/








































































































































































































































































































































































































































































































































Похожие




Текст
НЕ. СОКОЛОВ
Систематика млекопитающих
ОТРЯДЫ: КИТООБРАЗНЫХ, ХИЩНЫХ, ЛАСТОНОГИХ, ТРУБКОЗУБЫХ, , ХОБОТНЫХ, ДАМАНОВ, СИРЕН, ПАРНОКОПЫТНЫХ, МОЗОЛЕНОГИХ, НЕПАРНОКОПЫТНЫХ
ДОПУЩЕНО МИНИСТЕРСТВОМ ВЫСШЕГО
И СРЕДНЕГО СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР
В КАЧЕСТВЕ УЧЕБНОГО ПОСОБИЯ
ДЛЯ БИОЛОГИЧЕСКИХ СПЕЦИАЛЬНОСТЕЙ ВЫСШИХ
УЧЕБНЫХ ЗАВЕДЕНИИ
6
МОСКВА «ВЫСШАЯ ШКОЛА» 1979
С 59
Рецензенты:
кафедра охраны природы Казанского государственного университета (зав. кафедрой проф. В. А. Попов); проф. Б. А. Кузнецов (зав. кафедрой зоологии Московской сельскохозяйственной академии)
Соколов В. Е.
С 59 Систематика млекопитающих (китообразные, хищные, ластоногие, трубкозубые, хоботные, даманы, сирены, парнокопытные, мозоленогие, непарнокопытные): Учеб, пособие. — М.: Высш, школа, 1979. — 528 с., ил.
В пер.: 2 р.
Книга завершает 3-томное издание (первые два тома вышли в 1973 и 1977 гг.) сводки систематики мировой фауны современных млекопитающих. Это уникальное издание содержит все известные в мировой зоологической литературе сведения по систематике млекопитающих.
В книге приводятся характеристики отрядов, семейств, кратко — родов, полный перечень видов. В характеристике указываются их морфологические особенности, биология, распространение и хозяйственное значение. Указываются редкие виды и подвиды млекопитающих, включенные в «Красную книгу».
Предназначается для студентов университетов. Может быть использована преподавателями вузов, научными сотрудниками-биологами, охотоведами, работниками охраны природы и преподавателями биологии средней школы.
21008__291
С----------— 74—79 2005000000
001(01)—79
С59
ББК 28.6
Владимир Евгеньевич Соколов
СИСТЕМАТИКА МЛЕКОПИТАЮЩИХ
Редакторы Л. П. Петровская, Т. А. Рыкова. Художники: М. Никитин, В. Иванов, В. Козлов, А. Коленков. Переплет художника А. В. Алексеева. Художественный редактор Т. А. Коленкова. Технический редактор Н. В. Баранова. Корректор С. К. Завьялова.
ИБ № 1720
Изд. № Е—330. Сдано в набор 14.08.78. Подп. в печать 22.05.79. Т-11101. Формат 70X100’/ie. Бум. тип. № 2. Гарнитура литературная. Печать высокая. Объем 42.90 усл. печ. л., 42,89 уч,-
изд. л. Тираж 25 000 экз. Зак. № 671. Цеиа 2 руб.
Издательство «Высшая школа», Москва, К-51. Неглинная ул., д. 29/14
Ярославский полиграфном бипат Союзполиграфпрома при Государственном комитете СССР »о делам издательств, полиграфии и книжной торговли. 150014, Ярославль, ул. Свободы, 97.
от АВТОРА
Предлагаемая книга завершает трехтомное издание сводки мировой фауны современных млекопитающих. Первый том (издательство «Высшая школа», 1973) включает отряды однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых и ящеров. Второй том (издательство «Высшая школа», 1977) посвящен зайцеобразным и грызунам. В третьем томе дается описание отрядов китообразных, хищных, ластоногих, трубкозубых, хоботных, сирен, даманов, парнокопытных, мозоленогих и непарнокопытных. В связи с тем что общий очерк морфологии млекопитающих приведен в первом томе, во втором и третьем томах объяснение морфологическим терминам не дается.
Книга предназначена в основном для студентов, поэтому в диагнозах опущены некоторые детали, в большинстве случаев не даются ссылки на использованные источники, а в список литературы включены лишь основные сводки или работы, разрешающие спорные вопросы. Однако, учитывая, что публикация систематики современных млекопитающих в отечественной литературе предпринимается впервые и настоящее издание может представлять интерес и для научных работников, в книге приведен объем знаний, превышающий тот, который читается автором в университетском курсе «Систематика млекопитающих». Особенно это относится к морфологическим характеристикам и географическому распространению. В книгу включен перечень видов зарубежной фауны, отсутствующий в университетском курсе. Данные по числу семейств, родов и видов мировой фауны млекопитающих, принятых в настоящем издании «Систематики млекопитающих», и их распределение по отрядам сведены в таблицу. Эти материалы даются в сопоставлении с данными Симпсона (Simpson, 1945), Моррисона (Morrison, 1965) и Андерсона и Джонса (Anderson and Jones, 1967).
Иллюстрации для книги взяты в основном из Уокера и др. (Walker, 1964), Грассе (Grasse, 1955), Холла и Келсона (Hail and Kelson, 1959) и некоторых других.
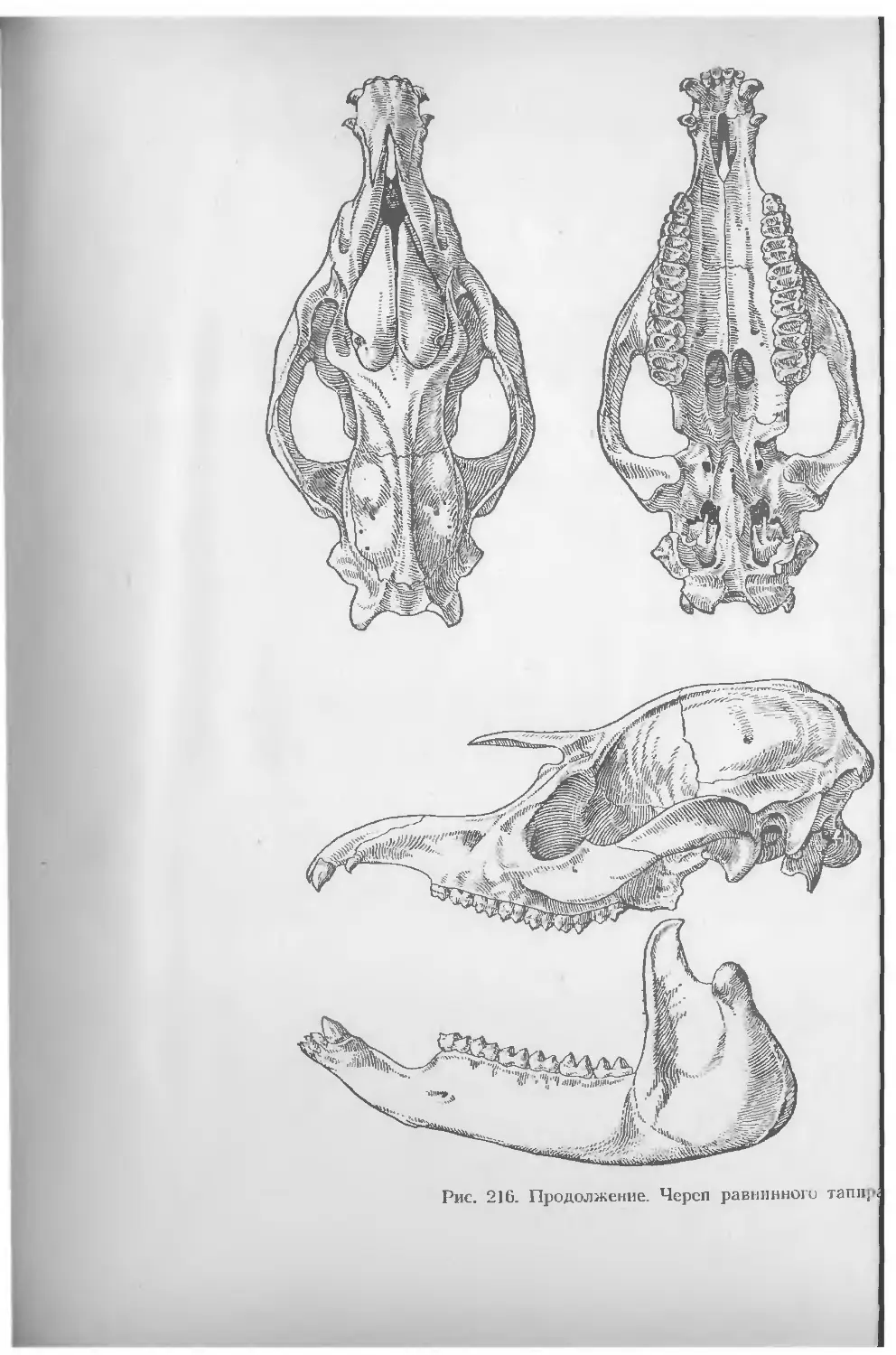
Ряд фотографий зверей с натуры сделаны автором (рис. 28, 55, А, 124, 130, 139, 171, 189, 210,Б, 211,А, 218); рисунки черепов по материалам Зоологического музея МГУ изготовлены М. Никитиным (рис. 3, 7, 16, 47, 64, 122, 134, 150, 151, 152, 195, 198), В. Ивановым (рис. 148, 165, 168, 190, 193, 202, 218), В. Козловым (рис. 124, 163, 180, 179, 206, 215) и А. Коленковым (рис. 58, 65, 127, 136, 167, 209, 212, 213, 214, 216).
Приношу глубокую благодарность за большую помощь в работе профессорам Б. А. Кузнецову, А. П. Кузякину и В. А. Попову и руководству Зоологического музея МГУ за предоставленную розможн^1к.жцользй'
Система класса млекопитающих к настоящему времени даже в его высших таксономических категориях окончательно не установлена и вызывает споры. Так некоторые систематики отряды хищных и ластоногих объединяют в один отряд, отряд китообразных разделяют на два отряда. Большинство ученых включают отряд мозоленогих в отряд парнокопытных. Естественно, разногласия возрастают с понижением рангов таксонов. Для иллюстрации цифровых расхождений в количестве семейств, родов и видов класса млекопитающих, подсчитанных разными систематиками, приводится следующая таблица.
Количество семейств, родов и видов в различных отрядах класса млекопитающих
Отряд Число
по Симпсону. 1945 по Моррисону, 1965 по Андерсону и Джонсу. 1967 по В. Соколову, 1973—1979
семейств родов семейств 1 родов ВИДОВ 1 семейств , родов ВИДОВ a S родов | ВИДОВ I
Monotremata Bonaparte, 1838 2 3 2 3 6 2 3 6 2 3 6
Marsupialia liliger, 1811 8 60 8 80 248 8 81 242 9 71 253
Insectivora Bowdich, 1821 8 68 8 62 374 8 77 406 7 61 376
Dermoptera liliger, 1811 1 I 1 I 2 1 1 2 1 1 2
Chiroptera Blumenbach, 1779 17 116 17 180 981 16 173 875 17 148 942
Primates Linnaeus, 1758 11 59 11 58 193 10 47 166 11 53 195
Edentata Gray, 1798 3 14 3 14 32 3 14 31 3 14 29
Pholidota Weber, 1904 I 1 1 1 7 I 1 8 I I 7
Lagomorpha Brandt, 1855 2 10 2 9 66 2 9 63 2 11 61
Rodentia Bowdich, 1821 32 339 32 363 1729 34 354 1687 35 355 1594
Cetacea Brisson, 1762 9 35 10 38 92 102 382 842 8 38 83
Carnivora Bowdich, 1821 7V 96H 7 95 252 7 96 253 7 90 232
Pinnipedia liliger, 1811 3J 16/ 3 21 32 3 20 31 3 21 34
Tubulidentata Huxley, 1872 1 1 1 I 1 1 1 I I I I
Proboscidea liliger, 1811 1 2 1 2 2 1 2 2 I 2 2
Hyracoidea Huxley, 1869 1 3 I 2 6 I 3 11 1 3 11
Sirenia liliger, 1811 2 2 2 2 4 2 3 5 3 3 5
Artiodactyla Owen, 1848 8f 84V 8 77 188 8V 73V 16711 8 68 159
Tylopoda liliger, 1811 If 2J I 3 6 И 2J 4J 1 2 4
Perissodactyla Owen, 1848 3 6 3 0 16 9 6 16 3 6 16
| 121 | 918 1122 |1017 |4237 122 | 1004 [ 4060 !124|952 |4012
1 Объединены в один отряд; .
г Андерсон и Джонс (1967) разделяют отряд Cetacea на два отряда: Mysticeti н Odontecetl.
ОТРЯД КИТООБРАЗНЫХ
ORDO CETACEA BRISSON, 1762
К отряду принадлежат высокоспециализированные млекопитающие, приспособившиеся к постоянной жизни в воде.
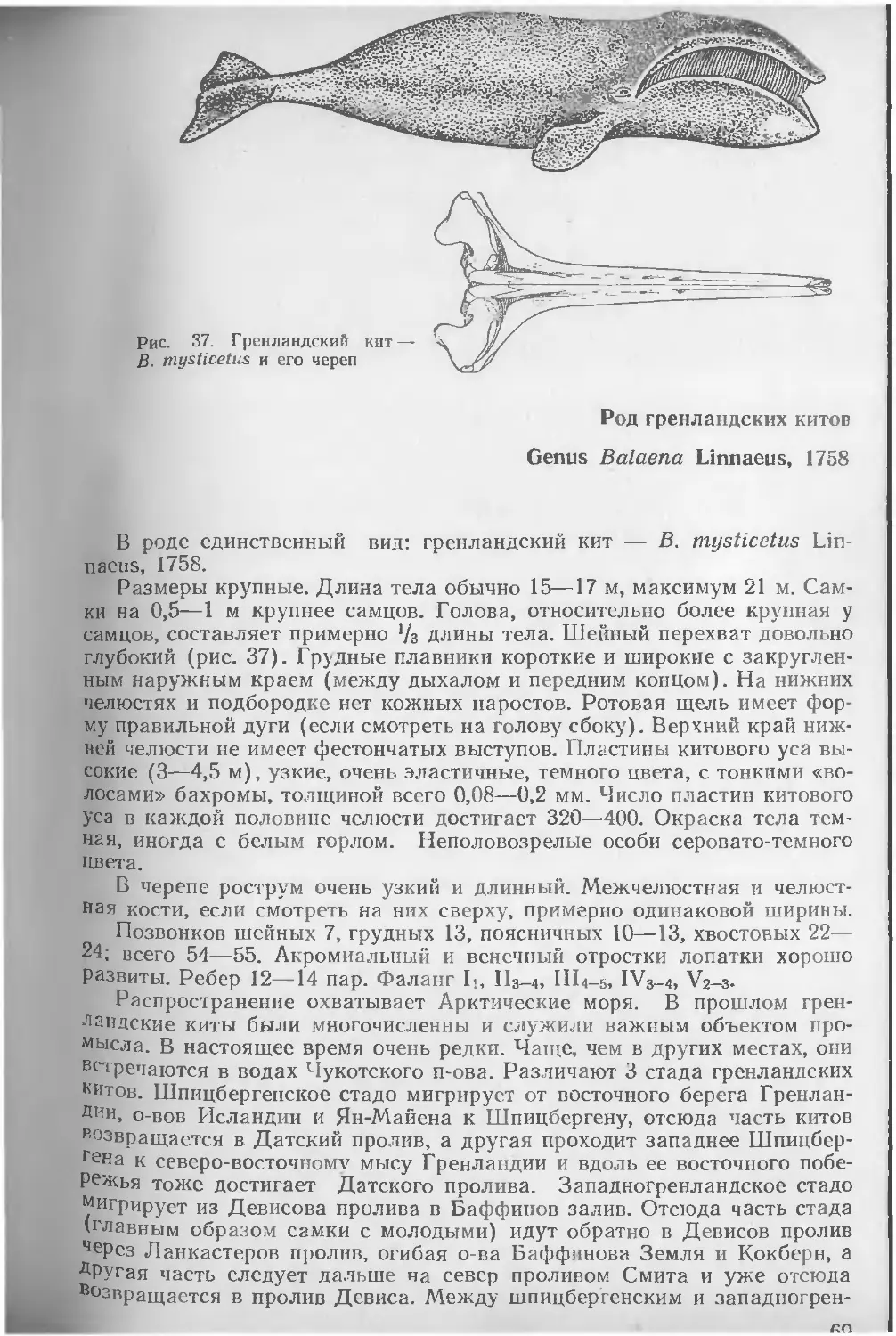
Размеры средние и крупные (наибольшие среди млекопитающих). Длина тела животных от 1,1 м, масса 30 кг у некоторых дельфинов, до 33 м и 150 т у голубого кита Наименьшее сопротивление воды при плавании обеспечивается хорошо обтекаемым телом, торпедообразным или каплевидным, вытянутым или несколько укороченным (рис. 1). Этому способствуют также редукция ушных раковин и расположение сосков в особых карманах и пениса в специальной сумке. Передние конечности превратились в ласты (грудные плавники), которые служат в основном рулями. Свободные задние конечности подверглись атрофии. На конце латерально уплощенного хвостового стебля имеются парные, горизонтально расположенные бесскелетные хвостовые лопасти. Хвостовой отдел туловища служит основным локомоторным органом. У большинства видов на спине находится непарный бесскелетный спинной плавник, который служит своего рода стабилизатором при плавании. Голова часто массивная, более или менее удлиненная. Ее терминальная часть кончается тупо, заострена или имеет вытянутый рострум-—«клюв». Почти без видимого шейного перехвата голова переходит в туловище, постепенно без резкой границы сужающееся в хвостовой стебель.
Характерно для китообразных, что у них нет сплошного волосяного покрова. У усатых китов на морде расположены отдельные щетинковидные волоски, обладающие типичным строением вибрисс наземных млекопитающих; по-видимому, они служат органом осязания и играют определенную роль при отыскании массовых скоплений планктона. Во взрослом состоянии у зубатых китов вибриссами обладают только Platanista gangetica и Inia goeffrensis, живущие в мутной речной воде; вибриссы у них наблюдаются не только на морде, но и на туловище. У большинства других зубатых китов (кроме белухи, нарвала) вибриссы имеют только эмбрионы. Кожные железы полностью отсутствуют, за исключением млечных желез. В коже на внутренней поверхности эпидермиса расположены многочисленные ячеи, в которые поднимаются дермальные сосочки. Некоторые ученые предполагают, что своеобразное строение кожи китов обусловливает возникновение ламинарного потока воды вокруг плывущего животного, что позволяет затрачивать минимум усилий при плавании и развивать большую скорость.
Окраска тела обусловлена пигментом, локализующимся в основном в эпидермисе. У некоторых китообразных она подвержена возрастной изменчивости (например, белуха, нарвал).
Лопасти хвостового плавника и спинной плавник относятся к кожным образованиям. Наряду с обычными артериями и венами в плавниках
* В отРяле можно выделить китообразных малых размеров с длиной тела 1,1—12 м, редких — с длиной тела 12,1—22 и крупных — 22,1—33 м.
Рис. 1. Относительные размеры и форма тела китообразных:
1 — дельфин-белобочка, 2 — грин да, 3 — косатка, 4 — северный плавун; 5 — кашалот самка, 6 — каталог самец» 7 — голубой кит, 8 — финвал, 9 — сейвал, 10 — горбатый кит. 11 — малый полосатик, 12 — серый кит, /3 — гренландский кит
проходят характерные только для них комплексные сосуды, каждый из которых состоит из крупной артерии с очень толстой средней оболочкой, окруженной 10—13 более мелкими тонкостенными венами. В основании спинного плавника дельфина-белобочки таких сосудов насчитывается свыше двух десятков, в хвостовом и грудном плавниках их значительно меньше (Томилин, 1957).
В черепе китообразных кости располагаются симметрично или асимметрично. В последнем случае кости правой стороны расширены и смещены влево, а левой — утолщены. Одни кости могут надвигаться на другие. Мозговой отдел черепа укорочен и округлен. Верхнезатылочная кость сильно развита и спереди причленяется к лобным, в то время как теменные смещаются в стороны. Носовые кости невелики по размерам, так что наружные костные носовые отверстия сдвинуты далеко назад и располагаются непосредственно перед мозговым черепом. Носовые ходы короткие и проходят более или менее вертикально. Верхнечелюстные и межчелюстные кости и сошник сильно вытянуты и образуют рострум. Костное небо удлинено за счет крупных крыловидных костей, вследствие чего хоаны оказываются смещенными назад. Нижние челюсти длинные и имеют рудиментарный венечный отросток, которого иногда нет вовсе.
Рис. 2. Скелет и контуры дельфина-белобочки — Delphinus delphis
Во взрослом состоянии зубы (гомодонтного типа) имеют только представители подотряда зубатых китов. У усатых китов закладки зубов имеются только у эмбрионов. Для взрослых усатых китов характерны особые роговые образования эпителия неба — так называемые «усы», функционально заменившие зубы. «Усы» служат для отцеживания мелких планктонных организмов, которыми питаются эти киты. Акт пережевывания пищи у китообразных утрачен.
Кости посткраниального скелета в большинстве своем губчатого строения с большим содержанием жира. В поясе передних конечностей отсутствует ключица (рис. 2). Лопатка широкая,' вееровидная со слабо развитым гребнем. Грудина сильно варьирует по форме и расчлененности. Плечевая кость чрезвычайно укорочена. Локтевая и лучевая кости также сильно укорочены, уплощены и расширены. В связи с тем что нагрузка на конечность происходит перпендикулярно к ней, конечность уплощается в том же направлении. Подвижное сочленение находится только в плечевом суставе. Все кости свободной конечности имеют общий кожный покров и сочленены между собой неподвижно, а иногда и срастаются друг с другом. Чаше, по сравнению с другими млекопитающими, наблюдается слияние карпальных костей. Пальцев 4 или 5. Средние обладают увеличенным количеством фаланг, крайние — уменьшенным. При сравнении числа фаланг пальцев кисти взрослых китообразных с эмбрионами отмечается их сокращение с возрастом.
Закладка задних конечностей появляется на ранней эмбриональной стадии, но вскоре исчезает. У взрослых китообразных от пояса задних конечностей сохраняются только рудименты таза, расположенные в мускулатуре по большей части в виде двух костных стерженьков (слившихся подвздошной, седалищной и лобковой костей). Связь с позвоночником эти кости утрачивают, но к ним прикрепляются мышцы мочеполовой системы, в связи с чем, очевидно, и не происходит их полная редукция.
Позвоночник состоит из четырех отделов: шейного, грудного, поясничного и хвостового. Крестцовые позвонки отсутствуют вследствие утраты задних конечностей. Шейный отдел представлен семью сильно Укороченными позвонками, которые могут сливаться между собой в одну или несколько групп. В остальных отделах количество позвонков непостоянно. В хвостовом отделе развиваются гемальные дуги, аналогичные дугам низших позвоночных. У быстроплавающих китов число хвостовых позвонков возрастает.
Число ребер варьирует от 10 до 17 пар. Нормальное сочленение с позвонками имеют передние 2—8-я пары; у задних ребер головка и шейка отсутствуют. К грудине присоединяются передние 1—8-я пары ребер. Для глубоконыряющих китов характерно уменьшенное .число ребси. при
крепляющихся к грудине (у кашалота — три) или развитие в ребрах хрящевых отделов (клюворылы). Грудная клетка в отличие от большинства наземных млекопитающих не сжата с боков, а округлая в поперечнике или несколько уплощена дорзо-вентрально.
Язык хорошо развит, мягкие губы отсутствуют. Слюнных желез нет или они рудиментарны.
Желудок сложный, состоит из 3—5 отделов. Кишечник достигает различной длины: он может быть в 5—6 раз длиннее тела (усатые киты, клюворылы) или в 15—16 раз (кашалот, афалина) и до 32 раз (дельфин Ла-Платы).
Печень относительно невелика, желчного пузыря нет. Поджелудочная железа удлиненная и слабо дольчатая, значительно реже разъединенная. Ее протоки открываются в желчный проток.
Наружные ноздри парные у усатых китов, непарные (одна ноздря) у зубатых. Они смещены на верх головы и обладают особыми клапанами, запирающими эти отверстия при нырянии. Трахея и бронхи укорочены, что способствует убыстрению акта дыхания. Легкие однодольчатые с сильно развитой гладкой мускулатурой. Число альвеол относительно больше и размеры их намного крупнее, чем у наземных млекопитающих.
Китообразные способны подолгу не дышать, находясь под водой от 2—10 до 30—40 мин (по некоторым данным, до 2 ч). Длительность ныряния у них обеспечивают значительная емкость легких, слабая чувствительность дыхательного центра к накоплению в организме углекислоты, повышенное содержание миоглобина. Кислородная емкость крови несколько увеличена за счет более высокого содержания гемоглобина в крови и повышения его концентрации в эритроцитах.
Процесс дыхания китообразных обычно можно подразделить на выдох после длительного ныряния, промежуточные короткие вдохи и выдохи и глубокий вдох перед длительным нырянием. Во время промежуточных вдохов и выдохов кит ныряет неглубоко, плывет почти всегда по прямой линии, дышит через правильные промежутки времени. Число промежуточных вдохов — выдохов различно у разных видов и тем больше, чем дольше остается животное под водой во время основного ныряния. Выдох может начинаться у поверхности под водой, в результате чего выдыхаемый с силой воздух образует водяной фонтан, форма и размеры которого в общем характерны для различных видов Ч
Некоторые китообразные способны нырять на значительную глубину (кашалоты — свыше 1000 м). Быстрый подъем нырнувшего животного с большой глубины на поверхность воды должен вызвать выделение из крови газообразного азота, растворенного в ней вследствие повышенного давления при нырянии. Пузырьки азота могут закупоривать кровеносные сосуды, вызывая кессонную болезнь и даже смерть. Однако у китообразных кессонной болезни не бывает. Вероятно, это объясняется тем, что количество азота в легких не намного превышает азотную емкость тканей кита, а воздух в процессе ныряния в легкие больше не поступает (в противоположность тому, что имеет место у водолазов).
Головной мозг китообразных во многих отношениях высоко дифференцирован. Однако по ряду показателей сохраняет весьма примитивные признаки, которые, видимо, были характерны для мозга млекопитающих исходного типа. Масса головного мозга в абсолютных цифрах (белуха—• 1,6—2,3 кг; кашалот — 7,8—9,2 кг) достигает наибольшей величины среди млекопитающих, но относительно массы всего тела она незначитель-
1 По-видимому, фонтан может образовываться и в результате конденсирования пара в теплом выдыхаемом воздухе (в высоких широтах), и в результате захватывания и распыления воды, которая остается при выныривании кита в ложбине, где располагается дыхало.
на (морская свинья — 0,8%, голубой кит — 0,007%). В укороченном, имеющем округлую форму головном мозгу крупные извилины и борозды располагаются в продольном направлении. Характерно сильное развитие коры полушарий переднего мозга. Происходит редукция Rhino-cephalon. Bulbus olfactorius у большинства китообразных отсутствует (bulbus olfactorius описана у бутылконоса, голубого кита и финвала).
Глаза небольшие. Роговица и склера достигают значительной толщины. Хрусталик имеет своеобразную округлую форму. Веки неразвиты. Зрение китообразных, по-видимому, монокулярное, они близоруки. Слезные железы редуцированы, отсутствует и слезно-носовой проход. Жирный секрет гардеровой железы предохраняет глаз от механического и химического воздействия воды. Имеются конъюнктивные железы, не встречающиеся у других млекопитающих.
Органы слуха сильно видоизменены. Ушная раковина редуцирована. Наружный слуховой проход открывается позади глаза маленьким отверстием. Интересно мнение, что рудиментарный слуховой проход может служить самостоятельным чувствующим органом, воспринимающим изменение давления. Барабанная перепонка выгнута наружу (усатые киты) или внутрь (зубатые киты). С внешней стороны барабанная перепонка усатых китов покрыта своеобразной ушной пробкой, состоящей из ороговевшего эпителия и ушной серы. Китообразные способны улавливать широкий диапазон звуковых волн от 150 до 120—140 тыс. Гц (Slijper, 1962), т. е. даже ультразвуковые колебания. Высокая степень развития слуховых отделов мозга зубатых китов свидетельствует об особой остроте их слуха, почти уникальной среди млекопитающих; у усатых китов слух хуже по сравнению с наземными млекопитающими. Китообразные способны к эхолокации, как и летучие мыши. В связи с тем что голосовые связки у китообразных отсутствуют, они не могут издавать звуки обычным для млекопитающих способом. Возможно, звуки производятся в результате вибрации нижней части перегородки между носовыми мешками или колебаниями складки наружного клапана в результате пропускания воздуха из дорзальных носовых мешков. Дельфины способны испускать серии коротких звуковых импульсов, длительность которых составляет 1 мс, а частота следования меняется от 1—2 до нескольких сот герц.
В кровеносной системе китообразных сильно развита артериальная чудесная сеть (retia mirabilia), находящаяся на вентральной стороне позвоночного стебля, в области шеи, грудной клетки, между ребрами, в основании головного и вокруг спинного мозга. Она образована огромным числом извилистых артерий мускульного типа, связанных между собой многими анастомозами. Имеется и венозная чудесная сеть, располагающаяся в основании черепа и в области живота.
Температура тела китообразных сходна с температурой наземных млекопитающих и колеблется в пределах от 35 до 40°С (верхняя граница отмечалась у раненых китов или пойманных после преследования дельфинов). Сохранение высокой температуры тела в воде, проводящей тепло во много раз лучше, чем воздух, осуществляется толстым слоем подкожной жировой клетчатки в коже.
Почки китообразных многодольчатые, относительно более крупные, чем у наземных млекопитающих. Мочевой пузырь небольшой и в шейке не имеет сфинктера, что способствует частому мочеиспусканию небольших порций мочи, которая, возможно, может служить сигналом для других китов. Китообразные, по-видимому, не пьют морской воды. Они получают необходимую влагу из пищи, используя и оксидазную воду.
Семенники располагаются в брюшной полости. Из дополнительных Желез имеется только простата. Os penis отсутствует. Матка двурогая.
У самок усатых китов половое и заднепроходное отверстия разделяются друг от друга значительным пространством, а у зубатых они расположены в едином углублении и окружены общим сфинктером. Самцы постоянно или очень подолгу в течение года способны к оплодотворению. Высказываются предположения, что овуляция у китообразных провоцируется половым актом. У самок в начале беременности в матке могут находиться два или три зародыша, из которых вскоре остается только один. Плацента диффузного типа.
Роды происходят под водой. Детеныш рождается вполне развитый, способный к самостоятельному передвижению. Пропорции его тела весьма сходны с пропорциями тела взрослых китов, а размеры достигают V2—’А длины тела матери. Самки некоторых китообразных могут быть оплодотворены вскоре после родов во время лактационного периода. Кормление детенышей совершается под водой, длительность каждого приема пищи -— несколько секунд. Молоко вбрызгивается в ротовую полость детеныша сокращением особых мышц самки. Млечные железы самки располагаются по бокам полового отверстия. Два соска (по одному с каждой стороны) залегают в щелевидных складках и только в период лактации выдаются наружу. Самки китов продуцируют различное количество молока в сутки: от 200—1200 г у дельфинов до 90—150 л у финвала и 200 л у голубого кита (Слепцов, 1955). Молоко густое, обычно кремового цвета. Характерно, что его поверхностное натяжение в 30 раз больше, чем у воды, что особенно важно, так как струя молока не расплывается в воде. Питательность китового молока весьма высока.
Рост детенышей во время вскармливания их молоком происходит быстро. Например, детеныш голубого кита за 7 месяцев жизни вырастает с 7 до 16 м, т. е. среднесуточный прирост в длину составляет 4,5 см.
Половой диморфизм проявляется главным образом в различной длине тела самцов и самок. Самки усатых китов крупнее самцов, а большинства зубатых, наоборот, меньше. Диплоидное число хромосом у Твидов зубатых китов и 4 видов усатых (сейвала, малого полосатика, финвала и серого кита) равно 44, а у кашалотовых — 42.
Распространены во всех океанах и в большинстве морей Земного шара. Факторы, определяющие размещение китообразных, — наличие пиши и температура воды. Некоторые виды распространены широко и встречаются как в теплых; так и в холодных морях (некоторые виды из семейства дельфинов), другие имеют меньший ареал (серые киты обитают в субтропических, умеренных и холодных водах северной половины Тихого океана и в Чукотском море), ареал третьих еще более ограничен (нарвал не покидает вод Арктики), наконец, ареал речных, озерных и эстуарных форм совсем незначителен.
Большинство видов — стадные животные; держатся группами от нескольких голов до сотен и тысяч особей Встречаются как возле побережий, так и в открытом море. Представители некоторых видов способны подниматься вверх по крупным рекам, впадающим в море, а отдельные виды постоянно живут в реках. У большинства китов питание специализировано, и среди них встречаются планктофаги, теутофаги, ихтиофаги и сакрофаги. Кормятся массовой или штучной добычей. Среди китообразных есть быстрые пловцы (косатки, многие дельфины) и относительно медленно передвигающиеся (серые киты). Большинство китов постоянно держатся в поверхностных водах. Некоторые, например кашалот, могут нырять на значительную глубину. Численность различных видов китообразных неодинакова. Многие из них весьма многочисленны и могут встречаться тысячными стадами (дельфин-белобочка), другие, напротив, очень редки и встречи с ними отмечены всего несколько раз (некоторые представители рода ремнезубов, карликовый кашалот, кар
ликовая косатка). Естественные колебания численности не изучены; вряд ли они могут быть значительными.
Чрезмерно интенсивный промысел пагубным образом сказывается на численности китов, заметно снижая ее, и в некоторых случаях может угрожать полным уничтожением этих животных. 'Гак, ничтожная в настоящее время численность гренландских китов — следствие хищнического выбоя этих многочисленных когда-то животных.
Для большинства видов характерны периодические миграции. У некоторых видов протяженность миграционных путей относительно невелика (азовочерноморская морская свинья —из Азовского моря в Черное и обратно); у других огромна (некоторые крупные киты — от тропических вод до высоких широт).
Китообразные — преимущественно моногамы. Периоды спаривания и щенки обычно растянуты во времени. Рождают одного, редко двух детенышей. Сильно развит материнский инстинкт.
Врагов, кроме человека и косатки, практически не имеют. Остатки дельфинов находили в желудках тигровых и гренландских акул.
Почти 100% особей заражены эндо- и эктопаразитами. Эктопаразиты: китовые вши — Cyamus, копепода — Penella, усоногие ракообразные— Conchoderma — свойственны особенно крупным китам.
Практическое значение в недалеком прошлом было довольно велико. Некоторых из китообразных и в настоящее время добывают ежегодно в больших количествах (кашалоты), других промышляют лишь случайно. Почти все органы кита используются для выработки ценных пищевых и технических продуктов. Промысел китообразных может вестись только при строгом соблюдении мер, необходимых для поддержания их численности. В настоящее время в целях сохранения китообразных целесообразно прекратить их промысел на ряд лет.
Ископаемых остатков китообразных известно относительно немного. Наиболее примитивные из известных нам китообразных (Protocetus, подотряд Archaeoceti) были найдены в среднем эоцене Северной Африки. Представители подотряда Archaeoceti образовали специализированную ветвь, дивергировавшую от ранних предковых форм и развивавшуюся самостоятельно до полного исчезновения в верхнем эоцене. Ископаемые остатки из позднего среднего и верхнего эоцена свидетельствуют, что Archaeoceti имели две основные группы, различающиеся по размерам и скелету. Представитель одной из них — Basilosaurus, имел змееподобное тело длиной 17—22 м, специализированный позвоночник, способный изгибаться, как у змеи, и горизонтально расположенный маленький хвостовой плавник. Представители другой группы — Dorudon, Prozeuglodon, были значительно меньших размеров и походили на дельфинов. Их позвоночник напоминает позвоночник современных дельфинов. О предках китообразных нет единого мнения. Ряд ученых высказываются в пользу происхождения китообразных от копытных, с которыми они имеют ряд общих черт: диффузную плаценту, двурогую матку, сложный желудок. Анализы числа хромосом и изучение реакции преципитации белка крови китообразных подтвердили эту гипотезу. Другие исследователи на основании сходства в строении скелета и зубной системы древних китообразных с креодонтами предполагают, что китообразные произошли от по-еледних. Высказывается предположение, что родоначальниками китообразных явились более древние формы, чем креодонты, а именно меловые насекомоядные. Некоторые ученые (например, Beddard, 1900; Клей-ненберг, 1958; Андерсон и Джонс, 1967) предполагают днфилитическое происхождение отряда китообразных. Эволюция усатых и зубатых китов, По их мнению, шла путем конвергенции, а не дивергенции, и, следовательно, правильнее считать эти подотряды отрядами. В подтверждение
И
своей гипотезы авторы приводят многочисленные морфологические различия между усатыми и зубатыми китами. Проблема дифилитического происхождения китообразных разработана еще недостаточно и требует дальнейшего изучения. Наиболее древние остатки усатых китов известны из верхнего олигоцена, а зубатых — из значительно более древних отложений — верхнего эоцена. Предки усатых китов имели зубы, о чем свидетельствуют не только палеонтологические, но и эмбриологические данные.
Систематика китообразных к настоящему времени изучена недостаточно полно. Как уже сказано выше, даже целостность всего отряда ставится некоторыми учеными под сомнение. Объемы некоторых семейств также не установлены окончательно.
В отряде китообразных обычно различают 2 современных подотряда: зубатых китов — Odontoceti, Flower, 1867;
усатых китов — Mysticeti Flower, 1864.
В фауне СССР представлены оба подотряда.
ПОДОТРЯД ЗУБАТЫХ КИТОВ
SUBORDO ODONTOCETI FLOWER, 1867
Размеры мелкие и средние. Длина тела 1,1—21 м.
Череп резко асимметричен, хотя на ранних стадиях эмбриогенеза он характеризуется всеми признаками, свойственными черепу наземных млекопитающих. Причины асимметрии до сих пор точно не установлены. Одни исследователи предполагают возможность ее возникновения вследствие неодинакового давления воды при плавании на разные участки черепа, другие — из-за редукции обонятельных нервов. Носовые кости развиты слабо и не прикрывают заднюю часть костного носового отверстия (рис. 3). Сами отверстия смещены влево и открываются в общую камеру. Верхнечелюстные, межчелюстные и носовые кости сильно надвинуты на лобные и почти целиком закрывают их. В черепе имеются примитивные черты строения, о чем свидетельствует нахождение в нем os parasphenoideum, tabularia и postfrontalia.
О 1 65
Зубов от — или -у до, примерно .
Зубы гомодонтные, монофиодонтные. Выделяют (Яблоков, 1958а) 3 типа зубов.
1. Простые колышковидиые зубы с сильно развитой полостью пульпы и тонкими слоями цемента и эмали у взрослых животных. Этот тип зубной системы встречается у дельфина-белобочки, обыкновенной морской свиньи, гринды и др. Кроме гринды, все они имеют большое количество зубов, равномерно распределенных.
2. Для зубов второго типа свойственно сильное развитие слоя цемента и отсутствие эмали на зубах взрослых особей (у молодых на коронке имеется тонкий слой эмали). Зубы простые колышкообразные, крупнее, чем у первого типа, их число достигает 30—50. Полость пульпы хорошо развита или отсутствует. Этот тип зубов имеют кашалот и, по-видимому, карликовый кашалот, белуха, серый дельфин, карликовая косатка, ира-вадийский дельфин, малая косатка.
3. Плоские клиновидные зубы третьего типа обладают сильно развитым слоем эмали и цементом, заполняющим полость и охватывающим таким образом весь зуб, за исключением коронки. Причем слой 12
Рис. 3. Череп дельфина-белобочки — Delphinus delphis сверху, снизу, сбоку:
1 — лобная кость, 2 — носовая, 3 — верхнечелюстная, 4 — межчелюстная, 5 — чешуйчатая, 6 —скуловая, ✓ — крыловидная, 8 — затылочная, 9 — небная, 10'— костное носовое отверстие, 11 — хоаны
цемента своеобразно налегает на слой эмали в средней части зуба. Число зубов невелико, и располагаются они только в нижней челюсти. Этот тип свойствен северному плавуну, клюворылу и, по-видимому, остальным представителям этого семейства, кроме тасманова клюворыла.
Зубы косатки занимают промежуточное положение между первым и вторым типами — они похожи на зубы первого типа, но отличаются от
них большими размерами, сравнительно небольшой пульпарной полостью и большим количеством дентина и напоминают зубы второго типа, но в противоположность последним имеют слабо развитый слой цемента. Для большинства представителей зубатых китов характерна индивидуальная изменчивость числа зубов. У некоторых китов количество зубов в верхней челюсти меньше, чем в нижней (северный плавун, кашалот, серый дельфин и Др.), у других, наоборот, в верхней челюсти число зубов больше, чем в нижней (нарвал, белуха, обыкновенная и бесперая морские свиньи и др.), наконец, у третьих число зубов в верхней и нижней челюстях одинаковое.
Пищеварительная система характеризуется полным и постоянным отделением от дыхательного тракта. Она имеет следующие основные черты. 1. Язык в отличие от наземных млекопитающих несет иную функцию. Будучи в целом очень подвижным, он ориентирует пойманную добычу в ротовой полости, проталкивает ее в глотку и препятствует попаданию туда воды. 2. Мягкое небо утрачено. 3. Начальные отделы пищеварительного тракта покрыты изнутри многослойным ороговевшим эпителием, что, очевидно, предохраняет их от повреждения твердыми частями целиком проглатываемой пиши. 4. Многокамерный мускулистый желудок имеет (Слепцов, 1955) в первом своем отделе у видов, питающихся преимущественно головоногими моллюсками (кашалот, северный плавун), выстилку из железистого эпителия, по другим сведениям (Яблоков, 19586) у кашалота первый отдел желудка имеет обычный многослойный эпителий кожного типа. Последние отделы желудка, по-видимому, участвуют в процессе всасывания пищи, так как их строение сходно со строением кишечника. 5. Слепая кишка отсутствует (она имеется у гангского дельфина), а отделы кишечника разграничены друг от друга нечетко.
В дыхательной системе непарная ноздря запирается плотными кожно-мышечными складками таким образом, что выступы одной заходят во впадину другой. Над черепом располагается ряд особых парных воздухоносных мешков: верхних — дорзальных, нижних — премаксиллярных, задних — назофронтальных с дополнительными полостями и боковых — антеролатеральных (рис. 4). Основная их функция состоит в запирании носового прохода при нырянии, причем чем больше давление воды, тем сильнее их действие. Кроме того, они, по-видимому, участвуют в издава-нии звуков. Гортань вдается в хоаны в виде трубки, образованной вытянутыми черпаловидными хрящами и надгортанником. Эту трубку охватывает особый мышечный сфинктер. В бронхиолях есть кольцевые гладкомышечные сфинктеры (у бутылконосов отсутствуют), развитые наиболее хорошо у мелких видов, характеризующихся частыми выныриваниями и, соответственно, частым дыханием и имеющих большую относительную емкость легких. Очевидно, система сфинктеров служит приспособлением к изменению давления в легких во время частых ныряний и сильных вдохов и выдохов.
В головном мозгу хорошо развиты вентральные ядра слухового нерва и передние оливы.
Половой диморфизм проявляется наиболее резко в размерах животных: самцы большинства видов крупнее самок. У некоторых видов имеются и другие отличия, так, у самцов косаток спинной плавник более высокий, чем у самок, для самцов нарвалов (главным образом) характерен бивень.
Распространены во всех океанах и почти во всех открытых морях Земного шара.
Почти все зубатые киты быстрые пловцы, что обусловлено особенностями их питания. Некоторые способны к глубокому и длительному по-
1 л
6
Рис. 4. Воздушные мешки у морской свиньи — Р. pho-саепа (А) и белухи — D. ieucos (Б) (схема расположения):
I — дорзальные, 2 — премаксиллярные, 3 — назофронтальиые, 4— актеролатеральные, 5— дополнительные полости назофрон-тальных мешков, 6 — надчерепной носовой проход, 7 —- костные носовые проходы
гружению. Держатся как у берегов, так и в открытом море. Некоторые заходят в реки, а представители семейства речных дельфинов постоянно обитают в реках (например, в Ганге, Амазонке). Все представители зубатых китов в отличие от усатых питаются штучной добычей. В зависимости от преимущественного питания тем или иным видом пищи выделяют ряд приспособительных групп (Томилин, 1957). 1. Ихтиофаги питаются главным образом стайной пелагической рыбой. Рострум у них длинный и узкий, зубы многочисленные. Сюда, в частности, относятся дельфины-белобочки и продельфины. 2. Бенто-ихтиофаги питаются в основном придонными рыбами и беспозвоночными. Рострум значительно укорочен, число зубов меньше. К этой группе принадлежат морские свиньи, белухи, афалины. 3. Теутофаги кормятся преимущественно головоногими моллюсками и имеют небольшое количество зубов, причем на верхней челюсти их нет вовсе. К ним принадлежат кашалоты, клюворылы, бутылконосы. 4. Теутоихтиофаги поедают головоногих моллюсков и рыб (гринды, малые косатки) и занимают промежуточное положение между 2-й и 3-й группами. Рострум у них укорочен, но зато расширен за счет межчелюстных костей. Хотя зубы и уменьшены в числе, но они хорошо развиты. 5. Сакрофаги (косатки) наряду с рыбами питаются и крупными морскими млекопитающими. Имеют короткие, но мощные челюсти с немногочисленными, но очень крепкими зубами.
Практическое значение зубатых китов довольно велико. Кашалотов ежегодно добывают в больших количествах. В то же время ряд редких и малочисленных видов практического значения не имеет. Многие виды зубатых китов нуждаются в охране.
Система зубатых китов изучена недостаточно хорошо. Существенны расхождения среди ученых даже по числу и объему семейств. Единствен-пое, в чем сходятся все систематики, — выделение двух семейств: речных Дельфинов — Platanistidae Gray, 1863, и клюворыловых —Ziphiidae Cray, 1865. Большинство исследователей объединяют кашалота и карликового кашалота в семейство кашалотовых —Physeteridae Gill, 1872, однако есть предложение выделить карликового кашалота в отдельное семейство Kogiidae (Nishiwaki, 1972). Наибольшие расхождения касаются дельфинов. Одни исследователи (например, Томилин, 1957) всех Пх .объединяют в одно семейство Delphinidae, другие (Hershkowitz, 1966;
Rice a. Scheffer, 1968; Яблоков и др., 1972) выделяют из этого семейства нарвалов и белух в отдельное семейство Monodontidae, третьи (Simpson, 1945) считают отдельным семейством также и морских свиней — Phocoe-nidae, четвертые (Fraser a. Purves, 1960) обособляют еще одно семейство—-длинномордых дельфинов — Stenidae. Предлагается (Nishiwaki, 1972) еще более дробная система: Monodontidae, Delphinidae, Orcaeiidae, Phocoenidae, Grampidae, Globicephalidae.
До получения более обоснованных данных представляется правильным объединение дельфинов в 2 семейства: Delphinidae Gray, 1821, и Monodontidae Gray, 1821.
Таким образом, подотряд зубатых китов объединяет 5 современных семейств (см. табл.):
речных дельфинов —-Platanistidae Gray, 1863 (4 рода);
дельфиновых — Delphinidae Gray, 1821 (19 родов);
нарваловых — Monodontidae Gray, 1821 (2 рода);
кашалотовых — Physeteridae Gray, 1821 (2 рода); клюворыловых — Ziphiidae Gray, 1865 (5 родов). В СССР все семейства, кроме речных дельфинов.
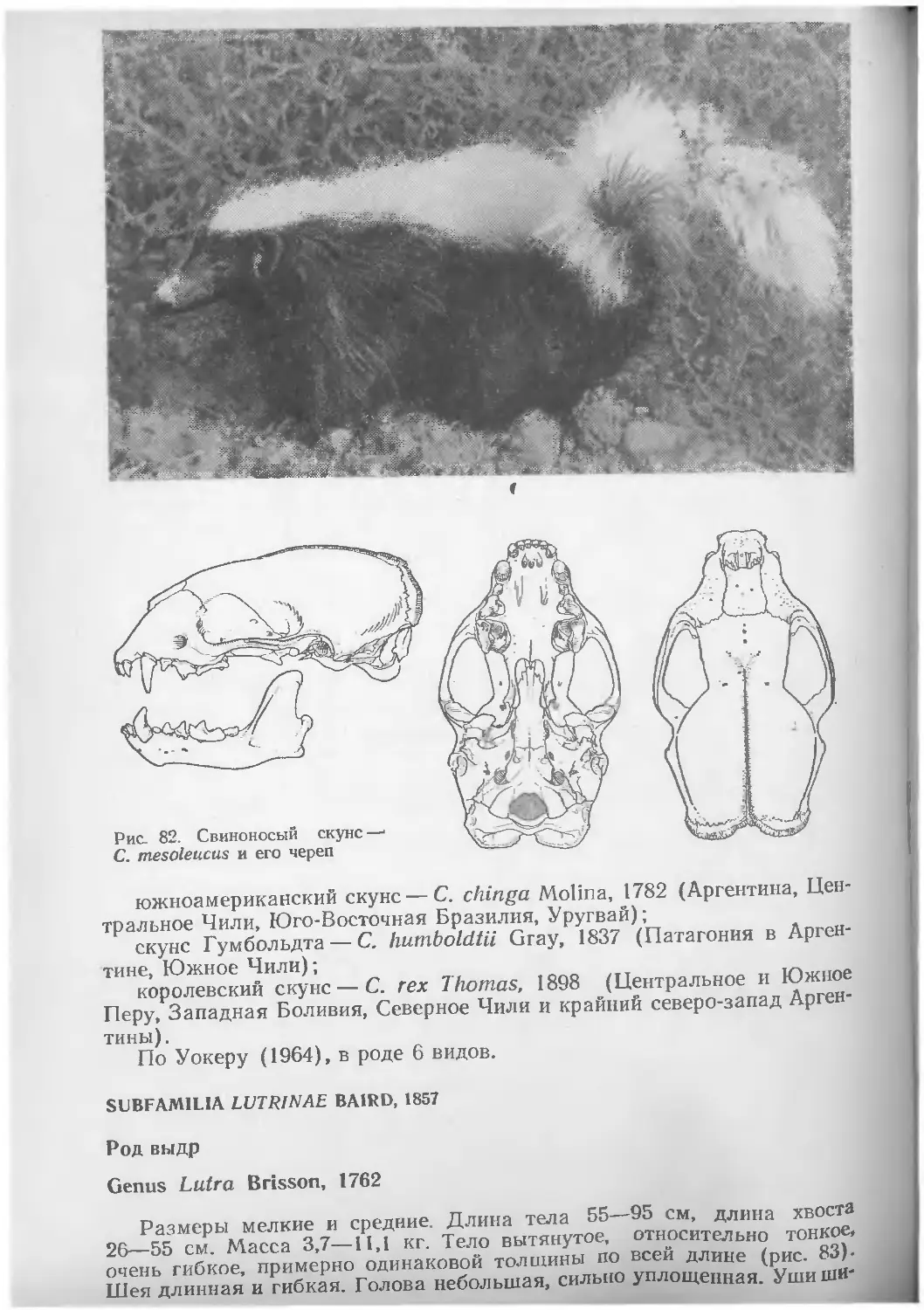
СЕМЕЙСТВО РЕЧНЫХ ДЕЛЬФИНОВ
FAMILIA PLATANISTIDAE GRAY, 1863
Размеры мелкие. Длина тела 1,5—2,9 м. Масса до 40—123 кг. Самки несколько крупнее самцов. «Клюв» тонкий и длинный (Че—V? длины тела), резко отделенный от лобного выступа. Лобный выступ иногда покатый или несколько более выпуклый. У амазонского и гангского дельфинов имеется шейный перехват. Грудные плавники широкие и короткие. Спинной плавник небольшой у амазонского и гангского дельфинов и относительно крупный у дельфина Ла-Платы и озерного. Окраска тела сильно варьирует от бурой или черноватой до почти белой. Зрение развито слабо или полностью отсутствует, например у гангских дельфинов, в глазах которых нет хрусталика.
Череп речных дельфинов почти симметричен. Половинки нижней че-люсти срастаются между собой (или по крайней мере тесно прилегают друг к другу) примерно на протяжении половины своей длины. Наружный край верхнечелюстной кости с продольным гребнем.
Позвонков от 41 до 45. Шейные не срастаются между собой.
Зубов у лаплатских дельфинов от 200 до 242 в обеих челюстях, у представителей других родов от 100 до 130.
Распространены в р. Инд, Ганг и Брахмапутра в Индии, Бангладеш и Пакистане; в оз. Дунтинху в Китае; в р. Амазонке и Ориноко; в р. Ла-Плате и вдоль атлантического побережья от центральной Бразилии до Центральной Аргентины в Южной Америке.
Обитают преимущественно в реках и их устьях. Питаются различными рыбами и пресноводными ракообразными. Держатся обычно поодиночке или маленькими группами по 2—12 особей. Менее активны, чем представители семейства дельфинов. Продолжительность ныряния невелика — не более нескольких минут. Размножение почти не изучено. Хозяйственного значения не имеют. В ряде мест нуждаются в охране.
В семействе 4 рода (4 вида):
лаплатских дельфинов — Pontoporia Gray, 1846 (1 вид);
амазонских дельфинов — Inia d’Orbigny, 1834 (1 вид);
озерных дельфинов — Lipotes Miller, 1918 (1 вид);
гангских дельфинов — Platanista Wagler, 1830 (I вид).
Рис. 5. Лаплатский дельфин — Р. blainvillei
Род лаплатских дельфинов
Genus Pontoporia Gray, 1846
В роде единственный вид: лаплатский дельфин — Р. blainvillei Gervais et d’Orbigny, 1844.
Длина тела 1,5—1,75 м. Масса от 29 до 61 кг. Характерен очень длинный (относительно более длинный, чем у других китообразных) и узкий «клюв», отделенный заметной бороздой от лобного возвышения (рис. 5). Голова округлая. Спинной плавник высокий, треугольный. Он слегка вырезан по заднему краю и расположен несколько кзади от середины туловища. Окраска серая, немного более светлая на брюшной стороне.
Зубов в каждой половине челюсти от 50 до 61.
Распространен вдоль Атлантического побережья Южной Америки от 30° ю. ш. в Бразилии до п-ова Вальдес в Аргентине (42°30' ю. ш.), а также в Ла-Плате. Отмечены сезонные миграции: зимой перемещается к северу вдоль побережья Бразилии.
Экология изучена слабо. Питается в основном рыбами (Clupeidae, Penaeus), а также кальмарами. В поисках пищи, вероятно, использует эхолокацию.
Рис. 6. Амазонский лельфпн — I. geoffrensis
Род амазонских дельфинов
Genus Inia d’Orbigny, 1834
В роде единственный вид: амазонский дельфин — /. geoffrensis Bla-inville, 1817.
Крупные речные дельфины с длиной тела до 3 м. Самка длиной 216 см имела массу 67,5 кг (Pilleri и Gihr, 1969). Длинный «клюв» слегка изогнут вниз. Лобное возвышение крутое, округлое (рис. 6). Овальное дыхало несколько смещено влево. На морде имеются редко расположенные уплощенные короткие щетинки. Спинной плавник длинный и низкий, гребневидный, полого спускающийся вперед и назад. Его вершина располагается примерно на уровне 2/3 длины тела, считая от головы. У амазонского дельфина довольно хорошо выражена шея, вследствие чего голова у него более подвижна, чем у других китообразных. Лопасти грудных и хвостового плавников крупные и широкие. Глаза маленькие, но полностью функционирующие.
Окраска тела, по-видимому, зависит от возраста. У молодых дельфинов спинная сторона сероватая или черная, брюшная светло-серая. Взрослые особи совсем светлые, иногда с розоватым оттенком.
Зубов 33—34 в каждой половине челюсти.
Хромосом в диплоидном наборе 44.
Распространение охватывает систему р. Амазонки с ее притоками в Бразилии, Боливии, северной части Перу, Эквадоре, Колумбии, Венесуэлы (верховье р. Риу-Негру), а также систему р. Ориноко.
Питается мелкой рыбой. Вероятно, использует эхолокацию для поисков добычи. Записанные у животных, содержавшихся в неволе, звуки состояли из отдельных щелчков и серий щелчков очень низкой интенсивности. Часто копает «клювом» ил, доставая пищу. Зрение развито достаточно хорошо, хотя ведущими органами чувств, по-видимому, являются слух и осязание. Держатся поодиночке, парами или мелкими группами до 6 особей. При плавании могут иногда выпрыгивать более чем на 1 м из воды. Под водой обычно остаются 0,5—2 мин. Плавают медленно со скоростью до 8—10 миль в час. В период наводнения могут покидать русло рек, перемещаясь в затопленные леса.
Род озерных дельфинов
Genus Lipotes Miller, 1918
В роде единственный вид: озерный дельфин — L. vexillifer Miller, 1918.
Длина тела до 2,5 м. «Клюв» немного загнут кверху. Средней высоты спинной плавник полого спускается вперед и назад и располагается немного позади середины тела. Грудные плавники закруглены на концах.
Окраска тела бледно-голубовато-серая сверху и бледноватая снизу. Глаза сильно редуцированы и, по-видимому, почти не функционируют.
В черепе рострум слегка изогнут концом кверху.
32 — 34 Зубная формула: •
Позвонков в отделах позвоночника: шейных 7, грудных 10, поясничных 7—8.
Обитает только в оз. Дунтинху в 1000 км от устья Янцзы (Китай).
Экология не изучена. Питается, по-видимому, рыбами. В поисках до бычи часто копается «клювом» в тине. Держится группами по 3—4, иногда до 10—12. Период размножения охватывает позднюю весну. Отмечены незначительные сезонные миграции: в поздне-весенний период (дождливый сезон) мигрирует из озера вверх по впадающим в него рекам.
Род гангских дельфинов
Genus Platanista Wagler, 1830
В роде единственный вид: гангский дельфин — Р. gangetica Lebeck, 1801.
Длина тела достигает 2,3 м. Самец длиной 199 см имел 84 кг (Kasu-уа, 1972). Голова отделена от туловища заметным шейным перехватом. Спинной плавник небольшой, гребневидный, расположен слегка позади середины тела (рис. 7). Грудные плавники большие, широкие, на концах тупо закруглены. От «клюва» круто вверх поднимается лобный выступ. Глаза не имеют хрусталика. Зрительный нерв очень тонкий (Pilleri, 1970). Верх тела темно-серый, иногда почти черный, брюхо несколько светлее.
Позвонков в отделах позвоночника: шейных 7, грудных 10—11, поясничных 7—8, хвостовых 24—25; всего 49—51. Фаланг в пальцах передней конечности К II4...5 11 К—5 IV4.-5
Распространен в реках Ганг и Брахмапутра (Индия, в том числе Ассам; Бангладеш),Хугли (Индия), Инд (Пакистан) и Сутледж (Северо-Западная Индия).
Питается рыбами и различными пресноводными беспозвоночными. Количество съедаемой ежедневно пищи в неволе составляет 8,5% массы
Рис. 7. Гангский дельфин—Р. gangetica и его череп: Л—вид снизу; Б— вид сверху и несколько спереди; Б — вид сбоку
тела животного. Способность к эхолокации хорошо развита. Среди звуков, которые производит гангский дельфин, наиболее часто (87%) слышны щелчки, которые, по-видимому, имеют эхолокационное значение. Период спаривания приходится на июль — сентябрь. Длительность беременности 8—9 месяцев. Детеныши рождаются с апреля по июль. Новорожденный имеет длину 45—70 см и массу примерно 7 кг. В годовалом возрасте детеныш достигает длины примерно 116 см. Держатся небольшими группами в 3—10 особей. Под водой остается от */2 до 2 мин. Часто плавает на боку (особенно на мелководье). Возможно, гангские дельфины осуществляют местные миграции.
Некоторые ученые (Pilleri a. Cihr, 1971) выделяют дельфинов, обитающих в р. Инд, в особый вид: Р. indi Blyth, 1859. Однако для достоверного решения этого вопроса необходим дополнительный материал.
Гангский дельфин из р. Инд может оказаться перед опасностью исчезновения, если не будет налажена его охрана. Он включен в «Красную книгу».
СЕМЕЙСТВО ДЕЛЬФИНОВЫХ
FAMILIA DELPHINIDAE GRAY, 1821
Китообразные мелких размеров. Длина тела 1,1—10 м *. Самцы обычно крупнее самок. Половой диморфизм может проявляться и в других признаках — форме и размере спинного плавника, форме грудных плавников. Хвостовой плавник с глубокой выемкой между лопастями. У большинства представителей имеется крупный спинной плавник (иногда он отсутствует), расположенный примерно посредине тела. На горле борозд нет. «Клюв» хорошо развит, длинный или полностью отсутствует. Окраска тела разнообразная: одноцветная (серая, черная), темная сверху и светлая снизу или с различными светлыми полосами или пятнами.
В черепе слабо развиты скуловая и височная кости. Проксимальные отделы лицевых костей не образуют гребней. Крыловидные и носовые кости относительно невелики по размерам. Имеются пред- и заглазнич-ные отростки. Симфиз нижней челюсти обычно не превышает '/з ее длины. Рострум длинный или короткий. Ширина нижней и верхней челюстей примерно одинакова.
0—65 Зубная формула; --•
2 — 58
Зубы гомодонтные, у большинства дельфинов небольших размеров, иногда крупные и массивные.
Шейные позвонки срастаются в различных комбинациях.
Распространены почти во всех морях Земного шара. Некоторые распространены очень широко, почти всесветно (дельфин-белобочка, косатки). другие более локально.
Подвижные животные, ведущие стайный образ жизни. Питание разнообразное. Среди представителей семейства можно выделить ихтиофагов, теутофагов, теутоихтиофагов и саркофагов.
Некоторые представители семейства имеют промысловое значение. Другие, вследствие своей малочисленности или по другим причинам, почти не добываются и практического значения не имеют.
1 В семействе можно выделить дельфиновых мелких размеров с длиной тела 1.1—4 м, средних — 4,1—7 и крупных 7,1—10 м.
Систематика семейства дельфинов трактуется разными учеными по-разному (см. с. 15, 16).
Семейство дельфиновых содержит 19 родов (46 видов):
крупнозубых дельфинов — Steno Gray, 1846 (1 вид);
длинноклювых дельфинов — Sotalia Gray, 1866 (8 видов);
продельфинов—Stenella Gray, 1866 (5 видов);
дельфинов-белобочек — Delphinus Linnaeus, 1758 (1 вид);
афалин — Tursiops Gervais, 1855 (1 вид);
китовидных дельфинов — Lissodelphis Gloger, 1841 (2 вида);
малайзийских дельфинов — Lagenodelphis Fraser,. 1956 (1 вид);
короткоголовых дельфинов — Lagenorhynchus Gray, 1846 (6 видов);
бесклювых дельфинов — Peponocephala Nishiwaki et Norris, 1966 (1 вид);
пестрых дельфинов — Cephalorynchus Gray, 1846 (4 вида);
иравадийских дельфинов — Orcaella Gray, 1866 (1 вид);
малых косаток — Pseudorca Reinhardt, 1862 (1 вид);
косаток — Orcinus Fitzinger, 1860 (1 вид);
серых дельфинов — Grampus Gray, 1828 (1 вид);
гринд — Globicephala, Lesson, 1828 (3 вида);
карликовых косаток — Feresa Gray, 1870 (1 вид);
морских свиней — Phocoena G. Cuvier, 1817 (4 вида);
белокрылых морских свиней — Phocoenoides Andrews, 1911 (1 вид); бесперых морских свиней — Neophocaena Palmer, 1899 (3 вида).
В водах СССР встречаются или могут быть встречены представители всех родов дельфинов, кроме длинноклювых, малайзийских, пестрых и иравадийских дельфинов.
Род крупнозубых дельфинов
Genus Steno Gray, 1846
В роде единственный вид: крупнозубый дельфин — S. bredanensis Lesson, 1828.
Размеры мелкие. Длина тела до 2,5 м. Взрослый самец с длиной тела 2,2 м был массой 102 кг. Спинной плавник высокий, расположен немного кзади от середины туловища, вершина его загнута назад (рис. 8). Вытянутый и несколько сжатый с боков «клюв» постепенно пёреходит в лобное возвышение. Длина грудного плавника составляет примерно */? длины тела, а размах хвостовых лопастей — 'Д Спинная сторона темносерого цвета, брюхо беловатое. «Клюв» белый.
Зубов в каждой половине челюсти по 20—27. Поверхность коронок покрыта вертикальными бороздками и валиками.
Позвонков шейных 7, грудных 13, поясничных 15—16, хвостовых 30— 31; всего 66. Фаланг в передней конечности I3 Ils—э Ше-? IV6 V2.
Распространен в Северной Атлантике от Голландии до Португалии, побережья Сенегала и Берега Слоновой кости на востоке и от Виргинии до Флориды и, возможно, Кубы на западе; в Южной Атлантике — от 1°14', ю. ш., 17°20' з. д. и берегов Бразилии и Аргентины на северо-восток до о-вов Тристан-да-Кунья (32° ю. ш., 2° з. д.), в северной части Тихого океана к югу от Калифорнии, Гавайских о-вов и Японии; в южной части Тихого океана у Галапагосских о-вов; в Индийском океане от Аденского до Бенгальского заливов и Явы.
Экология не изучена. Встречается иногда крупными стадами до 100 особей. Питается головоногими моллюсками и рыбой. Специально тренированный крупнозубый дельфин отказывался нырять глубже 30 м.
Род длинноклювых дельфинов
Genus Sotalia Gray, 1866
Размеры мелкие. Длина тела 1,1—2,5 м. Наименьший среди китообразных белый дельфин —1,1—1,2 м. Взрослый самец западноафриканского дельфина с длиной тела 2,5 м имел массу 166 кг. Спинной плавник средней высоты расположен примерно посредине тела и обычно имеет треугольную форму или выемку на заднем крае (рис. 9). Грудные плавники широкие. «Клюв» длинный, четко отделен от лобного возвышения. Окраска тела изменчива. Большинство видов бурые, серые или черные на спинной стороне тела и светлее на брюшной; калимантанский и китайский дельфины белого цвета.
В черепе характерен длинный симфиз челюсти и широкий промежуток между крыловидными костями.
Позвонков у белого дельфина: шейных 7, грудных 12, поясничных 11, Хвостовых 26; всего 56.
Фаланг пальцев передней конечности у этого же дельфина Б Ilg Ills IV4 V2.
Распространены в тропических морях и больших реках Южной Америки, по побережью Западной Африки от Сенегала до Камеруна, в Бенгальском заливе, Аравийском море, Персидском заливе, Красном море, Сиамском заливе и у берегов Калимантана и Юго-Восточного Китая.
Питаются в основном рыбами и в значительно меньшей степени ракообразными. Особенности размножения не известны. Держатся поодиночке или небольшими группами по 2—8 особей, и лишь иногда встречаются более крупные стада. Плавают медленно, хотя быстрее, чем речные дельфины. Иногда выпрыгивают над поверхностью воды. Дышат с интервалом от 5 до 85 с. Наибольшая активность дельфинов отмечается в ранне-Утренние и поздневечерние часы.
В роде 8 видов:
белый дельфин — S. fluviatilis Gervais et Deville, 1853 (p. Амазонка и нижнее течение ее притоков между Рио-Токангинс в Бразилии и Рио-Хуаллага в Перу);
ПО
Рис. 9. Западноафриканский дельфин — S. teuszi
гайанский дельфин —S. guianensis Р. J. Van Beneden, 1864 (побережье и реки Гайаны и Венесуэлы, включая оз. Маракайбо);
бразильский дельфин —S. brasiliensis Е. Van Beneden, 1875 (окрестности Рио-де-Жанейро, но ареал, очевидно, тянется вдоль побережья Атлантического океана к юго-западу по крайней мере до Сантос);
китайский дельфин — S. chinensis Osbeck, 1765 (моря Китая к северу от о-ва Калимантан; вдоль побережья Китая, а также в реках юго-запада Китая, встречается и в р. Янцзы на 750 миль вверх по ее течению);
калимантанский дельфин — S. borneensis Lydekker, 1901 (Южно-Китайское море от о-ва Калимантан до Таиланда);
крапчатый дельфин — S. lentiginosus Gray, 1866 (Бенгальский залив от Мадраса до Шри Ланка и на запад до Южной Африки; встречи в водах Австралии остаются под вопросом);
свинцовый дельфин — S. plumbed G. Cuvier, 1829 (от берегов о-ва Калимантан в Южно-Китайском море на запад через Малаккский пролив, Бенгальский залив, Аравийское и Красное моря до Суэцкого канала);
западноафриканский дельфин — S. teuszi Kukenthal, 1892 (вдоль берегов Западной Африки и в устьях рек от Сенегала до Камеруна).
Некоторые исследователи (например, Rice a. Scheffer, 1968; Nishiwa-ki, 1972) выделяют свинцового, китайского, калимантанского, крапчатого и западноафриканского дельфинов в род Sousa Gray, 1866, или белого дельфина (Nishiwaki, 1972) разделяют на 3 вида: S. fluviatilis Gervais et Deville, 1853; S. pallida Gervais, 1855 и S. tucuxi Gray, 1856. Вероятно, два последних вида — лишь возрастные стадии S. fluviatilis. Все перечисленные выше виды иногда объединяют всего в 3 вида: Sotalia fluviatilis, Sousa chinensis и Sousa teuszi {Marine mammal commission, 1977).
Иногда роды Sotalia, Sousa и Steno относят в особое семейство Ste-nidae Erases et Purves, 1960 ( Fraser a. Purves, 1960; Anderson a. Jones, 1967; Nishiwaki, 1972).
Род продельфинов
Genus Stenella Gray, 1866
Размеры мелкие. Длина тела до 2,7 м. Расположенный посредине тела спинной плавник загнут вершиной назад (рис. 10). Грудные плавники серпообразные. «Клюв» длинный. Нижняя челюсть немного длиннее верхней.
Окраска тела подвержена видовой и индивидуальной изменчивости. Так, у полосатого продельфина спина и верхние отделы боков тела тем-
Рис. 10. Полосатый продельфин — 5. coeruleoalbus и его череп
ные, нижняя часть боков тела и брюхо белые; от глаз к грудному плавнику тянется узкая темная полоса (одинарная или двойная), другая полоса проходит над плавником по боку тела по направлению к заднепроходному отверстию. У пятнистого дельфина темная спина покрыта белыми, а светлое брюхо — темными пятнами.
В черепе имеются черты строения, сближающие продельфинов, с одной стороны, с родом дельфинов-белобочек, с другой — с родом афалин. Общие признаки для продельфинов и дельфинов-белобочек следующие: длинный рострум, намного превышающий мозговой отдел черепа, большое число мелких зубов (34—65 в каждой половине челюсти) и обычно сливающиеся в ростральной части черепа межчелюстные кости. Однако, как и у афалин, продельфины не имеют продольных борозд на костном небе. Мозговой отдел черепа вздут в затылочной области и сплющен в лобной. Крыловидные кости граничат между собой по всей своей длине.
Позвонков шейных 7, грудных 15—16, поясничных 18—22, хвостовых 35—37; всего 77—79. Фаланг в передней конечности 1[_3 Пэ-ю Шт IV3-4 Vi-2.
Распространены от холодных до тропических морей в Тихом и Атлантическом (включая Средиземное море) океанах о г Гренландского и Берингова морей до прибрежных вод Мыса Доброй Надежды, мыса Горн и Австралии, а также в Индийском океане.
Экология изучена слабо. Объектами пищи служат рыбы и головоногие моллюски. Новорожденные в длину достигают около 120 см. Держатся группами от нескольких особей до сотен и даже тысяч голов. Длин-норылый продельфин хорошо приручается. Существенного промыслового значения не имеют. Полосатый продельфнн добывается местными жителями в прибрежных водах Японии.
Систематика продельфинов разработана недостаточно ввиду малочисленности добытых экземпляров. Hershkovitz (1966) в роде выделяет 10 видов: S. asthenops Соре, 1865; S. clymene Gray, 1846; S. coeruleoalbus Meyen, 1'833; S. crotaphiscus Cope. 1865; S. aubia G. Cuvier, 1812; S. frontalis G. Cuvier, 1829; S. graffmani Lonnberg, 1934; S. longirostris Gray, 1828; S. malayana Lesson, 1826; S. pernettyi Blainville, 1817.
Рис. 11. Малайский продельфин — S. dubia, вид снизу и сбоку
Рис. 12. Большелобый 5. frontalis и его череп
продельфин —
Примерно такую же систему предлагает и Nishiwaki (1972), различая: S. caeruleoalba (-S. coeriileoalbus); S. styx Gray, 1846 (выделяет из S. caeruleoalba); S. malayana; S. longirostris; S. microps Gray, 1971 (выделяет из S. longirostris); S. attenuata; S. plagiodon Cope, 1866 (=S. pernettyi); S. graffmani. Это число видов, вероятно, завышено. Некоторые исследователи насчитывают в роде 5 видов: S. caeruleoalbus, S. dubia, S. pernettyi S frontalis и 5. longirostris (Томилин, 1971) или 4 вида: X. caeruleoalba, S. dubia, S. longirostris, S. roseiventris Wagner, 1853 (Rice a. Scheffer, 1968).
Впредь до ревизии рода, по-видимому, правильнее объединять в роде лрсдельфинов 5 видов:
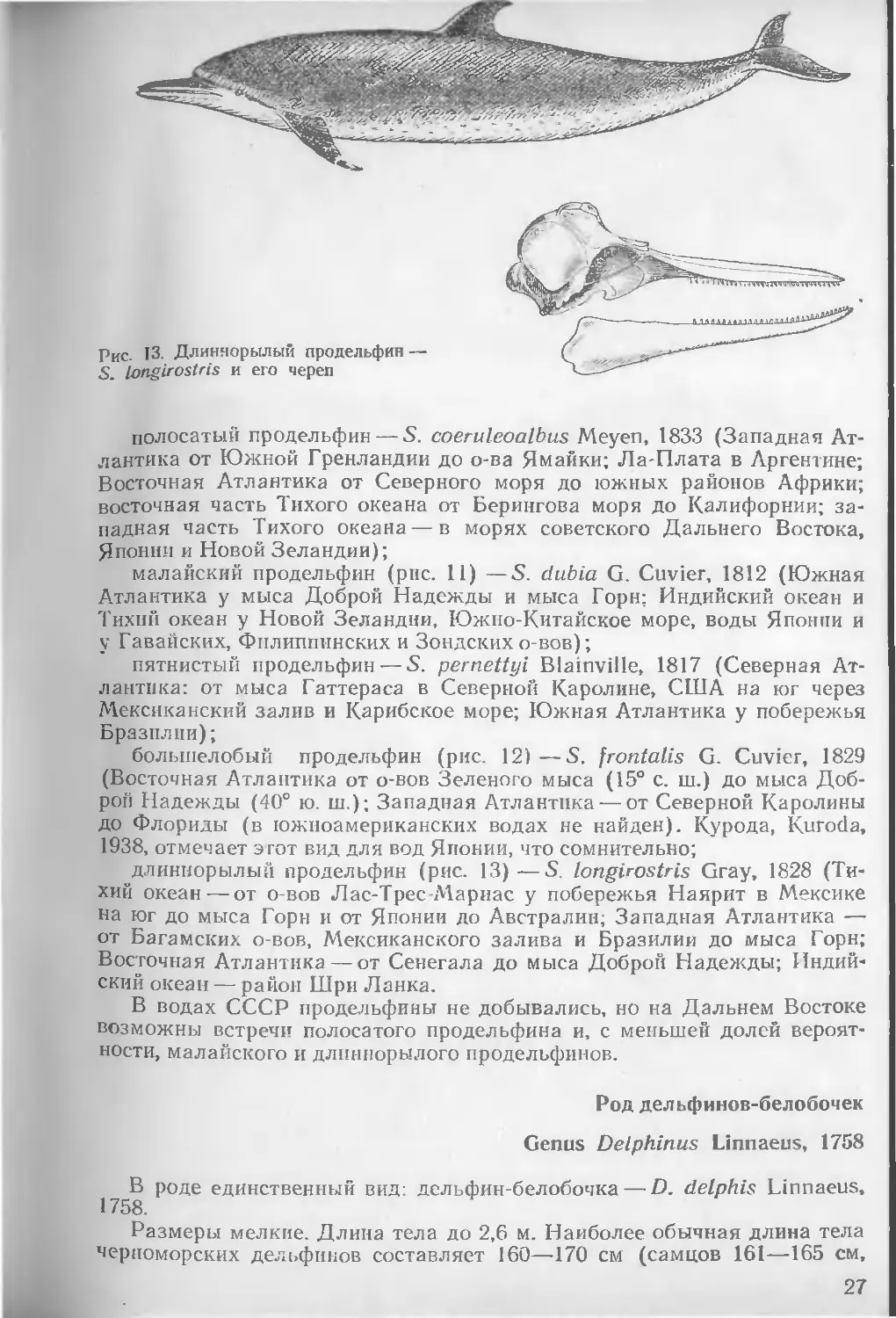
Рис. 13. Длиннорылый продельфин — S. longlrostris и его череп
полосатый продельфин — S. coeruleoalbus Меуеп, 1833 (Западная Атлантика от Южной Гренландии до о-ва Ямайки; Ла-Плата в Аргентине; Восточная Атлантика от Северного моря до южных районов Африки; восточная часть Тихого океана от Берингова моря до Калифорнии; западная часть Тихого океана — в морях советского Дальнего Востока, Японии и Новой Зеландии);
малайский продельфин (рис. 11) —S. dubia G. Cuvier, 1812 (Южная Атлантика у мыса Доброй Надежды и мыса Горн; Индийский океан и Тихий океан у Новой Зеландии, Южно-Китайское море, воды Японии и у Гавайских, Филиппинских и Зондских о-вов);
пятнистый продельфин — S. pernettyi Blainville, 1817 (Северная Атлантика: от мыса Гаттераса в Северной Каролине, США на юг через Мексиканский залив и Карибское море; Южная Атлантика у побережья Бразилии);
большелобый продельфин (рис. 12)—S. frontalis G. Cuvier, 1829 (Восточная Атлантика от о-вов Зеленого мыса (15° с. ш.) до мыса Доброй Надежды (40° ю. ш.); Западная Атлантика — от Северной Каролины до Флориды (в южноамериканских водах не найден). Курода, Kuroda, 1938, отмечает этот вид для вод Японии, что сомнительно;
длиннорылый продельфин (рис. 13)—S. longirostris Gray, 1828 (Тихий океан — от о-вов Лас-Трес-Мариас у побережья Наярит в Мексике на юг до мыса Горн и от Японии до Австралии; Западная Атлантика — от Багамских о-вов, Мексиканского залива и Бразилии до мыса Горн; Восточная Атлантика — от Сенегала до мыса Доброй Надежды; Индийский океан — район Шри Ланка.
В водах СССР продельфины не добывались, но на Дальнем Востоке возможны встречи полосатого продельфина и, с меньшей долей вероятности, малайского и длпннорылого продельфинов.
Род дельфинов-белобочек
Genus Delphinus Linnaeus, 1758
В роде единственный вид: дельфин-белобочка — D. delphis Linnaeus, 1758.
Размеры мелкие. Длина тела до 2,6 м. Наиболее обычная длина тела черноморских дельфинов составляет 160—170 см (самцов 161—165 см,
Рис. 14. Дельфин-белобочка — D. delphis
Рис 15. Изменчивость окраски дельфина-белобочки
самок 158 см), максимальная — 210—219 см; дельфины с длиной тела свыше 200 см встречаются крайне редко. Атлантические и северотихоокеанские дельфины-белобочки крупнее черноморских: их максимальная длина тела достигает соответственно 258 и 259 см. «Клюв» длинный. Высокий спинной плавник серпообразно изогнут назад. Узкие грудные плавники также изогнуты (рис. 14).
Окраска тела подвержена индивидуальной изменчивости, но обычно бывает черной на спине, серой на боках с темными и светлыми полосами, образующими сложный рисунок, брюхо светлое. От основания грудных плавников вперед, к подбородку, тянется по темной полосе. Различные комбинации окраски черноморских дельфинов показаны на рис. 15. Дальневосточные дельфины-белобочки отличаются от черноморских и атлантических отсутствием отчетливо выраженных боковой и диагональной полос и тем, что темная окраска верхней части боков тела резко отграничена от светлой нижней (Томилин, 1957). В общем же окраска в различных точках ареала сходна.
Рострум в 1,5—2 раза длиннее черепной коробки. Спереди он раздвоен. Характерно, что на костном небе есть два продольных желобка. Крыловидные кости граничат между собой по всей своей длине. Верхнезатылочный гребень асимметричен: его правая половина больше левой. Нижняя челюсть немного длиннее верхней. Возрастные изменения черепа характеризуются следующими особенностями. У новорожденных мозговая коробка не имеет гребней, кости черепа срастаются между собой неполностью, зубов еще нет. На первом году жизни швы между костями замыкаются, закладываются гребни и прорезываются зубы (45—48 в каждой челюсти). С переходом на самостоятельное питание усиление мышечного аппарата влечет за собой увеличение размеров костных гребней и выступов; округлость черепа утрачивается. Наиболее интенсивно рост черепа происходит в первые два года, причем увеличение общей длины черепа идет главным образом за счет его лицевого отдела. За это же время полностью развивается зубная система. К трем годам череп окончательно сформирован. Черепные гребни достигают максимального развития к 4—5 годам. Черепа атлантических и тихоокеанских дельфинов-белобочек имеют более крупные размеры по сравнению с черноморскими.
65 Конические зубы многочисленны — до ——- • 58
Ребер до 13—15 пар; число их может быть неодинаково на левой и правой сторонах. Описана находка рудиментов задних конечностей, расположенных по бокам мочеполовой щели у самки, пойманной в районе Ялты. Они представляли собой лопасти треугольной формы высотой 3,4 (правая) и 1,6 см (левая). Каждая из них содержала по 2 костных элемента (рудиментов бедра и голени) и 3 хрящевых справа и 2 хрящевых слева (рудименты предплюсны, плюсны и фаланг). Грудина цельная, Т-образная. Позвонков шейных 7, грудных 14, поясничных 21, хвостовых 31—32, всего 73—74. Общее число позвонков может варьировать от 70 до 75. Шейные позвонки часто срастаются между собой. Фаланг в передней конечности 12_3, Пв-э, Ш5-7, IV2-4, Vi-2. Наибольшую длину имеют II и III пальцы.
Хромосом в диплоидном наборе 44.
Распространен в умеренных и теплых водах всех океанов и большинства морей Земного шара: в Западной Атлантике — от Нью-Фаундленда и штата Массачусетс в США до Богамских о-вов, Мексиканского залива, о-ва Ямайка, побережья Южной Бразилии и Аргентины между 27—• 45° ю. щ„ о-ва Тристан-да- Кунья; в Восточной Атлантике-—от Исландии, Норвегии и Балтийского моря к северу, западу и на юг до южных частей Африки; в Индийском океане; в южной части Тихого океана —• от Новой Зеландии и Австралии на западе до Перу и Чили на востоке. Наиболее многочисленны дельфины-белобочки в умеренных водах.
В СССР — в Черном, Балтийском, Охотском, Японском и Беринговом морях.
Держатся дельфины-белобочки группами, иногда образуя тысячные стада. В различные сезоны скопления дельфинов дифференцируются по полу и возрасту. Зимой различают 2 типа косяков, состоящих из взрослых самок с детенышами и из взрослых самцов. Летом 6 типов: предщенные (беременные самки); детные (кормящие самки с детенышами); брачные (половозрелые особи обоего пола с детенышами, переходящими на самостоятельное питание); неполовозрелые; остаточные самцовые и остаточные самочные (два последних типа встречаются весной и в начале лета и представляют собой еще не распавшиеся зимние косяки).
Под водой обычно остается 0,5—3 мин, иногда до 5. Испуганное стадо
дельфинов-белобочек развивает скорость до 45—55 км/ч, причем звери делают огромные пологие прыжки в воздухе.
Дельфин-белобочка — пелагическая форма и питается главным образом рыбой. В Черном море поедает в основном шпроты и хамсу, в меньшей степени пелагических игл, пикшу, барабулю, ставриду и морского таракана (из ракообразных). В других местах ареала употребляет в пищу сельдей, мойву, сайру, анчоуса, макрель, сардину, кефаль и других рыб, а также головоногих моллюсков (кальмаров).
Детеныши рождаются в основном в летние месяцы. Размеры новорожденных 80—95 см. Лактационный период продолжается 4—6 месяцев. Спаривание происходит до его окончания. Длительность беременности составляет 10—11 месяцев. Рожают самки ежегодно, но значительная их часть остается яловыми. Половозрелость наступает не ранее чем в трехлетием возрасте. Продолжительность жизни до 30 лет.
До 1966 г. на Черном море ежегодно добывали большое количество дельфинов-белобочек. С 1966 г. в СССР введен запрет на добычу дельфинов. Болгария и Румыния также прекратили промысел дельфинов.
От добытых дельфинов-белобочек использовали главным образом подкожное сало, из которого вырабатывали медицинский и технический жир.
Некоторые исследователи (Nishiwaki, 1972) выделяют в роде помимо D. delphis еще 2 вида: D. capensis Gray, 1828, и D. bairdii Dull, 1873. По-видимому, правильнее считать их в лучшем случае подвидами.
Род афалин
Genus Tur slops Gervais, 1855
В роде единственный вид: афалина — Т. truncatus Montagu, 1821.
Размеры мелкие. Наибольшая длина тела 3,9 м. Средняя длина самцов, добытых в Черном море, составила 228 см, самок 222 см, наиболее крупным оказался самец с длиной тела 310 см. Афалины из других мест крупнее черноморских: в водах Японии длина самцов в среднем равна 2.7 м. самок 2,8 м, в Атлантике афалины достигают в длину 3,1—3,3 м, в Средиземном море—-3,2 м. У афалины средней длины «клюв» четко отделен бороздой от лобного возвышения. Высокий спинной плавник имеет глубокую вырезку по заднему краю (рис. 16). Грудные плавники относительно широкие. Нижняя челюсть немного длиннее верхней. Окраска тела подвержена значительным индивидуальным вариациям. Верх тела темно-серый (может быть и светло-серым), бока серые, обычно резко отграниченные от белого (иногда серого) брюха, но серая окраска боков тела может переходить в светлый тон брюха и постепенно. У афалин, обитающих в Черном море, выделяют 2 цветовые группы (Барабаш-Никифоров, 1940, 1960). Тип А характеризуется более или менее четкой границей между темной окраской спины и белой окраской брюха и тем, что в темном поле средней части тела расположен светлый угол, вершиной обращенный к спинному плавнику. У типа Б пигментированная верхняя поверхность тела не имеет резкой границы с нижней поверхностью; она представлена более или менее размытой прямой, волнистой или изломанной линией, не имеющей светлого угла у спинного плавника. Количественное соотношение особей типа А и Б колеблется по годам. У 50% афалин того и другого типов встречается лобно-грудная линия, соединяющая глаза и постепенно расширяющаяся от наружных углов глаз к грудным плавникам. У афалин Атлантики светлая вырезка, свойственная типу А, и лобно-грудная линия всегда отсутствуют. У афалин из Средиземного моря лобно-грудная линия может встречаться.
Рис. 16. Афалина — Т. truncatus и ее череп
В черепе рострум средней длины. Его размеры подвержены половому диморфизму: у самок он длиннее и уже, чем у самцов. На правой стороне черепа межчелюстная и носовые кости граничат между собой, на левой они не соприкасаются. Крыловидные кости с косым срезом по своей задней ча-сти соединяются друг с другом широкими выступами. Зубы относительно крупные (до 10 мм в диаметре), число их в верхней челюсти 19—28, а в нижней 19—25. Зубы часто стираются в той илз иной степени.
Позвонков шейных 7, грудных 12—14, поясничных 17, хвостовых 26— 30; всего 63—65. Шейные позвонки могут срастаться между собой в различных комбинациях. Грудина состоит из трех срастающихся сегментов. Ребер 12—13 пар. Фаланг в передней конечности К-2, II7-9, Ш5-8, IV2_4, Vj_2. Самый длинный палец второй, изредка — третий.
Хромосом в диплоидном наборе 44.
Распространена почти во всех теплых и умеренных водах: в Атлантическом океане — от Норвежского, Северного и Балтийского морей до Средиземного и Черного моря и Южной Африки на востоке и от Южной Гренландии до штатов Флориды и Техаса в США, Мексики и о-ва Барбадос на западе; в Тихом океане-—от Калифорнии до Чили на востоке и от Японии до Австралии и Новой Зеландии на западе; в Индийском океане — от Австралии до Африки.
В СССР: в Черном и Балтийском морях и на Дальнем Востоке.
Держатся обычно группами в несколько десятков голов. Наибольшая скорость при плавании 45—55 км/ч. Способна издавать звуки, лежащие в частотном пределе от 7 тыс. до 170 тыс. Гц. Исследователи описывают 6 типов таких звуков.
Относится к группе бентоихтиофагов. В Черном море основными объектами пищи служат пикша и камбала, а также кефаль, лобан, скат, умбрина, скорпена, барабуля, пеламида и хамса. В других местах поедает кефалевых, смариду, угрей, небольших акул и др. При содержании в океанарии в день съедает 20—32 кг рыбы. За пищей может нырять на глубину до 90—175 м, оставаясь под водой 5—7, иногда 15 мин. Детеныши рождаются в весенне-летнее время. Длина новорожденного составляет около 1 м. Лактационный период, по наблюдениям в океанариуме, продолжается у разных особей от 3,5 до 18 месяцев. Беременность длится примерно 12 месяцев, а весь половой цикл — 2 года. Половозрелость наступает в 6 лет (данные по океанариуму). Хорошо приручается и легко переносит неволю. Способность к дрессировке выше, чем у многих других видов дельфинов и собак, но ниже по сравнению с некоторыми человекообразными обезьянами.
Некоторые исследователи выделяют в роде 2 вида (Hershkovitz, 1966; Rice a. Scheffer, 1968): Т. truncatus и Т. gilli Dall 1873, или даже 7 (Nis-hiwaki, 1972): Т. truncatus; Т. gilli; Т. пииапи Andrews, 1911; Т. aduncus Ehrenberg, 1833; Т. parvimanus Reinhardt, 1888; Т. gephyreus Lahille, 1908; T. abusalam Ruppel, 1842. Для обоснования реальности существования всех этих видов (в частности Т. gephyreus, о чем пишет Pilleri а. Gihr, 1972) необходим дополнительный материал.
В водах СССР добывались в относительно небольшом числе в Черном море. С 1966 г. промысел запрещен.
Род китовидных дельфинов
Genus Lissodelphis Gloger, 1841
Размеры мелкие. Длина тела 1,8—2,4 м. Голова с низким лбом, челюсти относительно короткие и узкие. Грудные плавники серповидные (рис. 17). Спинного плавника нет. Лопасти хвостового плавника отно-
Рис. 17. Северный дельфии — L. borealis и его череп
сительно невелики. Хвостовой стебель тонкий. Окраска тела черная и только на брюшной стороне от горла до хвоста тянется узкая светлая полоса, образующая между грудными плавниками ромбовидное пятно (иногда все брюхо белое), и дорзальная сторона верхней челюсти, часть лба и конец нижней челюсти белые (северный дельфин) или белые все брюхо, бока тела и грудные плавники (южный дельфин).
В черепе узкий рострум лишь немного длиннее мозговой коробки. Крыловидные кости не соединяются друг с другом. Межчелюстные кости спереди уплощены. Симфиз нижней челюсти короткий.
Зубы мелкие (около 3 мм в диаметре); 40—47 в верхней челюсти и 42—49 в нижней.
Позвонков шейных 7, грудных 14—15, поясничных 29—30, хвостовых 37—39; всего 88—90. Почти полукруглая по форме лопатка имеет очень крупные коракоидный и акромиальный отростки. Фаланг в передней конечности 1,-2. П8, Ш9, IV3, V2_3.
Распространены в Южной Атлантике у Австралии, Тасмании, Новой Зеландии и Чили, а в северной части Тихого океана от Берингового моря До Японии и Калифорнии.
Экология изучена недостаточно. Держатся небольшими группами (по 3—10 особей), иногда объединяются в большие стада (северные дельфины до 200 и более голов). Стайки северных дельфинов обычно передвигаются медленно, хотя отдельные особи могут достигать скорости Зо км/ч. Численность популяции северных дельфинов определяется примерно в 100 000 особей (Nishiwaki, 1972). Питаются, по-видимому, в основном рыбой и головоногими моллюсками.
Китовидных дельфинов иногда промышляет местное население у берегов Японии.
В роде 2 вида: южный дельфин — L. peroni Lacepede, 1804 (южноамериканские воды от Южной Бразилии и побережья Патагонии в Атлантическом океане до Чили (37° ю. ш.) в Тихом океане; от Новой Зеландии 2—671 33
и Тасмании до Новой Гвинеи в западной части Тихого океана и у побережья Южной Африки в Атлантическом океане);
северный дельфин — L. borealis Peale, 1843 (северная часть Тихого океана от Берингова моря до Сан-Диего в Калифорнии на востоке и Японии на западе).
В водах СССР китовидные дельфины не добывались, но у берегов советского Дальнего Востока может быть встречен северный дельфин.
Род малайзийских дельфинов
Genus Lagenodelphis Fraser, 1956
В роде единственный вид: малайзийский дельфин — L. hosei Fraser, 1956.
Размеры мелкие. Длина тела примерно 2,1—2,4 м.
В черепе рострум по длине равен мозговой коробке, в основании широкий. Крыловидные кости крупные; они примыкают друг к другу или разделены небольшим промежутком. Предчелюстные кости срослись между собой дорзально по средней линии, напоминая таковое у дельфина-белобочки; у малайзийского дельфина оно не столь резко выражено.
Зубов в каждой половине челюсти по 40—44.
Обнаружен только в устье р. Лутонг на о-ве Калимантан в Южно-Китайском море.
Экология не изучена.
Род короткоголовых дельфинов
Genus Lagenorhynchus Gray, 1846
Размеры мелкие. Длина тела до 3 м. «Клюв» короткий, не более 7 см, у большинства хорошо отграниченный бороздкой от покатого лобного возвышения. Высокий спинной плавник серпообразно изогнут и расположен на середине спины или немного впереди от нее (рис. 18). Серповидная форма характерна и для грудных плавников. По дорзальной и вентральной сторонам хвостового стебля проходят кожные кили. Окраска на верхней части тела темная. На боках обычно проходят широкие и узкие продольные полосы. Брюхо белое.
В черепе широкий у основания рострум равен по длине мозговой коробке или чуть длиннее ее (рис. 18). Крупные крыловидные кости примыкают друг к другу или имеют между собой небольшое пространство.
Зубная формула:
20 — 45
20 — 45
Зубы мелкие.
Позвонков от 71 до 94. Так у атлантического дельфина позвонков шейных 7, грудных 15, поясничных 18—22, хвостовых 38—41; всего 78— 82. Тела их сильно уплощены. Позвонки поясничного отдела имеют очень длинные остистые и поперечные отростки.
Хромосом в диплоидном наборе у тихоокеанского дельфина 44.
Распространены почти во всех морях Земного шара, исключая полярные.
Экология изучена недостаточно. Для беломордого дельфина объектами пищи служат сельдь, треска, мерланг, мойва, навага, реже головоногие моллюски и ракообразные. Атлантический дельфин поедает сельдь,
макрель, лососевых, головоногих моллюсков, придонных раков-отшельников. Тихоокеанский дельфин питается мелкой стайной рыбой — сардиной, сельдью, анчоусами, сайрой и др., и головоногими моллюсками Щенка преимущественно летом. В это же время происходит и спаривание. Держатся как небольшими группами, так и стадами в тысячу и больше особен. У тихоокеанского дельфина в водах Японии отмечены миграции. Беломордый дельфин на зиму уходит в южные районы ареала.
В наших водах промысел не ведется. У берегов Норвегии добывают беломордого и атлантического дельфинов, а у японских берегов — тихоокеанского. Короткоголовые дельфины хорошо приручаются при содержании в неволе.
Система рода разными исследователями трактуется по-разному. Hershkovitz, 1966, включает в род: L. acutus Gray, 1828; L. albirostris Gray, 1846; L. cruciger Quoy et Gaimard. 1824; L. electra Gray, 1846 (большинство исследователей в настоящее время выделяют этот вид в особый род: Peponocephala Nishiwaki et Norris, 1966); L. obliquidens Gill, 1865; L. thicolea Gray, 1846. По Rice a. Scheffer, 1968 в роде следующие виды: L. acutus; L. albirostris; L. cruciger; L. australis Peale, 1948 (этот вид выделен из L. cruciger); L. obscurus Gray, 1828 (этот вид выделен из L. cruciger); L. obliquidens. По Nishiwaki, 1972, род содержит L- acutus; L. albirostris; L. cruciger; L. wilson Lillie, 1915 (выделяет этот вид из L. cruciger); L. fitzroyi Waterhouse, 1836 (выделяет из L. cruci-Sei), L. superciliosus Lesson et Gamot, 1826 (выделен из L. cruciger); L. obscurus; L. australis; L. obliquidens; L. tricola.
Впредь до ревизии рода короткоголовых дельфинов и учитывая выделение L. electra в особый род, целесообразно объединить в роде Lageno-chynchus следующие 6 видов:
беломордый дельфин — L. albirostris Gray, 1846 (Северный Ледовитый океан и Северная Атлантика от Баренцева моря, Гренландского моря и Девисова пролива на юг до Португалии на востоке и Ньюфаундленда на западе) ;
атлантический дельфин — L. acutus Gray, 1828 (Северная Атлантика От Гренландии до штата Массачусетс, США и от Шпицбергена до Британских о-вов и Голландии);
Рис. 19. Бесклювый дельфин — Р. electra и его череп
тихоокеанский дельфин — L. obliquidens Gibl, 1865 (северная часть Тихого океана от Аляски и Алеутских о-вов до Калифорнийского залива на востоке и от Курильских о-вов до Японии на западе);
южный дельфин — L. australis Peale, 1848 (умеренные воды южной части Южной Америки и Фолклендские о-ва);
L. cruciger Quoy et Gaimard, 1824 (южные части Тихого и Атлантического океанов до антарктических льдов);
темный дельфин — L. obscuras Gray, 1828 (умеренные воды у Южной Америки, Южной Африки, о-ва Кергелен, Южной Австралии, Новой Зеландии и южная часть Тихого океана).
В водах СССР: беломордый (Мурманское побережье, Балтийское море), атлантический (Балтийское море и, возможно, Мурманское побережье) и тихоокеанский дельфины (Японское море, воды Советского Приморья и Курильские о-ва).
Род бесклювых дельфинов
Genus Peponocephala Nishiwaki et Norris ,1966
В роде единственный вид: бесклювый дельфин — Р. electra Gray, 1846.
Размеры мелкие. Длина тела самцов до 280 см, размеры самок немного меньше. Тело удлиненное и стройное с длинным хвостовым стеблем. Треугольный спинной плавник расположен примерно посредине тела. «Клюва» нет. Передняя часть головы закруглена, плавно изгибаясь от ротовой щели к дыхалу (рис. 19). Нижняя челюсть немного короче верхней. Окраска однотонная темно-серая, немного светлее на брюхе. Губы и область заднепроходного и полового отверстий не пигментированы. Светлое пятно находится на груди между плавниками и спереди от пупка.
В черепе анторбитальная борозда больше и глубже, чем у короткоголовых дельфинов.
22__25
Зубная формула: _24 •
Позвонков шейных 7, грудных 14, поясничных 17, хвостовых 44; всего 82. Первых три шейных позвонка срастаются между собой. Фаланг в пальцах 13, Н9, Ш7, IV4, V3.
Рис 20. Пестрый дельфин — С. commersoni
Распространение охватывает теплые воды северной части Тихого и Атлантического океанов, побережья Индии, Японии, Индонезии, Гавайских о-вов, Сенегала, Гвинеи.
Экология не изучена. Образ жизни, по-видимому, сходен с образом жизни короткоголовых дельфинов. Изредка добывается местными жителями.
В систематическом отношении бесклювый дельфин близок к роду короткоголовых дельфинов и только в 1966 г. было предложено выделить этого дельфина в отдельный род (Nishiwaki a. Norris, 1966). Иногда бесклювых дельфинов вместе с гриндой, косаткой, малой косаткой и карликовой косаткой выделяют в особое семейство Globicephalidae (Nishiwaki, 1972).
Род пестрых дельфинов
Genus Cephalorhynchus Gray, 1846
Размеры мелкие. Длина тела 1,2—1,8 м. Морда удлиненная, хотя «клюв» выражен нерезко и незаметно переходит в лобное возвышение (рис. 20). Спинной плавник небольшой, расположен несколько кзади от середины тела и имеет небольшую серповидную вырезку по заднему краю. Окраска своеобразная: представляет комбинацию резко отграниченных друг от друга белых и черных полей. Так, у С. commersoni голова, полоса от нее к каждому грудному плавнику, спинной плавник, пятно в области заднепроходного отверстия и задняя часть хвостового стебля вместе с хвостовым плавником черные, остальная часть тела белая. У всех видов плавники черные.
Распространены в Южном полушарии в западной части Тихого океана и в Южной Атлантике.
Экология изучена слабо. Питаются в основном каракатицами и креветками. Иногда встречаются крупные стада из сотен голов.
В роде 4 вида:
пестрый дельфин — С. commersoni Lacepede, 1804 (Южная Атлантика От Санта-Крус до Огненной Земли, на восток ареал идет до Фолклендских о-вов, а на запад продолжается в южную часть Тихого океана вдоль южного побережья Чили; в южной части Индийского океана обнаружен у о-ва Кергелен);
белобрюхий дельфин — С. eutropia Gray, 1849 (южная часть Тихого океана у побережья Чили между 33 и 40° ю. ш.);
дельфин Хэвисайда — С. heavisidei Gray, 1828 (вдоль берегов Южной Африки в Атлантическом и Индийском океанах);
Рис 21. Иравадийский дельфин — О brevirostris
дельфин Гектора — С. hectori Van Beneden, 1881 (западные области Тихого океана от о-ва Калимантан в Южно-Китайском море до Новой Зеландии).
Некоторые исследователи (Nishiwaki, 1972) выделяют из С. hectori еше один вид — С. albifrons True, 1889.
Род иравадийских дельфинов
Genus Orcaella Gray, 1866
В роде единственный вид; иравадийский дельфин — О. brevirostris Gray, 1866.
Размеры мелкие. Длина тела достигает примерно 2,2 м. Передняя часть головы округлая с едва заметным «клювом». Небольшой спинной плавник серповидно вырезан по заднему краю и сдвинут кзади от середины тела (рис. 21). Грудные плавники широкие. Окраска темно-серая, иногда с голубоватым оттенком. Брюшная сторона тела может быть светлее спинной.
„ 15—17
Зубная формула: ——-—-- •
Позвонков шейных 7, грудных 14, поясничных 14, хвостовых 28; всего 63. Только первый и второй шейные позвонки срастаются между собой. Фаланг пальпев 12, Не, Шв. IV3, Vi- Иногда V палец не окостеневает.
Распространен в прибрежных водах и эстуариях Индийского океана и Южно-Китайского моря от северного Калимантана до Бенгальского залива; встречается также в р. Меконг, Ганг и Иравади.
Экология изучена слабо. Держится небольшими группами. Под водой остается обычно 1—2,5 мин. Питается в основном рыбой. На р. Иравади местные рыбаки использовали этого дельфина для загона рыб в сети.
Некоторые исследователи (например, Nishiwaki, 1972) предлагают выделить иравадийских дельфинов в особое семейство Orcaelidae.
Род малых косаток
Genus Pseudorca Reinhardt, 1862
В роде единственный вид: малая косатка — Р. crassidens Owen, 1846.
Размеры средние. Длина тела до 6 м. Самки на 60—100 см меньше самцов. Тело удлиненное. Небольшая округлая голова имеет крупную 38
Рис. 22. Малая косатка — Р. crassi-dens и ее череп
жировую подушку. Тупой «клюв» едва заметен. Верхние челюсти немного длиннее нижних (рис. 22). Спинной плавник средних размеров, расположен посредине тела и сзади глубоко вырезан. Грудные плавники узкие, заостренные на концах, в длину достигают 7io длины тела. Окраска черная, на брюшной стороне чуть светлее.
В черепе рострум короткий (не длиннее мозговой коробки), широкий. Характерно, что межчелюстные кости очень широки, причем одинаково по всей длине. Спереди их края закруглены. Укороченные крыловидные кости граничат между собой.
Зубов в верхней челюсти 8—11, в нижней 8—12. В высоту зубы достигают 8 см, в диаметре 1,5—2,7 см.
Позвонков шейных 7, грудных 10, поясничных 9—11, хвостовых 22— 24; всего 50—51. Передние, а иногда и все шейные позвонки у взрослых животных срастаются между собой. Грудина состоит из 4 сегментов. Ребер 10 пар, из них 4 пары причленяются к грудине. Фаланг в передней конечности В—г, Не—8, Ш5-6, IV2—4, Vi—2-
Распространена в умеренных водах Тихого, Индийского и Атлантического океанов: в Восточной Атлантике — от Северного и Балтийского морей до Средиземного моря и мыса Доброй Надежды; в Западной Атлантике — от шт. Северная Каролина, США до Карибского моря и Буэнос-Айреса в Аргентине; в восточной части Тихого океана — от Японии До Австралии, Тасмании и Новой Зеландии; в Индийском океане — у Шри Ланка и Южной Индии.
В водах СССР не добывалась, но встречи возможны у Курильских о-вов, в Японском и Балтийском морях.
Экология почти не изучена. Питается главным образом головоногими моллюсками и стайной рыбой — пикшей, треской, лососевыми. Спаривание и щенка растянуты во времени. В зимнее время в теплых водах Иногда встречаются стада малых косаток в несколько сотен (до тысячи) иолов. В неволе хорошо приручается. Промыслового значения не имеет.
череп
Иногда (например, Nishiwaki, став семейства Globicephalidae.
1972) малую косатку включают в со-
Род косаток
Genus Orcinus Fitzinger, i860
В роде единственный вид: косатка — О. orca Linnaeus, 1758.
Размеры крупные, наибольшие в семействе. Длина тела до 10 м. Самцы значительно крупнее самок. Масса тела до 8 т. Голова немного сжата в дорзо-вентральном направлении. «Клюва» нет. Широкие грудные
плавники овальной формы (рис. 23). Спинной плавник достигает в вы-соту больших размеров, особенно у самцов, — до 1,7 м. Его величина и относительные размеры грудных плавников у обоих полов и хвостового у самцов с возрастом увеличиваются. Тело сверху черное, брюхо и горло белые. Позади спинного плавника и на виске над глазом распространено по белому пятну (иногда нет вовсе). Белая окраска брюха разделяется в задней части тела на три языка, из которых два боковых (правый и левый) заканчиваются позади заднепроходного отверстия, а средний может заканчиваться там же, либо идти вплоть до хвостовых лопастей (тогда последние бывают снизу белого цвета), либо совсем отсутствовать. Иногда правое и левое белые пятна сливаются между собой позади спинного плавника. На Дальнем Востоке встречаются совершенно темные косатки лишь с белым пятном над глазом.
В черепе широкий и уплощенный рострум немного короче или равен по длине мозговой коробке. Верхнечелюстные кости сильно развиты. Затылочный гребень, которого нет у молодых животных, особенно сильно развит у старых самцов. Височные ямы достигают крупных размеров. В черепе хорошо проявляется половая и возрастная изменчивость: височная впадина и затылочный гребень с возрастом увеличиваются; у взрослых самцов индексы ширины рострума несколько больше, чем у молодых; у самцов нижние челюсти относительно немного длиннее, чем у самок; затылочный гребень взрослых самцов превышает гребень самок и совсем отсутствует у молодых животных. Зубы крупные: 10—14 в верхней челюсти и 8—14 в нижней. Все они сжаты спереди назад, имеют сильно развитые корни, покрыты эмалью и очень прочны. Наибольший диаметр зубов достигает 3—5 см, высота 13 см. При закрытой пасти зубы верхней челюсти заходят между зубами нижней, образуя прекрасный хватательный аппарат. Сжатые спереди назад зубы, видимо, специально адаптированы к питанию крупной добычей, из которой косатки способны вырывать большие куски мяса (Томилин, 1957). Передние зубы, отклоненные несколько вперед и наружу, выдерживают самые сильные рывки добычи, а средние и задние выполняют функцию окончательного ее удерживания. Коронки зубов иногда стачиваются до пульпы.
Позвонков шейных 7, грудных 11—12, поясничных 10, хвостовых 21— 24; всего 50—52. Первые 2—3 шейных позвонка обычно срастаются между собой. Грудина состоит из 3—4 сегментов. Ребер 11—12 пар, из них 5—7 передних соединены с грудиной. Фаланг в пальцах П, Пб-7> III4 5, IV3—4, V2-з- Характерно, что длина фаланг пальцев и пястных костей меньше их ширины.
Хромосом в диплоидном наборе 44.
Распространена почти во всех морях от Арктики до Антарктики. В плотные льды не идет.
В СССР: в Балтийском, Баренцевом и Белом морях и на восток до северо-западной и западной части Карского моря. В морях Лаптевых и Восточно-Сибирском не обнаружена. На востоке от побережья Чукотского п-ова на юг в Беринговом, Охотском и Японском морях и с восточной стороны Курильских о-вов.
Держатся группами, иногда собираясь стадами до 150—200 голов. Способны плыть с большой скоростью (до 55 км/ч), часто выстраиваясь при этом в шеренгу или колонну, в которой ряд (по 3—6 косаток) следует за рядом. Под водой обычно остается по 5—6 мин.
Косатка — саркофаг с широким диапазоном кормов. Питается в основном рыбами, а также головоногими моллюсками и морскими млекопитающими. Из рыб поедает сельдь, треску, корюшку, мойву, макрель. Палтуса, тунца, пеламиду, скатов, акул и др. Нападает на большинство Усатых китов, дельфинов и ластоногих. Охотятся за добычей всем ста-
дом и, окружив крупного кита, рвут его на части. Массовая щенка и спаривание приходится на летние месяцы (в Японских водах на май — июль). Новорожденные имеют длину тела 2,1—2,7 м. Годовалые детеныши достигают длины 3,5 м. Длительность беременности составляет более года (по некоторым данным до 16 месяцев). В неволе хорошо приручается.
Специальный промысел не ведется.
Иногда косаток вместе с гриндами, малыми косатками, карликовыми косатками и бесклювыми дельфинами выделяют в особое семейство Globicephalidae (Nishiwaki, 1972).
Род серых дельфинов
Genus Grampus Gray, 1828
В роде единственный вид: серый дельфин— G. griseus G. Cuvier, 1812.
Размеры мелкие и средние. Длина тела до 4,3 м. Вследствие сильного развития жировой подушки голова спереди округлая, «клюв» не выражен. Спереди на голове проходит продольная бороздка. Нижняя челюсть немного короче верхней. Спинной плавник, расположенный почти посредине тела, вырезан по заднему краю. Длинные и узкие грудные плавники серповидной формы и заострены на концах. Окраска тела подвержена изменчивости: индивидуальной (от серой до черно-серой) и возрастной (у молодых животных она более светлая); на спине более темная по сравнению с другими частями туловища, постепенно светлеет к брюху. Цвет всех плавников такой же, как и спины. Между грудными плавниками находится светлое якоревидное пятно. Другие светлые пятна и полосы возникают в результате различных повреждений кожи.
В черепе рострум широкий, его длина короче мозгового отдела. Проксимальные части широких межчелюстных костей образуют выпуклость. Крыловидные кости примыкают друг к другу. Носовые и лобные образуют задненосовой гребень. Симфиз нижней челюсти короткий.
Зубы у большинства имеются только в передней части нижней челюсти в количестве 2—7 пар; в верхней челюсти их число может колебаться от 0 до 2 пар. У взрослых дельфинов зубы могут достигать 1,5 см в диаметре и почти до 4 см в высоту (корень и коронка). Утрата зубов в верхней челюсти, по-видимому, филогенетически недавняя. Очевидно, это следствие питания головоногими моллюсками (Томилин, 1957).
Позвонков шейных 7, грудных 12—13, поясничных 18—19, хвостовых 30—31; всего 68—69. Фаланг в передней конечности К, IIs-io, HIs-s, IV3-5, Vi.
Распространен в теплых и умеренных водах обоих полушарий: в восточной части Тихого океана обнаружен у о-ва Стюарт, побережий Британской Колумбии, Монтерей и Калифорнии (37° с. ш.), о-ва Гваделупа, в Калифорнийском заливе и у берегов Чили; в западной части Тихого океана — от Курильских о-вов и Японии до берегов Китая, Австралии и Новой Зеландии; населяет Индийский океан, Красное море, Восточную Атлантику от Британских о-вов и Средиземного моря до мыса Доброй Надежды и Западную Атлантику от штата Массачусетс, США до штата Нью-Джерси, США. Везде малочислен.
В водах СССР не добывался, но встречи возможны в Японском море и у Курильских о-вов.
Экология изучена недостаточно. Держится небольшими группами по 4—6 особей; иногда объединяется в более крупные стада до 20 голов. Питается преимущественно головоногими моллюсками. Детеныши рож-42
даются, по-видимому, зимой, в относительно теплых водах. Длина новорожденного составляет более половины длины матери (140—166 см).
Иногда серых дельфинов выделяют в особое семейство Grampidae (Nishiwaki, 1972).
Род гринд
Genus Globicephala Lesson, 1828
Размеры средние. Длина тела до 6,5 м. Самцы крупнее самок примерно на 1 м. Крупная жировая лобная подушка придает голове округлую форму. «Клюв» едва заметен (рис. 24). Ротовая щель проходит наискось сверху вниз под углом 30—45° к продольной оси тела. Верхняя челюсть немного длиннее нижней. Дыхало смещено несколько влево от средней линии. Низкий спинной плавник расположен почти на границе передней трети туловища. Задний край плавника вогнут. Грудные плавники узкие и длинные (до Чл длины тела). С возрастом у самцов на хвостовом стебле образуются высокие продольные вентральный и дорзальный кили. У самцов жировая подушка головы и горб, расположенный между головой и спинным плавником, крупнее, чем у самок. Окраска тела темно-серо-голубая или черная. Па вентральной стороне тела может располагаться светло-серая полоса, резко расширяющаяся на груди и горле.
череп широкий, слегка сплющенный в дорзо-вентральном направлении. Рострум по длине примерно равен мозговой коробке. Межчелюстные кости очень широкие. Крыловидные кости примыкают друг к другу. Носовые кости сдвинуты на лобные и расположены выше уровня всех других костей.
о , 7—13
Зубы находятся в передних половинах челюстей: --— •
С возрастом зубы могут стираться и выпадать.
Грудина состоит из 3—4 сегментов. Из 11 пар ребер с грудиной сочленены 5 пар. Позвонков шейных 7, грудных 11, поясничных 12—14, хвостовых 28—29; всего 58—59. Первые 5—6 шейных позвонков срастаются между собой.
Фаланг 1з—4, Пэ—14, Illg—и, IV2—з, Vi—2.
Хромосом в диплоидном наборе у обыкновенной гринды 44.
Распространены очень широко, почти во всех морях, исключая полярные от южной части Гренландии, Норвегии, Аляски и Японии на севере до мыса Доброй Надежды, мыса Горн, Новой Зеландии и о-ва Кергелен на юге.
Держатся группами иногда по нескольку сотен голов. Не потревоженные гринды часто выстраиваются колонной по 2—4 в ряд, один ряд за другим. Могут достигать скорости 40 км/ч. Черная гринда, по-види-мому, способна нырять на глубину до 370 м. Гринды теутоихтиофаги — питаются преимущественно головоногими моллюсками, в меньшей степени стайной рыбой (сельдевыми и тресковыми). Спаривание и щенка растянуты, но в основном приходятся на лето. Беременность и лактационный периоды продолжаются примерно по году. Длина тела новорожденных обычно 137—210 см. Окраска новорожденного из серой вскоре превращается в темную. В неволе хорошо приручаются.
Регулярный промысел ведется у о-вов Фарерских и у о-ва Ньюфаундленд, где гринд промышляют, загоняя их на отмель у пологого берега, иногда несколько сотен, даже тысяч голов. Добывают также у Шетландских и Оркнейских о-вов и в морях Дальнего Востока, где промысел ведут японцы. В водах СССР промысла гринд нет.
В роде 3 вида:
обыкновенная гринда — G. melaena Traill, 1809 (Северная Атлантика от Южной Гренландии и Ньюфаундленда до Средиземного моря);
тропическая гринда — G. macrorhyncha Gray, 1846 (тропические воды Мирового океана);
черная гринда — G. scammoni Соре, 1869 (умеренные воды северной части Тихого океана на севере до Командорских и Алеутских о-вов).
Некоторые исследователи в род гринд объединяют лишь 2 вида: G. melaena и G. macrorhyncha (Rice a. Scheffer, 1968) или даже один (Томилин,1957).
В водах СССР не добывались, но возможны встречи обыкновенной гринды в Баренцевом море и черной гринды в водах Дальнего Востока. Иногда гринд включают в состав семейства Globicephalidae (Nishiwaki, 1972).
Род карликовых косаток
Genus Feresa Gray, 1870
В роде один вид: карликовая косатка — F. attenuata Gray, 1875.
Размеры мелкие. Длина тела самцов достигает 250 см, самок — 240 см. «Клюва» нет, голова спереди закруглена. Довольно высокий 44
Рис. 25. Карликовая косатка — F. attenuata спинной плавник расположен примерно посредине тела и серповидно вырезан по заднему краю (рис. 25). Длина грудных плавников составляет примерно ‘/в длины тела. Окраска тела темно-серая. Белые лишь губы и пятно в области заднепроходного отверстия. Якоревидное пятно на груди светлое.
В черепе рострум занимает примерно половину его длины. Передние отделы предчелюстных костей уплощены; их внутренние края отделены друг от друга. В области симфиза нижняя челюсть имеет киль.
Л 10—и
Зубная формула: 1J_ j1 ’
Позвонков шейных 7, грудных 12—13, поясничных 16—17, хвостовых 32—34; всего 68—71. Первых три шейных позвонка срастаются между собой.
Распространена в Атлантике (обнаружена у берегов Сенегала) и в Тихом океане (у о-вов Гавайских и Хонсю).
В водах СССР не обнаружена, однако может быть встречена у Курильских о-вов и в Японском море.
Экология не изучена.
Иногда карликовую косатку включают в состав семейства Globicep-balidae (Nishiwaki, 1972).
Род морских свиней
Genus Phocoena G. Cuvier, 1817
Размеры мелкие. Длина тела до 2 м. Наиболее крупные морские свиньи из Черного и Азовского морей достигают в длину самцы 167 см, самки 180 см; из северной части Тихого океана — самки 178 см; из северной части Атлантики — самцы 186 см. Средняя длина тела половозрелых особей из Черного и Азовского морей составляет у самцов 141 см, у самок 148 см. «Клюва» нет. Голова притуплена, лобный выступ спереди уплощен (рис. 26). Передние края невысокого, почти треугольного спинного плавника, коротких овальных грудных плавников и лопастей хвостового плавника имеют небольшие роговые бугорки (иногда они сохраняются только на спинном плавнике). У морских свиней из Черного и Азовского морей по переднему краю спинного плавника их насчитывается 12—16, иногда 18—20. Окраска тела на спине темная (от почти черной до серой), с боков светлее, брюхо белое или светло-серое.
В черепе широкий рострум равен или несколько короче мозговой ко-Р°бки. Задние отделы межчелюстных костей образуют перед костным но-
совым отверстием вздутие. Крыловидные кости разделены между собой промежутком. Границы межтеменных костей всегда хорошо заметны.
Зубная формула:
16 — 30
16 — 27
У черноморско-азовских морских свиней число зубов в верхней челюсти бывает от 44 до 60 (наиболее часто 54), а в нижней — от 38 до 55 (обычно 44—46). С возрастом зубы могут снашиваться или даже полностью выпадать. Коронка зубов несколько сплющенная, отделена от корня шейкой. Форма коронок изменчива: от почти конической до слабо выраженной трехбугорчатой. Передняя сторона коронки немного повернута внутрь ротовой полости. При закрытой пасти зубы верхней челюсти располагаются снаружи от зубов нижней челюсти.
Позвонков шейных 7, грудных 12—14, поясничных 14—17, хвостовых 27—32; всего 62—68. Шейные позвонки могут срастаться между собой. Ребер 12—14 пар. Грудина цельная. Фаланг в передней конечности Ii—з> Hs-io, Ills-8, IV2-6, Vl-3.
Хромосом в диплоидном наборе у обыкновенной морской свиньи 44.
Распространение охватывает почти все моря Земного шара, исключая полярные.
Морские свиньи ихтиофаги. В Черном море пищей служат в основном придонные рыбы: бычки, обыкновенная камбала, морской язык, лещ, в меньшей степени пелагические рыбы: атеринка, хамса, судак, кефаль и сельдь. В Северной Атлантике поедают мойву, сельдь, сардину, макрель, треску, мерланга, сайду, мелких лососевых и некоторых других, а также головоногих и пластинчатожаберных моллюсков, а в Баренцевом и Белом морях песчанку, сельдь, мойву, навагу. В северной части Тихого океана кроме трех последних видов объектами питания служат сайка, американская сельдь, сиги. За пищей могут нырять на глубину 50—75 м.
Детеныши рождаются в летние месяцы. Пик спаривания приходится на конец лета — начало осени. Беременность длится 9—10 месяцев. Новорожденные имеют длину тела 63—86 см (около половины длины матери). Лактация продолжается около 4 месяцев. Самки способны к оплодотворению через 2—3 месяца после родов, во время молочного
кормления. Щенка ежегодная. Обычно держатся небольшими группами, хотя иногда собираются большими стадами.
В Северной Атлантике у обыкновенной морской свиньи, по-видимому существуют сезонные миграции — осенью к югу, а летом к северу. Обыкновенные морские свиньи Азовского моря на зиму мигрируют в Черное море. Более медлительны, чем, например, дельфин-белобочка. Под водой обычно не задерживаются дольше 6 мин. Приручаются плохо Обыкновенные морские свиньи регулярно промышлялись в Черном море весной и осенью, когда они мигрируют из Азовского моря через Керченский пролив следом за хамсой. С 1966 г. промысел запрещен.
В роде 4 вида:
обыкновенная морская свинья — Р. phocoena Linnaeus, 1758 (Северный Ледовитый океан и Северная Атлантика от Баренцева моря, Исландии и Девисова пролива на севере до Черного, Азовского и Средиземного морей на юге и от берегов Сенегала на востоке до штата Делавэр в США на западе; в северной части Тихого океана от мыса Барроу на Аляске до Калифорнийского залива на востоке и Японии на западе);
калифорнийская морская свинья — Р. sinus Norris et Me Farland, 1958 (Калифорнийский залив);
очковая морская свинья — Р. dioptrica Lahille, 1912 (Южная Атлантика от Ла-Платы в Аргентине до Огненной Земли, о-вов Фолклендских и Южной Георгии);
черная морская свинья — Р. spinipinntis Burmeister, 1865 (побережье Южной Америки к югу от Ла-Платы в Аргентине (35° ю. ш.) на востоке и до Пайта в Перу (5° ю. ш.) на западе).
В СССР: обыкновенная морская свинья в Баренцевом, Белом, Балтийском, Северном, Черном, Азовском, Японском, Охотском, Беринговом морях и на юге Чукотского моря.
Некоторые исследователи объединяют морских свиней, белокрылых морских свиней и бесперых морских свиней в отдельное семейство Pho-coenidae Bravard, 1885.
Род белокрылых морских свиней
Genus Phocoenoides Andrews, 1911
В роде единственный вид: белокрылая морская свинья — Р. dalli True, 1885.
Размеры мелкие. Длина тела самцов до 207 см, самок 180 см. «Клюва» нет. Нижняя челюсть слегка выступает за конец верхней челюсти. Вдоль дорзальной и вентральной сторон хвостового стебля проходят горбовидные кили. Верхний киль достигает особой высоты и, по-видимо-му, увеличивается с возрастом. Контуры спинного плавника составляют почти равнобедренный треугольник (он лишь слегка вогнут по заднему краю). Располагается он немного впереди от середины тела (рис. 27). На переднем краю спинного плавника расположено несколько мелких роговых бугорков. Грудные плавники относительно малы, с широким основанием. Окраска тела темная (от темно-стальной до черной); молодые Животные пигментированы интенсивнее, а новорожденные слабее, чем взрослые. Окраска головы, спины, верха передней и всей задней асти хвостового стебля темная — от темно-стальной до черной. На боках ела расположено по крупному белому пятну, тянущемуся вперед до Уровня переднего края спинного плавника или почти до самого глаза. *<аУДально это пятно по бокам хвостового стебля идет дальше уровня Днепроходного отверстия, а на брюхе не доходит до него. Спинной Лавник частично или целиком белый.
В черепе широкий и уплощенный рострум короче мозговой коробки. Крыловидные кости не граничат между собой.
' 19 — 23 — 28
Зубная формула;; 20_24_2g-‘
Зубы маленькие, долотовидные, едва возвышающиеся над деснами. Часть зубов может оставаться непрорезывающимися, в деснах. Межаль-веолярные перегородки развиты слабо. Зубы окружены бугорками ороговевшего эпителия десны, которые могут возвышаться над коронками зубов и функционировать вместо них.
Позвонков большое количество: шейных 7, грудных 15—18, поясничных 23—27, хвостовых 39—49; всего от 92 до 98. Шейные позвонки срастаются между собой. Число шевроновых костей колеблется от 23 до 35. Ребер 15—18 пар, из которых 3—5-я передних сочленены с грудиной. Фаланг в передней конечности Б-г, Нб-7. ПЦ-ь IV2-3» V1-2.
Хромосом в диплоидном наборе 44.
Распространена в северной половине Тихого океана от Берингова моря до Калифорнийского залива на востоке и Японии на западе.
В СССР: Японское, Охотское, Берингово и Чукотское моря.
Экология изучена слабо. Держится небольшими группами по 2—4 особи, иногда поодиночке или большими стадами до 1000 голов. Под водой обычно не задерживается дольше 5 мин. Может развивать скорость до 37 км/ч. Питается, по-видимому, в основном головоногими моллюсками и рыбами из сем. Moctophidae, сем. Sudidae, треской, сайрой, морской щукой, ставридой, мойвой, сельдью, анчоусом. Промысел не ведется.
Род бесперых морских свиней
Genus Neophocaena
Palmer, 1899
Длина тела обычно 150 сМ. Максимальная длина тела не превышает 190 см. Голова округлая, имеется хорошо выраженный лобный выступ (рис. 28). Спинного плавника и «клюва» нет. Грудной плавник составляет в длину примерно */б длины тела. По средней линии спины от шейного отдели до начзла хвосто-вого стебля тянется полоса 3—6 см шириной из многочисленных роговых бугорков. Глаза маленькие. Окраска спины свинцово-черная, брюхо светлее. Молодые животные светлее взрослых. На груди и брюхе расположено серое пятно. Грудные и хвостовой плавники име
ют иногда светлые участки различных размеров.
Череп несколько уплощенный. Закругленный спереди рострум широкий и короткий (значительно короче мозговой коробки). Маленькие крыловидные кости далеко отстоят друг от друга. Межчелюстные кости широкие, почти не суживающиеся кпереди.
Зубная формула: - ;-•
Зубы имеют расширенную и уплощенную коронку.
Позвонков шейных 7, гРУДных 13—14, поясничных П—14, хвостовых 28^31; всего 60—63. Первые пять шейных позвонков срастаются между собой. Фаланг 12, Ц5..7>
IV3—4, V2.
Рис. 28 Японская морская свинья—N. sunameri и ее череп
распространены от Пакистана и Индии на восток до о-вов угва, Калимантан и Японии. Встречаются в некоторых реках, в частности поднимаются вверх по Янцзы до 1000 км от устья и обитают в оз. Дунтинху.
В водах СССР не обнаружены, но японская морская свинья может быть встречена на Дальнем Востоке.
Экология изучена слабо. Держатся преимущественно в прибрежной зоне, в эстуариях, поднимаются вверх по рекам. Плавают поодиночке или группами по 4—5 особей. Питаются придонными ракообразными из группы длиннохвостых раков, рыбой и головоногими моллюсками. Приручаются плохо.
Регулярный промысел не ведется.
В роде 3 вида:
бесперая морская свинья — N. phocaenoides G. Cuvier, 1829 (от Пакистана и Индии на восток до п-ова Малайзия, о-вов Ява и Калимантан);
восточноазиатская морская свинья — N. asiaeorientalis Pillery et Gihr, 1972 (побережье Китая, р. Янцзы);
японская морская свинья — N. sunameri Pilleri et Gihr, 1975 (воды Японии).
Иногда все виды объединяют в один N. phocaenoides (Marine Mammal Commission, 1977).
СЕМЕЙСТВО НАРВАЛОВЫХ
FAMILIA MONODONTIDAE GRAY, 1821
Китообразные мелких размеров. Длина тела до 6,1 м, масса примерно до 2 т. Самцы крупнее самок. Спинного плавника нет, хотя по спине может проходить низкая и длинная кожная складка. Голова округлая, небольшая, без «клюва», отделена от туловища слабо или хорошо выраженной шеей. Борозд на горле нет. Грудные плавники короткие, широкие, закругленные по свободному краю. Окраска тела однотонная, темная или светлая, иногда с темными пятнами, подвержена возрастной изменчивости: у молодых она темная, с возрастом светлеет.
В черепе рострум относительно короткий и широкий. Лобные и межтеменные кости образуют гребень. Каменистая кость прирастает к черепу. Широкие крыловидные кости причленяются к чешуйчатым костям.
„ 1 — Н
Зубная формула: --— •
У самцов левый верхний зуб может образовывать бивень длиной до 3 м, направленный вперед.
Позвонков 49—55. Первый и второй шейные позвонки между собой не срастаются, остальные шейные позвонки могут срастаться в отдельные группы. У локтевой кости нет локтевого отростка.
Распространены в Арктике, кругополярно. Заходят во льды.
В семействе 2 рода (2 вида):
белух — Delphinapterus Lacepede, 1804 (1 вид);
нарвалов — Monodon Linnaeus, 1758 (1 вид).
В водах СССР — оба рода. Белухи имеют промысловое значение.
Род белух
Genus Delphinapterus Lacepede, 1804
В роде единственный вид: белуха — D. leucas Pallas, 1776.
Максимальная длина самцов на Дальнем Востоке достигает 599 см, самок 499 см, в районе Гренландии — самцов 545 см, самок 449 см. 50
Рис. 29. Белуха — D. leucas и ее череп
Голова имеет сильно выдающийся лобный выступ. Относительно небольшая голова отделена от туловища хорошо выраженной шеей (рис. 29). Отсутствие спинного плавника, видимо, объясняется приспособлением к жизни во льдах (плавник мог бы мешать плаванию среди льдов) (Гентнер, 1930) или возможностью поворачиваться вдоль длинной оси тела, что важно при ловле рыб (Яблоков, 1959). У некоторых особей на месте отсутствующего спинного плавника развивается невысокий кожный гребень.
Окраска тела однотонная: темная у молодых, белая или желтая у взрослых. В кожном покрове сильно утолщенная (до 15 мм) наружная часть эпидермиса — так называемая «броня», предохраняет от порезов при соприкосновении со льдом, а относительно более толстый по сравнению с другими китами слой подкожной жировой клетчатки уменьшает теплоотдачу у этого северного вида. Интересна возрастная изменчивость окраски тела: светло-серая у новорожденных, через несколько дней или недель после рождения она переходит в почти черную у сосунков и затем с возрастом постепенно светлеет. Быстрее всего побеление происходит на спине, боках тела и брюхе. Темный край хвостовых лопастей не подвергается депигментации. Предполагают, что белая окраска белух уменьшает теплоотдачу или помогает им при охоте, пугая рыб и заставляя НХ плыть в нужном для белух направлении. Все эти сомнительные предположения нуждаются в серьезных подтверждениях. Поверхность тела большинства зверей покрыта многочисленными шрамами, пятнами и полосами.
Череп относительно узкий, уплощен в дорзо-вентральном направлении. Широкий у основания рострум примерно в 1,5 раза длиннее мозговой коробки. Верхнечелюстные кости ограничивают снаружи костные ноздри и идут далеко назад почти до затылочных костей. Крыловидные кости крупные. Характерно развитие на черепе мощных гребней.
5 — 11
Число зубов изменчиво: ------— (Клейненберг и др., 1964).
Для простых колышкообразных зубов белухи характерна своеобразная режуще-удерживающая форма, которая образуется в результате стачивания зубов. Максимальные размеры зубов в верхней челюсти: длина 5,8 см, толщина 1,2 см; в нижней челюсти: длина 5,0 см и толщина 1,8 см. На межчелюстных костях зубы, как правило, отсутствуют. Прорезываются зубы у годовалых детенышей.
Позвонков шейных 7, грудных 11—12, поясничных 6—12, хвостовых 21—26; всего 49—53. В шейном отделе атлас и эпистрофей остаются свободными, тогда как другие позвонки могут срастаться по два-три вместе. Из 11—12 пар ребер 5—7 срастаются с грудиной. Фаланг 12-4, П6_д, Ш6_7, IV5-6> V4-6. III и IV пальцы по длине почти равны между собой. У взрослых животных наблюдается уменьшение числа фаланг некоторых пальцев по сравнению с эмбрионами.
Желудок состоит из 5 отделов. Длина кишечника от 22,5 до 37 м. Двенадцатиперстная кишка сильно развита.
Распространена в северных морях, омывающих берега Северной Америки и Евразии от 81° с. ш. на юг до Японии и Аляски в Тихом океане и до Британских о-вов и Бискайского залива в восточной части Атлантики и штата Массачусетс в США в западной Атлантике.
В СССР: моря Баренцево, Белое, Карское. Лаптевых, Восточно-Сибирское, Чукотское, Берингово, Охогское, Японское и изредка Балтийское.
Держатся белухи стадами различной величины. Летом иногда собираются до тысячи и более голов. Зимой образуют небольшие группы по нескольку десятков особей. В стадах наблюдается группировка по полу и возрасту: впереди находятся взрослые самцы, сзади самки с детенышами. В летнее время белухи встречаются у берегов, в открытом море у кромки льда и в полыньях. Зиму проводят, по-видимому, в разводьях среди льдов и в незамерзающих участках моря. Часто поднимаются далеко вверх по течению рек (например, за 2000 км от устья Амура).
Белухи быстро не плавают — максимум до 22 км/ч. Под водой обычно остаются 1—1,5 мин (не больше 15 мин). Превосходно развит слух. Характерны разнообразные звуки. Целям гидролокации служат 2 типа звуков: длительные импульсы с переменной частотой колебаний (служат для общей ориентировки) и короткие импульсы переменной длительности на одной и той же частоте (для изучения близкорасположенных объектов).
В летнее время питаются в основном рыбой. В Белом море поедает навагу, треску, пикшу, камбалу, сельдь, мойву, корюшку, пинагора, миногу, а также креветок и двустворчатых моллюсков; в Карском море — сайку, навагу, сиговых, нельму, сибирского осетра, щуку, корюшку, сельдь; на Дальнем Востоке — кету, навагу, сельдь, а также горбушу, кунджу, охотского сига, камбалу, красноперку, бельдюгу, бычков, морских собачек, миногу, ракообразных и головоногих моллюсков. Зимнее питание не изучено.
Щенка и спаривание большинства самок приходятся на середину или конец лета. Длина тела новорожденного обычно 130—150 см. Длительность лактационного периода не установлена. Спариваются самки задолго до окончания лактации. Беременность продолжается 11—12 месяцев. Рождение детенышей происходит ежегодно. Самки достигают поло-возрелости в 6 лет, самцы — в 6—9 лет.
Неволю переносит хорошо и может подолгу жить в океанарии.
Промышляют белуху на Новой Земле, на Дальнем Востоке в Тугур-ском заливе, у берегов Гренландии, Шпицбергена, в заливах Св. Лаврентия и Гудзона. От добытых зверей получают прочную кожу и ценный жир.
Род нарвалов
Genus Monodon Linnaeus, 1758
В роде единственный вид: нарвал — М. monoceros Linnaeus, 1758.
Длина тела у самцов до 610 см, у самок до 420 см. Общая форма тела сходна с белухой. Округлая небольшая голова отделяется от туловища слабо выраженной шеей. Округлость голове придает спермацетовая по
душка, слегка нависающая над верхней челюстью. Ротовое отверстие небольшое. Верхняя губа несколько выступает вперед над мясистой нижней губой. Спинного плавника нет. Слегка выпуклая спина несет невысокую складку кожи — до 5 см высотой и 75 см длиной. Грудные плав-
ники короткие и широкие (рис. 30). Окраска тела у молодых одноцветная темная (голубовато-серая, шиферная или аспидная), у взрослых светлая с многочисленными темными пятнами неправильной формы не более 5 см в диаметре, располагающимися на голове, спине и боках. На верхних частях головы, шеи и хвостового стебля пятна иногда сливаются
имеете, образуя общий темный фон.
Череп имеет в общем такое же строение, как и у белухи, но асимметрия выражена резче. Она проявляется в более сильном развитии челюстной и межчелюстной костей левой стороны, тогда как мозговой отдел Черепа сильнее развит на правой стороне.
Зубов в нижней челюсти нет. В верхней челюсти — всего 2 зуба (у самок они обычно не прорезываются). У самцов левый зуб превращается
в винтообразно закрученный справа налево направленный вперед бивень до 3 м длиной. Правый же зуб чаще скрыт в десне. Иногда у самцов развиваются два бивня. Изредка бивни образуются и у самок. Значение бивня для нарвала не выяснено. По этому поводу высказывается много предположений. По Томпсону (1939), спирально закрученный бивень служит стабилизатором против вращения тела вдоль оси при круговых движениях хвостового плавника во время плавания. Правильнее, по-видимому, считать бивни вторичнополовым признаком самцов (Чапский, 1941; Слепцов, 1955), который может служить и органом защиты. Вероятно, они могут выполнять и функцию, приписываемую им Томилиным (1957), — пробивать во льду отдушины, через которые может дышать не только сам самец, но и все члены стада — лишенные бивней самки и молодые звери.
Позвонков шейных 7, грудных 11—12, поясничных 6—10, хвостовых 26—27; всего 50—55. Шейные позвонки обычно не срастаются между собой. Грудина более короткая и широкая по сравнению с белухой. Фаланг в передней конечности В-2, П5-8, П14_6, IV2-4, V2-3-
Хромосом в диплоидном наборе 44.
Распространен в водах Арктики и Северной Атлантики к северу от линии плавучих льдов. На юг идет (зимой) до Британских о-вов, Голландии и Лабрадора в Атлантике и до Берингова моря в Тихом океане.
В СССР: во всех арктических морях.
Держатся стадами, причем крупные косяки состоят из взрослых самцов, самок и молодняка, а мелкие — только из взрослых самцов или из самок с детенышами. Прежде встречались огромные стада по нескольку тысяч голов, теперь они не превышают сотни.
Добывает себе пищу в пелагической части моря. Питается в основном головоногими моллюсками, а также рйбой: скатами, камбалой, палтусом, сельдью, лососевыми, тресковыми. Сроки спаривания и щенки растянуты. Длина новорожденного около 1,5 м.
Специально промышляется лишь местными жителями Гренландии.
СЕМЕЙСТВО КАШАЛОТОВЫХ
FAMILIA PHYSETER1DAE GRAY, 1821
Китообразные мелких и средних размеров с длиной тела 2,1—21 м. Половой диморфизм выражен резко (самцы значительно крупнее самок) или слабо.
Голова очень крупная или относительно небольшая. Дыхало расположено посредине головы или на ее переднем конце с левой стороны. Спинной плавник высокий, серповидно вырезанный сзади или низкий, горбообразный.
В черепе рострум расширен и имеет на дорзальной поверхности большое углубление, в котором помещается крупная спермацетовая подушка. У самцов она выступает вперед по отношению к костям черепа. Затылочная кость образует высокий гребень. Слезная кость приращена к черепу. Узкая нижняя челюсть не доходит до переднего отдела головы; симфиз обеих половин составляет обычно не менее */3 ее длины.
Зубная формула:
0—10
0 — 28 ’
Зубы расположены в глубоких альвеолах, сохраняются у большинства животных только в нижней челюсти. В верхней челюсти они мелкие, непрочные или отсутствуют вовсе. Функционально их замещают ороговевшие выступы на небе.
Распространены главным образом в теплых и умеренных водах Ат-лантического, Тихого и Индийского океанов. Представители рода кашалотов обычны, многочисленны, а рода карликовых кашалотов встречаются очень редко.
Кашалотовые — теутофаги, питаются в основном головоногими моллюсками разных размеров, в том числе и крупными.
Практическое значение имеет только кашалот, который промышляется.
В семействе 2 рода (3 вида):
карликовых кашалотов — Kogia Gray, 1846 (2 вида) и
кашалотов — Physeter Linnaeus, 1758 (1 вид).
Некоторые ученые (например, Kellog, 1928; Nishiwaki, 1972) выделяют карликовых кашалотов в особое семейство Kogiidae.
В СССР: оба рода.
Род карликовых кашалотов
Genus Kogia Gray, 1846
Размеры наименьшие в семействе. Длина самцов в среднем 2,9 м (до 3,4 м), самок 2,4 м. Голова в отличие от головы представителя рода кашалотов небольшая и спереди закруглена (рис. 31). Спермацетовый мешок развит хорошо, однако слабее, чем у кашалота. Дыхало расположено почти симметрично на теменной части головы. Высокий спинной плавник находится примерно посредине тела. Грудные плавники относительно узкие. Окраска спинной стороны тела, хвостовых и грудных плавников темная, почти черная. Бока светло-серые, брюхо светлое. Позади глаз и впереди грудных плавников снизу вверх, в темноокрашенную область, проникают один или два светлых выступа. Светлые пятна могут встречаться и на вентральной поверхности хвостовых лопаетей.
В черепе широкий у основания рострум резко суживается кпереди. Углубление на его дорзальной поверхности неглубокое и имеет продольный гребень, образованный межчелюстными и верхнечелюстными костями. Носовые кости не срастаются с межчелюстными. Верхнечелюстные кости имеют очень крупные предглазничные отростки. Скуловая кость не причленяется к чешуйчатой. Симфиз нижней челюсти короче половины ее длины.
Зубов 8—16 пар на нижней челюсти и 1—3 пары в переднем отделе верхней (иногда здесь зубы полностью отсутствуют).
Позвонков шейных 7, грудных 12—14, поясничных 9—12, хвостовых 21—27. Шейные позвонки срастаются между собой. Уплощенная лопатка имеет тонкий акромиальный и крупный коракоидный отросток. Грудина короткая. Ребер 13—14 пар. Число фаланг в пальцах подвержено индивидуальной изменчивости: I2, Щ-я, III4-8, IV4-8, V2-7.
Хромосом в диплоидном наборе у карликового кашалота 42.
Распространены в умеренных и тропических водах Тихого, Атлантического и Индийского океанов.
Экология изучена слабо. Численность везде очень мала. Держатся поодиночке или небольшими группами до 5 голов. Питаются в основном головоногими моллюсками и в меньшей степени рыбами и ракообразными. Сроки спаривания и щенки растянуты. Самка спаривается задолго До окончания лактационного периода. Беременность длится примерно у месяцев. Самки размножаются, вероятно, ежегодно. Детеныш остает-Ся с матерью около года.
Рис. 31. Карликовый кашалот — К. breviceps и его череп
Хозяйственного значения не имеют и специально не промышляются.
В роде 2 вида:
карликовый кашалот — К. breviceps Blainviile, 1838 (умеренные и тропические воды Тихого, Атлантического и Индийского океанов);
малый кашалот -— К. simas Owen, 1866 (прибрежные воды Южной Африки, Южной Австралии, Индии, о-ва Шри Ланка, Гавайских о-вов, Японии, восточного побережья США).
В водах СССР: могут быть встречены оба вида в Японском море и у южной части Курильских о-вов.
