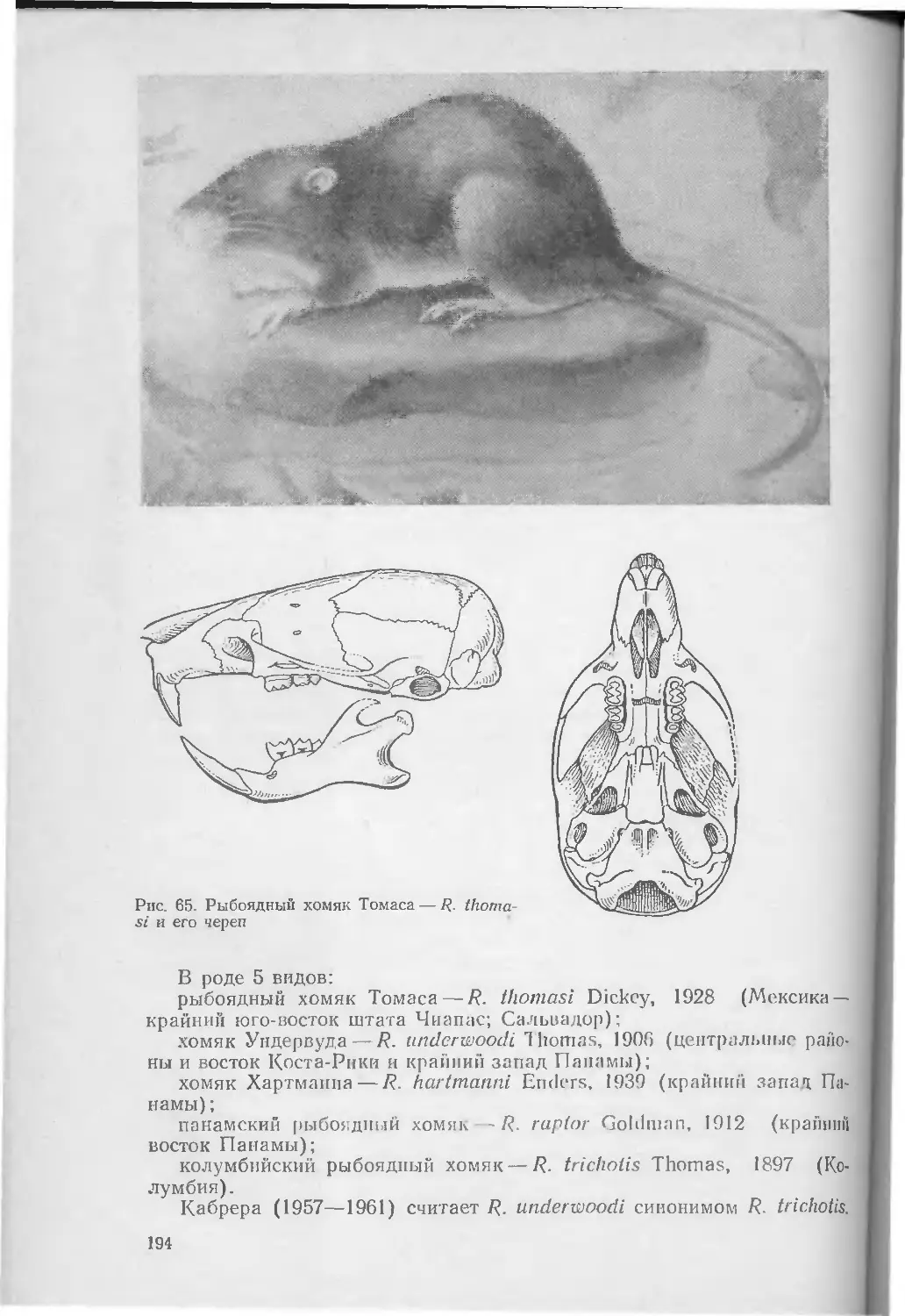
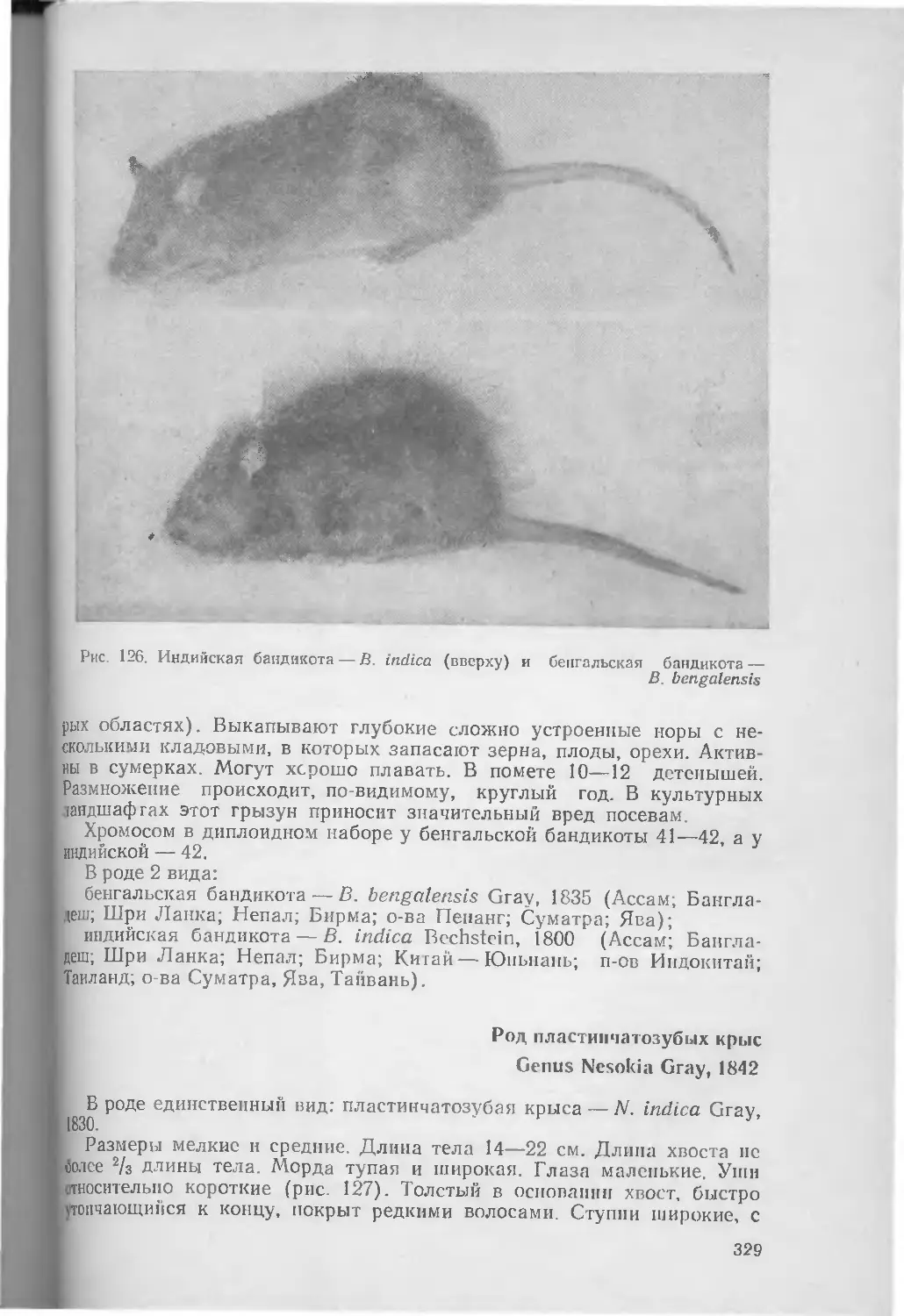
/










































































































































































































































































































































































































































































































Похожие




Текст
BE СОКОЛОВ
Систематика млекопитающих
Отряды: зайцеобразных, грызунов
ДОПУЩЕНО МИНИСТЕРСТВОМ ВЫСШЕГО И СРЕДНЕГО СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР В КАЧЕСТВЕ УЧЕБНОГО ПОСОБИЯ ДЛЯ СТУДЕНТОВ УНИВЕРСИТЕТОВ
596.5 С59
Рецензенты учебного пособия:
кафедра охраны природы Казанского университета (зав. кафедрой проф. В. А. Попов) и проф. Б. А. Кузнецов (зав. кафедрой зоологии Московской сельскохозяйственной академии)
60 000—059
С 001(01)—77
69—77
ОТ АВТОРА
Первая часть «Систематики млекопитающих» (1973) включала отряды однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых и ящеров. Предлагаемая читателям вторая часть «Систематики млекопитающих» посвящена отрядам зайцеобразных и грызунов. В третью часть войдут все остальные отряды млекопитающих. Описание морфологии млекопитающих дается в первой части, поэтому объяснений морфологических терминов в настоящем издании не приводится.
В связи с тем, что книга предназначена в основном для студентов, в диагнозах опущены некоторые детали. Так, в описании зубов даются лишь их основные особенности. По этой же причине в список литературы включены в большинстве случаев только основные сводки, посвященные рассматриваемым отрядам.
Большинство иллюстраций взято из книг Уокера (Walker, 1964) и Эллермэна (ЕПсгшап, 1940—1941), остальные рисунки изготовил художник-зоолог 11. 11. Кондаков. Фотографии 12, 15, 26, 26, 29, 32, 42, 67, 70, 79, 101, 112, 117, 137 сделаны М. Никитиным, 144 — П. П. Гамбаряном.
Критические замечания, сделанные по первой части «Систематики млскопитаю-щих» профессорами В. Г. Гептнером,
Н. Н. Калабуховым и А. П. Кузякиным, учтены при написании второй части. Большую помощь в подготовке рукописи к публикации оказали профессора |В. Г. Гептнер, Б. А. Кузнецов, А. П. Кузякин, В. А. Попов. Пользуюсь случаем принести всем им глубокую благодарность.
ОТРЯД ЗАЙЦЕОБРАЗНЫХ
ORDO LAOOMORPHA BRANDT, 1855
Млекопитающие мелких и средних размеров с длиной тела 12— 74 см'. У крупных видов задние конечности удлинены, уши сильно вытянуты. Они имеют хорошо всем известный «заячий» облик. Мелкие виды обладают почти одинаковыми по длине передними и задними конечностями и невысокими округлыми ушами. Хвост снаружи незаметен или очень короток, весь покрыт волосами. Передние конечности пятипалые, задние четырех- или пятипалые. На пальцах хорошо развиты когти. Подошвы лап покрыты волосами. Ноздри окружены полоской голой кожи. Лопасти верхней и нижней губ способны смыкаться позади резцов по средней линии. Волосяной покров мягкий и густой, лишь у немногих жесткий, щетиновидный, изреженный. У многих видов волосяной покров
изменяется по сезонам года как по структуре и густоте волос, так и по окраске. Окраска его у большинства серая, бурая или охристая. Полоска покрытой волосами кожи вдается внутрь рта вдоль зубных рядов — примерно на уровне нижних щечных зубов. Потовые железы в коже туловища отсутствуют; имеются только на подошвах лап. Обычны анальные и паховые кожные специфические железы. Сосков 2—5 пар.
Череп легкий с губчатой структурой отдельных костей. Боковые части верхнечелюстных костей имеют отверстия (рис. 1). Резцовые отверстия
крупные, овальной формы, вытянутые далеко назад; оканчиваются между щечными зубами. Костное небо короткое, располагается позади широких и длинных резцовых отверстий на уровне IV предкоренного или I коренного зубов. Подглазничное отверстие маленькое. Настоящий крылоклиновидный канал отсутствует. Сочленовная ямка для нижней че-
люсти вытянута в поперечном направлении. Нижняя челюсть с расширенной угловой частью и удлиненным сочленовным мыщелком. Характерны боковые движения нижней челюсти при пережевывании пищи. Перети-
рание пиши происходит попеременно то правым, то левым рядом зубов. Зубная формула современных видов: I с — р т —=26 — 28.
В верхней челюсти две пары резцов, причем одна из них располагается позади второй. Передние резцы более крупные, долотовидные, задние мелкие, в сечении округлой или овальной формы. Па па ружной поверхности передней пары резцов есть продольная бороздка. Для резцов характерен постоянный рост в течение всей жизни животного. Клыки от-
1 В пределах отряда можно выделить 3 размерные группы зайцеобразных: мелкие с длиной тела 12—33 см, средние — 34—54 см и крупные — 55—74 см.
Рис. 1. Череп беляка — L. timidus снизу и сбоку
сутствуют, между резцами и щечными зубами находится большой промежуток — диастема. Эмаль на фронтальной поверхности передних резцов значительно толще, чем на задней; это обеспечивает неравномерное стачивание зубов, причем их передний крап остается всегда острым. У второго (заднего) резца верхней челюсти такого режущего острого края нет. Щечные зубы гипселодонтного типа и у современных видов не имеют корней. Правый и левый ряды верхних щечных зубов расставлены шире, чем нижние. У верхних щечных зубов более высокий наружный край коронки, а у нижних щечных, наоборот, более высокий внутренний край.
Ключица имеется. Малая и большая берцовые кости срастаются в дистальной части. Пяточная кость длинная. Головной мозг продолговатой формы с относительно гладкими большими полушариями, не прикрывающими мозжечка. Желудок простой. Слепая кишка крупная, с развитой спиральной складкой. Семенники в период размножения располагаются спереди от основания полового члена, как у сумчатых. Матка двойная. Плацента дискоидальпая и гемоэндотелиальная.
Распространены на всем земном шаре, за исключением Антарктики, южной части Южной Америки, Австралийской области, Повой Зеландии, Индонезии (кроме о-ва Суматра), Мадагаскара, Филиппинских о-вов и о-вов Карибского моря. С успехом акклиматизированы в Австралии, Новой Зеландии, на юге Южной Америки, Яве и па различных островах Атлантического и Тихого океанов.
Населяют все зоны от арктических тундр до тропических лесов и пустынь. В горы поднимаются до альпийского и субальпийского поясов. Ведут наземный образ жизни. Некоторые колониальные, другие встречаются поодиночке. В спячку пе впадают. Активны днем (пищухи) или в сумерки и ночью (кролики и зайцы). Некоторые прекрасные бегуны. Убежищем служат выкопанные ими поры, расщелины екал, заросли травы и кустов. Питание исключительно растительноядное. Предпочитают зеленые части травянистых растений, поедают также кору кустов и молодых деревьев, иногда их веточки. Характерно поедание собственного кала, содержащего витамины (копрофагия). Самка приносит от 1 до
4 пометов в году, по нескольку детенышей в каждом. Виды, обитающие в норах или естественных укрытиях, приносят голых или покрытых шерстью, но слепых детенышей, а обитающие на поверхности земли — зрячих детенышей, покрытых шерстью, способных передвигаться сразу после рождения.
Некоторые виды отряда имеют охотничье-промысловое значение и добываются в больших количествах. Кролик стал одомашненным и лабораторным животным. Могут приносить известный вред сельскохозяйственным культурам. Кролик местами сильно вредит сельскому хозяйству (например, в Австралии). Некоторые виды служат хозяевами паразитам— переносчикам возбудителей ряда трансмиссивных заболеваний человека и домашних животных.
Зайцеобразные ведут происхождение, по-видимому, от примитивных насекомоядных (Тениус и Хофер, Thenius u. Hofer, 1960). Есть предположение о независимом развитии отряда зайцеобразных, начиная с позднемелового времени (Тобиен, 1974). Наиболее древние находки (представители сем. Eurymylidae) относятся к верхнему палеоцену. Семейство Eurymylidae представляет собой боковую ветвь центрального ствола зайцеобразных, который в эоцене разделился на зайцев и пищух. Исходными формами для этого разделения послужили представители семейства Palaeolagidae. Представители этого семейства, имевшие в верхней челюсти 6 щечных зубов (подсемейство Palaeolaginae), дали начало зайцам, а с 5 верхними щечными зубами (подсемейство Amphilaginac) — пищух (Гуреев, 1964). Некоторые исследователи предполагают и другие пути филогенеза зайцеобразных, считая, например, Eurymylus из палеоцена возможной исходной группой как для пищух, так и для зайцев (Симпсон, Simpson, 1945).
С 1811 г. зайцеобразных выделяют в особый подотряд Duplicidentata (Иллигер, liliger, 1811) в составе отряда грызунов — Glires. В 1855 г. Брандт (Brandt) дал этому подотряду название зайцеобразных — Lago-morpha. В 1912 г. Джидлей (Gidley, 1912) предложил выделить этот подотряд в самостоятельный отряд Lagomorpha. В настоящее время большинство систематиков рассматривает зайцеобразных как особый отряд, хотя, например, Б. А. Кузнецов (1965), Ландри (1974) считают зайцеобразных подотрядом отряда грызунов.
В отряде 2 семейства:
пищуховых — OchotonidaeThomas, 1897 (1 род);
Зайцевых —• Leporidae Gray, 1821 (10 родов).
Оба семейства представлены в фауне СССР.
СЕМЕЙСТВО ПИЩУХОВЫХ
F АМИЛА OCHOTONIDAE THOMAS, 1897
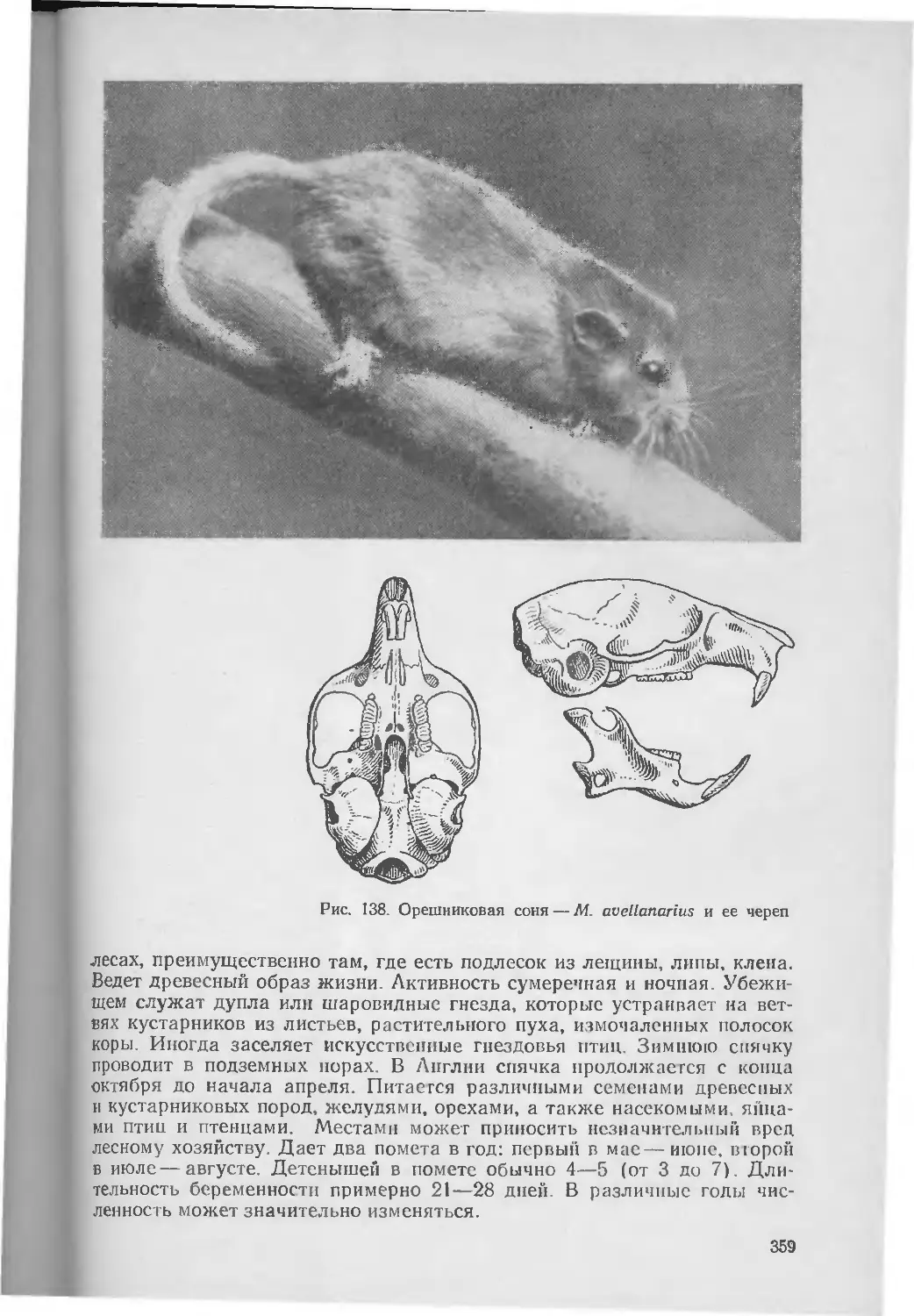
Размеры мелкие. Длина тела колеблется от 12 до 28 см-Масса 170— 240 г. Конечности относительно короткие, задние по длине почти равны передним, лишь-немного (па 20—25%) превышая их (рис. 2). Ступня относительно короткая. Хвост снаружи незаметен. Уши короткие, их длина лишь у некоторых видов больше половины длины головы. Вершины ушей округлые. Глаза относительно небольшие. Передние конечности имеют 5 пальцев, задние — 4. Подошвы конечностей покрыты длинными волосами. Волосяной покров высокий, мягкий и густой. Окраска его красноватая или буроватая, до серой, на брюшной стороне обычно более светлая. Линька может быть одна, две, возможно, три в течение года. Зимний мех более светлый, чем летний (обычно серый разных оттенков).
Рис. 2. Пика — О. princeps и череп алтайской пищухи — О. alpina
Молодые звери темнее старых. Характерны длинные вибриссы, у некоторых видов превышающие треть длины тела. Кожные подушки пальцев голые. Периапальных желез пег. Специфическая кожная апокриновая железа располагается в заднем отделе щек и служит, по-впдимому, для мечения территории пли имеет значение в размножении животных. Мошонки пет. Положение семенников снаружи незаметно. Сосков 2 пли 3 пары.
Череп удлиненной формы, уплощенный сверху, сильно сжатый в межорбитальной области (рис. 2). Надглазничных отростков пет. Скуловые кости длинные. Небные кости сильно развиты и занимают более
половины площади костного неба. Слуховые барабаны с губчатыми 2 0 3 2
стенками. Зубная формула: i• — с — р т — — 26. Седьмой шейный
позвонок не имеет артериального канала в поперечном отростке. Груд-ных позвонков 22; в отличие от зайцев они с короткими остистыми отростками. Ключица хорошо развита. Кисть сочленяется главным образом с локтевой костью и лишь отчасти с лучевой. Лобкового сочленения
в тазе нет.
Число хромосом в диплоидном наборе изменяется от 38 у палласовой пищухи, 40 у северной, 42 у алтайской до 62 у степной и красной и 68 у пика и воротничковой.
Ареал охватывает горы Северной Америки от южной части центральной Аляски до центральных районов Калифорнии и штата Ныо-Мекси-ко, Юго-Восточную Европу и Азию на север до Арктического побережья и на восток до Анадырского залива, Северной Японии (Хоккайдо) и Корейского п-ова, а на юг до северных частей Ирана (гор Копетдага), Афганистана, Пакистана, Индии и Бирмы.
Пищухи обитают в высокогорье, используя в качестве убежищ расщелины между камнями, или на открытых равнинах и предгорьях, где роют норы. Высотное распределение пищух — почти от уровня моря до выше уровня верхней границы леса, примерно на высоте 4100 м в Северной Америке или до 6000 м в Азии.
Даурская пищуха встречается в северных частях своего ареала в степях, реже в закрепленных песках. В полупустыне и пустыне обитает по более влажным понижениям и речным долинам, где она предпочитает селиться в зарослях ириса. Иногда даурская пищуха попадается в поясе горных лугов. Поселяется колониями, так что подчас трудно отличить границы одной норы от другой или одной колонии от соседней. Нора неглубокая со многими (до 15—20) выходами, может занимать площадь до 25—35 м2. Степная пищуха обитает преимущественно в степях, где выбирает для своих поселений густые заросли травы и кустарника на равнине, в балках. Встречается на сильно увлажненных участках, в долинах горных рек, изредка в лесу (в горах Улутау).
Палласова пищуха населяет разнообразные места — от пустынь до гор. Наиболее обычна на скальных обнажениях, каменистых россыпях. Роет сложные норы глубиной 20—50 см со многими выходами. В норе на глубине 30—50 см бывает от 1 до 3 гнездовых камер диаметром 15— 24 см, выстланных сухой травой, и несколько отнорков, служащих уборными. Общая длина норы достигает 4—10 м. Кроме описанных выше постоянных нор, устраивает несколько временных, неглубоких, с меньшим количеством выходов. Если зимой снег достаточно глубокий (не менее 20—30 см), палласова пищуха роет в нем норы и устраивает шаровидные гнезда.
Рыжеватая пищуха населяет горы, причем в предгорьях селится у подножья холмов, на полях, в глинобитных постройках. Выше в горах она населяет каменистые осыпи, скальные выходы. Обычно выбирает пологие склоны, но иногда держится и на почти отвесных обрывах. Нору устраивает под камнями, в расщелинах скал. Нора простая, длиной до 3 м и глубиной до 1 м с гнездовой камерон. Характерным местом обитания алтайской пищухи служат каменистые россыпи как па открытых горных склонах, так и на покрытых лесом или кустарником. Обычно выбирает склоны южной и западной экспозиции. Нередко встречается по берегам водоемов. Роет норы.
Красная пищуха, как и алтайская, предпочитает каменистые россыпи. Убежищем служат пустоты между камнями.
Большеухая пищуха встречается обычно в высокогорье среди каменистых россыпей. Предпочитает южные и юго-западные склоны гор. Гнезда устраивает в пустотах между камнями.
Пищухи живут поодиночке или колониями. Хорошо развита звуковая сигнализация, особенно в случае опасности. Крик пищух громкий, напоминает свист или циканье. Активны днем, иногда ночью. Большинство видов в спячку не впадают. В поисках пищи далеко от своих убежищ не отходят. Характерно запасание на зиму кормов, причем зверьки имеют своеобразные приемы сушки и хранения запасов. Во время сборов разнообразных растений для запасов пищухи многократно перекладывают и просушивают их, прячут под навесами каменных плит или собирают стожки. Иногда на стожки кладут камни, чтобы защитить от развеивания их ветром. Так, палласова пищуха на равнинах собирает стожки растений, предварительно высушивая их. Число видов растений в стожках более 60. Количество стожков и их размеры зависят от числа пищух, участвовавших в их устройстве. Масса стожка, собранного одной палла-совой пищухой, до 3—4 кг, а собранного группой — до 16—20 кг. Пика, алтайская пищуха собирают запасы сена под навесами камней. Большеухая пищуха питается практически почти всеми растениями (примерно 46 видов), встречающимися около ее нор. Запасать траву начинает с первых чисел июля. Высушенные растения прячет в щелях и под камнями. Степная пищуха приступает к запасанию пищи в июле — августе. Растения складывает в стожки. Копрофагия известна у северной пищухи. Период размножения растянут и не совпадает у различных видов и в различных местах обитания у одного и того же вида. Например, у пика период размножения приходится с мая по сентябрь, у северной пищухи в Японии — с мая по июль, у большеухой пищухи — с апреля по август и т. д. Длительность беременности примерно 30 дней. Глаза открываются на десятый день. В одном помете от 2 до 7 детенышей. Пометов в году, по-видимому, два. Новорожденные густо покрыты волосами, масса 8—9 г.
Местами пищухи могут вредить сельскому хозяйству, принося ущерб пастбищам.
В семействе 1 род пищух — Ochotona Link, 1795, и 18 видов.
Род пищух
Genus Ochotona Link, 1795
См. описание семейства.
В роде, по-видимому, 18 видов:
большеухая пищуха — О. roylei Ogilby, 1839 (О. macrotis Gunther, 1875) (СССР — Центральный и Восточный Тянь-Шань, Восточный Памир; Тибет, Непал, Пенджаб, Кашмир; Северная Бирма, Северо-Западный, Центральный и Южный Китай);
китайская пищуха-—О. erythrotis Buchner, 1894 (северо-восточный Тибет, Сычуань и Ганьсу);
камская пищуха — О. kamensis Argiropulo, 1948 (северо восточный Тибет, Кам);
непальская пищуха — О. nepalensis Hodgson, 1841 (Непал);
красная пищуха — О. rutila Severtzov, 1872 (СССР: Тянь-Шань. Гнс-саро-Алай, Западный Памир; горы Северо-Западного Китая);
даурская пищуха — О. daurica Pallas, 1776 (СССР — Южное Забайкалье и Чуйская степь на Алтае; МНР; провинции Китая — Шэньси, Шаньси, Ганьсу; Сикким; Иран);
черногубая пищуха — О. curzoniae Hodgson, 1858 (Тибет, Ганьсу, Непал, Сикким);
пищуха Козлова — О. koslovi Buchner. 1894 (северный Тибет);
рыжеватая пищуха — О. rufescens Gray, 1842 (СССР — горы Копет-даг и Большие Балханы в Туркмении; Иран, Афганистан, Пакистан);
ладакская пищуха — О. ladacensis Gunther, 1875 (Тибет, Кашмир);
палласова пищуха — О. pallasi Gray, 1867 (О. pricei Thomas, 1911) (СССР — Чуйская степь на Алтае и мелкосопочник Восточного Казахстана; МНР);
алтайская пищуха — О. alffina Pallas, 1773 (СССР — Центральный Казахстан, Алтай, Саяны, Прибайкалье, Забайкалье; МНР; Китай);
северная пищуха — О. hyperborea Pallas, 1811 (СССР — от Енисея до Камчатки и Сахалина включительно, на север до устья Лены и берегов Чукотского моря и изолированная популяция на Северном Урале);
воротничковая пищуха — О. collaris Nelson, 1893 (юго-восток Аляски, не включая, однако, Тихоокеанского побережья; южная половина провинции Юкон в Канаде, юго-запад провинции Маккензи и крайний северо-запад Британской Колумбии);
пика — О. princeps Richardson, 1828 (провинции Канады — южная половина Британской Колумбии, крайний юго-запад Альберта; отсюда ареал идет двумя языками на юг в северо-западные штаты США; один язык следует через центральные районы штатов Вашингтон и Орегон в северо-восточные части Калифорнии и северо-западные области Невады, а второй через северо-запад и центр Айдахо, запад и юг Монтаны и запад Вайоминга в центральную область Юты; два небольших изолированных участка ареала лежат в центре штата Невада, а третий, более крупный, — в центре Колорадо, юге Вайоминга, севере Ныо-Мсксико и востоке Юты);
тибетская пищуха — О. thibetana Milne-Edwards, 1872 (Тибет, Ганьсу, Шанси, Сычуань, Юньнань);
степная пищуха — О. pusilia Pallas, 1768 (СССР — степи Заволжья, Южного Урала и Северного Казахстана);
пищуха Томаса — О. thomasi Argyropulo, 1948 (северо-восточный Тибет и Ганьсу).
