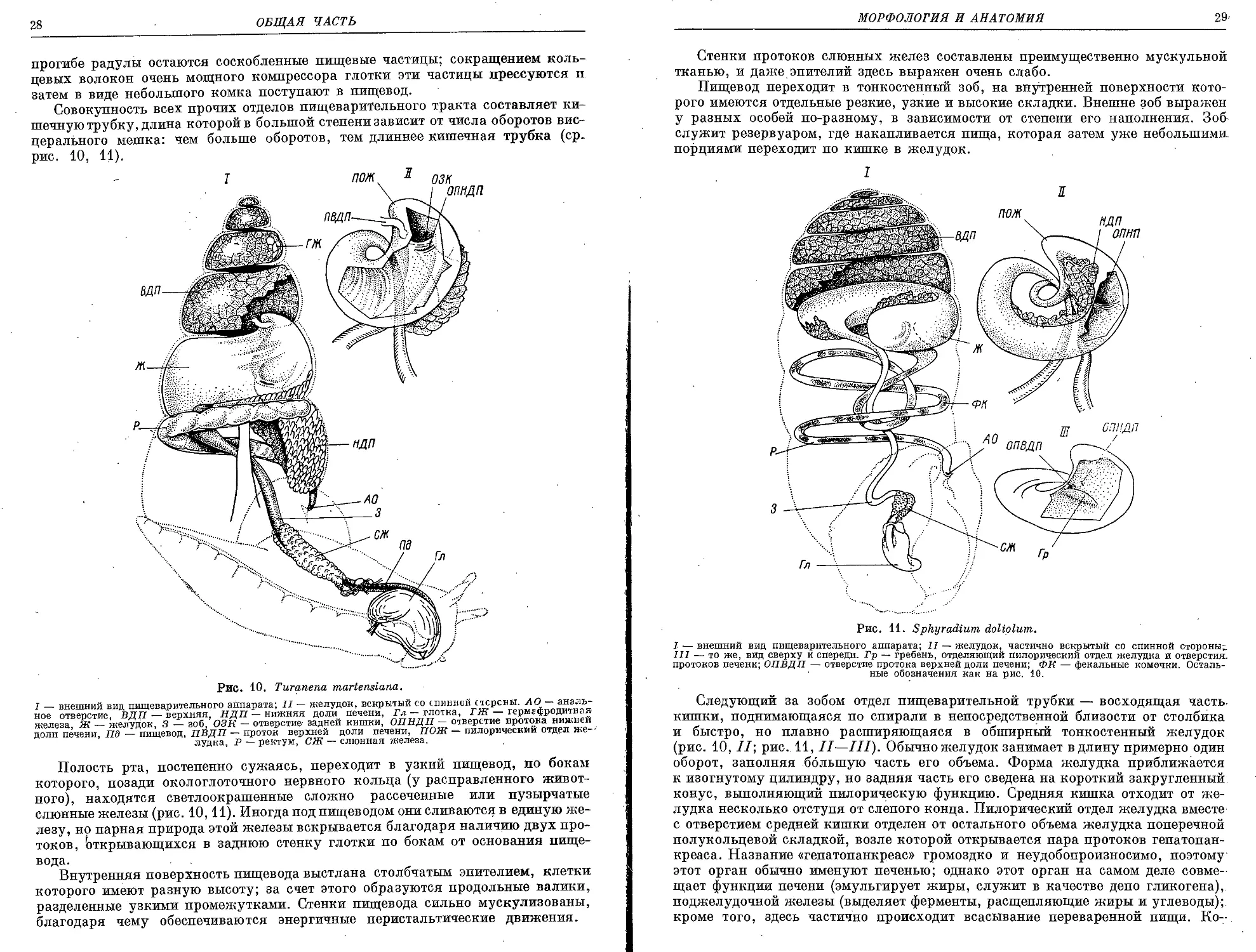
Рис. 10. Turanena martensiana.
I — внешний вид пищеварительного аппарата; II — желудок, вскрытый со спинной стсрсны. АО — аналь-
ное отверстие, ВДП — верхняя, НДП — нижняя доли печени, Гл — глотка, ГЖ — гермафродитная
железа, Ж — желудок, 3 — зоб, ОЗК — отверстие задней кишки, ОПНДП — отверстие протока нижней
доли печени, Пд — пищевод, ПВДП — проток верхней доли печени, ПОЖ — пилорический отдел же-
лудка, Р — ректум, СЖ — слюнная железа.
Полость рта, постепенно сужаясь, переходит в узкий пищевод, по бокам
которого, позади окологлоточного нервного кольца (у расправленного живот-
ного), находятся светлоокрашенные сложно рассеченные или пузырчатые
слюнные железы (рис. 10,11). Иногда под пищеводом они сливаются в единую же-
лезу, но парная природа этой железы вскрывается благодаря наличию двух про-
токов, Открывающихся в заднюю стенку глотки по бокам от основания пище-
вода.
Внутренняя поверхность пищевода выстлана столбчатым эпителием, клетки
которого имеют разную высоту; за счет этого образуются продольные валики,
разделенные узкими промежутками. Стенки пищевода сильно мускулизованы,
благодаря чему обеспечиваются энергичные перистальтические движения.
МОРФОЛОГИЯ И АНАТОМИЯ
29-
Стенки протоков слюнных желез составлены преимущественно мускульной
тканью, и даже эпителий здесь выражен очень слабо.
Пищевод переходит в тонкостенный зоб, на внутренней поверхности кото-
рого имеются отдельные резкие, узкие и высокие складки. Внешне зоб выражен
у разных особей по-разному, в зависимости от степени его наполнения. Зой
служит резервуаром, где накапливается пища, которая затем уже небольшими,
порциями переходит по кишке в желудок.
1
Рис. 11. Sphyradium doliplum.
I — внешний вид пищеварительного аппарата; II — желудок, частично вскрытый со спинной стороны;.
III — то же, вид сверху и спереди. Гр — гребень, отделяющий пилорический отдел желудка и отверстия,
протоков печени; ОПВДП —• отверстие протока верхней доли печени; ФК — фекальные комочки. Осталь-
ные обозначения как на рис. 10.
Следующий за зобом отдел пищеварительной трубки — восходящая часть-
кишки, поднимающаяся по спирали в непосредственной близости от столбика
и быстро, но плавно расширяющаяся в обширный тонкостенный желудок
(рис. 10, II; рис. 11, II—III). Обычно желудок занимает в длину примерно один
оборот, заполняя большую часть его объема. Форма желудка приближается
к изогнутому цилиндру, но задняя часть его сведена на короткий закругленный
конус, выполняющий пилорическую функцию. Средняя кишка отходит от же-
лудка несколько отступя от слепого конца. Пилорический отдел желудка вместе
с отверстием средней кишки отделен от остального объема желудка поперечной
полукольцевой складкой, возле которой открывается пара протоков гепатопан-
креаса. Название «гепатопанкреас» громоздко и неудобопроизносимо, поэтому
этот орган обычно именуют печенью; однако этот орган на самом деле совме-
щает функции печени (эмульгирует жиры, служит в качестве депо гликогена),
поджелудочной железы (выделяет ферменты, расщепляющие жиры и углеводы);
кроме того, здесь частично происходит всасывание переваренной пищи. Ко-
30
ОБЩАЯ ЧАСТЬ
ричневато-бурая или красноватая печень состоит из двух долей — верхней и
нижней. Более массивная и компактная верхняя доля занимает почти весь объем
верхних оборотов, нижняя располагается между петлями кишечника.
Весь отдел кишечника, лежащий позади желудка, имеет почти постоянный
диаметр и подразделяется на два отдела — среднюю и заднюю кишку. Морфо-
логически эти отделы между собой практически не различаются; границей между
ними служит задняя стенка легочной полости. Тот отдел, который расположен
рядом с легким и сопровождает его по всей длине, называется задней или прямой
кишкой (ректумом), а отдел кишечника, заключенный между желудком и зад-
ней стенкой легкого, именуется средней кишкой. Внутри средней и большей
части задней кишки имеется тифлозоль, начинающийся обычно еще в желудке.
Суммарная длина средней и задней кишки различна. Если длина ректума
коррелятивно связана с длиной легочной полости, то длина средней кишки
связана, возможно, с особенностями питания, о которых известно очень мало.
Ректум открывается на мантийном валике рядом с пневмостомом и снабжен
сфинктером, а также небольшим выростом прилежащих тканей мантийного
валика.
Опорно-двигательный аппарат
Все многообразие движений моллюска обеспечивается работой четырех мы-
шечных комплексов: колумеллярной мускулатурой; мощной системой мус-
кульных волокон, рассеянных в толще покровов ноги; совокупностью муску-
лов, управляющих движениями глотки; собственной мускулатурой органов
(особенно богата мускульная система в глотке и в дистальных участках поло-
вого аппарата). Кроме того, исключительно важна роль сильно развитой системы
синусов и лакун, залегающих в толще ноги и связанных с полостью тела.
Если наличие некоторого мускульного тяжа предполагает, как правило,
наличие тяжа-антагониста, то самый мощный и основной мускульный ствол —
колумеллярный мускул — лишен противодействующих мускульных элемен-
тов. Колумеллярный мускул со всеми ветвями служит общим ретрактором,
втягивающим внутрь раковины все выступающие при движении моллюска части.
Роль его антагонистов—протракторов—выполняется согласованным сокраще-
нием кольцевой мускулатуры ноги в совокупности с перераспределением по-
лостной жидкости внутри синусов и лакун.
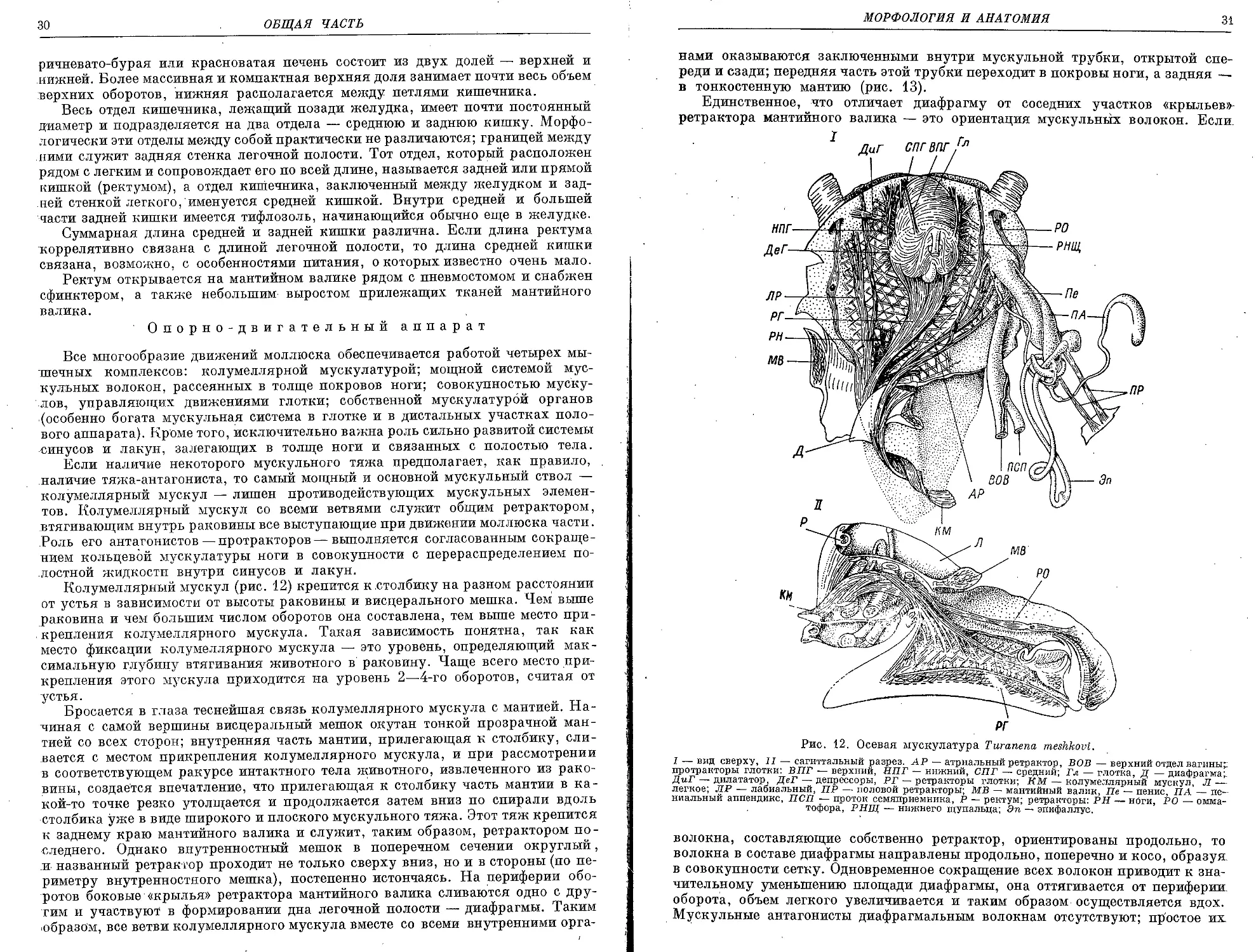
Колумеллярный мускул (рис. 12) крепится к столбику на разном расстоянии
от устья в зависимости от высоты раковины и висцерального мешка. Чем выше
раковина и чем большим числом оборотов она составлена, тем выше место при-
крепления колумеллярного мускула. Такая зависимость понятна, так как
место фиксации колумеллярного мускула — это уровень, определяющий мак-
симальную глубину втягивания животного в раковину. Чаще всего место при-
крепления этого мускула приходится на уровень 2—4-го оборотов, считая от
устья.
Бросается в глаза теснейшая связь колумеллярного мускула с мантией. На-
чиная с самой вершины висцеральный мешок окутан тонкой прозрачной ман-
тией со всех сторон; внутренняя часть мантии, прилегающая к столбику, сли-
вается с местом прикрепления колумеллярного мускула, и при рассмотрении
ъ соответствующем ракурсе интактного тела животного, извлеченного из рако-
вины, создается впечатление, что прилегающая к столбику часть мантии в ка-
кой-то точке резко утолщается и продолжается затем вниз по спирали вдоль
столбика уже в виде широкого и плоского мускульного тяжа. Этот тяж крепится
к заднему краю мантийного валика и служит, таким образом, ретрактором по-
следнего. Однако внутренностный мешок в поперечном сечении округлый,
и названный ретрактор проходит не только сверху вниз, но и в стороны (по пе-
риметру внутренностного мешка), постепенно истончаясь. На периферии обо-
ротов боковые «крылья» ретрактора мантийного валика сливаются одно с дру-
тим и участвуют в формировании дна легочной полости — диафрагмы. Таким
образом, все ветви колумеллярного мускула вместе со всеми внутренними орга-
МОРФОЛОГИЯ И АНАТОМИЯ
31
нами оказываются заключенными внутри мускульной трубки, открытой спе-
реди и сзади; передняя часть этой трубки переходит в покровы ноги, а задняя —
в тонкостенную мантию (рис. 13).
Единственное, что отличает диафрагму от соседних участков «крыльев»-
ретрактора мантийного валика — это ориентация мускульных волокон. Если
РГ
Рис. 12. Осевая мускулатура Turanena meshkovi.
I — вид сверху, II — сагиттальный разрез. АР — атриальный ретрактор, ВОВ — верхний отдел вагины-
протракторы глотки: ВНР — верхний, НПГ — нижний, СПГ — средний; Гл — глотка, Д — диафрагма;.
ДиГ — дилятатор ДеГ — депрессоры, РГ — ретракторы глотки; КМ — колумеллярный мускул, Л —
легкое; ЛР — лабиальный, ПР — половой ретракторы; МВ — мантийный валик, Пе — пенис, ПА — пе-
ниальный аппендикс, ПСП — проток семяприемника, Р — ректум; ретракторы: PH — ноги, РО — омма-
тофора, РНЩ — нижнего щупальца; Эп — эпифаллус.
волокна, составляющие собственно ретрактор, ориентированы продольно, то
волокна в составе диафрагмы направлены продольно, поперечно и косо, образуя,
в совокупности сетку. Одновременное сокращение всех волокон приводит к зна-
чительному уменьшению площади диафрагмы, она оттягивается от периферии
оборота, объем легкого увеличивается и таким образом осуществляется вдох.
Мускульные антагонисты диафрагмальным волокнам отсутствуют; простое их
-32
ОБЩАЯ ЧАСТЬ
расслабление не может привести к сокращению объема полости легкого. Оста-
ется допустить, что выдох осуществляется увеличением давления на диафрагму
со стороны полостной жидкости. Увеличение давления в свою очередь должно
быть связано с уменьшением объема тела, а уменьшение объема тела обеспечи-
вается главным образом сокращением собственной мускулатуры ноги. Детали
этих процессов нуждаются в. выяснении.
Периферическая поверхность мускульной трубки прикрыта мантийным
листком — крышей легочной полости; диафрагма заключена между линиями
прикрепления крыши легкого; пространство, заключенное между диафрагмой
Рис. 13. Осевая мускулатура Brephulopsis bidens (витки колумеллярного мускула сильно
раскручены).
К Л— крыша легкого (участок), ММ— мантийный мускул, РМВ—ретрактор мантийного валика.
Остальные обозначения как на рис. 12.
и крышей легочной полости, играет роль респираторного органа — легкого.
От диафрагмы отходит двуветвистый половой ретрактор (виды, у которых
отсутствует пениальный аппендикс, имеют одноветвистый половой ретрактор).
Не разбирая деталей происхождения отдельных мышечных пучков, можно го-
ворить о том, что вся описанная мышечная система представляет собой морфо-
логически единый комплекс колумеллярной мускулатуры: ретрактор мантий-
ного валика и задний ретрактор тела — педальные и головные стволы—диаф-
рагма—половой ретрактор. На это обстоятельство надо обратить внимание,
так как бытует представление о независимости диафрагмы и полового ретрак-
тора от колумеллярной мускулатуры: В этой связи уместно вспомнить, что
половой ретрактор Helicodonta (подотряд Helixina) настолько сильно сдвинут
к основному стволу колумеллярного мускула, что выглядит ветвью последнего
(Шилейко, 1978а). «У некоторых Euxininae (Clausiliidae) ретрактор пениса
не прикрепляется к диафрагме, а сливается с задним ретрактором колумел-
шярного мускула» (Лихарев, 1962, с. 29). (
На схеме (рис. 14) висцеральный мешок условно сильно раскручен, так как
МОРФОЛОГИЯ И АНАТОМИЯ
33
на тугой спирали, какую представляет собой внутренностный мешок на самом
деле, разобраться в описанных отношениях очень трудно.
Не вполне выяснена роль полового ретрактора. Наиболее заметна его функ-
ция, отображенная в названии этого мускульного тяжа: при спаривании пенис
и пениальный аппендикс выворачиваются наподобие пальцев перчатки и вво-
рачиваются затем обратно за счет сокращения полового ретрактора. Показа-
тельно, что места прикрепления ветвей этого мускула — та граница, до кото-
рой происходит выворачивание дистальных участков совокупительного аппарата.
Рис. 14. Схема строения осевой мускулатуры.
КЛП — крыша легочной полости, КМ — колумеллярный мускул (основной ствол), ЛП — легочная по-
лость, Ом — омматофор, ПВ — педальные ветви колумеллярного мускула. Остальные обозначения как
на рис. 12 и 13 (РМВ — ратрактор мантийного валика).
Однако возможно, что это не единственная функция полового ретрактора: его
основная роль очень эпизодична, так как она осуществляется всего несколько
раз в году — во время спаривания. В то же время очевидно, что при сокраще-
нии полового ретрактора, крепящегося к нижней поверхности диафрагмы, по-
следняя будет оттягиваться внутрь, в результате чего подость легкого будет
увеличиваться. Иными словами, есть основания предполагать, что половой
ретрактор принимает участие в вентиляции легочной полости. Понятно тем не
менее, что это не может быть основной функцией названного ретрактора, во-пер-
вых, потому, что он вплоть до достижения моллюском субдефинитивного состоя-
ния развит чрезвычайно слабо, и, во-вторых, потому, что известны легочные
моллюски, вообще лишенные полового ретрактора (например, некоторые
Euomphaliinae из Hygromiidae).
Вернемся к рассмотрению колумеллярного мускула. Как отмечено выше,
самая внутренняя периферическая часть колумеллярного мускула — ретрактор
мантийного валика — прикрепляется к внутренней части края мантии по всему
периметру, но в той части, которая прилегает к столбику, мышечные волокна
наиболее мощные. От той же исходной точки на столбике отходит центральный
ствол колумеллярного мускула, распадающийся вскоре на несколько стволов
второго порядка; эти стволы объединяются в две группы — правую и левую,
поскольку нога и голова обладают билатеральной симметрией, и каждая сто-
рона снабжается симметрично расположенными мускулами. Симметрия нару-
3 А. А. Шилейко
34
ОБЩАЯ ЧАСТЬ
шается только тем, что дистальные отделы полового аппарата располагаются на
правой стороне, и там же открывается отверстие полового атриума. Соответ-
ственно на правой стороне добавляется система атриальных ретракторов, не
имеющая аналогов на левой стороне.
Центральное место занимает очень мощный парный ретрактор глотки,
ветви которого крепятся к нижнебоковой поверхности задней части глотки
справа и слева от радулярного влагалища. Ветви глоточного ретрактора почти
на всем' протяжении слиты одна с другой и разделяются только в непосредст-
венной близости от глотки. Следовательно, оба ретрактора действуют как одно
целое и практически не участвуют в обеспечении боковой подвижности глотки.
Ниже глоточных ретракторов, т. е. ближе к ретрактору мантийного валика,
располагается парный педальный ствол, отчленяющийся от ретракторов глотки
сравнительно высоко. Входя в полость ноги, педальный ствол почти сразу же
начинает распадаться на отдельные мышечные пучки второго и следующего
порядков, и вся система крепится близ центра ноги. Таким образом, при сокра-
щении ретрактора ноги и всех его ветвей начинается втягивание центральной
части ноги, в результате чего нога сгибается поперек продольной оси.
Справа и слева от центрального ствола колумеллярного мускула отходят
боковые мускульные ленты, направляющиеся к голове (рис. 12). Каждая из
них примерно на уровне мантийного валика распадается на ретракторы ом-
матофоров и ветвь, лежащую ниже ретракторов омматофоров; эта ветвь расчле-
няется вдоль уже в пределах ноги: лента, занимающая более дорсальное поло- ;
жение, обеспечивает вворачивание нижних (губных) щупалец, а вентральная
лента несколькими пучками крепится изнутри к покровам головы в районе
околоротовых лопастей и осуществляет во взаимодействии с глоточным ретрак- ]
тором вворачивание головного конца животного. На той стороне, где располо-
жено половое отверстие, добавляется система атриальных ретракторов, отходя-
щих в пределах ноги от боковой мускульной ленты этой же стороны. Основное
назначение системы атриальных ретракторов заключается, вероятно, в обеспе-
чении симметричного и равномерного втягивания ноги в раковину, поскольку
дистальные отделы полового аппарата — относительно очень массивные обра-
зования, и при отсутствии специальных ретракторов для их втягивания та сто-
рона, к которой прилежат пенис и вагина, неизбежно отставала бы при сокраще-
нии и вворачивании животного в раковину от другой стороны.
В передней ча,сти ноги имеется богатая система мышечных пучков, управляю-
щих движениями головы и глотки. При рассматривании сверху видны две сим-
метрично расположенные мышечные ленты, начинающиеся на спинной стороне
глотки по бокам от начала пищевода и идущие по поверхности глотки вперед и
несколько в стороны; они прикрепляются к коже головы на уровне верхней
части глотки. Эти мышцы выполняют роль верхних протракторов и одновре-
менно леваторов глотки: при их сокращении задняя часть глотки приподнима-
ется и подается вперед.
Слабее развита пара средних протракторов глотки, которые можно увидеть
при рассматривании глотки сбоку: они прикрепляются на уровне передней
границы между передней и средней третью глотки.
Наконец, самая мощная пара глоточных протракторов — нижняя, она от-
ходит рядом с местом прикрепления ретрактора глотки и крепится к коже го-
ловы несколькими ветвями (рис. 12, /).
Таким образом, вертикальная подвижность задней части глотки при фикси- ;
рованной передней части обеспечивается верхними и нижними протракторами:
верхние выполняют также роль частично леваторов и поднимают задний конец j
глотки, а нижние, сокращаясь, выполняют роль антагонистов по отношению ]
к верхним. От центральной части нижней поверхности глотки отходят много-
численные мелкие депрессоры, связывающие глотку с дном полости тела. Нако-
нец, последняя группа мышц, обслуживающая глотку, — пара дилататоров-
(расширителей), которые прикрепляются к нижне-боковым поверхностям •
глотки и кожи головы в районе омматофоров; их антагонистом служит мышеч-
МОРФОЛОГИЯ И АНАТОМИЯ
35
ный узел, помещающийся внутри глотки между основаниями субрадулярных
хрящей.
Заслуживает упоминания также система мускульных тяжей, выстилающая
снизу полость тела; среди них хорошо видны поперечные и косые тяжи (рис. 12, Z).
Заканчивая характеристику опорно-двигательного аппарата, надо отме-
тить, что поступательное движение моллюска обеспечивается не столько мы-
шечными сокращениями подошвы ноги (эта часть ноги вообще относительно
бедна мускульными элементами), сколько скольжением по слизистой дорожке
с помощью мерцательного эпителия, выстилающего центральную часть по-
дошвы. В толще ноги залегает педальная железа, вырабатывающая слизь жид-
кой консистенции. При движении эта слизь поступает наружу через щелевидное
отверстие педальной железы, расположенное под ртом, и стекает вниз, под
подошву.
Нервный аппарат
Концентрация ганглиев, входящих в состав окологлоточного нервного кольца
Pupillina, не достигает такой степени, как у высших Geophila: даже в сезон
размножения, во время интенсивной нейросекреторной деятельности, всегда
можно различить каждый ганглий. Это обстоятельство было отмечено еще Стин-
бергом (Steenberg, 1925), изучившим внешнее строение ЦНС Chondrina similis
Рис. 15. Окологлоточное нервное кольцо Cochlicopa lubrica (коннективы сильно растянуты,
лабиальные нервы не изображены).
Штриховые линии — пищевод. А — аорта; ганглии: БГ — буккальный, ВГ — висцеральный, ЛПГ —
левый париетальный, лПлГ — левый плевральный, ППаГ — правый париетальный, ППлГ — правый
плевральный, ПЦГ — правый церебральный; нервы: АН — анальный, ИН — интестинальный, НИ —
пениса, ОбН — обонятельный, ОН — оптический, ПаН — паллиальный. ПН — педальный, ФН — фарин-
геальный; РЛО — ретрактор левого омматофора, Ст—статоцист, ЦПН — церебро-плевральная кон-
нектива.
и Vertigo moulinsiana. Тонкое строение ганглиев Achatinella fulgens освещено
Ван Молем (Van Mol, 1967). У расправленного животного окологлоточное нерв-
ное кольцо залегает позади глотки, охватывая пищевод. При втягивании глотки
окологлоточное кольцо за счет растяжения церебральных комиссур и церебро-
педальных коннектив пропускает глотку сквозь себя и оказывается впереди
нее.
У полностью вытянувшегося из раковины моллюска, вскрытого со спинной
стороны, легко рассмотреть самые крупные нервные узлы — церебральные
3*
36
ОБЩАЯ ЧАСТЬ
ганглии, лежащие на верхне-боковых поверхностях пищевода (рис. 15). Между
собой эти ганглии связаны двумя комиссурами: мощной верхней, идущей по
верхней поверхности пищевода, и чрезвычайно тонкой нижней (субцеребраль-
ной), охватывающей пищевод снизу. Передние части церебральных ганглиев
слегка вытянуты и представляют собой процеребрум. Процеребрум левого
ганглия посылает от себя 4 основных нервных ствола, процеребрум правого — 5.
Самые наружные стволы наиболее заметны — это оптические нервы. Они на-
правляются к ретракторам омматофоров и в составе последних доходят до кон-
цов щупалец, где располагаются глаза. Следующий, считая к сагиттальной
Рис. 16. Строение омматофора Cochlicopa lubrica.
I — сагиттальный срез через частично ввернутый омматофор, II — схема расположения рецепторов в рас-
правленном щупальце. Гз — глаз, ЗН — зрительный нерв, МВо — мышечные волокно, МС — мышечный
слой, ОбГ — обонятельный ганглий, ОбН — обонятельный нерв, ООН — окончания обонятельного нерва,
РО — ретрактор омматофора, СГз — сетчатка глаза, X — хрусталик, ПЭЩ — покровный эпителий
щупальца.
плоскости ноги, — нерв пениса, отходящий от правого церебрального ганглия
и не имеющий аналога на правой стороне. Нерв пениса иннервирует дисталь-
ную часть полового аппарата вместе с областью полового атриума.
Наиболее сложно организована система обонятельных нервов, также отходя-
щих от передней части процеребрума. Вначале это один относительно тонкий
нерв, идущий параллельно ретрактору омматофора. В апикальном отделе щу-
пальца, под глазом, этот нерв внезапно и резко расширяется в небольшой обо-
нятельный ганглий сложной конфигурации (рис. 16). От обонятельного ганг-
лия отходит несколько довольно толстых нервов, которые, подходя к стенкам
омматофора, интенсивно древовидно ветвятся. Щупальце, изображенное на
рис. 16, I, частично ввернуто; если оно будет полностью расправлено, глаз
займет терминальное положение, а участки покровов, иннервируемые ветвями
обонятельных нервов, расположатся вокруг глаза близ вершины щупальца,
как показано на схеме (рис. 16, II).
Следующая пара нервов, отходящих от процеребрума — фарингеальные (гло-
точные), уходящие под наружную мускульную оболочку глотки одной нитью и
разветвляющиеся в толще ее покровов.
МОРФОЛОГИЯ И АНАТОМИЯ
37
Наконец, самая внутренняя пара хорошо заметных нервных тяжей —
церебро-буккальные коннективы, подходящие к маленьким буккальным ганг-
лиям: последние располагаются на поверхности глотки по бокам от пищевода
и иннервируют глотку и переднюю часть пищевода. Иннервация глотки осуще-
ствляется также тонкими нервами, отходящими от дистальной части церебро-
буккальных коннективов. Как и церебральные ганглии, буккальные тоже свя-
заны короткой комиссурой, проходящей над пищеводом.
Помимо перечисленных нервов от процеребрумов отходят три пары очень
тонких и плохо заметных лабиальных нервов, идущих вдоль глотки к переднему
концу тела. Они обслуживают нижние щупальца, околоротовые лопасти, верх-
нюю губу и систему мышечных волокон, окаймляющую ротовое отверстие.
Под церебральными, прилегая справа и слева к нижне-боковым поверхно-
стям пищевода, располагается пара округлых педальных ганглиев. Соединяю-
щая их комиссура настолько коротка, что ганглии выглядят просто плотно
прижатыми один к другому. На верхней поверхности ганглиев, в задней их
части, хорошо видны относительно крупные статоцисты, нередко окруженные
кольцом пигмента. На срезах обычно можно рассмотреть многочисленные сфе-
рические и овальные статолиты, но ресничную выстилку внутренней поверх-
ности статоцистов рассмотреть не удалось. Основная роль педальных ганглиев —
иннервация ноги; в соответствии с этим от вентральных поверхностей ганглиев
отходит несколько коротких сильно ветвящихся педальных нервных тяжей.
В каудальном направлении от педальных ганглиев и несколько выше их
заметны левый плевральный и правый плевральный ганглии, имеющие округло-
треугольную форму. Комиссура между ними отсутствует, но каждый из них
связан коннективами с церебральным и педальным ганглиями той же стороны.
От левого плеврального ганглия отходит, насколько удается проследить,
единственный нерв, подходящий к ретрактору левого омматофора. Что же ка-
сается ретрактора правого омматофора, то он в значительной мере иннервиру-
ется нервом пениса, так как этот ретрактор находится в тесном контакте с пе-
нисом. Контакт обеспечивается соединительнотканно-мускульными перепон-
ками.
Основное назначение плевральных ганглиев — иннервация колумеллярной
мускулатуры, и, кроме того, Стинберг (Steenberg, 1925) отмечает, что от этих
ганглиев к переднему концу тела отходят очень тонкие нервы; однако соот-
ветствующих иллюстраций этих нервов Стинберг не приводит.
Далее на левой стороне пищевода расположен левый париетальный ганг-
лий, иннервирующий левую часть мантии и мантийного валика. Правый парие-
тальный ганглий сильно сдвинут назад, снабжает нервами правую сторону
мантии и мантийного валика и тесно сближен с непарным висцеральным (абдо-
минальным) ганглием, расположенным слева от правого париетального. От
висцерального ганглия отходят два нерва: анальный и интестинальный.
Половой аппарат
Репродуктивный тракт, начинающийся гонадой (гермафродитной железой),
спускается по спирали вдоль столбика в непосредственной близости от него,
переходит в полость ноги и заканчивается половым отверстием позади основа-
ния глазного щупальца (рис. 17). Петля, образованная дистальными участками
мужского отдела, вытянута вдоль женских путей. '
В общем, несмотря на своеобразие в деталях строения дистальных участков
мужского отдела, половой аппарат Pupillina построен по той же схеме, что
и у всех прочих Geophila.
Гонада залегает в толще тканей верхнего отдела печени и составлена не-
сколькими гроздями фолликулов. У животных большого и среднего размера
число гроздей может достигать 7—8, у мелких гонада состоит из двух гроздей.
В каждом фолликуле формируются как мужские, так и женские гаметы. Грозди
гермафродитной железы впадают в тонкий и почти прямой участок гермафро-
38
ОБЩАЯ ЧАСТЬ
дитного протока. Немного отступя от самой нижней грозди гермафродитный
проток Buliminidae окружен мутовчато расположенными семенными пузырь-
ками (vesiculae seminalis); у прочих семейств эта мутовка отсутствует и семен-
ные пузырьки входят в состав гермафродитного протока. Далее проток утол-
щается, образуя множество плотно сомкнутых извивов, и переходит в квад-
ривий, частично погруженный в ткани белковой железы. Строение квадривия
Pupillina сходно с таковым Helicoidea (Л. Шилейко, А. Шилейко, 1975), т. е.
Рис. 17. Положение полового аппарата Turanena in situ.
Отделы пениального аппендикса: А, — базальный, А6 — терминальный; ЕЖ — белковая железа; ВОВ —
верхний, НОВ — нижний отделы вагины; ГЖ — гермафродитная железа, ГП — гермафродитный проток,
ДСП — дивертикул семяприемника, Пе — пенис, ПО — половое отверстие, ПР — половой ретрактор,
РСп — резервуар семяприемника, Сп — семяпровод, СВд — спермовидукт, СП — семенные пузырьки,
Ц — цэкум, Эп — эпифаллус.
здесь имеется восходящий проток (самый дистальный участок протока гонады),
нисходящий проток (ведущий в просвет спермовидукта), сперматека (депо ал-
лоспермы) и камера оплодотворения, откуда оплодотворенные яйца поступают
в матку.
Среди Pupillina имеются как яйцекладущие, так и яйцеживородящие формы.
У последних в матке формируется молодь, которая сразу же по выходе из ро-
дительского организма уже ведет самостоятельное существование. Стенки
матки у яйцеживородящих видов могут чрезвычайно сильно растягиваться,
и, когда матка заполнена яйцами, вскрыть животное, не повредив стенок ор-
гана, очень трудно, тем более что они становятся почти прозрачными. У боль-
шинства мелких яйцекладущих форм яйца до момента откладки находятся
в слепых карманах матки, которые Стинберг (Steenberg, 1925) называет cul-
de-sac.
МОРФОЛОГИЯ И АНАТОМИЯ
39
К матке тесно прилегает простата, и этот объединенный отдел называется
спермовидуктом, что не совсем точно; однако термин «спермовидукт» прижился
в отечественной литературе, и нет смысла его заменять более точным. Для так-
сонов, составленных мелкими формами, характерно резкое уменьшение про-
статы; например, у многих Vertiginidae простата сравнительно с маткой кро-
шечная и представлена всего несколькими фолликулами, расположенными
у нижнего края белковой железы; у самых мелких форм (Truncatellina) про-
стата внешне вообще не выражена. Кроме того, слабое развитие простаты в не-
которых случаях связано с яйцеживорождением и носит в таком случае времен-
ный характер. Так, в период созревания эмбрионов в матке у Pupilia простата
выражена гораздо слабее, чем
тогда, когда матка яиц не содер-
жит.
Поскольку семяпровод начи-
нается от нижнего конца простаты,
место отхождения этого протока
у видов с короткой простатой
расположено выше, т. е. ближе
к белковой железе.
Матка обычно отделена от ци-
линдрической вагины кольцевой
перетяжкой; от вагины ответв-
ляется проток семяприемника.
Участок вагины, заключенный
между маткой и основанием про-
Рис. 18. Схема строения совокупитель-
ного аппарата Buliminidae:
At—Аъ — отделы пениального аппендикса,
Пе — пенис, ПО — половое отверстие, ПР —
половой ретрактор, Сп — семяпровод, Ф —
флагеллум (бич), Ц — цэкум, Эп — эпифал-
лус.
тока семяприемника, именуется верхним отделом вагины (превагина); нижний
отдел вагины располагается между основанием протока семяприемника и
половым атриумом.
Семяприемник состоит из протока, резервуара и дивертикула, отходящего
от протока на большем или меньшем расстоянии от резервуара. Дивертикул
семяприемника является наименее обязательным атрибутом, так как он может
быть развит в различной степени вплоть до полной редукции даже в пределах
одного вида (Hesse, 1933; Матёкин, 1959).
О значительной изолированности Pupillina от прочих Geophila и высокой
степени специализации подотряда свидетельствует в основном строение дисталь-
ных участков мужского отдела (рис. 18). Самая общая схема строения сово-
купительного аппарата такова. Пениальная трубка разделяется на два отдела:
проксимальный — эпифаллус, и дистальный — пенис. Единственное исклю-
чение составляют Achatinellidae, у которых эпифаллуса нет. Каждый из отделов
пениальной трубки снабжен придатком. На эпифаллусе расположен либо цэ-
кум в виде округлого бугорка, заключающего щелевидную полость (Bulimi-
nidae), либо придаток червеобразной, веретеновидной или конической формы,
строение которого достаточно разнообразно (Orculoidea, Pupilloidea, Chond-
rinoidea).
Близ атриальной области собственно пениса располагается аппендикс, со-
стоящий из пяти отделов: 1) базальный цилиндрический толстостенный отдел
(Л3); 2) шаровидный отдел, отчлененный перетяжкой от базального (А2); 3) ко-
роткий узкий цилиндрический отдел (А3); 4) длинная и очень тонкая трубка
40
ОБЩАЯ ЧАСТЬ
с мускулизованными стенками (Л4); 5) удлиненная булавовидная терминальная
ампула (Л5). Половой ретрактор двуветвистый; одна из его ветвей крепится
к пенису, другая — чаще всего на границе между и Л2, гораздо реже к сред-
ней части Аг.
Рассмотрим вначале органы, относящиеся к эпифаллусу (рис. 19). Как
отмечалось, у Buliminidae они представлены бугорком со щелевидной полостью
(рис. 19, 7). У всех прочих групп, имеющих дополнительный вырост на эпифал-
лусе, он (вырост) имеет различную форму и первично продольную внутреннюю
перегородку (рис. 19,77); характерно, что Гиттенбергер именует этот вырост
у Spelaeodiscidae слепым мешком пениса («Penisblindsack»), а не цэкумом (Git-
tenberger, 1975, S. 264). Поскольку нет уверенности, что описанные придатки —
гомологичные структуры (хотя это, конечно, не исключено), то название «цэ-
кум» лучше оставить для при-
датка, имеющегося у Buliminidae,
а для прочих таксонов использо-
вать название «придаток эпифал-
луса».
Рис. 19. Дополнительные органы на
эпифаллусе Pupillina.
1 — цэкум Bullminoidea, II — придаток эпи-
фаллуса Orculoidea и Pupilloidea.
Если цэкум возник в результате местной Дифференциации ткани эпифал-
луса, то строение придатка эпифаллуса наводит на мысль, что его формирование
связано со складыванием эпифаллуса; вспомним, что у Achatinellidae семяпро-
вод впадает в пенис апикально или субапикально; если проксимальная часть
пениса и дистальная часть семяпровода преобразуются в эпифаллус, то воз-
никновение придатка эпифаллуса с продольной перегородкой внутри будет за-
кономерным результатом этого процесса.
Роль цэкума Buliminidae легко понять, если вскрыть особь, содержащую
внутри эпифаллуса сперматофор: последний несет «шпору» (рис. 20), формирую-
щуюся в полости цэкума и точно отвечающую по форме этой полости (Schikow,
1978). Сперматофор, снабженный шпорой, неспособен двигаться внутри пе-
ниальной трубки, поскольку шпора играет роль якоря, закрепленного в по-
лости цэкума. Сперматофор способен покинуть мужской отдел лишь тогда,
когда последний будет вывернут в достаточной степени. Заслуживает также
внимания строение задней части сперматофора Buliminidae (рис. 20): в общей
схеме сперматофор представляет собой желоб, верх которого затянут тонкой
пленкой, а в задней части пленка отсутствует и сперматофор открыт. Функцио-
нальный анализ этих особенностей приведен ниже, а пока отметим, что при-
даток эпифаллуса у прочих таксонов к образованию сперматофора отношения
не имеет, о чем свидетельствует как внешний вид сперматофора (например, см.
рис. 85), так и его положение внутри эпифаллуса.
Интересные модификации полового аппарата наблюдаются у пацифических
Achatinellidae (Cooke, Kondo, 1960); в частности, у некоторых групп наблюда-
ется функциональная замена аппендиксом пениса, в связи с чем последний пре-
терпевает редукцию. Кук и Кондо нашли три варианта строения пениального
аппендикса: секреторный, секреторно-эйякуляторный и копуляторный. По-
нятно, что возникновение последнего варианта связано именно с редукцией
пениса (что касается первых двух вариантов, то участие базального отдела пе-
ниального аппендикса в выделении какого-либо секрета сомнительно; этот
вопрос рассматривается ниже, при функциональном анализе совокупительного
аппарата). Видимо, субституция аппендиксом пениса наблюдается и у Jaminia
quadridens (Buliminidae, Jaminiinae): достаточно сравнить тончайший ните-
МОРФОЛОГИЯ И АНАТОМИЯ
41
видный пенис и мощный базальный отдел пениального аппендикса у этого
вида, чтобы убедиться в справедливости такого предположения.
На том основании, что многие авторы гомологизировали (впрочем, бездо-
казательно) пениальный аппендикс Pupillina со стилофорами и слизистыми же-
лезами йысших Geophila (Albers, 1860; Semper, 1870; Ihering, 1892, 1909; Wieg-
mann, 1901; Forcart, 1940), принято считать, что аппендикс выполняет раздра-
жающую функцию при спаривании^ Существует также предположение, что
роль этого придатка заключается в восприятии сперматофора партнера.
Рис. 20. Строение сперматофора и его положение в эпифаллусе у Pseudonapaeus albiplicatus.
I — внешний вид сперматофора, II — эпифаллус со сперматофором внутри и .серия поперечных срезов
на разных уровнях. Зачерненная часть — стенки сперматофора, Ш — шпора. Остальные обозначения
как на рис. 18.
Функция эпифаллуса (и бича, нередко присутствующего у различных Pu-
pillina) такая же, как и у представителей всех других подотрядов, имеющих
сперматофор, — формирование этого сперматофора. Роль цэкума показана выше
(образование шпоры на поверхности сперматофора). Что касается пениального
аппендикса, то функция его своеобразна и будет рассмотрена в разделе, посвя-
щенном размножению (см. с. 4.7).
Изложенные материалы характеризуют половой аппарат Pupillina в самой
общей форме. На самом деле разнообразие в строении совокупительного ап-
парата таково, что здесь можно видеть наличие пяти вариантов строения (рис. 21),
характер связи между которыми неясен.
Варианты эти следующие.
1. Первичное и полное отсутствие эпифаллуса, и, следовательно, всех при-
датков этого отдела, а также сперматофора (рис. 21, II) (Achatinelloidea).
2. Наличие придатка эпифаллуса, близ основания которого крепится пе-
ниальная ветвь полового ретрактора — либо немного ниже придатка (Orcu-
loidea — рис. 21, III), либо несколько выше (Pupilloidea — рис. 21, IV).
Картину, изображенную на рис. 21, I (Gochlicopoidea), можно, вероятно, ис-
толковать как производное обсуждаемого варианта за счет редукции придатка
42
ОБЩАЯ ЧАСТЬ
эпифаллуса, резкого уменьшения Аг и, как следствие, редукции ветви полового
ретрактора, который всегда крепится к Аг,
3. Пенис и эпифаллус резко разделены чрезвычайно узким пережимом, при-
чем единственная ветвь полового ретрактора крепится к месту перехода семя-
провода в эпифаллус, чего не наблюдается ни в каких других случаях. Кроме
Рис. 21. Варианты строения совокупительного аппарата (Z— VII) к гермафродитного про-
тока (VIII—IX) у Pupillina.
I— Cochlicopoidea; 11 — Achatinelloidea; Ill — Orculoidea; IV — Pupllloidea; V — Vertiginoidea; VI —
Chondrinoidea; VII — Buliminoidea; VIII — гермафродитная железа и проток, свойственные всем над-
семействам, кроме Buliminoidea; IX — гермафродитная железа и проток, свойственные Buiimlnoidea.
ГЖ— гермафродитная железа, ГП — гермафродитный проток, Не — пенис, ПА — пениальный аппен-
дикс, ПР — половой ретрактор, ПЭ — придаток эпифаллуса, Сп — семяпровод, Ц — цэкум, ЭП — эпифал-
лус, СП — семенные пузырьки.
того, совокупительный аппарат здесь не несет никаких придатков (рис. 21, У)
(Vertiginoidea).
4. Пенис и эпифаллус образуют практически замкнутую петлю за счет
того, что место перехода семяпровода в эпифаллус плотно контактирует и срас-
тается с атриальным участком пениса. Исходно в месте контакта пениса и эпи-
фаллуса имеется вырост, гомологичный, вероятно, придатку эпифаллуса. Одна
из ветвей полового ретрактора крепится к дистальному отделу пениса, адругая —
к проксимальной части эпифаллуса. Существенно также, что мышечная лента,
„ представляющая собой половой ретрактор, расщепляется иначе, чем двуветвис-
тый ретрактор в прочих вариантах: здесь скорее можно говорить о расслоении
мышечной ленты (рис. 21, УГ) (Chondrinoidea).
МОРФОЛОГИЯ И АНАТОМИЯ
43
5. Подробно разобранный выше вариант, для которого характерно наличие
на эпифаллусе настоящего цэкума; цэкум всегда расположен значительно выше
(т. е. ближе к семяпроводу) места прикрепления пениальной ветви полового
ретрактора (рис. 21, VII) (Buliminoidea).
Достойно также внимания то, что при вариантах, представленных на рис. 21,
I—VII, семенные пузырьки входят в состав гермафродитного протока (рис. 21,
VIII), а у Buliminidae семенные пузырьки всегда морфологически обособлены
и расположены на проксимальной части этого протока (рис. 21, IX).
Перейдем к рассмотрению собственно пениса. У многих Pupillina пенис
представляет собой простую трубку с более или менее ясно выраженным узором
Рис. 22. Схема строения пениса Pseudonapaeinae (Buliminidae).
Общий вид и поперечные срезы через пенис; внизу — парасагиттальный срез пениса по линии а—б. ВВ —
внутренние, НВ — наружные (внешние) ветви V-образных складок, Кк — козырек, Не — пенис, Ск —
складки внутри пениса, Эп — эпифаллус.
на внутренней стенке, состоящим из складок, ориентированных в основном
продольно. Многочисленные усложнения оформления внутренней поверхности
пениса наблюдаются преимущественно у Buliminidae и сводятся к двум основ-
ным направлениям.
1. Складки на внутренней поверхности пениса образуют какой-либо харак-
терный узор. Чаще всего узор составлен одной складкой (пилястром) V-образ-
ной формы (или таких складок две); место сгиба складок всегда направлено
в сторону полового атриума (рис. 22).
2. Исходные продольные складки внутри пениса дифференцируются на ряды
бугорков характерной формы, которая здесь обозначается как призмоконичес-
кая. Это выражение означает, что основание каждого бугорка имеет форму овала
или круга, что свойственно конусу; вершинная же часть бугорка превращена
в гребень, что создает сходство с призмой.
В любом случае может формироваться папилла пениса — либо в виде откры-
того желоба (незамкнутая), либо в виде трубки (настоящая, или замкнутая).
Примитивные Buliminidae имели, вероятно, пенис в наименее специализи-
рованном варианте, т. е. пенис представлял собой простую трубку с системой
продольных складок на внутренней поверхности. При образовании замкнутой
папиллы, которая возникает за счет разрастания тканей, расположенных в месте
44
ОБЩАЯ ЧАСТЬ
перехода эпифаллуса в пенис, фактура внутренних стенок теряет значение и
почти исчезает (так как сперматофор со стенками пениса теперь не соприкаса-
ется). В противном случае специализация направлена на усложнение внутрен-
ней фактуры пениса, что нередко сопровождается возникновением незамкну-
той папиллы. Надо заметить, что замкнутая пениальная папилла характерна
преимущественно для высших Geophila и в обсуждаемом подотряде — явле-
ние редкое: это три подсемейства Buliminidae (Merdigerinae, Chondrulopsininae,
Multidentulinae). Первое из указанных подсемейств, по-видимому, монотипичес-
кое, два остальных по числу видов также очень небольшие.
Естественно, гипотетический исходный тип строения пениса среди рецепт-
ных форм не обнаружен; однако нечто близкое к нему имеется у некоторых
Pseudonapaeinae: при вскрытии Pseudonapaeus regelianus найдено, что от клоаки
вверх поднимается несколько параллельных высоких складок, а выше середины
пениса они вдруг резко сминаются, полностью теряют правильность формы и
расположения; здесь же имеется несколько бугорков и неправильной формы
складок, по-разному ориентированных. Вся эта система в совокупности образует
клапан, изолирующий отверстие эпифаллуса от дистального участка пениса
(рис. 157).
У Subzebrinus labiellus среди серии параллельных складок отчетливо вы-
деляются две, идущие рядом и уходящие в эпифаллус. Близ отверстия эпифал-
луса они образуют короткий тоннель, смыкаясь верхними кромками (рис. 185).
Для множества Pseudonapaeinae характерно, как отмечалось, наличие
одной, или чаще, двух V-образных складок (рис. 22). Поскольку складки удоб-
нее всего изучать на вскрытом пенисе, т. е. на плоскостной развертке, когда
эти складки лежат рядом, а не напротив, как на самом деле внутри пениса, то
для удобства изложения будем различать внутренние и внешние ветви этих
складок; внутренние — те, которые уходят в эпифаллус, наружные (внешние)
смыкаются над отверстием эпифаллуса, также нередко образуя тоннель, верх-
няя часть которого может выступать наподобие козырька. На базе такой струк-
туры за счет резкого разрастания козырька может формироваться стержне-
видная «папилла», как у Pseudonapaeus submucronatus (рис. 183).
Трехлопастная папилла, свойственная видам рода Laevozebrinus, образуется
также за счет разрастания козырька (верхняя лопасть) и примыкающих участ-
ков V-образных складок (боковые лопасти).
БИОЛОГИЯ И ЭКОЛОГИЯ
Питание
Почти все представители Pupillina питаются растительным материалом —
свежим, сухим или гниющим. Единственное исключение составляют тропичес-
кие Sagdoidea, включающие только хищные формы. Хищничество резко стиму-
лирует совершенствование экскреторного аппарата, поэтому у Sagdoidea на-
блюдается настоящая сигмуретрия; однако наличие пениального аппендикса
достаточно характерного строения (Pilsbry, 1940) свидетельствует о принад-
лежности Sagdoidea к Pupillina.
Потребление свежих частей высших растений наблюдается редко и отмечено
только для некоторых крупных представителей Buliminidae. Гораздо более
обычная пища — отмершие, сухие или гниющие части травянистых растений,
лиственный опад, разлагающаяся влажная древесина, грибы, лишайники.
Вероятно, заглатывание древесных частиц носит случайный характер, и они
поглощаются вместе с гифами грибов, которыми пронизана разлагающаяся дре-
весина. Опад хвойных пород не потребляется, и обитание некоторых мелких
видов в хвойных лесах определяется наличием там грибов.
Особенности питания отражаются на строении радулярных зубов; их мно-
гообразие можно свести к пяти основным типам (рис. 23), между которыми на-
блюдаются и промежуточные состояния.
БИОЛОГИЯ И ЭКОЛОГИЯ
45
I тип: центральный зуб с тремя зубцами, латеральные — двухзубцовые,
причем внутренний зубец, или эктокон, т. е. обращенный к средней линии ра-
дулы, больше наружного — энтокона. Маргинальные зубы несут по нескольку
тонких и длинных зубцов (рис. 23, I).
II тип: центральный зуб — однозубцовый, латеральные — как в типе I,
но зубцы более массивные. Латеральные постепенно переходят в маргинальные,
приобретая еще по 1—2 дополнительных зубца, располагающихся кнаружи
от основного (рис. 23, II).
III тип: как центральный, так и первые латеральные зубы — однозубцовые.
По мере приближения к краям радулярной мембраны латеральные становятся
Рис. 23. Основные типы радулярных зубов Pupillina.
Арабские цифры — порядковые номера зубов в ряду. I — Lauria cylindracea, II — Abida secale, III —
Chondrina avenacea, IV — Columella edentula, V — Vertigo moulinsiana (изображения зубов — из: Steen-
berg, 1925).
двухзубцовыми и переходят в маргинальные, несущие по 3—5 тонких зубцов
и напоминающие гребень (рис. 23, III).
IV тип: центральный зуб с тремя зубцами, из них средний лишь немного
больше боковых, латеральные — также трехзубцовые, причем два боковых
зубца примерно одного размера, а между ними располагается тонкий допол-
нительный зубец. На зубах, сидящих ближе к краям радулы, рядом с первым
появляется еще по 1—2 дополнительным, а справа и слева от боковых зубцов
возникают часто зачатки самых наружных зубцов. В этом типе граница между
латеральными и маргинальными зубами особенно сильно размыта (рис. 23, IV).
V тип: между тремя исходными зубцами центрального зуба появляются два
тонких дополнительных, в результате центральный зуб становится - пятизуб-
цовым. Латеральные зубы несут по три основных зубца, а между ними еще по
два дополнительных, иными словами, латеральные зубы отличаются от цент-
ральных в основном лишь тем, что они несколько асимметричны. Маргинальные
зубы похожи на латеральные, но число дополнительных зубцов увеличивается
до 5—6; кроме того, у них резко сужена базальная пластинка (рис. 23, V).
Часто выделяют еще один тип, характеризующийся наличием только двух
зубцов центрального зуба, т. е. срединный зубец редуцирован.
Рассмотрение вопроса о том, какой из типов радулы первичен, не входи®
в задачи настоящего исследования; однако надд отметить, что, по-видимому,
46
ОБЩАЯ ЧАСТЬ
I тип вообще характерен для растительноядных Geophila. От этого типа наме-
чаются две линии специализации: одна линия — к питанию лишайниками, про-
израстающими на скалах, для чего необходимо скоблить жесткие субстраты
(типы II и III); вторая линия — к употреблению рыхлого мягкого детрита и гри-
бов (типы IV и V). Таким образом, виды, переходящие к жизни среди скал, не-
зависимо от их систематического положения, приобретают радулу с более мас-
сивными зубами, позволяющими эффективно сдирать лишайники с шерохова-
той поверхности камней. Напротив, у видов, освоивших влажные, затененные,
богатые травянистой растительностью биотопы, развиваются дополнительные
зубцы, благодаря чему зубы превращаются в подобие миниатюрных щеточек.
Сходные тенденции наблюдаются и у других групп Geophila (Лихарев, 1962;.
Solem, 1973).
Примечательно, что у видов, населяющих широкий спектр биотопов и имею-
щих широкий ареал, у видов с гибкой экологией (Zeissler, 1973), радула отно-
сится к типу I.
В главе, посвященной строению пищеварительного аппарата, разобран
механизм работы глотки и радулы; здесь уместно добавить несколько слов по
поводу функций отдельных групп радулярных зубов. Поскольку при основном
трущем движении радулы о субстрат радулярная мембрана натянута между
крыльями одонтофора, весь комплекс зубов, соприкасающийся с поверхностью
пищевого объекта, выполняет единообразные движения. Однако в силу того,
что центральные и первые латеральные зубы длиннее и мощнее крайних лате-
ральных и маргинальных, основную работу выполняют именно они: борозды
на поверхности пищевого куска «пропахиваются» преимущественно срединным
зубцом центрального зуба и медиальными зубцами латеральных зубов. Не слу-
чайно сломанные зубцы наблюдаются среди центральных и первых латеральных
и никогда — среди маргинальных. Отсюда следует, что назначение маргиналь-
ных зубов иное, чем центральных и латеральных. К тому же строение мар-
гинальных зубов менее разнообразно.
Роль маргинальных зубов становится понятной, если проследить их переме-
щение по мере втягивания и продольного складывания радулярной мембраны.
По завершении скоблящего движения на центральных и первых латеральных
зубах остаются частицы пищевого материала; глубина погружения зубов в пи-
щевой объект и, следовательно, ширина «захвата» радулы определяются как
размерами моллюска, так и тем, насколько тверда поверхность пищевого
куска. При последующем втягивании радула складывается вдоль и пищевые
частички оказываются внутри узкого желоба, почти кольцевого на поперечном
разрезе, дно которого покрыто центральными и первыми латеральными, а боко-
вые поверхности — крайними латеральными и маргинальными зубами.
При последующем сокращении мощного компрессора глотки пищевые частицы
спрессовываются, и маргинальные зубы при этом погружаются в пищевой комок
всеми зубцами, не давая ему расползаться. Эта роль существенна, так как если
пищевые частицы не будут собраны в компактный комок, то при следующем
выдвижении радулы размазанные по ее поверхности частицы будут увлекаться
к ротовому отверстию; комок же удерживается за счет трения о верхний свод
ротовой полости. При завершении следующего скоблящего движения и втяги-
вания радулы этот комок передается дальше — по направлению к пищеводу.
Размножение
Все Pupillina,— гермафродитные животные, но, подобно большинству дру-
гих Geophila, у них наблюдается протерандия, т. е. вначале протекает преиму-
щественно мужская фаза, а затем женская. Внешне это выражается в относи-
тельных размерах гермафродитного протока и белковой железы. Во время про-
текания мужской фазы гермафродитный проток очень сильно развит, широкий,
набухший; белковая железа, напротив, небольшая. Когда в гонаде проте-
БИОЛОГИЯ И ЭКОЛОГИЯ
47
кает преимущественно оогенез, картина обратная: сильноразвитая белковая
железа при тонком гермафродитном протоке (Лившиц, Шилейко, 1978).
Протерандрия у улиток малого размера приводит к частичной или полной
дегенерации дистального участка мужского отдела гениталий. Так, имеется
прямая и легко заметная зависимость между степенью развития совокупитель-
ного аппарата и состоянием матки у Pupillidae: если матка не содержит яиц,
то пенис, эпифаллус и пениальный аппендикс хорошо развиты и все их отделы
хорошо развиты и ясно различимы; если же в матке имеется развивающаяся
Рис. 24. Схема хода спермиев в половых путях Buliminidae.
Справа (в кружке) — частично вывернутый пениальный аппендикс. Изображения, расположенные справа
от штриховой линии, относятся к одному из партнеров, а изображения слева от штриховой линии — к дру-
гому. А,—As — отделы пениального аппендикса, ДСП — дивертикул, РСп — резервуар семяприемника,
ПаА — папилла аппендикса, Сд5 — сперматофор, Ср — сфинктер, Ц — цэкум, Ш — шпора сперматофора,
РА — ретрактор аппендикса. Объяснения в тексте.
молодь, то протоки мужского отдела становятся тонкими, границы между от-
делами пениального аппендикса стираются (ср. два изображения на рис. 98).
Еще дальше этот процесс заходит у Valloniidae: дистальная часть мужского
отдела после выполнения его функции полностью редуцируется. Судя по тому,
что особи с развитым пенисом и пениальным аппендиксом встречаются исклю-
чительно редко (из более чем 100 исследованных мною особей различных видов
Vallonia развитый мужской отдел найден лишь у одного экземпляра V. costata),
можно быть уверенным, что редукция происходит очень быстро.
Наиболее своеобразные, сравнительно с другими легочными моллюсками,
особенности физиологии размножения Pupillina определяются наличием у них
сложного дополнительного органа — пениального аппендикса, функционально
связанного с ходом мужских половых продуктов и с процессом спаривания.
При гистологическом исследовании особей двух видов Buliminidae (Pse-
udonapaeus trigonochilus и P. albiplicatus), собранных непосредственно перед
массовым спариванием (в мае) в среднем течении р. Нарын на Тянь-ТПяно,.
внутри А5 были обнаружены спермин. Очевидно, что это не может быть алло-
сперма (д. е. сперма партнера), во-первых, потому, что спаривания еще не было
48
ОБЩАЯ ЧАСТЬ
(оно началось на следующий день), и, во-вторых, потому, что для восприятия
спермиев партнера существует семяприемник. Следовательно, обнаруженные
спермин есть аутосперма. Скопления сперматозоидов, покидая семяпровод,
проходят полость эпифаллуса и проксимальной части пениса и поступают
в пениальный аппендикс, где проходят, возможно, период физиологического
дозревания. Поскольку в названных протоках мерцательный эпителий отсутст-
вует, движение спермиев, очевидно, частично активное, а частично обеспечива-
ется перистальтикой протоков. После того как сперматозоиды собраны
внутри А5, внутри эпифаллуса (и бича, если он имеется) формируется спермато-
фор.
Рис. 25. Схема строения и действия пениального аппендикса.
I — состояние покоя; II — пениальный аппендикс частично вывернут, ретрактор сокращен. А,—А4 —
отделы пениального аппендикса; ПаА — папилла аппендикса, РА — ретрактор аппендикса.
Надо заметить, что скорость образования сперматофора очень высока.
Прямые наблюдения за скоростью секретирования сперматофора отсутствуют,
но косвенные данные заставляют предполагать, что весь процесс завершается
за считанные часы; один из наиболее важных косвенных показателей заключа-
ется в том, что обнаружить сперматофор внутри эпифаллуса удается исключи-
тельно редко — неизмеримо реже, чем найти его в семяприемнике (см. также
Breure, Eskens, 1977). Очевидно, скорость формирования сперматофора в эпи-
фаллусе многократно превышает скорость его разрушения в семяприемнике:
при вскрытии 72 экз. Р. albiplicatus, собранных в долине р. Нарын 1 июня 1972 г.
в утренние часы во время массового спаривания, у 34 экз. были найдены спер-
матофоры в семяприемнике и лишь у одной особи — в эпифаллусе.
Рассмотрим рис. 24, иллюстрирующий ход спермиев в половых путях. У жи-
вотного, вполне готового к спариванию, спермин собраны внутри As, а сперма-
тофор сформирован, но пуст (вспомним, что задний конец сперматофора открыт).
В процессе спаривания выворачивается пенис, а также два нижних отдела
пениального аппендикса (или только Л^. Пустой сперматофор начинает пода-
ваться в половые пути партнера, и когда большая часть его уже покинула пре-
делы пениса, в полость сперматофора .начинает нагнетаться сперма. При этом
первостепенную роль играют энергичные сокращения мощной мускулатуры
(главным образом кольцевой) А3 и А4. Регулятором поступления спермы в спер-
БИОЛОГИЯ И ЭКОЛОГИЯ
49
матофор служит А2, снабженный сфинктером и папиллой (рис. 25). Далее за-
полненный сперматофор втягивается в семяприемник партнера. Если имеется
дивертикул семяприемника, то передний конец сперматофора направляется
в дивертикул. Здесь сперматофор лизируется, а спермин собираются в резер-
вуар семяприемника; часть сперматозоидов здесь фагоцитируется, а часть, спус-
тившись по протоку семяприемника, поднимается затем по серозному каналу
спермовидукта в камеру оплодотворения.
Большая часть Pupillina — яйцекладущие животные, хотя известно немало
яйцеживородящих. Таковы Lauriinae (Orculidae), все Pupillidae, многие Acha-
tinellidae.
Откладывание яиц и рождение молоди у тропических видов растянуто во-
времени или, если год делится на сухие и влажные сезоны, происходит в начале
сезона дождей. В умеренных широтах цикличность размножения более четкая
и основная масса молоди появляется в начале лета, хотя у яйцеживородящих
видов всегда имеется определенный процент взрослых особей, содержащих
эмбрионы в течение почти всего сезона' активности. Только что вылупившаяся
молодь менее устойчива к различным неблагоприятным воздействиям, в первую,
очередь к засухе, и взрослые особи, носящие эмбрионы, служат резервом вос-
становления популяции, если суровые погодные условия года приведут к гибели
вылупившуюся весной молодь.
Половая активность регулируется не столько внутренними ритмами, сколько-
конкретной погодной обстановкой. Так, Vallonia costatav лабораторных условиях
размножается круглый год, если поддерживается достаточная влажность почвы.
Яйца откладываются в укромные сырые места: в дерновинки трав, под от-
ставшую кору пней и поваленных деревьев, под камни, в нижние слои листвен-
ного опада и в расщелины скал. Размеры кладки различны, число яиц у крупных
Buliminidae достигает 40 и даже более. Время развития яиц во внешней среде
различно и зависит как от систематической принадлежности вида, так и от
погодных условий, в первую очередь от температуры и влажности.
Среда и адаптации
Все многообразие признаков, за которыми признается адаптивное значение,
можно сгруппировать в несколько совокупностей в зависимости от того, чем
руководствоваться при выделении этих совокупностей. Так, если рассматривать
особенности, меняющиеся в ходе онтогенеза, то можно выделить ювенильные
и дефинитивные группы признаков; если оценивать признаки в историческом
аспекте, то на первый план выходит их таксономический вес и речь идет уже
о том, какие признаки или какие состояния признаков и почему обеспечили
становление данного таксона. Если же в основу поставить наблюдения за из-
менчивостью признаков, носящей экологический характер, то возникает
возможность связать появление различных признаков с воздействием каких-то
конкретных условий.
Нетрудно заметить, что «экологические» признаки представляют собой,
образно говоря, ассортимент того разнообразия изменчивости, которое вид спо-
собен проявить, адаптируясь к различным условиям (в этом отношении самым
ярким примером может служить Pseudonapaeus albiplicatus с поразительно
широким размахом изменчивости конхологических признаков). Наиболее пер-
спективные, «удачные» пункты этого ассортимента ложатся затем в основу
дальнейшей эволюции таксона, в процессе которой повышается таксономический
вес наиболее эффективно действующих признаков (состояний признака) —
от форм внутривидовой изменчивости до признака таксона ранга рода, подсе-
мейства и т. д.
Поскольку в адаптивном и эволюционном отношении ювенильные и дефинити-
ные признаки практически равноценны, то нельзя говорить о резких границах
между отмеченными категориями признаков, но рассматривать их удобнее
отдельно.
4 А. А. Шилейко
50
ОБЩАЯ ЧАСТЬ
Ювенильные адаптации. Наиболее яркие и существенные
признаки, отличающие молодых особей от взрослых, связаны с устьевой арма-
турой и наблюдаются у некоторых Orculidae.
По времени образования в онтогенезе устьевые образования бывают трех
типов: либо они появляются в раннем постэмбриогенезе и не испытывают су-
щественных изменений вплоть до завершения формирования раковины; либо
раковина.нарастает, не имея устьевой арматуры, до субдефинитивного состояния,
когда быстро достраивается завершающий участок устья вместе с зубным оформ-
лением; либо наконец, в процессе формирования раковины образуются какие-то
элементы устьевой арматуры, не связанные с оформлением дефинитивного
I
Рис. 26. Устьевая арматура Sphyradium doliolum на разных стадиях постэмбриогенеза.
Зачерненные части — париетальная и колумеллярная пластинки. Объяснения в тексте.
устья. Второй тип среди Pupillina самый обычный, но пока нас интересуют
первый и третий типы.
Рассмотрим вначале этапы формирования раковины Sphyradium doliolum,
принимая во внимание условия существования вида в течение постэмбриогенеза
(Шилейко, 1967а).
Этот вид размножается, откладывая по 10—20 яиц в различные укрытия.
Из яйца вылупляется моллюск с раковиной, составленной полутора оборо-
тами (рис. 26); впоследствии эмбриональные обороты легко отличить от дефини-
тивных благодаря спиральной скульптуре. Только что вылупившийся моллюск
имеет в устье маленькие, но хорошо различимые спиральную париетальную
и нитевидную колумеллярную пластинки; когда появляется первый дефинитив-
ный оборот, париетальная пластинка занимает около 3/4 его длины. Далее, по
сформировании 2—3 дефинитивных оборотов, париетальная пластинка занимает
почти весь последний оборот. Когда раковина насчитывает 3—4 дефинитивных
оборота, постепенно меняется облик париетальной пластинки: нижняя кромка
ее начинает волнообразно изгибаться. Наиболее развитая пластинка наблюда-
ется на стадии 5—6 дефинитивных оборотов; по мере формирования последнего
оборота кромка париетальной пластинки выравнивается. Длина ее на ранних
стадиях (1—4 оборота) относительно невелика: не более одного оборота. На
стадии 5—6 оборотов длина пластинки достигает иногда 2 целых оборотов,
а при дальнейшем росте длина ее вновь уменьшается. Наконец, у вполне сфор-
мированных раковин париетальная пластинка простирается обычно на 1—1.2 обо-
рота, редко больше. Говоря о длине париетальной пластинки, я имею в виду
только ту ее часть, которая вполне заслуживает это название, так как по мере
БИОЛОГИЯ И ЭКОЛОГИЯ
51
роста задние части пластинки резорбируются и остаются лишь в виде светлой
полоски.
Сопоставим приведенные материалы с условиями, в которых живет моллюск
на разных стадиях постэмбриогенеза. Поскольку яйца откладываются в сильно
затененные и укрытые места, то и вылупившаяся молодь попадает в те же условия.
Далее моллюск, по мере увеличения числа оборотов, поднимается в средние и
верхние слои лесной подстилки или в верхние горизонты осыпи. Устье в это
время относительно широкое, и по мере роста раковины сравнительное развитие
париетальной пластинки усиливается. В субдефинитивном состоянии животное
живет уже в тех же условиях, что и взрослый моллюск, а потому нуждается
И
Рис. 26 {продолжение).
в усилении защиты устья; кроме того, в это время он питается особенно интен-
сивно, зоб способен сильно растягиваться, а париетальная пластинка, играя
роль перегородки между зобом и задней кишкой, служит ограничителем, устра-
няющим давление на ректум со стороны зоба (подробнее о роли париетальной
пластинки сказано ниже). Соответственно именно в это время париетальная
пластинка достигает наиболее сильного развития.
Наконец, взрослые животные обитают преимущественно в средних, и особенно
верхних слоях подстилки, обладая уже дефинитивным устьем и суженным по-
следним оборотом.
Второй вид, подлежащий рассмотрению, — эндемик талышских лесов
Pagodulina lederi — еще более показателен в этом отношении. Молодой мол-
люск, вылупившийся в нижних горизонтах подстилки, вообще не имеет в уСтье
никаких зубов, пока не достигнет субдефинитивного состояния (6—оборотов).
На этой стадии формируется довольно мощное устьевое вооружение: колумел-
лярная и париетальная пластинки и удлиненно-бугорковидная палатальная
складка. По достижении полного числа оборотов (8—8х/2) упомянутые зубы
оказываются внутри раковины и с течением времени испытывают некоторую
редукцию. Иными словами, провизорный аппарат целиком формируется на
средних стадиях образования раковины и,,выполнив свою функцию, деградирует.
Третий случай — Lauria cyllndracea (рис. 27). Это живородящий вид, каж-
дая особь которого производит на свет от 4 до 12 молодых улиточек. Последние
появляются, имея уже сформированную раковину, составленную 2 оборотами
и снабженную одной слаборазвитой париетальной пластинкой. Когда нарастает
первый дефинитивный оборот, на базальном крае устья очень быстро формиру-
ется складка, параллельная плоскости устья, и к стадии 31/2 оборотов таких
складок насчитывается обычно 3. Когда раковина насчитывает 4 оборота, на-
чинается резорбция первой по времени образования, т. е. задней, складки.
д*
52
ОБЩАЯ ЧАСТЬ
Максимальное число базальных складок, присутствующих одновременно,
не превышает 5, и такое положение наблюдается при 41/2—5 оборотах раковины.
По мере дальнейшего роста новые складки начинают появляться менее регу-
лярно и по достижении полного числа оборотов их либо уже нет совсем, как
это бывает чаще всего, либо они присутствуют в начале последнего оборота и
-со временем резорбируются окончательно.
Париетальная пластинка появляется, как сказано выше, в эмбриогенезе и
«охраняется пожизненно. По мере роста животного верхние части пластинки
исчезают, оставляя след в виде тонкой светлой полоски. Сама пластинка при-
сутствует постоянно лишь в последнем обороте, имея почти постоянную высоту;
Рис. 27. Провизорные базальные складки Lauria cylindracea, просвечивающие сквозь стенку
раковины.
Цифры указывают число оборотов. Изображения приведены к одному размеру.
когда уже формируется губа, нижний конец пластинки утолщается, превращаясь
в зуб.
На фоне изложенных фактов рассмотрим, как и где протекает жизнь молоди.
Перед рождением потомства материнская особь спускается в самые нижние
горизонты лесной подстилки (возможно, что молодые улитки сами уползают вниз);
следовательно, ювенильные особи попадают в условия постоянного почти цолного
затенения и высокой влажности. В подобной обстановке закономерно слабое
развитие устьевой арматуры. Растущий моллюск по-прежнему ведет скрытный
образ жизни, но постепенно поднимается в более высоколежащие слои опада, и
проблема водного баланса несколько обостряется, но еще не является ведущей.
В связи с этим наблюдается появление базальных складок, функционально
аналогичных дефинитивной губе. По достижении 4 оборотов моллюск живет
уже вместе со взрослыми особями, но испытывает нужду в дополнительных
защитных приспособлениях, так как просвет устья у них относительно больше,
чем у взрослых, и наибольшее число базальных складок, а также развитие каж-
дой из них достигают в это время максимума.
Во всех трех рассмотренных случаях показательно, что наибольшего разви-
тия провизорные образования достигают тогда, когда молодь живет уже бок о бок
со взрослыми формами, но еще не защищена дефинитивными образованиями.
Хотелось бы обратить особое внимание на то, что все приведенные примеры
касаются Orculidae (вероятно, нечто аналогичное наблюдается и у Strobilopsi-
dae), т. е. семейства, наиболее архаичного и своеобразного по сравнению с про-
чими Pupillina, населяющими Палеарктику. У всех других семейств специаль-
ных устьевых морфологических адаптаций, которые носили бы провизорный
характер, не наблюдается.
БИОЛОГИЯ И ЭКОЛОГИЯ
53
С рассмотренным кругом явлений очень тесно смыкается одна особенность,
свойственная подавляющему большинству Pupillina: окраска раковин доста-
точно разнообразна — от одноцветно-темно-бурой до чисто-белой, нередко с ри-
сунком. Но при всем разнообразии окраски дефинитивных оборотов начальные
обороты всегда темные (см.: Матёкин, 1959, с. 1523; Стеклов, 1966, с. 74). Эту
закономерность также уместно поставить в связь с различиями в условиях су-
ществования молоди и взрослых животных. Лесные виды на любых стадиях
онтогенеза находятся в почти одинаковых и однородных условиях, и дефинитив-
ные обороты от эмбриональных окраской не отличаются. Но виды, населяющие
открытые пространства, имеют светлую
эмбриональных. Эти виды на разных
этапах роста живут в существенно
различных условиях: молодь оби-
тает в условиях сильного затенения,
в дерновинках трав, в припочвенном
•слое, а затем, по мере роста, подни-
мается на стебли растений, под пря-
мые лучи солнца. Б первую очередь
это касается ксеротермных предста-
вителей Buliminidae.
Дефинитивные признаки.
Устьевая арматура. В разных под-
Рис. 28. Устьевое вооружение Pupillina.
I — устье с губой и поверхностными зубами;
II — устье без губы с погруженными зубами.
Вверху — устье в нормальном положении, вни-
зу — вид справа; губа и зубы просвечивают сквозь
•стенку раковины, г — губа, Пз — погруженные
зубы.
окраску дефинитивных оборотов и темную
отрядах Geophila рассеяно несколько групп с хорошо развитым и специализи-
рованным вооружением устья, однако нет другого подотряда, для которого
в целом наличие складок, пластинок и бугорков в устье было бы столь характер-
ным, как для Pupillina.
В понятие устьевой арматуры включаются губа, имеющая вид складки,
параллельной краю устья, и система зубов разной величины и формы, располо-
женная внутри устья. Топографически зубы бывают двух типов, которые здесь
обозначаются как поверхностные и погруженные. Поверхностные зубы лежат
примерно в плоскости устьевого края и представляют собой местные утолщения
губы (рис. 28,I). Погруженные зубы с губой чаще всего не связаны и отодвинуты
в глубь устья на большее или меньшее расстояние от края (рис. 28, II).
Различия между поверхностными и погруженными элементами устьевого
края настолько серьезны, что эти категории заслуживают отдельного рассмот-
рения.
Губа образуется в результате того, что в субдефинитивном состоянии, перед
резким ослаблением секреторной деятельности в связи с завершением роста
раковины, железы мантийного валика в течение короткого времени очень ин-
тенсивно выделяют СаСО3, который откладывается по всему свободному краю
устья в виде кольцевого утолщения — губы. Если в крае мантийного валика
имеются зоны особо интенсивной секреции, то в таких зонах образуются утол-
щения губы — поверхностные зубы.
Ведущий фактор жизнедеятельности сухопутных моллюсков — водный ре-
жим; основная и постоянная опасность, которая им грозит, — это опасность
обезвоживания. Потеря влаги происходит через устье. Очевидно, совершенст-
вование средств, замедляющих испарение — едва ли не самая важная проблема,
которая обостряется тем сильнее, чем суше условия обитания. Имеется три спо-
соба замедлить скорость испарения: 1-й — сужение последнего оборота к устью,
54
ОБЩАЯ ЧАСТЬ
2-й — формирование губы и поверхностных зубов, 3-й — образование защитной
пленки — эпифрагмы — за счет усиления секреции слизи краем мантии.
3-й способ обычно дополняет один из первых двух.
Существует два основных способа переживать засушливое время. Одни виды
закапываются в почву, другие приклеиваются устьем к сухому листу, камню,
стебельку травы и т. п. Среди приклеивающихся форм выделяются в свою очередь
две группы: виды, обитающие в лесах, осыпях, трещинах скал, т. е. в мезофиль-
ных условиях, й виды, живущие открыто в степях и полупустынях.
Характер морфологических адаптаций у представителей этих экологических
групп существенно различается. Обратимся вначале к рассмотрению приспособ-
лений, свойственных видам, уходящим на засушливый период в почву.
Существование тесной связи между условиями влажности и степенью разви-
тия элементов устьевой арматуры у Chondrula tridens было показано Матёкиным
(1950): в общем чем суше условия,,тем интенсивнее развиты устьевые зубы.
В связи с этим был сделан вывод, что зубы служат для выжимания слизи из ноги
моллюска при его втягивании в раковину; эта слизь идет на построение эпи-
фрагмы. Существенно, что у названного вида элементы устьевой арматуры отно-
сятся к категории поверхностных. Такая же зависимость имеется и у других
видов, обладающих поверхностной арматурой устья, причем не только на меж-
популяционном уровне, но и на межвидовом. Например, сильно развитая губа
Chondrulopsina intumescens несет лишь слабые вздутия, а у близкого вида
Ch. fedtschenkoi эти вздутия превращаются в мощные бугорки, степень развития
которых, однако, сильнд варьирует в зависимости от условий, в которых обитает
конкретная популяция.
Надо уточнить, что слизь, идущая на построение эпифрагмы, на самом дел&
снимается не с ноги моллюска, а с края мантийного валика по той причине,
что при втягивании улитки в раковину нога с краем устья не соприкасается,
а с губой— тем более. В этом можно убедиться, если закрепить раковину ку-
сочком пластилина, ориентировав ее устьем к себе, и направить на моллюска
сильный пучок света от осветителя: тогда легко заметить, что когда улитка,
раздражаемая световым и тепловым потоком, будет втягиваться в раковину,
внутренние края устья вместе с губой и всем вооружением погружены в толщу
мантийного валика, и слизь с ноги отирается не устьевыми зубами (с которыми
нога, повторяю, не соприкасается), а лопастями мантийного валика. Лишь
когда нога полностью втянута, а центральное отверстие мантийного края,
куда убралась нога, затянуто тонкой пленкой прозрачной слизи, начинается
втягивание края мантии. Именно в этот момент с его поверхности с помощью
губы и поверхностных зубов снимается слизь, насыщенная гранулами СаСО3,
и эпифрагма образуется именно за счет этой слизи. Лишь самый центральный
участок эпифрагмы включает ту слизь, которая была предварительно снята
лопастями мантийного валика с поверхности ноги.
Трудно не согласиться с тем, что в аридных условиях эпифрагма, насыщен-
ная гранулами СаСО3, более эффективно задерживает испарение, чем эпифрагма,
бедная СаСО3. Следовательно, чем суше условия, тем более строгие требования
предъявляются к свойствам эпифрагмы и тем сильнее работают зоны секреции.
В результате возникают два независимых явления: усиление поверхност-
ной устьевой арматуры и интенсификация насыщения эпифрагмы солями каль-
ция. Поскольку слизь с поверхности ноги снимается лопастями мантийного
валика, то за счет этой слизи строится лишь срединная часть эпифрагмы,
а периферическая часть формируется за счет секрета, выделяемого краем мантии,
т. е. за счет того же источника, что и поверхностные устьевые зубы.
Таким образом, представляется вероятным, что интенсивность развития
устьевых зубов действительно зависит от степени сухости климата, но эта зави-
симость не прямая, а «сцепленная» с наличием в крае мантии зон интенсивной
секреции слизи, богатой СаСО3. Эта слизь нужна в первую очередь для усиления
эпифрагмы; но существование таких зон определяет и образование зубов на крае
устья, способствующих в свою очередь выжиманию слизи из мантийного края.
БИОЛОГИЯ И ЭКОЛОГИЯ
55
Иными словами, побочный эффект развития зон усиленной секреции приобрел
самостоятельное адаптивное значение.
Рассмотрение видов, которые на время засухи приклеиваются устьем к суб-
страту, а не закапываются, удобнее начать с наиболее ярко выраженных ксеро-
филов — форм, живущих открыто. Чрезвычайно существенно, что все виды,
относящиеся к этой категории, лишены не только зубов поверхностного типа,
но и губы (погруженные зубы могут присутствовать). Поэтому отмеченная выше
связь между развитием поверхностных элементов устьевой арматуры, качеством
эпифрагмы и условиями влажности касается только закапывающихся форм.
Каким же образом предотвращается потеря влаги у видов, живущих от-
крыто, т. е. в наиболее аридных условиях? С наступлением засухи моллюск
поднимается на стебли трав или камень. Пока животное ползет, оно удержи-
вается на вертикальной поверхности за счет клейкой слизи, выделяемой педаль-
ной железой. Затем улитка останавливается и начинает втягиваться, причем
базальный край устья обращен вверх, а вершина раковины вниз. Когда втянуты
голова и передняя часть ноги, базальный край устья входит в контакт с суб-
стратом, и в этом месте устье приклеивается за счет слизи, выделенной краем
мантии. Теперь, когда устье зафиксировано на вертикальной поверхности,
постепенно втягивается остальная часть ноги; по мере втягивания палатальный
и колумеллярный края устья опускаются на субстрат; в последнюю очередь
с поверхностью соприкасается париетальный край.
По мере увеличения площади контакта краев устья с поверхностью слизь
выделяется теми участками мантийного края, которые входят в соприкоснове-
ние с поверхностью камня или стебелька. В результате по окончании втягивания
моллюска устье оказывается приклеенным по всему периметру — частично
той слизью, которая выделена краем мантии, частично слизью, снятой с ноги
лопастями мантийного валика. Описание всех этих процессов основано на
прямых наблюдениях за втягиванием Brephulopsis cylindrica.
В дальнейшем, по мере втягивания моллюска все глубже в раковину, в устье
может образоваться еще несколько эпифрагм, но максимальный эффект защиты
от потери влаги все равно будет обеспечиваться самой первой эпифрагмой хотя
бы потому, что большая часть устья закрыта субстратом, и лишь зазоры между
ним и краем устья заполнены высохшей слизью.
Очевидно, что в таких условиях усиление эпифрагмы гранулами СаСО3,
равно как и образование поверхностных устьевых зубов, функционально
не оправдано. Более того, в этих условиях отсутствие дефинитивных признаков
оказывается, как мы увидим дальше, очень выгодной особенностью.
Однако путь все более точной пришлифовки поверхности устья к субстрату
(этот путь более характерен для видов, живущих в мезофильных условиях)
чреват серьезной опасностью: если сквозь эпифрагму газообмен продолжает
осуществляться, то при очень точном притирании края устья к поверхности
может быть достигнута полная герметизация устья, и тогда газообмен будет
прекращен. С этим, на мой взгляд, связано волнообразное изгибание края
устья: если устье по всему периметру лежит не в одной плоскости, какие-то
участки будут плотно прижаты к поверхности, а какие-то будут немного отста-
вать от нее (рис. 29, I). С тем же обстоятельством связано и возникновение
мелких складочек и бугорков, расположенных на самом краю устья и выступаю-
щих из плоскости устья: они обеспечивают некоторый зазор между устьем и
субстратом (рис. 29, II). Наиболее реальна опасность герметизации у видов
с цельным устьем, и не случайно оба примера, приведенные на рис. 29, каса-
ются Ghondrinidae — семейства, многие члены которого имеют цельное или
почти цельное устье.
Если взять любую раковину и приложить ее устьем к стеклу, на которое
нанесен слой пластилина, то мы получим отпечаток устья, позволяющий судить,
какие участки раковины, и насколько плотно прилегают к субстрату. Изучение
множества таких отпечатков параллельно с рассматриванием самого устья
показывают, что при максимально плотном прилегании устья к субстрату участок,
56
ОБЩАЯ ЧАСТЬ
сильнее всего отстоящий от субстрата, приходится на ангулярную область, т. е.
как раз на то место, где расположен пневмостом. Отсюда становится понятным
смысл особенности, нередко встречающейся в различных таксонах подотряда:
поднятый к устью последний оборот. Очевидно, чем сильнее поднят оборот,.
тем ангулярная область дальше отодвинута от субстрата, как бы тесно устье
к нему ни прилегало (рис. 30). Закономерно,
что поднятый к устью последний оборот встре-
чается только у видов, приклеивающихся к суб-
страту, и никогда не наблюдается у закапы-
вающихся форм.
Перейдем к рассмотрению устьевой арма-
туры погруженного типа. Говоря о погруженных
зубах, я имею в виду в первую очередь эле-
менты палатального края, поскольку длинные
париетальные и колумеллярные пластинки не-
Рис. 29. Два способа предотвращения герметизации
устья у Chondrinidae.
I — края устья образуют изгибы (Abida secale brauniopsisY,.
II — на устьевом крае имеются мелкие складки, выступающие-
из плоскости устья (Abida secale brongersmai) (контуры рако-
вин — из: Gittenberger, 1973).
связаны непосредственно с дефинитивным устьем и имеют иное происхождение и
иные функции. Зубы, расположенные на париетальном и колумеллярном краях
дефинитивного устья, не обязательно связаны с первичными колумеллярными
и париетальными пластинками, могут формироваться независимо и чаще всего-
относятся к категории поверхностных. За редким исключением (Truncatellina)
Рис. 30. Способ предотвращения герметизации устья у Euxinolauria superstructa.
I — вид справа; устье прижато к субстрату, ангулярная область поднята, пневмостом имеет контакт с внеш-
ней средой (показано стрелкой); II — устье и часть последнего оборота (зачерненные части — участки,,
контактирующие с субстратом). СИН — синулус.
погруженные элементы имеют вид более или менее удлиненных складок, ори-
ентированных вдоль, т. е. по ходу оборотов.
Нередки случаи, когда последний оборот к устью сужается. Тогда получа-
ется, что периметр субдефинитивного устья уменьшается, а периметр края
мантии остается прежним. В результате мантийный край сминается, и в образо-
вавшихся складках закладываются погруженные зубы. Это было показана
для Clausiliidae (Edlauer, 1941) и справедливо, вероятно, для Pupillina. Возни-
кает вопрос, насколько неизбежно появление погруженных палатальных скла-
док при сужении последнего оборота. Очевидно, эта связь обычна и законо-
мерна, но не обязательна: так, Pseudonapaeus otostomus обладает веретеновидной
БИОЛОГИЯ И ЭКОЛОГИЯ
57
раковиной, но погруженные зубы у этого вида отсутствуют; напротив, Р. еп-
toptyx башневидной формы, но внутри устья залегает продольная складка.
Погруженные зубы, возможно, играют некоторую роль средства ослабления
испарения (путем уменьшения площади устья), но эта функция не основная.
Удлиненные структуры в силу своей формы вряд ли эффективны при выжима-
нии слизи из мантийного края, так как для этой цели нужны образования,
ориентированные перпендикулярно оси втягивания, а при продольном положе-
нии складок мантия скользит по ним, как по рельсам. Следовательно, основная
роль погруженных складок иная, а именно обеспечение эффективности дыхания
и защита ректума, а также управление раковиной. Рассмотрим эти вопросы
подробнее.
Связь процессов дыхания с наличием устьевых
зубов. Если в раковине имеется парцетальный зуб, то в верхней части
палатального края, как правило, имеется тоже развитый в разной степени зуб.
Гребни этих зубов направлены один к другому. Исходя из положения зубов
относительно края мантии, можно уверенно предположить, что в совокупности
они образуют нечто вроде синулуса, в котором располагаются пневмостом, анус
и отверстие мочеточника.
У многих Vertigo, Euxinolauria и некоторых других форм синулус еще резче
обособляется, благодаря тому что палатальная стенка имеет вдавление. Следова-
тельно, названные зубы защищают органы, прилегающие к легочной полости
(преимущественно, конечно, ректум, ректальный синус и анальное отверстие),
от давления со стороны внутренних органов. Это важный момент, поскольку
во время интенсивного питания зоб, расположенный рядом с ректумом и лежа-
щий параллельно ему, значительно раздувается и давит на все окружающие
органы. Легко понять, что при расширенном зобе задняя кишка оказывается
так сильно прижатой к стенке раковины, что фекальные массы не могут поки-
нуть кишечник. Эта проблема особенно обостряется в том случае, когда пос-
ледний оборот устья сужается. Наличие жесткой перегородки между зобом и
ректумом устраняет эту опасность, и роль такой перегородки выполняет пари-
етальный зуб.
Если принять во внимание, что основное давление приходится именно сбоку,
то станет понятным, почему париетальный зуб «нужнее» палатального; этим
и объясняется тот факт, что один париетальный зуб (без палатального) встреча-
ется гораздо чаще, чем один палатальный (без париетального). Кстати, те ис-
ключительно редкие случаи, когда развит только палатальный зуб, не со-
провождаемый париетальным (Pseudochondrdla lederi, Imparietula schelkovni-
kovi), касаются только поверхностных зубов и никогда — погруженных.
Параллельно с описанными процессами наличие синулуса способствует
упорядочению процессов газообмена, защищая пневмостом и прилегающие
участки легочной полости.
Участие погруженных зубов в управлении рако-
виной. Раковину и висцеральный мешок с легочной полостью можно уподо-
бить двум конусам, вставленным один в другой. Если поверхности конусов глад-
кие, то внутренний может вращаться относительно наружного и наоборот.
Но любая жесткая складка, вдающаяся в полость устья, будет неизбежно вы-
полнять роль стопора, предотвращающего вращение висцерального мешка вну-
три раковины, а система таких складок создает очень жесткое сцепление рако-
вины с телом моллюска.
Поскольку висцеральный мешок завит в спираль, вращающий момент
может возникнуть только в области, прилегающей к устью, т. е. как раз там,
где и расположен комплекс погруженных зубов.
Облегчая управление раковиной, погруженные зубы несколько замедляют
скорость втягивания; думается, что именно с этим связано отсутствие погружен-
ных зубов у мелких форм, живущих в условиях резких температурных колеба-
ний. Паллиативный выход из положения наблюдается у Truncatellinat неболь-
шой палатальный зуб, сидящий глубоко внутри раковины, увеличивает сцепле-
58
ОБЩАЯ ЧАСТЬ
ние между телом животного и раковиной, не препятствуя в то же время быстрому
втягиванию. Аналогичное явление наблюдается у некоторых Buliminidae:
Pseudonapaeus entoptyx, Siraphoroides moltschanovi.
Заслуживает внимания в этой связи редко встречающаяся модификация ко-
лумеллярной пластинки, когда она поднимается вверх параллельно столбику,
одновременно расширяясь в виде лопасти. Подобное строение колумеллярной
пластинки наблюдается у Azeca, Vertilla, немногих Euxinolauria и некоторых
Achatinellidae. Сравнивая положение раковины относительно ноги у ползущих
Azeca и Cochlicopa, Пол (Paul, 1974) обратил внимание на то, что у Azeca рако-
вина вытянута параллельно ноге, а у Cochlicopa раковина на 20—30° отклонена
Рис. 32. Раковины тропических Pupillina.
I — Boisidia gereti', II — Aulacospira haloloma\ III — Hypselo-
stoma roebeleni (вид сверху и снизу).
в сторону. Эту разницу Пол связывает с тем, что Azeca имеет колумеллярную
пластинку описанного строения, a Cochlicopa не имеет. Это мнение справедливо:
когда животное ползет, вертикальная расширенная лопасть колумеллярной
пластинки лежит на спинной стороне ноги, не давая раковине отклоняться в сто-
рону; но Пол упустил из виду, что ту же роль еще эффективнее играет седло-
видная система, образованная париетальной и субпариетальной пластинками,
которые связаны между собой поперечными утолщениями: когда улитка ползет,
это седло охватывает сверху спинную сторону ноги и фиксирует раковину от-
носительно тела моллюска в определенном положении (рис. 31).
Такая жесткая система эффективна, но в то же время представляет собой
своего рода подводный камень, так как лишает раковину подвижности относи-
тельно ноги и затрудняет активное управление ею посредством колумеллярной
мускулатуры. Утратившие устьевую арматуру Cochlicopa в этом отношении
прогрессивнее, поскольку способны активно' управлять раковиной. Как Azeca,
так и Cochlicopa обитают преимущественно среди лесного опада, мелких сучьев,
перепутанных, травинок, и способность в таких условиях активно управлять
раковиной, бесспорно, ценное приобретение. Это одно из важных обстоятельств,
объясняющих, почему Azeca имеет относительно узкий ареал реликтового типа
и встречается в ограниченном наборе биотопов, тогда как Cochlicopa распростра-
нены почти повсеместно по всей Голарктике.
Поддержание раковины в определенном положении есть частный случай
управления ею, и важная роль погруженных зубов заключается во многих
случаях именно в поддержании раковины, ее более или менее постоянной ориен-
тации. Большим своеобразием отмечена раковина тропических Hypselostoma
БИОЛОГИЯ И ЭКОЛОГИЯ
59
(рис. 32, III): устье не опущено, как бывает чаще всего, а наоборот, поднято и
направлено вверх так резко, что оно видно при рассматривании раковины со
стороны вершины. Поэтому, когда животное ползет, вершина раковины направ-
лена вниз и, вероятно, не касается или почти не касается субстрата, т. е. все
время находится на весу. В результате обостряется проблема равновесия, наи-
более выгодного положения висцеральной массы и раковины относительно ноги.
4 мощных зуба внутри устья с притупленными гребневидными вершинами, упи-
раясь в дорсолатеральные поверхности ноги справа и слева, сильно ограничи-
вают свободу перемещения раковины и устойчиво удерживают ее в наиболее
выгодном положении. Параллельно сочетание париетального и верхнего пала-
тального зубов образуют синулус, т. е. эти зубы несут также и традиционную
функцию защиты ректума и пневмостома.
Как отмечалось, выше, возникновение погруженных зубов связано с суже-
нием последнего оборота; роль этого процесса и эволюционные последствия за-
служивают специального рассмотрения.
Сужение последнего оборота — дрейнее приобретение, возникавшее не раз;
поэтому вполне правдоподобно предположение, что первично это сужение возни-
кло вообще вне связи с какими-то внешними условиями, а для облегчения управ-
лением длинной раковиной, вернее, для ее удержания в наиболее выгодном по-
ложении относительно ноги.
Уменьшение диаметра последнего оборота обеспечивает более тесное взаим-
ное прилегание внутренних органов и, в частности, более тесный контакт колу-
меллярного мускула со столбиком. Прибегая к вольному сравнению, можно
сужение последнего оборота уподобить тугому корсету: танцовщик или гимнаст,
от которых требуются четкие строго скоординированные движения, туго затя-
гивают пояс, играющий при поворотах туловища роль наружной опоры. Выше
было показано, что длинная колумеллярная пластинка — архаичный признак,
исходно характерный для подотряда; наличие этой пластинки облегчает управ-
ление раковиной, и эта роль выполняется тем успешнее, чем туже колумел-
лярный мускул прилегает к колумеллярной пластинке; в свою очередь тугое
прилегание колумеллярного мускула обеспечивается сужением последнего обо-
рота.
Таким образом, у тех групп, развитие которых пошло по пути увеличения
относительной высоты раковины, начался процесс формирования погруженной
арматуры, отдельные элементы которой все более тонко адаптировались для осу-
ществления пространственной ориентации раковины при передвижении.
Дальнейшее развитие устьевого вооружения погруженного типа в этом на-
правлении зашло в эволюционный тупик: сложная система зубов так загромо-
здила устье, что существенно повлияла на скорость втягивания животного и
при усилении аридизации условий сыграла роковую роль, особенно для жи-.
вотных малого размера.
Убедиться в том, что улитки с сильно армированным устьем тратят на втя-
гивание больше времени, чем животные того же размера и с тем же числом обо-
ротов, обладающие простым устьем, несложно. Надо подобрать соответствую-
щую пару видов, прикрепить раковины на пластилине и, раздражая моллюсков
касаниями иглы, заставить их втянуться в раковину: моллюск с армированным
устьем затратит на втягивание больше времени. Это и понятно: если устье ли-
шено зубов, то для втягивания достаточно простого сокращения колумеллярного
мускула; если же в просвете устья имеются многочисленные жесткие выступы,
то для втягивания помимо сокращения осевой мускулатуры необходимо еще и
сложное перераспределение гемолимфы в туловищных синусах, на что тратится
дополнительное время.
Выход из этого эволюционного тупика был найден формами, еще не успев-
шими достичь необратимой и критической специализации сложного устьевого
аппарата. Их развитие пошло, по-видимому, по пути тонкой функциональной
дифференциации колумеллярной мускулатуры, что дало возможность управ-
лять раковиной, сохраняя в то же время способность быстро втягиваться.
60
ОБЩАЯ ЧАСТЬ
Высказанная гипотеза основана на следующих фактах.
Среди Vertigininae и Gastrocoptinae виды с самым мощным вооружением устья
обнаруживаются среди форм, вымерших к середине неогена (Стеклов, 1967;
Стеклов, Цытович, 1967). Далее, наиболее архаичный вид среди современных
Vertigininae — Vertilla angustior имеет сложное устье; напротив, наиболее про-
грессивный вид, обладающий колоссальным ареалом, очень широким размахом
изменчивости и встречающийся нередко массами, — Vertigo modesta проявляет
тенденцию к редукции зубов вплоть до полного их исчезновения.
Не менее показательны в этом отношении виды подсемейства Lauriinae.
Представители рода Euxinolauria, населяющие в основном западные районы
Кавказского перешейка, живут только в лесах (большей частью — в реликто-
вых лесах) и характеризуются сложной устьевой арматурой. Представитель
олиготипического рода Lauria cylindracea имеет обширный ареал, освоил широ-
кий спектр биотопов — от разреженных лиственных лесов до полупустынь и
имеет относительно простое устье: оно снабжено лишь небольшим париетальным,
зубом.
Объяснение изложенным фактам я склонен связывать в конечном счете со
скоростью втягивания моллюсков в раковину. Euxinolauria живут в лесах, в ус-
ловиях сильного затенения, им не грозит быстрое высыхание под прямыми лу-
чами солнца. На западном Кавказе бывают бездождевые периоды, когда леса
сильно просыхают, но этот процесс настолько медленный, что совершенно без-
различно, сколько времени потратит моллюск на втягивание — несколько се-
кунд или несколько минут.
Показательно также еще одно обстоятельство. Виды, обитающие в наиболее
влажных условиях — в сырых тенистых ущельях, на мокрых скалах (Е. recti-
dentata, Е. paulinae),—обладают только погруженными элементами устьевого
обрамления. В наиболее сухих условиях живет Е. pulchra". вид населяет широко-
лиственные леса северо-западного Кавказа, которые в бездождевые периоды
просыхают особенно сильно; и именно у этого вида развиваются в дополнение-
к погруженным зубам поверхностные бугорки и складки, выполняющие роль,
защиты от потери влаги.
На эту закономерность порой наслаиваются еще и особенности экологии
отдельных видов. Так, в Батумском ботаническом саду в зарослях бамбука оби-
тают совместно три вида — Е. tenutmarginata, Е. superstructa и Е. sinangula.
Судя по строению устья, наиболее влаголюбивый вид — Н. tenutmarginata,.
лишенный поверхностных зубов, но обладающий широким отворотом края
устья, что обеспечивает плотное прилегание к субстрату. Е.. sinangula, напро-
тив, обладает не только погруженными, но и поверхностными зубами, что по-
зволяет предполагать большую устойчивость к высыханию, чем у Е. tenuimargi-
nata. Промежуточное положение занимает Е. superstructa.
Совершенно иная ситуация складывается в условиях, среди которых обитает
Lauria cylindracea. Палящие лучи солнца выполняют роль жесткого инстру-
мента отбора на скорость втягивания. При малом размере животного проблема
стоит столь остро, что даже незначительное замедление процесса втягивания
грозит гибелью от обезвоживания. К тому же надо иметь в виду, что мезофиль-
ные виды сухопутных моллюсков подвержены тепловому шоку: например, некото-
рые виды слизней, вынесенные под яркое солнце в жаркий день, иногда гибнут
в течение считанных минут, хотя за такое время организм слизня не может по-
терять столько воды, чтобы можно было подозревать гибель от обезвожи-
вания.
Поэтому при почти полном отсутствии зубов у L. cylindracea развивается
сильный отворот губы, способный пришлифовываться к субстрату с минималь-
ным зазором. Вспомним, что провизорные базальные складки, периодически
появляющиеся у этого вида по мере роста раковины, носят в момент формирова-
ния поверхностный характер; поскольку сильно отвернутая губа может появи-
ться лишь на завершающем этапе строительства раковины, а потребность в за-
щите устья существует и у молоди, хотя и не в такой резкой форме, возникает
БИОЛОГИЯ И ЭКОЛОГИЯ
61'
нужда в каких-то структурах, способных на время заменить дефинитивный от-
ворот устьевого края.
Корреляция высоты раковины, степени скошенности устья и развития зубов-
прослеживается в морфологическом ряду Boisidia—Aulacospira—Vallonia. От-
носительно высокая коническая раковина В. gereti имеет умеренно скошенное
устье и два небольших зуба, ограничивающих подвижность раковины: париета-
льный и базально-палатальный (рис. 32,7). У А. haloloma гораздо более короткая:
(короткоконическая) раковина, устье сильнее скошено, и соответственно набор
устьевых зубов ограничен небольшой колумеллярной пластинкой (рис. 32, II).
Наконец, раковина многочисленных видов Vallonia еще более уплощена, устье
резко скошено, устьевые зубы отсутствуют.
Подведем основные итоги анализу устьевой арматуры.
1. Зубы в устье представлены двумя категориями — поверхностные и погру-
женные. Поверхностные представляют собой местные утолщения губы; погру-
женные зубы возникают как закономерный результат при сужении последнего-
оборота в конце постэмбриогенеза.
2. Поверхностные зубы и губа служат в основном для выжимания слизи и»
края мантии и образования плотной эпифрагмы, насыщенной гранулами угле-
кислого кальция.
3. Поверхностные зубы имеются у форм, закапывающихся на время сухого
сезона в почву.
4. Основное назначение погруженных зубов — обеспечение ориентации ра-
ковины относительно ноги и облегчение управлением высокой раковиной.
5. По мере усиления аридизации условий наблюдается в общем усиление-
поверхностных зубов и ослабление погруженных. Эта тенденция касается форм,
живущих открыто и переживающих засуху, приклеившись устьем к поверхности
субстрата.
6. Виды, живущие открыто в аридных степных, полупустынных и пустын-
ных ландшафтах, лишены устьевых зубов и хорошо развитой губы; у таких ви-
дов иногда развивается широкий отворот устьевого края, что позволяет им плот-
но притираться устьем к субстрату.
Окраска. Перейдем к рассмотрению адаптивных особенностей окраски рако-
вины. Как было отмечено, первично раковина одноцветно темная: коричнева-
тых, буроватых или красноватых тонов, с относительно слаборазвитым извест-
ковым слоем. Известно единственное исключение — Gastrocoptinae, у которых
раковина стекловидно-прозрачная при жизни моллюска и известково-белая
после его гибели. Однако в фауне СССР это подсемейство представлено всего
двумя видами, и от детального обсуждения группы в целом лучше воздержаться.
Стекловидно-прозрачные раковины характерны также и для живых особейг
относящихся к видам рода Vallonia, но другим родам Valloniidae свойственны
раковины темной окраски.
Изменение цвета раковины в большинстве таксонов начинается опять-таки
с освоением новой адаптивной зоны — открытых сухих пространств. Конечно,,
в первую очередь изменение окраски коснулось тех видов, которые именно пере-
шли в новую среду, а не в те мезофильные местообитания, которые имеются даже
в полупустынях. Скажем, условия жизни глубоко в трещинах скал, в нижних
горизонтах осыпей, под большими камнями, в отношении влажности и освещен-
ности не столь уж сильно отличаются от условий жизни в лиственных лесах.
Иное дело виды, подвергающиеся прямому воздействию солнечных лучей
и в полной мере заслуживающие названия ксерофилов. Эти виды имеют ярко
выраженную тенденцию к приобретению светлой окраски.
Матёкин (1959) считает, что интенсивность развития светлых штрихов на
темном фоне вплоть до формирования чисто-белых раковин определяется пре-
имущественно наличием в почве доступных карбонатов. Прямые наблюдения
показывают, что дело не столько в карбонатном режиме, сколько в том количе-
стве солнечной энергии, которое потребно моллюску для завершения жизнен- i
ного цикла. Об этом говорят следующие факты.
62
ОБЩАЯ ЧАСТЬ
На Тянь-Шане, в среднем течении р. Нарын, примерно в 50 км выше Ташку-
мыра, вдоль берега в изобилии встречаются сизовато-каштановые Pseudonapaeus
trigonochilus вместе с одной из форм Р. albiplicatus, в окраске которой, особенно
на нижних оборотах, преобладает снежно-белый цвет. Обе формы живут при
одинаковом карбонатном режиме, но в сухую погоду Р. trigonochtlus закапыва-
ются в почву, а Р. albiplicatus остаются на поверхности почвы, приклеиваясь
устьем к камням.
Другой пример. Имеющий обширный ареал Chondrula tridens заселяет боль-
шой набор биотопов, где, в частности, сильно колеблется содержание СаСО3;
тем не менее раковины живых моллюсков никогда не бывают белыми — их ок-
раска колеблется от темно-бурой до светло-коричневойили желтовато-коричне-
вой. Вместе с тем Ch. trtdens никогда во время засухи не остаются на поверхности
почвы — они всегда закапываются. Надо заметить, что при этом толщина стенок
раковин членов разных популяций заметно колеблется в зависимости от коли-
чества СаСО3 в данном местообитании.
Далее, в долине р. Аламедин на Киргизском хр. близ г. Фрунзе встречаются
коричневые вздутые приземистые Pseudonapaeus albiplicatus]. asiatica и строй-
ные полосатые Р. albiplicatus f. secalina, но первые населяют мелкообломочные
осыпи и способны закапываться в землю, а вторые образуют плотные колонии
под большими камнями и не закапываются. Поскольку названные формы живут
в условиях одинакового карбонатного режима, различия в окраске их раковин
приходится относить за счет каких-то иных воздействий.
Наконец, последний пример. В среднем течении р. Пскем на западном Тянь-
Шане под камнями на пойменных террасах встречаются вместе, т. е. в одних ми-
кробиотопах, каштановые гладкие Р. entoptyx, полосатые морщинистые Р. albi-
plicatus f. albiplicata и глянцево блестящие одноцветно белые или желтоватые
Р. stabilis. Очевидно, что различия в их окраске связаны не с карбонатным ре-
жимом, а с особенностями экологии видов. К сожалению, более конкретно го-
ворить пока нет возможности, так как время моего пребывания в указанном
месте было ограничено несколькими днями.
В отношении окраски раковины своеобразны Chondrinidae: несмотря на то
что отдельные виды живут в лесных биотопах, семейство в целом составлено ксе-
рофилами, связанными с горными скалистыми или каменистыми местностями.
Даже лесные виды населяют в основном горные леса, богатые скальными вы-
ходами. Днем представители семейства большей частью находятся на поверх-
ности скал, под прямыми лучами солнца. Несмотря на это, раковины белой
окраски среди Chondrinidae не встречаются, лишь у некоторых видов на поверх-
ности раковин возникают светлые радиальные штрихи. Окраска раковин явно
маскирующая, и для того, чтобы заметить сидящих на скале моллюсков, нужен
наметанный глаз. ।
То же касается и Pyramidulidae. Очевидно, оба семейства — исходно очень
теплолюбивые группы, не нуждающиеся в отражении солнечных лучей поверх-
ностью раковины. Существенно, что ювенильные особи обитают вместе со взро-
слыми.
Что касается степени кальцинированности субстрата, то в аридных зонах,
где живут Pupillina с белой раковиной,- дефицит карбонатов — явление редкое
и уровень содержания СаСО3 в почве может находить отражение в толщине сте-
нок раковины, а не в ее окраске.
Те редчайшие случаи, когда у палеарктических видов имеются цветовые
узоры иного характера (вроде широкой спиральной темной ленты у Caucasicola
raddei или Euxinolauria zonifera), связаны, вероятно, с достижением расчленяю-
щего зрительного эффекта, в результате чего отдельные светлые элементы окра-
ски в лесном полумраке заметны хорошо, а контуры раковины в целом теря-
ются. Расчленяющая окраска в животном мире распространена широко и слу-
жит главным образом для защиты от хищников, разыскивающих добычу с по-
мощью зрения.
Размеры и форма раковины. Как будет показано ниже, есть основания ду-
БИОЛОГИЯ И ЭКОЛОГИЯ
63
мать, что исходно раковина Pupillina имела средние размеры (порядка 5—10 мм
в высоту) и форму высокого цилиндра или конуса. Специализация по размерам
и форме пошла по четырем основным направлениям: 1) сохранение архаичного
облика с появлением различных мелких адаптаций; 2) увеличение размеров
(в первую очередь в высоту) и приобретение веретеновидной или башневидной
формы раковины; 3) уменьшение размеров с параллельным приобретением оваль-
ной формы; 4) уменьшение числа оборотов и появление микрохеликоидных
раковин.
Говоря о тенденциях в целом, нельзя забывать, что вообще разнообразие фор-
мы и размеров очень велико и во всех размерных группах имеются многочис-
ленные примеры параллельного развития по данным признакам. Так, высо-
кие цилиндрические раковины характерны для крошечных Truncatellina (Verti-
ginidae), средних размеров Orculella (Orculidae) и сравнительно крупных Lae-
vozebrinus (Buliminidae) (среди них L. ujfalvyanus, высота раковины которого
достигает 24 мм); овальные или короткие овально-цилиндрические раковины
обычны среди мелких Vertigo, среднего размера Multidentula и крупных Pupini-
dius.
Экологическая изменчивость формы и размеров раковины среднеазиатских
Buliminidae проанализирована Матёкиным (1959), и расхождения во взглядах
на таксономическую структуру семейства не меняют принципиальных положе-
ний. Поскольку в настоящей книге признается существование полиморфного
вида Pseudonapaeus albiplicatus, изменчивость формы раковины в пределах
этого вида толкуется с тех же позиций, с каких ее оценивал Матёкин; изменчи-
вость остальных форм, признаваемых здесь за самостоятельные виды, есть про-
явление параллельной изменчивости, т. е. фенотипические особенности близких,
а иногда и не очень близких видов примерно одинаковы в одинаковых условиях.
Формирование больших раковин светлой окраски отмечается для форм, на-
селяющих аридные местности и живущих открыто. В таких условиях увеличе-
ние объема тела означает прежде всего увеличение объема жидкости, заключен-
ной в теле моллюска, а «. . . увеличение запасов воды обеспечивает улиткам
не только возможность пассивного существования, но и возможность. . . актив-
ных передвижений даже днем» (Матёкин, 1959, с. 1524). Выше указывалось,
что у таких форм дефинитивные признаки, связанные с устьем, почти отсут-
ствуют: нет отворота края устья, нет поверхностных зубов, и даже губа обычно
выражена очень слабо (характерные примеры: Laevozebrlnus eremita, Brephulop-
sis cylindr ica, Napaeopsis hohenackeri,Zebrinadetritd). Отсутствие названных при-
знаков обеспечивает сохранение потенциальной способности к дальнейшему
росту, так как развитие отворота устьевого края означает неизбежное прекра-
щение роста раковины. Увеличение размеров, а следовательно, и объема рако-
вины является в свете изложенного важной адаптацией к открытому образу
жизни в сухих условиях.
В тех же аридных местностях нередко встречаются и мелкие формы, как, на-
пример, Geminula continens или Gibbulinopsis signata, но конкретные условия,
среди которых они живут, существенно отличны: мелкие моллюски не живут
открыто, а забираются глубоко в трещины скал, где температура ниже и, глав-
ное, не испытывает резких колебаний, а влажность выше. Существенно также,
что в укрытиях отсутствует столь важный иссушающий фактор, как ветер.
Виды, активно зарывающиеся в почву при наступлении сухого периода,
образовали жизненную форму с характерным обликом раковины: последний
оборот вздут, преобладает над остальными, раковина укороченная и по форме
приближается к овальной. При зарывании основную роль играет нога, и есте-
ственно, что у зарывающихся видов нога мощная и мускулистая. Увеличение
последнего оборота в данном случае как раз и связано с необходимостью разме-
щения объемистой ноги при втягивании животного (Матёкин, 1959); в то же
время для зарывающегося животного очевидны преимущества приземистой ра-
ковины перед удлиненной; пожалуй, среди закапывающихся форм нет ни одной
с высокой и узкой раковиной.
€4
ОБЩАЯ ЧАСТЬ
Напротив, моллюски, имеющие минимальную связь с грунтом, часто об-
ладают высокой раковиной конической, цилиндрической, веретеновидной
или башневидной формы. Когда животное поднимается на какую-нибудь верти-
кальную поверхность, такая раковина свободно свисает вниз, а при ползании
по горизонтальной плоскости волочится по субстрату.
Что касается размера закапывающихся видов, то они не бывают ни очень,
маленькими, ни очень большими (надо оговориться, что это касается только
Pupillina: такие крупные животные, как Achatina или Helix, при соответствую-
щих условиях успешно закапываются на значительную глубину). Частицы
почвы в сравнении с улитками такого размера, как Vertigo или Columella,
слишком велики, чтобы моллюск мог их раздвигать, а при большом размере
возникают трудности, связанные с необходимостью развивать большие мус-
кульные усилия для погружения в столь плотную среду, как почва.
Выше было показано, что у видов с высокой раковиной, часто поднимаю-
щихся на вертикальные поверхности, обычно наблюдается сужение последнего
оборота. Вначале эта особенность была связана, вероятно, с необходимостью
более эффективно управлять раковиной, но в дальнейшем повлекла за собой
целую цепь следствий.
Сужение последнего оборота приводит к появлению веретеновидной формы;
но центр тяжести веретена совпадает с его геометрическим центром, а центр
тяжести конуса сдвинут к его основанию. Если у конической раковины центр
тяжести приходится на верхнюю часть последнего оборота или на границу по-
следнего и предпоследнего оборотов, то у веретеновидных раковин центр тяже-
сти смещается к вершине, т. е. отодвигается от устья. Чем дальше от устья
расположен центр тяжести, тем резче обостряется проблема управления рако-
виной.
Иными словами, складывается ситуация замкнутого круга: сужение послед-
него оборота призвано облегчить управление раковиной, но оно же вызывает
сдвиг центра тяжести, и в результате вновь обостряется проблема управления
раковиной, но уже по иным причинам.
Существует, однако, возможность выйти из этого положения, реализован-
ная представителями нескольких семейств, особенно членами семейства Chon-
drinidae. Выход был найден в сдвигании устья влево, что приводит если не к пол-
ному совмещению, то во всяком случае к сильному сближению осей ноги и
столбика (рис. 33). Когда моллюск ползет вверх по вертикальной поверхности,
то чем ниже (т. е. в данном случае ближе к вершине) расположен центр тяжести,
тем сильнее выражен вращающий момент, возникающий в области устья и скру-
чивающий основание висцерального мешка, и тем большее усилие требуется от
колумеллярной мускулатуры для поддержания раковины в наиболее выгодном
положении относительно ноги. Очевидно, что чем сильнее в этих условиях бу-
дут сближены оси ноги и столбика, тем более эффективно будет гаситься вра-
щающий момент.
Можно предполагать, что сильное скручивание основания внутренностного
мешка нежелательно по трем причинам: во-первых, тем самым скручивается
длинное узкое легкое, на стенках его могут возникнуть пережимы и затруд-
нить газообмен; во-вторых, при скручивании пережимаются туловищные си-
нусы, чем затрудняется циркуляция гемолимфы; в-третьих, при движении мол-
люск обычно питается, а сильное перекручивание основания висцерального
мешка может привести к пережиманию кишечника в области устья.
Все эти рассуждения касались представителя Ghondrinidae — старой,
сравнительно архаичной и консервативной группы. Представители более мо-
лодых и прогрессивных таксонов встали на принципиально иной путь: те виды,
которые перешли к жизни на вертикальных поверхностях, резко усовершенст-
вовали в функциональном отношении колумеллярную мускулатуру, не прибе-
гая к тем способам, которыми воспользовались Chondrinidae. Такой способ,
конечно, прогрессивнее, хотя бы потому, что не сдерживаемая никакими жест-
кими стопорами в виде устьевых зубов деятельность колумеллярного мускула
БИОЛОГИЯ И ЭКОЛОГИЯ
65
со всеми его ветвями приводит к эффективному и активному управлению рако-
виной, к способности осуществлять тонкие и адекватные реакции (рис. 33,
ср. Z, II и III, IV). Поэтому в данном случае несущественно, насколько велик
будет угол между осью ноги и осью столбика; точнее, в данном случае постоян-
ный угол, строго говоря, отсутствует.
При анализе адаптивных тенденций неизбежна некоторая схематизация,
без которой разобраться в наблюдаемом разнообразии просто невозможно.
Так, здесь принимается, что виды живут либо в лесах, либо в открытых мест-
Рис. 33. Положение устья относительно оси раковины.
I — Granaria frumentum, устье сдвинуто влево; II — то же, животное ползет по вертикальной поверхности;
III— Turanena leptogyra, устье оттянуто вправо; IV — то же, животное ползет по вертикальной поверх-
ности. Ось: а—а' — раковины, b—Ь' — ноги. Объяснения в тексте.
ностях; в последнем случае это или закапывающиеся, или живущие
открыто формы. Однако в реальной ситуации в пределах какого-нибудь одного
биотопа, скажем осыпи, обитает порой целый комплекс видов, живущих по
существу в достаточно различных условиях: одни виды населяют самые нйжние
горизонты этой осыпи и живут практически в гигромезофильных условиях;
другие живут по краям осыпи, почти открыто и во время засухи закапываются
в почву; третьи дневное время проводят на поверхности камней, приклеив-
шись устьем. Экологическая пластичность многих видов еще более затрудняет
выявление общих закономерностей.
Наконец, известны случаи, когда раковины представителей таксона вообще
лишены видимых адаптивных черт. Наиболее показательно в этом отношении
семейство Pyramidulidae, составленное единственным родом с немногочислен-
ными видами. Это мелкие моллюски, облигатные термофилы и ксерофилы, от-
крыто сидящие на скалах. Поразительно поэтому полное отсутствие привычных
в такой ситуации конхологических адаптаций: ни скошенного устья, способ-
ного плотно прилегать к субстрату, ни губы, ни устьевой арматуры, ни какой-
либо специализированной формы раковины. Очевидно, надо допустить,что это
очень старая и очень консервативная группа с малой амплитудой фенотипи-
ческой изменчивости и высоко совершенными физиологическими,адаптациями.
Половой аппарат. Разбирая конхологические адаптации, мы видели, что
эволюция представителей подотряда направлена главным образом на приобрете-
5 А. А. ШилейкО
66
ОБЩАЯ ЧАСТЬ
ние разнообразных особенностей, обеспечивающих успешную борьбу с потерей
воды. Эти адаптации на самом деле более глубоки и отнюдь не ограничены воз-
никновением конхологических черт: приспособления затрагивают как особен-
ности внутреннего строения, так и физиологические свойства животных. От-
носительно последних известно очень немного, и то, что известно, касается
представителей других подотрядов. Поэтому остановимся лишь на адаптациях,
выражающихся в реорганизации полового аппарата.
В первую очередь надо вспомнить, что основной защитой спермиев от вы-
сыхания служит сперматофор, имеющийся у большинства семейств подотряда.
Понятно, что крошечный слизистый комок, который представляет собой скопле-
ние спермиев, высохнет гораздо скорее, чем само животное, и поэтому возникно-
вение сперматофора имело место даже в относительно влажных условиях —
у видов, населяющих мезофильные местообитания.
Как это ни парадоксально на первый взгляд, но сперматофор гораздо нужнее
животным, обитающим в мезофильных или ксеро-мезофильных условиях, чем
моллюскам, населяющим настоящие аридные области. На самом деле это не
парадокс: в районах с жарким и сухим климатом опасность высыхания грозит
и самим животным, тем более что процесс спаривания продолжается довольно
долго.
Можно представить себе два способа свести к минимуму подобную опасность,
оба способа сводятся к резкому сокращению времени спаривания, и оба спо-
соба были реализованы различными представителями подотряда.
Первый способ — сворачивание сперматофора в клубок внутри пениса,
причем по строению пениса можно судить, способен ли сперматофор сворачи-
ваться в клубок. Хороший пример — Bulimininae, ярко выраженные ксеро-
филы. Внутри пениса у членов этого подсемейства нет никаких направляющих
складок, и вся пениальная камера представляет собой объемистый мешок, спо-
собный растягиваться, где скапливаютря сперматофоры; о том, что они хорошо
развиты, можно судить по наличию длинного бича (рис. 239).
Второй способ сокращения времени спаривания — редукция сперматофора;
тогда весь акт передачи комка спермиев осуществляется резким сокращением
нижних отделов пениального аппендикса. В качестве наиболее яркого примера
можно указать на Jamlnia quadridens (Jaminiinae) (рис. 235). У этого вида пенис
настолько тонкий, что заставляет предполагать отсутствие сперматофора (вряд
ли хорошо развитый сперматофор способен пройти по столь узкому протоку);
в проксимальной части нитевидного пениса имеется накопительная камера,
4 ограниченная подкововидной складкой. Размеры этой камеры также наводят на
мысль об отсутствии сперматофора: здесь может поместиться крошечный сли-
зистый комок, содержащий скопления спермиев, но не склеротизованный сперма-
тофор. Следовательно, сперматозоиды концентрируются внутри пениальной
камеры и затем передаются в пениальный аппендикс, откуда они поступают в ва-
гину партнера. В этой связи надо обратить внимание на мощное развитие ниж-
них отделов пениального аппендикса, принявших на себя функцию пениса:
картина, аналогичная той, что наблюдается у некоторых Achatinellidae (Cooke,
Kondo, 1962). Что касается наличия дивертикула семяприемника у J. quadri-
dens, то о его функции можно только догадываться: возможно, он принял на
себя роль резервуара, так как сам резервуар семяприемника у этого вида раз-
вит слабо.
Надо добавить, что у видов, исходно живущих в местностях с жарким и влаж-
ным климатом (Achatinellidae), сперматофор первично отсутствует.
Все случаи, варианты и примеры, разбиравшиеся в настоящей главе, носят
характер частных адаптаций; но, кроме них, известны явления, которые можно
расценить как приспособления общего характера, «ad hoc адаптации», т. е.
адаптации на все случаи, не являющиеся ответом на некие конкретные воздей-
ствия. В частности, речь идет о путях и способах редукции придаточных орга-
нов полового аппарата.
В монографии по Helicoidea (Шилейко, 1978а) описаны чрезвычайно инте-
БИОЛОГИЯ И ЭКОЛОГИЯ
67
ресные случаи редукции придаточных органов женского отдела; в процессе
изучения Pupillina посчастливилось столкнуться с аналогичными картинами,
и теперь имеет смысл рассмотреть весь известный круг явлений.
Редукция придаточных органов может осуществляться не менее чем тремя
способами.
1. Наиболее ортодоксальный способ — постепенная редукция, основные
стадии которой можно проследить в цепи рецептных форм. Такой редукции
подвержены, например, такие коррелятивно связанные органы, как бич и ди-
вертикул семяприемника, длина кото-
рых может как варьировать в преде-
лах одной популяции (иногда вплоть до
полного исчезновения), так и обнару-
живать изменчивость клинального типа.
Постепенная редукция характерна так-
же для верхней пары стилофоров Hyg-
romiidae, где можно проследить все
этапы редукции — от хорошо выражен-
ных стилофоров (Trichiinae) до полного
их отсутствия (часть Hygromiinae) че-
рез слаборазвитые верхние стилофоры
Chilanodon (Hygromiinae), которые мож-
но обнаружить только на продольном
разрезе вагины (рис. 34, I).
2. «Мгновенная» и полная редукция
органов. Этот путь отмечен для Circas-
Рпс. 34. Редукция придаточных органов поло-
вого аппарата у Hygromiidae (подотряд Heli-
xina).
I — постепенная редукция верхнего стилофора у Hyg-
roniiinae; II — ступенчатая редукция стилофоров
п слизистых желез у Cireassina cireassica; III — мута-
генная односторонняя редукция у среднеазиатских
видов (Nanai о. cumulata—Arehaica heptapotamica');
IV — «мгновенная» редукция вагинальных придат-
ков у Euomphaliinae.
sina cireassica, у одних популяций которого имеются и стилофоры, и слизистые
железы, у других — только стилофоры, у третьих — только слизистые же-
лезы и у четвертых отсутствуют как стилофоры, так и слизистые железы
(рис. 34, II).
Явление того же порядка обнаружено у пары среднеазиатских Hygromiidae:
Nanaja cumulata и Arehaica heptapotamica', у первого вида имеются две пары сти-
лофоров, сидящих симметрично на вагине, у второго — одна пара таких же
стилофоров (рис. 34, III)', существенно, что иных отличий между этими видами
обнаружить не удалось.
Далее, у Euomphaliinae (Hygromiidae) имеются или два равноценных ва-
гинальных придатка, или один, или ни одного, но никогда не было замечено,
чтобы один придаток был развит лучше другого (рис. 34, IV).
Иными словами, во всех перечисленных примерах нет оснований подозре-
вать постепенную редукцию.
Среди Pupillina подобное явление обнаружено у Multidentulinae (Bulimi-
nidae): Multidentula обладают как дивертикулом семяприемника, так и пени-
альным аппендиксом, Improvisa — только пениальным аппендиксом, без ди-
вертикула и Senaridenta — только дивертикулом, без аппендикса (рис. 35).
По всем остальным признакам виды этих родов чрезвычайно близки.
Действительно ли это результат «мгновенной» редукции или она осуществля-
ется в цепи поколений, пока неясно, но совершенно очевидно, что скорость этой
5*
68
ОБЩАЯ ЧАСТЬ
Рис. 35. «Мгновенная» редукция придатков полового аппарата у Multidentulinae.
I — Multidentula, имеется пениальный аппендикс и дивертикул семяприемника; II — Improvisa, пениаль-
ный аппендикс имеется, дивертикул отсутствует; Ill — Senaridenta, дивертикул семяприемника имеется,
пениальный аппендикс отсутствует. Внизу — замкнутая папилла пениса, свойственная всем членам под-
семейства (из: Шилейко, 1978 г.).
Рис. 36. «Мгновенная» редукция базального отдела пениального аппендикса (Ат) у Gemi-
nula. isseliana f. callilabris из Горгана.
1 — половой аппарат, II — вскрытый пенис, III — пенис другой особи, А, отсутствует. А,—Л5 — отделы
пениального аппендикса; ДСП — дивертикул семяприемника, Пе — пенис, ППе — папилла пениса,.
Ц — цэкум.
БИОЛОГИЯ И ЭКОЛОГИЯ
69
редукции в любом случае неизмеримо выше, чем скорость эволюционных изме-
нений всех прочих признаков.
3. Способ частичной мутагенной редукции ранее известен не был и обнаружен
у двух видов Buliminidae: крымского Brephulopsis bidens и Geminula isseliana f. cal-
lilabris из окр. Горгана (Иран). У обоих видов среди особей с нормальным пе-
ниальным аппендиксом обнаружены экземпляры, у которых полностью отсут-
ствует базальный отдел аппендикса (А3), и И2, сохраняя обычное строение,
впадает непосредственно в пенис (рис. 36, 37). Всего было вскрыто около ЮОэкз.
В. bidens, и соотношение особей с нормальным аппендиксом с особями, лишен-
ными А1Г примерно составляет 1 : 1; G. isseliana f. callilabris вскрыто 9 экз.,
у 5 из них аппендикс обычного строения, а у четырех — без Av
Рис. 37. «Мгновенная» редукция базального отдела пениального аппендикса (Л,) у Бге-
phulopsis bidens из Крыма, с. Чистенькое.
I — пенис особи с аппендиксом обычного строения; II — пенис особи с аппендиксом, лишенным базаль-
ного отдела и соответствующей ветви полового ретрактора. Прочие признаки особей идентичны. А,—At —
отделы пениального аппендикса, ВА — ветвь аппендикса (полов'ой ретрактор), ВП — ветвь пениса (поло-
вой ретрактор), Пе — пенис.
О том, насколько быстро может проходить редукция и как стойко она может
удерживаться, дает некоторое представление работа Сориты (Sorita, 1978),
который исследовал обширный материал из разных местонахождений по двум
близким, но надежно различимым по раковине видам Bradybaenidae. У всех
изученных особей Aegista proba goniosoma стилофор, резервуар слизистых же-
лез и сами слизистые железы развиты нормально; большая часть исследован-
ных A. mikuriyensls была вовсе лишена придатков вагины, но у отдельных осо-
бей можно было заметить рудименты этих органов на разных этайах редукции.
В этой связи уместно вспомнить, что у Monacha orientalis (Hygromiidae, Euom-
phaliinae) вагина, как правило, лишена придатков, но изредка встречаются
особи, несущие рудименты вагинального придатка и (или) слизистых желез.
Из всего сказанного следует, что один из важных путей морфологической
эволюции наземных Pulmonata заключается в упрощении полового аппарата
за счет редукции тех или иных придаточных органов. Поскольку названные
органы играют ту или иную роль в совокуплении и связанных с ним процес-
сах, можно заключить, что основная тенденция направлена на упрощение этих
процессов и часто на сокращение времени спаривания. Особенно показательно,
что наиболее интенсивная редукция придатков, осуществляемая различными
70
ОБЩАЯ ЧАСТЬ
способами, свойственна видам, освоившим исторически молодые аридные типы
ландшафтов, либо видам, стоящим на пути к выходу из мезофильных условий
в ксерофильные.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ PUPILLINA
Первый шаг при изучении распространения таксона на какой-то территории
заключается в оценке трех основных показателей. .
1. Структура фауны, т. е. качественное соотношение таксонов разной зоо-
географической природы.
2. Степень (уровень) эндемизма, т. е. доля эндемиков в фауне данного ре-
гиона.
3. Ранг эндемизма, т. е. ранг наивысших из эндемичных для данного региона
таксонов (Старобогатов, 1970).
На основании этих трех показателей осуществляется следующий шаг —зоо-
географическое районирование. Однако такое районирование целесообразно
проводить на базе всей фауны (в данном случае — фауны наземных моллюс-
ков) , так как использование материалов по одной сравнительно небольшой группе
«таит в себе ту опасность, что какие-то биогеографические границы останутся
невыявленными или некоторым границам будет придаваться меньшее значение,
чем они имеют в действительности» (см. Старобогатов, 1970, с. 9). Поэтому от
районирования в данной книге лучше воздержаться.
Вначале рассмотрим общее распространение Pupillina, затем — особен-
ности пространственного распределения членов подотряда по территории СССР
и, наконец, проанализируем состав и степень эндемизма фауны Pupillina по
отдельным регионам.
Ареалы семейств и подсемейств Pupillina
В этом разделе специальное внимание уделяется особенностям распростра-
нения по территории нашей страны. Случаи изменения ареалов в результате
интродукции не учитываются.
Achatinellidae. О-ва Тихого океана и местами — прибрежные территории
материков в тропической и субтропической зонах. Наибольшее многообра-
зие на Гавайских о-вах.
Amastridae. Гавайские о-ва.
Ceriidae. О-ва Карибского бассейна и Флорида.
Cochlicopidae. Европа, о-в Мадейра, Азорские о-ва, Северная Африка,
Азия, Северная Америка. В СССР повсеместно.
Orculidae.
Ocrulinae. Средняя и южная Европа, Северная и Северо-Восточная Африка,
Малая, Средняя и Передняя Азия; кроме того, из Индии известна Orcula hima-
layana (Benson) (Rajagopal, Subba Rao, 1972). В СССР — Карпаты, Крым,
Кавказ, Средняя Азия, причем Крым и Средняя Азия включаются только за
счет Sphyradium doliolum — представителя монотипического рода. Таким об-
разом, если не считать названного вида, то члены подсемейства обитают в двух
регионах, причем на Карпатах живет единственный вид — Orcula dolium,
а остальные 4 вида (2 рода)— на Кавказе. Из этих 4 видов три (Pilorcula trifila-
rts, Orculella ruderalis, О. scyphus f. batumensis) имеют малоазиатское происхож-
дение, a О. bulgarica известна из Болгарии, Армении, Нахичевани и окр. Анапы.
Своеобразный род Pilorcula представлен на Кавказе и в Малой Азии; в Кире-
наике (Северная Африка) обитает несколько видов, удивительно напоминающих
Pilorcula уплощенной (примятой) вершиной, наличием складочек на столбике,
доходящих до края устья и присутствием хорошо выраженных кожистых реб-
рышек на поверхности раковины; правда, их отличает от Pilorcula отчетливо
прерванное устье (Brandt, 1956, 1958; Zilch, 1960).
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
71
Pagodulininae. Южные и восточные Альпы и прилежащие горные системы;
Талыш. В Талыше живет единственный представитель подсемейства Pagodu-
lina lederi, обитающий только в реликтовых лесах. Этот вид составляет моно-
типический подрод Crystallifera.
Lauriinae.Европа (кроме северной), о-ва Мадейра, Порту-Санту, Азорские
и Канарские, Северная Африка, Малая Азия, Крым, Кавказ. В фауне СССР
подсемейство представлено' 10 видами; один вид — Lauria cylindracea — имеет
обширный ареал, включающий большую часть территории Европы, Северную
Африку, Малую Азию, Крым и Кавказ. Остальные 9 видов (род Euxinolaurid)
— кавказские эндемики, тяготеющие преимущественно к юго-западному
Закавказью. На Центральном Кавказе встречается лишь один вид —Е. са-
ucasica, а на север вдоль черноморского побережья до юга Краснодарского
края проникает два вида — Е. zonifera и Е. pulchra.
Argninae. Южные и восточные Альпы, Карпаты, Балканский п-ов. Един-
ственный в СССР вид Argna bielzi населяет леса Карпат и Прикарпатья.
Strobilopsidae. Юго-Восточная Азия, Корейский п-ов, Япония, Филип-
пины; оба американских материка, Вест-Индия, Галапагосские о-ва. В СССР
один вид (Strobilops coreand) в заповеднике «Кедровая падь» на юге Приморско-
го края.
Spelaeodiscidae. Карпаты и горы Балканского п-ова, включая Албанию.
В нашей стране не исключено присутствие Spelaeodiscus triarius на Карпатах.
Valloniidae.
Acanthinulinae. Европа, Африка, Азия с Японией, о-в Ява, Америка, Ав-
стралия. В СССР 3 рода. Род Spermodea характеризуется реликтовым ареалом:
прибрежные области западной и северо-западной Европы; на территории нашей
страны обнаружен лишь в Калининградской обл. Единственный современный
представитель рода Acanthinula (A.aculeata) — типично европейско-кавказский
вид: на севере ареала он обитает в равнинных лесах, и чем южнее, тем выше под-
нимается в горы (до 1500 м над ур. м.). Наконец, ареал рода Zoogenetes (един-
ственный представитель — Z. harpd) может быть охарактеризован как циркум-
бореально-альпийский.
Valloniinae. Палеарктика. За единственным исключением, эндемизм на ви-
довом и подвидовом уровне, причем широкораспространенные виды имеют,
вероятно, европейское происхождение, а эндемики Дальнего Востока —амери-
канское. Исключение составляет монотипический подрод Planivallonia, встре-
чающийся в горах на севере Средней Азии; подрод известен также из миоцено-
вых отложений центрального Казахстана.
Pupillidае. Евразия, Африка с атлантическими островами, о-в Реюньон,
Северная и Центральная Америка, Гавайские о-ва, Австралия, Тасмания.
В СССР семейство встречается повсеместно, главным образом за счет единст-
венного представителя рода Pupilla — Р. muscorum. Вместе с тем как на Кав-
казе, так и в Средней Азии род представлен либо европейскими видами, либо
эндемиками, но близкими к европейским видам. На Дальнем Востоке имеется
эндемичный подрод Fragipuptlla с двумя видами, предположительно американ-
ского происхождения. Ареал рода Gibbulinopsls в нашей стране охватывает
Кавказ, Среднюю Азию и южное Приморье, причем один вид номинативного
подрода (G. interrupta') встречается на Кавказе, а другой (G. cryptodon) — на
Дальнем Востоке; подрод Primipupilla представлен тремя видами —G.(P.)sig-
nata (Кавказ и Средняя Азия), G. (Р.) gracilis и G. (Р.) nanosignata (Средняя
Азия). Наконец, тропический род Pupoides представлен единственным видом
в Таджикистане.
Vertiginidae.
" Gastrocoptinae. О-ва Зеленого Мыса, Кавказ, Урал, Сибирь, Средняя и Вос-
точная Азия, Шри-Ланка, Филиппины, Северная, Центральная и Южная Аме-
рика, о-ва Пацифики, Маскаренские и Бермудские о-ва, Австралия. В СССР
два вида: Gastrocopta theeli имеет спорадическое распространение — При-
морье, бассейн Енисея, Челябинская обл., Северный Кавказ, Дагестан, бас-
72 ОБЩАЯ ЧАСТЬ
сейн р. Риони; G. huttoniana живет в Индии и обнаружена в выбросах р. Ка-
фирниган в Таджикистане.
Vertigininae. Евразия, Северная Африка, Северная и Центральная Америка,
Вест-Индия, Бермудские о-ва. В СССР повсеместно, причем на большей части
территории встречаются европейские виды или виды европейского корня, а на
Дальнем Востоке и Курильских о-вах обитают три вида, общих с фауной Япо-
нии, и два эндемичных вида — Vertigo microsphaera и V. circumlabiata.
Truncatellininae. Евразия, Африка, Америка, Гавайи, о-ва Кермадек и
Маркизские. В СССР 2 рода. Род Columella встречается почти повсеместно.
С. edentula и С. columella — циркумбореально-альпийские виды, в южной
части СССР (на Кавказе, в Крыму и в Средней Азии) встречаются только в го-
рах, а на севере — в равнинных лесах и на влажных лугах. С. aspera преиму-
щественно в северной Европе, и С. acicularis — эндемик Заилийского хр. Виды
второго рода — Truncatelllna — характеризуются широкими ареалами и насе-
ляют степную зону европейской части, Предкарпатье, Крым, Кавказ, Сред-
нюю Азию. Дальше всего на север, в лесную зону, проникают Т. cylindrica
и Т. costulata, но здесь они играют роль интразональных элементов,, придержи-
ваясь сухих опушек, суходольных лугов.
Chondrinidae. Европа, кроме северных областей, Канарские о-ва, Северная
и Южная Африка, Передняя и Малая Азия, Кавказ, Копетдаг. В СССР два
рода. Род Granaria представлен только на Карпатах единственным видом —
G. frumentum; род Chondrina, широко распространенный по средиземноморским
странам и Европе, встречается как на Карпатах (Ch. clienta clienta), так и
в Крыму, на Кавказе и в западном Копетдаге, причем в Крыму обитают Ch. (Ch.)
clienta caucastca и Ch. (Granopupa) rhodta taurica, на Кавказе Ch. (Ch.) clienta
caucasica и Ch. (Ch.) amphorula, а в Закавказье и на Копетдаге — Ch. (Granopupa)
granum.
Pyramidulidae. Европа, средиземноморские страны, Передняя Азия, Сред-
няя Азия, Китай, Корейский п-ов, Япония. В СССР единственный вид Руга-
midula rupestris распространен в Европе от Ирландии до берегов Средиземного
моря, в Северо-Западной Африке, Передней Азии, Иране, северной Индии. На
территории нашей страны встречается в Подолии, в Крыму, на Кавказе и
в горных районах Средней Азии.
Buliminidae.
Pseudonapaeinae. Закавказье, Средняя, Малая и Центральная Азия, Афга-
нистан, Индия, ? Япония. В СССР подсемейство обильно представлено на Кав-
казе и особенно в Средней Азии; причем характерно, что единственный общий
для этих регионов вид Geminula isseliana обитает в юго-восточном Закавказье,
огибает с юга по Иранскому нагорью Каспийское море и доходит до юго-запад-
ных'районов Копетдага,- Кроме этого рода на Кавказе и в Средней Азии одно-
временно представлен только род Тигапепа. В Закавказье живет один вид
Т. scalaris, в Копетдаге — тоже один (Т. herzi)', в СССР вид не найден, но при-
сутствие его в западном Копетдаге не исключено; остальные 8 видов обитают
в горах восточнее Копетдага. Некоторое число видов рода Тигапепа обитает
в восточном Китае, Гималаях и Индии . В Средней Азии подсемейство представ-
лено 4 родами (не считая Geminula и Тигапепа), на Кавказе—-5, причем на Кав-
казе из них полностью эндемичны 4, а один род (Pseitdochondrula) связывает
Кавказ с Малой Азией и Балканами. В Средней Азии полностью эндемичны
2 рода (Subzebrinus и Mastoides)-, род Pseudonapaeus связывает Среднюю Азию
с Центральной, a Laevozebrinus включает 4 вида, 3 из которых встречаются
только в Средней Азии, а один (L. eremita) имеет широкий ареал — от Ирана
через Копетдаг до Тянь-Шаня и Гиссаро-Дарваза.
Chondrulopsininae. Все три известных вида (2 рода) встречаются только в го-
рах Средней Азии (на Копетдаге отсутствуют).
Jaminiinae. Южная Европа и Малая Азия.
Merdigerinae. Леса Западной и средней Европы и Северной Африки. В СССР —
лесная зона европейской территории, Карпаты и Крым. Многократные упоми-
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
73
нания о присутствии вида «Епа» obscura на Кавказе связаны с тем, что этот вид
по раковине чрезвычайно похож на кавказский вид Akramovskiella'umbrosa
{Pseudonap aeinae).
Bulimininae. Передняя и Малая Азия, южное Закавказье, Северная Аф-
рика. В СССР один вид Buliminus urmianus, известный из Ирана, Ирака, найден
в Нахичеванской АССР (окр. Джульфы) и в Азербайджанской ССР (Зангилан-
ский р-н). Род Bulimirius распространен преимущественно в Передней и Малой
Азии. Эндемиком Кавказа является монотипический род Adzharia.
Retowskiinae. Западная часть Кавказского перешейка и северо-восточная
Турция (бассейн р. Чорох).
Eninae. Европа (кроме северных районов), Канарские о-ва, Северная Аф-
рика, Малая Азия, Кавказ. Если в самых общих чертах распространение под-
семейства можно охарактеризовать как европейско-средиземноморское, то
для его представителей, обитающих в нашей стране, характерна высокая сте-
пень эндемизма на видовом и родовом уровнях: из 9 родов, присутствующих
в нашей фауне, только 4 {Ena, Zebrina, Napaeopsis, Chondrula) встречаются за
пределами СССР. Епа — типично европейский лесной род, Е. montana обитает
в нашей стране на Карпатах и местами — в лесной зоне европейской части.
То же можно сказать и о роде Zebrina, с той лишь разницей, что это род не лес-
ной, а степной, и у нас представлен европейским видом Z. detrita, живущим
у юго-западных границ СССР. Ареал рода Chondrula охватывает центральную
и южную Европу, Северо-Западную Африку и Переднюю Азию до Ирана.
В СССР род присутствует в степной зоне европейской части, на Карпатах,
в Крыму и на Кавказе. Наконец, вид, который здесь обозначен как Napaeopsis
hohenackeri, возможно, и не относится к роду Napaeopsis, поскольку строение
совокупительного аппарата типового вида этого рода неизвестно.
Остальные 5 родов распространены следующим образом. Роды Thoanteus
и Ramusdulus (оба монотипические) — эндемики горного Крыма. Род Peristoma
представлен в Крыму и на западном Кавказе; монотипический род Caucasi-
cola — эндемик лесов Кавказа. Наконец, дитипический род Brephulopsis ис-
ходно, вероятно, эндемичен для Крыма, и находки В. cylindrica за пределами
Крыма связаны с завозом человеком.
Multidentulinae. Кавказ, Малая и Передняя Азия, а также восточная часть
Балканского п-ова и о-ва Греческого архипелага.
Распространение Pupillina на территории СССР
и структура фауны
Приведенная характеристика распространения Pupillina исходит из ареа-
лов таксономических единиц; теперь рассмотрим распространение подчиненных
таксонов по отдельным регионам.
Если ориентироваться на распространение Pupillina, то территория СССР
достаточно определенно разбивается на следующие физико-географические
регионы, характеризующиеся определенным составом фауны: 1), лесная зона
европейской части (на приводимых в настоящей главе таблицах сокращается
как «Европа, лес»); 2) степная зона европейской части СССР (на таблицах сокра-
щается как «Европа, степь»); 3) Карпаты и Прикарпатье; 4) Крым (преимуще-
ственно горный); 5) Кавказ и Закавказье; 6) горные системы Средней Азии;
7) Сибирь и Алтай; 8) Дальний Восток.
Приводимое деление условно хотя бы потому, что сама территория СССР
не представляет собой естественного региона; строго говоря, из всех выделен-
ных пространственных единиц один лишь горный Крым может быть охаракте-
ризован как естественный регион. Некоторые пояснения будут приведены ниже,
а пока можно заметить, что без некоторой схематизации вообще никакое зоогео-
графическое деление невозможно.
В табл. 1 суммированы материалы по распространению родов; рядом с циф-
рой, указывающей число видов в роде, указано в скобках, какое число из этих
74
ОБЩАЯ ЧАСТЬ
Таблица 1
Название рода, число видов Европа, лес Европа, степь | Карпать Крым Кавказ Средняя Азия Алтай, Сибирь Дальний Восток
Cochlicopa, 4 3 (0) 2(0) 3 (0) 3 (0) 3 (0) 4(0) 4 (0) 3(0)
Pilorcula, 1 — — — — 1 (1) — — —
Orculella, 3 — — — — 3(2) — — —
Orcula, 1 — — 1 (1) — — — —
Sphyradium, 1 — — 1 (0) 1 (0) 1 (0) 1 (0) — —
Pagodulina, 1 — — 1 (1) — — —
Euxinolauria, 9 — — — — 9(9) — — —
Lauria, 1 — — — 1 (0) 1 (0) — — —
Argna, 1 — — 1 (1) — — —
Strobilops, 1 — — — — — — — 1 (1)
Zoogenetes, 1 1(0) — — — 1 (0) — 1(0) 1 (0)
Acanthinula, 1 1 (0) — 1 (0) 1 (0) 1 (0) — —
Spermodea, 1 1 (1) — — — — — —
Vallonia, 9 3(0) 4(0) 4(0) 3(0) 4(0) 6 (1) 4(0) 5 (2)
Gibbulinopsis, 5 2(1) 3(2) 1 (1)
Pupilla, 13 1(0) 2(0) 3(0) 2 (0) 6(2) 8(5) 2(0) 3(2)
Pupoides, 1 —. — — — 1 (1) — —
Gastrocopta, 2 — — — — 1 (0) 1 (1) 1 (01 1 (0)
Vertigo, 13 6(0) 4(0) 5 (0) 4(0) 6(1). 4(0) 3 (0) 6(5)
Vertilla, 1 1 (0) . — 1 (0) 1 (0) 1 (0) — — —
Columella, 5 3(1) 1 (0) 2(0) 1 (0) 2(0) 4 (2) 2(0) 2(0)
Truncatellina, 4 2(0) 3(0) 3(0) 4(0) 2(0)
Granaria, 1 — 1 (1) — —
Chondrina, 4 — — 1(0) 2(1) 3(1) 1 (0) — —
Pyramidula, 1 — 1(0) 1 (0) 1(0) 1 (0) 1 (0) — —
Pseudonapaeus, 14 — — — 14 (13) 1 (0) —
Subzebrinus, 1 — — — — — 1 (1) — —
Turanena, 9 — —. — — 1 (1) 8(8) — —
Pseudochondrula, 5 —i — — 5(5) — —
Mastoides, 2 —. — — — — 2?2) ' —- —
Laevozebrinus, 4 — — — — — 4(3) — —
Imparietula, 2 — — — — 2(2) — — —
Ljudmilena, 2 — — — — 2(2) — — —
Akramovskiella, 3 — — —— — 3(3) — — —
Differena, 1 — — — — 1 (1) — — —
Geminula, 2 — — — — 1(0) 2(1) — —
Chondrulopsina, 2 — — — — — 2(2) — —
Siraphoroides, 1 — - — — — 1 (1) — —
Merdigera, 1 1(0) — 1(0) 1(0) — — — —
Buliminus, 1 —. —. 1 (1) — — —
Adzharia, 1 — — — — — 1(1) — —. —
Andronakia, 1 — — — — 1 (1) — — —
Retowskia, 1 — — — — 1(1) — — —
Ena, 1 1 (0) —-. 1(0) — — — —
Peristoma, 4 — —— 2(2) 2(2) — — —
Caucasicola, 1 — — — 1(1) — — —
Zebrina, 1 — 1 (1) — — — — —
Brephulopsis, 2 — 1(0) — 2(1) 1(0) — — —
Napaeopsis, 1 — — — — 1 (1) — — —
Thoanteus, 1 — — — 1 (1) —. — — —
Ramusculus, 1 — — — 1 (1) —- — — —
Chondrula, 4 — 1 (0) 2(1) 1(0) 3(2) — — —
Multidentula, 1 — — 1(1) — —- —
Improvise, 1 — — — — 1 (1) — — —
Senaridenta, 1 — — — — 1(1) — —
видов являются эндемиками данной территории. Помимо действительных энде-
миков, т. е. видов, действительно не встречающихся за пределами данного ре-
гиона, имеется некоторое число видов, которые можно назвать условными эн-
демиками, или субэндемиками. Так, условным эндемиком лесной зоны европей-
ской части СССР можно назвать Spermodea lamellata, так как в других регио-
нах нашей страны он не встречается, но населяет леса прибрежных территорий
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
75
Западной Европы. То же касается карпатских эндемиков Orcula dolium, Argna
bielzi, Chondrula bielzi и Granaria frumentum: эти виды широко распространены
по территории всей обширной горной системы Карпат и сопредельных горных
Cochltcopidae
г Pyramidulidae ;
^Strobilopsidae
15 Е-
: Orcu.ltd.ae
10 г
5 г
Рис. 38. Соотношение общего числа видов и эндемиков по регионам и семействам.