Род кашалотов
Genus Physeter Linnaeus, 1758
В роде единственный вид: кашалот — Р. catodon Linnaeus, 1758.
Размеры наибольшие в семействе и среди зубатых китов. Ясно выражен половой диморфизм: длина самцов до 18—21 м, самок до 11—13 м. Форма тела каплевидная, значительно утолщенная в переднем отделе. Огромная выпуклая сверху голова, достигающая 7з длины тела, спереди оканчивается тупо — «лобовиной» (рис. 32). Размеры головы в основном определяются мощным спермацетовым мешком, расположенным над черепом и выступающим вперед за передний конец челюсти. Спермацетовый орган представлен двумя сообщающимися между собой полостями, заполненными маслянистой жидкостью — спермацетом и окруженными фиброзной и мышечной тканью. В состав спермацета входит в ос-
новном жировоск (цетин). От взрослых кашалотов удается получить от 1 до 5,7 т спермацета. Голова самцов относительно крупнее (до 1/з длины тела), чем у самок (немногим более ’Л длины тела). С возрастом головной отдел кашалота относительно увеличивается, а хвостовой соответственно укорачивается.
Непарное дыхало S-образной формы располагается в отличие от других китообразных в передней части головы слева от средней линии и достигает в длину от 20 до 60 см. Глаза находятся примерно посредине головы на уровне задних углов рта. Длина разреза глазной щели 6—12 см, Диаметр глазного яблока 15—17 см. Позади глаз и несколько ниже их открываются небольшие (до 1 см длиной) серповидные ушные отверстия. 1 олова от туловища отделяется слабо заметным шейным перехватом. В области грудных плавников туловище достигает наибольшей толщины, а затем постепенно утончается к хвосту. Грудные плавники короткие, широкие, с закругленными наружными краями. Крупные лопасти хвостового плавника разделены глубокой выемкой. Позади низкого горбообразного спинного плавника располагаются 2—6 более низких бугров. Вдоль нижней стороны хвостового стебля проходит невысокий кожный киль. Кожный покров (в частности, и дерма) достигает значительной толщины (у некоторых особей на брюхе до 50 см). На горле находятся 0 4Q продольных бороздок глубиной до 1 см и длиной от 5 до 50 см. озможно, благодаря им глотка может несколько расширяться при за-латывании крупной добычи. •
Окраска тела однородно темная (черная, черно-бурая, темно-серая) несколько более светлым брюхом, на котором расположено белое пят-
но неправильной формы с сильно изменяющимися у разных особей очертаниями.
В резко асимметричном черепе верхние боковые отделы верхнечелюстных костей образуют гребни, охватывающие с боков ложе для спермацетовой подушки. Правая носовая кость срастается с правой межчелюстной. Горизонтально расположенные небные кости сзади перекрываются крыловидными костями. Половинки нижней челюсти образуют между собой симфиз, составляющий до 50% (или несколько более) длины всей челюсти.
0 — 11 Зубная формула: ——-— •
Гомодонтные колышкообразные зубы кашалота, часто слегка изогнутые в средней части, могут достигать до 27 см в длину.. Наиболее крупные зубы находятся посредине челюсти. У самцов зубы крупнее, чем у самок. Зубы эмбрионов могут быть трехвершинными. Прорезываются зубы детенышей обычно к концу лактационного периода. Они прочно сидят в альвеолах, возвышаясь над уровнем десны лишь на Vs—Vs своей длины.
Позвонков шейных 7, грудных 11, поясничных 8—9, хвостовых 20— 24. В отличие от других китообразных у кашалотов атлант остается свободным, а все остальные шейные позвонки срастаются между собой. В пятипалой передней конечности плечевая кость иногда срастается с локтевой и лучевой. Узкая лопатка имеет большой коракоидный отросток. Ребер до 11 пар, из которых только 3 соединены с грудиной.
Хромосом в диплоидном наборе 42.
Распространение охватывает все моря от Северного Ледовитого океана до морей Антарктики. Предпочитают теплые и умеренные воды. В летнее время мигрируют к северу, а зимой — к югу. Северная граница летнего распространения в Тихом океане проходит примерно по 62° с. ш., а в Атлантике местами достигает и даже переходит 75° с. ш. Места зимовок в западной части Тихого океана располагаются в основном южнее Японских о-вов, а в Восточной Атлантике — в тропических водах у Западной Африки. В водах СССР обычны на Дальнем Востоке, возможны также встречи у Мурманского побережья.
Питается в основном головоногими моллюсками, преимущественно кальмарами, в значительно меньшей степени — рыбой. Может нападать на крупных кальмаров до 10 м и более в длину и поедать их. В поисках добычи может нырять на большую глубину (более чем на 1000 м) и оставаться подолгу под водой (15—60 мин). Кашалоты — полигамы, и в период размножения около одного половозрелого самца держатся 10— 15 самок. Растянутый во времени период спаривания приходится главным образом на весенне-летнее время и сопровождается драками между самцами. Продолжительность беременности точно не установлена. По данным различных исследователей, она составляет от 10—12 до 16— 17 месяцев. Новорожденные встречаются круглый год. Рождение обычно происходит в теплых водах более низких широт. Новорожденные имеют длину тела 3—3,5 м, массу около 1 т. Длительность лактации неизвестна; предполагают, что она длится от 5—6 до 12—13 месяцев. Спаривание у самок происходит еще до окончания лактации. Рождают детенышей, по-видимому, один раз в 3—4 года. Половозрелость наступает у самок в возрасте 4, у самцов — 4—5 лет.
Кашалот имел до недавнего времени важное промысловое значение. Так, в сезон 1966/67 г. и в календарный 1967 г. китобойными флотилиями 12 стран было добыто почти 26 тыс. особей (Берзин, 1971). В последние годы в связи с общим уменьшением численности кашалота (к 1972 г. 58
«мерная численность в Мировом океане составляла около 700 тыс.) ^обыча сокращена (до 23 тыс. голов в промысловом сезоне 1972/73 гг.)«
Желателен полный запрет промысла кашалота на ряд лет.
СЕМЕЙСТВО КЛЮВОРЫЛОВЫХ
FAMILIA ZIPHIIDAE GRAY, 1865
Мелкие китообразные с длиной тела до 6—12,5 м. Морда образует суженный «клюв». Полулунное по форме дыхало расположено выпуклостью назад и вперед. Спинной плавник находится на уровне заднепроходного отверстия или немного впереди от него. Хвостовые лопасти не имеют между собой выемки. Грудные плавники расположены низко на боках тела. На горле проходят борозды.
В черепе длинный и узкий рострум длиннее мозговой коробки. Верхнечелюстные, межчелюстные и лобные кости образуют гребни. Каменистые кости срастаются с черепом. Крупные слезные кости не сливаются со скуловыми. Большие крыловидные кости отстоят друг от друга на небольшом расстоянии. Нижняя челюсть шире верхней и немного выдается впереди нее.
„ 0—19
Зубная формула: --— •
У эмбрионов закладываются многочисленные зубы, позднее редуцирующиеся.
У взрослых особей большинства представителей семейства немногочисленные зубы имеются только на нижней челюсти.
Число позвонков не превышает 50. Ребер не больше 10 пар; их грудные отделы не окостеневают. У взрослых особей сохраняются рудименты обонятельных нервов.
Распространены в теплых, умеренных и холодных водах всех океанов.
Теутофаги; питаются в основном головоногими моллюсками. Способны к глубоким и продолжительным ныряниям.
Практическое значение клюворылов относительно невелико. В водах СССР они промышляются в небольшом количестве.
В семействе 5 родов (18 видов):
тасмановых клюворылов — Tasmacetus Oliver, 1937 (1 вид);
плавунов — Berardius Duvernoy, 1851 (2 вида);
ремнезубов — Mesoplodon Gervais, 1850 (12 видов);
клюворылов — Ziphius G. Cuvier, 1823 (1 вид), и бутылконосов — Hyperoodon Lacepede, 1804 (2 вида).
Разделение семейства на роды в настоящее время не вызывает сомнений. Однако система видов клюворылых нуждается в дальнейшей разработке (особенно рода ремнезубов).
В СССР: все роды, кроме тасмановых клюворылов.
Род тасмановых клюворылов Genus Tasmacetus Oliver, 1937 IggB роде единственный вид: тасманов клюворыл — Т. shepherdi Oliver, Длина тела около 6 м. Голова округлая с коротким и толстым «клю-59
вом». Глаза относительно крупные. Окраска спинной стороны тела черная, брюшной — белая.
19
Зуб°в — ;
всего 90.
На конце нижней челюсти располагается пара более крупных зубов с конической коронкой.
Распространен в водах Новой Зеландии. Очень редок (известно всего 3 экземпляра).
Экология не изучена.
Род плавунов
Genus Berardius Duvernoy, 1851
Длина тела 7,8—12,5 м. Самки крупнее самцов. Голова относительно мала. «Клюв» уплощен в дорзо-вентральном направлении. Лобный выступ на голове небольшой (рис. 33). У самцов он поднимается вверх и отграничен отчетливой поперечной бороздой. У самок лобное возвышение более пологое и поперечной борозды нет. Дыхало имеет полулунную форму и выпуклостью обращено вперед. На голове располагаются 2 (редко 3—4) глубокие продольные бороздки длиной около 60 см. Они хорошо видны у эмбрионов. Форма спинного плавника изменчива: от низкого, горбовидного до относительно высокого, более стройного. Его длина больше высоты в 2—3 раза. Грудные плавники расположены близко к брюшной поверхности тела. Они относительно короткие и широкие. Хвостовые лопасти имеют между собой небольшую выемку. Окраска тела с возрастом темнеет. У северного плавуна спинная сторона тела шиферно-серая, брюшная немного светлее, а у южного плавуна спина темноголубая, бока серовато-голубые, брюхо серое или светло-серое. У северного плавуна на брюхе могут быть белые пятна. Белые шрамы (вероятно, от укусов самцов) встречаются в большем числе на спине, чем на брюхе.
Асимметрия костей черепа выражена слабее, чем у других представителей семейства клюворыловых. Узкий рострум в два раза превышает длину мозговой коробки; он слегка сплющен в дорзо-вентральном направлении. Челюстные гребни низкие. Межчелюстные кости симметрич-
Ы ДРУГ ДРУГУ и имеют почти одинаковую ширину по всей своей длине. Лобные кости с широкими глазничными отростками. Крупные носовые кости сильно надвинуты на лобные и занимают наиболее высокое положение в черепе.
Зубы — от 2 до 4 пар, расположены в переднем отделе нижнеи челюсти Они имеют коронку треугольной формы и передние более крупные зубы при закрытой пасти остаются снаружи. В десне может быть скрыто еше несколько зубов. Передние зубы могут достигать в высоту 79—89, задние — 53—67 мм. У самок задних зубов, как правило, нет. У новорожденных и сосунков зубы еще не прорезываются. У молодых зверей передние зубы могут быть длиннее, чем у взрослых. С возрастом зубы несколько стачиваются, но становятся массивнее, а часть, находящаяся
в альвеоле, уплощается.
Позвонков шейных 7, грудных 10—И, поясничных 12, хвостовых 17— 19' всего 47—48. Тела трех передних шейных позвонков срастаются между собой. Грудина состоит из 5 сегментов.
Распространены в Тихом и Индийском океанах и Южной Атлантике.
Экология изучена слабо. Головоногие моллюски служат основным объектом питания, а рыбы — тресковые, морские ерши, скаты и др., второстепенным. Под водой остаются обычно 8—20 мин. Спаривание и щенка растянуты во времени, хотя спаривание происходит главным образом в феврале, а роды в декабре. Новорожденный достигает в длину 4,5— 4,8 м. Беременность длится около 10 месяцев. У северного плавуна отмечена сезонная миграция: весной половозрелые самцы, самки и молодые животные (обычно вместе) группами по 10—40 голов держатся на севере в Беринговом и Охотском морях, а осенью на юге у Японии и Калифорнии.
В СССР специальный промысел на плавунов не ведется и добывают их только случайно.
В роде 2 вида:
южный плавун — В. arnouxi Duvernoy, 1851 (примерно от 33° ю. ш. на юг до арктических льдов в Южной Атлантике, Тихом и Индийском океанах);
северный плавун — В. eairdi Stejneger, 1883 (северная часть Тихого океана от Берингова моря и далее на юг до Монтерей в Калифорнии на востоке и до Японии на западе).
В СССР: на Дальнем Востоке встречается северный плавун.
Род ремнезубов
Genus Mesoplodon Gervais, 1850
Длина тела до 6,7 м. Спинной плавник невысокий, его вершина загнута назад (рис. 34). Грудные плавники заострены на концах. Окраска тела темная (от черной до серой), но брюхо'светлое.
В черепе длинный и тонкий рострум сверху более узкий, чем у других Представителей семейства клюворыловых. Верхнезатылочная и межтеменная кости образуют высокий гребень. Лобные кости имеют небольшие глазничные отростки. Средняя обонятельная кость полностью окостене-вает. Симфиз нижней челюсти значительных размеров (1/5—’/з длины челюсти) .
От двух до четырех сильно сжатых с боков зубов располагаются в передней трети нижней челюсти и при закрытой пасти находятся вне ее. Корни зубов проходят наискось к продольной оси челюсти. У некоторых Пидов ремнезубов зубы резко увеличены в размерах. В десне верхней Челюсти скрываются многочисленные мелкие зубы.
Рис. 34. Командорский ремнезуб — М. stejnegeri и его череп
Позвонков шейных 7, грудных 9—11, поясничных 9—12, хвостовых 18—21; всего 46—49. Атлант и эпистрофей срастаются между собой (иногда к ним прирастает и третий шейный позвонок). Остистые отростки позвонков очень крупные. В грудине различают 5 (реже 4) отделов. Из пяти пальцев передней конечности самые длинные II и III.
Фаланг В-2, IU-6, ПЦ-в, IV3-4, V2-3.
Ареал охватывает Восточную Атлантику — от Норвегии и Британских о-вов до о-ва Мадейра, Средиземного моря и побережья Южной Африки; Западную Атлантику — от Ньюфаундленда, Канады до Карибско-го моря, о-ва Тринидад и от устья р. Чубут в Аргентине (42° ю. ш.) до Фолклендских о-вов; восточную часть Тихого океана — от Берингова моря до Ла-Хоя в Калифорнии и побережья Чили; западную часть Тихого океана — от Берингова моря до Японии и от Австралии до Новой Зеландии; Индийский океан.
Экология изучена крайне слабо. Питаются, очевидно, в основном головоногими моллюсками. Сроки спаривания и щенки сильно растянуты.
Специальный промысел за ремнезубами не ведется.
В роде 12 видов:
атлантический ремнезуб — М. bidens Sowerby, 1804 (Северная Атлантика на востоке от Норвегии, Балтийского моря и Британских о-вов до Средиземного моря и на западе от Ньюфаундленда до штата Массачусетс, США);
антильский ремнезуб — М. europaeus Gervais, 1855 (северная часть Западной Атлантики от Тринидада, Ямайки и Мексиканского залива до Нью-Йорка; одна находка известна из пролива Ла-Манш);
ремнезуб Тру — М. mirus True, 1913 (Северная Атлантика от Флориды и Новой Шотландии на западе до Британских о-вов на востоке; одна находка отмечена у Южной Африки);
австралийский ремнезуб — М. pacificus Longman, 1926 (известен по одной находке на побережье Квинсленда в Австралии);
ремнезуб Грея — М. grayi Haasl, 1876 (воды Южной Африки, Южной Австралии, Новой Зеландии, Патагонии, Фолклендских о-вов; одна находка известна близ Голландии);
ремнезуб Гектора — М. hectori Gray, 1871 (побережье Тасмании, Новой Зеландии и Фолклендских о-вов);
командорский ремнезуб — М. slejnegeti True, 1885 (северная часть
Тихого океана от Берингова моря на юг до Японии и штата Орегон в гША);
Ь ремнезуб Хабса — М. carlhubbsi Moore, 1963 (северная часть Тихого кеана от Японии на восток до штата Вашингтон в США и Калифорнии);
° новозеландский ремнезуб — М. bowdoini Andrews, 1908 (побережье Новой Зеландии, Тасмании и Западной Австралии);
японский ремнезуб — М. ginkgodens Nishiwaki et Kamiya, 1958 (побережье Японии и Калифорнии);
ремнезуб Лейарда — М. layardi Gray, 1865 (побережье Южной Африки, Южной Австралии, Новой Зеландии, Чатемских и Фолклендских ° В тупорылый ремнезуб — М. densirostris Blainville, 1817 (тропические и субтропические воды Мирового океана к северу и к югу от экватора).
Систематика рода ремнезубов разработана недостаточно четко. Ряд
видов описан всего по 1—2 экземплярам и отличия между некоторыми из них при более тщательном исследовании могут оказаться лишь индивидуальными вариациями. Так, можно предполагать, что М. stejnegeri True, 1885; М. bowdoini Andrews, 1908; Af. ginkgodens Nishiwaki et Kamiya, 1958 и M carlhubbsi Moore, 1963, населяющие Тихий океан, отно-
сятся к одному виду.
В водах СССР найден только один вид — командорский ремнезуб, а некоторые другие могут встречаться, например, в южной части Балтийского моря — атлантический ремнезуб, в морях Дальнего Востока — японский ремнезуб и ремнезуб Хабса.
Род клюворылов
Genus Ziphius G. Cuvier, 1823
В роде единственный вид: настоящий клюворыл — Z. cavirostris G. Cuvier, 1823.
Наибольшая длина тела 7,9 м. Самки крупнее самцов в среднем на 30 см. В отличие от бутылконосов и плавунов лобный выступ значительно более низкий и спереди без резкой границы переходит в короткий, конический «клюв» (рис. 35). Характерна небольшая ротовая щель (меньшая, чем у других представителей семейства клюворыловых). На горле проходят две продольные бороздки. Спинной плавник низкий, с сильно изменяющейся формой у разных особей — от серповидной до треугольной. Грудные плавники относительно длинные и узкие. Нижняя челюсть сильнее выдается вперед, чем верхняя. Дыхало выгнутой стороной обращено назад. Окраска тела сильно изменяется в зависимости от пола и возраста. Встречаются целиком серые (наиболее часто), темно- или голубовато-серые, бурые или черные животные, у других брюшная сторона может быть светлее спинной или наоборот. Могут быть и другие вариации. На коже много депигментированных пятен различной формы и величины.
Рострум резко асимметричного черепа наиболее широкий и короткий (лишь немного длиннее мозговой коробки) среди представителей семейка клюворыловых и слабо отграничен от мозговой коробки. Сошник °ора3ует крепкое мезоростральное окостенение. Продольные челюстные гребни развиты крайне слабо. На дорзальной стороне черепа между внешними краями верхнечелюстных костей находится крупная впадина, в Которой располагается спермацетовый мешок. Челюстные, межчелюстные и носовые кости образуют позади костных ноздрей большой выступ в виде свода, нависающего над ноздрями.
Единственная пара зубов располагается на переднем конце нижней
Рис. 35. Настоящий клюворыл — Z. cavirostris и его череп
челюсти и при закрытом рте выступает перед верхней челюстью. Зубы самцов крупнее и массивнее, чем у самок (диаметр зубов у самцов 25— 29 мм, у самок 10—14 мм). Молодые животные в деснах верхней и нижней челюстей имеют по 28—30 зачатков зубов, которые с возрастом редуцируются.
Позвонков шейных 7, грудных 9—10, поясничных 10—11, хвостовых 19—22; всего 46—49. Три передних шейных позвонка обычно срастаются между собой. Грудина состоит из 5 отделов. Ребер 9 пар. Фаланг в передней конечности: 1ь 113_6, Ш5-б, IV4, Vi-2.
Распространен в Северной Атлантике — от Северного и Балтийского морей до Средиземного моря и от побережья штата Массачусетс в США до Флориды и Вест-Инчии; в Южной Атлантике — у побережья Южной Африки и Аргентины; в северной части Тихого океана — от Берингова моря до Калифорнии на востоке и Дальнего Востока и Японии на западе, а также у Гавайских о-вов и острова Мидуэй; в южной части Тихого океана — от архипелага Бисмарка до Австралии и Новой Зеландии на западе и побережья Чили на востоке; в Индийском океане — от Южной Африки до Австралии.
В СССР: встречается в Балтийском море и в водах Дальнего Востока.
Экология изучена слабо. Пелагическая форма, питается, по-видимому, в основном головоногими моллюсками. За добычей ныряет обычно на 10—30 мин. Сроки спаривания и щенки растянуты. Держатся небольшими группами. Везде малочислен.
Род бутылконосов
Genus Hyperoodon Lacepede, 1804
Длина тела самцов высоколобого бутылконоса до 9,4 м, самок—-до 8,7 м. «Клюв» выражен более четко, чем у других представителей семейства клюворыловых, так как лобный выступ жировой подушки почти отвесно опускается к его основанию (рис. 36). С возрастом лобный выступ увеличивается в размерах. У самцов лобная жировая подушка развита сильнее. Небольшой спинной плавник с вырезом по заднему краю расположен позади середины тела. Грудные плавники также относительно Невелики. Окраска на спинной стороне темная (черно-серая), на брюшной более светлая. Плавники более темного цвета, чем туловище.
В черепе сильно развиты продольные челюстные гребни (особенно у взрослых самцов), образующие у основания рострума значительный выступ. Носовые кости крупные и сверху вогнуты. Вместе с межчелюстными костями они поднимаются позади костных носовых отверстий вертикально вверх и даже нависают над ними.
Зубов 2, реже 4 пзры. Самцы имеют более крупные зубы, чем самки. у высоколобого бутылконоса две пары зубов только у некоторых взрослых самцов. Самки и многие самцы задней пары зубов вообще не имеют, а если они прорезываются, то бывают значительно меньше передних.
3—671
65
Передняя пара зубов располагается на вершине нижней челюсти и при закрытой ротовой полости остается вне ее. Зубы округлые или слабо овальные в сечении. В длину достигают 5 см.
Позвонков шейных 7, грудных 8—9, поясничных 9—11, хвостовых 18—20; всего 43—46. Шейные позвонки срастаются между собой. Грудина состоит из трех отделов, фаланг 1,_2, П5_6, Ш5, IV4-5, V2-3-
Распространены в Северном Ледовитом и Атлантическом океанах и южных частях Индийского и Тихого.
Экология изучена недостаточно. Держатся высоколобые бутылконосы преимущественно небольшими группами по 10—20 особей — взрослых самцов, самок и детенышей или только самцов. Иногда образуют большие стада до нескольких сотен голов. За добычей могут нырять на большую глубину, оставаясь под водой 10—20 мин (иногда до часа или несколько дольше). Отмечены сезонные миграции. Так,-высоколобый бутылконос весну и лето проводит в Норвежском море, у Шпицбергена и Ньюфаундленда, а зиму — в более южных водах.
Питаются главным образом головоногими моллюсками и в значительно меньшей степени рыбой — сельдью, треской и другими и голотуриями. Сроки спаривания растянуты. Беременность продолжается год или даже 15 месяцев. Детеныши рождаются весной и летом. Новорожденный достигает в длину 3—3,5 м.
В наших водах специально не промышляется.
В роде 2 вида:
высоколобый бутылконос — И. ampullatus Forster, 1770 (Северный Ледовитый океан, Белое и Баренцево моря; Северная Атлантика от Денисова пролива и Шпицбергена до Балтийского и Средиземного морей на востоке и штата Род-Айленд в США на западе);
плосколобый бутылконос — Н. planifrons Flower, 1882 (от льдов Антарктики до Буэнос-Айреса и залива Баня-Бланка в Аргентине и Южной Атлантике до Вальпараисо, 33° ю. ш. и Новой Зеландии в южной части Тихого океана и до южных и западных районов Австралии в Индийском океане).
Описанный Лонгманном в 1926 г. Mesoplodon planifrons иногда рассматривается как синоним Hyperoodon planifrons (Hershkovitz, 1966) или как подвид Mesoplodon mirus Moore, 1960, или в качестве единственного вида отдельного рода Indopapacetus Moore, 1968 (Moore, 1968). Описание сделано всего по двум черепам, поэтому для окончательного решения необходим дополнительный материал.
В СССР: встречается только высоколобый бутылконос в морях Белом, Баренцевом, Балтийском.
ПОДОТРЯД УСАТЫХ китов
SUBORDO MYSTICETI FLOWER, 1864
Размеры от мелких до наибольших в отряде. Длина тела от 6,1 м У карликового кита до 33 м у голубого кита. Самки крупнее самцов. Характерна крупная голова, составляющая 1/3—'/s длины тела и огромная ротовая полость. Наружные ноздри парные, расположены в особом углублении в теменной области. На морде имеются небольшие щетинки, выполняющие тактильную функцию. К сумке одного такого волоса подходит до 400 нервных окончаний.
В черепе кости располагаются симметрично. В относительно небольшом мозговом отделе черепа характерны резко наклоненные вперед 66
„хнезатылочиые кости. Очень сильно развиты верхнечелюстные и ежчелюстные кости, образующие длинный и узкий рострум, а такж§ Мижнечелюстные кости. Обычно верхняя челюсть загибается дистальным ониом вниз, а нижняя челюсть вверх, причем средние отделы нижнече-юстных костей сильно выгнуты в наружные стороны. Сохраняются обо-ятельные раковины. У лобных костей имеются широкие глазничные от-остки. Носовые кости образуют передний отдел наружных костных носовых отверстий.
В процессе эмбрионального развития у усатых китов происходит закладка зубов, но вскоре рост их прекращается и они резорбируются, функционально отсутствующие зубы заменены так называемым китовым усом, образующим своеобразный цедильный аппарат. Он состоит из 130—-470 роговых пластин на каждой стороне верхней челюсти. Пластина китового уса имеет в общем форму треугольника, причем стороной, образующей наименьший катет, пластина укреплена в десне верхней челюсти, наибольший катет обращен наружу, а сторона, соответствующая гипотенузе, —• в ротовую полость. Эта последняя сторона пластины расщеплена на многочисленные и довольно длинные волосовидные щетинки — как бы бахрому. Бахрома соседних пластин, переплетаясь между собой, образует войлокообразный внутренний покров пластин. Пластины китового уса имеют неодинаковые размеры; наиболее высокие располагаются примерно посредине верхней челюсти, поэтому вперед и назад происходит уменьшение пластин. В каждой пластине различают сердцевинный, или мозговой, слой и два наружных, корковых. Мозговой образован многочисленными роговыми трубочками (полыми сосочками), проходящими по всей длине пластины. У внутреннего края пластины корковые слои исчезают, и роговые трубочки непосредственно переходят в бахрому. При плавании кита с открытой пастью вода проходит сквозь бахрому пластин китового уса, а все пищевые объекты, вплоть до очень маленьких (планктонных ракообразных), оседают на ней. Закрыв
рот, кит движениями языка отправляет пищу в глотку.
Грудина представлена одной костью, к которой причленяются только передние ребра.
Пищеварительная система, в противоположность зубатым китам, не отделена полностью и постоянно от дыхательного тракта, так как черпаловидный хрящ и надгортанник не образуют удлиненной трубки, входящей в хоаны. В этом проявляется сходство усатых китов с большинством наземных млекопитающих. Верхние части альвеолярных перегородок легких имеют большое число мускульных волокон. Глотка узкая. Желудок обычно трехкамерный. Слепая кишка имеется.
Усатые киты космополиты; обитают во всех океанах и большинстве открытых морей Земного шара.
Держатся как в открытом море (полосатики), так и у берегов (серые киты, горбачи). Больших стад не образуют. Ряд видов совершают дальние сезонные миграции. Подводные звуки, издаваемые усатыми кита-Ми, — это крики низкой частоты, обычно значительно более длительные, чем «щелчки» зубатых китов. Питаются массовой пищей —• зоопланктоном, стайной рыбой.
Самки размножаются ежегодно или через год-два. Беременность Длится около года, лактационный период — 4—7 месяцев.
Практическое значение усатых китов достаточно велико. Многие из них имели, а некоторые и в настоящее время имеют промысловое значение. Переопромышление некоторых видов привело к почти полному Их уничтожению (например, гренландского кита). Желательно полное запрещение промысла всех усатых китов.
В подотряде 3 семейства и 6 родов:
гладких китов — Balaenidae Gray, 1825 (3 рода);
серых китов — Eschrichtiidae Ellerman et Morrison-Scott, 1951 (1 род);
полосатиковых — Balaenopteridae Gray, 1864 (2 рода).
В СССР: все семейства.
СЕМЕЙСТВО ГЛАДКИХ КИТОВ
FAMILIA BALAENIDAE GRAY, 1825
Размеры изменчивы — от мелких (карликовый кит — 6,1—6,4 м) до средних и крупных (гренландский, южный киты — до 17'м, иногда 21 м)1.
Огромная голова составляет ’Д—*/з длины тела. Тело толстое. Спинной плавник имеется только у карликового кита. На брюшной стороне кожных складок нет. Грудные плавники короткие и широкие. Полость рта очень высокая. Пластины китового уса в ней достигают в высоту до 2—4,5 м. Число пластин в каждой половине челюсти 210—400. Левый и правый ряды пластин не соединены спереди роговыми стерженьками. Щель закрытого рта искривлена в виде дуги, обращенной выпуклостью вверх, и делит лицевой отдел на две неравные части: меньшую верхнюю и значительно большую нижнюю.
В черепе рострум узкий, резко изогнутый дистальным концом вниз. Передний край верхнезатылочной кости заходит далеко вперед за линию орбит. Носовые кости небольшие и не имеют спереди расширения. Задний край носовых и предчелюстные кости располагаются спереди по отношению к надглазничному отростку лобных костей. Нижнечелюстные кости сильно выгнуты в стороны, их венечный отросток рудиментарный. Костные слуховые барабаны имеют ромбовидную форму. Верхнечелюстные кости сзади не имеют лобно-носового отростка.
Все шейные позвонки срастаются между собой в единое образование.
Распространение охватывает арктические моря, Атлантику и Тихий океан.
Плавают медленно. Питаются планктонными ракообразными. Миграции выражены относительно слабо. Держатся в поверхностных слоях воды поодиночке или небольшими группами.
Биология размножения изучена слабо. Длительность беременности около 12 месяцев. Детеныши рождаются в Северном полушарии в конце зимы — с января по апрель.
Большого хозяйственного значения в настоящее время не имеют, так как в результате неумеренного выбоя в последние столетия их численность упала настолько, что возникла опасность полного уничтожения. Благодаря запрету промысла этих китов численность постепенно восстанавливается. Добыча небольшого числа гладких китов разрешена лишь местному населению.
В семействе 3 рода (3 вида):
гренландских китов — Balaena Linnaeus, 1758 (1 вид);
южных китов — Eubalaena Gray, 1864 (1 вид);
карликовых китов— Caperea Gray, 1864 (1 вид).
Иногда роды гренландских и южных китов объединяют в один род — Balaena Linnaeus, 1758 (например, см. Rice a. Scheller, 1968).
В СССР: 2 рода — гренландские и южные киты.
' В семействе можно выделить гладких китов мелких размеров с длиной тела 6,1 —11 м, средних— 11,1—15 и крупных—15,1—21 м.
В роде единственный вид: гренландский кит — В. mysticetus Linnaeus, 1758.
Размеры крупные. Длина тела обычно 15—17 м, максимум 21 м. Самки на 0,5—1 м крупнее самцов. Голова, относительно более крупная у самцов, составляет примерно */з длины тела. Шейный перехват довольно глубокий (рис. 37). Грудные плавники короткие и широкие с закругленным наружным краем (между дыхалом и передним концом). На нижних челюстях и подбородке нет кожных наростов. Ротовая щель имеет форму правильной дуги (если смотреть на голову сбоку). Верхний край нижней челюсти не имеет фестончатых выступов. Пластины китового уса высокие (3—4,5 м), узкие, очень эластичные, темного цвета, с тонкими «волосами» бахромы, толщиной всего 0,08—0,2 мм. Число пластин китового уса в каждой половине челюсти достигает 320—400. Окраска тела темная, иногда с белым горлом. Неполовозрелые особи серовато-темного цвета.
В черепе рострум очень узкий и длинный. Межчелюстная и челюстная кости, если смотреть на них сверху, примерно одинаковой ширины.
Позвонков шейных 7, грудных 13, поясничных 10—13, хвостовых 22— 24; всего 54—55. Акромиальный и венечный отростки лопатки хорошо развиты. Ребер 12—14 пар. Фаланг 1>, Пз-4. ПД-б, IV3-4, V2-3.
Распространение охватывает Арктические моря. В прошлом гренландские киты были многочисленны и служили важным объектом промысла. В настоящее время очень редки. Чаще, чем в других местах, они встречаются в водах Чукотского п-ова. Различают 3 стада гренландских китов. Шпицбергенское стадо мигрирует от восточного берега Гренландии, о-вов Исландии и Ян-Майена к Шпицбергену, отсюда часть китов возвращается в Датский пролив, а другая проходит западнее Шпицбергена к северо-восточному мысу Гренландии и вдоль ее восточного побережья тоже достигает Датского пролива. Западногренландское стадо мигрирует из Девисова пролива в Баффинов залив. Отсюда часть стада (главным образом самки с молодыми) идут обратно в Девисов пролив ЧеРез Ланкастеров пролив, огибая о-ва Баффинова Земля и Кокберн, а дРугая часть следует дальше на север проливом Смита и уже отсюда Б°звращается в пролив Девиса. Между шпицбергенским и западногрен
дп
ландским стадами существует постоянный обмен особями. Более изолировано третье стадо — берингово-чукотское, которое мигрирует из моря Бофорта вдоль Аляски в Чукотское море, а затем через Берингов пролив в Берингово море (где зимует) и обратно. В настоящее время первые два стада почти полностью уничтожены.
В водах СССР встречается у Новой Земли в Карском, Восточно-Сибирском, Баренцевом, Чукотском, Беринговом и Охотском морях.
Основными объектами пищи служат мелкие планктонные рачки Са-lanus finmarchicus и крылоногие моллюски Limacina helicina, в меньшей степени Clio borealis.
Период деторождения длится с февраля по май. Новорожденный имеет длину около 4,5 м. Период спаривания растянут. Продолжительность беременности, по данным различных авторов, от 9 до 14 месяцев. Находка в теле одного гренландского кита датированного гарпуна свидетельствует, что длительность жизни, видимо, составляет около 40 лет.
Держатся поодиночке или небольшими группами по 2—5 особи. Под водой остаются обычно 5—10, иногда 20 мин, раненые — до 40—80 мин. Плавают медленно со скоростью 7—8 км/ч, раненые до 13—17 км/ч. Фонтан гренландских китов высокий — до 4 м, двойной.
Гренландский кит хорошо приспособлен к жизни во льдах, превосходно ориентируется среди разводий и способен даже взламывать лед толщиной до 20—30 см.
Промысел гренландского кита разрешен только местному населению для собственного потребления. Весной и осенью чукчи и эскимосы Чукотского п-ова добывают китов из вельботов ручными гарпунами и ружьями. От одного кита получают до 30—36 т жира.
Гренландский кит включен в «Красную книгу» как вид, которому грозит исчезновение.
Род южных китов
Genus Eubalaena Gray, 1864
В роде единственный вид: южный кит — Е. glacialis Muller, 1776.
Размеры средние и крупные. Длина тела обычно 13—17 м, (максимум до 21 м). Самки на 0,3—1 м крупнее самцов. Самка длиной 17,1 м была массой 67,2 т. Голова составляет примерно 'Д Длины тела. Шейный перехват менее глубокий, чем у гренландского кита (рис. 38). На верху головы между концом рыла и дыхалом располагается крупный кожный нарост с утолщенным и плотным дермальным слоем и очень толстым эпидермисом. Такие же наросты, но меньших размеров расположены на подбородке, нижней челюсти и на верху головы позади большого нароста. На наростах растут редкие щетинковидные волосы. Наружная поверхность эпидермиса наростов изъедена китовыми вшами (Cyamus), которые поселяются здесь во множестве. Ротовая щель сбоку имеет сложную S-образную форму. Верхний край нижней челюсти фестончато вырезан. Пластины китового уса не такие длинные, как у гренландских китов (максимум до 2,6 м) и не так эластичны. Толщина «волосков» бахромы усов в средних отделах около 0,13 мм. Число пластин меньше, чем у гренландского кита, — 210—260 в каждой половине челюсти. Окраска тела черная или темно-голубоватая. Брюшная сторона несколько светлее. Иногда на брюхе расположены белые пятна разной величины; из-
редка встречаются южные киты с белым брюхом. Новорожденные имеют светло-серую окраску, с возрастом темнеют.
В черепе рострум уже, чем у гренландского кита, меньше загнут вниз и относительно короче. Если смотреть на череп сверху, то межчелюстная кость шире челюстной.
Позвонков шейных 7, грудных 14—15, поясничных 10—11, хвостовых 25; всего 56—57. Ребер 14—15 пар. Лопатка имеет только акромиальный отросток. Фаланг 13, П5, Ш6, IV4, V4.
Распространение охватывает Северную Атлантику — от берегов Северо-Западной Африки. Средиземного моря, о-ва Мадейры, Азорских, Бермудских о-вов и Флориды к северу до Шпицбергена, Кольского п-ова, о-ва Ян-Майен, Исландии, Ньюфаундленда и пролива Девиса. В тропические и высокоширотные воды Атлантического океана южные киты не идут. В Тихом океане южный кит встречается в Северном полушарии от Аляски и Алеутских о-вов вдоль побережья Северной Америки до штатов Орегон и Калифорния в США и от Анадырского залива, Охотского и Японского морей до Желтого и Восточно-Китайского морей. В Южном полушарии он населяет акваторию примерно от 15 до 60° ю. ш. (в основном между 30 и 50° ю. ш.): воды Австралии, Новой Зеландии, у берегов Южной Америки, Южной Африки, у о-вов Крозе, Кергелен, Амстердам, Сен-Поль и в открытые воды Антарктики. В соответствии с районами обитания среди южных китов можно выделить 3 подвида: бискайский кит, обитающий в Северной Атлантике, японский кит — из северной части Тихого океана и австралийский кит — из морей Австралии. Иногда эти подвиды рассматривают как отдельные виды: Е. glacialis; Е. sieb'oldi; Е. australis.
В водах СССР бискайский кит встречается у берегов Кольского п-ова, а японский кит в Анадырском заливе у восточного побережья Камчатки, в Охотском и Японском морях.
Держатся в большей части поодиночке, иногда образуют небольшие гРуппы по 2—3 особи. Хотя южные киты более подвижны, чем гренландские, их скорость плавания также невысока — обычно около 7 км/ч, а у испуганных китов — до 14 км/ч. Они не ныряют глубже 15—18 м и остаются под водой 10—20 мин. Потревоженный кит может пробыть под водой 30 и даже 50 мин. Фонтаны южного кита высокие — до 4,5 м и Двойные.
Рис. 39 .Карликовый гладкий кит — С. marginata
Основными объектами питания южного кита в Тихом океане служат различные виды морских ракообразных — Calanus и Euphasia, а в Атлантическом океане — ракообразные — Calanus finmarchicus и Thysae-nossa inermis, реже — крылоногие моллюски.
Детеныши рождаются раз в два года зимой и весной в теплых водах. Длина тела новорожденных достигает 4—4,5 м. Лактация продолжается около 6—7 месяцев. Оплодотворение самок происходит вскоре после родов. Длительность беременности около года.
На южном ките найдено большое количество эктопаразитов — усоногих раков Coronula и различных видов китовых вшей — Cyamus. Китовые вши обычно поселяются на кожных наростах на голове, а также вокруг дыхала, глаз и мочеполового отверстия.
Хозяйственного значения в настоящее время не имеет. Длительный промысел (бискайских китов промышляли еще в IX—X вв., причем от одного кита охотники получали до 15—18 т жира) подорвал популяции южного кита. Южный кит включен в «Красную книгу» как вид, которому угрожает исчезновение.
Род карликовых китов
Genus Caperea Gray, 1846
В роде единственный вид: карликовый кит—С. marginata Gray, 1846.
Размеры мелкие. Длина тела 6,1—6,4 м, причем голова составляет примерно четверть этой длины. Имеется небольшой спинной плавник с относительно глубокой вырезкой по заднему краю (рис. 39). Окраска тела черная с различно расположенными серыми пятнами разной формы. Иногда на брюхе проходит белая полоса. Резко выделяется белая окраска языка и слизистой оболочки ротовой полости. Китовый ус белого цвета с темными краями. В каждой половине челюсти находится до 230 пластин китового уса. Наибольшие из них в длину достигают 70 см, а в ширину 12 см. Бахрома усов тонкая.
Позвонков шейных 7, грудных 17, поясничных 2, хвостовых 14 15. всего 40—41. Шейные позвонки срастаются между собой. Ребер 17 пар-Для них характерны большая ширина и уплощенность.
Распространены в Южном полушарии. В Южной Атлантике — от Буэнос-Айреса (Аргентина) до Мыса Горн, Фолклендских о-вов и Юя<-ной Африки; в южной части Тихого океана — у берегов Новой Зеландии и индоокеанского побережья Южной и Юго-Западной Австралии.
Экология изучена слабо. Держатся, по-вндимому, поодиночке. Даль-..игпяпии нр установлены.
Рис. 40. Серый кит — Е. gibbosus и его череп
СЕМЕЙСТВО СЕРЫХ КИТОВ
FAMILIA ESCHRICHTIIDAE ELLERMAN
ET MORRISON-SCOTT, 1951
Длина тела взрослых самок обычно 12—15 м, взрослых самцов 11— 13 м. Наиболее крупная самка имела длину 15 м, самец — 14,6 м. Масса самки длиной 13,3 м была 31,46 т. По ряду признаков серые киты занимают промежуточное положение между полосатиковыми и гладкими китами. Тело менее стройное, чем у полосатиковых. Спинного плавника нет (рис. 40). На его месте на уровне заднепроходного отверстия находится низкое, сжатое с боков горбовидное возвышение. Позади этого возвышения тянется несколько (6—14) низких, закругленных выступов. На заднем краю хвостового плавника между хвостовыми лопастями расположена глубокая вырезка. Кожных складок на брюшной поверхности тела нет. только в области горла проходят 2 (реже 3—4) глубокие борозды. Начинаются они сразу за симфизом нижней челюсти и достигают В длину около 2 м.
Нижняя челюсть, килевидная снизу, имеет такую же длину, как и верхняя челюсть.
Ротовая щель серых китов в противоположность гладким китам и по-лосатиковым делит голову почти на две равные половины — верхнюю и нижнюю. Полость рта низкая и пластины китового уса тоже низкие (20— 30 см высотой), толстые, реже расположенные, чем у других усатых китов, и с более грубой бахромой. Отдельные «волоски» бахромы имеют в Длину 20—25 см и толщину у основания 0,9—1,1 мм, а в средних отделах — 0,4—1,0 мм. В каждой половине челюсти насчитывается 130—• 180 пластин китового уса белого цвета. Спереди правый и левый ряды пластин разделены широким промежутком.
На верху головы у взрослых китов находится около 60 вибрисс и еще Примерно 120 вибрисс — на нижней челюсти. Зародыши имеют на верху головы вдвое большее число вибрисс. Сильно развита подкожная жировая клетчатка, достигающая на груди толщины 30—45 см. Окраска тела серая с буроватым оттенком и многочисленными светлыми пятнами различного расположения и формы. Эти пятна образуются в результате деятельности эктопаразитов.
Череп более массивный, чем у представителей других семейств под-ОтРяда усатых китов. Он шире, чем у гладких китов, но уже, чем у по-лосатиковых. Рострум широкий и толстый, слегка загнутый вниз. Передний край верхнезатылочной кости далеко не доходит до линии, соединя-
ющеи ороиты. нижние челюсти массивные, широкие, почти прямые рудиментарным венечным отростком. Костные слуховые барабаны сжаты с боков. Носовые отростки предчелюстных костей и крупные носовые кости продолжаются назад дальше уровня передней границы надглазничных отростков лобных костей. Верхнечелюстные кости с носовыми отростками.
Позвонков шейных 7, грудных 14, поясничных 12—14, хвостовых 21 — 23; всего 56. Благодаря тому что шейный отдел позвоночника сильно удлинен и позвонки в этом отделе не срастаются, голова имеет некоторую подвижность. Ребер 14. Тазовые кости относительно крупные. Фаланг в четырехпалой передней конечности (нет I пальца) П3, III3, IV4, Vt.
Хромосом в диплоидном наборе 44.
Распространение охватывает субтропические, умеренные и холодные воды северной части Тихого океана и Чукотское море.
Серые киты — типичные прибрежные обитатели и без ущерба для себя могут даже обсыхать во время отлива. По-видимому, с целью избавления от эктопаразитов они часто трутся телом о дно или заходят в пресноводные лагуны. Во время миграции держатся поодиночке или небольшими группами до 10—12 китов. Б высоких широтах на местах кормежек собираются стадами до 150 голов. Серые киты тихоходны — скорость плавания 7—10 км/ч и даже у напутанных животных скорость плавания не превышает 14—18 км/ч. Спасаясь от нападения косаток, серые киты затаиваются или подплывают вплотную к берегу. Под водой при кормежке остаются обычно 3—4 мин. Длительность дыхательных интервалов 1—7, иногда до 20 мин. Высота раздвоенных фонтанов 1,5—3 м.
Серый кит питается у дна на небольших глубинах — 15—60 м. Нередко роется мордой в иле, захватывая ртом и процеживая илистые массы. В Беринговом и Чукотском морях питается в основном придонными ракообразными: Ampelisca macrocephala, Lembos arcticus, Anonyx nugax, Eusirus sp., Atylus sp., Mysius oculata. В Желтом море (в мае) поедает мелких рачков Nephros thompsonii, а в водах Северной Калифорнии в июле — Euphasia pacifica.
Период спаривания длится с декабря по март с пиком в январе — феврале. Самки приносят детенышей зимой (в январе — феврале) на мелководье у побережья Корейского п-ова и Калифорнии. Новорожденные имеют длину тела 3,6—-5,5 м. Вскоре после рождения детенышей самки спариваются. Лактация продолжается 6—12 месяцев. Длительность беременности около года. Половозрелоегь наступает в 8—9 лет.
В летнее время взрослые особи плывут в более северные районы ареала (Чукотское море — Берингов пролив), а молодые остаются южнее (у Корякского побережья). Распространены в летнее время в Охотском море (охотско-корейская популяция) и в Чукотском и Беринговом морях (чукотско-калифорнийская популяция). Из Охотского моря серые киты на зиму уходят к берегам Корейского п-ова и Южной Японии. Киты чукотско-калифорнийской популяции зимуют у берегов Калифорнии. Протяженность миграции составляет 5—10 тыс. км.
В СССР: серый кит в Беринговом, Чукотском, Охотском и Японском морях.
Серый кит сильно заражен эктопаразитами: усоногими ракообразными— Cryptolepas rhachianactei и китовыми вшами — Cyamus scammonii.
В 1853—1856 гг. число китов в водах Калифорнии составило примерно 30 тыс. Усиленный промысел серых китов с 1850 по 1925 г. почти полностью истребил их. К 1930 г. серых китов оставалось всего несколько сотен. В 1938 г. было введено ограничение промысла, а с 1947 г. полное его запрещение. К 1955 г. численность чукотско-калифорнийского стада возросла до 3—4 тыс. голов, к 1060 — до 5—6 тыс. голов, к 1970 г. пример-74
но Д° У—11 тыс. голов, ъ настоящее время серых кшив иришишлши> ли1ПЬ местные жители для своих нужд. Недавно правительство США разрешило добывать по 100 серых китов ежегодно с целью выяснения необходимости их дальнейшей охраны. Можно предполагать, что в будущем без ущерба для серых китов может быть организован разумный промысел.
В семействе один род серых китов — Eschrichtius Gray, 1864 (один вид)»
Род серых китов
Genus Eschrichtius Gray, 1864
См. описание семейства.
В роде единственный вид: серый кит — Е. gibbosus Erxleben, 1777 (ареал совпадает с ареалом семейства).
СЕМЕЙСТВО ПОЛОСАТИКОВЫХ
FAMILIA BALAENOPTERIDAE GRAY, 1864
Размеры от мелких до наибольших, не только в отряде китообразных, но и в классе млекопитающих. Длина тела от 6,7 до 33 м. Тело стройное с обтекаемыми контурами или более короткое, утолщенное. Голова в 3,5—5 раз короче длины тела. Грудные плавники относительно небольшие, узкие и заостренные или огромные, достигающие длины тела. Спинной плавник тонкий, серпообразный, вырезанный по заднему краю или невысокий, горбовидный. По нижней поверхности передней части тела проходит от 12 до 120 продольных кожных складок. Функция их не совсем ясна. Возможно, они имеют значение для облегчения скольжения тела кита в воде, а также, растягиваясь, способствуют увеличению ротовой полости, что важно при ловле добычи. Характерно необычайно сильное развитие сети эластиновых волокон и сухожильных пучков в коже в области складок. Полость рта относительно низкая. Соответственно пластины китового уса также невысоки — обычно не более 1 м. В каждой половине челюсти их насчитывается 270 -430. Спереди правый и левый ряды пластин соединяются между собой многочисленными роговыми стерженьками. Ротовая щель слабо изогнутая и делит лицевой отдел головы на неравные части — большую нижнюю и меньшую верхнюю.
В черепе рострум довольно широкий, слабо изогнутый. Верхнечелюстные кости имеют лобно-носовые отростки. Верхнезатылочная кость передним краем находится на уровне орбит или лишь немного выдается вперед дальше их уровня. Носовые кости небольшие, спереди резко расширены. Носовые кости и носовые отростки предчелюстных костей продолжаются назад дальше уровня переднего края надглазничных отростков лобных костей. Выгнутые наружу нижнечелюстные кости с развитым венечным отростком. Костные слуховые барабаны имеют яйцевидную форму.
Шейные позвонки обычно не срастаются между собой, а если и срастаются, то только первые 2—3 позвонка. Передние конечности четырехпалые (без третьего пальца), предплечье в них значительно длиннее тела.
Распространены во всех океанах Земного шара.
Питаются планктонными ракообразными и стайными рыбами. Самки приносят детенышей ежегодно или раз в два года. У большинства имеют место сезонные миграции.
Рис. 41. Голубой кит—В. musculus и его череп
Большинство представителей семейства имели, а некоторые имеют и сейчас важное хозяйственное значение, составляя значительный процент в ежегодной мировой добыче китов. Чрезмерный промысел поставил под угрозу существование некоторых видов полосатиков.
В семействе 2 рода (6 видов):
настоящих полосатиков — Balaenoptera Laccpede, 1804 (5 видов) и горбатых китов — Megaptera Gray, 1846 (1 вид).
В СССР: оба рода.
Род настоящих полосатиков
Genus Balaenoptera Lacepede, 1804
Тело удлиненное, стройное, хорошо обтекаемое. Длина тела от 6,7— 9 м (малый полосатик) до максимальных в отряде китообразных — 23— 33 м (голубой кит). Самка голубого кита с длиной тела 27,1 м имела массу 136,4. Голова составляет ’Л—'7s длины тела (рис. 41). Спинной плавник высокий, уплощенный, серповидно вырезанный по заднему краю. Грудные плавники умеренной длины — не более 1/7 длины тела, ланцетовидной формы с ровными краями. На голове и грудных плавниках нет шишковидных кожных наростов. Нет кожного нароста и на переднем-конце нижней -челюсти. По брюшной стороне тела проходит 40—120 продольных, относительно узких кожных складок.
Пластины китового уса имеют высоту до 18—21 см у малого полосатика, до 70—90 см у финвала и до 130 см у голубого кита. Число пластин в каждой половине челюсти у голубого кита 270—400, финвала — 260— 470, сейвала — 219—402, полосатика Брайда — 250—280, малого полосатика — 231—348. Окраска китового уса различна у разных видов настоящих полосатиков: у голубого кита пластины смоляно-черные; финвала — темно-голубые, причем в передней половине правого ряда целиком белые или белые с продольными серыми полосками; у сейвала черно-сероватые с белой или серой бахромой; полосатика Брайда — серовато-черные, в передней части белые; малого полосатика — белые и желтые. В ротовой полости на передней части неба проходит одна продольная срединная борозда.
Окраска тела у голубого кита темно-серая с голубоватым оттенком, с большим количеством серых и более светлых пятен, образующих своеобразный мраморный рисунок. У финвала окраска темно-серая на спине и более светлая или белая на брюхе, причем правая сторона нижней челюсти снаружи светлее левой, а в ротовой полости, наоборот, темнее.
Примерно такая же окраска у сейвала и малого полосатика. Полосатик Брайда на спинной стороне голубовато-черный, а брюшной — серый или светло-серый. Тело настоящих полосатиков, особенно в задней части на боках, покрыто многочисленными светлыми пятнами с радиально расположенными волнистыми линиями. Эти пятна появляются в результате
деятельности эктопаразитов.
При сравнении черепов настоящих полосатиков и горбатого кита можно отметить, что у первых скуловые дуги расставлены менее широко, верхнеглазничные отростки лобной кости слабее выдаются в стороны, но сильнее расширены спереди назад, граница между верхнечелюстными и
межчелюстными костями по верху черепа менее извилиста.
Позвонков шейных 7, грудных И—16, поясничных 12—16, хвостовых 17—27; всего 47—65. Лопатка имеет хорошо развитые акромиальные и коракоидные отростки. Фаланг 14_6, П5-9, IV5_7, V2-4-
Хромосом в диплоидном наборе у малого полосатика, сейвала и финвала по 42.
Распространены во всех океанах и большинстве морей Земного шара.
Настоящие полосатики больших стад обычно не образуют, а держатся поодиночке, попарно или по три вместе. Лишь иногда они скаплива-
ются в больших количествах на кормовых полях. Обычная скорость движения кормящихся китов 5—15 км/ч. Испуганный или раненый кит способен на короткой дистанции развить скорость до 37—55 км/ч. Голубой кит ныряет, по-видимому, не глубже 100 м; загарпуненный финвал достиг глубины 500 м (Kooyman a. Anderson, 1969). Под водой голубой кит остается 5—50 мин, финвал во время кормежки 5—8 мин, сейвал 2— 12 мин, малый полосатик — 2—3 мин. Форма и размер фонтанов специфичен для каждого вида: у малого полосатика он имеет высоту 1—2 м и форму опрокинутого конуса, у голубого кита — высоту 6—9 м и форму более вытянутого опрокинутого конуса.
Объектами питания служат в основном планктонные ракообразные и рыбы. Голубой кит поедает главным образом ракообразных и за рыбами специально не охотится. В Северной Атлантике это Thysanoessa inermis, Meganislephanes norvegica, Femora longirornis, на Дальнем Востоке — Thysanoessa inermis, T. raschii, T. longipes, T. spinifera, Calanus cristatus, C. tonsus, Euphasia pacifica, Nematoscelis megalops, в Антарктике — Euphasia superba, реже Thysanoessa macrura. Финвал употребляет в пищу кроме ракообразных различных видов Thysanoessa, Calanus, Euphasia и некоторых других, разнообразных рыб — сельдь, мойву, песчанку, сайку, навагу, минтая, треску и др., и некоторых головоногих моллюсков. Сейвал и малый полосатик также питаются смешанной пи-
Щей из ракообразных, рыб и головоногих моллюсков, полосатик Врай-Да — в основном стайными рыбами — сельдью, макрелью, анчоусом, сардинами, а также ракообразными. Преобладание того или иного пищевого объекта отмечается у разных видов настоящих полосатиков в зависимости от сезона и географического места. Например, сейвал в КБкном полушарии питается лишь рачками Euphasia superba и Parathes-г° guidichaudi, в Северном — разными видами ракообразных и рыбой, а У Курильских островов и Японии еще и головоногими моллюсками. Желудок настоящих полосатиков объемистый. У голубого кита он вмещает 1,5 т пищи. В желудке одного финвала было найдено 2800 (560 кг) кальмаров.
Период спаривания у настоящих полосатиков сильно растянут во времени, но пик его приходится на зимние месяцы; у голубого кита Северного полушария — на январь, Южного полушария — на июнь. Длительность беременности около года (примерно 11 месяцев у голубого кита и финвала, около 12 месяцев у сейвала, 10—12 месяцев у полосатика Брайда, от 10 до 18 месяцев, по данным разных исследователей, у малого полосатика). Длина тела новорожденных голубого кита обычно 7—• 8 м, финвала 6—6,4, сейвала 4,2—5,3, малого полосатика 2,1—2,7 м. Продолжительность лактации самок сейвала 5 месяцев, финвала 6 и голубого кита 7. У голубых китов обычно детеныши рождаются через каждые два года. Половозрелости достигают в 3—5-летнем возрасте.
Из эктопаразитов настоящие полосатики обычно бывают заражены веслоногими ракообразными — Penella balaenoptera, усоногими ракообразными — Coronula regina и морскими уточками Conckoderma auri-tum и С. virgatus, а также различными видами китовых вшей — Cyamus sp. Тело китов бывает покрыто зеленой пленкой диатомовых водорослей.
Настоящие полосатики представляют ценный объект промысловой охоты. Хотя в результате усиленного промысла в ряде мест численность их сильно сократилась, все же и теперь некоторых из них еще добывают.
Голубых китов в последние годы добывали в основном в Антарктике, а прежде — в проливе Девиса, у берегов Финмаркен, Северной Америки и Японии. В Антарктике за 50 лет с 1909 г. было добыто 324 338 голубых китов. В настоящее время промысел голубых китов прекращен, так как численность их стала угрожающе низкой. Несколько лучше обстояло дело с финвалом, промысел которого в последние годы еще продолжался, но сейчас прекращается. Наибольшее число финвалов промышляли в Антарктике, меньше в северной части Тихого океана и еще меньше в Северной Атлантике. В Антарктике финвалов с 1909 г. за 50 лет добыли около 500 тыс. голов. Сейвала раньше в основном промышляли в водах Японии, Бенинских о-вов, Южной Георгии, Южной Африки, Норвегии, теперь добывают по всему его ареалу. В связи с тем, что основными объектами промысла в последние годы стали финвал и сейвал, их численность заметно уменьшается. Целесообразно введение на ряд лет полного запрета на их промысел. Малый полосатик имел лишь ограниченное промысловое значение, а в настоящее время промышляется более интенсивно. Норвежцы за период с 1938 по 1956 г. ежегодно добывали от 537 до 4328 малых полосатиков (всего за этот срок добыто 45 655). Небольшое число полосатиков Брайда добывают в водах Японии, Южной и Экваториальной Африки. Примерная численность в Мировом океане к 1972 г. малых полосатиков 150—200 тыс., голубых китов 10 тыс. В северной части Тихого океана численность полосатиков Брайда составляла к 1965 г. примерно 10 тыс. голов.
В роде 5 видов:
малый полосатик — В. acutorostrala Lacepede, 1804 (Северная Атлантика от Шпицбергена, Новой Земли и Баффинова залива до Средиземного и Черного морей и Флориды, в Южной Атлантике от мыса Доброй Надежды на востоке и Ла-Плата (Аргентина) на западе на юг до льдов Антарктики; в северной части Тихого океана от Берингова моря до Калифорнии на востоке и Японского и Желтого морей на западе; в южной части Тихого океана от Австралии, Новой Зеландии и Чили на юг до льдов; в Индийском океане от Южной Африки до Бенгальского залива и Явы);
полосатик Брайда — В. edeni Anderson, 1878 (Индийский океан от Австралии до Южной Африки; в Атлантике от мыса Доброй Надежды до Сенегала на востоке и Карибского моря на западе; в Тихом океане известен у берегов Калифорнии, Японии и Новой Зеландии);
сейвал — В. borealis Lesson, 1828 (Северный Ледовитый океан и Северная Атлантика — от Новой Земли, Шпицбергена, пролива Девиса и Лабрадора до Испании, Средиземного моря и Северной Африки на востоке и Мексики на западе; Южная Атлантика — от Анголы и Южной Африки на востоке до 20° ю. ш. в Бразилии и Земли Грейама (65° ю. ш.) на западе; северная часть Тихого океана — от Восточной Сибири и Западной Аляски до Японии и Корейского п-ова, Сиамского залива на западе и Калифорнии на востоке; в южной части Тихого океана — у Австралии, Калимантана, Новой Зеландии, Эквадора, Перу и Чили; в Индийском океане — у берегов Индонезии);
финвал — В. physalus Linnaeus, 1758 (обширные пространства Атлантического, Тихого й Индийского океанов от экваториальных вод до льдов в Северном и Южном полушариях);
голубой кит — В. musculus Linnaeus, 1758 (Северный Ледовитый, Атлантический, Тихий и Индийский океаны; более обычен в водах к северу от 35° с. ш. и к югу от 40° ю. ш. до кромки льдов в обоих полушариях).
В СССР: малый полосатик (от Чукотского моря и Берингова пролива на юг до границы СССР; у Новой Земли, в Белом, Баренцевом морях, у Мурманского побережья, изредка заходит в Балтийское море и дважды отмечен в Черном море); полосатик Брайда (возможны встречи у Курильских о-вов); сейвал (на востоке от Курильских о-вов к северу до Берингова пролива, на западе — в Баренцевом, Белом и изредка Балтийском морях); финвал (Берингово, Чукотское, Охотское, Карское, Белое, Баренцево и Балтийское моря); голубой кит (Мурманское побережье, у Новой Земли, Балтийское море и от Курильских о-вов на север до Чукотки).
Иногда из малого полосатика выделяют вид В. bonaerensis Burmeister, 1867 (например, Nishiwaki, 1972), а из В. musculus — В. brevicauda (Zemsky et Boronin, 1964), что, по-видимому, неправильно.
От одного голубого кита можно получить до 15—20, даже 50 т жира, от финвала — 6—7, от сейвала — 2—4, от малого полосатика — около 1 т.
В «Красную книгу» внесен голубой кит как вид, находящийся под угрозой исчезновения, и финвал как вид, который может оказаться перед угрозой исчезновения, если не будет налажена его охрана.
Род горбатых китов
Genus Megaptera Gray, 1846
В роде единственный вид: горбатый кит — М. novaeangliae Borowski, 1781 ( = M. nodosa Bonnaterre, 1789).
В отличие от настоящих полосатиков тело укороченное и толстое. Длина тела не превышает 18 м; у самок до 17,4 м, у самцов до 16 м. Крупная притупленная голова занимает около 30% длины тела. Спинной плавник низкий и толстый, горбовидный с небольшой выемкой на пологом переднем крае (рис. 42). Грудные плавники очень длинные — до /з—’/< длины тела с бугристыми передними краями, несущими 10 крупных бугров. В переднем отделе верхней части головы располагаются в 3 5 рядов бородавчатые кожные наросты. Средний ряд насчитывает о—8 наростов, а боковые — по 5—15. По 10—15 таких же наростов проходит вдоль каждой половины нижней челюсти. По форме и размерам наросты напоминают половину апельсина; каждый нарост имеет обычно по одному волоску. На переднем крае нижней челюсти находится большой, до 30 см в диаметре, кожный нарост неправильной формы. Полосы
Рис. 42. Горбатый кит — М. novaeangliae и его череп
на брюхе крупные (в 2,5 — 3 раза шире и глубже, чем, например, у финвала) и немногочисленные — от 12 до 36. Пластины китового уса относительно высокие — до 1 м, в каждой половине верхней челюсти их до 270—400. Пластины пепельно-черного цвета, бахрома жесткая, бурая.
Окраска тела на спине и боках черная, иногда с коричневым оттенком, на брюхе варьирует от черной и пестрой до белой. В ротовой полости в передней части неба проходят две продольные борозды.
Позвонков шейных 7, грудных 14, поясничных 10—11, хвостовых 21— 22; всего 52—53. Лопатка не имеет акромиального и коракоидного отростков. Фаланг в передней конечности 12—з, II?, iV’e-z, V2-3.
Распространение всесветное — от Арктики до Антарктики. В северном полушарии, по-видимому, 4 стада горбатых китов: в северной половине Тихого океана одно мигрирует от Чукотского моря до Калифорнии и Мексики, другое — от Алеутских о-вов и Камчатки до о-вов Тайвань, Рю-Кю, Бонимских и Мариамских; в Северной Атлантике третье стадо — от Шпицбергена, Новой Земли и Финмаркена до Северо-Восточной Африки и о-вов Зеленого Мыса, четвертое — от Гренландии и Исландии до Антильских о-вов. В Южном полушарии выделяют 5 стад, совершающих миграции от Антарктики в меридианальном направлении: первое— к западным берегам Южной Америки, второе — к восточным берегам Южной Америки и западным берегам Южной Африки, третье — к восточным берегам Южной Африки и к Мадагаскару; четвертое — к западным берегам Австралии, пятое — к восточным берегам Австралии, Новой Зеландии и некоторым островам Меланезии и Полинезии. Между отдельными стадами существует обмен особями (например, между обоими северотихоокеанскими стадами, восточно- и западноавстралийскими и т. д.).
В СССР горбатый кит встречается у берегов Камчатки, в Беринговом проливе, в Анадырском заливе, в Чукотском, Охотском и Японских морях, а также в Баренцевом море и на востоке до о-вов Новой Земли; изредка заходит в Балтийское море.
Обычно держится недалеко от берегов. Более тихоходен, чем настоящие полосатики. Скорость не превышает 13—15 км/ч, а у раненых до 80
25 км/ч. Нередко полностью выпрыгивает из воды. Под водой обычно остается 5—6 мин, максимум 15—30 мин. Высота фонтанов 2—5 м.
Питается рыбой, придонными и пелагическими ракообразными, реже головоногими и крылоногими моллюсками. В Антарктике поедает преимущественно рачка Euphasia superba, на Дальнем Востоке ракообразных jYiysus oculata, Thysandessa inermis, T. longipes, T. spinifera, Euphasia pacifica, Ananyx nugax, Themisto sp., Eualus gaimardi и рыб — сельдь, корюшку, мойву, навагу, минтая, треску, сайку, кету, горбушу, песчанку, морского ленка, морского ерша. Питание головоногими моллюсками отмечалось у Курильских о-вов. В Северной ‘Атлантике пищей служат сельдь, мойва, рачки Thysanoessa inermis и Meganictiphanes norvegica, изредка головоногие и крылоногие моллюски. Желудок горбатого кита может вместить до 600 кг рыбы или 500 кг рачков.
Период спаривания у самок горбатых китов растянут на весь год с пиком в зимние месяцы. Горбатые киты способны производить частотно-модулированные звуки, особенно в низких широтах, что, вероятно, свидетельствует о их связи с размножением. Длительность беременности П—12 месяцев. Длина тела новорожденного 4—4,5 м. Лактация продолжается, по различным данным, от 5 до 10 месяцев. Самки приносят детенышей почти ежегодно (по данным других исследователей, раз в два года). Половозрелость наступает в 5-летнем возрасте. Физической зрелости достигает в 7,5—16,5 лет. Предельный возраст (определенный по ушным пробам) самцов 48 лет, самок 38 лет.
На поверхности тела горбатых китов поселяется относительно больше эктопаразитов, чем у настоящих полосатиков. Среди эктопаразитов усоногие раки — Coronula diadema и С. regina, морские угочки — Conchoderma auritum, веслоногие раки — Penella sp., китовые вши — Paracyamus boopis, Cyamus elongatus, жгутиковые — Hematophagus. Обычно все тело горбача обрастает диатомовыми водорослями.
В прошлом горбатые киты интенсивно промышлялись. Только в Антарктике с 1904 по 1956 г. их было добыто около 62 тыс. голов. В результате переопромышления численность резко снизилась (к 1972 г. примерно до 4,2—5,6 тыс. голов) и в 1963 г. промысел был запрещен.
Горбатые киты внесены в «Красную книгу» как вид, находящийся под угрозой исчезновения.
ОТРЯД хищных
ORDO CARNIVORA BOWDICH, 1821
К отряду хищных принадлежат разнообразные по строению тела и образу жизни млекопитающие. Некоторые из них высоко специализированы. Телосложение от легкого и стройного до тяжелого и неуклюжего. Размеры от мелких (у ласки длина тела до 26 см) до крупных (у бурого медведя длина тела до 300 см, у тигра — до 317 см) Г Половой диморфизм, как правило, отсутствует или выражен слабо. Голова различной формы: округлая у кошек, вытянутая у волков, уплощенная и тупомордая у выдр, остромордая у енотов. Ушные раковины очень крупные, остроконечные или короткие, округлые, иногда недоразвитые, с произвольно замыкающимся слуховым проходом. Глаза обычно средних размеров. Хвост у большинства видов длинный, у некоторых короткий, по всей длине покрытый волосами. Конечности стопоходящие, полустопоходящие или пальцеходящие. Пальцев обычно 5, реже 4 (I палец редуцирован); ни I, ни V не противопоставляются другим. Пальцы вооружены когтями; у некоторых видов они втяжные.
Тело всегда покрыто волосяным покровом. Волосы различных категорий: направляющие, остевые и пуховые. У большинства видов волосяной покров густой, высокий и мягкий, реже редкий и грубый. Характерен сезонный диморфизм: в зимнее время волосяной покров становится выше и гуще; иногда меняется его окраска. Линек две или одна. Окраска тела может быть одноцветной или с полосами и пятнами. Вибриссы хорошо развиты. Как правило, в коже расположены сальные и потовые железы. У некоторых видов в анальной области находятся специфические кожные железы, выделяющие секрет с резким, иногда очень неприятным запахом. Сосков от одной до 6—7 пар, в паховой области и на брюхе.
Череп длинный с вытянутой лицевой частью и укороченной мозговой или округлый с укороченным лицевым отделом и относительно крупной мозговой коробкой или промежуточной формы. Глазницы направлены вперед и у некоторых видов расположены близко друг к другу. Слуховые барабаны большие, окостеневающие (за исключением Nandinia). В связи с сильным развитием челюстной мускулатуры для черепа характерны крупные гребни и широко расставленные скуловые дуги. Костное небо продолжается кзади по крайней мере до уровня последнего коренного зуба. Слезные кости маленькие.
2 — 3 1 2 — 4 1—4
Зубная формула: i — — с — р — - m -=28 — 50.
2 — о I 2 — 4 1 — 5
Зубная система гетеродонтная и дифиодонтная. Резцы обычно не-
1 Представителей отряда можно разделить на 3 группы: мелкие с длиной тела 15—114 см, средние—115—215 и крупные — 216—317 см.
больших размеров. Клыки сильно развитые, остроконечные. Щечные зубы туберкулосекториального типа, четырех- или многобугорчатые, с острыми, реже тупыми вершинами. Четвертый верхний предкоренной и первый нижний коренной особенно велики и имеют острые высокие вершины. Эти зубы называются хищническими. Зубы всегда имеют корни (брахиодонтного типа).
Локтевая и-малая берцовые кости хорошо развиты и не срастаются с лучевой и большой берцовой. Запястные кости: ладьевидная, полулунная и центральная сливаются в одну кость. Ключицы нет или она рудиментарна. Обычно os penis имеется (кроме гиен).
Головной мозг относительно крупный, макросматического типа. Большие полушария развиты хорошо и обычно имеют 3 надсильвиевых борозды. Желудок простой, с многочисленными железами. Длина кишечника у большинства невелика. Слепой кишки нет или она слабо развита, или довольно длинная. Матка двойная или двурогая. Плацента отпадающая, зонарная. У самцов семенники всегда располагаются вне брюшной полости в мошонке или подкожно.
Распространение охватывает практически весь земной шар. Нет хищных лишь в Антарктике и на некоторых мелких островах. Нет аборигенных видов в Австралии. В ряде мест хищные акклиматизированы: например, европейская красная лиса интродуцирована в Австралии, индийский мангуст — на некоторых островах Вест-Индии, американская норка — в СССР. Динго, широко распространенные в Австралии, завезены сюда, по-видимому, предками современных австралийцев.
По образу жизни весьма разнообразны. Среди них есть обитатели открытых пространств, жители лесов, древесные, хорошо копающие землю, полуводные или почти водные. Большинство оседлые, другие широко мигрируют. Некоторые виды в холодных и умеренных широтах на зиму впадают в спячку или сон. Как правило, держатся поодиночке или парами, иногда объединяются в стаи. Активны преимущественно в сумерки и ночью. Большинство видов приспособлено к питанию животной пищей, значительно реже употребляют растительную, некоторые всеядны. В виде исключения могут быть энтомофаги. По характеру размножения некоторые виды монога.мы; большинство соединяются в пары лишь на один сезон размножения. Полигамии нет. Детенышей в помете от 1—2 до 20—22. Детеныши рождаются слепые, с закрытыми ушными отверстиями, не способные к самостоятельному передвижению; их развитие происходит медпенно.
Представители отряда имеют большое практическое значение. Многие — ценные объекты пушного промысла. Есть виды, истребляющие вредных для сельского хозяйства животных, некоторые относятся к вредителям сельского и охотничьего хозяйства. Отдельные виды используются для спортивной охоты. Среди хищных есть виды, распространяющие некоторые заболевания домашних животных и человека (например, бешенство).
Вследствие неумеренного промысла численность некоторых видов хищных сильно снизилась и в недалеком прошлом они были близки к Полному исчезновению (соболь, калан). Ряд видов и подвидов уничтожены полностью или почти полностью (фолклэндский волк, предкавказ-ский леопард, персидский и атласский львы и др.). Ряд хищных в различных странах охраняются законом, запрещены их охота или промысел. Другие, напротив, объявлены вне закона и за их добычу полагается определенная премия. Хищные послужили родоначальниками некоторых Домашних животных (например, произошедшая от волка собака — древнейшее из всех одомашненных животных). Ряд видов служат объектом звероводства и разводятся при клеточном содержании.
Классификация отряда хищных разными исследователями представляется по-разному. Одни разделяют отряд хищных на 2 подотряда: вымершие Creodonta Соре, 1875, с четырьмя семействами и современные Fissipeda Blumenbach ,1791, с одним вымершим и семью современными семействами. Некоторые ученые (например, Симпсон, 1945) считают, что ластоногие, близко примыкающие к хищным, также являются представителями этого отряда и выделяют их в качестве особого подотряда Pinnipedia liliger, 1811. Действительно, ряд признаков ластоногих сближает их с отрядом хищных, однако правильнее считать хищных и ластоногих отдельными отрядами.
Иногда современных хищных разделяют на 2 подотряда: Canoidea куда включают семейства волчьих, медвежьих, енотовых и куньих, и Feloidea с семействами виверровых, гиеновых, кошачьих. Ряд исследователей (например, Симпсон, 1945) считают Canoidea и Feloidea не подотрядами, а надсемействами.
Расхождения есть и в трактовке системы родов и видов. Некоторые исследователи склонны к дробному делению представителей хищных на многочисленные роды и виды. В последние годы эта точка зрения подвергается основательной критике.
В отряде 7 семейств (89 родов):
волчьих — Canidae Gray, 1821 (12 родов); медвежьих — Ursidae Gray, 1825 (4 рода); енотовых — Procyonidae Bonaparte, 1850 (8 родов); куньих — Mustelidae Swainson, 1835 (24 рода); виверровых — Viverridae Gray, 1821 (35 родов); гиеновых — Hyaenidae Gray, 1869 (3 рода);
кошачьих — Felidae Gray, 1821 (4 рода).
В СССР: представители всех семейств, кроме виверровых (енот-полоскун из Procyonidae акклиматизирован).
Свое происхождение хищные ведут от примитивных меловых насекомоядных. С ними тесно связаны примитивные хищные — Creodonta, составляющие особый вымерший подотряд хищных. Некоторые из мелких креодонт напоминали ласок, другие — волков или львов. Их череп, как правило, был большим и низким. Число зубов достигало 44. Коренные зубы были тритуберкулярного типа. /Мозг маленький, почти гладкий. Слуховые барабаны, за исключением Mesonyx, не окостеневали. Конечности стопоходящие, полустопоходяшие или пальцеходящие. Креодонты были представлены 4 семействами:
Arctocyonidae Murray, 1866, — всеядные креодонты с вытянутым туловищем и высокими конечностями, щечные зубы были с многочисленными тупыми вершинами на жевательной поверхности, конечности пятипалые, стопоходящие; 27 родов были распространены в Северной Америке, Европе и Восточной Азии с нижнего палеоцена по нижний олигоцен;
Mesonychidae Соре, 1875, — различного размера от величины куницы до очень большого Andrewsarchus, длина черепа которого достигала 1 м, конечности были параксонические, первый палец у примитивных форм несколько противопоставлялся другим пальцам, а позднее он становился рудиментарным, конечности претерпели переход от стопоходящих к пальцеходящим; примерно 8 родов были распространены в Голарктике со среднего палеоцена по верхний эоцен;
Oxyaenidae Соре, 1877 — напоминали кошек, череп широкий, нижняя челюсть высокая и массивная, тело и хвост длинные, конечности короткие, пятипалые, стопо- или полустопоходяшие; примерно 10 родов были распространены в Северной Америке, Азии и Европе с верхнего палеоцена по верхний эоцен;
Myaeodontidae Leidy, 1869, — череп и ряды щечных зубов удлинен-,1ижняя челюсть относительно слабая, тело менее массивное, чем у flbie’aenidae, конечности довольно длинные, пятипалые, стопо- или паль-лЯщие; примерно 24 рода были распространены в Северной Амери-цеХдзиИ, Европе и Африке с нижнего эоцена по верхний миоцен.
Ке’ Кпеодонты были многочисленными в палеоцене, достигли расцвета в т не сильно уменьшились в числе в олигоцене и исчезли в миоцене.
Э011Бчизкое к креодонтам семейство Miacoidea Simpson, 1931 (его иног-включают в подотряд Creodonta), по-видимому, было родоначальника современных хищных. Представители этого семейства были мелких КС"педних размеров, имели полное число зубов, длинный и низкий череп, И окостеневшие слуховые барабаны; мозговая коробка была большая оа3мерам, чем у креодонт; туловище, хвост и пятипалые конечности б тли очень длинные. Таким образом, миацоиды имели промежуточные признаки между креодонтами и современными хищными. Примерно 18 родов миацоид были распространены в Северной Америке, Европе и Азии со среднего палеоцена по верхний эоцен.
СЕМЕЙСТВО ВОЛЧЬИХ
FAMILIA CANIDAE GRAY, 1821
размеров в
К семейству принадлежат хищные мелких и средних большинстве случаев стройного телосложения. Длина тела от 40 см (фе-нек) до 160 см (волк) *. Хвост обычно довольно длинный от 11 до 55 см. Масса от 2 кг у фенека до 80 кг у волка. Голова удлиненная с вытянутой мордой. Уши, как правило, высокие, иногда короткие, вершина их заостренная и лишь изредка округлая. Конечности очень длинные (гривистый волк), менее длинные (волк), укороченные (лисица) или короткие (енотовидная собака, кустарниковая собака); передние и задние примерно одинаковой длины. Тело в области крестца немного ниже, чем в области лопаток. Конечности пальцеходящие. Передние — пятипалые, за исключением гиеновой собаки (4 пальца). Первый палец всегда небольших размеров и расположен выше остальных. Задние конечности четырехпалые. Когти не втяжные, тупые, слабо изогнутые.
Волосяной покров обычно высокий, густой и пушистый, у южных форм более грубый и редкий. Окраска его обычно одноцветная — серая, желтоватая, рыжая или черная, иногда с пятнами белыми, черными и рыжими. У одного вида (песец) имеет место резкая сезонная смена окраски. В течение года одна или две линьки. В кожном покрове находятся альные и потовые железы, причем последние, по-видимому, не участву-т в терморегуляции. Из специфических желез имеются небольшие нальные (иногда отсутствуют) и железы в подушечках лап. Некоторые наДЫ °бладают так называемой «фиалковой» железой, расположенной СосВерХНеЙ стоРоне основания хвоста. Семенники находятся в мошонке, сков 3—8 пар.
вые бРеП хаРактеризуется удлиненным лицевым отделом. Костные слухо-Разм аРа®аны небольшие, лишь в виде исключения достигают крупных сосц еР°в’ Всегда есть крылоклиновидный канал. Боковой затылочный и невидный отростки небольшие.
с ДлиДйеДСтави1елей семейства по размерам можно разделить на 3 группы: мелкие тела 40—79 см; средние — 80—119, крупные—120—160 см.
Зубная формула обычно
4 2
— m —- = 42, но может изменяться от
гп = 38 у кустарниковых собак — Speothos до
3 — 4
m ------=46 — 48 — 50 у большеухих лисиц.
Клыки большие, островершинные. Хищнические зубы сильно развиты. Щечные зубы относительно низкие, несколько уплощенные, с невысоки-ми тупыми буграми на жевательной поверхности.
Слепая кишка обычно длинная, спирально изогнутая. Иногда она развита слабо.
Распространены по всему Земному шару, исключая Антарктику и некоторые океанические острова — Новую Зеландию, Меланезию, Полинезию, Молуккские, Сулавеси, Тайвань, Мадагаскар.
Наземные формы, населяющие самые разнообразные ландшафты, преимущественно открытые: тундру, лесотундру, лесостепь, степь, пустыню. Большинство быстро бегающие, способные догонять добычу при преследовании. Некоторые хорошо роют и находят себе убежище в норах. Держатся поодиночке, иногда парами, некоторые — семейными группами и стаями, иногда собираясь в значительных количествах. Активны, как правило, в сумерки и ночью. Из органов чувств наиболее хорошо развиты слух и обоняние, в меньшей степени зрение. Питаются различными позвоночными, иногда падалью, реже беспозвоночными. В качестве дополнительного объекта питания могут служить растения. Большинство ведет оседлый образ жизни, некоторые совершают дальние, сезонные или нерегулярные кочевки. По характеру размножения моно-гамы, у некоторых видов пары соединяются на длительный период. Беременность от 51 дня у фенека до, вероятно, 80 у гиеновой собаки. В течение года дают один помет из 4—7 детенышей, иногда до 13—18, по некоторым данным, до 22—23 (песец). У большинства видов самец принимает участие в воспитании детенышей. Половой зрелости обычно до
стигают в конце первого года жизни.
Ряд видов имеет значение в пушном промысле и как объекты охотничьего спорта. Некоторые виды истребляют вредных грызунов; другие могут приносить известный ущерб сельскому и охотничьему хозяйствам; немногие виды — хранители заболеваний, опасных для человека и жи
вотных.
Систематика семейства окончательно не установлена. Некоторые исследователи (например, Андерсон и Джонс, 1967) насчитывают в нем 15 родов:
Alopex Каир, 1829;
Atelocynus Cabrera, 1940;
Canis Linnaeus, 1758;
Cerdocyon H. Smith, 1839;
Chrysocyon H. Smith, 1839;
Dasycyon Krumbiegel, 1953;
Dusicyon H. Smith, 1839;
Fennecus Desmarest, 1804;
Nyctereutes Temminck, 1839;
Urocyon Baird, 1858;
Vulyes Bowdich, 1821;
Cuon Hodgson, 1838;
Lycaon Brookes, 1827;
Speothos Lund, 1839;
Otocyon Miiller, 1836.
С другой стороны, предлагается уменьшить число родов в семействе до 7—8 (например, Гептнер, 1967): Canis, Nyctereutes, Chrysocyon,SP* othos, Cuon, Lycaon, Otocyon. Впредь до серьезной ревизии семеист можно, следуя Симпсону (1945), выделить в семействе 3 подсемейст и 12 родов (35 видов).
Subfamilia Caninae Gill, 1872
нотовидных собак — Nyctereutes Temminck, 1839 (1 вид);
о чков — Cants Linnaeus, 1758 (6 видов);
песцов — Alopex Каир, 1829 (1 вид);
лисиц — Vulpes Oken, 1816 (9 видов);
(Ьенеков — Fennecus Desmarest, 1804 (1 вид);
серых лисиц — Urocyon Baird, 1858 (2 вида);
южноамериканских лисиц — Dusicyon Н. Smith, 1839 (10 видов);
гривистых волков — Chrysocyon Н. Smith, 1839 (1 вид).
Subfamilia Simocyoninae Zittel, 1893
красных волков — Сиоп Hodgson, 1838 (1 вид);
гиеновых собак — Lycaon Brookes, 1827 (1 вид);
кустарниковых собак — Speothos Lund, 1839 (1 вид).
Subfamilia Otocyoninae Trouessart, 1885
большеухих лисиц — Otocyon Muller, 1836 (1 вид).
Некоторые исследователи (например, Фрешкоп, Frechkop, 1959) полагают, что енотовидная собака стоит ближе к Procyonidae, чем к Canidae.
В СССР 5 родов: енотовидных собак, волков, песцов, лисиц, красных волков.
SUBFAMILIA CANINAE GILL, 1872
Род енотовидных собак
Genus Nyctereutes Temminck, 1839
В роде единственный вид: енотовидная собака — N. procyonoid.es Gray, 1834.
Размеры мелкие. Длина тела 65—80 см. Длина хвоста 15—25 см. Масса до 10 кг. Туловище удлиненное, конечности относительно короткие, хвост опускается ниже скакательного сустава, но у стоящего животного земли не достигает. Голова относительно небольшая, с короткой заостренной мордой. Уши невысокие, слегка выдающиеся над волосяным покровом (рис. 43). Волосяной покров высокий, густой, довольно грубый, краска его зимой грязно-буровато-серая с черноватым или бурым оттенком. Вдоль средней линии спины проходит темная полоса, расширяю-Щзяся в области плеч. Брюхо желтовато-бурое, грудь буро-черноватая.
°Рда светлая, с характерным черным рисунком в виде маски, распо-чег>аЮ1ЦИМСЯ в области глаз, перед ними и на щеках. Задние части ушей сам Ые Летом окраска более светлая. Морда покрыта короткими воло-Хар11’ Сильно удлиняющимися позади глаз и образующими на щеках Лин КТеРНЫе «бакенбарды». Волосы на хвосте очень длинные и густые. СоиЬКЙ г°дна в течение года, весной, начинается с марта или апреля, сков 5 пар.
саги еРеп Довольно массивный, с сильно развитыми у старых животных ро Ттальным и затылочным гребнями. Скуловые дуги выдаются в сто-Реди СЛаб°' Длина лицевого отдела черепа почти равна мозговому. Впе-Га от углового отростка нижней челюсти на ее нижнем крае распола-Ка СЯ Свое°бразный массивный выступ, отделенный от углового отрост-глубокой впадиной. Такое строение нижней челюсти имеется еще
Рис. 43. Енотовидная собака — N. procyonoides и ес череп
о у серой лисицы. Угловой отросток очень высокий, не имеет населенного кверху зубца.
ПР 3 1 4 2
Зубная формула: i— с р m --- — О 1 О
Изредка в верхней челюсти 3 коренных зуба (и тогда общее число зубов 44).
у Зубы относительно небольшие. Клыки укороченные, хотя и сильные. R ихние хищнические зубы относительно слабые. Верхние резцы долото-биазной формы без боковых зубцов. Третий верхний резец отделен от второго значительным промежутком.
Диплоидное число хромосом 42.
Распространена в южной части Советского Дальнего Востока, Северном Вьетнаме, Китае, на Корейском п-ове и в Японии.
В СССР: естественный ареал охватывает Уссурийский край и южную часть Амурского, где общая численность достигала на 1974 г. 26— 29 тыс. особей.
Енотовидную собаку начиная с 1929 г. акклиматизировали во многих местах СССР. С 1929 по 1955 г. расселено 8850 енотовидных собак в 82 областях, краях и республиках. В европейской части СССР енотовидная собака в настоящее время образовала обширный сплошной ареал, на севере охватывающий всю Карелию, южную часть Кольского п-ова, Архангельскую область до лесотундры включительно с заходами в тундру, на юге распространяющийся до Черного, Азовского и Каспийского морей. Попытки акклиматизации в Сибири и Средней Азии, по-ви-димому, успеха не имели. В конце 50-х годов из европейской части СССР проникли в Польшу, Румынию, Финляндию, Швецию и ГДР.
Населяет пойменные леса, заросли кустарников и тростника по берегам рек и озер. Может самостоятельно вырывать норы или находить себе убежище среди камней, в расщелинах скал, в норах барсуков и лисиц, под буреломом, в крупных дуплах упавших деревьев. Норы иногда с несколькими выходами, обычно простые, длиной 1,5—3 м, оканчивающиеся гнездовой камерой. Пища очень разнообразная. Летом в основном питается мелкими позвоночными (мышевидными грызунами, птицами, пресмыкающимися, лягушками, рыбами), различными беспозвоночными (главным образом, насекомыми), а также плодами, ягодами, луковицами, корневищами. В осеннее время важное значение приобретает растительная пища. Имеет место географическая изменчивость питания, о северных и средних широтах среди животных кормов в первую очередь преобладают мышевидные грызуны, а на юге и западе — насеко-ые- Но мере продвижения к югу в питании возрастает значение птиц и моллюсков. Активна, как правило, в сумерки и ночью. В осеннее время сильно жиреет, накапливая под кожей до 18—23% к общей массе жира
** 5% внутреннего жира.
Сон северных районах в ноябре—феврале — марте впадает в зимний с ‘единственный случай в семействе). Общий обмен веществ может а аться во время сна примерно на 25%. Во время оттепелей бывает зимИВ1~а И брОДит в поисках пищи. В южных районах совсем не впадает в осе и„СОн или впадает только на короткое время. Енотовидная собака— ние^ко™ ЗВеРь’ ио в неблагоприятные по питанию годы могут быть даль-°сенью1°ГаМЬ1’ с°единяющиеся в пары на один сезон. Пары формируются Крае ’ °бычно в октябре или ноябре. Гон происходит в Уссурийском РедиНе 5еРедине маРта — начале апреля, на Украине и Татарии — в се->кИтед15)еВРаля’ на северо-западе — в начале — середине марта. Продол-
юсть гона 2—3 недели. Длительность беременности обычно
59 дней (иногда до 61—70). Детенышей в помете 6—7, но может быть до 15—16. Новорожденные имеют массу по 60—96 г. Они слепые, покрц. ты короткой темной, почти черной густой и мягкой шерстью. Глаза от-крываются на 9—10-й день, зубы прорезываются на 14—16-й. Длительность лактации 45—60 дней. Примерно с 3-недельного возраста щенки начинают есть лягушек, насекомых, которых приносят родители. В воз. расте около 5 месяцев щенки достигают размера взрослых и по внешнему виду почти от них не отличаются. Половозрелость наступает в 8—10 месяцев. Продолжительность жизни, по-видимому, около 8 лет, в неволе —. до 11 лет.
Хотя мех енотовидной собаки не очень высокого качества, тем не менее зверь имеет некоторое значение в пушном промысле. В ряде северо-западных и центральных областей СССР енотовидная собака стала од. ним из основных объектов пушного промысла. С 1960 по 1970 г. в СССР заготавливали 34,7—78,4 тыс. шкурок ежегодно. В охотничьих хозяйствах может наносить существенный урон, разоряя гнезда наземногнездя-щихся промысловых и охотничьих птиц. Особенно вредна енотовидная собака в плавнях и на побережьях лиманов, где она поедает в больших количествах яйца и птенцов водоплавающих птиц. Иногда вредит огородным и бахчевым культурам. Оценка вредности или пользы енотовидной собаки должна производиться для каждого конкретного места отдельно.
Род волков
Genus Cams Linnaeus, 1758
Размеры мелкие, средние и крупные, наибольшие в семействе. Длина тела 70—160 см. Длина хвоста 20—50 см. Масса 6—80 кг. Конечности высокие, туловище умеренно вытянутое (рис. 44). Хвост пушистый, никогда не достигающий земли у стоящего зверя и не опускающийся ниже скакательного сустава. Морда относительно широкая и короткая. Уши средней длины, стоячие, заостренные. Волосяной покров относительно грубый, высокий.. Окраска его сероватая, желтоватая или рыжеватая с примесью черных волос. В году 2 линьки. «Фиалковая» железа имеется, но развита слабо. Сосков 5 пар.
Череп массивный, скуловые дуги расставлены широко. Лицевой отдел длиннее мозгового. У взрослых и старых зверей сильно развиты гребни, особенно сагиттальный. Лобная часть черепа выпуклая с большими полостями. Надглазничные отростки тоже выпуклые с отогнутыми книзу вершинами.
„ . 3
Зубная формула: i --- с
3
1 4 2
— р — m — — 42.
14 3
Зубы крупные и сильные. Клыки мощные, слабо изогнутые, относительно короткие. Хищнические зубы хорошо выражены. Режущий кран резцов с небольшими дополнительными выступами (трехлопастные).
Диплоидное число хромосом у шакала, койота и волка по 78.
Распространены очень широко. Ареал охватывает всю Евразию с рядом арктических островов; всю Африку без Мадагаскара; всю Северну!0 Америку, включая Арктический архипелаг; Центральную Америку на юг до Коста-Рики включительно; встречаются на северном, восточной и западном побережьях Гренландии.
Представители рода обитают во всех ландшафтах от тундр до пус' тынь и высокогорий. Шакал обычно поселяется в густых зарослях, кус' тарнике, тростниках или в оазисах, не избегает соседства человека. BH'j
К
бор лей..
Зеравшан* 10 км?
рЫ L-НОСТЬ С чается i
места для поселения определяется наличием добычи, густых зарос-служащих убежищем и доступностью воды. В тугаях Амударьи, —. плотность населения шакала колеблется от 3 до 6 особей на
-.г Койот населяет самые различные места — тундру, лес, степь, го-^плоть до альпийской зоны. Предпочитает открытую холмистую мест-- - водоемами. Встречается рядом с поселками людей. Волк ветре-в самых разнообразных географических зонах и высотных поясах, ЧС*"иная от тундр, где преимущественно он приурочен к долинам рек и На ослям на возвышенных сухих участках. В лесной зоне избегает сплош-заР лесных массивов, однако придерживается укромных глухих мест в Н те глухих оврагов, болот, зарослей кустарников. В степях держится по ВИухим оврагам, балкам; в полупустынях и пустынях встречается там, где есть доступная для водопоя вода. Поднимается высоко в горы, до 3_-4 тыс. м над уровнем моря.
По характеру питания представители рода — плотоядные, преимущественно поедающие различных позвоночных. Питание шакала разнообразно — приближается к всеядности. Число видов растений и животных, входящих в состав пищи шакала в Средней Азии, более 90. Шакал охотится на средних и мелких млекопитающих, различных птиц, пресмыкающихся и земноводных, поедает разнообразных насекомых и падаль. Большое значение имеют и растения: костянки лоха, плоды груши, боярышника, винограда, арбузы, дыни и т. д. Койот в основном питается мелкими позвоночными; анализ желудков дает следующее соотношение (%): 33 — кролик и зайцы; 25 — падаль; 18 — мелкие грызуны и суслики; 4 — олени и антилопы; 2 — птицы; 2 — растительность; 1 — насекомые; 14—-домашние млекопитающие и птицы (Banfield, 1974). У волка основной объект питания копытные; важное значение имеют и сурки, зайцы, барсуки, лисы, а также и мелкие грызуны. Охотятся чаще всего, загоняя добычу. В летнем питании волка велика роль птиц и мелких грызунов. В это время, а также осенью они охотно поедают ягоды и различные плоды.
Как правило, держатся поодиночке, парами или семейными группами, состоящими из родителей, молодых этого года, и молодых прошлого года (переярки). Изредка объединяются в стаи при охоте за крупными копытными. Ведут оседлый, реже кочевой образ жизни. Так, волки в тундре совершают большие меридианальные кочевки вслед за оленьими стадами. Кочуют волки и следом за мигрирующими копытными: сайгаками в Казахстане, дзеренами в Монголии. Моногамы, у некоторых видов пары сохраняются пожизненно (волк, шакал). Течка происходит в разное время в зависимости от географического места. У шакала она приходится в Болгарии на середину января — середину февраля, в Закавказье и Средней Азии — на первую половину февраля — начало марта, У койота длится с конца января по конец марта, у волка протекает в декабре в Средней Азии, феврале— марте в Карельской АССР, на Южном рале, в Казахстане. Продолжительность течки отмечена для шакала примерно 26—28 дней, койота — 4—5 дней, волка — около 2 недель. Длительность беременности у койота 60—63 дня, у волка 62—75 дней. Дают один помет в год. Самка шакала приносит в среднем 5 детенышей, оиота 1—19, в среднем 5,7, волчица — от 1 до 9—14, в среднем 4—6. Шенка шакала на Кавказе и в Закавказье происходит обычно в норе gPpCToro устройства: она имеет в длину около 2 м и гнездовую камеру, вредней Азии чаще устраивает логово в густых зарослях. Койот У 1чно занимает нору сурка или барсука. Волчица щенится в логове, Раиваемом в естественных укрытиях: расщелинах скал, ямах, зарос-Те > изредка в простых неглубоких норах с 1—3 выходами. Продолжи-льность лактации у шакала 50—70 дней на Кавказе, до 90 дней в Тад-
t?' ,В°ЛК ~ 6. lupus (А), чепрачный шакал —С. mesomelas (Б) и койот — С. latrans (В) и череп волка
жикистане; у волка примерно 1,5 месяца. Самцы шакала и волка участвуют в воспитании щенков. Начиная с 2—3-недельного возраста родители подкармливают щенят отрыгнутым полупереваренным мясом. Половозрелость койотов и волков наступает к 2 годам, самок шакалов в возрасте около года, самцов — около 2 лет. Продолжительность жизни шакала 12—14 лет.
Волк послужил родоначальником распространенной всесветно домашней собаки — С. familiaris Linnaeus, 1758. Предполагавшееся ранее происхождение собаки от волка и шакала, по-видимому, исключается в частности, в связи с тем, что у волка и собаки сходное диплоидное число хромосом (78), отличающееся от шакала (74). Австралийский динго вероятно, — одичавшая собака.
Представители рода приносят существенный вред животноводству и охотничьему хозяйству. В последнее время, однако, ряд ученых доказывают положительную роль волков в естественных биоценозах как регуляторов численности растительноядных животных. Могут распространять бешенство. Имеют второстепенное значение в пушном промысле. В СССР ежегодно заготовлялось в 1960—1970 гг. по 17—31,4 тыс. волков и по 11,2—15,5 тыс. шакалов.
В роде, по-видимому, 6 видов:
эфиопский шакал — С. simensis Ruppel, 1835 (Эфиопия);
полосатый шакал — С. adustus Sundevall, 1846 (от Сенегала на северо-западе Африки до Эфиопии и Сомали на северо-востоке и до Намибии, Ботсваны, Южно-Африканской Республики, Родезии и Мозамбика на юге);
чепрачный шакал—С. mesomelas Schreber, 1775 (от Анголы, Родезии, Мозамбика до Танзании, Кении, Сомали, Эфиопии, Судана и Уганды);
шакал — С. aureus Linnaeus, 1758 (Европа — Греция, Албания, Югославия, Болгария, изредка заходит в Венгрию и Румынию, в СССР по Кавказскому побережью Черного моря от Новороссийска до Батуми и в восточной половине Кавказа за Главным хребтом на север до устьев Терека и Кумы, забегает в Молдавию; Азия — юго-западный Таиланд, Бирма, Индия, Бутан, Непал, Шри Ланка, Афганистан, Иран, Малая Азия, Аравийский п-ов, в СССР долины рек и оазисы Средней Азии; Африка — Кения, Эфиопия, Сомали, Судан, АРЕ, Ливия, Тунис, Алжир, Марокко, Рио-де-Оро, Сенегал, Гвинея-Бисау, Дагомея, Камерун);
койот—С. latrans Say, 1823 (почти вся Северная Америка, кроме Западной Аляски, — отсутствует на запад от линии, проходящей примерно от Коцебу на севере и залива Кука на юге; нет в Северо-Восточной Канаде, к северу от линии, тянущейся от устья Маккензи до залива Джемса; отсутствует в восточных штатах США к востоку от линии, проходящей по западным берегам озер Онтарио и Эри и на юг через штаты Индиана, юг Иллинойса, юго-восток АТиссури, Арканзас и запад Луизианы; вся Мексика, кроме п-ова Юкатан; Центральная Америка на юг до центральных районов Коста-Рики и кроме восточных и северо-восточных районов Гондураса и Никарагуа);
волк—С. lupus Linnaeus, 1758 (до недавнего времени почти вся Евразия — Индостан на юг примерно до 15° с. ш. и Китай до‘Ян-Цзы-Цзяна, Северная Америка, кроме крайнего юга, северное, западное и восточное побережья Гренландии, СССР — вся территория страны, крО' ме Соловецких о-вов, Земли Франца-Иосифа, Северной Земли, Кара гинского, Командорских и Шантарских о-вов. В настоящее время истреблен в Ирландии, Англии, Франции, Голландии, Бельгии, Дании, ШвеИ^ царии, во всей Центральной Европе, в Японии, в ряде областей евро пейской части СССР, южных районах Канады, почти по всей террИ'
Q4
ооии США, за исключением Аляски, штата Миннесота и национальных Тяпков Йеллоустон и Глэшиер, почти по всей территории Мексики, за Псключением нескольких глухих областей).
И Многие американские ученые (например. Холл и Келсон, Hall а. Kelson, 1959) выделяют, из С. lupus отдельный вид С. niger Bartram, 1?91 (юго-восточные штаты США). Андерсон и Джонс, 1967, насчитывают в Роде 9 видов-
В СССР: шакал и волк.
В «Крвсную книгу» включены эфиопский, шакал как исчезающий вид сохранившийся лишь в некоторых местах Симиенских гор в Эфиопии' волк как вид, который может попасть в категорию исчезающих, если ситуация с его охраной не изменится к лучшему; подвиды волка исчезающие: С. lupus irremolus Goldman, 1937, сохранившийся в США в национальных парках Йеллоустонском и Глэшиер общей численностью около 20 особей и С. I. rufus Audubon et Bachman, 1851, сохранившийся в США в восточном Техасе и Луизиане общей численностью около 50 особей.
Род песцов
Genus Alopex Каир, 1829
В роде единственный вид: песец — A. lagopus Linnaeus. 1758.
Размеры мелкие. Длина тела 46—73 см. Длина хвоста 25—52 см. Масса 2—8,8 кг. Общий вид напоминает обыкновенную лисицу, туловище удлинено, конечности относительно короткие, хвост длинный, опускается значительно ниже скакательного сустава конечности и у стоящего животного достигает земли. Голова с удлиненной мордой, уши средней высоты с закругленными вершинами (рис. 45). Волосяной покров высокий, очень густой и мягкий. Летом он более низкий, дымчато-серовато-бурый, на брюхе белесый. Зимой значительно более высокий и густой, чисто-белый или серовато-бурый. В году 2 линьки: весной и осенью. «Фиалковой» железы нет. Сосков 5—8 пар.
Череп относительно массивный с умеренно вытянутым лицевым отделом. Гребни на черепе, в частности сагиттальный гребень, развиты слабо. Надглазничные отростки невелики по размерам, сверху плоские.
3 14 2
Зубная формула: i с — р — m — = 42.
Клыки сравнительно короткие, но массивные, хищнические зубы хорошо развиты. Резцы не имеют дополнительных выступов.
Диплоидное число хромосом 48—50.
Распространен в лесотундре, летом в тундре, на арктических остро-Вах Евразии и Северной Америки, а также на о-вах Командорских и Алеутских. В период зимних миграций заходит в северную часть лесной зоны и на льды южной области Ледовитого океана.
Наиболее охотно поселяется в сильно пересеченной тундре с озерами среди возвышенностей, а также на возвышенных берегах внутренних °доемов и морей. Предпочитает открытые пространства и только в годы £!с°кой численности селится среди кустарниковой растительности.
качестве убежища, особенно в период размножения, использует норы, вг>Т°₽Ь1е занимает из года в год, при этом постоянно расширяет их и пре-Ращает в сложный многоярусный лабиринт. Иногда использует норы РУгих животных, например сурков. По характеру питания песец — все-
ное животное с тенденцией к плотоядности. Для СССР известно 125 дсв животных и 25 видов растений, поедаемых песцом. Для тундро-
Рис. 45. Песец —Д. lagopus и его череп
н
песца основными объектами питания служат мелкие грызуны, осо-н°г лемминги. Значительно реже он поедает других млекопитающих, ^еНГча даже оленей (больных или обессиленных длительным голода-йН°м) На побережье питается различными выбросами моря. Птиц по-кИС' ~ чаще всего весной и летом. Растительную пищу употребляет глав-е,да образом в летнее время и осенью. Вне периода размножения держится поодиночке.
Определенный участок обитания свойствен песцу только в период зМНожения и составляет от 2 до 30 км2 в зависимости от географиче-Ра" места и кормности года. В конце лета —начале осени песцы начи-С ют широко кочевать. Дальность миграций зависит от наличия кормов. В годы, бедные кормами, песцы могут уходить на сотни и даже более тысячи километров. Наиболее далекие миграции чаще всего совершают самцы. Большинство песцов при этом погибает от голода или болезней или вылавливается охотниками. Оставшиеся в живых звери в феврале— мапте начинают обратные кочевки. Активность песца не приурочена к определенному времени суток. Однако осенью и зимой он преимущественно активен ночью, а летом — в течение любого времени суток.
В природных условиях песцы обычно моногамы. Иногда встречаются случаи полигамии. При разведении в неволе применяется полигамия. Гон происходит с конца января по март—апрель. В одной и той же мест-
ности сроки гона могут изменяться по годам в зависимости от кормовых условий и возрастного состава популяции. Течка продолжается 12—14 дней, а в состоянии охоты самка находится около 4—5 дней. Длительность беременности составляет 49—56 дней, в среднем 52 дня. Массовое щенение приходится на май — июнь; оно совпадает с отелом у северных оленей и с прилетом птиц. Самки приносят в среднем 7—10 детенышей, максимально до 22 (или даже до 25; Банфилд, 1974) — это наибольшее число в семействе и отряде. Число щенков в помете сильно меняется в зависимости от кормовых условий года. Самка мечет щенков только в норе. Глаза у них открываются на 9—18-й день, на 12-й день намечается прорезывание зубов. Длительность лактации 1,5—2 месяца. В месячном возрасте щенки подкармливаются мясной пищей. Половая зрелость наступает к концу первого года жизни. Продолжительность жизни составляет 8—10 лет.
Отчетливо выражена динамика численности песцов в зависимости от обилия кормов (например, численность континентальных песцов зависит от леммингов — основного объекта их питания). Отмечены пики численности с колебаниями от 2 до 6 лет между ними. Общая численность песцов в СССР перед началом промысла в разные годы колеблется от нескольких десятков до нескольких сотен тысяч. Основные враги песцов — волк, а также ездовые и пастушьи собаки.
Песец — ценный пушной зверь. По размерам добычи песцов СССР занимает первое место в мире. Так, с 1960 по 1970 г. ежегодные заготовки шкурок песца составляли 51,9—95,9 тыс. С 30-х годов в СССР нача-то промышленное разведение песцов в зверосовхозах. Вольное разведение песцов осуществляется на Командорских о-вах. Отрицательное зна-ение песцов выражается в распространении и передаче бешенства.
Род лисиц
Genus Vulpes Oken, 1816
РО Размеры мелкие и средние. Длина тела 40—90 см. Длина хвоста до см. Масса 2—14 кг. Туловище удлиненное, хвост очень длинный—• егДа опускается ниже скакательного сустава и у стоящего животного ^71 97
Рис. 46. Продолжение. Череп обыкновенной лисицы
Достигает земли (рис. 46). Конечности относительно короткие, голова с вытянутой мордой. Уши относительно длинные с заостренной вершиной. ”а лапах пальцевые мозоли небольшие, пяточные мозоли неразвиты и п°крыты шерстью.
Волосяной покров высокий, густой и мягкий или более редкий и гру-Ь1Н- В окраске его преобладают рыжие, желтоватые и буроватые тона, иногда с серебристо-белыми и черными отметинами. Хорошо выражен сезонный и географический диморфизм. В зимнее время в северных часик ареала мех более густой и высокий. «Фиалковая» железа хорошо Рззвита. В году бывает 2 линьки. Сосков 4—6 пар.
”ереп вытянутый, стройный, относительно низкий, с широко расстав-
4*
ленными скуловыми дугами. Гребни на черепе развиты слабо или их нет вовсе. Надглазничные отростки большие, сверху слегка вогнутые, с острым краем. Межглазничная область уплощенная.
Зубная формула: i — с — р — гл — = 42. 3 14 3
У некоторых особей нижних коренных зубов может быть только 2 Резцы обычно не имеют дополнительных выступов. Клыки тонкие, но очень длинные и сильно изогнутые. Щечные зубы с очень острыми вер-шинами и ребрами.
Диплоидное число хромосом у лисицы 35—40, у песчаной лисицы 40-у бенгальской 60.
Распространены очень широко: вся Европа, вся Африка, кроме Мадагаскара, и большая часть Азии, на север до побережья Северного Ледовитого океана (кроме северной части Таймыра) и о-вов Колгуев, Южного из Новоземельских и Белого. На юг ареал идет до Тонкина Юньнаня, Сычуаня, Ассама и Индии — до м. Коморина; охватывает Аравийский п-ов; на востоке до о-вов Карагинского, Св. Лаврентия, Курильских, Сахалина, Шантарских и Японских; в Северной Америке — на юг до Северной Мексики и Калифорнии и на север до северного побережья Лабрадора, Гудзонова залива и линии Честертон (на западном берегу Гудзонова залива)-—Коппермайн (на Арктическом побережье).
Лисица населяет самые разнообразные ландшафты и все зоны и высотные пояса от тундры до пустыни и высокогорий. Сплошных равнинных лесных массивов избегает. Предпочитает селиться в открытых пространствах с разреженными лесами, перемежающимися полями, в лесостепях и степях с балками и оврагами. Корсак и американский корсак встречаются в сухих степях и пустынях. Убежищем служат норы барсуков, сурков, песцов. Иногда выкапывают норы сами на возвышенных местах и по склонам оврагов. Собственные норы не отличаются большой сложностью. Участок обитания лисицы может изменяться в зависимости от географического местоположения, сезона и кормности года, обычно его нлощадь составляет несколько квадратных километров. В центральной зоне европейской части СССР диаметр такого участка равен 6—10 км.
Состав нищи лисицы сильно изменяется в зависимости от географического местоположения, по годам и сезонам. По характеру питания лисица всеядная. В СССР насчитывается до 300 видов животных и несколько десятков видов растений, которыми она питается. Основной объект пищи — мелкие грызуны, чаше всего серые и лесные полевки, в меньшей степени — различные птицы, особенно наземногнездящиеся. В летнее время систематически поедает насекомых и плоды. В зимнюю бескормицу питается падалью, отбросами, может нападать на домашнюю птицу и на довольно крупных млекопитающих, в частности молодых косуль. Корсак в отличие от других представителей рода нашей фауны в меньшей степени использует растительные корма. Он приспособился охотиться на различных млекопитающих до молодых сурков и зайцев включительно, но главным образом на мелких грызунов. В летнее время в больших количествах он поедает мелких птиц, пресмыкающихся и насекомых. У американского корсака пища на 64% состоит из мелких млекопитающих — кроликов, сусликов, хомяков; на 9,6—из птиц, на 8,8 — из беспозвоночных, главным образом насекомых, и на 17,6% 113 растений.
Корсак активен преимущественно в сумерках. Лисица встречается в любое время суток, но наиболее активна ранним утром и поздним вечером. Вне периода размножения^исица держится поодиночке. Течка про* 100
кходит с января по март в зависимости от географического местоположения. Самка находится в охоте всего несколько дней. В период гона между самцами происходят сильные драки. Как правило, лисица моно-гам. Пара соединяется на один сезон. Длительность беременности — 49— 58 дней, чаще 52 дня. Детенышей в помете в среднем 4—6, максимум до И- Корсак образует пару, по-видимому, на всю жизнь. Беременность около 60 дней; Число детенышей у него бывает от 2 до 16, но чаще 3—6. Американский корсак —моногам. Пара соединяется, по-видимому, на всю жизнь. Течка происходит с конца декабря по начало февраля. Детеныши рождаются с марта по апрель. Детенышей в помете обычно 4—5. Продолжительность лактационного периода у лисицы 1,5 месяца. Глаза открываются у лисят на 13—15-й день. В этот же срок открываются ушные отверстия и прорезываются зубы на верхней челюсти. Около нор лисята остаются до 1,5—2-месячного возраста. К 6—7 месяцам они приобретают вид взрослого животного. В конце лета — начале осени лисята переходят к самостоятельной жизни. Половозрелость наступает примерно в годовалом возрасте. Длительность жизни в неволе до 15 лет. Врагами являются волк, рысь, росомаха, среди птиц — орлы и орланы.
Представители рода — важные промысловые пушные звери. В СССР добывается большое число лисиц и корсаков: с 1960 по 1970 г. лисиц заготовляли ежегодно 180,8—652,1 тыс., а корсака — 5,2—-34,0 тыс. Лисица приносит определенную пользу сельскому хозяйству, уничтожая большое количество вредных грызунов. Некоторый вред может наносить лисица охотничьим хозяйствам, поедая промысловых птиц, зайцев, ондатр, молодняк копытных. Однако это ее отрицательное значение, по-видимому, часто преувеличивается.
В роде 9 видов:
афганская лисица —V. сапа Blanford, 1877 (Афганистан, северо-запад Индии, Восточный Иран; в СССР — юго-западная Туркмения);
корсак—V. corsac Linnaeus, 1768 (Северо-Восточный Китай, Внутренняя Монголия; МНР; Афганистан и, по-видимому, Северо-Восточный Иран; в СССР — равнины Средней Азии, Казахстана и прилегающих территорий на запад изредка до северного побережья Азовского моря и востока Днепропетровской области и на север до Самарской луки, южных районов восточной половины Татарской АССР, южной оконечности Урала, Верхнеуральска, Троицка, Шадринска, Тюмени, Ишима, Тары, Омска, Барабинска и по левобережью Оби до Барнаула, граница распространения огибает Алтай с запада и по северному склону Зайсанской котловины идет до государственной границы; другой участок ареала в СССР — юг Забайкалья);
бенгальская лисица — V. bengalensis Shaw, 1800 (Индия, Непал),
лисица — V. vulpes Linnaeus, 1758 (Северная Африка — Марокко, Алжир, Ливия, АРЕ; вся Европа; Азия — Малая Азия, Аравийский п-ов, Ирак, Иран, Афганистан, Индия, Китай, МНР, Корейский п-ов, Япония; в СССР — вся европейская часть на север до побережья Северного Ледовитого океана и вся азиатская часть на север примерно до южной границы тундры, Сахалин и Курильские о-ва; Северная Америка — на север До побережья Берингова моря и северного побережья материка, кроме оеверо-западных территорий Канады, почти вся территория США на юго-восток до центральных районов штатов Северная и Южная Каролина, Ажорджия, крайнего северо-запада штата Флорида, на юг до побережья ексиканского залива и центральных районов Техаса; нет лисицы в ж ной половине провинции Альберта й юго-западе Саскачеван в Кана-Де> Далее почти по всей Монтане, кроме ее запада, восточном Вайомин-е> востоке Колорадо, востоке и западе Е1ью-Мексико, в западных рай-иах штатов Северная и Южная Дакота, Небраска, Канзас, Оклахома,
востоке штата Вашингтон, центральных районах штата Орегон, севере Невады, крайнем западе Юты, юге Калифорнии и почти на всем Тихоокеанском побережье США и Канады, кроме штатов Орегон и Аляски; самые южные точки ареала располагаются в Центральной Калифорнии^ на юге Невады, севере Аризоны, юге Нью-Мексико, центральных районах Техаса, по побережью Мексиканского залива);
тибетская лисица— V. ferrilata Hodgson, 1842 (Тибет и Непал);
американский корсак— V. velox Say, 1823 (Канада — юго-восток провинции Альберта, юг Саскачеван, юго-запад Манитобы; США — штаты Монтана, Северная и Южная Дакота, Вайоминг, Небраска, север Коло- 4 радо, запад Канзаса, крайний запад Миннесота и Айова, запад Оклахомы, северо-запад Техаса, Нью-Мексико, Юта, Аризона, Невада, крайний юго-восток Орегона, крайний юго-запад Айдахо, Калифорния; Мексика— п-ов Калифорния, штаты Сонора, Чиуауа, Коауила, Закатекас, Нуево-Леон);
песчаная лисица— V. riippeli Schinz, 1825 (Сомали, Эфиопия, Судан, Чад, Нигер, Алжир, Марокко, Ливия, АРЕ, Синайский п-ов, юг Аравийского п-ова, Иран, Афганистан);
южноафриканская лисица — V. chama A. Smith, 1833 (Намибия, Ботсвана, юг и юго-запад Анголы, Родезия, Южно-Африканская Республика) ;
африканская лисица—V. pallida Cretzschmar, 1826 (от Судана до Сенегала).
Американские ученые (например, Холл и Келсон, 1959) выделяют из V. velox особый вид V. macrotis Merriam, 1888 (южная и юго-западная часть ареала); лисицу с американского континента считают особым видом V. fulva Desmarest, 1820. Иногда (например, Гептнер, 1967) африканскую V. chama и африкано-переднеазиатскую V- ruppeli относят к Fennecus.
В «Красную книгу» включен подвид американского корсака — V. velox hebes Merriam, 1902, как исчезающий подвид, возможно, еще сохранившийся в Канаде на юго-западе провинции Саскачеван и в США на территории штатов Северная и Южная Дакота, Айова и на востоке Колорадо.
Род феиеков
Genus Fennecus Desmarest, 1804
В роде единственный вид: фенек — F. zerda Zimmermann, 1780.
Размеры мелкие. Длина тела 35—40 мм, длина хвоста 19—30 см. Масса около 1,5 кг. Внешне похож на лисицу. Характерны огромные уши, достигающие примерно 15 см в длину (рис. 47). Лицевой отдел головы относительно короткий. Глаза большие. Волосяной покров высокий, густой и мягкий. Окраска его на спине рыжеватая, бледно-желтоватая или почти белая. Брюхо белое. Кончик хвоста черный.
В черепе характерны умеренно расширенный мозговой отдел, огромные костные слуховые барабаны и узкий лицевой отдел. Межглазничное сужение выражено слабо. Скуловые дуги расставлены широко. Костное небо в области коренных зубов широкое.
Диплоидное число хромосом 64.
Распространен в Северной Африке (Марокко, Алжир, Ливия, АРВ и на юг до Нигера и Судана), на Синайском и Аравийском п-вах.
Населяет пустыни. Активен ночью. Убежище находит в норах, которые выкапывает сам. Питается разнообразной пищей: мелкими грызунами, ящерицами, птицами и их яйцами, насекомыми, некоторыми рас-
Рис. 47. Фенек — F. zerda и его череп
тениями. Может подолгу обходиться без воды. Длительность беременности примерно 50 дней. Детеныши рождаются обычно в марте или в начале апреля. В помете от 2 до 5 детенышей. Продолжительность жизни 10—12 лет.
Род серых лисиц
Genus Urocyon Baird, 1858
Размеры мелкие. Длина тела 48—68 см. Длина хвоста 11—44 см. Масса 2,5—7 кг. Внешне похожи на лисицу, но с укороченным хвостом (рис. 48). Уши средней высоты, остроконечные. Волосяной покров высокий и грубый. Окраска его на голове, спине, боках и хвосте серая. Брюхо, горло и внутренние части конечностей белые. По середине спины проходит черная полоса. Конец хвоста черный
Диплоидное число хромосом у серой лисицы 66.
Распространены на крайнем юге провинции Онтарио и Квебек в Ка-наде, в США (кроме некоторых северо-западных и центральных штатов), в Центральной и северной части Южной Америки и на некоторых островах у Калифорнии.
Населяют леса, участки поросшие кустарником, болота, скалы, аридные области. Активны ночью. Убежище находят в пещерах, расщелинах 104
скал, дуплах упавших деревьев; иногда выкапывают норы (в северных частях ареала). Хорошо лазают по деревьям (единственный случай в семействе) и спасаются на них в случае опасности, а иногда залезают на них и в поисках добычи. Питаются различными растениями, главным образом плодами и семенами, а также мелкими животными. Пища серой лисицы состоит на 28,6% из растений, 20,5 — кроликов, 30,5 — мелких грызунов, 1 —землероек, 26 — птиц, 2,5 — насекомых, 2,5% —рыб (Бан-филд, 1974). Моногамы. Гон у серой лисицы с конца января по март. Беременность длится примерно 63 дня. Детеныши рождаются весной от 2 до 7 (обычно 4) в помете. Масса новорожденного в среднем 115 г. Семья распадается поздним летом или ранней осенью, но самец и самка могут продолжать жить совместно. Половозрелость у самок наступает в возрасте около года.
В роде 2 вида:
серая лисица — U. cinereoargenteus Schreber, 1775 (крайний юг провинций Онтарио и Квебек в Канаде; США, кроме штатов Вашингтон, востока Орегона, севера Невады, Айдахо, Монтана, Вайоминг, севера Юты, востока Колорадо, западных районов штатов Северная Дакота, Южная Дакота, Небраска, Канзас и Оклахома и крайнего севера Техаса; вся Центральная Америка, кроме северных районов Гондураса и Никарагуа; запад Венесуэлы);
островная лисица — U. littoralis Baird, 1858 (острова, расположенные у штата Калифорния в США: Санта-Каталина, Сан-Клементе, Сан-Николас, Сан-Мигель, Санта-Крус).
Род южноамериканских лисиц
Genus Dusicyon Н. Smith, 1837
Размеры мелкие и средние. Длина тела 60—100 см. Длина хвоста 30—35 см. Общий вид напоминает лисиц. Конечности относительно высокие. Морда вытянутая. Уши крупные остроконечные или средней высоты округлые (рис. 49). Волосяной покров высокий, густой и мягкий. Его окраска может быть одноцветная рыжеватая, коричневатая или черноватая или на спине темно-серая, на брюхе рыжеватая. Хвост покрыт удлиненными волосами.
Диплоидное число хромосом у бразильской лисицы 74.
Распространены в Южной Америке.
Населяют различные ландшафты: от степей до лесов и от уровня моря до 4 тыс. м над его уровнем. Активны в основном в сумерки и ночью. Питаются разнообразными позвоночными: земноводными, пресмыкающимися, птицами и их яйцами, мелкими и средних размеров млекопитающими, а также различными беспозвоночными и плодами. Детеныши Рождаются в октябре — ноябре; в помете их от 3 до 6. Могут приносить вРед сельскому хозяйству.
В роде, по-видимому, 10 видов (список видов приводится по Кабрера, Cabrera, 1957):
Вая)^™^1*351 лисица— cuipaeolus Thomas, 1914 (юго-запад Уруг-андская лисица — D. culpaeus Molina, 1782 (Анды в Перу, Боливии, или, Аргентине, Эквадоре);
чилоэсская лисица — D. fulvipes Martin, 1837 (о-в Чилоэ около Чили); тин^)КНОаМеРИКанСКая лисица— griseus Gray, 1836 (Чили и Арген-Уп^1аРагвайская лисица — D. gymnocercus Fischer, 1814 (Парагвай, цугваи, юго-запад Бразилии, Аргентина);
Рис. 49. Андская лисица — D. culpaeus
перуанская лисица — D. inca Thomas, 1914 (юг Перу);
D. sechurae Thomas, 1900 (северо-запад Перу и юго-запад Эквадора); бразильская лисица — D. vetulus Lund, 1842 (Бразилия);
малая лисица — D. microtis Sclater, 1882 (бассейн Амазонки в Бразилии, Эквадор, Перу и Колумбия);
майконг — D. thous Linnaeus, 1766 (Колумбия, Венесуэла, Гайана, Суринам, Бразилия, Боливия, Парагвай, Уругвай, север Аргентины).
Некоторые исследователи (например, Кабрера, 1957) выделяют D microtis в особый род Atelocynus Cabrera, 1940; D. thous в род Cer-docyon H. Smith, 1839.
Малая лисица включена в «Красную книгу» как малочисленный вид, хотя и не находящийся в непосредственной опасности исчезновения.
Род гривистых ВОЛКОВ
Genus Chrysocyon Н. Smith, 1839
В роде единственный вид: гривистый волк — С. brachyurus, liliger, 1811.
Размеры крупные. Длина тела около 125 см. Длина хвоста примерно 30 см. Высота в плечах 75 см. Масса около 23 кг. Лицевой отдел удлинен Уши большие. Характерны высокие конечности (рис 50). Волосяной покров высокий и относительно мягкий. На шее и плечах волосы удлинены в небольшую гриву. Окраска его желтовато-рыжая. Проксимальные отделы конечностей темные или почти черные. Иногда спина, хвост и область рта черные. Конец хвоста и подбородок могут быть белыми.
Диплоидное число хромосом 76.
Распространен в центральной части Бразилии, на крайнем востоке Боливии, в Парагвае, Уругвае и Севере Восточной Аргентине.
Рис. 50. Гривистый волк — С. brachyurus
Населяет пампасы, окраины болот, а в Мато-Гросу — леса. Численность гривистых волков в Бразилии на площади 650 тыс. км2 в 1964— 1967 гг. была примерно 1,5—2 тыс. особей. Держатся поодиночке. Активен ночью. Питается мелкими и средними по размерам млекопитающими, птицами, пресмыкающимися, насекомыми, плодами. Самка приносит обычно 2 (иногда 3) детеныша.
Включен в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения, если не будут предприняты меры охраны.
SUBFAMILIA SIMOCYONJNAE ZITTEL, 1893
Род красных волков
Genus Сноп Hodgson, 1838
В роде единственный вид: красный волк—С. alpinus Pallas, 1811.
Размеры средние. Длина тела около 100 см. Длина хвоста примерно 50 см. Масса около 17 кг. Ноги высокие. Хвост относительно длинный, спускается ниже скакательного сустава, по у стоящего животного, однако. до земли не достает. Голова с укороченной, заостренной мордой (рис. 51). Уши большие, стоячие, с закругленными вершинами, высоко Посажены на голове; их основания сближены. Волосяной покров зимой очень высокий, густой и мягкий. Окраска его однотонная, рыжая. Летом полосы значительно короче, грубее, более темные. Окраска может меняться индивидуально и отчасти географически. Хвост покрыт длинными, пушистыми волосами. Сосков 6—7, редко 8 пар.
Череп массивный, укороченный (особенно в лицевом отделе), с широко расставленными скуловыми дугами. На черепе хорошо развитые
Рис. 51. Красный волк — С. alpinus и его череп
ебни, в частности сагиттальный. Лобная область выпуклая. Надглаз-Г ичные отростки относительно небольшие.
Н 3 14 2
Зубная формула: 1 — с — р — m — — 40.
Зубы крупные. На хищническом зубе нижней челюсти позади главно-зубца имеется всего один небольшой дополнительный зубец. Клыки массивные, слабо изогнутые, короткие. Резцы не имеют дополнительных выступов.
Распространен в Китае — в Синьцзян-Уйгурском автономном районе, СЫчуане, Юньане, Фуцзяне, на п-овах Индостан, Индокитай и Малакка, о-вах Суматра и Ява; на Корейском п-ове, в южных и восточных частях МНР; в СССР — в южных районах Дальнего Востока, на юге Восточной и Средней Сибири, а также в восточной части Средней Азии.
Обитает в различных ландшафтах: на равнинах, побережье моря и в горах, поднимаясь в субальпийский и альпийский пояса до 4 тыс. м над уровнем моря. Питается в основном крупными копытными, в пределах СССР особенно горными козлами. В летнее время в больших количествах употребляет и растительные корма. В периоды размножения красные волки держатся стаями, насчитывающими от 5 до 30, иногда и больше особей. Охотятся стаями, подолгу преследуя жертву. Активны преимущественно днем. Вне периода размножения широко мигрируют в поисках добычи. Размножение изучено плохо. По наблюдениям в Пекинском зоопарке, гон бывает в январе — феврале. Беременность длится около 60 дней. Щенков в помете 5—9. Вследствие своей малочисленности практического значения не имеет.
Включен в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
Род гиеновых собак
Genus Lycaon Brookes, 1827
В роде единственный вид: гиеновая собака — L. nidus Temminck, 1820.
Размеры средние. Длина тела 76—100 см. Длина хвоста 30—40 см. Высота в плечах около 60 см. Масса 16—23 кг. Внешне похожа на собаку. Морда широкая, челюсти мощные. Уши большие, глаза относительно маленькие (рис. 52). Конечности высокие, четырехпалые. Волосяной покрой низкий и редкий. Окраска его сильно изменчива, представлена комбинацией различных по величине и форме пятен темно-бурого, желтого, черного и белого цвета. Голова обычно темная, конец хвоста белый. Кожные железы продуцируют секрет с сильным мускусным запахом.
Череп мощный, с большими гребнями. Скуловые дуги широко рас-ставленные, крепкие, зубы массивные. Лицевой отдел короткий и ши-
Диплоидное число хромосом 78.
Распространена в Африке от Берега Слоновой Кости и Гвинеи на западе, Мали, Нигера, юга Алжира, Судана, Эфиопии и Сомали на евере и до Южно-Африканской Республики на юге. Сохранилась к на-тоящему времени в основном в национальных парках и кое-где в не споенных человеком ландшафтах.
Встречается на равнинах и в горах от уровня моря до верхней грани-леса несколько выше 2700 м над уровнем моря. Держится стаями Ычн° около 10 голов вместе (от 4 до 60). Активна круглосуточно. Пи-
Рис. 52. Гиеновая собака — L. pictus
тается разнообразными позвоночными, включая крупных антилоп. Обычно охотятся стаями, загоняя добычу. Сезонность в размножении, по-видимому, отсутствует. Продолжительность беременности 63—80 дней. В помете 6—8 детенышей. Продолжительность жизни около 10 лет.
Включена в «Красную книгу» как малочисленный вид, который в ближайшее время может оказаться под угрозой исчезновения.
Род кустарниковых собак
Genus Speothos Lund, 1839
В роде единственный вид: кустарниковая собака — S. venaficus Lund,
Размеры мелкие. Длина тела 57—75 см. Длина хвоста 12—15 см. Масса 5—7 кг. Телосложение тяжелое. Туловище удлиненное, конечности короткие. Голова большая, лицевой отдел укороченный. Уши ма^ ленькие, округлые. Глаза относительно большие (рис. 53). Волосяной покров высокий, грубый. Окраска туловища темно-бурая, иногда почти черная, голова и плечи желтоватые, оранжеватые или беловатые.
Характерно наименьшее число зубов в семействе:
.3 I 4 1
*тст рттт=38-
НО
Рис. 53. Кустарниковая собака—-«S. venaticus и ее череп
^БрСаП3РиОХНнеяНа В Панаме’ Гайане, на севере и в центральных облас-Вая, в Эквадоре СеВер°’Востоке ПеРУ> в°етоке Боливии, севере Параг-Актив2ЛночьюеСЛРпаВпНПЫ’ Песчаные Районы. Экология изучена слабо, мому Даями Ynn Проводит в ноРах броненосцев. Охотится, по-видн-гРызуиами R пом Р°₽ШО влавает и ныРяет. Питается преимущественно уиами. в помете до 4—5 детенышей.
ПанамыДваьшелХМпеР’ “ КеЛСеОН’ ’959) ^арниковую собаку из Включай^ п особый вид —S. panamensis Goldman, 1912.
Щее время и не iS₽nHyi° К'ТУ>> как малочисленный вид, хотя в настоя-Р я и не подвергающийся опасности исчезновения.
Рис. 54. Большеухая лисица — О. megalotis
SUBFAMILIA OTOCYONINAE TROUESSART, 1885
Род большеухих лисиц
Genus Otocyon Muller, 1836
В роде единственный вид: большеухая лисица — О. megalotis Desmarest, 1822.
Размеры мелкие. Длина тела 46—58 см. Длина хвоста 24—34 см. Высота в плечах 35—40 см. Масса 3—4,5 кг. Внешне похожа на лисицу. Уши очень большие, широкие в основании, остроконечные — до 11—13см в высоту (рис. 54). Волосяной покров средней высоты, желтоватый или желтовато-бурый. Проксимальные отделы конечностей, конец хвоста и вершины ушей черные.
„ 314 3—4
Зубная формула: i — с — р — m -=46 — 50.
4 1 4 4 — 5
Характерны многочисленные зубы — наиболее многочисленные среди наземных плацентарных млекопитающих. Зубы небольшие, слабые.
Диплоидное число хромосом 72.
Распространена в Южно-Африканской Республике, Ботсване, Намибии, на юге Анголы, возможно, на западе Родезии, а также в Замбии и от Танзании на север до юга Судана, Эфиопии и Сомали.
Населяет аридные земли. Активна преимущественно ночью. Держатся поодиночке, парами или небольшими группами (по-видимому, семьями). Питается насекомыми, в основном термитами, а также плодами, клубнями, мелкими млекопитающими, иногда падалью. Детеныши рождаются с декабря по апрель. Продолжительность беременности 60-^ 70 дней. Детенышей в помете 2—5.
СЕМЕЙСТВО МЕДВЕЖЬИХ
FAMILIA URSIDAE GRAY. 1825
Размеры средние и крупные. Длина тела ПО—302 см1. Высота в хол-ке до 135 см. Хвост очень короткий и обычно скрыт в мехе (снаружи не аметен). Масса от 50—100 до 500—700, возможно, до 1000 кг. Сложение тяжелое, неуклюжее. Туловище удлиненное, конечности относительно короткие, стопоходящие, пятипалые (рис. 55). Все пальцы касаются земли и вооружены длинными, изогнутыми, мощными, невтяжными когтями На передних конечностях они обычно значительно больше, чем на задних. На нижней поверхности кисти и ступни кроме обнаженных пальцевых подушек имеется поперечная голая подушка, а также и голые участки кожи позади иее. Шея толстая, обычно длинная. Голова крупная, чаше с короткой, реже удлиненной мордой. Глаза небольшие. Уши v некоторых видов маленькие, почти не выступающие над мехом, или умеренной длины, или большие и широкие.
Волосяной покров высокий, косматый; у северных и горных видов густой, у малайского медведя — низкий и редкий. Окраска его одноцветная, белая, черная или бурая с различными оттенками. У некоторых видов имеет место индивидуальная и географическая изменчивость окраски. На груди бывает белое пятно, иногда белый ошейник. У черного медведя морда светлая или коричневая. У очкового медведя глаза окружены светлыми кольцами. Сезонный диморфизм выражается в том, что высота и густота меха в зимнее и летнее время различны. Анальных желез нет или они развиты очень слабо. Функционирующих сосков одна пара — грудных.
Череп массивный, крупный с удлиненным лицевым отделом (у малайского медведя лицевой отдел сильно укорочен). Скуловые дуги большинства видов слабо расставлены в стороны (у малайского медведя — сильно). Гребни на черепе развиты сильно. Костные слуховые барабаны обычно уплощенные. Костный слуховой проход имеет вид удлиненной трубки. Во внутренней полости слухового барабана перегородки нет. Крылоклиновидный канал имеется. Глазницы черепа невелики, височная яма очень крупная. Челюсти мощные. На нижнем крае нижней челюсти под угловым отростком находится небольшой загнутый внутрь альвеолярный отросток.
2__з 1 2___4 2
Зубная формула: i —-— с — р —---— m —---— = 34— 38 — 40 — 42.
I
Часто имеет место индивидуальная и возрастная изменчивость зубной системы. Клыки очень сильно развиты, хищнические зубы выражены слабо. Крайние резцы крупнее внутренних. Коренные зубы с низкими широкими коронками и притупленными бугорками.
Распространены в Европе, включая Северную Исландию; в Северо-западной Африке; Азии, кроме Аравийского п-ова на юго-западе и о-вов Пулавеси и Филиппинских на юго-востоке; в Арктике; Северной Америке на юг до центральных районов Мексики; горных областях северо-запада Южной Америки. За последние десятилетия ареал сильно сокра-ился под влиянием деятельности человека (прямого уничтожения или зменения условий жизни). Медведи исчезли на значительных территориях Европы, Северной Америки, Северной Африки, Японии.
Населяют леса на равнинах и в горах, обитают и в безлесных горных
„ 1 Представителей длиной тела 110—173
семейства по размерам можно разделить на 3 группы: мелкие с см, средние — 174—238 и крупные — 239—302 см.
ИЗ
Рис. 55 Бурый медведь — U. arctos (А) чепный идТ (В) ' americanus (£) белый медведь — U. mariti-
Рис. 55. Продолжение. Череп бурого медведя (Г),
Районах, белый медведь встречается на побережье Ледовитого океана и во льдах Арктики. Все формы наземные; белый медведь полуводныи, губач, малайский медведь — довольно высоко специализированные др< весные формы. Держатся поодиночке или семьями, самка с молодыми и иногда с самцом. Активны главным образом ночью, отчасти днем, иног Да круглосуточно. По характеру питания всеядные, некоторые специализированы к поеданию растительной пищи, а другие — животной. Белый Медведь питается почти исключительно мясом млекопитающих. Большинство ведет оседлый образ жизни; самцы белых медведей широко кочуют в течение круглого года, а самки с молодыми — часть года. Бурый медведь, белогрудый и черный медведи впадают в зимний сон. У белого Медведя в зимний сон впадают только беременные самки. Некоторые ви Ды зимой совсем не спят. Медведи—моногамы, однако пары подолгу Ие Держатся и самец в заботах о потомстве участия не принимает. В по-Мете обычно 1—2, редко 3 детеныша. _
Практическое значение представителей семейства невелико. В неко
Рис 55. Продолжение. Череп белого медведя (Д)
торых местах они служат предметом спортивной охоты. В ряде мест медведи находятся под запретом, в частности белый медведь; в некоторых местах медведи вредят сельскому хозяйству (животноводству), поэтому в этих областях их истребление не ограничивается. В СССР в период с 1960 по 1970 г. заготовляли ежегодно по 200—300 шкур медведей.
В семействе, по-видимому, 4 рода (7 видов):
медведей — Ursus Linnaeus, 1758 (4 вида);
очковых медведей — Tremarctos Gervais, 1855 (1 вид);
медведей-губачей — Melursus Meyer, 1793 (1 вид);
малайских медведей — Helarctos Horsfield, 1825 (1 вид).
Покок (Рососк, 1941), Симпсон (1945) насчитывают в семействе 6 родов: Helarctos; Melursus; Selenarctos Heude, 1901; Tremarctos, Ursus, Thalarctos Gray, 1825. Андерсон и Джонс (1967) включают Thalarctos в род Ursus. Более правильно отнести сюда же и Selenarctos. По Гептне-ру и др. (1967) в семействе следует выделить всего 2 рода: Helarctos с одним видом и Ursus со всеми остальными видами.
В СССР: один род медведей — Ursus..
Род медведей
Genus Uгsus Linnaeus, 1758
Размеры мелкие, средние и крупные, наибольшие в семействе. Длина белогрудого медведя 130—200 см, черного— 150—180, бурого — белого— 160—302 см. Масса до 640 кг у бурого медведя и до 1Л60 кг-""У белого. Сложение массивное, тяжелое. Конечности средней 1J ы иЛи довольно высокие (белый медведь). На нижних поверхнос-Б лап расположены голые участки различного размера, иногда очень Тйбольшие (белый медведь). Шея длинная (особенно у белого медведя). Голова крупная или небольшая (белый медведь). Уши небольшие округ-ые глаза маленькие. Губы подвижные или малоподвижные (белый медведь). Хвост очень короткий. Когти на лапах очень крупные или относительно небольшие (белый медведь). Зимний мех очень густой, мягкий длинный или относительно короткий (белый медведь). В летнее время мех короче и более редкий или остается таким же густым (белый медведь).
Череп крупный и массивный. Лицевой отдел удлиненный или несколько укороченный. Скуловые дуги расставлены нешироко.
3 ] 2 4 2
Зубная формула: i — с — р —- ш — =34 — 42.
3 12 4 3
Число предкоренных зубов может изменяться индивидуально. Клыки сильно развиты, коренные зубы средних размеров или небольшие (белый медведь). У бурого медведя с возрастом зубы стираются, у белого медведя, даже у самых старых особей, зубы остаются нестертыми.
Диплоидное число хромосом бурого, черного, белогрудого и белого медведей по 74.
Распространение соответствует указанному в описании семейства, кроме Индостана, Малакки, Шри Ланка и Южной Америки.
Населяют лесные районы, отчасти безлесные горы и плоскогорья, белый медведь обитает на побережье Ледовитого океана и во льдах Арктики. Кроме белого медведя хорошо лазают по деревьям (бурый медведь в юности). Бурый медведь питается самой разнообразной пищей, главным образом растениями, их зелеными частями, ягодами, плодами; поедает много беспозвоночных, в меньшей степени мелких позвоночных; за крупными копытными охотится только в голодные периоды года; вылавливает рыбу; охотно ест падаль. У него хорошо прослеживается сезонная изменчивость в питании. Пища черного медведя на 76,7% состоит из растений, 7,4 — насекомых, 15,2 — падали и 0,7% —мелких млекопитающих. Белогрудый медведь в основном питается различными частями растений; в редких случаях поедает падаль и снулую рыбу. Основным объектом питания белого медведя служит нерпа, в меньшей степени — Морской заяц. Бурый медведь активен по большей части в сумерки и ночью, у белого медведя суточного ритма активности не наблюдается, а зиму бурый и белогрудый медведи, а также самки белого медведя впадают в зимний сон (в литературе имеются данные, что в зимний сон иногда впадают также и самцы белого медведя).
в Ьерлогу бурый медведь обычно устраивает в укромном сухом месте—• Углублении под вывернутыми корнями деревьев, в расщелине скалы. ®ЛогРУДЬ1й медведь на зиму ложится в дупла старого дерева, обычно ПОлей Максимовича и чозений, липы и дубов, а также в пещерах и злах. Белый медведь устраивает берлогу только на материке и остро-х в сугробах снега. В пределах СССР область залегания беременных ь»ок белого медведя в берлоге ограничивается островами Новая Зем
ля, Земля Франца-Иосифа, Северная Земля, Новосибирские, Медвежьи Врангеля и некоторыми другими. Наибольшее количество самок белое’ медведя залегает на о-ве Врангеля. Длина камеры берлоги достигае° 160—180 см, высота 100—120 см. Длина хода, ведущего в камеру беп логи, бывает до 2,5—3 м. Залегание в зимний сон происходит й" в одно и то же время у различных зверей одного вида даже в одном рай6 оне. Необходимым условием для зимнего сна служит накопление значд, тельных жировых запасов. Во время зимнего сна снижаются частот дыхания и пульсация сердца. Температура может уменьшаться до 29°с Бурые медведи могут находиться в зимнем сне от 75 до 195 дней. В юж.’ пых районах ареала, на Кавказе, в Южном Закавказье многие бурце медведи вообще не впадают в зимний сон.
Бурые и белогрудые медведи ведут, как правило, оседлый образ жизни, хотя в случае бескормицы могут уходить на большие расстояния, в частности белогрудые медведи — за 200—350 км. Более или менее регулярные сезонные миграции отмечаются у бурых медведей в горах. Миграция белых медведей обусловлена состоянием плавучих льдов и распределением тюленей. В летнее время белые медведи откочевывают к северу, а на зиму — к югу.
Самки медведей размножаются раз в 2 года. Гон у медведей в различных географических точках происходит в разные сроки. У бурых медведей обычно гон приходится на вторую половину мая, июнь, до середины июля; у черного медведя — с середины апреля по конец августа. Гон может сопровождаться драками самцов. Гон белых медведей растянут с апреля по август. Продолжительность беременности бурых медведей составляет 6—8 месяцев, черных медведей—около 220 дней, белых медведей— около 8 месяцев. Детенышей в помете 1—5, чаще 2. Самки бурых медведей рожают с декабря по апрель, белых—с января по июль, белогрудых — с конца декабря до середины февраля, изредка до начала марта. Медвежата рождаются очень небольшими; у бурого медведя их длина достигает примерно 23 см, масса 500 г, черного медведя — 20 см и 240—330 г. Новорожденные слепые, с закрытыми слуховыми отверстиями, покрыты короткой редкой шерстью. На 30—32-й день у них открываются глаза и слуховые отверстия. Длительность лактации 6—8 месяцев. У бурого медведя к 3-му месяцу жизни имеется полный состав молочных зубов. Медвежата переходят к самостоятельной жизни на 3-м году или в конце 2-го года. Половозрелость наступает в 3—4 года или несколько позднее. Продолжительность жизни 30—40 лет.
Врагов у медведей, кроме человека, очень немного. У белых медведей их нет вовсе, у бурого медведя может быть уссурийский тигр (в редких случаях), у белогрудого медведя — тигр и волки. Медведи местами служат объектом спортивной охоты, а местами промысловой. В СССР с 1956 г. добыча белого медведя запрещена.