Систематика пищух требует уточнений. Различные исследователи по-разному трактуют видовой состав рода. Так, Эллермэн и Моррисон-Скотт (1966) выделяют из О. roylei отдельный вид О. macrotis; О. erythrotis включают в О. rutila; О. nepalensis в О. roylei; О. curoniae в О. daurica. Гуреев (1964) объединяет алтайскую, воротничковую, северную пищух и пика в один вид — О. alpina. Холл и Келсон (Hall a. Kelson, 1959) считают О. collaris и О. princeps отдельными видами.
В СССР: большеухая, красная, даурская, рыжеватая, палласова, алтайская, северная и степная пищухи.
СЕМЕЙСТВО ЗАЙЦЕВЫХ
FAMILIA LEPORIDAE GRAY, 1821
Самые крупные представители отряда. Длина тела примерно 25— 74 см. Как правило, самки крупнее самцов. Хвост короткий, по хорошо заметен снаружи (за исключением бесхвостых кроликов, Romcrolagus). Уши длинные, их длина намного превышает половину длины головы. Конечности пятипалые с сильной редукцией первых пальцев па передней и задней конечностях. Задние конечности обычно значительно длиннее передних. Ступня относительно длинная. Нижняя поверхность когтей
и ступней имеет волосяные щетки. Подушки пальцев не бывают голыми. Когти почти прямые, заостренные. Волосяной покров обычно густой, высокий и мягкий, но у некоторых видов редкий и грубый. Окраска его сильно изменчива — от серого до грязно-желтого, бурого и белого. Некоторые формы имеют полосы. У некоторых видов при сезонной линьке изменяется морфология волос и их окраска. Линьки в течение года две или три. Сосков от 3 до 5 пар. Многочисленны специфические кожные железы, образованные сальными железами. Они находятся в перианаль-ной, паховой, препуциальной, клиторальной, подбородочной и у некоторых видов — в носовой областях. Паховые железы открываются в глубокие безволосые впячивания, подошвенные состоят из потовых желез.
Череп более или менее выгнут по верхнему профилю и умеренно сжат в межорбитальной области. Есть надглазничные отростки. Боковые стороны верхнечелюстных костей имеют решетчатое строение. Отросток скуловой кости не доходит дс/ слуховых барабанов. Скуловая кость относительно короткая. Стенки костно-слуховых барабанов не губчатые. Костное небо образовано главным образом небными отростками « о - , .2 0 3 2—3 по
верхнечелюстных костей. Зубная формула: 1~ с Р
У шейных позвонков поперечные отростки имеют канал для позвоночной артерии. Грудных позвонков 19. Ключицы обычно рудиментарные. Кисть сочленяется преимущественно с лучевой костью и лишь отчасти с локтевой. Лобковый симфиз хорошо развит.
Семенники располагаются в брюшной полости. В период размножения опускаются в слабо выраженную мошонку. Матка двойная.
Распространение соответствует ареалу отряда.
Зайцы широко использовались для акклиматизации. Хорошо известна успешная (с точки зрения акклиматизационных работ) интродукция кроликов в Австралию, приведшая к катастрофическому положению сельское хозяйство в некоторых районах этой страны в результате силь-
ного увеличения численности кроликов.
Населяют самые разнообразные природные зоны — от тундры до пустыни, а также окультуренные ландшафты. Поднимаются в горы, в альпийские луга на высоту до 4900 м.
Ведут, как правило, сумеречный и ночной образ жизии. Активны круглый год, в спячку не впадают. Передвигаются обычно прыжками. Могут достигать скорости до 70 км в час. Некоторые виды (Sylvilagus aquatints и S. palustris) ведут полуводный образ жизни, но не имеют специальных приспособлений для плавания. Держатся, как правило, поодиночке. Звуковой сигнализации нет. Обычно издают громкий крик только в случае крайней опасности. Медленно бегающие виды (представители рода Sylvilagus) выкапывают норы. Сложные поры устраивают кролики. Для быстро бегающих видов днсвпым убежищем может служить неглубокая впадина куста или пучок травы. В отличие от пищух зайцы не запасают пищи. Питаются в основном различными травянистыми растениями, а также корой молодых деревьев, веточками некоторых древесных и кустарниковых растений. Некоторые виды поедают собственный кал. Этот кал отличается от обычного сухого, более твердого, своей полужидкой консистенцией и богатым содержанием витамина В.
Самки большинства видов приносят несколько пометов в году. Овуляция может быть спонтанной или провоцированной. Число детенышей в помете колеблется от 2 до 8, иногда до 15 (кролик). При недостатке пищи и воды, заболеваниях, беспокойстве характерна резорбция развивающихся эмбрионов. У видов, живущих в порах, детеныши рождаются голыми, слепыми и беспомощными, а у быстро бегающих, не имею
щих хороших убежищ, детеныши рождаются зрячими, покрытыми шерстью и способными почти сразу после рождения к самостоятельному передвижению. Длительность беременности от 26 до 55 дней. Половозре-лость наступает в возрасте от 10 недель (Sylvilagus) до 2 лет (некоторые виды Lepus). Врагами зайцев служат многочисленные хищные птицы и млекопитающие. Мех зайцев идет в виде сырья для фетровальной промышленности, реже в качестве мехового товара.
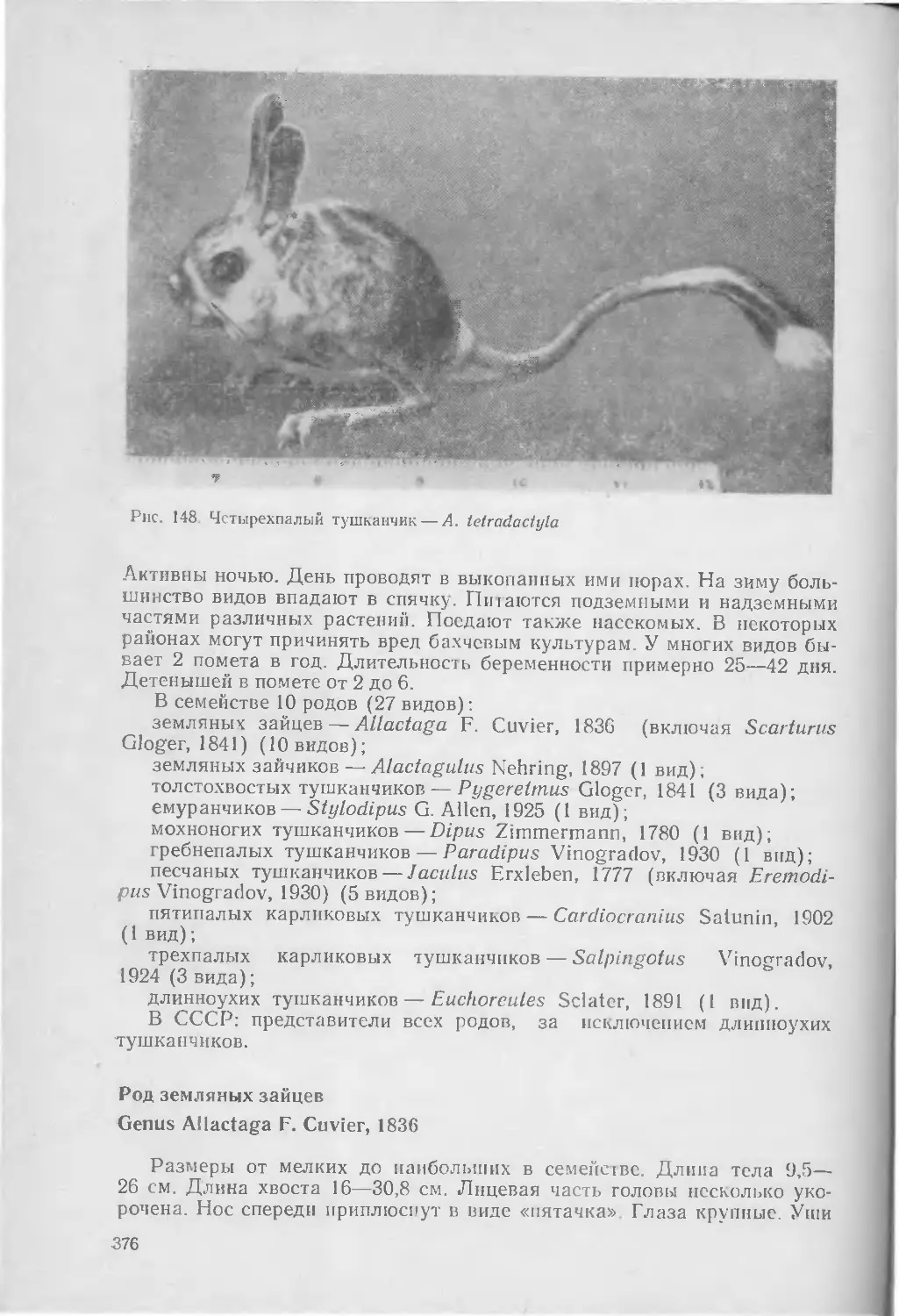
В семействе, по-видимому, 10 родов (43 вида):
лазающих зайцев — Pentalagus Lyon, 1903 (1 вид);
полосатых зайцев — Nesolagus Major, 1899 (1 вид);
бушменовых зайцев — Bunolagus Thomas, 1929 (1 вид);
щетинистых зайцев — Caprolagus Blyth, 1845 (1 вид); » африканских зайцев — Poelagus St. Leger, 1932 (1 вид), зайцев — Lepus Linnaeus, 1758 (23 вида);
бесхвостых кроликов — Romerolagus Merriam, 1896 (1 вид);
американских кроликов — Sylvilagus Gray, 1867 (включая Microla-gus Trouessart, 1897) (12 видов);
южноафриканских кроликов — Pronolagus Lyon, 1903 (1 вид); кроликов — Oryctolagus Liljeborg, 1874 (1 вид).
Система родов семейства Зайцевых окончательно не уточнена как по количеству, так и по родственным отношениям. Так, из рода Sylvilagus иногда выделяют род Brachylagus (Симпсон, 1945) или Microlagus (Гуреев, 1964); роды Bunolagus и Poelagus включают в род Lepus (Симпсон, 1945) или Poelagus в род Caprolagus (Гуреев, 1964).
В СССР: зайцы и кролики.
Род лазающих зайцев
Genus Pentalagus Lyon, 1903
В роде единственный вид: лазающий заяц — Р. furnessi Stone, 1900.
Размеры средние: длина тела 43—51 см. Длина хвоста 1,5 см (рис. 3). Уши небольшие или средних размеров, примерно 4,5 см в длину. Ноги короткие. Ступни короткие и широкие. Характерны очень длин-
Рис. 3. Лазающий заяц — Р. furnessi
ные (10—20 мм), толстые искривленные когти. Волосяной покров густой и мягкий, темно-коричневый с золотистым налетом. Брюхо светло-рыжевато-коричневое. Посредине груди и брюха проходит узкая белая полоска.
Череп массивный, уплощенный с крупными, широко расставленными скуловыми дугами. Орбиты небольшие, овальной формы. Костные слуховые барабаны относительно небольшие. Щечные зубы с многочисленными эмалевыми складками на жевательной поверхности. Плечевая и бедренная кости относительно укороченные, утолщенные.
Распространен на островах Рюкю (о-ва Амамиосима, Токуносима).
Экология не изучена. Населяет участки леса с густой растительностью. Активен ночью. Лазает по стволам деревьев. Убежище наводит в дуплах. Общая численность, вероятно, 500—900 животных.
Включен в «Красную книгу». Охраняется государством с 1921 г.
Род полосатых зайцев
Genus Nesolagus Major, 1899
В роде единственный вид: полосатый заяц — N. netscheri Schlegel et Jentink, 1880.
Размеры средние. Длина тела 36—40 см. Длина хвоста примерно 1,5 см. Уши короткие (рис. 4). Голова очень крупная. Задние конечности укороченные. Волосяной покров низкий, густой и мягкий. Общий тон ок
Рис. 4. Полосатый заяц — N netscheri
раски спины грязно-желто-ссрый. Брюхо белое. По телу проходят темпо-бурые полосы. Коричневая полоса-тянется от носа до хвоста по средней линии спины. По бокам тела вдоль них проходит по широкой полосе и еще одна полоса идет от крестца к задним конечностям. Вокруг глаз темные кольца. От заднего угла глаза к основанию уха тянется также темная полоса; тога же цвета боковые стороны ушей и ошейник, заканчивающийся на груди треугольным пятном, вершиной направленным вниз. Крестец н хвост рыжие.
Череп массивный, широкий, уплощенный. Скуловые дуги широко расставлены в стороны.
Распространен на о-ве Суматра.
Экология изучена слабо. Населяет влажные горные леса. Встречается в горах на высоте 600—1400 м над уровнем моря. Ведет преимущественно ночной образ жизни. Питается различными саккулентами. День проводит в норах. Как правило, занимает чужие норы, реже копает собственные.
Включен в «Красную книгу».
Род бушменовых зайцев
Genus Bunolagus Thomas, 1929
В роде один вид: бушменов заяц — В. monticularis Thomas, 1903.
Длина тела около 42 см, хвоста 9 см. Конечности короткие, покрытые густым мехом. Уши длинные, почти голые, желтовато-серого цвета. Волосяной покров мягкий, средней высоты, на спине и голове кофейно-коричневого цвета. Брюхо розовато-желтоватое. Вокруг глаз белые кольца. Хвост одноцветный, коричневый. В черепе характерна крупная межтеменная кость. Скуловые дуги не расставлены широко. Небо короткое. Межглазничное пространство широкое. Костные слуховые барабаны средних размеров. Верхние резцы с небольшим желобком.
Распространен в Капской провинции (Южно-Африканская Республика).
Экология не изучена.
Эллермэн (1953) включает бушменова зайца в состав рода зайцев. А. А. Гуреев (1964) отмечает неясное систематическое положение рода бушменовых зайцев и почему-то совсем не приводит видовое название этого зайца в своей монографии. Ангерманн (1966) подтвердила правильность выделения бушменовых зайцев в особый род.
Род щетинистых зайцев
Genus Caprolagus Blyth, 1845
В роде единственный вид: щетинистый заяц — С. hispidus Pearson, 1839.
Размеры средние. Длина тела примерно 47 см. Длина хвоста 2,5 см. Масса 2,5 кг. Уши очень короткие — около 55 мм, и широкие. Глаза маленькие. Задние конечности лишь незначительно длиннее передних. Когти длинные и острые. Остевые волосы грубые, щетинковидные, пух мягкий. Окраска спины темно-коричневая с примесью черных и коричневатобелых волос. Брюхо коричневато-белое. Хвост коричневый, сверху немного темнее.
Череп массивный, с уплощенной крышей. Костное небо удлиненное. Лицевая часть черепа укорочена. Носовые кости относительно небольшие. Межтеменной кости нет. Орбиты небольшие. Костные слуховые барабаны маленькие. Нижняя челюсть крупная, высокая в альвеолярной части и с укороченной дистальной. Щечные зубы и резцы мощные, причем резцы сильно загнуты внутрь.
Ареал охватывает Северо-Восточную Индию, восточную Бенгалию, Непал, Ассам.
Экология не изучена. Держится парами. Роет собственные норы. Питается в основном корой деревьев и корнями.
А. А. Гуреев (1964) включил в род щетинистых зайцев: С. nigricollis F. Cuvier, 1823: С. dayanus Blanford, 1874; С. ruficaudatus Geoffroy, 1826; C. sinensis Gray, 1832; C. peguinsis Blyth, 1855; C. siamensis Boncho-
te, 1902; C. hainanus Swinhoe, 1870; C. yarcandensis Gunther, 1875; C. brachyurus Temminck, 1845; C. marjorita St. Leger, 1929 и C. alleni Me-arns, 1890. По-видимому, более правильно всех их (за исключением предпоследнего вида) отнести к роду Lepus, а С. marjorita — к роду Poelagus (Ангерманн, 1966).
Включен в «Красную книгу». *
Род африканских зайцев
Genus Poelagus St. Leger, 1932
В роде один вид: африканский заяц — Р. marjorita St. Leger, 1929. Размеры средние. Длина тела примерно 45 см. Длина хвоста 5 см. Масса около 2,7 кг. Уши очень короткие. Задние конечности короткие. Когти острые. Волосяной покров жесткий. Остевые волосы грубые, щетинковидные. Пух мягкий, но редкий. Спина серовато-коричневая и желтая, на боках более желтая; затылок красновато-желтый. Брюхо белое. Хвост коричневато-желтый сверху и белый снизу.
Череп массивный, с укороченным лицевым отделом. Сильно развиты гребни скуловых отростков верхнечелюстных костей. Костное небо удлиненное. Нижняя челюсть крупная с высоким альвеолярным отделом и короткой диастемой. Лобные кости длинные. Костные слуховые барабаны относительно крупные. Орбиты относительно большие. Скуловые дуги без длинного каудального отростка. Верхние резцы с простой бороздкой. Передняя поверхность нижних резцов слегка уплощена.
Распространен в Уганде, Заире, Анголе, Руанде, Бурунди, Судане.
Экология не изучена. Встречается в саваннах и лесах. Активен ночью. Днем прячется в зарослях растений. Делает тропы в траве. Сезонности в размножении, по-видимому, нет. Детенышей приносит в норах. Длительность беременности около 35 дней.
А. А. Гуреев (1964) относит африканских зайцев к роду зайцев.
Род зайцев
Genus Lepus Linnaeus, 1758
Размеры средние и крупные. Длина тела 44—47 см. Длина хвоста 5— 11 см. Он хорошо виден снаружи. Нижняя поверхность хвоста белая. Уши длинные (рис. 5). Задние конечности значительно длиннее передних. Подошвы лап покрыты густыми волосами. Обычная окраска бурая или серовато-бурая сверху и более светлая или белая снизу. Зимой у большинства видов наблюдается посветление окраски, а у некоторых (беляк, американский заяц) зимний мех становится чисто белым. Линьки в течение года две. Некоторые виды линяют дважды в году, но не имеют белой зимней окраски. Волосяной покров густой и мягкий, иногда более грубый, но не бывает щетинистым, как у жесткошерстных или африканских зайцев.
Череп крупный, легкого строения, с удлиненным лицевым отделом (см. рис. 1). Верхнечелюстные чешуйчатые кости с резко выраженным сетчатым строением. Небо относительно короткое. .............с от-
ростки короткие и широкие. Костные слуховые барабаны крупные. Орбиты большие. Межтеменная кость срастается с соседними костями. Каудальный отросток скуловой дуги короткий. Заглазничное сужение значительно уже межглазничного. Скуловой отросток верхнечелюстной кости более или менее расширен.
Рис. 5. Беляк — L. titnidus в летнем (Д) и зимнем (Б) меху
У изученных видов (толай, русак, беляк, темношейный, китайский, американский, белохвостый, калифорнийский и антилоповый зайцы) хромосом в диплоидном наборе 48.
Распространение охватывает большую часть Евразии, Африки и Северной Америки. Зайцы успешно акклиматизованы в некоторых местах Южной Америки, Австралии, Новой Зеландии и других, в частности, pjcaK — на северо-востоке США и на юге провинции Онтарио в Канаде.
Населяют весьма различные ландшафты. Большинство живет в открытых травянистых равнинах, некоторые из них как в открытых, так и в антропогенных ландшафтах и в лесах. Американский заяц в основном обитает в вечнозеленых лесах Северной Америки. Капский заяц предпочитает открытые пространства. Беляк населяет преимущественно леса различных типов (в том числе горные), чаще всего их окраины, в тундре — заросли кустарников, а в степях, например в Казахстане, разнообразные биотопы: заросли камыша, березовые колки и т. д. Толай обитает в тугаях, степях, полупустынях и пустынях и иногда поднимается высоко в горы. Русак встречается в пустынно-степных, степных и лесостепных ландшафтах и на полях в лесной зоне. Активны преимущественно в сумерках и ночью. Летом беляк питается преимущественно травянистыми растениями, а зимой — древесно-кустарниковыми (в основном ивой, осиной, березой), толай — летом различными травянистыми растениями, а зимой кустарниками. Русак с ранней весны до поздней осени поедает травянистые, а зимой —- древесные и кустарниковые растения. Отмечена звуковая сигнализация ударами лапы о землю. Если зверек схвачен хищником, он громко кричит. Крик несколько напоминает плач ребенка.
В европейской части СССР беляк начинает размножаться с февраля в южных частях ареала, с марта в средней полосе и с апреля — па севере. Самка дает в течение года 2—3 помета. Па севере третьего помета не бывает. На юге третий помет приносят примерно 7з самок. В каждом помете бывает от 1 до 10, в среднем 3—4 детеныша. Продолжительность беременности примерно 47—55 диен. Зайчата рождаются зрячими, покрытыми шерстью, тотчас начинают сосать мать и способны после этого к самостоятельному передвижению. Половозрслость наступает в годовалом возрасте. Толай дает по 3—4 помета в год,- в каждом из которых по 3—6, реже 8—10 зайчат. Первый помет — во второй половине февраля— начале марта. У русака период размножения растянут с февраля по сентябрь, иногда и позже. За год бывает до 2 пометов на
севере и 3 или даже 4 на юге по 1—7 (обычно 3—4) зайчат в каждом. Американский заяц размножается с марта по сентябрь (иногда и зимой). Длительность беременности 36 дней. За год бывает 4 помета. Среднее число детенышей в помете 3,82 (провинция Альберта). В разные годы численность зайцев подвержена значительным изменениям. В европейской части СССР периоды пониженной численности беляков обычно длятся 5—7 лет, а на северо-востоке Сибири 9—11 лет. Подъем численности характеризуется быстрыми темпами, но коротким периодом. Для американского зайца в Северной Канаде отмечено возрастание численности примерно в 3400 раз.
Зайцы имеют существенное промысловое значение, особенно беляк и русак. В некоторых районах и в отдельные годы зайцы могут приносить известный вред сельскому хозяйству.
Система рода окончательно не установлена. По-видимому, в роде 23 вида
кустарниковый заяц — L. saxalitis F. Cuvier, 1823 (горные области юга Южной Африки);
1 капский заяц — L. capensis Linnaeus, 1758 (включая L. atlanticus de Winton, 1898, L. arabicus Ehrenberg, 1833, и L. salae Jentink, 1880) (широко распространен по Африке);
толай — L. tolai Pallas, 1778 (включая L. tibetanus Waterhause, 1841) (СССР — от северо-восточных берегов Каспийского моря на восток в Южном Казахстане, среднеазиатские республики, Чуйская долина на Алтае, Забайкалье; МНР; Китай, Северо-Западная Индия, Афганистан, Пакистан, Иран);
эфиопский заяц — L. habessinicus Hemprich cl Ehrenberg, 1832 (восточная часть Африки);
рыжебокий заяц — L. crawshay de Winton, 1899 (Африка от юго-востока ЮАР, юга Трансвааля, Ботсваны и севера Юго-Западной Африки на север до Кении и южного Судана; кроме того, северо-восточная Сахара) ;
заяц Уайта — L. whytei Thomas, 1894 (Малави, восток Замбии, север Мозамбика и Танзании);
бирманский заяц — L. peguensis Blyth, 1855 (включая L. hainanus Swinhoe, 1870) (Бирма, Индокитай, Хайнань);
русак — L. europaeus Pallas, 1778 (Европа на север до Англии, Южной Швеции, Южной Финляндии; СССР — европейская часть на север до северных побережий Ладожского и Онежского озер, района г. Архангельска, центральных частей Коми АССР, далее северная граница идет к г. Перми, огибает Уральский хребет с юга и в Азии проходит по линии Шадринск, Курган, Тара, Омск, на восток до линии Целиноград — р. Сырдарья к западу от г. Кзыл-Орда, на юг от северного берега Каспийского моря к северному берегу Аральского моря, постепенно происходит расширение ареала на восток; акклиматизирован в ряде районов Южной Сибири и Дальнего Востока);
сиамский заяц — L. siamensis Bonhotc, 1902 (Таиланд, Лаос, Бирма); темношейный заяц — L. nigricollis F. Cuvier, 1823 (включая L. daya-nus Blanford, 1874; L. ruficaudatus Geoffroy, 1826) (п-ов Индостан, Шри Ланка, акклиматизирован на о-вах Св. Маврикий, Суматра и Калимантан) ;
1 Перечисление дается не по филогенетическому принципу па основании работ Алле на, 1939; Эллермэна и др., 1953; Холла и Кслсопа, 1959; Петтера, 1959, 1963; А. А. Гуреева, 1964; Эллермэна и Моррисон-Скотта, 1966.
беляк — L. timidus Linnaeus, 1758 (включая L. arcticus Ross, 1819, и L. othus Merriam, 1900) (север Западной Европы; СССР — от государственной границы на западе До Тихого океана на востоке, включая Саха’ лин, на север до побережья Северного Ледовитого океана, южная граница проходит примерно от Беловежа через Киев, Харьков, Воронеж, Камышин, Оренбург, Актюбинск, Караганду, оз. Балхаш и на Джунгарский Алатау; МНР; Северо-Восточный Китай; о-в Хоккайдо; северные районы Северной Америки — Канадский Арктический архипелаг, запад и северо-запад Аляски в районах, прилегающих к Чукотскому морю, Берингову проливу и Берингову морю, провинции Канады: северо-восток Маккензи, Киватин, кроме крайнего юго-запада, северо-восток Манитобы, примыкающий к Гудзонову заливу, север Квебека, Ньюфаундленд, кроме юго-запада; южное и северное побережье Гренландии);
курчавый заяц — L. oiostolus Hodgson, 1840 (Тибет, Ганьсу, Сычуань, Юньнань, Кашмир, Непал, Сикким);
китайский заяц — L. sinensis Gray, 1832 (Корейский п-ов, Юго-Восточный Китай и Тайвань);
маньчжурский заяц — L. brachyurus Temminck, 1845 (Япония, Корейский п-ов, Северо-Восточный Китай; СССР — южные районы Дальнего Востока на север до Благовещенска, Малого Хингана, Хабаровска, р. Са-марги);
яркендский заяц — L. yarkandensis Gunther, 1875 (Яркенд в Китае);
антилоповый заяц — L. alleni Mearns, 1890 (США — крайний юг штата Аризона; штаты Мексики-—Сонора, Синалоа, север Наярит, а также о-ов Тибурон в Калифорнийском заливе у побережья Соноры);
американский заяц — L. americanus Erxlebcn, 1777 (север Северной Америки: Аляска, кроме ее севера — районов, примыкающих к морю Бофорта и Чукотскому морю, территории около залива Кускоквим и п-ова Аляска; почти вся Канада, кроме Канадского Арктического архипелага, северо-запада Маккензи, Киватина и северных районов Квебека и Ньюфаундленда; штаты США — Вашингтон, кроме юго-востока, Орегон, кроме северо-востока и крайнего юго-запада и юго-востока, северо-восток Калифорнии, крайний северо-запад Невады, западная половина Монтаны и крайний северо-восток Айдахо, кроме юго-запада, центральные и северо-восточные районы Юты, западная половина Вайоминга, Колорадо, кроме юго-запада и восточной ее трети, север Нью-Мексико, Северная Дакота, кроме юга и юго-запада, северная половина Миннесоты, север Висконсина, Мичиган, кроме крайнего юго-запада, все северо-восточные штаты на юг по границе, которая идет примерно от г. Кливленда на оз. Эри на юго-восток до пересечения с 40° с. ш., затем резко поворачивает на юг до востока и юго-востока Теннесси и запада Северной Каролины, откуда круто сворачивает на северо-восток, проходит через середину штата Виргиния и выходит на побережье Атлантического океана в штате Нью-Джерси).
бесхвостый заяц — L. townsendii Bachan, 1839 (юг Канады — край юга центральной области Британской Колумбии на границе со штатом Вашингтон, южная половина провинций Альберта и Саскачеван, крайний юг Манитобы и небольшой участок па юго-западе Онтарио; западные, центральные и северные штаты США — восточные половины Вашингтона и Орегона, Айдахо, узкая полоса вдоль границы штатов Калифорния и Невада на юг примерно до уровня горы Уитни, северо-восточный угол штата Невада, Юта, кроме юго-востока, Монтана, кроме узкой полосы вдоль западной границы штата, Северная Дакота, Южная Дакота, Миннесота, кроме северо-востока, Айова, кроме южных районов, небольшой участок на юго-западе Висконсина, Небраска, восточная поло-
вина Канзаса, кроме ее самой южной части, Вайоминг, Колорадо, кроме небольших территорий на юго-востоке и юго-западе); «
белобокий заяц — L. mexicanis Lichtenstein, 1830 (южная Мексика — юг штата Дуранго, южная половина Закатекас, юг Сан-Луис-Потоси, Идальго, Тласкала, Пуэбла, север Оахака, северо-восток Герреро, северная половина Мичоакан, Халиско, кроме зоны побережья Тихого океана);
желтоватый заяц — L. flavigularis Wagner, 1844 (крайний юг Мексики: небольшая территория на юго-востоке штата Оахака, примыкающая к Тихоокеанскому побережью);
калифорнийский заяц — L. californicus Gray, 1837 (штаты США — юг середины Вашингтона, примыкающий к штату Орегон, Орегон, кроме северо-востока и северо-запада, южная половина Айдахо, кроме ее восточной части, Калифорния, кроме горных районов, Невада, Юта, кроме северо-востока, Аризона, юго-западный угол и восточная половина Колорадо, Нью-Мексико, небольшая территория в юго-восточном углу Вайоминга, южная половина Южной Дакоты, Небраска, западная половина Миссури, Канзас, Оклахома, северо-запад Арканзаса, Техас, кроме небольшой территории на востоке; север Мексики — п-ов Калифорния, северная половина штата Сонора, Чиуауа, кроме ее западных районов, Коауила, север Дуранго, Нуэво-Леон, запад Тамаулипас, восточная половина Закатекас, Сан-Луис-Потоси, Керетаро, север Халиско, а также о-ва Кармен в Калифорнийском заливе и Маргерита и Магдалена в Тихом океане у п-ова Калифорния) ;
чернобурый заяц — L. insularis Bryant, 1891 (о-в Эспириту-Санто в Калифорнийском заливе);
заяц Гейлларда — L, gaillardi Mearns, 1896 (два небольших изолированных участка, расположенных: один в юго-западном углу штата Нью-Мексико в США, северо-восточном углу штата Сонора и северо-западном углу штата Чиуауа в Мексике и второй — в центральных районах штата Дуранго, примыкающих к границе со штатом Чиуауа);
Эллермэн и Моррисон-Скотт (1966) и Эллермэн и др. (1953) выделяют из L. capensis в особые виды L. atlanticus, L. arabicas, L. salae. Холл и Келсон (1959) и Банфилд (Banfield, 1974) из L. timidus— L. arcticus и L. othus, а С. И. Огнев (1940) и Н. А. Бобринский и др. (1965) из L. total—L. tibetanus. С. И. Огнев (1940) выделяет L. brachyurus в особый род Allolagus. А. А. Гуреев (1964) выделяет L. dayanus, L. rufi-caudatus из L. nigricollis и L. hainanus из L. peguensis и всех их вместе с L. sinensis, L. siamensis, L. yarcandensis, L. orachyurus и L. alleni объединяет в род Caprolagus.