По оси ординат — число видов. Зачерненные участки диаграмл^соответствуют. количеству эндемичных
видов.
территорий. К условным эндемикам можно причислить и несколько кавказских
видов, живущих и на территории Малой Азии.
Уже при первом взгляде на табл. 1 бросается в глаза обилие пустых клеток:
из 448 клеток в 314 имеются прочерки, т. е. заполнено лишь около 30% клеток.
Это свидетельствует о высокой дисперсности фауны, т. е. большинство родов
приурочено к одному или немногим регионам. Графически та же закономер-
ность иллюстрируется рис. 38.
76
' ОБЩАЯ ЧАСТЬ
Повсеместно (во всех 8 регионах) распространено только 4 рода из 55:
Cochlicopa, Vallonia, Pupilla, Vertigo (7% общего числа родов); в 7 регионах
представлен единственный род Columella (2%) — он отсутствует лишь в степ-
ной зоне европейской части, хотя в интразональных биотопах, таких как бай-
рачные леса, поймы рек с высоким травостоем, С. edentula имеется и на терри-
тории степной зоны. Родов, которые были бы представлены в 6 регионах, нет.
В 5 регионах имеется один род Truncatellina (2%): он отсутствует в лесной зоне
европейской территории Союза, в Сибири и на Алтае, а также на Дальнем Во-
стоке. Перечисленные роды можно назвать широкораспространенными, хотя
отнесение последнего рода к этой категории условно. Характерно, что все эти
роды древние, европейского происхождения, или роды, широко представленные
во всей Голарктике (происхождение неясно).
Распространение остальных 49 родов (89%) ограничено: в 4 регионах встре-
чается 8 родов (14%), в трех — 3 рода (7 %), в двух — 5 родов (8%), и основная
масса родов (33 рода, 60%) встречается лишь в одном каком-либо регионе.
Наиболее пестрый в систематическом отношении состав родов, распростра-
ненных в трех или четырех регионах: здесь и древние своеобразные Pyramiduli-
dae, приуроченные ко всем горным системам, кроме Алтая, Сибири и Дальнего
Востока, и лесные Acanthinula и Zoogenetes, и род Gastrocopta, распространение
которого в Палеарктике в настоящее время имеет реликтовый характер. Роды,
представленные в двух регионах, имеют или европейское распространение,
включающее Карпаты и Крым (Merdigera, Ena), или крымско-кавказское
(Lauria, Peristoma), или кавказско-среднеазиатское (Turanena, Geminula).
Самую большую долю составляют роды, представленные в каком-нибудь
одном регионе, т. е. эндемики. Поэтому имеет смысл специально рассмотреть
вопрос об эндемизме Pupillina и способах его оценки.
Степень эндемизма фауны
Степень, или уровень, эндемизма обычно выражают через отношение числа
эндемиков к общему числу видов в регионе; однако относительное богатство
фауны региона при этом никак не учитывается. Между тем ясно, что если в од-
ном регионе, допустим, всего 4 вида, из которых один эндемик, то это совсем
иное явление, чем если в регионе 100 видов, из которых эндемичны 25, хотя
показатель относительного эндемизма в обоих случаях одинаков — 25% .
Поэтому представляется желательным вместо уровня абсолютного энде-
мизма (отношение числа эндемиков к общему числу видов) ввести показатель
относительного эндемизма D, где доля эндемиков оценивалась бы через отно-
сительное богатство фауны. Попытка такой оценки была осуществлена Вилен-
киным й Шилейко (1979).
Для каждого из выделенных регионов известно число неэндемичных родов и
видов (C7g, U3) и число эндемичных родов, видов (Eg, Es). Чтобы получить срав-
нительную оценку степени эндемизма фаун регионов, принимаем, что по каж-
дому региону имеем дело не с выборочными оценками, а с генеральными сово-
купностями (в противном случае полученные далее выводы следует рассматри-
вать как предварительные).
Между величинами Ug и 1g [Eg-)-l ] существует тесная значимая корреляция:
Н=+0.943, р <; 0.01 (статистические расчеты проведены в соответствии с ру-
ководством Урбаха, 1964). Уравнение регрессии имеет вид 1g [^+1] =
= —0.585+0.085 Ug; выражение 1g [£^g+l] берется вместо Eg, так как в этом
случае получается более тесная корреляция.
Приведенное уравнение описывает среднюю тенденцию, существующую
на территории СССР, согласно которой происходит увеличение числа эндемич-
ных родов при переходе к регионам с большим числом неэндемичных родов.
Используя приведенное, уравнение, находим ожидаемое число эндемичных ро-
дов (Eg) в каждом регионе с известным Ug. Степень эндемизма фауны D есте-
ственно оценивать, сравнивая реальное число эндемиков Eg с ожидаемым Eg.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
77
Таблица 2
Регион us D Е,г j, ,+ • ioo% xL g —j- U g г • I г • IT
Европа, лес 10 1 1 0 9.1 5 6
Европа, степь 8 0 0 0 0.0 5 7.5
Карпаты 14 3 3 0 17.6 5 3
Крым 16 2 5 -3 11.1 8 4
Кавказ 21 18 15 3 46.1 1 1
Средняя Азия 15 6 4 2 28.6 2 2
Сибирь. Алтай 7 0 0 0 0.0 5 7.5
Дальний Восток 9 1 1 0 10.0 5 5
Приведем исходные данные и оценки эндемизма на родовом уровне (табл. 2).
Как следует из уравнения и табл. 2, если величины Eg и Eg равны, т. е. D —
=Eg—Ёё=0, то все регионы, где выполняется это условие, одинаково насыщены
эндемиками. Пригодной для сравнения оценкой степени относительного энде-
мизма фауны регионов будет величина разности D. В табл. 2 для сравнения при-
ведены также традиционные, предполагающие независимость величин Eg и
Ug оценки степени эндемизма в процентах:
Е„+ U„
8 1
• 100%..
Для суждения о том, в какой мере совпадают представления о степени энде-
мизма фаун, основанные на оценках разными способами, в табл. 2 приведены
ранги (здесь «ранг» — термин статистический, а не зоогеографический) энде-
мизма каждого региона в соответствии с величинами/) (г-I) и 100% •£,g/(£,g+C7g).
(г-II). Коэффициент ранговой корреляции Спирмена ps=+0.60, его отличие
от нуля недостоверно. Это значит, что разные способы оценки эндемизма приво-
дят к закономерно различным результатам.
Надо сделать еще одно замечание по табл. 2 и уравнению. Полученные по
уравнению величины Eg округляются до целого числа, поэтому суммы Eg и Eg
в таблице могут не совпадать. Из уравнения следует, что при Ug 94-Ю энде-
миков быть не может. Существенно, что при использовании оценки степени от-
носительного эндемизма D ее величина будет совпадать для региона, содержа-
щего £е=1и l/g=9, и для региона, в котором Е = 15 и ?7g=21: в обоих случаях
/)=0. При обычном же вычислении доли эндемиков от.общего списка родов мы
получим для этих же регионов соответственно 10 и 41.7%.
Рассмотрим аналогичным образом эндемизм на видовом уровне; исходные
данные и оценки эндемизма сведены в табл. 3.
Исходные обозначения в та!бл. 3 те же, что и в табл, 2.
Соответствующее уравнение имеет вид 1g [TJs+U^—1-492+0.086 Us.
Таблица 3
Регион us Es ®s D г • 1 г • II
Европа, лес 20 2 1 1 9.1 2.5 5
Европа, степь 20 0 1 —1 0.0 4.5 6.5
Карпаты 28 4 7 -3 12.5 6 4
Крым 26 6 5 1 18.7 2.5 3
Кавказ 39 42 72 -30 51.8 7 2
Средняя Азия 30 41 11 30 57.7 1 1
Сибирь, Алтай 20 0 1 —1 0.0 4.5 6.5
Дальний Восток 14 12 0 0 46.1 0 0
78
ОБЩАЯ ЧАСТЬ
Имеются два момента, на которые надо обратить внимание при анализе ма-
териалов, представленных в табл. 3; первое — фауна Дальнего Востока не
следует общей тенденции, второе — две последовательности, в которые выстраи-
ваются регионы в зависимости от того метода, которым проведен учет энде-
мизма. По процентам имеем следующую последовательность: Средняя Азия—
Кавказ—Крым и Карпаты—лесная зона европейской части; в двух остальных
регионах эндемики отсутствуют. Иной вид имеет последовательность, в которую
выстраиваются регионы по показателю относительного эндемизма: Средняя
Азия—лесная зона европейской части и Крым—степная зона европейской части
и Сибирь с Алтаем—Карпаты, Кавказ.
Какие же выводы можно сделать из приведенных материалов?
Повышенный сравнительно с общей тенденцией показатель относительного'
эндемизма (D >0) свидетельствует о том, что фауна рассматриваемого региона
слабо связана с фаунами прочих сравниваемых территорий либо вследствие
физической изоляции, либо за счет более тесных связей с иными физико-геогра-
фическими единицами (которые в обсуждение не входят).
Напротив, пониженный показатель относительного эндемизма (Р <С 0)
говорит о том, что обсуждаемая территория есть лишь участок более обширного
естественного региона. Так, рассматриваемый здесь участок Карпат — малая
часть горной страны; рассматриваемый участок, именуемый «степной зоной ев-
ропейской части СССР», — по существу произвольно выбранная часть степной
зоны средней и южной Европы и т. п.
Следовательно, избыток свидетельствует о связи изолированной территории
с какими-то иными регионами, не входящими в рассмотрение (или о полной
обособленности региона), а недостаток — о фаунистической (и, как правило,
физико-географической) принадлежности региона. В любом случае, конечно,
могут быть исключения, связанные с существованием каких-то иных законо-
мерностей.
Обратимся к рассмотрению конкретных примеров. Как видно из данных
табл. 3, соотношение Us и Es на Дальнем Востоке выходит за рамки закономер-
ности, ясно прослеживающейся во всех прочих регионах. Это естественно, так
как Дальний Восток принадлежит к другой иерархической системе, или (в тер-
минах зоогеографии) относится к Маньчжуро-Китайской подобласти Палеарк-
тики, тогда как прочие регионы составляют часть Средиземноморской зоогео-
графической подобласти Палеарктики.
Наиболее высок уровень Ds в фауне Pupillina Средней Азии. За исключением
Копетдага, фауна которого практически лишена эндемиков, горы Средней Азии
изолированы территориями, мало пригодными для жизни сухопутных моллю-
сков: на западе — пустынями, на юге и востоке — высокогорьями Гиндукуша,
Памира и Кунь-Луня, на севере — сухими степями восточного Казахстана
с резко континентальным климатом. Поэтому в обширной, замкнутой и очень
сложной системе хребтов с огромным набором биотопов видообразование идет
очень интенсивно, что и выражается высоким значением Ds: как следует из
табл. 3, здесь на 30 (!) видов больше, чем можно ожидать, исходя из общей тен-
денции.
Минимальный «избыток» видов (Ds=l) имеется в лесах европейской части
СССР и в Крыму. Столь незначительное отклонение вряд ли целесообразно
рассматривать специально, так как оно может быть вызвано и случайными фак-
торами, и субъективностью оценки видового и подвидового ранга отдельных
форм. То же касается фаун степной зоны европейской части, а также Сибири
с Алтаем, только здесь наблюдается минимальное отклонение от средней тен-
денции в отрицательную сторону (Ds=—1).
Небольшое отклонение значения Ds в фауне Карпат («нехватка» трех видов)
станет понятным, если вспомнить, что обсуждаемая часть Карпат составляет
небольшую часть обширной горной страны, фауна которой к тому же тесно свя-
зана с фауной Альп.
Самый яркий пример отрицательного отклонения дает фауна Pupillina
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
79
Кавказа (Ds=—30). Фауна Большого Кавказа, имея известную степень обо-
собленности, отчетливо и тесно связана с фауной Закавказья, а та в свою оче-
редь есть часть малоазиатской фауны. Именно резкая «несамостоятельность»
фауны Pupillina Кавказского перешейка и выражается столь ощутимым отри-
цательным значением показателя относительного эндемизма.
Формальный смысл отклонений от средней тенденции на уровне родов
тот же, что и на уровне видов, но зоогеографическая сущность этих отклонений
заслуживает отдельного рассмотрения. Небольшой избыток эндемичных родов,
как следует из данных табл. 2, имеет место на Кавказе и в Средней Азии.
Избыток родов, как и избыток видов, в горных районах Средней Азии объяс-
няется почти полной физической изоляцией региона; но повышенное число энде-
мичных родов свидетельствует, кроме того, и о достаточно давней изоляции;
здесь имеется даже эндемичное подсемейство Chondrulopsininae.
Для многих фаунистических элементов Кавказа характерен высокий уровень
реликтовости, иными словами, здесь имеется 9 монотипических родов (Pilor-
cula, Differena, Adzharia, Andronakia, Retowskia, Caucasicola, Multidentula,
hnprovisa, Senaridenta) и значительное число олиготипических. Больших родов
очень мало: самый большой (эндемичный, кстати) род Euxinolauria включает
9 видов; характерно, что род распадается на 4 подрода. За счет этой реликто-
вости и возникает небольшой «избыток» эндемичных родов на Кавказе.
Большая часть регионов (5 из 8) показывает точное соответствие ожидаемой
степени относительного эндемизма на уровне родов (Dg=0), из чего следует
представление о «взаимной сбалансированности» их фаун на родовом уровне.
Иными словами, на этом уровне фауны 5 регионов, обладая известной самостоя-
тельностью, тем не менее отчетливо взаимосвязаны.
Уместно заранее ответить на вопрос, возникший, вероятно, у читателя:
а разве без приведенных выкладок не ясно, что фауна, например, советской части
Карпат есть часть фауны горной страны в целом? Да, о связях фаун отдельных
регионов судить, конечно, можно путем простого сопоставления фаунистиче-
ских списков, но составить объективное представление о сравнительной сте-
пени их обособленности нельзя. Так, например, Средняя Азия занимает первое
место по эндемизму на видовом уровне как по оценке методом процентов, так и
по оценке величины Ds. Но, например, Кавказ по абсолютному эндемизму, вы-
раженному в процентах, занимает второе место, а по показателю Ds — послед-
нее. Из пятой слева колонки табл. 3 мы получаем сведения о том, что число
эндемичных видов на Кавказе по сравнению со всей фауной Pupillina этого ре-
гиона выше, чем где бы то ни было (кроме Средней Азии); однако по процентному
содержанию эндемичных видов еще нельзя сказать, много это или мало для дан-
ной фауны. Из четвертой слева колонки табл. 3 видно, что если учесть еще и
общее богатство фауны Кавказа, то эндемиков должно быть много больше.
Из этого, в частности, следует прогноз: поскольку фауна Кавказа есть часть
фауны Малой Азии, то при полной ревизии малоазиатской фауны Pupillina
число эндемичных видов резко возрастет.
Следовательно, вычисляя значение D, мы получаем объективную информа-
цию о том, какова ожидаемая степень эндемизма для отдельных регионов; со-
поставляя этот показатель с действительным, можно судить о степени насыщен-
ности фауны эндемиками. Любые отклонения вычисленного показателя от реаль-
ного, особенно если отклонения значительны, имеют объективные причины,
анализ которых составляет одну из важных задач зоогеографического иссле-
дования.
Имеется существенное ограничение изложенного метода, которое определя-
ется рамками применения термина «фауна». Так, была сделана попытка про-
анализировать тем же методом фауну Кавказа, сравнивая для этой цели малако-
фаунистические округа и подокруга, выделенные Лихаревым и Раммельмейер
(1952, с. 42—57). Здесь обнаружено отсутствие корреляции между U и Е как
на видовом, так и на родовом уровне. Очевидно, если можно говорить о фауне
Кавказа (пусть даже как части более обширного региона), то применение поня-
80
ОБЩАЯ ЧАСТЬ
тия «фауна» в том смысле, в котором это понятие употребляется здесь, к более
дробным единицам — округам и подокругам — неправомерно: это уже не
фауна физико-географической территориальной единицы во всем разнообразии
ее условий, но скорее сочетания видов, свойственные отдельным типам ланд-
шафта. Например, «фауна» армянского округа — это в основном набор ксеро-
термных видов, «фауна» талышского округа — преимущественно комплекс
видов субтропического леса и т. и.
ВОПРОСЫ СИСТЕМЫ И ФИЛОГЕНИИ
Развитие взглядов на систематическую
структуру и положение Pupillina
в отряде Geophila
История исследования Pupillina складывалась из трех основных направле-
ний, которые развивались в значительной мере независимо одно от другого:
это, во-первых, установление систематической обособленности Pupillina и выяс-
нение объема таксона; во-вторых, разработка системы самого крупного (во вся-
ком случае, в Палеарктике) семейства Buliminidae; в-третьих, исследование
внутреннего строения Pupillidae s. lat.
! Начало первому направлению было положено небольшой статьей Пилсбри
(Pilsbry, 1900), который обнаружил своеобразие в строении почки Partula и
Achatinella и установил подотряд Orthurethra; при всех дальнейших перестрой-
ках системы Geophila (Vasopulmonata, Stylommatophora) этот подотряд в от-
ношении объема оказался самым стабильным, и к этому таксону относили любой
вид, обладавший почкой ортуретрального облика. Существенно, что изолирован-
ное положение Orthurethra определяется двумя полностью независимыми при-
знаками: строением почки и наличием сложно устроенного пениального аппен-
дикса, первично имеющегося у всех членов подотряда и всегда отсутствующего
у любых других таксонов Geophila. При кардинальном пересмотре системы
Geophila (Шилейко, 1979а) самостоятельность Orthurethra была подтверждена,
лишь немного изменен объем подотряда (введены надсемейства; Cerioidea и
Sagdoidea, а также переведено в другой подотряд семейство Partulidae). Пред-
ложение названия Pupillina вместо Orthurethra вызвано тем, что противопостав-
ляемые Orthurethra таксоны (Mesurethra, Sigmurethra) признаны сборными,
a Heterurethra входят в состав другого отряда (Succineida).
Самый значительный этап в разработке системы подотряда в целом связано
именем Стинберга. После изучения Valloniidae (Steenberg, 1918) повилось исклю-
чительно тщательное исследование (Steenberg, 1925), посвященное преимуще-
ственно европейским видам Pupillidae, Vertiginidae, Orculidae и Chondrinidae;
на основании собственных и литературных материалов Стинберг суммировал
свои взгляды на систему следующим образом.
Orthurethra
1 fam. Chondrinidae (Abida, Sandhalia, Chondrina)
2 fam. Orculidae (Orcula, Orculella, Pagodulina)
3 fam. Pupillidae
1 subfam. Lauriinae (Lauria)
2 subfam'. Pupillinae (Pupilia)
4 fam. Vertiginidae
1 subfam. Nesopupinae (Nesopupa)
2 subfam. Vertigininae (Vertigo)
3 subfam. Truncatellininae (Truncatellina, Columella)
5 fam. Valloniidae
1 subfam. Valloniinae (Vallonia, Acanthinula, Spermodea, Zoogenetes)
2 subfam. Pyramidulidae (Pyramidula)
? 3 subfam. Spelaeodiscinae (Spelaeodiscus)
6 fam. Patulastridae (— Pleurodiscidae) [Patulastra (Pleurodiscus)]
7 fam. Strobilopsidae (Strobilops)
8 fam. Enidae
СИСТЕМА И ФИЛОГЕНИЯ
81
9 fam. Pachnodidae (или Pachnodinae как подсемейство Enidae) (Pachnodus и, вероятно.,
много других родов).
10 fam. Partulidae
11 fam. Cochlicopidae
12 fam. Amastridae-
13 fam. Achatinellidae
? 14 fam. Pacificellidae
15 fam. Tornatellinidae
16 fam. Auriculellidae
Через два года после опубликования исследования Стинберга Пилсбри
(Pilsbry, 1927, Man. Conch., v. 28, p. 8), придерживаясь более широкого толко-
вания Pupillidae, вводит в систему филогенетические связи:
Gastrocoptinae ' Vertiginidae
Pupillinae Oreulinae Nesopupinae
________________________________I___________________________:
I
Ancestral
---------------------Strobilopsidae
Pupillidae
Valloniidae I
Система Бейкера (Baker, 1935) представляет собой по существу синтез взгля-.
дов Стинберга и Пилсбри:
Pupillinae VaHoniinae Vertiigininae
Авторы последующих систем (Thiele, 1931; Wenz, Zilch, 1959—1960; Taylor,
Sohl, 1962) придерживаются в целом взглядов Стинберга, с учетом воззрений
Пилсбри и Вейкера; эти системы носят в большой степени компилятивный ха-
рактер.
Взгляды, развиваемые в настоящей работе, есть развитие и дополнение си-
стемы Стинберга на основании главным образом новых фактов, полученных
при исследовании анатомии видов, обитающих преимущественно ца территории
СССР.
Показательно развитие второго направления, т. е. разработка системы Buli-
minidae (=Enidae).
Первым, кто предпринял попытку внести какую-то упорядоченность в много-
образие моллюсков с булиминидным обликом раковины, был Пфайфер (Pfeif-
fer, 1846). Все 598 известных к тому времени видов «Bulimits» он по конхологи-
ческим признакам разбил на}32 категории, причем названия этим категориям
6 А. А. Шилейко
82 ОБЩАЯ ЧАСТЬ
предложены не были. Категории в свою очередь сгруппированы Пфайфером
в совокупности 4 рангов:
А — перистом расширенный или отвернутый,
В — перистом прямой.
А разделены на три категории более низкого ранга: I — пупок отсутствует,
II — пупок проколовидный, III — пупок в разной степени расширен.
В таким же образом включает две совокупности: I — край устья утолщен,
II — край устья простой.
Каждая из совокупностей разделяется еще на две категории, имеющие та-
кие же краткие описательные характеристики.
Ныне система Пфайфера представляет лишь исторический интерес, так как
сюда включены все виды, имеющие соответствующую форму раковины, в том
числе представители Partulidae, Bulimulidae и других похожих по раковине
семейств.
Примерно такой же характер имеет и система Кобельта (Kobelt, 1902), ко-
торый создал фундаментальное справочное руководство, но не пошел дальше
формальных описаний, снабженных иллюстрациями и материалами по синони-
мике. Заключительным этапом «конхологического» периода изучения системы
Buliminidae стала система Линдгольма (Lindholm, 1925а), представляющая
собой попытку внести упорядоченность и уточнить характер соподчинения уста-
новленных к тому времени таксонов, а также оформить систему номенклатурно.
Первая система груйпы, в основу которой были положены анатомические
признаки, принадлежит А. Вагнеру (Wagner А., 1927), который различал два
самостоятельных семейства: Chondrulidae (пениальный аппендикс отсутствует,
половой ретрактор одноветвистый) и Enidae (пенис с хорошо развитым аппен-
диксом, половой ретрактор двуветвистый). Здесь надо заметить, что оба при-
знака тесно скоррелированы: если нет пениального аппендикса, нет, естественно,
и соответствующей ветви полового ретрактора. Последнее семейство, по Ваг-
неру, включает два подсемейства: Spelaeoconchinae (систематическое положение
неясно) и Napaeinae (дивертикул семяприемника отсутствует). Очевидно, под-
разумевалось и третье — номинативное подсемейс^о, характеризующееся
альтернативным признаком: дивертикул семяприемника имеется.
Тиле (Thiele, .1931) включает оба вагнеровских семейства в качестве под-
семейств в Enidae, добавляет подсемейство Jaminiinae и расширяет диагнозы за
счет включения признаков раковины.
Анатомировав 58 видов и использовав литературные данные, Гессе (Hesse,
1933) разделяет Enidae на 4 подсемейства; Chondrulinae, Jaminiinae, Eninae и
Spelaeoconchinae; при этом он оперирует теми же признаками: отсутствие или
наличие пениального аппендикса и (или) дивертикула семяприемника. Удиви-
тельно, что Гессе не сделал никаких выводов из собственных материалов по вну-
треннему строению гениталий. Так, в более ранней работе (Hesse, 1885) он за-
метил и нарисовал призмоконические бугорки внутри пениса (видимые сквозь
покровы органа) у Buliminus detritus, В. hohenackeri, В. bidens var. B.etowskiana
и В. pupa (Hesse, 1885, Taf. 3, Figs. 2, 3, 5, 6); в работе 1933 г. Гессе отмечает
наличие незамкнутой папиллы у Mastus pupa (S. 156, Fig. 2) и у Chondrus
tournejortianus (S. 188, Fig. 25), а также замкнутую папиллу у Jaminia pupoides
(S. 163, Fig. 6). Далее, Гессе анатомировал 2 экз. Adzharia renschi и нашел, что
у одного экземпляра дивертикул семяприемника имеется, а у другого отсутствует,
и этот факт его не насторожил и не заставил пересмотреть таксономический вес
признака наличия—отсутствия дивертикула: он выделяет роды на основании
названного признака. Вероятно, здесь сказался гипноз авторитета Вагнера.
Очень тщательно выполненная работа Форкарта (Forcart, 1940), посвященная
богатой фауне Buliminidae Турции, сделана по существу на теоретической базе
Вагнера—Гессе: здесь семейство рассматривается как состоящее из двух тра-
диционных подсемейств (Eninae — пенис с аппендиксом и Chondrulinae —
пенис без аппендикса). Хотя на иллюстрациях в монографии Форкарта видны
СИСТЕМА И ФИЛОГЕНИЯ
83
основные детали внутреннего строения пениса, эти детали также не были ис-
пользованы в таксономических построениях.
В исследовании Матёкина (1959), касающемся среднеазиатских представи-
телей семейства, основной акцент сделан на выяснении размаха изменчивости
как конхологических, так и анатомических признаков. Не обсуждая сейчас част-
ные вопросы о числе видов и их границах, хотелось бы подчеркнуть, что работой
Матёкина ярко показана ограниченность «анатомического» метода, если ис-
пользуется только внешний вид полового аппарата. Даже если не углубляться
в рассуждения о неправомерности применения родового названия Jaminia
к среднеазиатским формам (типовой вид рода Jaminia резко отличается от всех
среднеазиатских видов даже внешним обликом полового аппарата), то чрез-
вычайно показателен вывод Матёкина о «несостоятельности выделения рода
Тигапепа, так как виды этого «рода» принадлежат.к роду Zebrina Held.» (см.
Матёкин, 1959, с. 1519). И действительно, если ограничиться рассмотрением
только внешнего вида полового аппарата, то виды рода Тигапепа очень похожи
на виды рода Zebrina. Однако об их резком различии свидетельствует органи-
зация совокупительного аппарата, причем внутреннее строение пениса Тига-
пепа свидетельствует о близости рода другим среднеазиатским таксонам,
а строение пениса Zebrina очень напоминает таковое видов других европейских
родов.
Очевидно, что при исследовании систематической структуры Buliminidae
знание деталей внутреннего строения мужского отдела полового аппарата при-
обретает особую важность. При пересмотре системы семейства (Шилейко, 19786,
1978в, 1978г) это обстоятельство было учтено; в результате было показана зна-
чительно большая сложность исторически сложившихся взаимоотношений
между различными группами внутри Buliminidae, чем это принималось ранее.
Разделение семейства на 8 подсемейств на основании как конхологических, так
и анатомических признаков призвано отразить сложность таксономической
структуры семейства и наметить естественные группировки внутри него.
Позже (Шилейко, 19796) выделение подсемейства Chondrulinae было при-
знано нецелесообразным (этот вопрос подробно обсуждается ниже). Существенно,
что выделенные подсемейства имеют достаточно четкий зоогеографический
облик.
Наконец, основы третьего направления, т. е. исследование внутренней ор-
ганизации прочих семейств и разработка вопросов их систематики, заложены
Стинбергом в уже цитированной работе (Steenberg, 1925), хотя сведения о вну-
треннем строении отдельных видов содержатся еще в трудах Мокец-Тандона
(Moquin-Tandon, 1855), Лемана (Lehmann, 1873) и некоторых других исследо-
вателей. В последние десятилетия были опубликованы исследования, посвя-
щенные полной или частичной ревизии отдельных семейств и подсемейств:
Achatinellidae (Cooke, Kondo, 1960), Spelaeodiscinae (Gittenberger, 1969, 1974,
1975), Chondrinidae (Gittenberger, 1973), Lauriinae (Шилейко, 19756), Orculidae
(Шилейко, 19766; Gittenberger, 1978).
Таксономическая и филогенетическая
структура подотряда Pupillina
«В целом проблема. . . находится в очень
здоровом состоянии, поскольку вопросов в ней
гораздо больше, чем ответов».
Р. X. Райт1..
Как будет видно из дальнейшего текста, палеонтологических материалов, по
которым можно судить об истории фауны Pupillina, а тем более о происхожде-
нии подотряда, удручающе мало. Поскольку эти материалы в отечественной ли-
1 Цит. по: С. А. Корытин. Запахи в жизни зверей. М., Знание, 1978, с. 127.
6*
84
ОБЩАЯ ЧАСТЬ
тературе уже обсуждались (Лихарев, Раммельмейер, 1952; Стеклов, 1966, 1968)
и ничего существенного добавить не представляется возможным, краткие сооб-
ражения по истории фауны помещены в настоящий раздел, где они излагаются
в связи с вопросами филогении подотряда и подчиненных таксонов.
Общ ие замечания
Представления зоологов о системе таксонов часто более совершенны, чем
представления о путях, приведших к современному положению. Причины этого
очевидны: палеонтолог оперирует лишь частями организмов, сохранившимися
в ископаемом состоянии, а неонтолог имеет возможность изучать многообразие
современных видов с привлечением богатого арсенала методов анатомии, физи-
ологии, биохимии и других, неприменимых к палеонтологическому материалу.
Трудности усугубляются неполнотой палеонтологической летописи.
По этой причине тесно взаимосвязанными понятиями «филогения» и «система»
в реальных условиях часто приходится оперировать по отдельности. Если пра-
вильно (адекватно) понятая филогения, опирающаяся на надежный ископаемый
материал, обычно приводит к столь же адекватной системе, то нельзя утверж-
дать обратного: система, разработанная с учетом всех признаков, имеющих таксо-
номический вес, может быть удовлетворительной в том смысле, что объясняет
известные факты и не вступает с ними в противоречия, но при этом пути разви-
тия таксонов и особенно время их становления могут оставаться в той или иной
мере предположительными.
Приведенные соображения в полной мере иллюстрируются ситуацией, наб-
людающейся в подотряде Pupillina, поскольку система, порядок соподчинения
таксонов, принимаемый здесь, удовлетворительно, как будто, отражает поло-
жение, сложившееся к настоящему времени, но недостаточность палеонтолог
гических материалов делает филогенетические положения лишь предположи-
тельными.
В этой связи наиболее существенное обстоятельство заключается в том, что
самые ранние ископаемые находки членов подотряда датируются мелом, и они
представлены большей частью уже вполне надежно определяемыми видами сов-
ременных родов, о чем дает представление табл. 4. Помимо рода Protornatel-
Ипа в таблицу включены только таксоны, представленные в рецентном состоя-
нии; кроме того, имеется некоторое число родов, вымерших в течение неогена.
В табл. 4 не включен также род Dendropupa, описанный из верхней перми —
карбона (обсуждение положения этого рода см.'ниже).
Как видно из данных табл. 4, наиболее древняя находка представителя под-
отряда сделана в верхнемеловых отложениях (Северная Америка): это Protor-
natellina isoclina (Achatinellidae), напоминающая современных Т ornatellina',
обитающих на островах Хуан Фернандес, т. е. на участке ареала семейства,
ближе всего расположенного к Северной Америке. Наиболее ранние ископаемые
находки представителей прочих таксонов обнаруживаются в палеоцене Европы
и состоят уже из вполне надежно определяемых видов современных родов
Azeca, Cochlicopa, Orcula, Acanthinula, Vallonia, Vertigo. Более того, Cochli-
copa, Vertigo и Vallonia найдены в палеоцене не только Европы, но и Северной
Америки. Отсюда с очевидностью следует, что подотряд в целом возник много
раньше — возможно, еще в раннем мезозое или даже в палеозое.
В этой связи многозначительна находка Dendropupa в верхней перми' — кар-
боне Северной Америки и Европы, которую Венц (Wenz, 1938—1944) и Мур
(Moore, 1960) относят к Cyclophoridae; такое толкование спорно, поскольку для
переднежаберных моллюсков, имеющих крышечку, длинные париетальные
пластинки нехарактерны, и трудно себе представить сочетание таких признаков,
как наличие крышечки и париетальной пластинки, вдающейся в'просвет устья.
Напротив, длинная париетальная пластинка — характерная особенность наи-
более архаичных Pupillina: Achatinelloidea и Orculoidea.
СИСТЕМА И ФИЛОГЕНИЯ
85
Таблица 41
Название рода Плей- стоцен Плио- цен Мио- цен Олиго- цен Эоцен Палео- цен 'Мел
Protomatellina Azeca
Cochlicopa
OrCI 11A
Pagodulina
'Leiostyla
Lauria
Strobilops
Acanthinula Spermodea
\7я 11 onia
Pupilia s. laL
Pupoides
Gastrocopta s. lat
Vertigo
VertiHa Columella
Trunc atelli na G ranaria 4
Chondrina Pyramidula
Zebrina Napaeus
Chondrula
Retowski a
Multid entula
1 Составлено по: Стеклов, 1966; Wenz, Zilch, 1959—1960.
Далее, в эоцене Европы найдены виды современных родов Strobilops, (?)
Pupotdes, Abida, Pyramidula; в олигоцене — Leiostyla, Spermodea, Papilla,
Gastrocopta, Truncatellina. Существенно, что и в это время связи между родами,
подсемействами и семействами также не прослеживаются. Отсутствие такой
связи можно частично объяснить неполнотой летописи, но это объяснение можно
86 ОБЩАЯ ЧАСТЬ
бы принять, если бы отсутствие видимых связей не было столь полным. Поэтому,,
даже учитывая, что в палеонтологических рядах имеются обширные лакуны,
надо допустить, что и эти роды также сформировались значительно раньше.
К обсуждению этих вопросов мы еще вернемся, пока' лишь отметим, что
палеонтологические материалы не дают прямых ответов на вопросы филогене-
тического характера. Поэтому в качестве основы для реконструкции филогене-
тических отношений внутри подотряда приходится опираться главным образом,
на анализ конхологических и анатомических признаков современных видов.
Проблемы филогении
В качестве отправного пункта для всех дальнейших рассуждений надо об-
судить два взаимосвязанных вопроса: какой тип раковины мыслим в качестве
исходного и раковины каких форм ближе всего к этому гипотетическому типу.
Несмотря на то что высота раковин различных Pupillina образует непрерыв-
ный ряд от 0.7 до более чем 30 мм, всю совокупность семейств можно довольно
определенно разбить на три группы (микрохеликоидные раковины в расчет не
принимаются, так как они несомненно вторичны): мелкие (1—4 мм), средние
(5—10 мм) и большие (более 10 мм). К первой группе принадлежат Vertiginidae
и Pupillidae, ко второй — Achatinellidae, Orculidae и Cochlicopidae и к третьей—
Buliminidae. Своеобразные Chondrinidae занимают промежуточное положение
между второй и третьей группами (рис. 39).
Наиболее примитивные, на мой взгляд, Achatinellidae и Orculidae относятся
ко второй размерной группе, а раковины их снабжены длинными париетальными
и колумеллярными пластинками, происхождение и функциональное назначе-
ние которых рассмотрено выше (с. 20 и 57). Такими признаками, кстати, харак-
теризуется и раковина Dendropupa. Исходя из этого и принимая во внимании
облик раковины Protornatellina, можно представить себе раковину анцестраль-
ных форм следующим образом.
Форма раковины высококоническая, высота 6—10 мм, число оборотов
5—7, устье с париетальной и колумеллярной пластинками, присутствующими
на всех стадиях постэмбриогенеза; эмбриональные обороты, вероятно, со спираль-
ной скульптурой, так как такая скульптура первично характерна и для Orcu-
lidae, и для Achatinellidae, и для наиболее архаичных Valloniidae, и для прими-
тивных Buliminidae — Retowskiinae.
Возможно, возникновение групп, относящихся как к первой, так и к тре-
тьей размерным группам, приведшее к обособлению таксономических единиц,,
связано с освоением двух основных типов лесных биотопов: подстилки и стволов.
Естественно, адаптация к жизни в слое лиственного опада привела к становле-
нию Vertiginidae и Pupillidae. Размеры Achatinellidae колеблются в заметных
пределах, что стоит в связи с характером биотопов, освоенных разными ви-
дами: самые большие формы обитают на стволах деревьев, самые маленькие —
в слое лиственного опада. Наиболее архаичные (по крайней мере среди пале-
арктических групп) Buliminidae — Retowskiinae — обитатели реликтовых влаж-
ных лесов юго-западного Закавказья,, причем примечательно, что крупные
Retowskia (взрослые!) живут на стволах деревьев, часто на мокрых скалах в те-
нистых ущельях, а значительно более мелкие Andronakia, как и молодь Retow-
skia, — в толстом слое подстилки и часто даже закапываются во влажную почву.
Способность к закапыванию также определяется размерами животных:
самые мелкие формы не закапываются активно, но могут лишь использовать
естественные трещины в земле. Это связано с тем, что отдельные частицы почвы
(песчинки) соизмеримы по размерам с такими раковинами, как, например,
у Vertigo, и моллюски не могут в силу малых размеров развить достаточные уси-
лия для раздвигания почвенных частиц. Редко и неглубоко могут закапываться
несколько более крупные Pupillidae и Cochlicopidae; многие вышедшие в откры-
тые пространства Buliminidae уже активно закапываются на глубину несколь-
ких сантиметров.
СИСТЕМА И ФИЛОГЕНИЯ
87
Уменьшение раковины достигалось не только уменьшением размеров как
таковых, с сохранением в целом пропорций раковины, но и приобретением спо-
собности размножаться раньше, чем раковина достигнет дефинитивного со-
стояния. Это вызвало раннюю остановку роста раковины и затем образование
постоянной микрохеликвидной формы. Естественно, что этот путь привел
к резко выраженной протерандрии: совокупительный аппарат полностью исче-
зает после выполнения его функции (Valloniidae). Вместе с тем афаллия, полная
или частичная, нередко встречается и в других группах, отличающихся в соче-
тании с малыми размерами яйцеживорождением (Lauria, Gibbulinopsis, Pupilla).
Формирование в матке крупных яиц, в которых развиваются эмбрионы, имею-
щие раковину, обостряет проблему размещения внутренних органов, и у осо-
бей различных видов нередко прослеживается ясная закономерность: чем
больше в матке яиц и чем старше эмбрионы, тем хуже развит мужской отдел;
в максимально выраженной ситуации он превращается в систему нитевидных
протоков, занимающих в совокупности ничтожно малый объем.
Исключение составляют некоторые Chondrinidae, у которых при сравнительно
больших размерах раковины наблюдается порой полная афаллия. В этом слу-
чае афаллия, вероятно, не связана с функциональным состоянием полового
аппарата, так как среди одной выборки Chondrina clienta caucasica, собранных
в один день, встречены как афалличные, так и эуфалличные особи. Таким об-
разом, афаллия у Chondrinidae имеет скорее всего иную природу, чем у разоб-
ранных выше групп; это еще один довод в пользу представления об изолирован-
ном положении семейства.
Чрезвычайно интересные соображения по поводу ранней эволюции Geophila
в целом и Pupillina в частности были высказаны Я. И. Старобогатовым в личной
беседе, который справедливо указал на способ питания как один из важных
факторов исторического развития наземных легочных моллюсков.
Всех Pulmonata, по Старобогатову, можно разделить по типам питания на
8 основных групп:
1) детритофаги (питаются бактериями и переработанной ими органикой),
2) микромикофаги (питаются гифами грибов), 3) макромикофаги (питаются
плодовыми телами макромицетов), 4) лихенофаги (питаются лишайниками),
5) фитофаги (питаются живыми тканями покрытосеменных растений), 6) оли-
гохетофаги (питаются почвенными олигохетами), 7) малакофаги (питаются
наземными моллюсками), 8) полифаги (едят все).
Чрезвычайно важно, что легочные моллюски не едят ни голосеменных ра-
стений, на папоротникообразных, ни мхов.
Детритофагия первична для легочных моллюсков, но она не могла обеспе-
чить становления наземных форм, во всяком случае в широких масштабах.
Поэтому формирование Geophila связано с переходом на питание грибами, вна-
чале, очевидно, гифами грибов. Но микромикофагия обеспечила формирование
мелких наземных форм; переход к макромикофагии повлек за собой увеличение
размеров (и увеличение плотности популяций).
Новый и очень перспективный пищевой ресурс появился с возникновением
покрытосеменных растений; что касается лихенофагии, то она всегда была
тупиковым направлением пищевой специализации.
Если эти положения верны, то возникновение Pulmonata надо датировать
примерно рубежом кембрия и ордовика. Geophila появились вместе с назем-
ным растительным покровом, т. е. в девоне—карбоне, и это могли быть древние
Achatinina или Pupillina мелких или средних размеров. В начале позднего
мела появились покрытосеменные растения — тогда же могли возникнуть фи-
тофаги.
Поскольку основная масса Pupillina — микро- или макромикофаги, их воз-
раст можно оценить как раннекарбоновый; многие Buliminidae — фитофаги,
и, следовательно, появились в верхнем мелу; лихенофагами являются преиму-
щественно Chondrinidae и Pyramidulidae, и о возрасте их судить трудно.
88
ОБЩАЯ ЧАСТЬ
Мне приятно выразить глубокую благодарность Я. И. Старобогатову за
разрешение изложить его неопубликованные данные.
Переходя к рассмотрению анатомических признаков, которые могут быть
использованы при анализе системы и филогении подотряда, надо в первую оче-
редь констатировать, что, за исключением полового аппарата, внутренняя
организация обнаруживает большое постоянство. Поэтому приходится опери-
ровать почти исключительно признаками, касающимися репродуктивного тракта.
Наиболее характерная диагностическая черта полового аппарата Pupil-
lina — первичное наличие сложного пениального аппендикса, который имеется
исходно у большинства Pupillina (исключение Chondrinidae) и полностью и
всегда отсутствует у всех прочих подотрядов Geophila. Поэтому вопрос об исто-
рическом развитии этого придатка заслуживает специального рассмотрения.
Вряд ли подлежит сомнению, что гермафродитный половой аппарат исход-
ных водных форм был прост и лишен черт специализации: от гермафродитной
железы отходил проток, не полностью разделенный на два отдела — мужской
и женский. В условиях жизни на суше возникает необходимость сократить по
возможности время спаривания. Предпосылкой для реализации этой тенденции
служит возникновение специализированной камеры, где происходило бы на-
копление аутоспермы, чтобы затем передать партнеру этот комок целиком. Та-
кая камера, вероятно, могла возникнуть как можно ближе к выходному отвер-
стию мужского протока, и формирование пениального аппендикса есть в ко-
нечном счете закономерный результат развития и дифференциации этой камеры.
Но если теперь сперма передается не постепенно, а сразу в виде плотной аг-
грегации, то необходимо и соответствующее помещение, куда может вместиться
этот комок целиком. С этим, видимо, связано развитие семяприемника в виде
резервуара, соединенного с вагиной протоком. Образование дивертикула семя-
приемника коррелятивно связано с появлением сперматофора, который на-
правляется вначале именно в дивертикул; у таксонов, исходно лишенных спер-
матофора (Achatinellidae), дивертикул семяприемника всегда отсутствует.
Параллельно с дифференциацией аппендикса, но в значительной мере не-
зависимо, дистальная часть семяпровода преобразовалась в орган, передаю-
щий скопления спермиев партнеру, т. е. в пенис. Появление Pupillina имело
место в условиях постоянно влажного и теплого климата, когда сперматофор
как средство защиты спермы от высыхания не нужен. Поскольку последний фор-
мируется в эпифаллусе, то первично отсутствует и эпифаллус. Achatinellidae,
до сих пор живущие в условиях влажного тропического климата без смены се-
зонов (наибольшее разнообразие — на Гавайских о-вах), не приобрели ни спер-
матофора, ни, следовательно, эпифаллуса и сохранили примитивное состояние
дистального отдела полового аппарата (рис. 21, II).
Группы, попавшие в условия чередования влажного и сухого сезонов, на-
против, имели мощный стимул к приобретанию сперматофора; однако строение
эпифаллуса, дополнительных органов на этом протоке (цэкума и придатка эпи-
фаллуса) и сперматофора у различных семейств Pupillina достаточно резко раз-
личается, что может свидетельствовать об их значительной взаимной изолиро-
ванности.
ВОПРОСЫ СИСТЕМЫ ПОДОТРЯДА
Рассмотрев ряд общих положений, обратимся к анализу схемы, представ-
ленной на рис. 39. В первую очередь надо обратить внимание еще раз на то,
что монофилетичность подотряда вряд ли может быть поставлена под сомнение:
все таксоны, включаемые в подотряд, исходно характеризуются двумя взаимно
независимыми признаками: ортуретральной организацией экскреторного ап-
парата и наличием однотипно устроенного пениального аппендикса.
Исключение составляют две группы. Во-первых, Sagdoidea с ярко выражен-
ной сигмуретрией; но это надсемейство составлено хищниками, а для всех хищ-
ных форм независимо от систематического положения характерна сигмуретрия
(Шилейко, 1979а). Еще Бейкер (Baker, 1935, р. 191) писал, что «мужской отдел
90
ОБЩАЯ ЧАСТЬ
этих Orthurethra (Pupillidae s. lat., — А. Ш.) строго параллелен сигмуретраль-
ным Sagdidae». Надсемейство Sagdoidea в схему на рис. 39 не включено из-за
того, что анатомия его представителей не изучена на должном уровне и связи
таксона неясны.
Второе исключение — семейство Chondrinidae, почка которых сохраняет
ортуретральный тип строения, но неизвестно пи одного вида, у которого был бы
пениальный аппендикс или орган, который можно было бы счесть гомологом
аппендикса. У некоторых родов, например Granaria, на мужском отделе име-
ется придаток, но это наверняка не гомолог аппендикса, так как половой рет-
рактор крепится ближе к половому атриуму, чего не наблюдается при редукции
или видоизменении аппендикса у любых других таксонов. Названный орган
скорее можно гомологизировать с придатком эпифаллуса Orculoidea, хотя та-
кой вывод надо рассматривать только как предварительный.
Постулируя естественность Pupillina, нельзя, однако, не признать значи-
тельного разнообразия, имеющего место внутри подотряда. Общепринято вы-
делять три надсемейства: Achatinelloidea (единственное семейство Achatinel-
lidae), Cochlicopoidea (два семейства — Amastridae и Cochlicopidae) и Pupil-
loidea. Внутри последнего надсемейства долгое время различали только три
семейства: Pupillidae, Valloniidae и Buliminidae. Гетерогенность, точнее раз-
нообразие, первого из перечисленных семейств была показана Стинбергом
(Steenberg, 1925), но материалы, полученные этим исследователем, привели
только к разделению Pupillidae s. lat. на несколько самостоятельных семейств.
Позже Бейкер (см. Baker, 1935, р. 191) пишет, что он склоняется к более ши-
рокому пониманию Pupillidae, т. е. семейства Стинберга включаются в Pu-
pillidae в качестве подсемейств. Причина того, что Стинберг не сумел убедить.
Бейкера в справедливости своих построений, заключается, вероятно, в том,,
что он не подчеркнул должным образом обособленность и резкую очерченность-
выделенных им семейств, не указал на взаимную несводимость особенностей
строения полового аппарата, свойственных разным таксонам.
За основу для дальнейших рассуждений возьмем три факта.
Первый факт: древность всех семейств и подсемейств (в любом понимании).
Второй факт: самые ранние находки членов основных семейств представлены,
уже надежно определяемыми видами современных родов. Иными словами,
роды Azeca, Cochlicopa, Orcula, Vallonia, Vertigo существуют не менее 60 млн. лет.
Третий факт: палеонтологический материал не дает ни одного (!) фактичес-
кого подтверждения связей между любыми семействами и подсемействами Pu-
pillina. Другими словами, как современный, так и ископаемый материал оп-
ределенно указывает на четкую взаимную разобщенность всех семейств, вклю-
чаемых в подотряд.
Здесь нужно сделать одну оговорку. Стеклов (1966) описал из миоцена
(караганский песчаник) Предкавказья род Pupilorcula и указал, что этот род
занимает промежуточное положение между Pupilia и Orcula и «является как бы
связующим звеном между подсемействами Orculinae и Pupillinae, совмещая
в себе признаки обоих подсемейств» (см. Стеклов, 1966, с. 162). Единственным
основанием для такого толкования Pupilorcula Стеклов считал проникновение
колумеллярной пластинки в глубь раковины; однако он сам подчеркивал, что
все остальные признаки Pupilorcula свойственны Pupillinae. Если принять, что
исходное состояние для всех Pupillina — наличие длинных колумеллярной и
париетальной пластинок, то придется признать, что Pupilorcula — один из
архаичных родов Pupillidae s. str., не имеющий отношения к Orculidae.
Первый и основной вывод, который следует из трех приведенных фактов,
заключается в том, что принимаемая система должна отражать эти факты, сви-
детельствующие в первую очередь о большой раздробленности подотряда. Именно
эту цель и преследует автор, выделяя в подотряде 9 надсемейств (в.схему на
рис. 39 не включены Sagdoidea и Cerioidea).
Диагнозы надсемейств, приведенные в систематической части, трактуют
различия, существующие между ними. Однако целесообразно рассмотреть,
СИСТЕМА ПОДОТРЯДА
91
существуют ли между ними общие черты (кроме ортуретральной почки и на-
личия пениального аппендикса), и если существуют, то таков их таксономичес-
кий вес. Если при сравнении семейств постоянно ставить перед собой вопрос —
что между ними общего? — то придем к выводу, что общего очень немного (при
любом сочетании семейств).
Детальное сопоставление признаков, используемых в систематике, приводит
к заключению, что основные критерии, которыми руководствовались старые
авторы при отнесении таксонов к Pupillidae s. lat., заключались, во-первых,
в малом размере животных, во-вторых, в «пупиллидной» форме раковины (кото-
рая, кстати, достаточно разнообразна), и, в-третьих, в наличии устьевой арма-
туры. Для понимания таксономической структуры подотряда этих признаков,
конечно, недостаточно, и главным образом в силу того, что они неконкретны.
Малый размер раковины как признак, имеющий филогенетическую ценность,
отпадает, так как уменьшение размеров могло происходить не один раз и члены
любого таксона, вставшие на путь освоения такого биотопа, как припочвенные
слои лиственного опада, неизбежно приобретут малые размеры. То же касается
и формы раковины, которая достаточно разнообразна и нередко вполне харак-
терна для отдельных семейств и подсемейств. Так, не видя устья, только по форме
и размеру раковины очень часто можно установить семейственную принадлеж-
ность того или иного вида. Наконец, очевидно, что само по себе наличие или,
отсутствие устьевой арматуры ничего не дает для суждений о филогенетической
или систематической природе таксона. Если же ставить вопрос более конкретно,
т. е. сравнивать отдельные элементы обрамления устья, то можно констатиро-
вать большое сходство в устьевой арматуре между Achatinellidae, Orculidae и
Strobilopsidap и одновременно видеть существенные различия между этими
таксонами и всеми прочими семействами.
Не менее резко различается характер устьевой арматуры у Pupillidae и
Vertiginidae, особенно если сравнивать наиболее архаичные виды. Устье Ver-
tiginidae формально имеет некоторое сходство с устьем Chondrinidae (в сочета-
нии с отсутствием пениального аппендикса это, очевидно, единственное основа-
ние для сближения названных семейств), но совокупность признаков иного по-
рядка убеждает в резких различиях, существующих между Pupillidae. и Ver-
tiginidae. В первую очередь эти различия проявляются в совершенно иной ор-
ганизации совокупительного отдела гениталий.
Существует, таким образом, альтернатива: или не выделять в подотряде
Pupillina вообще никаких надсемейств, или принять предложенную схему
(рис. 39). Если выделение надсемейств Cochlicopoidea и Achatinelloidea можно
считать обоснованным и общепризнанным, то не менее обоснованным выглядит
и такое разделение Pupilloidea auct., какое осуществлено в настоящей книге.
Тогда, может быть, отказаться от выделения надсемейств вообще, тем более что
15 выделяемых семейств (включая Ceriidae, Sagdidae и Thysanophoridae) при-
надлежат 9 надсемействам, из которых 5 монотипических, а остальные вклю-
чают всего по 2—3 семейства? Такое решение вряд ли целесообразно, потому что
повышение таксономического ранга группы свидетельствует о двух моментах:
о степени обособленности группы и косвенно о ее значительном возрасте. Выде-
ление надсемейств, в том числе и монотипических, призвано подчеркнуть как
взаимную обособленность, так и возраст таксонов.
Под «взаимной обособленностью» здесь понимается несводимость деталей
строения половых аппаратов, свойственных отдельным семействам, друг к другу.
Перейдем к обсуждению более частных вопросов, касающихся системати-
ческой структуры отдельных таксонов.
Надсем. ORCULOIDEA
Изменения в составе Orculidae и близких групп, предлагаемые в настоящей
книге, касаются двух пунктов: включение Lauriinae в состав Orculidae и вве-
дение семейства Spelaeodiscidae в состав Orculoidea.
92
ОБЩАЯ ЧАСТЬ
По первому пункту можно высказать следующие соображения.
С тех пор как Стинберг (Steenberg, 1925) предложил свою систему Pupil-
lidae s. lat., отнесение Lauriinae в качестве подсемейства к семейству Pupil-
lidae s. str. стало общепринятым. Однако ранее Пилсбри (Pilsbry, 1916, Man.
Conch., 2nd ser., v. 24 : X) пришел к выводу, что семейство Orculidae слагается
родами Orcula и Lauria. Настоящее исследование заставляет отдать должное
проницательности Пилсбри: из материалов, помещенных в систематической
части, следует, что длинные хорошо развитые париетальные и колумеллярный
пластинки — первичная, исходная особенность, и вероятность того, что они по-
являлись в рамках подотряда не один раз, крайне невелика. Это заключение
основано прежде всего на ранней закладке названных пластинок (еще в эмбрио-
Рис. 40. Внешний вид мужского отдела полового аппарата.
I — Spelaeodiscus triarius; II — Euxinolauria paulinae. Пе — пенис, ПА — пениальный аппендикс, ПЭ —
придаток эпифаллуса, Эп — эпифаллус. Объяснения в тексте.
генезе) и их, следовательно, нельзя расценивать как позднее и частное приобре-
тение, подобное зубам на крае дефинитивного устья, которые появлялись, ко-
нечно, неоднократно.
Существуют и другие признаки, указывающие на близость Lauriinae и Ог-
culinae: широко отвернутые края устья, лишенные губы; характерная радиаль-
ная морщинистость, иногда почти ребристость; приподнятость последнего обо-
рота к устью. Убедительным доказательством филогенетического родства слу-
жит наличие спиральной скульптуры на эмбриональных оборотах у наиболее
архаичных Lauriinae юго-западного Закавказья — Euxinolauria silicea и Е. te-
nuimarginata. Эта скульптура исчезающе слаба, но сам факт ее наличия лишний
раз говорит о близости Lauriinae к Orculinae, для которых спиральная скульп-
тура на эмбриональных оборотах очень характерна.
Наконец, очень важным доводом в пользу сближения двух обсуждаемых так-
сонов можно считать значительное сходство половых аппаратов, свойственных
их представителям/
Spelaeodiscinae принято относить к Valloniidae; правда, особой уверенности
в такой трактовке нет (Wenz, Zilch, 1959—1960). Что сближает Spelaeodiscidae
с Valloniidae? В основном — микрохеликоидная форма раковины и резкая ради-
альная скульптура. Но микрохеликоидная раковина не раз появлялась среди
разных таксонов Pupillina; что же касается радиальной скульптуры, то она не-
менее характерна и для Orculidae. В тоже время для всех Valloniidae характерна
СИСТЕМА ПОДОТРЯДА
резко выраженная протерандрия с последующей редукцией мужского отдела,
чего у Spelaeodiscidae не наблюдается. Чрезвычайно существенно, что у всех
трех родов Spelaeodiscidae {Spelaeodiscus, Virpazaria, Klemmia) отсутствует
цэкум, но имеется хорошо развитый придаток эпифаллуса (Gittenberger, 1975).
На рис. 40 приведены изображения совокупительного аппарата Spelaeodiscus
triarius и Euxinolauria paulinae'. бросается в глаза чрезвычайное сходство всех
основных признаков названных видов (рис. 40, /, II).
По всей вероятности, Spelaeodiscidae — старое семейство; однако члены его-
обитают в биотопах, не создающих хороших условий фоссилизации (влажные
Рис. 41. Строение париетальной пластинки.
I — Sircbilops согеапа, вид снизу; II — то же, вид сбоку; III — Sphyradium doliolum, вид снизу: IV —
то же, вид сбоку.
лиственные горные леса), либо живут, в пещерах, а потому шансы на их находку
в ископаемом состоянии незначительны. Поэтому в ископаемом состоянии се-
мейство известно только из четвертичных отложений, да и то встречается редко
(Lozek, 1964).
Отнесение Strobilopsidae к Orculoidea вряд ли может вызвать возражения:
у представителей семейства также имеется не цэкум, а придаток эпифаллуса
(Baker, 1935); сходный характер имеет скульптура раковины, и, что очень
существенно, наличествуют длинные париетальные пластинки. Обращает на
себя внимание еще одно обстоятельство: у Sphyradium doliolum (Orculidae)
нижняя кромка париетальной пластинки, особенно у не вполне взрослых эк-
земпляров, волнообразно изгибается (рис. 41, III, IV) и вершины «волн» за-
метно утолщены. Эта структура напоминает строение париетальных пластинок
Strobilopsidae, на кромке которых имеются узелки (утолщения), которые легко
расценить как дериваты утолщений на кромке пластинки Sphyradium (рис. 41,
Z, П).
Вопрос об исторических связях Valloniidae находится пока в области до-
гадок (рис. 39). Единственным доводом в пользу отнесения семейства к Orcu-
loidea служит тот факт, что эмбриональные обороты Некоторых Vallonia несут
тончайшую спиральную скульптуру, свойственную Orculidae, а дефинитивная.
94
ОБЩАЯ ЧАСТЬ
представлена нередко кожистыми ребрышками, также характерными для мно-
гих Orculidae, особенно на ранних дефинитивных оборотах.
Не менее сложен вопрос и о составе Valloniidae. Здесь не внесено никаких
изменений в объем и состав семейства; тем не менее надо указать на некоторые
обстоятельства, заставляющие подозревать гетерогенность группы. В первую
очередь обратим внимание на подсемейство Acanthinulinae, составленное не
менее чем четырьмя родами: Planogyra, Spermodea, Acanthinula и Zoogenetes,
причем два последних рода ныне монотипические. Ярко выраженная спираль-
ная скульптура на эмбриональных оборотах Acanthinula aculeata удивительно
напоминает таковую Orculidae; молодые особи Sphyradium doliolum имеют пе-
риостракальные шипы на периферии оборотов, как у A. aculeata, да и вообще
молодь S. doliolum похожа на Acanthinula. Наконец, неясно, что представляют
собой выросты на эпифаллусе A. aculeata: во-первых, их два, что вообще бес-
прецедентно, и, во-вторых, если говорить о гомологиях, то они (или один из них)
могут быть гомологами только придатка эпифаллуса, о чем говорит место при-
крепления пениальной ветви полового ретрактора.
Все это вместе взятое подтверждает, как будто, правомерность отнесения
Acanthinula к Orculoidea, но отсутствие достаточных материалов по организа-
ции остальных родов подсемейства затрудняет решение вопроса о том, насколько
они близки к Acanthinula: следует ли перевести в Orculoidea только Acanthi-
mula или Acanthinulinae auct.
В любом случае как систематическое положение, так и состав семейства
Valloniidae нуждаются в выяснении.
Надсем. COCHLICOPOIDEA
Поскольку внутреннее строение Amastridae должным образом пока не ис-
следовано, обсуждению подлежат главным образом Cochlicopidae. Тем не менее
объединение этих семейств в одно надсемейство, если ориентироваться на при-
знаки раковины, выглядит правомерным. Так, очень характерная «полирован-
ная» раковина Cochlicopa с сильно уплощенными оборотами, обрубленным стол-
биком и прямыми слегка утолщенными краями устья очень похожа на раковину
Leptachatina (Amastridae). С другой стороны, нельзя не обратить внимания на
чрезвычайно широкое конхологическое разнообразие Amastridae, что не ис-
ключает таксономической гетерогенности семейства.
Если раковина, свойственная членам надсемейства, своеобразна, то половой
аппарат сравним с таковым Orculoidea. У Cochlicopidae имеются две основные
особенности: отсутствие каких бы то ни было придатков на эпифаллусе и от-
сутствие ветви полового ретрактора, которая обслуживала бы пениальный ап-
пендикс.
Судя по тому, что у всех исследованных видов Cochlicopa обнаружен дивер-
тикул семяприемника, находящийся в явно рудиментарном состоянии, надо по-
лагать, что отсутствие придатка эпифаллуса есть не первичное состояние, а ре-
зультат редукции. Надо вспомнить, что как дивертикул семяприемника, так
и придаток эпифаллуса функционально связаны с наличием сперматофора.
Так как дивертикул имеется, надо думать, что сперматофор исходно присутст-
вует; но тогда исчезновение придатка эпифаллуса, равно как и явно рудимен-
тарное состояние дивертикула, наводит на мысль о том, что сперматофор в настоя-
щее время редуцирован.
Об этом же свидетельствует и строение пениального аппендикса. Аг очень
короткий, но зато чрезвычайно сильно развит А3, резкие сокращения мощной
мускулатуры которого способны быстро и эффективно передать партнеру скоп-
.ление спермиев. С резким укорочением Аг связано и исчезновение соответствую-
щей ветви полового ретрактора, что может быть уверено расценено как вторичное
состояние.
СИСТЕМА ПОДОТРЯДА
95
Надсем. PUPILLOIDEA
Единственное семейство, включаемое в состав Pupilloidea, имеет настолько
характерный и постоянный облик раковины, что одних только конхологических
признаков достаточно для признания четкой обособленности Pupillidae от всех
прочих Pupillina. Представлению об обособленности семейства не мешает и су-
ществование таких родов, v.a.v.Pupoides,Microstele, идругих, характеризующихся
конической раковиной и в среднем несколько большими размерами, чем Glb-
bulinopsis и Pupilla.
Основное и яркое отличие Pupilloidea от наиболее близких Orculoidea за-
ключается в том, что если в устье Pupilloidea имеются париетальная и колумел-
лярная пластинки, то они появляются только в субдефинитивном состоянии.
Тем не менее обсуждаемые пластинки Pupilloidea гомологичны таковым Orcu-
loidea: миоценовый род Pu.pilorcu.la характеризуется глубоким проникновением
колумеллярной пластинки внутрь устья (Стеклов, 1966) а у некоторых совре-
менных американских Pupilla значительной длины достигает париетальная
пластинка, например у Р. syngenes avus из Большого Каньона (Pilsbry, 1920,
Man. Conch., v. 26, р. 172, fig. 4).
Надсем. VERTIGINOIDEA
Возникновение таксона, как уже отмечалось, связано, по-видимому, с освое-
нием условий жизни в слое лесного опада; наиболее заметный результат пере-
хода в подобные условия — ярко выраженная миниатюризация животных: это,
самые мелкие представители подотряда.
Вопрос о составе и объеме надсемейства пока далек от разрешения, и здесь
уместно привести лишь некоторые материалы и соображения, не претендующие
на что-либо большее, чем рабочая гипотеза.
Тиле (Thiele, 1931) выделяет в подсемействе Vertigininae среди прочих
трибы Vertigineae и Nesopupeae, причем указывает, что анатомия большинства
представителей последней трибы неизвестна, но у Nesopnpa имеется пениаль-
ный аппендикс, и соответственно раздвоенный половой ретрактор. Сущест-
венно, что иных дифференциальных признаков трибы не имеют. Тем не менее
авторы следующей большой сводки (Wenz, Zilch, 1959—1960) пошли дальше
и возвели трибы в ранг подсемейства, причем если сличить диагнозы этих
подсемейств, окажется, что они почти совпадают. Недостаток материалов по
анатомии Nesopupinae не позволяет пока дать оценку подсемейству в целом,
но очень вероятно, что серьезных различий между подсемействами Vertigininae
и Nesopupinae нет. Единственное различие заключается в наличии или отсут-
ствии пениального аппендикса, но нет никаких доказательств, что редукция
аппендикса происходила один раз, а не два или больше.
Nesopupinae auct. — обитатели тропической зоны, где они живут главным
образом в сильно или умеренно увлажняемых местностях (о. Мадейра, о-ва Бо-
нин, Гавайи, Новая Каледония, Квинсленд, Новый Южный Уэльс, о-ва Паци-
фики, о-в Кокос, Индия, Шри-Ланка, Калимантан, Филиппинские о-ва, Африка,,
о-в Св. Елены, Средняя и Южная Америка, Вест-Индия). В то же время Verti-
gininae больше тяготеют к умеренным широтам северного полушария, хотя
и встречаются в Восточной Азии, Центральной Америке и Вест-Индии.
Не исключено, что именно отсутствие аппендикса позволило Vertigininae
заселить тайгу и достичь северного полярного круга: показательно, что наиболее
процветающий, наиболее изменчивый, наиболее крупный по размерам и наиболее
широко распространенный вид — Vertigo modesta — один из самых северных
представителей подсемейства. Причина связи между отсутствием аппендикса
и освоением высоких широт кроется, видимо, в том, что отсутствие аппендикса
означает совершенствование процесса обмена спермой; не исключено также, что»
эта связь частично имеет энергетический характер.
96
ОБЩАЯ ЧАСТЬ
Однако своеобразие совокупительного аппарата Vertiginidae отнюдь не
сводится к отсутствию пениального аппендикса, что само по себе — не такое
уж редкое явление. Пенис у членов семейства отделен от эпифаллуса очень
резким пережимом, а половой ретрактор крепится к месту соединения семяпро-
вода с эпифаллусом. Выведение этого варианта организации совокупительного
-аппарата от какого-либо другого при современном состоянии знаний по анато-
мии Pupillina (строение множества тропических видов неизвестно) выглядело
>бы чисто спекулятивным и совершенно бездоказательным; но перестройки
.дистального отдела полового аппарата в любом случае достаточно глубоки и
серьезны, чтобы дать основание наряду с конхологическими признаками выде-
лить Vertiginidae в отдельное надсемейство, подчеркнув тем самым их обособлен-
ность и своеобразие.
Если совокупительный аппарат Columella и Truncatellina легко сопостав-
ляется с таковым Vertigo и представляет собой укороченный вариант последнего,
т?о с Gastrocoptinae дело сложнее и положение этого подсемейства неясно. Оче-
видно одно: к Chondrinidae эта своеобразная группа отношения не имеет. Здесь
Gastrocoptinae оставлены в составе Vertiginidae потому, что я не располагал
достаточным материалом для решения вопроса о систематическом положении
•этого резко очерченного таксона.
Надсем. CHONDRINOIDEA
Выше не раз подчеркивалось своеобразие Chondrinidae во многих отноше-
ниях. В частности, это единственный из хорошо очерченных таксонов Pupillina,
у которого полностью и всегда отсутствует пениальный аппендикс; более того,
у большинства родов мужской отдел гениталий воббще лишен всяческих при-
датков.
Поскольку эволюция полового аппарата Pupillina направлена, насколько
удается проследить, в сторону упрощения, то надо принять, что самыми прими-
тивными членами любого подчиненного таксона являются носители наиболее
сложно устроенного совокупительного аппарата. Исходя из этого, самыми
архаичными родами среди Chondrinidae следует считать Granaria и Solatopupa,
у членов которых на дистальных протоках мужского отдела имеется вырост,
гомологичный, видимо, придатку эпифаллуса.
К другим своеобразным особенностям полового аппарата Chondrinidae
надо отнести тесную морфологическую связь проксимального участка эпифал-
луса (часто несущего рудиментарный бич) с тканями дистального конца пениса,
.а также необычное строение полового ретрактора, крепящегося одной лентой
к пенису, а другой — к эпифаллусу.
Все эти особенности в сочетании с яркими конхологическими признаками
и узким экологическим спектром (подавляющее большинство видов — облигат-
ные скаложилы, живущие открыто и питающиеся лишайниками) свидетельствуют
об узкой и далеко зашедшей специализации, что очень затрудняет выяснение
исторических связей семейства. Те признаки, на которые указывает Гиттенбер-
гер (Gittenberger, 1973), имея в виду возможную связь Chondrinidae с Gastro-
coptinae, вряд ли могут считаться надежными для решения проблемы.
Еще более трудны для понимания Pyramidulidae. Автор отчетливо осознает
всю шаткость аргументов в пользу помещения семейства в Ghondrinoidea, но
для выделения таксона в отдельное надсемейство мы не располагаем достаточ-
ным числом весомых признаков, а для объединения Pyramidulidae с любой дру-
гой группой Pupillina оснований еще меньше, чем для сближения семейства
с Chondrinidae. Что касается возможной связи Pyramidulidae с Pleurodiscidae,
то такая связь кажется весьма вероятной: допустимо также, что род Pyramidula
следует относить к Pleurodiscidae. Не исключено, что и род Toltecta, положение
которого неясно (пока его обычно относят к Endodontidae), также принадлежит
семейству Pleurodiscidae.
СИСТЕМА ПОДОТРЯДА
97
Условное отнесение Pyramidulidae к Chondrinoidea основано лишь на сходстве
раковины Pyramidula с начальными оборотами многих Chondrinidae как по
форме, так и по скульптуре, а также по окраске; мешковидный вырост на пенисе
Pyramidula, служащий, вероятно, для накопления семенной жидкости перед
спариванием, можно расценить как гомолог придатка на мужских половых путях
Granaria и Solatopupa.
Надсем. BULIMINOIDEA
Изолированное положение Buliminidae определяется тремя группами при-
знаков, не зависимых одна от другой: во-первых, весь богатый набор конхологи-
ческих особенностей; во-вторых, у всех членов семейства в отличие от прочих
таксонов семенные пузырьки морфологически обособлены и сидят мутовкой
на гермафродитном протоке немного ниже гонады; в-третьих, очень маловеро-
ятно, чтобы цэкум Buliminidae был гомологичен придатку эпифаллуса, имею-
щемуся у членов остальных групп, так как эти органы имеют различное строе-
ние, различное положение на эпифаллусе относительно места прикрепления
полового ретрактора и различные функции.
Поскольку пениальный аппендикс в высокой степени характерен для под-
отряда вообще, а для Buliminidae в особенности, наличию или отсутствию
этого органа принято придавать большой таксономический вес (признак ха-
рактеризует таксоны ранга не менее, чем подсемейство). Однако материалы,
как полученные в процессе настоящего исследования, так и содержащиеся
в литературе, заставляют пересмотреть традиционные взгляды.
Не обсуждая способов редукции пениального аппендикса (эти способы об-
суждены на с. 67—69), можно, вероятно, говорить о том, что в настоящее
геологическое время наиболее прогрессивные представители подотряда до-
стигли такого уровня физиологического совершенства, что пениальный аппен-
дикс часто перестает быть обязательным атрибутом. Если ранее, на заре станов-
ления таксона, наличие пениального аппендикса было исходной и важной осо-
бенностью всех видов, то сейчас мы видим, что пениальный аппендикс у ряда
семейств и подсемейств отсутствует. Отсутствию аппендикса принято придавать
большой таксономический вес, и в пределах Buliminidae установилась традиция
тотчас относить к подсемейству Chondrulinae любой вид, у которого установлено
отсутствие органа, поскольку эта негативная черта — единственная дифферен-
циальная особенность данного подсемейства. В результате к Chondrulinae
относят виды таких непохожих друг на друга родов, как Chondrula, Senaridenta
и Cyrenaeus, не обращая внимания на то, что по всем остальным признакам, как
анатомическим, так и конхологическим, Senaridenta ничем серьезным не отли-
чается от Multi dentula (Multidentulinae), a Cyrenaeus — от Paramastus (Buli-
mininae). В то же время род Chondrula обнаруживает совершенно отчетливую
связь с европейскими Eninae.
Остановимся на этом вопросе подробнее. Гудец (Hudec, 1972с), вскрывая
«Jaminia» pupoides из различных мест, обнаружил, что у особей, собранных
близ Степанакерта, отсутствует пениальный аппендикс. Это послужило поводом
для описания нового вида и отнесения его не только к роду Chondrula, но даже
к его номинативному подроду.
Конхологические признаки, особенно характер оформления устья, доста-
точно резко изолируют группу видов, относимых ранее к роду Jaminia {pupoides,
lamelliferus, nachicievanjensis, angustior, phasianus, ovularis) от всех прочих Buli-
minidae и настолько близки у всех этих видов, что, не зная анатомии и при
отсутствии совместных находок, их вполне можно принять за один вид; показа-
тельно, что их считали синонимами или очень близкими видами все крупные кон-
хиологи. Особое ударение надо сделать на том, что раковина всех перечисленных
форм специализирована и богата признаками; ведь когда обнаруживается почти
полная идентичность видов, раковина которых бедна признаками (например,
Merdigera obscura и A kramovskiella umbrosa), это можно счесть конвергентным сход-
7 А, А, Шилейко
98
ОБЩАЯ ЧАСТЬ
ством, но когда столь богатая и сложно устроенная устьевая арматура повторя-
ется в рамках одного семейства, это не может не свидетельствовать о действи-
тельном родстве. Показателен также тот факт, что если у прочих видов, имеющих
устьевые зубы, форма и число последних подвержены значительной изменчи-
вости, то у обсуждаемого конхологического комплекса все элементы зубной
арматуры отличаются замечательным постоянством. Сходство между перечис-
ленными таксонами подтверждается и другими признаками: форма раковины,,
число и степень выпуклости оборотов, форма устья, отворот его края, размеры
раковины, форма пупка.
Наконец, решающий признак, объединяющий все эти формы, заключается
в наличии замкнутой папиллы пениса, что среди Buliminidae вообще встреча-
ется редко (рис. 35).
Следовательно, родство обсуждаемых форм не вызывает сомнений. Другой
вопрос, какова степень этого родства. Два крайних решения заключаются в аль-
тернативе: либо это «анатомические» формы одного вида, либо разные роды;
однако в любом случае можно быть абсолютно уверенным, что они не имеют
прямого отношения ни к Chondrulinae auct., ни к Eninae, ни к Napaeinae auct.r
т. е. ни к одному из трех признававшихся ранее подсемейств.
В пользу первого решения говорит тот факт, что воообще известны аналогич-
ные случаи редукции отдельных придатков полового аппарата, приводящие
к тому, что несомненно близкие формы приобретают признаки родового и даже
подсемейственного уровня, например, анатомические формы Circasstna circas-
sica из Hygromiidae (Шилейко, 1978а). Но в данном случае различия между
pupoides, lamelliferus и nachicevanjensis не ограничиваются наличием или отсут-
ствием пениального аппендикса и дивертикула семяприемника: они отли-
чаются также относительной длиной пениса и дивертикула семяприемника^
а также положением цэкума на эпифаллусе. С учетом этих различий, пра-
вильно было бы придать каждой из анатомических форм ранг рода, выделив их
вместе с тем отдельным подсемейством Multidentulinae.
Не менее показателен случай с родом Cyrenaeus. Для рода Paramastus (Buli-
minidae) характерно наличие пениального аппендикса. Брандт (Brandt, 1958),.
анатомировав виды из Северной Африки (Киренаика), установил, что у всех
этих видов аппендикса нет (см. также Zilch, 1960, S. 59, Taf. 9, Fig. 4, 5). Эти
послужило Геллеру (Heller, 1971b) основанием выделить североафриканские
виды в отдельный род Cyrenaeus, и, что самое показательное, отнести новый род.
к Chondrulinae, хотя переднеазйатские и африканские виды обладают комплек-
сом общих признаков: очень сходной раковиной, своеобразной скульптурой
(Zilch, 1951), одинаковой внешней организацией пениса, в частности наличием
мешковидного выроста на проксимальном конце, и хорошо развитым бичом.
Таким образом, есть все основания для уверенности, что редукция пениаль-
ного аппендикса в семействе Buliminidae осуществлялась не менее трех раз; так-
сон, основанный на таком признаке, как отсутствие аппендикса, есть группа,
полифилетическая, и, следовательно, не имеет таксономического статуса.
Очевидно, если и выделять Chondrulinae в отдельное подсемейство, то только-
ограничив его родом Chondrula с подродами; однако этот род так близок другим
родам Eninae, что этот шаг вряд ли целесообразен.
Изложенные факты интересно рассмотреть в историческом аспекте. Если
принять, что семейство сформировалось в условиях умеренно влажных лесных
биотопов, то получается, что более архаичные группы Buliminidae, имеющие-
пениальный аппендикс, и поныне обитают в лесах или в биотопах, по условиям
сходных с лесными. Все без исключения случаи редукции аппендикса касаются
представителей, перешедших к обитанию в открытых пространствах, хотя при-
сутствие аппендикса не служит препятствием к освоению аридных ландшафтов.
Иными словами, все «лесные» группы имеют аппендикс, а среди «нелесных»-
наблюдаются формы, как имеющие аппендикс, так и лишенные его. В связи
с этим правдоподобно предположение, что стимулом к редукции органа послужил
переход в новую адаптивную зону.
ПРИНЦИПЫ И МЕТОДЫ РАЗРАБОТКИ СИСТЕМЫ
99
ПРИНЦИПЫ И МЕТОДЫ РАЗРАБОТКИ СИСТЕМЫ PUPILLINA
В результате настоящего исследования наибольшим перестройкам подверглось
семейство Buliminidae, поэтому, при рассмотрении методов и принципов раз-
работки адекватной системы целесообразно главное внимание уделить этому
семейству.
Таксономическая структура Pupillina вообще и Buliminidae в особенности
имеет несколько своеобразных черт, благодаря которым любые построения могут
вызвать разногласия; поэтому возникла потребность изложить соображения,
послужившие основой для выводов, сформулированных в терминах практической
таксономии и изложенных в систематической части.
Широко бытует мнение, что существуют два противоположных подхода
к разработке классификации зоологических таксонов: представители этих под-
ходов именуются либо «дробителями», либо «объединителями». Первые стре-
мятся выделить в отдельный род или подрод любой вид, чем-то отличающийся
от близких видов, и создают большое число моно- или олиготипических таксонов,
чем, по мнению Майра (1971), понижают информативную ценность системы или
затрудняют извлечение информации. Вторые, напротив, создают большие роды,
пренебрегая тем, что внутри родов обычно имеются подчиненные группировки.
Возможно, среди зоологов имеются «принципиальные» сторонники как той,
так и другой концепции, но при внимательном рассмотрении оказывается, что
большую часть серьезных систематиков нельзя отнести к категории «принци-
пиальных» поклонников одного или другого подхода. Действительно, любой,
кто признает существование эволюции, неизбежно допускает, что существо-
вал таксон, предковый по отношению к данному таксону, включавший опре-
деленное число подчиненных таксонов. Каждый из этих подчиненных таксонов
эволюционировал; одни таксоны менялись филетическим путем, другие ис-
пытали дивергенцию, третьи оказались тупиковыми и вымерли. Все эти процессы
реально имели место, моменты дивергенции на самом деле фиксированы во
времени и в пространстве, и деятельность систематика уподобляется решению
кроссворда, где связи между элементами задачи реально существуют, и дело
сводится к обнаружению этих связей, т. е. задача систематика состоит в том,
чтобы определить ход и результаты эволюционного процесса возможно коррект-
нее и изложить в форме, принятой в таксономической практике.
. Способ адекватного описания таксономической и филогенетической струк-
туры, исторически сложившихся отношений лишь один: написание системы
в виде списка таксонов, расположенных в определенном порядке и в опреде-
ленной иерархической форме. Изображение филогенетических связей в виде
ветвящихся «древес» — хорошее дополнение к систематическому списку.
. Естественно, чем сложнее структура таксона, чем больше внутри него уров-
ней, или ступеней, различий, тем сложнее будет и отражение этой структуры,
представленное в виде системы. Если система достаточно адекватно отражает
•сложившееся в природе положение и по возможности пути, приведшие к такому
положению, то она имеет высокую информативную ценность независимо от того,
объединил ли систематик в один род десяток прежних родов или, напротив,
разбил один род на много родов, одни из которых оказались монотипическими,
а другие состоящими из многих подродов.
Таким образом, разного рода советы и рекомендации относительно того,
делить или не делить, в каких случаях и как делить или объединять таксоны,
представляются лишь первичным и чисто формальным средством для внесения
какой-то упорядоченности в существующее многообразие. Эти советы можно
и даже нужно использовать в условиях острого дефицита информации по строе-
нию отдельных видов в плохо изученных группах.
Обилие монотипических таксонов в такой-то группе дли в. таком-то районе—
не произвол систематика, а факт, имеющий причинно-следственное объяснение.
Если исследователь не располагает пока материалом, достаточным для того,
чтобы дать такое объяснение, — это не повод для отрицания самого факта.
7*
100
ОБЩАЯ ЧАСТЬ
В качестве примера сравним среднеазиатскую фауну Buliminidae (без Ко-
петдага) с крымско-кавказской и среднеевропейской. В Средней Азии на тер-
ритории СССР семейство представлено 7 родами и 29 видами; монотипических
родов только два (Subzebrinus и Siraphoroides). Род Pseudonapaeus представлен
13 видами, один из которых обнаруживает крайнюю степень внутривидовой
изменчивости (Р. albiplicatus). В Крыму, на Кавказе и в средней Европе имеется
при почти том же числе видов 19 родов, т. е. почти втрое больше; из них 10 мо-
нотипических, причем ни один из них не содержит более 5 видов (картина,
правда, неполна из-за недостаточной изученности фауны прилегающих районов
Турции и Ирана).
Подобное соотношение свидетельствует об определенных качественных
различиях между фаунами регионов. В Средней Азии, главным образом на
Тянь-Шане и особенно в северо-восточной его части, процесс видообразования
в настоящее время идет гораздо интенсивнее, чем в западных регионах; отме-
ченное соотношение свидетельствует также о том, что в Крыму, на Кавказе
и в средней Европе основные экологические ниши давно заполнены, фауна в це-
лом прошла через сито отбора и имеет «устоявшийся» в общем облик, где каждый
или почти каждый вид четко отделен от ближайших видов; эти разрывы между
видами нередко настолько резки, что сплошь да рядом представляется целесо-
образным выделять отдельные виды в монотипические роды или подроды,
подчеркивая эту характерную черту фауны обсуждаемых регионов. Кстати,
подобную же картину получаем, сравнивая фауну Hygromiidae Средней Азии
и западных регионов (Шилейко, 1978а).
Надо подчеркнуть, что систематик не имеет иных способов выразить степень
обособленности между фаунистическими элементами, да они и не нужны: опери-
руя с названиями таксонов разного ранга, всегда можно выработать решение,
наиболее точно отражающее положение, сложившееся в природе.
Говоря иными словами, получается, что автор этих строк, изучая фауну
Средней Азии, стоит на позициях объединителя, а когда дело касается фауны
Кавказа, Крыма и средней Европы, выступает как дробитель. Это еще раз
свидетельствует о том, что создание для себя заранее определенной платформы
неизбежно приведет к тенденциозным построениям, и исследователь будет вы-
нужден затискивать в определенные избранные рамки любые ситуации, с кото-
рыми ему придется столкнуться. Из созданной На такой базе системы нельзя
будет извлечь никакой информации, кроме формальных сведений о том, какой
вид какому близок, причем о степени этой близости также, как правило, судить
будет весьма трудно.
В результате надо констатировать, что средствами, имеющимися в распоря-
жении систематика, мы излагаем определенные факты, и ничего не выигрываем,
если будем следовать советам — как «создавать» роды оптимального объема.
Много сложнее положение с анализом на уровне вида. Пока речь идет о за-
паднопалеарктических видах, особенно связанных с лесом, то ситуация таксо-
номических затруднений, как правило, не вызывает: каждый вид отделен от
близких видов более или менее ясным хиатусом и, если даже строго придержи-
ваться типологической концепции, почти всегда можно определить по морфоло-
гическим признакам, что есть вид, а что — внутривидовая форма изменчивости.
Например, конхологическое разнообразие крымских ксерофилов Brephulopsis
cylindrica и В. bidens уверенно можно определить как проявление локальной
изменчивости, а столь же широкий спектр изменчивости закавказского вида
Pseudochondrula seductilis укладывается в рамки понятия о подвидах.
Использование терминов «локальная раса» и «подвид» в данном случае свя-
зано с величиной ареала внутривидовых форм, многообразием физико-географи-
ческих условий на территории ареала и расстоянием между ареалами. Так,
в Крыму соседние достоверно различающиеся по признакам раковины популя-
ции могут занимать территорию, не превышающую 1 га, и быть разделенными
пространством в несколько десятков метров. Климат открытых местностей Крыма
повсеместно достаточно однороден. Что же касается Закавказья, то здесь,
ПРИНЦИПЫ И МЕТОДЫ РАЗРАБОТКИ СИСТЕМЫ
101
как правило, популяции занимают территории более обширные и разделены
пространствами в несколько километров или десятков километров. Сущест-
венно также, что климат открытых местностей в Закавказье при продвижении
с запада на восток заметно меняется в сторону нарастания аридности и конти-
нентальности; соответственно меняются и фитоценотические условия. В преде-
лах подвидовых ареалов Р. seductilis, границы которых установлены пока не-
достаточно четко, можно найти участки, где на ограниченной и однородной
в физико-географическом отношении территории имеется несколько изолиро-
ванных локальных рас.
Совершенно иное положение наблюдается в горных районах Средней Азии,
преимущественно на Тянь-Шане и Джунгарском хр. Показателем поразитель-
ного конхологического разнообразия, наблюдающегося в этих горных системах,
служит тот факт, что начиная с 1874 г. (работа Мартенса) для фауны Bulimi-
nidae этого региона было предложено 97 видовых и инфравидовых наз-
ваний, имеющих номенклатурную пригодность, и это при условии, что фауна
здесь собиралась и изучалась куда менее интенсивно, чем например, на Кавказе.
Существует единственная попытка оценить в широких масштабах характер
изменчивости среднеазиатских Buliminidae — это работы Матёкина (1959, 1966).
Установив чрезвычайно широкий, необычный для других регионов размах из-
менчивости конхологических признаков, Матёкин свел все разнообразие в 4 вида
(Jaminia potaniniana, J. labiella, J. fedtschenkoi, J. kasnakowi), подчеркнув тем
самым поразительную амплитуду изменчивости признаков раковины. Основа-
нием для такого вывода послужили два факта: во первых, возможность на боль-
шом материале проследить плавное изменение почти всех конхологических
признаков (пропорции, размеры и форма раковины, окраска, скульптура,
устьевая арматура); во-вторых, отсутствие видимых отличий внешнего вида
полового аппарата у различных форм, точнее, столь же плавное изменение в этих
признаках (относительная и абсолютная длина вагины, пениса, пениального
аппендикса и дивертикула семяприемника). Четыре вида, выделенные Матёки-
ным, имеют дифференциальную анатомическую характеристику.
Расхождения взглядов, развиваемых в данной книге, с воззрениями Матё-
кина основаны на трех обстоятельствах.
1. При исследовании внутреннего строения пениса обнаружены существен-
ные детали, свидетельствующие о качественных различиях между отдельными
группами видов.
2. Если две или более конхологически различимые формы обнаружены в пре-
делах одного микробиотопа, то это служит доказательством того, что они не
конспецифичны: «. . .морфологически различные популяции могут быть сим-
патрическими только в том случае, если они принадлежат к разным видам»
(Рауп и Стэнли, 1974, с. 104, 105). Разумеется, когда речь идет о таких
малоподвижных животных, как сухопутные улитки, совместное обитание по-
нимается в самом строгом смысле: обитают в пределах одной осыпи, на одном
однородном участке склона, встречены под одним камнем, в пределах радиуса
индивидуальной активности и т. п.
3. Если между двумя формами, различающимися по признакам раковины,
отсутствуют видимые анатомические отличия, то это не может явиться поводом
для категорического утверждения их конспецифичности. Изолирующие ме-
ханизмы достаточно разнообразны и отнюдь не сводятся к механической изоля-
ции, основанной на различиях в строении полового аппарата (см. Майр, 1968,
с. 86). Таким образом, виды, не имеющие видимых анатомических различий,
но отличимые по раковине, здесь расцениваются как таковые, если они встречены
вместе.
В статье 1959 г. Матёкин проанализировал связи между отдельными фено-
типическими значениями конхологических признаков и условиями, в которых
обитают животные, и показал зависимость определенных конхологических
форм от условий в конкретных типах биотопов. Сделанные выводы не противо-
речат излагаемым здесь взглядам, но я рассматриваю эту связь морфологии с эко-
102
ОБЩАЯ ЧАСТЬ
логическими условиями как проявление жизненных форм, т. е. сходных фено-
типических вариантов,, возникших на базе различных генотипов под воздейст-
вием сходных условий.
Рассматривая Bradybaenidae Средней Азии, Матёкин (1972) обнаружил
явление конхологического полиморфизма Bradybaena plectotropis и, обозначив
этот комплекс форм термином «надвид», разделил его на несколько полувидов
(semispecies). Термин «надвид» был введен Майром (Мауг, 1931); позже Майр
дает такое определение: «Надвид — это монофилетическая группа близкородст-
венных и в значительной мере или полностью аллопатрических видов» (см. Майр,
1971, с. 70). Ситуация, наблюдаемая в случае с В. plectotropis, больше отвечает
понятию «Rassenkreis» Ренша (Rensch, 1929), поскольку тут речь идет о внутри-
видовой изменчивости, а не о разных, хотя и очень близких, видах. Однако
термин «надвид» на практике удобен и взгляды Матёкина были мною приняты
(Шилейко, 1978а). Если придерживаться той точки зрения, что аллопатрические
формы, не различающиеся анатомически и связанные между собой промежу-
точными значениями признаков раковины, почти всегда конспецифичны, то
ситуацию, наблюдаемую в случае с Pseudonapaeus albiplicatus, приходиться
расценивать точно так же, т. е. считать Р. albiplicatus надвидом, состоящим
из некоторого числа полувидов (см. с. 247). Однако в данном случае применение
терминов «надвид» и «полувид» не всегда может быть обосновано достаточно
строго; поэтому здесь формы внутривидовой изменчивости обозначаются как
таковые (forma, f.).
Ситуация сильно осложняется еще одним обстоятельством, с которым мне
довелось ознакомиться лишь в сентябре 1980 г., когда при любезном содействии
К. К. Увалиевой я получил возможность собрать материал на Заилийском хр.
у Алма-Аты. В ущелье Тургень встречаются совместно и без каких-либо пере-
ходов четко различающиеся Bradybaena plectotropis и В. phaeozona, Pseudo-
napaeus subobscurus и Р. secalinus, Р. asiaticus и Р. secalinus, т. е. эти формы здесь
ведут себя как хорошие виды; однако эти же таксоны на Киргизском и Талас-
ском хребтах ведут себя совсем иным образом: на ограниченном участке склона
можно собрать серию, которая будет содержать «настоящих» Р. asiaticus, «на-
стоящих» Р. secalinus и всевозможные переходные между ними формы. Очевидно,
окончательное суждение о таксономическом статусе форм, объединенных здесь
в полиморфный вид Р. albiplicatus, можно будет составить после тщательных
сборов на южных склонах Заилийского хр. и на хребтах Кунгей и Терскей.
О методах, использованных в настоящей работе, можно сообщить следующее.
Если ограничиться использованием признаков, привлекавшихся до сих пор
для изучения системы, то трудно себе представить перспективы дальнейшего
совершенствования наших представлений о структуре Pupillina и в первую
очередь Buliminidae.
Действительно, за немногими исключениями виды в пределах Buliminidae
отличаются один от другого преимущественно тремя группами признаков:
наличием или отсутствием отдельных элементов полового аппарата (дивертикул
семяприемника, бич, пениальный аппендикс, цэкум), степенью развития этих
элементов, пропорциями различных участков дистального отдела репродуктив-
ного тракта.
Перетасовывание этих признаков иногда дает возможность понять объем
родов и в сочетании с признаками раковины интуитивно угадывать степень бли-
зости этих родов между собой, но перечисленные признаки совершенно беспер-
спективны для расшифровки таксономической и филогенетической структуры
семейства.
Таким образом, возникла проблема поиска качественно новых структур,
которые можно было бы использовать для целей систематики. Такие структуры
обнаружены при исследовании внутреннего строения мужского отдела репро-
дуктивного тракта. Материалы, приведенные в систематической части, свиде-
тельствуют о надежности и перспективности этих признаков. Тот факт, что эти
признаки, как и любые другие, подвержены изменчивости, не снижает их
МЕТОДЫ ИССЛЕДОВАНИЯ
103
ценности: дело лишь в том, чтобы знать пределы изменчивости и коррелятивные
связи.
Изложенные в определенной форме и одинаковым образом, все построения
автора любой системы выглядят одинаково достоверными и равнозначными.
На самом деле обычно это не так. Каждый автор ревизии представляет себе,
в какой степени надежны выводы по каждому из ревизованных таксонов. Поэтому
очень полезны соображения самого автора ревизии по поводу того, какие группы
или фауна каких районов «решены» достаточно достоверно, а какие — в той или
иной степени предположительно, и сделанные в их отношении выводы представ-
ляют лишь некоторый итог, не претендующий на полноту или законченность.
Думается, что система подотряда в целом, включая таксоны родовой группы,
недалека от истины; во всяком случае, она объясняет известные факты и не
противоречит им. То же касается таксономических решений на уровне видов
и родов, обитающих к западу от Каспийского моря, а также широко распро-
страненных видов. Исключение составляют некоторые из родов, ареал которых
охватывает не только Кавказ, но и соседние участки Ирана и Турции. Вероятно,
произойдут какие-то изменения, когда достаточно детально будут исследованы
типовые виды родов Sesteria, Euchondrus, Imparietula, Amphiscopus.
Бесспорно, далека от завершения (главным образом на видовом уровне)
система Buliminidae Средней Азии. Во-первых, предстоит уточнить статус
многих названий, которые провизорно включены в синонимы тех или других
видов. Во-вторых, для применения критерия совместной встречаемости нужны
дополнительные и абсолютно достоверные сведения о том, какие конхологи-
ческие формы обитают совместно в пределах одного микробиотопа.
Паллиативно решен вопрос о видах рода Тигапепа. Дело в том, что каждая
из 24 исследованных выборок достоверно отличается от любой другой; широкой
изменчивости подвержены все учитываемые признаки. В таких случаях
напрашивается вывод о существовании одного вида с широким диапазоном
изменчивости. Но в ущельи р. Некем на западном Тянь-Шане обнаружено сов-
местное обитание двух форм, четко различающихся как конхологически, так
и анатомически. Следовательно, приходится признать существование не менее
двух видов; но как их отличать в тех случаях, когда они встречаются порознь
в виде иных экологических форм? Строение гениталий у членов анатомически
исследованных популяций (7 выборок) идентично одному из видов, обитающих
в долине Пскема, при условии широкой изменчивости признаков раковины.
Картина еще более усложняется тем, что нельзя отвергать и такую возможность,
как существование кольцевых перекрываний, т. е. обе пскемские формы могут
оказаться двумя концами одной цепи. Известно к тому же, что близкие Виды,
почти не различимые при условии аллопатричности, в зоне контакта могут
хорошо различаться (Майр, 1968; Schindel, Gould, 1977). Для решения этих во-
просов нужны обширные дополнительные сборы и анатомирование массового
материала. Решение, предложенное в данной книге, призвано облегчить даль-
нейшие исследования тем, что здесь изложена вся синонимика и очерчены ос-
новные морфологические варианты, которым пока придается ранг самостоятель-
ных видов.
МЕТОДЫ ИССЛЕДОВАНИЯ
Моллюска, как и любой зоологический объект, можно исследовать со мно-
жества точек зрения. Биохимическое, например, исследование легочных мол-
люсков проводится теми же методами, что и изучение других животных; стра-
тиграфическая роль Pupillina выясняется теми же способами, что и других мел-
ких ископаемых групп. Поэтому здесь речь пойдет лишь о специфических мето-
дах или специфических модификациях методов, которые применимы именно
к рассматриваемому подотряду.
Сбор, фиксация и хранение материала. Особен-
ности сбора материала по Pupillina определяются главным образом тем, что
многие виды отличаются небольшими, а то и совсем крошечными размерами.
104
ОБЩАЯ ЧАСТЬ
Разные виды встречаются почти во всех природных условиях, имеющих место
на территории нашей страны. Представителей подотряда нет на участках с за-
соленными почвами, в зоне вечных снегов и в песчаных пустынях; во всех осталь-
ных зонах, биотопах и ландшафтах они есть.
Самая общая рекомендация — быть предельно внимательным в полевых
условиях. Собирая мелких моллюсков в лесу, в первую очередь надо обратить
внимание на отставшую кору пней, поваленных деревьев и на слой листвен-
ного опада. Укрытия осматриваются на месте, а часть лиственной подстилки
рекомендуется доставить в лабораторию, где поверхность листьев осматривается
с помощью лупы, дающей увеличение в 2—4 раза (более сильные лупы имеют
слишком малую площадь обзора). Далее осмотру подлежат нижние части дре-
весных стволов, на которые нередко поднимаются как мелкие формы, так и более
крупные Buliminidae; виды родов Vertigo, Acanthinula, Zoogenetes нередко встре-
чаются на грибах. По утрам и в пасмурные дни хорошие результаты дает ко-
шение энтомологическим сачком в траве по лесным опушкам и полянам; таким
образом можно получить материал по видам рода Vertigo и особенно Columella',
кошение сачком рекомендуется также среди кустов и травостоя в речных до-
линах и поймах.
Особенно внимательно надо осматривать древесные стволы в лесах Кавказа:
там обитает несколько видов Buliminidae, которые встречаются главным обра-
зом на деревьях; в Батумском ботаническом саду на стволах бамбука на высоте
человеческого роста и выше иногда сотнями сидят моллюски разных видов Lau-
riinae.
Более бедные сборы дают суходольные луга, лесостепные и степные участки.
Здесь моллюски встречаются в основном в дерновинках трав и возле корней
кустарников, но собрать хорошую серию на месте трудно. Поэтому раститель-
ную труху вместе с верхним слоем почвы надо насыпать в мешочек, сшитый из
плотного полотна, или полиэтиленовый пакет и разобрать пробу в лаборатории
с помощью оптики. Именно в таких местах чаще всего встречаются самые мелкие
моллюски — виды рода Truncatellina, а также несколько видов рода Pupilia.
В горных районах среди каменистых россыпей на склонах иногда массами
встречаются различные представители Buliminidae, Valloniidae и Pupillidae.
Здесь надо стремиться осмотреть не только верхний слой камешков и щебенки,
но и дойти до более глубоких горизонтов, где начинаются уже плотные слежав-
шиеся слои. Моллюски часто уходят в нижние горизонты осыпей, особенно
в жаркую сухую погоду. Кроме того, мелкие виды проникают в трещины скал
удивительно глубоко, поэтому есть смысл прибегнуть к помощи рычага, осто-
рожно расшатывая каменные плиты. На крутых склонах при этом возникает
риск обрушить вниз крупные камни, поэтому действовать надо с максимальной
осмотрительностью.
В горах нередко бывает так, что раковины какого-то вида встречаются в боль-
ших количествах, а живых моллюсков собрать не удается. В таких случаях
можно попробовать один из двух способов. Первый способ заключается в том,
чтобы отвернуть несколько больших камней и взрыхлить почву под ними на глу-
бину 8—10 см. Таким способом можно обнаружить некоторые виды закопав-
шихся Buliminidae. Второй путь можно использовать, если поблизости есть
источник воды: вылить среди россыпей в одном месте несколько ведер воды и
подождать несколько минут, затенив это место от прямых солнечных лучей.
Улитки нередко «поддаются на провокацию» и выползают на поверхность.
Если после нескольких дней засухи прошел дождь, то надо выйти на сборы
немедленно после дождя: некоторые виды спешат в укрытие сразу, как только
выглянет солнце.
В аридных районах некоторые крупные Buliminidae (Napaeopsis hohenackeri,
Zebrina detrita, Brephulopsis cylindrica) обычно массами поднимаются на стебли
растений, образуя плотные скопления, и их обнаружить не составляет труда.
Наконец, Находясь среди скал, необходимо тщательно осмотреть их по-
верхность: в таких биотопах живут Pyramidulidae и Chondrinidae. Несмотря на
МЕТОДЫ ИССЛЕДОВАНИЯ
105
то что они сидят открыто на поверхности скал, они отнюдь не бросаются в глаза
благодаря маскирующей окраске.
Выбросы рек и ручьев, оставленные после разливов, содержат богатый набор
раковин, причем основная масса принадлежит мелким видам: на 1 м береговой
линии в среднем течении р. Чегем на Северном Кавказе мною однажды была
собрана серия, содержавшая свыше 1000 раковин Vertigo, Truncatellina, Pupilla
и Vallonia.
Собранный материал сразу же, в поле, сортируют по размеру, так как хра-
нение мелких улиток вместе с крупными крайне нежелательно: мелкие животные
прилипают к телам крупных, обволакиваются их слизью, попадают в устье
больших раковин. Поэтому каждый сбор лучше всего хранить в двух коробоч-
ках или пробирках, заткнутых ватой (не пробкой!). Если сосуд будет плотно
закупорен, то животные могут погибнуть, особенно в жаркую погоду, от пере-
грева и недостатка кислорода.
Предметом особой заботы должно быть следующее. Каждый сбор должен
содержать животных, собранных в одном месте, а не просто «в долине реки та-
кой-то». В применении к данной группе этот вопрос ныне стоит, как отмеча-
лось выше, особенно остро: известно некоторое число родов, где границы между
видами улавливаются с трудом и без должной достоверности; анатомическое
исследование также не всегда дает однозначный ответ при решении вопроса о ви-
довой самостоятельности. В таких случаях крайне важно знать, какие формы
встречаются вместе и, следовательно, относятся к разным видам. .
Следующий этап подготовки материала к исследованию — фиксация. По-
скольку мелких моллюсков вскрывать трудно, желательно иметь лгатериал для
гистологического исследования, с тем чтобы в дальнейшем можно было приме-
нить метод графической реконструкции. Спиртовая фиксация для этой цели не
годится — срезы прокрашиваются слабо и неконтрастно, поэтому можно ре-
комендовать как самый простой способ фиксацию кипящим формалином. Этот
способ был описан ранее (Шилейко, 1978а) и сводится к тому, что на смочен-
ную водой лучинку сажают ползущего моллюска; выбрав момент, когда живот-
ное максимально вытянется из раковины, лучинку быстрым движением опус-
кают в кипящий 10%-ный формалин, после чего немедленно перекладывают
в 4%-ный формалин нормальной температуры. Для гистологических работ же-
лательны экземпляры, не втянувшиеся в раковину, а при фиксации холодным
раствором моллюски успевают сократиться и уйти в раковину.
После того как часть материала зафиксирована формалином (для этого доста-
точно 5—6 экз.), остальных моллюсков убивают выдерживанием в воде. Брат-
чик (1976) предлагает следующую модификацию этого способа: животных поме-
щают в банку с холодной водой и плотно закрывают крышкой; затем банку по-
мещают в водяную баню, где температуру в течение 40—50 мин доводят до
70 °C, после чего моллюсков перекладывают в спирт крепостью 70%. При этом
колумеллярный мускул чаще всего отстает от столбика, и впоследствии тело
моллюска легко извлечь из раковины.
Сухие раковины удобно хранить следующим образом. Материал размещается
по маленьким пробиркам, каждая из которых вкладывается в спичечный коро-
бок, туда же помещается полевая этикетка. Копию этикетки, аккуратно оформ-
ленную и написанную тушью, приклеивают на лицевую часть коробка, а на
торце его пишут только видовое название. Размещенный таким образом ма-
териал укладывают в однотипные коробки так, чтобы торец, на котором напи-
сано видовое название, был обращен вверх. Для семейства или рода отводят
отдельную коробку, а виды внутри нее располагают по алфавиту (рис. 42).
Этот способ дает возможность быстро отыскать в коллекции весь материал по
нужному виду.
Камеральная обработка. Изучать материал начинают с опре-
деления видовой принадлежности исследуемой формы. Для определения и изу-
чения раковин нужно иметь бинокуляр, тонкий пинцет, иглы, штангенциркуль,
106
ОБЩАЯ ЧАСТЬ
мягкую кисточку, пипетку, пузырек с ксилолом или толуолом и предметное сте-
кло, смазанное пластилином или воском.
Для начала раковину ориентируют устьем к себе и вершиной от наблюда-
теля (рис. 43, слева) и в таком положении закрепляют ее на стекле с пластили-
ном. Затем оценивают форму раковины, число и характер выпуклости оборотов
и проводят измерения (рис. 43). Ввиду того что многие виды отличаются малыми
размерами, непосредственное измерение их трудно, ошибка в измерениях бу-
дет относительно велика, и существует риск повредить раковину при зажима-
нии ее в штангенциркуле; в таких случаях можно использовать окуляр-микро-
Рис. 42. Удобный способ хранения коллекции сухих раковин.
метр или изготовить точный рисунок и, зная увеличение, измерять уже изобра-
жение на бумаге. Нарисовать контуры раковины можно с помощью рисовального
аппарата и фотоувеличителя: на чисто вымытом предметном стекле раковину за-
крепляют с помощью крохотного кусочка пластилина, а рядом на стекле тушью
проводят линию длиной 1 мм; затем это стекло помещают в увеличитель и обво-
дят контуры проекции раковины карандашом.
Если надо выяснить расположение элементов устьевой арматуры внутри
последнего оборота, на сухую и очищенную от почвенных частиц раковину на-
носят несколько капель ксилола: раковина смачивается и делается почти про-
зрачной. Таким путем можно, не повреждая раковины, обнаружить пластинки
и складки, расположенные внутри последнего оборота, которые не удается рас-
смотреть сквозь устье.
Специальное внимание надо обратить на скульптуру поверхности раковины
и, в частности, на скульптуру эмбриональных оборотов: там часто имеются спи-
ральные ребрышки, но они выражены подчас исчезающе слабо. Эмбриональные
обороты рекомендуется рассматривать при большом увеличении в узком ярком
пучке света, идущем под острым углом к поверхности предметного столика.
Раковину лучше держать при этом в руке и медленно поворачивать ее в пучке
света, по-разному наклоняя; если раковина очень мала, то можно ее закрепить.
МЕТОДЫ ИССЛЕДОВАНИЯ
107
базальным краем на пластилине в «стоячем» положении и менять ее положение
относительно пучка-света легкими касаниями иглы.
Радиальные элементы дефинитивной скульптуры всегда можно заметить
при некотором увеличении, но спиральные бороздки не во всех случаях выра-
жены хорошо. Для их выявления надо в первую очередь осмотреть заустьевую
часть последнего оборота, так называемый затылок раковины, а также склоны
пупка: на этих участках спиральные линии, если они имеются, часто выражены
лучше, чем в других местах; выявление спиральных бороздок облегчается также
узким пучком света, если направить его на раковину со стороны вершины почти
параллельно оси столбика.
Рис. 43. Схема основных промеров раковины.
Диаметр раковины: БД — большой, МД — малый; высота: ВПО — последнего оборота, ВР — раковины,
ВУ — устья; ШУ — ширина устья.
Многие виды плохо различимы по раковине, поэтому после тщательного изу-
чения раковины нельзя ограничиться только определением ее по ключу, по
изображениям или просматривая описания. Определение необходимо вести,
придерживаясь последовательности: определительный ключ—описание—изо-
бражение. Нередко для большей надежности животное нужно изучить анатоми-
чески.
Вскрытие проводится в ванночке, заполненной 70%-ным спиртом по обыч-
ной методике (Лихарев, 1962; Шилейко, 1978а), а при анатомическом иссле-
довании мелких видов работа облегчается применением способа, предложенного
Акрамовским (1976, с. 78): освобожденное от раковины тело моллюска надевают
на тонкую энтомологическую булавку (минуцию), так чтобы булавка прошла
сквозь полость столбика. Острый конец минуции вкалывают в кусок- пласти-
лина, прикрепленный к внутренней стенке ванночки, а тупой конец погружают
в дно. Тогда тело моллюска можно поворачивать вокруг его оси (это необхо-
димо при раскручивании висцерального мешка); кроме того, сдвигая моллюска
к тупому концу булавки, его можно закрепить в любом положении, зажав между
булавкой и дном ванночки. Альмухамбетова с успехом вскрывает мелкие виды
на куске влажной бумаги. Этот способ имеет то преимущество, что объект не
плавает и отпрепарированные органы легко расправляются.
При гистологическом исследовании материала, фиксированного формалином,
самые лучшие результаты дает бутаноловая проводка. При этом надо помнить,
что при слишком быстрой дегидратации и передерживании в бутаноле ткани
коробятся и становятся ломкими, особенно крыша легочной полости. Поэтому
для мелких форм рекомендуется постепенная проводка по следующей Цримерной
108
ОБЩАЯ ЧАСТЬ
схеме [спирт или бутанол (время, мин)]: 70° (не менее 1 сут) -> 70° (40—60)
->80° (20) ->90° (10) ->96° (10) -> смесь 96-градусного спирта с бутанолом
в соотношении 3 : 1 (10) -> то же 1 : 1 (10) -> то же 1 : 3 (10 мин) -> чистый
бутанол при нормальной температуре (15) -> чистый бутанол при 37 °C (5—7) ->
-> первый парафин (40) -> второй парафин (40) -> заливка. Эта схема рассчи-
тана для животных с высотой раковины 3—4 мм; при проводке объектов мень-
шего размера время экспозиции в каждом растворе уменьшается, а для мол-
люсков большего размера — увеличивается.
Для удобства ориентировки мелкого объекта его желательно покрасить,
добавив в первую из смесей спирта с бутанолом чуть-чуть эозина.
Препараты радул лучше всего готовить водно-бальзамным методом Целлера,
описанным в монографии Лихарева (1962, с. 99, 100).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Приводимые в настоящем разделе описания таксонов построены по единому
плану, куда, в частности, включены сведения о находках в ископаемом состоя-
нии; если таких данных нет, то соответствующая рубрика в описаниях опущена.
Равным образом рубрика «Внутреннее строение» приводится только для ана-
томически изученных видов.
Распространение монотипических родов и подродов указывается при опи-
саниях соответствующих видов.
Помимо традиционных таблиц для определения подсемейств, родов и видов
для семейства Buliminidae составлена еще одна (дополнительная) таблица, по
которой можно определить по конхологическим признакам вид, минуя опреде-
ления подсемейства и рода. Включение этой таблицы вызвано тем, что некоторые
роды, а тем более подсемейства, не имеют дифференциального конхологического
диагноза, и для точного определения часто необходимо анатомическое исследо-
вание. В тех случаях, когда пригодный для анатомирования материал отсут-
ствует, можно определить вид, пользуясь дополнительной таблицей. Если опре-
деление по этой таблице приводит к названию рода, это означает, что опреде-
ление следует продолжить, открыв таблицу для определения видов данного
рода. Однако надо иметь в виду, что определения, сделанные по этой таблице,
не всегда надежны ввиду того, что раковины видов из разных родов иногда чрез-
вычайно похожи (в частности, Р seudochondrula lederi и Imparietula brevior прак-
тически неразличимы); кроме того, имеются виды, отличающиеся очень широ-
ким размахом изменчивости.
УСЛОВНЫЕ СОКРАЩЕНИЯ И ОБОЗНАЧЕНИЯ
Вописаниях
БД — большой диаметр (ширина) раковины, ВР — высота раковины, МД — малый
диаметр раковины (приводится только для уплощенных раковин).
В рисунках к систематической части
А. — базальный отдел пениального аппендикса; А., — шаровидный отдел пениального
аппендикса; As — короткий мускулизованный отдел пениального аппендикса; — ните-
видный отдел пениального аппендикса; А., — терминальный отдел пениального аппендикса
(ампула); АР — атриальный ретрактор; Б Ж — белковая железа; ВОВ — верхний отдел
вагины; Г Ж — гермафродитная железа; ГII — гермафродитный проток; ДСП — диверти-
кул семяприемника; Кк — козырек, разрастание складки внутри пениса возле отверстия
эпифаллуса; КС — кольцеобразные складки на внутренней поверхности пениса; КЩ —
кольцевая щель в проксимальной части эпифаллуса у места впадения семяпровода; М —
матка; НОВ — нижний отдел вагины; ОВ — отверстие вагины; ОН — отверстие пениса;
Пе — пенис; ПаА — папилла аппендикса; ПКБ — призмоконические бугорки на внутрен-
ней поверхности пениса; ППе — папилла пениса; ПР — половой ретрактор; Пре — про-
стата; ПСП — проток семяприемника; ПЭ — придаток эпифаллуса; РПА — рудимент пе-
ниального аппендикса; РПрО — ретрактор правого омматофора; РСП — резервуар семя-
приемника; Сп — семяпровод; Ск — складки на внутренней поверхности пениса; Ф — фла-
геллум (бич); Ц — цэкум; П1 — шпора сперматофора; Э — эмбрионы, видимые сквозь по-
кровы матки; Эп — эпифаллус; Я — яйца, видимые сквозь покровы матки.
110
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ И НЕКОТОРЫХ ПОДСЕМЕЙСТВ
ПОДОТРЯДА PUPILLINA
1 (2). Нижний конец столбика косо обрублен; пупок запаян, пупочная впа-
дина отсутствует ................................ Cochlicopidae (с. 111).
2 (1). Нижний конец столбика не обрублен; пупок имеется хотя бы в виде
узкой щели; если запаян, то имеется пупочная впадина.
3 (10). Высота раковины меньше ее диаметра, равна ему или превышает
диаметр незначительно. Дефинитивные обороты без спиральных бороз-
док и без кожистых шипов на периферии.
4 (5). На париетальной стенке устья имеются длинные продольные пластинки
.............................................. Strobilopsidae (с. 154).
5 (4). Париетальная стенка без длинных продольных пластинок.
6 (7). Поверхность раковины тонко, густо и резко радиально исчерчена , . .
................................................ Pyramidulidae (с. 236).
7 (6). Скульптура иная.
8 (9). Раковина светлой окраски, гладкая или с кожистыми радиальными
ребрышками; края устья сильно сближены или устье цельное ....
....................................... Valloniidae: Valloniinae (с. 162).
9 (8). Раковина темной окраски, поверхность ее несет грубые редкие сглажен-
ные ребра, лишенные кожистой оторочки; места прикрепления устья
расставлены ................................... Spelaeodiscidae (с. 156).
10 (3). Высота раковины больше ее диаметра; если же раковина кубаревидная -
или уплощенная, то дефинитивные обороты с ясными спиральными бо-
роздками или кожистыми шипами на периферии.
11 (12). Раковина хрупкая, края устья неотвернутые, ломкие, острые. Поверх-
ность раковины с радиальными кожистыми ребрышками, присутствую-
щими на всех оборотах; иногда эти ребрышки вытянуты в шипы .....
..................................... Valloniidae: Acanthinulinae (с. 158).
12 (11). Раковина не хрупкая, края устья в той или иной мере отвернуты;
если же не отвернуты, то утолщенные, не ломкие. Кожистые ребрышки
отсутствуют или имеются только на верхних оборотах.
13 (14). В устье залегают 1—2 колумеллярные пластинки, прослеживающиеся
на значительной части длины столбика, и тонкая высокая париетальная
пластинка, уходящая в глубь раковины на разное расстояние, эти пла-
стинки присутствуют на всех стадиях развития; либо устье резко поднято,,
так что его ангулярная область почти касается нижнего шва................
...................................................Orculidae (с. 116).
14 (13). Совокупность признаков иная.
15 (16). Раковина коническо-цилиндрическая, высококоническая или вере-
теновидная, высота ее не менее 4 мм; в устье не менее 4 пластинчатых
зубов; затылочное утолщение отсутствует .... Chondrinidae (с. 224).
16 (15). Раковина от овальной до цилиндрической; если веретеновидная, то-
зубы в устье отсутствуют. Затылочное утолщение имеется, если же оно
отсутствует, то высота раковины не превышает 3.5 мм.
17 (18). Высота вполне сформированной раковины не превышает 2.6 мм;
иногда высота раковины достигает 3.5 мм, но в этом случае поверхность
ее почти гладкая, без правильной радиальной скульптуры, а устье без
губы, с ломкими краями ............................ Vertiginidae (с. 195).
18 (17). Высота вполне сформированной раковины не менее 3 мм; если меньше,,
то края устья с губой и затылочным утолщением.
19 (20). Высота раковины 2.7—5.5 мм; при высоте 5.0—5.5 мм форма ее ци-
линдрическая ......................................... Pupillidae (с. 171).
20 (19). Высота раковины не менее 6 мм; если несколько меньше, то форма ее
удлиненно-овальная ...................................Buliminidae (с. 238).
£
<6
1. РОД COCHLICOPA
111
I. Надсем. COCHLICOPOIDEA Pilsbry, 1900
Раковина средних размеров, исходно удлиненно-овальная, гладкая. Пупок
•запаян. Эмбриональные обороты гладкие. Устье без зубов или со сложной ар-
матурой. Края устья утолщены, не отвернуты, без губы.
Семенные пузырьки не обособлены от гермафродитного протока. Пенис без
характерных внутренних структур, пениальный аппендикс имеется, но лишен
ветви полового ретрактора. Эпифаллус без придаточных органов.
Сем. COCHLICOPIDAE Pilsbry, 1900
Pilsbry, 1900, Man. Conch., 22.
Раковина удлиненно-овальная или овально-коническая, сильно блестящая.
Простата лентовидная. Проток семяприемника тонкий, недлинный, резервуар
маленький, с собственным коротким протоком, дивертикул семяприемника
также короткий.
Распространение. Голарктика. -4 рода.
В ископаемом состоянии семейство известно начиная с палеоцена (Европа,
Северная Америка).
1. Род COCHLICOPA Ferussac, 1821
Ferussac (fils), 1821 : 28, 55; Risso, 1826 : 79; Jeffreys, 1830 : 347 (Cio-
nella, типовой вид Helix lubrica Miiller, 1774); Leach in Turton, 1831 : 82 (Zua,
типовой вид Helix lubrica Miiller, 1774); Pilsbry, 1908, Man. Conch., 19 : 308; Thiele,
1931 : 503; Лихарев, Раммельмейер, 1952 : 125; Wen?, Zilch, 1959—
1960 : 145; Акрамовский, 1976 : 128 (Cionella).
Типовой вид Helix lubrica Muller, 1774; по последующему обозначе-
нию (Pilsbry, 1908).
В связи с разноречивыми мнениями, имеющимися в литературе, необхо-
димы некоторые пояснения. Названия Cochlicopa Ferussac, 1821 и Cionella
Jeffreys, 1830 основаны на одном типовом виде — Helix lubrica Muller, 1774.
Обозначение типового вида для рода Cochlicopa было сделано, по-видимому,
Пилсбри (см. синонимику), а обозначение типового вида рода Cionella — при
его установлении.
Недоразумения начались со статьи Кеннерда (Kennard, 1942), который безос-
новательно решил, что Cochlicopa — замещающее название для Polyphemus
Montfort, 1810, non Muller, 1774. На самом деле в работе Ферюссака (Ferussac,
1821) сказано, что подрод Cochlicopa делится на Polypherrlus Montfort и Styloides
Ferussac; в той же работе Ферюссака, но несколько ниже (с. 55) приводится пе-
речень видов, принадлежащих подроду Cochlicopa, причем Helix lubrica Mul-
ler отнесен к Styloides вместе с 8 другими видами. Далее Риссо (Risso, 1826),
не обозначая типового вида, сузил объем Cochlicopa до одного вида — Helix
lubrica. Ошибочность толкования Кеннерда была показана Уотсоном (Watson,
1943), но в более поздней работе Пилсбри (Pilsbry, 1948) взгляд Кеннерда был
принят, и в последующей обзорной публикации Тейлора и Соля (Taylor, Sohl,
1962) принято название Cionella со ссылкой на Кеннерда и Пилсбри. Поэтому
правы Беквер и Миллер (Bequaert, Miller, 1973, р. 193), восстановившие назва-
ние Cochlicopa.
Устье без зубов. Столбик снизу обрублен.
Распространение. Голарктика. 10—15 видов.
В ископаемом состоянии род известен начиная с палеоцена (Европа, Север-
ная Америка).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ COCHLICOPA
1 (4). Большой диаметр раковины не менее 2.8 мм.
2 (3). Оборотов не менее 6V2, выпуклых........2. С. pseudonitens.
112
I. НАДСЕМ. COCIILICOPOIDEA
3 (2). Обороток no более 6, слабовыпуклых ....................1. С. nitens.
4 (1). Большой диаметр раковины но превышает 2.7 мм.
5 (6). Большой диаметр раковины по менее 2.2 мм, изгиб столбика слабо выра-
жен, окраска свежих раковин темно-роговая или красновато-роговая
........................................................... 3. С. lubrica.
6 (5). Большой диаметр раковины не превышает 2 мм, изгиб столбика обычно
ясный, окраска свежих раковин большой частью светло-роговая . . .
..................................................... 4. С. lubricella.
I
Рис. 44. Виды рода Cochlicopa.
I — С. pseudonitens Uvalieva, голотип; II — С. nitens (Gall.) из поймы р. Оки, с. Белые Колодези; III —
Ct lubrica (Mull.) из Москвы, Кунцево; IV — С. lubricella (Porro) из сухого склона на левом берегу р. Оки
у с. Белые Колодези.
1. Cochlicopa nitens (Gallenstein, 1852) (рис. 44, II; рис. 45).
Gallensteiu, 1852 : 75 (Achatina); Pilsbry, 1908, Man. Conch., 19 : 323; Л и-
харев, Раммельмейер, 1952 : 125 (lubrica var. nitens)-, Ehrmann, 1956 : 33,
t. I, fig. 2 (lubrica Lokalrasse nitens)-, Hudec, 1960 : 278, obr. 2; Zilch, Jaeckel,
1962 : 76 (lubrica nitens); Lozek, 1964 : 194, t. V, 5; Матёкин, 1966 : 109 (CionellaJ,ub-
rica var. nitens); Дамянов, Лихарев, 1975 : 164, фиг. 100.
Terra typica — Клагенфурт, Каринтия (Австрия).
Синтипы хранятся в Зенкенбергском музее (Франкфурт-на-Майне),
№ 166385/7 (Zilch, 1962, Arch. Moll., 91, S. 230).
Материал: около 60 экз. из 6 местонахождений в Московской области,
на Северном Кавказе и на Тянь-Шане.
Раковина овально-коническая, слегка вздутая, очень сильно блестящая,
с широко и плавно закругленной вершиной, просвечивающая, сравнительно
тонкостенная. Оборотов 5—51/2, слегка выпуклых, плавно нарастающих. Пос-
ледний оборот почти прямой, иногда очень плавно и незначительно поднимается
к устью. Окраска одноцветно каштановая или красновато-роговая. Поверх-
ность раковины выглядит полированной, иногда даже с зеркальным блеском,
скульптура в виде редких чрезвычайно плавных и легких радиальных морщи-
нок; при увеличении в 50—60 раз обычно видна также легкая спиральная струй-
чатость. Устье овальное, слегка скошенное, края его тупые, совершенно не от-
вернуты. Остаток колумеллярной пластинки хорошо заметен в виде вертикаль-
ного утолщения края устья.
Размеры: ВР 6.8—7.5, БД 2.8—3.1 мм; в первоописании размеры не ука-
заны, сообщается, что раковины больше, чем у Cochlicopa lubrica.
Внутреннее строение. Материал: 7 экз. из окр. с. Апаринки
Московской обл., 2 мая 1963 г.; 6 — из верховьев р. Чаткал, Тянь-Шань,
25 мая 1972 г.; сборы и определения автора.
1. РОД COCHLICOPA
ИЗ
Белковая железа пигментированная, голубовато-серая, с 1—2 глубокими
ложбинами на поверхности. Нижний отдел вагины немного длиннее верхнего.
Семяпровод впадает строго терминально; эпифаллус в дистальной части вере-
теновидно расширяется, затем следует небольшое сужение и далее идет дисталь-
ное расширение, также веретеновидной формы. Пенис длинный, тонкий, цилин-
дрический; пениальный аппендикс расположен у его проксимального конца.
Внутренние стенки пениса ниже места впадения аппендикса с неясными про-
дольными складками;внутри
веретеновидного вздутия рас-
полагаются 1—2 мощные гоф-
рированные складки, за ко-
торыми находится узкое от-
верстие эпифаллуса. слит
сЛ2; оба эти отдела вместе
имеют вид короткого закруг-
ленного конуса, обращенного
вершиной к пенису. А3 при-
мерно вдвое длиннее, чем
Л1+Л2. Нитевидный Ai уме-
ренно длинный, А5 нес-
колько короче. Внутри Л 2
имеется хорошо выраженная
папилла. Половой ретрактор
крепится к пенису на сто-
роне, противолежащей месту
впадения аппендикса. Про-
ток семяприемника короткий,
дивертикул в l1^—2 раза
длиннее, резервуар неболь-
шой, шаровидный, с собст- * У
Рис. 45. Cochlicopa nitens (Gall.)
из верховьев р. Чаткал, Тянь-
Шань, 25 V 1972 г. Внешний вид
полового аппарата.
венным относительно длинным и очень тонким протоком, длина которого пре-
вышает длину дивертикула. Резервуар семяприемника едва доходит до сере-
дины спермовидукта.
Распространение и местообитание. Вид широко, но
спорадически распространен по территории Палеарктики. Поскольку раньше
его часто указывали под названием С. lubrica, уточнить ареал пока не пред-
ставляется возможным; встречается в европейской части, в том числе на Карпа-
тах и в Крыму, на Кавказе, в горах Средней Азии (на Копетдаге отсутствует),
в Сибири и на юге Дальнего Востока. Обитает в подстилке лиственных и сме-
шанных лесов, во влажных каменистых россыпях, в высокотравных поймах.
2. Cochlicopa pseudonitens Uvalieva, 1967 (рис. 44, 7).
У валиева, 1967 : 215, рис. 2, 3/
Terra typica — Южный Алтай, пойма р. Малая Нарымка.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 11 экз. из типовой серии.
Раковина овально-коническая, очень блестящая, с узко закругленной верши-
ной. Оборотов 7, выпуклых, быстро нарастающих. Последний оборот прямой.
Окраска от темно-роговой до темно-коричневой. Поверхность раковины выгля-
8 А. А, Шилейко
114
I. НАДСЕМ. COCHLICOPOIDEA
дит полированной, с редкими вялыми морщинками. Устье овальное, с острым
углом на стыке палатального и париетального краев. Края устья прямые, не
отвернуты, тупые и слегка утолщены. Складкообразная пластинка на колумел-
лярном крае имеет плавные обводы и ясно заметна.
Размеры в первоописании: ВР 7—8.2, БД 3—3.5 мм; размеры голотипа не
указаны.
Внутреннее строение. Материал мною не исследован, приводи-
мый ниже текст составлен по первоописанию и оригинальному рисунку.
Рис. 46. Cochlicopa lubrica (Miill.) из
Москвы, Кунцево, 5 IX 1976 г. Внешний
вид полового аппарата.
Рис. 47. Cochlicopa lubricella (Рогго)
из Ботанического сада Еревана,
15 IV 1971 г. Внешний вид полового аппа-
рата.
Проксимальный конец эпифаллуса умеренно утолщен. Отделы пениального
аппендикса выражены неясно; половой ретрактор крепится к пенису возле
основания аппендикса. Проток семяприемника очень тонкий, дивертикул от-
сутствует.
Распространение и местообитание. Вид найден в трех
местах на Южном Алтае: в пойме р. Малая Нарымка, на берегу р. Большая На-
рымка и в окр. с. Верхне-Катунска. Встречен на высотах от 500 до 1800 м над
уровнем моря. Обитает в очень влажных условиях, в поймах рек, ручьев и по
берегам родников.
3. Cochlicopa lubrica (Muller, 1774) (рис. 44, III; рис. 46).
Muller, 1774 : 104 (Helix); С 1 е s s i n, 1875 : 41, t. 2, fig. 4a, b (Cionella columna);
Pilsbry, 1908, Man. Conch., 19 : 312, t. 49, figs. 33—35; Лихарев, P аммель-
мейер, 1952 : 125, рис. 35; Ehrmann, 1956 : 33, t. 1, fig. 1, la; Loze k, 1956 : 84,
t. II, 3; Hude c, 1960 : 287, obr. 1; Zilch, Jaeckel, 1962 : 76; L о z e k, 1964 :
193, t. V, 7, 8; M а т ё к и и, 1966 : 109 (Cionella); Gittenberger et al., 1970 : 45,
fig. 27, Kaart 8; Даминов, Лихарев, 1975 : 162, фиг. 97, 98; Акра м о в с к и й,
1976 : 129, рис. 51, табл. V, 39 (Cionella).
Terra typica — Фридриксдаль у Копенгагена.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 44 местонахождений в Венгрии,
европейской части СССР, на Кавказе, в Средней Азии, Западной Сибири и на
Дальнем Востоке. . -
1. РОД COCHLICOPA
115
Раковина удлиненно овально-коническая, с очень плавно закругленной
вершиной, сильно блестящая, умеренно твердостенная, от просвечивающей до
почти прозрачной. Оборотов —6, слабовыпуклых, очень плавно нарастаю-
щих. Последний оборот прямой. Окраска от желтовато-роговой до красновато-
роговой, одноцветная. Поверхность раковины очень гладкая; при увеличении
20 раз и более видны редкие вялые радиальные морщинки, а местами иногда
можно заметить тончайшие исчезающе слабые спиральные бороздки. Устье
немного косое, овальное, в ангулярной области заостренное, места его прикреп-
ления связаны тонкой прозрачной мозолью; после гибели животного парие-
тальная мозоль очень быстро мутнеет и становится белой, непрозрачной. Края
устья тупые, не отвернуты, слегка утолщены по краю. Складка на колумелляр-
ном крае, представляющая собой остаток колумеллярной пластинки, нерезкая,
но ясно заметная.
Размеры: ВР 5.3—6, БД 2.2—2.7 мм; в первоописании: ВР 2.5, БД 1 линия,
т. е. ВР около 5.5, БД около 2.2 мм.
Среди особей с обычным контуром раковины изредка встречается форма
pietfjerl Weinland, 1876, отличающаяся резко увеличенным шагом оборотов при
том же их числе. В результате высота раковины может достигать 8 и даже И мм.
Гудец (Hudec, 1960) предполагает, что это явление может быть вызвано парази-
тарной кастрацией..
Внутреннее строение. Материал: 15 экз. из Москвы, Кунцево,
7 мая 1976 г.; 4 — из того же места, 5 сентября 1976 г.; 4 — из Чегемского
ущелья на Северном Кавказе, 15 мая 1970 г.; 3 — из Талыша, окрестности
с. Бибионы, 25 июня 1964 г.; сборы и определения автора.
Белковая железа с выраженной в разной степени апикальной вырезкой,
которая часто переходит и на латеральную поверхность органа. Отделы вагины
примерно равной длины или верхний отдел немного длиннее. Семяпровод впа-
дает в эпифалдус терминально, с плавным и постепенным расширением. Четкой
границы между эпифаллусом и цилиндрическим пенисом нет, в общем петля,
образованная дистальными протоками мужского отдела, недлинная, без крутых
изгибов; ясно выраженных и постоянных вздутий не наблюдается. Поскольку
два базальных отдела пениального аппендикса слиты в короткий округлый ко-
нус, аппендикс составлен 4 отделами. А3 заметно длиннее, чем М1-|-Л2; степень
развития и особенно А5 сильно зависит от функционального состояния по-
лового аппарата. Половой ретрактор крепится несколько проксимальнее места
впадения пениального аппендикса. Проток семяприемника тонкий, дивертикул
немного длиннее протока семяприемника ниже места раздвоения; проток ре-
зервуара примерно такой, же длины, как и дивертикул. Резервуар семяприем-
ника тесно прилегает к спермовидукту в его нижней части, заметно ниже се-
редины.
Распространение и местообитание. Палеарктика. Вид
населяет любые типы биотопов с мезофильными условиями, за исключением
чистых хвойных лесов. В северной части ареала местами выходит в тундру,
где держится преимущественно в моховых подушках. В полупустынных облас-
тях находки вида приурочены к местам выхода грунтовых вод или к осыпям,
в нижних горизонтах которых влажно даже во время засушливого периода.
4. Cochlicopa lubricella (Porro, 1838) (рис. 44, IV; рис. 47).
Menke, 1828:29 (Achatina lubrica var. exigua, nom. nudum); Porro, 1838:
53 (Bulimus lubricus var. lubricella); Pilsbry, 1908, Man. Conch., 19 : 321 (lubrica
lubricella); Лихарев, Раммельмейер, 1952 : 125 (lubrica var. exigua);
W a 1 d e n,' 1955 : 403 (minima Siemaschko); Ehrmann, 1956 : 33, t. I, fig. 3
(lubrica Lokalrasse exigua); Lozek, 1956 : 85, t. II, 4;. Z i 1 c h, J a e c k e 1,
1962 : 77 (lubrica minima Siemaschko, 1847, = exigua Menke, 1830,=lubricella); Lozek,
1964 : 194, t. V, 6; Матёкин, 1966 : 109 (Cionella lubrica var. lubricella); Gitten-
berger et al., 1970 : 46, fig. 28—31, Kaart 9; Даминов, Лихарев, 1975 : 163,
фиг. 99; Акр амовский, 1976 : 129, рис. 51А, табл. V, 40 (Cionella).
8*
116
II. НАДСЕМ. ORCULOIDEA
Terra typica — Италия, провинция Комо (Ломбардия).
Место хранения типов не установлено.
Материал: более 40 экз. из 7 местонахождений в Подмосковье, на
Тянь-Шане, в Закавказье и Приморском крае.
Раковина удлиненно овально-коническая, с очень плавно закругленной
вершиной, умеренно блестящая, просвечивающая. Оборотов 4—5, сильно упло-
щенных (особенно верхние), плавно нарастающих. Последний оборот прямой.
Окраска бледно-роговая или светло-коричневая, в общем светлее, чем у любого
другого вида рода. Поверхность раковины практически гладкая, но без стекло-
видного блеска. Устье от овального до удлиненно-овального, с углом в ангулярной
области. Париетальная мозоль обычно хорошо заметна. Края устья прямые,
притупленные, слабо утолщены. Колумеллярный край устья почти отвесный,
складка выражена слабо или почти не выражена; в последнем случае ее можно
обнаружить при косом положении раковины.
Размеры: ВР 3.8—5.2, БД 1.8—2 мм; в первоописании размеры не указаны,
сообщается, что меньше, чем у С. lubrica.
Внутреннее строение. Материал: 5 экз. из окр. с. Ромашково
Московской обл. 15 июля 1974 г.; 3 — из Ботанического сада Еревана, 15 ап-
реля 1971 г.; сборы и определения автора.
Поверхность белковой железы с более или менее глубоким желобком. От-
делы вагины примерно равной длины. Семяпровод, плавно расширяясь, перехо-
дит в цилиндрический эпифаллус; между тонким цилиндрическим пенисом и
эпифаллусом имеется очень слабая перетяжка, у некоторых экземпляров она
полностью отсутствует. Все отделы пениального аппендикса сохраняют само-
стоятельность; в частности, между Аг и А2 заметна граница, хотя Аг чрезвы-
чайно короткий. А3 в П/з—2 раза длиннее, чем Аг и А2, вместе взятые. очень
короткий, А5 небольшой, хотя объем его в зависимости от сезона несколько ме-
няется: у особей, собранных в июле, он развит лучше, чем у животных, собран-
ных в апреле. Суммарная длина Л4иА’5 приблизительно равна длине А3. По-
ловой ретрактор крепится немного проксимальнее основания пениального ап-
пендикса. Проток семяприемника тонкий, длина его примерно равна длине ди-
вертикула. Резервуар небольшой, его проток не длиннее самого резервуара.
Семяприемник прилегает к спермовидукту ниже середины последнего.
Распространение и местообитание. Вид широко, но
спорадически распространен по территории Палеарктики. Населяет сравни-
тельно сухие, хорошо дренируемые участки в редколесье, зарослях кустарников,
на склонах оврагов.
II. Надсем. ORCULOIDEA Steenberg, 1925
Раковина средних размеров, исходно цилиндрическая или овально-цилиндри-
ческая, с радиальной скульптурой. Эмбриональные обороты со спиральными
тонкими шнурами на поверхности. Пупок точковидный, реже перспективный.
Среди устьевых зубов имеются длинные париетальные и колумеллярные плас-
тинки. Края устья тонкие или незначительно утолщены, отвернуты; губа раз-
вита слабо или отсутствует.
Семенные пузырьки не обособлены от гермафродитного протока. Пенис
с эпифаллусом, несущим различной формы (чаще всего конический) придаток.
Пениальная ветвь полового ретрактора крепится близ основания придатка
эпифаллуса. Внутренние стенки пениса с тонкими продольными немного спи-
ральными складками. Пениальный аппендикс имеется, половой ретрактор
двуветвистый.
I. Сем. ORCULIDAE Steenberg, 1925
Steenberg, 1925 : 201; Pilsbry, 1922, Man. Conch., 27.
Поверхность раковины с ясной и тонкой радиальной скульптурой, нередко
имеются кожистые ребрышки. Эмбриональные обороты тонкозернистые или со
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ
117
спиральной скульптурой. Устье с пластинчатыми зубами. Края устья по всему
свободному краю широко и плавно отвернуты. Пупок узкий, проколовидный.
Гермафродитный проток без обособленных семенных пузырьков. Простата
лентовидная. Папилла пениса отсутствует. Первично имеется как пениальный
аппендикс, так и придаток эпифаллуса, но оба органа подвержены сильным ви-
доизменениям или редукции. Бич отсутствует или рудиментарный, обнаружи-
вается лишь при вскрытии эпифаллуса. Дивертикул семяприемника имеется
или отсутствует.
Распространение. Средиземноморские страны, Крым, Кавказ,
средняя Европа, Средняя Азия. 4 подсемейства.
В ископаемом состоянии семейство известно начиная с палеоцена (Европа).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ ORCULIDAE
1 (2). Устье настолько резко поднято, что ангулярная область его почти ка-
сается шва. Элементы устьевой арматуры расположены в глубине рако-
вины и при нормальном положении не видны .... II. Pagodulininae.
2 (1). Устье большей частью поднято, но его ангулярная область заметно не
доходит до шва. При нормальном положении раковины в устье видна по
меньшей мере париетальная пластинка.
3 (4). Края устья умеренно отвернуты; на палатальной стенке устья, отступя
от края, расположены три складки, целиком видимые при нормальном
положении раковины ....................................IV. Argninae.
4 (3). Края устья широко отвернуты; на палатальной стенке раковины не более
двух складок.
5 (6). Раковина цилиндрическая или булавовидная; поверхность эмбриональ-
ных оборотов несет тонкие, но ясно различимые спиральные нити . . .
....................................................... I. Orculinae.
6 (5). Раковина овальная, овально-широко-коническая или овально-цилиндри-
ческая; поверхность эмбриональных оборотов без спиральных нитей или
они исчезающе слабые................................ III. Lauriinae.
I. Подсем. ORCULINAE Steenberg, 1925
Дефинитивные обороты тонко радиально исчерчены и часто несут редкие
кожистые ребрышки. Последний оборот почти прямой или плавно приподнят
к устью. Нижние концы колумеллярных и париетальной пластинок видны
при нормальном положении раковины.
Придаток эпифаллуса, даже если он рудиментарный, всегда ясно выражен.
Проток семяприемника без морфологически выраженных желез.
Распространение как у семейства.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ORCULINAE
1 (2). Устье цельное, на колумеллярном крае имеются 2—4 нитевидные скла-
дочки. Вершина раковины притуплена и слегка уплощена. Дивертикул
семяприемника развит ...................................1. Pilorcula.
2 (1). Устье не цельное, колумеллярный край без нитевидных складочек.
Вершина раковины куполовидная или коническая. Дивертикул семя-
приемника отсутствует.
3 (4). Вершина раковины коническая. Пениальный аппендикс отсутствует,
придаток эпифаллуса хорошо развит, удлиненно-конической формы
..........................................................3. Orcula.
4 (3). Вершина раковины куполовидная. Пениальный аппендикс имеется,
хотя бы рудиментарный. Придаток эпифаллуса развит в различной сте-
пени.
118
II. НАДСЕМ. ORCULOIDEA
5 (6). Пениальный аппендикс хорошо развит, но лишен ветви полового ре-
трактора ................................................2. Orculella.
6 (5). Пениальный аппендикс рудиментарный, представлен небольшим вы-
ростом удлиненно-овальной формы, к которому терминально крепится
половой ретрактор ...................................,4. Sphyradium.
1. Род PILORCULA Germain, 1912
Germain, 1912 : 448; Thiele, 1931 : 510 (род Orcula. секция Pilorcula}', Л и-
х а р е в, Р а м м е л ь м е й е р, 1952:158 (Orcula part.); Wenz, Zilch, 1959—
1960 : 155 [Orcula (Pilorcula)]', Шилейко, 19766 : 51.
Типовой вид Pupa raymondi Bourguignat, 1863; по монотипии.
Вершина раковины сильно притупленная, часто почти плоская, со слегка
эмбриональными оборотами. Радиальные кожистые ребрышки
выдающимися
1
хорошо развиты. Колумеллярный
край устья с 2—4 тонкими нитевид-
ными складочками. Край устья цель-
ный. Имеется очень тонкая, но длин-
ная палатальная складка, которая
сквозь устье не видна.
Пениальный аппендикс и прида-
ток эпифаллуса хорошо развиты.
Семяприемник с длинным диверти-
кулом.
Рис. 48. Pilorcula trifilaris (Mouse.) из-
долины р'. Техури, юго-западные склоны
Большого Кавказа.
I — раковина, II — положение париетальной и
колумеллярной пластинок внутри последнего обо-
рота.
Распространение. Передняя и Малая Азия, Кавказ. 2—3 вида.
Pilorcula trifilaris (Mousson, 1863) (рис. 48, 49).
M o u s s о n, 1863 : 391 (Pupa)', 1873 : 12, t. 8, fig. 8 [P. (Sphyradium) bifHaris part.];
0. Boettger, 1881:229 (P. raymondi Bourg.— part.); Rosen, 1911:120, t. I,
fig. 9a, b, c, t. 2, fig. 3 [P. (Orcula) raymondi f. bifilaris, I. bifilaris truncata nov., f. trifilaris,
f. quadrifilaris truncata nov.]; Retows k i, 1914 : 311 [P. (Orcula) raymondi part.]; R o-
s e n, 1914 : 191 [P. (Orcula) raymondi part.]; Pilsbry, 1922, Man. Conch., 27 : 38,
40, t. 4, figs. 1—6 (Orcula raymondi part.); Лихарев, Раммельмейер, 1952 :
161, рис. 75 (О. raymondi part.); H u d e c, L e z h a w a, 1969 : 99, Abb. 5, t. VI, fig. 13
(O. raymondi part.); Шилейко, 19766 : 51, рис. 2 (О. raymondi part.).
Terra typica—„Редуткале“ (немного севернее Поти, западный Кавказ).
Место хранения типов не установлено.
Как видно из материалов по синонимике, Беттгер свел название trifilaris
в синонимы названию raymondi, и с тех пор эта точка зрения укоренилась в ли-
тературе. Однако Pupa raymondi была описана из Ливана (окр. Бейрута) и най-
дена затем на Синайском п-ове (форма hebraica Tristram). Доктор Л. Форкарт
любезно прислал мне изображения лектотипа номинативной формы Р. raymondi
и формы hebraica, а также некоторые пояснения. Сравнивая эти изображения
с кавказскими раковинами, можно убедиться, что названия trifilaris и raymondi
относятся.к разным видам. Кроме того, о видовой самостоятельности Pilorcula
trifilaris свидетельствует то, что вид населяет влажные леса и поймы рек,,
a Pupa raymondi встречается в открытых аридных местностях.
Материал: более 40 экз. из 5 местонахождений на западном Кавказе.
Раковина от цилиндрической до булавовидной, с сильно уплощенной верх-
ней частью и слегка выступающей вершиной. Оборотов 8—9, довольно выпук-
1. РОД PILORCULA
119
лых. Последний оборот к устью слегка поднят, высота его немного более х/3 вы-
соты раковины. Окраска одноцветная, темно-роговая или красновато-роговая.
Дефинитивная скульптура в виде тонких радиальных периостракальных пластин-
чатых ребрышек, которые под швом превращены в шипы наподобие ресниц.
Эти ресницы хорошо выражены у молодых особей, а у старых чаще всего
стираются. Устье коротко овальное или округлое, цельное, немного раструбооб-
разное, края его светлые, широко и плавно отвернуты. На париетальной стенке
расположена высокая пластинка, которая вначале свисает в просвет устья
Рис. 49, Pilorcula trifilaris (Mouss.) из долины р. Техури, юго-западные склоны Большого
Кавказа, 30 IV 1971 г.
I — внешний вид полового аппарата; 11 — квадривий вместе с участком гермафродитного протока; III —
участок сперматофора, извлеченный из дивертикула семяприемника; IV — внутреннее строение придатка
эпифаллуса (из: Шилейко, 19766).
вертикально, затем в глубине последнего оборота ее нижний край отгибается
к периферии оборота почти под прямым углом, и кромка его направлена к кромке
палатальной складки (рис. 48, II). Вся париетальная пластинка уходит в глубь
раковины примерно на Р/а оборота. На расширенном колумеллярной крае видны
2—4 нитевидные складочки, из которых вторая сверху представляет собой
собственно колумеллярную пластинку; эта пластинка внутри последнего обо-
рота сильно увеличивается и направлена горизонтально, располагаясь парал-
лельно париетальной (рис. 48, II). Пупок маленький, каплевидный.
Размеры: ВР 3.8—5, БД 2 мм; в первоописании: ВР 4.2, БД 2 мм.
Внутреннее строение. Материал: 9 экз. из юго-западной части
Большого Кавказа, долина р. Техури,; 30 апреля 1971 г., сбор и определение
автора.
Апикальная вырезка на белковой железе развита слабо. Семяпровод плавно
переходит в недлинный эпифаллус, диаметр которого превышает диаметр
цилиндрического пениса. Чрезвычайно длинный пениальный аппендикс от-
120
II. НАДСЕМ. ORCULOIDEA
ходит дистальнее середины пениса, близ клоаки. Самый длинный из отделов-
аппендикса — Ар, проксимальная часть его слегка извита и утолщена. Л2-|~Л3;
удлиненно-грушевидной формы, отделен от А± кольцевой перетяжкой.
тонкий, нитевидный, извитой, очень плавно переходит в короткий Аъ. Стенки
Л2-|-Л3 утощены, внутри с тонкими продольными складками, при переходе в Ах
имеется глубокая кольцевая борозда. Внутренняя поверхность как эпифаллуса,,
так и пениса несет нерезкие продольные складки, при переходе семяпровода-
в эпифаллус имеется несколько тонких кольцевых складочек. Удлиненно-меш-
ковидный придаток эпифаллуса сидит на тонкой ножке, внутренняя его поверх-
ность с многочисленными очень тонкими кольцевыми складками. Половой,
ретрактор крепится к эпифаллусу проксимальнее цэкума. Семяприемник с длин-
ным дивертикулом, плотно прилегающим к спермовидукту и почти доходящим
до белковой железы. Резервуар семяприемника округлый, с коротким, но ясным
собственным протоком.
Распространение и местообитание. Колхидская низ-
менность, западная часть Большого Кавказа на север до Майкопа, по северным
склонам Большого Кавказа на восток до Орджоникидзе (Калитина, 1958).
Встречается в подстилке влажных лиственных лесов и в поймах с густой,
травянистой растительностью; живет также на задернованных склонах с ка-
менистым покрытием.
2. Род ORCULELLA Steenberg, 1925
Steenberg, 1925 : 176: Thiele, 1931 : 510: Wenz, Zilch, 1959—1960 :
156; Акрамовский, 1976:140; Шилейко, 19766:52.
Типовой вид Pupa orientalis L. Pfeiffer, 1861; по первоначальному
обозначению.
Вершина раковины не притупленная, куполовидная. Радиальные кожистые
ребрышки развиты слабо, раковина иногда резко радиально исчерчена. На колу-
меллярном крае устья нитевидных складочек нет, но на самом столбике имеется
1—3 сглаженные пластинки, уходящие глубоко внутрь раковины. Устье не
цельное. Палатальная складка большей частью отсутствует.
Пениальный аппендикс и придаток эпифаллуса хорошо развиты. Семяпри-
емник без дивертикула.
Распространение. Восточное Средиземноморье и Закавказье.
Около 10 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ORCULELLA
1 (2). Пениальный аппендикс состоит не менее чем из трех ясно выраженных
отделов (подрод Mesorculella) ..................1. О. (М.) ruderalis.
2 (1). Пениальный аппендикс на отделы практически не разделен, почти равно-
мерно цилиндрический по всей длине (подрод Orculella s. str.).