В роде 4 вида:
бурый медведь—Ursus arcios Linnaeus, 1758 (Евразия на север Д° границы леса; южная граница проходит по берегам Средиземного моря> по северным районам Аравийского п-ова, двумя языками вдается на территорию Ирана — на западе и юго-западе через Луристан до Шираза и на севере от Талыша до Копетдага, по горным системам Гиндукуш3 и Памира в Афганистане, по северным районам Индостана, связанный с Гималаями, охватывает с юга, востока и севера Тибет, проходит по границам пустыни Гоби, огибая ее с запада, севера и востока, выходя побережью Желтого моря к югу от Корейского п-ова; северо-запад Аф рики—горы Атласа; Северная Америка — к западу примерно от 90° з. Д-> на юг до 22° ю. ш. в штате Дуранго, Мексика и на север до линии, проходящей примерно от Честерфилда на Гудзоновом заливе на северо-запаД
северного побережья материка и далее на запад по побережью моря Ьорта или несколько отступая от него в глубь материка, до северо-b рядных оконечностей Аляски; в СССР — по всей лесной зоне частично заП тепной и степной, востока тундры, на Кавказе, в горах Средней леС к настоящему времени ареал бурого медведя, разбился на участ-А31 нНОгда небольшие, изолированные друг от друга; во многих местах ^И’оЫе медведи очень малочисленны. В СССР ареал по сравнению с дру-6 ми странами изменился меньше, хотя и сильно сократился в южных, ГИигральных и западных районах европейской части страны, на Кавказе, Ц Средней Азии и на юге Дальнего Востока. В Европе представители ро-В наиболее многочисленны в Скандинавии, Румынии, Югославии п Болгарии' в Испании, Франции, Италии, Польше, Чехословакии, Венгрии, Албании они очень малочисленны, а в других странах исчезли совершенно. В Северной Африке полностью истреблены. В Северной Америке сохранились на Аляске, в Северной и Западной Канаде, в штатах США —Монтана, Айдахо, Вайоминг и Колорадо; на Мексиканском плоскогорье) ;
черный медведь—U. atnericatius Pallas, 1780 (почти вся Северная Америка, кроме крайнего севера материка и беренгийского побережья Аляски на севере; южная граница проходит примерно от Хьюстона в Техасе на юго-западе и юг через штаты Мексики Нуево-Леон и Тамаулипас, далее на восток до Тихоокеанского побережья штата Наярит, затем на север вдоль восточных границ штатов Синалоа и Сонора, через центральные районы штатов Аризона и Юта, огибает с севера Неваду, языком вдается в центральные районы Калифорнии и выходит в Средней Калифорнии к Тихоокеанскому побережью);
белогрудый медведь—И. thibetanus G. Cuvier, 1823 (Белуджистан в Пакистане, Гиндукуш в Афганистане, Пенджаб, Кашмир, Гималаи в Непале, Сиккиме, Бутане и Тибете, Ассам, Бирма, п-ов Индокитай, п-ов Малакка к югу примерно до его центральных районов, Юго-Восточный, Восточный Китай, восток Северо-Восточного Китая, о-ва Хайнань, Тайвань и Японские, в СССР — Уссурийский край и область, прилежащая к Амуру с севера);
белый медведь—U. maritimus Phipps, 1774 (плавучие льды, острова и Азиатское и Американское побережья Северного Ледовитого океана, У Европейского побережья появляется случайно; на север примерно до 88° с. ш.; на материке встречен на расстоянии до 150 км к югу от берега).
Иногда (например, Эллермэн и Моррисон-Скотт, 1966) белогрудого медведя выделяют в особый род Selenarctos Heude, 1901, а белого — в inalarctos Gray, 1825. Более правильно включать их в род Ursus.
Некоторые исследователи насчитывают в роде большее число видов, например Холл и Келсон (1959) только для Северной Америки—78+белый медведь, которого они выделяют в особый род Thalarctos,—всего 79, что, конечно, ошибочно.
В СССР; бурый, белогрудый и белый медведи.
bpf «Красную книгу» включены: подвид белогрудого медведя— U. thi-<2QQnus gedrosianus Blanford, 1877, сохранившийся в количестве около в ОСОбей„в южной части Центрального Белуджистана как подвид, под-^Рга1°Щийся опасности исчезновения; подвид черного медведя—U. ате-Hoij^1S еттоп$Ц Dall, 1895, из Юго-Восточной Аляски и Северо-Запад-Как Ританской Колумбии, сохранившийся в количестве около 500 особей °пасМаЛ0ЧИСЛенный Подвид’ хотя в настоящее время не подвергающийся исчезновения; подвид бурого медведя — U. arctos nelsoni Мег-част' прежде обитавший в Мексике на территории от Северной п-ова Калифорния, Северо-Восточной Соноры, Чиуауа и Коауила
Рис 56. Очковый медведь — Т. ornaius
на юг до севера Дуранго, к наст05.Л тему времени исчезнувший полйо' I стью или, может быть, сохрани шийся в незначительном числе штате Сонора; подвид бурого Ме? ведя U. arctos richardsoni Swain son, 1838, обитающий в Канаде н' Арктическом побережье и в тунд/ между Гудзоновым заливом « р. Маккензи как малочисленный подвид (сохранилось от 500 д0 1000 особей), хотя в настоящее вре. мя и не подвергающийся опасности исчезновения; белый медведь как вид, который в ближайшее время может подвергнуться опасности ис-
чезновения, если ситуация с его охраной не изменится в лучшую сторону (общее число белых медведей по всему ареалу около 20 тыс. особей).
Род очковых медведей
Genus Tremarctos Gervais, 1855
В роде единственный вид: очковый медведь — Т. ornatus F. Cuvier, 1825.
Размеры мелкие и средние. Длина тела 150—180 см. Длина хвоста около 7 см. Масса около 140 кг.
Внешне сходен с бурым медведем. Волосяной покров высокий, лохматый. Окраска его одноцветная черная или черно-коричневая, за исключением крупных белых колец или полуколец, окружающих глаза, и белого полукольца в нижней части шеи, от которого на грудь спускаются две белые полосы (рис. 56). Степень развития белых колец и полос на голове, шее и груди у различных очковых медведей может существенно изменяться, а у некоторых форм совсем отсутствовать.
Диплоидное число хромосом 52.
Распространен в Западной Венесуэле, Колумбии, Эквадоре, Перу и Западной Боливии, возможно и в Панаме.
Населяет горные леса, однако часто встречается выше их верхней границы — до 30 тыс. м над уровнем моря, иногда спускается в саванны и кустарниковые равнины. Экология слабо изучена. Питается почти исключительно листьями, плодами и корнями растений. Хорошо лазает
по деревьям.
Включен в «Красную книгу» как малочисленный вид, который может подвергнуться исчезновению, если ситуация с его охраной не изменится
в лучшую сторону.
Рис. 57. Медведь-губач — М. ursinus
Род медведей-губачей
Genus Melursus Meyer, 1793
В роде единственный вид: медведь-губач, М. ursinus Shaw, 1791.
Размеры мелкие и средние. Длина тела 140—180 см. Длина хвоста 10—12 см. Масса 55—136 кг. Общий облик типично медвежий. Голова крупная, с сильно удлиненной лицевой частью (рис 57). Губы крупные, подвижные и голые, ноздри могут произвольно смыкаться. Волосяной покров высокий, лохматый. Окраска его одноцветная, бурая или серая, иногда светло-бурая или красноватая. На груди имеется V-образная белая или желтая полоса.
214 2 лп
Зубная формула: i — с — р — m — W.
Распространен на п-ове Индостан и о-ве Шри Ланка.
Населяет леса. Активен в основном ночью. В зимний сон не впадает. Всеяден: пища состоит из насекомых, их личинок, яиц, птиц, падали, а также разнообразных растений. Своеобразен способ добывания муравьев н термитов; медведь-губач раскапывает их гнезда, песок и пыль сдувает в сторону, затем сильно вытягивает губы и всасывает термитов. Период размножения в Индии приходится на июнь, а на о-ве Шри Ланка. по-видимому, продолжается круглый год. Длительность беременности около 7 месяцев. В помете 1—2, реже 3 детеныша. Через 2 3 месяца после рождения медвежата с матерью покидают берлогу. В течение двух Или трех лет медвежата остаются с матерью. Предполагают, что самец принимает участие в воспитании медвежат. Продолжительность жизни около 40 лет.
Рис. 58. Малайский медведь — Н. malayanus и его череп
Род малайских медведей
Genus Helarctos Horsfield, 1825
В роде единственный вид: малайский медведь — И. malayanus Raf-1321
^е5размеры мелкие — наименьшие в семействе. Длина тела ПО—140 см. м сса 27—65 кг. Туловище относительно укороченное, а конечности окие, с большими кистями и стопой. Когти очень крупные, изогну-вЫ щёя длинная, голова крупная, с короткой .мордой. Уши короткие, ТЬ1оуглые( рис. 58). Волосяной покров низкий редкий. Окраска его одно-ветная черная с беловатым или с оранжевым пятном на груди. Морда еповатая или оранжевая. Конечности иногда светлые. В черепе сильно укорочен лицевой отдел, широко расставлены скуловые дуги. Зубная система характеризуется уменьшенным количеством предкоренных зубов (редуцированы второй, иногда и третий предкоренные). Таким образом, общее число зубов уменьшается до 38 или 34.
Диплоидное число хромосом 74.
Распространен в Бирме, Таиланде, п-овах Индокитай и Малакка, о-вах Суматра и Калимантан и, возможно, в Южном Китае.
Населяет лесные районы на равнинах и в горах. Активен ночью. Превосходно лазает по деревьям, день проводит на деревьях, где строит себе подобие гнезда. Всеяден. Поедает различные растения, насекомых, мелких позвоночных. В зимний сон не впадает. В помете обычно 2 медвежонка.
СЕМЕЙСТВО ЕНОТОВЫХ
FAMILIA PROCYONIDAE BONAPARTE, 1850
Размеры мелкие и средние. Длина тела 31—180 см. Длина хвоста 12—69 см. Масса 0,8—160 кг. Внешний вид крайне разнообразен. Некоторые напоминают медведя (большая панда), другие кошку (малая панда), третьи лисицу или енотовидную собаку (еноты), или куницу (какомицли); кинкажу и носуха очень своеобразны. Голова обычно крупная, морда короткая и притупленная или вытянутая и заостренная, иногда очень длинная и тонкая. Уши большие, короткие и округлые. Глаза обычно большие. Хвост длинный, лишь у большой панды короткий. Y кинкажу хвост хватательный (единственный случай в отряде). Конечности пятипалые, стопо- и полустопоходяшие. Нижняя поверхность кисти и ступни обычно голые. Третий палец наиболее длинный. Когти короткие, изогнутые и сжатые с боков, у большинства видов невтяжные, у некоторых полувтяжные. Волосяной покров, как правило, густой и высо-ии. Окраска его очень изменчива: от однотонной, желтой или буроватой, до пестрой и яркой, с комбинациями ярко-рыжего и черного цвета; У некоторых видов хвост с темными поперечными кольцами. У большой У ДИ в анальной области находится специфическая кожная железа, кинкажу специфическая кожная железа расположена на брюхе.
р ЧеРеп весьма различного строения. Лицевой отдел укороченный, уме-Но ю вытянутый пли очень сильно удлиненный. Мозговая коробка обыч-чен«аЗВИТа иильно. Крылоклиновидный канал отсутствует (за исклюем малой и большой панды).
•Зубная формула: i — с -у Клыки крупные или небольшие,
3 — 4 2
р ---- m----— =36 — 40.
3 — 4 2 — 3
предкоренные зубы заостренные, но
маленькие. Жевательная поверхность коренных зубов тупобугорчатая или с острыми гребнями, или промежуточного типа. ’
Ключица рудиментарная.
Ареал семейства состоит из двух частей. Большая его часть располагается в Америке, меньшая — в Юго-Восточной Азии. Американская часть ареала охватывает почти всю Северную Америку, за исключение^ ее северных районов, Центральную Америку и большую часть Южной кроме ее самых южных частей. В Азии ареал включает горные области Гималаев в пределах Непала и Сиккима и провинции Китая, Юньнань Сычуань и на север до границ Ганьсу. 1
Населяют обычно тропические и субтропические районы. Лишь одщ вид (енот-полоскун) заходит далеко на север, где даже впадает в зимний сон. Большинство видов лесные, некоторые хорошо лазают по деревьям н живут в дуплах. Кинкажу приспособлен к древесному образу жизни Активны ночью, а один вид (носуха) — днем. Как правило, держатся поодиночке, иногда образуют семейные группы. Носуха встречается стаями. Большинство всеядные: питаются корнями, побегами, семенами, плодами, насекомыми, ракообразными, земноводными, пресмыкающимися, птицами, и мелкими млекопитающими, некоторые плотоядные или растительноядные. Органы чувств развиты не очень хорошо. Для самок большинства видов характерен моноэстральный цикл. Длительность беременности 54—77 или 210—270 дней. В помете 1—6, чаще 2—4 детеныша.
Некоторые систематики разделяли семейство енотовых на 2—3 или даже 7 семейств, выделяя каждый род в самостоятельное семейство. Большую панду часто помещали в отдельное семейство или присоединяли к семейству медвежьих; одну или вместе с малой пандой.
В семействе 8 родов (15 видов):
какомицли — Bassariscus Coues, 1887 (2 вида);
енотов — Procyon Storr, 1780 (2 вида);
носух — Nasua Storr, 1780 (Звида);
горных носух — Nasuella Hollister, 1916 (1 вид);
кинкажу — Potos Е. Geoffroy St.-Hilaire et G. Cuvier, 1795 (1 вид); олинго— Bassaricyon J. Allen, 1876 (4 вида);
малых панд — Ailurus F. Cuvier, 1825 (1 вид);
больших панд — Aluropoda Milne-Edwards, 1870 (1 вид).
Некоторые виды семейства — объекты охоты и промысла. Енот акклиматизирован в некоторых странах Европы и в южных районах СССР.
Род какомицли
Genus Bassariscus Coues, 1887
Длина тела 30—47 см. Длина хвоста 31—53 см. Масса 0,8—1,1 Конечности относительно короткие. Тело удлиненное. Хвост очень длинный. Голова широкая, с укороченным лицевым отделом. Уши крупные, округлые или заостренные (рис. 59). Подошвы кисти и стопы голые или покрыты волосами. Когти невтяжные или полувтяжные. Окраска спинной стороны тела темно-желтая с черным или темно-бурым оттенком. Брюхо белое, беловатое или желтоватое. Глаза окружены черными или темно-бурыми кольцами. На хвосте, покрытом удлиненными волосам! > чередуются белые и черные или темно-желтые кольца.
-3 1 4 А.лп
Зубная формула: 1 с —— р — m —
Диплоидное число хромосом североамериканского какомицли 38.
Распространены на юге и юго-западе США, в Мексике и Центральной Америке.
Населяют различные ландшафты, но чаще встречаются в скалистых каменистых местах, обычно около воды. Очень проворные, хорошо азают по деревьям. Ведут ночной образ жизни. Всеядны. Кормом слу-,)faJ Растения, беспозвоночные, птицы и мелкие млекопитающие. У севе-н мериканского какомицли летом и осенью основную часть пищи со-авляют насекомые, а зимой и весной — грызуны. Самки североамери-ве СКого какомицли приносят потомство обычно в мае или июне, а д тРальноамериканского— в январе. В помете бывает от одного или ног-Х Л° пяти Детенышей (обычно три или четыре). Масса новорожден-ра ° Около 30 г. В возрасте 4,5 месяцев молодые животные достигают
Меров взрослых. В неволе жили до 8 лет. Хорошо приручаются.
В роде 2 вида:
североамериканский какомицли—В. astutus Lichtenstein, 1830 (Мексика; штаты США — Орегон, Калифорния, Невада, Юта, Колорадо, Канзас, Оклахома, Техас, Нью-Мексико, Аризона);
центральноамериканский какомицли — В. sumichrasti Saussure, I860 (крайний юг Мексики и вся Центральная Америка).
Некоторые исследователи (например, Уокер, 1964) выделяют центральноамериканского какомицли в особый род Jentinkia Trouessarf 1904).
Род енотов
Genus Procyon Storr, 1780
Длина тела 41—60 см. Длина хвоста 20—40 см. Масса 1,5—22 кг. Туловище относительно короткое, конечности невысокие (рис. 60). Хвост укороченный, покрыт удлиненными волосами. Голова широкая, с короткой, заостренной лицевой частью. Уши крупные, несколько заостренные, глаза большие. Пальцы на лапах глубоко разделенные, хватательные. Кисти и ступни покрыты укороченными волосами, резко отличающимися от волос остальных отделов конечностей. Мозоли на подошвах передних и задних лап безволосые. Когти невтяжные. Волосяной покров густой высокий, с хорошо развитым пухом. Окраска его на спине буровато-серая, на брюхе несколько более светлая. Морда грязно-белая; от носа по направлению ко лбу тянется узкая черная полоска, соединяющаяся с темными пятнами вокруг глаз и на щеках. На хвосте от 5 до 7 резко отграниченных черных или бурых колец, чередующихся с сероватыми или беловатыми кольцами. На нижней поверхности хвоста кольца выражены менее резко, чем на верхней стороне. В году одна линька.
Череп с обширной уплощенной мозговой коробкой и укороченным лицевым отделом. Имеются небольшие гребни. Костные слуховые пузыри крупные, округлые.
Зубная формула:
3 14 2
i — с — р — ш — = 40.
3 1*4 2
Коренные зубы с широкими жевательными поверхностями и острыми бугорками. Верхние клыки прямые, короткие. Нижние клыки изогнутые Хищнические зубы слабо выражены.
Диплоидное число хромосом енота-полоскуна 38.
Распространены в Северной Америке, кроме ее северных районов, в Центральной Америке и в северных и центральных районах Южной Америки.
Населяют лесные районы или заросли кустарников, обычно недалеко от воды. Убежищем служат дупла деревьев пли расщелины скал. Активны в основном ночью. Хорошо лазают по деревьям и хорошо плавают. В центральных и южных частях ареала активны круглогодично, а в северных районах на холодное время года впадают в зимний сон. Длительность зимнего сна в Канаде длится около 4 месяцев. Всеядные: поедают различные растения, беспозвоночных и мелких позвоночных. Период гона растянут с января по июнь, в зависимости от географического места. Продолжительность беременности примерно 63 дня. В помете 2—8 детенышей. Новорожденные покрыты густой шерстью, слепые. Прозревают на 18—23-й день. Лактация длится около 7 недель. К концу лета детеныши становятся самостоятельными. Половозрелость наступает в возрасте около года. В неволе жили до 13 лет.
Основные враги енота — волки, рыси, филины.
А
126
Рис. 60. Енот-полоскун — Р. lotor и его череп
Енот-полоскун — ценный пушной зверь, имеющий промысловое значение. Некоторую пользу приносит енот-полоскун, поедая вредных для лесного и сельского хозяйства насекомых.
В роде 2 вида:
енот-полоскун — Р. lotor Linnaeus, 1758 (от Южной Канады через всю территорию США и Мексику, включая Центральную Америку);
енот-ракоед — Р. cancrivorus Cuvier, 1798 (Коста-Рика, Панама, Эквадор, Гвиана, Венесуэла, Бразилия, Парагвай, север Аргентины).
Некоторые систематики принимают в пределах рода 6 (например, Андерсон и Джонс, 1964) или 7 (например. Холл и Келсон, 1959) видов; кроме двух вышеперечисленных еще Р. insularis Merriam, 1898; Р. п^У~ ^rdi Bangs, 1898; Р. pygniaeus Merriam, 1901; Р. minor Miller, 1911;
• Bloveralleni Nelson et Goldman, 1930.
Вероятно, все они не более чем подвиды.
Енот-полоскун акклиматизирован в некоторых странах Европы и в СССР. В СССР опыты с акклиматизацией енота-полоскуна проводились начиная с 1936 г. и повторялись несколько раз по 1962* г. Попытки акклиматизировать енота-полоскуна в Средней Азии, по-видимому, потерпели неудачу. В Восточном Закавказье " (Азербайджане), на южном склоне Главного Кавказского хребта акклиматизация енотов-полоскунов прошла успешно. Видимо, успешно завершилась и попытка акклиматизации енота-полоскуна в Белоруссии.
Род носух
Genus Nasua Storr, 1780
Длина тела 41—67 см. Длина хвоста 32—69 см. Масса около 11 кг.
пдвише удлиненное, конечности средней высоты. Хвост очень длинный /ис 61)- Передние конечности короче задних. Голова с сильно вытянули лицевым отделом. Передняя часть верхней губы и нос вытянуты в большой подвижный хоботок. Волосяной покров короткий, мягкий пли ” лее высокий и тогда несколько более грубый. Окраска спины рыжева-°°.бурая, рыжевато-буро-серая пли черная, брюхо черноватое или темно-
ое Морда, щеки и горло обычно беловатые, а лапы черноватые. 11а морде могут быть черные пятна. Хвост в чередующихся белых и черных кольцах. _
3 1 4 2
Зубная формула: i — с — р —— ш — = 40.
Диплоидное число хромосом у обыкновенной носухи 38.
Распространены в штатах США — юго-восток Аризоны, юго-запад Нью-Мексико, крайний юг Техаса; почти по всей Мексике, кроме п-ова Калифорния и центральных районов страны; по всей Центральной Америке, почти по всей Южной Америке.
Населяют лесные районы. Активны круглосуточно, особенно днем. Всеядны. Обычно держатся группами от 5—6 до 40 особей. Продолжительность беременности около 77 дней. Ранним летом или весной самки приносят в помете от 2 до 6 молодых.
Местное население иногда охотится за носухами и использует их мясо в пищу.
В роде, по-видимому, 3 вида:
обыкновенная носуха — N. nasua Linnaeus, 1766 (Бразилия, Боливия, Парагвай, Аргентина, Колумбия, Северная Аргентина, Перу, Эквадор, Венесуэла, Гайана, Суринам, Гвиана);
коати — N. narica Linnaeus, 1766 (США — юго-восток Аризоны, юго-запад Нью-Мексико, крайний юг Техаса; вся Мексика, кроме п-ова Калифорния, запада штата Сонора, юго-запада Чиуауа, Дуранго, Закате-кас, Сан-Луис-Потоси, Гуанохуато, востока Халиско, востока Мичоакан, Мехико; вся Центральная Америка);
носуха Нельсона — N. nelsoni Merriam, 1901 (о-в Косумель в Юкатанском проливе).
Видовая самостоятельность коати и особенно носухи Нельсона сомнительна.
Род горных носух
Genus Nasuella Hollister, 1916
т>
“ роде единственный вид: горная носуха — A. olivacea Gray, 1843.
внешним видом напоминает носуху, но хвост более короткий. Голо-относительно меньше и сильнее вытянута.
в Р еРеп горной носухи в отличие от черепа носухи меньших размеров, теп еДНем отделе сильнее сжат с боков, его костное небо заходит относи-
рНо дальше назад.
аспространена в Андах Венесуэлы, Колумбии и Эквадора.
Дология не изучена. По-видимому, всеядна.
5~-б71
19Q
Рис. 62. Кинкажу — Р. flavus и его череп
Род кинкажу
Genus Potos Е. Geoffroy St.-Hilaire et G. Cuvier, 1795
В роде единственный вид: кинкажу — Р. flavus Schreber, 1774.
Длина тела 41—57 см. Длина хвоста 39—55 см. Масса 1,4—2,7 Туловище удлинено. Передние конечности значительно короче задШ • Голова округлая с укороченным лицевым отделом (рис. 62). БолыШ округлые уши широко расставлены. Глаза большие. Хвост длинный, хв тательный, весь покрыт шерстью; пальцы с короткими изогнутыми рыми когтями.
Волосяной покров низкий, плотный и мягкий. Окраска его на сШ рыжевато-коричневая или бурая. У некоторых особей вдоль спины пР ходит срединная черная полоса. Брюхо желтоватое или буроватое. М " I
3 2
—- m— = 36.
3 2
примерно к юго-востоку от
темно-бурая или черноватая. Хвост одноцветный. На брюхе находится кожная специфическая железа.
3 1
Зубная формула: i — с —— р 3 *
Диплоидное число хромосом 38.
Распространен в Южной Мексике, оо° з Д-, кРоме центральных районов и севера п-ова Юкатан; по всей Центральной Америке, в Бразилии, Гайане, Венесуэле, Эквадоре, Перу, Колумбии.
Обитает в лесах. Ведет древесный образ жизни. Активен ночью. День „поводит в дуплах. Держится поодиночке или парами. Питается в основном плодами, в меньшей степени насекомыми и мелкими позвоночными. Весной или летом самка приносит одного — редко двух детенышей. Глаза у них открываются на 10-й день. Продолжительность жизни около 19 лет. Хорошо приручается.
Род олинго
Genus Bassaricyon J. Allen, 1876
Длина тела 35—47 см. Длина хвоста 40—48 см. Туловище удлиненное. Конечности относительно короткие, передние короче задних. Голова широкая, несколько уплощенная. Морда укороченная, заостренная (рис. 63). Уши средней величины, округлые. Глаза большие. Подошвы лап покрыты шерстью, пальцы с острыми искривленными когтями. Хвост покрыт удлиненными волосами, не хватательный. Волосяной покров низкий, очень плотный и мягкий. Спина желтовато-бурая или золотистая с примесью черных и сероватых тонов. Брюхо бледно-желтоватое. На хвосте иногда намечаются темноватые кольца.
„ 3 14 2
Зубная формула: I — с —— р — m — = 40.
Диплоидное число хромосом у пушистохвостого олинго 38.
Распространены в Центральной Америке, Эквадоре, Перу, Гайане, Венесуэле, Бразилии, Колумбии.
Обитают в джунглях от уровня моря до 2 тыс. м над его уровнем, экология изучена слабо. Активны ночью. Превосходно лазают, ведут в основном древесный образ жизни. Встречаются обычно небольшими группами. Питаются преимущественно плодами. В помете, вероятно, только один детеныш.
В Роде, по-видимому, 4 вида:
пушистохвостый олинго — В. gabbii J. Allen, 1876 (Панама, Коста-Ика> Никарагуа, Колумбия);
чирикуиский олинго — В. pauli Enders, 1936 (Чирикуи в Панаме); Еен°Линго Аллена — В. alleni Thomas, 1880 (Эквадор, Перу и, возможно, Ч^°ЛИнго Беддарда — В. beddardi Pocock, 1921 (Гайана и, возможно, здор и Бразилия).
бый Ногда (например, Холл и Келсон, 1959) из В. gabbii выделяют в осока рВИД олинго> В- lasius Harris 1932, обитающего в Картаго, Коста-Ри-Ват ОЗМожн°> что В. pauli — лишь подвид В. gabbii. Некоторые исследо-вида (напРимеР’ Гептнер и др., 1967) выделяют в роде один или
Рис 63. Пушистохвостый олинго — В. gabbii и его череп
Род малых панд
Genus Ailurus F. Cuvier, 1825
Род представлен единственным видом малая панда — A. fulge,'s F. Cuvier, 1825.
Длина тела 51—63. Длина хвоста 28—48 см. Масса 3—4,5 кг. ТуЛ°' вище удлиненное, конечности короткие (рис. 64). Пальцы с длинным®
Рис. 65. Большая панда — A. melanoleuca
полувтяжными когтями. Подошвы лап покрыты волосами. Хвост длинный, равномерно покрыт удлиненными волосами, не хватательный. Голова большая округлая с укороченным лицевым и крупным мозговым отделом. Глаза относительно маленькие. Уши широко расставлены, большие округлые. Волосяной покров высокий, плотный и мягкий. Окраска одноцветная, рыжеватая до темно-бурой, более темная по средней линии спины. Кольца вокруг глаз, морда, губы, щеки и края ушей белые. Задняя часть ушей и брюшная сторона тела темно-рыжевато-бурые или черные. На хвосте имеются нерезко выраженные черные кольца, конец хвоста черноватый.
3 13 2
Зубная формула: i — с — р — m — = 38.
3 14 2
Диплоидное число хромосом 36.
Распространена в Юньнане и Сычуане в Китае, в Северной Бирме, Сиккиме и Непале.
Населяет горные бамбуковые леса от 2 до 4 тыс. м над уровнем моря. Активна ночью. Наземное животное. Питается проростками бамбука, травой, корнями, плодами, изредка поедает яйца и самих птиц или мелких млекопитающих. Держатся парами или семейными группами. Продолжительность беременности от 90 до 150 дней. Самка приносит детенышей в дупле дерева или в расщелинах скал от 1 до 4 в помете (обычно 1 или 2). У детенышей глаза открываются на 21—30-й день. Молодые остаются с родителями до появления следующего помета.
Род больших панд
Genus Ailuropoda Milne-Edwards, 1870
В роде единственный вид,- большая панда — A. melanoleuca David, 1869.
Размеры наибольшие в семействе. Длина тела 120—180 см. Длина
Рис. 65. Продолжение. Череп большой панды
I
хвоста около 12 см. Масса 75—160 кг. Внешний облик очень своеобраз-ный, медведеподобный (рис. 65). Тело укороченное. Конечности невысокие стопоходящие. Значительная часть подошв лап покрыта волосами однако в пяточной области имеется обнаженная мозоль; такие же некрупные мозоли расположены у каждого пальца и, кроме того, позади пальцевых мозолей находятся вытянутые поперек подошвы обнаженные мозоли на передних и задних конечностях. Когти не длинные, слабо изогнутые, но сильные. Голова массивная, медведеобразная, с укороченным лицевым отделом. Уши крупные, округлые, стоячие. Глаза небольшие. Волосяной покров высокий, густой и мягкий. Глазные пятна, уши, конечности и широкая поперечная полоса, проходящая вокруг тела на уровне плеч, черные, остальная часть тела белая. Имеется подхвостовая специфическая кожная железа.
В черепе лицевой отдел укорочен, скуловые дуги мощные, широко расставлены.
3 14 2
Зубная формула: i с --3 ш = 40.
О 1 о
Диплоидное число хромосом 42.
Распространена в провинциях Китая: Сычуане, Ганьсу и Тибете; в настоящее время, по-видимому, встречается лишь в Западном Сычуане на территории, протянувшейся с севера на юг примерно на 280 км.
Населяет горные леса на высоте 1500—2000 м над уровнем моря. Обычно держатся поодиночке. Может лазать по деревьям, однако в основном ведет наземный образ жизни. Убежищем служат дупла деревьев, расщелины скал и пещеры. Питается преимущественно проростками и корнями бамбука, но поедает также и другие растения, а также и животных, в частности мелких грызунов. В помете 1—2 детеныша массой каждый около 2 кг.
Включена в «Красную книгу» как малочисленный вид, который, однако, в настоящее время не подвергается опасности исчезновения.
СЕМЕЙСТВО КУНЬИХ
FAMILIA MUSTELIDAE SWAINSON, 1835
Размеры мелкие (наименьшие в отряде) или средние. Длина тела от 11 (ласка) до 150 см (калан)1. Туловище сильно удлинено, реже относительно короткое. Конечности короткие, пальцеходящие или полусто-походящие, иногда стопоходящие пятипалые, с невтяжными когтями. Подошвы кисти и стопы покрыты волосами, или имеют голые подушки, или целиком голые. У некоторых видов между пальцами находится плавательная перепонка. Конечности калана сильно изменены: задние превратились в ласты, а на передних пальцы укорочены и соединены между собой. Голова обычно небольшая. Глаза средних размеров, иногда большие. Уши чаще маленькие, изредка крупные с округлыми вершинами. У некоторых видов (водных) уши сильно редуцированы, слуховые проходы могут замыкаться. Длина хвоста варьирует от очень короткого до длинного. Волосяной покров может быть густым, высоким и мягким, реже грубым и щетинистым. Окраска его изменяется от одноцветной, темно-бурой, желто-рыжей, черной или белой до двух- или многоцветной, состоящей из вариаций черного, рыжего, желтого и бурого цветов. На голове или туловище могут проходить полосы и пятна. В году бывает одна или две линьки. Сосков от 1 до 4 пар. Хорошо развиты анальные
1 Представителей семейства по размерам можно разделить на 3 группы: мелкие с длиной тела 11—56 см, средние — 57—102 и крупные— 103—150 см.
железы, выделяющие секрет с резким запахом. Некоторые виды способ-ibi выбрызгивать этот секрет на расстояние.
Череп невелик по размерам, с укороченным лицевым отделом и большой вздутой мозговой частью. У крупных фор^и на черепе сильно развиты гребни. Костные слуховые барабаны крупные, но уплощенные. Крылоклиновидного канала нет. Глазницы, как правило, большие.
1 2—4
— р------ m
1 2 — 4
3
Зубная формула: i -----—
2 — 3
1
= 28 — 38.
1 — 2
Второй верхний коренной зуб отсутствует. Число зубов значительно изменяется в разных родах, главным образом за счет предкоренных и коренных зубов. Только у калана уменьшение зубов до 32 произошло за счет редукции пары нижних резцов. Хищнические зубы сильно развиты. Жевательная поверхность коренных зубов может иметь острые или притупленные вершины. Os penis имеется.
Распространены почти всесветно, за исключением Фолклендских и о-вов Галапагосских, Вест-Индии и некоторых о-вов Берингова моря, Исландии, большинства Арктических, Австралии, островов южной части Тихого океана и Мадагаскара. В результате активного преследования со стороны человека в ряде мест они уменьшили свой ареал или даже совсем исчезли.
Среди представителей семейства имеются наземные формы, полудре-весные и скальные, полуводные и почти исключительно водные. Соответственно населяют самые различные ландшафты и.встречаются от тундры до пустыни и от подножья гор до альпийских лугов. Как правило, представители семейства ведут одиночный образ жизни, иногда держатся семьями и лишь в виде исключения образуют небольшие группы. По характеру питания большинство хищники, но используют, однако, и другие корма. У некоторых имеется тенденция к всеядности. Как правило, ведут оседлый образ жизни. Обычно убежищем служат собственные подземные норы. Некоторые виды в холодный период года впадают в зимний сон. Активны преимущественно ночью, отчасти в сумерки. Из органов чувств лучше всего развит слух, у некоторых также и обоняние. В году дают один помет, в котором бывает 1—10 детенышей.
Практическое значение представителей семейства велико, так как многие виды принадлежат к ценным пушным промысловым или разводятся в зверосовхозах: куницы, илька, выдры, соболь, калан, американская норка.
Выдры приносят некоторый вред, так как поедают рыбу (в абсолютных величинах, однако, очень немного).
Система семейства окончательно не установлена. В семействе, по-видимому, 5 подсемейств и 24 рода (64 вида):
Subfamilia Mustelinae Gill, 1872
куниц — Martes Pinel, 1792 (6 видов);
росомах—Gulo Storr, 1780 (1 вид);
ласок и хорьков — Mustela Linnaeus, 1758 (13 видов);
Grammogale Cabrera, 1940 (1 вид);
перевязок— Vormela Blasius, 1884 (1 вид);
тайр — Eira Hamilton — Smith, 1842 (1 вид);
гризонов — Galictis Bell, 1826 (2 вида);
патагонских ласок — Lyncodon Gervais, 1844 (1 вид) ;
африканских хорьков — Ictonyx Каир, 1835 (1 вид);
североафриканских ласок — Poecilictis Thomas et Hinton, 1920 (1 вид);
африканских ласок — Poecilogale Thomas, 1883 (1 вид).
Subfamilia Mellivorinae, Gill, 1872
медоедов — Mellivora Storr, 1780 (1 вид);
Subfamilia Alelinae Burmeister, 1850
барсуков — Meles Brisson, 1762 (1 вид);
теледу — Arctonyx F. Cuvier, 1825 (1 вид);
американских барсуков—Taxidea Waterhouse, 1839 (1 вид);
хорьковых барсуков — Melogale I. Geoffroy, 1834 (3 вида);
Subfamilia Mephitinae Gill, 1872
полосатых скунсов — Mephitis G. Cuvier, 1800 (2 вида);
пятнистых скунсов — Spilogale Gray, 1865 (2 вида), свиноносых скунсов — Conepatus Gray, 1867 (7 видов);
Subfamilia Lutrinae Baird, 1875
выдр — Lutra Brisson, 1762 (12 видов);
гигантских выдр — Pteronura Gray, 1837 (1 вид);
бескоготных выдр — Aonyx Lesson, 1827 (2 вида);
африканских выдр — Paraonyx Hinton, 1921 (1 вид);
каланов — Enhydra Fleming, 1822 (1 вид);
В СССР 8 родов: куниц, росомах, ласок и хорьков, перевязок, медоедов, барсуков, выдр, каланов,
SUBFAMILIA MUSTELINAE GILL, 1872
Род куниц
Genus Martes Pinel, 1792
Размеры мелкие. Длина тела обычно 40—50 см, максимум до 80 см (харза). Длина хвоста 34—42 см. Сложение стройное. Туловище вытянутое, но не очень сильно. Голова относительно крупная. Уши большие, округленные. Морда у основания широкая, клинообразная, сужающаяся к носу (рис. 66). Конечности относительно короткие, пальцеходящие. Лапы широкие. Волосяной покров обычно высокий, густой, мягкий и пушистый, иногда грубый. Хвост покрыт удлиненными волосами. У большинства видов окраска одноцветная, бурая или коричневатая. На горле и груди обычно светлое пятно. У харзы окраска яркая и пестрая с комбинацией белого, черного, желтого и бурого. В году две линьки — весенняя и осенняя. Сезонный диморфизм в волосяном покрове проявляется в значительном укорочении его летом и сильным изреживании. Паховых сосков две пары.
Череп крупный, относительно узкий и вытянутый. Только у старых особей развиваются крупные гребни. Мозговой отдел большой. Лицевая часть удлинена. Скуловые дуги выражены слабо и нешироко расставлены в стороны. Надглазничные отростки небольшие. Костные слуховые барабаны крупные.
3 14 1
Зубная формула: i — с — р — гл — = 38. 3 14 2
Первые предкоренные невелики по размерам и иногда выпадают. о Диплоидное число хромосом у лесной, американской и каменной куниц, ильки и соболя по 38, у харзы — 40.
Распространены по всей Европе, большей части Азии — на юг до Ма лой Азии, Сирии, Палестины, Ирана, Пакистана, севера п-ова Индостан— Кашмира, Пенджаба, Гималаев (кроме того, небольшой участок ареала имеется на юге Индостана), п-вов Индокитай и Малакка, о-вов 138
Рис. 66. Соболь — М. zibellina и его череп
Суматра, Банка, Явы и Калимантана; в северной половине Северной Америки; северная граница везде примерно совпадает с границей леса.
Населяют равнинные и горные леса. Хорошо приспособлены к лазанию по деревьям. Для соболя характерна приуроченность к северным хвойным лесам таежного типа, особенно темнохвойным насаждениям имеющим в своем составе сибирский кедр. В северных хвойных лесах встречаются илька и американская куница. Лесная куница населяет самые разнообразные типы лесов. Каменная куница встречается преимущественно в горах; она предпочитает безлесные склоны, ущелья, каменистые россыпи, а из лесов — предгорные, широколиственные. На равнинах каменная куница превратилась в синантропный вид; поселяется предпочтительно в каменных сооружениях человека. В центральной лесостепи выбирает для поселения старые высокоствольные дубравы, особенно с пересеченным рельефом. Типичные места обитания харзы — нагорная хвойная тайга.
Ведут оседлый образ жизни. У большинства видов участок обитания постоянен и занимается зверьком в течение всей жизни. Его размер изменяется в зависимости от местоположения в ареале, кормиости участка, сезона и характера кормности года. У соболя на Камчатке размеры индивидуального участка редко превышают 10 км2, на острове Большой Шантар он в среднем равен 13 км2, в Якутии — от 12 до 30 км2, в Восточном Саяне — от 4 до 10 км2, на Северном Урале — от 4 до 6 км2. У лесной куницы размеры участка закономерно уменьшаются с севера на юг. На Кольском п-ове площадь участка составляет 15—50 км2, на Северном Урале — около 6 км2, в Жигулях — примерно 4 км2, на Северо-Западном Кавказе — 32—72 га. Индивидуальный участок самца американской куницы в Канаде примерно 0,9 кв. мили, самки — 0,27 кв. мили. Значительно менее постоянны и больше по размерам участки обитания харзы. За сутки харза обычно проходит 10—20 км.
По характеру питания представители рода хищники, однако в их корме значительную роль играют беспозвоночные и растения. Основной объект питания соболя — лесные полевки и другие мелкие грызуны, поедает также бурундуков, белок, землероек, зайцев, ондатр, глухарей, рябчиков и мелких птиц, рыбу. В теплое время года он питается также насекомыми, ягодами (в частности, брусникой) и в больших количествах поедает «орешки» кедрового стланца. При неурожае последних в питании соболя значительно увеличивается значение теплокровных животных.
У лесной куницы идут в пищу также весьма разнообразные корма: в основном мышевидные грызуны (главным образом рыжие полевки), а также белки, птицы, насекомые, ягоды и орехи. Поедает также зайцев, птичьи яйца, амфибий, пресмыкающихся. Существует определенная за-= кономерность географической изменчивости в питании: возрастание всеядности с севера на юг. Сезонная смена кормов заключается в преимущественном поедании белки в снежный период, а также тетеревиных птиц и рябины. Насекомые, черника и мышевидные грызуны используются главным образом в бесснежный период.
Разнообразен набор кормов у американской куницы: мышевидные грызуны составляют основу питания — иногда до 66% от всей пищи, белки и летяги — до 10,2, кролики и пищухи — в разные годы от 3 до 40, птицы — в Британской Колумбии до 12,3; ягоды — летом до 17, насекомые (жуки, осы) —около 5%, поедается также падаль.
Для каменной куницы характерно значительное содержание в пище растительных кормов, в частности плодов самых разнообразных деревьев и кустарников. Охотится за мышевидными грызунами, различными птицами, ящерицами, поедает большое количество насекомых.
Илька питается в основном млекопитающими (около 80% всей пищи) ' белками, летягами, полевками, кроликами, землеройками, а также птИцами — 11,7%. рыбой — 1,7, насекомыми — 3,3 и плодами — 23,4%.
Основным кормом для харзы служат кабарга и телята других копытных зверей (массой до 10—12 кг), также мелкие промысловые звери, а весной и летом — рябчик. В летнее время она поедает также мелких птии, мышевидных грызунов, сеноставок, моллюсков, орехи, различные плоды и насекомых. Осенью харза использует в пищу рыбу, плоды и опехи. Харзы часто охотятся группами до пяти особей. При погоне они сменяют друг друга, а если жертва бежит по дуге, то одна-две харзы перерезают ей путь.
1 убежищем для представителей рода служат главным образом дупла и расшелины в скалах или каменистых россыпях. Держатся поодиночке, иногда семейными группами. Активны в разное время: лесная куница в сумерки и ночью, соболь как в сумерки, ночью, так и в течение дня, каменная куница — преимущественно в сумерки и ночью, но также и
дне^, харза, по-видимому, днем.
Представители рода — моногамы. Гон у соболей растянут с середины июля до середины августа. Течка у самок продолжается 1—3 дня. После перерыва на 7—10 и до 20 дней течка повторяется. У лесной и каменной
куницы гон приходится на конец июня и продолжается до начала августа с максимумом в конце июля — начале августа. У американской куницы гон в июле — августе. У ильки — в марте и апреле. У харзы спаривание бывает, по-видимому, с первых чисел июня до середины июля. Продолжительность беременности соболя 245—298 дней, лесной куницы (по различным данным) —от 236 до 274 дней, американской куницы — 220— 275 дней, каменной куницы — от 236 до 275 дней, ильки — 338—358 (в среднем 352) дней.
Характерна длительная диапауза в развитии оплодотворенной яйцеклетки. У соболя плацентация эмбрионов происходит лишь в конце февраля — начале марта, у лесной куницы — в марте, у американской
куницы в феврале или позднее. С периодом имплантации связаны явления так называемого «ложного гона» (некоторые авторы полагают, что «ложный гон» — атавистический признак). После плацентации развитие зародышей происходит быстро и оканчивается в течение 27-—28 дней. Соболь дает в помете 1—7 детенышей (наиболее часто 3—-4), лесная куница 2—8 (чаще 3—5), американская куница 1—4 (в среднем 2,6), каменная куница — 3—7, илька—1—4 (в среднем 2,7), харза — обычно 2—3, редко 4. Детеныши рождаются беспомощные, слепые, с закрытыми ушными отверстиями, покрытые редким пухом.
Длина тела новорожденных соболят 11-—12 см, масса около 30 г. Йховые отверстия открываются на 23—24-й день, прозревают на 30— 36-й день. Резцы прорезываются на 38-й день, смена молочных зубов на постоянные начинается в 3,5 месяца. Из гнезда начинают выходить в г!?'месячном возрасте. Животные корма начинают использовать на 50— о2-й день. Лактация продолжается примерно 2 месяца. Масса месячного соболенка около 260 г, двухмесячного (к июлю—августу)—примерно JJ г; в этом возрасте онн почти достигают размера взрослых. Половой зрелости в неволе достигают в возрасте 2 лет 3 месяцев. В природе большинство самок становятся беременными на втором году жизни. Масса новорожденного американской куницы около 28 г. Уши открываются на "Ц день, а глаза прозревают на 39-й день. Лактация длится около ньНед?ль- В 3,5 месяца детеныши достигают размеров взрослых живот-их. у большинства половая зрелость наступает в 2-летнем возрасте. Летеныши каменной куницы прозревают на 30—36-й день. Лактация Длится 40—45 дней.
На территориях совместного обитания соболя и лесной куницц (Урал, Приуралье, Зауралье) они способны давать помеси — так называемых кидасов. Самки кидасов плодовиты при обратном скрещивании как с соболем, так и с лесной куницей. Самцы кидасов при таком скрещивании бесплодны. Не установлена плодовитость кидасов между собой.
Врагов у представителей рода немного, во всяком случае они не оказывают какое-либо серьезное влияние на изменение численности популяции. Наиболее серьезным фактором, воздействующим на численность представителей рода, является промысел, а также деятельность человека, изменяющая характер ландшафта.
Большинство представителей рода — ценные пушные звери, которых и промышляют, и разводят на фермах. С 1960 по 1970 г. в СССР заготовляли ежегодно 125—195 тыс. шкурок соболя и 35—95 тыс. шкурок куниц. Харза приносит вред охотничье-промысловому хозяйству.
Система рода разными исследователями представляется по-разному. По-видимому, в роде можно выделить 6 видов (Гептнер, 1967):
соболь — М. zibellina Linnaeus, 1758 (Азия — Монгольский Алтай; Северо-Восточный Китай; возможно, Корейский п-ов; Япония — о-ва Хоккайдо, Хонсю, Сикоку, Кюсю, и Цусима; в СССР в историческое время ареал занимал почти всю территорию Сибири, на север ареал шел до границы леса, на восток до побережья Тихого океана, на юг от Тихого океана до Южного Алтая, выходил за государственную границу, от Южного Алтая тянулся по зоне лесостепи на север и запад через Новосибирск, Ишим, Курган, Челябинск. В европейской части СССР северная граница ареала, видимо, совпадала с границей леса, южная шла примерно от 55—56° с. ш. на Урале к верховьям Уфы, устью Белой, верховью Вятки и на область Великого Устюга, затем поворачивала на запад в Южную Карелию в область Онежского озера; кроме того, изолированный участок ареала, возможно, располагался в Литве и Белоруссии. В XVI—XVII вв. в результате неумеренного промысла соболь исчез на значительных территориях, особенно по периферии ареала. К XX в. он сохранился на немногих небольших участках, изолированных один от другого, почти полностью исчез из европейской части страны. После Октябрьской революции были приняты меры охраны соболя: ограничение сроков добывания и запрет промысла с 1935 г. на 5 лет, организация заказников и заповедников, реакклиматизацпя — с 1949 по 1970 г. было расселено около 19 тыс. соболей. Все это благоприятно сказалось на увеличении численности соболя и на восстановлении его ареала. К настоящему времени западная граница ареала начинается примерно от 65°30' с. ш. на Урале и идет на юг по его западным склонам, захватывая Печоро-Илычский заповедник, до истоков Яйвы, откуда поворачивает на восток, проходит севернее г. Серова и направляется к верховьям Носки, впадающей в Иртыш ниже Тобольска; северная граница, — по-видимому, на большей своей части совпадает с границей леса, иногда отступая от него южнее — например, в междуречье Лены и Колымы, или, наоборот, поднимаясь к северу—по лесам речных долин, например по Анадыри и по кедровому стланику в Коряцкой земле; восточная граница проходит по Тихоокеанскому побережью, включая о-ва Шантарские и Сахалин, а из Курильских — Итуруп, Кунашири и Парамушир; южная граница по сравнению с южной границей восстановленного ареала, указанной выше, сдвинута несколько к северу. В очерченных границах ареал остается, однако, еще разбитым на участки, из которых самые крупные: урало-обский, занимающий Урал и левобережье Оби, енисейский — правобережье Енисея к северу от 57° с. ш., саянский- Саяны и Тува, ленский — территория между Леной и Вилюем, витимо-забайкальекий
пространство по верхней Лене, Витиму и вдоль восточного побережья райкала, алдано-зейский — территория между Алданом и нижним Аму-пом, камчатский — весь полуостров; более мелкие участки — сахалинский, уссурийский, южнозабайкальский, алтайский; много и других еще более мелких очагов);
лесная куница — М. rhartes Linnaeus, 1758 (Европа — на север до Гранины леса на крайнем севере Скандинавского п-ова, на запад, включая Днглию и Ирландию, на юг до Северной Испании — Пиренеев включительно и далее по Средиземноморскому побережью, включая о-ва Балеарские, Сардинию и Сицилию и Северную Грецию; Азия — Малая Азия, Северный Ирак, в Иране ареал образует 2 мыса: по горной системе Эльбурс и по Загросским горам. В СССР северная граница ареала начинается на западе от государственной границы на Кольском п-ове на широте Колы и идет на юго-восток по границе леса до юго-восточного побережья Белого моря, обходит Белое море по берегу и направляется на восток примерно по Полярному Кругу до Оби, далее сворачивает на юго-восток примерно к точке пересечения 63° с. ш. с 80-й параллелью и несколько восточнее от этой параллели круто сворачивает на юг, превращаясь в восточную границу ареала. Несколько южнее места пересечения 80-й параллелью Оби граница ареала сворачивает на юго-запад, становясь южной границей, выходит на Иртыш у устья Тары и направляется на запад севернее оз. Салтаим, проходит немного севернее 56° с. ш. и выходит на Урал через Сысерть к Нязепет-ровску, далее следует на юг, охватывает Урал с юга, захватывая долину р. Урал, и доходит до Уральска. Несколько севернее она переходит на Волгу, примерно от Саратова идет на запад до пересечения Хопром 52° с. ш., далее направляется по Хопру до Новохоперска, к Павловску на Дону, к устью Оскола на Донце и Изюму, Днепропетровску, Балте, переходит Днестр и охватывает с юга лесные области Молдавии в южной части этой республики; западная граница ареала совпадает с государственной границей нашей страны от Баренцева почти до Черного моря. Изолированная в пределах СССР от европейско-сибирской части ареала кавказская часть соединяется с ней на юго-западе вне пределов нашей страны; кавказский участок занимает леса Главного Кавказского хребта, доходя на восток до внутренних лесистых областей Дагестана, на западе до бассейна р. Пшехи, левого притока Белой, и на юг до государственной границы);
американская куница-—М. americana Turton, 1806 (северная часть Северной Америки — Аляска, кроме юго-западной части с п-овом Аляска и побережья моря Бофорта; провинции Канады — Юкон, Маккензи, кроме северо-востока, Британская Колумбия с архипелагом Александры, о-вами Королевы Шарлотты и о-вом Ванкувер, северная половина и узкая полоса юго-запада Альберта, Манитоба, Онтарио, Квебек, кроме северо-запада, Ньюфаундленд с о-вом Ньюфаундленд, Нью-Брансуик, Новая Шотландия, о-в Принца Эдуарда; штаты США —Мэн, Вермонт, Нью-Гэмпшир, запад Массачусетса, Нью-Йорк, север Пенсильвании, восток Огайо, Мичиган, крайний северо-восток Иллинойса, Висконсин, кроме Юго-запада, северная половина Миннесоты, крайний северо-восток Неверной Дакоты, западная треть Монтаны, северная половина и юго-восток Айдахо, северо-запад и юг Вайоминга, северо-восток Юты, западная половина Колорадо, север центральных районов Нью-Мехико, западная половина, север и юго-восток Вашингтона, западная треть и северо-восток Орегона, северная половина Калифорнии);
каменная куница — М. foina Erxleben, 1777 (Европа — на юг до Средиземного моря, включая о-ва Родос и Крит, на запад до Атлантическо-Го побережья, но в Англии и Ирландии отсутствует, на север до Балтий-
скоро моря, включая Данию; Азия — Малая Азия, Сирия, Палестина, горы Ирака, север и запад Ирана, Афганистан, Белуджистан в Пакистане, северный Кашмир, северный Пенджаб, Непал, Сикким, в Китае Тибет, возможно, Хэбэй, Шаньси и юг Северо-Восточного Китая; от центральноазиатской области ареал узким мысом вдается на северо-восток охватывая горы Кашгарии, запад Джунгарии, северо-запад и север' МНР. В СССР северная граница ареала проходит от государственной границы по Балтийскому побережью и берегу Финского залива примерно до района, расположенного к западу от Ленинграда, где поворачивает на юг, обходит Ленинградскую и Псковскую области с запада и проходит по центральным районам Белоруссии. Поворачивает на восток через Южную Белоруссию, Полесье и юго-запад Брянской области, далее направляется на Новосиль восточнее Орла и к Боброву юго-восточнее Воронежа, поворачивает на юг и затем на запад, превращается в южную границу ареала, проходит севернее Луганска, спускается на Донецк, следует к Мелитополю, выходит к Перекопу и дальше проходит по берегу Черного моря. В Крыму ареал образует изолированный участок в горной части полуострова. Кавказский участок изолирован в СССР от остального ареала, соединяясь с ним за пределами нашей страны,— его северная граница проходит от Кубани до Махачкалы, а южная совпадает почти на всем протяжении с государственной границей; отдельный участок имеется в Талыше. В Средней Азии и Южной Сибири участки ареала охватывают Копетдаг, Большой Балхан, горные хребты Памиро-Алтайской и Тянь-Шаньской систем, Тарбагатай и Саур, Кал-бинский Алтай и Южный Алтай);
илька — М. pennanti Erxleben, 1777 (в Северной Америке северная граница ареала проходит по побережью залива Св. Лаврентия провинции Квебек в районе г. Наташкуана и идет на север, захватывая юго-запад провинции Ньюфаундленд, и затем по центральным районам Квебека на запад к южной оконечности Гудзонова залива, проходит по южному берегу этого залива, северной границе провинции Манитоба, южным районам Маккензи и, пересекая северо-западный угол Британской Колумбии, выходит к Тихому океану, южная граница ареала начинается на Атлантическом побережье в южной части штата Мэн, идет на юго-запад через юго-восток Нью-Гэмпшира, середину Массачусетса и Коннектикута, северо-запад Нью-Джерси, запад Виргинии, северо-запад Северной Каролины, северо-восток Теннесси, поворачивает на северо-запад и проходит по восточной части Кентукки, северу Иллинойс, северо-востоку Иова, юго-западу Миннесота, крайнему северо-востоку Северной Дакоты, пересекает государственную границу Канады, проходит по южной трети провинции Саскачеван и затем круто сворачивает на юг, проходя через юго-восток Альберта, восточную треть штата Монтана, США, крайний северо-запад Вайоминга и по границе между этим штатом и Айдахо и Юта спускается до центральных районов Юты, затем резко сворачивает на север к южным районам Айдахо и вдоль границы этого штата с Орегоном и Вашингтоном идет на север до границы с Канадой, вновь резко поворачивает на юг, пересекая центральные районы штатов Вашингтон и Орегон, крайний северо-восток Калифор-нии и по границе этого штата с Невадой спускается языком до центральных районов Калифорнии, вновь поднимается к северу и несколько севернее Сан-Франциско выходит к Тихому океану; восточная и западная границы ареала проходят соответственно по Тихоокеанскому и Атлантическому побережьям);
харза — М. flavigula Boddaert, 1785 (Азия — восток и север Северо-Восточного Китая, Восточный Китай, на западе включая Ганьсу и Шень-си, на юге Сычуань и Юньнань, п-ова Индокитай и Малакка, о-ва Хай-
лань, Тайвань, Суматра, Банка, Ява, Калимантан, верхнюю Бирму, горные области п-ова Индостан: Ассам, Бутан, Сикким, Непал, на западе включая Джамму и Кашмир, изолированный участок расположен на западе южной оконечности полуострова; в СССР — Уссурийский край и область среднего и частью нижнего Амура: северная граница ареала на западе начинается на Амуре около 53°30' с. ш. и направляется к среднему течению Зеи, затем к среднему течению Бурен, на верховья Бид-жана и Биры, затем следует на средние течения левых притоков Амура __Кура и Урми и выходит к Амуру у устья Хунгари, пересекает Амур
и выходит к Тихому океану примерно на 48° с. ш., к югу от обозначенной линии ареала выходит к государственной границе).
Ряд исследователей (например, Эллермэн и Моррисон-Скотт, 1966) считают, что подвид соболя из Японии и с Корейского п-ова заслуживает выделения из М. zibellina в особый вид М. melampus Wagner, 1841; многие (например, Эллермэн и Моррисон-Скотт, 1966) выделяют из М. flavigula южноиндийскую харзу — М. gwatkinsi Horsfield, 1851; Хаг-мейер (Hagmeier, 1961) объединил соболя, лесную и американскую куниц в один вид, что неправильно.
Род росомах
Genus Gulo Storr, 1780
В роде единственный вид: росомаха — G. gulo Linnaeus, 1758.
Размеры крупные. Длина тела 70—105 см. Длина хвоста 18—23 см. Высота в плечах 35—45 см. Внешний облик своеобразен, общий вид несколько напоминает медведя. Сложение тяжелое. Туловище укороченное, широкое. Голова относительно небольшая, несколько вытянутая. Уши маленькие, широко расставленные (рис. 67). Глаза небольшие. Конечности умеренной высоты, полустопоходящие. Лапы относительно короткие, очень широкие. Когти крупные. Хвост короткий, по длине примерно равен голове и покрыт удлиненными густыми волосами. Зимний волосяной покров очень высокий, густой, лохматый и довольно грубый. Летний мех значительно более редкий и низкий. В году 2 линьки — осенняя и весенняя. Общий тон окраски бурый, у некоторых особей рыжевато-бурый. Вдоль спины проходит широкое пятно темно-коричневого или шоколадного цвета; оно занимает верх шеи, холку и всю среднюю часть спины до крестца. От боков шеи через лопатки по бокам туловища, по огузку, охватывая его, тянутся две светло-коричневые полосы. На середине шеи и на груди между передними лапами обычно расположены белые пятна неправильных очертаний. Концы лап почти черные. На брюхе перед половым отверстием у самцов и у самок имеется железистый участок, который, по-видимому, служит для маркировки территории. Кроме того, имеются парные анальные специфические кожные железы, секрет которых росомаха может выбрызгивать на расстояние до 3 м. Сосков 2 пары.
Череп крупный, широкий, с укороченным лицевым отделом и хорошо выраженными гребнями. Мозговая коробка относительно невелика. Глазницы небольшие. Скуловые дуги мощно развитые, сильно расставленные в стороны. Надглазничные отростки развиты слабо. Костные слуховые пузыри маленькие, уплощенные. Костное небо очень широкое.
3 14 1
Зубная формула: 1 — с — р m — =38.
Зубы мощные. Хищнические зубы большие, с острыми гранями. Первые предкоренные зубы иногда могут отсутствовать.
Рис. 67. Росомаха — G. gulo и ее череп
Диплоидное число хромосом 42.
Распространена в Северной Америке на север до побережья материка (кроме Арктического побережья Аляски), захватывая и Арктический архипелаг, кроме о-вов Банкс, Принс-Патрик, Свердруп и некоторых других в северо-восточной части архипелага; южная граница проходит по штатам Нью-Брунсуик, Мэн, Вермонт, Нью-Йорк, Пенсильвания, северу Огайо, югу Индианы, северу Миннесоты, Северная Дакота, далее к западу тремя мысами вдается к югу: один от Северной Дакоты и Монтаны до западной Небраски, второй от Монтаны и Айдахо до Юты и Колорадо, третий проходит по западным районам Вашингтона, Орегона и северо-западной Калифорнии и по горам Сьерра-Невада примерно до 35° с. ш.; в Европе — Скандинавский п-ов и Финляндия; в Азии — север МНР, Северо-Восточный Китай. В СССР ареал занимает большую часть территории страны — северная граница идет по арктическому побережью, включая о-ва Большой Ляховский и Новая Сибирь, восточная граница проходит по Тихоокеанскому побережью, включая Сахалин и Шантарские острова, южная граница на Дальнем Востоке проходит несколько севернее государственной границы и немного западнее нижней Бурей и Зеи уходит за пределы страны; от Зеи до Южного Алтая граница идет южнее территории СССР, кроме степей Забайкалья; да-' лее граница проходит западнее Кузнецкого Алатау и идет к Уралу на Ирбит — Егоршино—Пермь, проходит немного севернее Кирова, по 60° с. ш. пересекает Вологодскую область и в области Ленинграда выходит к Финскому заливу.
Населяет хвойную тайгу и лесотундру, встречается также в открытой тундре, хотя здесь росомаха немногочисленна. Иногда заходит в полосу смешанных лесов и даже в западносибирскую лесостепь. Обитает как в равнинных лесах, так и в горных. Логово устраивает в расщелинах скал или под вывернутыми корнями деревьев. В зимнее время питается в основном копытными (преимущественно падалью). На живых копытных росомаха нападает, если они больны или ранены1 или если это молодняк. Главным образом охотится за северными оленями и лосями. При малочисленности копытных зверей, а также в летний период основными кормами становятся заяц-беляк, тетеревиные птицы, лесные полевки. Весной и летом росомаха в большом количестве поедает птиц и яйца, личинок ос, а также ягоды: морошку, малину, чернику, бруснику, клюкву. В «демпинговые» годы большое значение приобретает питание мышевидными грызунами. Росомаха всегда предпочитает падаль свежему мясу.
Участок обитания у росомахи обширный. Его площадь в малокормных местах достигает 1600 км2. В более кормных местах, например в Финской Лапландии, охотничьи участки у самок были около 300 км2, в Британской Колумбии— 124—132 км2. У трупа крупного копытного собирается обычно несколько росомах, которые живут здесь несколько дней. Активны преимущественно ночью. Период гона, по-видимому, приходится на июнь — сентябрь. Детеныши рождаются, в зависимости от географического местоположения, с конца февраля по начало мая. Таким образом, длительность беременности составляет около 8—9 месяцев. В развитии яйцеклетки имеет место диапауза. Самка приносит приплод раз в два года. Обычно в помете бывает 2—3, редко 4 детеныша. Длительность лактации около 3 месяцев. Глаза у детенышей открываются на 5-й неделе. Половозрелость наступает, по-видимому, на 3-м году жизни. Основной враг росомахи, кроме человека, волк.
Росомаха приносит известный вред, поедая молодняк копытных и Похищая добычу и приманки из самоловов и капканов промысловых охотников. Поэтому промысел росомахи разрешен в течение круглого
Рис. 68 Ласка — М. nivalis (/I) и ее череп (5), американский хорек — М. nigripes (В) и его череп (Г)
года. Шкуры росомахи, хотя и красивые, ценятся невысоко из-за грубого волоса.
Род ласок и хорьков
Genus Mustela Linnaeus, 1758
Размеры мелкие. Длина тела от 11 см у ласки до 56 см у степного хорька. Туловище тонкое, вытянутое, у некоторых видов очень сильно. Конечности короткие, пальцеходящие. Лапы довольно широкие. Хвост различной длины; достигает половины длины тела. Покрыт удлиненными волосами. Голова небольшая, несколько уплощенная, узкая, тупомордая (рис. 68). Небольшие уши широко расставлены, вершины их за-кРуглены. Глаза относительно крупные. Волосяной покров густой, невысокий, у некоторых видов плотно прилегающий к телу, у других пушистый. Окраска его одноцветная, черно-желтая с темным брюхом или Двухцветная со светлым брюхом. Иногда на голове имеется узор темного или светлого цвета в виде маски. На груди и на нижней стороне бедер могут быть черные пятна. Обычно на холодный период года волосяной Покров становится гуще, выше и бледнее. У некоторых видов (ласка, горностай) на севере имеет место резко выраженный сезонный диморфизм: мех на зиму белеет. В году бывает 2 линьки: весной и осенью.
Сосков на брюхе и в пахах 3—5 пар. Хорошо развиты специфические анальные железы, продуцирующие пахучий секрет, который некоторые виды (например, лесной хорек) могут выбрызгивать на расстояние.
Череп у большинства видов небольших размеров, иногда маленький довольно узкий, уплощенный, со слабо развитыми гребнями. Мозговой отдел увеличен, вытянут, лицевой укорочен. Верхний профиль черепа прямой. Скуловые дуги тонкие, слабо расставленные в стороны. Над-глазничные отростки небольшие. Межглазничное и заглазничное сужения обычно слабые. Иногда череп несколько укорочен, мозговой отдел относительно уменьшен, а лицевой увеличен, скуловые дуги сильно развиты и шире расставлены в сторону, заглазничное сужение выражено сильно. Костные слуховые барабаны у всех сильно вздуты и имеют удлиненно-бобовидную форму.
„ 3 13 1
Зубная формула: i с — р — m — = 34.
Диплоидное число хромосом у ласки 38, горностая и длиннохвостой ласки — по 42, солонгоя — 44, американской норки — 30, лесного хорька — 40.
Распространены очень широко: по всей Европе, кроме Исландии, арктических островов и островов Средиземного моря; в Азии на север встречаются до побережья материка, включая Новосибирские о-ва, на юге ареал захватывает Палестину, Сирию, северную половину Ирака, северо-запад Ирана, мысом вдается на юго-восток по горам системы Загроса, но не доходит до берегов Персидского залива, включает Северный и Северо-Восточный Иран, систему Гиндукуша, Кашмир, Гималаи, Бирму, п-ова Индокитай и Малакку, о-ва Суматру, Яву и Калимантан, на востоке Азии ареал доходит до Тихого океана, включая о-ва Карагин-ский, часть Курильских, Шантарские, Сахалин, Японские, Рюкю, Тайвань и Хайнань; на Командорских о-вах представителей рода нет; в Африке ареал охватывает Марокко, горы Атласа в Алжире, Тунисе и АРЕ; по всей Северной Америке, включая северные и северо-восточные районы Гренландии; по северной части Южной Америки, охватывая с юга Венесуэлу, с юга и юго-запада Колумбию, с запада Перу, с запада и юга Боливийские Анды и на широте озера Титикака вплоть до Тихого океана; в ареал входят все о-ва Атлантического побережья Северной Америки и все о-ва Тихоокеанского побережья, включая Кадьяк, но исключая Вест-Индию и острова Берингова моря, кроме Унимана.
Населяют самые разнообразные биотопы и ландшафты всех зон от тундры до степей, пустынь и тропических лесов, поднимаются высоко в горы до альпийского пояса включительно. Ласка встречается в самых разнообразных биотопах и ландшафтах: в тундре, тайге, но наиболее обычна в лесостепи и степи; часто селится в полях, скирдах, хозяйственных постройках, по окраинам или даже в самих городах. Горностаи более многочислен в лесостепи, зоне леса и лесотундре; везде предпочитает держаться вблизи водоемов. Солонгой преимущественно обитает в горных местах, предпочитая открытые ландшафты. Колонок — житель горной и равнинной тайги; долин рек, каменистых россыпей, в глубь таи-ги проникает следом за распространением сельскохозяйственных угодий; обитает и в лесостепи — по березовым и осиновым колкам и в за рослях тростника по берегам рек и озер. Длиннохвостая ласка предпо читает степи; осиновое редколесье и поймы рек. Норки встречаются по берегам небольших рек. Лесной хорек — обитатель разреженных лесов и лесов с чередующимися полями и лугами, часто держится по опушкам леса, встречается в поселках, по окраинам городов. Степной хорек насе 150
пяет открытые ландшафты: луга, степи, полупустыни, американский хорек— сухие прерии.
1 Все представители рода — наземные хищники, однако большинство может лазать по деревьям. Все они могут плавать, а некоторые приспособлены к полуводному. образу жизни. Очень быстры в движениях, ловко передвигаются в кучах валежника, среди камней, в норах грызунов. Убежищем служат в основном норы грызунов. Иногда поселяются среди камней, в кучах валежника. Активны преимущественно в сумерки и ночью, хотя у большинства видов суточная активность выражена недостаточно четко и их можно встретить в течение любого времени суток.
По характеру питания все представители рода резко выраженные хишники. Ласка питается в основном полевками, мышами, хомячками, песчанками, иногда землеройками, рыбой; известны случаи нападения на крупную добычу: глухарей и зайцев. В питании горностая преобладают грызуны, причем в отличие от ласки ими могут быть довольно крупные: водяная полевка и обыкновенный хомяк, пищухи; второстепенное значение имеют птицы; еще меньшую роль играют земноводные, пресмыкающиеся и насекомые. Колонок поедает в основном грызунов мелких и средних размеров, а также птиц и в некоторых районах рыб, пресмыкающихся и земноводных; из растительных объектов — кедровый «орешек». Длиннохвостая ласка питается в основном мелкими млекопитающими: полевками, бурундуками, гоферами, сусликами, пищухами, луговыми собачками; в значительно меньшей степени птицами и насекомыми. Пищу норок составляют прибрежные, водные и лесные животные: полевки, земноводные, рыбы, раки, птицы, насекомые. Основными объектами питания лесного хорька служат мышевидные грызуны, а из них серые полевки; в больших количествах также земноводные, в меньшей степени — птицы. Степной хорек охотится за более крупной добычей: сусликами, хомяками, пищухами, но в летнее время и за беспозвоночными. Американский хорек питается почти исключительно луговыми собачками, а также сусликами, кроликами, полевками.
Все представители рода — оседлые формы. Размеры индивидуальных участков изменяются в зависимости от географического местоположения, сезона и кормности места и года.
Горностаи и хорьки — моногамы. У ласки время гона в разных местах ареала очень изменчиво; беременность, по данным разных авторов, от 35—37 дней до 7—8 недель, в помете от 3 до 10 (обычно 4—7) детенышей; детеныши рождаются слепыми, беспомощными, покрытыми редким белесым пушком, долго остаются при матери. Гон горностая длится с середины февраля по первую половину июня (Северный Казахстан); беременность — 9—10 месяцев с латентным периодом и 6 недель без него; детенышей в помете от 2 до 18 (обычно 4—8); новорожденные слепые, голые, беспомощные; самки становятся половозрелыми в 2—3 месяца, а самцы значительно позже. У солонгоя гон происходит в Казахстане со второй половины января до начала апреля, беременность — 30 41 день; детенышей в помете от 1 до 8; продолжительность лактации 34—41 день.
Сроки гона колонка в разных местах ареала различны: в Западной Сибири с начала февраля по март включительно, в Приморье с первой тРети марта до конца апреля, в зверосовхозе под Москвой с 25 апреля До 15 мая; беременность длится 38—41 день (по другим данным, 35— 42 дня), детенышей в помете 4—-10; детеныши рождаются слепые и Почти голые, прозревают на 38—39-й день, продолжительность лактации около 2 месяцев. У длиннохвостой ласки гон проходит в июле — августе; переменность длится 220—337 (в среднем 279) дней; латентный период в Развитии яйцеклетки — примерно .7 месяцев; в помете 3—9 (в сред
нем 6) детенышей; масса новорожденного около 3,1 г, глаза открываются на 31—37-й день; лактация — около 3,5 недель.
Течка европейской норки в Московском зоопарке происходила в конце апреля; беременность длится 42—46 дней, число детенышей в помете 3—7, масса новорожденного примерно 6,5 г; прозревают на 30—31-й день-самка лактирует 2—2,5 месяца; в возрасте 70—84 дня становятся самостоятельными; половозрелость наступает примерно в годовалом возрасте. У американской норки течка в СССР протекает с конца февраля до начала апреля; беременность — 36—80 дней, из них эмбриональная диапауза занимает от 1 до 7 недель; число детенышей в помете от 1 до 9 (в исключительных случаях до 16), масса новорожденного 6—11 г; прозревают на 30—31-й день; лактация длится 2—2,5 месяца; продолжительность жизни в неволе — 7—10 лет. Гон лесного хорька начинается со второй половины июня, беременность примерно 42 дня, детенышей в помете от 2 до 12 (обычно 4—6); масса новорожденного примерно 7 г; прозревают на 34—36-й день; половозрелости достигают в возрасте около года, а полного развития — к двум годам. У степного хорька гон идет с февраля по март, в Забайкалье — до конца мая, беременность длится 36—43 дня, латентный период 7—8 дней, детенышей в помете 3—18, масса новорожденного примерно 4,5—10 г; прозревают на 28—34-й день; половозрелости достигают в 10 месяцев, а полного развития — в 2 года.
Система рода окончательно не установлена. Можно выделить 13 видов (Гептнер, 1967):
ласка — 714. nivalis Linnaeus, 1766 (вся Европа, кроме Ирландии и, по-видимому, Кипра; Азия — Малая Азия, Северный Ирак, Северный и Юго-Восточный Иран, Северный Афганистан, Кашгария, Джунгария, МНР, Северо-Восточный Китай, Корейский п-ов, Япония; возможно, Северный Вьетнам; Африка — Алжир, Марокко; возможно, Тунис; АРЕ; Северная Америка — северная Гранина ареала проходит по Арктическому побережью материка, но к западу от Гудзонова залива отходит от берега и идет примерно по линии от Честерфилда на Бетерс-Инлет на северном побережье, к югу от о-ва Виктория и далее на восток вновь следует по побережью, восточная граница идет по р. Св. Лаврентия к восточной оконечности оз. Гурон, южному берегу оз. Эри, направляется на юго-восток, пересекает Пенсильванию и выходит к Атлантическому океану; южная граница следует от Джуно на Тихоокеанском побережье через Британскую Колумбию, юго-запад Альберты, север Монтаны, в ареал входят Северная Дакота, Южная Дакота, кроме запада, Небраска, Айова, северные части Иллинойса и Индианы, Огайо, почти вся Пенсильвания; Западная Вирджиния, отсюда небольшой мыс ареала выдвинут на юго-запад, достигая северо-западного угла Южной Каролины; в СССР — вся территория страны, но отсутствует на арктических островах, кроме Большого Ляховского. Нет ласки на Командорских островах, но водится, по-видимому, на Шантарских, обитает на Сахалине, из Курильских — на Кунашире);
горностай — М. erminea Linnaeus, 1758 (вся Европа на юг до Пиренеев и Альп включительно, в Югославии — лишь на севере, отсутствует в Болгарии, Албании, Греции и Турции; Азия — восток Афганистана, Кашмир и область к северо-востоку от Пешавара, МНР, Северо-Восточный Китай, возможно, Северная Корея, Япония — Хоккайдо и Хонсю, Северная Америка — вся северная половина материка, включая Арктический Архипелаг, северную и восточную Гренландию, южная граница проходит от Средней Калифорнии через Центральную Неваду, юг ЮтЫ, Колорадо, север Нью-Мексико, затем резко поднимается к юго-востоку Альберты и Южному Саскачевану, далее идет к Северной Дакоте, Северной Айове и через южную оконечность Мичигана и Эри выходит к
Атлантическому океану к Чесапикскому заливу; в СССР — большая |АСТЬ территории страны: северная граница проходит по Арктическому Побережью, включая Новосибирские о-ва; восточная граница следует по Тихоокеанскому побережью, включая Сахалин, Шантарские о-ва, а из Курильских — Парамушир, Шумшу и Кунашир; западная граница идет по западной государственной границе от Баренцева до Черного моря; южная граница протянулась от Дуная к устью Дона (нет горностая только в Крыму) и по узкой полосе степей, прилежащих к Черному и Азовскому морям, далее следует к устью Кумы и дельте Волги; обитает зверек на изолированном участке на Кавказе; граница идет от устья Волги вдоль побережья Каспия до устьев Урала и Эльбы, северо-западного побережья Аральского моря, проходит по его северному берегу и через низовья Сырдарьи до Кзыл-Орды, поднимается к северу через южные предгорья Улутау, идет примерно севернее средней части Балхаша и поворачивает на юг к западной части Алакульской впадины, оставляя ее в пределах ареала; в Средней Азии горностай распространен между Алаколем и Джунгарским Алатау и между последним и Заилий-ским Алатау, на Тянь-Шане, в Фергане и Памиро-Алтайской системе; от Алакульской впадины и до устья Уссури ареал уходит за государственную границу СССР, в Уссурийском крае горностай отсутствует по Уссури и нижнему течению ее притоков и по приморской полосе на север до района немного севернее устья Самарги);
солонгой — М. altaica Pallas, 1811 (Азия — Кашгария и Джунгария в Китае и МНР, в МНР Монгольский Алтай и восточная часть республики на юг примерно до 46° с. ш.; Корейский п-ов, северо-восток Китая и на восток до Шаньси и Хутея, на юг до Сычуаня, Юньнаня и Фуцзяна и включает Тибет; Гималаи от Сиккима до Кашмира, Ладака и Гиль-гита, Каракорум и восток Афганистана; в СССР имеется несколько участков северо-западных, северных и северо-восточных частей видового ареала, разобщенных между собой и соединяющихся на юге за пределами СССР: Памир, Тянь-Шань, Тарбагатай, Саур, Зайсанская котловина и окружающие ее горы, по Черному Иртышу, Калбинский Алтай, Алтай; по-видимому, отсутствует на Восточном Алтае, в Западных Саянах и в Туве; узкая полоса Прибайкалья к западу от Байкала; большой район Забайкалья, на север, по-видимому, до Лены; далее на востоке встречается на Алдане, в месте слияния Шилки с Аргунью, в западных частях Амурской области и в Приморье у впадения Бикина в Уссури и по долине ее и западным предгорьям Сихотэ-Алиня узкой полосой ареал идет на юг и выходит к океану у Находки);
индийский солонгой — М. kathiah Hodgson, 1835 (Индия — горы Каракорум, Кумаон; Непал и на восток до Ассама в Индии, Бирмы, п-ова Индокитай и Китая — Сычуаня, Юньнаня, Фуцзяня);
колонок — М. sibirica Pallas, 1773 (Азия — северная часть МНР, Северо-Восточный Китай и на юг, включая Гуандунь, Юньнань и Тибет; Гималаи, включая Кашмир и Непал; Верхняя Бирма, о-ва Японские, Тайвань и Ява; в СССР — на востоке европейской части СССР граница проходит примерно от 63° с. ш. на Урале на юго-запад на Среднюю Вет-ЛУГУ, вдоль Ветлуги и Волги к Казани, идет к юго-востоку, захватывая 1атарию, север и северо-восток Куйбышевской области и Башкирию и выходит к Южному Уралу; в Сибири северная граница идет от 63° с. ш. На Урале на восток, пересекает Обь, следует на верховья Пура, круто сворачивает к северу вдоль 80-й параллели, доходит до Полярного кру-га, идет на северо-восток, пересекает Енисей несколько выше Дудинки и Достигает 69° с. ш., затем спускается южнее, на 63° с. ш„ пересекает перховья Вилюя, идет до Лены вдоль Вилюя, несколько отступя к севе-РУ от него, затем по правобережью Лены до устья Алдана, по Алдану
примерно до его поворота на юг и отсюда выходит к устью Охоты у впадения в Охотское море; к югу от указанной границы колонок обитает по всей Сибири и на пространстве от Тихого океана до Южного Алтая ареал выходит за пределы страны; отсутствует колонок лишь на юге юго-западного Забайкалья; южная граница от Алтая идет по Иртышу Северному Казахстану — северным частям Павлодарской области, Северо-Казахстанской области, Кокчетавской, б. Акмолинской области Кустанайской области и выходит к Уралу в районе Орска; на Сахалин в 1932 г. завезен колонок из Японии и в настоящее время здесь (по крайней мере, на Южном Сахалине) существует его устойчивая популяция; для последнего столетия характерно расширение ареала колонка в Средней Сибири, Якутии, Дальнем Востоке и особенно на западе; так, еще в 70—80-е годы прошлого века его не было на Урале и весь европейский участок ареала был освоен колонком в текущем столетии) ;
длиннохвостая ласка — М. frenata Lichtenstein, 1831 (Северная, Центральная и Южная Америка — северная граница ареала начинается у Тихоокеанского побережья юго-западного угла провинции Британская Колумбия в Канаде, поднимается к северу, несколько отступя от берега, до центральных районов этой провинции, поворачивает на восток, проходя посредине Альберта, южной трети Саскачевана, крайнего юго-запада Манитобы, по северо-восточному углу штата Миннесота в США, южному и северо-восточному берегам оз. Верхнего, пересекает юго-восток Онтарио, крайний юг Квебека, север штата Мэн, в США и в южной части провинции Нью-Брансуик выходит к Атлантическому океану; в состав ареала входит вся Северная Америка, кроме территории, охватывающей в США крайний юг Невады, юго-восток Калифорнии, восток и север Аризоны, северо-запад Нью-Мексико, и в Мексике п-ов Калифорнию и западную половину штата Сонора; в ареал входит вся Центральная Америка, а в Южной Америке Колумбия, Венесуэла, Эквадор, Перу и Боливия);
белополосая ласка — М. strigidorsa Gray, 1853 (Непал, Индия — Сикким и Ассам, Бирма и п-ов Индокитай);
голостопая ласка — М. nudipes Cuvier, 1821 (о-ва Суматра и Калимантан, п-ов Малакка);
европейская норка — М. lutreola Linnaeus, 1761 (Европа — Финляндия, Польша, Венгрия, Чехословакия, Румыния, Австралия, Болгария, север и запад Югославии; в СССР северная граница проходит по Карелии примерно по 66° с. ш., затем по берегу Белого моря к Архангельску, обходит с юга тундру Канлыг, выходит к устью Печоры, огибает с юга Большеземельскую тундру, пересекает Урал в районе Полярного круга, резко сворачивает на юг и далее идет на восток, выходя на Обь немного ниже Тобольска, затем следует на восток несколько севернее Оби до нижнего течения Агана, правого притока Оби, откуда сворачивает на юг, становится восточной границей и, плавно изгибаясь на юго-запад, достигает Урала в районе Челябинска, следует на юг через Орск к Актюбинску, отсюда граница, становясь уже южной, тянется на запад к Уральску, огибает с востока бассейн Еруслана и выходит на Волгу, по ее долине узкой полосой ареал спускается до Каспия; в районе сближения Волги и Дона южная граница переходит на Дон, огибает Калмыцкие степи и выходит к устью Кумы, отсюда идет по побережью Кас-пия намного южнее Махачкалы и сворачивает на запад на Орджоникидзе, примерно на меридиане Туапсе пересекает Кавказский хребет и вдоль Черноморского побережья уходит на запад за пределы нашей страны. В Крыму европейской норки нет, на запад ареал везде от Карелии Д° Черного моря уходит за государственную границу);
американская норка—М. vison Schreber, 1777 (большая часть Северной Америки — северная граница проходит по Арктическому побережью материка, не захватывая п-ов Унгава, и отсекает часть материка от Эскимо-Пойнт на Гудзоновом заливе до пересечения 125° з. д. с северным побережьем: западная граница проходит по Тихоокеанскому, побережью на юг до Сан-Франциско; восточная граница образована Атлантическим побережьем на юг до Мексиканского залива; южная граница идет по берегу этого залива примерно до Корпус-Кристи, а отсюда через штаты Техас, Нью-Мексико, Юта, Невада и северную часть Калифорнии выходит к Тихому океану; в СССР с 1933 г. начали акклиматизировать американскую норку: с 1933 по 1948 г. на территории СССР было выпущено около 4 тыс. американских норок в 50 точках 22 областей, к 1964 г. в европейской части страны выпустили более 2 тыс. зверьков, а в Сибири и на Дальнем Востоке около 14 тыс; в ряде мест к настоящему времени сложились устойчивые популяции американской норки- так, в Татарии, частично в Башкирии, американские норки достигли промысловой численности, успешно прошла акклиматизация в Белоруссии. В тех местах, где американская норка поселяется вместе с европейской, она вытесняет последнюю. Две крупные популяции американской норки образовались в Уссурийском крае и части Приамурья, на Алтае и в смежных местах. Неясны еще результаты акклиматизации •на Кавказе, в Средней Азии, на Крайнем Севере. Американская норка имеет более крупную и ценную шкурку, чем европейская. Из отрица-
тельных качеств американской норки надо отметить, что она в значительных количествах поедает ондатру, особенно молодняк. Кроме того, в ряде мест она может стать конкурентом соболя);
лесной хорек-—М. putorius Linnaeus, 1758 (почти вся Европа — Финляндия к югу от 64° с. ш., крайний юг Скандинавского п-ова, на западе ареал включает Великобританию (в Ирландии нет); южная граница проходит по Средиземноморскому побережью, оставляя вне ареала Балеарские о-ва, Корсику, Сардинию и Сицилию, пересекает Балканский п-ов через север Югославии и идет по Дунаю, захватывая Добруджу; в СССР — северная граница проходит примерно от 62° 40' с. ш. на государственной границе с Финляндией к Спасской губе Онежского озера, а затем к Кеми на Белом море и по морскому побережью следует до пересечения его Полярным кругом; отсюда граница, становясь восточной, идет в верховья Вычегды и на Урал в район 58° с. ш., проходит по Уралу до широты Магнитогорска, здесь сворачивает на запад и уже как южная граница идет примерно по 53° с. ш. к Волге несколько южнее Самарской Луки, затем тянется по Волге несколько южнее Саратова, спускается на юг на Дон и следует вдоль него До станицы Цимлянской на Дону, откуда выгибается на юг к Ставрополю, идет к Краснодару и выходит к Азовскому морю, огибает его, проходит к устью Днепра, несколько отступя от Черноморского побережья (Крым в ареал не входит) и вдоль берега Черного моря через устье Дуная уходит за государственную границу; западная граница ареала в рССР совпадает с государственной границей от Черного моря до о2 40' с. ш.; за последние десятилетия происходит энергичное расселение лесного хорька на север, что связано отчасти с потеплением климата н вырубкой лесов и распашкой земли; так, в Карелии с 1930—1932 по 1Jo6 г. лесной хорек продвинулся на север на 250 км, по Северной Двине на 320 км);
американский хорек — М. nigripes Audubon et Bachman, 1851 (Канада— юг провинций Альберта и Саскачевана; штаты США—центр и восток Монтаны, западная половина Северной Дакоты, Южная Дакота, Роме северо-востока, Небраска, восточная половина Вайоминга, Коло-
радо, крайний юго-восток Юты, северо-восток Аризоны, Нью-Мексико, северо-запад Техаса, западные половины Оклахомы и Канзаса; в настоящее время встречается на западе Северной и Южной Дакоты, в Северной Монтане, в провинции Альберта и, возможно, на юг до Техаса н центра штата Нью-Мексико);
степной хорек — М. eversmanni Lesson, 1827 (Европа — Румыния Венгрия, запад Австрии, Чехословакия, север Болгарии, северо-восток Югославии; Азия —МНР; в Индии Кашмир, в Китае Тибет и восточные провинции страны от Хэйлунцзяна на севере до Сычуаня на юге, включая автономный район Внутренняя Монголия; в СССР изолированный участок находится в Закарпатье в районах Ужгорода и Мукачева, к северу от Карпат северная граница ареала начинается в районе Равы-Русской на польской границе севернее и северо-восточнее Львова и идет на Житомир и Киев и далее к Чернигову, затем поднимается на северо-восток к Брянску, потом на восток на Калугу, Тулу, Рязань и Горький с выгибом на север между двумя последними городами во Владимирскую область, перейдя Волгу в районе устья Камы, граница поднимается круто на северо-восток, охватывая Татарскую автономную республику и юг Кировской области, далее следует к Уралу и, обходя с востока Татарию, резко спускается к югу, идет по Уфе и Белой, на широте Магнитогорска резко поворачивает на восток и пересекает Урал, затем через Миасс выходит на Свердловск и далее на север к верховьям Сосьвы, далее на Тобольск и вдоль Иртыша до устья р. Шиш, потом выходит на Обь между устьями Васюгана и Кети, следует на Енисей между Красноярском и устьем Ангары, дальше на восток обходит Байкал с юга, тянется к Витиму, верховью р. Читы в Читинской области, опускается на юг на Аргунь и уходит за государственную границу; отдельный участок ареала расположен на Среднем Амуре в районе Благовещенска и на восток от него до Бурен; участок южной границы ареала расположен от Равы-Русской, включая Львов и Станислав, в северной Молдавии выходит на юг за пределы СССР; далее южная граница идет по берегу Черного и Азовского морей, включая Крым, и от Краснодара через Нальчик и Орджоникидзе следует до Махачкалы на Каспийском море; узкая полоса ареала тянется по Черноморскому побережью почти до Сухуми, а по Каспийскому побережью от Махачкалы до Дербента; от Дербента граница огибает Каспий с северо-запада, севера, северо-востока и востока вплоть до района Красноводска, отсюда, огибая Большой Балхан с севера, идет к низовьям Амударьи, далее на восток граница неопределенна, по-видимому, она проходит в некотором отдалении от Аральского моря, выходит на Сырдарью в районе Кзыл-Орды, идет вдоль Сырдарьи на юго-восток, потом удаляется от нее и следует на Бухару, откуда, делая изгиб на юг, юго-восток и восток, идет на Сурхандарью и через Алайскую долину уходит за государственную границу в Китай; между Алайской долиной и низовьями Аргуни в Забайкалье южная граница ареала проходит вне территории нашей страны; ареал за последнее столетие расширялся на запад и отчасти на север, так что сто лет назад степного хорька в Татарии не было; в 1928 г. он проник в Венгрию, в 1948 г. в Чехословакию, в 1962 г. в Австрию).
В СССР: ласка, горностай, солонгой, колонок, европейская норка, лесной и степной хорьки, американская норка (акклиматизирована).
Американские систематики (например, Холл и Келсон, 1959) выделяют из М. nivalis американскую ласку — М. rixosa Bangs, 1896. Японского колонка иногда (например, Строганов, 1962) считают особым видом М. itatsi Temminck, 1844, а не подвидом колонка М. sibirica. Возможно (Гептнер, 1967), американский хорек М. nigripes— лишь подвид степного хорька—М. eversmanni.
В «Красную книгу» включен американский хорек как вид, которому грозит исчезновение.
Представители рода имеют важное промысловое значение. В СССР в период с 1960 по 1970 г. ежегодно заготовляли (тыс. шкурок): 117,2— 334,3 горностая, 57,3—164,7 колонка, 11,7—34,8 норок, 1,8—9,4 солонгоя, 15 J—114,8 степного хорька и 17—407 лесного хорька.
Genus Grammogale Cabrera, 1940
В роде единственный вид: G. africana Desmarest, 1818 (Южная Америка — бассейн Амазонки, Восточный Эквадор, Северо-Восточное Перу).
Симпсон (1945) не включает Grammogale в свой список родов. Оставляет его в стороне и В. Г. Гептнер (1967). Андерсон и Джонс (1967) приводят его в своем перечне.
Род перевязок
Genus Vormela Blasius, 1884
В роде единственный вид: перевязка — V. peregusna Giildenstaedt, 1770.
Размеры мелкие. Длина тела 26—35 см, длина хвоста 11—20 см. Туловище сильно удлиненное и тонкое. Голова небольшая с притупленной мордой. Уши крупные округлые, широко расставленные и широкие в основании. Конечности короткие (рис. 69). Волосяной покров низкий и грубый. Окраска его пестрая и подвержена значительной индивидуальной изменчивости. Окраска головы состоит из участков белого и черного цветов. Верхняя и нижняя губы и подбородок белые. Через глаза про
ходит широкая поперечная черная полоса, переходящая у углов рта в черное поле горла. Позади этой полосы между глазами и ушами расположена тоже широкая поперечная белая полоса, идущая на бока головы ниже ушей и заканчивающаяся на боках шеи. Остальная часть головы черная. Уши у основания и сзади тоже черные, но по краю они чисто-белые. Шея сверху буровато-черная с тремя продольными светлыми полосами. Спина бурая с многочисленными разной формы мелкими и крупными светлыми пятнами или желтая с бурыми пятнами. В области лопаток спереди наискось вниз и назад по бокам проходит по белой полосе вплоть до последних ребер. Навстречу этим полосам от крестца вниз, вбок и вперед также идет по белой полосе. Весь низ тела черный с буроватым оттенком. Хвост в основании ржаво-коричневый или бурый. Посредине он палево-белесый с коричневатой примесью, а на конце черный. Имеются анальные кожные специфические железы. Сосков 5 пар.
Череп небольшой, с укороченным широким лицевым отделом и сильно выраженными гребнями. Мозговая коробка широкая и невытянутая. Скуловые дуги развиты сильно и поставлены широко; имеются большие заглазничные отростки. Костные слуховые барабаны небольшие, треугольной формы. Костное небо широкое.
3 13
Зубная формула: i — с — р —• 3 13
m = 34.
Зубы хорошо развиты, хищнические — крупные, клыки длинные, особенно верхние.
Распространена в Юго-Восточной Европе, части Передней Азии, Средней Азии, Казахстане, на севере Центральной Азии. В СССР западная граница проходит несколько к западу от Дона, а северная тя-
Рис. 69. Перевязка — V. peregusna и ее череп
нется от района Воронежа на восток на Хопер и далее через Волгу на средние течения Узеней в междуречье Волги и Урала, затем к верховьям Эмбы и, поворачивая на восток, захватывает Приаральские Каракумы и направляется к горам Улутау, верховьям Сарысу и Семипалатинску, входит в предалтайские степи и следует к Бийску, откуда поворачивает на юго-запад, проходит по предгорьям Алтая, включая район Усть-Каменогорска, Катон-Карагая и Зайсанскую котловину, и здесь выходит за пределы СССР; 100—-200 лет назад ареал простирался на запад значительно дальше и перевязка встречалась вплоть до Волыни, Подолии, Молдавии и Одессы; распашка целинных степей, их хозяйственное использование и исчезновение объектов питания, в частности сусликов, сделали здесь обитание для перевязок невозможным. В Европе вне СССР в ареал входит Добруджа в Румынии, Болгария, Восточная и Южная Югославия, европейская часть Турции, крайний восток Греции, в Азии — Монгольская Народная Республика, Автономный район Внутренняя Монголия в Китае, включая Шеньси, Афганистан, Белуджистан в Пакистане, Иран, Ирак, Сирия, Палестина и Малая Азия.
Населяет сухие степи, полупустыни и пустыни, иногда облесенные долины рек, окраины лесных массивов лесостепи, а также антропогенные ландшафты. Питается в основном песчанками (главным образом большой), сусликами, тушканчиками, хомяками, различными мышевидными грызунами. В качестве убежища обычно выбирает норы различных грызунов. Активна в вечерние и ранние утренние часы, а зимой в Туркмении и днем. Характерна поза устрашения, при которой перевязка высоко поднимается на ногах, вскидывает голову, заносит распушенный хвост на спину, рычит и скалит зубы. Биология размножения изучена слабо. На Северном Кавказе период гона приходится на март. Самка приносит в помете от 3 до 8 детенышей. Самец, по-видимому, в воспитании молодых участвует. Практическое значение невелико; шкур ка перевязки малоценная. Определенное значение зверек имеет как истребитель эпидемиологически опасных грызунов.
Род тайр
Genus Eira Hamilton-Shith, 1842, ( = Tayra Oken, 1816, jun. horn;
Galera Gray, 1843)
В роде единственный вид: тайра — Е. barbara Linnaeus, 1758.
Размеры средние. Длина тела 60—68 см, длина хвоста 30—47 см. Масса около 4,5 кг. Туловище удлиненное, конечности относительно высокие, хвост длинный (рис. 70). Голова широкая, на длинной шее, уши большие, широко расставленные, с округлыми вершинами. Подошвы лап безволосые. Когти сильные, изогнутые, невтяжные. Волосяной покров низкий и грубый, в хвосте волосы удлиненные. Окраска волос на туловище темно-бурая, голова и шея серые, бурые или черные, на груди желтые или белые пятна.
Диплоидное число хромосом 38.
Распространена в Южной Мексике, Центральной Америке, северной части Южной Америки до Парагвая и Аргентины и на о-ве Тринидад.
Населяет леса. Наземный хишник, однако хорошо лазает по деревьям и плавает. Держится парами или семейными группами. Активна ночью и ранним утром. Питается различными мелкими млекопитающими и птицами, а также плодами и медом. Биология размножения почти неизвестна. В помете от 2 до 4 детенышей. В неволе тайра жила до 12 лет.
Род гризонов
Genus Galictis Bell, 1826 ( = Grison Okean, 1816, jun. hom)
Размеры мелкие. Длина тела 40—55 см, длина хвоста 15—19 см. Масса от 1 до 3,2 кг. Туловище сильно вытянутое, конечности короткие (рис. 71). Шея длинная, голова небольшая, уши широко расставленные, маленькие, с закругленными вершинами. Волосяной покров средней высоты, довольно грубый. Хвост покрыт удлиненными волосами. Окраска волос на спине дымчато-серая, желтовато-серая или буроватая. По лбу и дальше по бокам головы и шеи проходит белая полоса. Морда кпереди от этой полосы, нижняя часть шеи, конечности и брюхо темнобурые или черные.
Диплоидное число хромосом у гризона 38.
Распространена в южной части Северной Америки, Центральной и Южной Америке.
Населяют как открытые пространства, так и леса. Поднимаются в горы до 1200 м над уровнем моря. В качестве убежища занимают норы других животных, иногда прячутся в камнях. Держатся обычно группами. Хорошо лазают по деревьям и хорошо плавают. Активны круглосуточно. По характеру питания хищники, хотя набор кормов достаточно обширный. Самка приносит в помете от 2 до 4 детенышей. Хорошо приручаются. Одно время использовались при охоте за шиншиллами.
В роде 2 вида:
гризон — G. vittatus Schreber, 1776 (юг Мексики — штаты юг Сан-Луис-Потоси, Идальго, восток Пуэбла, Веракрус, северо-восток и восток Оахака, Табаско, Чиапас, Кампече, Юкатан, Кинтано-Роо; вся Центральная Америка; Южная Америка —• Западная Колумбия, Эквадор, Перу, Венесуэла, Гайана, Суринам, Бразилия);
малый гризон — G. cuja Molina, 1782 (Южная Америка — Чили, Аргентина, Бразилия, Парагвай, Уругвай, Западная Боливия).
Иногда малого гризона выделяют в особый вид Grisonella Thomas, 1912.
Род патагонских ласок
Genus Lyncodon Gervais, 1844
В роде единственный вид: патагонская ласка — L. patagonicus Blain-ville, 1842.
Размеры мелкие. Длина тела 30—35 см, длина хвоста 6—9 см. Туловище сильно удлинено, конечности короткие. По внешнему виду напоминает гризона, но хвост относительно короче (рис. 72).
— = 28.
1
m
3 12
Характерно число зубов: i — с - р —
3 12
Редукция происходит за счет предкоренных и коренных.
Волосяной покров средней высоты, редкий и довольно грубый. Волосы на спине серовато-бурые, с белыми вершинами. Верх головы белый и от нее по бокам головы и по плечам проходит широкая белая полоса. Ьрюхо и конечности темно-бурые.
Распространена в Аргентине и Чили.
Населяет пампасы. Экология изучена слабо.
6—671
161
Рис. 72. Патагонская ласка — L patagonicus
1ЛО
Род африканских хорьков
Genus Ictonyx Каир, 1835
В роде единственный вид: африканский хорек-— /. striatus Perry, 1810.
Размеры мелкие. Длина тела 28—38 см, длина хвоста 28—30 см. Туловище удлиненное, конечности короткие, хвост длинный с длинными волосами (рис. 73). Голова широкая, уши небольшие, широко расставленные, округлые. Волосяной покров высокий, грубый и редкий. Окраска его яркая, состоит из комбинации белого и черного цветов. Позади глаз и между ними располагаются белые пятна, отделенные черной полосой, проходящей на уровне ушей от белого затылочного поля. Вершины ушей белые. Вдоль спины на белом фоне проходят две черные продольные полосы. Нижняя часть боков тела, брюшная сторона и вся морда, за исключением вышеупомянутых белых пятен, черные. Сильно развиты анальные железы.
3 13 1
Зубная формула: 1 — с — р —• гп = 34. 3 13 2
Распространен в Африке от Сенегала, Северной Нигерии, Судана, Эфиопии на юг до Южно-Африканской Республики включительно.
Встречается в различных биотопах.
Ведет наземный образ жизни, но хорошо лазает по деревьям и плавает. Держится обычно поодиночке. Активен ночью. День проводит в норах других животных или в собственных. В случае опасности длинная шерсть на теле поднимается дыбом и зверек выбрызгивает на врага неприятно пахнущий секрет анальных желез. Может также притворяться мертвым. Питается различными мелкими млекопитающими, а также насекомыми, пресмыкающимися и яйцами птиц. В помете—2—3 детеныша.
Род североафриканских ласок
Genus Poecilictis Thomas et Hinton, 1920
В роде единственный вид: североафриканская ласка — Р. lybica Hemprich et Ehrenberg, 1832 (рис. 74).
Размеры мелкие. Длина тела 22—28 см, длина хвоста 13—18 см. Масса около 250 г. Туловище вытянутое, конечности короткие, хвост Длинный. Морда с удлиненной лицевой частью. Волосяной покров высокий, косматый. В окраске характерно сочетание белого и черного цветов. Морда черная, лоб белый, затылок черный. Спина белая с черными пятнами различных размеров. Брюхо и конечности черные. Хвост белый, постепенно темнеющий к концу. В черепе характерны крупные костные. слуховые барабаны.
Распространена в Северной Африке от Марокко до АРЕ и на юг до Нигерии и Судана.
Населяет'- окультуренные ландшафты на границе с пустыней. Экология изучена слабо. Активна ночью. Питается, по-видимому, мелкими животными. Самка приносит в помете 1—3 детеныша с января по март. Беременность — 37 дней (по другим данным, 77). Легко приручается.
6* IfiQ
Рис. 74. Североафриканская ласка — Р. lybica и ее череп
Род африканских ласок
Genus Poecilogale Thomas, 1883
В роде единственный вид: африканская ласка — Р. albinucha Gray, 1864. Tv-
Размеры мелкие. Длина тела 25—35 см, длина хвоста 15—23 см. 1у' ловище и шея сильно удлинены, конечности короткие. Голова небольшая, уплощенная, с укороченным лицевым отделом. Внешним виде ~ напоминает ласку. Волосяной покров низкий, на хвосте волосы удлине ны. По верху головы и шеи проходит белая полоса. Вдоль спины расп лагаются четыре беловатые или оранжево-желтые и чередующиеся ними три черные полосы. Бока тела, брюхо и конечности черные. Сил но развиты анальные железы.
3 12 1
Зубная формула: i — с —— р — m —— = 22.
о 1 • А
1А4
Распространена в Южной и Центральной Африке: ЮАР, Мозамбике, Родезии, Малави, Замбии, Анголе, Заире, Бурунди, Руанде, Танзании й Юго-Западной Уганде.
Населяет различные биотопы. Ведет наземный образ жизни, но хорошо лазает по деревьям. Убежищем служат норы. Активность круглосуточная. Держатся обычно парами или семейными группами. Питается мелкими млекопитающими. Благодаря длинному, гибкому телу и коротким конечностям способна преследовать добычу в ее же норах. Поедает также птиц, пресмыкающихся, насекомых и их личинок. В помете обычно 2 детеныша.
SUBFAMILIA MELL1VORINAE GILL, 1872
Род медоедов
Genus Mellivora Storr, 1780
В роде единственный вид: медоед — М. capensis Schreber, 1776.
Размеры средние. Длина тела 68—75 см, длина хвоста 17—20 см. Внешним видом напоминает барсука. Туловище массивное, умеренно вытянутое. Голова относительно крупная и широкая. Лицевой отдел слабо вытянут, притуплен (рис. 75). Ушные раковины представлены только невысокими валиками. Конечности короткие, толстые. Лапы широкие, передние типичные роющие с очень длинными когтями. Когти на задних конечностях несколько короче. Подошвы голые. Волосяной покров грубый и редкий, пуховых волос нет. В зимнее время волосы более длинные и несколько более густые, чем летом. По верху головы располагается беловатое поле: оно начинается позади глаз, идет по верху шеи и по всей спине, спускаясь на бока и постепенно расширяясь от шеи к середине туловища или бедрам. Вся брюшная поверхность тела, конечности и нижняя часть боков черные. В летнее время характерны темные тона на белом поле. Имеются анальные железы, выделяющие секрет с сильным запахом. Сосков 2 пары.
Череп крупный, массивный с увеличенной длинной и широкой мозговой коробкой. Лицевой отдел относительно короткий и широкий. Костные слуховые барабаны очень крупные, угловато-овальной формы, вздутые. Заглазничное сужение длинное, хорошо выраженное. Скуловые Дуги сильные, широко расставленные в стороны. Затылочный гребень развит сильно, а сагиттальный слабо. Костное небо короткое и широкое.
3 13 1
Зубная формула: i —- с - р —— m — =32. 3 2 3 1
Щечные зубы с невысокими вершинами. Хищнические зубы развиты сильно; клыки массивные, но короткие. Второй нижний коренной зуб, как правило, отсутствует. Передний верхний предкоренной зуб крупный, имеет два корня.
Распространен в Африке от Южно-Африканской Республики на се-веР До Эфиопии, Северного Сомали, Судана и Южного Марокко; ® Азии — в Сирии, Палестине, на Аравийском п-ове, в Ираке, Иране, •Афганистане, Индии. В СССР южная граница ареала проходит по государственной границе от Каспийского моря до Амударьи, западная граница — по побережью Каспия на север немного севернее Красновод-ска, северная — от южного берега Кара-Богаз-Гола примерно к дельте Амударьи и по ее левому берегу до государственной границы.
Встречается в предгорьях, долинах рек и тугайных зарослях, реже песках. По характеру питания почти всеяден. Поедает мелких грызу-
Рис. 75. Медоед — М. capensis и его череп
лов, ежей, насекомых и их личинок, земляных пчел, ос, их личинок и мед. Активен, как правило, в сумерки и ночью, но иногда и днем. День проводит в вырытой им норе. Она обычно простая длиной 1—3 м с одним входом и гнездовой камерой. Беременность — 6—7 месяцев.
Практического значения не имеет. В СССР редок и охраняется законом.
„ SUBFAM1L1A MELINAE BURMEISTER, 1850
Род барсуков
Genus Meles Brisson, 1762
В роде единственный вид: барсук — М. meles Linnaeus, 1758.
Размеры средние. Длина тела 60-—90 см, длина хвоста 12—24 см. Масса 16—17 кг, иногда до 24 кг. Тело умеренно удлиненное, массивное, приземистое, расширенное в заднем отделе и резко сужающееся кпереди. Голова с вытянутой клиновидной мордой (рис. 76). Ушные раковины небольшие, округлые. Глаза маленькие. Шея короткая, почти незаметная. Конечности короткие массивные с широкими лапами, стопоходящие, пальцы удлиненные. Когти слабо изогнутые, сильные. Волосяной покров грубый, высокий и редкий; особенно изрежен на брюшной стороне тела. Зимой мех более длинный, густой с большим количеством пуховых волос. Спина и бока в зимнее время буровато-серые или серые с черноватой рябью. Голова белая; по бокам головы от верхней губы несколько спереди от угла рта и назад через глаз, все расширяясь дальше к уху и к плечу, проходит по черной полосе. Между этими черными боковыми полосами заключена широкая белая полоса, идущая от кончика носа через лоб, темя, затылок, переднюю часть шеи, иногда через всю шею и постепенно сливающаяся с окраской спины. Низ головы и горло черные. Между этой окраской и черными боковыми полосами располагаются широкие боковые белые поля, которые проходят назад примерно до уровня окончания верхней белой полосы, иногда и дальше, и незаметно переходят в окраску верха тела. Уши черные с широким белым краем. Хвост той же окраски, что и верх тела. Брюшная сторона тела и конечности черные с буроватым оттенком. Хвост той же окраски, что и верх тела. В течение года бывает только одна линька, которая растянута на весь летний период. Хорошо развиты анальные железы. Между заднепроходным отверстием и основанием хвоста имеется специфическая кожная железа, располагающаяся в особом кармане. Сосков три пары: две брюшных и одна паховая.
Череп крупный, массивный. Мозговой отдел округло-овальный, лицевая часть черепа относительно узкая, длинная. Сагиттальный и затылочный гребни хорошо развиты. Скуловые дуги сильные, широко расставленные сзади. Заглазничное сужение хорошо выражено. Костные слуховые барабаны невздутые, умеренной величины. Твердое небо узкое и Длинное.
Зубная формула: i — с -у- р 1 ш — =38 — 34.
Мелкий по размерам первый верхний предкоренной зуб часто выпадает, иногда отсутствует и первый нижний предкоренной зуб, и тогда общее число зубов может равняться 36 или даже 34. Характер жевательной поверхности щечных зубов весьма своеобразен и представляет собой комбинацию зубов с острыми краями режущего типа и бугорчато-пере-тирающего. Клыки крупные: верхний хищнический зуб небольшой, зяа-
Рис. 76. Барсук — М. meles и его череп
чительно меньше по размерам первого верхнего коренного зуба, который имеет широкую (перетирающую) жевательную поверхность. Все остальные щечные зубы с острыми режущими краями. Нижний хищнический зуб крупный удлиненный, с тремя острыми вершинами и удлиненной уплощенной и вогнутой задней частью.
J Диплоидное число хромосом 44.
Распространен в Европе на юг до побережья Средиземного моря /включая о-ва Родос и Крит), на север до центральных районов Норвегии, Швеции и Финляндии, на запад по побережью материка, включая Англию и Ирландию; в Азии в ареал входят Малая Азия, Сирия, Палестина, Ирак, Иран, Северный Афганистан, МНР, Северный, Восточный, Южный, Юго-Западный Китай, Северная Бирма, Северная часть п-ова Индокитай, Корейский п-ов, Японские о-ва. В СССР северная граница ареала проходит примерно от Кандалакши по южному берегу Белого моря на Архангельск, далее на Мезень (65° с. ш.), к истокам р. Пеши, впадающей в Чешскую губу, к г. Ижма, на р. Печору (65° с. ш.) и Урал. В Сибири эта граница идет от нижнего течения Оби к верховьям Пури и Таза (около 63° с. ш.), пересекает Енисей у 61° с. ш., устье Витима и затем тянется на юго-запад к западному берегу Байкала, обходит озеро с юга, огибает Витимское плоскогорье, следует на г. Зея, верхнюю Бурею, среднее течение Амгуни, пересекает Амур и на 52° с. ш. выходит к океану; к югу от указанной границы ареал барсука занимает всю территорию страны.
Населяет самые разнообразные биотопы во всех географических зонах, кроме тундры. Предпочитает уединенные овражистые места недалеко от водоемов. Поднимается в горы на высоту до 3000 м над уровнем моря. Больших сплошных лесных массивов избегает. Для поселения обычно выбирает сухие, легко поддающиеся рытью песчаные и супесчаные почвы с глубоким расположением грунтовых вод. Активен преимущественно в сумерки и ночью. День проводит в норах, которые, как правило, выкапывает сам. Основные работы по рытью нор (выкапывание новых, ремонт старых) происходят в августе — сентябре. В одной и той же норе барсуки могут жить в течение долгого времени и за этот срок выкапывают очень сложную систему ходов со многими выходами и подземными переходами. Количество выходов в норе может колебаться от 1 до 50. Гнездовая камера (иногда 2—3) располагается на расстоянии 5—10 м от входного отверстия. В камеру затаскивается подстилка из сухих листьев и травы, которая периодически обновляется. Камера обычно располагается на глубине не менее 1 и до 2—3 м от поверхности. Ее средние размеры 74X76 см при высоте 38 см. Общая длина ходов может быть от 35 до 81 м. На зиму выходные отверстия норы барсук забивает землей и листьями.
По характеру питания барсук всеяден. Пищей служат мышевидные грызуны, в основном полевки, реже мелкие птицы, земноводные (главным образом лягушки), пресмыкающиеся (в основном ящерицы), различные насекомые и их личинки, моллюски, земляные черви и растительные корма — плоды, орехи, ягоды, клубни, корневища и пр. Состав кормов значительно меняется в географическом аспекте и по сезонам. Так, процент встречаемости грызунов среди кормов, поедаемых барсуком в Тульских засеках, составляет 69,0; в Татарии 53,6; в Жигулях 56,6; в Бузулукском бору 17,1; в горном Крыму 29,8; в дельте Или 20,8; на Рыбинском водохранилище 14,5. В первой половине лета процент встречаемости мышевидных грызунов в кормах барсука в Молдавии составил 18,3, личинок жуков 33,3, а во второй половине лета — осенью первых 67,5, вторых 7,0.
К осени барсук очень сильно жиреет и на холодный период года
Рис. 77. Теледу — A. collaris
впадает в зимний сон. При этом существенного замедления физиологических функций и понижения температуры не происходит. В местах с мягким климатом (например, Англия, в СССР — Закавказье) барсук бывает активен в течение круглого года.
Гон и спаривание у барсуков могут происходить в весеннее и летнее время. Самец и самка образуют пару, по-видимому, на ряд лет, а возможно, и на всю жизнь. Продолжительность беременности при летнем спаривании составляет 271—284 дня, а при весеннем — до года. Имеет место эмбриональная диапауза. Самка приносит 1—6, чаше 2 детенышей. Новорожденные слепые, беспомощные, покрыты редкими белыми волосами, с массой 63—84 г. В возрасте 9—11 дней открываются ушные отверстия. На 35—42-й день детеныши прозревают. В месячном возрасте прорезаются молочные зубы, примерно с 3-месячного возраста переходят к самостоятельному питанию. Половая зрелость у самок наступаег примерно в 2-летнем, а у самцов — в 3-летнем возрасте.
Практическое значение невелико; шкура ценится невысоко. Широко используется сало как целебное снадобье. Полезен истреблением вредных сельскохозяйственных и лесных вредителей — насекомых и млекопитающих В СССР с 1960 по 1970 г. ежегодно заготовляли от 5 до 22 тыс. шкур барсука.
Род теледу
Genus Ar atonyх F. Cuvier, 1825
В роде единственный вид: теледу — A. collaris F. Cuvier, 1825.
Размеры средние. Длина тела 55—70 см, длина хвоста 12—17 см. Масса 7—14 кг. Внешним видом несколько напоминает барсука (рис. 77), однако морда сильнее вытянута, передние конечности имеют очень мощные, сильно искривленные светлые когти; хвост относительно 170
Рис. 78. Американский барсук— Т. taxus и его череп
длинный. Волосяной покров редкий и грубый. Спина желтоватая, сероватая или черноватая. На голове проходят белые и черные полосы. Уши и хвост белые. Брюхо и конечности черные, горло белое. Анальные железы развиты хорошо.
Распространен на большей части Китая, в Индии — в северо-восточной части и Ассаме, Бирме, п-ове Индокитай, Суматре.
Населяет различные биотопы на равнинах и в горах. Активен ночью. День проводит в расщелинах скал или в вырытых ими глубоких норах. Всеяден. В помете обычно 4 детеныша.
Род американских барсуков
Genus Taxidea Waterhouse, 1839
1778 Р°Де единственныв вид: американский барсук — Т. taxus Schreber,
Размеры средние и крупные. Длина тела 42—72 см, длина хвоста Ю—-15 см. Масса 3,5—11,4 кг. По внешнему виду напоминает барсука (рис. 78). Волосяной покров высокий и густой. Спина сероватая, ино-
171
гда рыжеватая. Белая узкая полоса проходит от носа по верху головы и шее до области между лопатками. Голова с боков белая с черными пятнами между глазом и ухом, вершины ушей белые, брюхо грязно-желтое; конечности темно-бурые или черные. Наиболее длинные волосы располагаются по бокам тела. Анальные пахучие железы хорошо развиты.
Зубная формула:
2 з
1 3 1
гртт т 34-
Диплоидное число хромосом 32.
Распространен почти по всей Северной Америке. Западная граница ареала начинается от Тихоокеанского побережья северной части штата Калифорния в США и на некотором расстоянии от берега тянется на север, пересекает западную четверть Орегона, западную треть штата Вашингтон, отсекает юго-восточный угол провинции Британская Колумбия в Канаде и поднимается до северных частей провинции Альберта, где переходит в северную границу ареала, которая следует на юго-восток через центральные районы Саскачевана, юг Манитобы, крайний юго-запад Онтарио и далее по южным берегам Великих озер до северо-восточной части штата Огайо. Отсюда идет на юг, юго-запад и запад, проходя по южным районам штатов Огайо, Индиана и Иллинойс, центр Миссури, где поворачивает на юг, отсекая юго-восточный угол Оклахомы, восточную треть Техаса и на крайнем юге этого штата выходит к ЛТексиканскому заливу, откуда уже в Мексике сворачивает на запад. Проходит по северу штата Тамаулипас и юг Нуэво-Леон, идет на юг через восток Сан-Луис-Потоси, Керетаро и в южной части Пуэбла поворачивает на северо-запад, пересекая юг Мексико, северо-восток Мичоакан, юго-запад Гаунохуато, восток Халиско, юг Закатекас, западную треть Дуранго, крайний юго-запад Чиуауа и по границе между штатами Синалоа и Сонора выходит к Калифорнийскому заливу. П-ов Калифорния входит в ареал.
Обычно населяет сухие открытые пространства. Держится, как правило, поодиночке. Активен в основном в сумерки, ранним утром и вечером. По образу жизни напоминает барсука. Убежищем служат сложно устроенные глубокие норы с гнездовой камерой. В северных частях ареала в холодный период года впадает в зимний сон. Питается сусликами, гоферами, луговыми собачками, мышевидными грызунами, птицами, беспозвоночными, падалью. Иногда запасает пищу, закапывая ее в землю. Период спаривания приходится на август и сентябрь. В развитии оплодотворенной яйцеклетки имеет место латентный период. Имплантация яйцеклетки обычно происходит не ранее февраля. Действительный период развития яйцеклетки равен примерно 6 неделям. В помете 1—5, чаще 2 детеныша. Длительность лактации около 6 недель. Молодые переходят к самостоятельной жизни в конце лета. В неволе жили до 13 лет.
Род хорьковых барсуков
Genus Melo gale 1. Geoffroy, 1834
Размеры мелкие. Длина тела 33—43 см, длина хвоста 15—23 см. Масса примерно 1,75 кг. По внешнему виду больше напоминает хорька, чем барсука. Тело удлиненное, конечности короткие (рис. 79). Передние конечности с большими, изогнутыми когтями. Хвост относительно длинный, покрыт густыми длинными волосами. Голова укороченная, широкая; уши широко расставлены, крупные с округленными вершина-172
мп. «ишна иледно-оурая или темно-бурая, вдоль нее по середине проходит белая или рыжеватая полоса, брюхо светлее спины, верх морды темный, а бока головы, горло и нижняя часть морды белые.
. 3
<5
.„141
Зубная формула: 1 -у с — р — m — = 38.
Диплоидное число хромосом у китайского барсука 38.
Распространены в Непале, в Индии — Ассаме, а также в Бирме, на п-ове Индокитай, в Китае, на о-вах Тайвань, Ява и Калимантан.
Населяют открытые пространства и леса. Активны в сумерки и ночью. День проводят в норах и естественных укрытиях. Китайский барсук с Тайваня хорошо лазает по деревьям. Всеядны: кормом служат мелкие позвоночные, насекомые, земляные черви, различные плоды. Самка приносит обычно 1—3 детенышей в мае или июне.
В роде 3 вида:
бирманский барсук — М. personata Geoffroy, 1831 (Непал, Ассам в Индии; Бирма, п-ов Индокитай);
китайский барсук — М. moschata Gray, 1831 (Китай от Сычуаня на юг до Юньнаня и Фуцзяня, о-ва Тайвань и Хайнань, Ассам в Индии, Бирма, п-ов Индокитай);
восточный барсук — М. orientalis Horsfield, 1821 (о-ва Ява и Калимантан).
Некоторые ученые (например, Чэйзен, 1940) относят китайского и восточного барсуков в особый род Helictis Gray, 1831. По-видимому, более правильно считать Helictis подродом Melogale.
SGBFAMILIA MEPHITINAE GILL, 1872
Род полосатых скунсов
Genus Mephitis G. Cuvier, 1800
Размеры мелкие. Длина тела 28—38 см, длина хвоста 18,5—43,5 см. Масса 0,75—2,5 кг. Туловище умеренно удлиненное, конечности невысокие, полустопоходящие. Хвост длинный, покрытый длинными волосами (рис. 80). Голова с укороченным лицевым отделом, широкая в мозговой части, сужающаяся к носу. Уши средней высоты, с широким основанием и округлыми вершинами. Подошвы лап голые. Когти передних лап длинные, сильные, а задних короткие. Волосяной покров высокий, густой и мягкий. Затылочная часть головы и верх шеи белые. В области плеч белое поле разделяется на две полосы, проходящие по бокам спины к основанию хвоста. Между полосами волосы имеют черную окраску. Все остальные части тела также черного цвета. Иногда конец и бока хвоста могут быть белыми. У некоторых животных белым остается только лоб. Очень сильно развиты анальные железы, продуцирующие секрет с отвратительным запахом.
В черепе хорошо выражен сагиттальный гребень. Костйые слуховые барабаны маленькие.
Зубная формула:
3 13 1
-- с - р - m -= 34
3 1 н 3 2
Диплоидное число хромосом у полосатого скунса 50.
Распространены от Южной Канады на юг через США, Мексику до Никарагуа.
Населяют разнообразные биотопы, в том числе леса и различного типа открытые пространства, включая пустыни. Наибольшая плот-174
ность полосатого скунса отмечена на сельскохозяйственных землях (13,5 скунса на 259 га). Встречаются, как правило, поодиночке. В зимнее время самки могут образовывать группы из нескольких животных. Активны ночью, но иногда в сумерках и даже днем. Хорошо плавают. По деревьям лазают плохо. В случае опасности зверек поворачивает заднюю часть тела в сторону врага, закидывает хвост на спину и сокращением сфинктеров выбрызгивает секрет анальных желез, метко попадая им в цель. Секрет имеет отталкивающий запах, а если попадает на слизистую оболочку глаза, вызывает сильное жжение и непродолжительную слепоту. Осенью зверьки сильно жиреют. В северных частях
_r__________ vnym. в лилидние время года впадает в зимний сон.
На севере Канады в конце октября — начале ноября полосатый скуцё собирает материал для своего зимнего гнезда, а в начале декабря впадает в сон. Взрослые самки и молодые редко просыпаются раньше конца марта, а взрослые самцы бывают активны во время случающихся зимой оттепелей. Убежищем служат норы или любые сухие укромные места Часто занимают норы кроликов, иногда выкапывают их сами. В норе устраивают подстилку из сухой травы и листьев. Всеядны. В Канаде осенью и зимой пища полосатого скунса состоит на 26,7% из плодов и ягод, 19,6% составляют млекопитающие (полевки, хомяки, кролики), 6,8 — насекомые (кузнечики, личинки жуков), 10,8 — трава, листья, почки, 13,5 — падаль, 11,3 — зерно и орехи; весной и летом 43% составляют насекомые, 27,5 — плоды, 16,2 — млекопитающие, 8,7%—зерна. Иногда также поедают яйца птиц и птенцов, рыб, моллюсков, лягушек, ящериц и змей.
Самцы полигамы. Самки моноэстральны. Течка длится примерно 3 дня. Период спаривания приходится на конец зимы — начало весны. Продолжительность беременности примерно 63 дня. В помете от 2 до 10, обычно 5—6 детенышей. Новорожденные появляются на свет слепые, с закрытыми ушами, покрытые короткой мягкой шерстью. В возрасте 2 недель открываются глаза. В возрасте 4 недель детеныши начинают принимать оборонительную позу, поднимая хвост. Длительность лактации 6—7 недель. В это же время они покидают убежище и следуют за матерью в поисках пищи в течение конца лета и осени. Вместе с матерью впадают в зимний сон. Половозрелость наступает примерно в годовалом возрасте. В неволе живут до 10 лет.
Одно время полосатых скунсов разводили на зверофермах, предварительно удалив анальные железы. В настоящее время мех полосатого скунса не находит спроса.
В 1933—1939 гг. делались неудачные попытки акклиматизировать полосатого скунса в СССР — в Воронежском заповеднике, на о-ве Петрова в Приморском крае, в Харьковской обл., Киргизской и Азербайджанской ССР, Дагестанской АССР. Одна из причин неудач заключалась в том, что выпускали зверьков с предварительно удаленными анальными железами — главным органом защиты зверьков.
Большую пользу приносят полосатые скунсы сельскому хозяйству, поедая огромное количество вредителей полей.
В роде 2 вида:
полосатый скунс — М. mephitis Schreber, 1776 (провинции Канады — крайний юго-запад Маккензи, восточная половина Британской Колумбии, Альберта, Саскачеван, кроме северо-востока, южная половина Манитобы, Онтарио, южная половина Квебека, Нью-Брансуик, о-в Принца Эдуарда, север Новой Шотландии; вся территория США, кроме юго-востока штата Калифорния, центральных районов Невады и юго-запада Юты; штаты севера Мексики — восточная половина Соноры, Чиуауа, крайний север Дуранго, северные половины штатов Коауила и Нуэво-Леон, крайний север Тамаулипас);
мексиканский скунс — М macroura Lichtenstein, 1823 (штаты США — юго-восток Аризоны, юго-запад Нью-Мексико, юго-запад Техаса; штаты Мексики: восточные две трети Соноры, Чиуауа, западные две трети Коауила, юг Нуэво-Леон, юго-запад Тамаулипас, Дуранго, Зака-текас, Наярит, Халиско, Агуаскальентес, Гуанохуато, Мичоакан, кроме юга, Мексико, Идальго, Тлакскала, Морелос, Пуэбла, центральные районы Веракрус, северная половина Герреро, северная половина Оахака, южная половина Гватемалы; Сальвадор; южные две трети Гондураса; юго-запад Никарагуа).
Род пятнистых скунсов
Genus Spilogale Gray, 1865
Размеры мелкие. Длина тела 11,5—34,5 см, длина хвоста от 7 до 22 см. Масса 0,2—I кг. Голова небольшая, с укороченным лицевым отделом, притупленная на конце (рис. 81). Глаза довольно крупные, iuin средней высоты, широкие в основаниях с округлыми вершинами. Когти на передних лапах примерно в два раза длиннее, чем на задних, волосяной покров мягкий, относительно высокий на туловище, низкий на голове. Хвост покрыт очень длинными волосами. Общий тон окраски тела черный. По спинной стороне и бокам проходят шесть продольных полос, часто разделенных на отдельные черточки или пятна. На лбу треугольное пятно. Конец хвоста обычно белый. Сосков 3—5 пар.
Череп небольшой, несколько уплощенный, округлый. Костные слуховые барабаны уплощены.
„ 3 13 1
Зубная формула: i — с - р —- m — = 34.
3 13 2
Диплоидное число хромосом у пятнистого скунса 64.
Распространены на крайнем юго-западе Канады, по всей территории США, кроме востока и северо-востока, Мексике и Центральной Америке на юг до Коста-Рики включительно.
Населяют заросли кустарников, каньоны, фермы. Избегают сплошных лесов и влажных мест. Активны ночью. В зимний сон не впадают однако в холодную погоду из убежища не показываются. Убежищем служат чужие норы, дупла деревьев, пространство под упавшими стволами, а также любые укромные и сухие места. Секрет, вырабатываемый анальными железами, имеет особо неприятный и сильный запах и если попадает в глаза, вызывает сильное жжение. Зверек способен выбрызгивать струю секрета на расстояние до 3,5 м. Близки к всеядности, но более плотоядны, чем полосатый скунс. В летнее время поедают различные корма и насекомых, иногда птиц и их яйца, а зимой и весной в основном млекопитающих (кроликов, полевок, крыс, мышей), а также зерна кукурузы. В течение круглого года охотно употребляют в пищу падаль, при случае поедают ящериц, змей и лягушек. Хотя пятнистые скунсы наземные животные, они довольно хорошо лазают по деревьям. В помете 2—6, обычно 4—5 детенышей. Беременность длится около 120 дней. В северных частях ареала гон происходит в конце зимы, а роды весной, в южных определенной сезонности в размножении нет. На юге самка может иметь два помета в течение одного года (второй помет — поздним летом). Новорожденный имеет длину (с хвостом) около 10 см и массу примерно 22,5 г. Тело покрыто редкими, короткими и тонкими волосами. Глаза и уши закрыты. К трем неделям они одеваются в густой мех, имеющий расцветку взрослых зверей. В 32-дневном возрасте открываются глаза, и детеныши начинают принимать позу угрозы. В 6 недель начинают функционировать анальные железы. В 3,5 месяца достигают размеров взрослых и переходят к самостоятельной жизни.
В роде, по-видимому, 2 вида:
пятнистый скунс —S. putorius Linnaeus, 1758 (северная граница ареала проходит от Тихоокеанского побережья крайнего юго-запада провинции Британской Колумбии в Канаде, спускается на юг через середину штата Вашингтон в США, почти достигая Орегона, поворачивает на северо-восток к северным частям Айдахо, захватывая передний юго-запад Монтаны, спускается на юг к юго-восточному углу Айдахо, следует на северо-восток к северо-восточной части Южной Дакоты, сворачивает на юг к юго-западному углу этого штата, далее идет к северо-восточному его углу и включает весь штат Миннесота, выходя к западной оконечности озера Верхнего, круто спускается на юг до северо-восточного угла штата Луизиана, так же круто поднимается к северу к юго-западной оконечности штата Индиана, следует на восток с небольшим уклоном к северу до центральных районов юга Пенсильвании, плавно проходит на юго-запад к западной трети штата Северная Каролина, пересекает западную часть Южной Каролины и далее выходит к Атлантическому океану в северо-восточной части Флориды; крайний юг Флориды, юго-восток штата Калифорния, все Тихоокеанское побережье от северных частей штата Соноры в Мексике до штата Оахака, Тихоокеанское побережье Сальвадора и Никарагуа, берег Мексиканского залива штатов Мексики Веракрус и Табаско, берег с прилегаЮ
щими к Карибскому морю районами Гондураса, Никарагуа и Коста-Рики в ареал не входят; на юг ареал тянется до центральных районов Коста-Рики включительно);
малый скунс — 3. pygmaea Thomas, 1898 (узкая полоса Тихоокеанского побережья Мексики от юга штата Синалоа на юг до центральных районов штата Оахака).
Некоторые исследователи объединяют оба вида в один. Другие (например, Холл и Келсон, 1959) насчитывают в роде еще 2 вида: S. gracilis Merriam, 1890, и 3. anguslifrons Howell, 1902.
Род свиноносых скунсов
Genus Conepatus Gray, 1837
Размеры мелкие. Длина тела 30—49 см, длина хвоста 16—41 см. Масса 2,3—4,5 кг. Общий вид напоминает представителей описанных выше родов скунсов, однако голова с удлиненной и расширенной мордой (рис. 82). Носовой отдел широкий, выступающий вперед, безволосый и внешне несколько напоминает пятачок свиньи. Когти на передних лапах очень большие. Волосяной покров ниже, чем у скунсов двух предыдущих родов. Характерны две цветовые вариации: от затылка по всей спине, расширяясь к хвосту, тянется белое поле или две белые полосы проходят по бокам спины от затылка до бедер. Хвост белый, все остальные части тела черные.
3 12 1
Зубная формула: i — с — р — m — = 32.
3 13 2
Ареал охватывает обширную территорию от юго-запада США на юг до Магелланова пролива.
Населяют леса и открытые ландшафты. Держатся поодиночке. Активны ночью. День проводят в расщелинах скал, среди камней или в норах других животных. Питаются в основном насекомыми и их личинками. Поедают также мелких позвоночных и некоторые растительные объекты, в частности плоды. Пищу обычно добывают, роясь в грунте когтями и носом. Самки приносят 2—5 детенышей. Беременность длится 42 дня.
Мех свиноносых скунсов использует местное население.
В роде 7 видов:
свиноносый скунс — С. mesoleucus Lichtenstein, 1832 (штаты США — юго-восток Юты, Нью-Мексико, кроме северо-запада и юго-востока, южная половина Техаса, центральные части запада Аризоны и ее юго-восток; Мексика, кроме штатов северо-запада Соноры, Тамаулипас, восточной половины Сан-Луис-Потоси, Веракрус, юга и севера Чиапас, Табаско, Кампече, Юкатан, Кинтано-Роо; центральные районы Гватемалы; Гондурас, кроме северных прибрежных районов; северо-запад Никарагуа);
восточномексиканский скунс — С. leuconotus Lichtenstein, 1832 (США — крайний юг штата Техас; штаты Мексики — Тамаулипас, восточная половина Сан-Луис-Потоси, Веракрус, кроме южной трети);
полуполосый скунс— С. semistriatus Boddaert, 1784 (Мексика — южная половина штата Веракрус, Табаско, Кампече, Юкатан, Кинтано-*°°, Белиз; север Гватемалы; север Гондураса; северо-восток Никара-гУа; Коста-Рика; Панама; северо-восток Бразилии; Западный Эквадор; к>г, север и центр Колумбии; Западная Венесуэла; Северное и Северо-Западное Перу);
аргентинский скунс—С. castaneus D’Orbigny et Gervais, 1847 (Аргентина) ;
<
южноамериканский скунс — С. chinga Molina, 1782 (Аргентина, Центральное Чили, Юго-Восточная Бразилия, Уругвай);
скунс Гумбольдта — С. humboldtii Gray, 1837 (Патагония в Аргентине, Южное Чили);
королевский скунс — С. rex Thomas, 1898 (Центральное и Южное Перу, Западная Боливия, Северное Чили и крайний северо-запад Аргентины).
По Уокеру (1964), в роде 6 видов.
SUBFAMILIA LUTRINAE BA1RD, 1857
Род выдр
Genus Lutra Brisson, 1762
Размеры мелкие и средние. Длина тела 55—95 см, длина хвоста 26—55 см. Масса 3,7—11,1 кг. Тело вытянутое, относительно тонкое, очень гибкое, примерно одинаковой толщины по всей длине (рис. 83)-Шея длинная и гибкая. Голова небольшая, сильно уплощенная. Ушиши-
у - Kuuny
роко расставлены, неоолыпие, округлые. Наружные слуховые проходы могут закрываться специальными клапанами. Глаза большие. Ноздри способны замыкаться при нырянии. Конечности короткие, стопоходящие. Подошвы лап голые. Между пальцами имеются плавательные перепонки. Когти небольшие. Хвост длинный, очень толстый у основания и постепенно сужающийся к концу, слегка уплощенный в дорзо-вентральном направлении. При медленном движении выдры в воде в качестве локомоторных органов служат лапы. При быстром движении выдра прижимает лапы к туловищу и двигается змееобразными движениями туловища и хвоста. Волосяной покров низкий, очень ровный по всему телу, густой, с многочисленными пуховыми волосами. На брюхе волосяной покров гуще, чем на спине. Окраска спинной части тела темно-бурая. Брюхо немного светлее спины с примесью серебристых желтоватых или белесых тонов. В отличие от остальных представителей семейства (кроме калана) хвост покрыт такими же, как и на туловище, короткими, плотно прилегающими волосами. Имеются анальные железы. Сосков 2—3 пары. Окраска хвоста такая же, как и спины.
Череп очень широкий и сильно уплощенный. Лицевой отдел укороченный, мозговая коробка крупная, широкая (ее ширина превышает длину). Скуловые дуги развиты сильно и далеко расставлены в стороны. Сагиттальный гребень маленький. Заглазничное пространство резко сужено. Заглазничные отростки крупные. Костное небо длинное и узкое. Костные слуховые барабаны небольших размеров, слабо вздутые, уплощенные.
„ 3 14 1
Зубная формула: i — с -— р —— m —— = 36. 3 13 2
Хищнические зубы развиты сильно. Клыки тонкие и длинные. Пред-коренные зубы с острыми режущими выступами, коренной зуб верхней челюсти с уплощенной коронкой и небольшими тупыми вершинами.
Диплоидное число хромосом у канадской и гладкошерстной выдры 38.
Распространены чрезвычайно широко: по всей Европе; в Азии (кроме крайнего севера и Аравийского п-ова), включая о-ва Шри Ланка, Ява, Суматра, Калимантан, Банка, Хайнань, Тайвань, Японские и Сахалин; в Африке: северо-западные районы и вся территория к югу от Сахары, кроме крайнего юга и юго-запада); в Северной Америке, за исключением некоторых областей крайнего севера и пустынь на юге; в Центральной и Южной Америке, включая Огненную Землю.
Встречаются почти во всех ландшафтах и всегда около водоемов, как правило, пресноводных. Активны в сумерки и ночью, а там, где их не беспокоят, и днем. Держатся обычно поодиночке или семейными группами (самка с молодыми). Убежищем служат постоянные норы и временные укрытия (по данным некоторых авторов, выдра не имеет постоянных нор, а только выводковые и временные). У постоянной норы вход открывается под водой, затем идет наклонный ход к берегу выше уровня воды длиной 1,5—2 м, оканчивающийся гнездовой камерой диаметром 40—50 см. Временными убежищами бывают подмывы берега, кучи плавника, подмытые корни деревьев. В питании выдры преобладает рыба. На Сахалине в основном выдра питается мелкими непро-мысловыми рыбами длиной 5—20 см. За сутки взрослый зверь съедает примерно 1 кг рыбы, редко отлавливая больше, чем может съесть. ПИ' щей служат также лягушки, птицы, млекопитающие, водные насекомые, речные раки, пресноводные моллюски. Течка и спаривание выдры про' исходят часто в марте. Гон сопровождается жестокими драками самцов-182
Беременность длится 10—12,5 месяцев. Длительность латентного периода в развитии яйцеклетки весьма неустойчива и зависит от воздействия внешней среды. После окончания латентного периода развитие зародыша заканчивается в течение 60 дней. Самка приносит от 2 до 4 детенышей, по-видимому, раз в два года. Молодые прозревают в возрасте примерно 35 дней. Самец в воспитании молодых участвует редко. Половая зрелость наступает у самок на 3-м году, у самцов в конце 3-го, начале 4-го года жизни. Постоянных врагов, кроме человека, у выдры нет, как нет и серьезных конкурентов.
Хотя выдра имеет ценный, прочный и теплый мех, существенного практического значения она не имеет ввиду малочисленности. В связи с этим выдра не может оказывать отрицательного влияния на рыбное хозяйство. Для Сахалина показан полезный экономический эффект выдры в рыбном хозяйстве (Вшивцев, 1972), поскольку она питается в основном мелкими непромысловыми рыбами, которые во множестве уничтожают икру и мальков ценных видов лососевых. Все выдры Сахалина (примерно 5500 особей) за год съедают около 100 млн. непромысловых рыб, которые могли бы уничтожить такое количество икры и мальков лососевых, которое равно производительности всех рыборазводных заводов острова. В СССР с 1960 по 1970 г. ежегодно заготовляли по 2,4—9,0 тыс. шкур выдры.
Система рода окончательно не установлена. По-видимому, в роде 12 видов:
выдра-—L. lutra Linnaeus, 1758 (вся Европа, включая Англию и Ирландию, однако отсутствует на Балеарских о-вах, Корсике, Сардинии, Крите и Кипре; Азия — ?Лалая Азия, Сирия, Палестина, север Ирака, Афганистан, север и юг Индии (в центральных районах нет), о-в Шри Ланка, Ассам, Бирма, п-ова Индокитай и Малакка, о-ва Суматра и Ява, ?ЛНР, Китай, о-ва Хайнань и Тайвань, Япония; Африка—Алжир и Марокко; в СССР — почти вся страна, северная граница ареала в европейской части проходит от государственной границы по морскому побережью до устья Печоры и отсюда к южной Обской губе, южному Таймыру, верховьям Вилюя и севернее от него выходит на Лену несколько ниже устья Вилюя, далее следует к истокам Индигирки, среднему течению Колымы, охватывает с севера бассейн Анадыря и в его устье выходит к Тихому океану, кроме того, в ареал входят Сахалин и Шантарские острова; южная граница идет от устья Дуная по Черноморскому побережью до устья Дона, исключая Крым, следует по Дону и в области сближения Дона и Волги переходит на Волгу и охватывает узкой полосой ее долину до дельты; в Калмыцких и Предкавказских степях выдры нет; изолированный с севера участок ареала охватывает весь Кавказ, Далее южная граница с Волги на р. Урал переходит, вероятно, на широте Иргиза, вдоль по Уралу выдра распространена до его устья; по Р- Урал южная граница огибает Уральские горы и следует на восток по границе лесной зоны и лесостепи до Иртыша и по нему вверх до его истоков, затем идет на Обь, южнее Новосибирска огибает Алтай с севера и выходит на Иртыш; в Средней Азии выдра распространена на Копет-Даге и по Атреку до устья, по р. Теджену, р. Кушке, есть на Западном Памире, в Джунгарском и Заилийском Алатау);
канадская выдра — L. canadensis Schreber, 1776 (почти вся Северная Америка, кроме юга: Аляска, кроме прибрежной зоны Моря Бофорта; Канада, кроме северо-востока Северо-Западных Территорий и Канадского Арктического архипелага; США, кроме южной половины штата Калифорния, центральных районов Невады, западной половины Юты, крайнего юга Аризоны, юга и юго-востока Нью-Мексико, юга и юго-запада Техаса);
островная выдра — l. mira uoioman, гули jo-ob прница олшъпищ в архипелаге Александра, о-ва США в Тихом океане у северной части Британской Колумбии);
мексиканская выдра — L. annectens Major, 1897 (юг Мексики, штаты — юг Синалоа, южная половина Дуранго, Наярит, Халиско, Колима, Герреро, Мексико, Веракрус, Пуэбла, Чиапас, Оахака, Табаско, Кампече, Юкатан, Кинтано-Роо; Белиз; Гватемала, Сальвадор, Гондурас; Никарагуа; Коста-Рика; Панама; Колумбия; Западная Венесуэла; Западный Эквадор);
гвианская выдра — L. enudris F. Cuvier, 1823 (Восточная Венесуэла, Гайана, Суринам, Гвиана, Северо-Восточная Бразилия);
кошачья выдра — L. felina Molina, 1782 (в недалеком прошлом Тихоокеанское побережье Южной Америки от Перу на юг до Огненной Земли включительно, в настоящее время главным образом на островах — от о-ва Чилоэ у центральной части Чили и к югу до Магелланова пролива; в водах Перу сохранилось всего 200—300 особей);
перуанская выдра — L. incarum Thomas, 1908 (Юго-Восточное Перу и, возможно, прилегающие районы Боливии) ;
лаплатская выдра — L. platensis Waterhouse, 1838 (Южная Бразилия, Парагвай, север Аргентины, Уругвай; везде крайне малочисленна, а в Аргентине, по-видимому, исчезла);
южная выдра — L. provocax Thomas, 1908 (Западная Патагония в Аргентине, Центральное и Южное Чили; везде крайне малочисленна, а в Аргентине, по-видимому, исчезла);
гладкошерстная выдра — L. perspicillata Geoiiroy, 1826 (Индия,втом числе Ассам; Непал; Бирма; возможно, Китай — восток Юньнаня; п-ова Индокитай и Малакка, о-в Суматра);
суматранская выдра — L. sumatrana Gray, 1865 (о-ва Суматра, Калимантан, Банка, п-ов Малакка и п-ов Индокитай на север до северной части Вьетнама);
белогорлая выдра — L. maculicollis Lichtenstein, 1835 (Южно-Африканская Республика, север Ботсваны, север Юго-Западной Африки, Ангола, Замбия, Малави, Мозамбик, на север до Южного Сомали, Эфиопии, Судана и на запад до Либерии).
Иногда L. perspicillata выделяют в особый род Lutrogale Gray, 1865. В. Г. Гептнер (1967) считает вероятным, что L. mira — лишь подвид L. canadensis.
В «Красную книгу» включены L. felina, L. platensis и L. provocax как виды, которым грозит исчезновение.
Род гигантских выдр
Genus Pteronura Gray, 1837
В роде единственный йид: гигантская выдра — Р. brasiliensis Gmelin, .1788.
Размеры средние и крупные. Длина тела 100—150 см, длина хвоста примерно 70 см. Масса около 24 кг. Внешним обликом не отличается от выдры, но имеет значительно более крупные размеры, сильнее уплощенный в дорзо-вентральном направлении хвост (рис. 84). Окраска шоколадно-бурая с беловатым пятном на груди и горле. Специфические анальные железы хорошо развиты. Зверь способен выбрызгивать их секрет на некоторое расстояние.
Распространена от Восточной Венесуэлы, Гвианы и Северо-Восточно Бразилии на юг до Южной Бразилии, Уругвая, Парагвая и Северо-Восточной Аргентины. Везде малочисленна, в ряде мест истреблена.
Рис. 84. Гигантская выдра — Р. brasiliensis
Населяет медленно текущие речки и реки. Образ жизни, по-видимому, мало чем отличается от образа жизни выдры. Активна днем. Убежищем служат пещеры в берегах рек. Держится обычно небольшими группами. Поедает в основном рыб, а также водных млекопитающих, птиц и их яйца. В помете 1—2 детеныша.
Включена в «Красную книгу» как вид, находящийся под угрозой исчезновения.
Род бескоготных выдр
Genus Aonyx Lesson, 1827
Размеры средние. Длина тела 61—100 см, длина хвоста 30—55 см. Масса 14—23 кг. По внешнему облику сходны с выдрой (рис. 85). На передних конечностях между пальцами у их основания расположены слабо развитые перепонки. Когтей на пальцах нет или они развиты очень слабо. На задних конечностях плавательная перепонка развита несколько сильнее и местами достигает частей первой фаланги. Окраска желтовато-бурая или бурая, иногда серая. Брюхо может быть светлее спины. На горле и груди иногда есть белые пятна.
Для щечных зубов характерны уплощенные, расширенные жевательные поверхности, приспособленные к раздроблению раковин моллюсков и панциря ракообразных.
Рис 85. Капская выдра — A. capensis. А — кисть; Б — стопа
Диплоидное число хромосом у серой выдры 38.
Ареал охватывает Африку от мыса Доброй Надежды к северу до Эфиопии, бассейна Конго и Либерии и Южную и Юго-Восточную Азию с о-вами Суматра, Ява, Калимантан и Палаван.