Петтер (1959, 1963) относит к nomina dubia L. aegyptius Desmarest, 1822 (APE и Судан); L. victioriae Thomas, 1893 (Танзания, Кения, Уганда); L. microtis Heuglin, 1865 (Судан); L. zechi Matschie, 1899 (Toro); L. chadensis Thomas et Wroughton, 1907 (окрестности оз. Чад); L. kaku-megae Heller, 1912 (Кения).
В СССР: толай, русак, беляк, манчжурский заяц.
Род бесхвостых кроликов
Genus Romerolagus Merriam, 1896
В роде единственный вид: бесхвостый кролик — R. diazi Diaz, 1893.
Размеры мелкие. Длина тела 28—31 см. Хвост снаружи незаметен (рис. 6). Уши относительно укороченные (4,0—4,4 см), округлые. Задние конечности сравнительно короткие. Волосяной покров мягкий, довольно 20
Рис. 6. Бесхвостый кролик — R. diazi и его череп
густой. Окраска его на спине серовато-бурая и черная с примесью желтых волос. На брюхе пепельно-серая, с грязно-желтым оттенком. Линька, по-видимому, один раз в году.
В черепе характерен укороченный лицевой отдел. Надглазничные отростки короткие. Скуловые дуги сравнительно массивные. Гребни скуловых отростков верхнечелюстных костей сильно развиты. Костные слуховые барабаны относительно крупные.
Распространен на очень ограниченной территории в Мексике — штат Мексике, гора Рио-Фрио, западный склон вулкана Попокатепетль, штат Пуэбла, гора Истаксиуатль, север штата Морелос. Весь ареал составляет в длину примерно 40 км.
Обитает в горных лесах на высоте 300—3600 м над уровнем моря. Держится в зарослях густой травы, где протаптывает хорошо видимые тропинки. Экология изучена слабо. Хотя активен бесхвостый кролик преимущественно ночью, его можно встретить также и днем, особенно в туманные дни и в период размножения. Убежищем служат выкапываемые им норы. Питается молодыми побегами трав. Период размножения приходится на период с марта по июнь — начало июля. Длительность беременности 38—40 дней. В помете от 1 до 4 детенышей (в среднем 3).
Включен в «Красную книгу».
Род американских кроликов
Genus Sylvilagus Gray, 1867
Размеры от мелких до средних. Длина тела 25—54 см, длина хвоста примерно 2,5—6 см. Уши варьируют по длине, но, как правило, они средних размеров (рис. 7). Волосяной покров обычно жесткий, щетинистый, редковатый, у некоторых видов волосы более или менее мягкие. Окраска его варьирует от коричневой до коричнево-серой, иногда на спине красновато-коричневая. Брюшная сторона обычно белая или грязно-желто-белая. Хвост может быть коричневым сверху и белым снизу или одноцветным, темным. Линька, как правило, бывает один раз в год. Зимой мех белым не бывает.
Череп массивный, лицевой отдел укороченный (рис. 7). Глазничные отростки узкие и длинные. Скуловые дуги крупные, с сильно развитыми гребнями скуловых отростков верхнечелюстных костей.
Хромосом в диплоидном наборе у флоридского и степного кроликов и кролика Нуталла по 42, у новоанглийского кролика 52.
Распространены на американском континенте: в Северной Америке от Южной Канады (на запад примерно 50° с. ш., на востоке 45° с. ш.) и далее на юг через Центральную и Южную Америку, до северных границ Патагонии).
Населяют кустарниковые заросли или влажные, тропические леса и болота морских побережий, причем некоторые виды (водяной кролик) приспособились к плаванию. Айдахский кролик живет в пустыне. Ряд видов хорошо приспособились к жизни в антропогенном ландшафте. Только айдахский кролик роет норы, другие виды обычно занимают норы различных животных, особенно в северных частях ареала, или находят убежище в любых укромных местах. Активны в сумерках и ночью. Питаются преимущественно травянистыми растениями. Веточки и кора деревьев поедаются главным образом в зимнее время. Индивидуальная территория невелика. Передвигаются обычно не спеша; наибольшая скорость может достигать 33—40 км в час. Сезон размножения в северной части ареала длится с февраля или марта по сентябрь. В помете от 2 до 7, обычно 4 детеныша. Длительность беременности 26—32 дней. Детеныши рождаются голыми и слепыми, с массой 25—35 г, но развиваются очень быстро. Самка помещает их обычно в гнездо, сделанное из растительных остатков и собственной шерсти в какой либо ямке. Мать обычно кормит детенышей один или два раза в день в течение 16—22 дней. За сезон бывает от 3 до 5 пометов. Глаза открываются у детенышей па 5—8-й день. Гнездо покидают спустя 2 подели после рождения. Сначала держатся вместе, но в возрасте примерно 7 недель семья распадается. Половозрелось наступает в 2—3 месяца. Максимальная продолжительность жизни 3 года.
Рис. 7. Флоридский кролик — S. floridanus и его череп
В США являются одним из важных объектов спортивной охоты.
В роде 12 видов:
айдахский кролик — S. idachoensis Merriam, 1891 (два изолированных участка на западе и юго-западе США — небольшая территория па юго-востоке штата Вашингтон л более крупная, занимающая юго-восток штата Орегон, крайний северо-восток Калифорнии, северную половину Невады, юг Айдахо, кроме ее западных районов, небольшой участок на юге Монтаны и северо-запад Юты);
бразильский кролик — S. brasiliensis Linnaeus, 1758 (юг Северной, Центральная и Южная Америка — Мексика вдоль побережья Мексиканского залива от юга штата Тамаулипас, на юг до штата Чиапас и п-ова Юкатан; Центральная Америка, кроме Тихоокеанского побережья; Ко
лумбия, Венесуэла, Эквадор, Бразилия, Перу, север и северо-восток Аргентины);
калифорнийский кролик — S,. bachmani Waterhouse, 1838 (США — юго-запад штата Орегон, западная часть штата Калифорния, протянувшаяся вдоль Тихоокеанского побережья; штат Нижняя Калифорния в Мексике, а также о-в Седрос);
болотный кролик — S. palustris Bachman, 1837 (штаты США — Флорида, крайний юго-восток Алабамы, южная половина Джорджии, юго-восток Южной Каролины, восток Северной Каролины и крайний юго-восток Виргинии);
флоридский кролик — S. floridanus J. Allen, 1890 (север и северо-запад Южной Америки — Колумбия, Венесуэла и о-ва Маргарита и Лос-Тестигос к северу от Венесуэлы; Центральная Америка — изолированный участок в срединных районах Гондураса и Никарагуа, не выходящих к побережью, крайний северо-запад Коста-Рики, примыкающий к побережью Тихого океана; второй изолированный участок занимает небольшую территорию, захватывая штат Мексики Юкатан и север штата Кампече; остальная часть ареала занимает юго-западный угол Гватемалы и захватывает почти всю Мексику от южной половины штата Чиапас и юго-востока штата Оахака на север через всю страну, кроме узкой полосы, вытянувшейся вдоль Тихоокеанского побережья и берега Мексиканского залива, п-ова Калифорния и центральных штатов Мексики, включающих почти весь штат Чиуауа, запад Коауила, восток Дуранго, и северо-запад Закатекас; в США небольшой «язык» ареала поднимается из Мексики на север, захватывая почти весь юг штата Аризоны, кроме его юго-западного угла, и крайний юго-запад Ныо-Мексико; изолированный небольшой участок находится на западе штата Техас; от восточных частей ареала в Мексике последний продолжается к северу и занимает восточную половину США от Атлантического побережья на востоке до примерно 105° з. д. на западе и на север до линии, проходящей от Атлантического побережья штата Массачузетс к восточной оконечности оз. Онтарио, отсюда поднимающейся к северу примерно до Монреаля и идущей на запад к восточной части оз. Гурон, далее следующей по южным берегам оз. Гурон, Мичиган и Верхнее и от г. Дулут в западной оконечности оз. Верхнее, тянущейся по северной части штата Миннесота, несколько отступя к югу от государственной границы, далее эта линия поворачивает на северо-запад в провинцию Манитоба и достигает южной части оз. Манитоба, откуда следует на юго-запад и переходит в западную границу ареала);
новоанглийский кролик — S. transitionalis Bangs, 1895 (США — узкая полоса, захватывающая юг штата Мэи, Нью-Гэмпшир, кроме севера, Вермонт, кроме северо-востока, восточную половину штата Нью-Йорк, Массачузетс, Коннектикут, Род-Айленд, Пенсильвания, кроме западной трети, Нью-Джерси, Мэриленд, восток Западной Виргинии, запад Виргинии, крайний восток Кентукки, восточная треть штата Теннесси, крайний север Джорджии и северо-восток Алабамы);
кролик Нуталла — S. nuttalli Bachman, 1837 (запал США — восточные половины штатов Вашингтон и Орегон, северо-восток Калифорнии, Айдахо, кроме северо-восточной части, Монтана, кроме крайнего северо-запада, крайний запад Северной Дакоты и крайний юго-запад Южной Дакоты, Вайоминг, кроме юго-востока, западная половина Колорадо, север Ныо-Мексико, северо-восток Аризоны, Юта, кроме крайнего юго-запада, Невада, кроме юга; от штата Вашингтон небольшой участок ареала вдается с юга по 120° з. д. в провинцию Канады Брит. Колумбия, а от шт. Монтана — к северу в провинции Альберта и Саскачеван);
степной кролик — S. audoboni Baird, 1858 (ареал имеет сложную конфигурацию; штаты США — южная половина Калифорнии, вдаваясь языком к северу по долине р. Сакраменто, юг Невады, Юта, кроме северо-запада и узкой полосы, проходящей отсюда по центральным районам на юг и пересекающей Аризону с севера на юго-восток, тогда как вся остальная территория Аризоны входит в состав ареала; юго-восток Монтаны, Вайоминг, кроме крайнего запада, юго-запад Северной Дакоты, запад Южной Дакоты, запад Небраски, Колорадо, кроме центральных районов, Нью-Мексико, Техас, кроме востока, западные половины Оклахомы и Канзаса; Мексика — п-ов Калифорния и штаты Сонора, кроме востока, север Синалоа, Чиуауа, кроме запада, восточная половина Дуранго, Нуэво-Леон, западная половина Тамаулипас, Закатекас, Сан-Луис-Потоси, Ачуаскальентес, север Халиско, северная половина Гуанахуато, Керетаро, Пуэбло, центр Веракрус) ;
водяной кролик — S. aquaticus Bachman, 1837 (юго-восток США — восток штата Техас, восточная половина Оклахомы, Арканзас, Луизиана, штат Миссисипи, Алабама, Джорджия, кроме севера и юго-востока, крайний северо-запад Южной Каролины, запад Теннесси, крайний юго-восток Канзаса, юг и юго-восток штата Миссури, юг Иллинойса);
амильтемский кролик — S. insonus Nelson, 1904 (Мексика — штат Герреро);
мексиканский кролик-—S. cunicularis Waterhouse, 1848 (штаты Мексики— юг Синалоа, западная половина Наярит, запад Халиско, Колима, Мичоакан, кроме крайнего севера, Герреро, западная половина Оахака, крайний запад центральных районов Веракрус, Пуэбла, кроме крайнего севера, Тласкала, Мексико, юг Хидальго);
островной кролик — S. graisoni J. Alien, 1877 (о-ва Лос-Трсс-Марияс у Тихоокеанского побережья Мексики).
Иногда из S. bachmani выделяют особый вид S. mansuetus Nelson 1907 (Холл и Келсон, 1959), а из S. brasilensis — S. gabbi J. Allen, 1877 (Гуреев, 1964). А. А. Гуреев (1964) объединяет калифорнийского и айдахского кроликов в особый род Microlagus Trouessart, 1897.
Род южноафриканских кроликов
Genus Pronolagus Lyon, 1903
В роде единственный вид: южноафриканский кролик — Р. crassicau-datus I. Geoffroy, 1832.
Размеры средние. Длина тела 35—50 см. Длина хвоста 4—13 см. Уши довольно короткие или средних размеров (5—11 см). Задние конечности относительно короче по сравнению с зайцами (рис. 8). Когти на конечностях короткие, тупые. Волосяной покров густой, мягкий, волосы слегка волнистые. Окраска его охристо-рыжая или охристо-бурая, наиболее яркая на голове, боках тела, конечностях и особенно на верхней стороне хвоста. Нижняя сторона тела, внутренняя поверхность пог белая или серовато-белая.
Череп относительно нс массивный. Лобные кости короткие. Надглазничные отростки широкие. Скуловые дуги массивные, с сильно развитыми гребнями скуловых отростков верхнечелюстных костей. Орбиты округлые. Носовые кости длинные.
Распространен в южной части Африки на север до южных окраин Анголы, Заира и Танзании.
Рис. 8. Южноафриканский кролик — Р. crassi
Обитатель в основном скалистых районов Встречается также по опушкам густых лесов. Активен ночью. В отличие от других представителей семейства характерна звуковая сигнализация между отдельными особями не только по поводу тревоги. Пптасюя в основном травянистыми растениями.
Эллермэп и др. (1953) объединяют в род южноафриканских кроликов 3 вида: Р. crassicaudatus, Р. randensis Jameson, 1907 и Р. rupcstris A. Smith, 1834, а Робертс (1954) 9 видов: /’. crassicaudatus; Р. rupcstris; Р melanurus Rflppel, 1842; Р. curryi Thomas, 1902; Р. barreth Roberts, 1948; P. caucinus Thomas, 1929; P. kobosetisis Roberts, 1938; P. ratideusis; P. whitei Roberts, 1938. По-видимому, более правильно объединить их в один вид (Гуреев, 1964).
Рис. 9. Дикий кролик — О. cuniculus и его череп
Род кроликов
Genus Oryctolagus Lilljeborp, 1874
В роде единственный вид: дикий кролик — О. cuniculus Linnaeus, 1758.
Размеры средние. Длина тела 36—44 см, длина хвоста 4—7 см Задние конечности длинные (рис. 9). Уши удлиненные — до 7 см. Глаза крупные. Подошвы лап густо покрыты волосами. Когти сильные и довольно длинные. Окраска мягкого и густого волосяного покрова буровато-серая, несколько светлее на брюхе. Хвост белый снизу и буровато-черный сверху.
Череп не массивный, с удлиненным лицевым отделом (рис. 9). Надглазничные отростки узкие и длинные. Скуловые дуги тонкие. Гребни скуловых отростков верхнечелюстных костей развиты слабо. Орбита округлая. Носовые кости длинные. Костные слуховые барабаны относительно крупные.
Хромосом в диплоидном наборе 44.
Распространен в Центральной и Южной Европе и Северной Африке.
Акклиматизирован в Великобритании, Новой Зеландии, Австралии, Южной Америке и в некоторых частях США. В Австралии в ряде районов размножился в таком количестве, что уничтожал пастбища и стал национальным бедствием.
В СССР акклиматизирован в Юго-Западной Украине, где встречается между реками Днестром и Днепром, под Одессой, Херсоном, Николаевом, а также в Каховском и ряде других районов Херсонской области.
Дикие кролики роют норы. Селятся колониями. Обитают в различных ландшафтах, но предпочитают кустарниковые заросли. Нора имеет одну гнездовую камеру и несколько выходов. Система нор иногда занимает площадь более гектара. От нор далеко не отходят и питаются на прилегающих участках. Поедают в основном различные травянистые растения. Ведут главным образом ночной образ жизни. Пометов от 3 до 5 в год. Число детенышей в помете 4—7, иногда до 9. Продолжительность беременности 28—33 дня. Детеныши рождаются голыми, слепыми, совершен--но беспомощными. Хотя кролики имеют высокую потенциальную скорость размножения, опыты показали, что каждая самка в среднем приносит за год всего 10—11,5 детеныша. По наблюдениям в Австралии показано большое значение в поведении диких кроликов пахучего секрета кожных специфических желез (паховых, анальной, подбородочной), в частности для мечения территории, маркировки субдоминантой и т. д.
Одомашненные кролики имеют существенное значение в народном хозяйстве: от них получают мясо, шерсть, пушнину, а также используют как лабораторных животных.
ОТРЯД ГРЫЗУНОВ
ORDO RODENTIA BOWDICH, 1821
К отряду относятся, как правило, мелкие, иногда средних размеров млекопитающие. Длина тела от 5 см у мышовок до 130 см у капибары Внешний вид в зависимости от образа жизни весьма разнообразен. Ушных раковин нет или они в разной степени развиты от едва заметного кожного валика до больших размеров, достигающих почти половины длины тела. Конечности стопоходящие или полустопоходящие. Передние конечности обычно пятипалые, иногда четырехпалые; число пальцев на задних конечностях изменяется от 5 до 3. Пальцы вооружены когтями разной величины и формы. Хвост снаружи может быть совсем незаметен (как у морских свинок) или очень длинный, раза в полтора превышающий длину тела (тушканчики, мышовки); между этими двумя крайностями можно наметить ряд переходов. Волосяной покров весьма разнообразен— от густого и мягкого, хорошо разделяющегося на категории волос (направляющие, остевые, промежуточные, пуховые) до изрежениого, щетинковидного или даже образующего иглы. Окраска крайне разнообразна. На туловище потовых желез нет, имеются только сальные. Потовые железы расположены на подошвах. Характерны многочисленные специфические кожные железы: в углах рта, анальные, среднебрюшные, поясничные, боковые и др. Сосков 2—12 пар.
Череп образован плотными костями без губчатых участков. Лицевой отдел, как правило, короче мозгового (рис. 10). Глазница открытая, позади соединяется с височной ямкой. Костные слуховые барабаны образованы барабанной костью. Предчелюстные кости имеют отросток, достигающий лобных костей. Сочленовная головка нижней челюсти обычно вытянута в продольном направлении, а сочленовная ямка имеет, соответственно, удлиненную форму, поэтому основное движение нижней челюсти при жевании передне-заднее. Правая и левая половины нижней челюсти соединены между собой подвижно. Сокращение transversus mandibulae может сближать их и отводить вершины нижних резцов в стороны. Зуб-
, -I 0 о — 2 1— 31ООО
пая формула: i — с — р -—- т —~ ~ — 12 — 22.
Клыков нет. Характерна большая диастема между резцами и щечными зубами. Резцы не имеют корней и растут в течение всей жизни животного. Эмаль покрывает только переднюю часть резцов. Такое расположение твердой эмали спереди, а мягкого дентина сзади обусловливает возмож-
1 В пределах отряда по размерам можно выделить 3 группы грызунов: мелкие — с длиной тела 5—46 см, средние — 47—88 см и крупные — 89—130 см.
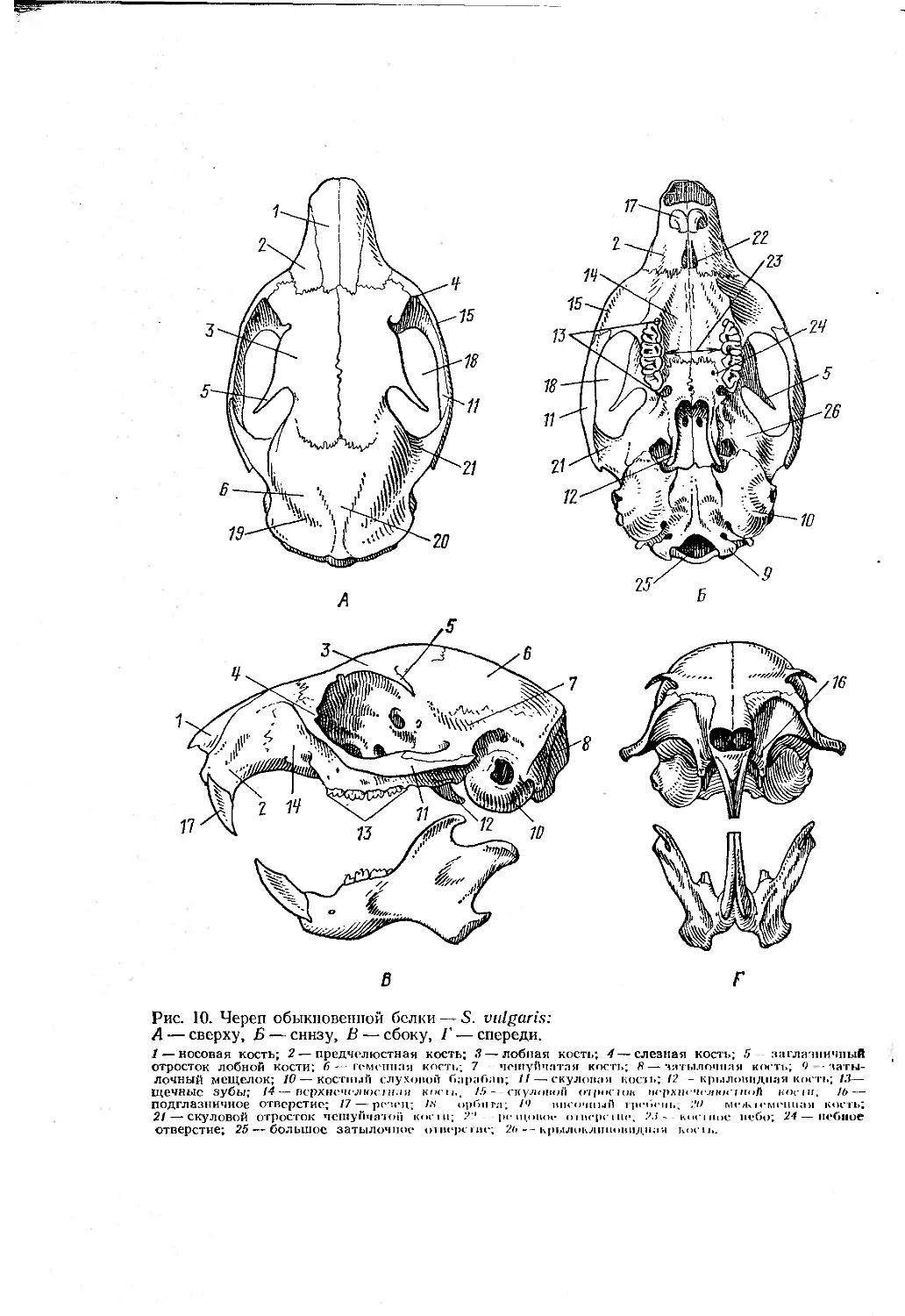
Рис. 10. Череп обыкновенной белки — S. vulgaris:
А — сверху, Б — снизу, В — сбоку, Г — спереди.
1 — носовая кость; 2— предчелюстная кость; 3— лобная кость; 4 — слезная кость; 5 заглачинчпый отросток лобной кости; 6— гомеппая кость; 7 чешуйчатая кость; 8—затылочная кость; <) -затылочный мещелок; 10 — костный слуховой барабан; 11—скуловая кость; 12 - крыловидная кость; 1.3— щечные зубы; 14— верхнечелюстная косы., /5--скуловой отроскж верхнечелюстной ноет, 1Ь— подглазничное отверстие; 17 — резец; М орбита; /Ч височный щебень, 2tf межи’менная кость; 21 — скуловой отросток чешуйчатой кооп; 2? ре иннии- <нвереiне, 2.1 - - к<м*|ное небо; 21— небное Отверстие; 25 — большое затылочное ошерсгие; 2ь— крылоклпиовиднаи к<>си>.
ность постоянного самозатачивания резцов, поэтому их грызущая поверхность всегда очень острая, долотовидная, скошенная назад. Расстояние между правым и левым щечными зубными рядами верхней челюсти обычно несколько меньше, чем между соответственными рядами щечных зубов нижней челюсти. Щечные зубы имеют, а иногда не имеют корней или корни появляются у взрослых (или старых) особей. Жевательная поверхность щечных зубов изменчива — от бугорчатой или гребенчатой до складчатой. Высота коронки щечных зубов также изменчива — от низкой до высокой.
Ключица обычно имеется; иногда она сильно редуцирована. Большая и малая берцовые кости не срастаются или срастаются в дистальной части между собой. Локтевая и лучевая кости свободны. Os penis обычно имеется. Головной мозг крупный. Поверхность полушарий обычно гладкая, и они не прикрывают мозжечок. Комплекс жевательных мышц увеличен в размерах и разделяется на три или большее число порций, имеющих различную функцию. Он может быть выдвинут в оральном направлении книзу от скуловой дуги у Sciuromorpha; проходить сквозь подглазничное отверстие между вентральной и дорзальной ветвями передней части скуловой дуги у Hystricomorpha или образовывать комбинацию этих двух модификаций у Myomorpha.
Желудок простой или многокамерный. Слепая кишка имеется (за исключением сонь); спиральной складки в ней нет. Семенники располагаются в брюшной полости, подкожно в паховой части или в мошонке. Матка двурогая. Плацента дискоидальная, отпадающая, алаптохори-альная.
Распространены почти по всему земному шару, за исключением некоторых арктических и океанических островов и Антарктиды. Расселились всюду, куда проник человек. Обитают почти по всей территории СССР.
Живут в самых разнообразных зонах, высотных поясах и ландшафтах от арктических тундр до пустынь и от равнин, расположенных ниже уровня океана, до субнивального пояса высоких гор. Приспособлены к жизни в разнообразных биотопах. Большинство — наземные, некоторые всю жизнь проводят под поверхностью земли. Есть среди грызунов полу-водные формы, превосходно плавающие и ныряющие. Некоторые виды живут только в кронах деревьев и, пользуясь «планерными» складками кожи на боках тела, могут «перелетать» с дерева на дерево на расстояние до сотни и более метров. Убежища грызунов крайне разнообразны (норы, дупла, расщелины скал и т. д.). Большинство активны круглогодично. Ряд видов в условиях холодных и умеренных зон впадают в спячку разной длительности.
Среди грызунов есть виды только ночные, кормящиеся только в светлую часть суток и активные в любое время суток. Питаются преимущественно растительными кормами: семенами, плодами, сочными зелеными частями растений, даже корой и древесиной; многие виды — также насекомыми и другими беспозвоночными. Некоторые стали исключительно насекомоядными или хищными, например ряд видов крупных крыс.
Плодовитость грызунов различна. Для большинства характерна высокая плодовитость: несколько (до 6—8) пометов в год с большим количеством (до 8—15) детенышей в каждом. У некоторых видов рождение детенышей раз в году (1—2). Многим свойственно раннее половое созревание— на 2-м — 3-м месяце жизни. Для высокоплодовнтых видов характерна неустойчивая численность: годы их крайнего обилия сменяются годами почти полного вымирания па больших территориях. Количество особей на единице площади по годам может различаться в десятки тысяч раз. Известны случаи и полного вымирания на обширных пространствах
за короткое время (желтая пеструшка в Казахстане и на севере Средней Азии).
Практическое значение грызунов при их почти повсеместном распространении, периодически высокой численности велико и разнообразно. Некоторые виды (ондатра, белка, тарбаган) имеют большое промысловое значение. Лесные полевки, лемминги и некоторые из серых полевок (узкочерепная, экономка и пр.), населяющие обширные пространства тайги, лесотундры и тундры, приносят очень большую пользу как основные кормовые объекты ценных пушных хищников (соболя, песца и др.). Есть массовые вредители полевых, садовых и лесных растений, а также запасов пищевых продуктов. Наконец, за счет некоторых грызунов в природе и в непосредственном окружении человека веками существуют очаги опасных для человека болезней (чумы, туляремии, риккетсиозов, спирохетозов, лептоспирозов, лейшманиозов и др.). Особенно большую эпидемиологическую опасность представляют синантропные грызуны — домовая мышь и крысы, из крыс наиболее опасна и вредна серая крыса.
Предками грызунов следует считать примитивных насекомоядных. Возникновение грызунов произошло, по-видимому, в дотретичное время. Во всяком случае, древнейшие из известных ископаемых остатков грызунов в палеоцене Нового Света были уже достаточно специализированными формами. Наиболее древние из известных грызунов Северной Америки и Европы принадлежат к семейству Paramyidae. Они возникли в позднем палеоцене Северной Америки и рапном эоцене Европы (Вууд, 1959). К концу нижнего эоцена появились два новых семейства: Pseudos-ciuridae в Европе и Sciuravidae в Северной Америке. Такое внезапное появление непарамиидных грызунов привело некоторых исследователей (Шауб, 19536) к заключению, что грызуны имеют значительно более длительную историю и населяли некоторые области нашей планеты, где их остатки еще не найдены, и что ранние парамииды не были предками современных грызунов, а просто сохранившейся ветвью предковых форм. Причем последние возникли, вероятно, в мелу и в палеоцене подверглись обширной радиации. Так ли это было на самом деле, сказать с достоверностью в настоящее время невозможно.
После появления грызунов в Европе и Северной Америке эволюция их на этих континентах не была сходной. В Европе псеудосциуриды в среднем эоцене дали начало Theridomyidae, которые просуществовали до нижнего олигоцена. В Северной Америке парамииды были многочисленны в эоцене и начале олигоцена. В то же время обычными были и Sciuravidae. От парамиид или сциуравид в позднем эоцене возникло несколько других семейств грызунов. К раннему олигоцену процесс радиации различных групп грызунов был закончен.