3 (4). Колумеллярный край с 1—2 сглаженными валикообразными пластин-
ками .............................................2. О. (О.) scyphus.
4 (3). Колумеллярные пластинки отсутствуют или еле угадываются ....
................................................3. О. (О.) bulgarica.
1. Подрод MESORCULELLA Schileyko, 1976
Шилейко, 19766 : 53.
Типовой вид Orculella ruderalis Akramowski, 1947; по первоначаль-
ному обозначению.
Отделы (не менее трех) пениального аппендикса хорошо заметны. Проток
внутри А2-{-Аз очень тонкий, извилистый. Половой ретрактор крепится близ
основания цэкума или чуть дистальнее его.
2. РОД ORCULELLA
121
1. Orculella (Mesorculella) ruderalis Akramowski, 1947 (рис. 50, 51).
Акр а м овский, 1947в : 179, рис. 1, 2 (scyphus ruderalis); Лихарев, P а м-
•мельмейер, 1952 : 160, рис. 74 (Orcula ruderalis); Акрамовский, 1971a : 109;
1976 : 140, рис. 60, табл. 5, 56; Шилейко, 19766 : 53, рис. 3.
Terra typica — Армения,, Ехегнадзорский р-н, с. Гнишик.
Г о л о т и п хранится в Зоологическом ин-те АН СССР.
Материал: 12 экз. из типового местонахождения.
Раковина цилиндрическая, с закругленной куполовидной вершиной, уме-
ренно твердостенная, матовая. Оборотов 8—9, слабовыпуклых, последний обо-
рот к устью плавно, но
отчетливо приподнят, вы-
сота его немного более У3
высоты раковины. Окраска
почти одноцветная, рого-
вая разных оттенков. Ни-
тевидные ребрышки на
эмбриональных оборотах
очень тонкие, но ясно раз-
личимы. Скульптура де-
финитивных оборотов в ви-
де редких неправильных
Рис. 50. Orculella (Me- Рис. 51. Orculella (Mesorculella) ruderalis Akr. из Армении,
sorculella) ruderalis Akr. с. Гнишик, 1941 г. Половой аппарат (из: Шилейко, 19766).
из Армении, с. Гнишик.
радиальных морщин, между которыми видны еще более тонкие морщинки. Устье
округлое, почти отвесное, края его широко и плавно отвернуты, места прикрепле-
ния не сближены, связаны светлой мозолью, на которой может присутствовать
очень слабый ангулярный бугорок. Париетальная пластинка вначале вертикаль-
ная, в глубине устья ее кромка несколько отвернута к периферии, заканчивается
эта пластинка примерно на границе предпоследнего и последнего оборотов.
Колумеллярная пластинка в виде бугорка, если рассматривать ее сквозь устье,
но в глубине последнего оборота она заметно усиливается и заканчивается в пред-
последнем обороте. Пупок чрезвычайно узкий,: проколовидный.
Размеры в первоописании: ВР 5—6, БД 2.25—2.5 мм.
Внутреннее строение. Материал: 5 экз. из типового местона-
хождения, сбор и определение Н. Н. Акрамовского.
На поверхности белковой железы хорошо заметен желобок, в котором рас-
полагается участок кишки. Семяпровод впадает в эпифаллус строго апикально.
Пенис и эпифаллус почти одинаковой длины и почти одинакового диаметра.
122
II. НАДСЕМ. ORCULOIDEA
Пениальный аппендикс умеренно длинный, отходит немного отступя от полового
атриума. А± цилиндрический, с неясными кольцевыми перетяжками; A2-f-A3
в общем веретеновидной формы, слабо утолщен, Ai слегка извит, недлинный,
Аъ выражен хорошо. Придаток эпифаллуса длинный, конический, внутри со
слабыми продольными гофрированными складками. Половой ретрактор кре-
пится к эпифаллусу на одном уровне с основанием придатка эпифаллуса, но
на противолежащей стороне протока (рис. 51). Проток семяприемника длинный,
резервуар объемистый, проток в нижней части образует один-два крутых изгиба.
Распространение и местообитание. Вид известен только
из типового нахождения в Армении (с. Гнишик). «Там обыкновенна и много-
численна на сухих пастбищах горностепного пояса; под камнями и щебнем,
а также в трещинах выходов горных пород и под подушковидными колючими
полукустарниками» (Акрамовский, 1976, с. 141).
2. Подрод ORCULELLA s. str.
Пениальный аппендикс почти не подразделен на отделы, с очень тонким
внутренним протоком. Половой ретрактор крепится проксимальнее придатка
эпифаллуса.
2. Orculella (Orculella) scyphus (L. Pfeiffer, 1848) (рис. 52, 53).
L. Pfeiffer, 1848a : 7 (Pupa); Retowski, 1889:254 [P. (Orcula) doliolum
var. batumensis]; Rosen, 1914:190, t. 2, fig. 1 [P. (Orcula) kasnakovi]; Pilsbry,
1922, Man. Conch., 27 : 34, t. 3, figs. 7—1,0 (Orcula scyphus batumensis); 1925, Man. Conch.,
28 : 92 (Orcula (Orculella)]; Hesse, 1924, t. I, figs. 1—6 (0. batumensis); Лихарев,
Раммель мейер, 1952 : 160 (О. batumensis); Ш и л е й к о, 19766 : 55, рис. 5.
Terra typica — «Brussa Natoliae» (Бруса, Турция).
Место хранения типов не установлено.
Материал: более 60 экз. из 7 местонахождений в Аджарии и Турции.
Раковина от цилиндрической до овально-цилиндрической, с короткой ку-
половидной верхней частью, твердостенная, слегка просвечивающая, матовая
или блестящая. Оборотов 9—11, уплощен-
ных, последний оборот в последней трети
плавно, но отчетливо приподнимается, высота
его заметно меньше половины высоты рако-
вины, но более У3 этой величины. Окраска
одноцветно светло-роговая. Спиральные ни-
тевидные ребрышки на эмбриональных обо-
ротах очень тонкие, но вполне отчетливые,,
они хорошо заметны .щэи увеличении 50—
60 раз в косом свете. Скульптура дефини-
тивных оборотов развита в разной сте-
Рис. 52. Orculella (Orculella) scyphus (L. Pfr.)
I — номинативная форма из Халкиса, Греция; II —форма
batumensis Ret. из Артвинского округа, Турция.
пени — от тонкой равномерной ребристости (причем ребрышки могут нести
слабую кожистую оторочку) до вялой неравномерной морщинистости, которая
чередуется с почти гладкими участками; в последнем случае поверхность вы-
глядит блестящей. Устье округлое, края его широко отвернуты, места при-
крепления устья связаны развитой в разной степени париетальной мозолью.
В случае сильного развития мозоли она может иметь на концах утолщения,
а в средней части небольшой выступ, направленный к переднему концу пари-
етальной пластинки. Последняя вначале свисает вертикально, в глубине по-
следнего оборота сгибается вдоль и ее нижняя часть отгибается кнаружи, при-
2. РОД ORCULELLA
123
нимая почти горизонтальное положение. Длина париетальной пластинки — от
1 до 2 оборотов. К олумеллярных'пластинок 2—3. Кроме того, в конце последнего
оборота залегает короткая расплывшаяся палатальная складка, которую можно
увидеть, просветлив раковину ксилолом и рассматривая ее в проходящем свете.
Пупок почти запаян или имеет вид очень узкой изогнутой щели.
Размеры ВР 6.5—10, БД 1.8—3 мм; в первоописании: ВР 9, БД 3 мм.
Широкая изменчивость вида дала основание Пилсбри (Pilsbry, 1922, Man.
Conch., 27: 31) объединить значительное число старых видов в полиморфный вид
•О. scyphus, включающий несколько подвидов; однако ареалы этих форм частью
Рис. 53. Orculella (Orculella) scyphus (L. Pfr.) из Турции, вилайет Ван, с. Шамунис,
, 22 IX 1956 г. Половой аппарат (из: Шилейко, 19766).
недостаточно ясны, частью заведомо перекрываются. Поэтому речь идет, веро-
ятно, большей частью не о подвидах, а об экологических формах. Оконча-
тельного заключения пока дать нельзя ввиду недостатка фактических сведений.
Внутреннее строение. Материал: 1 экз. из Турции, вилайет
Ван, с. Шамунис, 22 сентября 1956 г., сбор К. Линдберга, определение И. М. Ли-
харева (форма crassa Pilsbry).
Белковая железа с небольшой апикальной вырезкой. Семяпровод впадает
апикально в расширенный конец эпифаллуса. Пенис умеренно тонкий и уме-
ренно длинный. Пениальный аппендикс достаточно длинный, но несет явст-
венные признаки дегенерации: отделы аппендикса практически не выражены,,
а сам он тонкий, лишь слегка утолщенный в апикальной области. Придаток
эпифаллуса хорошо развит, конический, причем создается впечатление, что
пенис переходит в придаток, а эпифаллус отходит сбоку в виде тонкой в начале
и расширенной в конце трубки. Внутри придатка эпифаллуса имеются две
мощные противолежащие продольные складки. Внутренняя поверхность
проксимальной части пениса и дистальной части эпифаллуса покрыта неболь-
шими,; но четкими бугорками. Проток в проксимальном отделе эпифаллуса
124
II. НАДСЕМ. ORCULOIDEA
тонкий, со множеством кольцевых глубоко врезанных бороздок. Проток семя-
приемника в нижней части утолщен, почти прямой. Половой ретрактор крепится
на границе нижней и средней трети эпифаллуса.
Распространение и местообитание. Восточное Сре-
диземноморье, включая юго-западную часть Закавказья (Аджария). Населяет
открытые ландшафты, держится в осыпях.
3. Orculella (Orculella) bulgarica (Hesse, 1915) (рис. 54).
Hesse, 1915 : 62 (Orcula); Pilsbry, 1922, Man. Conch., 27 : 26, 256, t. 21, fig. It
(Orcula); Лихарев, Раммельмейер, 1952 : 159, рис. 72 (Orcula); Акрамов-
ский, 1968 : 26 (Orcula); Дамянов, Лихарев, 1975 : 114, фиг. 47 (Orcula); А к-
рамовский, 1976 : 139, табл. VI, 54 (Orcula).
Terra typica — «Gebedsche bei Varna» (Болгария, Варна).
Типы хранятся, вероятно, в Зоологическом музее Берлинского универси-
тета им. Гумбольдта вместе с другими материалами П. Гессе.
Материал: 2 экз. из Армении, берег Севана, и 1 экз. из Дарошама.
Раковины от цилиндрической до коротко овально-конической, с короткой
округлой верхней частью. Оборотов 8—10, довольно выпуклых; последние
три оборота почти равной ширины. Последний оборот
к устью энергично, но без перегиба, поднят. Окраска од-
ноцветная, темно-роговая, к устью немного светлее. Спи-
ральные нитевидные ребрышки на эмбриональных оборотах
хорошо заметны. Дефинитивная скульптура в виде тонких
радиальных морщинок или ребрышек, которые на ранних
оборотах и возле устья располагаются чаще, чем на послед-
них трех. Устье широкоовальное, с плавно й широко
отвернутыми краями. Париетальная пластинка представлена
Рис. 54. Orculella (Orculella) bulgarica (Hesse) из Армении, окр. оз. Се-
ван.
клиновидным зубом, расположенным близ края устья, и очень слабым продоль-
ным валиком, отходящим от этого зуба и быстро сходящим на нет. При рас-
сматривании раковины в нормальном положении колумеллярный край ровный
или с очень слабым утолщением; если раковину повернуть вокруг оси стол-
бика по часовой стрелке, то на столбике можно' заметить конической формы
припухлость — остаток колумеллярной пластинки. Пупок почти полностью
запаян.
Размеры: ВП 6.5—8, БД 2—3.2 мм; в первоописании: ВР 6.5, БД 3 мм.
До сих пор вид единодушно относили к роду Orcula, но, как видно из описа-
ния, раковина его гораздо ближе к некоторым видам рода Orculella, в частности
О. scyphus.
Распространение и местообитание. Вид описан из се-
веро-восточной Болгарии, затем найден в горном массиве Витоша (центральная
часть западной Болгарии), в окр. Анапы, в выбросах Аракса близ Дарошама
(Нахичеванская АССР) и на восточном берегу оз. Севан недалеко от устья
р. Масрик. Населяет, по-видимому, влажные леса и открытые влажные склоны.
3. Род ORCULA Held, 1837
Held, 1837:919; Pilsbry, 1922, Man. Conch., 27 : 1; Thiele, 1931:510;
Лихарев, Раммельмейер, 1952 : 158; W e n z, Zilch, 1959—1960 : 155;
Дамянов, Лихарев, 1975 : 113; Акрамовский, 1976 : 139; Шилейко,
19766 : 56; Gittenberger, 1978 : 18.
3. РОД ORCULA
125
Типовой вид Pupa dolium .Draparnaud, (1801; по обозначению Herr-
mannsen, 1847.
Вершина раковины большей частью заостренная. Радиальная ребристость
отсутствует, скульптура представлена неравномерной радиальной исчерчен-
ностью. Устье с одной париетальной и 1—2 колумеллярными пластинками.
Палатальных складок нет.
Пениальный аппендикс отсутствует, но хорошо развит конический придаток
эпифаллуса. Проток семяприемника длинный, лишен дивертикула.
Распространение. Средняя и частично южная Европа. Несколько
видов. Поскольку род достоверно отличается от прочих родов семейства глав-
Рис. 55. Orcula dolium.
(Drap.) из Австрии, Ка-
ринтия.
Рис. 56. Orcula dolium (Drap.) из Татр, Польша, 12 VIII
1952 г.
Половой аппарат; звездочка — перегородка внутри придатка эпифал-
луса.
ным образом по анатомическим признакам, родовая принадлежность ископаемых
видов неясна. Самые первые находки раковин Orculidae сделаны в палеоценовых
отложениях Европы.
Orcula dolium (Draparnaud, 1801) (рис. 55, 56).
Draparnaud, 1801 : 58 (Pupa); Draparnaud, 1805 : 62 (Pupa); M о q u i n-
T a n d о n, 1855 : 384, pl. 27, figs. 29—31 [Pupa (Sphyradium)]; Pilsbry, 1922, Man.
Conch., 27 : 7,1.1, figs. 1—4; Steenberg, 1925 : 53, pl. 8, figs. 1, 2, fig. 28 du texte; Л и-
харев, Раммельмейер, 1952 : 158, рис. 71; Ehrmann, 1956 : 50, fig. 27;
Lozek, 1956 : 103, t. 7, fig. 2; Gittenberger, 1978 : 25, Figs. 16—19.
Terra typica — Франция.
Место хранения типов не установлено. .
Материал: более 60 экз. из двух местонахождений в Венгрии, Чехосло-
вакии, Австрии и ФРГ.
Раковина от цилиндрической до коническо-цилиндрической, просвечиваю-
щая, шелковисто поблескивающая, сравнительно твердостенная. Верхняя часть
раковины в виде конуса, вершина слегка приостренная. Оборотов 9—10, плавно
нарастающих, последний оборот к устью медленно и плавно поднимается,
126
II. НАДСЕМ. ORCULOIDEA
высота его немного меньше половины высоты раковины, изредка (у высоких
раковин) — около 1/А этой величины. Окраска одноцветная, от желтовато-
роговой до бурой и красноватой. Эмбриональные обороты покрыты неясной зер-
нистостью, но близ границы с дефинитивными иногда можно заметить исчезаю-
ще слабые элементы спиральной скульптуры, заметные не на каждой раковине.
Дефинитивные обороты неравномерно покрыты сглаженными радиальными мор-
щинками, одни из них выражены резче, другие слабее, иногда несколько мор-
щинок располагаются вплотную одна к другой, а за ними следует почти глад-
кий участок. Устье округлое или широкоовальное, почти отвесное. Парие-
тальная пластинка умеренно развита, заканчивается на границе последнего
и предпоследнего оборотов. Колумеллярная пластинка слабее и короче парие-
тальной. Пупок цилиндрический; если рассматривать раковину строго снизу,
то видна лишь часть его, так как над пупком нависает колумеллярная часть
устья.
Размеры ВР 6.5—8.5, БД 2.6—3.6 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 7 экз. из Татр, 12 августа
1952 г., сбор и определение А. Риделя.
Белковая железа с хорошо развитой глубокой апикальной вырезкой. Се-
мяпровод впадает апикально в слегка расширенный конец эпифаллуса, длина
которого примерно вдвое больше прямого цилиндрического пениса. На границе
пениса и эпифаллуса крепится мощный половой ретрактор, а на противолежа-
щей стороне пениальной трубки располагается мощный придаток эпифаллуса
конической формы. То, что это придаток эпифаллуса, а не пениальный аппен-
дикс, доказывается как его положением (пениальный аппендикс располагается
близ полового атриума), так и пространственным отношением его к половому
ретрактору (у всех членов семейства половой ретрактор всегда более или менее
сближен с основанием придатка эпифаллуса). Стинберг (Steenberg, 1925, pl. 8,
figs. 1, 2) отмечает наличие у этого вида рудиментарного бича, отсутствующего
у исследованных мною экземпляров; однако наличие припухлости на одной
стороне проксимальной части эпифаллуса позволяет предположить, что эта
припухлость есть остаток бича, который, как и всякий рудиментарный орган,
подвержен повышенной изменчивости. Внутренние стенки пениса, эпифаллуса
и придатка эпифаллуса покрыты тонкими продольными гофрированными склад-
ками. Кроме того, как видно на продольном разрезе пениальной трубки (рис. 56),
полость пениса и эпифаллуса сообщаются не непосредственно, а через полость
придатка эпифаллуса, так как существует поперечная перемычка, помеченная
на рис. 56 звездочкой. Проток семяприемника очень мощный, прямой или в раз-
ной степени изогнутый, резервуар неясно отграничен от протока, прилегает
к белковой железе.
Распространение и местообитание. Горные области
средней Европы — от Альп до Карпат. Предположение Лихарева и Раммель-
мейер (1952) о том, что вид может быть встречен в Закарпатье на территории
СССР, вполне правдоподобно, хотя пока не подтвердилось. Вид обитает в под-
стилке лиственных и смешанных лесов, а также на 'влажных скалах.
4. Род SPHYRADIUM Charpentier, 1837
Charpentier, 1837 : 15 [Pupa (Sphyradium.)]; Moquin-Tandon, 1855 :
381 ГР. (Sphyradium) part.]; Thiele, 1931 : 510 (род Orcula, секция Sphyradium); W e n z,
Zilch, 1959—1960 :155 [Orcula (Sphyradium)]; Шилейко, 19766:56.
Типовой вид Bulimus doliolum Bruguiere, 1792; по обозначению
Мартенса (Martens in: Albers, 1860: 295).
Вершина раковины куполовидная. На дефинитивных оборотах имеются
хорошо выраженные редкие тонкие радиальные кожистые ребрышки. 1—2 колу-
меллярные пластинки видны через устье при нормальном положении раковины.
Палатальных складок нет.
4. РОД SPHYRADIUM
127
Пениальный аппендикс рудиментарный, представлен лишь слабо развитым
остатком Alt к которому крепится половой ретрактор. Придаток эпифаллуса
также рудиментарный, но всегда ясно выражен, расположен на границе первой
и второй трети эпифаллуса. Последний имеет собственный очень тонкий ретрак-
тор; кроме того, дистальная часть эпифаллуса связана с его проксимальной
частью посредством дополнительной мышечной ленты. Семяприемник без ди-
вертикула.
Монотипический род.
Sphyradium doliolum (Bruguiere, 1792) (рис. 57, 58).
Bruguiere, 1792 : 351 (Bulimus); Schneider, 1878 :27 {Pupa bifilaris Mousson,.
1873); 0. Boettger, 1879 : 32 (Pupa); 1889a : 23 (Pupa); Soos, 1917 : 112 (Orcula);
Pilsbry, 1922, Man. Conch., 27 : 17, t. 2, figs. 1, 2, 4 (Orcula); Лихарев, Рам-
мельмейер, 1952 :159, рис. 73 (Orcula); Ehrmann, 1956 :, 41, t. 1, fig. 21 [Orc ula (Scy-
phus)]; L о i, e k, 1956 : 103, t. 7, fig. 1 (Orcula); Zilch, Jaeckel, 1962 : 61 [Orcula
(Sphyradium, Scyphus)]; Шилейко, 1967a : 18, рис. 5, 6 (Orcula); Дамянов, Ли-
харев, 1975 : 116, фиг. 49, 50 (Orcula); Акраио вский, 1976 : 140, рис. 59, табл.
VI, 55, 57 (Orcula); Шилейко, 19766 : 56, рис. 6.
Terra tуpica — не указана.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 87 местонахождений в Закарпатье,
в Крыму, на Кавказе и в Средней Азии, а также в Южной Европе.
Раковина от цилиндрической до булавовидной, с более или менее куполовид-
ной верхней частью, слабо блестящая, слегка просвечивающая. Оборотов 8—
972, сильно уплощенных, мед-
ленно и плавно нарастающих.
Последний оборот к устью в пос-
ледней трети очень плавно, но
отчетливо поднимается,, высота
его у высоких раковин немного
больше Уз высоты раковины,
у низких раковин—немного
меньше У2 этой величины, со
всеми промежуточными значе-
Рис. 57. Sphyradium doliolum (Brug.)
из окр. Тбилиси.
I — вполне сформированная раковина,
II — раковина молодого экземпляра.
ниями. Окраска одноцветная, светло-роговая, реже буроватая. Спиральные нити
на эмбриональных оборотах видны вполне отчетливо. Дефинитивные обороты
покрыты отдельными редкими кожистыми ребрышками, которые у старых рако-
вин нередко стираются; на ранних оборотах они сохраняются дольше. Между
ребрышками видны очень вялые и неясные морщинки. Устье округлое или усе-
ченно овальное, с широко отвернутыми и слегка утолщенными белыми краями;
места .прикрепления устья расставлены. Париетальная пластинка по всей длине
вертикальная, обычно плавно широко гофрированная, уходит в глубь раковины
на 1—iy2 оборота. Колумеллярных пластинок 1 или 2; иногда чтобы их рассмот-
реть, раковину надо немного повернуть вокруг оси столбика по часовой стрелке.
Пупок почти запаян, от него остается очень узкая щель.
Размеры ВР 4—5, БД 2.2—2.4 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 4 экз. из Северной Осетии,
левый берег р. Ардон, 18 июля 1974 г., сбор Я. И. Старобогатова; 7 — из
Тянь-Шаня, заповедник Аксу-Джабаглы, 19 сентября 1973 г., сбор И. М. Хо-
хуткина; 13 — из Грузии, с. Коджори близ Тбилиси, 24 апреля 1971 г.; 9 —
128
II. НАДСЕМ. ORCULOIDEA
из Тянь-Шаня, Ферганский хр. 18 мая 1972 г.; 3 — из Армении, с. Агверан
близ Еревана, 25 сентября 1975 г.; сборы и определения автора; 2 — из Арме-
нии, Гегарт, 10 августа 1940 г., сбор и определение Н. Н. Акрамовского.
Поверхность белковой железы с глубоким изогнутым желобом. Семяпровод
впадает строго апикально в плавно расширенный конец эпифаллуса. Пенис
умеренно тонкий и умеренно длинный. Пениальный аппендикс представлен
только рудиментом Ат. Эпифаллус тонкий, длинный. Система половых ретрак-
торов крайне своеобразна. Самый мощный мускул прикрепляется терминально
к рудименту пениального аппендикса; тонкая мышечная лента соединяет район
Рис. 58. Sphyradium doliolum (Brug.) из Ферганского хр., Тянь-Шань, 18 V 1972 г.
Половой аппарат; звездочка — рудимент пениального аппендикса (из: Шилейко, 19766).
аппендикса с более проксимальной частью мужской петли; наконец, имеется
очень тонкий и слабый ретрактор, соединяющий диафрагму с эпифаллусом.
Придаток эпифаллуса рудиментарный,; тонкий, конический. Проток семяпри-
емника в нижней части слегка утолщен, далее постепенно истончается и внезапно
переходит в удлиненный резервуар, значительно не доходящий до белковой
железы.
Распространение и местообитание. Средиземномор-
ские страны,, южная, западная и средняя Европа,. Малая Азия, Крым, Кавказ,;
Северный Иран,; Средняя Азия. Встречается в большом наборе биотопов:
от умеренно увлажняемых разреженных лесов до влажных речных долин и
аридных открытых участков, где держится в осыпях. Нередко образует массо-
вые скопления.
II. Подсем. PAGODULININAE Pilsbry, 1924
Pilsbry, 1924, Маи. Conch., 27 : 166; Klemm, 1939 : 198.
Дефинитивные обороты ребристо исчерчены. Последний оборот к устью резко
поднят и почти достигает шва. Зубы расположены в глубине последнего оборота
и сквозь устье не видны.
1. РОД PAGODULINA
129
Придаток эпифаллуса имеется или отсутствует. На протоке семяприемника
имеется железа фасолевидной формы.
Монотипическое подсемейство.
Распространение. Южная и частично средняя Европа и горная
система Эльбурс с Талышом.
В ископаемом состоянии подсемейство известно из верхнего миоцена J Пред-
кавказья и верхнего плиоцена Италии.
1. Род PAGODULINA Glessin, 1876
Stabile, 1864 : 100 (Pagodina, non Van Beneden, 1852); Glessin, 1876 : 198;
Hesse, 1916b : 124 (Pagodula, non Monterosato, 1884); Pilsbry, 1924, Man. Conch.,
27 : 166; Thiele, 1931 : 510; Лихарев, Раммельмейер, 1952 : 162; W e n z,
Zilch, 1959—1960:156; Стеклов, 1966:165; Дамянов, Лихарев, ^1975:
117; Шилейко, 19766 : 56.
Рис, 59. Виды рода Pagodulina.
1 — Р. (Pagodulina) pagodula (Des Moulins) из северной Италии, близ г. Изео; II — то же, устьевая арма-
тура (верхние обороты удалены, раковина просветлена ксилолом); III — Р. (Crystallifera) lederi (Bttg.)
из Тальппа, с. Бибионы. ЦП — колумеллярная пластинка, ПаП — париетальная пластинка, ПаС — па-
латальная складка, У — устье раковины.
Типовой вид Pupa pagodula Des Moulins, 1830; по монотипии.
Признаки и распространение как у подсемейства. 3—5 ви-
дов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ PAGODULINA
1 (2). Ребристая исчерченность дефинитивных оборотов редкая. Придаток эпи-
фаллуса имеется (подрод Pagodulina s. str.) . . . . 1. Р. (Р.) pagodula.
2 (1). Ребристая исчерченность дефинитивных оборотов густая. Придаток
эпифаллуса отсутствует (подрод Crystallifera) .... 2. Р. (С.) lederi.
1. Подрод PAGODULINA s.^str.
Придаток эпифаллуса хорошо развит.
Распространение. Южная и частично средняя Европа.
1. Pagodulina (Pagodulina) pagodula (Des Moulins, 1830) (рис. 59, I, II\
рис. 60).
Des Moulins, 1830 : 158, figs. 1—3 (Pupa); Beck H., 1837 : 84 (Pupa); R о s s-
m ae s s 1 e r, 1837, Iconogr., Heft 5—6: 15, fig. 325 (Pupa); Mo quin-T andon,
1855 : 388, pl. 27. figs. 35—41 [P. (Odostomia)]; Westerland, 1887 : 90; Pilsbry,
9 А. А. Шилейко
130
II. НАДСЕМ. ORCULOIDEA
1924, Man. Conch., 27 : 169, t. 20, figs. 1—4; Germain, 1930 : 420, figs. 367—370, pl. 12,
fig. 354; Klemm, 1939 : 198; Ehrmann, 1956 : 52, t. 1, fig. 22; L о z e k, 1956 ::
104, t. 7, 4; 1964 : 209, t. 8, fig. la, b; Hude c, 1965b : 159, Abb. 6; A 1 z о n a. 1971 :
70; Дамянов, Лихарев, 1976 : 117, фиг. 51, 52.
Terra typica — Perigord (Перигё, юго-западная Франция, западная часть.
Центрального массива).
Место хранения типов не установлено.
Материал: 24 экз. из 8 местонахождений в Испании, Италии, Болгарии
и Австрии.
Раковина от овальной до короткоцилиндрической, слегка блестящая, уме-
ренно твердостенная. Оборотов?—8,выпуклых,медлецно нарастающих. Послед-
ний оборот, начиная с сере-
дины, плавно постепенно под-
нимается, а возле самого устья
поднимается так резко, что.
ангулярная область устья
располагается примерно на
уровне нижнего шва или
даже немного выше шва. Вы-
сота последнего оборота сос-
тавляет примерно 1/3 высоты
раковины. Окраска одноцвет-
но-роговая. Эмбриональные
обороты покрыты неясной зер-
нистостью, скульптура дефи-
нитивных оборотов в виде
тонких равномерно располо-
женных косых радиальных
Рис. 60. Pagodulina (Pagodulina^
pagodula (Des Moulins) из север-
ной Италии, Бус-дель-ке у гор.
Изео, 19 III 1977 г.
Половой аппарат (внутри вскрытого-
пениса видны тетрагональные кри-
сталлы); звездочка — бобовидная же-
леза на протоке семяприемника.
ребрышек, промежутки между которыми и 1Чг—3 раза шире самих реб-
рышек. Устье почти цельное, сильно сближенные места его прикрепления
соединены светлой мозолью. Ангулярная область слегка обособлена от стенки
раковины; благодаря наличию сглаженного зубовидного утолщения губы на
палатальном крае намечается синулус. Помимо палатального утолщения име-
ются вялая колумеллярная пластинка, видимая лишь при косом положении
раковины, и длинная продольная палатальная складка, лежащая в глубине
устья. Снаружи этой складке соответствует углубление в виде широкой ложбин-
ки на базально-боковой поверхности последнего оборота. Наконец, имеется
париетальная складка, начинающаяся на границе последнего и предпоследнего-
оборотов, которая лежит напротив палатальной и занимает примерно 1/3 длины
оборота. Элементы устьевой арматуры при нормальном положении раковины не
видны и их можно обнаружить только после удаления всех оборотов, кроме по-
следнего и части предпоследнего. Края устья широко отвернуты. Пупок запаян
за счет полного смыкания внутренних стенок столбика, но пупочная впадина
имеется.
Размеры: ВР 2.8—4, БД 1.8—2.5 мм; в первоописании: ВР 4, БД 2.5 мм.
Внутреннее строение. Материал: 2 экз. из северной Италии,,
центральные Альпы, Бус-дель-ке близ г. Изео, 19 марта 1977 г., сбор И. М. Ли-
харева, определение автора. Материал представлен формой subdola Gredler, 1856.
1. РОД PAGODULINA 131
Выемок и ложбин на поверхности белковой железы не обнаружено. Семя-
провод впадает в эпифаллус субтерминально; на продольном разрезе соответ-
ствующего участка эпифаллуса можно различить рудимент бича в виде щели,
расположенной в тканях проксимальной части эпифаллуса. Эпифаллус не-
сколько длиннее тонкого цилиндрического пениса; на границе между этими от-
делами располагается длинный конический червеобразно изогнутый или пря-
мой придаток эпифаллуса. Половой ретрактор крепится проксимальнее этого
придатка. Внутренние стенки пениса покрыты тонкими прямыми продольными
складками. В просвете пениса у обоих исследованных экземпляров обнаружены
прозрачные очень мелкие кристаллы тетрагональной формы, назначение и про-
исхождение которых неясно. Вероятно, что это не артефакт: во-первых, потому,
что такие же кристаллы обнаружены и у Другого вида этого рода, и, во-вторых,
потому, что ни у каких других видов ничего подобного никогда не отмечалось.
Проток семяприемника снабжен фасолевидной железой, назначение которой
скорее всего сводится к лизису сперматофора; не исключена также ее гаметоли-
тическая деятельность. Участок протока семяприемника выше железы заметно
короче протока резервуара. Дивертикул семяприемника длинный, извитой,
конец его лежит на базальной части белковой железы.
Распространение и местообитание. Западная и частично
средняя Европа и Балканский п-ов. Населяет лиственные леса, преимущест-
венно горные. Не исключено наличие вида в Закарпатье.
2. Подрод GRYSTALLIFERA Schileyko, 1976
Шилейко, 19766:57.
Типовой вид Pagodiria pagodula var. lederi Boettger, 1886; по перво-
начальному обозначению.
Придаток эпифаллуса отсутствует.
Монотипический подрод.
2. Pagodulina (Crystallifera) lederi (О. Boettger, 1886) (рис. 59, IIP, рис. 61).
О. Boettger, 1886b : 305, t. 3, fig. 8 (Pagodina pagodula var. lederi); Pilsbry,
1924, Man. Conch., 27 : 178, t. 20, figs. 11, 12 ; F о г с a r t, 1935 : 421 (Pagodulina pagodula
lederi); Лихарев, Раммельмейер, 1952 : 162, рис. 76; Шилейко, 19676 :
.25, рис. 2, 3; 19766 : 57, рис. 7.
Terra typica — Талыш.
Лектотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 3726 (Zilch, 1947, Arch. Moll., Bd 76 : 75, Abb. 5).
Материал. 18 экз. из 8 местонахождений в Талыше.
Раковина от короткоцилиндрической до овально-цилиндрической, сравни-
тельно твердостенная, с шелковистым блеском. Оборотов 8—8х/2, слабо выпук-
лых; последний оборот, начиная с последней трети, плавно, но энергично
поднимается, и ангулярная область устья лишь немного не доходит до шва. Окра-
ска одноцветная, роговая или светло-роговая. Эмбриональные обороты покрыты
очень неясной микроскопической зернистостью, дефинитивные несут многочислен-
ные тонкие острые равномерно расположенные ребрышки, промежутки между
которыми примерно равны ширине ребра или несколько уже. Устье овальное,
обращено немного вверх, края его умеренно отвернуты и слегка утолщены, места
прикрепления связаны четкой париетальной мозолью; если мозоль хорошо раз-
вита, устье выглядит почти цельным. Середина палатального края устья
имеет небольшое вдавление. Элементы устьевой арматуры при нормаль-
ном положении раковины не видны; если удалить часть стенки раковины на
последнем обороте, то можно заметить длинную невысокую продольную пала-
тальную складку, слабую колумеллярную и плохо различимую париетальную
пластинки. Палатальную складку у свежих раковин иногда можно обнаружить
9*
132
II. НАДСЕМ. ORCULOIDEA
в проходящем свете слева от устья, на базально-боковой стенке последнего
оборота. Пупок почти запаян: от него остается очень узкая изогнутая щель.
Размеры: ВР 3.2—3.8, БД 2—2.2 мм; в первоописании: ВР 3.7, БД 2 мм.
Внутреннее строение. Материал: 5 экз. из с. Бибионы, Талыш,
8 апреля 1965 г.; 3 — из с. Сейфидор, Талыш, 1 июля 1964 г.; 1 — из листвен-
ного леса в горах над Астрой, Талыш, 20 сентября 1966 г.; сборы и определения
автора.
На поверхности белковой железы никаких вырезок и желобков не наблюда-
ется. Семяпровод впадает в эпифаллус терминально; при вскрытии места перехода
Рис. 61. Pagodulina (Crystallifera') lederi (Bttg.) из Талыша, с. Бибионы, 8 IV 1965 г.
I — половой аппарат, II — квадривий с участком гермафродитного протока; звездочка — бобовидная же-
леза на протоке семяприемника (из: Шилейко, 19766).
семяпровода в эпифаллус можно обнаружить рудимент бича в виде тонкой щели,
слепой конец которой направлен в сторону семяпровода. Граница между пени-
сом и эпифаллусом не выражена и отмечена лишь местом прикрепления полового
ретрактора. Пенис состоит из двух отделов, внешне различающихся в разной
степени: проксимального, внутренние стенки которого покрыты очень тонкими
кольцевыми бороздками, и дистального, заключающего множество мелких
прозрачных тетрагональной формы кристаллов, химическая природа которых
не выяснена. Не исключено, что роль кристаллов заключается в раздражении
партнера при спаривании; во всяком случае участие кристаллов в процессе
совокупления доказывается тем, что они нередко обнаруживаются внутри семя-
приемника. Внутренние стенки дистального отдела покрыты тонкими глубокими
кольцевыми бороздками. Основание протока семяприемника в той или иной мере
расширено; несколько выше в колене протока располагается хорошо развитая
железа фасолевидной формы. Участок протока семяприемника, лежащий выше
железы, в 3—4 раза длиннее протока резервуара. Место раздвоения протока
1. РОД EUXINOLAURIA 13g"
семяприемника непостоянно и находится то выше, то ниже его середины..
Как проток, так и дивертикул семяприемника многократно изогнуты и часто
извиты. Дивертикул достигает белковой железы.
Распространение и местообитание. Хр. Эльбурс.,,
часть которого составляют Талышские горы. Вид населяет горные широколист-
венные леса, где держится в мощном слое опада и во влажной гниющей древе-
сине.
III. Подсем. LAURIINAE Steeuberg, 1925
Steeuberg, 1925 201; Pilsbry, 1926, Man. Conch., 27.
Скульптура дефинитивных оборотов — от редких вялых морщинок до- радик-
альной ребристости. Последний оборот к устью в той или иной мере поднят, но-
ангулярная область его далеко не доходит до нижнего шва. Устье с 2—7 зубами-,,
хорошо заметными при нормальном положении раковины; париетальная плат-
стинка всегда хорошо развита.
Придаток эпифаллуса разнообразной формы и строения. Проток семяприемз-
ника без морфологически выраженных желез.
Распространение. Средиземноморские страны, о. Мадейра^,
Канарские й Азорские о-ва, Европа, Крым, Кавказ.
В ископаемом состоянии подсемейство известно начиная с верхнего олиго-
цена Европы и о-ва Мадейра.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ LAURIINAE
1 (2). В устье не менее 4 зубов...............................1. Euxinolauria^
2 (1). В устье не более 3 зубов.....................................2. Lauria».
1. Род EUXINOLAURIA Lindholm, 1924
Lindholm, 1924:80; Thiele, 1931:509 (род Lauria, секция Euxinolauria)^
Wenz, Zilch, 1959—1960 : 171 [Leiostyla (Euxinolauria)]; Шилейко, 19756: 1768
Типовой вид Pupa (Charadrobid) pulchra Betowski, 1883; по первона-
чальному обозначению.
Раковина от овально-цилиндрической до овально-ширококонической, св'
слегка заостренной или умеренно сглаженной вершиной, от почти гладкой до-,
ребристой. Высота раковины от 2.5 до 6 мм при ширине 1.3—3 мм. Оборотов
6г/2—8, последний оборот к устью поднят. В наиболее полном наборе устьевая
арматура полностью сформированных раковин представлена 2 париетальными и
2 колумеллярными пластинками, 1 базальным зубом, 2 палатальными складками
и продольным утолщением палатального края, лежащим в плоскости устья..
Кроме того, край устья может нести серию бугорков. В случае ослабления усть-
евого аппарата могут исчезать колумеллярная пластинка (при сильном разви-
тии субколумеллярной), базальный зуб, одна из палатальных складок, утол-
щение палатального края и бугорки на устьевом крае.
Придаток эпифаллуса чаще всего конической или червеобразной формы,,
внутреннее его строение разнообразно. Яйцекладущие животные.
Распространение. Кавказ. 9 видов.
В ископаемом состоянии род известен начиная с миоцена (Европа).
Дифференциальный анатомический диагноз дать пока невозможно из-з®
крайне слабой изученности наиболее близкого по раковине рода Leiostyla,.
населяющего западное Средиземноморье, частично Среднюю Европу и о-в®
восточной Атлантики; наибольшего разнообразия род достигает на о. Ма-
дейра.
134
И. НАДСЕМ. ORCULOIDEA
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ EUXINOLAURIA
1 (4). Скульптура представлена отчетливыми сглаженными ребрышками (под-
род Euxinolauria s. str.).
2 (3). Париетальная пластинка сильно отогнута к палатальному краю устья;
палатальное утолщение продолжается в верхнюю палатальную складку
.................................................... 8. Е. (Е.) silicea.
3 (2). Париетальная пластинка вертикальная; палатальное утолщение огра-
ничено краем устья..................................7. Е. (Е.) pulchra.
4 (1). Скульптура в виде радиальной исчерченности или морщинистости, либо
раковийа почти гладкая.
5 (6). Париетальная и субпариетальная пластинки сближены одна с другой,
согнуты вдоль и лежат почти горизонтально (подрод Neolauria) ....
9. Е. (N.) zonifera.
6 (5). Париетальная и субпариетальная пластинки не сближены или умеренно
сближены, вертикально свешиваются в просвет устья.
7 (8). Синулус почти не выражен; большой диаметр раковины не превышает
1.5 мм (подрод Matschachelia) .................1. Е. (М.) rectidentata.
8 (7). Синулус выражен хорошо; если же слабо, то большой диаметр раковины
не менее 2 мм (подрод Caucasipupa).
9 (10). Палатальное утолщение слабое, поэтому синулус выражен нечетко
............................................ 2. Е. (С.) tenuimarginata.
10 (9). Палатальное утолщение хорошо развито, поэтому синулус вполне
отчетливый.
11 (12). Раковина удлиненно-овально-цилиндрическая......................
. ...........................................5. Е. (С.) caucasica.
12 (11). Раковина от овальной до удлиненно-овальной.
13 (14). Колумеллярная пластинка в глубине устья круто изгибается и под-
нимается почти вертикально, одновременно расширяясь; в результате
сквозь устье при нормальном положении раковины видна боковая поверх-
ность заднего участка пластинки, лежащая параллельно плоскости
устья ..............................................4. Е. (С.) paulinae.
14 (13). Задний конец колумеллярной пластинки не поднимается вертикально;
если же поднимается, то не образует расширения.
15 (16). Завиток куполовидный, палатальный край устья ниже утолщения
вдавлен, плоскость утолщения составляет острый угол с плоскостью
устья ....................................... 3. Е. (С.) superstructa.
16 (15). Завиток конический, со слегка выпуклыми сторонами, палатальный
край устья почти без вдавления, палатальное утолщение лежит в пло-
скости устья .......................................6. Е. (С.) sinangula.
1. Подрод MATSCHACHELIA Schileyko, 1975
Шилейко, 19756:1769.
Типовой вид Euxinolauria (Matschachelia) rectidentata Schileyko,
1975; по первоначальному обозначению.
Раковина удлиненно-овальная, тонко скульптурированная, хрупкая, ма-
ленькая. Париетальных и колумеллярных зубов по 2, базальный зуб 1, пала-
тальная складка 1. Париетальные пластинки не сближены, опускаются в про-
свет устья почти вертикально.
Семяприемник имеет короткий дивертикул. Эпифаллус при переходе в пенис
образует колено, в котором располагаются две продольные и одна поперечная
складки, благодаря чему форма просвета этого отдела очень сложная.
Монотипический подрод.
1. РОД EUXINOLAURIA
135'
1. Euxinolauria (Matschachelia) rectidentata Schileyko, 1975 (рис. 62, 63).
Шилейко, 19756 : 1769, рис. 1, 1—2, 6, 1.
Terra typica — долина p. Мачахели-цкали в Аджарии.
Г о л о т и и хранится в Зоологическом ин-те АН СССР.
Материал: 6 экз. из типового местонахождения.
Раковина удлиненно-овальная, тонкостенная, хрупкая, просвечивающая,
блестящая, со слегка выступающей вершиной. Оборотов 7, несильно выпуклых.
Окраска одноцветная, светло-роговая. На дефинитивных оборотах скульптура
на первый взгляд отсутствует, но при значительном увеличении заметны редкие
слабые неправильные радиальные морщинки, выраженные несколько яснее
у шва. Колумеллярный край устья отвесный, базальный
палатальный почти параллелен колумеллярному. Сину-
лус умеренно выражен. Все пластинки и складки прямые
или слабо изогнуты. Париетальная пластинка, связанная
с небольшим ангулярным утолщением, опускается почти
отвесно, параллельно субпариетальной; последняя начи-
нается глубже париетальной, но заканчиваются они
в глубине устья примерно на одном уровне. На правой
стороне париетальной пластинки может присутствовать
маленькое зубообразное вздутие. Колумеллярная пла-
стинка отсутствует или представлена небольшим бугорком.
Субколумеллярная пластинка развита хорошо, несколько
Рис. 62. Euxinolauria {Matschachelia} rectidentata Schileyko. Го-
лотип.
плавно закруглен,
отогнута вниз и видна со стороны пупка в виде тонкой светлой полоски. Базаль-
ный зуб небольшой, расположен заметно отступя от края устья. Палатальная
складка, начинаясь от края устья:, постепенно увеличивается в высоту, затем
резко обрывается и тонким шнуром тянется до половины последнего оборота;
в виде светлой полоски она видна снаружи. На палатальном крае устья име-
ется небольшое продольное вздутие, в этом месте наружный край устья имеет
очень слабое вдавление. Пупок открытый, цилиндрический, проколовидный.
Размеры: ВР 2.7—2.8, БД 1.3—1.5 мм; голотип: ВР 2.8, БД 1.5 мм.
Внутреннее строение. Материал: 2 экз. из долины р. Мача-
хели-цкали, Аджария, 23 июля 1973 г., сбор И. М. Лихарева, определение
автора.
Поверхность белковой железы без вырезок и ложбинок. Семяпровод терми-
нально впадает в плавно закругленный конец эпифаллуса, внутри которого
имеется ясно выраженная кольцевая щель. Пениальный аппендикс отходит
почти от полового атриума, очень длинный, сильно мускулизован, А2 очень
плавно переходит в длинный конический А3. Длина собственно пениса примерно
равна длине Аг. Полость пениса в проксимальной части посредством продоль-
ных складок разделяется на две продольные камеры. Здесь же имеется попереч-
ная складка, конец которой направлен в полость эпифаллуса. Вся эта конструк-
ция, будучи гомологом придатка эпифаллуса, представляет собой вместилище
аутоспермы, где она накапливается перед поступлением в пениальный ап-
пендикс. Эпифаллус примерно вдвое длиннее пениса, с узкой полостью, стенки
которой покрыты множеством кольцевых морщин. Одна ветвь полового ретрак-
тора крепится к А± сразу же под А2, вторая — к пенису чуть дистальнее опи-
санного выше расширения гомолога придатка эпифаллуса. Проток семяприем-
ника прямой, резервуар маленький, шаровидный, причем создается впечат-
ление, что продолжение протока составлено не резервуаром, а очень коротким
рудиментарным дивертикулом.
136
II. НАДСЕМ. ORCULOIDEA
. P ас пространение и местообитание. Вид известен только
из типового местонахождения, где был найден в очень влажных условиях в уз-
ком лесистом ущелье, выходящем к реке.
Рис. '63., Еuxinolauria (Matschachella) rectidentata Schileyko. Паратин.
3^—{половой аппарат, II — внутреннее строение пениса и пениального аппендикса (из: Шилейко, 19756).
2. Подрод GAUCASIPUPA Pilsbry, 1926
Caziot, Mergier, 1909 : 141 (Caucasica, типовой вид Pupa caucasica, non Cau-
easica- 0. Boettger, 1877); Pilsbry, 1926, Man. Conch., 27 : 260 (nom. nov.); Thiele,
1931:509 (род . Lauria, секция Caucasipupa);. Wenz, Zilch, 1959—1960:170 [Leio-
styla (Caucasipupa)]; Шилейко, 19756 : 1771.
Типовой вид Pupa caucasica L. Pfeiffer, 1857; по тавтонимии.
Раковина от удлиненно-овальной до овально-широко-конической, может
быть как хрупкой и тонкостенной, так и прочной, толстостенной; поверхность
ее блестящая или матовая. Скульптура почти отсутствует или представлена
слабыми радиальными морщинами. Париетальных зубов 2, колумеллярных 1—
-2, базальный зуб может отсутствовать, палатальная складка 1; кроме того, может
-присутствовать палатальное утолщение. Париетальные пластинки спускаются
«в просвет устья вертикально, лишь кромка их может отгибаться к палатальной
«стенке.
Семяприемник без дивертикула, проток его в нижней части в той или иной
«степени извит или даже скручен. Придаток эпифаллуса заключает внутри си-
стему четких бугорков и (или) продольную складку.
Распространение. Западная часть Большого Кавказа и Закав-
казья. Один вид встречается на южных склонах центральной и восточной части
^Главного Кавказского хр. 5 видов.
1. РОД EUXINOLAURIA
137
2. Euxinolauria (Caucasipupa) tenuimarginata (Pilsbry, 1922) (рис. 64, 65).
Pilsbry, 1922, Man. Conch., 27 : 76, t. 10, figs. 1—2 [Lauria (Leiostyla)]; Лиха-
рев, Раммельмейер, 1952 : 155, рис. 67 [L. (Leiostyla)]; Hudec, L ezhaw a,
1967 : 74, Abb. 2 [Z,. (Leiostyla)]; 1969 : 96, t. II, 5 [L. (Leiostyla)]; Шилейко, 49756 r
1771, рис. 1, 3—5 и 6, 2.
Terra typica — Нотанеби, в 40 км к северу от Батуми.
Голотип хранится в Филадельфийской Академии естественных наук,.
№ 130116, коллекция Беттгера.
Материал: около 100 экз. из Батумского ботанического сада.
Раковина овально-ширококоническая, тонкостенная, слабо или умеренно-
блестящая, с неширокой чуть приостренной вершиной. Завиток почти кониче-
ский, со слегка выпуклыми сторонами. Оборотов 7—ТЧ2, несильно выпуклых,,
плавно нарастающих. Последний оборот, начиная с последней трети, ииавнц. ж
постепенно поднимается, в результате чего ангулярная
область устья заметно придвинута к шву. Окраска от
зеленовато-роговой до светло-каштановой, над швом рас-
положена широкая спиральная светлая лента, кото-
рая на последнем обороте проходит по средней линии обо-
рота. Эмбриональные обороты очень тонко и неясно
зернистые, дефинитивные с неправильными тонкими
вялыми морщинками, усиливающимися на затылке. Устье
неправильно овальное, его вертикальная ось немного
отклонена вправо от оси раковины, причем устье слегка
Рис. 64. Euxinolauria (Caucasipupa) tenuimarginata (Pils.) из Ба-
тумского ботанического сада.
наклонено так, что плоскость его смотрит немного вверх. Края устья тон-
кие, широко отвернуты. Устьевая арматура сложная. Две тонкие париеталь-
ные пластинки, уходящие глубоко в просвет устья; субпариетальная короче и;
ниже париетальной. Между передним концом париетальной пластинки и ме-
стом прикрепления устья заметен небольшой ангулярный бугорок, развитый:
в разной степени (иногда отсутствует). Колумеллярная пластинка хорошо раз-
вита, слегка отклонена к базальной стенке. Базальный зуб маленький, пластин-
чатый, палатальная складка высокая и длинная, снаружи просвечивает в виде
светлой полоски, ее гребень направлен к гребню субпариетальной пластинки.
Кроме того, в верхней части палатального края имеется небольшое утолщение.
Париетальные и колумеллярные элементы растут вместе с раковиной, и их задние-
концы по мере роста раковины резорбируются. Базальные зубы формируются-
несколько раз в течение роста раковины, и более ранние зубы просвечивают
сквозь стенку, раковины. Пупок узкий, сквозной, цилиндрический.
Размеры: ВР 2.8—3.5, БД 2—2.2; в первоописании: ВР 2.8, БД 2.0 мм..
Внутреннее строение. Материал: 9 экз. из Батуми, Ботаниче-
ский сад, 7 апреля 1971 г., сбор и определение автора.
Приводимое описание представляет собой компиляцию из собственных на-
блюдений и материалов Гудеца и Лежавы (Hudec, Lezhawa, 1967, S. 74), по-
скольку все животные, бывшие в моем распоряжении, собраны в начале апреля
и их половой аппарат несет признаки ювенильности. К таким признакам отно-
сятся: относительно большая длина всех протоков сравнительно с их толщиной...
слабо развитая матка, не вполне сформированный придаток эпифаллуса и сла-
бая выраженность границы между семяпроводом и эпифаллусом. Единствен-
ный экземпляр, исследованный Гудецом и Лежавой, собран в середине авгу-
ста, и, судя по рисунку (Hudec, Lezhawa, 1967, S. 75, Abb. 2), гениталии его»
полностью сформированы.
.138
II. НАДСЕМ. ORCULOIDEA
Белковая железа без вырезки и без желобков на поверхности. Длина пени-
ального аппендикса превышает суммарную длину пениса и эпифаллуса, причем
с возрастом разница в длине между этими протоками увеличивается. А2 и А3
выражены слабо. Внутреннее строение придатка эпифаллуса взрослого экзем-
пляра неизвестно: у исследованных мною особей внутри него заметна высокая
продольная складка, направленная в полость эпифаллуса, который на конце
^слегка или сильно вздут. Кольцевая щель внутри этого вздутия выражена более
Рис. 65. Euxinolauria (Caucasipupa) tenuimarginata (Pils.) из Батумского ботанического сада,
7 IV 1971 г.
£ половой аппарат, II — внутреннее строение придатка эпифаллуса и проксимальной части эпифаллуса
(из: Шилейко, 19756).
или менее ясно. Ветви полового ретрактора к диафрагме подходят единым тя-
жем; .одна ветвь крепится на границе двух нижних отделов пениального аппен-
дикса, вторая — немного дистальнее придатка эпифаллуса. Проток семяприем-
ника у молодых особей прямой, у половозрелых образует легкие изгибы.
Распространение и местообитание. Вид обнаружен
в трех местах юго-западной Грузии: у станции Нотанеби в 40 км севернее Ба-
туми, у с. Чаквистави в 20 км восточнее Батуми и в Батумском ботаническом
саду. Населяет толстый слой лиственного опада; в Ботаническом саду массами
поднимается на стебли бамбука на высоту до 2.5 м.
3. Euxinolauria (Caucasipupa) superstructa (Mousson, 1876) (рис. 66, 67).
Mousson, 1876 : 37, pl. 2, fig. 1 [Pupa (Pupilla)]; 0. Boettger, 1879 : 404
(Charadrobia)]", 1881 : 229 [P. (Charadrobia)]; 1883 : 180 [P. (Charadrobia) superstructa
et var. lederi]; 1886a : 149, T. 1, Fig. 5 [P. (Charadrobia) superstructa var. unibasalis]; R e-
towski, 1914:310 [P. (Lauria)]; Rosen, 1914:189 [P. (Lauria)]; Pilsbry,
1922, Man. Conch., 27 : 77, t. 9, figs. 1—4 (Lauria (Leiostyla)]-, Лихарев, Раммель-
1. РОД EUXINOLAURIA
139
мейер, 1952 : 155, рис. 68 [L. (Leiostyla)]-, Hude с, Lezhawa, 1969 : 95, Abb. 1,
2, t. II, 4, t. VI, 15 [L. (Leiostyla) superstructa et f. lederi]-, Шилейко, 19756 : 1772, рис. 2,
1—6 и 6, 3.
Terra typica — Лайлаш, Сванетский хр.
Место хранения типов не установлено.
Материал: около 30 экз. из 6 местонахождений на западном Кавказе.
Раковина удлиненно-овальная, слабо блестящая, умеренно твердостенная,
с узкой чуть прпостренной вершиной. Завиток куполовидный. Оборотов 7—8,
слабовыпуклых. Последний оборот, начиная с последней трети, очень медленно и
плавно поднимается. Окраска роговая, под швом располагается коричневая лента,
почти неразличимая на верхних оборотах; на последнем обороте эта лента при-
ходится на периферию. Эмбриональные обороты неясно зернистые, дефинитив-
ные покрыты вялыми неправильно расположенными ра-
диальными сглаженными морщинками.. Устье неправильно
овальное, обращено немного вверх; края его отвернуты,
колумеллярный и палатальный несут вмятины. Париеталь-
ная пластинка высокая, тонкая, изогнутая: кромка ее
отклонена к палатальной стенке; субпариетальная ниже и
короче, отвесная, начинается она глубже париетальной.
Вплотную к париетальной, а иногда и на ее базальной
части, расположен маленький заостренный ангулярный
бугорок (может отсутствовать). Супраколумеллярная
Рис. 66. Euxinolauria (Caucasipupa) superstructa (Mouss.) из Ба-
тумского ботанического сада.
пластинка в виде маленького бугорка, колумеллярная почти горизонтальная,
длинная; базально-колумеллярная короткая, но сравнительно высокая. Пала-
тальная складка высокая, неровная, ее гребень направлен к гребню субпарие-
тальной пластинки. Базальный край устья в разной степени смят в короткие
морщинки. На палатальном крае видна мощная гребневидная складка, лежа-
щая почти в плоскости устья. Благодаря тому что ее гребень близко подходит
к кромке париетальной пластинки, формируется ясно выраженный синулус.
Пупок узкий, сквозной, проколовидный.
Размеры: ВР 4.5—5.3, БД 2.4—2.5 мм; в первоописании: ВР 5, БД 2.5 мм.
Внутреннее строение. Материал: 4 экз. из Батуми, Ботаниче-
ский сад, 7 апреля 1971 г., сбор и определение автора.
Поверхность белковой железы без вырезок. Семяпровод впадает терминально,
с плавным расширением; в толще стенок эпифаллуса в месте впадения семяпро-
вода залегают 1—2 узкие глубокие полукольцевые бороздки. Эпифаллус длин-
нее пениса, терминальное расширение его выражено слабее, чем у других видов
рода. Внутренние стенки эпифаллуса более или менее гладкие, проток узкий.
Внутренняя поверхность пениса бугорчатая. Строение пениального аппендикса
своеобразно: Аг и А2 относительно очень короткие, А3, напротив, чрезвычайно
длинный. Пенис немного короче суммарной длины Аг, А2 и А3. Придаток эпи-
фаллуса длинный, конический, изогнутый, внутри покрыт неясными бугорками,
расположение которых таково, что можно предполагать возникновение неко-
торых из них за счет распадения продольной,складки. Одна из ветвей полового
ретрактора крепится к границе пениса и эпифаллуса, которая наружно почти
не выражена, а вторая ветвь — к середине А3. Проток семяприемника при осно-
вании вздут, вслед за вздутием имеется резкий изгиб; резервуар слабо обособ-
лен от протока.
Распространение и местообитание. Западный Кавказ:
Сванетский и Сурамский хр., окр. Кутаиси, Сухуми, Нотанеби, Бакуриани,
Цагери, Боржоми, Аджарисцкали, Батумский ботанический сад. В последнем
пункте обнаружен в сыром понижении среди зарослей бамбука.
II. НАДСЕМ. ORCULOIDEA
4. Euxinolauria (Caucasipupa) paulinae (Lindholm, 1913) (рис. 68, 69).
liodholm, 1913:62 (Lauria); 1914:36 (Л. paulinae subsp. unicolumellaris); Ro-
se n, 1914 : 231 [Pupa (Dauria)]; Pilsbry, 1922, Man. Conch., 27 : 75 [Lauria (Leio-
Лихарев, Раммельмейер, 1952 : 154, рис. 66 [Lauria (Leiostyla)]; HI и-
Л«йко, 19756 : 1773, рис. 2, 3—4 и 6, 4.
Terra typtca — Нотанеби, в 40 км к северу от Батуми.
Лектотип иЗ паралектотипа хранятся в Зоологическом ин-те АН СССР,
эбозначаются впервые.
М а т е р и ал: 7 экз. из трех местонахождений в Аджарии и Турции.
Э?ис. 67. Euxinolauria (Caucasipupa.) superstructa (Mouss.) из Батумского ботанического сада,
7 IV 1971 г.
7— половой аппарат, II—детали строения мужского отдела (из: Шилейко, 19756).
Жаковина удлиненно-овальная, тонкостенная, просвечивающая, завиток
атонический, со слегка выпуклыми сторонами, вершина узкая, закругленная.
'Оборотов 7—7х/2, умеренно или слабо выпуклых, плавно нарастающих. Послед-
ний оборот к устью медленно и плавно поднимается. Окраска одноцветная,
окрасновато-бурая. Эмбриональные обороты покрыты очень неясной зернисто-
стью, дефинитивные тонко и неравномерно радиально исчерчены. Устье усеченно-
чэвальное, почти отвесное, края его слегка утолщены и отвернуты. Парие-
тальная пластинка высокая, отвесная, нижняя кромка ее слегка отклоняется
к палатальной стенке; субпариетальная ниже и короче париетальной. Колу-
-меллярная пластинка вначале невысокая, горизонтальная; с правой стороны
столбика в глубине устья она образует широкий изгиб — вначале вниз, с перело-
жим, затем вверх, и вертикальная часть расширяется в виде лопасти и утолща-
«зтся по гребню. Супраколумеллярная пластинка небольшая или отсутствует
>(f, unicolumellaris Lindholm). Базальная складка короткая и тонкая. Палаталь-
ная складка длинная, неровная, ее гребень обращен к гребню париетальной пла-
‘Феинки. Хорошо выраженный синулус образован за счет развития бугорко-
1. РОД EUXINOLAURIA
141
видного утолщения палатального края, близко подходящего к гребню париеталь-
ной пластинки. Пупок каплевидный, маленький.
Размеры: ВР 3.2—3.5, БД 1.75—1.9 мм; лектотип:
Внутреннее строение. Материал: 2 экз.
из долины р. Мачахели-цкали, Аджария, 23июля 1973 г.,
сбор И. М. Лихарева, определение автора.
Поверхность белковой железы лишена вырезок. Се-
мяпровод впадает терминально в хорошо развитое
плавное расширение эпифаллуса. Внутри этого расши-
рения содержится глубокая и узкая кольцевая щель.
Эпифаллус немного длиннее пениса, внутренняя его
поверхность несет множество высоких бугорков; подоб-
ная бугорчатость наблюдается и внутри пениса, но там
она выражена гораздо слабее. Пениальный аппендикс
очень длинный. 24X+A2 заметно длиннее пениса, А3
Рис. 68. Euxinolauria (Caucasipupa') paulinae (Lindh.) из долины
р. Мачахели-цкали, Аджария.
ВР 3.2, ВД 1.9 мм.
слабо выражен. У обоих изученных экземпляров Аа вдается в хорошо
выраженной папиллой (рис. 69, II), но у тех представителей подсе-
мейства, по которым изучен большой материал, эта папилла обычно выражена
I
Рис. 69. Euxinolauria (Caucasipupa) paulinae (Lindh.) из долины р. Мачахели-цкали, Аджа-
рия, 23 VII 1973 г.
I—половой аппарат, II— детали строения мужского отдела (из: ШиЛейко, 19756).
в разной степени, поэтому и здесь наличие столь сильно развитой папиллы может
не иметь постоянного таксономического значения. Придаток эпифаллуса мас-
сивный, конический, почти прямой, внутри заключает хорошо развитую про-
дольную перегородку, разделяющую полость придатка на две камеры. Проток
142
II. НАДСЕМ. ORCULOIDEA
семяприемника в общем конический: базальная его часть значительно шире ди-
стальной, заканчивающейся маленьким шаровидным резервуаром.
Распространение и местообитание. Обитает в прибреж-
ных районах Аджарии, в радиусе 40—50 км от Батуми. Населяет сырые листвен-
ные леса. Форма unicolumellaris описана из окр. Артвина (Турция, вилайет
Чорох).
5. Euxinolauria (Caucasipupa) caucasica (L. Pfeiffer, 1857) (рис. 70, 71).
L. Pfeiffer, 1857 : 88 (Pupa); 0. Boettger, 1879 : 30 [Pupa (Reinhardtia)];
Rosen, 1911 : 122 [P. (Charadrobia)]; 1914 : 190 [P. (Lauria)]; Pilsbry, 1922, Man.
Conch., 27 : 74, t. 5, figs. 15—17 [Lauria (Leiostyla)]; Лихарев, Рамме льмейер,
1952 : 153, рис. 65 [Lauria (Leiostyla)]; Ha de c, Lezhawa, 1967 : 73, Abb. 1, t. I,
1 [Lauria (Leiostyla)]; Шилейко, 19756 : 1773, рис. 3, 1—2 и 6,- 5.
Terra typica — Казбек.
Место хранения типов не установлено.
Материал: более 100 экз. из пяти местонахождений на южных склонах
Большого Кавказа.
Раковина удлиненно-овально-цилиндрическая, сравнительно твердостенная,,
свежие раковины просвечивают и слегка блестят. Вершина узкая, закруглен-
ная. Оборотов 7—8х/2, слабовыпуклых, последний обо-
рот приподнят только возле самого устья. Окраска ко-
ричневатых тонов, одноцветная. Эмбриональные обороты'
покрыты тончайшей зернистостью, дефинитивные — тон-
кими редкими вялыми сглаженными морщинками; в общем:
поверхность выглядит почти гладкой. Устье неправильно
овальное, отвесное, края его широко отвернуты и слегка
утолщены. Париетальная пластинка высокая, ее гребень,
отклонен к палатальному краю; субпариетальная короче
и ниже париетальной. Приостренный ангулярный бугорок
расположен при основании переднего края париетальной
пластинки или непосредственно на ее боковой поверх-
ности. Колумеллярная пластинка вначале горизонталь-
ная, в глубине устья ее гребень отклоняется книзу
Рис. 70. Euxinolauria (Caucasipupa) caucasica (L. Pfr.) из Закаталь-
. ского заповедника, хр. Печегель.
и затем вертикально поднимается по столбику. Супраколумеллярная пла-
стинка слабая, короткая, иногда едва заметна. Базальная складка корот-
кая, лежит отступя от устьевого края и может быть очень слабо выраженной.
Палатальная складка длинная, высокая, неровная, над ней может залегать еще
одна короткая маленькая (верхняя палатальная) складка. Гребень палатальной
складки обращен к гребню субпариетальной пластинки. На палатальном крае,
немного выше уровня, на котором расположен гребень париетальной пластинки,,
имеется зубовидное утолщение, замыкающее в совокупности с ангуляр-
ным бугорком и париетальной пластинкой круглый синулус. Пупок сквозной,,
узкий, цилиндрический.
Размеры: ВР 5—6.5, БД 2.5—3 мм; в первоописании: ВР 6, БД 3 мм.
Внутреннее строение. Материал: 3 экз. из Закатальского запо-
ведника, хр. Печегель, 20 мая 1953 г,, сбор и определение И. М. Лихарева;.
2 — из окр. Цагери, Грузия, 12 августа 1957 г., сбор Н. Н. Акрамовского,
определение автора.
На поверхности белковой железы вырезок и ложбин нет. Семяпровод впадает-
в эпифаллус с резким расширением, внутри которого имеется несколько коль-
цевых и полукольцевых бороздок. На поверхности этого вздутого отдела эпи-
1. РОД EUXINOLAURIA
143
фаллуса выделяется рисунок в виде темного удлиненного пятна, окруженного
еще более темной каймой. Эпифаллус заметно длиннее пениса, почти прямой.
Проток внутри эпифаллуса узкий, в дистальной части его имеется несколько
коротких продольных складок. Пенис неопределенной формы, изнутри покрыт
неясными складками и впадает в массивный придаток эпифаллуса, внутренняя
поверхность которого несет очень четкие ряды крупных конических бугорков.
Пениальный аппендикс длинный, впадает в пенис заметно отступя от полового
Рис. 71. Euxinolauria (Caucasipupa) caucasica (L. Pfr.) изЗакатальского заповедника, хр. Пе-
чегель, 20 V 1953 г.
I — половой аппарат, II — строение придатка эпифаллуса и проксимального участка эпифаллуса
(из Шилейко, 19756).
атриума. Аг вздутый, недлинный, не длиннее собственно пениса. А2 неясно от-
делен от Аг, заключает хорошо развитую папиллу. А3 выражен неясно. Одна из
ветвей полового ретрактора присоединяется к границе между Аг и А2, вто-
рая — к дистальной части эпифаллуса возле основания придатка эпифаллуса.
Основание протока семяприемника расширено и образует плавный изгиб. Проток
тонкий, плавно переходит в удлиненно-булавовидный резервуар.
Распространение и местообитание. Вид встречается
в широколиственных лесах центральных и восточных участков южных склонов
Большого Кавказа.
6. Euxinolauria (Caucasipupa) sinangula Schileyko, 1975 (рис. 72, 73).
Шилейко, 19756 : 1775, рис. 3, 3—4 и 6, 7.
Terra typica — Батумский ботанический сад.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 7 экз. из типового местонахождения.
144
II. НАДСЕМ. ORCULOIDEA
Раковина овальная, с уклоном в сторону овально-ширококонической,
умеренно твердостенная, слабо просвечивающая. Вершина слегка заостренная.
Оборотов 8, слабовыпуклых, медленно и плавно на-
растающих. Окраска свежих раковин роговая, вокруг
пупка имеется слабое светлое поле; один из экземпля-
ров типовой серии имеет бесцветную раковину. После
гибели животного раковина приобретает желтоватый
оттенок. Эмбриональные обороты неясно мелкозерни-
стые, скульптура дефинитивных оборотов — неравно-
мерные тонкие радиальные морщинки. Устье в общем
округлое, синулус ясно выражен. Париетальная пла-
стинка тонкая. На правой поверхности этой пластинки
Рис. 72. Euxinolauria (Caucasipupa) sinangula Schileyko. Голо-
тип.
Рис. 73. Euxinolauria (Caucasipupa) sinangula Schileyko из Батумского ботанического сада,
7 IV 1971 г. Паратип.
I — половой аппарат, II — строение мужского отдела (из: Шилейко, 19756).
имеется небольшой, но ясный бугорок. Субпариетальная пластинка парал-
лельна париетальной, но короче ее и меньшей высоты. Колумеллярная
пластинка короткая, субколумеллярная гораздо длиннее, заметно отогнута
к базальной стенке; задний ее конец слегка поднимается по столбику.
Базальный зуб отсутствует. Палатальная складка уходит в глубь раковины
немного меньше, чем на половину оборота, снаружи ей соответствует продольное
1. РОД EUXINOLAURIA
145
углубление. Края устья утолщены и широко отвернуты, на палатальном
крае имеется мощное продольное утолщение. Пространство между наруж-
ными концами палатальной складки и субколумеллярной пластинки несет
4—6 бугорков. Базальная поверхность раковины с тупым килем. Пупок
проколовидный.
Размеры: ВР 4.3—4.5, БД 2.5—2.8 мм; голотип: ВР 4.5, БД 2.8 мм.
Внутреннее строение. Материал: 3 экз. из типового место-
нахождения (Батумский ботанический сад), 7 апреля 1971 г., сбор и определение
автора.
Белковая железа без вырезок и ложбинок. Семяпровод впадает в эпифаллус
строго апикально, причем не постепенно, а с резким утолщением, поэтому гра-
ница между этими протоками хорошо заметна. Внутри утолщения в стенках
эпифаллуса залегает серия узких глубоких кольцевых бороздок. Эпифаллус
немного длиннее пениса, дистальная часть эпифаллуса связана с проксималь-
ной частью пениса несколькими тонкими мышечными лентами. Внутренняя
поверхность эпифаллуса покрыта множеством небольших, но ясных бугорков^
внутри пениса эти бугорки также имеются, но выражены гораздо менее ясно.
примерно одной длины с участком пениса, расположенным проксимальнее
основания пениального аппендикса. А 2 очень небольшой,Л3 тонкий, но явственно
обособлен от Л4. Придаток эпифаллуса в виде короткого округлого конуса,
внутренняя его поверхность несет комплекс очень четких конических бугор-
ков. Одна ветвь полового ретрактора крепится к Аг сразу же под А2, вторая
немного дистальнее придатка эпифаллуса. Проток семяприемника в нижней
части извит или даже скручен, удлиненный резервуар неясно отграничен от
протока.
Распространение и местообитание. Вид обнаружен
в Батумском ботаническом саду, в зарослях бамбука — в подстилке и на стеб-
лях.
3. Подрод EUXINOLAURIA s. str.
Раковина овальная или удлиненно-овальная, ребристая. Париетальных
зубов 2, колумеллярных 2, базальный зуб слабый или отсутствует, палаталь-
ные складки мощные. Париетальные пластинки не сближены, опускаются
в просвет устья почти вертикально или заметно отклонены к палатальной стенке.
Семяприемник без дивертикула. На границе пениса и эпифаллуса распо-
лагается тонкий цилиндрический придаток эпифаллуса, скрученный в клубок
и обтянутый тонкой соединительно-тканной пленкой.
Распространение. Западный Кавказ. 2 вида.
7. Euxinolauria (Euxinolauria) pulchra (Retowski, 1883) (рис. 74, 75).
R e t о w s k i, 1883b : 57 [Pupa (Charadrobia)]; 1887 : 35, T. 1, Fig. 1 [P. (Charadrobia)
pulchra f. bilabiata]; 0. Boettger, 1888 : 149 (Lauria pulchra var. nitens); Retowski,
1888 : 287 [Pupa (Charadrobia)]', 1889 : 253 [P. (Lauria) pontica]; Rosen, 1911 : 122
[P. (Charadrobia) pulchra et var. nitens]; Retowski, 1914 : 310 [P. (Lauria)]; Rosen,
1914 : 189 [P. (Lauria)]; Pilsbry, 1922, Man. Conch., 27 : 31, t. .9, figs. 11, 13, 14 [Lauria
(Leiostyla)]; Лихарев, Раммель мейер, 1952:156, рис. 69 [L. (Leiostyla)], 157
(var. nitens, var. pontica); Шилейко, 19756 : 1777, рис. 4, 1—2 и 6, 8.
Terra typica — морские выбросы в Крыму, возле Судака.
Место хранения типов не установлено.
Материал: 27 экз. из пяти местонахождений на западном Кавказе.
Раковина удлиненно-овальная, твердостенная, слабо просвечивающая,
с узкой закругленной вершиной. Завиток в виде высокого купола. Оборотов
7—8, уплощенных, последний оборот очень плавно, но явственно приподнят
к устью. Окраска одноцветная, роговая разных оттенков — от очень светлых
до почти каштановых. Эмбриональные обороты покрыты очень неясной зерни-
стостью, дефинитивные равномерно покрыты тупыми сглаженными ребрышками;
10 А- А. Шилейко
146
II. НАДСЕМ. ORCULOIDEA
на последнем обороте ребрышки ослаблены и теряют правильность расположе-
ния. Устье неправильно овальное, с утолщенными ишироко отвернутыми краями.
Париетальная пластинка длинная, высокая, изогну-
тая, слитая с ангулярным бугорком, поэтому при рас-
сматривании раковины в нормальном положении она
выглядит раздвоенной. Нижняя кромка париетальной
пластинки направлена вниз, к палатальной стенке не
отклонена. Субпариетальная пластинка ниже и ко-
роче париетальной. Очень мощная колумеллярная
пластинка вначале почти горизонтальная, лишь слегка
отклонена вниз, затем плавно изгибается и уходит
вверх по столбику. Супраколумеллярная короткая,
но ясно выраженная. Базальная складка небольшая,
лежит в глубине устья, иногда почти отсутствует.
Нижняя часть колумеллярного и базальный края не-
Рис. 74. Euxinolauria (Euxinolauria) pulchra (Ret.) из долины
р. У бин на северо-западном Кавказе.
сут несколько небольших складочек. Палатальный край устья несет мощное
гребневидное утолщение, лежащее в плоскости устья. Пупок открытый, очень
узкий, округлый.
Рис. 75. Euxinolauria (Euxinolauria) pulchra (Ret.) из долины р. Убин на северо-западном
Кавказе, 17 IX 1970 г.
I —'половой аппарат, II — строение пениса и придатка эпифаллуса (из:. Шилейко, 19756).
Размеры: ВР 3.4—4.5, БД 2—2.2 мм; в первоописании: ВР 4, БД 2 мм.
Внутреннее строение. Материал: 2 экз. из Краснодарского
края, берег р. Убин, 17 сентября 1970 г., сбор и определение автора.
1. РОД EUXINOLAURIA
147
Поверхность белковой железы гладкая. Семяпровод впадает в сильно рас-
ширенный терминальный участок эпифаллуса, на внутренних стенках кото-
рого легко видеть множество глубоких кольцевых бороздок. Эпифаллус корот-
кий, толстый, пенис несколько длиннее эпифаллуса, внутренняя поверхность
его бугорчатая, в дистальной части имеются продольные складки. Пениальный
аппендикс характеризуется чрезвычайно сильным развитием. At очень длин-
ный, почти равен суммарной длине пениса и эпифаллуса, Л2 плохо обособлен,
А3 почти не выражен, относительно очень короткий, Л5 сильно расширен.
Придаток эпифаллуса в виде тонкого цилиндрического отростка становится
заметным только после удаления окутывающего его тонкого прозрачного чехла;
в интактном же состоянии придаток эпифаллуса выглядит компактным бугор-
чатым наростом на границе между пенисом и эпифаллусом. Пениальная ветвь
полового ретрактора крепится к пенису дистальнее придатка эпифаллуса,
вторая ветвь — к границе между и А2. Проток семяприемника прямой или
слабо извит, удлиненно булавовидный резервуар неясно отграничен от про-
тока.
Распространение и местообитание. Юго-западная и
западная часть Большого Кавказа и западная часть Малого Кавказа. Насе-
ляет подстилку широколиственных лесов, а также влажные склоны с густым
травостоем. Форма pontica описана из Самсуна (Малая Азия).
8. Euxinolauria (Euxinolauria) silicea Schileyko, 1975 (рис. 76).
Hudec, Lezhawa, 1969: 96, Abb. 3, t. VI, 14 [Lauria (Leiostyla) pulchra]; ITT n-
л e й к o, 19756 : 1777, рис. 6, 8.
Terra typica — долина p. Техури в юго-западной части Большого Кавказа
выше с. Талери.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 1 экз. из типового местонахождения (голотип).
Раковина умеренно тонкостенная, прочная, удлиненно-овальная, слегка
блестящая. Оборотов 8, умеренно выпуклых. Скульптура эмбриональных обо-
ротов в виде тончайшей неясной пунктировки; кроме того, слегка поворачивая
раковину в пучке сильного света, при увеличении свыше 40 раз можно рас-
смотреть очень слабые элементы спиральной скульптуры. Дефинитивные обо-
роты покрыты равномерно расположенными сглаженными четкими радиальными
ребрйшками, сильно ослабленными на последнем обороте. Окраска одноцвет-
ная, темно-роговая. Устье уховидное, с очень четко обособленным синулусом,
отграниченным от остального просвета устья париетальной пластинкой и верх-
ней палатальной складкой. Париетальная пластинка необыкновенно высокая;
изгибаясь плавным полукругом и отгибаясь нижней кромкой к палатальной
стенке, она уходит в глубь последнего оборота. На наружной боковой поверх-
ности париетальной пластинки виден небольшой приостренный бугорок.
При нормальном положении раковины задняя часть париетальной пластинки
хорошо видна. На фоне этой задней части хорошо заметна субпариетальная
пластинка значительно меньшего размера, идущая параллельно париетальной.
Колумеллярная пластинка мощная, горизонтальная; над ней имеется очень
маленькая супраколумеллярная, не уходящая в глубь раковины. Базальный
зуб отсутствует, но на базальном крае устья имеется небольшой бугорок.
Нижняя палатальная складка хорошо развита, уходит в глубь раковины почти
на целый оборот (видно при просветлении раковины ксилолом). Верхняя
палатальная складка начинается от палатального вздутия устьевого края,
затем уходит в глубь устья, причем задний конец этой складки виден при нор-
мальном положении раковины. Переднему концу верхней палатальной складки
соответствует вмятина палатальной стенки. Базальная поверхность раковины
с тупым килем. Пупок открытый, проколовидный, располагается на дне пупоч-
ной впадины несколько эксцентрично.
10*
148
II. НАДСЕК. ORCULOIDEA
Размеры голотипа: BP 3.7, БД 2 мм.
Гудец и Лежава (Hudec, Lezhawa, 1969) приводят фотографию раковины и
описание внешнего вида полового аппарата Lauria pulchra; однако внешний
•облик раковины Euxinolauria silicea настолько отличен от Е. pulchra, что не
приходится сомневаться, что изученный названными авторами материал отно-
сится к Е. silicea. Надо добавить, что материал Гудеца и Лежавы происходит
из окр. с. Балда Гегечкорского р-на западной Грузии;
найденный мною экземпляр (пустая раковина пре-
красной сохранности) обнаружен неподалеку от этого
села. Следовательно, приводимое в указанной статье
описание деталей анатомии Lauria pulchra относится
к Euxinolauria silicea.
Внутреннее строение. Материал мною
не исследован. Судя по данным Гудеца и Лежавы,
основные отличия от предыдущего вида сводятся к сле-
дующему. Пенис и пениальный аппендикс разделены не
у полового атриума, а отступя от него; эпифаллус зна-
Рис. 76. Euxinolauria (Euxinolauria) silicea Schileyko. Голотип.
чительно длиннее пениса; на границе между этими отделами имеется ясное
вздутие, но природа его не выяснена; вероятно, это стиснутый в комок черве-
образный придаток эпифаллуса.
Распространение и местообитание. Обе находки вида
сделаны в долине р. Техури; раковина, найденная близ с. Талери (голотип),
обнаружена на влажном склоне под камнем вместе с Pilorcula trifilaris.
А. Подрод NEOLAURIA Schileyko, 1975
Шилейко, 19756 : 1778.
Типовой вид Lauria (Leiostyla) zonifera Pilsbry, 1934; по первоначаль-
ному обозначению.
Раковина овальная, небольшая, тонко скульптурированная, хрупкая.
Париетальная и субпариетальная пластинки сближены, отогнуты к палаталь-
ному краю. Остальные устьевые зубы развиты относительно слабо, базальный
зуб может отсутствовать.
Семяприемник без дивертикула, проток его почти прямой. На границе
пениса и эпифаллуса имеется сложный комплекс, состоящий из обширного
тонкостенного мешка, несущего на внутренних стенках бугорки, и тонкого
цилиндрического придатка эпифаллуса.
Монотипический подрод.
9. Euxinolauria (Neolauria) zonifera (Pilsbry, 1934) (рис. 77, 78).
О. Boettger, 1883 : 182, pl. 7, fig. 2 [Pupa (Charadrobia) superstructa var. zonata
non Pupa zonata Gassies, 1869]; 1889a : 22 (Lauria zonata et mut. albina); Rosen, 1911 :
122 (Pupa zonata)', 1914 : 190 (P. zonata)', Pilsbry, 1922, Man. Conch. 27 : 79,-pl. 9, figs. 8—
10 [Lauria (Leiostyla) zonata}', 1934, Man. Conch., 28 : 139 (L. zonifera, нот. nov. pro Pupa.su-
perstructa var. zonata Boettger); Лихарев, P а мме льмейер, 1952 : 155, рис. 68
[L. (Leiostyla) superstructa var. zonata}; Hudec, Lezhawa, 1969 : 95 [L. (Leiostyla)
superstructa f. zonata}; Шилейко, 19756 : 1778, рис. 4, 3—4 и 6, 10.
Terra typica — Цебельда, Военно-Сухумская дорога, Абхазия.
Место хранения типов не установлено.
Материал: более 100 экз. из 10 местонахождений на западном Кав-
казе.
1. РОД EUXINOLAURIA
149
Раковина овальная, тонкостенная, слегка просвечивающая, умеренно
блестящая, вершина узко закругленная, завиток куполовидный. Оборотов
6—7, слабовыпуклых, последний оборот плавно, но
довольно энергично поднят к устью. Окраска от светло-
роговой до каштановой, над швом проходит более
темная, чем фон, лента; на последнем обороте эта
лента занимает его середину. Эмбриональные обороты
с неясной зернистостью, дефинитивные несут очень
вялые сглаженные морщинки. Устье неправильно оваль-
ное. Париетальная пластинка высокая, неровная, на-
правлена почти горизонтально к палатальной стенке. Суб-
париетальная пластинка гораздо ниже и короче, тесно
Рис. 77. Euxinolauria {Neolauria} zonifera (Pils.) из Кавказского
заповедника, Краснодарский край.
придвинута к париетальной. Колумеллярная пластинка вначале почти гори-
зонтальная; в глубине устья, изогнувшись, она уходит вверх по столбику, при-
чем после изгиба ее высота уменьшается. Супраколумеллярная пластинка неболь-
Рис. 78. Euxinolauria {Neolauria) zonifera (Pils.) из Кавказского заповедника, близ Гузе-
рипля, 6 VIII 1956 г.
I — половой аппарат,
II — строение пениса и
эпифалпуса (из: Шилейко, 19756).
шая, почти вплотную подходит к париетальной стенке. Базальная складка корот-
кая или отсутствует. Палатальная складка развита в различной степени, гре-
бень ее направлен к гребню париетальной пластинки или отклонен к колумел-
лярному краю. Пупок очень узкий, щелевидный или каплевидный.
Размеры: ВР 3—4, БД 1.8—2 мм; в первоописании: ВР 4, БД 2 мм.
150
II. НАДСЕМ. ORCULOIDEA
Внутреннее строение. Материал: 2 экз. из Кавказского за-
поведника, близ Гузерипля, 6 августа 1956 г., сбор и определение И. М. Ли-
харева.
Белковая железа без вырезок и ложбинок. Семяпровод впадает терминально.
Эпифаллус в дистальной части сужен, затем расширяется и имеет в целом була-
вовидную форму. Внутренние стенки эпифаллуса покрыты правильно распо-
ложенными узкими кольцевыми бороздками, последняя (проксимальная) и»
этих бороздок расширена и образует кольцевую щель. Внутренние стенки
пениса покрыты вялыми продольными складками. В проксимальной части
пенис образует резкий изгиб и расширяется в объемистый тонкостенный ме-
шок, внутренние стенки которого покрыты четкими коническими бугорками,
которые видны даже без вскрытия мешка, просвечивая сквозь его покровы.
Рядом с местом впадения эпифаллуса в этот мешок имеется тонкий червеобраз-
ный придаток эпифаллуса с очень узким протоком. Пениальный аппендикс от-
ходит от атриальной области пениса; Ат немного длиннее пениса, А2 небольшой,
но ясно обособленный, А3 также хорошо развит, удлиненно-булавовидный.
Внутри А2 заметна папилла. Пениальная ветвь полового ретрактора крепится
к пенису немного дистальнее мешка. Другая ветвь ретрактора подходит к At
немного отступя от А2. Проток семяприемника в нижней части утолщен, по-
степенно сужается к резервуару, почти прямой; проток переходит в резервуар
очень плйвно.
Распространение и местообитание. Западная часть
Большого Кавказа и Аджария; населяет подстилку широколиственных лесов,,
встречается также в малоподвижных осыпях, заросших кустарником.
2. Род LAURIA Gray, 1840
G ray in : Turton, 1840 : 193; Lowe, 1852 : 275 (Gastrodon, non Rafinesque, 1815);
0. Boettger, 1879 : 29, 403 (Reinhardtia, типовой вид Turbo cylindracea Da Costa, обозна-
чение Pilsbry, 1922, Man. Concn., 27 :,43); Thiele, 1931 : 509; Лихарев, Рам-
мельмейер, 1952 : 151; Wenz, Zilch, 1959—1960 : 171; Шилейко, 19756 :
1779; Акрамовский, 1976 : 147.
Типовой вид Pupa umbilicata Draparnaud, 1801 (syn. Turbo cylind-
racea Da Costa, 1778).
Раковина овально-цилиндрическая или удлиненно-овальная, со сглаженной
вершиной, слабо скульптурированная. Высота раковины от 2.3 до 4.5 мм при
ширине 1.5—2 мм. Оборотов 5—7. Последний оборот к устью не поднят или
поднят незначительно. Устьевая арматура взрослых особей представлена
только двумя зубами: париетальной и колумеллярной пластинками. Кроме
того, нередко присутствует ангулярный бугорок.
На границе между пенисом и эпифаллусом имеется конический придаток,
несущий на внутренних стенках ясные бугорки. Яйцеживородящие животные.
Распространение. Средиземноморье, большая часть Европы,
Крым, Кавказ, Эфиопия. 1—2 вида.
В ископаемом состоянии представители рода известны из верхнего плиоцена
Европы; кроме того, Стеклов (1966) обнаружил дефектную раковину, близкую
к L. cylindracea, в неогене Предкавказья (назрановская свита, нерасчлененный
миоплиоцен).
Lauria cylindracea (Da Costa, 1778) (рис. 79, 80).
Da .Costa, 1778:89 (Turbo); Draparnaud, 1801:58 \(Pupa umbilicata}-,
O. Boettger, 1886a : 148 (Charadrobia)-, Pilsbry, 1922, Man. Conch., 27 : 47, pl. 6,
figs. 1-^8; Пузанов, 19256 : 116 (subsp. cymmetrica); Steenberg, 1925 : 58, pl. 9,
figs. 1, 2, fig. 29 du text; Лихарев, Раммельмейер, 1952 : 152, рис. 63, 64; Ш и-
л е й к о, 1967а : 14, рис. 1—3; Шилейко, 19756 : 1779, рис. 5, 1, 2 и 6, 11; Акр а-
м о в с к и й, 1976 : 147, рис. 65, табл. VI, 64, 66.
-2. РОД LAURIA
151
Terra typica — Англия.
Место хранения типов не установлено.
Материал: несколько сотен раковин из 53 местонахождений в Крыму,
на Кавказе и в Западной Европе.
Раковина от овально-цилиндрической до удлиненно-овальной, тонкостен-
ная, просвечивающая, блестящая, с широко закругленной вершиной. Завиток
от куполовидного до почти конического (у высоких ра-
ковин). Оборотов 6—7, слабовыпуклых, последний обо-
рот к устью почти не поднят. Окраска одноцветная, жел-
товато-роговая или (гораздо реже) светло-коричневая.
Эмбриональные обороты почти гладкие, дефинитивные
покрыты тонкими вялыми радиальными морщинками,
устье округло-треугольное, не отвесное, слегка обращено
вниз. Края устья тонкие, широко отвернуты. Устьевая
арматура несложная. Париетальная пластинка появляется
еще в эмбриогенезе и растет вместе с раковиной, причем
задний конец ее по мере роста раковины резорбируется.
Рис. 79. Lauria cylindracea (Da Costa) из Степанакерта, Нагорный
Карабах.
Таков же характер роста и колумеллярной пластинки, но в отличие от парие-
тальной рост ее прекращается до окончательного формирования дефинитивного
Рис. 80. Lauria cylindracea (Da Costa) из Чегемского ущелья на Северном Кавказе,
23 IX 1970 г.
I — половой аппарат; II — детали строения мужского отдела (из: Шилейко, 19756).
устья, и у вполне развитых раковин эта пластинка сквозь устье не видна или
обнаруживается в виде слабого утолщения на заднем крае столбика. У рако-
вин, закончивших рост, палатальных складок нет, но в течение роста периоди-
152
II. НАДСЕМ. ORCULOIDEA
чески появляются короткие поперечные палатальные складки. Кроме того,
в верхней части палатального края имеется слабое утолщение губы. Пупок
узкий, сквозной, цилиндрический.
Размеры: ВР 2.3—4.5, БД 1.5—2 мм; в первоописании размеры не указаны.
Для территории Кавказа отмечено присутствие второго вида этого рода —
Lauria sempronii (Charpentier, 1837) и его формы — var. caspia Pfeiffer, 1871.
Изучение обширного материала заставляет считать, что на территории СССР
присутствует лишь один широко изменчивый вид — L. cylindracea. Поскольку
L. sempronii описан из западной Европы (Тироль), лучше пока воздержаться
от сведения этого названия в синонимы к L. cylindracea, однако достойно вни-
мания, что Биггс указывает на находку L. cylindracea var. caspica (sic!) в север-
ном Иране и тут же сообщает, что Томлин, определивший эту форму, считает,
что «Вестерлунд полагает v. caspicaформой sempronii Ch., но последняя обычно
рассматривается как незначительная вариация cylindracea» (Biggs, 1971, р. 215;.
перевод А. Ш.).
Внутреннее строение. Материал: 6 экз. из окр. Степанакерта,
1 августа 1964 г., сбор Г. М. Овчинникова; 5 — из Чегемского ущелья на Се-
верном Кавказе, 23 сентября 1970 г.; 34 — из окр. Ленкорани, июнь—июль.
1964 г., сборы и определения автора.
> Поверхность белковой железы гладкая. Семяпровод впадает в резкое рас-
ширение булавовидного эпифаллуса, длина которого примерно равна длине
пениса или немного меньше. Просвет дистальной части эпифаллуса шире, чем
просвет проксимальной. Кольцевые складки, покрывающие внутреннюю по-
верхность эпифаллуса, по направлению от придатка эпифаллуса к семяпроводу
становятся все более узкими и тесно расположенными. Кольцевая щель вы-
ражена ясно. Внутренняя поверхность пениса почти гладкая. Придаток эпи-
фаллуса обширный, мешковидно-конический; конические бугорки, покрываю-
щие его внутреннюю поверхность, видны вполне отчетливо. Пениальный аппен-
дикс впадает в пенис на некотором расстоянии от полового атриума. Аг-|-42
почти слиты, немного длиннее пениса или равны ему. А3 хорошо обособлен.
Пениальная ветвь полового ретрактора крепится при основании придатка
эпифаллуса, другая ветвь подходит к А2. Проток семяприемника почти прямой,
заканчивается хорошо отграниченным маленьким шаровидным резервуаром.
Матка большинства исследованных особей содержала от 2 до 7 эмбрионов, ра-
ковина которых образована 2 оборотами.
Распространение и местообитание. Приморские об-
ласти Европы от о-ва Боргхольма и южной Норвегии вдоль атлантического и
средиземноморского побережий — до Малой Азии; Северная Африка, Крым,
Закавказье; возможно, Дагестан и Копетдаг. Населяет широколиственные леса,
осыпи, кустарниковые заросли с повышенным увлажнением; попадается также
на открытых участках в местах выхода грунтовых вод.
IV. Подсем. ARGN1NAE Hudec, 1965
Hudec, 1965b : 162; Pilsbry, 1924, Man. Conch., 27.
Дефинитивные обороты большей частью ребристые или ребристо исчерчен-
ные; ребрышки не имеют кожистых оторочек. Последний оборот приподнят
к устью. Нижние концы колумеллярной и париетальной пластинок видны при
нормальном положении раковины.
Придаток эпифаллуса отсутствует. Внутри пениса, на границе с эпифал-
лусом, имеется система мелких бугорков. Проток семяприемника без морфоло-
гически выраженных желез.
Распространение. Альпы, Карпаты. 2—3 рода.
В ископаемом состоянии подсемейство известно начиная с верхнего олиго-
цена (Европа).
1. РОД ARGNA
153
1. Род ARGNA Cossmann, 1899
Hartmann, 1840—1844 : 53 (Sphyradium, non Charpentier, 1837); Westerlund,
1887 : 78 (Coryna, non Billerg, 1833); Cossmann, 1889, Ann. geol. univ., 5 : 1104; Gude,
1911 : 361 (Agardhia, типовой вид Pupa ferrari Porro, 1838); Pilsbry, 1924, Man. Conch.,
.27 : 129 (A gardhia); Thiele, 1931 : 509 (Agardhia)-, Лихарев, Раммельмейер,
1952 : 157 (Agardhia)- W e n z, Zilch, 1959—1960 : 171; Hudec, 1965b : 157; Дами-
нов, Лихарев, 1975 : 105; Шилейко, 19756 : 1781.
Типовой вид Pupa ferrari Porro, 1838; по первоначальному обозна-
чению.
На палатальном крае устья имеются 2—4 складки, часть из которых видо-
изменена в бугорки.
Распространение. Южные и восточные Альпы, Карпаты. Около
10 видов.
В ископаемом состоянии род известен начиная с верхнего миоцена (Европа).
Argna bielzi (Rossmaessler, 1859) (рис. 81, 82).
Rossmaessler, 1859, Iconogr., 3, 109, fig. 942 (Pupa)-, Pilsbry, 1924, Man.
<Conch., 27 : 153, t. 16, figs. 12—14 (Agardhia)-, Лихарев, Раммельмейер, 1952 :
157, рис. 70 (Agardhia)-, Hudec, 1965b : 157, Abb. 1; Gross u, Negrea, 1968 : 723,
fig. 5 [Argna (Agardhia)]; Шилейко, 19756 : 1781, рис. 5, 3—5 иб, 12.
Terra typica — «Nagy Fain, unweit Bethlen» (Трансильвания).
Тип, вероятно, хранится в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: 22 экз. из А местонахождений в Закарпатье и Татрах.
Раковина высокоцилиндрическая, с широко закругленной короткой вер-
шиной, умеренно блестящая, твердостенная. Оборотов S1^—10, слабо или
умеренно выпуклых, очень медленно нарастающих. Пос-
ледний оборот возле самого устья довольно энергично;
с перегибом, поднят, высота его примерно равна 1/3 вы-
соты раковины или немногим больше. Окраска одноцвет-
ная, темно- или светло-роговая. Дефинитивная скульптура
в виде правильно расположенных сглаженных округлых
ребрышек. Устье усеченно-овальное, незначительно ско-
шенное, края его умеренно отвернуты, места прикрепле-
ния почти не сближены и соединены светлой размытой
мозолью. Зубов 5—7: длинная париетальная пластинка,
справа от которой в глубине устья может находиться
еще небольшой ангулярный бугорок; колумеллярная пла-
Рис. 81. Argna bielzi (Rssm.) из Закарпатья, с. Квасы близ
Рахова.
стинка, ниже которой также иногда располагается утолщение задней стенки
столбика; базальный бугорок, сидящий на некотором расстоянии от устьевого
края; две палатальные складки, из которых нижняя длинная, а верхняя в виде
округлого бугорока. Пупок узкий, сквозной, цилиндрический.
Размеры: ВР 4.5—5,5, БД 1.6—2 мм; в первоописании: ВР 5.5, БД 2 мм.
Внутреннее строение. Материал: 4 экз. из Закарпатья, Ра-
ховский р-н, с. Квасы, 14 сентября 1969 г., сбор и определение автора.
По выгнутой поверхности белковой железы проходит широкий и глубокий
желоб. Семяпровод впадает в утолщенную проксимальную часть эпифаллуса,
в стенках которой заключена глубокая кольцевая щель. Помимо этой щели
в утолщенной стенке проксимального конца эпифаллуса дистальнее терминаль-
ного расширения заключена продольная щелевидная полость, сообщающаяся с
протоком эпифаллуса. Пенис тонкостенный, цилиндрический, по его внутренней
154
II. НАДСЕМ. ORCULOIDEA
поверхности проходят многочисленные тонкие поперечные складки. Граница
между пенисом и эпифаллусом внешне не выражена, но при вскрытии пениаль-
ной трубки видно, что эта граница отмечена участком, поверхность которого
покрыта мелкими коническими бугорками; вероятно, этот участок гомологичен
придатку эпифаллуса Lauriinae. Пениальный аппендикс впадает в пенис не-
много отступя от полового атриума. A1-j-A2 значительно короче пениса, А3
хорошо выражен. Одна ветвь полового ретрактора присоединяется к дисталь-
ной части пениса, вторая — к границе между Аг и Л2. Дивертикул семяприем-
ника в несколько раз длиннее протока резервуара.
Ряс. 82. Argna bielzi (Rssm.) из Закарпатья, с. Квасы близ Рахова. 14 IX 1969 г.
I — половой аппарат; II — детали строения мужского отдела (звездочка — участок между пенисом и эпи-
фаллусом, несущий на внутренней поверхности конические бугорки); III — частично вскрытый участок
пениального аппендикса другого экземпляра (из: Шилейко, 19756).
Распространение и местообитание. Карпаты; в СССР
обитает в горных и предгорных районах западной Украины. Населяет под-
стилку широколиственных лесов, особенно в сырых ущельях; встречается также
в гниющей древесине и под корой старых пней.
II. Сем. STROBILOPSIDAE Pilsbry, 1918
Pilsbry, 1918, Man. Conch., 25.
Поверхность раковины с выраженной в разной степени радиальной скуль-
птурой — от исчерченности до грубой ребристости. Эмбриональные обороты
покрыты микроскопической зернистостью. Устье с пластинчатыми зубами,
уходящими глубоко внутрь раковины. Края устья отвернуты. Пупок воронко-
видный.
Гермафродитный проток без обособленных семенных пузырьков. Простата
короткая. Папилла пениса отсутствует. Пениальный аппендикс имеется, но
не все его отделы хорошо выражены. Бич, придаток эпифаллуса и дивертикул
семяприемника отсутствуют.
1. РОД STROBILOPS
155
Распространение. Юго-восточная Азия и Филиппинские о-ва;
в Америке от юго-восточной части Канады до Венесуэлы и северо-восточной
Бразилии; Галапагосские о-ва. 2 рода.
В ископаемом состоянии семейство известно начиная с эоцена (Европа) и
миоцена (Предкавказье). На Американском континенте ранее верхнего плио-
цена представители семейства не найдены.
1. Род STROBILOPS Pilsbry, 1892
Morse, 1864 : 26 (Strobila, non Sars, 1835); Sandberger, 1872—1875, Land-
п. Siisswasser-Conch. Vorwelt : 258 (Strobilus, non Anton, 1839); Pilsbry, 1892 (1893) :
403; 1927, Man.. Conch., 28: 12; Лихарев, Pa мме льмейер, 1952 : 169; Wen z,
Zilch, 1959—1960 : 177; Стеклов, 1966 : 169.
Типовой вид Helix labyrintica Say, 1817; по монотипии.
Поверхность раковины от почти гладкой до ребристой. Устье узкое, ско-
шенное, с утолщенными и плавно отвернутыми краями. Внутри устья до трех
париетальных пластинок, кромки которых снабжены узелками. Иногда имеется
колумеллярная пластинка и базально-палатальные складки.
Распространение. Юго-восточная Азия, Дальний Восток и Аме-
рика. 5 подродов, не менее 20 видов.
Представители рода найдены в эоцене—верхнем плиоцене Европы, верх-
нем миоцене Украины и Предкавказья.
Strobilops coreana Pilsbry, 1927 (рис. 83).
Pilsbry, 1927, Man. Conch., 28 : 43, pl. 10, figs. 8—10; Лихарев, P а мме ль-
мейер, 1952 : 170, рис. 83.
Terra typica — окр. Пхеньяна (КНДР).
Голотип хранится в Академии естественных наук Филадельфии (США).
Материал: 24 экз. из заповедника, «Кедровая падь», Приморский край.
Раковина прижатая, с куполовидно-коническим завитком, твердостенная,
слабоблестящая. Оборотов 5, слабовыпуклых, последний оборот с легким на-
меком на угол по перифе-
рии, к устью очень слабо
опущен. Высота завитка
примерно равна высоте
устья. Окраска одноцвет-
ная, коричневатая или кре-
Рис. 83. Strobilops coreana
Pils. из заповедника «Кедро-
вая падь», Приморский край.
мовая. Дефинитивная скульптура — тонкие радиальные морщинки, дающие впе-
чатление очень тонкой правильной исчерченности. Устье полулунное, косое, края
его отвернуты и утолщены. Две париетальные пластинки начинаются от края
устья и уходят в глубь раковины на 1/3—1/2 оборота. Кромки их передних кон-
цов гладкие, отступя от края снабжены серией узелков (рис. 41, I—II) и затем
плавно сходят на нет. В глубине устья не видные при прямом положении ра-
ковины располагаются маленькая колумеллярная пластинка, 2—3 базально-
палатальные складки и тонкая короткая сутуральная складка (может отсут-
ствовать). Пупок круглый, воронковидный, расположен на дне пупочной впа-
дины эксцентрично.
Размеры: ВР 1.8—2, БД 2.5—2.8, МД 2.6=2.7 мм; в первоописании:
ВР 1.85, БД 2.7 мм.
Внутреннее строение не исследовано; судя по тому, что из-
вестно по типовому виду рода S. labyrintica, для полового аппарата характерны
156
II. НАДСЕМ. ORCULOIDEA
следующие особенности. Бич отсутствует, семяпровод впадает строго терми-
нально. Придаток эпифаллуса притупленно-конический. Пениальный аппен-
дикс хорошо развит, но границы между его отделами сглажены. Одна ветвь
полового ретрактора крепится к нижней части аппендикса, вторая — к гра-
нице пениса и эпифаллуса, при основании придатка. Дивертикул семяприем-
ника отсутствует, проток семяприемника очень тонкий, резервуар маленький,
не доходит до белковой железы (Baker, 1935, pl. 26, figs. 4, 5, 7).
Распространение и местообитание. Вид найден только
в двух местах: в окр. Пхеньяна и в заповеднике «Кедровая падь». Встречен
в подстилке широколиственного и смешанного леса.
III. Сем. SPELAEODISCIDAE Steenberg, 1925
Steenberg, 1925:202 (FaZZoniidae, ? Spelaeodiscinae).
Поверхность раковины покрыта грубыми радиальными сглаженными реб-
рами. Эмбриональные обороты покрыты микроскопической зернистостью.
Устье без зубов или с поверхностными зубами, лежащими в плоскости устья
и не уходящими внутрь раковины. Края устья умеренно отвернуты. Пупок
в разной степени развернутый.
Гермафродитный проток без обособленных семенных пузырьков. Простата
недлинная. Папилла пениса отсутствует. Пениальный аппендикс нормально
развит, придаток эпифаллуса конический. Бич и дивертикул семяприемника
отсутствуют.
Распространение. Юго-восточная Европа. 1—2 рода.
В ископаемом состоянии семейство известно лишь из четвертичных отло-
жений Чехословакии.
1. Род SPELAEODISCUS Brusina, 1886
Brusin а, 1886 : 37; Р i 1 s b г у, 1926, Man. Conch., 27 : 180; Thiele, 1931 : 516;
Wenz, Zilch, 1959—1960 : 176; Gittenberger, 1969 : 288; Дамянов, Ли-
харев, 1975 : 136.
Типовой вид Helix Hauffeni F. Schmidt, 1855; по монотипии.
Устье без зубов, лишь на палатальном крае может присутствовать выражен-
ная в разной степени вмятина.
Распространение. Чехословакия, Румыния, Болгария, Югосла-
вия, Албания. Несколько видов.
Spelaeodiscus triarius (Rossmaessler, 1839) (рис. 84, 85).
R о ssm a essler, 1839, Iconogr., 2, 3—4 : 12, fig. 611 (Helix); P i 1 s b r y, 1926,
Man. Conch., 27 : 182, t. 22, figs. 1—3; G г о s s u, 1955 : 213, fig. 104; Urbanski, 1964 :
21; Gittenberger, 1969 : 290, t. I, 1, 2; Hude c, 1970 : 33, obr. 1 (f. triadis); Git-
tenberger, 1975:264, t. I, 1, t. II, 3; Дамянов, Лихарев, 1975:137,
фиг. 72, 73.
Terra typica — Банат.
Лектотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 5065 (Zilch, 1969, Arch. Moll., 99 : 227, Т. 3, Fig. 14).
Материал: 2 экз. из Болгарии, окр. Враца.
Раковина уплощенная, относительно твердостенная, слегка просвечиваю-
щая, со слегка куполовидным или почти коническим завитком, высота которого
превышает высоту устья примерно в полтора раза. Оборотов 6—672, выпуклых,
медленно нарастающих, последний оборот к устью резко, с перегибом опущен.
Окраска одноцветная, от темно-роговой до красновато-роговой. Эмбриональ-
ные обороты покрыты неясной зернистостью, дефинитивные с редкими грубыми
закругленными ребрами, расстояние между которыми превышает ширину
самого ребра. Кожистых оторочек на ребрах нет. Устье округло-треугольное,
1. РОД SPELAEODISCUS
157
сильно скошенное, края его острые, отвернутые, внутри с валикообразной
расплывшейся губой. На палатальном крае устья часто имеется развитая в раз-
личной степени вмятина, которая отсутствовала
земпляров. Пупок широкий, развернутый, пер-
спективный, сквозь него просматриваются все
обороты.
Размеры: ВР 2.4-3, БД 3.8-6.3, МД 3.5-
4.5 мм; в первоописании: ВР 12/5, БД 3 линии,
т. е. ВР около 2.9, БД около 6.3 мм.
Рис. 84. Spelaeodiscus triarius (Rssm.) из Болгарии,
окр. Враца.
у обоих исследованных эк-
Внутреннее строение. Материал: 1 экз. из Болгарии, пещера
Леденика близ Враца, 4 июля 1969 г., сбор и определение И. М. Лихарева.
Белковая железа с широким вялым апикальным желобком. Семяпровод
впадает терминально в расширенный конец эпифаллуса. На продольном срезе
Рис. 85. Spelaeodiscus triarius (Rssm.) из Болгарии, Стара Планина, пещера Леденика близ
Врана, 4 VII 1969 г.
I — половой аппарат; II — сперматофор, извлеченный из протока семяприемника; III — детали строения
мужского отдела.
эпифаллуса видно, что внутри него кроме канала, составляющего продолжение
семяпровода, имеется слепой канал (вероятно, гомолог бича). Диаметр эпифал-
луса по направлению к придатку эпифаллуса постепенно уменьшается: прит
даток хорошо развит, закругленно-конический, внутри него имеется неполная
перегородка, т. е. канал эпифаллуса образует внутри придатка очень крутой
изгиб. Придаток делит эпифаллус в соотношении примерно 1:2. Пенис подраз-
делен на длинный цилиндрический дистальный отдел и короткий проксималь-
ный, на внутренних стенках пениса залегает серия тонких продольных складок.
Пениальный аппендикс хорошо развит. А2 не обособлен от А1г А3 не выражен,
158
И. НАДСЕМ. ORCULOIDEA
Ai короткий, не длиннее, чем А5. Пениальная ветвь полового ретрактора кре-
пится немного ниже основания придатка эпифаллуса, ветвь аппендикса развита
слабее и подходит к нижней части А±, из чего можно заключить, что при спа-
ривании пениальный аппендикс почти не выворачивается. Проток семяприем-
ника в нижней части расширен, по направлению к резервуару постепенно су-
жается, резервуар небольшой, не доходит до белковой железы. Из нижней
части протока семяприемника извлечен сперматофор сложной и своеобразной
формы (рис. 85, II).
Распространение и местообитание. Юго-восточная Ев-
ропа: северо-западная Болгария, северо-восточная Сербия, Румыния, Венгрия;
в Татрах Польши и Чехословакии — подвид tatricus Hazay. В СССР не найден,
но не исключено, что со временем вид будет обнаружен у юго-западных границ.
Населяет преимущественно горные районы, где встречается в подстилке широко-
лиственных лесов и на влажных открытых склонах; подобно прочим предста-
вителям рода, нередко встречается в приустьевых частях пещер.
IV. Сем. VALLONIIDAE Morse, 1864 1
Morse, 1864; Pilsbry, 1892, Man. Conch., 8 : 247.
Поверхность раковины гладкая или с радиальными кожистыми ребрышками.
Эмбриональные обороты покрыты микроскопической зернистостью или несут
•спиральные нити, выраженные в разной степени. Устье без зубов. Края устья
•острые или широко отвернуты. Пупок от узкого, цилиндрического до очень
широкого, перспективного.
Простата в виде нескольких долек при основании белковой железы. Па-
пилла пениса отсутствует. Семяприемник с очень коротким протоком и малень-
ким резервуаром. Кроме пениального аппендикса, придатков на совокупитель-
ном аппарате, за редким исключением, нет. Характерна резко выраженная
лротерандрия, поэтому мужской отдел, как правило, обнаружить не удается.
Распространение. Почти весь земной шар. 2—3 подсемейства.
В ископаемом состоянии семейство известно начиная с палеоцена (Европа,
Северная Африка, Азия, Северная Америка).
I. Подсем. ACANTHINULINAE Steenberg, 1917
Steenberg, 1917 : 14.
Раковина от сильно уплощенной до кубаревидной, хрупкая, тонкостенная,
коричневатой или бурой окраски. Края устья тонкие, не отвернутые, без губы.
Как эмбриональные, так и дефинитивные обороты несут, как правило, тонкие
частые спиральные линии. На дефинитивных оборотах имеются, кроме того,
хорошо развитые радиальные кожистые ребрышки. Пупок перспективный, но
не развернутый.
Анатомический диагноз в настоящее время сформулировать невозможно
за недостатком материалов.
Распространение. Почти весь земной шар. Около 10 родов.
В ископаемом состоянии подсемейство известно начиная с палеоцена
(Европа).
х В сводке Лихарева и Раммельмейер (1952) среди представителей семейства указывается
Vallonia patens Reinhardt, 1883; позже этот вид отмечается Иззатуллаевым (1975) для фауны
Таджикистана; под названием Punctum microрleurum • lederi Bttg. его указывает Шилейко
(19676) для Талыша. Специально проведенное исследование показало, что Vallonia patens
в фауне СССР отсутствует, а обсуждаемый вид есть Toltecia pusilia (Lowe, 1831) (=Helix mi-
cropleuros Paget, 1854, =Patula lederi Boettger, 1880c), принадлежащий сем. Punctidae (см.:
Giusti, 1976, p. 175; Gittenberger et al., 1980, p. 12).
1. РОД ZOOGENETES
15»
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
ACANTHINULINAE
1 (2). Большой диаметр раковины превышает ее высоту или равен этой вели-
чине: раковина ширококоническая......................3. Spermodea.
2 (1). Большой диаметр раковины меньше ее высоты: раковина высококони-
ческая или кубаревидная..
3 (4). Спиральная скульптура на эмбриональных оборотах отчетливая. Ради-
альные ребрышки на периферии оборотов вытянуты в треугольные шипы
.................................................. 2. Acanthinula.
4 (3). Спиральная скульптура на эмбриональных оборотах чрезвычайно тонкая
и плохо различимая. Радиальные ребрышки не вытянуты в шипы . . .
.................................................... 1. Zoogenetes.
1. Род ZOOGENETES Morse, 1864
Morse, 1864 : 32; Pilsbry, 1926, Man. Conch., 27 : 196; Thiele, 1931 : 515
[Acanthinula (Zoogenetes)]; Wen z, Zilch, 1959—1960 : 176.
Типовой вид Helix harpa Say, 1824; по монотипии.
Раковина высококоническая, с невысокими радиальными кожистыми реб-
рышками на дефинитивных оборотах и слабой спиральной струйчатостью на
эмбриональных. Пупок узкий.
Монотипический род.
Zoogenetes harpa (Say, 1824) (рис. 86).
Say, 1824, Long’s Second Exped., appendix : 256, t. 15, fig. 1 (Helix)', L. Pfeiffer,
1847a : 147 (Bulimus); Gerstfeldt, 1859 : 517, t. 9, fig. 26 (Helix amurensis); Tryon,
1867 : 311, pl. 15, fig. 1 (Zoogenites); Линдгольм, 19226 : 309; Steenberg, 1925 :
102, pl. 28, tigs. 1, 2, 4, 5, pl. 29; P i 1 s b r y, 1926, Man. Conch., 27 : 196, t. 32, fig. 10; Л и-
харев, Раммельмейер, 1952:168, рис. 81; Ehrmann, 1956 : 57; Zilch,
J a e c k e 1, 1962 : 106, Abb. 50.
Terra typica — Северная Америка.
Место хранения типов не установлено.
Материал: свыше 100 экз. из 53 местонахождений в Западной Европе,
Архангельской и Мурманской обл., в Забайкалье, на Дальнем Востоке и на
Аляске.
Раковина высококоническая, тонкостенная, хрупкая, с почти коническим
контуром завитка и закругленной вершиной. Оборотов 4, быстро нарастающих,
выпуклых, последний оборот очень плавно и очень
слабо приподнят к устью. Высота завитка примерно
в ПД раза больше высоты устья. Окраска оливковая,
одноцветная. Самое начало эмбриональных оборотов
покрыто неясной микроскопической зернистостью,
остальная поверхность эмбриональных оборотов несет
характерную тончайшую скульптуру из неровных
спиральных нитей (видно при увеличении в 50—60 раз
Рис. 86. Zoogenetes harpa (Say) из Аяна, побережье Охотского
моря.
в сильном косом свете), создающих впечатление микроскопической струйча-
тости. Дефинитивная скульптура в виде очень тонкой радиальной исчерчен-
ности и редких неравномерно расположенных кожистых ребрышек, кромки
которых имеют нередко коричневый оттенок. Устье овальное, умеренно косое;
устьевые края, за исключением верхней части колумеллярного, не отвернуты,
острые, тонкие. Пупок узкий, сквозной, цилиндрический.
160
П. НАДСЕМ. ORCULOIDEA
Размеры: ВР 3—5, БД 2.5—3.5 мм; в первоописании: ВР 5, БД 3.5 мм.
Внутреннее строение. Исследовано 9 экз., но все они оказа-
лись афалличными, в матке содержались эмбрионы в числе 4—6.
Белковая железа с апикальной вырезкой. Простата заметна в виде несколь-
ких тонких фолликулов у нижнего края белковой железы. Проток семяприем-
ника очень плавно переходит в тонкий резервуар, не доходящий до белковой
железы.
Распространение и местообитание. Север европейской
части СССР, Сибирь, Камчатка, Командорские о-ва; по восточному побережью
Азии спускается до Сихотэ-Алиня; высокогорье Кавказа; Скандинавия, высо-
когорье Швейцарии; северные районы Северной Америки.
Населяет смешанные и хвойные леса, живет в подстилке, иногда поднимается
на стволы деревьев, часто встречается на грибах.
2. Род ACANTHINULA Beck, 1847
Н. В е с k, 1847 : 109; Pilsbry, 1926, Man. Conch., 27 : 188; Thiele, 1931 : 515;
Лихарев, P амме льмейер, 1952 : 166; Wenz, Zilch, 1959—1960 : 174;
Акр амо вс к ий, 1976 : 150.
Типовой вид Helix aculeata Muller, 1774; по монотипии.
Раковина кубаревидная, с радиальными кожистыми ребрышками, которые
на периферии оборотов вытянуты в треугольные шипики. На поверхности эм-
бриональных оборотов имеются ясно различимые спиральные нити. Пупок
узкий.
Распространение. Европа, Кавказ, Северная и Восточная Африка,
Малая Азия, Тянь-Шань (?). 2—3 вида.
В ископаемом состоянии род известен начиная с палеоцена (Европа).
Acanthinula aculeata (Muller, 1774) (рис. 87, 88).
Muller, 1774 : 81 (Helix)-, Moquin-Tandon, 1855 : 189, pl. 15, figs. 5—9
[Helix (Fruticicola)]; Hesse, 1916a : 116; В о у с о t t, 1917b : 221—226, fig.; Steen-
berg, 1918 : 2, figs. 1, 2; P i 1 s b r y, 1926, Man. Conch., 27 : 191, t. 32, figs. 4—6; Лиха-
рев, P амме льмейер, 1952 : 166, рис. 79; Ehrmann, 1956 : 56, fig. 29; L o-
zek, 1956:109, t. 9, 1; Zilch, J a e c k e 1, 1962 : 105; L о z e k, 1964 : 224, t. 9,
2a—с; M а т ё к и и, 1966 : 115; Giusti, 1968 : 246, fig. 4; Даминов, Лихарев,
1975 : 136, фиг. 70, 71; А к р а м о в с к и й, 1976 : 151, рис. 68, табл. VII, 70.
Tetra typica — Фридриксдаль (окр. Копенгагена).
Место хранения типов не установлено.
Материал: около 200 экз. из 107 местонахождений в Закарпатье, под
Москвой, на Кавказе и в Западной Европе.
Раковина кубаревидная, тонкостенная, хрупкая, со слегка куполовидным
контуром завитка и закругленной вершиной. Оборотов 4, сильновыпуклых,,
но не ступенчатых, в профиле плавно закругленных. Последний оборот к устью
почти не опущен. Высота устья немного меньше высоты завитка. Окраска светло-
или темно-роговая. Спиральные нити на эмбриональных оборотах расположены
густо и заметны иногда даже на старых вытертых раковинах. Дефинитивные
обороты покрыты множеством тончайших густо расположенных радиальных
морщинок и редкими кожистыми ребрышками, вытянутыми на периферии обо-
ротов в загнутые наподобие ресниц треугольные шипы. Эти шипы могут быть
развитыми в разной степени. Кроме того, имеется и спиральная исчерченность
дефинитивных оборотов, выраженная в разной степени. Устье округлое, косое,
края его тонкие, колумеллярный и базальный несколько отвернуты, иногда
заметна слабая светлая губа по линии перегиба. Пупок цилиндрический, круг-
лый, сквозной.
Размеры: ВР 1.7—-2.1, БД 2—2.3, МД 1.9—2.1 мм; в первоописании: «diam.
3/4lin.», т. е. БД около 1.6 мм; очевидно, Мюллер имел дело с молодым экземп-
ляром-
3. РОД SPERMODEA
161
Внутреннее строение. Материал: 3 экз. из Московской обл., с. Раз-
доры, 5 мая 1961 г., сбор и определение автора. Исследованные особи оказа-
лись афалличными, поэтому описание составлено по литературным данным
(Steeuberg, 1918; Germain, 1930; Giusti, 1968) и дополнено собственными на-
блюдениями афалличных животных.
Белковая железа с вялой широкой апикальной вырезкой. Семяпровод впа-
дает в короткий веретеновидный эпифаллус. В дистальной части протока видна
пара небольших цилиндрических
придатков. Пенис недлинный, в его
атриальный отдел впадает короткий
аппендикс: А± и А2 слиты в единый
почти шаровидный отдел; А3 не вы-
ражен; Ai очень короткий, перехо-
дит в короткобулавовидный Аа. По-
ловой ретрактор крепится к диа-
фрагме единой лентой, расщепляется
довольно низко. Короткая пениаль-
ная ветвь крепится возле придатков
Рис. 87. Acanthinula aculeata (Miill.)
из Подмосковья, с. Жаворонки.
Рис. 88. Acanthinula aculeata (Miill.). Половой ап-
парат (из Steeuberg, 1918).
эпифаллуса, ветвь аппендикса — к средней части Л1+М2. Семяприемник имеет
очень тонкий проток и маленький удлиненно-овальный резервуар, не доходящий
до белковой железы.
Распространение и местообитание. Большая часть
Европы, на восток примерно до 40° в. д., на север примерно до широты Петро-
заводска; Северная Африка; Крым, Северный Кавказ, Закавказье.
Обитает преимущественно в лесах, в южной части ареала поднимается
в горы. Держится в местах с повышенной влажностью, в укрытиях, в толстом
слое лиственного опада; нередко на плодовых телах грибов.
3. Род SPERMODEA Westerlund, 1902
Westerland, 1902 : 90; Pilsbry, 1926, Man. Conch., 27 : 187; Thiele,
1931 : 515 [Acanthinula (Spermodea)]; Wenz, Zilch, 1959—1960 : 175.
Типовой вид Helix lamellata Jeffreys, 1830; по монотипии.
Раковина ширококоническая, с низкими правильно расположенными кожи-
стыми ребрышками на дефинитивных и микроскопической зернистостью на
эмбриональных оборотах. Пупок узкий.
Распространение. Западная Европа. Несколько видов.
В ископаемом состоянии род известен начиная с верхнего олигоцена (Европа).
11 А. А. Шилейко
162
II.,НАДСЕМ. ORCULOIDEA
Spermodea lamellate (Jeffreys, 1830) (рис. 89).
J effreys, 1830 : 333 (Helix); Reeve, 1863 : 81, fig. (Helix); Boycott, 1917a :
157—1'78 (Acanthinula); Steenberg, 1918 : 6, figs. 3, 4 (Acanthinula); Pilsbry, 1926,
Man. Conch., 27 : 187, t. 32, figs. 1—3; Лихарев, Раммельмейер, 1952 : 167,
рис. 80; Ehrmann, 1956 : 56, fig. 30; Zilch, Jaeckel, 1962 : 105; Gitte n b e r-
ger et al., 1970 : 57, fig. 51.
Terra typica Англия («Scarborough, York»).
Место хранения типов не установлено.
Материал: 25 экз. из пяти местонахождений в Ирландии, Дании и ФРГ.
Раковина ширококоническая, довольно тонкостенная, просвечивающая,
с шелковистым блеском. Завиток ширококуполовидный, вершина плавно за-
кругленная. Оборотов 5х/2—6, выпуклых, ступенчатых, плавно нарастающих.
Высота завитка примерно в 1х/2 раза превышает вы-
соту устья. Последний оборот прямой. Окраска ро-
говая или красновато-роговая. Эмбриональные
обороты покрыты микроскопической зернистостью;
спиральная скульптура полностью отсутствует. На
поверхности дефинитивных оборотов располагаются
Рис. 89. Spermodea lamellata (Jeffr.) из Ирландии.
тонкие невысокие радиальные пластинчатые ребрышки. Устье полулунное,
отвесное, края его не отвернуты, кроме колумеллярного, места прикрепления
широко расставлены. Пупок узкий, сквозной, цилиндрический, иногда слегка
перекрыт отворотом колумеллярного края.
Размеры: ВР 1.9—2.3, БД 2—2.3, МД 1.9—2.1 мм; в первоописании: ВР 1.9,
БД 2.2 мм.
Внутреннее строение. Материала, пригодного для анатомиро-
вания, в моем распоряжении не было. Вид был исследован Бойкоттом (Boycott,
1917а) и Стинбергом (Steenberg, 1918), которые имели дело только с афаллич-
ными особями.
Белковая железа с глубоким желобом на боковой поверхности, матка со сле-
пыми карманами. Проток семяприемника тонкий, плавно переходит в удлинен-
ный резервуар, далеко не доходящий до белковой железы. Стинберг (Steenberg,
1918) отмечает наличие ретрактора семяприемника и верхнего отдела вагины
(«свободного овидукта»).
Распространение и местообитание. Прибрежные обла-
сти западной и северо-западной Европы; в СССР — Калининградская обл.
(Zilch, Jaeckel, 1962). Населяет подстилку лиственных и смешанных лесов.
II. Подсем. VALLONIINAE Morse, 1864
Раковина сильно уплощена, светлой окраски, часто стекловидно-прозрачная.
Края устья тонкие, сильно отвернуты, место перегиба усилено губой. Спираль-
ная скульптура отсутствует или выражена слабо на эмбриональных оборотах
Дефинитивные обороты гладкие или с редкими кожистыми радиальными реб-
рышками; в редких случаях радиальная скульптура представлена не столько
периостракальными элементами, сколько складками известкового слоя. Пупок
развернутый, широкий, перспективный.
Анатомический диагноз в настоящее время сформулировать невозможно, так
как для членов подсемейства в высокой степени характерна афаллия и найти
особь с развитым мужским отделом удается чрезвычайно редко.
Монотипическое подсемейство.
Распространение. Почти вся Голарктика.
В ископаемом состоянии подсемейство известно начиная с палеоцена (Европа,
Северная Африка, Передняя и Восточная Азия, Северная Америка).
1. РОД VALLONIA
163
1. Род VALLONIA Risso, 1826
R i s s о, 1826 : 101; S t e г к i, 1893 : 234; Thiele, 1931 : 515; Pilsbry, 1935,
Man. Conch., 28 : 174, 195; Лихарев, Раммельмейер, 1952 : 163; Стеклов,
1966 : 166; Gittenberger, 1972 : 63.
Типовой вид Vallonia rosalia Risso, 1826 (syn. Helix pulchella Miiller,
1774); по монотипии.
Признаки подсемейства.
Распространение. Голарктика. Около 20 видов.
В ископаемом состоянии род известен начиная с палеоцена примерно в пре-
делах современного ареала.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ПОДВИДОВ
VALLONIA
1 (2). Устье цельное и несколько обособлено от последнего оборота (подрод
Planivallonia) .......................<.............9. V. (Р.) zaru.
2 (1). Устье не цельное (подрод Vallonia s. str.).
3 (8). Эмбриональные обороты со спиральной скульптурой.
4 (7). Высота завитка не превышает половины высоты устья.
5 (6). Последний оборот с плавным углом выше средней линии. Пупок очень
мелкий..........................................2. V. (V.) amurensis.
6 (5). Последний оборот в профиле широко закруглен, без угла. Пупок углуб-
ленный ....................................... 1. V. (V.) costata.
7 (4). Высота завитка приблизительно равна высоте устья.............
...............................3. V. (V.) cyclophorella kamtschatica.
8 (3). Эмбриональные обороты без спиральной скульптуры или она едва
заметна.
9 (10). Раковина при рассматривания сверху (или снизу) выглядит оваль-
ной, последний оборот к устью расширяется в виде раструба...........
. . ...................................... 6. V. (V.) excentrica.
10 (9). Раковина в плане округлая, последний оборот почти цилиндрический.
11 (16). Последний оборот почти прямой, т. е. к устью не опущен или опущен
очень слабо.
12 (15). Последний оборот почти' вдвое шире предпоследнего.
13 (14). Большой диаметр раковины не менее 3 мм.....................
7. V. (V.) tenuilabris tenuilabris.
14 (13). Большой диаметр раковины не превышает 2.2 мм ..............
. . . .............:................ 7а. V. (V.) tenuilabris peteri.
15 (12). Последний оборот примерно в 1х/2 раза шире предпоследнего . . .
............................................5. V. (V.) pulchella.
16 (11). Последний оборот к устью более или менее ясно опущен.
17 (18). Губа внутри устья хорошо развита.......4. V. (V.) enniensis.
18 (17). Губа отсутствует или очень слабая......8. V. (V.) ladacensis.
1. Подрод VALLONIA s. str.
Устье не цельное, хотя места его прикрепления чаще всего сближены.
Распространение. Голарктика.
В ископаемом состоянии подрод известен с палеоцена в пределах современ-
ного ареала.
1. Vallonia (vallonia) costata (Muller, 1774) (рис. 90, 11; рис. 91).
Muller, 1774 : 31 (Helix)-, Risso, 1826 : 102, pl. 4, fig. 30 (rosalia); M о q u i n-Tan-
d о n, 1855 : 140, pl. 11, figs. 28—34 [Helix (Lucena) pulchella costata]; Pilsbry, 1892,
Man. Conch., 8 : 252, t. 32, figs. 18—22, 27, t. 35, fig. 54; S t e e n b e r g, 1918 : 9, figs. 5—7
(pulchella var. costata); Лихарев, Раммельмейер, 1952 : 164, рис. 77Б; Ehr-
mann, 1956 : 54, t. 2; fig. 25; L о z e k, 1956 : 107, t. 8, 1; Z i I в h, Jaeckel, 1962 :
11*
164
II. НАДСЕМ. ORCULOIDEA
103; Loze к, 1964 : 219, t. 10, За—с; Матёкин, 1966 : 115; Gittenberger et al.,
1970:55, fig. 45, Kaart 19; Дамянов, Лихарев, 1975:113, фиг. 68a—г, 69;
Акр амовский, 1976 :150; рис. 66, табл. VII, 68.
Terra typica — Фридриксдаль (окр. Копенгагена).
Место хранения типов не установлено.
Материал: несколько сотен экз. из 62 местонахождений в Венгрии,
Подмосковье, в Нижнем Поволжье, на Карпатах, в Крыму, на Кавказе и в За-
кавказье, в Средней Азии, Сибири и на Дальнем Востоке.
Раковина низкоконическая, тонкостенная, хрупкая, просвечивающая,
составлена Зх/4—33/4 слегка ступенчатыми оборотами. Последний оборот возле
I I
Рис. 90. Виды рода Vallonia (подрод Vallonia s. str.).
I — V. (Vallonia') amurensis Sterki из о-ва Два Брата в зал. Петра Великого; II — V. (Vallonia) eostata
(Miill.) из Москвы, Кунцево.
устья опущен с плавным перегибом, в профиле плавно закруглен, примерно
в 1х/2 раза шире предпоследнего. Высота завитка составляет примерно половину
высоты устья или меньше. Окраска серая, иногда с желтоватым оттенком. Спи-
ральная скульптура на эмбриональных оборотах ясно видна при 40—50-кратном
увеличении. Дефинитивные обороты несут редкие тонкие кожистые пластинча-
тые ребрышки, между которыми заметна тонкая, но резкая радиальная исчер-
ченность, создающая впечатление шелковистой струйчатости. Устье круглое,
косое, места его прикрепления сильно сближены. Края устья тонкие, резко
отвернуты, по линии перегиба усилены широкой белой губой. Пупок широкий,
развернутый, перспективный, углубленный.
Размеры: ВР 1.3—1.4, БД 2.2—2.7, МД 2.1—2.4 мм; в первоописании:
«diam. 1 lin.», т. е. БД примерно 2.1—2.2 мм.
Внутреннее строение. Материал: из нескольких десятков жи-
вотных, собранных в различных районах и в разные сезоны, обнаружен лишь
1 экз. с развитым мужским отделом полового аппарата: окр. с. Агверан близ
Еревана, 25 сентября 1975 г., сбор и определение автора.
Белковая железа с глубоким латеральным желобом. Простата не обнаружена.
Проксимальный отдел матки имеет плотные стенки и образует карман; дисталь-
ный отдел органа тонкостенный, многоскладчатый. Семяпровод впадает терми-
нально в слегка расширенный конец эпифаллуса, образующий крутое колено;
судя по месту прикрепления полового ретрактора, этим коленом отмечена гра-
ница между пенисом и эпифаллусом. Внутренняя поверхность этих протоков без
1. РОД VALLONIA
165
каких-либо специализированных структур. Пениальный аппендикс развит,
слит с А2; А3 заметно обособлен, короткий. Половой ретрактор отходит
от диафрагмы единой лентой, которая раздваивается только близ пениса. Ветвь
аппендикса крепится к верхней части Л1+Л2. Проток семяприемника прямой,
недлинный, резервуар слабо обособлен.
Распространение и местообитание. Голарктика. Насе-
ляет богатый набор биотопов с достаточным увлажнением. Встречается как
в лесах, так и на открытых местах под укрытиями, на равнине и в горах; в тун-
дре отсутствует. Завезен в Австралию и Тасманию.
2. Vallonia (Vallonia) amurensis Sterki, 1892 (рис. 90, 1).
Sterki in: Pilsbry 1892, Man. Conch., 8 : 254, pl. 33, figs. 50—52 (Vallonia
eostata nbs. amurensis); Лихарев, Раммельмейер, 1952 : 165 (V. eostata amu-
rensis).
Terra typica — окр. Казакевичево на Амуре.
Место хранения типов не установлено.
Материал: более 100 экз. из 14 местонахождений на юге Приморского
края и в долине Амура (с. Вятское ниже Хабаровска).
Раковина почти плоская, с едва возвышающимся завитком, тонкостенная,
хрупкая, просвечивающая, составлена оборотами. Последний оборот к устью
несильно, с легким перегибом, опущен,
в профиле имеет закругленный угол
выше средней линии; ширина его при-
мерно в 1х/2 раза больше ширины пред-
последнего оборота. Высота завитка
ценыпе половины высоты устья. Ок-
раска белая или слегка желтоватая; у жи-
вых животных раковина обычно почти
бесцветная. Эмбриональные обороты не-
много темнее дефинитивных, их поверх-
ность с ясно заметными спиральными
нитями. На дефинитивных оборотах
имеются тонкие периостракальные пла-
стинчатые ребрышки, в промежутках
между которыми видна тонкая резкая
радиальная исчерченность, местами соз-
дающая впечатление струйчатости. Устье
почти круглое, ‘косое, места его при-
Рис. 91. Vallonia (Vallonia) eostata (Mull.) из
с. Агверан близ г. Еревана, 25 IX 1975 г.
Половой аппарат.
крепления сильно сближены и соединены тонкой мозолью, так что можно гово-
рить о наклонности к образованию цельного устья. Устьевые края плавно, но
сильно отогнуты почти под прямым углом, тонкие, острые. Губа отсутствует или
исчезающе слабая. Пупок очень мелкий и очень широкий — мельче и шире,,
чем у любого другого представителя рода в фауне СССР.
Размеры: ВР 0.9—1.1, БД 2.2—2.5, МД 2—2.3 мм; в первоописании:
БД 2.2 мм.
Распространение и местообитание. Помимо двух место-
нахождений в долине Амура обнаружен Р. Я. Братчиком и мною на многих
островах в зал. Петра Великого, часто вместе с V. eostata. Обитает в лиственном
опаде, под камнями и в скоплениях растительных остатков.
166
II. НАДСЕМ. ORCULOIDEA
3. Vallonia (Vallonia) cyclophorella kamschatica Likharev, 1963 (рис. 92, 1).
Лихарев, 1963 : 70, рис. 2.
Terra typica — Камчатка, окр. Милькова.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: более 30 экз. из типового местонахождения и из окр. Аяна
на побережье Охотского моря.
Рис. 92. Виды рода Vallonia (подрод Vallonia s. str.).
I — V. (Vallonia) cyclophorella hamtschatica Likh., голотип; II— V. (Vallonia) enniensis Gredl. из Болгарии,
Кюстандил; III — V. (Vallonia) pulchella (Miill.) из Москвы, Кунцево; IV — V. (Vallonia) excentrica Sterki
из Москвы, Кунцево; У — V. (Vallonia) ladacensis (Nev.) из Гиссарского хр.; VI — V. (Vallonia) tenuilabris
peteri Schileyko, suhsp. nov., голотип.
Раковина низкоконическая, тонкостенная, просвечивающая, составлена
Зх/4—Зх/2 выпуклыми ступенчатыми оборотами. Последний оборот в 1% раза
шире предпоследнего, к устью незначительно, с легким перегибом, опущен.
Высота завитка приблизительно равна высоте устья или несколько меньше.
Окраска бледнотроговая или сероватая. Эмбриональные обороты с тончайшей
спиральной скульптурой, заметной только на свежих раковинах, и с тонкой
микроскопической зернистостью. Дефинитивные обороты тонко исчерчены и не-
1. РОД VALLONIA
16'7
сут тонкие пластинчатые ребрышки (на последнем обороте около 50 ребрышек
или меньше). Устье округлое, Места его прикрепления умеренно сближены, края
тонкие, коротко отвернуты, без губы или с очень слабой губой. Пупок округ-
лый, развернутый, перспективный.
Размеры: ВР 1.1—1.3, БД 2.2—2.5, МД 1.9—2.1 м; голотип: ВР 1.25, БД 2.4,
МД 2.1 мм.
Распространение и местообитание. Подвид известен
из типового местонахождения, где встречен в лесу с преобладанием белой березы
и тополя, а также из окр. Аяна — в трещинах скал на обрывистом берегу над
морем. Номинативный подвид распространен в горных районах запада Северной
Америки.
4. Vallonia (Vallonia) enniensis (Gredler, 1856) (рис. 92, II).
G r e d 1 e г, 1856 : 56 (Helix pulchella var.); Sandbergs r, 1875, Land. u. Suss-
wasser-Conch. Vorwelt. : 856 (costellata); Лихарев, P амме льмейер, 1952 :
164 (pulchella var.); Sparks, 1953 : 111, figs, le—h; Ehrmann, 1956 : 53; Lozek,
1956 : 108, t. 8, 2; Z i 1 c h, J a e c k e 1, 1962 : 102; Lozek, 1964 : 222, t. 10, 2a—с; Д a-
м я н о в, Лихарев, 1975 : 134, фиг. 68н—p (pulchella enniensis).
Terra typica — Тироль (Австрия).
Место хранения типов не установлено.
Материал: 12 экз. из пяти местонахождений в Подмосковье, на Карпа-
тах (Рахов), в Болгарии (Кюстандил), Франции и Греции.
Раковина низкоконическая, тонкостенная, просвечивающая, составлена
Зх/2—4 довольно выпуклыми оборотами. Последний оборот в профиле плавно за-
круглен, почти вдвое шире предпоследнего, к устью почти не опущен или опущен
очень слабо, с легким плавным перегибом. Высота завитка примерно равна
высоте устья или немного меньше. Окраска желтовато-серая или раковина почти
бесцветная, прозрачная. Поверхность эмбриональных оборотов, микрозерни-
стая, без спиральной скульптуры. Дефинитивные обороты покрыты тонкой, но
резкой радиальной исчерченностью и узкими морщинками; через более или
менее правильные промежутки располагаются более резкие морщинки (ребры-
шки), лишенные кожистой оторочки; слабая оторочка имеется у еще не вполне
сформированных свежих раковин. Устье почти круглое, места его прикрепления
сближены, края острые, умеренно отогнутые под прямым углом и слегка уси-
лены по линии перегиба тонкой губой. Пупок углубленный, перспективный,
развернутый.
Размеры: ВР 1.1—1.3, БД 2.1—2.8, МД 1.9—2.5 мм.
Распространение и местообитание. Южная, юго-
восточная и средняя Европа; Малая Азия; Латвия; Закарпатье и Прикарпатье;
Московская обл.; окр. Новороссийска. Обитает преимущественно на лугах и
по хорошо освещенным опушкам лиственных лесов, а также в кустарниковых
зарослях.
5. Vallonia (Vallonia) pulchella (Muller, 1774) (рис. 92, III).
KJ ii 11 e г, 1774 : 30 (Helix)-, Pilsbry, 1892, Man. Conch., 8 : 248, t. 32, figs. 1 —5;
Лихарев, Рам мель мейер, 1952 : 164, рис.' 77А; Sparks, 1954 : 110—121,
fig. li—1; Ehrmann, 1956 : 53, t. I, fig. 23; Lozek, 1956 : 109, t. 8, 3; Z i 1 c h, J a-
e c k e 1, 1962 : 102; Lozek, 1964 : 221, t. 10, fig. 5a—с; M а т ё к и н, 1966 : 115; G i t-
t enb er ger et al., 1970 : 56, figs. 4, 47, 50, Kaart 21; Д амянов, Лихарев, 1975 :
133, фиг. 68и—м; Акрамовский, 1976 : 149, рис. 67А, табл. VII, 67.
Terra typica — Фридриксдаль у Копенгагена.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 57 местонахождений в Подмосковье,
на Кольском п-ове, Нижнем Поволжье, на Карпатах, в Крыму, на Кавказе и
в Закавказье, в Средней Азии, на Алтае и на Дальнем Востоке.
168
II. НАДСЕМ. ORCULOIDEA
Раковина прижатая, низкоконическа'я, умеренно тонкостенная, просвечи-
вающая, составлена З’Д—4 умеренно выпуклыми оборотами. Последний оборот
в профиле плавно широко закруглен, в 1х/2 раза шире предпоследнего, к устью
почти не опущен. Высота завитка составляет примерно половину высоты устья.
Окраска белая, светло-серая или желтоватая; свежие раковины нередко стекло-
видно-прозрачные. Спиральная скульптура на эмбриональных оборотах отсут-
ствует, но имеется неясная микроскопическая зернистость. Дефинитивные обо-
роты выглядят гладкими, но при увеличении в 20—30 раз заметны тонкая нерав-
номерная радиальная исчерченность и легкие морщинки. Устье круглое, косое,
места его прикрепления сближены, края устья сильно отвернуты, с хорошо разви-
той губой по линии перегиба. Пупок широкий, перспективный, углубленный.
Размеры: ВР 1.3, БД 2.1—2.6, МД 1.9—2.4 мм; в первоописании: «diam.
1 lin.», т. е. БД примерно 2.1—2.2 мм.
Внутреннее строение. Строение мужского отдела неизвестно;
Уотсон (Watson, 1920) приводит изображение полового аппарата афалличной
особи; мною исследовано несколько десятков экземпляров, но все они также
оказались афалличными.
Распространение и местообитание. Голарктика. На-
селяет влажные микробиотопы почти во всех ландшафтных зонах: подстилку
лиственных'и смешанных лесов, дерновины на пойменных лугах, в сухих мест-
ностях живет под камнями близ рек и ручьев; среди мха на опушках. На Коль-
ском п-ове изредка встречается в тундре.
6. Vallonia (Vallonia) excentrica Sterki, 1892 (рис. 92, IV).
Sterki in: Pilsbry, 1892, Man. Conch., 8 : 249, t. 32, figs. 6—9; Sterki,
1893 : 252, pl. 8, figs. В, M; Pilsbry, 1948 : 1025, fig. 545; Sparks, 1953 : 115, fig.
Im—p; Ehrmann, 1956 : 54, t. 2, Abb. 24; Lozek, 1956 : 109, t. 8, 4; Z i 1 c h, Jae-
ckel, 1962 : 103; Lozek, 1964 : 222, t. 10, 4a—c; Gittenberger et al., 1970 : 55,
figs. 46, 49, Kaart 20; Дамянов, Лихарев, 1975 : 134, фиг. 68Д—3 (pulchella excen-
trica).
Terra typica — не указана; сообщается, что живые моллюски найдены только
у Квебека, а пустые раковины обнаружены во многих местах Европы и Север-
ной Америки.
Место хранения типов не установлено.
Материал: более 100 экз. из 24 местонахождений в окрестностях Мо-
сквы, Ужгорода, на Северном Кавказе и в Западной Европе.
Раковина низкоконическая, умеренно тонкостенная, слабо просвечивающая,
составлена 23/4—Зх/4 выпуклыми оборотами. Последний оборот в профиле широко
и плавно закруглен, вдвое или немного более чем вдвое шире предпоследнего,
к устью раструбообразно расширен, не опущен. Высота завитка больше поло-
вины высоты устья. При рассмотрении раковины снизу бросается в глаза нару-
шение привычной геометрии раковины: контур ее не округлый, а овальный,
причем этот овал обычно не совсем правильной формы. Окраска белая, с серова-
тым или желтоватым оттенком; свежие раковины нередко стекловидные. Повер-
хность эмбриональных оборотов лишена элементов спиральной скульптуры,
покрыта очень неясной микроскопической зернистостью. Дефинитивные обороты
почти гладкие, с сильно сглаженными радиальными морщинками. Устье округ-
лое, места его прикрепления умеренно сближены и соединены тонкой мозолью.
Края устья слегка утолщены, отвернуты под прямым углом. По линии перегиба
имеется мощная губа, лишенная, однако, четких границ. Форма пупка отра-
жает своебразный характер нарастания оборотов: он имеет овальную форму, и
шов, видимый сквозь пупок, представляет собой спираль, как бы слегка сплю-
щенную в плоскости нарастания.
Размеры: ВР 1.1—1.2, БД 2—2.3, МД 1.8—2 мм; в первоописании: ВР 1.1,
БД 2.3, МД 1.8 мм.
Распространение и местообитание. Вид широко, но
спорадически распространен по территории Голарктики; вопрос о присутствии
169
вида в Сибири нуждается в выяснении. Обитает в тех же местах, что и V. pul-
chella', иногда оба вида встречаются вместе.
7. Vallonia (Vallonia) tenuilabris tenuilabris (Al. Braun, 1842).
Braun Al., 1842, D. Naturf. Vers. Ber. : 143 (Helix); Pilsbry, 1892, Man. Conch.,
8 : 258; Лихарев, Раммельмейер, 1952 : 165, рис. 78; Ehrmann, 1956 :
54; Lozek, 1956 : 107; Zilch, Jaeckel, 1962 : 104; Lozek, 1964 : 223, t. 10, fig.
la—c.
Terra typica — плейстоценовые отложения Европы.
Место хранения типов не установлено.
Материал: 22 экз. из Западной и Восточной Сибири (4 местонахождения).
Раковина низкоконическая, тонкостенная, составлена Зх/2—4 выпуклыми
оборотами. Последний оборот в профиле плавно и широко закруглен, в базаль-
ной части вздут, почти вдвое шире предпоследнего, возле самого устья еле за-
метно опущен (почти прямой). Высота завитка равна высоте устья или немного
меньше. Окраска серая, желтоватая или желтовато-серая. Эмбриональные
обороты покрыты микроскопической зернистостью и тончайшей неясной спи-
ральной струйчатостью; дефинитивные чрезвычайно тонко радиально исчерчены;
на последнем обороте имеются отдельные тонкие морщинки. Устье округлое,
косое, места его прикрепления умеренно сближены. Края устья тонкие, коротко
отвернутые, кроме верхней части палатального края; губа отсутствует. Пупок
широкий, перспективный. Размеры: ВР 1.7—1.8, БД 3—3.4, МД 2.8—3.1 мм.
Распространение и местообитание. В Европе встре-
чается в миндель-рисских отложениях. Ныне: Сибирь, горные области Средней
Азии, бассейн Амура, Северный Китай. Встречается в подстилке смешанных и
хвойных лесов.
7а. Vallonia (Vallonia) tenuilabris peteri Schileyko, subsp. nov. (рис. 92, VT).
Terra typica — Приморский край, зал. Петра Великого, о-в Козлова.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 22 экз. на 5 о-вах в зал. Петра Великого.
От номинативного подвида устойчиво отличается очень тонкой стекловидной
раковиной и меньшими размерами: ВР 1.3, БД 1.9—2.1, МД 1.75—1.8 мм;
голотип: ВР 1.3, БД 2.0, МД 1.8 мм.
Внутреннее строение. Вскрыто 4 экз., все оказались афаллич-
нЫми.
Распространение и местообитание. Вид найден на сле-
дующих Островах: Козлова, Два Брата, Верховского, Пахтусова, Веры. Обна-
ружен в трещинах скал и на мшистых камнях.
8. Vallonia (Vallonia) ladacensis (Nevill, 1878) (рис. 92, V).
N e v i 1 1, 1878 : 4 [Helix (Vallonia)}; E. Martens, 1882a : 3, t. 3, fig. 3 [7/. (Vallonia)];
Линдгольм, 1931 : 42; Лихарев, Раммельмейер, 1952 : 166 (tenuilabris
var. ladacensis); Матёкин, 1966 : 116 (tenuilabris ladacensis); Иззату ллаев, 19706 :
81; R a j a g о p a 1, S u b b a Rao, 1972 : 200, text-figs. 1,2; Акрамовский, 1976 :
150, рис. 67Б, табл. VII, 69.
Terra typica — Индия, Кашмир, xp. Ладакх.
Лектотип хранится в Зоологическом управлении Индии, Калькутта.
Материал: 13 экз. из двух местонахождений в Армении и Таджики-
стане.
Раковина прижатоконическая, с коническим контуром короткого завитка,
тонкостенная, хрупкая. Высота завитка примерно равна высоте устья или не-
много меньше. Оборотов 4, выпуклых, последний оборот почти вдвое шире пред-
последнего, к устью несильно, с легким перегибом опущен. Окраска желтовато-
роговая. Эмбриональные обороты выглядят почти гладкими, дефинитивные
170
II. НАДСЕМ. ORCULOIDEA
покрыты очень тонкими радиальными морщинками и редкими низкими слабыми
кожистыми ребрышками, которые располагаются через разные интервалы и
легко утрачиваются; на некоторых раковинах их вовсе не видно. Устье уме-
ренно косое, места его прикрепления сильно сближены, края Слабо отвернуты,
почти не утолщены, со слабой губой. Пупок развернутый, открытый, вполне
перспективный. Размеры (размах изменчивости велик): ВР 1.6—3.4, БД 2.5—
4, МД 2.2—3.3 мм; лектотип (по обозначению Rajagopal et Subba Rao, 1972):
BP 2.0, БД 4.0, МД 3.25 мм. У популяций, обитающих на территории СССР,
несмотря на заметные различия в размерах, прочие признаки сохраняют
постоянство. Возможно, что в Армении и в Таджикистане вид представлен
разными подвидами, отличающимися размерами раковин.
Распространение и местообитание. Горные области
Закавказья, Афганистана, Индии, Средней и Центральной Азии. Населяет
осыпи и горные леса.
