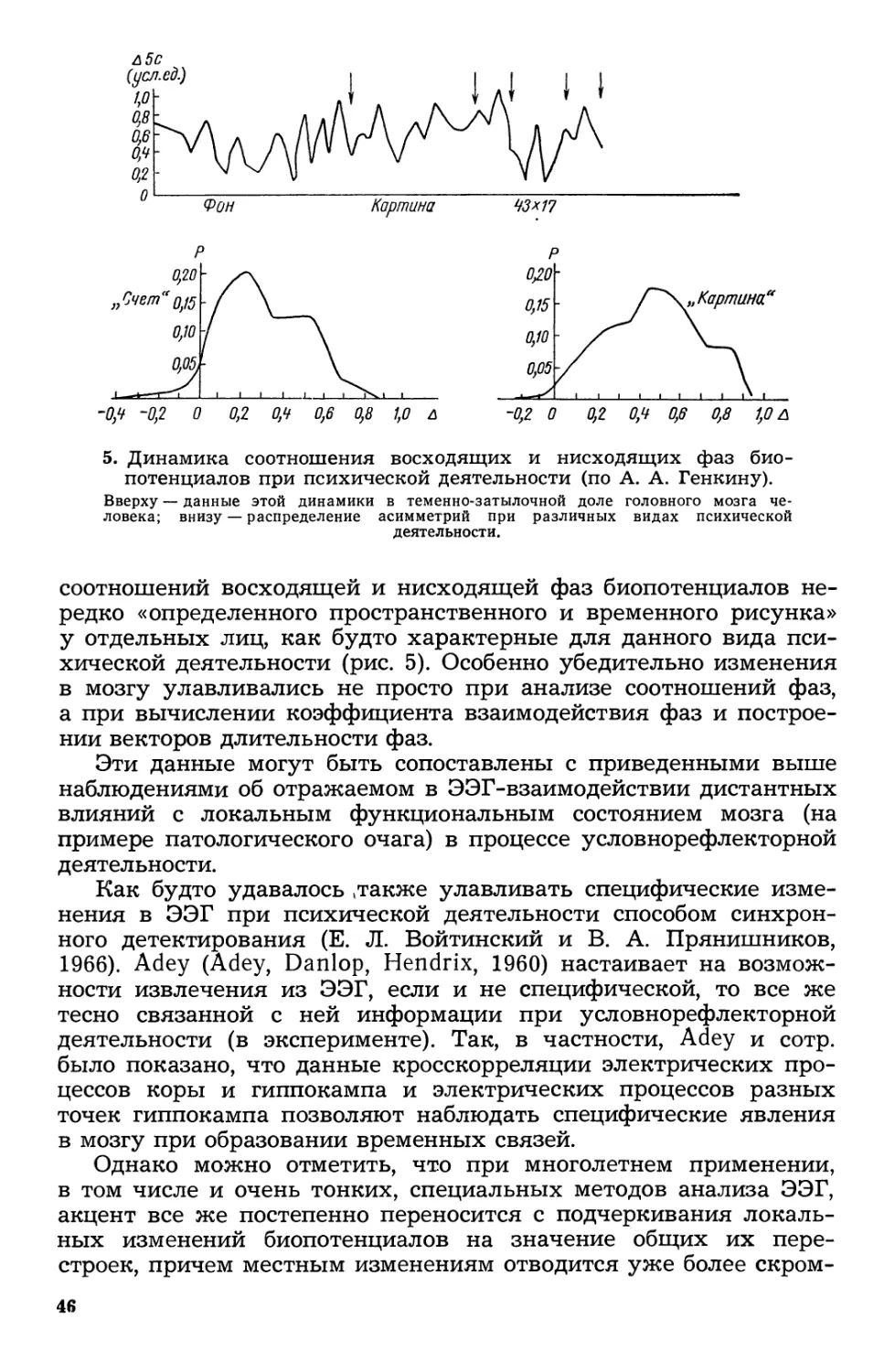
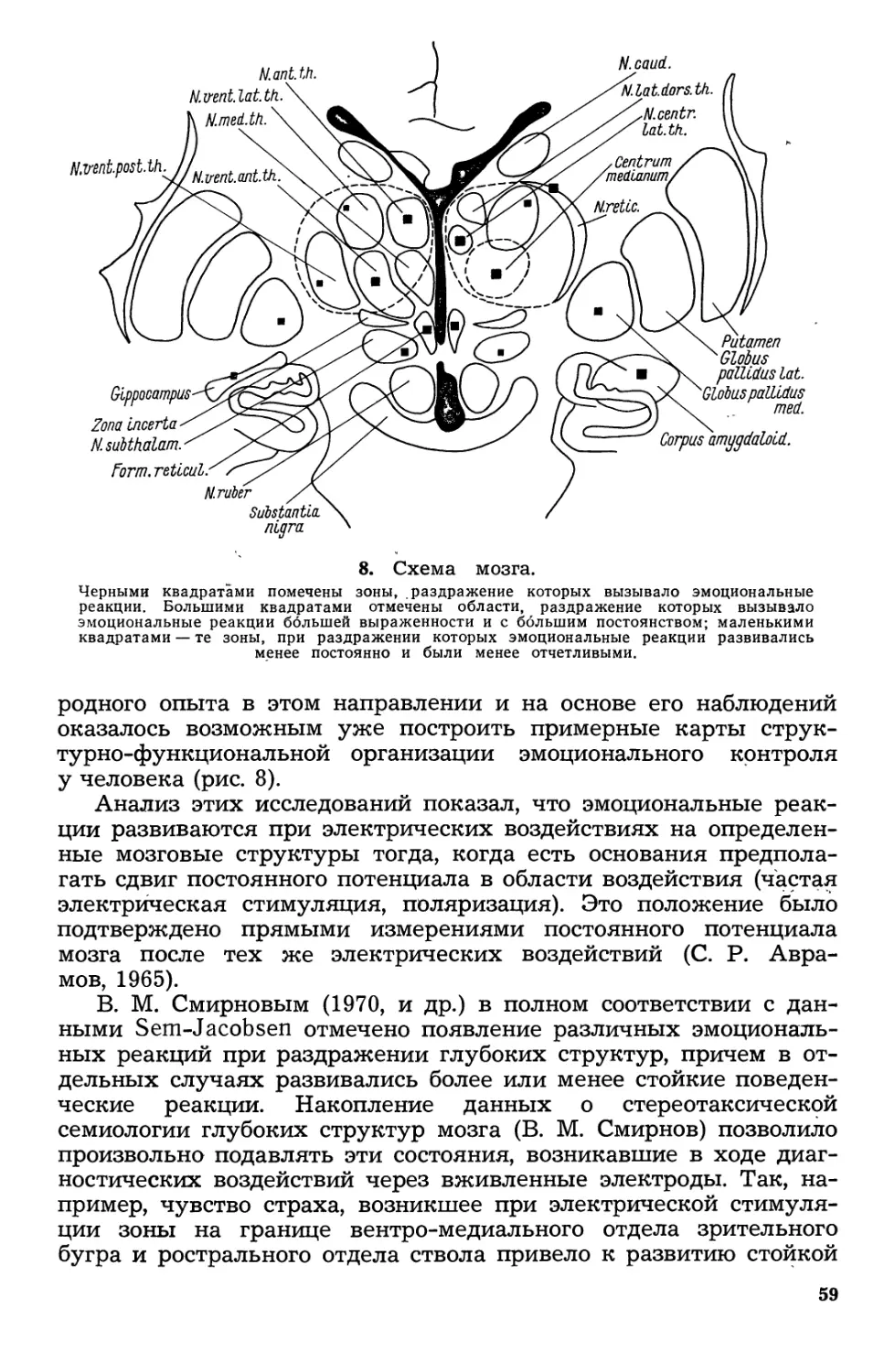
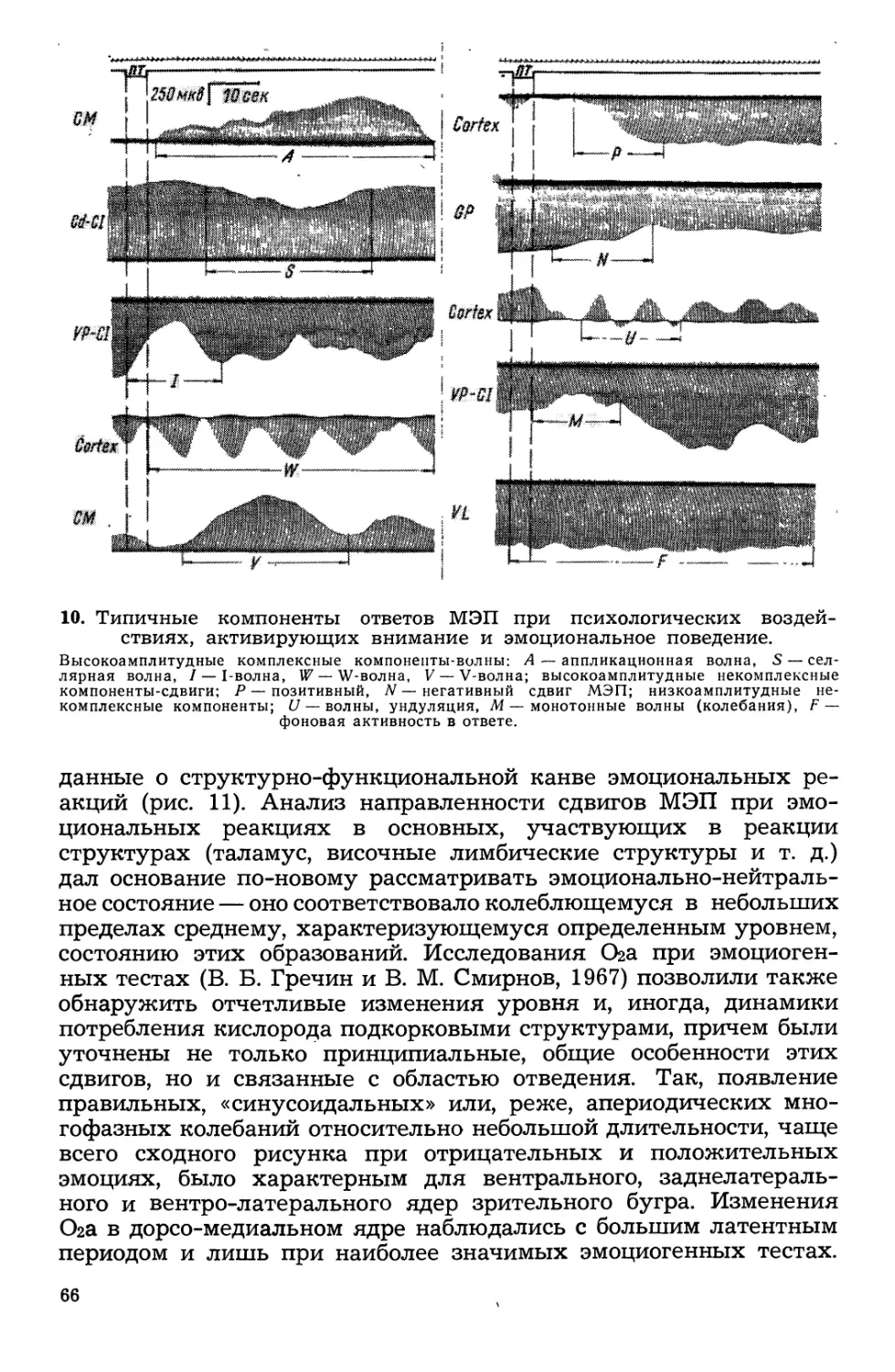
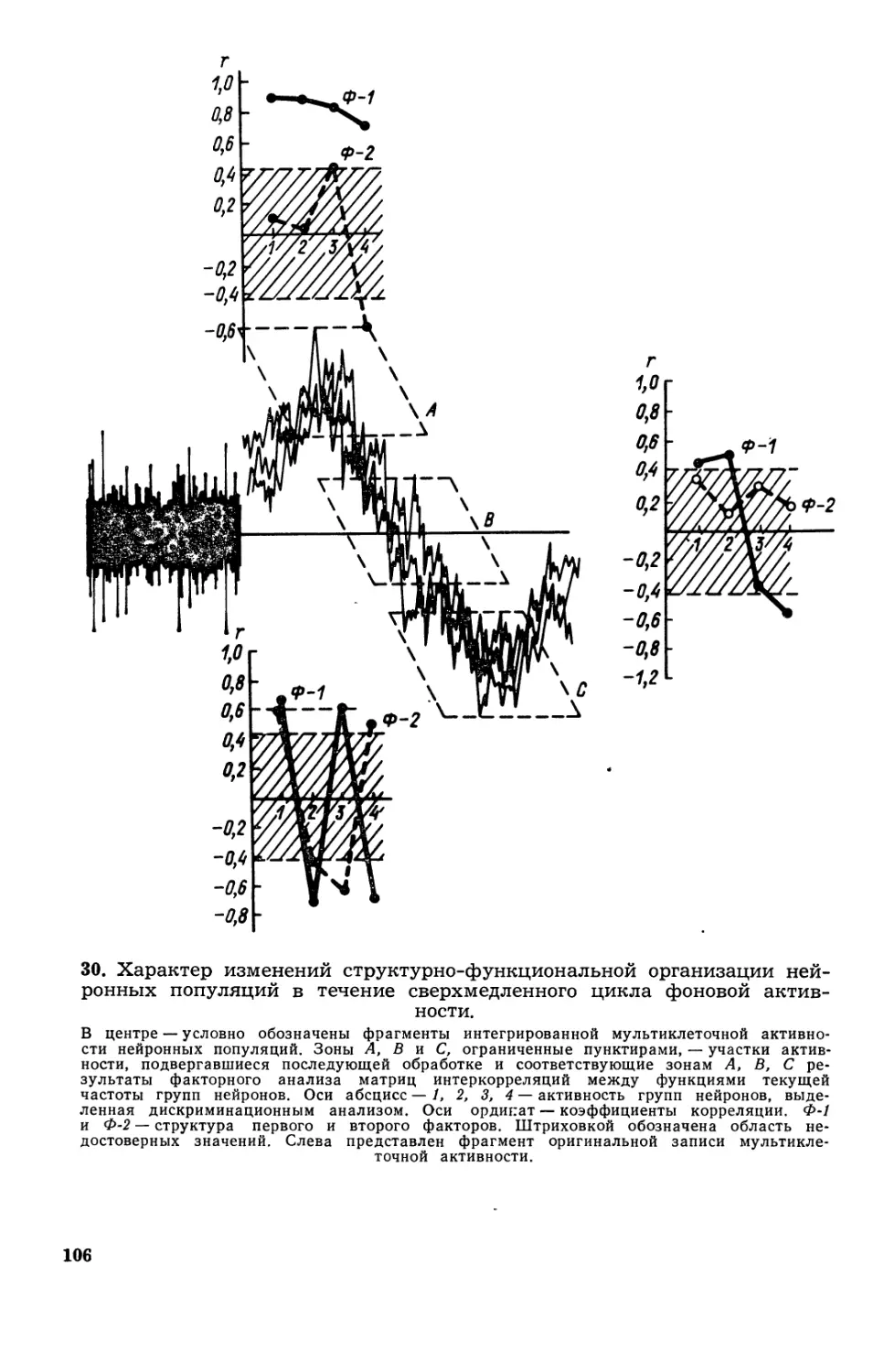
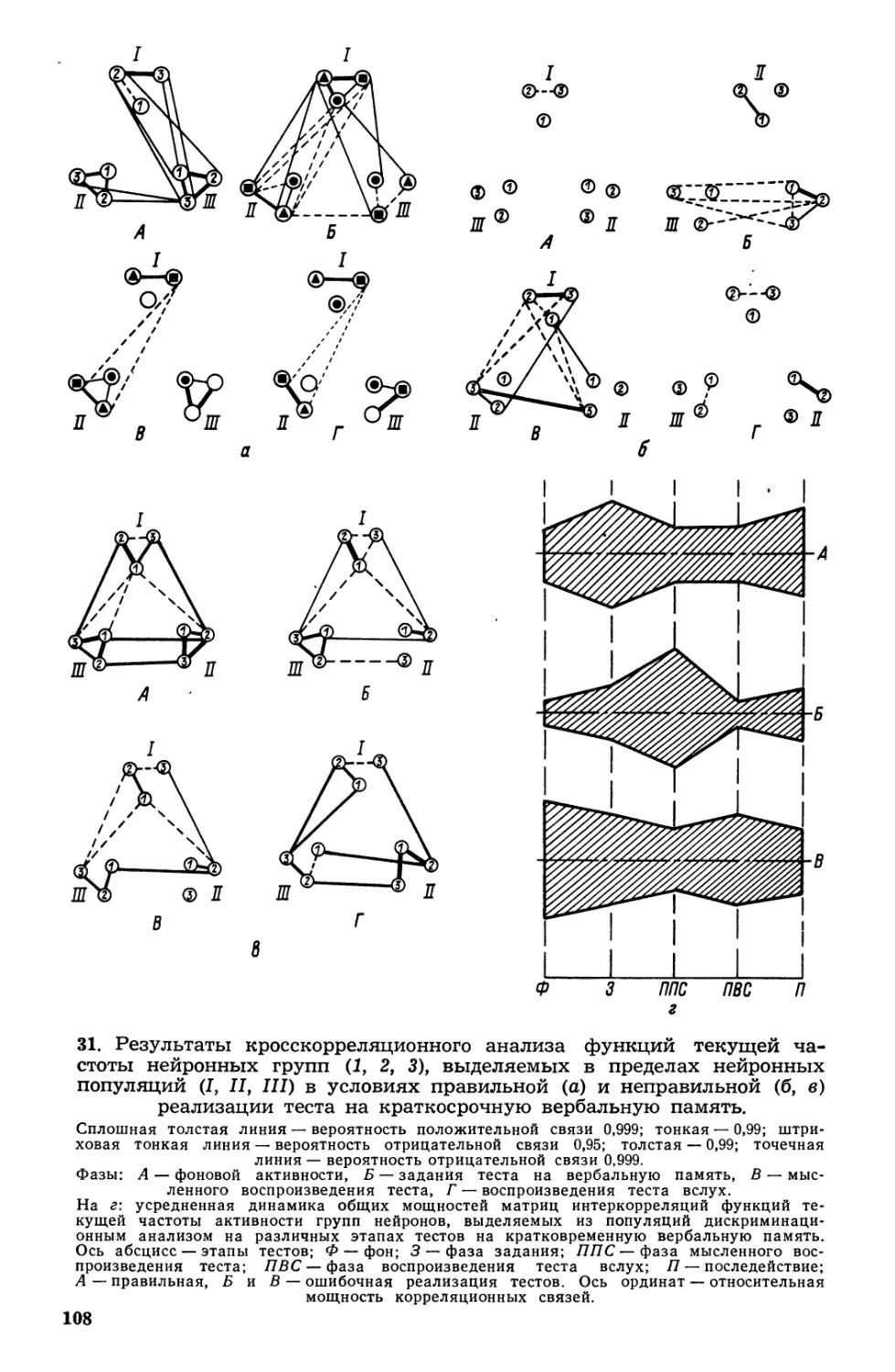
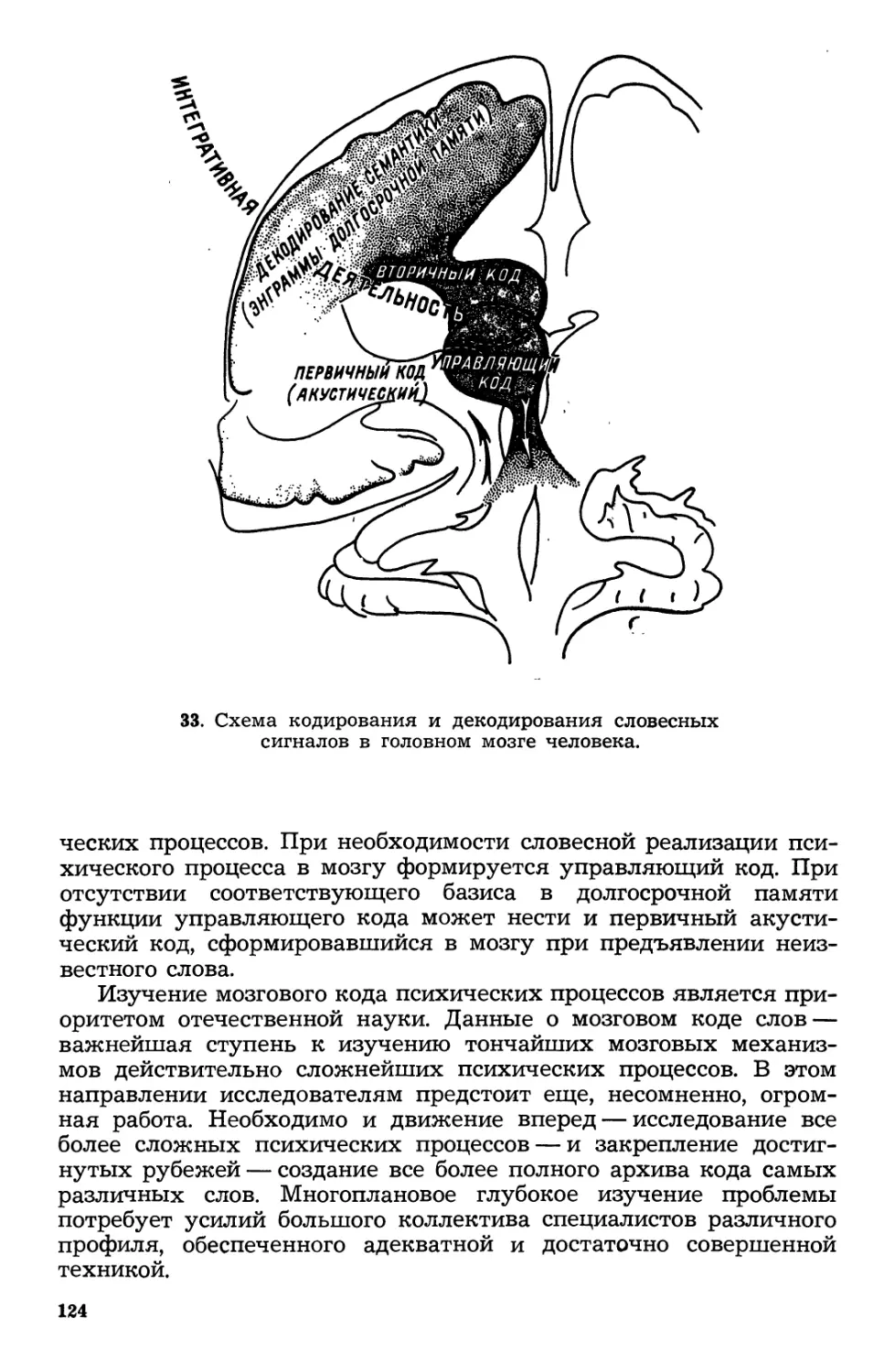
/




























































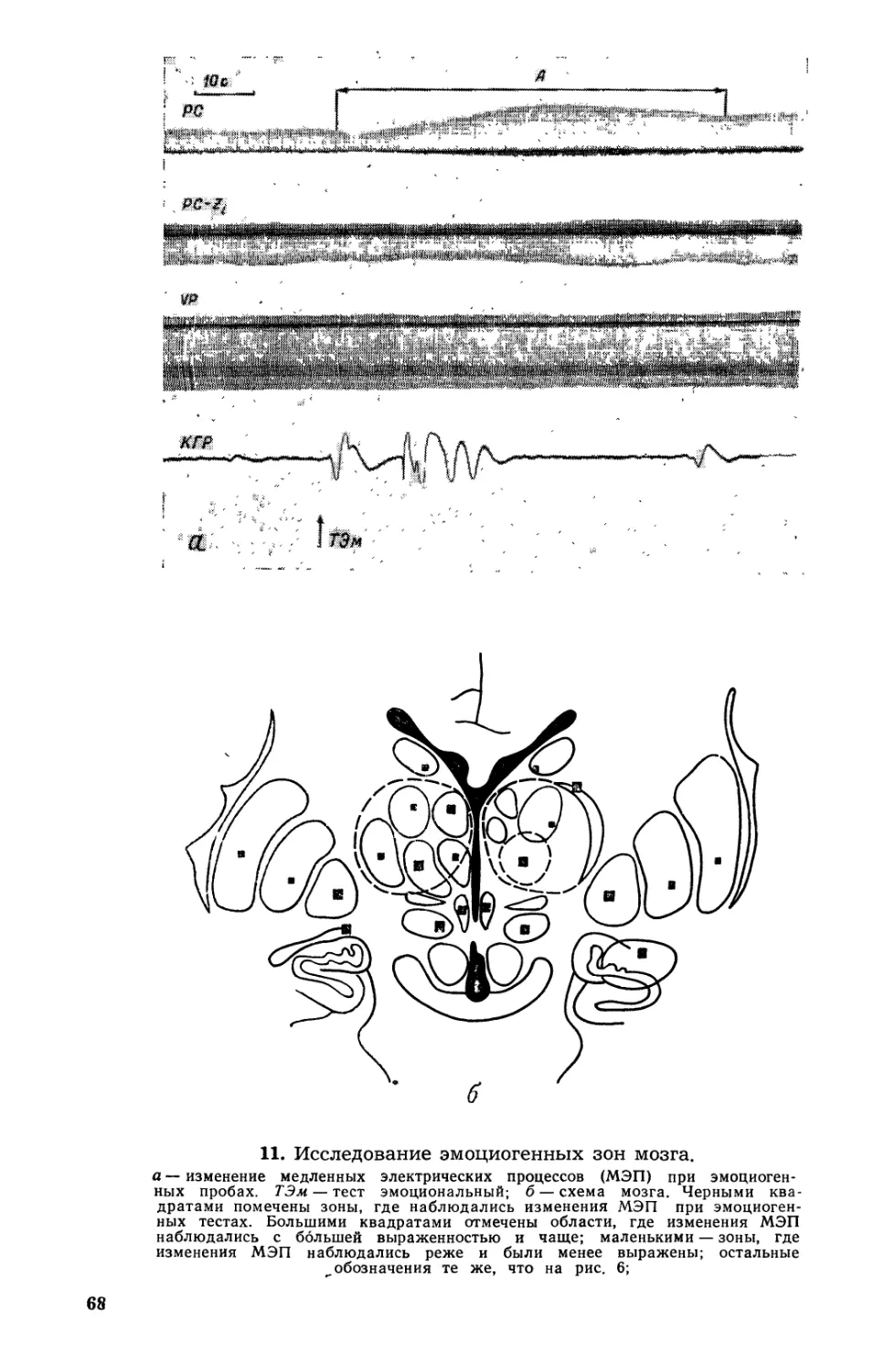
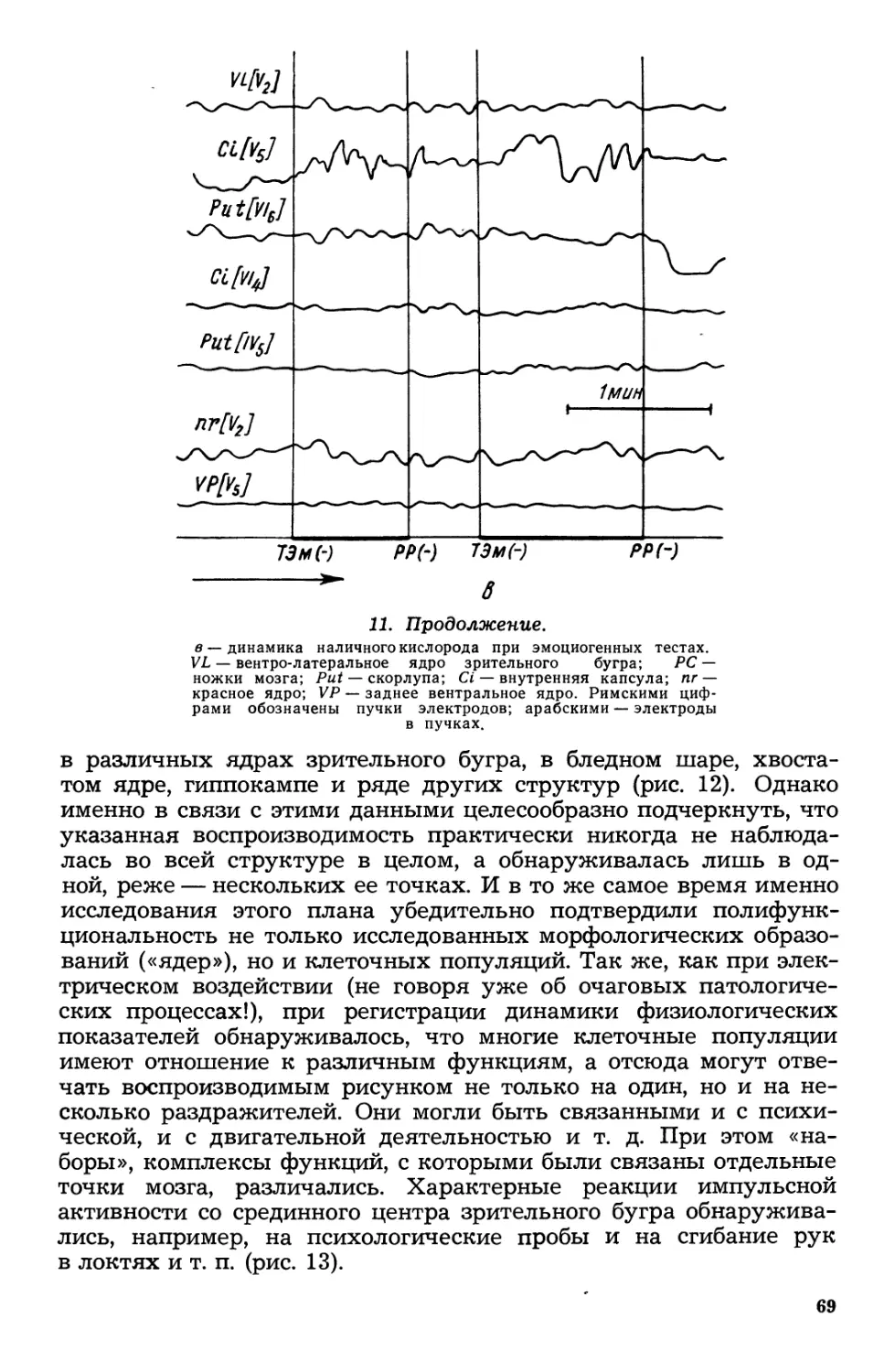
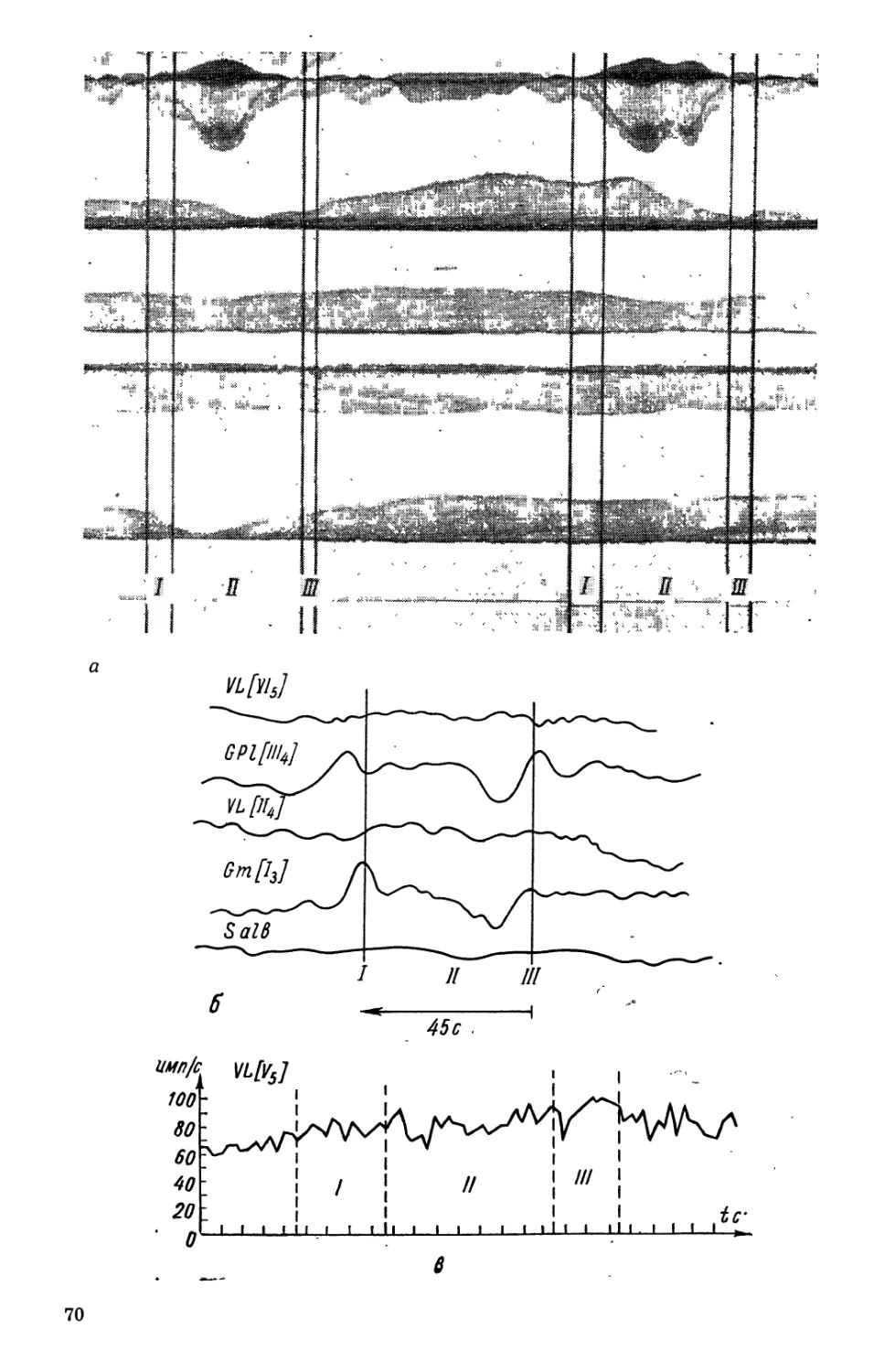
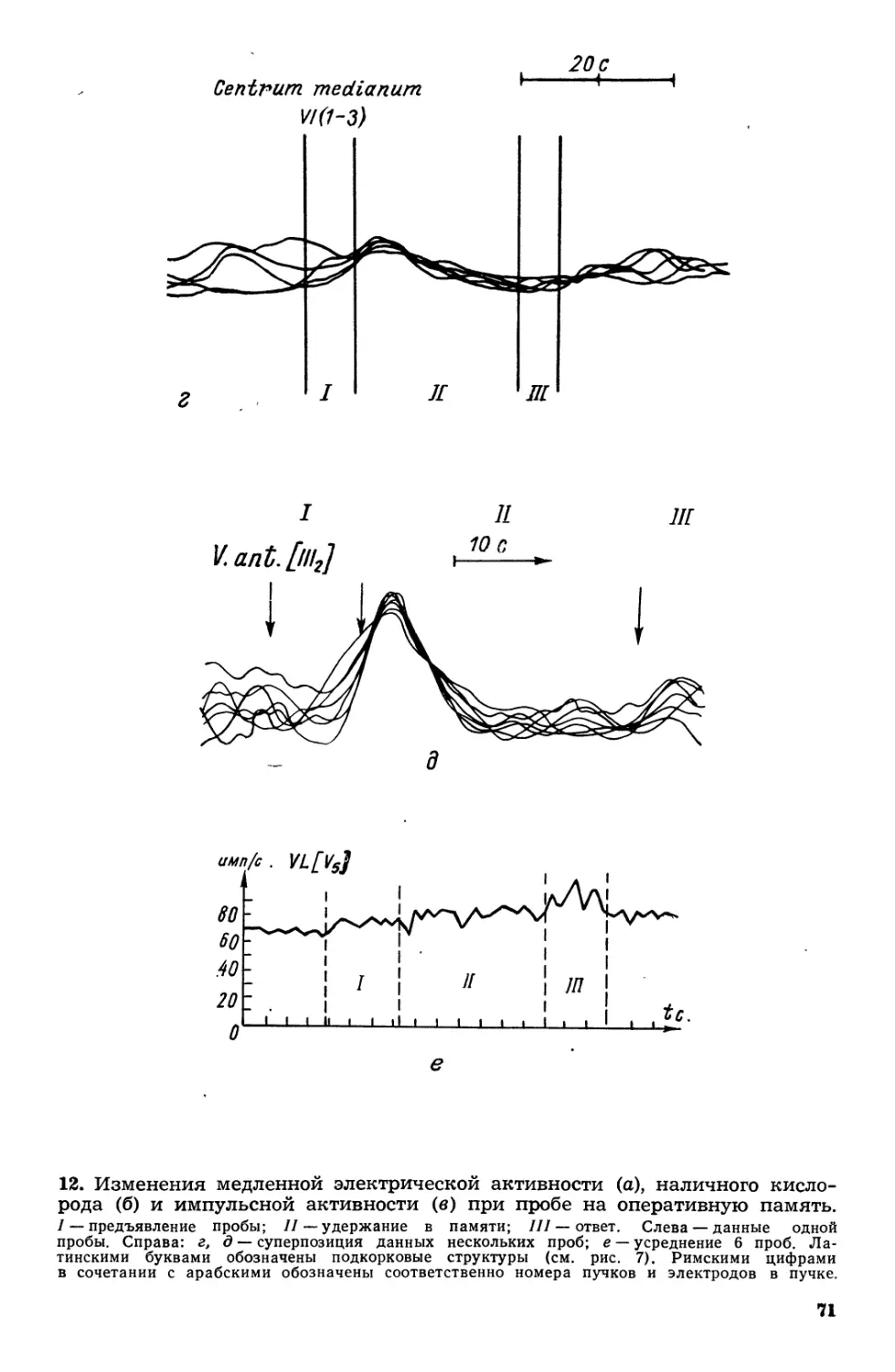
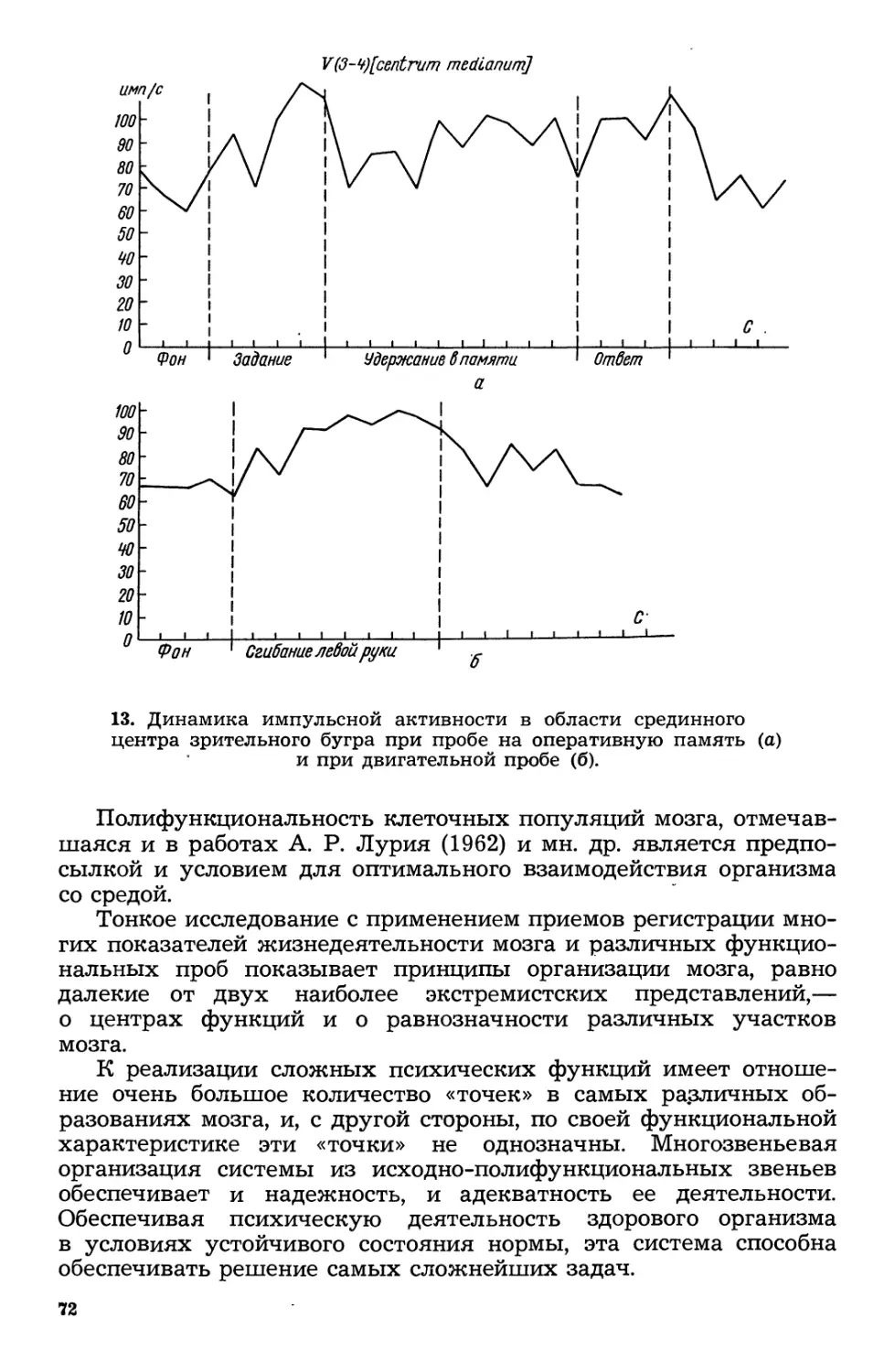

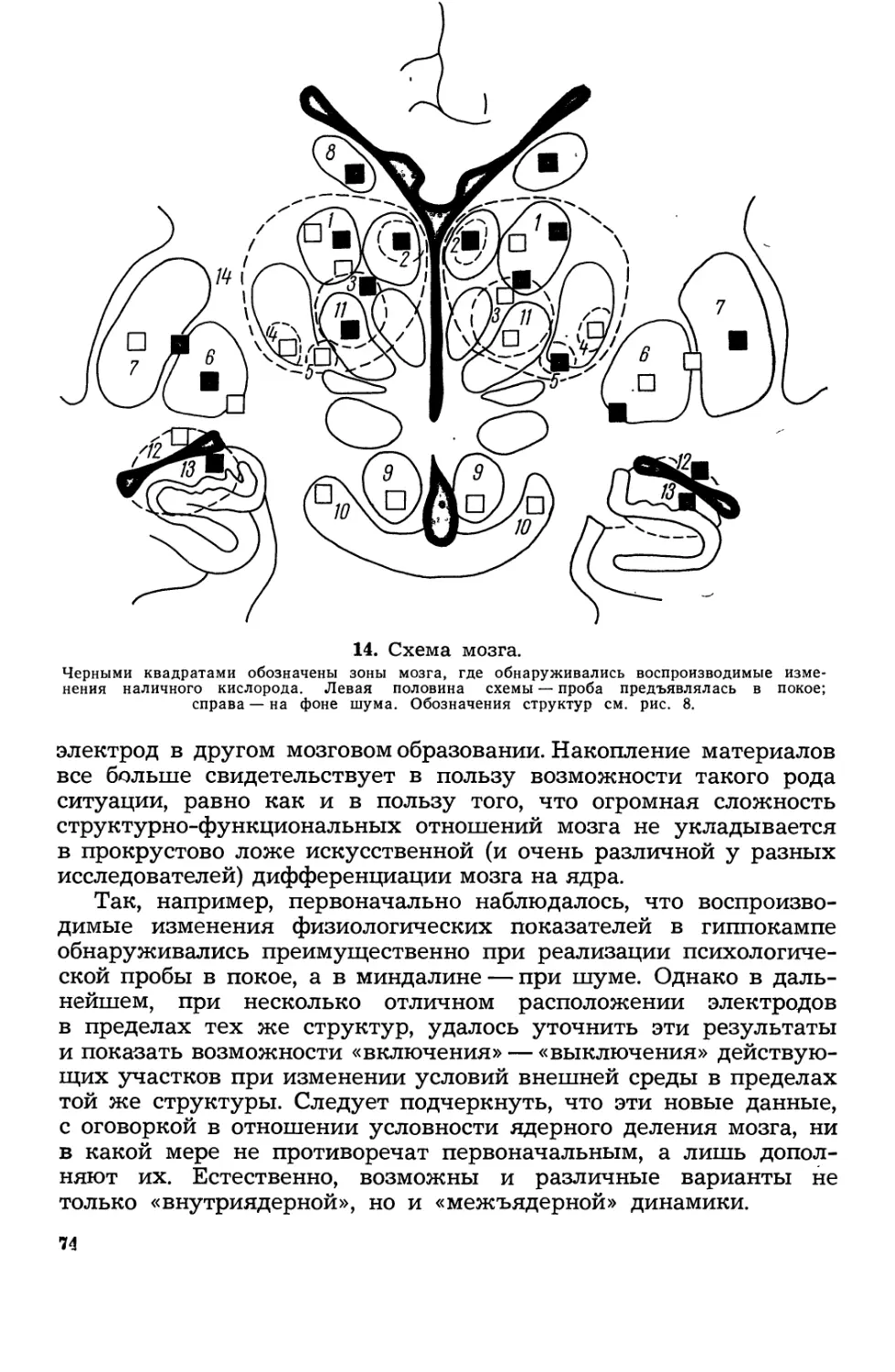



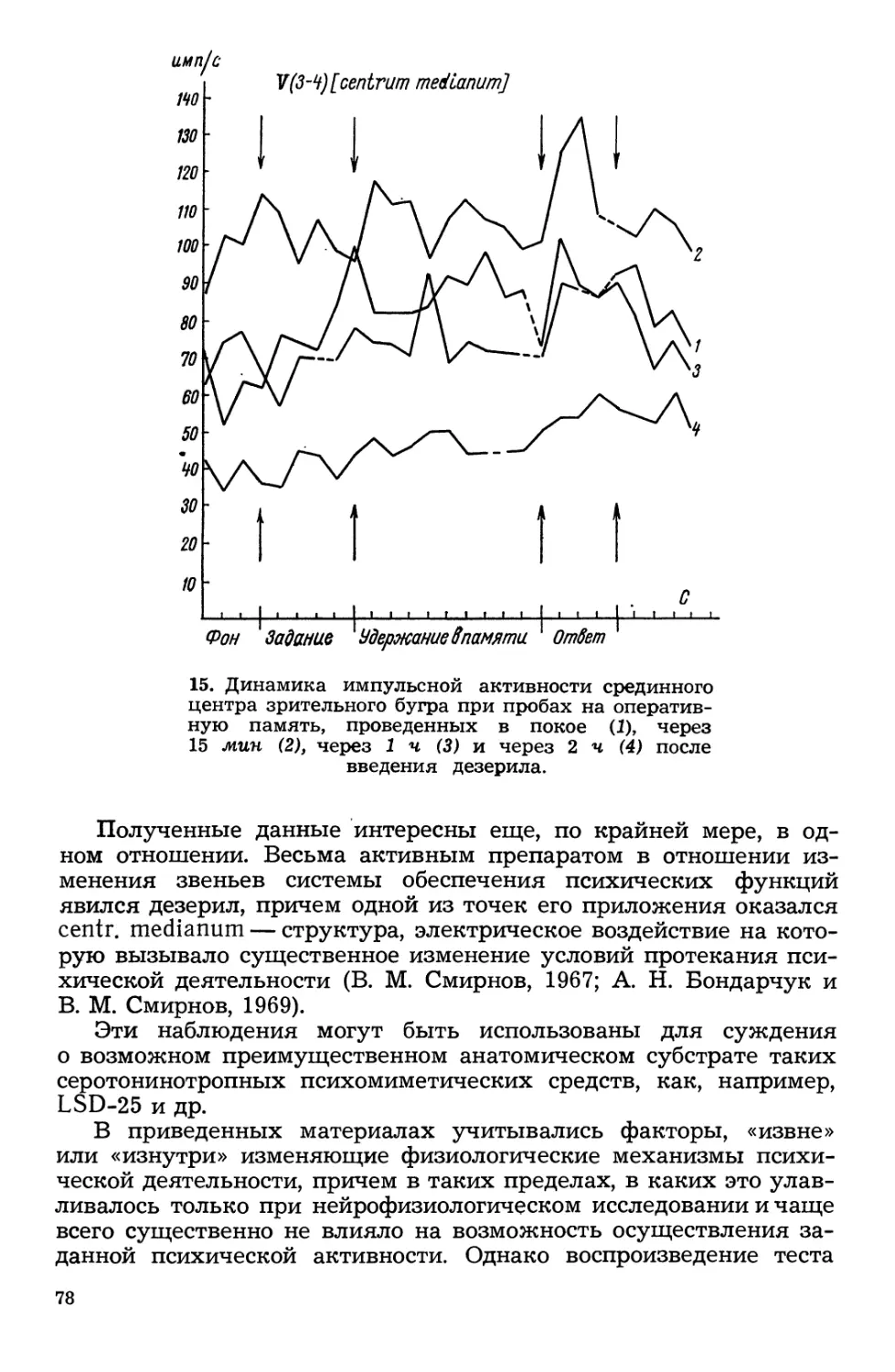
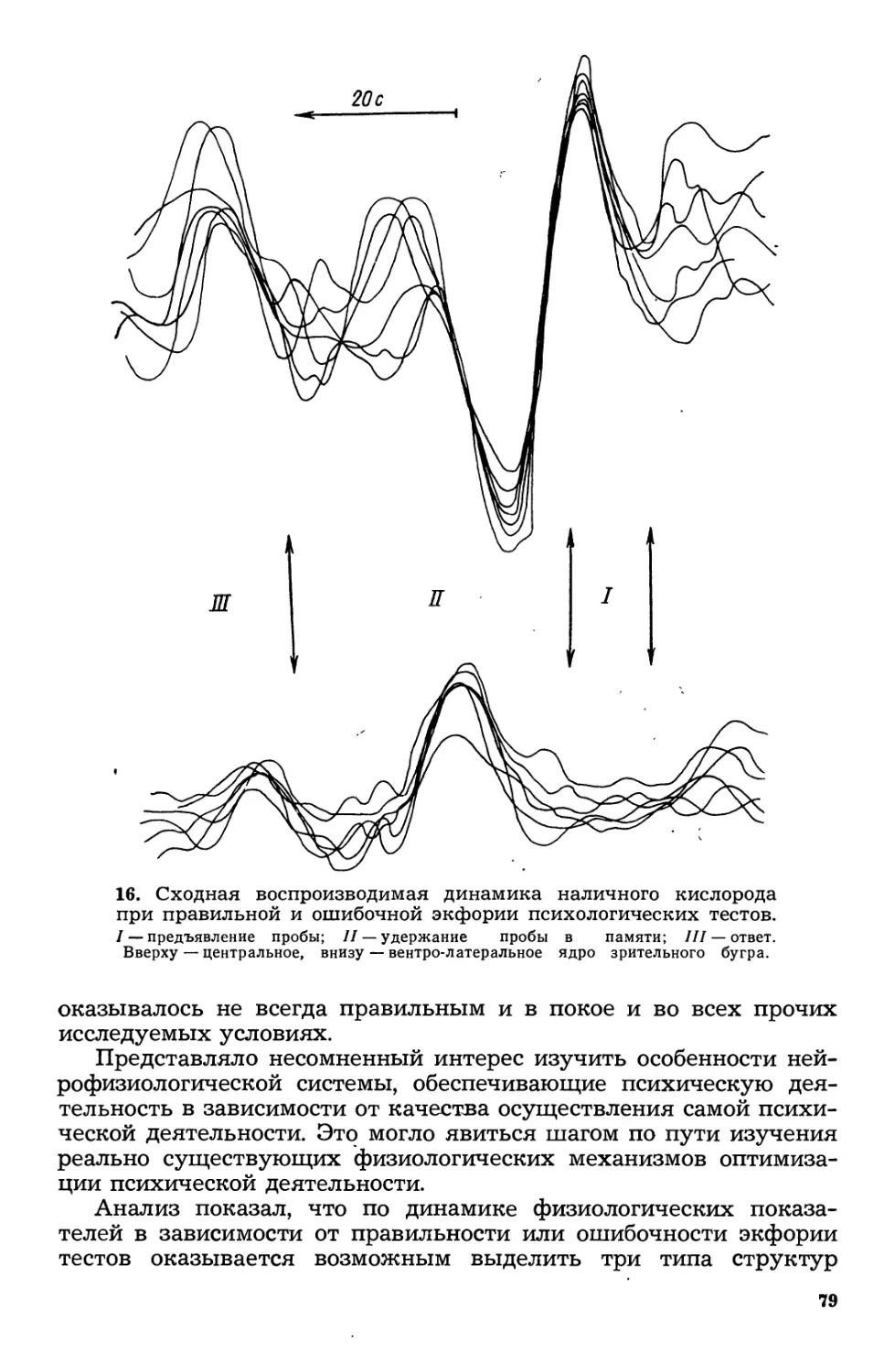
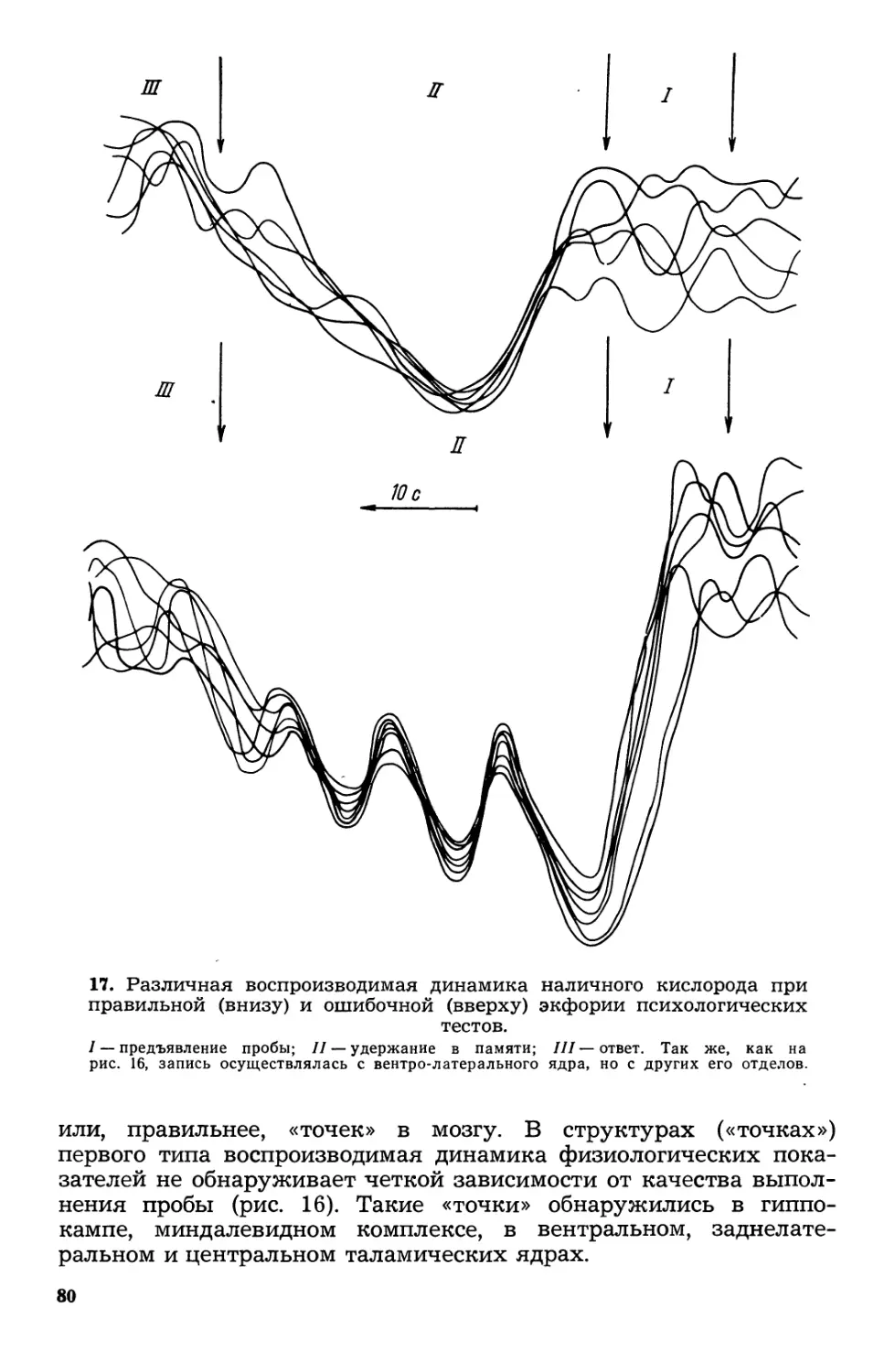



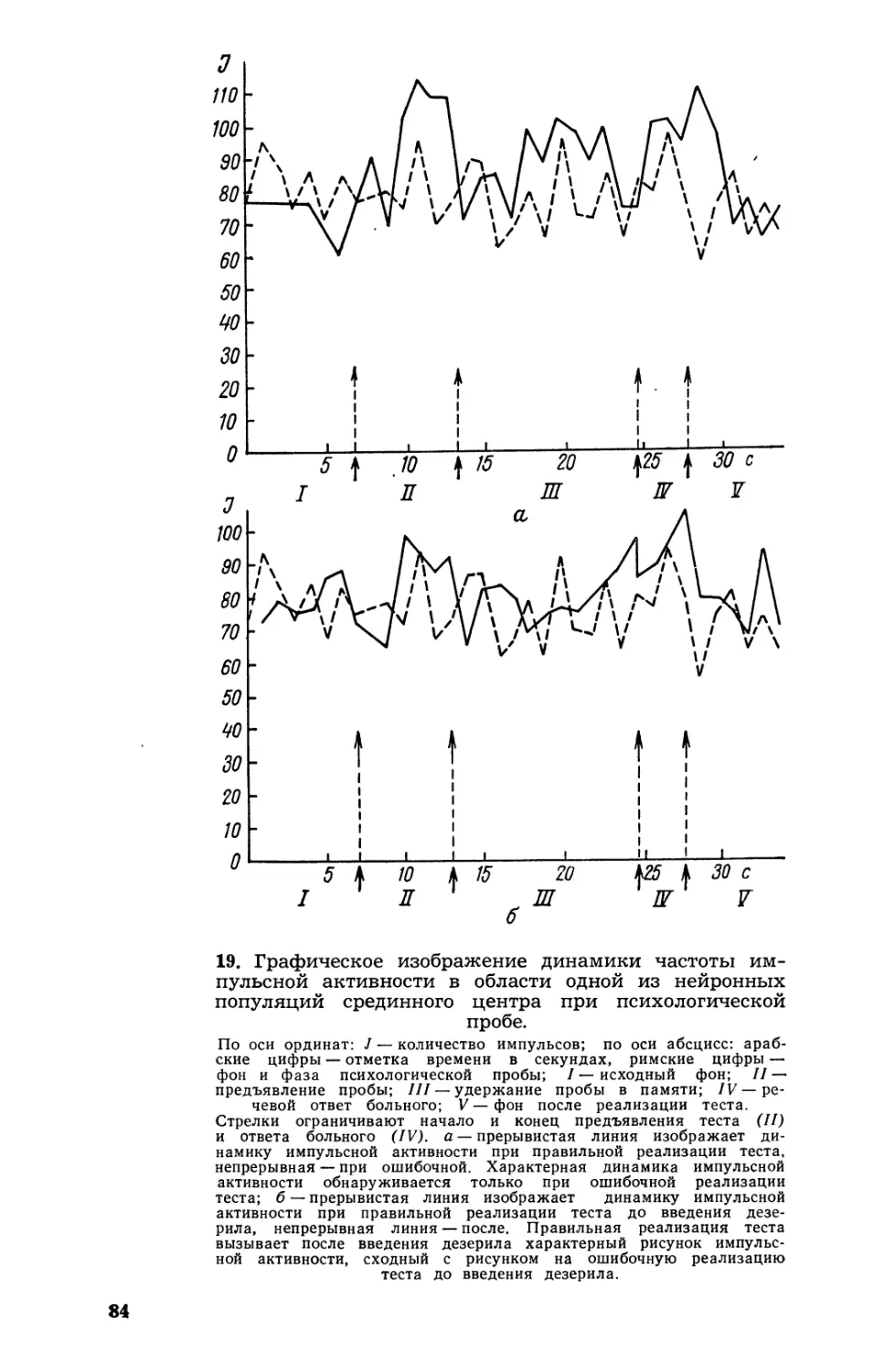








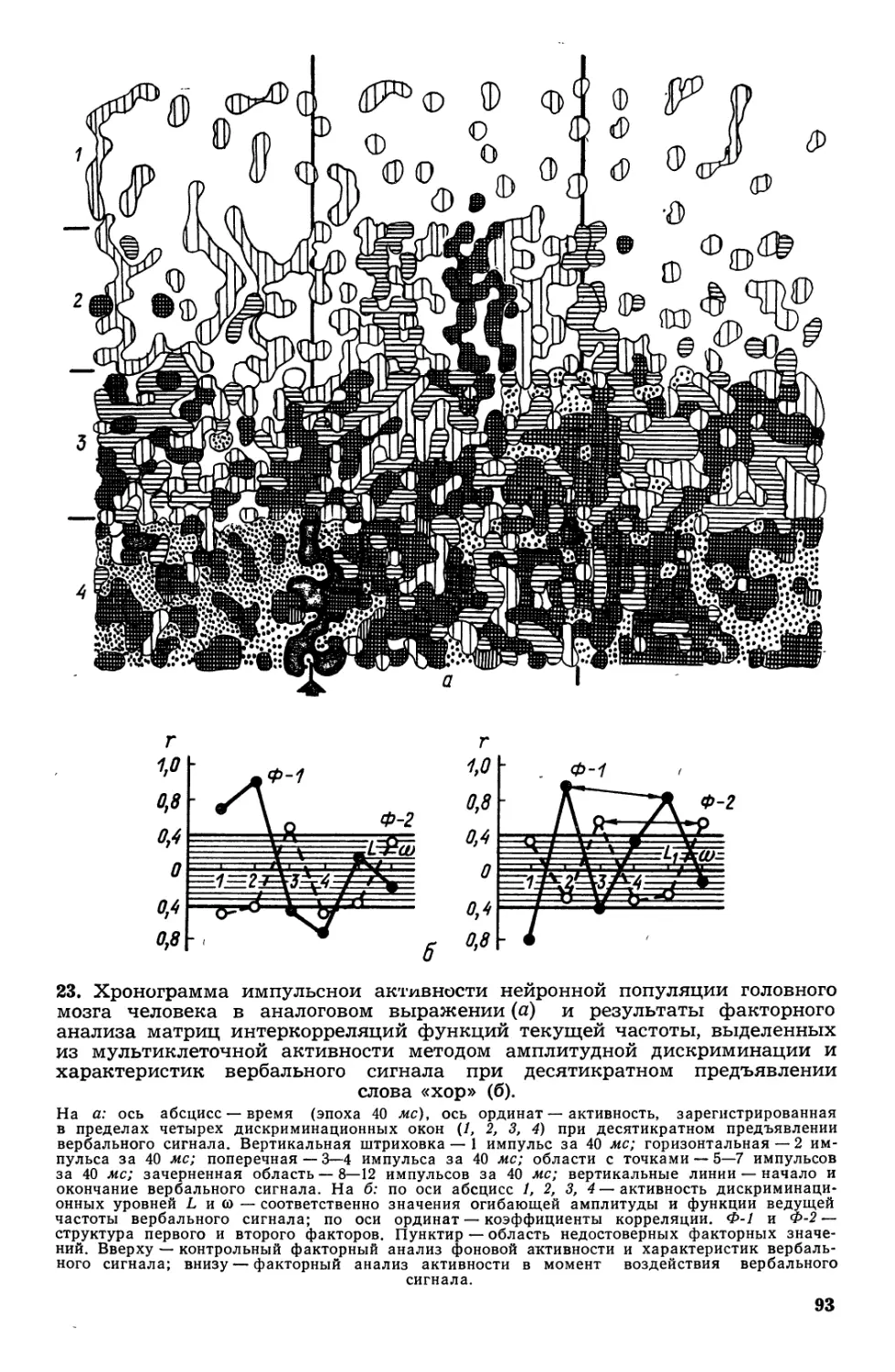


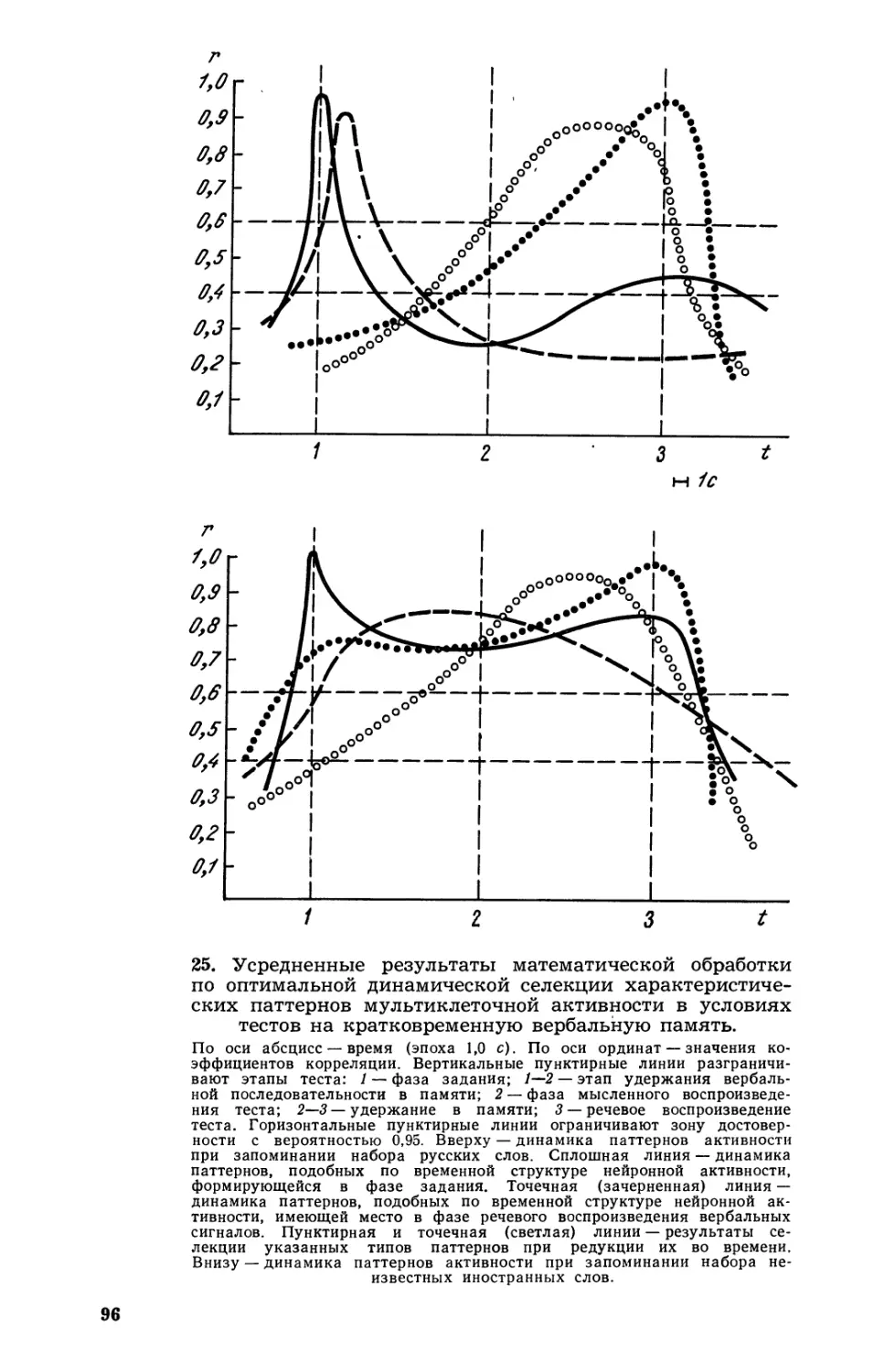





















































Текст
Н. п. БЕХТЕРЕВА
НЕЙРОФИЗИОАоrИЧЕСКИЕ АспЕктыI
ПСИХИЧЕСКОЙДЕЯТЕАЬНОСТИ
ЧЕАОВЕКА
Издание второе, переработаНRое
и дополненное
.,
/M
Л Е Н И Н r РАД. сеМ Е Д И Ц И Н А»
ЛЕНИнrРАДСКОЕ ОТДЕЛЕНИЕ. 1974
удк 612.821
Б е х т е р е в а Н. п.