Грызуны представляют собой наиболее обширный по числу семейств (35), родов (355) и видов (1597) отряд млекопитающих. Систематика отряда из-за многочисленности и разнообразия его представителей, иногда имеющих конвергентные признаки, очень трудна и сложна. До настоящего времени среди специалистов нет единого мнения даже относительно крупных таксономических категорий (подотрядов, надсемейств).
Брандт (Brandt, 1855) первым предложил разделение однопарпорез-цовых (Simplicidentata) па Sciuromorplia, Myomorplia и Hystricomorplia. Разделение базировалось па характере расположения жевательной мышцы и скуловой дуги.
Туллберг (Tullberg, 1899) объединяет грызунов по принципу наличия или отсутствия выгиба угла нижней челюсти в две крупные группы — трибы: Hystricognathi и Sciurognathi. В первой из них он выделяет: Ва-thyergomorphi, Hystricomorphi, а во второй — Myomorphi и Sciuromorphi.
Suborder Simplicidentati
Tribus I. Hystricognathi
Subtribus 1. Bathyergomorphi
Familia Bathyergidae
Subtribus 2. Hystricomorphi
Familia Hystricidae
Familia Caviidae
Familia Erethizontidae
Familia Chinchillidae
Familia Aulacodidae (Aulacodus-Thryonomys)
Familia Echinomyidae
Familia Petromyidae
Tribus II. Sciurognathi
Subtribus 1. Myomorphi.
Sectio 1. Ctenodactyloidei
Familia Ctenodactylidae
Sectio 2. Anomaluroidei
Familia Anomaluridae
Familia Pedetidae
Sectio 3. Myoidei
Subsectio Myoxiformes
Familia Myoxidae (Myoxus = Glis)
Subsectio 2. Dipodiformes
Familia Dipodidae
Subsectio 3. Muriformes
Familia Spalacidae
Familia Nesomyidae
Familia Cricetidae
Familia Lophiomyidae
Familia Arvicolidae
Familia Hesperomyidae
Familia Muridae
Familia Gerbillidae
Subtribus 2. Sciuromorphi
Sectio 1. Sciuroidei
Familia Haplodontidae (Haplodon = Aplodontia)
Familia Sciuridae
Sectio 2. Castoroidei
Familia Castoridae
Sectio 3. Geomyoidei
Familia Geomyidae
Вебер (Weber, 1904, 1928) разделяет подотряд Simplicidentata па 10 триб:
1. Haplodontoidea (семейство Haplodontidae);
2. Sciuroidea (семейства Sciuridae, Pteromyidae, Xcridac, Tamiidae, Marmotidae);
3. Castoroidca (семейство Casloridae);
4. Geomyoidca (семейства Ileleromyidae, Geomyidae);
5. Anomaluroidea (семейства Anomaluridae, Pedetidae);
6. Myoxoidea (семейства Myoxidae, Platacanlhoniyidae);
7. Dipodoidea (семейства Sicistidae, Dipodidae);
8. Myoidea (семейства Spalacidae, Nesomyidae, Muridae);
9. Bathyergoidea (семейство Bathyergidae);
10. Hystricoidea (семейства Hystricidae, Erethizontidae, Caviidae,
Chinchillidae, Capromyidae, Octodontidae, Ctenodactylidae, Thryonomyi-dae).
Миллер и Джидли (Miller a. Gidley, 1918) на основании характерных особенностей жевательной мускулатуры и скуловой дуги выделяют среди грызунов 5 надсемейств:
1. Sciuroidae (семейства Sciuridae, Geomyidae, Heteromyidae, Cas-toridae);
2. Muroidae (семейства Muscardinidae, Cricetidae, Platacanthomyi-dae, Rhizomyidae, Spalacidae, Muridae);
3. Dipodoidae (семейства Graphiuridae, Aplodontiidae, Anomaluri-dae, Idiuridae, Zapodidae, Dipodidae, Ctenodactylidae, Pedetidae);
4. Bathyergoidae (семейство Bathyergidae);
5. Hystricoidae (семейства Hystricidae, Erethizontidae, Echimyidae, Petromyidae, Myocastoridae, Thryonomyidae, Dinomyidae, Cuniculi-dae, Heptaxodontidae, Dasyproctidae, Chincillidae, Abrocomidae, Caviidae, Hydrochoeridae).
Винге (Winge, 1924) разделяет грызунов на 8 семейств:
Haplodontidae,
Anomaluridae,
Dipodidae,
Myoxidae,
Myridae,
Hystricidae,
Sciuridae,
Saccomyidae ( = Heteromyidae).
Эллермэн (Ellerman, 1940) возвращается к системе Туллберга (1899), несколько изменяя ее.
Hystricognathi
I. Bathyergomorph Series
1. Надсемейство Bathyergoidae (семейство Bathyergidae).
IL Hystricomorph Series
2. Надсемейство Hystricoidae (семейства Echimyidae, Dinomyidae, Erethizontidae, Dasyproctidae, Hystricidae, Cuniculidae, Chinchillidae) ;
3. Надсемейство Cavioidae (семейство Caviidae).
Sciurognathi
4. Надсемейство Aplodontoidae (семейство Aplodontiidae);
5. Надсемейство Sciuroidae (семейство Sciuridae);
6. Надсемейство Castoroidac (семейство Castoridae);
7. Надсемейство Gcomyoidac (семейства Heteromyidae, Geomyidae). III. Myomorpha Series
8. Надсемейство Anomaluroidac (семейство Anomaluridae);
9. Надсемейство Pcdetoidac (семейство Pedetidae);
10. Надсемейство Ctcnodactyloidac (семейство Ctenodactylidae);
11. Надсемейство Dipodoidae (семейство Dipodidae);
12. Надсемейство Muroidae (семейства Muscardinidae, I.opliiomyi-dae, Spalacidae, Rhizomyidae, Muridae).
Симпсон (1945) предлагает разделять грызунов па три группы, как и Брандт (1855):
Подотряд Sciuromoplia
Надсемейство Aplodontoidca (семейство Aplodontidae);
Надсемейство Sciuroidea (семейство Sciuridae);
Надсемейство Geomyoidea (семейства Geomyidae, Heteromyidae);
Надсемейство Castoroidea (семейство Castoridae):
Sciuromorpha incertae sedis;
Надсемейство Anomaluroidea (семейство Anomaluridae). Anomaluroidea incertae sedis (семейство Pedetidae).
Подотряд Myomorpha
Надсемейство Muroidea (семейства Cricetidae, Spalacidae, Rhizo-myidae, Muridae);
Надсемейство Gliroidea (семейство Gliridae, Platacanthomyidae, Se-leviniidae);
Надсемейство Dipodoidea (семейства Zapodidae, Dipodidae).
Подотряд Hystricomorpha
Надсемейство Hystricoidea (семейство Histricidae);
Надсемейство Erethizontoidea (семейство Erethizontidae);
Надсемейство Cavioidea (семейства Caviidae, Hydrochoeridae, Dinomyidae, Heptaxodontidae, Dasyproctidae);
Надсемейство Chinchilloidea (семейство Chinchillidae);
Надсемейство Octodontoidea (семейства Capromyidae, Octodon-tidae, Ctenomyidae, Abrocomidae, Echimyidae, Thryonomyidae, Pet-romyidae).
Hystricomorpha incertae sedis
Надсемейство Bathyergoidea (семейство Bathyergidae);
Hystricomorpha, или Alyomorpha incertae sedis;
Надсемейство Ctcnodactyloidca (семейство Ctcnodactylidae).
Штелин и Шауб (Stchlin и Schaub, 1951) и Шауб (Schaub, 1953а) предложили разделение грызунов в основном по принципу строения зубной системы. За ними пошли Грассе и Декейзер (Grasse et Dekeyser, 1955). Вводятся новые названия для подотрядов, инфраотрядов, некоторых надсемейств.
Подотряд Non-Pentalophodonta
Надсемейство Sciuroidea (семейство Sciuridae);
Надсемейство Aplodontoidea (семейство Aplodontidae);
Надсемейство Eomyoidea (семейство Anomaluridae);
Надсемейство Gliroidea (семейства Gliridae, Seleviniidae);
Надсемейство Geomyoidea (семейства Heteromyidae, Geomyidae) ;
Надсемейство Ctenodactyloidea (семейство Ctenodactylidae).
Подотряд Myodonta
Надсемейство Dipodoidea (семейства Zapodidae, Dipodidae);
Надсемейство Muroidea (семейства Cricetidae, Platacanthomyidae, Microtidae, Gerbillidae, Muridae).
Nonlophodonta incertae sedis
Надсемейство Pedetoidca (семейство Pedetidae).
Подотряд Penlalophodoiila
Инфраотряд Palaeotrogomorpha
Надсемейство Hystricoidea (семейство Hystricidae);
Надсемейство Castoroidea (семейство Castoridae);
Palaeotrogomorpha incertae sedis (семейства Thryonomyidae, Eupetauridae, Petromyidae, Bathyergidae, Spalacidae, Rhizomyidac).
Инфраотряд Nototrogomorpha
Надсемейство Erethizontioidea (семейство Erethizontidae);
Надсемейство Cavioidea (семейства Caviidae, Hydrochoeridae);
Надсемейство Dinomyoidea (семейства Cuniculidae, Chinchilli-dae, Dinomyidae);
Надсемейство Octodontoidea (семейства Capromyidae, Myocas-toridae, Echimyidae, Abrocomidae, Ctenomyidae, Octodontidae).
Nototrogomorpha incertae sedis, (семейство Dasyproctidae).
Вууд (Wood) в статье 1955 г. не только сохраняет три подотряда Брандта, но и вводит новые:
Подотряд Sciuromorpha
Надсемейство Aplodontoidea (семейство Aplodontidae);
Надсемейство Sciuroidea (семейство Sciuridae).
Sciuromorpha incertae sedis
Надсемейство Ctenodactyloidea (семейство Ctenodactilidae).
Подотряд Theridomyomorpha
Надсемейство Anomaluroidea (семейство Anomaluridae);
Sciuromorpha, или Theridomyomorpha incertae sedis (семейство Pedetidae).
Подотряд Castorimorpha
Надсемейство Castoroidea (семейство Castoridae).
Подотряд Myomorpha
Надсемейство Muroidea (семейства Cricetidae, Muridae);
Muroidea incertae sedis (семейства Spalacidae, Rhizomyidae);
Надсемейство Geomyoidea (семейства Heteromyidae, Georriyi-dae);
Надсемейство Dipodoidea (семейства Zapodidae, Dipodidae).
Myomorpha incertae sedis
Надсемейство Gliroidea (семейства Gliridae, Seleveniidae).
Подотряд Caviomorpha
Надсемейство Octodontoidea (семейства Octodontidae, Echimyidae, Ctenomyidae, Abrocomidae);
Надсемейство Chincilloidea (семейства Chinchillidae, Capromyidae);
Надсемейство Cavioidea (семейства Caviidae, Hydrochoeridae^ Dinomyidae, Heptaxodontidae, Dasyproctidae, Cuniculidae);
Надсемейство Ercthizontoidea (семейство Erethizontidae).
Подотряд Hystricomorpha
Надсемейство Hystricoidea (семейство Hystricidae);
Надсемейство Thryonomycidea (семейства Thryonoinyidae, Pet-romuridae).
Подотряд Bathyergomorpha
Надсемейство Bathyergoidca (семейство Bathyergidae).
В статье 1965 г. Вууд подчеркивает, что в настоящее время трудно с
определенностью утверждать, на какие подотряды следует разделить грызунов. Он предлагает следующую систему:
Подотряд Protrogomorpha
Надсемейство Aplodontoidea (семейство Aplodontidae).
Подотряд Caviomorpha
Надсемейство Octodontoidea (семейства Octodontidae, Echimyidae, Ctenomyidae, Abrocomidae, Capromyidae);
Надсемейство Chinchilloidea (семейства Chinchillidae, Dasyproctidae, Cuniculidae, Heptaxodontidae, Dinomyidae);
Надсемейство Cavioidea (семейства Caviidae, Hydrochoeridae); Надсемейство Erethizontoidea (семейство Erethizontidae).
Подотряд Myomorpha
Надсемейство Muroidea (семейства Cricetidae, Muridae);
Надсемейство Geomyoidea (семейства Geomyidae, Heteromyidae);
Надсемейство Dipodoidea (семейства Dipodidae, Zapodidae);
Надсемейство Spalacoidea (семейства Spalacidae, Rhizomyidae); Надсемейство Gliroidea (семейства Gliridae, Seleveniidae).
He входят в подотряды:
семейство Sciuridae;
Надсемейство Castoroidea (семейство Castoridae);
семейство Ctenodactylidae;
семейство Anomaluridae;
семейство Pedetidae;
семейство Hystricidae;
семейство Bathyergidae.
Учитывая значительное расхождение во мнениях ученых относительно деления грызунов на подотряды, надсемейства, а иногда и семейства, по-видимому, для наших целей целесообразно следом за Андерсоном и Джонсом (1967), принимая в основном систему Симпсона (1945), не выделять специально подотряды, однако условно объединить грызунов в три группы, название каждой из которых поставить в кавычки. Таким образом, мы принимаем в отряде грызунов 35 семейств и 355 родов:
«Белкообразные» — «Sciuromorpha» Brandt, 1855
Надсемейство Aplodontoidea Matthew, 1910;
семейство аплодонтовых Aplodontidae Trouessart, 1897 (1 род); Надсемейство Sciuroidea, 1872;
семейство беличьих—Sciuridae Gray, 1821 (39 родов);
семейство летяговых—Pteromyidae Brandt, 1855 (13 родов); Надсемейство Geomyoidea Weber, 1904;
семейство гоферовых — Geomyidae Gill, 1872 (8 родов);
семейство мешотчатых прыгунов — Heteromyidae Allen et
Chapman, 1893 (5 родов);
Надсемейство Castoroidea Gill, 1872;
семейство бобровых-—Castoridae Gray, 1821 (1 род);
Надсемейство Anomaluroidea Gill, 1872;
семейство шипохвостовых — Anomaluridae Gill, 1872 (Зрода);
семейство долгоноговых — Pedetidae Owen, 1847 (1 род);
«Мышеобразные» — «Myomorpha» Brandt, 1855
Надсемейство Muroidea Miller et Gidley, 1918;
семейство хомяковых — Cricetidae Rochebrune, 1883 (396 родов) ;
семейство слепышевых — Spalacidae Gray, 1821 (1 род);
семейство бамбуковых крыс — Rhizomyidae Miller et Gibley, 1918 (3 рода);
семейство мышиных — Muridae Gray, 1831 (100 родов);
Надсемейство Gliroidea Simpson, 1945;
семейство соневых — Gliridae Thomas, 1897 (7 родов);
семейство колючих сонь — Platacanthomyidae Miller et Gidley, 1918 (2 рода);
семейство селевиниевых — Seleviniidae Argyropulo et Vinogradov, 1939 (1 род);
Надсемейство Dipodoidea Weber, 1904;
семейство мышовковых Zapodidae Coues, 1875 (4 рода);
семейство тушканчиковых — Dipodidae Waterhouse, 1842 (10 родов);
«Дикобразные» — «Hystricomorpha» Brandt, 1855
Надсемейство Hystricoidea Gill, 1872;
семейство дикобразовых — Hystricidae Burnett, 1830 (4 рода);
Надсемейство Erethizontoidea Simpson, 1945;
семейство американских дикобразов — Erethizontidae Thomas, 1897 (4 рода);
Надсемейство Cavioidea Kraglievich, 1930;
семейство свинковых — Caviidae Waterhouse, 1839 (5 родов);
семейство водосвинковых —Hydrochoeridae Gill, 1872 (5 родов);
семейство пакорановых — Dinomyidae Alston, 1876 (1 род);
семейство семизубовых — Heptaxodontidae Miller et Gidley, 1918 (2 рода);
семейство агутиевых — Dasyproctidae Smith, 1842 (4 рода);
Надсемейство Chinchilloidea Kraglievich, 1940;
семейство шиншилловых — Chinchillidae Bennett, 1833 (3 рода);
Надсемейство Octodontoidea Simpson, 1945;
семейство хутиевых — Capromyidae Smith, 1842 (3 рода);
семейство нутриевых — Myocastoridae Miller et Gidley, 1918 (1 род);
семейство восьмизубовых — Octodontidac Waterhouse, 1839 (5 родов);
семейство тукотуковых — Ctenomyidae Tate, 1935 (1 род);
семейство шиншиллокрысовых — Abrocomidae Miller et Gidley, 1918 (1 род);
семейство колючих шиншилл — Echimyidae Miller el Gidley, 1918 (14 родов);
семейство тростниковых крыс — Thryonomyidae Pocock, 1922 (1 род);
семейство скальных крыс — Petromyidae Tullberg, 1899 (1 род).
Не входят в указанные выше 3 группы:
Надсемейство Bathyergoidea Osborn, 1910;
семейство землекоповых — Bathyergidae Waterhouse, 1841 (5 родов);
Надсемейство Ctenodactyloidea Simpson, 1945;
семейство гундиевых — Ctenodactylidae Zittel, 1893 (4 рода).
Белкообразные
Sciuromorpha Brandt, 1855
В черепе скуловая дуга образована в основном скуловой костью. Подглазничное отверстие небольшое. Его ширина редко превышает ширину первого коренного зуба. Жевательная мышца не проходит сквозь подглазничное отверстие. В нижней челюсти всегда больше трех щечных зубов. Малая и большая берцовые кости разделены.
СЕМЕЙСТВО АПЛОДОНТОВЫХ
FAMILIA APLODONTIDAE TROUESSART, 1897
Наиболее примитивные среди грызунов и наиболее древние из известных групп современных грызунов. Размеры мелкие. Длина тела 30— 46 см. Длина хвоста 1—3,5 см. Масса 900—1800 г. Сложение тяжелое (рис. 11). С виду это неуклюжие животные, с крупной головой, короткими стопоходящими ногами. Ушные раковины небольшие, округлые на вершинах, едва выступающие над уровнем меха. Глаза маленькие. Передние и задние конечности пятипалые, но на передних конечностях I палец очень короткий. На задних конечностях I и V пальцы короче средних. Пальцы с сильными когтями, на передних конечностях особенно крупными. Волосяной покров плотный, мягкий и относительно короткий. Окраска его на спинной стороне одноцветно-серая, серовато-или красновато-бурая, на брюшной слегка светлее. С возрастом животные сереют. Линька в течение года одна; начинается в июле или августе и продолжается 2—3 месяца. Сосков 3 пары. Имеется анальная железа, продуцирующая секрет с характерным мускусным запахом.
Череп сильно уплощен и резко расширен в задней части (рис. 11). Скуловые дуги широко расставлены. Нижняя челюсть очень большая и мощная. Угловой отросток нижней челюсти необычайно сильно завернут внутрь. Венечный отросток высокий и загнут назад. Костное небо широкое, продолжается позади последних щечных зубов. Посторбитальных 10 2 3
отростков нет. Зубная формула: t — с — р — т — = 22.
Щечные зубы с постоянным ростом. Большая и малая берцовые кости не срастаются. Os penis имеется.
Характерно очень сильное развитие слюнных желез. Кардинальная часть желудка вытянута в рогоподобный ио форме отросток. Семенники находятся в брюшной полости, а в период размножения образуется полумошонка.
Распространены в западной части Северной Америки, в Каскадных горах и горах Сьерра-Невада, включая крайний юго-запад провинции Британская Колумбия в Канаде, запад штатов Вашингтон и Орегон и крайний северо-запад штата Калифорния; кроме того, имеется три изолированных участка: один в виде узкой полосы проходит с севера па юг в северо-восточной части штата Калифорния, захватывая небольшую
Рис. 11. Аплодонтиа — A rufa и ее череп
территорию на западе штата Невада; два других располагаются в центральных районах штата Калифорния на его Тихоокеанском побережье.
Населяют лесные районы и территории, покрытые густой растительностью. Предпочитают хорошо оводненные места, районы, где бывают частые и сильные дожди.
Обычно селятся вдоль водоемов, где почва насыщена влагой. Встречаются почти от уровня моря до 2200 м над ним. Ведут преимущественно подземный образ жизни Выкапывают сложную систему пор, близко расположенных к поверхности земли, в диаметре 10—-25 см. Норы имеют гнездовую камеру и многочисленные входы. Гнездовая камера располагается примерно на глубине 1,5 м от поверхности земли. В качестве подстилки используются ветви деревьев и мягкие сухие растения. Отнорки гнездовой камеры служат уборными. Рядом находится кормовая камера. Обычно не уходят от гнездовой камеры далее 50—120 м. Подземные туннели часто чистят, а если они заполняются водой после дождя, то аплодонтии по ним плавают. Зимой делают туннели в снегу, иногда передвигаются и по поверхности снега. В спячку не впадают.
Питаются надземными и подземными частями различных растений, предпочитая сочные стебли. Обычно кормятся на-земле, но иногда, чтобы погрызть ветви, могут залезать и на деревья до 6 м высотой. Характерно запасание пищи в норах. Ведут преимущественно ночной образ жизни, но иногда активны и в дневное время.
Период спаривания приходится на конец зимы или раннюю весну и продолжается примерно 6 недель. За год самка дает 1 помет. Длительность беременности 28—30 дней. С февраля по апрель самка приносит 2—6 детенышей (обычно 2 или 3). Рожает в гнездовой камере, выстланной сухой травой. Новорожденные слепые. Глаза открываются примерно в 10-дневном возрасте. Лактация длится около 2 месяцев. К концу июня полувзрослые детеныши покидают гнездо. Половозрелости самки достигают в 2 года. Местами животные могут вредить сельскохозяйственным культурам.
В семействе 1 род аплодонтов —Aplodontia Richardson, 1829, и 1 вид.
Род аплодонтов
Genus Aplodontia Richardson, 1829
См. описание семейства.
В роде один вид: аплодонтиа — A. rufa Rafinesque, 1817 (ареал совпадает с ареалом семейства).
СЕМЕЙСТВО БЕЛИЧЬИХ
FAMILIA SCIURIDAE GRAY, 1821
Размеры изменчивы: от мелких до средних. Длина тела от 6 (мышиная белка) до 60 см (суркН) ’. По внешнему виду представителей семейства можно в общем разделить на 2 типа: сходных, с одной стороны, с хорошо известными всем белками, с другой — с сусликами. Глаза довольно крупные. Конечности хорошо развиты; задние обычно длиннее передних, но не более чем в 2 раза. Задние конечности пяти-, передние четырех- или пятипалые. I.V палец на передних и задних конечностях наиболее длинный. Пальцы с острыми когтями. Длина хвоста варьирует от короткого до длинного (длиннее тела). Хвост всегда
густо покрыт волосами, иногда длинными с кистью на конце. Волосяной покров густой и мягкий, относительно высокий или сильно изреженный, щетинковидный. Окраска его одноцветная или с полосами и пятнами,
варьирует от черного и белого до рыжего или темно-грязно-желтого. Сосков от 2 пар у некоторых тропических и древесных белок до 6 пар у некоторых неоарктических земляных белок.
Череп с хорошо развитыми посторбитальными отростками. Костные
слуховые барабаны относительно небольшие, округлые по форме. Резцовые отверстия обычно короткие. Межглазничное пространство широкое. Скуловые дуги расставлены сильно или слабо. Костное небо до-
вольно широкое, иногда слегка продолжается каудальнее последних
1 о 1 _____2 3
щечных зубов. Зубная формула:/ — с — р —-— т -у = 20 — 22.
Щечные зубы с корнями; брахиодонтного или гипселодонтиого типа. Малая и большая берцовые кости нс срастаются.
Распространены по всему земному шару, за исключением Австралийской области, Мадагаскара, южной части Южной Америки (Патагонии,
1 В пределах семейства можно выделить 3 группы беличьих: мелкие с длиной тела до 23 см, средние — 24—42 см, крупные — 43—60 см.
Чили, большей части Аргентины), полярных областей и некоторых пустынь п-ова Аравия и АРЕ.
Населяют самые разнообразные ландшафты: леса, открытые равнины, пустыни, тундры, горы, от тропиков до Арктики. Ведут наземный и древесный образ жизни. Активны главным образом днем. Питаются преимущественно различными растительными объектами, иногда насекомыми и мелкими позвоночными. Некоторые виды впадают в зимнюю спячку. Длительность беременности 22—45 дней. Самки приносят от 1 до 15 голых и слепых детенышей. Для некоторых видов отмечены дальние миграции. Ведут одиночный, иногда колониальный образ жизни.
Ряд видов (белка, сурок) имеют промысловое значение. В семействе есть вредители сельского хозяйства (ряд сусликов) и хранители опасных заболеваний человека (некоторые сурки, суслики).
В семействе 39 родов (228 видов) (в нижеследующем списке родов за основу принят порядок, предложенный Симпсоном, 1945, с незначительными изменениями):
белок-—Sciurus Linnaeus, 1758 (36 видов);
бороздчаторезцовых белок — Syntheosciurus Bangs, 1902 (2 вида);
карликовых белок-—Microsciurus J. Allen, 1895 (17 видов);
белок-пигмеев — Sciurillus Thomas, 1914 (1 вид);
карликовых сулавесских белок — Prosciurillus Ellerman, 1949 (3 вида);
рубиновых белок — Rubrisciurus Ellerman, 1954 (1 вид);
кистеухих белок — Rheithrosciurus Gray, 1867 (1 вид);
красных белок — Tamiasciurus Trouessart, 1880 (2 вида);
пальмовых белок — Funambulus Lesson, 1832 (5 видов);
гигантских белок — Ratufa Gray, 1867 (4 вида);
масличных белок — Protoxerus Major, 1893 (2 вида);
африканских пальмовых белок—Epixerus Thomas, 1909 (1 вид);
полосатых белок — Funisciurus Trouessart, 1880 (6 видов);
кустарниковых белок — Paraxerus Major, 1893 (11 видов);
солнечных белок — Heliosciurus Trouessart, 1880 (3 вида);
мышиных белок — Myosciurus Thomas, 1909 (1 вид);
прекрасных белок — Callosciurus Gray, 1867 (17 видов);
сулавесских белок — Hyosciurus Tate et Archbold, 1935 (1 вид);
сундасциурус — Sundasciurus Moore, 1958 (12 видов);
тамиопс -— Tamiops J. Allen, 1906 (2 вида);
многополосых белок — Menetes Thomas, 1908 (1 вид);
длинноносых белок-—Rhinosciurus Gray, 1843 (1 вид);
малайских белок — Lariscus Thomas et Wroughton, 1909 (2 вида);
дремомис — Dremomys Heude, 1898 (4 вида);
калимантанских белок — Glyphctes Thomas, 1898 (1 вид);
черноухих белок — Nannosclurus Trouessart, 1880 (5 видов);
крошечных белок — Exilisciurus Moore, 1958 (2 вида);
магрибских белок — Atlantoxerus Major, 1893 (1 вид);
земляных белок — Xerus Hemprich et Ehrenberg, 1832 (4 вида);
тонкопалых сусликов—Spermophilopsis Blasius, 1884 (1 вид);
бурундуков— Tamias liliger, 1811 (18 видов);
беличьих бурундуков — Sciurotamias Miller, 1901 (2 вида);
скалистых сусликов-—Olospermophilus Brandl, 1844 (8 видов);
пустынных сусликов — Xerospermophilus Merriam, 1892 (2 вида);
антилоповых сусликов — Ammospermophilus Merriam, 1892 (5 видов); иктидомис — Ictidomys Allen, 1877 (5 видов);
сусликов — Citellus Oken, 1816 (20 видов);
сурков — Marmota Frisch, 1775 (13 видов);
луговых собачек—Cynomys Rafinesque, 1817 (5 видов);
Эллермэн (1940) по-другому располагает роды в семействе Sciuridae:
Myosciurus Thomas, 1909
Ndnnosciurus Trouessart, 1880
Sciurillus Thomas, 1914
Microsciurus J. Allen, 1895
Syntheosciurus Bangs, 1902
Sciurus Linnaeus, 1758
Tamiasciurus Trouessart, 1880
Callosciurus Gray, 1867
Funambulus Lesson, 1832
Dremomys Heude, 1898
Ratufa Gray, 1867
Menetes Thomas, 1908
Lariscus Thomas et Wroughton, 1909
Glyphotes Thomas, 1898
Rheithrosciurus Gray, 1867
Rhinosciurus Gray, 1843
Hyosciurus Tate et Archbold, 1933
Heliosciurus Trouessart, 1880
Paraxerus Major, 1893
Funisciurus Trouessart, 1880
Protoxerus Major, 1893
Epixerus Thomas, 1909
Xerus Hemprich et Ehrenberg, 1832
Atlantoxerus Major, 1893
Spermophilopsis Blasius, 1884
Sciurotamias Miller, 1901
Tamias liliger, 1811
Citellus Oken, 1816
Marmota Blumenbach, 1779
Cynomys Rafinesque, 1817
Myyp (1959) предлагает иной порядок расположения родов в семействе Sciuridae (трибы и подтрибы в списке нами опущены):
Ratufa Gray, 1867
Protoxerus Major, 1893
Epixerus Thomas, 1909 z
Heliosciurus Trouessart, 1880
Funambulus Lesson, 1832
Funisciurus Trouessart, 1880
Paraxerus Major, 1893
Myosciurus Thomas, 1909
Callosciurus Gray, 1867 (включая Tamiops)
Sundasciurus Moore, 1958
Glyphotes Thomas, 1898
Xannosciurus Trouessart, 1880
Dremomys Heude, 1898
Lariscus Thomas et Wroughton, 1909
Menetes Thomas, 1908
Rhinosciurus Gray, 1843
Hyosciurus Archbold et Tate, 1935
Prosciurillus Ellerman, 1947
Rubrisciurus Ellerman, 1954
Exilisciurus Moore, 1958
Rheithrosciurus Gray, 1867
Sciurus Linnaeus, 1758 !
Guerlinguetus Gray, 1821
Microsciurus Allen, 1895
Syntheosciurus Bangs, 1902
Sciurillus Thomas, 1914
Tamias liliger, 1811
Eutamias Trouessart, 1880
Spermophilus Cuvier, 1825 ( = Citellus) (включая Otospermophilus, Ic-tidomys, Xerospermophilus)
Ammospermophilus Merriam, 1892
Cynomys Rafinesque, 1817
Marmota Frisch, 1775
Tamiasciurus Trouessart, 1880
Sciurotamias Miller, 1901
Xerus Hemprich et Ehrenberg, 1832
Atlantoxerus Major, 1893
Spermophilopsis Blasius, 1884
Многие исследователи (например, Симпсон, 1945) включают летяг в семейство беличьих в качестве подсемейства. Следуя Пококку (Ро-cock, 1923) и большинству отечественных систематиков, здесь они выделены в особое семейство.