Населяют долины и устья рек. Слабое развитие перепонок на лапах не мешает бескоготным выдрам превосходно плавать и нырять. Активны преимущественно ночью, хотя в местах, где их не беспокоят, зверей можно видеть в любое время суток. Держатся семейными группами. Как правило, собственных нор не копают, а находят себе убежище под навесом размытого берега, стволами плавника. Питаются в основном пресноводными ракообразными, а также моллюсками, черепахами, рыбами, лягушками, ящерицами, водными птицами и мелкими млекопитающими. Поедая добычу, берут ее передними лапами. В помете от 2 до 5 детенышей. Беременность длится около 63 дней. Чаще всего рождение детенышей происходит весной. Молодые остаются с родителями по крайней мере в течение одного года. Основные враги — крокодилы.
В роде 2 вида:
серая выдра — A. cinerea liliger, 1815 (Южный Китай — Юньнань, Фуцзянь, о-в Хайнань; Северная Бирма; Индия, в том числе Ассам; Непал; п-ова Индокитай и Малакка; о-ва Суматра, Ява, Калимантан, Палаван);
капская выдра — A. capensis Schinz, 1821 (Южно-Африканская Республика, Родезия, Ангола, Замбия, Мозамбик и далее на север до Кении и Эфиопии и на запад до Сенегала).
Некоторые исследователи (например, Уокер, 1964; Соколов, 1973) выделяют серую выдру в особый род АтЫопух Rafinesque, 1832. Иногда в род Аопух включают Paraonyx Hinton, 1921, в качестве подрода (например, Андерсон и Джонс, 1967; Харрис, Harris, 1968; И. Соколов; 1973; Розивир, Rosevear, 1974).
Род африканских выдр
Genus 'Paraonyx Hinton, 1921
В роде единственный вид: африканская выдра — Р. congica Lonnberg, 1910.
Размеры средние. Длина тела около 60 см, длина хвоста примерно 35 см. Масса около 7 кг. По внешнему облику сходна с выдрой, однако на передних конечностях пальцы имеют очень небольшие плавательные перепонки, тогда как на задних лапах они развиты сильно. Когти на пальцах маленькие и тупые. Окраска меха светло-бурая с сероватыми и беловатыми лицевыми и горловыми отметинами.
Зубы развиты слабо, брахиодонтного типа.
Распространена в бассейне р. Конго, в Руанде, Бурунди, Юго-Западной Уганде, Габоне, Камеруне и прилегающих районах Нигерии.
Встречается только в густых дождевых лесах, по берегам небольших горных потоков. Экология изучена крайне слабо. По-видимому, африканская выдра по сравнению с другими выдрами наименее приспособлена к водному образу жизни. Питается в основном мелкими наземными позвоночными, в меньшей степени рыбой. В помете 2—3 детеныша. Беременность длится около 2 месяцев.
Некоторые исследователи (например, Уокер, 1964) выделяют в роде еще 2 вида: Р. microdon Pohle, 1920, и Р. philippsi Hinton, 1921.
Подвид из Камеруна — Р. с. microdon Pohle, 1920, включен в «Красную книгу» как находящийся под угрозой исчезновения.
Род каланов
Genus Enhydra Fleming, 1822
В роде единственный вид: калан—Е. lutris Linnaeus, 1758.
Размеры средние и крупные. Длина тела 100—150 см, длина хвоста 30—36 см. Масса до 36—42 кг. Общий вид весьма своеобразен. Сложение тяжелое; туловище сильно удлиненное, цилиндрическое. Шея короткая и утолщенная. Голова округлая, притупленная (рис. 86). Ушные раковины очень малы, обычно скрыты в волосяном покрове. Щелевидный слуховой проход может смыкаться при нырянии. Ноздри также щелевидные и тоже способны закрываться. Глаза небольшие. Вибриссы очень длинные и многочисленные. Конечности сильно укороченные, причем задние отставлены далеко назад. Стопа превратилась в ласт, все ее пальцы одеты общим кожным покровом; V палец наиболее длинный, а I — самый короткий. Ласт с верхней и с нижней поверхности покрыт густыми волосами. Очень короткая передняя конечность не имеет расчлененных пальцев. На ее подошве расположены своеобразные кожные подушки: одна длинная поперечная и спереди от нее четыре небольших, из которых вторая от наружного края заключает два пальца (III и IV), остальные — по одному. Подушки не покрыты волосами. Когти на передних и задних конечностях очень небольшие.
Хвост относительно короткий, несколько уплощенный в дорзо-вент-ральном направлении. Волосяной покров по всему телу почти одинаково не очень высокий, но необычайно густой и мягкий с сильно развитыми пуховыми волосами. Общая окраска его темно-бурая (индивидуальные вариации от рыжего до почти черного), морда буровато-серая, голова
Рис. 86. Калаи— Е. lutris, его череп и нижняи поверхность кисти
почти белая. Нижняя поверхность тела намного светлее свины. Хвост покрыт короткими густыми волосами. Сезонной изменчивости в окраске и густоте меха почти нет. Линька происходит постепенно на протяжении всего года с интенсификацией у командорских каланов в апреле — июле. В зимнее время на брюхе волосы гуще, чем на спине. Характерно почти полное отсутствие подкожного жирового слоя. Сосков функционирует одна пара; иногда спереди от них имеется еще одна пара рудиментарных.
Череп массивный, укороченный и широкий. Особенно укорочен лицевой отдел, который как бы срезан спереди. Длина лицевого отдела примерно в два раза короче мозговой части. Бугры и гребни черепа резко выражены. Скуловые дуги сильно развиты и широко расставлены в стороны. Заглазничный перехват резко выражен. Костные слуховые барабаны крупные, расположены поперечно, уплощенные. Носовые кости короткие и широкие. Характерно очень крупное носовое отверстие. У многих особей наблюдается асимметрия черепа, имеющаяся уже у эмбрионов и новорожденных.
3 13 1
Зубная формула: 1 —- с — р —- m — =32. 2 13 2
Очень своеобразно отсутствие нижнего первого резца. Хищнические зубы не развиты. Характерны мощные верхние третий предкоренной и коренной, причем коренной больше предкоренного. Жевательные поверхности щечных зубов почти плоские с тупыми закругленными вершинами — они приспособлены к перетиранию пищи.
В скелете характерны широкая лопатка, узкий и длинный таз, поставленный параллельно позвоночнику, массивная укороченная скрученная по спирали плечевая кость и сильно укороченное бедро. Большая берцовая кость толстая, малая — очень тонкая.
Своеобразны слабо дифференцированный очень длинный кишечник, превышающий примерно в 10 раз длину тела животного, и очень большой желудок. Легкие относительно крупные, в связи с этим диафрагма косо поставлена и прикрепляется по спинной стороне дальше назад, чем на брюшной (признак настоящих водных животных). Семенники располагаются под кожей.
Диплоидное число хромосом 38.
Распространены в СССР у Командорских о-вов, южной оконечности Камчатки и Курильских о-вов; в прошлом обитали, кроме того, на Южном Сахалине, по восточному берегу Камчатки и, возможно, шли на север приблизительно до 60° с. ш. (или даже 64° с. ш.). Вне СССР встречались на Хоккайдо, севере Хонсю, на Алеутских и Прибыловых о-вах, по Тихоокеанскому побережью Северной Америки и прилежащим островам примерно от 62° с. ш. на юг до средних частей или даже южной оконечности п-ва Калифорния. Из-за неумеренного промысла каланы были почти полностью истреблены на всем ареале. В результате охранных мероприятий к настоящему времени ареал каланов в значительной степени восстановлен.
Обитает по морским побережьям с крутыми берегами, многочисленными рифами, обширными зарослями из водорослей. Как правило, держится недалеко от берега в пределах 18—30 км. В тихую погоду остается на воде и на ночь. Для отдыха, а также во время штормов вылезает на камни и каменистый берег. Летом может ночью спать в воде, иногда нарочно запутываясь в водорослях, чтобы не снесло течением. Активен преимущественно днем; в летнее время в основном утром и вечером, а зимой в течение всего дня. На берегу калан неуклюж, передвигается
медленно, сильно выгибая на ходу спину. В воде же он очень подвижен, легко плавает и ныряет, оставаясь под водой обычно 1-—3 мин, а в случае опасности — до 5 мин. Часто лежит брюхом кверху на поверхности воды с пойманной добычей, прижимая ее передними лапами к груди и поедая в такой позе. В этом же положении самка держит на груди своего детеныша. Животные встречаются обычно небольшими стадами. У курильской популяции каланов отмечена сезонная миграция — в зимнее время к югу в район о-вов Итуруп и Уруп, а летом к северу — к о-ву Па-рамушир, а может быть, и к побережью Камчатки. Пищей служат в основном морские ежи, в меньшей степени двухстворчатые моллюски, осьминоги, крабы, раки-отшельники и некоторые рыбы.
Определенных сроков гона нет. Однако наиболее часто самки с новорожденными встречаются на Командорах с апреля по июль, а в американской популяции — в апреле — мае. Длительность беременности 8— 9 месяцев. Роды происходят на суше. Самка приносит, как правило, одного детеныша. Новорожденный зрячий, с молочными зубами, в ювенильном меху. Имеет общую длину с хвостом около 55 см, массу 1,5 кг. Через 2,5—3 недели после рождения самостоятельно плавает на большие расстояния. У полугодовалого детеныша общая длина возрастает до 110— 115 см, масса до 11 кг. С матерью детеныш остается минимум один год. К этому времени общая длина тела достигает 125—130 см, масса 17— 19 кг. Половой зрелости каланы достигают, по-видимому, на 3-м году жизни (по некоторым данным, в 1,5—2 года). Основной враг каланов — косатка.
Калан имеет самый ценный мех. Промысел калана достигал больших размеров. Так, в 1798—1845 гг. Российско-Американской компанией было реализовано 118 тыс. шкур каланов, к 1867 г. русскими было вывезено 260 тыс. шкур. В 1871—1890 гг. американцы добыли на Аляске свыше 90 тыс. каланов. Неумеренный промысел привел примерно к середине XIX в. почти к полному уничтожению этого зверя. Первые попытки охраны каланов (хотя и безуспешные) были предприняты еще Российско-Американской компанией. В 1911 г. была заключена Вашингтонская международная конвенция по охране морского котика и калана; в 1913 г. на Алеутских о-вах был организован долгосрочный национальный заповедник. После Великой Октябрьской социалистической революции промысел каланов в СССР был запрещен законом и была организована охрана этого зверя, которая дала хороший результат, и численность каланов постепенно возросла. Если каланов не беспокоить, они успешно уживаются и по соседству с человеком. По учету 1963 г., в СССР курильское стадо достигало 3300—3500 голов, камчатское, по-видимому, — нескольких сотен голов, командорское — несколько более 1500 голов. К 1968 г. общее число каланов составляло не менее 40 тыс. голов, из которых примерно 1/5 часть обитала в СССР (Барабаш-Никифоров и др., 1968). В США численность каланов у берегов Аляски и Алеутских о-вов составляла 27—47 тыс., а у берегов Калифорнии — около 650 голов. В 1962 г. у американских берегов был проведен пробный промысел каланов.
Попытки искусственного переселения каланов по большей части заканчивались безуспешно. С 1937 по 1940 г. два калана, привезенные с о-ва Медного, жили в вольере на Мурманском побережье, а один из них, случайно оказавшийся на воле, прожил до 1944 г. По-видимому, успешно закончилось заселение каланами прибрежных вод о-ва Беринга в 1969 и 1970 гг.
СЕМЕЙСТВО ВИВЕРРОВЫХ
FAMILIA VIVERRIDAE GRAY, 1821
Размеры мелкие. Длина тела 17—100 см1, длина хвоста 12—90 см. Масса 0,7—15 кг. Внешний вид весьма своеобразен, но в общем напоминает внешний вид представителей семейства куньих, а иногда — кошачьих. Туловище и хвост длинные. У Arctictis хвост хватательный. Шея средней длины, голова небольшая с удлиненной, утончающейся к носу лицевой частью. Уши невысокие, обычно округлые, широко расставленные. Глаза большие. Конечности короткие, пятипалые, но I и V пальцы располагаются выше уровня других и не функционируют. Иногда на задних конечностях отсутствует I палец, реже его нет и на передних и задних конечностях. Когти втяжные или невтяжные. Конечности пальцеходящие или полустопоходящие. Волосяной покров низкий, довольно грубый, обычно с большим количеством полос или пятен, иногда одноцветный. Превалирует бурая расцветка. Хвост часто имеет цветные кольца. Сосков две или три, реже одна пара. У большинства в анальной области расположены специфические кожные железы.
Череп низкий с удлиненным лицевым отделом. Крылоклиновидный канал обычно имеется.
3 1 3—4 1—2
Зубная формула: 1 —- с — р —- m -— = 32 — 40.
3 13" ~ 4 1 2
Хищнические зубы относительно невелики. Первый нижний резец выше двух других. Клыки и предкоренные зубы маленькие. Коренные зубы крупные.
Распространены в Южной Европе, Африке, на Мадагаскаре, в Южной и Юго-Восточной Азии, включая Индонезию и Филиппины. Некоторые виды акклиматизированы в различных местах, например гималайская цивета в Японии, a Herpestes на Гавайских о-вах и в Вест-Индии.
Встречаются преимущественно в лесах, густых зарослях кустарника и высокой травы. Активны ночью или днем. Убежища находят в дуплах деревьев, пещерах, реже в норах (обычно занимают чужие норы). Некоторые живут в постройках человека. Держатся поодиночке, парами или группами. Некоторые (например, Cynictis) живут колониями в земляных норах, что вообще нетипично для хищных. Хорошо лазают по деревьям, и некоторые виды проводят на них большую часть своей жизни. Представители трех родов — Osbornictis, Atilax и Cynogale — полуводные. Большинство хишники: питаются различными мелкими позвоночными и беспозвоночными. Второстепенным объектом питания может служить растительная пища — плоды, орехи, луковицы и т. д. Некоторые виды поедают падаль. Органы чувств хорошо развиты. Гон приходится на весну и лето; у некоторых видов сезонности в размножении нет. Длительность беременности известна у немногих видов: примерно 60 дней. В помете от 1 до 6 слепых, но покрытых волосами детенышей. У отдельных видов в году бывает 2 помета.
В семействе 6 подсемейств, 35 родов и 71 вид:
Subfamilia Viverrinae Gill, 1872
африканских линзангов — Poiatia Gray, 1865 (1 вид);
генет—Genetta Oken, 1816 (9 видов);
малых цивет—Viverricula Hodgson, 1838 (1 вид);
водяных цивет — Osbornictis J. Allen, 1919 (1 вид);
1 Представителей семейства можно разделить на 3 группы: мелкие с длиной тела J7—44 см, средние — 45—72 и крупные—73—100 см.
цивет—Viverra Linnaeus, 1758 (4 вида);
пятнистых линзангов — Prionodon Horsfield, 1824 (2 вида).
Subfamilia Paradoxurinae Gill, 1872
пальмовых цивет — Nandinia Gray, 1843 (1 вид);
мелкозубых цивет — Arctogalidia Merriam, 1897 (1 вид);
мусангов — Paradoxurus F. Cuvier, 1821 (3 вида);
гималайских цивет — Pagutna Gray, 1831 (1 вид);
сулавесских цивет — Macrogalidia Schwarz, 1910 (1 вид);
бинтуронгов — Arctictis Temminck, 1824 (1 вид).
Subfamilia Hemigalinae Gill, 1872
фаналок — Fossa Gray, 1864 (1 вид);
полосатых цивет — Hemigalus Jourdan, 1837 (1 вид);
цивет Оустона — Chrotogale Thomas, 1912 (1 вид);
калимантанских цивет—Diplogale Thomas, 1912 (1 вид);
выдровых цивет — Cynogale Gray, 1837 (1 вид);
мелкозубых мунго — Eupleres Doyere. 1835 (2 вида).
Subfamilia Galidiinae Gill, 1872
кольцехвостых мунго — Galidia I. Geoffrey, 1837 (1 вид);
широкополосых мунго — Galidictis I. Geoffrey, 1839 (2 вида);
узкополосых мунго — Mungotictis Pocock, 1915 (2 вида);
бурохвостых мунго — Salanoia Gray, 1864 (2 вида).
Subfamilia Herpestinae Gill, 1872
сурикат — Suricata Desmarest, 1804 (1 вид);
мангустов — Herpestes liliger, 1811 (14 видов);
карликовых мангустов — Helogale Gray, 1862 (2 вида);
водяных мангустов — Atilax Cuvier, 1826 (1 вид);
полосатых мангустов — Mangos Е. Geoffrey et G. Cuvier, 1795 (1 вид);
кузиманз—Crossarchus F. Cuvier, 1825 (3 вида);
либерийских мангустов — Liberiictis Hayman, 1958 (1 вид);
белохвостых мангустов — Ichneumia I. Geoffrey, 1837 (1 вид);
черноногих мангустов — Bdeogale Peters, 1850 (3 вида);
умби — Rhynchogale Thomas, 1894 (1 вид);
желтых мангустов—Cynictis Ogilby. 1833 (1 вид);
серых мангустов — Paracynictis Pocock, 1916 (1 вид).
Subfamilia Cryptoproctinae Trouessart, 1885
фосс— Cryptoprocta Bennett, 1833 (1 вид).
Существенного практического значения не имеют, за исключением случаев, когда, будучи интродуцированными в некоторые области, они размножились в больших количествах (Herpestes) и уничтожают большое количество мелких млекопитающих и птиц.
SUBFAMILIA VIVERRINAE GILL, 1872
Род африканских линзангов
Genus Poiana Gray, 1865
В роде единственный вид: африканский линзанг — Р. richardsoiu Thomas, 1842.
Размеры мелкие. Длина тела около 33 см, длина хвоста примерно 38 см. Тело удлиненное, конечности средней высоты. Голова небольшая с вытянутой мордой. Уши средней высоты, слегка заостренные. Хвост очень длинный (рис. 87). Окраска меха светло-буровато-серая или ржаво-желтая с темно-бурыми или черными пятнами и кольцами на спине и боках. Хвост с темными узкими и широкими, иногда только широкими 192
Рис. 87 Африканский линзанг — Р. rlchardsoni и его череп
кольцами. Пятна на туловище не образуют полос и не располагают рядами. На шее и голове имеются темные полосы. п„Л„„ппй Кпгти
Распространен в Сьерра-Леоне, Либерии, на Береге ль гема-и от юга Камеруна до севера Заира, а также на о-ве Ьиного.
Экология изучена слабо. о няп
Населяет леса. Строит гнезда на деревьях на высоте о поме-землей. Активен ночью. Всеяден. Дает два помета в год. В Д
те 2 или 3 детеныша. пилР1,Яют в оо-
Некоторые исследователи (например, Розивир, 1974) в>д
Де еще один вид — Р. leightoni Pocock, 1908.
Рис. 88. Пятнистая генета — G. tigrina
Род генет
Genus Genetta Oken, 1816
Размеры мелкие и средние. Длина тела 42—58 см, длина хвоста 39— 42 см. Масса около 2 кг. Общий облик, типичный для виверр. Туловище удлиненное, конечности короткие, когти короткие, искривленные, втягивающиеся; хвост длинный, но короче тела. Морда средней длины, заостренная (рис. 88). Волосяной покров мягкий и густой. Окраска изменчива, но у большинства видов сероватая или желтоватая с бурыми или черными пятнами на спине и боках, образующими продольные ряды. Хвост в белых и черных кольцах, примерно одинаковой ширины. На хвосте волосы более длинные, чем на туловище. Среди представителей этого рода обычны меланисты. Брюшных сосков 2 пары.
Диплоидное число хромосом у малой генеты 52.
Распространены в Юго-Западной Европе, на Аравийском п-ове, в Израиле и Африке, за исключением Сахары.
Населяют леса, кустарники, заросли высокой травы. Активны преимущественно ночью. День проводят в расщелинах скал, в норах, выкопанных другими животными, в дуплах или ветвях деревьев. Обычно встречаются поодиночке или парами. Хорошо лазают по деревьям, но большую часть времени проводят на земле. Питаются мелкими млекопитающими, птицами, пресмыкающимися, а также насекомыми. В году бывает два помета (в Южной Африке весной и осенью), в каждом из которых 2—3 детеныша.
В неволе могут жить до 34 лет.
В роде 9 видов:
ложная генета — G. villiersi Dekevser, 1949 (от Сенегала и Гамбии на восток до области южнее озера Чад);
эфиопская генета — G. abyssinica Rflppell, 1836 (Эфиопия);
генета Джонстона — G. johnstoni Pocock, 1907 (Либерия);
гигантская генета — G. victoriae Thomas, 1901 (Заир — бассейн Р- Конго к северу от Эквадора);
обыкновенная генета — G. genetta Linnaeus, 1758 (Ливия, Алжир, Марокко и затем от Сенегала, огибая Сахару с юга, до Красного моря и от Эфиопии через Восточно-Центральную Африку до Мозамбика, Замбии, Анголы и Южно-Африканской Республики);
ангольская генета — G. angolensis Bocage, 1882 (Ангола; Замбия, Заир);
серваловая генета — G. servalina Pucheran, 1855 (юг Нигерии, Камерун, Габон, бассейн р. Конго и на восток до Кении);
лесная генета —G. pardina 1. Geoffrey, 1832 (Западная Африка от 1 амбии до Камеруна);
пятнистая генета — G. tigrina Schreber, 1778 (от Южно-Африканской Республики и Намибии на юге до Эфиопии на севере).
. Иногда из G. tigrina выделяют в качестве отдельных видов G. rttbi-ginosa Pucheran, 1855, и G. mossambica Matschie, 1902. По Уокеру (1964), в роде 6 видов, а по Андерсону и Джонсу (1967), — 5.
Род малшх цивет
Genus Viverricula Hodgson, 1838
В роде единственный вид: малая цивета — V. indica Desmarest, 1817
Размеры средние. Длина тела 45—63 см, длина хвоста 30—43 см" Масса 2—4 кг. Туловище удлиненное, конечности короткие, хвост длинный, но короче туловища. Голова небольшая с укороченной заостренной мордой (рис. 89). Уши средней высоты, широкие в основании, сближены между собой на верху головы, с закругленными вершинами. Волосяной покров редкий и грубый, на хвосте несколько удлиненный. Окраска его желто-бурая, буроватая или сероватая; на конечностях черная. По передней части тела и спине разбросаны мелкие черные пятна. На боках они крупнее и образуют продольные ряды. По спине могут проходить от б до 8 темных полос. Хвост с чередующимися 6—9 белыми и 6—9 черными кольцами. Хорошо развиты анальные железы.
Диплоидное число хромосом 36.
Распространена в Индии, на о-ве Шри Ланка, в Южном Китае, на о-вах Тайвань и Хайнань, в Юго-Восточной Азии до Суматры, Явы и Бали. Акклиматизирована на о-вах Сокотра, Коморо, Филиппинских и Мадагаскаре.
Населяет леса. Встречаются поодиночке и парами. Ведет наземный образ жизни. Активна ночью. Питается мелкими позвоночными, насекомыми и их личинками, плодами и корнями. Охотно поедает падаль. На о-ве Шри Ланка сезонности в размножении не отмечено. В помете 2— 5 детенышей. Местные жители иногда содержат малых цивет в неволе с целью получения от них секрета анальных желез. Этот секрет употребляют в качестве добавки к табаку для придания ему специфического запаха.
Род водяных цивет
Genus Osbornictis J. Allen, 1919
В роде единственный вид: водяная цивета — О. piscivora J. Allen, 1919.
Размеры средние. Длина тела примерно 50 см, длина хвоста около 41 см. Внешний вид, типичный для цивет. Морда средней длины, заостренная. Волосяной покров высокий, густой и мягкий. Окраска его одноцветная каштаново-красная или темно-рыжая. Хвост черный с удлиненными волосами. Между глазами два продольных небольших белых пятна. Передняя часть, а также бока морды и бока головы ниже глаз беловатые. Темных пятен и полос на туловище нет. Подошвы передних и задних лап, в отличие от представителей других родов, безволосые.
Череп удлинен. Зубы относительно невелики и слабые.
Распространена в Заире.
Экология не изучена. По-видимому, населяет берега лесных водоемов. Ведет полуводный образ жизни, питается рыбами.
Род цивет
Genus Viverra Linnaeus, 1758
Размеры средние и крупные. Длина тела 60—83 см, длина хвоста 30— 48 см. Масса 7-—11 кг. Туловище удлиненное, с боков несколько уплощенное, в крестце выше, чем в холке (рис. 90). Конечности средней высоты или короткие. Голова с вытянутой мордой; уши средних размеров, от-
<3
стоят далеко друг от друга (азиатские виды) или поставлены близко (африканская цивета). У двух видов этого рода (V. zibetha, V. tangalun-ga} III и IV пальцы передней конечности с кожными лопастями, в которые прячутся втяжные когти. Волосяной покров высокий, но редкий и грубый. По средней линии спины проходит гребень особо высоких черных волос. Удлиненные волосы имеет и хвост. Общий тон окраски рыжевато-коричневый или сероватый с черными пятнами. Шею охватывают чередующиеся черные и белые кольца (обычно три черных и два белых). Хвост также с черными и белыми кольцами. Конечности обычно черные. Сильно развиты анальные железы.
Диплоидное число хромосом у африканской и большой цивет по 38.
Распространены в Индии — Ассаме, Китае, Непале, на п-овах Индокитай, Малакка и на Филиппинских о-вах, а также в Африке.
Населяют разнообразные ландшафты. Встречаются обычно поодиночке или парами. Активны ночью. Убежищем часто служат норы других животных. В основном наземные животные, хотя могут хорошо лазать по деревьям и плавать. Питаются мелкими млекопитающими, птицами, пресмыкающимися, земноводными, насекомыми, а также плодами и корнями растений. В каждом из двух пометов, которые самка приносит в течение года, бывает от 1 до 4, обычно 2—3 детеныша. Имеют некоторое практическое значение, так как от них получают секрет анальных желез, употребляющийся в парфюмерии. Местные жители содержат африканских цивет в неволе и собирают несколько раз в неделю секрет анальной железы — от каждого зверя за неделю получают 3—4 г секрета.
В роде 4 вида: '
большая цивета — V. zibetha Linnaeus, 1758 (Китай от провинции Фуцзянь на запад до провинций Сычуань и Шэньси, о-в Хайнань и от Бирмы на запад через Ассам в Индии до Непала; п-ова Индокитай и Малакка);
крупнопятнистая цивета—V. megaspila Blyth, 1862 (Индия, Бирма, п-ова Индокитай и Малакка);
тангалунга— V. tangalunga Gray, 1832 (п-ов Малакка, Филиппинские о-ва);
африканская цивета— V. civetta Schreber, 1778 (Африка от Сенегала на восток до Сомали и на юг до Южно-Африканской Республики).
Многие исследователи (например, Симпсон, 1945; Уокер, 1964) выделяют африканскую цивету в особый род Civettictis Pocock, 1915. Следуя Эллермэну и др. (1953), здесь этот вид включен в род Viverra.
Подвид крупнопятнистой циветы—V. megaspila civettina Blith, 1862, включен в «Красную книгу» как находящийся под угрозой-исчезновения. Прежний ареал включал прибрежные области Малабара, Траванкура и Западного Гхата в Южной Индии; современный ареал неизвестен.
Род пятнистых линзангов
Genus Prionodon Horsfield, 1824
Размеры мелкие. Длина тела 37—43 см, длина хвоста 30—35 см. Масса около 0,7 кг. Сложение изящное. Туловище удлиненное, конечности короткие (рис. 91). Шея длинная. Морда удлиненная и узкая. Когти втяжные. Кожные защитные лопасти для когтей имеются на задних лапах. Волосяной покров низкий, густой и мягкий. У полосатого линзанга окраска беловато-серая или буровато-серая на. спинной стороне тела и кремовая на брюшной. Поперек спины располагаются четыре или пять широких черных и темно-бурых полос. Бока тела и конечности с темными пятнами; хвост с чередующимися светлыми и темными
кольцами. Пятнистый линзанг имеет оранжево-бурую или светло-бурую окраску. Черные пятна на верхней части туловища проходят более или менее правильными рядами. На хвосте 8 или 10 темных колец.
Диплоидное число хромосом у полосатого линзанга 34.
Распространены в Непале, Ассаме в Индии, Бирме, на п-овах Индокитай и Малакка, о-вах Суматра, Ява, Калимантан.
Экология изучена слабо. Населяют густые леса и заросли кустарников. Активны в основном ночью. Хорошо лазают по деревьям. Питаются преимущественно птицами, а также мелкими млекопитающими и насекомыми. Самка дает два помета в год, в каждом 2 или 3 детеныша. Размножение, вероятно, происходит в феврале и августе.
В роде 2 вида:
полосатый линзанг — Р. linsang Hardwicke, 1821 (юг Бирмы, п-ов Малакка, о-ва Суматра, Ява, Калимантан);
пятнистый линзанг — Р. pardicolor Hodgson, 1841 (Непал, Ассам в Индии, север Бирмы, п ов Индокитай).
Некоторые исследователи (например, Симпсон, 1945) выделяют пятнистого линзанга в особый род Parditis Thomas, 1925.
SUBFAMILIA PARADOXERINAE GILL, 1872
Род пальмовых цивет
Genus Nandinia Gray, 1843
В роде единственный вид: пальмовая цивета — N. binotata Gray, 1830.
Размеры средние. Длина гела 44—57 см, длина хвоста 46—62 см. Туловище удлиненное. Конечности короткие, с острыми искривленными ког-
I
hpipt" ?0Па И ЕИСТ!> кРУпные, подошвы их голые. Длинный хвост превы-поЛ ТеЛа' Голова широкая с укороченной мордой (рис. 92). Копт плп Расставленные уши с округлыми вершинами. Волосяной
mpvq о зкии, густой и мягкий. На хвосте волосы удлинены. Окраска МежпчГп1ительно варьирует, но в общем она сероватая или буроватая. имрк^ лечами обычно располагаются два кремовых пятна. На хвосте имеются темные кольца.
. Диплоидное число хромосом 38.
мячц2иаР°Странсна в Африке от Южно-Африканской Республики до Сомали на северо-востоке и Сенегала на северо-западе.
гг>О1л»ТСеЛЯеТ леса‘ Ведет наземный и древесный образ жизни. Активна ip имущественно ночью. Питается главным образом, по-видимому, пло-
ОЛП
Рис. 93. Мелкозубая цивета — A. trivirgata и ее череп
дами, но употребляет в пищу также и мелких млекопитающих и птип. В году, по-видимому, 2 помета, в каждом из которых от 2 до 3 детенышей. Длительность беременности около 64 дней.
Род мелкозубых цивет
Genus Arctogalidia Merriam, 1897
В роде единственный вид: мелкозубая цивета — A. trivirgata Gray, 1832.
Размеры мелкие и средние. Длина тела 43—53 см, длина хвоста 51— 66 см. Туловище удлиненное. Хвост превышает длину тела. Конечности средней высоты. Голова большая, широкая в мозговой части с относительно длинной мордой, сужающейся к носу (рис. 93). Уши небольшие, широко расставленные с закругленными вершинами. Верхняя часть туловища, проксимальный отдел хвоста и наружные части конечностей рыжевато-бурые; голова более темная. Дистальная часть хвоста буроватая. По средней линии верха морды проходит беловатая полоска. Вдоль спины тянутся три бурые или черные продольные полосы, причем средняя полоса, проходящая вдоль хребта, обычно сплошная, а боковые могут быть разбиты на отдельные пятна или совсем отсутствовать. Брюхо серовато-белое или кремово-рыже-бурое. На груди беловатое пятно. Самки имеют у мочеполового отверстия специфическую кожную железу.
Щечные зубы малы.
Распространена в Ассаме в Индии, в Бирме, Таиланде, на п-овах Индокитай и Малакка, о-вах Суматра, Ява и Калимантан.
Род мусангов
Genus Paradoxurus F. Cuvier, 1821
Размеры мелкие и средние. Длина тела 43—71 см, длина хвоста ^Л— 66 см. Масса 1,5—4,5 кг. По внешнему виду напоминает мелкозубых ни-вет. Туловище вытянутое. Конечности относительно короткие. Хвост длинный. Голова с удлиненной узкой мордой и широкой мозговой частью (рис. 94). Широко расставленные уши среднего размера. Основная окраска меха сероватая, изменяющаяся до буроватой. Длинные остевые волосы имеют черные кончики и иногда создают общий черноватый тон. По спине проходят черноватые полосы, а по бокам располагаются продольными рядами черные пятна. Хвост темный. Сосков 3 пары.
Диплоидное число хромосом у обыкновенного мусанга 42.
Распространены на о-ве Шри Ланка, в Индии, Юго-Восточной Азии, Южном Китае и на Филиппинских о-вах.
Населяют леса. Превосходно лазают по деревьям и в основном ведут древесный образ жизни. Активны ночью. Часто встречаются около чело-202
веческих построек. Питаются мелкими позвоночными, в том числе млекопитающими, а также насекомыми, плодами и семенами.
Вероятно, бывает более одного помета в году. В каждом помете от 2 до 4 детенышей.
В роде 3 вида:
обыкновенный мусанг—Р. hermaphroditus Pallas, 1777 (Индия, о-в Шри Ланка, Бирма, п-ова Индокитай и Малакка, Южный Китай, Суматра с малыми соседними островами, Ява, Калимантан, Сулавеси, Филиппинские о-ва, Тимор, Керам и о-ва Кай);
шриланкский мусанг — Р. zeylonensis Pallas, 1778 (о-в Шри Ланка); южноиндийский мусанг — Р. jerdoni Blanford, 1885 (Южная Индия).
Род гималайских цивет
Genus Pagutna Gray, 1831
В роде единственный вид: гималайская цивета — Р. larvata Hamilton-Smith, 1827.
Размеры средние и крупные. Длина тела 51-—76 см, длина хвоста 51-—63 см. Общий облик напоминает мусанга (рис. 95). Характерна одноцветная окраска тела и отсутствие полос и пятен. Волосяной покров серый, иногда с рыжеватым оттенком. От лба до носа проходит беловатая полоса. Белые отметины над глазом и под глазом тянутся до уха. Хвост обычно темнее туловища. Лапы черноватые. Анальные железы сильно развиты. Сосков 2 пары.
Диплоидное число хромосом 44.
Распространена в Китае от провинций Фуцзянь и Гуаньдун до Юньнани и на север до Сычуани, Шэньси и Хэбэй; о-ва Хайнань и Тайвань; от Бирмы, Ассама в Индии на запад до Кашмира: Андаманские о-ва; п-ова Индокитай и Малакка, о-ва Суматра и Калимантан; акклиматизирована в Японии.
Рис. 96. Бинтуронг — A bintu-rong и его череп
Населяет леса и заросли кустарников. Экология изучена слабо. Ведет древесный образ жизни. Активность ночная. Всеядна: поедает мелких позвоночных, насекомых, плоды и корешки. В помете от 3 до 4 детенышей. Примерно через три месяца после рождения детеныши достигают размера взрослых.
Род сулавесских цивет
Genus Macrogalidia Schwarz, 1910
В роде единственный вид: сулавесская цивета — М. musschenbroekl Schlegel, 1879.
Размеры крупные. Длина тела около 100 см, длина хвоста примерно 60 см. Туловище сильно вытянутое и тонкое. Конечности средней длины. Голова с длинной мордой. Уши средней высоты, закругленные, широко расставленные. Волосяной покров низкий и густой. Окраска его на спинной стороне тела от светло-буроватой до темно-бурой. Щеки и надглазничное пятно обычно грязно-желтые или сероватые. Брюхо рыжеватобеловатое. Хвост в чередующихся темно- и светло-бурых кольцах.
Распространена на северо-востоке о-ва Сулавеси.
Экология изучена слабо. Населяет леса. Питается мелкими млекопитающими, а также плодами.
Род бинтуронгов
Genus Arctictis Temminck, 1824
В роде единственный вид: бинтуронг — A. binturong Raffles, 1821.
Размеры средние и крупные. Длина тела 61—96 см, длина хвоста 56— 89 см. Масса 9—14 кг. Туловище удлиненное, конечности относительно короткие. Голова широкая в мозговой части и укороченная, суживающаяся к носу — в лицевой. Уши средней высоты, округлые, широко расставленные (рис. 96). Сильный хвост с хватательным кончиком (хватательный хвост среди хищных имеется еще только у Potos flavus). Характерен высокий, грубый и косматый волосяной покров. Наибольшей длины волосы достигают в основной части хвоста. На задних поверхностях ушей располагаются длинные кисточки. На теле волосы блестящие, черные, часто с серыми или желтоватыми кончиками. Края ушей белые. Сосков 2 пары.
Диплоидное число хромосом 42.
Распространен в Бирме (возможно, также в Ассаме в Индии, Бутане, Непале, Сиккиме), на п-овах Индокитай и Малакка и о-вах Суматра, Ява, Калимантан и Палаван.
Населяет густые леса, везде редок. Экология изучена слабо. Ведет преимущественно древесный образ жизни. Активен ночью. Медлительное животное. Питается, по-видимому, в основном плодами, а также падалью. Легко приручается.
SUBFAMILIA HEMIGALINAE GILL, 1872
Род фаналок
Genus Fossa Gray, 1864
В роде единственный вид: фаналока — F. fossa Schreber, 1777.
Размеры мелкие. Длина тела около 40 см, длина хвоста примерно 20 см. Туловище умеренно удлиненное. Голова с расширенной мозговой
частью и длинной и узкой лицевой. Хвост длинный, составляющий примерно половину длины тела (рис. 97). Конечности средней высоты, тонкие, вероятно, приспособленные к бегу. Волосяной покров низкий; на хвосте волосы незначительно длиннее, чем на туловище. Окраска спины и боков сероватая с рыжеватым оттенком. По спине и бокам проходят с каждой стороны по четыре продольных ряда черных пятен; кроме того, черные пятна имеются на бедрах. Брюхо сероватое или беловатое. Характерно отсутствие анальных желез.
Диплоидное число хромосом 42.
Распространена была в прошлом по всем лесам о-ва Мадагаскар, а в настоящее время встречается лишь в восточной и северо-западной части острова.
Экология изучена слабо. Населяет подлесок густых дождевых лесов. Питается пресмыкающимися и насекомыми. Активна ночью.
Включена в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
Род полосатых цивет
Genus Hemigalus Jourdan, 1837
В роде единственный вид; полосатая цивета — Н. derbyanus Gray, 1837.
Размеры мелкие и средние. Длина тела 41—51 см, длина хвоста 25 38 см. Масса 1,7—3 кг. Туловище удлиненное, тонкое (рис. 98). Хвост длинный и мускулистый. Характерна небольшая с вытянутым и тонким лицевым отделом голова. Конечности средней высоты, пятипалые с большими искривленными, втягивающимися когтями. Подошвы кистей и ступней безволосые. Волосяной покров беловатый или желтовато-оранжевый, на брюхе более светлый. Хвост обычно черный. Узкая срединная темная полоса проходит от носа до затылка, а по бокам от нее широкие темные полосы окружают глаза и продолжаются назад к основанию 206
Рис 99. Цивета Оустона— С. owstoni и ее череп
уха. Две широкие полосы, иногда разбитые на пятна, идут назад по шее и дугообразно спускаются к локтю. На спине позади плеч располагаются четыре или пять широких поперечных полос, кроме того, две слабо выраженные полосы имеются у основания хвоста. Характерно, что волосы на спине и шее направлены вершинами вперед. Анальные железы маленькие.
Диплоидное число хромосом 42.
Распространена в Таиланде, на п-ове Малакка, Суматре, некоторых островах к западу от нее и Калимантане.
Экология не изучена. Питается насекомыми и червями.
Род цивет Оустона
Genus Chrotogale Thomas, 1912
В роде единственный вид: цивета Оустона — С. owstoni Thomas, 1912.
Размеры средние. Длина тела 50—63 см, длина хвоста 38—48 см. Туловище удлиненное, тонкое. Конечности средней высоты. Голова широкая в мозговом отделе и сильно сужается к носу; морда длинная (рис. 99). Спина с резко контрастирующими чередующимися четырьмя темными и четырьмя светлыми поперечными полосами. Такие же полосы расположе
ны в основании хвоста. На шее имеются продольные полосы. Брюхо светло-грязно-желтое. Узкая оранжевая полоса проходит вдоль середины брюха и груди. Концевые две трети хвоста черные. Анальные железы хорошо развиты.
Коронки резцов очень широкие, плотно прилегающие друг к другу. Ряды резцов образуют полукруг (единственный случай среди хищных).
Распространена во Вьетнаме и Лаосе.
Экология не изучена. Поедает земляных червей.
Род калимантанских цивет
Genus Diplogale Thomas, 1912
В роде единственный вид: калимантанская цивета — D. hosei Thomas, 1892.
Размеры средние. Длина тела около 60 см. Длина хвоста примерно 30 см. Туловище удлиненное. Шея довольно длинная, голова небольшая, расширенная в мозговой части и сильно суживающаяся, удлиненная в лицевой. Уши средней высоты, довольно широко расставленные, округлые. Конечности короткие, довольно тонкие. Окраска спинной стороны тела темно-бурая или черная, брюшной — сероватая или желтовато-белая. Ушные раковины с внутренней стороны белые. Над глазами располагаются желто-серые пятна; они продолжаются на щеки и сливаются с белым полем, идущим от губ и горла. Темный хвост покрыт укороченными волосами и не имеет колец.
Распространена на о-ве Калимантан.
Встречается в горах от 600 до 1200 м над уровнем моря. Экология не изучена. По-видимому, хорошо лазает по деревьям. Всеядна.
Некоторые исследователи (например, Андерсон и Джонс, 1967) включают калим антанскую цивету в род Hemigalus.
Род выдровых цивет
Genus Cynogale Gray, 1837
В роде единственный вид: выдровая цивета — С. bennetti Gray, 1837. Размеры средние. Длина тела 57—67 см, длина хвоста 12—20 см. Масса 3—5 кг. По внешнему виду несколько напоминает выдру. Туловище удлиненное, массивное, шея укороченная. Конечности и хвост короткие (рис. 100). Уши едва возвышаются над мехом. Отверстия ушей и ноздрей могут смыкаться при нырянии. Ноздри располагаются на конце морды, но открываются вверх. Между пальцами развита плавательная перепонка. Волосяной покров низкий, но очень густой и мягкий. Общий тон окраски буро-серый или черноватый. Брюхо светло-бурое. Пятен и полос нет. Характерны длинные вибриссы, часть которых располагается под ушами. Около половых органов находятся специфические кожные железы, продуцирующие секрет с сильным запахом. Сосков 2 пары.
Распространена на п-овах Индокитай и Малакка и о-вах Суматра и Калимантан.
Держится около рек и в болотистых местах. Экология изучена слабо. По-видимому, ведет полуводный образ жизни, хотя и не является хорошим пловцом. Питается в основном рыбами, ракообразными, возможно, моллюсками, а также мелкими млекопитающими, птицами и, вероятно, плодами. Может хорошо лазать по деревьям.
Род мелкозубых мунго
Genus Eupleres Doyere, 1835
Размеры средние и крупные. Длина тела 60—90 см, длина хвоста 20— 30 см. Туловище и шея удлиненные. Конечности с низко расположенными первыми пальцами. Когти относительно длинные, невтяжные или полу-втяжные. Голова узкая и длинная, особенно в лицевом отделе (рис. 101). Уши полуокруглые, широко расставленные. Волосяной покров высокий, густой и мягкий. Его окраска одноцветная буроватая у самцов и сероватая у самок; на брюшной стороне тела более светлая, чем на спинной. Хвост покрыт удлиненными волосами. Анальных желез нет.
Зубы слабо гетеродонтные небольших размеров. Первый предкоренной отделен от второго большой диастемой.
Экология изучена слабо. Мелкозубый мунго населяет дождевые леса. Ведут наземный образ жизни. Активны ночью. День проводят в расщелинах скал и норах.
Пища разнообразная; состоит из насекомых, их личинок и других беспозвоночных, пресмыкающихся, земноводных и, вероятно, птиц и мелких млекопитающих.
Распространены на Мадагаскаре.
В роде 2 вида:
мелкозубый мунго — Е. goudoti Doyere, 1835 (Восточный Мадагаскар; в настоящее время встречается лишь во влажной прибрежной зоне);
большой мунго — Е. major Lavauden, 1929 (Северный Мадагаскар).
Мелкозубый мунго включен в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
Рис. 101. Мелкозубый мунго — Е. goudoti и его череп
SUBFAMILIA GALIDIINAE GILL, 1872
Род кольцехвостых мунго
Genus Galidia I. Geoffroy, 1837
В роде единственный вид: кольцехвостый мунго — G. elegans I. Geoffroy, 1837.
Размеры мелкие. Длина тела около 38 см, длина хвоста примерно 30 см. Туловище удлиненное, конечности средней высоты. Голова широкая в мозговом отделе с длинной мордой, сужающейся к носу. Уши средней высоты, закругленные, широко расставленные. Пальцы и когти короткие. Волосяной покров одноцветный, бурый. Опушенный длинными волосами хвост с чередующимися темно-бурыми и черными кольцами. У самок около половых органов имеются кожные специфические железы.
Клыки короткие. Второй верхний предкоренной зуб очень маленький.
Диплоидное число хромосом 44.
Распространен на Мадагаскаре.
Экология изучена слабо. Очень малочислен. Активен днем. Хорошо лазает по деревьям. Как правило, держатся поодиночке. Питается мелкими млекопитающими, птицами, пресмыкающимися и насекомыми.
Рис. 102. Широкополосый мунго — G. striata и его череп
Род широкополосых мунго
Genus Galidictis I. Geoffrey, 1839
Размеры мелкие. Длина тела и хвоста примерно по 35 см. Туловище удлиненное, конечности средней высоты (рис. 102). Пальцы и когти длинные. Уши средних размеров, широко расставлены. Волосяной покров светло-бурый или сероватый, на голове более темный. Вдоль спины проходит от 5 до 10 темно-бурых или черных полос. У самок в области мочеполового отверстия расположены специфические кожные железы. Матка двойная (это, по-видимому, единственный случай среди хищных).
Распространены на Мадагаскаре.
Экология не изучена.
По-видимому, всеядны.
В роде 2 вида:
широкополосый мунго — G. striata I. Geoffroy, 1826 (Мадагаскар); полосатый мунго— G. fasciata Gmelin, 1788 (Мадагаскар).
Род узкополосых мунго
Genus Mungotictis Pocock, 1915
Размеры от мелких до крупных. Длина тела 35—84 см, длина хвоста 35—60 см. Туловище удлиненное, конечности средней высоты (рис. 103). Голова вытянутая, широкая в мозговом отделе и постепенно сужающаяся к носу. Уши широкие в основании, округлые, широко расставленные. Волосяной покров на спине светло-серый или темно-оливково-серый, на
Рис. 103. Узкополосый мунго — М. subsriafa
брюхе темно-желтый. Вдоль спины и по бокам проходит 11 черноватокоричневых (М. lineata) или 10 рыжевато-коричневых (М. substriata) узких полос.
Распространены на Мадагаскаре.
Экология не изучена. Активны днем.
В роде 2 вида:
линейчатый мунго — М. lineata Pocock, 1915 (Мадагаскар);
узкополосый мунго — М. substriata Pocock, 1915 (Мадагаскар).
Род бурохвостых мунго
Genus Salanoia Gray, 1864
Размеры мелкие. Длина тела примерно 33 см, длина хвоста около 18 см. Туловище удлиненное. Конечности средней высоты. Голова широкая в мозговом отделе, вытянутая в длинную и узкую морду. Уши широкие, короткие, далеко расставленные друг от друга. Когти слабо искривлены. Волосяной покров рыжевато-бурый с темными пятнами или оливково-бурый с желтыми пятнами. Хвост покрыт удлиненными волосами того же цвета, что и на туловище. Колец на хвосте нет.
Экология не изучена.
Распространены на Мадагаскаре.
В роде 2 вида:
бурохвостый мунго — S. olivacea I. Geoffrey, 1839 (Мадагаскар);
одноцветный мунго — S. unicolor I. Geoffroy, 1837 (Мадагаскар).
a
Рис. 104. Суриката — S. suricata и ее череп
SUBFAM1L1A HERPESTINAE GILL, 1872
Род сурикат
Genus Suricata Desmarest, 1804
В роде единственный вид: суриката — 5. suricata Schreber, 1777.
Размеры мелкие. Длина тела 25—35 см, длина хвоста 17—25 см. Туловище умеренной длины. Конечности высокие четырехпалые. Подошвы лап голые. Когти длинные. Голова вытянутая с длинной узкой мордой, й'ши небольшие, широко расставленные, округлые (рис. 104). Волосяной покров высокий и мягкий, особенно на спине и боках. Окраска его светло-серая; по спине проходят неясно выраженные черные поперечные полосы. Голова почти белая, уши черные, вокруг глаз черные кольца. Хвост желтоватый с черным концом покрыт более короткими волосами, чем туловище. Сосков 3 пары.
Рис. 105 Египетский мангуст — Н ichneumon (Л), мангуст-крабоед— Н. urva (5)
Зубная формула: i — с
1 3 2
— р-------- m =36 — 38.
1 н 3 — 4 2
Диплоидное число хромосом 36.
Распространена в Южно-Африканской Республике, Ботсване и Намибии.
Живет обычно колониями. На равнинах копает норы, а в скалистых местах находит убежища среди камней. Активна ночью. Днем обычно сидит около норы и греется на солнце. Часто встает столбиком. Питается луковицами, а также насекомыми, в частности термитами, змеями, ящерицами, мелкими млекопитающими, птицами и их яйцами. В помете от 2 до 4 детенышей.
Рис. 105. Продолжение. Череп малого мангуста -Н. auropunctatus
Род мангустов
Genus Herpestes liliger, 1811
Размеры мелкие и средние. Длина тела 23—64 см, длина хвоста 23 51 см. Масса 0,5—3,2 кг. Туловище удлиненное, конечности короткие пятипалые (рис. 105). Хвост длинный, покрыт более длинными волосами, чем туловище. Голова относительно небольшая, расширенная в мозговом отделе, с укороченной мордой, постепенно сужающейся к носу. Нижняя поверхность кистей и стоп обычно безволосая. Когти на передних конечностях острые и искривленные. Волосяной покров мягкий низкий или высокий. Окраска его изменчива: зеленовато-серая, желтовато-бурая или серовато-бурая; иногда спина испещрена пятнами белого или желто
бурого цвета. Брюхо обычно светлее спины и боков тела и может быть белым.
Имеются небольшие анальные железы. Некоторые виды могут выбрызгивать неприятно пахнущий секрет этих желез, как это делают скуц. сы. Сосков 2 или 3 пары.
3 1 3—4 2
Зубная формула: i — с — р —-- га — = 36 — 40.
3 13 ~“ 4 2
Диплоидное число хромосом у самок египетского мангуста 44, самцов— 43; у самок стройного мангуста — 42; самцов — 41; у самок южноафриканского мангуста — 40, самцов — 39; у самок обыкновенного, малого, короткохвостого, бурого мангустов и мангуста-крабоеда — 36, самцов -— 35.
Распространены в Южной Европе, Африке, на Аравийском п-ове, в Малой Азии, Иране, Афганистане, Индии, на о-ве Шри Ланка, в Непале, Бирме, Южном Китае, на о-вах Тайвань и Хайнань, п-овах Индокитай и Малакка, о-вах Суматра, Ява, Калимантан. Мангусты акклиматизированы в Вест-Индии, на о-вах Гавайских, Фиджи. Акклиматизация была проведена с целью уничтожения крыс и змей. Однако сильно размножившиеся мангусты оказались вредными: они поедали большое количество различных мелких млекопитающих и птиц.
Населяют разнообразные биотопы: от густых лесов в холмах до песчаных пустынь. Периодичности в суточной активности нет. Обычно держатся поодиночке, иногда объединяются небольшими группами до 4— 12 особей. Хищники; пищей служат мелкие млекопитающие, пресмыкающиеся, в том числе ядовитые змеи, земноводные, рыбы, ракообразные, насекомые. Некоторые виды питаются плодами и другими частями растений. Мангусты не имеют иммунитета к яду и справляются даже с крупными ядовитыми змеями благодаря исключительно быстрой реакции. Сезонности в размножении не имеют. Длительность беременности примерно 60 дней. В помете от 2 до 4 детенышей. Хорошо приручаются. Длительность жизни 7—12,5 лет.
В роде 14 видов:
египетский мангуст—И. ichneumon Linnaeus, 1758 (Испания, Португалия, Палестина, Алжир, АРЕ и от Кении и Нигерии на юг до Южно-Африканской Республики);
яванский мангуст — И. javanicus Geoffroy, 1818 (п-ова Индокитай и Малакка и о-в Ява);
малый мангуст — Н. auropunctatus Hodgson, 1836 (северная часть Аравийского п-ова, Иран, Ирак, Афганистан, Индия, Непал, Бирма, Таиланд, о-в Хайнань, Малайзия);
обыкновенный мангуст—Н. edwctrdsi Geoffroy, 1818 (Аравийский п-ов, Ирак, Иран, Афганистан, Индия, Непал, о-в Шри Ланка);
мангуст Смита — Н. smithi Gray, 1837 (Индия, о-в Шри Ланка);
бурый мангуст— Н. fuscus Waterhouse, 1837 (Южная Индия и о-в Шри Ланка);
полосатошейный мангуст—Н. vilticollis Bennett, 1835 (Южная Индия и о-в Шри Ланка);
мангуст-крабоед — Н. urva Hodgson, 1836 (Южный Китай, о-ва Тайвань и Хайнань, Непал, Ассам в Индии, Бирма, п-ов Индокитай);
короткохвостый мангуст—Н. brachyurus Gray, 1837 (Малайзия, о-ва Суматра и Калимантан);
мангуст Хозе — Н. hosei Jentink, 1903 (Калимантан);
воротничковый мангуст — Н. semitorquatus Gray, 1846 (о-ва Суматра и Калимантан);
Рис. 106. Карликовый мангуст — Н. parvula
длинноносый мангуст — Н. naso de Winton, 1901 (от юго-восточной Нигерии на юг до устья р. Конго);
южноафриканский мангуст — И. pulverulentus Wagner, 1839 (Южно-Африканская Республика, Намибия, Южная Ангола);
стройный мангуст — Н. sanguineus Riippel, 1835 (Африка к югу от Сахары и о-в Занзибар).
По Андерсону и Джонсу (1967), в роде 13 видов. Некоторые исследователи (например, Симпсон, 1945; Андерсон и Джонс, 1967) выделяют длинноносого мангуста в особый род Xenogale J. Allen, 1919.
Род карликовых мангустов
Genus И elogale Gray, 1862
Размеры мелкие — наименьшие в семействе. Длина тела 17—24 см, длина хвоста 15—23 см. Туловище вытянутое. Конечности короткие, пятипалые. Голова широкая в мозговой части и суживающаяся клинообразно к носу (рис. 106). Морда средней длины. Уши небольшие, округлые, широко расставленные. Волосы на хвосте длиннее, чем на туловище. Волосяной покров низкий густой и мягкий. Окраска его на спине пятнистая, бурая или серая, иа брюхе рыжевато-серая или светло-бурая. Иногда попадаются совершенно черные особи. Сосков 3 пары.
„ 3 1 3 2
Зубная формула: i — с — р — m = 36.
3 13 2
Распространены от Эфиопии и Сомали на юг до Южно-Африканской Республики.
Населяют засушливые места, заросли акаций, горные и густые равнинные леса от уровня моря до 1800 м над ним. Обычно держатся группами по 4—12 особей. Активны днем. Питаются разнообразными растениями и животными, в том числе насекомыми, пресмыкающимися, мелкими млекопитающими, а также плодами, ягодами.
Рис. 107. Водяной мангуст—Л. paludinosus
В помете от 2 до 4 детенышей.
В роде 2 вида:
1846 (Южно-Африкан-
карликовый мангуст — Е. parvula Sundevall, 1846 (Южно-Африканская Республика, Ботсвана, Намибия, Ангола, Родезия, Мозамбик, Малави, Замбия, Заир, Танзания, Уганда, Кения, Эфиопия и Сомали);
мангуст Дыбовского — И. dybowskii Pousargues. 1894 (Заир, Судан, Уганда).
Иногда (Уокер, 1964) мангуста Дыбовского выделяют в особый род — Dologale Thomas, 1926.
Род водяных мангустов
Genus Atilax F. Cuvier, 1826
В роде единственный вид: водяной мангуст — A. paludinosus G. Cuvier, 1777.
Размеры средние. Длина тела 45—60 см, длина хвоста 30—40 см. Внешний вид сходен с остальными мангустами, но сложение более тяжелое, хотя конечности относительно несколько выше (рис. 107). Конечности пятипалые, с широко расставленными пальцами. Когти короткие и тупые. Подошвы лап голые. Голова массивная. Уши небольшие, слабо выдаются над мехом, округлые и далеко отстоят друг от друга. Между носом и верхней губой проходит узкая полоска безволосой кожи. Волосяной покров высокий и грубый, у основания хвоста волосы удлинены. Окраска одноцветная, буроватая с черным оттенком. Голова и брюхо светлее спины. Сосков 2 пары.
Диплоидное число хромосом у самок водяного мангуста 36, у самцов 35.
Распространен от Южно-Африканской Республики до Эфиопии на северо-востоке и Сьерра-Леоне на северо-западе, за исключением пустынных и полупустынных областей.
Населяет берега различных водоемов, часто устья рек. Держится поодиночке или парами. Прекрасно плавает и ныряет. Активен преиму-218
Рис. 108. Полосатый мангуст — М. mungo
щественно ночью. Питается водными и прибрежными животными: рыбами, лягушками, ракообразными, насекомыми, змеями, мелкими млекопитающими и птицами. В помете 2—3 детеныша.
Род полосатых мангустов
Genus Mangos Е. Geoffroy et G. Cuvier, 1795
В роде единственный вид: полосатый мангуст — М. mango Gmelin, 1788.
Размеры мелкие. Длина тела 30—45 см, длина хвоста 23—29 см. Туловище удлиненное, конечности короткие. Хвост довольно длинный, покрыт более высокими по сравнению с туловищем волосами. Голова умеренной длины, постепенно суживающаяся от мозгового отдела к кончику носа (рис. 108). Конечности пятипалые. Когти на передних лапах длинные. Подошвы лап голые. Волосяной покров грубый, пух развит слабо. Окраска буровато-серая. По спине и верхним отделам боков тела располагаются поперечные чередующиеся бурые и желтоватые или беловатые полосы. Соски брюшные — 3 пары.
Диплоидное число хромосом 36.
Распространен от Южно-Африканской Республики и Намибии на северо-восток до Сомали и Судана и на северо-запад до Гвинеи-Бисау и Гамбии.
Населяет скалистые участки, густые заросли кустов, обычно недалеко от воды. Держатся группами из 6—20 особей. Активен преимущественно утром и вечером. Питается насекомыми, моллюсками, пресмыкающимися, мелкими млекопитающими, яйцами птиц, плодами и ягодами. Детенышей в помете 3—6.
Иногда (например, Розевир, 1974) в роде выделяют еще один вид — Al. gambianus Ogilby, 1835.
Род кузиманз
Genus Crossarchus F. Cuvier, 1825
Размеры мелкие. Длина тела 30—41 см, длина хвоста 15—25 см. Масса 0,5—1,3 кг. Туловище вытянутое, приземистое. Конечности короткие. Хвост относительно короткий. Голова небольшая с вытянутым лицевым
отделом. Уши небольшие округлые, широко расставленные. Волосяной покров высокий и грубый, образован коричневыми, серыми и желтыми волосами. Окраска головы обычно светлее, а конечностей — темнее, чем на туловище. Полос и пятен на теле нет.
Диплоидное число хромосом у длинноносого кузиманзе 36.
Распространены в Западной и Центральной Африке.
Населяют леса. Держатся группами от 6 до 20 особей. Постоянно кочуют, редко оставаясь больше чем два дня на одном месте. Активны днем. Питаются насекомыми, их личинками, мелкими пресмыкающимися, ракообразными, различными плодами и ягодами.
В роде 3 вида:
ангольский кузиманзе — С. ansorgei Thomas, 1910 (Ангола);
длинноносый кузиманзе — С. obscurus F. Cuvier, 1825 (от Сьерра-Леоне до Камеруна);
заирский кузиманзе— С. alexandri Thomas et Wroughton, 1907 (север Заира).
Некоторые исследователи (например, Андерсон и Джонс, 1967) насчитывают в роде 4 вида.
Род либерийских мангустов
Genus Liberiictis Hayman, 1958
В роде единственный вид: либерийский мангуст—L. kuhni Hayman, 1958.
Описание дано только по черепам. По-видимому, сходен с представителями рода кузиманз. Отличается от них числом предкоренных зубов, которых по четыре в верхней и нижней челюсти по сравнению с тремя у кузиманз. Череп относительно крупнее, лицевой отдел более удлиненный, а зубы относительно слабее. Окраска волосяного покрова, по словам местных жителей, черная.
Распространен в Северо-Восточной Либерии. Населяет высокоствольные леса. Держатся группами по 3—5 особей.
Род белохвостых мангустов
Genus Ichneumia I. Geoffroy, 1837
В роде единственный вид: белохвостый мангуст — I. albicauda G. Cuvier, 1829.
Размеры средние. Длина тела 53—61 см, длина хвоста 42—47 см. Масса около 4,5 кг. Туловище удлиненное. Конечности относительно высокие, пятипалые. Нижняя поверхность кистей голая. Хвост длинный, покрыт длинными волосами. Голова небольшая с вытянутым лицевым отделом (рис. 109). Уши средних размеров, широко расставленные, округлые. От носа к верхней губе спускается полоска белой кожи. Волосяной покров высокий, грубый. Окраска его буроватая с белым или желтым оттенком. Конечности черные. Проксимальная четверть хвоста той же окраски, что и туловище, остальная часть белая. Сосков 2 пары. Общее число зубов 40.
Диплоидное число хромосом 36.
Распространен от Южно-Африканской Республики, Ботсваны и Намибии на северо-восток до Сомали, Судана, южной части Аравийского п-ова и на северо-запад до Сенегала.
Населяет густые леса и заросли по берегам водоемов. Обычно держится поодиночке, иногда парами. Активен в сумерки и ночью. День
Рис. ПО. Черноногий мангуст — Bdeogale sp.
проводит в норах дикобразов или в расщелинах скал. Может лазить по Деревьям. Питается мелкими млекопитающими, птицами и их яйцами, насекомыми, пресмыкающимися. В помете обычно 2 детеныша.
Род черноногих мангустов
Genus Bdeogale Peters, 1850
Размеры мелкие и средние. Длина тела 37—60 см, длина хвоста 17—• 37 см. Внешним обликом напоминают белохвостого мангуста (рис. НО). Туловище удлиненное, конечности короткие, четырехпалые (отсутствует 1 палец). Голова вытянутая, уши широкие, короткие, округлые, широко расставленные. От носа к верхней губе проходит полоска белой кожи. Волосяной покров густой, низкий. Общий тон окраски сероватый и буроватый. Конечности черные. Волосы на хвосте чуть длиннее, чем на туловище. Подошвы лап голые.
Распространены в Западной, Восточной и Южной Африке.
Экология не изучена. Питаются мелкими млекопитающими, пресмыкающимися и насекомыми.
В роде 3 вида:
пушистохвостый мангуст — В. crassicauda Peters, 1852 (Мозамбик Малави, Замбия, Танзания и Кения);
черноногий мангуст—В. nigripes Pucheron, 1855 (от Нигерии до Анголы и Заира);
мангуст Джексона — В. jacksoni Thomas, 1894 (Кения и Уганда).
Род умби
Genus Rhynchogale Thomas, 1894
В роде единственный вид: умби — R. melleri Gray, 1865.
Размеры средние. Длина тела 44—48 см, длина хвоста 30—40 см. Внешним обликом напоминает белохвостого мангуста, но отличается от него отсутствием большого пальца передней конечности и голой полоски кожи между носом и верхней губой. Нижняя поверхность стопы покрыта волосами. Волосяной покров грубый. Обшая окраска его сероватая или бледно-бурая. Голова и брюхо светлее спины, а конечности и хвост— темнее. Соски брюшные — 2 пары.
Общее число зубов 40—42.
Распространен в Южно-Африканской Республике, Мозамбике, Малави, Замбии, Анголе, Заире, Западной Танзании.
Экология не изучена. Ведет наземный образ жизни. Активен ночью. Встречаются поодиночке. Питается мелкими позвоночными, насекомыми, плодами.
Род желтых мангустов
Genus Cynictis Ogilby, 1833
В роде единственный вид: желтый мангуст— С. penicillata G. Cuvier, 1829.
Размеры мелкие. Длина тела 27-—38 см, длина хвоста 18—28 см. Внешним видом сходен с другими мангустами. Туловище удлиненное, конечности короткие. Голова небольшая с укороченной мордой (рис. 111). Передние конечности пятипалые, задние четырехпалые. Первый палец передних конечностей небольшой, расположен выше уровня других пальцев и не касается земли. Общий тон окраски волосяного покрова от темно-оранжево-желтого до светло-желтовато-серого. Брюхо и конечности светлее спины. Хвост на конце белый. Летний мех в феврале — январе рыжеватый, низкий, зимний — в июне — августе — желтоватый, высокий и густой. Соски брюшные — 3 пары.
Общее число зубов 40.
Диплоидное число хромосом 36.
Распространен в Южно-Африканской Республике, Ботсване, Намибии и Анголе.
Населяет засушливые районы, но может также встречаться в зарослях кустарников вдоль берегов реки и среди камней. Живут колониями (редкий случай среди хищных). Число особей в колонии может достигать 50 или даже несколько больше. Обычно выкапывают норы, но иногда занимают норы других животных. Колониальные норы имеют сложное устройство с многими входами и камерами. Для лучшего обзора встает на задние конечности. Охотятся поодиночке или парами. Обычно от нор не уходит далее чем на 1 км. Активен преимущественно днем.
Питается мелкими млекопитающими, птицами и их яйцами, пресмыкающимися, насекомыми и их личинками. В помете от 2 до 4 детенышей.
Некоторые ученые (например, Симпсон, 1945) включают в род желтых мангустов серого мангуста — Paracynictis selousi.
Рис. Ill. Желтый мангуст — С. peni-cillata
Род серых мангустов
Genus Paracynictis Pocock, 1916
В роде единственный вид: серый мангуст — Р. selousi de Winton, 1896.
Размеры мелкие и средние. Длина тела 39—47 см, длина хвоста 28—• 40 см. По внешнему облику сходен с другими мангустами. Конечности четырехпалые с длинными слегка искривленными когтями. Окраска спины желтовато-серая, брюха — темножелтая. Конечности черные. Хвост с
белым кончиком.
Распространен в Южно-Африканской Республике, на юге Мозамбика, в Родезии, Ботсване, Намибии, Анголе, Замбии.
Экология не изучена. По-видимому, живут по нескольку зверьков вместе. Роет сложные норы. Активен ночью. Питается различными мел-
кими позвоночными и насекомыми.
SUBFAMILIA CRYPTOPROCTINAE TROUESSART, 188-5
Род фосс
Genus Cryptoprocta Bennett, 1833
В роде единственный вид: фосса — С. ferox Bennett, 1833.
Размеры средние и крупные. Длина тела 61—76 см, длина хвоста около 66 см. Это наиболее крупный хищник, обитающий на Мадагаскаре. Внешний облик кошачий (рис. 112). Туловище удлиненное. Конечности высокие; задние выше передних. Шея длинная, голова короткая. Когти короткие, но очень острые втяжные. Хвост длинный; покрыт волосами такой же длины, как и на туловище. Волосяной покров низкий, гладкий мягкий и густой. Характерны очень длинные вибриссы. Имеются анальные и прескротальные железы, выделяющие сильно пахнущий секрет.
3 13 1
Зубная формула: 1 — с —— р — гп — =32.
Распространена на Мадагаскаре, кроме центрального горного плато.
Населяет лесные районы. Активна ночью. Ведет в основном древесным образ жизни. Прекрасно лазает. Пищей в основном служат лемуры, за которыми фосса охотится по деревьям. В помете 2—3 детеныша. Продолжительность жизни, по-видимому, около 17 лет.
Включена в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
СЕМЕЙСТВО ГИЕНОВЫХ
FAMILIA HYAEN1DAE GRAY, 1869
Общим обликом напоминают волков, но отличаются от них относительно укороченным туловищем, более массивной передней частью тела в слабее развитой задней.
Размеры мелкие и средние. Длина тела 55—165 см, длина хвоста 20—33 см. Масса 10—80 кг. Голова крупная, удлиненная. Иногда лицевая часть несколько укорочена. Морда может быть узкой или широкой, иногда укороченной. Уши большие округлые. Передние конечности выше задних, поэтому тело в холке выше, чем в крестце. Конечности четырехпалые (у земляного волка на передних лапах пять пальцев), пальцехо-224
дящие. Когти крупные тупые, слабо искривленные, невтягивающиеся. Волосяной покров низкий или более высокий, редкий и грубый. На верхней части шеи и загривке обычно располагаются удлиненные волосы, образующие гриву. Хвост также покрыт длинными волосами. Окраска меха преимущественно бурая с темными пятнами и полосами. Имеются специфические кожные анальные железы, продуцирующие секрет с резким запахом. Сосков 2 или 3 пары.
Череп крупный с мощными гребнями. Лицевой отдел удлиненный, мозговой невелик. Скуловые дуги массивные, широко расставленные в стороны. Крылоклиновидного канала нет. Костные слуховые барабаны сильно вздутые, с внутренней камерой, разделенной перегородкой у земляного волка и неразделенной — у гиен.
3 1 3 — 4
Зубная формула: i —- с — р--
3 11 — 3
1
гп ----=28—34.
1 — 2
Третий резец гиен крупнее двух других. Клыки мощные у гиен или длинные и слабые у земляного волка. Хищнические зубы гиен развиты очень сильно, а земляного волка — слабо. Предкоренные зубы небольшие и широко отставленные друг от друга у земляного волка или мощные, конической формы, с хорошо развитыми коронками, приспособленными к раздроблению костей, у гиен. Коренные зубы земляного волка небольшие, у гиен они крупные и сильные.
Характерно отсутствие os penis.
Распространены в Африке, Передней, Средней и Южной Азии.
Обычно населяют равнины или кустарниковые заросли, иногда прибрежные леса. Активны ночью, день проводят в норах других животных, пещерах, иногда в густых зарослях. Держатся поодиночке или парами, но к добыче могут собираться большими группами. По характеру питания трупояды, исключение составляет земляной волк, который питается в основном насекомыми. Длительность беременности 90—ПО дней. Число детенышей в помете от 1 до 6 (у земляного волка), чаще от 2 до 4. Как правило, дают один помет в год. Детеныши рождаются слепыми. Продолжительность жизни в неволе до 25 лет.
Большого практического значения гиены не имеют, однако, будучи «санитарами», быстро уничтожающими всякую падаль, полезны. Иногда нападают на домашний скот.
В семействе 2 подсемейства и 3 рода (4 вида):
Subfamilia Hyaeninae Mivart, 1882
пятнистых гиен — Crocuta Каир, 1828 (1 вид);
полосатых гиен — Hyaena Brisson, 1762 (2 вида).
Subfamilia Protelinae Mivart, 1882
земляных волков — Proteles Е. Geoffroy St.-Hilaire, 1823 (1 вад).
В СССР: один род полосатых гиен.
Иногда земляных волков выделяют в особое семейство — Protelidae.
SUBFAMILIA HYAENINAE MIVART, 1882
Род пятнистых гиен
Genus Croatia Каир, 1828
В роде единственный вид: пятнистая гиена—С. cr acuta Erxleben, 1777.
Размеры наибольшие в семействе. Длина тела 127—165 см, длина хвоста 25—33 см, высота в плечах 76—91 см. Масса 59—82 кг. Сложение 8—671 225
Рис. 113. Пятнистая гиена — С. crocuta и ее череп
массивное. Передняя часть тела развита значительно сильнее задней. Голова крупная с вытянутой, но притупленной мордой (рис. 113). Челюсти очень сильно развиты. Конечности четырехпалые. Волосяной покров низкий, мягкий. Основная окраска его желтовато-серая с многочисленными округлыми бурыми или черными пятнами. Грива на шее развита слабо.
„ 3 14 1
Зубная формула: i — с — р m — = 34,
Диплоидное число хромосом 40.
Распространена от Южно-Африканской Республики на север до Сахары и от Сомали на востоке до Сенегала на западе.
Населяет различные ландшафты от жарких пустынь до горных лесов, но предпочитает степи и саванны. В горы поднимается до 4 тыс. М
226
над уровнем моря. Большая плотность пятнистых гиен наблюдается вблизи концентраций диких копытных или около некоторых поселений человека (например, в Эфиопии и Сомали). Активна в основном ночью. Могут вести преимущественно оседлый образ жизни (в кратере Нгорон-горо) или кочевой (на равнине Серенгети). Охотятся и поедают гну, зебр, газелей, питаются также падалью. На равнине Серенгети и особенно в кратере Нгоронгоро пятнистые гиены кормятся в основном добываемыми ими копытными (Крук; Kruuk, 1972). Около поселков поедают преимущественно падаль и отбросы. Соотношение полов у взрослых зверей примерно 1 : 1. Сезонности в размножении нет. Однако в сезон дождей может быть пик рождаемости детенышей. Продолжительность беременности 98—99 или 109—111 дней (по данным разных исследователей). В помете обычно 2 детеныша (редко 1 или 3). Новорожденные беспомощные, с открытыми глазами, покрыты коричневато-черными мягкими волосами, масса каждого около 1,5 кг. Дней через 10 после рождения уже могут быстро двигаться. В 1,5—2 месяца приобретают волосяной покров, характерный для взрослых. Лактация длится около 6 месяцев, однако молоко матери продолжает играть важную роль в питании детенышей вплоть до достижения ими годовалого возраста. Половозрелость наступает примерно к 3-му году (у самок немного позже, чем у самцов). Продолжительность жизни около 25 лет.
Род полосатых гиен
Genus Hyaena Brisson, 1762
Размеры меньше, чем у пятнистых гиен. Длина тела 91—120 см, длина хвоста около 31 см. Масса 27—54 кг. Внешний вид, типичный для гиен (рис. 114). Голова менее массивная, чем у пятнистой гиены, и более узкая, уши большие, широкие и высокие. Конечности четырехпалые. Волосяной покров высокий, грубый. Окраска его сероватая, светло-желтовато-бурая, буровато-серая. Поперек тела и кольцеобразно окружая конечности, располагаются темно-бурые или черные полосы. По верху шеи и по ее бокам удлиненные волосы образуют гриву, она проходит по загривку и середине спины. Сосков 3 пары.
3 1
Зубная формула: i — с — р 3 1
4
3 ™
X—34,
Диплоидное число хромосом у полосатой гиены 40.
Распространены почти по всей Африке, в Передней, Средней и Малой Азии. В СССР встречаются в Закавказье и Средней Азии.
Населяют саванны, глинистые, реже песчаные пустыни, каменистые предгорья. Активны в основном ночью. Днем скрываются среди камней или в каких-либо пещерах. Вне периода размножения ведут бродячий образ жизни. Питаются преимущественно падалью. Иногда охотятся на различных позвоночных, в том числе на мелкий домашний скот. Могут поедать овощи, дыни, плоды деревьев.
Сезонности в размножении, как правило, нет; на севере ареала течка отмечена в январе—феврале. Беременность длится около 3 месяцев. В помете от 2 до 6 (обычно 2—4) детенышей. Длительность лактационного периода примерно 2 месяца. В воспитании детенышей принимают участие самка и самец. Половозрелость наступает в 3—4 года. Продолжительность жизни примерно 24 года.
8* 227
Рис. 114. Полосатая гиена — Н. hyaena (Л) и бурая гиена— Н. Ьгиппеа (Б)
В роде 2 вида:
полосатая гиена — 77. hyaena Linnaeus, 1758 (Африка от Марокко и Сенегала на западе до Танзании, Кении, Эфиопии, Сомали, Судана и АРЕ на востоке; Аравийский п-ов, Малая Азия, Палестина, Сирия, Ирак, Иран, Афганистан, Пакистан, почти весь Индостан; в СССР— Азербайджан, Восточная Грузия, Копетдаг в районе Бадхыза, южная часть Узбекистана и полупустыни Южного Таджикистана);
бурая гиена — И. Ьгиппеа Thunberg, 1820 (Южно-Африканская Республика, юг Мозамбика, Родезия, Ботсвана, Намибия).
114. Продолжение. Череп полосатой гиены
В СССР: полосатая гиена.
В «Красную книгу» включены:
бурая гиена как вид, который в ближайшее время может подвергнуться опасности исчезновения;
подвид полосатой гиены — Н. hyaena barbara Blainville, 1844, прежде широко распространенный в Алжире, Тунисе и Марокко, а в настоящее время сохранившийся лишь в горах — как подвид, находящийся под угрозой исчезновения.
Рис. 115. Земляной волк—
Р. cristatus и его череп
SUBFAMILIA PROTELWAE MIVART, 1882
Род земляных волков
Genus Proteles I. Geoffroy St.-Hilaire, 1824
В роде единственный вид: земляной волк — Р. cristatus Sparrman, 1783.
Размеры наименьшие в семействе, длина тела 55—80 см, длина хвоста 20—30 см, высота в плечах 45—50 см. Характерны высокие, относительно тонкие и слабые конечности. Шея длинная (рис. 115). Голова удлиненная с узкой мордой и слабыми челюстями. Пальцев на передних конечностях пять, а на задних четыре. Волосяной покров образован длинными грубыми остевыми и мягкими, извитыми пуховыми волосами. Начиная от затылка по верхней части шеи до спины располагаются удлиненные волосы, образующие своеобразную гриву. На хвосте волосы особо удлинены. Окраска тела желтовато-серая с белыми поперечными полосами на спине и боках. Конец хвоста и проксимальные отделы конечностей черные. Анальные железы хорошо развиты.
3 I
Зубная формула: i — с —— р 3 1
3
2 — 1
= 28 — 32 (обычно 30).
ш
Зубы слабые, отделенные друг от друга промежутками. Диплоидное число хромосом 40.
Распространен в Южно-Африканской Республике, Намибии, Родезии, Анголе, Замбии, Танзании, Кении, Эфиопии, Сомали, Судане.
Населяет открытые песчаные равнины или заросли кустарников. Держится поодиночке, иногда парами или семейными группами из пятишести особей. Активен ночью. День проводит в норе, которую выкапывает сам, или занимает норы трубкозубов. Питается в основном термитами и личинками насекомых, которых достает, разрывая землю. Поедает также мелких млекопитающих, мелких птиц и их яйца. Самка приносит 2—4, иногда до 6 детенышей. В южной части ареала рождение детенышей происходит в ноябре—декабре. В ряде мест, например в Кении, охраняется законом.
СЕМЕЙСТВО КОШАЧЬИХ
FAMILIA FELIDAE GRAY, 1821
Размеры тела от мелких до крупных. Длина тела 46—317 см, длина хвоста 15—114 см. Масса 2,5—390 кг. У всех представителей семейства внешний облик однообразный — «кошачий». Сложение стройное. Туловище гибкое, более или менее вытянутое. Шея укороченная. Голова округлая с короткой лицевой частью. Глаза крупные с вертикальным зрачком. Уши относительно невелики с закругленными или заостренными вершинами. Конечности высокие или не очень высокие, пальцеходящие с округлой лапой. Передние конечности пятипалые (первый палец расположен высоко и очень небольшой), задние — четырехпалые. Когти у всех видов, кроме гепарда, втяжные, большие, сильно изогнутые и острые. Комбинация подвижных пальцев и втяжных когтей делает лапу совершенным хватательным органом. Волосяной покров обычно низкий, но густой и мягкий. Окраска варьирует от серой, рыжеватой или желтовато-бурой до ярко-рыжей и коричневой, обычно с полосами и пятнами. Хвост покрыт равной длины волосами. Обычно имеются слабо развитые анальные железы. Сосков 2—4 пары.
Череп с сильно укороченной лицевой частью, округлый, широкий с мощными и короткими челюстями. Скуловые дуги сильные, широко расставленные. Крылоклиновидного канала нет. Костные слуховые барабаны сильно вздуты, округлой формы, внутри разделены перегородкой.
3 13 1
Зубная формула: i — с — р m — = 30.
Второй верхний предкоренной зуб (первого верхнего предкоренного У кошек вообще нет) небольших размеров, иногда не развивается или выпадает или всегда отсутствует (манул, рысь). Тогда зубная формула приобретает следующий вид.
„ 3 12
Зубная формула: i — с — р —
m— = 28.
1
Резцы небольших размеров. Клыки сильно развиты, удлиненные, острые. Позади клыков обычно имеется диастема, достигающая наибольшей величины у дымчатого леопарда (у гепарда ее нет). Коренные зубы с режущими высокими выступами на жевательных поверхностях. Хищнические зубы развиты сильно, причем нижний меньше по размерам, чем
верхний. Верхний хищнический зуб с тремя корнями. Коренные зубы сильно сжаты с боков.
Os penis иногда отсутствует.
Диплоидное число хромосом 38 и только у Felis pardalis и F. wiedi по 36.
Ареал обширен, охватывает весь земной шар, за исключением Антарктики, Австралии, Мадагаскара и некоторых океанических островов.
Населяют преимущественно леса, а также пустыни, горы, приречные долины. Наземные, хотя многие из кошек хорошо лазают по деревьям. Большинство представителей семейства ярко выраженные хищники, за исключением суматранской кошки, которая предпочитает плоды, и кошки-рыболова, которая поедает рыб и моллюсков. Ловят добычу, подстерегая или скрадывая ее. Держатся поодиночке или семейными группами. Для кошек характерен полиэстр а льный тип овуляции. Длительность беременности от 56 дней (бенгальская и дикая кошки) до более чем 100 (тигр, ягуар). Самка приносит от 1 до 6, обычно 2—3 детенышей. Большинство видов имеет по одному помету в год. Существенного хозяйственного значения не имеют. Некоторые виды служат объектом промысла или спортивной охоты. Ливийская кошка — Felis silvestris libyca была одомашнена в Египте около 4—3 тыс. лет до н. э.
Систематика семейства на уровне родов разработана недостаточно. Количество родов и их объем определяются по-разному. Так, Огнев (1935) насчитывает в семействе 22 рода:
Felis Linnaeus, 1758;
Lynx Kerr, 1792 (рысь);
Caracal Gray, 1843 (каракал);
Otocolobus Brandt, 1841 (манул);
Eremaelurus Ognev, 1927 (барханный кот);
Puma I ar dine, 1834 (пума);
Leptailurus Severtzov, 1858 (сервал);
Prionailurus Severtzov, 1858 (бенгальская кошка);
Pardofelis Severtzov, 1858 (мраморная кошка);
Profelis Severtzov, 1858 (золотая кошка);
Zibetailurus Severtzov, 1858 (кошка-рыболов);
Ictailurus Severtzov, 1858 (суматранская кошка);
Neofelis Gray, 1867 (дымчатый леопард);
Leopardus Gray, 1842 (оцелот);
Herpailurus Severtzov, 1858 (ягуарунди);
Dendrailurus Severtzov, 1858 (пампасская кошка);
Acinonyx Brookes, 1828 (гепард);
Leo Oken, 1816 (лев);
Tigris Oken, 1816 (тигр);
Pardus Fitzinger et Giebel, 1868 (леопард);
Uncia Gray, 1854 (снежный барс);
Jaguarius Severtzov, 1858 (ягуар).
Аллен (Allen, 1938), Новиков (1956, 1963) объединяют всех кошек в род Felis, кроме гепарда, которого выделяют в род Acinonyx. Возможно, это наиболее естественная система, поскольку между всеми кошками можно найти переходные формы.
Промежуточные позиции занимает, например, Уокер (1964), выделяющий в семействе 6 родов: Lynx, Felis, Panthera, Neofelis, Uncia и Acinonyx. По Андерсону и Джонсу (1967), в семействе 4 рода: Acinonyx, Felis, Neofelis и Leo Brehm, 1829 ( = Panthera). Симпсон (1945), Грассе (1955), Новиков (1971) выделяют в семействе 3 рода: Felis, Panthera и Acinonyx. Впредь до тщательной ревизии семейства представляется це-232
лесообразным вслед за Гептнером и Слудским (1972) принять в семей стве 4 рода (36 видов):
больших кошек—Panthera Oken, 1816 (4 вида);
снежных барсов — Uncia Gray, 1854 (1 вид):
кошек — Fells Linnaeus, 1758 (30 видов):
гепардов — Acinonyx Brookes, 1828 (1 вид).
Единого мнения нет и по поводу объединения родов в подсемейства. Большинство исследователей принимают в семействе 2 подсемейства: Felinae Trouessart, 1885, с родами Panthera, Uncia, Felis и Acinonychi-nae Pocock, 1917, с родом Acinonyx или 3 подсемейства: Felinae, Panthe-rinae Pocock, 1917 (роды Panthera и Uncia) и Acinonychinae. Иногда выделяют и подсемейство рысей — Lyncinae.
Поскольку четкого разграничения между родами современных кошек нет, по-видимому, нецелесообразно объединять современных кошек более чем в одно подсемейство—Felinae, как это и делает, например, Симпсон (1945).
В СССР: представители всех четырех родов.
Род больших кошек
Genus Panthera Oken, 1816
Размеры крупные — наибольшие в семействе. Длина тела 91—317 см, длина хвоста 70—114. Масса 91—390 кг. Общий облик, типичный для кошек (рис. 116). Туловище удлиненное, иногда сильно. Голова средних размеров или крупная с относительно вытянутым лицевым отделом. Уши небольшие, с закругленной вершиной. Конечности относительно невысокие, толстые. Когти втяжные. Волосяной покров у большинства видов по всему телу одинаковой высоты. У самцов льва в передней части тела удлиненные волосы образуют гриву. На конце хвоста может иметься кисть волос. Окраска тела одноцветная, или с черными поперечными полосами, или по светлому фону разбросаны черные пятна. Обычно на задней стороне уха расположено белое пятно. Сосков 2 или 3 пары.
Череп массивный с сильными скуловыми дугами и мощными гребнями и буграми. Лицевой отдел развит очень сильно, мозговой — слабо. Костные слуховые барабаны маленькие, не вздутые. Общее число зубов 30. Клыки относительно короткие, но мощные.
Диплоидное число хромосом у льва, тигра, барса и ягуара по 38.
До недавнего времени были широко распространены в Азии: на юг — по побережью материка, включая п-ов Малакку, о-ва Шри Ланка, Суматру, Яву, Бали и Кангеан (к северу от Бали), на восток — до побережья материка, исключая о-ва Хайнань, Тайвань, Японские и Шангарские, на севере включает Приамурье, Забайкалье, Алтай и Кашгарию, Восточный и Южный Казахстан и Среднюю Азию, Иран, Кавказ и Малую Азию; в Европе — южные части Балканского п-ова; вся Африка; юг Северной Америки, Центральная Америка, Северная и центральные районы Южной Америки на юг, включая Перу, Боливию, Парагвай, юг Бразилии, в Аргентине провинции Сан-Луис и Ла-Пампа. За последнее время ареал сильно сократился (исчезли в Европе, Казахстане и Средней Азии, Сибири, во многих районах Африки, Южной Азии и Америки).
Населяют различные ландшафты от полупустынь, степей и саванн (лев, леопард) до приречных зарослей, равнинных и горных лесов (тигр, ягуар, леопард) и высокогорий (леопард). Леопард может лазать по деревьям. Встречаются поодиночке, кроме льва, который обычно держится небольшими стаями (прайдами), вероятно представляющими семейные группы. В Серенгети состав 12 изученных прайдов льва был
Рис. 116. Лев — Р. 1ео (4), тигр — Р, tigrib (Б), леопард— Р. pardus (В)
Рис. 116. Продолжение. Ягуар — Р. опса (Г) и череп тигра
следующим: общее число 4—37 особей, из них взрослых самцов 2—4, полувзрослых самцов 1 (отмечен всего в одном прайде), взрослых самок 2—II, полувзрослых самок 1—5 (отмечены в 4 прайдах), больших (по размерам) детенышей 1—11 (отмечены в 5 прайдах), маленьких (по размерам) детенышей 1—22 (отмечены в 9 прайдах). Размеры участка, занятого одним прайдом, варьируют от 34 км2 в национальном парке Найроби в Кении до 400 км2 в Масаи. Участки соседних прайдов могут налегать друг на друга. Активны преимущественно ночью. У львов пищу в прайде добывают в основном самки. 48% охот совершается львами в одиночку, 20%—двумя зверями и 32%—3—14 (обычно 3—8) львами (Шаллер, Schaller, 1972).
Ярко выраженные хищники, питающиеся в основном крупными копытными. Львы в Африке охотятся за зебрами, антилопами, газелями, иногда нападают на домашний скот. Преимущественный состав корма львов значительно варьирует в разных местах: так, в национальных парках Маньяра, Найроби, Кафуе и Крюгера процент от общего состава пищи соответственно составлял: гну-—2,0; 48,3; 6,1; 23,6; зебр — 16,0; 20,7; 7,3; 15,8; импал—11,0; 2,7; 2,0; 19,7; буйволов — 62,0; 0; 30,5; 9,2. В Индии львы поедают кабанов, а также нильгау, замбаров и аксисов, изредка домашний скот. Тигры в Закавказье, Средней Азии, Казахстане питались в основном кабанами, а также косулями, оленями, сайгаками, джейранами, иногда домашним скотом. Употребляли в пищу саранчу, некоторых других насекомых, птиц, мелких млекопитающих; на Дальнем Востоке в Сихотэ-Алине объектом пищи служат главным образом кабан, а также изюбр, лось, пятнистый олень, косуля, кабарга, изредка медведь, рысь, барсук, волк, маньчжурский заяц, рябчик, рыба, из домашних животных тигр предпочитает собак. Леопард предпочитает нападать на мелких и средних копытных, поедает также мелких и среднего размера хищных (вплоть до молодых медведей), различных грызунов (в частности, дикобразов), зайцеобразных, птиц, нападает на домашний скот, собак.
Сезонности в размножении у льва и тигра в тропической зоне, очевидно, нет. Однако наибольшее число рождений у львиц в парке Крюгера (ЮАР) происходит с марта по июль, в Восточной Африке — с ноября по март. По-видимому, пики рождаемости в различных местностях и в разные годы варьируют. У леопарда гон на Кавказе происходит в январе, в Туркмении — в конце ноября — январе, в Приморье — в январе. Для льва характерна полигамия. Беременность длится у льва 100—119 дней, тигра — 98—112, обычно 105 дней, леопарда — 90—105, обычно 92—95 дней. Самки тигра рожают раз в 2—3 года, льва — раз в 2 года. В помете у львицы бывает 1—6, редко 7 детенышей. У тигрицы и леопарда 1—6 (обычно 2—4). Тигрята уссурийского тигра рождаются слепыми (иногда зрячими), беспомощными. Масса новорожденного 0,8—1 кг, длина тела 31—40 см. Они покрыты довольно густыми волосами, несколько более светлой окраски, чем у взрослых. Прозревают на 6—8-й день, ушные отверстия открываются на 4—5-й день, на 13-й день прорезаются первые зубы (два средних резца), ювенильная линька проходит в 3,5—5,5 месяцев. Продолжительность лактации до 5—6 месяцев жизни, но примерно с 2-месячного возраста тигрята регулярно подкармливаются мясом. С тигрицей детеныши держатся 2, иногда 3 года. Масса двухгодовалых около 100 кг, а трехлеток примерно 150 кг. Половозрелость наступает, как правило, на 4-м году жизни. Способность к размножению сохраняется до 20 лет. Продолжительность жизни 40—50 лет.
Котята леопарда рождаются слепыми, беспомощными, покрытые густым и высоким волосяным покровом. Окраска его светло-бурая с мелкими темно-бурыми и черными пятнами, низ тела и хвоста белый с круп-
ними черно-бурыми пятнами. Масса новорожденного 0,5—0,7 кг, длина тела около 15 см. Прозревают на 12—15-й день. В 2-месячном возрасте выходят из логова, начинают есть мясо, сменяют ювенильный мех. Лактация продолжается 5—6 месяцев. Детеныщи держатся с матерью, пока у нее не начнется течка. Половозрелость наступает в 2—3 года. Способность к размножению сохраняется 12—15 лет. Продолжительность жизни 21 год (в неволе).
Существенного практического значения не имеют. До последнего времени (а в некоторых местах и сейчас) это ценный трофей спортивной охоты. Иногда могут причинять вред скотоводству (как правило, незначительный). Известны случаи людоедства (тигр и леопард); людоедами становятся старые, больные или раненые звери.
Ввиду быстрого снижения численности повсеместно заслуживают охраны. В случае перенаселения можно искусственно регулировать численность.
В роде 4 вида:
лев — Р. leo Linnaeus, 1758 (в историческое время ареал охватывал всю Африку, кроме влажных лесов на западе; в Европе — Балканский п-ов и Армению; в Азии — Аравийский п-ов, Турцию, Ирак, Сирию, Иран, Афганистан, Индию; на север шел примерно до 28°30' с. ш., на юг немного южнее тропика и от Синда на западе до Бенгалии на востоке; к настоящему времени львы уничтожены в Южной и на большей части территории Северной Африки, сохранились, как правило, только на охраняемых территориях в Сомали, Судане и на запад от них к югу от Сахары до Сенегала и на юг до Юго-Западной Африки, Ботсваны, Родезии, Мозамбика; в Греции они исчезли к концу I в. н. э., в Закавказье — в X в., в Иране последний лев был убит в 1930 г.; в Индии есть небольшое число львов— в 1955 г. их было 250, в 1968 г. 177 особен сохранилось в лесу Гир на п-ове Катхиавар на северо-западе страны);
тигр — Р. tigris Linnaeus, 1758 (Северный Иран, Афганистан, п-ов Индостан, кроме пустынь северо-запада и о-ва Шри Ланка, Бирма, Таиланд, Индокитай, п-ов Малакка, о-ва Суматра, Ява, Бали, Джунгария, Синьцзян-Уйгурский автономный район, Восточный и Северо-Восточный Китай на запад до Большого Хингана включительно; в СССР обитал в Талыше и Ленкоранской низменности и отсюда проникал в различные районы Кавказа, но к середине XX в. на Кавказе исчез, последний заход из Ирана в Ленкоранскую низменность известен в 1966 г.; в Казахстане и Средней Азии держался главным образом по тугаям в долинах рек, однако к 20—30-м годам нашего века повсеместно исчез, хотя отдельные тигры еще встречались и позже, например в низовьях Сырдарьи в 1945 г., в низовьях Или в 1939 г., в Тигровой балке в Таджикистане в 1953 г., в низовьях Амударьи в 1968 г.; в Приморье и Уссурийском крае численность тигров в 1974 г. была 100—150 голов, что при соответствующей охране, по-видимому, обеспечивает сохранность вида в пределах СССР; с Дальнего Востока происходили заходы в Забайкалье, Прибайкалье и Восточную Сибирь);
леопард — Р. pardus Linnaeus, 1758 (Африка, кроме Сахары, Аравийский п-ов, Малая Азия, Иран, Афганистан, Пакистан, п-ов Индостан и о-в Шри Ланка, Бангладеш, Бирма, Таиланд, п-ов Малакка, о-в Ява, архипелаг Кангеан и, возможно, Суматра, п-ов Индокитай, восток Тибета, Восточный и Северо-Восточный Китай, Корейский п-ов; в СССР: горная система Кавказа, Средняя Азия — Копетдаг, Малый и Большой Балхан, Бадхыз, правобережье верхней Амударьи и Пянджа, Дальний Восток—крайний юг Уссурийского края. Встреченные в Забайкалье, Приамурье, в остальных частях Уссурийского края звери в большинстве случаев были бродячими; в последнее время ареал значительно сокра-
тился, особенно в Северной и Южной Африке; у нас на Кавказе сохранился только в Талыше, но, возможно, это не аборигенные звери, а заходящие из Ирана; исчез в северо-западной части Копетдага, почти истреблен в Большом Балхане);
ягуар — Р. опса Linnaeus, 1758 (обитал по всем тропическим и субтропическим лесам Северной, Центральной и Южной Америки; в настоящее время исчез из США, отсутствует на большей части территории Мексики и Аргентины и на освоенных человеком площадях Центральной и Южной Америки).
В «Красную книгу» включены: подвиды тигра с о-ва Суматра — Р. tigris sumatrae Pocock, 1929, и с юга Китая, Бирмы, п-ова Малакка, Таиланда, Кампучии, Лаоса, Вьетнама; леопард — Р. pardus Linnaeus, 1758, и ягуар — Р. опса Linnaeus, 1758, как подвиды и виды, которые в ближайшее время могут оказаться под опасностью исчезновения.
В «Красную книгу» включены также находящиеся под угрозой исчезновения: подвид льва из Индии — Р. leo persica Meyer, 1826; подвиды тигра из Индии (сохранилось на 1972 г. 1827 особей), Непала (на 1970 г. сохранилось 200), Бутана, Бангладеш (на 1970 г. сохранилось 20 тигров) и Западной Бирмы — Р. tigris tigris Linnaeus, 1758; из Северного Афганистана, Северного Ирана (на 1950 г. общее число достигало 50—80 особей) и Талыша— в СССР — Р. tigris virgata liliger, 1815; с Дальнего Востока СССР (на 1974г. сохранилось 100—150 тигров); Северо-Восточного Китая и северо-восточной части Корейского п-ова (сохранилось 40— 50 тигров) — Р. tigris altaica Temminck, 1844; с юга и востока о-ва Ява (численность очень низка: 3—7 в лесу Бетери и 5 — в Сукамати) — Р. tigris sondaica Temminck, 1845; из провинции Сычуань, долины р. Янцзы и провинции Фуцзянь Китая — Р. tigris amoyensis Hilzheimer, 1905; с западной части о-ва Бали (осталось, по-видимому, несколько особей) —Р. tigris balica Schwarz, 1912; подвиды леопарда из Алжира, Марокко (к 1964 г. сохранилось 50—100 особей) и Туниса — Р. pardus panthera Schreber, 1777; из южной части Аравийского п-ова — Р. pardus nimr Hemprich et Ehrenberg, 1833; из Турции, Сирии, возможно Ливана, Израиля, Туркменской ССР и Таджикской ССР — Р. padrus tulliana Valenciennes, 1856; с Дальнего Востока СССР, из Северо-Восточного Китая и северной части Корейского п-ова — Р. pardus orientalis Schlegel, 1857; с п-ова Синай и северной части Саудовской Аравии (численность очень низка, возможно уничтожен полностью) — Р. pardus jarvisi Pocock, 1932.
Род снежных барсов
Genus Uncia Gray, 1854
В роде единственный вид: снежный барс — V. uncia Schreber, 1775. Размеры крупные. Длина тела 103—130 см, длина хвоста 80—105 см. Масса 22—39 кг. Туловище сильно вытянуто и слегка приподнято в крестцовой области (рис. 117). Голова относительно небольшая, округлая. Глаза большие, зрачок круглый. Уши короткие с закругленной вершиной. Конечности относительно короткие, лапы широкие и массивные. Когти втяжные. Волосяной покров очень высокий, густой и мягкий. Гривы и удлиненных волос на щеках нет. Хвост покрыт высоким густым мехом одинаковой высоты. Окраска зимой на спине и верхней части боков сероватая, почти белая с дымчатым налетом, бока книзу, брюхо и внутренние части конечностей светлее спины. По этому фону разбросаны редкие крупные кольцевидные и мелкие сплошные пятна черного или темно-серого цвета. В задней части тела пятна часто сливаются в 238
Рис. 117. Снежный барс — U. tii'icia и его череп
короткие продольные полосы. Летний мех имеет общий фон светлее зимнего, а пятна более черные; волосы летом значительно короче и реже, чем зимой.
Череп массивный, но в меньшей степени, чем у представителей рода больших кошек (Panthera). Гребни и бугры на нем крупные, хотя и не столь развиты, как у последних. Лицевой отдел относительно короче, а мозговой сильнее вздут и крупнее, чем у больших кошек. По общему виду череп более сходен с черепом рода кошек (Felis). Костные слуховые барабаны относительно небольшие, слабо вздутые. Общее число зубов 30.
Распространен в Монгольском и Гобийском Алтае, Хангае, горах области оз. Хубсугул в МНР, Тибете до Алтынтага на севере и Кама — на востоке, Гималаях от восточной оконечности до Кашмира на западе, Гиндукуше в Афганистане. В СССР обитает в горах востока Средней Азии и юга Сибири, от Амударьи до Байкала, на западе в Памиро-Гис-сарской системе и Тянь-Шане, на Алтае — южном, центральном, восточном и северо-восточном, в Саянской системе и по хребту Танну-Ола и на восток до гор Прибайкалья по Ангаре и Верхней Лене; в последние годы ареал сократился, в частности, снежного барса уже давно нет в Прибайкалье и Саянах.
Держится в горах преимущественно в субальпийском и альпийском поясах, часто у границы вечного снега. Зимой следом за копытными спускаются к лесному поясу. Предпочитает скальные участки. Активен в сумерках-— вечером и утром, изредка днем. Питается преимущественно сибирскими горными козлами, реже горными баранами, косулями, молодыми кабанами, иногда сурками, сусликами, зайцами, мышевидными грызунами, уларами, кекликами. Изредка нападает на домашний скот. Течка в неволе в марте — мае; в Узбекистане в конце февраля — марте. Беременность 93—ПО дней. В помете 1—5 (обычно 2—3) котят. Новорожденные слепые, беспомощные, с длиной тела около 30 см, массой около 0,5 кг. Они покрыты густым, ио грубым волосяным покровом, на спине светло-буроватого цвета, испещренным темно-бурыми пятнами, на брюхе — сливочно-белого. Половозрелость у самок наступает в 2—3 года. Продолжительность жизни до 15 лет.
Существенного практического значения не имеет. Промышляется в незначительном количестве.
Включен в «Красную книгу» как вид, находящийся под угрозой уничтожения.
Род кошек
Genus Felis Linnaeus, 1758
Размеры мелкие, средние и крупные. Длина тела 46—197 см, длина хвоста 15—91 см. Масса 2,5—105 кг. Туловище умеренно удлиненное или относительно укороченное, покатое от крестца к плечам, реже линия спины прямая (рис. 118). Голова округлая, иногда вытянутая (пума, дымчатый леопард), крупная. Уши большие, иногда очень большие (оцелот), округлые или с острыми вершинами, иногда с кисточкой волос на них (рысь, каракал). Уши могут отстоять далеко друг от друга или почти сходиться внутренними краями (оцелот). Глаза большие, иногда очень большие, зрачок вертикальный, изредка круглый. Конечности короткие или средней длины, иногда высокие (рысь). У большинства лапы небольшие, изредка широкие и мощные (пума, рысь, дымчатый леопард). Когти полностью втяжные (кроме суматранской кошки). Волосяной покров высокий, густой и мягкий. Окраска его разнообразна: одноцветная, 240
Рис. 118. Дикая кошка — F. silvestris (Л), коптка рыболов— F. viverrma (Б), ягуарунди— F yagouarundi (В), кошка Темминка — F, temmincki (Г), рысь — F. lynx (Д1
Рис. 118. Продолжение Селилп___г ,
кал — Л caracal (М). nvMa — F concolo^rix леопард—< tiebulosa (И\. ' сопсо1°' (3),
(^), кара-дымчатый
Рис. (18. Продолжение. Череп дикой кошки
обычно желтых тонов, одноцветная со следами пятен, у большинства пятнистая или полосатая. Пягна бывают разных размеров, сплошные и кольцевые, они образуют ряды или сливаются или почти сливаются в продольные, реже поперечные полосы. У некоторых видов (дымчатый леопард) на тыльной стороне уха находится белое пятно. Сосков 2—4 пары.
Череп относительно крупный, но не массивный. Гребни и выступы развиты слабо или их нет вовсе. Мозговая коробка большая, вздутая. Лицевой отдел короткий. Скуловые дуги слабые. Глазница большая, у некоторых видов замкнутая или почти замкнутая. У немногих видов (дымчатый леопард, пума) череп сходен с черепом представителей рода больших кошек. Он массивный с сильными гребнями, выступами и ску--элшыы.. мммй»» л-гип.ктсп|.ил пытятиын Костные слуховые баоабаны
очень большие, вздутые. Общее число зубов 30. Размеры второго верхнего предкоренного зуба очень малы. У манула, большинства каракалов, рысей он отсутствует и общее число зубов составляет 28.
Диплоидное число хромосом у бенгальской, золотой, дикой, черноногой, барханной кошек, кошки Темминка, ягуарунди, сервала, рыси, красной рыси, каракала и пумы по 38, у оцелота, пампасской и американской кошек по 36.
Распространение соответствует ареалу семейства. В Европе и Северной Америке местами истреблены.
Населяют различные ландшафты от пустынь до северной границы леса. Держатся поодиночке. Активны, как правило, в сумерках и ночью, некоторые (барханная кошка) и днем. Хорошо лазают по деревьям. Некоторые плавают (камышовая кошка). Убежищем служат дупла деревьев, расщелины скал, пещеры, густые заросли, норы дикобразов, лисиц, барсуков.
Питаются преимущественно мелкими позвоночными, главным образом грызунами и птицами. Так, амурская кошка летом охотится за мышевидными грызунами, бурундуками, белками, различными птицами и их птенцами, зайцами, молодыми косулями, ящерицами, змеями и рыбой, а зимой ловит маньчжурских зайцев, белок, фазанов, рябчиков и рыб. Камышовая кошка питается водяными крысами, мышевидными грызунами, зайцами-русаками, акклиматизированными нутриями, сусликами, поросятами кабана, различными водными и прибрежными птицами, черепахами, змеями, ящерицами, рыбой. Лесная дикая кошка кормится в основном грызунами, в том числе серыми крысами, водяными полевками, ондатрами, различными мелкими и средней величины птицами, реже насекомыми, рыбой. Степная дикая кошка питается в основном песчанками, полевками, тушканчиками, зайцами-песчаниками, реже птицами и пресмыкающимися, изредка жуками, саранчой, растениями. Основным кормом каракала служат, по-видимому, зайцы-толаи, а также различные грызуны, поедает он также птиц, пресмыкающихся и насекомых. Рысь охотится за зайцами — беляком и русаком, самками и молодью лося и северного оленя, косулей, пятнистым оленем, различными грызунами и птицами; основным кормом в большинстве мест служит заяц-беляк и (или) копытные. Красная рысь питается преимущественно кроликом и зайцами, а также мышевидными грызунами, белками, иногда оленями. Барханная кошка поедает зайца-песчаника, тонкопалого суслика, большого песчаного тушканчика и других грызунов, в меньшей степени птиц, пресмыкающихся и членистоногих. Манул кормится пищухами, полевками, сусликами, хомячками. Кошка-рыболов поедает в основном рыбу, суматранская кошка — плоды.
Гон у амурского кота в Приморье приходится на март, у камышового кота — с января по первую половину апреля, у дикой лесной кошки —-в основном на январь— март, у дикой степной кошки и рыси — январь — февраль. Беременность у большинства видов длится около 60 дней (у рыси 67—74 дня, пумы — 96 дней). Детенышей в помете 1—7 (обычно 3—4). В году иногда бывает 2 помета (камышовая кошка, дикая лесная кошка). Пума приносит один помет в два, иногда три года. Детеныши рождаются слепыми, беспомощными, покрытыми густым и высоким волосяным покровом. Основная окраска его обычно светло-бурая с темными продольными полосами на спине. Масса новорожденного камышовой кошки 45—55 г. Прозревают котята на 10—12-й день. Лактация длится около 2 месяцев. Масса новорожденного лесной дикой кошки 38—45 г. Прозревают на 9—12-й день. Лактация продолжается 3—4 месяца (у степной дикой кошки 2—2,5 месяца). Масса новорожденного рыси 250—300 г. Котята прозревают на 12-й день. Лактация тянется около 244
85 дней. Масса новорожденного красной рыси 283—368 г; прозревают на 9-й день, лактация — около 2 месяцев. Половозрелость наступает у дикой степной кошки в возрасте 21—22 месяцев, рыси — около 2 лет, пумы — в 2—3 года. Продолжительность жизни дикой лесной кошки до 21 года, рыси — до 25 лет.
Хозяйственное значение невелико. Большинство — промысловые виды, дающие малоценную пушнину, но мех рыси ценится высоко. В СССР с 1960 по 1970 г. ежегодно заготовляли по 1,8—5,8 тыс. шкур рысей и по 5 2—34,0 тыс. — остальных кошек. Могут причинять вред охотничьему хозяйству, уничтожая охотничью фауну (особенно камышовый кот).
Каракал и манул, имеющие низкую численность, нуждаются в охране.
Ливийская форма дикой кошки — F. silvestris libyca Forster, 1780 послужила предком домашней кошки. Одомашнение произошло в Египте примерно 4—3 тыс. лет до н. э. Отсюда домашняя кошка была вывезена в другие страны: в Китай не ранее VI в. до н. э., в Европу — в Причерноморье в VI—I вв. до н. э., в Рим в IV в. н. э., в Среднюю и Западную Европу — в средние века, в Древнюю Русь — в VII—XIV вв. н. э.
Система рода окончательно не установлена. Ниже приводится перечень видов по В. Г. Гептнеру (1972). В роде, по-видимому, 30 видов:
бенгальская кошка — F. bengalensis Кегг, 1792 (Южный Китай и Тибет, п-ова Индокитай и Малакка, Бирма, Индия, Непал, Пакистан, о.-ва Суматра, Ява, Бали, Калимантан и Филиппинские);
пятнисторыжая кошка — F. rubiginosa I. Geoffroy, 1831 (Южная Индия и Шри Ланка);
кошка-рыболов — F. viverrina Bennett, 1833 (Индия, Шри Ланка, Непал, п-ов Индокитай, о-ва Суматра и Ява);
ириомотейская кошка — F. iriomotensis Imaizumi, 1967 (о-в Ириомоте из группы о-вов Рюкю);
амурская кошка — F. euptilura Elliot, 1871 (Корейский п-ов, о-ва Чечжудо и Сайсю, Восточный и Северо-Восточный Китай; СССР: южная часть Дальнего Востока, где ареал разделен небольшим разрывом на 2 участка — один ограничен с севера линией, начинающейся на западе несколько восточнее г. Благовещенска и тянущейся вдоль Амура на расстоянии 30—50 км к северу от реки до 131° в. д., откуда граница резко сворачивает на восток, затем, образуя дугу на юго-восток и, пересекая Амур, уходит в Китай; северная граница второго участка ареала вновь входит в пределы СССР у устья Уссури и по правому берегу Амура идет до устья Хунгари, где немного отходит к востоку и затем резко поворачивает на юг по западному склону Сихотэ-Алиня на расстоянии 120—150 км к востоку от Уссури до 45° или 45° 15' с. ш., здесь граница сворачивает на северо-восток, следуя по восточному склону Сихотэ-Алиня до устья Ботги на 48° с. ш., где выходит к морю; к югу от этой границы ареал занимает всю территорию Уссурийского края);
суматранская кошка — F. planiceps Vigors et Horsfield, 1827 (о-ва Суматра, Калимантан, п-ов Малакка, Южный Таиланд);
мраморная кошка — F. marmorata Martin, 1837 (Индия — Сикким и Ассам; Непал; Северная Бирма; п-ова Индокитай и Малакка, о-ва Суматра и Калимантан);
калимантанская кошка — F. badia Gray, 1874 (о-в Калимантан);
кошка Темминка — F. temmincki Vigors et Horsfield, 1827 (Южный Китай, Тибет; Непал; Индия — Ассам; Бирма; п-ова Индокитай и Малакка, о-в Суматра);
золотая кошка — F. aurata Temminck, 1827 (Африка, узкая полоса к югу от Сахары от р. Гамбии ца северо-западе до Кении на востоке; северная граница ареала идет от Гамбии через север Сьерра-Леоне, Либерию, центр Берега Слоновой Кости, Гану, центр Нигерии, Северный Ка
мерун, юг Центральноафриканской Республики, Северную Уганду; южная граница ареала проходит через Северную Анголу, юг Заира и до Руанды и Бурунди; по Танзании сведений нет);
оцелот — F. pardalis Linnaeus, 1758 (юг Северной Америки: узкая западная полоса ареала начинается в США в центре штата Аризона, проходит на юг через его юго-восточный угол и далее в Мексике по штату Сонора, вдоль границы этого штата со штатом Чиуауа и затем такой же узкой полосой идет по Тихоокеанскому побережью, захватывая штаты Синалоа, Наярит, западные половины Халиско, Колима, запад Мичоакан, Герреро, юг Оахака, где эта полоса ареала сливается с восточной полосой, которая начинается в США, захватывая весь штат Техас, восточную половину штата Коауила в Мексике, Нуэво-Леон, Тамаулипас, крайний восток Сан-Луис-Потоси и Веракрус; далее к юго-востоку ареал включает штаты Мексики: Чиапас, Табаско, Кампече, Юкатан, Кинтано-Роо, всю Центральную Америку и переходит в Южную Америку с запада от Колумбии, Эквадора и Перу, на восток идет до устья Амазонки и Восточной Бразилии и с севера от Венесуэлы на юг до Парагвая, Боливии и Северной Аргентины; о-в Тринидад);
американская кошка —К. wiedi Schinz, 1821 (юг Северной Америки — в своей северной части ареал напоминает ареал оцелота: узкая западная полоса ареала начинается в Мексике в южной половине штата Синалоа и идет на юг, включая западные половины штатов Наярит, Халиско и Мичоакан, Колима, Герреро и Оахака, где сливается с восточной полосой ареала, которая начинается в США на юго-западе штата Техас и идет в Мексику, захватывая штаты Нуэво-Леон, Тамаулипас, крайний восток Сан-Луис-Потоси, восток Идальго и Веракрус, далее к юго-востоку ареал включает штаты Табаско, Чиапас, Кампече, Юкатан, Кинтано-Роо и всю территорию Центральной Америки; в Южной Америке ареал охватывает большую площадь — от Колумбии на юг до Боливии, Северной Аргентины, Восточного Парагвая и Северного Уругвая);
пампасская кошка — F. colocolo Molina, 1782 (Бразилия, Эквадор, Анды в Перу, Западная Боливия, Аргентина, Чили);
кошка Жоффруа — F. geoffroyi D’Orbigny et Gervais, 1844 (Боливия, Парагвай, Аргентина);
онцилла — F. pardinoides Gray, 1867 (~F. tigrina Schreber, 1775) (юго-восточная половина Коста-Рики, Панама, Гвиана, Венесуэла, Колумбия, Эквадор, Бразилия, Парагвай и Аргентина);
чилийская кошка — F. guigna Molina, 1782 (Чили и Аргентина);
аргентинская кошка — F. paieros Desmarest, 1816 (Аргентина);
ягуарунди — F. yagouarundi Geoffroy, 1803 (юг Северной Америки: США—крайний юго-восток штата Аризона и далее на юг узкой полосой через восточную часть штата Сонора; Мексика — юго-запад Чиуауа, запад Дуранго, юг Синалоа, Наярит, западные половины штатов Халиско и Мичоакан, Колима, Герреро, южная половина Оахака, куда подходит восточная часть ареала, тянущаяся полосой вдоль Мексиканского залива, от крайнего юга штата Техас в США и далее на юг, включая штаты Мексики Тамаулипас, крайний восток Сан-Луис-Потоси, Веракрус; в ареал входят также штаты Мексики Чиапас, Табаско, Кампече, Юкатан, Кинтано-Роо; вся Центральная Америка и почти вся Южная Америка: Колумбия, возможно Эквадор, Венесуэла, Гвинея, Бразилия, Перу, Парагвай, Аргентина);
камышовая кошка — F. chans Gflldenstaedt, 1776 (Африка: дельта Нила и его нижнее течение до Кепа на 28° с. ш. и по Средиземноморскому побережью на 250 км на запад от Александрии; Азия: Ирак, часть Сирии, Палестина, западная и южная части Малой Азии, Иран, Афганистан, Пакистан, Индия, Шри Ланка, Непал, Бирма, п-ов Индо-246
китай, Китай — провинция Юньнань; в СССР на юге в двух районах, которые соединяются между собой вне территории государства: кавказский район — от дельты Волги узкой полосой к югу по западному берегу Каспийского моря до устья Кумы, включая ее нижнее течение, далее до устья Терека и вверх по его течению до Моздока и по Сунже до Грозного, к югу от Терека ареал распространяется до государственной границы с Ираном, захватывая Восточное Закавказье на запад до предгорий Большого и Малого Кавказа; среднеазиатский район представлен несколькими изолированными друг от друга участками — на юго-востоке Туркмении по Атреку и его притокам; по Теджену и Мургабу; по Амударье и ее правым притокам Кызылсу, Вахшу, Кафирнигану; Гис-сарская долина на востоке с заходом несколько за Душанбе и долина Сурхандарьи и нижний Тупаланг; по р, Зеравшан вверх до Педжикен-та; вся Сырдарья);
дикая кошка — F. silvestris Schreber, 1777 (включая F. libyca Forster, 1780) (Африка: вся территория континента, кроме центральных районов Сахары и западноафриканской лесной области; Европа: вся территория включая Англию, на север до Балтийского моря, на юг до Южной Испании, Италии с Сицилией и Балкан; в Ирландии отсутствует; Азия: Малая Азия, Аравийский п-ов, Ирак, Сирия, Палестина, Иран, Афганистан, Пакистан, Индия — Пенджаб, Синд, Кач, Раджпутана, центральные районы; Южная и Юго-Западная МНР; Северо-Западный Китай; СССР: запад и север Волынской, север Ровенской и Житомирской, некоторые места Каменец-Подольской области, леса Карпат, Закарпатская, Станиславская, Дрогобычская, Черновицкая, южная часть Львовской и юго-восток Тернопольской областей, Молдавия, леса Большого и Малого Кавказа и предкавказских равнин, юг и юго-восток Кавказа, Казахстан и Средняя Азия, на север до линии, проходящей от устья Эмбы к северной оконечности Аральского моря, устью Сарысу, на р. Чу и захватывающей южную часть Бетпак-Дала, следующей к западному берегу Балхаша, вдоль его южного берега и охватывающей дугой Алакульскую впадину и уходящей за государственную границу в Китай);
китайская кошка — F. bieti Milne-Edwards, 1892 (Китай — Ганьсу и Сычуань; МНР);
сервал — F. serval Schreber, 1776 (Африка — Алжир и вся территория к югу от Сахары);
рысь — F. lynx Linnaeus, 1758 (Северная Америка: вся северная часть материка, включая Баффинову Землю, но без остальных о-вов Канадского Арктического архипелага, обитает на о-ве Ньюфаундленд; южная граница ареала проходит по территории США от юго-западной части штата Орегон на юго-восток к северной части Невады, югу Айдахо, откуда резко сворачивает к югу к центральным районам Юты, так же круто поднимается к северу до северо-западного угла Колорадо, выгибаясь дугой к центральным районам этого штата, опускается к его юго-западному углу, затем поворачивает на северо-восток и север и доходит до центральных районов юга Северной Дакоты, откуда спускается к югу до юго-востока Небраски, поворачивает на юго-восток к южным частям Индианы, сворачивает на северо-восток и в районе штата Коннектикут выходит к Атлантическому океану; Европа: (СССР см. ниже) в недалеком прошлом вся территория Европы, на север до Северной Скандинавии, на юг до Южной Испании, возможно Сардинии, Апеннинского п-ова и Балкан, на Британских о-вах и Сицилии не обитала; к настоящему времени рысь сохранилась местами в Испании, Швеции, Норвегии, Финляндии, Польше, Чехословакии, Румынии и на Балканах; Азия: Малая Азия, Северный и Восточный Иран, Северо-Западный и Северный Иран, Афганистан, Индия — Кашмир, Китай, Северо-Западный, Северный и
Северо-Восточный Тибет, МНР — Монгольский и Гобийский Алтай, Хан-гай, Хентей и отроги Большого Хингана; СССР: почти вся территория страны — северная граница ареала проходит несколько южнее северной границы леса, Сахалин, на Шантарских и Курильских о-вах нет; на Камчатке появилась, по-видимому, с 1930—1937 гг., южная граница идет от Карпат по югу Полесья, через Брянские леса, Рязанскую Мещеру, Мордовию, Татарию, Южный Урал, Барабинскую лесостепь и Алтай, где уходит за государственную границу, далее к востоку южная граница ареала в нашей стране совпадает с государственной границей; кроме того, в ареал входят горы Кавказа, Закавказья и Средней Азии);
красная рысь — F. rufus Schreber, 1777 (Северная Америка: Канада— южные районы провинций Британская Колумбия, Альберта, Саскачеван, Манитоба, Онтарио, Квебек, а также провинции Нью-Брунсуик и Новая Шотландия; вся территория США; Северная и Центральная Мексика — южная граница ареала проходит от юго-западной части штата Халиско на юго-восток через середину Мичоакан, северо-восток Герреро, север Оахака и в северо-восточной части этого штата сворачивает на северо-запад, проходя на некотором расстоянии от берега Мексиканского залива по западным границам штата Веракрус и центральным районам Тамаулипас, на севере этого штата граница ареала выходит к Мексиканскому заливу);
каракал — F. caracal Schreber, 1776 (Африка: весь материк, кроме сплошных лесов Западной и Центральной Африки бассейнов Нигера и Конго и прибрежных областей Мозамбика и Наталя в Южно-Африканской Республике; Азия: Афганистан, Иран, Малая Азия, Ирак, Сирия, Палестина, Аравийский п-ов, Пакистан, северо-запад Индии; СССР — территория Средней Азии, ограниченная с запада Каспийским морем, с севера линией, проходящей от Мангышлака и северной части п-ова Буза-чи на восток по северному чинку Устюрта к северо-западной части Аральского моря, с востока западным берегом Аральского моря и Амударьей, с юга — государственной границей);
черноногая кошка — F. nigripes Burchell, 1882 (центральные и восточные части Юго-Западной Африки, Центральная и Южная Ботсвана, Южно-Африканская Республика);
барханная кошка — F. margarita Loche, 1858 (Африка: Алжир, Южная Ливия, Северный Нигер, Юго-Западный Судан; Азия: Синайский п-ов, восток и юг Аравийского п-ова; СССР — песчаные пустыни Средней Азии на территории, ограниченной с юга южной границей Каракумов, с востока — берегом Каспийского моря на север до Мангышлака, с севера — северным чинком Устюрта, западным, южным и восточным берегами Аральского моря и далее Сырдарьей, с востока — восточными окраинами пустынь Кызылкумы и Каракумы; ареал окончательно не установлен: вероятно, в него включаются Ирак, Иран, Афганистан, вся Сахара и весь Аравийский п-ов);
манул — F. manul Pallas, 1776 (Азия: Северо-Восточная Турция, возможно север Ирака, Иран, Афганистан, Пакистан, Индия—Кашмир; Китай — Тибет, Ганьсу, Сычуань и крайний северо-восток; МНР; в СССР 4 отдельных участка соединяются друг с другом за пределами страны: первый на крайнем юге Центрального Закавказья; второй в Средней Азии, Казахстане, по южносибирским хребтам включает Копетдаг, Большие Балханы, возвышенную область восточной половины Туркмении, обрывы Устюрта, среднее течение Эмбы, южные оконечности Мугоджар, юго-запад Таджикистана, возможно Зеравшанский и Туркестанский хребты и их отроги, мелкие останцы Кызылкумов, западные отроги и предгорья Тянь-Шаня, Каратау, Киргизию, Чу-Илийские горы, предгорья Заилийского Алатау, Джунгарский Алатау, Саурские горы, Тарбагатай, 248
Зайсанскую котловину, Черный Иртыш, Калбинский Алтай, Чингизтау, Казахскую складчатую страну на юг до Балхаша, вероятно, останцы Бедпак-Далы, далее на восток — Южный Алтай, Чуйскую и Курайскую степи, Тувинскую АССР; третий в степной части Западного Забайкалья; четвертый в Восточном Забайкалье — Ачинская и Ьорзинская степи и по р. Окон);
пума — F. concolor Linnaeus, 1771 (Северная Америка: вся территория, кроме северных районов, северная граница ареала начинается на западе на Тихоокеанском побережье в районе о-вов Королевы Шарлотты, поднимается к северо-востоку и юго-восточному углу провинции Юкон, откуда плавной дугой спускается к юго-востоку, пересекая северо-восточные районы Британской Колумбии, южную треть провинции Альберта, юго-запад Саскачевана, северо-восток штата Северная Каролина, США, затем проходит по северо-западному углу штата Миннесота и вновь поднимается к северу в Канаду, поворачивает на восток и проходит севернее Великих озер по южным частям Онтарио и Квебека, захватывая Нью-Брансуик и Новую Шотландию, в северных районах которой граница ареала выходит к Атлантическому океану; вся территория Центральной и Южной Америки);
дымчатый леопард — F. nebulosa Griffith, 1821 (Южный Китай — Фуцзянь и соседние районы, о-ва Тайвань и Хайнань; п-ова Индокитай и Малакка; Южный Таиланд; Бирма; Непал; Индия—-Сикким; о-ва Суматра и Калимантан).
Предлагаются и другие трактовки системы рода, например считают F. euptilura подвидом F. bengalensis (например, Эллермэн и Моррисон-Скотт, 1966), из F. silvestris в качестве особого вида выделяют F. libyca Forster, 1780 (например, Эллермэн и Моррисон-Скотт; 1966; Новиков, 1963; Бобринский и др., 1965), из F. lynx выделяют F. canadensis Kerr, 1792 (Холл и Келсон, 1959); выделение различных видов в отдельные роды см. выше в описании семейства.
В «Красную книгу» включены: каракал из Туркменской ССР — F. caracal michaelis Heptner, 1945, как малочисленный подвид, который, однако, в настоящее время не находится под угрозой исчезновения; оцелот— F. pardalis и дымчатый леопард — F. nebulosa как виды, которые в ближайшее время могут исчезнуть; подвид рыси, прежде населявший Пиренейский п-ов, а в настоящее время лишь горы Южной Испании,—• F. lynx pardina Temminck, 1824; подвид оцелота из южного Техаса в США и восточного Коауила в Мексике — F. pardalis albescens Pucheran, 1855; подвиды пумы — из Канады и северных штатов Восточных США—• F. concolor cougar Kerr, 1792, и из Флориды — F. concolor coryi Bangs, 1896 (всего 150—300 особей); подвид дымчатого леопарда из горной центральной части о-ва Тайвань — F. nebulosa brachyurus Swinhoe, 1862, как подвиды, которые находятся под угрозой исчезновения.
Род гепардов
Genus Acinonyx Brookes, 1828
В роде единственный вид: гепард —A. jubatus Schreber, 1775.
Размеры крупные. Длина тела 123—150 см. Длина хвоста 63—-75 см. Высота в плечах примерно 100 см. Масса обычно 50—65 кг. Туловище несколько удлинено. Конечности очень высокие, тонкие, но сильные (рис. 119). Лапы небольшие. Характерны невтяжные когти. Голова небольшая, округлая. Глаза большие, с круглым зрачком. Уши короткие, закругленные, кисточек волос на них нет. Хвост длинный и тонкий. Волосяной покров низкий, редкий (даже зимой) и грубый. Общий тон
Рис. 119. Гепард — A. jubatus и его череп
окраски желтоватый или рыжеватый. Спина, бока и наружные отделы конечностей покрыты мелкими темными пятнами. По средней линии спины пятна иногда располагаются полосами. Кончик хвоста черный, перед ним располагается белое кольцо, а еще кпереди — черное.
Брюшная часть туловища и внутренние поверхности конечностей беловатые. От затылка, по верхней части шеи и до лопаток тянется гребень удлиненных волос, образующих невысокую гриву. На щеках и конце хвоста удлиненных волос нет. Сосков 4 пары.
Череп высокий, относительно небольшой, легкого строения, гребней и выступов нет или они развиты слабо, за исключением крупного затылочного гребня. Мозговая коробка большая, вздутая, лицевой отдел укорочен. Глазницы большие, открытые сзади. Твердое небо укорочено. Костные слуховые барабаны небольшие, но вздутые.
3 13 1
Зубная формула: i — с — р — гп — = 30,
но иногда второй верхний предкоренной зуб отсутствует.
Клыки небольшие. Диастемы после них нет.
Диплоидное число хромосом 38.
Был распространен до недавнего времени в Африке, кроме центральных районов Сахары, сплошных лесов и гор; в Азии ареал охватывал Аравийский п-ов, равнины Ирака, Ирана, Белуджистан, Афганистан (кроме Гиндукуша), Индию к югу от Ганга и от Раджпутаны до Бенгалии, Пенджаб, Синд, Центральную Индию, Северный Декан, в СССР пустыни Южного Казахстана и Средней Азии, а до средних веков гепард обитал и на равнинах Восточного Закавказья вплоть до Западной Грузии. К настоящему времени ареал сильно сократился. Гепард сохранился в Африке фактически лишь в глухих местах или на охраняемых территориях (общая численность 10—15 тыс. особей), исчез в Индии и в СССР (некоторые авторы предполагают, что возможно обитание гепарда в небольшом числе в Мангышлаке, Бузачи, Устюрте, на северной окраине Каракумов).
Населяет саванны, глинистые, реже песчаные пустыни. Предпочитает слабопересеченную местность. Держится часто парами. Активен днем. Приспособлен к очень быстрому бегу; на коротких дистанциях может развивать скорость до 112 км/ч. Добычу сначала скрадывает, а подкравшись метров за 100—150, бросается на нее и на дистанции до 300 м догоняет. Дальше 400—500 м добычу не преследует. Питается преимущественно различными копытными — газелями, антилопами, в СССР джейранами, а также сайгаками и горными баранами. Иногда нападает на домашний скот. Беременность 84—95 дней. Детенышей в помете 2—4. Котята рождаются слепыми и беспомощными. Семья подолгу не распадается. Самка ходит с молодыми, почти равными ей по размерам.
Практического значения не имеет. В отличие от многих других кошек гепард хорошо приручается и во взрослом состоянии. В прежние времена феодалы и правители Египта, Индии и многих других стран (в том числе Киевской Руси и Московского княжества) использовали прирученных гепардов для охоты.
Включен в «Красную книгу» как вид, который в ближайшее время может оказаться под угрозой исчезновения.
ОТРЯД ЛАСТОНОГИХ
ORDO PINNIPEDIA I LLIGER, 1811
К ластоногим относятся млекопитающие крупных размеров, приспособившиеся к жизни в воде, но в разной степени связанные с сушей или льдами. У кольчатой нерпы — Pusa hispida — одного из самых мелких ластоногих, взрослый самец в длину достигает 150 см и массы 100 кг. У наиболее крупного среди ластоногих южного морского слона — Miro-unga leonina взрослый самец имеет в длину до 650 см, а массу до 3630 кг. Размеры самцов и самок могут быть одинаковы (большинство Phoci-dae), или самки могут быть немного крупнее самцов (роды Monachus, Lobodon, Hydrurga, Leptonychotes), или, наконец, самцы могут быть значительно крупнее самок (все Otariidae).
Для ластоногих характерны многие признаки, свидетельствующие о глубокой адаптации к водной среде. Форма тела у них обтекаемая, в общем веретеновидная, округлая в поперечном сечении, лишенная каких-либо выступов, которые мешали бы плаванию. Передние и задние пятипалые конечности превратились в ласты. Проксимальные отделы конечностей (плечо и бедро) в противоположность наземным млекопитающим не свободны, а включены в общий мешок покровов туловища. Свободная передняя конечность начинается с середины предплечья (у тюленевых свободной остается лишь кисть). Задние конечности свободны дистальнее нижней трети голени или только с предплюсны. Кисть одета общим кожным покровом и пальцы снаружи не заметны. На задних ластах пальцы видны, хотя они и соединены между собой плавательной кожной перепонкой.
Небольшой хвост тюленевых и ушастых тюленей находится в выемке между задними конечностями. Хвост моржей включен в широкую складку кожи, соединяющую задние конечности, и снаружи незаметен. Ушные раковины ушастых тюленей имеют небольшие размеры, у моржей и тюленевых их нет вовсе. Щелевидные ноздри могут замыкаться во время ныряния.
Кожный покров у ластоногих имеет значительную толщину, превышающую в абсолютных цифрах толщину кожи большинства наземных млекопитающих. Эпидермис сильно развит, у моржей на внутренней поверхности его расположены ячеи. В дерме есть специфические образования— пучки гладких мышечных волокон, не связанные с корнями волос. Мощная подкожная жировая клетчатка, образованная сплошным скоплением жировых клеток и редкими пучками коллагеновых волокон, образует надежный термоизолирующий слой, одинаково хорошо функционирующий и в воздушной среде и в воде. Сальные и потовые железы имеются. Волосы большинства ластоногих можно разделить на остевые, промежуточные и пуховые (последние своей формой обычно напоминают остевые). Волосяной покров густой или сильно изрежен-252
ный. У многих ластоногих имеет место возрастная смена волосяного покрова. Волосы не имеют mm. arrectorus pilorum. Как правило, волосы растут пучками, причем пуховые и промежуточные волосы располагаются каудальнее остевого.
Самки ушастых тюленей, моржа, морского зайца и тюленя-монаха имеют по две пары сосков, остальных представителей тюленевых —
одну.
Череп ластоногих с большим, несколько уплощенным мозговым отделом. Лицевая часть черепа укорочена. Глазницы крупные, незамкнутые. Для большинства характерен узкий межглазничный промежуток и широко расставленные скуловые дуги. Слезная кость у взрослых особей отсутствует. Межтеменной кости нет и у молодых зверей.
1 —3 1
Зубная формула: i ——— с —
3—4 0—2
р ---- гл ---= 18 — 36.
3—4 0—1
Молочная смена зубов развита слабо. Предшественники постоянных зубов имеются у резцов, клыков и второго, третьего и четвертого коренных зубов. Молочные зубы выпадают в возрасте 4—5 месяцев. У большинства видов зубы служат только для схватывания добычи. Тем не менее, за исключением моржа, зубы ластоногих хорошо дифференцированы на резцы, клыки и щечные. Все щечные (кроме первого) более или менее однотипны, обычно уплощены с боков. Их форма несколько варьирует в зависимости от видовой принадлежности. Щечные зубы имеют по 2 корня. Разработана методика определения возраста ластоногих по шлифам зубов.
Позвонков у ластоногих обычно шейных 7, грудных 14—15, поясничных 5—6, крестцовых 3, хвостовых 10—12. В скелете свободных конечностей характерно сильное укорочение костей (особенно плечевой и бедренной). Ключицы нет. Лопатка широкая. Грудина состоит из 8— 9 частей. Сильно развит дельтовидный гребень плечевой кости. Локтевая и лучевая кости латерально уплощены. Локтевой отросток сильно развит. В запястье ладьевидная, полулунная и центральная кости слиты. Таз ластоногих по сравнению с наземными млекопитающими имеет укороченные подвздошные и удлиненные седалищные и лобковые кости. Лобковый симфиз отсутствует. Бедренные кости широкие и уплощенные в передне-заднем направлении. Малая и большая берцовые кости сращены между собой в проксимальных отделах и соединяются связкой в дистальных. Метатарзальная кость и проксимальная фаланга I пальца развиты сильнее, чем эти же элементы других пальцев.
Головной мозг крупный, более сферической формы, чем у Наземных хищных. Большие полушария с многочисленными извилинами. Мозжечок относительно крупных размеров. Масса головного мозга от 250— 300 г у обыкновенного тюленя, 375 г у калифорнийского морского льва, до 1 кг у моржа.
Желудок простой. Слепая кишка развита слабо. Печень обычно шестилопастная, имеет желчный пузырь. Желчный проток и проток поджелудочной железы открываются в небольшую полость в стенке двенадцатиперстной кишки. Почки многодольчатые.
У многих ластоногих в желудке находят камни различных размеров. Для объяснения функций этих проглоченных камней существует много теорий. Ряд ученых полагают, что эти камни служат своеобразным балластом, помогающим животным при нырянии. Другие считают, что камни необходимы для сохранения активности мышц желудка при длительном голодании в период размножения. Имеются высказывания, что камни способствуют перетир'анию рыбьих костей или освобождению от паразитических червей.
. „«vjvnviiii имеег типичное для большинства млекопитаю-
щих строение. Однако для Phoca и Halichoerus характерны сплошные хрящевые кольца трахеи, а для Hydrurga, напротив, слабо развитые вентрально расположенные небольшие хрящи — остатки этих колец. Легкие тюленевых симметричны, а у ушастых тюленей правое легкое значительно больше левого. В бронхах ластоногих находятся многочисленные мышечио-эластиновые клапаны. В терминальных отделах бронхиального дерева мышцы развиты сильнее, чем у наземных млекопитающих, ио слабее, чем у китообразных. Диаметр альвеол ушастых тюленей от 100 до 300 мкм, моржа — от 75 до 125, тюленевых — от 50 до 150 мкм.
Строение сердца и артериальной системы не обнаруживает у ластоногих существенных отклонений по сравнению с наземными млекопитающими. Число сердцебиений колеблется у взрослых животных от 55 до 122 в минуту. Резко уменьшается число сокращений сердца во время ныряния: до 4—15 ударов в минуту.
Венозная система ластоногих имеет ряд особенностей. V. jugularis internus сильно редуцирована, так как венозная кровь от головы проходит по двум v. hypocondylar, которые соединяются в большой синус. Вены, отходящие от этого синуса, расположены по бокам спинного мозга, соединяются между собой анастомозами и каудальнее, в основании шеи образуют крупную v. extradural. Эта вена проходит дорзальнее спинного мозга — между твердой мозговой оболочкой и периостеумом спинномозгового канала. Спереди v. extradural получает кровь от спинной мускулатуры и межреберных вен, а сзади от почечного и тазового сплетений и вен брюшной стенки. В крестцовом отделе v. extradural раздваивается и продолжается каудальнее в тазовое сплетение. Характерно отсутствие клапанов в этой вене. Почечное сплетение соединено также, помимо v. extradural, с задней полой веной (парной в области почек) посредством почечных вен. Печеночный синус, образованный расширением печеночных вен и расположенный каудальнее диафрагмы, получает кровь от задней полой вены. Спереди от диафрагмы задняя полая вена имеет мощный мышечный сфинктер. Кпереди от сфинктера в заднюю полую вену открываются многочисленные вены перикардиального сплетения, связанные между собой анастомозами и расположенные в основании перикардиума.
Венозные сплетения, синусы и крупная задняя полая вена служат депо венозной крови. Сфинктер задней полой вены, по-видимому, препятствует попаданию венозной крови в сердце и затем в мозг. Эти приспособления, а также уменьшение потока крови к мышцам и повышенное содержание в мышцах миоглобина позволяет ластоногим на длительное время задерживать дыхание.
Матка двурогая. Семенники моржей и настоящих тюленей располагаются в паховой области под кожей, а у ушастых тюленей в мошонке. Имеется os penis.
Температура тела ластоногих с относительно редким волосяным покровом обычно составляет 36,5—37,5°С. Наружные покровы имеют значительно более низкую температуру, близкую к температуре окружающей среды, превышая ее всего на 1—2°. Таким образом предотвращается излишняя теплоотдача с поверхности тела. На глубине около 4 см от поверхности тела температура сохраняется постоянной. У всех ластоногих и особенно ушастых тюленей ласты служат местом интенсивной отдачи, избытка тепла.
Распространены ластоногие в основном в холодных и умеренных водах Атлантического и Тихого океанов, в Северном Ледовитом океане и морях Антарктики. Летняя изотерма 20°С в тех местах, где она под-254
ходит к побережью, служит примерной границей их распространения. В более теплых водах встречается обычно только тюлень-монах. Обитают ластоногие и во внутренних водоемах, как пресноводных (Ладожское озеро, Байкал), так и соленых (Каспийское море).
Ластоногие в разной степени связаны с сушей. Бее они размножаются (приносят детенышей) и линяют только на земле или на льду. Однако некоторые (например, обыкновенный тюлень) проводят на суше много времени, тогда как другие (северный морской котик) могут не вылезать на сушу в течение 6—8 месяцев в году. Ластоногие — превосходные пловцы, способные оставаться под водой от 5—6 до 20—40 мин и нырять на глубину до 180 (сивуч, северный морской слон) — 275 м (гренландский тюлень) или даже до 350—600 м (тюлень Уэдделла).
Ряд ластоногих ведет оседлый образ жизни, тогда как другие совершают миграции, причем иногда очень дальние: например, северный морской котик проплывает в год путь длиной около 10 тыс. км.
Ластоногие, как правило, редко держатся поодиночке, особенно в период размножения, когда некоторые из них образуют так называемые «залежки» из тысяч особей. Так, северные морские котики концентрируются в период размножения на Прибыловых о-вах на участке радиусом около 50 км в количестве более миллиона голов.
Питаются ластоногие животной пищей: рыбой, ракообразными, моллюсками, морскими птицами, другими ластоногими. Слух развит хорошо. Звуки, производимые ластоногими, очень разнообразны — от коло-колоподобиых звуков и интенсивных щелчков моржа до своеобразных, очень продолжительных серий (до 42 с) резонантных звуков тюленя Уэдделла и подводных рычаний, лая и щелчков калифорнийского морского льва. Ряд ластоногих, вероятно, способны к гидролокации, помогающей в поисках добычи. Обоняние острое, а зрение слабое.
Самки рожают на льду или на берегу, как правило, одного, редко двух детенышей не чаще 1 раза в год. По-видимому, у всех ластоногих (возможно, исключая моржей) имеет место 2-месячная или более задержка имплантации бластоцисты в стенку матки. Половозрелости достигают не ранее 3 лет. Продолжительность жизни до 41 года (серый тюлень, в неволе). Естественные враги некоторых ластоногих — крупные акулы (в умеренных и теплых водах), косатки, белый медведь, а также морской леопард, реже морж (в холодных водах).
Многие виды ластоногих до сих пор служат объектами промысла. Добываются в основном из-за жира и шкуры.
У некоторых видов в результате неконтролируемого промысла настолько снизилась численность, что им угрожает исчезновение.
В отряде 3 семейства и 21 род:
ушастых тюленей — Otariidae Gill, 1866 (7 родов);
моржовых — Odobenidae J. Allen, 1880 (1 род);
тюленевых — Phocidae Brooker, 1828 (13 родов).
Систематическое положение как самого отряда, так и отдельных групп внутри него вызывает некоторые разногласия среди ученых. Так, одни из них (Грей, Gray, 1874; Вебер, Weber, 1928; Симпсон, Simpson, 1945; Смирнов, 1908, 1929, 1935, и др.) считают ластоногих за подотряд, входящий вместе с хищными в отряд Carnivora. Другие (Шеффер, Scheffer, 1958; Огнев, 1935; Бобринский, 1944; Чапский, 1961, 1963, 1975, и др.) выделяют ластоногих в ранг отряда. Несмотря на ряд сходных признаков ластоногих и хищных, до сих пор не удается найти их общих предков, послуживших начальным звеном дивергенции. Эта дивергенция произошла, безусловно, очень давно, и за долгий срок эволюционного развития ластоногие настолько резко обособились от хищных, что выделение их в особый отряд представляется более правильным.
Наиболее древние ластоногие известны из миоцена. К этому времени они почти не отличались от современных форм и их распространение во многих отношениях совпадало с распространением ныне живущих ластоногих. Поэтому предков ластоногих, очевидно, следует искать где-то в эоцене или даже в верхнем мелу. Отсутствие к настоящему времени находок ископаемых остатков ластоногих на предмиоценовых скалах, которые содержат кости китообразных, можно объяснить тем, что предки ластоногих образовывали небольшие и малочисленные популяции.
Поскольку о предках ластоногих ничего не известно, высказывается ряд предположений, реконструирующих происхождение этой группы млекопитающих. Обсуждают возможность происхождения ластоногих от еще не дифференцированной насекомоядно-креодонтной группы в позднем мелу — примерно в то время, когда обособились китообразные и сирены. Хотя современные ластоногие сохраняют задние конечности и типичную эутерийную зубную систему и их волосяной покров менее модифицирован по сравнению с китообразными и сиренами, возможно, что эти три группы имели в мелу общих предков.
Другое предположение говорит о происхождении ластоногих от общего ствола с хишными в позднем эоцене. Современные ластоногие наиболее близко стоят к группе собак — енотов — медведей — куниц (надсемейство Canoidea, по Симпсону, 1945) и менее близко к группе виверр— гиен — кошек (надсемейство Feloidea, по Симпсону, 1945). Миварт (Mivart, 1885) подчеркивал сходство ушастых тюленей и тюленевых, соответственно, с медведями и выдрами. Келлог (Kellog, 1922) замечает, однако, что предков ушастых тюленей следует искать среди хищных задолго до того, как появились настоящие медведи и волки. Многие авторы (особенно Хауэлл, Howell, 1930) указывают на анатомическую близость современных ластоногих и хищных. Эади и Кирк (Eadie and Kirk, 1952) показали, что по концентрации катионов Na и К кровь морского слона более близка к крови кошки и собаки, нежели к любым другим млекопитающим. Леон и Виенс (Leone a. Wiens, 1956) на основании серологических данных заключили, что ластоногих как систематическую группу можно рассматривать как семейство в пределах отряда хищных. Следует, однако, заметить, что делать подобные заключения только на основании сравнительноанатомических и серологических исследований современных форм было бы не совсем правильно. Дальнейшее изучение морских или даже пресноводных предмиоценовых отложений должно прояснить вопрос происхождения ластоногих.