2. Подрод PLANT VALLONIA Schileyko,. subgen. nov.
Типовой вид Vallonia zaru Almuhambetova, 1979.
Устье цельное, слегка обособлено от стенки предпоследнего оборота.
Ра спространение. Заилийской хр. Ныне — один вид.
В ископаемом состоянии подрод известен из среднего (?) миоцена Централь-
ного Казахстана.
9. Vallonia (Planivallonia) zaru Almuhambetova, 1979 (рис. 93).
Альмухамбетова, 1979 : 32, рис. 3.
Terra typlca — Средняя Азия, Заилийский хр., ущелье р. Тургень, санато-
рий Тау-Тургень.
Рис. 93. Vallonia (Planivallonia) zaru Almuhambetova. Голотип.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 1 раковина (голотип).
Раковина почти плоская, тонкостенная, слегка шелковисто поблескивающая.
Оборотов 4, сильновыпуклых, разделенных глубоко врезанным швом. Контур
последнего оборота в пришовной части плавно закруглен, затем на небольшом
отрезке почти прямой, горизонтальный, и далее на периферии оборота и при
переходе на базальную поверхность вновь образует широкое закругление. Близ
устья последний оборот плавно, но энергично, с легким перегибом, опущен.
Поверхность эмбриональных оборотов единственной раковины слегка корро-
дирована, но спиральной скульптуры, по-видимому, лишена. Дефинитивные
III. НАДСЕМ., PUPILLOIDEA
171
обороты несут многочисленные нерегулярно расположенные тонкие радиальные
морщинки, которые на базальной части раковины выражены значительно сла-
бее. Устье цельное, отстает от стенки раковины; края его тонкие, сильно отвер-
нутые, с исчезающе слабой губой. Позади устьевого края имеется кольцевая
перетяжка, резче всего выраженная на базальной части. Пупок чрезвычайно
широкий, очень мелкий.
Размеры голотипа: ВР 1.0, БД 2.8, МД 2.2 мм.
Вид наиболее близок Vallonia gigantea Steklov, 1967 (см.: Стеклов, Цытович,
1967), описанной из среднего (?) миоцена в долине Костанкол (центральный
Казахстан); сходство между этими видами определяется наличием цельного,
заметно отстающего устья'. V. zaru отличается от V. gigantea гораздо более мел-
кими размерами и резко уплощенной раковиной.
Распространение и местоообитание. Единственный эк-
земпляр обнаружен на территории санатория Тау-Тургень на Заилийском хр.,
в траве.
III. Надсем. PUPILLOIDEA Turton, 1831.
Раковина небольшая, от удлиненно-овальной до цилиндрической, с радиаль-
ной скульптурой. Эмбриональные обороты покрыты микроскопической зер-
нистостью. Пупок точковидный. Устье с зубами или без зубов, однако длинные
париетальные и колумеллярные пластинки отсутствуют; они могут быть только
в приустьевой части и формируются в субдефинитивном состоянии. Края устья
слегка отвернуты, с губой и затылочным утолщением.
Семенные пузырьки не обособлены от гермафродитного протока. Пенис
с эпифаллусом, внутри гладкий или с вялыми продольными складками. На про-
ксимальной части эпифаллуса, близ границы с пенисом, имеется небольшой
цилиндрический придаток; пениальная ветвь полового ретрактора крепится
дистальнее придатка эпифаллуса. Пениальный аппендикс имеется, половой
ретрактор двуветвистый.
Сем. PUPILLIDAE Turton, 1831
Turton, 1831 : 8; Р i 1 s b г у, 1921, Man. Conch., 26: 1; Steenberg, 1925 : 169.
Устье с 1—7 зубами, реже без зубов. Края устья в равной степени отвернуты,
очень редко прямые. Пупок узкий — щелевидный или проколовидный.
Простата представлена небольшим числом долек у нижнего края белковой
железы. Папилла пениса и бич отсутствуют. Дивертикул семяприемника имеет
наклонность к редукции.
Распространение. По всему земному шару. 1—2 подсемейства.
В ископаемом состоянии семейство известно начиная с палеоцена (Северная
Америка: род Albertanella) и верхнего олигоцена (Северная Америка, Европа,
Северная Африка, Восточная Азия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ PUPILLIDAE
1 (2). Раковина коническая ............................3. Pupoides.
2 (1). Раковина от овальной до цилиндрической.
3 (4). Зубов в устье не более 4..........................2. Pupilia.
4 (3). Зубов в устье не менее 5.....................1. Gibbulinopsis.
1. Род GIBBULINOPSIS Germain, 1919
Germain, 1919 : 265 [Pupilla (Gibbulinopsis)]; Thiele, 1931 : 508 (род Pupilia,
секция Gibbulinopsis); W e n z, Zilch, 1959—1960 : 166 [Pupilla (Gibbulinopsis)].
Типовой вид .Orthogibbus pupula Deshayes, 1863; по первоначаль-
ному обозначению.
В устье обычно 6 зубов: ангулярный, париетальный, колумеллярный и три
палатальных; верхний палатальный может отсутствовать. Ангулярный в виде
172
III. НАДОЕМ. PUPILLOIDEA
пластинки, лежащей почти в плоскости устья, а париетальный расположен
перпендикулярно плоскости устья.
Распространение. Кавказ, Средняя и Центральная Азия, Даль-
ний Восток и Япония; Африка; о-в Реюньон (типовое местонахождение типо-
вого вида); Австралия. 10—15 видов.
В ископаемом состоянии род известен начиная с верхнего олигоцена (Ев-
ропа, восточная Азия).
таблица ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ GIBBULINOPSIS
1 (6). Нижняя палатальная складка удлиненная. Устье цельное, реже прер-
вано на париетальной стенке (подрод Primipupilla).
2 (5). Палатальных складок две. Высота последнего оборота больше 1/3 высоты
раковины.
3 (4). Ширина раковины 1.8—2 мм.....................4. G.' (Р.) signata.
4 (3). Ширина раковины 1.2—1.4 мм...............5. G. (Р.) nanosignata.
5 (2). Палатальная складка одна. Высота последнего оборота меньше V3 высоты
раковины.......................................3. G. (Р.) gracilis.
6 (1). Нижняя палатальная складка в виде округлого бугорка. Устье прервано
на париетальной стенке (подрод Gibbulinopsis s. str.)
7 (8). Обороты выпуклые. Вершина ангулярного зуба направлена вниз и лишь
слегка отклонена к палатальной стенке. Вид обитает на Кавказе . . .
................................................ 2. G. (G.) interrupta.
8 (7). Обороты слабовыпуклые. Вершина ангулярного зуба направлена к верх-
ней части палатального края. Вид обитает на Дальнем Востоке . . .
................................................1. G. (G.) cryptodon.
1. Подрод GIBBULINOPSIS s. str.
Раковина от овально-цилиндрической до короткоцилиндрической, слабо
скульптурированная. Край устья прерван на париетальной стенке.
Распространение как у рода.
1. Gibbulinopsis (Gibbulinopsis) cryptodon (Heude, 1880) (рис. 94, I).
H e u d e, 1880 : 77, pl. 18, fig. 20 (Pupa); M oellen dorf f, 1884, Jahrb. dtsch.
Malak. Ges., XI : 176 [Pupa [Pupilla) heudeana]; 1901 : 381 (Pupilla heudeana et subsp. grandis);
Pilsbry, 1921, Man. Conch., 26 : 200, pl. 22, figs. 1—-5 (P. heudeana); Yen, 1939 : 74,
t. 6, fig. 31, 32 (P. cryptodon et subsp. grandis); Kuroda, 1963 : 26 [Pupilla).
Как видно из синонимики, Меллендорф заменил название cryptodon на том
основании, что счел его гомонимом Pupa cryptodus Al. Braun, 1842, и Пилсбри
использовал название, предложенное Меллендорфом. Однако эти названия раз-
личаются двумя буквами, и, следовательно, не конкурируют.
Terra typica — Цзянси, юго-восточный Китай.
Место хранения типов не установлено.
Материал: более 100 экз. из 8 местонахождений на островах залива
Петра Великого.
Раковина от овально-цилиндрической до короткоцилиндрической, слегка
просвечивающая, шелковисто поблескивающая, с широко закругленной вер-
шиной. Оборотов 5х/2—6, выпуклых, последние три оборота (у коротких рако-
вин — последние два) почти одной ширины. Последний оборот к устью медленно
и плавно поднимается, высота его немного более г/3 высоты раковины, у корот-
коцилиндрических форм — немного меньше половины высоты раковины.
Окраска одноцветно роговая. Скульптура эмбриональных оборотов в виде тон-
чайшей сетки, слегка напоминает соты. Дефинитивные обороты покрыты тон-
кими правильными притупленными ребрышками или морщинками. Устье округ-
лое, края его широко отвернуты и утолщены. Снаружи свободный край устья
1. РОД GIBBULINOPSIS
173
окаймлен хорошо развитым желтоватым затылочным утолщением. Зубов
обычно 6: в глубине устья располагается пластинчатая, слегка изогнутая про-
дольная париетальная пластинка; почти на одной линии с нею, чуть ближе
к палатальной стенке, расположен ангулярный зуб, отклоненный вправо.
Колумеллярная пластинка начинается у заднего края столбика и тут же уходит
в глубь раковины. Два расположенных рядом бугорковидных палатальных
зуба, из которых нижний развит лучше
верхнего, лежат в глубине устья. Наконец,
имеется зубовидное палатальное утолщение
губы, лежащее в плоскости устья. Пупок
открытый, проколовидный, круглый или
слегка овальный.
Размеры: ВР 2.3—3, БД 1.6—1.8 мм.
Рис. 94. Виды рода Gibbulinopsis (подрод Gibbu-
linopsis s. str.).
I — G. (Gibbulinopsis) cryptodon (Heude) из о-ва Пастухова,
зал. Петра Великого; II — G. (Gibbulinopsis) interrupta
(Reinh.) из с. Коджори близ Тбилиси.
I
п
Внутреннее строение. Вскрыто более 30 экз. из острова Два
Брата в зал. Петра Великого; у всех особей в матке содержались яйца (время
сбора август), а мужской отдел был настолько редуцирован, что представлял
собой систему тончайших нитевидных трубочек; можно лишь установить, что
семяпровод впадает, видимо, терминально, и пениальный аппендикс, а также
придаток эпифаллуса присутствуют.
Распространение и местообитание. Восточный Китай,
Япония, Корея; В СССР обнаружен на о-вах в зал. Петра Великого, где встре-
чается большими колониями в трещинах скал.
2. Gibbulinopsis (Gibbulinopsis) interrupta (Reinhardt, 1876) (рис. 94, II).
Reinhardt in : E. Martens, 1876b : 367 [Bulimus (Pupilla)]; Reinhardt,
1877a : 79, t. 3, fig. 4 (Pupa)-, О. В о e t t g e r, 1880a : 237 [P. (Pupilla)]- 1881 : 228 [P. (Pu-
pilla)]; Rosen, 1911 : 120 [P. (Pupilla)]; 1914 : 193 [P. (Pupilla)]; Pilsbry, 1921, Man.
Conch., 26 : 196, pl. 21, figs. 5—8 (Pupilla); Акрамовский, 1949 : 144 (Pupilla); Л и-
харев, Раммельмейер, 1952:148 (Pupilla); Акрамовский, 19716:6
(Pupilla); 1976 : 146, рис. 64Д, табл. VI, 62 (Pupilla).
Terra typica — Боржоми.
Место хранения типов не установлено.
Материал: более 40 экз. из 5 местонахождений в Грузии, Армении,
Азербайджане и горном Дагестане.
Раковина короткоцилиндрическая, реже овально-цилиндрическая, слабо
блестящая, с широко закругленной вершиной. Оборотов 5х/2—бУз, выпуклых,
последний оборот к устью слегка и плавно, без перегиба, поднят; высота его
заметно меньше половины высоты раковины. Окраска одноцветная, от светло-
роговой до светло-серой. Скульптура в виде тонкой радиальной исчерченности,
которая на нижних оборотах становится неясной и размытой. Устье округлое,
места его прикрепления слегка сближены и соединены тонкой мозолью. Заты-
лочное утолщение мощное, той же окраски, что и фон раковины, или чуть свет-
лее. Зубов 5: чуть отступя от края располагается приостренный бугорок, кото-
рый можно толковать и как ангулярный, и как переднюю часть париетальной
пластинки. Вершина бугорка направлена вниз и лишь слегка отклоняется
к палатальной стенке. На одной линии с названным бугорком, значительно
глубже в устье лежит короткая париетальная пластинка. Бугорковидная
колумеллярная пластинка занимает заднюю часть столбика. Значительно
174
III. НАДСЕМ. PUPILLOIDEA
отступя от края устья расположены два палатальных бугорка: нижний, более
вытянутый, которому на затылке соответствует короткая вмятина, и более
округлый верхний. Кроме того, на середине палатального края имеется выра-
женное в разной степени утолщение губы. Пупок сквозной, короткокаплевид-
ный. 1
Размеры: ВР 3—3.5, БД 1.5—1.8 мм; в первоописании: ВР 3, БД 1.5,
высота и ширина устья 1 мм.
Внутреннее строение. Исследовано 12 экз. из окр. Тбилиси,
но у всех изученных особей матка содержала крупные яйца, а мужской отдел
был недоразвитым.
Распрост.ранение и местообитание. Центральная часть
Северного Кавказа, Дагестан, центральное и восточное Закавказье. Обитает
в подстилке редколесья, под камнями в степной и полупустынной зонах.
2. Подрод PRIMIPUPILLA Pilsbry, 1921
Pilsbry, 1921, Man. Conch., 26 : 153 (род Pupilla, секция Primipupilla).
Типовой вид Pupa signata Mousson, 1873; по первоначальному обо-
значению.
Раковина высокоцилиндрическая, с резкой радиальной скульптурой. Устье,
как правило, цельное.
Распространение. Закавказье, Средняя Азия, Афганистан, се-
веро-западный Китай. Несколько видов.
В ископаемом состоянии подрод известен начиная , со среднего миоцена
(Предкавказье).
3. Gibbulinopsis (Primipupilla) gracilis (Izzatullaev, 1970) (рис. 95, I).
Иззатуллаев, 1970a : 53, рис. 1 (Pupilla); 19706:81 (Pupilla).
Terra typica — Таджикистан, Гиссарский xp., выбросы p. Сардай-Мион
близ пос. Ромит.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 1 экз. (голотип).
Раковина высокая, стройная, почти строго цилиндрическая, с короткой
широко закругленной вершиной, довольно твердостенная. Оборотов 8х/2, уме-
ренно выпуклых. Последний оборот к устью очень слабо приподнят, высота его-
немного менее трети высоты раковины. Об окраске судить трудно, так как един-
ственный известный экземпляр представлен пустой раковиной, утратившей пер-
воначальный цвет. Похоже, что раковины живых моллюсков имеют одноцветно-
роговую окраску. При увеличении более чем в 50 раз заметно, что поверхность
эмбриональных оборотов мелкозернистая. Дефинитивные обороты покрыты
правильными сглаженными радиальными ребрышками. Устье цельное, края
его широко отвернуты и образуют манжету. Ангулярный зуб имеет вид корот-
кого округлого гребня, лежащего почти в плоскости устья. Париетальная
пластинка высокая, задний конец ее сквозь устье не просматривается. Колу-
меллярная пластинка при нормальном положении устья не видна, но, повора-
чивая раковину, можно заметить ее в виде бугорковидного утолщения на задней
поверхности столбика. На нижней части палатальной стенки видна невысокая'
расплывшаяся, слегка изогнутая складка. Чуть выше середины палатального
края губа образует утолщение, лежащее в плоскости устья. Позади устьевого
края лежит хорошо выраженное затылочное утолщение, за которым имеется
продольная вмятина, отвечающая палатальной складке. Пупок округлый.
Размеры голотипа: ВР 5.2, БД 1.8 мм.
Вид описан по единственному экземпляру, поэтому нет уверенности в его
самостоятельности. Не исключено, что это высокая форма G. signata, так как
у некоторых популяций этого вида наблюдаются стадии редукции верхней
1. РОД GIBBULINOPSIS
175
палатальной складки. Архангельский (1933) сообщает, что в окр. Самарканда
по берегам р. Сиаб, в Ургуте и Сары-Мазаре не раз была найдена форма, кото-
рую он обозначил как Pupilla sp. и которая «отличается от других Pupilla
размерами — до 5 мм, коричневым цветом и косой, хорошо заметной исчерчен-
ностыо» (loc. cit., с. 22). Причем Архангельский отличал эту форму от G. sig-
nata, поскольку этот вид упоминается отдельно. Возможно, что Архангельский
имел дело с G. gracilis.
Рис. 95. Виды рода Gibbulinopsis (подрод Primipupilla),
I — G. (Primipupilla) gracilis (Izzatullaev), голотип; II — G. (Primipupilla) signata (Mousson) из Ферган-
ского xp.; Ill — то же, из Гиссарского xp., правый берег р. Искандердарья; IV — G. (Primipupilla
nanosignata Schileyko et Izzatullaev, голотип.
Распространение и местоообитание. Вид описан по
одной раковине, найденной в выбросах р. Сардай-Мион. Если подтвердится
предположение,' что упоминаемая Архангельским Pupilla sp. относится к этому
виду, то ареал помимо высокогорья Гиссарского хр. охватывает также окр, Са-
марканда.
4. Gibbulinopsis (Primipupilla) signata (Mousson, 1873) (рис. 95, II, III;
рис. 96).
? I s s e 1, 1865 : 423, t. 2, figs. 15—17 (Pupa armeniaca); Mousson, 1873 : 211»
pl. 8, fig. 7 (Pupa); Мартенс, 1874 : 23, табл. 2, фиг. 19, табл. 3, фиг. 40 (Р. cristata);
Е. Martens, 1882а : 28 (Pupa); G г е d 1 е г, 1889, Nachrbl. dtsch. malak. Ges., 21:162
(P. diecki); Westerlund, 1893, Nachrbl. dtsch. malak. Ges., Aug. : 121 (P. signata var.
cyclostoma et var. debilis); Pilsbry, 1921, Man. Conch., 26 : 194, pl. 21, figs. 1—3 (Pupilla);
Лихарев, Рамме ль мейер, 1952 : 150, рис. 61 (Pupilla); Матёкин, 1966 :
111, табл. 2, рис. 7 (Pupilla); Акрамовский, 1976 : 146, рис. 54Е, табл. VI, 63 (Pu-
pilla).
Terra typica — выносы Аракса.
Место хранения типов не установлено.
Материал: более 100 экз. из 32 местонахождений на Кавказе и в Сред-
ней Азии.
Раковина цилиндрическая, с широко закругленной вершиной, умеренно
твердостенная; свежие раковины шелковисто поблескивают. Оборотов 6—8,
выпуклых, последний оборот близ устья заметно, с плавным перегибом, поднят,
высота его немного больше трети высоты раковины. Окраска одноцветная:
176
III. НАДСЕМ. PUPILLOIDEA
светло-роговая, коричневая или (реже) каштановая. Эмбриональные обороты
с микроскопической, но ясной зернистостью. Скульптура дефинитивных обо-
ротов в виде тонкой радиальной ребристости; обычно ребрышки в той или иной
мере сглажены, некоторые из них вильчато расщеплены. Устье цельное, причем
иногда встречаются раковины, у которых устьевой край отчленен от стенки
раковины по всему периметру. Края устья широко отвернуты, образуют ман-
жету. Помимо утолщения на середине палатального края, лежащего в плоскости
устья, имеется 5 зубов: париетальная изогнутая пластинка, нередко связанная
с ангулярный бугорком; глубоко
лежащий колумеллярный зуб; две
палатальные складки, причем ниж-
няя длинная, при нормальном по-
ложении раковины ее задний ко-
нец не виден, так как закрыт
столбиком (снаружи этой складке
соответствует вмятина), а верхняя
палатальная заметно короче, часто
бугорковидная. Нередко эти склад-
ки взаимосвязаны припухлостью
палатальной стенки. Затылочное
утолщение хорошо выражено; при
рассматривании раковины снизу
видно, что в базальной части оно
переходит в притупленный киль,
частично обходящий проколовид-
ный пупок.
Размеры: ВРЗ.З-5.4, БД 1.8-
2 мм; в первоописании: ВР 4,
БД 2 мм.
Рис. 96. Gibbulinopsis (Primipupilla)
signata (Mouss.) из долины р. Нарын,
Тянь-Шань, 1 VI 1972 г. Половой ап-
парат.
Внутреннее строение. Материал: 4 экз. из окр. Еревана,
15 сентября 1939 г., сбор и определение Н. Н. Акрамовского; 7 — из склонов
на правом берегу р. Нарын близ Таш-Кумыра, 1 июля 1972 г., сбор и определе-
ние автора.
Белковая железа с развитой в разной степени вырезкой на боковой стороне.
Семяпровод впадает с эпифаллус терминально; эпифаллус относительно длин-
ный, равномерно цилиндрический. Хорошо развитый удлиненно-конический
придаток эпифаллуса на конце закруглен, располагается ниже места прикрепле-
ния полового ретрактора и делит эпифаллус в отношении 1 : 3—1 : 4. Дисталь-
ный отдел пениса цилиндрический, проксимальный утолщенный, веретеновид-
но-цилиндрический, короче дистального. и А2 неясно отделены один от
другого, А3 плавно, без резкой границы переходит в А4. Половой ретрактор кре-
пится к диафрагме единой лентой, которая раздваивается, подходя к эпифал-
лусу; пениальная ветвь заметно короче ветви аппендикса, прикрепляющейся
примерно к середине Кроме того, выше середины эпифаллуса имеется еще
один тонкий дополнительный ретрактор. Дивертикул семяприемника тонкий
и длинный, немного не доходит до белковой железы; резервуар семяприемника
небольшой, округлый, с собственным коротким протоком. Простата в виде
нескольких долек, расположенных на верхней части матки. У особей, собранных
в сентябре под Ереваном, матка была наполнена развивающимися эмбрионами,
2. РОД PUPILLA
ПТ
а совокупительный аппарат был представлен очень тонкими протоками с явными
признаками дегенерации; экземпляры, собранные 1 июня на берегу р. Нарын,
имели толстостенную матку, эмбрионы отсутствовали, мужской отдел хорошо
развит.
Распространение и местообитание. Восточное Закав-
казье, Северный Иран, юг Средней Азии, Афганистан, северо-западный Китай,
юго-восточный Казахстан. Населяет степные, полупустынные и горностепные
участки, где держится в мелкообломочных осыпях, под камнями и в скоплениях
отмерших растительных остатков. Нередко образует огромные скопления,
плотность которых достигает нескольких сотен'экз. на 1м2.
5. Gibbulinopsis (Primipupilla) nanosignata Schileyko et Izzatullaev, 1980
(рис. 95, IV).
Шилейко, Иззатуллаев, 1980 : 283, рис.
Terra typica — Гиссарский хр., правый берег р. Искандердарья.
Голотип и 12 паратипов хранятся в Зоологическом ин-те АН СССР,
48 паратипов — в Институте зоологии и паразитологии АН ТаджССР.
Материал: 60 экз. из 5 местонахождений на Туркестанском и Гиссар-
ском хребтах.
Раковина цилиндрическая, с широко закругленной вершиной, сравнительно
тонкостенная, с шелковистым блеском. Оборотов 5—6 (у голотипа 6), выпуклых,
последний оборот к устью незначительно приподнят, высота его больше 1/3 вы-
соты раковины, но меньше х/2 этой величины. Окраска коричневая. Эмбриональ-
ные обороты с неясной зернистостью. Скульптура дефинитивных оборотов
в виде тончайшей ребристости. Край устья на париетальной стенке у одних
раковин прерван, у других имеется развитая в разной степени париетальная
мозоль; когда эта мозоль развита сильно, создается впечатление цельного устья.
Устьевые края широко отвернуты, образуют манжету. Кроме утолщения пала-
тального края, лежащего в плоскости устья, имеется 5 зубов: ангулярный
бугорок, глубоко лежащая париетальная пластинка, колумеллярный зуб и
две палатальные бугорковидные складки примерно одинаковой величины и
формы. На затылке вмятина, соответствующая нижней палатальной складке,
развита слабо или отсутствует. Пупок проколовидный.
Размеры: ВР 2.3—3.1, БД 1.2—1.4 мм; голотип: ВР 2.9, БД 1.3 мм.
Распространение и местообитание. Северные предгорья
восточной части Туркестанского хр. — с. Ворух; там же обнаружен в выносах
реки; Гиссарский хр. — окр. озер Искандеркуль и Варенкуль, а также по
берегам р. Искандердарьи. В окр. с. Ворух обнаружен вместе с G. signata;.
на Гиссарском хр. встречен как вместе с G. signata, так и в виде изолированных
популяций.
2. Род PUPILLA Turton, 1831
Turton, 1831 : 99; Pilsbry, 1921, Man. Conch., 26 : 152; Thiele, 1931 : 508;.
Лихарев, Раммельмейер, 1952 : 145; Wenz, Zilch, 1959—1960 : 165;
Стеклов, 1966 : 152; Акрамов ский, 1976 : 143.
Типовой вид Pupa marginata Draparnaud, 1801 (syn. Turbo muscorum
Linnaeus, 1758); по монотипии.
В устье не более 4 зубов: париетальный, колумеллярный и два палаталь-
ных; однако элементы устьевой арматуры непостоянны, зубы развиты отно-
сительно слабо и подвержены редукции вплоть до полного исчезновения всех
зубов. Ангулярный зуб если имеется, то выражен очень слабо и имеет вид ма-
ленького округлого бугорка.-
Распространение. Голарктика. Не менее 30 видов.
В ископаемом состоянии род известен начиная с верхнего олигоцена (Ев-
ропа, Северная Африка, Северная Америка).
12 А. А. Шилейко
178
Ш. НАДСЕМ. PUPILLOWEA
1
2
3
4
5
6
7
8
9
10
И
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ PUPILLA
(24). Раковина твердостенная или умеренно хрупкая. Имеется губа и раз-
витое в разной степени затылочное утолщение. Устьевые зубы большей
частью имеются (подрод Pupilla s. str.).
(3). Раковина левозавитая ...........................4. Р. (Р.) gallae.
(2). Раковина правозавитая.
(5). На палатальном крае устья, немного отступя от сильно развитой губы,
расположена удлиненная складка, продольная ось которой почти па-
раллельна краю устья; эта складка имеет тенденцию расщепляться вдоль
на два округлых бугорка.........................7. Р. (Р.) inequidenta.
(4). Палатальная стенка устья без складок; если складки имеются, они
имеют иной облик и направлены по ходу оборотов либо представлены
округлыми бугорками.
(7). На палатальной стенке устья имеются два бугорковидных зуба одина-
ковой формы и размеров..........................1. Р. (Р.) bipapulata.
(6). На палатальной стенке устья не более одного зуба; если же зубов два,
то один из них бугорковидный, другой вытянутый, в виде складки.
(17). Раковина цилиндрическая.
(12). Радиальная скульптура в виде тончайших правильно расположенных
ребрышек, часто имеющих кожистую оторочку (особенно на ранних
дефинитивных оборотах).
(11) . Устье без зубов или имеется слабый париетальный зуб. Затылочное
утолщение слабое или умеренно развито .... 10. Р. (Р.) turcmenica.
(10)^ Устье с тремя зубами, реже с двумя — тогда имеются колумеллярный
и париетальный зубы. Затылочное утолщение хорошо развито . . . .
................................................ 9. Р. (Р.) sterrii.
(9) . Раковина почти гладкая или с радиальными морщинками; кожистые
оторочки в любом случае отсутствуют.
(14) . В устье не менее трех ясно выраженных зубов...................
.............................................. 2. Р. (Р.) triplicate.
(13). Зубов в устье не больше двух.
(16). Раковина высокая, стройная, устье без зубов ............... . . .
............................................ И. Р. (Р.) anzobica.
(15). Раковина приземистая, короткоцилиндрическая, устье с одним или
двумя зубами.........................................6. Р. (Р.) inops.
(8). Раковина овально-цилиндрическая или удлиненно-овальная.
(19). Дефинитивные обороты покрыты плавными равномерно расположен-
ными радиальными морщинками. На линии перегиба устьевого края
располагается мощная порожистая губа. Зубы в устье отсутствуют . . .
............................................ 8. Р. (Р.) striopolita.
(18) . Дефинитивные обороты гладкие или покрыты неравномерно располо-
женными морщинками. Губа имеется, умеренно развита. Зубы в устье
имеются или отсутствуют.
(21) . Колумеллярный зуб хорошо развит, часто приостренный...........
..............................................2. Р. (Р.) triplicate.
(20) . Колумеллярный зуб отсутствует или слабый, в виде небольшой при-
пухлости на задней части столбика.
(23) . При 6 оборотах большой диаметр раковины не менее 1.6 мм . . .
..............................................5. Р. (Р.) muscorum.
(22). При 6 оборотах большой диаметр раковины не превышает 1.5 мм
........................;.....................3. Р. (Р.) bigranata.
(1). Раковина хрупкая. Губа, затылочное утолщение и устьевые зубы отсут-
ствуют (подрод F г agi pupilla).
(26). Раковина стройная, отношение ее высоты к большому диаметру не
менее 1.8 . ............................ '......12. Р. (F.) alabiella.
(25). Раковина' приземистая, отношение ее высоты к большому диаметру
не более 1.7 ..................................13. Р. (F.) limata.
г. РОД PUPILLA
179
1. Подрод PUPILLA s. str.
Раковина относительно твердостенная, с затылочным утолщением; устье
обычно с зубами.
Р аспространение как у рода.
Рис. 97. Виды рода Pupilla (подрод Pupilla s. str.).
I — P. (Pupilla) bipapulata Акт. из Армении, с. Гнишик; II — Р. (Pupilla) triplicata (Stud.) из ущелья
р. Фиагдон на Северном Кавказе; III — Р. (Pupilla) bigranata (Rssm.) из долины р. Оки у с. Белые Коло-
дези; IV — то же, из Загреба, Югославия; V — С. (Pupilla) gallae Tzv. из Джунгарского хр.; VI —
Р. (Pupilla) muscorum (L.) из Подмосковья, с. Ромашково; VII — то же, из Ужгорода; VIII — то же,
из долины р. Поной на Кольском полуострове; IX — то же, из Зеравшанского хр. (форма asiatica Mlldff);
X — то же, из окр. Ульяновска (форма milaschevitschi landh.).
1. Pupilla (Pupilla) bipapulata Akramowski, 1947 (рис. 97, Г, рис. 98).
Акрамовский, 1947а (1943) : 255; 1947в : 182, рис. 3, 4; 1949 : 144; Лихарев,
Раммельмейер, 1952:149; Акрамовский, 19716:6; 1976:146, рис. 64Б,
табл. VI, 61.
12*
180
III. НАДСЕМ. PUPILLOIDEA
Terra typica — Армения, Абовянский р-н, Гегард.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: более 30 экз. из типового местонахождения и из окр. с. Гни-
шик, Армения.
Раковина цилиндрическая, с правильно закругленной вершиной, слабо-
блестящая. Оборотов 6—б^з, сильновыпуклых. Последний оборот к устью
очень плавно и незначительно поднят, высота его немного более чем вдвое
меньше высоты раковины. Окраска одноцветная, светло-роговая. Дефинитивная
скульптура в виде тончайших радиальных ребрышек, которые на верхних
Рис. 98. Pupilla (Pupilla) bipapulata Akr. из Армении, с. Гнишик. Половой аппарат двух
экземпляров.
оборотах располагаются более правильно и тесно, чем на нижних. Устье округ-
лое, края его отвернуты; места прикрепления слегка сближены и связаны тон-
кой мозолью, часто совсем прозрачной. Зубов 4: слегка изогнутая париеталь-
ная пластинка, колумеллярный и два палатальных. Если удалить часть задней
стенки последнего оборота, то можно заметить, что задний конец париетальной
пластинки круто понижается, затем эта пластинка продолжается еще на неко-
торое расстояние в виде светлого нитевидного утолщения, очень плавно сходя-
щего на нет. Колумеллярный зуб бугорковидный, занимает задний край стол-
бика. Две палатальные складки хорошо развиты, причем нижней соответствует
размытый желобок на затылке. Затылочное утолщение явственное. Пупок про-
коловидный.
Размеры: ВР 2.2—2.7, БД 1.25 мм; голотип: ВР 2.25, БД 1.25 мм.
Внутреннее строение. Материал: 5 экз. из Армении, с. Гни-
шик, 1941 г., сбор Э. А. Давтяна, определение Н. Н. Акрамовского.
Белковая железа с резким и глубоким латеральным желобком. Семяпровод
впадает в эпифаллус терминально. Эпифаллус цилиндрический, слабо извит.
Придаток эпифаллуса тупоконечный, почти цилиндрический, хорошо развит.
2. РОД PUPILLA 181
Граница между пенисом и эпифаллусом внешне практически не выражена.
Дистальная часть пениса короткая, поэтому пениальный аппендикс отходит
от пениса в непосредственной близости с половым атриумом. Аг и А2 внешне
почти не отделены один от другого, но при исследовании внутреннего строения
можно установить, что А2 сохраняет нормальное строение, т. е. имеется вну-
тренняя полость, в которую коротким сосочком вдается Л3, снабженный тонким
каналом и терминальным отверстием. А3 хорошо развит и ясно отделен от А4.
Пениальная ветвь полового ретрактора очень короткая, может быть раздвоен-
ной; ветвь ретрактора крепится к середине Аг. Резервуар семяприемника
очень тонкий, короткий, примерно того же размера и той же формы, что и
дивертикул; в результате складывается впечатление, что проток семяприемника
на конце вильчато раздвоен. Среди исследованного материала оказались особи
с развивающимися яйцами в матке (рис. 98, слева), а также экземпляры, матка
которых не содержала яиц (рис. 98, справа). У первых мужской отдел в значи-
тельной степени редуцирован (редукция особенно коснулась пениального ап-
пендикса), у вторых все участки мужского отдела хорошо развиты.
Распространение и местообитание. «Южная и юго-
восточная Армения: от бассейна реки Азат через Даралагез до Кафанского
и Мегринского районов: горностепной пояс и сухие пастбища нижнеальпий-
ской полосы; фригана и горная степь, в сухих растительных остатках под колю-
чими подушковидными полукустарниками, под кустами и в трещинах дерна»
(Акрамовский, 1976, с. 146). Самадов (1954, 1960) указывает, что вид живет
также в Ферганской долине; это сообщение вызывает сомнение.
2. Pupilla (Pupilla) triplicata (Studer, 1820) (рис. 97, IT, рис. 99).
Studer, 1820 : 89 [Glischrus (Pupa)]; Reinhardt, 1877a : 79, pl. 3, fig. 2 (Pupa
triplicata luxurians); 0. Boettger, 1879 : 26 [P. (Pupilla) suboviformis]; Pilsbry, 1921,
Man. Conch., 26 : 189, t. 21, figs. 11—13; Steenberg, 1925 : 66, pl. 11, figs. 1—3; A k-
p амовский, 1949 : 144; Лихарев, Раммельмейер, 1952 : 148; Ehr-
mann, 1956 : 48, t. I, fig. 19; L о z e k, 1956 : 102; Zilch, Jaeckel, 1962 : 99; L o-
zek, 1964 : 214, t. 9, fig. 5; M а т ё к и н, 1966 : 111, табл. 2, рис. 6; Ш и л е й к о, 1969 :
30, рис. 2 (triplicata f. luxuriana); Дамянов, Лихарев, 1975 : 100, фиг. 33; Акра-
мовский, 1976 : 145, рис. 63, 64В, табл. VI, 60, 65.
Terra typica — «Kt. Waadt bei Bex-Vieux» (Швейцария, кантон Во).
Лектотип хранится в Естественноисторическом музее в Берне (Forcart,
1957: 195, Taf. 3, Fig. 15).
Материал: более 150 экз. из 34 местонахождений в Западной Европе,
в Московской обл., в Крыму, на Кавказе, в Средней Азии, на Байкале.
Раковина от удлиненно-овальной до почти цилиндрической, умеренно бле-
стящая, с закругленной слегка суженной вершиной. Оборотов 51/2—6, уме-
ренно выпуклых или уплощенных. Последний оборот к устью плавно, но до-
вольно энергично приподнят, высота его всегда заметно меньше половины вы-
соты раковины, а у относительно высоких цилиндрических раковин — немного
более трети этой величины. Окраска одноцветно-роговая, разных оттенков;
в нескольких пунктах восточного Кавказа встречены серовато-белые раковины.
Эмбриональные обороты покрыты неясной зернистостью. Скульптура дефини-
тивных оборотов представлена тонкой радиальной морщинистостью, иногда
морщинки тонкие и густо расположенные, иногда расплывшиеся и сравнительно
редкие; встречаются и слабо скульптурированные раковины. Устье округлое,
большей частью с тремя зубами: короткой париетальной пластинкой, колумел-
лярным бугорковидным утолщением и палатальной складкой, постепенно схо-
дящей на нет в глубине устья. Иногда добавляется еще один палатальный буго-
рок (f. luxurians Reinhardt, 1877). Степень развития устьевой арматуры раз-
лична; колумеллярный зуб может отсутствовать, палатальная складка также
выражена в разных популяциях по-разному. Затылочное утолщение хорошо
развито, обычно светлее фона. Пупок проколовидный.
182
III. НАДСЕМ. PUPILLOIDEA
Размеры: BP 2—3.2, БД 1.3—1.6 мм; лектотип: ВР 2.6, БД 1.4 мм (Forcart,
1957).
Внутреннее строение. Материал: 4 экз. из Еревана, 29 июня
1969 г., сбор и определение Н. Н. Акрамовского, 3 — из долины р. Чегем на
Северном Кавказе, 17 мая 1970 г., сбор и определение автора.
Матка всех исследованных экземпляров содержала от 2 до 5 относительно
очень крупных яиц, в связи с чем мужской отдел был в значительной степени
редуцирован; у одного экземпляра вообще не удалось найти мужских протоков.
Поэтому приводимое ниже описание основано
на изучении этих особей и на литературных
данных (Steenberg, 1925; Матёкин, 1966; Ак-
рамовский, 1976).
Белковая железа с глубоким и резким ла-
теральным желобком. Семяпровод впадает
в утолщенный конец эпифаллуса, который
плавно переходит в пенис. Придаток эпифал-
луса недлинный, конический. Пениальный ап-
пендикс у особей с нормально развитым муж-
ским отделом имеет обычное строение, а у жи-
вотных, в матке которых развиваются яйца, раз-
вит слабо и отделы его едва угадываются.
Пениальная ветвь полового ретрактора корот-
кая, ветвь аппендикса крепится к А х сразу же
под А2- Резервуар семяприемника с коротким
Рис. 99. Pupilla (Pupilla) trtplicata (Stud.) из ущелья
р. Чегем, Северный Кавказ, 17 V 1970 г. Половой ап-
парат.
собственным протоком, дивертикул очень тонкий, лишь слегка длиннее ре-
зервуара.
Распространение и местообитание. Южная Европа,
местами южные районы центральной Европы; восточные Карпаты, Крым,
Новочеркасск; отдельные популяции в бас. Волги и Оки; Северный Кавка»
и Закавказье; Малая, Передняя и Средняя Азия, южный Алтай; наиболее
восточная находка сделана 18 сентября 1981 г. Е. И. Компанцевой на восточ-
ном берегу Байкала у мыса Крестовского. Вид населяет почти все типы био-
топов с умеренным увлажнением — от опушек лиственных лесов до россыпей
камней в полупустынях и в горах.
3. Pupilla (Pupilla) bigranata (Rossmaessler, 1839) (рис. 97 , 77/, IV).
Rossmaessler, 1839, Iconogr., II, 10 : 27, fig. 645 (Pupa); Pilsbry, 1921,
Man. Conch., 26 : 182, t. 20, figs. 22, 23; S t e e n b e г g, 1925 : 67, fig. 16A, В du texte; Л и-
x a p e в, Раммельмейер, 1952:147; Ehrmann, 1956:47, t. I, fig. 18; Lo-
z e k, 1956 : 100, t. 6, fig. 2; Zilch, Jaeckel, 1962 : 99 (trtplicata f. bigranata); L o-
z e k, 1964 : 214, t. 9, fig. 6 (trtplicata f. bigranata).
Terra typica — не указана; сообщается, что типовой материал получен от
проф. Хазе из Дрездена.
Типы, вероятно, хранятся в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: более 60 экз. из 8 местонахождений в Югославии, в Москов-
ской обл., Средней Азии (Зеравшанский, Киргизский и Заилийский хребты).
Раковина от удлиненно-овальной до овально-цилиндрической, с округло-
конической верхней частью, умеренно твердостенная, слегка блестящая. Обо-
ротов 6—7, несильно выпуклых; последний оборот к устью слабо и очень плавно
2. РОД PUPILLA
183
приподнят, высота его меньше половины высоты раковины. Окраска одно-
цветно-роговая, затылочное утолщение светлее фона. Дефинитивные обороты
слабо скульптурированы: имеются отдельные редкие тонкие радиальные мор-
щинки. Устье округлое, с плавно, отвернутыми краями. Затылочное утолщение
массивное, отделено от края устья кольцевой перетяжкой. В устье два зуба:
небольшой округлый бугорковидный париетальный и по-разному развитый
палатальный; в минимальном выражении он может иметь вид «наплыва»; но
чаще палательный зуб имеет вид бугорка или удлиненной складки. Кроме того,
у некоторых экземпляров заметно слабое колумеллярное утолщение. Пупок
маленький, овальный.
Размеры: ВР 2—2.9, БД 1.5 мм; в первоописании: ВР линии, БД 3/4
линии, т. е. ВР около 3, БД около 2 мм.
Внутреннее строение. Материал мною не исследован; судя
по изображениям, помещенным в монографии Стинберга (Steenberg, 1925),
вид отличается от Р. trtplicata более длинным эпифаллусом и более четко обо-
собленным базальным отделом пениального аппендикса.
Бесспорно, Р. trtplicata и Р. bigranata — близкородственные формы. Цильх
и Эккель, а также Ложек (см. синонимику) считают Р. bigranata экологической
формой от Р. trtplicata', действительно, встречаются раковины Р. trtplicata
с ослабленной зубной арматурой и расплывшимся палатальным зубом. Однако
эти виды, во-первых, встречаются вместе (например, на берегу р. Оки близ
с. Белые Колодези, на однородном участке сухого склона), во-вторых, эти виды
различаются контуром раковины: если Р. trtplicata имеет цилиндрическую
раковину, то контур раковины Р. bigranata в большей или меньшей степени
овальный.
Распространение и местообитание. Юго-западная и
Западная Европа, западные районы Украины, долины рек Ока, Волга, Москва;
южные отроги Урала с прилегающими территориями Казахстана и Западной
Сибири; горные местности Средней Азии; отдельные находки на восточном
и Северном Кавказе. Населяют открытые склоны, хорошо прогреваемые солн-
цем; в осыпях, среди щебенки, под корнями кустарника.
4. Pupilla (Pupilla) gallae Tzvetkov, 1940 (рис. 97, V; рис. 100).
Цветков, 1940 : 384; Лихарев, Раммельмейер, 1952 : 150, рис. 62;
Матёкин, 1966 : 111.
Terra typica — Заилийский хр., Иссыкское ущелье, в,50 км к востоку от
Алма-Аты.
Голотип хранится в Зоологическом музее Московского ун-та.
Материал: 17 экз. из трех местонахождений в юго-восточном Казах-
стане (берег р. Нарынкол близ устья р. Текес, ущелье Аксай на Заилийском
хребте и типовое местонахождение).
Раковина левозавитая, удлиненно-овальная, слегка блестящая, с закруглен-
ной вершиной. Оборотов 5—б1^, умеренно выпуклых. Последний оборот к устью
плавно, но явственно приподнят, высота его немного меньше половины высоты
раковины. Окраска одноцветно светло-роговая или серовато-роговая. Эмбрио-
нальные обороты неясно микрозернистые. Дефинитивная скульптура в виде
радиальных довольно правильных нерезких сглаженных морщинок. Устье
округлое, края его умеренно отвернуты, не утолщены, места прикрепления
устья не сближены. Затылочное утолщение хорошо развито, обычно немного
светлее фона. Зубов 3: короткая гребневидная париетальная пластинка, на-
против которой в глубине устья расположен округлый па латальный бугорок,
и колумеллярная пластинка, обычно слаборазвитая и не всегда видная при
нормальном положении раковины. Эту пластинку можно рассмотреть, слегка
повернув раковину вокруг оси столбика. Пупок короткокаплевидный.
Размеры ВР 2.7—3, БД 1.5—1.6 мм; голотип: ВР 3.0, БД 1.6 мм.
184
III. НАДСЕМ. PUPILLOIDEA
Внутреннее строение. Материал: 3 экз. из ущелья р. Аксай
на Заилийском хребте, июль 1977 г., сбор и определение С. Альмухамбетовой.
Все исследованные животные находились в мужской фазе, поэтому белковая
железа была развита слабо, и с этим, возможно, связано отсутствие вырезок
и желобков на поверхности органа. Семяпровод впадает терминально; эпифаллус
очень плавно расширяется и, образовав колено, впадает в пенис, длина которого
Рис. 100. Pupilla (Pupilla) gallae Tzv. из
ущелья р. Аксай на Заилийском хр.,
VII 1977 г. Половой аппарат.
Рис. 101. Pupilla (Pupilla)%muscorum (L.)
из побережья Княжой губы Белого моря,
28 VIII 1961 г. Половой ^аппарат.
в 2—3 раза меньше длины эпифаллуса. Придаток эпифаллуса расположен на
границе с пенисом, короткий, небольшой, цилиндрический. Внутренние стенки
мужских протокдв без каких-либо специализированных структур. Пениальный
аппендикс хорошо развит. А± и А2 слиты вместе, тонкостенные, сквозь про-
зрачные покровы легко заметить длинную остроконечную папиллу аппендикса,
занимающую большую часть длины A1-j-A2. А3 выражен хорошо, но переходит
в плавно, без резкой границы. Пениальный аппендикс располагается очень
низко, непосредственно у полового атриума. Пениальная ветвь полового ре-
трактора крепится к эпифаллусу значительно выше придатка, ветвь аппендикса
подходит к границе между А1-|-А2 и А3. Нижний отдел вагины относительно
очень длинный, проток семяприемника отходит, следовательно, высоко. Резер-
вуар семяприемника у исследованных особей был пуст и говорить о форме его
трудно. Дивертикул семяприемника тонкий и короткий, короче резервуара.
Семяприемник располагается примерно на уровне средней части матки.
Распространение и местообитание. Горные системы
юго-восточного Казахстана: хребты Заилийский, Терскей и, вероятно, Кунгей.
Населяет подстилку лиственных лесов, доходит до верхней границы субальпий-
ского пояса.
2. РОД PUPILLA
185
5. Pupilla (Pupilla) muscorum (Linnaeus, 1758) (рис. 97, VI—X; рис. 101).
Linnaeus, 1758 : 767 (Turbo); Draparnaud, 1801 : 58 (Pupa marginata);
C. Pfeiffer, 1821 : 59, pl. 3, figs. 21, 22 (P. unidentata); Hanley, 1855 : 353, pl. 4,
fig. 6 (Pupa); Moquin-Tandon, 1855 : 392, pl. 28, figs. 5—15 (P. muscorum et f. al-
bina, i. edentula); S 1 a v i к in: M ar t i n i, Chemnitz, 1868, Conch.-Cab., I, 15 : 12, t. 2,
figs. 1—5 (var. edentula); С 1 e s s i n, 1871, Fauna von Augsburg: 101 (var. pratensis); 1876 : 202
(var. elongata); Westerlund, 1876 : 98 (var. lundstrbmi); Moellendorff, 1901 : 381
(subsp. asiatica); Lindholm, 1911 : 39 (var. milaschevitschi); Steenberg, 1925 : 62,
figs. 30, 31, pl. 10, figs. 1—3; Pilsbry, 1927, Man. Conch., 28.: 173, pl. 20, figs. 1—7; Л и-
xa p ев, Раммельмейер, 1952 : 146, рис. 59; E h r m a n n, 1956 : 46, t. I, fig. 16;
Lozek, 1956 : 99, t. 6, fig. 1; 1964 : 215, t. 9, 1, 7; M а т ё к и н, 1966 : 111, табл. 2, рис. 4;
Gittenberger et al., 1970 : 53, fig. 41, Kaart 18; Акрамовский, 1976 : 145,
рис. 64A, табл. VI, 58.
Terra typica — Швеция.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 63 местонахождений в Европе и
европейской части СССР, на Кавказе, в Средней Азии, в Сибири и на Дальнем
Востоке.
Раковина овально-цилиндрическая, в разной степени вытянутая, обычно
твердостенная, но в условиях дефицита кальция может быть тонкостенной,
сильно просвечивающей. Оборотов 6—8, слабо или умеренно выпуклых; послед-
ний оборот к устью плавно приподнят, причем иногда настолько сильно, что
верхний край устья прикрепляется выше середины последнего оборота (var.
lundstromi Westerlund). Высота последнего оборота почти равна трети высоты
раковины (у высоких экземпляров), но чаще — несколько меньше половины
этой величины. Окраска одноцветная от роговой до темно-каштановой или
красноватой. Скульптура в виде тонкой радиальной исчерченности, правиль-
ность которой местами нередко нарушается. Устье округлое, края его утолщены
и слегка отвернуты. Затылочное утолщение развито в различной степени:
обычно у более коротких раковин это утолщение более мощное и светлее ок-
рашено. В максимальном суммарном наборе в устье 4 зуба: париетальный,
колумеллярный, палатальный и утолщение губы па палатальном крае; однако
мне неизвестно ни одного случая, чтобы у экземпляров из одной выборки при-
сутствовало одновременно все 4 зуба. Чаще всего одновременно имеется не
более 2 зубов: париетальный и палатальный бугорки. Весьма обычны также
раковины с каким-нибудь одним зубом; реже всего встречаются раковины со
слабым колумеллярным утолщением и палатальным зубовидным наплывом
на губе (такая раковина изображена в работе Цильха и Эккеля — см. синони-
мику). Нередко устье совсем без зубов, причем Хенлей (Hanley, 1855, р. 352)
указывает, что единственный сохранившийся экземпляр в коллекции Линнея
как раз характеризуется полным отсутствием устьевых зубов. Пупок коротко-
каплевидный.
Размеры: ВР 3—4, БД 1.6—1.8 мм; экземпляр из типовой серии, изображен-
ный в работе Хенлея и измеренный мной по линии рядом с рисунком, которая
показывает натуральную величину, имеет высоту 3.4 мм.
Внутреннее строение. Материал: 8 экз. из побережья Княжой
губы Белого моря, 28 августа 1961 г., 5 — из окрестностей с. Белые Колодези
на правом берегу р. Оки, 7 июля 1974 г.; 4 — из с. Ромашково Московской обл.,
1 мая 1959 г; 3 — из с. Квасы близ Рахова, Закарпатье, 14 сентября 1969 г.;
сборы и определения автора.
Белковая железа с глубоким желобком, проходящим по боковой поверхности
вдоль или поперек железы. Семяпровод впадает терминально в слабое расшире-
ние конца эпифаллуса. Придаток эпифаллуса хорошо развит. Граница между
пенисом и эпифаллусом наружно не выражена. Все отделы пениального ап-
пендикса нормально выражены, но у особей, в матке которых содержатся яйца,
отделы слабо обособлены один от другого. Пениальная ветвь полового ретрак-
тора короткая, крепится несколько выше придатка эпифаллуса, ветвь аппен-
186
III. НАДСЕМ. PUPILLOIDEA
дикса подходит-к Аг сразу под А2. Дивертикул семяприемника почти такой же
длины, как и резервуар, или немного длиннее.
Распростр ан ение и местообитание. Голарктический
вид, распространен почти повсеместно, в том числе и в зоне тундры. Населяет
разнообразные типы биотопов с умеренным увлажнением — как на равнине,
так и в горах. В связи с широким спектром биотопов вид образует множество
форм межпопуляционной изменчивости.
Рис. 102. Pupilla (Pu-
pilla) Inops (Reinh.) и?.
Ленинакана, Армения.
Рис. 103. Pupilla (Pu-
pilla) inops (Reinh.) из
Армении, Мегринский р-н,
с. Личк, 1-3 VIII 1947 г.
Половой аппарат.
6. Pupilla (Pupilla) inops (Reinhardt, 1877) (рис. 102, 103).
Reinhardt, 1877a : 79, t. 3, fig. 3 (Pupa triplicata var. inops); 0. Boettger,
1879 : 399 (P. muscorum var. caucasica); Pilsbry, 1921, Man. Conch., 26 : 180 (muscorum-
inops); Акрамовский, 19476 : 269 (armeniaca Issel, 1865); 1949 : 144; Л и x a p e в,
Раммельмейер, 1952 : 149; К а л и т и н a, 1958 : 172, Акрамовский, 19716
6; 1976 : 145, рис. 64Г, табл. VI, 59.
Terra typica — Боржоми.
Место хранения типов не установлено.
Материал: более 40 экз. из 5 местонахождений на Кавказе.
Раковина от короткоцилиндрической до овально-цилиндрической, с плавно
закругленной вершиной, матовая. Оборотов 6, умеренно выпуклых, последний
оборот к устью очень плавно и очень слабо приподнят. Высота последнего оборота
равна половине высоты раковины или немного меньше. Окраска одноцветно-
роговая. Скульптура — тонкая, очень вялая радиальная морщинистость, вы-
раженная подчас настолько слабо, что раковина выглядит почти гладкой.
Устье округлое, места его прикрепления не сближены. Затылочное утолщение
хорошо развито, белое или светло-серое, всегда светлее фона. Зубов один или
два: небольшой париетальный бугорок и выраженный в разной степени колу-
меллярный зуб, расположенный на задней поверхности столбика (может от-
сутствовать). Пупок сквозной, каплевидный.
Размеры: ВР 2.5—3.3, БД 1.5—1.7 мм; в первоописании Рейнхардт огра-
ничился указанием, что эта форма мельче, чем «настоящая» Р. triplicata.
2. РОД PUPILLA
187
Внутреннее строение. Материал: 5 экз. из Армении, Мегрин-
ский р-н, окр. с. Личк, 13 августа 1947 г., сбор и определение Н. Н. Акрамов-
ского.
Белковая железа с глубоким латеральным желобком. Семяпровод впадает
терминально или чуть сбоку. Эпифаллус цилиндрический, граница между
ним и пенисом не выражена. Придаток эпифаллуса хорошо развит, конический,
расположен близ места прикрепления пениальной ветви полового ретрактора.
Все отделы пениального аппендикса хорошо выражены, кроме42, который почти
слит с Аг. Ветвь полового ретрактора, подходящая к пенису, очень короткая,
иногда раздвоенная; ветвь аппендикса крепится в том месте, где должна быть
граница между Аг и А2. Проток семяприемника недлинный, прямой. Диверти-
кул рудиментарный, примерно той же длины, что и резервуар.
Уникальная особенность вида заключается в том, что мужской отдел хорошо
развит одновременно с развитием яиц в матке.
Распространение и местообитание. Восточное Пред-
кавказье и Закавказье. Населяет высокогорные и среднегорные участки, по
долинам рек спускается в предгорья. По-видимому, типовое местонахождение —
один из наиболее западных пунктов ареала. Встречается в лиственной под-
стилке лесов и кустарниковых зарослей и в малоподвижных, заросших травой
осыпях.
7. Pupilla (Pupilla) inequidenta Schileyko et Almuhambetova, 1979 (рис. 104,
105).
Альмухамбетова, 1979 : 30, рис. 1.
Terra typica — Пойма p. Аксай в ущелье Аксай, Заилийский хребет.
Г о л о т и п хранится в Зоологическом ин-те АН СССР.
Материал: 25 экз. из типового местонахождения.
Раковина овально-цилиндрическая, умеренно твердостенная, слегка про-
свечивающая, слабо блестящая. Оборотов 5т/2—6, умеренно выпуклых, послед-
ний оборот к устью плавно, но вполне явственно поднимается, высота его за-
метно меньше половины высоты раковины. Окраска одноцветно-роговая.
Поверхность эмбриональных оборотов тонко, но отчетливо мелкозернистая.
Скульптура дефинитивных оборотов в виде тонкой и не очень равномерной
радиальной исчерченности; степень выраженности этой скульптуры в пределах
типовой серии колеблется от слабой до сравнительно резкой. Устье округлое,
места его прикрепления не сближены, края устья коротко, но сильно отогнуты.
Чуть отступя от линии перегиба располагается высокая мощная губа, которой
отвечает резкое светлое затылочное утолщение. Зубов 2: небольшой бугорко-
видный париетальный и очень своеобразный палатальный — он слегка удлинен
и располагается позади губы почти параллельно ей; у большинства раковин
намечается его разделение на два округлых бугорка. У голотипа такое деление
лишь слегка обозначено, но у нескольких паратипов палатальный зуб отчет-
ливо двувершинный. На затылке палатальный зуб просвечивает, но соответст-
вующего углубления нет. Пупок очень маленький, овальный.
Размеры: ВР 2.8—3.5, БД 1.5—1.8 мм; голотип: ВР 3.5, БД 1.8 мм.
Внутреннее строение. Материал: 4 экз. из типового местона-
хождения, июль 1977 г., сбор С. Альмухамбетовой.
Латеральная поверхность белковой железы сложнорассеченная: помимо
глубокого желобка имеется еще 1—2 выемки. У всех исследованных мной эк-
земпляров в матке находились эмбрионы на поздних стадиях развития, а муж-
ской отдел характеризовался явными признаками дегенерации. Семяпровод
впадает в небольшое расширение проксимальной части эпифаллуса. Придаток
эпифаллуса конический, небольшой. Пениальный аппендикс впадает в пенис
далеко от полового атриума, отделы его у анатомированных экземпляров вы-
ражены плохо. Одна из ветвей полового ретрактора крепится близ основания
188
III. НАДСЕМ. PUPILLOIDEA
пениального аппендикса, другая — немного проксимальней придатка эпифал-
луса. Семяприемник не обнаружен, что связано, вероятно, с гипертрофирован-
ным состоянием матки.
Распространение и местообитание. Вид пока известен
только из типового местонахождения на Заилийском хр., где он обнаружен на
камнях и в растительной трухе.
Рис. 104. Pupilla (Pupilla)
inequidenta Schileyko et Al-
, muhambetova. Голотип.
Рис. 105. Pupilla (Pupilla)
inequidenta Schileyko et Almu-
hambetova. Паратип: пойма
p. Аксай на Заилийском хр.,
VII 1977 г. Половой аппарат.
8. Pupilla (Pupilla) striopolita Schileyko, sp. nov. (рис. 106).
Terra typica — Тянь-Шань, Чаткальский хр., берег р. Касансай над с. Ала-
Бука, 29 мая 1972 г., сбор, и определение автора.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 21 экз. из 5 местонахождений на Тянь-Шане.
Раковина овально-цилиндрическая, тонкостенная, просвечивающая, с шел-
ковистым блеском и сильно сглаженными радиальными морщинками. Вершина
раковины ширококуполовидная. Оборотов 6, умеренно выпуклых, последний
оборот к устью очень плавно и постепенно поднимается, высота его заметно
меньше половины высоты раковины. Окраска светло-роговая. Микроскульптура
эмбриональных оборотов в виде неправильной зернистости, хорошо заметна
при увеличении приблизительно в 50 раз. Поверхность дефинитивных оборотов
покрыта плавными радиальными морщинками, правильность расположения
которых местами нарушается, особенно на последнем обороте. Устье округлое,
места его прикрепления умеренно сближены за счет закругленного угла в верх-
ней части палатального края. Края устья широко отвернуты, острые, по линии
перегиба усилены мощной порожистой губой цвета слоновой кости. Сущест-
венно, что отогнутая часть устьевого края переходит в губу очень плавно,
т. е. губа не обособлена кольцевой ложбинкой, как это обычно бывает у других
2. РОД PUPILLA
189
представителей рода. Устьевые зубы отсутствуют. Затылочное утолщение сильно
развито. Пупок маленький, округлый.
Размеры: ВР 2.8—3.3, БД 1.9—2 мм; голотип: ВР 3.2, БД 2.0 мм.
Вид ближе всего к Р. muscorum; стойко отличается характерной губой, не
отделенной от края устья. Видовая самостоятельность подтверждается совмест-
ным нахождением обоих видов в пойме верхнего течения р. Чаткал.
Рис; 106. Pupilla (Pupilla} strlopolita Рис. 107. Pupilla (Pupilla} sterrii (Voith).
Schileyko, sp. nov. Голотип. I — из Регенсбурга, Бавария, ФРГ; II — из с. Код-
жори близ Тбилиси.
Распространение и местообитание. Вид известен из
следующих местонахождений: Чаткальский хр. — склоны по берегам р. Касансай
над с. Ала-Бука и верхнее течение р. Чаткал; Таласский хр. — ущелье Беш-Таш
близ гор. Таласа; Киргизский хр. — ущелье Карабалты и склоны над с. Лу-
говое. Кроме того, по свидетельству С. Альмухамбетовой, вид живет на Заилий-
ском хр.
Встречается преимущественно в мелкообломочных осыпях; в высокогорье
Чаткальского хр. найден в пойменных зарослях ольхи.
9. Pupilla (Pupilla) sterrii (Voith, 1840) (рис. 107).
V о i t h in: Fiirnrohr, 1840 : 469 (Pupa); Pilsbry, 1921, Man. Conch., 26 : 185,.
t. 23, figs. 1—4 (Pupilla сира non Jan); Steenberg, 1925 : 70, pl. 12, figs. 1, 2, pl. 13,
fig. 4, pl. 26, figs. 3, 4 [P. сира Jan var. (?) sterri]; Линдгольм, 1931 : 44; Л и x a p e в,
Раммельмейер, 1952 : 147, рис. 60; Ehrmann, 1956 : 47, t. 1, fig. 17; Loze k,
1956 : 101, t. 6, fig. 3; Z i 1 c h, J a e c k e 1, 1962 : 99; L о z e k, 1964 : 216, t. 9, fig. 2;
Матёкин, 1966 : 111, табл. 2, рис. 5; Д а и я н о в, Лихарев, 1975 : 101, фиг. 34.
Terra typica — Регенсбург, Бавария.
Место хранения типов не установлено.
Материал: более 50 экз. из 6 местонахождений в окр. Регенсбурга,
на Кавказе и в долине р. Оки.
Раковина цилиндрическая, слабо блестящая, с закругленной вершиной,
сравнительно тонкостенная, шелковисто поблескивающая. Оборотов —6х/2,
сильновыпуклых. Последний оборот к устью очень слабо и очень плавно при-
поднят, высота его всегда заметно меньше половины высоты раковины. Окраска
одноцветная, от светло-роговой до красновато-роговой. Дефинитивная скульп-
тура в виде тончайших тесно расположенных ребрышек, гребни которых на
верхних оборотах несут чрезвычайно слабую кожистую оторочку. Устье ок-
руглое, с тонкими отвернутыми краями; места его прикрепления слегка сбли-
жены и связаны тонкой мозолью. Зубов в максимальном наборе три: короткие
бугорковидные париетальный и колумеллярный и удлиненный палатальный,
которому снаружи соответствует углубление. Однако степень выраженности
элементов устьевой арматуры изменчива: встречаются раковины совсем без
зубов, либо имеется один колумеллярный, или только париетальный. Затылоч-
ное утолщение обычно хорошо развито. Пупок сквозной, [проколовидный.
190
III. НАДОЕМ. PUPILLOIDEA
Размеры: BP 3—3.3, БД 1.7—1.8 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 6 экз. из окр. с. Коджори
близ Тбилиси, 14 июля 1973 г., сбор И. М. Лихарева; 2 — из того же места,
27 апреля 1970 г., сбор и определения автора.
В матке у всех исследованных животных содержались яйца, поэтому совоку-
пительный аппарат претерпел редукцию. Белковая железа с глубоким латераль-
ным желобком. Семяпровод впадает терминально в небольшое утолщение на
конце эпифаллуса. Придаток эпифаллуса относительно длинный, цилиндриче-
ский. Пенис также цилиндрический, внешне не отделен от эпифаллуса. Отделы
пениального аппендикса выражены неясно. Семяприемник с коротким диверти-
кулом.
Рис. 108. Pupilla (Pu-
pilla) turcmentca (Bttg.)
из Гиссарского хр., пра-
вый берег р. Сари-Тач.
Рис. 109. Pupilla (Pu-
pilla) anzobica Izzatul-
laev. Голотип.
Рис. 110. Виды рода Pupilla (под-
род Fragipupilla).
I — Р. (Fragipupilla) limata Schileyko, sp.
nov., голотип; II — P. (Fragipupilla) ala-
biella Schileyko, sp. nov. Голотип.
Распространение и местообитание. Горные области
средней и частично южной Европы; возможно, Карпаты. Одна находка сделана
на Кавказе (окр. Тбилиси). Указания на присутствие вида в Средней Азии
требуют подтверждения, так как там живет близкий вид — Р. turcmentca.
Населяет ущелья и долины горных рек; реже — сухие склоны, где держится
в осыпях и под отдельно лежащими крупными камнями.
10. Pupilla (Pupilla) turcmenica (О. Boettger, 1889) (рис. 108).
О. Boettger, 1889b : 958, pl. 26, fig. За—c (Pupa (Pupilla) сира var. turcmenica];
Pilsbry, 1921, Man. Conch., 26 : 188, pl. 23, figs. 6—9 (сира turcmenica); Лихарев,
Раммельмейер, 1952 : 148 (sterri var. turcmenica).
Terra typica — Копетдаг, гора Аг-даг.
Место хранения типов не установлено.
Материал: 23 экз. из 4 местонахождений в Средней Азии.
Раковина цилиндрическая, тонкостенная, просвечивающая, матовая. Эмбри-
ональные обороты вместе с первыми дефинитивными образуют притупленную
короткокуполовидную вершину. Оборотов 6, выпуклых, последний оборот
к устью плавно приподнят, высота его заметно меньше половины высоты рако-
вины. Окраска одноцветная, роговая или коричневая. Микроскопическая зер-
нистость поверхности эмбриональных оборотов хорошо заметна. Дефинитивные
обороты покрыты тончайшими радиальными ребрышками, каждое из которых
несет кожистую оторочку. Эта оторочка лучше выражена на верхних оборотах.
Устье округлое, места его прикрепления слегка сближены. Колумеллярный,
2. РОД PUPILLA
191
базальный и нижняя часть палатального краев умеренно отвернуты, не утол-
щены или незначительно утолщены. Иногда имеется слабый глубоко сидящий
париетальный бугорок, но чаще зубы полностью отсутствуют. Затылочное
утолщение развито в различной степени, но всегда выражено. Пупок очень
узкий, округлый.
Размеры: ВР 3—3.2, БД 1.1—1.4мм; в первоописании: ВРЗ—Зг/2, БД l5/g мм.
Распространение и местообитание. Помимо типового
местонахождения на восточном Копетдаге вид найден 3. Иззатуллаевым на
Гиссарском хр., правый берег р. Сари-Тач, С. Альмухамбетовой на Киргиз-
ском хр. над с. Луговое и мной на Киргизском хр. в ущелье Карабалты. Встре-
чается также в западной части Китая и Тибете. Обитает в осыпях на сухих
склонах.
11. Pupilla (Pupilla) anzobica Izzatullaev, 1970 (рис. 109).
Иззатуллаев, 1970а : 54, рис. 2 (Pyptlla laps, cal.); 19706 : 81.
Terra typica — окр. перевала Анзоб на Гиссарском хр. на высоте около
3500 м над уровнем моря.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 9 экз. из типовой серии.
Раковина высокоцилиндрическая, с короткой закругленно-конической вер-
шиной, довольно тонкостенная, просвечивающая, поверхность ее имеет шелко-
вистый блеск. Оборотов б1^—8 (у голотипа 8), умеренно выпуклых. Последний
оборот к устью очень плавно и незначительно поднимается, высота его со-
ставляет примерно треть высоты раковины или, у более коротких раковин, не-
много больше. Окраска одноцветная, роговая. Эмбриональные обороты тонко-
зернистые, дефинитивные неравномерно покрыты вялыми радиальными мор-
щинками. При увеличении более чем в 50 раз при сильном косом свете видно,
что поверхность раковины несет тончайшую неопределенно ориентированную
струйчатость, создающую впечатление, что поверхность слегка ирризирует.
Устье округлое, лишенное зубов, с плавным углом в верхней части палатального
края. Устьевые края незначительно утолщены, слегка отвернуты, затылочная
борозда слабая, затылочное утолщение нерезкое. Пупок маленький, овальный,
не сквозной.
Размеры в первоописании: ВР 4.0 + 1.05, БД 1.7+0.09 мм; голотип: ВР 4.1,
БД 1.7 мм.
Распространение и местообитание. Вид пока известен
только из типового местонахождения близ перевала Анзоб на Гиссарском хр.,
на высоте 3450—3500 м над уровнем моря. Обнаружен на сухих склонах под
камнями, у корней колючих подушковидных полукустарников.
2. Подрод FRAGIPUPILLA Schileyko, subgen. nov.
Типовой вид Pupilla (Fragipupilla) alabiella Schileyko, sp. nov.
Раковина тонкостенная, без затылочного утолщения. Устье без зубов.
Распространение. Дальний Восток. 2 вида.
12. Pupilla (Fragipupilla) alabiella Schileyko, sp. nov. (рис. 110, II, рис. 111).
Terra typica — о. Верховского в зал. Петра Великого.
Г о л о т и п хранится в Зоологическом ин-те АН СССР.
Материал: свыше 100 экз. из о-вов Верховского и Два Брата в зал.
Петра Великого.
Раковина от удлиненно-овальной до почти цилиндрической, тонкостенная,
хрупкая, просвечивающая, с шелковистым блеском и тонкими радиальными
ребрышками. Вершина притупленная, плавно закругленная. Оборотов 5—6,
умеренно выпуклых, разделенных узким глубоким швом. Последний оборот
192
III. НАДСЕМ. PUPILLOIDEA
к устью слегка и плавно приподнят, высота его всегда меньше половины вы-
соты раковины, но больше трети этой величины. Окраска роговая, одноцвет-
ная. Эмбриональные обороты покрыты тончайшей, но ясно различимой «сеточ-
кой». Дефинитивные обороты несут характерную скульптуру в виде тонких
радиальных ребрышек, промежутки между которыми шире самих ребрышек.
Эти скульптурные элементы часто теряют правильность расположения, распо-
лагаясь то гуще, то реже; одни не доходят до шва, другие не совсем параллельны
соседствующим. На старых вытертых раковинах ребрышки тоже видны, но они
Рис. 111. Pupilla (Fragipupilla) alabiella Schileyko, sp. nov. из острова Верховского в зал.
Петра Великого, 26 VI 1971 г., паратип.
I — половой аппарат, II — передний конец тела животного с просвечивающими внутренними органами.
значительно менее острые, чем на свежих, из чего можно заключить, что верхняя
кромка каждого ребрышка сформирована только периостракумом. Устье слабо
скошено, неправильно округлое, со слегка отвернутыми и почти не утолщенными
краями. Места прикрепления устья расставлены и связаны тонкой прозрачной
мозолью, которая после гибели моллюска мутнеет и становится белой. От ангу-
лярной области край устья отходит почти горизонтально, слегка наклонно вниз,
далее круто изгибается и спускается почти отвесно; базальный край плавно
закруглен. Зубы полностью отсутствуют. Пупок сквозной, узкий, каплевидный.
Размеры ВР 2.2—3.3, БД 1.4—1.6 мм; голотип: ВР 3.2, БД 1.6 мм.
Вид напоминает североамериканский Pupilla hebes (Ancey, 1881), описанный
из высокогорья Невады и найденный затем во многих горных районах Запада
США; отличается меньшими размерами, более резкой скульптурой и отсутст-
вием зубов (у Р. hebes имеется легкое утолщение на столбике).
В н у т р е н,н ее строение. Материал: 19 экз. из типового место-
нахождения: зал. Петра Великого, о. Верховского, под маяком, 26 июня 1971 г.,
сбор и определение автора.
2. РОД PUPILLA
193
Белковая железа с глубоким латеральным желобком. Семяпровод впадает
терминально; между семяпроводом и эпифаллусом имеется резкая граница.
Придаток эпифаллуса слабо выражен и располагается в месте перехода эпифал-
луса в пенис. Эпифаллус недлинный, равномерно цилиндрический; пенис также
цилиндрический, его дистальная часть значительно короче проксимальной.
Пениальный аппендикс хорошо развит, At слит с Л2; А3 не выражен или выра-
жен слабо, А4 большего диаметра, чем у прочих видов рода. Пениальная ветвь
полового ретрактора крепится на уровне придатка эпифаллуса, на противоле-
жащей придатку стороне, ветвь аппендикса — возле места отхождения А3.
Проток семяприемника недлинный, резервуар небольшой, с коротким, но ясно
выраженным собственным протоком. Дивертикул семяприемника тонкий, не до-
ходит до белковой железы.
Распространение и местообитание. Вид найден только
на двух островах в зал. Петра Великого (о-ва Верховского и Два Брата).
Обитает в трещинах скал, среди растительной трухи. В обоих случаях обнару-
жены массовые скопления.
13. Pupilla (Fragipupilla) limata Schileyko, sp. nov. (рис. 110, I; рис. 112).
Terra typica — Чукотский п-ов, окр. Марково.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: около 100 экз. из 2 местонахождений на Чукотке.
Раковина овально-цилиндрическая, сравнительно тонкостенная, слабо
блестящая, с закругленной чуть приостренной вершиной. Оборотов —6,
умеренно выпуклых, последний обо-
рот к устью очень слабо приподнят,
высота его немного меньше половины
высоты раковины. Окраска одно-
цветная, от светло-роговой до корич-
невой. Эмбриональные обороты по-
крыты неясными тончайшими грану-
лами и плохо различимой «сеточкой».
Дефинитивные обороты с очень тон-
кими кожистыми ребрышками, пра-
вильность расположения которых
местами нарушена. Устье округлое,
места его прикрепления не сбли-
жены, края слегка отвернуты, не
утолщены. Зубы и затылочное утол-
щение отсутствуют. Пупок очень уз-
кий, короткощелевидный.
Рис. 112. Pupilla (Fragipupilla) limata
Schileyko, sp. nov. из леса близ 163-ro км
дороги Эгвекинот—Иультин на Чукотском
полуострове,х 29 VII 1974 г., паратип. По-
ловой аппарат.
Размеры: ВР 2.8—3.1, БД 1.6—1.7 мм; голотип; ВР 3.0, БД 1.7 мм.
Вид близок предыдущему, отличается главным образом более приземистой
раковиной; видимые анатомические различия сводятся в основном к отсутствию
дивертикула семяприемника у данного вида.
Внутреннее строение. Материал: 3 экз. с Чукотского п-ова,
берег р. Амгуэмы, 163-й км дороги Эгвекинот—Иультин, 29 июля 1974 г.,
сбор Р. Я. Братчика, определение автора.
Белковая железа с глубоким латеральным желобком. Семяпровод впадает
терминально; между семяпроводом и эпифаллусом имеется резкая граница.
13 А. А. Шилейко
194
III'. НАДСЕМ. PUPILLOIDEA
Эпифаллус в начале цилиндрический, к пенису сужается. Хорошо развитый
конический придаток эпифаллуса расположен примерно на границе пениса
и эпифаллуса. Дистальная и проксимальная части пениса почти равной длины.
Пениальный аппендикс хорошо развит, Ах и А2 почти слиты, As очень плавно-
переходит в А4. Пениальная ветвь полового ретрактора крепится к эпифаллусу
заметно выше придатка, ветвь аппендикса крепится к А1 чуть ниже А2. Диверти-
кул семяприемника отсутствует, резервуар овальный, проток недлинный.
Несмотря на то что в матке у исследованных особей обнаружены яйца в коли-
честве 1—3, совокупительный аппарат развит нормально, без признаков деге-
нерации. Эта особенность помимо данного вида отмечена для кавказского
Pupilla inops.
Распространение и местообитание. Вид обнаружен-
в двух местах на Чукотском п-ве: в березовой роще близ Марково и в тундровом
редколесье на берегу р. Амгуэмы вдоль дороги Эгвекинот—Иультин.
3. Род PUPOIDES L. Pfeiffer, 1854
L. Pfeiffer, 1854 (1854—1860) : 192; Е. Martens in: Albers, 1860 : 296 (Leu-
cochila, типовой вид Pupa jallax, syn. P. albilabris C. R. Adams); L. Pfeiffer, 1878, No-
mencl. Helic. Vivent. : 292 (Leucochiloides, типовой вид Bulimus lardeus Pfr., обозначение Con-
nolly, 1912, Ann. S. Afr. Mus., XI, p. 176); Pilsbry, Vanatta, 1900 : 585; Pilsbry,
1921, Man. Conch., 26 : 108; Thiele, 1931 : 508; Wenz, Zilch, 1959—1960 : 167.
Типовой вид Bulimus nitidulus L. Pfeiffer, 1854; по обозначению-
Гюда (Gude, 1914, Fauna, of British India, Moll., II : 259).
Устье без зубов или имеется один ангулярный бугорок.
Распространение. Африка, Южная Азия, Америка, Австралия
(Квинсленд). Не менее 20 видов.
В ископаемом состоянии род известен из эоцена и нижнего миоцена Север-
ной Америки, а также из плейстоцена Египта, Бермудских о-вов и Северной
Америки.
Pupoides coenopictus (Hutton, 1834) (рис. ИЗ).
Hutton, 1834, J. Asiat. Soc. Bengal., 3 : 85 (Pupa coenopicta); Pilsbry, 1921, Man.
Conch., 26 : 123, pl. 13, figs. 1—3; Ray, Mukherjee, 1963 (1969): 412, fig. 2; Л и x a-
рев, Старобогато в, 1967 : 180; Иззатуллаев, 1970а : 81; 19706 : 82, рис.;
М i е n i s, 1974 : 99.
Terra typica — Индия, Бомбей.
Место хранения типов не установлено.
Материал: 4 экз. из выносов р. Вахш близ заповедника «Тигровая
балка», 1 из Афганистана, Кандагар, 5 из Ирана, Ахваз, 12 из о. Харат,
Красное море, архипелаг Дахлак.
Раковина высококоническая, просвечивающая, с узкозакругленной вер-
шиной. Оборотов 51/2—6, выпуклых, последний оборот близ устья плавно,,
но довольно энергично поднят, высота его немного больше половины высоты
раковины. Окраска одноцветная, светло-роговая. Эмбриональные обороты по-
крыты мельчайшей неясной зернистостью; кроме того, при увеличении около»
60 раз можно рассмотреть исчезающе слабые радиальные морщинки (видны
при сильном косом свете, если раковину постепенно поворачивать вокруг оси).
Скульптура дефинитивных оборотов в виде радиальных вялых, тонких, не-
правильно расположенных морщинок; в общем раковина выглядит почти глад-
кой. Устье овальное, с широко отвернутыми краями, места его прикреп-
ления слегка сближены и соединены тонкой мозолью. В ангулярной. области,
ниже верхней кромки париетальной мозоли, расположен небольшой при-
остренный бугорок, вершина которого несколько отклонена к палатальной
стенке. На колумеллярной крае зубы отсутствуют, кроме очень слабого колу-
меллярного утолщения; это утолщение сквозь устье не видно, и, чтобы его об-
IV. НАДСЕМ. VERTIGINOIDEA
195
нарушить, надо удалить часть задней стенки последнего оборота. Пупок очень
.узкий, каплевидный.
Размеры: ВР 4.2—5.2, БД 2—2.5 мм; в первоописании: ВР 5.1, БД 2.4 мм.
Внутреннее строение. Материал: 4 экз. из о. Харат, Красное
море, архипелаг Дахлак, 6 февраля 1980 г. Сбор и определение автора.
Белковая железа с глубоким продольным желобком и апикальной вырез-
кой. Верхний и нижний отделы вагины длинные. Мужской отдел полового
аппарата у всех исследованных жи-
вотных обнаруживает признаки се-
зонной дегенерации, что сказы-
вается в очень малых размерах его
(ср. Baker, 1935, pl. 27, fig. 1 —
изображение полового аппарата
Р. marginatus nitidulus лс нормально
развитым мужским отделом), слабом
развитии эпифаллуса и в отсутствии
ясного деления пениального аппен-
дикса на отделы. Семяприемник без
дивертикула, не обнаруживает де-
ления на проток и резервуар.
Рис. 113. Pupoides coenopictus (Hut-
ton).
I — из заповедника «Тигровая балка», выносы
•р. Вахш, раковина; II — из о-ва Харат, Красное
море , архипелаг Дахлак, 6 II 1980 г. Половой
аппарат.
Распространение и местообитание. Индия, Шри-Лан-
ка, южный Иран, Афганистан, Аравийский п-ов, Северо-Восточная Африка
южный Таджикистан (бассейн р. Вахш); завезен в Южную Америку и Вест-
Индию. Населяет открытые каменистые склоны^
IV. Надсем. VERTIGINOIDEA Pilsbry, 1918 |
Раковина маленькая или очень маленькая, от овальной до цилиндрической,
гладкая или с радиальной скульптурой. Эмбриональные обороты гладкие.
Пупок точковидный. Устье с зубами или без зубов; если колумеллярные и
париетальные пластинки имеются, то они формируются в субдефинитивном со-
стоянии. Края устья прямые или отвернутые, тонкие или в разной степени
утолщенные; за редким исключением, без губы.
Животные без губных щупалец, за исключением Gastrocoptinae.
Семенные пузырьки не обособлены от гермафродитного протока. Пенис
с эпифаллусом, резко обособленным очень четкой кольцевой перетяжкой.
Пениальная трубка внутри с нерезкими продольными складками. Придаточ-
ные органы мужского отдела отсутствуют, за исключением Nesopupinae, у чле-
нов которого имеется пениальный аппендикс. Половой ретрактор нерасщеп-
ленный, крепится к месту перехода семяпровода в эпифаллус.
Сем. VERTIGINIDAE Pilsbry, 1918J
Pilsbry, 1918, Man. Conch., 25 : 68;JS t e e n b e r g, 1925 : 179. @
Признаки надсемейства.
Распространение. По^всему земному шару. 3 или 4 подсемей-
ства.
В ископаемом состоянии семейство известно начиная с палеоцена (Европа,
Северная Африка, Северная Америка, Бермудские о-ва).
13*
196
IV. НАДСЕМ. VERTIGINOIDEA
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ VERTIGINIDAE
1 (2). Раковина цилиндрическая, покрыта резкой ребристой скульптурой,
либо края устья тонкие, ломкие, почти не отвернуты, лишенные губы
............................................ III. Truncatellininae.
2 (1). Раковина от овальной до овально-цилиндрической, гладкая или относи-
тельно тонко скульптурированная. Края устья отвернуты, обычно с губой.
3 (4). Раковина белая или стекловидно прозрачная, с мощной париетальной
пластинкой, нередко раздвоенной ..................I. Gastrocoptinae.
4 (3). Раковина от светло-роговой до . красновато-бурой; если париетальная
пластинка имеется, то она умеренно развита и не раздвоена.........
.............................................II. Vertigininae.
I. Подсем. GASTROCOPTINAE Pilsbry, 1918
Pilsbry, 1918, Man. Conch., 24 : X.
Раковина овальная, белая или стекловидно-прозрачная. Ангулярная и
париетальная пластинки слиты или соединены мозолью; при рассматривании
раковины в нормальном положении создается впечатление, что передняя часть
париетальной пластинки раздвоена.
Пенис недлинный, с одним коротким придатком; эпифаллус внешне не вы-
ражен.
Монотипическое подсемейство.
1. Род GASTROCOPTA Wollaston, 1878
Wollaston, 1878 : 515; О. В о е 11 g е г in : Е. М ar t е n s, 1880—1881 : 64 (Беисо-
chilus, типовой вид Pupa armifera Say); Sterki in : P i 1 s b г у, 1891, Proc. Acad. Nat.
Sci. Philad. : 315 (Bifidaria, типовой вид Pupa servilis Gould); Pilsbry, 1916, Man. Conch.,
24:6; Thiele, 1931:512; Лихарев, Раммельмейер, 1952:128; Wenz,
Zilch, 1959—1960 : 160; Стеклов, 1966 : 130.
Типовой вид Pupa acarus Benson, 1856.
Признаки подсемейства.
Распространение. О-ва Зеленого мыса, Кавказ, южный Урал,
Сибирь, Средняя и восточная Азия, Шри-Ланка, Филиппины; оба американ-
ских материка; о-ва Тихого и Индийского океанов, Бермудские о-ва; Австра-
лия. 9 подродов, несколько десятков видов. Оба современных вида, представ-
ленных в фауне нашей страны, принадлежат подроду Sinalbinula Pilsbry, 1916
(типовой вид Pupa contracta Say, 1822).
В ископаемом состоянии род известен-начиная с верхнего олигоцена (Европа,
Северная Америка); в неогене был обширно представлен на Кавказе.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ GASTROCOPTA
1 (2). Субпариетальная пластинка имеется. Палатальных складок чаще всего
две ..............................................2. G. huttoniana.
2 (1). Субпариетальная пластинка отсутствует. Палатальных складок обычно
три............................................... . 1. G. theeli.
1. Gastrocopta theeli (Westerlund, 1877) (рис. 114, /; рис. 115)
Westerlund, 1877 : 102, fig. 4 (Pupa}', Mousson, 1887 : 19, t. I, fig. 6 [P. (Ver-
tigo) denudata]; Westerlund, 1887 : 129 (Pupa); Pilsbry, 1916, Man. Conch., 24 :
109, t. 21, figs. 2—4 (coreana); F о г с a г t, 1935 : 422, Abb. 3; Лихарев, Раммель-
мейер, 1952 : 128, рис. 37; Lozek, 1964 : 209, t. 8, fig. 9; Матёкин, 1966 : 110,
табл. 2, рис. 8; Крестьянинов, 1973 : 53—54, рис.
Terra typica — Микулино, окр. Енисейска.
Лектотип (№ 1657) и 20 паралектотипов хранятся в Естественно-
историческом музее Стокгольма, 5 паралектотипов — в Естественноисториче-
1. РОД GASTROCOPTA
197
ском музее Гётеборга и 1 паралектотип — в Зоологическом ин-те АН СССР
(выделены И. М. Лихаревым и мною).
Материал: более 100 экз. из 9 местонахождений в Приморском крае,
в Грузии, Дагестане и на Северном Кавказе.
Раковина от удлиненно-овальной до овально-конической, с притупленной
вершиной, прозрачная, бесцветная; после гибели животного раковина нередко
мутнеет и становится известково-белой, непрозрачной. Оборотов 5—ЕПД,
сильновыпуклых, последний оборот к устью слегка приподнят, высота его со-
ставляет около половины высоты раковины. Поверхность эмбриональных обо-
ротов неясно микрозернистая, дефинитивные пок-
рыты тончайшей радиальной исчерченностыо,
нередко почти совсем гладкие. Устье овальное
или округло-треугольное, края его тонкие,
умеренно отвернуты; сильно развитая парие-
Рис. 114. Виды рода Gastrocopta. Рис. 115. Gastrocopta theeli (West.)
I — G. theeli (West.) из Уссурийского заповедника, Пример- из с' тРеА?Ула Приморского края,
ский край; II — G. huttoniana (Benson) из выносов р. Ка~ 4 VII 1971 г. ПОЛОВОЙ аппарат,
фирниган, Гиссарский хр.
тальная мозоль создает впечатление почти цельного устья. Зубов 6. Передняя
вершина двувершинной париетальной пластинки (эту пластинку, вероятно,
правильнее называть ангулярно-париетальной, так как передняя вершина
соответствует, по-видимому, ангулярному зубу) отклонена к палатальной
стенке, задняя (собственно париетальная пластинка) направлена вертикально
вниз. Субпариетальная пластинка отсутствует. Колумеллярная пластинка хо-
рошо развита, горизонтальная. Базально-колумеллярный бугорок округлый,
расположен немного отступя от устьевого края. Палатальных складок три,
самая длинная нижняя, самая короткая супрапалатальная; нижней палаталь-
ной складке на затылке соответствует слабая вмятина, две верхних представ-
ляют собой утолщения губы. Имеется небольшое затылочное утолщение. Пу-
пок в виде узкой и короткой щели.
Размеры: ВР 1.7—2.2, БД 0.9—1.2 мм; в первоописании: ВР 2.25,
БД 1.25 мм; лектотип: ВР 2.1, БД 1.05 мм.
Внутреннее строение. Материал: 7 экз. из с. Пейшула в При-
морском крае, 4 июля 1971 г., сбор и определение автора.
Белковая железа с латеральным желобком. Семяпровод терминально впа-
дает в очень короткий эпифаллус, который при переходе в пенис образует ко-
лено. Пенис недлинный, цилиндрический. Никаких придатков совокупитель-
ного аппарата не обнаружено. Половой ретрактор крепится немного прокси-
мальнее середины пениса. Семяприемник состоит из очень тонкого протока
и маленького веретеновидного резервуара, прилегающего к срединной части
матки.
198 IV. НАДСЕМ. VERTIGINOIDEA
Распространение и местообитание. Ареал вида носит
.реликтовый характер: Южное Приморье, окр. Енисейска, Челябинска, южный
Алтай (Увалиева, 1965), выбросы рек Куры и Риони, высокогорье Дагестана
г(Ботлих) и Северного Кавказа (среднее течение р. Чегем).
Населяет широколиственные и смешанные сильно увлажненные леса, где
держится в нижних слоях опада; иногда встречается во влажной гниющей дре-
весине.
2. Gastrocopta huttoniana (Benson, 1849) (рис. 114, II).
В е n s о п, 1849, Ann. Mag. Nat. Hist., 2, IV : 126 (Pupa)- Westerlund, 1898 :
181 (P. theeli)-, Pilsbry, 1917, Man. Conch., 24 : 137, t. 21, figs. 15—17; Лихарев,
P а м м e л ь м e й e p, 1952 : 129, рис. 38; И з з а т у л л а е в, 19706 : 80.
Terra typica — Симла (северная Индия, Гималаи).
Место хранения типов не установлено.
Материал: более 100 экз. из выносов р. Кафирниган.
Раковина удлиненно-овальная, свежие раковины стекловидно-прозрачные,
бесцветные, блестящие; после гибели животного мутнеют, становятся белыми
и утрачивают блеск. Оборотов 5, сильновыпуклых, последний оборот близ
устья плавно приподнят, высота его примерно равна половине высоты раковины.
Эмбриональные обороты с неясной тончайшей зернистостью. Поверхность де-
финитивных оборотов выглядит почти гладкой; при увеличении в 20'—30 раз
заметны неправильно расположенные тонкие радиальные морщинки. Устье
от усеченно-овального до округло-треугольного, края его тонкие, умеренно
отвернуты, места прикрепления связаны отчетливой, но тонкой мозолью.
Зубов 7—8. Ангулярно-париетальная пластинка двувершинная, причем перед-
няя вершина отклонена немного к палатальному краю, а задняя — отвесная; при
рассмотрении спереди пластинка напоминает формой ласточкин хвост. Суб-
париетальная пластинка в виде небольшого бугорка, иногда развитого слабо.
Колумеллярная пластинка горизонтальная. Базально-колумеллярное утол-
щение бугорковидной формы, немного приостренное. Палатальных складок
.две, нижняя длиннее верхней, снаружи ей соответствует слабое углубление;
кроме того, у некоторых раковин намечается супрапалатальная складка
в виде легкого утолщения губы в самой верхней части палатального края.
Пупок маленький, овальный.
Размеры: ВР 2—2.3, БД 1.1—1.2 мм; в первоописании: ВР 2, БД 1.1мм.
Распространение. Вид описан из Индии; неоднократно найден
в выносах р. Кафирниган близ Душанбе. Очевидно, моллюски обитают в до-
лине этой реки между Гиссарским и Каратегинскйм хребтами.
II. Подсем. VERTIGININAE Pilsbry, 1918|
Раковина овальная, коричневатых или красноватых тонов. Ангулярный бу-
горок отсутствует, париетальная пластинка не раздвоена.
Пенис различной длины, резко отделен от эпифаллуса очень узким пере-
жимом.
Распространение. Голарктика и Вест-Индия. 6—8 родов.
В ископаемом состоянии подсемейство известно начиная с палеоцена
{Европа, Северная Африка, Северная Америка, Бермудские о-ва).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ VERTIGININAE
1 (2). Палатальная складка длинная, ее задний конец при нормальном поло-
жении раковины не виден. Колумеллярная пластинка поднимается вер-
тикально, образует расширенную лопасть..............2. Vertilla.
2 (1). Задний конец любой из палатальных складок виден при нормальном
положении раковины. Колумеллярная пластинка горизонтальная, ло-
пастей не образует .................................1. Vertigo.
1. РОД VERTIGO 19&
1. Род VERTIGO Muller, 1774
Muller, 1774 : 124; Gray, 1821 : 239; Jeffreys, 1830 : 357 (Alaea, типовой
вид A. palustris, syn. Vertigo antivertigo)', Pilsbry, 1919, Man. Conch., 25 : 69; Thiele,
1931:505; Лихарев, Раммельмейер, 1952:133; W e n z, Zilch, 1959—
1960 : 148; Стеклов, 1966 : 143, Акрамовский, 1976 : 134.
Типовой вид Vertigo pusilia Muller, 1774; по монотипии.
Раковина право- или левозавитая. Колумеллярная пластинка почти гори-
зонтальная. Задний конец верхней палатальной складки виден через устье или
эта складка отсутствует.
Распространение. Голарктика. Не менее 70 видов.
В ископаемом состоянии род известен начиная с палеоцена (Европа, Север-
ная Африка, Северная Америка).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ПОДВИДОВ
VERTIGO
1 (12). На париетальном крае устья 2—3 зуба (подрод Vertigo s. str.).
2 (3). Раковина левозавитая .......................4. V. (V.) pusilla^
3 (2). Раковина правозавитая.
4 (5). Края устья притупленные, утолщенные .... 6. V. (V.) hydrophife„
5 (4). Края устья тонкие, губа отсутствует или расположена отступя от края
устья.
6 (9). Устьевой край сильно отвернут; имеется выраженный в разной степени,-
базально-колумеллярный зуб.
7 (8). Дефинитивные обороты, кроме последнего, радиально ребристо исчер-
чены ...........................................2. V. (V.) substriata»
8 (7). Дефинитивные обороты слабо скульптурированы....................
...........................................1. V. (V.) antivertigo.
9 (6). Устьевой край незначительно отвернут; базально-колумеллярный зуб'
отсутствует.
10 (11). Последний оборот почти гладкий............5. V. (V.) sieversL.
11 (10). Последний оборот с отчетливой радиальной скульптурой .........
...................................... 3. V. (V.) pseudosubstr iata.
12 (1). На париетальном крае не больше одного зуба (подрод Isthmia).
13(14). Края устья массивные, утолщенные губой ........................
......................................... 9. V. (I.) circumlabiata.,
14 (13). Края устья тонкие, губа отсутствует или расположена отступя от края.
15 (18). На базально-колумеллярном крае устья имеется зуб.
16 (17). Окраска раковины красновато-коричневая, большой диаметр не менее
1. 3 мм ...................................7. V. (I.) moulinsiana.
17 (16). Окраска раковины роговая, большой диаметр не превышает 1.2 мм;
. . . .........................................8. V. (I.) pygmaea.
18 (15). Базально-колумеллярный зуб отсутствует.
19 (20). Поверхность дефинитивных оборотов покрыта тонкими резкими ра-
диальными ребрышками......................... 12. V. (I.) microsphaera.
20 (19). Дефинитивные обороты слабо скульптурированы.
21 (24). Палатальный край устья прямой или слегка вдавлен.
22 (23). Последний оборот к устью плавно приподнимается, начиная с середины
или с последней трети; затылочное утолщение слабое, но всегда выражено
...................................................11. V. (I.) japonica.
23 (22). Последний оборот прямой или слегка приподнят у самого устья;
затылочное утолщение отсутствует...............10. V. (I.) hirasei.
24 (21). Палатальный край плавно и равномерно выгнут.
25 (26). Раковина цилиндрическая, устье с 4 зубами ............ . . „
.............1 . . . ’.............. 13а. V. (I.) modesta alpestris»
26 (25). Раковина от овальной до овально-цилиндрической, число зубов в устье
от 0 до 4............................. 13. V. (I.) modesta mo desta^
200
IV. НАДСЕМ. VERTIGINOIDEA
1. Подрод VERTIGO s. str.
На париетальной стенке устья от 2 до 3 зубов.
Р а сп р остр анение. Голарктика.
В ископаемом состоянии подрод известен начиная с палеоцена (Европа,
Северная Африка, Северная Америка).
1. Vertigo (Vertigo) antivertigo (Draparnaud, 1801) (рис. 116, 117).
Draparnaud, 1801 : 57 (Pupa); Montagu, 1803 : 337, t. 12, fig. 8; (Turbo sex-
dentatus); Draparnaud, 1805 : 60, t. 3, figs. 32, 33 (Pupa); Studer, 1820 : 89 (P. 8-
dentata); H. В e c k, 1837 : 85 (Alaea); Moquin-Tandon, 1855 : 407, pl. 29, figs. 4—
7 (Pupa); Mousson, 1873 : 213, t. 8, fig. 10 (sinuata); Pilsbry, 1919, Man. Conch.,
25 : 163, t. 16, figs. 4—6; Steenberg, 1925 : 79, pl. 18, figs. 1—3, pl. 19, fig. 1; Лиха-
рев, Раммельмейер, 1952 : 135, рис. 45; Ehrmann, 1956 : 40, t. I, fig. 10;
Loz ek, 1956 : 92, t. 4, fig. 3; Zilch, Jaeckel, 1962 ; 82; L о z e k, 1964 : 202, t. 7,
fig. 1; Матёкин, 1966 : 109; Стеклов, 1966 :145, табл. Ill, 51;Акрамовский,
1976 : 137, рис. 56В, табл. V, 49.
Terra typica — Франция.
Место хранения типов не установлено.
Материал: свыше 80 экз. из 19 местонахождений в европейской части '
СССР, на Кавказе и в Средней Азии.
Раковина овальная, тонкостенная, просвечивающая, блестящая, с чуть
приостренной вершиной. Оборотов 5, выпуклых; последний оборот возле устья
слегка приподнят, высота его нем-
ного больше половины высоты
раковины. Окраска каштановая,
иногда с вишневым оттенком,
верхние обороты обычно не-
сколько светлее нижних. Эмбри-
ональные обороты почти гладкие,
аппарат.
Рис. 116. Vertigo (Vertigo) antiver-
tigo (Drap.) из Подмосковья, дер.
Костино.
Рис. 117. Vertigo (Vertigo) antivertigo (Drap.) из
Подмосковья, дер. Костинр, 1 VII 1958 г. Половой
дефинитивные — тоже, но здесь имеются сильно сглаженные неправильно распо-
ложенные радиальные морщинки, не нарушающие, однако, впечатления гладкой
поверхности. Устье треугольно-овальное, с хорошо выраженным вдавлением на
палатальном крае. Затылочное утолщение светлое, ясно заметное. Края устья
слегка отвернуты, места прикрепления устья расставлены и связаны тонкой
прозрачной мозолью. Губа отсутствует. Зубов 7—10. На париетальной стенке
2—3 пластинки (субпариетальная иногда отсутствует); 1—2 колумеллярные
пластинки, короткая базально-колумеллярная складка и 2—3 палатальных;
1. РОД VERTIGO
201
если их две, то отсутствует самая верхняя. Нижней палатальной складке соот-
ветствует продольная вмятина на затылке. Пупок почти запаян.
Размеры: ВР 2—2.2, БД 1.2—1.3 мм; в цервоописании размеры не указаны.
Внутреннее строение. Материал: 3 экз. из кишлака Буралик
на Кураминском хр., 2 июня 1977 г., сбор и определение 3. Иззатуллаева;
5 — из дер. Костино в Подмосковье, близ Пушкино, 1 июля 1958 г., сбор и
определение автора.
Белковая железа с глубоким латеральным желобком. При основании же-
лезы видна простата в виде нескольких светлых долек, от которых отходит
тончайший семяпровод. Последний впадает в эпифаллус с резким изгибом,
к которому крепится половой ретрактор. Эпифаллус в l1/^—2 раза короче
пениса, цилиндрический или веретеновидный. Пенис тонкостенный, граница
между ним и эпифаллусом обозначена очень четко благодаря наличию узкого
пережима (сфинктера). Проток семяприемника очень тонкий, длинный, без
резкой границы переходит в маленький резервуар, прилегающий к верхней
части матки на границе с белковой железой.
Распространение и местообитание. Большая часть тер-
ритории Палеарктики: на восток до Забайкалья включительно, на север до
зоны тундры. Чаще всего встречается на влажных лугах с высоким травостоем
и в долинах горных рек; живет также в подстилке лиственных и смешанных
лесов, особенно в оврагах и понижениях, где почва всегда влажная. В сырую
погоду поднимается на стебли трав и кустарников.
2. Vertigo (Vertigo) substriata (Jeffreys, 1830) (рис. 118, 119).
Jeffreys, 1830 : 515 (Alaea); Held, 1837 : 304 (Pupa curta); Reeve, 1863 : 119,
fig. (striata); Mousson, 1876 : 143, t. 10, tig. 6, 7 (Pupa pygmaea var. nitidula); О. В о e t-
t g e r, 1880c : 381 (Pupa substriata var. mitis); Lindholm, 1910 : 35 (Alaea substriata
f. viridana); Pilsbry, 1919, Man. Conch., 25 : 172, t. 17, fig. 10; Steenberg, 1925 :
83, pl. 16, fig. 4, pl. 20, fig. 1; Лихарев, Раммельмейер, 1952 : 135, рис. 44;
Ehrmann, 1956 : 41, t. 2, fig. 13; L о z e k, 1956 : 94, t. 4, fig. 4; Zilch, Jaeckel,
1962 : 85; L о zek, 1964 : 204, t. 7, fig. 4; Gittenberger et al., 1970 : 51, fig. 37,
Kaart 16.
Terra typica — «In a marshy piece of ground near Rawleigh House, ... Barn-
staple» (Англия, Барнстапл).
Место хранения типов не установлено.
Материал: около 70 экз. из 26 местонахождений в европейской части
СССР, в Крыму и на Кавказе.
Раковина овальная или удлиненно-овальная, просвечивающая; вершина
закругленная. Оборотов 4х/2, довольно выпуклых, последний оборот возле устья
едва заметно приподнят, высота его немного больше половины высоты раковины.
Окраска роговая или светло-роговая. Эмбриональные обороты покрыты микро-
скопической зернистостью, дефинитивные несут ясную скульптуру — от чет-
кой равномерной морщинистости до тонкой ребристости; на последнем обороте
эта скульптура ослабляется. Устье округло-треугольное, вдавлено на палаталь-
ном крае. Затылочное утолщение относительно слабое. Зубов 6: ангулярная,
париетальная и колумеллярная пластинки, базальная и две палатальных
складки. Пупок очень маленький, каплевидный.
Размеры: ВР 1.5—1.8, БД 1—1.1 мм; в первоописании: ВР 0.06, БД 0.04
дюйма, т. е. ВР 1.5, БД 1 мм.
Внутреннее строение. Материал: 1 экз. из с. Ромашково Мос-
ковской обл., 1 мая 1961 г.; 7 — из Чегемского ущелья на Северном Кавказе,
15 мая 1969 г.; сборы и определения автора.
Белковая железа с очень резким латеральным желобком. Простата хорошо
заметна у нижней кромки белковой железы. На границе семяпровода и эпи-
фаллуса имеется резкий изгиб. Пенис и эпифаллус примерно равной длины,
разделены очень резкой перетяжкой. Половой ретрактор крепится к месту
202
IV. НАДСЕМ. VERTIGINOIDEA
перехода семяпровода в эпифаллус. Проток семяприемника очень тонкий,
округлый резервуар прилегает к нижней кромке белковой железы.
Распространение и местообитание. Северная, средняя
и восточная Европа, Северный Кавказ. Обитает во влажных высокотравных
лугах и в подстилке лиственных и смешанных лесов. Изолированный участок
ареала на Алтае.
3. Vertigo (Vertigo) pseudosubstriata Lozek, 1954 (рис. 120).
Lozek, 1954 : 327, t. I, fig. 1; 1964 : 204, t. 8, fig. 6; Матёкин: 1966 : 109; У ва-
ли e в a, 1967 : 213, рис. 1 (7. laevis).
Terra typica — Чехословакия, Моравия, раскопки позднепалеолитического
апоселения в Дольни Вестонице над р. Дый.
Рис. 118. Vertigo (Ver-
tigo) substriata (Jeffr.) из
Чегемского ущелья на
Северном Кавказе.
Тис. 119. Vertigo (Ver-
tigo) substriata (Jeffr.) из
‘Чегемского ущелья на
Северном Кавказе,
Ж5 V 1969 г. Половой ап-
парат.
Рис. 120. Vertigo
(Vertigo) pseudosub-
striata Lozek из Юж-
ного Алтая (голотип
V. laevis UValieva).
Голотип хранится в малакологической коллекции Центрального гео-
•аттического института в Праге.
Материал: 6 экз. из одного местонахождения на Южном Алтае (голо-
тип и паратипы V. laevis).
Раковина удлиненно-овальная, тонкостенная, просвечивающая, с шелко-
вистым блеском. Вершина узко закругленная. Оборотов 5, выпуклых, плавно
нарастающих. Последний оборот к устью приподнят, высота его чуть больше
половины высоты раковины. Окраска одноцветная, от светло- до темно-роговой.
• Эмбриональные обороты покрыты очень неясной тончайшей зернистостью,
дефинитивные местами почти гладкие, с отдельными тонкими неправильно рас-
положенными морщинками.' Устье неправильно округлое, со слегка отверну-
тыми тонкими краями; затылочное утолщение отсутствует. Места прикрепления
устья не сближены. Зубов 5: две париетальные пластинки (субпариетальная
.длиннее и выше париетальной); колумеллярная пластинка мощная, зубовид-
аая; две хорошо развитые палатальные складки начинаются слегка отступя от
гкрая устья. Пупок узкий, сквозной, овальный.
Размеры: ВР 2.1—2.2, БД 1.2 мм; в первоописании: ВР 2.15, БД 1.2 мм.
Исследование типового материала по V. laevis и сопоставление его с описа-
нием и хорошей фотографией голотипа V. pseudosubstriata (описание сделано
1. РОД VERTIGO
203
на чешском языке и повторено на русском и немецком) показало, что все при-
знаки обеих форм совпадают настолько, что трудно говорить даже о подвидах.
Распространение и местообит ан и я. Вид найден в полу-
ископаемом состоянии в Моравии; затем Матёкин (1966) указывает присутствие
его на Тянь-Шане (точнее не указано), и, наконец, под названием V. laevis
отмечается для Южного Алтая, хребтов Терскей, Заилийский и Джунгарский
(Увалиева, 1967; Альмухамбетова, Увалиева, 1979). Обитает в лесной под-
стилке, на альпийских лугах, по берегам ручьев; поднимается до высоты;
3500 м над уровнем моря.
4. Vertigo (Vertigo) pusilia Muller, 1774 (рис. 121, 122).
Muller, 1774 : 124; Mo qui n - T an d on, 1855 : 409, pl. 29, figs. 12—14 [Ver-
tigo (Vertilla)]; Reeve, 1863 : 120, fig.; Pilsbry, 1919, Man. Conch., 25 : 161, t. 16,
figs. 1—3; Steenberg, 1925 : 86, pl. 21, figs. 1, 2, pl. 26, fig. 2; Л и x a p e в, Рам-
мельмейер, 1952 : 134, рис. 43; Ehrmann, 1956 : 39, T. I, Fig. 9; L о z'e k, 1956 ;
91, t. 4, fig. 2; Zilch, Jaeckel, 1962 : 82; Lozek, 1964 : 202, t. 4, fig. 12; С т e k-
л о b, 1966 : 143, табл. Ill, 52; Gittenberger et al., 1970 : 51, fig. 35, Kaart 14; S zi-
g e t h y, 1973 : 21, figs. 3, 9; А к p а м о в с к и й, 1976 : 136, рис. 56А, табл. V, 46; Аль-
мухамбетова, Увалиева, 1979 : 38.
Terra typica — не указана; -сообщается лишь, что вид живет на гниющих
стволах («in truncis putridis»); вероятно, типовая серия происходит из окр. Ко-
пенгагена, так как многие виды Мюллером описаны из этой местности.
Рис. 121. Vertigo (Vertigo) pu-
silia . Miill. из долины р. Те-
хури на юго-западных скло-
нах Большого Кавказа.
Рис. 122. Vertigo (Vertigo)
pusilia Mull. Половой аппарат
(из: Steenberg, 1925).
Место хранения типов не установлено.
Материал: несколько сотен экз. из 52 местонахождений в западной
Европе, европейской части СССР и на Кавказе.
Раковина левозавитая, коротко- или удлиненно-овальная, блестящая,
сильно просвечивающая, с плавно закругленной вершиной. Оборотов 5, вы-
пуклых, последний оборот к устью почти не поднят, высота его заметно больше
половины высоты раковины. Окраска одноцветная, от роговой до каштановой.
Поверхность эмбриональных оборотов покрыта неясной зернистостью, дефини-
тивные обороты очень тонко радиально исчерчены; эта исчерченность иногда
переходит в морщинистость — на верхних оборотах и на затылочном утолще-
204
IV. НАДСЕМ. VERTIGINOIDEA
нии. Устье округлое, с палатальным вдавлением, края его тонкие, слегка от-
вернутые. Затылочное утолщение массивное. Зубов 6, по два на каждом крае;
кроме того, могут присутствовать небольшой базальный бугорок и легкая
супрапалатальная складка. Пупок открытый, очень маленький, каплевидный.
Размеры: ВР 2.0—2.2 , БД 1.0—1.1 мм; в первоописании: ВР 1, БД 1/i ли-
нии, т. е. ВР 2.1—2.2, БД примерно 0.25 мм; указание большого диаметра,
по-видимому, ошибочно.
Внутреннее строение. Материал мной не исследован, описание
составлено по данным Стинберга (Steenberg, 1925).
Белковая железа с латеральным желобком. На границе семяпровода и тон-
кого эпифаллуса имеется изгиб, к которому подходит половой ретрактор. Пере-
тяжка между пенисом и эпифаллусом ясная. Пенис в 2—22/2 раза длиннее эпи-
фаллуса и отличается большим диаметром. Резервуар семяприемника удли-
ненно-овальный .
Распространение и местообитание. Европа, Кавказ,
Малая Азия, Алтай, северный Казахстан; в европейской части СССР к востоку
до Волги, на север примерно до 60° с. ш. Населяет умеренно увлажняемые лист-
венные и смешанные леса, где держится в подстилке и на обомшелых стволах.
Встречается также на открытых луговинах: в дерновинках злаков и под кам-
нями .
5. Vertigo (Vertigo) sieversi (О. Boettger, 1879) (рис. 123).
О. Boettger, 1879 : 407, t. 10, figs. 6, 7 [Pupa (Vertigo)]; 1880a : 141 (var. punctum
et var. subalpestris); Rosen, 1914:195 {Papa); Лихарев, Раммельмейер,
1952 : 135 {substriata var. sieversi).
Terra typica — у оз. Табацкури (южная Грузия, Триалетский хр.).
Место хранения типов не установлено.
Материал: 18 экз. из 6 местонахождений на Кавказе.
Раковина удлиненно-овальная или овально-цилиндрическая, тонкостенная,
сильно или умеренно просвечивающая. Оборотов 4^2, умеренно выпуклых,
последний оборот плавно и незначительно приподнят, высота его составляет
около половины высоты раковины. Окраска одноцветная, светло-роговая.
Эмбриональные обороты почти гладкие, первые дефинитивные тонко и равно-
мерно исчерчены; на последнем обороте эта исчерченность выражена заметно
«слабее или почти отсутствует. Устье в общем округлое, палатальное вдавление
«слабое, места прикрепления устья не сближены, связаны тонкой, но отчетливой
мозолью. Затылочное утолщение развито в разной степени, но всегда имеется.
Края устья незначительно утолщены или совсем не утолщены, почти не отвер-
нуты. Зубов 5. На париетальном крае два небольших зуба, париетальный раз-
вит лучше супрапариетального; последний иногда развит настолько слабо, что
представляет собой небольшую припухлость париетальной стенки. Колумелляр-
ная пластинка в виде уплощенного бугорка, лежит в задней части столбика.
Нижняя палатальная складка развита сильнее верхней, на затылке нижней
палатальной складке соответствует вмятина; значительно более слабая вмятина
может соответствовать и верхней палатальной. Пупок очень узкий, в виде
короткой округлой щели.
Размеры: ВР 1.9—2.1, БД 1 мм; в первоописании: ВР 17/8, БД 1 мм.
Акрамовский (1976) считает, что данный вид является подвидом V. sub-
striata, живущим на Северном Кавказе и в Закавказье. Доводом в пользу пред-
ставления о видовой самостоятельности V. sieversi служит указание Беттгера
(Boettger, 1880а, S. 141), что Pupa sieversi var. punctum и var. subalpestris встре-
чены на Казбеке вместе с Р. substriata. Кроме того, оба вида вместе собраны
мною в верхнем течении р. Чегем.
Распространение и местообитание. Вид широко, но
•спорадически распространен по центральной и западной частям Северного Кав-
1. РОД VERTIGO
205
каза и Закавказью. Населяет подстилку лиственных и смешанных лесов,
встречается также под корой старых пней. В высокогорье может жить в осыпях
и среди щебенки у подножья скал.
6. Vertigo (Vertigo) hydrophila (Reinhardt, 1877) (рис. 124).
Reinhardt, 1877b : 323, t. 11, fig. 6 [Pupa [Vertigo')]; Pilsbry, 1919, Man.
Conch., 25 : 152, pl. 14, figs. 4, 5; Лихарев, 1957 : 78.
. Terra typica — «Hakodate, Yesso» (Япония, Хакодате).
Место хранения типов не установлено.
Материал: 1 экз. с о. Итуруп.
Раковина овально-коническая, с закругленной вершиной, тонкостенная,
слегка просвечивающая. Оборотов 4х/2, умеренно выпуклых, разделенных глу-
Рис. 123. Vertigo (Vertigo) sieversi (Bttg.) из
Кавказского заповедника, Краснодарский
край.
Рис. 124. Vertigo (Vertigo) hydrophila
(Reihn.) с о. Итуруп, Курильские о-ва.
боким швом. Последний оборот к устью слегка и плавно, но с легким переги-
бом, поднят, высота его примерно равна половине высоты раковины. Окраска
одноцветно-роговая. Эмбриональные обороты неясно зернистые, дефинитивные
покрыты слабыми сглаженными радиальными морщинками, резче выражен-
ными на ранних оборотах. Устье округлое, с плавным углом на верхней части
палатального края. Устьевые края слегка утолщены, палатальный почти пря-
мой, колумеллярный слегка отвернут. Зубов 6, по два на каждом крае. Парие-
тальная пластинка в виде короткого округлого гребня. Колумеллярная пла-
стинка горизонтальная, субколумеллярная зубовидная, округлая. Нижняя
палатальная складка заметно длиннее верхней, со слегка утолщенным греб-
нем. Палатальным складкам снаружи соответствуют чрезвычайно слабые вмя-
тины. Затылочная борозда еле выражена, затылочное утолщение слабое. Пупок
точковидный.
Размеры в первоописании: ВР 1.75, БД 1 мм; единственный исследованный
экземпляр имел такие же размеры.
Распространение. Северная Япония и о-в Итуруп.
2. Подрод ISTHMIA Gray, 1821
Gray, 1821 : 239.
Типовой вид Helix (Isthmia) cylindrica Draparnaud, 1801 (syn. Vertigo
pygmaea Draparnaud, 1801).
На париетальной стенке устья 1 зуб или париетальные зубы отсутствуют.
Распространение. Палеарктика; ареал состоит из европейско-
кавказского и дальневосточного участков, связанных между собой циркум-
бореально-альпийским ареалом Vertigo modesta.
206
IV. НАДСЕМ. VERTIGINOIDEA
В ископаемом состоянии подрод известен начиная с палеогена (Европа,.
Северная Америка).
7. Vertigo (Isthmia) moulinsiana (Dupuy, 1849) (рис. 125, 126).
Dupuy, 1849 : 284 (Pupa)- К okeil in : G aliens t ein, 1852: 80 (P. laevigata)^
Hey nemann, 1862 : 11, t. I, figs. 6—8 (ventrosa)-, Wes t erlun d, 1868, Coll. Typ.
moll. Siiecia : 60 (Pupa lilljeborgi)', 1875, Malak. Bl., XXII : 133 (P. kiisteriana)- Pilsbry,-'
1919, Man. Conch., 25 : 178, pl. 17, figs. 1—3; Steenberg, 1925 : 73, pl. 14, fig. 1, pl. 15,
figs. 1—3, pl. 16; Лихарев, Раммельмейер, 1952 : 136, рис. 46; Ehrmann,
1956 : 40, t. 2, fig. 11; Lozek, 1956 : 92; Z i 1 c h, J a e c k e 1, 1962:83; Lozek,
1964 : 203, t. 7, fig. 2; G i 11 en b erger et al., 1970 : 50, fig. 39; S z i g e t h y, 1973 :
24, fig. 5, 11; А к p а м о в с к и Й, 1976 : 137, рис. 56Д, табл. V, 50.
Terra typica — окр. Лиона, Франция.
Место хранения типов не установлено.
Материал: 40 экз. из 19 местонахождений в европейской части СССР^
Финляндии, Швеции, Дании, Чехословакии, Болгарии и Италии.
Рис. 125. Vertigo (Isthmia)
moulinsiana (Dupuy) из
Подмосковья, окр. За-
горска.
Рис. 126. Vertigo (Isth-
mia) moulinsiana (Dupny)
из Подмосковья, окр. За-
горска, 2 VI 1959 г. По-
ловой аппарат.
Раковина овальная, блестящая, прозрачная, с узко закругленной верши-
ной. Оборотов 41 /2, выпуклых, последний оборот к устью слабо поднят, высота
его заметно больше половины высоты раковины, ^иногда достигает почти
2/3 этой, величины. Окраска одноцветная,"(красновато-коричневая или темно-
вишневая. Микроскопическая зернистость на эмбриональных оборотах слабо
заметна. Дефинитивные обороты почти гладкие, местами с очень вялыми сгла-
женными радиальными морщинками. Устье округлое, немного сердцевидное,
края его отвернуты, особенно сильно отвернут базальный край. Несколько
отступя от палатального края залегает светлая расплывшаяся губа, выражен-
ная в различной степени. Затылочное утолщение массивное. На париетальном
и колумеллярной краях по одной пластинке, на палатальном — две складки.
Кроме того, нередко присутствует базальный зуб. Пупок точковидный.
Размеры: ВР 2—3, БД 1.3—2 мм; в первоописании: ВР 2.5—3, БД 2 мм.
Вальден (Walden, 1966) считает V. moulinsiana и V. lilljeborgi разными ви-
дами: он приводит таблицу, где сравниваются основные признаки этих форм.
Из таблицы следует, что наиболее существенные различия носят количествен-
ный характер и касаются в первую очередь признаков, связанных с устьевой
1. РОД VERTIGO
207
арматурой: у V. lilljeborgi устьевые зубы развиты слабее. Существенно, что
Вальден сравнивал V. moulinsiana из Дании, ФРГ, Франции, Британских о-вов
и Италии с V. lilljeborgi из Швеции и Финляндии. Поэтому представляется,
что f. lilljeborgi — это «ослабленный» вариант V. moulinsiana, встречающийся
^лиз северных (и восточных) границ ареала вида.
Внутреннее строение. Материал: 1 экз. из окр. Загорска, Мос-
ковская обл., 2 июня 1959 г., сбор и определение автора. .
Белковая железа с очень резким и глубоким желобком на латеральной по-
верхности. Семяпровод довольно плавно расширяется в эпифаллус, не образуя
резкого изгиба; к проксимальной части эпифаллуса крепится половой ретрак-
тор. Резкий пережим между эпифаллусом и пенисом, столь характерный для
других видов рода, у данного вида отсутствует: имеется лишь плавное сужение
протока. Пенис примерно вдвое длиннее эпифаллуса. Резервуар семяприем-
ника в виде короткой булавы, тесно прилегает к нижней кромке белковой же-
лезы.
Распространение и местообитание. Центральная и
частично южная Европа; в СССР — европейская часть, на север до Литвы,
па восток до Волги; южная граница ареала не установлена. Местами встреча-
-ется в Закавказье. В Армении — по берегам стоячих водоемов на травянистых
растениях, почти не спускаясь на почву (Акрамовский, 1976).
8. Vertigo (Isthmia) pygmaea (Draparnaud, 1801) (рис. 127, 128).
Draparnaud, 1801 : 57 {Pupa); 1805 : 60, pl. 3, figs. 30, 31 {Pupa); J e f f г e у s,
1830 : 359 {Alaea vulgaris); Reeve, 1863 : 118, fig.; Pilsbry, 1919, Man. Conch., 25 :
174, pl. 17, figs. 15—17; Steenberg, 1925 : 77, pl. 14, fig. 3, pl. 16, fig. 5, pl. 17; Л и-
x ар ев, Раммельмейер, 1952 : 136, рис. 47; E h r m a n n, 1956 : 40, t. I, fig. 12;
Lozek, 1956 : 93, t. 4, fig. 5; Z i 1 c h, J a e c k e 1, 1962 : 85; L о i e k, 1964 : 203, t. 7,
fig. 3; Матёкин, 1966 : 109; Gittenberger et al., 1970 : 51, fig. 38, Kaart 15;
S zi get hy, 1973 : 21, fig. 6, 12; А к p а м о в с к и й, 1976 : 137, рис. 56Д, табл. V, 51.
Terra typica — Франция.
Место хранения типов не установлено.
Материал: около 100 экз. из 34 местонахождений в европейской части
СССР, на Кавказе и на Тянь-Шане.
Раковина удлиненно-овальная, матовая, просвечивающая. Оборотов 5,
слабо- или умеренно выпуклых, последний оборот к устью почти не поднят,
высота его всегда больше половины высоты раковины. Окраска одноцветно
«светло-роговая. Эмбриональные обороты с тончайшей зернистостью, дефини-
тивные выглядят почти гладкими, но при увеличении порядка 50 раз видна тон-
чайшая струйчатая радиальная исчерченность. Устье неправильно округлое,
края его слабо отвернуты, тонкие, палатальное вдавление очень слабое, заты-
лочное утолщение, напротив, мощное. Места прикрепления устья не сближены,
«связаны очень тонкой мозолью. На середине париетального края располага-
ется короткая гребневидная пластинка. Колумеллярная пластинка развита
хорошо; на стыке колумеллярного и базального краев расположен небольшой
пластинчатый зуб. На палатальном крае обычно 2 складки, нижняя длиннее
верхней, на затылке ей соответствует продольный желобок. Иногда имеется
«еще маленькая супрапалатальная складка. Пупок почти запаян.
Размеры: ВР 2—2.2, БД 1.2 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 3 экз. из Москвы, Кунцевский
парк, 29 августа 1978 г.; сбор и определение автора; 3 — из окрестностей Сте-
панавана, 30 мая 1956 г., сбор и определение Н. Н. Акрамовского.
Белковая железа с резким латеральным желобком. Семяпровод впадает
в эпифаллус с резким изгибом, скорее даже изломом, к которому крепится
половой ретрактор. Эпифаллус тонкий, почти цилиндрический, кольцевой
пережим, отделяющий его от пениса, не очень резкий, хотя выражен вполне
ясно. Пенис в проксимальной части плавно изогнут, и этот изгиб, видимо,
208
IV. НАДОЕМ. VERTIGINOIDEA
постоянен, потому что отмечен у всех исследованных животных. Резервуар
семяприемника небольшой, шаровидный, прилегает к границе спермовидукта
и белковой железы.
Распространение и местообитание. Центральная, за-
падная, южная и восточная Европа, Кавказ, Сибирь до Забайкалья, горные
Рис. 127. Vertigo (Isth-
mia) pygmaea (Drap.) из
Подмосковья, с. Ромаш-
ково.
Рис. 128. Vertigo (Isth-
mia) pygmaea (Drap.) из
окр. Степанавана, Армения,
30 V 1956 г. Половой ап-
парат.
районы Средней Азии; Малая Азия, северный Иран. Завезен в Новую Зелан-
дию и северо-восточную часть Северной Америки. Встречается во всех типах
биотопов с умеренным увлажнением (сырых мест избегает), кроме тундры
и чисто хвойных лесов. Живет на равнине и в горах.
9. Vertigo (Isthmia) circumlabiata Schileyko, sp. nov. (рис. 129).
Terra typica — о. Кунашир, окр. Серноводска.
Голотип хранится в Зоологическом ин-те АН СССР.
Материи л: 4 экз. из окр. Серноводска на о. Кунашир, и 1 экз. из Аяна
(берег Охотского моря).
Раковина овальная, с почти гладкой поверхностью, умеренно блестящая,
относительно твердостенная. Оборотов 4х/2—43/4, умеренно выпуклых, в про-
филе плавно закругленных. Последний оборот к устью слегка и очень плавно
поднимается, высота его несколько превышает половину высоты раковины.
Эмбриональные обороты гладкие, дефинитивные также в общем лишены ре-
гулярной правильной скульптуры и лишь местами заметны одиночные тонкие
радиальные линии. Окраска роговая, эмбриональные обороты почти бесцвет-
ные, первые дефинитивные желтоватые, два последних коричневые. Устье почти
правильно округлое, места его прикрепления широко расставлены и связаны
тонкой светлой мозолью. Края устья почти не отвернуты, но сильно утолщены
по всему свободному краю; если рассматривать раковину справа, то на середине
палатального края заметен небольшой выступ, направленный вперед. В устье
4 зуба, все они имеют вид коротких пластинок и сидят довольно глубоко: па-
1. РОД VERTIGO
20»
риетальный, колумеллярный и два палатальных, из которых нижний больше
верхнего. На затылке палатальные зубы просвечивают в виде коротких желто-
ватых пятен, но соответствующие вдавления почти не выражены. Пупок почти
полностью запаян, от него остается еле заметная щель.
Размеры: ВР 2—2.4, БД 1.3—1.6 мм; голотип: ВР 2.2, БД 1.5 мм.
Вид похож на некоторые формы из группы V. modesta, отличается почти
полным отсутствием скульптуры, более овальной раковиной и сильно утол-
щенными краями устья.
clrcumlabiata Schileyko, sp. nov.
11 — паратип из Аяна на побережье Охотского моря.
Рис. 130. Vertigo (Isth-
mia) hirasei Pils. с о-ва
Шикотан, Курильские
о-ва.
Рис. 129. Vertigo (Isthmia)
I — голотип,
Распространение и местообитание. Вид найден авто-
ром в двух местах: 4 экз. в подстилке смешанного леса на о. Кунашир близ
Серноводска и 1 экз. примерно в тех же условиях в окр. Аяна на берегу Охот-
ского моря.
10. Vertigo (Isthmia) hirasei Pilsbry, 1901 (рис. 130).
Pilsbry, 1901, Nautilus, 14 : 128; 1919, Man. Conch., 25 : 154, pl. 14, figs. 13, 14.
Terra typica — «Yanagawa, Chikugo, in Klushin» (Япония, о. Кюсю, Яна-
гава близ г. Омута).
Голотип хранится в Академии естественных наук Филадельфии (США),
№ 79738.
Материал: 27 экз. из окр. бухты Крабовая на о. Шикотан.
Раковина овально-цилиндрическая, т. е. последние два оборота почти одной
ширины, а более ранние обороты образуют закругленно-коническую вершину.
Стенки раковины тонкие, сильно просвечивающие, поверхность с шелковистым
блеском. Оборотов 41/2, выпуклых, разделенных глубоким швом. Последний
оборот прямой или чуть приподнят у самого устья, высота его несколько пре-
вышает половину высоты раковины. Окраска одноцветная, желтовато-роговая.
Эмбриональные обороты отчетливо микрозернистые, дефинитивные покрыты
тонкой ребристой исчерченностью, более резкой на ранних оборотах; на послед-
нем обороте местами заметна исключительно тонкая струйчатость, видная при
увеличении более чем в 50 раз. Устье округлое, края его не утолщены, умеренно
отвернуты, светлые. Затылочная бороздка и затылочное утолщение отсутствуют.
Зубов 4. Париетальная пластинка округло-треугольная, колумеллярная почти
горизонтальная, располагается у заднего края столбика. Палатальных складок
две, нижняя длиннее верхней и выше нее. Пупок в виде округлой щели.
Размеры: ВР 1.5—1.7, БД 1 мм; в первоописании: ВР 1.65, БД 1 мм.
Распространение и местообитание. Япония и о. Шико-
тан. Обитает в подстилке лиственных лесов.
14 А. А. Шилейко .....
210
IV. НАДСЕМ. VERTIGINOIDEA
И. Vertigo (Isthmia) japonica Pilsbry et Hirase, 1904 (рис. 131).
Pilsbry, Hirase, 1904 : 118; Pilsbry, 1919, Man. Conch., 25 : 155, t. 15,
figs. 1, 2 (V. japonica subsp. coreana et tosana)', Лихарев, Раммельмейер, 1952 :
137, рис. 49.
Terra typica — «Ikusagawa, Ojima» (Япония, о-ва Осима, Икусагава).
Голотип хранится в Академии естественных наук Филадельфии (США),
№ 85746.
Материал: более 100 экз. из 17 местонахождений на юге Приморского
края, в том числе на о-вах зал. Петра Великого, в окр. Ванино и Аяна, а также
на Курильских о-вах.
Рис. 131. Vertigo (Isth-
mia) japonica Pils. et
Hir. из Приморского
края, берег зал. Восток.
Рис. 132. Vertigo (Isthmia) microsphaera
Schileyko, sp. nov.
I — голотип; II — паратип из о. Шикотан,
Курильские о-ва.
Раковина овально-цилиндрическая, тонкостенная, слегка блестящая. Обо-
ротов 41/2—5, выпуклых. Последний оборот к устью плавно приподнят, высота
его составляет около половины высоты раковины. Окраска одноцветная, светло-
роговая. Эмбриональные обороты почти гладкие, дефинитивные покрыты до-
вольно правильными сглаженными тонкими радиальными морщинками. Устье
имеет немного сердцевидную форму из-за того, что палатальный край посре-
дине несильно и плавно, но отчетливо вдавлен. Затылочное утолщение плохо
заметно отчасти оттого, что оно развито умеренно, отчасти из-за окраски, не от-
личающейся от окраски остальной поверхности раковины. Края устья тонкие,
незначительно отвернуты. Зубов 4: париетальный, колумеллярный и два па-
латальных. Все зубы выражены примерно в одинаковой степени, имеют вид
тонких полукруглых пластинок. Пупок чрезвычайно узкий.
Размеры: ВР 1.5—1.7, БД 0.7—1 мм; в первоописании: ВР 1.7, БД 1 мм.
Распространение и местообитание. Япония, Корей-
ский п-ов, Южное Приморье, южные Курильские о-ва, найден также в окр. Ва-
нино и в Аяне на берегу Охотского моря (Аян, очевидно, северная граница
ареала). Населяет преимущественно опушки лиственных лесов и редколесье;
держится обычно на стеблях трав.
12. Vertigo (Isthmia) microsphaera Schileyko, sp. nov. (рис. 132).
Terra typica — о-в Беринга, окр. бухты Подутесной.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 1 экз. с о-ва Беринга (голотип) и 1 экз. с о. Шикотан,
окр. бухты Крабовой.
Раковина короткоовальная, с четкими радиальными ребрышками на всех
дефинитивных оборотах, почти шаровидная, тонкостенная, шелковисто побле-
скивающая. Оборотов 4у4, сильно выпуклых. Последний оборот возле самого
устья еле заметно приподнят (практически прямой), высота его немного больше
1. РОД VERTIGO
211
половины высоты раковины. Эмбриональные обороты почти гладкие, дефини-
тивные покрыты тонкими, но резкими правильными радиальными ребрышками.
Окраска коричневатая, верхние обороты немного светлее нижних. Устье округ-
лое, места его прикрепления не сближены, связаны тонкой мозолью, причем
у голотипа участок мозоли, прилегающий к колумеллярному краю, несколько
утолщен. Края устья тонкие, незначительно отвернуты. Имеется 4 глубоко
сидящих умеренно развитых коротких пластинчатых зуба: париетальный,
колумеллярный и два палатальных, из которых нижний развит сильнее верх-
него. На затылке палатальные зубы немного просвечивают в виде желтоватых
удлиненных пятнышек, нижнему палатальному соответствует неглубокая вмя-
тина. Пупок у голотипа точковидный, у паратипа в виде узкой щели.
Рис. 133. Vertigo (Isthmia) modesta (Say).
I — V. modesta alpestris (Aid.) из леса близ г. Говерлы в Закарпатье; II—IV — V. modesta modesta: II — ns
Ораниенбаума под Ленинградом (f. ronnebyensis West.); Ill — из с. Покур Тюменской обл.; IV — из до-
лины р. Амгуэмы, Чукотский п-ов (f. hoppit Moll.). ,
Размеры голотипа: ВР 1.8, БД 1.1 мм; паратип: ВР 1.4, БД 0.8 мм.
Благодаря резкой радиальной скульптуре вид слегка напоминает V. sub-
striata', отличается меньшими размерами, наличием только одного париеталь-
ного зуба, относительно более широкой раковиной, а также тем, что радиальные
ребрышки выражены не только на средних оборотах, но и на последнем.
Распростр ан ениеи местообитание. Вид найден в двух
местах: на о-ве Беринга (сбор Р. Я. Братчика) в трещинах скал вместе с V. mo-
desta и на о-ве Шикотан (сбор О. Г. Кусакина) в зарослях бамбука на склоне
сопки вместе с V. hirasei.
13. Vertigo (Isthmia) modesta modesta (Say, 1824) (рис. 133, II—IV;
рис. 134). Ц’
Say, 1824, Long’s Second Exped., appendix : 259, t. 15, fig. 5 (Pupa); Moller, 1842 :
4 (P. hoppii); L. Pfeiffer, 1847a : 148 (P. shuttleworthiana); Gould, 1848 : 263 (P. de-
cora); G r e d 1 e r, 1856 : 122, t. 2, fig. 3 (P. genesii); M о r e 1 e t, 1858 : 9 (P. borealis);
Wallenberg, 1858, Malak. BL, 5 : 32, 99, t. 1, fig. 3, 4 (arctica); Westerlund,
1871, Nova Acta Reg. Soc. Sci. Uppsal. 3, VIII : 94 (Pupa ronnebyensis); Westerlund,
1877 : 103 (P. inermis, non Deshayes, 1863); Pilsbry, 1919, Man. Conch., 25 : 123, t. 10,
figs. 1, 2, 192, t. 17, fig. 14 (ronnebyensis); 204, t. 18, figs. 10—12, 17 (genesii); Lindholm,
1925b,: 242 (genesii subsp. geyeri); Лихарев, Раммельмейер, 1952 : 138, рис. 50
(ronnebyensis), рис. 51; 139, рис. 52 (genesii et var. geyeri); Ehrmann, 1956 : 42, fig. 20
(ronnebyensis), fig. 21 (arctica), 43, fig. 22 (genesii); Loz e k, 1956 : 96, t. 5, fig. 1 (arctica);
Zilch, Jaeckel, 1962 : 89 (genesii), 90 (genesii ssp. geyeri); Лихарев, 1963 : 68
(modesta hoppii); Lozek, 1964 : 205, t. 7, fig. 7 (arctica), t. 7, figs. 9, 10 (genesii); Walden,
1966 : 58 (subsp. arctica).
Terra typica — «Inhabits the Northwest Territory» (северо-запад Северной
Америки).
Место хранения типов не установлено.
Материал: несколько сотен экз. из 36 местонахождений в окр. Ленин-
града, в западной Сибири (Тюменская обл., Сургутский р-н, с. Покур), на
14*
212
IV. НАДСЕМ. VERTIGINOIDEA
восточном побережье Сибири (Аян), на Чукотке, Камчатке и Командор-
ских о-вах).
Раковина от удлиненно-овальной до овальной, тонкостенная, просвечива-
ющая, хрупкая, блестящая. Верхняя часть более или менее коническая или
куполовидная. Оборотов 47г—5, выпуклых или сильновыпуклых, последний
оборот к устью почти не поднят или поднят непосредственно возле самого
устья. Высота последнего оборота составляет около половины высоты раковины
(иногда немного больше или немного меньше). Окраска одноцветная, роговая,
красноватая, каштановая или бурая. Эмбриональные обороты с микроскопиче-
ской зернистостью, скульптура дефинитивных оборотов — от правильной очень
реп
тонкой морщинистости до редкой сла-
бой исчерченности, в последнем случае
раковина выглядит почти гладкой. Устье
овальное, с тонкими слегка отверну-
тыми краями, места его прикрепления
не сближены, вдавления на палатальном
крае устья нет, затылочное утолщение
очень слабое или отсутствует. Зубов
в наиболее полном наборе 4: париеталь-
ный, колумеллярный и два палатальных;
все они имеют вид тонких коротких по-
лукруглых пластинок. Однако редукция
нескольких или даже всех зубов — яв-
ление обычное. Так, в сборах Братчйка
на о. Беринга среди многих раковин
с 4 зубами наблюдались раковины только
с одной нижней палатальной складкой,
причем можно было проследить все ста-
дии редукции верхней палатальной. На
Чукотке (сборы в 8 местонахождениях)
Рис. 134. Vertigo {Isthmia) modesta modesta
(Say) из окр. Ачайваяма, Корякский нац.
округ, 7 VII 1960. Половой аппарат.
большая часть раковин лишена зубов, но попадаются экземпляры с одной
только париетальной и (или) одной колумеллярной пластинками. Материал,
собранный мной в Аяне, обнаруживает еще более пеструю картину: 4 зуба,
3 зуба (отсутствует верхний палатальный), 1 зуб (колумеллярный). Мерч
(Morch, 1868) отмечает, что среди V. hoppit в Гренландии в пределах одной по-
пуляции встречаются наряду с «зубатыми» раковинами экземпляры без устьевых
зубов. Пупок очень узкий, но открытый, сквозь него видна часть предпослед-
него оборота.
Размеры: ВР 1.7—2.8, БД 1.1—1.6 мм; в первоописании: «length less than
7io inch».1 Пилсбри (Pilsbry, 1919, Man. Conch., 25) указывает: BP 2.6,
БД 1.3 мм; в этой же работе на с. 124 приведены изображения основных форм
изменчивости.
Первым, кто высказал мысль о полиморфизме V. modesta, был Пилсбри
(Pilsbry, 1948); позже Лихарев (1963) поддерживает эту точку зрения, но за-
мечает, что в комплекс форм V. modesta включаются популяции, отличающиеся
слабым развитием зубов (не более трех) или их отсутствием; он говорит о при-
сутствии на Камчатке V. modesta modesta с 4 зубами (т. е. форма, отличающаяся
от V. alpestris только контуром) и V. modesta hoppi, у которого зубов 3, 1 или
зубы отсутствуют. Очевидно, эти формы подвидами признать нельзя, и они
1 Высота менее 41» дюйма.
2. РОД VERTILLA
213
представляют собой в одних случаях экологические формы, а в других — ва-
рианты индивидуальной изменчивости. Существенно также, что в литературе
не удалось найти указаний на совместную встречаемость двух форм этого вида,
которые не были бы связаны между собой промежуточными значениями при-
знаков.
Внутреннее строение. Материал: 4 экз. из окр. Ачайваяма,
Чукотский п-ов, 7 июля 1960 г., сбор и определение И. М. Лихарева.
Белковая железа с четким латеральным желобком. Семяпровод впадает
апикально в тонкий длинный цилиндрический эпифаллус, диаметр которого
не отличается от диаметра семяпровода; граница между этими протоками отме-
чена только резким изгибом, к которому подходит половой ретрактор. Пенис
примерно в 1У2 раза длиннее эпифаллуса и значительно толще его, внутри с про-
дольными складками, которые просвечивают сквозь стенки пениса. На границе
пениса и эпифаллуса наблюдается очень резкий изгиб. Проток семяприемника
в базальной части слегка расширен, далее тонкий, цилиндрический; резервуар
маленький, прилегает к нижнему краю белковой железы.
Распространение и местообитание. Циркумбореальный
ареал охватывает северную часть Евразии и Северной Америки. Населяет
широкий спектр хорошо увлажняемых биотопов — подстилка смешанных
и лиственных лесов, тундровое редколесье, россыпи камней на склонах и в тун-
дре, высокотравные поймы. •
13а. Vertigo (Isthmia) modesta alpestris Alder, 1839 (рис. 133, /).
Alder, 1839, Trans, Nat. Hist. Soc. Northumberland, II : 340 (V. alpestris); M i d den-
do r f f, 1853, Bull. Acad. St. Petersb., 11 : N 263, 264 (Pupa pygmaea var. alpestris); Pil-
sbry, 1919, Man. Conch., 25 : 197, t. 18, figs. 1, 2 (alpestris); Лихарев, Раммель-
мейер, 1952 : 137, рис. 48 (alpestris).
Terra typica — Англия, графство Нортумберленд.
Материал: 16 экз. из трех местонахождений в Альпах и на Карпатах.
Отличается от номинативного подвида, а также от всех форм экологической
изменчивости несколько меньшими размерами и более цилиндрической формой
раковины при наличии всех четырех устьевых зубов.
Размеры: ВР 1.8—2, БД 1.1 мм; в первоописании: ВР 0.07, БД 0.04 дюйма.
Распространение и местообитание. Скандинавия и вы-
сокогорные области средней Европы, Крым, Сибирь, Сихотэ-Алинь с пред-
горьями. Неселяет биотопы, сходные с теми, в которых обитает номинативный
подвид.
2. Род VERTILLA Moquin-Tandon, 1855
Moquin-Tan don, 1855 : 408 [ Vertigo (Vertilla)]; Th i e 1 e, 1931 : 505 [7. (Ver-
tilla)]; Лихарев, Раммельмейер, 1952 : 139 [V. (Vert'illa)]; Wenz, Zilch,
1959—1960 : 149 [V. (Vertilla)]; Стеклов, 1966 : 147 [V. (Vertilla)].
Типовой вид Vertigo (Vertilla) plicata A. Muller, 1838 (syn. Vertigo
angustior Jeffreys, 1830); по обозначению Pilsbry, 1920, Man. Conch., 25 : 210.
Раковина левозавитая. Колумеллярная пластинка длинная, почти верти-
кальная. Верхняя палатальная складка очень длинная, задний конецее сквозь
устье не виден.
Распространение. Европа, Кавказ. 1—2 вида.
В ископаемом состоянии род известен начиная со среднего миоцена (Европа,
Предкавказье).
Vertilla angustior (Jeffreys, 1830) (рис. 135, 136).
J effreys, 1830 : 361 (Vertigo); Charpentier, 1837 : 18, t. 2, fig. И (V. venetzii);
A. Miillerin: Wiegmann, 1838, Arch. Naturg., I : 210, t. 4, fig. 6 (V. plicata); Held,
1837 : 304 (V. hamata); Moquin-Tandon, 1855 : 408, pl. 29, figs. 8—11 [V. (Vertilla)
plicata]; Reeve, 1863 : 121, fig. (V. vertigo); Pilsbry, 1919, Man. Conch., 25 : 211, t. 5,
figs. 13, 16—18 (Vertigo); Steenberg, 1925 : 88, pl. 22, figs. 1—3 (Vertigo); Лихарев,
214
IV. НАДОЕМ. VERTIGINOIDEA
Раммельмейер, 1952 : 139, рис. 53 [V. (Vertilla)]; Ehrmann, 1956 : 39, fig. 19'
(Vertigo); Lozek, 1956 : 91, t. 4, fig. 1 (Vertigo); Zilch, Jaeckel, 1962 : 81 [V. (Ver-
tilla)]; Lozek, 1964 : 201, t. 6, fig. И (V. (Vertilla)]; Стеклов, 1966 : 148, табл. Ill,
58, 59 [V. (Vertilla)]; Gittenberger et al., 1970 : 52, fig. 34, Kaart 17 [7. (Vertilla)];
Акрамовский, 1976 : 138, рис. 58, табл. V, 47 (Vertigo).
Terra typica — «Marino near Swansea» (Англия, западное побережье, окр. Су-
онси).
Место хранения типов не установлено.
Материал: около 100 экз. из 45 местонахождений в Московской обл.,
в Закарпатье, на Северном Кавказе и Средней Европе.
Раковина левозавитая, овальная, тонкостенная, сильно просвечивающая,
умеренно блестящая. Оборотов 4—41/2, выпуклых; последний оборот к устью
Рис. 135. Vertilla angustior
(Jeffr.) из Чегемского ущелья на
, 4 Северном Кавказе.
Рис. 136. Vertilla angustior
(Jeffr.) из Чегемского ущелья на
Северном Кавказе, 16 V 1969 г.
Половой аппарат.
не поднят, высота его составляет примерно половину высоты раковины. Окраска
от роговой до каштановой, иногда с легким багровым оттенком. Поверхность
эмбриональных оборотов покрыта тончайшей неясной зернистостью, дефини-
тивные несут вялые, широкие и очень невысокие морщинки, которые на послед-
нем обороте часто почти сглаживаются. Устье сердцевидное, края его тонкие,
отвернутые, на середине палатального края видно, хорошо выраженное вдавле-
ние. Места прикрепления устья слегка сближены. На париетальной стенке две
почти одинаковые пластинки. Колумеллярная пластинка в виде тонкой лопа-
сти, почти вертикальная. Базальный зуб небольшой, бугорковидный, может
отсутствовать. Нижняя палатальная складка бугорковидная, верхняя пала-
тальная длинная, пластинчатая, в глубине последнего оборота крючковидно
изогнута; задний конец ее можно рассмотреть только на просвет сквозь стенку
последнего оборота. Гребень верхней палатальной складки близко подходит
к гребню париетальной пластинки, за счет чего образуется округлый синулус.
На затылке верхней палатальной складке соответствует глубокий желобок,
прорезающий хорошо развитое затылочное утолщение. Пупок почти запаянный.
Размеры: ВР 1.5—1.8, БД 0.8—0.9 мм; в первоописании: ВР 0.06,
БД 0.035 дюйма, т. е. ВР около 1.5, БД около 1 мм.
Внутреннее строение. Материал: 6 экз. из Чегемского ущелья .
на Северном Кавказе, 16 мая 1969 г.; 3 — из с. Апаринки Московской обл.,
4 мая 1963 г.; сборы и определения автора.
1. РОД COLUMELLA
215
Белковая железа с узким и глубоким латеральным желобком. Семяпровод
впадает терминально в очень легкое утолщение эпифаллуса. Постоянный изгиб
на границе семяпровода и эпифаллуса отсутствует, к месту контакта этих про-
токов подходит половой ретрактор. Эпифаллус тонкий, почти цилиндрический.
Перетяжка между эпифаллусом и пенисом очень резкая. Пенис в 1х/2—2 раза
длиннее эпифаллуса и значительно толще его. Овальный резервуар семяприем-
ника плотно вжат в ткани нижней части белковой железы и верхней части матки.
Расп.ространение и местообитание. Средняя, частично
северная и южная Европа; Англия; на восток — до Урала; Крым, Северный
Кавказ и Закавказье, северный Иран. Населяет влажные луга и подстилку
лиственных, реже смешанных лесов. В тайге и тундре отсутствует.
III. Подсем. TRUNCATELLININAE Steenberg, 1925
Steeuberg, 1925 : 201; Pilsbry, 1921, Man. Conch., 26.
Раковина цилиндрическая, реже удлиненно овально-цилиндрическая, от
светло-роговой до красноватой. Ангулярный бугорок отсутствует, париеталь-
ная пластинка не раздвоена, бугорковидная, часто отсутствует.
Пенис короткий, без придатков; эпифаллус короткий или очень короткий;
кольцевая перетяжка между пенисом и эпифаллусом широкая.
Распространение. По всем континентам, кроме Австралии. 9 родов.
В ископаемом состоянии подсемейство известно начиная с верхнего олиго-
цена (Европа, о-в Мадейра).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ TRUNCATELLININAE
1 (2). Устье с губой или с утолщенными краями. Поверхность раковины ребри-
стая .......................................... 2. Truncatellina.
2 (1). Устье без губы, края его тонкие. Поверхность раковины гладкая или
тонко исчерченная ..................................1. Columella.
1. Род COLUMELLA Westerlund, 1878
Charpentier, 1837 : 15 {Sphyradium part.); С 1 e s s i n, 1876 : 208 {Edentulina,
non L. Pfeiffer, 1855); Westerlund, 1878 : 193; Pilsbry, 1926, Man. Conch., 27 :
232; Thiele, 1931 : 504; Лихарев, Раммельмейер, 1952 : 143; F о г с a г t,
1959а : 7—18; W е n z, Zilch, 1959—1960 : 146; Акрамовский, 1976 : 131.
Типовой вид Pupa inornata Clessin, 1872 (syn. Pupa edentula Dra-
parnaud, 1805); по обозначению Вестерлунда (Westerlund, 1887).
Раковина хрупкая, темных тонов, слабо скульптурированная. Устье без
зубов, без губы и без затылочного валика, края устья острые, почти не отвер-
нуты.
Распространение. Голарктика. 5—6 видов.
В ископаемом состоянии род встречается .начиная с верхнего плиоцена
(Европа, Африка, Северная Америка).
таблица для определения видов columella
1 (4). Последний оборот несколько вздут.
2 (3). Скульптура в виде радиальной морщинистости, яснее выраженной на
первых дефинитивных оборотах . . . . ............1. С. columella.
3 (2). Скульптура в виде тонкой радиальной ребристости, ясно выраженной .
на всех дефинитивных оборотах....................5. С. intermedia.
4 (1). Последний оборот не вздут.
5 (б). Раковина покрыта тонкой частой правильной радиальной исчерченностью
................................................4. С. aspera.
6 (5). Раковина почти гладкая или с редкими сглаженными морщинками.
216
IV. НАДСЕМ. VERTIGINOIDEA
7 (8). Раковина башневидно-цилиндрическая. Высота устья превышает его
ширину ............................................... . 3. С. acicularis.
8 (7). Раковина в виде высокого купола или короткоцилиндрическая. Высота
устья примерно равна его ширине..........................2. С. edentula.
1. Columella columella (G. Martens, 1830) (рис. 137, Г).
G. Martens, 1830, Correspondenzbl. wiirttemberg. landwirtsch. Vereins, 17 : 171
(Pupa)-, С1 e s s i n, 1872:57, t. 4, fig. 8 (Pupa gredleri); Лихарев, Рам-
мельмейер, 1952:144 (edentula var. columella); Акрамовский, 1976:132,
табл. V, 45.
Terra typica — северная Европа.
Лектотип хранится в Естественноисторическом музее в Базеле,
№ 5900-а (Forcart, 1959а: И, АЬЬ. 4).