Нейрофизиолоrические аспекты
психической деятельности
человека
Издание второе, переработанное и дополненное, 1974.
в работе излаrаются основные этапы изучения физиолоrии мозrовоrо
обеспечения психических процессов и современные данные о нейрофизио
лоrических механизмах этих процессов, полученные в результате прямоrо
изучения физиолоrии мозrа человека.
Мноrолетние исследования по ходу диаrностики и лечения больных
методом вживления электродов с помощью комплексноrо метода, вклю
чающеrо наблюдение динамики физиолоrических показателей мозrа при
реализации психической деятельности и динамики спонтанных и вызван
ных психических процессов при локальных электрических воздействиях
на мозr, позволили накопить большое количество новых данных о физио
лоrических механизмах психических явлений. В результате анализа этих
данных выдвинуто предположение, что мозrовое об спечение психической
деятельности осуществляется корково подкорковой структурно функцио"
нальной системой со звеньями различной степени жесткости.
Рассматриваются основные теоретические перспективы развития проб
лемы и возможности расширения лечебных воздействий в психиатриче
ской и нервной клинике на основе новых данных о мозrе человека.
2 e издание дополнено материалами о наиболее тонких характеристи
ках импульсной активности нейронных ансамблей мозrа человека при
психической деятельности.
Моноrрафия рассчитана на физиолоrов, невропатолоrов, психиатров
и врачей друrих специальностей.
Книrа содержит 33 рисунка. Библиоrрафия 337 названий.
For Summary see page 151.
Б 51700 102
36 74
039(01 ) 7 4
@ Издательсl'ВО «Медицина», Москва, 1974 r..
с изменениями
СОДЕРЖАНИЕ
Предисловие ко второму изданию
Введение .'.
Функциональное состояние мозrа человека при реализации условно
рефлеI ТОРНЫХ реакций и психолоrических проб
Электроэнцефалоrрамма при условнорефлекторных реакциях
Электроэнцефалоrрамма при психолоrических пробах
Электросубкортикоrрамма при психолоrических пробах. 3наче
ние ее изменений при психической деятельности
Структурно функциональная и нейрофизиолоrическая орrанизация моз
rOBoro обеспечения психической деятельности .
Принципы исследования элементов системы мозrовоrо обеспече
ния психической деятельности
Физиолоrическое значение и особенности различных звеньев си
стемы мозrовоrо обеспечения психических функций ....
Нейрофизиолоrическая сущность некоторых изменений, развива
ющихся в звеньях системы мозrовоrо обеспечения психической
деятельности . .
Динамика взаимодействия звеньев мозrовой системы обеспечения
психической деятельности .
Некоторые принципиальные ВОПRОСЫ и перспективы"развития проблемы
Изучение физиолоrии мозrа человека и эксперимент на живот
ных
Указатель литературы
4
5
19
19
41
47
57
57
73
85
101
110
132
. 135
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
в области изучения мозrовоrо обеспечения психической дея
тельности очень трудными были первые шаrи. Сейчас, косда
получение данных о мозrовой структурно функциональной opra
низации различных видов деятельности в условиях примене
ния метода вживленных электродов становится обычной диаrно
стической процедурой, открылись исключительные возможности
дальнейшеrо развития проблемы. Клинико физиолоrические pa
боты показали жизненность и перспективность методических
лечебных разработок на основе rипотезы об устойчивом патоло
rическом состоянии и поддерживающих ero реакциях, о роли
этоrо состояния при так называемых хронических заболеваниях
мозrа и лечебном значении ero преодоления, пере стройки мозrа
на новый L уровень функционирования. Доказана целесообраз
ность комплексноrо хирурrическоrо и медикаментозноrо лече
ния, множественных микролизисов, лечебной электрической сти
муляции мозrа и клиническоrо применения так называемоrо
управляемоrо эксперимента, адаптивноrо биоуправления. Особые
возможности представляет сочетание адаптивноrо биоуправления
с л ечебной электрической стимуляцией, позволяющее cTporo
целесообразно формировать новые функциональные системы
в условиях именно Toro режима работы мозrа, при котором ocy
ществимо оптимальное использование ero резервов.
Основным теоретическим направлением развития вопроса
явилось изучение тонкой структуры нейрофизиолоrических изме
нений в звеньях мозrовой системы обеспечения психических про
цессов, в первую очередь........... тех изменений, которые лежат в oc
нове «зашифровки» И «расшифровки» В мозrу словесных раздра
жителей.
Результаты исследований показали, что в мозrу развиваются
перестройки импульсной активности ансамблей нейронов, завися
щие от акустических характеристик слов, от частоты их употреб
ления и от наличия или отсутствия соответствующеrо базиса
энrрамм долrосрочной памяти. Эти исследования оказались Ha
столько значимыми, чтО их, по видимому, правомерно расцени--
вать как начало изучения HepBHoro кода психических процес
сов........... импульсноrо кода слов в первую очередь.
Важность новых материалов и целесообразность компактноrо
представления лоrики развития вопроса о физиолоrических Mexa
низмах психической деятельности и определила целесообразность
BToporo издания данной книrи.
,.
ВВЕДЕНИЕ
ПреДПОЛО:Нtение о связи мозrа с «разумом», «управляющим
духом» всем тем, что теперь называют психической деятель
ностью и центральной реrуляцией функций орrанизма заслуrа
мыслителей, живших мноrие сотни лет до нас (rиппократ и др.).
Абсолютная невозможность экспериментальноrо изучения
принципов и конкретных форм внутренней орrанизации актив
ности мозrа в те далекие времена порождала подчас неожидан
ные мнения. Так, около пятнадцати веков авторитетом rалена
поддерживалось представление о важнейшей роли ликворной си
стемы в психической деятельности. Ведущая роль в орrанизации
и реализации психической деятельности на разныIx этапах разви--
тия естественных наук в достаточно катеrоричной форме при
писывалась различным rлубинно расположенным образованиям
(Декарт, Виллис, Ланцизий и, MHoro позднее, Пенфильд) или,
наоборот, коре. Практически без сколько--нибудь серьезных фак
тических оснований эта, вторая точка зрения оказывается пред
ставлен ной в.более или менее схематичной форме во мноrих paH
них работах и достиrает cBoero апоrея в хорошо известной френо
лоrии rалля (XVIII в.).
Эти представления вновь активизируются, но уже на основе
большоrо клинико анатомическоrо материала с конца первой чет
верти XIX в., бурно развиваются во второй половине XIX в. и
формируют теорию локализационизма в учении о CTPYKTypHO
функциональной орrанизации мозrа (Буйо, 1825; Брока, 1861; Bep
нике, 1874, и мн. др.).
Каждодневная практика невропатолоrии, психиатрии и oco
бенно нейрохирурrии продолжала подтверждать тезис о связи
мозrа с психической деятельностью (с. С. Корсаков, 1890;
В. М. Бехтерев, 1900; Foersten, Gagel, 1933; Alpers, 1937; Griinthal,
1939; Buseh, 1940; М. М. Александровская и др., 1947; Gless и Grif
fith, 1952; Williams а. Pennybacker, 1954; Orthner, 1957; r. Б. Абра--
Мович и В. В. Захарова, 1961; А. Л. Абашев Константиновский,
1961, 1964; Victor et al., 1961; Barbizet, 1963; Milner, 1962, 1967;
Levita et al., 1967, и мн. др.).
ПО существу, тенденции cTporo связывать высшие ПСИХJ1че
ские функции с определенными областями коры, такие удобные
в невролоrической и нейрохирурrической клинике, нередко так
MHoro позволяющие решать в судьбе больноrо, живут в клинике
в более или менее удобно «оrоворенной» форме и по сие время.
Однако очаrовые патолоrические процессы «естественная
5
модель», воспроизводящая эксперимент с экстирпацией и отчасти
раздражением мозrа, сравнительно мало дали для физиолоrии
мозrа, понимания орrанизации системы мозrа, ответственной за
реализацию различных сложных психических функций. Анализ
же нейрофизиолоrических механизмов обеспечения этих функций
в связи с клинико--анатомическими наблюдениями оставался
чисто вербальным.
Массивность изменений структуры при очаrовых поражениях
мозrа не давала ключа и к пониманию феномена нарушения MHO
rих функций при изолированном очаrе.
Наиболее адекватным подходом к оценке данных, представ
ляемых очаrовыми патолоrическими процессами, являлась, и до
настоящеrо времени, по видимому, в значительной мере яв
ляется, позиция Хьюrлинса Джексона о неправомерности пред
положения о локализации центров функций на основе их Hapy
шений при повреждениях какой--либо (одной) области мозrа.
Оставаясь в пределах своеобразноrо невролоrическоrо «под
спорья» И не претендуя на объяснение общих принципов слож
нейшей деятельности мозrа, локализационистическое направле--
ние было весьма полезным практически, что как бы компенсиро
вало ero теоретическую неполноценность.
При развитии этоrо направления все, однако, дошло до лоrи
ческоrо абсурда, коrда мозr, и прежде Bcero кору, попытались
превратить в лоскутное rосударство карликовых суверенных
княжеств, каждое из которых «решало» свои, более или менее
сложные вопросы от контроля движений до релиrиозноrо
чувства, от контроля речи до социальноrо «я» И т. д.
Особенно серьезный удар локализационизму, как это ни пара
доксально, нанесло именно ero развитие.
Схематизированные карты локализации мозrовых функций,
особенно в первой половине ХХ в., переходят из учебника в учеб
ник, однако та же практика невролоrии накапливает теперь уже
все болыпе противоречий в этих представлениях. Противоречий
оказывается в клинико анатомических и просто клинических
данных такое множество, что они, по существу, создают реаль
ную предпосылку для друrой крайности в учении о функциях
мозrа почву для трудности безоrоворочноrо о"рицания нали--
чия, хотя и небольшоrо, но ощутимоrо рациональноrо «зернышка»
В своеобразных (мяrко rоворя) позициях Lashley (1958).
Можно отметить при этом, что, несмотря на известную
(в определенных пределах) практическую ценность локализацио
низма, именно теоретические представления эквипотенциализма
в более или менее трансформированном виде постоянно оживают
и в наши дни.
Одним из наиболее ярких примеров модернизации эквипотен--
циализма являются, пожалуй, взrляды Pribram (1969), по кото--
рым сложная психическая деятельность представлена в мозrу
полностью в каждом ero микрообъеме, а ее реализация связана
с вовлечением в деятельность макрообъема мозrа.
6
В известноЙ мере в противовес локализационистическим
воззрениям в физиолоrической и психолоrичuеской литературе
нередко все еще поддерживается, на первыи взrляд, вполне,
казалось бы, приемлемая точка зрения о том, что весь мозr
принимает участие в психической деятельности, психическая
деятельность обеспечивается мозrом «в целом». Действительно,
сложная психическая деятельность есть результат активности
или rотовности к ней очень мноrих отделов мозrа. И в то же
самое время подобная научная установка является, по существу,
демобилизующей, обесценивая или, в лучшем случае, не стиму
лируя поиск структурно--функциональной орrанизации обеспече--
ния мозrом психической деятельности.
Оrромный по объему и ценности, но жестко сфокусированный
по направленности опыт Penfield (1958) приводит к трансформи--
рованному оживлению некоторых представлений Meyer (1779)
о связи коры с более или менее сложными видами памяти и о pe
шении всех сложнейших вопросов реализации центральной инте--
rрации центрэнцефалической системой.
Обсуждению и критике локализационизма, эквипотенциа
лизма, холизма и центрэнцефалической системы посвящено очень
большое число работ. С известным правом можно сказать, что на
критический анализ полностью или частично ошибочных пред
ставлений в физиолоrии мозrа человека потрачено, пожалуй, осо--
бенно в последние десятилетия, едва ли не больше сил, чем на
создание чеrо--то позитивноrо, пусть даже и не всеrда полностью
адекватно отражающеrо реальные отношения :в мозrу.
Однако в равной мере, а может, и больше, чем в друrих об--
ластях естествознания, в связи с запросами практики меди
цины, теории физиолоrии, а также философским аспектом вопроса
в учении о сложнейших функциях человеческоrо мозrа особенно
нужны и методы изучения мозrа, и факты о ero функциях, и по--
строенные на этой основе rипотезы и теории. Культивизация
«критическоrо направления» привела к появлению в биолоrиче
ской и философской литературе высказываний, rде под тонкой
пленкой вербальноrо материализма выпукло. контурируются идеи
отрыва психических функций от материальноrо субстрата. Это
проявляется в форме утверждений о наличии одноrо эквивалента
мноrих конкретных психических процессов, в форме прямоrо OT
рицания необходимости и возможности поисков физиолоrических
коррелятов психических явлений и т. д. Вряд ли эти взrляды по
своей внутренней суrцности так уяt далеки от oTKpoBeHHoro дуа--
лизма.
Объективный анализ мноrих современных «теоретических по--
строений» erцe раз подчеркивает не возможность реальноrо про
rpecca в этой проблеме без адекватноrо метода ее изучения.
Как хорошо известно, исследование закономерностей высшей
нервной деятельности человека чрезвычайно широко ocyrцe
Ствлялось и осуrцествляется методами психолоrии. Началом соб--
ственно физиолоrическоrо изучения высшей нервной деятель
'7
ности человека, по видимому, следует считать сеченовское pac
смотрение механизмов психическоrо и павловский период анализа
патолоrии высшей нервной деятельности человека на основе
данных, полученных в условнорефлекторных экспериментах на
животных.
rениальность основополаrающеrо павловскоrо эксперимента
неоспорима. Она прочно обосновала принцип, на котором бази
руются исключительные возможности мозrа животных и че
лове ка.
Оrромной заслуrой И. П. Павлова и ero школы явилось по
строение вероятных схе:м: структурно функциональноrо узора
в мозrу, лежащеrо в основе условнорефлекторной деятельности,
а также физиолоrических механизмов, определяющих разные ее
проявления, уточнение особенностей этих механизмов в зависи
мости от свойств среды и орrанизма.
Экспериментальный подход к изучению физиолоrии высшей
нервной деятельности у человека был вначале реализован исполь
зованием тех же методических приемов, что и у животных
(оборонительная и слюнная методики), а затем неско лько модифи
цирован с помощью более «человеческоrо» варианта условнореф
лекторноrо метода двиrательной методики на речевом подкреп
лении и использованием далее разноrо рода «эквивалентов»
условнорефлекторной ситуации. С помощью варианта, обозначае
Moro чаще Bcero как речедвиrательная методика, или двиrатель
ная методика на речевом подкреплении (А. r. Иванов Смолен
ский, 1933), были проведены исследования на детях и взрослых,
больных и здоровых испытуемых.
Большим количеством наблюдений была подтверждена уни--
версальность принципиальных положений высшей нервной дея--
тельности и уточнены типовые варианты ее нарушений при изме
нениях функциональноrо состояния rоловноrо мозrа человека.
Прошедшие rоды показали, однако, что применение условно--
рефлекторных методик у человека принесло в науку несравненно
меньше, чем их использование. в экспериментах на животных.
Для TOHKoro физиолоrическоrо исследования здоровоrо чело
века с колоссальными возможностями ero мозrа методики оказы
вались слишком примитивными, а примененные изолированно,
не давали возможность судить о конкретных центральных Mexa
низмах реакции.
При исследовании психической деятельности человека необ
ходим о было использование и условнорефлекторных, и пси
холоrических приемов. етодической попыткой преодоления
сложностей в изучении нейрофизиолоrических механизмов пси--
хических явлений явил ось объединение классических условно--
рефлекторных и психолоrических приемов с возможностями
более или менее прямоrо (или опосредованноrо) наблюдения ней
родина мики мозrа.
Реrистрация физиолоrических показателей жизнедеятельности
мозrа человека оказалась облеrчена в конце двадцатых rодов Ha
8
стоящеrо столетия, коrда был реализован прием относительно
прямоrо наблюдения за динамикой функциональноrо состояния
мозrа человека электроэнцефалоrрафия (ээr). Этапы ее BHe
дрения в различные клиники широко известны она заняла
прочное место в ряду локально диаrностических и функцио
нально диаrностических приемов. Она широко применяется в ней
рохирурrической, невролоrической, психиатрической и друrих
клиниках, признана необходимейшим приемом в анестезиоло--
rии и т. д.
Уже в 30--х rодах делались попытки наблюдения динамики
ээr человека при условнорефлекторных реакциях (Durup а. Fes
sard, 1935; Loomis et al., 1935а, Ь; 1936а, Ь; Knott, 1939).
Исследования этоrо рода на человеке подтвердили, уrлубили
и расширили мноrие положения условнорефлекторной теории.
Они позволили предельно полно изучить также меняющийся
в этих условиях сравнительно монотонный биоэлектрический
узор здоровоrо мозrа человека и более боrатый узор больноrо.
Было показано, что биоэлектрическая активность мозrа типа
электроэнцефалоrраммы меняется при реализации условных реф
лексов. Изменения MorYT быть различными при осуществлении
положительных и тормозных реакций. Конкретный тип разви--
вающихся изменений в наибольшей степени зависит от исходноrо
биоэлектрическоrо фона. При исходно умеренно синхронизиро
ванном фоне ээr здоровоrо человека осуществление положитель
ных условных реакций вызывает изменения типа десинхрониза
ции, а упроченных тормозных увеличение синхронизации. При
друrом исходном фоне и в норме MorYT наблюдаться любые дpy
rие соотношения, вплоть до обратных. Изменения в ээr перво--
начально развиваются во всех случаях очень диффузно, что paB
новероятно отражает и «rенер лизацию возбуждения», и участие
мноrих образований мозrа в орrанизации условнорефлекторной
реакции. Эти изменения затем оrраничиваются, что может с раз
ных позиций рассматриваться и как отражение концентрирова
ния нервных процессов, и как постепенное формирование peaK
ции на оптимальной структурной основе, оптимизацию условий
реализации реакции.
При исходно устойчивой десинхронизации биопотенциалов
реализация условных реакций практически не вызывала, по
крайней мере, видимых сдвиrов ээr, что в значительной мере
может быть связано прежде Bcero с опосредованностью (через
оболочки мозrа, кости черепа, апоневроз, кожу) реrистрации био
потенциалов здоровоrо человека и, таким образом, неполнотой
наблюдения. Эти же условия реrистрации и в тех случаях, rде
наблюдались отчетливые изменения ээr, определили далеко не
Полное выявление составляющих ээr и отсюда их динамики.
В этом отношении наблюдения в эксперименте на живот
Ных с записью электрокортикоrраммы и субкортикоrраммы с по
МОЩью вживленных электродов непосредственно с мозrа дали
значительно больше сведений о ero динамике и состоянии и по--
9
зволили уже более десяти лет назад осуществить исследование
не только ээr, но и нейронной активности при классической
условнорефлекторной деятельности.
Исследователи не удовлетворились, однако, скудными резуль
татами, полученными на человеке: были предприняты дополни--
тельные шаrи в изучении вопроса в трех основных направлениях:
1. Исследования были предприняты у больных с измененной
в связи с поражением мозrа ээr, исходная полиморфность KOTO
рой позволяла наблюдать при любой, в том числе и условно--
рефлекторной, активности значительно более «боrатую» дина...
мику, чем в норме.
2. Разнообразились и усложнялись функциональные пробы.
Вместо простой системы формирующих «классический» условный
рефлекс световых, звуковых, проприоцептивных и т. п. раздра
жителей при реrистрации ээr применялись и различноrо рода
психолоrические пробы.
3. При сохранении стандартизированных «классических»
условнор.ефлекторных проб или при применении более слож
ных психолоrических проб ээr субъекта подверrались специаль
ной обработке с целью извлечения максимальной информации из
данных исследований при использовании анализаторов, электрон
новычислительной техники и специально разработанных, в том
числе и именно для этой цели, приемов математическоrо aHa
лиза ээr.
Можно отметить, что при сочетании метода условных реф
лексов и ээr у больных и в наблюдениях с записью ээr при.
предъявлении психолоrических проб было получено MHoro инте
pecHoro, в частности, о роли различных областей коры мозrа
в осуrцествлении определенноrо вида психической активности и
о принципах взаимодействия больших областей мозrа при реали
зации психической деятельности.
Известным этапом в изучении нейрофизиолоrии психической
деятельности явилось открытие феномена ожидания, Е волны,
условноrо неrативноrо отклонения CNV (Walter, 1965; Walter
и др., 1964), возникающеrо в лобных областях мозrа человека
в тех условиях, коrда появление KaKoro To одноrо раздражителя
определяет или делает вероятным появление BToporo. Феномен
обнаруживал существенные различия в зависимости от психиче
cKoro и эмоциональноrо состояния субъекта и степени тренировки
испытуемоrо, отражая, по rрею Уолтеру, «субъективную вероят--
ность события». Надежность появления феномена в определен
ных условиях делала ero практически идеальным явлением для
изучения процесса формирования rотовности к действию, актив
ности субъекта в ситуации взаимодействия человека со средой.
В настоящее время получены четкие доказательства подкорко
Boro эквивалента этоrо первоначально рассматривавшеrося как
чисто KopKoBoro явления (В. А. Чернышева в работе: Н. П. Бех
терева, В. А. Чернышева, 1969; В. А. Илюхина, Ю. В. Хон, 1973)
и изучены корково подкорковые механизмы ero реализации.
10
#i
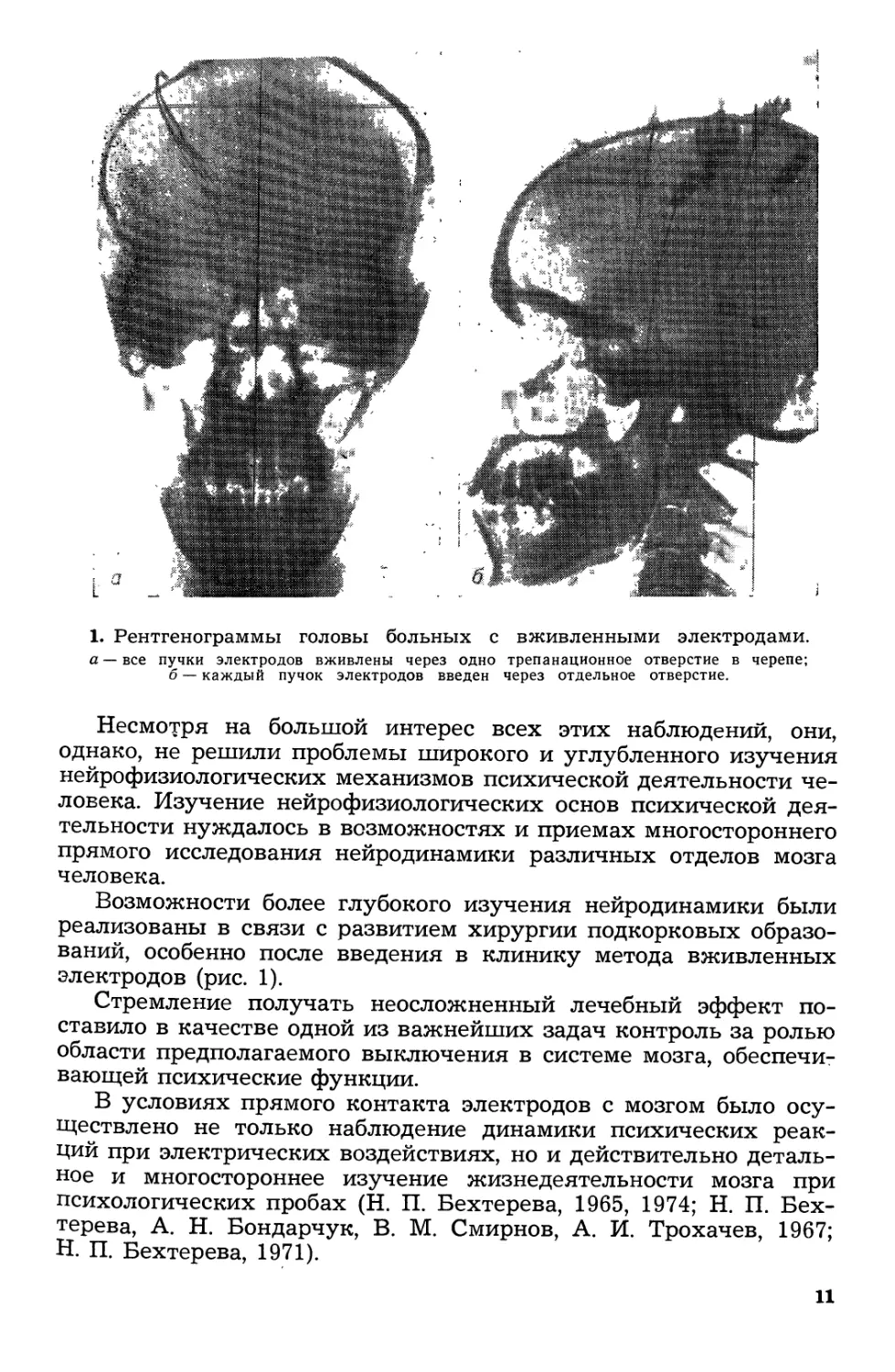
1. PeHTreHorpaMMbI rоловы больных с вживленными электродами.
а все пучки электродов вживлены через одно трепанационное отверстие в черепе;
б каждый пучок электродов введен через отдельное отверстие.
Несмотря на большой интерес всех этих наблюдений, они,
однако, не решили проблемы широкоrо и уrлубленноrо изучения
нейрофизиолоrических механизмов психической деятельности че--
ловека. Изучение нейрофизиолоrических основ психической дея
тельности нуждалось в возможностях и приемах MHorocTopoHHero
прямоrо исследования нейродинамики различных отделов мозrа
человека.
Возможности более rлубокоrо изучения нейродинамики были
реализованы в связи с развитием хирурrии подкорковых образо
ваниЙ, особенно после введения в клинику метода вживленных
электродов (рис. 1).
Стремление получать неосложненный лечебный эффект по--
ставило в качестве одной из важнейших задач контроль за ролью
области предполаrаемоrо выключения в системе мозrа, обеспечи-:-
вающей психические функции.
В условиях прямоrо контакта электродов с мозrом было ocy
Ществлено не только наблюдение динамики психических peaK
ций при электрических воздействиях, но и действительно деталь
ное и MHorocTopoHHee изучение жизнедеятельности мозrа при
Психолоrических пробах (Н. П. Бехтерева, 1965, 1974; Н. П. Бех
терева, А. Н. Бондарчук, В. М. Смирнов, А. И. Трохачев, 1967;
Н. П. Бехтерева, 19,71).
11
заr
""" .............
Выз8анные от6еть/
СНУ
3лектростuмуляцuя
3лектрОЛUЗlJС
3лектрополнризация
мап · + J ........
.11111111111111111 1.11111111111111111111111111111_
OpqW'''..
Кл,тfJIIНQR акти8ность
. . .1..
Функцuональные пробы
flсuzолоеuческuе тесты
"7+J, 15 2.......... .... .... п1:.т "
"1 7. -4. . . . . N . . . . . . . . . . 17 7. .. . .. н-+"
, 1 1 , JY
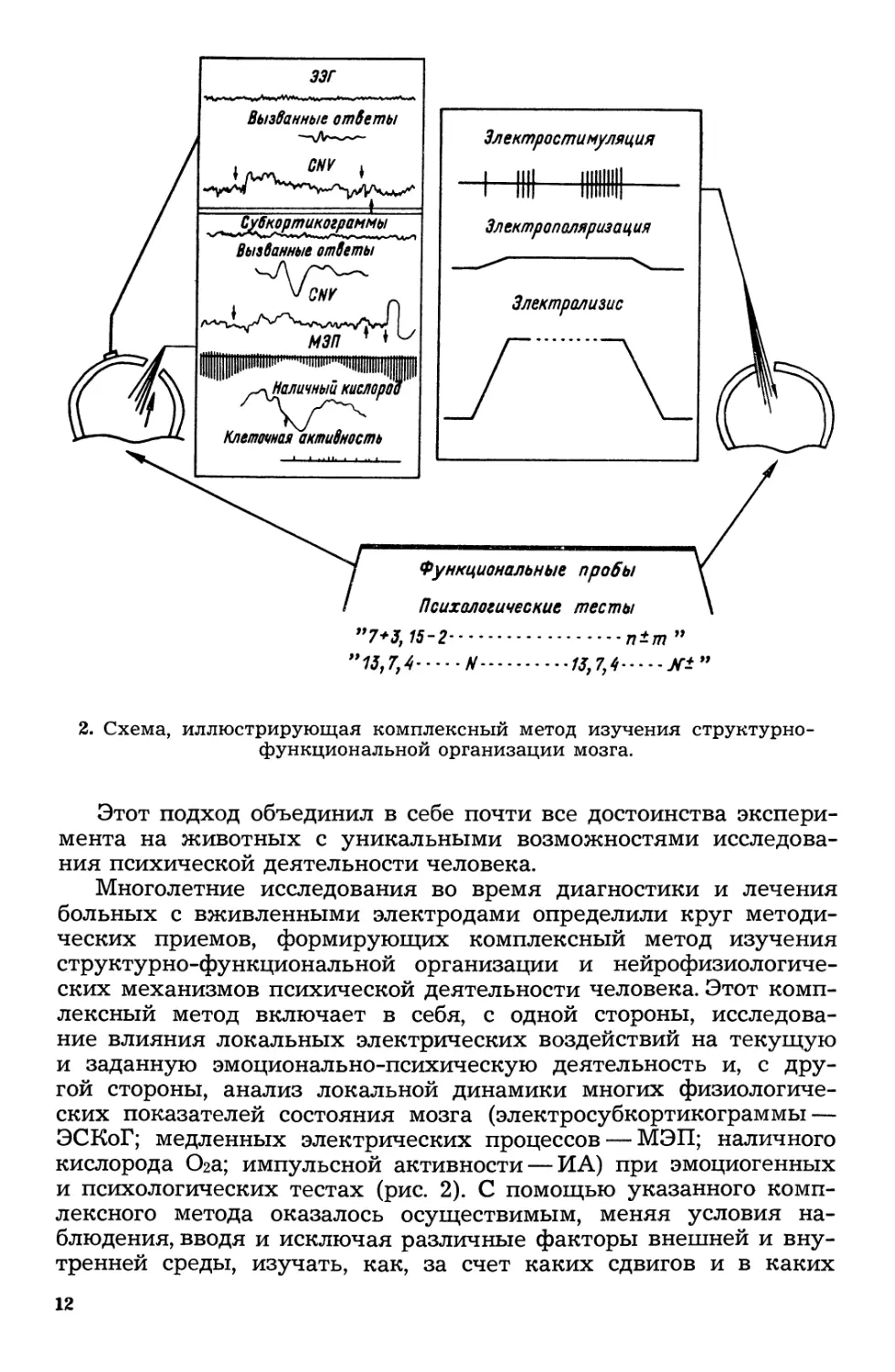
2. Схема, иллюстрирующая комплексный метод изучения CTpYKTypHO
функциональной орrанизации мозrа.
Этот подход объединил в себе почти все достоинства экспери
мента на животных с уникальными возможностями исследова
ния психической деятельности человека.
ноrолетние исследования во время диаrностики и лечения
больных с вживленными электродами определили Kpyr методи
ческих приемов, формирующих комплексный метод изучения
структурно функциональной орrанизации и нейрофизиолоrиче
ских механизмов психической деятельности человека. Этот комп
лексный метод включает в себя, с одной стороны, исследова
ние влияния локальных электрических воздействий на текущую
и заданную эмоционально--психическую деятельность и, с дpy
rой стороны, анализ локальной динамики мноrих физиолоrиче
ских показателей состояния мозrа (электросубкортикоrраммы
эскоr; медленных электрических процессов ЭП; наличноrо
кислорода 02а; импульсной активности ИА) при эмоциоrенных
и психолоrических тестах (рис. 2). С помощью указанноrо комп
лексноrо метода оказалось осуществимым, меняя условия Ha
блюдения, вводя и исключая различные факторы внешней и BHY
тренней среды, изучать, как, за счет каких сдвиrов и в каких
12
структурах мозrа решается любая, реализуемая мозrом психоло
rическая задача. Можно попутно упомянуть, что разработка
дан ноrо комплексноrо приема изучения мозrа оказалась исклю
чительно полезной для изучения мозrовых механизмов двиrа
тельной активности и для реlllения ряда практических, диаrно
стических и лечебных задач (Н. П. Бехтерева, А. Н. Бондарчук,
В. М. Смирнов, А. И. Трохачев, 1967; Н. П. Бехтерева, А. Н. Бон
дарчук, 1968; Н. П. Бехтерева, А. Н. Бондарчук, В. М. Смир
нов, 1969).
Уже то, что получено к настоящему времени в результате
обследования около 2000 зон мозrа в пределах различных тала--
мических ядер, стриопаллидарной системы, верхних отделов
ствола и медиобазальных отделов височных долей, позволило
сформулировать и подтвердить rипотезу о принципах обеспече--
ния мозrом психической деятельности (Н. П. Бехтерева, 1966).
Основные материалы по проблеме нейрофизиолоrических
аспектов психической деятельности получены сейчас в отноше
нии rлубоких структур мозrа, причем изучен и находится в про
цессе изучения ряд важнейших аспектов этой проблемы.
Проведено исследование так называемых общих изменений
в мозrу, фона, развивающеrося в процессе реализации психиче
ской деятельности и, воз ожно, определяющеrо оптимальность
условий ее протекания. Обнаружены и продолжают исследо
ваться зоны, «точки» мозrа, наиболее тесно связанные с психи
ческими функциями, по видимому, являющиеся элементами моз'"
rовой системы, обеспечивающей психические функции. Исследо
ваны и продолжают изучаться степень важности и роль этих
различных элементов нейронных (или точнее нейронноrли
альных) популяций в системе обеспечения психических функций.
Изучается нейрофизиолоrическая сущность изменений, разви
вающихся в процессе психической деятельности. Исследуются
механизмы, ;или, правильнее, объективные признаки объедине
ния исследуемых элементов в процессе психической деятель
ности в систему.
В специально психолоrических исследованиях, как известно,
нередко значительно разнообразятся психолоrические пробы и
именно характер выполнения их подверrается преимуществен
ному анализу. При изучении нейрофизиолоrических аспектов
Психической деятельности сейчас еще наиболее целесообразно
выбирать психолоrические тесты из наиболее стандартных и изу--
ченных и преимущественно анализировать динамику физиолоrи
ческих показателей. Исследование различных физиолоrJ1ческих
показателей жизнедеятельности мозrа при психолоrических про
бах оказалось приемом, значительно более тонким, чем наблю
дение изменений психической деятельности при очаrовых пато
лоrичеСRИХ процессах, хирурrических локальных разрушениях
и электрическом воздействии на мозr.
Использование именно психолоrических тестов, с друrой CTO
роны, явилось значительно более адекватным при изучении пси
13
хической деятельности человека, в том числе и ее нейрофизио
лоrическоrо аспекта, чем применение классических условнореф
лекторных проб.
Комплексный подход позволил значительно полнее, чем это
делалось до сих пор, изучить структурно функциональные и фи--
зиолоrические основы психическоrо, показать участие в пс хиче
ской деятельности и тех образований, которые моrли в резуль
тате данных выключения и раздражения несправедливо счи
таться «немыми» зонами.
Результаты, получаемые при электрических воздействиях на
мозr через вживленные электроды и при реrистрации различных
параметров жизнедеятельности мозrа, нередко были вполне сопо--
ставимыми или, наоборот, не были полностью сравнимыми. По
лучаемые данные в этом последнем случае характеризовали как
бы разные стороны явления, отвечая избирательно или преиму--
щественно лишь на некоторые из сформулированных вопросов,
и моrли, пожалуй, наиболее адекватно, хотя и с известным orpa
ничением, рассматриваться как показатели взаимодополняющие
(в смысле, приданном этому термину квантовой механикой).
Оrраничение в использовании этоrо термина определялось прежде
Bcero тем, что в отличие от «истинных» взаимодополняющих
явлений некоторые параметры жизнедеятельности мозrа моrли
быть записаны одновременно, хотя, естественно, очень сложно
ответить на вопрос об уровне «потерь» В этом случае. Различные
физиолоrические показатели оказались избирательно или пре
имущественно полезными в отношении разных аспектов изучае
мых явлений.
Вряд ли правомерно rоворять сейчас о том, что полностью
возможно наложение психическоrо на физиолоrическое. Мы все
еще относительно мало знаем о том, что происходит в мозrу и oco
бенно в совершеннейшем мозrу человека при психической дея
тельности, сложнейшей из всех возможных видов деятельности
живоrо орrанизма.
Долrая и кропотливая работа предстоит исследователям этой
области, работа, которая хоть HeMHoro может быть облеrчена под
ведением этапных итоrов.
Данная работа и представляет собой в первую очередь KpaT
кое обобщение ряда важнейших аспектов нейрофизиолоrиче--
ских исследований психической деятельности человека. На части
и rлавы работа разделена по принципу основных направлений
в изучении данной проблемы. В работе представлены материалы
об изм нениях ээr при условнорефлекторных реакциях и прак--
тически не высказано никаких новых соображений о механизмах
и уровнях замыкания условных связей у человека. В задачу на--
стоящей работы ни в коей мере не входил и анализ всех мате'7'
риалов об электроэнцефалоrрафическом выражении условнореф
лекторных реакций, этому посвящены обзоры В. с. Русинова
и М. я. Рабиновича (1958), Р. С. Мнухиной (1964) и мноrих дpy
rих. Такая же попытка была предпринята ранее и нами (1959).
14
Данный материал приведен потому, что он является важным для
уточнения общих изменений состояния мозrа в этих условиях,
изменений, отражающих процесс оптимизации состояния мозrа
ДЛЯ деятельности. Можно, однако, утверждать, что д я суждения
об уровнях и механизмах замыкания условных связеи у человека
ни одна лаборатория мира не обладает достаточно полноценным
фактическим материалом. Анализ этих механизмов не входил
и в задачу автора. В то же самое время именно этот вопрос
исключительно плодотворно изучается и может изучаться в экс--
перименте (П. К. Анохин, М. Н. Ливанов, В. С. Русинов, Н. Ю. Бе--
ленков, М. М. Хананашвили, Н. Н. Василевский и мноrие друrие
в нашей стране и различных странах мира).
С друrой стороны, в работе обсуждаются доступные на данном
уровне знаний аспекты структурно функциональных и нейрофи--
зиолоrических механизмов психических явлений человека. По
вполне понятным причинам ни одна экспериментальная лабора
тория мира в свою очередь не располаrает подобными данными.
Использованное построение работы ни в коей мере не должно
быть поводом к предположению о существенных различиях Me
жду условнорефлекторным и психическим у человека (реальных
или обусловленных взrлядами автора). Однако, не отрицая зна
чения изучения механизмов классических (или близких к ним)
условнорефлекторных реакций у человека, надо признать, что
именно специально психическое, фундаментом KOToporo является
условнорефлекторный принцип, представляет собой важнейшую
задачу в изучении мозrа человека и задачу TaKoro рода, KOTO
рая должна и может быть решена только в условиях изучения
человека.
Техническая революция текущеrо столетия, явившаяся pe
зультатом реализации работы мозrа человеческой мысли,
предъявила в свою очередь совершенно особые требования
к мозrу человека, потребовала обеспечения взаимодействия opra--
низма с практически совершенно новой средой. Большие CKO
рости, а отсюда необходимость срочной адаптации мозrовых си
стем реrуляции функции к новому поясу времени, резко OT
личным климатическим условиям) новому языковому, а иноrда
и социальному окружению, ВХCiДЯТ в практику повседневной
жизни все возрастающеrо количества людей. Резко изменились
требования к быстроте реакций и их точности во мноrих отраслях
народноrо хозяйства, причем в целом современная система чело
век........... машина привела во мноrих случаях и к большему услож--
нению необходимых операций.
Рассмотрению этих аспектов проблемы посвящено большое
КОличество психолоrических и физиолоrических исследований,
мноrопланово изучающих вопросы реакций человека и их опти--
Мизации в мире все усложняющейся техники (Б. Ф. Ломов, 1966;
r. М. Зараковский, В. И. Медведев, В. П. Зинченко, 1970).
Однако не только запросы медицинской практики и теорети
ческий интерес вопроса, но и технический скачок, выявивший
15
orpoMHbIe резервы мозrа, поставил и при этом в качестве одной
из первоочередных проблему изучения физиолоrии мозrа чело
века, физиолоrических принципов и конкретных механизмов,
обеспечивающих исключительные резервы мозrа и ero надеж
ность.
Каждая следующая ступень в изучении мозrовоrо обеспече
ния психической деятельности, несомненно, важна сама по себе,
как этап развития естественных наук, как этап развития боль
шой теории. Даже очень маленький шаr вперед в науке о мозrе
человека исключительно важен для клиники, как необходи
мейшее условие для усовершенствования лечения нервных и
психических заболеваний, расширения спектра излечимых забо
леваний.
Кроме Toro, успехи в изучении физиолоrии мозrа исключи
тельно важны для дальнейшеrо развития философии и социоло
rии. Современные социолоrи обеспокоены тем, как мозr человека
справится с обилием информации, с неизмеримо возросшими и
продолжающими возрастать требованиями, предъявляемыми
к нему. Научно техническая революция, подrотовленная мыслью
rениев и талантов и обеспеченная трудом миллионов, предъявила
в свою очередь orpoMHbIe требования к мозrу. Через rлаза и
уши хочет этоrо человек или не хочет к нему поступает
оrромный объем сведений. Ero мозr хочет он этоrо или нет
реаrирует на оrромный поток информации. Существует ли pe
альная уrроза, что мозr человека может не справиться с этой
сложностью?
Теоретики экспериментальной физиолоrии показали предпо
ложительные основные принципы, по которым во взаимодей
ствии с внешней средой, под влиянием этой внешней среды, при
способления к ней, развивался мозr. Физиолоrи мозrа человека
должны попытаться ответить на вопросы: каким образом оказы
вается возможным не только колоссальное индивидуальное yco
вершенствование, выявление возможностей мозrа, но и резкий
переход на _ новые ступени взаимодеЙ:ствия со средой во все
усложняющейся обстановке; как мозr человека меньше, чем за
два поколения, оказался способным адаптироваться в практически
совершенно новом мире?
Ч то будет с мозrом человечества, если и дальше с orpoMHbIM
ускорением будет увеличиваться наrрузка на мозr человека?
Существует ли в мозrу механизм самосохранения, самозащиты?
В сложнейшем взаимодействии со средой, при сочетании по
степенноrо развития и скачка, сформировался мозr cOBpeMeHHoro
человека с ero поистине колоссальными возможностями. Как на
протяжении мноrих веков он не только сохранил, но и умножил
свои удивительные возможности?
На мцожество важных вопросов должны ответить физиолоrи
мозrа. Большинство из этих вопросов требует дальнейших
исследований и раздумий; на некоторые из них, по видимому,
можно ответить и сейчас.
16
Известно, что в мозrу есть механизм, обеспечивающий избы
точные возможности при встрече с каждой новизной. Те, кому
удалоСЬ «подсмотреть», что п оисходит В мозrу в момент, коrда
обстановка оказывается новои, коrда неожиданно совершается
переход к старой обстановке, коrда есть хоть какие нибудь OCHO
ваниЯ для Toro, чтобы «удивиться», MorYT сказать, что мозr
в этих случаях как бы «проиrрывает» массу rотовностей к новой
ситуации. В это время активируется orpoMHoe количество HepB
НЫХ элементов, включается масса связей между различными
участками и элементами мозrа. Не исключено, что этот же меха--
низм, хотя бы частично, леr в основу сохранения возможностей
мозrа, возможностей вида.
Весьма вероятно, что реакция на новизну и является чем то
вроде естественной тренировки мозrа, вроде механизма, который,
обеспечивая избыточную rOToBHoCTb к каждой конкретной, даже
маленькой новизне данной минуты, на долrие века сохранил бес--
конечно большие возможности мозrа.
Чем больше количество новизны, чем больше раз за короткие
отрезки времени «удивляется» мозr, чем больше сведений посту--
пает через сенсорные входы, тем быстрее развивается мозr pe
бенка, тем полнее выявляется потенциал мозrа человечества, тем
большие возможности открываются перед ним. Может быть, не
будет большим преувеличением сказать, что научно техническая
революция сеrодняшнеrо дня есть результат взаимодействия
по принципу положительной обратной связи мозrа человечества
и внешней среды, изменяемой этим мозrом. Научно технический
проrресс, таким образом, с одной стороны принятые человече
ством решения личностей и, с друrой CTOpOHЫ, массовый подъем
возможностей человечества, массовая rенерация, столкновение,
взаимообоrащение идей. rде--то, сами Toro не заметив, мы пере
шли ступень, за которой наступила общая активация мозrа чело
вечества, после чеrо произошел «взрыв» В форме научно--техни--
ческой революции.
Но научно техническая революция это и orpoMHoe увеличе
ние возможностей изучения caMoro мозrа, решения задачи что
же такое наш мозr? И здесь вновь мы сталкиваемся с социаль
ным аспектом проблемы.
Социальные запросы пробуждают и активируют интерес
к мозrу. Успехи в изучении физиолоrических основ психической
деятельности приобретают в свою очередь социальное значение
потому, что именно они MorYT и должны дать ключи к управле
нию возможностями мозrа. Не в наших силах и не в наших инте
ресах замедлить научно--технический проrресс в целом и проrресс
в изучении мозrа человека в частности, но в нашей власти при
дать желаемое направление этому проrрессу.
СеrОДНЯlllНИЙ уровень знаний по физиолоrии мозrа человека
позволяет надеяться, что в самом ближайшем будущем мы полу--
чим действительно Реальную Власть над Мозrом. Эта власть
должна быть использована только в интересах Человека!
2 н. п. Бехтерева
17
Современный технический проrресс создал предпосылки для
использования в диаrностике и лечении заболеваний мозrа TOH
чайших методик физиолоrическоrо эксперимента, обеспечив тем
самым в свою очередь уникальные возможности диаrностики,
лечения и исследования механизмов мозrа человека. П рименение
математических методов, анализирующей адаптированной аппа
ратуры, а также аналоrовой и цифровой вычислительной Tex
ники для исследования физиолоrических явлений, наряду с pe
шением ряда практических задач, позволило поставить задачу
изучения физиолоrическоrо кода нервных явлений человека.
В свою очередь проблема изучения нейрофизиолоrии чело
века может расцениваться как реальная только сейчас, коrда она
оказалась обеспеченной всем проrрессом медицины, биолоrии и
техники.
Вполне понятно, что скорейшее приближение к полному pe
шению всей этой сложнейшей проблемы в обозримый срок Bpe
мени возможно лишь при концентрации усилий мноrих научных
rрупп, изучающих механизмы психической деятельности чело
века. Поэтому целью данной работы являлось не только подведе
ние этапноrо итоrа, но и стимуляция дальнейших исследований
в области нейрофизиолоrических аспектов психической деятель--
ности человека.
ФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ мозrА ЧЕЛОВЕКА
ПРИ РЕАЛИЗАЦИИ
УСЛОВНОРЕФЛЕКТОРНЫХ РЕАКЦИЙ
И психолоrИЧЕСКИХ ПРОБ
ЭЛЕКТРОЭНЦЕФАлоrР АММА ПРИ УСЛОВНОРЕФЛЕКТОРНЫХ
РЕАКЦИЯХ
Исследование физиолоrических характеристик мозrа при yc
ловнорефлекторной деятельности с помощью электроэнцефало--
rрафии (ээr) насчитывает почти тридцатилетнюю историю экспе
риментов на животных и, как это ни парадоксально, более чем
тридцатилетнюю историю наблюдений на человеке.
Изучение ээr при условнорефлекторных реакциях в экспери...
менте и у человека проводилось в течение этих десятилетий
с четко очерченной целью,........... предполаrалось, что в ээr можно
было найти отражение свойств и динамики основных нервных
процессов. В большинство обзоров данноrо вопроса вошла одна
из ранних работ М. Н. Ливанова и К. Л. Полякова (1945), rде при
использовании двух ритмических раздражителей (световое и
электрическое) были исследованы фазы становления условноrо
рефлекса у кролика. Аналоrичные фазовые изменения в ээr
были затем прослежены в эксперименте Morrell и Jasper (1956)
и др. В дальнейших работах М. Н. Ливанова и сотр., а также
orpoMHoM количестве друrих отечественных и зарубежных иссле...
дований были показаны закономерности изменений ээr живот
ных В зависимости от характера раздражителей и типа условной
связи, области отведения биопотенциалов и вида животноrо
(М. Н. Ливанов и А. М. Рябиновская, 1947; А. Б. KoraH, 1949, 1967;
М. Н. Ливанов и Т. А. Королькова, 1949; Maikowski, 1955;
И. Н. Книпст, 1955; Angyan, Hosznos, 1951; r. Т. Сахиулина,
1955 1957; Л. r. Трофимов, Р. Н. Лурье, Н. Н. Любимов и
М. Я. Рабинович, 1955; А. И. Ройтбак, 1956; Jouvet и Hernandez
Peon, 1957; В. С. Русинов, 1953, 1955, 1957; П. К. Анохин, 1957,
1968; Ioshii, Pruvot и Gastaut, 1956; Morrell и Jasper, 1956;
М. М. Хананашвили, 1972).
Проведенные исследования позволили изучить мноrие KOH
кретные нейрофизиолоrические механизмы условнорефлекторной
деятельности, а также ряд общебиолоrических закономерностей
функционирования мозrа (П. К. Анохин, 1968).
Данные, полученные у различных животных, по биоэлектри--
ческому компоненту исследуемых .реакций, были, однако, трудно
СОпоставимыми между собой и не только по конкретным фор--
мам биопотенциалов, но и по направленности и степени распро
страненности наблюдаемых перестроек.
2*
19
Уже эти факты делают вполне понятными трудности исполь
зования результатов подобноrо эксперимента для суждения об
аналоrичных механизмах мозrа человека. Дополнительные слож
ности взаимодействия «экспериментальной» И «человеческой»
физиолоrии мозrа возникли и на уровне самой условнорефлек
торной методики. Механическое перенесение экспериментальных
приемов в исследования человека далеко не всеrда давало ОjКи
даемые результаты и, как правило, приводило к результатам
значительно менее общ rо значения, чем в эксперименте. Важ
нейший фактор существенное усложнение механизмов мозrа
на уровне человека наряду с несопоставимостью некоторых
физиолоrических показателей мозrа животных и человека опре
делил необходимость проведения практически Bcero спектра
исследований на человеке.
Изучение вопроса о динамике ээr при условно рефлекторных
актах или реакциях, сходных с условнорефлекторными, в иссле
дованиях на человеке было начато уже в 30 x rодах настоящеrо
Столетия (Durup и Fessard, 1935; Loomis, Harvey а. Hobart, 1935,
1936; Knott, 1939, и др.).
Основн й задачей этих и последующих исследований, иноrда
почти примитивных, применительно к возможностям человече
cKoro мозrа было, так же как в эксперименте, изучение с по
мощью ээr динамики нервных процессов мозrа в модельных
условиях воспроизведения простейшей схемы психическоrо.
В первых и в подавляющем большинстве последующих pa
бот в качестве подкрепления использовалось световое или про
приоцептивное раздражение или и световое, и проприоцептивное
(<<подкреплением» служил свет, на включение ROToporo испытуе
мый должен был осуществить обычно простую двиrательную
реакцию). В качестве условноrо раздражителя чаще Bcero исполь
зовался звук, ориентировочная реакция на который предвари
тельно обычно уrашалась (Loomis, Harvey, Hobart, 1935; Knott,
1939, и др.). в зависимости от частных целей исследования варьи
ровала относительная сложность задачи (абсолютно они, как
правило, были очень просты), вводилось или исключалось KaKoe
либо условие исследования, вырабатывались положительные или
тормозные реакции при совпадении или большем или меньшем
несовпадении раздражителей.
Результаты уже первых исследований в достаточной мере
подчеркнули роль сложности предъявляемой задачи в характере
и динамике ээr проявлений. В дальнейшем эти факты рассма--
триваются в свете представлений о значении фактора ориентиро
вочно исследовательских составляющих для стойкости не только
самой условнорефлекторной реакции, но и связанных с ней ээr--
изменений (Л. r. Воронин и Е. Н. Соколов, 1955; П. К. Анохин,
1957, и др.).
При использовании специальных методических приемов, rде
ориентировочно исследовательский компонент не Mor yracHYTb,
реакция депрессии альфа ритма в ээr была очень прочной.
20
с друrой стороны, при упрощении и стандартизации условий
исследования можно было наблюдать постепенное уменьшение
выраженности реакции депрессии альра ритма на повторные
сочетанные предъявления раздражителеи.
При повторении однообразных сочетаний звука со светом
в наблюдениях Knott и Henry (1941) депрессия альфа ритма ста--
новилась все менее отчетливой не только на звук, но и на свет,
что объяснялось в работе механизмами центральноrо торможе
ния и адаптации. Этот факт в дальнейшем получил множество
подтверждений, а в отдельных исследованиях был подверrнут
и специальному изучению (Е. Н. Соколов, 1956). Наиболее чет
кое объяснение этоrо феномена приведено в работах П. К. AHO
хина (1962, и др.), rде показано значение биолоrически незначи--
Moro «подкрепления», формирования условной реакции в этих
случаях не на базе истинноrо безусловноrо рефлекса. Условная
реакция, сформированная на основе совпадающеrо и несовпадаю
щеrо предъявления раздражителей при использовании сочетания
звука со светом, на которое испытуемому предлаrалось нажимать
на ключ, исследовалась Jasper и Shagass (1941). При совпадении-
раздражителей условный рефлекс образовывался леrко и также
леrко уrасал после 3 5 rо'неподкрепления. Исследование дина
мики ээr при совпадающих, запаздываюЩИХ и следовых услов .