Территория, где возникли ластоногие, по-видимому, — Северное полушарие, в котором в настоящее время обитает 2 семейства из 3 и 12 родов из 20.
Что касается филогенетического взаимоотношения семейств, то нельзя сказать, что какое-нибудь из них предок другого. Каждое, по-видимому, развивалось обособленно. Все же ряд общих признаков сближает семейство ушастых тюленей и семейство моржовых в одну общую группу, выделяемую в надсемейство Otarioidea, которому проти-вопоставляется надсемейство Phocoidea с одним семейством тюленевых. Несходство между ними дало повод некоторым ученым утверждать о независимом происхождении этих двух групп. Предполагается, что среди наземных Creodonta выделились 2 группировки животных, которые независимо одна от другой позднее приспособились к водной жизни (Huber, 1934). Против этого мнения высказываются паразитологи, обнаружившие у всех представителей отряда совершенно специфичных паразитов (Anoplura), что свидетельствует о монофилитичности ластоногих (Hopkins, 1949). С другой стороны, некоторых носовых клещей, свойст-
венных ластоногим, можно разделить на 2 рода: Orthohalarchne, паразитирующих только у ушастых тюленей и моржей, и Halarchne, встречающихся только у тюленевых (Newell, 1947).
Представители семейства Otariidae более близки по ряду признаков к наземным млекопитающим. Однако современные данные не позволяют утверждать, что какое-то из трех семейств ластоногих более древнее, чем’ любое другое.
СЕМЕЙСТВО УШАСТЫХ ТЮЛЕНЕЙ
FAMILIA OTARIIDAE GILL, 1866
Тело стройное, вытянутое, с крупными передними ластами —- примерно до ‘А длины тела. Размеры средние или крупные: длина тела от 150 до 380 см, масса до 1100 кг. Самцы значительно крупнее самок. Хвост короткий, но хорошо заметный. Ушные раковины с хрящами, небольшие, покрытые волосами. Шея длинная, гибкая, мускулистая, и ее сильное движение взад и вперед придает некоторую движущую силу телу при передвижении по суше. Полагают, что если бы морские львы обладали только половиной длины своей шеи, они не смогли бы двигаться по земле. Во время плавания голова и шея подвижны при поисках пищи, задний конец тела действует как руль.
Передние конечности на суше поддерживают тело, сгибаясь лате-ральио в кисти под прямым углом. При движении по земле животное переставляет по очереди каждый передний ласт или при быстром движении опирается на оба передних ласта сразу. Задние ласты при движении по земле подгибаются под тело в пяточном сочленении и перемещаются оба вместе. В воде передние конечности служат локомоторными органами. Их гребущая поверхность увеличивается благодаря тому, что наружный край ласта продолжается за кончики пальцев и поддерживается предпальцевыми хрящами. Когти при движении по земле не могут функционировать и редуцируются до небольших утолщений.
Передний край ластов усилен утолщением плотной соединительной ткани. Задние конечности используются при плавании главным образом как рули, причем они соприкасаются друг с другом подошвами. Гребущая поверхность задних ластов увеличена за счет кожной оторочки, которая поддерживается предпальцевыми хрящами, как и у передних ластов. Эта оторочка может подгибаться, и три длинных когтя внутренних пальцев функционируют. Когти двух наружных пальцев очень малы.
Внутренняя поверхность ластов лишена волосяного покрова и, вероятно, служит местом отдачи избытка тепла в случае перегрева организма. Для кожного покрова характерно более сильное, чем у других ластоногих, развитие сети эластиновых волокон. Сальные железы относительно невелики, а потовые очень крупные. В противоположность остальным ластоногим в остевых волосах ушастых тюленей хорошо развит сердцевинный слой. Пуховые волосы могут быть многочисленными.
В черепе относительно удлиненный лицевой отдел. Надглазничные отростки крупные. Костные слуховые барабаны небольшие и плоские. Крылоклиновидный канал имеется. Передние отделы лобных костей входят выступами между носовыми костями.
Зубная формула: i с -J- р m —j—=34 — 36.
Первый и второй верхние резцы малы, каждый имеет по две вершины — переднюю и заднюю, отделенные друг от друга глубокой бороздой.
Третий (наружный) верхний резец клыкоподобный. Клыки крупные, конически заостренные и несколько изогнутые. Предкорениые и коренные простые, с одной основной вершиной. Предкорениые, а иногда и верхние коренные зубы с одним корнем.
Распространены в арктических, умеренных и субтропических водах Тихого океана, Южной Атлантики и южной части Индийского океана: по побережьям Южной и запада Северной Америки, Южной Африки, Австралии, Новой Зеландии и некоторых океанических островов.
Держатся стадами, особенно в период размножения. Залежки устраивают в тихих заливах, на скалистых изолированных островах. Хорошие пловцы: скорость плавания калифорнийского морского льва под водой может достигать 17 км/ч, а северного морского котика — 26 км/ч. Для двух родов — северных и южных морских котиков — характерны периодические миграции.
Активны ночью и днем. Объектами питания в основном служат рыбы, головоногие моллюски и ракообразные.
Для большинства характерна полигамия. В период размножения один самец имеет в гареме от 3 до 50 самок. Самка рожает на берегу. Вскоре после этого происходит спаривание. Длительность беременности колеблется от 250 до 365 дней. По-видимому, для всех представителей семейства свойственна задержка в имплантации яйцеклетки. Самец в воспитании детеныша участия не принимает. Новорожденный начинает плавать обычно не ранее 2-недельного возраста. Лактация продолжается обычно 3—4 месяца.
Ушастые тюлени служат объектом промысла. От них получают меховое и кожевенное сырье и жир.
Семейство насчитывает 7 родов (14 видов):
южных морских львов — Otaria Регоп, 1816 (1 вид);
новозеландских морских львов — Phocarctos Gray, 1844 (1 вид);
австралийских морских львов — Neophoca Gray, 1866 (1 вид); калифорнийских морских львов — Zalophus Gill, 1866 (1 вид);
сивучей — Eumetopias Gill, 1866 (1 вид);
южных морских котиков — Arctocephalus Е. Geoffroy et F. Cuvier, 1826 (8 видов);
северных морских котиков — Callorhinus Gray, 1859 (1 вид).
В водах СССР (на Дальнем Востоке) 2 рода: сивучей и северных морских котиков.
Род южных морских ЛЬВОВ
Genus Otaria Peron, 1816
В роде единственный вид: южный морской лев — О. byronia Blain-ville, 1820 (по Райс и Шефферу, Rice a. Scheffer, 1968, правильнее— О. flavescens Shaw, 1800).
Длина тела самца до 245 см, самки — до 195 см. Масса самца достигает 317 кг (по другим данным, 520 кг), самки— 136 кг. Окраска волосяного покрова преимущественно темно-бурая, хотя у отдельных особей может быть серой, светло-золотистой или желтоватой. Брюхо темножелтое. Самцы имеют на шее небольшую гриву, которая достигает плеч (рис. 120). Новорожденные черного цвета, ио вскоре становятся бурыми или темно-бурыми. На 4-м году жизни окраска самцов темнеет и коричневеет, а самок светлеет и сереет.
Костное небо очень длинное: расстояние от его заднего края до резцов превышает 55% длины черепа. Ширина черепа на уровне клыков превышает 25% длины черепа.
Рис. 120. Южный М07 лев — О. byronia (Л австралийский мош лев — N. cinerea
Распространен по Атлантическому и Тихоокеанскому побережью Южной Америки (на север до Рио-де-Жанейро в Бразилии и Зорритос в Перу — примерно 4° ю. ш.), в Магеллановом проливе, у Фолклендских о-вов.
Иногда южные морские львы заходят в крупные реки. Однажды они были встречены в р. Ла-Плате на расстоянии 300 км от устья. Истинных миграций не отмечено, хотя для некоторых популяций описаны передвижения в южном направлении.
Пища состоит главным образом из кальмаров, ракообразных (Muni-da), мелких рыб, иногда пингвинов. Вне периода размножения южные морские львы держатся крупными смешанными стадами. Период размножения приходится на конец декабря — середину января. В это время о*
размножающиеся и неразмножающиеся звери разделяются. Стада размножающихся южных морских львов состоят из половозрелых самцов и самок и новорожденных детенышей. Самцы охраняют свои гаремы от посягательств других самцов. Даже во время прилива, когда вода почти покрывает их, самцы не покидают своих территорий. Южные морские львы для размножения выбирают песчаный берег и плоские скалы. Обычное число самок в гареме на Фолклендских о-вах 9, у уругвайской популяции—-7,6. Длительность беременности примерно 330 дней. Самки рождают одного детеныша, редко — двух с длиной тела примерно 80 см. Волосяной покров новорожденного черного цвета. Рождение детенышей происходит с конца декабря до середины января (у большинства самок— в начале января). Лактация продолжается примерно 6 месяцев. Спариваются самки спустя несколько дней после родов. Во время размножения (примерно 2 месяца) самцы ничего не едят и почти не спят. Самки достигают половозрелости к концу 4-го года жизни, а самцы — в 6 лет. Продолжительность жизни, по-видимому, составляет около 20 лет. Естественных врагов мало: на молодых зверей иногда нападают морские леопарды. Численность определяется примерно в 700 тыс. голов (Scheffer, 1958) или в 800 тыс. (King, 1964). Из них на Фолклендских о-вах насчитывается 400 тыс. (к 1964 г.).
Хотя южных морских львов промышляли еще в XVI в., серьезного промыслового значения они не имеют из-за трудности добывания их на малодоступных прибрежных скалах. В настоящее время промысел ведется по лицензиям.
Род новозеландских морских львов
Genus Phocarctos Gray, 1844
В роде единственный вид: новозеландский морской лев — Р. hookeri Gray, 1844.
Взрослый самец достигает в длину примерно 2,8—3 м, а самка — 1,8 м. Масса самца примерно 400 кг, самки — 230 кг. Самцы черноватобурые с хорошо развитой гривой из длинных темных волос. Окраска самок значительно светлее, рыжевато-коричневая или тускло-желтая. Новорожденные имеют нежный волосяной покров каштанового цвета.
Расстояние от заднего края костного неба до резцов составляет примерно 45% длины черепа. Ширина черепа на уровне клыков примерно равна 23% длины черепа. Костное небо в области клыков имеет глубокую впадину.
Распространен в течение всего года у побережья Оклендских о-вов (у Новой Зеландии).
Экология изучена слабо. Залежки устраивают на песчаных берегах, а также в кустах и траве вдали (до 800 м) от берега. В начале октября взрослые самцы вылезают на берег и захватывают территорию для гарема. Самки появляются месяцем позже. В гареме насчитывается в среднем 12 самок. Детеныши рождаются в последнюю неделю декабря— в январе. Лактационный период продолжается примерно 7 месяцев. Спаривание происходит вскоре после родов. К концу января — началу февраля большая часть самок оказывается покрытыми, большинство детенышей уже могут плавать, гаремы начинают распадаться и самцы уходят в море на кормежку. Линька взрослых и молодых животных происходит в апреле. Пищей новозеландских морских львов служат мелкая камбала и другие мелкие рыбы, крабы и двустворчатые моллюски, иногда поедают пингвинов. Численность составляет 10—50 тыс. голов. Новозеландских морских львов промышляли в XIX в. В настоящее
время лежбища этих зверей на Оклендских о-вах объявлены резерватом Некоторые ученые (например, Scheffer, 1958) включают новозеланд ского морского льва в род Neophoca Gray, 1866.
Род австралийских морских льво»
Genus Neophoca Gray, 186f
В роде единственный вид: австралийский морской лев — N. cinerec Peron, 1816 (рис. 120).
Длина тела самца достигает 270—360 см, масса — 410 кг. Длина теля самки 180—300 см, масса — до 230 кг. У самцов окраска тела и головь темно-коричневая, а гривы желтоватая. У молодых самцов шея светлая гривы нет. Спина у самок ярко-коричневая, брюхо желтоватое или цели ком темно-желтое.
Череп сходен с черепом новозеландского морского льва, однако ши рина его меньше.
Распространение ограничено южным побережьем Австралии о: Хутман Рокс на западе до о-ва Кенгуру'на востоке.
Экология изучена недостаточно. Ведет оседлый образ жизни и мо жет в течение всей жизни держаться у места своего рождения. Питается в основном рыбой, крабами и пингвинами. Хорошо лазает по скалам иногда уходит далеко от воды (до 10 км). В желудке всегда находя: камни. Рождение детенышей приходится на октябрь — начало декабря Гаремы состоят из 4—5 самок. В период размножения отмечено поеда ние самцами пингвинов.
Численность невелика и к 1958 г. не превышала 2—10 тыс. голов Промыслового значения эти ластоногие не имеют.
Род калифорнийских морских львом
Genus Zalophus Gill, 1861
В роде единственный вид: калифорнийский морской лев — Z. califor nianus Lesson, 1828.
Длина тела самца достигает 200—250 см, масса — 200—300 кг. .Дли на тела самки 150—200 см, масса 50—100 кг. Окраска варьирует от светло- до темно-шоколадно-коричневой. У взрослых самцов на шее име ется грива. Лобный отдел головы у них выступает благодаря сильнс развитому сагиттальному гребню (рис. 121). С возрастом верх головь У самцов может светлеть. Ласты по наружной поверхности покрыть шерстью приблизительно наполовину, а на внутренней стороне ее неновее.
Череп удлиненный, с относительно узким лицевым отделом. Носовы» кости длинные и узкие. Носовые отростки межчелюстных костей равно мерно сужаются в заднем отделе.
Крайние резцы крупные, клыкоподобные. Коронки щечных зубов ко иусовидные, без дополнительных вершин. Пространство между четвер тым и пятым щечными зубами мало или его нет совсем.
Хромосом в диплоидном наборе 36.
Распространен в субтропических и умеренных водах Тихого океана Три локальных стада образуют отдельные подвиды: на западном побе режье Северной Америки.от Сан-Франциско на юг до центральных рай онов Калифорнийского залива, на Галапагосских о-вах и у берегов Япо «ии (в южной части Японского моря по обеим сторонам о-ва Хонсю) Калифорнийские морские львы — прибрежные обитатели, как правиле
не встречающиеся в открытом море далее десятка миль от берега. У берегов Калифорнии и Мексики обычно придерживаются мелких островов и скал.
Слух и обоняние развиты хорошо. Способны нырять на глубину до 83 м. Питаются преимущественно головоногими моллюсками и различными рыбами (главным образом, сельдью, мерлузой). Взрослый самец в неволе за день съедает до 30 кг рыбы. Сезон размножения и, соответ
ственно, пребывание на лежбищах длится 3—4 месяца, на юге американской части ареала — с мая по август, а на севере — с июня до октября. Взрослые самцы занимают в период размножения береговые участки, образуя залежки вместе с самками. Самцы удерживают свою территорию агрессивным поведением, которое обычно не переходит в схватку (Peterson a. Bartholomew, 1967). Самки не обращают внимания на границы индивидуальных территорий самцов и свободно передвигаются от одного участка к другому. Самцы не делают попыток удержать самок, так что гаремы фактически не существуют. Период беременности продолжается 342—365 дней (считая вместе с латентным периодом). Детеныши рождаются с конца мая до конца июня, а на Галапагосских о-вах — с октября по декабрь. Новорожденные имеют золотистобурую окраску разных оттенков. Они достигают в длину примерно 75 см, массы •—5,5—6,3 кг. Через 10 дней после рождения детеныши начинают плавать. Первые несколько дней самка остается с новорожденным почти все время. Примерно, в 2-недельном возрасте детеныши группируются в наиболее тихом месте лежбища. Они проводят много времени, играя на суше и в воде. С возрастом они все больше держатся в воде. Самки кормят только своих детенышей, распознавая их по голосу и запаху (Peterson a. Bartholomew, 1967). Лактация длится 5— 6 месяцев. Детеныши остаются с матерью примерно в течение года. Спаривание происходит вскоре после родов. После периода размножения калифорнийские морские львы покидают лежбища и все остальное время проводят в море, хотя дальних миграций не совершают и держатся не далее 2—10 миль от берега. Многие самцы зимой уплывают к северу до штатов Орегон и Вашингтон (США), о-ва Ванкувер и Британской Колумбии.
Половозрелости самки достигают в 3 года, самцы — в 5 лет. Враги калифорнийских морских львов — косатки и акулы. Длительность жизни в неволе до 30 лет.
Численность калифорнийских морских львов у берегов Калифорнии и Мексики определяется в 50 тыс. голов, а у Галапагосских о-вов — около 20 тыс., в водах Японии — всего 200—500 особей (Scheffer, 1958). Поданным Nishiwaki (1972), японский подвид/, californianus japonicus Peters, 1866, по-видимому, исчез полностью. Он включен в «Красную книгу».
Калифорнийские морские львы хорошо содержатся в неволе. Легко тренируются для различных представлений в цирках. Специального промысла не ведется, и их хозяйственное значение невелико.
Род сивучей
Genus Eumetopias Gill, 1866
В роде единственный вид: сивуч Е. jubatus Schreber, 1776.
Размеры наиболее крупные в семействе. Самец достигает длины 385 см и массы 1120 кг, самка — соответственно 270 см и 350 кг. Телосложение тяжелое. Морда широкая и тупая с чуть вздернутым носом (рис. 122). Молодые животные темно-коричневой окраски, взрослые — темно-желтой или желтоватой. Волосяной покров состоит из прямых грубых остевых и очень редких, тонких пуховых волос.
Волосяные пучки содержат один остевой и один-два пуховых волоса. На шее самцов развита грива из удлиненных и грубых волос. Волосы покрывают сверху значительную часть кистей (не менее 2/3 длины).
Рис. 122. Сивуч — Е. juba-tus и его череп
Череп, особенно взрослых самцов, массивный. Лицевой отдел удлинен. Костное небо имеет почти прямой задний край, иногда с небольшим выступом. Носовые отростки межчелюстных костей равномерно суживаются кзади.
Крайние резцы крупные клыкообразные. Последний заднекоренной зуб верхней челюсти отделен от первого предкоренного промежутком, величина которого больше длины первого из этих зубов. Коронки всех коренных зубов одновершинные и приблизительно равны между собой по величине.
Хромосом в диплоидном наборе 36.
Распространен в субарктических и холодных умеренных водах Тихого океана от северо-восточной части Берингова моря, Алеутских о-вов и западного побережья Северной Америки на юг до Южной Калифорнии и на запад до Командорских о-вов, Камчатки, Курильских о-вов и Японии. Иногда поднимаются вверх по рекам; один сивуч был добыт в р. Колумбии в 150 км от устья. В СССР распространены от Берингова пролива на юг до государственной границы. Наиболее крупные залежки отмечены на о-вах Курильских, Ионы, Ямских, Командорских, северной оконечности Сахалина, по восточному побережью Камчатки.
Обычно встречаются у скалистых берегов. Держатся стадами, которые занимают летом, как правило, из года в год одни и те же участки на берегу. Лежбища бывают различные по величине и составу. В период размножения обычны гаремные лежбиша, состоящие из крупных самцов и группирующихся около каждого из них самок. В стороне от гаремных лежбищ собираются звери, не участвующие в размножении,— от нескольких десятков до нескольких сотен. В южных районах, где не бывает льдов, лежбища существуют почти круглый год. На севере с появлением льдов сивучи уходят с мест лежбищ, хотя, по-видимому, далеких миграций не делают.
Питается в основном рыбами и головоногими моллюсками. Поедает треску, минтая, мойву, навагу, сайку, камбалу, палтуса, терпуговых, корюшку, сельдь, мяконькую, акул, лососей, из головоногих моллюсков кальмаров, осьминогов, каракатиц. В водах Курильских островов и, видимо, Камчатки, основная пища сивуча — минтай. Лишь очень редко нападает на тюленей. Характерно заглатывание камней.
В начале мая взрослые самцы захватывают территорию на гаремных залежках. В каждом гареме обычно по 10—20, иногда более самок. Детеныши (обычно один в помете) рождаются с середины мая до начала июля. Самка вскоре после родов спаривается с самцом— хозяином гарема. Имплантация бластоцисты задерживается до октября. Гаремы разбиваются в июле—-августе. Новорожденный имеет длину тела около 1 м и массу 15—23 кг. Лактационный период продолжается не менее 3 месяцев, возможно, до года. Иногда можно видеть годовалых сивучей, держащихся с матерью. Хотя молодые сивучи способны плавать сразу после рождения, они не могут пробыть долго в воде, так как не имеют достаточного слоя подкожной жировой клетчатки, уменьшающей теплоотдачу. Только по достижении возраста нескольких недель они много времени проводят, плавая и играя в воде. Среди молодняка отмечается большая смертность. Многие падают в воду и тонут, так как не могут вскарабкаться на обрывистые скалы, других давят самцы во время частых драк. Половозрелости самки достигают в 3 года, а самцы в 5 лет, но обладателями гаремов становятся не ранее 7-летнего возраста. Мировая численность сивучей составляет 240—300 тыс. голов, причем в восточной части ареала их насчитывают примерно 200 тыс.
Естественный враг — косатка.
Промысловое значение невелико.
Рис. 123. Новозеландский морской котик—A. forsteri
Род южных морских котиков
Genus Arctocephalus Е. Geoffroy et F. Cuvier, 1826
По облику южные морские котики напоминают северных морских котиков, но имеют другую форму головы с несколько более вытянутым лицевым отделом (рис. 123). Длина тела самца от 152—183 см у керге-ленского морского котика, до 244 см у австралийского морского котика. Масса соответственно от 112 до 315 кг. Длина тела самок этих двух видов от 120 до 180 см, масса от 35 до 90 кг. Окраска коричневая, черная, серая или желтовато-серая; волосяной покров с нежным и плотным пухом. Самцы имеют густую гриву, образованную длинными остевыми волосами, черными в основании и середине и бурыми на вершине. Мех южных морских котиков по качеству несколько хуже, чем северных.
Диплоидное число хромосом у южноамериканского и фернандесско-го морских котиков по 36.
Распространены по берегам Южной Африки, Западной и Южной Австралии, Новой Зеландии, южной части Южной Америки, Галапагосских о-вов и ряда островов к северу от Антарктиды.
Придерживаются скалистых берегов, причем гуадалупский морской котик находит себе убежище в пешерах.
Питаются мелкой рыбой, головоногими моллюсками и ракообразными. В желудках обычно находят небольшое количество камней.
Южноамериканский морской котик, по-видимому,— единственный вид рода, которому свойственны сезонные миграции. На зиму он откочевывает от Фолклендских о-вов к Уругваю. В период размножения в октябре — ноябре самцы южных морских котиков захватывают на залежках территорию и образуют гаремы. В прежние времена число самок в гареме одного самца доходило до 50, а в настоящее время составляет 3—5, редко 15. Длительность беременности у южноафриканского морского котика в среднем 259 дней, а у южноамериканского морского котика 330. Детеныши рождаются в конце ноября — начале декабря. Но
ворожденный достигает в длину примерно 45—62 см, имеет черную окраску. Сеголеток южноамериканского морского котика линяет в марте— апреле (в 4—5-месячном возрасте). Новый волосяной покров имеет оливково-серые остевые и бурые пуховые волосы. Через год снова происходит линька, и молодой зверь приобретает серебристо-серую окраску. Следующая линька приходится на январь—февраль. Самки оплодотворяются через 5—10 Дней после рождения детенышей. Хар-актерна задержка имплантации бластоцисты. Лактационный период может продолжаться до года. Примерно в 5-месячном возрасте детеныши начинают подкармливаться ракообразными и мелкими рыбами. Самки достигают половозрелости в 2—-3-летнем возрасте, а самцы — в 3-летнем, хотя гаремы образуют значительно позже. Продолжительность жизни в неволе до 20 лет.
Главные враги южных морских котиков, кроме человека, косатки и акулы. У одной крупной акулы в желудке было найдено 5 молодых южноамериканских морских котиков.
В роде 8 видов:
южноафриканский морской котик — A. pusillus Schreber, 1776 (включая A. tasmanicus) (юго-западное и южное побережья Африки со многими прибрежными островами, от мыса Кросс на севере — 21°14/ ю. ш. до залива Алгоа на юге —30° ю. ш., причем зимой доходит до Анголы, а также побережье Австралии от Сиднея до западной границы Виктории) ;
кергеленский морской котик — A. gazella Peters, 1875 (Южные Шетландские, Южные Оркнейские и Южные Сандвичевы о-ва, Южная Георгия, Буве, Кергелен и Херд);
новозеландский морской котик — A. forsteri Lesson, 1828 (включая A. doriferus) (о-в Южный в Новой Зеландии и прилегающие острова от о-вов Чатам до о-ва Макуори, а также побережье Юго-Западной Австралии от о-ва Эклипс, Западной Австралии и Южной Австралии до о-ва Кенгуру);
субтропический морской котик — A. tropicalis Gray, 1872 (острова к северу от антарктической конвергенции (пояса смешения холодных антарктических и теплых субантарктических вод) —от Тристан-да-Кунья и Гоф в Южной Атлантике на западе до Амстердама и Сен-Поль в Индийском океане на востоке);
южноамериканский морской котик —A. australis Zimmermann, 1783 (побережье и близлежащие острова Южной Америки от Рио-де-Жанейро в Бразилии, на юг вокруг Мыса Горн и по западному берегу на север до Лимы в Перу, а также Фолклендские о-ва);
галапагосский морской котик — A. galapagoensis Heller, 1904 (Галапагосские о-ва);
фернандесский морской котик — A. philippii Peters, 1866 (о-ва Хуан-Фернандес к западу от Чили);
гуадалупский морской котик — A. townsendi Merriam, 1897 (о-в Гуа-далупа к востоку от Калифорнии).
Некоторые исследователи (King, 1964) разделяют южноафриканского морского котика на 2 вида: A. pusillus (Африка) и A. tasmanicus Scott et Lord, 1926 (Тасмания и юг Австралии); новозеландского морского котика на 2 вида: A. forsteri (Новая Зеландия) и A. doriferus Wood Jones, 1925 (Юго-Западная, Западная и Южная Австралия); южноамериканского и галапагосского морских котиков объединяют в один вид — A. australis; кергеленского морского котика и субтропического морского котика в один вид —A. tropicalis; гуадалупского и фернан-десского морских котиков в один вид — A. philippii. Иногда A. tasmanicus и A. doriferus объединяют в один вид A. doriferus (Rice a. Scheffer,
1968). Таким образом, Кинг (1964) насчитывает в роде южных морских котиков 7 видов: A. pusillus, A. australis, A. doriferus, A. tropicalis, A. tasmanicus, A. philippii, A. forsteri, а Райс и Шеффер (1968) 6 видов: A. australis, A. doriferus, A. forsteri, A. pusillus, A. tropicalis, A. philippii. Гуадалупский морской котик, возможно, заслуживает выделения в особый подрод или даже род Arctophoca Peters, 1866 (Rice a. Scheffer, 1968).
Общая численность южных морских котиков к 1964 г. определялась примерно в 735—885 тыс. голов (в основном по King, 1964): южноамериканских морских котиков на Атлантическом побережье более 100 тыс. голов и на Фолклендских о-вах примерно 20 тыс. голов; галапагосских морских котиков к 1972 г. 1000 голов; южноафриканских морских котиков (африканская популяция) примерно 500 тыс. голов; кергеленских и субтропических морских котиков от 28 до 50 тыс. голов; фернандес-ских морских котиков к 1971 г. примерно 450 голов; гуадалупских морских котиков к 1972 г. более 500, но не превышает 600 голов; новозеландских морских котиков на австралийском континенте около 35 тыс. голов и в Новой Зеландии примерно 50 тыс. голов, но в связи с резким увеличением численности за последние годы, возможно, последняя популяция достигла 100 тыс. голов.
В «Красную книгу» включены галапагосский морской котик — A. ga-lapagoensis Heller, 1904, фернандесский морской котик — A. philippii Peters, 1866, и гуадалупский морской котик — A. townsendi Merriam, 1897, как виды, которые в ближайшее время могут оказаться под угрозой исчезновения, если условия их охраны не улучшатся.
Южных морских котиков промышляют. В Уругвае ежегодно добывают 4—5 тыс. южноамериканских морских котиков. Южноафриканских морских котиков начали добывать с открытием Южной Африки и вскоре их запасы были подорваны. После восстановления численности история с переопромышлением повторилась в XVIII столетии. В настоящее время ведется промысел южноафриканских морских котиков у берегов Африки — летом (в октябре — ноябре) в основном на самцов и зимой (в июле — августе) на 7—10-месячных детенышей. За кергеленским морским котиком наиболее интенсивный морской промысел велся в 1800-х годах. У о-ва Южная Георгия, например, этим занимался 31 корабль; известно, что один из них добыл за сезон 57 тыс. котиков. В настоящее время на ряде островов промысел кергеленского морского котика запрещен, а на о-ве Буве и островах, принадлежащих Англии, его добыча разрешена лишь по лицензиям.
Фернандесский и гуадалупский морские котики до конца XVIII столетия занимали обширный ареал по всему Тихоокеанскому побережью Америки от Фаралонских о-вов у Сан-Франциско до Галапагосских и далее на юг до о-ва Масафуэра и чилийского побережья. Промысел велся на истребление этих зверей. В 1805 г. у берегов Калифорнии за 3 недели было добыто 8 тыс. котиков. На о-ве Масафуэра в течение 7 лет добыли 3 млн. зверей. К концу XIX столетия эти морские котики уже не имели промыслового значения.
В начале XX столетия полагали, что гуадалупский морской котик исчез полностью и только в 1954 г. была обнаружена небольшая колония котиков на о-ве Гуадалупе. Новозеландских морских котиков (в районе Новой Зеландии) в конце XVIII — начале XIX в. добывали по нескольку сотен тысяч голов ежегодно. К середине XIX в. из-за резкого снижения численности эти котики уже не представляли интереса для промысла.
В течение последних 50 лет правительство Новой Зеландии ввело запрет на промысел новозеландских морских котиков, устраивая за этот 268
срок несколько экспериментальных сезонов, открытых для добычи. Морских котиков из австралийской популяции разрешено добывать лишь по лицензиям.
Род северных морских котиков
Genus Callorhinus Gray, 1859
В роде единственный вид — северный морской котик — С. ursinus Linnaeus, 1758.
Половой диморфизм резко выражен: длина тела самца до 2,1 м, масса до 300 кг, самки — соответственно до 1,5 м и 65 кг. Телосложение самцов массивное, самок — изящное (рис. 124). Ласты очень длинные, лишенные волос. Хрящевые окончания пальцев, поддерживающие края ластов, сильно развиты. Когти на передних ластах едва заметны или их нет вовсе. Морда укороченная, заостренная. Глаза небольшие, широко расставленные. Наружные ушные раковины длиной до 5 см.
Окраска взрослых самцов на спинной стороне тела от темно-серой до коричневой, на брюшной красновато-коричневая, у взрослых самок серовато-коричневая или серая. Молодые самцы и самки серебристо-серые, более светлые на брюшной стороне. Новорожденные черного цвета, затем приобретают серую окраску. Для кожного покрова характерны мощная сеть эластиновых волокон и хорошо развитые гладкие мышцы. По сравнению с другими ластоногими относительно более сильно развиты потовые железы, особенно сильно на ластах. Вероятно, оголенные поверхности крупных ластов служат участками отдачи тепла как во время пребывания на суше, так и в воде. Волосы морских котиков растут пучками, состоящими из одного остевого, двух-трех промежуточных и 10—30 пуховых волос. В пуховых волосах нет сердцевинного слоя и по форме они отличаются от остевых.
Череп вытянут в длину относительно слабо. Его лицевой отдел укорочен. Скуловые дуги слабо расставлены в стороны. Носовые отростки межчелюстных костей в задних отделах (на границе с носовыми костями) расширены. Носовые кости относительно короткие и широкие.
Щечные зубы мелкие и, как правило, одновершинные. Верхний первый коренной зуб имеет два корня.
Хромосом в диплоидном наборе 36.
Северные морские котики в летний период концентрируются на побережье Прибыловых о-вов в Беринговом море, на Командорских о-вах и о-ве Тюленьем в Охотском море. Небольшая популяция обитает на Курильских о-вах. В зимнее время северные морские котики широко рассеиваются в южных частях Берингова моря, в Охотском, северных областях Тихого океана, Японском море. На юг они спускаются до Сан-Диего в Калифорнии и Японии. Взрослые самцы, по-видимому, не предпринимают дальних миграций и зиму проводят поблизости от летних лежбищ. Так, в командорской популяции северных морских котиков самцы старших возрастов проводят зиму частично в Беринговом море, частично несколько южнее Алеутской гряды. Большинство же самок и неполовозрелых самцов зимуют в открытых водах Тихого океана; по-ви-Димому, около берегов Японии.
Взрослые самцы северных морских котиков с о-ва Тюленьего проводят зиму на севере Японского моря, а самки и остальные самцы — в южной части моря, частично выходя за его пределы к востоку от побережья Японии. В течение зимы котики ведут чисто пелагический образ жизни.
Большую часть жизни северные морские котики проводят в воде.
Рис. 124. Северный морской котик — С. ursinus и его череп
Они быстро плавают и глубоко ныряют. Известны случаи, когда морские котики ныряли на глубину до 73 м.
Объектами питания служат около 60 видов морских организмов, главным образом головоногие моллюски, рыбы и ракообразные. Ежедневно северному морскому котику необходимо съедать пищи примерно 7% от собственной массы. Кормятся северные морские котики главным образом вечером, ночью и рано утром. Днем обычно спят. Основной сезон нагула — с осени до конца весны. У берегов Японии северные морские котики поедают главным образом светящихся и обычных анчоусов и головоногих моллюсков, в меньшей степени скумбрию, сайру, сельдь, иногда лососей; у американских берегов питаются в основном мойвой, мерлузой, минтаем, морским ершом, сайрой, анчоусами, сельдью, лососевыми, головоногими моллюсками; у Командорских о-вов поедают треску, терпуговых, бычков, головоногих моллюсков; у берегов Камчатки большое значение в питании имеет песчанка, а в Японском море — минтай. В период размножения половозрелые самцы, имеющие гаремы, совсем не питаются.
Весной половозрелые самцы (так называемые секачи) первыми приплывают к местам размножения (в конце апреля — начале мая) и лишь после них в июне — июле здесь же появляются самки. Самцы занимают каждый свой участок берега и охраняют его от других самцов. Самцы группируют гаремы, число самок в которых может достигать 40—50. Из-за самок между самцами часто возникают драки. Спустя несколько дней после прибытия к местам размножения самка рожает одного детеныша и через несколько дней спаривается. Таким образом, длительность беременности составляет около года, причем на латентный период в развитии оплодотворенной яйцеклетки приходится 3,5—4 месяца. Половозрелость у самок наступает в 3—4-летнем возрасте, способность к размножению они сохраняют до 21 года, однако наиболее интенсивное размножение происходит в возрасте от 5 до 9 лет. У большинства самцов половозрелость наступает в 5-летнем возрасте, но обладателями гаремов они становятся в возрасте не ранее 7—12 лет. Продолжительность жизни примерно до 30 лет.
Новорожденные северные морские котики достигают длины 55— 65 см и массы около 5 кг. В возрасте 6—8 недель детеныши начинают плавать. Лактация продолжается около 3—4 месяцев. Самка кормит только своего детеныша, находя его среди тысяч других, по-видимому, при помощи слуха и обоняния. К концу лактационного периода черные котики линяют, превращаются в так называемых серых котиков. Смертность детенышей очень высока — до 50% в первый год жизни. С конца июля до поздней осени звери линяют — сначала неполовозрелые, а потом взрослые животные. В сентябре—.декабре северные морские котики начинают мигрировать на юг.
Одно время считали, что американская и азиатская популяции северных морских котиков мигрируют в зимние месяцы в разные места. В настоящее время установлено, что северные морские котики от берегов Аляски могут мигрировать в воды Японии и возвращаться обратно к Аляске или оставаться в азиатских водах. Вообще же, как правило, «инстинкт дома» у северных морских котиков очень сильно развит и звери обычно возвращаются к тем местам, где родились. Наиболее стабильна популяция северных котиков с о-ва Тюленьего, почти не пополняющаяся за счет котиков из других стад. Северные морские котики по сравнению с другими ластоногими совершают наиболее далекие миграции. В течение 6—7 месяцев в году они проплывают путь более чем в 10 тыс. км. Во время миграций они держатся небольшими группами или даже поодиночке.
Общая численность северных морских котиков к 1970 г. определялась в 1580—1920 тыс. голов. Численность популяции северных морских котиков Прибыловых о-вов к настоящему времени, по-видимому, достигла максимума, составляя примерно 1,2—1,5 млн. голов. Численность стада котиков на Командорских о-вах составляет около 200 тыс. голов, на о-ве Тюленьем— 150—160 тыс. голов и на Курильских о-вах— 15— 20 тыс. голов. Недавно обнаружена небольшая популяция северных морских котиков (300—400 животных), размножающихся на о-ве Сан-Мигуель у берегов Калифорнии.
Хозяйственное значение северных морских котиков велико — они дают превосходный ценный мех. Интенсивный промысел северных морских котиков ведется издавна. С момента организации Российско-Американской К° (1780 г.) он принял особенно крупные масштабы. Так, в период с 1799 по 1867 гг. на о-вах Командорских и Прибыловых было добыто свыше 2,5 млн. северных морских котиков. К началу XX столетия в результате хищнического промысла американских, японских и русских зверобоев численность северных морских котиков резко упала, не превышая к 1910 г. 132 тыс. голов. В 1911 г. была подписана конвенция между США, Великобританией (представлявшей Канаду), Россией и Японией о предотвращении дальнейшего уничтожения северных морских котиков. Пелагический промысел был запрещен, а США и Россия выплачивали по 15% Японии и Великобритании от дохода, получаемого от берегового промысла котиков. Япония платила 10% своего дохода остальным трем странам. Это соглашение существовало до 1941 г., когда было расторгнуто Японией, и было возобновлено в 1957 г. Главное мероприятие новой конвенции — запрещение морского промысла котиков, наносящего виду большой ущерб.
О-ва Прибылова объявлены специальной правительственной резервацией США. Ежегодно около 80% 3—4-летних холостяков убивают здесь на мех, жир и мясо. Общее количество добычи составляет около 60 тыс. холостяков и 30 тыс. самок различных возрастов.
Заповедны лежбища котиков на о-ве Тюленьем и Командорских о-вах (СССР). Здесь также производится выборочный забой котиков; в 1961 г.— 12 тыс. голов, в 1962 г.— 13,5; в 1963 — 14,7; в 1964—18,9; 1965 — 20 тыс. голов.
СЕМЕЙСТВО МОРЖОВЫХ
FAMILIA ODOBENIDAE J. ALLEN, 1880
Размеры крупные: длина тела самцов до 4,1 м по поверхности тела или до 3,7 м — по горизонтали, масса около 2 т. Самки меньше самцов — длина до 3,4 м, масса до 900 кг (разница между ними не столь велика, как у ушастых тюленей).
Тело толстое, неуклюжее, голова округлая, относительно небольшая, с короткой, очень широкой мордой (рис. 125). Хвост небольшой-—в виде едва заметного кожного выроста. Шея короткая. Глаза небольшие, высоко расположенные, широко расставленные. На верхней губе в 13— 14 рядов расположены толстые — диаметром 1,5—2 мм — вибриссы. Длина вибрисс до 30 см, они богато иннервированы. Общее число их около 400. Ушная раковина отсутствует; наружное слуховое отверстие защищено складкой кожи. Конец языка в противоположность другим ластоногим не имеет выемки.
Кожа очень толстая. Эпидермис достигает наибольшей толщины среди ластоногих и по всей внутренней поверхности имеет многочисленные неправильной формы ячеи. Роговой слой эпидермиса очень сильно
Рис. 125. Морж — О. rostnarus и его череп
развит. В дермальном слое кожи сеть эластиновых волокон и пучки гладких мышечных волокон развиты слабо. Подкожная жировая клетчатка очень толстая. Сальные железы маленькие, а потовые крупных Размеров. Кожа покрыта редкими волосами; у старых особей кожа почти голая. Волосы растут пучками, которые обычно содержат по одному остевому волосу и 1—2 пуховых. В зимнее время в коже откладываются большие запасы жира, причем многочисленные жировые клетки заполняют даже эпидермальные ячеи. У половозрелых самцов кожа на шее, груди и плечах образует шишкообразные утолщения, которые служат своеобразным вторичнополовым признаком. Эти утолщения отличаются необычайно прочной вязью пучков коллагеновых волокон, толстым эпидермисом и особенно плотным роговым слоем. Такая *броня» защищает животных во время драк от ударов клыков против-
ника. Окраска тела желтовато-серая. У новорожденных волосяной покров густой, но без пуха. С возрастом волосяной покров все более редеет.
Пятипалые конечности моржей во многом напоминают конечности ушастых тюленей, но относительно меньше по размерам, и предпальце-вые хрящи не такие длинные. Передние ласты достигают в длину менее 25% длины тела, несколько крупнее задних и имеют пять маленьких когтей. Передние и задние ласты утолщены по переднему краю. Ладонная и подошвенная поверхности ластов безволосые, с очень грубой кожей. Тыльная сторона ластов (по крайней мере у молодых зверей) покрыта волосами. Задние ласты участвуют в передвижении по твердому субстрату, сгибаясь в пяточном сочленении и подворачиваясь вперед под тело. При плавании конец туловища двигается латерально, как у тюленевых, но в противоположность последним передние ласты также участвуют в движении наподобие весел. Скорость плавания достигает 24 км/ч.
Сосков 2 пары. Семенники располагаются под кожей. Боковые стенки у взрослых моржей, преимущественно самцов, образуют большие парные мешки. Каждый из них тянется назад между мышцами шеи до переднего края лопатки. Воздух поступает в мешки из легких. Вокруг отверстий мешков имеются сфинктеры. Мешки, по-видимому, несут гидростатическую функцию и являются резонаторами.
Череп очень массивный, примерно одинаковой ширины в мозговом и ростральном отделах (у взрослых животных). Это связано с чрезвычайно сильным развитием верхних клыков, сидящих в глубоких и широких альвеолах. Скуловые кости короткие и высокие. Костные слуховые барабаны уплощены. Имеется крылоклиновидный канал. Заднеглазничные отростки отсутствуют, но предглазничные хорошо выражены. Граница между лобными носовыми костями проходит по прямой линии или лобные кости вдаются выступом между носовыми костями. Сагиттального гребня нет. Половинки нижней челюсти у взрослых особей срастаются между собой.
Зубная формула индивидуально изменчива:
1 —2 ---- с
О
3 — 4 О
р ------ tn — — 18 — 24.
3 — 4 О
Верхние клыки имеют постоянный рост и превратились в огромные бивни, иногда прямые, иногда дугообразно изогнутые. Они достигают в длину у самцов 82 см, а у самок 70 см. Клыки имеют длину примерно 2,5 см у 1-летних моржей, 10 см у 2-летних и 27 см у 5-летних. Первоначально клыки обладают небольшим колпачком эмали, который стирается уже у двухлетнего животного, так что клыки состоят только из тонкого наружного слоя цемента и плотного дентина. Все остальные зубы примерно однотипные, округлые в поперечнике, с низкой коронкой, плоской или слабоконической жевательной поверхностью, приспособленной к дроблению и перетиранию пищи. У взрослых особей коронки зубов обычно сточены почти до края альвеол. Зубы имеют по одному корню.
Позвонков обычно 14 грудных и 6 поясничных. Лопатка высокая и узкая.
Os penis больших размеров (до 64 см длины).
Хромосом в диплоидном наборе 32.
Распространены моржи циркумполярно в прибрежных водах и неглубоких участках Северного Ледовитого океана, смежных районах Атлантического океана и Берингова моря. В настоящее время они встречаются, кроме большей части побережья Восточно-Сибирского моря, вдоль всего побережья Северного Ледовитого океана в пределах СССР
и в Беринговом море до мыса Наварин на юге, по побережью Аляски до Британского залива на юге и мыса Барроу на востоке, по берегам Западной Гренландии, арктическим берегам Канады.
Большинство моржей в, зимнее время мигрируют к югу, а летом к северу, однако не уходят далеко от кромки льда. Тихоокеанские моржи зиму проводят на мелководье северной части Берингова моря. В апреле _мае они начинают движение к северу вдоль Южной и Восточной
Чукотки, появляясь и у п-ова Стюарт. К началу июня основная часть популяции моржей проходит Берингов пролив и большинство направляется в западные районы Чукотского и восточные Восточно-Сибирского морей, а остальные — к северному побережью Аляски. Осенью миграция проходит в обратном направлении. Незначительная часть самцов остается летом в Беринговом море.
Моржи обитают в прибрежной зоне и, как правило, не встречаются над глубинами более 90 м. Обычно держатся смешанными стадами — самки, самцы и детеныши—до сотни и более голов вместе (иногда до нескольких тысяч). Весной и летом располагаются группами на плавучих льдах, осенью большими стадами устраивают лежбища на берегах. Иногда половозрелые самцы образуют отдельные лежбища на берегу. Чтобы выбраться на лед или взобраться на скалы, моржи пользуются своими клыками. Активны днем и ночью. Органы обоняния и слуха развиты плохо. Вибриссы служат тактильными органами чувств, помогающими найти пищу в иле дна океана.
Объектами питания в основном служат донные животные: пластинчатожаберные (Му a runcata, Saxicava arctica, Cardium groenlandicum и др.) и реже брюхоногие (Buccium) моллюски, значительно реже ге-фиреи и голотурии и еще более редко черви и ракообразные. Масса содержимого желудка одного моржа может достигать 55 кг. Известны случаи нападения отдельных моржей на нарвалов, белух и тюленей.
Моржи, по-видимому, моногамы и в период размножения образуют семейные группы из 3—6 животных. Такая группа состоит из самца, самки и разновозрастных детенышей. Имеются предположения и о полигамное™ моржей (например, Nishiwaki, 1972). Период спаривания приходится на апрель — май (самки спариваются вскоре после рождения детеныша). В это время самцы дерутся между собой. Беременность длится 11—12 месяцев. Задержки в имплантации яйцеклетки, по-видимому, нет. Самка рожает одного, редко двух детенышей раз в два года. Детеныши появляются на свет с апреля по июнь. Длина новорожденного около 125 см, масса 45—68 кг. У него нет зубов, тело покрыто довольно густым волосяным покровом серого цвета. Детеныш сразу может плавать. Он держится около матери до 2—3-летнего возраста. Самка очень привязана к своему детенышу и активно защищает его. Лактационный период длится около года, иногда до двух лет. Половозрелое™ У самок наступает в 4—5-летнем возрасте, а у самцов примерно в 6— ' лет. Длительность жизни моржей, по-видимому, может достигать 30—40 дет.
Врагов у моржей немного: косатка и отчасти белый медведь.
Промысел моржей ведется издавна. Так, в конце XIX столетия ежегодная добыча тихоокеанского моржа в течение многих лет находилась на Уровне 15—20 тыс. голов, не считая потерь (раненые и утонувшие животные), составлявших примерно 40% (Ивашин и др., 1972). В результате неумеренной охоты численность моржей сильно снизилась и в ряде мест они были полностью истреблены. В результате охранных мер время популяция моржей в районах Чукотского моря, в проливе и прилежащих районах моря вновь стала довольно
многочисленной. Общее число зверей здесь достигает 45—90 тыс. осо-
в настоящее Беринговом
бей. Численность на побережье Аляски определяется примерно в 10— 15 тыс. голов; у Западной Гренландии— 10 тыс.; у северного побережья Европы моржи очень малочисленны.
Ежегодно на Аляске добывают примерно 1300 моржей, у Гренландии — примерно 500. Для местного населения Чукотки установлен ежегодный лимит добычи моржей. В СССР с 1956 г. введен запрет на судовой промысел. От моржей получают мясо, шкуру, жир и клыки.
Подвид моржа моря Лаптевых, восточной части Карского моря и западной части Восточно-Сибирского моря — О. rosmarus laptevi Chapskii, 1940, включен в «Красную книгу».
В семействе один род:
моржей — Odobenus Brisson, 1762 (1 вид).
Род моржей
Genus Odobenus Brisson, 1762
В роде единственный вид: морж — О. rosmarus Linnaeus, 1758. См. описание семейства.
СЕМЕЙСТВО ТЮЛЕНЕВЫХ
FAMILIA PHOCIDAE BROOKER, 1828
Длина тела и масса сильно колеблются: от 125 до 650 см и от 90 кг до 3,5 т. Кольчатая нерпа наименьшая среди ластоногих, а морские слоны имеют наибольшие размеры. Самцы по величине примерно равны самкам (Pagophilus, Pusa), или немного меньше их (Phoca, Hydrurga, Leptonychotes, Lobodon, Monachus, Ommatophoca), или немного крупнее (Halichoerus, Histriophoca), или значительно больше (Mirounga).
Голова не резко суживается кпереди. Наружные ушные раковины полностью отсутствуют. Хвост короткий, но хорошо выраженный. Передние ласты меньше задних и составляют значительно менее 'Д длины тела. По сравнению с ушастыми тюленями и моржами они расположены на туловище ближе к голове.
У тюленевых локомоторным органом служит задняя часть тела, в связи с чем исходя из принципов механики длинная шея была бы бесполезной; шейный отдел у тюленевых выражен слабо. Задние конечности вытянуты назад и не сгибаются в пяточном сочленении. Они не могут подгибаться под тело и не используются при передвижении по суше. Главный способ движения по твердому субстрату заключается в том, что масса тела попеременно переносится то на грудину — и тогда подтягивается задняя часть тела вперед, то на область таза — и тогда передний отдел тела двигается вперед. Передние ласты у различных тюленей в разной степени принимают участие в движении по твердому субстрату. Тюлень Уэдделла передвигается совсем без помощи передних ластов. Серый тюлень использует их в качестве своеобразных балансов при переползании по скалистым участкам или цепляется за грунт сильно развитыми когтями. Морской слон держит передние ласты под прямым углом, используя их в качестве опоры для своего тяжелого тела. Крабоед, передвигаясь по суше, употребляет волнообразное «плавательное» движение и может двигаться очень быстро. При плавании передние конечности прижимаются к телу и используются только при поворотах. Передние и задние ласты покрыты шерстью с обеих сторон.
У большинства тюленевых когти на ластах развиты хорошо, но подвергаются редукции на задних ластах у тюленей-монахов и тюленей 276
Уэдделла и на передних и задних ластах у морских леопардов. Когти тюленя Росса на всех ластах редуцированы до небольших бугорков. Предпальцевые хрящи, поддерживающие края ластов, сильно развиты, но среди настоящих тюленей это исключение.
Плавание осуществляется за счет последовательных медиально направленных ударов задних ластов, причем сами ласты расправляются во время толкательного движения и сжимаются при возвращении в исходное положение. Движению вперед помогают боковые изгибы задней части тела, которые осуществляются сильно развитой спинной мускулатурой. Достаточно точных сведений о скорости плавания тюленей в естественных условиях нет, но известно, что на коротких участках они в случае необходимости могут развивать скорость до 12—15 миль/ч.
Абсолютная толщина кожи у большинства представителей семейства меньше, чем у других ластоногих. В эпидермисе по большей части зернистый и блестящий слои не выделяются или они развиты слабо. В дерме сосочковый слой толще сетчатого. По степени развития в дерме сети гладких мышечных пучков и эластиновых волокон тюленевые стоят на втором месте после ушастых тюленей. Относительная толщина подкожной жировой клетчатки (в % к толщине кожи) у обыкновенных тюленей больше, чем у остальных ластоногих. У тюленя Уэдделла масса подкожного жира составляет около 113 кг —более чем '/4 от общей массы животного. Сальные железы очень крупные. Потовые железы развиты слабее, чем у других ластоногих. Их нижние отделы идут лишь немного глубже в кожу, чем луковицы волос.
Волосяной покров грубый и содержит остевые, промежуточные и пуховые волосы. Пуховые волосы по форме напоминают остевые с их уплощенным стержнем. Взрослые особи морских слонов почти полностью лишены волосяного покрова. Окраска меха разнообразная. Некоторые имеют пятнистую окраску, а представители родов Pagophilus и Histri-ophoca имеют на теле окрашенные полосы, для них характерен и половой диморфизм в окраске. Имеет место возрастная изменчивость окраски волосяного покрова. Во время сезонной линьки сменяются не только волосы, но и роговой слой эпидермиса, который слущивается целыми пластами. Вибриссы на морде обычно хорошо развиты, образуя 6—10 рядов.
Мошонки нет. Семенники абдоминальные. Имеется os penis. Сосков одна или две пары.
В черепе крупный (округлый или бочкообразный) мозговой отдел. Лицевая часть относительно короткая и узкая. Для большинства видов характерно очень узкое межглазничное пространство. Заднеглазничные отростки небольшие или их нет вовсе. Костные слуховые барабаны большие. Крылоклиновидного канала нет. Задние отделы носовых костей образуют выступ, расположенный между передними частями лобных костей.
2 — 3
£ 1
Зубная формула: i
5 — 6
р 4-m -----= 26—36.
5
Коронки коренных зубов обычно с дополнительными боковыми вершинами. Предкоренные (кроме первого) в большинстве случаев, а заднекоренные всегда с двумя корнями.
Хромосом у обыкновенного тюленя и кольчатой нерпы 32, у морского зайца, тюленя Уэдделла и северного морского слона 34.
Распространены в прибрежных и открытых водах преимущественно полярных, а также тропических областей всех океанов, кроме Индийского. Населяют и некоторые пресноводные озера — Байкал, Ладожское И др.
Устраивают залежки на скалах, песчаных берегах или на льду. Некоторые из них, например кольчатые нерпы, проводят на суше много времени. Обычно держатся группами, немногие поодиночке {Hydrurga}, Активны днем и ночью.
Питаются рыбой, головоногими моллюсками и ракообразными.
Большинство тюленевых во время размножения держатся парами, полигамия отмечена только у серого тюленя и морского слона. Длительность беременности 270—350 дней. У самок некоторых родов известна задержка имплантации яйцеклетки. Лактация обычно продолжается несколько недель. Половозрелость наступает в возрасте от 2 до 8 лет.
Тюленевых промышляют в больших количествах, получая кожу, мех, сало и мясо.
В семействе 13 родов (19 видов):
обыкновенных тюленей — Phoca Linnaeus, 1758 (2 вида);-
нерп — Pusa Scopoli, 1777 (3 вида);
полосатых тюленей — Histriophoca Gill, 1873 (1 вид);
гренландских тюленей — Pagophilus Gray, 1844 (1 вид);
длинномордых тюленей — Halichoerus Nilsson, 1820 (1 вид);
морских зайцев — Erignathus Gill, 1866 (1 вид);
тюленей-монахов — Monachus Fleming, 1822 (3 вида);
крабоедов — Lobodon Gray, 1844 (1 вид);
тюленей Росса — Ommatophoca Gray, 1844 (1 вид);
морских леопардов — Hydrurga Gistel, 1848 (1 вид);
тюленей Уэдделла — Leptonychotes Gill, 1872 (1 вид);
хохлачей — Cystophora Nilsson, 1820 (1 вид) и
морских слонов — Mirounga Gray, 1827 (2 вида).
Систематика семейства окончательно не установлена и число родов в нем разными исследователями принимается неодинаковое. Например, Чапский (1975) насчитывает в семействе 10 родов: Phoca, Halichoerus, Erignathus, Monachus, Leptonychotes, Ommatophoca, Hydrurga, Cystophora, Mirounga.
Род обыкновенных тюленей
Genus Phoca Linnaeus, 1758
Длина тела колеблется у самцов от 1,5 до 1,9 м, а у самок от 1,2 до 1,5 м, длина хвоста — от 75 до 102 мм, масса — от 50 до 150 кг. Окраска волосяного покрова обыкновенного тюленя в основном серебристая, серая, серовато-коричневая или почти черная; на этом основном фоне разбросаны многочисленные мелкие темные пятна (рис. 126). На боках и брюхе пятен значительно меньше. Обычно окраска спины темнее, чем брюха. У взрослых островных тюленей окраска тела обычно черная со светлыми пятнами и кольцами на спинной стороне. Эмбриональный волосяной покров высокий, чисто-белый. Сосков одна пара.
В черепе костные слуховые барабаны треугольной формы. Костный слуховой проход не образует колена. На переднем крае носовых, костей три выступа. Задний край костного неба глубоко вырезан. Скуловые дуги расставлены относительно широко.
Q Ж .314 1
Зубная формула: i с -j— р т — =34.
Верхние резцы имеют уплощенные с боков корни. Добавочные вершины на коренных зубах развиты слабо.
У обыкновенного тюленя хромосом в диплоидном наборе 32.
Распространены по побережью Атлантического и лихого океанов умеренных и субарктических зон Северного полушария.
Обыкновенные тюлени — преимущественно прибрежные обитатели. Они предпочитают эстуарии и области, где песчаные отмели обсыхают при отливах.
Питаются в основном рыбами, поедая наиболее многочисленных, встречающихся в. данный момент. Наиболее обычными объектами питания служат сельди, корюшка, мойва, навага, песчанка, лососевые. На втором месте — бычки, камбала, треска. Из беспозвоночных употребляют в пищу креветок, реже бокоплавов и головоногих моллюсков, изредка крабов, геферей, донных моллюсков. Ныряют, по-видимому, не глубже 90 м.
Самки обыкновенных тюленей, населяющих Атлантику и восточную часть Тихого океана, приносят детенышей в мае — июне на берегу, песчаных отмелях. Эмбриональный белый волосяной покров спадает у детенышей еще в матке или же сразу после рождения, поэтому их волосяной покров не отличается от волосяного покрова взрослых. Они способны плавать очень скоро после появления на свет. Длина тела новорожденного составляет примерно 86 см, масса 11 кг. Продолжительность лактации около 3 недель. После ее окончания взрослые звери линяют, затем спариваются. Латентный период в развитии оплодотворенной яйцеклетки длится примерно 2 месяца. В западной части Тихого океана размножение обыкновенных тюленей носит несколько иной характер. Самки приносят детенышей на льду с февраля по апрель. В зависимости от холодной погоды эмбриональный мех детенышей может не спадать 2—3 недели; в это время молодой не идет в воду. Длительность лактации такая же, как и у обыкновенного тюленя, населяющего Атлантику (для островного тюленя характерна трехмесячная лактация). Линька взрослых приходится на май — июнь, а спаривание — на июнь — июль. Половозрелость наступает, по-видимому, к концу третьего года жизни. Продолжительность жизни в неволе составляет 14—18 лет.
Общая численность обыкновенных тюленей 150—450 тыс. (Scheffer, 1958).
Охота за обыкновенными тюленями сложна и регулярного промысла в большинстве мест не ведется. Постоянно добывают обыкновенных тюленей у берегов Исландии, о-ва Хоккайдо. Шкуру детенышей употребляют как пушное сырье, а взрослых — как кожевенное. Обыкновенные тюлени могут причинять известный вред рыбному хозяйству, поедая большое количество рыб, в том числе ценных лососевых, и портя сети.
Враги обыкновенного тюленя — белые медведи, косатки, акулы. На детенышей могут нападать даже орланы.
В роде 2 вида:
обыкновенный тюлень — Р. vitulina Linnaeus, 1758 (побережье Атлантического и Тихого океанов умеренных и субарктических зон Северного полушария; часто заходит в крупные реки, поднимаясь далеко вверх против течения; в водах СССР — на Мурмане, в южных районах Балтийского моря, на Дальнем Востоке обычен вдоль всего побережья Японского моря; в Татарском проливе, в Охотском и Беринговом морях и в Чукотском— до мыса Шмидта);
островной тюлень — Р. kurilensis Inukai, 1942 (восточный берег о-ва Хоккайдо, о-ва Курильские, Командорские и Прибылова).
В водах СССР: оба вида.
Островной тюлень включен в «Красную книгу».
Система рода не установлена окончательно. Некоторые исследователи (например, Nishiwaki, 1972; Чапский, 1975) выделяют в нем всего один вид — Р. vitulina-, другие (Rice a. Scheffer, 1968)—3 вида: Р. vitu-
Рис. 126. Обыкновенный тюлень—Р. vitulina (Л), островной тюлень — Р. kurilensis (Б)
Рис. 126. Продолжение. Череп обыкновенного тюленя
Нпа (Атлантика и Американское побережье Тихого океана), Р. kurilen-sis и Р. largha Pallas, 1811 (Азиатское побережье Тихого океана) или 2 вида: Р. vitulina и Р. largha (Чапский, 1966).
Иногда (Чапский, 1975) в род включают на правах подродов Pusa, Histriophoca и Pagophilus.
Род нерп
Genus Pusa Scopoli, 1777
Нерпы — наиболее мелкие представители семейства. Размеры самцов и самок примерно одинаковы: максимальная длина тела 150 см, масса 90—100 кг. Темная спина обычно покрыта светлыми пятнами с темными центрами или темными пятнами (рис. 127). Иногда спина одноцветная оливково-коричневая или коричнево-серебристо-серая. Брюхо светлое, беловатое или желтоватое, без пятен. Для кольчатой нерпы характерен в окраске спины сложный светлый кольчатый рисунок на темном фоне. У байкальского тюленя спина серовато-серебристая, а пятнистый рисунок полностью отсутствует у всех возрастных групп. Для взрослого каспийского тюленя типичны многочисленные неправильной формы и разной величины темные (буроватые или почти черные), часто сливающиеся пятна, между которыми просвечивает основной сизовато-серый на спине и сизовато-белый на брюхе фон. На боках и брюхе пятна редки или полностью отсутствуют. Взрослые самки окрашены более тускло, с менее контрастными пятнами. Еще менее четкие пятна у молодых зверей.
Байкальский тюлень отличается от двух других представителей рода относительно более длинными передними ластами с мощными длинными когтями треугольной в поперечном сечении формы, которыми он пользуется при передвижении по твердому субстрату, заметно выдвигая их вперед. Задние ласты байкальского тюленя, наоборот, несколько слабее и когти на них менее развиты, чем у кольчатой нерпы и каспийского тюленя. Каспийский тюлень имеет несколько более вытянутую морду, чем Другие виды рода. Сосков одна пара.
Рис. 127. Каспийский тюлень—Р. caspica (4 — самец, Б — са,мка с бельком)
Череп с расширенным мозговым и укороченным лицевым отделами. Скуловая кость сильно наклонена передним концом вниз. Носовые косги относительно узкие и длинные. Костные слуховые барабаны имеют треугольную форму. Костный слуховой проход не согнут в виде колена.
3 Зубнан формула: i с
1 4
Г ртт
— = 34. 1
У верхних резцов корни с боков уплощены. Добавочные вершины щечных зубов развиты сильно.
Хромосом в диплоидном наборе у всех трех видов по 32.
Распространены в Северном Ледовитом океане, на севере Атлантики и Тихого океана, в Балтийском и Каспийском морях, в озерах Байкал, Ладожское. Сайменское.
Питаются преимущественно рыбой. Кольчатая нерпа в арктических морях поедает в основном сайку, затем навагу и корюшку. В Охотском море она ест весной планктонных ракообразных, а осенью рыбу — мойву, корюшку; в Балтийском море — салаку, кильку, треску, угря, судака, сига; в Ладожском озере—корюшку, ряпушку, ерша, сига. Байкальский и каспийский тюлени питаются в основном малоценными рыбами: первый— бычками и голомянкой, второй — бычками, килькой, атериной. а также креветками, бокоплавами, обыкновенными раками, реже поедает сельдь и воблу. Кольчатая нерпа ныряет на глубину, по-видимому, до 90 м.
Рис. 127. Продолжение. Череп каспийского тюленя
Щенка у самок кольчатой нерпы приходится на середину марта — середину апреля. Самка рожает детеныша на прочном не дрейфующем льду в норе, вырытой в снегу, или в естественной пещере торосов. Самка байкальского тюленя приносит детеныша в феврале — марте обычно в логовищах, выкопанных в наносах снега у торосов. Шенка каспийских тюленей происходит в конце января и не позднее первых чисел февраля на льду в северной, замерзающей части Каспийского моря. Новорожденный достигает в длину 60—70 см, масса его 3—4,5 кг. Он покрыт длинной и густой шерстью белого, соломенно-белого или сероватого цвета. Детеныши кольчатой нерпы и каспийского тюленя линяют спустя 10— 20 дней после рождения. Лактационный период продолжается 1—3 месяца (наиболее долго у байкальского тюленя). Спаривание происходит, видимо, до окончания лактации (у кольчатой нерпы пик спаривания при
ходится на середину апреля, у каспийского тюленя —на двадцатые числа февраля). Для развивающейся яйцеклетки характерна задержка имплантации в течение 3—3,5 месяцев (она установлена у кольчатой нерпы, но, по-видимому, имеет место и у двух других видов). После спаривания у взрослых зверей начинается линька: у кольчатой нерпы — в июне — июле, байкальского тюленя — в мае, каспийского — с конца февраля по май. Половозрелость самок наступает в 4—6, самцов—-в 5— 7 лет.
Продолжительность жизни, по-видимому, до 43 лет. Врагами могут быть белый медведь, косатка, лиса, иногда морж.
Для каспийских тюленей характерны сезонные перемещения. С поздней осени до весны они концентрируются в северных районах Каспийского моря, сначала образуя залежки на отмелях, а с января — на льдах, где они размножаются и линяют. Примерно с мая каспийские тюлени начинают откочевывать на юг, где в менее прогреваемых водах Среднего и Южного Каспия большая часть популяции проводит лето.
Существует 2 гипотезы по поводу появления тюленей в двух внутренних водоемах — Байкале и Каспийском море. Согласно одной из них в плейстоцене во время третьего оледенения представители северной фауны, в частности нерпы, были вынуждены мигрировать к югу от наступающего западносибирского ледника. Тюлени обитали в то время в гигантских озерах вдоль южной границы ледника. Более правдоподобной представляется вторая гипотеза, по которой байкальский и каспийский тюлени имеют более древнее (дочетвертичное) происхождение и представляют собой предков одной из ветвей тюленей, обитавших в Сарматско-Понтическом бассейне.
В роде 3 вида:
кольчатая нерпа — Р. hispida Schreber, 1775 (распространение очень широкое — кругополярное в Северном Ледовитом океане; наиболее многочисленна в морях, омывающих берега Евразии и Северной Америки; значительно меньше их в Центральном полярном бассейне с его приполюсными районами; населяет также субарктические воды Атлантики и северную часть Тихого океана, Балтийское море и Ладожское и Сай-менское озера);
байкальский тюлень — Р. sibirica Gmelin, 1788 (Байкал, преимущественно его северная часть; иногда заходит в реки Ангару, Селенгу и Баргузин);
каспийский тюлень — Р. caspica Gmelin, 1788 (Каспийское море и лишь изредка низовья Волги и Урала).
Иногда нерп включают в качестве подрода в род Phoca (например, Чапский, 1975).
Кольчатая нерпа служит важным объектом промысла. Ежегодно ее добывают много тысяч голов, получая от зверей кожу, мех, жир и мясо. У нас промысел кольчатой нерпы ведется на Дальнем Востоке, а также в Белом и Баренцевом морях. Б Охотском море в 1947—1954 гг. ежегодно добывали в среднем по 20 тыс. голов. С 1969 г. введен лимит на добычу кольчатой нерпы судовым промыслом в Охотском море в размере 32 тыс. голов, в 1970 г. — 26 тыс., в 1972 г. — 25 тыс. В Беринговом море судовая добыча запрещена. С 1970 г. на Охотском и Беринговом морях ограничивается и береговая добыча. Байкальского тюленя в настоящее время промышляют в небольшом количестве зимой с лимитом добычи примерно 2—3 тыс. голов.
Каспийский тюлень издавна служит важным промысловым видом. В конце XVIII — начале XIX столетий его ежегодно добывали в среднем по 160 тыс. голов. В последнее время добыча каспийских тюленей резко сократилась, что связано с уменьшением их общей численности. Добы
вали каспийского тюленя на плаву, осенью на островных залежках в северо-восточной части моря и зимой на льду. С 1966 г. запрещены промысел взрослых самок в зимний период и бой тюленей на воде, а с 1967 г. запрещен осенний промысел на береговых лежбищах. Ежегодно разрешается добывать зимой детенышей 60—65 тыс. голов.
В СССР: все 3 вида нерп. Кольчатая нерпа обитает во всех наших морях Северного Ледовитого океана, включая Баренцево и Белое, в Балтийском море и Ладожском озере, в Тихом океане от северных частей Берингова моря на юг до Татарского пролива и южной присахалинской области Охотского моря; у Командорских и Курильских о-вов, за исключением самой северной части последних, кольчатая нерпа обычно не встречается. Байкальский тюлень —в оз. Байкал, каспийский тюлень — в Каспийском море.
Примерная численность кольчатой нерпы составляет в Охотском море около 800 тыс., в Беринговом — 350—400 тыс., байкальского тюленя— 40—100 тыс., каспийского тюленя — 500—600 тыс.
Подвид кольчатой нерпы, обитающей в озере Сайма (Р. hispida saimensis Nordquist, 1899), включен в «Красную книгу» как малочисленный, но в настоящее время не находящийся под угрозой исчезновения.
Род полосатых тюленей
Genus Histriophoca Gill, 1873
В роде единственный вид: полосатый тюлень — Н. fasciata Zimmermann, 1783.
Средняя длина тела 155—165 см, масса до 150 кг. Самцы несколько крупнее самок. Для самцов характерен темный основной фон окраски от аспидно-бурого до почти черного с несколькими светлыми полосами шириной 10—12 см, проходящими в виде ошейника вокруг шеи, каждого грудного ласта и заднего отдела туловища в области крестца (рис. 128). Самки и молодые звери имеют светлую окраску от палевой до темносерой с неясно выраженными полосами. Сосков одна пара.
Мозговая часть черепа сильно увеличена, а лицевая укорочена. Костные слуховые барабаны вытянуты и несколько выгнуты. Костный слуховой проход изогнут. Передний край носовых костей образует три выступа, из которых боковые развиты сильнее среднего. Носовые отростки межчелюстных костей глубоко вклиниваются между носовыми и верхнечелюстными костями.
о 3 14 1
Зубная формула: 1 — с у р у гл у =34.
У верхних резцов корни уплощены с боков. Щечные зубы небольшие, без передних добавочных вершин, маленькая задняя дополнительная вершина появляется лишь с третьего предкоренного зуба.
Хромосом в диплоидном наборе 32.
Распространение охватывает Берингово и Охотское моря. Встречаются также вдоль Курильских о-вов, восточного побережья Камчатки и в южных районах Чукотского моря от Колючинской губы на западе До мыса Барроу на востоке. Отдельные особи отмечены у Корейско-го п-ова и о-ва Хоккайдо. До последнего времени не установлены с Достоверностью осеннее и зимнее местопребывание полосатых тюленей. Возможно, весенняя локализация тюленей в двух разобщенных частях океана в Охотском и Беринговом морях соответствует двум популяциям этого вида.
Полосатый тюлень тесно связан со льдами. Обычно залежки устраи-
Рис. 128. Полосатый тюлень — Н. fasciata и его череп
ваются вдали от берега на дрейфующих торосистых льдах с разводьями и трещинами. Держатся разрозненно: поодиночке или небольшими группами. По-видимому, мигрируют: на юг со льдом зимой и на север — летом.
Питание изучено слабо. Основными кормовыми объектами в Охотском море, по-видимому, служат минтай, треска, мойва, головоногие моллюски и ракообразные. В Беринговом море — креветки, сайка, навага, сельдь, изредка головоногие моллюски. Молодые животные питаются в основном мелкими ракообразными. Более активно кормятся ночью.
Размножаются на дрейфующих льдах. Беременность длится около 9 месяцев. С конца марта до первых чисел мая самка приносит прямо на льду, без убежища, детеныша длиной в среднем 85 см и массой около 86 кг с густой и длинной белой шерстью. Лактационный период продолжается около месяца. Линька ювенильного волосяного покрова происходит в конце апреля. Период спаривания приходится на конец апреля— май (конец июля—август; King, 1964). В развитии оплодотворенной клетки, по-видимому, имеет место задержка имплантации на 2,5 месяца. Половозрелость у самок наступает в 4—5, а у самцов в 5—6 лет. Взрослые звери линяют с конца апреля до середины июля.
Примерная численность полосатых тюленей составляет, по данным авиаучета, в Беринговом море 80—90 тыс. голов (1964) и в Охотском море__80 тыс. голов (1968) (Ивашин и др., 1972). Враги полосатых тюле-
ней — полярная акула, косатка и белый медведь.
Полосатых тюленей промышляют со зверобойных судов в Охотском и Беринговом морях, однако их хозяйственное значение невелико. С 1965 по 1968 г. ежегодно судовым промыслом добывали от 15,6 до 23 7 тыс. голов. Введен ежегодный лимит на судовой промысел полосатых тюленей: 3 тыс. в Беринговом море и 7 тыс. в Охотском.
Полосатого тюленя иногда (например, Чапский, 1975) включают на правах подрода в род Phoca.
Род гренландских тюленей
Genus Pagophilus Gray, 1844
В роде единственный вид: гренландский тюлень — Р. groenlandica Erxleben, 1777.
Максимальная длина тела достигает 205 см, масса 164 кг (рис. 129). Длина пальцев на передних конечностях постепенно убывает от почти одинаковых I и II к V. Когти на пальцах передних ластов крупные. Полового диморфизма в размерах тюленей практически нет. Верх тела взрослого самца обычно желтоватый или почти белый, низ — серебристый. Темно-бурая или почти черная полоса начинается в области лопаток и разделяется на две ветви, идущие назад и вниз по обеим сторонам тела и постепенно сужающиеся и сходящие на нет в крестцовой области. Передняя часть головы черно-бурая или черная. У самок полосы на теле более светлые, размытые по наружному краю и могут прерываться. Гренландские тюлени, как и полосатые тюлени, — единственные представители ластоногих, имеющие полосы на теле, с половым диморфизмом в окраске.
В черепе носовые кости спереди имеют 3 выступа, нависающие над наружным носовым отверстием. Носовые отростки межчелюстных костей могут вклиниваться между носовыми и верхнечелюстными костями. Задний край костного неба образует почти прямую линию. Костные слуховые барабаны треугольной формы с костными слуховыми проходами в виде мощных толстостенных трубок, коленообразно загнутых вперед.
о 3 1 4
Зубная формула: i ~ z — р —
m — = 34.
1
Коронки верхних предкоренных зубов имеют по крайней мере одну Дополнительную вершину позади основной, а коронки нижних предкоренных — минимум две дополнительные вершины (по одной спереди и сзади). Третий и четвертый нижние предкорениые имеют чаще по три Добавочные вершины (одну спереди от основной вершины и две сзади), предкорениые (кроме первого) и коренные зубы с двумя корнями.
Хромосом в диплоидном наборе 32.
Распространен в Северной Атлантике, морях Гренландском, Норвежском, Баренцевом, Карском, Белом.
Ареал гренландских тюленей, учитывая миграционные пути, ограничивается на юге 50° с. ш., на севере 75° с. ш. в западной части ареала и с. ш. в восточной, на западе — устьем Маккензи и на востоке—Но-вой Землей (в отдельные годы Северной Землей).
Гренландский тюлень — обитатель холодных вод. Держится у кромки
Рис. 129. Гренландский тюлень—Р. groenlandica и его череп. А — взрослая самка, Б — двухлеток
льда и в районах устойчивых дрейфующих льдов. Во время размножения и линьки встречается в глубине ледовых массивов.
В питании в течение года происходит смена кормов. Летом, когда гренландский тюлень держится у кромки льда, он питается в основном крупным зоопланктоном - эуфаузиидами, бокоплавами и головоногими моллюсками, а также сайкой. Осенью и в начале зимы переходит на питание рыбой: сайкой, мойвой, сельдью, в меньшей степени треской, морским окунем; значительно реже поедает ракообразных.
Сезонные перемещения характерны. К концу зимы тюлени концентрируются в местах размножения на льду в трех районах: 1) в районе Ньюфаундленда (здесь два центра размножения: в проливе Св. Лаврентия и у восточного побережья Ньюфаундленда, к северу от о-ва Белл), 2) в Гренландском море между Исландией и Шпицбергеном — к северу от Ян-Майена, примерно на 79° с. ш., 3) в Белом море. Эти три района размножения соответствуют трем популяциям гренландских тюленей, по-видимому, значительно изолированным друг от друга. Предполагается, что 1% ньюфаундлендской популяции смешивается с ян-майенской и 10% последней — с беломорской.
Гренландские тюлени беломорской популяции проводят летнее время у края льдов Баренцева и Карского морей, около Шпицбергена, Новой Земли, Земли Франца-Иосифа и Северной Земли. Поздней осенью взрослые звери мигрируют к Канинскому побережью, а в январе — феврале появляются в Белом море. Неполовозрелые тюлени появляются здесь не раньше апреля. Для размножения тюлени выбирают прочные льдины,
устраивая на них так называемые детные залежки, иногда тянущиеся на многие десятки километров и насчитывающие десятки тысяч особей. Самки приносят детенышей с конца января до конца апреля, причем большинство детенышей рождается с 20 февраля по 5 марта. В этот период самки почти не питаются. В помете, как правило, один детеныш. Лактация продолжается около 3 недель. Самка обычно кормит детеныша 2—3 раза в течение светлого времени суток, иногда и ночью. За сутки детеныш прибавляет в массе до 2,5—3 кг. После окончания лактации происходит спаривание — с середины до конца марта. Только в это время самцов гренландских тюленей можно видеть на льду; самцы жестоко Дерутся между собой. Длительность беременности составляет примерно 11 месяцев, включая задержку имплантации в течение 2—2,5 месяцев.
Новорожденный достигает в длину 80—90 см и массы примерно 8 кг. Он покрыт густой и длинной шерстью, белой, с зеленоватым оттенком, поэтому его называют «зеленец». Через 3—5 дней зеленоватый оттенок исчезает и детеныш становится чисто-белым — «белёк». Через 3—4 недели после рождения белый мех начинает заменяться обычным для тюленей коротким и жестким волосяным покровом серого цвета, более темным на спине, с темно-серыми или черными пятнами — «серка». Эта окраска сохраняется до 5—8-летнего возраста. Когда линька у детенышей завершается, они становятся способными плавать и переходят к самостоятельному существованию.
После размножения беломорские гренландские тюлени мигрируют к северу — к границе Баренцева моря, где образуют линные залежки, с- конца марта до конца мая линяют сначала взрослые самцы, затем неполовозрелые самки и последними взрослые самки. С первой половины мая гренландские тюлени начинают покидать Белое море, мигрируя в арктические районы Баренцева и Карского морей.
Ньюфаундлендская популяция лето и раннюю осень концентрируется в водах западной Гренландии и в восточном секторе Канадской Арктики. Зимой тюлени передвигаются к югу, в пролив Св. Лаврентия, и в фев
рале взрослые животные образуют здесь, а также к северо-востоку от Ньюфаундленда детные залежки. Самки приносят детенышей примерно в те же сроки, что и в Белом море, и после спаривания тюлени перемещаются на льды восточного берега Ньюфаундленда для линьки. Затем постепенно двигаются к северу вдоль побережья Лабрадора.
В районе Ян-Майена миграции гренландских тюленей сходны с миграциями двух описанных выше популяций, но самки приносят детенышей несколько позже — большинство между 18 и 20 марта. Льды в это время дрейфуют к югу, и после периода спаривания тюлени опять плывут к северу от Ян-Майена на линные залежки. Линька происходит в апреле, а к середине мая они перемещаются к северу на места кормежек между Шпицбергеном и Гренландией.
Половозрелость у самок наступает в 4—5, а у самцов в 5 лет. Самки способны размножаться до 16—20-летнего возраста. Длительность жизни может, по-видимому, достигать 30 лет.
Численность беломорской популяции способом аэрофотосъемки в 1928 г. была определена в 3—3,5 млн. голов. В 1952 и 1953 гг. аэрофотосъемка линных залежек показала, что тюленей 722 и 514 тыс. (общее число 1,2 и 1,5 млн. голов). В 1962, 1963 и 1967 гг. на линных залежках насчитывалось 179, 165 и 334 тыс. тюленей. Примерная численность гренландских тюленей в районе Ньюфаундленда составляет около 3— 4,5 млн. голов и у Ян-Майена 0,5—1 млн. Некоторыми исследователями (Чапский, 1975) суммарная численность гренландских тюленей определяется в 3—3,5 млн. голов.
Гренландский тюлень издавна служит важным объектом зверобойного промысла. В основном добывают детенышей в 2—10-дневном возрасте. После первой линьки, когда молодой тюлень покрывается густым и мягким мехом серого цвета, шкурка его ценится еще выше. От добытых взрослых тюленей используют главным образом шкуру как кожевенное сырье и подкожное сало.
В районе Ньюфаундленда промысел гренландских тюленей ведется более 200 лет и наибольшая добыча (687 тыс. голов) датируется 1831 г. В настоящее время судовой промысел ежегодно дает около 180 тыс. детенышей и 80 тыс. неполовозрелых и взрослых тюленей, а береговой промысел — еще 60—65 тыс. тюленей. Промысел в Ньюфаундлендских водах ведется главным образом судами Канады и Норвегии. В 1961 и 1963 гг. в районе Ньюфаундленда промышляло советское судно, взявшее за 2 года около 29 тыс. тюленей.
Первое упоминание о промысле этих тюленей у Ян-Майена относится к 1720 г. До 1890 г. промысел вели многие страны, но после этой даты — главным образом Норвегия, а с 1955 по 1956 г. и СССР. В 1874 г. было добыто 200 тыс. тюленей, но с 1900 г. численность добычи сильно сократилась и средняя ежегодная добыча Норвегии за период 1951—1960 гг. составила около 32 тыс. тюленей, а с 1961 по 1965 г.— 18,6 тыс., а СССР — с 1958 по 1961 гг. примерно 6 тыс. тюленей, с 1962 по 1965 гг.— около 4 тыс.
В Белом море наиболее высокая добыча тюленей пришлась на 1925 г. — она составила 500 тыс. голов, из которых 343 тыс. добыли норвежские суда. После Отечественной войны 1941—1945 гг. в Белом море промышляли только советские суда. В 1954—1964 гг. средняя ежегодная добыча составляла примерно 100 тыс. тюленей. С 1965 г. судовой промысел гренландских тюленей в Белом море запрещен на 5 лет. Добыча белька разрешается только местному населению в размере не более 20 тыс. голов при полном запрете выбоя взрослых самок на детных залежках. Ограничение промысла беломорского гренландского тюленя благотворно сказалось на увеличении его численности.
Гренландского тюленя иногда включают на правах подрода в род Phoca (например, Чапский, 1975).
Род длинномордых тюленей
Genus Halichoerus Nilsson, 1820
В роде единственный вид: длинномордый тюлень — Н. grypus Fabri-cius, 1791.
Длина тела самца достигает 3,3 м, самки — 2,3 м; масса соответственно 315 и 250 кг. Морда у этих тюленей относительно более длинная, чем у других ластоногих, с ровным, даже несколько выпуклым межглазничным пространством (рис. 130). Расстояние от конца морды до глаза в два раза больше, чем от глаза до уха. Ноздри крупные. Грудные ласты обладают значительной подвижностью и имеют длинные, узкие искривленные когти. Окраска сильно варьирует. На спинной стороне она светло- или темно-серая, иногда почти черная, обычно с темно-бурыми или черноватыми пятнами разной формы и размеров. Брюхо светлее спины, иногда почти белое с яркими черными пятнами. Самцы немного темнее самок. Сосков одна пара.
Череп массивный. Лицевая часть черепа удлинена. Короткие и широкие носовые кости спереди образуют три крупных примерно одинаковых размеров выступа, нависающих над костным носовым отверстием. Узкие носовые отростки межчелюстных костей в отличие от большинства тюленей не вклиниваются между носовыми и верхнечелюстными костями. Костное небо с дугообразной глубокой выемкой по заднему краю. Костные слуховые барабаны треугольной формы, причем их ширина больше длины. Костный слуховой проход не изогнут в виде колена. Своеобразны крупные передние небные отверстия, от которых почти до резцов тянутся глубокие корытообразные выемки. Костное небо имеет по заднему краю глубокую дугообразную выемку.
3 1 4 1—2 „
Зубная формула: 1 — с — р — ш —j— =34 — 36.
В большинстве случаев верхнего второго коренного зуба нет. Три первых нижних и верхних предкоренных зуба и верхний заднекоренной имеют по одной вершине. Четвертый нижний н верхний предкоренные и нижний коренной либо с двумя — передней и задней, либо с одной — задней дополнительными вершинами. Верхние резцы с уплощенными с боков корнями. Предкоренные зубы (из верхних по крайней мере три передних) имеют по одному корню.
Хромосом в диплоидном наборе 32.
Ареал распадается на 3 участка. Одна популяция (западноатлантическая) распространена у Американского побережья в Западной Атлантике в районе залива Св. Лаврентия и до Южной Гренландии. Вторая популяция (восточноатлантическая) занимает прибрежные районы островов Великобритании, Ирландии, Исландии, Шпицбергена, Скандинавского п-ова, Мурманского побережья, а также южные и восточные районы Северного моря. Третья балтийская популяция встречается в Балтийском море, главным образом в Финском и Ботническом заливах и у Аландских о-вов и, особенно в зимнее время, в самой северной части собственно Балтийского моря.
В СССР длннномордый тюлень встречается в Балтийском море, у Мурманского побережья, у Земли Франца-Иосифа.
Длинномордые тюлени не мигрируют, но после размножения широко распределяются по всему ареалу.
Рис. 130. Длинномордый тюлень — Н. grypus и его череп
292
Держатся главным образом у скалистых берегов. Пищей служат преимущественно пелагические крупные рыбы — треска, камбала, лососи, реже более мелкие — бычки, сельдь, мойва и др., еще реже ракообразные и моллюски. Может нырять за пищей на глубину до 128 м.
Детеныши рождаются в Балтийском море и в Западной Атлантике поздней зимой или ранней весной (большей частью в середине марта), а в Восточной Атлантике — осенью (пик в конце ноября — начале декабря) . Период деторождения сильно растянут. Самки первых двух популяций рожают на льду, а третьей — на берегу. В помете один, очень редко два детеныша. Длина тела новорожденного 85—100 см, масса 9—15 кг. Детеныш ежедневно прибавляет в массе примерно 0,5 кг (за первые три дня лактации по 1,8 кг). Высокий белый эмбриональный волосяной покров сохраняется 3 недели после рождения, после чего заменяется коротким голубовато-серым на спине и более светлым на брюхе. После линьки детеныши уходят в воду. Смертность детенышей в возрасте до 1 года достигает 60%- Лактационный период продолжается около 2-— 3 недель, причем в это же время происходит и спаривание. В развитии оплодотворенной яйцеклетки имеет место примерно 3-месячная задержка имплантации. Самка рожает обычно ежегодно.
Длинномордые тюлени — полигамы, самец собирает около себя гарем, насчитывающий до 10—20 самок. Между самцами бывают драки. Интересно, что на льдах у Канадского побережья полигамия не отмечена: с самкой и детенышем всегда держится лишь один самец. Вскоре после спаривания начинается линька. Тюлени балтийской популяции образуют на льду линные залежки. В британских водах самки линяют в конце января— начале февраля, самцы — в марте.
Половозрелость у самок наступает в 3-летнем, а у самцов в 6—7-лет-нем возрасте. Продолжительность жизни в неволе примерно 40 лет. На длинномордых тюленей могут нападать акулы.
Общая численность длинномордых тюленей определяется в 25—• 50 тыс. голов (Scheffer, 1958) или в 55—60 тыс. голов (Чапский, 1975). Хозяйственное значение невелико и специального промысла вследствие малочисленности этого вида не ведется.
Род морских зайцев
Genus Erignathus Gill, 1866
В роде единственный вид: морской заяц — Е. barbatus Erxleben, 1777.
Морской заяц—один из наиболее крупных представителей семейства тюленевых. Длина тела самца достигает 2,7 м, масса 360 кг. Длина тела самки немного меньше. Бросается в глаза непропорциональность телосложения: крупное туловище н относительно небольшая голова. Морда умеренно вытянута, глаза относительно небольшие (рис. 131). Передние ласты сильно укорочены. В отличие от других тюленей они расположены ближе к голове и наиболее длинный палец — третий. Когти на передних ластах крупные черные, с хорошо заметной поперечной поясковой структурой, а на задних слабые и заостренные. Характерны многочисленные Длинные толстые н гладкие вибриссы, крючкообразно изогнутые на концах. Вибриссы находятся на несколько вздутой верхней губе, цвет их белый. Волосяной покров у взрослых особей редкий. У взрослых животных окраска его однотонная, на спинной стороне буровато-оливковая до черновато-серой, на брюшной светло-серая, но бывают и пятна: мелкие темные на боках и 2—3 крупных, белесоватых на спине. На темени и над глазами обычно располагается по белесому пятну. Сосков две пары.
В черепе скуловые кости широкие и короткие. Костные слуховые ба-
Рис. 131. Морской заяц— Е. barbaius
и его череп
рабаны трапециевидной формы. Костный слуховой проход не изогнут коленообразно. Межглазничное пространство широкое и уплощенное. Костное небо с неглубокой дугообразной выемкой по заднему краю. Носовые кости спереди образуют широкий, иногда слегка раздвоенный средний выступ и два боковых, которые длиннее среднего, однако не выступают в виде зубцов над носовым проходом.
3 14 1
Зубная формула: i — с -у- р — m — = 34.
Верхние щечные зубы обычно только с одной дополнительной задней вершиной, а нижние (кроме первого и второго) с двумя — передней и задней. Зубы морского зайца слабее, чем у других тюленей, с тонкими корнями. Верхние резцы имеют конусовидные, округлые в поперечнике корни. Уже у молодых зверей вершины зубов стираются, у старых же от зубов сохраняются одни корни.
Хромосом в диплоидном наборе 34.
Распространен морской заяц циркумполярно по побережью материков и островов вдоль Северной Америки и Евразии. Южная граница ареала достигает Хоккайдо, Татарского пролива, восточной части Берингова моря, Ньюфаундленда, Франции, Британских о-вов и Норвегии. На север изредка проникает в Центральный Полярный бассейн (даже до района полюса). В водах СССР встречается вдоль всего северного побережья страны от Мурмана до Чукотки, а в бассейне Тихого океана — в северных, западных и восточных областях Берингова моря, в большей части Охотского моря (кроме юго-восточного района) и в северной части Японского моря.
Ведет относительно оседлый образ жизни; регулярные миграции отмечены у некоторых популяций. Так, весной морской заяц, по-видимому, мигрирует вдоль всего западного побережья Новой Земли к архипелагу Земли Франца-Иосифа и на восток — в Карское море. На Дальнем Востоке часто заходит в реки. Под лежбище занимает галечные берега или песчаные отмели. С появлением льда ложится на лед. Наиболее обычен в дрейфующих льдах.
Обычно держится поодиночке или небольшими группами (в арктических областях), хотя иногда и образует большие скопления до 1000 и более голов (на Дальнем Востоке в конце лета и осенью). Питается придонными животными, в основном беспозвоночными, а также рыбой. В связи с этим морской заяц встречается главным образом на мелководье— не глубже 30—50 м. Из беспозвоночных в пищу употребляет креветок, крабов, равноногих раков, реже бокоплавов, брюхоногих и пластинчатожаберных моллюсков, червей, геферий. Из рыб поедает главным образом сайку. За пищей может нырять на глубину 90—100 м.
Самки приносят детенышей с апреля до конца мая на прочных дрейфующих льдах. Больших залежек они не образуют. В высоких арктических широтах размножение начинается позже — с марта. Специальных убежищ, как правило, не делают. При рождении детеныш достигает длины 120 см и массы 36 кг. Эмбриональный коричневато-серый волосяной покров сбрасывается еще в утробе матери, и детеныш рождается с вторичным, густым и мягким, но коротким волосом темного, буроватооливкового цвета. Лактация продолжается примерно 4 недели, после чего наступает период спаривания. Самки рождают детенышей через год, что не типично для настоящих тюленей. Задержка в имплантации оплодотворенной яйцеклетки составляет 2—2,5 месяца. Линька происходит с мая по август с разгаром в нюне (в Арктике позднее, чем на Дальнем Востоке). Половозрелость, по-видимому, наступает в 6-летнем возрасте У самок и в 7-летнем — у самцов.
Общая численность морских зайцев разными авторами в 50-х годах определялась в 75—150 тыс. голов. Естественный враг—белый медведь.
Морские зайцы служат важным промысловым объектом для местного населения. Шкура — ценное кожевенное сырье. Кроме того, от каждого зверя получают до 100 кг подкожного сала и столько же мяса, которое употребляется в пищу и человеком. В водах СССР судовой промысел морского зайца запрещен.
Род тюленей-монахов
Genus Monachus Fleming, 1822
Длина тела самца тюленя-монаха 210—250 см (по некоторым данным, может достигать 3 м), масса до 300 кг. Длина тела самки несколько меньше — по 2.7 м. Размеоы двух доугих видов меньше. У гавайского
Рис. 132. Продолжение. Гавайский тюлень-монах —•
М. schauinslandi (Б)
тюленя-монаха самки крупнее самцов. Когти на передних ластах большие, на задних ластах очень маленькие. Вибриссы уплощенные, гладкие. Спинная поверхность тела сероватая или желтоватая с черными пятнами или почти одноцветная серо-бурая или темно-бурая; брюшная белесая или желтовато-белая (рис. 132). Подпуши у тюленей-монахов нет. Сосков две пары.
Череп с широко расставленными скуловыми дугами (особенно у старых особей) и несколько расширенным носовым отделом. Носовые отростки межчелюстных костей обычно не вклиниваются между носовыми и верхнечелюстными костями. Передний край носовых костей образует два выступа, разделенных выемкой. Костное небо имеет дугообразный задний край со срединной угловой вырезкой. В отличие от других тюленевых у тюленей-монахов мощно развит задний отдел нижней челюсти. Имеются крупные предглазничные отростки. Костные слуховые барабаны небольшие, треугольной формы. Костный слуховой проход не согнут в виде колена.
„ 2 1 4 1
Зубная формула: 1 — с -у р у m -у = 32.
Щечные зубы тесно прилегают друг к другу и, как правило, не имеют дополнительных вершинок (если они имеются, то очень малы). Все щечные зубы, кроме первого предкоренного, с двумя корнями. Внутренние верхние резцы имеют уплощенные корни.
Хромосом в диплоидном наборе 34.
Ареал рода разорван на 3 изолированных области: 1) Средиземное, Адриатическое, Эгейское и Черное моря и прилежащие к Северной Аф
рике районы Атлантического океана; 2) Мексиканский залив и Кариб-ское море; 3) Гавайские острова. За исключением калифорнийского морского льва, тюлени-монахи — единственные тюлени, обитающие в тропиках.
Несмотря на то что тюлень-монах известен людям с глубокой древности (упоминают Гомер, Плиний, Плутарх, а Аристотель даже исследовал его анатомию), экология этого вида изучена крайне недостаточно. Характер питания почти не известен; по-видимому, поедает рыб и моллюсков. В Черном море пищей служит главным образом камбала, в меньшей мере— макрель и анчоус. Гавайский тюлень-монах поедает различных рифовых рыб, а также головоногих моллюсков. Питаются в основном ночью.
Период размножения, очевидно, растянут во времени и приходится на конец лета — начало осени. У тюленя-монаха беременность длится 11 месяцев. Самка приносит детеныша в сентябре — октябре в расщелинах и выемках скал или на песчано-галечных берегах. Новорожденный достигает в длину около 120 см. Он имеет темно-бурую или почти черную окраску спины, постепенно переходящую на брюхе в светло-бурую с крупным белесым пятном. Лактация длится примерно 6—8 недель. После этого у детеныша проходит линька и он может сойти в воду. Спаривание у самок, по-видимому, происходит спустя примерно 2 месяца после рождения детеныша. Самки гавайского тюленя-монаха имеют растянутый период деторождения — с января по июнь; наибольшее число самок приносит детенышей с середины марта до конца мая. Длина новорожденного около 125 см, масса 16 кг. Черный мягкий волосяной покров спустя 3— 5 недель после рождения заменяется серебристо-серо-голубым на спине и серебристо-белым на брюхе. Самки приносят детенышей, по-видимому, раз в два года. Линька тюленей происходит с мая по ноябрь, в основном в июле.
Ввиду малочисленности никакого хозяйственного значения не имеют.
В роде 3 вида:
тюлень-монах — М. monachus Hermann; 1779 (Черное и Средиземное моря, за исключением большей части Средиземноморского побережья Африки, а также Африканское побережье Атлантического океана от Гибралтара до мыса Бланк);
карибский тюлень-монах — М. tropicalis Gray, 1850 (ранее встречался по берегам мелких островов у восточного Гондураса, Юкатана, Вест-Индии, Багамских о-вов и Флориды; в настоящее время, если еще не полностью уничтожен, то сохранился в небольшом количестве; последнее сообщение об этом виде известно из Ямайских вод и относится к 1943 г.);
гавайский тюлень-монах — М. schauinslandi Matschie, 1905 (держится у небольших изолированных островов и рифов западной половины Гавайских о-вов между 20 и 30° с. ш.).
Численность везде очень мала и определяется для тюленя-монаха разными авторами в 500—5000 голов, а для гавайского тюленя-монаха 700—1000 голов. В наших водах (Черное море) тюлень-монах встречался до конца прошлого столетия единичными особями и небольшими группами у юго-западного побережья Крыма. В настоящее время небольшое число их обитает в Черном море у побережья Турции, Болгарии н Румынии.
Необходимы решительные меры для предотвращения полного исчезновения тюленей-монахов. Все три вида включены в «Красную книгу»: тюлень-монах и карибский тюлень-монах как виды, находящиеся под угрозой исчезновения (последний, возможно, уже не существует), а га-, вайский тюлень-монах как вид, который в ближайшее время может оказаться на грани исчезновения.
Род крабоедов
Genus Lobodon Gray, 1844
В роде единственный вид: крабоед — L. carcinophagus Hombron et Jacquinot, 1842.
Длина тела 2,3—2,6 м, масса 200—250 кг. Самки немного крупнее самцов. Взрослый крабоед имеет серебристо-буровато-серую спину и светлое брюхо, на боках и плечах различные по величине бурые отметины. Мех быстро выцветает, особенно летом, приобретая кремово-белую окраску (отсюда произошло другое название крабоеда — «белый тюлень»). В январе белый мех линяет и заменяется новым, темным. С возрастом волосяной покров светлеет.
2 1 5
Зубная формула: i — с — р + m — - =32.
Зубы с характерными выступами (см. рис. 133,В), которые, возможно, помогают отфильтровывать при питании планктон — основную пищу крабоеда.
Распространение циркумполярное и приурочено к дрейфующим льдам антарктических морей. Осенью вместе со льдами тюлени могут передвигаться к северу до 55—65° ю. ш. Зиму, весну и часть лета они обычно проводят в северной части ареала. Летом мигрируют в антарктические прибрежные воды — к западу от Земли Грейама и в южную часть моря Росса.
Залежки обычно устраивает на льду или на снегу и лишь изредка на скалах. Зимой отмечались залежки до 3 тыс. голов. На суше может передвигаться быстрее других тюленей — до 24 км/ч. Питается главным образом пелагическими ракообразными, преимущественно Euphausia superba, реже мелкой рыбой. Кормится в основном в темное время суток во время подъема рачков в верхние слон воды.
Детеныши рождаются ранней весной, в середине сентября — начале ноября на паковом льду. Новорожденный имеет в длину примерно 135 см и покрыт мягким, пушистым мехом серовато-коричневого цвега. Продолжительность лактации по разным данным от 2 до 5 недель. После ее окончания детеныш линяет. Спаривание приходится на середину лета. Длительность беременности составляет около 9 месяцев. Линька взрослых крабоедов длится с января по март. Самки достигают половозрелос-ти в 2-летнем возрасте. Продолжительность жизни до 29 лет.
Естественный враг крабоедов — косатка. 4
Крабоед — один из наиболее обычных и многочисленных представителей отряда ластоногих. Общая численность достигает 2—5 млн. Специального промысла не ведется.
Род тюленей Росса
Genus Ommatophoca Gray, 1844
В роде единственный вид: тюлень Росса — О. rossi Gray, 1844.
Длина тела достигает 2,8 м, масса 230—250 кг. Самки, по-вндимому, немного мельче самцов. Шея короткая н толстая, с большими складками кожи. Голова укороченная и может втягиваться в жировые складки шеи. Глаза очень большие, ротовая шель относительно невелика. Грудные ласты длинные. Волосяной покров низкий и грубый. Окраска его на спин
ной стороне тела темно-серая, wa брюшной беловатая с косыми светлосерыми полосами по бокам.
2 1 5
Зубная формула: i —— с —— р + m —- = 32.
Л 1 о
Зубы относительно мелкие. Резцы и клыки тонкие, острые.
Встречаются тюлени Росса по кромке льда в антарктических водах. В секторе, обращенном в сторону Индийского океана, находок тюленя Росса не было.
Экология изучена плохо. Держатся поодиночке или небольшими группами. Плавание отличается большой скоростью и высокой степенью маневренности. Пищей служат в основном головоногие моллюски, в меньшей степени рыбы. О размножении ничего не известно. В начале декабря встречали новорожденных детенышей. Линька происходит в январе — феврале; в это время тюлени не питаются. Половозрелость у самок наступает в 3—4-летнем возрасте, у самцов несколько позже. Продолжительность жизни около 12 лет.
Общая численность тюленя Росса составляет 20—50 тыс. голов.
Род морских леопардов
Genus Hydrurga Gistel, 1848
В роде единственный вид: морской леопард-—Н. leptonyx Blamville, 1820.
Длина тела 2,5—3,5 м, масса до 380 кг. Самки в среднем на 10% крупнее самцов. Бросается в глаза непропорционально крупная голова с большой пастью (рис. 133). Существует 2 основных типа окраски: темно-серая или черная на спинной стороне тела и светло-серая на брюшной, и, соответственно, светло-серая сверху и снизу с темными пятнами. Подпуши нет.
2 1 5
Зубная формула: i — с — р + m — = 32. Я 1 и
Зубы приспособлены к плотоядному характеру питания. Высокие коронки коренных зубов имеют по три острые вершины, загнутые назад.
Распространение охватывает моря вокруг Антарктиды. В летнее время морские леопарды держатся у паковых льдов, а зимой поднимаются к северу к свободным от льда островам. Есть предположение, что некоторые звери проводят во льдах круглый год.
Морской леопард — единственный представитель семейства, постоянно питающийся теплокровными животными. Он нападает на пингвинов, а также на других птиц, молодых ластоногих и рыб. Птиц морской леопард ловит, подплывая к ним под водой. На море обычно держатся поодиночке, но на берегу иногда собираются группами до 250 особей. Самка приносит детеныша в ноябре — январе на паковом льду (отмечены случаи щенки и на берегу в Южной Георгии). Новорожденный имеет длину тела около 150 см, массу 27 кг. Волосяной покров его густой, мягкий. Окраска его темно-серая на спине и почти белая на брюхе с неправильными черными пятнами. Полагают, что лактация продолжается до 2 месяцев. Половозрелости самки достигают в 3 года, а самцы в 4 года. Спаривание, а затем линька приходится у большинства особей на февраль.
Естественный враг, вероятно, косатка.
Общая численность морских леопардов составляет 100—300 тыс. голов. Специального промысла не ведется.
Рис. 133. Морской леопард — //, leptonyx (А), его зубы (Б) и голова и зубы крабоеда— L. carcinophagus (В)
Род тюленей Уэдделла
Genus Leptonychotes Gill, 1872
В роде единственный вид: тюлень Уэдделла — L. weddelli Lesson, 1826.
Длина самки достигает примерно 3,3 м, масса 426 кг. Самец имеет меньшие размеры: длину тела до 2,9 м. Окраска спинной стороны тела темно-серая, часто почти черная, брюшной — светлее, с желтовато-белыми пятнами (рис. 134). Летом мех выцветает, становясь серовато-бурым. Молодые звери светло-серые с более светлой слабопятнистой брюшной стороной. Сосков одна пара.
_ „ 2 13 2
Зубная формула: 1 — с — р — m — = 32.
Верхние резпы не одинаковы по размерам: наружный примерно в 4 раза крупнее внутреннего. Щечные зубы небольшие, с одной вершиной умеренной величины.
Хромосом в диплоцдном наборе 34.
Рис. 134. Тюлень Уэдделла — L. weddelli И его череп
Распространен тюлень Уэдделла в антарктических водах циркумполярно, встречаясь южнее всех остальных антарктических тюленей. Наиболее северные находки тюленей Уэдделла относятся к о-вам Фолклендским и Кергелен, Южной Австралии и Новой Зеландии.
Тюлени Уэдделла приурочены к берегам Антарктического материка и прилежащим островам и лишь изредка встречаются на дрейфующих льдах. Ведут оседлый образ жизни. Местные кочевки могут быть связаны с изменением ледовой обстановки. Зимой большую часть времени проводят в воде, постоянно поддерживая открытыми лазки во льду, обгрызая клыками нарастающий лед. Ориентация и отыскание лазок полярной ночью во время плавания подо льдом является серьезной навигационной проблемой для этих тюленей. Объектами питания служат главным образом мелкие рыбы, а также головоногие моллюски, ракообразные и голотурии. Тюлень Уэдделла, по-видимому, обладает способностью иметь относительно больший запас кислорода, чем другие представители семейства. Это обусловливает, в частности, возможность нырять на глубину до 600 м. Держатся поодиночке или небольшими группами до 40 голов (в период размножения). Беременность длится от 9 до 10 месяцев. Детеныши рождаются прямо на льду в сентябре-октябре. Новорожденный достигает почти половины длины матери и массы около 29 кг. Волосяной покров его густой, мягкий, сероватой окраски. В 2-недельном возрасте детеныши начинают линять, и примерно через 3—4 недели молодые имеют волосяной покров, сходный с волосяным покровом взрослых тюленей. Продолжительность лактации 6—7 недель. Самки остаются на льду в течение недели до родов и неделю после них. Взрослые звери линяют летом — с декабря по март, причем во время линьки плавают и питаются. Половозрелости самки достигают в 3-летнем возрасте. Самцы способны к оплодотворению с 6—8 лет. Длительность жизни до 16 лет.
Естественных врагов тюлени Уэдделла, по-видимому, не имеют, так как обычный враг ластоногих — косатка, не подплывает близко к берегу, где держатся эти тюлени. Численность тюленей Уэдделла определяют в 200—500 тыс. голов (Scheffer, 1958).
Род хохлачей
Genus Cystophora Nilsson, 1820
В роде единственный вид: хохлач — С. cristata Erxleben, 1777.
Длина тела самца 2,2—2,4 до 3,0 м, масса от 260—300 до 407 кг. Размеры самки несколько меньше. Взрослые самцы обладают своеобразным носовым мешком, который начинается позади глаз и тянется вперед, представляя собой разрастание носовой полости, разделенное продольной перегородкой (рис. 135). В возбужденном, а иногда и в спокойном состоянии хохлач раздувает носовой мешок, так что вся верхняя часть •лицевого отдела головы вздувается в виде нароста, который может достигать 25 см в высоту, 30 см в длину и 18 см в поперечнике. В неразду-'Ом состоянии мешок выглядит морщинистым снаружи, передний конец J0 свисает свободно перед ртом. Характерны очень крупные ноздри, охлач имеет также своеобразный дополнительный перепончатый мешок, образованный эластичной межносовой перегородкой, который выдается из левой ноздри, если животное надувает его воздухом. Этот мешок может достигать размера страусиного яйца. Функция его неясна, •эксперименты, проведенные на мертвом животном, показали, что хохлач, по-видимому, сначала наполняет желудок и пищевод воздухом, которым затем при закрытых ноздрях заполняет носовой мешок. При сокра-
Рис. 135. Хохлач — С. cristata (А—самец, Б — самка) и его череп
шении мускулатуры в стенках мешка воздух переходит в дополнительный носовой мешок. Окраска волосяного покрова на теле самца обычно сверху голубоватая или темно-серая, а по бокам и снизу более светлая. Как правило, имеются темные или светлые разных размеров и очертаний пятна, наиболее крупные в средней части тела. Морда черноватая. Окраска самок светлее, и пятна выражены менее резко. Сосков одна пара.
Череп очень массивный, с широко расставленными скуловыми дугами. Носовые кости сильно расширены спереди, где они образуют крупный выступ, и клиновидно сужены сзади. Задний край костного неба
имеет вид двух дуг, сходящихся посредине на конце межнебного шва, где имеется небольшой выступ. Костные слуховые барабаны имеют трапециевидную форму. Носовые отростки межчелюстных костей далеко не доходят до задне-верхнего края наружного костного носового отверстия.
Зубная формула:
2 1 4
2 1
30.
— с — р — m
1 1 4
Корни верхних внутренних резцов уплощены с боков. На коронках несточенных коренных и некоторых предкоренных зубов имеется по очень маленькому зубчику спереди и сзади от основной слабо выраженной вершины. Продольная ширина коронки щечных зубов больше ее высоты. Корни щечных зубов часто срастаются между собой. Большинство щечных зубов имеет по одному корню, коренной и четвертый предкорен-ной — по 2 корня.
Хромосом в диплоидном наборе 34.
Распространение связано с паковым льдом Субарктики Атлантического и Северного Ледовитого океанов. Отдельные странствующие особи отмечены у Юкона, Флориды и Португалии. В воды СССР заходит в северные районы Белого моря. Однажды отмечен у архипелага Земли Франца-Иосифа.
Хохлач — пелагическое животное и обычно встречается на глубоководье у кромки дрейфующих льдов. Питается главным образом рыбой — сельдью, морским окунем, тресковыми, камбалой, также головоногими моллюсками, а возможно и пелагическими ракообразными. За пищей может нырять на значительную глубину. В период лактации и линьки, по-видимому, не питается.
Существует, вероятно, 2 района размножения хохлача: один к северу от о-ва Ян-Майен (размножающиеся здесь весной хохлачи мигрируют затем к северу в сторону Свалбарда) и второй около Ньюфаундленда (после весеннего размножения тюлени мигрируют отсюда к берегам Гренландии и в Датский пролив). Для размножения выбирают прочные многолетние паковые льды со многими разводьями. Большинство детенышей рождается в конце марта — начале апреля. Самка приносит одного детеныша (возможно, в редких случаях 2). Масса новорожденного 11—14 кг, длина около 79—112 см. Эмбриональный высокий и мягкий светлый волосяной покров сбрасывается еще в утробе матери. Новорожденный имеет короткий и довольно грубый волосяной покров — пепельный или сизо-серый на спине и белый на брюхе. Пятна отсутствуют; они появляются на втором году жизни. Спустя 2—3 недели после рождения, когда заканчивается лактационный период, матери покидают детенышей, которые, однако, остаются на льду еще в течение примерно двух недель.
В.период размножения держатся семейными группами: самец, самка и детеныш. Период спаривания приходится на конец лактационного периода и продолжается примерно 2 недели. Беременность около 11 месяцев. Имплантация бластоцисты задерживается на 3—4,5 месяца. Линька происходит в июне — первой половине июля, главным образом в Датском проливе. Половозрелости самки достигают примерно в 3—4-летнем возрасте, а самцы — в 5—6 лет. Самцы и самки держатся отдельными стадами, за исключением периода спаривания.
Численность хохлачей определяют в 300—500 тыс. Естественные враги— полярная акула, косатка и белый медведь. Продолжительность жизни примерно 35 лет.
Промысловое значение имеют главным образом новорожденные, шкурки которых служат ценным меховым сырьем. От взрослых зверей получают подкожный жир и шкуру, которая используется в качестве
кожевенного сырья. Норвегия и Канада с 1960 по 1967 г. ежегодно добывали от 28 до 65 тыс. хохлачей. Зверобойные суда СССР вели промысел хохлачей с 1955 по 1966 г. в районе о-ва Ян-Майен и добыли за этот срок около 25 тыс. хохлачей, в основном детенышей. Численность хохлача в результате промысла уменьшается. В последние годы введена некоторая регуляция промысла, например запрещена добыча на линных залежках.
Род морских слонов
Genus Mirounga Gray, 1827
Наиболее крупные представители семейства. Самцы имеют длину тела 4,5—6,5 м и массу до 3,5 т. Самки значительно меньше: длина 3— 3,5 м, масса до 900 кг. Взрослые особи имеют одноцветную желтоватую, серовато-коричневую или голубовато-серую окраску. Брюхо светлее спины. Самки темнее, чем самцы. Волосяной покров низкий, грубый, без пуха. Конец морды удлинен в хобот, достигающий 35 см длины (рис. 136). Он направлен вниз, свисая спереди перед ротовым отверстием. В период размножения хобот может подниматься немного кверху, частично благодаря действию мышц, а частично за счет наполнения его сосудов кровью. Он действует как резонирующая камера, и рев крупных самцов слышен благодаря этому за несколько километров. Хобот начинает развиваться у двухлетних животных. Шея у самцов имеет утолщенную кожу, защищающую от укусов соперника во время драк в период размножения, и обычно покрыта многочисленными шрамами.
4 1
гга т=30-
2 1
Зубная формула: i — с -у- р
Хромосом в диплоидном наборе 34.
Распространены в субантарктических Тихоокеанскому побережью Северной Америки от Калифорнии на север до о-ва Ванкувер.
Питаются рыбами и головоногими моллюсками. За добычей могут нырять на глубину до нескольких сот метров. Кормятся зимой и в течение нескольких месяцев летом. Во время размножения и линьки голодают. Самки северных морских слонов во время лактационного периода питаются в море. Размножение и линька происходят на земле или на льду. Морские слоны образуют крупные залежки на берегах островов. Для них характерна полигамия. Самец имеет гарем, состоящий у южных морских слонов из 10—30, иногда до 100 самок, а у северных—10— 12 самок. Самцы достигают половозрелости в 4 года, но образуют гаремы не ранее 6—8 лет. Период деторождения у северных морских слонов приходится на декабрь—-январь, а у южных —на октябрь. Самки начинают спариваться с 2-летнего возраста. Беременность длится примерно 350 дней. Новорожденный имеет длину тела около 1,2 м и массу примерно 50 кг. Он покрыт коричневато-черным пушистым мехом. Смертность детенышей высока — составляет около 50%. Приблизительно в 10-дневном возрасте детеныш теряет эмбриональный волосяной покров. Линька заканчивается в возрасте 34 дней. Когда спустя 3 недели он переходит к самостоятельному образу жизни, его масса составляет примерно 175 кг. К этому времени самка вновь спаривается. Самка приносит детенышей ежегодно в течение 7 лет. Линька взрослых животных приходится на декабрь — февраль у южных морских слонов и на май — июль у северных; продолжается она 30—40 дней.
водах циркумполярно и по
Рис. 136. Южный морской слои — М. leonina и его череп
После размножения и линьки южные морские слоны уходят в море и, по-видимому, проводят зиму на краю пакового льда. Северные морские слоны ведут оседлый образ жизни. Продолжительность жизни самок около 12 лет. Самцы могут жить дольше — до 20 лет, но вследствие промысла их предельный возраст также обычно не превышает 12 лет. Отмечены случаи поедания морских слонов косатками. На детенышей морских слонов нападает морской леопард.
В роде 2 вида:
южный морской слон — М. leonina Linnaeus, 1758 (субантарктические воды циркумполярно на север до 16° ю. ш. и на юг до антарктических паковых льдов—-78° ю. ш.; размножается у Пунта-Норте и Огненной Земли в Аргентине и на о-вах Фолклендских, Южных Шетландских, Южных Оркнейских, Южной Георгии, Южных Сандвичевых, Гоф, Марион, Принца Эдуарда, Крозе, Кергелен, Херд, Макуори, Окленд, Кемпбелл);
северный морской слон — М. angustirostris Gill, 1866 (острова у берегов Мексики и Калифорнии на север до о-вов Ванкувер и Принца Уэльса; размножается на о-вах Сан-Николас, Сан-Мигуэль, Гуадалупе и Сан-Бенито).
Северный морской слон еще недавно был близок к уничтожению чрезмерным промыслом, но в последнее время благодаря запрещению промысла его численность значительно возросла и продолжает увеличиваться.
Общее число южных морских слонов определяется в 600—700 тыс. голов, а северных — всего в 10—15 тыс. голов.
Южных морских слонов промышляют на береговых залежках, причем существуют ограничения промысла по сезонам, размерам добываемых тюленей не менее 3,5 м длиной и их числу. Например, в 1951 г. было разрешено добыть 8 тыс. морских слонов; добыли 7877. От добытых зверей получают жир и кожу.
ОТРЯД ТРУБКОЗУБЫХ
ORDO TUBULIDENTATA HUXLEY, 1872
Длина тела 100—158 см, длина хвоста 44—61 см, высота в плечах СО—65 см. Масса взрослого животного обычно 50—70 кг, редко до £2 кг.
Тело массивное, с удлиненной головой, заканчивающейся пятачком, напоминающим пятачок свиньи (рис. 137). Длинный язык может высовываться из рта на 30 см. Ноздри округлые, способные смыкаться. Уши длинные (15—21 см), трубчатые. Они загибаются назад, когда животное копает нору, и могут двигаться независимо одно от другого. Шея короткая. Спина дугообразно выгнута. Сильный мускулистый хвост утолщен v основания (около 40 см в окружности) и утончается к концу. Конечности короткие и сильные, пальцеходящие. Передние конечности короче задних, четырехпалые (отсутствует I палец), задние — пятипалые. Пальцы сращены между собой у основания. Длинные, прямые, сильные, но тупые когти, напоминающие одновременно ногти и копыта, приспособлены для копания земли. Толстая розовато-серая кожа покрыта редким грубым волосяным покровом. Окраска кожи варьирует от тускло-коричневато-серой до желтовато-серой. В пахах расположены специфические кожные железы, продуцирующие желтоватый пахучий секрет. На конце морды и около глаз располагается большое число вибрисс. Два соска находятся в брюшной области и два — в паховой.
Череп сильно вытянут в длину. Предчелюстные кости небольшие. Скуловая дуга тонкая, но полная. Нижняя челюсть прямая, пластиновидная с длинным симфизом.
Зубная формула обычно: i - с
0 2
3
т=20-
— р — m 0 2
Иногда в небе находятся непрорезавшиеся маленькие клыки.
Зубная система гетеродонтная, дифиодонтная. Многочисленные зубы эмбрионов пронизаны параллельно расположенными вертикальными каналами, заполненными пульпой. Молочные зубы не прорезаются. У взрослых зубы имеются только в задней части челюстей. Зубы не имеют эмали и покрыты снаружи цементом. Каждый зуб состоит из большого числа трубочек, содержащих пульпу и окруженных гексагональными призмами дентина. Второй коренной зуб имеет около 1500 таких призм. Зубы лишены корней. Вершины предкоренпых зубов конические, а коренных — битуберкулярные.
Характерны большие ключицы, что связано с копательными функциями передних конечностей. Плечевая кость короткая, сильная. Локтевая кость несколько сплющена, локтевой отросток большой. Лучевая кость короткая, сильная. Бедренная кость массивная. Малая и большая
Рис. 137. Трубкозуб—О. afer и его череп берцовые кости срастаются в проксимальных отделах. Таз массивный, седалищная кость не соединяется с позвонками.
Мозг с большими обонятельными долями: большие полушария бедны извилинами. Желудок простой, крупный. Желчный пузырь имеется. Слепая кишка большая. Слюнные железы сильно развиты. Матка двойная, плацента зональная. Семенники располагаются в пахах под кожей.
Ископаемые остатки известны из плиоцена Европы (Франция, Греция) и Азии (Ирак, Индия), миоцена Африки, плейстоцена Мадагаскара. Они принадлежат к современному роду Orycteropus (или близким ему формам), поэтому дают мало сведений о происхождении всей группы. Сравнительноанатомические исследования современных форм трубкозубых обнаруживают сходство их с примитивными копытными.
Распространены по большей части Африки к югу от Сенегала, Центрального Судана и Эфиопии до мыса Доброй Надежды. Во многих мес-
тах своего прежнего ареала в настоящее время истреблены человеком.
Встречаются в различных ландшафтах, предпочитая, однако, поросшие кустарником степи; густых лесов избегают. В горы поднимаются до 2000 м над уровнем моря. Ведут одиночный образ жизни. Активны ночью. Придерживаются индивидуальных участков. День проводят в норе, которую способны выкопать с поразительной быстротой. Норы длиной около 2—3 м, диаметром примерно 40 см, уходят на глубину 1,2—1,5 м и обычно заканчиваются гнездовой камерой, выстланной сухими листьями. В поисках пищи трубкозуб может проходить за ночь 10—12, а то и 30 км. Питается муравьями (Formicidae) и термитами (преимущественно Macroterm.es). Благодаря толстой коже не чувствителен к их укусам. Самка приносит в течение года одного, редко двух детенышей
Рис. 137. Продолжение
(в Эфиопии в мае — июне, в Центральной Африке в октябре — ноябре). В возрасте двух недель детеныш начинает сопровождать свою мать в ночных походах. Полугодовалый зверь вырывает собственную нору, но продолжает ходить с матерью. В течение следующего сезона размножения детеныш покидает мать, если он самец, и остается с ней, если это самка. Поэтому взрослую самку часто можно видеть с двумя разновозрастными детенышами. В последние годы численность сократилась. В неволе жили до 10 лет.
Отряд включает одно семейство: трубкозубовых — Orycteropodidae Bonoparte, 1850 (1 род).
СЕМЕЙСТВО ТРУБКОЗУБОВЫХ
FAMILIA ORYCTEROPODIDAE BONOPARTE, 1850
См. описание отряда.
В семействе один род трубкозубов — Orycteropus Geoffroy, 1795 (и один вид).
Род трубкозубов
Genus Orycteropus Е. Geoffroy, 1795
В роде единственный вид: трубкозуб — О. ajer Pallas, 1766.
См. описание отряда.
ОТРЯД ХОБОТНЫХ
ORDO PROBOSCIDEA ILLIOER, 1811
Размеры наибольшие среди современных наземных млекопитающих. Самцы крупнее самок. Наиболее крупные представители отряда достигают длины тела 7,5 м, высоты 4 м, массы 7,5 т. Тело массивное, удлиненное (рис. 138). Шея короткая. Голова огромная с большими веерообразными ушами, маленькими глазами (с мигательной перепонкой) и длинным мускулистым хоботом, на конце которого располагаются ноздри. Хобот образован вытянутыми носом и верхней губой: он представляет собой полое внутри мускулистое образование, разделенное по всей длине продольной перегородкой. На конце хобота имеются только дорзальные или дорзальный и вентральный хватательные пальцевидные отростки. Функция хобота многообразна. Он служит для дыхания, обоняния, осязания, помогает при питье и еде. Хоботом слон срывает траву, ветви деревьев, плоды и отправляет их в рот, всасывает в хобот воду и затем впрыскивает в рот. Конечности высокие, колоннообразные, пятипалые. Пальцы покрыты общим кожным покровом, но заметны снаружи. На передних конечностях 5, иногда 4 копытца, на задних — 3 или 4.
Кожа слона сероватой окраски, имеет значительную толщину. Ее наружная поверхность неровная, покрыта эпидермальными буграми различной толщины. Эпидермис имеет ячеистую внутреннюю поверхность. В ячеи поднимаются дермальные сосочки. Сальных и потовых желез нет. Волосы у взрослых редкие, щетинковидные. У новорожденных волосяной покров довольно густой. В височной области находится кожная специфическая железа, продуцирующая в период течки обильный секрет жидкой консистенции с неприятным запахом. Небольшое (около 4 мм в диаметре) выводное отверстие этой железы помешается на расстоянии около 20 см кзади от наружного угла глаза. Выход секрета из резервуара железы осуществляется сокращением гл. glandulae temporalis.
Сосков одна пара — в области груди, между передними ногами.
Череп слона огромных размеров, но несколько укороченный. В связи с развитием хобота и бивней увеличивается весь передний отдел черепа за счет утолщения костей и приобретения ими пневматично-губчатой структуры, которая особенно сильно развита в крыше черепа. Скуловые кости маленькие, расположены в самой середине скуловой дуги.
„ 1 0 3 3
Зубная формула: 1 — с — р — m — = 26.
В постоянной смене зубов сохраняются только верхние третьи резцы. Они обладают постоянным ростом и превратились в большие бивни, которые у самок азиатских слонов, как правило, отсутствуют. Бивни образованы сплошным дентином, а их концы покрыты эмалью, стираю-312
Рис. 138. Индийский слон — Е. tnaximus
щейся с возрастом. Щечные зубы представлены вторым, третьим и четвертым предкоренными и первым, вторым и третьим коренными зубами. Из щечных зубов одновременно функционирует всего по одному зубу в каждой половине челюсти. По мере изнашивания сработанные зубы замещаются новыми, которые прорезаются сзади и продвигаются вперед на место своих стершихся и выпавших (часто по кускам) предше-2
ственников. У индийского слона р сменяются на втором году жизни, 3 4 1
Рз---на пятом, р — —на девятом, m — начинают функционировать
только в возрасте 15 лет. Полная серия щечных зубов бывает израсходована к 50—60 годам (существуют индивидуальные колебания). Щечные зубы массивные, удлиненные спереди назад, с высокой коронкой (гипселодонтные) и сложной структурой, полилофодонтного типа. Каждый зуб состоит из большого числа поперечных пластин дентина, покрытых эмалью. Пространство между пластинками заполнено цементом.
Кости скелета массивные, составляют 12—15% общей массы тела. Кости предплечья и голени разделены. Фаланги пальцев уменьшены в размерах. Ключица отсутствует.
Мозг по массе наибольший среди наземных млекопитающих. Однако большие полушария относительно невелики и не закрывают мозжечка. Желудок простой, слепая кишка большая, толстая кишка длинная — превышает половину длины тонкой кишки. Желчного пузыря нет. Дыхание почти исключительно диафрагмальное. Семенники находятся в брюшной полости. Матка двурогая. Плацента смешанного типа: часть ее диффузная и неотпадающая, а часть зональная, котилодонтная и отпадающая.
Распространены в Южной Азии и Африке.
Населяют леса и саванны, иногда высокотравье. Обычно не уходят
далеко от воды. Самки, детеныши и молодые самцы образуют стада д0 30—400 голов. Взрослые самцы держатся обычно поодиночке, временами присоединяясь к стадам. Размеры стада зависят от наличия пищи воды и фактора беспокойства. Активны в светлое время суток; в жаркие часы отдыхают. Питаются исключительно растениями, включая листья плоды, кору, корни. Имеют место кормовые миграции. Обычно передвигаются шагом и бегать могут только на короткие дистанции. Хорошо плавают. Слух развит хорошо, обоняние превосходно, зрение относительно слабое. Хорошо представлена звуковая коммуникация.
Беременность от 20 до 22 месяцев. Самка приносит одного, редко двух детенышей. Масса новорожденного около 100 кг. Вскоре после рождения детеныш следует за матерью. Молоко сосет ртом. Лактация длится около двух лет. Половозрелость наступает примерно на 9—-20-й год. Продолжительность жизни обычно 50—60 лет.
Слоны были предметом интенсивного промысла из-за ценимых на рынке бивней. В результате прямого уничтожения и косвенного воздействия деятельности человека численность резко упала и, как правило, в настоящее время слоны многочисленны лишь на охраняемых территориях. Азиатские слоны издавна использовались в качестве рабочих животных.
Хоботные, по-видимому, имели общих предков с сиренами и даманами. Но уже с палеоцена каждая из этих групп развивалась самостоятельно.
Наиболее древние находки хоботных известны из эоцена Северной Африки (АРЕ). Они принадлежат к роду Moeritherium Andrews, 1901, семейства Moeritheriidae Andrews, 1906, подотряда Moeritherioidae Osborn, 1921. Это были животные размером с крупную свинью, без хобота и настоящих бивней. Щечные зубы отличались от щечных зубов современных слонов, но пара резцов была удлинена.
Древнейший представитель подотряда Mastodontoidae Osborn, 1921,— Palaeomastodon Andrews, 1900, из нижнего олигоцена АРЕ, хотя и сильно отличался по строению черепа и зубов от современных хоботных, был первым настоящим слоном. В его верхней и нижней челюстях з
было всего по одной паре резцов (у Moeritherium i у ). Эти резцы были настоящими бивнями. Бивни верхней челюсти более поздних мастодонтов из верхнего третичного периода (Mastodon Cuvier, 1817; Zy-golophodon Vacek, 1877; Stegamastodon Pohlig, 1912) становятся силь-. нее и длиннее, а бивни нижней челюсти (и сама нижняя челюсть) постепенно укорачиваются, усложняются щечные зубы.
Представители подотряда Elephantoidea Osborn, 1921 (сюда относятся и современные слоны) появляются в плейстоцене, например род Archidiskodon Pohlig, 1885 (Северная и Южная Африка и от Южной Англии до Индии).
Мамонты — род Mammanthus Burnett, 1830, были широко распространены в плейстоцене Европы, Азии и Северной Америки. Представители родов Elephas и Loxodonta известны с плейстоцена.
В отряде 1 семейство слоновых — Elephantidae Gray, 1821, и 2 рода.
СЕМЕЙСТВО СЛОНОВЫХ
FAMILIA ELEPHANTIDAE GRAY, 1821
См. описание отряда.
В семействе 2 рода (2 вида):
индийских слонов — Elephas Linnaeus, 1758 (1 вид);
африканских слонов — Loxodonta F. Cuvier, 1827 (1 вид).
Род индийских слонов
Genus Elephas Linnaeus, 1758
В роде единственный вид: индийский слон — Е. maxitnus Linnaeus, 1758 (рис. 138).
Размеры наименьшие в семействе. Длина тела 5,5—о,4 м, высота в плечах 2,5—3 м, длина хвоста 1,2—1,5 м. Масса около 5 т. Наиболее высокая точка тела — вершина головы. Бивни имеются, как правило, только у самцов; они достигают 1,5 м и массы 20—25 кг. Размеры ушей значительно меньше, чем у африканского слона. На конце хобота один дорзальный отросток. На передних конечностях 5 копытец, на задних — 4. Окраска тела от темно-серой до бурой.
Поперечных пластин дентина в каждом щечном зубе от 6 до 27 (больше, чем у африканского слона). Ребер 19 пар. Хвостовых позвонков 33. Сердце часто имеет двойную вершину.
Распространен в Индии, Бирме, Непале, Бутане, Таиланде, Кампучии, Лаосе, Вьетнаме, Малайзии, о-вах Калимантан, Суматра, Шри Ланка.
Населяет различные ландшафты от густых лесов до высокотравных равнин. Держатся стадами в 15—30 особей; обычно предводительствует старая самка. Пища — преимущественно трава, листья, молодые побеги, плоды. Сезонности в размножении нет. Беременность 20—22 месяца. Самка приносит 1—2 детенышей. Новорожденный имеет высоту в плечах примерно 1 м, массу 90 кг. Половозрелость наступает в 9— 12 лет. В течение жизни самка дает в среднем 4 помета.
Индийский слон используется как домашнее животное для различных тяжелых работ, транспортировки и охоты.
Прямое истребление индийских слонов и в большей степени изменение условий существования резко снизили их численность и сократили ареал. Ареал превратился в ряд отдаленных друг от друга участков, приуроченных к глухим или охраняемым местам. Общее число диких индийских слонов в Индии составляет 7—8 тыс., включая 2 тыс. в Ассаме, 4 тыс. в Мисоре, Тамилнаду и Каррака, 800 в Бихаре, Ориссе и Западной Бенгалии и 400 в Уттар-Прадеш. В Таиланде их примерно 500, на Суматре около 1500, п-ове Малакка 500, Северном Калимантане 2 тыс., о-ве Шри Ланка 3 тыс.
Индийский слон включен в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения. •
Род африканских слонов
Genus Loxodonta F. Cuvier, 1827
В роде единственный вид: африканский слон — L. africana Blumen-bach, 1797 (рис. 139).
Крупнее азиатского слона. Длина тела достигает 6—7,5 м, высота в плечах до 4 м, длина хвоста 1—1,3 м. Средняя масса самки 3 т, самца 5 т (до 7,5 т). Наиболее высокая точка тела — область плеч. Бивни крупнее у самцов и значительно меньших размеров у самок — до 18 кг. Наиболее крупный из известных бивней достигал в длину 3,5 м и массы 107 кг. Уши большие: от основания до вершины иногда достигают 1,5 м. На конце хобота есть дорзальный и вентральный отростки. На передних конечностях копыт 5, реже 4, на задних — 3. Окраска тела коричневатосерая.
.415
Рис. 139. Африканский слон — L. africana
Поперечных пластин дентина в каждом щечном зубе от 5 до 14 — меньше, чем у индийского слона. Ребер 21 пара — больше, чем у индийского слона. Хвостовых позвонков меньше — максимум 26.
Распространен по всей территории Африки к югу от Сахары. Однако почти непрерывный в прошлом ареал в настоящее время разорван. Африканский слон не встречается на большей части Южной Африки, Намибии, Ботсваны, Эфиопии; полностью исчез из Северного Сомали. От Судана на запад граница ареала теперь примерно совпадает с 12° с. ш., однако отдельные изолированные участки ареала сохраняются и севернее (у оз. Чад, в Мали, Мавритании).
Населяют разнообразные ландшафты (за исключением полупустынь и пустынь )от уровня моря до 3660 м над его уровнем (в виде исключения, даже до 4570 м). Держится стадами, которые в прежнее время достигали 400 голов. Питается преимущественно ветвями, побегами, корой и корнями деревьев и кустарников. Размножение с определенным сезоном не связано. Беременность длится примерно 22 месяца. Самка обычно приносит раз в 4 года одного детеныша. Масса новорожденного около 100 кг, высота в плечах примерно 1 м. Половозрелость наступает в 12—20 лет. Продолжительность жизни 60—70 лет.
Африканские слоны хорошо одомашниваются. В походе Ганнибала на Рим участвовали африканские слоны. В начале XX в. в Заире успешно проводилось одомашнивание африканских слонов. Практического применения домашние африканские слоны не получили.
До сих пор ведется (теперь лицензионный) отстрел африканских слонов. В прежние годы их промысел из-за ценных бивней был сильно развит: в конце XIX столетия ежегодно убивали 60—70 тыс. африканских слонов. Такой интенсивный промысел резко снизил численность слонов и в 1920—1928 гг. ежегодно добывали всего по 6 тыс. Меры по охране восстановили численность (сейчас их насчитывают около 350 тыс. голов) и в ряде мест привели даже к перенаселению и подрыву кормовой базы (например, в Национальном парке Тсаво в Кении).
ОТРЯД ДАМАНОВ
ORDO HYRACOIDEA HUXLEY, 1869
Млекопитающие мелких размеров: длина тела 30—60 см. Хвост или отсутствует, или имеет длину 1—3 см. Масса взрослого животного до 3 кг. По внешнему виду напоминает бесхвостого сурка или крупную пищуху с выгнутой дугообразно спиной и туловищем, сжатым с боков (рис. 140). Морда укорочена. Верхняя губа раздвоена посредине. Уши небольшие. Глаза относительно крупные. Конечности короткие, сильные, стопоходящие. Передние конечности четырехпалые с уплощенными ногтями, напоминающими копыта. На задних лапах по три пальца (отсутствуют I и V). Внутренний (II) палец с раздвоенной терминальной фалангой имеет длинный искривленный коготь, который служит для расчесывания волос. Два других пальца с короткими, уплощенными, копытообразными ногтями. Ось конечностей проходит через III палец. Подошвы лап голые, с многочисленными протоками потовых желез, открывающимися на поверхности кожи. Во многих литературных источниках сказано об уникальной способности специальных мышц в подошвах лап втягивать центральную часть свода подошвы при опоре ее на субстрат и создавать таким образом вакуум. При этом лапа «присасывается» к поверхности камня или стволу дерева. Даже застреленный, зверек будто бы остается висеть на одной ноге на совершенно гладкой отвесной скале. Последние исследования, однако, не обнаружили никаких особых мышц в подошвах даманов, а многочисленные наблюдения не подтвердили способности «присасываться» к гладкой отвесной поверхности. Лазание даманов почти по отвесным скалам и стволам деревьев объясняется большой ловкостью этих на первый взгляд несколько неуклюжих зверьков и тем, что крупные подошвы их лап, обладающие относительно большой площадью, покрыты резиноподобным толстым, постоянно увлажненным эпидермисом, обеспечивающим большое сопротивление при трении.
Волосяной покров густой, образован мягким пухом и грубыми остевыми волосами. Окраска его обычно коричнево-серая. По всему телу разбросаны отдельные вибриссы, выступающие над уровнем меха. Особенно многочисленны они на морде и в несколько меньшей степени— на лапах. На середине спины расположена крупная кожная специфическая железа, образованная гипертрофированными сальными и потовыми железами. Она покрыта светлыми волосами, отличающимися по окраске от окружающих волос. У молодых животных железа неразвита или развита слабо; у самцов сильнее, чем у самок. Когда животное раздражено или испугано, волосы на железе поднимаются вертикально, открывая железистое поле. Сосков одна пара грудных и две — паховых или одна пара подмышечных и одна (или две) паховых. Некоторые исследова-
Рис. 140. Капский даман — Р. capensis
тели считают возможным теплоотдачу испарением пота с поверхности подошв, когда зверьки лежат на камнях, подставив их под лучи солнца.
Череп с укороченным лицевым отделом и уплощенной крышей. Глазницы сдвинуты вперед. Заглазничные отростки хорошо выражены к иногда смыкаются со скуловой дугой. Межтеменная кость относительно крупная. Костные слуховые барабаны маленькие. Скуловые дуги сильные.
10 4 3
Зубная формула: i с — р m — - 34.
Зубная система дифиодонтная и гетеродонтная. Длинные полукруглые и остроконечные верхние резцы обладают постоянным ростом и похожи на резцы грызунов. Поперечное сечение резцов треугольное; их внутренняя уплощенная сторона не имеет эмали. Нижние резцы долотообразные. В верхней и нижней челюстях резцы отделены от щечных зубов небольшой диастемой. Коренные зубы имеют по 4 корня и по 2 гребня на жевательной поверхности; они брахиодонтные или гппсело-донтные, лофодонтные. Напоминают коренные зубы носорога.
Позвоночник выгнут дорзально. Два крестцовых позвонка срастаются с подвздошной костью. В запястье развита центральная косточка (примитивный признак). Ключицы нет.
Мозг макросматический. Для глаз характерно, что часть радужной оболочки над зрачком слегка вдается в переднюю камеру и предотвращает попадание на сетчатку лучей света, падающих сверху. В конечностях имеется чудесная сеть артериальных сосудов. Желудок простой. Желчного пузыря нет. Матка двурогая. Плацента сначала диффузная, затем зональная гемихориальная, отпадающая. Семенники в брюшной полости.
Рис. 140. Продолжение. Его череп, кисть (Л) и стопа (Б)
Распространены почти по всей Африке от Южно-Африканской Республики на север до линии, проходящей от Сенегала через Южный Алжир, Ливию и АРЕ, а также на Синайском и Аравийском п-овах, в Сирии и Палестине.
Обитают в горных областях среди скальных обнажений, в засушливых степях и лесах. Поднимаются в горы до 4,5 тыс. м над уровнем моря. Имеются наземные и древесные формы. Очень подвижны, способны быстро бегать и прыгать по крутым скалам и хорошо лазить по деревьям. Наземные формы образуют колонии в 5—60 особей. Активны днем (преимущественно утром и вечером) или ночью. Хорошо развиты зрение и слух. Питаются в основном различными частями растений, иногда поедают насекомых и их личинки. В поисках пищи могут уходить на расстояние до 1—3 км. В питьевой воде не нуждаются. Сезонности в размножении, по-видимому, нет. Беременность 7—7,5 месяцев. Самка приносит 1—3, иногда до 6 детенышей, по-видимому, один раз в год. Детеныши рождаются с открытыми глазами, покрытые волосами, способные быстро бегать.
На основании ископаемых остатков можно считать, что даманы произошли от примитивных Condylathra— первичнокопытных, послуживших основным стволом, от которого в дальнейшем произошли многочисленные копытные (Thenius und Hofer, 1960). Этим объясняются сходные признаки с непарнокопытными и то, что серологические исследования показывают сходство даманов со слонами. В начале плиоцена даманы распространились из Африки в Европу и Азию. Сначала даманы были описаны как самостоятельный род отряда грызунов. Затем их относили к непарнокопытным, а со второй половины XIX столетия выделили в самостоятельный отряд.
В отряде одно семейство дамановых — Procaviidae Thomas, 1892, и 3 рода.
СЕМЕЙСТВО ДАМАНОВЫХ
EAMILIA PROCAVIIDAE THOMAS, 1892
См. описание отряда.
В семействе 3 рода (11 видов):
скалистых даманов — Procavia Storr, 1780 (5 видов);
горных даманов — Heteroxyrax Gray, 1868 (3 вида);
древесных даманов — Dendroxyrax Gray, 1868 (3 вида).
Иногда (Roche, 1972) горных даманов включают в качестве подрода в род Dendroxyrax.
Род скалистых даманов
Genus Procavia Storr, 1780
Длина тела 30—55 см, масса 1,4—2 кг. Хвост снаружи незаметен (см. рис. 140). Волосяной покров относительно низкий и грубый. Окраска его на спинной стороне тела корнчневато-серая, на брюшной — кремовая. Окраска волос спинной железы черная, бледно-желтая или оранжевая. Сосков одна пара грудных и две пары паховых.
Коренные зубы гипселодонтные. Первый нижний предкоренной иногда отсутствует. Длина ряда верхних предкоренных значительно меньше ряда верхних коренных зубов. Размеры верхних щечных зубов увеличиваются спереди назад до второго коренного зуба, самого большого по величине.
Распространение совпадает с распространением отряда.
Населяют полупустыни, степи, саванны и горы. Держатся небольшими группами и колониями до 50 особей. Активны в светлую часть суток, особенно утром и вечером, а иногда и теплыми ночами nphi лунном свете. Убежище находят под камнями, в расщелинах скал или в выкопанных ими норах. Пища преимущественно растительная — трава, мох, листья, побеги кустарников и их кора. Поедают также и животную пищу, особенно саранчовых. Беременность длится примерно 214—232 дня. Детенышей в помете 2, реже 3, иногда до 6 (капский даман).
Б роде 5 видов:
капский даман — Р. capensis Pallas, 1766 (Южная Африка — Южно-Африканская Республика, Намибия, северо-восток Ботсваны и Родезия);
каокефелдский даман — Р. welwitschii Gray, 1868 (Каокефелд в Намибии и юго-западная часть Анголы);
рыжий даман — Р. ruficeps Hemprich et Ehrenberg, 1832 (Гамбия, Сенегал, Дагомея, Нигерия, Южный Алжир, Камерун, Чад, Судан, Центральноафриканская Республика);
даман Джонстона — Р. johnstoni Thomas, 1894 (Малави, Северная Танзания, Руанда, Бурунди, Южная Уганда, Кения, Северо-Восточный Заир);
сирийский даман — Р. syriacus Schreber, 1784 (Кения, Эфиопия, Сомали, Судан, юг АРЕ, Синайский и Аравийский п-ова, Сирия, Палестина) .
Некоторые исследователи (Roche, 1972) считают род скалистых даманов монотипическим и объединяют все указанные выше виды в один — Р. capensis Pallas, 1766.
Род горных даманов
Genus Heteroxyrax Gray, 1868
Длина тела 30—38 см, хвост снаружи неразличим. Масса 0,5—4,5 кг. Волосяной покров плотный, низкий и довольно грубый. Окраска его на спинной стороне тела коричневато-белесая, с темной рябью, на брюшной — белая. Окраска волос спннной железы желтая или не отличается от волос спины.
Сосков одна пара грудных и две пары паховых, иногда соски только паховые.
Коренные зубы брахиодонтные. Увеличение размеров верхних щечных зубов спереди назад выражено слабее, чем у скалистых даманов. Первый нижний предкоренной зуб всегда имеется.
Длина ряда верхних предкоренных зубов немного меньше длины ряда верхних коренных или равна ей.
Распространены в Центральной и Южной Африке.
Населяют преимущественно скальные выходы на равнинах и в горах. Поднимаются в горы до высоты 3800 м над уровнем моря. Живут колониями от нескольких десятков до сотен зверьков. Активны днем, иногда и в лунные ночи. Убежищем служат расщелины скал и пустоты между камнями. Питаются преимущественно корнями и луковицами. Охотно поедают саранчу. Беременность 225 дней. Самка приносит двух детенышей (иногда одного или трех) обычно в феврале—марте.
В роде 3 вида:
ахаггарский даман — Н. antinae Heim de Balsac et Begouen, 1932 (нагорье Ахаггар на юге Алжира, Центральная Сахара);
Рис. 141. Западный даман— D. dorsalis и его череп
даман Брюса — Н. brucei Gray, 1868 (АРЕ, Эфиопия, Сомали, юг Судана, Восточный Заир, Уганда, Кения, Руанда, Бурунди, Танзания, Малави, Замбия, Северный Мозамбик, Ангола, северо-восток Южно-Африканской Республики, северо-восток Ботсваны);
лоадийский даман — И. chapini Hatt, 1933 (холмы Лоади в восточной части Центрального Заира).
Иногда в роде выделяют всего один вид (Roche, 1972).
Род древесных даманов
Genus Dendroxyrax Gray, 1868
Длина тела 40—60 см, длина хвоста 1—3 см. Масса 1,5—2,5 кг. Волосяной покров выше и мягче, чем у представителей других родов (рис. 141). Окраска коричневая. Волосы на спинной железе белые, желтовато-белые или желтовато-коричневые. Под подбородком может быть белое пятно. Сосков, как правило, только одна пара паховых, иногда одна пара подмышечных и одна или две пары паховых.