Род белок
Genus Sciurus Linnaeus, 1758
Размеры мелкие и средние. Длина тела 20—31 см. Длина хвоста 20—31 см. Хвост всегда густо покрыт волосами (рис. 12). Длина его может быть больше длины тела, примерно равна ей или значительно короче. Задние конечности заметно длиннее передних. На передних и задних конечностях самый длинный палец четвертый. Уши обычно крупные, иногда с кисточкой волос на вершине, как у S. vulgaris и S. aberti. Волосяной покров у большинства форм в зимнее время довольно высокий, густой и мягкий, а летом более грубый, редкий и короткий. Линька два раза в году; хвост линяет один раз. У обыкновенной белки весенняя линька происходит непосредственно вслед за течкой. У самцов она начинается раньше и растягивается до середины лета. Самки линяют позднее самцов, но более интенсивно — линька у них заканчивается к рождению молодых. Волосы весной сначала начинают линять на голове (вокруг глаз) и постепенно линька передвигается по спине к основанию хвоста, а потом спускается на бока и конечности. Осенью первыми также начинают линять самцы, после окончания гона. Взрослые самки, не имевшие второго выводка, линяют раньше самок, принесших второй помет. Осенняя линька начинается у основания хвоста и продвигается по спине к голове, одновременно линяют задние конечности. Линька молодых примерно совпадает со сроками линьки взрослых самок. Обычная окраска спинной стороны тела одноцветная серая, сероватобурая, черновато-бурая с различными оттенками рыжего цвета, брюшной— белая, грязно-желтая, желтая или оранжевая. Хорошо выражены географическая изменчивость окраски меха и полиморфизм окраски в пределах одной популяции. Обычно выделяют пять форм окраски волосяного покрова (Кузнецов, 1965):
1. Краснохвостка; зимний мех светло-серый, летний ярко-рыжий; хвост и кисточки на ушах рыжие; остевые волосы на хвосте у основания серые, на вершинах рыжие. 2. Бурохвостка: зимний мех серый сред-
Рис. 12. Обыкновенная белка—5. vulgaris
ней темноты или светло-серый, летний рыжий или буроватый, хвост и кисточки на ушах бурые; остевые волосы на хвосте у основания серые, посередине коричневые, на вершинах черные. 3. Темнохвостка: зимний мех темно-серый, летний бурый или черноватый; кисточки на ушах черноватые; хвост черный; остевые волосы на хвосте с сероватыми основаниями. 4. Чернохвостка: зимний мех темно-серый, летний черный; хвост и кисточки на ушах черные. 5. Серохвостка: зимний мех очень светлый, серебристо-серый; хвост серый, седоватый, кисточки па ушах обычно рыжие.
Череп с крупной мозговой частью, резко снижающейся в заднем направлении (см. рис. 10). Сагиттального гребня пет, теменные гребни развиты слабо. Заглазпичиые отростки средних размеров Костные слуховые барабаны обычно относительно крупные. Костное небо широкое, оканчивается сразу позади зубных рядов Зубная формула-. 1 0 1—2 .3 on оо
t — с — р-----т — = 20 — 22.
10 1 3
Распространены очень широко; в Европе от Ирландии па восток и от Северной Скандинавии до Средиземного моря; большая часть европейской части СССР, на юг до Крыма, Кавказа, Закавказья; Малая Азия, частично Сирия, Иран; Сибирь на восток до Анадырского залива и на юг до Алтая, Трансбайкала, Амура и Уссури; северная часть МНР;
Северный Китай; Корейский п-ов; Сахалин; Япония; юго-восточная Канада, большая часть США, Мексика; на юг через Центральную Америку до Колумбии, Эквадора, Перу, Боливии, Северной Аргентины, Бразилии, Гвианы, Венесуэлы; о-ва Тринидад и Тобаго.
Населяют различные типы лесов: от северных до тропических. Большую часть жизни проводят на деревьях, превосходно лазая и прыгая с ветки на ветку. Наибольшая активность приходится на раннеутренние и вечерние часы. Питаются преимущественно различными растительными объектами. Обыкновенная белка в основном питается семенами хвойных пород: ели, сосны, кедра, лиственницы, пихты, а также желудями, орехами, грибами, почками и побегами лиственных деревьев, ягодами, иногда и животными кормами — такими, как насекомые, яйца птиц, мелкие птицы, их птенцы. На зиму обыкновенная белка делает небольшие запасы желудей, орехов, грибов.
Убежищами белок служат гнезда, которые они устраивают в дуплах или на ветвях деревьев. Гнездо-гайно шарообразной формы с боковым входом, сделано из сучьев различной величины. У обыкновенной белки тайно часто выстлано лыком.
В спячку не впадают, но в штормовую или очень холодную погоду могут долго оставаться в гнезде. У большинства представителей рода, как правило, два помета в году. У обыкновенной белки весенняя течка приходится на конец января — начало марта. Вторая течка — летом. Течка продолжается около двух недель. Длительность беременности 38— 39 дней у обыкновенной белки, примерно 44 дня — у серой. В помете у обыкновенной белки от 3 до 10 детенышей, а у серой от 1 до 5, обычно 2—3. Новорожденные детеныши обыкновенной белки имеют длину тела 5—5,6 см, массу 7,5—8,5 г. Они голые и слепые. Волосяной покров у них появляется на 14-е сутки, резцы нижней челюсти прорезаются на 21 — 23-и сутки, резцы верхней челюсти — на 37—41-е сутки. Глаза открываются на 30—32-е сутки. К этому времени бельчата уже могут лазать.
Для ряда видов белок характерны дальние миграции. Обыкновенные белки во время миграции передвигаются в большом количестве, достигая по фронту иногда 100—300 км. Скорость передвижения во время миграции составляет 3—4 км/ч. Во время миграций они могут переплывать реки, большие озера (как Телецкое на Алтае) и даже морские заливы. Продолжительность миграции и число вовлеченных в нее особей может быть значительным. Так, в 1917 г. белка шла через Северную Двину непрерывно в течение 45 дней. Многие гибнут в пути. Во время миграции обыкновенная белка иногда заходит в тундру и проникла (примерно в 1920 г.) на Камчатку. Для белок характерны значительные изменения численности по годам; резкое уменьшение численности обычно следует на следующий год после неурожая кормов. На численность белок влияют и метеорологические факторы: ранние или поздние заморозки, дождливые весна, лето и осень, засуха летом, суровая зима и т.д. Серая белка в неволе жила до 15 лет.
Большинство видов рода не имеет какого-либо существенного хозяйственного значения для человека, за исключением обыкновенной белки, которая в СССР занимает важное место в заготовках пушнины (в отдельные годы заготавливали до 9—21 млн. шкурок).
Число видов в роде окончательно не установлено. На территории Сев. и Юж. Америки Холл и Келсоп (1959) и Кабрера (Cabrera, I960) насчитывают 34 вида. Вместе с 2 видами Евразии это составляет 36 видов:
обыкновенная белка — S. vulgaris Linnaeus, 1758 (Британия и Ирландия, Франция, Испания, Италия, Швейцария, Норвегия, Швеция, Дания, Финляндия, ФРГ, ГДР, Нидерланды, Венгрия, Польша, Чехо
Словакия, Австрия, Югославия, Румыния, Болгария, Греция; СССР — в лесной и лесостепной зонах от западных границ на восток до Сахалина и Тихоокеанского побережья, Сахалина и Камчатки включительно; северная граница ареала в СССР проходит от г. Печенги на севере Кольского п-ова, южнее Мурманска, севернее оз. Имандра к Терскому берегу, проходит восточнее горла Белого моря и далее от устья р. Мезени по р. Суле, северным притокам р. Усы к Северному Уралу, затем следует несколько севернее г. Салехард, пересекает р. Таз примерно в 250 км выше устья, идет к г. Дудинка, устью р. Хатанга, пересекает реки Лену, Яну и Индигирку примерно по 70° с. ш., р. Колыму — у г. Нижне-Ко-лымска, р. Анадырь — в ее среднем течении и выходит к Пенжинской губе; Камчатку заселила в 20-х годах текущего столетия; Япония; южная граница проходит по Северо-Восточному и Северному Китаю, Северной Монголии и Калбинскому Алтаю, пересекает р. Иртыш у г. Омска, следует на г. Курган, г. Магнитогорск, Южный Урал и через города Бузулук— Куйбышев —Саратов-— Воронеж — Купянск— Артемовск — Харьков — Полтава — Днепропетровск — Тирасполь к лесам Центральной Молдавии; имеются островные участки распространения, изолированные от основного ареала, как к югу, так и к северу от него, а также на крайнем северо-востоке; акклиматизирована в 1940 г. в лесах Южного Крыма, в 1937 г.— в горных лесах Главного Кавказского хребта, в Тебердинском заповеднике, в 1951—1954 гг. — в сосновых островных лесах Северного и Центрального Казахстана;
кавказская белка — S. anomalus Gmelin, 1778 (Иран; Восточная Турция; Сирия; Палестина; Израиль; СССР — южный склон Главного Кавказского хребта от Абхазии на восток до р. Герды-ман-Чая, северный склон Малого Кавказа от р. Чороха до восточных склонов Карабаха, окрестности Дилижана, Кировакана, оз. Гель-Гель, хребет Муров-Даг; на юге Армении и Азербайджана отсутствует);
каролинская белка — S. carolinensis Gmelin, 1788 (юго-восток Канады •—юг провинции Манитобы, юго-восток Онтарио, юг Квебека; по всей восточной половине США — на восток до Атлантического океана, кроме небольшого участка центральной части штата Флорида, примыкающей к Мексиканскому заливу, на западе граница проходит с севера через центральные районы штата Северная Дакота, пересекает штат Миннесота в его юго-западном углу, далее проходит в западной части штата Иова, в юго-восточном углу Небраски, центральных районах Канзаса и Оклахомы, пересекает почти посредине Техас и выходит к побережью Мексиканского залива);
золотистобрюхая белка — S. aureogaster Cuvier, 1829 (запад Мексики— штаты Тамаулипас, Веракрус, запад Сан-Луис-Потоси, запад Керетаро, запад Идальго, крайний запад Оахака, юг Табаско, крайний север Чияпас; неб. территория на крайнем северо-западе Гватемалы;
мексиканская серая белка — S. poliopus Fitzingcr, 1867 (юг Мексики— штаты Халиско, Колима, Герреро, Мичоакан, Пуэбла, Оахака, Наярит);
белка Нельсона — S. nelsoni Merriam, 1893 (Центральная Мексика — штаты Мехико и Морелос);
S. colliaei Richardson, 1839 (западная часть нейтральных районов Мексики — области, примыкающие к тихоокеанскому побережью штатов Наярит и Халиско);
синалоасская белка — S. sinaloensis Nelson, 1899 (Мексика — крайний юг штата Синалоа);
сонорская белка — S. truei Nelson, 1899 (крайний юго-восток штата Сонора в Мексике);
общественная белка — S. socialis Wagner, 1837 (юг Мексики — юго-восток штата Герреро, юг Оахака, юг Чиапас; крайний запад Гватемалы) ;
серожелтая белка — S. griseoflavus Gray, 1867 (юг Мексики — штат Чиапас; юг Гватемалы, однако без тихоокеанского побережья);
юкатанская белка — S. yucatanensis J. Allen, 1877 (юго-запад Мексики— штаты Кампече и Юкатан; северная половина Гватемалы и Британский Гондурас);
пестрая белка—S. variegatoides Ogilby, 1839 (юг Мексики — штат Чиапас; юг и юго-запад Гватемалы; Сальвадор; Гондурас; Никарагуа; северная половина Коста-Рики, тихоокеанская половина Панамы);
S. deppei Peters, 1863 (юг Мексики — штаты Сан-Луис-Потоси, Веракрус, Пуэбла, Табаско, Кампече, Юкатан, Кинтана-Роо, Чиапас, северо-восток Оахака; Британский Гондурас; Гватемала, кроме тихоокеанского побережья; крайний север Сальвадора, центральные районы Гондураса; центральные районы Никарагуа; крайний север Коста-Рики);
серая белка — S. griseus Ord, 1818 (узкая полоса на западе США— центральные районы штата Вашингтон, запад — кроме тихоокеанского побережья штата Орегон, штат Калифорния, кроме северо-востока, юго-востока и центральных районов; крайний запад штата Невада);
белка Аберта — S. aberti Woodhouse, 1853 (пять изолированных участков на юго-западе США — юго-восток штата Юта, севернее г. Гал-лап на границе штатов Аризона и Нью-Мексико, узкая полоса, проходящая через центральные районы штата Аризона, северо-западный угол штата Нью-Мексико и занимающая всю середину штата Колорадо; северо-запад штата Чиуауа в Мексике; крайний юго-запад штата Чиуауа* северо-запад штата Дуранго и северо-восток штата Синалоа);
кайбабская белка — S. kaibabensis Merriam, 1904 (Кайбабское плато, северная сторона Гранд-Каньон реки Колорадо в штате Аризона, США);
черная белка—S. niger Linnaeus, 1758 (центральные и восточные штаты США — восточная граница проходит по тихоокеанскому побережью; северная граница идет от штата Делавэр, через южные и затем западные районы штата Пенсильвания и крайний юго-запад штата Нью-Йорк, по южным берегам озер Эри, Гурон и Мичиган, вдоль северо-восточных районов штата Висконсин граница выходит к западной оконечности оз. Верхнее, идет вдоль его северо-западного берега примерно до г. Тандер-Бей на юго-западе провинции Онтарио в Канаде, далее следует на запад чуть севернее государственной границы между Канадой и США; пересекает крайний север штата Миннесота к югу от государственной границы и идет параллельно ей по северу штата Северная Дакота, затем на границе со штатом Монтана резко поворачивает на юг и переходит в западную границу, которая вдается языком с запада в штат Южная Дакота — до ее центральных районов, пересекает северо-западный угол штата Небраска и юго-восточный угол штата Вайоминг, захватывает северо-восток штата Колорадо и весь штат Канзас, кроме его юго-запада, проходит через западные районы штата Техас и уходит в Мексику, где захватывает северо-восток штата Коауила и север штата Нуэво-Леон, после чего поворачивает па восток и в юго-восточном углу штата Техас выходит к побережью Мексиканского залива);
S. oculatus Peters, 1863 (центральные районы Мексики — штаты Керетаро, Веракрус, Пуэбла, Идальго, Сан-Луис-Потоси, Гуанахуато, Мехико);
белка Аллена — S. alleni Nelson, 1898 (северо-восток Мексики — юго-восток штата Коауила, центральные районы штата Нуэво-Леон*
юго-запад штата Тамаулипас и северо-восток штата Сан-Луис-Потоси) ;
наяритская белка — S. nayaritensis J. Allen, 1889 (штаты Мексики — запад Наярит, юг Дуранго, юго-запад Закатекас и центр. Халиско);
S. apache J. Allen, 1893 (штаты Мексики — восток Соноры, запад Чиуауа, северо-восток Синалоа, западная половина Дуранго);
чирикауасская белка — S. chiricahuae Goldman, 1933 (горы Чири-кауа, на крайнем юго-востоке штата Аризона, США);
аризонская белка — S. arizonensis Coues, 1867 (три изолированных участка: узкая полоса, проходящая с востока на центральные районы штата Аризона, США и захватывающая небольшую территорию штата Нью-Мексико к югу от 34-й параллели; горы Санта-Рита на юго-востоке штата Аризона; крайний юг штата Аризона, северо-восток штата Сонора в Мексике и крайний северо-запад штата Чиуауа);
S. granatensis Humboldt, 1811 (Коста-Рика, Панама, Колумбия, Эквадор, Венесуэла);
белка Ричмонда — S. richmondi Nelson, 1898 (Никарагуа);
S. aestuans Linnaeus, 1766 (Гайана, Суринам, Бразилия, Венесуэла);
S. Ignitus Gray, 1867 (Аргентина, Боливия, Перу, Бразилия);
S. pucheranii Fitzinger, 1867 (Колумбия);
S. sanborni Osgood, 1944 (Перу);
S. stramineus Eydoux et Souleyet, 1841 (Перу, Эквадор);
S. flammifer Thomas, 1904 (Венесуэла);
S. igniventris Wagner, 1842 (Бразилия, Венесуэла, Колумбия, Перу);
S. langsdorffi Brandt, 1835 (Бразилия, Боливия);
S. pyrrhinus Thomas, 1898 (Перу);
S. pyrrhonotus Wagner, 1842 (Боливия, Бразилия).
После ревизии рода число видов, вероятно, будет сокращено. Эл-лермэн (1940) кроме указанных выше выделяет следующие виды: S. negligens Nelson, 1898; S. durangi Thomas, 1893; S. hoffmani Peters, 1863; S. miravallensis Harris, 1931; S. griseogena Gray, 1867; S. chaptna-ni Allen, 1899; S. nesaeus G. Allen, 1902; S. griseimembra Allen, 1914; S. candelensis Allen, 1914; S. argentinus Thomas, 1921; S. ferminae Cabrera, 1917; S. gerrardi Gray, 1861; S. splendidus Gray, 1842; S. pyrrhinus Thomas, 1898; S. alphonsei Thomas, 1906; S. ingrami Thomas, 1901; S. bo-liviensis Osgood, 1921; S. rhoadsi Allen, 1914; S. tricolor Poeppig, 1844; S.nigratus Pinto, 1931; S. duida Allen, 1914.
Ряд исследователей (Myyp, 1959; Андерсон и Джонс, 1967), следуя за Греем, выделяют S. granatensis, S. richmondi и некоторые другие виды в особый род Guerlinguetus Gray, 1821. По-видимому, правильнее рассматривать его в качестве подрода Sciurus, как это делают, например, Симпсон (1945) и Холл и Келсон (1959) и др.
В СССР обитают обыкновенная и кавказская белки.
S. kaibabensis и 2 подвида черной белки: S. niger cinereus Linnaeus, 1758, и S. п. vulpinus Gmelin, 1788, включены в «Красную книгу».
Род бороздчаторезцовых белок
Genus Syntheosciurus Bangs, 1902
Внешний облик беличий. Размеры мелкие. Длина тела 15—17 см. Длина хвоста 14—15 мм. Волосяной покров густой и мягкий. Окраска верхней части тела оливковая, иногда с примесью черного цвета. Брюшная сторона, особенно вдоль средней линии, оранжево-рыжая (S. Ьго-chus).
В черепе лицевой отдел относительно удлиненный. Профиль крыши черепа выпуклый. Костные слуховые барабаны относительно небольшие, 5 а заглазничные отростки средних размеров. Крупных щечных зубов— .
4
По передней поверхности верхних резцов S. brochus проходит продольная бороздка. У S. poasensis бороздки нет.
Распространены в Панаме и Коста-Рике.
Экология не изучена. Встречаются высоко в горах.
В роде 2 вида:
бороздчаторезцовая белка — 5. brochus Bang, 1902 (крайний запад Панамы);
поэзская белка — S. poasensis Goodwin, 1942 (центральный район Коста-Рики);
Эллермэн (1949) насчитывает в роде 1 вид.
Возможно, бороздчаторезцовые белки являются лишь подродом рода белок.
Род карликовых белок
Genus Microsciurus J. Allen, 1895
За исключением рода Sciurillus, представители этого рода являются наиболее мелкими среди американских древесных белок. Длина тела 12—16 см. Длина хвоста 9—15 см. Пышность и мягкость волосяного покрова значительно изменяются в зависимости от местообитания. Карликовые белки, живущие па равнине, имеют низкий и грубый мех, а высоко в горах — густой и высокий. Хвост покрыт относительно короткими волосами. Окраска спинной части тела варьирует от серой и оливковобурой до темно-рыжевато-бурой; брюшная сторона обычно более светлого тона. Уши маленькие с округлыми вершинами, густо покрытые волосами.
Череп значительно сжат сзади, лобные кости очень широкие, носовые короткие. Скуловая дуга широкая. Костные слуховые барабаны небольших размеров.
Распространены от южной части Никарагуа на юг через Эквадор до Перу и Риу-Негру в Южной Америке.
Экология почти не изучена. Населяют леса от равнин до 1950 м над уровнем моря в горах. Питаются плодами пальм и орехами. Передвигаются очень быстро, делают длинные прыжки. Держатся обычно парами.
В роде 17 видов (Эллермэн, 1940):
М. alfari Allen, 1895 (крайний юго-восток Никарагуа, юг Коста-Рики, юго-запад и восток Панамы);
бокуетская белка — М. boquetensis Nelson, 1903 (запад Панамы);
М. similis Nelson, 1899 (горный хребет Западная Кордильера на юге Колумбии);
М. otinus Thomas, 1901 (горный хребет Центральная Кордильера па севере Колумбии);
М. isthmius Nelson, 1899 (северо-восток Колумбии и крайний восток Панамы);
М. mimulus Thomas, 1898 (северо-восток Эквадора и юго-восток Колумбии) ;
М. palmeri Thomas, 1909 (Колумбия);
М. simonsi Thomas, 1900 (центральные области Эквадора);
перуанская белка — М. peruanus Allen, 1897 (северо-восток Перу);
Рис. 13. Белка-пигмей — S. pusillu&
напосская белка — М. napi Thomas, 1900 (граница между Эквадором и Колумбией);
красномордая белка — М. rubrirostris Allen, 1914 (центральные области Перу);
флоренциасская белка — AL Jlorencial Allen, 1914 (северо-западная Колумбия);
М. avunculus Thomas, 1914 (Восточный Эквадор);
М. septentrionalis Anthony, 1920 (Никарагуа);
сабаниллская белка — S. sabanillac Anthony, 1922 (Южный Эквадор);
М. manarius Thomas, 1920 (р. Риу-Пегру);
желтобрюхая белка — М. flaviventer Gray, 1867 (Бразилия).
По-видимому, число видов должно быть сокращено. Андерсон и Джонс (1967) указывают для этого рода 7 видов. Кабрера (1960) для всей Южной Америки приводит лишь 1 вид: М. flaviventer Gray, 1867.
Род белок-пигмеев
Genus Sciurillus Thomas, 1914
В роде единственный вид: белка-пигмей — S. pu.sillus Desmarest, 1822.
Размеры мелкие (рис. 13). Длина тела 9—11 см. Длина хвоста примерно 10 см. Самцы крупнее самок. Окраска спинной стороны тела серовато-бурая или бурая, брюшной обычно светло-бурая. Голова может быть рыжей и наружные края ушей черные. Уши небольшие, с округлой вершиной. Хвост покрыт короткими волосами.
В черепе костное небо широкое. Скуловые дуги расставлены очень широко. Заглазничные отростки небольшие.
Распространена в Южной Америке от Гайаны, Суринама и Гвианы на юг через северо-восточную часть Бразилии до Амазонки и в бассейне этой реки в Перу.
Экология изучена слабо. Населяет высокоствольные леса. Держится на вершинах деревьев. Питается главным образом плодами, орехами и корой деревьев. Спаривание приходится на май — август. Большинство самок приносит детенышей в июне.
Эллермэн (1940) объединяет в роде 3 вида: S. pusillus, S. murinus Muller et Schlegel, 1844; S. evidens Miller et Hollister, 1921. Правильнее к роду белок-пигмеев относить лишь один вид (S. pusillus), а 2 других выделить в род карликовых сулавесских белок.
Род карликовых сулавесских белок
Genus Prosciurillus Ellerman, 1949
Сходны с белками-пигмеями. Размеры мелкие. Длина тела 10— 13 см. Длина хвоста 7—11 см. Окраска спинной стороны изменяется от буроватой до оливковой, а брюшная обычно серая.
Распространены на Сулавеси и мелких прилегающих островах.
Экология не изучена.
В роде 3 вида:
Р. murinus Forsten, 1844 (включая S. evidens Miller et Hollister, 1921) (северо-восток Сулавеси);
P. leucomus Muller et Schlegel, 1839 (Сулавеси);
P. abstrurus (Сулавеси).
Род рубиновых белок
Genus Rubrisciurus Ellerman, 1954
В роде единственный вид: рубиновая белка — R. rubrlventer Forsten, 1844.
Размеры относительно крупные. Длина черепа между затылочной и носовыми костями (63,8—68,3 мм) больше, чем у представителей родов Callosciurus и Sciurus, напоминая черепа Ratufa и Rheithrosciurus. От Ratufa рубиновая белка отличается тем, что в черепе имеется длинный сагиттальный гребень, а также более узкими лобпыми костями и более длинным костным небом.
Распространена в Северном Сулавеси.
Экология не изучена.
Род кистеухих белок
Genus Rheithrosciurus Gray, 1867
В роде единственный вид: кистеухая белка — /?, macrotis Gray, 1856.
Размеры средние и крупные. Длина тела 28—53 см. Длина хвоста 28—36 см. Масса до 2 кг. Уши большие с длинными кисточками волос на вершинах. Хвост густо покрыт длинными волосами, длина которых к концу его возрастает; волосы грубые, слегка волнистые. Окраска спинной стороны светло-бурая или бурая. По бокам тела проходит продольная полоса грязно-желтого или белого цвета, тянущаяся от района лопатки до бедра, а ниже ее проходит темно-бурая полоса. Боковые части морды серые. Передние лапы черные, задние ярко-бурые. Брюшная часть тела белая. Волосы на хвосте, особенно около его вершины, имеют длинные серые или белые концы. Череп в отличие от большинства видов семейства более широкий и уплощенный. Рострум черепа удлинен. Лобные кости широкие. Заглазничные отростки хорошо развиты. Задняя часть нижней челюсти более округла, чем у других белок. Угловой отросток высокий. Венечный отросток не редуцирован. Резцы сильно утолщены спереди назад, их передняя поверхность имеет многочисленные (7—10) продольные бороздки. Щечные зубы уменьшены, предкоренные зубы как в верхней, так и в нижней челюстях меньше по размерам, чем коренные. Все зубы брахиодонтного типа.
Распространена на о-ве Калимантан.
Населяет леса, ведет в основном наземный образ жизни. Бегает быстро по земле, длинный хвост держит загнутым над спиной. Питается различными плодами, местами может вредить садам.
Род красных белок
Genus Tamiasciurus Trouessart, 1880
Размеры средние. Длина тела 16—23 см. Длина хвоста 9—16 см. Внешним обликом напоминают обыкновенных белок (рис. 14), но хвост относительно короче — составляет примерно 40% длины тела. Кисточки волос на ушах и подошвах лап имеют сезонный характер. Окраска спинной стороны варьирует от золотисто-бурого до буроватого или оливково-бурого цвета, в зимнее время принимающего сероватый оттенок. В летнее время обычно по бокам идет черноватая полоса. Брюшная сторона белая. Окраска хвоста имеет золотисто-бурый, желтый или белый оттенок.
Череп в общем напоминает таковой обыкновенной белки, однако он не так сильно уплощен в заднем направлении. Костное небо слегка продолжено позади последних щечных зубов. Третий предкоренной зуб имеется или может отсутствовать.
Распространены в Канаде и США от Аляски, Квебека и Британской Колумбии на юг до Нью-Мексико, Южной Каролины и Калифорнии.
Древесные животные, хотя значительную часть времени проводят на земле, хорошо плавают. Дальние миграции совершают редко. Активны в основном днем, а также в лунные ночи. В спячку не впадают, но остаются в своем гнезде в случае плохой иогоды. Каждая особь обычно имеет несколько гнезд. Зимнее гнездо преимущественно устраивают в дуплах, а летние — среди ветвей деревьев. Иногда выкапывают норку под камнями. Питаются широким набором растительных объектов, поедают также птенцов и яйца птиц. Основным кормом служат семена
Рис. 14. Белка Дугласа — Т dnuglasi и ее череп
хвойных деревьев. Делают запасы шишек. Период размножения приходится на конец зимы, весну. Беременность длится 36— 10 дней. Самка приносит от 1 до 7, обычно 4—6 детенышей. В течение года 2 помета. Половозрелость наступает примерно в годовалом возрасте.
В роде 2 вида:
красная белка — Т. hudsonicus Erxleben, 1777 (ареал охватывает север Северной Америки и спускается языком через западные штаты США к югу по направлению к Мексике: Аляска, кроме прибрежной зоны Чукотского моря и моря Бофорта, приустьевых областей рек Юкон и Кускоквим и п-ова Аляска; почти вся территория Канады — северная граница идет с запада южнее г. Гордон в штате Аляска на границе с Канадой примерно к месту впадения р. Пил в р. Маккензи, к южной части Большого Медвежьего озера и к г. Черчилл на западном берегу Гудзонова залива, огибает Гудзонов залив по его южному берегу, севернее г. Пост-де-ла-Баден поворачивает на северо-восток и идет к г. Форт-Шимо в южной части залива Унгава, отсюда сворачивает на восток и выходит к морю Лабрадор южнее г. Нейн, южная граница ареала имеет сложную конфигурацию: от атлантического побережья США в районе г. Норфола в штате Виргиния она идет на запад севернее границы со штатом Северная Каролина, затем сворачивает на юго-запад, пересекая Северную Каролину восточнее ее границы с Теннесси, доходит до штата Георгия и сворачивает на северо-восток к юго-западному углу штата Пенсильвания, затем следует на запад, пересекая юг штата Огайо, проходит вдоль границы штата Индиана со штатом Кентукки, поворачивает на север вдоль границы штата Индиана с Иллинойс, сворачивает на запад и пересекает штат Иллинойс в его центральных районах, проходит вдоль границы штата Иова с Миссури до юго-западного угла штата Иова, сворачивает на север и проходит вдоль границ штатов Иова с Небраска и Миннесота, Южная и Северная Дакота, доходит до государственной границы с Канадой, круто сворачивает на юго-запад, образуя небольшой выступ к центральным районам штата Северная Дакота, опять выходит к государственной границе и пересекает ее, направляясь к южной оконечности оз. Манитоба, проходит по восточному берегу озер Манитоба и Виннипегосис и от северной оконечности последнего поворачивает на запад, пересекая центральные районы провинций Саскачеван и Альберта и в окрестностях г. Джаспер-Плейс сворачивает на юго-восток, проходя несколько восточнее границы между провинциями Альберта и Британская Колумбия, выходит за государственную границу, поворачивает на восток и идет несколько южнее границы между провинцией Альберта, Канада и штатом Монтана, США, до юго-западного угла провинции Саскачеван, откуда сворачивает круто к югу, пересекает центральные районы штата Монтана, восточные районы штата Вайоминг, центральные районы Колорадо и восточные — штата Ныо-Мексико, где доходит до границы со штатом Техас к югу от г. Карлсбад, откуда сворачивает на северо-запад к юго-западному углу штата Юта, далее поворачивает на север, проходя восточнее вдоль границы между штатами Невада и Юта, пересекает штат Айдахо в его юго-западном углу и штат Орегон в северо-восточном углу, проходит наискось с юго-востока на северо-запад через штат Вашингтон, примерно от г. Уолла-Уолла к месту восточнее г. Сиэтл, пересекает государственную границу п выходит к тихоокеанскому побережью па уровне северной оконечности о-ва Ванкувер, в ареал входят острова архипелага Александра, о-в Ванкувер и три небольших изолированных участка на юго-востоке штата Аризона, юго-востоке штата Юта и на границе между штатами Южная Дакота н Вайоминг);
белка Дугласа — Т. douglasi Bachman, 1838 (узкая полоса па западе Северной Америки, вдоль тихоокеанского побережья: граница ареала идет по юго-западу провинции Британская Колумбия, Канада примерно от уровня северной оконечности о-ва Ванкувер на юг к центральным
Рис. 15 Северная пальмовая белка — F. pennanti
районам штата Вашингтон, США; пересекает этот штат примерно по 100° з. д., следует далее на юг, делая небольшой выступ к востоку, и выходит к общей точке на границе трех штатов: Орегон, Невада и Калифорния, двигается к югу примерно по границе между двумя последними штатами, достигает центральных областей штата Калифорния и сворачивает на север, достигая района к северу от г. Чико, откуда опять поворачивает на юг вдоль р. Сакраменто и достигает ее устья).