ных рефлексах показало предшествование депрессии альфа ритма
подкреплению. В полном соответствии с данными павловскоrо
эксперимента обе эти ситуации отставление и запаздывание
моrли приводить к развитию cOHHoro состояния. После -выработки
дифференцировочной реакции дифференцировочный раздражи '
тель постепенно переставал оказывать влияние на электроэнце
фалоrрамму.
Изучение динамики биоэлектрической реакции во времени
в различных областях коры по той же методической схеме, что
у J asper и Shagass, было предпринято в ряде работ и, в частности,
в исследованиях Н. А. Мушкиной (1956). В то время как характер
изменений в слуховом и зрительном анализаторах оказывался
одинаковым при наличных условных связях, при отставлении
света обнаруживалась фазовость изменений. На первом этапе
выработки отставленных условных рефлексов реакция уrнетения
альфа ритма начиналась одновременно в слуховой и зрительной
областях, что рассматривалось в качестве ээr выражения фазы
rенерализации нервных процессов. На втором этапе уrнетение
альфа ритма в слуховой области наступало при действии звука,
а в зрительной области к моменту cBeToBoro подкрепления
(фаза постепенной концентрации). И, наконец, при продолжении
сочетаний реакция уrнетения альфа ритма в обеих корковых
областях начала развиваться перед моментом cBeToBoro под
крепления.
Так же подробно была прослежена динамика ээr при Bыpa
ботке дифференцировки. Первоначально реакция уrнетения
альфа ритма реrистрировалась и в зрительной, и в слуховой
21
.,
областях одновременно. Вторая фаза характеризовалась yraca
нием реакции уrнетения альфа ритма в зрительной области. Затем
наступала третья фаза........... полное уrасание реакции уrнетения
альфа--ритма на применение данноrо сочетания раздражителей и
в области представительства «безусловноrо» раздражителя, и
в области представительства условноrо сиrнала. Эти данные pac
сматривались как развитие торможения первоначально в области
представительства «безусловноrо» раздражителя и последующее
распространение ero на область представительства условноrо
сиrнала.
ОтраЯ-tение в ээr процесса запаздывания при двиrательном
условнорефлекторном акте изучали В. Е. Майорчик, В. С. Руси
нов и r. д. Кузнецова (1954). В их наблюдениях при изменении
времени отставления в ээr появлялись диффузные медленные
волны, постепенно концентрирующиеся в теменной области.
Анализ данных (ээr, электромиоrраммы эмr и др.) и
варьирование условий исследования явились поводом для заклю
чения об отражении медленными волнами в данном случае про--
цесса торможения. Сходные ээr изменения наблюдали при OT .
ставлении cBeToBoro «подкрепления» А. Jus и К. Jus (1954).
Исследователи так же, как и J asper и Shagass, отмечали, что
в процессе выработки этих условных реакций моrло наблю..
даться развитие cOHHoro состояния у испытуемых.
Как известно, отсутСтвие медленных волн в ээr в «тормоз
ных» ситуациях в большинстве исследований, проведенных у че
ловека, в 50 x rодах рассматривалось не как отсутствие связи
определенноrо ээr рисунка и HepBHoro процесса, а как отраже
ние концентрирования процесс а торможения, оrраничения ero
небольшим участком, не улавливаемым при ээr. Это предполо
жение казалось очень лоrичным прежде Bcero на основе всей
истории клинической электроэнцефалоrрафии весь клиниче
ский опыт ээr делал предположение о связи медленных волн
с торможением, пусть даже и не прямой, весьма заманчивым.
При анализе исследований, проведенных с записью ээr при
условнорефлекторных реакциях у человека, и рассмотрении xa
рактера наблюдавшейся ээr--динамики наиболее вероятным сей
час кажется, что появление медленной активности в подобных
условиях в первую очередь было связано именно с развитием
COHHoro состояния. У здоровых лиц в достоверных условиях ис
следования практически не наблюдалось появления медленных
волн в «тормозной» ситуации вне засыпания.
Условнорефлекторное изменение альфа ритма прослежива--
лось и в друrих методических вариантах опытов (Shagass, 1942;
И. С. Беритов и А. О. Воробьев, 1943, и мн. др.).
Частных вариантов исследований ээr изменений при условно--
рефлекторной активности было очень MHoro (r. В. rершуни и
и. И. Короткин, 1947; r. В. rершуни, в. А. Кожевников, А. М. Ма--
русева, Л. А. Чистович, 1948; В. Е. Майорчик и Б. r. Спирин,
1951; Р. Н. Лурье и В. С. Русинов, 1955; Ю. r. Кратин, 1955;
22
П. о. Макаров, 1956, 1957; Л. А. Новикова и Е. Н. Соколов, 1957;
Lansing 1957, и мн. др.). Широко проводилась запись ээr во
вреl\IЯ условнорефлекторных реакций, выработанных на основе
двиrательной методики с речевым подкреплением А. r. Иванова
Смоленскоrо.
К середине 50 x rодов во всем мире накопилось очень боль
тое количество данных исследований динамики ээr при yc
ловнорефлекторной активности. Предполаrалось, что возбужде--
ние отражается в ээr десинхронизацией это казалось весьма
вероятным. Предполаrалось, что торможение может (или точ
нее должно) отраj-каться медленными волнами или синхрониза
цией. В то же calVloe время сходство, но не полная однозначность
lетодических условий и в связи с этим трудность сопостав
ления результатов не позволяли провести достаточно. полно
ценный анализ данных. В связи с этим, а также неуrасающей
верой исследователей в возможность обнаружения ээr корре
лятов нервных процессов Gastaut после личных исследований
-в данном направлении (1955 1956) орrанизует широкое изучение
ээr при условнорефлекторной деятельности большой rруппы
исследователей в ряде стран Европы (Gastaut, Jus, Morrell, Storm
van Leewen, Bekkering, Каmр, Werre, 1957; Gastaut и др., 1957).
Работа проводилась с caMoro начала с введением элемента OT
бора испытуемых, а потому любые полученные закономерности
оказывались хотя и общими, но далеко не исчерпывающими.
Известная условность результатов прежде Bcero заключалась
в том, что исследования были предприняты не у случайноrо
континrента лиц с различными ээr, а у лиц с хорошо выражен
ными альфа и роландическим ритмами. С друrой стороны, было
проведено выделение и последующее описание лишь наиболее
часто повторяющихся проявлеНИЙ, все то, что наблюдалось не
всеrда, отбрасывалось как несущественное.
Методически данное исследование было осуществлено сле
дующим образом. Реrистрировалась ээr, электромиоrрамма
(эмr), электрокардиоrрамма (экr), пневмоrрамма и электродер
MorpaMMa (Krp). ээr оценивалась визуально и с помощью ча
стотно амплитудноrо анализатора. Звуковое раздражение соче
талось со светом или с пассивным (или активным) движением
руки. Звуковое раздражение, таким образом, в зависимости от
условий опыта приобретало способность вызывать депрессию за
тылочноrо или роландическоrо ритмов. И, наконец, в третьем
варианте опыта звуковое раздражение сочеталось с комплекс
ным, сложным подкреплением........... звук подкреплялся светом, на
который осуществлялось активное или пассивное движение руки.
Этот вариант опыта Mor приводить к депрессии на звук и ролан--
Дическоrо ритма, и альфа ритма (причем последнеrо билате
рально) или только роландическоrо ритма.
В результате проведенных наблюдений и их анализа Gastaut
и ero коллеrи констатировали, что область преимущественноrо
подавления синхронизированноrо ритма типа альфа--ритма или
23
Схема основной пространственной Динамики явлений синхронизации
(по G а s
Порядковый номер
реакции
Раздражители
условный
безусловный
1
2
3
1
2
3
1
2
3
Звук +
Свет
Звук +
Движение руки
Звук +
Свет, движение руки
N < м * Звук + Свет, движение руки
N<M+1 Звук + Свет, движение руки и сло
N < М + к + 1 ** во «хорошо»
Звук Свет
Звук Движение руки
Звук Свет, движение руки
* N KaKoe TO повторное сочетание раздражителей (обычно меньше два
телей, коrда ээr выражение положительных условных реакций становится
** К сочетание, при котором в связи с каким то внешним или внутрен
роландическоrо ритма зависит прежде всеrо от характера под
крепления. При двиrательных условных рефлексах удалось
обнаружить фазу, коrда блокирование роландическоrо ритма вы--
являлось ТОЛЬКО В двиrательной области противоположноrо по
лушария. Данные Gastaut в отношении корреляции возбужде--
нил и блокады альфа-- и роландическоrо ритмов в ээr оказались
принципиально теми же, что и в проведенных ранее исследо--
ваниях.
При предъявлении испытуемым сочетания: звук (тормоз
ной) + С'вет, на который давалась инструкция сжима.ть кисть, OT
24
идесинхронизации при условнорефлекторных реакциях у человека
t а u t и др.)
Характер реакции
Область проявления реакции
Блокада альфа ритма
Блокада роландическоrо ритма
Блокада альфа ритма и роланди
ческоrо ритма
Блокада роландическоrо ритма
То же
Увеличение выраженности альфа
I ритма
Увеличение выраженности ролан
дическоrо ритма
в зависимости от характера пред
шествовавшей положительной peaK
ции
1) увеличение выраженности
роландическоrо ритма и YBe
личение выраженности альфа
ритма или .
2) увеличение выраженности
роландическоrо ритма
Теменно височно затылочные области
обоих полушарий
Центральная область полушария,
контралатеральноrо руке, осуществля
ющей движение
Теменно височно затылочные области
обоих полушарий н центральная
область полушария, контралатерально
ro руке, осуществляющей двиrатель
ную реакцию
Центральная область полушария,
контралатеральноrо руке, осуществляю
щей двиrательную реакцию
Центральные области обоих полуша-
рий
'Теменно височно затылочные области
обоих полушарий
Центральная область полушария,
контралатеральноrо руке, осущест-
вляющей двиrательную реакцию
То же
Теменно височно затылочные области
обоих полушарий, центральная область
полушария, контралатеральноrо руке,
осуществляющей двиrательную реак-
цию
дцатоrо, но индивидуально существенно варьирует); М сочетание раздражи
неотчетливым.
ним фактором происходит «растормаживание» ээr реакции.
мечалось первоначальное усиление роландическоrо ритма и лишь
затем депрессия ero, только на фоне которой и проявлялось дви
жение. Усиление роландическоrо ритма расценивалось как OTpa
жение в ээr торможения. Как проявление развития тормозных
процессов запредельноrо типа рассматривалось, с друrой CTO
роны, самопроизвольное исчезновение реакции блокировани я
при положительных реакциях, которое при повторениях соче
таний сменялось повышением амплитуды и замедлением ритмов
коры, вплоть до развития картины, характерной для леrкоrо сна
(появление TeTa и дельта ритмов). И в этом случае в ситуации,
25
rде можно было предположить развитие торможения, в ээr Ha
блюдались синхронизация ритмов и их замедление (таблица).
Gastaut и ero коллеrи полаrали, что подтвердили связь TOp
можения в условнорефлекторной ситуации у человека с синхро
низацией и развитием медленных волн в ээr.
Надо отметить, что трактовка явлений, наблюдаемых в ээr
в плане первичноrо развития торможения запредельноrо типа
и затем сна, базируется в исследованиях Gastaut и соавт., так же
как и в друrих исследованиях, в первую очередь на желании
увидеть сходное с павловскими данными. .
Gastaut и соавт., как и подавляющее большинство исследова
телей до них, леrко оперировали известными и неизвестными
величинами в наблюдениях развивавшееся в ээr рассматрива
лось как отражение Toro, что должно было бы развиваться по
классическим законам павловской физиолоrии. На основе соб
ственных допущений далее определенным биоэлектрическим яв
лениям приписывалась однозначная связь с основными HepB
ными процессами, и затем уже появление какоrо либо из этих,
«привязанных» К нервным процессам биоэлектрических призна
ков расценивалось как свидетельство развития соответствующеrо
HepBHoro процесса.
Gastaut было выделено 4 основных типа явлений: два, свя
занных с возбуждением не зависящим и зависящим от услов--
норефлекторной деятельн сти, и два, связанных с процессом TOp
можения.
Развитие распространенной блокады альфа--ритма при воз--
буждении объяснялось на основе активации меззнцефалическоrо
уровня ретикулярной формации, развитие локальной блокады
(также связанной с возбуждением) за счет активации неспе
цифических таламических образований, причем в появлении
этой второй ээr реакции известное место отводится уrнетению
мезэнцефалической системы при повторении стимулов. Для Toro,
чтобы обеспечить стройность своей rипотезы, Gastaut оперировал
в этом случае очень укладывающимися в ero схему и не очень
соответствующими реалЬНОСТf1 представлениями об антаrонизме
этих двух уровней срединных структур (Bremer, 1962). По види
мому, отсутствие к тому времени достаточно убедительных дaH
ных о синхр(' :изирующих образованиях мозrа (Morruzzi, 1962)
в значительной мере определило использование Gastaut для
объяснения уrашения реакции блокады альфа ритма l1 увеличе
ния синхронизации в ээr лишь механизмов развития торможе
ния в мезэнцефалической и таламокортикальной системах.
Проводя такое территориально сложноорrанизованное, KOM
плексное исследование, Gastaut обсудил и вопрос об уровнях
замыкания условной связи у человека. Он представил ряд схем,
на которых центр тяжести не только биоэлектрических, но и
условнорефлекторных событий уверенно приписан таламиче--
скому уровню. Не споря с этим положением в данном случае по
существу, следует все же подчеркнуть, что, приведя вполне
полноценный, хотя и частный, фактический материал, Gastaut
в определении уровня замыкания условной связи допустил недо--
пустимое: он идентифицировал узловые пункты, «ритмоводи
тели» , наблюдавшихся электроrрафических реакций с облаСТЯМvI
высшей интеrрации. И это осуществляется в то время, коrда
принцип включения активирующих влияний неспецифической
системьr при самых различных внешних воздействиях был
в достаточной мере уже изучен. Gastaut (1957, 1958) настаивал,
что роль коры больших полушарий при такой трактовке физио
лоrических данных никак «не умаляется», так как таламическое
замыкание рефлекторной дуrи всеrда сопровождается актива
цией соответствующеrо таламокортикальноrо звена и т. д. (Ga--
staut и Roger, 1962). Следует подчеркнуть, что эти взrляды
Gastaut очень близки, по существу, представлениям Penfield
о центрэнцефалической системе. И замыкание условной связи
происходит в области срединных образований мозrа, и кора при
нимает участие в реакции, допустимо себе представить, лоrично
продолжая эту точку зрения, что кора в этом случае фиксирует
опыт происходящеrо или представляет прошлый опыт (память)
в распоряжение формирующейся реакции.
Возвращаясь к оценке нейрофизиолоrическоrо аспекта иссле--
дованных в rоловном мозrу сдвиrов, видно, что Gastaut, не при--
бавив ничеrо существенно HOBoro к известному ранее, подтвер
дил, уточнил и, в связи с детальностью обработки большоrо и
сравнимоrо исходноrо материала, утвердил в положении законо
мерностей развитие у лиц с исходно хорошим альфа--ритмом
распространенной или несколько более оrраниченной десинхро
низации или синхронизации (в том числе и с замедлением ритма)
в тех условиях, коrда соответственно классическим законам
павловской физиолоrии были основания ожидать возникновения
распространенноrо или оrраниченноrо возбуждения или TOpMO
жения.
Анализируя эти исследования, приходится признать, что, если
для корреляции состояния возбуждения с десинхронизацией
в ээr все же имелись хоть какие--то основания, то с торможе
нием дело обстояло сложнее. Дело осложнялось, во первых, ero
электроэнцефалоrрафическим выражением оно оказывалось
уже очень «неспецифичным». Как известно, исходным призна
ком, положенным в основу подбора испытуемых, был хороший
альфа--ритм покоя; покой, таким образом, электроrрафически
оказывался полностью или почти полностью идентичным TOp
можению.
BO--BTOpЫX, и это rораздо важнее, связь возбуждения
с десинхронизацией и торможения с синхронизацией в такой
форме моrла быть утверждена лишь для частноrо континrента
лиц с исходно имеющимся альфа ритмом. В подобной си:rуации
у достаточно большоrо континrента здоровых лиц без четкоrо
альфа ритма в ээr покоя не только не были выраженными
проявления возбуждения в виде десинхронизации, но и просто
27
отсутствовали ээr признаки торможения в виде синхронизации.
Таким образом, даже если и признать значение обнаруженных
корреляций, связь нервных процессов и ээr--проявлений оказы
валась и после этоrо исследования в ЛУЧlllем случае удовлетво
ряющей ЛИlllЬ принципу условной вероятности (роль условия
исходно ХОрОlllИЙ альфа ритм и роландический ритм). Вопросы
о нейрофизиолоrическом выра ении основных нервных про
цессов и о тонких нейрофизиолоrических механизмах психиче
ской деятельности человека для реlllения НУjКдались в дальней
ших исследованиях, новых материалах и, rлавное, НОВЫХ под
ходах.
Совершенно закономерно мысль исследователей направилась
прежде Bcero по трем основным путям, нередко очень тесно пе--
реплетавшимся между собой. Пути эти определялись стремле
нием: 1) обнаружить более мноrоrранную, менее монотонную
динамику ээr; 2) получить материалы об изменениях ээr
в условиях уже специально человеческой психической деятель
ности и, наконец, 3) выявить в ээr с помощью математики и
аппаратуры то, что ускользало от невооруженноrо rлаза иссле
дователя.
Наблюдение полиморфной динамики ээr у человека было
возможным при использовании для исследования вариаций фи
зиолоrическоrо состояния испытуемых и проведении исследова
ний у больных. Извлечение из ээr дополнительной. информации
требовало применения не только стандартных инструменталь
ных, но и некоторых специальных, в том числе и разработанных
для этой цели, приемов. Исследование ээr в условиях специ--
ально человеческой психической деятельности предполаrало
проведение электроэнцефалоrрафии при предъявлении различ
ных психолоrических тестов (что, естественно, никак не исклю
чало также разнообразной обработки данных).
Изучение динамики полиморфной ээr при условнорефлек
торной деятельности было проведено у здоровых лиц при засы--
пании и у больных споражениями мозrа.
Исследования, I1Iроведенные в период засыпания и развития
сна, в целом обнаруjКИЛИ в ээr пространственную динамику,
сходную с наблюдавшейся у здоровых лиц- в бодрствующем со--
стоянии. Сходство увеличивалось, если испытуемые пробужда
лись от первых раздражителей. Если же пробуждение было He
полным, конкретный тип реакций Mor быть прямо обратным
тому, что отмечалось в бодрствующем состоянии, на положи
тельные раздражители наблюдались Iреакции оживления альфа--
ритма (Roger, Sokolov, Voronin, 1957). В этих же работах была
показана леrкость развития уrашения подкрепляемой реакции
и появления или усиления медленных волн одновременно
с уrлублением сна. Эти наблюдения, а также исследования, про--
веденные с использованием ээr при условнорефлекторной дея--
тельности, осуществляемой на фоне фармаколоrических проб
(М. Н. Ливанов, Н. А. rаврилова, А. С. Асланов, 1966), убеди
28
тельно показали роль исходноrо ээr фона в отношении KOH
RретноЙ формы развивающейся реакции и неправомерность KOH
статации uднозначной связи между нервными процессами и
определенными электроrрафическими реакциями.
Исследование динамики полиморфной ээr больных с пора
жениями мозrа при условнорефлекторной деятельности обнару
жило, . что обычно распространенная или оrраниченная в связи
с характером раздражителей и фазой выработки условной связи
динамика биопотенциалов может оказаться в исключительно
четкой форме «подчиненной» корковому патолоrическому очаrу.
Эти наблюдения не только еще раз подтвердили неоднознач
ность соотношения: нервные процессы ээr, но и в очень Ha
rлядной форме у человека показали взаимодействие распростра
ненных дистантных влияний с функциональным состоянием от--
дельных корковых зон. На ээr больных моrли наблюдаться
и обычные перестройки альфа ритма, если он сохранялся, но
rораздо более отчетливо, вне прямой зависимости от локализа
ции полушарноrо патолоrическоrо очаrа в мозrу и типа раздра
жителя обнаруживалась, как одна из возможных и в данном
случае........... наиболее интересных реакций, динамика медленных
волн в области KopKoBoro очаrа, а также и различные . друrие,
связанные с корковым очаrом перестройки так называемых па
толоrических форм активности.
При этом усиление или уменьшение выраженности медлен
ных волн в области очаrа моrло развиваться сразу по предъяв
лении условноrо сиrнала, спустя некоторое время, во время
предъявления подкрепляющеrо раздражителя и т. п. И сохра--
няться лишь короткое время или задерживаться в ээr и после
реализации условной реакции. Эти факты в том, что касалось
значения патолоrическоrо очаrа в динамике биоэлектрической
реакции, оказывались вполне объяснимыми на основе представ
лений А. А. YXToMcKoro (1945) о доминанте, ориrинально разви
ваемых применительно к новым возможностям cOBpeMeHHoro
эксперимента В. С. Русиновым (1953, 1955, и др.). В том, что Ka
салось caMoro характера изменений в области очаrа, развитие
медленных волн на первый взrляд представлялось убедительной
иллюстрацией павловских представлений о запредельном тормо--
жении. Действительно, эти изменения моrли рассматриваться
именно таким образом и особенно, если считать увеличение BЫ
раженности или появление медленных волн свидетельством
развития торможения. Но по ряду проявлений условнорефлек
торной деятельности для TaKoro рода суждений все же не всеrда
были прямые основания. Медленные волны в области очаrа,
в том числе и расположенноrо в области одноrо или обоих пред
ставительств предъявленных раздражителей, развивались не--
редко наиболее отчетливо при первой условнорефлекторной ре--
.акции. Понятно, что это несколько Пlротиворечило высказанной
rочке зрения, хотя допустимо себе представить, что в дальней--
тем ээr--динамика «осложнялась» фактором повышения воз--
29
1
2
3
'1
5
6
7
V WWVtY
8
8
1 && "' ."'& . t .8L....... ь-....
2
3
1.}
5
6 V
7
8
9
а
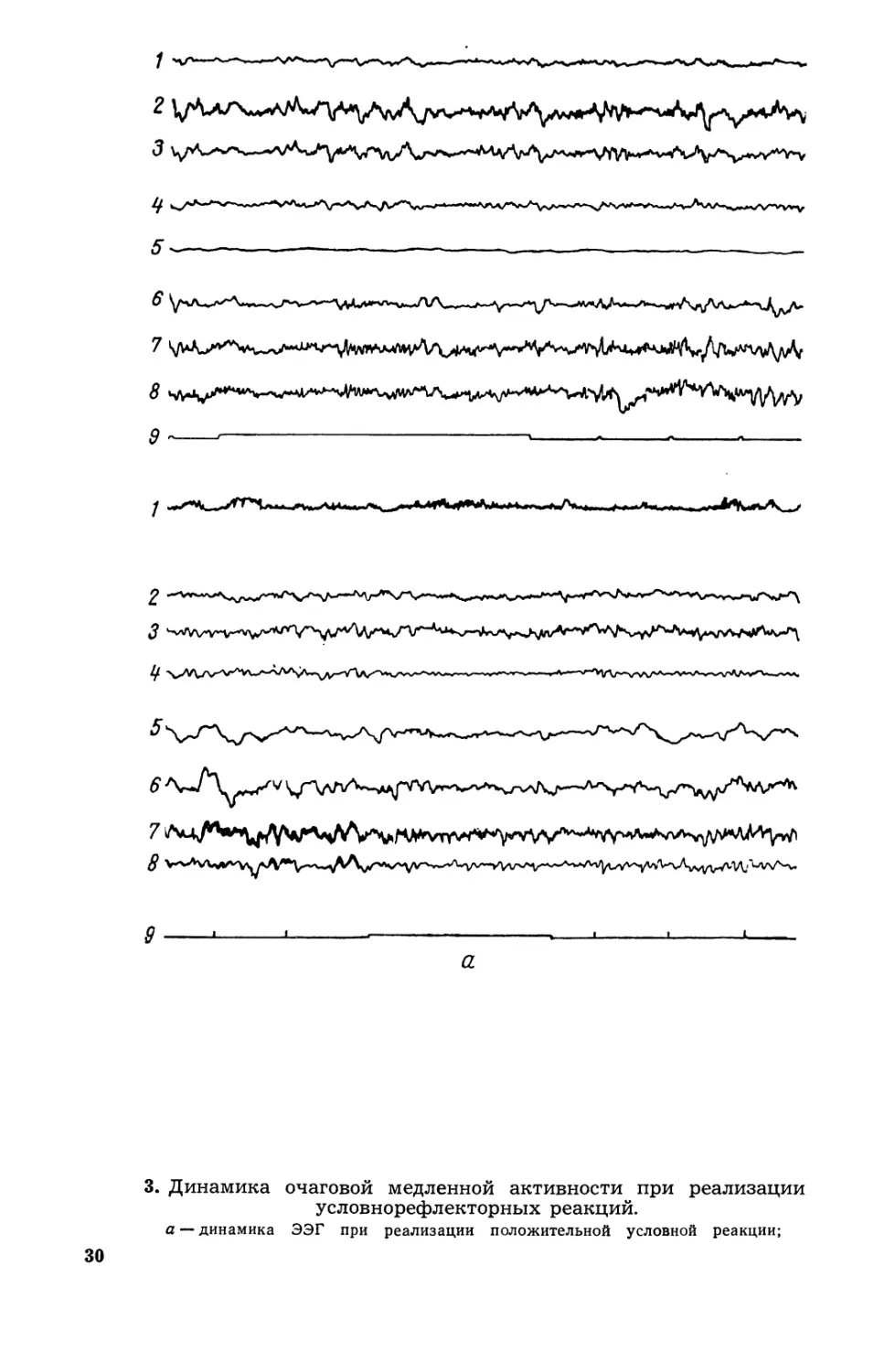
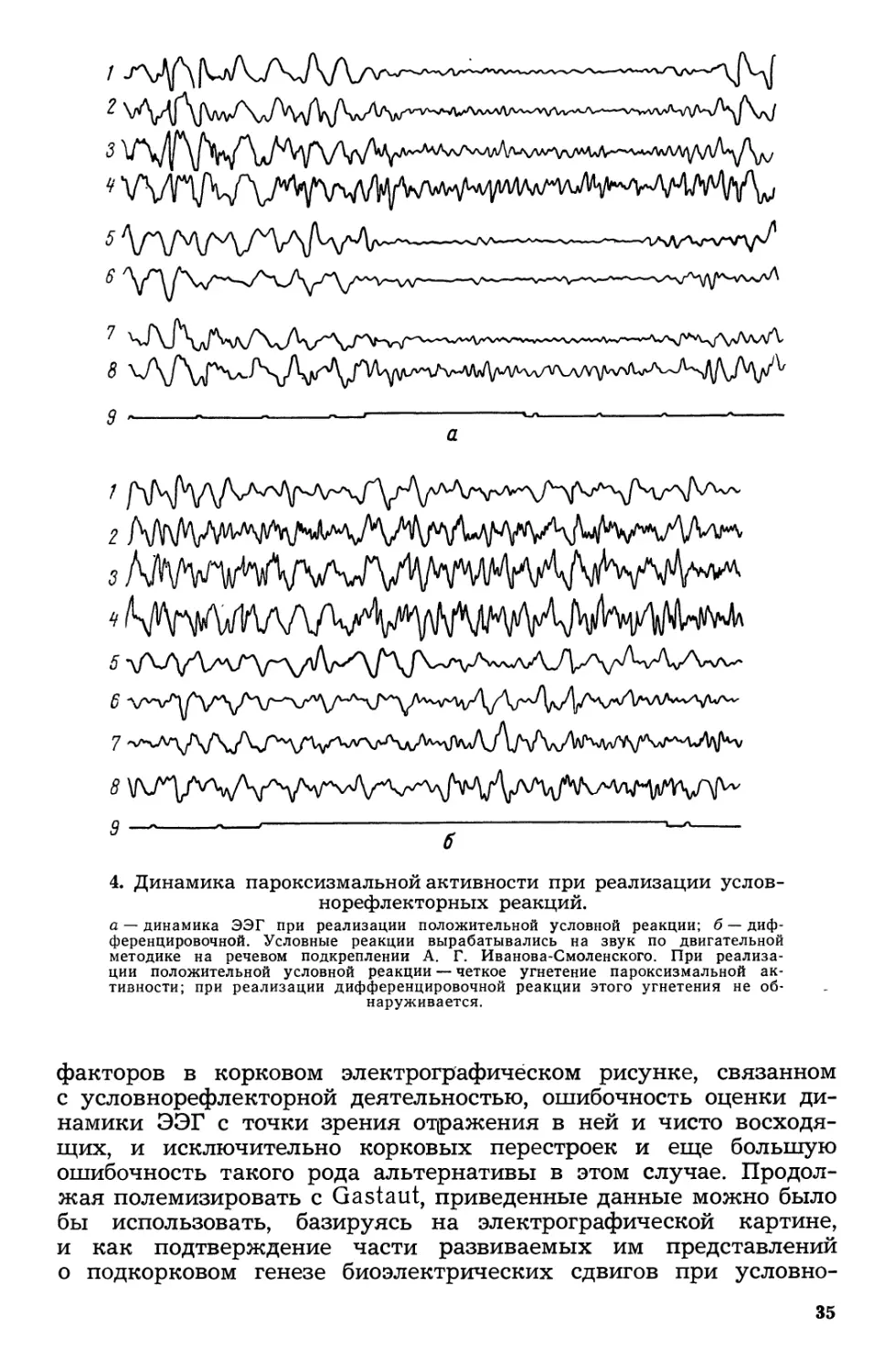
3. Динамика очаrовой медленной активности при реализации
условнорефлекторных реакций.
а динамика ээr при реализации положительной условной реакции;
30
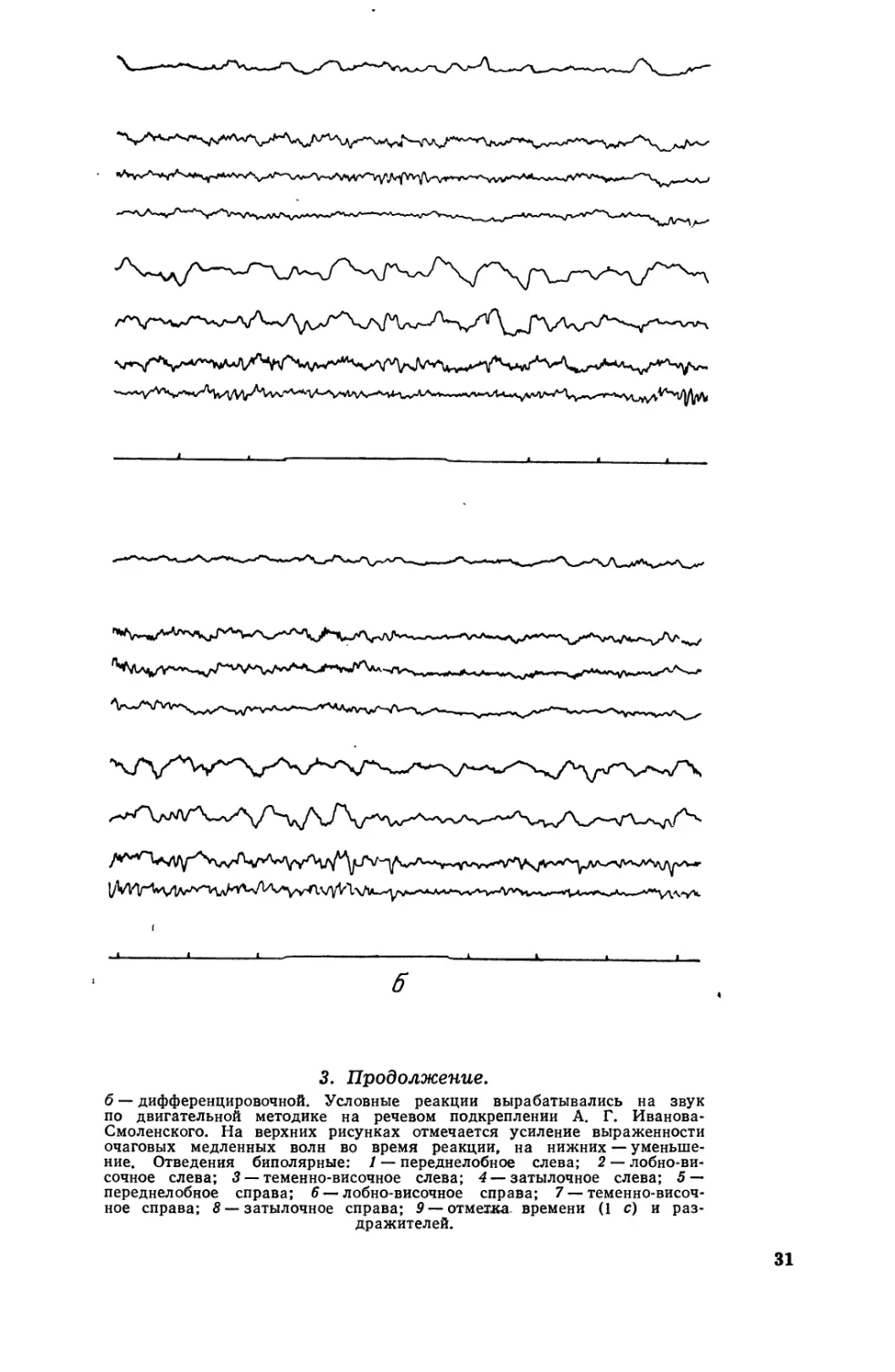
""'У"
б
3. Продолжение.
б дифференцировочной. Условные реакции вырабатывались на звук
по двиrательной методике на речевом подкреплении А. r. Иванова.
Смоленскоrо. На верхних рисунках отмечается усиление выраженности
очаrовых медленных волн во время реакции, на нижних уменьше-
ние. Отведения биполярные: 1 переднелобное слева; 2 лобно ви-
сочное слева; 3 темен но-височное слева; 4 затылочное слева; 5
переднелобное справа; 6 лобно височное справа; 7 теменно височ-
ное справа: 8 затылочное справа; 9 oTMe:rxa времени (1 с) и раз.
дражителей.
31
. будим ости В rоловном мозrу и, в частности, в коре в связи с yc
ловнорефлекторной деятельностью. Так называемое «уrашение»
подкрепляемых реакций ситуация, также предположительно
связанная с торможением, далеко не четко коррелировала с раз
витием медленных волн у тех же больных. И, наконец, даже
при обширном и увеличивающемся по выраженности и ПlPотя
женности очаrе медленных волн в наиболее значимых для реак--
ции корковых зонах сама условная реакция, как правило, реа--
лизовалась.
Оставалось предположить, что: 1) вследствие раЗБИТИЯ очаrо
Boro патолоrическоrо процесса произошли уже компенсаторные
перестройки, «перераСПlPеделения контроля» функций в мозrу, и
именно поэтому даже увеличение очаrа уrнетения (торможения?)
в коре не вызывает существенноrо дефекта в условнорефлектор--
ной деятельности, орrанизованной на взаимодействии звуковых,
световых и проприоцептивных сиrналов, или 2) то, что проявля
лось в виде медленных волн, не отражало торможения в коре,
ведущеrо к ее выключению из деятельности, а было отражением
возбуждения, развивающеrося на KaKOM TO ином исходном
YIPoBHe функционирования ,мозrа. Такая возможность была дo
статочно вероятна на основе непредвзятоrо анализа клинических
данных, обнаруживающих сохранность иноrда довольно широ
Koro Kpyra типично «корковой» деятельности у больных не
только с очаrовыми, но и распространенными медленными вол--
нами. Неоспоримыми, однако, в наблюдениях с реrистрацией
ээr при условнорефлекторной деятельности у больных, с ис
ходно полиморфной ээr оставались факты о возможном
полиморфизме электроrрафическоrо выражения функциональ"
ных изменений в мозrу при условнорефлекторной деятельности
t1 неправомерности оценки наблюдаемых в ээr явлений исклю,
чительно с точки зрения их дистантной обусловленности. Те
электроrрафические изменения, которые развиваются в мозrу
в процессе условнорефлекторной деятельности, происходят при
взаимодействии дистантных влияний с корой. Свидетельством
этоrо, более убедительным, чем «иrра» синхронизации десин
хронизации в коре, является ээr выражение взаимодействия
«вызванной» ситуации (дистантно обусловленной ээr реакциw)
с наличным корковым патолоrическим очаrом и, в частности.
ээr изменения, развивающиеся в этих условиях в области KOp
J(oBoro патолоrическоrо очаrа. Это положение принципиально
важно при решении вопроса о возможности извлечения из ээr
информации о дробных, преимущественно локально корковых
событиях, в частности при собственно психической деятельности.
Исследование ээr больных с очаrовыми поражени.я.ми rолов
Horo мозrа при тормозных условных реакциях показало возмож
ность наблюдать у них разные типы реакций, в том числе уве'"
личеНJlе и уменьшение ВЫlраженности очаrовых медленных волн.
Таким образом, и на тормозные, раздражители обнаруживалась
определенная динамика первично корковых медленных волн.
32
причем она моrла выражаться их перестройками разной на..
правленности (рис. 3).
Исследования, проводимые в этих, cBoero рода «модельных»
ситуациях исходноrо наличия патолоrическоrо очаrа в коре, под
черкнуЛИ сложность вопроса о сущности электроrрафических
феноменов при условнорефлекторной деятельности и особенную
противоречивость материалов о взаимоотношениях медленной
активности и торможения. Принципиальную важность положе
нил о необязательности связи медленных волн с торможением
и, наоборот, малую вероятность такой связи подчеркивал
и П. К. Анохин (1968). И все же клинико физиолоrичеСRие
исследования показали, что очаrовые медленные волны, разно
направленная динамика KOTOp IX обнаруживалась при реализа--
ции и положительных и тормозных условных с язей, действи--
тельно обладают, по крайней мере, одной из функций тормозноrо
процесса охранительной (как известно, специально в отноше
нии медленных волн это свойство было предположено Walter,
1953).
в ходе совместных исследований с нейрохирурrами (Н. п. Бехтерева
и А. Н. Орлова, 1957) была убедительно показана относительная безопас
ность хирурrических манипуляций в зоне очаrа медленной активности и,
наоборот, резкое повышение частоты патолоrических реакций при слабой
выраженности или практическом отсутствии в области патолоrическоrо
очаrа медленной активности.
Усиление медленных волн не было единственной формой
взаимодействия патолоrическоrо очаrа с приходящими в кору
В процессе условнорефлекторной деятельности стволовыми, Ta
ламическими, лимбическими и друrими влияниями. Взаимоот
ношение очаrа и этих явлений моrло складываться таким обра
зом, что биоэлектрическим эффектом оказывалось значительное
уменьшение выраженности медленных волн или усиление ча
стых колебаний типа заостренных бета волн и т. д.
Учет этих реакций оказался полезным в предоперационной оценке co
стояния больных. Если наличие и усиление медленных волн при раздра
жителях в зоне патолоrическоrо очаrа являлись признаком, проrности
чески блаrоприятным в отношении реакции на операцию, то усиление BЫ
раженности частых колебаний в зоне очаrа оказывалось признаком
неблаrоприятным. Предотвращение неблаrоприятных реакций во время опе
рации осуществлялось с помощью специальной предоперационной подrо
тnвки больных (Н. П. Бехтерева и А. Н. Орлова, 1957), однако это уже co
B eM особый вопрос, не связанный с темой данной книrи.
Природный эксперимент болезни мозrа позволил иссле--
Д(Jвать те же явления и при первичном поражении ствола.
Как известно, одним из частых проявлений стволовоrо пато
лоrическоrо очаrа является развитие более или менее выражен
ной пароксизмальной активности, обнаруживающейся в виде
вспышек бета , альфа--подобноrо, TeTa и дельта или смешанноrо
ритма. Исследования, проведенные у больных с вживленными
электродами, показали, что вспышки эти MorYT обнаруживаться
не только в коре больших полушарий, но и как бы «пронизы
вать» все вещество мозrа, быть действительно распространен--
33
ными (что, конечно, не исключает возможности появления и чи--
сто локальных вспышек). .
Динамика раСПРОС'I)раненных вспышек в ээr при условно
рефлекторной деятельности во MHoroM повторила все то, что
можно было наблюдать у здоровых лиц в отношении динамики
альфа ритма. Распространенные вспышки подавлялись при пер
вом предъявлении индифферентных раздражителей и при по
вторных предъявлениях положительных условных сиrналов. Они
сохранялись при повторяемых индифферентных сиrналах, при--
чем при прочных дифференцировочных сиrналах медленная
активность во вспышках моrла не только сохраняться, но и YBe
личиваться по периоду (рис. 4).
Таким образом, в одних и тех же условиях, коrда в связи
с характером условнорефлекторной реакции можно было пред--
полаrать развитие или усиление возбудительноrо процесса,
у здоровых людей в бодрствующем состоянии наиболее частой
(но отнюдь не обязательной!) реакцией была десинхронизация,
блокада альфа ритма. Изменение физиолоrическоrо состояния
здоровых лиц засыпание моrло в тех же условиях и с близ
кими пространственными закономерностями вызвать реакцию
синхронизации альфа ритма. Обе эти электроrрафические peaK
ции моrли отражать четко дистантные электроrрафические
сдвиrи. «Вмешательство» KopKOBoro патолоrическоrо очаrа в эти
отношения привело к большему полиморфизму ва,риантов peaK
ций и обнаружило возможность уже чисто корковых сдвиrов
в этих условиях. Развитие патолоrическоrо процесса в области
срединных структур мозrа, сопровождавшееся пароксизмальной
активностью, в подчеркнутой, «преувеличенной» форме вновь
позволило наблюдать развитие десинхронизации, дистантную pe
акцию при положительных условных рефлексах.
В этом, последнем, суrубо специальном случае внутреннее
торможение как бы оказывалось «связанным» С медленными
волнами в rоловном мозrу, и в коре в частности, причем в этой
ситуации медленная активность в коре является действительно
следствием событий на стволовом уровне. Таким образом, при
поражен иях мозrа, при всех оrраничениях TaKoro рода данных
удавалось полнее, чем в норме, изучить и роль местных, и роль
общих (дистантных) влияний в происхождении ээr--картины при
реализации условных реакций.
Дальнейшая детализация относительной роли MecTHoro и об
щеrо фактора, а также некоторое уточнение возможноrо физио
лоrическоrо значения наблюдаемой электроrрафической дина
мики при условнорефлекторной деятельности оказались возмож
ными при сочетании «физиолоrическоrо И патолоrическоrо», при
ээr у больных с очаrовыми поражениями мозrа, осуществлен--
ной в процессе условнорефлеКТОIРНОЙ деятельности, реализуемой
на фоне резкоrо оrраничения зрительной импульсации (Н. П. Бех
терева, 1961). Проведенные в этих условиях исследования еще
раз подчеркнули относительность значения общих и местных
34
з
'1
5
6
7
8
9 ...r
а
1
Lf
5
7
8
9
.......,.
5
4. Динамика пароксизмальной активности при реализации услов
норефлекторных реакций.
а динамика ээr при реализации положительной условной реакции; 6 диф
ференцировочной. Условные реакции вырабатывались на звук по двиrательной
методике на речевом подкреплении А. r. Иванова Смоленскоrо. При реализа
ции положительной условной реакции четкое уrнетение пароксизмальной aK
тивности; при реализации дифференцировочной реакции этоrо уrнетения не об-
наруживается.
факторов в корковом электроrpафическом рисунке, связанном
с условнорефлекторной деятельностью, Оlllибочность оценки ди
намики ээr с точки зрения ОТlPажения в ней и чисто восходя--
щих, И исключительно корковых перестроек и еще большую
ошибочность TaKoro рода альтернативы в этом случае. Продол--
жая полемизировать с Gastaut, приведенные данные можно было
бы использовать, базируясь на электроrрафической картине,
и как подтверждение части развиваемых им представлений
о подкорковом rенезе биоэлектрических сдвиrов при условно
35
рефлекторной деятельности, и как антитезу основной линии ero
анализа материала о физиолоrическом значении этоrо подкор
KOBoro уровня.
Исследования, проведенные с записью ээr при условнореф
лекторных реакциях, прежде Bcero с исключительной убедитель--
ностью свидетельствуют о непраВОМ1ерности утверждения об
однозначной связи биоэлектрических явлений мозrа с основными
нервными ПlPоцессами. Локальные и распространенные измене
ния ээr при условнорефлекторных реакциях зависели от исход
Horo биоэлектрическоrо и, таким образом, функциональноrо фона
мозrа. Лоrичным следствием всех проведенных наблюдений яв
ляется неправомерность идентификации ведущих, узловых пунк
тов В мозrу, ответственных за биоэлектрическую перестройку,
с местом OCHoBHoro, определяющеrо, замыкания условной связи.
Важно подчеркнуть, что хотя источником суммарных сдвиrов
биоэлектрической активности, реrулируемых в ээr при условно
рефлекторных реакциях, служат явления, разыrрывающиеся,
по--видимому, на мезеНЦiефальном и диэнцефальном уровнях, это
не свидетельствует ни за, ни против замыкания условной связи
на тех же уровнях.
И в то же самое время еще более неправомерно было бы
впасть в друrУIО крайность, пренебреrать данными о биоэлектри
ческих перестройках в мозrу в этих условиях. При учете исход
Horo биоэл,ектрическоrо фона эти данные MorYT быть использо
ваны для оценки условий, развивающихся при взаимодействии
со средой в сложнейшей из известных на земле структур мозrу
человека, для оптимальноrо протекания условнорефлекторной
деятельности. Биоэлектрические явления свидетельствуют о BO
влечении обширных территорий мозrа в это состояние оптими
;зации условий протекания исследуемой деятельности. Однако,
наблюдая этот феномен, всеrда следу,ет помнить, что «вся кора»,
«весь мозr» и т. д. В этом случае обеспечивают только возмож
ность деятельности, а не конкретную деятельность, которая имеет
свои, rораздо БОЛ1ее дробные механизмы. Отражение в ээr
прежде Bcero «rотовности К функционированию» отмечает и
А. Б. KoraH (1967). Иошии, Мацумото, Orypa, Симокоти, Лмаrути
и Лмасаки (1962) и др. настаивают на том, что ээr--эффекты при
условнорефлекторной деятельности следует связывать не с aK
тивностью образований (центров), действительно ответственных
за реакцию, и не с динамикой нервных процессов, а с динамикой
фоновой возбудимости мозrа.
Изменения в ээr наблюдались всеrда на больших «площадях» мозrа,
в пределах долей и областей коры. Эти данные о закономерности обшир
ных изменений в коре в этих условиях были получены в середине и
в конце 50 x rодов, коrда осуществлялись уже микроэлектродные иссле
дования условнорефлекторных реакций (Джаспер, Риччи и Доун, 1958,
1962, и др.), показавшие исключительную дробность нервных явлений на
клеточном уровне. Можно указать, что понимание также достаточно слож
ных явлений на клеточном уровне стало в условнорефлекторной ситуации
возможным лишь значительно позднее, после методическоrо вклада в проб
36
лему, сделанноrо в первую очередь Н. Н. Василевским (1965 1968). Следует
подчеркнуть, что соrласование Toro, что наблюдалось на нейрональном
уровне и в ээr, было (и в известной мере остается и сейчас!) 9чень слож
НЫМ. И однако, ни исследование закономерностей клеточноrо уровня, ни
исследование ээr в одной и той же ситуации не отрицали друr друrа,
а дополняли, как всеrда дополняли и будут дополнять друr друrа любые
исследования микро и макроуровней любой проблемы.
Можно ли все же на сеrодняшний день принять какую--либо
или какие либо точки зрения в вопросе о соотношении нервных
процессов и биоэлектрических явлений? По видимому, HeKOTO
рые стороны зтоrо вопроса MorYT быть поняты на основе пред--
ставлений В. С. Русинова (1953) о важности учета направления
изменений биоэлектрической активности. В этом случае разви
вающееся преобладание торможения, сдвиr в сторону преоблада
ния торможения в мозrу MorYT отражаться в ээr синхронизацией
и замедлением активности и, наоборот, сдвиr в сторону преоб
ладания возбуждения повлечет за собой учащение биоэлектри--
ческой активности и e де синхронизацию, наиболее отчетливой
формой которой является блокада альфа ритма при исходном
присутствии ero в ээr. Эта очень" ..частная схема, действующая
при соблюдении целоrо ряда условий (исходный фон и Т. д.),
однако, отражает лишь один из аспектов действительных отно--
шений и прежде Bcero TaKoro рода соотношение нервных про
цессов и биоэлектрической активности, rде биоэлектрическая aK
тивность изменяется вслед за перестройкой нервных процессов.
Наблюдение ээr в различных физиолоrических и патолоrи--
ческих ситуациях заставляет с равной вероятностью учитывать
и еще одну возможность измен,ения в ээr по отношению к ди
намике нервных процессов, в частности связанных с условно
рефлекторной деятельностью, MorYT быть не только вторичными,
но и первичными, облеrчая или, наоборот, затрудняя эту дина
мику. Применительно к данному конкретному случаю взаимо
действию нервных процессов и ээr в наиболее четкой форме
это положение рассмотрено М. Н. Ливановым (1957, 1962а, б),
считаюIЦИМ, что исходная био лектрическая динамика является
фоном, который может облеrчать распространение нервных про
цессов. 'Представления John (1967), полаrающеrо, что синхрони--
зация в ээr облеrчает механизмы «считывания», различаются
с приведенными представлениями М. Н. Ливанова rлавным об--
разом по форме.
Две эти стороны взаимоотношений ээr и деятельности, KOH
тролируемой мозrом, в еще более общем виде формулируются
как свойства ээr не только отражать то, что происходит в мозrу,
но и определять ero текущее функциональное состояние, обеспе--
чивать функциональный уровень, - оптимальный для протекания
любой данной деятельности (М. Н. Ливанов, 1962б; Е. Н. Соко--
лов, 1962; Н. П. Бехтерева, 1966). Следует, однако, подчеркнуть,
что при анализе этих явлений, при уrлублении в проблему,
37
исследователи сталкиваются сейчас с все большими сложно
стями. Так, тонкие экспериментальные нейрофизиолоrические
исследования (П. К. Анохин, 1968; r. А. Вартанян, 1966, 1970,
и др.) представляют все больше убедительных данных о непра
вомерности утверждений об иррадиации торможения в мозrу, по
крайней мере, в той трактовке этоrо явления, которая предлаrа--
лась и. П. Павловым. Новые данные о физиолоrической сущ
ности активирующих и тормозных реакций в мозrу получены
сейчас уже и в прямых наблюдениях на человеке (Н. П. Бехте
рева, А. Н. Бондарчук, В. М. Смирнов, 1969). Даже если бы
не возражать по существу против предположения о том, что по
ээr можно rоворить не только о предпосылках к иррадиации
нервных процессов, но и о движении их, коррелирующем с дви
жением KaKoro To вида биопотенциалов, приходится признать,
что очень MHoroe, используемое в качестве доказательств TaKoro
движения, методически спорно.
При специальном изучении этоrо вопроса одним из важней--
ших условий исследования движения биопотенциалов должно
быть проведение расчетов на максимально мономорфном виде
биоэлектрической активности и тончайшее изучение фазовоrо
СДБиrа волн во мноrих соседних областях мозrа. TaKoro рода
условиям удовлетворяют лишь отдельные работы и, в частности,
некоторые исследования Ю. В. Дубикайтиса (1964, 1968) и Petsche
(1967, 1968). Pet che с помощью известноrо собственноrо варианта
топоскопии на примере активности пик волны было показано,
что во вспышках движутся по коре позитивные волны. Однако
именно Petsche и Ю. В. Дубикайтис сравнительно мало интере
совались соотношением биоэлектрической дина ики с динамикой
нервных процессов.
Рассматривать злектроrрафическую динамику явлений можно
и в несколько ином аспекте. Можно рассматривать десинхрони
зацию при положительных условных реакциях и особенно re
нерализованную как отражение первоначально очень неэконом
Horo механизма вовлечения в работу и рабочую rOToBHoCTb боль
шинства областей мозrа при существенном общем повышении
уровня возбудимости коры (А. Б. KoraH, 1962; Chang, 1962;
М. Н. Ливанов, 1962). Оrраничение реакции при повторениях
сочетаний есть внешне проявление уменьшения количества
структур, rOTOBbIx к деятельности, возможное при условнореф
лекторном исследовании исключительно вследствие стереотипии
ситуации, резкоrо снижения фактора новизны и ero вероятности.
Синхронизация биопотенциалов при тормозных реакциях,
с этой точки зрения, отражает оrраничение зон, находящихся
в состоянии деятельности или оптимальной rотовности к вовле
чению именно в данную деятельность. Это состояние может
развиться принципиально за счет выключения активирующих
влияний или активации синхронизирующих влияний любоrо
уровня. Однако биолоrически выrодным (экономным) может
быть включение этих механизмов при простейших условнореф
38
лекторНЫХ реакциях на «узловых» уровнях. Не исключено, Ta
ким образом, и не только в связи с данным положением, что,
если реализация положительной условной связи, особенно перво
начально, приводит в рабочее состояние (по крайней мере, до
«фазы падения вероятности новизны») обширные области мозrа,
то тормозные реакции MorYT реализоваться и обычно в норме
реализуются в этих простейших модельных условиях на основе
значительно более экономичных механизмов.
Далее. Если реакция десинхронизации, однообразная биоэлек
трически в ээr, и, если судить по данным прямоrо наблюдения
мозrа, электрокортикоrрафии (экоr), не однозначная реально, MO
жет рассматриваться как отражение оптимизации функциональ
Horo состояния мозrа для деятельности, то как можно представить
значение синхронизации биопотенциалов коры при реализации
тормозных реакций?
у здоровоrо человека с хорошо выраженным альфа ритмом
картина ээr в этих условиях нередко не отличима от покоя или
проявляется как бы yr лублением рисунка покоя, увеличением
размахов колебаний и (иноrда) замедлением их периода. Иссле
дуя в течение ряда лет ээr человека с точки зрения взаимоот
ношения ее ритмов с динамикой возбудимости мозrа (Н. П. Бех
терева и соавт., 1958 1965, и др.), удалось показать, что, хотя
соотношения волн ээr и колебаний возбудимости не всеrда
просты, само их наличие, по--видимому, не должно вызывать
сомнений (Brazier, 1962). Отсюда, если не пытаться «примыс
лить» что либо к видимому в ээr, придется признать, что при
данных простых условных реакциях в ситуации «торможение»
восстанавливаются, развиваются или усиливаются колебания
возбудимости, резко уменьшающиеся в ситуации «возбуждение
десинхронизация». Это состояние синхронизации может явиться
следствием действительноrо развития торможения в области
диэнцефальных и мезенцефальных активирующих структур или
активации тормозящих подкорковых механизмов и синхрони
зирующих влияний (или взаимодействия этих механизмов). Top
можение в исследуемых условиях может Iразвиться и на уровне
коры больших полушарий, реальность чеrо можно было просле
дить в конкретных специальных условиях и предположить при
дробных пространственно механизмах сложной психической дея
тельности. Функциональные перестройки этоrо рода, развиваю
щиеся на различных уровнях срединных структур мозrа, MorYT,
как показано выше, повлечь за собой не только диффузные, но
и локальные функциональные изменения в коре больших полу
шарий, связанные с применяемым раздражителем и местным
состоянием мозrа (патолоrический очаr). Статистически наиболее
вероятными в норме, однако, являются ээr изменения, разви--
вающиеся на сравнительно больших площадях коры и являю
щиеся скорее Bcero не отражением наличия торможения в коре,
а следствием возникновения перестроек в нижележащих CTPYK
турах. Эти представления не очень далеки от точки зрения
39
Adrian (1954) и др. о возможности блокирования потока инфор
мации в определенных случаях на разных уровнях нервной
системы. TaKoro рода возможность не противоречит также пред--
ставлениям п. К. Анохина (1968) об орrанизации функциональ--
ных систем по пейсмекерному типу. Колебания возбудимости
в коре в этих условиях оказываются состоянием, оптимальным
для восстановления ее воЗможностей при сохранении, хотя в этом
случае и дискретной, rотовности к деятельности вообще, а не
к какой 'rо данной, как в ситуации «возбуждение». Ритмические
смены одноrо состояния друrим являются одним из наиболее
общих биолоrических законов, на основе KOToporo в живом opra
низме происходит длительное сохранение и восстановление воз
можностей opraHa и орrанизма в целом. Отсюда то, что наблю
дается часто в ээr здоровоrо человека при тормозных простых
условных реакциях и в друrих аналоrичных состояниях, яв
ляе'r.СЯ скорее Bcero обусловленным дистантно развившимся TOp
можением (?) или активацией «тормозноrо» механизма, выrодным
для орrаниз а состоянием действительно активноrо покоя, rде
восстановление возможностей протекает при сохранении rOToB
ности к включению в деятельность, но в дискретной форме.
Безусловно, такой трактовке может быть противопоставлен
ряд возражений. Так, у ряда лиц синхронный альфа ритм не об
наруживается ни в покое, ни в ситуации «торможение»; В HeKO
торых физиолоrических состояниях (засыпание) именно положи
тельная условная реакция наблюдается при синхронизированной
ээr и т. п. В первом случае можно предположить, что колебания
возбудимости MorYT быть связаны и не только с альфа ритмом,
а и с более частыми колебаниями, однако наблюдение динамики
этих ритмов по ээr затруднено; второй случай, скорее под--
тверждение, чем возражение данным представлениям, первой
фазой выхода из состояния дремотыI (или сна) оказывается, если
судить по ээr, фаза отчетливых колебаний возбудимости коры
со всеми ее свойствами и возможностями (хорошо известно, что
при усилении раздражителей или частом повторении их проис
ходит «пробуждение» со всеми ero поведенческими и биоэлек
трическими характеристиками).
rенерализованная синхронизация биопотенциалов, как указы--
валось выше, может быть понята с точки зрения блокады BOCXO
дящих активирующих влияний ретикулярной формации и (или)
активации синхронизирующих структур ствола (Morruzzi, 1962).
Однако сейчас уже есть данные, полученные непосредственно
у человека, о механизмах более BbIcoKoro уровня, способных BЫ
зывать тормозные эффекты. Их значение должно быть учтено
в понимании реакции синхронизации более локальной. Эти явле--
нил у человека MorYT быть связаны с деятельностью, в частности,
срединноrо центра зрительноrо буrlPа, xBocTaToro ядра и, воз--
можно, друrих структур (Н. П. Бехтерева, Н. И. Моисеева,
В. М. Смирнов, А. Н. Орлова, 1964; Н. п. Бехтерева, А. Н. Бон
дарчук, В. М. Смирнов, 1969).
40
При оценке всех этих явлений не следует забывать также
возможность и первично корковых механизмов синхронизации
(М. Н. Ливанов, 1962), в данном случае, однако, несколько менее
вероятных, хотя и не невозможных, и в то же самое время
в самом возникновении десинхронизации, обусловленной влия
ниями ствола или подкорки, чрезвычайно важно учитывать роль
кортико--фуrальных влияний (Morruzzi, 1962).
Приведенные наиболее вероятные возможности взаимодей
ствия синхронизации и торможения не исключают и прямой
связи этих явлений, т. е. развития торможения непосредственно
в зоне синхронных колебаний в коре; они лишь показывают
необязательность этой прямой связи и проливают свет еще на
один возможный структурный и физиолоrический механизм ox
ранительноrо свойства торможения.
Конечно, весь опыт изучения ээr человека утверждает не
строrую обязательность появления синхронной активности при
тормозных реакциях. Следует, по видимому, признать, что по
явление синхронной активности и связанноrо с ней усиления
ритмических колебаний возбудимости есть, если и не обязатель
ная, то одна из наиболее выrодных для орrанизма реакций, peaK
ция охранительноrо типа, способствующая сохранению или BOC
становлению рабочеrо потенциала мозrа.
Следует, однако, напомнить, что наиболее серьезным дефек
том всех проведенных наблюдений, с точки зрения ценности
именно физиолоrических данных, является суждение о состоя
нии мозrа на основе одноrо параметра ээr и, кроме Toro, ээr,
в значительной мере деформированной в связи с непрямым xa
рактером наблюдения ее, отведением через покровы мозrа.
ЭЛЕКТРОЭНЦЕФАлоrРАММА ПРИ психолоrИЧЕСКИХ
ПРОБАХ
Изменения ээr человека были прослежены не только на про
стейшей модели психическоrо........... условнорефлекторных реакциях,
но и при изменениях сознания, и в зависимости от особенности
личности, и при психолоrических пробах.
С точки зрения анализируемоrо в данной работе вопроса
нейрофизиолоrических аспектов психической деятельности
наибольший интерес представляет динамика ээr при различных
психолоrических тестах. Условнорефлекторный принцип лежит
в основе всех психических явлений человека. Однако при
изучении нейрофизиолоrическоrо аспекта психических явлений
необходимо было использование в сочетании с нейрофизиолоrиче
скими приемами не только наиболее распространенных, простей
тих условнорефлекторных проб, но и возможностей психолоrи
ческоrо исследования. Именно в этом и только в этом случае
исследователь наиболее прямо адресовался к тому, что явилось
rиrантским скачком в живой природе, к психической деятель
ности человека.