Материал: 27 экз. из 5 местонахождений в Мурманской обл., Закав-
казье, на Чукотке и на побережье Охотского моря (Аян).
Рис. 137. Виды рода Columella.
1 — С. columella (G. Mart.) из Аяна,
берег Охотского моря; II — С. eden-
tula (Drap.) из Курска.
Рис. 138. Columella edentula
(Drap.) из Курска, VII 1967 г.
Половой аппарат.
Раковина цилиндрическая, высокая, тонкостенная, хрупкая, блестящая,
с короткой закругленной вершиной. Оборотов 6х/2—8, выпуклых или сильно-
выпуклых. Последний оборот заметно вздут, к устью не поднят, высота его со-
ставляет треть высоты раковины или немного больше. Окраска от роговой до
каштановой, вершина несколько светлее. Скульптура верхних дефинитивных
оборотов в виде очень тонких правильных радиальных морщинок, которые
на нижних оборотах расплываются, и расположение их теряет упорядоченность.
Устье округлое, умеренно скошенное, с тонкими острыми прямыми краями,
лишь верхняя часть колумеллярного края немного отвернута. Пупок сквозной,
цилиндрический, округлый.
Размеры: ВР 2.8—3.5, БД 1.3—1.5 мм.
Распространение и местообитание. Северная Европа,
высокогорье центральной Европы, Сибирь, горный Крым, Малый Кавказ (гора
Капут-джух); Тянь-Шань; Северная Америка. Населяет пойменные луга с вы-
соким травостоем, влажное редколесье, опушки широколиственных лесов,
в горных районах — влажные осыпи, трещины скал. Часто поднимается на
стебли трав.
1. РОД COLUMELLA
217
2. Columella edentula (Draparnaud, 1805) (рис. 137, II\ рис. 138).
Draparnaud, 1805 : 59, t. 3, figs. 28, 29 (Pupa); Beck, 1837 : 85 (Alaea); Held,
1837 : 307 (Vertigo lepidula); Mo quin - T and on, 1855 : 402, pl. 28, figs. 28—30 (Ver-
tigo); Reeve, 1863 : 122, fig. (Vertigo); Steenberg, 1925 : 98, fig. 20 du texte, pl. 27,
figs. 1—4, pl. 28, fig. 3; Pilsbry, 1926, Man. Conch., 27 : 236, t. 30, figs. 9—11; Лиха-
рев, Раммельмейер, 1952 : 143, рис. 56; Ehrmann, 1956 : 45, fig. 26; L o-
z e k, 1956 : 98, t. 5, fig. 3; Zilch, Jaeckel, 1962 : 45, Abb. 26 (Columella edentula
et f. turritella Westerlund); Lozek, 1964 : 197, t. 6, fig. 4; M а т ё к и н, 1966 : НО; Git-
tenberger et al., 1970 : 47, fig. 29, Kaart 10; А к p а м о в с к й й, 1976 : 131, рис. 53,
табл. V, 44.
Terra typica — Франция.
Синтипы хранятся в Естественноисторическом музее Вены (коллекция
Драпарно).
Материал: более 100 экз. из 31 местонахождения в Московской и Кур-
ской обл., в Закарпатье, на Кавказе, в Средней Азии, на побережье Охотского
моря (Аян) и на Сахалине.
Раковина от высококуполовидной до короткоцилиндрической, но даже у ци-
линдрических раковин заметно, что ширина оборотов сверху вниз постепенно
увеличивается; тонкостенная, хрупкая, блестящая, с правильно закругленной
вершиной. Оборотов 6—б1^ выпуклых или умеренно выпуклых, последний
оборот не вздут, к устью не поднят или поднят еле заметно; высота его немного
меньше половины высоты раковины. Окраска от светло-роговой до каштановой,
иногда со слабым зеленоватым оттенком, вершина обычно светлее. Скульптура
дефинитивных оборотов очень слабая, представлена редкими неправильно рас-
положенными сильно расплывшимися морщинками; близ устья эти морщинки
расположены чаще. Устье округлое, скошенное, края его тонкие, хрупкие,
не отвернуты, кроме верхней части колумеллярного. Пупок сквозной, узкий,
цилиндрический, совершенно круглый.
Размеры: ВР2.2—2.7, БД 1.2—1.4 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 10 экз. из окр. Курска,
18 июля 1966 г., сбор Ю. И. Чернова; 1 — из Закарпатья, с. Квасы близ Ра-
хова, 20 сентября 1969 г.; сбор и определения автора.
Белковая железа с глубоким желобом, направленным почти перпендику-
лярно длинной оси органа, т. е. пересекающим белковую железу поперек.
Семяпровод впадает в тонкий почти цилиндрический эпифаллус, к которому,
заметно отступя от семяпровода, подходит половой ретрактор. Эпифаллус впа-
дает в пенис не апикально; эти отделы примерно равной длины или пенис
немного короче. Проток семяприемника отходит очень высоко, поэтому нижний
отдел вагины в несколько раз длиннее верхнего. Как проток, так и резервуар
семяприемника тесно прилегают к женским путям; резервуар от шаровидного
до коротко веретеновидного, прилегает к матке ниже ее середины. Простата
не обнаружена.
Распространен и е и местообитание. Европа, кроме самых
южных районов, Кавказ с Закавказьем, Средняя Азия, Северная Америка.
Живет примерно в тех же местах, что и предыдущий вид, иногда оба вида
встречаются вместе.
3. Columella acicularis Almuhambetova, 1979 (рис. 139, Г).
Альмухамбетова, 1979 : 31, рис. 2; Альмухамбетова, Увалиева,
1979 : 36, рис. 16.
Terra typica — Заилийский хр., ущелье Тау-Тургёнь.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал. 3 экз. из типовой серии.
Раковина башневидно-цилиндрическая, умеренно тонкостенная, матовая.
Оборотов б^—7, несильно выпуклых. Первые 2—2х/2 оборота образуют плавно
закругленную куполовидную вершину, диаметр последующих оборотов очень
218
IV. НАДСЕМ. VERTIGINOIDEA
плавно и постепенно увеличивается. Последний оборот незначительно вздут,
к устью практически не поднят, высота его несколько превышает треть высоты
раковины. Окраска буроватая или темно-роговая, причем верхние 2—2х/2 обо-
рота светлее остальных. Скульптура как эмбриональных, так и дефинитивных
оборотов в виде тончайшей микроскопической зернистости; на дефинитивных
могут, кроме того, присутствовать отдельные очень вялые радиальные мор-
щинки, которые в конце пос-
леднего оборота, близ устья,
выражены немного лучше. Устье
небольшое, удлиненно-оваль-
ное, высота его приблизи-
тельно в IJ/g раза превышает
ширину. Места прикрепления
устья слегка сближены, края
Рис. 139. Виды рода Columella,.
I — С. acicularis Almuhambetova, голотип;
II — С. aspera WaldSn, паратип; III —
С. intermedia Schileyko et Almuhambetova,
sp. nov., голотип.
не отвернуты, острые или незначительно утолщены. Колумеллярный край
почти отвесный, в нижней части очень слабо, но Явственно выгнут влево (от
наблюдателя). Базальный край вместе с палатальным образует плавную дугу,
а в верхней части палатального края имеется слабый закругленный угол. Пупок
почти правильно круглый, расположен чуть эксцентрично относительно центра
пупочной впадины.
Размеры: ВР 2.6—2.9, БД 1.3—1.4 мм; голотип: ВР 2.8, БД 1.3 мм.
Распространение и местообитание. Вид собран С. Аль-
мухамбетовой в ущелье Тау-Тургень на Заилийском хр., у подножья склона
в растительной трухе.
4. Columella aspera Walden, 1966 (рис. 139, II).
Walden, 1966 : 53, Abb. 1; Schmid, 1967 : 157; Gittenberger et al.,
1970 : 48, fig. 30, Kaart 10.
Terra typica — «VSstmanland, Linde Kirchspiel, im Nadelwald 6. Siggebohyt-
tan» (Швеция, лен Вестманланд).
Голотип хранится в Естественноисторическом музее Гётеборга, Швеция.
Материал: 2 экз. из типового местонахождения (паратипы).
Раковина в виде высокого купола, тонкостенная, сильно просвечивающая.
Оборотов 5—61/2, выпуклых, каждый последующий оборот несколько шире
предыдущего. Последний оборот прямой, высота его меньше половины высоты
раковины. Окраска коричневая, иногда с легким зеленоватым оттенком. Скульп-
тура в виде тонкой и густой радиальной исчерченности, одинаково хорошо вы-
раженной как на верхних, так и на нижних оборотах. Устье округлое, с тон-
кими ломкими краями; отвернут лишь самый верхний участок колумеллярного
края. Пупок точковидный.
Размеры голотипа: ВР 2.30, БД 1.37 мм.
Распространение и местообитание. Вид широко распро-
странен в северной и северо-западной Европе. В СССР найден на Валдайской
возвышенности (Шиков, 1979). Населяет влажные, преимущественно лесные,
биотопы.
2. РОД TRUNCATELLINA
219
5. Columella intermedia Schileyko et Almuhambetova, sp. nov. (рис. 139, 727).
Альмухамбетова, Увалиева, 1979 : 36, рис. 2 (Columella sp.).
Terra typica — Таласский хр., ущелье Бахраусу.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 1 экз. (голотип).
Раковина цилиндрическая, высокая, умеренно тонкостенная, слабо блестя-
щая, с короткой закругленной вершиной. Оборотов 7, сильно выпуклых, по-
следний оборот несильно, но явственно вздут, прямой, высота его больше трети
высоты раковины. Окраска одноцветно светло-роговая. Поверхность эмбрио-
нальных оборотов тонкозернистая; дефинитивные обороты несут частые тонкие
ребрышки, которые на двух нижних оборотах располагаются несколько гуще,
чем на верхних. Устье удлиненное, высота его больше ширины. Устьевые края
тонкие, прямые, кроме верхней части колумеллярного края. Пупок точковид-
ный.
Размеры голотипа: ВР 2.9, БД 1.2 мм.
Формой раковины вид более всего напоминает С. columella, но четко отлича-
ется ребристой скульптурой, похожей на ту, которая имеется у представителей
рода Truncatellina. От последних отличается большей величиной и тонкими лом-
кими устьевыми краями.
Распространение и местообитание. Единственный эк-
земпляр (пустая раковина очень хорошей сохранности) обнаружен в ущелье
Бахраусу на Т^аласском хр., под стелющейся арчой.
2. Род TRUNCATELLINA Lowe, 1852
Lowe, 1852 : 275; Reinhardt,. 1879 : 133 (Isthmia, non. Gray, 1821); Hesse,
1915, Nachrichtenbl. dtsch. malak. Ges., 47 : 53 (Laurinella, типовой вид Pupa minutissima
auct., syn. Vertigo cylindrtca Ferussac, 1807, обозначение Pilsbry); Pilsbry, 1920, Man.
Conch., 26 : 58; Thiele, 1931 : 504; Лихарев, Раммельмейер, 1952 :
140; Wenz, Zilch, 1959—1960 : 148; Стеклов, 1966 : 150; Акр амо в ский,
1976 : 132.
Типовой вид Pupa linearis Lowe, 1852; по монотипии.
Раковина относительно прочная, обычно не темная, с резкой радиальной
скульптурой. Устье с губой или с утолщенными отвернутыми краями, обычно
с затылочным валиком. Зубы чаще всего имеются, бугорковидные. В наиболее
полном наборе имеются париетальный, колумеллярный и палатальный бугорки,
последний лежит глубоко внутри устья.
Распространение. Европа, Канарские о-ва, о-ва Зеленого Мыса,
Средняя Азия и Кавказ, о-ва Лиукиу, Восточная и Южная Африка. Несколько
десятков видов.
В ископаемом состоянии род известен начиная с верхнего олигоцена (Ев-
ропа, о-в Мадейра).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ TRUNCATELLINA
1 (6). Устье раковины с зубами.
2 (3). Палатальный зуб целиком виден при нормальном положении раковины
...................................................1. Т. callicratis.
3 (2). При нормальном положении раковины палатальный зуб полностью или
частично заслонен столбиком.
4 (5). Палатальный край устья широко отвернут и усилен мощной губой . . .
................................................... 2. Т. costulata.
5 (4). Палатальный край устья незначительно отвернут, с тонкой губой . .
............................................ 3. Т. claustralis.
6(1). Устьевые зубы отсутствуют . ..................4. Т. cylindrica.
220
IV. НАДОЕМ. VERTIGINOIDEA
1. Truncatellina callicratis (Scacchi, 1833) (рис. 140, III', рис. 141).
Scacchi, 1833 : 11 (Turbo); G r e d 1 e r, 1853, Programm d. k. k. Gymnasium z. Bo-
zen : 114 (Pupa strobeli); Benson, 1854 : 97 (P. rivierana); Pilsbry, 1921, Man. Conch.,
26 : 71, t. 8, figs. 10—12 (rivierana); Steenberg, 1925 : 91, pl. 23, figs. 1—3 (rivierana);.
Лихарев, Раммельмейер, 1952 : 141 (strobeli); Ehrmann, 1956 : 44 (stro-
beli); Lozek, 1964 : 199, t. 6, fig. 10 (strobeli); F о r c a r t, 1965 : 84; M а т ё к и н, 1966:
109 (strobeli); Д а м я н о в, Лихарев, 1975 : 128, фиг. 63 (strobeli); Акрамовский,.
1976 : 133, рис. 54, 55А, табл. V, 41; Альмухамбетова, Увалиева, 1979 : 37, рис. За.
Terra typica — Неаполь, Ботанический сад.
Место хранения типов не установлено.
Материал: около 40 экз. из 15 местонахождений на Северном Кавказе,,
в Закавказье и в Средней Азии.
I Л Ш IV
• Рис. 140. Виды рода Truncatellina.
1 — Т. eostulata (Nilss.) из с. Голубинка, Крым; II — Т. cylindrica (Гёг.) из Армении, с. Агверан близ--
г. Еревана; III — Т. callicratis (Scacchi) из Талыша, с. Космальян; IV — Т. claustralis (Gredl.) из с. Го-
лубинка, Крым.
Раковина цилиндрическая, реже овально-цилиндрическая, умеренно про-
свечивающая, с широко закругленной вершиной, которая часто несколько упло-
щена. Оборотов 5—6V2, сильно выпуклых, последний оборот к устью почти
не поднят, высота его изредка составляет половину высоты раковины, но обычно
заметно меньше. Окраска светло-роговая, вершина обычно несколько светлее.
Дефинитивные обороты несут резкие радиальные ребрышки, расстояние между
которыми больше ширины ребра. Устье округлое, без затылочного утолщения
или с очень слабым утолщением, края устья слегка отвернуты, места прикреп-
ления почти не сближены и соединены тонкой прозрачной мозолью, выраженной
в разной степени. Зубов обычно три: короткая полукруглая париетальная пла-
стинка, бугорковидный колумеллярный зуб и округлый палатальный бугорок,
иногда слегка вытянутый по ходу оборота. Палатальный бугорок при нормаль-
ном положении раковины располагается примерно на пересечении вертикальной
линии, проведенной от париетальной пластинки, и горизонтальной линии, про-
веденной от колумеллярной пластинки. Иногда зубы в той или иной мере реду-
цированы: в первую очередь исчезает париетальный, затем палатальный; реже
всего отсутствует колумеллярный зуб. Пупок проколовидный.
Размеры: ВР 1.6—2.2, БД 0.8—0.95 мм. В Цейском ущелье на Северном Кав-
казе Т. А. Тавасиева собрала серию очень крупных раковин: ВР до 2.5, БД 1 мм.
Внутреннее строение. Материал: 4 экз. из Талыша, с. Космальян
4 июля 1964 г.; 3 — из Армении, с. Агверан близ Еревана, 26 сентября 1975 г.;
сборы и определения автора.
Белковая железа с чрезвычайно резкой и широкой косой бороздой на лате-
ральной поверхности. Тонкий семяпровод, образовав крутой изгиб, впадает
в короткий вздутый эпифаллус, отделенный от цилиндрического или веретено-
видного пениса отчетливой кольцевой перетяжкой. Половой ретрактор крепится.
2. РОД TRUNCATELLINA
221
к самой дистальной части эпифаллуса. Проток семяприемника короткий, за-
канчивается маленьким овальным резервуаром, прилегающим к нижней части
матки.
Распространение и местообитание. Южная Европа, ме-
стами — южные районы центральной Европы, Северный Кавказ и Закавказье,
Передняя Азия, Иран, Копетдаг, горные районы Средней Азии, южный Алтай.
Встречается обычно в сухих растительных остатках на открытых склонах
и в редколесье; сплошных лесов избегает.
Рис. 141. Truncatellina, callicratis (Scacchi) Рис. 142. Truncatellina costulata (Nilss.) из
из, Талыша, с. Космальян, 4 VII 1964£г. Армении, с. Агверан близ г. Еревана,
Половой аппарат. 26 IX 1975 г. Половой аппарат.
2. Truncatellina costulata (Nilsson, 1822) (рис. 140, Г, рис. 142).
Nilsson, 1822 : 51 \Рира)‘, Schmidt, 1849 : 141 (Р. ascaniensis)-, Pilsbry,
1921, Man. Conch., 26 : 78, t. 8, figs. 17, 18; S t e e n b e r g, 1925 : 94, pl. 24, figs. 1, 2; Л n-
харев, Раммельмейер, 1952:141, рис. 54; Ehrmann, 1956:44, fig. 24;
Lozek, 1956 : 97, t. 5, fig. 6; Zilch, Jaeckel, 1962 : 80, Abb. 31; Lozek, 1964 :
200, t. 6, fig. 9; M а т 6 к и h, 1966:109; Wirth, 1970:328, Abb. I, 1; Акрамов-
ский, 1976 : 133, рис. 55Б, табл. V, 42.
Terra typica — Швеция.
Место хранения типов не установлено.
Материал: более 100 экз. из 7 местонахождений на Северном Кавказе,
в Закавказье, в Крыму и на северном Тянь-Шане.
Раковина цилиндрическая, реже овально-коническая, относительно тонко-
стенная, слабо блестящая, с широко закругленной вершиной. Оборотов 5г/2—
6х/2, выпуклых, последний оборот к устью очень плавно и едва уловимо поднят,
высота его всегда меньше половины высоты раковины. Окраска светло-роговая.
Дефинитивные обороты несут резкие притупленные ребра, промежутки между
которыми равны ширине ребра или немного больше. Устье неправильно округ-
лое, с небольшим расплывшимся затылочным утолщением. Края устья сильно
утолщены, слегка отвернуты, с плавным палатальным вдавлением и широкой
губой. Зубов 3: невысокая слегка изогнутая париетальная пластинка, бугорко-
видный колумеллярный зуб и округлый палатальный бугорок, лежащий глу-
222
IV. НАДСЕМ. VERTIGINOIDEA
боко в устье. При нормальном положении раковины палатальный зуб распола-
гается в левом (от наблюдателя) углу устья. Пупок округлый.
Размеры: ВР 1.7—2, БД 0.8—1 мм.
Внутреннее строение. Материал: 2 экз. из Армении,, с. Агверан
близ Еревана, 26 сентября 1975 г., сбор и определение автора.
Внешний вид полового аппарата очень похож на таковой предыдущего вида.
Отличается немного более длинным эпифаллусом и более резко выраженной гра-
ницей между семяпроводом и эпифаллусом.
Распространение и местообитание. Вид спорадически
распространен по территории центральной и южной Европы, на восток до Волги „
на север примерно до линии Эстония—Москва—Ульяновск; Закавказье, Сред-
няя Азия. Населяет лесную подстилку в разреженных широколиственных лесах
и кустарниковых зарослях, реже — под укрытиями на открытых сухих лугах.
3. Truncatellina [claustralis (Gredler, 1856) (рис. 140, IV).
G г е d 1 е г, 1856 : 116, t. 2, fig. la—с {Pupa claustralis et var. anodus); Reinhardt,
1877a : 82, t. 3, fig. 6 {P. clavella); Pilsbry, 1921, Man. Conch., 26 : 79, pl. 11, figs. 1, 2;
Лихарев, Раммельмейер, 1952 : 141; Ehrmann, 1956 : 45, L о z e k, 1956 :
97, t. 5, fig. 5; Zilch, Jaeckel, 1962 : 80, Abb. 32; Lozek, 1964 : 200, t. 6, fig. 5, 6;
Да мянов, Лихарев, 1975 : 126, фиг. 61.
Terra typica — «Salegg» (австрийский Тироль).
Место хранения типов не установлено.
Материал: 3 экз. из Крыма, с. Голубинка.
Раковина цилиндрическая, умеренно тонкостенная,, слегка просвечива-
ющая. Оборотов 5У2—6У2, выпуклых, последний оборот к устью почти не под-
нят, высота его меньше половины высоты раковины. Окраска одноцветная,,
светло-роговая. Дефинитивные обороты тонко и равномерно ребристые. Устье
округло-треугольное, места его прикрепления слегка сближены, края слегка
утолщены и умеренно отвернуты. Палатальный край почти прямой, отвесный.
Париетальная пластинка сравнительно длинная, но невысокая; колумеллярная
пластинка в виде удлиненного бугорка, расположена вдоль оси столбика. Па-
латальный зуб сидит так глубоко, что сквозь устье, как правило, не просматри-
вается; обнаружить его можно, просветлив раковину ксилолом. Если палаталь-
ный зуб просматривается в устье, то не целиком: он частично заслонен столби-
ком. Пупок узкий, короткощелевидный.
Размеры: ВР 1.2—1.8, БД 0.5—0.8 мм; в первоописании: ВР 3/5—3/4,- БД
около */4 линии, т. е. БД примерно 1.25—1.56, БД примерно 0.5 мм.
Распространение и местообитание. Южная и частично
средняя Европа, Предкарпатье, Крым, Центральное Закавказье. Встречается
в|подстилке лиственного редколесья и в осыпях. |
4. Truncatellina cylindrica (Ferussac, 1807) (рис. 140, II; рис. 143).
Draparnaud, 1805 : 59, t. 3, figs. 26, 27 {Pupa muscorum, non Linnaeus, 1758);
Ferussac, 1807 : 52 {Vertigo); Hartmann, 1821 : 220; t. 2, fig. 5 {Pupa minutissima);
Held, 1837 : 308 {Vertigo pupula); Mo quin-T an don, 1855 : 399, t. 28, figs. 20—24
[17. {Isthmia) muscorum]; Reeve, 1863 : 123, fig. {Pupa minuta Studer, 1820); Mousson,
1876 : 40 [P. {Pupilla) micula]; S t e e n'b erg, 1925 : 96, pl. 25, fig. 1, pl. 26, fig. 1; L i n d-
h о 1 m, 1926 : 176 {cylindrica var. costigerella), 177 {tauricola); Лихарев, Раммель-
мейер, 1952 : 142, рис. 55; Ehrmann, 1956 : 44, fig. 23; Lozek, 1956 : 96, t. 5,
fig. 4; Zilch, Jaeckel, 1962 : 79 {cylindrica et var. odontostoma Westerlund); Lozek,
1964 : 199, t. 6, figs. 7, 8; Gittenberger et al., 1970 : 48, fig. 33, Kaart 12; Дами-
нов, Лихарев, 1975 : 127, фиг. 62; Акрамовский, 1976 : 134, рис. 55В, табл. V,
43.
Terra typica — Франция.
Место хранения типов не установлено.
Материал: около 80 экз. из 7 местонахождений в Поволжье,- в Крыму,
на Северном Кавказе и в Закавказье.
2. РОД TRUNCATELLINA
223
Раковина цилиндрическая, тонкостенная, просвечивающая, матовая,
с несколько уплощенной вершиной. Оборотов 5—6х/2, сильновыпуклых, послед-
ний оборот к устью очень плавно, несильно, но явственно приподнят, высота
его меньше половины высоты раковины. Окраска светло-роговая, реже красно-
ватая или красновато-коричневая. Скульптура в виде тонких ребрышек, рас-
стояние между которыми примерно равно ширине ребра или чуть больше.
Реже встречаются раковины с густо расположенными ребрышками, в резуль-
тате чего поверхность раковины выглядит ребристо исчерченной. Устье оваль-
ное, без затылочного утолщения, края его тонкие, слегка коротко отвернуты,
почти без губы. Колумеллярный край устья слегка расширен. Места прикреп-
Рис. 143. Truncatellina cylindrica (Fer.) из Армении, с. Агверан близ г. Еревана,
25 IX 1975 г. Половой аппарат.
ления устья незначительно сближены, париетальная мозоль отсутствует или
выражена слабо. Устьевые зубы отсутствуют. Пупок маленький, овальный.
Размеры:} ВР 1.6—2.4, БД 0.8—1 мм.
Внутреннее строение. Материал: 5 экз. из с. Агверан близ Ере-
вана, 25 сентября 1975 г.; 3 — из Крыма, долина р. Бельбек, 4 августа 1962 г.;
сборы и определения автора.
Белковая железа с очень широким и глубоким косым желобком. Топкий
семяпровод впадает в эпифаллус с крутым изгибом, к которому крепится полов ой
ретрактор. Эпифаллус короткий, в виде изогнутого конуса, отделен от пениса
широкой кольцевой перетяжкой, на внутренней поверхности этой перетяжке
соответствует-кольцевой валик. Пенис в 3—4 раза длиннее эпифаллуса, круто
изогнут в дистальной части. Обширная полость проксимальной части пениса
.гладкостенная, с двумя противолежащими широкими продольными складками.
Внутренняя поверхность значительно более узкой дистальной части пениса
покрыта мелкими овальными бугорками. Нижний отдел вагины немного длин-
нее верхнего, проток семяприемника короткий, резервуар небольшой, капле-
видный,, вжат в нижнюю часть многоскладчатой матки.
Распространение и местообитание. Центральная и юж-
ная Европа; в европейской части СССР до Эстонии, Московской обл. и Южного
Урала; Северный Кавказ и Закавказье; Северо-Западная Африка, Малая
Азия. Населяет преимущественно степные и полупустынные участки, держится
в скоплениях сухих растительных остатков; иногда в сухом редколесье и в за-
рослях кустарников.
224
V. НАДСЕМ. CHONDRINOIDEA
V. Надсем. CHONDRINOIDEA Steenberg, 1925
Раковина маленькая или средних размеров, башневидная или веретеновид-
ная, в виде исключения может быть кубаревидной, с радиальной скульптурой.
Эмбриональные обороты покрыты микроскопической зернистостью. Пупок
точковидный, изредка почти запаянный. Устье со сложной арматурой, состав-
ленной пластинчатыми зубами, которая иногда может упрощаться вплоть до
полного исчезнования всех зубов. Края устья не утолщены, отвернуты, обычно
без губы.
Семенные пузырьки не обособлены от гермафродитного протока. Пенис
с эпифаллусом, внутри с системой складок, достигающей иногда значительной
сложности. Граница между пенисом и эпифаллусом отмечена коническим при-
датком, нередко отсутствующим. Эпифаллус плавно переходит в семяпровод^
причем место контакта между этими протоками обычно в той или иной мере по-
гружено в ткани атриальной области пениса. Половой ретрактор одноветви-
стый, крепится близ середины пениса, но нередко от ретрактора отходит допол-
нительная лента, фиксирующая эпифаллус. Пениальный аппендикс отсутствует,
I. Сем. CHONDRINIDAE Steenberg, 1925
Steenberg, 1925 : 201; Gittenberger, 1973 : 3.
Раковина удлиненная. Устье с зубами, края его отвернуты. Придаток эпи-
фаллуса конический или отсутствует, половой ретрактор крепится к пениальной
трубке заметно ниже ее середины.
Распространение. Средиземноморские страны, Центральная и За-
падная Европа, Крым, Кавказ, Копетдаг; Южная Африка. 6—8 родов.
В ископаемом состоянии семейство известно начиная со среднего эоцена
(Европа).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ПОДРОДОВ
CHONDRINIDAE 1 2 3 4 5
1 (2). Пенис с хорошо развитым коническим придатком, длина которого не
менее х/3 длины пениса.................................1. Granaria.
2 (1). Придатка пениса нет или он рудиментарный.
3 (4). Половой ретрактор крепится к пенису рядом с половым атриумом . . .
.................................................... 2. Abida.
4 (3). Половой ретрактор отсутствует; если имеется, то крепится к пенису
заметно отступя от атриума (род Chondrina).
5 (6). Внутри пениса залегает подкововидная складка; в месте изгиба этой
складки пенис более или менее заметно утолщен.....................
.......................................3. Chondrina (Chondrina).
6J5). Пенис-цилиндрический, без подкововидной складки.............
.......................................3. Chondrina (Granopupa).
1. Род GRANARIA Held, 1837
Held, 1837 : 918; Swains on, 1840 : 334 (Pupella, типовой вид Pupa variabilis
Draparnaud, 1801); Herrmannsen, 1846—1849 : 488; Gittenberger,
1973 : 17, 41.
Типовой вид Pupa frumentum Draparnaud,; 1801; по обозначению
Геррманнсена (Herrmannsen, 1847 : 488).
Пениальная трубка с придатком, гомологичным, вероятно,; придатку эпи-
фаллуса; длина придатка не менее 1/3 длины пениса. Половой ретрактор кре-
пится примерно к середине пениса. Резервуар семяприемника достигает белко-
вой железы, проток семяприемника на большей части протяжения вжат в про-
стату.
1. РОД GRANARIA
225
Распространение. Европа и Малая Азия. 7 видов.,
В ископаемом состоянии род известен начиная со среднего эоцена (Европа).
Granaria frumentum (Draparnaud, 1801) (рис. 144, 145).
Draparnaud, 1801 : 59 (Pupa); Held, 1837 : 918; Pilsbry, 1918, Man.
Conch., 24 : 297, t. 42, figs. 1—4 (Abida); Steenberg, 1925 : 43, figs. 2—10, 14, 15 du
texte (Abida); Лихарев, Раммельмейер, 1952:130, рис. 39 (Abida); Ehr-
mann, 1956 : 37, t. I, fig. 6 (Abida); Lozek, 1956 : 88, t. 3, fig. 1 (Abida); Zilch, J a-
e c k e 1, 1962 : 96 (A. frumentum et f. minor Rossmaessler); Lozek, 1964 : 211, t. 8, fig. 7
(Abida); Hudec, 1965b : 161, fig. 7 (Abida); Gittenberger, 1973 : 45, text-fig. 12,
t. I, 1; Д а м я н о в, Лихарев, 1975 : 109, фиг. 42, 43 (Abida).
Terra typica — Франция.
Лектотип хранится в Естественноисторическом музее в Вене,, № 77703.
Материал: 12 экз. из трех местонахождений в северной Италии и в Бол-
гарии.
Раковина от коническо-цилиндрической до веретеновидной, сравнительно
тонкостенная, слегка просвечивающая, верхняя часть ее представляет собой
довольно высокий конус, вершина узкая, закруглен-
ная. Оборотов 8—9, несильно выпуклых, последний
оборот практически прямой, но в ангулярной области
резко поднят за счет губы; высота его немного больше
трети высоты раковины. Окраска одноцветная, светло-
роговая; за устьем, на затылке, имеется светлое поле.
Микроскопическая зернистость эмбриональных оборотов
хорошо заметна, дефинитивные обороты покрыты тонкими
сглаженными правильно расположенными морщинками,,
которые на верхних оборотах выражены резко, но посте-
пенно расплываются, и поверхность последнего оборота
на некоторых раковинах местами уже почти гладкая.
Устье округлое, места его прикрепления слегка сбли-
жены и соединены тонкой мозолью; устьевые края отвер-
Рис. 144. Granaria frumentum (Drap.) из Италии, окр. Вероны.
нуты и незначительно утолщены. Устьевая арматура сложная: хорошо развитая
ангулярная: пластинка короткая, изогнутая, почти сливается с верхним концом
палатального края; париетальная пластинка высокая, слегка изогнутая, гре-
бень ее иногда слегка отклонен к палатальной стенке. В глубине устья имеется
короткая спиральная пластинка, которую можно увидеть, рассматривая рако-
вину несколько снизу. Колумеллярных пластинок две, верхняя развита сильнее
нижней. На палатальном крае 5 складок, из которых верхняя (сутуральная)
лежит очень глубоко и при нормальном положении раковины не видна. Осталь-
ные 4 примерно равной длины и заканчиваются в пределах верхней половины
последнего оборота. Пупок имеет характерный облик: так как колумеллярный
край устья расширен, он на большом протяжении спаян с базальной стенкой
последнего оборота, образуя косой рубчик, и только в самой нижней части от-
стает, за счет чего возникает очень маленькое пупочное отверстие в виде изогну-
той капли.
Размеры: ВР 7—10, БД 2.7—3.7 мм; в первоописании размеры не указаны.
Внутреннее строение. Материал: 2 экз. из Болгарии, Широка
Планина, с. Мировцы, 30 июня 1969 г., сбор и определение И. М. Лихарева;
2 — из Италии, Бус-дель-ке у г. Изео, 19 марта 1977 г., сбор И. М. Лихарева;
.2 — из Италии, окр. Вероны, 6 сентября 1974 г., сбор А. Виктора; определения
автора.
15 А. А, Шилейко
226
7. НАДСЕМ. CHONDRINOEDEA
Белковая железа с нерезкой субапикальной вырезкой. Место перехода семя-
провода в эпифаллус слито с тканями атриальной области и дистального конца
пениса, так что мужские протоки образуют замкнутую петлю. Эпифаллус пря-
мой, цилиндрический, длина его равна длине пениса. На границе между этими
отделами располагается хорошо развитый конический придаток, внутри кото-
рого имеется несколько продольных складок. Две соседствующие складки осо-
бенно хорошо развиты, возле отверстия эпифаллуса они образуют пару лопастей,,
которые тут же обрываются. Внутри пениса напротив отверстия эпифаллуса
Рис. 145. Granaria frumentum (Drap.) из Италии, окр. Вероны, 6 IX 1974 г. Половой аппарат
и вскрытый пенис.
и чуть ниже его расположено подушковидное утолщение, прорезанное рядами
резких глубоких и узких поперечных бороздок. Ниже этого утолщения внутрен-
ние стенки пениса несут несколько слабых продольных складок. Пенис цилинд-
рический или веретеновидный, диаметр его в 2—3 раза превышает диаметр эпи-
фаллуса. Половой ретрактор в нижней части расслаивается и двумя тонкими,,
но широкими лентами крепится к эпифаллусу и пенису. Проток семяприемника
длинный, почти прямой; массивный резервуар, ясцо отделенный от протока,
прилегает к верхней части спермовидукта и к нижней части белковой железы.
Между протоком семяприемника и верхним отделом вагины натянута тонкая
прозрачная соединительно-тканная перепонка.
Распространение и местообитание. Горные области
Средней, Южной и Западной Европы, включая советскую часть Карпат с При-
карпатьем. Обитает на степных участках с коротким травяным покровом,,
а также на камнях и скалах.
2. Род ABIDA Turton, 1831
Studer, 1820:86 (Torqullla, non Bruennich, 1771); Turton, 1831:101; Mer-
m e t, 1843 : 45 (Stomodonta, типовой вид Pupa secale Draparnaud, 1801); Pilsbry, 1918,
Man. Conch., 24 : 262; Thiele, 1931 : 511; Лихарев, Раммельмейер, 1952
129; W e n z, Zilch, 1959—1960 : 156; Gittenberger, 1973 : 19, 82.
2. РОД ABIDA
227
Типовой вид Pupa secale Draparnaud, 1801; по монотипии.
Придаток эпифаллуса отсутствует или рудиментарный; в последнем случае
длина его не превышает у7 длины пениса. Половой ретрактор крепится к дисталь-
ному отделу пениса. Слабо обособленный от протока резервуар семяприемника
доходит или почти доходит до нижней границы белковой железы. Проток семя-
приемника не вжат в простату.
Распространение. Горные районы южной и средней Европы. 9 видов.
В ископаемом состоянии род известен начиная со среднего эоцена (Европа).
Рис. 146. Abida Рис. 147. Abida secale (Drap.) из Франции, окр. Дижона, 23 VIII
secale (Drap.) из 1969 г. Половой аппарат и детали строения пениальной трубки.
Франции, окр.
Дижона.
Abida secale (Draparnaud, 1801) (рис. 146, 147).
Draparnaud, 1801 : 59 (Pupa}', Steeuberg, 1925 : 40, pl. 5, fig. 1; вид очень
изменчив, поэтому к настоящему времени описано множество его форм, которым часто при-
давали ранг вида. Не имея достаточного материала для суждений о внутривидовой структуре,
отсылаю читателя к чрезвычайно обстоятельной работе Гиттенбергера (Gittenberger, 1973,
S. 84, Text.-Fig. 36, 39, Karte 5), где приведена полная синонимика и подробно обсуждаются
направления изменчивости вида; см. также Alzona, 1971, р. 77.
Terra typica — Франция.
Лектотип хранится в Естественноисторическом музее в Вене, № 77709.
Материал: 14 экз. из двух местонахождений во Франции и Италии.
Раковина коническо-цилиндрическая, нижняя короткая цилиндрическая
часть переходит в коническую вершину. Оборотов &/2—9уа, слабовыпуклых,;
последний оборот возле самого устья приподнят, высота его не превышает
Уз высоты раковины. Окраска от светло-роговой до коричневой, иногда с сизо-
ватым налетом. Эмбриональные обороты с ясной микроскопической зерни-
стостью, дефинитивные покрыты тонкими правильными радиальными морщин-
ками, иногда переходящими в легкие ребрышки. Устье округлое или овальное,
места его прикрепления слегка сближены и иногда связаны тонкой прозрачной
мозолью. Края устья отвернуты и слегка утолщены. Ангулярная пластинка
длинная, изогнутая, возле ее верхнего конца имеется небольшой бугорок.
Париетальная пластинка начинается глубоко в устье. Колумеллярных пласти-
15*
228
F. НАДСЕМ. CHONDRINOIDEA
нок две, верхняя длиннее нижней. Базальный бугорок слабый, часто отсут-
ствует. На палатальной стенке 4 складки, сутуральная очень короткая, лежит
глубоко и сквозь устье плохо заметна. Три палатальных складки длинные, но.
задние их концы просматриваются через устье. Пупок цилиндрический, круглый.
Размеры: ВР 6—9.5, БД 2—2.9 мм.
Внутреннее строение. Материал: 3 экз. из Франции, окр. Ди-
жона, 23 августа 1969 г., сбор Н. К. Верещагина, определение автора.
Белковая железа без ясно выраженной вырезки. Семяпровод плотно сра-
стается с нижним отделом пениса; несколько отступя отходит эпифаллус, обра-
зуя в месте отхождения маленький и очень крутой изгиб с сомкнутыми сторо-
нами. Далее диаметр эпифаллуса быстро увеличивается. Эпифаллус переходит
в пенис без видимой границы, дистальная часть мужского протока образует
правильную замкнутую петлю. Внутренняя поверхность эпифаллуса вначале-
неясно бугорчатая, затем появляются 1—2 продольные складки, каждая из ко-
торых резко гофрирована. В пределах пениса внутренние стенки несут несколько
винтообразно направленных тонких складок. Половой ретрактор, отходит от
диафрагмы единой лентой, затем расслаивается и подходит двумя широкими,
но очень тонкими тяжами к дистальной части пениса и проксимальной части
эпифаллуса. Семяприемник не подразделен на проток и резервуар, представ-
ляет собой длинный рукавообразный орган, прилегающий к спермовидукту;
вершина его не достигает белковой железы или слегка касается ее нижнего края.
Распространение и местообитание. Франция, Италия,
и Пиренейский п-ов. Населяет главным образом горные районы.
3. Род CHONDRINA Reichenbach, 1828
Reichenbach, 1828 : 93; L i n d s t г б m, 1868 : 18 (Alloglossa, типовой видНгШ-
mus avenaceus Bruguiere, 1792); Pilsbry, 1918, Man. Conch., 25 : 1; Л и x a p e в, Рам-
мельмейер, 1952 : 130; Wenz, Zilch, 1959—1960 : 157; Gittenberger,
1973 : 22, 158; Акрамовский, 1976 : 142.
Рис. 148. Виды рода Chondrina (подрод Chondrina s. str.).
I — Ch. (Chondrina) avenacea (Brug.) из Австрии, Кайзербрюнн; II — Ch. (Chondrina) clienta caucasica Ehrm-
из Дагестана, с. Акуша; III — Ch. (Chondrina) amphorula Schileyko, sp. nov., голотип.
Типовой вид Bulimus avenaceus Bruguiere, 1792 (рис. 148, I); обозна-
чение Райхенбаха (Reichenbach, 1836 : 152).
Придаток эпифаллуса отсутствует. Близ середины пениальной трубки может
присутствовать расширение, содержащее подкововидную складку. Половой
ретрактор крепится несколько дистальнее середины пениса или отсутствует.
3. РОД CHONDRINA
229
Резервуар семяприемника не доходит до белковой железы, проток семяприем-
ника в простату не вжат или вжат частично.
Распространение. Южная и частично центральная Европа, Ка-
нарские о-ва, Северо-Западная Африка, Крым, Кавказ, Передняя Азия, Иран,
Копетдаг. Около 30 видов.
В ископаемом состоянии род известен начиная с верхнего плиоцена (Европа).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ,
ВИДОВ И ПОДВИДОВ chondrina
1 (6). Ангулярная пластинка отчетливо выражена, удлиненная. Края устья
сильно отвернуты (подрод Chondrina s. str.).
2 (3). На палатальной стенке устья 2 складки; третья (нижняя палатальная),
если имеется, то очень слабая'.................2. Ch. (Ch.) amphorula.
3 (2). На палатальной стенке устья не менее 3 отчетливо выраженных складок.
4 (5). Радиальная скульптура в виде более или менее правильной морщини-
стости или даже ребристости. Подвид обитает на Карпатах . ,
...................................... 1. Ch. (Ch.) clienta clienta.
5 (4). Радиальная скульптура в виде вялых морщинок, правильность распо-
ложения которых местами нарушена. Подвид обитает в Крыму и на
Кавказе.................................la. Ch. (Ch.) clienta caucasica.
6 (1). Ангулярная пластинка отсутствует или представлена слабым округлым
бугорком. Края устья незначительно отвернуты (подрод Granopupa).
7 (8). Раковина высококоническая, обороты сильновыпуклые. Радиальные
ребрышки на поверхности раковины резкие............................
........................................... 4. Ch. (G.) rhodia taurica,
8 (7). Раковина коническо-цилиндрическая, обороты умеренно выпуклые.
Радиальные ребрышки на поверхности раковины в разной степени
размыты......................................3. Ch. (G.) granum,
1. Подрод CHONDRINA s. str.
Ангулярная пластинка удлиненная, начинается от верхней кромки пала-
тального края и уходит в глубь устья. Края устья отвернуты.
Внутри пениса залегает подкововидная складка; там, где она образует ко-
лено, пенис более или менее заметно утолщен.
Распространение. Южная и частично Центральная Европа; Северо-
Западная Африка; Крым, Цавказ, Закавказье.
В ископаемом состоянии подрод известен начиная с плейстоцена (Европа),
1. Chondrina (Chondrina) clienta clienta (Westerlund, 1883).
Westerlund., 1883 : 60 (Pupa avenacea var.. clienta)-, W esterlund, 1887 ;
97 (P. avenacea var. paucidens), 98 (P. avenacea var. subhordeum); Лихарев, Раммель-
мейер, 1952 : 130, рис. 40 (Chondrina avenacea part.); Nordsieck, 1962 : 15, t, I,
fig. 16, Abb. 12; Nordsieck, 1970 : 257, Abb. 21—22; Gittenberger, 1973 : 209;
Дамянов, Лихарев, 1975 : 112, фиг. 46.
Terra typica — «Choc im Tatra Gebirge» (Татры).
Лектотип хранится в Естественноисторическом музеё Гетеборга,
№ 2174.
Материал: 4 экз. из одного местонахождения в Австрии.
Раковина высококоническая, тонкостенная, слегка просвечивающая. Оборо-
тов 7—8, выпуклых, последний оборот возле самого устья поднят — плавно
или с легким перегибом, высота его несколько превышает х/3 высоты раковины.
Окраска одноцветная, темно-роговая, красновато-коричневая или каштановая,
иногда с сизым налетом. Поверхность эмбриональных оборотов с ясно выраженной
микроскопической зернистостью г скульптура дефинитивных оборотов подвер-
230
V. НАДСЕМ. CHONDRINOIDEA
жена значительной изменчивости: от вялой морщинистости до ребристости,
но в любом случае скульптурные элементы сохраняют правильность расположе-
ния. Устье неправильно овальное, слабо скошенное, края его тонкие, умеренно
отвернуты, места прикрепления устья сближены. Устьевая арматура: длинная
ангулярная пластинка может быть цельной или прерванной, в первом случае
ее задний конец находится рядом с передним концом париетальной. Колумел-
лярных пластинок две, верхняя развита лучше нижней. Палатальных складок
3—4, самая верхняя развита слабее прочих. Пупок маленький, несквозной,
каплевидный.
Размеры: ВР 6—7, БД 2.0—2.4 мм; в первоописании: ВР 7, БД 2 мм. Лекто-
тип: ВР 6.48, БД 2.32 мм.
Внутреннее строение неизвестно; Гиттенбергер (Gittenberger,;
1973 : 209) сообщает, что все вскрытые им экземпляры оказались афалйичными.
Распространение и местообитание. Средняя и южная
Европа, включая Балканский п-ов. Обитает на скалах.
la. Chondrina (Chondrina) clienta caucasica Ehrmann, 1931 (рис. 148, 77;
рис. 149).
E hran ann, 1931 : 19; Лихарев, Раммельмейер, 1952 : 131, рис. 41; Git-
tenberger, 1973 : 207, Text.-Fig. 112; Акрамовский, 1976 : 142.
Terra typica — Лечхумский хр. (южные склоны Большого Кавказа).
Лектотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 44091а.
Материал: несколько сотен экз. из 12 местонахождений в Крыму и
на Кавказе.
От номинативного подвида устойчиво отличается более темной окраской,
более вытянутой и стройной раковиной, менее резкой и менее правильной скуль-
птурой.
Размеры: ВР 6—7, БД 2.3—2.8 мм; в первоописании размеры не указаны,
но отмечено, что на приводимой фотографии увеличение 7х/2 раз; размеры
раковины на фотографии 49x17 мм, следовательно, размер сфотографирован-
ного экземпляра (очевидно, лектотипа): ВР около 6.6, БД около 2.3 мм.
Гиттенбергер (Gittenberger, 1973) подтвердил, что кавказские популяции
действительно составляют иной подвид, нежели европейские. Указания на то,;
что в СССР присутствует Ch. avenacea (рис. 148, I) нужно, вероятно, считать
ошибочными (типовое местонахождение Ch. avenacea — окр. Парижа). Вместе
с тем в указанной работе Гиттенбергера отсутствуют сведения о крымских
представителях вида. Акрамовский (1976) в ареал подвида Крым не включает,
но считает, что в Крыму обитают популяции, относящиеся к этому виду. Срав-
нение раковин из 12 местонахождений в Крыму и на Кавказе привело меня
к представлению, что в этих регионах обитает одна и та же форма.
Внутреннее строение. Материал: 9 экз. из Куртатинского
ущелья, долина р. Фиагдон на Северном Кавказе, 10 мая 1970 г.; 7 — из уще-
лья р. Чегем на Северном Кавказе, 21 сентября 1970 г.; 7 — из Дарьяльского
ущелья у с. Чми, 8 мая 1970 г., сборы и определения автора; 4 — из Армении,
Иджеванский р-н, урочище Ланидзор, сбор и определение Н. Н. Акрамовского.
Белковая железа с глубоким желобом, лежащем на верхне-боковой поверх-
ности органа. Семяпровод срастается с тканями дистальной части пениса и
здесь внешне незаметен. Несколько отступя от места, куда впадает семяпровод,
отходит короткий эпифаллус, образующий вместе с пенисом короткую петлю.
Если эту петлю вытянуть вдоль женских путей, то верхний конец ее будет
прилегать к нижней части спермовидукта. На внутренней поверхности эпифал-
луса лежат несколько продольных складок, которые при переходе в пениаль-
ный отдел становятся более слабыми, образуют беспорядочные изгибы и сходят
на нет. Дистальная часть пениса внутри почти гладкая, затем возникает мощ-
3. РОД CHONDRINA
231
ный продольный валик, который вскоре раздваивается; его ветви, подходя
к эпифаллусу, снова смыкаются, образуя подкововидную или петлевидную
складку. В том месте, где залегает эта петля, пенис более или менее явственно
вздут; это вздутие видно и на рисунке в работе Гиттенбергера (loc. cit., S. 208,
Text.-Fig. 112), где изображен внешний вид полового аппарата животного,
собранного в Чегемском ущелье. Половой ретрактор слабый, крепится не-
много ниже середины пениса. Резервуар семяприемника неясно отделен от про-
тока, заканчивается немного выше середины спермовидукта и значительно не
доходит до белковой железы.
Рис. 149. Chondrina (Chondrina) clienta caucasica Ehrm. из Северной Осетии, Куртатинское
ущелье, 10 V 1970 г. Половой аппарат и вскрытый пенис.
Из 27 исследованных экземпляров 11 оказались афалличными.
Распространение и местообитание. Большой Кавказ
с Закавказьем и горный Крым. Обитает на скалах — как в безлесных участках,
так и в редколесье; реже на скальных выходах в лесу. Часто образует плот-
ные колонии.
2. Chondrina (Chondrina) amphorula Schileyko, sp. nov. (рис. 148, III',
рис. 150).
Terra typica — Дагестан, окр. с. Верхнее Инхело близ пос. Карата.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 62 экз. из 5 местонахождений в Дагестане, на Северном
Кавказе и в Армении.
Раковина высококоническая, умеренно тонкостенная, с легким шелковистым
блеском, слегка просвечивающая. Оборотов 7—8, выпуклых; верхние обороты
с правильными радиальными морщинками, которые на нижних оборотах лежат
более редко; последний оборот возле самого устья заметно, с перегибом, поднят,
высота его слегка превышает х/3 высоты раковины. Окраска каштановая или
темно-коричневая, однотонная. Микроскопическая зернистость на эмбриональ-
ных оборотах отчетливая. Поверхность дефинитивных оборотов покрыта тон-
кими косыми морщинками, которые на верхних оборотах очень тонки, распо-
ложены правильно и тесно; на нижних оборотах расстояние между морщин-
ками увеличивается, сами они делаются более резкими, а правильность их
232
V. НАДСЕМ. CH0NDRIN01DEA
расположения местами может нарушаться. Устье овальное, слегка скошено,,
края его тонкие, белые, умеренно отвернуты. Париетальная пластинка слабая,
йевысокая; ангулярная и субпариетальная также развиты относительно слабо.
Колумеллярная пластинка не вполне горизонтальная: ее задний конец заметно
поднимается по столбику; субколумеллярная бугорковидная, заметно меньше
колумеллярной; субпалатальная складка отсутствует или очень маленькая,
округлая. Лежащие выше верхняя и нижняя, палатальные складки тонкие, ко-
роткие; супрапалатальная большей частью отсутствует или очень слабая,
имеет вид небольшой припухлости палатальной стенки. Пупок несквозной,;
Маленький, каплевидный.
Рис. 150. Chondrina (Chondrina) amphorula Schileyko, sp. nov. из Дагестана, с. Карата,
2 VII 1967 г. Паратип. Половой аппарат и вскрытый пенис.
Размеры: ВР 6—6.5, БД 2.2—2.3 мм; голотип: ВР 6.4, БД 2.3 мм.
Вид близок к Ch. clienta caucasica; отличается тем, что с первого взгляда
у Ch. amphorula видны только две складки на палатальной стенке (субпалаталь-
ная, если имеется, видна лишь при тщательном рассмотрении), а у Ch. clienta
caucasica всегда хорошо заметны три складки на палатальной стенке. Известно,
что палатальные складки, особенно самая верхняя и самая нижняя, часто по-
разному выражены даже в пределах одной выборки, но в данном случае видо-
вая самостоятельность Ch. amphorula подтверждается тремя обстоятельствами:
во-первых, совместной встречаемостью обоих видов; во-вторых, отсутствием
промежуточных форм; в-третьих, наличием стойких анатомических отличий:
у Ch. amphorula относительно более длинная и тонкая мужская петля (пенис-}-
эпифаллус), чем у Ch. clienta caucasica, что показано на большом материале
из разных мест.
Внутреннее строение. Материал: 7 экз. из Дагестана, окр.
с. Верхнее Инхело близ пос. Карата, 2 июля 1967; 9 — из с. Чми на правом
берегу р. Терек, 8 мая 1970 г.; 5 — из с. Верхний Чегем в Чегемском ущелье,
21 сентября 1970 г.; сборы автора; 2 — из Армении, Иджеванский р-н, урочище
Ланидзор, сбор Н. Н. Акрамовского; определения автора.
Белковая железа с резким косым желобком на верхней части. Семяпровод
плотно контактирует с покровами дистального отдела пениса, но полностью
с ними не сливается. Эпифаллус постепенно расширяется и при переходе в пе-
3. РОД CHONDRINA
233
нис образует отчетливое вздутие. Если всю петлю, образуемую дистальными
протоками мужского отдела, вытянуть вдоль женских путей, то верхний конец
этой петли окажется значительно выше середины спермовидукта. На внутрен-
ней поверхности эпифаллуса лежат 1—2 вялые продольные складки, которые,
не доходя до пениального отдела, исчезают. Дистальная часть пениса внутри
гладкая, несколько выше формируется продольная складка, которая вскоре
раздваивается и внутри вздутия на границе пениса и эпифаллуса образует
петлю за счет того, что ветви ее сливаются концами; между ветвями может при-
сутствовать еще одна маленькая продольная складка. Половой ретрактор сла-
бый, крепится примерно возле середины пениса. Небольшой резервуар семя-
приемника ясно выражен, проток семяприемника тонкий и короткий: резер-
вуар, если распрямить проток, окажется возле средней части спермовидукта.
Распространение и местообитание. Вид найден в Да-
гестане, в ущелье р. Чегемир. Терек на Северном Кавказе, а также в Иджеван-
ском р-не Армении. Населяет те же биотопы, что и Ch. clienta caucasica, часто
встречаясь вместе с нею.
2. Подрод GRANOPUPA О. Boettger, 1889
О. Boettger, 1889с : 249; М о n't е г о s a t о, 1894 : 170 (Rupestrella, типовой вид
Pupa rupestns Ph.); Pilsbry, 1918, Man. Conch., 24 : 332; Лихарев, Раммель-
мейер, 1952 : 131; Wenz, Zilch, 1959—1960 : 157; Gittenberger, 1973 : 36;
Акрамовский, 1976 : 141.
Типовой вид Pupa granum Draparnaud, 1801; по монотипии.
Подкововидной складки внутри пениса нет; пениальная трубка на внутрен-
них стенках несет плавные широкие продольные складки, в месте перехода
пениса в эпифаллус могут присутствовать мелкие бугорки.
Ангулярная пластинка отсутствует или представлена округлым слабораз-
витым бугорком. Края устья почти не отвернуты.
Распространение. Средиземноморские страны, Копетдаг. 2—
3 вида.
В ископаемом состоянии подрод известен начиная с верхнего плиоцена
(Европа).
Данный подрод принято рассматривать в качестве самостоятельного рода;
однако обширное диагностическое описание Chondrina и Granopupa, помещен-
ное в работе Гиттенбергера (loc. cit., S. 22, 36, 37), показывает, что различия
между этими таксонами очень невелики и значительно меньше, чем отличия
любого из этих таксонов от других родов семейства. Сравнение приводимых
в настоящей книге диагнозов Chondrina и Granopupa также свидетельствует
о тесной близости таксонов. Поэтому представляется более правильным счи-
тать Chondrina s. str. и Granopupa подродами рода Chondrina.
3. Chondrina (Granopupa) granum (Draparnaud, 1801) (рис. 151, 7; рис. 152),
Draparnaud, 15)01 : 59 (Pupa)', Westerlund, 1898 : 165 [P. (Torquilla.) pro.-
fuga]; Pilsbry, 1918, Man. Conch., 24 : 335, pl. 47, figs. 3, 12 (Granopupa)', Л и x a p е в,
Раммельмейер, 1952 : 132, рис. 42 (Granopupa)', Ehrmann, 1956 : 38 (Granopupa)’,
M а т ё к и н, 1966 : 110, табл. 6, рис. 7 (Granopupa)-, Gittenberger, 1973 : 36, figs. 6—
8 (Granopupa)', Акрамовский, 1976 : 142, рис. 61, табл. IV, 52 (Granopupa).
Terra typica — Франция.
Лектотип хранится в Естественноисторическом музее в Вене, № 77707,
Материал: около 60 экз. из двух местонахождений на Большом Бал-
хане и Копетдаге.
Раковина коническо-цилиндрическая, стройная, умеренно тонкостенная^
матовая или слабо блестящая, с узко закругленной вершиной. Оборотов
53/4—83/4, умеренно выпуклых, последний оборот к устью не ноднят или не-
значительно поднят за счет устьевого края, высота его немногим более */3 вы-
234
V. НАДСЕМ. CHONDRINOIDEA
соты раковины. Окраска одноцветная, от светло-роговой до коричневой. Микро-
скопическая зернистость на эмбриональных оборотах очень четкая, дефинитив-
ные покрыты тонкими сглаженными довольно правильно расположенными
ребрышками. Устье округлое или овальное, с легким углом в верхней части
палатального края. Места прикрепления устья незначительно сближены и
соединены слабой, но отчетливой мозолью, усиленной на концах; за счет уси-
ления мозоли в ангулярной области образуется небольшой округлый бугорок.
Зубов 5—7. В наиболее полном наборе имеются париетальная и две колумел-
лярные пластинки, из которых
верхняя массивнее нижней, а так-
же одна базальная и три пала-
тальных складки, из которых
самая длинная — нижняя, а са-
мая короткая — верхняя. Ба-
зальная и верхняя палатальная
РСП
Рис. 152. Chondrina (Granopupa) granum (Drap.)
из Копетдага, 28 IV 1953 г. Половой аппарат.
Рис. 151. Виды рода Chondrina (под-
род Grano pupa).
I — Ch. (Granopupa) granum (Drap.) из
Копетдага; II — Ch. (Granopupa) rhodia
taurica (Kessler) из Крыма, с. Батилиман.
складки подвержены редукции и могут иногда отсутствовать. Пупок очень
узкий, короткощелевидный.
Размеры: ВР 4—6, БД 1.5—1.8, мм; в первоописании размеры не указаны.
Для некоторых южноевропейских популяций характерны более мелкие раз-
меры: ВР может составлять 3.1 при БД 1.4 мм.
Внутреннее строение. Материал: 3 экз. из Копетдага, 28 ап-
реля 1953 г., сбор и определение И. М. Лихарева.
Белковая железа с вялым желобком, занимающим апикальное положение.
Помимо этого желобка другой такой же желобок присутствовал у всех трех
исследованных экземпляров и на поверхности спермовидукта. Насколько этот
признак постоянен, трудно сказать, так как у изученных особей матка была
в активном состоянии, с набухшими стенками; возможно, что, когда падает
активность женской фазы и стенки Матки спадаются, отмеченный желобок
исчезает. Вагина относительно короткая. Семяпровод плотно контактирует
с дистальным отделом пениса и притянут к нему тонкими соединительно-ткан-
ными тяжами, но ткани семяпровода и пениса сохраняют обособленность. Эпи-
фаллус от пениса внешне не отделен, внутренние стенки его не несут каких-
либо отчетливых и характерных структур; на границе с пенисом на внутрен-
3. РОД CHONDRINA
235
ней поверхности видны небольшие округлые бугорки, которые можно заметить
и без вскрытия пениальной трубки: они просвечивают в виде светлых пятны-
шек. Пенис тонкий, цилиндрический, внутри с несколькими широкими нерез-
кими продольными складками. Проток семяприемника довольно длинный,
заканчивается небольшим округлым резервуаром, заметно не доходящим до
белковой железы.
Распространение и местообитание. Южная Европа,
Северо-Западная Африка, Передняя и Малая Азия с восточным Закавказьем,
Иран, Копетдаг, Большой Балхан. Встречается преимущественно на известко-
вых скалах и на сухих каменистых склонах, открытых или поросших кустар-
ником.
4. Chondrina (Granopupa) rhodia taurica (Kessler, 1860) (рис. 151, II;
рис. 153).
Кесслер, 1860 : 226 {Pupa taurica n. sp.?); Пузанов, 1925a : 87, рис. V, 12, 13,
рис. VII, 5 (Modicella taurica)-, Цветков, 1939 : 181; Лихарев, Раммельмейер,
1952 : 132 (Granopupa).
Terra typica — Крым, окр. Ялты.
Место хранения типов не установлено.
Материал: около 50 экз. из трех местонахождений в горном Крыму.
Раковина высококоническая, стройная, относительно хрупкая, матовая,
слегка просвечивающая, с закругленной вершиной. Оборотов 6—7, сильно
выпуклых, последний оборот у вы-
соких раковин к устью почти не
поднят, у низких приподнят с лег-
ким перегибом; высота его всегда
заметно меньше половины высоты
раковины, а иногда составляет даже
почти Уз этой величины. Окраска от
коричневой до каштановой, иногда
с легким сизым налетом. Микрозер-
нистая скульптура эмбриональных
оборотов очень четкая. Дефинитив-
ные обороты несут правильно рас-
положенные закругленные четкие
радиальные ребрышки. Устье оваль-
ное, с легким углом в верхней части
палатального края. Места прикре-
пления устья слегка сближены, свя-
заны очень тонкой мозолью, иногда
слегка усиленной на концах. Края
Рис. 153. Chondrina (Granopupa) rhodia
taurica (Kessler) из Крыма, с. Батилиман,
V 1930 г. Половой аппарат.
устья тонкие, отвернутые, без губы. Зубов 5. Короткая, но высокая полу-
круглая париетальная пластинка занимает примерно середину париетального
края. Колумеллярная пластинка несколько сильнее субколумеллярной. Верх-
няя палатальная складка немного длиннее и выше нижней палатальной.
Пупок короткоовальный.
Размеры: ВР 4.4—5.8, БД 1.7—2.2 мм: в первоописании: ВР 5.8, БД 2.1 мм.
Кесслер описал эту форму предположительно как Самостоятельный вид,
отличающийся от Pupa rhodia Roth, 1839 (Terra typica — о-в Родос, Греция)
большими размерами и почти равной длиной палатальных складок (у Р. rho~
236
V. НАДСЕМ. CHONDRINOIDEA
dia верхняя палатальная складка почти втрое длиннее нижней). Тем не менее
Ретовский (Retowski, 1883а) полагает, что крымские популяции должны быть
отнесены к номинативной форме. Пузанов справедливо указывает, что «. . . от-
личия крымских экземпляров от эгейских настолько очевидны и определенны,
Географическая их разобщенность столь велика, что не может быть и речи об их
Идентичности; вопрос лишь в том, толковать ли крымскую форму как вид или
подвид. Лично я склоняюсь к первому. . .» (Пузанов, 1925а, с. 87). Лихарев
и Раммельмейер (1952) считают крымскую форму подвидом. Характеризуя
ареал Granopupa (Rupestrella) rhodia, Клемм (Klemm, 1962, S. 223) не называет
Крым, считая, очевидно, что в этом регионе обитает самостоятельный вид;
Пагет (Paget, 1976, S. 709) определенно указывает, что в Крыму живет G. rho-
dia taurica. Это решение представляется самым приемлемым, так как различия
между крымскими и Эгейскими популяциями заключается лишь в относитель-
ной длине палатальных складок. Что касается размеров, то, промерив несколько
десятков раковин из Крыма, я нашел вполне сформированные раковины высо-
той не более 4.4 мм, т. е. того же размера, что и раковины номинативного под-
Рида.
Внутреннее строение. Материал: 4 экз. из Крыма, окрестности
Батилимана, май 1930 г., сбор В. А. Линдгольма, определение автора.
Белковая железа с очень вялой апикальной вырезкой. Семяпровод сли-
вается с тканями атриального отдела и затем, плавно и постепенно расширяясь,,
переходит в эпифаллус. Если считать границей между пенисом и эпифаллусом место
прикрепления полового ретрактора, то пенис приблизительно в 11/2 раза короче
эпифаллуса. На внутренних стенках протоков, составляющих мужскую петлю,
специализированных структур не найдено, кроме 1—2 очень широких и плав-
ных продольных складок, идущих по очень пологой спирали. Резервуар семя-
приемника неясно отграничен от прямого протока, заметно не доходит до бел-
ковой железы.
Распространение и местообитание. Горный Крым.
Ареал номинативного подвида: Адриатическое побережье Балканского п-ва,
о-ва Ионического и Эгейского моря, южная Греция, Ливан, Малая Азия.
Обитает на открытых прогреваемых солнцем скалах.
?П. Сем. PYRAMIDULIDAE Kennard et Woodward, 1914
Раковина микрохеликоидная. Устье без зубов, с прямыми краями. Прида-
ток эпифаллуса отсутствует; на пенисе имеется мешковидный вырост. Половой
ретрактор крепится к пениальной трубке возле ее середины.
Монотипическое семейство.
Распространение. Южная часть Палеарктики от Европы до Япо-
нии.
В ископаемом состоянии семейство известно начиная с эоцена (Европа).
1. Род PYRAMIDULA Fitzinger, 1833
Fit zinger, 1833 : 95; Т h i e 1 e, 1931 : 503; Pilsbry, 1935, Man. Conch., 28 : 182;
Лихарёв, Раммельмейер, 1952 : 168; Habe, 1956 : 117 (подрод Pyramidulops,
типовой вид Pyramidula conica Pilsbry et Hirase); Wenz, Zilch, 1959—1960 : 145; A k-
p а м о в с к и Й, 1976 : 130.
Типовой вид Helix rupestris Draparnaud, 1801; по монотипии.
Признаки семейства.
Распространение как у семейства. 2—3 вида.
Pyramidula rupestris (Draparnaud, 1801) (рис. 154, 155).
Draparnaud, 1801 : 71 (Helix)', Fitzinger, 1833 : 95; Beck, 1837 : 9 (Euryom-
phalia umbilicata); Held, 1837 : 916 (Patula)', M oquin-Tan don, 1855 : 192, pl. 15,
figs. 10—13 [Helix (Hygromane) rupestris et f. saxatilis et f. trochoides)]; Watson, 1920 : 6,
1. РОД PYRAMIDULA.
237
pl. II, fig. 4; P i 1 s b г у, 1935, Man. Conch., 28 : 183, t. 25, fig. 9; Лихарев, Рамме ль-
мейер, 1952 : 169, рис. 82; Ehrmann, 1956 : 57, fig. 31; Lozek, 1956 : 111, t. 9,
fig. 2; 1964 : 196, Abb. 38; Матёкин, 1966 : 115, табл. 6, рис. 6; Акрамовский,
1976 : 130, рис. 52, табл. V, 38.
Terra typica — Кастельно и Арбуа, Франция.
Место хранения типов не установлено.
Материал: более 100 экз. из 23 местонахождений на Карпатах, в Крыму,
яа Кавказе и на Тянь-Шане.
Раковина кубаревидная, тонкостенная, слабо просвечивающая, с кониче-
ским контуром короткого завитка и закругленной вершиной. Оборотов 4—
4х/2, сильно выпуклых, ступенчатых; участки шва,: разделяющие соседние обо-
Рис. 154. Pyramidula гире-
stris (Drap.) из Дагестана,
с. Акуша.
Рис. 155. Pyramidula rupe-
stris (Drap.) из Армении, Ид-
жеванский р-н, 6 V 1953 г.
Половой аппарат.
роты, часто не вполне параллельны один другому. Последний оборот к устью
слегка и плавно,- без перегиба, опущен. Высота завитка примерно в 1х/2 раза
больше высоты устья, но подвержена значительной изменчивости. Окраска
одноцветная, от красновато-роговой до каштановой и темно-бурой. Дефини-
тивные обороты тонко, но глубоко и резко радиально исчерчены. Устье округлое,,
края его тонкие, острые, не отвернуты, кроме верхней части колумеллярного
края. Места прикрепления устья слегка сближены. Пупок относительно широ-
кий, перспективный.
Размеры: ВР 1.7—2.1, БД 2.2—2.7 мм; в первоописании: ВР 1.8, БД 2.3 мм.
Внутреннее строение. Материал: 4 экз. из Армении, Иджеван-
ский р-н, с. Агран, сбор и определение Н. Н. Акрамовского; 5 — из Дагестана,;
каменистое плато у с. Акуша, 27 июня 1967 г., сбор и определение автора.
Поверхность белковой железы без ясных желобков. Проксимальный отдел
эпифаллуса вытянутый, веретеновидный, переходит в тонкий нитевидный
отдел, который постепенно расширяется; перед впадением в пенис эпифаллус
вновь сужается, а немного проксимальнее этого сужения к эпифаллусу кре-
пится половой, ретрактор. Далее пениальная трубка вновь расширяется и
переходит в цилиндрический пенис, на середине которого сидит округлый
мешковидный придаток, полость которого широко сообщается с полостью пе-
ниса. Каких-либо специализированных структур на внутренней поверхности
пениса и его придатка не обнаружено. Проток семяприемника тонкий,- отходит
примерно от середины вагины, заканчиваается небольшим округлым резервуа-
ром, заметно не доходящим до белковой железы.
238
VI. НАДСЕМ. BULIMINOIDEA
Распространение и местообитание. Ирландия, южная
Европа и юг Центральной Европы; северо-западная Африка, Передняя Азия,.
Иран, северная Индия; в СССР — Волыно-Подольская возвышенность, При-
карпатье, Крым, Кавказ, Средняя Азия. Горный вид, обитает на известковых
скалах и в осыпях, часто образует плотные скопления.
VI. Надсем. BULIMINOIDEA Kobelt, 1880
Раковина средних размеров или довольно большая, от овальной до верете-
новидной и башневидной, часто с радиальной и (или) спиральной скульптурой.
Эмбриональные обороты гладкие, или, редко, со спиральными нитями (Retow-
skiinae). Пупок точковидйый или щелевидный. Устье простое или с зубами,,
в последнем случае они обычно присутствуют близ края устья (поверхностные).
Устьевые края прямые или отвернутые, тонкие или утолщенные, нередко с гу-
бой.
Семенные пузырьки обособлены от гермафродитного протока и мутовчато
расположены вокруг проксимальной части протока. Пенис с эпифаллусом,,
внутренние его стенки гладкие или с системой складок; нередко присутствует
папилла пениса. Имеется пениальный аппендикс (иногда вторично редуциро-
ван) и цэкум, сидящий на эпифаллусе. Половой ретрактор двуветвистый, пе-
ниальная ветвь крепится на границе пениса и эпифаллуса дистальнее цэкума.
Сем. BULIMINIDAE Kobelt, 1880
Kobelt, 1880, Iconogr., 7; Kobelt, 1902 : 397; Hesse, 1933 : 145.
Проток семяприемника длиннее резервуара. На пенисе, кроме пениального-
аппендикса, придатков нет. Цэкум эпифаллуса отсутствует лишь в виде ред-
чайшего исключения.
Вероятно, роды, представленные в Африке, на Новой Гвинее и в северной
Австралии (Cerastua, Amimopind), составляют отдельное семейство, характе-
ризующееся чрезвычайно коротким протоком семяприемника, двумя придат-
ками пениса и отсутствием цэкума (см. Solem, 1964).
Распространение. Европа, кроме северных областей, о-ва восточ-
ной Атлантики, Северная Африка, Передняя и Малая Азия, Кавказ, Средняя,,
Центральная и Юго-Восточная Азия с Японией. 8 подсемейств.
В ископаемом состоянии семейство известно начиная с миоцена (Европа,.
Кавказ, Алжир).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ
BULIMINIDAE
1 (14). Эмбриональные обороты гладкие.
2 (3). Бич не короче эпифаллуса ........................Bulimininae (с. 330).
3 (2). Бич отсутствует либо значительно короче эпифаллуса.
4 (9). Папилла пениса замкнута, т. е. с внутренним каналом.
5 (6). Раковина без зубов и с очень слабой губой . . . Merdigerinae (с. 328).
6 (5). Раковина с зубами и (или) хорошо развитой губой.
7 (8). В устье не менее 5 зубов.....................Multidentulinae (с. 371).
8 (7). В устье не более 3 зубов...................Chondrulopsininae (с. 321).
9 (4). Папилла пениса отсутствует или незамкнутая, т. е. с бороздой на по-
верхности.
10 (11). Пенис чрезвычайно тонкий, нитевидный .... Jaminiinae (с. 326).
11 (10). Пенис нормально развит, не нитевидный.
12 (13). Внутренняя поверхность пениса несет призмоконические бугорки,,
развитые иногда только в Проксимальной или только в дистальной части
органа..............................................Eninae (с. 338).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ
239
13 (12). Внутренняя поверхность пениса гладкая или несет рельефный узор
из складок ................................... Pseudonapaeinae (с. 243).
14 (1). Эмбриональные обороты с ясной спиральной скульптурой...........
............................................. Retowskiinae (с. 334).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ НЕКОТОРЫХ РОДОВ И ВИДОВ
BULIMINIDAE (ДОПОЛНИТЕЛЬНАЯ')
1 (16). Раковина левозавитая.
2 (7). Виды обитают в Средней Азии.
3 (6). Окраска коричневая или каштановая, нередко с сизоватым налетом.
4 (5). В глубине устья залегает удлиненная палатальная складка . . . .
. . . ........................... Pseudonapaeus entoptyx (с. 269).
5 (4). Палатальная стенка устья без складки .........................
.................................. Pseudonapaeus goldfussi (с. 267).
6 (3). Раковина белая или серая, иногда с темными радиальными пестринами
......................................Laevozebrinus urgutensis (с. 303).
7 (2). Виды обитают в Крыму или на Кавказе.
8 (13). Устье раковины с 3—5 зубами.
Я (10). Нижний колумеллярный зуб не связан с верхним; оба зуба лежат
в плоскости устья...........................Jaminia quadridens (с. 326).
10 (9). Нижний (наружный) колумеллярный зуб связан тонкой перемычкой
с пластинчатым верхним (внутренним), т. е. нижний лежит в плоскости
устья, а верхний составляет его продолжение и уходит по столбику вглубь
раковины.
11 (12). Устье овальное, палатальный зуб в виде приостренного бугорка рас-
полагается в плоскости устья . . . Pseudochondrula seductilis (с. 289).
12 (11). Устье округло-треугольное, палатальный зуб треугольной формы,
нижний край его располагается в плоскости устья, а верхний заметно
отодвинут вглубь устья ......................Ljudmilena tricollis (с. 312).
13 (8). Устье раковины без зубов.
14 (15). Раковина овально-коническая, белая или сероватая. Вид обитает
в Крыму..................................... . Thoanteus gibber 1 (с. 358).
15 (14). Раковина от коническо-цилиндрической до почти башневидной, желто-
ватая, роговая или каштановая. Вид обитает на Кавказе...................
.............................. Pseudochondrula sinistrorse (с. 294).
16 (1). Раковина правозавитая.
17 (18). Раковина от овально-цилиндрической до цилиндрической, почти
гладкая. Устье с 4 короткими гребневидными зубами: париетальным,
колумеллярный и 2 палатальными.........................Geminula (с. 317).
18 (17). Раковина не обладает всей совокупностью указанных признаков.
19 (20). Раковина слабо скульптурирована, от почти кубаревидной до высоко-
конической, в любом случае контур завитка имеет отчетливо конические
очертания. Обороты сильновыпуклые. Места прикрепления устья сильно
сближены; устье без зубов, края его отвернуты .... Turanena (с. 274).
20(19). Набор признаков иной.
21 (78). Виды обитают к западу от Каспийского моря.
22 (39). Устье с 1—7 отчетливыми зубами либо с глубоко лежащей удлиненной
палатальной складкой.
23 (26). Зуб один, занимает середину палатального края.
24 (25). Оборотов не больше 7, высота раковины не превышает 10 мм ...
...................................Pseudochondrula tuberifera (с. 294). .
25 (24). Оборотов не меньше 9, высота раковины не меньше 11 мм.........
. .............................Imparietula schelkovnikovi (с. 308).
1 Изредка встречаются правозавитые, особи.
240
VI. НАДСЕМ. BULIMINOIDEA
26 (23). Зубов не менее двух, либо имеется одна удлиненная палатальная
складка, лежащая глубоко в устье.
27 (28). В глубине устья залегает длинная палатальная складка, развитая
иногда слабо ................................Brephulopsis bidens (с. 354).
28 (27). Палатальная складка указанного типа отсутствует.
29 (30). Раковина от овальной до почти короткоцилиндрической, сложена
6—7 оборотами. Зубов 6—7: небольшой ангулярный, мощный париеталь-
ный, бугорковидный колумеллярный, небольшой базально-колумелляр-
ный и 2—3 округлых палатальных; высота раковины 4—6,5 мм ....
............................................. Multidentulinae 1 (с. 371).
30 (29). Раковина не обладает'совокупностью перечисленных признаков.
31 (32). Устье с двумя бугорковидными округлыми зубами — колумеллярный
и палатальным..................................Chondrula bicallosa (с. 367).
32 (31). Устье с 3—6 зубами.
33 (36). Колумеллярный зуб состоит из двух частей, соединенных перемычкой:
бугорковидной наружной части и пластинчатой внутренней, уходящей
вверх по столбику.
34 (35). Раковина от удлиненно-овальной до коническо-цилиндрической; устье
овальное, палатальный край его равномерно выгнут ........................
................................... Pseudochondrula tetrodon (с. 290).
35 (34). Раковина овально-цилиндрическая, цилиндрическая или булавовид-
ная; устье округло-треугольное, с легким вдавлением на середине пала-
тального края ................................Ljudmilena sieversi (с. 309).
36 (33). Колумеллярный зуб одиночный.
37 (38). Раковина белая, одноцветная или с темными радиальными пестринами.
На поверхности дефинитивных оборотов заметны следы спиральной
скульптуры ......................................Chondrus zebra (с. 351).
38 (37). Раковина от серой до коричневой, без темного рисунка. Элементы
спиральной скульптуры отсутствуют .......................................
.......................................Chondrula (Chondrula) (с. 363).
39 (22). Устье без зубов.
40 (59). Поверхность дефинитивных оборотов со спиральной скульптурой,
которая может быть тонкой или выраженной местами, особенно на ба-
зальной части; либо скульптура тонкобугорчатая.
41 (46). Раковина белая, нередко с темными радиальными пестринами или
рыжеватыми размытыми радиальными полосами.
42 (43). Устье внутри желтое..............Napaeopsis hohenackeri (с. 356).
43 (42). Устье внутри белое или серое.
44 (45). Форма раковины удлиненно-овальная . . . Zebrina detrita (с. 349).
45 (44). Форма раковины от веретеновидной до цилиндрической .....
.........................................Brephulopsis cylindrica (с. 353).
46 (41). Раковина коричневая, каштановая, роговая, реже желтая с темной
спиральной лентой; радиальные пестрины или полосы отсутствуют.
47 (48). В результате пересечения радиальных и спиральных элементов скульп-
туры поверхность раковины тонкобугорчатая; вид обитает в западных
районах Украины, Прибалтике и местами в средней полосе европейской
части................................................Ena montana (с. 339).
48 (47). Тонкобугорчатая скульптура отсутствует; виды обитают на Кавказе
и в Крыму.
49 (50). Вид обитает в Крыму...................Peristoma rupestre (с. 343).
50 (49). Виды обитают на Кавказе.
51 (52). Высота раковины 21—27 мм,' с темно-бурой широкой спиральной
лентой, реже одноцветно желтая .... Caucasicola raddei (с. 347).
52 (51). Высота раковины не превышает 16 мм, спиральная лента отсутствует,
окраска коричневых тонов.
1 Роды и обитающие в СССР виды, принадлежащие этому подсемейству, достоверно»
различаются только по анатомическим признакам.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ
241
53 (56). Края устья притупленные, утолщены мощной губой.
54 (55). Внутри пениса залегает незамкнутая папилла ...................
.................................... Imparietula brevior (с. 307)
55 (54). Внутри пениса имеется пара V-образных складок, папилла
отсутствует .............................Pseudochondrula lederi (с. 292).
56 (53). Края устья острые, губа лежит отступя от устьевого края.
57 (58). Спиральные бороздки выражены ясно. Большой диаметр раковины
не менее 6 мм ...............................Peristoma boettgeri (с. 345).
58 (57). Спиральные бороздки выражены слабо. Большой диаметр раковины
не более 5 мм................................Peristoma lanceum (с. 343).
59 (40). Поверхность дефинитивных оборотов без спиральной скульптуры
или эта скульптура настолько слаба, что лишь местами еле угадывается.
60 (61). Раковина цилиндрическая, очень стройная: при 8—11 оборотах высота
раковины 9—14, большой диаметр 2—3 мм..................................
.................................... Ramusculus subulatus (с. 360).
61 (60). Раковина менее стройная, оборотов не более 7.
62 (63). Раковина белая, овально-коническая, с темными радиальными пестри-
нами. Края устья почти не отвернуты .... Adzharia renschi (с. 332).
63 (62). Набор признаков иной.
64 (65). Раковина удлиненно-овальная, с широкой притупленной вершиной.
Устье большое, овальное, места его прикрепления широко расставлены.
Края устья сильно отвернуты, с широкой расплывшейся губой . . .
......................................... Buliminus urmianus (с. 331).
65 (64). Раковина не обладает совокупностью указанных признаков.
66 (69). Виды обитают в Крыму.
67 (68). Высота раковины не менее 13, большой диаметр не менее 5 мм . . .
......................................Peristoma merduenianum (с. 341).
68 (67). Высота раковины не более 11, большой диаметр не более 4 мм . . .
.............................................Merdigera obscura (с. 328).
69 (66). Виды обитают на Карпатах, на Кавказе и в средней полосе европейской
части СССР.
70 (73). Высота раковины не менее 16, большой диаметр не менее 6 мм.
71 (72). Устье овально-четырехугольное. Вид обитает в западной части Кавказа
Chondrula caucasica (с. 369).
72 (71). Устье овальное. Вид обитает на Карпатах.......................
............................................Chondrula bielzi (с. 370).
73 (70). Высота раковины не более 14, большой диаметр не более 5.5 мм.
74 (75). Последний оборот несколько оплывший, т. е. максимальный диаметр
его ниже средней линии. Края устья немного отвернуты и утолщены за
счет сильно расплывшейся, но довольно массивной белой губы ....
......................................... Differena leucostoma (с. 316).
75 (74). Последний оборот не оплывший, максимальный диаметр его примерно
совпадает со средней линией. Края устья если и отвернуты, то не утол-
щены; если имеется губа, она располагается отступя от устьевого края.
76 (77). Виды обитают на Кавказе . ..............Akramovskiella (с. 312).
77 (76). Вид обитает в Крыму, на Карпатах и в средней полосе европейской
части........................................Merdigera obscura (с. 328).
78 (21). Виды обитают к востоку от Каспийского моря.
79 (92). Устье раковины с 1—3 зубами, либо в глубине устья залегает удли-
ненная складка.
80 (81). Устьевая арматура представлена одной продольной палатальной склад-
кой, лежащей в глубине устья . . Siraphoroides moltschanovi (с. 325).
81 (80). Складка указанного типа отсутствует, зубы бугорковидные или в виде
коротких пластинок.
82 (83). На париетальной стенке два четких приостренных бугорковидных
зуба — ангулярный и париетальный ................'.....................
....................................Triangustoma dentatum (с. 296).
16 А. А. Шилейко
242
VI. НАДСЕМ. BULIMINOIDEA
83 (82). На париетальной стенке один зуб (ангулярный если есть, то малень-
кий, представляет собой утолщение париетальной мозоли).
84 (87). В устье три отчетливых зуба: париетальный, колумеллярный и пала-
тальный.
85 (86). Все три зуба лежат в плоскости устья и представляют собой утолщения
губы и париетальной мозоли ..............................................
................................. Chondrulopsina fedtschenkoi (с. 323).
86 (85). Зубы не лежат в плоскости устья: париетальный углублен, колумел-
лярный уходит вверх по столбику .........................................
....................... Pseudonapaeus albiplicatus f. retrodens (с. 255).
87 (84). Зуб только один — париетальный.
88 (89). Скульптура составлена грубыми редкими ребрами .................
.................. Pseudonapaeus albiplicatus f. dissimilis 1 (с. 256).
89 (88). Скульптура в виде исчерченности или морщинистости.
90 (91). Раковина белая или серая, обычно с темными радиальными пестринами,
развитыми в различной степени ...........................................
...................... Pseudonapaeus albiplicatus f. secalina (с. 252).
91 (90). Раковина одноцветная или почти одноцветная, темно-роговая или
серовато-роговая ........................................................
...................... Pseudonapaeus albiplicatus f. aptycha (с. 254).
92 (79). Устье без зубов; могут присутствовать лишь небольшие утолщения
устьевого края или ангулярный бугорок в виде слабой припухлости
париетальной мозоли.
93 (94). Устье внутри желтое или коричневое .............................
..................................... Laevozebrinus eremita (с. 301).
94 (93). Устье внутри белое или серое.
95 (102). Края устья отвернуты и утолщены за счет хорошо развитой губы.
96 (97). Скульптура в виде резких грубых белых ребер....................
................................................. Mastoides (с. 297).
97 (96). Ребра на поверхности раковины отсутствуют; если имеются, то нерез-
кие, расплывшиеся.
98 (99). Устье округло-треугольное .......................................
.............................. Pseudonapaeus trigonochilus (с. 265).
99 (98). Устье овальное или округлое.
100 (101). На устьевом крае имеются 2—3 слабых вздутия, имеющих вид мест-
ных припухлостей: париетальный (в виде расширения париетальной
мозоли), колумеллярный (может отсутствовать) и палатальный в виде
легкого продольного утолщения' губы......................................
..................................Chondrulopsina intumescens (с. 322).
101 (100). 'Край устья без вздутий, порожисто утолщен за счет узкой, но вы-,
сокой губы, лежащей вплотную к краю .....................................
........................................Subzebrinus labiellus (с. 273).
102 (95). Края устья прямые или отвернуты, но губа, если имеется, распола-
гается отступя от устьевого края.
103 (104). Раковина тонкая, стройная, цилиндрическая, сложена 8—11 обо-
ротами. Окраска белая, серая или светло-роговая, с многочисленными
темными радиальными пестринами. Высота раковины 14—24, большой
диаметр 5—8 мм..........................Laevozebrinus ujfalvyanus (с. 303).
104 (103). Набор признаков иной.
105 (106). Раковина от высококонической до почти башневидной, с широкой
тупой вершиной; составлена 71/3—8 оборотами. Окраска первых оборотов
коричневато-розовая или серовато-роговая; 2—3 последних оборота серые
или почти белые, нередко с темными нерезкими радиальными штрихами.
Высота раковины 11—14.5, большой диаметр 4.5—6 мм........................
...................................... Laevozebrinus lenis (с. 305).
1 Париетальный зуб иногда развит очень слабо.
I. ПОДСЕМ. PSEUDONAPAEINAE
243
106 (105). Раковина не обладает всей совокупностью перечисленных признаков
...............................................Pseudonapaeus (с. 244).
I. Подсем. PSEUDO NAPAEINAE Schileyko, 1978
Шилейко, 1978 г : 843.
Раковина почти гладкая или с радиальной скульптурой, выраженной в раз-
ной степени. Зубы в устье, как правило, отсутствуют; в виде редкого исключе-
ния может присутствовать 1—3 зуба или палатальная складка. Эмбриональные
обороты гладкие.
Эпифаллус без бича (кроме родов Serina и Тигапепа'), на конце широко за-
круглен. Семяпровод впадает эксцентрично. Цэкум хорошо развит. Пенис
внутри с продольными складками, которые могут группироваться в 1—2 V-
образных складки либо в 2—3 кольцевидных. Иногда за счет разрастания и
дифференцировки складок формируется незамкнутая папилла. Призмокониче-
ские бугорки и проксимальный вырост пениса отсутствуют. Пениальный яппен-
дикс имеется, дивертикул семяприемника чаще всего присутствует.
Распространение. Закавказье, прилежащие районы Турции и
Ирана, Афганистан, Индия, Средняя и Центральная Азия, Корейский п-ов, Япо-
ния. Не менее 20 родов.
Ареал нуждается в уточнении, так как важные диагностические признаки
подсемейства связаны с внутренним строением пениальной трубки; в то же
время организация пениса исследована в основном у видов, обитающих в пре-
делах нашей страны. Вероятно, род Luchuena, живущий в Японии, также
относится к этому подсемейству (см. Minato, 1977).
В ископаемом состояции подсемейство известно начиная с плейстоцена
(Средняя и Центральная Азия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ПОДРОДОВ
PSEUD ONA РА EINA Е
1 (20). Дивертикул семяприемника имеется.
2 (7). Раковина коническая, обороты сильновыпуклые, места прикрепления
устья сближены, устье без зубов.
3 (6). Пениальный аппендикс нормально развит, половой ретрактор двувет-
вистый.
4 (5). Папилла пениса имеется, дистальный конец пениса заключает камеру,
ограниченную двумя сфинктерами. Эпифаллус на конце широко закруглен
..............................................Akramovskiella (с. 312).
5 (4). Папилла пениса отсутствует, пениальная трубка до самого атриума
ровная, без камер. Эпифаллус переходит в короткий конический бич
................................................... Тигапепа (с. 274).
6 (3). Пениальный аппендикс рудиментарный, половой ретрактор одноветви-
стый ..............................................Differena (с. 315).
7 (2). Раковина не обладает полным набором перечисленных черт.
8 (9). В устье раковины 4 зуба...................... Geminula (с. 317)
9 (8). В устье раковины не более 3 зубов.
10 (11). Папилла пениса трехлопастная: по поверхности центральной стержне-
видной лопасти проходит продольная борозда, охваченная в нижней части
парой коротких лопастей.........................Laevozebrinus (с. 301).
11 (10). Папилла пениса иного строения или отсутствует.
12 (13). На париетальной стенке устья 2 бугорковидных зуба ..........
......................................... . Triangustoma (с. 296).
13 (12). На париетальной стенке не более одного зуба или устье без зубов.
14 (15). Резервуар семяприемника сидит на протоке всем широким основанием,
т. е. лишен собственного протока................Mastoides (с. 297).
15 (14). Резервуар семяприемника с собственным протоком.
16*
244
VI. НАДСЕМ. BULIMINOIDEA
16 (17). Раковина левозавитая; в глубине устья на палатальной стенке имеется
продольная складка..................Pseudonapaeus (Siraphorus) (с. 269).
17 (16). Раковина право- или левозавитая; палатальных складок нет.
18 (19). Раковина удлиненно-овальная, верхняя часть ее вытянутая, узкоко-
ническая. Пенис со стержневидной папиллой, зажатой между двумя про-,
дольными складками ...................Pseudonapaeus (Aridenus) (с. 271).
19 (18). Раковина иной формы. Папилла пениса отсутствует, на вйутреннёй
поверхности пениса имеются 1 или 2 V-образные складки.................
.....................Pseudonapaeus (Pseudonapaeus s. str.) (с. 246).
20 (1). Дивертикул семяприемника отсутствует.
21 (22). Участок эпифаллуса между цэкумом и семяпроводом сильно мускули-
зован. Род представлен в Средней Азии..............Subzebrinus (с. 272).
22 (21). Эпифаллус по всей длине равномерно и умеренно мускулизован. Роды
представлены на Кавказе.
23 (26). Папилла пениса имеется.
24 (25). Устье с париетальными и колумеллярными зубами ...............
............................................. Ljudmilena (с. 309).
25 (24). Париетальная и колумеллярная стенки устья лишены зубов . . .
.................................................. Imparietula (с. 306).
26 (23). Папилла пениса отсутствует . .........Pseudochondrula (с. 288).
1. Род PSEUDONAPAEUS Westerlund, 1887
Westerlund, 1887 : 66; там же: (Chondrulopsis, типовой вид Buliminus sogdianus
Е. Martens, 1874); Kobelt, 1888, Iconogr., N. F., IV : 40 (Sewertzowia, типовой вид Buliminus
dlssimilisE. Martens, 1882); Westerlund, 1896 : 189 (Pseudopetraeus, типовой вид Buli-
minus albiplicatus E. Martens, 1874); Kobelt, 1904, Iconogr., N. F., XI: 133 (Severtzovia nom.
emend, pro Sewertzowia Kobelt, 1888); Lindholm, 1925a : 27 [Ena (Pseudopetraeus), секция
Cauliculus, типовой вид Buliminus schnitnikovi Lindholm, 1922]; там же : [Ena (Pseudona-
paeus), секция Laeonapaeus, типовой вид Buliminus goldfussi Kobelt, 1893], 28 [Zebrina (Sub-
zebrinus), секция Oedichilus, типовой вид Buliminus merzbacheri Weber, 1913]; там же : [Zeb-
rina (Subzebrinus), секция Styloptychus, типовой вид Buliminus kasnakovi Westerlund, 1898],
30 [Sewertzowia (Parachondrula), типовой вид Buliminus retrodens Martens, 1879]; Thiele,
1931 : 519 (Sewertzowia), 520 [Zebrina (Pseudonapaeus)]; Лихарев, Раммельмейер,
1952 : 189 (Sewertzowia), 195 (Subzebrinus part.), 201 [Subzebrinus (Styloptychus), 202; W e n z,
Zilch, 1959—1960 : 180 (? Sewertzowia), 184; M а т ё к и н, 1966 : 112 [Jaminia (Chondru-
lopstna) part.]; Шилейко, 1977 : 40; 1978 г : 844.
Типовой вид Buliminus asiaticus Е. Martens, 1881 (=Pseudonapaeus
albiplicatus f. asiatica E. Martens); обозначение Линдгольма, 1922a.
Раковина различной формы, в разной степени скульптурирована. Устье
без зубов или с 1—3 зубами, края устья отвернуты.
Бич отсутствует. Внутри пениса имеется одна или две V-образных складки.
Папилла отсутствует (кроме подрода Aridenus). Дивертикул семяприемника
нормально развит.
Распространение. Малая Азия, Иран, Афганистан, Средняя и
Центральная Азия, северо-западная Индия. Не менее 30 видов.
В ископаемом состоянии род известен начиная с плейстоцена (Средняя и
Центральная Азия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ФОРМ PSEUDONAPAEUS
1 (4). Раковина левозавитая.
2 (3). На палатальной стенке имеется продольная складка, уходящая в глубь
раковины......................................... 13. Р. (S.) entoptyx.
3 (2). Палатальная стенка без складок..............И. Р. (Р.) goldfussi.
4 (1). Раковина правозавитая.
5 (12). На париетальной стенке имеется развитый в разной степени зуб или
раковина с резкими радиальными ребрами.
1. РОД PSEUDONAPAEUS
245
6 (7). Поверхность раковины с резкими радиальными ребрами...............
............................... 2з. Р. (Р.) albiplicatus f. dissimilis.
7 (6). Поверхность раковины гладкая или слаборебристая.
8 (9). Раковина тонкая, высокая, цилиндрическая.........................
................................... 2в. Р. (Р.) albiplicatus f. secalina.
9 (8). Раковина короткоцилиндрическая или овально-цилиндрическая.
10 (11). В устье имеется лишь один париетальный зуб ......................
................................... 2е. Р. (Р.) albiplicatus f. aptycha.
11 (10). Кроме париетального, имеются выраженные в разной степени колу-
меллярный и палатальный зубы ..............................................
. . . ......................... 2ж. Р. (Р.) albiplicatus f. retrodens.
12 (5). Париетальный зуб отсутствует. Если имеются радиальные ребра, то они
сглаженные, нерезкие.
13 (14). Раковина удлиненная, веретеновидная, с широко отвернутыми устье-
выми краями, на столбике имеется косая складка.............................
..............................................5. Р. (Р.) otostomus.
14 (13). Совокупность признаков иная.
15 (32). Раковина слабо скульптурированная, одноцветная, роговая, кашта-
новая или коричневая, но не белая и не желтоватая.
16 (23). Последний оборот сильно вздут и явно преобладает над остальными;
раковина короткоконическая, веретеновидно-коническая или удлиненно-
овальная.
17 (18). Верхняя часть раковины сужена, на колумеллярной крае никаких
складок нет.....................................14. Р. (A.) submucronatus.
18 (17). Верхняя часть раковины не сужена, на колумеллярном крае чаще
всего имеется выраженная в разной степени складка.
19 (22). Края устья усилены очень мощной губой.
20 (21). Раковина овально-коническая, большой диаметр не менее 5.5 мм . .
.................................................... 1. Р. (Р.) regelianus.
21 (20). Раковина веретеновидно-коническая, большой диаметр не превы-
шает 5 мм ..............................................9. Р. (Р.) galinae.
22 (19). Губа развита умеренно или отсутствует ...........................
................................... 2д. Р. (Р.) albiplicatus f. asiatica.
23 (16). Последний оборот не вздут; раковина удлиненно-овальная, цилиндри-
ческая, башневидная или булавовидная.
24 (25). Устье округло треугольное . ..............10. Р. (Р.) trigonochilus.
25 (24). Устье округлое или овальное.
26 (27). Позади палатального края устья имеется желтоватое пятно . . . .
.................................... 2г. Р. (Р.) albiplicatus f. subobscura.
27 (26). Желтого пятна на затылке нет.
28 (29). Раковина башневидная......................8. Р. (Р.) schnitnikovi.
29 (28). Раковина удлиненно-овальная или овально-цилиндрическая.
•30 (31). Внутри пениса залегают две мощные продольные складки . . . .
...................................................3. Р. (Р.) miser.
81 (30). Внутри пениса залегают 2—3 кольцеобразные складки ......
...........................................12. Р. (Р.) chodschendicus.
32 (15). Раковина не одноцветная; если одноцветная, то белая или желтоватая.
Поверхность гладкая или ребристая.
33 (38). Раковина стройная — цилиндрическая или башневидная.
34 (35). На столбике имеется нерезкая косая складка; раковина башневидная
....................................................4. Р. (Р.) kasnakowi.
35 (34). Косой складки на столбике нет; раковина цилиндрическая.
36 (37). Раковина белая, с темными штрихами, морщинистая или тонкоребри-
стая ..................................2а. Р. (Р.) albiplicatus f. albiplicata.
37 (36). Раковина одноцветно-белая или желтоватая, очень гладкая (как поли-
рованная) ..............................................7. Р. (Р.) stabilis.
38 (33). Раковина вздутая — овально-цилиндрическая или овально-коническая.
246
VI. НАДСЕМ. BULIMINOIDEA
39 (40). Раковина почти гладкая, овально-цилиндрическая ....................
................................... 26. Р. (Р.) albiplicatus f. sogdiana.
40 (39). Раковина с сильно сглаженными ребрами, овально-коническая . .
.....................................................6. Р. (Р.) diplus.
1. Подрод PSEUDONAPAEUS s. str.
Раковина право- или левозавитая, в разной степени скульптурирована,
одноцветная или с радиальными пестринами. Зубы в устье развиты или отсут-
ствуют, но продольной палатальной складки нет.
Папилла пениса отсутствует, внутри пениса имеется пара или, реже, одна
V-образная складка; изредка рельеф внутренних стенок пениса составлен
множеством бессистемно расположенных складок.
Распространение как у рода.
1. Pseudonapaeus (Pseudonapaeus) regelianus (Ancey, 1886) (рис. 156, 157).
Ancey, 1886a : 26 (Bulimlnus asiaticus var. Regeliana)', Лихарев, Раммель-
мейер, 1952 : 203 (Pseudonapaeus asiaticus part.).
Terra typica — окр. Алма-Аты («Wjernoje»).
Место хранения типов не установлено.
Материал: 7 экз. из окр. Алма-Аты и из Джунгарского хр.
Раковина овально-коническая, умеренно твердостенная, слегка блестящая,
составлена 6—7 умеренно выпуклыми оборотами. Последний, оборот непосред-
ственно возле устья несильно и плавно, но явственно
приподнят, высота его приблизительно равна половине
высоты раковины или немного больше. Окраска почти
одноцветная, серовато-роговая или роговая с легким
желтоватым оттенком; последний оборот за устьем белый
или светло-серый. Скульптура — тонкая неравномерная
радиальная морщинистость; возле устья, особенно на
базальной поверхности раковины, иногда заметны спи-
ральные бороздки. Устье слабо скошено, округлое, края
его соединены слабой мозолью. Колумеллярный и базаль-
ный края отвернуты сильно, палатальный слабо, но все они
Рис. 156. Pseudonapaeus (Pseudonapaeus) regelianus (Ancey) из Джун-
гарского хр.. Саркандский р-н.
утолщены за счет губы, лежащей вплотную к' краю устья. Если раковину по-
вернуть по часовой стрелке примерно на 45° против ее нормального положения,
то видно, что колумеллярный край сильно расширен и уплощен и задняя его
кромка усилена расплывшейся валикообразной складкой. Эта складка видна
и при нормальном положении устья в виде вялого утолщения колумеллярного
края. Пупок узкий, щелевидный.
Размеры: ВР 11—14, БД 5.5—7 мм; в первоописании: ВР 13, БД 6г/3 мм.
Внутреннее строение. Материал: 3 экз. из Джунгарского хр.
Саркандский р-н, июнь 1970 г., сбор К. К. Увалиевой, определение автора.
Белковая железа с нерезкой апикальной вырезкой. Нижний отдел вагины
в несколько раз короче верхнего. Семяпровод впадает в эпифаллус эксцен-
трично и чуть отступя от вершины последнего. Хорошо развитый цэкум рас-
полагается примерно на середине эпифаллуса или чуть выше. Эпифаллус рав-
номерно цилиндрический, отделен от пениса ясной кольцевой перетяжкой.
Внутренние стенки пениса покрыты мощными складками: в дистальной части
они тонкие и продольные, в проксимальном отделе утолщаются, сминаются и
1. РОД PSEUDONAPAEUS
247
превращаются в систему четких, по-разному ориентированных коротких
валиков и бугорков, которые в интактном пенисе почти полностью заполняют
его просвет близ отверстия эпифаллуса. Все отделы пениального аппендикса
хорошо заметны; сильно развитый А3 плавно, без резкой границы переходит
в нитевидный Л4. Проток семяприемника длинный, у исследованных экземпля-
ров образует 2 резких изгиба в нижней части. Проток резервуара очень корот-
кий, дивертикул развит необыкновенно сильно, верхний конец его лежит на
поверхности белковой, железы, достигая ее середины.
Пос
Рис. 157. Pseudonapaeus {Pseudonapaeus) regelianus (Ancey) из Джунгарского хр., Сарканд-
ский р-н, VI 1970 г.
I — половой аппарат; II — внутреннее строение пениса.
Распространение и местообитание. Джунгарский и
Заилийский хр. и предгорья. Обитает в осыпях и зарослях кустарников.
2. Pseudonapaeus (Pseudonapaeus) albiplicatus (Е. Martens, 1874) (рис. 158—
164).
Мартенс, 1874 : 20, табл. 2, рис. 15 (Buliminus).
Чрезвычайно изменчивый политипический вид, представляющий собою
обширный комплекс конхологических форм, различающихся порой исклю-
чительно резко. Обоснование излагаемой здесь таксономической структуры
приведено в общей части. Ниже дается описание анатомических особенностей
вида и характеристика изменчивости черт внутреннего строения, затем —
описание основных форм изменчивости признаков раковины и синонимика.
Внутреннее строение. Материал.:
f. albiplicata,'. 9 экз. из окрестностей оз. Сары-Чилек, 22 сентября 1972 г.;
2 — из Чаткальского хр., 20 сентября 1972 г., сборы В. В. Горохова; 3 — из
Джунгарского хр., ущелье Талды-Булак, июнь 1970 г., сбор К. К. Увалиевой;
248
VI. НАДСЕМ. BULIMINOIDEA
5 — из Кетмень-Тюбинской долины, 12 июня 1973 г., сбор Р. Я. Бутылина;
14 — из долины р. Ала-бука-сай на Чаткальском хр., 20 мая 1972 г.; 2 — из-
ущелья р. Аламедин на Киргизском хр., 14 июня 1972 г.; 11 — из прирусло-
вого вала р. Пскем, 17 мая 1972 г.; 72 — из прируслового вала правого берега
р. Нарын ниже Токтогула, 1 июня 1972 г.; сборы и определения автора;
f. sogdiana: 4 экз. из Душанбе, 4 апреля 1968 г.; 5 экз. из с. Зидды, Гиссар-
ский хр., 31 июля 1968 г., сбор и определение 3. Иззатуллаева; 6 — из Зерав-
шанского хр., окр. оз. Маргузор, 8 августа 1973 г., сбор Ю. А. Мешкова; 7 —
из Большого Балхана, 9 мая 1972 г.; 4 — из долины р. Касансай, 29 мая 1972 г.;
сборы и определения автора;
Рис. 158. Основные формы Pseudonapaeus (Pseudonapaeus) albiplicatus (Mart.).
I — f. albiplicata (Mart.) из Ташкента; II — f. sogdiana (Mart.) из Душанбе; Ill — f. secalina (Mart.) из
ущелья p. Аламедин, Киргизский хр.; IV — f. subobscura (Ancey) из долины p. Чу; V — f. asiatica (Mart.)
из Фрунзе; VI — t. aptycha (Ancey) из Узун-агача близ Алма-Аты; VII — f. retrodens (Mart.) из Ферган-
ского хр., урочище Ак-Терек; VIII — f. dissimilis (Mart.) из Арасан-булака, Алма-Атинская обл.
f. secalina: 4 экз. из Джамбульской обл., Курдайский р-н, июнь 1969 г.;:
5 — из Таласского хр., заповедник Аксу-Джабаглы, 1 июня 1969 г.; сборы
К. К. Увалиевой; 3 — из окр. оз. Иссык близ Алма-Аты, 5—16 мая 1963 г.,
сбор А. Гусева; 18 — из Киргизского хр., ущелье р. Аламедин, 14 июня 1972 г.;
7 — из Киргизского хр., ущелье над с. Луговое, 6 июня 1972 г.; 7 —. из Кир-
гизского хр., ущелье Карабалты, 11 июня 1972 г.; 6 — из Таласского хр.^
ущелье Беш-Таш, 4 июня 1972 г.; сборы и определения автора;
f. asiatica: 2 экз. из Гиссарского хр., перевал Анзоб, 30 июня 1956 г., сбор
К. Я. Грунина; 12 — из Киргизского хр., ущелье р. Аламедин, 14 июня 1972 г.;
5 — из Киргизского хр., ущелье Карабалты, 11 июня 1972 г.; сборы и опре-
деления автора;
f. aptycha: 2 экз. из окр. Алма-Аты, 1907 г., сбор Шнитникова, определение
автора;
f. subobscura: 7 экз. из южного берега оз. Иссык-куль, 12 июня 1970 г.,
сбор Л. В. Славошевской; 2 — из ущелья Чим-Тургень, 12 октября 1976 г.;
сбор И. М. Лихарева; определения автора;
1. РОД PSEUDONAPAEUS
249
f. retrodens: 10 экз. из предгорий Ферганского хр., урочище Ак-Терек,
14 июля 1937 г., сбор А. Н. Кириченко, определение автора.
Белковая железа с вырезкой, занимающей апикальное положение или вы-
раженной также и на латеральной поверхности органа. Семяпровод впадает
эксцентрично и обычно слегка сдвинут вниз по эпифаллусу, в результате чего
возникает очень короткий широко закругленный бич; впрочем, чаще можно
говорить лишь о том, что закруглен задний конец эпифаллуса, и семяпровод
впадает субапикально. Положение хорошо развитого цэкума варьирует, но
в общем он располагается близ середины эпифаллуса и никогда не бывает резко
сдвинутым ни к верхнему, ни к нижнему концам эпифаллуса. На границе
между эпифаллусом и пенисом имеется легкий «перелом», а проксимальный
конец пениса в разной степени утолщен. Пенис изредка почти цилиндрический,
но чаще с широкой перетяжкой посредине. Внутреннее строение пениса доста-
точно разнообразно, но похоже, что это разнообразие того же свойства, что и
разнообразие раковин, т. е. определенные детали строения совокупительного
аппарата свойственны отдельным популяциям или группам популяций, но
в пределах этих совокупностей сохраняют относительное постоянство. Суще-
ственно, что имеет место преемственность между разными вариантами строения
пениса; образно говоря, наблюдаемое разнообразие — вариации на одну тему.
Так, серия продольных складок, проходящих снизу доверху, увенчанная
коротким придатком возле отверстия эпифаллуса обнаружена внутри пениса
у f. aptycha из окр. Алма-Аты (рис. 163), f. sogdiana из Большого Балхана (фор-
мально определяется как oxiana), f. secalina из окр. оз. Иссык близ Алма-Аты
(рис. 161) и f. asiatica из ущелья Карабалты и на Киргизском хр. (рис. 162).
Упорядочение этой серии в две V-образные складки, внутренние ветви которых
уходят в просвет эпифаллуса, а наружные смыкаются над отверстием этого
протока, наблюдается у f. secalina из ущелья р. Аламедин на Киргизском хр.,
из окр. с. Луговое на том же хребте; у f. albiplicata из окр. оз. Маргузор на
Зеравшанском хр., из прибрежной зоны р. Нарын и из ущелья р. Пскем
{рис. 159), а также у f. asiatica из ущелья Беш-Таш на Таласском хр. и из не-
скольких мест Киргизского хр. У f. retrodens картина примерно та же, но между
внутренними ветвями V-образных складок имеется небольшой бугорок (рис. 164).
Наличие нескольких продольных гофрированных складок, неясно группирую-
щихся в подобие пары V-образных складок, обнаружено у f. sogdiana из
с. Зидды на Гиссарском хр. и из Душанбе (рис. 160), а также у f. subobscura
из южного берега оз. Иссык-куль. В остальном исследованном материале наблю-
даются многочисленные промежуточные варианты. Ветви полового ретрактора
в разной степени сближены на диафрагме, чаще всего располагаются вплотную
-одна к другой; одна ветвь крепится к выше его середины, часто непосред-
ственно под А2, а вторая — к середине пениса, немного выше или немного
ниже нее. Большой изменчивостью отмечена длина протока семяприемника —
от прямого, вытянутого вдоль вагины (например, f. secalina из ущелья р. Ала-
медин), до многократно извитого (например, f. asiatica из перевала Анзоб).
Резервуар семяприемника всегда имеет ясно выраженный собственный корот-
кий проток, длина которого равна или немного меньше длины резервуара.
Дивертикул семяприемника всегда хорошо развит, верхний конец его либо
немного не доходит до белковой железы, либо слегка касается ее нижнего края.
Распространение. Вид широко распространен по горным и пред-
горным районам Средней Азии. Поскольку конхологические формы представ-
ляют собой, по существу, жизненные формы, свойственные определенным ти-
пам биотопов, то характер местообитаний указывается после описания каждой
из форм.
2а. Pseudonapaeus (Pseudonapaeus) albiplicatus f. albiplicata (E. Martens, 1874)
(рис. 158, Z; рис. 159).
Мартенс, 1874 : 20, табл. 2, рис. 15 (Buliminus)', Ancey, 1886a : 30 (Buliminus)',
Ancey, 1886b : 329 (B. przevalskii): Westerland, 1887 : 67 (Buliminus)', К о b e 1 t,
250
VI. НАДСЕМ. BULIMINOIDEA
1890, Iconogr., N. F., 4 (B. ferghanensis); Ancey, 1893 : 36 (B. annenkovi); West erlu nd,
1896 : 191 [5. albiplicatus var. narynensis, B. (Pseudopetraeus) biformis], 193 (B. errans); R o-
s e n, 1897 : 170 (B. retteri); Wester lun d, 1898 : 163 (B. albiplicatus f. implicate.); К o-
b e 1 t, 1902 : 507, t. 82, figs. 4, 5 [5. (Subzebrinus)]; Lindholm, 1927a : 101 (Subzebri-
nus jlammulatus); Lindholm, 1927b : 258 (?Subzebrinus leucopleurus); Лихарев,
Раммельмейер, 1952 : 199, рис. 110 [(Subzebrinus (Subzebrinus)]; Матёкин, 1966 :
114 [Jaminia (Chondrulopsina) potaniniana subsp. albiplicata'. Шилейко, 19786 : 353,
рис. 4, I.
Terra typica — Ташкент и Шахимардан.
Лектотип происходит из Ташкента, хранится в Зоологическом музее
Берлинского ун-та им. Гумбольдта, № 22399а (Kilias, 1971).
Материал: несколько сотен экз. из 43 местонахождений на Тянь-Шане,.
Памиро-Алае и в Джунгарии.
Раковина от коническо-цилиндрической до цилиндрической и почти башне-
видной, толстостенная, часто блестящая. Оборотов 7—8, слабовыпуклых или
почти плоских, последние
2—4 оборота почти одина-
ковой ширины, последний
оборот к устью очень слабо
поднят, высота его нес-
колько меньше половины
высоты раковины. Окраска
состоит из темного фона
(чаще всего коричневого) и
светлых радиальных пест-
рин, развитых в различ-
ной степени; изредка эти
пестрины развиты так силь-
но, что можно скорее го-
ворить о белой раковине
с темными пестринами.
Скульптура только ра-
Рис. 159. Pseudonapaeus
(Pseudonapaeus) albiplicatus
f. albiplicata (Mart.) из ущелья
p. Пскем, 20 V 1972 г. Поло-
вой аппарат и вскрытый пе-
нис (из: Шилейко, 19786).
диальная, от тонкой исчерченности до грубой морщинистости, иногда даже
почти ребристости. Если скульптура развита хорошо, то положительные эле-
менты рельефа белые, а промежутки между ними темные. Устье слабо ско-
шено, усеченно-овальное, края его снабжены широкой губой и умеренно от-
вернуты. Изредка в глубине устья на столбике может присутствовать бугор-
ковидный зуб.
Размеры: ВР 8—15, БД 3—5 мм; в первоописании: ВР 11.5—12, БД 5
(Ташкент) и ВР 8, БД 3 мм (Шахимардан). Килиас (Kilias, 1971) указывает
следующие размеры лектотипа: ВР 11.4, БД 4.6 мм.
Местообитание. Населяет горные и предгорные открытые участки
с мезофильными или даже гигрофильными условиями; осыпей избегает, пред-
почитает участки с рыхлой почвой. Встречается также на склонах, поросших
кустарником.
26. Pseudonapaeus (Pseudonapaeus) albiplicatus f. sogdiana (E. Martens, 1874)
(рис. 158, IT, рис. 160).
Мартенс, 1874 : 19, табл. 2, рис. 14а—с (Buliminus); Е. М а г t е n s, 1876а : 335,
pl. 12, fig. 8 (В. oxianus, var. elatior et var. brevior); 1882a : 21 (B. labiellus var. kokan-
1. РОД PSEUDONAPAEUS
251
densis); 0. Boettger, 1889b : 955, t. 26, fig. 1 (B. walteri); Ancey, 1893 : 38 (B. turanicus,
nom. nov. pro B. komarovt Kobelt, non Boettger, 1880); 1902 : 47 (B. larvatus); Линдгольм,
1922a : 273 [/?. (Subzebrinus) korshinskii]; Лихарев, Раммельмейер, 1952 : 197,
рис. 108 [Subzebrinus (Subzebrinus) sogdianus]; Матёкин, 1966:131 [Jaminia (Chondru-
lopsina) potaniniana subsp. potaniniana]; Шилейко, 19786 : 355, рис. 5, I.
Terra typica — верховья долины Зеравшана, Кули-Калан.
Один синтип хранится в Зоологическом музее Берлинского ун-та им. Гум-
больдта, № 22406 (Kilias, 1971).
Материал: несколько сотен экз. из 29 местонахождений на южном
Тянь-Шане, Памиро-Алае, Копетдаге и на Большом Балхане.
Рис. 160. Pseudonapaeus (Pseudonapaeus) albiplicatus f. sogdiana (Mart.) из Душанбе,
28 IV 1968 г. Половой аппарат и вскрытый пенис (из: Шилейко, 19786).
Раковина от овальной до овально-цилиндрической или удлиненно-овальной,
блестящая или умеренно блестящая. Оборотов 6.5—8, слабовыпуклых, послед-
ний оборот к устью слегка приподнят, высота его сильно варьирует в зависи-
мости от высоты завитка, но в среднем составляет около половины высоты ра-
ковины. Окраска от светло-роговой до чисто белой, однако оба этих крайних
варианта встречаются сравнительно редко; чаще всего на белом фоне имеются
многочисленные радиальные пестрины, иногда носящие характер правильных
радиальных полос различной ширины. Скульптура в виде нерезких и нерегу-
лярно расположенных радиальных морщин. Устье слабо скошено, места его
прикрепления слабо или умеренно сближены, края отвернуты и усилены белой
губой.
Размеры: ВР 11—24, БД 6—9 мм; в первоописании: ВР 11.5, БД 6 мм; од-
нако Килиас (Kilias, 1971) указывает следующие размеры для синтипа, изо-
брженного Мартенсом на рис. 14а: ВР 10, 2, БД 5.3 мм.
Местообитание. Населяет крупнообломочные осыпи, заросли ку-
старников; нередко на степных участках предгорий среди травянистой расти-
тельности.
252
VI. НАДСЕМ. BULIMINOIDEA
2в. Pseudonapaeus (Pseudonapaeus) albiplicatus f. secalina (E. Martens, 1881)>
(рис. 158, JU; рис. 161).
E. Martens, 1881 : 27, t. 6, figs. 8, 9 (Buliminus secalinus); 1882b : 106 (B. entodon)*
Ancey, 1886b : 337 (B. entodon var. dextroversa); 1893 : 36 (B. semenovi); Rosen, 1893b :
179 [5. (Subzebrinus) varenzovi]; Скворцов, 1940:368 (Ena secalina var. maibulaki)*
Лихарев, Раммельмейер, 1952 : 198, рис. 109 [Subzebrinus (Subzebrinus)], 191,
рис. 101 [Sewertzovia (Parachondrula) entodon]; M а т ё к и н, 1966 : 131 [Jaminia (Chondrulop-
sina) potaniniana subsp. albiplicata part.].
Terra typica — Кульджа (Западный Китай).
Место хранения типов не установлено.
Материал: несколько сотен экз. из 19 местонахождений на северной
и северо-восточном Тянь-Шане и в Джунгарии.
Раковина от башневидной до коническо-цилиндрической и почти правильно
цилиндрической, в разной степени блестящая. Оборотов 7—9, умеренно вы-
пуклых или уплощенных, пос-
ледний оборот к устью почти
не поднят, высота его меньше-
половины высоты раковины,
часто не более трети этой ве-
личины. Окраска от красно-
вато-роговой до белой с тем-
ными радиальными пестри-
нами. Скульптура в виде-
тонкой неравномерной ра-
диальной исчерченности, ко-
торая никогда не переходит
в ребристость. Устье слабо
скошено, места его прикре-
пления не сближены или
сближены незначительно,,
края устья слабо отвернуты
и соединены париетальной
мозолью, на которой в ангу-
Рис. 161. Pseudonapaeus (Pseudo-
napaeus) albiplicatus f. secalina-
(Mart.) из окр. оз. Иссык близ.
Алма-Аты, 5—16 V 1963 г. По-
ловой аппарат и вскрытый пенис _
лярном районе может располагаться небольшой бугорок. Кроме того, еще-
один зуб может присутствовать на париетальной стенке. В пределах некоторых
популяций париетальный зуб может быть развит в разной степени — от кро-
шечного, обнаружить который можно лишь, если провести концом тонкой
иглы по париетальной стенке, до хорошо заметного, клиновидного. Нередко»
оба названных зуба отсутствуют вовсе. Пупок в виде широкой щели.
Размеры: ВР 6—14, БД 3—5 мм; в первоописании: ВР 9, БД 3.5 мм.
Местообитание. Населяет преимущественно открытые склоны, где-
держится в дерновинках трав, в тонких мелкообломочных осыпях и под кам-
нями. Нередко образует огромные скопления под каменными плитами.
2г. Pseudonapaeus (Pseudonapaeus) albiplicatus f. subobscura (Ancey, 1886)>
(рис. 158, IV).
Ancey, 1886b : 330 (Buliminus subobscurus); Лихарев, Раммельмейер,.
1952 : 204 (Pseudonapaeus subobscurus); Матёкин, 1966 : 131 [Jaminia (Chondrulopsina}
potaniniana subsp. asiatica part.].
1. РОД PSEUDONAPAEUS
253
Terra typica — Дарати-булак в долине р. Текес.
Место хранения типов не установлено.
Материал: более 40 экз. из 5 местонахождений на северном Тянь-
Шане и в Джунгарии.
Раковина от удлиненно-овальной до почти цилиндрической и овально-
конической, матовая или слегка блестящая, умеренно твердостенная. Оборотов
7—8, слабовыпуклых, разделенных светлым швом. Последний оборот к устыо
почти не поднят, высота его обычно меньше половины высоты раковины. Ок-
раска бурая, темно-роговая или темно-серая, за устьем обычно имеется желто-
Рие. 162. Pseudonapaeus (Pseudonapaeus) albiplicatus f. asiatica (Mart.) из ущелья Карабалты
на Киргизском хр., 11 VI 1972 г.
I — половой аппарат и вскрытый пенис, 11 — вскрытый пенис другого экземпляра.
ватоё пятно. Скульптура в виде слабой радиальной исчерченности. Устье оваль-
ное, очень слабо скошено, края его незначительно отвернуты, со светлой губой,
развитой иногда очень интенсивно. Пупок в виде овальной щели.
Размеры: ВР 9—12, БД 4—5 мм; в первоописании: ВР 9.5, БД 4.3 мм.
Местообитание. Форма встречается относительно нечасто на щеб-
нистых слабо залесенных склонах, населяет малоподвижные осыпи и прикор-
невые участки под кустарником. Придерживается участков с рыхлой почвой.
2д. Pseudonapaeus (Pseudonapaeus) albiplicatus f. asiatica (E. Martens, 1881)
(рис. 158, V; рис. 162).
E. M a r t e n s, 1881 : 29, t. 6, fig. 12—14 (Buliminus asiaticus); 1882a : 22, t. 3, fig.^5
(B. sogdianus var. Kuldshanus), 25, t. 3, fig. 6, 7 (Bullminus); Ancey, 1886a : 23 (B. herzen-
steini), 24 (B. herzensteini var. pellucens), 28 (B. Vamberyi); 1886b : 333 (B. asiaticus var. cy-
lindroconus); Westerlund, 1887 : 68 (Buliminus); Kobelt, 1902 : 505, t. 82, fig. 1, 2
[B. (Pseudopetraeus) asiaticus]; Лихарев, Раммельмейер, 1952 : 203, рис. 116
(Pseudonapaeus asiaticus); Матёкин, 1966:114 [Jaminia (Chondrulopsina) potaniniana
subsp. asiatica]; Шилейко, 19786 : 357, рис. 5, II.
Terra typica — Кульджа (Западный Китай).
M e сто хранения типов не установлено.
254
VI. НАДСЕМ. BULIMINOIDEA
Материал: свыше 100 экз. из 18 местонахождений в Средней Азии.
Раковина с трудом отличима от раковины Р. regelianus. Отличия заключа-
ются главным образом в том, что у данной формы отсутствует валикообразная
складка на колумеллярной крае, а сам устьевой край не так сильно расширен.
Размеры: ВР 7—15, БД 4—6 мм; в первоописании: ВР 8—15, БД 4—5.5 мм.
Местообитание. Щебнистые склоны с кустарником либо редким
лесом; иногда (особенно мелкие формы) — на открытых склонах, под камнями.
Рис. 163. Pseudonapaeus (Pseudonapaeus) albiplicatus f. aptycha (Ancey) из окр. Алма-Аты,
VII (?) 1907 г. Половой аппарат и вскрытый пенис.
2е. Pseudonapaeus (Pseudonapaeus) albiplicatus f. aptycha (Ancey, 1886)
(рис. 158, VP, рис. 163).
Ancey, 1886b : 334 (Buliminus aptychus et var. capusiana)', Westerland, 1887 : 68
(Buliminus); К о b e 1 t, 1902 : 958 [Buliminus (Chqndrulopsis)]; Лихарев, Раммель-
мейер, 1952 : 191 [Sewertzowia (P arachondrula) aptycha]; Матёкин, 1966 : 130 [Jaminia
(Chondrulopsina) potaniniana subsp. retrodens part.].
Terra typica — Алма-Ата.
Место хранения типов не установлено.
Материал: 17 экз. из двух местонахождений в окр. Алма-Аты.
Раковина короткоцилиндрическая или овально-цилиндрическая, твердо-
стенная, слабо блестящая. Оборотов 6—7, умеренно или слабо выпуклых, раз-
деленных узким довольно глубоким швом. Последний оборот к устью плавно
и несильно, но вполне заметно поднят, высота его приблизительно равна поло-
вине высоты раковины. Окраска одноцветная, темно-роговая или серовато-
роговая, или со светлыми слабо выраженными радиальными пестринами. Скуль-
птура в виде тонкой неравномерной радиальной исчерченности или вялой мор-
щинистости. Устье слабо скошено, овальное или округлое, с притупленными
краями, слегка отвернутыми и снабженными светлой расплывшейся губой.
На париетальной стенке в глубине устья расположен развитый в различной
степени зуб. Пупок в виде широкой щели.
Размеры: ВР 9—11, БД 5 мм; в первоописании: ВР 10, БД 5 мм.
1. РОД PSEUDONAPAEUS
255
Местообитание. Форма встречена только на Заилийском хр., где
широко распространена в предгорьях и населяет большей частью сухие склоны
со степной растительностью.
2ж. Pseudonapaeus (Pseudonapaeus) albiplicatus f. retrodens (E .Martens, 1879)
(рис. 158, VII; рис. 164).
E. Martens, 1879, Sitzungsb. Ges. naturf. Fr. Berlin : 126 {Buliminus retrodens)-,
Westerlund, 1887 : 40 {B. retrodens)- 1894, Nachrbl. dtsch. malak. Ges., 26, 9/10 : 169
[5. {Chondrulopsis) retroplicatus]; 1896 : [B. {Chondrulopsis) drymaeus]; К о b e 1 1, 1902 : 514,
t. 82, fig. 19, 20 [5. {Chondrulopsis) retrodens], 705, t. 105, fig. 9—11 [5. {Chondrulopsis) dry-
maeus]; Лихарев, Раммельмейер, 1952 : 190, рис. 100 [Sewertzowia {Parachond-
rula) retrodens, S. {P.) drymaeus]; Матёкин, 1966 : 114 [Jaminia {Chondrulopsina) potani-
niana subsp. retrodens]; Шилейко, 19786 : 352, рис. 3, III.
Рис. 164. Pseudonapaeus {Pseudonapaeus) albiplicatus f. retrodens (Mart.) из Ферганского хр.,
урочище Ак-Терек, 14 VII 1937 г. Половой аппарат и вскрытый пенис.
Terra typica — Кульджа (Западный Китай).
Лектотип хранится в Зоологическом музее Берлинского ун-та им. Гум-
больдта, № 34878а (Kilias, 1971).
Материал: около 70 экз. из 12 местонахождений на Тянь-Шане и
в Джунгарии.
Раковина удлиненно-овальная или овально-цилиндрическая, блестящая или
матовая, твердостенная или умеренно твердостенная, в разной степени просве-
чивающая, иногда сильно просвечивающая. Оборотов 7—8, слабовыпуклых,
плавно нарастающих. Последний оборот к устью слабо поднят, высота его
приблизительно равна половине высоты раковины. Окраска светло-роговая,
коричневая, красноватая или каштановая, чаще всего однотонная, но нередко
видны светлые узкие радиальные штрихи (у скульптурированных раковин),
последний оборот за устьем светлый. Скульптура — от тончайшей шелкови-
стой исчерченности, когда раковина выглядит полированной, до морщини-
стости и до ребристости, причем в последнем случае ребра более светлые, чем
256
VI. НАДСЕМ. BULIMINOIDEA
промежутки между ними. Устье овальное, в базальной части сужено, края его
слабо отвернуты. В типичном случае имеются три хорошо развитых зуба:
париетальный, колумеллярный в виде косой складки, уходящей в глубь ра-
ковины, и палатальный, представляющий собой расширенный участок губы.
При ослаблении устьевой арматуры в первую очередь исчезает палатальный
зуб, затем колумеллярный — и такие формы уже по существу есть f. aptycha.
Пупок в виде широкой или узкой щели.
Размеры: ВР 9—13, БД 3.5—5 мм; в первоописании: ВР 8—12, БД 4—5 мм.
Лектотип имеет следующие размеры: ВР 10, БД 4.3 мм (Kilias, 1971).
Местообитание. Кустарниковые заросли и лиственные леса, реже
на открытых степных участках и в осыпях.
2з. Pseudonapaeus (Pseudonapaeus) albiplicatus f. dissimilis (E. Martens, 1882).
(Рис. 158, VIII).
E. M ar t en s, 1882b : 106 (Buliminus dissimilis); A n c e y, 1886b : 336 (B. leucoptychus);
Westerlund, 1887 : 70 (B. dissimilis); С 1 e s s i n, 1894 : 64 [2?. (Chondrulopsis) costatus];
Kobelt, 1902 : 506, t. 82, fig. 3a, b [5. (Severtzovia) dissimilis]; Weber, 1913 : 16 (B. merz-
bacheri); Лихарев, Раммельмейер, 1952 : 189, рис. 99 (Sewertzowia (Sewertzowia)
dissimilis], 201, рис. 113 [Subzebrinus (Subzebrinus) costatus]; M атёкин, 1966 : 130 [Jaminia
{Chondrulopsina) potaniniana subsp. albiplicatus part.].
Terra typica — Арасан-булак (Алма-Атинская обл.).
Лектотип хранится в Зоологическом музее Берлинского ун-та им. Гум-
больдта, № 101575а (Kilias, 1971).
Материал: более 80 экз. из трех местонахождений в юго-восточном
Казахстане.
Раковина от почти кубаревидной до овальной и цилиндрической, относи-
тельно очень твердостенная. Оборотов 6—8, выпуклых, разделенных глубоким
неровным швом. Последний оборот заметно или резко поднят к устью, высота
его, в зависимости от формы раковины, может быть больше половины высоты
раковины и составлять почти треть этой величины. Окраска от одноцветно-
белой до красновато-роговой, причем в последнем случае положительные эле-
менты рельефа раковины все равно остаются белыми. Скульптура в типичном
случае в виде резких тупых массивных ребер, однако провести резкую грань
по этому признаку между данной формой и номинативной невозможно, поскольку
у последней также нередко имеются развитые ребра. Устье отвесное, с отвер-
нутыми светлыми краями, без зубов или с париетальным зубом, который, осо-
бенно у коротких раковин, бывает хорошо развитым. Пупок воронковидный:
в начале развернутый, но затем становится проколовидным, так что сквозь
него видно не более половины предпоследнего оборота.
Размеры: ВР 5.5—10, БД 3.5—4.5 мм; в первоописании: ВР 5.5—8, БД 4 мм.
Килиас (Kilias, 1971) указывают следующие размеры лектотипа: ВР 7.3, БД 4.1 мм.
Местообитание. Форма встречается только в юго-восточном Казахстане, где
населяет сухие ущелья и степные участки с кустарником и многочисленными
осыпями; нередко на отвесных скалах.
3. Pseudonapaeus (Pseudonapaeus) miser (Е. Martens, 1874) (рис. 165, 166).
Мартенс, 1874 : 21, табл. 2, рис. 17 (Buliminus)', Westerlund, 1896 : 194 (В. mi-
ser var. misellus); 1897 : 71 (Buliminus); Kobelt, 1902 : 522, t. 83, fig. 12 (Buliminus);
Лихарев, Раммельмейер, 1952 : 204, рис. 117; M а т ё к и н, 1966 : 130 [Jaminia
{Chondrulopsina) potaniniana subsp. asiatica part.].
Terra typica — Ура-Тюбе (предгорья Туркестанского хр.). '
Место хранения типов не установлено.
Материал: более 70 экз. из 13 местонахождений на Тянь-Шане.
Раковина от удлиненно-овальной до овально-цилиндрической и почти башне-
видной, с притупленной вершиной, гладкая, слабо блестящая или матовая.
Оборотов 5—6, медленно нарастающих, умеренно или слабо выпуклых. Послед-
1. РОД PSEUDONAPAEUS
257
ний оборот к устью почти не поднят, высота его чаще всего немного больше
половины высоты раковины, но встречаются почти башневидные раковины,
у которых высота последнего оборота лишь немного более трети высоты рако-
вины. Окраска одноцветная, серая, светло-роговая или светло-коричневая.
Скульптура в виде редких неравномерно расположенных сильно сглаженных
радиальных морщинок, но в целом раковина по первому впечатлению почти
гладкая. Устье овальное, слабо скошено, края его слегка отвернуты, с тонкой
светлой губой; колумеллярный край почти отвесный, палатальный и базальный
плавно и равномерно выгнуты. Пупок в виде узкой щели, слегка прикрыт отво-
ротом колумеллярного края.
Рис. 165. Pseudonapaeus Рис. 166. Pseudonapaeus (Pseudonapaeus) miser (Mart.) из ущелья
(Pseudonapaeus) miser p. Пскем, 17 V 1972 г. Половой аппарат и вскрытый пенис.
(Mart.) с перевала Ан-
зоб^на Гиссарском хр.
Размеры: ВР 5—11, БД 2.2—5 мм; в первоописании: ВР 10, БД 5 мм.
Внутреннее строение. Материал: 3 экз. из Чаткальского хр.,
перевал Орток-бель, 20 мая 1973 г., сбор В. В. Горохова; 8 — из Зеравшан-
ского хр., окр. оз. Маргузор, 8 августа 1973 г., сбор Ю. А. Мешкова; 17 —
из долины р. Пскем на западном Тянь-Шане, 17 мая 1972 г., сбор и определения
автора.
Белковая железа с нерезкой апикальной вырезкой. Нижний отдел вагины
примерно вдвое короче верхнего. Семяпровод впадает эксцентрично, но эпи-
фаллус бывает как закругленным на конце, так и слегка приостренным. Эпи-
фаллус недлинный, прямой, цилиндрический. Хорошо развитый цэкум рас-
положен заметно выше середины эпифаллуса. Пенис цилиндрический или ди-
стальная часть его цилиндрическая, а проксимальная в разной степени рас-
ширена, овальная. Внутри пениса у особей из долины р. Пскем залегают две
широкие продольные складки, почти не связанные между собой, а у особей из
Чаткальского и Зеравшанского хребтов наблюдались обычные для видов рода
две V-образные складки; у одного экземпляра из Чаткальского хр., однако
одна из складок оказалась редуцированной до одиночного гребневидного ва-
лика, входящего в промежуток между ветвями противолежащей складки.
17 А, А. Шилейко
258
VI. НАДСЕМ. BULIMINOIDEA
Все отделы пениального аппендикса нормально выражены; ветви полового
ретрактора сливаются непосредственно перед диафрагмой, одна ветвь крепится
ги несколько дистальнее, другая — около сере-
дины А1 или немного выше. Проток семяпри-
емника прямой, резервуар почти лишен собст-
венного протока, дивертикул семяприемника
тонкий, заметно не доходит до белковой же-
лезы.
Распространение и местоо-
битание. Вид широко, но спорадически
распространен по всей Средней Азии и юго-
западному Алтаю. Населяет разные типы био-
топов, но избегает лесов.
Рис. 167. Pseudonapaeus (Pseudonapaeus) kasnakowi
(West.) из ущелья Хушкак-дере на Гиссарском хр. (I) и
Р. (Pseudonapaeus) otostomus (West.) из окр. Душанбе
(II).
4. Pseudonapaeus (Pseudonapaeus) kasnakowi
(Westerlund, 1898) (167, /; рис. 168).
[Buliminus (Brephulus)]; К о b e 1 1, 1902 : 702, t. 105,
fig. 3, 4 [.B. (Subzebrinus)]; Лихарев, Раммельмейер, 1952 : 201, рис. 114 [Subzebri-
nus (Styloptychus)]; Матёкин, 1966 : ИЗ, табл. 3, рис. 4 [faminia (Chondrulopsina)]; Ilf и-
л e й к о, 19786 : 349, рис. 2, II.
Рис. 168. Pseudonapaeus (Pseudonapaeus) kasnakowi (West.) из ущелья Хушкак-дере на Гис-
сарском хр., 9 VIII 1967 г. Половой аппарат и вскрытйй пенис (из: Шилейко,|1978б).
Terra typica — «Turkestan. Talbar, Thai Ak-su, Totkaul, Thai Waksch,
alluv. Schichten des Fl. Ssurchab in der Nahe von Diuvan, Sarypul», т. e. все
типовые серии происходят из бас. Вахша.
1. РОД PSEUDONAPAEUS
259
Лектотип1 хранится в Естественноисторическом музее Гётеборга,
№ 1806; 36 паралектотипов хранятся в Зоологическом ин-те АН СССР.
Материал: более 40 экз. из 5 местонахождений из Дарвазского и Гис-
сарского хребтов, и из хр. Хазрати-шах.
Раковина высокая, тонкая, стройная, башневидная, слегка блестящая,
умеренно твердостенная. Оборотов 9—10, немного выпуклых, разделенных
мелким швом. Последний оборот прямой или слегка приподнят к устью, высота
его всегда гораздо меньше половины высоты раковины, обычно около трети
этой величины или немногим больше. Окраска от серой до роговой, со светлыми
радиальными пестринами. Скульптура в виде вялых неравномерно располо-
женных радиальных морщинок, спиральные бороздки отсутствуют. Устье
округлое, края его утолщены за счет широкой расплывшейся губы и слегка
отвернуты. На колумеллярном крае имеется спиральная складка, которая
при рассматривании спереди видна не всегда, но хорошо заметна, если рако-
вину повернуть вокруг оси по часовой стрелке. Пупок в виде щели, может быть
сильно прикрыт отворотом колумеллярного края.
Размеры: ВР 9—12, БД 2.5—3 мм; в первоописании: ВР 9—11, БД 2.5 мм;
лектотип: ВР 9.5, БД 3.2 мм.
Внутреннее строение. Материал: 5 экз. из хр. Хазрати-шах,
правый берег р. Яхсу, 15 июня 1974 г.; 4 — из Дарвазского -хр., окр. Голу-
бого оз., 11 июля 1975 г.; 4 — из Гиссарского хр., 25 мая 1969г.; 3 — из ущелья
Хушкак-дере близ с. Зидды на Гиссарском хр., 9 августа 1967 г.; сборы и
определения 3. Иззатуллаева.
Белковая железа имеет, помимо апикальной вырезки, глубокий желоб
на боковой поверхности. Нижний отдел вагины в ИД—2 раза короче верхнего.
Эпифаллус заканчивается широко закругленным вздутием, сбоку от которого
впадает семяпровод. Эпифаллус короткий и толстый, цэкум расположен в са-
мом низу его, по соседству с пенисом, длина которого значительно меньше
длины эпифаллуса. Складки внутри пениса образуют две V-образные фигуры,
их внутренние ветви уходят в эпифаллус, образуя короткие лопасти. Вторые
ветви складок сливаются на противолежащей стенке пениса; вся эта конструк-
ция на интактном пенисе образует клапан, способный полностью изолировать
дистальную часть пениса от проксимальной. Отделы пениального аппендикса
развиты в разной степени; случай наиболее сильной редукции аппендикса
обнаружен Матёкиным (1959). Соответственно и половой ретрактор может быть
одноветвистым. В исследованном материале не обнаружено особей со следами
редукции ветви аппендикса, а сам аппендикс был нормально развитым, все
его отделы были хорошо выражены. Пениальная ветвь полового ретрактора
крепится к середине пениса или немного дистальнее, а ветвь аппендикса к верх-
ней части Ат заметно отступя от А2. Проток семяприемника относительно
короткий и прямой, проток небольшого резервуара очень короткий; дивертикул
нормально развит, но не доходит до нижнего края белковой железы.
Распространение и местообитание. Ареал вида огра-
ничен системой хребтов Гиссаро-Дарваза. Основные биотопы — умеренно
увлажняемые открытые или поросшие кустарником щебнистые склоны или
малоподвижные осыпи.
5. Pseudonapaeus (Pseudonapaeus) otostomus (Westerlund, 1898) (рис. 167, II;
рис. 169).
Westerlund, 1898 : 164 [Buliminus (Brephulus)]; Розен, 1901 : 10 (В. otostomus
var. servatus); К о b e 11, 1902 : 703, t. 105, fig. 5, 6 [5. (Subzebrinus)]; Лихарев, Рам-
мельмейер, 1952 : 202, рис. 115 [Subzebrinus (Styloptychus)]; Матёкин, 1966 : 113
[Jaminia (Chondrulopsina)].
Terra typica — «Totkaul» (Тоткуль, долина Вахша).
Лектотип1 хранится в Естественноисторическом музее Гётеборга,
№ 1797; 9 паралектотипов хранятся в Зоологическом ин-те АН СССР.
1 Обозначается здесь И. М. Лихаревым.
17*
260'
VI. НАДСЕМ. BULIMINOIDEA
Материал: более 30 экз. из трех местонахождений на хребтах Хазрати-
шах и Гиссарском.
Раковина высокая, стройная, веретеновидная или, реже, веретеновидно-
цилиндрическая. Оборотов 8—9, слабовыпуклых, разделенных мелким швом.
Последний оборот непосредственно возле устья энергично приподнят. Окраска
роговая или каштановая, обычно с узкими радиальными светлыми пестринами.
Скульптура в виде сильно сглаженных неравномерно расположенных радиаль-
ных морщин, которые, изредка становятся довольно резкими. Спиральная
скульптура отсутствует. Форма устья оправдывает видовое название — «ухо-
Рис. 169. Pseudonapaeus {Pseudonapaeus) otostomus (West.) из окр. Душанбе, 4 V 1968 г. По-
ловой аппарат и вскрытый пенис.
видная», края устья сильно и плавно отвернуты, на стыке колумеллярного
и базального краев имеется ясный перегиб. На почти отвесном колумеллярном
крае залегает косая спиральная складка, заметная при нормальном положении
раковины. Пупок прикрыт отворотом колумеллярного края, от него остается
узкая щель.
Размеры: ВР 7—12.5, БД 2.5—3.3 мм; в первоописании: ВР 9—11,
БД 2.5 мм; лектотип: ВР 12.5, БД 3.3 мм.
Внутреннее строение. Материал: 4 экз. из хр. Хазрати-шах,
кишлак Сиёфарк на правом берегу р. Яхсу, 15 июня 1974 г.; 3 — из окр. Ду-
шанбе, 4 мая 1968 г.; сборы и определения 3. Иззатуллаева.
Белковая железа с апикальной вырезкой, которая продолжается и на ла-
теральную поверхность органа. Нижний отдел вагины приблизительно вдвое
короче верхнего. Семяпровод впадает резко эксцентрично, конец эпифаллуса
не только широко закруглен, но и заметно расширен; эпифаллус короткий,
толстый, неизвитой. Цэкум развит в различной степени, располагается близ
середины эпифаллуса, иногда цэкум плохо зам:етен. На границе между пенисом
и эпифаллусом имеется резкое сужение, эпифаллус впадает в пенис не терми-
нально, но несколько сбоку. Пенис внутри с парой V-образных складок, форма
1. РОД PSEUDONAPAEUS
261
которых подвержена изменчивости: чаще всего редуцирована одна половина
одной складки, в результате получается одна V-образная складка, напротив
которой помещается одиночный продольный валик, входящий в пространство
между ветвями V-образной складки. Все отделы пениального аппендикса нор-
мально выражены и ясно различимы. Половой ретрактор крепится к диафрагме
единой лентой, которая распадается на два тяжа заметно отступя от диафрагмы.
Одна ветвь крепится примерно к середине пениса, другая — немного выше
середины Аг. Проток семяприемника прямой, вытянут вдоль вагины, резер-
Рис. 170. Pseudonapaeus Рис. 171. Pseudonapaeus (Pseudonapaeus) diplus из окр. Кызыл-
(Pseudonapaeus) diplus Унгура, Ферганский хр., 9 июля 1961 г. Половой-аппарат и
изокр. Арсланбоба, Фер- вскрытый пенис.
ганский хр.
вуар с собственным коротким протоком, дивертикул немного не доходит до
белковой железы.
Распространение и местообитание. Как ареал, так и
основные типы биотопов примерно совпадают с тем, что известно о Р. kasnakowi.
6. Pseudonapaeus (Pseudonapaeus) diplus (Westerlund, 1896) (рис. 170, 171).
Westerlund, 1896 : 191 [Buliminus (Pseudopetraeus)].
Terra typica — Арсланбоб, Ферганский xp.
Лектотип1 хранится в Естественноисторическом музее Гётеборга,
№ 2018; 6 паралектотипов — в Зоологическом ин-те АН СССР.
Материал: 12 экз. из одного местонахождения на Ферганском хр.
Раковина овально-коническая, умеренно твердостенная, с почти кониче-
ским завитком. Оборотов 6, умеренно выпуклых, разделенный неровным швом.
Последний оборот непосредственно возле устья несильно, но явственно при-
поднят, высота его больше половины высоты раковины. Окраска роговая, со
светлыми радиальными штрихами, которыми отмечены положительные эле-
менты скульптуры, представляющие собой сглаженные, не очень равномерно
расположенные, но довольно частые ребра. На верхних оборотах ребра слабые
или вообще почти не выражены; лучше всего ребра развиты на последнем
обороте. Устье овальное, с отвернутыми белыми краями, усиленными широкой
1 Обозначен здесь И« М. Лихаревым.
262
VI. НАДСЕМ. BULIMINOIDEA
губой. Колумеллярный край расширен, без складок. Пупок в виде короткой
широкой щели.
Размеры: ВР 8—11, БД 4—5 мм; в первоописании: ВР И, БД 4.5—5 мм;
лектотип: ВР 11, БД 4.9 мм.
Внутреннее строение. Материал: 5 экз. из Ферганского хр.,
окр. Кызыл-Унгура, 9 июня 1961 г., сбор И. М. Лихарева, определение автора.
Белковая железа с очень резкой апикальной вырезкой, переходящей и на
боковую поверхность железы. Нижний отдел вагины не более чем в полтора
раза короче верхнего. Семяпровод впадает эксцентрично, но совершенно не
сдвинут вниз по эпифаллусу. Цэкум хорошо развит, расположен выше сере-
дины цилиндрического эпифаллуса. Пенис, веретеновидный, на внутренней его
поверхности легко заметить две чрезвычайно мощные продольные складки,
расщепленные лишь в непосредственной близости от эпифаллуса. Все отделы
пениального аппендикса ясно выражены. Половой ретрактор отходит от диаф-
рагмы единым тяжом, который вскоре раздваивается: одна ветвь крепится
немного дистальнее середины пениса, вторая — к верхней части Ат слегка
отступя от А2. Проток семяприемника прямой, резервуар небольшой, с соб-
ственным коротким протоком, длина которого меньше длины самого резервуара.
Дивертикул недлинный, заметно не доходит до нижней кромки белковой железы.
Распространение и местообитание. Вид найден в двух
точках на Ферганском хр., в зарослях кустарников среди камней.
7. Pseudonapaeus (Pseudonapaeus) stabilis Schileyko, sp. nov. (рис. 172, 173).
Terra typica — западный Тянь-Шань, долина p. Пскем, в 15 км выше с. На-
най.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: более 50 экз. из типового местонахождения.
Раковина пулевидная, почти одноцветно светлая, с блестящей полирован-
ной поверхностью, довольно твердостённая, с конической короткой верхней
частью и широкой закругленной вершиной; цилиндрическая часть раковины
составлена тремя последними оборотами. Свежие раковины блестят, как по-
лированные. Оборотов 6—7, слабовыпуклых; последний оборот занимает более
трети, но менее половины высоты раковины; лишь у немногих раковин высота
его составляет примерно половину высоты раковины. К устью последний обо-
рот плавно, без перегиба, поднят. Окраска на первый взгляд почти однотонная,
светло-роговая, желтовато-роговая или белесовато-желтоватая;' однако при
внимательном рассмотрении видно, что имеются группы более светлых, чем
фон, штрихов. Верхние обороты чаще всего с явственной желтизной или рыже-
ватые. Скульптура в виде очень редких и очень сильно сглаженных радиальных
морщин, местами имеются отдельные слабые вмятины и намеки на спиральную
исчерченность, но в целом блеск поверхности раковины обеспечивается именно
отсутствием ясных скульптурных элементов. Устье без зубов, места его при-
крепления слабо сближены, соединены толстой светлой мозолью, благодаря
чему овальное устье часто выглядит почти цельным. Устьевые края сильно
расширены, отвернуты и усилены расплывшейся белой губой. Колумеллярный
край почти отвесный, на границе его с базальным имеется плавно закруглен-
ный угол. Базальный и палатальный края образуют широкую равномерно вы-
гнутую дугу. Пупок узкощелевидный.
Размеры: ВР 11.5—16, БД 5—6 мм; голотип: ВР 15.0, БД 6.0 мм.
Внутреннее строение. Материал: 12 экз. из типовой серии
(долина р. Пскем, 20 мая 1972 г.), сбор и определение автора.
Белковая железа с вялой апикальной вырезкой и слабым латеральным же-
лобком. Нижний отдел вагины в 11/2—2 раза короче верхнего. Семяпровод впа-
дает эксцентрично, задний конец эпифаллуса закруглен или очень слабо при-
острен. Эпифаллус короткий, цилиндрический, не извит; хорошо развитый цэ-
кум располагается близ его середины. Пенис длиннее эпифаллуса или эти от-
1. РОД PSEUDONAPAEUS
263
делы примерно одной длины. Эпифаллус впадает в пенис чуть сбоку, после чего
следует вздутие проксимального конца пениса, поэтому граница между этими
протоками хорошо заметна. Внутри пениса залегают две обычные V-образные
складки, однако у некоторых экземпляров
они трансформируются , в систему из одной
V-образной складки и одного продольного ва-
лика, который входит в просвет между ветвя-
ми двойной складки (см. поперечный разрез
пениса и вывернутый пенис на рис. 173). Все
отделы пениального аппендикса хорошо за-
метны. Ат длинный, сужается к концу, Л2
правильно шаровидный, четко выражен,
А3 относительно длинный, тонкий, незна-
чительно шире, чем Л4. Проток семя-
Рис. 172. Pseudonapaeus (Pseudonapaeus) stabilis.
I — голотип, 11 — паратип.
приемника почти прямой, тесно прилегает к вагине. Проток резервуара почти
отсутствует и представлен только тем коротким участком, который отделяет
Рис. 173. Pseudonapaeus (Pseudonapaeus) stabilis из долины р. Пскем выше с. Нанай, 20 мая
1972 г. Паратипы.
I— половой аппарат, II —-вскрытый пенис, III — другая особь (пенис и вагина вывернуты).
овальный резервуар от общего протока. Дивертикул семяприемника недлинный,
не достигает белковой железы.