Эллермэн (1940) выделяет еще 2 вида: Т. regalis Howell, 1936, и F. fremonti Audubon et Bachman, 1854.
Род пальмовых белок
Genus Funambulus Lesson, 1832
Размеры мелкие. Длина тела II—18 см. Хвосг имеет примерно такую же длину. Внешним видом напоминает бурундука (рис. 15). Волосяной покров мягкий, густой, низкий. Хпост покрыт более длинными волосами, чем туловище. Окраска спинной стороны варьирует от светло-серовато-бурого до почти черного. Голова серая или рыжевато-бурая, у некоторых видов с декабря по май рыжеватая. Обычно на спине проходят три широкие светлые полосы, а у некоторых видов дополнительно
я короткая полоса по бокам тела. Брюшная сторона у F. layardi рыжая, у всех других видов беловатая, грязно-желтая или светло-бурого цвета.
Череп с более или менее удлиненным лицевым отделом. Заглазнич-ные отростки невелики. Подглазничные отверстия средних размеров. Костное небо не широкое. Венечные отростки обычно низкие.
Распространены на о-ве Шри Ланка, в Индии, Пакистане.
Населяют различные биотопы. Некоторые встречаются в пальмовых или кустарниковых зарослях, в то время как другие (редко встречающиеся и слабо изученные) населяют густые джунгли и высокоствольные леса. Активны днем. Питаются на земле и на деревьях семенами, орехами, побегами, листьями, цветками, насекомыми и их личинками. Местами могут вредить посевам. Самки строят из растительных волокон шаровидное гнездо, где приносят потомство. В году бывает до трех пометов, в каждом от 2 до 4, обычно 3 детеныша. Длительность беременности 40—45 дней. Половозрелости самки достигают в возрасте 6—8 месяцев.
В роде 5 видов:
пальмовая белка—*F. palmarum Linnaeus, 1766 (о-в Шри Ланка и юг Индии на север до штатов Мадхья-Прадеш и Бикар);
северная пальмовая белка — F. pennanti Wroughton, 1905 (Белуджистан и Синд в Пакистане и Северный Пакистан; штаты Индии — Пенджаб, северо-запад Уттар-Прадеш, Раджстхан, Гуджарат, Мадхья-Прадеш, Западная Бенгалия, и на юг примерно до г. Хубли в штате Карнатака; Непал);
трехполосая белка — F. tristriatus Waterhouse, 1837 (Южная Индия на север, примерно до Бомбея и Мадраса);
белка Лайарда — F. layardi Blyth, 1849 (о-в Шри Ланка, Южная Индия).
F. sublineatus Waterhouse, 1838 (о-в Шри Ланка, Южная Индия).
Род гигантских белок
Genus Ratufa Gray, 1867
Размеры средние и крупные. Длина тела 25—47 см. Хвост иногда короче, чаще значительно длиннее. Масса до 1,5—3 кг. Уши короткие, широкие, с округлой вершиной; у R. тасгоига — с кисточкой. Стопа и особенно кисть широкие. Когти крупные, мощные. Хвост покрыт очень длинными и густыми волосами (рис. 16). Окраска волосяного покрова яркая, значительно варьирующая у разных видов. У некоторых из них верхняя часть тела ярко-черная, брюхо светло-желтовато-бурое. У других спинная сторона рыжая, у третьих от темно-бурой до серой, а брюшная от грязно-желтой до белой. Лапы приспособлены к древесному образу жизни. На конечностях наиболее длинный IV палец.
Череп тяжелый, широкий, с сильно выдающимися заглазпичпыми отростками. Задняя часть мозговой коробки сильно снижается в каудальном направлении. Скуловые дуги широкие. Рострум короткий п шнро-4
кий. Лобные кости очень широкие. Число щечных чубов—; брахподонт-лого типа.
Распространены в Индии, на о-ве Шри Лапка, в Непале, Бирме, Индокитае, Малайзии, Суматре, Яве, Калимантане и некоторых мелких прилегающих островах.
Ведут древесный образ жизни. На землю спускаются редко. Очень проворно лазают по деревьям, делают длинные прыжки до 6 м длиной.
Рис. 16. Индийская белка — /?. indica
Держатся поодиночке или парами. Питаются плодами, орехами, корой различных деревьев, насекомыми, а также япцами птиц. В качестве убежищ используют дупла. Во время размножения самки строят большие гнезда Беременность составляет примерно 28 дней. В помете обычно 1—2 детеныша. По-видимому, у каждой самки в течение года бывает несколько пометов. В неволе жили до 16 лет.
В роде 4 вида:
гигантская белка — R. macroura Pennant, 1769 (о в Шри Ланка, Южная Индия);
индийская белка — R. indica Erxleben, 1777 (Индия — от южной оконечности полуострова на север до штатов Орисса и Мадхья-Прадеш);
двухцветная белка — R. bicolor Sparrman, 1778 (о-ва Натуна, Ява, Бали, Суматра; Малайзия, Таиланд, Индокитай, Хайнань, Юньнань, Бирма, Ассам, Непал, а также мелкие острова по соседству с Суматрой и Малайзией);
кремовая белка — R. affinis Raffles, 1822 (Южный Таиланд, Малайзия, о-ва Суматра и Калимантан).
Род масличных белок
Genus Protoxerus Major, 1893
Средних размеров белки. Длина тела 23—33 см. Длина хвоста 25— 38 см. Масса 540—730 г. Конечности приспособлены к древесному образу жизни, очень сильные с мощными сильно изогнутыми и острыми когтями. Хвост густо покрыт длинными волосами (рис 17). На брюхе волосы редкие, иногда оно почти голос (редкий случай в отряде грызунов). Уши небольшие, округлые. Волосяной покрои короткий, грубый, па брюшной стороне редкий; иногда она почти голая Окраска спинной стороны варьирует от беловато-оливковой до почти черной Шеки беловатые или серо-белые. Бока тела буровато-серые, брюшная сторона белая или грязно-желтая. Вдоль середины спины может проходить широкая черная полоса. Хвост иногда обрамлен черными и белыми пли черными и рыжими волосами. Сосков 4 пары.
Череп массивный. Подглазничные отверстия крупные, округлой формы. Лобные кости широкие. Заглазничныс отростки развиты средне.
Рис. 17. Масличная белка Р. stangeri
У взрослых особей хорошо выражен сагиттальный гребень. Рострум короткий, широкий. Небо нормальное. Носовые кости короче, чем наименьшая межглазничная ширина. Костные слуховые барабаны не умень-
4 шены. Резцы очень сильные. Число брахиодонтных щечных зубов —.
4
Распространены в Африке от Либерии, Ганы и о-ва Фернандо-По до Кении и Анголы.
Экология изучена слабо. Населяют густые, высокоствольные леса. Убежищем служат дупла или гнезда на ветвях деревьев. Подвижные, быстро прыгающие белки. Обычно держатся на вершинах деревьев. Кормятся как на деревьях, так иногда и на земле. Питаются различными семенами, орехами, плодами, в некоторых районах в основном плодами масличной пальмы. В помете обычно от 3 до 4 детенышей.
В роде 2 вида:
тонкохвостая белка — Р. aubinii Gray, 1873 (от Либерии до Верхней Вольты);
масличная белка — Р. stangeri Waterhouse, 1842 (от Сьерра-Леоне до Кении и на юг до Северной Анголы, а также Фернандо По).
Эллермэн, 1940, выделяет Р. aubinii в отдельный род Myrsilus Thomas, 1909, что, видимо, неверно.
Род африканских белок
Genus Epixerus Thomas, 1909
В роде единственный вид: африканская белка — Е. ebii Tennninck, 1853.
Размеры средние. Длина тела 25—30 см. Длина хвоста 28- 30 см. Хвост покрыт длинными густыми волосами — очень пушистый.
Рис. 18. Funisciurus pyrrhopus и ее череп
Брюхо покрыто волосами слабо, часто почти голое, как у масличных белок. Уши довольно широкие, округлые. Окраска спинной стороны тела рыжая или рыжая с черным. На внешней стороне конечностей преобладает рыжий цвет. Внутренние части конечностей, бока головы и уши желтовато-рыжие или ярко-рыжие. Горло, грудь и брюхо желтовато-рыжие. Хвост часто окаймлен белой полоской.
Череп относительно длинный и узкий. Лобные кости узкие. Носовые кости длиннее, чем наименьшая межглазничная ширина. Подглазничные отверстия сходны с таковыми у масличных белок. Костные слуховые барабаны относительно уменьшены в размерах. Костное небо простирается немного далее назад задних щечных зубов. Сагиттального гребня у взрослых особей нет. Резцы мощные.
Распространена в Западной Африке — Гане, Камеруне, Габоне.
Экология не изучена. Населяет леса. Встречается редко. Часто спускается на землю. Питается плодами, орехами.
Включена в «Красную книгу».
Некоторые ученые (например, Эллермэн, 1940) выделяют в роде и второй вид: Е. wilsoni du Chaillu, 1860.
Род полосатых белок
Genus Funisciurus Trouessart, 1880
Мелких размеров белки (рис. 18). Длина тела 31 см. Длина хвоста 10—20 см. Масса 70—700 г. Хвост густо опушен длинными волосами у одних видов и слабо — у других. Уши короткие, округлые. Волосяной покров мягкий. У одних видов имеются продольные полосы па синие, у других они отсутствуют, некоторые имеют беловатые полосы на боках. Основная окраска спинной стороны у всех видов представлена различной комбинацией рыжего и черного, желтовато-бурого или оливково-бурого цветов. Брюшная сторона беловатая или кремовая. Сосков 2 пары.
Череп со слабо выраженными гребнями. Заглазничные отростки средних размеров. Рострум слегка удлинен. Скуловая дуга укорочена. Небо узкое. Щечные зубы полугнпселодонтные.
Распространены в Африке от Сьерра-Леоне и Берега Слоновой Кости на восток до Заира и Танзании и на юг до Анголы и Юго-Западной Африки.
Населяют заросли пальм, саванны и леса до 2000 м над уровнем моря. Держатся обычно поодиночке или парами. Наибольшая активность ранне-утренняя или вечерняя. Ведут в основном древесный образ жизни, но часто ищут пищу и на земле. Питаются различными семенами, орехами, плодами, поедают также насекомых, яйца птиц и рептилий. Строят гнезда из сухих листьев и растительных волокон. В году 2—3 помета.
В роде, по-видимому, 6 видов:
конголезская белка — F. congicus Kuhl, 1820 (северная часть Юго-Западной Африки на север до р. Конго; область Катанга в Заире; возможно, в Восточной и Северо-Восточной Анголе);
F. bayonii Bocage, 1890 (Северо-Восточная Ангола и к югу от р. Ка-сан в Заире);
F. substriatus de Winton, 1899 (Южная Гана, Того, Дагомея и Юго-Восточная Нигерия);
оранжевоголовая белка-—F. leucogenys Waterhouse, 1842 (от Ганы до юго-востока Центрально-Африканской Республики), а также о-в Фернандо-По) ;
F. pyrrhopus F. Cuvier, 1833 (от Гамбии на восток до Уганды и на юг до Северной Анголы);
F. anerythrus Thomas, 1890 (от юго-запада Нигерии до Центрально-Африканской Республики и Уганды и на юг до юго-запада Заира).
Некоторые ученые (например, Эллермэн, 1940) насчитывают в роде 15 видов.
Род кустарниковых белок Genus Paraxerus Major, 1893
j Размеры мелкие и средние. Длина тела 15—25 см. Длина хвоста при-I мерно такая же. В отличие от полосатых белок кустарниковые белки I имеют более высокие и слегка заостренные уши, 6 сосков вместо четы-I рех. Хвост пушистый — покрыт длинными волосами. Волосяной покров I: на туловище мягкий, короткий, густой. Окраска существенно варьирует. ! По спинной стороне тела могут проходить 4 продольные полосы, пере-I межающиеся тремя светлыми полосами. Верхняя сторона может быть В черной, черновато-бурой, зеленовато-серой или ярко-рыжей со слабо В намеченными желтовато-серыми пятнами. Брюшная сторона белая, бу-| рая, рыжевато-бурая или ярко-рыжая.
I Череп в отличие от полосатых белок с относительно укороченным I рострумом. Щечные зубы — полугипселодоптпые.
I Распространены в Восточной и Южной Африке от Судана па юг до i I Юго-Западной Африки, Ботсваны и юго-восточной части Южпо-Афри-} Г канской Республики.
;. [ Населяют леса. Активны, по-видимому, и днем и ночью, наибольшая 1- | активность приходится на сумеречное время. Большую часть времени 1 проводят на вершинах деревьев, но часто их можно видеть и на земле в
поисках пищи. Питаются различными семенами и плодами. Поедают
также луковицы и корни различных растений, насекомых, яйца птиц. Делают небольшие запасы семян. Убежищем часто служат дупла. В помете обычно 2 детеныша. Кустарниковые белки служат объектом охоты для местного населения; их употребляют в пищу.
В роде 11 видов:
белка Александра — Р. alexandri Thomas et Wrougthon, 1907 (Заир и Уганда);
Р. boehmi Reichenow, 1881 (север и восток Заира, юг Судана, север Замбии, Уганды, запад Кении, северо-запад Танзании);
полосатая кустарниковая белка — Р. fluvivittis Peters, 1852 (север Мозамбика, Танзания, юг Кении);
чернорыжая белка — Р. lucifer Thomas, 1897 (плато Ниика — 10°311 ю. ш., 33О42* в. д. и гора Мисуку — 9'э43| ю. ш., 33°31* в. д.);
рыжая белка —Р. palliatus Peters, 1852 (Южно-Африканская Республика, Южная Родезия, Малави, Юго-Восточная и Восточная Танзания, Восточная Кения, Южное Сомали);
Р. vexillarius Kershaw, 1923 (Центральная и Восточная Танзания); белка Купера —Р. cooperi Hayman, 1950 (Камерун);
зеленая белка — Р. poensis A. Smith, 1834 (от Сьерра-Леоне до р. Вольта и от Дагомеи до Заира; также о-в Фернандо-По);
белка Винсента — Р. Vincenti Haymann, 1950 (Северный Мозамбик);
Р. ochraceus Huet, 1880 (Танзания, Кения, Судан);
Р. cepapi A. Smith, 1836 (Мозамбик, Трансвааль, Южная Родезия, Ботсвана, Юго-Западная Африка, Юго-Западная Ангола, Юго-Восточная Катанга, Замбия, Малави, Юго-Западная Танзания).
Эллермэн, 1940, выделяет Р. poensis, Р. lucifer и Р. vexillarius в род Heliosciurus; из Р. boehmi выделяет Р. vulcanorum Thomas, 1918, из Р. palliatus Р. bridgemani Dollman, 1914. Кроме того, в составе кустарниковых белок отдельными видами он считает: Р. byatti Kershaw, 1923; Р. sponsus Thomas Thomas et Wroughton, 1907; P. emini Stuhlmann, 1894; P. antoniae Thomas et Wroughton; 1907.
Род солнечных белок
Genus Heliosciurus Trouessart, 1880
Мелкие и средних размеров белки с длиной тела 15—25 см. Длина хвоста 15—30 см. Масса 205—403 г. Волосяной покров довольно густой. Окраска его значительно варьирует. На спинной стороне он может быть зеленовато-серым, буроватым или черным и обычно имеет многочисленные беловатые или светло-серо-бурые отметины. Окраска на брюхе изменяется от беловатой до желтоватой и рыжевато-бурой. Сосков 3 пары.
В черепе заглазничные отростки хорошо развиты, костные слуховые барабаны средних размеров. Передняя поверхность резцов оранжевая.
4
Щечных зубов— - Резцы утолщены.
4
Распространены в Африке примерно от 15° с. ш. до 15“ ю. ш.
Населяют различные биотопы от густых лесов до открытых саванн. Питаются в основном семенами, орехами, плодами, а также насекомыми и яйцами птиц. Наиболее активны утром и вечером. Хороню лазают но деревьям, но много времени проводят и на земле. Держатся семьями. Убежищем для них служат дупла деревьев, а также гнезда, в которых они приносят детенышей. Рождение детенышей происходит главным образом в июле — августе и в декабре — январе.
Рис. 19. Мышиная белка — М. pumilio н ее череп
В роде 3 вида:
рувензорская белка — Н. ruwenzorii Schwann, 1907 (восток Заира, Руанда, Бурунди, юго-восток Уганды);
гамбийская белка — Н. gambianus Ogilby, 1822 (от Сенегала на восток до Дарфура в Судане и Кении и па юг до Анголы и севера Замбии);
красноногая белка — И. rufobruchium Waterhouse, 1842 (от Сенегала до юго-восточного Судана; Копия, юг Мозамбика и юго восток Южной Родезии);
Эллермэн (1940) относит к роду солнечных белок: // gambianus. Н. ruwenzorii, Н. poensis, Smith, 1830; Н. lucifer Thomas, 1897; // vexil-larius Kershaw, 1923, и H. bayonii Bocage, 1890; II riifobrachium он счи тает подвидом H. gambianus. Правильнее II. poensis, II. lucifer и H. vchil larius включить в род кустарниковых белок, а 11. bayoni — в род полосатых белок. Н. rufobrachium, по-видимому,— самостоятельный вид (Ro-sevear, 1969).
Род мышиных белок
Genus Myosciurus Thomas, 1909
В роде один вид: мышиная белка — М. pumilio Le Conte, 1857.
Размеры очень небольшие — наименьшие среди белок. Длина тела 6—7,5 см Длина хвоста около 5 см. Хвост покрыт относительно короткими волосами и не пушистый. Уши небольшие, округлые. Глаза относительно крупные (рис. 19). Окраска спинной стороны грязно-желто-зеле-новатая, а брюшной — оливково-белая. Края ушей белые. Вокруг глаз светло-оранжевые кольца.
В черепе характерны очень широкие лобные кости. Заглазничные отростки едва заметны. Костное небо относительно узкое. Скуловые кости широкие. Щечных зубов —. Предкоренной зуб в верхней челюсти 4
всего один. Резцы светло-желтые, относительно слабые.
Распространена в Камеруне, Габоне и Западной Африке.
Экология не изучена. Встречается в высокоствольных лесах. Подвижный, быстрый зверек.
Род прекрасных белок
Genus Callosciurus Gray, 1867
Мелких и средних размеров белки с длиной тела 13—28 см. Длина хвоста 8—25 см. Внешним видом напоминают обыкновенных белок (рис. 20). Волосяной покров довольно грубый, но окраска очень красивая. Некоторые виды почти чисто-белые или светло-кремового цвета с розоватым оттенком; у других окраска различных оттенков серого с бурым, па брюхе ярко-рыжевато коричневая, причем передние конечности могут быть коричневыми, а задние белыми У некоторых видов окраска трехцветная: верх головы, спина, бока тела и хвост ярко-черные или
Рис. 20. Белка Превоста — С. prevosti
бурые; брюхо от светлого до темного рыжевато-бурого и на боках, где два поля разной окраски соприкасаются друг с другом, идет чисто-белая полоска или узкая серая. Белка Финлансона может быть полностью черной или совершенно белой, или ее брюшная сторона— белой, рыжей или черной. Если на брюхе окраска рыжая, то все животное также рыжего или рыжеватого цвета Если же брюхо не рыжее, то основание хвоста обычно бывает бледно окрашено. Другой вид — С. eryth-racus, обычно оливкового цвета сверху и ярко-рыже-бурого или белого снизу.
Череп напоминает череп рода Sciurus. У некоторых видов имеется тенденция к удлинению рост-рума. Лобные кости очень широкие, а посторбитальпый отросток сильно развит.
Распространены в Юго-Восточной Азии, Восточной Индии, на Филиппинах и Тайване. Золотоспинная белка акклиматизирована в Японии.
Населяют различные ландшафты, в том числе антропогенные, леса, сады. Держатся на деревьях, но часто спускаются на землю. Активны днем. Питаются различными семенами, орехами, плодами, цветками, проростками, вероятно, поедают насекомых и яйца птиц. Убежищем в основном служат дупла, но также устраивают гнезда из сучьев и листьев. Обычно 3—4 детенышей в помете.
В роде 17 видов (Тэйлор, 1934; Эллермэн, 1940; Лори и Хилл, 1954; Эллермэн и Моррисон-Скотт, 1966; Медвэй, 1969):
С. erythraeus Pallas, 1779 (Ассам; Бирма; провинции Китая — Сычуань, Юньнань, о-в Хайвань, Фуцзянь, о-в Тайвань; п-ов Индокитай, Таиланд, п-ов Малакка);
желтолапая белка —С. flavimanus I. Geoffroy 1831 (п-ов Индокитай, Бирма);
белка Финлайсона — С. finlaysoni Horsfield, 1823 (Таиланд, п-ов Индокитай, Бирма);
золотоспинная белка — С. caniceps Gray, 1842 (Сикким, Бирма, Таиланд, о-в Тайвань, п-ов Малакка и мелкие соседние о-ва);
ирравадийская белка — С. pygerythrus Geoffroy, 1831 (Непал, Ассам, Бирма; п-ов Индокитай, по-видимому, Юго-Восточный Китай);
С. quinquestriatus Anderson, 1871 (Юньнань в Китае и Восточная Бирма);
С. jentinki Thomas, 1887 (северный Калимантан);
белка Превоста — С. prevosti Desmarest, 1822 (п-ов Малакка, о-ва Суматра, Калимантан, Сулавеси, Натуна);
С. nigrovittatus Kloss, 1908 (о-ва Суматра, Калимантан, Ява);
С. notatus Boddaert, 1785 (п-ов Малакка, о-ва Ява, Бали, Калимантан, Суматра);
С. albescens Bonhote, 1901 (о-в Калимантан);
С. leucomus Forsten, 1844 (о-в Сулавеси);
С. mowewensis Roux, 1910 (юго-восточная часть о-ва Сулавеси);
С. elbertae Schwarz, 1911 (о-в Восточный Кабаена у юго-западного побережья Сулавеси);
С. sarasinorum Meyer, 1898 (о-в Сулавеси);
краснобрюхая белка —С. rubriventer Forsten, 1839 (Северный Сулавеси) ;
белка Алстона — С. alstoni Anderson, 1879 (о-в Калимантан);
По Уокеру (1964), в роде 20 видов; по Андерсону и Джонсу (1967)—13.
Муур (1959) включает в род прекрасных белок род Tamiops. Чэйзен (1940) объединяет род прекрасных белок с родом Sciurus.
Род сулавесских белок
Genus Hyosciurus Tate et Archbold, 1935
В роде единственный вид: сулавесская белка — II. heinrichi Tate et Archbold, 1935.
Размеры мелкие. Длина тела 20—24 см. Длина хвоста 9—13 см. Характерен короткий хвост и необычайно сильно развитые когти, особенно на передних конечностях. Волосяной покров густой и мягкий. Характерна сильно вытянутая лицевая часть головы. Кисти и ступни сильно удлинены, узкие. Окраска верхней части тела темная — рыжевато
коричневая. Ниже располагается неправильной формы белая полоса шириной примерно 30 мм.
Череп обладает чрезвычайно удлиненным лицевым отделом, маленькими заглазничными отростками. Подглазничные отверстия образуют длинный канал. Число щечных зубов — .
4
Распространена на о-ве Сулавеси.
Экология не изучена. Населяет горные области, встречаясь на высоте 1700—2300 м над уровнем моря. Роет норы в земле, большую часть времени проводит среди низкой растительности.
Род сундасциурус
Genus Sundasciurus Moore, 1958
Мелкого и среднего размера белки с длиной тела примерно 15— 25 см. Длина хвоста 10—27 см. Морда относительно удлиненная. Уши небольшие, округлые. Хвост опушен не очень длинными волосами. Спинная сторона тела темно-буроватая с черными пестринами, брюшная ярко-каштанового, грязно-желтого или белого цвета.
Распространены на о-вах Калимантан, Суматра, Филиппинских и п-ове Малакка.
Экология не изучена. Населяют густые леса равнин и гор. Больше наземные, чем древесные формы. Насекомые и мелкие позвоночные животные преобладают в питании в большей степени, чем у других белок.
В роде 12 видов:
стройная белка — S. tenius Horsfield, 1824 (от Южного Таиланда до Суматры, о-ва Натуна);
белка Лови — S. lowi Thomas, 1892 (о-ва Калимантан, Суматра, Натуна, п-ов Малакка, о-ва Ментавай);
S. hippurus Geoffrey, 1832 (п-ов Малакка, о-ва Суматра, Калимантан);
чернобрюхая белка — S. melanogaster Thomas, 1895 (о-ва Ментавай);
S. brookei Thomas, 1892 (о-в Калимантан);
самарская белка — S. samarensis Steere, 1890 (о-в Самар);
минданаосская белка — S. mindanensis Steere, 1890 (о-в Минданао);
филиппинская белка — S. philippinensis Waterhouse, 1839 (о-в Минданао);
балабакская белка — S. steeri Gunter, 1876 (о-в Балабак);
палаванская белка — S. juvencus Thomas, 1908 (о-в Палаван);
белка Моллендорфа — <8. tndllendorffi Matschie, 1898 (о-ва Кала-миан);
белохвостая белка — S. albicauda Matschie, 1898 (о-ва Каламиан).
Андерсон и Джонс (1967) насчитывают в роде 7 видов, а Уокер (1964) — 2.
Род тамиопс
Genus Tamiops J. Allen, 1906
Размеры мелкие. Длина тела 10—16 см. Длина хвоста 8—11 см. Масса от 40 до 85 г. Обычно по спине проходят пять темных продольных полос, чередующихся со светлыми. Основная окраска спины желтовато-коричневая. Брюхо белое, бледно-коричневое или ярко-рыжее. Вер-
Рис. 21. Многополосая белка—М. berdmorei
шины ушей обычно с небольшими белыми кисточками. Хвост не пушистый. В течение года бывает, по-видимому, две линьки. Сосков 3 пары.
Распространены в Южном Китае, на севере Индии, в Бирме, Таиланде, на п-ове Малакка, в Индокитае.
Населяют леса и кустарники от уровня моря до 1500 м над уровнем моря. Ведут древесный образ жизни. Убежищем служат дупла. Держатся поодиночке и парами. Активность дневная, преимущественно раннеутренняя. Питаются орехами, плодами, семенами, насекомыми.
В роде 2 вида:
гималайская белка — Т. maclellandi Horsfield, 1839 (п-ов Малакка; Таиланд; п-ов Индокитай; Бирма; Ассам; Непал; Юньнань в Китае);
Т. swlnhoe Milne-Edwards; 1874 (Китай от Ганьсу и Шаньси на юг до Сычуаня, Юньнаня, Фуцзяня, о-вов Хайнань и Тайвань; Северная Бирма, п-ов Индокитай).
Андерсон и Джонс (1967) выделяют в роде 4 вида.
Эллермэн (1940), Эллермэн и Моррисон-Скотт (1966), Муур (1959) включают тамиопс в род прекрасных белок.
Род многополосых белок
Genus Menetes Thomas, 1908
В роде единственный вид: многополосая белка — TH. berdmorei Blyth, 1849.
Размеры мелкие. Длина тела 17—20 см. Длина хвоста 15—17 см. Внешним видом напоминает бурундука. Мордочка вытянутая и заостренная (рис. 21). Волосяной покров густой и мягкий. На спине он Сероваго-бурый, на голове сероватый, по бокам светло-серый, па брюхе беловатый или грязно-желтый. Вдоль тела по спине и бокам проходят чередующиеся черные и белые или грязно-желтые полосы. Хвост покрыт густыми, довольно длинными пушистыми волосами. Сосков 3 нары.
Череп удлиненный с узким и вытянутым рострумом. Заглазпичиые отростки средних размеров или относительно небольшие. Передняя поверхность резцов оранжевого цвета.
Распространена в Таиланде, Аммане, Кампуче, Бирме.
Населяет скалистые участки, окруженные густыми зарослями кустарников. Часто встречается в антропогенных ландшафтах, в полях, а
также в лесах. Ведет в основном наземный образ жизни, лишь иногда залезая на деревья.
Эллермэн и Моррисон-Скотт (1966) включают в ареал многополосой белки Малаккский п-ов. Медвэй (1969), однако, не приводит этот вид в списке малайских млекопитающих.
Род длинноносых белок
Genus Rhinosciurus Gray, 1843
В роде единственный вид: длинноносая белка — /?. laticaudatus Muller et Schlegel, 1839.
Размеры мелкие. Длина тела 18—24 см. Длина хвоста 10—15 см. Масса 187—225 г. Морда сильно удлинена, уши короткие, округлые (рис. 22). Язык очень длинный, способный высовываться далеко изо
tus и ее череп
рта. Стопа очень узкая. Хвост покрыт длинными и густыми волосами. Окраска волосяного покрова на спине рыжевато-бурая до черноватобурой. Брюхо обычно беловатого цвета. Хвост серый, с белым кончиком.
Череп характеризуется очень вытянутым лицевым отделом (рис. 22). Верхние резцы сильно редуцированы, очень узкие, иногда едва различимы. Посторбитальные отростки короткие. Костные слуховые барабаны увеличенных размеров.
Распространена в южной части Малаккского п-ова, на Суматре, Калимантане и на прилегающих островах. Держится в долинах и на холмах в первичном и вторичном лесах. Ведет в основном одиночный наземный образ жизни. Питается преимущественно мелкими насекомыми: крупными муравьями, термитами, жуками. Поедает также плоды и земляных червей. По-видимому, пользуется языком при ловле насекомых. Самка приносит 1—2 детенышей.
Род малайских белок
Genus Lariscus Thomas et Wroughton, 1909
Размеры мелкие. Длина тела 16—20 см. Длина хвоста 7—11 см. Масса 120—130 г. Морда удлинена, но не в такой степени, как у длинноносых белок. Волосяной покров довольно изреженный. По окраске имеется две резко отличные формы, связанные, однако, между собой переходами. У одних верх головы и спина имеют темно-серую окраску, бока бурые, брюхо белое или грязно-желтое. По спине и вдоль каждого бока проходит по черной полосе. Хвост черноватый с серыми вершинами волос. Вторая цветовая форма совсем не имеет полос, причем спина шоколадно-бурая, а брюхо почти полностью белое. Бока имеют окраску, переходную между спиной и брюхом. Хвост покрыт относительно редкими, хотя и длинными волосами. Сосков 3 пары.