41
Попытка корреляции ээr и психических явлений была пред
принята еще Berger (1933). Изменения ээr были в дальней
шем прослежены при использовании в качестве раздражителей
психолоrических проб различной сложности (Travis, 1937;
П. И. Шпильберr, 1947; Е. Н. Семеновская и Р. Н. Лурье, 1948,
и др.) и, в чаСТНОСТИ, словесных сиrналов. В этих работах было
показано, что ээr эффект (и прежде Bcero ero выраженность) на
словесные раздражители находится в зависимости от смысловоrо
значения слов и состояния испытуемоrо. В обычных условиях во
время «леrкоrо» разrовора реrистрировалось лишь небольшое
снижение амплитуды альфа--волн (самонаблюдения Adrian и Mat
thews, 1934). Travis и Eg4an (1938) наблюдали небольшое учащение
альфа ритма при прослушивании читаемоrо вслух текста (с 10,15
до 10,75). Эффект этот в целом был так невелик, что Gastaut
(1957) считал возможным пренебречь реакцией на словесный
приказ при оценке условнорефлекторных изменений ээr в тех
исследованиях, rде он включался в комплекс раздражителей.
В то же время словесное смысловое условие моrло опреде
лить реакцию в ээr на любой друrой конкретный раздражитель
(Н. А. Мушкина, 1956) и, с друrой стороны, словесный раздра--
житель в соответствии с ero смыслом Mor вызвать эффект KOH
KpeTHoro раздражителя (Р. Н. Лурье и В. С. Русинов, 1952). При
подкреплении словесноrо раздражителя конкретным (слово «блок»
в наблюдениях Shagass, 1942) оно само начинало вызывать реак--
ЦИЮ, характерную для использованноrо подкрепления.
Важнейшую роль в динамике ээr на словесные раздражи--
тели имело их эмоциональное значение. Если слово оказывалось
для испытуемоrо эмоционально значимым, депрессия альфа--
ритма оказывалась очень выраженной и длительной (А. И. Ройт--
бак и Н. А. Саванели, 1953; Bagchi, Howell, Schmall, 1945, и др.).
Проведение таких исследований у больных (И. А. Пеймер, 1954;
Е. М. Яковлева, 1952, 1956) моrло приводить к появлению или
усилению в ээr и патолоrической биоэлектрической активности.
Однако повторение и эмоциоrенно значимых слов приводило
обычно к уrасанию ээr реакции на них.
Реакция на словесный раздражитель была прослежена также
в процессе засыпания и в rипнозе. При засыпании (Jung, 1954)
словесный раздражитель вызывал реакцию активации в ээr
появлялись альфа и бета--волны и уменьшались медленные KO
лебания. Исследования с использованием rипнотическоrо внуше
нил показали возможность и уменьшения, и увеличения ээr--
эффекта на словесные раздражители (М. Н. Ливанов, И. В. Стрель
чук, А. М. Мелехова, 1963; М. П. Невский, 1954; Loomis, Harvey
и Hobart, 1936).
ээr изменения в виде депрессии альфа--ритма наблюдались
при показе рисунков (Jasper, Shagass, 1941; Williams, 1939; Golla,
Hutton and Walter, 1943), мысленном воспроизведении изображе
ний (Walter а. Yeager, 1956). Мысленное воспроизведение движе
ний вызывало блокаду роландическоrо ритма (Gastaut, 1952;
42
Magnus, 1954). Снижение амплитуды волн ээr наблюдалось при
запоминании цифр (Л. А. Новикова, 1955).
Депрессию альфа ритма оказалось возможным наблюдать
в самых различных ситуациях связанных с просмотром кино
фильма (Gastaut а. Bert, 1954; Faure и Cohen--Seat, 1954; Lelord,
1957; Verdeaux а. Verdeaux, 1955), при воспроизведении одноrо из
компонентов искусственно созданной мысленной ассоциации
(Laufberger, 1950) и т. п. В тех случаях, коrда у испытуемых, про
сматривающих кинофильм, исходно в ээr реrистрировался po
ландический ритм, а поведение актеров на экране было очень
динамичным, можно было наблюдать не только подавление, но
и усиление этоrо ритма.
Большое количество исследований проведено с записью ээr
при счете в уме и при решении арифметических задач (Mundy
Castle, 1957; И. А. Пеймер, 1960; А. А. rенкин, 1963; М. Н. Лива
нов, 1962; М. Н. Ливанов, Н. А. rаврилова, А. С. Асланов, 1966;
Glass, 1964, 1965; Е. Ю. Артемьева и Е. д. Хомская, 1966; Volavka
et al., 1967; Vogel et al., 1968, и др.). Наиболее постоянно в этих
условиях наблюдалась депрессия альфа ритма, причем в ряде
работ (но не всеrда!) отмечалась прямая зависимость трудности
выполняемоrо задания и выраженности депрессии альфа ритма
(И. А. Пеймер, 1960). Исследование ээr у здоровых лиц и
у больных в процессе опознавания образов при различной дли
тельности экспозиции подтвердцло зависимость изменений ээr
от концентрации внимания, от эмоциональноrо и психическоrо
напряжения (Е. А. Жирмунская, Е. С. Бейн, В. Н. Волков, 1969).
В условиях психическоrо напряжения нередко отмечалась
отчетливая динамика бета волн увеличение их выраженности
(Kennard, Fister, Rabinovitsh, 1955; Mundy Castle, 1957; Е. С. Бейн
и др., 1967; Volavka et al., 1967) или, наоборот, уменьшение бета
индекса. В сравнительно большом количестве исследований OT
мечалось и усиление тета волн у ряда лиц при интеллектуальной
деятельности (Arellano и Schwab, 1950; Brazier а. Casby, 1952;
Mundy Castle, 1957; И. А. Пеймер и А. А. Фадеева, 1957; Liu
Schih yih и др., 1964; А. Н. Брендстед, 1966; Е. С. Бейн и др.,
1967; Vogel и др., 1968, и др.), а также появление «каппа--ритма»
(Kennedy, Gottsdanker, Arnington, Gray, 1948; Liu Schih yih и др.,
1964). И в то же самое время в ряде проведенных исследо
ваний указывается, что некоторые изменения ээr при YMCTBeH
ной деятельности MorYT в значительной мере определяться apTe
фактами, а во мноrих работах отмечается, что у некоторых лиц
в условиях интеллектуальной деятельности ээr, по крайней
мере, видимо, не перестраивается (Е. Ю. Артемьева, Е. д. XOM
ская, 1966, и мн. др.). Однако во всех случаях, если изменения
ээr обнаруживались при психолоrических пробах, они были
распространенными, хотя и моrли быть в каких--то определен
ных областях преимущественно выраженными (Adey, 1969).
Известное внимание было уделено изменениям не только
спонтанной ээr, но и вызванных потенциалов при психической
43
деятельности (И. А. Пеймер, В. А. EropoB, М. Л. Модин, Э. В. Бон
дарев, Н. А. rOBopoBa, Э. Ф. Васильева, 1966; Chapman, 1966;
Shagass, Canter, 1966, и др.). Особенно убедительно и дeMOHCTpa
тивно разница вызванных потенциалов в зависимости от связи
стимула с каким либо из видов интеллектуальной активности
показана в работе Сhарmап вызванный ответ на значимые
раздражители был несравненно более отчетливым, чем на н зна
чимые. Grey Walter (1967), реrистрировавший вызванные потен
циалы с различных областей мозrа человека с помощью вжив--
ленных электродов, подчеркивает, что множественность зон
неспецифических ответов свидетельствует о вовлечении (и воз
можности вовлечения) обширных областей мозrа в процесс BЫC
шей интеrрации.
Болыпая часть исследований ээr при психической деятель
ности, равно как и изучение ээr при некоторых психических
СОСТОЯНИЯХ и условнорефлекторных реакциях, были проведены
с использованием лишь визуальноrо анализа. Однако в некоторых
работах с целью получения данных о количественной стороне раз
вивающихся перестроек для выявления изменений ээr, усколь
зающих при визуальном анализе, а также для уточнения области
преимущественных изменений использовались приемы MaTeMa
тическоrо и инструментальноrо анализа (Shipton а. Walter, 1957;
Mundy--Castle, 1957; Kennard, 1953, Kennard et al., 1955).
С использованием мноrоканальноrо MHorocTopoHHero анализа
биопотенциалов мозrа человека при ассоциативном эксперименте,
решении арифметических задач, тестов Роршаха, Крепелина,
Бурдона и др. было проведено исследование Storm van Leeuwen
с соавт. (1963).
Данные ээr оценивались с учетом субъективной реакции
исследуемоrо лица. Это исследование, однако, скорее продемон--
стрировало возможности анализа, чем ero результаты, и не шло
ни в какое сравнение с последующими, действительно практи
чески исчерпывающими вопрос работами Storm van Leeuwen
и др. (1965) по сопоставлению поведенческих и ээr lреакций,
проведенных в экспериментах на собаках.
Использование тех приемов анализа биопотенциалов мозrа,
rде представлялись результаты частотной или частотно--ампли
тудной обработки ээr и не делалось попытки извлечения из
ээr каких либо особых сведений, позволило уточнить данные
визуальноrо наблюдения и оценить не только качественную, но
и количественную сторону динамики биопотенциалов мозrа
(Creutzfeldt, 1969). Этот анализ не внес чеrо либо принципиально
HOBoro в изучение связи ээr динамики и нервных процессов,
развиваюIЦИХСЯ при условнорефлекторных реакциях и психоло
rических пробах.
Данные, которые было действительно невозможно получить
лишь при визуальном анализе ээr, представило изучение COOT
ношения биопотенциалов в различных отделах мозrа при пси
хической деятельности. Извлечение этой информации из ээr
44
приемом кросскорреляции (Walter, 1953; Darrow, Wl1son, Vieth,
Маllе, 1960) показал о, что при арифметическом счете синфаз
ность биопотенциалов в ээr центральной и теменной области
выше, чем синфазность теменной и затылочной областей. В по--
кое наблюдались обратные отношения. Особый интерес в этом
плане представляют работы, проведенные с использованием кросс--
корреляционноrо анализа биопотенциалов 50 и более отведений
(М. Н. Ливанов, 1962б, М. Н. Ливанов, Н. А. rаврилова, А. с. Ac
ланов, 1966). В этих исследованиях показаны общие и местные
изменения синхронности биопотенциалов в покое, при интеллек
туальной деятельности у здоровых лиц, специфика динамики
биопотенциалов лобных отделов мозrа в этих условиях и особен
ности взаимоотношений биопотенциалов в покое, при бреде и
интеллектуальной активности у психически больных. .В этом
случае, по существу, изучались также особенности функцио--
нальных изменений больших областей мозrа, причем даже так
называемые местные изменения являлись перестройками биопо--
тенциалов в пределах долей мозrа.
Обобщение мноrолетних исследований (М. Н. Ливанов, 1972)
показало, что в покое у здоровоrо человека 50 О/о точек коры
работают независимо, 45 О/о слабо синхронизированы, 2,2 О/о ........... вы--
сокосинхронизированы, причем в этом случае синхронизированно
работают соседние участки мозrа. При умственной и физической
работе наблюдается резкое увеличение количества участков, pa
ботающих синхронизированно, а число независимо работающих
участков сокращается., При этом отмечается не только общее
увеличение сильных и слабых корреляций, но и перераспреде--
ление корреляций, причем изменения зависят от сложности за--
дачи. На фоне приема аминазина уменьшается число корреляций
и увеличивается время решения задачи. Число корреляций при
неврозах выше, чем в норме, «стимуляция больноrо пункта»
(предъявление эмоциоrенноrо слова) вызывает обычно увеличе
ние корреляций, хотя может и уменьшить их. По этим данным
создается впечатление, что в увеличении (изменении) корре--
ляций значительную роль иrрает развитие эмоциональных со--
стояний. М. Н. Ливанов вновь подчеркивает, что корреляци
онные отношения отражают не связи, а условия, делающие
возможными их реализацию. На основе своих, теперь уже mhoro--
численных, данных исследований нейрофизиолоrии здоровоrо
и больноrо человека М. Н. Ливанов подчеркивает и развивает
представление о роли взаимодействия мноrочисленных корковых
пунктов в мозrовом обеспечении функций. .
В некоторых исследованиях были предприняты специальные
попытки извлечения из ээr информации об изменениях, разви
вающихся в коре в области соответствующих «представительств»
высших функций при ПСИХОJIоrических пробах. В исследованиях
А. А. rенкина (1961, 1964, 1966) и использовавших предложен--
ный им способ анализа ээr Е. ю. Артемьевой и Е. д. Хом--
ской (1966) была показана возможность установить изменения
45
fj 5с
(усл.ед.)
1,0
0;8
D,B
0,'1
0/2
О
ФОН
Картина
'1ЗХ17
р
Ц20
r.
".Jчет /5
р
0;'0
o,'! 2 О 0,2 '! 6 0;8 1,0 fj
2 О 42 О, Ч 0,0 48 O ./j
5. Динамика соотношения восходящих и нисходящих фаз био
потенциалов при психической деятельности (по А. А. rенкину).
Вверху данные этой динамики в теменно затылочной доле rоловноrо мозrа че
ловека; внизу распределение асимметрий при различных видах психической
деятельности.
соотношений восходящей и нисходящей фаз биопотенциалов He
редко «определенноrо пространственноrо и BpeMeHHoro рисунка»
у отдельных лиц, как будто характерные для данноrо вида пси
хической деятельности (рис. 5). Особенно убедительно изменения
в мозrу улавливались не просто при анализе соотношений фаз,
а при вычислении коэффициента взаимодействия фаз и построе
нии векторов длительности фаз.
Эти данные MorYT быть сопоставлены с приведенными выше
наблюдениями об отражаемом в ээr взаимодействии дистантных
влияний с локальным функциональным состоянием мозrа (на
примере патолоrическоrо очаrа) в процессе условнорефлекторной
деятельности.
Как будто удавалось ,также улавливать специфические изме
нения в ээr при психической деятельности способом синхрон--
Horo детектирования (Е. Л. Войтинский и В. А. Прянишников,
1966). Adey (Adey, Danlop, Hendrix, 1960) настаивает на возмож
ности извлечения из ээr, если и не специфической, то все же
тесно связанной с ней информации при условнорефлекторной
деятельности (в эксперименте). Так, в частности, Adey и сотр.
было показано, что данные кросскорреляции электрических про--
цессов коры и rиппокампа и электрических процессов разных
точек rиппокампа позволяют наблюдать специфические явления
в мозrу при образовании временных связей.
Однако можно отметить, что при мноrолетнем применении,
в том числе и очень тонких, специальных методов анализа ээr,
акцент все же постепенно переносится с подчеркивания локаль--
ных изменений биопотенциалов на значение общих их пере
строек, причем местным изменениям отводится уже более CKpOM
46
ная роль части общих (К. А. rенкин, 1966; Е. Ю. Артемьева и
Е. Д. Хомская, 1966).
Надо признать, что современные методы анализа позволили
довольно тонко дифференцировать различные состояния по ээr
человека (Adey, 1970; Berkhout, Walter, Adey, 1969; Э. С. Бейн,
Е. А. Жирмунская, В. Н. Волков, 1971).
Инструментальный и специальный математический анализы,
таким образом, подтвердили наличие изменений ээr у человека
при психической деятельности. Они в то же время не опро
верrли, а скорее (за редким исключением) подчеркнули распро
страненность развивающихся изменений в коре, закономерность
перестроек ээr не только при простых условнорефлекторных
реакциях, но и при психолоrических пробах на больших пло--
щадях коры. В нейрофизиолоrическом плане подавляющее боль
шинство сказанноrо по поводу сущности изменений ээr при
условнорефлекторной деятельности может быть отнесено и к из
менениям ээr при психолоrических пробах. Известным разли
чием явились большая дробность и большая полиморфность из
менений ээr при собственно психолоrических пробах. Однако
инструментальный и математический анализы подчеркнули, что,
хотя при психолоrических пробах и удается отмечать локальные
изменения ээr, развитие этих локальных изменений биопотен
циалов всеrда является частью общих перестроек биоэлектри
ческой активности. Таким образом, и ээr изменения при психи
ческой активности также отражали не только и не столько
деятельное состояние отдельных структур, сколько общие изме
нения в мозrу, связанные, по видимому, с оптимизацией условий
для деятельности.
Это положение определялось, по видимому, не только особен
ностями изучаемой деятельности, но и свойствами избранноrо
физиолоrическоrо параметра ........... ээr.
Нельзя полностью исключать и еще одноrо возможноrо Mexa
низма, лежащеrо в основе массивности изменений ээr, а следо
вательно, и в основе перестроек функциональноrо состояния
мозrа. Эта динамика моrла бы быть объяснимой, если бы дей
ствительно один из «языков мозrа» (Pribram, 1971) и, в част
ности, язык долrосрочной памяти был бы основан на принципе,
сопоставимом с rолоrрафическим.
ЗЛЕКТРОСУБкортикоrРАММА ПРИ психолоrИЧЕСКИХ
ПРОБАХ
ЗНАЧЕНИЕ ЕЕ ИЗМЕНЕНИЙ ПРИ ПСИХИЧЕСКОЙ
ДЕЯТЕЛЬНОСТИ
Анализ данных прямоrо отведения ээr с поверхности мозrа
человека при психолоrических пробах проведен в отдельных
исследованиях (А. А. rенкин, 1964; Chapman, 1966).
Действительно, систематическое изучение динамики электро--
субкортикоrраммы при психической деятельности в условиях ее
4'7
прямоrо отведения от мозrа человека оказалось возможным после
введения в практику метода вживленных электродов, получив
шеrо доступ в клинику после почти столетнеrо использования
в эксперименте (Л. П. Симонов, 1866). Применение вживленных
электродов для диаrностики и лечения rиперкинезов, эпилепсии,
психических болезней и некоторых друrих заболеваний насчи--
тывает сейчас уже историю более двух десятилетий (Bickford
и др., 1955, 1958; Heath, 1954; Meyers и др., 1959; Bastes, 1961;
Sem Jacobsen, 1961, 1968; Walter а. Grow, 1961; Н. П. Бехтерева,
К. В. rрачев, А. Н. Орлова, С. Л. Лцук, 1963; Н. п. Бехтерева,
А. Н. Бондарчук, В. М. Смирнов, А. и. Трохачев, 1967; Н. П. Бех--
терева, 1972, и мн. др.).
Прошедшие rоды позволили не T05IbKO использовать в кли--
нике уже известные из эксперимента orpoMHbIe возможности
вживленных электродов, но и определить Kpyr оrраничений ме--
тода применительно к новым условиям ero использоваtIия. Осо--
бые требования клиники явились стиму лом К существенному
усовершенствованию самой методики. Эти усовершенствования
оказались наиболее значительными в двух аспектах в аспекте
облеrчения, ускорения и уточнения расчетов координат введения
электродов во время самой операции (что было осуществлено
при использовании ЭВМ Ю. r. Иванников, В. В. Усов, 1967)
и в аспекте послеоперационноrо уточнения индивидуальных
структурно--функциональных отношений мозrа (Н. П. Бехтерева,
А. Н. Бондарчук, В. М. Смирнов, А. и. Трохачев, 1967). Возмож
ности в этом последнем плане значительно возросли тоrда, коrда
желаемоrо результата научились достиrать не только электри--
ческими воздействиями через вживленные электроды, но и более
тонким и значительно более щадящим способом исследованием
различных параметров жизнедеятельности мозrа при COOTBeT
ствующих функциональных пробах.
Этот прием в варианте реrистрации электросубкортикоrраммы
(ЭСКоТ), медленных электрических процессов (МЭП), отражающих дина
мику постоянноrо потенциала, сопротивления в ткани мозrа и процесса
поляризации золотых электродов, условно иноrда обозначаемых в форме
так называемоrо постоянноrо потенциала (ПП), наличноrо кислорода
(О2а) и импульсной активности (ИА) с вживленных электродов при дви
rательных пробах пассивных и активных движениях, оказался исклю
чительно эффективным для уточнения отношения различных структур
к мозrовой системе орrанизации движений (В. Б. Тречин, 1966; А. И. Tpo
хачев, 1965, 1966). Наиболее выrодными параметрами для решения этоrо
вопроса оказались МЭП, 02а, ИА. При записи динамики этих показателей
при повторных двиrательных пробах наблюдалась воспроизводим:ость
развивающейся реакции, что очень отчетливо выявлял ось при супер
позиции ряда последовательных отрезков кривых. (Вполне понятно,
что сравнение реакции ИА осуществлялось после соответствующей об
работки. )
Используя принципиально такой же подход, оказалось возможным
изучить и связь различных образований мозrа с психической деятель
Ностью. Различные физиолоrические показатели оказывались избира
тельно или преимущественно полезными в отношении разных аспектов
изучаемых явлений.
48
Таким образом, наблюдения за изменениями жизнедеятель
ности мозrа и Bcero орrанизма в целом при электрических воз
действиях на мозr через вживленные электроды и анализ проте--
кающих в мозrу процессов, реrистрируемых с помощью тех же
электродов, в сочетании составили комплексный метод, схема KO
Toporo представлена на рис. 2.
Наиболыпее количество исследований проведено в наСТОЯIЦее
время с записью электросубкортикоrраммы при психолоrических
пробах и при развитии спонтанных и вызванных психических
и психопатолоrических явлений. Появление спайков в rлубине
.1I0бной доли при возбуждении и rаллюцинациях и спайков в rи--
поталамусе при возбуждении наблюдали Sem Jacobsen .и соавт.
(1953, 1956), Hodes и соавт. (1954) и др. Sem Jacobsen наблюдал
также подавление определенных видов ритма, близкоrо к альфа
ритму, при чтении книrи. Penfield и Jasper (1954) наблюдали раз
витие амнезии при эпилептическом разряде в одной из миндалин.
Сочетание биоэлектрических эффектов в области крючка с раз
витием спутанности сознания наблюдала Brazi r (1966). Анализ
электрических явлений коры, обонятельноrо мозrа и базальных
rанrлиев человека провел с помощью ЭВМ Chapman (1966). Элек
трокортикоrрамма (экоr) и эскоr изучались во время обучения,
эмоций, тестов на память и друrих проб у больных с эпилептиче
скими rиперкинезами, болями и хроническими психозами.
Каmр, Schriger, Storm van Leeuwen (1972) наблюдали вспышки
бета ритма в нижних отделах фронтальной коры у больных
с вживленными электродами, причем эти вспышки усиливались
в ситуации, коrда больной должен был «предсказать» какие либо
события. Исследователи высказали rипотезу о том, что вспышки
бета--ритма отражают кратковременное внезапное снятие напря
жения.
в отличие от исследований с «психолоrическим уклоном», rде, как
правило, весьма разнообразились психолоrические пробы, а потому физио
лоrические данные трудно поддавались сравнению и анализу, в наших
работах центром тяжести явился физиолоrический рспект исследований.
Анализу подверrались преимущественно физиолоrические показатели, коли
чество используемых психолоrических тестов оrраничивалось, а результаты
их выполнения учитывались лишь в той мере, в какой это было необхо
димо для анализа динамики физиолоrических параметров.
Основным из использованных психолоrических тестов в проводимых
исследованиях был тест на краткосрочную оперативную память типа
пробы Бине. В этом случае физиолоrическая динамика мозrа (эскоr,
МЭП, 02а, ИА) реrистрировалась до пробы, во время предъявления циф
pOBoro ряда, известных или неизвестных слов, или квази слов, или ряда
триrрамм, во время удержания их в памяти, повторения задачи (по при
казу «повторить») И В период последействия. В некоторых случаях проба
усложнял ась (вариант В. М. Смирнова, 1968) произносились однознач
ные цифры, которые больному надо было вычитать или складывать в уме,
и воспроизводился (через те же 10 30 + 2 3 с) лишь конечный результат
ряда таких операций. Реже применялась задача прибавления в уме He
четноrо однозначноrо числа (например, 3) к KaKOMY TO заданному при Tpe
бовании произнесения вслух либо только четноrо, либо только нечетноrо
результата. Мозrовые механизмы эмоциональных реакций изучались
в целом прИнципиально тем же путем, но при использовании специально
3 н. п. Бехтерева
49
эмоциоrенных тестов (В. М. Смирнов, 1963 1968). Основными пробами яв
лялись предъявление эмоциоrенных слов, карт Роршаха, воспроизведение
эмоционально значимых ситуаций и т. п.
Тесты на оперативную память использовались как прошедшие боль
шую проверку временем стандартные психолоrические пробы. Это в из
вестной мере позволило в связи с задачами чисто физиолоrическоrо плана
отвлечься на данном этапе от собственно психолоrическоrо аспекта иссле
дования.
Изменения электросубкортикоrраммы при психолоrичеСКJ1Х
пробах были более или менее отчетливы частота и амплитуда
ее составляющих на разных этапах проб более или менее отли
чались от исходных. Эти изменения моrли быть очень существен
ными, и тоrда создавалось впечатление, что динамика эскоr
выявляла образования мозrа, имеющие первостепенное значение
для данноrо вида деятельности. Наоборот, в друrих образова
ниях мозrа изменения эскоr при психолоrических пробах визу
ально практически не улавливались, что моrло бы явиться OCHO
ванием для суждения об отсутствии связи этих вторых образова
ний с психической деятельностью (рис. 6).
Однако так же, как и в отношении ээr, при исследовании
эскоr обнаружилось, что выраженность изменений в какой то
области значительно чаще была связана с состоянием структуры,
чем с использованной пробой.
Это положение, как известно, широко используется в клинической
электроэнцефалоrрафии самые различные функциональные пробы MorYT
способствовать обнаружению очаrовоrо патолоrическоrо процесса по ЭЭr
вне прямой зависимости от ero локализации (сказанное, естественно, не
исключает и не умаляет избирательноrо локальнодиаrностическоrо значе
ния некоторых функциональных проб).
Сравнение эскоr, полученных при применении различных
функциональных проб, показало, что изменение эскоr моrло
быть выраженным вне прямой связи с характером раздражителя
и не обязательно быть наиболее выраженным при психолоrиче
ских пробах.
Рисунок эскоr при 'психолоrической пробе обнаруживал, Ta
ким образом, прямую связь с исходной эскоr и не был одинако
вым, повторяющимся при повторении проб не обладал чертами
убедительной воспроизводимости. Использование приемов MaTe
матической обработки данных способом измерения относительной
длительности восходящих и нисходящих фаз волн по А. А. reH
кину (Н. П. Бехтерева, А. А. rенкин, Н. И. Моисеева, В. М. Смир"
нов, 1965) подтвердило лишь достоверность отличия эскоr при
реализации психолоrических проб от фона. Условия, искусственно
созданные в виде отбора лиц со сходными ээr при исследовании
динамики ээr при условнорефлекторных реакциях (Gastaut и co
авт., 1957), привели, таким образом, как видно из этих данных,
действительно к результату, имеющему суrубо частное зна
чение.
Обработка эскоr с помощью получающеrо сейчас все боль--
шее распространение приема анализа, OCHoBaHHoro на принципе
50
.::t-
't";) со
f l
с::::
:I:
:s:
E-t
ro
Q)
r::
о
ro
:I:
Q)
\о
О
r::
):S:
о
(,)
Q)
:;rt
:s:
о
t:: 1=:
О
><
:s:
(,)
r::
:s:
r::
ro
;:))
ro
о
:s:
E-t
о.
О
\о
()
о
О.
E-t
Q)
1=:
ст)
.
ф
э.
t-...
QQ
использования возможностей специалиста и ЭВМ (Н. И. Моисеева
и В. А. Орлов, 1965), позволила показать наличие изменений
в эскоr в процессе реализации психолоrической пробы и в тех
отведениях, rде рассмотрение одной эскоr не обнаруживало
отчетливой динамики.
Сдвиrи в эскоr в различных ядерных образованиях наблюда
лись в форме перестроек рисунка биоэлектричес ой активности
в пределах Bcero спектра биопотенциалов. При эr;rом в зависи
мости от исходноrо фона очень отчетливыми моrли быть разли--
чия в рисунке эскоr на предъявление пробы и во время ответа.
Так, например, в отведении миндалина скорлупа на предъявле
ние пробы моrло происходить значительное усиление дельта
активности, при ответе доминировала альфа активность и т. д.
При друrих количественных соотношениях изменений эскоr
составляющих пр нципиально такая же картина наблюдалась
в области rиппокампа. В случае, коrда требовалось мысленно от--
вечать на задание (не произнося ответа вслух), характерным для
вышеприведенных образований моrло быть и усиление бета
активности. При непра ильном ответе в э:rих же CTpYKT pax бо
лее выраженной становилась тета активность и :r. д. Статисти
ческая обработка эскоr обнаружила, что при различии каждоrо
KOHKpeTHoro рисунка ее при одном из. использованных видов пси
холоrических проб (например, при прибавлении нечетноrо числа
к любому исходному и произнесении только четных или нечет
ных ответов) суммарная тенденция эскоr--изменений моrла OT
ражать отчетливые отличия в зависимости от характера ответа
произнесения или непроизнесения ero.
Факт этот привлекает внимание в свете связи характера изме
нений друrих физиолоrических показателей мозrа (Н. П. Бехте
рева, А. И. Трохачев, 1966б) с качеством ответа, хотя, естественно,
эти различия MorYT быть связаны и с сущностью физиолоrиче
ских механизмов, лежащих в основе реализации различных вари
антов пробы, и с наличием или отсутствием MOTopHoro компо
нента в реакции и т. д.
Как указывалось ыше, конкретный рисунок изменений
эскоr не был воспроизводимым при повторных пробах.
В то же самое время повторение использованных психолоrиче--
ских проб с большим постоянством приводило К развитию изме
нений эскоr, что вполне понятно, так как в этой психолоrиче
ской пробе всеrда присутствовал фактор новизны, стимулирую
щий активацию внимания больноrо (это последнее положение
никоим образом не выделяет использованную пробу из ряда пси
хических деятельностей человека, так как очень мноrие из них,
если не все, всеrда содержат этот элемент новизны). Этот факт
нередко обнаруживался уже и при реrистрации ээr при психо
лоrических пробах, однако именно в условиях прямоrо отведения
биопотенциалов от мозrа человека и соответствуюrцей обработки
данных он приобрел значение закономерности и Mor явиться
поводом к рассмотрению ero физиолоrическоrо значения.
52
Это рассмотрение представляется целесообразным прежде
Bcero в ряде основных аспектов. Так, зависимость изменений
эскоr в этом случае от исходноrо фона является еще одним под
тверждением Toro, что биоэлектрические явления, реrистрируе
мые в ээr, по видимому, не только (и не столько) отражают про
исходящее в мозrу, но и служат поддержанию определенноrо
Функциональноrо состояния структур мозrа может быть, управ
ляют этим функциональным состоянием (см. выше), устанавливая,
.а затем поддерживая ero на необходимом для протекания какой
то данной деятельности уровне хотя, может быть" и не в cTporo
индивидуальном, а в KaKOM TO типовом плане (Н. П. Бехтерева,
В. В. Зонтов, 1961). Вполне естественно на основе подобной точки
зрения рассматривать данные об изменениях эскоr при психо
лоrической пробе, полученные при статистической обработке, как
отражающие количественно измеренный сдвиr, характеризующий
оптимизацию системы (мозrа) применительно к данным вызван
ным (созданным) условиям.
Естественно, что приведенная трактовка на сеrодняшний день
все еще больше возможность, чем действительность, хотя и воз
можность весьма вероятная. Необходима дальнейшая -детальная
математическая обработка эскоr как при самых различных, так
и при сходных деятельностях, равно как и разработка приемов
анализа эскоr, позволяющая сравнивать результаты исследова
ний у разных больных с поправкой на характеристику фоно
ВОЙ эскоr. ·
с друrой стороны, приведенная трактовка в общем плане
не нова, так как является еще одним аспектом известноrо поло
жения о необходимости определенноrо функциональноrо СОСТОЯ--
ния центральной нервной системы для различных видов деятель
ности. Таким образом, если рассматривать вопрос с точки зрения
развития при психической деятельности определенноrо, опти
мальноrо для данной активности при данных исходных условиях,
отражаемоrо в эскоr функциональноrо состояния, представле
ния эти принимают уже и совсем знакомый (<<приемлемый») вид.
Как же будут происходить события в мозrу, если должна раз
виться друrая, новая деятельность? Вполне естественным на oc
нов е рассмотрения этоrо вопроса будет ответ о том, что в этом
случае функциональное состояние мозrа (отражаемое в эскоr)
должно измениться. А отсюда вряд ли с достаточными OCHOBa
ниями можно возражать против предположения о том, что в Ka
кой то данный минимальный отрезок времени, коrда мозr Ha
строен на одну психическую деятельность, коrда мноrие ero
структуры вовлечены в эту деятельность, а друrие rOToBbI обес
печить ее развитие (и именно ее!), друrая сложная психическая
деятельность невозможна. Таким образом, одномоментно проис
ходит и может происходить в мозrу только одна сложная психи
ческая деятельность. Известный еще со времен Юлия Цезаря
феномен одновременности осуществления ряда деятельностей
реален, но он не происходит по принципу одномоментноrо ocy
53
F
А
8
:
I
I
,......., ,...........,,..........,
I . I I I
. _',.
I I I I
L-...., :
. : t t I .
1: I
I I I
I I I I
,..........J ,..........
I t . t
1: I I
I I I I
: t : I
I ,........, 1"""""'1 ,
I I I I I
: : : : I :
:........; I I I I '
I I '
I I I I I
I I I I I
I I I I .
t I I I
J --..... ----
. I
I I
I .
:
с
I
I I
I :
т
.. . t n
Аt 1 Аt 2 Аt з ...
а
А
I
,
т
I
I
.
I
.
I
.
,
.. ...
с
"1. Схема, иллюстрирующая динамику изменения функцио
нальных состояний мозrа, оптимальных для разных видов
деятельности.
а процесс представлен в двумерной системе координат; б то же
в цилиндрической системе. А.. В, С различные деятельности. По оси
абсцисс динамика процессов во времени. По оси ординат на верхнем
rрафике и на поверхности цилиндра на нижнем rрафике условно пред-
ставлены уровни функциональноrо состояния моэrа.
ществления всех этих деятельностей. В каждый данный момент
реализуется ЛИll1Ь одна из них, а одновременность ряда актив
ностей обеспечивается быстрым и оптимальным (в смысле мини
мальных потерь!) пере ключе ни е м с одной деятельности на дpy
rую. В павловском смысле TaKoro рода переключение, по види
мому, может происходить у лиц С подвижным и сильным типом
ВЫСll1ей нервной деятельности. Для наrлядности эти представле
ния про иллюстрированы rрафически (рис. 7).
- Вполне понятно, что уровни функциональноrо состояния мозrа
приведены на верхнем rрафике условно. В действительности они
представляют собой интеrральные величины, элементами KOTO
рых являются состояния всех участвующих в деятельности (и, Be
роятно, rOToBbIx к ней) структур мозrа. Отсюда уровни А, В, С
ЛИll1Ь схематично MorYT рассматриваться в. представленном виде
на оси ординат. Работая на основе возможностей MHoroMepHoro
пространства, мозr, естественно,. «заслуживает» И использования
ординат этоrо пространства при rрафическом изображении ero
состояний, rде время представлено одной из ординат, а деятель--
ность, отнесенная к состоянию, друrими. Попыткой изобразить
rрафически данную ситуацию в менее далекой от реальности
форме является нижний рисунок.
Приведенные положения об одномоментности одной деятель
ности леrRО MorYT быть проверены каждым при условии Toro, что
54
хоть одна из реализуемых деятельностей происходит в - связи
с объективно, независимо от субъекта, развивающимися собы--
тиями, постоянство связи субъекта с которыми может быть
в дальнейшем проконтролировано им самим или любым друrим
лицом. TaKoro рода деятельностью, жестко «привязанной» К собы
тиям, развивающимся вне субъекта, может быть мозrовая актив
ность, возникающая в связи с просмотром фильма, слушанием
радиопередачи и т. п.
В интересах автодорожников, а не для изучения принципов физиоло
rическоrо обеспечения психической деятельности несколько лет назад
в Анrлии был проделан следующий эксперимент: в определенный час,
коrда по радио передавали увлекательный рассказ, был установлен KOHT
роль за реакциями водителей на одном из сложных отрезков пути. Подав
ляющее большинство опрошенных в конце пути водителей были уверены,
что ни на секунду не отвлекались от слушания текста. Однако все, кто
правильно решил те дорожные задачи, для решения которых было недо--
статочно автоматических реакций, полностью «потеряли» куски текста,
звучавшие по радио в эти мrновения. Те же, которые оказались в TPYД
ной дорожной ситуации не вполне адекватными, значительно полнее BOC
производили текст. (Эти данные были сообщены мне в 1967 r. в Бристоле
rpeeM Уолтером, Реем Купером и др. при обсуждении вопроса об OДHO
моментности только одной психической деятельности.) Этот эксперимент,
по видимому, свидетельствует об опасности слушания увлекательных пе
редач при вождении машин; в очень доходчивой форме он иллюстрирует
и особенности реализации мозrом психической деятельности.
Чем можно объяснить, однако, тот факт, что прерываемая
(и прерывистая объективно) психическая деятельность может
восприниматься субъективно, как непрерывная? Вероятно, пони
мание этоrо феномена лежит в принятии правильности высказы
вания математика А. И. Колмоrорова (1969) о дискретности мыш
ления. Исходно всеrда дискретное мышление, субъективно вос--
принимаемое как процесс непрерывный, является основной
предпосылкой для такой же субъективной оценки событий при
попеременной реализации двух и более деятельностей. (Идея
о дискретности психических процессов находит косвенное под
тверждение в данных Stevens (1968) о дискретности выделения
медиаторов.) Сейчас проведены исследования, rде предприняты
попытки доказать возможность протекания не одной, а мноrих
деятельностей одновременно (Gomes, .1966). Эти исследования не
исключают, однако, возможности одномоментноrо протекания
только одной сложной деятельности, а использованные в них ме--
тодические приемы не сравнимы с использовавшимися нами.
Исследование динамики друrих показателей жизнедеятель
ности мозrа также подтвердило изменение функциональноrо со--
стояния ero различных структур при психической деятель
ности обнаруживались изме ения уровня МЭП, 02а, ИА. Изу
чение общих изменений в rоловном мозrу было дополнено этими
материалами, однако в целом для решения именно вопроса
о функциональном состоянии мозrа основные материалы были
получены с помощью эскоr, более динамичной, чем МЭП и 02а.
Понятно, что весьма плодотворным в тонкой оценке сущности
55
общих изменений в мозrу может оказаться дальнейший анализ
динамики ИА показателя, также динамичноrо.
Исследования с записью ээr с поверхности кожи черепа и He
посредственно с мозrа при простой (условные реакции) и более
сложной (психолоrические пробы) условнорефлекторной дея--
тельности rармонически дополнили друr друrа. Представленный
спектр исследований показывает некоторые нейрофизиолоrиче--
ские аспекты психическоrо в предельно простых, модельных и во
все более усложняющихся условиях. Естественно, что очень MHO
roe в физиолоrическом анализе БО-?Iее простых психических явле
ний может быть 'отнесено и к явлениям более сложным. В то же
самое время именно усложнение условий исследования и получе
ние все более полноценных физиолоrических данных при прямом
отведении биопотенциалов от мозrа позволили не остановиться на
рассмотрении только самых общих сторон вопроса.
Проделанный путь исследований определил место электроэн--
цефалоrраммы в ряду нейрофизиолоrических параметров как
приемлемоrо и, по--видимому, адекватноrо показателя общих из
менений в мозrу, развивающихся с более или менее дробной про
странственной мозаикой, зависящей от исходноrо фона и реали
зуемой деятельности. Настоящий этап исследований, и прежде
Bcero в связи с не воспроизводимостью полиморфноrо рисунка
ээr, зареrистрированной в условиях прямоrо отведения при по
вторе нии тестов, заставляет рекомендовать максимальную OCTO
рожность в суждении по ней о структурно функциональной opra
низации изучаемой деятельности. Математические приемы ана--
лиза, часто первоначально подтверждавшие локальные изменения
в мозrу, как правило, при продолжении исследований с большим
постоянством подтверждали значение этих локальных изменений
как части общих перестроек и, таким образом, преимуществен--
ную ценность ээr для изучения именно функциональноrо фона
мозrа.
Функциональный фон мозrа является частным проявлением
общих rомеостатических реакций, rде варьирующее в сравни--
тельно небольших пределах «постоянство» внутренней среды
орrанизма является тем соматическим базисом, который опреде--
ляет возможность осуществления всех поведенческих реакций
от обеспечивающих существование вида до сложнейших психи
ческих реакций, связанных с индивидуальной жизнью человека
в социальной среде.
СТРУКТУРНО ФУНКЦИОНАЛЬНАЯ
И НЕйРоФизиолоrИЧЕСКАЯ оРrАНИЗАЦИЯ
мозrовоrо ОБЕСПЕЧЕНИЯ ПСИХИЧЕСКОЙ
ДЕЯТЕЛЬНОСТИ
ПРИНЦИПЫ ИССЛЕДОВАНИЯ ЭЛЕМЕНТОВ СИСТЕМЫ
мозrовоrо ОБЕСПЕЧЕНИЯ ПСИХИЧЕСКОЙ
ДЕЯТЕЛЬНОСТИ
CBoero рода «классическим» приемом исследования тонких
структурно функциональных отношений мозrа в эксперименте до
caMoro последнеrо времени является наблюдение динамики про
явлений спонтанной и заданной деятельности орrанизма во время
локальных электрических воздействий на rоловной мозr. Этим
способом изучается центральная реrуляция различных функций
и делаются попытки выяснения мозrовоrо контроля эмоциональ
ных реакций животных в условиях острых и хронических экс
периментов, а в последние rоды, в связи с усовершенствованием
телеметрии, и в условиях свободноrо поведения.
Метод локальных электрических воздействий на rоловной
мозr человека используется в нейрохирурrической практике rлав--
ным образом для уточнения эпилептоrенноrо очаrа и области
предполаrаемоrо разрушения при rиперкинезах. Он применяется
TaKjКe в условиях длительноrо обследования больных при лече
нии сцособом множественных вживленных электродов. Резуль
таты, полученные при локальных' электрических воздействиях
на мозr человека, дополнили, детализировали и уrлубили CBeдe
нил, представленные анализом нарушений высших психических
функций при очаrовых патолоrических процессах мозrа. Элек
трические воздействия на мозr позволили получить большой ма--
териал о мозrовой структурной орrанизации эмоциональноrо
контроля.
Хорошо известно, как MHoro лет данные Olds (Olds et Milner,
1954, 1966), по существу, оставались исключительно интересным
набором фактов, трактовка которых была в лучшем случае rипо--:
тетической. Действительное понимание субъективной суrцности
«зон наказания», «зон поощрения» и т. д. оказалось возможным
после Toro, как при раздражении мозrа человека стали наблю
даться эмоциональные реакции, в том числе и сопровождавшиеся
развитием «влечения» К получению повторных раздражений
(В. М. Смирнов, 1967) и являвшиеся иноrда основой сложных по
веденческих перестроек (Н. П. Бехтерева, К. В. rрачев, А. Н. Op
лова, С. Л. Лцук, 1963; В. М. Смирнов, 1967).
Развитие эмоциональных реакций при электрических воздей
ствиях на rлубокие структуры мозrа человека описано в работах:
57
Heath (1954, 1963), Peacock (1954), Monroe, Heath (1954), King и др.
(1954), Heath и Mickle (1960), Sen1--Jacobsen, Torki1dsen (1960), Ange
leri, Ferro Milone, Parigi (1961), Spiegel, Wycis (1961), Delgado
(1963, 1970), Delgado, Hamlin (1962), Sem J acobsen (1964, 1968),
Н. П. Бехтеревой, К. В. rрачева, А. Н. Орловой, С. Л. Лцук (1963),
van Buren (1963), В. М. Смирнова (1963 1969), Walter, Chapman,
Porter (1964), Chapman (1966), Umbach (1966).
Изменения эмоциональноrо состояния различноrо типа и раз
личной степени выраженности приведены в обобщающем труде
Sem Jacobsen (1968), который делит эти изменения на девять
типов:
1. Больной спокоен, у Hero появилось чувство «блаrополучия».
Он может быть при этом немножко сонливым положительная
реакция 1 степени.
2. Состояние больноrо отчетливо изменено, настроение xopo
шее и появляется ощущение хорошеrо состояния. Он спокоен и
доволен собой, часто улыбается. При этом наблюдается леrкая
эйфория, но поведение остается в пределах нормы. Он хочет по
вторения стимуляций........... положительная реакция 11 степени.
3. Эйфория четко выходит за пределы нормы. Больной
rpoMKo смеется, доволен собой, ему очень нравится стимуляция,
)1 он хочет повторных стимуляций положительная реакция
111 степени.
4. Больной становится беспокойным, напряженным или пе
чальным отрицательная реакция 1 степени.
5. Раздражительность может сочетаться со слабой депрессией.
Больной чувствует себя несчастным, ему неуютно отрицатель
ная реакция 11 степени.
6. Больной подавлен, раздражителен или даже сердит, боится,
иноrда обижен, иноrда плачет отрицательная реакция 111 CTe
пени. ,
7. Внезапные эмоциональные вспышки позитивноrо или Hera
тивноrо типов.
8. Амбивалентность. Больному иноrда нравится стимуляция,
иноrда нет.
9. Реакция типа орrазма с соответствующими изменениями
настроения. Больной сначала положительно реаrирует на стиму
ЛЯЦИЮ, затем внезапно наступает такое полное удовлетворение,
что он более или менее долrий период отказывается от стимуля
ций. Продолжение стимуляций в этот период определенно He
приятно.
Sem J acobsen подчеркивает необходимость учета и анализа
поведенческих реакций, так как далеко не во всех случаях сло
весный отчет больноrо оказывается достаточно надежным. Oco
бенно показательным является стремление R повторным стиму
ляциям.
Эмоциональные реакции, развивающиеся в этих условиях,
подробно, rлубоко и систематично изучались В. М. Смирновым
(в нашей лаборатории, ИЭМ АМН СССР). В развитие междуна
58
1
'"
Gippocaтpl1$
20ла iпcerta
N. subthalaт.
forт. reticlJl.
N. ruber
SuЬstапtiа.
лigrа
РlJtатеп
Globu$
p(]lli(ius lat.
GloblLS pallidlJs
. тed.
, ,
8. Схема мозrа.
Черными квадратами помечены зоны, раздражение которых вызывало эмоциональные
реакции. Большими квадратами отмечены области, раздражение которых вызыв!\ло
эмоциональные реакции большей выраженности и с б6льшим постоянством; маленькими
квадратами те зоны, при раздражении которых эмоциональные реакции развивались
M Hee постоянно и были менее отчетливыми.
родноrо опыта в этом направлении и на основе ero наблюдений
оказалось возможным уже построить примерные карты струк--
турно Функциональной орrанизации эмоциональноrо К9НТРОЛЯ
у человека (рис. 8).
Анализ этих исследований показал, что эмоциональные peaK
ции развиваются при электрических воздействиях на определен
ные мозrовые структуры тоrда, коrда есть основания предпола
raTb сдвиr постоянноrо потенциала в области воздействия (част я
электрИческая стимуляция, поляризация). Это положение было
подтверждено прямыми измерениями постоянноrо потенциала
мозrа после тех же электрических воздействий (С. Р. Авра--
мов, 1965).
В. М. Смирновым (1970, и др.) в полном соответствии с дан--
ными Sem--J acobsen отмечено появление различных эмоциональ--
ных реакций при раздражении rлубоких структур, причем в от--
дельных случаях развивались более или менее стойкие поведен
ческие реакции. Накопление данных о стереотаксической
семиолоrии rлубоких структур мозrа (В. М. Смирнов) позволило
произвольно подавлять эти состояния, возникавшие в ходе диаr
ностических воздействий через вживленные электроды. Так, Ha
пример, чувство страха, возникшее при электрической стимуля--
ции зоныI на rранице вентро медиальноrо отдела зрительноrо
буrра и ростральноrо отдела ствола привело к развитию сто кой
59
фобии. Фобия была полностью
снята последующей стимуля
цией зоны в области средин
Cd Horo центра зрительноrо буrра.
Ci Развитие фобии в данном
случае моrло рассматриваться
Pиt как возникновение устойчи
Boro патолоrическоrо состоя
СР ния (Н. П. Бехтерева и
А. Н. Бондарчук, 1968), пре
одоление KOToporo оказалось
Ат! возможным немедленно при
изменении режима работы
мозrа, вызвавшеrо дезинтеrра
Не цию системы, обеспечившей
появление данной поведенче
ской реакции.
С помощью стимуляции
rлубоких структур мозrа
удается достиrать и дезинте
rрации систем, обеспечиваю
щих поведенческие реакции,
ВОЗНИКIllие на эмоциональной
основе в условиях «eCTeCTBeH
ной патолоrии». Так, лечеб--
ная электрическая стимуля
ция области подушки зри
тельноrо буrра вызвала дe
фраrментацию, «побледнение» и последующее исчезновение фан
томно болевоrо синдрома давностью в несколько десятилетий
(В. М. Смирнов в работе Н. П. Бехтеревой, А. Н. Бондарчук,
Л. А. Мелючевой, В. М. Смирнова, 1972) (рис. 9).
А Н! УА
ИВ
СМ
/l
Yl
VP
Hthl
HF
Но
SN
РС
9th! zi
(l)1 (//}2 фs (t4/5 /6
9. Схематическое изображение подкор
ковых структур, электростимуляция
которых вызывала стойкие эмоцио
нальные реакции или дезинтеrрацию
систем, обеспечивающих поведенче
ские реакции.
1 при электростимуляции возникала фобия;
2 фобия исчезала; 3 возникновение навяз-
чивоrо смеха; 4 ликвидация навязчивоrо сме-
ха; 5 исчезновение фантомной боли и дефраr-
ментация фантома; 6 уrашение фантома.
Электрические воздействия на ряд подкорковых структур
срединный центр, переднее вентральное ядро, ретикулярное ядро
таламуса, субталамическую область, свод, внутренний членик
бледноrо шара и друrие образования (Jung, 1954; Hassler, 1961;
Umbach, 1961; В. М. Смирнов, 1967; А. Н. Бондарчук и В. М. Смир
нов, 1969) вызывали изменения Toro типа, которые в экспери
менте оцениваются как изменения уровня бодрствования и из
менения сознания. Эти наблюдения позволили показать, что
стимуляция одних подкорковых структур (элементов xBocTaToro
ядра vап Buren, 1963; Н. П. Бехтерева, Н. И. Моисеева, А. Н. Op
лова, В. М. Смирнов, 1964, 1966; срединноrо центра зрительноrо
буrра В. М. Смирнов, 1967; А. Н. Бондарчук, В. М. Смирнов,
1969) наиболее часто приводит к уrнетению возможностей BЫ
полнения, по крайней мере, некоторых видов интеллектуальной
деятельности. Так, раздражение xBocTaToro ядра в наблюдениях
van Buren вызывало торможение речи, затруднение восприятия
и даже помрачнение сознания. Раздражение xBocTaToro ядра и не--
60
которых структур передней ядерной rруппы таламуса моrло при
водить к повышению числа ошибок при выполнении психолоrи
ческих тестов (Н. П. Бехтерева и соавт., 1964). Наоборот, уrнетаю
IЦие электрические воздействия (частая электрическая стимуля
ция, поляризация) в области Toro же срединноrо центра, а также
вентро латеральноrо ядра моrли приводить к увеличению психи
ческих возможностей, что было объективизировано в виде повы
шения пороrов распознавания изображений с различными стати
стическими характеристиками (Л. И. rорелик, в. м. Смирнов,
И. М. Тонконоrий, 1969). В. М. Смирнов описывает также различ
ные степени этоrо повышения «уровня бодрствования» при элек
трических воздействиях на rлубокие структуры вплоть до
KpaTKoBpeMeHHoro развития rипоманиакальноrо состояния.
Изменения речи при электрических воздействиях наблюда
лись как один из элементов изменения уровня бодрствования.
При электрических воздействиях на rлубокие структуры (раз
личные таламические ядра, мезенцефальные отделы ствола,
хвостатое ядро и друrие образования) наблюдались также явле
ния типа остановки речи, ее замедления, ускорения, изменения
rолоса, тональности речи, афазии (Sem Jacobsen, 1965, 1968; Oje..
тапп et al., 1968Ь; В. М. Смирнов и А. Н. Шандурина, 1969; Дель--
rадо, 1970). Воздействия на rлубокие структуры левоrо полуша
рин в наблюдениях Sem J acobsen вызывали и нарушения письма
(аrрафию).
При раздражении области, расположенной в 1,5 СМ от средней
линии, вблизи лобной части мозолистоrо тела Sem--J acobsen (1968)
отмечал вокализацию.
Как известно, Penfield связывал наблюдавшиеся им явления
активации памяти при стимуляции верхнелатеральных отделов
височной доли с вовлечением в реакцию rиппокампа. Эти пред
ставления были подтверждены Walter и Crow (1964), наблюдав
шими воспроизведение картин прошлоrо опыта при стимуляции
миндалины и rиппокампа. В исследованиях Mahl и Delgado (цит.
по Sperry, 1966) уточнено, что внешнее окружение может моди
фицировать эти вызванные переживания. Несмотря на то, что все
же не исключена связь наблюдавшихся феноменов или, по
крайней мере, леrкости их выявления с болезненным состоя
нием мозrа, трудно, по существу, возразить против Toro, что, про
изводное от нормальных возможностей коры или ее болезнен
Horo состояния, это явление отражает некоторые стороны мозrо--
вых механизмов памяти. Существенные нарушения краткосроч
ной вербальной памяти при стимуляции латеральноrо таламуса,
теменной и височной области и подушки зрительноrо буrра слева
наблюдали Ojemann и Fedio (1968а).
В настоящее время накоплен большой материал не только об
активации, но и уrнетении памяти при воздействии на ПОДRОрКО
вые структуры. t
Так, в работах Gerard (1961), Sem Jacobsen (1965), Brazier
(1966), Chapman (1966), В. М. Смирнова (1968) приводятся
61
данные о том, что электрическое воздействие на покрышку' cpeд
Hero мозrа, некоторые ядра таламуса, медиобазальные отделы
височной доли и друrие образования может вызывать развитие
более или менее длительной амнезии.
В наблюдениях Sem Jacobsen (1968) при раздражении rлубокJ.1Х
структур мозrа были воспроизведены два типа феноменов, опи
санных Penfield типа воспроизведения картин прошлоrо и BHe
запноrо. «переноса В прошлое». Феномены приобрели, таким об
разом, более общее значение, так как в этом случае не моrли
быть отнесены за счет эпилептической ауры стимуляция про--
водилась у больных паркинсонизмом.
Электрическое воздействие на rлубокие структуры мозrа по
казало, что в этих условиях MorYT наблюдаться также мноrие
психопатолоrические феномены. При стимуляции и поляризации
rлубоких структур мозrа описано появление разноrо рода иллю
зий, rаллюцинаций, бредовых идей, аrрессивных реакций, взры--
вов ярости, феноменов типа deja vu и др. (Umbach, 1961; Chapman,
1966; Jasper, Rasmussen, 1958; В. М. Смирнов, 1967; Delgado,
1970, и др.).
Понимание приведенных выше результатов лечебной элек
трической стимуляции rлубоких структур мозrа также невоз--
можно без учета роли памяти в развитии и поддержании устой
чивых патолоrических состояний и модуляции не только KpaTKO
срочной, но и долrосрочной памяти при воздействии на rлубокие
структуры. Дезинтеrрация фобий, фантомно болевых синдромов
и друrих состояний, по видимому, возникает за счет воздействия
стимуляций преимущественно на долrосрочную память.
В исследованиях В. М. Смирнова (1967), В. М. Смирнова и
А. Н. UUандуриной (1969) специальное внимание уделено pac
стройствам схемы тела при воздействиях на подкорковые ядра.
Наступавшие кратковременные расстройства схемы тела, равно
как и некоторые иллюзии, наиболее адекватно, по видимому,
моrли бы быть поняты С точки зрения BpeMeHHoro нарушения
функциональноrо состояния детекторов ошибок.
Большой интерес представили данные Albe..Fessard (1954),
Spiegel, Wycis, Orchinick, Freed (1956), Penfield (1958), Chapman
(1958), Bickford, Mulder, Dodge, Svien, Rome (1958) и В. М. Смир--
нова (1969), наблюдавших факты, которые можно было бы TpaK
товать как изменение и нарушение «шкалы времени» при элек
трических воздействиях на мозr. Нарушения временной шкалы
чаrце Bcero проявлялись развитием более или менее продоляси--
тельной дезориентировки во времени, но моrло наблюдаться и
резкое ускорение и замедление различных действий. Весьма
демонстративно это обнаруживалось в форме ускорения произне
сения вслух цифровоrо ряда при раздражении медиобазальныIx
структур височной доли (наблюдение В. М. Смирнова; см. ра--
боту Н. П. Бехтерева, А. И. Бондарчук, В. М. Смирнов, 1969)
и в форме ускорения речи при стимуляции лобной доли (Sem
J acobsen, 1968).
62
Эти материалы показывают, что мозrовой контроль психиче--
ских функций обеспечивает различные их стороны и в том числе
развитие явлений во времени и масштаб этоrо времени. Эти дaH
ные свидетельствуют также о возможности обеспечения мозrом
не одноrо, а мноrих различных масштабов времени.
Реальность возможности жизни мозrа в разных системах времени,
протекания психических явлений в иной системе времени, чем обычная,
доказывается не только приведенными, хотя и объективными, но пока еще
едиНичными наблюдениями, и также не только субъективно известной
способностью нашей памяти в сложных, «стрессовых» жизненных ситуа..
циях «пробеrать» в короткие отрезки времени по следам длинноrо пути,
и наконец, не только сходными явлениями при сновидениях. Два послед
них случая всеrда MorYT быть подверrнуты критике с позиций избира
тельной отрывочности памяти в этих случаях, использования следов не
полной цепи событий, а цепи представительных «отрезков» И, таким обра
30М, оперированию в соизмеримом с обычным масштабе времени.
Более убедительной возможностью перехода в друrие системы' Bpe
мени представляется наличие у отдельных людей исключительной спо
собности за минуты или даже секунды оперировать умножением MHoro
значных (шестизначных!) чисел, извлечением квадратных корней из дe
вятизначных чисел и т. п. В том случае, если это никак не связано с YMe
лым шарлатанством, это действительно поддается одному и только одному
объяснению счет осуществляется по обычным законам математических
операций, но в друrой системе времени. Мноrочисленные записи очевид
цев, в том числе и медицинских специалистов, свидетельствовавшие о том,
как известный такими способностями Aparo (18. . 1949) уставал за KOpOT
кие (полчаса) сеансы TaKoro счета, дают некоторые основания полаrать,
что в эти моменты не только структуры мозrа, обеспечивающие счетные
операции, но и весь мозr, а может быть, и весь орrанизм, жили по дpy
rOMY времени. Пересчеты в отношении цифровых операций показывают,
что «полчаса Aparo» равнялись в лучшем случае непрерывной обычной
работе в течение мноrих часов.
TaKoro рода способность «выходить за рамки обычноrо масштаба Bpe
мени», по видимому, реальна и в то же самое время в связи с ее биоло
rической опасностью, при вовлечении Bcero мозrа или орrанизма в суще
ствование по ускоренной (или замедленной) проrрамме она, по видимому,
обычно прочно заторможена. Растормаживание ее возможно как электри"
ческими воздействиями на мозr, так и, вероятно, фармаколоrическими.
Интересным при решении этой проблемы явля тся получение ключа
к эпизодической возможности ускорения времени в одной двух функцио
нальных системах. Это может оказаться при разумном подходе к вопросу
не так опасно, как общее ускорение или замедление мозrовоrо отсчета
времени при жизни в обычных условиях. (Понятно, что общее ускорение
может привести к быстрому износу особенно индивидуально малоустой
чивых систем, а замедление резко понизит социальную приспособляемость
человека. )
Приведенные (и мноrие аналоrичные) данные подтверждают
важнейшую роль подкорковых структур в обеспечении нормаль
Horo протекания эмоциональных и психических реакций и их зна--
чение в разноrо рода психических нарушениях, подчеркивав--
шихся уже и в тех работах, которые базировались на данных,
получаемых при очаrовых поражениях мозrа (Nielsen, 1958;
Roberts, 1958, и др.).
в плане изучения мозrовых механизмов эмоциональных и
психических реакций электрическое раздраjКение мозrа предста
63
вило: 1) сравнительно большой (но, как видно будет позднее,
далеко не полный!) материал о зонах мозrа, имеющих отноше
ние к центральному контролю эмоций; 2) сведения о наличии
в мозrу структур, изменяющих возможности осуществления пси
хической деятельности, и 3) очень интересные и разнообразные
наблюдения различных психических феноменов, их изменений
и изменений элементов этих феноменов. Этот последний MaTe
риал представляется, однако, скорее сейчас еще в виде отдель--
ных фактов, чем данных Toro рода, по которым моrли бы быть
построены хотя бы суrубо ориентировочные карты мозrовоrо
контроля психических функций.