Коренные зубы брахиодонтные. Как и у горных даманов, увеличение размеров верхних щечных зубов спереди назад выражено слабее, чем у скалистых даманов. Первый нижний предкоренной зуб всегда имеется. Длина ряда верхних предкоренных зубов превышает длину ряда верхних коренных.
Распространены в Экваториальной и Южной Африке, включая о-ва Фернандо-По, Занзибар и Пембу.
Населяют леса. Поднимаются в горы до высоты 4,5 тыс. м над уровнем моря. Ведут древесный образ жизни, но часто спускаются на землю. Очень подвижны; бегают вверх и вниз по наклонным стволам деревьев, перепрыгивают с ветки на ветку. Активны ночью. День проводят в дуплах или в густой кроне деревьев. Держатся поодиночке. Характерны громкие крики, которые древесные даманы издают после захода солнца. Питаются листьями, побегами, почками и насекомыми. Размножение не приурочено к определенному времени года. Беременность около 7 месяцев. В помете 1—2, редко 3 детеныша. Половозрелость наступает в 2 года. Продолжительность жизни в неволе 6—7 лет.
В роде 3 вида:
западный даман — D. dorsalis Fraser, 1854 (о-в Фернандо-По, прибрежные леса от Гамбии на юг до северной оконечности Анголы, Центральный и Северо-Восточный Заир, Северная Уганда);
древесный даман — D. arboreus A. Smith, 1827 (Южно-Африканская Республика, Замбия, Восточный Заир, Танзания, Руанда, Бурунди, Западная Уганда, Центральная и Южная Кения);
восточный даман — D. valtdus True, 1890 (Восточная Танзания, о-ва Занзибар и Пемба).
ОТРЯД СИРЕН
ORDO SIRENIA ILLIGER, 1811
Вторичноводные млекопитающие (морские или пресноводные), приспособившиеся к постоянной жизни в воде. Длина тела 2,5—5,8 м (у вымершей морской коровы до 7,2—10 м). Масса до 650 кг (у морской коровы до 4 т). Тело массивное веретеновидное (рис. 142). Шея короткая и толстая, но подвижная. Голова сравнительно небольшая, округлая с относительно маленьким ртом. Сильно развитая верхняя губа образует мягкий «губной диск» — своеобразный хобот, снабженный органами тактильного чувства. Ротовое отверстие находится на нижней поверхности головы. Наружные носовые отверстия открываются наверху головы и способны замыкаться. Глаза небольшие, с подвижными веками без ресниц; мигательная перепонка хорошо развита. Ушных раковин нет, ушные отверстия очень малы. Передние конечности пятипалые, превращены в ласты. Задние конечности редуцированы. Ласты свободно двигаются в плечевом суставе и в отличие от китообразных подвижны в локтевом и кистевом сочленениях. Пальцы одеты общим кожным покровом и снаружи незаметны. Бесскелетный горизонтальный хвостовой плавник треугольной или округлой формы; он служит локомоторным органом. Кожа толстая, покрытая редкими щетинистыми волосками. Подкожная жировая клетчатка сильно развита. На губах расположены многочисленные толстые вибриссы. Два соска находятся в грудной области.
Череп с сильно развитым рострумом, образованным большими предчелюстными костями. Костное носовое отверстие сильно сдвинуто кверху. Носовые кости очень малы или отсутствуют. Слезная кость маленькая, без слезного канала. Хотя костное небо длинное, небольшие небные кости принимают малое участие в его образовании. Нижняя челюсть массивная с косо наклоненным вниз симфизом.
„ 1
Зубы представлены резцами —
(у взрослых зверей они могут отсутствовать) и отделенными от них широкой диастемой коренными. Число коренных зубов колеблется от 3 до 10 в каждой половине челюсти. Их жевательная поверхность несет два поперечных гребня, образованных тремя буграми каждый. При снашивании жевательная поверхность уплощается. Передняя часть неба и противолежащий ей отдел нижней челюсти (область симфиза) покрыты грубыми роговыми пластинами, служащими для перетирания пищи. Такие же пластины имеются и на верхней поверхности небольшого языка.
Кости скелета плотные и тяжелые; длинные кости без срединной мозговой полости. Передние конечности с сильно укороченными плечом и предплечьем (рис. 142). Ключицы нет. Кости задней свободной конеч-
Рис. 142. Ламантин — Т. manat us (виден хвост сверху),
его скелет и череп
ности отсутствуют. Таз находится в редуцированном состоянии и представлен одной или двумя парами костей. Лобковых костей нет. Позвонки не срастаются между собой. В шейном отделе 6 (Trichechidae), 7 (Dugongidae) или 6 или 7 (Hydrodamalidae) позвонков. Os penis нет.
Желудок сложный, с резкой перетяжкой между кардиальной и пилорической частями, слепым железистым выростом в кардиальном отделе и двумя слепыми мешками — в пилорическом. Кишечник очень длинный. Он превышает длину тела в 13—20 раз. Слепая кишка хорошо развита, у некоторых видов с двумя дополнительными придатками. Легкие простые, длинные и узкие, не разделяющиеся на доли. Мозг небольшой с малочисленными извилинами; обонятельные доли хорошо развиты. Почки дольчатые. Матка двурогая. Плацента зональная, неотпадающая. Семенники располагаются в брюшной полости.
Распространены в тропических водах Индийского, Атлантического и восточной части Тихого океанов, а также в бассейнах рек Амазонки, Ориноко, в реках тропической Западной Африки. Вымершая морская корова обитала в Беринговом море.
Местные жители охотятся за сиренами из-за их вкусного мяса и прочной кожи.
Ископаемые представители отряда известны со среднего эоцена АРЕ и Ямайки. Наиболее древние из них, хотя и имели ряд примитивных признаков (полная зубная система, отсутствие роговых пластин, довольно хорошо развитый таз, рудиментарные задние конечности), были настоящими водными животными. В строении их черепа и зубов прослеживается сходство с примитивными хоботными и даманами. По-видимому, предками сирен были наземные звери, близкие к исходным формам хоботных, даманов и копытных.
В отряде 3 семейства и 3 рода:
ламантиновых — Trichechidae Gill, 1872 (1 род);
дюгоневых — Dugongidae Gray, 1821 (1 род);
морских коров—Hydrodamalidae Palmer, 1895 (1 вымерший род).
Многие исследователи (например, Симпсон. 1945) включают морских коров в ранге подсемейства в семейство Dugongidae.
СЕМЕЙСТВО ЛАМАНТИНОВЫХ
FAMILIA TRICHECHIDAE GILL, 1872
Размеры наименьшие в отряде. Длина тела до 4,5 м (обычно от 2 до 4 м). Масса около 200 кг. Верхняя губа глубоко расщеплена и каждая половина подвижна независимо от другой (рис. 143). Хвостовой плавник округлый, без вырезки (см. рис. 142). Грудные плавники у двух видов имеют небольшие когтевидные копытца (на 2—5 пальцах, рис. 143). Окраска туловища одноцветная — темно-серая или черноватая.
В черепе предчелюстные кости относительно невелики. Имеются небольшие толстые носовые кости, причленяюшиеся сбоку к лобным костям. Скуловые кости тонкие и не достигают предчелюстных костей. Орбита замкнута сзади или почти замкнута. Лобная часть черепа вытянутая. Обонятельные камеры удлинены и содержат две эктотурбиналь-вые кости. Симфиз нижней челюсти скошен не очень сильно, составляя угол примерно в 45° к продольной оси черепа. Венечный отросток хорошо развит и направлен вперед. Сочленовные поверхности нижнеи челюсти удлинены в поперечном направлении.
У взрослых особей функционирующих резцов нет (в молочной смене они имеются по 2 в каждой половине челюсти). Коренные зубы мно-326
гочисленные, непостоянного числа (до 10, обычно не более 6). Они обладают низкой коронкой, покрытой эмалью, без цемента. Жевательные поверхности с двумя острыми, поперечно расположенными гребнями. Корни у зубов замкнутые. По мере снашивания и выпадения передних коренных зубов они замещаются передвигающимися вперед задними. Всего в течение жизни в каждой половине челюсти сменяется до 30 зубов, а возможно и несколько больше. Замещение зубов сзади наперед осуществляется благодаря тому, что костная перегородка, прилегающая к зубу, спереди облитерируется, а находящаяся сзади зуба — нарастает. Такое горизонтальное замещение зубов происходит и у слонов. Необходимость постепенной смены зубов обусловлена, по-видимому, поеданием пищи с примесью грунта, способствующего стиранию зубов.
Позвонков шейных 6, грудных 17, поясничных 2, крестцовый 1, хвостовых 22—24. Остистые отростки на позвонках развиты хорошо. Подвздошная кость сильно уменьшена в размерах. Седалищная кость пластинчатая, треугольная.
Распространены в прибрежных водах и реках от штата Северная Каролина в США на юг через Мексиканский залив и о-ва Карибского моря до 20 южной широты на восточном берегу Южной Америки, в бассейнах рек Амазонка и Ориноко, в прибрежных водах и реках в тропической Западной Африке от р. Сенегал (16° с. ш.) на юг до р. Кванза (10° ю. ш.).
Населяют мелкие прибрежные воды морей, заливов, проливов, устья Рек. Поднимаются вверх по течению Рек и встречаются в некоторых озерах. Trichechiis inunguis обитает только в системах рек Амазонки и Ориноко.
Держатся поодиночке или небольшими группами (до 15—20 вместе е Детенышами). В холодную погоду со-ираются иногда до 40 и более особей в каком-либо теплом месте.
Рис. 143 Морда ламантина — Т. гпа-natus спереди и его грудной плавник (видны копытца)
Питаются морскими и пресноводными растениями, а также наземными, свисающими над водой. За сутки съедают до 40 кг пищи. В Гайане ламантинов иногда используют для очистки каналов от водорослей. В настоящее время обсуждается вопрос о более широком использовании ламантинов для этой цели. Иногда совершают миграции (в водах Мексики — протяженностью до 100 км). Активны преимущественно ночью. Отдыхают у поверхности воды, погрузив хвост глубже головы и выгнув спину. Иногда отдыхают на дне. Под водой остаются до 16 мин. Характерна значительная чувствительность к изменениям температуры воды. Подводные звуки, издаваемые ламантинами, служат им, по-видимому, для связи.
Сезонности в размножении нет. Беременность длится 152—180 дней. Детеныш (как правило, один, редко два) рождается под водой и немедленно поднимается матерью к поверхности воды. Новорожденный имеет длину около 1 м и массу 18—27 кг. Лактация продолжается примерно 18 месяцев. В течение двух лет после рождения молодое животное остается с матерью. Половозрелость наступает в возрасте 3— 4 лет.
От ламантинов получают мясо, жир и кожу. Интенсивный промысел значительно снизил в некоторых областях их численность.
В семействе один род ламантинов — Trichechus Linnaeus, 1758 и 3 вида.
Род ламантинов
Genus Trichechus Linnaeus, 1758
См. описание семейства.
В роде 3 вида:
ламантин — Т. manatus Linnaeus, 1758 (Атлантическое побережье Северной Америки от Бофорта в штате Северная Каролина на юг вдоль берегов Центральной Америки и Южной Америки до 20° ю. ш., берега о-вов Карибского моря; численность в США около 1000 зверей, в Гайане — несколько тысяч, обычен в Белизе);
бразильский ламантин — Т. inunguis Natterer, 1883 (бассейны рек Ориноко и Амазонки);
африканский ламантин— Т. senegalensis Link, 1795 (прибрежные воды и реки, в частности Нигер, Конго тропической Западной Африки от р. Сенегал — 16° с. ш. на юг до р. Кванза в Анголе— 10° с. ш.).
Все 3 вида включены в «Красную книгу»; ламантин и африканский ламантин как виды, не находящиеся под непосредственной угрозой исчезновения, но нуждающиеся в охране, а бразильский ламантин как вид, находящийся на грани исчезновения. В Бразилии и Перу бразильский ламантин, а в США, на Кубе, Ямайке, в Пуэрто-Рико, Тринидаде, Гондурасе и Гайане ламантин охраняется законом.
СЕМЕЙСТВО ДЮГОНЕВЫХ
FAMILIA DUGONGIDAE GRAY, 1821
Наиболее крупные современные представители отряда с длиной тела до 5,8 м (самец из Красного моря). Обычно же размеры меньше: длина тела самцов около 3 м, самок 2,7 м. Верхняя губа расщеплена относительно слабо (сильнее у молодых животных). Ноздри сдвинуты на верхнюю сторону головы в большей степени, чем у других представителей отряда. Хвостовой плавник с глубокой выемкой между правой и левой половина-
39Я
Рис. 144. Дюгонь — £>. dugon и его череп
ми (рис. 144). Грудные ласты не имеют копытца. Окраска спинной стороны туловища зеленоватая, сероватая, буроватая или черноватая, брюшной — розоватая или беловатая.
В черепе сильно увеличены в размерах предчелюстные кости. Носовых костей нет. Скуловые кости соединяются с предчелюстными. Ширина лобного отдела черепа превышает его длину. Костное небо узкое. Обонятельные камеры укороченные. Эктотурбиналии отсутствуют. Симфиз нижней челюсти сильно скошен вниз — почти вертикален. Венечные отростки развиты хорошо и направлены вверх. Сочленовные поверхности нижней челюсти почти сферической формы.
• Зубная формула молодых зверей: i — с
О О
ш — = 26, 4
— Р — 0 3
2 о
а взрослых, i
0 0 2 — 3
с — р — m ---=10— 14.
0 0 2—3
Резцы взрослых самцов крупные, бивнеподобные, а у самок небольшие, иногда не прорезывающиеся сквозь десну. Коренные зубы столбчатые, покрытые цементом, без эмали, имеют простые, открытые корни.
Позвонков шейных 7, грудных 19, поясничных 4, крестцовый 1, хвостовых от 28—29 до 59—60. Остистые отростки на позвонках небольшие. Седалищная и подвздошная кости палочковидные.
Распространение охватывает прибрежную зону и эстуарии Индийского и западной части Тихого океанов. Вдоль Африканского побережья
встречается от залива Делагоа в Мозамбике на север до Красного моря включительно. От Африки ареал простирается на восток до о-вов Рюкю на севере и берегов Северной Австралии и Соломоновых о-вов на юге. В ареал входят также берега Мадагаскара и о-ва Индо-Австралийского архипелага.
Обитают на мелководье в тропических морях с обильной водной растительностью. Изредка встречаются в устьях рек. Держатся поодиночке, за исключением периода размножения. Иногда наблюдаются крупные скопления. Питаются преимущественно ночью. Кормом служат морские водоросли, главным образом зеленые, и другие растения, а возможно и мелкие беспозвоночные. В течение дня обычно отдыхают на глубокой воде, а ночью приплывают к берегам на мелководье кормиться. Часто дюгонь собирает целые растения в одну кучу и съедает их позднее. Перед тем как съесть растение, дюгонь обычно тщательно прополаскивает его в воде, чтобы удалить песок. Под водой остается от одной до 10 мин. Зрение развито слабо, слух — хорошо. Сезонности в размножении, по-видимому, нет. Длительность беременности 11—12 месяцев. Самка приносит одного детеныша в тихих прибрежных водах.
Дюгоней промышляют из-за мяса и жира. От зверя массой 200— 300 кг получают 24—56 л жира. Во многих местах численность дюгоня сильно снизилась. Так, зверь стал очень редок в Красном море, в Персидском заливе, около Шри Ланка, Малайзии, у восточного берега Африки; у Австралии еще достаточно многочислен.
В семействе один род дюгоней — Dugong Lacepede, 1799, и один вид.
Род дюгоней
Genus Dugong Lacepede, 1799
См. описание семейства.
В роде единственный вид: дюгонь — D. dugon Muller, 1776 (распространение см. в описании семейства).
Включен в «Красную книгу» как вид, который в ближайшее время может оказаться под угрозой исчезновения.
СЕМЕЙСТВО МОРСКИХ КОРОВ л
FAMILIA HYDRODAMALIDAE PALMER, 1895
Вымершие животные. Длина тела 7,2—10 м. Масса до 4 т. У самки длиной 741 см окружность шеи у затылка составляла 204 см, окружность туловища на уровне плеч 367 см, наибольшая окружность туловища посредине 622 см, окружность хвостового стебля в месте отхождения лопастей 143 см, расстояние между концами хвостовых лопастей 199 см. Самцы были немного крупнее самок. Голова небольшая, укороченная, незаметно переходившая в шею, которая в свою очередь постепенно продолжалась в туловище (рис. 145). Грудные плавники заканчивались, по описанию Стеллера, чем-то напоминающим лошадиное копыто. Грудные плавники служили для медленного плавания и передвижения по дну на мелководье.
В кожном покрове эпидермис и подкожная жировая клетчатка достигали очень большой толщины. Окраска тела была от темно- до серо-коричневой.
В черепе массивные межчелюстные кости перегнуты вниз (сильнее, чем у ламантинов, но слабее, чем у дюгоней). Соответственно нижняя челюсть отогнута вниз, занимая по степени изгиба среднее положение 330 ___
между ламантинами и дюгбнями. Имеются маленькие носовые кости; они частично перекрываются лобными костями.
Зубов нет. Заменяющие их функционально роговые пластины сильно развиты.
Шейных позвонков, по данным разных ученых, 6 или 7.
Морские коровы были распространены в прибрежных водах о-вов Беринга, Медного и мелких о-вов Командорского архипелага.
Описаны эти животные Стеллером, участником экспедиции Витуса Беринга в 1741 г.
Пищу составляли в основном водоросли, которые растут в затишных местах. За зиму звери сильно худели. Очень медлительные, они совершенно не боялись людей. Обычно самка и самец держались вместе с сеголетком и молодым прошлого года. Раненому животному другие морские коровы пытались оказать помощь.
Морских коров хищнически промышляли из-за жира, кожи и мяса. К 1754 г., по-видимому, они были полностью истреблены на о-ве Медном, а к 1768 г. — на о-ве Беринга. Все современные упоминания о встречах морских коров не заслуживают доверия.
Многие исследователи (Симпсон, 1945; Андерсон и Джонс, 1967, и др.) относят морскую корову в ранге подсемейства Hydrodamalinae к семейству дюгоневых — Dugongidae. Следуя Веберу (Weber, 1928), Мор (Mohr, 1957) и Гептнеру, 1967, здесь морские коровы выделены в особое семейство, хотя такое выделение и не бесспорно (например, Тениус, Thenius, 1972, с позиций палеонтологии считает его неправильным).
В семействе 1 род морских коров — Hydrodamalis Retzius, 1794 (1 вид).
Род морских коров
Genus Hydrodamalis Retzius, 1794
См. описание семейства.
В роде единственный вид: морская корова — И. gigas Zimmermann, 1780 (распространение см. в описании семейства).
ОТРЯД ПАРНОКОПЫТНЫХ
ORDO ARTIODACTYLA OWEN, 1848
Размеры от мелких до крупных. Карликовая антилопа имеет длину тела до 52 см, высоту в холке 25 см, массу 2—3 кг (у большинства видов) . Жираф имеет длину тела почти до 500 см, общую высоту до 600 см, высоту в холке около 350 см. Бегемот с длиной тела до 450 см, высотой в холке до 150 см, массой до 3200 кг.
У большинства видов тело стройное, на высоких тонких конечностях, иногда толстое, неуклюжее, приземистое, на толстых ногах. Шея длинная, иногда очень длинная. Морда вытянутая. Хвост сильно редуцирован, у некоторых снаружи почти незаметен или отсутствует, лишь у немногих он длинный. У большинства видов один или оба пола имеют пару рогов. Рога костные, ежегодно сменяющиеся или роговые, на костных стержнях, постоянные. Волосяной покров, как правило, хорошо развит. В большинстве случаев он образован в основном остевыми волосами, имеющими сильно развитую воздухоносную сердцевину. У некоторых видов волосяной покров щетинистый; иногда он почти редуцирован. Для большинства видов характерно значительное число разнообразных кожных специфических желез. Одна или две пары сосков располагаются в области паха; у некоторых видов соски в количестве 5—6 пар находятся и на брюхе, и в паху.
Имеется тенденция к пневматизации черепных костей. У ряда видов орбита и височная яма черепа отделены друг от друга смыкающимися между собой орбитальными отростками лобной и скуловой костей, у других между этими отростками имеется более или менее широкий промежуток, у третьих, наконец, орбита сзади совсем открытая. Слезная кость хорошо развита. Теменные кости обычно уменьшены в размерах; лобные — крупные. Носовая кость не заходит далеко назад. Крылоклиновидный канал отсутствует.
0 — 3
Зубная формула: 1 ------—
1 — 3
Зубная система гетеродонтная и дифиодонтная. Число зубов значительно варьирует. Верхние резцы, клыки и первые предкоренные зубы уменьшены в размерах или отсутствуют. У некоторых видов клыки достигают полного развития или даже гипертрофированы. Щечные зубы тупобугорчатые; у немногих бунодонтные или селенодонтные, обычно гипселодонтные (иногда брахиодонтные).
Отличительная особенность — число пальцев: у большинства на передних и задних конечностях по 4 пальца (редуцируется I). У немногих видов конечности имеют по два пальца (III и IV). Лишь у некоторых видов семейства пекари передние конечности четырехпалые, а задние —
трехпалые. Ось конечности парнокопытных проходит между III и IV пальцами. Боковые пальцы меньше средних и расположены выше их. Терминальные фаланги средних пальцев сильно увеличены и одеты копытами. Кости конечностей удлинены, что связано с приспособлением к быстрому бегу. Утрачена возможность супинации и пронации конечностей. Отделы конечностей двигаются в суставах в сагиттальной плоскости. Ключицы нет. У бедренной кости нет третьего трохантера. Локтевая кость у большинства видов редуцирована и обычно срастается с лучевой. Малая берцовая кость сильно уменьшена и часто срастается с большой берцовой. Пояснично-грудных позвонков обычно 19.
Желудок у представителей семейств свиных, пекариевых и бегемотовых двух- и трехкамерный «нежвачный». У представителей семейства оленьков — трехкамерный «жвачный», представителей семейств оленевых, жирафовых, вилороговых и полорогих — четырехкамерный «жвачный». Во время питания жвачное животное слабо пережевывает пищу и заглатывает ее в первый отдел желудка, где пища подвергается процессу размягчения. Затем она отрыгивается в ротовую полость, где снова пережевывается, перемешивается со слюной и опять проглатывается. Слепая кишка парнокопытных различного строения, иногда отсутствует. Матка двурогая. Плацента диффузная или поликотиледонная.
Семенники находятся в мошонке (у бегемотов под кожей).
Предки парнокопытных, по-видимому, развились из примитивных плацентарных верхнего мезозоя, имевших непосредственную связь с примитивными хищными Creodonta. Расхождение стволов хищных и копытных произошло где-то в конце мезозоя. В начале третичного периода в палеоцене уже существовали многочисленные «копытные», относящиеся к вымершим ныне отрядам Condylarthra, Amblypoda, Notoungula-ta. Очевидно, именно Condylarthra послужили исходными формами для современных парнокопытных, которые появляются в нижнем эоцене Северной Америки и Европы.
Распространены по всему Земному шару, исключая Австралию, Новую Зеландию, Антарктиду и некоторые изолированные острова.
Населяют преимущественно открытые ландшафты, но также лес, горы, берега водоемов. Большинство видов — быстробегающие животные. Некоторые лазают по скалам. Есть и амфибиальные виды. Большинство парнокопытных ведут оседлый образ жизни или совершают незначительные вертикальные перемещения. Для некоторых характерны значительные сезонные миграции. Обычно держатся небольшими или более крупными стадами. Питаются растениями. Только некоторые виды используют наряду с растительной и животную пищу. Характерна полигамия. Спаривание обычно происходит осенью, а рождение молодых— весной. Детенышей в помете у большинства видов 1—2 (иногда до 10— 14). Детеныши вскоре после рождения способны к самостоятельному передвижению.
В году бывает 1—2 линьки. У большинства выражен половой диморфизм, у некоторых сезонный.
В отряде 2 подотряда и 8 семейств: нежвачные — Suiformes, Jaeckel, 1911 (3 семейства) и жвачные—Ruminantia Scopoli, 1777 (5 семейств).
Многие парнокопытные имеют существенное хозяйственное значение. Некоторые послужили родоначальниками ценнейших домашних пород животных, от которых человек получает животные белки и шерсть. Некоторые содержатся в полудомашнем состоянии или находятся в различных стадиях одомашнения.
Почти везде парнокопытные служат важными объектами промысла, дающими мясо, кожу, сырье для фармацевтических препаратов. В последние годы все более увеличивается значимость парнокопытных как 334
объектов спортивной охоты. Вследствие этого некоторые из них полностью истреблены, численность других сильно уменьшена и они нуждаются в охране.
ПОДОТРЯД НЕЖВАЧНЫХ
SUBORDO SUIFORMES JAECKEL, 1911
Размеры мелкие, средние или крупные. Длина тела от 70 до 450 см. Туловище массивное, неуклюжее. Голова вытянутая, часто клиновидная. Половой диморфизм выражен слабее, чем у представителей подотряда жвачных. Кожный покров обычно утолщенный, с мощно развитой подкожной жировой клетчаткой. Волосяной покров изреженный, иногда он почти редуцирован. В нем характерны грубые щетины. Рогов не бывает. Конечности короткие, четырехпалые (у пекари задние конечности трехпалые). Размеры боковых пальцев почти такие же, как и средних. Метакарпальные и метатарзальные кости не срастаются или почти не срастаются между собой. Os canon не образуется. Локтевая, а иногда малая берцовая кости развиты хорошо и не срастаются первая с лучевой, вторая — с большой берцовой костями.
В черепе нежвачных, за исключением бегемотов, орбиты сзади открыты. Обычно имеется крупный сагиттальный гребень. Сочленовная поверхность сочленовного отростка нижней челюсти вытянута в продольном направлении.
1—3 I 3 — 4 3
Зубная формула: i —-— с —— р —-— m — = 32 — 44.
Клыки обычно развиты сильно, часто выдаются из ротовой полости по бокам головы. Нижние клыки с открытыми корнями, обладают постоянным ростом. Щечные зубы бунодонтные, лишь иногда гипселодонтные.
«Нежвачный» желудок разделяется на 1—3 отдела и не приспособлен к отрыгиванию пищи. Слепая кишка средних размеров или ее нет вовсе. Плацента диффузная.
Ареал обширный: в Евразии северная граница проходит по югу Скандинавского п-ова, Финскому заливу, р. Оке, Южному Уралу, Алтаю, Саянам, северной оконечности Байкала, Нижнему Амуру; южная граница тянется по Тихоокеанскому и Индоокеанскому побережьям Азиатского материка, включая Японские о-ва и о-ва Юго-Восточной Азии вплоть до Австралии; на Аравийском п-ове проходит в его северо-западной части и продолжается по Средиземноморскому побережью Европы; охватывает всю Африку и Мадагаскар, за исключением Сахары; включает южные районы Северной Америки на север до штатов Техас, Нью-Мексико и Аризона, Центральную и Южную Америку на юг до Уругвая и Центральной Аргентины.
Населяют различные биотопы тропической й умеренной зон, придерживаясь преимущественно густых зарослей поблизости от водоемов. Представители рода бегемотов ведут полуводный образ жизни. Обычно Держатся небольшими группами, реже поодиночке.
По характеру питания растительноядны с тенденцией к всеядности. Самки приносят обычно большое число детенышей — до 10—12.
Служат объектом охоты. В некоторых местах вредят посевам, огородам и бахчам.
Подотряд включает 3 семейства и 8 родов:
свиных — Suidae Gray, 1821 (5 родов);
пекариевых — Tayassuidae Palmer, 1897 (1 род);
бегемотовых — Hippopotamidae Gray, 1821 (2 рода).
СЕМЕЙСТВО СВИНЫХ
FAMILIA SUIDAE GRAY, 1821
Среднего размера парнокопытные. Характерны длинная заостренная голова, короткие шея и конечности, сжатое с боков массивное туловище. Длина тела 70—205 см, хвоста 3,5—32 см. Масса до 275 кг. Вытянутая морда усечена на конце и оканчивается небольшим, малоподвижным хоботком с дисковидным терминальным хрящом. Хрящевидный конец морды, употребляемый для вскапывания земли, усилен необычной предносовой костью, расположенной ниже вершины носовых костей черепа. Ноздри терминальные, глаза маленькие, уши большие, часто с пучком волос на вершине. Некоторые члены семейства имеют складки и гребни кожи на морде, причем костных образований в этих местах нет. Конечности имеют по 4 пальца, средние из которых (III и IV) крупные, а два других расположены выше, сдвинуты назад и не касаются земли в обычных условиях. Опирается на них животное лишь при ходьбе по мягкому грунту.
Некоторые свиньи совсем лишены волос, у других тело покрыто грубой щетиной. У некоторых видов есть грива, а хвост имеет кисточку на конце. Взрослые животные обычно одноцветные, а молодые имеют полосатую окраску (за исключением бабируссы и домашней свиньи). Сосков от 3 до 6 пар (у бабируссы только одна). Желудок двухкамерный простой нежвачный с небольшим дополнительным кардиальным отделом. Имеются простая и короткая слепая кишка и желчный пузырь. Матка двурогая. Плацента диффузная.
Для черепа характерна постепенно поднимающаяся спереди назад крыша, сходящаяся под острым углом с затылочным отделом. Спереди череп не расширен. Лобные, теменные и верхнезатылоч'ные кости пневматизированы. Орбита сзади незамкнутая. Слезная кость крупная, обычно с двумя отверстиями слезного канала. Боковая затылочная и чешуйчатые кости прикрывают собой мастоидную, которая снаружи незаметна. Костное небо продолжается кзади от задней границы щечных зубов. Костные слуховые барабаны вздутые, грушевидной формы с многочисленными костными перегородками во внутренней полости. В противоположность другим парнокопытным крыловидная ямка хорошо выражена.
1—3 1 3 — 4 3
Зубная формула: 1— - с —- р - ш —=32 — 44.
— о 1 ~~ т: О
Верхние резцы уменьшаются от первого к третьему, а нижние длинные, узкие, плотно прилегают друг к другу и расположены почти горизонтально. Большие верхние клыки растут наружу и назад, а нижние клыки — вверх и назад, имея тенденцию образовывать полный круг. Обычно, однако, верхний и нижний клыки растут друг против друга, образуя острый угол. Клыки сильнее развиты у самцов. У бабируссы верхние клыки направлены вверх, прободая кожу, выходят наружу, искривляются назад и вниз и часто касаются лба. Щечные зубы бунодонт-ного и брахиодонтного типа; верхние предкоренные более простые по структуре, чем коренные. С возрастом эмаль стирается, и все зубы исчезают, за исключением клыков и последних коренных. Третий коренной зуб бородавочника часто бывает единственным коренным зубом у этого вида и сходен с подобными зубами других млекопитающих. Он состоит из значительного числа плотно расположенных цилиндров дентина, скрепленных цементом.
Пястные и плюсневые кости не срастаются.
Распространены в Старом Свете, в Евразии к югу от 58° с. ш., в Аф
рике, на Мадагаскаре, в Японии, на Тайване, Филиппинах и в Индонезии. Свиньи были акклиматизированы в Австралии, Тасмании, на Новой Гвинее, в Новой Зеландии, на Британских о-вах и в ряде мест Северной и Южной Америки.
По большей части обитают в густых зарослях лесов, лесостепей, степей, гор, речных долин. Предпочитают увлажненные места. Обычно держатся группами до 40—50 особей, хотя самцы бородавочника, например, живут в одиночку. Бегают не быстро и на небольшие расстояния. Хорошо плавают. Любят лежать в грязевых ваннах. Активны в основном ночью. Всеядны, но предпочитают растительные корма. Характерна полигамия. Пометов в году один, реже два. Детенышей в помете 1—14. Длительность беременности 112—175 дней. Лактационный период длится 2—3 месяца. Половозрелость наступает в 9—15 месяцев. Длительность жизни почти 20 лет.
Почти все представители семейства промысловые животные, которых добывают из-за мяса.
В семействе 5 родов (7 видов):
кистеухих свиней — Potamochoerus Gray, 1854 (1 вид);
кабанов — Sus Linnaeus, 1758 (3 вида);
бородавочников — PhacochoerusF. Cuvier, 1817 (1 вид);
лесных свиней — Hylochoerus Thomas, 1904 (1 вид); бабирусс — Babyrousa Perry, 1811 (1 вид).
В СССР: род кабанов.
Род кистеухих свиней
Genus Potamochoerus Gray, 1854
В роде единственный вид: кистеухая свинья — Р. porcus Linnaeus, 1758.
Длина тела 100—150 см, длина хвоста 30—45 см, высота в холке 58—• 96 см. Масса 60—80 кг. Голова удлиненная, туловище сжато с боков (рис. 146). Уши длинные, заостренные, с пучками длинных волос на вершинах. Средней длины грубые волосы равномерно покрывают тело. Подшерстка нет. Окраска туловища варьирует от красновато-коричневой до черной, часто с примесью белых и желтоватых тонов. По спине иногда проходит белая срединная полоса. Молодые с бледно-желтыми или темно-желтыми продольными полосами на темно-коричневом фоне. На верху шеи и спины имеется хорошо выраженная грива светлого цвета. Есть предглазничные, межпальцевые и карпальные кожные специфические железы. Перед глазами у самцов располагаются кожные бугры высотой до 40 мм; они покрыты волосами и поэтому бывают незаметны. Сосков 3 пары.
3 1 4 3
Зубная формула: i — с —— р — m —- = 42. О 1 О О
Средняя длина верхнего клыка 76 мм, нижнего 165—190 мм.
Распространение охватывает Африку к югу от Сахары и о-ва Мадагаскар, Занзибар, Мафия и Майот из группы Коморских о-вов (возможно, на этих островах кистеухая свинья была акклиматизирована).
Населяет леса и кустарниковые заросли по берегам рек на равнинах и в горах. Активна ночью; день отдыхает в высокой густой траве, тростнике, в которых проделываются тропинки и туннели. Держатся стадами от 4 до 20, иногда до 100 особей. Питается сочной травой, корнями, клубнями, луковицами, плодами, мелкими наземными животными. Часто причиняет вред посевам и посадкам. Самка приносит детенышей в ок-
Рис. 146. Кистеухая свинья — Р. porctts
тябре-марте. Беременность длится около 4 месяцев. Самка имеет один или два помета в году. Детенышей в помете бывает от 3 до 6. Продолжительность жизни 15—20 лет.
Род кабанов
Genus Sus Linnaeus, 1758
Длина тела у карликовой свиньи 50—65 см, а у наиболее крупных особей кабана до 205 см у самцов и 170 см у самок. Длина хвоста кабана около 30 см. Высота в холке карликовой свиньи 25—30 см, кабана — до 120 см. Масса тела кабана 75—250 кг (изредка до 300 кг) у самцов и 35—150 кг у самок. Передняя часть уплощенного с боков туловища выше задней и значительно крупнее ее (рис. 147). Длинная голова, занимающая до трети длины всего тела, без резкого шейного перехвата переходит в туловище. Уши длинные и широкие. Глаза маленькие, глубоко сидящие. Хвост тонкий короткий, не достигающий скакательного сустава. Пальцы на ногах способны сильно раздвигаться, что важно при ходьбе по болотистому грунту. В последнем случае кабан для опоры использует и боковые пальцы, причем массовая нагрузка на 1 см2 сокращается приблизительно в 1,5 раза — с 500 до 320—390 г.
Волосяной покров обычно хорошо развит, иногда изрежен. Он состоит из щетины, остевых и пуховых волос. Многие особи имеют баки из удлиненных волос по бокам головы (особенно сильно развиты у бородатой свиньи) и гриву на загривке. Кабаны линяют один раз в году в весенне-летние месяцы. Окраска волосяного покрова темно-серая до бурой или черной. У молодых в первое время после рождения на светло-буром или рыжевато-буром фоне проходят светлые желтовато-палевые полосы. Взрослый наряд у кабана появляется в возрасте около 4 месяцев. Брюшных и паховых сосков 3—6 пар.
В черепе сагиттального гребня нет.
Зубная формула полная, что отличает представителей этого рода от других родов семей-.314 3
Рис. 147. Кабан — S. scroja и его череп
Крайние резцы и первый предкоренной часто уже у молодых животных выпадают. Первый предкоренной не имеет молочного предшественника. Средние резцы крупнее крайних. Клыки с незамкнутыми корнями, постоянно растущие. Нижний клык трехгранный. Клыки самцов значительно крупнее, чем самок. Щечные зубы брахиодонтного типа.
Диплоидное число хромосом у кабана 36.
Распространены в умеренных и южных районах Евразии, Северной Африке, Японии, на Тайване, Хайнане, Малайском архипелаге до Филиппин, Сулавеси, Молуккских о-вов, Новой Гвинеи и Меланезии.
Образ жизни примерно сходен у всех видов. Держатся в самых разнообразных зонах и биотопах. Обычно встречаются в густых тростниковых и кустарниковых зарослях по берегам водоемов, на склонах гор. Предпочитают сырые, болотистые места. Любят купаться в лужах с грязью. Активны преимущественно ночью.
Кабаны, за исключением взрослых самцов и самок с маленькими поросятами, в течение круглого года держатся стадами — обычно по 3— 10 голов. Число животных в стаде колеблется по сезонам, по годам и в зависимости от места обитания. Правильные сезонные переходы отмечены у кабанов в горах, где они совершают вертикальные миграции в зависимости от урожая кормов и состояния снегового покрова. Так, в средние и нижние пояса гор кабаны спускаются осенью с созреванием здесь фруктов и орехов. Здесь же они остаются на зимовку или уходят в многоснежные зимы еще ниже, в предгорья.
По характеру питания всеядны. Основные корма кабана можно разбить на 4 группы (Гептнер и др., 1961): 1) подземные части растений (корни, корневища, луковицы, клубни) — потребляются на протяжении большей части года; 2) плоды фруктовых деревьев, орехи, ягоды и семена — поедаются после созревания и, в случае хорошего урожая, зимой добываются из-под снега; 3) зеленые вегетативные надземные части растений — потребляются в период вегетации; кора, ветки и побеги — вынужденные корма — используются зимой; 4) животные корма (земляные черви, насекомые и их личинки, моллюски, рыбы, лягушки, ящерицы, змеи, яйца птиц, грызуны, насекомоядные, падаль и т. д.) •—служат кормом преимущественно в теплый период года. Обычно кабан достает пищу в поверхностных слоях почвы, взрывая ее. Значимость тех или иных кормовых объектов существенно меняется по сезонам и в различных местах ареала. Бородатая свинья питается плодами, личинками насекомых, корнями растений, сельскохозяйственными культурами.
Гон у кабанов бывает в ноябре-январе. У бородатой свиньи сезонности в размножении нет. Характерна ограниченная полигамия: на одного самца приходится 1—3 самки. Между самцами происходят жестокие драки. К началу гона у взрослых самцов в коже боков передних отделов тела развивается чрезвычайно плотное переплетение соединительной ткани — калкан, предохраняющий зверя от глубоких ран, наносимых клыками соперника. За время гона самцы теряют до 20% своей массы. Длительность беременности самок кабана 112—140 дней (у размножающихся впервые короче по сравнению со старыми свиньями), бородатой свиньи — примерно 120 дней. Опорос у кабана происходит с марта по май. Число поросят в помете кабана от 3 до 12, в среднем 4—6, бородатой свиньи — от 4 до 8, карликовой свиньи — 3—4. Масса новорожденного кабана составляет 0,6—1 кг. В малокормные годы число поросят в помете значительно снижается по сравнению с хорошим по кормовым условиям годом. Лактация длится 2,5—3,5 месяца. Физиологическая половозрелость у кабана наступает на первом году жизни, но самки, как правило, впервые принимают участие в гоне в возрасте 18—20 месяцев, самцы — на 4-м или 5-м году жизни. Продолжительность жизни кабанов
и карликовых свиней в природе 10—12 лет, в неволе — до 20 лет. Враги кабанов — волки и тигры.
Хозяйственное значение велико; большое число кабанов ежегодно добывают на мясо. В некоторых местах кабаны причиняют вред сельскому хозяйству, особенно посадкам картофеля, огородам и бахчам. Кабан— один из родоначальников домашней свиньи.
Классификация рода кабанов разработана недостаточно хорошо. По-видимому, наиболее правильно выделять 3 вида:
бородатая свинья — S. barbatus Muller, 1838 (о-ва Калимантан, Суматра, Ява, Сулавеси, Филиппинские и Молуккские, п-ов Малакка);
кабан — Sus scrofa Linnaeus, 1758 (Северная Африка — Марокко и Северный Тунис; Евразия от Атлантического до Тихого океанов, Средняя, Южная и Восточная Европа, Малая, Средняя, Центральная и Восточная Азия и некоторые острова Японии. В СССР: прибалтийские республики, Белоруссия, Западная и Центральная Украина, Калининградская, Ленинградская, Новгородская, Псковская, Смоленская, Калининская, Московская, Калужская, Брянская, Орловская, Воронежская, Кировоградская области, Молдавская ССР, Кавказ и Закавказье; низовья рек Кумы, Волги и Урала; долины рек и горные районы Казахстана и Средней Азии; Алтай, Саяны, Прибайкалье, Южное Забайкалье, Амур вниз до г. Комсомольска, Уссурийский край);
карликовая свинья — S. salvanius Hodgson, 1847 (южные предгорья Восточных Гималаев в Северной Индии, Сиккиме, Непале и Бутане).
Иногда бородатую свинью разделяют на 2 вида: бородатая и яванская— S. verricosus Muller et Schlegel, 1842 (Хальтенорт, Haltenorth, 1963). По И. И. Соколову (1959), в роде 5 видов: S. barbatus, S. cristatus, S. vittatus, S. scrofa, S. salvanius.
В СССР: только кабан.
Карликовая свинья включена в «Красную книгу» как малочисленный вид, сведения о котором крайне незначительны.
Род бородавочников
Genus Phacochoerus F. Cuvier, 1817
В роде единственный вид: бородавочник—Р. aethiopicus Pallas, 1767.
Длина тела 105—150 см, длина хвоста 35—50 см, высота в холке 55— 85 см. Масса 60—150 кг. Самцы значительно крупнее самок, но с более коротким хвостом. Тело бочковидное, голова очень большая, с значительным расширением в передней части морды (рис. 148). Уши средней длины, заостренные, снаружи голые, изнутри покрыты белыми волосами. Волосяной покров состоит из длинной редкой гривы тонких волос на загривке, которая тянется до середины спины, затем прерывается и опять появляется на крестце, и редкой щетины, покрывающей все остальное тело. Кроме того, имеются удлиненные волосы в виде баков на щеках. На конце хвоста — кисть из черных волос. Окраска кожи серая, а волос— от желто- до темно-коричневой. Молодые имеют рыжевато-коричневый Цвет. Длинные, высокие складки на щеках покрыты белыми волосами. Между глазом и углом рта с каждой стороны морды имеются своеобразные кожные выросты — «бородавки». У самцов их по две (задняя бородавка может достигать в высоту 15 см), а у самок по одной и они меньших размеров. Запястье с роговыми мозолями. Имеются подглазничные железы. Сосков две пары.
1 1 3 3
Зубная формула: i —- с — р — m — =34. 3.1 3
Рис 148. Бородавочник — Р. aethiopicus
Резцы слабые. Клыки очень мощные, верхние значительно длиннее передних, и у старых самцов могут достигать 60 см длины и быть изогнуты на три четверти окружности. Последний коренной зуб сильно развит и в процессе роста постепенно вытесняет расположенные перед ним зубы. У старых бородавочников обычно всего 16 зубов, так как часть резцов, предкоренных и коренных зубов выпадает.
Диплоидное число хромосом 34.
Распространен в Африке к югу от Сахары (за исключением крайнего юга континента).
Населяет саванны и светлые леса на равнинах и в горах. Обычно активен днем. Встречаются поодиночке, парами или небольшими группами— 1—2 самки с детенышами. Иногда попадаются стада до 30— 300 особей Самцы обычно держатся в одиночестве. Часто эти животные находят себе убежище в норах трубкозубов, куда обычно залезают, пятясь задом. При беге способны развивать скорость до 47 км/ч. Всеядны. В поисках пищи для подкапывания земли используют клыки. Во время добывания пищи животное часто опускается на запястные сочленения и. может передвигаться в таком положении.
Беременность длится 125—175 дней. В году часто бывает 2 помета: в феврале—апреле и в августе — ноябре. Самка приносит детенышей в норе. Детенышей в помете от 2 до 4, иногда до 7. Новорожденные одноцветные, серовато-розовые, без полос. В недельном возрасте детеныши начинают выходить вместе с матерью из норы. Остаются с матерью до года. Половозрелость наступает в 9—18-месячном возрасте. Продолжительность жизни 15—20 лет.
Род лесных свиней
Genus Hylochoerus Thomas, 1904
В роде единственный вид: лесная свинья — Н. meinertzhageni Thomas, 1904.
Длина тела 155—18Q см. ллмна хвоста 25—35 см. высота в холке
^0—НО см. Масса 100—250 кг. Самцы значительно крупнее самок, а особи из Восточной Африки крупнее, чем из Западной. Туловище округлое, голова длинная и широкая (рис. 149). Уши средней длины, широкие, слабо заостренные. Носовой диск достигает в диаметре 13 см. Волосы длинные, грубые, черные, редкие, с возрастом изреживаются. На шее и лбу обычна грива. На конце хвоста находится небольшая кисть черных волос. На бегу животное поднимает хвост вертикально вверх. Кожа чер-иозато-серого цвета. Ниже и позади каждого глаза с каждой стороны Морды расположен продольный кожный вырост (особенно крупный у самщ.ъ). богатый кожными железами. • Лицевых кожных бугров до в
отличие от бородавочника. Перед глазом имеется щелевидное отверстие предглазничной железы. Есть препуциальная железа. Сосков 2 пары.
1
2 — 3
Зубная формула: i
с
1 3 3
— Р — m — = 32 — 34.
Верхние резцы слабые, нижние расположены горизонтально и направлены вперед. Верхние клыки мощные, длиной до 31 см, изогнуты назад.
Распространена в Африке от Либерии на восток до Заира, Кении, Танзании и юга Эфиопии.
•Населяет большие лесные массивы и их опушки на равнинах и в го-
рах. Обычно держатся семейными группами по 4—6 особей. Убежищем иногда служат выкопанные ими ямы в земле. Активна утром и вечером. При добывании пищи землю не вскапывает. Питается вегетативными
частями растений и плодами. Беременность длится примерно 125 дней.
Самка рождает обычно 4 детеныша. Сезонности в размножении нет.
Род бабирусс
Genus Babyrousa Perry, 1811
В роде единственный вид: бабирусса — В. babyrussa Linnaeus, 1758.
Длина тела 90—110 см, длина хвоста 20—30 см, высота в холке 65— 80 см. Масса 60—-100 кг. Голова относительно небольшая, заостренная кпереди. Спина арковидно выгнута. Конечности относительно высокие (рис. 150). Пятачок маленький. Уши короткие, заостренные. Хвост длинный, без пучка волос на конце. Кожа грубая гладкая, коричневато-серого или черно-серого цвета, покрыта редкими и короткими беловато-серы-
Р«с. 150. Бабирусса — В. Ьа-byrussa и ее череп
ми или желтоватыми волосами. Нижняя поверхность тела и внутренние поверхности ног иногда светлее остальных частей тела. Кожа обычно висит складками. Сосков одна пара.
2 12 3
Зубная формула: i — с —— р — m —- = 34.
О 1 Л о
У самцов верхние клыки очень большие (длиной до 31 см); они растут вверх, прободают верхнюю губу, загибаются дугообразно назад и вниз к надглазничной области; у старых самцов их концы могут проникать в верхнечелюстную кость. Функция этих клыков неясна. У самок верхних клыков нет или они развиты слабо. Нижние клыки (также загнутые вверх и назад), как и верхние, не могут быть использованы в качестве оружия, потому что они не затачиваются друг о друга. По-видимому, сильное развитие клыков — вторичный половой признак.
Распространена на о-вах Сулавеси, Буру (Северные Молуккские о-ва) и Сула.
Населяет влажные леса, густые заросли тростника по берегам рек и озер. Держатся поодиночке или небольшими группами. Активна ночью. При добывании пищи обычно только самцы вскапывают землю, а самки и молодые следуют за ними сзади. Питаются в основном растительными кормами. Быстро бегают и хорошо плавают. Плавать могут даже в море, переплывая небольшие проливы между островами. Гон у большинства приходится на октябрь. Продолжительность беременности около 5 месяцев. Детеныши рождаются в феврале. Новорожденные поросята не имеют полосатой окраски. В помете обычно 1—2 детеныша. Новорожденный имеет длину тела 15—20 см. Половозрелости достигают примерно в возрасте года. Живут до 20 лет. Местные жители часто держат бабирусс в неволе.
СЕМЕЙСТВО ПЕКАРИЕВЫХ
FAMILIA TAYASSUIDAE PALMER, 1897
Внешне сходны со свиньями (см. рис. 151). Длина тела 75—НО см, длина хвоста 1,5—5,0 см (в хвосте всего 6—9 позвонков по сравнению с 20—23 у свиней). Относительно коротконоги; высота в холке 44— 57 см. Масса 18—50 кг. Передние конечности с 4 пальцами, задние — с 3 (у некоторых ископаемых видов было 2 пальца). Морда относительно короче, чем у свиней, с подвижным хрящевым, почти голым пятачком, на котором открываются ноздри. Окостенений в пятачке нет. Оба пола имеют на крестце специфическую кожную железу. Глаза и уши маленькие. Уши округлые, покрыты редкими волосами. Туловище сжато с боков. Хвост снаружи почти незаметен. Все тело покрыто шетиноподобны-ми волосами. Окраска волосяного покрова от темно-коричневой до серокоричневой; иногда на шее от плеч до горла проходит белый ошейник или на нижней челюсти и горле располагается белое пятно. Сосков одна пара.
Череп укорочен в ростральной части. Костные слуховые барабаны сердцевидной формы. Лицевая часть слезной кости и слезное отверстие отсутствуют. Резцовые отверстия маленькие, округлой формы.
2 13 3
Зубная формула: i — с — р — m — = 38.
О 1 О О
Верхние клыки длиной до 4 см направлены вниз (а не вверх и наружу, как у свиней). Их передний край остро затачивается нижними клыками. Третьего верхнего резца нет. Нижние резцы расположены гори-346
Рис. 151. Ошейниковый пекари — Т. tajacu и его череп
зонтальио и направлены вперед. Коронки коренных зубов лофодонтного типа. V палец задней конечности представляет только небольшой остаток от проксимального отдела плюсны. У ошейникового пекари срастаются проксимальные отделы плюсневых костей III и IV пальцев, а у белогубого пекари —эти же отделы плюсневых и пястных костей III и IV пальцев. С возрастом у белогубого пекари срастаются и дистальные части пястья. Желудок подразделяется на 3 отдела, из которых передний имеет два колбасовидных выроста. Желчного пузыря нет.
Диплоидный набор хромосом у ошейникового пекари 30.
Распространены в Северной и Южной Америке от штатов Техас, Нью-Мексико и Аризона на юг до Средней Аргентины, Парагвая и Уругвая.
Населяют засушливые места (ошейниковый пекари) или влажные тропические леса (белогубый пекари). Держатся небольшими группами по 5—15 (ошейниковый пекари, рис. 151) или крупными стадами — до 50—100 и более голов (белогубый пекари). В одном стаде собираются и самцы и самки. Активны ночью и в прохладные утренние и вечерние часы. Зрение развито слабо, слух и обоняние — хорошо. Всеядны, но предпочитают растения; не пренебрегают и мелкими наземными животными. Пекари полигамы. Период спаривания не приурочен к определенному времени. Длительность беременности 140—150 дней. Детенышей в единственном в течение года помете 1—4 (обычно 2). Половозрелость наступает в 9—18-месячном возрасте.
За пекари охотятся из-за мяса (чтобы мясо было съедобным, удаляют крестцовую железу). В 1965 г. только через г. Икитое в Перу было экспортировано 129 тыс. шкур ошейникового и 30 тыс. шкур белогубого пекари.
В семействе единственный род: пекари — Tayassu Fischer, 1814 (2 вида).
Род пекари
Genus Tayassu Fischer, 1814
См. описание семейства.
В роде 2 вида:
ошейниковый пекари — Т. tajacu Linnaeus, 1758 (ареал соответствует ареалу семейства);
белогубый пекари — Т. albirostris liliger, 1811 (от Южной Мексики до Парагвая).
СЕМЕЙСТВО БЕГЕМОТОВЫХ
FAMILIA HIPPOPOTAM1DAE GRAY, 1821
Размеры средние н крупные. Длина тела 170—450 см, длина хвоста 15—50 см, высота в холке 70—165 см. Масса 250—3200 кг. Тело бочковидное (рис. 152). Шея короткая, конечности короткие, столбовидные. Морда широкая с большим ртом. Глаза и уши маленькие, располагаются, как и ноздри, высоко на голове. Ноздри могут смыкаться. Волосы щетинковидные, очень редкие. Крупные кожные железы бегемота продуцируют красноватый секрет — «кровавый пот». Он защищает поверхностные слои кожи от набухания в воде. У карликового бегемота пот бесцветный. Кожа содержит слой жира до 5 см толщиной. На конечностях по 4 пальца, между которыми имеется слабо развитая плавательная перепонка. Средние пальцы больше боковых. Сосков одна пара.
Рис. 152. Бегемот — Н, amphibius, пасть бегемота и его череп
Череп массивный, особенно его лицевой отдел. Кости не пневматизированы. В слезной кости нет отверстия слезного протока. Костные слуховые барабаны маленькие.
Зубная формула:
2
1—2
1 4 3
с — р — m — = 38 — 40.
14 3
Резцы и клыки бивнеподобные с постоянным ростом. Нижние клыки достигают особенно больших размеров. Щечные зубы сменяются в течение жизни спереди назад.
Желудок крупный, трехкамерный, «нежвачный». Слепая кишка и желчный пузырь отсутствуют. Плацента диффузная, эпителио-хореаль-ная. Семенники расположены в пахах под кожей.
Распространены в Африке к югу от Сахары.
Населяют берега водоемов. Хорошо плавают и ныряют. Карликовый бегемот приспособлен к жизни на суше в большей степени, чем бегемот. Держатся поодиночке, парами и группами по 6, иногда до 30 особей. День проводят в воде, ночью кормятся на суше. Питаются различными водными и наземными растениями. Самцы метят территорию мочой и экскрементами, разбрасывая их в стороны движениями хвоста. Длительность беременности 200—240 дней. Самка рождает 1 детеныша массой 27—45 кг. Продолжительность лактационного периода 4—5 месяцев. Половозрелость наступает примерно в 3-летнем возрасте.
За бегемотами охотятся из-за жира, мяса и кожи.
В семействе 2 рода (2 вида);
бегемотов — Hippopotamus Linnaeus, 1758 (1 вид);
карликовых бегемотов — Choeropsis Leidy, 1853 (1 вид).
Род бегемотов
Genus Hippopotamus Linnaeus, 1758
В роде единственный вид: бегемот — Н. amphibius Linnaeus, 1758.
Длина тела 400—450 см, длина хвоста 35—50 см, высота в холке 140—165 см. Масса 2000—3200 кг. Самцы значительно крупнее самок. Волосы на теле настолько редки, что животные кажутся безволосыми. Толщина кожи 2—5 см. На шее и груди имеются глубокие складки (рис. 152). Окраска от медно- до темно-коричневой.
В черепе орбита сдвинута назад и вверх.
214 3
Зубная формула: 1 — с —j— р — ш — = 40.
Нижние клыки достигают 64,5 см длины и массы 4 кг.
Распространен в Африке к югу от Сахары и от Гамбии на западе до Эфиопии на востоке.
Населяет различные водоемы, предпочтительно с заводями и болотистыми берегами с зарослями тростников. Может долго оставаться в воде, выставляя над ее поверхностью глаза, уши и ноздри. Во время ныряния бегемот закрывает ноздри и уши. Под водой остается обычно 3 5 мин (детеныши около 20 с), но может до получаса (по другим данным, максимум 6 мин). Благодаря специфическому удельному весу, способен ходить по дну. Держатся поодиночке (обычно старые самцы), парами или небольшими группами до полудюжины голов; лишь иногда встречаются по 20—30 особей вместе. Активен ночью. В поисках пищи обычно не удаляется далее 300 м от воды, но иногда уходит и за 30 км. Пища в основном состоит из травы. В желудках 122 добытых в Уганде бегемотов было зарегистрировано 27 видов трав. Иногда посещает поля. Су-
Рис. 153. Карликовый бегемот — С. liberiensis (самка с детенышем)
точная потребность в корме составляет около 40 кг. Размножается в любое время года, но массовый гон приходится на июнь — сентябрь. Между самцами происходят жестокие драки. Беременность 225— 257 дней, в среднем 233 дня. В помете бывает один, редко два детеныша. Новорожденный имеет более многочисленные волосы на теле, чем взрослые звери. Самка рожает детеныша на земле (в зоопарках имели место роды и в воде). Масса новорожденного 35—40 кг. Он способен сразу ходить и плавать. Самка кормит детеныша как на суше, так и в воде. Длительность лактации 4—6 месяцев. К концу первого года жизни масса детеныша примерно 250 кг. Самки достигают половозрелости примерно в 6 лет. Продолжительность жизни 40—50 лет.
Род карликовых бегемотов
Genus Choeropsis Leidy, 1853
В роде единственный вид: карликовый бегемот — С. Uberiensts Morion, 1844.
Длина тела 170—185 см, длина хвоста 15—17 см, высота в холке 70— 80 см. Масса 250—275 кг. Тело бочкообразное, приземистое, задняя часть ги° немного приподнята. Голова сравнительно маленькая и короткая. Шея короткая и толстая (рис. 153). Глаза и уши не расположены так высоко на голове, как у бегемота. Ноздри крупные и почти округлые. Ко-практически лишена волос, за исключением ушей, губ и хвоста. Спинная сторона тела зеленовато-черная, бока более серые, а брюхо серовато-белое или желтовато-зеленое. Секрет, выделяемый кожными железами, прозрачный.
В черепе, по сравнению с черепом бегемота, сильнее развит мозговой отдел.
„ ‘ 2 1 4 3
Зубная формула: 1 — с -j— р — m — = 38.
Резцы и клыки относительно не столь крупные, как у бегемота.
Распространен в Сьерра-Леоне, Либерии, Гвинее, на западе Берега Слоновой Кости и в Нигерии.
Населяет влажные леса, болота, берега водоемов. Экология изучена слабо. В меньшей степени приспособлен к водному образу жизни, чем бегемот. При испуге спасается бегством в лес. Обычно встречаются поодиночке или парами. Активен ночью. Питается различными растениями. Самка после 200—210 дней беременности рожает на земле одного детеныша. Масса новорожденного 4,5—7 кг. Детеныш остается с самкой до 3-летнего возраста. Продолжительность жизни около 35 лет.
Включен в «Красную книгу» как малочисленный вид, который, однако, в настоящее время не находится под непосредственной опасностью исчезновения.
ПОДОТРЯД ЖВАЧНЫХ
SUBORDO RUMINANTIA SCOPOLI, 1777
Размеры мелкие, средние и крупные. Телосложение у большинства стройное, конечности длинные, четырех- или двупалые. Концевые фаланги пальцев несут настоящие копыта. Копытоходящие животные. Боковые пальцы (если конечность четырехпалая) недоразвиты, и при ходьбе, как правило, не касаются почвы. Половой диморфизм обычно хорошо выражен. У большинства видов есть рога. За малым исключением (Тга-guhdae), все жвачные имеют специфические кожные железы на голове, ь паху, на конечностях. Одна или две пары сосков располагаются в паху.
В черепе характерно сильное или умеренное развитие лицевого отдела, а иногда, наоборот, мозговой части. Орбита сзади замкнутая. Сагиттального гребня нет. Этмоидальные отверстия обычно большие. Сочленовная головка нижней челюсти и соответствующая ей суставная ямка вытянуты в поперечном направлении. На лицевой поверхности слезной кости у многих видов имеется ямка для пр ед глазничной железы.
О 0—1 2—3 3 „
Зубная формула обычно: i —- с - р — — m -— = 28— 34.
3 12 — 3 3
Резцы верхней челюсти отсутствуют, и их заменяет валик сильно ороговевшего эпителия. Верхних клыков у большинства видов нет, но у безрогих форм они имеются и у самцов развиты сильно. Клыки нижней челюсти имеют уплощенные расширенные коронки и по форме напоми-ют резцы. Между клыками и щечными зубами находится диастема. Коренные зубы селенодонтного типа, а у некоторых видов — гипсело-донтного. Первый предкоренной верхней и нижней челюстей не развивается. Пястные и плюсневые кости боковых пальцев в четырехпалой конечности у большинства видов сильно редуцированы или полностью отсутствуют (за исключением Tragulidae). Имеется os canon (только у Hyemoschus из Tragulidae ее нет). Локтевая кость сильно редуцирована в дистальном и среднем отделах и часто срастается с лучевой (у Tragu-1ns она не редуцирована и самостоятельна). От малой берцовой кости сохраняется дистальный отдел в виде лодыжковой кости — os malleola-ге> функционально входящий в предплюсну (у Tragulidae малая берцовая кость развита нормально). Основания поперечных отростков шейных позвонков прободены каналом позвоночной артерии.
Желудок сложный, в большинстве случаев состоящий из 4 отделов, «жвачный» (у Tragulidae в желудке 3 отдела). Первые отделы желудка — рубец, сетка и книжка — относятся к преджелудку и не имеют железистого эпителия. Четвертый отдел — сычуг является железистым отделом. Животные при приеме пищи жуют ее поверхностно, быстро заканчивая кормление. Корм попадает в рубец, там набухает и затем отрыгивается в полость рта для дополнительного пережевывания, которое животное может делать не спеша в каком-либо укромном месте. В Рубце жвачных имеется богатая бактериальная флора, обеспечивающая
бактериальное брожение проглоченной пищи, и очень много простейших, роль которых в основном сводится к химической обработке корма. Бактерии и простейшие, кроме того, синтезируют белки и витамины; затем они усваиваются животными при переваривании этих микроорганизмов. Слепая кишка хорошо развита. Плацента котиледонная, иногда диффузная (Tragulidae).
Ареал подотряда совпадает с ареалом отряда.
Населяют самые различные биотопы. Обычно держатся стадами иногда очень значительными. Лишь представители Tragulidae — одиночные звери. Питаются различными растениями, преимущественно травами. В помете 1—2 детеныша, и только у водяного оленя 4—7.
В подотряде 5 семейств и 60 родов:
оленьковых — Tragulidae Milne-Edwards, 1864 (2 рода);
оленевых — Cervidae Gray, 1821 (12 родов);
жирафовых — Giraffidae Gray, 1821 (2 рода);
вилороговых — Antilocapridae Gray, 1866 (1 род);
полорогих — Bovidae Gray, 1821 (43 рода).
СЕМЕЙСТВО ОЛЕНЬКОВЫХ
FAMILIA TRAGULIDAE MILNE-EDWARDS, 1864
Наиболее мелкие представители отряда. Длина тела 45—85 см, длина хвоста 2—15 см, высота в холке 20—40 см. Масса 2—15 кг. Сложение стройное. По внешнему виду похожи на маленьких оленей. Спина округло-выпуклая, шея короткая. Голова небольшая, рогов не бывает. Конец морды голый. Глаза большие, уши маленькие и заостренные или средних размеров, округлые. Конечности относительно короткие, четырехпалые. Средние пальцы—III и IV значительно больше боковых — II и V.
Волосяной покров густой. На спинной стороне тела окраска его от рыже-буро-черной до серо-черной; имеются белые пятна или полосы. Брюшная сторона тела белая или желтоватая. Предглазничная и межпальцевые кожные железы отсутствуют. Возрастного диморфизма в окраске нет. У Tragulus пари и Т. javanicus на подбородке имеется специфическая кожная железа. Сосков две пары.
Лицевой и мозговой отделы черепа примерно равной длины. Глазницы большие, их края в стороны не выдаются. Сагиттального гребня нет. Слезные кости с одним отверстием слезного канала. Костное небо широкое. Нижняя челюсть со слабо развитым венечным отростком.
. 0 1 3 3
Зубная формула: 1 — с — р — гп = 34.
О 1 О о
Верхние клыки самцов сильно развиты, изогнуты, заострены и выдаются из-под губ. Нижние клыки похожи на резцы. Щечные зубы брахио-донтного типа.
Лучевая и локтевая кости разделены между собой. Малая и большая берцовые кости срастаются в дистальном отделе. Плюсневые кости III и IV пальцев, а также пястные кости III и IV пальцев (исключая Hyemoschus) срастаются в столбчатую кость — os canon.
Желудок трехкамерный (книжка развита очень слабо). Желчный пузырь имеется. Слепая кишка простая. Плацента диффузная.
Распространены в Западной Африке и Юго-Восточной Азии, включая о-ва Шри Ланка, Суматру, Яву, Калимантан и прилежащие мелкие острова.
Обитают в густых лесах, по опушкам леса, в зарослях кустарников и мангровых зарослях. Некоторые прекрасно плавают. Ведут ночной оди-
ночный образ жизни. Питаются различными растениями, их плодами, иногда и животными. Продолжительность беременности 150—172 дня. В помете 1—2 детеныша. Спустя полчаса после рождения он способен подняться на ноги. В 2-недельном возрасте начинает питаться твердой пищей. Размеров взрослого достигает в 5 месяцев.
Служат объектом промысла для местных жителей.
Б семействе 2 рода (4 вида):
водяные оленьки — Hyemoschus Gray, 1845 (1 вид);
азиатские оленьки — Tragulus Brisson, 1762 (3 вида).
Род водяных оленьков
Genus Hyemoschus Gray, 1845
В роде единственный вид: водяной оленек — Н. aquaticus Ogilby, 1841.
Длина тела 75—85 см, высота в холке 35—40 см, длина хвоста 10— 15 см. Масса 10—15 кг. Уши длинные, округлые (рис. 154). Хвост с наружной стороны покрыт более длинными волосами, чем с внутренней. Боковые пальцы развиты хорошо. Спина темно-буро-оливковая или рыже-бурая с мелкими светлыми пятнами. Брюхо желтовато-белое или белое. Специфическая кожная железа на подбородке отсутствует.
Межчелюстные кости не соприкасаются с носовыми.
Рис. 155. Малый оленек — Т. javanicus (Л); пятнистый оленек — Т. memitma (Б) и его череп
Пястные кости не образуют os canon, а плюсневые сливаются в нее лишь у взрослых зверей.
Распространен в Африке от Сьерра-Леоне до Заира и Уганды.
Обитатель густых зарослей кустарников и подроста леса, берегов рек. Экология изучена слабо. Хорошо плавает и ныряет. От опасности спасается в воду. Помимо различных растений поедает насекомых, крабов, рыб, мелких млекопитающих. Держится поодиночке, за исключением периода размножения. Продолжительность беременности 4—6 месяцев. В помете один детеныш. Местное население активно охотится за водяными оленьками. Неволю оленьки переносят очень плохо.
Род азиатских оленьков
Genus Тragulus Brisson, 1762
Длина тела 40—75 см, длина хвоста 2—10 см, высота в холке 20— 35 см. Масса 2,2—8 кг. Конечности тонкие, с маленькими боковыми пальцами (рис. 155). Волосы короткие, плотно прилегающие.
Окраска с рядами продольных пятен (пятнистый оленек) или одноцветная (большой и малый оленьки). Подбородочная железа отсутствует (пятнистый оленек) или имеется (два других вида).
Межчелюстная кость соприкасается с носовой. Пястные кости образуют os canon.
Распространены в Юго-Восточной Азии.
Населяют засушливые места, первичные леса, мангровые заросли. Экология изучена слабо. Одиночные, ночные и сумеречные, очень осто-356
рожные звери. Плавают и ныряют. У пятнистого оленька гон приходится на июнь — июль. Беременность продолжается примерно 150— 155 дней. В помете 1, редко 2 детеныша. Служат объектом промысла для местного населения.
В роде 3 вида:
пятнистый оленек — Т. meminna Erxleben, 1777 (Южная Индия до центральных провинций, о-в Шри Ланка);
большой оленек — Т. пари F. Cuvier, 1822 (от Таиланда и Аннама до Суматры и Калимантана с прилегающими островами);
малый оленек — Т. javanicus Osbeck, 1765 (от Таиланда, Кампучии и Тенассерима до Суматры, Явы и Калимантана с мелкими соседними островами).
СЕМЕЙСТВО ОЛЕНЕВЫХ
FAMILIA CERV1DAE GRAY, 1821
Размеры мелкие, средние, реже крупные. Длина тела 90—310 см, высота в холке 35—233 см. Aiacca 7—825 кг. Тело стройное с длинными ногами. У большинства представителей (за исключением Moschus и Hydropotes) самцы имеют рога (у Rangifer рога имеют и самки). В развитом состоянии рога представляют костные, обычно ветвящиеся образования, сидящие на особых выростах (апофизах) лобных костей. Только у оленей родов Elapodus, Mazama и Pudu рога не ветвятся, у всех же остальных они состоят из основного ствола (штанги) и отростков, число которых бывает различно у разных родов и видов оленей и у разных возрастных групп. Самый нижний, отходящий над розеткой отросток рога — первый надглазничный, расположенный над ним —• второй надглазничный, третий отросток — средний, а все отростки над ним — верхушечные или венечные. Концы отростков, как и вершина основного ствола, могут вторично ветвиться или уплощаться. Рога один раз в году сбрасываются и отрастают вновь. Растущий рог последовательно проходит стадии соединительнотканную, хрящевую и костную. Снаружи в период роста рога покрыты кожей с короткими волосами: на этой стадии их называют пантами. После окостенения рогов кожа на них отмирает и спадает. В основании рога находится поясок («розетка») с неровной поверхностью, по которому происходит будущая резорбция кости и отпадение рога.
Окраска взрослых особей обычно однотонная, темная, иногда со светлыми пятнами. У примитивных форм и у новорожденных большинства видов оленей окраска пятнистая. У специализированных видов (например, северного оленя) и молодых одноцветная окраска. У многие оленей на задней стороне ляжек расположено светлое пятно — «зеркало». Для волос характерен мощно развитый сердцевинный воздухоносный слой. В кожном покрове имеются разнообразные специфические железы: предглазничные, межпальцевые, плюсневые, хвостовые и др. Сосков обычно 2 пары.
Пальцев четыре, реже два. Боковые пальцы—II и V четырехпалой конечности сильно уменьшены и при ходьбе по твердому грунту не каса ются его (исключение представляет северный олень). Боновые пальцы < небольшими копытами.
В черепе пневматизация костей слабая (отмечается лишь у верхне челюстной, слезной'и переднего отдела лобной). На наружной поверхности слезной кости обычно располагается предглазничная ямка специфи
ческой кожной железы. В слезной кости два (в виде исключения одно) отверстия слезного канала. Сагиттальный гребень не развит. Этмоидальные отверстия большие. У видов, имеющих рога, лобные кости простираются далеко назад и теменные кости сильно уменьшены. Межчелюстная кость относительно невелика. Костные слуховые барабаны маленькие и соединены с черепом неплотно.
„ 0 0—1 3 3
Зубная формула: i — с -- р —m—=32 — 34.
3 1 3 3
Щечные зубы селенодонтные, брахиодонтные. Верхние клыки обычно редуцированы, иногда же развиты хорошо или даже гипертрофированы (у безрогих). Форма нижних клыков сходна с формой резцов.
Плюсневые кости боковых пальцев четырехпалой конечности редуцируются, а от пястных сохраняются лишь их верхние или нижние части; иногда исчезают и они.
Желудок сложный, четырехкамерный. Желчный пузырь, за редким исключением, отсутствует. Плацента котиледонтная (число котиледонов меньше, чем у Bovidae) или диффузная.
Ареал охватывает Северную, Центральную и Южную Америку (кроме Огненной Земли и Вест-Индии); береговую полосу Гренландии, Северо-Западную Африку; Европу и Азию с прилежащими островами: Шпицберген, Новая Земля, Северная Земля, Новосибирские о-ва; острова Восточной Азии, кроме Курильских и островов Берингова моря; все острова между Азией и Австралией и Шри Ланка.
Населяют в основном леса на равнинах, в предгорьях и горах. Заходят в альпийский пояс и лесостепь. Один вид обитает в тундре. Держатся обычно группами. Питаются различными растениями. Для некоторых оленей характерны миграции. Самки обычно моноэстральные, хотя тропические виды могут быть полиэстральными. Обычно период гона приходится на осень — зиму. Длительность беременности примерно от 5 до 10 месяцев. Число детенышей в помете варьирует от 1 до 4—7.
В семействе 5 подсемейств и 12 родов (32 вида);
подсемейство Moschinae Zittel, 1893;
род кабарог — Moschus Linnaeus, 1758 (1 вид);
подсемейство Hydropotinae Trouessart, 1898;
род водяных оленей — Hydropotes Swinhoe, 1870 (1 вид); подсемейство Cervulinae Sclater, 1870;
род мунтжаков — Muntiacus Rafinesque, 1815 (1 вид);
род хохлатых оленей — Elaphodus Milne-Edwards, 1872 (1 вид);
подсемейство Odocoileinae Pocock, 1923;
род косуль — Capreolus Gray, 1821 (1 вид);
род американских оленей — Odocoileus Rafinesque, 1832 (6 видов);
род мазам — Mazama Rafinesque, 1817 (4 вида);
род пуду — Pudu Gray, 1852 (2 вида);
род лосей — Alces Gray, 1821 (1 вид);
род северных оленей — Rangif er Н. Smith, 1827 (1 вид); подсемейство Cervinae Baird, 1857;
род оленей Давида-—Elaphurus Milne-Edwards, 1866 (1 вид);
род оленей — Cervus Linnaeus, 1758 (12 видов).
Систематика семейства окончательно не установлена. Ряд исследователей (например, Симпсон, 1945) помимо указанных выше родов выделяют также Blastocerus Wagner, 1844; Ozotoceras Ameghino, 1891 (=Blastoceros Fitzinger, 1860); Hippocamelus Leuckart, 1816; Axis FL Smith, 1827; Damn Frisch, 1775.
Все представители семейства служат объектами охоты: от них получают мясо, кожу и сырье для фармацевтической промышленности.
SVBFAMILIA MOSCHINAE ZITTEL, 1803
Размеры мелкие. Рогов нет. Из кожных специфических желез имеются только хвостовая и мускусная в области препуциума.
В черепе костные слуховые барабаны очень маленькие. На слезных костях по одному отверстию слезных каналов и нет вдавления для предглазничной железы.
0 13 3
Зубная формула: i —- с — р m —- =34. О 1 О о
Верхние клыки у самцов очень длинные, а у самок короткие.
Боковые пальцы развиты хорошо и имеют нормальный скелет фаланг. От метакарпальных и метатарзальных костей II и V пальцев сохраняются лишь дистальные отделы. Желчный пузырь имеется.
Род кабарог
Genus Moschus Linnaeus, 1758
В роде единственный вид: кабарга — М moschiferus Linnaeus, 1758. Длина тела 85—100 см, длина хвоста 4—6 см, высота в холке 55— 67 см, высота в крестце 67—80 см. Масса 10—17 кг. Задние конечности длиннее передних, поэтому задняя часть животного значительно выше передней (рис. 156). Голова относительно небольшая, с большими глазами. У самцов сильно развиты верхние клыки, выступающие из-под верхней губы и направленные вниз, — они достигают 7 см в длину. У самок верхние клыки короткие и за пределы губы не выступают. На конце морды расположено большое голое поле, окружающее ноздри. Уши длинные и широкие с округлыми вершинами. Хвост очень мал, скрыт в волосах, у самцов голый — только на конце находится пучок волос. Конечности четырехпалые с хорошо развитыми боковыми пальцами, копыта которых достигают основания копыт средних пальцев. У стоящего животного они касаются почвы.
Общая окраска туловища от рыжеватой до черно-коричневой. По спине и бокам разбросаны светлые буровато-охристые или рыжеватожелтые пятна. Обычно они расположены продольными рядами. С возрастом пятнистость уменьшается и может исчезнуть вовсе. По нижней стороне головы и шеи тянутся парные белые полосы. Брюхо светло-бурое или сероватое. Волосяной покров состоит в основном из плотно прилегающих друг к другу, слегка извилистых крупных остевых волос. Зимой В них чрезвычайно сильно развит сердцевинный слой. Пуховые волосы немногочисленны.
Предглазничная кожная железа отсутствует. Хвостовая и большая мускусная железа (расположенная в особом мешочке на брюхе в области препуциума) функционально, по-видимому, связаны с половой деятельностью кабарог. Сосков одна пара.
В черепе сильно развит мозговой отдел, а лицевая часть короткая. Слезные кости широкие, без углубления для предглазничной железы. Этмоидальные отверстия небольшие. Носовой отросток верхнечелюстной кости тянется далеко назад, достигая или почти достигая лобной кости.
Верхние клыки у самцов с постоянным ростом, имеют значительную длину — до 7—10 см и острый режущий задний край. Нижние клыки напоминают по форме резцы; коронки их уплощены и лопатообразно расширены. Позвоночник сильно изогнут. Грудная клетка короткая и узкая.
Рис. 156. Кабарга — Л4. то-schiferus и ее череп
Распространена в Восточной Азии почти от северной границы леса до Южного Китая, Ассама, Бирмы и в Гималаях; в СССР — на Алтае, в Саянах, в Восточной Сибири, на Дальнем Востоке (кроме Камчатки).
Обитатель горной тайги. Прекрасно приспособлена к лазанию и прыганию по скалам. Держатся, за исключением периода размножения, как правило, поодиночке. Ведет оседлый образ жизни, строго придерживаясь своего индивидуального участка размером обычно в 200—300 га. Активна в сумерки и ночью. В пище важное значение имеют древесные и в меньшей степени наземные лишайники. Кроме того, круглый год использует в пищу древесные корма: тонкие ветки и молодые побеги, хвою и кору (главным образом зимой), листья и почки. Летом основным кормом служат травянистые растения. Изредка посещает солонцы. Гон приходится на ноябрь — декабрь. Беременность длится 185—195 дней. Латентного периода в развитии зародыша нет. Самка приносит в мае — июне двух, реже одного и еще реже трех детенышей. Масса новорожденного 300—500 г. Половозрелость наступает в возрасте 15—17 месяцев.
Враги — росомаха, рысь, на Дальнем Востоке харза, в меньшей степени волк, тигр, медведь.
Кабаргу промышляют главным образом из-за мускусной железы, секрет которой идет в парфюмерную промышленность в качестве закрепителя запаха при изготовлении высококачественных духов.
Иногда в роде насчитывают 3 вида: М. chrysogaster Hodgson, 1839; М. berezovskii Flerov, 1928; М. moschiferus Linnaeus, 1758, илг M. moschiferus, М. berezovskii и М. sibiricus Pallas, 1779 (Флеров 1952; Соколов, 1959; Уайтхэд, Whitehead, 1972), что, по-видимому, недо статочно обосновано; изредка кабарог выделяют в особое семействе (например, И. И. Соколов, 1959).
SUBFAMILIA HYDROPOTINAE TROUESSART, 189?
Размеры мелкие. Рогов нет. Из кожных специфических желез име ются предглазничная, паховая и на задних конечностях — межпальцевая
В черепе костные слуховые барабаны очень большие. На слезны? костях по два отверстия слезных каналов.
0 13 3
Зубная формула: i —- с —— р - m -= 34.
3 13 3
Верхние клыки у самцов очень длинные, у самок короткие. От мета карпальных и метатарзальных костей II и V пальцев сохраняются липп дистальные отделы.
Желчного пузыря нет.
Некоторые исследователи (например, Симпсон, 1945) включают во дяных оленей в состав подсемейства лосей.
Род водяных олене»
Genus Hydropotes Swinhoe, 187(
В роде единственный вид: водяной олень-—М. inermis Swinhoe, 1870 Длина тела 75—97 см, длина хвоста 5—8 см, высота в холке 45— 55 см. Масса 12—15 кг. Тело приземистое с относительно короткими конечностями. Профиль спины несколько округлый. Шея недлинная. Го лова небольшая с укороченной мордой (рис. 157). Глаза средних раз меров. На конце морды большой голый участок кожи. Уши короткие округлые, покрыты густыми волосами. Задние конечности выше перед них. Боковые пальцы развиты слабо и расположены высоко. Коротки»
Рис. 157. Водяной олень — 11. inermis и его череп
хвост густо покрыт волосами. В волосяном покрове пуховые волосы редки. Окраска желто-бурая до светло- или темно-рыже-бурой летом и темно-бурая зимой. Брюхо светлее спины. Из специфических кожных желез имеются небольшие предглазничные, маленькие паховые (единственный случай среди оленей) и на задних конечностях — межпальцевые. Сосков 2 пары.
В черепе этмоидальные отверстия средних размеров. Слезные кости с ямками для предглазничных желез. Ямки у самцов более глубокие. Верхние клыки самцов достигают длины 6 см; они слегка изогнуты назад, с острым задним краем, подвижные. Клыки самок небольшие. Резцы с длинными лопатообразными коронками.
Распространен по берегам и островам р. Янцзы, в восточных районах Центрального Китая и на Корейском п-ове.
Населяет преимущественно заросли по берегам рек и озер на равнинах и возвышенностях. Питается в основном травянистыми растениями. За исключением периода размножения, держатся в большинстве случаев поодиночке. Характерен тревожный крик в виде своеобразного лая. Гон приходится на декабрь. Продолжительность беременности около 6 месяцев. Самка рождает от 2 до 7 (максимальное число в семействе) дете
нышей в июне — июле. Спинная сторона новорожденного в белых пятнах. Половозрелость наступает в 6—18 месяцев. Продолжительность жизни 10—12 лет.
SUBFAMILIA CERVVLINAE SCLATER, 1870
Размеры мелкие. У самцов есть рога. Из кожных специфических желез имеются предглазничная, лобная, подбородочная и на задних конечностях — межпальцевая.
В черепе костные слуховые барабаны очень маленькие. На слезных костях по два отверстия слезных каналов.
013 3
Зубная формула: i —- с — р — m — =34, 3 13 3
Верхние клыки самцов очень длинные, самок — короткие. От метакарпальных костей II и V пальцев сохраняются лишь проксимальные отделы.
Желчного пузыря нет.
Род мунтжаков
Genus Muntiacus Rafinesque, 1815
В роде единственный вид: мунтжак — М. muntjak Zimmermann, 1780. Длина тела 89—135 см, длина хвоста 13—23 см, высота в холке 40— 65 см, масса 40—50 кг. Тело приземистое на относительно коротких ногах, спина округлая (рис. 158). Шея короткая. Профиль головы прямой. На конце морды расположен безволосый участок кожи. Глаза и уши средних размеров. Вершины ушей округлые. У самцов имеются простые (с очень небольшим надглазничным и иногда двумя концевыми отростками) рога длиной 4—25 см. Пеньки рогов очень длинные и далеко выступают за задний край черепа, рога же короткие с 1—3 отростками. Задние конечности длиннее передних. Боковые копыта маленькие. Волосяной покров почти без пуха, низкий и редкий у особей, обитающих в тропиках и более высокий и густой у особей из северных частей ареала. Окраска спины от желтовато- или серовато-охристой до бурой и черно-бурой; брюхо беловатое. Самки светлее самцов. Молодые звери пятнистые. Предглазничные железы очень сильно развиты. Имеются также лобная, парная подбородочная и на задних конечностях — межпальцевая железы. Плюсневой железы нет.
Диплоидное число хромосом 46.
Распространен на о-ве Шри Ланка, в Восточной Индии, Юго-Восточном Тибете, Бирме, на п-овах Индокитай и Малакка, в Китае (на север до 32° с. ш.), на о-вах Тайвань, Хайнань, Калимантан, Суматра, Ява, Бали и прилежащих мелких островах.
Населяет густые заросли в лесах и антропогенных ландшафтах преимущественно недалеко от воды. Поднимается в горы до верхней границы леса (примерно до 4 тыс. м над уровнем моря). Активен в сумерках и ночью. Держатся поодиночке и парами, иногда семьями. Питается различными травами, листьями кустарников, плодами, грибами т. д. Вовремя гона или будучи испуганными издают громкие лающие звуки. Если опасность не исчезла, мунтжак может «лаять» в течение часа и более. Сезонности в размножении нет. Наибольшее число рождений на Яве и Суматре приходится на вторую половину года. Длительность беременности около 6 месяцев. В помете один, редко два детеныша. Масса олененка при рождении составляет 550—650 г. Молодой олень переходит к
самостоятельной жизни в 6-месячном возрасте. Половозрелость наступает у самок в 7—8 месяцев, а у самцов в годовалом возрасте. Продолжительность жизни 12—15 лет. Основные враги — тигр и леопард.
Местные жители охотятся за мунтжаком из-за мяса и шкуры. Пойманные мунтжаки хорошо переносят неволю.
Некоторые исследователи насчитывают в роде от 4 видов (Андерсон и Джонс, Anderson a. Jones, 1967) до 6 (Уокер, Walker, 1964). По Эл-лермэну и Моррисон-Скотту (Ellerman a. Morrison-Scott, 1966) и Уайт-
хэду (1972), в роде 5 видов: М. muntjak Zimmermann, 1780 (п-ов Индостан, о-в Шри Ланка, Бирма, Таиланд, п-ов Индокитай, п-ов Малайка, о-ва Хайнань, Калимантан, Суматра, Ява); М. rooseveltorum Osgood, 1932 (п-ов Индокитай); М. reevesi Ogilby, 1839 (Восточный Китай и о-в Тайвань); М. crinifrons Sclater, 1885 (Восточный Китай) и М. feae Thomas et Doria, 1889 (Таиланд). Следуя Хальтенорту (Haltennorth, 1963), более правильно объединить их в один вид.
Подвиды из Тенассерима — М. т. feae Thomas et Doria, 1889, и Юго-Восточного Китая — М. т. crinifrons Sclater, 1885, включены в «Красную книгу»: первый как находящийся под угрозой исчезновения, а второй как малочисленный подвид, состояние которого неизвестно.
Род хохлатых оленей
Genus Elaphodus Milne-Edwards, 1872
В роде единственный вид: хохлатый олень — Е. cephalophus Milne-Edwards, 1872.
Длина тела НО—160 см, длина хвоста 7—15 см, высота в холке 52— 72 см. Масса около 50 кг. По внешнему виду напоминает мунтжака, лишь хвост относительно короче, и неразветвленные рога скрыты в длинных волосах, покрывающих лоб. Пеньки рогов короткие. Волосяной покров густой, грубый, зимой темнее и гуще, чем летом. Окраска спинной стороны тела бурая до черно-бурой или серо-черная до черной. Голова и шея более светлые. Брюхо слегка светлее спины. Самцы с темной шапкой длинных (до 17 см) волос на лобной части головы. Лобной кожной железы нет. Предглазничная железа крупная.
В черепе этмоидальные отверстия маленькие или их нет вовсе. Ямки на слезных костях сильно развиты. Верхние клыки у самцов удлинены, изогнуты, но не обладают подвижностью.
Распространен в Северной Бирме, Юго-Восточном Тибете и Китае между 22 и 33° с. ш.
По образу жизни сходен с мунтжаком. Держатся поодиночке, иногда парами. Питается травянистыми растениями. Период спаривания в Бирме приходится на апрель — май. И самец, и самка «лают» во время гона или при испуге. Длительность беременности около 6 месяцев. Детенышей в помете один, редко два. Олененок имеет такую же окраску, как и взрослые, но с каждой стороны от средней линии спины тянется по ряду светлых пятен.
SUBFAMILIA ODOCOILEINAE POCOCK, 1923
Размеры мелкие, средние или крупные. У самцов есть рога (у северных оленей их имеют и самки). Рога имеют различную форму — от не-разветвленной спицы до сложноразветвленных; надглазничный отросток обычно отсутствует. Из кожных специфических желез имеются (в разной комбинации у разных видов): предглазничная, лобная, предплюсневая, плюсневая, тарзальная и на задних конечностях — межпальцевая.
В черепе костные слуховые барабаны маленькие или средней величины. Сошник, как правило, разделяет хоаны. На слезных костях по два отверстия слезных каналов. На этих костях имеется впадина для предглазничной железы (отсутствует у косули и одного вида пуду).
о 0 — 1 3 3 „„ „
Зубная формула: i с —;— р m — = 32— 34,
3 10 0
Вепхние клыки маленькие или их нет вовсе.
Рис. 159. Косуля — С. capreolus и типы рогов косуль, обитающих в СССР (верхний ряд: европейская косуля с Карпат — С. с. capreolus, остальные: сибирская косуля — С. с. pygargus из Прибайкалья)
От метакарпальных костей II и V пальцев сохраняются лишь дистальные отделы. Cuneiforme отделено от naviculo-cuboideum или срастается с ним.
Желчного пузыря нет.
Род косуль
Genus Capreolus Gray, 1821
В роде единственный вид: косуля — С. capreolus Linnaeus, 1758.
Размеры мелкие. Длина тела 100—150 см, длина хвоста 1—2 см, высота в холке 65—100 см. Масса 20—59 кг. Телосложение стройное, шея длинная, голова небольшая (рис. 159). Ноги тонкие и высокие, задние конечности немного длиннее передних. Хвост короткий, прикрыт мехом и снаружи почти незаметен. Безволосое пятно на конце морды большое, занимает пространство между ноздрями и охватывает их снаружи. Уши длинные и широкие. Рога относительно небольшие; их стволы почти прямые, округлые в сечении, без надглазничного отростка. Обычно ствол рога на вершине разделяется на три отростка: один направлен вперед («средний» отросток), один вверх (конец ствола) и один назад («задний» отросток). Ствол рога от самого основания усажен большим числом удлиненных выступов и бугорков. Окраска волосяного покрова од
ноцветная, летом рыжая с более светлым брюхом, а зимой серая с различными оттенками от бурой до рыжеватой. Околохвостовое «зеркало» обычно белое. Новорожденные телята пятнистые. Предглазничные железы маленькие. Межпальцевые железы сильнее развиты на задних конечностях. Предплюсневые железы крупные. Сосков 2 пары.
Череп с несколько укороченным лицевым отделом. Ямка предглаз-ничной железы на слезной кости выражена очень слабо. Этмоидальные отверстия большие.
0 0 3 3
Зубная формула: i —- с —- р —- m — = 32. 3 13 3
Верхние клыки, как правило, отсутствуют (иногда бывают у молодых самцов).
Диплоидное число хромосом у европейской косули 70, сибирской — 68.
Распространение охватывает умеренный пояс Евразии, всю Западную, Южную и Среднюю Европу (включая Англию, южную часть Скандинавского п-ова, Финляндию и Сицилию), Малую Азию, Сирию и Палестину, Северный Ирак, Северный Иран; в СССР — в европейской части на восток до Ленинградской, Новгородской, Калининской, Московской, Брянской, Орловской, Харьковской, Воронежской, Днепропетровской областей и Молдавии, Кавказ, Крым, горы Средней Азии, Казахстан, Сибирь до 58—60° с. ш., Дальний Восток; северные и восточные районы Центральной Азии, Корейский п-ов.
Населяет преимущественно несплошные смешанные и широколиственные леса, перемежающиеся прогалинами, лужайками и вырубками в лесной и лесостепной зонах. Встречается также в зарослях по берегам степных и даже полупустынных озер; хорошо уживается в условиях антропогенного ландшафта. Держатся в летнее время обычно поодиночке. Осенью иногда сбиваются в смешанные стада, насчитывающие до 20— 30 голов. К зиме эти группы разбиваются на более мелкие — по 2— 4 особи. Активны летом утром, вечером и с перерывами ночью, а зимой — круглосуточно. Во многих частях ареала имеют место сезонные кочевки. Так, в лесостепной зоне косули уходят осенью в малоснежные районы, а весной — на летние пастбища. В горах наблюдаются вертикальные миграции.
Питается летом в основном разнообразными травами, листвой и побегами кустарников и деревьев, а зимой главным образом тонкими ветвями и почками кустарников и подроста деревьев, засохшей травой Охотно посещает солонцы. Самцы «лают» при тревоге.
Период гона приходится на июль — октябрь, иногда ноябрь (в разных участках ареала по-разному). Косулям свойственна ограниченная полигамия. Самец в течение нескольких дней держится с одной самкой, а потом нередко гоняется за второй, а иногда и за третьей. Длительность беременности варьирует от 6 до 10 месяцев, обычно около 9 месяцев. Характерна латентная стадия в развитии оплодотворенной яйцеклетки (у самок, спаривающихся в ноябре — декабре, латентной стадии, как правило, не бывает). Телята рождаются обычно во второй половине мая — июне. В помете бывает 2, реже 3 детеныша. Новорожденные первые 6—8 дней остаются на месте, затаившись в траве; в возрасте двух недель уже не отстают в беге от матери. Продолжительность лактации 2—3 месяца. В месячном возрасте молодые уже начинают поедать траву. Самки живут до 11 —12, самцы — до 16 лет. Главные враги — волк, росомаха и рысь.
Косуля —объект охоты; используются мясо и кожа.
Рис. >60. Белохвостый олень — О. virginiana.
А — самец, J5 — самка с теленком
Рис. 160. Продолжение.
Череп белохвостого оленя
Род американских оленей
Genus Odocoileus Rafinesque, 1832
Размеры мелкие и средние. Длина тела ПО—200 см, длина хвоста 10—35 см, высота в холке 70—120 см. Масса 22—205 кг. Телосложение стройное, конечности высокие (рис. 160). Шея средней длины, изящная. Туловище удлиненное. Голова относительно небольшая. На конце морды имеется голое пятно. Уши средней величины или большие (у О. hemionus и О. dichotomus до 20 см длиной), заостренные или слегка округлые. Хвост короткий или средней длины. Рога в большинстве случаев средних размеров (от 25 до 79 см длиной) с небольшим или большим числом отростков. У рогов первый — надглазничный — отросток расположен низко (О. antisensis и О. bisulcus) или высоко (остальные виды). Он маленький у О. virginianus и О. hemionus или мощный у остальных видов, раздвоенный (О. hemionus и О. dichotomus) или цельный (прочие виды). Главный ствол над надглазничным отростком изгибается вперед и внутрь; очень слабо у О. antisensis и О. bisulcus, слабо у О. bezoarticus и О. dichotomus или сильно у О. hemionus и О. virginianus. Остальные отростки отсутствуют или немногочисленны и коротки у О. antisensis и О. bisulcus или коротки и немногочисленны у О. bezoarticus или длинны и многочисленны у остальных видов, причем в большинстве случаев они дихотомически ветвятся (особенно у О. dichotomus и О. hemionus).
Волосяной покров летом низкий, плотно прилегающий к телу, а зимой густой и относительно высокий. Пуховые волосы всегда редкие и короткие. Окраска спины от серо-желтой и серо-бурой до желто-рыжей, рыжей и темно-рыжей. Брюшная сторона белая. У чернохвостого оленя линька два раза в году: осенью и весной. Зимний мех светлее летнего. Молодые олени имеют белые пятна, образующие продольные ряды. У О. dichotomus, О. antisensis и О. bisulcus телята одноцветные.
Предплюсневые и предглазничные специфические кожные железы имеются у всех представителей рода, а плюсневые развиты только у td. virginianus (за исключением южноамериканских форм) и у О- hemionus-, у О. dichotomus их нет вовсе. Межпальцевые железы у О. virginianus и О. hemionus имеются на передних (здесь они развиты слабо) и задних конечностях, а у О. bezoarticus — только на задних.
Череп с прямым профилем черепной коробки. Глазницы средней величины. Ямка предглазничной железы на слезной кости у большинства видов хорошо выражена. Этмоидальные отверстия большие. Носовые и
предчелюстные кости не соприкасаются (за исключением О. antisensis н О. bisulcus). Костные слуховые барабаны обычно маленькие.
0 13 3
Зубная формула: i —— с — р — m -=32(34).
3 13 3
Верхний клык небольшой или его нет вовсе.
Диплоидный набор хромосом у белохвостого и чернохвостого оленей по 70.
Распространены в Северной, Центральной и Южной Америке от 62° с. ш. до 48° ю. ш.
Обитатели равнин и гор (поднимаются на высоту до 4,5 тыс. м над уровнем моря), болот, глухих и изреженных лесов, антропогенных ландшафтов, высокогорных степей. Обычно держатся небольшими семейными группами, иногда объединяются в крупные стада (особенно О. vir-ginianus и О. hemionus зимой при глубоком снеге). Питаются различными травами, листьями и побегами кустарников, грибами, орехами и т. д.
Гон и смена рогов происходят в определенные сезоны года или не приурочены к ним. Чернохвостый олень сбрасывает рога с января по март; новые рога становятся твердыми и очищаются от кожи у взрослых самцов в конце августа — начале сентября, у белохвостого оленя эти даты, соответственно, январь — февраль и сентябрь. В США гон у чернохвостого оленя происходит с конца сентября до середины ноября, а у белохвостого — с октября по начало декабря. В Южной Америке гон белохвостого оленя не приурочен к определенному сезону, хотя в Перу наиболее активно происходит в феврале — марте. Беременность продолжается 6,5—12 месяцев. Телята чернохвостого оленя рождаются с апреля по июль, но большинство -— в июне, белохвостого оленя в Северной Америке — в основном в мае и начале июня. Детенышей в помете 1—4. Олененок начинает подкармливаться твердой пищей уже в возрасте нескольких дней. Молодая самка обычно следует за матерью до 2-летнего возраста, а самец покидает ее в годовалом возрасте.
Продолжительность жизни 12—15 лет, а в неволе до 20 лет. В США численность чернохвостых оленей в 1969 г. была около 3 млн. голов, а белохвостых — 8 млн. (по сравнению с 40 млн. в 1818 г.).
Белохвостый и чернохвостый олени служат объектами спортивной охоты в США. В 1969 г. в США по лицензиям было добыто около 600 тыс. голов чернохвостых оленей.
В роде 6 видов:
белохвостый олень — О. virginianus Zimmermann, 1780 (Северная Америка к югу примерно от 55° с. ш., кроме следующих штатов США: Калифорнии, почти всей Невады, юго-востока Орегона, юго-запада Айдахо, почти всей Юты, западной половины Колорадо, северо-запада Нью-Мексико, северной половины Аризоны; вся Мексика, но отсутствует на п-ове Калифорния; Центральная Америка; Южная Америка на юг примерно до 12° ю. ш.);
чернохвостый олень — О. hemionus Rafinesgue, 1817 (запад Северной Америки: Канада — крайний юг провинции Юкон, Британская Колумбия, Альберта, Саскачеван, юг Манитобы; западные штаты США на восток, до линии, проходящей через восточные районы штата Миннесота, восток Небраски, центральные районы Канзаса, запад Оклахомы, запад Техаса; южная часть ареала двумя языками вдается в Мексику: на западе— на п-ове Калифорния и в провинции Сонора и в центральной части страны в провинциях Коауила, северо-западе Чиуауа, северо-западе Дуранго и севере Закатекас);
болотный олень — О. dichotomus liliger, 1811 (Бразилия и на юг до Уругвая, на запад до Парагвая и Северной Аргентины);
пампасный олень — О. bezoarticus Linnaeus, 1766 (центральные части Южной Америки: Бразилия, Уругвай, Аргентина, Боливия, Парагвай);
перуанский олень — О. antisensis D’Orbigny, 1834 (Эквадор, Перу Боливия, Северная Аргентина, Чили, Уругвай);
южноандский олень — О. bisulcus Molina, 1782 (Чили, Южная Аргентина).
Некоторые исследователи (например, Симпсон, 1945; Кабрера, Cabrera, 1957—1961; Андерсон и Джонс, 1967; Уайтхэд, 1972) в роде Odocoi-teas оставляют только 2 вида: белохвостого и чернохвостого оленей; болотного оленя выделяют в род Blastocerus Wagner, 1844 (по Андерсону и Джонсу, Blastocerus Gray, 1850); пампасного оленя в род Blasloceros Fitzinger, 1860 (= Ozotoceras Ameghino, 1891); перуанского и южноандского оленей — в род Hippocatnelus Leuckart, 1816. По-видимому, более правильно объединить их всех в один род так, как это делает Хальтенорт (1963). Иногда (например, Холл и Келсон, 1959) название рода «Odo-coileus» заменяют названием «Dama Zimmermann, 1780».
В «Красную книгу» включены:
подвиды, находящиеся под опасностью исчезновения, — подвид белохвостого оленя из низовий р. Колумбия, США, О. virginianus leucurus Douglas, 1829 (численность к 1972 г. составила 300—500 голов) и подвид чернохвостого оленя с о-ва Цедрос у западного побережья п-ова Калифорния, О. hemionus cerrosensis Merriam, 1898 (численность к 1972 г. очень низка; отмечена группа в 7 голов);
малочисленные виды, которые в ближайшее время могут оказаться под угрозой исчезновения:
перуанский олень (общая численность неизвестна; в Чили к 1972 г. их было не более 50 голов);
болотный олень (общая численность неизвестна; в Бразилии к 1972 г. их было около 2 тыс. голов, в Уругвае они почти полностью исчезли, в Перу 150—200 голов, в Боливии еще многочислен);
пампасный олень как малочисленный вид, состояние которого неясно подвид белохвостого оленя из Южной Флориды — О. virginianus cal-vium Barbour et G. Allen, 1922, как подвид, находившийся под угрозок исчезновения и ныне переставший быть таковым в результате предпринятых мер по его охране (численность к 1972 г. возросла до 600 голов пс сравнению с 300 голов в 1960 г.).
Род мазав
Genus Mazama Rafinesque, 181
Размеры мелкие. Длина тела 80—135 см, длина хвоста 7—17 см, вы с°та в холке 30—75 см. Масса 8—25 кг. Шея и голова коротки (рис. 161). На конце морды имеется безволосый участок кожи. Глаза средней величины, верхнее веко с густыми длинными черными ресница ми. Уши укороченные округлые. Конечности тонкие или несколько утол Щенные. Копыта небольшие, боковые копыта очень малы. Рога коротки
СМ длины)’ неветвящиеся, прямые, направлены наискось назад.
Окраска спины от желто-рыжей до темно-рыже-бурой или черноватс рыжей или от светло-серо-бурой до темно-серо-бурой. Брюхо желто-рь желтоватое или белое. На лбу имеется шапка удлиненных воло< С-езонная изменчивость волосяного покрова невелика или отсутствуе новорожденного на спине расположены белые пятна. Из специфши ских кожных желез у всех видов имеются межпальцевые железы, a nrej
глазничных и предплюсневых иногда нет. Плюсневых желез нет. Сосков 2 пары.
Череп короткий с утолщенным лицевым отделом. Лоб вогнут. Глазница большая. Ямка на слезной кости небольшая или глубокая. Этмоидальные отверстия крупные. Костные слуховые барабаны маленькие.
„ , о 0—1 3 3
Зубная формула: i — с - р — m -=32—34.
3 13 3
Верхний клык обычно отсутствует, иногда имеется, но небольших размеров.
Диплоидный набор хромосом большой мазамы 68.
Распространены от Центральной Мексики до Центральной Аргентины.
Обитают в глухих лесах, или в светлых лесах с кустарниковыми зарослями, или в саваннах на равнинах и в горах до 5 тыс. м над уровнем
моря. Обычно держатся около водоемов, хорошо плавают. Встречаются поодиночке или нарами. Активны рано утром, вечером или ночью. Питаются травянистыми растениями, зелеными побегами. У тропических Форм нет сезонности в размножении. Беременность длится около 7 месяцев. Детенышей в помете один, редко два. Половозрелость наступает в 6 18 месяцев. Продолжительность жизни 10—15 лет. Основные враги — ягуар, пума и крупные змеи (анаконда, боа). Местные жители охотятся на мазам: эти олени часто уничтожают посевы на полях, мясо их идет в пищу.
В роде 4 вида:
большая мазама—М. americana Erxleben, 1777 (от Центральной Мексики на юг до Южной Боливии, Парагвая и Северной Аргентины);
серая мазама — М. gcuazoubira Fischer, 1814 (от Юкатана на юг до Центральной Аргентины);
Рис. 163. Лось — A. alces
малая мазама — М. папа Lesson, 1842 (от Колумбии и Венесуэлы на юг до Северной Аргентины, Северного Парагвая и Боливии);
карликовая мазама — М. bricenii Thomas, 1908 (Западная Венесуэла, Северо-Западная Боливия и Юго-Восточное Перу).
Некоторые исследователи (например, Хальтенорт, 1963) включают род пуду в род мазам. По Уокеру (1969), в роде 10 видов. Иногда (например, Кабрера, 1957—1961) малую и карликовую мазам объединяют на правах подвидов в вид М. rufina Bourcier et Pucheran, 1852.
Род пуду
Genus Pudu Gray, 1852
Размеры мелкие. Длина тела 80—93 см, длина хвоста 5—10 см, высота в плечах 30—35 см. Масса 7—11 кг. По внешнему виду похожи на мазам (рис. 162). Спина дугообразно выгнута. Конечности, шея и голова короткие. Уши невысокие, округлые, покрыты густыми волосами. Хвост 374
можег отсутствовать (северный пуду). Рога неветвящиеся, короткие. Длиной 7—10 см. Волосяной покров густой. Окраска его на спине темно-рыже-бурая или темно-серо-бурая, на брюхе рыжеватая. Имеются пред-глазничные и межпальцевые кожные железы. Предплюсневые и плюсневые железы отсутствуют.
В черепе предчелюстные кости не достигают носовых. Ямка предглаз-ничной железы на слезной кости мелкая или глубокая.
„ 0 0 3 3
Зубная формула: 1 — с - р —- m -— = 32.
3 13 3
Верхних клыков нет.
Распространены в Южной Колумбии, Эквадоре, Северном Перу, Чили и Западной Аргентине.
Населяют густые заросли кустарников в лесах от уровня моря до 4 тыс. м над его уровнем. Питаются травой, плодами, семенами, листьями кустарников. Держатся поодиночке, парами или семейными группа-
Рис. 164. Некоторые типы рогов лося — A. alces. А — среднерусский лось — A. a. alces; Б — рога лося из Брянской области; В—Г — рога лося — A. a. alces из Западной Сибири; Д — рога уссурийского лося — А. а. canieloides
ми. Продолжительность беременности примерно 7 месяцев. В помете один детеныш. Вдоль спины у него от плеч до хвоста проходят 3 ряда белых пятен.
В роде 2 вида:
южный пуду — Р. pudu Molina, 1782 (Чили и Западная Аргентина);
северный пуду — Р. mephistophiles De Winton, 1896 (Южная Колумбия, Эквадор, Северное Перу).
Южный пуду очень редок и включен в «Красную книгу».
Род лосей
Genus Alces Gray, 1821
В роде единственный вид: лось — A. alces Linnaeus, 1758.
Размеры крупные, наибольшие в семействе. Длина тела 240—310 см, длина хвоста 5—10 см. Масса 300—820 кг. Телосложение тяжелое. Конечности длинные, туловище короткое, голова большая, удлиненная. Тело в холке выше, чем в крестце (рис. 163). Хвост короткий, но ясно видный снаружи. Верхняя губа очень большая, свисающая над нижней. Конец морды покрыт волосами, за исключением небольшого ромбического или треугольного участка посредине верхней губы. Ноздри большие, направлены вниз. Уши длинные, широкие. На горле имеется свисающий вниз кожный вырост, сильнее развитый у самцов (иногда достигает в длину 40 см), функция его не ясна, специфические железы в нем отсутствуют.
Форма рогов изменчива от оленьего типа с толстым коротким стволом и отростками, направленными вперед, в стороны и назад и загибающимися кверху, до более характерных для лосей рогов с коротким стволом и большим плоским расширением — так называемой лопатой, от которой вперед, наружу и назад отходят отростки (рис. 164). Наблюдается географическая изменчивость в типе и размерах рогов. Лоси с наибольшими по размерам рогами — в Восточной Сибири и на Аляске. Взрослые лоси сбрасывают рога ежегодно с ноября по декабрь, а рост новых рогов начинается в апреле—мае. К сентябрю рога уже очищаются от кожи. Конечности лосей с хорошо развитыми боковыми пальцами, обладающими относительно крупными копытами, которые при ходьбе по болотистой почве участвуют в опоре на грунт.
Окраска тела одноцветная темная, черно-бурая или почти черная, ноги светлее туловища, «зеркала» на ляжках нет. Молодые рыжеватые. В зимних волосах лосей сильно развит воздухоносный сердцевинный слой. На загривке и горле волосы удлинены. Предглазничные и межпальцевые железы маленькие. Имеются небольшие плюсневые железы. Сосков 2 пары.
Череп с удлиненным лицевым отделом, укороченными носовыми костями и большим носовым отверстием. Межчелюстные кости сильно удлинены. Ямка предглазничной железы на слезной кости небольшая. Этмоидальные отверстия крупные. Характерно расположение роговых отростков лобных костей — они направлены в стороны и находятся в плоскости лобной поверхности черепа.
£ 3
Зубная формула:
0 3 3
с — р ------- m — = 32.
1 Р 3 3
Верхних клыков нет ни у самцов, ни у самок. Щечные зубы массивные и широкие. Нижние клыки сходны по форме с резцами.
Диплоидный набор хромосом 68—70.
Распространен в Северной Америке, Евразии (включая Северный Кавказ).
Населяет лесную зону, частично лесотундру, лесостепи и степи (участки леса). Обитатель лесов. Предпочитает сырые и заболоченные места. Встречается и в горах, но избегает сильно пересеченных участков. Питается летом в основном травянистыми растениями, особенно водными и полуводными, а также побегами и листьями деревьев и кустарников. Зиг-мой переходит на питание главным образом веточками деревьев и кустарников, хвоей и корой деревьев. Охотно посещает солонцы. Держатся поодиночке или небольшими группами по 3—5 особей. Обычно старые самцы вне периода гона живут обособленно. Зимой стадность выше, чем летом. Летом активен в дневные часы, но с появлением большого количества слепней и оводов переходит к ночной активности. Днем часто заходит в воду. Наблюдаются сезонные миграции лосей, особенно далекие на севере ареала в многоснежных областях (с глубиной снегового покрова 70 см и больше), откуда на зиму они уходят в районы с менее глубоким снегом. Подобные миграции имеют место и в горах.
Гон приходится на сентябрь — октябрь. Обычно с самцом держится одна самка. Между самцами могут происходить драки. Беременность длится 225—237 дней. Лосиха приносит 1—2 лосят в мае — июне. Масса новорожденного 6—16 кг. В 3-дневном возрасте лосенок уже хорошо ходит, а на 10-й день не отстает от матери. Лактация продолжается 3,5— 4 месяца. Половой зрелости достигает в 3-летнем возрасте. Продолжительность жизни около 20 лет. Основные враги — волк и бурый медведь. Служит объектом промысловой и спортивной охот*ы.
Проведены работы по одомашниванию лосей с целью использования
Рис. 165. Северный олень—. tarandus
их как транспортного животного (под вьюком и в санях) в условиях лесного бездорожья. Местами при высокой численности лоси вредят подросту деревьев, особенно в лесопитомниках (например, посадкам сосны).
В результате неконтролируемого промысла к началу XX столетия численность лосей в России сократилась. После решительных мер по их охране в СССР в настоящее время ареал почти полностью восстановлен. Он охватывает с запада на восток всю лесную и лесостепную зоны страны, за исключением Камчатки и Сахалина.
Род северных оленей
Genus Rangifer Н. Smith, 1827
В роде единственный вид: северный олень — R. tarandus Linnaeus, 1758.
Размеры средние. Длина тела 150—220 см, длина хвоста 7—20 см, высота в холке 80—150 см. Масса 100—220 кг. Туловище удлиненное, ноги относительно короткие, шея относительно длинная (рис. 165). Голову держит низко и кажется менее стройным, чем остальные олени. Хвост короткий, но снаружи заметен. Уши невелики, с притупленными вершинами. На конце морды голого участка кожи нет. Средние пальцы с большими, широкими и уплощенными копытами. Боковые пальцы имеют копыта, которые при ходьбе животного касаются грунта. Это обстоятельство, а также то, что пальцы северного оленя подвижны и могут широко раздвигаться, позволяет ему передвигаться по топкой почве и по снегу. Нагрузка на 1 см2 опорной поверхности конечности значительно ниже, чем у других оленей (140 г на 1 см2 по сравнению, например, с 560 г у лося). Северные олени — единственные представители семейства, у которых и самцы, и самки имеют рога. Длинный основной ствол ро-378
Рис. 165 Продолжение. Череп северного оленя
гов идет, дугообразно изгибаясь, сначала назад и вверх, а затем вверх и вперед (рис. 166). Вершина главного ствола уплощена и надглазничные отростки образуют на концах лопатообразные расширения. Обычно правый и левый рога несимметричны. Рога самцы сбрасывают после гона (ноябрь—декабрь), а самки после отела (май—июнь). Начало роста рогов у самцов — в апреле. Кожа спадает с рогов в конце августа — начале сентября.
Окраска летнего меха однотонная, бурая с различными оттенками (сероватым, пепельным, кофейным). Зимой мех со светлыми и темными пестринами, иногда совсем темный или почти белый. На задних отделах ляжек имеется небольшое, более светлое «зеркало». Для зимнего меха
Рис. 166. Некоторые типы рогов северного оленя — /?. ta-randus из европейской части СССР
характерно чрезвычайно сильное развитие в волосах воздухоносной сердцевины, что делает его очень ломким. На шее волосы сильно удлинены— образуется своеобразная грива. Межпальцевые и предглазничные железы имеются. Линька одна в году: начинается в конце зимы и тянется все лето.
Череп с сильно развитым лицевым отделом и укороченным мозговым. Слезные кости удлиненные с хорошо выраженной ямкой предглазничной железы. Этмоидальные отверстия крупные. Носовые кости не укороченные. Костные слуховые барабаны маленькие.
„ 0 13 3
Зубная формула: 1 —- с - р — гл — =34.
3 13 3
Верхние клыки малы, но имеются и у самцов, и у самок. Щечные зубы небольшие с низкими коронками, причем нижние слабее верхних. Нижние резцы и клыки сходны по форме между собой.
Диплоидный набор хромосом 70.
Распространен в арктической и таежной зонах Старого и Нового Света, включая ряд островов Полярного бассейна; в СССР южная граница ареала проходит от Онежского озера через Вологодскую область к истокам Печоры, в Западной Сибири примерно по 58—59° с. ш., на Алтае к востоку от Телецкого озера и верхнего течения Оби, в Восточной Сибири — по Северному Забайкалью и на Дальнем Востоке по северным окраинам Амурского края.
Населяет тундру, лесотундру, тайгу и высокогорье некоторых горных систем (например, Алтая, Саян). Несколько условно северных оленей можно разделить на тундровых, лесных и горных. Однако у всех северных оленей отмечается тяготение к открытому ландшафту. Лесные олени и лето и зиму проводят в лесу, предпочитая разреженные участки и заболоченные места. Горные олени летом держатся высоко в горах, а зимой обычно спускаются в лесной пояс. Тундровые олени летнее время проводят в тундре, где, спасаясь от гнуса, выбирают открытые, обдуваемые ветром места. К зиме они уходят на юг в лесотундру и северную тайгу. Местами северные олени совершают сезонные кочевки на расстояние в сотни километров. Особенной дальностью отличаются миграции северных оленей в Арктике и Субарктике: на полярных островах до 300—500 км, на границе тундры и тайги — до 200—700 км.
Северные олени иногда сбиваются в огромные стада, насчитывающие многие тысячи голов. Такие стада непостоянны и легко распадаются на меньшие группы. В летнее время в условиях полярного дня периоды активности и последующего отдыха чередуются несколько раз в сутки и их 380
длительность варьирует в зависимости от погодных условий и наличия гнуса.
Характерно приспособление к питанию в зимнее время лишайниками: наземными (ягелем) и древесными. Летом главным кормом служат разнообразные травы, листья и побеги ив и карликовой березы. Охотно кормится грибами. Недостаток белковых и минеральных веществ в рационе заставляет при случае поедать леммингов, яйца птиц и птенцов.
Гон на территории СССР приходится на сентябрь—ноябрь. При одном взрослом самце в это время держатся 3—13 самок, несколько телят и 2—3 молодых самца. Между самцами имеют место не очень ожесточенные драки. Длительность беременности 192—246 дней. Отел происходит в мае— июне. Самки приносят по одному детенышу бурой или буроватосерой окраски. В первую же неделю после рождения теленок хорошо бегает и даже плавает. Молоко северного оленя чрезвычайно богато белками. Лактация продолжается до осени, а иногда и зимой. Половозрелость наступает на 2-м году жизни, но самцы в размножении раньше 3-го года участия не принимают. Главные враги— волк и в меньшей степени медведь и росомаха.
Ценный промысловый зверь, дающий мясо и шкуру. Родоначальник домашнего северного оленя, широко разводимого в северных районах.
Подвид северного оленя с Новой Земли — /?. tarandus pearsoni Ly-dekker, 1903 — включен в «Красную книгу».
SUBFAMILIA CERV1NAE BAIRD, 1857
Размеры мелкие, средние или крупные. У самцов есть рога — большие, обычно разветвленные, всегда с надглазничным отростком. Из кожных специфических желез имеются предглазничная, плюсневая, тарзальная, метатарзальная, подхвостовая и на задних конечностях межпальцевая.
В черепе костные слуховые барабаны маленькие или средней величины. Сошник короткий и не разделяет хоан. На слезных костях по два отверстия слезных каналов. На этих костях имеется большая впадина для предглазничной железы.
Верхние клыки маленькие или их нет вовсе.
От метакарпальных костей II и V пальцев сохраняются лишь проксимальные отделы. Cuneiforme отделено от naviculo-cuboideum.
Желчного пузыря нет.
Род оленей Давида
Genus Elaphurus Milne-Edwards, 1866
В роде единственный вид: олень Давида — Е. davidianus Milne-Ed-wards, 1866.
Размеры средние. Длина тела около 150—215 см, длина хвоста 50 см, высота в холке 115—140 см. Масса 150—200 кг. Туловище удлиненное, конечности высокие. Шея относительно короткая, голова длинная и узкая (рис. 167). Профиль верхней части головы прямой. Уши короткие заостренные. Конец морды голый. Хвост длинный с удлиненными концевыми волосами. Копыта средних пальцев крупные, боковые развиты хорошо и касаются почвы при ходьбе по мягкому грунту. Рога, достигающие в длину 87 см, весьма своеобразны (такого типа единственные среди
Рис. 167 Олень Давида — Е. davidianus
оленей): отростки основного ствола направлены только назад; самый нижний и самый длинный из них ответвляется от главного ствола, отступая всего несколько сантиметров от черепа, и может ветвиться сам (иногда имеет до 6 концов). В летнее время окраска спины желто-серая, брюха — светло-желто-бурая. Имеется небольшое околохвостовое «зеркало». Зимой окраска серо-бурая. Молодые светло-рыже-бурые со слабыми желто-белыми пятнами. Межпальцевые и плюсневые кожные железы отсутствуют. Предглазничные железы очень крупные.
Череп длинный и узкий. Лобный отдел немного вогнут. Слезные кости с большими ямками предглазничных желез. Этмоидальные отверстия длинные и узкие. Костные слуховые барабаны маленькие.
О 1 3
Зубная формула: i — с —— р m ole
Диплоидный набор хромосом 68.
По-видимому, населял болотистые районы Северного и Центрального Китая. К середине XIX столетия сохранился только в императорском охотничьем парке в окрестностях Пекина, где и обнаружен в 1865 г. французским миссионером Давидом. Был вывезен в 1869 г. в Европу и в настоящее время имеется во всех больших зоопарках мира в количестве примерно 450 голов. Последний экземпляр в Китае погиб во время боксерского восстания в 1920 г. В 1960 г. в Китае был реакклиматизи-рован.
Образ жизни в естественных условиях не известен, но, по-видимому, обитал по берегам водоемов в заболоченных местах. Питается водными болотистыми травянистыми растениями. Держится стадами разного размера. Спаривание приходится на июнь — июль. Беременность про-
Рис. 167. Продолжение. Череп оленя Давида
Рис. 168. Благородный олень — С. elaphus (Д), пятнис-тий ЛЛЛИк-- с гипппп /Б1
Рис. 168. Продолжение. Череп благородного оленя
должается 250—270 дней. Самки приносят 1—2 оленят в апреле — мае.
Половозрелость наступает в 27, редко в 15 месяцев.
Род оленей
Genus Cervus Linnaeus, 1758
Размеры от мелких до крупных. Длина тела 105—265 см, длина хвоста 10—55 см. Телосложение стройное, пропорциональное (рис. 168, 169). Конечности тонкие, задние лишь немного длиннее передних. Хвост длинный, средней длины или короткий. На конце морды имеется обширный безволосый участок. Уши длинные. Конечности четырехпалые. Боковые пальцы развиты слабее средних, однако сохраняют скелет фаланг. Рога различных размеров — от средних до очень больших с разным количеством отростков (одним надглазничным и еще минимум двумя кроме него). Отростки направлены вперед (кроме лани). Рог в сечении округлый и овальный (у лани на конце уплощенный) (рис. 169). Зимой в волосах сильно развит сердцевинный слой. Окраска взрослых животных круглый год одноцветная или пятнистая (только в летнее время). Как правило, имеется белое «зеркало» на задней поверхности ляжек. В году 2 линьки. Молодые в первые месяцы жизни имеют пятнистую окраску. Многочисленны специфические кожные железы: предглазничные, межпальцевые, плюсневые (иногда они малы или отсутствуют), хвостовые (имеются у немногих видов).
В черепе слезные кости с глубокими ямками для предглазничных желез. Этмоидальные отверстия большие. Лобные кости крупные, а теменные уменьшены. Костные слуховые барабаны средних размеров.
0
Зубная формула: i с
0—1 3 3
----- р — m — = 32 — 34.
1 и 3 3
Верхний клык у самцов обычно имеется, но развит слабо, у некоторых видов его нет вовсе, у самок бывает редко.
Диплоидный набор хромосом у пятнистого оленя 64—68, лани — 68, барасинги — 56, индийского замбара — 58, благородного оленя — 68.
Распространены в Евразии на север примерно до 60° с. ш., в Северо-Западной Африке и Северной Америке от 60° с. ш. на севере до Мексики на юге.
Населяют леса разных зон от лесостепей до гор. Держатся небольшими группами, чаще не больше 10 голов. Взрослые самцы большую часть года проводят отдельно от самок. Активны утром и вечером. Во многих местах совершают сезонные миграции. Летом поедают разнообразные травы, листву и молодые побеги деревьев и кустарников, а зимой основу витания составляют древесно-веточные корма. Охотно посешают солонцы. Самцы благородного оленя сбрасывают пога в марте — апреле, пятнистого оленя — в апреле — мае. Через несколько дней начинается рост новых рогов. В августе — сентябре происходит чистка рогов от кожи. Период спаривания у благородного оленя приходится на сентябрь—октябрь, пятнистого оленя — сентябрь — ноябрь, лани — октябрь. Самцы собирают вокруг себя гаремы из нескольких самок. Гон сопровождается ревом и драками самцов. Беременность длится около 8 месяцев. Телята рождаются в конце мая — июне. Самки приносят по одному теленку. Половозрелость наступает в 2-летнем возрасте, но самцы приступают к размножению значительно позже.
Главный враг оленей-—волк, изредка на них нападают тигр, рысь, медведь. Вследствие неумеренного преследования в ряде мест ареала численность оленей значительно сократилась или они исчезли.
Рис. 169. Аксис — С. axis (А) и лань С. llama (Б)
Олени — промысловые животные. Некоторых (пятнистого и марала)' успешно разводят в условиях полувольного содержания для получения молодых, еще неокостеневших рогов — пантов, из которых изготовляют лечебный препарат—пантокрин.
В роде 12 видов:
свиной олень — С. porcinus Zimmermann, 1780 (Северная Индия, Бирма, Таиланд, Кампучия, Лаос, Вьетнам, о-ва Филиппинские и Бавеан к северу от Явы, Австралия; акклиматизирован на о-ве Шри Ланка и в Виктории);
аксис—С. axis Erxleben, 1777 (Индия, о-в Шри Ланка, Северный Андаман, Пакистан, Бангладеш, Бирма, Таиланд, п-ов Индокитай, о-ва Каламиан из группы Филиппинских и о-в Бавеан к северу от Явы; акклиматизирован на Новой Зеландии, Гавайских островах, в Австралии и Аргентине);
пятнистый олень —С. nippon Temminck, 1838 (Восточный Китай, о-в Тайвань, Корейский п-ов, в СССР южная часть Дальнего Востока, Япония; успешно акклиматизирован во многих местах, например в Новой Зеландии, в СССР — в ряде заповедников и охотничьих хозяйствах европейской части страны);
беломордый олень — С. albirostris Przewalski, 1883 (Восточный, Средний и Северный Тибет, Западный Китай);
лань—С. dama Linnaeus, 1758 (Центральная, Западная и Южная Европа, Северная Африка, Малая Азия; акклиматизирована в США, Канаде, Чили, Аргентине, Южной Африке, на Мадагаскаре, в Австралии, Тасмании, Новой Зеландии, а также некоторых районах Украины, Белоруссии и прибалтийских республик СССР);
иранская лань — С. mesopotamicus Brooke, 1875 (Юго-Западный Иран и прилегающие районы Ирака);
барасинга — С. duvaticeli Cuvier, 1823 (Центральная и Северо-Восточная Индия, Непал; акклиматизирован в Австралии);
олень-лира — С. eldi M’Clelland, 1842 (Ассам в Индии, Бирма, Таиланд, Кампучия, о-в Хайнань);
индийский замбар — С. unicolor Kerr, 1792 (Индия, о-в Шри Ланка, Бангладеш, Бирма, Таиланд, Лаос, Кампучия, Вьетнам, Сычуань и Юньнань в Китае, о-ва Хайнань, Тайвань, Суматра, Калимантан; акклиматизирован в Новой Зеландии, Австралии и Флориде в США);
гривистый замбар — С. timorensis De Blainville, 1822 (о-ва Зондские и Молуккские и северо-запад Новой Гвинеи; акклиматизирован в Австралии, Новой Зеландии, на о-вах Мадагаскар, Новая Каледония, Маврикия, Коморских);
филиппинский замбар — С. mariannus Desmarest, 1822 (Филиппинские о-ва; был завезен испанцами на о-в Гуам из группы Марианских о-вов, где хорошо акклиматизировался);
благородный олень — С. elaphus Linnaeus, 1758 (Европа—Англия, Португалия, Испания, Франция, Швейцария, от Нидерландов до Дании, Норвегия, Швеция, Польша, от ГДР и ФРГ на юг до Румынии, Болгарии и Греции, Корсика, Сардиния, Италия. В СССР — Прибалтика, Белоруссия, Карпаты, Крым, Воронежский заповедник, Северный Кавказ, Закавказье, Копетдаг, долины рек Пяндж и Амударья, Восточный Тянь-Шань, Алтай, Салаирский кряж, Кузнецкий Алатау, Саяны, Присаянье, Прибайкалье, Забайкалье, Южная Якутия, Амурский и Уссурийский края; Малая Азия; Иран; Афганистан; МНР; Китай — Хэйлунцзян, Ганьсу, Шаньси, Сычуань, Синьцзян-Уйгурский автономный район, Тибет; Индия — Сикким и Кашмир; Бутан; Африка—Алжир и Тунис; Северная Америка — провинции Британская Колумбия, Альберта, Саскачеван, Манитоба, юг Онтарио, вся территория США, за исключением край
него востока, юго-востока и юго-запада; Чиуауа в Мексике; акклиматизирован в Австралии, Новой Зеландии, Аргентине, Чили, в ряде охотничьих хозяйств и заповедников СССР).
В СССР: пятнистый олень, лань и благородный олень.
Некоторые исследователи (например, Эллермэн и Моррисон-Скотт, 1966) выделяют свиного оленя и аксиса в особый род Axis Н. Smith, 1827 (Уайтхэд, 1972, указывает для этого рода еще два вида: A. kuhlii Miiller et Schleger, 1844, и A. calamianensis Heude, 1888), а лань и иранскую лань — в род Dama Frisch, 1775 ( = Platyceros Zimmermann, 1780). Холл и Келсон (Hall a. Kelson, 1959) американские подвиды благородного оленя выделяют в особые виды: С. canadensis Erxleben, 1777, и С. nannodes Merriam, 1905; Эллермэн и Моррисон-Скотт (1966) исчезнувший подвид барасинги из Таиланда — в вид С. schomburgki Blyth, 1863.
В «Красную книгу» включены:
виды и подвиды, находящиеся под угрозой исчезновения: иранская лань, барасинга (численность в 1972 г. 4—4,6 тыс.), подвиды оленя-лиры из штата Манипур в Индии С. eldi eldi M’Clelland, 1842 (численность в 1972 г. около 50 голов) и из Таиланда, Вьетнама, Кампучии, Лаоса и с о-ва Хайнань С. е. siamensis Lydekker, 1915, подвиды пятнистого оленя— с о-ва Тайвань С. nippon taiouanus Blyth, 1860 (численность к 1967 г. около 300 голов), с о-вов Рюкю С. п. keramae Kuroda, 1924, из Северного Китая С. п. tnandarinus Milne-Edwards, 1871 (в диком состоянии, вероятно, исчезли), из провинции Шэньси в Китае С. п. grassianus Heude, 1884 (в диком состоянии, вероятно, исчезли), из Южного Китая С. п. kopschi Swinhoe, 1873 (в диком состоянии, вероятно, исчезли), подвиды благородного оленя — с Корсики и Сардинии С. elaphus corsicanus Erxleben, 1777 (на Корсике, по-видимому, исчезли; на о-ве Сардиния в 1968 г. было примерно 140 голов), с юга Тибета из долины Чумой С. е. wallichi Cuvier, 1823 (к настоящему времени, по-видимому, исчезли), из Северной Африки С. е. barbarus Bennett, 1833 (к 1972 г. в Алжире было примерно 150, а в Тунисе 250 голов), из Северной Индии, штат Кашмир С. е. hangul Wagner, 1844 (к 1972 г. сохранилось около 1000 голов), из окрестностей озера Лобнор С. е. yarkandensis Blanford, 1892 (к настоящему времени, вероятно, исчез), с Амударьи и из Северного Афганистана С. е. bactrianus Lydekker, 1900,(численность к 1972 г. 500— 600 голов);
малочисленный подвид, в настоящее время не находящийся под угрозой исчезновения, — аксис с о-ва Бавеан, С. axis kuhli Miiller et Schlegel (к 1969 г. его численность достигала примерно 500 голов);
малочисленный вид и подвид, сведения о состоянии которых неизвестны, — беломордый олень и подвид благородного оленя с Тибета, С. elaphus macneilli Lydekker (в 1940 г. численность около 5 тыс. голов);
подвид, ранее находившийся в опасности и теперь переставший быть таковым — благородный олень с гор Сьерра Невада, США (к 1971 г« численность достигла примерно 470 голов).
СЕМЕЙСТВО ЖИРАФОВЫХ
FAMILIA GIRAFFIDAE GRAY, 1821
Размеры крупные. Длина тела 200—400 см, высота в холке 150—• • 30 см. Туловище слабо (окапи) или сильно (жираф) покатое от холки к крестцу и у жирафа укороченное. Шея и конечности длинные (окапи), или очень длинные (жираф). Голова относительно невелика с удлиненным лицевым отделом, подвижными губами, крупными глазами, неболь-
1пими (жираф) или большими (окапи)' ушами. Язык длинный и очень подвижный. На лобной части головы у обоих полов небольшие рожки покрытые кожей: 2 у окапи или 2—5 у жирафы. Передние конечности немного длиннее задних. Боковые (II и V) пальцы отсутствуют: иногда остаются лишь проксимальные остатки их пястных и плюсневых элементов. Копыта низкие и широкие. Волосяной покров низкий, но плотный. Окраска его от бледно-желтой и белой до черно-коричневой. Могут быть крупные темные пятна. На конце хвоста пучок удлиненных волос. Большинство специфических кожных желез отсутствует. Сосков 1—2 пары у жирафы и 2 пары у окапи.
Череп вытянутый с пневматизированными костями крыши. Слезные кости с одним, реже двумя отверстиями слезных каналов. Ямок для предглазничных желез на слезных костях нет. Этмоидальные отверстия у окапи и молодых жираф средних размеров, а у взрослых жираф очень малы или совсем исчезают. Костное небо короткое, не достигает уровня щечных зубов. Костные слуховые барабаны узкие и массивные.
„ 0 0 3 3
Зубная формула: i — с — р — m — = 32.
3 13 3
Верхние клыки всегда отсутствуют. Щечные зубы брахиодонтного типа.
Желчного пузыря нет или он имеется только у эмбрионов (жираф). Слепая кишка большая. Желудок четырехкамерный, «жвачный».
Ареал охватывает Африку к югу от Сахары.
Населяют саванны и леса. Питаются в основном листьями и побегами деревьев, фруктами. Активны днем или ночью. Держатся группами или поодиночке. Период размножения не приурочен к определенному времени года. Длительность беременности 14—15.5 месяцев, лактации — 6—10 месяцев. Половозрелость наступает в 3 года. Продолжительность жизни 20—30 лет.
В семействе 2 рода (2 вида):
окапи — Okapia Lankester, 1901 (1 вид) и
жираф — Giraffa Brisson, 1762 (1 вид).
Род окапи
Genus Okapia Lankester, 1901
В роде единственный вид: окапи — О. johnstoni Sclater, 1901.
Длина тела около 210 см, длина хвоста 30—40 см, высота в холке 150—170 см. Масса около 250 кг. Шея относительно не очень длинная; составляет около 2/з длины туловища. Спина слабо поката от холки к крестцу (рис. 170). Конечности относительно короче, чем у жираф. Посредине верхней губы проходит небольшая вертикальная полоска голой кожи. Уши большие, широкие и трубчатые. Язык очень длинный, может доставать до уха. Имеется два небольших рога. Костные выросты рогов самца располагаются на лобных костях; они имеют в длину 10—12 см и направлены наискось назад. Вершины их обнажены или имеют небольшой роговой чехол. У многих самок рогов нет. Если они имеются, то меньше по размерам, чем у самцов, и полностью покрыты кожей. Общая окраска тела каштаново-бурая, на плечах и крестце черноватая. Взрослые самцы светлее самок. Ноги белые, их проксимальные отделы в поперечных темно-бурых кольцевидных полосах.
В черепе лобная часть и небно-челюстной отделы костного неба сильно пневматизированы. Лобный отдел между глазницами не очень широк. 390
В конечностях обычно имеются остатки проксимальных отделов пястья и плюсны II и V пальцев.
Желчного пузыря нет.
Диплоидный набор хромосом у самцов 45, у самок 46.
Распространен в Заире.
Населяет дождевые леса с высохшей, жесткой почвой. Питается побегами и листьями преимущественно молочаевых, а также фруктами. Держатся поодиночке. Из органов чувств лучше других развиты обоняние и слух. Активен преимущественно ночью. Длительность беременности 14—15 месяцев. Период лактации 6 месяцев.
Рис. 171 Жираф—G. Camelopardalis
Род жираф
Genus Giraffa Brisson, 1762
В роде единственный вид: жираф — G. Camelopardalis Linnaeus, 1758.
Длина тела 300—400 см, высота в холке 270—330 см, высота до темени 450—580 см, длина хвоста 90—110 см. Масса 500—750 кг. Туловище узкое, на высоких ногах, спина сильно покатая от загривка к крестцу. Шея очень длинная, составляет примерно 4/з длины туловища (рис. 171). Лицевой отдел головы удлинен. Уши небольшие, заостренные. Рога имеются у самцов и у самок; парные располагаются на теменных костях, а непарные—на передних частях лобных и задних отделах носовых костей. Их вершины округлы и покрыты кожей. Число рогов от 2 до 5. Окраска тела сильно варьирует, обычно на изменяющемся от беловатожелтого до бурого фона расположены бледно-желтые или темно-бурые, трех- или шестиугольные пятна. Самцы темнее самок и молодых животных.
В черепе пневматизированы лобная и теменная области. Лобный отдел между глазницами широкий.
Остатки пясти II и V пальцев, а также метатарзалий V пальца, а иногда и II пальца полностью отсутствуют.
Желчный пузырь имеется у эмбрионов.
Диплоидный набор хромосом 30.
Распространен в Африке к югу от Сахары.
Населяет главным образом саванны, поросшие акацией. Питается в основном листьями и побегами деревьев и кустарников. Держатся небольшими группами, состоящими обычно из одного самца и нескольких самок с их детенышами. Иногда эти группы объединяются в крупные стада до сотни голов. Основные органы чувств — зрение и слух. Активны преимущественно днем. Во время гона между самцами происходят схватки; самцы бьют друг друга боковыми ударами головы, иногда и ногами. Продолжительность беременности 14—15,5 месяцев, лакгации -Ю месяцев. Жирафенок способен вставать на ноги спустя 20 мнн после рождения. Молодые начинают поедать зеленый корм в возрасте 3 не-
дель. Половозрелость наступает в 3-летнем возрасте. Продолжительность жизни 20—30 лет. Основной враг — лев; на молодых животных нападает леопард.
СЕМЕЙСТВО ВИЛОРОГОВЫХ
FAMILIA ANTILOCAPRIDAE GRAY, 1866
Размеры средние. Телосложение изящное. Длина тела 113—140 см, длина хвоста 8—15 см, высота в холке 88—105 см. Масса 36—64 кг. Самцы несколько крупнее самок. Туловище вытянутое. Конечности тонкие, высокие. Шея средней длины. Голова небольшая, удлиненная (рис. 172). Безволосое пространство на конце морды маленькое. Глаза очень большие с длинными ресницами. Уши средних размеров. Рога имеются как у самцов, так и (меньших размеров) у самок; иногда самки безрогие. Рога длиной до 25 см; они сжаты с боков и имеют короткий отросток, отходящий вперед. Вершины рогов загнуты назад и несколько внутрь. Рога имеют тонкое, неразветвленное костное основание и роговой чехол, который ежегодно спадает и отрастает вновь. Процесс отрастания длится около 4 месяцев. Копыта небольшие, заостренные. Волосяной покров средней высоты, плотный, с небольшим количеством пуховых волос. Окраска спины от ржавчато-рыжей до коричневато-желтой. Брюхо и нижняя половина боков белые. Хорошо выражено белое околохвостовое «зеркало». У самцов на горле черный полуошейник и черный верх морды. Имеются специфические кожные железы: подушные, хвостовые, предплюсневые и межпальцевые. Предглазничных желез нет. Сосков 2 пары.
Череп вытянутый. Носовые кости соприкасаются с предчелюстными. Этмоидальные отверстия крупные. Слезные кости без ямок предглазничных желез, с двумя отверстиями слезных каналов. Глазницы большие. Лобные кости короткие, массивные, не пневматизированы.
0 0 3 3
Зубная формула: i — с - р — т — = 32.
3 13 3
Верхний клык всегда отсутствует, нижний сходен с третьим нижним резцом. Щечные зубы гипселодонтные, селенодонтные.
Характерно крупное сердце и большой поперечник гортани, что связано с адаптацией к быстрому бегу. Желчный пузырь имеется.
Диплоидный набор хромосом 56—58.
Распространен вилорог на западе центральных областей Северной Америки; в Канаде в южной половине провинций Альберта и Саскачеван, на юго-западе Манитобы; в западных штатах США, кроме западной половины и крайнего востока штата Вашингтон, запада Орегона, запада Калифорнии, восточная граница проходит по линии, идущей через западные районы штата Миннесота, запад Айовы, восток Канзаса, восток Оклахомы, восток Техаса; в северной части Мексики на юг до линии, проходящей через юг штата Сонора, юго-запад Чиуауа, центральные районы Дуранго, север Закатекас и север Идальго, восточная граница проходит на некотором расстоянии от побережья Мексиканского залива.
Населяет степи и полупустыни на равнинах и сглаженных возвышенностях с доступной водой для питья. Хорошие бегуны со скоростью бега до 65 км/ч; на коротких расстояниях способны достигать 80—95 км/ч. Летом держатся небольшими группами, а зимой объединяются в стада до 50—100 и более голов. До того, как численность вилорогов была уменьшена человеком, их стада насчитывали до 1000 голов. Дневной кормовой путь составляет примерно 5 км с дополнительными маршрутами на водопой. Питается вилорог в течение круглых суток различными 394
Рис. 172. Вилорог — Д. атепсапа и его череп
травами, особенно шалфеями. Активен днем и ночью. Из органов чувств наиболее хорошо развито зрение. В зимнее время стадо мигрирует в более защищенные места. В период размножения стада разбиваются и один самец ходит с небольшим гаремом до 15 самок. Гон приходится на середину августа — середину сентября. В это время самцы дерутся между собой. Длительность беременности 230—240 дней. Самка рожает в феврале—марте или в конце мая двух детенышей. Масса новорожденного 2—2,5 кг. На 4-й день после рождения детеныш способен убежать от преследующего его человека. Лактационный период длится 5—6 месяцев. Половозрелость наступает в 15—16 месяцев у самок и в 24 месяца у самцов. Продолжительность жизни до 8—10 лет.
За вилорогом охотятся из-за мяса и охотничьих трофеев. Их численность была сильно снижена (примерно с 40 млн. голов в 1800 г. до 19 тыс. в 1900 г.), но в последнее время в результате защитных мероприятий число их увеличивается (в 1966 г. — около 400 тыс. голов).
Подвиды вилорога с п-ова Калифорния — A. a. peninsularis Nelson, 1912 (численность в 1925 г. около 500 голов), из штата Сонора в Мексике (численность в 1959 г. около 1000 голов) и штата Аризона в США — Л. a. sonoriensis Goldman, 1945, включены в «Красную книгу» как подвиды, находящиеся под угрозой исчезновения.
В семействе 1 род вилорогов — Antilocapra Ord, 1818, и 1 вид.
Род вилорогов
Genus Antilocapra Ord, 1818
См. описание семейства.
В роде единственный вид: вилорог — A. americana Ord, 1815 (распространение см. в описании семейства),
СЕМЕЙСТВО ПОЛОРОГИХ
FAMILIA BOVIDAE GRAY, 1821
Размеры от мелких до больших. Так, Neotragus pygmaeus имеет высоту в холке около 25 см и массу 2—3 кг, а зубр — высоту в холке до 200 см и массу до 1000 кг. Общее сложение от легкого и стройного до тяжелого и массивного. Конечности обычно высокие.
Самцы, а у многих видов и самки имеют пару неветвящихся рогов (у Т etracer us их две пары). Рога представляют собой постоянные несменяемые костные выросты лобных костей, покрытые снаружи роговым чехлом эпидермального происхождения. Рост рога, в противоположность оленям, происходит от его основания. Таким образом, вершина рога представляет наиболее старую его часть. Характерно периодическое^усиление и замедление роста рогов, в результате чего на его роговой поверхности образуются своеобразные кольца. Форма рогов чрезвычайно разнообразна — от совершенно прямых, длинных и тонких до коротких, толстых и сильно изогнутых или спирально закрученных. Если направление изгиба или закручивание рога происходит внутрь, навстречу рогу противоположной стороны, то такие рога называются гомонимными, если же правый рог свернут или изогнут вправо, а левый влево — гетеро-нимными. В поперечном разрезе рога бывают круглые, овальные или треугольные. На поверхности их часто бывают выступы, поперечные складки и кольца или продольные ребра.
Окраска очень разнообразна — от белой до почти черной, обычно без резких цветовых узоров. У многих видов на ляжках имеется белое по
ле — «зеркало». В кожном покрове обычно много специфических желез: предглазничные, межроговые, паховые, межпальцевые, хвостовые ит. д. Сосков 1—2 пары.
Пальцев на конечностях по 4 (редко 2), но боковые (II и V) сильно укорочены и, хотя имеют небольшие копытца, при ходьбе по твердому грунту обычно не касаются его. От пястных костей боковых пальцев сохраняются только проксимальные и дистальные части.
В черепе сильно развиты лобные кости. Теменные кости сдвинуты назад. В слезной кости сильно развита лицевая часть с ямкой для предглазничной железы или без нее. Обычно имеется всего одно отверстие слезного канала. Этмоидальные отверстия отсутствуют или развиты слабо. Кости черепа сильно пневматизированы. Межчелюстная кость обычно относительно невелика, верхнечелюстная — очень крупная.
О
Зубная формула: i —-
О
О 2—3 3
с — р ---— m — = 28 — 32.
1 2—3 3
Иногда вторые предкоренные в нижней, а изредка и в верхней челюстях не развиваются или рано выпадают. Щечные зубы гипселодонтные и тетраселенодонтные (четырехлунчатые).
Желудок сложный, ясно разделенный на 4 отдела: рубец, сетку, книжку и сычуг. Желчный пузырь обычно имеется. Плацента поликоти-ледонная.
Распространены широко по земному шару. Восстановленный ареал охватывает Африку (без Мадагаскара), Европу (кроме Британских о-вов), на север идет до юга Скандинавского п-ова, Финского залива, Верхней Волги, Самарской Луки и юга Урала. За Уралом в ареал входит юг Западной Сибири и большая часть Средней и Восточной Сибири и Дальний Восток. На юго-востоке и юге Азии ареал охватывает всю южную часть Азиатского материка с большинством прилегающих островов. В Новом Свете ареал занимает большую часть Северной Америки на юг до Калифорнии, Флориды и северной Мексики, Арктический архипелаг, северное и восточное побережье Гренландии. В результате неумеренного промысла или по другим причинам ареалы большинства видов значительно сократились.