Лицевая часть черепа удлинена. Костные слуховые барабаны небольшие. Щечные зубы гипселодонтные.
Распространены на п-ове Малакка, Суматре, Калимантане, Яве и мелких прилегающих островах.
Населяют разные типы лесов в горах от 900 до 1500 м над уровнем моря. Некоторые живут на равнинах. Часто встречаются по берегам рек в джунглях. Большую часть времени проводят на земле. Гнезда строят в дуплах упавших деревьев.
В роде 2 вида:
трехполосая белка — L. insignis F. Cuvier, 1821 (п-ов Малакка, Суматра, Ява, Калимантан и мелкие прилегающие острова);
малайская белка — L. hosei Thomas, 1892 (север и северо-запад о-ва Калимантан).
Андерсон и Джонс (1967) насчитывают в роде 4 вида.
Малайская белка включена в «Красную книгу».
Род дремомис
Genus Dremomys Heude, 1898
Размеры мелкие. Длина тела 16—24 см. Длина хвоста 12—18 см. Масса 210—270 г. Стопа узкая. Мордочка сильно вытянута в длину и заострена (особенно у D. rufigenis) (рис. 23). Волосяной покров мягкий и густой. Подшерсток густой, серого цвета- Хвост покрыт пышными, длинными, густыми волосами. Окраска разнообразная. У некоторых ви-
Рис. 23. Краспощекая бел <а — D. rufigenis
дов спина оливкового цвета с легким буроватым тоном, у других основной цвет рыжевато-бурый. Брюхо белое или оранжевое. Нижняя часть головы и лоб ярко-рыжевато-коричневые. Могут быть беловатые пятна позади ушей и белые кольца вокруг глаз. Верхняя поверхность хвоста может быть почти черной, но вершинки волос обычно сероватого цвета; нижняя часть хвоста коричневая, на конце черная. Сосков 3 пары.
Лицевой отдел черепа удлинен. У некоторых видов он почти достигает длины лицевого отдела длинноносых белок. Характерно более каудальное расположение слезной кости по отношению к щечным зубам, чем у других представителей семейства. Костные слуховые барабаны часто относительно малы. Лобные кости широкие. Заглазпичные отростки средних размеров.
Распространены в Китае — Тибете, Южном Китае и па Тайване; Непале; Индии: Ассаме, Манипуре; Бирме; Таиланде, па п-овах Индокитай, Малакка и о-ве Калимантан.
Населяют густые леса, поднимаясь в горы па высоту до 3400 метров. Значительную часть времени проводят па земле, но часто поднимаются и на деревья. Питаются различными плодами, орехами, другими растительными объектами, а также насекомыми.
В роде 4 вида:
оранжевобрюхая белка — D. lokriah Hodgson, 1836 (Непал, Ассам, Западная Бирма и юго-восточный Тибет в Китае);
белка Перни — D. pernyi Milne-Edwards, 1867 (юго-восток Китая, о-в Тайвань; штат Индии Манипур; Ассам; Бирма);
D. everetti Thomas, 1890 (о-в Калимантан);
краснощекая белка — D. rufigenis Blanford, 1878 (п-ова Малакка, Индокитай; Таиланд; Бирма; Ассам; юго-восток Китая);
Андерсон и Джонс (1967) насчитывают в роде 5 видов.
Род калимантанских белок
Genus Glyphotes Thotnas, 1898
В роде единственный вид: калимаптапская белка — G. sitnus Thomas, 1898.
Размеры мелкие. Длина тела около 13 см. Длина хвоста примерно 10 см. Хвост покрыт короткими волосами. По бокам тела проходит белая или черная полоса. Дорзальные поверхности пальцев, бока носа,
область вокруг глаз и края ушей грязно-желтого цвета. Грудь и брюхо желтоватые. Кончик хвоста черный.
Лицевой отдел черепа сильно укорочен, широкий. Носовые кости короткие. Заглазничные отростки небольшие. Нижняя челюсть слабая, венечный отросток низкий. Верхние резцы очень широкие, не утолщенные в передне-заднем направлении, их концы имеют тенденцию выгибаться в стороны друг от друга. Нижние резцы сходны с верхними, а их вершины еще более расставлены в стороны. Число щечных зубов .
Распространена на горе Кинабалу в северной части о-ва Калимантан.
Экология не изучена. Встречается в лесу, в горах на высоте примерно 1200 м над уровнем моря.
По Андерсону и Джонсу (1967), род включает 2 вида.
Род черноухих белок
Genus Nannosciurus Trouessart, 1880
Размеры мелкие. Длина тела 7,5—11,5 см. Длина хвоста 5—8 см. Волосяной покров очень короткий, мягкий. Узкий уплощенный хвост покрыт короткими волосами. Уши небольшие, округлые с удлиненными волосами по задней кромке. Окраска спинной стороны варьирует от сероватой до рыжеватой или темно-рыже-бурой, а брюшной — от рыжеватой до серой или грязно-желтой. Хвост имеет окраску, сходную с телом. У N. melanotis белые полосы проходят от носа по направлению к ушам и заканчиваются в районе плеч.
В черепе подглазничное отверстие образует короткий канал. Венечный отросток сильно редуцирован. Костное небо обычно узкое. Щечных „ 4
зубов —.
Распространены на Яве, Калимантане, Суматре, Филиппинских и мелких прилегающих островах.
Населяют в основном густые леса от уровня моря до примерно 1650 м над его уровнем. Обычно встречаются по две или три вместе. Активность дневная. Большую часть времени проводят на земле.
В роде 5 видов:
N. coticinnus Thomas, 1888 (Филиппинские о-ва);
черноухая самарская белка —У. samaricus Thomas, 1897 (о-в Самар в группе Филиппинских о-вов)
N. surrutilus Hollister, 1913 (о-в Минданао в группе Филиппинских о-вов);
N. luncefordi Taylor, 1934 (о-в Минданао);
A', melanotis Muller et Schlegel, 1844 (о-ва Ява, Суматра, Калимантан) .
Андерсон и Джонс (1967) насчитывают в роде 7 видов.
Род крошечных белок
Genus Exilisciurus Moore, 1958
Мелких размеров белки. Длина тела примерно 7—10 см. Длина хвоста 5—8 см. Окраска спинной стороны темно-рыжевато-бурая или темно-бурая, иногда с черноватым оттенком. Брюшная сторона грязно-желтая, изредка с серым оттенком. Хвост темно-оранжево-бурый или чер
ный. Уши почти голые у Е. exilis или имеют длинные кисточки волос (Е. whitheadi) серого и черного цвета.
Распространены на о-ве Калимантан.
Населяют высокоствольные леса.
В роде 2 вида:
Е. exilis Muller et Schlegel, 1838 (Северный и Восточный Калимантан);
Е. whitheadi Thomas, 1887 (Северный Калимантан).
Андерсон и Джонс (1967) насчитывают в роде 3 вида.
Род магрибских белок
Genus Atlantoxerus Major, 1893
В роде единственный вид: магрибская белка — A. getulus Linnaeus, 1758.
Средних размеров белка с длинным хвостом, покрытым довольно длинными, редкими волосами. Волосяной покров жесткий, короткий, редкий. По бокам проходят белые полосы, иногда такая же полоса тянется вдоль середины спины. Брюхо покрыто очень редкими волосами.
Череп относительно уплощенный. Слезные кости не увеличены. Верхние резцы часто с небольшими бороздками.
Распространена в Северо-Западной Африке: в Марокко и Алжире.
Ведет наземный образ жизни. Убежищем служат норы. Наиболее обычна в скалистых участках. Активна преимущественно утром и вечером. Днем, во время жары находится в норе. Питается плодами и семенами Argania sideroxylon.
Род земляных белок
Genus Xerus Hemprich et Ehrenberg, 1832
Размеры средние. Длина тела 23—46 см. Длина хвоста 18—27 см. Масса 300—945 г. Внешним видом напоминают сусликов, но с длинным хвостом (рис. 24). Уши настолько малы, что снаружи почти не заметны. Ступни сильно удлинены. Три средних пальца длиннее боковых. Когти слегка изогнутые, длинные. Хвост покрыт длинными, но не густыми волосами. Волосяной покров жесткий, сильно изреженный, не имеет подшерстка. Щетинковидные волосы плотно прилегают к туловищу. Окраска спинной стороны буроватая до рыжевато-серой и желтовато-серой или бледно-рыжевато-бурая или серовато-бурая с легкими черными пестринами, по бокам может проходить белая полоса. Брюшная сторона белая или грязно-желтая. Хвост обычно более темный, чем тело. У неполово
зрелых волосы на хвосте имеют светлые вершинки, а у половозрелых светлые вершинки почти исчезают, и хвост становится значительно более темным, особенно по средней части. Конечности белые. Сосков от 1
до 3 пар.
Костное небо сдвинуто далеко назад по отношению к уровню последних щечных зубов. Костные слуховые барабаны округлые, крупных размеров. Посторбитальные отростки короткие, направлены назад. Белые или желтые резцы не имеют бороздок. Число щечных зубов — - Корен
ные зубы гипселодонтного типа.
Распространены в Северо-Восточной, Восточной, Южной и Юго-Западной Африке.
Рис. 24. Красная земляная белка — X. rutilus и ее череп
Населяют преимущественно открытые места, но обитают также и в Кеах. Ведут наземный образ жизни, копают норы, иногда достигающие । удину 1,7 м; Нора имеет несколько выходов. Часто живут колониями. Держатся вблизи нор. Активны днем. В спячку не впадают и запасов лящи не делают. Питаются главным образом сочными подземными чачи растений, поедают также семена и другие растительные корма, -оме того, насекомых и их личинок, ящериц, яйца и птенцов. Воду не гют; влага поступает в организм с пищей. В помете от 2 до 6 детенышей, обычно 4. Половозрелость наступает примерно в 6 месяцев. Хра- -чди возбудителей некоторых болезней человека.
В роде 4 вида:
красная земляная белка — X. rutilus Crctzsclmiar, 1826 (север Эфи-.п,Сомали, Кения);
X. erythropus Е. Geoffrey, 1803 (от Сенегала и Южной Мавритании области Голубого Нила в Судане; Кения и Заир);
X. inauris Zimmerman, 1780 (Южно-Африканская Республика, Ботсвана, запад Южной Родезии и Юго-Западная Африка);
X. princeps Thomas, 1929 (Юго-Западная Африка и юго-запад Ангелы).
Некоторые, например Уокер (1964), выделяют X. inauris и X. princeps в отдельный род Geosciurus Smith, 1834. что, по-видимому, неправильно.
Рис. 25. Тонкопалый суслик — S /ер-todactylus и его череп
Род тонкопалых сусликов
Genus Spermophilopsis Blasius, 1884
В роде единственный вид: тонкопалый суслик — S. leptodactylu Lichtenstein, 1823.
Размеры средние и крупные. Длина тела 20—30 см. Длина хвост примерно 10 см. Задние конечности длинное передних (рис. 25). Кисть стопа сильно удлинены, пальцы также длинные с очень длинными слаб изогнутыми, но сильными когтями. Па передних конечностях наиболс длинные II и III пальцы. Па задних конечностях самый длинны IV палец, слегка превышающий по длине второй и третий. Подошвы ла (особенно задних) покрыты волосами. Ушные раковины представлен лишь небольшой кожной складкой. Волосяной покров резко изменят по сезонам (линька бывает дважды в году). В зимнее время он выс кий, довольно мягкий и густой, в то время как в летнее время это коро кие относительно редкие, грубые щетинкоподобные волосы. У основ
ния пальцев волосы собраны в пучки и образуют оторочку по краю ступней. Окраска спины однотонная, светло-песчаная, иногда с более темными пестринами, брюхо белое. Хвост на конце снизу и приблизительно на */з сверху черный с оторочкой из светлых волос.
Лицевой отдел черепа несколько укорочен, а мозговой крупный, слегка уплощенный сверху. Заглазничные отростки хорошо развиты. Кост-5
нце слуховые барабаны крупные. Число щечных зубов . Зубы гипсе-лодонтного типа. Их жевательная поверхность бугорчатоскладчатая. Хромосом в диплоидном наборе 38.
Характерна черная пигментация твердой оболочки мозга.
Распространен в Северном Иране, Северном Афганистане; в СССР — в пустынях Южного Казахстана и Средней Азии: Каракумы, Кызылкумы, Муюнкумы, Забалхашье, Бадхыз, Карабиль.
Характерный обитатель песчаных пустынь. Особенно часто встречается в слабо закрепленных бугристых песках. Норы устраивает, как правило, в понижениях между барханами, предпочитая места с богатой туковичной растительностью. Селятся поодиночке, довольно далеко друг от друга. Активен днем. Летом наибольшая активность наблюдается в утренние часы, но животное можно встретить иногда и в самые жаркие дневные часы. В спячку не впадает, однако в холодную погоду по нескольку дней не появляется на поверхности земли. В поисках пищи может уходить далеко, за несколько сотен метров от своей норы. Питается различными травянистыми и кустарниковыми растениями, а также насекомыми. Особенно предпочитает сочные клубеньки мятлика, корневища песчаной осоки, луковицы тюльпанов и дикого лука. Без сочного корма существовать не может. В зимнее время поедает в большом количестве плоды коллигонума. Самка приносит один помет в год. Гон приходится на вторую половину февраля — начало апреля. Число детенышей в помете от 3 до 7 (обычно 5).
Второстепенный промысловый вид. Природный носитель возбудителей чумы, лихорадки папатачи, лейшманиоза, бруцеллеза, одной из форм клещевых сыпнотифозных лихорадок.
Род бурундуков
Genus Tamias liliger, 1811
Размеры мелкие. Длина тела 8—18 см. Длина хвоста 6—14 см. Масса 80—111 г. Как задние, так и передние конечности относительно короче, чем у белок. Задние длиннее передних (рис. 26). На передних конечностях III и IV пальцы — самые длинные. На задних IV палец длиннее других. Подошвы частично покрыты волосами. Уши небольшие, слабо опушенные. Волосы на хвосте не отчетливо расчесаны на две стороны. Волосяной покров короткий, с довольно грубой остью. Основная окраска на спине от серовато-бурой до рыжевато-бурой. Обычно вдоль спины проходит пять продольных темных полосок, чередующихся со светлыми. Брюхо беловатое. В течение года две линьки. Имеются довольно хорошо развитые защечные мешки.
Череп относительно крупный, вытянутый, несколько уплощенный сверху в мозговой части. Гребни па черепе не развиты. Посторбитальные отростки небольшие, слабые. Костные слуховые барабаны относительно крупные. Костное небо широкое. Инфраорбитальные отверстия крупные, | округлой формы. Число щечных зубов. Коронки низкие. Хромосом в диплоидном наборе у 9 изученных видов бурундуков (малого, сосново-
Рис. 26. Азиатски» бурундук — Т. sibiricus и его черен 76
го, Таунсенда, скалистого, длинноухого, Т. speciosus, уинтасского и азиатского) равно 38.
Распространены на большей части Северной Америки: на западе от рек Юкона и Маккензи на юг до Калифорнии и штатов Дуранго и Сонора в Мексике, а на востоке — юго-восточные районы Канады и восточные штаты США. В Старом Свете обитают в лесных районах Дальнего Востока (кроме Камчатки), в Сибири, в северо-восточном Казахстане, на северо-востоке европейской части СССР, а также в Среднем, Северном и Северо-Восточном Китае, на Корейском п-ове, на некоторых островах Японии и в северной части МНР.
Обитают в хвойных, смешанных и частично широколиственных лесах Старого и Нового Света. В горах поднимаются до верхней границы леса. Селятся часто по каменистым россыпям. Ведут в основном наземный образ жизни, хорошо лазают и по деревьям. Убежищем служат норы, которые животные сами выкапывают в земле. Активны днем. На зиму впадают в спячку. Питаются в основном различными растениями. Для азиатского бурундука основной пищей служат семена хвойных и лиственных деревьев и различных травянистых растений, особенно осок и зонтичных, поедает также почки и ягоды, весной вегетативные части растений, употребляет в пищу насекомых, моллюсков. Делает запасы пищи, масса которых может достигать 3—4 кг. Период размножения азиатского бурундука приходится на апрель — май. В течение года, как правило, один помет из 4—10 детенышей. Лишь в некоторых частях ареала бывает второй помет. Размножение восточного бурундука начинается в марте и продолжается в течение всего лета. За сезон бывает 2 помета. Длительность беременности примерно 31 день. Самка приносит от 2 до 9 детенышей массой около 3 г каждый. Детеныши рождаются голые и слепые. Глаза открываются примерно на 31 день. Половой зрелости достигает в возрасте 1 года.
Азиатский бурундук имеет небольшое промысловое значение. Может причинять вред лесному и сельскому хозяйству. Природный носитель одной из форм клещевых сыпнотифозных лихорадок.
В роде, по-видимому, 18 видов:
восточный бурундук— Т. striatus Linnaeus, 1758 (юго-восток Канады и северо-восток и восток США; северная граница ареала идет от берега залива Св. Лаврентия в окрестностях г. Минган на запад к г. Ист-мейн в южной части залива Джемса, отсюда к оз. Нипигон и далее на восток к южной оконечности озер Виннипег и Манитоба, поднимается по западному берегу оз. Манитоба до южных частей оз. Виннипегосис в круто сворачивает на юг, становясь западной границей ареала, выходит южнее государственной границы с США, поворачивает на запад вдоль нее, вновь сворачивает на юг и идет западнее границы между штатами Северная Дакота и Миннесота, пересекает северо-восточный угол штата Южная Дакота, заходит на территорию Миннесоты и пересекает ее в юго-западной части штата, переходит в штат Айова, пересеки его в северо-западном углу с северо-востока па юго-запад, в районе г. Омаха переходит в штат Небраска, идет на юг, включая в ареал юго-восточный угол этого штата и северо-восточный угол штата Канзас, затем резко сворачивает на восток к центральным районам штата Миссури, поворачивает на запад севернее от границы между штатами Миссури п Арканзас, заходит в юго-восточный угол штата Канзас, сворачивает д юг и следует западнее границы между штатами Оклахома и Арканзас, немного не доходит до штата Техас и поворачивает на восток, становясь южной границей ареала, следует по югу штата Арканзас несколько к северу от границы со штатом Луизиана, проходит по границе между
штатами Арканзас и Миссисипи, Арканзас и Теннесси; поворачивает на юго-восток, пересекая Теннесси в юго-западном углу, заходит в северо-восточную часть штата Алабамы, поворачивает на запад, достигая крайней северо-восточной части в штате Луизиана, затем идет на юг вдоль границы между этим штатом и Миссисипи, немного не доходит до берега Мексиканского залива, полого поворачивает на северо-восток и идет примерно до г. Мейкон в штате Джорджия, откуда более круто поворачивает к северу к району г. Шарлотт в штате Северная Каролина, отсюда идет на восток примерно до г. Роли, поворачивает на запад и следует к крайней северо-восточной точке штата Теннесси, откуда вновь поворачивает на восток и выходит к Атлантическому океану несколько южнее г. Норфолк);
альпийский бурундук—Т. alpinus Merriam, 1893 (небольшой участок в штате Калифорния в США вдоль 119° з. д. и между 36 и 38° с. ш.);
малый бурундук—Т. minimus Bachman, 1839 (почти вся южная половина Канады — юго-восток провинции Юкон, юг Маккензи, северо-восток Британской Колумбии, Альберта, кроме крайнего юго-востока, Саскачеван, кроме крайнего юго-востока, Манитоба, кроме небольшой территории на севере, Онтарио и небольшой участок на юго-западе Квебека; штаты США — северная половина Висконсина, север и северо-восток Миннесоты, крайний северо-восток и юго-запад Северной Дакоты, почти вся Монтана, кроме северо-западного угла, Вайоминг, запад Южной Дакоты и запад Небраски, южная половина Айдахо, восточная половина Орегона, кроме северо-восточного угла, центральные районы Вашингтона, северо-восток Калифорнии и северо-запад Невады, а также север и северо-восточные районы Невады, северные, центральные и восточные районы Юты, западные две трети штата Колорадо, север штата Нью-Мексико; три небольших изолированных участка ареала расположены на севере и востоке штата Аризона и на юге штата Нью-Мексико) ;
сосновый бурундук — Т. amoenus Allen, 1890 (юго-запад Канады — южная половина провинции Британская Колумбия и узкая полоса на юго-западе провинции Альберта, проходящая вдоль границы с Британской Колумбией примерно от г. Джаспер-Плейс до г. Летбридж; северо-запад США— восточные две трети штата Вашингтон, восточные три четверти штата Орегон, кроме юго-восточного угла, север Калифорнии, кроме северо-запада, Айдахо, за исключением юго-западного и юго-восточного углов, западная половина Монтаны и северо-западный угол Вайоминга; кроме того, имеются два небольших изолированных участка на северо-западе штата Вашингтон и в северо-западном углу штата Невада),
бурундук Таунсенда — Т. townsendi Bachman, 1839 (крайний юго запад провинции Британская Колумбия и юг о-ва Ванкувер в Канаде северо-запад США — западные половины штатов Вашингтон и Орегон север Калифорнии, запад Невады около г. Карсон-Сити) ;
сономский бурундук—Т. sonomac Grinnell, 1915 (США — Калифор ния: от Саи-Фрапциско узкая полоса, несколько отступя от берега па се вер почти до границы со штатом Орегон);
бурундук Мерриама — Т. mcrriami J. Allen, 1889 (США—южна: часть штата Калифорния; Мексика — два небольших изолированны участка на север и посередине п-ова Калифорния);
скалистый бурундук — Т. dorsalis Baird, 1855 (юго-запад США-восток и северо-восток штата Невада, юго-запад Орегона, запад штат Юта, северо-запад Колорадо, полоса, проходящая от северо-западног угла штата Аризона к ее юго-восточному углу, юго-запад Ныо-Мексик<
узкая полоса в Северной Мексике, захватывающая северо-восточный угол штата Сонора, запад штата Чиуауа и крайний северо-восток Дуранго) ;
колорадский бурундук — Т. quadrivittatus Say, 1823 (штаты США—-юго-запад Колорадо, юго-восток Юты, крайний северо-запад Оклахомы, северо-восток Аризоны, север Ныо-Мексико);
рыжехвостый бурундук — Т. ruficaudus Howell, 1920 (Канада — крайний юго-запад Альберта и юго-восток Британской Колумбии; штаты США—-запад Монтаны, север Айдахо и крайний северо-восток Вашингтона) ;
сероворотничковый бурундук—Т. cinereicollis J. Allen, 1890 (юго-запад США — два изолированных участка: один на юго-востоке Аризоны и юго-западе Ныо-Мексико, а второй на юге Ныо-Мексико);
длинноухий бурундук — Т. quadrimaculatus Gray, 1867 (юго-запад США — небольшая территория на северо-востоке штата Калифорния, вытянувшаяся узкой полосой от оз. Альманор к югу примерно до 37°ЗО' с. ш., захватывающая небольшой участок штата Невада к востоку от оз. Тахо);
Т. speciousus Merriam, 1890 (два изолированных участка на юго-западе США — один вытянулся узкой полосой в штате Калифорния, с севера на юг от оз. Игл примерно до г. Сан-Бернардино, захватывая небольшой участок штата Невада к востоку от оз. Тахо, а второй небольшой расположен у горы Пинос на юго-запад от пересечения 35° с. ш. и 119° з. д.);
панаминтский бурундук — Т. panamintinus Merriam, 1893 (юго-запад США — узкая полоса, тянущаяся по штату Невада вдоль границы между Невадой и Калифорнией примерно от г. Йерингтон на юго-восток до пересечения 116°30' з. д. с границей штатов Невада и Калифорния, откуда небольшой язык ареала вдается в Калифорнию, два других узких выступа спускаются из Невады на юг в Калифорнию в районе оз. Моно; небольшой изолированный участок лежит на северо-востоке графства Сан-Бернардино в штате Калифорния);
уннтасский бурундук — Т. umbrinus J. Allen, 1890 (юго-запад США — семь изолированных участков, расположенных: один — на западе штата Калифорния, в центральных районах и северо-востоке Невады и крайнем северо-западе Юты; второй — в северо-западном углу штата Вайоминг, юго-западе Айдахо и юге Монтаны; третий на юге Невады; четвертый в центральных районах и северо-востоке штата Юта; пятый в центральных районах штата Юта; шестой на севере Аризоны; седьмой на северо-западе Колорадо, востоке Юты и юге Вайоминга);
бурундук Палмера — Т. palmeri Merriam, 1897 (небольшая территория на юге штата Невада в США);
бурундук Буллера — Т. bulleri J. Allen, 1889 (север Мексики — два изолированных участка: один на юго-западе штата Закатекас, на западе Дуранго и крайнем севере Чиуауа, а второй на севере штата Коауила);
азиатский бурундук—Т. sibiricus Laxmann, 1769 (северо-восток европейской части СССР-—на запад до линии Архангельск -Верхняя | Тойма — Вологда и на юг до Ветлуги— Казани — Уфы — Южного Урала; Северо-Восточный Казахстан, Сибирь, Дальний Восток, кроме Камчатки, Сахалин, Шантарские о-ва; Средний, Северный и Северо-Восточ-। ный Китай, северная часть МНР, Корейский п-ов, некоторые острова Японии).
В СССР один вид: азиатский бурундук.
Некоторые исследователи (Симпсон, 1945; Бурлиер, 1955; Муур, 1959;
[ Уокер, 1964; Андерсон и Джонс, 1967) оставляют в роде Tamias только
T. striatus, а все остальные виды относят в род Eutamias Trouessart, 1880.
Видимо, более правильно объединить их в один род, как это делают Эллермэн (1940) и И. М. Громов (1965).
Количество видов бурундуков Нового Света после ревизии, вероятно, будет уменьшено.
Род беличьих бурундуков
Genus Sciurotamias Miller, 1901
Размеры мелкие и средние. Длина тела 19—25 см. Длина хвоста 13— 18 см. IV палец на задних лапах немного длиннее третьего. Волосяной покров густой и мягкий. Окраска его на спине одноцветная — серая или серовато-коричневая с узкой беловатой боковой линией, ниже которой проходит темная линия от плеча до бедра. Вокруг глаз может располагаться светлое кольцо. Пушистый хвост и конечности той же окраски, что и спина, или конечности рыжеватые. Брюхо бледно-рыжевато-коричневое. Грудь и горло белые. У S. davidianus подошвы лап покрыты густой шерстью, а у S. forresti подошвы задних лап почти голые. Сосков 3 пары. Имеются слабо развитые защечные мешки.
Череп вытянутый, его скуловые дуги расставлены нешироко. Носовой отдел сверху выгнут. Подглазничные отверстия имеют форму узких и коротких вертикальных щелей. Подглазничный канал хорошо развит, но короткий. Заглазничные отростки сильно редуцированны. Щечные зубы такие же, как у бурундуков.
Распространены в Южном (Юньнань) и Юго-Восточном Китае.
Населяют широколиственные и жестколиственные субтропические леса гор и равнин, а также скальные участки в пределах умеренного пояса и субтропической зоны. По образу жизни сходны с бурундуками, но в спячку не впадают. Убежищем служат расщелины и пустоты под камнями.
В роде 2 вида:
бурундук Давида — 3. davidianus Milne-Edwards, 1867 (Юго-Восточный Китай);
бурундук Форреста — 3. forresti Thomas, 1922 (Юньнань).
Род скалистых сусликов
Genus Otospermophilus Brandt, 1814
Размеры мелкие и средние. Длина тела 18—29 см. Длина хвоста 14—25 см. Внешне сходны с белкой (рис. 27). Уши относительно хорошо развиты, покрыты волосами. Хвост густо опушен. Подошвы задних лап голые или покрыты волосами. По бокам или в заднем отделе спины могут располагаться полосы, крупные или отчетливые мелкие пятна, иногда здесь неясная светлая пестристость. Имеются защечные мешки.
Череп с удлиненным лицевым отделом. Скуловые дуги расставлены нешироко. Мозговая коробка округлая, сравнительно низкая. Теменные гребни развиты. Заглазничное и межглазничное пространство широкое. Заглазничные отростки хорошо развиты. Костные слуховые барабаны умеренной величины. Щечные зубы с низкими коронками. У скалистого и калифорнийского сусликов по 38 хромосом в диплоидном наборе.
Распространены по узкой полосе запада Северной Америки от крайнего юго-запада Канады до центральной Мексики включительно.
Рис. 27. Калифорнийский суслик — О Ьсе-cheyi и его череп
Населяют равнины и предгорные степи и лесостепи. Встречаются также на скальных участках и в разреженных горных лесах.
В роде 8 видов:
скалистый суслик—О. variegatus Erxleben, 1777 (юго-запад США — штаты: южная половина Невады, Юта, кроме севера, Колорадо, кроме северо-запада и северо-востока, Аризона и примыкающий к этому штату небольшой участок юго-востока Калифорнии, Ныо-Мексико, крайний запад Оклахомы, юго-запад Техаса; север и центр Мексики па юг до штатов Мичоакан, Мексике и Пуэбло включительно, в ареал не включен штат Веракрус и территория штата Тамаулипас, прилегающая к Мексиканскому заливу);
калифорнийский суслик—О. beecheyi Richardson, 1829 (запад и юго апад США: крайний юго-запад штата Вашингтон, западная половина штатт Орегон, Калифорния, кроме юго-восточной части, крайний запад штата Невада в районе оз. Тахо, о-в Сапта-Каталнна у Южной Калифорнии; Мексика — северная часть п-ова Калифорния);
черноголовый суслик—О. atricapillus Bryant, 1889 (п-ов Калифорния в Мексике).
кольцехвостый суслик—О. annulatus Audubon et Bachman, 1842 (западные районы Центральной Мексики — штаты: юго-запад Наярит, запад Халиско, Колима, юго-запад Мичоакан, крайний запад Герреро);
тропический суслик — О. adocetus Merriam, 1903 (западные районы Центральной Мексики — штаты: центр Мичоакан и северо-запад Герреро) ;
золотистый суслик—О. lateralis Say, 1823 (юго-запад Канады — юго-восток провинции Британская Колумбия и юго-запад Альберта; запад США — штаты: крайний юго-восток Вашингтона, Айдахо, запад Монтаны, Орегон, кроме тихоокеанского побережья, север и северо-восток Калифорнии, северная половина Невады, Юта, кроме юго-запада и юго-востока, запад и юг Вайоминга, Колорадо, кроме восточной трети территории, крайний северо-восток Аризоны, север Ныо-Мексико; кроме того, три небольших изолированных участка ареала: один на юге Калифорнии, второй на юге Невады и третий на севере и в центре Аризоны и на западе Нью-Мексико);
суслик Каскадных гор — О. saturatus Rhoods, 1895 (Канада—-крайний юг провинции. Британская Колумбия; США — центральные и северные районы штата Вашингтон);
сьеррамадресский суслик—О. madrensis Merriam, 1901 (горы Запад: ной Сьерра-Мадре в штате Чиуауа в Мексике).