В целом, однако, полученные с помощью электрических воз
действий данные существенно расширили представление о CTPYK
турно функциональной орrанизации мозrа. Эти данные явились
cBoero рода опорным материалом и были именно таким образом
использованы в дальнейших, значительно более тонких исследо
ваниях, проводимых с помощью оценки нейродинамики мозrа
при реализации соответствующих видов деятельности.
При изучении центральных механизмов эмоциональных и .
более дифференцированных психических реакций значительно
более полные данные в условиях электрическоrо раздражения
были получены в отношении эмоциональноrо контроля. Создава
лось впечатление, что чем сложнее оказывалась психическая дея
тельность, тем меньшей полноты данные об ее орrанизации моrли
быть получены при локальных электрических воздействиях.
Исследование структур, «точек» мозrа, наиболее тесно связан
ных с реализацией психических функций, уrлубление изучения
мозrовоrо контроля эмоциональных и психических реакций OKa
залось возможным способом исследования воспроизводимости фи--
зиолоrических показателей состояния мозrа при повторных реа--
лизациях психолоrических (в том числе эмоциоrенных) проб.
В связи с особенностями эскоr и отсутствием в настоящее время
достаточно совершенных приемов анализа реакции с учетом ее
зависимости от исходноrо фона и, таким образом, невозможностью
получения в этом случае убедительно сравнимых результатов,
для решения этой задачи использовались rлавным образом дaH
ные динамики медленных электрических процессов (МЭП или
так называемоrо постоянноrо потенциала), наличноrо кисло
рода a и импульсной активности ИА. Особые возможности
представили мноrоканальные записи этих показателей. Исследова
ния показали, что МЭП и 02а являются весьма демонстративными
показателями развития эмоциональных состояний (В. М. Смир
нов, 1967; В. Б. rречин, В. М. Смирнов, 1967). На основании aHa
лиза МЭП и 02а при эмоциоrенных пробах (воспроизведение эмо
циоrенных ситуаций способом эмоциоrенных слов, вопросов,
предъявлением карт Роршаха и т. п.) наблюдалось два основных
типа изменений.
Изменения первоrо вида развивались спустя 0,3 с (от момента
окончания действия раздражителя) и длились около 10 15 с. Они
64
характеризовались изменением рисунка колебаний МЭП, наблю
давшихся при спокойном бодрствовании, и иноrда умеренным
сдвиrом уровня МЭП. Эти изменения не были связаны собственно
с развитием эмоциональноrо состояния и отражали состояние
активации внимания, предшествующее развитию эмоций. Они
наблюдались преимущественно в неспецифических ядрах зри
тельноrо буrра (переднее, медиальное, срединный центр), в мин
далине, rиппокампе и менее отчетливо в вентро латеральном
ядре, бледном шаре, красном ядре, некоторых отделах коры. Если
эмоция не развивалась, появившиеся изменения первоrо вида
исчезали. Наоборот, при развитии эмоции эти изменения перехо
дили в изменения BToporo вида, причем в большинстве случаев
при сильных эмоциональных переживаниях направленность
СДВиrа МЭП «коррелировала» с эмоциональной окраской peaK
ции: положительный сдвиr МЭП наблюдался при отрицательных
эмоциях, отрицательный, наоборот, при положительных, хотя
в ряде структур при отрицательных эмоциях наблюдался OДHO
значный сдвиr МЭП. Эти сдвиrи оказывались наиболее отчетли--
выми в неспецифических ядрах зрительноrо буrра (переднем, Me
диальном, срединном центре), в некоторых отделах вентро лате
ральноrо ядра, в субталамической области, rиппокампе, минда
лине, бледном шаре, хвостатом ядре, в клеточных образованиях
ножек мозrа, в некоторых отделах коры (В. М. Смирнов и
М. М. Сперанский, 1970).
Результаты динамики МЭП при эмоциоrенных пробах не
всеrда моrли быть пред ставлены в виде суперпозиции одна и
та же эмоциоrенная проба при повторении моrла терять эффек
тивность, данные предъявления различных эмоциоrенных проб
моrли давать неполностью сопоставимые результаты. В этих
исследованиях чаще Bcero можно было rоворить о принципиаль
ной воспроизводимости результатов на основе характера сдвиrов
и их направленности.
Однако анализ мноrолетних наблюдений медленных электри--
ческих процессов, их формы и амплитуды позволил В. М. Смир
НОВУ И М. М. Сперанскому (1972) создать своеобразную «азбуку
эмоций» по динамике МЭП (рис. 10).
Специальный интерес результаты реrистрации сдвиrов МЭП
при эмоциоrенных тестах приобретают, если сопоставить их
с эмоциональными реакциями, развивающимися при вызванном
внешним электрическим воздействием сдвиrе МЭП в мозrу. Pe
rистрация физиолоrических показателей при эмоциоrенных про
бах представила более полный материал о структурах мозrа, свя
занных с развитием эмоциональной реакции. Однако зона наи--
больших сдвиrов МЭП при эмоциоrенных тестах совпадала
с областью, электрическое воздействие на которую вызывало раз
витие эмоциональной реакции. Можно утверждать, что использо
ванные приемы электрических воздействий на мозr и исследова
ние физиолоrических показателей мозrа позволили получить
в отношении эмоциональных реакций взаимоподтверждающие
65
"
.444 t r i :Io"t t t " f 6.& 3' '-4 l' #.1.1 $е.;......
1i Z5flMfi6 j::'.cs«
С'1 r , ' < i; { '
i
t 01 I
I
ШвСl
Сдltех
I I
1. I
'014 +
,
! ИР"Сl
I
i
!
; Vt
i
t
I
I
I
t "tti ' A:A j St \)tttt J.4I*' t t 4 );J
еР
Curtex
10. Типичные компоненты ответов МЭП при психолоrических воздей
ствиях, активирующих внимание и эмоциональное поведение.
Высокоамплитудные комплексные компонеlIТЫ ВUЛНЫ: А аппликационная волна, S сел
лярная волна, 1 I волна, W W волна, V V волна; высокоамплитудные некомплексные
компоненты сдвиrи; Р позитивный, N неrативный сдвиr МЭП; низкоамплитудные He
комплексные компоненты; и волны, ундуляция, М монотонные волны (колебания), F
фоновая активность в ответе.
данные о структурно функциональной канве эмоциональных pe
акций (рис. 11). Анализ направленности сдвиrов МЭП при эмо
циональных реакциях в основных, участвующих в реакции
структурах (таламус, височные лимбические структуры и т. д.)
дал основание по новому рассматривать эмоционально нейтраль
ное состояние оно соответствовало колеблющемуся внебольших
пределах среднему, характеризующемуся определенным уровнем,
состоянию этих образований. Исследования 02а при эмоциоrен
ных тестах (В. Б. rречин и В. М. Смирнов, 1967) позволили также
обнаружить отчетливые изменения уровня и, иноrда, динамики
потребления кислорода подкорковыми структурами, причем были
уточнены не только принципиальные, общие особенности этих
сдвиrов, но и связанные с областью отведения. Так, появление
правильных, «синусоидальных» или, реже, апериодических мно--
rофазных колебаний относительно небольшой длительности, чаще
Bcero сходноrо рисунка при отрицательных и положительных
эмоциях, было характерным для вентральноrо, заднелатераль
Horo и вентро латеральноrо ядер зрительноrо буrра. Изменения
02а в дорсо--медиальном ядре наблюдались с большим латентным
периодом и лишь при наиболее значимых эмоциоrенных тестах.
66
Изменения 02а в ядрах зрительноrо буrра наблюдались и при
повторении тестов. Наоборот, в красном ядре, в ряде случаев
в черной субстанции и в покрышке, реакции при повторении проб
становились менее отчетливыми. Различие направленности сдви--
rOB a или их динамики (рисунка) в зависимости от знака эмо--
циональной реакции отмечено в покрышке, rиппокампе, минда--
лине, а также хвостатом ядре.
Понимание физиолоrических и в первую очередь нервных
механизмов этих перестроек в мозrу, равно как и самих механиз
мов развития эмоциональных реакций при изменениях внеlllней
и внутренней среды орrанизма, по видимому, наиболее правильно
на основе учета фактора полифункциональности нейронных по
пуляций и, отсюда, большоrо количества «афферентных приво--
дов» К различным подкорковым структурам.
Исследования МЭП и 02а при психолоrических пробах типа
проб Бине и др. оказалось возможным уже почти всеrда осуще--
ствлять способом суперпозиции отрезков кривых, т. е. собственно
изучением воспроизводимости реакции. Исследование сравни
мости данных импульсной активности осуществлялось на основе
данных интеrратора (Ю. К. Матвеев, 1971) после предваритель
Horo подсчета и соотнесения частоты импульсации в разные фазы
пробы к фоновой (Н. П. Бехтерева и А. И. Трохачев, 1967),
а та же на основе rрафиков, составленных в результате машин
Horo (ЭВМ В. А. Орлов) подсчета количества импульсов за
определенные отрезки времени (0,3 с, 0,9 с, 1 с и др.) и друrих
более сложных обработок этих процессов. Сейчас обработка дaH
ных TaKoro рода осуществляется с помощью аналоrо цифровоrо
вычислительноrо комплекса.
Существенной выrодой использования тестов на оперативную -
память являлась возможность повторения проб принципиально
Toro же типа по нескольку раз в течение данноrо исследования,
в том числе и при произвольном варьировании условий наблю
дения.
Первоначально более или менее значительные отклонения
физиолоrических показателей от исходноrо состояния отмечались
в различных rлубоких структурах (и в исследованных точках
коры).
Запись МЭП, 02а и ИА при повторном выполнении психоло
rических проб обнаружила, что динамика этих показателей во
мноrих точках мозrа оказывалась весьма сходной, а иноrда почти
идентичной, воспроизводимой. .
Устойчивая воспроизводимость динамики в данных условиях
наблюдения рассматривалась как подтверждение связи данных
образований мозrа с психической деятельностью. В целях удоб
ства изложения при описании обнаруженных структурно функ
циональных отношений, так же как и в отношении эмоциоrенных
тестов, приходится обычно rоворить о морфолоrических образо
ваниях, в которых наблюдалась эта воспроизводимость физио
лоrических показателей. Эта воспроизводимость наблюдалась
67
p
! +,
" 100.: "
+"'т'r9:'j!\ '1Jfi : 1"'"
>'(> -(1 " )I 'IoW H:j\ , >,1 o!.\I < 1'1I\ i ... > 1 IJ> ' ша \ IWHia '! "H"t I\1i ' 111ft 1(,-'I :1 j . ' IIUЩI > 'illl1Ш t .НПi. !IJ' 'tt\j .I&! 'JA)t.\. >]' ' l ) 'all!l:Y
д
, P/OYfl'
ffi ;;( ;;;f't;; jri' tw [ жi\i\; !;< l( t1i J\$ :' й i tl\ ] (i i; 1 !1 I L ;: i ; ri;j , t;k;; ;M j <;:,« 4i '
.. ".. :'::0(' '\,
. ив
... tw
, ::/1t, { \ ; f\ {\
Р"'\ Д: .. .,.1\ w.m
'с:.. , \11,,: j' '
" . у1 /1/'. f'v'
.
$ '
.1(Т!1й
;.::"
,О." ".
".:.; ...
.J:: ..
. ;,
:.:.- "
1 . 1 .:.:': 1 .:::..:.:........"
......... /11 .
;.;. .:..:
. ':':'. ..: ....
;,;;.;
":;. .;;. l' V
::; .. ,
i
.
6
11. Исследование эмоциоrенных зон мозrа.
а изменение медленных электрических процессов (МЭП) при эмоциоrен
ных пробах. ТЭм. тест эмоциональный; 6 схема мозrа. Черными KBa
дратами помечены зоны, rде наблюдались изменения МЭП при эмоциоrен
ных тестах. Большими квадратами arмечены области, rде изменения МЭП
наблюдались с б6льшей выраженностью и чаще; маленькими зоны, rде
изменения МЭП наблюдались реже и были менее выражены; остальные
обозначения те же, что на рис. 6;
68
VL[V 2 ]
Ci [VIJ
Put /iVsl
VP[VsJ
ТЗ м ( )
рр(-)
тэм( )
8
рр (--)
11. П родОJl,жен ие.
8 динамика наличноrо кислорода при эмоциоrенных тестах.
VL вентро латералъное ядро зрителъноrо буrра; РС
ножки мозrа; Put скорлупа; Ci внутренняя капсула; n,
красное ядро; V Р заднее вентральное ядро. Римскими циф-
рами обозначены пучки электродов; арабскими электроды
в пучках.
в различных ядрах зрительноrо буrра, в бледном шаре, хвоста--
том ядре, rиппокампе и ряде друrих структур (рис. 12). Однако
именно в связи с этими данными целесообразно подчеркнуть, что
указанная воспроизводимость практически никоrда не наблюда
лась во всей структуре в целом, а обнаРУjКивалась ЛИlllЬ в од--
ной, реже нескольких ее точках. И в то же самое время именно
исследования этоrо плана убедительно подтвердили полиФунк
циональность не только исследованных морфолоrических образо
ваний (<<ядер»), но И клеточных популяций. Так же, как при элек
трическом воздействии (не rоворя уже об очаrовых патолоrиче--
ских процессах!), при реrистрации динамики физиолоrических
показателей обнаруживалось, что мноrие клеточные популяции
имеют отношение к различным функциям, а отсюда MorYT OTBe
чать воспроизводимым рисунком не только на один, но и на He
сколько раздражителей. Они моrли быть связанными и с психи
ческой, и с двиrательной деятельностью и т. д. При этом «Ha
боры», комплексы функций, с которыми были связаны отдельные
точки мозrа, различались. Характерные реакции импульсной
активности со срединноrо центра зрительноrо буrра обнаружива--
лись, например, на психолоrические пробы и на сrибание рук
в локтях и т. п. (рис. 13).
69
*1..
;,
:-:, .;"
;':'\1:
{; % 2 +,
;
:-;: .;.:-
<.Х
i 1::i i,
f.? .. ;:;;:.:: : :::
... ;';';:,
.:;::: ;:;:;:::.;::.)
'Н
rL
t I
.:(.
':.:
'. '1 =<-:.: " III ... ,
< :: ; : :: :r; :, ' , , ,J 1'
.. , .....' ....:
....;..':::: (. .,;:;:-;,;"...........,.;.,.:::'.;.x;:":::;";:';:; oOOC< .;.:.
::. ' "
а
VL [У/51
S alB
1 1/ /1/
5 ...."
..
45с.
IJИЛ/С VL[ ]
100
80
60
40 1 11 111
20 t С'
О
6
70
Ceпt'Pum тediaпuт
VI(1 3)
20с
.
i
z
1
J[
DI
1
ал t. [1//21
I
+
11
10 С
l/l
.
J
д
омn/с. VL[V 5 J
80
50
.40
20
О
1
Jl
10
tc.
е
12. Изменения медленной электрической активности (а), наличноrо кисло
рода (6) и импульсной активности (в) при пробе на оперативную память.
/ предъявление пробы; // удержание в памяти; /// ответ. Слева данные одной
пробы. Справа: 2, д суперпозиция данных нескольких проб; е усреднение 6 проб. Ла-
тинскими буквами обозначены подкорковые структуры (см. рис. 7). Римскими цифрами
в сочетании с арабскими обозначены соответственно номера пучков н электродов в пучке.
71
v ((J Lf)[сслtr(jт теdiапит]
имп/с
100
80
80
70
60
50
'10
30
20
10 С.
О Удержание 6 памяти
а
100
90
80
70
80
50
'10
30
20
10 С.
О
о
13. Динамика импульсной активности в области срединноrо
центра зрительноrо буrра при пробе на оперативную память (а)
и при двиrательной пробе (6).
Полифункциональность клеточных популяций мозrа, отмечав
шаяся и в работах А. Р. Лурия (1962) и мн. др. является предпо
сылкой и условием для оптимальноrо взаимодействия орrанизма
со средой.
Тонкое исследование с применением приемов реrистрации MHO
rих показателей жизнедеятельности мозrа и различных функцио
нальных проб показывает принципы орrанизации мозrа, равно
далекие от двух наиболее экстремистских представлений,
о центрах функций и о равнозначности различных участков
мозrа.
К реализации сложных психических функций имеет отноше--
ние очень большое количество «точек» В самых ра;зличных об--
разованиях мозrа, и, с друrой стороны, по своей функциональной
характеристике эти «точки» не однозначны. Мноrозвеньевая
орrанизация системы из исходно полифункциональных звеньев
обеспечивает и надежность, и адекватность ее деятельности.
Обеспечивая психическую деятельность здоровоrо орrанизма
в условиях устойчивоrо состояния нормы, эта система способна
обеспечивать решение самых сложнейших задач.
72
и в то же самое время именно мноrозвеньевой, сложный характер
этой системы может определять сложность борьбы с различными патоло
rическими процессами, характеризующимися развитием устойчивоrо па
толоrическоrо состояния (Н. п. Бехтерева и А. Н. Бондарчук, 1968). В этом
случае все боrатство поддерживающих устойчивое состояние реакций при
обретает обратную направленность, поддерживрет ставшее устойчивым па
толоrическое состояние. Компенсаторные реакции, основанные на MHO
жестве звеньев любой системы реrуляции в центральной нервной системе,
имеющей при поддержании устойчивоrо состояния здоровья orpoMHoe по
ложительное значение, в условиях общей перестройки орrанизма в ходе
формирования так называемоrо хроническоrо заболевания, смены устой
чивоrо состояния нормы устойчивым патолоrическим состоянием, MorYT
приобретать, хотя бы частично, и отрицательное значение для орrанизма.
ФизиолоrИЧЕСКОЕ ЗНАЧЕНИЕ И ОСОБЕННОСТИ
РАЗЛИЧНЫХ ЗВЕНЬЕВ СИСТЕМЫ
мозrовоrо ОБЕСПЕЧЕНИЯ ПСИХИЧЕСКИХ ФУНКЦИЙ
Исследование роли различных образований системы мозrа,
обеспечивающей психическую деятельность, целесообразно для
изучения принципов и конкретных механизмов, создающих пред
посылки для реализации психической деятельности в eCTeCTBeH
ных условиях меняющейся внешней среды орrанизма и BHYTpeH
ней среды мозrа. Это исследование необходимо и для изучения
механизмов оптимизации самой психической деятельности.
Исследование роли факторов внешней среды в исследуемых
механизмах было проведено следующим образом: предъявление
психолоrических тестов проводилось в условиях оrраничения дo
полнительных (к основному) раздражителей при закрытых rла--
зах субъекта, при открытых rлазах в условиях умеренной OCBe
щенности, а также на фоне «шума» фотостимуляции неритми
ческоrо случайноrо или триrrерноrо вариантов (Н. П. Бехте--
рева, 1965).
Оказалось, что некоторые «точки» (области) мозrа, обнаружи
вавшие четкую воспроизводимость рисунка исследуемых по
казателей в покое, переставали обнаруживать ее в условиях
«шума», и, наоборот, в этих новых условиях воспроизводимость
моrла появляться в тех участках мозrа, rде она ранее OTCYT
ствовала (рис. 14). При этом новые «точки» С воспроизводимым
рисунком моrли появиться в пределах той же или друrой MOp
фолоrической структуры (ядра) мозrа. Эти последние данные, по
видимому, MorYT быть связаны не только с реально существую
щими отношениями, но и с неполнотой возможностей обсле
дования мозrа в условиях лечебноrо применения вживленных
электродов. Возможно, что при исчезновении «точек» С воспроиз
водимым рисунком В пределах одной структуры и появлении их
в друrой определяющим является не факт «выключения» одноrо
ядра и «включения» друrоrо. Речь идет о «выключении» одной
из зон ядра и отсутствии электродов в друrой точке этоrо же
ядра, которая моrла бы «включиться» в деятельность в этих
же условиях, и, наоборот, обратных соотношений CTpYKTypa
73
14. Схема мозrа.
Черными квадратами обозначены зоны мозrа, rде обнаруживались воспроизводимые изме-
нения наличноrо кислорода. Левая половина схемы проба предъявлял ась в покое;
справа на фоне шума. Обозначения структур см. рис. 8.
электрод в друrом мозrовом образовании. Накопление материалов
все бl)льше свидетельствует в пользу возможности TaKoro рода
ситуации, равно как и в пользу Toro, что оrромная сложность
структурно--функциональных отношений мозrа не укладывается
в прокрустово ложе искусственной (и очень различной у разных
исследователей) дифференциации мозrа на ядра.
Так, например, первоначально наблюдалось, что воспроизво--
димы е изменения физиолоrических nоказателей в rиппокампе
обнаруживались преимущественно при реализации психолоrиче
ской пробы в покое, а в миндалине при шуме. Однако в даль--
нейшем, при несколько отличном расположении электродов
в пределах тех же структур, удалось уточнить эти результаты
и показать возможности «включения»........... «выключения» действую
щих участков при изменении условий внешней среды в пределах
той же структуры. Следует подчеркнуть, что эти новые данные,
с оrоворкой в отношении условности ядерноrо деления мозrа, ни
в какой мере не противоречат первоначальным, а лишь допол--
няют их. Естественно, возможны и различные варианты не
только «внутриядерной», но И «межъядерной» динамики.
74
Со всеми приведенными дополнениями первостепенно важ
ным остается факт о постоянстве активности одних и о непо--
стоянстве мноrих друrих звеньев структурно функциональной
системы мозrа, обеспечивающих психическую деятельность в ме--
няющихся условиях нешней среды. Естественно отсюда rOBo
рить, что система имеет обязательные для данной деятельности
образования мозrа, работающие независимо от изменений внеш--
ней среды (по крайней мере в данных пределах), жесткие
звенья и звенья, обязательные для данной деятельности
лишь в каких--то определенных условиях внешней среды, звенья
rибкие.
Вполне понятно, что. термин «жесткие» не используется
в смысле жесткоrо замыкания реакции через один нейрон.
И внутри rибкоrо и жесткоrо звеньев системы деятельность ней--
ронной популяции может осуществляться на основе вероятно--
стно--статистическоrо принципа (А. Б. KoraH, 1962). Имен,НО Ta
кое сочетание жестких звеньев (или, вероятно, точнее........... относи--
тельно жестких!) и rибких обеспечивает экономичность и
исключительную rибкость, адекватность системы мозrа, обеспе--
чивающей психическую деятельность.
Опыт практической нейрохирурrии, в том числе и стереотак--
сической, показывает, что разрушение мноrих структур мозrа,
в частности и тех, связь которых с психической деятельностью
показана, не всеrда ведет к видимым психическим нарушениям.
Это может быть связано с самим психолоrическим исследова--
нием недостаточное варьирование условий, в которых прово
дится психолоrическое исследование, может не выявить развив--
шийся дефицит.
Это, более вероятно, связано с оrромными компенсаторными
возможностями мозrа и замещением дефекта, в том числе и пар--
ным образованием интактноrо полушария. Последнее положение
в наблюдениях на человеке подтверждается увеличением ча--
стоты психических осложнений при двусторонних операциях на
подкорковых структурах мозrа. Можно себе представить, что
двустороннее разрушение жестких звеньев системы физиолоrиче
cKoro обеспечения психических функций особенно опасно, а по
тому выявление их приобретает все большую практическую
важность.
Исследованные явления имеют еще ряд аспектов. Так, напри--
мер, как указывалось, реализация психической деятельности на
фоне «шума» не только нуждае'I' Я в деятельности дополнитель
ных образований мозrа, но и происходит при «выключении»
каких то друrих «точек», активных в состоянии покоя. Иными
словами, внешний «шум» В чем то усложняет систему (являясь
«помехой»), но В чем то ее и упрощает (оказываясь источником
необходимоrо тонуса?). Отсюда то, что наблюдается по показате
лям МЭП, 02а, ИА, может быть использовано и в дальнейшей pac
шифровке положения об оптимальном фоне для деятельности,
эти наблюдения показывают, что одной и той же деятельности,
75
но реализуемой в разных условиях, соответствует свое, опреде--
ленное, функциональное состояние мозrа, складывающееся из
фУНКЦИОI:Iальноrо состояния ero различных структур. Фоновое co
стояние мозrа, на котором реализуется деятельность, в частности
психическая, интеrральное функциональноrо состояния отдель
ных структур, отнесенное к структурно функциональной орrани
зации, обеспечивающей данную деятельность системы, и состоя
нию всех друrих образований мозrа.
Как можно себе представить конкретный механизм, вызываю
щий «включение» И «выключение» каких то элементов струк",
турно функциональной системы, обеспечивающей психическую
деятельность? Понимание этоrо вопроса возможно при учете
уже упоминавшеrося выше положения ополифункциональности
нейронных популяций, имеющих отношение к психической дея
тельности.
Изменение и пульсации в связи с изменившимися условиями
внешней среды может «включить» одни элементы системы
(структуры мозrа) и «выключить» друrие. Наоборот, исключение
каких то внешних раздражителей оказывается оптимальным для
выявления активности друrих структур мозrа, элементов CTPY
турно функциональной системы, обеспечивающих реализацию
психической деятельности при отсутствии определенноrо вида
(или количества!) раздражителей. Не исключено, однако, что He
постоянство активности мозrовых структур (<<rибких звеньев»)
отражает также обеспечение психической деятельности в изме--
нившихся условиях среды за счет резервных возможностей
мозrа.
ЭТИ положения удалось существенно дополнить r В условиях
изучения физиолоrической орrанизации психической деятель
ности при изменениях внутренней среды мозrа. Наблюдения этоrо
рода были проведены во время применения нейротропных пре--
паратов, активных в отношении различных видов синаптической
передачи в мозrу адренерrических, холинерrических и cepOTO
нинерrических синапсов (А. Д. Аничков, 1968; д. К. Камбарова,
1969; В. А. Чернышева, 1971). Применение нейротропных средств
не только изменяло фоновые показатели физиолоrической актив
ности мозrа, но и ее динамику при деятельности и, в частности,
рисунок МЭП и клеточной активности при психолоrических про
бах. Так, применение дезерила привело к исчезновению воспро
изводимых изменений МЭП при психолоrических пробах в об
ласти nucl. centralis thalami; педифена в nucl. ruber и globus
pallidus medialis; применение метамизила, напротив, в значитель
ной мере усилило воспроизводимые изменения МЭП при психо
лоrических пробах в nucl. ruber и globus pallidus medialis
(В. А. Илюхина, 1972).
Особый интерес предстаВI1ЛИ изменения импульсной актив
ности при применении нейротропных средств (прослеженные
нами совместно с Ю. К. Матвеевым и Д. К. Камбаровой, 1970).
Запись импульсной активности не только в покое, но и в условиях
7С
реализации психолоrических и двиrательных проб позволила
проследить и общие, дистантно обусловленные изменения, и из
менения, развивающиеся непосредственно в области реrистрации.
Общие изменения проявлялись изменением уровня нейронной
активности; при этом динамика рисунка изменений импульсной
активности при соответствующих пробах изменялась вторично.
Действие нейротропных средств непосредственно в области отве--
дения импульсной активности проявлялось четким изменением
рисунка динамики импульсной активности при соответствующих
пробах. Таким способом удавалось различать не только местные
и общие изменения, но и прослеживать фазовые перестройки
в мозrу. Так, например, применение дезерила вызывало перво
начально некоторое увеличение фоновой импульсации в средин--
ном центре зрительноrо буrра, обусловленное скорее Bcero дис
тантными активизирующими влияниями. Рисунок динамики
импульсации при психолоrической пробе в этих условиях лишь
незначительно отличался от исходноrо. Через 1 'Ч, фоновая
импульсация вернулась к исходной; характер динамики разрядов
при психолоrической пробе в это время изменился более значи
тельно. Он еще более существенно перестроился на фоне суще
cTBeHHoro падения фоновой импульсации через 2 11, (рис. 15). На--
блюдения этоrо ряда позволяют rоворить о том, что пере стройка
нейрофизиолоrической системы обеспечения психической дея--
тельности, изменение звеньев этой системы возможны под влия'"
нием не только внешней среды орrанизма, но и внутренней среды
мозrа. В этом, втором, случае перестройки развиваются в связи
с изменением в определенных синаптических образованиях
адренерrических, холинерrических, серотонинерrических. А от--
сюда эти же наблюдения MorYT быть использованы как свидетель
ство Toro, что нейрофизиолоrическая система обеспечения психи--
ческих функций состоит из звеньев с различной медиацией, т. е.
эта система полибиохимична. Подавление KaKoro To вида биохи--
мической передачи по крайней мере в той степени, в какой об
этом можно судить по исследуемым показателям, выключает
данную нейронную популяцию как звено этой системы (хотя хо--
рошо известно, что на очень мноrих клетках (Jung, 1963; Bradley,
1965; П. К. Анохин, 1968, и др.) располаrаются синапсы разных.
типов, не rоворя уже о возможности полибиохимичности клеточ
ной популяции).
Естественно, до Toro как делать какие либо решающие BЫ
воды, исследования должны быть продолжены и расширены, но
если полученные данные будут подтверждаться, это окажется
не только путем к дальнейшему нейрофизиолоrическому анализу
звеньев системы и системы в целом, но и будет иметь четкий
практический результат. Окажется возможным избирательно, на--
правленно и дозированно выключать определенные звенья си
стемы, не изменяя существенно условий деятельности друrих
ее элементов и тех же структур мозrа, как звеньев друrих функ
циональных систем.
77
им п/ (;
1110
130
120
110
V(З 'f) [ceпtruт тe iaпuтJ
!
80
70
3
'1
30
20
10
f
с
15. Динамика импульсной активности срединноrо
центра зрительноrо буrpа при пробах на оператив
ную память, проведенных в покое (1), через
15 мин (2), через 1 ч (3) и через 2 ч (4) после
введения дезерила.
Полученные данные 'интересны еще, по крайней мере, в oд
ном отношении. Весьма активным препаратом в отношении из
менения звеньев системы обеспечения психических функций
явился дезерил, причем одной из точек ero приложения оказался
centr. medianum структура, электрическое воздействие на KOTO
рую вызывало существенное изменение условий протекания пси
хической деятельности (В. М. Смирнов, 1967; А. Н. Бондарчук и
В. М. Смирнов, 1969).
Эти наблюдения MorYT быть использованы для суждения
о возможном преимущественно м анатомическом субстрате таких
серотонинотропных психомиметических средств, как, например,
LSD--25 и др.
В приведенных материалах учитывались факторы, «извне»
или «изнутри» изменяющие физиолоrические механизмы психи
ческой деятельности, причем в таких пределах, в каких это улав--
ливалось только при нейрофизиолоrическом исследовании и чаще
Bcero существенно не влияло на возможность осуществления за
данной психической активности. Однако воспроизведение теста
78
20с
....
ш
л
1
16. Сходная ВОСПРОИЗВОДимая динамика наличноrо кислорода
при правильной и ошибочной экфории психолоrических тестов.
1 предъявление пробы; // удержание пробы в памяти; /// ответ.
Вверху центральное, внизу вентро латеральное ядро зрительноrо буrра.
оказывалось не всеrда правильным и в покое и во всех прочих
исследуемых условиях.
Представляло несомненный интерес изучить особенности ней--
рофизиолоrической системы, обеспечивающие психическую дея
тельность в зависимости от качест..ва осуществления самой психи
ческой деятельности. Эт моrло явиться шаrом по пути изучения
реально существующих физиолоrических механизмов оптимиза
ции психической деятельности.
Анализ показал, что по динамике физиолоrических показа
телей в зависимости от правильности или ошибочности экфории
тестов оказывается возможным выделить три типа структур
79
ш
л
1
ш
1
j
л
17. Различная воспроизводимая динамика наличноrо кислорода при
правильной (внизу) и ошибочной (вверху) экфории психолоrических
тестов.
1 предъявление пробы; 11 удержание в памяти; /1/ ответ. Так же, как на
рис. 16, запись осуществлял ась с вентро латеральноrо ядра, но с друrих ero отде.т.ов.
или, правильнее, «точек» В мозrу. В структурах (<<точках»)
первоrо типа воспроизводимая динамика физиолоrических пока--
зателей не обнаруживает четкой зависимости от качества выпол
нения пробы (рис. 16). Такие «точки» обнаружились в rиппо--
кампе, миндалевидном комплексе, в вентральном, заднелате--
ральном и центральном таламических ядрах.
80
,/
ш!
л
! 1 !
..
а
ш I
л
! 1 !
18. Динамика наличноrо кислорода при правильных и ошибочных
выполнениях тестов.
Наличие воспроизводимых изменений наличноrо кислорода при правильно выпол
ненной психолоrической пробе и отсутствие воспроизводимых изменений в области
дорсо медиальноrо ядра зрительноrо буrра (а) при ошибках. Отсутствие воспроиз
водимой динамики наличноrо кислорода при правильном выполнении психолоrиче
ских проб и наличие воепроизводимой динамики при ошибочном выполнении проб
в хвостатом ядре (6).
В структурах (<<точках») BToporo типа функциональный ypo
вень воспроизводимо перестраивался и при правильной и при
ошибочной экфории тестов, но рисунок физиолоrических пока--
зателей в обоих этих случаях различался (рис. 17). К третьему
типу моrли быть отнесены, по крайней мере, некоторые точки
в пределах xBocTaToro ядра, срединноrо центра и друrих ядер,
rде воспроизводимые изменения физиолоrической динамики об
\
наруживаются только при ошибочном воспроизведении пробы
(В. Б. rречин, 1968) (рис. 18).
Обнаруженные факты интересны, в частности, и как свиде
тельство большей или меньшей дифференциации звеньев изу
чаемой физиолоrической системы. Это положение сближает моз
rовые механизмы психической деятельности с аналоrичными
механизмами друrих видов активности орrанизма.
С точки зрения проблемы оптимизации психической деятель--
ности, естественно, наибольший интерес представляют «точки»,
обнаруживающие воспроизводимые изменения при ошибочном
выполнении пробы. Весьма заманчиво было бы полаrать, что эти
точки представляют собой что--то вроде «детектора ошибок», aHa
лизатора правильности действий, cBoero рода реальный элемент
«акцептора действия» Анохина (1968). Впечатление о правомер
ности TaKoro суждения основано на взаимодополняющих фактах.
Действительно, если в хвостатом ядре реrистрируются воспроиз
4 Н. п. Бехтерева
81
водимые реакции именно при ошибках, то раздражение хвоста--
Toro ядра моrло приводить к повышению числа ошибок
(Н. П. Бехтерева, Н. и. Моисеева, А. Н. Орлова, В. М. Смирнов,
1964; Н. П. Бехтерева, А. А. rенкин, Н. И. Моисеева, В. М. Смир--
нов, 1965, и др.).
Во мноrих экспериментальных наблюдениях, несмотря на двой
ственность общих выводов, показаны четкие факты уrнетающей
(тормозящей) роли XBocTaToro ядра в отношении различных видов
деятельности. Как можно себе представить механизм развития
воспроизводимых отклонений физиолоrических параметров в XBO
статом ядре 11 друrих структурах при ошибках? По какому меха--
низму развивается «включение» этих образований? При поверх
ностном подходе к вопросу может создаться впечатление, что
«мозr умнее человека», что мозr «знает» об ошибке, в том числе
и тоrда, Rоrда о ней «не знает» человек. Дело, конечно, обстоит
иначе.
В качестве одноrо из вероятных объяснений MO:rKHO, по види
мому, принять следующее.
По крайней мере, краткосрочная память (а может быть, и
долrосрочная!) основана на почти «запоминании», сохранении
полноrо следа Bcero происшедшеrо события, в данном случае на
полном сохранении в памяти предъявленноrо теста. Однако, Ha
ряду с процессом запоминания, одновременно и в подавляющем
большинстве случаев целесообразно включается механизм «за--
бывания», который представляет собой не «стирание», а затор
маживание: перевод следа в форму, при которой «считывание»
оказывается затрудненным.
Преждевременное или неадекватное включение этоrо, BTO
poro, механизма может само вызывать реакцию kakoro--то эле--
мента акцептора действия.
Более вероятно, однако, что неадекватное включение затор
маживания создает диссоциацию между имеющимися следами
и возможностью их воспроизведения, результатом чеrо и яв"
ляется включение структуры, реrистрирующей эту диссоциацию,
рассоrласование со всеми возможными последствиями общей,
в том чи е и rуморальной, эмоционально детермированной aK
тивации. \
Продолжающееся накопление наблюдений дает все больше
оснований считать, что действительно обнаружены «детекторы
ошибок», но в решении этоrо вопроса, по крайней мере при
анализе физиолоrических показателей, необходимо специально
исследовать роль эмоциональноrо фактора, особенно значимоrо
именно при ошибочном ответе. При получении дополнитель--
ных результатов в этом плане следует, однако, учитывать, что
эти две точКи зрения включение структуры первично в связи
с ошибкой или в связи с эмоциональной реакцией ни в коей
мере не противоречат друr друrу, а скорее вторая возможность
(если она подтвердится!) дополняет первую. В исследовании
зтоrо вопроса важно учитывать латентный период, форму peaK
82
ции и динамику последействия, так как эти показатели в peaK
циях на психолоrическую пробу типа оперативной памяти и
эмоциоrенную чаще Bcero различаются.
Интересно отметить, что применение нейротропных средств
позволило вскрыть некоторые нейрофизиолоrические механизмы,
лежащие в основе свойств нейронной популяции реаrировать
rrолько на ошибочные реакции психолоrJ1ческих тестов. Так, при
применении дезерила в отдельных точках мозrа (в области cpe
динноrо центра и вентральноrо ядра зрительноrо буrра) отмеча
лось появление не наблюдавшеrося ранее воспроизводимоrо
рисунка при психолоrических пробах или своеобразная «rehepa--
лизация эффекта» появление xapaKTepHoro эффекта и на поло
жительные реализации теста там, rде до применения фармако
лоrических препаратов реакция отмечалась только при Оlllибках
(рис. 19). Можно попутно OTMeTJ1Tb, что в этой же точке после
введения дезерила характерные изменения импульсной актив
ности стали наблюдаться не только при движениях контралате
ральных конечностей (как это наблюдалось до введения препа
рата), но и rомолатеральных. Наоборот, в друrих точках мозrа
именно применение препарата приводило к появлению «специ
фических» свойств нейронная популяция, до применения ней
ротропноrо ср,едства обнаруживавшая характерный рисунок и на
правильные, и на ошибочные реализации проб, начинала изби
рательно реаrировать лишь при ошибочных реализациях. Эти
данные свидетельствуют о нейрофизиолоrических механизмах
формирования свойств нейронных популяций, о первостепенной
роли относительной или абсолютной активности различных био
химических медиаторных систем в проявлении этих свойств.
rипотеза, выдвинутая в 1966 r. (Н. П. Бехтерева), об обеспе
чении психической деятельности человека корково подкорковой
структурно функциональной системой со звеньями различной
степени жесткости получает сейчас все болыпее подтверждение
и, таким образом, является основой расшифровки, по крайней
мере, одной из сторон физиолоrических механизмов психических
явлений и прежде Bcero структурно функциональноrо их обес
печения. Проведенные исследования показали значительное пре...
обладание rибких звеньев в системе мозrовоrо контроля психи
ческой деятельности, что, по--видимому, и является одним из
основных отличий этой системы по сравнению с друrими си--
стемами центральной реrуляции функций. Соотношение жестких
и rибких звеньев в системах центральной реrуляции функций
может, вероятно, рассматриваться как принципиальный крите
рий их сложности, В основе усложнения систем лежит абсо--
лютное и относительное увеличение количества rибких эле...
ментов.
Вполне понятно, что им.енно это положение может быть уч
тено при расшифровке данных электрической стимуляции мозrа.
Обнаружение жестких звеньев любой системы возможно при
наблюдении исследуемоrо показателя жизнедеятельности мозrа
4*
83
:J
110
100
80
80
70
60
50
110
30
20
10
О
;;
100
эо
80
70
60
50
"о
80
20
10
О
5
1
1
,
t
10
Л
t
15 20
Ш
б
,
I
I
I
I
I
I
I
tlБдт t
+
I
I
I
I
I
I
I
5
1
19. rрафическое изображение динамики частоты им
пульсной активности в области одной из нейронных
популяций срединноrо центра при психолоrической
пробе.
По оси ординат: J количество импульсов; по оси абсцисс: араб-
ские цифры отметка времени в секундах, римские цифры
фон и фаза психолоrической пробы; / исходный фон; //
предъявление пробы; /// удержание пробы в памяти; /V ре-
чевой ответ больноrо; V фон после реализации теста.
Стрелки оrраничивают начало и конец предъявления теста (//)
и ответа больноrо (/V). а прерывистая линия изображает ди
намику импульсной активности при правильной реализации теста,
непрерывная при ошибочной. Характерная динамика импульсной
активности обнаруживается только при ошибочной реализации
теста; 6 прерывистая линия изображает динамику импульсной
активности при правильной реализации теста до введения дезе-
рила, непрерывная линия после. Правильная реализация теста
вызывает после введения дезерила характерный рисунок импульс-
ной активности, сходный с рисунком на ошибочную реализацию
теста до введения дезерила.
84
при стимуляции и при изучении показателей нейродинамики
мозrа в условиях предъявления соответствующих тестов. Иссле--
дование rибких звеньев практически реально лишь при изучении
нейродинамики мозrа при предъявлении функциональных проб
и варьировании условий наблюдения.
Исследование различных физиолоrических показателей жиз
недеятельности мозrа при психолоrических пробах оказалось
приемом, значительно более тонким, чем наблюдение изменений
психической деятельности при очаrовых патолоrических процес
сах, хирурrических локальных разрушениях и электрическом
воздействии на мозr. Оно позволило значительно полнее изучить
структурно функциональные и физиолоrические основы психи--
ческоrо, показать участие в психической деятельности и тех
образований, которые моrли в результате данных выключения
и раздражения несправедливо считаться «немыми» зонами.
Таким образом, было уточнено, что изучение наиболее слож--
ных мозrовых систем, и прежде Bcero системы обеспечения
психических функций, возможно только при использовании всех
приемов комплексноrо изучения физиолоrии мозrа, при сочета
нии электрических и фармаколоrич.ескиXJ воздействий на мозr
с прямым наблюдением нейродинамики мозrа.
НЕйроФизиолоrИЧЕСКАЯ СУЩНОСТЬ
НЕКОТОРЫХ ИЗМЕНЕНИЙ, Р АЗВИВАЮЩИХСЯ В ЗВЕНЬЯХ
СИСТЕМЫ мозrовоrо ОБЕСПЕЧЕНИЯ ПСИХИЧЕСКОЙ
ДЕЯТЕЛЬНОСТИ
Естественно, что одним из наиболее интриrующих вопросов
именно в данной проблеме является анализ сущности изменений,
развивающихся в различных образованиях мозrа в процессе
обеспечения психической деятельности. Ответ на этот вопрос
может быть разделен на две неравные по значению и объему
части: 1) какие физиолоrические изменения развиваются в эле--
ментах системы, обеспечивающей психическую деятельность
в процессе реализации этой деятельности? 2) Какие изменения
в этих структурах мозrа обеспечивают специфический характер
каждой конкретной психической деятельности? Иными словами,
можно ли в KaKOM TO виде физиолоrической активности OTЫ
скать рисунок, который будет характерен для KaKoro To опреде
ленноrо слова, фразы и т. п.?
Ответ на первый вопрос возможен сейчас.
Деятельное состояние структур, участвующих в психической
активности, связано с изменением всех физиолоrических пока
зателей. Изменяются уровни медленных электрических процес
сов и наличноrо кислорода. При этом изменения уровня МЭП
не только развиваются при включении структуры в деятель--
ность, но и наоборот изменение уровня МЭП (вероятно, преи--
мущественно постоянноrо потенциала) может «включить» CTPYK
туру В деятельность по крайней мере, эмоциональную. Далее:
85
активность структуры, развившаяся как следствие изменения
МЭП (также, вероятно, постоянноrо потенциала) в определенных
образованиях мозrа, может явиться основой сложных и очень
стойких поведенческих реакций (Н. П. Бехтерева, К. В. rрачев,
А. Н. Орлова, С. Л. Лцук, 1963; В. М. Смирнов, 1967). В экспе
рименте явления этоrо рода наиболее подробно изучены В. С. Py
синовым (1957), а затем и друrими, исследовавшими процесс
временной связи при создании искусственных очаrов в мозrу
слабой поляризацией. Эти факты четко обнаруя{ивают важней--
шую роль изменений МЭП и, в частности, постоянноrо потен
циала в функциональной активности мозrа.
Показатель 02а может свидетельствовать об изменении и по--
требления, и доставки кислорода. Отсюда снижение ero уровня
может свидетельствовать и о повышении ero потребления и об
уменьшении доставки. Вполне понятно, что в условиях деятель--
ности и в связи с особенностями мозrовых сосудов уменьшение
доставки кислорода в деятельные (а об этом можно судить по
друrим физиолоrическим показателям!) структуры маловероятно.
Хотя общеизвестно, что при паркинсонизме возможна rипоксия
мозrа, снижение уровня 02а наблюдалось в исследуемых усло
виях пространственно неоднозначно: в вентро латеральном ядре,
срединном центре, дорсо--медиальном ядре зрительноrо буrра
значительно, менее отчетливо в медиальном членике бледноrо
шара, хвостатом ядре и изредка в rиппокампе и миндалевидном
комплексе.
Более сложно обстоит вопрос при повышении уровня 02а
именно это состояние может быть связано и с уменьшением по--
требления, и с повышением доставки кислорода. Однако вновь
и в этом случае понимание физиолоrической СУIЦности явлений
оказывается возможным при сопоставлении различных физио
лоrических показателей. Свидетельствуя о деятельном состоянии
структуры, эти дополнительные (по отношению к данному) по
казатели делают маловероятным предположение о снижении
потребления кислорода и более вероятным превышение ero
потребления увеличившейся поставкой. Повышение уровня 02а
при психической деятельности изредка наблюдалось в отдель--
ных участках срединноrо центра и дорсо медиальноrо ядра та--
ламуса, в вентро латеральном таламическом ядре, медиальном
членике бледноrо шара. При этом имела значение не только
область отведения 02а, но и фаза выполнения психолоrической
пробы (В. Б. rречин, 1966). в отношении изменений эскоr
суждение более сложно, так как они, как указывалось, были
весьма разнообразны и очень существенно зависели от исходноrо
фона. Можно лишь утверждать, что изменения эскоr в обнару
женных пределах (развитие медленных волн, острых волн и т. п.
В структурах, имеющих отношение к обеспечению психической
деятельности) не препятствовали ее реализации. Полученный
в условиях прямоrо отведения от мозrа этот факт должен быть
учтен при расшифровке материалов о связи проявлений ээr
86
(ЭСI\оr) с основными нервными процессами. С любыми желае--
мыми оrоворками вновь приходится признать, что наличие мед--
ленных волн в какой то области мозrа не является cTporo
обязательным показателем наличия в ней торможения, хотя, ко--
нечно, и не свидетельствует против такой возможности. Пере--
стройки эскоr, МЭП, 02а в процессе психической деятельности
свидетельствуют об изменениях функциональноrо состояния
структур в связи с данной деятельностью. Воспроизводимые
сдвиrи МЭП и 02а, как показано выше, являются надежными
показателями тесноты связи структуры с деятельностью. Однако
расшифровка сущности этой связи оказывается реальной лишь
на основе анализа данных импульсной активности. Только ана--
лиз импульсной активности может ответить на вопрос, что проис
ходит в структуре при ее включении в деятельность и являются
ли друrие, закономерно развивающиеся физиолоrические сдвиrи
(МЭП, 02а) результатом включения структуры в деятельность
или столь же закономерноrо, необходимоrо для данной деятель--
ности выключения.
Изменения ИА в структурах, по друrим показателям обнару
живающих деятельное состояние во время психо оrической
пробы, были самыми различными, причем различия эти опре
делялись и направленностью изменений (учащение, урежение
ИА), и динамикой их. Реакция ИА моrла различаться в зависи
мости от области (<<точки») отведения, фазы выполнения пробы,
внешних факторов (<<помех» А. И. Трохачев, 1966) и от пра
вильности или ошибочности выполнения пробы.
Так, например, учащение импульсации при выполнении
пробы Бине моrло наблюдаться в различных ядрах зрительноrо
буrр,а, причем в большинстве образований оно обнаруживалось
в фазе удержания пробы в памяти. В фазе ее воспроизведения
(речевой ответ) моrло наблюдаться как дальнейшее учащение
импульсации, так и отсутствие реакции учащения импульсов или
даже их урежение. Четкие реакции импульсной активности об
наруживались в верхних .отделах ствола, причем на уровне по
крышки моста была обнаружена зона, rде именно речевой ответ
давал учащение импульсов, значительно более отчетливое, чем
друrие, в том числе и двиrательные, пробы (рис. 20).
Как уже указывалось выше, динамика реакций различалась
не только в зависимости от анатомически контурируемой CTPYK
туры (ядра), но и от точки отведения в пределах каждой CTPYK
туры, хотя В результате проведения большоrо количества иссле--
дований (А. И. Трохачев, 1966; Н. П. Бехтерева и А. и. Трохачев,
1967; Ю. К. Матвеев, 1971) и удалось обнаружить определенные
типовые варианты этой динамики. Естественно, оrромный инте--
рее представило уrлубление в нейрофизиолоrические механизмы
этоrо явления.
ю. К. Матвеевым (1971) при использовании методическоrо
приема, предложенноrо М. Н. Ливановым (1965), были проанали--
зированы процессы, которые внешне про.являлись как учащение
87
Имп /с
90
80
70
80
50
'10
30
20
10
20. Динамика импульсной активности в области покрышки моста при
пробе на оперативную память (реrистрация проведена с помощью инте
rpaTopa).
ИА. На примере динамики ИА при психолоrических пробах
и при двиrательных пробах показано, что учащение разрядов
происходит (или, осторожнее, может происходить!) при YMeHЬ
шении количества активных нейронов (рис. 21). Понимание этоrо
явления возможно прежде Bcero на основе фактора латеральноrо
торможения (Jung, Тбппiеs, 1950; Creutzfeldt, 1969; Baumgartner,
1961). Допустимо представить себе, что на основе латеральноrо
торможения активация тех элементов нейронной популяции, KO
торые имеют отношение к обеспечению контроля какой то функ
ции, происходит при уrнетении остальных элементов нейронной
популяции. Можно также предположить, что из нейронной по
пуляции в данную деятельность включаются не все адекватные
данной деятельности элементы, но KaKoe TO оптимальное коли
чество их. И, однако, трудно полностью oTBeprHYTb и еще одно
предположение, противоречащее таким заманчивым первым
двум. Если представить себе, что в любой данной полифункцио
нальной нейронной популяции полифункциональными являются
и составляющие ее нейроны, то тоrда весь процесс может pac
сматриваться не как активация определенноrо количества спе
цифических элементов, а просто как активация оптимальноrо
для заданной деятельности количества полифункциональных
нейронов. В первых случаях импульсная активность оказывается
как--то связанной со специфическим характером деятельности.
В последнем случае ей принадлежит более скромная роль типа
обеспечения функциональноrо состояния субстрата. Данные
о значцтельных изменениях количества импульсов при ошибоч
ных реализациях тестов подтверждают положение об оптималь
ном количестве «работаюIЦИХ» нейронов в данной деятельности,
никак не свидетельствуя в пользу специфическоrо или неспеци
фическоrо характера этих работающих нейронов. Исследования
с применением нейротропных препаратов, rде применение oд
Horo препарата направленноrо нейротропноrо действия изменило
88
DОЛЬ данной структуры в си--
TeMe обеспечения психических
функций, как будто свидетель
ствуют в пользу возможной
правильности первоrо предполо--
жения по крайней мере, при
менительно к некоторым под
корковым структурам. Учет
факторов полифункционально
сти мноrих подкорковых ней
ронных популяций требует, oд
нако, большей осторожности
в выводах по этим данным. По
нимание роли наблюдаемоrо яв
ления в целостном механизме
обеспечения какой--либо дея
тельности возможно лишь на
основе дальнеЙlllеrо анализа
мноrих данных количественноrо
исследования активности мозrа.
И все же проведенные на ней
ронном уровне исследования уже
приоткрывают завесу над «таин
ственными, по сравнению со
свойствами одноrо нейрона, свой
ствами нейронных популяций» (Mountcastle, 1966), активность
которых обеспечивает все виды деятельности мозrа.
Важнейшим результатом, который утверждают проведенные
нейрофизиолоrические исследования, является положение о един
стве нейрофизиолоrических механизмов различных видов дея--
тельности, реализуемой мозrом, в том числе и деятельности пси
хической, и об усложнении физиолоrии психической деятельности
уже лишь на уровне самой системы.. Тем самым уже не вер--
бально, а фактически поколеблен известный дуалистический
тезис UUерринrтона и реализуется rениальное предвидение
И. П. Павлова о возможности «систему беспространственных
понятий психолоrии наложить на материальную конструкцию
мозrа» 1.
IIo--видимому, все более детальный, поэтапный ответ на BO
прос о физиолоrических изменениях, развивающихся в элемен
тах системы, обеспечивающей психическую деятельность в про
цессе ее реализации, все более полный по мере совершенствова
ния техники физиолоrических исследований, может явиться
предпосылкой к исследованию BToporo вопроса. Ответ на второй
вопрос о физиолоrических механизмах конкретных психиче
ских явлений сложен еще и сейчас, однако отрицать прин
ципиальную возможность ero решения, как это, по существу,
8
7
6
5
1f
:з
2
1
I
I .
I
I
I
I
I
I
I
I
I
с
ФОН
21. Схема, представляющая дина
мику импульсной активности и ди
намику количества активных ней
ронов нейронной популяции при
пробе на оперативную память.
По оси ординат: J количество импуль
сов; N количество активных нейронов.
1 С '
Павлов И. п. обр. соч., т. 111. M. Л., 1951 1952, с. 203.
89
делается в ряде физиолоrических работ и в ряде философских
исследований, значит базироваться только на данных вчераш
Hero дня в науке о мозrе человека. Отрицание возможности
исследования физиолоrических коррелятов конкретных психи--
ческих явлений человека неизбежно ведет к разрыву между
материальным и идеальным, к отрыву психической деятельности
от процессов, протекающих в мозrу. Можно себе представ:ить, что
так же, как электронный микроскоп позволил уточнить морфо--
лоrическую основу так называемых функциональных изменений
в центральноЙ: нервной системе, проrресс в методичес их воз
можностях физиолоrии проложит пути к решению этоrо важней--
шеrо вопроса физиолоrии человеческоrо мозrа.
За те немноrие rоды, которые прошли со времени публикации
первоrо издания книrи, проводилась интенсивная разработка
именно этоrо вопроса о наиболее тонких коррелятах импульс--
ной активности с «внешними» (акустическими) и «внутренними»
семантическими характеристиками предъявляемых словесных
раздражителей. Вполне понятны поиски TaKoro рода коррелятов
именно в импульсной активности, а не в друrих, более медлен
ных электрических процессах. ;По видимому, именно импульсная
активность нервных клеток может рассматриваться как один из
рабочих механизмов передачи, кодирования и декодирования,
в том числе и словесных сиrналов в мозrу. Однако даже в элек--
трофизиолоrическом аспекте этот механизм не должен рассмат--
риваться как единственный, учитывая orpoMHoe значение био
химических механизмов обработки и хранения информации.
Целенаправленный поиск коррелятов словесных сиrналов в им--
пульсной активности нейронных популяций потребовал разра--
ботки специальноrо аппарата анализа, что и было осуществлено
под руководством П. В. Бундзена им самим совместно с Ю. л. ro--
rолицыным и А. С. Каплуновским (1972 1973). Для этой цели
дополнительно были использованы приемы анализа, применяе
мые в 1 Физиолоrическом институте Эрланrена (Фрr).
Запись мультиклеточной активности осуществлялась на четырехка
нальный маrнитописец мrЧ l при скоростимаrнитноrо носителя 10,5 .м.м/с.
На одном из каналов маrнитописца реrистрировались речевые команды
исследователя и соответствующие ответные реакции больных.
Анализ мультиклеточной активности нейронных популяций прово
дился в два этапа на аналоrо цифровом комплексе (ABM MH 14, ЭЦВМ
«Проминь 2», ЭЦВМ «Минск 32») в Институте экспериментальной меди
цины АМН СССР и ЭЦВМ «Линк 8» в 1 Физиолоrическом ИНСТИТУ1'е
Эрланrен Нюрнберr (ФРr).
Задачей первоrо этапа обработки экспериментальноrо материала
являлся поиск характерных изменений, связанных с выполняемым тестом
в популяциях, отобранных на основе данных суперпозиции (см. выше).
С этой целью использовалась непрерывная амплитудная дискриминация
мультиклеточной активности, в условиях которой область выходных зна
rrений полезноrо сиrнала разбивалась на несколько пропорциональных интер
валов. В каждом дискриминационном интервале (дискриминационное окно)
вычислялась функция истинной текущей частоты по количеству потен
циалов действия, оканчивающихся в дискриминационном окне. Эпоха
нализа составляла 20 .мс. С выходов дискриминаторов сиrналы поступали
90
на мноrоканальный маrнитописец M 168. В дальнейшем для поиска сле
довых процессов использовался метод динамической селективной Koppe
ляции.
Применение динамической корреляции имело целью выделение по
функциям текущей частоты паттернов активности, которые по распреде
лению импульсных разрядов статистически подобны пространственно вре
менному рисунку мультиклеточной активности, развившемуся в фазе
предъявления тестов или рельефам динамических спектров вербальных
тест стимулов. Основу метода динамической селективной Rорреляции co
ставляет вычисление кросскорреляционной функции при непрерывном oд
нонаправленном сдвиrе анализируемых рядов с целью определения зон
корреляционноrо сходства опорноrо (психолоrо специфическоrо) и анализи
pyeMoro рядов. На основании данных динамической корреляции на запись
мультиклеточной активности наносились синхросиrналы, отмечающие зоны
достоверной (99,9 О/о) корреляции, что позволило с большой точностью Bыдe
лять фраrменты активности, содержавшие искомые паттерны. Эти фраr
менты подверrались последующей математической обработке на системе
«Линк 8».
Основу цифровой обработки составляла проrрамма, которая позволила
со временем квантования биоэлектрическоrо процесса в 10 МС обеспечить
анализ и запоминание амплитуд экстремальных значений процесс а и ин
тервалов, характеризующих временное разнесение экстремумов. Эта про
rpaMMa включала две подпроrраммы: 1) обеспечивающую построение
амплитудных rистоrрамм максимумов, минимумов и абсолютных значе
ний потенциалов действия, а также построение интервальных rистоrрамм
по максимумам и минимумам процесс а, и представляющую информацию
о длительности восходящих и нисходящих фаз биопотенциалов; 2) обеспе
чивающую построение связанных амплитудно фазовых rистоrрамм раз
дельно по длительности восходящеrо и НИСХОДЯIЦеrо фронтов в сочетании
с абсолютной амплитудой спайка и по общей длительности фронтов
и абсолютной амплитуде спайка.
Данная проrрамма представляет возможность двупараметрическоrо раз
деления потенциалов действия по амплитуде, длительности и (или) дли
тельности фаз. Получаемые пространственные паттерны фактически OTpa
жали степень неопределенности процесса.
При анализе данных все шире используются возможности мноrофак
TopHoro анализа.
В исследованиях, проведенных П. В. Бундзеном с Ю. л. ro
rолицыным и А. С. Каплуновским (1973) и далее с Э. Э. Давидом,
удалось существенно дополнить полученные ранее данные о ха--
рактерном спектре импульсной активности при психолоrических
тестах и обнаружить прежде Bcero зависимость паттерна им
пульсной активности в ряде нейронных популяций от акустиче--
ских характеристик словесноrо раздражителя (1 тип паттерна).
В методику исследования больноrо были внесены в соответствии с за
дачей некоторые изменения. Больным, которым с диаrностическими и ле
чебными целями были вживлены множественные золотые электроды,
предъявлялись тесты на краткосрочную память в виде ДBYX Tpex (и более)
коротких русских слов, бессмысленных сочетаний букв (фонем), COCTa
ляющих эти слова, иностранных слов, неизвестных больному, и тех же
иностранных слов, значение которых затем заучивалось больным. Таким
образом, в первом случае запоминались слова, раЕ:ее хорошо известные
больному, и процесс запоминания Mor быть связан с обращением в долrо
срочную память, во втором и третьем случае раздражители, не несущие
для больноrо семантической информации, но разные по звучанию, заве
домо не имеющие смысла и имеющие неизвестный еще БОЛЬНQМУ смысл,
и В третьем и четвертом случае слова, одинаковые по звучанию, но
в одном случае незначимые, а в друrом имеющие уже определенное
91
оУ
[:J
/(0 I1A/f
ОФ
АНН
1l8Y
НА;;
ТКС
знп
22. Обобщенная блок схема управляемоrо экспери
мента.
БJl биоусилители (Disa); MAF 1 четырехканальный маrнито
писец; ОФ оrраничитель формирователь активности; АВМ
аналоrовая вычислительная машина MH 7; ЛВJl проrраммно
временное устройство; MAF 2 маrнитофон для воспроизведения
фоноrрамм; ТКС табло контрольных сиrналов; ЭЛЛ caMO
писец; КО контрольный осциллоrраф. Вверху и справа: микро-
фон, автоматический проектор, динамик.