Вид близок к номинативной форме Р. albiplicatus, отличается гладкой ра-
ковиной, мощным развитием губы и наклонностью к образованию цельного
264
VI. НАДСЕМ. BULIMINOIDEA
устья. Из числа анатомических признаков следует указать на короткий срав-
нительно с пенисом эпифаллус, и на относительно длинный А3. Подтвержде-
нием видовой самостоятельности служит то, что оба вида найдены вместе;
Р. albiplicatus представлен номинативной формой, имеющей резко исчерчен-
ную раковину.
Распространение и местообитание. Вид найден только
в типовом местонахождении, но на значительной площади: сборы проводились
на нескольких участках нижней и верхней пойменных террас правого берега
р. Некем. Кроме того, две пустые раковины найдены на расстоянии около 2 км
от реки в одном из узких боковых ущелий. Живые особи в большом числе об-
Рис. 174. Pseudonapaeus (Pseudonapaeus)
schnitnikovi (I) И P. (Pseudonapaeus) ga~
linae (II). Лектотипы.
ff
Рис. 175. Виды рода Pseudonapaeus (подрод
Pseudonapaeus s. str.).
I — P. (Pseudonapaeus) trigonochilus из поймы p. Ha-
рын; II — P. (Pseudonapaeus) goldfussi из окр. с. Джи-
лису, Алайский хр-.
наружены под камнями на открытой пойме, заросшей травянистой раститель-
ностью, а также в малоподвижных крупнообломочных осыпях вместе с тремя
другими видами: Р. albiplicatus (номинативная форма), Р. miser и Р. entoptyx.
8. Pseudonapaeus (Pseudonapaeus) schnitnikovi (Lindholm, 1922) (рис. 174, I).
Линдгольм, 1922 : 274 [Buliminus (Pseudonapaeus)]; Лихарев, Раммель-
мейер, 1952:205; Матёкин, 1966 : 131 [Jaminia (Chondrulopsina) subsp. asiatica].
Terra typica — окр. Алма-Аты.
Лектотип хранится в Зоологическом ин-те АН СССР.
Материал: 6 экз. из 4 местонахождений в окр. Алма-Аты.
Раковина башневидная, стройная, тонкостенная, слегка блестящая. Вер-
шина притупленная. Оборотов 7х/2—8, медленно нарастающих; верхние более
выпуклые, чем нижние, разделены довольно глубоким швом. Последний обо-
рот к устью плавно приподнят, высота его несколько меньше половины высоты
раковины. Окраска одноцветная, темно-роговая. Скульптура — тонкая не-
равномерная радиальная исчерченность. Устье небольшое, усеченно-овальное,
слабо скошено, высота его в 21!3—3 раза меньше высоты раковины. Места при-
крепления устья слабо сближены и соединены тонкой мозолью. Края устья
слабо отвернуты, слегка утолщены, со светлой губой. Палатальный край
вначале почти отвесный, далее вместе с базальным образует широкую дугу.
Колумеллярный край наверху расширен. Пупок в виде короткой щели, ча-
стично прикрыт отворотом колумеллярного края.
2. РОД SUBZEBRINUS
265
Размеры: ВР 9.5—11, БД 3.5—3.6 мм; в первоописании: ВР 10.75, БД 3.6 мм
(размеры голотипа).
Распространение и местообитание. Вид известен только
из Заилийского хр. близ Алма-Аты, встречен на открытых склонах.
9. Pseudonapaeus (Pseudonapaeus) galinae (Tzvetkov, 1940) (рис. 174, II).
Цветков, 1940 : 386 (Епа); Лихарев, Раммельмейер, 1952 : 206, рис. 120;
Матёкин, 1966 : 130 [Jaminia (Chondrulopsina) potaniniana subsp. asiatica].
Terra typica — предгорья Заилийского xp. близ Алма-Аты (оригинальная эти-
кетка Б. Н. Цветкова с пометкой «cotypes»: «Алма-Ата. Сбор на склоне речки
у совхоза НКВД. 1 X 1936 г.».
Лектотип и 6 паралектотипов хранятся в Зоологическом музее Москов-
ского ун-та, лектотип обозначается здесь.
Материал: свыше 200 экз. из Заилийского хр.
Раковина веретеновидно-коническая, умеренно вздутая, слабо блестящая.
Вершина узко закругленная. Оборотов 7—7х/2 (у лектотипа 7), слабовыпуклых,
разделенных неглубоким швом. Последний оборот к устью плавно приподнят,
высота его приблизительно равна половине высоты раковины. Окраска светло-
роговая. Скульптура в виде неравномерной тонкой радиальной морщинистости.
Устье округло-треугольное, слегка скошенное, высота его около 1/3 вы-
соты раковины. Края устья слабо отвернуты, утолщены массивной губой,
расположенной почти у самого устьевого края. Чуть выше середины палаталь-
ного края губа образует легкое утолщение. Имеется слабый ангулярный буго-
рок. Пупок очень узкий, щелевидный.
Размеры: ВР 12.0—14.5, БД 4.7—6.0 мм; лектотип: ВР 14.0, БД 5.8 мм.
Распространение и местообитание. Предгорья Заилий-
ского хр., живет на сухих открытых склонах.
10. Pseudonapaeus (Pseudonapaeus) trigonochilus (Ancey, 1886) (рис. 175, I;
рис. 176).
Ancey, 1886a : 28 (Buliminus); Westerlund, 1887 : 67 (Buliminus); 1896 : 189
LB. (Pseudopetraeus) trigonochilus, f. teres, f. expansilabris, f. planicollis, f. pachychila]; 1897 :10,
figs, la, 2a (B. castaneus, syn. nov.); Розен, 1901 : 9 (В. maracandensis); Kobelt, 1902 :
516, t. 82, figs. 22, 23 [B. (Subzebrinus)]; Lindholm, 1927a : 101 [Ena (Pseudonapaeus)
latilabris, syn. nov.]; Лихарев, Раммельмейер, 1952 : 205, рис. 118, 206, рис. 119
(latilabris); Матёкин, 1966 : 131 [Jaminia (Chondrulopsina) potaniniana subsp. albiplicata];
Иззатуллаев, 1975:41, рис. 16, в (castaneus).
Terra typica — Самарканд.
Место хранения типов не установлено.
Материал: свыше 100 экз. из 19 местонахождений на западном и цен-
тральном Тянь-Шане.
Раковина твердостенная, блестящая; форма ее подвержена значительной
изменчивости: она может быть цилиндрической, овально-цилиндрической или
булавовидной, с полным спектром переходов, причем булавовидные райовины
обычно встречаются на открытых незатененных склонах. Оборотов 8—Ю1^,
очень медленно и плавно нарастающих. Верхние 4—5 оборотов образуют ку-
половидную вершину и навиты очень тесно; следующие обороты, формирую-
щие цилиндрическую часть раковины, нарастают настолько медленно, что
ширина и высота почти одинаковы у смежных оборотов. Последний оборот в по-
следней четверти плавно поднимается к устью — иногда более, иногда менее
заметно; непосредственно перед устьем поднимается круче, с плавным переги-
бом. Высота последнего оборота в 2—3 раза меньше высоты раковины. Окраска
одноцветно роговая или каштановая, нередко с сизоватым оттенком, иногда
на затылке имеется неясное желтоватое пятно. Скульптура в виде тонкой шелко-
вистой довольно правильной радиальной исчерченности. Устье округло-тре-
угольное, края его расширены, утолщены и отвернуты, места прикрепления не
266
VI. НАДСЕМ. BVLIMIN01DEA
сближены, соединены светлой мозолью. Пупок сильно сжат, почти проколо-
видный.
Размеры: ВР 8—13, БД 4—5 мм; в первоописании: ВР 12—12х/3, БД 4х/4—
41/2 мм.
Внутреннее строение. Материал: 4 экз. из предгорьев Ферган-
ского хребта, Базар-Курганский р-н, урочище Ак-Терек, 1937 г., сбор А. Н. Ки-
риченко; 26 — из долины р. Нарын близ Токтогула, 1 июня 1972 г., сбор и
определения автора.
Белковая железа с апикальной вырезкой, которая не распространяется на
боковую поверхность железы. Нижний отдел вагины в 2—3 раза короче верх-.
Рис. 176. Pseudonapaeus (Pseudonapaeus) trigonochilus из предгорьев Ферганского хр., Ак-
Терек, июль (?) 1937 г. Половой аппарат и вскрытый пенис.
него. Семяпровод впадает резко эксцентрично, у некоторых экземпляров даже
заметно отступя от конца эпифаллуса, в результате чего формируется явствен-
ный почти шаровидный бич. Цэкум расположен возле середины эпифаллуса или
немного проксимальнее ее. Пенис цилиндрический, но нередко имеется широ-
кий кольцевой пережим, и пенис приобретает, таким образом, форму гантели.
Внутри пениса залегает пара V-образных складок, причем их нижние концы
сильно развиты, гребневидные; в интактном пенисе, располагаясь одна про-
тив другой, они заполняют почти весь просвет органа и изолируют дистальную
часть полости пениса от проксимальной. Все отделы пениального аппендикса
нормально развиты, характерной чертой этого вида является относительно
небольшая длина нитевидного Ait приблизительно равная длине А5. У всех
исследованных животных половой ретрактор единой лентой крепился к диа-
фрагме и распадался на два тяжа значительно отступя от нее. Пениальная
ветвь крепится близ середины пениса, ветвь аппендикса подходит к Ат немного
отступя от А2. Проток семяприемника длинный, образует не менее двух резких
изгибов и значительно превышает длину вагины. Небольшой овальный резер-
вуар имеет ясно выраженный собственный проток, длина которого приблизи-
тельно равна длине самого резервуара. Дивертикул семяприемника умеренной
1. РОД PSEUDONAPAEUS
267
длины, но так как резервуар располагается высоко, достигает белковой железы
и часто накладывается на ее нижнюю часть.
Распространение и местообитание. Южные склоны
Чаткальского хр., Ферганский и Алайский хребты, окр. Самарканда. Населяет
леса, заросли кустарников, малоподвижные сырые осыпи; очень плотная по-
пуляция обнаружена на глинисто-валунном откосе правого берега р. Нарын
в непосредственной близости от воды, где вид живет совместно с гладкостен-
ными блестящими яркоокрашенными Р. albiplicatus f. albiplicata.
ДСП
Рис. 177. Pseudonapaeus (Pseudonapaeus) goldfussi из окр. с. Джи- Рис. 178. Pseudonapaeus
лису, Алайский хр., 16 июня 1974 г. Половой аппарат и вскрытый' (Рseudonapaeus) chodschen-
пенис (из: Шилейко, 19786). dicus. Паратип.
,11. Pseudonapaeus (Pseudonapaeus) goldfussi (Kobelt, 1893) (рис.^175, II;
рис. 177).
Kobelt, 1893, Iconogr., N. F., 7 : 82, t. 171, fig. 1102 [Buliminus (Chondrula?)]; Л n-
харев, Раммельмейер, 1952 : 205 (trigonoahilus i.var. goldfussi); ШиЛейко,
19786 : 352, рис 3, IV. .
Terra typica — Алайский xp.
Голотип хранится в ЗенкенбергскохИ музее, Франкфурт-на-Майне,
№156686.
Материал: 16 экз. из двух местонахождений на Алайском и Ферган-
ском хребтах.
Раковина левозавитая, является зеркальным отражением раковины Р. tri-
gonochilus.
Внутреннее строение. Материал: 4 экз. из окр. Джилису на
Алайском хр., 16 июня 1974 г., сбор и определение 3. Иззатуллаева.
Отличия от Р. trigonochilus заключаются в том, что у всех четырех анато-
мированных особей ветви полового ретрактора присоединялись к диафрагме
независимо, хотя и рядом одна с другой; кроме того, проток семяприемника
почти прямой, проходит вдоль вагины и не образует сколько-нибудь крутых
изгибов.
268
VI. НАДСЕМ. BULIMINOIDEA
Распространение и местообитание. Известно несколько
изолированных популяций на Алайском и Ферганском хребтах, обитающих
в тех же условиях, что и Р. trigonochilus. Обоснование, видовой самостоятель-
ности левозавитых популяций — см.: Шилейко, 1975в; Александров, Сергиев-
ский, 1979.
12. Pseudonapaeus (Pseudonapaeus) chodschendicus (Mukhitdinov, 1976)
(рис. 178, 179).
Мухитдинов, 1976 : 39, рис. 1А, 2А (Chondrulopsina).
Рис, 179. Pseudonapaeus (Pseudonapaeus) chodschendicus из окр. с. Ашт на Кураминском хр.,
19 мая 1972 г. Паратип.
I — половой аппарат; II, III — вскрытый пенис; IV — вскрытый пенис другого экземпляра.
Terra typica —|Кураминский хр., окр. с. Ашт (место находки голотипа)
и пос. Чорух.
Г о л о т и п ^(’хранится в Зоологическом ин-те АН СССР.
Материал: 14 экз. из типовой серии.
Раковина с трудом отличима от раковины Р. miser, особенно если учесть
диапазон изменчивости этого вида. Основные отличия заключаются в том, что
если раковина Р. miser ближе к удлиненно-овальной, то форма раковины Р. chod-
schendicus скорее башневидная, с более широко, чем у Р. miser, закругленной
вершиной. В деталях внутреннего строения отличия между этими видами вполне
определенные.
Размеры голотипа: ВР 7.8, БД 3.4 мм.
Внутреннее строение. Материал: 7 экз. из окр. с. Ашт на Ку-
раминском хр., 19 мая 1972 г., сбор и определение А. Б. Мухитдинова.
Белковая железа с глубокой апикальной вырезкой. Нижний отдел вагины
незначительно короче верхнего или отделы почти равной длины. Семяпровод
впадает эксцентрично, бича нет. Очень резко выраженный цэкум располага-
ется выше середины цилиндрического почти прямого эпифаллуса. Пенис вере-
теновидный или цилиндрический, тонкостенный. Внутри дистальной части
1. РОД PSEUDONAPAEUS
269
его имеется несколько вялых продольных складок, а в проксимальной части
залегают две складки, также не очень резкие, но имеющие характерную форму
(рис. 179, II—IV), которую очень приблизительно можно охарактеризовать
как кольцевидную. Между этими складками имеется приостренный бугорок.
Пениальный аппендикс сложен всеми пятью отделами; лишь А3 плохо заметен
и иногда практически не выражен. Половой ретрактор крепится к диафрагме
единой лентой и распадается на две ветви заметно отступя от диафрагмы. Одна
ветвь подходит к пенису близ его середины, а вторая — к Alf также близ цен-
тральной его части. Проток семяприемника прямой, небольшой резервуар
имеет собственный проток такой же длины, как и резервуар, или несколько
больше; дивертикул семяприемника тонкий, короткий, рудиментарный, длина
его приблизительно равна длине резервуара вместе с его протоком или чуть
больше.
От Р. miser отличается коротким дивертикулом и внутренним строением
пениса: складки внутри этого отдела у Р. miser всегда ориентированы про-
дольно и не образуют кольцеобразных фигур.
Распространение и местообитание. Вид найден пока
только в двух местах на Кураминском хр. Встречен на открытых каменистых
склонах предгорий на высоте 1000—1500 м над уровнем моря. Живет под кам-
нями и корнями растений. Наиболее активен в мае—апреле, в это время пи-
тается эфемерными растениями (Мухитдинов, 1976).
2. Подрод SIRAPHORUS Lindholm, 1925, stat. nov.
L i n d h о 1 m, 1925 : 29, 32 (род Siraphorus); Thiele, 1931 : 522 [Ena (Siraphorus)};
Лихарев, Раммельмейер, 1952 : 206 (род Siraphorus)- Wenz, Zilch, 1959—
1960 : 188 (род Siraphorus); M атёкин, 1966 : 114 [Jaminia (Chondrulopsina) part.]; Ill и-
л e й к о, 1977 : 41.
Типовой вид Buliminus entoptyx Lindholm, 1925; по первоначаль-
ному обозначению.
Раковина левозавитая, почти гладкая, одноцветная. Устьевая арматура
составлена единственной продольной палатальной складкой, лежащей глубоко
внутри устья.
Папилла пениса отсутствует, внутри пениса имеется пара хорошо развитых
V-образных складок.
Монотипический подрод.
13. Pseudonapaeus (Siraphorus) entoptyx (Lindholm, 1925) (рис. 180, 181).
Lindholm, 1925a : 32 (Buliminus); Лихарев, Раммельмейер, 1952 : 207
(Siraphorus); Матёкин, 1966 : 114 [Jaminia (Chondrulopsina) potaniniana entoptyx}; Ши-
лейко, 1977 : 41, рис. 1, 1—4.
Terra typica — Кара-су (Ферганский xp.).
Место хранения типов не установлено.
Материал: более 70 экз. из трех местонахождений на Таласском, Пскем-
ском и Ферганском хребтах.
Раковина от почти цилиндрической до башневидной, слегка просвечиваю-
щая, составлена 7—8 слабовыпуклыми, плавно и медленно нарастающими обо-
ротами, разделенными мелким светлым швом. Вершина притупленная, широко
закругленная. Последний оборот прямой или чуть приподнят к устью, базаль-
ная его стенка при рассматривании сбоку явственно приподнята. Окраска от
светло-коричневой до каштановой и красновато-роговой, нередко с более свет-
лыми, чем фон, радиальными штрихами. Скульптура представлена сильно
сглаженными редкими неправильными радиальными морщинами; однако на
первый взгляд поверхность почти гладкая, поблескивающая. Устье слегка
скошено. В результате интенсивного развития палатальной мозоли устье цель-
ное или почти цельное, края его плавно и широко отвернуты, образуя белую
270
VI. НАДСЕМ. BULIMINOIDEA
или желтоватую манжету. В верхней части палатального края обычно при-
сутствует в разной степени выраженное утолщение. В глубине устья при нор-
мальном положении раковины видна нижняя часть палатальной складки, ухо-
дящей в глубь раковины на 3/4 оборота; снаружи эта складка видна как желто-
ватая спиральная полоска. Имеется также очень слабая колумеллярная складка,
видная при косом положении раковины. Пупок очень узкий, имеет овальные
контуры и слегка прикрыт отворотом колумеллярного края устья.
Рис. 180. Pseudonapaeus
(Siraphorus') entoptyx из
долины р. Пскем (из:
Шилейко, 1977).
Рис. 181. Pseudonapaeus (Siraphorus) entoptyx (Lindh.) из долины
р. Пскем, 20 V 1972 г.
1 — половой аппарат, II— вскрытый пенис (из: Шилейко , 1977).
Размеры: ВР 7.5—10, БД 2.5—3 мм; в первоописании: ВР 8.5—9, БД 3 мм.
Внутреннее строение. Материал: 14 экз. из долины р. Пскем
на западном Тянь-Шане, 20 мая 1972 г., сбор и определение автора.
Белковая железа с резкой апикальной вырезкой, переходящей на боковую
поверхность органа. Верхний отдел вагины в 1112—2 раза длиннее нижнего.
Семяпровод впадает эксцентрично, задний конец эпифаллуса широко закруглен.
Цэкум развит хорошо, напротив него на эпифаллусе заметен бугорок, заклю-
чающий Г—2 щелевидных полости (внутри цэкума такая полость одна). Цэ-
кум расположен приблизительно на середине эпифаллуса, отделенного от пе-
ниса ясно выраженным сужением. Длина пениса примерно равна расстоянию
от конца эпифаллуса до цэкума. Внутри пениса залегает пара очень мощных
V-образных складок, каждая из которых раздваивается только близ отверстия
эпифаллуса. Пениальный аппендикс слагается всеми пятью отделами: Ат 1 —
2 раза резко изогнут, Л2 небольшой, правильно шаровидный, А3 очень плавно
переходит в Л4, А5 сравнительно длинный и тонкий. Пениальная ветвь полового
ретрактора крепится близ середины пениса, вторая ветвь — к Аг близ А2.
Перед диафрагмой ветви сливаются. Проток семяприемника образует 2—
4 крутых изгиба, резервуар снабжен собственным коротким протоком. Дивер-
тикул верхним концом прилегает к нижней части белковой железы.
1. РОД PSEUDONAPAEUS
27f
Распространение и местообитание. Вид найден в трех
местах. Типовая серия происходит из долины р. Кара-су на Ферганском хр.;
кроме того, большое число экземпляров найдено в припойменной террасе
р. Пскем под камнями и среди корней кустарников, а также на западе Талас-
ского хр. в заповеднике Аксу-Джабаглы.
3. Подрод ARIDENUS Schileyko, subgen. nov.
Типовой вид Епа submucronata Lindholm, 1927.
Раковина правозавитая, почти гладкая, с легкой морщинистостью, одно-
цветная. Устье без зубов.
Внутри пениса имеется одна V-образная складка и стержневидная папилла,
лежащая между ветвями этой складки.
Монотипический подрод.
14. Pseudonapaeus (Aridenus) submucronatus (Lindholm, 1927) (рис. 182, 183).
Lindholm, 1927a : 100 [Ena (Pseudonapaeus)]; Лихарев, Раммельмейер,
1952 : 204.
Terra typica — Чаткальский xp., среднее течение p. Узун-ахмат, высота
1200—1400 м над уровнем моря.
Лектотип иЗ паралектотипа хранятся в Зоологическом ин-те АН СССР,
обозначается впервые.
Материал: свыше 200 экз. из двух местонахождений на Чаткальском и
Ферганском хребтах.
Раковина удлиненно-овальная или высококоническая, блестящая, относи-
тельно тонкостенная. Верхняя часть завитка суженная и вытянутая. Оборотов
6х/2—7, умеренно выпуклых. Последний оборот к
устью постепенно и незначительно поднимается, вы-
сота его немного больше половины высоты ракови-
ны. Окраска от светло-роговой до коричневой, од-
ноцветная или с очень слабыми радиальными свет-
лыми штрихами. Скульптура представлена вялыми
неравномерными радиальными морщинами. Устье
усеченно овальное, почти отвесное, края его умерен-
но отвернуты, тонкие, по линии перегиба лежит ши-
рокая светлая губа. Париетальная мозоль слабая.-
Пупок в виде широкой короткой щели, почти не при-
крыт отворотом колумеллярного края.
Рис. 182. Pseudonapaeus (Aridenus) submucronatus (Lindh.).
Лектотип.
Размеры: ВР 13—17, БД 7—9 мм; лектотип: ВР 15, БД 7 мм.
Внутреннее строение. Материал: 4 экз. из долины р. Нарыннад
Таш-Кумыром, 31 мая 1972 г., сбор и определение автора.
Белковая железа с резкой и глубокой апикальной вырезкой, которая не
заходит на латеральную поверхность железы. Нижний отдел вагины немногим
короче верхнего. Семяпровод впадает немного отступя от конца эпифаллуса, за
счет чего возникает шаровидный бич. Цэкум развит очень хорошо, почти шаро-
видный, располагается немного дистальнее середины эпифаллуса. Пенис в 2—
2х/2 раза короче эпифаллуса, внутри его проксимальной части залегает хорошо
развитая V-образная складка, ветви которой соединены внизу лишь тонкой и
невысокой перемычкой; у одного из анатомированных экземпляров удалось
обнаружить слабые следы второй такой же складки. Между ветвями основной
272
VI. НАДСЕМ. BULIMINOIDEA
складки залегает стержневидная папилла, по поверхности которой проходят
два винтообразных желоба и одна узкая глубокая борозда, по которой при спа-
ривании скользит сперматофор. На вывернутом пенисе папилла видна очень
хорошо (рис. 183, II, III). Отделы пениального аппендикса нормально развиты,
особенно четко выражен обычно малозаметный А3. Ветви полового ретрактора
отходят от диафрагмы порознь, одна из них присоединяется к пенису немного
выше основания пениального аппендикса, другая — к границе между и
Рис. 183. Pseudonapaeus (Aridenus) submucronatus (Lindh.) из долины р. Нарын выше Таш-
Кумыра, 31 V 1972 г.
I — половой аппарат, II — вскрытый пенис, III — особь с вывернутыми дистальными участками поло-
вого аппарата.
А2 или непосредственно к А2. Проток семяприемника почти прямой или обра-
зует 1—2 небольших изгиба в верхней части. Резервуар с собственным корот-
ким протоком, дивертикул почти касается нижней кромки белковой железы.
Распространение и местообитание. Вид найден в двух
местах бассейна р. Нарын: вдоль одного из правых притоков Нарына в преде-
лах Чаткальского хр. и в предгорьях Ферганского хр. вдоль русла той же реки,
ниже по течению. Обитает на открытых, совершенно безлесных склонах с арид-
ными условиями, в крупнообломочных осыпях. Очевидно, во время засухи за-
рывается глубоко в почву, так как 31 мая 1972 г. в жаркий сухой день среди
более 200 раковин найдены только 4 живых моллюска, и те под камнями весом
более 100 кг.
2. Род SUBZEBRINUS Westerlund, 1887
Westerlund, 1887 : 66; Thiele, 1931 : 520 [Zebrina (Subzebrinus)]; Лихарев,
Раммельмейер, 1952 : 195; Wenz, Zilch, 1959—1960 : 192; Матёкин, 1966 :
113 [Jaminia (Chondrulopsina) part.]; Шилейко, 1978:844.
Типовой вид Buliminus labiellus Martens, 1881; по обозначению Мел-
лендорфа (Moellendorff, 1901 f 327).
2. РОД SVBREBRINUS
273
Раковина цилиндрическая или овально-цилиндрическая, слабо скульптури-
рованная. Устье без зубов, но с порожистой губой.
Бич отсутствует. Внутри пениса имеется серия продольных гофрированных
складок. Пениальной папиллы нет. Дивертикул семяприемника отсутствует,
семяприемник представляет собой длинный рукав с терминальным расшире-
нием.
Монотипический род.
Рис. 184. Subzebrinus la- Рис. 185. Subzebrinus labiellus (Mart.) из верховьев р. Чарса-
biellus (Mart.) из Заилий- чай на Джунгарском хр., 12—15 VI 1969 г. Половой аппарат,
ского хр., Ортотау. вскрытый пенис и участок эпифаллуса (из: Шилейко, 19786).
Subzebrinus labiellus (Е. Martens, 1881) (рис. 184, 185).
Е. М а г t е n s, 1881 : 24, t. 6, fig- 1, 2 (Buliminus)-, 1882а : 21, t. 3, fig. 4 (Buliminus);
Ancey, 1886a : 22 (B. labiellus), 23 (var. strophostoma); 1886b : 338 (B. pullaster)-, Wester-
lund, 1887 : 66 (Buliminus); Kobelt, 1902 : 503, t. 81, fig 21 [5. (Subzebrinus?)]; Цвет-
ков, 1940 : 386 [Ena (Subzebrinus) eleonorae, syn. nov.]; Лихарев, Раммельмейер,
1952 : 197; Матёкин, 1966 : 113, табл. 3, рис. 2 [Jaminia (Chondrulopsina)]; Шилейко,
19786 : 346, рис. 2, .
Terra typica — Тарбагатай.
Лектотип хранится в Зоологическом музее Берлинского ун-та им. Гум-
больдта, № 101591а (Kilias, 1971).
Материал: более 60 экз. из 7 местонахождений на Тарбагатае, Заилий-
ском и Джунгарском хребтах.
Раковина более или менее удлиненная, от овально-цилиндрической до почти
строго цилиндрической, иногда бочонковидная, твердостенная. Оборотов 6—8,
несильно выпуклых, разделенных мелким беловатым швом. Последний оборот
к устью еле заметно приподнят, отношение его к высоте раковины непостоянно
из-за сильной изменчивости относительной высоты раковины. Окраска может
быть почти одноцветно белой, светло-серой или роговой, нередко с сизоватым
оттенком; чаще, однако, на относительно темном фоне видны более светлые, часто
18 А. А. Шилейко
Tilt
VI. НАДСЕМ. BULIMINOIDEA
белые, радиальные пестрины, оптически более плотные, чем фон. Если эти
пестрины развиты сильно и располагаются густо, то раковина выглядит белой
с темными пестринами. В соответствии с этим темные раковины просвечивают
сильнее, чем сильно кальцинированные белые. Скульптура в виде неправильно
расположенных умеренно развитых радиальных морщин и очень слабо выражен-
ных спиральных бороздок, которые местами лишь угадываются. Устье слабо
скошено, округлое, не оттянуто в сторону, места его прикрепления не сближены
и соединены тонкой прозрачной мозолью, которая после гибели животного быстро
мутнеет и становится белой. Устьевые края незначительно отвернуты, но поро-
жисто утолщены за счет узкой, но высокой губы, лежащей вплотную к краю.
В ангулярной области может присутствовать небольшое утолщение париеталь-
ной мозоли. Пупок очень узкий, слегка овальный.
Размеры: ВР 8—16, БД 4—6 мм; в первоописании: ВР 14, БД 6 мм.
Внутреннее строение. Материал: 7 экз. из Джунгарского хр.,
верховья р. Чарса-чай, 12—15 июня 1969 г., сбор И. С. Даревского; 3 — из Заи-
лийского хр., Ортотау, 3 июля 1968 г., сбор К. К. Увалиевой; 3 — из Джунгар-
ского хр., верховья р. Текели, 9 июля 1916 г.; 2 — из того же хребта, долина
р. Теректы, июль 1936 г., сборы Шнитникова; определения автора.
Вырезка на белковой железе нерезкая, занимает апикальное положение.
Нижний отдел вагины приблизительно вдвое короче верхнего. Семяпровод впа-
дает эксцентрично; бича нет, задний конец эпифаллуса широко закруглен. При-
мерно посредине эпифаллуса или немного ниже расположен бугорковидный
цэкум. Стенки эпифаллуса между семяпроводом и цэкумом сильно мускулизо-
ваны, и это заметно даже без гистологического исследования, так как ткани
этого участка на спиртовом материале слегка ирризируют. На границе эпифал-
луса и цилиндрического пениса имеется небольшое, но постоянное тонкостенное
вздутие. Пенис лишь немного короче эпифаллуса. Внутри пениса залегает се-
рия продольных гофрированных складок, местами аностомозирующих. У от-
верстия эпифаллуса две из этих складок сливаются верхними кромками, обра-
зуя небольшой козырек; прочие складки сходят на нет. Пениальный аппендикс
расположен близ полового атриума; А1 умеренно развит, зато массивный А2‘,
А3 недлинный, но ясно выражен; А5 очень мощный. От диафрагмы ветви поло-
вого ретрактора отходят независимо, одна из них крепится немного ниже сере-
дины пениса, другая — к А± чуть ниже А2. Дивертикул семяприемника отсут-
ствует, длина протока семяприемника подвержена значительной изменчивости:
у особей из верховьев р. Чарса-чай он длинный, образует 1—2 петли, и мешко-
видный резервуар касается белковой железы; у экземпляров из Ортотау проток
почти прямой, и резервуар заметно не доходит до белковой железы.
Распространение и местообитание. Встречается на ска-
листых кустарниковых склонах в поясе лиственных лесов Тарбагатая, Заилий-
ского и Джунгарского хребтов (Матёкин, 1966).
3. Род TURANENA Lindholm, 1922
Kobelt, 1902 : 1021 (Pseudonapaeus, non Westerlund, 1887); Линдгольм, 1922a:
275 (Ena (Turanena)]; Lindholm, 1925a : 27 [Ena (Turanena)]; Thiele, 1931 : 520—521
[Ena (Turanena)]; Wenz, Zilch, 1959—1960 : 185; Лихарев, Раммельмейер,
1952 : 192; Матёкин, 1966 : 111 [Zebrina (Turanena)]; Шилейко, 1978г : 845.
Типовой вид Buliminus herzi Boettger, 1889; по первоначальному обо-
значению.
Раковина от почти кубаревидной до высококонической, слабо скульптури-
рована, обороты сильновыпуклые. Устье без зубов, с отвернутыми краями,
места его прикрепления сильно сближены.
Бич ясно выражен, короткий, конический или притупленный. Внутри пениса
имеются 1—2 V-образных складки, способных к некоторой трансформации.
Папилла пениса отсутствует. Дивертикул семяприемника развит нормально.
3. РОД TURANENA
275
Распространение. Закавказье, Северный Иран, Копетдаг, Гима-
лаи, Памиро-Алай, Тянь-Шань, Джунгарский хр., Гиссаро-Дарваз (Зерав-
шанский хр.); Западный Китай. Около 15 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ TURANENA
1 (18). Виды обитают в Средней Азии и на Копетдаге.
2 (3). Раковина темная: коричневая, каштановая или бурая, просвечиваю-
щая. Устье с белой губой.............................7. Т. albolimbata.
3 (2). Раковина от белой до светло-роговой, слабо просвечивающая или не
просвечивающая. Белая губа не бросается в глаза на фоне светлой ра-
ковины.
4 (5). Раковина маленькая, низкокубаревидная, составлена 5—5У2 оборотами
........................................................ 8. Т. conicula.
5 (4). Раковина средних размеров или крупная; если маленькая, то не куба-
ревидная, а коническая. Оборотов не менее 6.
6 (7). Вид обитает на Копетдаге............................2. Т. herzi.
7 (6). Виды обитают на Памиро-Алае, Тянь-Шане и Джунгарском хр.
8 (9). Раковина большая: высота не менее 16 мм, большой диаметр не менее
10 мм ..............................................10. Т. stschukini.
9 (8). Раковина меньше: высота раковины не более 15, большой диаметр не
более 7 мм.
10 (15). Завиток стройный, с коническим контуром.
11 (12). Завиток тонкий, стройный, узкий..................5. Т. leptogyra.
12 (11). Завиток не узкий, «развалистый».
13 (14). Последние два оборота почти одной ширины. Эпифаллус примерно
вдвое длиннее пениса............................,.........4. Т. meshkovi.
14 (13). Предпоследний оборот несколько уже предпоследнего. Длина эпифал-
луса примерно равна длине пениса..........................6. Т. tenuispira.
15 (10). Контур завитка не конический, стороны его несколько выпуклые.
16 (17). Большой диаметр раковины не менее 6 мм...........9. Т. cognata.
17 (16). Большой диаметр не более 5 мм ................3. Т. martensiana.
18 (1). Вид обитает в Закавказье .........................1. Т. scalaris.
1. Turanena scalaris (Naegele, 1902) (рис. 186, XXT, рис. 187, 188).
N а е g е 1 е, 1902:6 \Buliminus (Pseudonapaeus)]; Kobelt, 1903, Iconogr., N. F.,
X : 21, fig. 1782 [B. (Pseudonapaeus)]; Акрамовский, 1949 : 149 (Ena); Л и x a p e в,
Раммельмейер, 1952 : 194, рис. 105; Акрамовский, 19716 : 7; 1976 : 160,
рис. 74, табл. VIII, 81.
Terra typica — окр. гор. Резайе в северо-западном Иране.
Место хранения типов не установлено.
Материал: 3 экз. из одного местонахождения в Армении.
Раковина почти кубаревидная, слегка блестящая. Оборотов 5х/2—6, быстро
нарастающих, выпуклых. Последний оборот к устью не поднят, высота его за-
метно превышает половину высоты раковины. Окраска одноцветно роговая.
Скульптура в виде тонкой слегка косой радиальной исчерченности. Устье оваль-
ное, сильно косое, места его прикрепления сближены умеренно. Колумелляр-
ный край отвесный, при переходе в базальный широко закруглен, палатальный
плавно и широко дугообразно выгнут. Губа развита относительно слабо. Пупок
открытый, проколовидный.
Размеры: ВР 6.5—7, БД 3.5—4 мм; в первоописании: ВР 7, БД 4 мм.
Внутреннее строение. Материал: 3 экз. из Армении, Даралагез,
с. Гнишик, 1941 г., сбор Э. А. Давтяна, определение Н. Н. Акрамовского.
Белковая железа с резкой седловидной апикальной вырезкой и глубокими
желобками на боковых поверхностях. Нижний отдел вагины в 2—2х/2 раза ко-
роче верхнего. Семяпровод впадает в эпифаллус под острым углом несколько
18*
216
VI. НАДСЕМ. BULIMINOIDEA
Рис. 186. Виды рода Тигапепа.
I— Т. stschukini (Lindh.), паралектотип; 11 — Т. cognata (Lindh.), паралектотип; III — то же,Гиз пере-
вала Орток-бель на Чаткальском хр.; IV — то же, из долины р. Касансай на Чаткальском хр.; V— Г. ,te-
nuispira Schileyko, sp. nov., голотип; VI — то же, из с. Арсланбоб, Ферганский хр.; VII — Т. meshkovl
Schileyko, sp. nov., голотип; VIII — то же, паратип; IX — Т. leptogyra (Lindh.), лектотип; X —тоже,
из поймы р. Пскем; XI — то же, из бокового ущелья в долине р. Пскем; XII — Т. albolimbata (Lindh.),
паралектотип; XIII — то же, из с. Чардоварна Киргизском хр.; XIV — то же, издолиныр. Ала-бука-сай
на Чаткальском хр.; XV — Т. martensiana (Ancey) из окр. оз. Искандер-куль на Гиссарском хр.; XVI —
то же, из с. Бахмир, Заалайский хр. [лектотип Т. liostraca (West.)l; XVII — Т. herzi (Bttg.) из Шахруда,
Иран; XVIII — Т. conicula (Mart.), голотип; XIX — то же, из окр. оз. Маргузор на Зеравшанском хр.;
XX — то же, из с. Арсланбоб, Ферганский хр.; XXI — Т. scalaris (Naeg.) из Армении, с. Гнишик.
3. РОД TURANENA
277
отступя от конца, в результате чего образуется короткий притупленный (по
определению Н. Н. Акрамовского, 1976, «шпателевидный») бич. Диаметр эпи-
фаллуса от места впадения семяпровода к пенису постепенно уменьшается,
приблизительно на середине его длины расположен небольшой цэкум. Пенис
в дистальной части тонкий, цилиндрический, в проксимальной — коротковере-
теновидный. Внутри пениса имеется система складок, сильно напоминающая
V-образные складки среднеазиатских представителей подсемейства и представ-
ляющая собой модификацию последних. Хорошо заметна одна V-образная
складка, рядом с которой заметны следы второй такой же складки. Все отделы
пениального аппендикса хорошо развиты; Аъ непривычно мал и короток. Про-
Рис. 187. Тигапепа sea- Рис. 188.§ Тигапепа scalaris (Naeg.) из Армении, с. Гнишик,
laris (Naeg.) из Армении, 1941 г. Половой аппарат и вскрытый пенис.
с. Гнишик.
токТсемяприемника прямой, резервуар маленький, шаровидный, проток резер-
вуара примерно вдвое длиннее самого резервуара. Дивертикул хорошо развит,
достигает белковой железы.
Как признаки раковины, так и анатомические черты свидетельствуют о без-
условной близости этого вида к среднеазиатским видам. Однако Т. scalaris
имеет и ряд отличий: размер и форма резервуара семяприемника, форма бича,
пропорции и сравнительное развитие отделов пениального аппендикса. По-
скольку неизвестно внутреннее строение типового вида рода Тигапепа, который
к тому же занимает промежуточное положение между закавказским и средне-
азиатскими видами и по конхологическим признакам и по географическому по-
ложению, следует пока воздержаться от выделения Т. scalaris в отдельный под-
род. Обсуждать таксономическую структуру рода в целом можно будет только
после анатомирования как типового вида, так и многих видов, обитающих в Ги-
малаях и в Индостане.
Распространение и местообитание. Вид известен из двух
мест: окр. г. Резайе в Северо-Западном Иране и с. Гнишик в Армении, где оби-
тает в трещинах скал в ксерофильном редколесье на южном склоне в урочище
Джафаригюх (Акрамовский, 1976).
2. Тигапепа herzi (О. Boettger, 1889) (рис. 186, XVII', рис. 189).
О. Boettger, 1889b : 950, t. 17, fig. 14 [Buliminus (Pseudonapaeus)].
278
ГЛ НАДСЕМ. BULIMINOJDEA
Terra typica — Иран, Эльбурс, окр. Шахруда.
Лектотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 156687.
Материал: 2 не вполне сформированных раковины из типового место-
нахождения.
Раковина высококоническая с переходом к удлиненно-овальной, умеренно
твердостенная, слегка блестящая. Оборотов 51/2—6, выпуклых. Последний обо-
Рис. 189. Тигапепа
herzi (Bttg.) из Шах-
руда, Иран. Раковина
не вполне сформирован-
ная.
Рис. 190. Тигапепа mar-
tenslana (Ancey) из окр.
оз. Искандер-куль на
Гиссарском хр.
рот к устью не поднят, высота
его заметно превышает половину
высоты раковины. Верхние обо-
роты рыжеватые, нижние —свет-
ло-сррые. Поверхность эмбрио-
нальных оборотов с настолько-
слабой кососпиральной струй-
чатостью, что обнаружить ее
удается лишь при значитель-
ном увеличении, поворачивая
раковину в сильном косом пуч-
ке света. Дефинитивная скульп-
тура в виде довольно правиль-
ных, но нерезких радиальных
Морщинок, окрашенных чуть
светлее фона. Устье с тонкими
отвернутыми краями. Фотогра-
фию лектотипа см.: Wenz, Zilch,.
1959-1960, S. 185, Abb. 641.
Размеры; BP 8—9, БД 4—5; лектотип: ВР 7, БД 4 мм.
Распространение и местообитание. Вид известен из ти-
пового местонахождения на юго-восточных склонах Эльбурса и найден на Иран-
ском плато (Biggs, 1962). Не исключено, что вид живет и на территории юго-
западной части Туркмении. Обитает на скалах.
3. Тигапепа martensiana (Ancey, 1886) (рис. 186, XV, XVГ, рис. 190, 191).
Мартенс, 1874 : 21, табл. 2, рис. 16 (Buliminus segregates Reeve var. minor);. A n c e y,
1886a : 45 (Buliminus)-, Westerlund, 1887 : 71 (Buliminus)-, 1896 : 192 (Buliminus (Ma-
stoides) liostracas]; Kobelt, 1902 : 522, t. 83, fig. 11 (Buliminus); Л и x a p e в, Раммель-
мейер, 1952 : 193; M а т ё к и.н, 1966 : 112, табл. 3, рис. 6 [Zebrina (Тигапепа)].
Terra typica — горы близ Шахимардана, предгорья Алая.
Место хранения типов не установлено.
Материал: более 50 экз. из 6 местонахождений на Зеравшанском, Турке-
станском, Гиссарском, Алайском, Заалайском хребтах и на хр. Кунгей.
Раковина коническая с уклоном в сторону овально-конической, умеренно
твердостенная, слабо блестящая. Оборотов 5—6, умеренно выпуклых, довольно
быстро нарастающих, разделенных глубоким швом. Последний оборот не-
сколько приподнят к устью, его высота составляет заметно больше половины
высоты раковины, но меньше 2/3 этой величины. Окраска от желтовато-белой до-
светло-коричневой. Дефинитивная скульптура в виде вялых косорадиальных
морщинок, но в целом раковина выглядит почти гладкой. Устье овальное, края
его умеренно и равномерно отвернуты, места прикрепления сближены и сое-
динены тонкой, часто прозрачной мозолью. Пупок в виде короткой округлой
щели.
Размеры: ВР 8—12, БД 4—5 мм; в первоописании: ВР 9, БД 4 мм.
Внутреннее строение. Материал: 4 экз. из окрестностей оз. Ис-
кандер-куль на Гиссарском хр., 23 июля 1968 г.; 3 — из окр. с. Танги-сурх,
берег р. Исфаринка, 14 июня 1977 г.; сборы и определения 3. Иззатуллаева.
Белковая железа с вялой апикальной вырезкой, без желобков на боковых
поверхностях. Верхний отдел вагины незначительно короче нижнего. Семя-
3. РОД TURANENA
279
провод подходит под прямым углом к эпифаллусу, бич конический и изогнутый.
Эпифаллус цилиндрический, недлинный, цэкум делит его на отрезки 1 : 2—
1:3. Пенис подразделяется на дистальный цилиндрический и проксимальный
веретеновидный отделы. Внутри проксимального отдела залегает высокая тон-
кая V-образная складка, ветви которой уходят в просвет эпифаллуса; гребень
складки неправильно гофрирован. Аг и А2 слиты в один булавовидный отдел,
длина которого не превышает длину пениса; остальные отделы пениального
аппендикса ясно выражены, но сравнительно короткие. Проток семяприемника
прямой, короткий; неболь-
шой овальный резервуар
с собственным протоком,
длина которого примерно
равна длине самого ре-
Рис. 191. Тигапепа martensiana (Ancey) из окр. оз. Иская-
дер-куль на Гиссарском хр., 23 VII 1968 г. Половой аппарат
и вскрытый пенис.
Рис. 192. Тигапепа те-
shkovi Schileyko, sp. nov.
Голотип.
зервуара. Дивертикул короткий, заметно не доходит до белковой же-
лезы.
Распространение и местообитание. От Туркестанского,
Зеравшанского и Гиссарского хребтов через Алайский, Заалайский и Ферган-
ский хребты до юго-западных склонов хребта Кунгей. Обитает на каменистых
открытых склонах и на сухих скалах.
4. Тцгапепа meshkovi 1 Schileyko, sp. nov. (рис. 186, VII, VIIT, рис. 192,
193).
Terra typica — долина p. Пскем в 15 км выше с. Нанай, западный Тянь-
Шань.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: более 60 экз. из двух местонахождений в долине р. Пскем.
Раковина удлиненно овально-коническая, сравнительно стройная, умеренно
блестящая, умеренно твердостенная, слегка просвечивающая. Оборотов 6, вы-
пуклых, предпоследний оборот почти одной ширины с последним или незначи-
тельно уже. Последний оборот к устью слегка плавно приподнят, высота его
немного больше половины высоты раковины. Окраска верхних оборотов каш-
1 Вид назван именем Юрия Андреевича Мешкова, плодотворно помогавшего мне в сборе
материала в Средней Азии.
280
VI. НАДСЕМ. BULIMINOIDEA
тановая, она может сохраняться и на нижних оборотах, но чаще два последних
оборота светлее за счет развития размытых белесоватых пестрин, штрихов и
пятен. Дефинитивная скульптура в виде тонких косорадиальных морщинок;
местами имеются редкие легкие вмятины. Устье овальное, места его прикреп-
ления сильно сближены и соединены тонкой мозолью. Края устья умеренно и
равномерно отвернуты, по линии отгиба располагается невысокая и неширокая
белая губа. Колумеллярный край расширен, отвесный, базальный широко
закруглен, палатальный плавно выгнут и немного оттянут в сторону. Пупок
узкий, открытый, овальный.
Рпс. 193. Turanena meshkovi Schileyko, sp. nov.
I — голотип, половой аппарат и вскрытый пенис; II — паратип, вскрытый пенис.
Размеры: ВР 11—14, БД 5—7.5 мм; голотип: ВР 12.7, БД 5.9 мм.'1г1
Внутреннее строение. Материал: 15 экз. из узкого ущелья^па
правом берегу р. Пскем, 21 мая 1972 г., сбор и определение автора.
Белковая железа с глубокой седловидной апикальной вырезкой. Отделы
вагины приблизительно одной длины. Эпифаллус составляет естественное про-
должение семяпровода, бич тонкий, коническо-цилиндрический, располага-
ется под разным углом к семяпроводу. Эпифаллус тонкий и относительно очень
длинный, хорошо развитый бугорковидный цэкум располагается примерно на
его середине. Пенис веретеновидный или булавовидный, в разной степени изог-
нут или прямой, заметно толще эпифаллуса. Пара V-образных складок на вну-
тренних стенках пениса имеется, но маскируется наличием дополнительных
складок и аностомозами; существенно, что нижние их концы не прерываются
резко, как у всех других видов, но плавно сходят на нет (рис. 193,1, II). Пени-
альный аппендикс хорошо развит. Аг тонкий и длинный, не короче пениса,
небольшой, но ясно обособлен, А3 короткий, выражен в разной степени, у не-
которых экземпляров почти неотличим от Л4. Ветви полового ретрактора тесно
сближены на диафрагме, одна из них подходит к середине пениса, другая к верх.
3. РОД TURANENA
281
ней части Аг. Проток семяприемника прямой; удлиненно-овальный или груше-
видный резервуар плавно переходит в проток, длина которого примерно равна
длине резервуара. Дивертикул почти доходит до белковой железы, на конце
иногда расширен.
По раковине вид чрезвычайно похож на Т. tenutspira, но резко отличается
относительно гораздо более длинным и тонким эпифаллусом и расположением
и формой складок на внутренней поверхности пениса: у Т. tenuispira имеется
одна V-образная складка, резко прерванная спереди. От другого близкого
вида — Т. leptogyra — отличается относительно более широким завитком и вну-
тренним строением пениса; вообще по последнему признаку вид отличается от
всех других видов рода.
Распространение и местообитание. Вид найден пока
только в долине р. Пскем в двух типах биотопов: в осыпях и на скалах, располо-
женных на припойменных террасах правого берега, и в узком безлесном ущелье,
сплошь заваленном камнями; стены этого ущелья почти целиком представляют
собой подвижные осыпи. В обоих типах биотопов вид встречен вместе с Т. lepto-
gyra.
5. Turanena leptogyra (Lindholm, 1927) (рис. 186, IX, X, XT, рис. 194, 195).
Lindholm, 1927а : 99 [Ena (Turanena)}', Лихарёв, Раммельм е'й е р, 1952 :
194 (Turanena martensiana part.).
Terra typica — Чаткальский хр., с. Балгут между перевалами Март и Кум-
бель.
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
здесь.
Материал: более 70 экз. из 6 местонахождений на Чаткальском, Пскем-
ском, Таласском и Алайском хребтах.
Раковина от высококонической до высококубаревидной; в любом случае
завиток образует правильный стройный конус; последний оборот может быть рас-
ширенным. Оборотов 6—6V2. У первого дефинитивного
оборота периферия совпадает со средней линией, у после-
дующих периферия постепенно сдвигается все ближе к
нижнему шву (это лучше видно на левой стороне ракови-
ны). Последний оборот к устью слегка приподнят, высо-
та его немного больше половины высоты раковины или
почти равна ей. Окраска от роговой до белесовато-корич-
невой, обычно со светлыми штрихами; верхние обороты ча-
ще всего темнее нижних. Скульптура в виде сильно размы-
тых тонких неравномерно расположенных морщинок. Устье
овальное, его длинная ось может быть почти параллельной
Рис. 194. Turanena leptogyra (Lindh.). Лектотип.
оси столбика, но может образовывать с ней заметный угол. На границе колумел-
лярного и базального краев имеется слабый закругленный угол, остальные края
образуют плавную дугу. Края устья прямые или слегка отвернуты, в послед-
нем случае по линии перегиба располагается очень тонкая, часто нитевидная
белая губа. Если устье не оттянуто в сторону, пупок проколовидный, если оття-
нуто — в виде короткой узкой щели.
Размеры: ВР 6—13, БД 3.5—6.5 мм; лектотип: ВР 10.0, БД 4.4 мм.'
Внутреннее строение. Материал: 2 экз. из ущелья Беш-Таш на
Таласском хребте близ гор. Талас, 2 июня 1972 г.; 9 — из узкого ущелья на пра-
вом берегу р. Пскем выше с. Нанай, 21 мая 1972 г.; 8 — из припойменной тер-
расы на правом берегу р. Пскем, 17 мая 1972 г.; сборы и определения автора.
282
VI. НАДСЕМ. BULIMINOIDEA
Белковая железа с очень вялой апикальной вырезкой. Нижний отдел вагины
почти равен верхнему или немного короче. Семяпровод впадает в эпифаллус под
различным углом — от прямого угла до очень острого, когда эпифаллус со-
ставляет прямое продолжение семяпровода, а бич почти лежит на семяпроводе.
Цэкум расположен выше середины эпифаллуса. Эпифаллус немногим тоньше
пениса, если последний имеет цилиндрическую форму; чаще пенис булавовид-
ный, внутри него залегает ясно выраженная V-образная складка, причем место
слияния ветвей этой складки расширено и образует короткий, но мощный гре-
бень. Все отделы пениального аппендикса выражены. Ветви полового ретрак-
тора могут крепиться к диафрагме как независимо одна от другой, так и единой
Рис. 195. Тигапепа leptogyra (Lindh.) из долины р. Пскем, 21 V 1972 г.
I — половой аппарат, II — вскрытый пенис, III — другой экземпляр (пенис и пениальный аппендикс
вывернуты).
лентой. Одна ветвь подходит к пенису немного ниже его середины, вторая —
к верхней части Аг. Проток семяприемника прямой или слегка изогнутый, ре-
зервуар небольшой, овальный, его проток такой же длины, как и сам резер-
вуар. Дивертикул развит в различной степени, его конец часто расширен.
Распространенней местообитание. Вид найден на Пскем-
,ском, Таласском, Чаткальском и Алайском хребтах. Населяет сухие открытые
осыпи и склоны.
6. Тигапепа tenuispira Schileyko, sp. nov. (рис. 186, V, VI; рис. 196, 197).
Terra typica — ущелье p. Кара-Костек близ Алма-Аты.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 19 экз. из двух местонахождений: Алма-Ата и Арсланбоб.
Раковина высококоническая, относительно стройная, твердостенная, слабо
просвечивающая, завиток имеет почти правильно конический контур, вершина
узкая, закругленная. Оборотов 61/2 выпуклых, плавно нарастающих, шов глу-
3. РОД TURANENA
283
бокий. При нормальном положении раковины контуры отдельных оборотов,
расположенные справа от оси раковины, правильно закруглены, а контуры, ле-
жащие слева, немного оплывшие, т. е. стенка оборота отлого спускается, а за-
тем к шву широко закругляется. Верхние обороты желтовато-роговые, два
нижних белые, но цветовая граница отсутствует, и на нижних оборотах также
имеются отдельные размытые пятна и штрихи желтовато-рогового цвета.
Эмбриональные обороты полированные, дефинитивные покрыты тонкими, но
резкими радиальными морщинками, расположенными без видимой системы,
причем морщинки выражены по-разному даже в пределах одного оборота.
Помимо морщинок на пос-
леднем обороте, а часто и
на предпоследнем, имеются
отдельные грубые вмятины.
Устье округлое, с широ-
ко отвернутыми краями;
Рис. 196. Тигапепа tenui-
spira Schileyko, sp. nov.
Голотип.
Рис, 197. Тигапепа tenuispira Schileyko, sp. nov. Паратип.
Половой аппарат и вскрытый пенис.
места перегиба слегка усилены расплывшейся губой. Места прикрепления устья
сильно сближены. Пупок не сквозной, в виде изогнутой щели.
Размеры: ВР 12—14, БД 5.5—7 мм; голотип: ВР 13.7, БД 6.7 мм.
Внутреннее строение. Материал: 3 экз. из типовой серии: окр.
Алма-Аты, ущелье р. Кара-Костек, июнь 1969 г., сбор С. Д. Карабаевой, опре-
деление автора.
Белковая железа со слабой апикальной вырезкой. Нижний отдел вагины
незначительно короче верхнего. Семяпровод впадает в эпифаллус под углом, бич
конический, изогнутый. Цэкум резко сдвинут к бичу. Эпифаллус недлинный,
цилиндрический, неясно отграничен от пениса. Тонкий дистальный конец пе-
ниса значительно длиннее слегка вздутого проксимального, содержащего V-
образную складку. неясно отграничен от А2, А3 короткий, четко обособлен.
Половой ретрактор отходит от диафрагмы двумя самостоятельными лентами,
одна из которых подходит к границе между отделами пениса, а другая— к гра-
нице между А у и А2. Проток семяприемника относительно очень короткий, почти
такой же длины, как суммарная длина резервуара и его протока. Дивертикул
значительно длиннее, доходит до белковой железы. Внутри дивертикула, изоб-
раженного на рис. 197, находится сперматофор.
S. г Вид близок к Т. leptogyra, отличается большими размерами и заметно более
отвернутым устьевым краем. Из анатомических отличий надо указать на поло-
284
VI. НАДСЕМ. BULIMINOIDEA
жение цэкума (очень близко к бичу), значительно более короткий А3 и также
гораздо более короткий проток семяприемника.
Распространение и местообитание. Кроме типового место-
нахождения в окр. Алма-Аты, материал из которого изучен анатомически, в Зоо-
логическом ин-те АН СССР хранится сбор Л. Молчанова, сделанный в зоне ело-
вых лесов близ Арсланбоба (Ферганский хр.), содержащий 14 пустых раковин,
практически неотличимых от раковин типовой серии. В типовом местонахожде-
нии моллюски собраны на скалах.
7. Turanena albolimbata (Lindholm, 1927) (рис. 186, XII, XIII, XIV;
рис. 198, 199).
Lindholm, 1927а : 98 [Ena (Turanena)]-, Лихарев, Раммельмейер, 1952 :
193—194 (martenslana var. albolimbata)-, Матёкин, 1966:112, табл. 3, рис. 5 [Zebrina
(Turanena)].
Terra typica — верховья p. Турдук (бассейн p. Нарын), Чаткальский хр.
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
здесь.
Материал: 29 экз. из трех местонахождений на Чаткальском и Киргиз-
ском хребтах.
Раковина высококоническая, сравнительно тонкостенная, блестящая, с узко'
закругленной вершиной. Завиток имеет не строго конические, слегка выпуклые
контуры. Оборотов 5х/2—6, умеренно выпуклых; послед-
ний оборот более или менее заметно оплывает, и самая
выпуклая его часть расположена много ниже периферии.
К устью последний оборот плавно и незначительно'
поднят, высота его заметно превышает половину вы-
соты раковины. Окраска темная, коричневая, каштановая,
иногда с легким лиловатым оттенком, за устьем имеется раз-
мытое светлое пятно. При увеличении около 50 раз
на эмбриональных оборотах заметна тончайшая слабо
различимая спиральная исчерченность; дефинитив-
Рис. 198. Turanena albolimbata (Lindh,). Паралектотип.
ные обороты несут нерезкие радиальные морщинки, расположенные то чаще, то
реже; иногда соседствующие морщинки не вполне параллельны одна другой.
Кроме того, особенно на двух последних оборотах, имеются вмятины. Устье
овальное, слегка оттянуто в сторону, края его слегка утолщены, белые, губа
очень слабая. Пупок узкий, расположен на дне довольно широкой пупочной
впадины.
Размеры: ВР 9—И, БД 5—6 мм; в первоописании: ВР 9.33—11.0, БД 5—
5.75 мм; лектотип: ВР 9.7, БД 5.0 мм.
Внутреннее строение. Материал: 4 экз. из Киргизского хр., Ма-
линовая щель над с. Чардовар, 9 июня 1972 г.; 1 — из Киргизского хр., ущелье
Карабалты, 11 июня 1972 г.; сборы и определения автора.
Белковая железа с заметной вырезкой, занимающей не только вершину ор-
гана, но и часть боковой поверхности. Нижний отдел вагины в ПД—2 раза ко-
роче верхнего. Семяпровод впадает в эпифаллус под почти прямым углом. Четко
выраженный цэкум расположен несколько выше середины цилиндрического эпи-
фаллуса, внешне слабо отграниченного от пениса. V-образная складка внутри
пениса очень резкая, высокая, ветви ее сближены так, что между ними остается
очень узкое пространство. На вывернутом пенисе эта складка хорошо заметна
(рис. 199, II). Все отделы пениального аппендикса хорошо выражены. А1 от-
носительно длинный и массивный, отделен от небольшого А2 вялым пережимом;
3. РОД TURANENA
285
А3 плавно, без резкой границы переходит в А4. Половой ретрактор крепится
к диафрагме независимыми ветвями, одна из которых присоединяется к средней
части пениса, а другая — к А4 заметно отступя от А2. Проток семяприемника
прямой, длиннее дивертикула, резервуар небольшой, с собственным коротким
протоком. 1
Рис. 199. Turanena albolimbata (Lindh.) из окр. с. Чардовар на Киргизском хр., 9 VI 1972 г.
1 — половой аппарат и вскрытый пенис, II — другой экземпляр (пенис и часть пениального аппендикса
вывернуты).
Распространение и местообитание. Вид найден на Чат-
кальском и Киргизском хребтах, в осыпях и на скалах.
8. Turanena conicula (Е. Martens, 1882) (рис. 186, XV111—XX', рис. 200, 201).
Е.“»М а г t е n s, 1882а : 23, t. 3, fig. 3 (Buliminus}.
Terra typica — Кульджа (Западный Китай).
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 16 экз. из трех местонахождений: Кульджа; предгорья
Алайского хр.; Зеравшанский хр.
Раковина короткоконическая, почти кубаревидная, умеренно твердостен-
ная, матовая. Оборотов 5—выпуклых, последний оборот к устью не поднят
или поднят едва заметно, высота его немного больше половины высоты раковины.
Верхние обороты рыжеватые, нижние — известково-белые. Скульптура в виде
довольно грубой неравномерной радиальной исчерченности. Устье округлое,
края его не отвернуты или слабо отвернуты, губа почти не развита, места при-
крепления устья относительно слабо сближены. Пупок проколовидный.
Размеры: ВР 5—6.5, БД 4—4.5 мм; голотип: ВР 6, БД 4 мм, то же в перво-
описании.
Внутреннее строение. Материал: 4 экз. из Зеравшанского хр.,
берег оз. Маргузор, 8 августа 1973 г., сбор Ю. А. Мешкова, определение автора.
286
VI. НАДСЕМ. BULIMINOIDEA
Белковая железа с очень плавной апикальной вырезкой. Нижний отдел ва-
гины незначительно короче верхнего. Семяпровод впадает в эпифаллус под не-
которым углом, бич развит хорошо, притупленно-конический. Цэкум располо-
жен выше середины цилиндрического эпифаллуса, диаметр которого меньше
диаметра пениса. На внутренней стенке пениса залегает мощная V-образная
складка, нижняя часть которой слегка обособлена, т. е. отстает от стенки пениса.
Отделы пениального аппендикса хорошо развиты, граница между As и не-
резкая. Ветви полового ретрактора отходят от диафрагмы независимо одна от
другой; одна ветвь крепится к пенису близ его середины, вторая — к слегка
отступя от А2. Проток семяприемника короткий, прямой, резервуар с собствен-
Рис. 200. Тигапепа coni-
cula (Mart.). Голотип.
Рис. 201. Тигапепа conicula (Mart.) из окр. оз. Маргузор
на Зеравшанском хр., 8 VIII 1973 г. Половой аппарат и вскры-
тый пенис.
ным коротким протоком, длина которого приблизительно равна длине самого ре-
зервуара, дивертикул хорошо развит, почти доходит до белковой железы.
Распространение и местообитание. Вид найден только
в трех местах: Кульджа, Алайский хр. (с. Чаувай) и Зеравшанский хр. (окр.
оз. Маргузор). Населяет очень сухие каменистые участки.
9. Тигапепа cognata (Lindholm, 1927) (рис. 186, II—IV; рис. 202, 203).
L i n d h о 1 m, 1927а : 99 [Ena (Тигапепа)]; Лихарев, Раммельмейер, 1952 :
193 (Тигапепа stschukini part.).
Terra typica — нижнее течение р. Турдук, Цаткальский хр.
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается здесь.
Материал: более 60 экз. из трех местонахождений на Ферганском и
Чаткальском хребтах.
Раковина высококоническая, слегка вздутая, с узко закругленной вершиной.
Контур завитка не вполне конический за счет того, что предпоследний оборот
лишь немного уже последнего. Оборотов 5х/2—6, выпуклых, последний оборот
к устью почти не поднят, высота его всегда заметно превышает половину высоты
раковины. Окраска от светло-роговой и рыжеватой до светло-серой, верхние
обороты немного темнее нижних. На поверхности эмбриональных оборотов при
увеличении более чем в 50 раз можно рассмотреть исключительно тонкую и
3. РОД TURANENA
287
частую спиральную исчерченность (заметна только на свежих раковинах), хотя
с первого взгляда обороты выглядят полированными. Скульптура дефинитивных
оборотов в виде множества косорадиальных неправильно, расположенных мор-
щин и многочисленных вмятин. Устье овальное, с умеренно отвернутыми тон-
кими краями; по линии перегиба заметна слабая губа. Пупок маленький, не
сквозной, овальный.
Размеры: ВР 11—15, БД 5—8 мм; лектотип: ВР 13.0, БД 6.6 мм.
Внутреннее строение. Материал: 3 экз. из перевала Орток-бель
на Чаткальском хр., 18 мая 1972 г., сбор Р. Я. Бутылина, определение автора.
Рис. 202. Тигапепа cog-
nata (Lindh.). Паралек-
тотип.
Рис. 203. Тигапепа cognata (Lindh.) с перевала Орток-бель на
Чаткальском хр., 18 V 1972 г. Половой аппарат и вскрытый пе-
нис.
Белковая железа с глубокой вырезкой, занимающей апикально-латеральное
положение. Отделы вагины приблизительно равной длины. Семяпровод впадает
в эпифаллус под прямым углом, бич короткий, конический. Эпифаллус сравни-
тельно длинный, цилиндрический; граница между ним и пенисом внешне не
выражена. Цэкум сильно сдвинут к бичу. Внутри цилиндрического пениса, не
подразделенного на отделы, залегает четкая V-образная складка. Все отделы
пениального аппендикса хорошо выражены. А1 не короче пениса; Аг небольшой,
обособленный; А3 плавно переходит в Л4. Ветви полового ретрактора на диаф-
рагме разделены, одна ветвь крепится к срединной части пениса, вторая —
к At заметно отступя от М2. Проток семяприемника почти прямой, резервуар не-
большой, овальный; дивертикул не доходит до белковой железы и заканчивается
расширением, формой и размером напоминающим резервуар.
Распространение и местообитание. Ферганский и Чат-
кальский хр. Встречается на открытых сухих каменистых склонах.
10. Тигапепа stschukini (Lindholm, 1927) (рис. 186, Г, рис. 204).
Lindholm, 1927а : 100 [Епа (Тигапепа)]; Лихарев, Ра мм ельмейер,
1952 : 193, рис. 104; Матёкин, 1966 : 112 [Zebrina (Тигапепа)].
288
VI. НАДСЕМ. BULIMINOIDEA
Terra typica — долина р. Джуван-Сюгет, правый приток р. Узун-Ахмат'
(бассейн р. Нарын).
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
здесь.
Материал: 14 экз. из типовой серии и 3 из Ферганского хр. Раковина
высококоническая,
твердостенная, блестящая. Оборотов 6—б1^, последний
оборот к устью довольно сильно, но плавно поднят,,
высота его заметно больше половины высоты рако-
вины. Окраска верхних оборотов рыжеватая, за-
тем постепенно светлеет, и последние два оборота бе-
лые, иногда со слабым лиловатым оттенком. Поверх-
ность эмбриональных оборотов с крайне тонкой
спиральной струйчатостью, скульптура дефинитив-
ных — тонкие радиальные морщинки, среди кото-
рых встречаются более грубые. Местами имеются
вмятины. Устье овальное, слабо скошено, внутри
темно-желтое. Париетальная мозоль слабая. Края
устья тонкие, широко отвернуты, губа сильно раз-
мыта. Пупок узкощелевидный.
Рис. 204. Тигапепа stschukini (Lindh.). Паралектотип.
Размеры в типовой серии: ВР 17—19.5, БД 10.5—11.8 мм; лектотип: ВР 18.0,
БД 10.2 мм. Раковины, собранные на Ферганском хребте, несколько мельче:
ВР 15-16, БД 8-9.5 мм.
Распространение и местообитание. Помимо типового
местонахождения на Чаткальском хр. сделана находка на Ферганском хр.
(Майли-сай), на открытом склоне (сбор 3. Иззатуллаева 15 УП 1979 г.).
4. Род PSEUDOCHONDRULA Hesse, 1933
Hesse, 1933 : 167; Лихарев, Раммельмейер, 1952 : 179, Wenz, Zilch,
1959—1960 : 183 (Jmparietula part.); Шилейко, 1978г : 845.
Типовой вид Buliminus florenskii Rosen, 1914 (syn. Pupa seductilis
Rossmaessler, 1837); по первоначальному обозначению.
Раковина цилиндрическая или овально-цилиндрическая, слабо исчерчена,
обороты слабовыпуклые. Устье без зубов или с 1—4 зубами. Края устья отвер-
нуты, места его прикрепления расставлены.
Бич почти отсутствует, представлен лишь самой терминальной частью эпи-
фаллуса, на конце притуплен. Кнутри пениса имеется пара V-образных скла-
док. Папилла пениса отсутствует. Проток семяприемника рукавообразный,
без резко выраженного резервуара и без дивертикула.
Распространение. Юго-восточная Европа, Малая Азия, Кавказ.
5—6 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ PSEUDOCHONDRULA
1 (4). Раковина левозавитая.
2 (3). Устье раковины с зубами .................................1. Р. seductilis.
3 (2). Устье раковины без зубов .................................4. Р. sinistrorsa.
4 (1). Раковина правозавитая.
5 (6). На париетальной стенке устья имеется развитый в разной степени зуб
................................................... 2. Р. tetrodon.
6 (5). Париетальная стенка устья лишена зубов.
4. РОД PSEUDOCHONDRULA
289
7 (8). В устье имеется палатальный зуб................... . 5. Р. tuberifera.
8 (7). Устье без зубов.........................................3. Р. lederi.
1. Pseudochondrula seductilis (Rossmaessler, 1837) (рис. 205, 7; рис. 206).
Rossmaessler, 1837, Iconogr., I, 4 : 10, t. 23, fig. 306 (Pupa)', Mortillet,
1854 : 8 (Buliminus seductilis var. armeniaca); Mousson, 1873 : 206 (Chondrus scapus var.
destitutus); 1876 : 42 (Ch. sagas var. destituta); O. Boettger, 1880b : 154, t. 5, fig. 5 (Buli-
minus komarovi); Retowski, 1883b : 55 \B. (Chondrula) incertus]; R о s.e n, 1914 : 188,
t. 2, fig. 10 (B. florenskii); Ретовский. 1915 : 45 [5. (Chondrula) lindholmi]; Hesse,
1933 : 171, fig. 13 (florenskii), 172, fig. 15 (seductilis); F о r c a r t, 1940 : 209, t. 2, fig. 42 (Im-
parietula armeniaca florenskii), 216, t. 2, figs. 48—50 (Imparietula); Hudec, Vasa t ko,
1971 : 3, Abb. 1, pl. I, 2 (Imparietula (Imparietula)}; Акрамовский, 1976 : 158, рис. 71Г,
табл. VII, 78 (Imparietula); Шилейко, 1978в : 521, рис 5, II.
Terra typica — Далмация (Югославия).
Типы, вероятно, хранятся в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: более 70 экз. из 6 местонахождений в Армении, Грузии
и северной Турции.
Раковина левозавитая, цилиндрическая, реже овально-цилиндрическая или
булавовидная, умеренно твердостенная или очень твердостенная, блестящая;
Рис. 205. Виды рода Pseudochondrula.
I — Р. seductilis (Rssm.) из окр. Ленинакана, Армения; II — Р. tetrodon (Mort.) из Армении, Даралагез,
урочище Хортых; III — Р. tuberifera (Bttg.) из Северной Осетии, Зругское ущелье; IV — Р. lederi (Bttg.)
из Северной Осетии, Зругское ущелье; V — Р. sinistrorsa Kokotschashvili et Schileyko, sp. nov., голотип.
свежие раковины иногда сильно просвечивающие. Оборотов 9—11, уплощен-
ных, разделенных мелким неровным швом. У цилиндрических раковин диа-
метр последних 4—5 оборотов почти одинаковый. Высота последнего оборота
в 2—3 раза меньше высоты раковины; к устью последний оборот плавно и слегка
приподнят. Окраска одноцветно-серая или светло-роговая, реже почти белая,,
верхние обороты часто темнее нижних. Эмбриональные обороты гладкие, дефи-
нитивные покрыты вялыми сглаженными тонкими неравномерно расположен-
ными радиальными морщинками; элементы спиральной скульптуры отсутствуют.
Устье овальное, края его слабо отвернуты, утолщенные, белые. Места прикреп-
ления устья расставлены и связаны тонкой мозолью. Зубов 3—4, степень их
развития подвержена сильной индивидуальной и биотопической изменчивости.
На Ширакском нагорье близ Ленинакана мной была собрана серия из 43 рако-
вин, в пределах которой находились особи с сильно развитыми зубами, и ра-
ковины, у которых колумеллярные зубы были развиты крайне слабо. В наибо-
лее полном наборе устьевая арматура выглядит следующим образом. Короткая
гребневидная париетальная пластинка заметно сдвинута к ангулярной об-
ласти, бугорковидный наружный колумеллярный зуб отделен ложбинкой от
расплывшейся колумеллярной пластинки, уходящей по столбику на несколько
19 А. А. Шилейко
290
VI. НАДСЕМ. BULIMINOIDEA
оборотов. Палатальный зуб поверхностного типа, расположен почти на крае
устья в виде приостренного бугорка. Пупок открытый, маленький, округло-
щелевидный.
Размеры: ВР 7.5—13.5, БД 3.5—4.2 мм; в первоописании: ВР 4, БД 13,4
линии, т. е. ВР около 8.5, БД около 3.7 мм.
Внутреннее строение. Материал: 10 экз. из Армении, Ширак-
ское нагорье, близ Ленинакана, 18 апреля 1971 г., сбор и определение автора.
Белковая железа без вырезки. Нижний отдел вагины в 2—3 раза короче
верхнего. Семяпровод впадает в эпифаллус не под углом, эксцентрично. Бич
рудиментарный, приострен-
ный, соединен с семяпрово-
дом тонкой прозрачной пе-
репонкой. Эпифаллус срав-
нительно длинный, резко
выраженный цэкум рас-
положен близ его середи-
ны, выше цэкума эпифал-
лус несколько утолщен. Пе-
пис цилиндрический, була-
вовидный или веретеновид-
ный, внутри него распо-
ложена пара V-образных
складок, внутренние ветви
которых уходят в эпифал-
лус, а наружные смыкают-
ся над отверстием эпифал-
луса и образуют козырек.
Рис. 206. Pseudochondrula sedu-
ctilis (Rssm.) из Ширакского на-
горья близ Ленинакана, Армения,
8 IV 1971 г. Половой аппарат и
вскрытый пенис (из: Шилейко,
1978в).
Все отделы пениального аппендикса нормально развиты и хорошо выражены.
Ветви полового ретрактора крепятся на диафрагме порознь или слиты, одна
ветвь подходит к пенису ниже его середины, вторая — к А± сразу же под А2.
Проток семяприемника почти прямой, неясно отграниченный резервуар отно-
сительно объемистый.
Сравнение материала, который мне удалось просмотреть, с описаниями и
изображениями раковин и анатомических признаков форм, указанных в сино-
нимике, приводит к представлению, что это вид, имеющий широкий ареал и
образующий множество локальных популяций, многие из которых принято рас-
сматривать как подвиды (Forcart, 1940). Вероятно, действительно имеется не-
сколько подвидов, но их морфологическая определенность сильно маскируется
биотопической изменчивостью. Рассмотрение внутривидовой структуры пред-
ставляется преждевременным из-за недостаточной изученности популяций из
разных местонахождений.
Распространение и местообитание. Югославия, Бол-
гария, Малая Азия, Армения, местами Грузия (Батуми). Населяет открытые
места, держится в основном в осыпях, цод камнями.
2. Pseudochondrula tetrodon (Mortillet, 1854) (рис. 205, II; 207).
Mortillet, 1854 : 11, t. 1, fig. 3 (Bulimus); Mousson, 1876 : 36, t. 2, fig. 6 [Buli-
minus (Chondrus) diffusus]; Westerlund, 1887 : 43 (Buliminus)-, Retowski, 1889 : 249
4. РОД PSEUDOCHONDRULA
291
[Buliminus (Chondrula) kollyi]; Kobelt, 1902 : 595, t. 91, figs. 21, 22 (Chondrula)', Rosen,
1914 : 186 [Buliminus (Chondrulus)]; F о г с a r t, 1940 : 215, t. 2, figs. 56, 57 (Imparietula);
Лихарев, Раммельмейер, 1952 : 174, рис. 86 (Chondrula); Акрамовский,
1976 : 156, рис. 71Б, табл. VII, 74, 75 (Imparietula).
Terra typica — Испир (Турция, вилайет Эрзурум).
Синтипы хранятся в Женевском естественноисторическом музее и
в Зенкенбергском музее, Франкфурт-на-Майне (Forcart, 1940).
Материал: более 70 экз. из двух местонахождений в Армении.
Раковина от удлиненно-овальной до коническо-цилиндрической, твердостен-
ная, блестящая, с притупленной вершиной. Оборотов l1/^—9, умеренно вы-
Рис. 207. Pseudochondrula tetrodon (Mort.) из Армении, с. Гнишик, 1941 г.
I — половой аппарат, II— вскрытый пенис, III— продольный разрез бича.
пуклых; последние 2—3 оборота почти одной ширины (у коническо-цилиндри-
ческих форм). Последний оборот возле самого устья чуть приподнят, высота
его примерно равна половине высоты раковины, иногда немного больше или
немного меньше. Окраска одноцветно светло-роговая, реже светло-серая.
Эмбриональные обороты слегка желтоватые, гладкие. Скульптура дефинитив -
ных оборотов представлена радиальными неравномерно расположенными сла-
быми сглаженными морщинками, более четкими и правильными на ранних оборо-
тах. Устье овальное, очень слабо скошено, края его слабо отвернуты, места
прикрепления расставлены и связаны гладкой блестящей мозолью. Устьевая
арматура представлена 4 зубами, степень развития которых подвержена силь-
ной индивидуальной и биотопической изменчивости. На склоне у с. Агверан
близ Еревана мною была собрана серия из 56 раковин, в пределах которой можно
проследить все градации развития зубов от малозаметных припухлостей до мощ-
ных бугорков и складок. В максимальном выражении устьевая арматура выгля-
дит следующим образом. Париетальная пластинка слегка изогнута,: постепенно
понижается в глубине устья. Наружный колумеллярный зуб в виде короткой
19*
292
VI. НАДСЕМ. BULIMINOIDEA
гребневидной складки, плоскость которой лежит под углом к плоскости устья.
Внутренняя колумеллярная пластинка отделена от наружного зуба ложбин-
кой и служит его естественным продолжением; она уходит вверх по столбику.
Палатальный зуб занимает середину палатального края и направлен концом
к париетальной пластинке. Пупок открытый, овально-щелевидный, маленький.
Размеры: ВР 9—16, БД 4—6 мм; в первоописании: ВР 14—16, БД 6 мм.
Внутреннее строение. Материал: 4 экз. из Армении, Даралагез,.
урочище Хортых, 13 июля 1942 г., сбор и определение Н. Н. Акрамовского.
Белковая железа без ясно выраженной вырезки и без желобков на боковой
поверхности. Нижний отдел вагины в —2 раза короче верхнего. Семяпро-
вод впадает в эпифаллус не под углом и не терминально. Короткий рудиментар-
ный конический бич располагается параллельно семяпроводу. Эпифаллус
толстый, короткий, прямой, цилиндрический; мешковидный цэкум распола-
гается примерно на середине его длины или незначительно сдвинут к пенису;
пенис внешне почти не выражен, точнее, мало чем отличается от эпифаллуса.
На внутренней поверхности пениса две ясно выраженных, хотя немного рас-
плывшихся V-образных складки обычного строения. Кроме того, присутствует
небольшой придаток в виде высокого бугорка, расположенный близ отверстия
эпифаллуса. Отделы пениального аппендикса ясно выражены, особенно хо-
рошо развит Alf и создается впечатление, что пенис отходит от основания этого
отдела, а не аппендикс отходит от пениса. Ветви полового ретрактора на диа-
фрагме располагаются рядом одна с другой. Пениальная ветвь ретрактора кре-
пится к середине пениса или немного дистальнее середины, вторая ветвь —
к Аг заметно отступя от А2. Семяприемник практически не разделен на проток
и резервуар, почти прямой, немного не доходит до белковой железы.
Распространение и местообитание. Вид найден в не-
скольких местах Грузии (окр. Тбилиси, Кутаиси, Боржоми), в южной Армении;
восточная Турция, Северо-Западный Иран. Встречается почти во всех зонах,,
но лесов избегает; преимущественно на открытых склонах и в осыпях.
3. Pseudochondrula lederi (О. Boettger, 1883) (рис. 205 IV; рис. 208, I—III).
О. Boettger, 1883 : 177, t. 7, fig. 1 {Buliminus); Westerlund, 1887 : 37 {В. le-
deri nbx. limis); Kobelt, 1902 : 578, t. 90, fig. 3 {Chondrula).
Terra typica — окр. с. Эшери немного севернее Сухуми, Сванетия.
Место хранения типов не установлено; в Зоологическом ин-те АН СССР
хранится один синтип.
Материал: 27 экз. из 4 местонахождений в Грузии и Северной Осетии.
Раковина от коническо-цилиндрической до почти башневидной, довольно
твердостенная, слегка блестящая. Оборотов 7—8, умеренно выпуклых, послед-
ний оборот к устью незначительно поднят, высота его составляет половину вы-
соты раковины или немного меньше. Окраска одноцветная, желтоватая, роговая
или каштановая. Скульптура в виде тонких неравномерно расположенных, но-
в целом довольно частых морщинок и тончайших спиральных бороздок, выра-
женных не по всей поверхности раковины; местами спиральная скульптура пред-
ставлена тонкими короткими штрихами. Наконец, в нижней части последнего
оборота эти спиральные штрихи несколько смяты и дают впечатление тончай-
шей неправильной бугорчатости. Устье овальное, очень слабо скошено, края
его заметно отвернуты и утолщены.
Размеры: ВР 10—13, БД 3.7—5 мм; в первоописании: ВР 10.25—10.75,
БД 3.75—4 мм.
В первоописании указано, что вид близок к Buliminus brevior Mousson, и
отличается следующими особенностями: мельче, чем brevior, менее вздут; ра-
ковина высокая, удлиненно-цилиндрическая. Исследование большого материала
показывает, что эти виды настолько похожи, что Р. lederi достоверно отличить
от Imparietula brevior по раковине практически невозможно, хотя они резко
отличны по анатомии. Имеется, однако, хорошее экологическое различие:
4. РОД PSEUDOCHONDRULA
293
Р. lederi обитает в лесах или, по крайней мере, в тенистых местах, a I. brevior
живет в открытых сухих биотопах.
Рис. 208. Pseudochondrula lederi (Bttg.) (I—III) из Северной Осетии, Зругское ущелье,
17 VII 1974 г., и Р. sinistrorsa Kokotschashvili et Schileyko, sp. nov. (7I7) из Западной Грузии,
с. Лаше, паратип.
1 — половой аппарат; II— вскрытый пенис; 111 — вскрытый пенис экземпляра из Цейского ущелья,
Северная Осетия, 18 VII 1974 г.; IV — половой аппарат и вскрытый пенис.
Внутреннее строение. Материал: 2 экз. из Северной Осетии,
Зругское ущелье близ с. Нар, 17 июля 1974 г., сбор Я. И. Старобогатова; 2 —
из Грузии, окр. Бакуриани, 30 июня 1974 г., сбор И. М. Лихарева; определе-
ния автора.
294
VI. НАДСЕМ. BULIMINOIDEA
Белковая железа с малозаметной апикальной вырезкой. Отделы вагины при-
мерно равной длины. Бич рудиментарный, округло-конический. Эпифаллус не
извит, цэкум в виде небольшого сглаженного бугорка занимает середину эпи-
фаллуса или несколько сдвинут к семяпроводу. Пенис внешне не отграничен от
эпифаллуса, внутренняя его поверхность с двумя V-образными складками.
Аг тонкий и длинный, А2 крошечный, А3 почти не отграничен от Ai и слабо выра-
жен. Ветви полового ретрактора отходят от диафрагмы независимо, пениальная
ветвь фиксирована в нижней части пениса, другая подходит к Аг заметно от-
ступя от Л2. Семяприемник почти не разграничен на проток и резервуар, почти
прямой, немного не доходит до белковой железы.
Распространение и местообитание. Отдельные находки
на Большом Кавказе, кроме восточной его части, и на северных склонах Ма-
лого Кавказа (Бакуриани). Живет на горных лугах с высоким травостоем и
в постилке смешанных и лиственных лесов.
4. Pseudochondrula sinistrorsa Kokotschashvili et Schileyko, sp. nov.
(рис. 205, V; рис. 208, /К).
Terra typica — западная Грузия, с. Лаше.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 98 экз. из типового местонахождения.
Раковина левозавитая, от коническо-цилиндрической до почти башневидной;
это единственная особенность, отличающая вид от правозавитого Р. lederi.
В организации полового аппарата имеются заметные отличия.
Размеры голотипа: ВР 12.8, БД 4.2 мм.
Внутреннее строение. Материал: 5 экз. из типового местонахож-
дения (паратипы): западная Грузия, юго-западные отроги Сурамского хр.,
с. Лаше, 25 сентября 1977 г., сбор Э. А. Гордадзе, определение Г. В. Кокочаш-
вили и автора.
Белковая железа без апикальной вырезки. Нижний отдел вагины значи-
тельно короче верхнего. Бич очень короткий, приостренный. Эпифаллус длин-
ный, извитой; удлиненный почти цилиндрический цэкум расположен немного
ниже середины эпифаллуса, ближе к пенису. Эпифаллус относительно длин-
ный, вначале цилиндрический, но в нижней части, несколько отступя от цэ-
кума, его диаметр заметно уменьшается. Пенис небольшой, формой напоминает
песочные часы; на его внутренней поверхности имеется несколько неупорядо-
ченных гофрированных складок. Отделы пениального аппендикса хорошо видны;
в частности, ясно выражен А3, четко отделенный от At. Семяприемник разгра-
ничен на проток и резервуар, хотя граница между этими отделами нерезкая;
в нижней части протока имеется резкий изгиб. Резервуар почти касается белко-
вой железы.
От наиболее близкого вида Р. lederi отличается большей длиной эпифаллуса,
более дистальным положением цэкума, ясно обособленным А3 и отсутствием
внутри пениса пары V-образных складок, вместо которых имеется несколько
мелких складок, расположенных без видимой системы.
Распространение и местообитание. Вид известен только
из типового местонахождения в юго-западной части Сурамского хр., где обна-
ружен в подстилке лиственного леса.
5. Pseudochondrula tuberifera (О. Boettger, 1879) (рис. 205, ПР, рис. 209).
О. Boettger, 1879 : 22, t. 1, fig. 9 [Buliminus {Chondrula)}', Kobelt, 1902 : 570,
t. 89, figs. 10, 11 {Chondrula)-, Rosen, 1914 : 184 {Buliminus)', Лихарев, Раммель-
мейер, 1952:179 {Pseudochondrula brevior var. tuberifera).
Terra typica — «Коби и Казбек».
Место хранения типов не установлено,
4. РОД PSEUDOCHONDRULA
295
Материал: 4 экз. из двух местонахождений в Северной Осетии и на
Сурамском хр.
Раковина удлиненно овально-коническая, матовая, слабо просвечивающая,
умеренно тонкостенная. Оборотов 7, верхние слегка выпуклые, нижние упло-
щенные, последний оборот к устью приподнят, высота его немного меньше по-
ловины высоты раковины. Окраска одноцветно роговая. Скульптура в виде
тонкой радиальной морщинистости, лучше выраженной на ранних оборотах.
Элементы спиральной скульптуры полностью отсутствуют. Устье овальное,
места его прикрепления расставлены, края утолщены, не отвернуты, кроме
колумеллярного. На середине палатального края в плоскости устья распола-
гается бугорковидный зуб; за этим зубом, несколько отступя от него, лежит
слабая продольная расплыв- i /
шаяся белая складка, замет-
ная лишь при косом положе-
нии раковины, так как при
нормальном положении она
полностью заслонена пала-
тальным зубом. При внима-
тельном рассмотрении видно,
что на колумеллярной крае
имеется очень вялая косая
складка, уходящая по стол-
бику вверх. Пупок коротко-
щелевидный, полуприкры-
тый.
Размеры: ВР 8—9.5, БД
3—3.5 мм; в первоописании:
ВР8.5-9.5, БД 3.5 мм.
Видовая самостоятель-
ность подтверждается тем,
Рис. 209. Pseudochondrula tube-
rifera (Bttg.) из Северной Осетии,
Зругское ущелье, 14 VII 1974 г.
Половой аппарат и вскрытый пе-
нис.
что вид найден в Северной Осетии (Зругское ущелье) вместе с наиболее близким
видом Р. lederi.
Внутреннее строение. Материал: 1 экз. из Северной Осетии,
Зругское ущелье, близ с. Нар, 17 июля 1974 г., сбор Я. И. Старобогатова, опре-
деление автора.
Белковая железа с плавной апикальной вырезкой. Отделы вагины примерно
равной длины. Задний конец эпифаллуса притуплен, семяпровод впадает не
терминально. Эпифаллус прямой, цилиндрический; слаборазвитый цэкум в виде
небольшой припухлости, заметной близ середины эпифаллуса. Пенис неболь-
шой, отделен от эпифаллуса кольцевой перетяжкой. Внутри пениса у единст-
венного изученного экземпляра располагаются три складки, одна из которых
круто изогнута в виде клюшки, две другие спереди упираются в изгиб изогну-
той складки, а сзади, не доходя до отверстия эпифаллуса, сходят на нет. Возле
отверстия эпифаллуса изогнутая складка образует маленький козырек. Отделы
пениального аппендикса развиты нормально. Ветви полового ретрактора на
диафрагме слиты, одна ветвь крепится к середине пениса, вторая — к Аг чуть
отступя от А2. Семяприемник почти прямой, не подразделен на проток и ре-
зервуар, верхняя часть его прилегает к поверхности белковой железы.
296
VI. НАДСЕМ. BULIMINOIDEA
Распространение и местообитание. Кроме Казбека и
окр. Коби, вид найден на Сурамском хр. и в Северной Осетии. Обитает на лугах
с хорошо развитым травяным покровом.
5. Род TRIANGUSTOMA Schileyko, gen. nov.
Типовой вид Chondrulopsina dentatum Likharev, 1967.
Раковина башневидная или коническая, тонко скульптурированная, обо-
роты уплощенные. Устье с палатальным зубом или вздутием; на колумелляр-
ном и париетальном краях также могут присутствовать зубы. Места прикрепле-
ния устья не сближены, края устья утолщены и отвернуты.
Конический бич хорошо выражен, небольшой. Внутри тонкостенного пениса
имеется незамкнутая папилла с боковыми лопастями. Дивертикул семяприем-
ника хорошо развит.
В настоящее время трудно судить о близости рода к какому-либо из извест-
ных, поскольку материалы по анатомии представителей семейства из северо-вос-
точного Ирана, Афганистана, Пакистана и северной Индии в литературе от-
сутствуют. Наличие папиллы своеобразного строения в сочетании с тонкостен-
ным пенисом и своеобразной раковиной ставит род в изолированное положение
относительно среднеазиатских Pseudonapaeinae.
Распространение. Северо-восточный Афганистан. Ареал и объем
рода нуждаются в уточнении.
Triangustoma dentatum (Likharev, 1967) (рис. 210, 211).
Лихарев in: Лихарев, Старобогато в, 1967 : 183, рис 12, 13 IChondru-
lopsina').
Terra typica — Афганистан, Джалала, долина р. Кунар.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 3 экз. из типовой серии.
Раковина от башневидной до конической, твердостенная, слегка блестящая.
Оборотов 6х/4—772, верхние обороты слегка выпуклые, нижние уплощенные,
разделены тонким врезанным швом. Последний оборот к
устью приподнят, снизу сдавлен, высота его меньше поло-
вины высоты раковины. Окраска верхних оборотов темно-
роговая, нижние обороты белые с многочисленными нерав-
номерными темными радиальными пестринами, имеющими
неправильные четкие контуры. Скульптура в виде тонкой
неясной радиальной исчёрченности. Устье округло-треуголь-
ное, места его прикрепления расставлены и соединены
тонкой мозолью. Края устья сравнительно сильно отвернуты,
расширены и утолщены. В устье 4 зуба: бугорковидный ан-
гулярный, расположенный на париетальной мозоли; несколь-
ко глубже лежит хорошо развитый париетальный округ-
ло-конической формы; колумеллярный и палатальный пред-
Рис. 210. Triangustoma dentatum (Likh.). Голотип.
ставлены легкими расширениями соответствующих участков устьевого края.
Пупок в виде узкой изогнутой щели.
Размеры: ВР 5.9—7.1 БД 2.5—2.9 мм; голотип: ВР 7.1, БД 2.8 мм.
Внутреннее строение. Материал: 2 экз. из Афганистану, до-
лина р. Кунар, Джалала, 2 февраля 1958 г., сбор К. Линдберга, определение
И. М. Лихарева.
Белковая железа без апикальной вырезки. Нижний отдел вагины незна-
6. РОД MASTOIDES
297
чительно короче верхнего. Семяпровод впадает в эпифаллус под почти прямым
углом; эпифаллус заканчивается хорошо развитым, хотя и недлинным удли-
ненно-коническим бичом. Бугорковидный цэкум занимает на эпифаллусе сре-
динное положение; на противоположной стороне эпифаллуса также имеется
небольшое вздутие. Пенис с очень тонкими прозрачными стенками, без продоль-
ных складок. Сквозь стенки пениса просвечивает незамкнутая папилла своеоб-
разного строения. Основу папиллы составляет стержень, заполненный рыхлой
соединительной тканью; отверстие эпифаллуса лежит при основании этого стер-
жня. От стороны папиллы, противолежащей той, у которой открывается отвер-
стие эпифаллуса, отходят справа и слева две широкие складки, окутывающие
опорный стержень в проксимальной части и почти смыкающиеся на противо-
Рис. 211. Triangustoma dentatum (Likh.). Паратип. Половой аппарат и вскрытый пенис.
положной стороне стержня; в дистальной части папиллы эти складки сходят
на нет, и апикальная треть длины папиллы лишена складок. Отделы пениаль-
ного аппендикса хорошо заметны. Ветви полового ретрактора на диафрагме не
слиты одна с другой. Пениальная ветвь крепится к месту перехода эпифаллуса
в пенис, ветвь аппендикса — к Аг непосредственно под А2. Проток семяприем-
ника образует петлю; резервуар с очень коротким собственным протоком, длина
которого меньше длины самого резервуара. Дивертикул длинный, накладывается
на белковую железу.
Как отмечено в первоописании, на дивертикуле семяприемника одного из
паратипов видно резкое вздутие (рис. 211). В этом вздутии располагается
шпора сперматофора, находящегося в дивертикуле.
Распространение и местообитание. Вид известен только
из типового местонахождения, где обнаружен на дне пещер, под камнями в пе-
регное.
6. Род MASTOIDES Westerlund, 1896
Westerlund, 1896 : 192 [Buliminus (Mastoides}]; Thiele, 1931 : 520 (Subzebrinus
part.); Лихарев, Раммельмейер, 1952 :195 (Subzebrinus part.); Wen z, Zilch,
1959—1960:192 [(Subzebrinus (Subzebrinus} part, j; Шилейко, 1978г : 845.
298
VI. НАДСЕМ. BULIMINOIDEA
Типовой вид Buliminus albocostatus Westerlund, 1896; по монотипии.
Раковина почти башневидная, с радиальными ребрами, выраженными в раз-
ной степени, обороты выпуклые. Устье без зубов, с отвернутыми и утолщенными
краями, места его прикрепления не1 сближены.
Бич отсутствует. Внутри пениса имеется стержневидная папилла, на вну-
тренних стенках пениса залегают складки примерно кольцевидной формы. Ди-
вертикул семяприемника нормально развит.
Распространение. Тянь-Шань. 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ MASTOIDES
1 (2). Раковина покрыта резкими острыми и массивными радиальными ребрами
............................................................. 2. М. albocostatus.
2 (1). Раковина без ребер или ребра притупленные, сглаженные.........................
......................................................... 1. М. orloffensis.
1. Mastoides orloffensis (Kobelt, 1905) (рис. 212, I; рис. 213).
Kobelt, 1905, Iconogr., N. F., XII : 19, fig. 1972 [B. (Mastoides) albocostatus orloffen-
sis]; Lindholm, 1927b : 259 [Subzebrinus (Oedichilus)]; Лихарев, Раммель-
мейер, 1952 : 200, рис. 112 [Subzebrinus (Subzebrinus)].
Terra typica — долина p. Талас.
Типовой материал, вероятно, хранится в Зенкенбергском музее, Франк-
фурт-на-Майне.
Материал: 5 экз. из Ферганского хр. и 2 из долины р. Талас.
Раковина от коническо-цилиндрической до почти башневидной, с широко
закругленной вершиной,: умеренно твердостенная. Оборотов 8—9, плавно на-
растающих, последний оборот к устью почти
не поднят, высота его больше одной трети, но
меньше половины высоты раковины. Окраска
роговая; скульптура в виде сглаженных широ-
ких ребер, развитых в разной степени, иногда
они почти не выражены, но прослеживаются
в виде светлых радиальных пестрин, слегка
приподнятых над поверхностью раковины.
Устье короткоовальное или почти круглое,
слабо скошенное, места его прикрепления сое-
динены мозолью, степень развития которой стоит
в прямой связи со степенью развития радиаль-
Рис. 212. Виды рода Mastoides.
I — М. orloffensis lILdb.) из Ферганского хр. Джаз-ичу; II —
М. albocostatus (West.) из с.Арсланбоб, Ферганский хр.
ных ребер. Края устья умеренно отвернуты, с белой расплывшейся губой. Пу-
пок в виде узкой или овальной щели.
Размеры: ВР 8—12.5, БД 3—4 мм; в первоописании: ВР 8.2—9.5, БД 3 мм.
Раковины из долины р. Талас (топотипы) заметно отличаются от раковин,
собранных на Ферганском хр.: первые имеют высоту раковины 8—8.3 мм и рез-
кие ребра на поверхности, вторые крупнее (ВР 11—12.5 мм) и почти гладкие,
но со светлыми пестринами. Хотя размеры и степень развития скульптуры —
это признаки вообще пластичные, нет уверенности в конспецифичности экзем-
пляров из указанных местонахождений. Однако поскольку внутреннее строение
«настоящих» М. orloffensis (из долины р. Талас) неизвестно, предлагать новое
6. РОД MASTOIDES
299
видовое название для экземпляров из Ферганского хр. представляется прежде-
временным.
Внутреннее строение. Материал: 3 экз. из Ферганского хр.,:
долина р. Джаз-ичу (бассейн р. Кара-Унгур), 16 июня 1961 г., сбор И. М. Ли-
харева, определение автора.
Белковая железа с глубоким и широком желобом, проходящим по ее боковой
поверхности и формирующим апикальную вырезку. Верхний отдел вагины не-
сколько длиннее нижнего. Бич отсутствует, эпифаллус цилиндрический, к пе-
нису плавно сужается. Хорошо развитый цэкум расположен заметно дисталь-
нее середины эпифаллуса. Проксимальная часть пениса овальная или фасоле-
видная; дистальная часть примерно такой же длины, цилиндрическая.
Рис. 213. Mastoides orloffensis из Ферганского хр., Джаз-ичу (бассейн р. Кара-Унгур),
16 VI 1961 г. Половой аппарат и вскрытый пенис.
Внутри проксимального отдела имеется очень сильно развитая стержневидная
незамкнутая папилла и пара складок, в совокупности напоминающая кольцо
или прямоугольник с закругленными углами; картина маскируется наличием
еще нескольких мелких складок. А± очень короткий, такой же длины, как ша-
ровидный А2; зато Л3 длинный — длиннее первых двух отделов, вместе взятых.
Ветви полового ретрактора отходят от диафрагмы единой лентой; пениальная
ветвь крепится к границе дистального и проксимального отделов пениса,
ветвь аппендикса — к А2. Проток семяприемника прямой, округло-треуголь-
ный резервуар сидит на нем всем широким основанием. Дивертикул недлин-
ный, заметно не доходит до белковой железы.
Распространение и местообитание. Вид найден в двух
местах: долина р. Талас и долина р. Джаз-ичу на Ферганском хр. Встречен
на скалах и в осыпях.
300
VI. НАДСЕМ. BULIMINOIDEA
2. Mastoides albocostatus (Westerlund, 1896) (рис. 212, II; рис. 214).
Westerlund, 1896 : 192 [Buliminus (Mastoides) albocostatus et var. distortus]; Лиха-
рев, Раммельмейер, 1952 : 200 [Subzebrinus (Subzebrinus)]; Шилейко, . 9786 :
351, рис. 3, II.
Terra typica; — Арсланбоб (Ферганский хр.).
Г о л о т и и хранится в Зоологическом ин-те АН СССР.
Материал: 7 экз. из типового местонахождения.
Раковина башневидная, высокая, стройная, с тонкой вершиной, умеренно
твердостенная. Оборотов 10—11, медленно нарастающих, умеренно выпуклых,
разделенных довольно глубоким швом. Последний оборот к устью энергично,
но довольно плавно поднят, высота его втрое или даже больше чем втрое
меньше высоты раковины. Окраска от светло-роговой до каштановой; скульп-
Рис. 214. Mastoides albocostatus из Арсланбоба, Ферганский хр., 20 VI 1961 г. Половой ап-
парат и вскрытый пенис (из: Шилейко, 19786).
тура в виде очень резких массивных радиальных ребер, промежутки между кото-
рыми большей частью не меньше ширины ребра. Устье короткоовальное, слабо
скошено,; места его прикрепления расставлены и соединены хорошо развитой
светлой мозолью, так что создается впечатление, что устье почти цельное. Края
устья умеренно отвернуты, с белой широкой губой; колумеллярный край пти-
роко отвернут и расширен. Пупок широкощелевидный.
Размеры: ВР 12—14, БД 3.5—4; голотип: ВР 12, БД 3.5 мм.
Внутреннее строение. Материал: 5 экз. из Ферганского хр.,
Арсланбоб,; правый берег р. Арал-сай, 20 июня 1961 г., сбор и определение
И, М« Лихарева.
По латеральной поверхности белковой железы проходит очень широкий и
глубокий желоб, формирующий также и апикальную вырезку. Отделы вагины
примерно равной длины. Семяпровод впадает в эпифаллус эксцентрично, бич
отсутствует, эпифаллус на конце слегка утолщен и широко закруглен. Цэкум
чрезвычайно резко выражен, короткоцилиндрический, слегка сдвинут вниз
относительно середины эпифаллуса. Пенис разделен на два отдела: проксималь-
ный, имеющий фасолевидную форму, и более длинный дистальный цилиндри-
ческой формы. Внутри дистального отдела залегают вялые продольные складки;
внутренняя поверхность проксимального отдела несет 2—3 характерные складки
примерно кольцеобразной формы с неровными краями; здесь же расположена
незамкнутая хорошо развитая стержневидная папилла с глубокой продольной
7. РОД LAEVOZEBRINUS
301
бороздой. Перед отверстием эпифаллуса небольшие лопасти, окаймляющие
борозду, немного расширяются. Строение пениального аппендикса очень ха-
рактерно: Аг и А2 слиты, образуют короткую булаву; А3 длиннее, чем Aj+Aa.
Ветви полового ретрактора отходят от диафрагмы порознь, одна крепится
к Лл+Лз несколько выше середины, вторая — к дистальному отделу пениса
возле места перехода его в проксимальный. Проток семяприемника повторяет
все изгибы вагины и спермовидукта, резервуар вовсе лишен собственного про-
тока и в виде округло-треугольного придатка сидит на основном протоке всем
основанием. Дивертикул немного не доходит до нижнего края белковой железы.
Распространение и местообитание. Вид найден дважды
приблизительно в одном месте на Ферганском хр., близ Арсланбоба; на скалах
и в зарослях кустарников.
7. Род LAEVOZEBRINUS Lindholm, 1925
Lindholm, 1925а : 28 [Zebrina {Subzebrinus), секция Laevozebrinus); Thiele, 1931 :
520 [Z. {S.), секция Laevozebrinus); Wenz, Zilch, 1959—1960 : 192 [Subzebrinus {Laevo-
zebrinus)]; Шилейко, 1978г : 845.
Типовой вид Buliminus urgutensis Kobelt, 1902; по первоначальному
обозначению.
Раковина от удлиненно-овальной до цилиндрической, нередко с радиальными
складками и морщинами. Устье без зубов, края его умеренно отвернуты, места
прикрепления не сближены. \
Бич отсутствует или очень слабо развит. Пенис внутри со складками и с трех-
лопастной папиллой. Дивертикул семяприемника имеется.
Распространение. Иран, Афганистан, горные области Средней
Азии; возможно, северо-восточная Индия. 4 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА
LAEVOZEBRINUS
1 (2). Раковина левозавитая ..........................2. L. urgutensis.
2 (1). Раковина правозавитая.
3 (4). Раковина цилиндрическая или коническо-цилиндрическая, высота ее
не менее 18 мм .....................................3. L. ujfalvyanus.
4 (3). Раковина меньше; если той же высоты, то иной формы.
5 (6). Высота раковины не менее 14 мм...................1. L. eremita.
6 (5). Высота раковины не более 13 мм...................... 4. L. lenis.
1. Laevozebrinus eremita (Benson, 1849) (рис. 215, Г, рис. 216).
Benson in: Reeve, 1849, Conch. Iconica, 5, pl. 78, fig. 573 {Bulimus); Мартенс,
1874 : 18, табл. 2, фиг. 13 {Buliminus)', Ancey, 1886a : 31 {Buliminus kuschakewitzi), 32
(B. kuschakewitzi var. candisata), 33 {B. bonvallotianus), 44 {B. potaninianus); 1887 : 6 {B. ere-
mita var. hepatica et var. procera); 0. Boettger, 1889b : 952 {B. eremita var. germabensis);
Westerlund, 1896 : 189 {B. eremita var. solivagus); Лихарев, Раммельмейер,
1952 : 196, рис. 107 [Subzebrinus {Subzebrinus) potaninianus]; Лихарев, Старобога-
тов, 1967 : 182 {Subzebrinus); Шилейко, 19786 : 353, рис. 4, II.
Terra, typica — Афганистан, Кандагар, перевал Болан.
Место хранения типов не установлено.
Материал: свыше 200 экз. из 19 местонахождений в Иране, на Копетдаге
и прилегающих районах Алая.
Раковина от удлиненно-овальной до овально-цилиндрической, твердостен-
ная, блестящая. Оборотов 7—8, реже до 9, умеренно или слабо выпуклых,
последний оборот к устью слабо поднят, высота его не менее половины высоты
раковины, часто заметно больше. Окраска чисто белая, однотонно желтоватая
или серая, обычно с развитыми в разной степени радиальными темными штри-
302
VI. НАДСЕМ. BULIMINOIDEA
хами и пестринами; верхние обороты чаще всего темнее нижних. Скульптура
в виде тонкой или грубой радиальной морщинистости, которая, однако, никогда
не переходит в ребристость. Устье овальное, почти отвесное, внутри желтое или
коричневатое, края его умеренно плавно отвернуты. Колумеллярный край от-
весный, базальный и палатальный образуют широкую плавную дугу. Губа
светлая, сильно расплывшаяся. Пупок в виде широкой щели.
Размеры: ВР 14—28, БД 7.5—12 мм; в первоописании: ВР 22—24, БД 9.5 мм.
Внутреннее строение. Материал: 6 экз. из Ургута близ Самар-
канда, сбор II. П. Архангельского, определение автора; 5 — из Мешхеда,
Иран, 28 октября 1974 г., сбор И. С. Даревского, определение И. М. Лихарева;
4 — из предгорьев Туркестанского хр., окр. Карасоколбобо близ Ура-Тюбе,
10 ноября 1969 г., сбор А. Б. Мухитдинова; 10 — из долины р. Шинг на Зерав-
шанском хр., 6 августа 1973 г., сбор Ю. А. Мешкова; определения автора.
Рис. 215. Виды рода Laevozebrinus.
1 — L. eremita из Ирана, окр. Мешхеда; II — L. urgutensis из Ургута близ Самарканда; III — L. lenis из
хр. Каратау, голотип; IV — L. ujfalvyanus из Арсланбоба, Ферганский хр.
Белковая железа с апикальной вырезкой. Нижний отдел вагины в 1У2—
2 раза короче верхнего. Семяпровод впадает в эпифаллус немного отступя от
конца последнего, за счет чего возникает короткий цилиндрический широко и
плавно закругленный бич. Эпифаллус длинный, цилиндрический, в разной сте-
пени извитой; хорошо выраженный цэкум расположен заметно ниже середины
эпифаллуса. Пенис наружно выражен у разных особей по-разному, форма его
от цилиндрической до булавовидной; в последнем случае видно разделение его на
расширенный проксимальный и цилиндрический дистальный отделы. Внутри
дистального отдела залегают тонкие вялые продольные складки; проксималь-
ная часть заключает незамкнутую массивную папиллу очень характерного
строения: поверхность ее складчатая и бугорчатая, в базальной части папиллы
имеются две отчетливые небольшие лопасти, ограничивающие борозду1 на по-
верхности органа; эта борозда продолжается на основную лопасть, н о, не до-
ходя до ее конца, сходит на нет. Аг тонкий и очень длинный, извитой и может
даже образовывать петли. А2 небольшой, но отчетливо заметный; Аа очень корот-
кий и тонкий, ненамного толще нитевидного Л4. Ветви полового ретрактора
тесно сближены на диафрагме, одна ветвь крепится к середине дистального от-
дела пениса, вторая — к Аг заметно отступя от Аг. Проток семяприемника очень
7. РОД LAEVOZEBRINUS
303
длинный, резко извит, образует петли. Резервуар имеет короткий, но ясно вы-
раженный проток, дивертикул недлинный, но за счет большой длины общего
протока семяприемника доходит до белковой железы.
Распространение и местообитание. Иран, Афганистан,
Индия (Джамму и Кашмир), Копетдаг, Алай. Населяет открытые предгорные
и межгорные пространства, незалесенные склоны, встречается в осыпях и кус-
Рис. 216. Laevozebrinus eremita из Ургута близ Самарканда. Половой аппарат и вскрытый пе-
нис (из: Шилейко, 19786).
тарниковых зарослях, нередко совместно с похожим на него Pseudonapaeus
albiplicatus f. sogdiana.
2. Laevozebrinus urgutensis (Kobelt, 1902) (рис. 215, II).
Kobelt, 1902 : 21, t. 248, fig. 1606 (Buliminus?)' Лихарев, Раммельмейер,
1952 : 196 (Subzebrinus (Subzebrinus) potaninianus var. urgutensis].
Terra typica — Ургут (Самаркандская обл.).
Лектотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 150811.
Материал: 2 экз. из Ургута и 1 экз. из северного Афганистана.
Раковина левозавитая, представляет собой зеркальное отражение раковины
L. eremita.
Внутреннее строение неизвестно, но есть все основания пред-
полагать, что оно идентично или очень похоже на таковое предыдущего вида,
от которого L. urgutensis возник, видимо, мутагенным путем.
Распространение. Известны две находки в типовом местонахож-
дении и несколько находок в северном Афганистане (Лихарев, Старобогатов,
1967).
1 Апсей назвал вид именем французского путешественника, но допустил ошибку в напи-
сании; в работе 1893 г. он эту ошибку исправил. В 1928 г. Вагнер по непонятной причине свя-
304
VI. НАДСЕМ. BULIMINOIDEA
3. Laevozebrinus ujfalvyanus (Ancey, 1886) 1 (рис. 215, IV', рис. 217).
Ancey, 1886a : 38 (Buliminus ufjalvyanus); Westerlund, 1887 : 66 (B. ufjalvyanus);
Kobelt, 1890, Iconogr., N. F., IV : 39, fig. 579 (B. alaicus); Ancey, 1893 : 35 (Buliminus);
Kobelt, 1902 : 502, t. 81, figs. 19, 20 [B. (Subzebrinus) ufjalvyanus]; Wagner, 1928 : 130
(B. ujfalvyanus nom. emend.); Lindholm, 1927b : 257 (Subzebrinus prochorovi); Л и x a-
рев, Раммельмейер, 1952:199, рис. Ill [5. (Subzebrinus)]; Hudec, 1963:24,
Obr. 1, photo (Subzebrinus); Матёки н, 1966 : 131 [Jaminia (Chondrulopsina) potaniniana
subsp. albiplicata part.}.
Terra typica — Узген (Ферганская долина).
Место хранения типов не установлено.
Материал: более 60 экз. из 5 местонахождений на Ферганском и
Алайском хребтах.
Раковина стройная, цилиндрическая или башневидная, реже коническо-
цилиндрическая, твердостенная, блестящая, с узкой притупленной вершиной.
Рис. 217. Laevozebrinus ujfalvyanus из предгорьев Ферганского хр., урочище Ак-Терек близ
Арсланбоба, 14 VII 1937 г.
I — половой аппарат; II — вскрытый пенис; III — вскрытый пенис другого экземпляра.
Оборотов 8—И, слабовыпуклых. Последний оборот к устью слабо поднят, вы-
сота его всегда значительно меньше половины высоты раковины, иногда со-
ставляет треть этой величины. Окраска белая, серая, реже светло-роговая,
с многочисленными темными радиальными пестринами. Скульптура в виде вя-
лых сглаженных морщин, но при первом взгляде раковина выглядит почти глад-
кой. Устье овальное, почти не скошено. Колумеллярный край отвесный, базаль-
ный и палатальный плавно равномерно выгнуты. Места прикрепления устья
зывает название вида с наименованием венгерских егерей, которое пишется так же, как
и фамилия исследователя, в честь которого назван вид (Ujfalvy).
7. РОД LAEVOZEBRINUS
305
связаны развитой в разной степени мозолью. Края устья умеренно отвернуты,
с сильно расплывшейся белой губой. Пупок широкощелевидный.
Размеры ВР 14—24, БД 5—8 мм; в первоописании: ВР 24, БД 7.5 мм.
Внутреннее строение. Материал: 12 экз. из предгорий Ферганского
хр., урочище Ак-Терек, 1937 г.; 2 — из окр. Арсланбоба близ того же уро-
чища, 14 июля 1937 г.; сборы А. Н. Кириченко, определения автора.
Белковая железа с вялой апикальной вырезкой, не заходящей на боковую
поверхность железы. Нижний отдел вагины приблизительно втрое короче
верхнего. Семяпровод впадает в эпифаллус заметно отступя от конца, в резуль-
тате чего образуется короткоцилиндрический чуть приостренный бич. Эпи-
фаллус длинный, неизвитой, цэкум располагается несколько выше его се-
редины. Пенис цилиндрический, заметно шире эпифаллуса. На внутренних
стенках пениса залегает серия хорошо выраженных продольных складок,
гофрированных или образующих многочисленные аНостомозы (рис. 217, ср. II
и III). Между этими крайними вариантами имеются многочисленные промежу-
точные состояния. Кроме того, пенис заключает незамкнутую папиллу с бороз-
дой и сильно изрезанной поверхностью. Боковые лопасти в базальной части
папиллы выражены слабо, но заметить их большей частью можно. Все отделы
пениального аппендикса хорошо выражены, особенно мощно развит А1г длина »
которого превышает длину пениса. Половой ретрактор крепится к диафрагме
двумя полностью независимыми одна от другой лентами. Одна ветвь ретрактора
присоединяется к средней части пениса, другая — к верхней части Аг. Проток
семяприемника длинный, образует 1—2 изгиба. Объемистый резервуар имеет
собственный проток, длина которого меньше длины резервуара. Дивертикул
семяприемника хорошо развит, немного не доходит до белковой железы.
i Распространение и местообитание. Ферганская долина,
Ферганский и Алайский хребты. Населяет открытые и заросшие кустарником
; участки с умеренным увлажнением.
s
: 4. Laevozebrinus lenis Schileyko, sp. nov. (рис. 215, III; рис. 218).
1 Terra typica — хр. Каратау, в 50 км к северо-востоку от г. Туркестан.
Голотип и паратипы хранятся в Зоологическом ин-те АН СССР.
Материал: 10 экз. из типового местонахождения.
Раковина высокая, от овально-конической до почти башневидной, но
и в последнем случае контуры раковины слегка овальные; умеренно твердо-
стенная, блестящая. Вершина широкая, притупленная. Оборотов 7—71/2
(у голотипа 7), медленно и равномерно нарастающих. Последний оборот у са-
мого устья очень слабо и плавно приподнят, высота его немного меньше поло-
вины высоты раковины. Окраска первых оборотов серовато-роговая или розо-
вато-коричневая, более .поздние обороты светлее: серые или почти белые, почти
гладкие. На поверхности видны неяркие радиальные темные штрихи и пе-
стрины. Устье округлое, внутри коричневатое или розоватое, слабо скошено.
Края устья незначительно отвернуты, белые, со слаборазвитой валикообраз-
ной губой. Париетальная мозоль умеренно развита, в ангулярной области не-
/ значительно утолщена и образует слабый бугорок. Пупок в виде очень узкой
и короткой щели.
Размеры: ВР 11.5—12.5, БД 4.3—4.5 мм; голотип: ВР 11.8, БД 4.5 мм.
Вид близок к L. eremita, отличается более мелкими размерами и более цилин-
дрической формой раковины.
Внутреннее строение. Материал: 3 экз. из хр. Каратау, в 50 км
; в северо-востоку от г. Туркестан, 25 мая 1966 г., сбор М. К. Пастухова.
; Белковая железа с плавной, но глубокой апикальной вырезкой. Нижний
Г отдел вагины в ТЧ2—2 раза короче верхнего. Семяпровод впадает эксцентрично,
но бича не образуется. Хорошо развитый цэкум расположен немного ниже сере-
дины короткого цилиндрического эпифаллуса. Пенис состоит из длинной цилин-
дрической дистальной части и короткой расширенной проксимальной. Внутри
20 А- Шилейко
306
VI. НАДСЕМ. BULIMINOIDEA
дистальной части залегают вялые продольные складки, заходящие и в прокси-
мальную часть, где располагается папилла. Пениальная папилла трехлопаст-
ная, притупленная или остроконечная, с глубокой продольной бороздой, не
доходящей до конца папиллы. Поверхность папиллы с многочисленными попе-
речными морщинами; при выравнивании морщин может, вероятно, значи-
тельно удлиняться. Отделы пениального аппендикса хорошо выражены: А!
к концу сильно истончается и равен в этом месте по диаметру А 3; А2 маленький.
Проток семяприемника значительно длиннее вагины, поэтому он образует
1—2 широкие петли. Резервуар маленький, с очень коротким протоком. Дивер-
Рис. 218. Laevozebrinus lenis из хр. Каратау, в 50 км к северо-востоку от г. Туркестана,
25 V 1966 г. Голотип. Половой аппарат и вскрытый пенис.
тикул короткий, немногим длиннее резервуара, заметно не доходит до белковой
железы.
Распространение и местообитание. Вид найден пока лишь
в типовом местонахождении —-на хр. Каратау. Населяет сухие открытые склоны.
8. Род IMPARIETULA Lindholm, 1925
”~| Lindholm, 1925а : 30 {Jaminia, секция Chondrula, подсекция Imparietula)', Kor-
ea г t, 1940 : 205; Wenz, Zilch, 1959—1960 : 183; Акрамов с кий, 1976 : 154.
Типовой вид Bulimus leucodon L. Pfeiffer, 1846; по первоначальному
обозначению..
Раковина овально-коническая или цилиндрическая, тонко скульптуриро-
ванная. Устье без зубов или присутствует палатальный бугорковидный зуб,
представляющий собой местное утолщение губы. Края устья отвернуты, места
его прикрепления не сближены.
Бич практически отсутствует. Пенис заключает незамкнутую папиллу
с неровной бугристой поверхностью. Дивертикул семяприемника отсутствует.
Распространение. Закавказье с прилегающими территориями
Турции и Ирана. В ископаемом состоянии род известен, начиная с верхнего
плиоцена (апшерон Предкавказья). 3—4 вида.
8. РОД IMPARIETULA
307
Отнесение двух видов, обитающих в СССР, к данному роду диктуется их
конхологическим сходством с типовым видом рода Imparietula, а также сход-
ством во внешнем виде полового аппарата (см.: Hesse, 1933, S. 169, fig. И).1
Вместе с тем надо иметь в виду, что анатомически род Imparietula диагносци-
руется главным образом отсутствием дивертикула семяприемника; однако из-
вестно, что в некоторых случаях этот негативный признак на родовом уровне
может быть ненадежным. Поэтому вопрос об объеме рода Imparietula нельзя
считать решенным. Подобным же образом остается открытым вопрос о том,
какой ранг следует придать названию Imparietinia Lindholm, 1925, S. 30,
поскольку анатомия типового вида Buliminus schelkovnikovi Rosen неизвестна.
Этот вид условно включен в род Imparietula.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА IMPARIETULA
1 (2). Раковина левозавитая; в верхней части палатального края имеется зуб
в виде бугорка, сидящего на губе...................2. I. schelkovnikovi.
2 (1). Раковина правозавитая; на палатальном крае устья может присутство-
вать небольшое утолщение, однако оно не может быть названо зубом
....................................................... 1.1. brevior.
1 . Imparietula brevior (Mousson, 1876) (рис. 219, Z; рис. 220).
Mousson, 1876 : 34, pl. 2, fig. 5 (Buliminus)', Schneider, 1878 : 22 (Bulimus)',
0. Boettger, 1881 : 222 (Buliminus)-, Westerlund, 1897 : 44 (Chondrula brevior var.
viator)', Kobelt, 1902 : 599, t. 92, figs. 7, 8 (Chondrula)', Hesse, 1933 i 172, fig. 14 (Pseudo-
chondrula)', Давтян, 1940 : 10, 16 (Jaminia)', Акрамовский, 1949 : 146 (Jaminia)',
Лихарев, Раммельмейер, 1952 : 179 (Pseudochondrula)', С в а д ж я и, 1953 : 48
(Pseudochondrula)', Hudec, Lezhawa, 1967 : 75, Abb. 3, t. I, 2; Акрамовский,
19716 : 7; 1976 : 156, рис. 71A, табл. VII, 76.
Terra typica — верхнее течение p. Араке, выносы.
Место хранения типов не установлено.
Материал: свыше 90 экз. из 9 местонахождений в Закавказье.
Раковина достоверно не отличается от раковины Pseudochondrula lederi-
возможно, единственным видимым отличием является немного больший диаметр
эмбриональных оборотов у данного вида;
однако это можно будет уточнить лишь
пос ле сравнений больших серий раковин,
определенных по анатомическим признакам.
Размеры: ВР 10—12, БД 4.5—5 мм;
в первоописании: ВР 11, БД 5 мм.
Внутреннее строение. Матери-
ал: 3 экз. из Армении, окр. с Личк, сбор и
определение Н. Н. Акрамовского; 9 — из
Ширакского нагорья близ Ленинакана, 19 ап-
реля 1971 г., сбор и определение автора.
Рис. 219. Виды рода Imparietula.
I — I. brevior из. окр. Ленинакана (ориг.); II — I. schelkov-
nikovi из Евлаха, гора Боз-даг, западный Азербайджан (из:
Rosen, 1914).
Белковая железа с узкой и глубокой апикальной вырезкой. Верхний отдел
вагины в l1/^—2 раза длиннее нижнего. Семяпровод впадает эксцентрично или
субтерминадьно. Цэкум хорошо развит, располагается близ середины эпифал-
' 1 У Гессе вид определен как Pseudochondrula lasistanica, но Форкарт (Forcart, 1940)
показал, что Гессе имел дело с Bulimus leucodon L. Pfeiffer.
20*
308
VI. НАДСЕМ. BULIMINOIDEA
луса или немного ниже. Эпифаллус равномерно цилиндрический, пенис расши-
ренный, большей частью коротковеретеновидный. Внутренние стенки пениса
несут вялые бессистемно расположенные мелкие складки. Примерно посредине
пениса на его внутренней поверхности залегает кольцевая складка. Пениальная
папилла хорошо развита, рельефный узор на ее поверхности образован корот-
кими извилистыми бороздками, расположенными без видимой системы. При
основании папиллы у нескольких экземпляров замечен небольшой придаток,
неясно отчлененный от внутренней поверхности пениальной трубки. У некото-
рых особей на внутренней поверхности пениса можно различить две слабые про-
дольные складки, которые, несомненно, есть гомологи V-образных складок,
характерных для многих других родов подсемейства. Пениальный аппендикс
Хорошо развит, длина его колеблется у членов одной популяции, но все его
отделы всегда ясно различимы. Ветви полового ретрактора на диафрагме сбли-
жены в разной степени, иногда полностью изолированы одна от другой. Одна
Рис. 220. Imparietula brevior из окр. Ленинакана, 19 V 1971 г. Половой аппарат и вскрытый
пенис.
из ветвей^крепится к сразу же под А2, другая — к нижней части
пениса. Проток семяприемника длинный, резервуар иногда почти не обособ
лен, конец семяприемника накладывается на нижнюю часть белковой
железы.
S& Распространение и местообитание. В связи с тем что
вид до сих пор мало кто отличал от Р. lederi, ареал его недостаточно ясен. Веро-
ятно, указания на находки в Турции и в южном Закавказье относятся к данному
виду, а на Большом Кавказе — к Р. lederi. Что касается северных склонов
Малого Кавказа, то возможно, что там встречаются оба вида. Характерный
вид лугостепи и нижнеальпийской полосы; встречается в горах, главным об-
разом в почве и под камнями.
9. РОД LJUDMILENA
309
2 . (?) Imparietula schelkovnikovi (Rosen, 1914) (рис. 219, II).
R о sen, 1914 (10.VII) : 188, t. 2, fig. 9 (Buliminus); Retowski, 1914 (28.X) : 307
IB. (Chondrulus) konigi]; Лихарев, Раммельмейер, 1952 : 179, рис. 91 (Pseudo-
chondrula).
Terra typica — гора Боз-даг близ Евлаха, западный Азербайджан.
Место хранения типов не установлено.
Материал мной не исследован, описание составлено по оригинальному
тексту и рисунку.
Раковина левозавитая, цилиндрическая, твердостенная, слабо блестящая,
просвечивающая. Оборотов 91/2—11, уплощенных, высота последнего оборота
немного больше одной трети высоты раковины. Окраска темно-роговая или
беловато-желтая, одноцветная. Скульптура в виде тонкой радиальной ис-
черченности. Устье заметно скошено, усеченно-овальное, места его прикрепления
расставлены. Устьевые края незначительно отвернуты, кроме колумеллярного,
который отвернут довольно сильно; устье окаймлено широкой белой губой.
Середину палатального края занимает бугорковидный зуб, представляющий
собой утолщение губы. Пупок широкощелевидный.
Размеры: -ВР 12.25-14, БД З^-Э3/* мм.
Как систематическое положение, так и таксономический статус дан-
ной формы неясны. Известно, что степень выраженности палатального зуба
у многих Buliminidae сильно варьирует, а основное отличие I. schelkovnikovi
от I. brevior— именно наличие палатального зуба; к тому же у последнего почти
всегда имеется палатальное утолщение на губе. Таким образом, не исключено,
что I. schelkovnikovi есть левозавитые I. brevior; если они образуют постоянные
популяции, их действительно можно считать самостоятельным видом; однако
вид описан только по двум раковинам,1 найденным в разных местах. Поэтому
возможно, что мы имеем дело со случайными мутациями.
Распространение. Вид известен из гор Боз-даг близ Евлаха и из
окр. Худата (северо-восточные отроги Большого Кавказа).
9. Род LJUDMILENA Schileyko, gen. nov.
Типовой вид Chondrus sieversi Mousson, 1873.
Раковина от овально-цилиндрической до булавовидной, слабо скульпту-
рированная. В устье три зуба: париетальный, колумеллярный и палатальный,
причем колумеллярный иногда расщеплен на две пластинки. Края устья уме-
ренно отвернуты и утолщены.
Бич чрезвычайно короткий. Пенис заключает стержневидную папиллу.
Дивертикул семяприемника отсутствует.
Распространение. Закавказье. 2 вида.
По строению устьевой арматуры виды данного рода более всего напоминают
некоторых представителей рода Pseudochondrula (особенно Р. seductilis и Р. te-
tr odon); анатомически род близок роду Geminula, отличается отсутствием ди-
вертикула семяприемника, очень длинным эпифаллусом и положением цэкума:
если у Geminula цэкум сильно сдвинут к семяпроводу, то у Ljudmilena цэкум,
напротив, располагается ниже середины эпифаллуса, ближе к пенису.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ LJUDMILENA
1 (2). Раковина правозавитая................................1. L. sieversi.
2 (1). Раковина левозавитая........................... . . 2. L. tricollis.
1. Ljudmilena sieversi (Mousson, 1873) (рис. 221, Г, рис. 222).
Mousson, 1873 : 207, t. 7, fig. 6 (Chondrus); Westerlund, 1887 : 48 (Bullminus);
Kobelt, 1902 : 572, t. 89, fig. 13 (Chondrula); Retowski, 1914 : 308 [Buliminus (Chond-
1 Кроме того, по трем раковинам описан Buliminus konigi (см. синонимику). Этот ниц
также описан из горы Боз-даг.
310
V/. НАДСЕМ. BULIMINOIDEA
rulus) adjaricus]; Hesse, 1933 : 170, fig. 12 (Pseudochondrula adjarica); F о г с а г t, 1940 :
208 (Imparietula adjarica); Акрамовский, 1949 : 147 [Jaminia (Chondrula)]; Лиха-
рев, Раммельмейер, 1952 : 178, .рис. 90 (Jaminia); Акрамовский, 1968 : 26
(Imparietula); 1976 : 157, рис. 71В, табл. VII, 77 (Imparietula).
Terra typica — выносы Аракса.
Место хранения типов не установлено.
Материал: более 60 экз. из трех местонахождений в Армении и север-
ной Турции.
Раковина овально-цилиндрическая, цилиндрическая или булавовидная,
слегка блестящая, иногда сильно блестящая. Верхние обороты образуют купо-
ловидную вершину, у булавовидных раковин эмбриональные обороты выда-
ются. Оборотов 8—12, верхние — выпуклые, последние 3—4 — уплощенные. По-
следний оборот к устью немного и плавно поднят, в базальной части сжат, вы-
сота его много или значительно меньше половины высоты раковины. Окраска
одноцветно светло-роговая, иногда имеются отдельные участки с восковой по-
лупрозрачностью. Скульптура дефинитивных оборотов в виде неравномерных
I
радиальных размытых сглаженных морщи-
нок. Устье округло-треугольное: наружный
край его от ангулярной области полого от-
клоняется вправо, затем, плавно изогнув-
шись, спускается отвесно, причем на сере-
дине палатального края имеется широкое, но
неглубокое вдавление; базальный край об-
разует короткую полукруглую дугу и пере-
ходит в прямой колумеллярный край, рас-
Рис. 221. Виды рода Ljudmilena.
1 — L. sieversi из Армении, с. Гнишик; II — L; tricollis
из Грузии, Боржоми.
положенный под углом к оси столбика. Края устья отвернуты и несколько утол-
щены, места прикрепления широко расставлены и связаны очень тонкой прозрач-
ной мозолью. Париетальная пластинка в виде округлого немного треугольного
бугорка, расположена глубоко в устье. Почти в плоскости устья, чуть ниже
середины колумеллярного края располагается округлый наружный колумел-
лярный зуб, связанный перемычкой с внутренним колумеллярный, имеющим
вид изогнутой пластинки, уходящей вверх по столбику, усиливая и утолщая его.
На палатальном крае лежит самый мощный округло-треугольный зуб, плоскость
которого составляет некоторый угол с плоскостью устья; ближе к наблюдателю
лежит нижняя кромка зуба. На затылке палатальному зубу соответствует
желобок. Наконец, иногда присутствует небольшое ангулярное утолщение.
Пупок открытый, имеет форму капли.
Размеры: ВР 8—11, БД 2.5—4 мм; в первоописании: ВР 8, БД 2.7 мм.
Buliminus adjaricus Retowski, 1914 (terra typica — окр. Артв.ина и выносы
р. Чорох возле Батуми) представляет собой либо экологическую морфу, либо
подвид, однако для обозначения названной формы в качестве подвида необхо-
димо точнее знать ареалы и характер изменчивости обеих форм. В. adjaricus
отличается большими размерами при большем числе оборотов (в первоописа-
нии: ВР 9—10.6, БД 3.75—4.5 мм при 10—12 оборотах), менее резкой взаимной
обособленностью наружного и внутреннего колумеллярных зубов, более резким
ангулярным бугорком и наличием такого же бугорка в левом углу париеталь-
ного края.
Внутреннее строение. Материал: 3 экз. из Армении, Дарала-
гез, с. Гнишик, 1942 г., сбор Э. А. Давтяна, определение Н. Н. Акрамовского.
Белковая железа с очень слабой апикальной вырезкой. Верхний отдел ва-
9. РОД LJUDMILENA
311
гины в l1^—2 раза длиннее нижнего. Бич чрезвычайно короткий, в виде ко-
роткого закругленного конуса, семяпровод впадает эксцентрично, но рядом
с бичом, составляет естественное продолжение эпифаллуса. Цилиндрический эпи-
фаллус отличается значительной длиной, образует 1—2 изгиба и (или) петли.
Высокий сильно развитый цэкум располагается заметно ниже середины эпифал-
луса. Внутреннее строение цэкума таково, что создается впечатление, что он
возник за счет разкого изгиба эпифаллуса, оба колена которого срослись вну-
тренними соприкасавшимися участками. Пенис маленький, четко подразде-
лен на дистальный цилиндрический и проксимальный вздутый отделы; первый
с неясными продольными складками на внутренней поверхности, второй внутри
почти гладкий, но близ основания папиллы присутствуют!—2 широкие размы-
тые складки, имеющие почти кольцевую ориентацию. Папилла тонкая, черве-
образная, с кольцевыми складками и короткой, но хорошо заметной бороздой;
Рис. 222. Ljudmilena sieversi из Армении, Даралагез, с, Гнишик, 1941 г. Половой аппарат и
вскрытый пенис.
отверстие эпифаллуса открывается на вершине широкого короткого выроста,
боковой червеобразный придаток которого и представляет собой папиллу. Пе-
ниальный аппендикс отходит близ полового атриума, все его отделы выражены
четко, особенной длиной отличается АЦ; он превышает длину пениса примерно
в П/а раза. Ветви полового ретрактора фиксированы на диафрагме одна подле
другой, но вполне самостоятельно. Одна ветвь крепится на границе дистального
и проксимального отделов пениса, вторая — к чуть ниже А2. Семяприемник
практически не разделен на проток и резервуар, почти прямой, рукавообразный,
немного не доходит до белковой железы.
Распространение и местообитание. Турция: долина
Чороха и по долине Аракса до Кагызмана; южная Армения; Нахичеванская
АССР; Иран; бассейн Аракса вниз до западных отрогов Карадага, окр. оз.
Резайе. Встречается в полупустынной зоне и в нижней части горностепного
пояса, под камнями и полукустарниками (Акрамовский, 1976).
312
VI. НАДСЕМ. BULIMINOIDEA
2. Ljudmilena tricollis (Mousson, 1876) (рис. 221, II).
Mortillet, 1854 : 12 (Bulimus cespttus part.); Mousson, 1876 : 141, t. 5, fig. 2.
(Chondrus)- Westerlund, 1887 : 54 (Buliminus)-, Kobelt, 1902 : 598, t. 92, figs. 4—6
(Chondrula)-, Lindholm, 1923 : 221 (Chondrula)-, Forcart, 1940 : 236, t. 3, figs. 83, 84
(Chondrula (Parachondrula)]; Лихарев, Раммельмейер, 1952 : 178 (Jaminia).
Terra typica — долина Куры, Ацкури.
Место хранения типов не установлено.
Материал: 4 экз. из трех местонахождений на Малом Кавказе.
От предыдущего вида отличается только тем, что раковина данного вида ле-
возавитая, а раковина L. sievirsi правозавитая. Размеры в первоописании:
ВР 8, БД 3.8 мм.
Распространение и местообитание. Вид спорадически
распространен по северо-западной части Малого Кавказа. Населяет открытые
каменистые склоны.
10. Род AKRAMOVSKIELLA Schileyko, gen. nov.
Типовой вид Buliminus umbrosus Mousson, 1873.
Раковина в общем удлиненно-коническая, со слаборазвитой радиальной
скульптурой. Устье без зубов. Края устья тонкие, слегка отвернутые, места
прикрепления устья слегка сближены.
Бич очень короткий, закругленный. Проксимальная часть пениса со стер-
жневидной незамкнутой папиллой, дистальная часть пениса содержит допол-
нительную камеру. Дивертикул семяприемника имеется.
Распространение. Кавказ, Закавказье и северная Турция. 3 вида.
Конхологически род почти не отличается от рода Merdigera; резко отлича-
ется от последнего, как и от всех других родов семейства, наличием незамкнутой
папиллы в сочетании с дополнительной камерой в нижней части пениальной
трубки, которая ограничена двумя сфинктерами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА AKRAMOVSKIELLA
1 (2). Обороты сильновыпуклые, вершина узкая, чуть приостренная; места
прикрепления устья заметно сближены............... . 3. A. andronakii.
2 (1). Обороты умеренно или слабо выпуклые, вершина широко закруглена;,
места прикрепления устья лишь слегка сближены.
3 (4). Поверхность дефинитивных оборотов густо и тонко радиально исчерчена.
.................................................. 2. A. schuschaensis.
4 (3). Поверхность дефинитивных оборотов тонко и неравномерно радиально»
исчерчена,............................................1. A. umbrosa-
1 . Akramovskiella umbrosa (Mousson, 1873) (рис. 223, Z; рис. 224).
Mousson, 1873 : 205, pl. 8, fig. 1 [Buliminus (Napaeus)]; 0. Boettger, 1880c : 381
[B. (Napaeus) talyschanus]; Розен, 1901 : 9 [В. (Napaeus) obscurus var. Humbert!]; К O-
b e 1 t, 1902 : 457, t. 77, fig. 3 [B. (Napaeus)]; Lindholm, 1915 : XLII [B. (Ena) hyrcanus];
Лихарев, Раммельмейер, 1952 : 181, 182 (Ena obscura var. umbrosa et var. ta-
lyschana).
Terra typica — Боржоми.
Место хранения типов не установлено.
Материал: более 30 экз. из 17 местонахождений в Закавказье и на
Северном Кавказе.
Раковина удлиненно-коническая, со слегка овальными контурами, тонко-
стенная, хрупкая. Оборотов б1^—8 , слабовыпуклых, медленно нарастающих.
Последний оборот к устью очень слабоприподнят, высота его приблизительно
равна половине высоты раковины. Окраска одноцветная, роговая, желтова-
тая или темно-оливковая, иногда с узкими радиальными штрихами, более
10. РОД AKRAMOVSKIELLA
313
светлыми, чем фон. Скульптура в виде тонкой неравномерной неясной ис-
черченности. Устье в общем овальное, слабо скошено, места его прикрепления
яезначительно сближены, париетальная мозоль очень слабая или отсутствует.
Края устья тонкие, острые,
слегка отвернуты; по линии
перегиба проходит светлая,
иногда с розоватым оттенком,
сильно расплывшаяся губа.
Колумеллярный край почти
отвесный, слегка прикрывает
узкий щелевидный пупок.
;Рис. 223. Виды рода Akramovski-
ella.
I — A. umbrosa из Армении, с. Гни-
шик; II — A. sehuschaensis из Армении,
с. Агдак; III,— A. andronakii из Арт-
вйна, Турция.
Размеры: ВР 7—13, БД 3—5.5 мм; в первоописании: ВР 7, БД 3 мм.
Внутреннее строение. Материал: 2 экз. из Армении, Даралагез,
с. Гнишик, 1940 г., сбор Э. А. Давтяна, определение Н. Н. Акрамовского;
Рис. 224. Akramovskiella umbrosa (Mouss.) из Армении, с. Гнишик, 1940 г. Половой аппарат
и вскрытый пенис.
2 — из окр. Ставрополя, 5 июля 1971 г., сбор В. Труфанова; 1 — из окр. Лен-
корани, 10 июля 1964 г.; сбор и определения автора.
Белковая железа с небольшой апикальной вырезкой. Нижний отдел вагины
в 2—3 раза короче верхнего. Семяпровод впадает немного сбоку, в результате
314
VI. НАДСЕМ. BULIMINOIDEA
чего образуется очень короткий широко закругленный, бич. Длина эпифаллуса
изменчива: у особей из Ставрополя эпифаллус очень короткий, у экземпляров
из Армении он в 2—21/2 раза длиннее, у первых цэкум располагается лишь
слегка выше середины эпифаллуса, у вторых—заметно выше. Пенис формой
напоминает песочные часы; в дистальном вздутии располагается камера, огра-
ниченная сверху и снизу сфинктерами, а снизу, кроме того, разрастанием тка-
ней, образующих нечто вроде замкнутой чрезвычайно короткой папиллы с ши-
роким каналом. В проксимальном расширении пениса видна нормально раз-
витая длинная папилла с бороздой на поверхности, выраженной по-разному.
Строение пениального аппендикса у ставропольских и армянских особей раз-
личается: у первых очень короткий, примерно такой же длины, как 42,
зато А3 очень мощный, длиннее первых двух отделов, вместе взятых; у вторых
все отделы развиты пропорционально: самый длинный А1У заканчивается неболь-
шим А2; А3 примерно втрое короче, чем Аг. Что касается единственного экзем-
пляра, собранного близ Ленкорани, то это не вполне зрелое животное,' но стро-
ение его ближе к строению армянских представителей, хотя длина эпифаллуса
занимает промежуточное положение между охарактеризованными вариантами.
Ветви полового ретрактора на диафрагме расставлены или слиты задолго до
прикрепления к ней. Одна из ветвей подходит к дистальной части пениса в рай-
оне камеры, другая — Аг (у ставропольских — к верхней его части, у армян-
ских — к нижней, у талышского — немного ниже середины). Проток семяпри-
емника прямой, короткий; резервуар с собственным коротким протоком, дивер-
тикул немного не доходит до белковой железы.
Как явствует из описания, различия между ставропольскими и армянскими
представителями вида довольно значительны; однако придавать ранг разных
видов этим популяциям представляется преждевременным, так как раковины
их практически неразличимы, размах изменчивости анатомических признаков
неясен из-за недостатка материала, а материал из типового местонахождения
анатомически вообще не изучен.
Распространение и местообитание. Северный Кавказ и
Закавказье. Обитает в подстилке смешанных и лиственных лесов, иногда под-
нимается на стволы деревьев.
2 . Akramovskiella schuschaensis (Kobelt, 1902) (рис. 223, II).
Kobelt, 1902 : 727, t. 107, fig. 9, 10 {Buliminus (Ena)]; Лихарев, Раммель-
мейер, 1952 : 194, рис. 106 (Turanena); Акрамовский, 1955 : 150 (Ena); 1968 : 26
(Ena); 19716 : 7 (Ena); 1976 : 160, рис. 73В, табл. VIII, 80 (Ena).
Terra typica — окр. Шуши (Нагорно-Карабахская АО).
Типы хранятся, вероятно, в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: 7 экз.’ из двух местонахождений в Закавказье.
Раковина от удлиненно-конической до почти башневидной, тонкостенная,
блестящая. Оборотов —1Ч3, выпуклых, разделенных глубоким швом.
Последний оборот к устью не поднят, высота его примерно равна половине
высоты раковины или немного меньше. Окраска одноцветно роговая. Скуль-
птура в виде густой и тонкой радиальной исчерченности. Устье овальное, сильно
скошено, места его прикрепления заметно сближены и соединены тонкой про-
зрачной мозолью. Края устья почти не отвернуты, кроме верхней части колу-
меллярного края. Губы нет или она очень слабая. Пупок проколовидный.
Размеры: ВР 6.5—8.5, БД 3.2—4 мм; в первоописании: ВР 8, БД 3.25 мм.
Муссон (Mousson, 1861) сообщает, что Дюбуа нашел в Закавказье (Сомхетия-
«Somketh») Bulimus benjamiticus Benson, 1859 — вид, описанный из Перу,
салимаи чрезвычайно похожий на A kramovskiella schuschaensis, хотяМуссон (Mo-
usson, 1873) подчеркивает его сходство с A. umbrosa. Геллер (Heller, 1971а),
обсуждая распространение Ena benjamitica и ссылаясь на Муссона, включает
в ареал вида Закавказье; причем надо подчеркнуть, что со времени описания
Buliminus benjamiticus было найдено не более 10 экз. этого вида — и все только
11. РОД DIFFERENA
315
на Синайском п-ове. Не исключено, что этотвид относится к роду A kramovskiella
(это можно установить, только изучив его анатомию), но очень сомнительно,
чтобы он встречался в Закавказье.
Внутреннее строение. Материал: один не вполне взрослый
экз. из Иджеванского р-на Армении, с. Агдак, урочище Лани-дзор, сбор и
определение Н. Н. Акрамовского.
Белковая железа с вялой апикальной вырезкой. Нижний отдел вагины
незначительно короче верхнего. Семяпровод впадает эксцентрично, бич чрез-
вычайно короткий, приостренный. Цэкум расположен заметно выше середины
эпифаллуса. Внешний вид пениса и внутреннее его строение, по-видимому,
почти такое же, как у предыдущего вида. Отделы пениального аппендикса
нормально и пропорционально развиты. Ветви полового ретрактора слиты
перед диафрагмой, одна ветвь крепится к А1 несколько ниже его середины, вто-
рая — к заднему отделу пениального вздутия, содержащего камеру. Проток
семяприемника прямой, овальный резервуар с собственным протоком, длина
которого не превышает длины резервуара, дивертикул немного не доходит до
белковой железы.
Распространение и местообитание. Восточная часть
Малого Кавказа: Армения, окр. Шуши и гора Иландаг в Нахичеванской АССР;
возможно, южная Грузия. Встречается в трещинах влажных скал, покрытых
редким лесом или кустарником, иногда также в высокогорном поясе (Акра-
мовский, 1976).
3 . Akramovskiella andronakii (Lindholm, 1913) (рис. 223, III).
Lindholm, 1913 : 20 [Buliminus (Ena)].
Terra typica — Артвин и Ломашен (северо-восточная Турция).
। Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
I здесь.
; Материал: 14 экз. из типовой серии.
Раковина в виде высокого и узкого конуса, умеренно тонкостенная, с узкой
вершиной. Оборотов 8, довольно выпуклых, разделенных глубоким швом, рав-
номерно и плавно нарастающих. Последний оборот, непосредственно возле
устья чуть поднят, высота его заметно меньше половины высоты раковины.
!, Окраска одноцветная, от темно-коричневой до роговой. Скульптура в виде
; сглаженных и неправильно расположенных радиальных морщинок. Устье не-
большое, почти правильно овальное, края его равномерно и несильно отвер-
1 нуты, места прикрепления заметно сближены и соединены тонкой, но отчетливой
париетальной мозолью. Верхняя часть колумеллярного края еле заметно на-
' клонена внутрь устья. Пупок в виде короткой и узкой щели.
Размеры в первоописании: ВР 11—14, БД 4.5—5 мм; лектотип: ВР 12.0,
БД 4.5 мм.
На сходство с Buliminus schuschaensis указывал еще Линдгольм в первоопи-
сании.
Распространение. Пока вид известен из северо-восточной Турции
близ границы с юго-восточной частью Грузии; поэтому не исключено присут-
ствие вида в Аджарии.
11. Род DIFFERENA Schileyko, gen. nov.
Типовой вид Differena leucostoma Schileyko, sp. nov.
Раковина высококоническая, с нерезкой и неправильной радиальной скуль-
птурой. Устье без зубов. Края устья утолщены широкой, но невысокой губой
и слабо отвернуты. Места прикрепления устья сближены.
Бич как таковой почти не выражен, представляет собой широкое закругле-
ние конца эпифаллуса. Необычайно длинный дистальный отдел пениса вну-
три с многочисленными складками и бугорками, в маленьком проксимальном
316
VI. НАДСЕМ. BULIMINOIDEA
отделе содержится незамкнутая папилла. Дивертикул семяприемника хорошо
развит.
Монотипический род.
Конхологически род ближе всего напоминает представителей рода Akramov-
skiella, но внутреннее строение его настолько своеобразно, что в настоящее время
трудно судить о близости его к какому-либо из существующих родов. Помимо
отмеченных в описании рода признаков надо указать еще на сильное видоизме-
нение пениального аппендикса (Лх и А2 слиты и представлены маленьким шаро-
видным отделом, а А3 крупный, удлиненно-булавовидный) и наличие лишь од-
ной ветви полового ретрактора, обслуживающей пенис.
Differena leucostoma Schileyko, sp. nov. (рис. 225, 226).
Terra typica — гора Кяпаз, северные отроги Мровдагского хр. в восточной
части Малого Кавказа.
Г о л о т и п хранится в Зоологическом ин-те АН СССР.
Материал: более 40 экз. из типового местонахождения.
Раковина высококоническая, с немного оплывшим контуром, умеренно
твердостенная, слабо блестящая. Оборотов —6, слабовыпуклых, разделен-
ных мелким светлым швом. Последний оборот к устью по-
чти не поднят, высота его равна половине высоты ракови-
ны или немного больше. Окраска одноцветная, светло-ро-
говая или серовато-желтая. Скульптура представлена тон-
кими радиальными неравномерно расположенными мор-
щинками, элементы спиральной скульптуры отсутствуют.
Устье овальное или округлое, места его прикрепления
умеренно сближены и соединены слабой мозолью. Устье-
вые края немного отвернуты, утолщены за счет чрезвы-
чайно расплывшейся, но довольно массивной белой губы,
просвечивающей сквозь стенку раковины: затылок по
всему периметру окаймлен широкой светлой полосой с раз-
мытыми границами. Отворот колумеллярного края.не при-
крывает сквозного круглого проколовидного пупка.
Рис. 225. Dijferena leucostoma Schileyko, sp. nov. Голотип.
Размеры: ВР 6.5—8.2, БД 3.3—4 мм; голотип: ВР 8.0, БД 3.6 мм.
Внутреннее строение. Материал: 2 экз. из типового местона-
хождения на горе Кяпаз в западной части Азербайджана, 24 июня 1954 г.,
сбор И. М. Лихарева, определение автора.
Белковая железа с седловидной вырезкой, занимающей апикальное поло-
жение. Нижний отдел вагины приблизительно вдвое короче верхнего. Половой
атриум глубокий, короткоцилиндрический. Семяпровод впадает В эпифаллус
сбоку чуть отступя от конца последнего. Эпифаллус на конце притуплен, слегка
утолщен и широко закруглен. Эпифаллус относительно недлинный, почти пря-
мой; ясно выраженный бугорковидный цэкум немного сдвинут вверх относи-
тельно середины эпифаллуса. Пенис ясно разделен на необыкновенно длинный
дистальный цилиндрический отдел, длина которого слегка превышает длину
эпифаллуса, и маленький слегка вздутый проксимальный, длина которого
в 10—12 раз меньше длины дистального отдела. Внутренняя поверхность ди-
стального отдела покрыта моногочисленными резкими бугорками и складками,
почти полностью заполняющими его просвет. Внутренняя поверхность прокси-
мального отдела пениса гладкая, здесь имеется маленькая коническая папилла.
Борозда на поверхности папиллы ясно заметна, но очень мелкая (см. попе-
речный срез папиллы на рис. 226, I). Пениальный аппендикс очень своеобразен:
12. РОД GEMINULA
317
и А2 слиты вместе и имеют почти шаровидную форму; папилла аппендикса
отсутствует. А3 примерно втрое длиннее, чем Л1+^42> мощный, булавовидный.