Обитают в самых разнообразных местах —от густых лесов до степей, полупустынь и пустынь на равнинах, в предгорьях и высоко в горах — выше почти всех остальных млекопитающих (до 5500 м над уровнем моря). Однако наибольшее число видов населяет открытые пространства. Держатся стадами, иногда очень крупными — до нескольких тысяч голов. Значительно реже встречаются маленькими группами или поодиночке. Питаются растениями, преимущественно травами.
Большинство видов — полигамы, хотя есть и моногамы. Самцы некоторых полорогих в период размножения имеют гарем самок. У обитателей тропиков, как правило, сезонности в размножении нет. Продолжительность беременности 4—11 месяцев. В помете от одного до 4—5 детенышей.
Многие виды полорогих имеют существенное значение как промысловые животные, от которых получают мясо и кожу. Ряд видов послужили родоначальниками главнейших домашних животных.
Систематика семейства разработана недостаточно. В семействе 43 ро-Да (109 видов) (система приводится в основном по Хальтенорту, 1963):
подсемейство Cephalophinae Brooke, 1876;
хохлатых дукеров — Cephalophus Н. Smith, 1827 (15 видов);
кустарниковых дукеров — Sylvicapra Ogilby, 1837 (1 вид);
подсемейство Neotraginae Sclater et Thomas, 1894;
карликовых антилоп — Neotragus Н. Smith 1827 (3 вида);
дикдиков — Madoqua Ogilby, 1837 (5 видов)’;
антилоп-прыгунов — Oreotragus A. Smith, 1834 (1 вид);
бейр — Dorcatragus Noack, 1894 (1 вид);
стенбоков — Raphicerus Н. Smith, 1827 (Звида);
ориби — Ourebia Laurillard, 1841 (1 вид);
подсемейство Antiiopinae Baird, 1857;
четырехрогих антилоп — Tetracerus Leach, 1825 (1 вид);
нильгау — Boselaphus De Blainville, 1816 (1 вид);
лесных антилоп — Tragelaphus De Blainville, 1816 (8 видов);
ориксов — Oryx De Blainville, 1816 (1 вид);
аддаксов — Addax Rafinesque, 1815 (1 вид);
водяных козлов — Kobus A. Smith, 1840 (5 видов);
тростниковых козлов — Redunca Н. Smith, 1827 (3 вида);
косульих антилоп — Pelea Gray, 1841 (1 вид);
импал — Aepyceros Sundevall, 1847 (1 вид);
лошадиных антилоп — Hippotragus Sundevall, 1846 (2 вида);
лиророгих бубалов — Damaliscus Sclater et Thomas, 1894 (3 вида);
'Конгони — Alcelaphus De Blainville, 1816 (1 вид);
гну — Connochaetes Lichtenstein, 1814 (2 вида);
подсемейство Gazellinae Cones, 1889;
гарн — Antilope Pallas, 1766 (1 вид);
спрингбоков — Antidorcas Sundevall, 1847 (1 вид);
дзеренов — Procapra Hodgson, 1846 (2 вида);
газелей — Gazella De Blainville, 1816 (12 видов);
дибатагов — Ammodorcas Thomas, 1891 (1 вид);
геренуков — Litocranius Kohl, 1886 (1 вид);
подсемейство Caprinae Gill, 1872;
сайгаков — Saiga Gray, 1843 (1 вид);
оронго — Pantholops Hodgson, 1834 (1 вид);
горалов — Nemorhaedus H. Smith, 1827 (1-вид);
cepoy— Capricornis Ogilby, 1837 (1 вид);
снежных коз-—Oreamnos Rafinesque, 1817 (1 вид);
серн — Rupicapra De Blainville, 1816 (1 вид);
такинов — Budorcas Hodgson, 1850 (1 вид);
овцебыков — Ovibos De Blainville, 1816 (1 вид);
таров — Hemitragus Hodgson, 1841 (1 вид);
горных козлов— Capra Linnaeus, 1758 (8 видов);
голубых баранов — Pseudois Hodgson, 1846 (1 вид);
гривистых баранов — Ammotragus Blyth, 1840 (1 вид);
горных баранов — Ovis Linnaeus, 1758 (2 вида);
подсемейство Bovinae Gill, 1872;
буйволов — Bubalus Н. Smith, 1827 (Звида);
настоящих быков-—Bos Linnaeus, 1758 (5 видов);
зубров — Bison Н. Smith, 1827 (2 вида).
SUBFAMILIA CEPHALOPHINAE BROOKE, 1876
Размеры мелкие. Рога имеют самцы, а иногда и самки. Рога короткие, не превышают длины ушей, прямые или слегка изогнутые кверху. Средние копыта низкие, узкие, остроконечные или овальные. Боковые копыта средних размеров или маленькие, уплощенные. Из кожных специфических желез у всех видов имеются предглазничные, почти у всех видов — межпальцевые, у ряда видов — паховые, и лишь у немногих — запястные и предплюсневые. Сосков 2 пары.
В черепе длина лицевого отдела и мозговой коробки примерно одинаковы. Основная затылочная кость кпереди резко сужается.
0 0 3 3
Зубная формула: i —- с — р — ш — =32.
О 1 О о
Ширина коронки первого нижнего резца равна ширине коронок двух остальных резцов и клыка вместе взятых.
Род хохлатых дукеров
Genus Cephalophus И. Smith, 1827
Размеры мелкие. Длина тела 55—145 см, высота в холке 30—85 см, длина хвоста 7—17 см. Масса 4—80 кг. Самки обычно крупнее самцов. Туловище округлое в сечении. Конечности средней длины, тонкие. Контур спины полуовальный, в крестце тело более высокое, чем в загривке (рис. 173). Шея короткая или средней длины. Голова небольшая, узкая. Уши короткие и округлые или средней длины, овальные. На конце морды безволосый участок кожи. Глаза относительно большие. Хвост короткий, с небольшой кистью волос на конце. Основание хвоста на внутренней стороне голое. Рога имеют самцы и в большинстве случаев самки. Размеры рогов самок меньше, чем у самцов. Рога конические, прямые или слегка изогнутые назад и немного вверх; они служат как бы продолжением верхнего профиля головы. Форма поперечника рогов округлая или овальная. Окраска копыт и рогов буро-черная или черная.
Волосяной покров короткий, плотный, прилегающий к телу и мягкий. Верх головы с шапкой удлиненных волос, полностью или частично скрывающих рога. Окраска спины рыже-бурая, желто-бурая, иногда буро-серая, черно-серая или черная, часто с темно-бурыми или черными узкими поперечными полосами. Брюхо светлее спины. У С. sylvicultor на ляжках треугольное оранжево-желтое пятно. Предглазничные железы сильно развиты. Межпальцевые железы имеют большинство видов. Паховые железы имеют все, кроме С. nigrifrons и иногда С. niger, С. maxwelli и С. monticola. Тарзальные и карпальные железы встречаются лишь изредка.
Череп средней длины, с расширенной мозговой коробкой и суженным лицевым отделом. Последний ненамного длиннее мозгового отдела. Глазница средних размеров и в стороны не выдается. Этмоидальных отверстий нет. Слезная кость с глубокой или очень глубокой ямкой пред-глазничной железы. Носовые кости резко расширены в задней части.
Щечные зубы брахиодонтные с квадратными коронками.
Отделы желудка слабо дифференцированы.
Диплоидное число хромосом у желтоспинного, черного, черноспинного, рыжебокого дукеров и дукера Максвелла по 60.
Распространены в Африке к югу от Сахары.
Обитают в разных типах лесов с густым подростом, в том числе в Дождевых лесах, зарослях кустарников, саваннах. Поднимаются в горы До 4 тыс. м над уровнем моря. Держатся поодиночке и только в период размножения парами. Сумеречные и ночные животные. Метят тропы секретом щечных желез. Питаются листьями, побегами, фруктами, ягодами и в меньшей степени травянистыми растениями. Известны случаи поедания насекомых, птиц и падали. Своеобразный свист служит сигналом тревоги. Размножение к определенному сезону не приурочено. Продолжительность беременности около 4 месяцев. В помете обычно один детеныш. Длительность жизни до 8—10 лет. Местные жители охотятся за хохлатыми дукерами из-за мяса.
Рис. 173. Черный дукер — С. niger (Д), зебровый дукер— С. zebra (Б), желтоспинный дукер — С. sylvicultor (В) и череп желтоспинного дукера
В роде, по-видимому, 15 видов:
голубой дукер —С. monticola Thunberg, 1789 (от Восточной Нигерии на западе до юга Судана, Кении и Танзании на востоке и Южно-Африканской Республики на юге);
дукер Максвелла — С. maxwelli Н. Smith, 1827 (от Нигерии и Северо-Западного Камеруна на востоке до Гамбии и прилегающих к ней районов Сенегала);
чернолобый дукер — С. nigrifrons Gray, 1871 (от Южного Камеруна и Габона на восток через Заир, Руанду и Бурунди до Уганды и Кении);
рыжебокий дукер — С. rufilatus Gray, 1846 (от Сенегала и Гамбии на западе до Юго-Западного Судана, Северо-Западной Уганды и Севе-
ро-Восточной Кении на востоке через Нигерию и Центральноафриканскую Республику);
чепрачный дукер — С. jentinki Thomas, 1892 (Либерия, Сьерра-Леоне, Берег Слоновой Кости);
желтоспинный дукер — С. sylvicultor Afzelius, 1815 (от Гвинеи-Бисау на восток до Западной Уганды и Кении; на юг до Анголы и Замбии;
танзанийский дукер — С. spadix True, 1890 (Танзания, Руанда и Бурунди) ;
зебровый дукер — С. zebra Gray, 1838 (Сьерра-Леоне, Либерия и Берег Слоновой Кости);
черный дукер — С. niger Gray, 1846 (от Сьерра-Леоне до Юго-Западной Нигерии);
занзибарский дукер — С. adersi Thomas, 1918 (о-в Занзибар);
красный дукер — С. natalensis A. Smith, 1834 (Заир, Южный Судан, Уганда, Кения, Сомали, Танзания, Мозамбик, Южно-Африканская Республика) ;
конголезский дукер — С. callipygus Peters, 1876 (Конго, Габон, Южный Камерун);
черноспинный дукер — С. dorsalis Gray, 1846 (от Сьерра-Леоне до Камеруна, Заира и Анголы);
белобрюхий дукер — С. leucogaster Gray, 1873 (от Камеруна до Конго);
дукер Оджильби — С. ogilbyi Waterhouse, 1838 (о-в Фернандо-По).
Некоторые исследователи (например, Хальтенорт, 1963) выделяют из красного дукера в качестве особого вида дукера Гарвея — С. harveyi Thomas, 1893, рассматривают занзибарского дукера в качестве подвида С. harveyi (что представляется очень вероятным); относят (Симпсон, 1945) С. monticola в особый род Philantomba Blyth, 1840. Хальтенорт (1963) включает в род Cephalophtis на правах подрода кустарниковых дукеров Sylvicapra.
Чепрачный дукер включен в «Красную книгу» как вид, находящийся под угрозой исчезновения.
Род кустарниковых дукеров
Genus Sylvicapra Ogilby, 1837
В роде единственный вид: кустарниковый дукер — S. grimmia Linnaeus, 1758.
Размеры мелкие. Длина тела 85—115 см, длина хвоста 8—18 см, высота в холке 45—70 см. Масса 10—15 кг. Самки крупнее самцов. У самок обычно рогов нет, а если имеются, то они значительно меньше, чем У самцов. Ось рогов служит продолжением верхнего профиля головы. Уши крупные, узкие, заостренные (рис. 174). Окраска спины сероватая, Желтоватая или рыжевато-желтая. Брюхо светлее спины. От носа к основанию лба проходит темная полоса. Имеются паховые железы.
Распространен почти по всей Африке к югу от Сахары.
Населяет разреженные леса, саванны и кустарниковые равнины. Поднимается высоко в горы и встречается на альпийских лугах. Держатся поодиночке или парами. Днем отдыхает в кустарниковых или травянистых зарослях. Активен в сумерках. Питается различными растениями, по-видимому, отчасти и животными. Сезонности в размножении нет. В период гона самцы дерутся между собой. Продолжительность беременности около 4 месяцев. Самка приносит одного, редко двух детенышей.
Рис 174 Кустарниковый дукер — S. grimmia и его череп
SUBFAMILIA NEOTRAGINAE SCLATER ET THOMAS, 1894
Размеры мелкие. Рога имеют только самцы (у одного из подвидов антилопы-прыгуна рогами обладают и самки, но меньших размеров). Они короткие или средней длины, прямые или слегка выгнутые вперед, гладкие или частично гофрированные, широко расставленные. Уши средней длины или длинные. Хвост короткий или средней длины. Боковые копыта большие, маленькие или отсутствуют полностью. Из кожных специфических желез обычно имеются предглазничная и межпальцевые железы; у некоторых видов есть запястная и паховая железы. Сосков одна, чаще две пары.
Череп с широким лбом, коротким или длинным лицевым отделом. На слезных костях ямки предглазничных желез, как правило, большие и глубокие. Носовые кости короткие или очень короткие.
о о з з
Зубная формула: i —- с — р — m — = 32. 3 13 3
Ширина коронки первого нижнего резца равна или превышает в два раза ширину коронок второго и третьего нижних резцов и клыка вместе взятых.
Ряд исследователей (например, Симпсон, 1945) объединяют подсемейство карликовых антилоп и подсемейство газелей в подсемейство Ап-tilopinae Baird, 1857. Следуя Хальтенорту, 1963, видимо, следует считать их отдельными подсемействами.
Род карликовых антилоп
Genus Neotragus Н. Smith, 1827
Одни из самых маленьких представителей семейства. Длина тела 50—62 см, длина хвоста 5—13 см, высота в холке 25—38 см. Масса 2,5— 6 кг. Сложение изящное, хрупкое, туловище округлое в поперечнике, спина дугообразно выгнута; в крестце она выше, чем в холке. Конечности средней высоты, тонкие. Шея короткая. Голова относительно крупная. Глаза большие. На конце морды имеется участок безволосой кожи. Уши средней длины, овальные. Хвост короткий, покрыт длинными волосами. Рога короткие, достигают длины 2,5—12,5 см, направлены наискось назад. Вершины рогов загнуты немного вверх и внутрь. Форма поперечника рогов округлая. Копыта узкие, заостренные. Боковых копыт нет. Волосяной покров низкий и гладкий. Спина от рыже-бурой до темно-серо-бурой. Брюшная поверхность тела, кисточка на хвосте и низ хвоста белые. Верхняя часть шеи отделяется от нижней темной полосой. Имеются предглазничные и межпальцевые железы. Паховые железы отсутствуют. Сосков 2 пары.
Череп с широкой мозговой коробкой и узкой и укороченной лицевой частью. Глазница большая. Этмоидальные отверстия небольшие или их нет вовсе. Костные слуховые барабаны маленькие.
Распространены в Западной, Центральной, Восточной и Южной Африке.
Обитают в густых лесах на равнинах и в горах, поднимаясь на высоту до 2 тыс. м над уровнем моря. Питаются в основном листьями и побегами, а также плодами и различными травами. Держатся поодиночке, парами и небольшими семейными группами. Сумеречные и ночные животные. Период гона приходится в Южной Африке на ноябрь — декабрь. Биология размножения изучена плохо.
В роде 3 вида:
карликовая антилопа—• N. pygmaeus Linnaeus, 1758 (от Сьерра-Леоне до Ганы);
антилопа-крошка — N. batesi de Winton, 1903 (от Нигерии и Камеруна на восток до Уганды и на юг до границы лесной зоны);
суни — N. moschatus von Dueben, 1846 (Южно-Африканская Республика, Мозамбик, Танзания, Кения, о-ва Мафия и Занзибар).
Некоторые исследователи (например, Симпсон, 1945) выделяют суни в особый род Nesotragus Dflben, 1847. Правильнее считать его подродом.
Подвид суни из Занзибара — N. moschatus moschatus Diiben, 1846, включен в «Красную книгу» как подвид, находящийся под угрозой исчезновения.
Рис. 175. Рыжебрюхий дикдик — Л1. phillipsi. голова и череп обыкновенного дикдика — М. kirki
Род дикдиков
Genus Madoqua Ogilby, 1837
Размеры наименьшие в семействе. Длина тела 45—80 см, длина хвоста 3—6 см, высота в холке 30—45 см. Масса 2—0,5 кг. Самки крупнее самцов. Сложение стройное. Конечности тонкие. Спина дугообразно выгнута. Тело в крестце выше, чем в холке. Шея короткая. Голова короткая с узкой мордой (рис. 175). Нос подвижен. У М. guentheri и М. kirki нос образует небольшой хоботок. На конце морды участка голой кожи нет. Глаза большие. Уши средней длины, овальные. Хвост очень корог-
кий. Длина рогов достигает 11 см. Они стоят далеко друг от друга и направлены наискось назад и вверх, причем их вершины выгнуты немного вверх и наружу. Поперечник рогов овальный и округлый. Копыта узкие, длинные и заостренные. Боковые копыта очень маленькие и уплощенные.
Волосяной покров низкий, нежный (хотя не имеет пуховых волос), гладкий, удлиненный на передней части груди и на фронтальном отделе головы. Спинная поверхность тела серо-белая, серо-желтая, серо-красная и почти ржаво-красная. Внутренняя поверхность ушей, кольца вокруг глаз, губы, подбородок, горло, грудь, брюхо, внутренние части ног белые или желто-белые. Предглазничные железы крупные. Имеются межпальцевые железы. Паховых желез нет. Сосков 2 пары.
Череп самцов короткий и широкий, а самок более вытянутый. Мозговой отдел округлый, вздутый. Глазницы большие. Ямки предглазничных желез на слезных костях очень большие, но не глубокие. Этмоидальные отверстия узкие, треугольной формы. Носовые кости очень короткие и широкие. Костные слуховые барабаны большие. Длинные предчелюстные кости соприкасаются не только с носовыми, но иногда и со слезными костями.
Распространены в Восточной и Юго-Западной Африке.
Обитают в густых зарослях кустарников в саваннах, на каменистых равнинах. Поднимаются в горы до 3 тыс. м над уровнем моря. Дневные и сумеречные животные. Держатся обычно парами. Самцы метят занятую территорию секретом предглазничных желез и кучками экскрементов. Характерен крик «зик-зик» или «дик-дик». Питаются листьями и побегами кустарников и различными травами. Могут подолгу обходиться без питьевой воды. У самок полиэстральный цикл. Размножение не приурочено к определенному сезону. Беременность продолжается примерно 6 месяцев. В помете обычно один детеныш. Как правило, самки приносят потомство дважды в году. Половозрелость наступает в 6 месяцев. Продолжительность жизни 3—5 лет; в неволе — до 10 лет.
В роде 5 видов:
горный дикдик — М. saltiana Desmarest, 1816 (Эфиопия);
малый дикдик — М. swaynei Thomas, 1894 (Сомали и Эфиопия);
рыжебрюхий дикдик — М. phillipsi Thomas, 1894 (Сомали и Эфиопия);
дикдик Гюнтера — М. guentheri Thomas, 1894 (Сомали, Эфиопия, Кения. северо-восток Уганды и юго-восток Судана);
обыкновенный дикдик — At. kirki Gunther, 1880 (Сомали, Кения, Уганда, Танзания и изолированный участок: Ангола и Намибия).
Некоторые исследователи (например, Симпсон, 1945) выделяют дик-Дика Гюнтера и обыкновенного дикдика в особый род Rhynchotragus Neumann, 1905. Более правильно рассматривать его в виде подрода.
Род антилоп-прыгунов
Genus Oreotragus A. Smith, 1834
В роде единственный вид: антилопа-прыгун — О. oreotragus Zimmermann, 1783.
Размеры мелкие. Длина тела 75—115 см, длина хвоста 8—13 см, высота в холке 50—60 см. Масса 14—18 кг. Самки немного меньше самцов. Спина дугообразно выгнута и крестец расположен выше загривка. Конечности средней высоты, относительно толстые. Шея короткая. Голова небольшая, короткая с широким лбом (рис. 176). На конце морды имеется участок безволосой кожи. Глаза большие. Уши длинные и широкие.
Рис 176 Антилопа-прыгун — О orpotragus
с закругленными вершинами. Хвост короткий с голой внутренней поверхностью. Длина рогов достигает 16 см; они прямые, слегка выгнуты вперед, далеко отстоят друг от друга. Самки одного из подвидов также имеют рога.
Характерны длинные фаланги пальцев; пястные кости короткие. Средние копыта длинные, сжатые с боков, со стертыми концами (являются опорными точками конечностей). Животные стоят, бегают и прыгают только на кончиках копыт, поэтому сухожильный аппарат конечностей сильно развит. Боковые копыта расположены высоко, они средних размеров, широкие, длинные и плоские. Волосяной покров без пуховых волос, грубый, но плотный. Спинная поверхность тела зологис-то-желтая или светло-желто-бурая. Спинка носа, середина лба, задние поверхности ушей и пальцы над средними копытами буро-черные или черные. Конечности буро-серые или серебристо-серые. Брюшная сторона тела, а также подбородок, губы, круги вокруг глаз белые или желто-белые. Предглазничные железы больших размеров. Межпальцевых и паховых желез нет. Сосков 2 пары.
Череп короткий и широкий. Лобный отдел очень широкий. Лицевая часть черепа узкая, короткая. Глазницы большие, трубкообразные, вы-
ЛЛЛ
ступающие в стороны. Этмоидальные отверстия крупные, маленькие или их нет вовсе. Костные слуховые барабаны небольшие
Распространена от Северной Нигерии до Эфиопии и Сомали, на юг через Восточную Африку до мыса Доброй Надежды и отсюда на север до Южной Анголы.
Обитает в зарослях кустарников на горных склонах или на скальных обнажениях в горах, на холмах и равнинах. Скалы — необходимое условие для жизни антилопы-прыгуна. Активна утром и вечером. Питается листьями, побегами, различными травами. Водопой регулярно не посещает. Размножение не приурочено строго к определенному сезону. Длительность беременности около 7 месяцев. Самка приносит одного детеныша, который остается с родителями до рождения следующего. Половозрелость наступает, по-видимому, в 1,5 года. Продолжительность жизни до 10— 12 лет.
Местное население охотится за антилопами-прыгунами из-за мяса и кожи.
Рис. 177. Бейра — D. megalotis
Род бейр
Genus Dorcatragus Noack, 1894
В роде единственный вид: бейра — D. megalotis Menges, 1894.
Размеры мелкие. Длина тела 80—90 см, длина хвоста 10—12 см, высота в холке 55—60 см. Масса 28 кг. Самцы и самки примерно одинаковой величины. Тело в крестце выше, чем в загривке. Шея короткая. Голова укороченная с широким лбом и заостренной мордой (рис. 177). На конце морды имеется очень маленький участок голой кожи. Глаза большие. Уши овальные, крупные. Хвост короткий, но заканчивается длинными волосами. Длина рогов достигает 14 см; они отстоят друг от Друга на значительном расстоянии, проходят параллельно один другому и немного выгнуты вперед. Форма поперечника рогов округлая. Средние копыта широкие и высокие, заостренные. Боковые копыта небольших размеров. Волосяной покров плотный, высокий и грубый. Окраска спинной стороны тела от рыжевато-серой до голубовато-серой. По нижним отделам боков тела проходит темно-бурая кайма, которая спереди через локти спускается к копытам. Брюшная сторона тела белая. Глаза окружены белым полем. Предглазничных и паховых желез нет. Имеются межпальцевые железы. Сосков одна пара.
Череп укорочен. Лобные кости широкие. На слезных костях ямок предглазничных желез нет. Носовые кости очень короткие, а наружное костное носовое отверстие очень большое. Межчелюстные кости граничат с носовыми. Костные слуховые барабаны большие.
Распространена в Сомали и прилегающих районах Эфиопии.
Обитает в предгорьях, засушливых местах и полупустынях и по склонам гор в зарослях кустарников со скальными обнажениями. Активна утром и вечером. Держится обычно парами или семейными группами. Сигнал тревоги — громкий свист. Питается листьями и побегами кустарников, кроме того, различными травами. При зеленом корме в водопое не нуждается. Самки приносят в апреле по одному детенышу.
Включена в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения (численность в 1905 г. в Сомали была примерно 1 тыс. голов).
Род стенбоков
Genus Raphicerus Н. Smith, 1827
Размеры мелкие. Длина тела 62—90 см, длина хвоста 5—10 см, высота в холке 45—60 см. Масса 8—16 кг. Сложение изящное. Конечности тонкие, средней длины или длинные. Туловище со слегка выгнутой спиной. Тело в крестце выше, чем в загривке. Шея средней длины. На конце морды имеется участок безволосой кожи. Голова узкая. Глаза большие. Уши крупные овальные. Рога достигают длины 5—19 см; они гладкие, прямые и далеко отстоят друг от друга. Форма поперечника рогов округлая. Средние копыта узкие, заостренные. Боковые копыта очень малы (R. melanotis} или отсутствуют (Р. sharpei и R. campestris). Волосяной покров низкий, гладкий и грубый. Окраска спинной стороны тела варьирует от рыже-бурой до оранжево-рыжей и темно-рыже-бурой. У стенбо-ка Шарпа имеются белые пятна на спине и боках. Вокруг глаз располагаются темные или белые кольца. Брюшная сторона тела белая или желто-белая, а у R. melanotis буро-желтая. Старые самцы темнее самок и молодых. Предглазничные железы хорошо развиты. Имеются межпальцевые железы. Паховых желез нет. Сосков 2 пары.
Череп с широкой лобной частью и узким лицевым отделом. Его верхний профиль прямой. Мозговая коробка черепа длинная. Глазницы большие. Ямки предглазничных желез на слезных костях маленькие, но глубокие. Этмоидальные отверстия большие. Межчелюстные кости сочленяются с носовыми. Костные слуховые барабаны большие. На лобной поверхности довольно сильно развит срединный гребень, проходящий вдоль сагиттального шва.
Диплоидное число хромосом у обыкновенного стенбока 30.
Распространены в Африке к югу от Центральной Кении и Центральной Анголы.
Обитают в открытых местах, поросших травой или кустарником, а также в холмистых районах, на горных плато. Держатся поодиночке или парами. В случае опасности часто затаиваются, плотно прижимаясь к земле и вытянув шею. Активны ранним утром и поздним вечером. Питаются листьями, побегами, почками кустарников и травянистыми растениями. Могут подолгу обходиться без воды. Периоды гона и рождения детенышей различны в зависимости от района ареала. Так, в Восточной Африке самки приносят детенышей с мая по декабрь, а в Южной Африке— с сентября по декабрь. Длительность беременности около 7 месяцев. Детенышей в помете один, редко два. Самка может приносить потомство два раза в год. Длительность жизни 10—12 лет.
В роде 3 вида:
обыкновенный стенбок — R. campestris Thunberg, 1811 (Южная Ангола, Юго-Западная Замбия, Намибия, Ботсвана, Южно-Африканская Республика, Мозамбик, Танзания, Кения, Южное Сомали);
Рис. 178. Орибп — О. ourebi
грисбок-—R. melanotis Thunberg, 1811 (Южно-Африканская Республика, Мозамбик, Родезия);
стенбок Шарпа — R. sharpei Thomas, 1897 (Южно-Африканская Республика, Ботсвана, Замбия, Мозамбик, Малави, Юго-Восточный Заир, Танзания).
Некоторые исследователи (например, Хальтенорт, 1963) включают стенбока Шарпа в качестве подвида в вид R. melanotis.
Род ориби
Genus Ourebia Laurillard, 1841
В роде единственный вид: ориби — О. ourebi Zimmermann, 1783.
Размеры мелкие. Длина тела 92—-НО см, высота в холке 50—67 см, Длина хвоста 6—10 см. Сложение изящное, конечности высокие. Спина прямая, в крестце чуть выше, чем в холке. Шея средней длины (рис. 178). Голова небольшая с прямым верхним профилем. На конце морды имеется безволосый участок кожи. Глаза большие. Уши удлиненные, овальной формы. Ниже основания ушей с каждой стороны головы находится по небольшому безволосому округлому участку кожи, не содержащего Желез (функция неизвестна). Хвост короткий с безволосой внутренней поверхностью и кистью волос на конце. Длина рогов 8—18 см; они вы-[нуты немного вперед, вершинами направлены круто вверх. Верхняя половина рогов гладкая, нижняя сильно гофрирована. Форма поперечника Рогов круглая. Средние копыта узкие и заостренные. Боковые копыта маленькие, короткие и широкие. Волосяной покров низкий, густой. Пудовых волос нет. Окраска спинной стороны тела коричнево-желтая, брюшной — белая. Пятна вокруг глаз, губы, подбородок и горло белые.
Хвост черный. Имеются паховые, межпальцевые, запястные и крупные предглазничные железы. Сосков 2 пары.
Череп широколобый с узким лицевым отделом. Глазницы большие. Этмоидальные отверстия длинные; иногда они отсутствуют. Костные слуховые барабаны маленькие. Межчелюстные кости не достигают носовых.
Иногда имеется рудиментарный верхний клык.
Распространена от Судана и Эфиопии, на юг через Восточную Африку до Южной Африки и затем к северу до Анголы.
Держится в равнинных, холмистых и горных открытых или заросших кустарником участках; поднимается на высоту до 3 тыс. м над уровнем моря. Встречаются поодиночке, парами и семейными группами. Часто объединяются в стада с антилопами. Активна утром и вечером. При испуге издает громкий свист. Питается в основном различными травами, в меньшей степени листьями и побегами кустарников и деревьев. Продолжительность беременности около 7 месяцев. В помете один детеныш. Рождение детенышей происходит в Южной Африке чаще в сентябре—• декабре. Лактационный период 4—5 месяцев. Продолжительность жизни 8—12 лет.
За ориби активно охотятся из-за их вкусного мяса.
SUBFAMILIA ANT1LOPINAE BAJRD, 1857
Размеры мелкие, средние или большие. Рога только у самцов или у самцов и самок (у последних меньших размеров). Рога короткие или длинные, тонкие или толстые, гладкие или гофрированные, прямые или по-разному изогнутые. Хвост длинный. На конце морды имеется большой участок безволосой кожи (кроме Aepyceros). Из специфических кожных желез в разных комбинациях имеются предглазничные, паховые, межпальцевые, предкопытные, плюсневые. Сосков одна или две пары.
В черепе лицевой отдел длинный или очень длинный. Мозговая коробка укороченная или длинная. Носовые кости хорошо развиты. Костные слуховые барабаны большие.
о о 3 3
Зубная формула: i —- с — р —- m -— = 32. 3 13 3
Ширина коронки первого нижнего резца примерно такая же, как и ширина коронок второго и третьего резцов и клыка вместе взятых.
Состав подсемейства окончательно не установлен. Некоторые ученые (например, Симпсон, 1945), называя это подсемейство Hippotraginae Brooke, 1876, не включают в него роды Tetracerus, Baselaphus, Tragelap-hus и Aepyceros, из которых трех первых они относят к подсемейству Bovin ае, а последний — к Antilopinae.
Род четырехрогих антилоп
Genus Tetracerus Leach, 1825
В роде единственный вид: четырехрогая антилопа — Т. quadricornis De Blainville, 1816.
Размеры мелкие. Длина тела 90—ПО см, длина хвоста 10—15 см, высота в холке 55—65 см. Масса 15—25 кг. Самцы немного крупнее самок. Телосложение легкое. Конечности средней высоты, тонкие. Голова удлиненная, узкая, с прямым верхним профилем (рис. 179). Глаза большие. Уши средней длины, широкие, овальной формы. На конце морды имеется большой участок безволосой кожи. Хвост средней длины, без конце-410
бой кисти волос. Рога имеют только самцы. Их длина невелика — всего 5—10, редко 12 см. Рога отстоят друг от друга на значительном расстоянии, слегка изогнуты и направлены вершинами вверх и внутрь. Форма их поперечника в основании треугольная (одной из вершин треугольник направлен вперед), а к концу округлая. Часто имеется дополнительная пара коротких рогов (длиной 2—4, редко 7 см), возвышающихся в передней части лобного отдела, непосредственно перед основными рогами. Дополнительные рога расположены друг к другу близко, стоят вертикально и имеют коническую форму со слегка загнутыми назад вершина
ми. Средние копыта узкие, остроконечные. Боковые копыта уплощенные и широкие.
Волосяной покров низкий, редкий и довольно грубый. Спина светло-желто-бурая или темно-бурая, брюхо беловатое или белое. Спинка носа и лоб, наружные части ушей и передние отделы ног темно-бурые. На щеках под глазами обычно расположено по два беловатых пятна. Над боковыми копытами и под ними проходят беловатые кольцевидные полосы. Имеются предглазничные железы. На задних конечностях под боковым копытом на внутренней стороне возвышается безволосый округлый участок кожи, содержащий железы. Межпальцевых и паховых желез нет. Сосков 2 пары.
Череп удлиненный. Его мозговой отдел составляет примерно половину длины лицевого отдела, который сильно сужен кпереди от глазниц. Глазницы большие, их края почти не выдаются в стороны. Носовые кости длинные и узкие. Этмоидальных отверстий нет. Костные слуховые барабаны средних размеров.
Щечные зубы крупные с высокими коронками.
Диплоидное число хромосом 38.
Распространена в Западной Индии.
Обитает на равнинах и в холмистых участках с разреженными лесами или зарослями кустарников, перемежающихся открытыми пространствами. Обычно держимся вблизи водоемов. Активна утром и вечером. Встречается по большей части поодиночке или парами, а также семьями: самка и 1—3 молодых. По-видимому, самец остается с одной и той же самкой в течение всей жизни. Питается листьями, почками, побегами, фруктами кустов и деревьев, а также травянистыми растениями. Обычно ежедневно посещает водопой, причем в жаркое время года два раза в день. Период спаривания приходится на летнее время — июнь—август. Беременность продолжается 6 месяцев (по некоторым данным, 7,5 месяцев). Детенышей в помете 1—3. Продолжительность жизни 10— 12 лет.
Род нильгау
Genus Boselaphus De Blainville, 1816
В роде единственный вид: нильгау—В. tragocamelus Pallas, 1766.
Размеры средние. Длина тела 180—200 см, длина хвоста 40—55 см, высота в холке 120—150 см. Масса 100—200 кг. Самцы крупнее самок. Передняя часть тела значительно массивнее задней (рис. 180). Линия спины полого спускается от загривка к крестцу (у самцов более круто, чем у самок). Шея короткая и у самцов более толстая, чем у самок. Конечности высокие, тонкие. Голова у самок удлиненная и узкая, у самцов более короткая. Ее верхний профиль прямой. На конце морды имеется широкий участок голой кожи. Глаза большие. Уши сравнительно короткие, ланцетовидно-заостренной формы. Хвост с кистью волос на конце. Рога имеются только у самцов, короткие и толстые, концы их слабо загнуты вверх и внутрь. Форма поперечника рогов в основании треугольная (один из углов треугольника направлен вперед), а в верхней части— округлая. Окраска рогов черная. Средние копыта узкие и заостренные; боковые копыта короткие, широкие и уплощенные. Окраска копыт бурая или черная.
Волосяной покров низкий, гладкий. От затылка до холки идет грива длиной 5—10 см. Спереди посредине шеи на горле имеются удлиненные волосы. Спина голубовато-серая у самцов и серо-рыжая у самок. Передние отделы конечностей с продольными темными полосами. Брюхо серо-
л 1 о
белое. Спинка носа, затылок, грива на загривке, кольца на проксимальных отделах ног темно-бурые. Грива на шее бело-серая или беловатобурая. Губы, подбородок, горловое пятно, два щечных пятна под глазами, вершины ушей, паха, кольцевидные полосы в нижних отделах ног и два овальных пятна около копыт белые. Имеются небольшие предглазничные железы и железистое поле между боковыми копытами задних конечностей. Межпальцевых и паховых желез нет. Сосков 2 пары.
Череп удлиненный с прямым верхним профилем. Мозговой отдел составляет в длину примерно половину лицевой части черепа. Его зарого-вая часть уплощена, теменные гребни сильно развиты. Носовые кости длинные и узкие. Глазница большая с выдающимся в стороны нижним и задним краем. На слезной кости нет ямки предглазничной железы. Этмоидальные отверстия очень малы или отсутствуют. Костные слуховые барабаны большие.
Щечные зубы большие с высокими коронками.
Диплоидное число хромосом 46.
Ареал охватывает Западную Индию к югу от Гималаев.
Обитает в открытых равнинных или холмистых местностях, поросших кустарником, отдельными деревьями или рощами. Активна в основном утром и вечером. Встречаются обычно небольшими группами, состоящими из самок с молодыми. Взрослые самцы держатся поодиночке или объединяются в маленькие группы. Изредка попадаются небольшие стада в 15—20 голов. Питается в основном листьями и побегами кустарников и деревьев, а также травянистыми растениями. На юге ареала размножение не приурочено к определенному времени года, а на севере гон приходится в основном на март—апрель. Самцы в это время сражаются между собой. Продолжительность беременности около 8 месяцев. Самка приносит одного, чаще двух детенышей. Половозрелость наступает примерно в 1,5 года. Длительность жизни 12—15 лет, в неволе 21 год.
Основные враги — тигр и леопард.
За нильгау охотятся редко, так как в качестве охотничьего трофея она не представляет интереса, а мясо ее невкусно.
Род лесных антилоп
Genus Tragelaphus De Blainville, 1816
Размеры мелкие, средние и большие. Длина тела ЮЗ—345 см, длина хвоста 18—100 см, высота в холке 65—180 см. Масса 35—800 кг, по некоторым данным 1000 кг (канна). Телосложение разнообразное: от легкого и изящного до массивного (рис. 181, 182, 183). Спина прямая или-немного выше в крестце, или, наоборот, выше в холке. Конечности длинные и тонкие или средней длины и довольно толстые. Шея укороченная. Голова сравнительно с телом крупная с прямым верхним профилем. Глаза большие или очень большие. На конце имеется участок голой кожи. Уши большие, широкие с округлой вершиной. Хвост средней длины, покрыт косматыми волосами или средней высоты волосами, но с кистью на конце. Рога имеют как самцы, так и самки (у последних они, как пра_; вило, меньше по размерам) или только самцы (Т. scriptus, Т. spekti, Т. angasi, Т. buxtoni, Т. imberbis, Т. strepsiceros). Длина рогов достигает 30—170 см. Они лирообразные или почти прямые, проходят наискось спереди назад и вверх, причем делают очень пологие 0,5—2,5 поворота вдоль длинной оси. Основания рогов стоят близко друг к другу. Форма поперечника рогов круглая или овальная с килем; их поверхность глад-
Рис. 181. Ситатунга—Т. spekii (.4). бушбок—Т scrip-tu& (i>) и его череп
л
большой куду —
Рис 182. Ньяла — Т. angasi (А) и J. strepsiceros (6)
кая или имеет в нижней части небольшие кольцевидные выступы. Копыта узкие и заостренные или уплощенные и округлые. Боковые копыта обычно развиты сильно, иногда их размеры малы.
Волосяной покров гладкий или частично взъерошенный. Вдоль средней линии спины от затылка до основания хвоста тянется полоса удлиненных волос, которые особой длины достигают на шее и загривке. Удлинены волосы на лбу и на подбородке у Т. oryx-, небольшая грива на верху шеи имеется у Т. spekii и Т. buxtoni, длинная грива проходит по низу шеи от горла до брюха у Т. strepsiceros и Т. angasi. Окраска спины от песчано-серой и серо-голубой до черно-серой или от песчано-желтой и рыже-бурой до черно-бурой. Фронтальная часть морды обычно более темная. Брюхо в большинстве случаев немного светлее, реже значительно светлее или темнее спины. Конечности обычно пестрые, трехцветные: с наружной стороны они имеют окраску спины, на внутренней белые, в верхней части с бурой или черной задней поперечной полосой, а в дистальных отделах спереди со светлой или белой кольцевидной полосой (чаще двойной), сзади белые или черные. На морде между глазами проходит светлая или белая V-образная полоса; под глазами располагаются светлые пятна; подбородок и губы белые. Бока тела одноцветные или по ним проходят 3—15 узких поперечных белых полос. Если их число невелико, они располагаются в плечевой области, если же многочисленны, то доходят до крупа и основания хвоста. Самки и молодые животные окрашены светлее самцов.
Предглазничных и межпальцевых желез нет. Имеются околокопыт-ные железы (Т. strepsiceros, Т. angasi, Т. imberbis). Одна или две пары паховых желез встречаются у Т. scriptus и Т. spekii, две пары у Т. imberbis, Т. angasi и Т. buxtoni-, у Т. strepsiceros, Т. oryx и Т. euryceros они отсутствуют. Сосков 2 пары.
Череп массивный, удлиненный. Мозговой отдел относительно развит и составляет не менее половины длины лицевой части черепа. Глазницы выступают в стороны слабо. Этмоидальные отверстия и костные слуховые барабаны большие. Носовые кости длинные, массивные.
Диплоидное число хромосом у самок большого куду 32 и самцов 31, ситатунги — 30, ньялы — 55, самок канны — 31 и самцов — 32, самцов бонго — 33.
Распространены в Африке к югу от Сахары.
Обитают в глухих лесах и в болотах, заросших тростником, саваннах, кустарниковых степях, полупустынях, по склонам гор, горным лугам выше границы леса на высоте до 4,5 тыс. м над уровнем моря. Предпочитают держаться около водоемов. В засуху могут мигрировать на большие расстояния. Встречаются поодиночке, парами или семейными группами, реже крупными стадами. Питаются листьями, почками, побегами кустарников и деревьев, фруктами, корнями, луковицами, клубнями, травянистыми растениями. Активны ранним утром, вечером и ночью. Тревожный крик напоминает лай собаки. Сезонности в размножении в Ряде районов нет. Беременность продолжается 7—9 месяцев. Самка рождает одного, редко двух детенышей. Продолжительность жизни 10— 25 лет. Некоторые виды хорошо одомашниваются (например, канна в Аскания-Нова, СССР).
Основные враги — леопард, лев, гиеновая собака,
В роде 8 видов:
бушбок—Т. scriptus Pallas, 1766 (Африка к югу от Сахары; Эфиопия и Сомали);
ситатунга — Т. spekii Sclater, 1864 (Ботсвана, Намибия, Ангола, Габон, Экваториальная Гвинея, Конго, Заир, Руанда, Бурунди, Замбия, 1 анзания, Кения, Уганда, юг Судана, Эфиопия, Центральноафриканская 14—671 417
Рис. 183. Бонго — Т euryceros (Д)
Республика, юг Чада, Нигерия, Того, Берег Слоновой Кости, Гвинея, Гамбия, Гвинея-Бисау);
ньяла — Т. angasi Gray, 1849 (Южно-Африканская Республика, Мозамбик, Малави);
горная ньяла — Т. buxtoni Lydekker, 1910 (юг Эфиопии);
малый куду—Т. imberbis Blyth, 1869 (Сомали, Эфиопия, юго-восток Судана, Уганда, Кения, Танзания);
большой куду — Т. strepsiceros Pallas, 1766 (Южно-Африканская Республика, Намибия, Ботсвана, Ангола, Замбия, юго-восток Заира, Родезия, Мозамбик, Малави, Танзания, Кения, Сомали);
канна—Т. oryx Pallas, 1766 (аридные районы Африки к югу от Сахары);
бонго — Т. euryceros Ogilby, 1837 (от Сьерра-Леоне на восток до юга Судана, Уганды, Кении и Северной Танзании).
Рис. 133. Продолжение. Канна — Т. oryx (Я)
Некоторые исследователи выделяют канну в особый род Тaurotragus Wagner, 1855 (например, Эллермэн и др., Ellerman et aL, 1953), а бонго— в особый род Boocercus Thomas, 1902 (например, Уокер, 1964) ; из Т. oryx выделяют Т. derbianus Gray, 1847 (например, Андерсон и Джонс, 1967). Следуя Хальтенорту, 1963, здесь эти разделения не приняты.
Канна из Западной Африки— Т. oryx derbianus Gray, 1847 включена в «Красную книгу» как подвид, находящийся под угрозой исчезновения.
Род ор иксов
Genus Oryx De Blainviile, 1816
В роде единственный вид: орикс — О. gazella Linnaeus, 1758.
Размеры средние. Длина тела 160—235 см, длина хвоста 45—90 см, высота в холке 90—140 см. Масса 100—120 кг. Самцы крупнее самок. 14* 419
Рис. 184. Орике — О. gazella
Туловище массивное. Конечности высокие, сильные. Загривок чуть выше крестца (рис. 184). Шея средней длины, толстая. Голова большая. На конце морды имеется маленький участок голой кожи. Глаза сравнительно невелики. Уши средней длины, ланцетовидно-овальной формы с закругленной вершиной. Хвост длинный с большой кистью на конце. Рога имеют и самцы, и самки; у самок они тоньше, но немного длиннее. От основания рога отходят назад и вверх, слабо или сильно саблеобразно изгибаясь назад. В поперечном сечении рога округлые, концы их очень острые. В нижних отделах они имеют крупные поперечно-кольцевые выступы. При рождении у детенышей уже есть рога 2—5 см длиной. Длина рогов взрослых животных 60—120 см. Средние копыта узкие и заостренные. Боковые копыта большие. Окраска копыт и рогов бурочерная или черная.
Волосяной покров низкий густой. У ориксов из южных и северных частей ареала в зимнее время волосы длиннее, чем летом. Посредине шеи до загривка проходит короткая стоячая грива. У разных подвидов окраска варьирует: спинная сторона тела беловатая, бело-серая или белая до серо-бурой, серо-буро-рыжей или рыжевато-бурой, брюшная — белая или буровато-белая, иногда с темно-бурым полем на груди. Шея и нижняя часть плечей может быть темнее туловища или от горла до груди может проходить бурая полоса. Вдоль нижних частей боков проходит светло-бурая, темно-бурая или черная полоса. Она может продолжаться вперед к области груди, где раздваивается и поднимается к основанию ушей, но может также подходить к предплечью передних конечностей, сливаясь здесь с широкими темными кольцевидными полосами. Чаще эти кольцевидные полосы не связаны с боковыми и вы-
АПГ\
ряжены четко или не очень резко. Передние и задние конечности могут быть белыми или белыми остаются только передние, задние — снаружи темные, а изнутри белые или и передние, и задние конечности целиком бурые или черно-бурые с белыми кольцами в проксимальных частях. Спереди на запястных отделах расположено слабо или сильно выраженное черно-бурое продольное пятно или полоса. На передней части плюсны может проходить аналогичная полоса. Голова белая или беловатая. На лбу перед рогами находится пятно; удлиненное пятно располагается на спинке носа, и узкая полоса проходит от основания рогов через глаз к подбородку; все они могут быть соединены между собой; их окраска буро-серая, буро-черная или черная. Молодые животные светло-желтобурые или светло-буро-рыжеватые на спинной стороне тела и белые на брюшной.
Предглазничные специфические кожные железы очень малы, чаще их нет вовсе. На передних и задних конечностях есть межпальцевые железы. Паховые и запястные железы отсутствуют.
Сосков 2 пары.
Череп узкий и длинный. Верхний профиль черепа почти прямой. Задний и нижний края глазниц сильно выдаются в стороны. Мозговой отдел черепа составляет примерно четверть длины лицевого. Носовые кости короткие или длинные. Этмоидальные отверстия средних размеров или крупные. Костные слуховые барабаны большие и узкие.
Щечные зубы, особенно коренные, с широкими четырехугольными коронками.
Диплоидное число хромосом 56—58.
Распространен в Африке до северной границы Сахары и на юг до мыса Доброй Надежды (за исключением густых лесов), на п-овах Аравийском и Синайском, в Ираке и Сирии.
Обитает в травянистых и кустарниковых степях, песчаных и каменистых полупустынях и пустынях. Питается травянистыми растениями, луковицами, клубнями, плодами, в меньшей степени листьями, почками и побегами кустарников и деревьев. В засушливых местах может подолгу обходиться без воды. Отмечены ежегодные дальние кормовые миграции: в дождливый сезон в июне — июле на север, в центральные части Сахары, а в засушливый период в марте — мае на юг к границе Судана.
Активен утром и вечером, иногда и ночью. Держатся парами или небольшими группами в 6—12 голов, состоящими по большей части из самца, нескольких самок и молодых, реже встречаются более крупные стада в 40—60, даже сотню или тысячу голов. Часто объединяются в группы вместе с антилопами и зебрами. Размножение не приурочено к определенному времени. Самцы во время гона дерутся между собой. Беременность продолжается 240—300 дней. В помете один, редко два детеныша.
Продолжительность жизни до 18 лет.
Ценный объект спортивной охоты.
Возможна доместикация.
Некоторые исследователи (например, Андерсон и Джонс, 1967) выделяют в роде 3 вида: О. gazella, О. leucoryx Pallas, 1777, и О. dammah Cretzschmar, 1826.
Ориксы из Омана — О. gazella leucoryx Pallas, 1777, и из Судана, Чада, Нигера и Мали — О. g. dammah Cretzschmar ( = 0. g. tao H. Smith, 1827) включены в «Красную книгу», первый как находящийся под угрозой исчезновения (численность в 1969 г. около 200 особей), второй как малочисленный подвид, который в ближайшее время может подвергнуться исчезновению.
Рис. 185. Алдакс — A. nasomaculatus
Род аддаксов
Genus Addax Rafinesque, 1815
В роде единственный вид: аддакс— A. nasomaculatus De Blainville, 1816.
Размеры средние. Длина тела 150—170 см, длина хвоста 25—35 см, высота в холке 95—115 см. Масса 60—125 кг. Самцы крупнее самок. Внешним сложением напоминает северного оленя. Туловище удлиненное. Шея короткая. Конечности средней высоты, сильные. Загривок немного выше крестца (рис. 185). Голова крупная, ее верхний профиль прямой. На конце морды имеется очень маленький участок голой кожи. Глаза мелкие. Уши средней длины ланцетовидной формы. Хвост длинный, тонкий, с небольшой кистью волос на конце. Рога имеют и самцы, и самки: у самцов они крупнее, достигают в длину 60—109 см (у самок 55— 80 см) и делают 2,5—3 изгиба (у самок 1,5—2). Рога проходят наискось спереди назад и вверх и немного наружу, винтообразно закручиваясь. Форма поперечника рогов округлая. Почти по всей своей длине (кроме вершины) рога имеют поперечные кольцевые выступы. Средние копыта сходны с копытами северных оленей’ низкие, уплощенные, по форме полулунные с округлыми концами. Боковые копыта очень большие. Копыта и рога серо-черные.
Волосяной покров летом более низкий и менее густой, чем зимои. На лбу между рогами и глазами находятся удлиненные волосы в виде шапки. На нижней стороне шеи длинные волосы образуют гриву. Голова и шея серо-бурые, зимой более темные, чем летом. Лобная шапка волос рыже-бурая или черно-бурая, затылок темно-бурый. Поперечная по-422
леса, пересекающая морду сверху, пятно позади глаза, губы и подбородок белые. Плечи, спина и бока зимой серо-бурые, а летом беловато-серо-бурые. Брюшная сторона тела бело-серая или белая. Предглазничные железы снаружи незаметны. Межпальцевые железы имеются на передних и задних конечностях. Паховых желез нет. Сосков 2 пары.-
Череп средней длины, широкий. Задние и нижние края орбит выдаются далеко в стороны. Мозговой отдел составляет всего 3/з длины лицевого отдела. Этмоидальные отверстия маленькие. Костные слуховые барабаны очень крупные.
Диплоидное число хромосом 58.
Распространен в Северной Африке, в Сахаре — от Западной Сахары, Сенегала и Гамбии на западе до АРЕ и Судана на востоке.
Обитает в пустынях и полупустынях, песчаных, каменистых или глинистых. Обычно держится небольшими семейными группами в 5—15 голов, состоящими из одного самца и самок с молодыми. Иногда объединяются в крупные стада, насчитывающие сотни голов. Отмечены кормовые миграции, связанные с поисками новых пастбищ: в Южной Сахаре в засушливое время года (июль — сентябрь) к югу до границы с саваннами Судана, а в зимний дождливый сезон (ноябрь — февраль) на север, в пустыню. Может подолгу обходиться без водопоя. Питается в основном травянистыми растениями, а также листьями и побегами кустарников. Активен утром, вечером и ночью. Период размножения не приурочен строго к определенному времени года. Беременность продолжается 10—12 месяцев. В помете, как правило, один детеныш. Уже в первые дни после рождения он начинает подкармливаться подножным кормом. Продолжительность жизни до 18 лет.
Аддаксы служат объектом спортивной охоты из-за мяса и кожи. Охрана аддакса не налажена и ему может угрожать уничтожение.
Включен в «Красную книгу» как малочисленный вид, который в ближайшее время может подвергнуться опасности исчезновения.
Род водяных козлов
Genus Kobus A. Smith, 1840
Размеры мелкие и средние. Длина тела 125—220 см, длина хвоста 18—45 см, высота в холке 70—130 см. Масса 50—250 кг. Самцы значительно крупнее самок. Телосложение от легкого до тяжелого. Тело в крестце немного выше, чем в загривке (рис. 186). Голова большая. На конце морды имеется средних размеров или большой участок голой кожи. Глаза большие. Уши средней длины с заостренными или закругленными вершинами. Конечности тонкие. Хвост средней длины с кистью волос на конце. Рога длиной 30—100 см; имеют их только самцы. Прямые или лирообразные рога расходятся у основания под углом друг к ДРУгу и направляются наискось назад и вверх. Вершины рогов S-образ-но изгибаются вперед и вверх. В поперечнике рога округлые. Почти по всей длине рога имеют поперечные кольцевидные выступы. Средние копыта длинные или очень длинные, заостренные. Боковые копыта боль-ШиС удлиненные. Окраска копыт и рогов серо-бурая или черно-бурая.
Волосяной покров грубый, низкий или средней высоты. На шее имеется грива. Верхняя часть тела желтовато-серая, черно-серая, желтова-Тая, серо-бурая, буро-рыжая, буро-черная или почти черная. Обычно спина темнее боков. Наружные стороны ног с бурыми или черными продольными полосами. В их проксимальных отделах часто расположены елые кольца. Губы, подбородок, участок вокруг глаз, кольцо около но-
Рис. 186. Личи — К. leche
са и основания ушей светлые или белые. Брюшная сторона тела и око-лохвостовое «зеркало» серые, желто-белые или белые. Предглазничная железа маленькая или ее нет вовсе. Паховые железы отсутствуют или имеются в количестве двух или четырех пар. Запястных и межнальцевых желез нет. Сосков 2 пары.
Череп узкий, нижние и задние края крупных глазниц выступают в стороны. Мозговая коробка составляет примерно треть длины лицевой части. Носовые кости длинные и узкие. Этмоидальные и надглазничные отверстия большие.
Диплоидный набор хромосом водяного козла и коба по 50, а нильского личи — 52.
Распространены в Африке к югу от Сахары.
Обитают в саваннах, по опушкам лесов, или в галерейных лесах вблизи водоемов, или в болотистых местах на равнинах и холмах. Держатся по большей части небольшими группами, состояшими из самца, нескольких самок и молодых. Могут хорошо плавать. Питаются травянистыми наземными и водными растениями, a A. ellipsiprymnus и
К. kob — также листьями и побегами кустарников. Активны утром, вечером и ночью. Период размножения обычно не приурочен к определенному времени года. Размножающиеся самцы занимают небольшую территорию, которую охраняют.. Длительность беременности 7—8 месяцев. В помете один, редко два, очень редко три детеныша. Половозрелость наступает, по-видимому, в 1,5 года. Продолжительность жизни около 12 лет, в неволе до 17 лет.
' Из-за красивых рогов служат объектом спортивной охоты.
В роде 5 видов:
водяной козел — К. ellipsiprymnus Ogilby, 1833 (Африка к югу от Сахары, от Сенегала на западе до Сомали на востоке);
коб — К. kob Erxleben, 1777 (от Сенегала и Гамбии на восток до Юго-Западной Эфиопии и на юг примерно до 17° ю. ш.);
пуку К. vardoni Livingstone, 1857 (север Ботсваны, Замбия, Малави, Танзания, Заир);
личи—A. leche Gray, 1850 (Ботсвана, Ангола, Замбия, юго-восток Заира);
нильский личи — К. megaceros Fitzinger, 1855 (район р. Нил в Южном Судане и Западная Эфиопия).
Некоторые исследователи (например, Эллермэн, 1953) выделяют из К. ellipsiprymnus особый вид К. defassa Rilppel, 1835; включают (например, Хальтенорт, 1963) К. vardoni в A. kob-, выделяют (Симпсон, 1945) A. kob в особый род Adenota Grav, 1847, a A- megaceros в род Onotragus Heller, 1913.
Личи включен в «Красную книгу» как малочисленный вид, который в ближайшее время может подвергнуться опасности исчезновения.
Род тростниковых КОЗЛОВ
Genus Redunca Н. Smith, 1827
Размеры мелкие или средние. Длина тела 110—160 см, длина хвоста 15—30 см, высота в холке 60—105 см. Масса 20—95 кг. Самцы по величине не отличаются от самок или чуть крупнее. Телосложение легкое (рис. 187). Спина почти прямая (крестец чуть выше загривка). Конечности высокие тонкие. Голова небольшая. На конце морды имеется довольно большой участок голой кожи. Глаза большие. Уши средней длины, заостренные. Под ухом находится небольшой, округлый по форме, лишенный волос железистый участок кожи желто- или черно-серой окраски. Хвост на всем протяжении покрыт длинными волосами. Только самцы обладают рогами; длина их 12—45 см. От основания они идут наискось назад и вверх, а затем выгибаются вверх и несколько вперед. В поперечнике рога округлые; только верхняя их треть не имеет поперечных выступов. У основания рогов имеются узкие или широкие (до 5 см) мясистые складки, которые с возрастом становятся жесткими. Средние копыта узкие, низкие и заостренные (обыкновенная и большая редунки) или широкие и высокие (горная редунка). Боковые копыта короткие и широкие. Копыта и рога черно-бурые или черные.
Волосяной покров низкий, мягкий. На шее, груди и брюхе волосы несколько длиннее, чем на спине. Спинная сторона тела буро-серая или буро-охристая, брюшная бело-серая, желто-белая или белая. Запястных, предглазничных и межпальцевых желез нет. Паховых желез одна или Две пары. Сосков 2 пары.
Череп с сильно выдающимися в стороны нижними и задними краями крупных глазниц. Мозговая коробка составляет примерно треть длины лицевого отдела черепа. Межчелюстные кости не соприкасаются с длин-
Рис. 187. Г орная редунка — R. fulvorufula
ними и узкими носовыми. Этмоидальные и надглазничные отверстия большие. Костные слуховые барабаны очень большие.
Второй верхний коренной зуб небольших размеров или его нет совсем.
Распространены в Африке к югу от Сахары.
Обыкновенная и большая редунки обитают в открытых травянистых или поросших кустарником местах, иногда заболоченных, на равнинах и холмах или горах, но всегда вблизи воды. Горная редунка предпочитает травянистые с отдельными кустами, каменистые или скальные склоны холмов или гор; может подолгу обходиться без воды. Активны в основном утром и вечером. Питаются преимущественно травянистыми растениями. Держатся поодиночке, парами или небольшими группами, обычно состоящими из одного самца и нескольких самок с молодыми. Период спаривания не ограничен каким-либо временем года. Длительность беременности 6—7 месяцев. Детенышей в помете один, редко два. Половозрелость наступает в 1,5 года. Продолжительность жизни 8 10 лет.
В роде 3 вида:
большая редунка — /?. arundinum Boddaert, 1785 (от Южно-Афри' канской Республики на северо-запад до Габона и на северо-восток Д° Руанды, Бурунди и Юго-Восточной Танзании);
обыкновенная редунка — R. redunca Pallas, 1777 (от Сенегала до Северо-Восточного Заира, Южного Судана и Эфиопии; Уганда, Кения, Руанда, Бурунди и Танзания);
Рис. 188. Косулья антилопа — Р. capreolus
горная редунка — R. fulvorufula Alzeiius, 1815 (три изолированных участка: Северный Камерун; Танзания, Кения, Уганда, юго-восток Судана и юго-запад Эфиопии; Южно-Африканская Республика и Ботсвана).
Род косульих антилоп
Genus Pelea Gray, 1841
В роде единственный вид: косулья антилопа — Р. capreolus Forster, 1790.
Размеры мелкие. Длина тела 115—125 см, высота в плечах 70—80 см, Длина хвоста 15—30 см. Масса 20—30 кг. Самцы примерно таких же размеров, как и самки. Телосложение легкое. Тело в крестце немного выше, чем в загривке. Конечности высокие и тонкие (рис. 188). Голова средних размеров, удлиненная. На конце морды имеется большой участок голой кожи. Хвост средней длины. Рога имеют только самцы. Округлые в сечении, рога далеко отстоят друг от друга и поднимаются вертикально вверх, располагаясь почти параллельно друг другу.. Длина рогов 15—25 см, их нижняя часть несет 10—15 кольцевидных выступов.
Средние копыта короткие, треугольные; боковые копыта тоже короткие и широкие. Окраска копыт и рогов черно-бурая. Волосяной покров низкий, но плотный и мягкий. Спинная сторона тела бледно-серая до серобурой, брюшная белая. Подбородок с поперечной черной полосой. Запястных, паховых и предглазничных желез нет. На передних и задних конечностях имеются межпальцевые железы. Сосков 2 пары.
Череп длинный и узкий. Глазницы большие. Их задние края сильно выступают в стороны. Мозговая коробка короткая, составляет примерно четверть длины лицевого отдела. Этмоидальные отверстия узкие. Носовые кости длинные и узкие. Ямки предглазничных желез на слезных костях отсутствуют. Костные слуховые барабаны маленькие.
Щечные зубы гипселодонтные. Второй верхний предкоренной очень маленький или его нет вовсе.
Распространена в Южно-Африканской Республике и Ботсване.
Обитает в скалистых, покрытых кустами горных или холмистых районах вблизи водоемов. Активна утром и вечером. Питается в основном травянистыми растениями, в меньшей степени листьями и побегами кустарников. В сумерках спускается со скал вниз на водопой. Держатся небольшими семейными группами, состоящими по большей части из одного самца и нескольких самок с молодыми. Иногда они соединяются по 20—30 особей вместе. Старые самцы держатся поодиночке. Период спаривания с конца ноября по конец декабря. Самцы в это время дерутся друг с другом. Беременность продолжается, вероятно, 6—7 месяцев. В помете обычно два детеныша. Половозрелость наступает, по-видимому, в 1,5 года. Продолжительность жизни до 8—10 лет.
Род им пал
Genus Aepyceros Sundevall, 1847
В роде единственный вид: импала—A. melampus Lichtenstein, 1812.
Размеры мелкие и средние. Длина тела 130—180 см, длина хвоста 25—42 см, высота в холке 75—100 см. Масса 40—90 кг. Самцы значительно (примерно на четвертую часть) крупнее самок. Телосложение изящное. Туловище в крестце немного выше, чем в холке (рис. 189). Конечности высокие и стройные. Голова удлиненная, узкая. На конце морды имеется небольшой участок голой кожи. Глаза большие. Уши длинные, узкие и заостренные. Только у самцов имеются рога: они лировидной формы, выгнуты назад, в стороны и вверх. Длина рогов 50—92 см. В их нижних отделах располагаются многочисленные кольцевидные выступы. Копыта средних пальцев узкие и заостренные, боковых копыт нет. Окраска рогов и копыт черно-бурая или черная. Волосяной покров низкий, плотно прилегающий к телу. Спинная сторона тела светло- или темно-рыже-бурая; а брюшная и пятно над глазом белые. Внутренние и наружные поверхности ушей, тонкая полоска вдоль спииы от ее середины до вершины хвоста, полоса на плюсне имеют черную окраску. Хвост покрыт длинными волосами. Предглазничных, паховых и межпальцевых желез нет, имеются только плюсневые. Сосков 2 пары.
Череп длинный и узкий с сильно выступающими в стороны краями глазниц. Мозговая коробка короткая, составляет примерно четверть длины лицевого отдела. Носовые кости длинные. Этмоидальные отверстия очень малы или их нет вовсе. Ямок предглазничных желез на слезных костях нет.
Щечные зубы гипселодонтные.
Распространена в Южно-Африканской Республике, Намибии, Ьот-
Рис. 189. Импала— A. melampus
сване, Родезии, Мозамбике, Малави, Замбии, Анголе, Заире, Танзании, Кении, Руанде, Бурунди, Уганде.
Обитает в разреженных лесах, саваннах с кустарниками и деревьями, галерейных лесах на равнинах и холмах.
Питается листьями, побегами и травянистыми растениями. Ежедневно ходит на водопой. Обычно активна утром и вечером. Держится небольшими группами по 5—10 особей, обычно самец и самки с молодыми, или стадами в 30—50 голов; изредка (особенно в засушливый сезон) собираются более крупными стадами в 100—200 животных. В одном стаде с импалами часто бывают слоны, зебры и другие антилопы. Испуганная импала способна делать огромные прыжки до 3 м в высоту и 10 м в длину. Размножение не приурочено строго к определенному времени года, однако в Юго-Западной и Южной Африке период спаривания приходится в основном на февраль—май, а в Восточной Африке — на октябрь — ноябрь. В этот период самцы сильно дерутся друг с другом. Длительность беременности 6,5 месяцев. В помете один, реже два детеныша. Половозрелость наступает, по-видимому, в 1,5 года. Продолжительность жизни около 10—12 лет. Основные враги — лев, леопард и гиеновая собака.
Импала из Намибии — А. т. petersi Bocage, 1879, включена в «Красную книгу» как подвид, находящийся под угрозой исчезновения (численность в Намибии в 1971 г. около 1 тыс. голов).
Род лошадиных антилоп
Genus Hippotragus Sundevall, 1846
Размеры средние и большие. Длина тела 188—267 см, длина хвоста 37—76 см, высота в холке 100—160 см. Масса 150—300 кг. Самцы крупнее самок. В холке животные немного выше, чем в крестце. Конечности
ЛОО
Рис. 190. Черная антилопа — Н niger
высокие, относительно тонкие, но сильные (рис. 190). Шея короткая и массивная. Голова большая. На конце морды имеется средних размеров участок голой кожи. Уши большие заостренные. Глаза относительно невелики. Хвост длинный с концевой кистью. Рога имеют оба пола, но у самок они примерно на треть меньше по размерам. От оснований рога описывают плавный полукруг вверх и назад, достигая в длину 50— 175 см. Форма их поперечника округлая. Почти по всей длине рогов, за исключением вершины, располагаются крупные кольцевидные выступы. Средние копыта узкие и заостренные. Боковые копыта крупных размеров. Окраска копыт и рогов от серо-бурой до черно-бурой. Волосяной покров редкий и низкий, плотно прилегающий к телу. По верху шеи до середины спины проходит грива с вертикально стоящими волосами длиной 5—15 см. Верхняя часть тела серо-голубая, серая, темно-бурая или черная. У глаз, на подбородке и на губах светлые пятна. Передние отделы ног со светло- или темно-бурыми полосами. Брюхо и околохвостовОе «зеркало» белые. Имеются слаборазвитая предглазничная железа и межпальцевые железы на передних и задних конечностях. Паховых и запястных желез нет. Сосков 2 пары.
Череп узкий, удлиненный. Сильно развит зароговый отдел черепа. Края орбит сильно выдаются в стороны. Лобные кости короткие. Носовые кости длинные и узкие. Костные слуховые барабаны маленькие (черная антилопа) или большие (лошадиная антилопа). Этмоидальные отверстия узкие, вытянутые. Надглазничные отверстия маленькие.
Жевательные поверхности коронок щечных зубов широкие, четырехугольные.
Диплоидный набор хромосом черной антилопы 60.
Рис. 190 Продолжение.
*тереп черной антилопы
Распространены в Африке к югу от Сахары и в Эфиопии, за иск ченнем густых лесов, горных массивов и пустынь.
Обитают в саваннах, разреженных лесах на равнинах и холм иногда поднимаются в горы на высоту до 2 тыс. м над уровнем М°Р Питаются травянистыми растениями, листьями и побегами кустарник и деревьев. Ежедневно ходят на водопой — обычно утром и вечером, к тнвны утром, поздним вечером и ночью. Держатся небольшими гру ми по 3—20, изредка 40—60 голов. Обычно с одним самцом ходят не сколько самок и молодых. Вместе с ними в одном стаде бывают ангнло пы других видов, зебры и страусы. В случае опасности могут бежать ci
скоростью до 50—70 км/ч. Период размножения не приурочен строго к определенному времени года. Беременность 9 месяцев. В помете один детеныш. Продолжительность жизни до 12—15 лет, в неволе до 17 лет. Хорошо переносит неволю.
В роде 2 вида:
лошадиная антилопа — //, equinus Desmarest, 1804 (Южно-Африканская Республика, Ботсвана, Намибия, Родезия, Ангола, Замбия, Малави, Мозамбик, Танзания, Руанда, Бурунди, Уганда, Кения, юг Судана, запад Эфиопии и от Заира на запад до Сенегала);
черная антилопа — //. niger Harris, 1838 (от Южно-Африканской Республики на северо-восток до Кении и Танзании и на северо-запад до Анголы и Заира).
Род лиророгих бубалов
Genus Damaliscus Sclater et Thomas, 1894
Размеры мелкие и средние. Длина тела 140—205 см, высота в холке 85—130 см, длина хвоста 30—60 см. Масса 85—150 кг. Самцы немного крупнее самок. Сложение тяжелое. Тело в холке значительно выше, чем в крестце. Конечности высокие, относительно тонкие. Голова большая — длинная, узкая. На конце морды имеется большой участок голой кожи. Глаза маленькие. Уши большие заостренные. Шея короткая. Средние копыта узкие с острыми концами. Боковые копыта короткие и широкие.' Хвост длинный и тонкий с пучком волос на конце. Рога имеют и самцы, и самки; у самцов они крупнее и достигают в длину 72 см. Форма рогов лировидная, вершины их направлены назад и вверх. В поперечнике рога округлые. Волосяной покров низкий, гладкий. Окраска его на туловище светло-желтая, желто-бурая, или рыже-бурая. Морда рыже-бурая с черным верхом. Проксимальные отделы конечностей и кисть на хвосте черные. Имеются небольшие предглазничные железы, крупные межпаль-цевые железы на передних конечностях и маленькие — на задних. Паховых желез нет. Сосков одна пара.
Череп удлиненный, узкий с прямым верхним профилем. Мозговая коробка составляет 'А—'/з длины лицевого отдела. Лицевая поверхность слезных костей очень велика. Ямки предглазничных желез на них большие. Края глазниц сильно выдаются в стороны. Этмоидальных отверстий нет. Носовые кости длинные и узкие. Костные слуховые барабаны крупные.
Верхние щечные зубы с широкими коронками. Коронки резцов длинные и широкие. Нижний второй предкоренной зуб часто отсутствует.
Диплоидное число хромосом у беломордого бубала 38.
Распространены от Сенегала на западе до Сомали на востоке и Южно-Африканской Республики на юге.
Обитают в саваннах на равнинах и холмах, в степях, полупустынях, влажных травяных зарослях и кустарниковых. Держатся небольшими группами, состоящими обычно из самца, нескольких самок и молодых. Питаются в основном травянистыми растениями. Размножение можег быть сезонным. В Серенгети у беломордого бубала гон происходит с декабря по март. Размножающиеся самцы занимают небольшую территорию (беломордый бубал — диаметром 200—400 м), которую охраняют. Самки могут по нескольку дней оставаться на территории самца. Беременность продолжается 7—8 месяцев. Детенышей в помете один, редко два. Продолжительность жизни 12—15 лет, в неволе 17 лет.
В роде 3 вида:
топи__D. lunatus Burchell, 1823 (Южно-Африканская Респуолика,
i t
Рис. 191. Конгони— A. buselaphu1.
Мозамбик, Родезия, Ботсвана, Намибия, Ангола, Замбия, Танзания, Кс-ния, Уганда, запад Эфиопии, юг Судана, Руанда, Бурунди, Заир, Цент ральноафриканская Республика, Чад, Камерун, Нигерия, Нигер, Верхня Вольта, Мали, ААавритания, Сенегал, Гамбия; заходят в Дагомею, Тоги и Гану);
бубал Хантера — D. hunteri Sclater, 1889 (между реками Тана в Ке нии и Джуба в Сомали);
беломордый бубал — D. dorcas Pallas, 1766 (Южно-Африканская Ped публика, Ботсвана). I
Некоторые исследователи присоединяют бубала Хантера к топи (ня пример Хальтенорт, 19ЬЗ) или относят топи в особый род Beatragus 1 lei 1ег, 1912. J
Беломордый бубал из Южно-Африканской Республики — D. dorcd dorcas, Pailas, 1766, включен в «Красную книгу» как подвид, недавч! находившийся под угрозой исчезновения, а в настоящее время в связ] с предпринятыми мерами по его охране переставший быть таковым. I
Род КОНГ он
Genus Alcelaphus De Blainville, 181
В роде единственный вид: конгони—A. buselaphus Pallas, 1766. I Размеры средние. Длина тела 175—245 см, длина хвоста 45—70 с высота в холке ПО—150 см. Масса 120—225 кг. Самцы немного крулн самок. Сложение тяжелое. В холке туловище значительно выше, чем крупе (рис. 191). Шея короткая. Голова узкая и длинная, верхний п, филь прямой. На конце морды имеется участок голой кожи. Глаза ср< нительно невелики. Уши ланцетовидной формы, довольно длинные. Хв<
средней длины, тонкий с удлиненными волосами. Рога имеют и самцы и самки; у самцов они длиннее и толще. В поперечнике рога округлые' но у основания сплющены в передне-заднем направлении. От основания рога изогнуты вперед и в стороны, а затем внутрь, а вершины направлены назад. Нижние две трети рогов несут 1—25 поперечных кольцевидных выступов. Длина рогов достигает 70 см. Средние копыта узкие и заостренные. Боковые копыта короткие и широкие. Окраска рогов и копыт от светло-желтой до темно-бурой. Волосяной покров низкий, гладкий, плотно прилегающий к телу. Окраска его от желто-бурой до рыжебурой. Брюшная сторона тела светлее спинной; иногда она белая. Подбородок, лоб и верхняя часть носа, а также волосы на хвосте темно-бурые или черные. У глаз могут быть темные полосы. Проксимальные отделы конечностей с темными и черными продольными полосами. Имеется светлое околохвостовое «зеркало». Взрослые животные темнее молодых. Предглазничиые железы слабо развиты. Межпальцевые железы большие на передних конечностях и маленькие на задних. Паховых желез нет. Сосков одна пара.
Череп удлиненный. Мозговая коробка составляет 71—’/з длины лицевого отдела. Зароговый отдел черепа укорочен. Края орбит сильно выдаются в стороны. Носовые кости узкие и очень длинные. Костные слуховые барабаны большие. Слезные кости с очень большой лицевой частью. Ямки предглазничных желез на них крупные. Этмоидальных отверстий нет.
Верхние щечные зубы с широкими коронками. Коронки резцов длинные и широкие.
Диплоидное число хромосом 39—40.
Распространен в Африке к югу от Сахары (за исключением лесных районов). В недалеком прошлом встречался по всей Африке и на Аравийском п-ове.
Обитает на равнинах и холмах, в степях и саваннах. Держатся небольшими группами, по большей части состоящими из самца, нескольких самок и молодых. Иногда собираются (в засушливый сезон) крупными стадами по нескольку сот голов. Образуют смешанные стада с другими антилопами, зебрами и страусами. Питается травянистыми растениями, в меньшей степени листьями и побегами кустарников и деревьев. Обычно ходит на водопой дважды в день. Активен утром и вечером. Отмечены сезонные кормовые миграции. Период размножения не приурочен строго к определенному времени года. Беременность длится около 8 месяцев. В помете один, редко два детеныша. Продолжительность жизни до 11—15, в неволе до 19 лет.
Некоторые исследователи (например, Андерсон и Джонс, 1967) выделяют в качестве особого вида A. lichtensteini Peters, 1849.
Два подвида конгони из Судана и Эфиопии — A. buselaphus tora Gray, 1873, и из Сомали и Эфиопии — A. buselaphus swaynei Sclater, 1892, включены в «Красную книгу» как подвиды, находящиеся под угрозой исчезновения. Численность первого из иих в Судане в 1965 г. была примерно 200—300 голов, а второго, сохранившегося только в Эфиопии, около 100 голов.
Род гну
Genus Connochaetes Lichtenstein, 1814
Размеры средние. Длина тела 170—240 см, длина хвоста 80 100 см, высота в холке 90—145 см. Масса 145—270 кг. Самцы крупнее самок. Сложение тяжелое, туловище в поперечнике округлое, спина прямая
Рис. 192. Голубой гну—С. tau-rinus и его череп
или покатая назад (рис. 192). Конечности тонкие, средней высоты. Ше короткая. Голова большая, морда широкая. На конце ее имеется боль шой участок голой кожи. Глаза относительно маленькие. Уши ланцете видно расширены. Рога имеются у обоих полов. От основания они изы баются в стороны и вниз, а затем дугообразно вверх, внутрь и немнот вперед. У самцов рога длиннее (до 85 см) и толще, чем у самок. Окрач ка рсюв серо-черная или черная; они гладкие. Средние копыта сравп! тельно небольшие, узкие; боковые копыта большие. Хвост длинный, ч конце есть кисть удлиненных волос. Волосяной покров низкий, гладки! плотно прилегающий к поверхности тела. На лбу, затылке, шее, горл!
передней части груди удлиненные волосы образуют своеобразную гриву. Окраска тела желто-буро-серая до темно-бурой. Брюхо иногда бело-серое. Хвост бело-серый или черный. Молодые животные светло-желто-серые или рыже-бурые. Имеются железы: предглазничные и межпальцевые передних конечностей. Паховых желез нет. Сосков одна пара.
Череп' вытянутый. Мозговой отдел составляет 'Л—7з длины лицевого. Края орбит сильно выступают в стороны. Носовые кости длинные и очень узкие.
Верхние щечные зубы с широкими коронками. Нижний второй пред-коренной зуб часто отсутствует.
Диплоидное число хромосом белохвостого и голубого гну по 58.
Распространены в Восточной, Южной и Юго-Западной Африке: от Кении на юго-запад до Южно-Африканской Республики и отсюда на северо-восток до Анголы.
Обитают в травянистых и кустарниковых степях на равнинах и холмах. Держатся стадами различной величины от 3—5 до нескольких десятков особей. Иногда объединяются в огромные стада, насчитывающие сотни и даже тысячи животных. Часто в одном стаде с гну находятся другие антилопы, зебры, страусы. Питаются в основном травянистыми растениями, реже листьями и побегами кустарников. Охотно используют водопой. Активны в основном утром и вечером. Размножение не приурочено строго к определенному времени года. Длительность беременности 8—8,5 месяцев. В помете один, редко два детеныша. Лактация продолжается 7—8 месяцев. Молодые в возрасте одной недели начинают подкармливаться травой. Половозрелость наступает в 12—15 месяцев. Продолжительность жизни до 16—20 лет.
В роде 2 вида:
белохвостый гну — G. gnou Zimmermann, 1780 (Южно-Африканская Республика);
голубой гну — С. taurinus Burchell, 1823 (Южно-Африканская Республика, Намибия, Ангола, Ботсвана, Родезия, Замбия, Малави, Мозамбик, Танзания, Кения).
Некоторые исследователи, например Симпсон, 1963, выделяют голубого гну в особый род Gorgon Gray, 1850.
Белохвостый гну включен в «Красную книгу» как вид, недавно находившийся под угрозой исчезновения, а в настоящее время в связи с предпринятыми мерами по его охране переставший быть таковым. Численность в 1965 г. 2117, в 1970 — 3100 голов.
SUBFAMILIA GAZELUNAE COUES, 1889
Размеры мелкие и средние. Рога имеют только самцы или самцы и самки (у последних меньших размеров). Рога различной формы, средней длины или длинные, тонкие или толстые, гладкие или с поперечными кольцевидными выступами. Участок голой кожи на конце морды маленький или отсутствует. Из специфических желез в разной комбинации имеются предглазничные, запястные, межпальцевые, спинная, паховые, препуциальная. Сосков одна или две пары.
Мозговая коробка черепа и лицевой отдел длинные. Носовые кости короткие или средней длины/ Костные слуховые барабаны большие.
° 0 2 — 3 3 Ой ЙО
Зубная формула: 1 —- с р — — m — = 28— <52.
3 1 Л — о о
Ширина коронки первого нижнего резца такая же или больше, чем ширина коронок второго и третьего нижних резцов и клыка вместе взятых.
Некоторые исследователи (например, Симпсон, 1945) объединяют газелей вместе с карликовыми антилопами в подсемейство Antilopinae Baird, 1857, включая сюда и род Aepiceros.
Род гарн
Genus Antilope Pallas, 1766
В роде единственный вид: гарна — A. cervicapra Linnaeus, 1758.
Размеры мелкие и средние. Длина тела 100—150 см, длина хвоста Ю—17 см, высота в холке 60—85 см. Масса 25—45 кг. Самцы значительно крупнее самок. Телосложение изящное. Тело в загривке немного ниже, чем в крестце. В поперечнике туловище округлое. Конечности длинные и стройные. Шея средней длины. Голова небольшая с узкой мордой, крупными глазами, довольно длинными узкими ушами (рис. 193). На конце морды имеется маленький участок голой кожи. Средние копыта небольшие, узкие, заостренные. Боковые копыта короткие и широкие. Рога длиной 35—73 см имеют только самцы. В основании рога толстые и расположены рядом друг с другом. Они спирально закручены, начиная от основания назад, вверх и в стороны. Почти по всей длине рога имеют кольцевидные выступы. Волосяной покров густой, гладкий. Его окраска изменяется в зависимости от возраста, пола и времени года. Окраска спинной стороны тела самок и молодых животных светло- или темно-желто-бурая, брюшной—.белая. Самцы на спинной стороне тела с возрастом меняют окраску от желто-бурой, черно-бурой до черной с голубоватым оттенком. Взрослые самцы летом бурые, а зимой черно-бурые или черные. У самцов и самок имеются белое хвостовое «зеркало» и белые кольца вокруг глаз. У молодых таких колец около глаз нет. Имеются специфические кожные железы: запястные, предглазничные и межпальцевые на передних и задних ногах. Сосков одна пара.
Череп средней длины, расширенный в стороны за счет выступающих краев глазниц. Последние имеют средние размеры. Поверхность лба сильно вдавлена между глазницами. Мозговой отдел черепа короткий; он составляет примерно треть длины лицевого отдела. Носовые кости средней длины. Ямки предглазничных желез на слезных костях крупные, но не глубокие. Этмоидальные отверстия узкие.
Щечные зубы гипселодонтные. Второй предкоренной зуб в нижней челюсти часто, а в верхней реже отсутствует. Ширина коронки первого нижнего резца в два раза превышает ширину коронок второго и третьего нижних резцов и клыка вместе взятых.
Диплоидное число хромосом 30.
Распространена в Западной Индии к югу от Гималаев почти до мыса Кумари и от Пенджаба на восток до Ассама. Во многих местах численность вследствие охоты и нарушения естественной среды обитания снижена или гарны исчезли совсем.
Обитает на равнинах и холмах с низкой травянистой растительностью и участками сельскохозяйственных культур. Держится небольшими стадами в 10—50 голов, состоящими по большей части из самок с молодыми и одного взрослого самца. Иногда объединяются в крупные стада по многу сотен и даже тысяч животных. Активна в основном утром и вечером. Во время игр часто делают высокие прыжки друг через Друга. Скорость бега очень велика. Питается травянистыми растениями, в меньшей степени листьями и побегами кустарников. Может подолгу обходиться без воды. Период спаривания у большинства особей приурочен к весне (февраль—март). Длительность беременности 6 -месяцев.
Рис. 194. Спрингбок — A. marsuniatts
Детенышей в помете 1—2. Самка способна приносить 2 помета в год. Половозрелость наступает в 1—1,5 года. Продолжительность жизни да 8—10 лет, в неволе до 15.
Род спрингбоков
Genus Antidorcas Sundevall, 18471
В роде единственный вид; спрингбок — A. marsupialis Zimmermann! 1780. I
Размеры мелкие и средние. Длина тела 120—150 см, длина хвоста 20—32 см, высота в холке 70—90 см. Масса 18—45 кг. Самцы незначи! тельно крупнее самок. Телосложение легкое. Конечности тонкие и высо| кие, но пропорциональные. Шея средней длины (рис. 194). Голова на большая с узкой мордой. На конце морды имеется маленький участо! голой кожи. Глаза крупные. Уши небольшие заостренные. Рога есть | самок и самцов. Их длина у самцов до 48 см, у самок до 28 см. В оспа вании рога сближены. Форма их лирообразная; они изогнуты назад I вверх, а концы обычно направлены внутрь. В поперечном сечении роп округлые. Поперечные кольцевидные выступы на них крупные. Цвет р« гов черно-бурый. Средине копыта узкие, заостренные. Боковые копг и хорошо развиты. Хвост средней длины, снизу голый, на конце с кисто] кой волос. Волосяной покров низкий, плотно прилегающий к телу. Спи| пая сторона тела желто-коричневая, брюшная белая. По бокам прох| дит темпо-бурая полоса. Голова беловатая или белая с темной полосг| идущей от основания рогов к углам рта; небольшая, тоже темная полс| ка проходит перед рогами. Имеется белое околохвостовое «зеркала Специфические кожные железы: большие предглазничные, межпальа вые передних и задних конечностей и характерная для этого вида спи
Рис. 195 Монгольский дзерен — Р. gutturosa зимой
ная железа, расположенная на средней линии спины у хвоста. Запястных желез нет. Сосков одна или две пары.
Череп небольшой. Длина мозговой коробки составляет примерно треть длины лицевого отдела. Носовые кости длинные и широкие. Этмоидальные отверстия отсутствуют или очень малы. Ямки предглазничных желез на слезных костях хорошо выражены. Глазницы средних размеров; их края сильно выдаются в стороны.
Второй верхний предкоренной зуб рудиментарный или, как и второй нижний предкоренной, отсутствует. Ширина коронки первого нижнего резца равна ширине коронок второго и третьего нижних резцов и клыка вместе взятых. Диплоидное число хромосом 56.
Распространен в Намибии, Ботсване, Анголе и Южно-Африканской Республике.
Обитает в полупустынях и пустынях с небогатой травянистой растительностью и редкими кустарниками. В прежние годы отмечались грандиозные миграции, когда более миллиона спрингбоков передвигались в одном направлении. Питается травой, листьями и побегами кустарников. При зеленом корме обходится без водопоев. Держится группами по 5—50 особей. Способен делать прыжки по 3—3,5 м в высоту и 15 м в длину. Гон с февраля по июль, в основном в мае. Беременность продолжается около 6 месяцев. В помете один, редко два детеныша. Продолжительность жизни до 10—15 лет.
В результате неумеренного уничтожения эти многочисленные еше недавно звери сохранились теперь только в некоторых национальных парках. Хорошо переносят неволю.
Рис 195. Продолжение. Череп монгольского нзерена
Род дзеренов
Genus Procapra Hodgson, 1846
Размеры мелкие и средние. Длина тела 95 148 , Д самок.
12 см, высота в холке 54—84 см. Масса 20—40 кг. Самць РУ ль_
Сложение плотное, но стройное (рис. 195). Конечности про р
ные, тонкие. Тело в холке немного выше, чем в крестце. Голова небольшая на тонкой шее, в нижней части ее у самцов сильно выступает гортань. Ноздри крупные, по форме напоминающие букву S. На конце морды участка голой кожи нет (тибетский дзерен) или она имеет вид узкой поперечной полоски (монгольский дзерен). Уши средней длины. Глаза не очень большие. Хвост на нижней поверхности голый. Рога имеют только самцы. Цвет их темно-серый или черный. Форма рогов лирообразная. В основании они сближены, затем расходятся, изгибаясь назад и в верхней части внутрь, так что вершины их сходятся. Нижние отделы рогов с поперечными кольцевыми утолщениями имеют в поперечнике овальную форму. Средние копыта маленькие, заостренные, боковые копыта короткие и широкие. Цвет копыт черный.
Окраска волосяного покрова в летнее время желтовато-песчаная, на груди и брюхе белая с грязно-желтым налетом. Зимой окраска более светлая. Околохвостовое белое «зеркало» заходит на круп выше хвоста. По бокам верхней губы удлиненные волосы образуют подобие усов. По-видимому, у дзеренов две линьки: весной и осенью. Предглазничные, запястные и паховые железы развиты слабо или их нет вовсе, межпальцевые железы небольшие. Имеется зароговая железа. Сосков одна пара.
В черепе несколько удлинен лицевой отдел. Ямки предглазничных желез на слезных костях небольшие или отсутствуют. Этмоидальные отверстия имеются.
Первый верхний предкоренной зуб иногда появляется поздно. Зубы брахиодонтного типа.
Для самцов характерны очень крупные гортанные хрящи.
Диплоидное число хромосом монгольского дзерена 58.
Распространены в МНР, в Китае—Ганьсу, Внутренней Монголии, Сычуане, Синьцзяне, Тибете, Хэйлунцзяне и в СССР (до недавнего времени) — в Чуйских степях, на юге Тувинской АССР и в Восточном Забайкалье.
Населяют степи, полупустыни и пустыни равнин и гор. Питаются различными злаками и разнотравьем. Если достаточно зеленых кормов, на водопой не ходят. Период спаривания с середины ноября по конец января. Продолжительность беременности около 6 месяцев. Самка приносит одного — редко двух детенышей с мая по июль. Половозрелость наступает примерно в 1,5 года. В летнее время дзерены активны утром и вечером, зимой в течение всего светлого времени суток. Держатся стадами; летом они насчитывают обычно по 20—30 голов, осенью стадо в среднем состоит из 60—80, а зимой из 100—120 голов. Самки с детенышами собираются в стада до 1 тыс. и более голов. Прекрасные бегуны, развивающие скорость до 60—65 км/ч, способны бежать с такой скоростью по 12—15 км. В ряде районов имеют место сезонные миграции, обусловленные поисками лучших пастбищ и оптимальных мест для размножения. Враг дзеренов — волк.
В СССР охота на дзеренов запрещена, а в МНР и Китае они служат объектом промысла.
В роде 2 вида:
монгольский дзерен — Р. gutturosa Pallas, 1777 (АШР, в Китае • Внутренняя А4онголия, Хэйлунцзян, Ганьсу, в СССР — Чуйские степи, юг Тувинской АССР и Восточное Забайкалье);
тибетский дзерен — Р. picticaudaia Hodgson, 1846 (Тибет, Синьцзян, Сычуань).
В СССР: встречался (может быть, встречается в Забайкалье и теперь) монгольский дзерен.
Род Ргосарга очень близок к роду Gazetla и, возможно, это лишь его подрод.
Род газелей
Genus Gazella De Blainville, 1816
размеры мелкие и средние. Длина тела 85—170 см, длина хвоста 15—30 см, высота в холке 50—110 см. Масса 12—85 кг. Самцы крупнее самок. Сложение легкое и стройное (рис. 196). Конечности высокие и тонкие. Тело в крестце немного выше, чем в загривке. Шея относительно длинная, голова небольшая. На конце морды между ноздрями имеется участок голой кожи. Глаза большие. Уши невелики, заостренные. Хвост средней длины, полностью оволоснен, кроме внутренней части основания. На конце хвоста небольшая кисть волос. Средние копыта небольшие заостренные; боковые копыта короткие широкие. Рога имеют самцы и самки (у джейрана самки безрогие). Рога самцов крупнее, чем у самок, и достигают 25—80 см длины. Рога лировидной формы; как правило, каждый в нижней части несколько выгнут вперед, в средней назад и наружу, а его вершина направлена внутрь или внутрь и вперед. Почти по всей длине рога расположены кольцевидные поперечные выступы. В поперечнике рога овальные или округлые.
Волосяной покров низкий густой. Пуховые волосы развиты слабо. Окраска его на спине и боках серовато-желтая или коричневая. Брюхо белое. Околохвостовое «зеркало» белое; обычно оно заходит выше основания хвоста. По боку тела иногда проходит продольное темное поле, изредка вместе с более светлой полосой. От уха к носу обычно тянется светлая полоса, в некоторых случаях темная. Возрастного и полового диморфизма в окраске нет. Хорошо развиты специфические кожные железы: предглазничные, паховые, запястные, межпальцевые. Сосков одна
пара.
Череп средней длины. Длина его мозговой коробки составляет Уз—7г длины лицевого отдела. В черепе слезные кости с крупными ямками предглазничной железы. Носовые кости широкие. Этмоидальные отвер-
стия имеются.
Диплоидное число хромосом газели-дамы 38, газели доркас — у самцов 31 и самок 30, газели Томсона — 58, джейрана — 30.
Распространены в Северной Африке (на западе примерно до 10° с. ш., на востоке примерно до 5° ю. ш.), на Аравийском п-ове, в Иране, Восточном Закавказье, Афганистане, МНР, в Китае — Внутренней ААонголии, Северном Тибете; Северо-Западной, Северной и Средней Индии; в СССР — Восточном Закавказье, Средней Азии.
Обитают в пустынях, степях и лесостепях, иногда поднимаются в горы до 2—3,5 тыс. м над уровнем моря. Держатся поодиночке, парами, семейными группами, а в некоторые периоды года (осенне-зимний) образуют крупные стада в сотни и даже тысячи голов. Питаются различными травянистыми растениями. При достаточном количестве зеленого, сочного корма в водопоях не нуждаются. В ряде мест отмечены сезонные миграции. Активны в летнее время в прохладные часы — преимущественно утром и вечером, зимой — в течение светлого времени суток.
Размножающийся самец занимает небольшую территорию (у газели Гомсона 100—200 м в диаметре), которую он активно охраняет. Период спаривания в умеренных широтах и субтропиках приходится на позднюю осень — начало зимы, а в тропиках очень растянут. Беременность продолжается 5—7 месяцев. Самки приносят 1—2, редко 3, очень редко 4 детенышей. Период лактации 6 месяцев. Самка газели Томсона спаривается спустя 2—4 недели после рождения детеныша и может приносить помет дважды за год. Половозрелость наступает в 1—1,5 года.
Рис. 196 Джейран — G. subguttarosa
Продолжительность жизни в природе у мелких видов до 6—8, а у крупных до 10—12 лет.
В ряде мест служат объектами охоты. В последние годы численность их сильно снизилась и охота в некоторых странах запрещена (например, на джейрана в СССР).
В роде, по-видимому, 12 видов.
газель Гранта — G. granti Brooke, 1872 (от Южного Судана, Южной Эфиопии и Южного Сомали на юг через Уганду и Кению до Танзании включительно);
сомалийская газель—G. soemmerringi Cretzschmar, 1826 (Судан, Эфиопия, Сомали);
Рис. 196. Продолжение. Черед джейрана
газель-дама — G. dama Pallas, 1766 (от Южного Марокко на юг до Сенегала и на восток до Белого Нила);
обыкновенная газель — G. gazella Pallas, 1766 (Северная Африка, Аравийский п-ов, Иран, Пакистан, Западная Индия);
газель доркас—G. dorcas Linnaeus, 1758 (Северная Африка к югу до 13° с. ш. и Аравийский п-ов);
краснолобая газель — G. rufifrons Gray, 1846 (полоса к югу от Сахары от Сенегала до Северной Эфиопии);
газель Томсона—G. thomsoni Giinther, 1884 (Кения, Танзания и отдельно Южный Судан);
суданская газель — G. tilonura Heuglin, 1868 (юго-восток Судана и север Эфиопии);
газель Спика — G. spekei Blyth, 1863 (Сомали и Эфиопия);
газель Пельцельна — G. pelzelni Kohl, 1886 (Сомали);
песчаная газель — G. leptoceros F. Cuvier, 1842 (Центральная Сахара и Аравийский п-ов);
джейран — G. subgutturosa Gilldenstaedt, 1780 (Аравийский п-ов, Иран, Афганистан, Пакистан, южная часть МНР, Северо-Западный Китай; в СССР — пустыни Азербайджана, Южного Казахстана, Узбекистана, Киргизии и Туркмении).
В СССР: джейран.
Некоторые исследователи предлагают и другие систематические построения рода. Например, Ланге (Lange, 1972) включает газель Пельцельна в качестве подвида в G. dorcas; суданскую газель в качестве подвида в G. rufifrons; песчаную газель в качестве подвида в G. subgutturosa; из краснолобой газели выделяет особый вид G. rufina Thomas, 1894.
Включены в «Красную книгу» как находящиеся под опасностью исчезновения: газели доркас из Северо-Восточного Марокко — G. dorcas massaesyla Cabrera, 1928; с Аравийского п-ова, из Ирана, Иордании и Сирии— G. d. saudiya Carruthers et Schwarz, 1935; обыкновенные газели с Аравийского и Синайского п-вов — G. gazella arabica Lichtenstein, 1827, из Марокко, Алжира и Туниса — G. g. cuvieri Ogylbi, 1841 (численность к 1968 г. в Марокко около 400, в Алжире 200—300, в Тунисе менее того); песчаная газель — в «Красной книге» отдельно включен также ее подвид с Аравийского п-ова — G. I. marica Thomas, 1897, он указан под названием G. subgutturosa marica Thomas, 1897; газель Пельцельна; газель-дама из А4арокко— G. dama mhorr Bennett, 1833; из Западной Сахары — G. d. lozanoi Morales Agacino, 1834 (численность в 1968 г. была около 50 голов).
Род дибатагов
Genus Ammodorcas Thomas, 1891
В роде единственный вид: дибатаг — A. clarkei Thomas, 1891.
Размеры средние. Телосложение изящное. Длина тела 152—168 см, длина хвоста 30—36 см, высота в плечах 80—88 см. М.асса 22—32 кг. Самцы немного крупнее самок. Туловище удлиненное. Конечности и шея очень длинные и тонкие. Тело в крестце чуть выше, чем в холке. Голова небольшая, вытянутая, губы очень подвижные. На конце морды имеется маленький участок голой кожи. Глаза большие. Уши длинные, ланцетовидно расширенные. Средние копыта небольшие, заостренные. Боковые копыта очень маленькие. Хвост, достигающий пяточного сочле-
Рис. 197. Геренук— L. walleri
нения, весь покрыт волосами. Рога длиной 25—34 см имеют только самцы. От основания рога изгибаются вверх и назад, а затем вперед и вверх и немного в стороны. Форма их поперечника в основании округлая. Нижняя половина рогов покрыта крупными поперечными кольцевидными выступами. Цвет рогов и копыт черно-бурый. Волосяной покров низкий, гладкий, плотно прилегающий к телу. Спина коричневая, брюхо белое. Белое околохвостовое «зеркало'» имеется. Есть запястные и крупные предглазничные специфические кожные железы. Д4ежпаль-цевые и паховые железы отсутствуют. Сосков 2 пары.
Череп узкий и низкий, черепная коробка составляет примерно 3/ю длины лицевого отдела. Лицевая часть черепа узкая, конусообразная. Глазницы большие, их края слабо выступают в стороны. Носовые кости длинные и широкие. Слезные кости с глубокими ямками предглазничных желез. Межчелюстные кости на значительном расстоянии сочленены с носовыми. Этмоидальные отверстия средних размеров.
Щечные зубы слабо гипселодонтные. Ширина коронки первого нижнего резца больше ширины коронок второго и третьего нижних резцов и клыка вместе взятых.
Распространен в Северном Сомали и Восточной Эфиопии.
Обитает в песчаных пустынях или степях с зарослями кустарников и отдельными деревьями. Питается в основном листьями и побегами кустарников и деревьев, в меньшей степени травянистыми растениями. Часто достает пшцу с деревьев, поднимаясь вертикально на задние ноги- Держатся поодиночке, парами или небольшими группами. Активен преимущественно утром и вечером. Размножение не приурочено строго к определенному времени, но разгар гона приходится на март—май. Беременность, по-видимому, продолжается 6—7 месяцев. В помете один Детеныш. Продолжительность жизни, по-видимому, до 8—10 лет. Основной враг — леопард.
Вод геренуков
Genus Litocranius Kohl, 1886
В роде единственный вид: геренук — L. walleri Brooke, 1878.
Размеры средние. Длина тела 140—160 см, длина хвоста 25—35 см, высота в холке 90—105 см. Масса 35—52 кг. Телосложение изящное. Характерны очень длинные и тонкие конечности и шея. Тело в крестце несколько выше, чем в холке (рис. 197). Голова небольшая, удлиненная. Губы очень подвижные. Глаза крупные. На конце морды имеется маленький участок голой кожи. Уши длинные и широкие. Хвост средней длины тонкий с небольшой кистью волос на конце. Только самцы имеют рога длиной 25—43 см. Они лирообразные, с вершинами, загнутыми вверх, вперед и внутрь. Форма их поперечника в основании округлая. В нижних отделах рогов расположены крупные поперечные кольцевые выступы. Цвет рогов черно-бурый. Средние копыта небольшие, узкие, заостренные. Боковые копыта очень маленькие. Волосяной покров низкий, плотно прилегающий к телу. Спинная сторона тела темно-бурая или коричневая, брюшная белая. Имеется небольшое околохвостовое белое «зеркало». Из специфических кожных желез имеются запястные, межпальцевые (на передних и задних конечностях) и крупные предглаз-ничные. Паховых желез нет. Сосков 2 пары.
Череп небольших размеров, низкий и длинный. Черепная коробка несколько уплощенная, широкая, достигает в длину примерно 2/3 лицевого отдела. Этмоидальные щели хорошо выражены. Глазницы очень большие, их края выступают в стороны. Слезные кости с мелкими ямками предглазничных желез.
Щечные зубы с очень низкими коронками и непропорционально по отношению к черепу малы. Ширина коронки первого нижнего резца равна ширине коронок второго и третьего нижних резцов и клыка вместе взятых.
Распространен в Восточной Африке от Сомали и Эфиопии до Кении.
Обитает в засушливых или относительно влажных степях с зарослями кустарников, на равнинах и холмах; поднимается в горы до 1800 м над уровнем моря. Питается листьями, побегами и веточками кустарников и деревьев. Часто встает при кормежке вертикально на задние ноги. Держится небольшими, обычно семейными группами: самец с несколькими самками и молодыми. Активен в основном утром и вечером. Период гона не приурочен строго к определенным месяцам. Беременность продолжается 7 месяцев. В помете один, редко 2 детеныша. Поло-возрелость наступает в 1—1,5 года. Продолжительность жизни до 8—10 лет.
SUBFAMILIA CAPRINAE GILL, 1872
Размеры мелкие и средние. Рога только у самцов или у самцов и самок, причем у последних они могут быть меньше по размерам. Рога обычно по-разному изогнутые, гладкие или поперечными кольцевидными выступами. Хвост короткий или средней длины. Из специфических желез в разных комбинациях имеются предглазничные, паховые, запястные, межпальцевые, зароговая, подхвостовая. Сосков одна или две пары.
Мозговая коробка черепа примерно одинаковой длины с лицевым отделом. Носовые кости хорошо развиты илн почти полностью редуцированы. Костные слуховые барабаны маленькие или средних размеров.
о 002—33
Зубная формула: 1 —- с - р —-— m —- — 28—32.
3 12 — 3 3
Ширина коронок от первого к третьему нижнему резцу и далее к клыку отчетливо или слабо уменьшается или не уменьшается вовсе.
Состав подсемейства окончательно не установлен. Некоторые исследователи (например, Хальтенорт, 1963) выделяют сайгаков в особое подсемейство Saiginae Pocock, 1910; оронго — в подсемейство Pantholopinae Pocock, 1910; такинов—в подсемейство Budorcatinae Knottnerus-Meyer, 1907; а овцебыков — в подсемейство Ovibovinae Lonnberg, 1900. Следуя Симпсону, 1945, здесь они приводятся в состав подсемейства козлов.
Род сайгаков
Genus Saiga Gray, 1843
В роде единственный вид: сайгак — S. tatarica Linnaeus, 1758.
Размеры мелкие. Длина тела ПО—146 см, длина хвоста 8—12 см, высота в холке 60—79 см. Масса 23—40 кг. Телосложение легкое, но несколько неуклюжее (рис. 198). Туловище удлиненное. Конечности тонкие, сравнительно короткие. Шея тонкая, а голова непропорционально большая со вздутым в передней части лицевым отделом, нависающим в виде мягкого подвижного хоботка над ртом. Ноздри округлые, тесно сближены и направлены вниз. Уши короткие с округлой вершиной. Хвост короткий. Средние копыта крупные, боковые небольшие. Рога имеют только самцы. Они расположены на голове почти вертикально, средних размеров (по длине примерно равны длине головы), неправильной лирообразной формы. Рога полупрозрачные, желтовато-белого цвета. В нижних двух третях рога имеют поперечные кольцевые валики. Летний мех низкий и относительно редкий, желтовато-рыжий, более темный по средней линии спины и постепенно светлеющий к брюху. Околохвостового «зеркала» нет. Зимний мех значительно выше и гуще; общий тон окраски очень светлый, глинисто-серый. В течение года бывает две линьки: весной и осенью. Имеются небольшие предглазничные, паховые, запястные и межпальцевые специфические кожные железы. Сосков 2 пары.
Череп весьма своеобразен: кости лицевого отдела сильно изменены и укорочены. Носовые кости очень короткие и направлены вперед и немного вверх. В связи с этим костные наружные носовые отверстия очень велики. В образовании их передне-бокового края принимают участие слезные кости (единственный случай в отряде). Швы между носовыми, лобными и, частично, слезными костями рано исчезают. Ямка для пред-глазничной железы на слезной кости выражена слабо. А4ежчелюстные кости очень малы и также, как и верхнечелюстные, не соприкасаются с носовыми костями. Края глазниц трубкообразные. Этмоидальных отверстий нет. Кости черепа слабо пневматизированы.
0 0 3 3
Зубная формула: i — с — р т = 30, 3 12 3
Диплоидное число хромосом 60.
Распространен в степях и полупустынях Восточной Европы, Средней и Центральной Азии. К началу XX столетия в результате неумеренного промысла численность сайгаков стала очень невысокой, они обитали в отдельных участках, подчас удаленных друг от друга на большие расстояния. Охранные мероприятия восстановили численность сайгаков и за последние десятилетия ареал сайгаков в СССР сильно увеличился. Теперь он занимает обширную территорию в калмыцких, нижневолжских и 15—671 449
Рис. 198. Сайгак — S. tatarica
казахстанских степях. Северная граница распространения сайгаков проходит примерно от устья р. Терек через Сальские степи к Волгограду и Уральску, южнее Урала и тянется через Казахстан примерно на уровне Атбасара и Караганды к хребту Чпнгизтау и к котловине оз. Зайсан, а затем уходит в Китай. На юг ареал доходит до южной границы Устыор-та, отрогов Тянь-Шаня. Вне пределов СССР сайгаки обитают в Джунгарии у подножья Восточного Тянь-Шаня и у хр. Байтаг-Богдо и южных склонов Монгольского Алтая, а также в котловине Больших Западных озер в .МНР.
Приспособлен к жизни на равнинах — обитает в степях и полупустынях. Бегает пиоходыо и способен развивать скорость до 70—80 км/ч, изредка делая «смотровые» прыжки вверх. Питается в основном злаками, солянками, разнотравьем, эфемерами, эфедрой, полынью, степными лишайниками. Зимой поедает солянки, полынь, ветошь злаков. Период спаривания приходится на конец ноября — конец декабря. В это время с одним самцом держится но 5—10, иногда до 40—50 самок. Между самцами происходят ожесточенные драки, иногда кончающиеся смертью одного из соперников. Продолжительность беременности 5 месяцев. Ягнение происходит в разных районах с конца апреля до начала июня. В помете 1—2 детеныша. В 10-диевном возрасте сайгачонок начинает следовать за самкой. В возрасте 4—8 дней начинает щипать траву. Лактационный период длится 4 месяца. Самки начинают приносить потомство в возрасте одного года. Самцы достигают половозрелости на втором году жизни. Летом активен утром и вечером, зимой в течение всего дня. Сайгак — типичное стадное животное. Обычно образует стада по нескольку десятков (наиболее часто 10—50) или сотен голов. Иногда скапливается более тысячи особей вместе, но и в этом случае сайгаки держатся отдельными табунками. Ведут постоянно кочевую жизнь. В ряде мест отмечены сезонные миграции — зимние, весенние и летние. Они происходят в зависимости от доступности и обилия кормов, наличия водопоев и глубины выпадающего снега.
Рис. 198. Продолжение. Череп сайгака
Рис. 199. Горал—N. goral
В настоящее время стал объектом промысла. В МНР сайгак нуждается в охране; подвид 5. t. mongolica Bannikov, 1946, заслуживает занесения в «Красную книгу».
Род оронго
Genus Pantholops Hodgson, 1834
В роде единственный вид: оронго — Р. hodgsoni Abel, 1826.
Размеры мелкие. Длина тела 120—130 см, высота в холке 90—100 см, длина хвоста 18—30 см. Масса 25—35 кг. Самцы немного крупнее самок. Телосложение легкое. Туловище удлиненное. Конечности длинные и тонкие. Копыта узкие, удлиненные. Шея средней длины, стройная. Голова короткая с приподнятым лбом. Глаза большие. Уши маленькие заостренные. На конце морды расположен очень маленький участок голой кожи. Рога имеют только самцы. В длину они достигают 70 см. Форма поперечника рогов эллипсовидная. Они слабо S-образно изогнуты, поставлены вертикально. Нижняя часть рогов имеет поперечные валики. Средние копыта узкие, длинные, боковые маленькие округлые. Волосяной покров высокий, очень густой с большим количеством пуховых волос. Линька одна •— в июле—августе. Окраска на спинной стороне тела серо-желтая, брюшная сторона белая или светло-серая. У старых самцов передние части ног и верх морды буро-черного или черного цвета. Предглазничных, хвостовых, межпальцевых специфических кожных желез нет. Имеются небольшие паховые железы. Сосков одна пара.
В черепе лицевой отдел удлинен. Орбиты и наружное костное носовое отверстие очень большие. Края глазниц выдаются в стороны. Этмоидальные отверстия отсутствуют. Костные слуховые барабаны маленькие.
„ О О 2 3
Зубная формула: i - с - р —- ш -= 28,
3 12 3
Рис. 199. Продолжение. Череп горала
Щечные зубы гипселодонтного типа.
Распространен в высокогорье Тибета от Ладака до области Дзерниг-Нор и Оронг-Нор на востоке.
Обитает на высокогорных равнинах между 4,5 и 4,7 тыс. м над уровнем моря. Держатся небольшими группами, зимой часто собираются в крупные стада. Активен в основном утром и вечером. Имеют место сезонные миграции. Гон приходится на ноябрь—декабрь. В это время между самцами происходят драки; они отбивают себе гаремы из 10—20 самок.
Длительность беременности примерно 6 месяцев. В помете обычно один детеныш. Половозрелости достигают в 1,5—2,5 года. Продолжительность жизни 10—15 лет.
Род горалов
Genus Nemorhaedus Н. Smith, 1827
В роде единственый вид: горал — N. goral Hardwicke, 1825.
Размеры мелкие. Длина тела 05—130 см, высота в холке 10—45 см, масса 25—35 кг. Телосложение плотное (рис. 199). Конечности короткие и крепкие. Ноздри направлены вперед и окружены участком голой кожи. Верхняя губа покрыта волосами, за исключением небольшой срединной вертикальной полоски голой кожи. Уши длинные. Хвост очень длинный, почти достигающий пяточного сустава, с большой кистью удлиненных волос. Рога имеют и самцы и самки. Рога короткие (короче головы), направлены назад, продолжая линию профиля лицевого отдела, слегка изогнуты и в основании имеют поперечные кольца. Поперечное сечение рогов округлое. Волосяной покров высокий, равномерный по всему туловищу. «Бороды» и околохвостового «зеркала» нет. Окраска зимнего меха серая, рыже-бурая и белая. Летний мех темнее, более редкий и низкий. Из специфических кожных желез имеются межпальцевые и небольшие предглазничные.
В черепе профиль предрогового отдела прямой или выпуклый. Носовые кости имеют границу только с лобными и слезными костями и не соприкасаются с верхнечелюстными и предчелюстными. Края глазниц не трубкообразные. Ямок для предглазничных желез на слезных костях нет. Этмоидальные отверстия отсутствуют.
0 0 3
Зубная формула: i с — р — 3 13
ш -|-=32, О
Диплоидное число хромосом у самок 55.
Распространен на Гималаях, в Восточном Тибете, Бирме, Юго-Восточном и Восточном Китае, Хэйлунцзяне, на Корейском п-ове, в СССР в Амурском и Уссурийском краях Дальнего Востока — в юго-западной части Буреинских гор, в горах Сихотэ-Алиня и на массиве Синий Утес в Хасанском р-не.
Обитатель скалистых участков среди леса или сухих открытых склонов гор от уровня моря до 4 тыс. м над его уровнем. Правильного ритма в суточной активности нет. Обычно держатся небольшими группами по 4—6 голов или в одиночку. Местами отмечены сезонные кочевки, связанные с поисками удобных пастбищ. Летом поедает различные травянистые растения, осенью — листья кустов, деревьев и желуди. Зимой большое значение в питании имеет веточный корм. Гон происходит с середины сентября до конца октября в восточных частях ареала и с ноября до начала декабря — в западных. Самки и самцы держатся парами. Самцы дерутся друг с другом лишь изредка. Длительность беременности 250—260 дней. В помете один, реже 2 и лишь иногда 3 детеныша. Первый месяц жизни ягненок горала больше лежит в укромном месте. Половозрелость наступает в 1,5 года.
Продолжительность жизни до 12—-15 лет.
Основной враг — волк.
В СССР охота на горалов ввиду их малочисленности запрещена.
Рис. 200. Сероу — С. sumatraensis
Род сероу
Genus Capricornis Ogilby, 1837
В роде единственный вид: сероу — С. sumatraensis Bechstein, 1799. Размеры мелкие и средние. Длина тела 120—180 см, длина хвоста 8—18 см, высота в холке 70—105 см. Масса 55—140 кг. Самцы и самки почти одинаковой величины. Тело приземистое. Конечности средней высоты, толстые. Шея средней длины. Голова удлиненная (рис. 200). На конце морды имеется маленький участок голой кожи. Глаза небольшие. Уши длинные, обычно превышающие длину рогов. Рога имеются и у самцов, и у самок. Их длина у самцов достигает 30 см, у самок они чуть меньше. Форма рогов в поперечном сечении округлая. Они слегка изогнуты и вершинами направлены вверх и назад. Поверхность рогов, кроме верхней трети, покрыта многочисленными поперечными кольцевидными . Средние копыта узкие, заостренные, боковые маленькие, волосяного покрова — густота, высота, наличие пуховых во-
ЛОс—существенно изменяются в различных точках ареала. Окраска спины и боков бурая с различными оттенками до черной. Брюшная сторона тела светло-серая. Молодые животные такой же окраски, что и взрослые, или немного светлее.
Подхвостовая и паховые специфические кожные железы отсутствуют, есть лишь межпальцевые на передних и задних конечностях и пред-глазпичные.
Череп сходен с черепом горала. На слезных костях имеются большие и глубокие ямки предглазничных желез. Края глазниц не выдаются в стороны.
О
О 3
Зубная формула:
— с
3
3 m —=32.
3
Распространен в Китае: Южном Цинхае, Сычуане, Юньнане и на восток до Фунцзяня; Непале; Бирме; в Индии: Ассаме и на запад до Пенджаба и Кашмира; на п-овах Индокитай и Малакка и на о-ве Суматра. Акклиматизирован в Техасе, США.
Образ жизни такой же, как у горала: обитатель обычно верхней границы леса, зарослей кустарников, перемежающихся скальными выходами. Питается листьями и побегами кустов, различными травянистыми растениями. Держатся поодиночке или парами, зимой небольшими группами. Активен в основном утром и вечером. Гон происходит с конца сентября по конец ноября. Длительность беременности 7—8 месяцев. Детенышей в помете один, редко два. Половозрелость наступает, по-видимому, в 2,5 года.
Продолжительность жизни 12—15 лет. Местами численность в результате охоты сильно снизилась и животные находятся под охраной закона (например, в Японии).
Сероу с о-ва Суматры —• С. sumairaensis sumatraensis Bechstein, Г/99 включен в «Красную книгу» как подвид, находящийся под угрозой исчезновения.
Рис. 201. Продолжение. Череп снежной козы
Род снежных коз
Genus Oreamnos Rafinesque, 1817
В роде единственный вид: снежная коза — О. americanus De Blainvil-le, 1816.
Размеры средние. Длина тела 150—175 см, длина хвоста 12—17 см, высота в холке 90—105. /Масса 80—135 кг. Телосложение крепкое. Туловище массивное. Конечности средней длины, утолщенные. Шея короткая. Голова удлиненная (рис. 201). На конце морды имеется маленький участок голой кожи. Уши короткие, узкие и заостренные. Рога у самцов и самок почти одинаковой длины — до 30 см. Они слабо изогнуты вершинами вверх и назад, расходятся от основания примерно под углом 45°. Форма их поперечника округлая. Поверхность нижней трети рогов имеет слабо выраженные поперечные утолщения. Средние копыта узкие, боковые крупные. Волосяной покров очень густой, высокий, с многочисленными пуховыми волосами. На крупе, загривке, проксимальных отделах конечностей, по низу шеи располагаются особо длинные волосы. Окраска волосяного покрова от чисто-белой до желтовато-белой. Хвостовых, паховых и предглазничных кожных специфических желез нет. Имеются за-роговые и слаборазвитые межпальцевые железы (иногда последние отсутствуют) .
Череп сходен с черепом горала, отличаясь от него некоторыми деталями в строении и пропорциях. Лицевой отдел черепа удлинен и сильно сужен перед глазницами. Этмоидальных отверстий нет.
£
1
Зубная формула:
3 3
р — m — = 32.
3 3
Диплоидное число хромосом 42.
Распространена на западе Северной Америки от Южной Аляски до западной части штата Монтана, центральных районов штата Айдахо и юга штата Орегон.
Обитает в горах выше границы леса по обрывам и склонам. Зимой в случае больших снегопадов спускается вниз. Питается различными травами, побегами и веточками кустов, деревьев и мхом. Охотно посещает солонцы. Держится поодиночке или небольшими группами. Гон происходит в ноябре. Беременность продолжается около 6 месяцев. Самка приносит 1—2 детенышей, которые спустя уже 10 мин после рождения способны встать на ноги. Половозрелость наступает в 2,5 года. Продолжительность жизни до 12—15 лет.
Служит объектом спортивной охоты.
Род серн
Genus Rupicapra De Blainville, 1816
В роде единственный вид: серна — R. rupicapra Linnaeus, 1758.
Размеры мелкие. Длина тела 125—135 см, длина хвоста 5—8 см, высота в плечах 70—80 см, масса самцов 30—50 кг, а самок—25—42 кг. Телосложение плотное, но стройное (рис. 202). Туловище укороченное, ноги высокие, но относительно толстые. Шея тонкая и длинная. Голова небольшая с маленькой вертикальной полоской голой кожи на верхней губе. Ноздри щелевидные, направлены вперед. Уши удлиненные. Хвост короткий с безволосой нижней поверхностью. Средние копыта большие, но узкие. Боковые копыта сильно развиты. Своеобразные по Форме рога
имеют и самцы, и самки (у последних они меньших размеров). Рога отходят от головы вертикально вверх, а их вершины загнуты крючком назад и вниз. Поверхность рогов гладкая. Форма поперечного сечения округлая. Летом окраска волосяного покрова шеи и спины ржаво-желтая или желтая, отграниченная от боков широкой черно-бурой или серо-бурой полосой. Грудь темно-бурая. Голова значительно светлее туловища. На морде от основания уха через глаз к углу рта тянется темно-бурая полоса. Ноги темные; задние светлее-передних. Околохвостово-го «зеркала» и «бороды» на морде нет. Зимний черно-бурый мех серн значительно темнее летнего. Имеется зароговая кожная специфическая железа (по-видимому, функционирует в период размножения) и межпальцевые железы. Предглазничные, паховые и запястные железы отсутствуют. Сосков 2 пары.
Кости черепа слабо пневматизированы. Края орбит трубкообразные. Слезная кость без ямки для предглазничной железы. Этмоидальные отверстия узкие, с возрастом обычно исчезающие. Костные слуховые барабаны маленькие.
0 0 3 3
Зубная формула: i - с — р — m -— = 32.
3 13 3
Диплоидное число хромосом 58.
Распространена в горах Центральной и Южной Европы от Пиренеев до Балкан и Карпат, Малой Азии и Кавказа; в СССР на Главном Кавказском хребте и в горах Малого Кавказа.
Обитает в горах обычно у верхней границы леса, там, где имеются крутые скалистые участки. Питается различными травянистыми растениями, в первую очередь злаками и разнотравьем. Зимой в состав кормов входят злаки, листья, побеги и ветви кустарников. Суточный ритм выражен не очень четко, хотя летом наиболее активна утром и вечером. Держатся обычно смешанными стадами. Гон в разных областях происходит с середины октября до начала января с разгаром в ноябре. В период гона с одним самцом держится несколько самок, иногда же встречается 2—3 взрослых самца и 5—6 самок. Длительность беременности 160— 190 дней. Самки рождают одного, реже двух и очень редко трех детенышей с конца апреля по начало июня. На 2-й или 3-й день после рождения детеныш следует за своей матерью. Лактация длится около 6 месяцев. Половозрелости самки достигают в 1,5—-2,5, а самцы в 2,5—3,5 года. Продолжительность жизни до 12—20 лет.
Основные враги — волк и рысь.
Серна служит объектом главным образом спортивной охоты.
Род танинов
Genus Budorcas Hodgson, 1850
В роде единственный вид: такин — В. taxicolor Hodgson, 1850.
Размеры средние. Длина тела 170—220 см, длина хвоста — 15—20 см, высота в плечах 100—130 см. Масса до 350 кг. Самцы значительно крупнее самок. Телосложение плотное. Туловище удлиненное, массивное. Спина полого спускается от загривка к крестцу (рис. 203). Конечности средней длины, утолщенные (особенно передние). Шея короткая, сильная. Голова большая, удлиненная с широкой мордой. На конце морды имеется участок голой кожи. Глаза небольшие. Уши маленькие, широкие, слегка заостренные на вершине. Хвост короткий, покрыт длинными волосами. Рога имеются и у самцов, и у самок. Они близко соприкасаются основаниями. Форма поперечника в основании овальная, а в верх-
Рис. 202. Серна — R. rupicapra
них отделах округлая. В основании рога расходятся в стороны, а затем направляются вверх, назад и вершинами немного внутрь. Нижние отделы рогов в поперечных кольцевых выступах, остальная часть их гладкая. У самцов длина рогов достигает 50 см; у самок они значительно меньше. Средние копыта широкие и округлые, боковые сильно развиты. Волосяной покров высокий, густой, косматый. Зимой сильно развиты пуховые волосы. На нижней стороне тела волосы удлинены. На подбородке и горле они образуют небольшую «бороду». Окраска волосяного покрова от бело-желтой до буро-рыжей, причем зимой она светлее, чем летом. Молодые животные темно-буро-рыжие. Специфические кожные железы отсутствуют. Сосков 2 пары.
Череп с укороченным мозговым отделом. Верхний профиль черепа от основания рогов до вершины носовых костей слегка выгнут. Лобный отдел черепа между небольшими, с выступающими в стороны краями глазниц, широкий. Затылочная кость узкая, направлена вертикально вниз сразу за рогами. Носовые кости укорочены и соприкасаются только с лобными, которые сильно пневматизированы. Наружное носовое костное отверстие очень велико. Ямки предглазничных желез на слезных костях отсутствуют. Костные слуховые барабаны маленькие.
„ 0 0 3 3
Зубная формула: i —- с — р — m — = 32.
3 13 з
Распространен от Бутана на восток через Северный Ассам, Северную Бирму до Юньнаня, Сычуаня, Шэньси и, возможно, Ганьсу.
Обитает в горах от 2 до 5 тыс. м над уровнем моря, над верхней границей леса в субальпийских и альпийских лугах со скальными участ-
Рис. 202. Продолжение. Череп серпы
ками. Зимой при многоснежье спускается в лес. Активен утром и вечером. Питается травянистыми растениями, побегами и листьями кустарников. Держатся небольшими группами, старые самцы — поодиночке. В конце июня собираются более крупными стадами в несколько десятков голов; предводителем обычно бывает наиболее сильный самец. Период гона приходится на западе ареала на конец августа — начало сентября, на востоке на конец июля — начало августа. Беременность продолжается 7—8 месяцев. В помете один, редко два детеныша. Прловозрелости Достигают примерно в 2,5 года. Продолжительность жизни до 12— 15 лет.
Такины из Западного Китая—В. taxicolor tibetana Milne-Edwards,
dai
Рис. 203. Такин — В. taxicolor
1874, и из Шэньси—В. t. bedfordi Thomas, 1911, включены в «Красную книгу» как малочисленные подвиды, состояние которых, однако, неизвестно.
Род овцебыков
Genus Ovibos De Blainville, 1816
В роде единственный вид: овцебык—О. moschatus Zimmermann, 1780.
Размеры средние. Длина тела 180—245 см, длина хвоста 7—10 см, высота в плечах НО—145 см. Масса 200—300 кг. Самки примерно на четверть меньше самцов. Телосложение неуклюжее, удлиненное. Тело приземистое на относительно коротких и толстых ногах. Шея короткая. Голова большая, морда широкая (рис. 204). На конце ее имеется участок голой кожи. Уши средней длины, заостренные. Короткий хвост покрыт очень длинными волосами. Средние копыта короткие и широкие, боковые развиты сильно. Рога имеются и у самцов, и у самок: у самцов они имеют длину до 73 см, самок — до 40 см. Уплощенные, но сильно утолщенные основания рогов располагаются посредине лобного отдела черепа и у старых особей на лбу смыкаются между собой. От своего основания каждый рог загибается в сторону между глазом и ухом вниз, близко к боковой части головы, а затем поднимается вверх. Форма поперечника 462
Рис. 204. Овцебык — О. moschatus и его череп
рогов круглая. Волосяной покров очень высокий (на спине до 16 см, а на шее, груди и в задней части тела — до 90 см)и густой с многочисленными пуховыми волосами. Зимой окраска его от темно- до черно-бурой летом темно-бурая. Спина старых самцов желто-коричневая. Зимний мех более длинный и.густой. Предглазничные железы имеются. По данным одних исследователей, есть и межпальцевые железы, по сведениям других, они отсутствуют. Паховых желез нет. Сосков 2 пары.
Череп массивный. Лицевой и мозговой отделы примерно равны по длине. Профиль крыши черепа слегка выгнут. Края глазниц трубчатые и выдаются в стороны. Слезная кость с крупной ямкой предглазнпчной железы. Межчелюстные кости не соприкасаются с носовыми. Костные слуховые барабаны небольшие.
„ 0 0 3 3
Зубная формула: i —с — р — m —=32.
3 13 3
Диплоидное число хромосом 48.
Распространен в тундре Северной Америки от Аляски до Гудзонова залива, а также на арктических островах, включая Гренландию. Акклиматизирован в Норвегии и Шпицбергене. Сделана попытка акклиматизации в СССР на Таймыре и о-ве Врангеля.
Обитает на равнинных и скальных участках арктической тундры. Избегает влажных мест. Питается различными травянистыми растениями, листьями и побегами кустарников, а также мхами и лишайниками. В светлое время года активен утром и вечером. В этот период держатся небольшими группами, состоящими из одного самца и 2—4 самок с молодыми. В темное время года соединяется большими стадами по 40—50 и до 100 голов. Гон происходит с конца июля по начало сентября. Беременность продолжается около 9 месяцев. Самка приносит одного детеныша массой около 7 кг. Длительность лактации 3—6 месяцев. Половозрелость наступает в 3—4 года. Продолжительность жизни до 18—23 лет.
Род таров
Genus Hemitragtis Hodgson, 1841
В роде единственный вид: тар —Я. jetnlahicus Н. Smith, 1826.
Размеры мелкие и средние. Длина тела 130—170 см, длина хвоста 10—20 см, высота в холке 60—100 см. Масса до 105 кг. Самки значительно (примерно на четверть) меньше самцов. Телосложение тяжелое, туловище вытянутое, узкое и высокое. В холке тело немного выше, чем в крестце. Шея короткая. Голова средней длины (рис. 205). Профиль ее верхней части прямой. На конце морды имеется маленький участок голой кожи. Уши небольшие, заостренные. Глаза довольно крупные. Хвост короткий с голой нижней поверхностью, на которой открываются многочисленные железы. Рога имеются и у самцов (длиной до 43 см), и у самок (длиной до 35 см). В основании рога тесно соприкасаются друг с другом, а затем идут вверх параллельно друг другу или расходятся под меньшим или большим углом и несколько загибаются назад. В поперечном сечении рога полукруглые. Вся их поверхность в мелких поперечных кольцевидных возвышениях. Волосяной покров густой с хорошо развитым пухом. На плечах, груди и шее удлиненные волосы обычно образуют своеобразную гриву в виде воротника. Общая окраска волос от светло-бурой и рыже-бурой до черно-бурой. Брюшная сторона более светлая. Предглазничных и паховых специфических кожных желез нет. Межпальцевые железы иногда имеются только на задних ногах. Сосков одна или две пары.
Рис. 205. Тар — Н. jemlachicus
В черепе лицевой отдел длинный, мозговой короткий.
.0 0 3 3
Зубная формула: 1 — с — р — m — = 32.
3 13 3
Диплоидное число хромосом 48.
Распространен на юго-востоке Аравийского п-ова, в Гималаях и южной части Западной Индии.
Обитает в скалах лесного пояса гор или выше границы леса. Держатся маленькими или крупными стадами в 30—40 голов. Активен в основном утром н вечером. Питается различными травянистыми растениями. Гон в Гималаях приходится на декабрь. Длительность беременности 180—240 дней. Самка приносит одного, редко двух детенышей. Лактационный период продолжается 6 месяцев. Половозрелость наступает в 1,5 года. Продолжительность жизни 10—14, редко 16 лет. Местные жители охотятся за таром. ,
Включены в «Красную книгу»: подвид с Аравийского п-ова — Н. j. ja-yakari Thomas, 1894, как находящийся под угрозой исчезновения и под; вид из Южной Индии — Н. j. hylocrius Ogilby, 1838, как малочисленный подвид, который в ближайшее время может подвергнуться угрозе исчезновения (численность к 1972 г. была примерно 1500 голов).
Род горных козлов
Genus Capra Linnaeus, 1758
Размеры мелкие и средние. Длина тела 100—170 см, длина хвоста 10—20 см, высота в холке 65—115 см. Масса 35—150 кг. Самки меньше самцов (рис. 206). Телосложение тяжелое, конечности относительно ко-
Рис. 206. Сибирский козел —а С. sibirica (А) и его череп
A*
Рис. 206. Продолжение. Винторогий козел — С falconerl (Б), бородатый козел— С. aegagrus (В)
роткие и толстые^ Шея короткая и сильная. Голова короткая. На верхней губе имеется узкая вертикальная полоска голой кожи. Ноздри сближены и направлены вперед. Уши короткие заостренные. Хвост короткий с голой нижней поверхностью. Рога имеют и самцы, и самки; у последних они небольших размеров. Рога самцов большие, обычно саблевидно изогнутые назад. У некоторых видов они изогнуты спиралеобразно — очень полого или довольно круто. В поперечном сечении рога округлые, треугольные, линзообразные или каплевидные. На передней поверхности нога могут быть большие бугры. Волосяной покров низкий, но густой. Пуховые волосы многочисленны. На нижней стороне морды обычно расположен пучок удлиненных волос — «борода». Линька бывает раз в году в весенне-летнее время. Окраска меха почти одноцветная, коричневая или серая. Летом она светлее, чем зимой. Межпальцевых, предглазничных, паховых и запястных кожных специфических желез нет. У самцов на нижней голой поверхности хвоста открываются протоки хвостовой железы, имеющей резкий («козлиный») запах. У некоторых видов имеется зарого^ вая железа. Сосков одна пара.
Череп с сильно пневматизированными костями, удлиненным зарого-вым отделом и изогнутой лобнотеменной областью. Этмоидальные отверстия узкие. Слезные кости не имеют ямок предглазничных желез.
„ „ .0 0 3 3
Зубная формула: i —~ с — р — m — = 32.
3 13 3
Диплоидное число хромосом альпийского, бородатого и винторогого козлов по 60.
Распространены в горах Южной Европы, Северо-Западной Африки, Передней, Средней и Центральной Азии и Северо-Западной Индии.
Держатся в скалистых участках гор на различной высоте от уровня моря до 5000 м. Приспособлены к лазанию по отвесным скалам. Питаются в летнее время различными травянистыми растениями; особенно злаками, некоторыми осоками, сложноцветными и другими, иногда побегами деревьев и кустарников. Зимой употребляют в корм высохшую траву, ветви, кору, почки деревьев и кустарников, хвою. Активны летом утром и вечером. Зимой пастьба и отдых чередуются более часто. В отдельных местах отмечены значительные суточные перемещения горных козлов — с мест отдыха на пастбища и обратно. Держатся обычно группами по 3—40 голов. В осеннее и зимнее время могут объединяться в крупные стада, состоящие из более чем сотни особей. Однако такие стада непостоянны и легко распадаются на более мелкие. Во многих местах отмечены сезонные кочевки, обусловленные степенью доступности корма и высотой снегового покрова. Гон приходится на октябрь—январь. Между самцами нередки ожесточенные драки. Беременность продолжается 21—25 недель. Самки приносят детенышей с марта по май. Козленок уже на 2-й день после рождения следует за матерью. Лактационный период длится около полугода, хотя травой козлята начинают подкармливаться примерно с месячного возраста. Половозрелости достигают примерно в 1,5 года. Живут до 12—18, редко 20 лет. Враги — барс, волк, рысь.
Козлы — охотничьи и промысловые животные. Некоторые из них в связи с малочисленностью охраняются законом (например, винторогий козел в СССР).
В роде, по-видимому, 8 видов:
пиренейский козел — С. pyrenaica Schinz, 1838 (Испания);
альпийский козел — С. ibex Linnaeus, 1758 (Альпы в Северной Италии) ;
бородатый козел — С. aegagrus Erxleben, 1777 (в литературе часто называют С. hircus Linnaeus, 1758, что правильнее относить к домашнему козлу) (Передняя Азия от островов Греции на восток через Малую Азию и Иран до Кавказа, Южной Туркмении, Белуджистана и Синда в Индии; в СССР: Главный Кавказский хребет, Закавказье, Копетдаг и Большие Балханы);
нубийский козел — С. nubiana F. Cuvier, 1825 (АРЕ, Эфиопия, п-ова Синайский и Аравийский);
кубанский тур — С. caucasica Giildenstaedt et Pallas, 1783 (СССР — западная часть Главного Кавказского хребта);
дагестанский тур — С. cylindricornis Blyth, 1841 (СССР — центральная и восточная части Главного Кавказского хребта);
сибирский козел-—С. sibirica Pallas, 1776 (горы Афганистана, Северо-Западной Индии, Средней Азии, Южной Сибири — на восток до Байкала, Северо-Западного Китая и Тибета; в СССР: Тянь-Шань, Памиро-Алай, Тарбагатай, Саур, Калбинский Алтай, Алтай, Саяны);
винторогий козел — С. falconeri Wagner, 1839 (горы Северной и Северо-Западной Индии, Белуджистана, Афганистана, в СССР: в Таджикистане в верховье Амударьи и по правобережью Нижнего Пянджа).
В СССР: бородатый, кубанский, дагестанский, сибирский и винторогий козлы.
Систематика рода окончательно не установлена. Некоторые исследователи (например, Хальтенорт, 1963 ) насчитывают в роде 4 вида: С. aegagrus, С. ibex (включая С. nubianus, С. caucasica, С. cylindricornis и С. sibirica), С. pyrenaica и С. falconeri, другие (например, Эллермэг и Моррисон-Скотт, 1966)—5 видов: С. hircus (=С. aegagrus), С. ibex, С. caucasica (=С. cylindrocornis), С. pyrenaica и С. falconeri.
Бородатый козел (возможно, также С. ibex и С. falconeri) послужи; предком домашнего козла. Одомашнивание произошло предположительно около 7 тыс. лет до нашей эры. В настоящее время по всему земному шару распространен домашний козел — С. hircus Linnaeus, 1758
В «Красную книгу» включены как находящиеся под угрозой исчезновения подвид пиренейского козла из Испании — С. pyrenaica pyrenaicc Schinz, 1838 (численность в 1965 г. не более 20 голов), подвид нубий ского козла из Эфиопии С. nubiana walie Riippel, 1835 (численность i 1968—1969 гг. около 150 голов), подвиды винторогого козла из Афгани стана и Пакистана С. falconeri megaceros Hutton, 1842 (численность i 1972 г. в Пакистане 50—80 голов) и С. f. jerdoni Hume, 1875 (числен ность в 1972 г. в Белуджистане 700—1000 голов), из Пакистана — С. fal coneri megaceros Hutton, 1913 (численность в 1971 г. около 200 голов) и винторогий козел как вид, который в ближайшее время может ока заться под угрозой исчезновения.
Род голубых баранш
Genus Pseudois Hodgson, 184
В роде единственный вид: голубой баран — Р. nayaur Hodgson, 183: Размеры мелкие и средние. Длина тела 110—165 см, длина хвост 10—20 см, высота в холке 75—90 см. Масса до 80 кг. Самцы значительн крупнее самок. Телосложение довольно тяжелое, сходное с телосложен! ем баранов. Туловище удлиненное. Конечности средней высоты или к< роткие (рис. 207). Тело в крестие немного выше, чем в холке. Шея к< плткяя Голова копоткая с прямым верхним профилем. На конце морд
Рис. 207. Голубой баран — Р. пауаиг
имеется маленький участок голой кожи. Глаза крупные. Уши небольшие, заостренные. Хвост короткий, с голой нижней поверхностью, где располагаются специфические железы. Рога имеют и самцы, и самки. Основания рогов почти соприкасаются, а затем рога расходятся V-образно вверх, назад и несколько внутрь. Форма рогов в поперечнике округлая. Вдоль внутренней стороны рогов проходит киль. Поверхность рогов покрыта небольшими кольцевидными выступами. У самцов длина рогов достигает 45—80 см, у самок 15—20 см. Волосяной покров низкий густой, в зимнее время имеет многочисленные пуховые волосы. «Борода» или грива отсутствует. Общая окраска тела серо-бурая летом и более светлая зимой. Самцы темнее самок. Предглазничных, межпальцевых и паховых специфических кожных желез нет. Сосков одна пара.
В черепе лицевой отдел средней длины, мозговая коробка короткая, причем зароговая часть круто обрывается вниз. Лобная часть черепа широкая. Края глазниц трубчатые, направлены в стороны. Имеются небольшие этмоидальные отверстия.
Зубная формула: i
£ 1
3 3
р — m -— = 32.
3 3
Диплоидное число хромосом 54.
Распространен в Китае — Тибет, Сычуань, Шэньси и Ганьсу; Непале и Индии— Кашмир.
Обитает на высокогорье между 2,5 и 5,5 тыс. м над уровнем моря, придерживаясь скалистых хребтов, осыпей и травяных склонов. В заросли кустарников или к границе леса опускается только зимой в случае глубокого снега и малой доступности корма. Питается травянистыми растениями и лишайниками. Держится небольшими группами, собираясь иногда зимой в крупные стада, насчитывающие по нескольку сотен голов. Активен в основном утром и вечером. Гон приходится на октябрь—ноябрь. Длительность беременности 6—7 месяцев. В помете один, редко два детеныша. Лактация продолжается 6 месяцев. Половозрелость наступает примерно в 1,5 года. Продолжительность жизни 12—15, редко до 18 лет.
Местные охотники добывают голубых баранов на мясо.
Род гривистых баранов
Genus Ammotragus Blyth, 1840
В роде единственный, вид: гривистый баран — A. lervia Pallas, 1777.
Размеры средние. Длина тела 150—165 см, высота в холке 75— 100 см, длина хвоста 15—25 см. Масса 40—140 кг. Самцы значительно крупнее самок. Телосложение тяжелое. Тело несколько приземистое, туловище удлиненное, узкое и высокое. Шея короткая. Голова вытянутая с прямым верхним профилем (рис. 208). Глаза довольно большие. Голый участок кожи на конце морды очень маленький. Уши небольшие заостренные. Рога имеют и самцы, и самки. У самцов длина рога достигает 80 см, у самок 40 см. Форма поперечника основания рогов треугольная (вершиной назад). По всей поверхности они имеют мелкие кольцевидные выступы. Правый и левый рога основаниями почти соприкасаются друг с другом, затем изгибаются назад, вверх и в стороны, а вершинами — внутрь. Зимний волосяной покров выше и более густой, чем летний, содержит много пуховых волос. Нижняя часть шеи, передняя часть груди и верхние отделы передних ног покрыты длинными волосами, образующими своеобразную гриву, скрывающую большую часть передних конечностей. Грива особенно сильно развита у самцов. Общая окраска волосяного покрова от темно-желтой до рыже-бурой, на брюшной стороне тела светлая или белая. Из специфических кожных желез имеется лишь подхвостовая; предглазничных, межпальцевых и паховых нет. Сосков одна пара.
Череп сходен с черепом представителей рода Capra, однако лицевой отдел гривистого барана более вытянут, а мозговой укорочен. Этмоидальные отверстия очень узкие, иногда отсутствуют. Носовые кости не соприкасаются с предчелюстными.
о о
Зубная формула: i —- с — р 3 1
3 3
— m —- = 32.
3 3
Диплоидное число хромосом 58.
Распространен в Северной Африке от Атлантического побережья до Красного моря и на юг до Судана и р. Нигер. В 1950 г. акклиматизирован в США в штатах Нью-Мексико и Калифорния.
Обитатель каменистых горных пустынь. Превосходно лазает по скалам и обрывам. Питается травянистыми растениями и побегами кустарников. Может подолгу обходиться без воды. Держатся небольшими группами, состоящими обычно из самца, самки и молодых различного возраста. Активен утром, вечером и ночью. Гон происходит в ноябре. Самцы имитируют драку, однако настоящих столкновений между ними почти не происходит. Длительность беременности 150—165 дней. Детенышей в помете 1—2, часто 3. Продолжительность лактации 6 месяцев. Полово-зрелость наступает в 1,5 года. Длительность жизни до 10—12 лет.
Местные жители промышляют гривистых баранов.
Род горных баранов
Genus Ovis Linnaeus, 1758
Размеры мелкие и средние. Длина тела ПО—200 см, длина хвоста 3—13 см, высота в холке 65—125 см. Масса 20—230 кг. Телосложение плотное. Туловище удлиненное, конечности высокие и тонкие (рис. 209). Морда средней длины, ноздри направлены вперед. На конце морды посредине верхней губы и по краю ноздрей проходит узкая полоска голой
Рис. 208. Гривистый баран—A. lervia
кожи. Короткий хвост снизу покрыт волосами. Самцы имеют крупные рога длиной 50—190 см. У самок они меньших размеров — длиной 10— 30 см или реже их нет вовсе. У самцов величина и форма рогов весьма изменчивы. В основании рога массивны и постепенно сужаются к вершине. Поперечное сечение их обычно треугольной формы. По характеру изгиба различают три типа рогов. Наиболее обычны гомонимные рога, загибающиеся сначала вверх, назад и наружу, а затем вниз, вперед и наружу. Реже встречаются первертированные рога с изгибом вверх, назад и наружу, а затем в концевой половине загибающиеся вниз и внутрь, к шее. Еще реже встречаются гетеронимные рога, направляющиеся сначала вверх, назад и наружу, а в концевом отделе — более или менее горизонтально внутрь, к шее. Средние копыта узкие, боковые копыта маленькие.
Волосяной покров обычно относительно низкий, лишь у некоторых форм в зимнее время по низу шеи и части груди имеется подвес из удлиненных волос. Окраска волосяного покрова одноцветная, от белой, белосерой и желтой до коричневой и черной. Брюхо светлее спины. Линька происходит один раз в году — весной. Предглазничные, паховые и межпальцевые железы хорошо развиты. Иногда имеется подхвостовая железа. Запястных желез нет. Сосков одна пара.
Череп с сильно пневматизированными костями. Зароговая часть черепа укорочена. Этмоидальных отверстий чаше нет или они развиты слабо. Слезная кость с хорошо выраженной ямкой предглазничной железы. Костные слуховые барабаны небольшие.
„ к .о о з з
Зубная формула; 1 — с — р — m — = 32
3 13 3
Рис. 208. Продолжение. Череп гривистого барана
Рис 209 Гориый баран — О аттоп
Резцы долотовидной формы.
Диплоидное число хромосом у горных баранов группы vignei — уриалов— 58, у горных баранов группы polii — архаров — 56 и у горных баранов группы musimon — муфлонов —54.
Распространены на некоторых островах Средиземного моря, в Передней, Средней, Центральной, Северо-Восточной и частично Северной Азии и на западе Северной Америки.