значение для больноrо. В четвертом случае вновь можно было предполо
жить обращение в долrосрочную память. Понятно, что в связи с этим
тесты и в первом, и четвертом случае моrли иметь известную общность
нейрофизиолоrических механизмов. Больной повторял про себя и вслух
тесты по команде врача соответственно через 15 и 30 с после предъявле
ния. Импульсная активность реrистрировалась с 30 40 точек мозrа пер
воначально с помощью интеrратора.
Методом суперпозиции отрезков кривых обнаруживались зоны мозrа,
связанные с обеспечением данной психолоrической (мнестической) пробы.
Воспроизводимые изменения обнаружены в centrum medianum, nucl. ventr.
latera1is thalami, nucl. dorsa1is superfic. thalami и др.
В дальнейшем исследовалась активность только этих зон мозrа, запись
которой осуществлялась на четырехканальном маrнитописце.
В работе использовался метод автоматизированноrо биоуправляемоrо
эксперимента (рис. 22) и ряд тестов предъявлялся, таким образом, син
хронно с определенным функциональным состоянием нейронной попу
ляции.
На предъявляемые словесные раздражители обнаружились
реакции как тоническоrо, так и фазическоrо типов. Фазический
компонент активности обнаруживал функциональную связь
с динамикой амплитудно частотных характеристик акустических
раздражителей (рис. 23).
Эта нейродинамика, представляющая собой пространственно
временные паттерны в виде изменения функций текущей ча
стоты отдельных rрупп н.ейронов, статистически соответствовала
временной структуре максимумов амплитудной характеристики
акустическоrо сиrнала и в ряде популяций (в области п. venfralis
92
1
\D
ф
d)
(1)
)
(JJ
LD
2
r r
1,0 1,0 Ф 1
0,8 0,8
0,4 4
О О
0,4 О,"
0,8 5 0,8
23. XpoHorpaMMa импульснои аК'!'иВНtlСТИ нейронной популяции rоловноrо
мозrа человека в аналоrовом выражении (а) и результаты факторноrо
анализа матриц интеркорреляций функций текущей частоты, выделенных
из мультиклеточной активности методом амплитудной дискриминации и
характеристик вербальноrо сиrнала при десятикратном предъявлении
слова «хор» (6).
На а: ось абсцисс время (эпоха 40 МС), ось ординат активность, зареrистрированная
в пределах четырех дискриминационных окон (1, 2, 3, 4) при десятикратном предъявлении
вербальноrо сиrнала. Вертикальная штриховка 1 импульс за 40 МС; rоризонтальная 2 им
пульса за 40 МС; поперечная 4 импульса за 40 МС; области с точками 5 7 импульсов
за 40 МС; зачерненная область 8 12 импульсов за 40 МС; вертикальные линии начало и
окончание вербальноrо сиrнала. На 6: по оси абсцисс 1, 2, 3, 4 активность дискриминаци.
онных уровней L и <.о соответственно значения оrибающей амплитуды и функции ведущей
частоты вербальноrо сиrнала; по оси ординат коэффициенты корреляции. Ф l и Ф 2
структура первоrо и BToporo факторов. Пунктир область недостоверных факторных значе
ний. Вверху контрольный факторный анализ фоновой активности и характеристик вербаль
Horo сиrнала; внизу факторный анализ активности в момент воздействия вербальноrо
сиrнала.
93
л ("НО /нсек)
ft
1\ J.
, , l'
t 2 J 4
t (сек)
24. Пример опережения паттерна ответа в одной из областей
латеральноrо членика бледноrо шара при анализе МКА ней
ронной популяции на краткосрочную вербальную память.
Представлена текущая частота MI(A (t==20 .ме) в разных областях лате-
ральноrо членика бледноrо шара. На верхней кривой отмечены момент
задания и момент ответа; на нижней момент возникновения опережаю-
щеrо паттерна. По координатным осям n (количество импульсов за 20 .ме)
и t (время в е).
lateralis thalami, centrum medianum thalami и др.) отражала COOT
ношение частот спектральных максимумов вербальноrо раз--
дражителя. По видимому, в данном случае удалось обнаружить
cBoero рода первичное кодирование словесных сиrналов по «aKY
стическому» типу. оно, вероятно, является необходимой сту--
пенью для дальнейшеrо взаимодействия с энrраммами долrосроч--
ной памяти.
В фазе удержания в памяти обнаруживается более или менее
стойкое воспроизведение возникшеrо паттерна, причем в течение
3 6 с (от этапа задания) развивается характерный процесс сжа
тия во времени ero последовательных элементов. Дальнейшая
динамика возникшеrо паттерна существенно связана с друrими
особенностями предъявляемоrо раздражителя.
Паттерн, возникший на хорошо известные слова, в течение
первых секунд затухает, причем к моменту мысленноrо ил
реальноrо воспроизведения заданных слов формируется новыи
пространственно временной паттерн импульсной активности, xa
рактеристики KOToporo приближаются к характеристикам BЫXOД
Horo сиrнала (слова, произносимые больным). Этот пат терн об
наруживается в импульсной активности нейронных популяций
еще до произнесения слова больным (примерно за 38 и более JИс)
и, по--видимому, может рассматриваться как управляющий сиr--
нал (рис. 24).
94
На иностранные слова, т. е. вербальные раздражители, имею--
щие ранее незнакомое акустическое звучание инеизвестное зна--
чение, обнаруживается наибольшая длительность реверберации
паттернов активности, сформированных в фазе задания вплоть
до фазы воспроизведения теста больным (рис. 25). Анализ им
пульсной активности при предъявлении тех же слов после заучи--
вания их значения, осуществленный методами классификацион--
Horo анализа (ю. л. rоrолицын, 1973), показал, что процесс
обучения новым вербальным раздражителям приводит к законо--
мерному уменьшению продолжительности времени обнаружения
возникшеrо паттерна.
При этом формируется динамика паттернов импульсной актив--
ности, подобная той, которая обнаруживалась ПрJlI предъявлении
известных слов (рис. 26).
Следовательно, речевой ответ в процессе выполне:ния теста
на вербальную память может наблюдаться при двух основных
вариантах нейродинамики в области зон мозrа, rде выявляется
первичный код:
1) при рекомбинации паттерна активности, образующеrося
в фазу задания, и развитии HOBoro статистически отличающеtося
HepBHoro кода (случаи краткосрочной оперативной памяти, бази--
рующейся на обращении к долrовременной памяти);
2) на фоне устойчивоrо сохранения характеристик паттерна
активности, возникающеrо в фазе задания иностранных слов,
имеющих незнакомую акустику и несущих неизвестную для
исследуемоrо семантическую наrрузку (случай кратковременной
памяти при отсутствии следов в долrовременной памяти). По
всей вероятности, в последнем случае имеет место мотивационно
обусловленная (подкрепленная) максимальная мобилизация ме--
ханизмов краткосрочной памяти, что выражается, в частности,
в ультрастабильности реверберации следовых процессов, сфор
мировавшихся в фазе задания.
Таким образом, математический анализ динамики Иl\iПУЛЬСНОЙ
активности работающих нейронных ансамблей показал различия
их функциональной орrанизации при оперативной памяти с Ha
личием базиса долrосрочной памяти и оперативной кратковре--
менной вербальной памяти без соответствующеrо базиса.
Кратковременная память характеризуется более устойчивой
реверберацией следовых процессов, которую, очевидно, право--
мерно рассма'-rривать как адаптивное автоподкрепление, необ--
ходимое для обучения, т. е. фОрl\iирования устойчивых струк--
турных (биохимических) изменений в нейронных системах,
с целью создания дополнительноrо базиса долrовременной па
мяти. На коротком временном интервале мнестической пробы
устойчивый следовой процесс обеспеЧJ1вает, по--видимому, такя<е
формирование необходимой ответной реакции.
Прямым подтверждением высказанной точки зрения явля--
ются результаты исследований по обучению больных ранее не--
известным иностранным словам.
95
96
r
1,0
1l,.9
8
1
IJ,G
IJ,S
4
IJ,J
2
!
I I
I .+
I ·
\ OOOOOO I \
I 00 . О ·
\ 00.. 00 \
I 00,..
\ О.. .
I 00 ... :
\ Ь О . ' .
........ ........ ........ ........ .......l1I
о. то .........
· \ 00 I ... I % :
О , .. О .
О ·
.
1
2
3
t
н fc
r
1,0
.9
1J,8
/J,7
O
tJ,S
fJ,4
З
11,2
9,1
I
.. .
I ...
.. 00 I ·
· о .
.
.
.
.
1
2
3
t
25. Усредненные результаты математической обработки
по оптимальной динамической селекции характеристиче
ских паттернов мультиклеточной активности в условиях
тестов на кратковременную вербальную память.
По оси абсцисс время (эпоха 1,0 с). По оси ординат значения ко-
эффициентов корреляции. Вертикальные пунктирные линии разrраничи-
вают этапы теста: 1 фаза задания; 1 2 этап удержания вербаль-
ной последовательности в памяти; 2 фаза мысленноrо воспроизведе-
ния теста; 2 3 удержание в памяти; 3 речевое воспроизведение
теста. rоризонтальные пунктирные линии оrраничивают зону достовер-
ности с вероятностью 0,95. Вверху динамика паттернов активности
при запоминании набора русских слов. Сплошная линия динамика
паттернов, подобных по временной структуре нейронной активности,
формирующейся в фазе задания. Точечная (зачерненная) линия
динамика паттернов, подобных по временной структуре нейронной ак-
тивности, имеющей место в фазе речевоrо воспроизведения вербальных
сиrналов. Пунктирная и точечная (светлая) линии результаты се-
лекции указанных типов паттернов при редукции их во времени.
Внизу динамика паттернов активности при запоминании набора не-
известных иностранных слов.
.,...
а
3
ппс
ПНС
1
9
7
8
5
4
6
10
1 СУ
11 /;)
Z б
До
обучения
1
После
оОучения
До
обучения
11
После
обучения
До
05учения
///
После
обучения
3 ппс пас
26. Результаты классификационноrо анализа паттернов динамики
мультиклеточной активности, полученных при обучении новым Bep
бальным сиrналам (иностранным).
а схема устойчивости паттерна. I(ривая [ показывает ультрастабильную структуру
паттерна активности, возникшеrо в момент задания, при реализации теста на заПО
минание неизвестных иностранных слов. После Toro, как смысл слов был выучен
испытуемым, в момент задания возникал тот же паттерн (l /)1 но В последующих фа
зах теста он с достоверностью изменялся ([[/). По оси абсцисс фазы теста: 3 за
дание, ППС повторение про себя, ПЕС повторение вслух. По оси ординат ДOCTO
верность сходства. 6 вид выводимых из ЭВМ результатов классификации. Слева
номера анализируемых фраrментов, по оси абсцисс мера сходства паттернов.
Справа сводная таблица результатов классификации. [1 [[1 [[[ пара слов, предъ
являемая по 2 раза до и после обучения. По rоризонтали фаза теста. Рамками
обведены фраrменты, имеющие достоверно сходную (99,9%) динамику мультиклеточ
ной активности. Фазы ППС после обучения в отдельные классы не выделены, но
сходны (достоверность 95%) с соответствующими фазами ПВС после обучения.
Наряду с таким типом перестройки, метод динамической кор--
реляции позволил выделить еще один тип паттернов, характери
стики которых ,не обнаруживали связи с амплитудно частотным
рельефом словесноrо сиrнала (Н. П. Бехтерева, П. В. Бундзен,
В. д. Кайдел, Э. Э. Давид, 1973). При предъявлении словесных
раздражителей в нейронных популяциях ассоциативных структур
(п. dorsomedialis, п. lateralis posterior thalami и др.) появлялась
и в дальнейшем более или менее устойчиво сохранялась про--
странственно временная перестройка импульсной активности OT
носительно более сложноrо рисунка по сравнению с паттернами
первоrо типа. В тестах использовались слова с высоким и низким
индексами' частоты и квази слова. Динамика этих паттернов,
условно названных П. В. Бундзеном «автономными», обнаружи--
вала зависимость от вероятности употребления вербальных сиr--
налов в речевом поведении человека.
97
При применении в тестах на краткосрочную память слов,
имеющих высокий частотный индекс, устойчивые автономные
паттерны наблюдаются только в первые 2 3 с ретенции теста
в памяти и в конце ретенции. При запом:инании слов, xapaK
теризующихся низким индексом частоты, автономные паттерны
обнаруживались значительно дольше, на протяжении 18 30 с
(при общей длительности ретенции 30 с). В случаях предъявле--
ния больному квази--слов отмечается ультраустойчивая перио--
дическая реверберация автономных паттернов, которая к концу
фазы ретенции обнаруживается в работе всех нейронных попу
ляций (рис. 27). Эту динамику можно рассматривать, как дистант
ный результат формирования энrрамм долrосрочной памяти при
воздействии на них «акустических» паттернов.
Динамический rистоrраммный анализ автономных паттернов
позволяет думать о том, что поздние высокочастотные компо--
ненты паттернов имеют пачечную структуру. В период удержа--
ния теста в памяти структура автономных паттернов изменялась
в соответствии с индексом частоты слов, по видимому, В зависи
мости от возможности семантическоrо деКОДYIрования.
Структура паттернов мультиклеточной активности при коди--
ровании слов с высоким индексом частоты уже в фазе задания
отличается тонкой дифференциацией, которая через 2 3 с после
фазы задания значительно реорrанизуется. Использование при
веденных выше приемов анализа позволило предположить, что
паттерны этоrо типа характеризуются не только пачечной струк--
турой, но И rенерацией потенциалов действия определенной
формы. Подобные по структуре паттерны активности возникают
вторично только после инструкции на воспроизведение теста.
При предъявлении слов с низким индексом частоты разви--
вается значительное изменение импульсной активности при OT
сутствии выраженных элементов структурирования импульсных
потоков. В начальных фазах ретенции тестов наблюдается резкое
уменьшение количества низко и высокоамплитудных потенциа--
лов действия при возрастании активности rрупп нейронов cq
среднеамплитудными значениями потенциалов действия. Пере
стройка ИМПУ.:JIЬСНОЙ активности оказывалась более устойчивой.
По мере удержания вербальных сиrналов в памяти происходило
постепенное незначительное усиление пространственной синхро
низации элементов популяции и стабилизация по форме потен
циалов действия. Дифференциация популяций достиrала макси--
мума на 6 12 й секундах ретенции слов в памяти.
Характерным для перестройки импульсной активности при
предъявлении квази слов является значительное возрастание
высокоамплитудных разрядов и нарастающая синхронизация
элементов популяции. Дифференциация популяций может на--
блюдаться только в конце фазы ретенции, а структурирования
импульсных потоков, которое наблюдается при предъявлении
семантически значимой информации в условиях запоминания
ква2И СЛОВ, не ОТl\1:ечается (рис. 28).
98
\
:\
: \
: \
I I
J....
t\
: \
: \
: . \
I }\
о I
t\
I
I
I
I
I
I
I
I
I
(Q
I
:r
:s:
ct$
04
:s:
Ef
(1)
:s:
==,
=
::a
D:::x:: 1
= :х::
Ф
:x:: .
(';j . In
(l')Q)=0
f-t:x::t;:
(';jQ)U
....cY) =
Q)('t)
(';j . о. (';j
-8< In
:х::= ,
f-tОU
52 O
(';jQ)(';j
:X:::x::=
g;t::=(';j
:x::t::=U
(';j ::f::t:
0.0.::a:::<J)
ф
о о:х::
:<0.=
=t::
(';j:x:: о
:< :<
5. =g
O=b
;;;jU f-t
=U
o==(';j
f-t...,.
Ou....
:x::°D:: 0
:s: :x::
,... ==0.0
t;: f-t О. ::а:::
1--4 ::t:o('t)
(';j::t:;
I I
с) :х:: ь: r 1'\
О =f-tQ)'"""I
::r: =.
CQ...; = ::f (';j
.I::i=и
........ 0.-9< ::а:::
Е-с Е-с -::.. о -9<
о m :х::
ct$ t ==0=
,w = ::а:::
):S: \J ::f U ::а о
o D::u:x::
::r: С) t:: = = о
'"" и ф::! tI" :х::
tJ 1""04 o.u=f-t
О (1) О. \О t;: О
Е-с 1::( о (';j Q) f-t
(1) ::r:::t: In U
=
:::::: :s: In ::а
о ::t: О
) Ь: Q) Q)
::a Q)....:з'О
= (';j 5
::f,.Q U
=t::0....
11:::1 Q)\O,.Q
F==i "'о" t:: О In
-8<t::
(';jD::
ct$ ::а::: о. = t;:
gjи
О (';j :Х::ф
Е-с :x::(';j ::a:::
О =><:Q)1n
1--4 tI" = f-t О
1--4 = О. (';j О.
t;:f-t('t)=
a('t)
, e ::t:
f-tО==
(';j ..".o.
:x::f-t....t::
=O(';j
0.::t:a:1
о.О::Ео
О U
o.U
,.Q:S::::t:
U:S:: ....
o :s::8.
1=; t::
. :s::
........ ><:
(';j::a
f-t::t:1n
D::U:S::O
ф:х::
Q)f-t
o.t>;:0u
In:s::
I ..,... (';j
@ ::t:
U f-t:S::
UQ)
:s:: I!:I CtI
::f('t):S:::x::
U:S::
\OO(';j
(';j o.::t:
C
,.QU
u°c.,
ol!:l;:::::
CQ
о
с)
:s:
:s:
::r:
ct$
::r:
:s:
о
ct$
CQ
CQ
О
с)
с)
(1)
::f
о
04
10-4
CQ
О
1::(
(1)
с)
ct$
:s:
ct$
::r:
:s:
.
.
. .
1000Р F80 0005 1000Р FTO 0005 1000Р {МЗ0 0005 1000Р (Н50 0005 1000Р {ио 0005
1 А1
, JATH А , =2000ту T=5тs
..
.
.....
.#
. #. .
,. :. ...
. ,:. . -.."":... .
. . fj. ,. ..
#". .
,.
,.
. .' . .
. . . . "'...:.. ,
. . .- .
. е. .
..
..
. .
. """,..;... "-.t. . :
.:-1. ', ) ."
.....", ..
о.Т _.. ..
". " .
.
. .
. ..
I' . .
'.
.' . .
. ,
.
,
а
, .,,' .
ь, -:.. .--.
, '. .... . . .
'. .
. .
. .:;.,..... .
1:': , . ,. . о.
? '. . ..:.
,.
A 1 =2000тv
T=5тs
1000Р С8РО 0005 1000Р СТО' О 005 1000Р СМ80 0005 1000Р СМ 50 0005 1000Р сио 0005
.
. .
.. .
'. .
.
i · ·
. :8.. .
.. .
" "
. е..",.". .
р'. .
.
:', .
,1.. '.
.,.:,. .' . .
r .......
.'
.. .,
..
; .:: .
.
.
L ..
., .
.. " .
, .
..
б
.
е,
.
. ,
, "
; ' ': . .' . .'
.,..... .
. ,. ... ,
" .
.
. .
,'. I
. .
" .. .' . ..
. '. .
. .'. , .. t
. ., .
, .
. "
. .
j' , .
l ' · ......
, ,
. ',,' ,
, . .
'.
,
'.
1000Р КВО 0005 1000Р КТО 0005 1000Р /(МЗ0 0005 1000Р КМ50 0005 1000Р кио 0005
.
. .. I
.-
. . .
. .. .
. ..:, .J'.
4.- · , .. ·
,. .. .
100
.'
. '
, .
.
. . .
, ..... . .
. .,
. .
.... .
. .
. .
. .
.
. .
) : :: .
, ':
, . I
, . ", .
, .'., .'
,...", . '. ..
r .. · · '.
"
.
,
. .
. ,. .
...
.' ,
# . , . .
...\:. -.' . ,
.,. r". .." , .
;-,.
Применение мноrофакторноrо анализа позволило выделить
новую высокоспецифичную форму кодирования словесных сиr
налов, обнаруживающуюся и одновременно с акустическим
кодированием, и в отсутствие акустическоrо кода. Этот код про--
является перестройками взаимодействия близлежащих и, по ви
димому, отдаленных rрупп нейронов динамической простран
ственно временной реорrанизацией нейронных ансамблей. Весьма
вероятным кажется, что определенные типы пространственной
реорrанизации нейронных ансамблей MorYT отражать смысловое
кодирование в мозrу. Высокая специфичность этоrо кода позво
лила опознавать в импульсной активности нейронных ансамблей
появление коррелятов слова еще до произнесения ero больным
и появление коррелятов отдельных фонем. Основываясь на этом
типе кода, удается исследовать уже и течение простейших соб
ственно мыслительных процессов в мозrу.
Таким образом, кодирование словесных сиrналов может ха--
рактеризоваться пере стройкой текущей частоты импульсной
активности нейронных ансамблей, появлением пачечной CTPYK
туры в импульсации, импульсов определенной формы и про
странственным паттерном взаимоотношений нейронных ансам--
блей.
ДИНАМИКА ВЗАИМОДЕЙСТВИЯ ЗВЕНЬЕВ мозrовой
СИСТЕМЫ ОБЕСПЕЧЕНИЯ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ
Исследование механизмов, или, точнее, пока получение объек--
тивных признаков объединения исследуемых структурно функ
циональных элементов в систему, является лоrическим элемен
том, без KOToporo не может быть действительной полноты сужде
ния об изучаемом вопросе даже и на настоящем уровне знаний.
Результаты электрическоrо воздействия и данные наблюдения
физиолоrических показателей состояния мозrа при психической
деятельности показали, что в обеспечении этой деятельности
участвует очень MHoro звеньев, с очень мноrими полифункцио--
нальными клеточными популяциями связаны и психические
функции. Лоrично и естественно предс;тавить себе, что все это не
разрозненные элементы, а звенья одной системы. Это слово очень
часто употребляется в биолоrии по существу бездоказательно;
как указывалось выше, основой для суждения о наличии системы
нередко является лишь результат. Наблюдение конечноrо инте
28. Структуры паттернов мультиклеточной активности, выделяемых при
кодировании слов, имеющих различный индекс частоты.
Верхние ряды на а, 6 и 8 rистоrраммы JATH. Нижние ряды двумерные распределения
центров тяжести JATH, вычисленные с помощью проrраммы JSCHW (см. текст). Ось абс
цисс rистоrрамм амплитуда спайков (A t ), ординат длительность спайков (Т). Под каж.
дой rистоrраммой представлено общее количество спайков, образующих rистоrрамму (Р).
индекс кодируемых вербальных сиrналов и размерность оси ординат. rруппа F rистоrрамм
соответствует кодировке слов, имеющих высокий индекс частоты; rруппа С кодировке
слов низкоrо индекса частоты, и rруппа К соответствует кодировке квази слов. Во всех
рядах динамических rистоrрамм: ВО фраrмент фоновой активности; ТО фаза предъяв
ления вербальных сиrналов; М 30 и М 50 фазы удержания вербальных сиrналов в па-
мяти; ио фаза воспроизведения вербальных сиrналов.
101
.....
f"J
б 1
ot::...
6C't:1
00.
t::: f-o
::СО
Ф
. &:0\0
ЕЗI=
\с:) 1:>::...... 3
::t>J..... о
('\3t:>
= .
00
g р.::С
>.1::i::C
Ji ::cl:>::
I:Q f-o
::S:Q)U
f-oОQ)
Q') '" I.t"') ('у) "" '"' ('\3::cg
t:::) (;::) t::::;) (:;::) t;::) 0.,.Q>.
ct$ U
"i..:) .......:) 00.
'i..;) f-oU
"'..;) ;::) ('\3 т I
;:::) I
I I ::С::!
:r:\01 .
CQ о н ('\3
:s: 0.;:: g f-o
Е-с = ,.Q
ct$ t:: f-o
04 1:>:: =::S: о
==I:Q
:c::f('\3Q)
Q) 1:>:: 0.1:;:
О ::C =U
t::Q) о
ct$ 0g:('\3t::
8 0 ::СI
:r: ,.Q::a:::Q)
I:QU::CC't:l
U
:<00
ct$ ('\30.==
\о C'I)::a::::af-o
О ('\3 I:Q 1:>::
04 -9< I:Q
0('\3=
:<f-o\О
....::c01:Q
:s: o.
04 C'I)::f=Q)
('\3=1=
0.-9< ::с
-9<"" ('\3
:s: ('\3m......;E
:s: ::co. o.
::а:::('\3Ф
:r: o.
Q) =('\3>,
s 0 31
=
о U::t>JOC\:a
:r: о) ('\3
Е-с ::z::0::c
О == O
I:t Q) -9<
f-o t::1
1--4 О:а\О
с.о \О (-о ......
с':::) :r: ('\3::с::а:::
('\3 = ::s::
:r: О=::СО
О
:s: Q)1:Q::r\O
::f f-o =
о: ('\3ф а
:a:aO
Q) I:Q::C
04 >.O,.QO
04 a
. о o.(-оо.,.Q
Iфl:;:
(-00
I:Q ('\3\0
ct$ 'О ф
. Q) 1::C
:s: :ан
::с =..... U
::c=Q..
ct$ Q)::fo.o
,:r: ::rl:>::
>. .
"'t....:) :s: t::Q)0
..з 00.0.
=0.1:t
I ::::s t:j I:>::
...... Q)
c\t ,.Q(-оО
"'t...;) ::t>J::c::c
::t>JQ)U
('\3=
o.::fo.
t:: =::a:::
t::: 0-9<1
-9<
, o.mlo...
0.0;::
О::а:::
. 5 I Qj"
U(-00
0('\3::С
t5 0.::c,.Q
а.. ::a:::=
rl::i
о.ф
Of-o
'
'-t 1/
//
t:::S
...........
t:::
..:t-
&
ё
;
1/
&
1;
'l
"*
......... I'
'-..
t:::: с::::
(\.)
cи
:;Т-::S:I!I
::S:S'1::i
-&0 U
('\3 a>
0.1=;::S:
ro
' b
Uf--o
gc
. f--o
('\3.........0
О. ..... I
('\3
a 5
001=;
=a>
0=0.
=Q)E-<
I::iI=;U
Q)т:м
:=::01:>::
:s::1::::s::
=:s::=
Q) ::S:
I=;
.........1:>::
:::::: ('\3
:Доf--o
= о. U
..aE-<:S::
I=; ::!
('\3..с:..а
о. \о о.
Q) Q)
Е-<Оо.
('\31:::1:::
1=;
:s:: .
I
('\3
о.а)
. >. g
OE-<Q
0.:::S::('\3
I::i >. I!I
1:>:: :Д
Q,) Q) u I::i
о м
:I::S:: ('\3 .
Q,) U I::::S::
('\3.... ('\3 :I:
(1) о...а (1') Q)
с::::
о I E.
('t:) C'I':) о .... I!I
О :::s::..a0
O I!I
""-'4о.l:::f--oC\j
I::i :::S::
",.: 1:>:: >.
'W<l I::i .....
Q) E-<
О;>;;с.>
:I:Q)
..a
1=;
('\3:s::Q)
o.:I:g
Q)Q)Q
('\3
1=; ?E
'!:QQ)
о о.
oa>
:I:1!It::
Q) О
I!I 1:>::
I f--o
C\1t)0
. :д
01::i:I:
о.оа)
I::i
D:::s::
Q)('\30.
О м 1:::
:I:!:Q('\3
..а ::::
::S:
('\3:::S:::s::
0.:S::
E-< t::
:I:('\3Q)
Q):I:0.
::f':S::E-<
I I::i U
i
":I:
('\3Q)('\3
o.
MI!I
0('\30
E-<t::
UQ)
('\31::ic.
Q)o.
Q)0.0
><:t:::::S::
u U
I
o.
CX)U
103
rральноrо эффекта используется как основание для постулирова
ния наличия центральной интеrрации и в том числе как свиде
тельство деятельности системы.
Современные возможности анализа физиолоrических данных
позволяют исследовать этот вопрос. Одним из адекватных при
емов ero изучения является корреляционный анализ, уже более
десяти лет завоевавший прочное положение в изучении взаимо
связи различных образований мозrа по их биоэлектрическим
характеристикам (Brazier, Barlow, 1956; О. М. rриндель, 1965;
В. В. Беляев, 1968).
Способом корреЛЯЦионноrо анализа эскоr Brazier (1967) изу--
чена динамика взаимодействия подкорковых структур человека
в бодрствующем состоянии и при засыпании.
Допустимо представить себе, что, если верно высказанное
выше предположение об отражении в ээr (эскоr) состояния,
оптимальноrо для какой то данной деятельности, складывающе--
rося из состояний мноrих отдельных структур мозrа, то именно
в эскоr (ээr) в первую очередь можно и нужно искать
признаки объединения отдельных структур в системы, обеспечи
вающие различные виды деятельности.
Блестящие иллюстрации этоrо положения имеются в работах
М. Н. Ливанова (1972) и ero сотр. (М. Н. Ливанов, Н. А. rаврилова,
А. С. Асланов, 1966), проведенные, однако, у человека лишь при
анализе ээr, реrистрируемой от покровов черепа.
Понятно отсюда, что способом корреляционноrо анализа иссле
довалась при изучении вопроса о системе, обеспечивающей дея
тельность в условиях прямоrо контакта с мозrом, прежде Bcero
эскоr.
Исследование фоновых электросубкортикоrрамм способом
корреляционноrо анализа (В. В. Беляев, 1968, 1970) показало Ha
личие характерных закономерностей СООТНОlllений биопотенциа--
лов в различных подкорковых структурах мозrа в условиях спо
койноrо бодрствования.
Применение фармаколоrических препаратов нейротропноrо
ряда наряду с уменьшением выраженности паркинсонических
проявлений моrло изменять эти отношения вплоть до разви
тия обратных фоновым (В. В. Беляев, А. д. Аничков; цит.
В. В. Беляев, 1968). На основе суrубо предварительных данных,
полученных в этом направлении при изучении психической дея
тельности, можно уже rоворить, что при ее реализации в под
корковых структурах удается обнаружить отчетливые изменения
взаимосвязи между различными образованиями. Изменения тес...
ноты связи на разных этапах выполнения пробы на оператив
ную память в наиболее четкой форме обнаруживались по пара--
метру времени различия состояний высокой корреляции между
структурами, условно........... времени «запаздывания» или «опереже
ния» биоэлектрических явлений в одной структуре по othollle--
нию к друrой (В. В. Беляев, 1968). При этом различия в зависи
мости от фазы пробы, ее характера и качества выполнения пробы
104
моrли обнаруживаться как по быстрой, так и по медленной co
ставляющей кросскоррелоrрамм (рис. 29). Кросскорреляционный
анализ эскоr, таким образом, обнаружил, что в процессе психи--
ческой деятельности взаимодействие подкорковых структур MO
жет перестраиваться вплоть до существенноrо отличия их от фо .
новых. Обращает внимание значительность синхронизации во
времени биоэлектрических процессов в различных rлубоких
структурах мозrа после ответа, как правильноrо, так и ошибоч--
Horo, что может быть связано, вероятно, с включением эмоцио
нальноrо компонента ( «самооценка» ) И сопутствующих ему
нейро rуморальных распространенных сдвиrов в мозrу.
Что же касается caMoro коэффициента корреляции биопотен
циалов, ero отклонения между исследованными структурами, как
видно (см. рис. 16, в), были в целом невелики и прослеживались
в трех основных вариантах сохранении на высоком или низ
ком уровнях по всем этапам пробы или падении в фазу удержа
ния в памяти с подъемом вновь после ответа. Отношения этоrо
последнеrо рода, как вполне понятно, характеризующие умень--
шение тесноты связи по данному параметру, отмечались между
красным ядром и центральным ядром зрительноrо буrра.
Значительно более детально процесс перестройки взаимоотно
шений между элементами нейронной популяции, реrистрируемой
от одноrо электрода, и между различными нейронными популя
циями, реrистрируемыми от тех электродов, в области располо
жения которых обнаруживались воспроизводимые изменения фи
зиолоrических параметров мозrа при психолоrических пробах,
выявился при анализе импульсной активности (Н. П. Бехтерева,
п. В. Бундзен, А. С. Каплуновский, Ю. К. Матвеев, 1972; Н. П. Бех
терева, П. В. Бундзен, 1974).
Исследования обнаружили, что в состоянии спокойноrо бодр
ствования (фон) активность нейронных популяций характери
зуется вариациями частоты импульсов, их rруппировки и xapaK
тером функциональной орrанизации. Способом факторноrо aHa
лиза (А. С. Каплуновский, 1974) было показано, что при
максимальной частоте импульсных разрядов орrанизация попу--
ляции отличается выраженной синерrичностью работы большин
ства rрупп нейронов. В периоды минимальной частоты разрядов
нейронных популяций, наоборот, обнаруживается значительная
дифференцированность популяции. Таким образом, в нейронных
популяциях «спонтанно» формируются ансамбли, в которых уча
ствует различное количество морфофункциональных rрупп ней--
ронов (рис. 30).
Предъявление психолоrических тестов вызывает пере стройку
«спонтанных» флюктуаций. В этих условиях формируются рабо--
чие нейронные ансамбли, в данном случае функционально объ
единенные в соответствии с реализуемой задачей.
При этом, так же как и в периоды максимальной дифферен
цированности популяции, в фоновой активности в рабочем состоя
IIИИ чаще Bcero обнаруживаются реципрокные взаимоотношения
10:;
r
110 . .
0,8
0,6
0,"
0,2
o.2
7
o."
1
o.6
,
r
1,0
0,8
0,6
0,"
0,2
\
\
\
ф 2 \.... ....
\
\С
................
O'2
,
ll"
I
076
O'8
12
7
r
1,0
0,8
0,6
0,4
0,2
ф 2
30. Характер изменений структурно функциональной орrанизации ней
ронных популяций В течение сверхмедленноrо цикла фоновой актив
ности.
В центре условно обозначены фраrменты интеrрированной мультиклеточной активно-
сти нейронных популяций. Зоны А, В и С, оrраниченные пунктирами, участки актив-
ности, подверrавшиеся последующей обработке и соответствующие зонам А, В, С ре-
зу льтаты факторноrо анализа матриц интеркорреляций между функциями текущей
частоты rрупп нейронов. Оси абсцисс 1, 2, 3, 4 активность rрупп нейронов, выде-
ленная дискриминационным анализом. Оси ОРДИI:ат коэффициенты корреляции. Ф-l
и Ф-2 структура первоrо и BToporo факторов. Штриховкой обозначена область не-
достоверных значений. Слева представлен фраrмент ориrинальной записи мультикле-
точной активности.
106
нейронов близлежащих rрупп, однако конкретная орrанизация
нейронной популяции может быть друrой. Физиолоrической
основой этоrо явления следует считать, по видимому, процесс
латеральноrо торможения (Ю. К. Матвеев, 1971).
Одним из важных условий, предопределяющих правильность
выполнения тестов на краткосрочную память, оказался уровень
текущей частоты и характер структурно функциональной opra
низации популяций в момент предъявления теста. Предъявление
словесных раздражителей в периоды экстремальных значений
функции текущей частоты и, таким образом, максимальной си
нерrичности или, наоборот, дифференцированности состояния ней
ронных популяций сопровождалось снижением качества выпол
нения тестов в виде появления перестановки словесных сиrна
лов и ошибочноrо их воспроизведения.
Отсюда, по видимому, можно предположить, что оптималь
ными исходными условиями для переработки информации на
уровне нейронных популяций rоловноrо мозrа, по всей вероят
ности, являются временные интервалы (фазы), коrда орrаниза--
ция нейронных популяций характеризуется определенной CTe
пенью индетерминизма.
Изучение динамики мультиклеточной активности ранее иден
тифицированных нейронных популяций показывает, что, BO
первых, в период удержания вербальноrо теста в памяти, как
и в фазе задания, наблюдается сокращение общеrо числа меж
популяционных связей с преимущественным сохранением (же
сткие компоненты) структуры связей, сформировавшихся в фазе
задания. BO BTOpЫX, наряду с проrрессивным снижением частоты
импульсных разрядов, возрастает мощность отрицательных KOp
реляционных связей, что свидетельствует об усилении реципрок--
ности работы нейронных ансамблей, прИНИМающих участие
в обеспечении психической деятельности. При" этом в нейродина--
мической системе стабильно сохраняются профили функцио
нальных отношений между нейронными ансамблями, в которых
с наибольшей вероятностью обнаруживаются следовые про--
цессы (рис. 31).
Анализ процессов самоорrанизации нейронных систеrVI в фазе
сохранения следовых реакций позволяет прийти к заключению,
что в данных условиях находит свое выражение не при:нцип TO
тальной минимизации взаИМЬдействия звеньев системы, а скорее
принцип оптимальной минимизаци:и взаимодействия функцио--
нально значимоrо нейродинамическоrо комплекса с нейронными
элементами, не определяющими непосредственно результат TeKY
щей деятельности (П. В. Бундзен, см. Н. П. Бехтерева,
П. В. Бундзен, 1974).
Вероятно, в этих условиях происходит минимизация возмож--
ностей взаимодействия работающей функциональной системы
с «посторонними факторами», со «средой». Реализация данноrо
принципа самоорrанизации способна обеспечить формирование
зкономичноrо режима работы системы наряду с активным повы
107
ф ф ФФ
01..;0 -= -- ::-а=:"
ш ф ФЛ .... .... ... ...
А Ш ф- .... -:.. -...:'" ...-- ....
А 6
1
1 @--:"-ф
I <D
/ I
/ I
I
I
I
/ ф ф ф
I
Л ш Ф Q)Л
п 8 Ш В r
а 5
в
r I I
в I I I
8 I
I I ,
ф ппс п
z
I
1
А
1
1
ф- -ф
(i)
л
@
А
ш 2 .....ф П
Б
Б
31. Результаты кросскорреляционноrо анализа функций текущей ча..
стоты нейронных rрупп (1, 2, 3), выделяемых в пределах нейронных
популяций (1, 11, 111) в условиях правильной (а) инеправильной (6, в)
реализации теста на краткосрочную вербальную память.
Сплошная толстая линия вероятность положительной связи 0,999; тонкая 0,99; штри.
ховая тонкая линия вероятность отрицательной связи 0,95; толстая 0,99; точечная
линия вероятность отрицательной связи 0,999.
Фазы: А фоновой активности, Б задания теста на вербальную память, В MЫC
ленноrо воспроизведения теста, r воспроизведения теста вслух.
На 2: усредненная динамика общих мощностей матриц интеркорреляций функций те.
кущей частоты активности rрупп нейронов, выделяемых из популяций дискриминаци.
онным анализом на различных этапах тестов на кратковременную вербальную память.
Ось абсцисс этапы тестов; Ф фон; 3 фаза задания; ППС фаза мысленноrо вос.
произведения теста; ПВС фаза воспроизведения теста вслух; П последействие;
А правильная, Б и В ошибочная реализация тестов. Ось ординат относительная
мощность корреляционных связей.
108
шением устойчивости информационно значимоrо структурно--
функциональноrо комплекса в условиях «шумовоrо» окружения.
Формирование функционально детерминированных нейрон
ных ансамблей является, по видимому, одним из важнейших
условий «настройки» полифункциональной нейронной популяции
на участие в обеспечении определенноrо вида деятельности.
Именно на этой основе уже в пределах ансамблей развиваются
изменения, соотносимые не только с фазовой структурой психо--
лоrическоrо теста, но и с характеристиками предъявляемых раз--
дражителей.
Направленное изучение нейрофизиолоrических механизмов
психической деятельности человека, проведенное способом ана--
лиза физиолоrических показателей мозrа в условиях их непо--
средственной реrистрации, позволило получить данные о HeKOTO
рых принципах работы мозrа. Исследовано около 2000 точек
мозrа. Избирательно изучались физиолоrические явления специ--
ально при очень малом количестве сравнительно простых видов
психической деятельности (чтобы уменьшить количество неизве--
стных при решении сложных «уравнений» И т. д. И т. п.), но это
было необходимо на данной фазе ИСQледований.
Результаты исследований, в которых впервые использовался
комплексный метод изучения физиолоrических коррелятов пси
хическоrо, уже позволяют судить, по крайней мере, о некоторых
физиолоrических механизмах........... основах психической деятель--
ности человека.
Эти исследования показали возможность изучать мозrовую
канву психическоrо, исследовать, как, за счет каких реакций
обеспечивается адекватность психической деятельности меняю--
щимся условиям среды, ее rибкость в этих условиях и в то же
время максимальная экономичность в работе мозrа. Начат новый
этап изучение, кода психических явлений.
Вполне понятно, что данные, полученные на больных, не
MorYT быть абсолютно однозначными для здоровоrо мозrа.
Однако установленные принципы, по видимому, имеют общую
ценность, и можно надеяться, что накопление дальнейших pe
зультатов не только подтвердит эту общность, но и позволит
постепенно ответить на мноrие, до сих пор еще не ясные во--
просы о механизмах самой сложной функции мозrа человека...........
психической деятельности.
НЕКОТОРЫЕ ПРИНЦИПИАЛЬНЫЕ ВОПРОСЫ
И ПЕРСПЕКТИВЫ РАЗВИТИЯ ПРОБЛЕМЫ
Проrресс исследований в самых различных областях науки
связан с запросами практики, методическими возможностями
и движущими идеями. Плодотворная, перспективная идея может
далеко осветить трудную дороrу в неведомое. Возникшее в мозrу
исследователя, несмотря на недостаточную информацию, верное
представление о предмете может быть «отложено» и забыто пер--
воначально в связи с отсутствием инструмента получения допол
нительной информации, проверки и разработки идеи. Оно мо--
жет быть иноrда через мноrие rоды вновь «открыто», может
«ожить» тоrда, коrда новое методическое решение позволит
поставить необходимый эксперимент, который явится оконча--
тельным судьей в оценке идеи.
В физиолоrии высшей нервной деятельности ярким примером
сочетания плодотворных идей и метода явился павловский этап
ее экспериментальноrо изучения.
ПОПЫТКИ изучения физиолоrии психических явлений чело
века с использованием модельных условнорефлекторных peaK
ций подтвердили, что условнорефлекторный принцип распро
страняется и на деятельность мозrа человека. Эти модельные
ситуации, однако, сравнительно мало дали для понимания нейро--
физиолоrических механизмов сложных психических явлений
человека. При сопоставлении данных, полученных при примене
нии условнорефлекторных и простейших психолоrических проб,
уже на Основе анализа только одноrо прямоrо показателя разви
тия изменений в мозrу электроэнцефалоrраl\IМЫ видно, что
ээr в этих двух принципиально сходных ситуациях может быть
отчетливо различной. Она различается и по распространенности
развивающихся биоэлектрических перестроек, и по преимуще--
ственному вовлечению уровня неспецифических и специфиче--
ских мозrовых структур, а также по ряду друrих факторов.
Отсюда следовал естественный лоrичеСI{ИЙ вывод о необходи
мости для изучения нейрофизиолоrических механизмов психиче
ских явлений использования в сочетании с самыми различными
нейрофизиолоrическими методиками не только стереотипных
условнорефлекторных проб, 1-10 и психолоrических приемов. При
изучении механизмов психических явлений человека aдeKBaT
ным оказывалось, таким образом, сочетание классических при
емов павловской физиолоrии и возможностей «старой», прошед
шей долrий путь развития, психолоrии с новейшими, наиболее
современными возможностями нейрофизиолоrии.
110
Среди orpoMHoro количества разноrо рода экстремистских
представлений психолоrов и клиницистов о физиолоrических
механизмах наблюдавшихся ими нормальных и патолоrических
психических явлений человека, несомненно, встречались «золо--
тые зерна». Так и сейчас трудно возразить что либо по существу
в отношении некоторых ранних взrлядов Jackson (см. выше).
Основные представления о принципах мозrовой орrанизации пси--
хической деятельности, развивавшиеся И. Н. Филимоновым
(1940), поддержанные и обоrащенные А. Р. Лурия (1962) и др. и во
MHoroM перекликающиеся с этими ранними взrлядами, проходят
проверку самыми тонкими из возможных сейчас исследований
мозrа человека. Успехи физиолоrии мозrа человека подтверж
дают сложность локализации функций и множественность обра--
З0ваний различных уровней мозrа, участвующих в обеспечении
психической деятельности. Функциональная мноrозначность моз
rOBbIx структур не только подтверждается на уровне клеточных
популяций, но и оказывается основой возможности адекватной
реализации слояснейших видов психической деятельности в из
менчивых условиях обычной жизни человека. Показанный на
тонком нейрофизиолоr:ическом уровне мноrозвеньевой характер
системы обеспечения психической деятельности убедительно под
тверждает, что нарушение одной и той же психической функции
действительно может развиваться при различных локализациях
патолоrическоrо очаrа, причем нередко лишь очень частные oco
бенности психопатолоrическоrо синдрома зависят от Toro, rде раз
вился очаr.
Правильность этих преД,ставлений, стройных и лоrичных, но
возникших на основе недостаточной информации и поэтому не
поднявшихся выше rипотезы, оказалось возможным подтвердить
в условиях применения комплексноrо метода исследования
структурно функциональной и нейрофизиолоrической орrаниза
ции мозrа.
По ходу диаrностики и лечения больных способом вживлен
ных электродов и, таКИIVI образом, в условиях прямоrо контакта
с мозrом с помощью данноrо комплексноrо метода удалось, нако--
нец, получить убедительные факты, позволившие исследовать
принципы, на основе которых происходит усложнение мозrовых
систем реrуляции функций. Одновременно удалось обнаружить
и некоторые особенности мозrовой системы обеспечения психи
ческой активности.
В развитие плодотворных идей прошлых лет и на основе
J1спользованноrо комплексноrо метода была высказана rипотеза
об обеспечении психической деятельности человека корково под
корковой структурно--функциональной системой со звеньями раз
ной степени жесткости, жесткими, обязательно участвующими
в реакции, обеспечиваIОЩИМИ экономичность работы мозrа, и
звеньями rибкими, включающимися в зависимости от вариаций
внешней среды, внутренней среды мозrа и качества выполняемой
психической деятельности.
111
Отражение внешнеrо мира с ero варьирующими в небольших
пределах, сравнительно жесткими закономерностями и множе
ством «случайных», нереrулярно возникающих явлений, приоб
ретающих доминирующее значение на уровне специально
человеческоrо взаимодействия, и вызвало к жизни, создало
(сформировало) как наиболее адекватную систему мозrовоrо
психическоrо контроля из жестких и rибких звеньев. Перефрази
руя мысль С. И. Вавилова (1950) о зависимости строения rлаза
от характеристик солнечноrо спектра, можно предположить, что
мозrовые аппараты высших форм отражения необходимо pac
сматривать с учетом основных. характеристик среды, в том числе
и факторов специально человеческой, общественной среды.
Весь опыт невролоrической и нейрохирурrической клиник
подтверждает заменяемость не только rибких, но и, при OДHOCTO
роннем поражении, жестких элементов. Можно представить
себе, что формирование системы, морфолоrически предопреде--
ленное опытом видовой эволюции, есть все же всеrда преимуще
ственно результат индивидуальноrо развития. На нашем исто--
рическом отрезке структурная орrанизация системы есть прежде
Bcero проявление статистическоrо влияния видовых характери
стик на индивидуальные.
С друrой стороны, сама эта структурная орrанизация или
анатомическое «предопределение» есть накопившийся и закре--
пившийся в процессе эволюции оптимальный опыт индивиду
умов. Как же лоrичнее Bcero, на основе имеющеrося опыта изу
чения мозrа, представить себе влияние индивидуальноrо пути
на характеристики вида? Что закрепляется в непрерывно иду--
щем процессе взаимодействия со все усложняющейся средой сей
час? Правомерно ли rоворить опереходе условнорефлекторных
реакций в безусловные (на уровне человека!) и целесообразен ли
этот переход?
По видимому, изменчивость внешней среды, определившая
формирование rибкоrо CTPYKTypHoro аппарата мозrовой деятель
ности (rибкие звенья системы) и rибкоrо физиолоrическоrо аппа
рата (условнорефлекторных реакций), предопределяет и преиму--
щественную целесообразность закрепления облеrчения формиро
вания некоторых, наиболее частых и адекватных, а rлавное,
биолоrически выrодных реакций. В этом случае закрепляется
(или, точнее, может закрепляться!) не сама реакция, а леrкость
ее формирования при необходимости. Влияние индивидуальноrо
опыта на видовой, таким образом, может проявляться в виде за
крепления основы реакций, что вновь характеризует примат фак
тора rибкости на данной фазе эволюции. Убедительные примеры
именно TaKoro пути, как известно, представляет эколоrическая
физиолоrия. Сходную точку зрения на данный вопрос можно
найти и в работе В. М. Бехтерева «Общие основы рефлексолоrии
человеКа».
Резкое абсолютное и относительное увеличение количества
rибких звеньев в системе контроля психической деятельности по
112
сравнению с друrими системам:и является механизмом, обеспе
чивающим исключительные возможности работы этой системы.
Именно этот фактор появление и далее абсолютное и OTHO
сительное увеличение количества rибких звеньев в системе ...........
является и основным принципом усложнения мозrовых систем.
Можно попутно отметить, что, с точки зрения соотношения же
стких и rибких элементов, существенно различаются между co
бой не только системы обеспечения психических и, например,
двиrательных функций, но и системы обеспечения сложных пси
хических и эмоциональных реакций, что, безусловно, ни в коей
мере не исключает теснейшеrо взаимодействия между ними.
Обнаружение преобладания rибких звеньев в мозrовой си
стеме обеспечения психических функций раскрыло и причины
Toro, почему физиолоrическое изучение этой системы так долrо
не давалось в руки исследователей, пытавшихся изучать ее на
основе данных выключения и раздражения мозrа (являвшихся
у человека, естественно, приемами лечения и диаrноза). Выявле
ние rибких звеньев системы в условиях раздражения мозrа прин
ципиально моrЛо бы быть осуществлено, однако практически это
моrло быть реализовано только в том случае, если бы были най--
дены условия, в которых Mor бы выявиться развившийся дефект.
Так, например, дефект KaKoro To вида психической деятельности
Mor бы быть выявлен при проведении стимуляции на фоне дo
полнительноrо «шума», наоборот, в условиях, сенсорной депри
вации и т. д. Однако, как известно, TaKoro рода специальных на--
блюдений у человека не производилось. Кроме Toro, метод элек
трическоrо раздражения имеет и принципиальные оrраничения
при изучении структурно функциональной орrанизации мозrа
в силу большой вероятности в условиях раздражения наблюдать
не ТОЛЬ1\.0 локально обусловленные, но и самые различные про--
стые и сложные дистантные эффекты.
Только объединение Bcero, что давал «естественный экспери
мент» очаrовые болезни мозrа, что давали приемы выключения
и раздражения участков мозrа, с действительно MHorocTopoH
ними исследованиями ero нейродинамики позволило показать,
что и каким образом «включается» и выключается в мозrу в про
цессе обеспечения психической деятельности.
Комплексный метод изучения нейрофизиолоrии мозrа позво
лил уже ,сделать ощутимый шаr вперед в познании принципов
построения мозrовых систем, подтвердив одновременно исклю--
чительную плодотворность самой идеи о функциональных систе
мах (П. К. Анохин, 1962).
Он, этот метод, оказался весьма выrодным для изучения TOH
ких механизмов работы нейронных популяций. Важнейшим
результатом исследования нейронных популяций исследова--
нил, проведенноrо при использовании различных, в том числе и
не только физиолоrических, математических, но и фармаколоrи
ческих приемов, оказалась убедительная демонстрация обrцности
исследованных нейрофизиолоrических механизмов различных
11/25 Н. п. Бехтерева
113
видов деятельности и усложнения этих механизмов лишь на
уровне системы. Тем самым на основе строrих фактов был суще
ственно поколеблен (если не опроверrнут!) известный дуалисти
ческий тезис UUерринrтона о принципиальных различиях HepB
Horo и психическоrо и созданы предпосылки для реализации
rениальноrо павловскоrо тезиса о необходимости наложения пси
хическоrо узора на физиолоrическую канву.
Принципиальная возможность, однако, означает лишь нали
чие пути. Наложение психическоrо узора на физиолоrическую
Канву в настоящее время действительно становится возможным,
однако каждый шаr на этом пути очень нелеrок. Полное изуче
ние именно этих сторон проблемы является сейчас прежде Bcero
вопросом техники, времени и рациональных клинико физиоло
rических симбиозов, обеспечивающих возможность исследования
максимально большоrо количества областей мозrа на различных
ero уровнях. Первоочередной задачей этоrо плана является изу
чение коры больших полушарий, что не только осуществимо, но
и очень важно по клиническим соображениям при некоторых
формах эпилепсии и при некоторых психических заболеваниях.
Вполне понятно, что данные, полученные у больных, не MorYT
быть конкретно однозначными для здоровоrо мозrа. Однако YCTa
новленные принципы, по видимому, имеют общую ценность,
а отсюда получение большоrо количества данных у различных
больных будет способствовать и накоплению данных, «прило
жимых» К здоровому мозrу.
Общую ценность наряду с рядом приведенных выше положе
ний MorYT иметь данные о принципиальных особенностях Jиеха
uизмов uaдe HOCTи мозrовоrо обеспечения психической деятель
ности.
В этом плане первостепенную роль иrрает мноrозвеньевая
структура системы психическоrо контроля (1) и наличие в ней
не только постоянных, жестких, но и переменных, rибких эле
ментов (2).
Свойство «включения» различных звеньев системы с по
мощью определенных видов биохимической медиации, обеспечи
вающей возможность функционирования полифункциональной
нейронной популяции, как элемента системы психическоrо KOH
троля, может привести и реально приводит к нарушению актив
ности этоrо звена при повреждении биохимическоrо медиатор
Horo механизма. Динамика биохимической медиации существенно
меняет свойства полифункциональной нейронной популяции,
«уrашая» или, наоборот, активируя различные ее возможности,
сужая или расширяя ее функциональный спектр. В то же самое
время неодинаковость биохимической медиации в различных
звеньях мноrозвеньевой системы обеспечения контроля психиче
ских функций и, таким образом, возможность ее функционирова
ния при нарушении KaKoro To одноrо вида биохимической медиа--
ции является, несомненно, еще одним важным механизмом ее
надежности (3).
114
Эти механизмы создают предпосылки к реализации психиче--
ской деятельности при изменениях внешней среды орrанизма
и внутренней среды мозrа.
Однако рассматривать механизмы надежности обеспечения
любой деятельности и, в частности, психической активности He
возможно без учета механизма ее оптимизации механизма, по
нижающеrо общую вероятность ошибок, определяющеrо качество
выполнения деятельности. Важнейшим механизмом оптимизации
психической деятельности является детекция ошибок, осуще
ствляемая в мозrу нейронными популяциями, избирательно или
исключительно реаrирующими на ошибочное выполнение задан
ной деятельности, в данном случае психолоrических проб. Co
трудником нашей лаборатории В. Б. rречиным было показано,
что в момент детекции ошибки нейронная популяция, осуще--
ствляющая функцию детектора ошиб и, становится «ведущей» по
отношению ко мноrим друrим нейронным популяциям, ее физио
лоrические процессы опережают аналоrичные физиолоrические
процессы, развивающиеся в ряде друrих нейронных популяций
(данные кросскорреляционноrо анализа динамики наличноrо кис
лорода). В свете этих данных приобретает все большую деталь
ность представление о роли детектора Оlllибок как образования,
вызывающеrо реакцию активации, определяющую возможность
осуществления последующей мозrовой деятельности на более
высоком энерrетическом уровне. В мозrу человека сейчас уже
обнаружен ряд зон, исключительно или избирательно реаrирую
щих на ошибочное выполнение психолоrическоrо задания, что по
зволяет rоворить не о детекторах ошибок, а об аппарате детекции
ошибок. Не исключено, что обнаруженный аппарат детекции
ошибок в целом или, что более вероятно, хотя бы частично яв
ляется общим для различных деятельностей.
Наличие в мозrу нейронных популяций, тонко реаrирую--
щих на ошибочное действие, а также разная степень выражен--
ности реакций на Оlllибку в психолоrической пробе дает извест
ные основания предполаrать, что значение различных элементов
аппарата детекции ошибок неодинаково, и, таким образом сам
аппарат детекции ошибок имеет известную иерархию. Возможно,
что эта иерархия динамична и определяется в большей мере xa
рактером выполняемой деятельности, однако этот вопрос, eCTe
ственно, нуждается в дальнейшем изучении. Можно с известной
уверенностью лишь сказать, что при реализации психической
деятельности важное значение имеют детекторы ошибок нейрон
ных популяций xBocTaToro ядра.
Таким образом, к механизмам надежности мозrовоrо обеспе
чения психической деятельности может быть отнесен и аппарат
детекции ошибок, способствующий оптимизации этой деятель
ности (4).
Аппарат детекции ошибок заслуживает, однако, рассмотрения
и еще в одном плане в плане роли ero нарушений в патоло
rии психической сферы. Наблюдавшееся нами нарушение дeTeK
11/25*
115
ции ошибок в нейронной популяции при применении нейротроп--
ных средств в виде появления одинаковой или весьма сходной
реакции и при ошибочных, и при правильных реализациях пси
холоrических проб характеризует первый возможный вид ero
переСТРОЙRИ (этот вид ero нарушений, по видимому, и лежит
в основе эмпирически обнаруженной опасности вождения aBTO
машин на фоне приема некоторых транквилизаторов.) Разруше
ние исключительных или избирательных реакций элементов
аппарата детекции ошибок и, таким образом, частичное или пол--
ное наРУlllение условий оптимизации психической деятельности
может лежать и в основе некоторых собственно психических
расстройств.
Изменение аппарата детекции ошибок, как показали наши на--
блюдения, может, однако, развиться и в друrой форме. Стимуля
ция тех структур, в нейронных популяциях которых представ--
лены элементы аппарата детекции ошибок, приводила к детерми
нации ошибок. Увеличение количества ошибок в психолоrических
пробах при стимуляции нейронных популяций XBOCTaToro ядра
показывает принципиальную возможность детерминации ошибок
при активации нейронных популяций этой области. По видимому,
как проявление нарушений аппарата детекции ошибок (типа ero
стойкой активации) можно рассматривать описанное В. М. Смир
новым (1967) появление нарушений схемы тела при электриче
ском воздействии на rлубокие структуры.
Можно себе представить, что детектор ошибок приобретает
как бы самостоятельное, самодовлеющее значение и в друrих,
более «естественных» условиях патолоrии и превращается из
детектора оптимизатора деятельности в детерминатор ее на--
. рушений. Такой процесс может также лежать в основе психопа
толоrических синдромов и, в частности, тех, которые прояв--
ляются навязчивым повторением каких--то действий, неадекват--
ностью поведения и т. п. Постоянная, не детерминированная
ошибкой, первичная по отношению к KaKOMY TO действию актив
ность структуры, иrрающей обычно роль детектора ошибок, все
время сиrнализирует о рассоrласовании осуществляемоrо дей
ствия (или любой друrой реальности) с планом, независимо от
правильности или ошибочности действия.
Таким образом, если изучение структурно--функциональной
основы и некоторых механизмов деятельности системы мозrо
Boro контроля психических функций уже осуществляется сейчас
достаточно широко, изучение конкретных физиолоrических KOp
релятов KOHKpeTHoro психическоrо нуждалось в новом методи
ческом «скачке».
Предпосылки к этому скачку созданы разработкой специаль
Horo метода анализа импульсной активност.и, позволяющеrо BЫ
являть наиболее тонкие ее изменения.
Результатом Bcero комплекса новых методических возможно
стей, стимулирующеrо взаимовлияния 'конкретных результатов
и «работающих» rипотез, явились новые возможности исследова--
116
ний мозrа человека, позволившие поставить принципиально но--
вые задачи и приступить к их разрешению. Очень схематизиро
ванно эти новые возможности характеризуются сравнительной
пеrкостью ответа на вопрос «rде?», определения зон мозrа (ней
ронных ансамблей), связанных с обеспечением деятельности, и,
отсюда, реальностью постановки вопроса, «как»,' «каким обра
зом?» KaKoro рода общие и местные перестройки в мозrу не
только происходят в процессе обеспечения психической деятель
ности, но и лежат в ее основе; прежде Bcero........... как происходит
восприятие, узнавание, воспроизведение слов и использование
их: и отсюда каким образом мозr оперирует специально чело
веческой информацией. Решение этих проблем предполаrает изу--
чение Toro, как физиолоrически осуществляется в мозrу к о Д и --
р о в а н и е и Д е к о Д и р о в а н и е словесных сиrналов, формиро
вание Toro важнейшеrо базиса, без KOToporo невозможно осуще
ствление собственно мыслительных процессов.
Важно подчеркнуть, что если рассматривать физиолоrические
аспекты изучения психических процессов, то именно этот вопрос
является на данном этапе развития науки по--настоящему клю--
чевым. Только ero решение позволит пере бросить необходимый
мост между, казалось бы, такими далекими друr от друrа биохи--;
мическими и психолоrическими данными о мозrе, создав предпо
сылки для построения действительно стройной системы представ--
лений о том, как мыслит человек при помощи мозrа. Общей Teo
ретической предпосылкой к поискам тонких физиолоrических
коррелятов нейрофизиолоrических механизмов психических про
цессов является представление о том, что в физиолоrической
активности определенных морфофизиолоrических единиц может
содержаться достаточно специфическое соответствие психическим
процессам их код и, в частности, прежде Bcero........... код словесных
сиrналов, что процесс кодирования словесных сиrналов осуще
ствляется, хотя бы частично, с помощью биоэлектрических про
цессов или хотя бы адекватно в них отражается.