Л4 и Аъ развиты нормально. Половой ретрактор представлен одной ветвью,
которая крепится на границе третьей и последней четверти дистального отдела
пениса, считая от атриальной области. Существенно также и то, что пениаль-
ный аппендикс отходит от пениса не вблизи атриума, а на границе дистального
Рис. 226. Differena leucostoma Schileyko, sp. nov. Паратипы.
I — половой аппарат и вскрытый пенис, II — вскрытый пенис другого экземпляра.
и проксимального отделов. Проток семяприемника утолщен, прямой, проток
резервуара тонкий, резервуар овальный или фасолевидный. Дивертикул длиннее
протока до места раздвоения, доходит до белковой железы.
Распространение и местообитание. Известна лишь
одна находка в типовом местонахождении на горе Кяпаз. Моллюски найдены
в трещинах скал и осыпях.
12. Род GEMINULA Lindholm, 1925
Lindholm, 1925а : 30 (род Jaminia, секция Chondrula, подсекция Geminula)-, Thi-
ele, 1931 : 518 (род Jaminia, секция Chondrula, подсекция Geminula)-, Wenz, Zilch,
1959—1960:179 -[Chondrula (Geminula)]-, Шилейко, 1978г : 848.
Типовой вид Buliminus didymodus О. Boettger, 1880 (syn. Bulimus
isselianus Bourguignat, 1865); по первоначальному обозначению.
Раковина от овально-цилиндрической до цилиндрической, поверхность ее
почти гладкая. Устье с четырьмя зубами, имеющими вид коротких гребневид-
ных бугорков: париетальным, колумеллярный и двумя палатальными. Края
устья слегка или значительно отвернуты, могут быть утолщенными.
318
VI. НАДСЕМ. BULIMINOIDEA
Бич отсутствует. Пенис с папиллой. Дивертикул семяприемника имеется.
Яркий признак рода — очень короткий проток семяприемника: он раздва-
ивается в непосредственной близости от места своего отхождения, распадаясь
на дивертикул и относительно длинный собственный проток резервуара.
Распространение. Закавказье, северный Иран, Копетдаг. 2 вида.
таблица для определения видов рода geminula .
1 (2). Края устья широко отвернуты и утолщены. Большой диаметр раковины
3—4 мм .................................................1. G. isseliana.
2 (1). Края устья слегка отвернуты, почти не утолщены. Большой диаметр
раковины 2.2—2.9 мм ....................................2. G. continens.
1. Geminula isseliana (Bo.urguignat, 1865) (рис.. 227, Z; рис. 228).
Bourguignat in:Issel, 1865 : 421, t. 2, fig. 37—40 (Bulimus), 422, t. 2, figs. 41 —
44 (B. Ghilanensis); Mousson, 1873 : 208 (Chondrus nucifragus, non Pfeiffer, 1848); О. В o-
et t ger, 1880c : 380 (Buliminus didymodus); Westerlund, 1887 : 43 (Buliminus)', 1896 :
189 [B. (Chondrula) tardtgyrus]; Kobelt, 1902 : 940, t. 132, figs. 7—9 (Chondrula); F о r-
cart, 1935:423 [7. (Multidentata) ghilanensis]; Акрамовский, 1949:147 [Jaminia
(Chondrula, Multidentula)]; Лихарев, Раммельмейер, 1952 : 177, рис. 89 (Jaminia);
Матёкин, 1966; 112, табл. 2, рис. 10а, б [Jaminia (Multidentata) didymoda]; Акрамов-
ский, 1976 : 154, рис. 70, табл. VII, 72 (Jaminia); Шилейко, 1977 : 46, рис. ЗА.
Terra typica — окр. оз. Севан.
Место хранения типов не установлено.
Материал: более 40 экз. из 4 местонахождений в Армении, Талыше и
северном Иране.
Раковина от почти цилиндрической до коническо-цилиндрической и удли-
ненно овально-конической, твердостенная, блестящая. Оборотов 7—8, слабо-
выпуклых; последний оборот непосредственно возле устья слегка приподнят,
высота его немного меньше половины высоты раковины. Если раковина овально-
цилиндрическая, то последние 2 оборота почти одинаковой ширины, если ци-
линдрическая, то одинаковую ширину имеют последние 3 оборота. Окраска
одноцветная, от коричневой до желтоватой; иногда наблюдается чередование
чуть более светлых радиальных полос с более темными. Скульптура в виде
редких очень тонких неравномерно расположенных радиальных морщинок,
отдельные участки раковины совсем гладкие. Кроме того, в нижней части по-
следнего оборота иногда заметны слабые спиральные бороздки и штрихи, осо-
бенно на затылке. Устье овальное, с отвернутыми утолщенными белыми краями,
образующими плавную широкую дугу от ангулярной области до стыка базаль-
ного и колумеллярного краев, где намечается сглаженный угол. Зубы распола-
гаются отступя от края устья. К изогнутой короткой париетальной пластинке
справа вплотную прилегает маленький бугорковидный зуб, который иногда
полностью слит с париетальной пластинкой. Колумеллярная пластинка слегка
изогнутая, горизонтальная, в виде козырька; резко изогнувшись в глубине
устья, она уходит вверх по столбику. Иногда в нижней части колумеллярного
края имеется еще один небольшой бугорок. Нижний палатальный зуб пластин-
чатый, верхний палатальный — бугорковидный; в самом верху палатального
края нередко можно заметить еще один небольшой ступенчатый бугорок.
Палатальные зубы просвечивают сквозь стенку раковины в виде светлых пятен.
Пупок открытый, овальный, очень маленький.
Размеры: ВР 6.5—10, БД 3—4 мм; в первоописании: ВР 8, БД 4 мм.
Внутреннее строение. Материал: 4 экз. из Армении, Дарала-
гез, с. Гнишик, 1940 г., сбор Э. А. Давтяна, определение Н. Н. Акрамовского;
9 — из северного Ирана, окр. Горгана, июнь 1914 г., сбор Е. М. Филипповича,
определение В. А. Линдгольма.
Белковая железа с узкой и глубокой латеральной бороздой. Относительная
длина отделов ватины непостоянна: у представителей армянской популяции
12. РОД GEMINVLA
319
верхний отдел вагины в 2—5 раз короче нижнего, у североиранских эти отделы
примерно равной длины. Семяпровод впадает в эпифаллус почти терминально,
в луковицеобразное расширение, выраженное по-разному. Цэкум заметно сдви-
нут к семяпроводу относительно середины эпифалллуса. Эпифаллус недлинный,
резких изгибов не образует. Пенис небольшой, часто разделен на цилиндриче-
ский дистальный и короткий расширенный проксимальный отделы; правда, гра-
ница между этими отделами подчас почти отсутствует. Внутренняя поверхность
дистального отдела несет серию продольных гофрированных складок, сходя-
щих на нет в проксимальном отделе. Папилла пениса тонкая, червеобразная,
довольно длинная, с ясной продольной бороздой, поверхность ее гладкая или
с кольцевыми и короткими
продольными складками.
Отделы пениального ап-
пендикса нормально и про-
порционально развиты, но
среди 9 экз. из Горгана об-
наружено явление, отмечен-
ное и у крымского вида
Brephulopsts bidens', у 4>осо-
Рис. 227. Виды рода Ge-
minula.
I — G. isseliana (Bourg.) из Та-
лыша, верховья p. Ленкоран-
чай; II — G. continens (Rosen)
из Большого Балхана, Туркме-
ния.
Рис. 228. Geminula isseliana (Bourg.) из Армении, с. Гнишик,
1940 г. Половой аппарат и вскрытый пенис (из: Шилейко,
19786).
бей уЦ отсутствовал, и шаровидный Л2 располагался на середине пениса (рис. 36);
соответствующая ветвь полового ретрактора в этом случае отсутствовала (ма-
териал из Горгана представлен формой callilabris О. Boettger). Если строение
пениального аппендикса нормальное, то ветви полового ретрактора крепятся
к диафрагме самостоятельно или в разной степени сближены. Одна ветвь при-
соединяется к А± слегка отступя от А 2. Пениальная ветвь крепится к прокси-
мальному отделу пениса близ эпифаллуса. Проток семяприемника чрезвычайно
короткий. Резервуар овальный, длина его протока превышает длину самого
резервуара в 1Ч2—2 раза. Дивертикул длинный, прилегает верхним концом
к нижней части белковой железы.
Распространение и местообитание. Северные склоны
Малого Кавказа на запад до Ханлара, долина среднего течения Араксн па за-
пад до Джрвежа; Талыш, северный Иран, северный Ирак (?), Копетдаг. Встре-
чается в зарослях фриганы и в осыпях полупустынных районов, включая лу-
гостепную полосу (Акрамовский, 1976).
320
VI. НАДСЕМ. BULIMINOIDEA
2. Geminula continens (Rosen, 1892) (рис. 227, IT, рис. 229).
Rosen, 1892 : 125 [Buliminus (Amphiscopus)}', 1893a : 173 [B. (Amphiscopus)}', К o-
b e 11, 1901, Iconogr., N. F., 9 : 48, t. 257, fig. 1661 (Buliminus)', F о г с а г t, 1959b : 315,
figs. 1—3 [Jaminia (Euchondrus)]', С а м а д о в, 1960 : 8 (Chondrula natalensis); Матёкин,
1966 : 112, табл. 2, рис. 11 [Jaminia (Multidentata)}.
Terra typica — Казанджик, северо-западные отроги Копетдага.
Паратипы хранятся в Зенкенборгском музее, Франкфурт-на-Майне,
№ 64121 (Forcart, 1959b); место хранения голотипа не установлено.
Материал: 24 экз. из 4 местонахождений на Большом Балхане, Копет-
даге и Нуратинском хр. (северо-западные отроги Алая).
Раковина цилиндрическая или почти цилиндрическая, умеренно твердостен-
ная, просвечивающая, блестящая, с широко закругленной вершиной. Оборотов
Рис. 229. Geminula continens (Rosen) из окр. с. Джут на Нуратинском хр., 14 V 1976 г. По-
ловой аппарат и вскрытый пенис.
6V2—71/2; верхние выпуклые, нижние уплощенные. Последний оборот возле
самого устья слегка, но явственно приподнят, высота его заметно меньше по-
ловины высоты раковины. Окраска одноцветно светло-роговая. Шелковистая
скульптура в виде тончайших неравномерно расположенных радиальных мор-
щинок, местами имеются почти гладкие участки. Элементы спиральной скульп-
туры полностью отсутствуют. Устье округлое, места его прикрепления связаны
тончайшей прозрачной мозолью, края устья несильно плавно отвернуты и не-
значительно утолщены. Гребневидная короткая париетальная пластинка не
совсем отвесная: ее вершина немного отклонена вправо. Горизонтальная колу-
меллярная пластинка в глубине устья резко изгибается и уходит вертикально
вверх. Базальный зуб отсутствует. Нижний палатальный зуб короткий,
округло-треугольный, верхний палатальный — бугорковидный. Кроме того, у не-
которых раковин в ангулярной области имеется очень слабый бугорок. Пупок
открытый, проколовидный, почти круглый.
Размеры: ВР 6—7, БД 2.2—2.9 мм; паратип (по: Forcart, 1959b): ВР 7,
БД 2.2 мм.
Внутреннее строение. Материал: 3 экз. из Нуратинского хр.,
окр. Джуша, 14 мая 1976 г., сбор и определение 3. Иззатуллаева.
1. РОД CHONDRULOPSINA
321
Белковая железа тупоконечная, с широким латеральным желобком. Отделы
вагины приблизительно равной длины. Семяпровод впадает в сильно расширен-
ный широко закругленный конец эпифаллуса несколько сбоку. Цэкум неболь-
шой, приостренный, сильно сдвинут к семяпроводу. Эпифаллус короткий,
диаметр его от семяпровода к пенису постепенно уменьшается. Пенис верете-
новидный, на его внутренних стенках заметны нерезкие кольцевые складки.
Папилла удлиненная, форма ее у всех трех экземпляров была различной, так
как на поверхности папиллы имеются многочисленные широкие вмятины и
складки, между которыми, однако, можно различить широкий желоб, соответст-
вующий продольной борозде. Кроме того, поверхность папиллы несет кольцевые
складочки. Папилла заполнена очень рыхлой паренхиматозной тканью. Ветви
полового ретрактора на диафрагме слиты, одна ветвь крепится к А± сразу же
под А.2, вторая ветвь — к пенису на границе его с эпифаллусом. Отделы пени-
ального аппендикса нормально и пропорционально развиты. Семяприемник
характерного для рода строения.
Распространение и местообитание. Большой Балхан,
Копетдаг и прилежащие районы Ирана; Зеравшанский, Нуратинский и Гис-
сарский хребты. Населяет открытые аридные участки, встречается в осыпях
и под каменными плитами.
II. Подсем. CHONDRULOPSININAE Schileyko, 1978
Шилейко, 1978г : 845.
Раковина слабо скульптурированная. Устье раковины с зубами, которые
могут быть представлены только одним палатальным вздутием губы или пала-
тальной глубоко лежащей складкой. Эмбриональные обороты гладкие.
Эпифаллус без бича либо с очень коротким шаровидным бичом. Семяпровод
впадает почти апикально или сбоку. Цэкум нормально развит. Пенис гладко-
стенный, заключает замкнутую папиллу. Призмоконические бугорки и прок-
симальный вырост пениса отсутствуют. Пениальный аппендикс имеется, ди-
вертикул отсутствует или короткий, рудиментарный.
Распространение. Средняя Азия. 2 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
CHONDRULOPSININAE
1 (2). В глубине устья на палатальной стенке залегает длинная продольная
складка ........................................2. Siraphoroides.
2 (1). Палатальная складка отсутствует; на палатальной стенке имеется разви-
тый в разной степени бугорок, имеющий иногда вид небольшого утолще-
ния губы........................................1. Chondrulopsina.
1. Род CHONDRULOPSINA Lindholm, 1925
Lindholm, 1925а : 30 [Sewertzowia (Chondrulopsina)]; Thiele, 1931 : 518
[S. (Chondrulopsina)]; Hesse, 1933 : 166 [Jaminia (Chondrulopsina)]; Лихарев, Pa M-
ме льмейер, 1952 : 191 [Sewertzowia (Chondrulopsina)]; Wenz, Zilch, 1959—1960 :
182 [Jaminia (Chondrulopsina)]; Матёкин, 1966 : 113 [7. (Chondrulopsina)]; Шилейко,
1978г : 845.
Типовой вид Buliminus haberhaueri Ancey, 1886 (syn. Buliminus
intumescens var. fedtschenkoi Ancey, 1886); по первоначальному обозначению.
Устье с губой, на которой располагаются 3 бугорковидных зуба, развитых
в разной степени; от них может оставаться лишь палатальное вздутие губы.
Просвет папиллы округлый или в виде прямой щели на поперечном разрезе.
Дивертикул семяприемника отсутствует или рудиментарный, в последнем слу-
чае не длиннее резервуара. Семяпровод по эпифаллусу не сдвинут.
Распространение. Средняя Азия. 2 вида.
21 А. А. Шилейко
322
VI. НАДСЕМ. BULIMINOIDEA
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CHONDRULOPSINA
1 (2). Все три устьевых зуба хорошо развиты............2. Ch. fedtschenkoi.
2 (1). Париетальный зуб отсутствует, палатальный и колумеллярный зубы
развиты слабо, имеют вид небольших утолщений губы; колумеллярный
часто совсем незаметен ................................1. Ch. intumescens.
1. Chondrulopsina intumescens (E. Martens, 1874) (рис. 230, I; рис. 231).
E. Martens, 1874, Sitzungsb. Ges. naturf. Fr. Berlin : 46 [Buliminus (Chondrula)]; W e-
st erlun d, 1887 : 69 (Buliminus); 1897 : 61 [B. (Chondrula) anomphalus f. infans]; Розен,
1901 : 10 (В. intumescens var. suitanus); Kobelt, 1902 : 524, t. 83, fig. 14 [B. (Chondru-
lopsis)]; Hesse, 1933 : 166, fig. 9 [Jaminia (Chondrulopsina)]; Лихарев, Раммель-
мейер, 1952 : 191, рис. 102 [Sewertzowia (Chondrulopsina)]; Матёкин, 1966 : 130
[Jaminia (Chondrulopsina) potaniniana subsp. albiplicata part.]; Лихарев, Старо-
го г а т о в, 1967:182 (Subzebrinus); Мухитдинов, 1976:41, рис. IB, 2В (Subze-
brinus likharevi).
Terra typica — Чупаната близ Самарканда.
Г о л о т и и хранится в Зоологическом музее Берлинского ун-та им. Гум-
больдта, № 22459 (Kilias, 1971).
Материал: более 60 экз. из 14 местонахождений на Тянь-Шане и на
Алае.
Раковина от цилиндрической до коническо-цилиндрической, твердостенная,
с тупой вершиной, умеренно блестящая. Оборотов 7—8, слабовыпуклых, разде-
ленных светлым швом. Последний оборот к
устью слабо приподнят, высота его немного
меньше половины высоты раковины или по-
чти равна ей. Окраска от роговой до красно-
ватой или коричневатой, неодноцветная, име-
ются то более, то менее светлые участки,
особенно на нижних оборотах. Скульптура
в виде тонкой, иногда почти шелковистой
исчерченности. Устье . в целом овальное,
Рис. 230. Виды рода Chondrulopsina.
1 — Ch. intumescens (Mart.) с перевала Орток-бель на Чат-
кальском хр.; II — Ch. fedtschenkoi (Ancey) из окр. Самар-
канда.
места его прикрепления расставлены и соединены развитой в разной степени
мозолью. Края устья отвернуты и утолщены расплывшейся, но мощной губой.
Париетальный бугорок развит обычно слабо и представлен местным расширением
париетальной мозоли. Колумеллярный зуб отсутствует или очень слабый,
в виде еле заметной припухлости; палатальный чаще всего хорошо виден,
но развит слабо, имеет вид продольного вздутия губы. Пупок узкий, щеле-
видный.
Размеры: ВР 8—11, БД 3—4.5 мм; в первоописании: ВР 8.5, БД 4 мм.
Внутреннее строение. Материал: 2 экз. из Чупанаты близ
Самарканда, 29 марта 1929 г., сбор П. П. Архангельского; 3 — из Алайского хр.,
Джылису, 16 июля 1974 г., сбор 3. Иззатуллаева; 3 — из Ферганского хр.,
берег р. Джаз-ичу, бассейн р. Кара-Унгур, 16 июня 1961 г., сбор И. М. Лиха-
рева; 2 — из Туркестанского хр., окр. с. Чорку Исфаринского р-на, 10 сен-
тября 1973 г., сбор А. Б. Мухитдинова; определения автора.
Белковая железа с плавной апикальной вырезкой. Отделы вагины прибли-
зительно равной длины или верхний немного длиннее. Семяпровод впадает
1. РОД CHONDRULOPSINA
323
почти терминально, лишь слегка сдвинут на сторону. Длина эпифаллуса из-
менчива, но в общем этот проток недлинный, лишен резких изгибов. Цэкум
сильно сдвинут к семяпроводу;, на противоположной стороне эпифаллуса также
имеется вздутие. Пенис тонкостенный, иногда сквозь его покровы просвечи-
вает папилла, имеющая вид трубки. Отверстие папиллы в виде длинной щели.
Отделы пениального аппендикса ясно различимы. Ветви полового ретрактора
Рис. ,231. Chondrulopsina intumescens (Mart.).
I— экземпляр из Алайского хр., с. Джилису, 16 VII 1974., половой аппарат и вскрытый пенис; II —
вскрытый пенис экземпляра из окр. Самарканда, с. Чупаната, 29 III 1929 г.
тесно сближены на диафрагме, одна ветвь крепится к /Ц немного выше его сере-
дины, вторая — к нижней части пениса. Проток семяприемника прямой, дивер-
тикул слабо развит, иногда отсутствует.
Распространение и местообитание. Горные области
Средней Азии, кроме северной части Тянь-Шаня и Джунгарского хр.; западный
Китай (Кульджа); северный Афганистан. Встречается в различных типах
биотопов; густых зарослей кустарника и лесов избегает.
2. Chondrulopsina fedtschenkoi (Ancey, 1886) (рис. 230, II; рис. 232).
А п с е у 1886а : 51 (Buliminus intumescens var. fedtschenkoi), 52 (В. haberhaueri); 1893 :
37 (В. intumescens var. inermis); Westerlund, 1897 : 61 (Chondrulus anomphalus); 1898 :
164 [Buliminus (Chondrulus) .bisinuatus]; Розен, 1901 : 10 [В. (Chondrulus) Rennenkamft];
Лихарев, Раммельмейер, 1952 : 192, рис. 103 [Sewertzowia (Chondrulopsina)]-,
Матёкин, 1966 : 113, табл. 3, рис. 3 [Jaminia (Chondrulopsina)]; Мухитдинов,
1976 : 40, рис. 1Б, 2Б (Chondrulopsina tristnuata); Шилейко, 19786 : 349, рис. 3, I.
Terra typica — окр. Самарканда.
Место хранения типов не установлено.
Материал: более 100 экз. из 10 местонахождений на Тянь-Шане и на
Алае.
Раковина от удлиненно-овальной до веретеновидной и почти цилиндрической,
блестящая, умеренно твердостенная, просвечивающая. Оборотов 6—9, слабо
выпуклых, разделенных мелким беловатым швом. Последний оборот к устью
заметно приподнят, высота его всегда меньше половины высоты раковины;
324
VI. НАДСЕМ. BULIMINOIDEA
у высоких многооборотных раковин высота последнего оборота может составлять
1/з высоты раковины. Окраска роговая, серая или каштановая, верхние обороты
обычно темнее нижних. Скульптура в виде тонкой неясной радиальной исчер-
ченности. Устье почти отвесное, овальное или округло-треугольное, места его
прикрепления слегка сближены и соединены мозолью, на которой располага-
ется париетальный зуб или только его передняя часть. Колумеллярный зуб
удлиненный, палатальный — бугорковидный; степень развития зубов подвержена
сильной биотопической изменчивости. Пупок маленький, в виде короткой
и узкой щели.
Размеры: ВР 7—9, БД 2.4—3.5 мм; в первоописании: ВР 8, БД 3.3 мм.
Внутренннее строение. Материал: 12 экз. из предгорьев
Алайского хр., окр. с. Валакиш, 27 сентября 1972 г., сбор Р. Я. Бутылина;
10 — из долины р. Нарын ниже
Токтогула, 31 мая 1972 г.; сбор
и определения автора.
Белковая железа с узкой и
глубокой апикальной вырезкой
и латеральным желобком. Отде-
лы вагины равной длины или
верхний отдел незначительно ко-
роче нижнего. Семяпровод впа-
дает почти терминально, конец
эпифаллуса слегка утолщен.
Ясно выраженный цэкум сильно
смещен к семяпроводу. На проти-
Рис. 232. Chondrulopsina jedtsdien-
koi (Ancey) из предгорьев Алайского
хр., 27 IX 1972 г. Половой аппарат и
вскрытый пенис (из: Шилейко,
19786).
волежащей цэкуму стороне тонкого цилиндрического эпифаллуса имеется не-
большое вздутие, расположенное почти симметрично цэкуму. Пенис тонко-
стенный, булавовидный или цилиндрический, его внутренняя поверхность
гладкая. Имеется замкнутая папилла с узким щелевидным или округлым
, просветом. Эпифаллус способен частично вворачиваться внутрь пениса, за
счет чего папилла удлиняется. Все отделы пениального аппендикса нормально
развиты; А3 переходит в Ai плавно, без резкой границы. Половой ретрактор
отходит от дифрагмы единой лентой, затем распадается; одна ветвь крепится
к границе между Ау и А2, вторая — к средней части пениса. Проток семяприем-
ника короткий, прямой и тонкий. Дивертикул рудиментарный или отсутствует.
Резервуар овальный, изогнутый, не всегда доходит даже до середины спермови-
дукта.
Распространение и местообитание. Горные и предгор-
ные районы Самаркандской обл.; Ферганский, Алайский, Кураминский, Тур-
кестанский, Заалайский, Гиссарский хребты. Обитает на щебнистых открытых
участках, среди камней и на стеблях полыни (Матёкин, 1966). Во время летней
засухи животные способны закапываться в почву на глубину 35—40 см (Мухит-
динов, 1976).
2. Род SIRAPHOROIDES Schileyko, 1977
Шилейко, 1977 : 43.
Типовой вид Siraphorus moltschanovi Likharev et Rammelmeyer,
1952; по первоначальному обозначению.
2. РОД SIRAPHOROIDES
325
Устье с широкой и мощной губой. В глубине устья залегает продольная
палатальная складка.
Просвет папиллы на поперечном разрезе серповидно изогнут. Дивертикул
семяприемника короткий, но несколько длиннее резервуара. Семяпровод сдви-
нут вниз по эпифаллусу, в результате чего образуется шаровидный бич.
Монотипический род.
Siraphoroides moltschanovi (Likharev et Rammelmeyer, 1952) (рис. 233).
Лихарев, Раммельмейер, 1952 : 207, рис. 121, 122 (Siraphorus); Матё-
кин, 1966 : 115 [Jaminia (Chondrulopsina)]; Шилейко, 1977 : 43, рис. 1, 5, 6.
Terra typica — Арсланбоб, Ферганский хр.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 6 экз. из двух местонахождений на Ферганском хр. (типо-
вая серия — 4 экз., и 2 экз., собранные близ типового местонахождения).
Рис. 233. Siraphoroides moltschanovi (Likh. et Ramm.) из урочища Ак-Терек близ с. Арслаи-
боб, Ферганский хр., VIII—IX 1937 г.
1 — раковина, II— половой аппарат и вскрытый пенис (из: Шилейко, 1977).
Раковина овально-цилиндрическая, слегка просвечивающая, составлена 6—
7 слабовыпуклыми оборотами. Вершина закругленная. 2—3 нижних оборота
почти одинаковой ширины, последний оборот к устью еле заметно поднят,
высота его составляет приблизительно половину высоты раковины. Окраска
от серой до серовато-роговой, верхние обороты немного темнее нижних
(видно на свежих раковинах). Скульптура в виде неравномерно расположенных
сильно сглаженных радиальных морщинок и складок; некоторые из складок
окрашены немного светлее, чем фон. Устье слегка скошено, усеченно-полуоваль-
ное, с белыми краями. Места прикрепления устья расставлены, края плавни
широко отвернуты. Париетальная мозоль почти не выражена, лишь в ангуляр-
ной области имеется небольшой светлый бугорок. В глубине устья залегает
палатальная складка, уходящая в глубь раковины на полоборота. Вздутый
задний конец складки образует округло-треугольный бугорок, против которого
326
VI. НАДСЕМ. BULIMINOIDEA
находится вздутие колумеллярной пластинки. Снаружи палатальная складка
слабо просвечивает в виде светлой полосы. Пупок узкий, полуприкрыт отворо-
том колумеллярного края.
Размеры: ВР 6.7—8, БД 3—3.2 мм; гОлотип: ВР 7.7, БД 3.0 мм.
Внутреннее строение. Материал: 2 экз. из Ферганского хр.,
Ак-Терек в окр. Арсланбоба, август-сентябрь 1937 г., сбор А. Н. Кириченко,
определение автора.
На поверхности белковой железы имеется лишь очень вялый латеральный
желоб. Нижний отдел вагины в 2х/2—3 раза длиннее нижнего. Семяпровод впа-
дает отступя от конца эпифаллуса, в результате чего образуется короткий почти
шаровидный бич. Цэкум резко выражен почти по всему периметру эпифаллуса,
заметно сдвинут к бичу. Пенис резко разделен на дистальную узкую цилиндри-
ческую и проксимальную веретеновидную части. Внутренняя поверхность
дистальной части пениса покрыта продольными складками, проксимальная
часть внутри гладкая, причем стенки в этом отделе очень тонкие и полупрозрач-
ные, позволяющие видеть контуры папиллы без вскрытия пениального чехла.
Папилла массивная, с широким внутренним каналом, имеющим на поперечном
разрезе серповидную форму. Наружная поверхность папиллы с поперечными
широкими складками. Пениальный аппендикс нормально развит, все 5 его
отделов ясно различимы. Ветви полового ретрактора отходят от диафрагмы
порознь, одна из них крепится к границе йежду дистальным и проксимальным
участками пениса, вторая — к Аг выше его середины. Проток семяприемника
почти прямой, резервуар на коротком протоке прилегает к середине короткого
спермовидукта; дивертикул немногим длиннее резервуара, значительно не
доходит до белковой железы.
Распространение и местообитание. Обе известные на-
ходки сделаны примерно в одном месте на Ферганском хр. близ Арсланбоба,
в плодово-ореховых лесах.
III. Подсем. JAMINIINAE Thiele, 1931
Thiele, 1931 : 517.
Раковина слабо скульптурированная. В устье 4—5 зубов: 2 колумеллярных,
1 палатальный и 1—2 париетальных. Эмбриональные обороты гладкие.
Бич чрезвычайно короткий, семяпровод впадает в эпифаллус эксцентрично.
Цэкум отсутствует. Пенис нитевидный, без папиллы, в проксимальной части
с подкововидной складкой. Призмоконические бугорки и проксимальный вырост
пениса отсутствуют. Пениальный аппендикс и дивертикул семяприемника
нормально развиты.
Монотипическое подсемейство.
Распространение. Северное и восточное Средиземноморье.
В ископаемом состоянии подсемейство известно начиная с плейстоцена
(Европа).
1. Род JAMINIA Risso, 1826
Risso, 1826 : 88; Moquin-Tandon, 1855 : 298 (Gonodon, non Gray, 1847);
Monterosato in: Gecconi, 1908 : 7 (Chondritortus, типовой вид Helix quadridens
Miiller, 1774); Hesse,: 1933 : 160; F о г с a г t, 1940 : 190; Лихарев, Раммель-
мейер, 1952 : 176; W e n z, Zilch, 1959—1960 : 182; Шилейко, 1978г : 846.
Типовой вид Jaminia heterstropha Risso, 1826 (syn. Helix quadridens
Muller, 1774); по монотипии.
Признаки подсемейства.
Распространение как у подсемейства. Несколько видов.
Jaminia quadridens (Muller, 1774) (рис. 234, 235).
М ii 11 е г, 1774 : 107 (Helix)', Germain, 1930 : 377, fig. 314, pl. 13, fig. 379 (Chondrula
(Jaminia)]; Mermod, 1930 : 356, fig. 79 (Chondrula); Hesse, 1933 : 161, fig. 5; Al zona,
1971:81; Шилейко, 1978в : 516, рис. 3, 1.
1. РОД JAMINIA
327
Terra typica — «in Italia».
Место хранения типов не установлено.
Материал: более 50 экз. из 15 местонахождений в Швейцарии, Франции,
Италии и Греции.
Раковина левозавитая, от овально-цилиндрической до коническо-цилиндри-
ческой, твердостенная, слабо просвечивающая, блестящая. Оборотов 8—10,
плавно нарастающих, уплощенных. Последний оборот к устью чуть поднят,
половины, высоты раковины, у высоких раковин
высота его несколько меньше
составляет около х/3 этой ве-
личины. Окраска одноцвет-
ная, роговая, белесова-
тая или светло-коричневая.
Скульптура в виде шелко-
вистой радиальной исчер-
ченности. Устье овальное,
места его прикрепления
расставлены и связаны тон-
кой прозрачной или бело-
ватой мозолью, на кото-
рой иногда имеется неболь-
Рис. 234. Jaminia quadri- Рис. 235. Jaminia quadridens (Mull.) из Швейцарии, с. Сьон.
dens (Mull.) из Швейцарии, Половой аппарат и вскрытый пенис (из: Шилейко, 1978в).
с. Сьон.
шое ангулярное утолщение. Париетальный зуб в виде короткой высокой пла-
стинки, лежащей перпендикулярно плоскости устья. Нижний колумеллярный
зуб короткий, верхний представляет собой конец пластинки, спирально уходя-
щей по столбику вверх. Палатальный зуб приостренный, представляет собой
утолщение мощной губы, лежащее слегка отступя от края устья. Губа значи-
тельно усиливает устьевые края, которые почти не отвернуты или отвернуты
незначительно. Пупок почти запаян: от него остается чрезвычайно узкая и ко-
роткая щель.
Размеры: ВП 6—12, БД 3—4 мм; в первоописании: ВР 5 lin., БД 1.5 lin.,
т. е. ВР около 10.5, БД около 3.1 мм.
Внутреннее строение. Материал: 3 экз. из Швейцарии, кан-
тон Вале, Сьон, сбор и определение Л. Форкарта.
Белковая железа с косой вырезкой, занимающей вершину и участок ла-
теральной поверхности органа. Нижний отдел вагины приблизительно втрое
короче верхнего. Семяпровод впадает в эпифаллус не под углом, но эксцентрично;
конец эпифаллуса образует очень короткий прямой конический бич. Эпифал-
лус прямой, вначале цилиндрический, к пенису постепенно сужающийся.
328
VI. НАДСЕМ. BULIMINOIDEA
Цэкум отсутствует. Пенис в проксимальной части образует крошечное расшире-
ние, затем нитевидно сужается и в дистальной части, близ полового атриума
вновь слегка расширяется. Внутренняя поверхность пениса несет несколько
слабых неясных продольных складок; в проксимальном расширении пениса
две из этих складок усиливаются и сливаются возле отверстия эпифаллуса,
образуя единую дугообразную складку. Между крыльями этой дуги заметна
еще одна тонкая одиночная складка. Все отделы пениального аппендикса хорошо
выражены, особенно мощно развит Аг и А3, отделенный от четкой границей.
Места прикрепления ветвей полового ретрактора на диафрагме слегка расстав-
лены, одна из них подходит к А± слегка отступя от Л2, вторая — к дистальной
слегка расширенной части пениса. Проток семяприемника почти прямой, не-
большой овальный резервуар снабжен собственным коротким протоком, длина
которого не превышает длину самого резервуара. Дивертикул семяприемника
заметно не доходит до белковой железы.
Распространение. Средиземноморско-западноевропейский вид. От
южной Испании к востоку до островов, лежащих у побережья Малой Азии;
Пиренеи, почти вся Франция; в южных Альпах от Пьемонта к востоку до Ти-
роля; Швейцария; несколько находок в южной части ФРГ. Населяет открытые
горные участки.
IV. Подсем. MERDIGERI NAE Schileyko, subfam. nov.
Раковина слабо скульптурирована. Зубы в устье отсутствуют. Эмбриональ-
ные обороты гладкие.
Эпифаллус с коротким закругленным бичом. Цэкум хорошо развит. Пенис
короткий, тонкостенный, заключает замкнутую короткую тонкостенную па-
пиллу. Призмоконические бугорки и проксимальный вырост пениса отсутствуют.
Пениальный аппендикс и дивертикул семяприемника нормально развиты.
Распространение. Европа, Северо-Западная Африка.
В ископаемом состоянии подсемейство известно, по-видимому, только из
четвертичных отложений (Европа).
Монотипическое подсемейство.
1. Род MERDIGERA Held, 1837
Held, 1837 : 917; Lindholm, 1925а : 141.
Типовой вид Helix obscura Muller, 1774; обозначение Геррманнсена
(Herrmannsen, 1847).
Признаки подсемейства.
Монотипический род.
Merdigera obscura (Muller, 1774) (рис. 236, 237).
Muller, 1774:103 {Helix); Moquin-Tandon, 1855:291, pl. 21, figs. 5—10
[Buliminus (Ena)]; Westerlund, 1887 : 31 {Buliminus); S t u г a n y, 1897 : 116 {Buli-
mus); Kobelt, 1902 : 934, t. 131, figs. 11, 12 [Buliminus {Ena)]; S teen berg, 1911 : 221;
Лихарев, Раммельмейер, 1952 : 181, рис. 93 (Ena); Ehrmann, 1956 : 59,
fig. 33 (Ena); Lozek, 1956 : 113, t. 10, fig. 4 (Ena); Zilch, J aeckel, 1962 : 108 (Ena);
Hudec, 1963 : 24—28, obr. 2 (Ena); Lozek, 1964 : 228, t. 11, fig. 4 (Ena); Gitten-
berger et al., 1970 : 58, fig. 32, 53 (Ena); Дамяно в, Лихарев, 1975 : 139, фиг. 74,
75 (Ena).
Terra typica — «in Truncis fagi (praefertim Gallici)», т. e. указано, что живет
на стволах буков, главным образом во Франции.
Место хранения типов не установлено.
Материал: более 100 экз. из 19 местонахождений в Подмосковье,
в Крыму, на Карпатах, в Чехословакии и Венгрии.
Раковина овально-коническая, умеренно твердостенная, слегка просвечи-
вающая, матовая, с узкой сглаженной вершиной. Оборотов 61/2—8, верхние
1. РОД MERDIGERA
329
слегка выпуклые, нижние уплощенные. Последний оборот к устью не поднят,
высота его составляет около половины высоты раковины. Окраска одноцветная,
от желтовато-роговой до каштановой. Скульптура в виде не-
равномерных сглаженных радиальных морщинок; спираль-
ные элементы скульптуры, как и зернистость, полностью
отсутствуют. Устье овальное, незначительно скошено, места
его прикрепления расставлены, края умеренно отвернуты,
с широкой расплывшейся светлой губой. Палатальный и ба-
зальный края образуют широкую дугу, при переходе в ко-
лумеллярный имеется закругленный угол; колумеллярный
край почти отвесный. Пупок широкощелевидный.
Рис. 236. Merdigera osbcura (Miill.) из Калининской обл.
Рис. 237. Merdigera obscura'(Miill.) из окр. г. Тутаев Ярославской обл., 3 X I960 г.
I — половой аппарат, II — вскрытый дистальный отдел пениса и пениального аппендикса, III — особь
с вывернутыми дистальными участками половых протоков.
Размеры: ВР 6.5—11, БД 3—4 мм; в первоописании: ВР 3—4 lin., БД l1/^ lin.,
т. е. ВР 6.5—8.5 БД около 3 мм.
Внутреннее строение. Материал: 2 экз. из Крыма, Красно-
330
VI. НАДСЕМ. BULIMINOIDEA
лесье близ Симферополя, 29 июля 1962 г.; 5 — из окр. г. Тутаев, 3 октября
1960 г.; сборы и определения автора.
Белковая железа с нерезкой апикальной вырезкой. Нижний отдел вагины
приблизительно вдвое короче верхнего. Семяпровод впадает в эпифауллус
под углом к ^короткому закругленному слегка утолщенному бичу. Эпифаллус
недлинный, цилиндрический, резких изгибов не образует; хорошо развитый
цэкум' расположен несколько выше его середины. Пенис булавовидный, очень
тонкостенный; сквозь его покровы можно рассмотреть короткую округлую
папиллу. Канал папиллы на поперечном разрезе щелевидный. Строение пени-
ального аппендикса отмечено некоторым своеобразием: очень короткий;
шаровидный А2 содержит папиллу, очень похожую на пениальную; А3 длиннее,
чем Аг и А2, вместе взятые; резкая граница между А3 и Ai отсутствует. Ветви
полового ретрактора на диафрагме тесно сближены, одна из них крепится к про-
ксимальной части пениса, вторая — кЛ2. Проток семяприемника прямой, шаро-
видный резервуар с собственным протоком, длина которого немного превышает
длину резервуара. Дивертикул семяприемника хорошо развит, хотя немного
не доходит до белковой железы.
Распространение и местообитание. Западная и сред-
няя, частично южная Европа, Балканский п-ов, Северо-Западная Африка;
Калининская, Смоленская, Витебская, Ярославская обл., окр. Ленинграда
и Москвы; горный Крым. Населяет лиственные леса, обитает в подстилке и
у корней деревьев и кустарников, нередко поднимается на древесные стволы.
V. Подсем. BULIMININAE Kobelt, 1880
Поверхность раковины от почти гладкой до бугорчатой. Зубы в устье от-
сутствуют. Эмбриональные обороты гладкие.
Эпифаллус с длинным бичом. Цэкум отсутствует. Пенис внутри со складками,
без папиллы и без призмоконических бугорков; его проксимальная часть меш-
ковидно расширена или вытянута в конический придаток. Пениальный аппен-
дикс и дивертикул семяприемника имеются или отсутствуют.
Распространение. Северная Африка, Передняя и Малая Азия,
северный Иран, Закавказье. 4 рода.
В ископаемом состоянии подсемейство неизвестно.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ BULIMININAE
1 (2). Вершина раковины широкая, притупленная, края устья широко отвер-
нуты .............................' ..............1. Buliminus.
2 (1). Вершина раковины узкая, заостренная, края устья слабо или незначи-
тельно отвернуты..................................2. Adzharia.
1. Род BULIMINUS Beck, 1837
Beck, 1837 : 68; Albers, 1850 : 183 {Petraeus, типовой вид Buliminus labrosus Oli-
vier, 1804); Lindholm, 1925a : 27; Thiel e, 1931 : 522; Hesse, 1933 : 191 {Petraeus);
Forcart, 1940 : 162 {Petraeus); Лихарев, Раммельмейер, 1952 : 187 {Petra-
eus); Wenz, Zilch, 1959—1960 : 190; Heller, 1975 : 7; Шилейко, 1978г : 843.
Типовой вид Buliminus labrosus Olivier, 1804; по обозначению Грея
(Gray, 1847).
Раковина удлиненно-овальная, светлоокрашенная, одноцветная. Края об-
ширного устья широко отвернуты.
Пенис с коротким мешковидным проксимальным расширением, на его внут-
ренней поверхности две широкие продольные складки. Пениальный аппендикс
имеется, половой ретрактор нормально развит, двуветвистый.
Распространение. Малая и Передняя Азия, Иран, южное За-
кавказье. Несколько видов.
1; РОД BULIMINUS
331
Buliminus urmianus 0. Boettger, 1898 (рис. 238, 239).
0. Boettger, 1898 : '26 [Buliminus (Petraeus') halepensis var. urmiana}', Лихарев,
Раммельмейер, 1952 : 187, рис. 97 (Petraeus halepensis)', Асадов, АкраМов-
c к и й, Джаббаров, 1975 : 103; Акрамовский, 1976 : 152.:
Terra typica — окр. города Резайе, северо-западный
Иран.
Место хранения типов не установлено.
Материал: 3 экз. из одного местонахождения близ
Джульфы (Нахичеванская АССР).
Раковина удлиненно-овальная, блестящая, сильно просве-
чивающая, твердостенная, с широкой притупленной вершиной.
Оборотов 7—8, сильно уплощенных; последний оборот к устью
плавно, но сильно поднят; высота его превышает половину вы-
соты раковины. Окраска свежих раковин мутно-белая с ро-
Рис. 238. Buliminus urmianus Bttg. из окр. Джульфы.
зоватым оттенком; однако вскоре после гибели моллюска раковина становится
непрозрачной и теряет розоватый оттенок. Скульптура в виде редкой и неравно-
мерной радиальной исчерченности. Устье овальное, очень слабо скошенное,
Рис. 239. Bullminus labrosus Olivier из Шефарама, Аравийский п-ов. Половой аппарат и
вскрытый пенис (из: Шилейко, 1978в).
внутри белое или желтоватое, места его прикрепления широко расставлены и
связаны мозолью. Устьевые края сильно отвернуты, с широкой расплывшейся
белой губой. На колумеллярном крае мощно заметить широкую вялую спираль-
ную пластинку. Пупок щелевидный.
332
VI. НАДСЕМ. BULIMINOIDEA
Размеры: ВР 18—24, БД 8.5—10 мм; в первоописании: ВР 18.5—20.5,
БД 9 мм.
Внутреннее строение. Пригодного для анатомирования ма-
териала в моем распоряжении не было; однако благодаря любезности д-ра
Дж. Геллера удалось анатомировать типовой вид рода В. labrosus Olivier, 1804
(см. Шилейко, 1978в). большое сходство раковин позволяет предполагать, что
и анатомически эти виды близки'.' Ниже дается описание полового аппарата
В. labrosus, составленное на основании изучения 3 экз. из Шефарама, Аравий-
ский п-ов, сбор и определение Дж. Геллера.
Белковая железа с небольшой апикальной вырезкой. Верхний отдел ва-
гины в 2—3 раза длиннее нижнего. Семяпровод впадает в эпифаллус под прямым
углом; длина бича заметно превышает длину слабоизвитого цилиндрического
эпифаллуса. Пенис мешковидный, проксимальная часть его образует неболь-
шое мешковидное расширение, направленное вверх, т. е. в направлении бича.
Внутренняя поверхность пениса несет две очень широкие плавные складки;
благодаря огромному числу микроскопических ворсинок, покрывающих внут-
реннюю поверхность пениса, она выглядит бархатистой. Короткая цилиндри-
ческая дистальная часть пениса отделена от проксимальной кольцевой перетяж-
кой. Пениальный аппендикс хорошо развит. Аг длинный и тонкий, А2 неболь-
шой, А3 внешне не выражен. Половой ретрактор отходит от диафрагмы единым
тяжем, который вскоре раздваивается: одна ветвь подходит к Ах заметно
выше его середины, вторая — к средней части пениса. Проток семяприемника
длинный, образует 2—3 крутых изгиба или даже петли. Проток резервуара
тонкий, резервуар удлиненно-овальный. Дивертикул заметно не доходит до
белковой железы.
Распространение и местообитание. Помимо типового
местонахождения, вид найден в Ираке; в Нахичеванской АССР близ устья
р. Алинджа-чай в 5 км западнее Джульфы; в Зангиланском р-не Азербайджана.
Населяет ущелья среди пустынных холмов в резко ксеротермных биотопах.
2. Род ADZHARIA Hesse, 1933
Hesse, 1933 : 158; Лихарев, Раммельмейер, 1952 : 175; Wen z, Zilch,
1959—1960:181.
Типовой вид Adzharia renschi Hesse, 1933; по монотипии.
Раковина овально-коническая, белая, с темными радиальными пестринами.
Края устья почти не отвернуты.
Пенис веретеновидно-цилиндрический, без проксимального выроста. Пе-
ниальный аппендикс и половой ретрактор отсутствуют.
Монотипический род.
Adzharia renschi Hesse, 1933 (рис. 240, 241).
Hesse, 1933:158; Лихарев, Раммельмейер, 1952:176.
Terra typica — у перевала Хуло на Аджаро-Имеретинском хр.
Лектотип хранится в Зоологическом музее Берлинского ун-та
им. Гумбольдта, № 75980а (Kilias, 1971).
Материала в моем распоряжении не было, описания раковины и полового
аппарата составлены но. оригинальному тексту и иллюстрациям.
Раковина овально-коническая, тонкостенная, слабо блестящая. Оборотов 8,
слабовыпуклых, высота последнего оборота значительно больше высоты рако-
вины. Окраска белая, со светло-коричневыми радиальными пестринами. Скуль-
птура в виде тонкой радиальной исчерченности. Устье усеченно-овальное, в ан-
гулярной области заостренное; места его прикрепления не сближены, соединены
слабой мозолью. Края устья не отвернуты, острые, тонкие; лишь верхняя часть
колумеллярного края расширена и сильно отвернута. Пупок в виде узкой щели.
2. РОД ADZHARIA
333
Размеры: ВР 19—20,5, БД 10 мм; лектотип: ВР 20.5 мм (по Kilias, 1971).
Внутреннее строение. Вагина короткая, отделы ее примерно
равной длины. Бич длинный, тонкий, заостренный; длина его лишь немногим
меньше суммарной длины пениса и эпифаллуса. У одного из исследованных
Гессе экземпляров граница между пенисом
и эпифаллусом вообще незаметна, у другого
эта граница в виде кольцевой перетяжки,
расположенной чуть отступя от места впа-
дения семяпровода; т. е. эпифаллус как та-
ковой почти отсутствует. Пенис веретеновид-
но-цилиндрический, хорошо развит. У од-
ной из особей обнаружен дивертикул семя-
приемника, длина которого примерно вдвое
Рис. 240. Adzharia renschi Hesse.
I — лектотип,. II — паралектотип. Контуры раковин сде-
ланы по фотографиям, помещенным при первоописании
(Hesse, 1933).
меньше длины протока резервуара; проток семяприемника в этом случае корот-
кий. У другой особи дивертикул семяприемника отсутствовал. Резевуар семя-
приемника небольшой, овальный.
Рис. 241. Adzharia renschi Hesse. Синтипы. Половой аппарат (из: Hesse, 1933).
Внутреннее строение пениса неизвестно, но внешний вид полового аппарата
настолько своеобразен, что этот монотипический род в пределы подсемейства
Bulimininae помещен провизорно, благодаря наличию хорошо развитого бича.
Уточнить систематическое положение можно будет после исследования строе-
ния совокупительного аппарата.
Распространение. Известна лишь одна находка (2 экз.) в типовом
местонахождении.
334
VI. НАДСЕМ. BULIMINOIDEA
VI. Подсем. RETOWSKIINAE Schileyko, 1978
Шилейко, 1978г : 849.
Поверхность раковины покрыта бугорчатой скульптурой, развитой в раз-
ной степени. Устье с более или менее резкой вмятиной на столбике; на палаталь-
ном крае также может присутствовать вмятина. Эмбриональные обороты несут
ясно выраженные спиральные нитевидные ребрышки.
Бич отсутствует. Внутренняя поверхность пениса с чрезвычайно сильно раз-
витыми призмоконическими бугорками. Папилла пениса отсутствует, как и
проксимальный вырост пениса. Цэкум имеется или отсутствует. Пениальный
аппендикс и дивертикул семяприемника нормально развиты.
Распространение. Кавказ. 2 рода.
В ископаемом состоянии подсемейство известно начиная с верхнего миоцена
(Предкавказье).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ RETOWSKIINAE
1 (2)., Палатальный край устья с резкой вмятиной, поверхность раковины
покрыта крупными округлыми бугорками, высота раковины не превы-
шает 10 мм...........................................1. Andronakia.
2 (1). Палатальный край устья без вмятины, поверхность раковины покрыта
мелкими бугорками, высота раковины не менее 13 мм .................
................................................ 2. Retowskia.
1. Род ANDRONAKIA Lindholm, 1914
Lindholm, 1914 : 38; 1925а : 30; Wenz, Zilch, 1959—1960 :181 [? Andronakia
(Chondrulinae)]. 1
Типовой вид Chondrula (?) catenulata Lindholm, 1913; по монотипии.'
Раковина высококубаревидная. Излом колумеллярного края слабо выражен,
палатальный край с резкой вмятиной. Спиральные ребрышки на эмбриональ-
ных оборотах параллельны шву, скульптура дефинитивных оборотов грубая.
Цэкум имеется. А3 очень длинный; А4, напротив, чрезвычайно короткий.
Монотипический род.
Andronakia catenulata (Lindholm, 1913) (рис. 242, 243).
Lindholm, 1913 : 22 {Chondrula ?); H u d e c, L e z h a w a, 1969 : 101, Abb. 6. t. 1.
Terra typica — перевал возле с. Кварцхан близ Васрия (вилайет Чорох,
северо-восточная Турция).
Лектотип хранится в Зоологическом ин-те АН СССР,, обозначается
здесь.
Материал: 3 экз. из окр. Батуми и 5 экз. из северо-восточной Турции.
Раковина высококубаревидная, хрупкая, тонкостенная, просвечивающая,
составлена l1/^—8х/2 выпуклыми оборотами, разделенными узким глубоким
швом. Последний оборот к устью не поднят, высота его приблизительно равна
половине высоты раковины. Окраска одноцветная: роговая, коричневая или
каштановая. Скульптура эмбриональных оборотов в виде тонких четких спи-
ральных ребрышек, которые резко обрываются на границе эмбриональной и де-
финитивной раковины. Дефинитивные обороты покрыты правильными косыми
радиальными ребрами, разбитыми на ряды бугорков, которые на первых дефи-
нитивных оборотах имеют вид утолщенных чешуек, а далее приобретают ок-
ругло-коническую форму. В промежутках между рядами бугорков при увеличении
1. РОД ANDRONAKIA
335
порядка 50 раз можно заметить тончайшую радиальную струйчатость. Устье
неправильной формы, места его прикрепления не сближены, края умеренно
отвернуты, тонкие. На середине палатального края расположена резкая вмя-
тина; другая вмятина, но очень слабая, имеется в ниж-
ней части колумеллярного края. Пупок при рассмат-
ривании строго снизу проколовидный, сквозной, почти
не прикрыт колумеллярный краем.
Размеры: ВР 5.5—8.5, БД 3.3—4.5 мм; лектотип:
8.5, БД 2.5 мм.
Внутреннее строение. Мною не ис-
следовано, описание составлено по материалам Гу-
деца и Лежавы (Hudec, Lezhawa, 1969). .
Белковая железа без вырезки. Половая клоака
очень мелкая. Вагина длинная, отделы ее примерно
равной длины. Семяпровод впадает терминально, пе-
Рис. 242. Andronakia catenulata (Lindh.) из окр. Батуми.
реходит в эпифаллус без резкой границы. Немного выше середины эпифал-
луса расположен бугорковидный цэкум. Пенис отделен от эпифаллуса кольце-
вой перетяжкой. Аппендикс отходит примерно от середины пениса. очень
Рис. 243. Andronakia catenulata (Lindh.) из окр. Батуми, гора Мтирала, 20 VIII 1966 г. По-
ловой аппарат (из: Hudec, Lezhawa, 1969).
короткий, А2 небольшой, но ясно заметный, А3 длинный, необыкновенно
короткий, выглядит как сужение между А3 и нормально развитым Аь.
Проток семяприемника ниже места раздвоения короткий, проток резервуара
336
VI. НАДСЕМ. BULIMINOIDEA
длиннее как самого резервуара, так и протока семяприемника. Дивертикул
длинный.
Распространение и местообитание. Кроме типового
местонахождения в северо-восточной Турции (бассейн р. Чорох) вид найден на
территории нашей страны на горе Мтирала в 28—30 км восточнее Батуми, на
высоте 1700 м над уровнем моря, среди влажного растительного детрита (Hu-
dec, Lezhawa, 1969).
2. Род RETOWSKIA О. Boettger, 1881
О. Boettger, 1881 : 219 (род Bullminus, секция Retowskia); Lindholm, 1925а : 29;
Thiele, 1931 : 523 [Buliminus (Retowskia)]; Hesse, 1933 : 203; Л и x a p e в, Раммель-
мейер, 1952 : 188; Wen z, Zilch, 1959—1960 : 190; Стеклов, 1966 : 179; Ш и-
л e й к о, 1978г : 849.
Типовой вид Buliminus Schlaeflii Mousson, 1863; по монотипии.
Раковина коротко- или высокоцилиндрическая. Излом колумеллярного
края ясно выражен, палатальный край без вмятины. Спиральные ребрышки
на эмбриональных оборотах направлены под острым углом к шву, скульптура
дефинитивных оборотов относительно тонкая.
Цэкум отсутствует. Отделы пениального аппендикса выражены пропорцио-
нально.
Распространение. Западная часть Кавказского перешейка. 1 вид.
В ископаемом состоянии род известен начиная с верхнего миоцена (Пред-
кавказье) (Стеклов, 1962а).
Retowskia schlaeflii (Mousson, 1863) (рис. 244, 245).
Mousson, 1863 : 390 [Buliminus (Chondrus)]; Westerlund, 1887 : 21 (Buliminus)',
O. Boettger, 1888 : 152 (B. Schlaeflii var. ingens); Kobelt, 1902 : 415, t. 72, fig. 17, 18
[B. (Retowskia)]; Лихарев. Раммельмейер, 1952 : 188, рис. 98; Шилейко,
1978г : 840, рис. 3, II.
Terra typica — «Redutkale», окр. Поти.
Место хранения типов не установлено.
Материал: 22 экз. из 8 местонахождений в западном Закавказье.
Раковина цилиндрическая, с узкозакругленной куполовидной верхней частью,
умеренно твердостенная, слегка просвечивающая. Оборотов 8, слабовыпуклых,
разделенных мелким швом. Последний оборот непосред-
ственно возле устья слегка поднят, высота его прибли-
зительно равна половине высоты раковины. Спиральные
нити на эмбриональных оборотах четкие, различимы даже
невооруженным глазом; поскольку они направлены под
углом к шву, длина каждого ребрышка составляет при-
мерно половину оборота. Скульптура дефинитивных обо-
ротов в виде косых радиальных морщин, разбитых на ряды
мелких бугорков причем на верхних оборотах эти бугорки
ориентированы вдоль морщин, на средних оборотах бугорки
большей частью округлые, на последнем и частично пред-
Рис. 244. Retowskia schlaeflii (Mouss.) из Аджаро-Имеретинского хр.,
близ с. Адигени.
последнем оборотах бугорки вновь становятся удлиненными, но ориентированы
параллельно шву. Благодаря этому создается впечатление, что местами имеется
спиральная исчерченность. Между морщинами при увеличении 30—40 раз
2. РОД RETOWSKIA
337
можно заметить тончайшую косую струйчатость. Устье овальное, косое, места
его прикрепления почти не сближены. Палатальный и базальный края широко
отвернуты, желтовато-розоватые, образуют плавную дугу. При переходе в почти
отвесный колумеллярный край имеется бухточка — излом устьевого края.
Пупок почти полностью запаян за счет смыкания внутренних стенок послед-
него оборота; пупочная впадина слегка прикрыта отворотом колумеллярного
края.
Размеры; ВР 13.5—23.5, БД 7—9.5 мм; в первоописании: ВР 13.5, БД 7 мм.
Внутреннее строение. Материал: 2 экз. из Аджаро-Имеретин-
ского хр., 12 июля 1973 г., сбор И. М. Лихарева, определение автора.
Рис. 245. Retowskia schlaeflii (Mouss.) из Аджаро-Имеретинского хр., близ с. Адигени,
21 VII 1973 г. Половой аппарат и вскрытый дистальный отдел (из: Шилейко, 1978 г.).
Белковая железа с очень глубокой апикальной вырезкой, на конце двулопаст-
ная. Половая клоака очень мелкая, мужское и женское отверстия расположены
рядом на дне неглубокой ямки. Вагина длинная, цилиндрическая, ее отделы
примерно равной длины. Семяпровод впадает строго терминально, и граница
между семяпроводом и эпифаллусом определяется лишь постепенным расшире-
нием протока, поскольку диаметр эпифаллуса в 4—5 раз больше диаметра семя-
провода. Цэкум отсутствует, но примерно на середине дистальной части мужского
отдела (пенис-|-эпифаллус) имеется постоянный изгиб, возможно, гомо-
логичный цэкуму. Пенис внешне не отделен от эпифаллуса. Папилла отсут-
ствует; на внутренних стенках пениса залегают ряды складок, разбитых па мощ-
ные призмоконические бугорки. При переходе в эпифаллус эта структура
сменяется системой коротких складок, ориентированных без видимой системы.
Продольные складки внутри эпифаллуса также несут бугорки, но гораздо бо-
лее мелкие. Все отделы пениального аппендикса имеют обычное строение и
легко различимы. Ветви полового ретрактора отходят от диафрагмы независимо
одна от другой, пениальная ветвь крепится к дистальной части пениса, ветвь
аппендикса — к Аг немного выше его середины. Проток семяприемника не-
22 А- А. Шилейко
.338
VI. НАДСЕМ. BULIMINOIDEA
длинный, прямой; объемистый резервуар с собственным тонким протоком,
длина которого превышает длину самого резервуара. Дивертикул почти до-
ходит до белковой железы.
Распространение и местообитание. Вид встречается
в горных районах западной части Большого Кавказа (на востоке включая вер-
ховья р. Белой) и Закавказья (на востоке включая Сурамский хр.). Живет на
влажных скалах среди леса и в подстилке, особенно в сырых ущельях.
VII. Подсем. Е N I N А Е Woodward, 1903
Поверхность раковины гладкая или слабо скульптурированная, изредка
мелкозернистая. Устье без зубов или с 1—3 зубами. Эмбриональные обороты
гладкие.
Эпифаллус с цэкумом и коротким коническим бичом, изредка отсутствующим.
Пенис внутри с призмоконическими бугорками, в подавляющем большинстве
случаев ясно выраженными. Имеется незамкнутая папилла пениса., прокси-
мальный вырост пениса отсутствует. Дивертикул семяприемника нормально
развит, пениальный аппендикс большей частью имеется (отсутствует только
у рода Chondrula).
Распространение. Европа и средиземноморские страны.
В ископаемом состоянии подсемейство известно начиная со среднего и верх-
него миоцена (Алжир, Европа).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ПОДРОДОВ ENINAE
1 (18). Пениальный аппендикс имеется.
2 (3). По краю устья располагаются 3 бугорковидных зуба . . .5. Chondrus.
.3 (2). Устье без зубов или имеется только одна палатальная складка.
4 (7). Раковина одноцветная, роговая, каштановая или коричневая, но не
белая.
5 (6). Радиальные и продольные элементы скульптуры, пересекаясь, образуют
зернистую фактуру поверхности. Места прикрепления устья не сближены
............................................................. 1. Епа.
6 (5). Зернистая скульптура отсутствует. Места прикрепления устья сбли-
жены .....................................................2. Peristoma.
7 (4). Раковина с радиальными пестринами или двуцветная: у шва проходит
широкая темная лента; если одноцветная, то белая или желтоватая.
8 (9). Раковина левозавитая. Верхние отделы пениального аппендикса отходят
от базальной части Аг....................................8. Thoanteus.
9 (8). Раковина правозавитая. Пениальный аппендикс обычного строения,
т. е. верхние его отделы отходят от вершины Аг.
10 (13). Раковина высокая, стройная, цилиндрическая или коническо-цилиндри-
ческая.
11 (12). Большой диаметр раковины не превышает 3 мм, края устья с очень
слабой губой.............................................9. Ramusculus.
12 (И). Большой диаметр раковины не менее 4 мм, края устья изнутри окайм-
лены широкой расплывшейся губой..........................6. Brephulopsis.
13 (10). Раковина слегка вздутая, высококоническая или овально-коническая.
14 (15). Раковина желтая, с широкой темной лентой под швом; редко одно-
цветно желтая ......................................3. Caucasicola.
15 (14). Раковина одноцветно белая или с темными радиальными пестринами
или полосами.
16 (17). Устье внутри желтое ...................... 7. Napaeopsis.
17 (16). Устье внутри белое или серое........................4. Zebrina.
18 (1). Пениальный аппендикс отсутствует (род Chondrula).
19 (20). В устье не менее 3 зубов .... 10. Chondrula (Chondrula s. str.).
20 (19). В устье не более двух зубов или эубы отсутствуют.
1. РОД ENA
339
21 (22). В устье залегают колумеллярный и палатальный зубы . •......
................................... 10. Chondrula (Eubrephulug).
22 (21). Устье без зубов или с небольшим ангулярным бугорком........
....................................... 10. Chondrula (Mastus).
1. Род ENA Turton, 1831
Turton, 1831 •. 80; Ehrenberg, 1831:2, 4 (Bulimina, non d’Orbigny, 1826);
Lindholm, 1925a : 26; T h i e 1 e, 1931 : 520; Лихарев, Раммельмейер, 1952 :
180; W e n z, Zilch, 1959—1960 : 183; Шилейко, 1978г : 847.
Типовой вид Bulimus montanus Draparnaud, 1801; обозначение Гер-
рманнсена (Herrmannsen, 1847).
Раковина коническая и слегка овальная, скульптура мелкобугорчатая.
Места прикрепления устья не сближены. Окраска одноцветная, темная. Устье,
без зубов.
Эпифаллус и проток семяприемника умеренно длинные. Цэкум не сдвинут,
к семяпроводу. Отделы пениального аппендикса нормально выражены. Приз-
моконические бугорки на внутренней поверхности пениса слабые, имеются
только в проксимальном отделе пениса. Папилла гладкостенная.
Распространение. Европа, преимущественно горные районы;
в связи с новыми материалами по организации членов подсемейства, сильно ска-
завшимися на системе, неясно, какие виды, кроме типового, должны включаться
в род Епа s. str. Поэтому ареал рода нуждается в уточнении, равно как и его
объем.
В ископаемом состоянии род известен начиная с верхнего миоцена (Европа).
Ena montana (Draparnaud, 1801) (рис. 246, 247).
Draparnaud, 1801 : 65 (Bulimus); Moquin-Tandon, 1855 : 289, pl. 21,
figs. 1—4 [Bulimus (Ena)]; W e s t e r 1 u n d, 1887:29 (Buliminus); Geyer, 1927:127,'
t. VI, 8, [Ena (N apaeus)]; A. Wagner, 1927 : 316; Лихарев, Раммельмейер,
1952 : 181, рис. 92; E h г m a n n, 1956 : 59, fig. 32; L о z e k, 1956 : 113, t. 10, fig. 5; Zilch,
Jaeckel, 1962:107; Hudec, 1963:24—28, Abb. 2; L о i e k, 1964:227, Abb. 18;
Дамянов, Лихарев, 1975 : 140, фиг. 76, 77; Шилейко, 1978в : 512, рис. 1, I.
Terra typica — Франция.
Место хранения типов не установлено.
Материал: более 70 экз. из 5 местонахождений во Франции, Закар-
патье и в Москве.
Раковина от высококонической до овально-конической, тонкостенная, блес-
тящая, с приостренной вершиной. Оборотов 7—8, умеренно выпуклых, разде-
ленных беловатым швом. Последний оборот к устью незначительно поднят, вы-
сота его обычно слегка превышает половину высоты раковины. Окраска моно-
тонная; роговая, каштановая или темно-коричневая. Скульптура представлена
неравномерно расположенными радиальными сглаженными морщинками, каж
дая из которых состоит из множества мелких бугорков. Ясно выраженные спи-
ральные бороздки отсутствуют. Устье от овального до удлиненно-овального,
незначительно скошено, края его слегка отвернуты, светлые. Места прикрои
ления устья не сближены. Чуть отступя от устьевого края лежит сильно распл ыв
шаяся плохо развитая губа. Колумеллярный край расширенный, почти относ
ный, слегка перекрывает широкий щелевидный пупок.
Размеры: ВР 13—16, БД 5—6.5 мм.
Внутреннее строение. Материал: 21 экз. из Закарпатья, с. Квасы
близ Рахова, 14 сентября 1969 г., сбор и определение автора.
Белковая железа с резкой и глубокой апикальной вырезкой. Верхний отдел
вагины в 1х/2—2 раза длиннее нижнего. Бич очень короткий, конический;
цилиндрический неизвитой эпифаллус составляет естественное продолжен ио
семяпровода. Цэкум занимает середину эпифаллуса или слегка сдвинут к со-
22*
340
VI. НАДСЕМ. BULIMINOIDEA
.мяпроводу. Пенис в 2—3 раза короче и немного толще эпифаллуса, цилиндри-
ческий или веретеновидный. Внутренняя поверхность дистальной части пениса
несет нерезкие продольные складки; в проксимальной части эти складки распа-
даются на ряды тесно сближенных бугорков. Папилла пени-
са цилиндрическая, к концу сужается; по ее поверхности
проходит мелкая борозда. Все отделы пениального аппендик-
са ясно выражены; обращает на себя внимание А3, длина
которого почти равна суммарной длине Ах и Л2. Половой ре-
трактор отходит от диафрагмы двумя самостоятельными
ветвями; пениальная ветвь крепится на границе пениса и
эпифаллуса, ветвь аппендикса — к Аг непосредственно под
А2. Проток семяприемника длинный, извитой; резервуар объе-
мистый, овальный или грушевидный, с собственным коротким
протоком. Дивертикул немного не доходит до белковой железы.
Рис. 246. Ena montana (Drap.) из с. Квасы близ Рахова, Закарпатье.
Распространение и
Бельгия, Нидерланды, Пиренеи,
местообитание. Европа: Англия,
северная Франция. Балканский п-ов, сред-
Рис. 247. Ena montana (Drap.) из с. Квасы близ Рахова, Закарпатье, 14 IX 1969 г. Половой
аппарат и детали его строения (из: Шилейко, 1978в).
неевропейские страны, западные районы Украины, Латвия, Эстония, Валдайская
возвышенность, окр. Москвы и Ленинграда. Населяет подстилку лиственных,
реже смешанных лесов; иногда поднимается на стволы Деревьев.
t
2. РОД PERISTOMA
341
2. Род PERISTOMA Krynicki, 1833
Krynicki, 1833 : 420; Lindholm, 1925a : 27 [Ena (Ena), секция Peristoma];
W e n z, Zilch, 1959—1960 : 183 [Ena (Peristoma)]; Шилейко, 1978г : 834.
Типовой вид Peristoma merduenianum Krynicki, 1833; по монотипии.
Раковина высококоническая, скульптура тонкая, радиальная; нередко име-
ются спиральные бороздки. Места прикрепления устья в той или иной мере
•сближены. Окраска одноцветная, от светло-роговой до коричневой. Устье без
зубов.
Эпифаллус и проток семяприемника короткие. Цэкум расположен вплотную
к семяпроводу, т. е. резко сдвинут вверх. Отделы пениального аппендикса хо-
рошо развиты. Призмоконические бугорки на внутренней поверхности пениса
крупные, четкие, разбросанные. Папилла многобугорчатая.
Распространение. Горный Крым и западный Кавказ. 4 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ PERISTOMA
1 (4). Поверхность раковины с ясными спиральными бороздками.
2 (3). Контур завитка почти строго конический. На последнем обороте бо-
роздки частые: промежутки между ними примерно равны ширине бороздок
...................................................... 4. Р. boettgeri.
3 (2). Контур завитка несколько куполовидный. На последнем обороте бо-
роздки редкие: промежутки между ними заметно больше ширины бо-
роздок ................................................3. Р. rupestre.
4 (1). Поверхность раковины без спиральных бороздок или эти бороздки очень
слабые.
5 (6). Большой диаметр раковины не менее 6 мм. Высота последнего оборота
приблизительно равна половине высоты раковины .......................
.............................................1. Р. merduenianum.
6 (5). Большой диаметр раковины не превышает 4.5 мм. Высота последнего
оборота заметно меньше половины высоты раковины . . . 2. Р. lanceum.
1. Peristoma merduenianum Krynicki, 1833 (рис. 248, I; рис. 249).
Krynicki, 1833 : 421, t. 9, fig. 7a—d; Rossmaessler, 1837, Iconogr., 1,6: 47,
fig. 388 (Bulimus tener); Anton, 1839 : 42 (B. fragilis); Westerlund, 1887 : 27 (Buli-
minus); Kobelt, 1902 : 463, t. 77, figs. 13, 14 [5. (Napaeus)]; A. Wagner, 1922 : 120,
pl. 5, fig. 331 (Ena); Пу завов, 1925a : 85, рис. V, 7, 8 [Buliminus (Ena)]; Hesse, 1933 :
176, fig. 17 [Ena (Peristoma)]; Лихарев, Раммельмейер, 1952 : 182 [Ena (Ena)];
Шилейко, 1978г : 834, рис. 1, I.
Terra typica — горный Крым («Мердвень и по Айле»).
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
здесь.
Материал: 160 экз. из 16 местонахождений в Крыму.
Раковина высококоническая, с почти правильно коническими обводами за-
витка и узкой притупленной вершиной. Оборотов 7—8, верхние более выпук-
лые, чем нижние. Высота последнего оборота равна половине высоты раковины
или немного больше. Окраска от желтовато-роговой до коричневой. Поверхность
раковины слабо скульптурирована: имеются легкие неравномерно расположен-
ные радиальные морщинки, и на некоторых раковинах можно различить исче-
зающе слабые намеки на элементы спиральной скульптуры. Устье овальное,
иногда почти цельное в результате того, что места его прикрепления сильно
сближены и соединены хорошо выраженной мозолью, на концах которой могут
присутствовать небольшие наплывы. Края устья тонкие, незначительно отвер-
нуты, по линии перегиба имеется слабая расплывшаяся губа. Наружный край
устья плавно выгнут. Колумеллярный край короткий, почти отвесный, умеренно
отвернут и почти не прикрывает пупок, имеющий вид широкой открытой щели.
342
VI. НАДСЕМ. BULIMIN01DEA
Размеры: ВР 13—16, БД 5—7; в первоописании: ВР 6 линий, БД 2.5 ли-
нии; лектотип: ВР 13.0, БД 5.1 мм.
Внутреннее строение. Материал: 4 экз. из юго-западной части
Крыма, Батилиман, май 1930 г., сбор В. А. Линдгольма, определение автора.
Рис. 248. Виды рода Peristoma.
I — Р. merduenianum Kryn. из Крыма, с. Батилиман; II — Р. rupestre (Kryn.)' из Крыма, Ялта;:
III — Р. boettgeri (CI.) из окр. Кутаиси.
Массивная белковая железа с вырезкой, занимающей вершину органа и час-
тично его боковую поверхность. Отделы вагины примерно равной длины. Цэ-
кум настолько сдвинут вверх, что вместе с бичом образует характерную, фигуру
Рис. 249. Peristoma merduenianum Kryn. из Крыма, с. Батилиман, V 1930 г. Половой аппарат
и вскрытый пенис (из: Шилейко, 1978 г.).
в форме сапожка, в центр «подошвы» которого впадает семяпровод. Эпифаллус
короткий, неизвитой, цилиндрический. Пенис массивный, короткобулавовид-
ный, слегка короче эпифаллуса, на его внутренней поверхности располагаются
высокие призмоконические бугорки, не сгруппированные в ряды. Незамкнутая
2. РОД PERISTOMA
343
папилла занимает значительную часть объема пениса, поверхность ее бугрис-
тая, борозда широкая и неглубокая. Напротив борозды на стенке пениса имеется
участок, ограниченный полукольцевой складкой, поверхность которого несет'
несколько бессистемно расположенных складок. Отделы пениального ап-
пендикса хорошо выражены, в том числе А3, который обычно плохо заметен.
Ветви полового ретрактора на диафрагме тесно сближены; одна ветвь кре-
пится к заметно выше его середины, вторая — к проксимальной части
пениса, возле эпифаллуса. Проток семяприемника короткий, прямой. Проток
резервуара не длиннее самого резервуара, имеющего овальную форму. Дивер-
тикул короткий, немного длиннее резервуара с его протоком, значительно
не доходит до белковой железы.
Распространение и местообитание. Горный Крым. На-
селяет влажные участки лиственных лесов, держится в подстилке, иногда под-
нимается на стволы деревьев; встречается также в трещинах скал.
2. Peristoma lanceum Schileyko, sp. nov. (рис. 250).
Terra typica — долина p. Бзыбь у Пицунды.
Голотип хранится в Зоологическом ин-те АН СССР.
Материал: 2 экз. из типового местонахождения.
Раковина высококоническая, стройная, с переходом к цилиндрической,
тонкостенная, матовая. Первые 3—4 оборота нарастают чуть медленнее ос-
тальных, в результате чего контур верхней части раковины
не строго конический, но очертания его едва вогнутые. Вер-
шина закругленная, относительно широкая. Оборотов 8—9,
слабовыпуклых; высота последнего оборота немногим пре-
вышает х/3 высоты раковины. Поверхность дефинитивных
оборотов покрыта неравномерно расположенными слабыми
морщинками; местами заметны очень слабые следы спи-
ральной исчерченности, но на базальной части раковины из-
вилистые спиральные бороздки видны ясно. Устье овальное,
места его прикрепления сильно сближены и соединены сла-
бой мозолью. Края устья умеренно отвернуты и по линии
отворота незначительно усилены слабой размытой губой. Пу-
пок в виде короткой, широкой щели.
Рис. 250. Peristoma lanceam Schileyko, sp. nov. Голотип.
Размеры голотипа: ВР 12.2, БД 4.2 мм; паратип: ВР 11.0, БД 4.1 мм.
Вид ближе всего к Р. merduenianum, отличается более высокой и стройной ра-
ковиной и относительно меньшей высотой последнего оборота.
Распространение. 2 экз. собраны д-ром Э. Клауссом в июне 1973 г.
в долине р. Бзыбь близ Пицунды; возможно, что указания на присутствие
Р. merduenianum в ряде пунктов западного Кавказа — Боржоми, Кутаиси (Mous-
son, 1873; О. Boettger, 1881 — под названием Buliminus tener Rossmaessler,
1837) — относятся к данному виду.
3. Peristoma rupestre (Krynicki, 1833) (рис. 248, II; рис. 251).
К г у n i с k i, 1833 : 413, t. 3, fig. 4 (Bulimus); Rossmaessl er, 1837, Iconogr., I,
6 : 46, fig. 385 (B. assimilis, non Potiez et Michaud, 1838); Westerlund, 1887 : 27 (Buli-
minus); Kobelt, 1902 : 464, t. 77, fig. 17, 18 [B. (Napaeus) assimilis]; 1907 : 167 [B. (Ena)
kusnetzowl]; Lindholm, 1908 : 230 [B. (Ena) braunert]; Пузанов, 1925a : 84, рис. V,
4—6 [В. (Ena)]; Лихарев, Раммельмейер, 1952:182 [Ena (Ena)]; Hudec,
1972a : 76, Abb. 1, 25 [Ena (Peristoma)].
344
VI. НАДСЕМ. BULIMINOIDEA
Terra typica — горный Крым, деревня Мердвень.
Место хранения типов не установлено.
Материал: 200 экз. из 33 местонахождений в Крыму.
Раковина от высококонической до овально-конической, в любом случае
контур завитка более или менее выпуклый. Стенки раковины относительно тон-
кие, просвечивающие. Оборотов 7—8, разделенных светлым швом, верхние
обороты более, выпуклые, чем нижние. Последний оборот к устью совершенно не
поднят, высота его заметно превышает половину высоты раковины. Окраска
Рис. 251. Peristoma rupestre (Кгуп.) из Крыма, Ангарский перевал на Чатыр-даге-,.
1 VIII 1962 г.
I — половой аппарат, II — вскрытые пенис и пениальный аппендикс, III — квадривий, IV — проксималь-
ный конец эпифаллуса.
одноцветно светло-роговая или темнее — до каштановой; надо иметь в виду,,
что после гибели животного раковина очень быстро выцветает и светлеет.
Скульптура в виде крайне тонких радиальных морщинок и частых волнистый
спиральных бороздок. Устье овальное, немного косое, места его прикрепления
умеренно сближены. Париетальная мозоль слабая. Края устья умеренно от-
вернуты, тонкие, с расплывшейся светлой губой по внутреннему периметру
свободного края. Колумеллярный край немного расширен, не прикрывает от-
крытого широкощелевидного пупка.
Размеры: ВР 15—20, БД 6—9 мм; в первоописании: ВР 7, БД 23/4 линии,
т. е. ВР около 15, БД около 6 мм.
Внутреннее строение. Материал: 2 экз. из Ялты, 8 июня 1975 г.,
сбор С. Д. Чистякова; 7 — из Ангарского перевала через Чатыр-даг, 1 августа
1962 г.; сбор и определения автора.
Белковая железа с глубокой апикальной вырезкой. Нижний отдел вагины
в 1х/2 раза короче верхнего. Бич удлиненный, конический, на конце притуплен.
Цэкум располагается чуть отступя от места впадения семяпровода. Эпифаллус
недлинный, изгибов не образует, цилиндрический, такой же длины, как и не-
2. РОД PERISTOMA
345
нис, или немного длиннее. Дистальная часть булавовидного пениса сужена,
проксимальная вздута, поэтому граница между пенисом и эпифаллусом обо-
значена ясно. Отделы пениса отделены один от другого сфинктером, заметным
на продольном разрезе пениальной трубки. Внутренняя поверхность прокси-
мального отдела покрыта небольшими, но ясно различимыми призмоконическими
бугорками. Папилла чрезвычайно объемистая, с бугристой поверхностью, за-
нимает почти весь объем проксимального отдела пениса. Борозда на поверх-
ности папиллы узкая и глубокая; в стенках папиллы заключена система синусов.
Отделы пениального аппендикса развиты, особенно А3: его длина примерно
равна суммарной длине Аг и А2. Половой ретрактор отходит от диафрагмы не-
зависимыми ветвями, одна из которых крепится к проксимальной части пениса,
другая — к границе между Аг и Л2. Проток семяприемника прямой, резервуар
объемистый, с коротким собственным протоком, дивертикул заметно не дохо-
дит до белковой железы.
Распространение и местообитание. Горный Крым. По
сравнению с другими видами рода наиболее гигрофильный вид: встречается
в тенистых лесах, особенно по берегам ручьев, в заболоченных местах, в тени-
стых ущельях.
4. Peristoma boettgeri (Clessin, 1883) (рис. 248, III; рис. 252).
Mousson, 1873 : 204 (Buliminus tener, non Rossmaessler, 1837); Clessin, 1883 : 49,
t. 2, fig. 15 (Buliminus); 0. Boettger, 1883 : 176, t. 6, Fig. 7, 8 [5. (Ena)]; 1886a : 144,
t. 3, Fig. 2a, b [B. (Ena) boettgeri var. connivens]; Kobelt, 1902 : 458, Fig. 4, 5 \B. (Napaeus)];
Rosen, 1914 : 183 [B. (N apaeus)]; Лихарев, Раммельмейер, 1952 : 183 [Ena
(Ena)].
Terra typica — не указана; no. Boettger, 1883 — Боржоми и Кутаиси.
Место хранения типов не установлено.
Материал: 63 экз. из 14 местонахождений в западной Грузии.
Раковина высококоническая, с почти строго коническим контуром завитка,
умеренно твердостенная, матовая. Оборотов 61/,—7х/2, умеренно выпуклых,
разделенных мелким беловатым швом. Последний оборот к устью не поднят,
высота его равна половине высоты раковины или немного больше. Окраска
одноцветно роговая, иногда с оливковым оттенком. Скульптура в виде тонкой
радиальной неравномерной морщинистости и хорошо выраженных тонких
частых слегка волнистых бороздок. Устье овальное, места его прикрепления
слегка сближены. Палатальный и базальный края устья тонкие, светлые, слабо
отвернуты; почти отвесный колумеллярный край отвернут немного сильнее,
но почти не прикрывает широкощелевидного пупка.
Размеры: ВР 12—15, БД 6.5—7.5 мм; в первоописании размеры не указаны,
характеристика вида дана в плане сравнения с Р. merduenianum.
Внутреннее строение. Материал: 2 экз. из окрестностей Ку-
таиси, 30 апреля 1971 г., сбор и определение автора.
Белковая железа с широкой апикальной вырезкой. Нижний отдел вагины
примерно такой же длины, как и верхний. Цэкум настолько резко сдвинут
к верхнему концу эпифаллуса, что вместе с закругленно-коническим бичом
образует подобие сапожка, в «подошву» которого впадает семяпровод. Начало
семяпровода отличается очень малым диаметром, но после изгиба у полового
атриума семяпровод плавно утолщается, и эта утолщенная часть тянется вдоль
пениса и эпифаллуса, до впадения в эпифаллус. Вообще мужской отдел отли-
чается очень малой длиной; эпифаллус изогнут, широкий, цилиндрический;
пенис в дистальной части узкий, но эта дистальная часть очень короткая, а про-
ксимальная почти шаровидная. На внутренней поверхности толстостенного
пениса сидят многочисленные призмоконические бугорки, расположенные без
видимой системы. Папилла мешковидная, занимает почти весь объем пениса,
поверхность ее бугристая. Форму папиллы можно охарактеризовать как дву-
лопастную: по поверхности большей лопасти проходит борозда, прорезающая
углубление, в котором помещается меньшая лопасть. Пениальный аппендикс
346
VI. НАДСЕМ. BULIMINOIDEA
отходит практически от полового атриума. шаровидно утолщен, Л2 тоже
шаровидный или уплощенный, А3 длиннее, чем два первых отдела, вместе взятые.
Проток семяприемника прямой, дивертикул заметно не доходит до основания
белковой железы.
Распространение и местообитание. Вид обнаружен
в нескольких пунктах Грузии: Кутаиси, долина р. Техури, Боржоми, Цагери,.
Рис. 252. Peristoma boettgerl (Cl.) из окр. Кутаиси, 30 VI 1971 г.
7 — половой аппарат, II — вскрытая дистальная часть мужского отдела, III — вскрытый проксималь-
ный конец эпифаллуса.
Тебердинский заповедник. Обитает в лиственных и смешанных лесах и в мало-
подвижных крупнообломочных осыпях. Нередко поднимается на стволы де-
ревьев.
3. Род CAUCASICOLA Hesse, 1917
О. Boettger, 1883 : 174 (Medea, non Eschscholtz, 1829 : 38); Hesse, 1916b : 124
(Helle, non Osten-Sacken, 1896); 1917 : 122 [Ena (Caucasicola)]; Lindholm, 1925a : 27
[Ena (Caucasicola)}', Thiele, 1931 : 520 [Ena (Caucasicola)]; Wen z, Zilch, 1959—1960 :
184 [Ena (Caucasicola)]; Шилейко, 1978г : 847.
Типовой вид Buliminus raddei Kobelt, 1880; по монотипии.
Раковина высококоническая; тонкая скульптура представлена радиальными
морщинками и спиральными линиями. Места прикрепления устья не сближены.
Окраска одноцветно желтая или имеется широкая спиральная темная лента.
Устье без зубов.
Эпифаллус и проток семяприемника недлинные. Цэкум расположен вплот-
ную к месту впадения семяпровода. Отделы пениального аппендикса нормально
развиты. Призмоконические бугорки на внутренней поверхности пениса четкие,
расположены густо. Папилла несет многочисленные острые бугорки, ложко-
видная.
Монотипический род.
3. РОД CAUCASICOLA
347
Caucasicola raddei (Kobelt, 1880) (рис. 253, 254)
K o belt, 1880, Iconogr., VII : 50, fig. 2008, 2009 (Buliminus); O. Boettger, 1883 :
175 [B. (Medea)]; Westerlund, 1887 : 21 (Buliminus)-, Kobelt, 1902 : 415,t. 72,
fig. 19—21 [B. (Medea)]-, Simroth, 1912 : 90, t. 10, fig. 47 [B. (Medea)]-, Retowski,
1914 : 303 [B. (Medea)]; Rosen, 1914 : 183 [B. (Medea)]; Л и x a p e в, P а м м e л ь м е й е р,
1952 : 183 [Ena (Caucasicola)]; Шилейко, 1978г : 834, рис. 1, II.
Terra typica — «? Avarien» (горный Дагестан — см. ниже).
Лектотип хранится, вероятно, в Зенкенбергском музее, Франкфурт-на-
Майне.
Материал: 132 экз. из 44 местонахождений на Кавказе.
Раковина высококоническая, со строго коническим контуром завитка, твер-
достенная, слабо просвечивающая. Оборотов 8—9, уплощенных, разделенных
мелким светлым зазубренным швом. Последний оборот непосредственно возле
устья слабо, но явственно приподнят, высота его не-
сколько превышает половину высоты раковины. Окраска
состоит из светло-желтого фона и спиральной ленты бу-
рого, коричневого или лиловатого цвета, которая чаще
всего занимает большую часть поверхности раковины; в
этом случае фон остается в виде узкой ленты под швом
и светлого поля вокруг пупка. Однако встречаются рако-
вины, лишенные темной ленты и имеющие одноцветно-
желтую окраску. Относительная ширина темной ленты
подвержена индивидуальной изменчивости; на ней не-
редко заметны светлые узкие радиальные пестрины.
Скульптура в виде неравномерной радиальной нерезкой
морщинистости и тонких волнистых спиральных бороз-
док. Устье овальное, слабо скошено, края его несильно
отвернуты, белые; рисунок на поверхности раковины ви-
ден и внутри устья. Губа слабо выражена. Палаталь-
Рис. 253. Caucasicola raddei (Kob.) из окр. Хосты (Сочи).
ный и базальный края устья плавно выгнуты, при переходе в колумелляр-
ный край заметен закругленный угол. Колумеллярный край почти отвесный,
отвернут сильнее прочих и прикрывает пупочную щель.
Размеры: ВР 21—27, БД 9—13 мм; в первоописании: ВР 26, БД 13 мм.
Внутреннее строение. Материал: 1 экз. из окр. Хосты,
20 сентября 1956 г., сбор и определение И. М. Лихарева; 3 — из ущелья р. Агуры,
14 октября 1955 г., сбор Е. С. Кирьяновой; 1 — из окр. Ходжоха в 40 км к югу
от Майкопа, сбор'Я. И. Старобогатова; 2 — из тисо-самшитовой рощи близ
Хосты, 20 апреля 1956 г., сбор И. И. Малевича; 1 — из долины р. Техури близ
с. Талери, 18 апреля 1971 г.; сбор и определения автора.
Белковая железа с апикальной вырезкой. Нижний отдел вагины в 2—3 раза
короче верхнего. Цэкум резко сдвинут вверх, образует в совокупности с бичом
фигуру, напоминающую сапожок; в центр «подошвы» сапожка впадает семя-
провод. Пенис массивный, булавовидный, в 2—3 раза короче эпифаллуса. На
внутренней поверхности пениса бросается в глаза множество высоких резких
.густо расположенных призмоконических бугорков; похожие бугорки покрывают
и поверхность папиллы. Борозда на поверхности папиллы очень широкая и глу-
бокая, поэтому папилла в целом формой напоминает ложку. Все отделы пениаль-
ного аппендикса выражены очень четко; относительно слабо развит А2- Обра-
щает на себя внимание необыкновенная длина Аа. Ветви полового ретрактора на
диафрагме в разной степени сближены, одна из ветвей крепится к выше его
середины, вторая — к пенису близ его границы с эпифаллусом. Проток семя-
348
VI. НАДСЕМ. BVLIMINOIDEA
приемника прямой, удлиненный резервуар имеет довольно тонкий проток, не
длиннее самого резервуара. Дивертикул не доходит до белковой железы.
Распространение и местообитание. Подкововидный
ареал начинается на Северном Кавказе примерно от Майкопа (отдельная на-
ходка у Нальчика), далее на запад до Новороссийска и Геленджика; западные
районы южных склонов Большого Кавказа на восток до р. Кодор; Розен пишет,
Рис. 254. Caucasicola raddei (Kob.) из окр. с. Ходжох близ Майкопа, 12 VII
1974 г.
I — половой аппарат, II — вскрытый проксимальный конец эпифаллуса, III — вскрытые пенис и часть-
пениального аппендикса (из: Шилейко, 1978 г.).
что «эта . . . улитка далеко не так редка в Кубанской обл., как в Черноморской
губернии. . .» (Розен, 1901 : 10). Что касается присутствия вида на восточном
Кавказе («Авария»), то Кобельт сам не был уверен в правильности указания;
во всяком случае, достоверных сведений о находках вида в Дагестане пока нет.
4. Род ZEBRINA Held, 1837
Risso, 1826:78 (Bulimulus, non Leach, 1815); H e Id, 1837 : 917; Westerlund,
1887 : 3 (Zebrinus, типовой вид Helix detritus Muller, 1774); Kobelt, 1902 : 1021 (Rhabdoena,
типовой вид Buliminus caesius Kobelt, 1902); Hesse, 1933 : 181; F о г c a r t, 1940 : 134;
Лихарев, Раммельмейер, 1952 : 184; W e n z, Zilch, 1959—1960 : 188; III n-
л e й к о, 1978г : 847.
Типовой вид Helix detritus Muller, 1774; по обозначению Грея (Gray, 1847).
Раковина удлиненно-овальная, слабо скульптурирована радиальными мор-
щинками и спиральными бороздками. Места прикрепления устья не сближены.
Окраска светлая, обычно с темными радиальными пестринами. Устье без зубов.
Эпифаллус и проток семяприемника недлинные. Цэкум расположён вплот-
ную к месту впадения семяпровода. Пениальный аппендикс развит хорошо,
и А2 слиты. Призмоконические бугорки внутри пениса четкие, разбросан-
4. РОД ZEBRINA
349'
ные. Папилла с гладкой поверхностью и двумя продольными бороздами,
стержневидная.
Распространение. Европа, кроме северных областей, Малая Азия,.
Ближний Восток. Несколько видов.
В ископаемом состоянии род известен начиная с нижнего плиоцена (южная
Европа). П. М. Асланян (1971) описал из конкских отложений близ г. Раздан
в Армении два вида, относящиеся предположительно к роду Zebrina (Z. коп-
kensis и Z. hrazdanica). Если родовая принадлежность определена верно, то воз-
раст рода датируется нижним—средним миоценом.
Zebrina detrita (Muller, 1774) (рис. 255, 256).
Muller, 1774 : 101 (Helix)' В г u g и i ere, 1789 : 312 (Bulimus radiatus); Rossmaes-
sler, 1835, Iconogr., I, 1 : 86, fig. 42 (B. radiatus); Moquin-Tandon, 1855 : 294,.
pl. 21, figs. 11 a 24 [2?. (Bulimulus)]; Mousson, 1859 : 171 (B. detritus var. tumidus Parreys);
Kobelt, 1902 : 418, t. 73, figs. 1—3 [Bullminus (Zebrina)]; K. Beck, 1912 : 47 (Bulimi-
nus); Geyer, 1927 : 128, t. 6, fig. 6a, b; F о r c a r t, 1940 : 134, t. I, fig. 10; С. В о e 11 g e r,
1953 : 151, 152; Ehrmann, 1956 : 58, t. 2, Abb. 27; Lozek, 1956 :112, t. 10, fig. 1;
Zilch, Jaeckel, 1962 : 108; L о i e k, 1964 : 228, t. 11, fig. 5; Hudec, 1967 : 19,.
obr. 3, 4,1.1, 3; Д а м я н о в, Л и х а р е в, 1975, 142, фиг. 78, 79; Ш и л е й к о, 1978г : 514г
рис. 1, II.
Terra typica — «Италия и Саксония».
Место хранения типов не установлено.
Материал: более 60 экз. из 6 местонахождений в Чехословакии, Венг-
рии и Болгарии.
Раковина удлиненно-овальная, с закругленно-коническим контуром за-
витка, твердостенная, блестящая. Оборотов 6—8, несильно выпуклых. Послед-
ний оборот к устью не поднят, высота его немного превы-
шает половину высоты раковины. Окраска известково-
белая, чаще всего с темными радиальными пестринами.
Эмбриональные обороты светло-серые или рыжеватые. Де-'
финитивные обороты покрыты вялыми неравномерно
расположенными морщинами, которые местами расчле-
нены редкими спиральными бороздками. Устье оваль-
ное, внутри светлое, места его прикрепления не сближе-
ны, соединены расплывшейся мозолью. Палатальный край
равномерно выгнут, базальный — коротко закруглен, колу-
меллярный — почти отвесный, прямой. Края устья белые,
.немного утолщены, притупленные, очень слабо отвер-
Рис. 255. Zebrina detrita (Miill.) из Венгрии, окр. г. Печ.
нуты, кроме расширенного колумеллярного, прикрывающего своим отворо-
том пупочную щель.
Размеры; ВР 17—27, БД 9—12 мм; в первоописании: ВР 9.5, БД 3.5 линии,,
т. е. ВР около 20 мм, БД около 8 мм.
Внутреннее строение. Материал: 7 экз. из Венгрии, окр. г.
Печ, 30 мая 1970 г., сбор Д. А. Сахарова, определение автора.
Белковая железа с очень вялым, порой едва заметным латеральным желоб-
ком, иногда занимающим и апикальную область. Верхний отдел вагины при-
близительно в РД—2 раза длиннее нижнего. Цэкум тесно сближен с утолщен-
ным округлым очень коротким бичом. Из-за этого создается впечатление, что
проксимальный конец эпифаллуса утолщен и расплющен и в центр образован
шейся площадки впадает семяпровод. Эпифаллус относительно короткий, ров-
ный, цилиндрический. Пенис вдвое толще эпифаллуса и слегка короче послед-
него, цилиндрический. Внутри . пениса находится хорошо развитая папилла
350
VI. НАДСЕМ. BULIMINOIDEA
с двумя противолежащими продольными узкими и глубокими бороздками. Вну-
тренние стенки пениса несут отдельные четкие призмоконические бугорки, не
обнаруживающие тенденции к группировке в ряды. внешне слит с А2, хотя
при вскрытии этих отделов обнаруживаются структуры, характерные для каж-
дого из этих отделов. А3 хорошо развит, длинный, постепенно, без ясной гра-
ницы, переходит в относительно короткий А4. Аа заметно короче, чем А4, или
эти отделы примерно равной длины. Ветви полового ретрактора на диафрагме
тесно сближены, одна ветвь прикреплена к верхней части А1+А2, вторая —
несколько выше середины пениса. Проток семяприемника почти прямой; оваль-
Рис. 256. Zebrina detrita (Miill.) из Венгрии, окр. г. Печ, VIII 1970 г.
I — половой аппарат; II — вскрытая дистальная часть мужского отдела; III — вскрытая проксималь-
ная часть эпифаллуса; IV — призмоконический бугорок — сильно увеличен (из: Шилейко, 1978в).
яый резервуар сидит на тонком протоке; дивертикул немного не доходит до бел-
ковой железы.
Распространение и местообитание. Средняя Европа, Бал-
канский п-ов, южные Альпы, Тироль, южная Франция, Малая Азия; возможно,
юго-западное Закавказье. Здун (1960) сообщает о находке моллюсков этого вида
в равнинной части Закарпатья (с. Мала Копаня). Типичный обитатель степных
ландшафтов, часто поднимается на стебли кустарников и травянистых растений,
может образовывать плотные скопления.
5. Род CHONDRUS Cuvier, 1817
Cuvier, 1817, Regne animal, II : 408; Westerlund, 1887 : 9 (Brephulus, non
Beck, 1837); Lindholm, 1925a : 29 (род Chondrus, секция Antichondrus, типовой вид
Helix tournefortianus Ferussac, 1821); Thiele, 1931 : 519; Hesse, 1933 :187; Forcart,
1940 : 176; Wenz, Zilch, 1959—1960 : 190; Шилейко, 1978г ; 847.
Типовой вид Bulimus zebra Olivier, 1801; по обозначению Грея (Gray,
1847, p. 175).
5. РОД CHONDRUS
35£
Раковина от цилиндрической до башневидной, скульптура тонкая, пред-
ставлена радиальными морщинками и элементами спиральной исчерченности..
Места прикрепления устья не сближены. Окраска белая, чаще всего с темными
радиальными пестринами. Устье с 3—4 поверхностными зубами.
Эпифаллус длинный, проток семяприемника различной длины. Цэкум рас-
положен близ середины эпифаллуса. Отделы пениального аппендикса выражены,,
пропорциональные. Призмоконические бугорки внутри пениса разбросанные...
Папилла тонкая, длинная, гладкостенная.
Рис. 257. Chondrus zebra Рис. 258. Chondrus zebra (Oliv.). Половой аппарат (из: Hesse,.
(Oliv.) из Греции, Ко- 1933).
ринфский перешеек.
Распространение. Балканский п-ов, Греция, Малая Азия. Не-
сколько видов.
Chondrus zebra (Olivier, 1801) (рис. 257, 258).
Olivier, 1801, vol. 2 : 14, Atlas, t. 17, fig. 10a, b (Bulimus)-, Ferussac, 1821 : 58'
[Helix (Cochlogena) zebriola]; Kobelt, 1877 : 266, t. 5, fig. 8 (Buliminus olimpicus); Gal-
land, 1884 : 281 (Brephulus bithynicus), 282 (B. zebra var. anatolica), 283 (B. bradus), 284
[B. zebropsis (Bourguignat)] — fide Forcart, 1940; Розен, 1901 : 10 (Buliminus)-, R o-
s e n, 1914 : 183 (Buliminus)-, Hesse, 1933 : 189; Forcart, 1940 : 177, Abb. 5, t. I, fig. 24;
Hudec, Vasa t ko, 1971 : 14, Abb. 2, pl. I, 4 (Ch. zebra tantalus)-, Д а м я н о в, Л и х а-
р е в, 1975 : 146, fig. 82, 83.
Terra typica — окр. Гемлика, северо-западная Турция.
Место хранения типов не установлено.
Материал: 19 экз. из четырех местонахождений в Болгарии, Турции и Греции..
Раковина высокая, тонкая, стройная, коническо-цилиндрическая или почти
башневидная, твердостенная, блестящая. Оборотов 8—12, уплощенных или
умеренно выпуклых, разделенных довольно глубоким, но узким швом. Послед-
ний оборот к устью едва заметно поднят, высота его заметно меньше половины
высоты раковины, часто составляет х/3 этой величины, а иногда и еще меньше.
Окраска известково-белая, чаще всего с темными радиальными пестринами.
Скульптура в виде тонкой косорадиальной морщинистости и слабых спираль-
ных бороздок. Устье удлиненно-овальное, места его прикрепления связаны
мозолью, на которой может присутствовать ангулярное утолщение. Края устья
немного утолщены и слабо отвернуты. Кроме ангулярного бугорка, имеется
короткая, но высокая париетальная пластинка, вялая складкообразная колу-
352
VI. НАДСЕМ. BULIMINOIDEA
меллярная пластинка и округло-треугольный палатальный зуб. Пупок в виде
очень узкой и короткой щели.
Размеры: ВР 12—19, БД 4—5.5 мм.
Внутреннее строение мною не исследовано, описание составлено
по литературным данным (Hesse, 1933; Forcart, 1940; Gittenberger, 1967; Hudec,
Vasatko, 1971).
Вырезка на белковой железе вялая, занимает латеральное положение. Ва-
гина довольно длинная, верхний отдел ее несколько длиннее нижнего. Бич от-
сутствует или очень слабо развит, соответственно семяпровод впадает терми-
нально или несколько сбоку. Цэкум занимает на эпифаллусе срединное поло-
жение. Эпифаллус относительно длинный, в разной степени извит. Насколько
можно видеть на просветленных препаратах, изображения которых помещены
в работе Форкарта (loc. cit., 178, Abb. 5, и 183, Abb. 6), на внутренней поверх-
ности пениса имеются призмоконические бугорки; папилла цилиндрическая, за-
нимает большую часть проксимального отдела пениса. Все отделы пениального
аппендикса ясно выражены. Ветви полового ретрактора на диафрагме в разной
степени Сближены, иногда слиты; одна ветвь крепится к вторая — к пенису,
к срединной или проксимальной его части. Однако Виттенбергер (loc. cit.,
S. 138—139, Fig. 7) обнаружил у Ch. zebra tantalus заметную степень редукции
аппендикса: отделы его тонкие, не все выражены, сам аппендикс впадает в про-
ксимальную часть пениса; соответствующая ветвь полового ретрактора отсут-
ствует. Проток семяприемника резких изгибов не образует, небольшой оваль-
ный резервуар с собственным коротким протоком, дивертикул почти касается
белковой железы.
Распространение и местообитание. Балканский п-ов,
о-ва Эгейского моря, Малая Азия. Розен (1901, с. 10) сообщает о находке
«вполне типичных экземпляров» в Поти на берегу моря между устьями Риони и
речки, вытекающей из оз. Палеостом. Вероятно, эти раковины были выброшены
морем. Обитает на открытых прогреваемых солнцем участках с низкой травя-
нистой растительностью.
6. Род BREPHULOPSIS Lindholm, 1925
Lindholm, 1925а : 28 [Zebrina. (Zebrina), секция Brephulopsis, nom. nov. pro Brephulus
Westerlund, 1902, non H. Beck, 1837]; Wen z, Zilch, 1959—1960 : 189 [Zebrina (Brephulop-
sis)]; Шилейко, 1978г : 848.
Типовой вид Buliminus attenuatus Krynicki, 1833 (syn. Buliminus bidens
Krynicki, 1833); по первоначальному обозначению.
Раковина высокая, большей частью цилиндрическая. Скульптура тонкая,
представлена, радиальными морщинками и нерезкими спиральными борозд-
ками. Окраска белая, чаще всего с темными радиальными пестринами и штри-
хами. Устье без зубов или имеется одна глубоколежащая палатальная складка.
Эпифаллус и проток семяприемника длинные, цэкум сдвинут к пенису от-
носительно середины эпифаллуса. Отделы пениального аппендикса, как пра-
вило, выражены нормально. Резкие призмоконические бугорки внутри пениса
сгруппированы в ряды. Папилла длинная, гофрированная.
Распространение. Крым, западное Закавказье, южные и юго-за-
падные области европейской территории СССР. 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ BREPHULOPSIS
1 (2). В глубине устья на палатальной стенке имеется продольная складка.
Пениальная папилла состоит из одной удлиненной лопасти.................
................................................... 2. В. bidens.
2 (1). Палатальная складка отсутствует. Папилла пениса двулопастная . .
............................................... 1. В. cylindrica.
6. РОД BREPHULOPSIS
353
1. Brephulopsis cylindrica (Menke, 1828) (рис. 259, I; рис. 260).
Menke, 1828 : 77 (Bulimus)^ Krynicki, 1833 : 395, t. 6, fig. 1 (Chondrus lineatus),
398 (Ch. turgidus), 398, t. 6, fig. 2a—d (Ch. fuscilabris), 401 (Ch. concolor); L. Pfeiffer,
1848b : 226 (Bulimus tauricus); Westerlund, 1887 : 7 (Buliminus); Пузанов, 1926 :
92, рис. 3 [В. (Zebrina)]; Лихарев, Раммельмейер, 1952 : 184 (Zebrina); Hudec,
1972a : 77, Abb. 2, t. 2, fig. 29 [Zebrina (Ramusculus?)]; 1972b : 102, Abb. 2, 6 [Z. (Ramuscu-
lus)] ; Арутюнова, 1975 : 104, рис. 1A (Z. (Ramusculus)].
Terra typica «Patriam ignoro» — неизвестна.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 17 местонахождений в Крыму,
окр. Одессы и черноморского побережья Кавказа.
Раковина от веретеновидной до цилиндрической, твердостенная, блестящая,
с узко закругленной вершиной. Оборотов 8—10, верхние несколько более вы-
пуклые, чем нижние, но в общем тоже уп-
лощенные. Шов мелкий. Последний оборот
к устью не поднят, высота его всегда меньше
половины высоты раковины, у высоких рако-
вин составляет почти х/3 этой величины. Ок-
раска подвержена индивидуальной и межпо-
пуляционной изменчивости, в общем состо-
ит из белого фона и коричневых или почти
черных радиальных пестрин. Скульптура
сложная. Во-первых, имеется тонкая ра-
диальная неравномерная морщинистость;
во-вторых, местами заметны вмятины;
в-третьих, обычно присутствуют редкие
Рис. 259. Виды рода Brephulopsis.
I — В. cylindrica (Menke) из Чатыр-дага, II — В. bidens
(Kryn.) из с. Краснолесье близ Симферополя.
радиальные бороздки, рассекающие радиальные морщинки, но не затрагиваю-
щие собственно поверхность раковины. Устье усеченно-овальное или заостренно-
овальное, незначительно скошено, места его прикрепления не сближены, сое-
динены тонкой мозолью. Края устья почти не отвернуты, чуть отступя от края
лежит чрезвычайно расплывшаяся широкая губа. На колумеллярном крае не-
редко можно заметить слабую валикообразную пластинку. Пупок узкощеле-
видный.
Размеры: ВР 20—30, БД 7—10 мм; в первоописании: ВР 10, БД 3 линии,
т. е. ВР около 21, БД около 6.5 мм.
Внутреннее строение. Материал: 16 экз. из Крыма, окр. Судака,
4 августа 1962 г., сбор и определение автора.
Белковая железа с широкой плавной апикальной вырезкой. Вагина срав-
нительно длинная, длина ее отделов одинакова. Бич тонкий, конический, ко-
роткий, семяпровод впадает в эпифаллус почти под прямым углом. Эпифаллус
длинный, извитой, образует несколько крутых изгибов и петель. Цэкум распо-
лагается значительно ниже середины эпифаллуса. Граница между пенисом и
эпифаллусом часто совсем внешне не выражена. Призмоконические бугорки на
внутренней поверхности пениса большие, высокие, собраны в продольные ряды;
эти бугорки очень хорошо заметны на вывернутом пенисе (рис. 260, III). Пе-
ниальная папилла длинная, гофрированная, складчатая, двулопастная, при-
чем одна из лопастей меньше другой. Аг немного длиннее пениса, у некоторых
экземпляров скручен, А2 небольшой, А3 сравнительно длинный, плавно пере-
ходит в Л4. Ветви полового ретрактора отходят от диафрагмы порознь, пениаль-
23 А- А- Шилейко
354
VI. НАДСЕМ. BULIMINOIDEA
ная ветвь подходит к верхней части пениса, ветвь аппендикса — к Ах заметно
отступя от Л2. Проток семяприемника сильно извит, длинный; овальный резер-
вуар с собственным коротким протоком, длина которого меньше длины резер-
вуара. Дивертикул почти касается белковой железы.
Распространение и местообитание. Крым; представля-
ется очень вероятным, что многочисленные находки в Молдавии, в окр. Одессы,
Рис. 260. Brephulopsis cylindrica (Menke) из Крыма, Судак, 4 VIII 1962 г.
I — половой аппарат и детали его строения, II — призмоконический бугорок — сильно увеличен, III —
особь с вывернутыми дистальными участками половых протоков.
Новороссийска, Анапы и Сухуми связаны с завозом этого вида. Ксерофильный
вид, образует плотные скопления на открытых степных участках, нередко вдоль
дорог. В жаркую сухую погоду образует грозди на стеблях трав и кустарниках.
Молодь живет в почве, в прикорневых частях трав.
2. Brephulopsis bidens (Krynicki, 1833) (рис. 259, II; рис. 261).
Krynicki, 1833:401, t. 3, fig. За—е (Bulimus); 404 (В. attenuatus); Bourgui-
gnat, 1876 : 80, t. 2, fig. 30 (В. theodostanus); Clessin, 1881 : 140 [B. (Zebrina) Retow-
skianus]; Retowski, 1883a : 18 (Buliminus); Hesse, 1885 : 51, t. 3, fig. 5 [2?. bidens
Kryn. var. Retowskiana Cl. (=B. theodostanus Bourg.)J; Kobe I t, 1902 : 428, t. 74, figs. 15—18
[B. (Brephulus)]; Пузанов, 1925a : 80, рис. 4 [В. (Brephulus)]; 1926 : 93, рис. 3 [В. (Brephu-
lus)]; Лихарев, Раммельмейер, 1952:186 (Chondrus); Hudec, 1972a : 77, Abb. 3—
5, 27, 28, 30 (Chondrus (Chondrus)]; Шилейко, 1978г : 838, рис. 2, II; Лившиц, Ш и-
л е й к о, 1978 : 77—83, рис. 2.
Terra typica — Крым.
Лектотип хранится в Зоологическом ин-те АН СССР (место сбора —
Симферополь), обозначается здесь.
Материал: несколько сотен экз. из 106 местонахождений в Крыму.
Раковина от цилиндрической до удлиненно-овальной, веретеновидной и
почти башневидной, твердостенная, блестящая. Оборотов 7—11, сильно упло-
6. РОД BREPHULOPSIS
355
щенных, разделенных мелким швом. Последний оборот к устью не поднят, вы-
сота его заметно меньше половины высоты раковины. Окраска белая, с темными
радиальными пестринами,, иногда одноцветно белая. Скульптура представлена
неравномерной тонкой радиальной морщинистостью, у членов некоторых популя-
ций можно рассмотреть вялые спиральные бороздки, выраженные местами;
как и у предыдущего вида, они прорезают радиальные элементы скульптуры и
не затрагивают собственно поверхности раковины. Устье усеченно-овальное,
слегка скошено, места его прикрепления не сближены, связаны тонкой мо-
золью. В устье два зуба: слабая косая валикообразная колумеллярная пла-
стинка и палатальная складка, расположенная в глубине устья. Края устья
почти не отвернутые, слегка утолщенные, тупые. Пупок узкий, щелевидный.
Рис. 261. Brephulopsis bidens (Kryn.) из окр. с. Партизаны близ Симферополя, 10 X 1971 г.
Половой аппарат и вскрытый пенис (из: Шилейко, 1978 г.).
Размеры: ВР 19—22, БД 5—6 мм; в первоописании: ВР 9, БД 23/4 липин;
лектотип: ВР 14.5, БД 5.0 мм. '
Внутреннее строение. Материал: 96 экз. из окр. с. Чистенькое
близ Симферополя, И февраля, 7 марта, 10 апреля, 13 мая, 17 июля, 20 августа
и 16 октября 1975 г.; 9 — из с. Партизаны близ Симферополя, 10 октября 1974 г.;
сборы Г. М. Лившица; 14 экз. из Краснолесья близ Симферополя, 2 августа
1962 г.; сбор и определения автора.
Белковая железа с неглубокой апикальной вырезкой. Верхний отдел на
гины не более чем в 1х/2 раза длиннее нижнего, нередко отделы почти равной
длины. Семяпровод подходит к эпифаллусу почти под прямым углом, имеете»
очень тонкий и короткий конический бич. Эпифаллус длинный, образует не
сколько изгибов или 2—4 петли. Сильно развитый цэкум располагается заметно
ниже середины эпифаллуса. Диаметр эпифаллуса ниже цэкума явственно умень-
шается и вновь увеличивается при переходе в пенис. Граница между этими от
делами отмечена внешне только местом прикрепления пениальной ветви поло-
вого ретрактора. На внутренней поверхности пениса имеются высокие реэкио
2.4*
356
VI. НАДСЕМ. BULIMINOIDEA
призмоконичесцие бугорки, собранные в правильные продольные ряды. Па-
пилла пениса длинная, в проксимальной части бугорчатая, ниже середины гоф-
рированная. По строению пениального аппендикса вид обнаруживает четко вы-
раженный диморфизм. Первый вариант характеризуется нормальным строением
аппендикса, т. е. все его .отделы нормально и пропорционально развиты. Для
второго варианта характерно полное отсутствие А1; и А2 в виде шаровидного
утолщения располагается непосредственно на поверхности срединной части
пениса (рис. 37); остальные отделы сохраняют традиционное строение. В типич-
ном случае не короче пениса, А2 небольшой, Л3 относительно длинный, не ко-
роче половины Лх. Половой ретрактор в этом случае крепится к Лх заметно от-
ступя от А2; если Аг отсутствует, половой ретрактор одноветвистый, подходит
к проксимальному отделу пениса. Численное соотношение этих вариантов при-
мерно 1 : 1 (из 96 экз., собранных близ с. Чистенькое, 46 имели редуцирован-
ный Ах, а 50 сохраняли нормальное строение аппендикса); вместе с тем найдено
3 экз., у которых обнаружен рудимент в виде короткой подставки, располо-
женной, однако, не в нижней части пениса, а на его середине. Сведения по сте-
пени половой изоляции этих форм отсутствуют, но есть основания полагать, что
отмеченные различия вряд ли служат препятствием к свободному спариванию
между представителями этих форм. Проток семяприемника образует несколько
изгибов или петель, резервуар небольшой, овальный, длина его протока при-
мерно равна длине самого резервуара. Дивертикул почти касается белковой
железы.
Распространение и местообитание. Крым. Населяет степ-
ные участки, как на равнине, так и в горах. На зиму моллюски закапываются
в почву, летом иногда образуют разреженные грозди на ветвях низкорослых
кустарников и стеблях трав.
7. Род NAPAEOPSIS Sturany et Wagner, 1914
Stu ran у, Wagner, 1915 (1914) : 41 [Napaeus (Napaeopsis)]; Thiele, 1931 : 523;
Hesse, 1933 : 153; F о г с a r t, 1940 : 157; Wenz, Zilch, 1959—1960 : 189 [Zebrina
(Napaeopsis)-, Акрамовский, 1976 : 161.
Типовой вид Bulimus cefalonicus Mousson, 1859; по обозначению Гессе
(Hesse, 1933, S. 153).
Раковина не имеет дифференциальных отличий от рода Zebrina.
Эпифаллус недлинный, проток семяприемника лишен дивертикула, до-
вольно длинный. Цэкум сдвинут к пенису относительно середины эпифаллуса.
.Отделы пениального аппендикса развиты нормально, кроме А3, который внешне
не выражен. Призмоконические бугорки внутри пениса четкие, высокие, не-
ясно сгруппированы в продольные ряды. Папилла короткая, с глубокой бо-
роздой.
Распространение. Балканский п-ов, Малая Азия, Кавказ.
Таксономическую структуру и объем рода в настоящее время трудно оце-
нить, поскольку внутреннее строение совокупительного аппарата типового вида
неизвестно. Bulimus hohenackeri провизорно включен в этот род ввиду большого
сходства его с В. cefalonicus.
Napaeopsis hohenackeri (L. Pfeiffer, 1848) (рис. 262, 263).
К г у n i c k i, 1837 : 53 (Bulimus, nom. nudum); L. Pfeiffer, 1848b : 223 (Bulimus)-,
Rossmaessler, 1859, Iconogr., Ill : 91, fig. 912, 913 (Bulimus); Mousson, 1863 : 381
(B. hohenackeri Kryn. var. intermedius); Schneider, 1878:22 (B. interfuscus Mouss.);
Hesse, 1885 : 50, t. 3, fig. 3, 3a (Buliminus); Westerlund, 1887 : 6 (Buliminus); К o-
b e 1 1, 1902 : 425, t. 73, figs. 17, 18 [2?. (Zebrina)]; Retowski, 1914 : 302 [R. (Zebrinus)];
Rosen, 1914 : 182 (Buliminus); Lindholm, 1922 : 358 [B. (Zebrina) hohenackeri f. leu-
colaemus]', F о г с a r t, 1940 : 158; Лихарев, Раммельмейер, 1952 : 184, рис. 94
(Zebrina); Hudec, Lezhawa, 1967, : 77, Abb. 4, t. I, 5 [N apaeopsis (Napaeopsis)]; Акра-
мовский, 1976:161, рис. 75, табл. VIII, 82.
7. РОД NAPAEOPSIS
357
Terra typica — Грузия.
Место хранения типов не установлено.
Материал: несколько сотен экз. из 122 местонахождений на Кавказе.
Раковина от высококонической до овально-конической,
твердостенная, блестящая. Оборотов 8—9, умеренно вы-
пуклых, разделенных узким швом. Последний оборот к устью
едва заметно приподнят или прямой, высота его заметно
превышает половину высоты раковины. Окраска молочно-
белая, часто с радиальными светло-коричневыми или ры-
жеватыми размытыми пестринами. Скульптура в виде гру-
бой очень неравномерной морщинистости и редких неболь-
ших вмятин, на многих раковинах имеются слабые намеки на
спиральную исчерченность. Устье овальное, наверху заост-
ренное, слабо скошено. Края устья тупые, незначительно
отвернуты. Чуть отступя от устьевого края лежит широ-
Рис. 262. Napaeopsis hohenackeri (L. Р£г.) из Грузии, Ахалцихе.
кая расплывшаяся желтая или бурая губа, внутри устье темно-желтое.' Пала-
тальный край вначале плавно и несильно выгнут, переходит в базальный с ши-
Рис. Z&S..Napaeopsis hohenackeri (L. Pfr.) из долины p. Куры близ Ахалцихе, 19 IV 1971 г.
1 — половой аппарат и детали его строения, 11 — вскрытый пенис.
роким закруглением, и при переходе в колумеллярный вновь следует более y:t
кое закругление. Колумеллярный край сильно отвернут, почти закрыиает пу-
пок, оставляя от него узкую щель.
358
VI. НАДСЕМ. BULIMINOIDEA
Размеры: ВР 20—30, БД 9—14 мм; в первоописании: ВР 24, БД 10-11 мм.
Внутреннее строение. Материал: 5 экз. из Армении, Ведин-
ский р-н, Хосровский лес, 18 июля 1969 г., сбор А. Погосовой, определение
Н. Н. Акрамовского; 15 — из Талыша, Зуванд, окр. с. Космальян, 4 июля
1964 г.; 7 — из долины р. Куры между Боржоми и Ахалцихе, 19 апреля 1971 г.,
сборы и определения автора.
Белковая железа неправильной формы, с желобками, вмятинами и широкой
апикальной вырезкой. Верхний отдел вагины несколько длиннее нижнего.
Семяпровод впадает в эпифаллус под незначительным углом к короткому чуть
приостренному бичу. Длина эпифаллуса подвержена значительной изменчивости,
однако он никогда не бывает длинным, всегда слабо извит или вообще не извит.
Цэкум небольшой, расположен примерно на границе нижней и средней трети
эпифаллуса, отделенного кольцевой перетяжкой от цилиндрического пениса.
Призмоконические бугорки на внутренней поверхности пениса выражены ясно,
но упорядоченность их расположения многократно нарушается. Пениальная па-
пилла короткая, коническая, остроконечная, на ее поверхности заметны неболь-
шие бугорки, похожие на те, которыми покрыта внутренняя поверхность чехла
пениса. Борозда на поверхности папиллы очень глубокая и узкая, стенки па-
пиллы насыщены множеством синусов. Пениальный аппендикс хорошо развит,
4Х не длиннее пениса, 42 маленький, А3 и А4 большей частью не обособлены
один от другого, лишь у некоторых особей намечается граница между этими
отделами. Половой ретрактор отходит от диафрагмы единой лентой, которая
тут же распадается на две ветви, одна из которых крепится к проксимальной
части пениса, а другая — к середине 4Х. Проток семяприемника, лишенный
дивертикула, прямой или немного извит, заканчивается слабо отграничен-
ным удлиненным резервуаром, прилегающим к нижнему краю белковой
железы. i
Распространение и местообитание. Северные области Си-
рии и Ирака (Курдистан), северный Иран, северо-восточная и восточная Тур-
ция, Закавказье, центральная и восточная часть Северного Кавказа. Населяет
степные участки, часто образует плотные колонии. В засуху моллюски массами
поднимаются на стебли трав и колючих полукустарников.
8. Род THOANTEUS Lindholm, 1925 '
Lindholm, 1925а : 28 [Zebrina (Zarina), секция Thoanteus]; Thiele, 1931 : 520
[Zebrina (Zebrina}, секция Thoanteus]', Wen z, Zilch, <n'9—1960 : 189 [Zebrina (Thoanteus}];
Шилейко, 1978г : 848.
Типовой вид Bulimus gibber Krynick, 1833; по первоначальному обо-
значению.
Раковина удлиненная, овально-коническая. Скульптура вялая, присут-
ствуют как радиальные, так и спиральные скульптурные элементы. Окраска
белая, почти монотонная, но верхние обороты темнее нижних, рыжеватые или
светло-коричневые. Устье без зубов.
Эпифаллус и проток семяприемника короткие. Цэкум сильно сдвинут к бичу.
Ах и А2 слиты в единый цилиндрический отросток, от базальной части которого
отходят остальные отделы аппендикса. Призмоконические бугорки очень ясные,
разбросанные. Папилла бугорчатая, длинная. Дивертикул семяприемника име-
ется.
Монотипический род.
Thoanteus gibber (Krynicki, 1833) (рис. 264, 265).
Krynicki, 1833 : 416, t. 8, fig. 6a—e (Bulimus gibler, laps, cal.); 1837 : 53 (B. gibber};
Rossmaessler, 1837, Iconogr., I, 6 : 47, fig. 389 (B. revolutus); Bourguignat,
1855 : 115, t. 15, fig. 3 (Buliminus phorcus); Westerlund, 1887 : 32 (Buliminus); К o-
b e It, 1902 : 465, t. 77, fig. 19—22 [B. (Napaeus?)]; Пузанов, 1925a : 84, рис. 5 [2?. (Pet-
8. РОД THOANTEUS
359
raews)]; Лихарев, Раммельмейер, 1952 : 186, рис. 96 (Zebrina); Hudec, 1972а :
102, Abb. 3, 7; Арутюнова, 1975 : 104, рис. 1Б; Шил ейк о, 1978г : 836, рис. 2, I.
Terra typica — горный Крым, «между Мердвенью и дер. Скелыо».
Лектотип хранится в Зоологическом ин-те АН СССР (место сбора —
«Крым»), обозначается здесь.
Материал: свыше 100 экз. из 36 местонахождений в горной части Крыма.
Раковина левозавитая, овально-коническая, верхняя часть завитка имеет
строго конические или слегка вогнутые очертания. Оборотов 7—8, умеренно
выпуклых, разделенных узким швом. Обороты правильно и равномерно нара-
стают, высота последнего оборота приблизительно равна половине высоты ра-
ковины. К устью последний оборот не поднят. Окраска белая или сероватая,
верхние обороты светло-коричневые или рыжеватые; на некоторых раковинах
заметны тонкие размытые и очень слабые радиальные полосы. Эмбриональные
обороты гладкие, блестящие; при увеличении порядка 100 раз видно, что они
покрыты скульптурой в виде неясной пунктировки, бугорков и складочек.
Скульптура дефинитивных оборотов представлена нерав-
номерной слабой морщинистостью и тонкими волнистыми
спиральными бороздками. Устье овальное, немного ко-
сое, края его заметно сближены, париетальная мозоль
слабая или отсутствует. Края устья незначительно отвер-
нуты, за исключением колумеллярного. Над узким
щелевидным пупком нависает отворот колумеллярного
края.
Размеры: ВР 19—22, БД 7—9 мм; в первоописании:
ВР 9.5, БД 4х/4 линии, лектотип: ВР 19.2, БД 8.2 мм.
В. А. Николаев нашел на горе Кош-Кая в Красно-
лесье близ Симферополя несколько правозавитых осо-
бей, 3 из которых он любезно передал мне; в настоя-
Рис. 264. Thoanteus gibber (Кгуп.) из Крыма, Чатыр-даг.
щее время они хранятся в Зоологическом ин-те АН СССР. Внутреннее строение
правозавитых особей оказалось идентичным таковому левозавитых.
В н у гр еннее строение. Материал: 4 экз. из Чатыр-дага, 17 ав-
густа 1891 г., сбор А. Силантьева, определение В. А. Линдгольма; 2 — из окр.
Краснолесья близ Симферополя, гора Кош-Кая, 19 июля 1978 г., сбор В. А. Ни-
колаева (правозавитые особи); 2 — из Чатыр-дага, скалы над с. Перевальным,
1 августа 1962 г.; сбор и определения автора.
Белковая железа с нерезкой апикальной вырезкой. Нижний отдел вагины
почти отсутствует, так как проток семяприемника отходит очень низко. Семя-
провод впадает в эпифаллус сбоку на заметном расстоянии от его конца, так что
остается хорошо выраженный короткий конический бич. Чуть ниже моста впа-
дения семяпровода располагается округлый цэкум. Эпифаллус немного длин-
нее вздутого мешковидного или веретеновидного пениса, внутренняя поверх-
ность которого несет сильно развитые беспорядочно расположенные призмоко-
нические бугорки. Папилла массивная, на ее поверхности хорошо заметна, уз-
кая глубокая борозда, доходящая почти до конца папиллы. Строение пениаль-
ного аппендикса необычайно своеобразно. 4Х и А2 слиты в единую слепо замкну-
тую и закругленную на конце трубку. А3 в виде длинного тонкого булавовид-
ного придатка отходит от нижней части Ах+А2. Л4 и Л5 размером и формой на-
поминают А3 и отходят от его верхнего конца. Одна ветвь полового ретрактора
крепится к проксимальному концу пениса, вторая — терминально к Л, | /1а.
Проток семяприемника толстый, сильно мускулизованный, почти прямой. Ди-
вертикул несколько короче протока семяприемника, резервуар объемистый,
360
VI. НАДСЕМ. BULIMINOIDEA
мешковидный, с собственным толстостенным протоком. Дивертикул, проток
резервуара и резервуар животного, изображенного на рис. 265, содержат сперма-
тофор.
Распространение и местообитание. Встречается отдель-
ными колониями на скалах и в осыпях горной части Крыма.
Рис. 265. Thoanteus gibber (Kryn.) из Крыма, Чатыр-даг, 17 VIII 1891 г. Половой аппарат и
детали его строения (из: Шилейко, 1978 г.).
9. Род RAMUSCULUS Lindholm, 1925
Lindholm, 1925а : 28 [Zebrina (Zebrina'), секция Ramusculus]; Thiele, 1931 : 520
[Zebrina (Zebrina), секция Ramusculus]; Wenz, Zilch, 1959—1960 : 189 [Zebrina (Ramus-
culus)]; Шилейко, 1978г : 848.
Типовой вид Bulimus subulatus Rossmaessler, 1837; по первоначальному
обозначению.
Раковина тонкая, стройная, цилиндрическая, тонко скульптурированная ра-
диальными морщинками. Окраска белая, с темными радиальными пестринами.
Устье без зубов.
Эпифаллус и проток семяприемника умеренно длинные. Цэкум слегка сдви-
нут к пенису относительно середины эпифаллуса. Пениальный аппендикс обыч-
ного строения. Призмоконические бугорки на внутренней поверхности малень-
кие, редкие, выражены только в дистальной части пениса. Папилла стержне-
видная, гладкостенная. Дивертикул семяприемника хорошо развит.
Монотипический род.
Ramusculus subulatus (Rossmaessler, 1837) (рис. 266, 267).
Rossmaessler, 1837, Iconogr., 1,6 : 48, fig. 393 (Bulimus)-, Retowsk i, 1883a : 21
[Buliminus (Zebrina)]-, Westerlund, 1887:26 (Buliminus)-, Kobelt, 1902:903,
t. 128, figs. 24—26 (B. (Brephulus)]; Пузанов, 1926 : 96, рис. 3,. 31 [Buliminus (Zebrina)];
Лихарев, Раммельмейер, 1952 : 185, рис. 95 (Zebrina); Hudec, 1972b : 101,
Abb. 1, 5 [Zebrina (Ramusculus)]; Шилейко, 1978г : 838, рис. 3, I.
9. РОД RAMUSCULUS
361
Terra typica — «? wahrscheinlich Taurien» («? вероятно, Крым»).
Типы хранятся, вероятно, в Зенкенбергском музее, Франкфурт-на-Майне.
Материал:' более 170 экз. из 8 местонахождений в Крыму.
Раковина очень стройная, цилиндрическая, тонкостенная, слегка блестя-
щая. Оборотов 8—11, верхние слегка выпуклые, нижние уплощенные; 3—4 по-
следних оборота одинаковой ширины. Последний оборот к устью не поднят,
высота его составляет приблизительно У3 высоты раковины. Окраска белая,
светло-серая или светло-роговая, с более или менее частыми темными радиаль-
ными пестринами. Иногда эти пестрины развиты настолько интенсивно, что
создается впечатление, что раковина темная со светлыми штрихами. Поверх-
Рис. 266. Ramusculus su-
bulatus (Rssm.) из Крыма,
яйла Чатыр-дага.
Рис. 267. Ramusculus subulatus (Rssm.) из Крыма, яйла Ча-
тыр-дага над с. Перевальным, 28 VII 1962 г. Половой аппарат
и вскрытый пенис (из: Шилейко, 1978 г.).
ность раковины покрыта вялыми неравномерно расположенными радиальными
морщинками; спиральные бороздки отсутствуют, лишь на некоторых ракови-
нах можно с трудом заметить чрезвычайно слабые следы этих бороздок. Устье
маленькое, овальное, косое, края его хрупкие, ломкие, не отвернуты, кроме 'ко-
лумеллярного, который почти полностью прикрывает короткощелевидный пу-
пок.
Размеры: ВР 9—14, БД 2.1—3 мм; в первоописании: ВР 4.5—5, БД 1 линия,
т. е. ВР 10—10.5, БД около 2.1 мм.
Внутреннее строение. Материал: 7 экз. из яйлыЧатыр-дага, над
с. Перевальным, 28 июля 1962 г., сбор и определение автора.
Белковая железа с очень вялой апикальной вырезкой. Вагина длинная, ее
нижняя часть несколько короче верхней. Бич хорошо выражен, короткий, ко-
нический, притупленный. Эпифаллус не извит, равномерно цилиндрический,
ясно выраженный округлый цэкум располагается близ середины эпифаллуса.
Пенис очень тонкий, граница его с эпифаллусом отмечена только местом прикреп-
ления полового ретрактора. Внутренняя поверхность пениса выглядит гладкой,
но в дистальной части можно рассмотреть маленькие разбросанные призмоко-
нические бугорки. Слабое развитие бугорков, характерных для всех членов под-
семейства, связано, вероятно, с малыми размерами животного. Папилла пениса
362
VI. НАДСЕМ. BULIMINOIDEA
стержневидная, с ясно заметной неглубокой бороздой. Пениальный аппендикс
хорошо развит, особенно ясно выражен As, который у других видов часто мало-
заметен. Пениальная ветвь полового ретрактора крепится к проксимальному
отделу пениса на границе его с эпифаллусом, ветвь аппендикса крепится к Аг
немного выше его середины. Проток семяприемника извит, резервуар небольшой,
дивертикул немного не доходит до белковой железы.
Распространение и местообитание. Горный Крым. Оби-
тает в трещинах скал, под известняковыми плитами. Благодаря тому что мол-
люски населяют укрытые биотопы, они образуют порой очень плотные изоли-
рованные колонии, плотность в которых достигает несколько сотен особей
на 1 м2.
10. Род CHONDRULA Beck, 1837
Н. Beck, 1837 : 87; Charpentier, 1837 : 15 (Eucora, non Hiibner, 1816); Held,
1837 : 918 (Gonodon, типовой вид Helix tridens Muller, 1774); Kimakowicz, 1890 : 228
(D'entistomus, типовой вид Helix tridens Muller, 1774); Lindholm, 1925a : 29 (Jaminia,
секция Chondrula)’, Thiele, 1931 : 518 (Jaminia, секция Chondrula)’, Hesse, 1933 : 153;
Лихарев, Раммельмейер, 1952 : 173; Wen z, Zilch, 1959—1960 : 179; Стек-
лов, 1966 : 181; Акрамовский, 1976 : 152; Шилейко, 1978г : 848.
Типовой вид Helix tridens Muller, 1774; по обозначению Мартенса
(Е. Martens, in: Albers, 1860, S. 236).
Раковина от удлиненно-овальной до почти цилиндрической, поверхность
ее покрыта радиальной скульптурой, выраженной в разной степени, но не реб-
ристой. Окраска от жедтовато-роговой до темно-бурой, одноцветная. Устье без
зубов или имеется 1—6 зубов поверхностного типа.
Эпифаллус умеренно длинный, проток семяприемника различной длины.
Цэкум занимает срединную часть эпифаллуса. Пениальный аппендикс, отсут-
ствует. Призмоконические бугорки внутри пениса хорошо выражены, раз-
бросанные. Папилла стёржневидная или в виде желоба, бугорчатая. Диверти-
кул семяприемника нормально развит.
Распространение. Европа, Средиземноморье, Кавказ. Около 15 ви-
дов.
В ископаемом состоянии представители рода известны начиная с нижнего
плиоцена (средняя и восточная Европа, Северная Африка, Предкавказье).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ
И | ВИДОВ CHOND HULA
1 (6). В устье не менее трех зубов, один или два из них занимают париеталь-
ную стенку (подрод Chondrula s. str.).
2 (3). Вершины колумеллярного и палатального зубов находятся почти на
одной горизонтальной линии.............3. Ch. (Chondrula) microtraga.
3 (2). Колумеллярный зуб находится заметно ниже палатального.
4 (5). Гребень париетальной пластинки направлен отвесно вниз или слегка
отклонен к палатальной стенке устья . . . 2. Ch. (Chondrula) tridens.
5 (4). Гребень париетальной пластинки отклонен к колумеллярной стенке
устья ...................................1. Ch. (Chondrula) sunzhica.
6 (1). В устье не более двух зубов, на париетальной стенке зубов нет, лишь
иногда имеется париетальный бугорок.
7 (8). Устье с двумя бугорковидными округлыми зубами — колумеллярный
и палатальным (подрод Eubrephulws) ...............................
................................. 4. Ch. (Eubrephulus) bicallosa.
8 (7). Устье без зубов, лишь иногда имеется ангулярный бугорок (подрод
Mastus).
9 (10). Края устья широко отвернуты. Вид обитает на Кавказе.......
..................................... 5. Ch. (Mastus) caucasica.
10 (9). Края устья очень слабо отвернуты. Вид обитает на Карпатах . . .
.............................................. 6. Ch. (Mastus) bielzi.
10. РОД CHONDRULA
363
1. Подрод CHONDRULA s. str.
Устье раковины с 3—6 зубами.
Половой ретрактор крепится к срединной части пениса.
Распространение. Европа, восточное Средиземноморье, Кавказ.
4—6 видов.
В ископаемом состоянии подрод известен начиная со среднего плиоцена (Ев-
ропа, Предкавказье).
1. Chondrula (Chondrula) sunzhica Steklov, 1962 (рис. 268).
Стеклов, 19626 : 141, табл. II, фиг. 3, 4 [Ch. (Chondrula) microtraga sunzhica}', 1966 :
187, рис. 68, табл. VI, 114, 115 [Ch. (Chondrula) microtraga sunzhica].
Terra typica — Акчагыльские и «подакчагыльские» отложения р. Сунжа у
с. Алды, Предкавказье.
Голотип хранится в Палеонтологическом ин-те АН СССР.
Материал: 16 экз. из трех местонахождений в восточной Грузии.
Раковина удлиненно овально-коническая, иногда почти цилиндрическая,
умеренно твердостенная, слабоблестящая. Оборотов 7—8, слабовыпуклых,
медленно и правильно нарастаю-
щих; последний оборот прямой,
высота его приблизительно рав-
на половине высоты раковины.
Окраска свежих раковин коричне-
вых тонов, губа и зубы ярко белые,
снаружи губа видна в виде светлой
Рис. 268. Chondrula (Chondrula) sunz-
hica Steklov из Грузии, Кварелский
р-н, с. Греми, гора Чрдили.
I — раковина в нормальном положении; 11 —
то же,. вид сзади, часть последнего оборота
удалена.
полосы. Скульптура дефинитивных оборотов представлена вялыми неравно-
мерно расположенными морщинками; в целом раковина с первого взгляда вы-
глядит почти гладкой. Устье овальное, края его утолщены и слегка отвернуты.
В устье 6 зубов. Ангулярный бугорок хорошо развит, короткогребневидный,
иногда с прогибом посредине; относительно плоскости устья он расположен
немного косо и соединяет место прикрепления устья с передним концом парие-
тальной пластинки, расположенным немного отступя от плоскости устья.
Париетальная пластинка высокая, изогнутая, кромка ее утолщена и отогнута
в сторону столбика. Верхняя колумеллярная пластинка расположена глубоко,
но хорошо видна при нормальном положении раковины; она почти вертикально
поднимается по столбику и быстро, но плавно сходит на нет. Нижняя колумел-
лярная пластинка мощная, горизонтальная, сидит вплотную к базальному краю.
Нижняя палатальная складка подобна короткому гребню, расположена под
углом к плоскости устья. Верхняя палатальная складка маленькая, приострен-
ная. Пупок в виде короткой изогнутой щели.
Размеры: ВР 11—13.5, БД 5—5.5 мм; голотип: ВР 11.9, БД 5.0 мм.
Неясно, почему Стеклов рассматривает данный вид как разновидность
Ch. microtraga-. он сам указывает, что «некоторые разновидности из группы
Ch. tridens легко могут быть смешаны с описываемой формой» (Стеклов, 1966,
с. 188). Действительно, раковины Ch. sunzhica гораздо больше похожи на
Ch. trtdens, чем на Ch. microtraga.
364
VI. НАДСЕМ. BULIMINOIDEA
Распространение и местообитание. Вид описан из плио-
ценовых отложений Предкавказья на р. Сунжа; найден М. Г. Нацвлишвили
в рецентном состоянии в трех местах восточной Грузии: Телавский р-н, с. Ал-
вани; Кварелский р-н, с. Енисели-Тарсагалавани и с. Греми (гора Чрдили).
Моллюски обнаружены в почве, среди кустарников.
2. Chondrula (Chondrula). tri dens (Muller, 1774) (рис. 269, I~ IV', рис. 270).
Muller, 1774 : 106 (Helix); L. Pfeiffer, 1848b : 129 (Buliminus albolimbatus),
240 (B. Bayeri); Moquin-Tandon, 1855:297, pl. 21, figs. 25—30 [B. (Chondrula)];
I s s e 1, 1865:421 (B. tridens var. attenuatus); 0. Boettger, 1879:23 [5. (Chondrula)
tridens var. Kubanensis]; С 1 e s s i n, 1879, Malak. Bl., N. F., I : 7, t. I, fig. 5 (Chondrus gali-
Рис. 269. Виды рода Chondrula (подрод Chondrula s. str.).
I— Ch. (Chondrula) tridens (Mull.) из Грузии, с. Самадло; II — то же, окр. Грозного; III — то же, окр.
Хасавюрта; IV — то же, окр. Симферополя, V — Ch. (Chondrula) microtraga (Bssm.) из Болгарии, Бургас.
ciensis); Е.Martens, 1880 : 147 [Buliminus (Chondrula) tridens var. caucasicus]; 0. Boettger,
1886c : 251, t. 8, fig. 6, a, b [B. (Chondrula) tridens var. major Kryn. f. marcida]; Wester-
lund, 1887 : 38 (Buliminus); Lindholm, 1901 : 172 (tridens var. tenutlabiata); Kobelt,
1902 : 914, t. 129, figs. 21—27; Retowski, 1914 : 304 [B. (Chondrulus) tridens, f. elon-
gata W., f. minor W., var. eximius Bssm., f. marcidus Bttg., var. exiguus et var. terkensis]; Hesse,
1933 : 153, fig. 1; Forcart, 1940 : 229, t. 3, fig. 76; Лихарев, Раммельмейер,
1952 : 173, рис. 85; Clauss, 1965 : 173 (сообщение о левозавитых раковинах); Hudec,
L е z h a w а, 1969 : 105, Abb. 8 [Chondrula (Chondrula) tridens kubanensis]; Шилейко,
1969 : 26—31, рис. 1; Gittenberger et al., 1970 : 57; L о i e k, 1970 : 73—79; H u-
d e c, 1972a : 73; Дамянов, Лихарев, 1975 : 152, фиг. 89, 90; А к p а м о в с к и й,
1976 : 153, рис. 69,. табл. VII, 71; Шилейко, 1978в : 518, рис. 4, I.
Terra typica — Италия.
Место хранения типов не установлено.
Материал: несколько сотен экз. из более чем 300 местонахождений
в европейской части СССР, в Чехословакии, Венгрии, Италии, Болгарии и на
Кавказе. >
Раковина в общем удлиненно-овальная, и, хотя форма ее сильно изменчива,
контур завитка не.бывает строго коническим: обводы его всегда в большей или
меньшей мере выпуклые. Оборотов 6—8, плавно нарастающих, умеренно или
слабо выпуклых. Последний оборот к устью не поднят или поднят едва заметно.
Высота последнего оборота лишь в редких случаях меньше половины высоты
раковины, обычно равна этой величине или немного превышает ее. Окраска
одноцветная, от желтовато-серой до темно-бурой и интенсивно каштановой.
10. РОД CHONDRULA
365
Скульптура в виде радиальных морщинок, выраженных по-разному; спираль-
ные бороздки отсутствуют. Устье овальное, незначительно скошено, места его
прикрепления расставлены. Устьевые края слегка отвернуты, немного утолщены,
тупые. Наиболее полный набор элементов устьевой арматуры составлен 5 зу-
бами. Небольшой ангулярный бугорок, представляющий собой местное вздутие
париетальной мозоли, не связан с коротким пластинчатым париетальным, гре-
бень которого направлен вниз или слегка отклонен к палатальной стенке. Колу-
мёллярный зуб округлый, занимает нижнюю часть колумеллярного края. Пала-
тальный зуб, представляющий собой утолщение губы, развит в различной сте-
пени; выше него имеется супрапалатальный, гораздо более слабый. При редук-
ции устьевого вооружения в первую очередь исчезают ангулярный и супрапала-
Рис. 270. Chondrula (Chondrula) tridens (Miill.) из Чегемского ущелья на Северном Кавказе,
14 V 1970 г.
I — половой аппарат; II — вскрытый пенис; III — фрагмент сперматофора, извлеченный из протока
семяприемника (из: Шилейко, 1978в). . , , , , , j , j
тальный бугорки. Пупок щелевидный, слегка прикрыт отворотом колумелляр-
ного края.
Размеры обычно укладываются в следующие значения: ВР 10—15, БД 4—
6 мм; однако размерные показатели сильно изменчивы: самые мелкие из про-
меренных мною раковин имели ВР 7.7, БД 3.5 мм, эта популяция обитает в уро-
чище Ара-Боран между Чегемским и Баксанским ущельями на Северном Кав-
казе; самая большая из промеренных раковин, собранная на южной окраине
г. Грозного: ВР 25.2, БД 9.1 мм.
Размеры в первоописании: ВР 4.5, БД 12/3 линии, т. е. ВР около 9.5, БД
около 3.5 мм.
Внутреннее строение. Материал: 5 экз. из Еревана, 26 сентября
1975 г., сбор и определение Н. Н. . Акрамовского; 2 — из северной Италии,
окр. Вероны, 6 сентября 1974 г., сбор А. Виктора; 4 — из Хасавюрта, Дагестан,
366
V7. НАДСЕМ. BULIMINOIDEA
2 июля 1968 г.; 5 — из Чегемского ущелья близ водопадов, 14 мая 1970 г.; 6 —
из с. Самадло близ Тбилиси, 25 апреля 1970 г.; 3 — из Закарпатья, Ужгород,
6 сентября 1969 г.; сборы и определения автора.
Белковая железа с плавной апикальной вырезкой. Отделы вагины примерно
равной длины. Бич тонкий, недлинный, конический, часто червеобразно изо-
гнут. Семяпровод впадает в эпифаллус под углом. Эпифаллус не образует рез-
ких изгибов, цэкум всегда располагается выше его середины. Пенис короче эпи-
фаллуса, веретеновидный или булавовидный; дистальный его конец тонкий,
внутри с продольными складками, проксимальный отдел пениса расширен, на
его внутренней поверхности сидят многочисленные бугорки, большей частью
утратившие исходную призмоконическую форму. Массивная папилла занимает
весь проксимальный отдел, бугорчатая, с глубокой узкой бороздой. Половой
ретрактор крепится на границе дистального и проксимального отделов пениса.
Проток семяприемника извит, нередко образует петли. Резервуар с собственным
коротким протоком. Дивертикул почти касается белковой железы.
Распространение и местообитание. По определению
Цильха и Эккеля, континентально-восточноевропейский вид (Zilch, Jaeckel,
1962, S. 106). От юго-западной Франции и Балеарских о-вов на восток до северо-
западного Ирана и Урала; Крым, Кавказ. По атлантическому берегу на север
доходит до Нидерландов; в более восточных областях северная граница распро-
странения примерно совпадает с северной границей степной зоны. Границы
ареала в европейской части СССР нуждаются в уточнении. Один раз вид был
найден на Оке близ Серпухова; на восток доходит до р. Урал. Населяет степные
и полупустынные участки, в засушливое время закапывается в почву.
3. Chondrula (Chondrula) microtraga (Rossmaessler, 1839) (рис. 269, V;
рис. 271).
Rossmaessler, 1839, Iconogr., 2, 3/4 : 30, Fig. 651 (Pupa); Mousson, 1859 : 288;
Forcart, 1940 : 231, t. 3, Fig. 71 (microtraga microtraga, m. tricuspidata); Hudec, 1967 :
19, obr. 2, t. 2, fig. 5; Дамянов, Лихарев, 1975 : 154, фиг. 21 (microtraga microtraga,
m. tricuspidata).
Terra typica — Греция.
Типы хранятся, вероятно, в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: 3 экз. из двух местонахождений в Болгарии.
Раковина удлиненно-овальная, слегка вздутая, твердостенная, последний
оборот составляет около половины высоты раковины. Оборотов 6—7, верхние
более выпуклые, чем нижние. Последний оборот к устью еле заметно поднят.
Окраска одноцветная, серая, желтоватая или роговая. Скульптура в виде тон-
кой неравномерной радиальной морщинистости. Устье немного скошено, ши-
рокоовальное или округлое, места его прикрепления широко расставлены, сое-
динены мозолью. Края устья светлые, умеренно отвернуты и утолщены за счет
невысокой, но широкой губы. Зубов 3—4: маленький ангулярный бугорок
(может отсутствовать), пластинчатый отвесный париетальный; бугорковидный
колумеллярный и мощный палатальный. Отворот колумеллярного края слегка
прикрывает щелевидный пупок.
Размеры: ВР 8—11, БД 3.5—4.5 мм; в первоописании: ВР 4—5, БД 12/3—
13/4 линии, т. е. ВР 8.5—10.5, БД 3.5—3.7 мм.
Внутреннее строение. Материал: 2 экз. из Болгарии, парк в Бур-
гасских минеральных водах, 14 июня 1967 г., сбор и определение И. М. Лиха-
рева.
Белковая железа с нерезкой апикальной вырезкой. Нижний отдел вагины
заметно короче верхнего. Бич тонкий, конический. Цэкум расположен заметно
ниже середины эпифаллуса. Пенис состоит из цилиндрического дистального и
расширенного проксимального отделов; дистальный внутри несет продольные
складки; внутренняя поверхность проксимального отдела покрыта разбросан-
ными четкими призмоконическими бугорками. Папилла длинная, бугорчатая,
10. РОД CHONDRULA
367
на конце тонкая. Борозда на поверхности папиллы широкая, напротив борозды
располагается высокий гребень, который на интактном пенисе способен запи-
рать отверстие эпифаллуса, заполняя собой борозду. Проток семяприемника
недлинный, зато собственный проток резервуара относительно длинный и тон-
кий; дивертикул очень длинный, в нижней части образует 2—3 крутых изгиба,
верхний его конец накладывается на белковую железу.
Рис. 271. Chondrula (Chondrula) microtraga (Rssm.) из Болгарии, парк в Бургасских минераль-
ных водах, 14 VI 1967 г. Половой аппарат и вскрытый пенис.
Распространение и местообитание. Балканский"?п-ов,
Румыния, западная часть Малой Азии. В СССР не найден, но не исключено его
присутствие близ границы с Турцией. Населяет главным образом горное ред-
колесье, живет в лесной подстилке, под корнями кустарников и в дерновинках
трав.
2. Подрод EUBREPHULUS Wagner, 1927
A. Wagner, 1927 : 310 [Brephulus (Eubrephulus)]; W е n z, Zilch, 1959—1960 : 180
(род Eubrephulus)-, Дамянов, Лихарев, 1975 : 159 (род Eubrephulus); Шилейко,
1978г : 849.
Типовой вид Bulimus bicallosus L. Pfeiffer, 1847; по обозначению Фор-
карта (Forcart, 1940).
В устье раковины 2 зуба — колумеллярный и палатальный.
Половой ретрактор крепится к границе пениса и эпифаллуса.
Распространение. Балканский п-ов, Северная Африка и Перед-
няя Азия. 2—3 вида.
4. Chondrula (Eubrephulus) bicallosa (L. Pfeiffer, 1847) (рис. 272, 273).
L. Pfeiffer, 1847b : 192 (Bulimus); Kobelt, 1902 : 433 (Buliminus); Wagner,
1927 : 310, t. 15, fig. 119, t. 18, fig. 154; Forcart, 1940 : 250 (Eubrephulus); Pinter I.,
Pinter L.,. 1970 : 89, fig. 2 (Eubrephulus); Hudec, Vasatko, 1973 : 5, Abb. 2, pl. I,
fig. 1 (Eubrephulus); Дамянов, Лихарев, 1975:160, фиг. 95, 96 (Eubrephulus);
Шилейко, 1978в : 518, рис. 4, П.