Приспособлены к жизни в открытых горных ландшафтах со сглаженным рельефом, встречаются на разной высоте: от уровня моря до 5000 м над его уровнем. В противоположность горным козлам, избегают скалистых участков, однако часто выбирают местами обитания обширные пологие пастбища, перемежающиеся обрывистыми ущельями и скалами, где они спасаются от жары летом и от ветра зимой, а в случае нужды и от врагов. В питании основное значение имеют злаки, некоторые виды разнотравья, в высокогорных районах осоки, в полупустынях солянки, полыни. Зимой поедают высохшую траву, ветки кустарничков и кустарников. Летом активны, как правило, в раннеутреннее и вечернее время. Зимой пасутся в светлые часы суток. Осенью и в конце зимы образуют крупные смешанные стада до сотни и более голов. Местами свойственны значительные сезонные кочевки, часто обусловленные недостатком пищи или выпадением глубокого снега.
Сроки гона несколько различны в разных географических районах и приходятся на октябрь—январь. Длительность гона 3—6 недель. В этот
Рис. 209. Продолжение. Череп горного барана
период горные бараны держатся группами, состоящими из одного или 2—3 самцов и 5—25 самок. Между самцами происходят драки, однако не очень ожесточенные. Беременность продолжается около '5 месяцев. Самки приносят 1—2, иногда 3 ягненка с конца марта до середины июня. Перед ягнением самка уходит из стада и рожает где-либо в укрытом месте. С 4-го дня после рождения ягненок сопровождает мать. С месячного возраста он начинает подкармливаться травой. Лактация продолжается до конца лета — начала осени.
Половой зрелости достигают в 1,5—3 года. Самцы начинают участвовать в размножении не раньше 4—5 лет, так как до этого их отгоняют от самок более сильные соперники.
Главный враг горных баранов — волк.
Важные промысловые животные. Родоначальником домашней овцы, по-видимому, были средиземноморские и переднеазиатские горные бараны, так как именно у них и домашних овец оказалось одинаковое диплоидное число хромосом — 54. Успешно используются для гибридизации с домашними породами овец и выведения новых пород (горный меринос, архаромеринос).
В роде, по-видимому, 2 вида:
горный баран — О. аттоп Linnaeus, 1758 (о-ва Корсика, Сардиния, Кипр; горы Передней, Средней и Центральной Азии; в СССР — Южное Закавказье, Копетдаг, Малые и Большие Балханы, Южный Чинк, Западный Устюрт, Мангышлак, скалы восточного побережья Каспия, останцы в Кызылкумах, Памир, Тянь-Шань, мелкосопочник Казахстана, Тарбага-тай, Сауры, Алтай, Танну-Ола; акклиматизирован в Крыму, а также в Швейцарии, Италии, Чехословакии, Югославии и т. д.);
снежный баран — О. canadensis Shaw, 1804 (Аляска, кроме крайнего северо-востока и запада; провинции Канады — Юкон, запад Маккензи, Британская Колумбия; штаты США — Вашингтон, Орегон, Айдахо, Монтана, Северная и Южная Дакота, Небраска, Вайоминг, Колорадо, Юта, Невада, Калифорния, Аризона, Нью-Мексико, юго-запад Техаса; штаты Мексики — Нижняя Калифорния, север Соноры, север Чиуауа, северо-запад Коауила; СССР — Северо-Восточная и Южная Якутия, север Дальнего Востока, горы Путоран к югу от п-ова Таймыр).
В СССР оба вида.
Систематика рода разными исследователями трактуется неодинаково. Так, И. И. Соколов (1959) включает в род Ovis в качестве подродов голубых и гривистых баранов; из вида О. аттоп выделяет: О. musimon Pallas, 1811, и О. orientalis Gmelin, 1774. Эллермэн и Моррисон-Скотт (1966), насчитывают в роде 5 видов: О. canadensis, О. аттоп, О. orientalis, О. laristanica Nasonov, 1909, и О. musimon. Холл и Келсон (1959) разделяют баранов Северной Америки на 2 вида: О. canadensis и О. da-lli Nelson, 1884. По Хальтенорту (1963), в роде всего один вид: О. аттоп. Воронцов и др. (1972) на основании цитогенетического анализа выделяют 7 видов: О. musimon, О. orientalis, О. vignei Blith, 1841, О. аттоп, О. nivicola Eschscholtz, 1829; О. canadensis, О. dalli. Следует отметить, что несмотря на то что среди горных баранов Евразии имеются формы с диплоидным набором хромосом 54, 56 и 58, их гибриды плодовиты.
По-видимому, более правильно, следуя Цалкину (1951), Гептнеру и др. (1961), считать в роде 2 вида.
Снежные бараны Северо-Американского континента включены н «Красную книгу», так как в ближайшее время они могут подвергнуться опасности исчезновения (общая численность в Северной Америке в 1960 г. была 18—20 тыс. голов),
SUBFAMILIA BOVINAE GILL, 1872
Размеры мелкие, средние и большие. Рога имеют самцы и самки, причем у последних они обычно не меньше. Рога изогнутые, гладкие. На конце морды имеется большой участок голой кожи. Хвост короткий или длинный. Специфических кожных желез нет. Сосков 2 пары.
Лицевой отдел черепа длинный, мозговая коробка короткая. Лобные кости очень большие. Этмоидальные отверстия отсутствуют или развиты очень слабо. Костные слуховые барабаны вздутые.
0 0 3 3
Зубная формула: i — с —- р —- m — = 32.
3 10 о
Ширина коронок от первого нижнего резца к клыку не изменяется или возрастает, очень незначительно.
Состав подсемейства окончательно не установлен. Ряд исследователей (например, Симпсон, 1945) включают сюда рода Tragelaphus, Taurotragus, Boselaphus и Tetracerus. Следуя Хальтенорту (1963), здесь они отнесены к подсемейству Antilopinae.
Род буйволов
Genus Bubalus Н. Smith, 1827
Размеры мелкие, средние или крупные. Длина тела 100—290 см, длина хвоста 15—90 см, высота в холке 62—180 см. Масса 150—1200 кг. Телосложение тяжелое (рис. 210). Спина прямая. Шея средней длины, у большинства массивная и сильная. Голова крупная с удлиненной мордой. Глаза сравнительно небольшие или крупные. Уши широкие, короткие, средней длины или длинные. Хвост короткий или длинный. Рога имеют разнообразный вид. Ими обладают и самцы, и самки (у последних они меньше). Рога средней длины— 15—40 см, прямые и направлены назад. Рога азиатского буйвола достигают в длину 35—195 см, широкие в основании и изгибаются серповидно назад и внутрь; в поперечнике они имеют треугольную форму, причем вершина треугольника направлена внутрь. Рога африканского буйвола очень изменчивы; их основания могут отстоять на значительном расстоянии друг от друга или сливаться между собой, они могут быть короткими и изогнутыми серповидно назад или очень длинными и выгнутыми в стороны, назад и вверх или назад и внутрь; в основании поперечник рогов треугольный. Средние копыта большие, широкие и округлые, причем боковые развиты сильно (азиатский и африканский буйволы) или средние копыта короткие и округлые, а боковые маленькие (аноа).
Волосяной покров низкий или средней длины, мягкий или грубый, иногда частично отсутствует. Окраска его на спинной стороне тела у аноа от бурой от черно-бурой, брюшной светло-бурая. У азиатского буйвола спина и брюхо имеют окраску от темно-серой до черной, а у африканского буйвола от рыже-бурой и темно-бурой до черной.
Череп массивный; у аноа короткий и широкий, азиатского и африканского буйволов — удлиненный. Глазницы большие. Их края выдаются в стороны. Лобная часть широкая.
Диплоидный набор хромосом у аноа 48, азиатского буйвола 50, африканского буйвола 52—54.
Распространены в Африке, Индии, о-ве Шри Ланка, Непале, Бирме, Таиланде, на юге Китая, в Кампучии, Лаосе, на о-вах Калимантан, Сулавеси, Филиппинских.
буйвол — £^°iuffe^ (E)PreSSlCOrn‘S И аФРиканский
Обитают в лесах и густых зарослях, в том числе на равнинах и в горах, поднимаясь до 3 тыс. м над уровнем моря. Питаются в основном травянистыми растениями наземными, болотными и водными. Активны утром, вечером и ночью. Держатся парами и семейными группами (аноа) или более или менее крупными стадами (азиатский и африканский буйволы). Как правило, сезонности в размножении нет. Все же рождение молодых обычно приурочено к концу засушливого периода. Длительность беременности 9,5—10 месяцев (аноа и азиатский буйвол) или 10— 11 месяцев (африканский буйвол). Самка приносит одного, редко двух детенышей. Лактационный период продолжается 6—9 месяцев. Полово-зрелость наступает в 1,5—2,5 года. Продолжительность жизни до 20— 25 лет.
В роде 3 вида:
аноа — В. depressicornis Н. Smith, 1827 (о-в Сулавеси);
азиатский буйвол — В. атее Кегг, 1792 (Индия, о-в Шри Ланка, Непал, Бирма, Таиланд, юг Китая, Кампучия, Лаос, о-ва Калимантан и Филиппинские);
африканский буйвол —В. caffer Spaceman, 1779 (Африка).
Систему рода разные исследователи представляют по-разному. Так, Симпсон (1945), И. И. Соколов (1953) и Гептнер (1961) все перечисленные выше виды относят к отдельным родам: Апоа Н. Smith, 1827, Buba-lus Н. Smith, 1827 и Syncerus Hodgson, 1847. Филиппинского буйвола из В. агпее иногда выделяют в особый вид В. mindorensis Heude, 1888, а карликового буйвола из В. caffer в особый вид В. nanus Boddaert, 1875.
Азиатский буйвол одомашнен и встречается во многих странах.
В «Красную книгу» включены:
как находящиеся под угрозой исчезновения подвид азиатского буйвола с о-ва Миндоро (из группы Филиппинских островов) В. агпее mindorensis Heude, 1888 (численность к 1971 г. около 150 голов); подвиды аноа — с северного полуострова Сулавеси — В. depressicornis depressicornis Н. Smith, 1827, и из горных областей о-ва Сулавеси В. d. quarlesi Owens, 1910 (численность очень мала и они близки к полному исчезновению), и как малочисленный вид, который в ближайшее время может оказаться под угрозой исчезновения, азиатский буйвол (численность к 1972 г. в Индии и Непале около 2 тыс. голов).
Род настоящих быков
Genus Bos Linnaeus, 1758
Размеры средние или большие. Длина тела 180—325 см, высота в холке 130—210 см, длина хвоста 70—140 см. Масса 325—1200 кг. Самки значительно меньше самцов (например, самки достигают высоты в холке 145 см). Туловище длинное (рис. 211). Передняя часть тела особой массивностью не отличается. Спина в области холки с небольшим горбом. Конечности относительно короткие и сильные. Шея короткая; обычно имеется хорошо выраженный подгрудок. Голова большая. Глаза относительно маленькие. Уши средних размеров или крупные овальные. Хвост длинный с кистью волос на конце. Рога имеют и самцы, и самки, но у самок они меньше. Рога небольшие или крупные (длина от 15— 68 см у бантенга до 60—115 см у гаура), расположены по бокам черепа, у основания отходят в стороны, затем загибаются вверх и немного вперед; вершины направлены вверх и несколько назад и внутрь. В поперечнике рога округлые, их поверхность гладкая. Волосяной покров изменчив: он может быть низким и изреженным или высоким и густым. По брюху, груди, низу боков и конечностям могут располагаться очень длин-
1ГС/К>П„ Гаурт В %aurus <А)- гаял — В. gaurus fronta-(Б) и як—В. mutus (В)
ные и густые волосы. Окраска волосяного покрова у разных видов отличается мало; от рыже-бурой до темно-бурой и черной. Специфических кожных желез нет. Сосков 2 пары.
Череп крупный с укороченным мозговым отделом и удлиненным лицевым. Кости черепа сильно пневматизированы. Лобные кости очень крупные. Их задний край образует между рогами валик. На слезных костях ямок для предглазничных желез нет. Этмоидальные отверстия отсутствуют или очень малы.
Диплоидное число хромосом у бантенга 60, гаура 58, яка 60.
Распространение охватывает Европу, Северную Африку, Переднюю, Среднюю и Южную Азию, включая Тибет, о-ва Яву, Бали, Суматру и Калимантан.
Обитают в разных типах лесов и джунглей, вплоть до разреженных, болот избегают, в горы поднимаются до 2 тыс. м над уровнем моря. Питаются преимущественно травянистыми растениями, в меньшей степени листьями и побегами кустарников. Активны в основном ранним утром и вечером. Держатся небольшими стадами преимущественно из одного самца и 2—30 самок. У яков самки с молодыми иногда собираются в стадо до 2000 особей вместе. Сезонности в размножении нет (бантенг, гаур) или гон приходится на апрель—май (купрей) или на сентябрь—октябри (як). Длительность беременности примерно 270—280 дней. В помета один, реже два детеныша. Самки яков размножаются раз в два года] Половозрелость наступает в 2—3 года. Продолжительность жизни 20—1 25 лет. I
В роде по-видимому 5 видов: I
бантенг — В. javanicus D’Alton, 1823 (Бирма, п-ова Индокитай и Mai лакка, о-ва Ява и Калимантан); |
гаур — В. gaurus Н. Smith, 1827 (Индия, Непал, Бирма, п-ова Индо] китай и Малакка); I
купрей — В. sauveli Urbain, 1937 (Кампучия); I
тур — В. primigenius Bojanus, 1827 (обитал в Северной Африке, поч| ти по всей Европе, на север до 60° с. ш., на Кавказе, в Крыму, Мало! Азии, на Южном Урале, Туркмении, Зауралье, юге Западной Сибири, 1 районе Красноярска, Забайкалье, Китае от 50 до 40° с. ш.; в Африке бы] истреблен примерно в 2400 г. до н. э., в Месопотамии к 600 г. до н. э| в Центральной и Западной Европе к 1400 г., в Каменском округе Кулун динской степи жил еще в XVI или XVII вв., а под Кузнецком-] в XVIII в.); I
як — В. mutus Przewalski, 1883 (Тибет, а в историческое время, па видимому, Алтай и Саяны). I
Систематика рода окончательно не установлена. Так, Симпса (1945), Гептнер и др. (1961) включают тура и яка в род Bos, а гаур! бантенга и купрея — в род Bibos Hodgson, 1837. И. И. Соколов (1958 предлагает следующую систему: род Bos с одним видом — туром, pd Poephagus Gray, 1843, с одним видом — яком и род Bibos с 3 видами-гауром, бантенгом и купреем.
Тур послужил предком европейского крупного рогатого ско’ (В. primigenius taurus Linnaeus, 1758). Одомашнивание тура произот ло, вероятно, в Греции около 2000 лет до нашей эры. Почти все остал ные представители рода также были одомашнены. Известны домашн формы бантенга на о-вах Бали и Яве — В. javanicus domesticus Gai 1917, гаура — гаял, В. gaurus frontalis Lambert, 1804, яка — домашн як, В. mutus grunniens Linnaeus, 1766.
В «Красную книгу» включены: подвергающиеся опасности исчезновения купрей (численность 1970 г. 30—70 юлов по сравнению с 500 в 1964 г.) и як;
Рис. 212. Зубр- в. bonasus (Л), бизон - В. bison (В)
Рис. 212. Продолжение. Череп зубра
малочисленные виды, которые в ближайшее время могут подвергнуться опасности исчезновения: бантенг (к 1972 г, очень малочислен; на большей части ареала исчез совершенно и полагают, что в дикой природе сохранился только на о-вах Калимантан и Ява) и гаур (сохранился только в удаленных местах и охраняемых парках).
Род зубров
Genus Bison Н. Smith, 1827
Размеры средние и большие. Телосложение тяжелое (рис. 212). Длина тела зубра достигает 220—300 см, длина хвоста 50—НО см, высота в холке 140—200 см. Масса 430—1000 кг. Длина тела самца бизона достигает 304—308 см, самки 213—289 см; длина хвоста самца 33—91 см, самки 30—50 см; высота в холке самца 167—186 см, самки 152 см. Масса самца 460—720 кг, самки 360—460 кг. Самки значительно меньше самцов. Передняя часть тела мощно развита, а задняя несколько ослаблена. Холка высокая горбовидная и спина сильно поката вниз, к хвосту. Сравнительно короткая, массивная голова с широким лбом посажена низко. Уши короткие и широкие, глаза небольшие. Рога имеют самцы и самки (меньших размеров). Рога относительно небольшие, толстые у основания, с загнутыми внутрь вершинами, в поперечном сечении круглые. Поверхность рогов гладкая. Копыта крупные. Цвет копыт и рогов серобурый или буро-черный. Волосяной покров густой, в передней части тела высокий, в задней низкий. Окраска его варьирует от серо-бурой до рыжебурой и темно-бурой; летом она более светлая. На подбородке и по низу шеи растут удлиненные волосы. Линька у зубра происходит весной, раз в году, а у бизона два раза — весной и осенью. Зимний мех более высокий и густой, чем летний. На морде и на конечностях специфических кожных желез нет.
Череп широкий, несколько укороченный с сильно пневматизированными костями. Края орбит сильно выдаются в стороны, особенно у старых животных. Слезные кости без ямок для предглазничных желез. Этмоидальных отверстий нет. Межчелюстные кости не достигают носовых костей.
Верхние щечные зубы гипселодонтные с дополнительной призмой на внутренней стороне.
Остистые отростки грудных и седьмого шейного позвонков очень сильно удлинены.
Диплоидное число хромосом у зубра и бизона по 60.
Распространен в Северной Америке и местами в Европе.
Обитают в лесах равнин и гор, в лесостепи и степи. Питаются травянистыми, а также древесно-кустарниковыми кормами. Зубры в Беловежской пуще летом в основном питаются свежей и сочной травой и в значительно меньшей степени тонкими ветками деревьев, кустарничками и т. д. Зимой кормом для них служат сено, тонкие ветви и кора ясеня, ивы, осины и др., древесный лишайник. Бизоны питаются травянистыми растениями прерий. В жаркое время года зубры ходят на водопой минимум два раза в сутки. Активны в основном утром и вечером. Держатся стадами. Самки зубров образуют группы по 6—8 голов, реже по 10—-15; самцы встречаются поодиночке или по 3—4 особи вместе. Бизоны держатся группами по 4—20 особей, а иногда объединяются в крупные стада, в прежние времена насчитывающие тысячи голов. У зубров в Беловежской пуще сезонные миграции почти отсутствуют, а на Кавказе довольно четко выражены вертикальные перемещения: летом выше по склонам тор, вплоть до верхней границы леса, а зимой — вниз. У бизонов 484
отмечены осенние миграции на 200—300 км к югу и весенние — возвращение на летние пастбища.
Гон у зубров приходится на август — первую половину сентября. Сампы в это время иногда устраивают схватки между собой. При одном самце в этот период находится от 2 до 6 самок. С ними остаются и телята. Продолжительность беременности около 9 месяцев. В мае — начале июня происходит отел. Самки приносят, как правило, по одному теленку, масса которого колеблется от 22 до 23 кг. В недельном возрасте теленок уже повсюду следует за матерью. В 19—22-дневном возрасте теленок начинает подкармливаться травой. Лактация продолжается от 5 месяцев до года, а иногда и несколько дольше. Теленок остается с матерью до р__2 лет. В оптимальных природных условиях самки размножаются
ежегодно. Половозрелость наступает в 2—3-летнем возрасте. Гон у бизонов приходится на июль — сентябрь; самцы в это время держатся с самками. Брачное поведение самца может быть определено как «временная моногамия»: в течение нескольких дней он держится около самки, находящейся в эструсе, и отгоняет от нее других самцов, иногда устраивая с ними схватки. Беременность 270—300 дней. Рождение детенышей в Канаде происходит с середины апреля до начала июня с пиком в первой половине мая. Как правило, рождается один теленок, двойни бывают редко. Лактационный период около 7 месяцев. Половозрелость самцов наступает в 2—3 года, а самок — в 2 года.
Продолжительность жизни до 20—25, реже до 30 лет.
В роде 2 вида:
зубр — В. bonasus Linnaeus, 1758 (в историческое время обитал в лесах Западной, Центральной и частично Юго-Восточной Европы, в лесной, лесостепной и отчасти степной зонах европейской части СССР и на Кавказе; примерно к середине XIX столетия в Западной и Центральной Европе был уничтожен, в настоящее время в результате работ по восстановлению зубров они сохраняются в Беловежской пуще — на территории Польши и СССР, в Хоперском, Мордовском, Приокско-Террас-ном и Кавказском заповедниках);
бизон — В. bison Linnaeus, 1758 (был широко распространен в Северной Америке, но к 1825 г. на востоке, а к 1890 г. на западе материка был почти полностью уничтожен — из примерно 50 млн. голов к этому времени оставалось немногим более 500; в настоящее время численность бизонов в результате охранных действий достаточно высока, чтобы не бояться его вымирания).
В СССР: зубр.
Некоторые исследователи (например, Хальтенорт, 1963) включают зубра на правах подвида в вид В. bison.
Хозяйственного значения не имеют.
Зубр включен в «Красную книгу» как вид, прежде находившийся «од угрозой исчезновения, а в настоящее время переставший быть таковым в результате предпринятых мер по его охране (к 1971 г. общая численность достигла 1166 голов, из них в Польше 3 стада — 242 зубра и в СССР Н стад — 258 зубров, кроме того, в 28 государствах Европы, Азии и Америки зубры содержатся в зоопарках и на охраняемых территориях) .
ОТРЯД МОЗОЛЕНОГИХ
ORDO TYLOPODA ILLIGER, 1811
Размеры средние и крупные: длина тела 120—300 см, длина хвоста 12—55 см, высота в холке 72—210 см. Масса 45—650 кг.
Телосложение неуклюжее (рис. 213). Шея длинная. Голова относительно небольшая с удлиненной мордой. Рогов нет. Верхняя губа раздвоенная. Уши небольшие или длинные заостренные. На спине имеется один или два горба или горбов нет. Хвост короткий или длинный с пучком волос на конце. Конечности двупалые (сохраняются III и IV пальцы), пальцеходящие. Проксимальные кости пальцев расширены дистально. Средние кости пальцев широкие и уплощенные, внедряются в широкую эластичную парную или непарную кожную подушку, которая образует подошву. Дистальные кости пальцев невелики, не уплощены и не обрамлены копытом, как у представителей отряда парнокопытных, а несут на верхней поверхности ногти. Боковые пальцы полностью редуцированы вместе с соответствующими пястными и плюсневыми отделами. Проксимальный отдел бедра свободен. Волосяной покров высокий, густой и мягкий. Могут быть затылочная и плюсневая кожные специфические железы. Сосков одна пара.
В черепе лицевой отдел сильно удлинен, а мозговая коробка укорочена. Орбита сзади замкнутая. Имеется сагиттальный гребень. Этмоидальные отверстия большие. Половины нижней челюсти в отличие от парнокопытных сросшиеся. Как и в нижней челюсти хищных, имеется угловой отросток; сочленовная головка сферическая.
1 1
Зубная формула: i — с —— р О *
2 — 3
1 -2(3)
3
— = 30—34(36).
Предчелюстные кости молодых животных имеют по 3 резца, а у взрослых сохраняется по одному, имеющему клыкообразную форму. Третий нижний резец тоже может иметь форму клыка. Клыки конические, заостренные. Нижний клык отделен от резцов пространством. Верхний первый предкоренной зуб может напоминать по форме клык и отделяться диастемой от других щечных зубов; иногда он редуцируется. Второй нижний предкоренной часто отсутствует. Иногда встречается третий нижний предкоренной зуб. Щечные зубы селенодонтные (лунчатые) , гипселодонтные.
Локтевая и лучевая кости сращены между собой в различной степени. Имеется двураздельная на дистальном конце os canon.
Желудок сложный четырехкамерный, «жвачный». Слепая кишка короткая. Желчного пузыря нет. Плацента диффузная. Форма эритроцитов овальная — единственный случай среди всех млекопитающих. Их число очень велико: 14—19 млн. в мм3.
Распространены в Южной Америке и в МНР в пустыни Гоби.
Обитают в степях, пустынях и горах. Питаются различными растениями. Активны днем. Держатся группами, обычно состоящими из самца (вожака) и нескольких самок с молодыми. Могут встречаться и более крупные стада. Бегают иноходью — передняя и задняя конечности одной стороны тела двигаются одновременно в одну сторону — со скоростью до 55 км/ч (гуанако). При отдыхе ложатся на землю так, что тело опирается на запястья, локти, грудь и колена, причем все эти участки безволосы и покрыты кожными мозолями. Течка бывает несколько раз в году. Продолжительность беременности 300—440 дней. Детенышей Б помете один, редко два. Продолжительность жизни до 15—45 лет.
Дикие формы послужили родоначальниками домашних животных: двугорбого и одногорбого верблюда, ламы и альпака.
В отряде одно семейство верблюдовых — Camelidae Gray, 1821, и 2 рода.
До последнего времени мозоленогих обычно включали в качестве подотряда в отряд парнокопытных. Своеобразие эмбрионального развития мозоленогих и их филогенез (предполагаемая близость с палеоценовыми предками Macraucheniidae), по-видимому, подтверждают необходимость выделения мозоленогих в особый отряд (Хавесон, Шмидт, 1974).
Наиболее древние представители Camelidae известны из верхнего эоцена Северной Америки (Poebrodon Gazin, 1955). Они были величиной примерно с косулю и обладали незамкнутой сзади орбитой, полной зубной системой и рудиментарными боковыми пальцами. В плейстоцене предковые формы лам мигрируют из Северной в Южную Америку, а предки верблюдов — в Азию.
СЕМЕЙСТВО ВЕРБЛЮДОВЫХ
FAMILIA CAMELIDAE GRAY, 1821
См. описание отряда.
В семействе 2 рода (4 вида):
верблюдов — Camelus Linnaeus, 1758 (2 вида);
лам — Lama G. Cuvier, 1800 (2 вида).
Род верблюдов
Genus Camelus Linnaeus, 1758
Размеры средние и крупные. Длина тела 220—340 см, длина хвоста 55—75 см, высота в холке 180—210 см. Масса 450—650 кг.
Передние конечности имеют неразделенную подошву (см. рис. 213).
Длинные задние конечности с сильно развитыми коленными мозолями. Хвост длинный с пучком волос на конце. Шея изогнутая. Глаза большие с тяжелыми веками. Уши маленькие, округлые, полностью покрытые волосами. Щелевидные ноздри способны смыкаться. Верхняя губа глубоко разделена. От ноздрей к верхней губе идет желобок. На спине располагается один или два кожных горба, образованных жировой тканью. Когда верблюд упитан, его горб торчит вверх, а у истощенного свисает набок. Окраска волосяного покрова варьирует от темно-бурой до серой. Удлиненные волосы имеются на голове, шее, горбах, бедрах и кончике хвоста; все остальное тело покрыто плотным волосяным покровом око-
I*-
Рис. 213. Дикий двугорбый верблюд — С. bactrianus в Заалтайской Гоби в МНР (Л), домашний верблюд (Б)
Рис. 213. Продолжение. Череп и передняя нога домашнего верблюда
Рис. 214. Гуанако — L. guanicoe
ло 50 мм высотой. На затылке у самцов и самок расположена пара специфических кожных желез примерно 6 см в поперечнике.
„ 113 3
Зубная формула: 1 — с — р —— га — = 34(36). о 1 Z(оу о
Распространены в диком состоянии в Заалтайской Гоби в МНР и, возможно, в прилегающих районах Китая. Домашних верблюдов разводят в Африке, на Аравийском п-ове, в Передней, Малой и Средней Азии, на юго-западе США.
Обитают в степях и полупустынях, но предпочитают кустарниковые и щебнистые пустыни. Кормятся, по-видимому, утром и вечером, а в середине дня и ночью отдыхают.
Дикие верблюды встречаются поодиночке или парами, но чаше группами по 4—6. Стада в 12—15 голов редки. Максимальная скорость бега около 16 км/ч. Хорошо переносит жару и холод. При высоких температурах и сухом корме долгое время без водопоя оставаться не может. Пьет солоноватую и даже соленую воду. Может выпить до 57 л воды за один прием. Ест практически все, что растет в пустыне. Нуждается в галофитах, без которых теряет массу. Гон бывает зимой — в январе— феврале. Единственный детеныш рождается в марте. Беременность длится 370—440 дней. К концу первого дня после рождения детеныш передвигается почти свободно. Лактационный период составляет около года.
Дикие верблюды МНР прежде служили объектами охоты для местных жителей. В настоящее время численность диких верблюдов чрезвычайно мала, и охота на них запрещена.
Рис. 214. Продолжение. Череп гуанако
Дикий верблюд включен в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
В роде 2 вида:
одногорбый верблюд — С. drotnedarius Linnaeus, 1758 (за историческое время известен только в домашнем состоянии в Африке, на Аравийском п-ове, в Передней, Малой и Средней Азии, на юго-западе США);
двугорбый верблюд—С. bactrianus Linnaeus, 1758 (в диком состоянии еще встречается на юго-западе МНР, в Заалтайской Гоби, домашняя форма населяет Малую Азию, Калмыкию, Казахстан, Среднюю Азию).
В СССР: домашние одногорбые и двугорбые верблюды.
Род лам
Genus Lama G. Cuvier, 1800
Размеры мелкие и средние. Длина тела 125—225 см, длина хвоста 15—25 см, высота в холке 70—130 см. Масса до 75 кг. Шея тонкая, почти прямая. Профиль морды прямой (рис. 214). Глаза большие, верхнее веко с длинными ресницами. Уши длинные. Губы не особенно крупные. Хвост короткий, на нижней поверхности почти не имеет волос. Волосяной покров плотный мягкий. У викуньи на передней части груди удлиненные волосы образуют своеобразную гриву. Окраска волосяного покрова варьирует от рыжевато-желтой до буро-рыжеватой. Брюхо беловатое. На внутренней и наружной сторонах плюсны обычно имеются обнаженные ланцетовидной формы участки кожи, богатые железами с ороговевшей поверхностью черного или серо-бурого цвета, — «каштаны». Ногти серо-черного цвета.
„ .112 3
Зубная формула: i с -j- р — m — = 30.
Нижние резцы викуньи имеют открытые корни, постоянный рост и эмаль только на наружной поверхности.
Распространены в Южной Америке от Южного Эквадора до Ла-Платы и Огненной Земли.
Обитают в степях, полупустынях и горах от предгорий до границы снега (до 5 тыс. м над уровнем моря). Питаются травянистыми растениями, мхами, охотно посещают солонцы, водопои, причем могут употреблять солоноватую или соленую воду. Любят стоять или лежать в горных потоках, хорошо плавают. Гон в ноябре — феврале у гуанако и с апреля по июнь у викуньи. Продолжительность беременности 10 месяцев у викуньи и И у гуанако. Обычно рождается один детеныш, который очень быстро после рождения способен следовать за матерью. Лактационный период около 4 месяцев. Подкармливаться травой начинают уже на второй неделе жизни. Половозрелость наступает в 1,5 или 2,5—3 года. Продолжительность жизни до 15—30 лет.
Держатся обычно небольшими группами. У гуанако стада состоят из одного самца и 4—10 самок, а у викуньи из одного самца и 5—15 самок. Способны быстро бегать со скоростью до 50—55 км/ч.
В роде 2 вида:
гуанако — L. guanicoe Miiller, 1776 (от Северного Перу до Ла-Платы и Огненной Земли);
викунья — L. vicugna Molina, 1782 (от Южного Эквадора до Центрального Чили, Западной Боливии и Северо-Западной Аргентины).
Домашние формы рода: лама — L. guanicoe glama Linnaeus, 1758, и альпака — L. guanicoe (или vicugna) pacos Linnaeus, 1758, в диком со-492
стоянии неизвестны. Они были, по-видимому, одомашнены задолго до того, как испанцы впервые завоевали Южную Америку. По мнению одних ученых (например, Херре — Негге, 1952), как лама, так и альпака произошли от гуанако, по мнению других (например, Капурро и Сильва— Саригго у Silva, 1960), предком альпаки была викунья.
Альпака по' размерам меньше ламы и имеет чрезвычайно длинный и тонкий пух, который по качествам (как сырье для текстильной промышленности) превосходит пух любых других млекопитающих.
Все представители (дикие и домашние) рода лам дают при скрещивании между собой плодовитое потомство. Молодые гуанако хорошо приручаются.
Викунья включена в «Красную книгу» как вид, который в ближайшее время может подвергнуться опасности исчезновения.
ОТРЯД НЕПАРНОКОПЫТНЫХ
ORDO PERISSODACTYLA OWEN, 1848
Размеры средние и крупные. Длина тела от 180 до 500 см1, высота в холке от 73 до 200 см. Телосложение легкое и стройное или тяжелое. Конечности короткие и толстые, средней длины или длинные, приспособленные к быстрому бегу, неспособные к супинации и пронации и двигающиеся только в сагиттальной плоскости. Ось конечностей проходит через III палец, наиболее длинный по сравнению с другими пальцами. I палец всегда отсутствует. Число пальцев изменчиво: четыре на передних конечностях (нет I пальца) и три на задних (нет I и V пальцев) — у тапиров; по три на передних и задних конечностях (нет I и V) — у носорогов; по одному на всех четырех конечностях (есть лишь 111 палец) —у лошадей. Концы пальцев защищены роговыми копытами. Волосяной покров низкий и грубый, иногда сильно изреженный или почти полностью редуцированный. В году две линьки. В носовой или в носовой и лобной частях могут находиться один или два рога длиной до 155 см эпидермального происхождения. Соски паховые, одна пара.
Череп массивный с большим лицевым отделом. Кости могут быть пневматизированы. Орбиты открытые или замкнутые. Носовые кости хорошо развиты, сзади расширены. Этмоидальных отверстий нет.
0—3 0—1 3 — 4 3
Зубная формула: i —-— с —-р р —-— m — =24 — 44.
Зубная система гетеродонтная, дифиодонтная.
Щечные зубы лофодонтные (с гребнями) или селенодонтные (лунчатые), брахиодонтные или гипселодонтные.
Локтевая и малая берцовые кости могут быть сильно уменьшены и сливаться соответственно с лучевой и большой берцовой костями. У астрагала лишь одна (проксимальная) сочленовная поверхность. Пяточная кость не сочленяется с малой берцовой.
Желудок простой. Слепая кишка большая. Желчного пузыря нет. Матка двурогая. Плацента диффузная.
Распространены в Центральной и Южной Азии, на п-ове Малакка, о-вах Суматра, Ява, Калимантан, в Африке (кроме Сахары), южной части Северной Америки, Центральной Америке и северной части Южной Америки.
Обитают в пустынях, степях, лесостепях, некоторые во влажных и болотистых тропических лесах. Как правило, держатся поодиночке или небольшими группами, реже более крупными стадами. Активны в светлое или темное время суток. Питаются различными растениями, глав
1 В пределах отряда можно выделить три группы непарнокопытных: мелкие с длиной тела 180—286 см, средние — 187—393 и крупные — 394—500 см.
ным образом травами. У некоторых имеют место сезонные миграции. Полигамы. Самки приносят в помете по одному детенышу, который вскоре после рождения способен следовать за матерью.
Хозяйственное значение невелико. Некоторые виды служат до сих пор объектами охоты. В прошлом их значение в этом отношении было значительно больше. Ряд видов стоит на грани исчезновения, а некоторые вымерли.
Тарпан и африканский осел являются родоначальниками двух важных домашних животных — лошади и осла.
В отряде 3 семейства и 6 родов:
лошадиных — Equidae Gray, 1821 (1 род);
тапировых — Tapiridae Burnett, 1830 (1 род);
носороговых — Rhinocerotidae Owen, 1845 (4 рода).
Предками непарнокопытных были примитивные хищные — Creodonta. В нижнем эоцене появляются представители семейства лошадей (например, Hyracotherium, Eohippus в Северной Америке и Европе) и тапиров; со среднего эоцена начинают встречаться представители семейства носороговых (например, Prohyracodon в Европе). В семействе лошадиных известно 24 вымерших рода, в семействе тапировых — 6 родов, в семействе носороговых — 46 родов.
СЕМЕЙСТВО ЛОШАДИНЫХ
FAMILIA EQUIDAE GRAY, 1821
Размеры мелкие. Длина тела у диких видов 200—280 см, длина хвоста 40—50 см, высота в холке 110—146 см. Масса 120—350 кг. У домашних лошадей высота в холке от 104 до 168 см, а масса — от 136 до 1000 кг. Телосложение легкое и стройное (рис. 215). Конечности высокие и тонкие с одним функционирующим пальцем — III. От II и IV пальцев сохраняются лишь рудименты в виде так называемых «грифельных» косточек. Волосяной покров низкий, густой, плотно прилегающий к телу. Конец морды без волос. На шее короткая прямостоящая грива из удлиненных волос (у домашних форм грива свисающая). Хвост по всей длине покрыт длинными волосами или длинные волосы образуют пучок только на его конце. Окраска волосяного покрова одноцветная, желтая или серая или на светлом фоне туловища (иногда только его передней части) располагаются черно-коричневые поперечные полосы. По средней линии спины может проходить темная полоса. Семенники находятся в мошонке.
В черепе сильно увеличен лицевой отдел. Носовые кости длинные и узкие. Орбиты маленькие замкнутые. Заглазничные отростки широкие.
3 0—1 3—4 3
Зубная формула: i —- с ---- р —---- m - = 36 — 44.
3 0 — 1 3 — 4 3
В верхней и нижней челюстях (в последней реже) иногда имеются первые предкоренные зубы. Щечные зубы с очень высокими коронками (крайняя степень гипселодонтности) и сложной системой эмалевых петель на жевательной поверхности. Клыки самцов сходны по размерам с резцами, сжатые с боков. У самок клыки маленькие или их нет вовсе. Между резцами и щечными зубами находится диастема.
Локтевая кость сильно уменьшена в размерах и срастается с лучевой. Малая берцовая кость рудиментарна и срастается с большой бердовой.
Распространены в Африке от Сомали, Эфиопии, Судана на юг до Южно-Африканской Республики, в Иране, Афганистане, в СССР —
Рис. 215. Бурчеллова зебра — Е. burchelli (Д). лошадь Пржевальского — Е. cabciUus przewalskii (Б), кулан — Е. hemionus (В) и его череп
17—G71
, в Туркмении, МНР, Северо-Западном Китае, Тибете, Непале, Западной Индии.
Населяют степи, саванны, полупустыни и пустыни, предгорья и горные плоскогорья. Приспособлены к быстрому бегу и способны довольно долго бежать со скоростью 50—60 км/ч, временами до 75 км (кулан). Держатся небольшими косяками в 3—5—10 голов, иногда большими стадами. Питаются травянистыми растениями. У кулана наибольшее значение в питании имеют злаки, полыни и солянки. В период дождей, когда растения содержат много воды, обходятся без водопоя. В остальное время года периодически ходят на водопой. Активны в основном в светлое время суток. У некоторых видов имеют место сезонные миграции. Гон у кулана с июня по август. У зебры сезонности в размножении нет. Продолжительность беременности у кулана 331—374 дня, зебры — 336—350 дней. В помете один, редко два жеребенка. Вскоре после рождения он способен следовать за матерью. Самки куланов, особенно молодые, приносят потомство ежегодно. Половозрелость у самок кулана наступает в 2—3 года, у самца в 3 года, но размножаться он начинает в возрасте не ранее 4—5 лет. У зебр половозрелость наступает в 1— 1,5 года.
Численность большинства видов в последние годы сильно снизилась и некоторые (например, лошадь Пржевальского) находятся на грани полного исчезновения. Тарпан и квагга вымерли в недалеком прошлом.
Хозяйственного значения в диком состоянии не имеют.
В семействе единственный род лошадей — Equus Linnaeus, 1758 (7 видов).
Род лошадей
Genus Equus Linnaeus, 1758
См. описание семейства,
В роде 7 видов:
квагга — Е. quagga Gmelin, 1788 (обитала в юго-западной части Южной Африки; в природе встречалась, по-видимому,-до 1878 г., в зоопарках последние квагги умерли: в 1872 г. в Лондоне, в 1875 г. в Берлине и в 1883 г. в Амстердаме);
горная зебра — Е. zebra Linnaeus, 1758 (от Анголы через Намибию до юго-западной и южной частей Южно-Африканской Республики);
бурчеллова зебра — Е. btirchelli Gray, 1824 (Сомали, Эфиопия, Судан, Кения, Уганда, Руанда, Бурунди, Танзания, юго-восток Заира, Малави, Замбия, Мозамбик, Родезия, Ангола, Ботсвана, Намибия, Южно-Африканская Республика);
зебра Грэви— Е. grevyi Oustalet, 1882 (Сомали, Восточная Эфиопия, Северная Кения);
дикий осел — Е. asinus Linnaeus, 1858 (по-видимому, чистокровные дикие ослы сохранились в небольшом числе только в Северном Сомали и Северной Эфиопии);
кулан — Е. hemionus Pallas, 1775 (Иран, Афганистан, СССР — юг Туркмении; МНР, Северо-Западный Китай, Тибет, Непал, Западная Индия); возможно, сохранился в Сирии и на Аравийском п-ове; акклиматизирован на о-ве Барса-Кельмес, Аральское море);
тарпан — Е. caballus Linnaeus. 1758 (в исторический период был распространен в степи и лесостепи Европы — примерно до 55° с. ш., Западной Сибири, Казахстана, Средней и Центральной Азии, в настоящее время, возможно, сохранился в МНР; последний южнорусский степной тарпан, Е. с. gmelini Antonins, 1912, в природе был убит недалеко от Ас-
кания-Нова в 1879 г., в неволе жил на конном заводе в 1914—1918 гг.; лесной тарпан — Е. с. silvaticus Vetulani, 1928, населял Центральную Европу, Белоруссию, Литву, Польщу, последние лесные тарпаны были истреблены в Центральной Европе в раннем средневековье, а на востоке в XVI—XVIII вв,; последний был убит в 1814 г. в Калининградской области; лошадь Пржевальского — Е. с. przewalskii Poljakov, 1881, была распространена в Западной Сибири, Казахстане, Средней Азии, Забайкалье, Центральной Азии; в начале XIX столетия еще встречалась в Казахстане: в настоящее время, может быть, в небольшом числе сохранилась в Джунгарской Гоби в Юго-Западной части МНР, хотя, по последним данным, она исчезла и там.
В СССР: кулан.
Систематика рода окончательно не установлена. Бурчеллову зебру иногда считают не видом, а подвидом (например, Гептнер и др., 1961), лошадь Пржевальского и степного тарпана выделяют в разные виды (например, Банников и Флинт, 1971).
Лошадь была одомашнена в Азии примерно 5—6 тыс. лет назад, и около этого же времени в Египте и Эфиопии произошло одомашнивание ослов.
В «Красную книгу» включены:
как находящиеся под угрозой исчезновения дикий осел; подвиды кулана— из Северо-Западной Индии и из Пакистана Е. h. khur Lesson, 1827; из Сирии, Палестины, Аравийского п-ова и Ирака Е. h. hemippus G. Geoffrey, 1855 (вероятно, этот подвид уже вымер); лошадь Пржевальского;
виды и подвиды, которым в ближайшее время может угрожать исчезновение, — горная зебра, остальные подвиды кулана (кроме указанных выше) из МНР Е. h. hemionus Pallas, 1775, из Туркмении Е. h. onager Boddaert, 1785, из Ладага, Непала, Сиккима и Тибета Е. h. kiang Moorcroft, 1841.
СЕМЕЙСТВО ТАПИРОВЫХ
FAMILIA TAPIRIDAE BURNETT, 1830
Размеры мелкие. Длина тела 180—200 см, длина хвоста 5—10 см, высота в холке 75—120 см. Масса 225—300 кг. Телосложение тяжелое (рис. 216). Тело округло сверху и суживается кпереди. Конечности короткие и толстые. Передние конечности четырех-, а задние трехпалые. III палец самый большой. Копыта небольшие овальные. Глаза маленькие. Уши короткие, округлые, малоподвижные. Морда оканчивается небольшим подвижным хоботком, образованным носом и верхней губой. Ноздри открываются на конце хобота. Кожа толстая. Волосы очень редкие, прямые, щетиновидные. Иногда по средней линии верха шеи проходит короткая грива. Окраска тела одноцветная—темно-коричневая или рыжеватая на спинной стороне и светлая на брюшной. У азиатских тапиров передняя часть тела и задние конечности черноватые, остальная часть тела белая. Молодые тапиры темные с белыми и желтыми продольными полосами и пятнами; в 6—8-месячном возрасте они приобретают окраску взрослых животных. Мошонки нет.
Череп удлиненный с высокой мозговой коробкой. Носовые кости короткие. Наружные костные носовые отверстия большие.
3 14 3
Зубная формула: i с -j— р m -у = 44.
Тпетий верхний резец клыкоподобный и крупнее клыка. Клыки хоро-
тапир — r^etrestris(£™Р Lndicus (А)> равнинный
S4X
Рис. 216. Продолжение. Череп равнинного тапир,
шо развиты. Щечные зубы с низкой коронкой (брахиодонтные) с серией поперечных гребней и выступов на жевательной поверхности. Последних три предкоренных зуба сходны с коренными.
Лучевая и локтевая кости развиты примерно одинаково и не сливаются между собой.
Распространены в Бирме, Таиланде, на п-ове Малакка, на о-ве Суматра и в Центральной и Южной Америке.
Обитают в лесных и кустарниковых зарослях рядом с водоемами или в заболоченных местах. В горы поднимаются до 4500 м над уровнем моря. Способны быстро двигаться в густых зарослях. Хорошо плавают и ныряют. Встречаются поодиночке или парами. Активны в сумерках и ночью. Держатся на индивидуальных участках. Питаются различными растениями, в том числе водными. Иногда делают набеги на поля. Сезонности в размножении нет. Продолжительность беременности 390— 400 дней.- Самка приносит одного, редко двух детенышей в среднем каждые 15 месяцев. Половозрелость наступает у самок в 3—4 года, у самцов на год позже. Продолжительность жизни около 30 лет. Тапиры хорошо приручаются.
В семействе 1 род: Tapirus Brisson, 1762, и 4 вида.
Род тапиров
Genus Tapirus Brisson, 1762
См. описание семейства.
В роде 4 вида:
равнинный тапир — Т. terrestris Linnaeus, 1766 (от бассейна Амазонки до Парагвая и Северной Аргентины);
горный тапир — Т. pinchaque Roulin, 1829 (Колумбия, Эквадор, Перу и Венесуэла);
центральноамериканский тапир — Т. bairdi Gill, 1865 (от Мексики на юг до Северной Колумбии и Эквадора);
чепрачный тапир — Т. indicus Desmarest, 1819 (Юго-Восточная Бирма, Южный Таиланд, п-ов Малакка, о-в Суматра).
В «Красную книгу» включены горный, центральноамериканский и чепрачный тапиры как виды, находящиеся под угрозой исчезновения.
СЕМЕЙСТВО НОСОРОГОВЫХ
FAMILIA RHINOCEROTIDAE OWEN, 1845
Размеры от мелких до крупных. Длина тела 200—500 см, длина хвоста 60—76 см, высота в холке 100—200 см. Масса 1000—3600 кг. Телосложение тяжелое. Конечности трехпалые короткие и массивные. Хвост тонкий. Шея короткая и толстая. Голова большая удлиненная. Глаза маленькие, располагаются по бокам головы посредине между ушами и ноздрями. Верхняя губа сильно развита, подвижная. Уши длинные, торчащие, овальные с небольшим пучком волос на вершине. На морде в носовом отделе располагается рог до 158 см длиной и иногда позади него в лобном отделе второй рог меньшей длины. Кожа толстая, в некоторых местах образует складки. Волосы редкие, щетиновидные, иногда почти полностью отсутствуют. Окраска тела от сероватой до коричневатой и черной. На конце хвоста кисть волос. Мошонки нет.
.ЧП9
0 — 2 0 3 — 4
Зубная формула: i —----р с 0_ । Р 3______4 га
Череп с удлиненным лицевым отделом, небольшой мозговой коробкой. Затылочный гребень сильно развит. Орбиты сзади не замкнуты.
3 — -24-34. 3
Резцы и клыки небольших размеров или их нет вовсе. Предкоренные зубы сходны с коренными. На жевательной поверхности щечных зубов располагаются поперечные гребни эмали. Щечные зубы с высокими коронками у белого носорога и с низкими у других видов.
Локтевая и лучевая, малая и большая берцовые кости хорошо развиты и не срастаются между собой.
Распространены в тропических областях Африки и Юго-Восточной Азии, включая п-ов Малакку и о-ва Яву, Суматру и Калимантан.
Обитают в саваннах, в кустарниковых зарослях по опушкам леса г в лесу, в местах, где имеется водопой. Держатся поодиночке, а в периох размножения парами. Белые носороги встречаются группами до 18 особей. Активны вечером, ночью и ранним утром. День проводят в укромных местах. Питаются различными растениями — в основном саккулен-тами. Периодически ходят на водопой. Строгой сезонности в размножении нет. Самка приносит обычно одного, редко двух детенышей раз i 2—3 года. Продолжительность беременности 510—570 дней. Детеньш. вскоре после рождения способен следовать за матерью и остается с ней до рождения следующего детеныша. Половозрелость наступает от 3— 4 до 7-—9 лет. Продолжительность жизни до 50—60 лет.
Численность носорогов за последние годы вследствие неумеренной охоты сильно упала, и он сохраняется практически только ных парках. Настойчиво преследовались охотниками, а
коньерами, главным образом из-за рогов, высоко ценимых в восточной медицине.
В семействе 4 рода (5 видов):
суматранских носорогов — Didermocerus Brookes, 1828 (=Dicerorchi nus Gloger, 1841) (1 вид);
индийских носорогов — Rhinoceros Linnaeus, 1758 (2 вида);
черных носорогов — Diceros Gray, 1821 (1 вид);
белых носорогов — Ceratotherium Gray, 1861 (1 вид).
в националь теперь бра
Род суматранских носо]
Genus Didermocerus Brookes,
В роде единственный вид: суматранский носорог — D. sumatrensh Fischer, 18Г4.
Размеры наименьшие в семействе. Длина тела 200—280 см, высота i холке 100—150 см. Масса до 1000 кг. Наиболее древний из ныне жпву; щих носорогов. Тело покрыто негустыми щетиновидными волосами. Ушг окаймлены волосами. Окраска тела от сероватой до черноватой. На морде два рога, передний из которых достигает в длину примерно 15—45 см а задний имеет вид выступа. У самки рог значительно короче, чем 5 самца.
Распространен в Ассаме, Бирме, на п-овах Индокитай и Малакка, о-вах Суматра и Калимантан. Везде малочислен.
Обитает в глухих лесах на холмах около водоемов. Держатся поо Диночке. Активен в сумерках. День проводит в укромных местах, в гря зевых ямах. Питается листьями, веточками, побегами бамбука, иногда плодами. Беременность 7—8 месяцев. В помете один детеныш.
Включен в «Красную книгу» как вид, находящийся под угрозой ис
Рис. 217. Индийский носорог — R. unicornis
чезновения. Суматранских носорогов в 1968 г. в Бирме было 24 особи; в 1972 г. в Таиланде 10—20; в 1965 г. в Малакке 10—30; в 1972 г. на Суматре 45—85.
Род индийских носорогов
Genus Rhinoceros Linnaeus, 1758
Длина тела 210—420 см, длина хвоста 60—75 см, высота в холке 110—200 см. Масса 2000—4000 кг. Кожа лишена волос, за исключением участков вокруг ушей и кончика хвоста (рис. 217). У индийского носорога кожа, особенно в задних частях тела, покрыта большими шишковидными буграми, а у яванского — полигональными выступами. Кожа образует своеобразные складки, придающие ей вид панциря. Так, в области плеч проходит поперечная складка, загнутая назад. Еще одна складка располагается перед задними конечностями. Окраска тела от серой до черноватой с розоватым оттенком по краям кожных складок. В носовой области находится единственный рог, достигающий длины 60 см (обычно около 20 см).
Распространены в Непале, Северо-Восточной Индии, Ассаме и на о-ве Ява.
Обитают в болотистых джунглях, поросших высокой травой и камышом. Держатся по большей части поодиночке. Метят индивидуальную территорию кучами помета. Активны преимущественно утром и вечером. Много времени проводят, лежа в грязевых ваннах. Питаются различными травянистыми растениями. Самка приносит одного детеныша в период между концом февраля и концом апреля. Продолжительность беременности у индийского носорога около 570 дней, у яванского — около 510 дней. Новорожденный имеет в длину 100—120 см, высоту в холке около 60 см, массу 34—75 кг. Длительность лактации около двух лет.
В роде 2 вида:
индийский носорог — /?, unicornis Linnaeus, 1758 (Непал, Ассам, Северная Бенгалия);
яванский носорог — /?. sondaicus Desmarest, 1822 (в недалеком про
шлом встречался в Индии, Бирме, Бангладеш, на п-овах Индокитай и Малакка, о-вах Суматра и Ява; в настоящее время сохранился лишь на Яве и, возможно, на Северной Суматре).
Оба вида включены в «Красную книгу» как виды, находящиеся под угрозой исчезновения. Индийских носорогов в 1971 г. насчитывали 550 в Индии и 70 в Непале, а яванских в 1972 г.-—48 на Западной Яве и, возможно, 20—25 на Северной Суматре.
Род черных носорогов
Genus Diceros Gray, 1821
В роде единственный вид: черный носорог — D. bicornis Linnaeus, 1758.
Длина тела 300—375 см, длина хвоста около 70 см, высота в холке 140—150 см. Масса 1000—1800 кг. На морде два рога, иногда три и даже пять (рис. 218). Длина переднего рога обычно около 50 см, максимум до 13-8 см. Верхняя губа спереди заострена и свисает хоботком над нижней. Этим хоботком черный носорог захватывает листья и веточки кустарников. Окраска тела от темно-желто-коричневой до темно-коричневой.
Распространен в Судане, Эфиопии, Сомали, Чаде, Нигерии, Камеруне, Центральноафриканской Республике, Заире, Уганде, Кении, Танзании, Л1озамбике, Малави, Замбии, Намибии, Ботсване, Южно-Африканской Республике. Во всех этих странах сохранился лишь местами, в основном в национальных парках.
Обитает в разреженных сухих лесах, саваннах, степях и даже полупустыне. В горы поднимается до 2700 м над уровнем моря. Держатся поодиночке. Активен преимущественно утром и вечером. Питается побегами и листьями кустарников. Ежедневно ходит на водопой. Сезонности в размножении нет. Продолжительность беременности 530—550 дней. Новорожденный имеет массу 20—35 кг. Лактационный период длится примерно 2 года. Половозрелость наступает примерно в 5 лет.
Включен в «Красную книгу» как вид, который в ближайшее время может оказаться под угрозой исчезновения. В 1961 г, общее число черных носорогов достигало 11—13,5 тыс. голов.
Род белых носорогов
Genus Ceratotherlum Gray, 1861
В роде единственный вид: белый носорог — С. simutn Burchell, 1817.
Размеры наибольшие не только в семействе, но и в отряде. Длина тела 360—500 см, высота в холке 160—200 см. Масса 2300—3600 кг. На морде два рога (рис. 219); из них передний достигает в длину 60—• 158 см. Верхняя губа спереди без хоботка (иногда белого носорога называют квадратногубым). Волосы на теле отсутствуют, кроме краев ушей и конца хвоста. Окраска тела несколько светлее, чем у черного, шиферно-серая.
Распространен в Южно-Африканской Республике, Южном Судане,, Уганде и соседних районах Заира, а также, возможно, в Центральноаф^ риканской Республике и Чаде.
Обитает в саванне, зарослях кустарников. Держится небольшим^] группами, иногда собирается в стада, насчитывающие до 16—18 особейЛ Активен утром и вечером, а в прохладную погоду и днем. Питаете^ травянистыми растениями. Нуждается в водопое и грязевых ваинахЛ
ДОЧ
гл
Рис. 219. Белый носорог—С. sima
в размножении нет, хотя массовый гои, по-видимо
Строгой сезонности
му, приходится на период с июля по сентябрь. Беременность околс 540 дней. Лактационный период длится примерно год. Самка приноси детеныша раз в 2,5—3 года. Половозрелость наступает в 4—5 лет.
Подвид белого носорога из Судана, Уганды и Заира — С. simti -cottoni Lydekker, 1908, включен в «Красною книгу» как подвид, нахо
дящийся под угрозой исчезновения.
ЛИТЕРАТУРА
Агарков Г. Б., Хоменко Б. Г., Хаджинский В. Г. Морфология дельфинов. — Киев: Наукова думка, 1974.
Барабаш-Никифоров И. И., Мараков С. В., Николаев А. М. Калаи, морская выдра. — Л.: Наука, 1968.
Берзин А. А. Кашалот. — М.: Пищевая промышленность, 1971.
Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Определитель млекопитающих СССР. Изд. 2-е. — М.: Просвещение, 1965.
Бромлей Г. Ф. Уссурийский кабан. —М.: Наука, 1964.
Бромлей Г. Ф. Медведи юга Дальнего Востока СССР. — М. — Л.: Наука, 1965.
Гентнер В. Г., Насимович А. А., Банников А. Г. Млекопитающие Советского Союза. Т. I. Парнокопытные и непарнокопытные/Под ред. В. Г. Гептнера и Н. П. Наумова.— М.: Высшая школа, 1961.
Гентнер В. Г., Наумов Н. П., Юргенсон П. Б., Слудский А. А., Чиркова А. Ф., Банников А. Г. Млекопитающие Советского Союза. Т. II, ч. 1. Морские коровы и хищные, Под ред. В. Г. Гептнера и Н. П. Наумова. — М.: Высшая школа, 1967.
Гептнер В. Г. и Слудский А. А. Млекопитающие Советского Союза. Т. II., ч. 2. Хищные (гиены и кошки)/Под ред. В. Г. Гептнера и Н. П. Наумова. — М.: Высшая школа, 1972.
Гептнер В. Г., Цалкин В. И. Олени СССР. — М.: МОИП, 1947.
Гептнер В. Г., Чапский К. К., Арсеньев В. А., Соколов В. Е. Млекопитающие Советского Союза. Т. 2, ч. 3. Ластоногие и зубатые киты/Под ред. В. Г. Гептнера и Н. П. Наумова. — М.: Высшая школа, 1975.
Громов И. М„ Гуреев А. А., Новиков Г. А., Соколов И. И., Стрелков П. П., Чапский К. К. Млекопитающие фауны СССР. Ч. II. — М. — Л.: Изд-во АН СССР, 1963.
Ивашин М. В., Попов Л. А., Цапка А. С. Морские млекопитающие. — М.: Пищевая промышленность, 1972.
Клейненберг С. Е. Млекопитающие Черного и Азовского морей. — М.: Изд-во АН СССР, 1956.
Клейненберг С. Е., Яблоков А. В., Белькович В. М., Тарасевич М. Н. Белуха — М.; Наука, 1964.
Морфология, физиология и акустика морских млекопитающих/Под ред. Соколова В. Е. — М.: Наука, 1974.
Морфология и экология морских млекопитающих (дельфины)/Под ред. Соколова В. Е. — М.: Наука, 1971.
Насимович А. А. Африканский слон. — М.: Наука, 1975.
Новиков Г. А. Хищные млекопитающие фауны СССР. -— М. — Л.: Изд-во АН СССР, 1956.
Огнев С. И. Звери СССР и прилежащих стран. Т. III. Хищные н ластоногие. — М. — Л.: Гос. изд. биологической и медицинской лит., 1935.
Слепцов М. М. Китообразные дальневосточных морей. — Владивосток, 1955.
Смирнов Н. А. Очерк русских ластоногих. «Записки Императорской Академии Наук», 1908, т. 23, № 4.
Смирнов Н. А. Определитель ластоногих (Pinnipedia) Европы н Северной Азии.— Изв. Леи. НИ пхт. ин-та, 1931, т. XII, вып. 1.
Смирнов Н. А, Морские звери арктических морей. — В кн.: Звери Арктики. М., 1935.
Соколов В. Е. Систематика млекопитающих. Отряды: однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых, ящеров. — М.: Высшая школа, 1973.
Соколов В. Е. Систематика млекопитающих. Отряды: зайцеобразных и грызунов.— М.: Высшая школа, 1976.
Соколов И. И. Копытные звери. Фауна СССР. Млекопитающие. Т. I, вып. 3.—М.— Л.: Изд-во АН СССР, 1957.
Тимофеев В. В., Надеев В. Н. Соболь. — М.: Изд-во технической и экономической литературы по вопросам заготовок, 1955.
Тимофеева Е. К. Лось. —Л.: Изд-во ЛГУ, 1974.
Томилин А. Г. Звери СССР и прилежащих стран. Т. IV. Китообразные. — М.: изд-во АН СССР, 1957.
Томилин А. Г. Китообразные фауны морей СССР. — М.: Изд-во АН СССР, 1962.
Флеров К. К. Кабарги и олени. Фауна СССР. Млекопитающие. Т. I, вып. 2. — М.— Л.: Изд-во АН СССР, 1952.
Хавесон Я. И., Шмидт Г. А. Уточнения положения группы Tylopoda в системе млекопитающих. Первый международный конгресс по млекопитающим. Т. И. М., 1974.
Чалкин В. И. Горные бараны Европы и Азии. — М.: МОИП, 1951.
Чапский К. К. Современное состояние и проблемы систематики ластоногих. — Тр. совет- яхт. комисс. АН СССР, 1961, вып. 12.
Чапский К. К. Отряд ластоногих. — В кн.: Млекопитающие фауны СССР. Ч. 2. — М.—Л.: Изд-во АН СССР, 1963.
Чапский К. К. Отряд ластоногих. — В ки.: Гептнер В. Г., Чапский К. К-, Арсеньев В. А., Соколов В. Е. Млекопитающие Советского Союза. Т. II., ч. 3. Ластоногие и зубатые киты/Под ред. В. Г. Гептнера и Н. П. Наумова. — М.: Высшая школа, 1975.
Экология и морфология белого медведя/Под ред. Соколова В. Е. — М.: Наука, 1973.
Яблоков А. В. О строении зубной системы и типах зубов у китообразных. — Бюлл. МОИП, отд. биол., 1958, вып. 3.
Яблоков А. В. К морфологии пищеварительного тракта зубатых китообразных. — Зоол. журн., 19586, т. 37, вып. 4.
Яблоков А. В., Белькович В. М., Борисов В. И. Киты и дельфины.—М.: Наука, 1972.
Allen G. М. A checklist of African mammals. Bull. Mus. Comp. Zool. Harv. College, 83. Cambridge, 1939.
Allen G. M. The mammals of China and Mongolia. Nat. Hist, of Centra. Asia, 11, pt I. N. Y.
Andersen H. T. Ed. The biology of marine mammals. New York a. London, Acad. Press, 1969.
Anderson S. and Jones Knox Jr. Recent mammals of the world. The Ronald Press Co. New York, 1967.
Beddard F. E. A book of whales. London. 1900.
Cabrera A. Catalogo des los Mamiferos de America del Sur. Revista del Museo Ar-hentino de Ciencias Naturales “Bernardino Rivardavio” e Institute Nacional de Investigation de las Ciencias Naturales. Ciencias Zoologicas. Tome IV. Buenos Aires, 1957— 1961.
Capurro S. L. у Sulva G. Estndios chromotograficos у electroforeticos en camelidos sudamericanos. Invest. Zool. Chilenas. Santiago de Chile. 6. 49—64.
Eadie J., Kirk R. L. The sodium and potassium concentration in the blood cells and plasma of the elephant seal. Austr. J. Sci, 1952, 15, p. 26—27.
Ellerman I. R. and Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. 1758 to 1964. II Ed. London, 1966.
Ellerman J. R., Morrison-Scott T. C. S. and Hayman R. W. Southern African Mammals. British Museum. London. 1953.
Fraser F. C. and Purves P. E. Hearing in Cetacea. Bull. Brit. Mus. Nat. Hist., Zool., 1960, v 7, 1—40.
Gray J. E. Hand-list of seals, morses, sea-lions and sea-bears in the British Museum, London, Brit. Mus., 1874.
Hahn H. Baumschliefer, Buschliefer, Klippschliefer. Die Neue Brehm-Bficherei. Wittenberg Lutherstadt, A. Zeimsen Verlag, 1959.
Hall E. R. and Kelson K. R. The Mammals of North America, vol. II. The Ronald Press. Co. New York, 1959.
Haltenorth Th. Klassifikation der Saugetiere: Artiodactyla. Handbuch der Zoologie. 8 B, 32 Lieferung, Berlin, 1963.
Harris C. J. Otters a study of the recent Lutrinae. London. Weidenfeld and Nicol-son, 1968.
Harrison R. J., Ed. Functional Anatomy of marine mammals, v. 1—III. London and New York. Acad. Press, 1972—1974.
Harrison R. J., Hubbard R. C., Peterson R. S., Kice С. E.. Schusterman R. J. Ed. The behavior and physiology of pinnipeds. New York, Appleton — Century—Grafts. 1968.
Herre IT. Studien fiber wilden und domestizierten Tylopden Sfidamerikas. Zool. Garten, Leipzig, N. F. 19, 70—98, 1952.
Hershkowitz P. Catalog of living whales. Smith Inst. Bull, v 246, 1—259. 1966.
Hopkins G. H. E. The host-associations of the life of mammals. Proc. Zool. Soc. London, 1949, 119, p. 387—604.
Howell A. B. Aquatic mammals, their adaptations to life in the water. Baltimore, Charles C. Thomas, 1930.
Hubar E. Anatomical notes on Pinnipedia and Cetacea. Publ. Carneg. Inst, 1934, 447, part 4. p. 105—136.
Kellog R. Pinnipeds from Miocene and Pleistocene deposits of California and a resume
of current theories regarding origin of Pinnipedia. Bull. Dep. Geol. Univ. Calif., 1922, 13, p. 23—132
Kelsall J: P. The caribou. Canadian wildlife service. 1968.
King J. E. Seals of the world. London. British Museum (Natural History), 1964.
Frank H. The spotted hyena. Chicago and London. The University of Chicago Press 1972.
Lange J. Studien an Gazellenschaden. Ein Beitrag zur Systematik der kleineren Ga-zellen, Gazella (De Blainville, 1816). Saugetiere. Mitt. 1972. 20 N 3: 193—199, 242—249.
Leone C. A. and Wiens A. L. Comparative serology of carnivores. J. Mammal., 1956 37, p. 11—23.
Meester I. and Setzer H. W. The Mammals of Africa. An identification manual. Washington. Smithonian Institution Press. 1971.
Miwart St. G. Notes on the Pinnipedia. Proc. Zool. Soc., London, 1885, p. 484—500.
Mohr E. Sirenen oder Seekfihe. Die Neue Brehm — Bucherei. Wittenberg Lutherstadt. A. Ziemsen Verlag. (957.
Newell J. M. Studies on the morphology and systematics of the family Halarachnidae Oudemans 1906 (Acari, Parasitoidea) Bull. Bingham Oceanogr. Coll., 1947, 10, p. 235— 266.
Nishiwaki H. in: Ridgway S. H., Ed. Mammals of the sea. Springfield, Illinois, USA. Ch. C. Thomas, 1972.
Nishiwaki M. and Norris K- A new genus, Peponocephala, for the Odontoceti cetacean species Electra electra. The scientific reports of the whales research institute, 1966, N 20 95—100.
Norris K. S. Ed. Whales, dolphins and porpoises. Berkely and Los Angeles. University of California Press, 1966.
Pilleri G., Ed. Investigations on Cetacea, v I—II. Berne, Switzerland. 1969—1972.
Rice D. W. and Scheffer V, B. A list of the marine mammals of the world. US Fish and Wildlife Service. Special scientific report — Fisheries No 579. Washington. 1968.
Ridgway S. H., Ed. Mammals of the sea. Springfield, Illinois, USA, Ch. C. Thomas. 1972.
Roche J. Systematique du genre Procavia et des damans en general. Mammalia 1972 v. 36, N 1: 22—49.
Rosevear D. R. 1974. The Carnivores of West Africa. The British Museum (Natural History) London.
. Schaller G. B. The Serengeti lion. Chicago and London. The University of Chicago Press. 1972.
Scheffer V. B. Seals, sea-lions and walruses. Stanford, Calif. Stanford Univ. Press, 1958.
Sikes S. K. The natural history of African elephants. New York, American Elsevier Publ. Co. Inc.
Simpson G. C. The principles of classification and a classification of mammals. Bull. Amer. Mus. Nat. Hist, v 85, 1945.
Slijper E. I. Whales. London. Hutchinson. 1962.
The scientific reports of the whales research institute, N I—27. Tokyo, Japan. 1948— 1975.
Thenius E. und Hofer H. Stammesgeschichte der Saugetiere. Berlin. Springer — Verlag, I960.
Walker E. P., Warnick F., Lange К. I., Uible H. E., Hamlet S. E. Davies M. A., Wright P. F. Mammals of the world, v II. Baltimore. The John Hopkins Press, 1964.
Weber M. Die Saugetiere v II. Systematischer Teil. Jena, Gustav Fischer, 1928.
Whitehead G. K. Deer of the world. Constable. London. 1972.
Young S. P. and Goldman E. A. The puma, p. 1, II. New York, Dover Publications, Inc, 1964.
Young S. P. and Goldman E. A. The wolves of North America p. I, II. New York, Dover Publications, Inc., 1964.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Аддакс 398, 422*, 423
Аксис 236, 387, 389
Альпака 487, 492, 493
Аноа 477—479
Антилопа 91, 110, 236, 251, 410, 421,
429, 431, 434, 436
— карликовая 333, 398, 403, 437
— косулья 398, 427
— лесная 398, 414
— лошадиная 398, 429, 430, 432
— четырех рога я 398, 410, 411
— черная 430, 431, 432
Антилопа-крошка 403
Антилопа-прыгун 398, 402, 405—407
Архар 474
Архаромеринос 476
Астрагал 494
Афалина 8, 15, 22, 25, 30, 31
Бабирусса 336, 337, 344—346
Бантенг 481, 484
Баран 476
• — голубой 469, 470, 476
—• горный 240, 251, 398, 471, 474—476
— гривистый 398, 471—473, 476
— снежный 476
Барасинга 386, 388, 389
Барс 233, 468
— снежный 232, 233, 238, 239
Барсук 89, 91, 100, 138, 167, 168, 170—
172, 236, 244
— американский 138, 171
— бирманский 174
— восточный 173, 174
— китайский 174
— хорьковый 138, 172
Бегемот 333, 334, 348—350, 351, 352
— карликовый 348, 350, 351
Бегемотовые 334, 335, 348
Бейра 398, 407
Белка 140, 141, 244
Белуха 5, 9, 12, 14, 15, 16, 50—53, 275
Бизон 482, 485
Бинтуронг 192, 204, 205
Бонго 417—419
Бородавочник 336, 337, 341, 342, 344
Броненосец 111
Бубал беломордый 432, 433
— лпророгий 398, 432
— Хантера 433
* Полужирным шрифтом даются ссылки на описание рода.
Буйвол 236, 398, 477
— азиатский 477, 479
— африканский 477, 478
— карликовый 478
Бутылконос 9, 14, 15, 59, 63, 65
— высоколобый 65, 66
— плосколобый 66
Бурундук 140, 151, 244
Бушбок 415, 417
Бык настоящий 398, 479
Верблюд 487, 490, 492
— двугорбый 487, 488, 492
— домашний 489
— одногорбый 487, 488, 492
Верблюдовые 487
Виверровые 84
Виверры 256
Викунья 492, 493
Вилорог 395, 396
Вилороговые 334, 354
Волк 82—85, 87, 90—95, 97, 101, 118, 126, 147, 236, 256, 341, 361, 367, 381, 386, 442, 454, 468, 476
— гривистый 85, 87, 106, 107
— земляной 224, 225, 230
— красный 87, 107, 108, 109
Волчьи 84
Выдра 82, 137, 138, 180—183, 185, 187, 256
— африканская 138, 187
— белогорлая 184
- — бескоготная 138, 185, 186
— гвианская 184
— гигантская 138, 184, 185
— гладкошерстная 182, 184
- — канадская 182, 183
— капская 186
— кошачья 184
— лаплатская 184
— мексиканская 184
— островная 184
— перуанская 184
— серая 186
— суматранская 184
— южная 184
Газель 227, 236, 251, 403, 437, 443
— Гранта 444
— доркас 443, 446
— краснолобая 446
— обыкновенная 446
— Пельцельна 446
— песчаная 446
Газель сомалийская 444
— Спика 446
— суданская 446
— Томсона 443, 446
Газель-дама 443, 446
Гарна 398, 437, 438
Гаур 480, 481
Гаял 480
Генета 191, 194
— ангольская 195
— гигантская 195
— Джонстона 195
— лесная 195
— ложная 195
• — малая 194
— обыкновенная 195
— пятнистая 194, 195
— серваловая 195
— эфиопская 195
Гепард 231—233, 249—251
Геренук 398, 447, 448
Гиена 225
— бурая 228
— полосатая 225, 227—229
— пятнистая 225—227
Гиеновые 84, 224
Гну 227, 236, 398, 434, 436
— белохвостый 436
— голубой 435, 436
Горал 398, 452—454, 456, 458
Горбач 67, 81
Горностай 149—152, 156, 157
Гофер 151, 172
Гризон 137, 160, 161
— малый 161
Грин да 6, 12, 15, 22, 37, 42, 43
— обыкновенная 44,
— тропическая 44
• — черная 43, 44
Грисбок 409
Гуанако 490, 491
Даман 317, 320, 326
— ахаггарскпй 321
— Брюса 322
— восточный 323
— горный 320, 321, 323
— Джонстона 321
— древесный 320, 323
— западный 322, 323
— каокефелдский 321
— капский 318, 319, 321
— лоаднйский 322
- — рыжий 321
— сирийский 321
— скалистый 320, 321, 323
Дельфин 5, 9, 10, 11, 22, 30, 32, 41
— амазонский 16, 18, 23
— атлантический 35, 36
— белобрюхий 37
- — беломордый 35, 36
— белый 23, 24
— бесклювый 22, 36, 37, 42
• — бразильский 24
• — гайанский 24
— гангскпй 14, 16, 19, 20, 21
— Гектора 38
— длинноклювый 22, 23
Дельфин длинномордый 15
— западноафриканский 23, 24
— нравадийскпй 12, 22, 38
— калимавтанский 23, 24
— китайский 23, 24
— китовидный 22, 32
— короткоголовый 22, 34, 35, 37
• — крапчатый 24
— крупнозубый 22, 23
— лаплатскпй 16, 17
— малайзийский 22, 34
— озерный 16, 19
— пестрый 22, 37
— речной 15, 16, 18
— свинцовый 24
— северный 33, 34
— серый 12, 14, 22, 42, 43
— темный 36
— тихоокеанский 35, 36
— Хэвисайда 37
— южный 33, 36
Дельфин-белобочка 6, 7, 11, 12, 13,
15, 21, 22, 24, 25, 27—30, 34, 47
Дельфин Ла-Платы 8
Дельфиновые 21
Джейран 236, 251, 443—446
Дзерен 91, 398, 441, 442
— монгольский 440—442
— тибетский 442
Днбатаг 398, 446
Дпкдпк 398, 404
— горный 405
— Гюнтера 405
— малый 405
— обыкновенный 404, 405
- — рыжебрюхий 404, 405
Дикобраз 236, 244
Динго 83
— австралийский 94
Дукер белобрюхий 401
— Гарвея 401
— голубой 400
- — желтоспниный 399, 400, 401
— занзибарский 401
— зебровый 400, 401
• — конголезский 401
— красный 401
— кустарниковый 397, 401, 402
— Максвелла 399, 400
• — Оджильби 401
— рыжебокий 399, 400
— танзанийский 401
— хохлатый 397, 399
— чепрачный 401
— чернолобый 400
— черноспинный 399, 401
•— черный 399, 400, 401
Дюгоневые 328, 332
Дюгонь 329, 330, 332
Енот 82, 123, 126, 256
Енот-полоскун 84, 124, 126, 127, 128
Енот-ракоед 127
Енотовые 84, 123, 124
Жвачные 334, 335, 353
Жираф 333, 389, 390. 392, 393
Жирафовые 334, 354, 389
Замбар 236
__ гривистый 388
— индийский 386, 388
— филиппинский 388
Заяц 91, 101, 140, 151, 240, 244
— маньчжурский 236, 244
— морской 117, 253, 277, 278, 293,
294, 295, 296
Заяц-беляк 147, 244
Заяц-песчаник 244
Заяц-русак 244
Заяц-толай 244
Зебра 227, 236, 421, 429, 431, 434, 436,
498
— бурчеллова 497—498
— горная 498, 499
— Грэви 498
Землеройка 105, 140, 141, 151
Зубр 396, 398, 482, 483, 484, 485
Изюбр 236
Илька 137, 138, 140, 141, 144
Импала 236, 398, 428, 429
Кабан 236, 240, 244, 337, 338, 339, 340, 341
Кабарга 141, 236, 359, 360
Какомицли 123, 124
— североамериканский 124—126
— центральноамериканский 125, 126
Калан 83, 136—138, 182, 187—190
— командорский 189
Канна 414, 417—419
Каракал 232, 240, 242, 244, 245, 248, 249
Кашалот 6, 8, 9, И, 12, 14, 15, 55, 56—59
— карликовый 11, 12, 15, 55, 56
— малый 56
Каша лотовые 54, 55, 61
Квагга 498
Кидасы 142
Кинкажу 123, 124, 130
Кит 8, 10, 14, 42, 67
— австралийский 71
— бискайский 71, 72
-— гладкий 68, 73
— голубой 5, 6, 9, 10, 76—79
— горбатый 6, 76, 79, 80, 81
— гренландский 6. 11, 67, 68, 69—71
— зубатый 5, 7, 9, 10, 12, 13, 14—16, 67
— карликовый 66, 68, 72
---гладкий 72
— серый 6, 8, II, 67, 68, 73—75
— усатый 5, 7—10, 12, 41, 66, 67. 73
— южный 68, 70—72
-— японский 71
Китообразные 5—12, 17, 18, 21, 50, 54, 57—59, 75, 76
Клюворыл 8, 13, 15, 59, 63
•— настоящий 63, 64
— тасмаиов 13, 59
Клюворылые 59, 63, 65
Коати 128, 129
Коб 424, 425
Койот 90—94
Коза снежная 398, 456, 457, 458
Козел 398, 449, 468
— альпийский 468
— бородатый 467—469
— винторогий 467—469
— водяной 398, 423—425
— горный 109, 465
— дагестанский 469
— домашний 469
— кубанский 469
• — нубийский 469
— пиренейский 468. 469
— сибирский 466, 469
— — горный 240, 468
— тростниковый 398, 425
Колонок 150, 154—157
— японский 156
Конгони 398, 433, 434
Корова морская 324, 326, 330, 331, 332
Корсак 100, 101
— американский 100—102
Косатка 6, 11, 15, 21, 22, 37, 40, 41, 74, 190, 255, 265, 267, 275, 279, 284, 287, 299, 303, 305, 308
— карликовая 11, 12, 22, 37, 42, 44, 45 — малая 12, 15, 22, 37, 38, 39, 40, 42 Косуля 100, 236, 240, 244, 358, 365, 366, 367, 487
— европейская 366, 367
— сибирская 366, 367
Кот барханный 232
— камышевый 244, 245
Котик морской 269, 271, 272
--- австралийский 266
---галапагосский 267, 268
---гуадалупский 266, 267, 268
---кергеленский 266, 267, 268
---новозеландский 266, 267, 268
---северный 255, 258, 266, 269—272
--- субтропический 267, 268
---фернандесскпй.266, 267, 268
---южноамериканский 266—268
---южноафриканский 267, 268
— — южный 258, 266—268
Кошка 82, 84, 123, 231, 232, 233, 240, 251
— американская 244, 246
— амурская 244, 245
— аргентинская 246
-— барханная 244, 248
— большая 233, 240, 243
— бенгальская 232, 244, 245
— домашняя 245
— дикая 232, 240, 243—245, 247
— — лесная 244, 245
— — степная 244, 245
— Жоффруа 246
— золотая 232, 244, 245
— ириомотейская 245
— калимантанская 245
• — камышевая 244, 246
- — китайская 247
— ливийская 232
— мраморная 232, 245
— пампасская 232, 244, 246
- — пятннсторыжая 245
— суматранская 232, 240, 244, 245
— Темминка 241, 244, 245
— черноногая 244, 248
Кошка чилийская 246
Кошка-рыболов 232, 240, 244, 245
Кошачьи 84, 191
Крабоед 278, 299
Креодонт 84, 85
Кролик 91. 100, 105, 140, 141, 178, 244
Куду большой 416—418
— малый 418
Кузиманзе 192, 219, 220
— ангольский 220
— длинноносый 220
— заирский 220
Кулан 497—499
Куница 84, 123, 137, 138, 141, 256
— американская 138, 140, 141, 143,
145
— каменная 138, 140, 141, 143
— лесная 138, 140—143, 145
Куньи 84, 136, 191
Купрен 481
Лама 487, 492, 493
Ламантин 32о, 328, 330, 332
— африканский 328
— бразильский 328
Ламантнновые 326, 327
Лань 386, 387, 388, 389
— иранская 388, 389
Ласка 82, 84, 136, 137, 138, 149—152, 156, 164
— американская 156
— африканская 137, 164
— белополосая 154
— голостопая 154
— длиннохвостая 150, 151, 154
— патагонская 137, 161, 162
— североафриканская 137, 163. 164
Ластоногие 41. 84, 252—257, 261, 269, 276, 277, 287, 299, 300, 303
Лев 84, 232—234, 236, 237, 238, 394, 417, 429
— атласский 83
— морской 257
---австралийский 258, 259, 261
---калифорнийский 253, 255, 258, 261—263, 298
---новозеландский 258, 260, 261
---южный 258—260
— персидский 83
Лемминг 97, 381
Лемур 223
Леопард 232—234, 237, 238, 364, 394, 414, 417, 429, 447
— дымчатый 231, 232, 240, 242, 243, 249
— морской 255, 260, 278, 300, 301
— предкавказский 83
Летяга 140
Линзанг африканский 191, 192, 193
— полосатый 199
— пятнистый 192, 198, 199
Лисица 85, 87, 89, 91, 95, 97, 100, 101, 102, 104, 105, 112, 123, 244, 284
— андская 105, 106
— афганская 98, 101
— африканская 102
— африкано-переднеазиатская 102
— бенгальская 100, 101
Лисица большеухая 86, 87, 112
— бразильская 105, 106
— европейская красная 83
— обыкновенная 95, 98, 99
— островная 105
— парагвайская 105
— перуанская 106
— песчаная 100, 102
— серая 87, 89, 104, 105
— тибетская 102
— уругвайская 105
— чилоэсская 105
— южноамериканская 87, 105
— южноафриканская 102 •
Личи 425
— нильский 424. 425
Лось 147, 236, 244, 361, 374, 375, 376, 377, 378
— среднерусский 376
— уссурийский 376
Лошадиные 495
Лошадь 494, 495, 498, 499
— домашняя 495
— Пржевальского 497—499
Мазама 358. 371, 373, 374
— большая 372, 373
— карликовая 374
— малая 374
— серая 373
Майкоиг 106
Мамонт 314
Мангуст 192, 215, 216. 218. 222, 223
• — белохвостый 192, 220—222
— бурый 216
— водяной 192. 218
— воротиичковый 216
— Джексона 222
— длинноносый 217
— Дыбовского 218
— египетский 214, 216
— желтый 192. 222, 223
— индийский 83
— карликовый 192, 217, 218
— короткохвостый 216
— либерийский 192, 220
— малый 215, 216
— обыкновенный 216
— полосатошейиый 216
— полосатый 192, 219
— пушистохвостый 222
- — серый 192, 223
— Смита 216
• — стройный 216, 217
— черноногий 192, 221, 222
— Хозе 216
•— южноафриканский 216, 217
— яванский 216
Мангуст-крабоед 214, 216
Манул 231, 232, 244, 245, 248
Медведь 113, 115—117, 118, 123, 145, 236, 256, 361, 381, 386
— белогрудый 115, 117—119
— белый 114—120, 255, 275, 279, 284, 287, 296, 305
— бурый 82, 114, 115, 117—120
-— малайский 113, 115, 116, 122, 123
— очковый 116. 120
-—черный 113—115, 117—119
Медведь-барибал 115
Медведь-губач 115, 116, 121
Медвежьи 84, 113, 124
Медоед 138, 165, 166
Меринос горный 476
Миацоид 85
Мозоленогпе 486, 487
Морж 252—255, 257, 273, 274, 275, 276, 284
Моржовые 255, 256, 272
Мунго большой 209
— бурохвостый 192, 212
— кольцехвостый 192, 210
— линейчатый 212
— мелкозубый 192, 209, 210
— одноцветный 212
— полосатый 211
— узкополосый 192, 211, 212
• — широкополосып 192, 211
Мунтжак 363—365
Мусанг 192, 202, 203
— обыкновенный 202, 203
— шрнланкскнй 203
— южнонндпйскпй 203
Муфлон 474
Мышь 151, 178
Нарвал 5, 10, 14, 16, 50, 53, 275
Нарваловые 50
Нежвачные 334, 335
Непарнокопытные 494, 495
Нерпа 278, 281. 284, 285
—• кольчатая 252, 277, 278, 281—285
Ннльгау 236, 398, 412—414
Норка 150, 151
— американская 83, 137, 150, 152, 155—157
— европейская 154—156
Носорог 494, 503
— белый 503, 505, 507
— индийский 503, 504, 505
— суматранский 503, 504
— черный 503, 505, 506
— яванский 504
Носороговые 495, 502
Носуха 123, 124, 128, 129
— горная 124, 129
— обыкновенная 129
— Нельсона 129
Нутрия 244
Ньяла 416—418
— горная 418
Овцебык 398, 449, 462, 463
Окапи 389, 390, 391
Оленевые 334, 354, 357
Оленек 356
— азиатский 355, 356
— большой 356, 357
— водяной 355
— малый 356, 357
— пятнистый 356, 357
Олень 91. 97, 236, 354, 357, 362. 363, 369, 370, 373, 378, 380, 386, 388, 396
— американский 358, 369
— беломордып 388, 389
— белохвостый 368—371
— благородный 384—386, 388, 389
Олень болотный 370, 371
—- водяной 354, 361, 362
— Давида 358, 381—383
— пампасный 371
— перуанский 371
— пятнистый 236, 244, 384, 386 388 389
— свиной 388, 389
— северный 97, 147, 244, 357, 358, 365 378, 379, 380, 381, 422
— хохлатый 358, 365
— чернохвостый 369—371
— южноандскнй 371
Олень-лнра 388, 389
Оленьковые 354
Олинго 124, 131
— Аллена 131
— Беддарда 131
— пушистохвостый 131, 132
— чирикунскин 131
Ондатра 101, 140, 244
Онцилла 246
Орибн 398, 409, 410
Орнкс 398, 419, 420, 421
Оронго 398, 449, 452
Осел 495
- — африканский 495
— - дикий 499
Оцелот 232, 240, 244, 246, 249
Панда большая 123, 124, 134, 135
— малая 123, 124, 132, 133
Парнокопытные 333, 334, 487
Пекари 333, 348
— белогубый 348
— ошейннковый 347, 348
Пекариевые 334, 335, 346
Перевязка 137, 157, 158, 159
Песец 85, 86, 95, 96, 100
- — тундровый 97
Песчанка 151
Пингвин 259, 260, 261
Пищуха 140, 151, 317
Плавун 59, 60, 63
- — северный 6, 13, 14, 60, 61
— южный 60, 61
Полевка 141, 151, 169, 178, 244
— водяная 151, 244
— лесная 100, 140
— рыжая 140
— серая 100, 151
Полорогие 334, 354, 396, 397
Полосатик 67, 76
— Бранда 76—79
— малый 6, 10, 76—79
— настоящий 76—81
Полосатнковые 68, 73, 75
Проделифпн 15, 22, 24, 26
— большеголовый 27
— большелобый 26, 27
— длиниорылый 25, 27
— малайский 26, 27
— полосатый 24, 25, 27
— пятнистый 24, 27
Пуду 358, 365, 374
— северный 375, 376
— южный 373, 376
Пума 232, 240, 243, 244, '249, 373
Редуика большая 425, 426
— горная 425, 426, 427
— обыкновенная 425, 426
Ремнезуб 10, 59, 61
— австралийский 62
— антильский 62
— атлантический 62, 63
— Гектора 62
— Грея 62
— командорский 62, 63
— Ленарда 63
— новозеландский 63
— Тру 62
— тупорылый 63
— Хабса 63
— японский 63
Росомаха 101, 137, 145—147, 149, 361, 367, 381
Рысь 101, 126, 231, 232, 233, 236, 240, 241, 244, 245, 247, 249, 361, 367, 386, 459, 468
— красная 244, 248
Сайгак 91, 236, 251, 398, 449—452
Свиные 334, 335, 336
Свинья 179, 309, 314, 336, 340, 346
— бородатая 338, 340, 341
— домашняя 336, 341
— карликовая 338, 340, 341
— кистеухая 337, 338
— лесная 337, 342, 344
— морская 9, 15, 22, 45, 46
— — белокрылая 22, 47, 48
----бесперая 14, 22, 47, 49, 50
•— — восточноазиатская 50
---калифорнийская 47
----обыкновенная 12, 14, 46, 47
— — очковая 47
---- черная 47
---- яванская 341
— — японская 49, 50
Сейвал 6, 10, 76—79
Сервал 232, 242, 244, 247
Серна 398, 458, 459, 460, 461
Cepov 398, 455
Сивуч 258, 263, 264, 265
Сирена 256, 324, 326
Ситатунга 415, 417
Скунс 175, 179, 216
— аргентинский 180
— восточномексиканскнй 179
— Гумбольдта 180
— королевский 180
— малый 179
— мексиканский 176
— полосатый 138, 174, 175, 176, 179
— пятнистый 138, 177, 178
— свиноносый 138, 179, 180
— южноамериканский 180
Слон 312, 314, 320, 429
— азиатский 312, 314, 315
— африканский 315, 316
— индийский 313, 315
— морской 256, 276, 277, 278, 306, 308
----северный 277, 306—308
----южный 252, 306, 307, 308
Слоновые 314
Собака 32. 82, 88, 89, 97, 256, 417
— гиеновая 85—87, 109, ПО, 417, 429
— домашняя 94
— енотовидная 85, 87, 90, 123
— кустарниковая 85—87, ПО, 111
Собачка луговая 151, 172
Соболь 83, 137—142, 145
Солонгой 150, 151, 156, 157
— индийский 155
Спрингбок 439
Стенбок 398, 408
— обыкновенный 408
— Шарпа 408, 409
Сунн 403
Суриката 192, 213
Сурок 91, 95, 100, 240
— бесхвостый 317
Суслик 100, 151, 159, 172, 240, 244
— тонкопалый 244
Тайра 137, 159, 160
Такнн 398, 449, 459, 461, 462
Танга лунга 197, 198
Тапир 494, 502
— азиатский 499
— горный 502
— равнинный 500, 501, 502
— чепрачный 500, 502
— центральноамериканский 502
Таппровые 495, 499
Тар 398, 464, 465
Тарпан 495, 498
— лесной 499
— степной 499
Теледу 170
Тигр 118, 232, 233, 234, 235, 236, 238, 341, 361, 364, 386, 414
— уссурийский 118, 236
Топи 432
Трубкозуб 231, 310, 311, 342
Трубкозубовые 311
Трубкозубые 309, 310
Тур 481
— дагестанский 469
— кубанский 469
Тушканчик 159, 244
— большой песчаный 244
Тюленевые 252, 254, 274
Тюлень II8, 265, 275, 278, 285, 287, 289, 290, 293, 298, 303
— байкальский 281—285
— длппномордый 278, 291—293
— гренландский 255, 278, 287—291
— каспийский 281—285
— настоящий 254, 277
— обыкновенный 253, 255, 277, 278— 281
— островной 280
•— полосатый 278, 285—287
— Росса 277, 299, 300
— серый 255, 276, 278
— ушастый 252—254, 256—258, 272, 276, 277
— Уэдделла 255, 276—278, 301, 302
Тюлсиь-монах 253, 255, 276, 278, 296, 297
•— гавайский 297, 298
— карибский 298
Умби 192, 222
Урнал 474
Фаналока 192, 205
Фенек 85—87, 102—103
Финвал 6, 9, 10, 76—79
Фосса 192, 223, 224
Харза 138, 140, 141, [44, 361
— южнопндийская 145
Хищные 82—85, 191
Хоботные 326
Хомяк 100, 151, 159
— обыкновенный 151
Хорек 137, 149, 151, 172
— американский 149, 151, 155—157
— африканский 137, 151, 162, 163
— лесной 150—152, 155—157
— степной 149—152, 156, 157
Хохлач 278, 303—306
Цивета 192, 196
— африканская 197, 198
Цивета большая 198
— водяная 191, 196
— выдровая 192, 208, 209
— гималайская 191, 192, 203
— калимаитаиская 192, 208
— крупнопятнпстая 198
— малая 191, 196
— мелкозубая 192, 201, 202
— Оустоиа 192, 207
—• пальмовая 192, 199, 200
— полосатая 192, 206
— сулавесская 192, 204
Шакал 90, 91, 94, 95
— полосатый 94
— чепрачный 92—94
— эфиопский 94
Шиншилла 161
Ягуар 232, 233, 235, 238, 373
Ягуаруиди 232. 241, 244, 246
Як 480, 481
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Acinonychinae 233 Acinonyx 232, 233, 249 — jubatus 249, 250 Addax 398, 422 — nasomaculatus 422 Adenota 425 Aepiceros 437 Aepyceros 398, 410, 428 — melampus 428, 429 -----petersi 429 Ailurus 124, 132 — fulgens 132, 133 Alcelaphus 398, 433 — buselaphus 433 ----- tora 434 -----swaynei 434 — lichtensteini 434 Alces 358, 376 — alces 374, 376 •----alces 376
-----cameloides 376
Alopex 86, 87. 95 — lagopus 95, 96 Aluropoda 124, 134 — melanoleuca 134 Amblonyx 186 Amblypoda 334 Ammodorcas 398, 446 — clarkei 446
Ammotragus 398, 471 — lervia 471, 472 Andrewsarchus 84 Anoa 479
Anoplura 256
Antidorcas 398, 439
— marsupia lis 439 Antilocapra 396 •— americana 395, 396 -----peninsularis 396 ----sonoriensis 396
Antilocapridae 354, 394 Antilope 398, 437 — cervicapra 437, 438 Antilopinae 398, 403, 410, 437
Aonyx 138, 185, 186
— capensis 186
— cinerea 186 Archaeoceti 11 Archidiskodon 314 Arctictis 191, 192, 205 — binturong 204, 205 Arctocephalus 258, 266 — australis 267, 268 — doriferus 267, 268 — forsteri 266—268 — galapagoensis 267, 268 •— gazella 267 — philippii 267, 268 — pusillus 267, 268 — tasmanicus 267, 268 — townsendi 267, 268 — tropicalis 267, 268 Arctocyonidae 84
Arctogalidia 201
— trivirgata 201
Arctonyx 138, 170
— collaris 170
Arctophoca 268
Artiodactyla 4, 333
Atelocynus 86, 106
Atilax 191, 192, 218
— paludinosus 218
Axis 358
Babyrousa 337, 344
— babyrussa 344, 345
Balaena 68, 69
— mysticetus 69
Balaenidae 68
Balaenoptera 76
— acutorostrata 78
— bonaerensis 79
— borealis 79
— brevicauda 79
— edeni 78
— musculus 79
— physalus 79
Balaenopteridae 68, 75
Baselaphus 410
Bassaricyon 124, 131
— alleni 131
— beddardi 131
— gabbii 131, 132
— lasius 131
— pauli 131
Bassariscus 124
— astutus 125, 126
— sumichrasti 126
Bdeogale 192, 221
— crassicauda 222
— jacksoni 222
— nigripes 221, 222
Beatragus 433
Berardius 59, 60
— arnouxi 61
— bairdi 60, 61
Bibos 481
Bison 398, 484
— bison 482, 485
—• bona sus 482, 485
Blastoceros 371
Blastocerus 358, 371
Boocercus 419
Bos 398, 479, 481
— gaurus 480, 481
— — frontalis 480, 481
— javanicus 481
— — domesticus 481
— primigenius 481
—- — taunts 481
— mutus 480, 481
— -— grunniens 481
— sauveli 481
Boselaphus 398, 412, 477
— tragocamelus 412, 413
Bovidae 354, 358, 396
Bovinae 398, 477
Bubalus 398, 477, 479
— arnee 479
— — mindorensis 479
— caffer 478, 479
Bubalus depressicornfs
478, 479
----depressicornis 479
----tjuarlesi 479
— mindorensis 479
— nanus 479
Budorcatinae 449
Budorcas 398, 459
— taxicolor 459, 462
---- bedfordi 462
----tibetana 461
Callorhinus 258, 269
— ursinus 269, 270
Camelidae 487
Camelus 487
— bactrianus 488, 492
— dromedarius 492
Canidae 84, 85, 87
Caninae 87
Canis 86, 87, 90
— adustus 94
— aureus 94
— familiaris 94
— latrans 93, 94
— lupus 93—95
irremolus 95
---- rufus 95
— mesomelas 93, 94
— niger 95
• — simensis 94
Canoidea 84
Caperea 68, 72
— marginata 72
Capra 398, 465, 471
— aegagrus 467, 469
- — caucasica 469
— cylindricornis 469
— falconeri 467, 469
----jerdoni 469
----megaceros 469
— hircus 469
— ibex 468, 469
— nubiana 469
— — walie 469
— pyrenaica 468, 469
— — pyrenaica 469
— sibirica 466, 469
Capricornis 398, 455
— sumatraensis 455
----sumatraensis 456
Caprinae 398, 448
Capreolus 358, 366
— capreolus 366
----capreolus 366
---- pygargus 366
Caracal 232
Carnivora 4. 82, 255
Cephalophinae 397, 398
Cephalophus 397, 399, 401
— adersi 401
— callipygus 401
— dorsalis 401
— harveyi 401
— jeutinki 401
— leuccgaster 401
Cephalophus maxwelli 400
— monticola 400, 401
— natalensis 401
— niger 401
- — nigrifrons 400
— ogiibyi 401
— rufilatus 400
.— spadix 401
— sylvicultor 400, 401
— zebra 400, 401
Cephalorynchus 37
— albifrons 38
— commersoni 37
— eutropia 37
. — heavisidei 37
— hectori 38
Ceratotherium 503. 505
— siinum 505, 507
----eottoni 507
Cerdocyon 86, 106
Cervidae 354, 357
Cervinae 358, 381
Cervulinae 358, 363
Cervus 358, 386
— albirostris 388
.— axis 387, 388
— canadensis 389
— dama 387, 388
— duvauceli 388
— eldi 388
----eldi 389
— — siamensis 389
— elaphus 384, 388
----bactrianus 389
---- barbarus 389
----corsicanus 389
— — hangul 389
----inacneilli 389
— — wallichi 389
----yarkandensis 389
— mariannus 388
— kuhli 389
— mesopotamicus 388
— nannodes 389
— nippon 384, 387
— — grassianus 389
----к eram a e 389
— — kopschi 389
— — mandarinus 38Э
---taiouanus 389
— porcinus 388
— schomburgki 389
— timorensis 388
— unicolor 388
Cetacea 4
Chiroptera 4
Choeropsis 350, 351
— liberiensis 351
Chrotogale 192, 207
— owstoni 207
Chrysocyon 86, 87. 106
— brachyurus 106, 107
Civettictis 198
Condylarthra 334
Conepatus 138, 179
— castaneus 179
— chinga 180
— humboldtii 180
— Icuconotus 179
Conepatus mesoleucus 179, 180
— rex 180
— semistriatus 179
Connochaetes 398, 434
— gnou 436
— taurinus 434. 436
Creodonta 84, 256, 334
Crocuta 225
— crocuta 225, 226
Crossarchus 192, 219
— alexandri 220
— ansorgei 220
— obscurus 220
Cryptoprocta 192, 223
— ferox 223, 224
Cryptoproctinae 192
Clion 86, 87. 107
— alpinus 107, 108
Cynictis 191, 192, 222
— penicillata 222, 223
Cynogale 191, 192, 208
— bennetti 208, 209
Cystophora 278, 303
— cristata 303, 304
Dama 358
Damaliscus 398, 432
— dorcas 433
-----dorcas 433
— hunteri 433
— lunatus 432
Dasicyon 86, 87
Dasycyon 86
Delphinapterus 50
— leucas 15, 50. 51
Delphinidae 15, 16, 21
Delphinus 22, 27
— bairdii 30
— capensis 30
— delphis 7, 13, 27, 28, 30
Dendrailuriis 232
Dendroxyrax 320, 323 — arboreus 323 — dorsalis 322, 323 — validus 323
Dermoptera 4
Diceros 503, 505
— bicornis 505
Didermocerus 503
— sumatrensis 503
Diplogale 192, 208
— hosei 208
Dologale 218
Dorcatragus 398, 407 — megalotis 407 Dorudon 11
Dugong 330 — dugon 329, 330
Dugongidae 326, 328, 332
Dusicyon 86, 87, 105
— culpaeolus 105
— culpaeus 105, 106
— fulvipes 105
• — griseus 105
— gymnocercus 105
—• inca 106
— microtis 106
— sechurae 106
Dusicyon thous 106
— vetulus 106
Edentata 4
Eira 137, 159
— barbata 159, 160
Elaphodus 357, 358, 365
— cephalophus 365
Elaphurus 358, 381
— davivianus 381, 382
Elephantidae 314
Efephantoidea 314
Elephas 314, 315
— maj;imus 315
Enhydra 138, 187
— lutris 187, 188
Eohippus 495
Equus 498
— asinus 498
— burchelli 497, 498
— caballus 498
----gmelini 498
----przewalskii 497, 499
----silvaticus 499
— grevyi 498
— hemionus 497, 498
------- hemionus 499
•---hemippus 499
---- khur 499
----kiang 499
----onager 499
— quagga 498
- r- zebra 498
Equidae 495
Eremaelurus 232
Erignathus 278, 293
— barbatus 293, 294
Eschrichtiidae 68, 73
Eschrichtius 75
— gibbosus 73, 75
Eubalaena 68, 70
— australis 70
— glacialis 70, 71
— sieboldi 70, 71
Eumetopias 258, 263
— jubatus 263, 264
Eupleres 192, 209
— goudoti 209, 210
— major 209
Felidae 84, 231
Felinae 233
Felis 232, 233, 240
— aurata 245
• — badia 245
— bengalensis 245, 249
— bfeti 247
— caracal 242, 248
----michaelis 249
— chairs 246
— colocolo 246
— concolor 242, 249
---- coryi 249
— — cougar 249
— euptilura 245, 249
— geoffroyi 246
— guigna 246
— iriomotensis 245
— iagouarundi 246
— libyca 247, 249
Felis lynx 241, 247, 249
----pardina 249
— manul 248
— margarita 248
— marmorata 245
— nebulosa 242, 249
— — brachyurus 249
— nigripes 248
— pajeros 246
— planiceps 245
— pardalis 232, 246, 249
albescens 249
— pardinoides 246
— rubiginosa 245
— ruins 248
— serval 242, 247
— Silvestris 241, 247, 249
------- libyca 232, 245
•— temmincki 241, 245
— tigrina 246
— viverrina 241, 245
— wiedi 232, 246
— yagouarundi 241
Feloidea 84, 256
Fennecus 86, 87, 102
— zerda 102, 103
Feresa 44
— attenuata 44, 45
Fissipeda 84
Fossa 192, 205
— fossa 205
Gal era 159
Galictis 137, 161
— cuja 161
— vittatus 160, 161
Galidia 192, 210
— elegans 210
Galidictis 192, 211
— fasciata 211
— striata 211
Galidiinae 192, 210
Gazella 398, 442, 443
— dama 446
----lozanoi 446
— — mhorr 446
— dorcas 446
----massacsyla 446
---- saudiya 446
— gazella 446
•---arabica 446
----cuvieri 446
— granti 444
— leptoceros 446
•---marica 446
— pelzelni 446
— rtififrons 446
— rufina 446
— soemmerringi 444
— spekei 446
— subgutturosa 444, 446
• — — marica 446
— tbomsoni 446
• — tilonura 446
Gazellinae 398, 436
Genetta 191, 194
— abyssinica 195
— angolensis 195
— genetta 195
Genetta johnstoni 195 — mossambica 195 — pardina 195 — rubiginosa 195 — servalina 195 — tigrina 194, 195 — victoriae 195 — villiersi 194 Giraffa 390, 392 — Camelopardalis 392 Giraffidae 354, 389 Globicephala 43 — macrorhyncha 44 — melaena 44 — scammoni 43, 44 Globicephalidae 16, 40, 44, 45
Gorgon 436
Grammogale 137, 157
— africana 157 Grampidae 16 Grampus 42 — griseus 42 Grison 161 — cuja 161 — vittatus 160, 161 Grisonella 161 Gulo 137, 145 — gulo 145, 146
Halarchne 257
Halichoerus 254, 276, 278, 291
— grypus 291, 292 Helarctos 116, 123 — malayanus 122, 123 Helictis 174 Helogale 192, 217 — dybowskii 218 — parvula 217, 218 Hemigalinae 192 Hemigalus 192, 206 —• derbyanus 206 Hemitragus 398, 464 — jemlachicus 464, 465 — — hylocrius 465 -----jayakari 465 Herpaihirus 232 Herpestes 191, 192, 215 . — auropunctatus 215, 216 •— brachyurus 216 — edwardsi 216 — fuscus 216 — hosei 216 — ichneumon 214, 216 — javanicus 216 — naso 217
•— pulverulentus 217 — sanguineus 217 — semitorquatus 216 — smithi 216 .— urva 214, 216 — vitticollis 216 Herpestinae 192, 213 Heteroxyrax 320, 321 — antinae 321 — brucei 322 — chapini 322 Hippocamelus 358, 371
Hippopotamidae 335, 348
Hippopotamus 350
— amphibius 349, 350
Hippotraginae 410
Hippotragus 398, 429
— equinus 432
•— niger 430, 432
Histriophoca 276, 277,278, 281, 285
— fasciata 285, 286
Hyaena 225, 227
— brunnea 228
•— hyaena 228
---barbara 229
Hyaenidae 88, 224
Hyaeninae 225
Hydrodamalidae 326, 330
Hydrodamalinae 332
Hydrodamalis 332
— gigas 331, 332
Hydropotes 357, 358, 361
— inermis 361, 362
Hydropotinae 358, 361
Hydrurga 252, 254, 276,
' 278, 300
— leptonyx 300, 301
Hyemoschus 353, 354, 355
— aquaticus 355
Hylochoerus 337, 342
— meinertzhageni 342, 344
Hyperoodon 59, 65
•— ampullatus 65, 66
— planifrons 65
Hyracoidea 4
Hyracotherium 495
Ichneumia 192, 220
— albicauda 220, 221
Ictailurus 232
Ictonyj; 137, 163
— striatus 162, 163
Indopapacetus 66
Inia 16, 18
— geoffrensis 5, 18
Insectivora 4
Jaguarius 232
Jentinkia 126
Kobus 398, 423
— defassa 425
• — ellipsiprymnus 424, 425
— kob 425
— leche 424, 425
— megaceros 425
— vardoni 425
Kogia 55
— breviceps 56
• — simus 56
Kogiidae 15
Lagenodelphis 34
— hosei 34
Lagenorhynchus 34, 35
— acutus 35
- — albirostris 35
• — australis 35, 36
— cruciger 35
Lagenorhynchus electra 35
— fitzroyi 35
— obliquidens 35, 36
— obscurus 35, 36
— superciliosus 35
— thicolea 35
Lagomorpha 4
Lama 487, 492
— guanicoe 490, 492
---glama 492
--- pacos 492
------- vicugna 492
Leo 232
Leopardus 232
Leptailurus 232
Leptonychotes 252, 278, 301
— weddelli 301, 302
Liberiictis 192, 220
— kuhni 220
Lipotes 16, 19
— vexillifer 19
Lissodelphis 32 — borealis 33, 34 — peroni 33
Litocranius 398, 448
— walleri 447, 448
Lobodon 252, 278, 299
— carcinophagus 299, 301
Loxodonta 314, 315
— africana 315, 316
Lutra 138, 180
— annectens 184
— canadensis 183, 184
— enudris 184
— felina 184
— incarum 184
— lutra 181, 183
— maculicollis 184
— mira 184
— perspicillata 184
— platensis 184
— provocax 184
— sumatrana 184
Lutrinae 138
Lutrogale 184
Lycaon 86, 87, 109
— pictus 109, 110
Lyncinae 233
Lyncodon 137, 161
— patagonicus 161, 162
— Lynx 232
Macrogalidia 192, 204
— musschenbroeki 204
Madoqua 398, 404
— guentheri 404, 405
— kirki 404, 405
— phillipsi 404
— salticana 405
— swaynei 405
Marsupialia 4
Martes 137, 138
— americana 143
— flavigula 144, 145
— foina 143
- — gwatkinsi 145
— martes 143
Martes melampns 145
— pennanti 144
— zibellina 139, 142, 145
Mastodon 314
Mastodontoidae 314
Mazama 357, 358, 371
— americana 372, 373
— bricenii 374
— gouazoubira 373
— папа 374
— rufina 374
Megaptera 76, 79
• — nodosa 79
— novaeangliae 79, 80
Meles 138, 167
— meles 167, 168
Melinae 138
Mellivora 138, 165
— capensis 165, 166
Mellivorinae 138, 165
Alelogale 138, 172, 174
— moschata 174
— orientalis 173, 174
— personata 174
Melursus 116, 121
— nrsinus 121
Mephitinae 138, 174
Mephitis 138, 174
— macronra 176
— mephitis 175, 176
Mesonychidae 84
Mesonyx 84
Alesoplodon 59, 61
— bidens 62
— bowdoini 63
— carlhnbbsi 63
— densirostris 63
— europaens 62
— ginkgodens 63
— grayi 62
— hectori 62
— layardi 63
— minis 62, 66
— pacificus 62
— planifrons 66
— stejnegeri 62, 63
Miacoidea 85
Mironnga 278, 306
— angnstirostris 308
— leonina 252, 307, 308
Moeritheriidae 314
Moeritherinm 314
Monachns 278, 296
• — monachns 296, 298
— schauinslandi 297, 298
— tropicalis 298
Monodon 50, 53
— monoceros 53
Monodontidae 16, 50
Monotremata 4
Moschinae 358, 359
Moschus 357, 358, 359
— berezovskii 361
— chrysogaster 361
— moschiferus 359—361
— sibiricus 361
Mungos 192, 219
— gambianus 219
— mungo 219
Mnngotictis 192, 211
— lineata 212
— substriata 212
Munida 259
Muntiacus 358, 363
— crinifrons 365
— feae 365
— mnntjak 363—365
---crinifrons 365
---feae 365
— rooseveltorum 365
— reevesi 365
Mnstela 137, 149
— altaica 153
— erminea 152
— eversmanni 156
— frenata 154
— itatzi 156
— kathiah 153
— lutreola 154
— nigripes 149, 155, 156
— nivalis 149, 152, 156
— nudipes 154
— pntorins 155
— rixosa 156
— sibirica 153, 156
— strigidorsa 154
— vison 155
Mnstelidae 84, 136
Mnstelinae 137. 138
Myaeodontidae 85
Mysticeti 4, 12, 66
Nandinia 192, 199
— binotata 199, 200
Nasua 124, 129
— narica 128, 129
— nasua 128, 129
— nelsoni 129
Nasnella 124, 129
— olivacea 129
Nemorhaedns 398, 454
— goral 454
Neofelis 232
Neophoca 258, 261
— cinerea 259, 261
Neophocaena 49
— asiaeorientalis 50
— phocaenoides 50
— sunameri 49, 50
Neotraginae 397, 402
Neotragus 403
— batesi 403
— moschatus 403
---moschatus 403
— pygmaeus 396, 403
Notoungulata 334
Nyctereutes 86, 87
— procyonoides 87, 88
Odobenidae 255, 272
Odobenus 276
— rosmarus 273, 276
---laptevi 276
Odocoileinae 358, 365
Odocoileus 358, 369, 371
— antisensis 369—371
— bezoarticus 369, 371
— bisulcus 369—371
— dichotomus 369, 370
Odocoileus hemionus 369, 370
•------ cerrosensis 371
— virginianus 369—371 -----calvium 371
-----leticiirus 371
Odontoceti 4, 12
Okapia 390
— johnstoni 390, 391 Ommatophoca 276, 278,
299
— rossi 299
Onotragus 425 Orcaelidae 16 Orcaella 38 — brevirostris 38 Orcinus 40 — orca 40
Orcamnos 398, 458
— americanus 456, 458 Oreotragus 398, 405 — oreotragus 405, 406 Orthohalarchne 257 Orycteropodidae 311 Orycteropus 311 — afer 310, 311
Oryx 398, 419 — dammah 421 — gazella 419—421 -----dammah 421 ----- leucoryx 421 -----tao 421 — leucoryx 421 Osbornictis 191, 196 — piscivora 196 Otaria 258 — byronia 258, 259 — flavescens 258 Otariidae 252, 255, 257 Otarioidea 256 Otocolobus 232 Otocyon 86, 87, 112 — megalotis 112 Otocyoninae 87, 112 Ourebia 398, 409 — ourebi 409 Ovibos 398, 462 •— moschatus 462, 463 Ovibovinae 449 Chis 398, 471, 476 ' — ammon 474, 476 — canadensis 476 — dalli 476 — laristanica 476 — musimon 476 — nivicola 476 — orientalis 476 — vignei 476 Oxyaenidae 84 Ozotoceras 358, 371
Pagophilus 276, 277, 278, 281, 287
— groenlandica 287, 288 Paguma 192, 203 — larvata 203 Palaeomastodon 314
Panthera 232, 233, 249 — leo 234, 237 •-----persica 238
— onca 235, 238 — pardus 234, 237, 238 ------ jarvisi 238
. nimr 238 — — orientalis 238 ------- panthera 238 --------tulliana 238 — tigris 234, 237, 238 ---------------- altaica 238 ---------------- amoyensis 238 — -z-~ balica 238 --------sondaica 238 ---------------- sumatrae 238 --------tigris 238 --------tulliana 238 —. — virgata 238 Pantherinae 233 Pantholopinae 449 Pantholops 398, 452 — hodgsoni 452 Paracynictis 192, 223 — selousi 223 Paradoxurinae 192 Paradoxurus 192, 202 — hermaphroditus 202 — jerdoni 203 •— zeylonensis 203 Paraonyx 138, 186, 187 — congica 187 --------microdon 187 — microdon 187 — phelippsi 187 Parditis 199 Pardofelis 232 Pardus 232 Pelea 398, 427 — capreolus 427 Peponocephala 35, 36 — electra 36 Perissodactyla 4, 494 Phacochoerus 337, 341 — aethiopicus 341, 342 Phoca 254, 276, 278, 287 — kurilensis 279—281 — largha 281 — vitulina 279—281 Phocarctos 258, 260 — hookeri 260 Phocidae 252, 255 Phocoena 15, 45 — dioptric a 47 -— phocoena 15, 46, 47 -— sinus 47 — spinipinnus 47 Phocoenidae 16 Phocoenoides 47 — dalli 47, 48 Phocoidea 256 Pholidota 4
Physeter 55, 56 — catodon 56, 57 Physeteridae 15. 16, 54 Pinnipedia 4, 84, 252 Platanista 16, 19 — gangetica 4, 19, 20 — indi 21
Platanistidae 15, 16
Platyceros 389
Poecilictis 137, 163
— lybica 163, 164
Poecilogale 138, 164
— albinucha 164
Poephagus 481
Poebrodon 487
Poiana 191, 192
— leightoni 193
— richardsoni 192, 193
Pontoporia 16, 17
— blainvillei 17
Potamochoerus 337
— porcus 337, 338
Potos 124, 130
— flavus 130, 205
Primates 4
Prionailurus 232
Prionodon 192, 198
— linsang 199
— pardicolor 199
Proboscidea 4, 312
. Procapra 398, 441, 442
— gutturosa 440, 442
— picticaudata 442
Procavia 320
— capensis 318, 321
— johnstoni 321
— ruficeps 321
— syriacus 321
— welwitschii 321
Procaviidae 320
Procyon 124, 126
— cancrivorus 127
— gloveralleni 127
— insularis 127
— lotor 127
— maynardi 127
— minor 127
— pygmaeus 127
Procyonidae 84, 87, 123
Profelis 232
Prohyracodon 495
Proteles 225, 230 — cristatus 230
Protelidae 225
Protelinae 225, 230
Protocetus 11
Prozeuglogon 11
Pseudois 398, 469
— nayaur 469, 470
Pseudorca 38
— crassidens 38, 39
Pteronura 138, 184
— brasiliensis 184. 185
Pudu 357, 358, 374
— mephistophiles 376
— pudu 373, 376
Puma 232
Pusa 276, 278, 281, 284
— caspica 282, 284
— hispida 252, 284
— — saimensis 285
— sibirica 284
Rangifer 358, 378
— tarandus 378, 380
— — pearsoni 381
Raphicerus 398, 408 — campestris 408 __ melanotis 408, 409 .— sharpei 408, 409 Redunca 398, 425 — arundinum 426 __ fulvorufula 426, 427 — redunca 426 Rhinoceros 503, 504 — sondaicus 504 — unicornis 504 Rhinocerotidae 495, 502 Rhynchogale 192, 222 — melleri 222 Rhynchotragus 405 Rodentia 4 Ruminantia 334, 353 Rupicapra 398, 458 — rupicapra 458, 460
Saiga 389, 449 — tatarica 449, 450 -----mongolica 452 Saiginae 449 Salanoia 192, 212 — olivacea 212 — unicolor 212 Selenarctos 116, 119 Simocyoninae 87, 107 Sirenia 4, 324 Sotalia 22, 23, 24 — borneensis 24 — brasiliensis 24 — chinensis 24 — fluviatilis 23, 24 — guianensis 24 — ientiginosus 24 — plumbea 24 — teuszi 24 Sousa 24 — chinensis 24 — fluviatilis 24 — pallida 24 — teuszii 24 — tucuxi 24
Spcothos 86, 87, 110 — panamensis 111 — venaticus 110, 111 Spiiogale 138, 177 — angustifrons 179 — gracilis 179 — pugmaea 179 — putorius 177, 178 Stegamostodon 314 Stenella 24 — asthenops 25 — attenuate 26 — caeruleoalba 26 — coeruleoalbus 25—27 — ciymene 25 — crotaphiscus 25 — dubia 25—27 — frontalis 25—27 •— gralfmani 25, 26 -— longirostris 25—27 — malayana 25, 26 — rmcrops 26 — pernettyi 25—27
Stenella plagiodon 26
— roseiventris 26
— styx 26
Stenidae 16, 24
Steno 22, 24
— bredanensis 22, 23
Suidae 335, 336
Suiformes 334, 335
Suricata 192, 213
— suricata 213
Sus 337, 338
— barbatus 341
— cristatus 341
— salvanius 341
— scrofa 339, 341
— verricosus 341
— vittatus 341
Sylvicapra 397, 401
— grimmia 401, 402
Syncerus 479
Tangasi 417
Tapiridae 495, 499
Tapirus 502
— bairdi 502
— indicus 500, 502
— pinchaque 502
— terrestris 500, 502
Tasmacetus 59
— shepherdi 59
Taurotragus 419, 477
Taxidea 138, 171
— taxus 171
Tayassei 348
— albirostris 348
— tajacu 347
Tavassuidae 335, 346
Tayra 159
Tetracerus 396, 398, 410,
477
— quadricornis 410, 411
Thalarctos 116, 119
Tigris 232
Tragelaphus 398, 410, 414, 477
— angasi 414, 416—418
— buxtoni 414, 417, 418
— derbianus 419
— euryceros 417, 418
— imberbis 414, 417, 418
— oryx 417—419
----derbianus 419
— scriptus 414, 415, 417
— spekii 414, 415, 417
— strepsiceros 414, 416—
418
Tragulidae 353, 354
Tragulus 354—356
— javanicus 356, 357
• — meminna 356. 357
— napu 357
Tremarctos 116, 120
• — ornatus 120
Trichechidae 326
Trichechus 328
— inunguis 328
— manatus 325, 327, 323
Trichechus selegalensis
328
Tubulidentata 4, 309
Tursiops 30
— abusalam 32
— aduncus 32
— gephyreus 32
— gilli 32
— nuuanu 32
— parvimanus 32 — truncatus 30—32
Tylopoda 4, 486
Uncia 232, 233, 238
— uncia 238, 239
Urocyon 86, 87, 104 — cinereoargenteus 104, 105
— littoralis 105
Ursidae 113
LJrsus 117
— americanus 114, 119
-----emmonsii 119
— arctos 1-14, 118
----nelsoni 119
•----richardsoni 120
• — maritimus 114, 119
• — thibetanus 119
-----gedrosianus 119
Vuverra 192, 196
— civetta 197
—----megaspila 198 ------ civettina 198
- — tangalunga 197, 198
— zibetha 198
Viverricula 191, 196
• — indica 195, 196
Viverridae 88, 191
Viverrinae 191, 192
Vormela 137, 157
— peregusna 157. 158
Vulpes 86, 87, 97
— bengalensis 101
— сапа 101
— canus 98
— chama 102
— corsac 101
— ferrilata 102
— fulva 102
— macrotis 102
• — pallida 102
— ruppeli 102
— velox 102
— — nebes 102
— vulpes 98, 101
Xenogale 217
Zalophus 258. 261
— californianus 261, 262
-----japonicus 263
Zibetaiiurus 232
Ziphiidae 15, 16, 59
Ziphius.59, 63
— cavirostris 63, 64
Zygolophodon 314
СОДЕРЖАНИЕ
От автора..................................................................... 3
Отряд китообразных Ordo Cetacea Brisson, 1762 ................................. 5
Подотряд зубатых китов Subordo Odontoceti Flower, 1867 ................ ... 12
Семейство речных дельфинов Familia Platanistidae Gray, 1863 .............. 16
Род лаплатских дельфинов Genus Pontoporia Gray, 1846 .................... 17
Род амазонских дельфинов Genus Inia d'Orbigny, 1834 ..................... 18
Род озерных дельфинов Genus Lipotes Miller, 1918 ... ............. 19
Род гангских дельфинов Genus Platanista Wagler, 1830 .................... 19
Семейство дельфиновых Familia Delphinidae Gray, 1821...................... 21
Род крупнозубых дельфинов Genus Steno Gray, 1846 ........................ 22
Род длп.иноклювы.х дельфинов Genus Sotalia Gray, 1866 . ................. 23
Род продельфинов Genus Stenella Gray, 1866 .............................. 24
Род дельфинов-белобочек Genus Delphinus Linnaeus, 1758 .... . 27
Род афалин Genus Tursiops Gervais, 1855 ............................ . 30
Род китовидных дельфинов Genus Lissodelphis Gloger, 1841 ... . . 32
Род малайзийских дельфинов Genus Lagenodelphis Fraser, 1956 ............ 34
Род короткоголовых дельфинов Genus Lagenorhynchus Gray, 1846 .... 34
Род бесклювых дельфинов Genus Peponocephala Nishiwaki et Norris, 196G 36
Род пестрых дельфинов Genus Cephalorhynchus Gray, 1846 .................. 37
Род иравадийских дельфинов Genus Orcaella Gray. 1866 .................... 38
Род малых косаток Genus Pseudorca Reinhardt, 1862 ....................... 38
Род косаток Genus Orcinus Fitzinger, 1860 ............................ 40
Род серых дельфинов Genus Grampus Gray, 1828 ........................ 42
Род гриид Genus Globicephala Lesson, 1828 ............................. 43
Род карликовых косаток Genus Feresa Gray, 1870 ........................ 44
Род морских свиней Genus Phocoena G. Cuvier, 1817........................ 45
Род белокрылых морских свиней Genus Phocoenoides Andrews, 1911 ... 47
Род бесперых морских свиней Genus Neophocaena Palmer, 1899 .............. 49
Семейство нарваловых Familia Monodontidae Gray, 1821................ .. 50
Род белух Genus Deiphinapterus Lacepede, 1804 ........................... 50
Род нарвалов Genus Monodon Linnaeus, 1758 ............................... 53
Семейство кашалотовых Familia Physeteridae Gray, 1821..................... 54
Род карликовых кашалотов Genus Kogia Gray, 1846 ........................ 55
Род кашалотов Genus Physeter Linnaeus, 1758 ............................. 56
Семейство клюворыловых Familia Ziphiidae, Gray, 1865 ..................... 59
Род Тасмановых клюворылов Genus Tasmacetus Oliver, 1937 ..... 59
Род плавунов Genus Berardius Duvernoy, 1851 . ................... ... 50
Род ремнезубов Genus Mesoplodon Gervais, 1850 ........................... 61
Род клюворылов Genus Ziphius G. Cuvier, 1823 ............................ 63
Род бутылконосов Genus Hyperoodon Lacepede, 1804 ........................ 6a
Подотряд усатых китов Subordo Mysticeti Flower, 1864 ........................
Семейство гладких китов Familia Balaenidae Gray, 1825 ...................
Род гренландских китов Genus Balaena Linnaeus, 1758 ....................
Род южных китов Genus Eubalaena Gray, 1864 .............................
Род карликовых китов Genus Caperea Gray, 1846 .....................
Семейство серых китов Familia Eschrichtiidae Ellerman et Morrison-Scott, 1951......................................................................
Род серых китов Genus Eschrichtius Gray, 1864 ..........................
Семейство полосатиковых Familia Balaenopteridae Gray, 1864 ..............
Род настоящих полосатиков Genus Balaenoptera Lacepede, 1804 ............
Род горбатых китов Genus Megaptera Gray, 1846 ..........................
Отряд хищных Ordo Carnivora Bowdich, 1821 ......................................
Семейство волчьих Familia Canidae Gray, 1821.............................
Subfamilia Caninae Gill, 1872 .................................
Род болков Genus Canis Linnaeus, 1758 ..................................
Род песцов Genus Alopex Каир, 1829 .....................................
Род лисиц Genus Vulpes Oken, 1816 ......................................
Род фенеков Genus Fennecus Desmarest, 1804 .............................
Род серых лисиц Genus Urocyon Baird, 1858 ..............................
Род южноамериканских лисиц Genus Dusicyon H. Smith, 1837 ...............
Род гривистых волков Genus Chrysocyon Н. Smith, 1839 ...................
Subfamilia Simocyoninae Zittel, 1893 .....................................
Род красных волков Genus Cuon Hodgson, 1838 ............................
Род гиеновых собак Genus Lycaon Brookes, 1827 ..........................
Род кустарниковых собак Genus Speothos Lund, 1839 ......................
Subfamilia Otocyoninae Trouessart, 1885 ..................................
Род большеухих лисиц Genus Otocyon Mijller, 1836 ................. . .
Семейство медвежьих Familia Ursidae Gray, 1825 ...........................
Род медведей Genus Ursus Linnaeus, 1758 ...................... . . .
Род очковых медведей Genus Tremarctos Gervais, 1855 .... ...
Род медведей-губачей Genus Melursus Meyer, 1793 ........................
Род малайских медведей Genus Helarctos Horsfield, 1825 .................
Семейство енотовых Familia Procyonidae Bonaparte, 1850 ...................
Род какомицли Genus Bassariscus Coues, 1887 ............................
Род енотов Genus Procyon Storr, 1780 ...................................
Род носух Genus Nasua Storr, 1780 ......................................
Род горных носух Genus Nasuella Hollister, 1916 . ......................
Род кинкажу Genus Potos E. Geoffroy, St.-Hilaire et G. Cuvier, 1795 . . .
Род олннго Genus Bassaricyon J. Allen, 1876 ............................
Род малых панд Genus Ailurus F. Cuvier, 1825 ...........................
Род больших панд Genus Ailuropoda Milne-Edwards, 1870 ..................
Семейство куньих Familia Mustelidae Swainson, 1835 .......................
Subfamilia Mustelinae Gill, 1872 .........................................
Род куниц Genus Martes Pinel, 1792 ................................ ...
Род росомах Genus Gulo Storr, 1780 .....................................
Род ласок и хорьков Genus Mustela Linnaeus, 1758 .......................
Cenus Grammogale Cabrera, 1940 ...........................................
Род перевязок Genus Vormela Blasius. 1884 ..............................
Род тайр Genus Eira Hamilton-Shith, 1842 ...............................
Род гризонов Genus Gaiictis Bell, 1826 .................................
Род патагонских ласок Genus Lyncodon Gervais, 1844 .....................
Род африканских хорьков Genus Ictonyx Каир, 1835 .......................
Род североафрнкаиских ласок Genus Poecilictis Thomas et Hinton, 1920 . .
Род африканских ласок Genus Poecilogale Thomas, 1883 ...................
Subfamilia Mellivorinae Gill, 1872 .......................................
Род медоедов Genus Mellivora Storr, 1780 ...............................
Subfamilia Melinae Burmeister, 1850 ......................................
Род барсуков Genus Meles Brisson, 1762 .................................
Род теледу Genus Arctonyx F. Cuvier, 1825 ..............................
Род американских барсуков Genus Taxidea Waterhouse, 1839 ...............
Род хорьковых барсуков Genus Melogale I. Geoffroy, 1834 ................
Subfamilia Mephitinae Gill, 1872 .........................................
Род полосатых скунсов Genus Mephitis G. Cuvier, 1800 ...................
Род пятнистых скунсов Genus Spilogale Gray, 1865 .......................
Род свиноносых скунсов Genus Conepatus Gray, 1837 ......................
Subfamilia Lutrinae Baird, 1857 ..........................................
Род выдр Genus Lutra Brisson, .1762 ....................................
Род гигантских выдр Genus Pteronura Gray, 1837 .........................
Род бескоготных выдр Genus Aonyx Lesson, 1827 ..........................
Род африканских выдр Genus Paraonyx Hinton, 1921........................
Род каланов Genus Enhydra Flemming, 1822 ...............................
Семейство виверровых Familia Viverridae Gray, 1821........................
Subfam ilia Viverrinae Gill, 1872 ......................................
Род африканских липзангов Genus Poiana Gray, 1865 ......................
Род генет Genus Genetta Oken, 1816......................................
Род малых цивет Genus Viverricula Hodgson, 1838 ........................
Род водяных цивет Genus Osbornictis J. Allen. 1919......................
Род цивет Genus Viverra Linnaeus, 1758 .................................
Род пятнистых линзангов Genus Prionodon Horsfield, 1824 ................
Subfamilia Paradoxurinae Gill, 1872 ......................................
Род пальмовых цивет Genus Nandinia Gray, 1843 ..........................
Род мелкозубых цивет Genus Arctogalidia Merriam, 1897 ..................
Род мусаигов Genus Paradoxurus F. Cuvier, 1821..........................
Род гималайских цнвет Genus Paguma Gray, 1831........................
Род сулавесских цивет Genus Macrogalidia Schwarz, 1918 . ................ 204
Род бинтуроигов Genus Arctictis Temminck, 1824 .......................... 205
Subfamilia Hemigalinae Gill, 1872 ........................................ 205
Род фаналок Genus Fossa Gray, 1864 ...................................... 205
Род полосатых цивет Genus Hemigalus Jourdan, 1837 ....................... 206
Род цивет Оустона Genus Chrotogale Thomas, 1912.......................... 207
Род калимаитанскпх цивет Genus Diplogale Thomas, 1912.................... 208
Род выдровых цивет Genus Cynogale Gray, 1837 ............................ 208
Род мелкозубых мунго Genus Eupleres Doyere, 1835 ........................ 209
Subfamilia Calidiinae Gill, 1872 ......................................... 210
Род кольцехвостых муиго Genus Galidia I. Geoffroy, 1837 ................. 210
Род широкополосых мунго Genus Galidictis I. Geoffroy, 1839 .............. 211
Род узкополосых мунго Genus Mungotictis Pocock, 1915..................... 211
Род бурохвостых мунго Genus Salanoia Gray, 1864 ............' ... . 212
Subfamilia Herpestinae Gill, 1872 ........................................ 213
Род сурикат Genus Suricata Desmarest, 1804 .............................. 213
Род мангустов Genus Herpestes liliger, 1811.............................. 215
Род карликовых мангустов Genus Helogale, 1862 ........................... 217
Род водяных мангустов Genus Atilax F. Cuvier, 1826 ...................... 218
Род полосатых мангустов Genus Mungos E. Geoffroy et G. Cuvier, 1795 . . 219
Род кузиманз Genus Crossarchus F. Cuvier, 1825 .......................... 219
Род либерийских мангустов Genus Liberiictis Hayman. 1958 ............... 220
Род белохвостых мангустов Genus Ichneumia I. Geoffroy, 1837 ............. 220
Род черноногнх мангустов Genus Bdeogale Peters, 1850 .................. 221
Род умби Genus Rhynchogale Thomas, 1894 ................................. 222
Род желтых мангустов Genus Cynictis Ogilby, 1833 ........................ 222
Род серых мангустов Genus Paracynictis Pocock, 1916...................... 223
Subfamilia Cryptoproctinae Trouessart, 1885 .............................. 223
Род фосс Genus Cryptoprocta Bennett, 1833 ........................ ... 223
Семейство гиеновых Familia Hyaenidae Gray, 1869 .......................... 224
Subfamilia Hyaeninae Mivart, 1882 ........................................ 225
Род пятнистых гиен Genus Crocuta Каир, 1828 ............................. 225
Род полосатых гиен Genus Hyaena Brisson, 1762 ........................... 227
Subfamilia Protelinae Mivart, 1882 ....................................... 230
Род земляных волков Genus Proteles I. Geoffroy St.-Hilaire, 1824 .... 230
Семейство кошачьих Familia Felidae Gray, 1821 ............................ 231
Род больших кошек Genus Panthera Oken, 1816.............................. 233
Род снежных барсов Genus Uncia Gray, 1854 . ............................. 238
Род кошек Genus Felis Linnaeus, 1758 .................................... 240
Род гепардов Genus Acinonyx Brookes, 1828 ............................... 249
Отряд ластоногих Ordo Pinnipedia liliger, 1811.............................
Семейство ушастых тюленей Familia Otariidae Gill, 1866 ................
Род южных морских львов Genus Otaria Peron, 1816......................
Род новозеландских морских львов Genus Phocarctos Gray, 1844 ....
Род австралийских морских львов Genus Neophoca Gray, 1866 ............
Род калифорнийских морских львов Genus Zalophus Gill, 1866 ...........
Род сивучей Genus Eumetopias Gill, 1866 ..............................
Род южных морских котиков Genus Arctocephalus Е. Geoffrov et F. Cuvier, 1826 ................................................'..............
Род северных морских котиков Genus Callorhinus Gray. 1859 ............
Семейство моржовых Familia Odobenidae J. Allen, 1880 ..................
Род моржей Genus Odobenus Brisson, 1762 ..............................
Семейство тюленевых Familia Phocidae Brooker, 1828 ....................
Род обыкновенных тюленей Genus Phoca Linnaeus, 1758 ..................
Род нерп Genus Pusa Scopoli, 1777 ....................................
Род полосатых тюленей Genus Histriophoca Gill, 1873 ..................
Род гренландских тюленей Genus Pagophilus Gray, 1844 .................
Род длинномордых тюленей Genus Halichoerus Nilsson, 1820 .............
Род морских зайцев Genus Erignathus Gill. 1866 . .....................
Род тюленей-моиахов Genus Monachus Fleming, 1822 .....................
Род крабоедов Genus Lobodon Gray, 1844 ...............................
Род тюленей Росса Genus Ommatophoca Gray, 1844 .......................
Род морских леопардов Genus Hydrurga Gistel. 1848 ................. .
Род тюленей Уэдделла Genus Leptonychotes Gill, 1872 ..................
Род хохлачей Genus Cystophora Nilsson, 1820 ..........................
Г* __________ —____ --------------rz.—ж. 1QO7
252
257 258
260 261
261 263
266 269 272 276 276
278 281 285 287 291 293 296 2ОД
299
300 301 303 члл
Отряд трубкозубых Ordo Tubulidentata Huxley, 1872 ............................ 309
Семейство трубкозубовых Familia Orycteropodidac Bonoparte, 1850 .... 311
Род трубкозубов Genus Orycteropus E. Geoffroy, 1795 ..................... 311
Отряд хоботных Ordo Proboscides liliger, 1811................................. 3J2
Семейство слоновых Familia Elephantidae Gray, 1821........................ 314
Род индийских слонов Genus Elephas Linnaeus, 1758 ....................... 315
Род африканских слонов Genus Loxodonta F. Cuvier, 1827 .................. 315
Отряд даманов Ordo Hyracoidea Huxley, 1869 ................................... 317
Семейство дамановых Familia Procaviidae Thomas, 1892 ..................... 320
Род скалистых даманов Genus Procavia Storr, 1780 ........................ 320
Род горных даманов Genus Heteroxyrax Gray, 1868 ......................... 321
Род древесных даманов Genus Dendroxyrax Gray, 1868 ...................... 323
Отряд сирен Ordo Sirenia liliger, 1811........................................ 324
Семейство ламаитнновых Familia Trichecliidae Gill, 1872 .................. 326
Род ламантинов Genus Trichechus Linnaeus, 1758 .......................... 328
Семейство дюгоневых Familia Dugongidae Gray, 1821 ........................ 328
Род дюгоней Genus Dugong Laccpede, 1799 ................................ 330
Семейство морских коров Familia Hydrodamalidae Palmer, 1895 ..... 330
Род морских коров Genus Hydrodamalis Retzius, 1794 ...................... 332
Отряд парнокопытных Ordo Artiodactyla Owen, 1848 ............................. 333
Подотряд нежвачных Subordo Suiformes Jaeckel, 1911............................ 335
Семейство свиных Familia Suidae Gray, 1821 ............................... 336
Род кнстеухих свиней Genus Potamochoerus Gray, 1854 ..................... 337
Род кабанов Genus Sus Linnaeus, 1758 .................................... 338
Род бородавочников Genus Phacochoerus F. Cuvier, 1817.................... 341
Род лесных свиней Genus Hylochoerus Thomas, 1904 ........................ 342
Семейство пекарневых Familia Tayassuidae Palmer, 1897 ...................... 34o
Род пекари Genus Tayassu Fischer, 1814..................................... 348
Семейство бегемотовых Familia Hippopotamidae Gray, 1821 .................... 348
Род бегемотов Genus Hippopotamus Linnaeus, 1758 ........................... 3^0
Род карликовых бегемотов Genus Choeropsis Leidy, 1853 ..................... 351
Подотряд жвачных Subordo Ruminantia Scopoli, 1777 .............................. 353
Семейство оленьковых Familia Tragulidae Milne-Edwards, 1864 ................ 354
Род водяных оленьков Genus Hyemoschus Gray, 1845 .......................... 355
Род азиатских оленьков Genus Tragulus Brisson, 1762 ....................... 356
Семейство оленевых Familia Cervidae Gray, 1821 ............................ 357
Subfamilia Moschmae Zittel, 1893 ........................................... 3u9
Род кабарог Genus Moschus Linnaeus, 1758 .................................. 359
Siibfamilia Hydropotinae Trouessart, 1898 .................................. 361
Род водяных оленей Genus Hydropotes Swinhoe, 1870 ......................... 361
Subfamilia Cervulinae Sclater, 1870 ........................................ 363
Род мунтжаков Genus Muntiacus Rafinesque, 1815............................. 363
Род хохлатых оленей Genus Elaphodus Milne-Edwards, 1872 ................... 365
Subfamilia Odocoileinae Pocock, 1923 ....................................... 365
Род косуль Genus Capreolus Gray, 1821 ..................................... 366
Род американских оленей Genus Odocoileus Rafinesque, 1832 ................. 369
Род мазам Genus Atazama Rafinesque, 1817................................... 371
Род пуду Genus Pudu Gray, 1852 ........................................... 374
Род лосей Genus Alces Gray, 1821 ......................................... 376
Род северных оленей Genus Rangffer H. Smith, 1827 ......................... 378
Subfamilia Cervinae Baird, 1857 ............................................ 381
Род оленей Давида Genus Elaphurtis Milne-Edwards, 1866 .................... 381
Род оленей Genus Cervus Linnaeus, 1758 .................................... 386
Семейство жирафовых Familia Giraffidae Gray, 1821 .......................... 389
Род окапи Genus Okapia Lankester, 1901 .................................... 390
Род жираф Genus Giraffa Brisson, 1762 ..................................... 392
Семейство вилороговых Familia Antilocapridae Gray, 1866 .................... 394
Род вилорогов Genus Antilocapra Ord, 1818 ................................. 396
Семейство полорогих Familia Bovidae Gray, 1821 ............................. 396
Sublamilia Cephalophinae Brooke, 1876 ...................................... 398
Род хохлатых дукеров Genus Cephalophus H. Smith, 1827 ....... 399
Род кустарниковых дукеров Genus Sylvicapra Ogilby, 1837 ...... 401
Subfamilia Neotraginae Sciater et Thomas, 1894 ............................. 402
Род карликовых антилоп Genus Neotragus H. Smith, 1827 ..................... 403
Род дикдиков Genus Madoqua Ogilby, 1837 ................................... 404
Род антилоп-прыгунов Genus Oreotragns A. Smith, 1834 ...................... 405
Род бейр Genus Dorcatragus Noack, 1894 .................................... 407
Род стенбоков Genus Raphicerns H. Smith, 1827 ............................. 408
Род ориби Genus Ourebia Laurillard, 1841 .................................. 409
Subfamilia Antilopinae Baird, 1857 ......................................... 410
Род четырехрогих антилоп Genus Tetracerus Leach, 1825 ...................... Ho
Род нильгау Genus Boselaphus De Blainville, 1816........................... 412
Род лесных антилоп Genus Tragelaphus De Blainville, 1816................... 414
Род ориксов Genus Oryx De Blainville, 1816................................. 419
Род аддаксов Genus Addax Rafinesque, 1815.................................. 422
Род водяных козлов Genus Kobus A. Smith, 1840 ............................. 423
Род тростниковых козлов Genus Redunca H. Smith, 1827 ...................... 425
Род косульих антилоп Genus Pelea Gray, 1841 ............................... 427
Род импал Genus Aepyceros Sundevall, 1847 ................................. 428
Род лошадиных антилоп Genus Hippotragus Sundevall, 1846 ................... 429
Род лиророгих бубалов Genus Damaiiscus Sciater et Thomas, 1894 . . 432
Род конгонп Genus Alcelaphus De Blainville. 1816........................... 433
Род гну Genus Connochaetes Lichtenstein, 1814.............................. 434
Subfamilia Gazellinae Coues, 1889 .......................................... 43S
Род гарн Genus Antilope Pallas, 1766 ...................................... 437
Род спрингбоков Genus Antidorcas Sundevall, 1847 .......................... 439
Род дзеренов Genus Ргосарга Hodgson, 1846 ................................. 441
Род газелей Genus Gazella De Biainville, 1816.............................. 443
Род дибатагов Genus Ammodorcas Thomas. 1891 ............................... 446
Род геренуков Genus Litocranius Kohl, 1886 ................................ 448
Subfamilia Caprinae Gill, 1872 ...................................... .... 448
Род сайгаков Genus Saiga Gray, 1843 ....................................... 449
Род оронго Genus Pantholops Hodgson, 1834 ................. . . ... 452
Род горалов Genus Nemorhaedus H. Smith, 1827 ........................ 454
Род cepoy Genus Capricornis Ogilby, 1837 ............................... 455
Род снежных коз Genus Oreamuos Rafinesque. 1817.......................... 458
Род серн Genus Rupicapra De Blainville, 1816............................. 458
Род такпнов Genus Budorcas Hodgson, 1850 ................................ 459
Род овцебыков Genus Ovibos De Blainville, 1816........................... 462
Род таров Genus Hemitragus Hodgson, 1841 ................................ 464
Род горных козлов Genus Capra Linnaeus, 1758 ............................ 465
Род голубых баранов Genus Pseudois Hodgson, 1846 ........................ 469
Род гривистых баранов Genus Ammotragus Blyth, 1840 ..................... 471
Род горных баранов Genus Ovis Linnaeus, 1758 ...................... . . 471
Subfamilia Bovinae Gill, 1872 .............................................. 477
Род буйволов Genus Bubalus H. Smith, 1827 ........................... ... 477
Род настоящих быков Genus Bos Linnaeus, 1758 .............................. 479
Род зубров Genus Bison H. Smith, 1827 ..................................... 484
Отряд мозоленогих Ordo Tylopoda liliger, 1811.................................... 486
Семейство верблюдовых Familia Camelidae Gray, 1821............... . • 487
Род верблюдов Genus Camelus Linnaeus, 1758 ........................... 487
Род лам Genus Lama G. Cuvier, 1800 ................................... 492
1 Отряд непарнокопытных Ordo Perissodactyla Owen, 1848 .................
Семейство лошадиных Familia Equidae Gray, 1821...........................
Род лошадей Genus Equus Linnaeus, 1758 .................................
Семейство тапировых Familia Tapiridae Burnett, 1830 .....................
Род тапиров Genus Tapirus Brisson, 1762 ................................
Семейство носороговых Familia Rhinocerotidae Owen, 1845 .................
Род суматранских носорогов Genus Didermocerus Brookes, 1828 ............
Род индийских носорогов Genus Rhinoceros Linnaeus, 1758 ................
Род черных носорогов Genus Diceros Gray, 1821 ..........................
Род белых носорогов Genus Ceratotherium Gray, 1861......................
Литеоатуоа ..............................................................
494
495
498
499
502
502
503
504
505
505
508
511