Только такое уrлубление в нейрофизиолоrические корреляты
психических процессов действительно и окончательно решит во--
прос о нейрофизиолоrических основах психическоrо во всей ero
полноте и сложности.
Были ли, однако, достаточные основания для поисков кода
ПСИХИЧеских явлений в биоэлектрических процессах? Может
быть, поиски этоrо кода на нейрофизиолоrическом уровне, и, по
существу, Bcero лишь поrоня за химерой?! Окончательный от--
вет на вопрос о правомерности исследований в этом направлении
заключался, как это ни парадоксально, лишь в ero решении.
Это не единственный случай в науке, коrда rолоса скептиков
моrли звучать очень rpoMKo. Известные основания для поисков
TaKorQ кода все же, пожалуй, были.
Это, во первых, данные Lettvin, Matl1rana, McCulloch, Pitts
(цит. по Н. Н. Василевскому, 1969), обнаруживших в экспери
менте избирательную реакцию определенных нейронов зритель--
117
ной системы ляrушки на характерные свойства предмета, и MHO
rие аналоrичные данные, в том числе и полученные у человека,
rде обнаруживалось изменение нейронной активности, связанное
с определенным фактором раздражителей (Jasper, 1966; Mountca..
stle et Granit; цит. по Jasper, 1966, и др.). Это, BO BTOpЫX, те дaH
ные эксперимента, rде применение меченоrо раздражителя,
в частности cBeToBoro раздражителя определенной структуры
(Н. Н. Василевский, С. И. Сороко, 1969), приводило к воспроизве--
дению в рисунке нейронной активности той же структуры, co
хранению следов раздражения в деятельности определенных
нейронов. Это, в третьих, некоторые данные, полученные в на--
блюдениях на больных с вживленными электродами (наблюдение
А. Н. Бондарчук в книrе Н. П. Бехтеревой, А. Н. Бондарчук,
В. М. Смирнова, А. И. Трохачева, 1967): при подаче триrrерной
световой стимуляции при отведении биопотенциалов от областей
мозrа, по данным электрическоrо воздействия, тесно связанных
с реrуляцией артериальноrо давления, наблюдался существен
ный и продолжавшийся более суток ero подъем! Это, наконец,
данные об очень характерном спектре импульсной активности
в фазу удержания в памяти при реализации использовавшихся
нами психолоrических проб (с воспроизведением случайноrо ряда
чисел) спектре, существенно отличном и от фоновоrо, и от
Toro, который наблюдается при «неудержании» пробы в памяти
с последующим ошибочным ее воспроизведением. Вполне по
нятно, что все эти данные в плане поставленной проблемы имели
косвенное значение. Однако принципиальная общность нейрофи--
зиолоrических механизмов различных видов деятельности по
зволяла надеяться, что в нейрональной активности элементов
системы обеспечения психическоrо можно найти и корреляты
конкретных психических явлений, хотя в данном случае про--
блема сиrнал шум исключительно усложнилась. На каком
этапе решения вопроса мы находимся сейчас?
Можно без преувеличения отметить, что первые шаrи в об--
ласти расшифровки импульсноrо кода психических явлений ока--
зались весьма обнадеживающими. В rлубоких структурах мозrа
обнаружены зоны, перестройка импульсной активности которых
зависела от акустических характеристик слова. Можно себе пред--
ставить, что TaKoro рода изменения принципиально моrли бы
быть обнаружены и на любой, менее специфический акустиче
ский сиrнал, и не только в мозrу человека, но и животных.
В мозrу животных rодом позже, чем это показано у человека,
действительно обнаружены весьма сходные явления на словесные
сиrналы (Walker, Halas, 1972). Эти данные показали, что в мозrу
идет первичный анализ словесных сиrналов, позволяющий уже
далее «обратиться» в долrовременную память мозrа за ero ceMaH
тическим значением, для ero декодирования. Правомерность Ta
ких представлений была подтверждена обнаружением BToporo
типа паттернов в мозrу человека, по своим характеристикам
не связанноrо с акустическими свойствами сиrнала. Этот, условно
118
названный «автономным», паттерн, по--видимому, имеет отноше
ние уже к семантическим свойствам сиrнала (Н. П. Бехтерева
и др., 1973).
Достаточно характерные пере стройки в виде изменения формы
потенциалов отмечались при предъявлении слов с высоким
индексом частоты. При предъявлении слов с низким индексом
частоты наблюдающиеся изменения были менее отчетливы, что
связано, по видимому, с зашумлением паттерна перестройками,
имеющими относительно неспецифический характер и связан
ными с процессами, направленными на активацию энrрамм долrо--
срочной памяти, процессами энерrетическоrо характера. И все же
микроструктура. паттернов, подобная той, которая обнаружи
вается при предъявлении слов с высоким индексом частоты на
6...........12 й секунде, обнаруживалась и в данном случае. Второй тип
паттерна должен. рассматриваться как уже один из элементов
биоэлектрическоrо кода специально человеческих словесных сиr--
налов, важнейший элемент дальнейшей интеrрации собственно
психической деятельности.
Третий вид кода характеризовался динамической простран
ственной реорrанизацией взаимодействия нейронных ансамблей,
что позволило обозначить ero как пространственно временной
код. Он наблюдался и в тех нейронных популяциях, rде обнару
живалась акустически зависимая пере стройка текущей частоты
разрядов нейронов и там, rде TaKoro рода перестройка не обнару
живалась. Именно этот вид кода, выделяемый с помощью фак
TopHoro анализа, позволил обнаружить биоэлектрические про
странственно--временные корреляты отдельных фонем и тем ca
мым создал новые возможности контроля мозrовоrо кода слов.
Понятно, что пространственно временной код фонем являлся
элементом комплекса развиваюIЦИХСЯ при акустическом к диро
вании реорrанизаций активности нейронных ансамблей.
Способом выявления пространственно временноrо кода yдa
лось улавливать появление в мозrу нейрофизиолоrических KOp
релятов слова еще до ero произнесения больным в тех нейронных
популяциях, rде акустический код не обнаруживался ни в MO
мент восприятия, ни к моменту воспроизведения слов. Именно
с помощью обнаружения в мозrу пространственно временноrо
кода оказалось возможным перейти от исследования кодирования
слов в мозrу к изучению механизмов выявления мозrом их
смысловой общности. Так, например, способом опознания кода
различных слов удалось после предъявления больному слов
«стул», «стол», «шкаф» выявить В мозrу код слова «мебель» еще
до ero произнесения больным и т. д. (рис. 32).
Пространственно временной код может быть, таким образом,
весьма значимым элементом и смысловоrо кода. Косвенным, хотя
и достаточно веским доказательством положения о наличии
в мозrу смысловоrо кода является выявление в мозrовых зонах
сходных перестроек на слова, далекие по акустическим свой
ствам и близкие по семантическим (Ю. Л. rоrолицын, 1973). Мно--
119
СР}
Зиионuе п м/'Ое.ll6"
I
I
1
I
, .
l'
IJmd"m п м"оеЛ6 П
. I
I
ь
'" I
J' 1 f!.9s(jJ)= 88
, I
- I .
(t)
SIJ/J ме
I \\ V. HW"tV» *
....
2IJме
f v.. ""VI\H\ 1.1.1-1.1.1. 1. VVV\A\t\'\'
т(ш)
о
@
отВет
отВет
/4,Dоелl.J "
,.1 ,
32. Изменение функций текущей частоты трех нейронных популяций при
тесте на индукцию.
а /1 21 3 моменты предъявления слов: «стол», «стул», «шкаф», 4 ответ испытуемоrо
«мебель»; 6 результаты селекции пространственно временной формы орrанизации нейрон
пой популяции, специфичной для обработки вербальноrо сиrнала обобщающеrо значения
(<<мебеJ1Ь» ); 8 профили взаимодействия нейронных ансамблей, выделенные методом фак
TopHoro анализа и специфичные для обработки вербальноrо сиrнала «мебель»; 2 селекция
специфической формы ансамблевой орrанизации по результатам анализа мультиклеточной
активности в тесте на индукцию. Ось абсцисс время; ось ординат степень сходства Te
кущей ансамблевой орrанизации популяции со специфической формой. Звездочкой обозна
чены периоды максимальноrо сходства.
rолетний опыт психолоrии детскоrо возраста по взаимодействию
сиrнальных систем в процессе обучения, а также данные М. Н. Ли
ванова (1972) о наличии пространственной биоэлектрической ди--
намики в мозrу при психической деятельности, дают веские OCHO
вания к развитию исследований пространственно временноrо кода
специально для изучения процессов смысловоrо кодирования. При
этом важным является изучение пространственных взаимоотно--
шений не только в одной нейронной популяции, но и между ди
стантно расположенными популяциями.
Проведенные исследования позволили получить также HeKO
торые материалы о механизмах вербальной памяти.
Устойчивость связанных с акустическими характеристиками
паттернов и автономных паттернов при предъявлении редких
слов, слов, семантическое значение которых н:еизвестно, и квази--
слов связана с особенностями механизмов краткосрочной памяти,
120
одной из сторон которых являются и относительно большие
энерrетические затраты на биоэлектрическом уровне. Устойчи
вость паттернов при краткосрочной памяти связана, вероятно, не
только с активационными механизмами, но и с отсутствием TOp
мозящеrо влияния четкой энrраммы долrосрочной памяти на био
электрический реверберационный процесс. На основании имею--
щихся литературных сведений можно предположить, что устой
чивые реверберационные процессы, характеризующиеся высокой
синхронизацией активности нейронов и, таким образом, резким
оrраничением «степеней свободы» нейронных систем, ведут R из
менениям синаптической проводимости и структурно конформа
ционным изменениям в субсинаптических мембранах нервных
клеток, т. е. являются механизмом, обеспечивающим консолида
цию новых энrрамм долrосрочной памяти (п. В. Бундзен, цит. по:
Н. П. Бехтерева, П. В. Бундзен, В. Д. Кайдель, Э. Э. Давид, 1973).
Естественно, первые исследования собственно биоэлектрическоrо
кода не в состоянии дать объяснения всем биоэлектрическим про
явлениям памяти и также вполне естественно не претендуют
на это.
Приведенные положения вполне соrласуются с представле
ниями П. В. Бундзена (1972) о иерархической орrанизации памяти
и MorYT быть сопоставлены с некоторыми аспектами rипотезы
Н. Ю. Беленкова (1973) о взаимодействии долrосрочной и KpaTKO
срочной памяти. Вовлечение тормозных влияний энrрамм долrо
срочной памяти особенно отчетливо при пробах с использованием
слов с высоким индексом частоты. Автореrуляция устойчивости
HepBHoro кода вербальных сиrналов осуществляется, по--види
мому, ПО принципу обратной связи в зависимости от степени
функциональной активности энrрамм долrосрочной памяти. Pery
ляция устойчивости следовых паттернов осуществляется путем
не только BpeMeHHoro, но и пространственноrо оrраничения HepB
Horo кода, причем последнее осуществляется, очевидно, за счет
процессов латеральноrо торможения (Ю. К. Матвеев, 1971).
По видимому, сейчас уже ясны не только некоторые частные
закономерности кодирования словесных сиrналов в мозrу чело
века, но и некоторые общие механизмы TaKoro рода, позволяю
щие наметить четкие пути дальнейшеrо изучения вопроса.
Понятие «код» В физиолоrии сейчас, пожалуй, несколько pac
ширено. Нередко о кодировании и декодировании сиrналов внеш
Hero мира в центральной нервной системе rоворят уже тоrда,
коrда дело идет, по существу, о передаче с нейрона на нейрон
через биохимическую медиацию, через высокоспецифические про
цессы в мембране и в самой нервной клетке более или менее
сложных сиrналов в «нерасшифрованном» виде, нейрофизиолоrи--
ческие характеристики которых неизвестны (Keidel, 1972). В ши
роком смысле слова это правомерно, тем более, что в TaKoro рода
работах не всеrда и ставится вопрос о принципах и конкретных
особенностях биоэлектрической и биохимической шифровки сиr--
налов в мозrу.
121
По отношению к специально человеческой (психической) дея
тельности задача неизмеримо усложняется. Дело идет не только
о «шифровке» физических свойств сиrнала, не только о необхо
димости использовать этот «зашифрованный» сиrнал как пуско
вой для более или менее сложной поведенческой реакции, но
и о шифровке тех сиrналов, каждый из которых при дешифровке
не только восстанавливает свое значение (нередко приобретая
в то же время «индивидуальную окраску»), но и ложится в oc
нову мыслительных процессов через бесчисленное множество
взаимодействий и взаимоисключений. Для Toro чтобы иметь право
рассматривать задачу как решимую, необходимо не только
использовать адекватные подходы, но и определить хотя бы при
близительно реальные сеrодняшние лимиты возможностей каж--
дой специальности в ее исследовании. По видимому, рассмотре--
ние вопроса о законах сложнейшей интеллектуальной деятель
ности на сеrодня и завтра является задачей психолоrов, хотя
не исключено, что послезавтра к этому вопросу MorYT найти под
ходы и нейрофизиолоrи.
Задача нейрофизиолоrии в данной проблеме это прежде
Bcero исследование законов шифровки и дешифровки, кодирова
ния и декодирования словесных сиrналов на различных уровнях
центральной нервной системы там, rде осуществляется «вход»
И «выход», В структурах, ответственных за шифровку сиrнала по
ero акустическим характеристикам, и тех, rде происходит непо
средственное взаимодействие с долrосрочной памятью и, наконец,
там, rде формируется семантический код. Вполне понятно, что
успех в значительной мере связан с возможностями и aдeKBaT
ностью аппарата извлечения (и изучения!) биоэлектрических
характеристик мозrа, наибо ее тесно связанных именно с этими
.явлениями. Можно считать, что изучение вопроса начато в 1971 r.
(Н. П. Бехтерева, П. В. Бундзен, А. С. Каплуновский, Ю. К. Мат--
веев, 1971).
Поиск HepBHoro кода, несомненно, следует вести в плане изу
чения импульсной активности коры больших полушарий (1\lilnet",
1972), прежде Bcero височной доли, причем возможно, что ЗД,есь
будет обнаружено множество активных зон, выявлено разделен-- /
ное на элементы. и функционально--объединенное представитель-.
ство энrрамм памяти.
В полном соответствии с усредненными современными пред
ставлениями о долrосрочной памяти и в данном случае предпола
rается, что ее основой являются конформационные изменения
белковых молекул. Однако возможность улавливания биоэлек
трическоrо паттерна в области хранения энrрамм долrосрочной
памяти, по видимому, реальна, так как механизм считывания,
видимо, связан с биоэлектрически детерминированной активацией
мембран.
Большинство современных представлений о биохимической
медиации в мозrу трудно совместимо с данными о возможности
обнаружения в удаленных от периферическоrо конца анализатора
122
паттернов, в частности типа первичноrо (акустическоrо) паттерна,
коррелирующеrо с характеристиками TaKoro сложноrо сиrнала
на входе, как слово. Если представить себе движение импульсов
через множество синапсов, rде электрические явления проходят
через промежуточную биохимическую фазу типа адреналиновой,
норадреналиновой, ацетилхолиновой и т. д. медиации, то центр
тяжести кодирования в связи с неизбежной оrромной потерей спе
цифичности характеристик биоэлектрическоrо потока должен был
бы уже на самых первых фазах кодирования быть перенесенным
с биоэлектрическоrо паттерна на структурную специфичность
нейронов (rлии) и обнаружение коррелятов биоэлектрических
мозrовых явлений с характеристиками сиrнала на входе было бы
принципиально нереальным. Однако исследования показали Ha
личие такой специфичной связи. . .
Наиболее адекватное объяснение TaKoro рода фактам моrло бы
быть дано с позиций, развиваемых Bogoch (1968) и сходных, под
держиваемых Adey (1970) о роли rликопротеинов и мукополиса
харидов в передаче информации в центральной нервной системе.
Adey (1970) подчеркивает роль мукополисахаридов мембраны кле--
ток, леrко соединяющихся с ионами, в высокой чувствительности
центральной нервной системы к электрическим импульсам. Еще
более заманч вы, казалось бы, представления Bogoch (1968, 1972),
предположившеrо, что rликопротеины обеспечивают быструю (без
задержки) и очень высоко избирательную передачу электриче
ских импульсов между нейронами центральной нервной системы
и такой же специфический и быстрый контакт нейронов и rлии.
Подобные представления моrли бы адекватно объяснить возмож
ность сохранения специфических соотношений входноrо сиrнала
в активности мозrа. Однако эти и друrие сходные rипотезы не по
лучают пока lllирокоrо распространения в связи с отсутствием
достаточно полноценноrо подтверждения.
Полученные непосредственно в исследованиях на человеке
прямые (биоэлектрический код) и косвенные (ero динамика
в связи с базисом долrосрочной памяти) данные позволяют уже
построить примерную схему ходuрованuя и деходuрованuя сло
весных сиrналов в rоловном мозrу человека (рис. 33).
Соrласно этой схеме, первоначальное кодирование слов осуще
ствляется по законам кодирования сложных звуковых (акустиче
ских) сиrналов и является независимым от смысловоrо содержа
ния этих слов. Возникший акустический код адресуется затем
к долrосрочной памяти, накопленной в результате индивидуаль--
Horo опыта человека, активирует ее или формирует при OTCYT
ствии соответствующеrо базиса. На уровне акустическоrо кодиро
вания в зависимости от ряда дополнительных факторов (эмоцио
нальная окраска ситуации, друrие побудительные мотивы и т. д.)
возможен предварительный отбор, «фильтрация» информации.
Активация долrосрочной памяти приводит к формированию
в мозrу новой оперативной единицы смысловоrо кода, моrущеrо
служить основой уже друrих, значительно более сложных психи
123
33. Схема кодирования и декодирования словесных
сиrналов в rоловном мозrе человека.
ческих процессов. При необходимости словесной реализации пси
хическоrо процесса в мозrу формируется управляющий код. При
отсутствии соответствующеrо базиса в долrосрочной памяти
функции управляющеrо кода может нести и первичный акусти
ческий код, сформировавшийся в мозrу при предъявлении неиз
BecTHoro слова.
Изучение мозrовоrо кода психических процессов ЯВЛяется при--
оритетом отечественной науки. Данные о мозrовом коде слов
важнейшая ступень к изучению тончайших мозrовых механиз
мов действительно сложнейших психических процессов. В этом
направлении исследователям предстоит еще, несомненно, orpom--
ная работа. Необходимо и движение вперед........... исследование все
более сложных психических процессов и закрепление достиr--
нутых рубежей создание все более полноrо архива кода самых
различных слов. Мноrоплановое rлубокое изучение проблемы
потребует усилий БОЛЫllоrо коллектива специалистов различноrо
профиля, обеспеченноrо адекватной и достаточно совершенной
техникой.
124
Проблема HepBHoro кода имеет, однако, не только методоло",
rическую и методическую сложность. Безусловно, rоловокру--
жительно интересно проникнуть в нервный код, действительно
разrадать воплощенную в осязаемые знаки тайну психическоrо.
Исключительно важно решение этой задачи и для физиолоrии,
и для психолоrии, и для невролоrии, психиатрии, философии.
Но как отзовется решение этой, чисто «человеческой» задачи
на человечестве?! Ведь, по существу, именно расшифровка HepB
Horo кода психических процессов может сделать эти процессы до
конца управляемыми,........... rораздо более управляемыми, чем это
принципиально возможно уже сейчас приемами нейрофармаколо
rии; более, чем это возможно, в частности, способом изменения
постоянноrо потенциала в отношении друrих, более простых
функций животноrо и человека.
Надо ли, отсюда, стремиться к этому успеху, к успеху в рас--
шифровке HepBHoro кода? Не выпустят ли ученые и в этом слу--
чае (и в который раз!) «джина из бутылки», KOToporo уже не
удастся упрятать обратно? Безусловно, надо, как нужно развитие
всех без исключения друrих областей науки, в том числе и тех,
проrресс в которых невольно явился уrрозой человечеству, так
как хорошо известно, что опасность никоrда не содержится в са--
мом знании.
Решение заrадки HepBHoro кода, дав в руки Человека истинную
власть над ним самим, сделает управляемым болезни HepBHoro
кода, те ero нарушения, которые обрекают сейчас еще десятки
и сотни тысяч людей во всем мире на длительное пребывание
в палатах психиатрических больниц.
Все это........... пока вопрос будущеrо. Но если обратиться к клини--
ческому, чисто прикладному плану, если внимательно проанали--
зировать уже имеющийся запас знаний о мозrе с точки зрения
их клинической ценности, видно, что MHoroe при ero заболева--
ниях решимо уже сейчас.
Так, например, определенные перспективы в лечении психи--
ческих расстройств может открыть использование данных о на--
рушениях аппарата детекции ошибок.
На основе полученных результатов об активности нейротроп--
ных средств в отношении детектора ошибок понятно, что спо--
собом коррекции ero нарушений как типа десnе'Цuалuза'Цuu, так
и типа превращения в детерминатор ошибок MorYT быть рацио
нально подобранные фармаколоrические средства. В тяжелей--
ших случаях устойчивой активации детектора ошибок может
быть, по видимому, разумным лечебное разрушение этоrо пора--
женноrо элемента аппарата детекции ошибок. Далее убедительно
показано, что эмоциональные реакции связаны со сдвиrами в оп--
ределенных структурах мозrа медленных электрических процес
сов и, по видимому, преимущественно постоянноrо потенциала.
Сдвиr постоянноrо потенциала наблюдается в мозrу при развитии
эмоции; вызванный искусственно, он приводит к развитию эмо
циональной реакции.
125
На основе этих сдвиrов МЭП (постоянноrо потенциала) при
сочетании их (по принципу условноrо рефлекса) с каким--то фак--
тором среды MorYT развиться исключительно устойчивые пове--
денческие реакции больноrо. Врачи, в Kpyr задачи которых не
входит коррекция эмоциональных расстройств, расценивают Ta
кую возможность как опасность, которой они всемерно стремятся
избежать. Но задача коррекции эмоциональных расстройств MO
жет быть поставлена как самостоятельная проблема и, как сви--
детельствуют данные уже сеrодняшнеrо дня, она частично pe
шена и прежде Bcero при использовании Toro же способа вжив
ленных электродов.
Проникновение в клинику метода вживленных э.цектродов и
нейротропных средств принципиально сравняло методический
уровень эксперимента и наблюдений на человеке. Более Toro,
уравнивание методических возможностей клиники и экспери
мента, по существу, подняло возможности клиники над экспери--
ментом, так как при исследовании человеI{а ОСУlцествляется сло--
весный контакт с больным, позволяющий уточнять и субъектив
ный компонент реакции, а не только ее объективное выраjf{ение.
В наибольшей мере это обоrатило Иl\1:енно изучение сенсорной
сферы, эмоциональных и психических явлений, rде субъективный
словесный отчет позволил буквально в секунды дать ответ на
мноrие вопросы, которые и за долrие rоды в эксперименте моrли
бы быть решены лишь приблизительно. Так, хорошо известно,
KaKJ1M сложным было до получения данных у человека объясне--
ние субъективной сущности явления, наблюдавшеrося в экспери--
менте Olds.
На сравнительно очень узком Kpyre больных, диаrностика и
лечение которых осуществлялись способом вживленных электро
дов, при объединении возможностей этой методики и возможно
стей нейротропных средств удалось, таким образом, получить
целый ряд данных, имеющих весьма общее значение не только
для физиолоrии, но и для широкой медицинской практики. В этих
условиях уже оказалось возможным построение карт эмоциоrен
ных зон (см. выше) и карт структурно--функциональной орrаниза--
ции контроля друrих функций, и, по--видимому, В ближайшем бу
дущем окажется реальным создание уточненных карт централь
ной биохимической медиации (д. К. Камбарова, 1969). Наложение
карт биохимической медиации на карту орrанизации централь
Horo контроля любой функции обеспечит в условиях нервной,
психиатрической и друrих клиник возможность воспроизведения
уже не только с помощью вживленных электродов, но и с по--
мощью нейротропных peДCTB желаемой реакции, хотя, конечно,
при применении нейротропных средств и не всеrда в такой изби
рательной форме, как это возможно при использовании вживлен--
ных электродов. Можно предполаrать, что с помощью все более
чистых, «направленных» нейротропных средств будут вызы--
ваться все более локальные изменения в мозrу. В этих условиях,
на основе развития заданных эмоциональных реакций и при
126
сочетании их с избранным фактором среды, будут воссоздаваться
необходимые, лечебно целесообразные поведенческие пере--
стройки. В случае эмоциональных реакций, на основе карт био
химической медиации и с учетом карт центральноrо эмоциональ--
Horo контроля, направленно и на научной основе по условнореф--
лекторному принципу будут вызываться поведенческие сдвиrи
как бы Toro типа, которые иноrда так умело вызывали по жела
нию в далекие прошлые rоды знатоки нейротропноrо действия
вытяжек из растений способом «приворотных И отворотных» зе--
лий и т. п. Вполне понятно, однако, что здесь правомерно лишь
чисто формальное сравнение. То, что на основе большоrо Hapoд
Horo опыта и нередко с существенной опасностью не только для
жизни «жертвы», но И для жизни «врача» проводилось чисто
эмпирически, а потому с вq можностью вероятностноrо решения
очень узкоrо Kpyra задач, окажется возможным на научной
основе, в условиях соблюдения необходимых мер обеспечения
безопасности и с rотовностью к коррекции возникающих ослож
нений у больных с самыми различными эмоциональными pac
стройствами.
Приложение знаний, полученных на узком континrенте боль--
ных, к широкой клинической практике, значительно расширит
ее возможности, расширит власть Врача над Болезнью.
Следует, однако, попутно подчеркнуть, что и при лечении эмо
циональных расстройств в этом «случае будущеrо» и при вполне
современном, хотя во MHoroM чисто эмпирическом, применении
нейротропных средств следует учитывать возможную фазовость
развития сдвиrов МЭП при эмоциях и также возможную фазо
вость развития эмоций. Выше было показано, что одни и те же
структуры как бы MorYT участвовать в эмоциях прямо противо
положноrо знака при изменении электрическоrо знака (или TeH
денции сдвиrа) уровня так называемоrо постоянноrо потенциала.
Физиолоrический анализ этоrо вопроса был бы не полным, если
бы не рассматривать и друrую пространственно временную воз
можность возможность друrоrо, более сложноrо рисунка физио
лоrических явлений в пространстве и во времени. Действительно
«активным», свидетельствующим о вовлечении структуры в ре--
акцию, может быть сдвиr медленных электрических процессов
какой--то одной, наиболее вероятно, отрицательной направлен
ности. Отсюда фазовая динамика МЭП в какой то структуре MO
жет не прямо, а лишь косвенно отражать фазовое развитие ЗМО--
циональных состояний. Структура оказывается тесно связанной
с эмоциональной реакцией при развитии в ней, скажем, отрица--
тельноrо сдвиrа медленных электрических процессов, а следую--
щий, например, положительный сдвиr может свидетельствовать
не об обеспечении ею же эмоций противоположноrо знака, а о He
обходимом «отключении» ее и тесно с этим во времени связанным
«включением» структур, действительно активно обеспечивающих
развитие новой эмоции. При этом «включение» этих, вторых
структур может быть связано с развитием в них также отрица
127
тельноrо сдвиrа МЭП. С учетом данных эксперимента на живот--
ных 'эта вторая возможность более вероятна.
Развитие новой эмоции, таким образом, по видимому, оказы
вается связанным с «включением» новых мозrовых структур при
«отключении» первых. Следовательно, уrлубление в мозrовые
механизмы эмоций позволяет предположи:ть, что развитие, Ha
пример, отрицательной эмоции зависит не только от активности
зон мозrа, прямо связанных с их обеспечением, но и от инакти
вации друrих зон, «работающих» при положительных эмоциях.
Отсюда не ИСКЛЮЧЕНО, что эти процессы отражают своеобразную
«слоЖность» развития отрицательных эмоций и, таким образом,
защитную роль оптимальноrо уровня положительных эмоций.
Если эт так, то, к сожалению, и отрицательное эмоциональное
состояние по тем же механизмам затруднит развитие положи
тельной эмоции. Такую возможность нужно знать и учитывать
и в лечебной, и педаrоrичесн:ой практике. Нейрофизиолоrические
исследования приоткрывают, таким образом, и завесу над MaTe
риальной сущностью инертности эмоций. ·
Следует, однако, попутно подчеркнуть, что и при лечении эмо
циональных расстройств в «случаях будущеrо», и при вполне
современном, хотя во MHoroM чисто эмпирическом, применении .
нейротропных средств следует учитывать возможную фазовость
развития сдвиrов МЭП при эмоциях и также возможную фазо
вость развития эмоций. Так, например, развитие сильноrо Hera
тивноrо отклонения МЭП, коррелирующеrо с положительным
эмоциональным состоянием, может вести к своеобразному «эф--
фекту последействия» в виде последующеrо развития позитивноrо
отклонения МЭП, что может блаrоприятствовать развитию отри
цательноrо эмоциональноrо состояния. Это, по видимому, осо--
бенно значимо в случаях эмоциональноrо дисбаланса.
При формировании лечебных поведенческих реакций следует
учитывать эти данные, так как вполне понятно, что фазовость
реакций может привести не к желаемой, а прямо противополож
ной ей поведенческой перестройке. Этот факт фазовости процес--
сов разъясняет, по видимому, и механизмы явления,обобщенноrо
в народном опыте крылатой фразой «от любви до ненависти
ОДИ:Н шаr». Эту же возможность фазовых сдвиrов обязательно
надо учитывать при назн чении и применении нейротропных
средств, так как MHoroKpaTHoe, длительное инеконтролируемое
применение этих препаратов может повлечь за собой не «сбалан
сирование», а, наоборот, «расшатывание» всей сложной динами
ческой системы контроля эмоций, обеспечить леrкость развития
фазовых эмоциональных реакций. Не исключено, что существен
ный рост количества психических расстройств в различных CTpa
нах сейчас в какой--то мере связан и с принявшим поистине ro--
мерические размеры и, отсюда, практически неконтролируемым
применением транквилизирующей терапии и особенно «caMOTepa
пии». Сказанное, естественно, отнюдь не должно рассматриваться
как призыв к отмене транквилизаторов и друrих нейротропных
128
средств, а лишь как пожелание к контролю над их применением
и учету реальности, а иноrда и неизбежности TaKoro рода ослож--
нений.
Выше рассматривались основные принципы и механизмы на--
дежности работы системы обеспечения психической деятельности.
В то же самое время уже само формирование даже очень надеж
ной и динамичной системы есть всеrда одновременно «невключе--
ние» в ту же систему мноrих нейронных популяций эволю--
ционно потенциальных «претендентов» на участие. Любая, даже
самая динамичная, система предполаrает известную стабилиза
цию, более или менее прочно закрепляющееся оrраничение степе
ней свободы в оперировании возможностями мозrа.
Функциональные системы, являющиеся структурно--функцио--
нальными элементами устойчивоrо состояния здоровья, MorYT
(в известных пределах) перестраиваться при изменениях внешней
среды, в процессе обучения (в том числе и эксперименталь--
Horo) и т. Д.
Однако именно устойчивость функционирования мозrовых си--
стем, хотя и относительная, приводит к тому, что при патолоrи
ческих изменениях мозrа, особенно у взрослоrо человека, далеко
не всеrда происходит использование всех ero резервов. Очень
важно учитывать в лечебной тактике и особенно стратеrии, что
если повреждение достаточно существенно, не устранено и все же
совместимо с жизнью, существование орrанизма в среде обеспечи--
вается за счет формирования HOBoro устойчивоrо состояния
устойчивоrо состояния болезни, устойчивоrо патолоrическоrо co
стояния, пришедшеrо на смену вариациям нормальноrо устойчи--
Boro состояния (Н. П. Бехтерева, А. Н. Бондарчук, 1968). KOHKpeT
ные механизмы патолоrии и поддержания устойчивых состояний
различны при различных заболеваниях, что и определяет типовые
и индивидуальные различия приемов лечения. И все же в лече--
нии хронических заболеваний мозrа целесообразно и необходимо
считаться с общим фактором устоuчuвЪLМ nаТОЛО2uчесхuм CO
стоянuем, поддерживаемым реакциями типа rомеостатических,.НО
направленных на поддержание не здоровья, а болезни. Этот фак--
тор нередко определяет малую эффективность хирурrическоrо и
фармаколоrическоrо лечения мноrих заболеваний и особенно ча--
стую «недолrовечность» первоначальноrо положительноrо эф--
фекта. Знание и учет этоrо фактора помоrают наметить новые и
нередко достаточно общие пути лечения, естественно, чаще Bcero
лишь дополняющие применяемое типовое и индивидуализирован
ное лечение KOHKpeTHoro заболевания.
Хорошо известны попытки вызвать общие перестройки мозrа
и орrанизма, уже давно применяющиеся в психиатрии (электро
шок и др.). Способом, целенаправленно и дозированно изменяю--
щим условия функционирования мозrа, а ПОТОМУ и значительно
более перспективным, является предлоЯ\енная недавно лечебная
электрическая стимуляция, вызывающая изменение режимов pa
боты мозrа с помощью воздействия на ero модулирующие образо
129
вания. Их стимуляция, наряду с подавлением проявлений бо
лезни, вызывает значительный общий эффект (Н. П. Бехтерева
и соавт., 1972). Так, в частности, только в результате электрости
муляции удалось наблюдать сохранившееся в последующем «pac
членение» и резкое уменьшение выраженности фантомно боле
Boro синдрома, стойкую активацию психических функций у тяже
лейшеrо больноrо эпилепсией и улучшение состояния здоровья
больных rиперкинезами. Электростимуляция мозrа снимала остро
раЗБившиеся и хронические фобические синдромы, вызывала и
подавляла друrие эмоциональные реакции, перестраивала эмо
циональные состояния. Вполне понятно, что сфера применения
лечебной электрической стимуляции........... приема, значительно более
щадящеrо, чем разрушение мозrа, должна и может быть pac
ширена, особенно при тех заболеваниях, при которых домини--
рует фактор устойчивоrо патолоrическоrо состояния. Вероятно,
ее целесообразно применять при целом ряде не поддающихся ле
чению; обычными методами навязчивых состояний и друrих
психкcJеских и невролоrических заболеваний.
В Jlечебную практику все шире входит сейчас еще один прием,
направленный на перестройку условий функционирования мозrа.
Речь идет о так называемом «эксперименте С обратной связью»,
известном также под названием адаптивноrо биоуправления. За--
дачами ero применительно к физиолоrии и патолоrии мозrа яв
ляются изучение реакций мозrа с точным учетом ero состояния
или заданная перестройка мозrа, ero «переобучение» и создание
новых устойчивых состояний с помощью воздействия на opra
низм (мозr) в прямой зависимости от какоrо либо изменения
в нем (Walter, Shipton, 1949); Н. П. Бехтерева и В. В. Усов, 1960;
п. В. Симонов и Ф. Е: Темников, 1965; М. Н. Ливанов и соавт.,
1966; Н. Н. Василевский, 1972; п. В. Бундзен и А. В. Чубаров, 1969).
Сочетание воздействий, технически основанных на принципе
обратной связи (а в общем плане на принципе условноrо реф
лекса), с новыми данными об орrанизации мозrа открывает еще
одну возможность в лечении ero заболеваний. «Разрушение» устой
чивоrо патолоrическоrо состояния, формирование новых отноше
ний в мозrу может быть осуществлено на основе электрическоrо
воздействия на зоны, активация которых в заданном направлении
изменяет (подавляет) болезненную симптоматику, в четкой зави
симости от появ ения или непоявления этой симптоматики (при
автоматизированном включении подкрепляющих воздействий).
Формирование новых устойчивых состояний принципиально ocy
ществимо при использовании эмоционально--активных зон тех
областей мозrа, активация которых вызывает положительные
или, наоборот, отрицательные эмоциональные реакции. Повторное
подкрепление желаемой реакции (состояния) воздействием, вызы--
вающим положительные эмоции, и, наоборот, болезненной peaK
ции воздействием, вызывающим отрицательную эмоцию, может
оказаться исключительно активным приемом переобучения мозrа.
Эффект может развиться необычайно быстро, быть прочным и
130
достиrаться за счет «вынужденноrо» И направленноrо использо
вания резервов, компенсаторных возможностей мозrа и, таким
образом, заданноrо формирования новых функциональных систем.
С усовершенствованием стереотаксической техники внутри
мозrовые лечебные воздействия становятся реальностью при ши
роком Kpyre заболеваний мозrа. В то же время следуе'J; подчерк
нуть, что они являются лишь одним из возможных приемов,
«привязанных» К какому либо болезненному проявлению. В по--
давляющем большинстве случаев возможно и показано проведе
ние сеансов «обучения» с применением информационной и моти
вационной техники без прямоrо воздействия на мозr.
Используя информационные и мотивационные стимулы,
удается перестраивать электрическую активность мозrа, частоту
проявления тремора (Н. В. Черниrовская и соавт., 1972а, б, в), co
стояние сознания (Caarder и Chas, 1971а, Ь), мышечный тонус
(Jacobs и Fenton, 1969; Budzinsky и соавт., 1970; Whatmore и
Kohli, 1968) и др.
Описанные новые пути лечения MorYT в отдельных случаях
применяться изолированно. Принципиально же их следует pac
сматривать как элемент комплексноrо лечения, в котором спе--
цифическое воздействие на .данную болезнь сочетается с воздей
ствием на общие механизмы, свойственные болезненным процес--
сам и прежде Bcero хроническим заболеваниям мозrа. Представ--
ление об устойчивом патолоrическом состоянии, дополнившее
старую концепцию «порочноrо Kpyra» и лоrично развивающее
теорию rомеостаза применительно к болезни, позволяет понять
общие механизмы целоrо класса болезней. Таким образом, в ле--
чении заболеваний мозrа имеются сейчас новые разноплановые
возможности. Однако следует отметить, что и новые данные
о мозrе, и новые лечебные возможности, полученные в резуль
тате развития физиолоrии мозrа, еще сравнительно мало исполь--
зуются в лечении ero заболеваний. Не исключено, что в психиат
рии в этом в значительной мере повинна память о ранних и
иноrда, как известно, траrических неудачах психохирурrии, с чем
современное положение вопроса имеет уже достаточно мало
общеrо.
Сейчас уже очевидно, что павловский принцип условных реф--
лексов лежит в основе сложнейших активностей мозrа не только
животных, но и человека. «Человеческий близнец» условнореф
лекторной методики, создававшийся по ее образу и подобию, так
и не достиr, однако, роста cBoero экспериментальноrо брата. Под
черкнув универсальность условнорефлекторноrо принципа, он
не только не позволил понять тонкостей психическоrо, но и не по
Mor создать в клинике Toro фундамента, на котором можно было
бы перестраивать здание пораженноrо человеческоrо разума. При
«переходе К человеку», при проникновении в клинику нужны
были при сохранении OCHoBHoro принципа новые конкретные
формы. «Возврат» В клинику условнорефлекторноrо метода не
только возможен, но и необходим, однако он должен и может
131
быть поднят на новый уровень, модифицирован на основе совре--
менных данных изучения механизмов мозrа человека. Принцип
условных рефлексов безусловно может уже сейчас, при исполь
зовании успехов современной «человеческой» нейрофизиолоrии,
дать MHoro в коррекции болезней и страданий человека. Некото--
рые возможные пути применения принципа условных рефлек
сов в клинике и были рассмотрены в данном разделе книrи.
Расширение клинической нейрофизиолоrии в свою очередь
будет способствовать дальнейшему быстрому накоплению Teope
тически важных сведений о мозrе человека. Анализ данных о ме--
ханизмах нормальных и патолоrических психических реакций
мозrа человека, полученных в условиях расширенноrо примене--
ния возможностей клинической нейрофизиолоrии в сочетании
с уrлубленным изучением мозrа в условиях наилучших возмож
ностей TaKoro изучения при прямом контакте с мозrом,
а также дальнейшее совершенствование в извлечении информа
ции из нейрофизиолоrических данных будут несомненно исполь
З0ваны как фундамент для создания достаточно общей теории
о нейрофизиолоrических основах психической деятельности
человека.
ИЗУЧЕНИЕ Физиолоrии мозrА ЧЕЛОВЕКА
И ЭКСПЕРИМЕНТ НА ЖИВОТНЫХ
Успехи в изучении нейрофизиолоrическоrо обеспечения раз
личных функций мозrа в значительной мере связаны с COBpeMeH
ными техническими возможностями и использованием вживлен
ных электродов в клинике. По данным электрических воздействий
начали создаваться мозrовые карты обеспечения эмоций, запол--
нение которых кажется лишь вопросом количества наблюдений
и разнообразия исследуемых образований мозrа. Заполнение моз
rовых карт эмоций осуществляется сейчас уже не только на
основе данных электрических воздействий, но и реrистрации мед--
ленных электрических, а затем и ряда друrих процессов мозrа.
Наблюдения на человеке в данной области, казалось бы, при
обрели обязательную для эксперимента cTporocTb, mhoroctopoh--
ность методических подходов при rораздо больших, чем в экспе
рименте, возможностях выявления субъективноrо компонента
реакции. Казалось бы, полная физиолоrия мозrовоrо обеспече.ния
эмоций человека вопрос caMoro ближайшеrо будущеrо. . .
Однако дело в действительности обстоит не так просто. На
уровне «человеческих» исследований при принципиальной OДHO
значности физиолоrических методических возможностей с экспе
риментом и, естественно, больших возможностей психофизиоло
l"'ическоrо комплекса, «законной преrрадой» в изучении мозrа
являются медицинские и этические проблемы. Выбор области
ВjКивления электродов всеrда осуrцествляется исходя из интере
сов больноrо. Оправданное задачами лечения и техническими воз--
132
можностями расширение спектра заболеваний медленно расши
ряет и Kpyr структур мозrа, являющихся стереотаксическими
мишенями. При сложнейших формах эпилепсии TaKoro рода ми--
шенью в ряде клиник стала уже и одна из опасных для введения
инородных тел областей rипоталамус. Правда, все еще очень
немноrие рискуют внедряться у человека в эту зону, хорошо зная
вероятность развития при этом разноrо рода дистрофических
осложнений. По вполне понятным причинам оправдана и OCTO
рожность В отношении стволовых структур. Электроды в HeKOTO
рые области коры вводятся ПрJ1 эпилепсии и начинают вводиться
при зрительном и слуховом протезировании. И все же, учитывая
специфику этих заболеваний, нет оснований полаrать, что. в бли
жайшее время удастся, не пренебреrая оrраничениями медико
этическоrо порядка, войти в дл:ительное соприкосновение с ДOCTa
точно большим количеством" зон всех областей коры. Таким об--
разом, по сравнению с экспериментом на животных исследования
мозrовоrо обеGпечения эмоций человека существенно страдаIqТ
в отношении в зможной полноты обследования мозrа.
Вполне понятно, что уточнение наблюдавшеrося при диаrно",
стической стимуляции эмоциональноrо эффекта требует e o по,,:
BTopHoro воспроизведения. Мноrократная стимуляция этих З9Н
В диаrностических целях непозволительна. Лечебная электрости
муляция эмоциоrенных зон дает как будто несколько больше про
стора для наблюдения динамики результатов. Но лечебная стиму
ляция должна проводиться лишь по показания м эмоциональных
расстройств, т. е. при исходном нарушении активности системы
обеспечения эмоций, и повторение стимуляции в блаrоприятном
случае должно вести к изменению качества реакции. Не безrра
ничны в случае изучения мозrовоrо обеспечения эмоциональных
реакций и возможности, открывающиеся за счет реrистрации фи
зиолоrических показателей при эмоциоrенных тестах. Животное
можно и иноrда нужно (в интересах человека!) поставить в слож--
нейшую эмоциоrенную ситуацию. Э'/и'ОЦUО2е1i1iЪLе теСТЪL для бол ъ
'ИО20 дОЛЖ1iЪL uзбuратъся nредеЛЪ1iО береЖ1iО, u особе1i1iО те, X TO
рЪLе вЪL3ъtвают отрuцател Ъ1iУЮ эмоцuю.
Возможности прямоrо контакта с мозrом в условиях лечеб
Horo применения метода вживленных электродов трудно пере--
оценить и особенно по сравнению с друrими физиолоrическими
приемамJi изучения мозrа человека. Однако, как известно, целый
ряд приемов может быть использован и у больноrо, и у здоровоrо
человека. Вживление электродов осуществляется исключительно
по медицинским показаниям, а потому и во всех случаях полу--
ченный результат должен рассматриваться с учетом этоrо обст() ""
тельства. Исследование во всех случаях вживления электродов
больноrо мозrа является самым серьезным оrраничением этих
наиболее тонких возможностей изучения мозrа человека.
Радикальные меры для устранения этоrо оrраничения в фи
зиолоrии мозrа человека предложить трудно. Одной из паллиа
тивных рекомендаций является исследование у каждоrо больноrо
133
и пораженной и менее пораженной системы и построение пред
ставлений о физиолоrии мозrа, прежде Bcero с учетом данных
менее пораженной системы. Накопление материала, полученноrо
у разных rрупп больных, может представить более или менее
близкие к норме данные о физиолоrии эмоций. Друrим приемом
преодоления оrраничений, связанных с исследованием больноrо
мозrа, является проведение их на фоне компенсирующей или
замещающей фармаколоrической терапии.
Таким образом, современные технические возможности, по
множенные на мноrоплановый проrресс в диаrностике и лечении
заболеваний мозrа, хотя и предопределили решающий скачок
в изучении нейрофизиолоrии человека и, в частности, наиболее
СЛожноrо ее раздела нейрофизиолоrическоrо обеспечения пси
хических функций, не сняли и не должны снять медико этиче--
ские оrраничения.
Перечисление оrраничений в изучении физиолоrии высшей
нервной деятельности человека, если пойти по частным вариан--
там, моrло бы занять, вероятно, больше печатных страниц, чем
занимает вся эта книrа. В этом, безусловно, нет необходимости.
Важно подчеркнуть, чт() возможности физиолоrии мозrа человека
cyrцecTBeHHo возросли и особенно за последние два десятилетия.
Сочетание объективных наблюдений с «человеческим» контактом
помоrает проникнуть в rлубь тех механизмов, суждение о KOTO
рых на основе эксперимента может быть только косвенным.
И все же всеrда следует помнить, что в решении широкоrо Kpyra
вопросов физиолоrии мозrа человека разумно определенное место
эксперимента на животных никоrда не будет занято исследова
нием caMoro человека.
УКАЗА ТЕЛЬ ЛИТЕРА ТУРЫ
А б а ш е в К о н с т а н т и н о в с к и й А. л. О психопатолоrических син
дромах очаrовоrо значения. Вопр. нейрохир., 1961, NQ 5, с. 45 47.
А б а ш е в К о н с т а н т и н о в с к и й А. л. Проблема локализации и
клиническая психиатрия. Журн. невропатол. и психиатр., 1964, NQ 1,
с. 43 49.
А б р а м о в и ч r. Б., Зах а р о в В. В. О психических нарушениях при
поражении мамиллярных тел и системно связанных с ними образова
ний. Труды научн. исслед. психоневролоrическоrо ин та им. В. М. Бех
терева, т. 21. л., 1961, с. 125 181.
А в Р а м о в с. Р. Динамика уровня постоянноrо потенциала в rлубоких
структурах мозrа человека в состоянии спокойноrо бодрствования и
при некоторых воздействиях. В сб.: Роль rлубоких структур rолов
Horo мозrа человека в механизмах патолоrических реакций. Л., 1965,
с. 3 8.
Александровская М. М., Невзорова Т. А., Шпир Е. Р. Слу
чай опухоли мамиллярных тел, психопатолоrия и соматические симп
TOMЫ. Журн. невропатол. и психиатр., 1947, NQ 2, с. 30 37.
А н и ч к о в А. д. Влияние центральных холинолитиков на биоэлектри
ческую активность миндалины и rиппокампа человека. В кн.: Физи
олоrия и патолоrия лимбико ретикулярноrо комплекса. М., 1968,
с. 225 226.
А н о х и н п. К. О физиолоrическом субстрате сиrнальных реакций.
Журн. высш. нервн. деят., 1957а, NQ 1, с. 39 48.
А н о х и н п. К. Значение ретикулярной формации для различных форм
высшей нервной деятельности. Физиол. журн. СССР, 1957б, NQ' 11,
с. 1072 1085.
А н о х и н П. К. Соотношение коры и подкорковых аппаратов в услов
ном рефлексе. В кн.: Электроэнцефалоrрафическое исследование
высшей нервной деятельности. М., 1962, с. 241 254.
А н () х и н п. К. Биолоrия и нейрофизиолоrия условноrо рефлекса. М.,
1968.
А Р т е м ь е в а Е. Ю., Х о м с к а я Е. д. О периодических колебаниях
асимметрии а волны при различных функциональных состояниях
мозrа. Материалы XVIII Междунар. KOHrpecca по психолоrии, симп. 6.
М., 1966, с. 39 46.
(Вейн Е. С., Жирмунская Е. А., Волков В. Н.) В е у n Е. 5., Z 11 i r m u n
s k а у а Е. А., V о 1 k о v У. N. Electroel1cephalographic investigations in the
process of recognizing ill1ages of objects during their tachistoscopic presenta
tion 1 (Normal subjects). Neuropsychologia, 1967, N 5, р. 203 217.
Б е й н э. С., Ж и р м у н с к а я Е. А., В о л к о в В. Н. Электроэнцефало
rрафические исследования в процесс е узнавания предметных изобра
жен ий при тахистоскопическом их предъявлении. Журн. невропатол.
и психиатр., 1971, NQ 9, с. 1288 1296.
Б е л е н к о в Н. ю. Энrрамма как эквивалент пусковой афферентации.
Успехи физиол. наук, 1973, NQ 1, с. 53 64.
Б е л я е в В. В. Взаимоотношения биоэлектрических процессов HeKOTO
рых структур rоловноrо мозrа человека. Автореф. дисс. канд. Л., 1968.
Б е л я е в В. В. Некоторые аспекты структурно функциональной орrани
зации центральной нервной системы человека по данным корреля
ционноrо анализа биоэлектрических процессов. В сб.: Нейрофизиоло
135
rические основы нормальных и патолоrических реакций мозrа чело
века. Л., 1970, с. 60 62.
Б е р и т о в И. С., В о р о б ь е в А. О. О происхождении блаrоприятствую
щеrо действия закрывания' rлаза на а волны у человека. Труды
Института физиолоrии АН rрузинской сеР, т. 5. Тбилиси, 1943,
с. 369 386.
Б е р и т а ш в и л и И. С. О нейронной и биохимической орrанизации HepB
Horo субстрата памяти в коре большоrо мозrа. Труды 5 й Всесоюзн.
конф. по нейрохимии. Тбилиси, 1970, с. 11 21.
(Бехтерев В. М.) В е с h t е r е v W. М. Уоп Demonstrationes Gehirns mit Zersto
rung der vorderen und inneren Theile der Hirnrinde beider Schlafenlappen.
Neurol. Zbl., 1900, Bd. 19, s. 990 991.
Б е х т е р е в а Н. П. О биоэлектрической активности больших полушарий
при супратенториальных опухолях. Автореф. дисс. докт. Л., 1958.
Б е х т е р е в а Н. П. О значении применения различных видов прерывистой
световой стимуляции в локальной диаrностике опухолей rоловноrо моз
ra. Журн. невропатол. и психиатр., 1961, N2 11, с. 1608 1614.
Б е х т е р е в а Н. П. rлубокие структуры мозrа человека и некоторые
механизмы патолоrических реакций. В сб.: Роль rлубоких структур
мозrа человека в механизмах патолоrических реакций. Л., 1965,
с. 25 30.
Б е х т е р е в а Н. П. Некоторые принципиальные вопросы изучения нейро
физиолоrических основ психических явлений у человека........... В сб.:
rлубокие структуры мозrа человека в норме и патолоrии. Л., 1966,
с. 18 21.
Б е х т е р е в а Н. П. Нейрофизиолоrические аспекты психической деятель
ности человека. Л., 1971.
Б е х т е р е в а Н. П., Б о н Д а р ч у к А. Н. Об оптимизации этапов хирур
rическоrо лечения rиперкинезов. Вопр. нейрохир., 1968, Н2 3, с. 39
44.
Б е х т е р е в а Н. П., Б о н Д а р ч у к А. Н., С м и р н о в В. М. Физиолоrи
ческое обоснование операций на подкорковых структурах при rипер
кинезах. Вопр. нейрохир., 1969, N2 1, с. 1 7.
Б е х т е р е в а Н. П., Б о н Д а р ч у к А. Н., С м и р н о в В. М., Т р о х а
ч е в А. И. Физиолоrия и патофизиолоrия rлубоких структур мозrа че
.ловека. Л. М., 1967.
Б е х т е р е в а Н. П., Б о н Д а р ч у к А. Н., М е л ю ч е в а Л. А., С м и p
н о в В. М. Лечебные электростимуляции rлубоких структур rоловноrо
мозrа. Вопр. нейрохир., 1972, Н2 1, с. 7 12.
Б е х т е р е в а Н. П., Б У н Д з е н П. В., М а т в е е в Ю. К., К а п л y
н о в с к и й А. С. Функциональная реорrанизация активности нейрон
ных популяций мозrа человека при кратковременной вербальной па
мяти. Физиол. журн. СССР, 1971, N2 12, с. 1745 1761.
Б е х т е р е в а Н. П., Б у н Д з е н П. В., К а п л у н о в с к и й А. С., М а T
в е е в Ю. К. К расшифровке нейрофизиолоrическоrо кода краткосроч
ной памяти у человека. Материалы XXIII совещ. по проблемам BЫC
шей нервной деятельности, т. 1. rорький, 1972, с. 95 96.
Б е х т е р е в а Н. П., Б У н Д з е н П. В., К а й Д е л В. Д., Д а в и Д э. э.
Принципы орrанизации структуры пространственно временноrо кода
краткосрочной вербальной памяти. Физиол. журн. СССР, 1973, N2 12,
с. oo oo.
(Бехтерева Н. П., fенкин А. А., Моисеева Н. И., Смирнов В. М.)
В е с h t е r е v а N. Р., G е n k i n А. А., М о i s е е v а N. 1., S m i r по v У. М.
Electrographic evidence оп participation of deep structures of the human brain
in certain mental processes (computer analysis data) In: 6th Intern. Congr.
EEG Сliп. Neurophysiol., Vienna, 1965, v. 15, р. 8.
(Бехтерева Н. П., fенкин А. А., Моисеева Н. И., Смирнов В. М.).
Bechtereva N. Р., Genkin А. А., Moiseeva N. 1., Smirnov У. М.
Electrographic evidence оп participation of deep structures of the human in
centrain mental processes (computer analysis of data). EEG СНп. Neurophys.,
Suppl., 1967, v. 25, р. 153 166.
136
Бехтерева Н. П., rенкин А. A , Смирнов В. М. rлубокие
структуры мозrа человека и психические функции. В сб.: Структура
и функция архипалеокортекса. rаrрские беседы. М., 1968, с. 338 354.
Б е х т е р е в а Н. П., 3 о н т о в В. В. К вопросу об электроrрафической
характеристике основных нервных процессов. Физиол. журн. СССР,
1961, N2 12, с. 1463 1470.
Б е х т е р е в а Н. П., r р а ч е в К. В., О р л о в а А. Н. Я Ц у к С. Л.
Использование множественных электродов, вживленных в подкорковые
образования rоловноrо мозrа человека для лечения rиперкинеза.
Журн. невропатол. и психиатр., 1963, N2 1, с. 3 8.
Б е х т е р е в а Н. П., К а м б а р о в а Д. К., М а т в е е в Ю. К. Функцио
нальная характеристика звеньев мозrовых систем контроля психиче
ских и двиrательных функций у человека. Физиол. журн. СССР,
1970, N2 8, с. 1081 1097.
Б е х т е р е в а Н. П., М о и с е е в а Н. И., О р л о в а А. Н., С м и p
н о в В. М. Некоторые данные о нейрофизиолоrии и функции ПОДКор
ковых структур мозrа человека. Материалы Х Всесоюзн. съезда фи
зиолоrов, биохимиков, фармаколоrов, т. 1. Ереван, 1964, с. 124 125.
Б е х т е р е в а Н. П., М о и с е е в а Н. И., О Р л о в а А. Н., С м и р
н о в В. М. О нейрофизиолоrии и функции подкорковых структур моз
ra человека. В сб.: Физиолоrия в клинической практике. М., 1966,
с. 111 121.
(Бехтерева Н. П., Моисеева Н. И., Смирнов В. М.) В е с h t е r е v а N. Р.,
М о i s е е v а N. 1., S m i r по v V. М. Complex reiterative patterns in patholo
gic Ьиmап electrosubcorticograms and operative memory. In: 6th Intern.
Congr. EEG СНп. Neurophysiol. Communication, Vienna, 1965, р. 241 244.
Б е х т е р е в а Н. П., О р л о в а А. Н. Клинико электрофизиолоrические
наблюдения при опухолях rоловноrо мозrа в условиях применения
различных лечебных мероприятий. В сб. трудов l й межобл. конф.
нейрохирурrов 3ап. Сибири и Урала. Новосибирск, 1957, с. 113 117.
Б е х т е р е в а Н. п. Т р о х а ч е в А. И. Некоторые данные об электри
ческой активности rлубоких структур мозrа при изучении оперативной
памяти. Вопр. психол., 1966а, N2 3, с. 44 48.
Б е х т е р е в а Н. П., Т р о х а ч е в А. И. Динамика электросубкротико
rpaMMbI уровня постоянноrо потенциала и клеточной активности в rлу
бок их структурах мозrа человека при оперативной памяти. Труды
XIII Междунар. психолоrическоrо KOHrpecca, 6 й симп. М., 1966б,
с. 5 7.
Б е х т е р е в а Н. П., Т р о х а ч е в А. И. Изменение электрических явле
ний в rлубоких структурах мозrа и оперативная память. В сб.: COB
ременные проблемы электрофизиолоrии центр. нервной' системы. М.,
1967, с. 31 40.
Б е х т е р е в а Н. П., У с о в В. В. Методика пр рывистой фотостимуля
ции В ритме собственных потенциалов мозrа при реrистрации электро
энцефалоrраммы. Физиол. журн. СССР, 1960, N2 1, с. 108 111.
Б е х т е р е в а Н. П., Ч е р н ы ш е в а В. А. Основные этапы изучения
вопроса о физиолоrических механизмах психической деятельности че
ловека. Вестник «Научная мысль», 1969, N2 3, с. 28 37.
Б о н Д а р ч у к А. Н., С м и р н о в В. М. Эффекты электростимуляции
срединноrо центра зрительноrо буrра у человека. Физиол. журн.
СССР, 1969, N2 4, с. 408 413.
Б Р е н Д с т е Д А. Н. Особенности биоэлектрической активности мозrа при
сложных формах деятельности ребенка. Материалы 21 ro совещ. по
проблемам высшей нервной деятельности. М. Л., 1966, с. 52 53.
Б у н Д з е н П. В. Нейрокибернетический подход к анализу процессов адап
тивной самоорrанизации функциональных систем rоловноrо мозrа.
Материалы симпозиума: Самореrуляция нейрофизиолоrических меха..
низмов интеrративной и адаптивной деятельности мозrа. Л., 1972,
с. 7 12.
Б у н Д з е н П. В., Ч у б а р о в А. В., Шиш к и н Б. М. Нейрокибернети
ческие аспекты изучения механизмов оптимальноrо управления в дея
6 Н. П. Бехтерева
137
тельности rоловноrо мозrа. В кн.: Очерки прикладной нейр()киберне
тики. Л., 1973, с. 35 120.
Б у н Д з е н П. В. r о r о л и Ц ы н ю. Л., Д а в и Д Э., к а п л у н о B
С К И Й А. С., Пер е п е л к и н П. Д. Структурно системный подход
к анализу процессов функциональной реорrанизации нейронных попу
ляций. Физиол. журн. СССР, 1973, NQ 12, с. oo oo.
В а в и л о в С. И. Солнце и rлаз. М., 1950.
В а р т а н я н r. А. Ионные механизмы возбуждения и торможения.
В кн.: М ханизмы деятельности центральноrо нейрона. М. Л., 1966,
с. 58 100.
В а р т а н .я н r. А. Взаимодействие возбуждения и торможения в НЕ:Йро
не. Л., 1970.
В а с и л е в с к и й Н. Н. О связи фоновой активности нейронов с OMaTO
сенсорной коры с особенностями их функциональной орrанизации.
Физиол. журн. СССР, 1965, NQ 6, с. 711 716.
В а с и л е в с к и й Н. Н. Нейрональные механизмы коры больших полуша
рий. Л., 1968.
В а с и л е в с к и й Н. Н. Нейрофизиолоrические механизмы реrуляции адап
тивной деятельности мозrа. В сб.: Эволюция, эколоrия и мозr. Л.,
1972, с. 3 29.
Василевский Н. Н., Климова Черкасова В. И., BapTa
н я н r. А. о структурно функциональных соотношениях возбужде
ния и торможения в центральной нервной системе. Физиол. журн.
СССР, 1965, NQ 4, с. 424 430.