368
VI. НАДСЕМ. BULIMINOIDEA
Terra typica — «Habitat prope Szlivno» (Болгария, Сливен).
Место хранения типов не установлено.
Материал: 3 экз. из двух местонахождений в Болгарии.
Раковина от удлиненно-конической до веретеновидной, твердостенная,
слегка блестящая, с узко закругленной вершиной. Оборотов 10—11, плавно
нарастающих, слабовыпуклых. Последний оборот к устью слегка поднят, вы-
сота его составляет значительно меньше половины высоты раковины. Окраска
одноцветно роговая. Скульптура в виде тонкой радиальной морщинистости,
местами имеются почти гладкие участки. Устье овальное, с широко отвернутыми
краями. На колумеллярном крае залегает хорошо развитая валикообразная
Рис. 272. Chondrula
(Eubrephuliis) bicallosa
(L. Pfr.) из Болгарии,
горы Странджа.
Рис. 273. Chondrula (Eubrephulus) bicallosa (L. Р£г.) из Болгарии,
горы Странджа, 15 VI 1967 г. Половой аппарат и вскрытый пе-
нис (из: Шилейко, 1978в).
пластинка, уходящая вверх по столбику. На палатальном крае, немного выше
его середины, залегает высокий бугорковидный зуб. Пупок узкий, проколо-
видный.
Размеры: ВР 16—21, БД 4.8—5 мм; в первоописании: ВР 20—21, БД 5 мм.
Внутреннее строение. Материал: 2 экз. из Странджи, Болгария,
15 июня 1967 г., сбор и. определение И. М. Лихарева.
Белковая железа с очень вялой апикальной вырезкой. Отделы вагины при-
мерно равной длины. Семяпровод впадает не под углом, но составляет естествен-
ное продолжение эпифаллуса. Нерезко выраженный цэкум расположен близ
середины неизвитого цилиндрического эпифаллуса. Пенис булавовидный, вну-
тренняя его поверхность усеяна неплотно прижатыми один к другому крупными
бугорками, форма которых не имеет геометрической правильности. Папилла
стержневидная, бугорчатая, массивная; по большей части ее длины проходит
неглубокая широкая борозда. Половой ретрактор крепится к проксимальному
отделу пениса у места перехода его в эпифаллус. Проток семяприемника обра-
зует петлю, длинный; овальный резервуар с собственным протоком, длина
которого почти равна длине самого резервуара. Дивертикул очень длинный,
верхний конец его накладывается на белковую железу.
10. РОД CHONDRULA
369
Распространение и местообитание. Юго-восток Болга-
рии, турецкая часть Фракии и северо-западное побережье Малой Азии. В СССР
вид не найден, но не исключено его присутствие близ границы с Турцией. Насе-
ляет открытые склоны и редкие дубовые леса при умеренной влажности. Живет
в почве, в лиственном опаде, в трещинах скал.
3. Подрод MASTUS Beck, 1837
Н. Beck, 1837 : 78; Hesse, 1933 : 155 (род Mastus); F о г с a г t, 1940: 241; Л и х а-
рев, Раммельмейер, 1952 : 175 (род Mastus); Wen z, Zilch, 1959—1960 : 157
(род Mastus); Даиядов, Лихарев, 1975 : 157 (род Mastus), см. также Hudec,
1972d : 324, Abb. 1, 2.
Типовой вид Helix pupa Linnaeus, 1758; по обозначению Геррманнсена
(Herrmannsen, 1847, S. 24).
Устье без зубов или со слабым ангулярным бугорком.
Половой ретрактор крепится к срединной или проксимальной части пениса.
Распространение. Восточное Средиземноморье, Карпаты, запад-
ный Кавказ. Около 10 видов.
В ископаемом состоянии подрод известен начиная с нижнего плиоцена (сред-
няя и восточная Европа, Северная Африка).
5. Chondrula (Mastus) caucasica (L. Pfeiffer, 1854) (рис. 274, II; рис. 275).
L. P f e i f f e r, in: M ar t i ni, Ch emni t z, 1854, Conch.-Cab. : 121, t. 36, fig. 14,
15 (Bulimus); Kobelt, 1880, Iconogr., VII : 51, fig. 2011 (Buliminus); O. Boettger,
1883 : 175 [B. (Ena)]; Retowski, 1887 : 28, t. I, fig. 9 [B. (Petraeus) ponticus]; Wester-
lund, 1887 :-30 (Buliminus); Kobelt, 1902 : 456 [B. (Napaeus) ponticus]; Rosen, 1914 :
183 [B. (Napaeus)]; Hesse, 1933 : 156, fig. 3 (Mastus); Лихарев, Раммельмейер,
1952 : 175, рис. 87 (Mastus).
Terra typica — Кавказ.
Место хранения типов не установлено.
Материал: 26 экз. из Нового Афона (Абхазия).
Раковина высококоническая, почти башневидная, тонкостенная, блестящая,
просвечивающая. Вершина узкая, но не острая. Оборотов 7—8, почти плоских,
разделенных мелким светлым швом. Последний оборот к устью почти не поднят,
высота его приблизительно равна половине высоты раковины. Окраска одно-
цветно роговая. Поверхность раковины тонко и густо радиально исчерчена, но
при рассматривании невооруженным глазом выглядит гладкой, слегка шелко-
вистой. Устье овально-четырехугольное, умеренно косое, места его прикреплег
ния слегка сближены, соединены мозолью, слегка утолщенной на концах. Края
устья острые, тонкие, светлые, широко отвернуты. Палатальный край в верх-
ней части отклонен вправо, затем образует широкую дугу, причем при переходе
палатального края в базальный имеется намек на слабый угол, затем переходит
в колумеллярный край, где угол явственный. Колумеллярный край отвесный,
на его внутреннем крае, в глубине устья, иногда заметна вялая валикообраз-
ная складочка. Пупок открытый, широкощелевидный.
Размеры: ВР 17—20, БД 6—7.5 мм; в первоописании: ВР 18, БД 7 мм.
Внутреннее строение мною не исследовано, описание составлено
по материалам Гессе (Hesse, 1933) и Форкарта (Forcart, 1940), в работе которого
помещены изображения просветленных препаратов гениталий близкого вида —
Ch. rossmaessleri (L. Pfeiffer).
Белковая железа с развитой в разной степени апикальной вырезкой. Отделы
вагины примерно равной длины или нижний немного короче верхнего. Бич,ко-
роткий, конический. Цэкум хорошо заметен, расположен примерно посредине
цилиндрического эпифаллуса. Внутреннее строение пениса неизвестно, но
у Ch. rossmaessleri ясно видна папилла, занимающая проксимальный отдел пе-
ниса, и бугорки на внутренней поверхности пениального чехла (Forcart, 1940,
S. 243, Abb; 12); у типового вида рода — Ch. pupa, согласно Гессе (Hesse, 1933,
24 А- Шилейко
370
VI. НАДСЕМ. BULIMINOIDEA
S. 156, fig. 2), имеется хорошо развитая удлиненная коническая папилла с очень
неровной поверхностью и широкой бороздой на поверхности. Половой ретрак-
тор крепится на границе дистального и проксимального отделов пениса. Проток
семяприемника прямой, овальный резервуар с собственным протоком, длина
которого примерно равна длине резервуара. Дивертикул немногим длиннее
резервуара с его протоком, значительно не доходит до белковой железы.
Рис. 274. Виды рода Chondrula (под-
род Mastus).
I — Ch. (Mastus) bielzi (Kim.) из Ясинской
котловины, Закарпатская обл.; II — Ch.
(Masius) caueasica (L. Р£г.) из Нового Афона.
Рис. 275. Chondrula (Mastus) caueasica (L. PI’r.)
из Нового Афона. Половой аппарат (из: Hesse,
1933).
Распространение. В первоописании местонахождение указано
неопределенно — Кавказ. Остальные находки сделаны у Нового Афона (Аб-
хазия), последняя — в 1912 г. Линдгольмом; кроме того, одна раковина най-
дена в морских выбросах на крымском побережье между Феодосией и Судаком.
Не исключено, что в настоящее время этот вид принадлежит к числу исчезнув-
ших в связи с разрушением природных биотопов.
6. Chondrula (Mastus) bielzi (Kimakowicz, 1890) (рис. 274, I).
Kimakowicz, 1890:88 [Dentistomus (Amphitrorsus)]; Kobelt, 1899, Iconogr.,
N. F., 9 : 19, fig. 1062 [Buliminus (Dentistomus)]; 1902 : 699, t. 104, fig. 25, 26 [B. (Dentisto-
mus)]; G г о s s u, 1955 : 228, fig. 117 (Mastus); Пелевина, 1959 : 67 (Mastus); Loiek,
1964 : 226, t. 11, fig. 3 (Mastus).
Terra typica — «Siebenbiirgen» (Трансильвания, западная Румыния).
Место хранения типов не установлено.
Материал: 8 экз. из трех местонахождений в Румынии и Закарпатье.
Раковина коническая, слабо блестящая, умеренно тонкостенная, с узкой
закругленной вершиной. Оборотов 7—8, слабовыпуклых. Последний оборот
к устью почти не поднят, высота его приблизительно равна половине высоты
раковины. Окраска одноцветно роговая. Скульптура в виде грубой и неравно-
мерной радиальной морщинистости. Устье овальное, слабо скошено, места его
прикрепления расставлены, соединены тонкой мозолью, на которой в ангуляр-
ной области может присутствовать слабый бугорок. Края устья слегка отвер-
1. РОД MULTIDENTULA
371
нуты и слабо утолщены, отступя от края устья имеется широкая сильно раз-
мытая губа. Палатальный и базальный края образуют плавную дугу; при пере-
ходе в отвесный колумеллярный край заметен закругленный угол. Пупок почти
проколовидный.
Размеры: ВР 16—24, БД 7—10 мм; в первоописании: ВР 22, БД 10 мм.
Распространение и местообитание. Лесистые Карпаты
(Закарпатская обл.) и Трансильвания (западная Румыния). А. А. Полевина
(1959) указывает присутствие вида в Закарпатской обл., не называя точно места
находок; в Зоологическом ин-те АН СССР хранится ее сбор с этикеткой: «Ра-
ховский район, с. Кузя»; там же хранятся 2 экз., определенные И. М. Лихаре-
вым, с этикеткой: «Закарпатье, Ясинская котловина, в орешнике, под камнем,
12 августа 1959 г., сбор С. Высоцкой». Моллюски встречаются в горных широко-
лиственных лесах.
VIII. Подсем. MULTIDENTULINAE Schileyko, 1978
Шилейко, 1978г : 846.
Раковина слабо скульптурйрована. В устье не менее 6 зубов поверхностного
типа. Эмбриональные обороты гладкие.
Бич отсутствует, цэкум хорошо развит. Внутренняя поверхность пениса глад-
кая. Имеется замкнутая пениальная папилла, проксимальный вырост пениса
отсутствует. Дивертикул семяприемника и пениальный аппендикс могут отсут-
ствовать, хотя исходно имеются.
Распространение. Балканский п-ов, Передняя и Малая Азия,
Кавказ. 3 рода.
В ископаемом состоянии подсемейство известно начиная с верхнего плиоцена
(Румыния, Предкавказье).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ MULTIDENTULINAE
1 (2). Дивертикул семяприемника и пениальный аппендикс нормально развиты
............................ . . ............... 1. Multidentula.
2 (1). Либо аппендикс, либо дивертикул отсутствуют.
3 (4). Пениальный аппендикс развит, дивертикул семяприемника отсутствует
.............................................. 2. Improvisa.
4 (3). Пениальный аппендикс отсутствует, дивертикул семяприемника имеется
................................................ . 3. Senaridenta.
1. Род MULTIDENTULA Lindholm, 1925
Lindholm, 1925а : 30 (род Jaminia, секция Chondrula, подсекция Multidentula)',
W е n z, Z i 1 c h, 1959—1960 : 182 [Jaminia (Multidentula)]', Шилейко, 1978г : 846.
Типовой вид Bulimus ovularisOlivier, 1801; по первоначальному обозна-
чению.
Конхологические признаки подсемейства.
Дивертикул семяприемника и пениальный аппендикс хорошо развиты.
Распространение. Балканский п-ов, Передняя и Малая Азия.
Несколько видов.
Форкарт (Forcart, 1940, S. 194) предложил подрод Bollingeria с типовым ви-
дом Chondrus pupoides Krynicki; однако при этом он имел в виду Buliminus la-
melliferus, который достоверно отличается от Ch. pupoides только по анатомиче-
ским признакам. Это обстоятельство было выяснено Гудецом (Hudec, 1972с),
который предложил подродовое название Tokatta nom. .emend, (род Jaminia) и
типовым видом обозначил В. lamelliferus', вместе с тем В. lamelliferus настолько
близок Bulimus ovularis, что трудно сомневаться в .принадлежности этих форм
24*
372
VI. НАДСЕМ. BULIMINOIDEA
одному роду (рис 276, I, II). Следовательно ^названия Bollingeria и Tokatia
есть младшие синонимы названия Multidentula.
I I Ш ИГ
Рис. 276. Виды подсемейства Multidentulinae.
I — Multidentula lamellifera (Rssm.) из окр. с. Ломашен, Турция; II — М. ovularis (Oliv.) из окр. г. Сам-
сун, Турция; III — Improvisa pupoides (Кгуп.) из Чегемского ущелья на Северном Кавказе; IV — Sena-
ridenta naehicevanjensis (Hudec) из окр. Степанакерта, Нагорный Карабах, паратип.
Multidentula lamellifera (Rossmaessler, 1859) (рис. 276, /; -»рис. 277).
Rossmaessler, 1859, Iconogr., Ill : 95, fig. 919 (Buliminus)', Retowski, 1883b :
56, t. 2, fig. 2 [jB. (Chondrula) clessini]; 1887 : 32 [S. (Chondrula) lamelliferusfvax. angustatus]',
I — половой аппарат, II — пенис другого экземпляра (из: Hesse, 1933).
Westerlund, 1887 : 45 (Buliminus)' Kobelt, 1902 : 601, t. 92, figs. 10, 11 (Buliminus)',
Lindholm, 1922 : 358 (Chondrula acutior); Hesse, 1933 : 162, fig. 6 [Jaminia (Multi-
dentula) pupoides part.]; Forcart, 1940 : 194, t. 3, fig. 62 [7. (Bollingeria)pupoides part.];
Haas, 1955 : 434, fig. 103 [7. (Chondrula)].
Terra typica — Сирия.
Типы хранятся, вероятно, в Зенкенбергском музее, Франкфурт-на-Майне.
Материал: 2 экз. из одного местонахождения в Турции.
2. РОД IMPROVISA
373
Раковина от овальной до почти короткоцилиндрической, с широкой закруг-
ленной вершиной, слабо блестящая, твердостенная. Оборотов 6—7, сравнительно
выпуклых. Последний оборот к устью явственно приподнят, высота его не-
сколько превышает половину высоты раковины. Окраска серовато-роговая или
желтоватая, верхние обороты иногда темнее нижних. Скульптура в виде тонкой
неравномерной морщинистости, морщинки расположены довольно редко, раз-
мытые. Устье овальное, места его прикрепления расставлены. Края устья утол-
щенные, тупые, слабо отвернуты. Устьевая арматура составлена следующими
элементами. В ангулярной области имеется небольшой бугорковидный зуб; по
соседству с ним, на середине париетальной стенки располагается зуб в виде ко-
роткой толстой пластинки, плоскость которой почти перпендикулярна плоскости
устья. Колумеллярный зуб развит хорошо, вначале горизонтальный, затем, резко
ослабляясь, поднимается вверх по столбику, Базально-колумеллярное поло-
жение занимает бугорок, отодвинутый в глубь устья сильнее прочих. На пала-
тальной стенке сидят три зуба, самый крупный — нижний, самый маленький —
верхний; последний может отсутствовать. Пупок короткощелевидный.
Размеры: ВР 4—6.5, БД 2.2—4 мм; в первоописании: ВР 6.5, БД 4 мм.
Внутреннее строение. Материал мной не исследован, описание
составлено по материалам Гессе (Hesse, 1933).
Отделы вагины примерно равной длины. Семяпровод впадает строго терми-
нально, ясно выраженный цэкум сдвинут к пенису относительно середины длин-
ного цилиндрического эпифаллуса. Пенис короткий, в проксимальной части
расширен, заключает короткую папиллу. Лх относительно короткий, А2 иногда
не обособлен от Аг. А3 длиннее первых двух отделов, вместе взятых. Л4 сравни-
тельно короткий, очень плавно и постепенно переходит в Л5. Ветви полового ре-
трактора тесно сближены на диафрагме, одна из них крепится к верхней части
Аг, вторая — к пенису возле границы его с эпифаллусом. Проток семяприем-
ника не извит, небольшой овальный резервуар с собственным протоком, длина
которого примерно равна длине резервуара. Дивертикул хорошо развит, длин-
ный.
Распространение и местообитание. Передняя и Малая
Азия. Достоверных находок, подтвержденных анатомическим исследованием,
на территории СССР не сделано (по раковине вид практически неотличим от
Improvisa pupoides), но не исключено его присутствие в Аджарии. Живет на
открытых местах с аридными условиями, под камнями и в осыпях.
2. Род IMPROVISA Schileyko, 1978
Шилейко, 1978г : 846.
Типовой вид Chondrus pupoides (Krynicki, 1833); по первоначальному
обозначению.
Конхологические признаки подсемейства.
Пениальный аппендикс нормально развит, дивертикул семяприемника от-
сутствует.
Распространение. Северный Кавказ и Закавказье. Вероятно, 1 вид.
Improvisa pupoides (Krynicki, 1833) (рис. 276, III; рис. 278).
Krynicki, 1833 : 410 (Chondrus); Mousson, 1863 : 389 (Ch. lamellijerus var. pha-
sianus); Rosen, 1914 : 186 [Buliminus (Chondrulus) lamellijerus var. phasianus], 187 (var.
angustior Ret.); Лихарев, Раммельмейер, 1952 : 176, рис. 88 (Jaminia); Hudec,
Lezhawa, 1969 : 103, Abb. 7, 19 [/. (Bollingeria)); Hudec, 1972c : 211—219 [imparietula
(Bollingeria)]; Акрамовский, 1976 : 157, рис. 72, табл. VII, 73 (Imparietula).
Terra typica — гора Машук близ Пятигорска.
Лектотип хранится в Зоологическом ин-те АН СССР, обозначается
здесь.
374
VI. НАДСЕМ. BULIMINOIDEA
..Материал: несколько сотен экз. из 31 местонахождения на Северном
Кавказе и в Закавказье.
Раковина не имеет постоянных и достоверных отличий от предыдущего вида.
Размеры те же; в первоописании: ВР 23/4, БД 1т/2 линии, т. е. ВР около 4.5,
БД около 3.2 мм; лектотип: ВР 5.4, БД 2,7 мм.
Внутреннее строение. Материал: 3 экз. из Армении, Даралагез,
с. Гнишик, сбор и определение Н. Н. Акрамовского; 3 — из Грузии, с. Бенара,
Рис. 278. Improvisa pupoides (Кгуп.) из Грузии, Аджария, с.'Бенара, левый берег р. Аба-
• стуманки, 10 VI 1949 г. Половой аппарат и вскрытый пенис.
берег р. Абастуманки, 10 июня 1949 г., сбор А. Н. Кириченко; 7 — из Чегем-
ского ущелья на Северном Кавказе, 17 мая 1970 г.; сбор и определения автора.
Белковая железа с широкой плавной апикальной вырезкой. Нижний отдел
вагины в 2—4 раза короче верхнего. Семяпровод очень плавно переходит в от-
носительно длинный цилиндрический эпифаллус; цэкум сильно сдвинут к семя-
проводу, либо располагается немного выше середины эпифаллуса. Пенис от почти
шаровидного до веретеновидного. Папилла толстостенная, иногда с продоль-
ными складками. Ветви полового ретрактора крепятся на диафрагме независимо,
но неподалеку одна от другой. Одна ветвь присоединяется к границе и А2,
вторая — к проксимальной части пениса. Аг иногда почти слит с А2, А3 очень
плавно сужается, переходя в относительно короткий А4. Проток семяприемника
прямой или образует нерезкие изгибы, плавно переходит в резервуар, плотно
прижатый к спермовидукту ниже белковой железы.
Распространение и местообитание. Северный Кавказ и
Закавказье. Населяет открытые склоны или плоскогорья, где встречается в осы-
пях, трещинах скал и под камнями; иногда в редких кустарниковых зарослях,
в почве и среди опада под колючими подушковидными полукустарниками (Ак-
рамовский, 1976).
3. РОД SENARIDENTA
375
3. Род SENARIDENTA Schileyko, 1978
Шилейко, 1978г : 846.
Типовой вид Chondrula nachicevanjensis Hudec, 1972; по первоначаль-
ному обозначению.
Конохологические признаки подсемейства.
Рис. 279. Senaridenta nachicevanjensis (Hudec) из окр. Степанакерта, Нагорный Карабах,
31 VII 1964 г. Паратип.
I — половой аппарат, вскрытый пенис и поперечный разрез папиллы; 77 — эпифаллус, вскрытый в об-
ласти цэкума; III — квадривий.
Пениальный аппендикс отсутствует, дивертикул семяприемника нормально
развит.
Монотипический род.
Senaridenta nachicevanjensis (Hudec, 1972) (рис. 276, IV; рис. 279).
Hudec, 1972с : 217, Abb. 2, 5 [Chondrula (Chondrula)]]
Terra typica — окр. Степанакерта, Нагорно-Карабахская АО.
Голотип хранится в Зенкенбергском музее, Франкфурт-на-Майне,
№ 225190.
Материал: 41 экз. из типовой серии.
Раковина не имеет постоянных и достоверных отличий от двух предыдущих
видов; те особенности, которые приведены в первоописании, основаны на недо-
статочном материале, так как Гудец при описании вида имел в распоряжении
несколько экземпляров из довольно большой серии, переданной мною в Зооло-
гический ин-т АН СССР.
Размеры: ВР 5—6.3, БД 3—3.5 мм; голотип: ВР 6.0, БД 3.3 мм.
376
INCERTAE SEDIS.
Внутреннее строение: И паратипов: Нагорно-Карабахская АО,
окр. Степанакерта, 31 июня 1964 г., сбор Г. М. Овчинникова, определение
В. Гудеца.
Белковая железа с глубокой апикальной вырезкой. Длина нижнего отдела
вагины непостоянна: она может быть почти равной длине верхнего отдела или
быть короче его в 2—4 раза. Семяпровод впадает в эпифаллус терминально или
почти терминально. Цэкум незначительно сдвинут вниз относительно середины
довольно длинного эпифаллуса. Пенис небольшой, почти шаровидный, папилла
округлая, короткая, толстостенная, со щелевидным просветом. Половой ретрак-
тор крепится выше середины пениса. Проток семяприемника длинный, извит или
образует петлю, резервуар с собственным очень коротким протоком. Диверти-
кул длинный, беспорядочно извитой, верхний конец его накладывается на бел-
ковую железу.
Распространение и местообитание. Вид известен только
из типового местонахождения близ Степанакерта (в первоописании ошибочно
указано, что Степанакерт находится в Нахичеванской АССР). Найден на сугли-
нистых почвах в довольно сырых условиях в редком лесу: грабинник, граб, дуб.
. INCERTAE SEDIS
Stenogyra retteri Kobelt, 1904.
Kobelt, 1904 : 87, 1906; Iconogr., N. F., 12 : 11, sp. 1953.
Terra typica — «Aulie-Ata im Syr-Darja-Gebiet», Аулие-Ата, ныне г. Джамбул
КазССР.
Голотип хранится в коллекции О. Розена (Геологический ин-т АзССР,
Баку).
Материала в моем распоряжении не было, здесь приводится несколько
сокращенный перевод первоописания.
Раковина без пупка, почти цилиндрическая, тонкостенная, просвечивающая,
блестящая, гладкая, при сильном увеличении заметна тонкая исчерченность,
янтарно-коричневая. 9 слабовыпуклых оборотов, медленно и постепенно нара-
стающих, последний едва больше предпоследнего, в базальной части слегка
сжат. Устье маленькое, овальное, сверху и снизу слегка сжатое, внутри'окра-
шено так же, как снаружи. Края устья отвесные, практически не отвернуты и
не утолщены, ломкие. Колумеллярный край короткий, едва выгнут.
Размеры: ВР 14, БД 4.25, высота устья 4 мм.
Вероятно, этот своеобразный вид не относится к роду Stenogyra и вообще
к семейству Subtilinidae; скорее всего, это представитель Pupillina, нб отнесение
его к какому-нибудь семейству затруднительно. Такие признаки, как запаян-
ный пупок, гладкая блестящая поверхность и янтарно-коричневая окраска, сбли-
жают вид с членами семейства Cochlicopidae, но раковины Cochlicopidae ни-
когда не бывают таких размеров и число оборотов никогда не превышает у них 6;
кроме того, устьевой край представителей Cochlicopidae слегка утолщен и
никогда не бывает ломким. Размер, форма раковины и число оборотов харак-
терны для Buliminidae, но отнесению вида к этому семейству препятствуют пере-
численные выше «кохликопидные» признаки. Решение о систематическом поло-
жении вида можно будет сделать после новых сборов и исследования внутрен-
него строения животных.
ЛИТЕРАТУРА
Акрамовский Н. Н. Заметка о наземных моллюсках долины р. Гарни-чая у Гегарта (Арме-
ния). — Научи, тр. Ереван, гос. ун-та, 1947а (1943); с. 255—258.
Акрамовский Н. Н. Наземные моллюски лавового плато в окрестностях гор. Еревана. —
Научи, тр. Ереван, гос. ун-та, 19476 (1943), с. 259—273.
Акрамовский Н. И. Новые и мало известные формы наземных раковинных моллюсков из
Вайка (Даралагеза) (Gastropoda, Vertiginidae). — Докл. АН АрмССР, 1947в, т. 7,
с. 179—183.
Акрамовский Н. Н. Наземные моллюски территории селения Гнишик в Советской Армении. —
В кн.: Зоол. сб. Вып. 6. Ереван, 1949, с. 127—183.
Акрамовский Н. Н. Новые и редкие наземные моллюски из Армении (Gastropoda—Stylom-
matophora). — Докл. АН АрмССР, 1955, т. 20, с. 149—153.
Акрамовский Н. Н. Некоторые дополнения к фауне моллюсков Армении, включая новости
для фауны Закавказья и СССР. — В кн.: Моллюски и их роль в экосистемах. Л.,
1968, с. 26—27.
Акрамовский Н. Н. Некоторые итоги изучения современной фауны моллюсков Армении. —
В кн.: Моллюски. Пути, методы и итоги их изучения. Л., 1971а, с. 108—111.
Акрамовский Н. Н. Краткий каталог современной фауны моллюсков Советской Армении. —
Биол. журн. Армении, 19716, т. 24, вып. 6, с. 3—12.
Акрамовский Н. И. Моллюски (Mollusca). — В кн.: Фауна Армянской ССР. Ереван, 1976,
с. 1—268.
Александров Д. А., Сергиевский С. О. Об одном варианте симпатрического видообразования
у брюхоногих моллюсков. — В кн.: Моллюски. Основные результаты их изучения.
Л., 1979, с. 153-154.
Альмухамбетова С. К. Новые виды Pupilloidea с Заилийского Алатау. — Изв. АН КазССР-
Сер. биол., 1979, вып. 3, с. 30—32.
Альмухамбетова С. К., Увалиева К. К. Моллюски семейства Vertiginidae (Mollusca, Gastro-
poda) юга и юго-востока Казахстана. — Изв. АН КазССР. Сер. биол., 1979, вып. 4,
с. 35—40.
Арутюнова Л. Д. Заметка о некоторых наземных моллюсках южнобережья Крыма. —
Биол. журн. Армении, 1975, т. 28, вып. 10, с. 104—109.
Архангельский П. П. К изучению моллюсков УзССР. — В кн.: Материалы Центр, гос. муз.
УзССР. Вып. 1. Самарканд, 1933, с. 1—32.
Асадов С. М., Акрамовский Н. Н., Джаббаров Д. Г. К изучению фауны наземных моллюсков
Малого Кавказа в пределах Азербайджанской ССР. — Изв. АН АзССР. Сер. биол.
наук, 1975, вып. 2, с. 100—106.
Асланян П. М. Конкские моллюски близ г. Раздан (Армянская ССР). — Изв. АН АрмССР.
Науки о Земле. 1971, вып. 4, с. 3—13.
Братчик Р. Я. Метод быстрой фиксации наземных моллюсков. — Зоол. журн., 1976, т. 55,
вып. 7, с. 1078—1079.
Виленкин Б. Я., Шилейко А. А. Способ оценки уровня эндемизма фауны. — Зоол. журн.,
1979, т. 58, вып. И, с. 1720—1727.
Давтян Э. А. Цикл развития Cystocaulus nigrescens. — Тр. Научи.-исслед. ин-та Нарком-
зема АрмССР, 1940, т. 3, с. 5—31.
Дамянов С., Лихарев И. М. Gastropoda terrestria. — В кн.: Фауна на България. Т. IV.
София, 1975, с. 1—425 (болг.).
Здун В. I. До фауни моллюсюв Закарпаття. — Наукови зап. науково-природов. муз.
АН УССР. Т. VIII. Зоология, 1960. с. 83—95.
Иззатуллаев 3. Новые виды наземных моллюсков рода Pupilla Turton, 1831 Таджикистана
(Gastropoda, Pulmonata, Pupillidae). — Докл. АН ТаджССР, 1970а, т. 13, вып. 3,
с. 53—55.
Иззатуллаев 3. Материалы к изучению наземных моллюсков Гиссарского хребта и сопре-
дельных районов Таджикистана. — Изв. АН ТаджССР. Отд-ние библ, наук, 19706,
№ 239, с. 79—86.
Иззатуллаев 3. Малоизвестные наземные моллюски (Mollusca, Gastropoda) фауны Средней
Азии. — Изв. АН ТаджССР. Отд-ние биол. наук, 1975, № 2 (59), с. 39—44.
378
ЛИТЕРАТУРА
Калитииа 3. И. К изучению зонального распределения наземных моллюсков северных скло-
нов Центрального Кавказа и Восточного Предкавказья. — В кн.: Здравоохранение
и медицина в Сев. Осетии. Вып. VII, ч. 1. М.; Л., 1958, с. 157—181.
Кесслер К. Ф. Путешествие с зоологической целью к северному берегу Черного моря и в Крым
в 1858 г. Киев, 1860, с. 1—248.
Крестьянинов Ю. С. Наземный моллюск Gastrocopta theeli (Westerlund, 1877) на Южном
Урале. — В кн.: Вопросы зоологии. Вып. 3. Челябинск, 1973. с. 53—54.
Лившиц Г. М., Шилейко А. А. Жизненный цикл моллюска Brephulopsis bidens. — Экология,
1978, вып. 5, с. 77—83.
Линдгольм В. А. Описание двух видов р. Buliminus из Русской Центральной Азии. —
Ежегод. Зоол. муз., 1922а, т. 23, с. 273—275.
Линдгольм В. А. Заметки о палеарктических наземных и пресноводных моллюсках. —
Ежегод. Зоол. муз., 19226, т. 23, с. 304—320.
Линдгольм В. А. Моллюски. — В кн.: Тр. Памир, экспедиции 1928 г. Т. VIII. 1931, с. 29—64.
Лихарев И. М. Материалы к фауне наземных моллюсков островов Итуруп и Шикотан (Южные
Курильские острова). — Тр. Ленингр. о-ва естествоиспыт., 1957, т. 73, вып. 4,
с. 77—80.
Лихарев И. М. Моллюски клаузилииды. — В кн.: Фауна СССР. Моллюски. Т. 3, вып. 4.
Нов. сер., № 83. М.; Л., 1962, с. 1—317.
Лихарев И. М. К фауне наземных моллюсков Камчатской области. — В кн.: Фауна Камчат-
ской обл., Тр. Камчатск. комплексной экспедиции, 1963, с. 65—81.
Лихарев И. М., Раммельмейер Е. С. Наземные моллюски фауны СССР. — В кн.: Опред.
по фауне СССР. Вып. 43. М.; Л., 1952, с. 1—511.
Лихарев И. М., Старобогатов Я. И. Материалы к фауне моллюсков Афганистана. — Тр.
Зоол. ин-та АН СССР, 1967, т. 42, с. 159—197.
Майр Э. Зоологический вид и эволюция. М., 1968, с. 1—597.
Майр Э. Принципы зоологической систематики. М., 1971, с. 1—454.
Мартенс Э. Слизняки (Mollusca).— В кн.: А. П. Федченко. Путешествие в Туркестан.
Вып. I, т. 2, ч. 1. 1874, с. 1—64.
Матёкин П. В. Фауна наземных моллюсков Нижнего Поволжья и ее значение для представ-
ления об истории современных лесов района. — Зоол. журн., 1950, т. 29, вып. 3,
с. 193—205.
Матёкин П. В. Приспособительная изменчивость и процесс видообразования у среднеазиат-
ских наземных моллюсков семейства Euidae. — Зоол. журн., 1959, т. 33, вып. 10,
с. 1518—1536.
Матёкин П. В. Определитель раковинных моллюсков Средней Азии — промежуточных хо-
зяев гельминтов. — В кн.: Гельминты животных Киргизии и сопредельных террито-
рий. Фрунзе, 1966, с. 97—137.
Матёкин П. В. Наземные моллюски семейства Bradybaenidae Средней Азии. — Сб. тр. Зоол.
муз. Моск. гос. ун-та, 1972, т. 12, с. 112—144.
Миничев Ю. С. Положение Soleolifera в системе Gastropoda. В кн.: Моллюски. Пути, методы
и итоги их изучения. Л., 1971, с. 8—10.
Миничев Ю. С., Старобогатов Я. И. К построению системы эвтиневральных брюхоногих мол-
люсков. В кн.: Моллюски, их система, эволюция и роль в природе. Л., 1975, с. 8—11.
Мухитдинов А. Б. Новые виды наземных моллюсков северного Таджикистана. — Изв.
АН ТаджССР. Отд-ние биол. наук, 1976, № 1 (62), с. 39—43.
Полевина А. А. К фауне наземных моллюсков Закарпатья. — Докл. и сообщ. Ужгор. гос.
ун-та. Сер. биол., 1959, т. 3, с. 65—68.
Пузанов И. И. Материалы к познанию наземных моллюсков Крыма. Ч. I. Моллюски горного
Крыма. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол., 1925а, т; 33, с. 48—104.
Пузанов И. И. Наземные моллюски Симферопольского естественно-исторического музея. —
Зап. Крымск, общ. естествоиспыт. Т. VIII, 19256, с. 112—120.
Пузанов И. И. Материалы к познанию наземных моллюсков Крыма. Ч. II. Моллюски степ-
ного Крыма. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол., 1926, т. 35, с. 84—101.
Рауп Д., Стэнли С. Основы палеонтологии. М., 1974, с. 1—390.
Ретовский О. Описание нового кавказского вида Buliminus (Mollusca Pulmonata). — Ежегодн.
Зоол. муз., 1915, т. 20, с. 45—46.
Розен О. В. Материалы к познанию фауны слизняков Кавказа и Средней Азии. — В кн.:
Дневник Зоол. отд-ния О-ва любит, естествозн., антропол. и этногр., 1901, т. III,
вып. 2, с. 7—12.
Самадов К. С. К изучению малакофауны Ферганской долины. — Тр. Ин-та зоол. и паразитол.
АН УзССР, 1954, т. 3, с. 127—134.
Самадов К. С. К познанию моллюсков Узбекистана. — Учен. зап. Ташкенток, вечерн. пед.
ин-та, 1960, вып. 9, с. 5—26.
Сваджян П. К. Данные о биологии наземных моллюсков — главнейших промежуточных
хозяев ланцетовидного сосальщика в АрмССР. — Изв. АН АрмССР. Биол. и сель-
скохоз. науки, 1953, т. 6, вып. 8, с. 47—61.
Скворцов А. А. К фауне моллюсков Южного Казахстана. — Тр. Каз. науч.-исслед. вет.
ин-та, 1940, т. 3, с. 361—378.
Старобогатов Я. И. Фауна моллюсков и зоогеографическое районирование континентальных
водоемов. Л., 1970, с. 1—372.
ЛИТЕРАТУРА
379
Стеклов А. А. Неогеновые виды кавказского рода Retowskia (Mollusca, Pulmonata). — Па-
леонтол. журн., 1962а, вып. 2, с. 71—75.
Стеклов А. А. Роль наземных брюхоногих моллюсков в стратиграфии неогеновых конти-
нентальных отложений Северного. Кавказа. — В кн.: Геология Центрального и За-
падного Кавказа. Т. 3. М., 19626, с. 141—157.
Стеклов А. А. Наземные моллюски неогена Предкавказья и их стратиграфическое значение. —
Тр. Геол, ин-та АН СССР, 1966, вып. 163, с. 1—262.
Стеклов А. А. Наземные моллюски неогеновых отложений Тувы. — Тр. Зоол. ин-та АН СССР,
1967, т. 42, с. 269—279.
Стеклов А. А. Этапы развития неогеновых наземных моллюсков Предкавказья. — Malaco-
logia, 1968, т. 6 (3), с. 243—251.
Стеклов А. А., Цытович М. В. О находке неогеновых наземных моллюсков в долине Костан-
кол в Центральном Казахстане. — Бюл. Моск, о-ва испыт. природы. Отд-ние геол.,
1967, т. 42, вып. 3, с. 108—119.
Увалиева К. К. Зоогеографический анализ малакофауны Южного Алтая и некоторые во-
просы ее происхождения. — В кн.: Моллюски. Вопросы теоретической и прикладной-ма-
лакологии, 2. Второе совещ. по изуч. моллюсков. М.—Л., 1965, с. 52.
Увалиева К. К. Новые виды наземных моллюсков с Южного Алтая. — Тр. Зоол. ин-та
АН СССР, 1967, т. 42, с. 213-220.
Урбах В. Ю. Биометрические методы. М., 1964, с. 1—416.
Цветков Б. Н. Новые данные о моллюсках Крыма. — Сб. тр. Гос. зоол. муз. при Моск. гос.
ун-те, 1939. т. 5, с. 171—182.
Цветков Б. Н. К познанию фауны моллюсков пастбищ Алма-атинской области. — Тр. Каз.
науч.-исслед. вет. ин-та, 1940, т. 3, с. 379—419.
Шиков Е. В. Моллюски хвойных лесов Валдайской возвышенности й сопредельных террито-
рий. — В кн.: Фауна Верхневолжья, ее охрана и использование. Калинингр. ун-т,
1981, с. 28-45.
Шилейко А. А. О факторах изменчивости некоторых наземных моллюсков. — Вести. Моск,
гос. ун-та. Сер. VI, 1967а, вып. 2, с. 14—21.
Шилейко А. А. Некоторые новые и малоизвестные моллюски Талыша. — Научи, докл. высш,
школы. Биол. науки, 19676, № 4, с. 24—32.
Шилейко А. А. Эколого-фаунистический обзор наземных раковинных моллюсков Даге-
стана. — Вести. Моск. гос. ун-та. Сер. VI, 1969, вып. 1, с. 26—31.
Шилейко А. А. Особенности строения экскреторного аппарата Pulmonata в связи с разработ-
кой системы подкласса. — В кн.: Моллюски, их система, эволюция и роль в природе.
JL, 1975а, с. 12-15. , •
Шилейко А. А. Моллюски подсемейства Lauriinae фауны СССР (Pulmonata, Pupillidae). —
Зоол. журн., 19756, т. 54, вып. 12, с. 1767—1782.
Шилейко А. А. Явление синистральности у брюхоногих моллюсков и его роль в процессе
видообразования. — В кн.: Фауна и ее охрана в республиках Закавказья. Ереван,
1975в, с. 171-173.
Шилейко А. А. Пути эволюции экскреторного аппарата Pulmonata и его филогенетическое
значение. — Зоол. журн., 1976а, т. 55, вып. 2, с. 215—225.
Шилейко А. А. Особенности организации и система Orculidae (Gastropoda). — Научн. докл.
высш, школы. Биол. науки, 19766, № 4, с. 47—58.
Шилейко А. А. Строение и систематическая принадлежность видов рода Siraphorus Lindholm,
1925 (Gastropoda, Enidae). — Научн. докл. высш, школы. Биол. йауки, 1977, № 9,
jz. 40-46.
Шилейко А. А. Наземные моллюски надсемейства Helicoidea. — В кн.: Фауна СССР. Мол-
люски. Т. III, вып. 6. Нов. сер., № 117. Л., 1978а, с. 1—384.
Шилейко А. А. Исследование типовых видов некоторых таксонов родовой группы в семействе
Buliminidae (=Enidae) (Gastropoda). I. Виды Средней и Центральной Азии. — Зоол.
журн., 19786, т. 57, вып. 3, с. 344—358.
Шилейко А. А. Исследование типовых видов некоторых таксонов родовой группы в семействе
Buliminidae I--Enidae) (Gastropoda). II. Виды Европы, Передней и Малой Азии. —
Зоол. журн., 1978в, т. 57, вып. 4, с. 512—522.
Шилейко А. А. Исследование типовых видов некоторых таксонов родовой группы в семействе
Buliminidae (=Enidae) (Gastropoda). III. Виды Крыма и Кавказа. Вопросы системы
семейства. — Зоол. журн., 1978г, т. 57, вып. 6, с. 834—850.
Шилейко А. А. Система и филогения отряда Geophila (=Helicida). —Тр. Зоол. ин-та АН СССР,
1979а, т. 80, с. 44-69.
Шилейко А. А. Вопросы системы и филогении подотряда Pupillina (Gastropoda : Geophila). —
В кн.: Моллюски. Основные результаты их изучения. Л., 19796, с. 14—16.
Шилейко А. А., Иззатуллаев 3. Таксономическая структура наземных моллюсков семейства
Pupillidae фауны СССР и описание нового вида из Средней Азии. — Докл. АН ТаджССР,
1980, т. 23, вып. 5, с. 282—285.
Шилейко Л. В., Шилейко А. А. К морфологии проксимального отдела полового аппарата
Stylommatophora (Gastropoda, Pulmonata). — Научн. докл. высш, школы. Виол,
науки, 1975, № 1, с. 7—13.
Albers С. Die Heliceen, nach naturlicher Verwandtschaft systematisch geordnet. Berlin, 1850,
S. 1—262.
380
ЛИТЕРАТУРА
Albers С. Die Heliceen, nach natiirlicher Verwandtschaft systematise)! geordnet. 2-te Ausgabe,
herausgegeb en von E. Martens. Leipzig, 1860, S. 1—359.
Alzona C. Malacofauna Italica. Catalogo e bibliografia dei molluschi viventi, terrestri e d’acqua
doce. — Atti della Soo. Ital. di Sei. Nat. e del Museo Civico di Storia Nat. di Milano,
1971, v. Ill, p. 1—433.
Ancey C. F. Essai monographique sur les Buliminus de 1’Asie centrale russe et de 1’Afghani-
stan. — Bull. Soc. malac. France, 1886a, v. 3, p. 15—64.
Ancey C. F. Supplement 1’essai monographique sur les Buliminus de 1’Asie Centrale Russe
et de 1’Afghanistan. — Bull. Soc. malac. France, 1886b, v. 3, p. 329—339.
Ancey C. F. A catalogue of the Bulimini found in Central Asia. — Conchologist’s Exchange,
1887, v. 2, N 1, p. 5-6.
Ancey C. F. Remarques sur quelques especes du genre Buliminus, avec les descriptions de plu-
sieurs especes nouvelles de Qe genre. — Bull. Soc. zool. France, 1893, v. 18, p. 35—40.
Ancey C. F. Two new Bulimini from Central Asia. — Nautilus, 1902, v. 16, N. 4, p. 47—48.
Anton H. E. Verzeichnis der Conchylien welche sich in der Sammlung von H. A. Anton befinden.
Hall, 1839, S. 1—110.
Baker H. B. Review of the anatomy of Pupillidae and related groups. — In: Pilsbry H., Manual
of Conchology, 1935, v. 28, p. 191—209.
Baker H. B. Heterurethrous and aulacopod. — Nautilus, 1955, v. 68, N 4, p. 109—112.
Beck H. Index molluscorum praesentis aevi musei principis augustissimi Christiani Frederici.
Hafniae, 1837, p. 1—100.
Beck H. Verzeichniss der Naturaliensammlung, welche auf Befehl Sr. Majestat des Konigs aus
verschiedenen koniglichen Musaen in Copenhagen. . . zur 24. Versammlung Deutscher
Naturforscher und Arzte nach Kiel gesandt war. — Amtl. Ber. Versdtsch. Naturf.
Arzte Kiel 1846, 1847, S. 109—110.
Beck К. O. Anatomic deutscher Buliminus-Arten. — Jenaische Zeitschr. f. Naturwissenschaft,
1912, Bd 48, Heft 3 (N. F., Bd 41), S. 187—262.
Benson W. H. Characters of a new European Pupa, and of a new Australian Bulimus. — Ann.
Mag. Nat. Hist., 1854, ser. II, v. 13, p. 97—99.
Bequaert J. C., Miller W. B. The mollusks of the arid southwest with an Arizona check list.
Univ. Arizona Press, Tucson, 1973, p. 1—271.
Biggs H. E. J. Mollusca of the Iranian Plateau — II. — J. Conch., 1962, v. 25, p. 64—72.
Biggs H. E. J. Mollusca of the Iranian Plateau — III.-,— J. Conch., 1971, v. 27, p. 211—220.
Boettger С. R. Reisenwuchs der Landschnecke Zebrina (Zebrina) detrita (Muller) als Folge pa-
rasitarer Kastration. — Arch. Moll., 1853, Bd 82, N 4—6, S. 151—152.
Boettger O. Kaukasische Mollusken gesammelt von Herrn Hans Leder. — Jahrb. dtsch. malak.
Ges., 1879, Bd 6, S. 1-42, 388-412.
Boettger O. Kaukasische Mollusken gesammelt von Herrn Hans Leder. — Jahrb. dtsch. malak.
Ges., 1880a, Bd 7, S. 109—150.
Boettger O. Armenische und transcaucasische Mollusken, aus einer Sendung des Hrn. Dr.
G. Sievers. — Jahrb. dtsch. malak. Ges., 1880b, Bd 7, S. 151—161.
Boettger O. Diagnoses molluscorum novorum ab ill. Hans Leder in regione caspia Talysch dicta
lectorum. — Jahrb. dtsch. malak. Ges., 1880c, Bd 7, S. 379—383.
Boettger O. Sechstes Verzeichniss transcaucasischer, armenischer und nordpersischer Mollu-
sken. — Jahrb. dtsch. malak. Ges., 1881, Bd 8, S. 167—261.
Boettger O. Siebentes Verzeichniss von Mollusken der Kaukasuslander. — Jahrb. dtsch. malak.
. Ges., 1883, Bd 10, S. 138—198.
Boettger O. Neuntes Verzeichniss (IX) von Mollusken der Kaukasuslander. — Jahrb. dtsch.
malak. Ges., 1886a, Bd 13, S. 121—156.
Boettger O. Die Binnenmoilusken des Talysch-Gebietes. — In: Radde G. Fauna und Flora des
siidwestlichen Caspi-Gebietes, 1886b, S. 256—350.
Boettger O. Abbildungen und Beschreibungen von Binnenmoilusken aus dem Talysch-Gebiet
im Siidwesten des Caspisees (XI). — Jahrb. dtsch. malak. Ges., 1886c, Bd 13, S. 241—258.
Boettger O. Diagnosen neuer kaukasischer Arten. — Nachrichtenbl. dtsch. malak. Ges., 1888,
Bd 20, S. 149-155.
Boettger O. Zehntes Verzeichniss (XII) von Mollusken der Kaukasuslander. — Ber. Senckenb.
Nat. Ges., 1889a, S. 3—37.
Boettger O. Die Binnenmoilusken Transcaspiens und Chorossans. — Zool. Jahrb., 1889b, Abt. 3,
Bd 4, S. 925—992.
Boettger O. Die Entwicklung der Pupa-Arten des Mittelrheingebietes in Zeit und Raum. —
Jahrb. nassau. Ver. Naturk. Wiesbaden, 1889c, Bd 42, S. 225—327.
Boettger O. Bemerkungen fiber einige Buliminus aus Kleinasien, Syrien und Cypern, nebst
Beschreibung neuer Arten. — Nachrichtenbl. dtsch. malak. Ges., 1898, Bd 30, S. 19—27.
Bourguignat J.-R. Amenites malacologiques. § 29. Catalogue des coquilles recueillies en Cri-
mee et dans 1’Empire ottomane. Paris, 1855, t. 1, p. 1—255.
Bourguignat J.-R. Species novissimae molluscorum in Europaeo systemati detectae, notis
succinctus breviter descriptiae. Lutetiae, 1876, p. 1—80.
Boycott A. E. Preliminary note on the genitalia of Acanthinula lamellata Jeffr. — J. Conch.,
1917a, v. 15, p. 157—178.
Boycott A. E. The genitalia of Acanthinula aculeata. — Proc. Malac. Soc. London, 1917b,
v. 12, p. 221—226.
ЛИТЕРАТУРА
381
'Brandt В. A. Zur Orculidenfauna der Cyrenaika. — Arch. Moll., 1956, Bd 85, S. 69—82.
Brandt R. A. Uber neue und wenig bekannte Binnenmoilusken der Cyrenaika. — Arch. Moll.,
1958, Bd 87, S. 1-20.
Breure A. S. H., Eskens A. A. C. Observations on the formation of spermatophores in a buli-
mulid land snail, Drymaeus canaliculatus (Pfeiffer, 1845) (Mollusca, Gastropoda, Pulmo-
nata). — Netherlands J. Zool., 1977, v. 27 (3), p. 271—276.
Bruguiere J. G. Histoire naturelie des Vers in Encyclopedic methodique. Paris, 1792, part II.
Brusina S. Ueber die Mollusken-Fauna Oesterreich-Ungarns. — Mitt. nat. Ver. Seiermark.,
1886, Bd 22 (1885), S. 29-56.
Carriker M. R. Morphology of the alimentary system of the snail Lymnaea stagnalis appressa
Say. — Trans. Wisconsin Acad. Sci., 1946, v. 38, p. 1—88.
Caziot E., Margier E. Etude historique de la classification des Pupa du systeme europeen. —
Bull. Soc. Zool. France, 1909, v. 34, p. 134—147.
Cecconi G. Contribute alia fauna delle Isol Tremiti. — Boll. Mus. Zool. Anat. comp. Univ.
Torino, 1908, v. 23 (583), p. 1—53.
Charpentier J. Catalogue des mollusques terrestres et fluviatiles de la Suisse. — Neue Denkschr.
allg. schweiz. Ges. Naturw-, 1837, Bd 1 (2), S. 1—28.
Clauss E. Chondrula tridens (0. F. Miiller), linksgewunden. — Malak. Abhandl. staatl. Mus.
Tierk. Dresden, 1965, Heft 2, N 10, S. 173—174.
Clessin S. Pupa edentula Drap.—inornata Mich.—columella Bens. — Malak. BL, 1872, Bd 20,
S. 50-61.
Clessin S. Mollusken des Wolgagebietes. — Jahrb. dtsch. malak. Ges., 1875, Bd 2, S. 36—42.
Clessin S. Deutsche Excursions-Mollusken-Fauna. Niirnberg, 1876, S. 1—288.
Clessin S. Mollusken aus Taurien. — Malak. BL, 1881, N[ F., Bd 3, S. 136—141.
Clessin S. Anhang zur Molluskenfauna der Krim. — Malak. BL, 1883, N. F., Bd 6, S. 37—52.
Clessin S. Beschreibung neuer Arten aus Umgebung des Issykkul-Sees. — Nachrichtenbl. dtsch.
malak. Ges., 1894, Bd 24, S. 64—66.
Cooke С. M., Kondo Y. Revision of Tbrnatellinidae and Achatinellidae (Gastropoda, Pulmo-
nata). — Bernice P. Bishop Mus., Bull. 221, 1960, p. 1—303.
Crampton D. M. The anatomy and method of functioning of the buccal mass of Testacella mangei
Ferussac. — Proc. Malac. Soc. London, 1975, v. 41, № 6, p. 549—570.
Da Costa E. M. Historia naturalis Testaceorum Britanniae. London, 1778, p. 1—254.
Des Moulins. Description d’une nouvelle espece vivante de Pupa du Perigord. — Actos Soc.
Linn. Bordeaux, 1830, t. 4, p. 158—166.
Draparnaud J. P. R. Tableau de Mollusques terrestres et fluviatiles de la France, Paris, 1801,
p. 1—116.
Draparnaud J. P. R. Histoire naturelle des mollusques terrestres et fluviatiles de la Franco.
Paris, 1805, p. 1—164.
Dupuy D. Catalogue extramarinorum Galliae Testaceorum brevioribus specierum nondum des-
criptorum diagnosibus. Paris, 1849, p. 1—4.
Edlauer E. Die ontogenetische Entwicklung der Verschlussapparates der Clausiliiden, untor-
sucht an Herilla bosniensis. — Zeitschr. wiss. Zool., 1941, Bd 155, Heft 2, S. 129—158.
Ehrenberg C. G. Symbolae Physicae, seu icones et descriptiones Animalium Evertebratorum
sepositis Insectis. Berlin, 1831, I Dec., 10 Tabl.
Ehrmann P. Zur Kenntnis von Chondrina avenacea (Brug.) und ihre nachsten Verwandton. —
Arch. Moll., 1931, Bd 63, Heft 1, S. 1-28.
Ehrmann P. Mollusken. — In: Tierwelt Mitteleuropas. Leipzig, 1956, Bd II, N 1, S. 1—204.
Ferussac J. B. L. d’A. [perej. Essai d’une methode conchyliologique appliquee aux Mollusques
terrestres et fluviatiles. Paris, 1807, p. 1—142.
Ferussac A. E. J. d’A [fils]. Tableaux systematiques des animaux mollusques. . . Tableaux
systematique de la famille LimaQons, Cochleae. Paris, 1821, p. 1—114.
Fitzinger L. Systematisches Verzeichniss der im Erzherzogthume Oesterreich vorkommondon
Weichtier. — In: Beitrage zur Landeskunde Oesterreich’s, 1833, Bd 3, S. 88—122.
Forcart L. Die Mollusken der nordpersischen Provinz Mazenderan und ihre tiergeographischo
Bedeutung. — Arch. Naturg., 1935, N. F., Bd 4, Heft 3, S. 404—447.
Forcart L. Monographie der tiirkischen Enidae (Moll., Pulm.). — Verhandl. naturforsch. Gos.
Basel, 1940, Bd 51, S. 106—263.
Forcart L. Ipsa Studeri Conchylia. — Mitt. Naturforsch. Ges. Bern, 1957, N. F., Bd 15, S. 157—
210.
Forcart L. Die palaearktischer Arten des Genus Columella. — Verhandl. Naturforsch. Gos.
Basel, 1959a, Bd 70, S. 7—18.
Forcart L. Two new subspecies of Jaminia (Euchondrus) continens (Rosen) from Iran. — J. conch,
1959b, v. 24, № 9, p. 315-317-
Forcart L. Rezente Land- und Siisswassermollusken der siiditalienischen Landschafton Apu-
lien, Basilicata und Calabrien. — Verhandl. Naturforsch. Ges. Basel, 1965, Bd 78, № 1,
S. 59—184.
Fiirnrohr A. E. Naturhistorische Topographie von Regensburg. 1840, Bd 3, S. 1—478.
Galland J. Histoire des Brephulus de 1’Asie-Mineure. — Bull. Soc. malac. Franco, 1884, v. 1,
p. 280—285.
Gallenstein H. R. Karnthens Land- und Siisswasser-Conchylien. — Jahrb. naturhist. Landes-
Mus. Karnthen, Klagenfurt, 1852, Bd 1, S. 57—134.
382
ЛИТЕРАТУРА
Germain L. Mollusques terrestres et fluviatiles de 1’Asie anterieure, 5. Catalogue des gastero-
podes de la Syrie et de la Palestine. — Bull. Mus. Hist. nat. Paris, 1912, t. 7, p. 440—452.
Germain L. Contributions a la faune malacologique de 1’Afrique Equatoriale. LVII. Sur quel-
ques genres et especes de Pulmones de 1’Afrique Orientale. — Bull. Mus. Hist. nat. Pa-
ris, 1919, t. 25, p. 258—265.
Germain L. Mollusques terrestres et fluviatiles. — In: Faune de France, 1930, fasc. 21, p. 1—897.
GerstfeldtG. Ueber Land- und Siisswasser-Mollusken Sibiriens und des Amur-Gebietes. — Mem.
Savants etrangers, 1859, Bd 9, S. 507—548.
Geyer D. Unsere Land- und Siisswasser-Mollusken. Stuttgart, 1927, S. 1—224.
Gittenberger E. Die Enidae (Gastropoda, Pulmonata) gesammelt von der niederlandischen bio-
logischen Expedition in die Tiirkei in 1959. — Zool. meded., 1967, Bd 42, N 13, S. 125—
141.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. I. Die Spelaeodiscinae. — Zool. meded.,
1969, Bd 43, N 22, S. 287—306.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. II. Die Gattung Planogyra (Valloniidae)
in Europa. — Basteria, 1972, v. 36, N 2—5, p. 63—74.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. III. Chondrinidae. — Zool. verhandl.,
1973, N 127, S. 1-267.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. IV. Erganzungen der Gattung Argna. —
Basteria, 1974, v. 38, N 1-2, S. 1—12.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. V. Die Spelaeodiscinae, erster Nachtrag. —
Zool. meded., 1975, Bd 48, N 23, S. 263—277.
Gittenberger E. Beitrage zur Kenntnis der Pupillacea. VIII. Einiges fiber Orculidae. — Zool.
vemandl., 1978, N 163, S. 1—44.
Gittenberger E., Backhuys W., Ripken Th. E. J. De landslakken van Nederland. Amsterdam—
Zuid, 1970, S. 1—177.
Gittenberger E., Menkhorst H. P. M. G., Raven J. G. M. New data on four European terrestrial
gastropods. — Basteria, 1980, v. 44, N 1—4, p. 11—16.
Giusti F. Notulae malacologicae. IV. L’isola di Montecristo e lo Scoglio d’Affrice. — Atti Soc.
Toscana Sci. Nat. Mem., 1968, set. B, v. 75, p. 239—254.
Giusti F. Notulae malacologicae. XVIII. I molluschi terrestri e salmastri dell Isole Eolie. —
Lav. Soc. Italiana Biogeogr. (n. ser.), 1973, v. 3, p. 113—306.
Giusti F. Notulae malacologicae. XXIII. I molluschi terrestri, salmastri e di acqua dolce dell’-
Elba, Giannutri e scogli minori dell’Arcipelago Toscano. — Lav. Soc. Italiana Bio-
geogr. (n. ser.), 1976, v. 5, p. 99—355.
Gould. — Proc. Boston Soc. Nat. Hist., 1848, v. 2, 1 Dec. 1847, p. 262—263.
Gray J. E. A natural arrangement of Mollusca, according to the internal structure. — London
med. Rep., 1821, v. 15, p. 229-239.
Gray J. E. A list of the genera of recent Mollusca, their synonyma and types. — Proc. zool.
Soc. London, 1847, v. 15, p. 129—219.
Gredler V. Tirol’s Land- und Siisswasser-Conchylien. I. Die Landconchylien. — Verhandl.
zool.-bot. Ver. Wien, 1856, v. 6, Abh., S. 25—162.
Grossu A. V. Gastropoda Pulmonata. Fauna Rep. Pop. Romine. — Mollusca, 1955, v. 3, fasc. 1,
p. 1—518.
Grossu A. V., Negrea A. Revision des especes du genre Argna Cossmann (Gastropoda, Pulmo-
nata) de Roumanie et description de quelques nouvelles unites taxonomiques. — Trav.
Mus. hist, natur. «Gr. Antipa», 1968, v. 8, N 2, p. 721—734.
Gude G. K. Note on some preoccupied molluscan names (generic and specific). — Proc, malac.
Soc. London, 1911, v. 9, p. 269-273, 361, 362.
Haas G. Palestinian species and races of Jaminia Risso (Mollusca: Gastropoda). — Fieldiana:
Zoology, 1955, v. 37, p. 415—443.
Habe T. Anatomical studies on the Japanese land snails (7). — Venus, 1956, v. 19, N 2, p. 109—
117.
Hanley S. Ipsa Linnaei conchylia. London, 1855, p. 1—556.
Hartmann J. D. W. System der Erd- und Flussschnecken der Schweiz mit vergleichender Auf-
zahlung alter in den benachbarten Landern Deutschland, Frankreich und Itaiien sich
vorfindenden Arten. — Steinmiiller’s «Neue Alpina», 1821, Bd 7, S. 194—268.
Hartmann J. D. W. Erd- und Siisswasser-Gastropoden der Schweiz. Mit Zugabe einiger merk-
wiirdigen exotischen Arten, St. Gallen, 1840, S. 1—37, 38—60.
Held F. Notizen fiber Weichtiere Bayerns. — Isis (Oken), 1837, v. 4, p. 303—309; v. 12, p. 901 —
919.
Heller J. Ena benjamiticus (Benson) in Galilee. — Argamon, 1971a, v. 2, N 3—4, p. 95—96.
Heller J. Revision of the genus Paramastus Hesse (Enidae). — Arch. Moll., 1971b, Bd 101,
N 5/6, S. 263—273.
Heller J. The taxonomy, distribution and faunal succession of Buliminus (Pulmonata: Enidae)
in Israel. — Zool. J. Linn. Soc., 1975, v. 55, N 1, p. 1—57.
Hembrow D. Observations of the structure and function of the buccal mass of Planorbarius cor-.
neus (L.). — Proc, malac. Soc. London, 1973, v. 40, p. 505—521.
Herrmannsen A. N. Indicus Generum Malacozoorum primordia, nomina, subgenerum, generum,
familiarum, tribuum, ordinum, classium, adjectis auctoribus, temporibus, locis, syste-
ЛИТЕРАТУРА
383
maticis atque litterariis, etymis, synonimis. Cassel, 1: 1846, p. 1—232,1847, p. 233—637;
2: 1847, p. 1-352, 1848, p. 353-492, 1849, p. 493-717.
Hesse P. Ueber einige Arten der Genus Buliminus. — Jahrb. dtsch. malak. Ges., 1885, Bd 12,
S. 48-52.
Hesse P. Beschreibungen neuer Arten. — Arch. Moll., 1915, Bd. 47, N 2, S. 58—63.
Hesse P. Zur Kenntnis der Molluskenfauna von Ostrumelien. — Nachrichtenbl. dtsch. malak.
Ges.,1 1916a, Bd 48, S. 113—122.
Hesse P. Kritische Fragmente. XVI. Zur Nomenklatur. — Nachrichtenbl. dtsch. malak. Ges.,
1916b, Bd 48, S. 122-124.
Hesse P. Kritische Fragmente. XVII. Nochmals Nomenklaturfragen. — Nachrichtenbl. dtsch.
malak. Ges., 1917, Bd 49, S. 122—124.
Hesse P. Die Anatomie einiger Orcula-Arten. — Arch. Moll., 1924, Bd 56, N 1, S’. 1—13.
Hesse P. Zur Anatomie und Systematik der Familie Enidae. — Arch. Naturg., 1933, N. F.,
Bd 11, S. 145—224.
Heude P.-M. Notes sur les mollusques terrestres de la vallee du Fleuve Bleu. — Mem. d’Hist.
Nat. 1’Emp. Chinois, 1880, p. 1—87.
Heynemann D. F. Zur Anatomie der Gattung Vertigo. — Malak. Bl., 1862, Bd 9, S. 11—13.
Hudec V. Kriticke hodnoceni druhu rodu Cochlicopa Risso 1826 (Mollusca) z Ceskoslovenska. —
Prace Brnenske zakladny Ceskoslov. Akad. ved, 1960, t. 32, N 7, s. 277—300.
Hudec V. К anatomii stredoasijskeho plze Subzebrinus ujfalvianus (Anc.) a jemu pribuznych
druhti Ena montana (Drap.), E. obscura (Mull.) a Zebrina detrita (Mull.). — Casopis
Nar. muz., odd. prirod., 1963, v. 132, N 1, s. 24—28.
Hudec V. Poznamky к anatomii nekterych druhti р1£й z Bulharska. — Casopis Nar. muz., odd.
prirod., 1965a, v. 134, N 1, s. 11—16.
Hudec V. Neue Erkenntnisse liber die Anatomie von Argna bielzi (Rossmassler) und Bemer-
kungen zur systematischen Stellung der Gattung Argna Cossmann. — Arch. Moll.,
1965b, Bd 94, N 3/4, S. 157—163.
Hudec V. Poznamky к anatomii nekterych druhu plzti z Bulharska. II, III. — Casopis Nar.
muz., odd. prirod., 1967, v. 136, N 1, s. 17—24; N 2, s. 81—89.
Hudec V. Poznamky к anatomii nekterych plgCi z Madarska. — Casopis Nar. muz., odd. prirod.,
1970, v. 137, N .3/4, s. 33—43.
Hudec V. Poznamky к anatomii nekterych druhti plzti z Krymu. — Casopis Nar., muz., odd.
prirod., 1972a, v. 141, N 1/2, s. 73—91.
Hudec V. Dodatek к anatomii nekterych druhti plzti z Krymu. — Casopis Nar. muz., odd.
prirod., 1972b, v. 141, N 1/2, s. 101—104.
Hudec V. Was ist eigentlich Chondrus pupoides Krynicki, 1833? — Arch. Moll., 1972c, Bd 102,
N 4—6, S. 211—219.
Hudec V. Bemerkungen zur Anatomie einiger Schneckenarten (Gastropoda) aus Rumanien. —
Ann. Zool. PAN, 1972d, Bd 29, N 10, S. 323—348.
Hudec V., Lezhawa G. I. Bemerkungen zur Erforschung der Landmollusken der Grusinischen
Sozialistischen Sowietrepublik. — Sbor. Narodn. musea Praze, 1967, Bd 23B, N 3,
S. 69—100.
Hudec V., Lezhawa G. I. Bemerkungen zur Erforschung der Landmollusken der Grusinischen
Sozialistischen Sowietrepublik. (II). — Sbor. Narodn. musea Praze, 1969, Bd 25B, N 3,
S. 93—155.
‘ Hudec V., Vasatko J. Beitrag zur Molluskenfauna Bulgarians. — Acta Sc. Nat. Brno, 1971,
n. s., t. V, Fasc. 2, ,S. 1—38.
Iconographie der Land- und Siisswasser-Mollusken. 1835—1920. Bd. 1—3 (1835—1859) von
Rossmaessler E. A.; fortgesetz von Kobelt W.: Bd 4—7 (1876—1880) und N. F.,
Bd 1—23 (1884-1920) und Suppl. Bd (1895—1897).
Ihering H. Morphologie und Systematik des Genitalapparatus von Helix. — Z. wiss. Zool.,
1892, Bd 54, H. 1/2, S. 386—433; H. 3, S. 425—520.
Ihering H. System und Verbreitung der Heliciden. — Verhandl. zool.-bot. Ges. Wien, 1909,
Bd 59, N 10, S. 420—455.
Issel A. Catalogo dei molluschi raccolti della missione italiana in Persia agguntavi la descri-
zione delle spezie nuove о poco note. — Mem. R. Acad. Sci. Torino, 1865, ser. 2, t. 23,
p. 387—439.
Jeffreys J. G. A synopsis of the testaceous pneumonobranchous mollusca of Great Britain.
Trans. Linn. Soc. London, 1830, v. 16 (2), p. 323—392; Suppl.: p. 505—523.
Kennard A. S. The Histoire and Prodromus of Ferussac. — Proc, malac. Soc. London, 1942,
v. 25, p. 12—17, 105—118.
Kilias R. Die Typen und Typoide der Mollusken-Sammlung des Zoologischen Museums in Ber-
lin (6). II. Euthyneura, Stylommatophora, Orthurethra, Pupillacea (Enidae). — Mill.
Zool. Mus. Berlin, 1971, Bd 47, S. 215-236.
Kimakowicz M. Beitrag zur Molluskenfauna Siebenbiirgens. — Verh. u. Mitt. Siebonbilrgons.
Ver. Naturw. Hermannstadt, 1890, Bd 11, S. 1—113.
Klemm W. Zur rassenmassigen Gliederung des Genus Pagodulina Clessin. — Arch. Nutiirg.,
193'9, N. F., Bd 8 (2), S. 198—262.
Klemm W. Zoologische Studien in West-Griechenland. X Teil. Die Gehauseschnockon. — Sit-
zungsb. Oesterr. Akad. Wiss. Math.-naturw. KI., Abt. I, 1962, Bd 171, H. 6/7, S. 203—258.
384
ЛИТЕРАТУРА
Kobelt W. См. Iconographie. . . 1876—1920.
Kobelt W. Ueber einige griechische Buliminus. — Jahrb. dtsch. malak. Ges., 1877, Bd 4,
S. 265—270.
Kobelt W. Die Familie Buliminidae. — In: Martini, Chemnitz. Systematisches Gonchylien-
Cabinet, 1902, II Theil, S. 397-1051.
Kobelt W. Eine Stenogyra aus Turkestan. — Nachrichtbl. dtsch. malak. Ges., 1904, Bd 36,
H. 2, S. 87.
Kobelt W. Diagnosen neuer Arten. — Nachrichtbl. dtsch. malak. Ges., 1907, Bd 39, H. 3,
S. 167—169.
Krynicki I. Novae species aut minus cognitae e Chondri, Bulimi peristomae helicisque generibus
praecipue Rossiae meridionalis. — Bull. Soc. Nat. Moscou, 1833, v. 6, p. 391—436.
Krynicki I. Conchylia. — Bull. Soc. Nat. Moscou, 1837, v. 10, p. 50—64.
Kuroda T. A catalogue of the non-marine mollusks of Japan, including the Okinawa and Oga-
sawara Islands. — In: A Congratulatory publication on the 77th Birthday of Dr. Tokubei
Kuroda, publ. Malac. Soc. Japan, 1963, p. 1—71.
Lehmann R. Die lebenden Schnecken und Muscheln der Umgegend Stettins und im Pommern
mit besonderer Beriicksichtigung ihres anatomischen Baues. Cassel, 1873, S. 1—238.
Lindholm W. A. Beitrage zur Kenntniss der Weichthierfauna Siid-Russlands. — Nachrichtbl.
dtsch. malak. Ges., 1901, H. 11—12, S. 161—186.
Lindholm W. A. Materialen zur Molluskenfauna von Westrussland, Polen und der Krim. —
Зап. Новоросс. о-ва естествоиспыт., 1908, 31, с. 199—232.
Lindholm W. A. Einige fur die Fauna des St.-Petersburger Gouvernements neue Landschnecken.—
Nachrichtbl. dtsch. malak. Ges., 1910, Bd 42, S. 34—35.
Lindholm W. A. Zur Molluskenfauna des mittleren Wolga-Gebietes. — Nachrichtbl. dtsch.
malak. Ges., 1911, Bd 43, S. 33—34.
Lindholm W. A. Beschreibung neuer Arten und Formen aus den Kaukasus-Gebiete. — Nach-
richtenbl. dtsch. malak. Ges., 1913, Bd 45, S. 17—23, 62—69.
Lindholm W. A. Beschreibung vier neuer Landschnecken und einer neuen Untergattung aus dem
siidwestlichen Transkaukasien. — Nachrichtbl. dtsch. malak. Ges., 1914, Bd 46, S. 33—
38.
Lindholm W. A. Diagnoses of two new Bulimini from Persia. — Ежегод. Зоол. муз., 1915,
т. 20, с. 41—43.
Lindholm W. A. Beschreibung einiger kaukasischer Landschnecken (Gastropoda Pulmonata). —
Ежегод. Зоол. муз., 1922, T. 23, с. 355—361.
Lindholm W. A. Ueber einige Chondrula-Arten aus dem Kaukasus-Gebiete. — Arch. Moll.,
1923, Bd 55, H. 6, S. 218—225.
Lindholm W. A. A revised systematic list of the genera of the Clausiliidae, recent and fossil,
with their sybdivision, synonymy, and types. — Proc, malac. Soc. London, 1924, Bd 16,
p. 53—80.
Lindholm W. A. Beitrag zur Systematik und Nomenklatur der Familia Enidae (Buliminidae). —
Arch. Moll., 1925a, Bd 57, S. 23—41, 140-142.
Lindholm W. A. Studien an palaearktischen Vertigo-Arten. — Arch. Moll., 1925b, Bd 57,
S. 241-251.
Lindholm W. A. Ein Beitrag zur Kenntnis der Molluskenfauna der Krim. — Arch. Moll.,
1926, Bd 58, S. 161—177.
Lindholm W. A. Diagnosen neuer Landschnecken aus dem Ferghana-Gebiete. — Zool. Anz.,
1927a, Bd 74, N 7/9, S. 97—102, 195-202.
Lindholm W. A. Neue Landschnecken aus den Russischen Zentralasien. — Arch.
Naturg., 1927b, Bd 92, H. 2, Abt. 2, S. 257-269.
Lindstrom G. Om Gotlands nutida Mollusker. Wisby, 1868, p. 1—48.
Linnaeus C. Systema naturae, ed. 10, reformata 1, Holmiae, 1758, p. 1—823.
Lowe R. T. Brief diagnostic notices of new Maderan land shells. — Ann. Mag. Nat. Hist.,
set. 2, 1852, Bd 9, p. 112—120, 275—279.
Lozek V. Novi mekkysi ceskoslovenskeho pleistocenu. Vertigo pseudosubstriata sp. n., Pu-
pilla muscorum densegyrata ssp. n. a Pupilla loessica sp. n. — Anthropozoikum, III,
Praha, 1954, s. 327—342. '
Lozek V. Klic ceskoslovenskych mekkysfi. — Slov. Akad. Vied. Bratislava, 1956, s. 1—437.
Lozek V. Quartarmollusken der Tschechoslowakei. Praha, 1964, S. 1—374.
Lozek V. Stepni plzi Chondrula tridens (Muller) a Helicella obvia (Hartmann) v jiznich-Ce-
chach. — Sb. Jihogesk. muz. Cesk. Budejov. Prir. vedy, 1970, t. 10, N 2, s. 73—79.
Markel K. Bau und Funktion der Pulmonaten-Radula. — Zeitschr. Wissenschaft. Zool., 1958,
Bd 160, № 3—4, S. 213—289.
Martens E. Binnen-Mollusken von Chiwa. — Jahrb. dtsch. malak. Ges., 1876a, Bd3, S. 334—337.
Martens E. Transkaukasische Mollusken von Dr. O. Schneider gesammelt, bestimmt von E.
v. Martens. — Jahrb. dtsch. malak. Ges., 1876b, Bd 3, S. 364—370.
Martens E. Aufzahlung der von Alexander Brandt in Russischen-Armenien gesammelten Mol-
lusken. —. Bull. Acad. St.-Petersb., 1880, Bd 26, S. 142—158.
Martens E. Conchologische Mittheilungen als Fortsetzung der Novitates conchologicae. Kas-
sel, 1880—1881, Bd 1, S. 1—101.
Martens E. Ueber Centralasiatische Mollusken. — Mem. Acad. Sci. St.-Petersb., 7 ser., 1882a,
Bd 30, N 11, S. 1-65.
ЛИТЕРАТУРА
385
Martens Е. Ueber von Herrn Apollo Kuschakewitz gesammelte centralasiatische Land- und
Siisswasserschnecken. — Sitzungsber. Ges. naturf. Fr., Berlin, 1882b, S. 103—107.
Mayr E. Birds collected during the Whitney South Sea expedition. XII. Notes on Halcyon chlo-
ris and some of its subspecies. — Amer. Mus. Novitates, 1931, N 469, p. 1—10.
Menke С. T. Synopsis methodica Molluscorum generum omnium et specierum earum quae in
Museo Menkeano adservantur, cum synonymia critica et novarum specierum diagnosibus,.
Pyrmonti, 1828, p. 1—91.
Mermet C. Histoire des mollusques terrestres et fluviatiles vivant dans les Pyrenees-Occiden-
tales. — Act. Soc. sci. Pau, 1843.
MermodG. Gasteropodes. — In: Catalogue des Invertebres de la Suisse, 1930, Fasc. 18, p. 1—583.
Mienis H. K. Pupoides coenopictus (Hutton, 1834) in Israel. — Basteria, 1974, v. 38, p. 99—100.
Minato H. Description of Luchuena ulva n. sp. from Okinoerabu Island, with systematic re-
view of the Genus Luchuena (Gastropoda : Enidae). — Veliger, 1977, v. 36, N 1, p. 14—18.
Moellendorff O. Binnen-Mollusken aus Westchina und Centralasien. — Ежегод. Зоол. муз.,
1901, T. 3, S. 299-412.
Moller H. P. C. Index molluscorum Groenlandiae. Hafniae, 1842, p. 1—24.
Montagu G. Testacea Britannica or natural History of British Shells. London, 1803, pl. 1—l(k
Monterosato T. A. Conchiglie terrestri viventi e fossili di Monte Pellegrino. — Natura. Sicil..
1894, v. 13, p. 165—173.
Moore R. C. (ed.). Treatise on invertebrate paleontology. Part I, Mollusca, 1. — Geol. Soc,
Amer, and Univ. Kansas Press, New York, 1960, p. 1—351.
Moquin-Tandon A. Histoire naturelle des Mollusques terrestres et fluviatiles de France, part. 2.
Paris, 1855, p. 1—646.
Morch O. A. L. On the land and fresh-water Mollusca of Greenland. — Amer. J. Conch., 1868,
N. S., v. 4, N 1, p. 25—40.
Morelet A. Coquilles terrestres du Kamtchatka. — J. Conchyl., 1858, v. 7, p. 7—21.
Morse E. S. Observations on the terrestrial Pulmonifera of Maine, including a catalogue of all
the species of terrestrial and fluviatile mollusca known to inhabit the State. — J. Port-
land Soc. nat. Hist., 1864, v. 1, N 1, p. 1—63.
Mortillet G. Descriptions de quelques coquilles nouvelles d’Armenie, et considerations malaco-
statiques. — Mem. Inst. Nat. Geneve, 1854, t. II, p. 1—15.
Mousson A. Coquilles terrestres et fluviatiles recueillies dans 1’Orient par M. le Dr. Alexandre.
Schlafli. — Vierteljahrschr. Nat. Ges. Zurich, 1859, Bd 4, S. 12—253, 283—297.
Mousson A. Coquilles terrestres et fluviatiles recueillies par Mr. le Prof. J. R. Roth son dernier,
voyage en Palestine. — Vierteljahrschr. Nat. Ges. Zurich, 1861, Bd 6, S. 1—34, 124—
156.
Mousson A. Coquilles terrestres et fluviatiles recueillies dans 1’Orient par M. Io Dr. Alox.
Schlafli (2eme Partie). — Vierteljahrschr. Nat. Ges. Zurich, 1863, Bd 8, S. 275—320,'
368—426.
Mousson A. Coquilles recueillies par M. le Dr. Sievers dans la Russie meridionale et asiatiquo. —
J. Conchyl., 1873, t. 21, p. 193—230.
Mousson A. Coquilles recueillies par M. le Dr. Sievers dans les contrees Transcaucasiqucs. —
J. Conchyl., 1876, t. 24, p. 24—51.
Mousson A. Coquilles terrestres et fluviatiles recueillies par M. Louis Graeser dans le bassin,
de 1’Amour. — J. Conchyl., 1887, t. 35, p. 10—32.
Muller O. F. Vermium terrestri um et fluviatilium sen animalium infusoriorum, helminticorum
et testaceorum/non marinorum succincta historia, 2. Havniae et Lipsiae, 1774, p. 1 - -214.'
Naegele G. Einige Nenheiten aus Vorderasien. — Nachrichtenbl. dtsch. malak. Ges., 1902,
Bd. 34, N 1—2, S. 1—9.
Nevill G. Mollusca from Kashmir and neighbourhood of Mari (Murree) in the Punjab. — In:
Sci. Res. Second Yarkand Mission, Mollusca. London, 1878, p. 4—21.
Nilsson G. Historia Molluscorum Sueciae terrestrium et fluviatilium brevitor delineate. Lundae,
1822, p. 1-124.
Nordsieck H. Die Chondrinen der Siidalpen. —Arch. Moll., 1962, Bd 91, N 1/3, S. 1—20.
Nordsieck H. Die Chondrind-Arten der dinarischen Lander. — Arch. Moll., 1970, Bd 100,
N 5/6, S. 243—261.
Olivier G. A. Voyage dans 1’Empire Othoman, 1’Egypte et la Perse, fait par ordre du Gouver-
nement, pendant les six premieres annees de la Republique. 6 v., Atlas, Paris, 1801.
Paget О. E. Die Molluskenfauna der Insel Rhodos. — Ann. Naturhislor. Mus. Wien, 1976,
Bd 80, S. 681—780.
Paul C. R. C. Azeca in Britain. — J. Conch., 1974, v. 28, p. 155—172.
Pfeiffer C. Systematische Anordnung und Beschreibung deutscher Land- und Wasser-Schnecken,
mit besonderer Riicksicht auf die bisher in Hessen gefundenen Arten. Gassel und Berlin,'
1821, S. 1—134.
Pfeiffer L. Methodische Anordnung der bekanntenBulimusarten. — Z. Malak., 1846, S. 135—149.
Pfeiffer L. Diagnosen neuer Landschnecken. — Z. Malak., 1847a, S. 145—151.
Pfeiffer L. Diagnosen neuer von Frivaldsky gesammelter Landschnecken. Z. Malak., 1847b,
S. 191-192.
Pfeiffer L. Diagnosen neuer von Frivaldsky gesammelter Landschnecken. — Z. Malak., 1848a,
S. 6-12. '
Pfeiffer L. Monographia Heliceorum viventium sistens descriptiones systematicas et criticas.
25 А. А. Шилейко
386
ЛИТЕРАТУРА
omnium huius familiae generum et specierum hodie cognitarum. Lipsiae, 1848b, v. 2,
p. 1—594.
Pfeiffer L. Novitates Conchologicae. Serie prima. Mollusca extramarina. Beschreibung und
Abbildung neuer oder kritischer Land- und Siisswasser-Mollusken. Cassel, 2, 1854—1860,
g, <139________303.
Pfeiffer L. Neue Landschnecken. — Malak. Bl., 1857, Bd 4, S. 85—89.
Pilsbry H. A. (in Tryon-Pilsbry). Manual of Conchology. 2-nd set., 1886—1935, v. 1—28.
Pilsbry H. A. Preliminary outline of a new classification of the helices. — Proc. Acad. Nat.
Sci. Philadelphia, 1892 (1893), v. 44, p. 387—404.
Pilsbry H. A. On the zoological position of Partula and Achatinella. — Proc. Acad. Nat. Sci.
Philadelphia, 1900, p. 561—567.
Pilsbry H. A. Land Mollusca of North America (north of Mexico), v. I, part. 2. Philadelphia,
1940, p. <575-994.
Pilsbry H. A. Land Mollusca of North America (north of Mexico), v. II, part. 2. Philadelphia,
1948, p. 521—1113.
Pilsbry H. A., Hirase Y. Descriptions of new Japanese land shells. — Nautilus, 1904, v. 17,
p. 114—126.
Pilsbry H. A., Vanatta E. G. A partial revision of the Pupae of the United States. — Proc. Acad.
Nat. Sci. Philadelphia, 1900, p. 582—611.
Pinter I., Pinter L. Mollusken aus Bulgarien. — Malak. Abh. Staatl. Mus. Tierk. Dresden,
1970, Bd 3, N 1, S. 81—98.
Porro C. Malacologia terrestre e fluviale italiana. Provincia Comasca. Guglielmini e Redaelli,
Milano, 1838, p. 1—136.
Rajagopal A. S., Subba Rao N. V. Some land mollusks of Kashmir, India. — Rec. Zool. Surv.
India, 1972, v. 66, N 1—4, p. 197—212.
Ray H. C., Mukherjee A. Fauna of Radjasthan, India. Part 3. Mollusca. — Rec. Zool. Surv.
India, 1963 (1969), v. 61, N 3—4, p. 403—436.
Reeve L. The land and freshwater mollusks indigenous to,-or naturalized in the British Isles.
London, 1863, p. 1—275.
Reichenbach H. G. L. Allgemeine Taschenbibliothek der Naturwissenschaften. 5 Theil. Zoologie
oder Naturgeschichte des Thierreiches. Dresden, 1828.
Reichenbach H. G. L. Allgemeine Taschenbibliothek der Naturwissenschaften, 5. Zoologie
oder Naturgeschichte des Thierreichs. Zweites Bandchen. Zittau; Leipzig, 1836.
Reinhardt O. Bemerkungen uber einige transcaucasische Pupa-Arten. — Jahrb. dtsch. malak.
Ges., 1877a, Bd 4, S. 76—87.
Reinhardt O. Diagnosen japanischer Landschnecken. — Jahrb. dtsch. malak. Ges., 1877b,
Bd 4, S. 320—325.
Reinhardt O. Die Isthmia-Arten und ihre geographische Verbreitung. — Sitzungsber. Ges.
naturf. Fr., Berlin, 1879, S. 133—139.
Rensch B. Das Prinzip geographischer Rassenkreise und das Problem der Artbildung. Berlin,
1929 S. 1__________206
Retowski O. Die Molluskenfauna der Krim. — Malak. BL, 1883a, N. F., Bd 6, S. 1—34.
Retowski O. Am Strande der Krim gefundene, angeschwemmte transcaucasische (?) Binnencon-
chylien. — Malak. BL, 1883b, N. F., Bd 6, S. 53-61.
Retowski O. Am Strande der Krim gefundene angeschwemmte Binnenconchylien. — Malak.
BL, 1887, N. F., Bd 9, S. 22—42.
Retowski O. Beitrage zur Molluskenfauna des Kaukasus. — Bull. Soc. Nat. Moscou, 1888,
N. S., t. 2; S. 277—288.
Retowski O. Liste der von mir auf meiner Reise von Konstantinopel nach Batum gesammelten
Binnenmoilusken. — Ber. Senckenb. Nat. Ges., 1889, S. 225—265.
Retowski O. Materialen zur Kenntnis der Molluskenfauna des Kaukasus. — Изв. Кавк, муз.,
1914, т. 6, с. 271—347.
Risso A. Histoire naturelle des principales productions de 1’Europe meridionale et particuliere-
ment.de celles des environs de Nice et des Alpes Maritimes. Paris, 1826, p. 1—439.
Rosen O. Beitrag zur Kenntnis der Molluskenfauna Transcaspiens und Ghorossans. — Nach-
richtbl. dtsch. malak. Ges., 1892, Bd 24, S. 121—126.
Rosen O. Essai d’une description de la faune malacozoologique de la region Transcaspienne
russe. — In: 2nd Congr. intern. Zool., 1893a, t. 2, p. 171—178.
Rosen O. Descriptio Bulimini novi Transcaspiae rossicae. — In: 2nd Congr. intern. Zool.,
1893b, t. 2, p. 179.
Rosen O. Contribution a la faune malacologique terrestre du Turkestan (description de deux
especes nouvelles). — Feuille Jennes Natur., 1897, p. 170.
Rosen O. Die Mollusken Ciskaukasiens und speciell des Kuban-Gebietes. — Ежегод. Зоол.
муз., 1911, т. 16, с. 86—142.
Rosen О. Katalog der Schalentragenden Mollusken des Kaukasus. — Изв. Кавк, муз., 1914,
т. 6, с. 141—152.
Rossmaessler Е. А. См. Iconographie. . . 1835—1859.
Scacchi A. Osservazioni zoologiche (Testacei). Napoli, 1833.
Schikow E. W. Beschreibung der Spermatophore von Ena montana (Draparnaud). — Arch.
Moll., 1978, Bd 109, H. 1/3, S. 59—61.
ЛИТЕРАТУРА
387
Schindel D. E., Gould S. J. Biological interaction between fossil species: character displace-
ment in Bermudian land snails. — Paleobiology, 1977, v. 3, N 3, p. 259—269.
Schmid G. Columella aspera Walden auf dem Spitzberg bei Tiibingen (Vertiginidae). — Mitt,
dtsch. malak. Ges., 1967, Bd 9, S. 157—160.
Schmidt A. Ueber eine bei Aschersleben vorkommende Pupa. — Z. Malak., 1849, Bd 9, p. 140—
144.
Schneider O. Naturwissenschaftliche Beitrage zur Kenntnis der Kaukasuslander, auf Grund
seiner Sammelbente. — Sitzungsber. Naturwiss. Ges. «Isis.», Dresden, (1877) 1878,
S. 1—160.
Semper C. Reisen im Archipel der Philippinen, 2. Theil 3: Landmollusken (1—7). Wiesbaden,.
1870, S. 1-327.
Simroth H. Neue Beitrage zur Kenntnis der kaukasischen Nacktschneckenfauna. — Изв. Кавк.,
муз., 1912, т. 6, с. 1—140.
Solem A. Amimopina, an Australian enid land snail. — Veliger, 1964, v. 6, N 3, p. 115—120.
Solem A. Convergence in Pulmonate Radulae. — Veliger, 1973, v. 15, N 3, p. 165—171.
Soos L. Zur systematischen Anatomie der ungarischen Pulmonaten. — Ann. Mus. Nat. Hung.,.
1917, Bd 15, S. 1-165.
Sorita E. On the genitalia of Aegista mikuriyensis Pilsbry, 1902 and Aegista proba goniosoma
Pilsbry et Hirase, 1904 (Bradybaenidae) in southwestern Kanto District, Honshu. —
Venus, 1978, v. 36, N 4, p. 181—190.
Sparks B. W. Fossil and recent English species of Vallonia. — Proc, malac. Soc. London,
1963, v. 30, N 4—5, p. 110—121.
Stabile G. Mollusques terrestres vivants du Piemont. — Atti Soc. Ital. Sci. Nat., 1864, v. 7,
p. 1—141.
Steenberg C.-M. Bloddyr. I. Landsnegle. — In: Danmarks fauna. Kobenhavn, 1911, p. 1—221.
Steenberg C.-M. Anatomie des Acanthinula et des Vallonia. — Vidensk. Medd. Dansk. Naturh,
Forening, 1918, v. 69 (1), p. 1—15.
Steenberg C.-M. Etudes sur 1’anatomie et la systematique des maillots (fam. Pupillidae s. lat.). —
Vidensk. Medd. Dansk. Naturh. Forening, 1925, v. 80, p. 1—215.
Sterki V. Observations on Vallonia. — Proc. Acad. Nat. Sci. Philadelphia, 1893, p. 234—279,
Studier S. Kurzen Verzeichniss der bis jetzt in unseren Vaterlande entdeckten Conchylien. —
Meissners Naturw. allg. Anz. schweiz. Ges. Naturw. Bern, 1820, Bd 3, S. 83—90, 91—94.
Sturany R. Uber die von Dr. H. Rebel in Bulgarien 1896 gesammelten Gehauseschnocken. —
Ann. Naturhist. Hofmuseums, 1897, Bd 12, S. 111—118.
Sturany R., Wagner A. J. Uber schalentragende Landmollusken aus Albanien und Nachbarge-
bieten. — Denkschr. k. Akad. Wiss. Wien, Math.-nat. KI., 1915 (1914), Bd 91, S. 19—138.
Sullivan J. T., Cheng T. C. Structure and function of the mantle cavity of Biomphalaria glab-
rata (Mollusca: Pulmonata). — Trans. Amer. Micros. Soc., 1974, v. 93, N 3, p. 416—420,
Swainson W. A treatise on Malacology; or natural classification of shells and shell fish. 1840,
p. 1—419.
Szigethy A. A Vertigo O. F. Muller, 1774 genus magyarorszagi fajai (Mollusca, Gastropoda). —
Soosiana, 1973, Bd 1, S. 19—30.
Taylor D. W., Sohl N. F. An outline of gastropod classification. — Malacologin, 1962,
v. 1, N 1, p. 7—32.
Thiele J. Handbuch der systematischen Weichtierkunde. Band I. Jena, 1931, S. 1- -778.
Trappmann W. Die Muskulatur von Helix pomatia L. — Z. wiss. Zool. A, 1916, Bd 105, N 4,
S. 489—585.-
Tryon G. W. Monograph of the terrestrial Mollusca of the United States. Amor. .1. Conch.,
1867, N. S., v. 3, N 4, p. 298—324.
Turton W. A manual of the land and fresh-water shells of British Islands, arranged according
to the more modern systems of classification: and described from perfect specimens in the
author’s cabinet: with coloured plates of every species. London, 1831, p. I 150.
Turton W. A manual of the land and freshwater shells of the British Islands, kd. 2 (revised by
J-. E. Gray), London, 1840, p. 1—324.
Urbanski J. Beitrage zur Kenntnis balkanischer Stylommatophoren (Systonuitische, zoogeo-
graphische und okologische Studien fiber die Mollusken der Balkan-llalbinsel. VII). —
Bull. Soc. amis Sci. et Lettr. Poznan, ser. D, 1964, Bd 4, S. 19 -50.
Van Mol J.-J. Etude morphologique et phylogenetique du ganglion corobrw'ido des Gasteropo-
des Pulmones (Mollusques). — Mem. Acad. roy. Belgique, 1967, t. 37, fuse. 5, p. 1 — 168.
Wagner A. Beitrage zur Molluskenfauna Zentraleuropas. — Ann. Zool. Mus. Polonici Hist.
Nat., 1922, Bd 1, H. 1/2, S. 112—123.
Wagner A. Studien zur Molluskenfauna der Balkanhalbinsel mit bosondoror Beriicksichtigung
Bulgarians und Thraziens, nebst monographischer Bearbeitung olnzolner Gruppen. —
Prace Zool. Pol. Panstw. Muz. Przyr., 1927, Bd 6, H. 4, S. 203 399.
Wagner H. Malakozoologische Ergebnisse der Reise Dr. A. Lendl's In Klcinasien. — Zool.
Anz., 1928, Bd 76, H. 1/2, S. 129—132.
Walden H. W. The land Gastropoda of the vicinity of Stockholm. Ark. Zool., 1955, v. 7,
N 21, p. 391—448.
Walden H. W. Einige Bemerkungen zum Erganzungsband zu Ehrmann’s «Mollusca» in «Tier-
welt Mitteleuropas». — Arch. Moll., 1966, Bd 95, H. 1/2, S. 41) 08.
2I>*
•'388
ЛИТЕРАТУРА
Walter Н. J. Illustrated biomorphology of the «angulata» lake form of the basommatophoran
snail Lymnaea catascopium Say. — Malac. Review, 1969, v. 2, p. 1—102.
Watson H. The affinities of Pyramidula, Patulastra, Acanthinula, and Vallonia. — Proc. Ma-
lac. Soc. London, 1920, v. 14, N 1, p. 6—30.
Watson H. Notes on a list of the British non-marine Mollusca. — J. Conch., 1943, v. 22, p. 13—
22, p. 25—47, 53-72.
Weber A. Mollusken. Wissensch. Ergebnisse der Reise Prof. Merzbacher im zentralen und ost-
lischen Tian-Schan 1907—1908. — Abhandl. Bayer. Akad. Wiss., math.-phys. KI.,
1913, Bd 5, S. 1-26.
Wenz W. Gastropoda., Band 6, Teil 1. — In: Schindewolf, Handbuch der Palaozoologie, Ber-
lin—Zehlendorf, 1938—1944, S. 1—1639.
Wenz W., Zilch A. Euthyneura. In: Handbuch d. Palaozoologie. Gastropoda, Th. 2. Berlin,
1959—1960, S. 1—834.
Westerlund C. A. Neue Binnenmollusken aus Sibiriens. — Nachrichtbl. dtsch. malak. Ges.,
1876, Bd 8, S. 97-104.
Westerlund C. A. Sibiriens Land- och Sotvatten-Mollusken. — Svenska Vetensk. Acad. 1877,
N. F., t. 14, N 12, p. 1—111.
Westerlund C. A. Fauna Europaea Molluscorum Extramarinorum. Prodromus. 2 fasc. 1878,
p. 1—320.
Westerlund C. A. Malakologische Miscellen. — Jahrb. dtsch. malak. Ges., 1883, Bd 10, S. 51—72.
Westerlund C. A. Fauna der in der palaarctischen Region lebenden Binnenconchylien. III. Gen.
Buliminus, Sesteria, Pupa, Stenogyra et Cionella. Berlin, 1887, S. 1—198.
Westerlund C. A. Neue centralasiatische Mollusken. — Ежегод. Зоол. муз., 1896, т. 1, с. 181—
198.
Westerlund С. A. Synopsis molluscorum extramarinorum regionis Palaearcticae. Fasc. I. Ge-
nera et species ex typus Bulimi et Pupae. Lundae, 1897, p. 1—124.
Westerlund C. A. Novum Specilegium Malacologicum. — Ежегод. Зоол. муз., 1898, т. 3,
с. 155—183.
Westerlund С. A. Methodus dispositionis conchyliorum extramarinorum in regione palaearctica
viventium, familias, genera, subgenera et stirpes sistens. — Rad. jugosl. znanci unicta,
1902, v. 151, p. 82—139.
Wiegmann F. Binnen-Mollusken aus Westchina und Centralasien. II. Die Buliminiden. — Еже-
год. Зоол. муз., 1901, т. 6, с. 220—298.
Wirth U. Zur Verbreitung und Okologie von Truncatellina costulata (Nilsson) (Gastropoda,
Pulmonata). — Faun.-Okol. Mitt., Zool., Inst. u. Mus. Univ. Kiel, 1970, Bd 3, H. 9/10,
S. 328—333.
Wollaston T. V. Testacea Atlantica or the land and freshwater shells of the Azores, Madeiras,
Salvages, Canaries, Cape Verdes, and Saint Helena. London, 1878, p. 1—588.
Yen T. C. Die chinesischen Land- und Siisswasser-Gastropoden des Natur-Museums Sencken-
berg. — Abhandl. Senckenb. Naturf. Ges., 1939, N 444, S. 1—234.
Zeissler H. Zum okologischen Verhalten von Orcula doliolum (Bruguiere). — Malak. Abhandl.
Staatl. Mus. Tierk. Dresden, 1973, Bd 4, N 6, S. 53—54.
Zilch A. Uber einige Pagodulina des Senckenberg-Museums. — Arch. Moll., 1947, Bd 76,
N 1/3, S. 73—75.
Zilch A. Ergebnisse zoologisch-geologischer Sammelreisen H. Kaltenbach’s in NO-Afrika, 5:
Familie Enidae. — Arch. Moll., 1951, Bd 80, N 1/3, S. 33—46.
Zilch A. Neue Landschnecken aus der Cyrenaika. — Arch. Moll., 1960, Bd89, N 1/3, S. 57—60.
Zilch A., Jaeckel S. G. A. Mollusken. — In: Tierwelt Mitteleuropas. Erganzung zu II Bd,
Lief. 1. Leipzig, 1962, S. 1—294.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ МОЛЛЮСКОВ1
Abida 8, 80, 85, 224, 226
Acanthinula 6, 74, 76, 80, 84, 85, 94, 104, 159,
160
Acanthinulinae 6, 71, 81, 94, 110, 158 '
acarus, Gastrocopta, Pupa 196
Achat ina 64
Achatinella 80
Achatinellidae 13, 19, 21, 39, 40, 49, 58, 66,
70, 81, 83, 84, 86, 88, 89*, 90, 91
Achatinelloidea 41, 42*, 84, 89*, 90, 91
Achatinina 87
acicularis, Columella 8, 72, 216, 217, 218*
aculeata, Acanthinula, Helix 6, 71, 94, 160,
161*
acutior, Multidentula, Chondrula 372
adjarica, Ljudmilena, Buliminus, Imparie-
tula, Pseudochondrula 309, 310
Adzharia 10, 73, 74, 79, 330, 332
Agardhia 153
Akramovskiella 10, 74, 241, 243, 312, 313*,
315, 316
alabiella, Pupilla 7, 178, 190*, 191, 192*
Alaea 199
alaicus, Laevozebrinus, Buliminus 304
Albertanella 171
albilabris, Pupoides, Pupa 194
albina, Euxinolauria, Lauria 148
albina, Pupilla, Pupa 185
albiplicata, Pseudonapaeus albiplicatus f.,
Jaminia 247, 248*, 249, 252, 265, 267,
304
albiplicatus, Pseudonapaeus, Buliminus, Sub-
zebrinus 3, 9, 41*, 47—49, 62, 63, 100,
102, 244, 247, 250*, 263, 264
albocostatus, Mastoides, Buliminus, Subze-
brinus 9, 242, 298*, 300*
albolimbata, Turanena, Ena, Zebrina 9, 275
276,* 284,* 285*
albolimbatus, Chondrula, Buliminus 364
Alloglossa 228
alpestris, Vertigo modesta 8, 199, 211*, 212,
213
Amastridae 70, 81, 89,* 90, 94
Amastrinae 13
Amimopina 238
Amphiscopus 103
amphorula, Chondrina 8, 72, 228*, 229,
231, 232*
amurensis, Zoogenetes, Helix 159
amurensis, Vallonia 6, 163, 164,* 165
anatolica, Chondrus, Brephulus 351
Andronakia 10, 74, 79, 86, 334
andro'nakii, Akramovskiella, Buliminus 10,
312, 313*, 315
angustatus, Multidentula, Buliminus 372
angustior, Multidentula, Buliminus 97, 373
angustior, Vertilla, Vertigo 8, 60, 213, 214*
annenkovi, Pseudonapaeus, Buliminus 250
anodus, Truncatellina, Pupa 222
anomphalus, Chondrulopsina, Buliminus, Chon-
drulus 323
Antichondrus 350
antivertigo, Vertigo, Alaea, Pupa 7, 199,
200*
anzobica, Pupilla 7, 178, 190*, 191
aptycha, Pseudonapaeus albiplicatus f., Bu-
liminus, Sewertzowia 9, 242, 244, 248*, 249,
254*
arctica, Vertigo 211
Argna 6, 74, 85, 153
Argninae 6, 20, 71, 117, 152
Aridenus 9, 244, 271
armeniaca, Pseudochondrula, Buliminus 289
armeniaca, Gibbulinopsis, Pupa, Pupilla 175,
186
armifera, Gastrocopta, Pupa 196
ascaniensis, Truncatellina, Pupa 221
asiatica, Pseudonapaeus albiplicatus 1., Bu-
liminus, Jaminia, Pseudonapaeus 9, 62,
102, 244—246, 248*, 249, 252*
asiatica, Pupilla, Pupa 179, 185
aspera, Columella 8, 72, 215, 218*
assimilis, Peristoma, Buliminus, Bulimus 343
attenuatus, Brephulopsis, Buliminus, Bulimus
352, 354
attenuatus, Chondrula, Buliminus 364
Aulacospira 61
Auriculellidae 81
avenacea, Chondrina, Bulimus .45*, 228*,
229, 230
avus, Pupilla syngenes 95
Azeca 58, 84, 85, 90
Basommatophora 20
batumensis, Orculella, Orcula, Pupa 70,
122*
bayeri, Chondrula, Buliminus 364
benjamiticus, ? Akramovskiella, Bulimus, Ena
314
1 Курсивом выделены синонимы, неверные определения и прочие названия, не-
соответствующие принятой в книге номенклатуре; жирным шрифтом — названия так-
сонов групп рода и семейства Pupillina, а также номера страниц, на которых начи-
нается описание таксона; звездочкой (*) отмечены страницы с рисунками.
390
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
bicallosa, Chondrula, Buliminus, Bulimus,
Eubrephulus 11, 240, 362, 367, 368*
bidens, Brephulopsis, Buliminus, Bulimus,
Chondrus 11, 25*, 32*, 69*, 100, 240,
352, 353*, 354, 355*
bielzi, Argna, Agardhia, Pupa. 6, 71, 75,
153,* 154*
bielzi, Chondrula, Buliminus, Dentistomus,
Mastus 11, 75, 241, 362, 370*
Bijidarta. 196
bifilaris, Pilorcula, Pupa 118, 127
Ы/formis, Pseudonapaeus, Buliminus 250
bigranata, Pupilla, Pupa 7, 178, 179*, 182,
183
bilabiata, Euxinolauria, Pupa 145
bipapulata, Pupilla 7, 178, 179*, 180*
bisinuatus, Chondrulopsina, Buliminus 323
bithynicus, Chondrus, Brephulus 351
boettgeri, Peristoma, Buliminus, Ena 11,
241, 341, 342*, 345, 346*
Boisidia 61
Bollingeria 371, 372
bonvallotianus, Laevozebrinus, Buliminus 301
borealis, Vertigo, Pupa 211
Bradybaenidae 69, 102
brardus, Chondrus, Brephulus 351
brauneri, Peristoma, Buliminus 343
brauniopsis, Abida secale 56*
Brephulopsis 11, 73, 74, 338, 352, 353*
Brephulus 350, 352
brevior, Imparietula, Buliminus, Bulimus,
Pseudochondrula 10, 109, 241, 292, 293,
307*, 308*, 309
brevior, Pseudonapaeus, Buliminus 250
brongersmai, Abida secale 56*
bulgarica, Orculella, Orcula '5, 70, 120, 124*
Bulimina 339
Buliminidae 3, 4, 9, 13, 14, 17, 38, 39*, 40,
43, 44, 47*, 49, 53, 58, 63, 67, 69, 72, 75*,
80—83, 86, 87, 89*, 90, 97—104, 109, 110,
238, 309, 376
Buliminmae 10, 66, 73, 97, 238, 330, 333
Buliminoidea 9, 42*, 43, 89*, 97, 238
Buliminus 10, 73, 74, 330, 336
Bulimulidae 82
Bulimulus 348
Bulimus 81
caesius, Zebrina, Buliminus 348
callicratis, Truncatellina, Turbo 8, 219,
220*, 221*
callilabris, Geminula isseliana f. 68*, 69,
319
candisata, Laevozebrinus, Buliminus 301
capusiana, Brephulopsis, Buliminus 254
caspia, Lauria, Pupa 152
caspica, Lauria, Pupa 152
castaneus, Pseudonapaeus, Buliminus 265
catenulata, Andronakia, Chondrula 10, 334,
335*
Caucasica 136
caucasica, Chondrina clienta 8, 22*, 72,
87, 228*, 229, 230, 231*, 232
caucasica, Chondrula 364
caucasica, Chondrula, Buliminus, Bulimus,
Mastus 11, 241, 362, 369, 370*
caucasica, Euxinolauria, Lauria, Pupa 6, 71,
134, 136, 142*, 143*
caucasica, Pupilla, Pupa 186
Caucasicola 11, 13, 73, 74, 79, 338, 346
Caucasipupa 6, 136
Cauliculus 244
cephalonicus, Napaeopsis,. Bulimus 35&
Cerastua 238
Ceriidae 70, 91
Cerioidea 80, 90
cespitus, Ljudmilena, Bulimus 312.
Chilanodon 67
chodschendicus, Pseudonapaeus, Chandrulop-
sina 9, 245, 267*, 268*
Chondrina 8, 72, 74, 80, 85, 224, 228, 233
Chondrina (subg.) 229
Chondrinidae 8, 19, 20, 55, 56*, 62, 64, 72,
75, 80, 83, 86-89*, 90, 91, 96, 97, 104,
110, 224
Chondrinoidea 8, 39, 42*, 89*, 96, 97, 224
Chondritortus 326
Chondrula 11, 73, 74, 85, 97, 98, 240, 306,
317, 338, 362, 363, 364*, 371
Chondrula (subg.) 362
Chondrulidae 82
Chondrulinae 82, 83, 97, 98
Chondrulopsina 10, 74, 321
Chondrulopsininae 10, 44, 72, 79, 238, 321
Chondrulopsis 244
Chondrus 11, 338, 350
Cionella 111
circassica, Circassina 67*, 98
circumlabiata, Vertigo 8, 72, 199, 208, 209*
Clausiliidae 32, 56
claustralis, Truncatellina, Pupa 8, 219, 220*,
222
clavella, Truncatellina, Pupa 222
clessini, Multidentula, Buliminus 372
clienta, Chondrina 229
clienta, Chondrina clienta 8, 72, 229
Cochlicopa 5, 58, 74, 76, 84—86, 90, 94, 111
Cochlicopidae 5, 21, 70, 75, 81, 89, 90, 94,
110, 111, 376
Cochlicopoidea 5, 41, 42, 89—91, 94, 111
coenopictus, Pupoides, Pupa 7, 194, 195*
cognata, Turanena, Ena 9, 275, 276*, 286,
287*
Columella 8, 20, 64, 72, 74, 76, 80, 85, 96,
104, 215, 219
columella, Columella, Pupa 8, 72, 215, 216*
columna, Cochlicopa, Cionella 114
concolor, Brephulopsis, Chondrus 353
conica, Pyramidula 236
conicula, Turanena, Buliminus 9, 275, 276*,
285, 286*
connivens, Peristoma 345
continens, Geminula, Buliminus, Jaminia 10,
63, 318, 319*, 320*
contracta, Gastrocopta, Pupa 196
coreana, Gastrocopta 196
coreana, Strobilops 6, 71, 93*, 155*
coreana, Vertigo japonica 210
Coryna 153
costata, Vallonia, Helix 6, 47, 49, 163, 164,*
165*
costatus, Pseudonapaeus, Buliminus, Subzeb-
rinus 256
costellata, Vallonia 167
costigerella, Truncatellina 222
costulata, Truncatellina, Pupa 8, 72, 219,
220*, 221*
crassa, Orculella 123*
cristata, Gibbulinopsis, Pupa 175 1
cryptodon, Gibbulinopsis, Pupa 7, 71, 172,
173*
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
391
cryptodus, Pupa 172
Crystallifera 5, 71, 129, 131
cumulate, Nanaja 67
сира auct., Pupilla 189
curta, Vertigo, Pupa 201
Cyclophoridae 84
cyclostoma, Gibbulinopsis 175
cylindracea, Lauria, Charadrobia, Turbo 6,
45*, 51, 52*, 60, 71, 150, 151*, 152
cylindrica, Truncatellina, Vertigo 8, 72, 219,
220*, 222, 223*
.cylindrica, Vertigo, Helix 205
cylindricus, Brephulopsis, Bulimus, Zebrina
11, 19, 55, 63, 73, 100, 104, 240, 352,
353*, 354*
cylindroconus, Pseudonapaeus 253
cymmetrica, Lauria cylindracea 150
Cyrenaeus 97, 98
debilis, Gibbulinopsis 175
decora, Vertigo, Pupa 211
Dendropupa 84, 86
Dendropupidae 14
dentatum, Triangustoma, Chondrulopsina 9,
241, 296*, 297*
Dentistomus 362
denudata, Gastrocopta, Pupa 196
destituta, Pseudochondrula, Chondrus 289
detrita, Zebrina, Buliminus 11, 19, 63, 73,
82, 104, 240, 348, 349*, 350*
dextro versa, Pseudonapaeus 252
didymoda, Geminula, Buliminus, Jaminia
317, 318
diecki, Gibbulinopsis, Pupa 175
Differena 10, 74, 79, 243, 315
diffusus, Pseudochondrula, Buliminus 290
diplus, Pseudonapaeus, Buliminus 9, 246,
261*
dissimilis, Pseudonapaeus albiplicatus f.,
Buliminus 9, 242, 244, 245, 248*, 256
distortus, Mastoides 300
doliolum, Sphyradium, Bulimus, Orculi,
Pupa, Scyphus 5, 16*, 21*, 29*, 50*,
70, 93*, 94, 126, 127*, 128*
dolium, Orcula, Pupa 5, 70, 75, 125*
dryinaeus, Pseudonapaeus, Huliminus, Se-
wertzowia 255
edentula, Columella, Alaea, Pupa, Vertigo
. 8, 45*, 72,_ 76, 215, 216*, 217
edentula, Pupilla 185
Edentulina 215
elatior, Pseudonapaeus 250
eleonorae, Subzebrinus, Ena 273
Ellobiidae 20
elongata, Chondrula 364
Ena 11, 73, 74, 76, 85, 274, 338, 339
Endodontidae 96
Enidae 3, 81
Eninae 11, 73, 82, 97, 238, 338
enniensis, Vallonia 7, 163, 166*, 167
entodon, Pseudonapaeus, Buliminus, Sewert-
zovia 252
entoptyx, Pseudonapaeus, Buliminus, Jami-
nia, Siraphorus 9, 57, 58, 62, 239, 244,
264, 269, 270*
eremita, Laevozebrinus, Buliminus, Bulimus, ‘
Subzebrinus 10, 63, 72, 242, 301, 302*,
303*, 305
errans, Pseudonapaeus, Buliminus 250
Eubrephulus 11, 339, 362, 367
Euchondrus 103
Eucora 362
Euomphaliinae '33, 67, 69
Euxininae 32
Euxinolauria 6, 13, 57, 58, 60, 71. 74, 79,
133, 134
Euxinolauria (subg.) 145
excentrica, Vallonia 7, 163, 166*, 168
exigua, Cochlicopa, Achatina, Bulimus, Cio-
nella 115
exiguus, Chondrula 364
eximius, Chondrula 364
expansilabris, Pseudonapaeus 265
fallax, Pupoides, Pupa 194
fedtschenkoi, Chondrulopsina, Jaminia, Se-
wertzowia 10, 54, 101, 242, 322*, 323, 324*
jerghanensis, Pseudonapaeus, Buliminus 250
ferrari, Argna, Pupa 153
jlammulatus, Pseudonapaeus, Subzebrinus 250
florenskii, Pseudochondrula, Buliminus, Im-
parietula 288, 289
fragilis, Peristoma, Bulimus 341
Fragipupilla 7, 71, 191
frumentum, Granaria, Abida, Pupa 8, 65*,
72, 75, 224, 225*, 226*
fulgens, Achatinella 35
fuscilabris, Brephulopsis, Chondrus 353
galiciensis, Chondrula, Chondrus 364
galinae, Pseudonapaeus, Ena 9, 245, 264*,
265
gallae, Pupilla 7, 178, 179*, 183, 184*
Gastrocopta 7, 20, 74, 76, 85, 196
Gastrocoptinae 7, 20, 60, 61, 71, 81, 96, 195,
196
Gastrodon 150
Gastropoda 14
Geminula 10, 72, 74, 76, 239, 243, 309, 317
genesii, Vertigo, Pupa 211
Geophila 14, 17, 20, 23, 37, 39, 41, 44, 46,
53, 80, 87, 88
gereti, Boisidia 58*
germabensis, Laevozebrinus 301
geyeri, Vertigo 211
ghilanensis, Geminula, Bulimus, Jaminia
318
gibber, Thoanteus, Buliminus, Bulimus, Ze-
brina 11, 239, 358, 359*, 360*
Gibbulinopsis 7, 71, 74, 87, 171, 172
Gibbulinopsis (subg.) 172
gibler (laps, cal.), Thoanteus, Bulimus 358
gigantea, Vallonia 171
goldfussi, Pseudonapaeus, Buliminus 9, 239,
244, 264*, 267*
goniosoma, Aegista proba 69
Gonodon 326, 362
goodalli, Azeca 58*
gracilis, Gibbulinopsis, Pupilla 7, 71, 172,
174, 175*
Granaria 8, 72, 74, 85, 90, 96, 97, 224
grandis, Gibbulinopsis, Pupilla 172
Granopupa 8, 72, 224, 229, 233
granum, Chondrina, Granopupa, Pupa 8, 72,
229, 233, 234*
gredleri, Columella, Pupa 216
392
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
haberhaueri, Chondrulopsina, Buliminus 321,
323
halepensis, Buliminus, Petraeus 331
haloloma, Aulacospira 58*, 61
hamata, Vertilla, Vertigo 213
harpa, Zoogenetes, Bulimus, Helix 6, 71, 159*
hauffeni, Spelaeodiscus, Helix 156
hebes, Pupilla 192
hebraica, Pilorcula raymondi f., Pupa 118
Helicodonta 32
Helicoidea 38, 66
Helix 64
Helixina 32, 67
Helle 346
hepatica, Laevozebrinus 301
heptapotamica, Archaica 67*
herzenstetni, Pseudonapaeus, Buliminus 253
herzi, Turanena, Buliminus 9, 72, 274—276*,
277, 278*
heterstropha, J aminia 326
Heterurethra 23, 80
heudeana, Gibbulinopsis, Pupa 172
himalayana, Orcula 70
hirasei, Vertigo 8, 199, 209*, 211
hohenackeri, Napaeopsis, Buliminus, Buli-
mus, Zebrina 11, 63, 73, 82, 104, 240,
356, 357*
hoppii, Vertigo, Pupa 211, 212
hrazdanica, Zebrina 349
humberti, Akramovskiella 312
huttoniana, Gastrocopta, Pupa 7, 72, 196,
197*, 198
hydrophila, Vertigo 8, 199, 205*
Hygromiidae 33, 67, 69, 98, 100
Hygromiinae 67
Hypselostoma 58
hyrcanus, Akramovskiella, Buliminus 312
Imparietinia 307
Imparietula 10, 74, 103, 244, 288, 289, 306,
307 *
implicata, Pseudonapaeus 250
Improvise 12, 67, 68*, 74, 79, 371, 373
incertus, Pseudochondrula, Buliminus 289
inequidenta, Pupilla 7, 178, 187, 188*
tnermis, Chondrulopsina 323
inermis, Vertigo, Pupa 211
infans, Chondrulopsina 322
ingens, Retowskia 336
inops, Pupilla, Pupa 7, 178, 186*, 194
inornate, Columella, Pupa 215
interfuscus, Napaeopsis, Bulimus 356
intermedia, Columella 8, 215, 218*, 219
intermedius, Napaeopsis 356
interrupta, Gibbulinopsis, Bulimus, Pupilla
7, 71, 172, 173*
intumescens, Chondrulopsina, Buliminus, Ja-
minia, Sewertzowia, Subzebrinus 10, 54,
242, 322, 323*
isoclina, Protomatellina 84
isseliana, Geminula, Bulimus, Chondrula,
Jaminia, Multidentula 10, 72, 317, 318,
319*
Isthmia 8, 199, 205
Jaminia 10, 83, 244, 306, 317, 321, 326, 362,
371
Jaminiinae 10, 40, 72, 82, 238, 326
jdponica, Vertigo 8, 199, 210*
kamtschatica, Vallonia cyclophorella 6, 163,
166*
kasnakovi, Orculella, Orcula, Pupa 122
kasnakovi, Pseudonapaeus, Buliminus, Ja-
minia 9, 101, 244, 245, 258*
Klemmia 93
kokandensis, Pseudonapaeus 250
kollyi, Pseudochondrula, Buliminus 291
komarovi, Pseudochondrula 289
komarovi auct., Pseudonapaeus, Buliminus
251
kbnigi, Imparietula, Buliminus 309
konkensis, Zebrina 349
korschinskii, Pseudonapaeus, Buliminus 25f
kubanensis, Chondrula 364
kuldshanus, Pseudonapaeus 253
kuschakewitzi, Laevozebrinus, Buliminus 301
kusnetzowi, Peristoma, Buliminus 343
ktisteriana, Vertigo, Pupa 206
labiellus, Subzebrinus, Buliminus, Jaminia.
9, 44, 101, 242, 272, 273*
labrosus, Buliminus 330, 331*
labyrinthica, Strobilops, Helix 155
ladacensis, Vallonia 7, 163, 166*, 169
Laemodonta 20
Laeonapaeus 244
laevigata, Vertigo, Pupa 206
laevis, Vertigo 202, 203
Laevozebrinus 10, 44, 63, 72, 74, 243, 301,
302 *
lamellata, Spermodea, Helix 6, 74, 161, 162*
lamellifera, Multidentula, Buliminus 12, 97,
98, 371, 372*
lanceum, Peristoma 11, 241, 341, 343*
lardeus, Pupoides, Buliminus 194
larvatus, Pseudonapaeus, Buliminus 251
lasistanica, Pseudochondrula 307
latilabris, Pseudonapaeus, Ena 265
Lauria 6, 74, 76, 80, 83, 85, 87, 92, 133, 136,
150
Lauriinae 6, 20, 49, 60, 71, 80, 91, 92, 117, 133
Laurin'ella 219
lederi, Euxinolauria, Pupa 138, 139
lederi, Pagodulina 5, 51, 71, 129*, 131, 132*
lederi, Pseudochondrula, Buliminus, Chond-
rula 9, 57, 109, 241, 289*, 292, 293*, 294,
295, 307
lederi, Punctum micropleurum, Pdtula 158
Leiostyla 85, 133, 136
lenis, Laevozebrinus 10, 242, 301, 302*,
305, 306*
lepidula, Columella, Vertigo 217
Leptachatina 94
leptogyra, Turanena, Ena 9, 65*, 275, 276*,
281*, 282*, 283
Leucochila 194
Leucochiloides 194
Leucochilus 196
leucodon, Imparietula, Bulimus 306, 307
leucolaemus, Napaeopsis, Buliminus 356
leucopleurus, Pseudonapaeus, ?Subzebrinus 250
leucoptychus, Pseudonapaeus, Buliminus 256
leucostoma, Differena 10, 241, 315, 316*,
317*
likharevi, Chondrulopsina, Subzebrinus 322
lilljeborgi, Vertigo, Pupa 206, 207
limata, Pupilla 7, 178, 190*, 193*
limis, Pseudochondrula 292
lindholmi, Pseudochondrula, Buliminus 289
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
.linearis, Truncatellina, Pupa 219
lineatus, Brephulopsis, Chondrus 353
.liostraca, Turanena, Buliminus 276*, 278
Ljudmilena 10, 74, 244, 309, 310 *
lubrica, Cochlicopa, Cionella, Helix 5, 22,
24*, 35*, 36*, 111, 112*, 114*, 116
lubricella, Cochlicopa, Bulimus, Cionella 5,
112*, 114*, 115
Luchuena 243
lundstromi, Pupilla, Pupa 185
luxuriana, Pupilla 181
.luxurious, Pupilla 181
matbulaki, Pseudonapaeus 252
major, Chondrula 364
mangei, Testacella 26
maracandensis, Pseudonapaeus, Buliminus 265
marcida, Chondrula 364
marginata, Pupilla, Pupa 177, 185
martensiana, Turanena, Buliminus, Zebrina
9, 15*, 28*, 275, 276*, 278, 279*, 281
Mastoides 9, 72, 74, 243, 297, 298*
Mastus 11, 339, 362, 369
Matschachelia 6, 134
.Medea 346
Melampus 20
Merdigera 10, 76, 312, 328
Merdigerinae 10, 44, 72, 238, 328
merduenianum, Peristoma, Buliminus, Ena
10, 241, 341, 342*, 343, 345
merzbacheri, Pseudonapaeus, Buliminus 244,
256
meshkovi, Turanena 9, 31*, 275, 276*, 279*,
280*
Mesorculella 5, 120
Mesurethra 23, 80
micropleuros, Toltecia, Helix 158
microsphaera, Vertigo 8, 72, 199, 210*
Microstele 95
microtraga, Chondrula, Pupa 11, 362—364*,
366, 367*
micula, Truncatellina, Pdpa 222
mikuriyensis, Aegista 69
milaschevitchi, Pupilla 179*, 185
minima, Cochlicopa 115
minor, Chondrula 364
minor, Granaria 225
minor, Turanena 278
minuta, Truncatellina, Pupa 222
minutissima, Truncatellina, Pupa 219, 222
misellus, Pseudonapaeus 256
miser, Pseudonapaeus, Buliminus 9, 245,
256, 257*, 264, 268, 269
mltis, Vertigo 201
modesta, Vertigo, Pupa 20, 60, 95, 205, 209,
211*, 212
modesta, Vertigo modesta 8, 199, 211*, 212*
moltschanovi, Siraphoroides, Jaminia, Si-
raphorus 10, 58, 241, 325*
montana, Ena, Buliminus, Bulimus 11, 73,
240, 339, 340*
moulinsiana, Vertigo, Pupa 8, 35, 45*, 199,
206*, 207
Multidentula 11, 63, 67, 68*, 74, 79, 85, 97,
37
Multidentulinae 11, 20, 44, 67, 68*, 73, 97, 98,
238, 240, 371, 372 *
muscorum, Pupilla, Pupa, Turbo 7, 71,
177—179*, 184*, 185, 189
muscorum, Truncatellina, Vertigo 222
nachicevanjensis, Senaridenta, Chondrula 12,
97, 98, 372*, 375
nanosignata, Gibbulinopsis 7, 71, 172, 175*,
177
Napaeinae 82, 97
Napaeopsis 11, 73, 74, 338, 356
Napaeus 85
narynensis, Pseudonapaeus 250
natalensis, Geminula, Chondrula 320
Neolauria 6, 148
Nesopupa 80, 95
Nesopupeae 95
Nesopupinae 80, 81, 95, 195
nitens, Cochlicopa, Achatina 5, 112*, 113*
nitens, Euxinolauria 145
nitidMa, Vertigo 201
nitidulus, Pupoides marginatus, Bulimus 194,
195
nucifragus, Geminula, Chondrus 318
obscura, Merdigera, Buliminus, Ena, Helix
10, 73, 97, 241, 328, 329*
[octodentata], 8-dentata, Vertigo, Pupa 200
odontostoma, Truncatellina 222
Oedichilus 244
olimpicus, Chondrus, Buliminus 351
Orcula 5, 74, 80, 84, 85, 90, 92, 117, 118, 124
Orculella 5, 63, 74, 80, 118, 120, 124
Orculella (subg.) 5, 122
Orculidae 4, 5, 13, 19, 20, 49, 50, 52, 63, 70,
75*, 80, 83, 86, 89*, 91—94, 110, 118
Orculinae 5, 70, 81, 90, 92, 117
Orculoidea 5, 39, 41, 42*, 84, 89*—91, 93—95,
118
orientalis, Monacha 69
orientalis, Orculella, Pupa 120
orloffensis, Mastoides, Buliminus, Subzebri-
nus 9, 298*, 299*
Orthurethra 23, 80, 90
otostomus, Pseudonapaeus, Buliminus, Jami-
nia, Subzebrinus 9, 56, 245, 258*, 259, 260*
ovularis, Multidentula, Buliminus, Jaminia
97, 371, 372*
oxianus, Pseudonapaeus, Buliminus 249, 250
Pachnodidae 81
Pachnodinae 81
Pachnodus 81
pachychyla, Pseudonapaeus 265
Pacijicellidae 81
Pagodina 129
Pagodula 129
pagodula, Pagodulina, Pupa 5, 129*, 130*
Pagodulina 5, 74, 80, 85, 129
Pagodulina (subg.) 129
Pagodulininae 5, 71, 117, 128
palustris, Vertigo, Alaea 199
Parachondrula . 244
Paramastus 97, .98
Partula 80
Partulidae 80—82
patens, Vallonia 158
Patulastra 80
Patulastridae 80
paucidens, Chondrina 229
paulinae, Euxinolauria, Eaurla 6, 60, 92*,
93, 134, 140, 141*
pellucens, Pseudonapaeus 253
Peristoma 11, 73, 74, 76, 338, 341, 342 *
394
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
peteri, Vallonia tenuilabris 7, 163, 166*, 169
Petraeus 330
pfeifferi, Cochlicopa lubrica f. 115
phaeozona, Bradybaena 102
phasianus, Multidentula 97, 373
phorcus, Thoanteus, Buliminus 358
Pilorcula 5, 70, 74, 79, 117, 118
planicollis, Pseudonapaeus 265
Planivallonia 7, 71, 163, 170
Planogyra 94
plectotropis, Bradybaena 102
Pleurodiscidae 80, 81, 96
Pleurodiscus 80
plicata, Vertilla, Vertigo 213
polyodon, Abida 20
Polyphemus 111
pomatia, Helix 26
pontica, Euxinolauria, Pupa 145, 147
ponlicus, Chondrula, Bhliminus 369
potaninianus, Laevozebrinus, Bullminus, Ja-
minia, Subzebrinus 101, 251, 301
pratensis, Pupilla 185
Primipupilla 7, 71, 172, 174
procera, Laevozebrinus 301
prochorovi, Laevozebrinus, Subzebrinus 304
profuga, Chondrina, Pupa 233
Protornatellina 84—86
przevalskii, Pseudonapaeus, Buliminus 249
Pseudochondrula 9, 72, 74, 244, 288, 289*,
309
Pseudonapaeinae 9, 43*, 44, 72, 73, 239, 243
seudonapaeus 9, 72, 74, 100, 243, 244, 274
Pseudonapaeus (subg.) 9, 246
pseudoriitens, Cochlicopa 5, 111, 112*, 113
Pseudopetraeus 244
pseudosubstriata, Vertigo 8, 199, 202*
pulchella, Vallonia 7, 163, 166*, 167, 169
pulchra, Euxinolauria, Lauria, Pupa 6, 60,
71, 133, 134, 145, 146*, 148
pullaster, Subzebrinus, Buliminus 273
Pulmonata 14, 69, 87
Punctidae 158
punctum, Vertigo- 204
pupa, Chondrula, Buliminus, Helix, Mastus
82, 369
Pupella 224
Pupilla 7, 39, 71, 74, 76, 80, 85, 87, 90, 94,
104, 105, 171, 177
Pupilla (subg.) 179
Pupillidae 3, 4, 7, 13, 47, 49, 71, 75, 80, 81, 86,
89*, 90—92, 94, 104, 110, 171
Pupillina 3, 13—15, 17, 18, 20, 23, 37—40*,
41, 43—45*, 46, 47, 49, 50, 52, 53*, 56,
62—64, 67, 70, 73, 76, 78—80, 83, 84,
86—89*, 90—92, 94, 96, 99, 102, 103, 376
Pupillinae 80, 81, 90
Pupilloidea 7, 39, 41, 42*, 89*, 90, 91, 95,
171
Pupilorcula 90, 95
Pupinidius 63
Pupoides 7, 74, 85, 94, 171, 194
pupoides, Improvisa, Chondrus, Imparietula,
Jaminia 12, 82, 97, 98, 371, 372*, 373
pupula, Gibbulinopsis, Orthogibbus 171
pupilla, Truncatellina, Vertigo 222
pusilia, Toltecia 158
pusilia, Vertigo 8, 199, 203*
pygmaea, Vertigo, Pupa 8, 199, 205, 207,
208*
Pypilla (laps, cal.) 191
Pyramidula 8, 74, 80, 85, 97, 236
Pyramidulidae 8, 13, 20, 62, 65, 72, 75*, 76,
80, 87, 89*, 96, 97, 104, 110, 236
Pyramidulinae 81
Pyramidulops 236
quadri dens, Jaminia, Chondrula, Helix 10,
40, 66, 239, 326, 327*
quadrifilaris truncata, Pilorcula 118
raddei, Caucasicola, Buliminus, Ena 11, 62
240, 346, 347*; 348*
radiata, Zebrina, Bulimus 349
Ramusculus 11, 73, 74, 338, 360
raymondi, Pilorcula, Orcula, Pupa 118
rectidentata, Euxinolauria 6, 60, 134, 135*
136*
regelianus, Pseudonapaeus, Buliminus 8, 44
245, 246*, 247*, 254
Reinhardtia 150
rennenkamji, Chondrulopsina, Buliminus 323
renschi, Adzharia 3, 10, 82, 241, 332, 333*
Retowskia 11, 74, 79, 85, 86, 334, 336 &>
retowskianus, Brephulopsis, Buliminus&Bu-
limus 82, 354
Retowskiinae 10, 73, 86, 239, 334
retrodens, Pseudonapaeus albiplicatus f., Bu-
liminus, Jaminia, Sewertzowia 9, 242,
244, 245, 248*, 249, 254, 255*
retroplicatus, Pseudonapaeus, Buliminus [255
retteri, Pseudonapaeus, Buliminus 250
retteri, Stenogyra 12, 376
Rhabdoena 348
rhodia, Chondrina, Granopupa, Pupa 235, 236
rivierana, Truncatellina, Pupa 220
roebelini, Hypselostoma 58*
ronnebyensis, Vertigo, Pupa 211
rosalia, Vallonia 163
rossmaessleri, Chondrula 369
ruderalis, Orculella, Orcula 5,-70, 120, 121*
Rupestrella 233
rupestre, Peristoma, Bulimus, Ena 11, 240,
341, 342*, 343, 344*
rupestris, Chondrina, Pupa 233
rupestris, Pyramidula, Helix, Patula 8, 21*,
72, 236, 237*
Sagdidae 90, 91
Sagdoidea 44, 80, 88, 90
Sandhalia 80
saxatilis, Pyramidula 236
scalaris, Turanena, Buliminus 9, 72, 275,
276*, 277*
scapus, Pseudochondrula, Chondrus 289
schelkovnikovi, Imparietula, Buliminus, Pse-
udochondrula 3, 10, 57, 239, 307*, 309
schlaeflii, Retowskia, Buliminus 11, 336*,
337*
schnitnikovi, Pseudonapaeus, Buliminus 9,
244, 245, 264*
schuschaensis, Akramovskiella, Buliminus,
Ena, Turanena 10, 312, 313*, 314, 315
scyphus, Orculella 5, 120, 122*, 123*, 124
secale, Abida, Pupa 8, 45*, 226, 227*
secalina, Pseudonapaeus albiplicatus f., Bu-
liminus, Subzebrinus 9, 62, 102, 242, 245,
248*, 249, 252*
seductilis, Pseudochondrula, Pupa 9, 100,
101, 239, 288, 289*, 290*, 309
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
395
semenovi, Pseudonapaeus, Buliminus 252
sempronii, Lauria 152
Senaridenta 12, 67, 68*, 74, 79, 97, 371, 375
Serina 243
seroatus, Pseudonapaeus, Buliminus 259
servilis, Gastrocopta, Pupa 196
Sesteria 103
Severtzovia 244
Sewertzowia 244, 321
sexdentatus, Vertigo, Turbo 200
shuttleworthiana, Vertigo, Pupa 211
sieversi, Ljudmilena, Chondrula, Chondrus,
Imparietula, Jaminia 10, 240, 309, 310*,
311*
sieversi, Vertigo, Pupa 8, 196, 204, 205*
Sigmurethra 23, 80
signata, Gibbulinopsis, Pupa, Pupilla 7,
25*, 63, 71, 172, 174, 175*, 176*, 177
silicea, Euxinolauria 6, 92, 134, 147, 148*
similis, Chondrina 35
Sinalbinula 196
sinangula, Euxinolauria 6, 60, 134, 143, 144*
sinistrorsa, Pseudochondrula 9, 239, 288,
289*, 293*, 294
sinuata, Vertigo 200
Siraphoroides 10, 74, 100, 321, 324
Siraphorus 9, 244, 269
sogdiana, Pseudonapaeus albiplicatus f., Bu-
liminus, Subzebrinus 9, 246, 248*, 249,
250, 251*, 303
Solatopupa 96, 97
solivagus, Laevozebrinus 301
Spelaeoconchinae 82
Spelaeodiscidae 6, 40, 71, 89*,• 91—93, 110,
156
Spelaeodiscinae 80, 83, 92, 156
Spelaeodiscus 6, 80, 93, 156
Spermodea 6, 71, 74, §0, 85, 94, 159, 161
Sphyradium 5, 74, 93, 118, 126, 153, 215
stabilis, Pseudonapaeus 9, 62, 245, 262, 263*
Stenogyra 376
sterrii, Pupilla, Pupa 7, 178, 189*
Stomodonta 226
striata, Vertigo 201
striopolita, Pupilla 7, 178, 188, 189*
strobeli, Truncatellina, Pupa 220
Strobila 155
Strobilops 6, 74, 80, 85, 155
Strobilopsidae 6, 13, 20, 52, 71, 75*, 80, 81,
91, 110, 154
Strobilus 155
strophostoma, Subzebrinus, Buliminus 273
stschukini, Turanena, Ena, Zebrina 9, 275,
276*, 286, 287, 288*
Styloides 111
Stylommatophora 23, 80
Styloptychus 244
subalpestris, Vertigo, Pupa 204
subdola, Pagodulina pagodula f. 130*
subhordeum, Chondrina, Pupa 229
submucronatus, Pseudonapaeus, Ena 9, 44,
245, 271*, 272*
subobscura, Pseudonapaeus albiplicatus f.,
Buliminus 9, 102, 245, 248*, 249, 252
suboviformis, Pupilla, Pupa 181
substriata, Vertigo, Alaea 7, 199, 201, 202*,
211
subulatus, Ramusculus, Buliminus, Bulimus,
Ena, Zebrina 11, 241, 360, 361*
Subulinidae 376
Subzebrinus 9, 72, 74, 100, 244, 272, 297, 301
Succineida 80
suitanus, Chondrulopsina, Buliminus 322
sunzhica, Chondrula- 11, 362, 363*
superstructa, Euxinolauria, Lauria, Pupa 6,
56*, 60, 134, 138, 139*, 140*
talyschanus, Akramovskiella, Buliminus, Ena
312
tantalus, Chondrus zebra 351, 352
tardigyrus, Geminula, Buliminus 318
tatricus, Spelaeodiscus triarius 158
taurica, Chondrina rhodia, Granopupa, Mo-
dicella, Pupa 8, 72, 229, 234*, 235*
tauricola, Truncatellina 222
tauricus, Brephulopsis, Bulimus 353
tener, Peristoma, Buliminus, Bulimus 341,
343, 348
tenuilabiata, Chondrula 364
tenuilabris, Vallonia 169
tenuilabris, Vallonia tenuilabris 7, 163, 169
tenuimarginata, Euxinolauria, Lauria 6, 60, i
92, 134, 137*, 138*
tenuispira, Turanena 9, 275, 276,* 281, 282,
283*
teres, Pseudonapaeus 265
terkensis, Chondrula 364
tetrodon, Pseudochondrula, Bulimus,_ Chond-
rula, Chondrulus, Imparietula 9, 240, 288,
289*, 290, 291*, 309
theeli, Gastrocopta, Pupa 7, 71, 196, 197*,
198
theodostanus, Brephulopsis, Bulimus 354
Thoanteus 11, 73, 74, 338, 358
Thysanophoridae 91
Tokatia 371, 372
Toltecia 96
Tornatellina 84
Tornatellinidae 81
Torquilla 226
tosana, Vertigo 210
tournefortianus, Chondrus, Helix 82, 350
triadis, Spelaeodiscus 156
Triangustoma 9, 243, 296
triarius, Spelaeodiscus, Helix 6, 71, 92*, 93,
156, 157*
Trichiin ae 67
tricollis, Ljudmilena, Buliminus, Chondrula,
Chondrus, Jaminia 10, 239, 309, 310*, 312
tricuspidata, Chondrula 366
tridens, Chondrula, Buliminus, Helix 11, 54,
62, 362, 363, 364*, 365*
trifilaris, Pilorcula, Orcula, Pupa 5, 70,
118*, 119*, 148
trigonochilus, Pseudonapaeus, Buliminus,
Subzebrinus 9, 47, 62, 242', 245, 264*,
265, 266*
triplicata, Pupilla, Glischrus 7, 178, 179*,
181, 182*, 183, 186
trisinuata, Chondrulopsina 323
trochoides, Pyramidula, Helix 236
truncata, Pilorcula, Pupa 118
Truncatellina 8, 17, 39, 56, 57, 63, 72, 74, 76,
80, 85,' 96, 104, 105, 215, 219, 220 *
Truncatellininae 8, 72, 80, 196, 215
tuberifera, Pseudochondrula, Buliminus, Chond-
rula 9, 239, 289*, 294, 295*,
tumidus, Zebrina, Buliminus 349
Turanena 9, 38*, 72, 74, 76, 83, 103, 239,
243, 274, 276*, 277
turanicus, Pseudonapaeus, Buliminus 251
396
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
turcmenica, Pupilla, Pupa 7, 178, 190*
turgidus, Brephulopsis, Chondrus 353
turritella, Columella 217
ujjalvyanus, Laevozebrinus, Buliminus 304
ujfalvyanus, Laevozebrinus, Subzebrinus 10,
63, 242, 301, 302*, 304*
umbilicata, Pyramidula, Euryomphalia 236
umbilicata, Lauria, Pupa, Turbo 150
umbrosa, Akramovskiella, Buliminus, Ena
10, 73, 97, 312, 313*
unibasalis, Euxinolauria 138
unicolumellaris, Euxinolauria 140, 142
unidentata, Pupilla, Pupa 185
urgutensis, Laevozebrinus, Buliminus, Sub-
zebrinus 10, 239, 301, 302*, 303 .
urmianus, Buliminus 10, 73, 241, 331*
Vallonia 6, 18, 47, 61, 74, 80, 84, 85, 90,
93, 105, 163
Vallonia (subg.) 163
Valloniidae 4, 6, 13, 20, 47, 61, 71, 75*, 80,
81, 86, 87, 89, 90, 92—94, 104, 110, 156,
158
Valloniinae 6, 71, 80, 81, 110, 162
vamberyi, Pseudonapaeus, Buliminus 253
varenzovi, Pseudonapaeus, Buliminus 252
variabilis, Granaria, Pupa 224
Vasopulmonata 80
venetzii, Vertilla, Vertigo 213
ventrosa, Vertigo 206
Vertigineae 95
Vertiginidae 4, 7, 13, 15, 20, 39, 63, 71, 75*,
80, 81, 86, 89*, 91, 96, 110, 195
Vertigininae 7, 60, 72, 80, 81, 95, 196, 198
Vertiginoidea 7, 42*, 89*, 95, 195
Vertigo 7, 17, 57, 63, 64, 74, 76, 80, 84—86,
90, 96, 104, 105, 198, 199
Vertigo (subg.) 200
vertigo, Vertilla, Vertigo 213
Vertilla 8, 58, 74, 85, 198, 213
viator, Imparietula, Chondrula 307
viridana, Vertigo, Alaea 201
Virpazaria 93
vulgaris, Vertigo, Alaea 207
walteri, Pseudonapaeus, Buliminus 251
zaru, Vallonia 7, 163, 170*
zebra, Chondrus, Buliminus, Helix 11, 240,
350, 351*
Zebrina 11, 73, 74, 83, 85, 244, 274, 301, 338,
348, 356
Zebrinus 348
zebriola, Chondrus, Helix 351
zebropsis, Chondrus, Brephulus 351
zonata, Euxinolauria,. Lauria, Pupa 148
zonifera, Euxinolauria 6, 62, 71, 134, 148,
149*
Zoogenetes 6, 71, 74, 76, 80, 94„ 104, 159
Zoogenites 159
Zua 111
СОДЕРЖАНИЕ
Предисловие ..............'................................................ 3"
Систематический указатель видов............................................... 5
Общая часть ................................................................ 13
Общая характеристика моллюсков подотряда Pupillina........................... 13
Морфология и анатомия представителей Pupillina.............................. 15
Внешний вид и раковина.................................................... 15
Реноперикардиальный комплекс и легкое..................................... 21
Пищеварительный тракт ................................................ . 23
Опорно-двигательный аппарат .............................................. 30
Нервный аппарат........................................................... 35
Половой аппарат.......................................................... 37
Биология и экология.......................................................... 44
Питание................................................................... 44
Размножение............................................................... 46
Среда и адаптации......................................................... 49
Ювенильные адаптации .................................................. 50
Дефинитивные признаки.................................................. 53
Устьевая арматура.................................................. 53
Окраска.......................................................... 61
Размеры и форма раковины ........................................... 62
Половой аппарат .................................................... 65
Географическое распространение Pupillina..................................... 70
Ареалы семейств и подсемейств Pupillina..............................1 . 70
Распространение Pupillina на территории СССР и структура фауны ....... 73
Степень эндемизма фауны................................................... 76
Вопросы системы и филогении.................................................. 80
Развитие взглядов на систематическую структуру и положение Pupillina в отряде
Geophila.................................................................. 80
Таксономическая и филогенетическая структура подотряда Pupillina......... 83'
Общие замечания.......................................................... 84
Проблемы филогении ....................................................... 85
Вопросы системы подотряда ................................................... 88
Надсем. Orculoidea........................................................ 91
Надсем. Cochlicopoidea.................................................... 94
Надсем. Pupilloidea ..................................................... 95
Надсем. Vertiginoidea .................................................... 95
Надсем. Chondrinoidea.................................................... 96
Надсем. Buliminoidea.................................................... 97
Принципы и методы разработки системы Pupillina.............................. 99
Методы исследования ........................................................ ЮЗ
Сбор, фиксация и хранение материала....................................... ЮЗ
Камеральная обработка ................................................... 105
Систематическая часть ...................................................... 109
Условные сокращения и обозначения........................................... 109
Таблица для определения семейств и-некоторых подсемейств подотряда Pupillina 110
I. Надсем. Cochlicopoidea................................................ 111
Сем. Cochlicopidae ................................................... Ill
1. Род Cochlicopa Ferussac........................................ Ill
II. Надсем. Orculoidea...................................................-. lift
398
ОГЛАВЛЕНИЕ
I. Сем. Orculidae....................'........................................ 116
I. Подсем. Orculinae........................................................ 117
1. Род Pilorcula Germain................................................. 118
2. Род Orculella Steenberg............................................... 120
3. Род Orcula Held....................................................... 124
4. Род Sphyradium Charpentier............................................ 126
II. Подсем. Pagodulininae.................................................. 128
1. Род Pagodulina Clessin................................................. 129
III. Подсем. Lauriinae..................................................... 133
1. Род Euxinolauria Lindholm . . . . !.................................... 133
2. Род Lauria Gray........................................................ 150
IV. Подсем. Argninae........................................................ 152
1. Род Argna Cossmann.................................................... 153
II. Сем. Strobilopsidae...................................................... 154
1. Род Strobilops Pilsbry ................................................ 155
III. Сем. Spelaeodiscidae . ............................................... 156
1. Род Spelaeodiscus Brusina..............................,............... 156
IV. Сем. Valloniidae........................................................ 158
I. Подсем. Acanthinulinae.................................................. 158
1. Род Zoogenetes Morse .................................................. 159
2. Род Acanthinula Beck................................................... 160
3. Род Spermodea Westerlund . . . :....................................... 161
II. Подсем. Valloniinae................................................... 162
1. Род Vallonia Risso.................................................... 163
III. Надсем. Pupilloidea......................... . ........................ 171
Сем. Pupillidae............................................................... 171
1. Род Gibbulinopsis Germain..........................................' 171
2. Род Pupilla Turton..................................................... 177
3. Род Pupoides L. Pfeiffer............................................... 194
IV. Надсем. Vertiginoidea ................................................ • 195
Сем. Vertiginidae........................................................... 195
I. Подсем. Gastrocoptinae.................................................. 196
1. Род Gastrocopta Wollaston...........................................•• • 196
II. Подсем. Vertigininae................................................ • 198
1. Род Vertigo Miiller.................................................... 199
2. Род Vertilla Moquin-Tandon.......................................... 5 213
III. Подсем. Truncatellininae................................................ 215
1. Род Columella Westerlund.............................................. 215
2. Род Truncatellina Lowe............................................. • 219
V. Надсем. Chondrinoidea................................................... • 224
I. Сем. Chondrinidae ................................................. • • 224
1. Род Granaria Held..................................................... 224
2. Род Abida Turton...................................7'............. • 226
3. Род Chondrina Reichenbach.............................................. 228
II. Сем. Pyramidulidae ...................................................... 236
1. Род Pyramidula Fitzinger.............................................. 236
IV. Надсем. Buiiminoidea........................................................ 238
Сем. Buliminidae ............................................................. 238
I. Подсем. Pseudonapaeinae.................................................. 243
1. Род Pseudonapaeus Westerlund..................................... • 244
2. Род Subzebrinus Westerlund ............................................ 272
3. Род Turanena Lindholm................................................. 274
4. Род Pseudochondrula Hesse.............................................. 288
5. Род Triangustoma gen. nov.............................................. 296
6. Род Mastoides Westerlund............................................... 297
7. Род Laevozebrinus Lindholm............................................. 301
8. Род Imparietula Lindholm.......................................... • 306
9. Род Ljudmilena gen. nov................................................ 309
10. Род Akramovskiella gen. nov.......................................... 312
11. Род Differena gen. nov.............................................. 315
12. Род Geminula Lindholm................................................. 317
II. Подсем. Chondrulopsininae . . ......................................... 321
ОГЛАВЛЕНИЕ
.’III!)'
1. Род Chondrulopsina Lindholm .......................................... 321
2. Род Siraphoroides Schileyko .......................................... A24
Ш. Подсем. Jaminiinae ............................................ 32(>-
1. Род Jaminia Risso..................................................... 326
IV. Подсем. Merdigerinae................................................... 32S
1. Род Merdigera Held.................................................... 328
V. Подсем. Bulimininae . ;.............................................. 330
1. Род Buliminus Beck................................................. 330
2. Род Adzharia Hesse ............................................... 332
VI. Подсем. Retowskiinae .................................................. 334
1. Род Andronakia Lindholm ............................................. 334
2. Род Retowskia 0. Boettger............................................ 336
VII. Подсем. Eninae...................................................... 3.38
1. Род Ena Turton . ..................................................... 339
2. Род Peristoma Krynicki................................................ 341
3. Род Caucasicola Hesse................................................. 346
4. Род Zebrina Held.................................................... 348
5. Род Chondrus Cuvier................................................ 350
6. Род Brephulopsis Lindholm............................................ 352
7- Род Napaeopsis Sturany et Wagner..................................... 356
8. Род Thoanteus Lindholm............................................... 358
9. Род Ramusculus Lindholm............................................. 360
10. Род Chondrula Beck . . . '.......................................... 362
VIII. Подсем. Multidentulinae ............................................ 371
1. Род Multidentula Lindholm............................................ 371
2. Род ImproVisa Schileyko............................................. 373'
3. Род Senaridenta Schileyko ........................................... 375
Incertae sedis.................................................................... 376
Литература............................................................... ; . . . 377
Алфавитный указатель латинских названий моллюсков................................. 389
Анатолий Алексеевич Шилейко
НАЗЕМНЫЕ МОЛЛЮСКИ ПОДОТРЯДА PUPILLINA
ФАУНЫ СССР
(Фауна СССР. Моллюски. Т. III, вып. 3)
Утверждено к печати
Зоологическим институтом Академии наук СССР
Редактор издательства И. Л. Песенко
Технический редактор М. Э. Карлайтис
Корректоры О. И. Буркова и С. В. Феофанова
ИВ № 20678
Сдано в набор 08.08.83. .Подписано к печати 14.06.84. . М-30767.
Формат 70xl087ie. Бумага книжно-журнальная. Гарнитура
обыкновенная. Печать высокая. Усл. печ. л. 35.00. Усл. кр.-
отт. 35.00. Уч.-изд. л. 38.91. Тираж 1000. Тип. зак. 666.
Цена 6 р. 30 к.
Издательство «Наука». Ленинградское отделение.
199164, Ленинград, В-164, Менделеевская лин., 1.
Ордена Трудового Красного Знамени
Первая типография издательства «Наука».
199034, Ленинград, В-34, 9 линия, 12.