(Василевский Н. Н., Вартанян r. А.). V а s s i 1 е v s k у N. N., V а r t а n i i а n
G. А. Relations between ,phases of the electrocorticogram, firing and excHa
bility of cortical neurons. In: 6th Intern. Congr. EEG сliп. Neurophysiol.
Vienna, 1965, v. 193, р. 81.
Василевский Н. Н., Вартанян f. А. Реакции и механизмы дея
тельности центральных нейронов. В кн.: Проблемы физиолоrии и па
толоrии высшей нервной деятельности, в. з. Л., 1966, с. 52 71.
В а с и л е в с к и й Н. Н., С о р о к о с. И. Следовые процессы и клеточ
ные механизмы памяти. Физиол. журн. СССР, 1969, NQ 8, с. 957 965.
(Войтинский Е. Л., Прянишников В. А.). V о i t i n s k i Е. L., Р r у а n i s с h
n i k о v С. А. The study of correlation between psychological and electroen
cephalographic data Ьу the method of synchronous detection. In: XVIII
Intern. Congr. of Psychol., v. 1. Abstract, 1966, р. 217.
В о р о н и н Л. r., С о к о л о в Е. И. О взаимоотношении ориентировочноrо
и условноrо рефлексов у человека. Вестн. Моск. YH Ta, 1955, NQ 9,
с. 39 54.
r е н к и н А. А. О выявлении в электроэнцефалоrрамме специфической
локальной реакции при сложной деятельности. Вопр. психол., 1961,
NQ 6, с. 114 126.
r е н к и н А. А. Об асимметрии длительностей фаз ээr при умственной
активности. Докл. АН СССР, 1963, NQ 6, с. 460 1463.
r е н к и н А. А. Длительность восходящих и нисходящих фаз электроэн
цефалоrраммы, как источник информации о нейрофизиолоrических
процессах. Автореф. дисс. канд. Л., 1964.
f е н к и н А. А. Статистические характеристики длительностей фаз коле
баний электроэнцефалоrраммы и некоторые механизмы произвольной
активности человека. Материалы XVIII Междунар. KOHrpecca психо
лоrии, симп. 6. М., 1966, с. 80 85.
r ерш у н и r. В., к о р о т к и н И. И. о субсензорных условных рефoJ ек
сах на звуковые раздражения. Докл. АН СССР, 1947, NQ 2, с. 417 480.
r ерш у н и r. В., к о ж е в н и к о в В. А., М а р у с е в а А. М., Ч и
с т о в и ч Л. А. Об особенностях образования временных связей на He
ощущаемые звуковые раздражения у человека. Бюлл. экспер. биол.
и мед., 1948, NQ 3, с. 205 209.
r о r о л и Ц ы н ю. Л. Метод объективной классификации паттернов муль
тиклеточной активности нейронных популяций rоловноrо мозrа чело
века. Физиол. журн. СССР, 1973, NQ 6, с. 960 962.
138
r о r о л и Ц ы н ю. л. Исследование методами классификационноrо aHa
лиза паттернов мультиклеточной активности нейронных популяций ro
ловноrо мозrа человека при краткосрочной памяти. Материалы Bce
союзн. конф. по нейрокибернетике. POCTOB Ha ДOHY, 1973, с. 68.
r о р е л и к л. И., С м и р н о в В. М., Т о н к о н о r и й И. М. Об участии
rлубоких структур rоловноrо мозrа человека в восприятии элементов
формы и изображений, различающихся по своим статистическим xa
рактеристикам. В сб.: Психолоrический эксперимент в невролоrиче
ской и психиатрической клинике, в. 46. Л., 1969, с. 31 51.
r р е ч и н В. Б. Некоторые данные о динамике кислорода в подкорковых
образованиях rоловноrо мозrа человека. В сб.: rлубокие структуры
rоловноrо мозrа человека в норме и патолоrии. М. л., 1966, с. 50 53.
r р е ч и н В. Б. Наличный кислород rлубоких структур мозrа человека и
динамика ero при некоторых функциональных пробах. Автореф. дисс.
канд. л., 1968.
r р е ч и н В. Б. Целесообразна ли калибровка оrоленноrо кислородноrо Ka
тода в практике интрацеребральных электродов. Физиол. журн.
СССР, 1969, NQ 9, с. 1180 1182.
r р е ч и н В. Б., С м и р н о в В. М. Изменение окислительноrо метаболизма
rлубоких структур мозrа человека при эмоциональных реакциях и
в rипнотическом сне. Труды ин та им. Бехтерева, т. 44. Л., 1967,
с. 52 67.
r р и н Д е л ь о. М. Значение корреляционноrо анализа для оценки ээr че
ловека. М., 1965.
Д у б и к а й т и с Ю. В. Оценка электроэнцефалоrрамм с помощью потенци
альных полей у больных с орrаническими поражениями rоловноrо
мозrа. Материалы Х съезда Всесоюзн. физиол. об ва им. И. П. Пав
лова, т. 1. М. Л., 1964, с. 17.
Д У б и к а й т и с ю. В. Электрофизиолоrическая локализация энилепто
rенной зоны при хирурrическом лечении очаrовой эпилепсии. Автореф.
дисс. л., 1968.
(Жирмунская Е. А., Вейн Е. С., Волков В. Н.). Z h i r m u n s k а у а Е. A ,
В е у n Е. S., V о 1 k о v У. N. Electroencephalographic investigations in the
process of recogniz'ing pictures of obiects during their tachistoscopic presenta
tion II. Neuropsychologia, 1969, v. 7, р. 167 178.
3 а р а к о в с к и й П. М., м е Д в е Д е в В. И., З и н ч е н к о В. П. Эрroно
мика. М., 1970.
И в а н н и к о в ю. r., у с о в В. В. Предоперационная и операционная
стереотаксические проrраммы для ЭВМ «Минск l». В кн.: Проблемы
клинической и экспериментальной физиолоrии rоловноrо мозrа. М.,
1967, с. 21 25.
И в а н о в С м о л е н с к и й А. r. Методика исследования условных реф
лексов у человека. М., 1933.
J1 л ю х и н а В. А. Медленные электрические процессы rоловноrо мозrа че
ловека при реализации психической и двиrательной деятельности
(в связи с состоянием центральной биохимической медиации). Автореф.
дисс. л., 1972.
И л ю х и н а В. А., Х о н ю. В. К вопросу о корково подкорковой opra
низации мозrовых систем обеспечения rотовности к действию у че
ловека. Физиол. журн. СССР, 1973, NQ 12, с. oo oo.
К а м б а р о в а Д. К. Биоэлектрическая активность мозrа человека при воз
действии на серотонинерrические CTPYKTYpы. В сб.: Механизмы HepB
ной деятельности. л., 1969, с. 56 65.
К а м б а р о в а Д. К. Центральные биохимические системы и патолоrия.
В сб.: Проблемы физиолоrии и патолоrии высшей нервной деятельно
сти. л., 1970, с. 185 205.
К н и п с т И. Н. Электрическая активность разных уровней коры rолов
Horo мозrа при выработке оборонительноrо условноrо рефлекса. Tpy
ды- ин та высш. нервн. деятельности АН СССР, т. 1. М., 1955, с. 294.
К о r а н А. Б. Электрофизиолоrическое исследование центральных Mexa
НИзмов некоторых сложных рефлексов. М., 1949.
6*
139
к о r а н А. Б. Выражение процессов высшей нервной деятельности элект
рических потенциалах коры мозrа при свободном поведении живот
Horo. В сб.: Электроэнцефалоrрафическое исследование высшей
нервной деятельности. М., 1962, с. 42 53.
К о r а н А. Б. Проблема движения нервных процессов в коре мозrа. Po
CTOB Ha ДOHY, 1967.
К о ж е в н и к о в В. А. Принципы метода анализа биотоков rоловноrо мозrа
как сложноrо колебательноrо процесса. В кн.: Проблемы физиолоrи
ческой акустики, т. 3. М. Л., 1955, с. 102 116.
К о л м о r о р о в а А. И. Автоматы и жизнь. В кн.: Возможное и невоз
можное в кибернетике. М., 1969, с. 10 29.
К о Р с а к о в С. С. Болезненные расстройства памяти и их диаrностика. М.,
1890.
(Ливанов М. Н.). L i v а n о v М. N. Spatial analysis of the bioelectric activity of
the brain. Proc. Intern. Union Sci. XXII Congr., v. 1, р. 2. Leiden, 1962а,
р. 899 907.
Л и в а н о в М. Н. О замыкании условных связей (по материалам электро
физиолоrических исследований. В сб.: Электроэнцефалоrрафические
исследования высшей нервной деятельности. М., 1962б, с. 174 185.
Л и в а н о в М. Н. Нейрокинетика. В сб.: Проблемы современной нейро
физиолоrии. М. Л., 1965, с. 37 72.
Л и в а н о в М. Н. Пространственная орrанизация процессов rоловноrо моз
ra. М., 1792.
Л и в а н о в М. Н., r а в р и л о в а Н. А., А с л а н о в А. С. Об отраже
нии некоторых психических состояний в пространственном распреде
лении биопотенциалов коры rоловноrо мозrа человека. Материалы
XVIII психолоrическоrо KOHrpecca, симп. 6: Электрофизиолоrические
корреляты поведения. М., 1966, с. 31 33.
Ливанов М. Н., Королькова Т. А. О быстрых колебаниях в элект
роэнцефалоrрамме и некоторых условиях, их усиливающих. rаrрские
беседы, т. 1. Тбилиси, 1949, с. 301 307.
Л и в а н о в М. Н., П о л я к о в К. Л. Электрические процессы в коре ro
ловноrо мозrа кролика при выработке оборонительноrо условноrо реф
лекса на ритмический раздражитель. Изв. АН СССР, отд. биол.
наук, 1945, N2 3, с. 286 305.
Л и в а н о в М. Н., Р я б и н о в с к а я А. М. К вопросу о локализации изме
нений в электрических процессах коры rоловноrо мозrа кролика при
становлении оборонительноrо условноrо рефлекса на ритмический раз
дражитель. Физиол. журн. СССР, 1947, Ng 5, с. 523 534.
Л и в а н о в М. Н., С т р е л ь ч у к Н. В., М е л и х о в а А. М. Электрофизио
лоrические исследования взаимодействия первой и второй сиrнальных
систем при rипнотическом состоянии. Материалы XVI совещания по
проблемам высшей нервной деятельности. М. Л., 1953, с. 123 125.
Л о м о в Б. Ф. Человек и техника. В сб.: Вопросы инженерной психолоrии.
М., 1966, с. 25 35.
Л У Р и я А. Р. Высшие корковые функции человека и их нарушение при
локальных поражениях мозrа. М., 1962.
Л У Р ь е Р. Н., Р У с и н о в В. С. Изменения в ээr человека при световых
раздражениях и следовых процессах. Пробл. физиол. оптики, 1952,
N2 8, с. 80 92.
Л у р ь е Р. Н., Р У с и н о в В. С. Изменения в ээr человека при образова
нии условноrо рефлекса на комплексный раздражитель. Пробл.
физиол. оптики, 1955, N2 11, с. 113 122.
l\fайорчик В. Е., Русинов В. С., Кузнецова r. Д. Динамика KOp
ковых процессов при выработке отставленных условных рефлексов
у человека. В сб.: К физиолоrическому обоснованию нейтрохирурrи
ческих операций. М., 1954, с. 48 59.
М а й о р ч и к В. Е., С пир и н Б. Т. Электрофизиолоrический анализ KOp
ковых процессов при образовании временной связи у человека.
Вопр. нейрохир., 1951, N2 3, с. 3 11.
140
М а к а р о в П. О. Условно рефлекторные изменения функциональноrо co
стояния оптическоrо анализатора и электроэнцефалоrраммы человека.
В сб.: Вопросы физиолоrии нервной и мышечной систем. Тбилиси,
1956, с. 361 371.
М а к а р о в П. о. Условно рефлекторные сдвиrи электроэнцефалоrраммы
человека в микроинтервалах времени, как показатель функциональной
подвижности коры rоловноrо мозrа. Материалы конф. по вопросам
электрофизиолоrии центр. нервной системы. Л., 1957, с. 84.
М а т в е е в Ю. К. Латеральное торможение в нейронных популяциях.
В кн.: Импульсная активность мозrа человека. М., 1971, с. 132.
М е Щ е р с к ий Р. М. Изменение электрической активности в корковом
конце зрительноrо анализатора кролика .при образовании условноrо
оборонительноrо рефлекса на световые раздражители. Труды ин та
высш. нервной деятельности АН СССР, т. 1. М., 1955, с. 265 278.
М о и с е е в а Н. И., О р л о в В. А. Прием статистической обработки данных
биоэлектрической активности rоловноrо мозrа с использованием ЭВМ.
В сб.: Роль rлубоких структур rоловноrо мозrа человека в механизмах
патолоrических реакций. Л., 1965, с. 98 100.
М о Р Д в и н о в Е. Ф., Е r о р о в А. С. Об одном ээr корреляте отношения
человека к пред стоящей деятельности. Вопр. психол. 1971, J'fg 3,
с. 121 125.
М н у х и н а Р. С. Электроэнцефалоrpафические исследования условнореф
лекторных реакций и их анализ в свете теории Н. Е. Введенскоrо.
Л., 1964.
М у ш к и н а Н. А. Динамика выработки наличных и следовых условных
рефлексов уrнетения альфа ритма и дифференцировок к ним. Журн.
высш. нервн. деят., 1956, J'fg 1, с. 157 163.
Н е в с к и й М. П. О биоэлектрической деятельности мозrа в rипнотическом
CHe. Журн. невропатол. и психиатр., 1954, J'fg 1, с. 26 32.
Н о в и к о в а Л. А. Электрофизиолоrическое исследование речевых кино
стезий. Вопр. психол., 1955, J'fg 5, с. 84 94.
Н о в и к о в а Л. А., С о к о л о в Е. Н. Исследование электроэнцефалоrраммы,
двиrательных и кожно rальванических реакций при ориентировочных
и условных рефлексах у человека. Журн. высш. нервн. деят., 1957,
J'fg 3, с. 363 373.
П а в л о в И. П. Собр. соч., т. 111. M. Л., 1951 1952, с. 203.
Пей м ерИ. А. Опыт комплексноrо электрофизиолоrическоrо исследования
больных с отдаленными последствиями закрытой травмы черепа.
Труды ВМА им. Кирова, т. 57. Л., 1954, с. 118 153.
Пей м ерИ. А. Электроэнцефалоrрафическое исследование высшей HepB
ной деятельности человека. Автореф. дисс. канд. Л., 1960.
Пей м ерИ. А., Ф а Д е е в а А. А. Электроэнцефалоrpафия при выработке
условных рефлексов у больных с последствиями закрытых травм
мозrа. Вопр. невропатол. и психиатр., 1957, J'fg 1, с. 51 60.
Пей м ерИ. А. и др. Электрофизиолоrическое и психофизиолоrическое
исследование механизмов передачи информации в центральной нервной
системе человека. Материалы XVIII Междунар. психолоrическоrо KOH
rpecca, симп. 2. М., 1966, с. 184 187.
Рой т б а к А. И., Дендриты и процесс торможения. rаrрские беседы, т. 11.
Тбилиси, 1956, с. 165 187.
Рой т б а к А. И., С а в а н е л л и Н. Д. Электроэнцефалоrрафическое иссле
дование шизофрении. Труды ин та физи_ л-оrии АН rруз. сер, т. 9.
Тбилиси, 1953, с. 201 220.
Р У с и н о в В. С. Учение И. П. Павлова о высшей нервной деятельности
и физиолоrические исследования. Вопр. нейрохир., 1953, J'fg 1, с. 3 8.
Р у с и н о в В. С. Доминантный очаr, вызванный в разных этажах цент--
ральной нервной системы и ero влияние на ход рефлекторной peaK
ции. Материалы VIII Всесоюзн. съезда физиолоrов, биохимиков,
фармаколоrов. Тбилиси, 1955, с. 523 525.
Р У с и н о в В. С. Электрофизиолоrические исследования высшей нервной
деятельности. Журн. высш. нервн. деят., 1957, J'f2 6, с. 854 867.
141
(Русинов В. С., Рабинович М. Н.). R u s i по v V. S., R а Ь i по v i с h М. 1. Ele
ctroencephalographic researches iп the la baratories and c1inics af the Soviet
Union. EEG СНп. N europhysiol., 1958, Suppl. 8. р. 1.
Р у с и н о в В. с. Общие и локальные изменения в электроэнцефалоrрамме
при выработке условных рефлексов у человека. В кн.: Электроэнце
фалоrрафическое исследование высшей нервной деятельности. М., 1962,
с. 288 297.
С а х и у л и н а r. т. Изменение в ээr собак в процесс е образования и уп
рочения условноrо рефлекса. Докл. АН СССР, 1955, NQ 1, с. 153 156.
С а х и у л и н а r. т. Изменение в электроэнцефалоrрамме собак в связи
с образованием условных рефлексов. Журн. высшей нервн. деят., 1957,
NQ 5, с. 741 753.
С е м е н о в с к а я Е. Н., Л у р ь е Р. Н. Изменения ээr зрительных и лоб
ных областей при напряжении внимания. Пробл. физиол. оптики, 1948,
NQ 6, с. 299 304.
С и м о н о в Л. Н. Опытное доказательство существования центров, задер
живающих рефлексы у млекопитающих животных. BoeH. Meд. журн.,
1866, т. 44, кн. XCVII, NQ 1, с. 47.
(Симонов п. В.) S i m о по v Р. V. Emotions Greativity. In: Psychology Today.
1970 р. 51 56.
С и м о н о в П. В. К вопросу о нарастающей тенденции к синтезу наук
(о некоторых проблемах современной психофизиолоrии). В сб.: MeTO
долоrические вопросы физиолоrии высшей нервной деятельности. Л.,
1970, с. 19 32.
С и м о н о в п. В., т е м н и к о в Ф. Е. Адаптивные биоэлектронные системы
восприятия, обучения и управления. В сб.: Бионика. М., 1965, с. 77 83.
С м и р н о в В. М. Психопатолоrическое исследование больных с rиперки
незами, леченных путем воздействия через вживленные электроды на
подкорковые структуры rоловноrо мозrа. Материалы 11 съезда обще
ства психолоrов, в. 4. М., 1963, с. 43 44.
С м и р н о в В. М. Исследование психической деятельности при электриче
ских воздействиях на rлубокие структуры rоловноrо мозrа человека.
В сб.: Роль rлубоких структур rоловноrо мозrа человека в механизмах
патолоrических реакций. Л., 1965, с. 100 111.
С м и р н о в В. М. Исследование механизмов эмоций в процессе лечения
методом долrосрочных интрацеребральных электродов. В сб.: Вопросы
современной психоневролоrии. Труды ин та им. Бехтерева, т. 38. Л.,
1966, с. 120 132.
С м и р н о в В. М. Изучение клинических эффектов при стереотаксических
воздействиях на rлубокие структуры rоловноrо мозrа. В сб.: Проб
лемы локализации и психоневролоrии. Труды ин та им. Бехтерева,
т. 43. Л., 1967, с. 22 48.
(С м и р н о в В. М.) S 111 i r n о v V. М. N europsychologicaI investigations of the
thalamus in mап. Science in progress. October 1968, р. 21 31.
С м и р н о в В. М. Нейропсихолоrия и вопросы изучения rлубоких структур
rоловноrо мозrа человека. Вестн. АМН СССР, 1970, NQ 1, с. 35 42.
С м и р н о в В. М., Ш а н Д у р и н а А. Н. О роли диэнцефально стволовых
и отриопаллидарных структур rоловноrо мозrа в механизмах речи и еЕ
нарушений. Труды 22 ro совещания по проблемам высш. нервнои
деятельности, посвящ. 120 летию со дня рожд. И. П. Павлова. Рязань,
1969, с. 220.
С м и р н о в В. М., С пер а н с к и й М. М. Способ формализованноrо описа
ния постоянноrо потенциала. Физиол. журн. СССР, 1970, NQ 8, с. 1172
1178.
С м и р н о в В. М., Ш а н Д у Р и н а А. Н., r о р е л и к Л. И. О роли под
корковых образований мозrа в механизмах самореrуляции высшей
нервной деятельности человека. Труды XXIII совещ. по проблемам
высшей нервной деятельности, в. 2. rорький, 1972, с. 175.
С о к о л о в Е. Н. Природа фоновой ритмики коры больших полушарий.
В кн.: Основные вопросы электрофизиолоrии центр. нервной системы.
Киев, 1962, с. 157.
142
т р о Ф и м о в Л. r., Л у р ь е Р. Н., Л ю б и м о в Н. Н., Р а б и н о в и ч М. я.
Исследование электрических явлений в области корковых концов раз
.,'ТIИЧНЫХ анализаторов при выработке оборонительных и пищевых yc
ловных рефлексов. Материалы VIII Всесоюзн. съезда физиолоrов,
биохимиков, фармаколоrов. Тбилиси, 1955, с. 613 615.
Т Р о х а ч е в А. и. Динамика клеточной активности подкорковых структур
м:озrа человека в процессе лечения методом долrосрочных rлубинных
интрацеребральных электродов. В сб.: Роль rлубоких структур rолов
Horo мозrа человека в механизмах патолоrических реакций. Л., 1965,
с. 115 117.
Т Р о х а ч е в А. И. Динамика клеточной активности подкорковых структур
м:озrа человека при некоторых моторных и психолоrических пробах.
В сб.: rлубокие структуры rоловноrо мозrа человека в норме и патоло
rии. Л., 1966, с. 157 160.
У х т о м с к и й А. А. Учение о доминанте. Собр. соч., т. 1. Л., 1950.
Ф и л и м о н о в И. Н. Локализация функций в коре большоrо мозrа.
Невропатол. и психиатр., 1940, N2 1 2, с. 14 25.
Х а н а н а ш в и л и М. М. Механизмы нормальной и патолоrической услов
норефлекторной деятельности. Л., 1972.
Ч е р н и r о в с к а я Н. В., Ц у к е р м а н А. С., М о в с и с я н Ц с. А. Опыт
применения метода адаптивноrо автореrулирования в невролоrической
клинике. Материалы симпозиума: «Самореrуляция нейрофизиолоrиче
ских механизмов интеrративной и адаптивной деятельности l\'!озrа».
Л., 1972а, с. 58 60.
Ч е р н и r о в с к а я Н. В., Ц у к е р м а н А. С., М о в с и с я н Ц С. А. Исполь
зование принципа адаптивноrо автоуправляемоrо эксперимента для ле
чения больных с центральными двиrательными нарушениями. MaTe
риалы Междунар. союза теоретической и прикладной биофизики
АН СССР. М., 1972б, с. 142.
Ч е р н и r о в с к а я Н. В., В а с и л е в с к и й Н. Н., Ц у к е р м а н А. С.,
М о в с и с я н Ц с. А. Метод адаптивноrо автореrулирования в изучении
управления двиrательной активностью больных с rиперкинезами.
Материалы 2 ro Междунар. симпозиума по реrуляции движений.
V arna . BuIgaria, Abstracts, 1972Ь, р. 48.
Ч е р н ы ш е в а В. А. Динамика медленных сдвиrов мозrовых потенциалов
в подкорковых образованиях rоловноrо мозrа человека при реализации
психической деятельности в условиях направленноrо изменения BHYT
ренней среды мозrа. Физиол. журн. СССР, 1971, N2 2, с. 150 158.
Ш п и л ь б е р r П. и. Токи действия сетчатки и мозrа человека и живот
ных В изучении зрительной функции. Материалы VII Всесоюз
Horo съезда физиолоrов, биохимиков и фармаколоrов. М., 1947,
с. 280 284.
Я к о в л е в а Е. К. О некоторых особенностях электрической активности
rоловноrо мозrа при наркозе навязчивых состояний. Журн. HeBpo
патол. и психиатр., 1952, N2 6, с. 20 23.
Я к о в л е в а Е. К. 00 отражении в ээr словесных воздействий при неврозе
навязчивых состояний. В сб.: Вопросы теории и практики ээr. Л.,
1956, с. 217 234.
А d е у W. R. Neural information processing; Windows without and the citade 1
within. In: Biocybernetics of the Central N ervous System. Ed. L. D. Proc.
tor. Boston, 1969, р. 1 27.
А d е у W. R. Spontaneous Electrical Brain Rhythms Accompanying Learned Res.
ponses. In: The Neurosciences. New York, 1970, р. 224 243.
А d е у W. R., D u n 1 о р С. W., н е n d r i х С. Е. Hippocampal slow waves,
distribution and phase relationships in tlle course of approach learning.
AN\A Arch. Neurol., Psychiat., 1960, v. 3, р. 74 90.
А d r i а n Е. D. The physiological basis of perception. In: Brain mechanislns
and consciousness. Ed. J. 1. Delafresnaye, Ch. С. Thomas. Springfield, 1954,
р. 237 243.
А d r i а n Е. D., М а t t h е w s В. Н. The interpretation of potential waves in the
cortex. J. Physiol., 1934, v. 81, р. 440.
143
А 1 Ь е F е s s а r d А. Е. Mechal1isms of llervous integration and conscious expe
rience. In: Brain mechanisms and consciousness. Ed. J. F. Delafresnaye,
Ch. С. Thomas. Springfield, 1954, р. 200 236.
А 1 р е r s В. J. Relation of the hypothalamus to disorders of persona1ity. Report
of а case. Arch. Neurol. Pshychiat., 1937, v. 38, р. 291 303.
А n g е 1 е r i F., F е r r о М i 1 о n е F., Р а r i g i S. Electric activity and reactivity
of the rhinencephalic, pararhinencephalic and thalamic structures. Trial of
plolonged ilnplantation of electrodes in human beings. Excerpta Med. Intern.
Congr. series, 1961, v. 37, р. 84 85.
А n g у а 11 А., Н а s z 11 О S Т Цит. по Р. М. Мещерскому, 1955.
А r е 11 а n о А. Р., S с h w а Ь R. S. Scalp and basal recordings during mental
activity. Proc. 1st Intern. Congr. Psychiatr. Paris, 1950.
В а g с h i В. К. The adaptatioll and variability of responses of the human
brain rhythm. J. Physiol., 1937, v. 3, р. 463 485.
В а g с h i В. К., Н о w е 11 R. W., S с h m а 11 Н. Т. The electroencephalogr. and
c1inical effectg of electricaIly induced convulsions in the treatment of menthal
disorders. Аm. J. Pshychiat., 1945, v. 102, N 1, р. 49 50.
В а r Ь i z е t J. Defect of memorizing of hippocampal.mamillary origin. J. Neu.
rol. Neurosurg. Psychiat., 1963, v. 26, р. 127 135.
В а r Ь i z е t J. Etudes sur la memoire. Paris, 1966.
В а t е s J. А. V. Depth eletrodes in the human subject. Basic technical, interpl"e..
tative and ethical considerations. Excerpta Med. Intern. Congr. series, 1961,
v. 37, р. 62 64.
В а u m g а r t h е r G. Die Reaktionel1 dell Neurone des zentralen visuellen Systems
der Katze im simultanen Helligkeitskontrast. In: R. Jung, Н. Kernhuber.
(Red.). N europhysiologie und Psychophysik des vesuellen Systems. Symposium
Freiburg. Berlin, 1961, S. 296 311.
В е r g е r Н. ОЬеl. das Elektronkephalogramm des Menschen. Arch. Psychiat.,
1929, Bd. 87, S. 527.
В е r k h о u t J., W а 1 t е r D. О., А d е у R. AIterations of the human electroen.
cephalogram induced Ьу stressful verbal activity. EEG clin. N europhysiol.,
1969, v. 27, р. 457 469.
R i с k f о r d R. G. а. oth. Changes in memory function produced Ьу electrica 1
stimulatioll of the temporal lobe in mап. In: The brain and human behavior.
Baltimore, 1958, р. 227 243.
В о g о с h S. Brain gIycoprotein 10B: Further evidence of the "Sign pose'. Role
of brain g1icoprotein in cell recognition, its change in brain tumor and the
presence of а "Disfance factor". 111: Structural and functional proteins of
the nervoys system. Ed. А. N. Davison, 1. G. Morgan, Р. Magel. N. У.,
1972, р. 39 54.
В о g о с h S. The Biochemistry of lnemory, with ап inquiry into the function of
the brain mucoids. New York, 1968.
В о u i 11 а u d J. Recherches c1iniques propres а dimontrer que Ia perte de lа pa
role correspond а lа lesioll des lebules anterieurs du cerveau. Arch. Gen.
et Med., 1825, v. VIII.
В о u i 11 а u d J. Traite c1inique et physiologique de l'encepha1ite. Paris, 1825.
В r а d 1 е у Р. В. The pharmacology of the EEG. In: Second Advance Course
in EEG. Salzburg, 1965, р. 157 161.
(Brazier М.). Б рей ж е М. Длительно сохраняющиеся электрические сле
ды в rоловном мозrе человека и их возможное отношение к высшей
нервной деятельности. В кн.: Электроэнцефалоrрафические исследо
вания высшей нервной деятельности. М., 1962, с. 341 353.
В r а z i е r М. А. В. The Contributioll of anestesiology to electroencephalography.
In: Anesthesiology and the Nervous System. Ed. J. В. Dillon а. С. М. Ballin
ger. N. У., 1966, р. 165 187.
В r а z i е r М. А. В. Absence of Dreaming or Failure to Recall from Experimental
Neurology. Suppl. 4; Physiological Correlates of Dreaming. Ed. С. D. Cle
mente. New York London, 1967, р. 91 98.
В r а z i е r М. А. В., В а r 1 о w 1. S. Some appIications of correlation analysis
to clinical problems in electroencephalography. EEG Clin. NeurophysioI.,
1956, v. 8, N 2, р. 325 331.
144
В r а z i е r М. А. В., С а s Ь у J. U. Cross Correlation and autocorrelation studies
of electroencephalographic potentiaIs. EEG CIin. Neurophysiol., 1952, v. 4,
р. 201 211.
(Bremer F.). Б р е м ер Ф. Анализ корковых процессов побуждения. В кв.:
Электроэнцефалоrрафические исследования высшей нервной деятель
ности. М., 1962, с. 119.
Broca Р. Цит. по А. Р. Лурия, 1962.
В u d z у n s k i Т., S t о У v а J., А d 1 е r С. Feedback induced muscle relaxation:
AppIication to tensicJn headache, J. Behav. Ther. Ехр. Psychiat., 1970, v. 1,
р. 205 211.
В u r е n J. М., van. Sentory, motor and autonomic effects of medial temporaI
stimulation in mап. J. Neurosurg., 1961, v. 18, р. 273 288.
В u r е n J. М., van. Confusion and disturbance of speech from stimulation in
vicinity of the head of the caudate nucleus. J. Neurosurg., 1963, v. 20,
N 2, р. 148 157.
В u s е h Е. Psychial symptoms in neurosurgicaI disease. Acta psychiat. neurol.,
1940, v. 15, р. 257 290.
(Chang Н. Т.). Ч а н r Х. Т. Некоторые данные об изменениях возбудимости
корковых и подкорковых нейронов и возможная роль этих изменений
в процессе выработки условных рефлексов. В кн.: Электроэнцефало
rрафЙческие исследования высшей, нервной деятельности. М., 1962,
с. 69.
С h а р m а n W. Р. Studies of the periamygdaloid area in relation to human
behavior. In: The brain and human behavior. BaItimore, 1958, р. 258
277.
С h а р m а n R. М. Ниmап evoked responses to meaningful stimuIi. XVIII In
tern. Congr. of psychology, symp. 6. М., 1966, р. 53 59.
С r е u t z f е 1 d t о. Attention as а Concept in Neurophysiology. Science, 1969,
v. 158, р. 495 496.
D а r r о w С., W i 1 s о n 1., V i е t h Р., 1\'1 а 1 1 е 1. Acceleration and momentum
incerebrbal function reflected in EEG phase relations. In: Recent Advances
BioI. Psychiatry. N. Y. London, 1960.
D е 1 g а d о J. М. R. Effect of Brain stimulation оп Tasn free stimulations.
EEG сliп. N europhysiol., 1963, Suppl. v. 24, р. 260 280.
D е 1 g а d о J. М. R. Modulation of Emotion Ьу CerebraI Radio Stimulation.
In: PhysiologicaI Correlates of Emotion. New York а. London, 1970, р. 189
204.
(DeIgado J. М. R.). д е л ь r а Д о Х. Мозr и сознание. М., 1970.
D е 1 g а d о J. М., Н а m 1 i n Н. Depth electrography. Confin. NeuroI., 1962,
v. 22 (3/5), р. 228 237.
D u r u р G., F е s s а r d А. L'electro encephalogramme de l'homme: Observations
pshychophysiologique relative а l'action des stimuli visuel е! auditif. Апп.
Psychol., 19'35, v. 36, р. 1 35.
F а u r е J., С о h е n S е а t. Responses to sensory stimuli, produced Ьу movie
activation. EEG CIin. NeurophysioI., 1954, v. 6, N 4, р. 523.
F о е r s t е r О., G а g е 1 о. Ein FaII von Ependymcyte des 111 VentrikeIs Eiu
Beitrag zur Frage der Beziehungen psychis cher Storungen zum Hirnstaum.
Zschr. ges N euroI. Psychiat., 1933, Bd. 149, S. 312 344.
G а а r d е r К., С h а s е Ch. ControI of States of Consciousness. 1 AItainment
Through ControI of PsychophysiologicaI VariabIes. Arch. Gen. Psychiat.,
1971а, v. 25, р. 429 435.
G а а r g е r К., С h а s е Ch. ControI of States of Consciousness. 11. Attainment
Through ExternaI Feedback Augmenting ControI of PsychophysiologicaI Va-
riabIes. Arch. Gen. Psychiat., 1971 Ь, v. 25, р. 436 41.
G а s t а u t Н. Etude electrocorticographique de la reactivite des rythmes rolan-
diques. Rev. Neurol., 1952, v. 87, N 2, р. 176 182.
G а s t а u t Н. The role of the reticulal. forll1ation in esta bIishing conditioned
reactions. In: Reticular formation of the brain. Ed. Н. Н. Jasper et al.
Boston, 1958а, р. 561 579.
G а s t а u t Н. Some aspects of the neurophysiologicaI basis of conditioned ref
lexes and behaviour. In: Ciba foundation symposium оп the neurologicaI
145
basis of behaviour, Ed. G. Е. W. Wolstenholme а. С. М. O'Connor. Al11ster-
dam London N. У., 1958Ь, р. 255 272.
G а s t а u t Н. J., В е r t J. EEG changes during cinematographic presentation.
EEG Clin. Neurophysiol., 1954, v. 6, N 3, р. 433 444.
(Gastaut Н. et. aI). r а с т о А. и др. Электроэнцефалоrрафическая -xapaKTe
ристика образования условных рефлексов у человека. Журн. высш.
нервн. деят., 1957, NQ 1, с. 25 38.
G а s t а u t Н. et з1. Etude topographigue des reactions Electroencephalographi
ques conditionees chez l'homme. EEG Clin. Neurophysiol., 1957, v. 9, N 1,
р. 1 34.
(Gastaut Н. et aI.). r а с т о А. и др. Нейрофизиолоrическое объяснение yc
ловных электроэнцефалоrрафических реакций. Журн. высш. нервн.
деят., 1957, М 2, с. 203 213.
(Gastaut N., Roger А.). r а с т о А., Р о ж е. А. Участие основных функцио
нальных структур rоловноrо мозrа в механизмах высшей нервной дея
тельности. В кн.: Электроэнцефалоrрафические исследования высшей
нервной деятельности. М,. 1962, с. 18 41.
G е r а r d R. W. The НхаНоп of experience. In: Brain mechanisms and learning.
Ed. J. F. Delafresnaye. Oxford, 1961, р. 21 32.
G 1 а s s А. Mental arithmetic and blocking of the occipital alp11a rhythlll. EEG
Clin. Neurophysiol., 1964, v. 17, N 6, р. 595 603.
G 1 а s s А. Evidence of the habituation of the EEG to mental arithmeiic. EEG
СIiп. NeurophysioI., 1965, v. 19, N 5, р. 531 533.
G 1 е s s Р., G r i f f i t h Н. В. Bilateral destruction of the hippocampus (corl1u
Ammonis) in а case of dementies. Mschr. Psychiat. Neurol., 1952, Bd. 123,
S. 19 204.
G о 11 а F. L., Н u t t о n Е. L., W а 1 t е r W. Grey. The objective study of menta 1
imagery. J. Ment. Sci., 1943, v. 89, р. 375.
С о m е s А. о. Brain consciousness problem in contemporary scientific research.
In: Brain and conscious experience. Ed. J. Eccles. Berlin Heidelberg Ne\v
York, 1966, р. 446 469.
G r ii n t h а 1 Е. Ober das Corpus mammi1Iare und den Korsakowschen sympto
mokomplex. СопНп Neurol., 1939, Bd. 2, S. 64 95.
Н а s s 1 е r R. Motorische und sensible Effekte umschriebener Reitungen und
AusschaItungen im menschlichen Zwischenhirn. Dtsch. Z. N ervenheilk., 1961,
Bd. 183, S. 148 171.
Н а s s 1 е r R., R i е с h е r t Т., М u n d i n g е r F., U m Ь а с h W., G а n g 1 Ь е r .
g е r Т. Physiological observations in streotaxic operations in extrapyramidaI
motor disturbances. Brain, 1960, v. 83, р. 337 350.
Н е а t h К. Physiological data.electrical recordings. In: Studies in Schizophrenia.
Cambridge', 1954, р. 151 156.
Н е i t h R. G. Closing Remarks with commentary оп depth electroencephalography
in epi1epsy and schizophrenia. In: EEG а. Behavior. N. У., 1963, р. 377
394.
Н е а t h R. G., М i с k 1 е W. А. Evolution оУ 7 years experience with dept11
electrodes studies in human patients. In: Electrical Studies оп the Una.
nesthetized Brain. Е. R. Ramey а. D. S. O'Doherty. Ed. Hoeber. New York,
1960, р. 214 242.
Н о d е s R., Н е а t h R. G., lVl i 1 1 е r W. Н. Еlеktrоепсерhаl0grаms and Subcorti
cograms made before stimulation. In: Studies in Schizophrenia. Cambridge,
1954, р. 157 195.
(Ioshii N. et aI.). И о ш и и Н. и др. Условный рефлекс и электроэнцефало
rрафия. В кн.: Электроэнцефалоrрафические исследования высшей
нервной деятельности. М., 1962, с. 187 198.
1 о s h i i N., Р r u v о t Р., G а s t а u t Н. Electrographi activity of the unsen
cephalic reticular formation during conditioning in the cat. EEG Clin. Neuro
physiol., 1957, v. 9, N 4, р. 595 08.
J а с о Ь s А., F е n t о n С. S. Visual feedback of myoelectric output to facilitate
muscle relaxation in normal persons and patients with neck injuries. Arch.
Physiol. Med., 1969, v. 50, р. 34 39.
J а s р е r Н. Н. PathophysiologicaI Studies of Brain Mechanisms in different
146
states of consciousl1ess. 111: Brail1 al1d cOl1scious experience. Ed. J. Eccles.
Berlil1 Heidelberg Ne\v York, 1966, р. 257 282.
J а s р е r Н. Н., R а s m u s s е 11 Т. Studies of c1il1ical al1d electrical respol1ses to
deep temporal stimulation il1 mа11 \vith some cOl1siderations of fUl1ctiol1al
a11atomy. Res. РиЫ. Ass. l1erv. ment. Dis., 1958, v. 36, р. 316 334.
(J asper Н., Ricci G. F., Doane В.). д ж а с пер r., р и ч ч и Д Ж., Д О У н Б.
Микроэлектродный анализ разрядоJ3 корковых клеток при выработке
оборонительных рефлексов у обезьян. В кн.: Электроэнцефалоrрафи
ческие исследования высшей нервной деятельности. М., 1962, с. 129.
J а s р е r Н., S h а g а s s С. COl1ditioning the occipital alpha rhythm in ma11.
J. Ехр. Psychol., 1941, v. 26, р. 373 388.
J о h 11 Е. R. Mechal1isms of memory. New York, London, 1967.
J о u у е t М., Н е r 11 а n d е z Р е о n R. l\lechanisrnes l1europhysiologiques соп
centrant l'atte11tion, l'habituatiol1 et le conditionnement. EEG Сliп. Neuro
physiol., 1957, Suppl., N 6, р. 39. .
J u 11 g R. Correlatiol1 of bioelectrical al1d autol1omic phel10mena with alteratiol1s
of cOl1sciousness and arousal in ma11. 111: Brail1 lnechal1isms and cOl1scious
11ess. Ed. J. F. Delafresnaye. Springfield, ]954, р. 310 339.
J u 11 g R. Brain processes and learning. In: Brain mechal1isms al1d learl1il1g,
v. 1. Ed. М. А. В. Brazier. Amsterdam, 1963, р. 189 201.
J u 11 g R., Н а s s 1 е r R. The extrapyramidal lnotor system. 111: Handbook of
physiology, sect. 1, v. 11. Washington, 1960, р. 863 927.
J u 11 g R., Т о n n i е s J. D. Hirnelektrische Ul1tersuchungel1 iiber Entstehul1g ul1d
Erha1tung VOI1 Кrаmрfеl1t1апduпgеп: Die Vorgange аm Reizort und dir
Bremsfahigkeit des Gehirl1s. Arch. Psychiatr. Nervel1kr., 1950, Bd. 185, S.
701 735.
(Jus. А., Jus К.) Ю с А., Юс К. Попытка электроэнцефалоrрафическоrо aHa
лиза процессов BHYTpeHHero торможения (запаздывания и ero перехода
в COH.) Журн. невропатол. и психиатр., 1954, NQ 9, с. 715 721.
К а m р А., S с h r i j е r С. F. М., S t о r m v а n L е е u w е n W. Occurance of
"beta bursts" in human frontal cortex related to psychological parameters.
EEG СНп. Neurophysiol., 1972, v. 33, N 3, р. 257 268.
К е i d е 1 W. D. Sinnesphysiologie, Т. 1. Allgemeine Sinnesphysiologie Visualles
System. Ber1in, Heidelberg, N. У., 1971.
К е n 11 а r d М. А., F i s t е r W. Р., R а Ь i n о v i t s h М. S. The use of frequency
analysis in the interpretation of the EEG of patients with psychological
disorders. EEG СНп. N europhysiol., 1955, v. 7, N 3, р. 29 38.
К е n n е d у 1. L., G о t t s d а n k е r R. М., А r m i n g t о n 1. G., G r а у F. Е.
А new EEG associated with hirurnd. Science, 1948, v. 108, р. 527 529.
К i n g Н. Е., J о u n g К. М., С о r r i g а n R., В е r s а d s k у L. Psychological ob
servations before and after stimulations. lп: Studies in Schizophrenia. СаП1
bridge, 1954, р. 309 344.
К 11 о t t J. R. Some effects of "mental set" оп the electrophysiological processes
of the human cerebral cortex. J. Ехр. Psychol., 1939, v. 24, р. 384 405.
К n о t t J. R., Н е n r у Ch. Е. The conditioning of the blocking of the alpha
rhythm of the human EEG. J. Ехр. Psychol., 1941, v. 28, р. 134 144.
(Lal1ge G., Storm van Leeuwen W., Werre J.). Л а н r е r., ш т о р м в а н Л е y
в е н В., В е р р е Д. Корреляция между психолоrическими и электро
энцефалоrрафическими явлениями. В кн.: Электроэнцефалоrрафиче
ские исследования высшей нервной деятельности. М., 1962, с. 369 377.
L а 11 s i n g R. W. Relation of brain and tremor rhythms to visual reaction time.
EEG СНп. Neurophysiol., 1957, v. 9, N 3, р. 497 504.
L а 11 s i s i 1. М. Dissertation de sode cogitandis anima. Ve11esia, 1939.
L а s h 1 е у К. S. Cerebral organization and behavior. In: The Brain and human
behavior. BaItimore, 1958, р. 1 18.
L а u f Ь е r g е r V. La reponse conditionnee par imagination dans l'encephalo
gramme. С. R. Soc. Biol., 1950, v. 144, р. 467 468.
L е 1 о r d G. Different moda1ities of reaction of the central and anterior rhythms
at 10 c/sek. EEG СНп. Neurophysiol., 1957, v. 2, N 3, р. 561.
L е t t v i п, М а t u r а n а, М с С u 11 о с h, Р i t t s. Цит. по Н. Н. Василевскому и
с. и. Сороко, 1969.
147
L е v i t а Е., R i k 1 а n М., С о о р е r S. Psychological comparison of unilateral
and bilateral thalamic surgery. J. Abnormal Psychol., 1967, v. 72, N 3,
р. 251 254.
L i n S h i k у i Ь, W u С h i n е Ь, S u n S h i k h е Ь, К о с h i n у u i i, L i n
J u n g t s а i Влияние умственной деятельности (счет в уме) на ээr лю..
дей различноrо возраста. Acta psychol. Sinica, 1964, v. 3, р. 281 289.
L о о m i s А. L., Н а r v е у Е. N., Н о Ь а r t G. Potencial rhythms of the cerebral
cortex during sleep. Science, 1935а, v. 81, р. 187.
L о о m i s А. L., Н а r v е у Е. N., Н о Ь а r t G. Further observations оп the
potential rhythms of the cerebral cortex during sleep. Science, 1935Ь, v. 82,
р. 198 200.
L о о m i s А. L., Н а r v е у Е. N., Н о Ь а r t G. Brain potentials during hypno
sis. Science, 1936а, v. 83, р. 239 241. . .
L о о m i s А. L., Н а r v е у Е. N., Н о Ь а r t G. Electrical potentials of the
Ьиmап brain. J. Ехр. Psychol., 1936Ь, v. 19, р. 249 279.
М а g n u s О. ТЬе central alpha rhythm ("rhythms еп arceau"). EEG Сliп
Neurophysiol., 1954, v. 6, р. 349 350.
М а h 1 G. F. ЦИТ. по Sperry R. W., 1966.
М а i k о w s k i J. Dynamika powstawania odruchow wyrunkowych w obrazic eeg.
(Doniesienie wstepl1e). Neurol. neurochir. i psychiatr. polska, 1955, r. 5,
N 3, s. 229 241.
М е у е r L. СЬ. А. Anastomisch physiologische Abhandlungen vom Gehirn. Ber
liп Leipzig, 1779.
М е у е r s R., К n о t t J. К., Н а у k е R. А. ,S w е е n е у D. В. ТЬе surgery oi
epilepsy. J. Nel1rosurg., 1950, v. 7, р. 337 346.
М i 1 n е r Н. Laterality effects in Audition. In: Interhemispheric Relations and
Cerebral Dominance. Ed. V. В. Mountcastle, Baltimore, 1962, р. 177 198.
М i 1 n е r В. Brain Mechanisms Suggested Ьу Studies of Temporal Lobes.
In: Brain Mechanisms Underlying Speech and Language. N. У., London, 1967,
р. 122 145.
М i 1 n е r В., Т а у 1 о r L. Right hemisphere superiority in tactile pattern recogni
tion after cerebral commissiurotomy: evidence for nonverbal memory. N euro.
psychologia, 1972, v. 10, р. 1 15.
М о n r о е R. R., Н а е t h R. G. Description of the patient group. In: Studies
in scgizophrenia, сЬар. 8, Sect. 111. Cambridge, 1954, р. 123 130.
(Morruzzi G.). М о р у ц ц и Д ж. Синхронизирующее влияние ствола мозrа
и тормозные механизмы, лежащие в основе возникновения сна под
влиянием сенсорных раздражителей. В кн.: ЭлеRтроэнцефалоrрафи
ческие исследования высшей нервной деятельности. М., 1962, с. 216
240.
М о r r е 11 F., J а s р е r Н. Н. Electrographic studies of the formation of tempo
rary connections in the brain. EEG Сliп. N europhysiol., 1956, v. 8, N 2,
р. 201 214.
М о u n t с а s t 1 е V. В. ТЬе neural replication of sensory events in the somatic
afferent system. In: Brain and Conscious experience. Ed. J. Eccles. Ber
1in Heidelberg New York, 1966, р. 85 115.
М u n d у С а s t 1 е А. С. EEG and mental activity. EEG C1in. Neurophysiol.,
1957, v. 2, N 4, р. 643.
N i е 1 s е n J. М. Cerebral 10calization and the psychoses. In: ТЬе brain and
Ьиmап behavior. Baltimore, 1958, р. 467 477.
О j е m а n n G., F е d i о Р. Effect of Stimulation of the human thalamus and
parietal and temporal white matter оп short term memory. J. Neurosurg,
1968, v. 29, N 1, р. 51 59.
О j е m а n n G. А., F е d i о Р., В u r е m J. М., van. Anemia from pulvinar and
subcortical parietal stimulation. Brain, 1968Ь, v. 91 (1), р. 99 116.
О 1 d s J. а. М i 1 n е r В. Positive reinforcement produced Ьу electrical stimula
tion od septal area and other regions of rat. In: Brain Physiology and
psyhychology. Ed. С. R. Evans а, А. D. J. Robertson. London, 1966, р. 51 64.
О r t h n е r Н. Anatomic pathologiques des trobles de la conscience d'origine by
pothalamique. Premiere Congr. Int. Sci. N eurol., Sec. J. Сотm. Wien, 1957,
р. 77 96.
148
Р е а с о с k S. М. Physiological responses to subcortical stimulation. In: Stu
dies in Schizophrenia. Cambridge, 1954, р. 235 250.
Реп f i е 1 d W. The role of the temporal cortex in recal1 of past experience and
interpretation of the present. In: Ciba foundation symposium оп the Neurolo
gical Basis of Behaviour. London, 1958, р. 149 172.
Реп f i е 1 d W. G., J а s р е r Н. Epi1epsy and the functional anatomy of the
human brain. Boston, 1954.
Р е t s с h е Н. Die Effassung von Form und VerhaIten der Potential felder ап
der HirnoberfHiche durch eine Kombinierte EEG toposkopische Methode. In:
\Veiner Zeitschrift fiir Nervenhei1kunde und deren Grenzgebiete. Wien. N. У.,
1967, S. 373 387.
Р е t s с h е Н. Epi1eptischer Anfall und Kortikale Neuronenpopulation. Wien. Z.
Nervenhei1k., 1968, Bd. 26, S. 46 55.
Р r i Ь r а m К. Н. The neurophysiology of remembering. Sci. Аm. J., 1969, v. 220,
N 1, р. 73 86.
Р r i Ь r а m К. Languages of the brain. New Jersey, 1971.
R о Ь е r t s L. Functional plasticity in cortical speech areas and the integration
of speech. In: The brain and human behavior. Baltimore, 1958, р. 449
466.
R о g е r А., S о k о 1 о w Е. N., V о r о n i n L. G. The motor conditioning during
wakefulness and during sleep. EEG Clin. Neurophysiol., 1957, v. 9, N 3,
р. 561 562.
S е m J а с о Ь s е n С. W. Depth electrographic recording in mап. Excerpta Med.,
1961, v. 37, р. 81 92.
S е m J а с о Ь s е n С. W. Electrical stimulation of the human brain. EEG C1in.
Neurophysiol., 1964, v. 17, р. 211.
S е m J а с о Ь s е n С. W. Depth electrographic stimulation and treatment of pa
tients with parkinson's disease including neurosurgical technique. Acta Neu
rol. Scand., 1965, v. 41, Suppl. 13, р. 365 377.
S е т . J а с о Ь s е n С. W. Depth electrographic observations related to parkin
son's disease. 2nd Symp. Parkinson's Dis. N. У., Part 2, 1966, р. 388.
S е m . J а с о Ь s е n С. W. Depth electrographic studies of the human brain and
behavior. Springfield, 1968.
S е m . J а с о Ь s е n С. W., в i с k f о r d R. G., D о d g е Jr. Н. W., Р е t е r s е n
М. С. Ниmап olfactory responses recorded Ьу depth electrography. Proc.
Мауо Clin., 1953, v. 28, р. 166 170.
S е m . J а с о Ь s е n С. W., Р е t е r s е n М. С., D о d g е Н. W., L а z а r t е J. А.,
Н о 1 m а n С. В. Elektroencephalographic rhythms from the depth of the
parietal, occipital and temporal lobes in тап. EEG Clin. N europhysiol., 1956,
v. 8, р. 263 278.
S е m . J а с о Ь s е n С. W., т о r k i d s е n А. Depth recording and electrical sti
mulation in the human brain. In: Electrical studies оп the inasthetized bra
in. N. У., 1960, р. 275.
S h а g а s s Ch. Conditioning the human occipital alpha rhythm to а voluntary
stymulus. J. Ехр. Psychol., 1952, v. 31, р. 367 379.
S h а g а s s С., С а n t е r А. Some personality correlates of cerebral evoked
response characteristics. XVIII. Intern. Congr. of Psychology, Symp. 6. М.,
1966, р. 47 52.
S h е r r i n g t о n С. S. Мап and his nature. London, 1942.
S h е r r i n g t о n С. S. The Integrative action of the N ervous System. London,
1947.
S h i Р t о n J., W а 1 t е r W. G. Les relations entre les activites alpha, les modes
de pensee et les affinites sociales. EEG Clin. Neurophysiol., 1957, Suppl. 6,
р. 185 202.
S 1 а t t е r К. Н. Alpha rhythms and mental imagery. EEG Clin. Neurophysiol.,
1960, v. 12, N 4, р. 851 859.
S Р е r r у R. W. Brain bisection and mechanisms of consciousness. In: Brain
and conscious experience. Ed. J. Eccles. Berlin Heidelberg New York,
1966, р. 298 313.
S Р i е g е 1 Е. А., W у с i s Н. Т., О r с h i n i k С. а. F r е е d Н. Thalamic chrono
taraxis. Аm. J. Psychiat., 1956, v. 113, р. 97 1 05.
149
S р i е g е 1 Е. А., W u с i s Н. Т. Stimulatiol1 of Ьrаil1stеП1 and basa1 ganglia il1
mап. 1п: Electrical stimulation of t11e brain. Austin, 1961, р. 487 497.
S t е v е n s Ch. F. Synaptic Physiology. Inst. Electr. Electron. Engin., 1968.
June, v. 56, N 6, р. 916 930.
S t о r m v а n L е е u w е n W., к а m р А., 1< о k М. L., Т i е 1 е n А. 1\'1. R е 1 а t i оп
entre l'activite electrique de certaines structures cerebrales et lе comportement
du chien. Rev. Neurol., 1963, v. 109, N 3, р. 258 259.
Т r а v i s L. Е. Brain potentials and the temporal course of consciousness. J.
Ехр. Psychol., 1937, v. 21, р. 302 309.
Т r а v i s L. Е., Е g а n 1. Р. Increase in frequency of t11e alpha rhythm Ьу yerbal
stimulation. J. Ехр. Psychol., 1938, v. 23, р. 384 393.
U m Ь а с h W. Cortica1 responses to subcortical stimulation of the diffuse projec
ting system in 662 stereotaxic operations in mап. Excerpta Nled. Intern.
congr. series, 1961, v. 37, р. 76.
U m Ь а с h W. Vegetative Reactionen bei elektrisc11er Reizung und Ausschaltung
in subcorticalen Hirnstrukturen des Menschen. Acta neuroveget. (Wien),
1961, Bd. 23, S. 225 245.
U m Ь а с h W. Elektrophysiologische uпd vegetative Pharomena bei stereotaktis
chell Hirnoperationen. Berlil1, Heidelberg, N. У., 1966.
V е r d е а u х G., V е r d е а u х J. Il1teret de l'enregistrement des 1.hythmes biolo
giques dans 1а recherche radiophonique (electroencephalographie polygrap
hie). Cahiers studes radio telev., 1955, v. 5, р. 78 81.
V i с t о r М. а. oth. Memory 10ss with 1esions of hippocampal formation. Arch.
Neurol., 1961, v. 5, N 3. р. 244 263.
V о g е 1 W., В r о v е r m а n D. М., К 1 а i Ь е r Е. L. EEG and mental abilities.
EEG Сliп. Neurophysiol., 1968, v. 24, р. 166 175.
V о 1 а v k а J., М а t о u s е k .L\1., R о u Ь i с ek J. Menta1 arithmetic and еуе ope
ning. Ап. EEG frequency analysis and GSP study. EEG Сliп. Neurophysiol.,
1967, v. 22, р. 174 176.
W а 1 k е r J., Н а 1 а s Е. Neuronal coding at subcortica1 auditory l1uclei. Physiol.
Behav., 1972, v. 8, N 6, р. 1099 1105.
W а 1 t е r \V. G. The Iiving brain. London, 1953.
W а 1 t е r W. G. Brain mechanisms and perception. Brit. J. Physio1. Optics,
1965, v. 22, N 1, р. 1 9.
W а 1 t е r W. G. The analysis, synthesis and identification of evoked responses
and CNV. EEG Сliп. Neurophysiol., 1967, v. 23, р. 489.
W а 1 t е r W. G. а. oth. The interaction od responses to semantic stimuli in the
human brain. EEG CIin. Neurophysio1., 1965, v. 18, р. 513 533.
Wa I t е r W. G., С о о р е r R., А I d r i d g е V. J., М с С а 11 u m W. С., W i n
t е r А. L. Contingent negative variation: ап electric sign of sensorY Inotor
association and expectancy in the human brain. Nature, 1964, v. 203, р. 380.
W а I t е r W. G., С r о w Н. J. Depth recording from the human brain. Excerpta
Med., 1961, v. 37, р. 64 65.
W а I t е r W. G., G r о \v N. S. Depth recording from the human brain. EEG
Сliп Neurophysio1., 1964, v. 16, р. 68 79.
W а 1 t е r W. G., S h i Р t о n Н. W. The effect of synchl'onizing light and sound
stimuIi with various components of the electro.encopha logram. J. Physiol.,
1949, v. 108, N 3, р. 50.
W а 1 t е r R. D., С h а р m а n L., Р о r t е r R., С r а n d а 1 Р., R а n d R. Beha
vioral responses to stimulation of human l1ippocampus, hippocampal gyrus
and amugda1e. EEG Сliп. Neurophysiol., 1964, v. 17, р. 451 472.
W а 1 t е r R. D., У е а g е r С. L. Visual imagery and elektroencephalograp11ic
changes. EEG C1in. Neurophysiol., 1956, v. 8, N 2, р. 193 199.
W е r n i с k е С. Der aphasische Sympomen.komplex. Breslau, 1874.
W h а t m о r е G. В., К о h I i D. R. Dysponesis: а neurophysiological factor in
functional disorders. Behav. Sci., 1968, У. 13, р. 102 124.
W i 1 I i а m s D. The abnormal cortical potentials associaied with high intracra
nial pressure. Brain, 1939, v. 62, N 3, р. 321 334.
W i 11 i а m s М., Р n n у Ь а с k е r S. Memory disturbances in third ,, entricle
tumours. J. Neu ol. Neurosurg. Psychiat., 1954, v. 17, р. 115 123.
W i 11 i s Т. Cerebral anatomy. London, 1964.
150
Bechtereva N. Р.
The neurophysiological aspects of mental activity in тап. The second edition,
remaked and supplemented, 1974.
In the monograph, the 111ain stages of the investigation into the physiology
of cerebral sustaining of mental processes, and the moderl1 data оп the l1europhy
siological mechanisms of these processes obtained in t11e direct study of the
human brain physiology, are presented.
Мапу years of observati0n of the patients during their diagnosis al1d treat
ment with the aid of implanted electrodes Ьу means of а complex method invol
ving examination of the brain physiological paran1eters during mental activity
and of the spontaneous and evoked mental processes after local electrical effects
оп the brain, made it possible to accumulate vast amount of ne\v data 011 the
physiological mechanisms of mental phenon1ena. The analysis of these data
suggests that the cerebral sustаiпiпg of mental activity is carried out Ьу ihe
cortical subcortical structural functional system with links of different grades of
rigidity.
The main theoretical perspectives of development of the problem and the
possibilities for enlargement of the range of therapeutic means in the psychiatric
and neurological cIinics, are considered оп the ground of the new evidence соп.
cerning t11e human brain.
The 2nd edition is completed with the data оп the finest characteristics of
the impulse activity of neuronal assemblies of the human brain during mental
processes.
The monograph is meant for physiologists, neurologists, psychiatrists, and
physicians of other specialities.
The book contains 33 figures. The reference amounts to 337 titles.
Наталья Петровна Бехтерева
НЕйРоФизиолоrИЧЕСКИЕАСПЕКТЫ
ПСИХИЧЕСКОй ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
Издание второе, переработанное и дополненное
Редактор Т. И. Трекова
Переплет художника В. В. Белякова
Художественный редактор А. И. Прuймак
Технический редактор Т. Т. Лебедева
Корректор Р. И. rольдuна
Сдано в набор 26/11 1974 r. Подписано к печати 7 jVI 1974 r. Формат бумаrи 60Х90 1 /16. Бу.
l\1ara машиномелованная. Печ. л. 9,5. Бум. л. 4,75. Уч.
изд. л. 11,06. ЛН
71. Тираж 5000 ЭК3.
Заказ N2 544. Цена 1 р. 32 к.
Ленинrрад, «Медицина», Ленинrрадское отделение. 192104, Ленинrрад, ул. Некрасова, 10.
Ленинrрадская типоrрафия N2 4 Союзполиrрафпрома при rосударственном комитете Совета
Министров СССР по делам издательств, полиrрафии и книжной торrовли, 196126, Ленинrрад,
Ф
126, Социалистическая ул., 14.