Многие исследователи (например, Эллермэн, 1940; Симпсон, 1945; Холл и Келсон, 1959; Муур, 1959, и др.) включают род скалистых сусликов в род сусликов.
Следом за И. М. Громовым (1965) здесь они выделены в особый род.
Род пустынных сусликов
Genus Xerospermophilus Merriam, 1892
Размеры мелкие. Длина тела около 15 см. Длина хвоста 6—11 см. Внешне сходны с бурундуками. Наружное ухо имеет вид небольшого кожного валика. Хвост равномерно, но слабо опушен. Подошвы задних лап покрыты волосами. Окраска волосяного покрова темно-песчаная. Нижняя поверхность хвоста может быть белой. Имеются слабо развитые защечные мешки.
Череп короткий и широкий. Заглазничная область шире межглазничной. Заглазничные отростки длинные. Костные слуховые барабаны крупные. Щечные зубы с низкими коронками. У голохвостого суслика в диплоидном наборе 36 хромосом.
Распространены на юго-западе Северной Америки.
Населяют пустыни.
В роде 2 вида:
мохавесский суслик — X. mohavensis Merriam, 1889 (центр штата Калифорния в США, в частности пустыня Мохаве);
голохвостый суслик — X. tereticaudus Baird, 1858 (крайний юго-запад США: крайний юг штата Невада, юго-восток Калифорнии и юго-запад Аризоны; Мексика — северо-восточная часть п-ва Калифорния и северо-западные районы штата Сопора; кроме того, небольшой изолированный участок находится в северо-восточной части н-на Калифорния к западу от границы основного ареала).
Некоторые исследователи (например, Эллермэн, 1940; Холл и Келсон, 1959; Муур, 1959, включают эти виды в род atelias).
Следом за И. М. Громовым (1969) здесь они выделены в особый род.
Род антилоповых сусликов
Genus Ammospermophilus Merriam, 1892
Размеры мелкие. Длина тела 14—16 см. Длина хвоста 6—10 см. Внешне сходны с сусликами. Волосяной покров редкий и грубый. Окраска его на спине варьирует от очень светлой серовато-коричневой до светло-рыжевато-коричневой с мелкой темной штриховатостью. Зимний более серый. По бокам туловища проходит по светлой полосе. Брюхо белое. Хвост уплощенный с темным концом; низ хвоста белый. Защечные мешки развиты относительно хорошо.
Верхний профиль черепа выпуклый. Мозговой отдел округлый. Ростральная часть черепа удлинена. Заглазничные отростки тонкие, направлены назад. Костные слуховые барабаны крупные, уплощенные с боков. Щечные зубы с низкими коронками.
У суслика Харриса и белохвостого суслика в диплоидном наборе по 32 хромосомы.
Распространены в юго-западной части Северной Америки.
Населяют равнинные и предгорные пустыни. Убежищем служат норы. Активны днем, круглогодично. В участках ареала с наиболее низкими температурами могут не выходить из нор в холодную погоду, однако в настоящую спячку не впадают. Питаются семенами, плодами, стеблями растений и их корнями, насекомыми и даже падалью. Могут делать запасы корма в норах или под камнями. В год бывает 1—2 помета, в каждом по 6—8 детенышей.
Местами могут приносить вред сельскому хозяйству.
В роде 5 видов:
суслик Харриса — A. harrisii Audubon et Bachman, 1854 (юго-запад США — южная половина штата Аризона и крайний юго-запад штата Нью-Мексико; север Мексики — северо-запад штата Сонора);
белохвостый суслик-—A. leucurus Merriam, 1889 (юго-запад США — крайний юго-восток Орегона, юго-запад Айдахо, крайний северо-восток, юго-восток и юг Калифорнии, Невада, кроме северо-востока, Юта, кроме северо-запада и северо-востока, север Аризоны, запад Колорадо и северо-запад Нью-Мексико; Мексика — п-ов Калифорния);
техасский суслик — A. interpres Merriam, 1890 (штаты США — центральные районы и юг Ныо-Мексико, крайний запад Техаса; Мексика — штат Коауила);
островной суслик—A. insularis Nelson et Goldman, 1909 (о-в Эспириту-Санто у юго-восточной части п-ва Калифорния);
суслик Нельсона — A. nelsoni Merriam, 1893 (США — центральные районы штата Калифорния).
Некоторые исследователи (например, Эллерман, 1940; Симпсон, 1945) включают этих сусликов в род Citellus.
Следом за Холлом и Келсоном (1959), Андерсоном и Джонсом (1967), И. М. Громовым (1969) и другими здесь они выделены в особый род.
Род иктидомисов
1 Genus Ictidomys Allen, 1877
! Размеры мелкие или средние. Длина тела 11—25 см. Длина хвоста 6—15 см. Внешне сходны с сусликами. Наружное ухо едва заметно. Хвост умеренно или хорошо опушен. Подошвы задних лап покрыты во-юсами. Спина покрыта белыми пятнами. Иногда по спине проходят про
дольные чередующиеся темные (буроватые или черноватые) или светлые полосы. По каждой темной полосе тянется ряд почти квадратных светлых пятен. На боках туловища полосы и пятна выражены слабее. У некоторых видов спина может быть одноцветно-буроватая. Защечные мешки средних размеров.
Череп яйцевидной формы, выпуклый сверху, реже уплощенный. Межглазничное и заглазничное пространства узкие, причем первое из них лишь незначительно меньше второго. Заглазничные отростки короткие и тонкие, направлены назад и вниз. Костные слуховые барабаны различных размеров и формы. Коронки щечных зубов средней высоты.
Диплоидное число хромосом у тринадцатиполосового суслика — 34, а у мексиканского и пятнистого по 32.
Распространены в центральных частях Северной Америки от Южной Канады на севере до Северной Мексики на юге.
Населяют степные и пустынно-степные ландшафты равнин и предгорий. Встречаются также и на каменистых участках и в светлых сосновых лесах.
В роде 5 видов:
тринадцатиполосый суслик — I. tridecemlineatus Mitchill (юг Канады— провинции: юго-восток Альберта, южная половина Саскачевана, юг и юго-запад Манитобы; северные, центральные и южные штаты США — север и восток Монтаны, Вайоминг, кроме крайнего запада, Северная Дакота, Южная Дакота, Небраска, Мшнесота, Иова, Висконсин, Мичиган, Огайо, кроме востока и юго-востока, Индиана, кроме юга, Иллинойс, кроме юга, Миссури, кроме юго-востока, Канзас, Оклахома, кроме крайнего юго-востока, Техас, кроме востока и юго-запада, северная половина Ныо-Мексико, небольшой участок иа западе Аризоны на границе с Ныо-Мексико, Колорадо, крайний северо-восток Юты);
мексиканский суслик — I. mexicanus Erxleben, 1777 (два изолированных участка: один на юго-западе США — юго-восток штата Нью-Мексико и юг, юго-восток и центральные районы штата Техас, а также в северо-восточной Мексике — штаты: восточная половина Коауила, Нуэво-Леон и Тамаулипас, кроме юга; второй — в центральных районах Мексики— штаты: восток Халиско, Гуанохуато, юг Керетаро, юг Хидальго, север Мексике, запад Пуэбло);
пятнистый суслик — I. spilosoma Merriam, 1893 (юг и центр США — штаты: крайний юг Южной Дакоты, крайний юго-восток Вайоминга, западная половина Небраски, восточная половина и юго-западный угол Колорадо, западная половина Канзаса, запад Оклахомы, западная половина Техаса, Нью-Мексико, восток Аризоны, юго-восточный угол Юты; Северная и Центральная Мексика — штаты: восток Чиуауа, запад Коауила, восток Дуранго, Закатекос, западная половина Сан-Луис-Потоси, север Тамаулипас, юг Нуэво-Леон);
перотесский суслик — I. perotensis Merriam, 1893 (восток Центральной Мексики — запад центра штата Веракрус и северо-восток штата Пуэбла);
суслик Франклина — /. franklinii Sabine, 1822 (юг Канады — центральные районы провинции Альберта, центр и юго-восток Саскачевана, юг Манитобы, крайний юго-запад Онтарио; северные и центральные штаты США: Миннесота, восток Северной Дакоты и Южной Дакоты, центр и восток Небраски, Айова, юг Висконсина, Иллинойс, кроме юга, крайний северо-запад Индианы, север Миссури, северо-восток Канзаса),
Многие исследователи (например, Эллермэн, 1940; Холл и Келсоп, 1959; Муур, 1959, и др.) включают эти виды в род Citellus.
Следом за И. М. Громовым (1965) здесь они выделены в особый род.
Рис. 28. Малый суслик — С pygmaeus и его череп
Род сусликов
Genus Citellus Oken, 1816 ( = Spermophilus F. Cuvier, 1825)
Размеры от мелких до средних. Длина тела 14—10 см. Длина хвое га 4-25 см. Задние конечности, как правило, немного длиннее передних. Четвертый палец на обеих конечностях длиннее третьего. Внутренние пальцы на передних конечностях имеются. Уши короткие, слабо опушен ные, обычно лпшьнемного выступающие над уровнем меха, покрывающего голову (рис. 28). К зиме отрастает мягкий и густой волосяной покров; в летнее время он становится более редкий, грубый и короткий. Окраска его разнообразная, обычно желтовато-серая, более темная сверху. Спина может быть испещрена светлыми пятнышками грязно желтого, светло-желтого или беловатого цвета. Изредка окраска может быть черноватой. Наконец, окраска сверху может быть очень светлая серовато-бу
рая с легким рыжеватым оттенком. По бокам туловища могут проходить белые полосы, ограниченные по сторонам более темной окраской. Эти полосы могут быть белые или грязно-желтые с более резко выраженными черными полосами по краям этой полосы. Наконец, вдоль спины могут располагаться многочисленные продольные темные буровато-серые полосы, перемежающиеся со светлыми полосами или вытянутыми в продольный ряд светлыми пятнами. Эти пятна, как и брюшная сторона тела, обычно грязно-желтого или беловатого цвета. Защечные мешки имеются. Сосков от 4 до 6 пар.
Череп с крупной округлой мозговой коробкой, несколько уплощенной сверху. Хорошо развит ламбдоидальный гребень и заглазничные отростки; последние в меньшей степени, чем у сурков. Костное небо продолжается немного кзади от последних щечных зубов. Подглазничное отвер-5 стие относительно крупное, треугольной формы. Щечных зубов у . Коронки высокие или средней высоты (сильно или средне выраженная гипселодонтность). Жевательная поверхность коренных зубов бугорчатогребенчатая.
Хромосом в диплоидном наборе от 30 у суслика Белдинга и 32 у суслика Франклина, длиннохвостого и колумбийского до 34 у американского, уинтского сусликов и 36 у желтого, краснощекого, реликтового, малого, даурского сусликов и суслика Ричардсона.
Распространены очень широко по степным, лесостепным, лугостепным и лесотундровым ландшафтам умеренных широт Евразии и Северной Америки.
Населяют степные, лесостепные и лугостепные ландшафты. По луговым участкам речных долин заходят за Полярный круг, а по остепнен-ным участкам в полупустыни, иногда в пустыпи; с горными степями поднимаются в горы до высот, превышающих 3000 м над уровнем моря. Ведут наземный образ жизни. Убежищем служат норы, которые зверьки выкапывают сами. Длина нор и их устройство могут изменяться у различных видов сусликов и в различных географических районах. Например, у длиннохвостого суслика норы могут достигать протяженности до пятнадцати м и находиться на глубине до 3 м. Такие норы суслик строит на песчаных почвах. На более тяжелых глинистых почвах глубина нор, как правило, не превышает 2 м, а протяженность 5—7 м. У крапчатого суслика норы по длине обычно не бывают более 3 м, а в глубину они достигают примерно 130 см. В норах обычно имеется гнездовая камера, куда суслик притаскивает сухую траву. Как правило, суслики поселяются колониями. Питаются они поблизости от нор. В пищу идут различные зеленые части растений, а также их подземные части и, особенно осенью, семена. Некоторые виды сусликов употребляют в пищу значительное количество животных кормов, в основном насекомых. Некоторые суслики могут делать значительные запасы пищи. Например, крапчатый суслик — семена лютика, фиалки, василька, осоки весом около 200 г или зерпа хлебных злаков весом до 700 г. Длиннохвостый суслик — семена хлебных злаков до 6 кг.
Наибольшая активность приходится на утренние часы, в мспынсй степени — на вечерние. Дневные часы суслики проводят преимущественно в норах. На холодный период года суслики впадают в спячку. Длительность ее, начало и конец сильно колеблются в зависимости от географического положения. Так, длиннохвостый суслик пробуждается в первых числах апреля, а последние длиннохвостые суслики выходят из нор в начале мая. Осенью первыми в спячку уходят старые самцы — в первой декаде сентября, во второй декаде сентября — старые самки, а
в начале октября — молодые звери текущего года рождения. Общая продолжительность спячки у них составляет примерно 6,5—7 месяцев. Американский суслик просыпается в конце марта — начале апреля.
Массовое залегание в спячку приходится на октябрь. Крапчатый суслик в Белоруссии выходит на поверхность в конце марта — начале апреля, в Чувашии обычно в середине апреля. В спячку впадает в начале октября, в Белоруссии — несколько позже, чем в Чувашии и на Украине. У ряда видов сусликов наряду с зимней спячкой имеет место и летняя, связанная с недостатком пищи. Например, у малого суслика летняя спячка наступает обычно в конце лета, однако не каждый год и с понижением температуры переходит в зимнюю. Самка суслика дает один помет в год. Число детенышей в помете от 2 до 13. Длительность беременности примерно 23—28 дней. Гон обычно начинается через несколько дней после пробуждения.
Суслики имеют второстепенное значение в пушном промысле. Многие виды вредят сельскому хозяйству. Малый и крапчатый суслики приносят серьезные убытки в зоне интенсивного земледелия, в степных частях Украины, Предкавказья, Поволжья, питаясь основными сельскохозяйственными культурами. В эпизоотологии чумы велика роль малого суслика.
В роде 20 видов (ниже приводится система и распределение по И. М. Громову, 1965):
длиннохвостый суслик — С. undulatus Pallas, 1778 (от северных хребтов Тянь-Шаня и гор Южной Сибири до Приамурья, МНР, Северного и Северо-Восточного Китая; в СССР — Джунгарский Алатау, Южный и Юго-Западный Алтай; восточнее идет по степным участкам в тайге и по безлесным склонам гор иа север до Бийска, Красноярска, Капска, Усть-Уды на Ангаре, Качуга на Лене, в Забайкалье до государственной границы; изолированные участки лежат в южной части Амурской области и в юго-западной Якутии);
американский суслик—С. parryi Richardson, 1825 (Северо-Восточная Сибирь, Аляска, кроме Юконского п-ова и долины р. Юкон, на вос- ток примерно до 150° в. д., а также п-ов Кенай; Канада и некоторые о-ва Берингова моря, Аляскинского залива и западной части Алеутской гряды; в СССР — Якутия к востоку от Лены: Верхоянский хребет, бассейны рек Яны, Индигирки, Колымы, к востоку от Колымы по всему материку; о-в Айон в Чаунской губе, побережья Чукотского и Охотского морей; Камчатка);
колумбийский суслик— С. columbianus Ord, 1815 (провинции Канады—восток Британской Колумбии, юго-запад Альберта; штаты США — запад Монтаны, север и центр Айдахо, северо-восток Орегона, восток Вашингтона);
реликтовый суслик — С. relictus Kaschkarov, 1923 (СССР — изолированные участки горных степей хребтов Тянь-Шаня и Памиро-Алая: Кегенского, Кетменьского, Кунгей и Терскей Алатау, Киргизского и Таласского Алатау, западного Тянь-Шаня, Ферганского, Гиссарского и Зеравшанского);
суслик Ричардсона — С. richardsoni Sabine, 1822 (провинции Канады—юг и юго-восток Альберта, южная половина Саскачевана, юг и юго-запад Манитобы; штаты США — Монтана, кроме северо-запада и юго-востока, Северная Дакота, кроме юго-запада, крайний запад Миннесоты, северо-восток Южной Дакоты, Вайоминг, кроме северо-востока, северо-запад Колорадо, крайний северо-восток Юты, северо-восток Айдахо: небольшой изолированный участок занимает юго-восточный угол Орегона, юго-западный угол Айдахо и северо-восток Невады);
суслик Белдинга — С. beldingi Merriam, 1888 (штаты США — восточная половина и юг Орегона, юго-восток Айдахо, крайний северо-запад Юты, северная половина Невады, северо-восток и восток Калифорнии);
уинтский суслик—С. armatus Kennicott, 1863 (штаты США — крайний юго-запад Монтаны, восток Айдахо, запад Вайоминга, север и центр Юты);
алашанский суслик—С. alaschanicus Bachner, 1888 (Гобийский Алтай в МНР; Алашань и Нань-Шань в Западном Китае);
суслик Таунсенда — С. townsendi Bachman, 1839 (штаты США—юг Вашингтона, восточная половина, кроме северо-востока Орегона, юг Айдахо, запад Юты, кроме крайнего юго-запада, Невада, кроме крайнего юга, два небольших участка на востоке Калифорнии);
айдахский суслик—С. brunneus Howell, 1928 (США — запад штата Айдахо);
крапчатый суслик — С. suslicus Giildenstaedt, 1770 (Европейская равнина на запад до левобережья Дуная и Прута, западных районов Львовской области, Люблянского воеводства Польши, на восток до Волги от Казани до Саратова, на север до линии Ровно — Чернигов — Почеп — Карачев — Кашира •— Рязань — Арзамас — на юг до низовьев Дуная, берег Черного моря вплоть до устья Днепра, правобережья Днепра, правобережья р. Ворсклы и р. Коломак, левобережья р. Северный Донец до Луганска, кроме того, имеются два изолированных участка на крайнем юго-западе Волынской области и в Белоруссии в районе Капыль-ской гряды и Новогрудско-Слонимской возвышенности);
европейский суслик—С. citellus Linnaeus, 1776 (Центральная и Юго-Восточная Европа на восток до западных районов Украины и Молдавии; Средне- и Нижне-Дунайская низменности, западная часть Трансильвании, Нижняя Австрия, ГДР, Чехословакия к западу от Западных Карпат,. юго-восточная часть Великопольской низменности; с территории Нижне-Дунайской низменности проникает в Болгарию, в юго-западные районы Югославии, на территорию восточной Греции и европейской Турции; в СССР встречается к северу до линии, проходящей южнее Черновиц на Заставна, Бор, Жмеринку, Тульчин, Могилев-Подольский, Сороки, затем северная граница переходит на правый берег Днестра, огибает с востока Бельцкую степь до с. Галерканы — севернее Кишинева, поворачивает к западу р. Прут у с. Пырлица);
малоазиатский суслик—С. xanthoprymnus Bennet, 1835 (предгорья и горы Закавказья до 2700 м над уровнем моря; Турция: Карское плоскогорье, горы по долине Ардакса в районе Эрзрума, западная часть Понтийских гор; в СССР — Западная Армения, долина Аракса, верховья р. Памбак и реки ее бассейна, западные и северные склоны горы Арагац);
даурский суслик — С. dauricus Brandt, 1843 (Юго-Восточное Забайкалье; Восточная часть .МНР, Северный и Северо-Восточный Китай; в СССР — южная часть Юго-Восточного Забайкалья к северу до линии, проходящей через Нерчинский Завод, южное Александровского Завода на ст. Оловянную па р. Ононе и по ее правому берегу до государственной границы с МНР);
горный суслик—С. musicus Menelrie, 1832 (центральная часть Кавказского хребта на высотах 1500—-3100 м над уровнем моря, между Черным, Пастбищным и Скалистым хребтами с севера н северным склоном Главного хребта с юга, а также к востоку от Грозного, между Терским и Сунженским хребтами);
малый суслик — С. pygmaeus Pallas, 1778 (от нижнего Днепра на западе до района Караганды и западной окраины Бетпак-Далы на востоке; северная граница проходит по левобережьям Ворсклы и Коломака,
правобережью Северного Донца, поворачивает к востоку на Саратов, по левому берегу Волги немного не доходит до Куйбышева, идет на крайний юго-восток Башкирской АССР, через Кустанай, Атбасар, Шад-ринск, Целиноград к Караганде; южная граница идет по черноморскому и азовскому побережью Крыма до Ставрополя, Пятигорска, Моздока, Маглобека, Грозного и Гудермеса, Дагестана, Каспийского побережья, Устюрта (43° с. ш.), берега Аральского моря до устья Сырдарьи, по ее правобережью до района истоков Сарысу и по западной окраине Бетпак-Далы к Караганде);
большой суслик—С. major Pallas, 1778 (СССР — Заволжье, Южный Урал, Западная Сибирь, Центральный и Восточный Казахстан);
красиощекий суслик—С. erythrogenys Brandt, 1843 (МНР, Китай-— Синьцзян, СССР — северная граница от района г. Карсакпая на восток обходит с юга горы Улутау, а с севера — Казахское нагорье, южнее Семипалатинска выходит к Иртышу, по его правобережью поднимается до района Тюкалинска, 'Гатарска и оз. Чаны, по Оби идет до Барнаула, выходит к р. Томь, являющейся восточной границей ареала, по ее левому берегу следует к Усть-Каменогорску, обходит с запада горы Южного Алтая, выходит к устью р. Кучум, северному берегу оз. Зайсан и по правому берегу Черного Иртыша уходит за пределы СССР; на западе я юго-западе до восточной окраины песков Муюнкумы, южной окраины Бетпак-Далы, песков Таукум и отрогов Чу-Илийских гор; предгорные и низкогорные районы Джунгарского Алатау, южное Прибалхашье и Ала-кольская котловина);
желтый суслик—С. fulvus Lichtenstein, 1823 (СССР — от Южного Заволжья до Центрального и Юго-Восточного Казахстана, к югу до Северного Ирана и Афганистана; в СССР на север до линии район г. Ча-паевска — верховье р. Ишим — среднее течение Сарысу — низовье р. Чу — вся равнинная часть Средней Азии, кроме песков Каракум и Кызылкум и Ферганской долины).
В СССР: суслики длиннохвостый, американский, реликтовый, крапчатый, европейский, малоазиатский, даурский, горный, малый, желтый, большой, краснощекий.
Б. А. Кузнецов (1965) считает американского суслика подвидом C.undulatus (с этим согласны и многие другие ученые), реликтового, ма-лоазийского, даурского сусликов — подвидами С. citellus, горного суслика—подвидом С. pygmaeus, краснощекого суслика— подвидом С. major.
Многие ученые (Эллермэн, 1940; Симпсон, 1945, и др.) включают роды скалистых сусликов, пустынных сусликов, антилоповых сусликов и l иктидомисов на правах подродов в род Citellus. Следуя И. М. Громову (1965), в настоящем руководстве всем им придан ранг рода.
Род сурков
Genus Marmota Frisch, 1775
Размеры от средних до максимальных в семействе. Длина тела от 30 до 60 см. Относительно короткий хвост достигает длины 10—25 см (рис. 29). Задние конечности лишь немного длиннее передних. Па обеих конечностях, как правило, третий палец длиннее четвертого. На передних конечностях первый палец сильно укорочен, иногда редуцирован. Уши короткие. Стопа широкая. Когти обычно хорошо развиты, мощные. Сосков 4 или 5 пар. Хвост равномерно и густо опушен. Защечные мешки рудиментарные и не функционируют. Характер волосяного покрова варь-
Г
ирует у разных видов от высокого, густого и мягкого до сильно изрежен-ного и значительно более грубого. Окраска волосяного покрова также изменчива. Спинная сторона может быть от ярко-буровато-желтого, темно-бурого, рыжевато-бурого цвета до смеси черных и белых волос. Брюшная сторона от беловатой до темно-бурой, желто-серой или рыжеоранжевой. По-видимому, в течение года бывает только одна линька, которая приходится на раннее лето.
Череп имеет относительно небольшую округлую мозговую коробку с сильно развитыми гребнями. Посторбитальные отростки очень крупные. Лобные кости сжаты. Подглазничные отверстия небольшие. Подглазничный канал хорошо развит. В отличие от всех других представителей семейства верхнечелюстная кость у взрослых животных не образует площадки над корнями верхних коренных зубов. Число щечных зубовЛ- .
Коронки у них средней высоты, жевательная поверхность бугорчато-гребенчатая; гребни и бугорки средней высоты. По сравнению с другими представителями семейства кости конечностей относительно более массивные с хорошо развитыми гребнями и буграми. Характерен мощный локтевой отросток.
Хромосом у седого сурка подвида broweri 36, у лесного, альпийского, длиннохвостого, серого и сурка Мензбира 38, у олимпикского и ванкуверского 40, у седого и желтобрюхого 42 (Воронцов и др., 1969).
Ареал охватывает в Новом Свете большую часть Канады, Соединенных Штатов от Аляски до Лабрадора и атлантического побережья США, на юг до Калифорнии, Нью-Мексико и Северной Оклахомы и Алабамы. В Евразии — Альпы, Карпаты, Польшу, Северную Румынию, Буковину, европейскую часть СССР (степи Дона, Донца, Средней и Нижней Волги, Среднего Урала), Северный Казахстан, Фергану, Памир, Семиречье, Алтай, При- и Забайкалье, область Верхоянска, Анадырскую область, Камчатку, Тибет, Китайский Туркестан, Ганьсу, северную часть МНР, Северо-Восточный Китай, Афганистан, Кашмир, Непал и от Сиккима до Юньнаня.
Населяют открытые ландшафты гор и равнин от холодных тундр за Полярным кругом до южных степей; в горы поднимаются до высот, превышающих 4500 м над уровнем моря. Убежищем служат глубокие норы, которые звери выкапывают сами. Выброшенная из норы почва образует так называемую сурчину, иногда достигающую (например, у байбака) в диаметре до 20—25 м и более 0,5 м, иногда до 1—1,5 м высоты. В норе имеется гнездовая камера и различное число входов (до 3, 4 и даже 7), в зависимости от возраста норы. Длина хода нередко достигает 15—20 м и в глубину может уходить на 2,5—3 м и даже несколько более. Кроме гнездовых нор имеются и более мелкие просто устроенные кормовые или защитные. В них зверь прячется во время кормежки, если вдруг появилась какая-нибудь опасность.
Питаются различными растительными объектами, в основном злаками, осоками, разнотравьем. Так, байбак в Стрелецкой и Каменной степях в основном поедает шалфей, клевер, понтийскую полынь, люцерну, овсюг. типчак, пырей, птичью гречиху, цикорий, вьюнок, одуванчик. В Казахстане наибольшее значение имеют злаки (типчак и пырей), бобовые (люцерна, клеверы, астрагалы), различные полыни, зонтичные, лебедовые, губоцветные. В поисках корма от поры далеко по отходит. При появлении опасности сурок встает на задние лапы столбиком и пронзительно и громко кричит, затем прячется в нору. Активность преимущественно раннеутренняя и менее четко выраженная — во второй половине дня Жаркие часы суток сурки проводят в норе.
На зиму впадают в спячку. Длительность спячки варьирует в зависимости от географического положения. Байбак в Стрелецкой и Каменной степях проводит в спячке 5,5—6 мес. В Саратовских степях и Волжско-Камском крае 7—7,5 мес., а на юге Целиноградской области около 8 мес. Сурки зимуют семьями в одной норе, куда осенью собирается до 12— 15 зверьков. В местах малой численности сурков в одной норе зимует меньшее количество особей, иногда даже одиночки. В последнем случае отмечается часто гибель зверей. Гон у сурков происходит почти сразу после того, как они появляются на поверхности земли после спячки. Длительность беременности примерно 35—42 дня. Иногда спаривание, по-видимому, происходит и до появления зверьков из зимовальных нор. В пом:ете от 2 до 10, обычно 4—5 голых и слепых детенышей. Масса новорожденного около 26 г. На 26—27-й день он прозревает. Примерно спустя месяц после рождения детеныши переходят на растительную пищу. Половозрелость наступает в 3 года (у лесного сурка в 2 года). Продолжительность жизни лесного сурка до 6 лет в естественных условиях и до 10 лет в неволе.
Сурки имеют существенное хозяйственное значение: их жир и мясо идут в пищу, шкурка используется как пушнина. Местами сурки могут вредить сельскохозяйственным посевам, однако вред их обычно незначителен. Более важно, что сурки участвуют в эпизоологии чумы в природе.
В роде (И. М. Громов, 1965) 13 видов:
гималайский сурок — М. himalayana Hodgson, 1841 (Центральная Азия от западного Куньлуня до восточного Напьшапя, северо-восточной Сычуани и Юиьнаня, к югу во всем Тибете и Гималаях; здесь и ниже распространение приводится по И. М. Громову, 1965);
байбак — М. bobak Muller, 1776 (СССР — Воронежская обл. — Каменная степь, Луганская обл. — Стрелецкая и Деркульская степи, Донецкая обл. — Провальская степь, Харьковская обл. — Великобурлук-ский р-н, Ростовская обл. — Кашарский р-н, юго-восток Чувашской АССР, юго-восток Татарской АССР, юг Ульяновской обл., юг и юго-восток Башкирии, Саратовская, Уральская, Оренбургская, Челябинская, Северо-Казахстанская, Павлодарская, Актюбинская, Кустанайская, Целиноградская, Кокчетавская, Тургайская области);
серый сурок — М. baibacina Kastschenko, 1899 (МНР, Северо-Западный Китай; в СССР — Алтай на восток до юга Телецкого озера, Чулыш-манского хребта, оз. Кындыктыколь и р. Бурхей-Муреи; Западный Саян; отдельно Томская и Кемеровская области, окрестности г. Новосибирска; к югу — до государственной границы и хребтов Южного Алтая, Саур, Тарбагатай, Чингизтау, Казахский мелкосопочник, Джунгарский, За-илийский и Киргизский Алатау, Центральный Тянь-Шань, па запад до северных склонов хребта Джумголтау, Сонкольского нагорья, восточных склонов Ферганского хребта, долины р. Арпы и хребта Джамантау; на восток и юго-восток — до государственной границы);
тарбаган — М. sibirica Raddc, 1862 (МНР, прилежащие к лей районы СССР, Северо-Восточный и Северо-Западный Китай; СССР -Забайкалье до Гусиного озера но правобережью р. Селенги и до горы Тологай по левобережью, далее - • по степям Юго-Восточного Забайкалья; юго-восток Тувинской АССР; юго-восточные Саяны);
черношапочный сурок — М. camtschatica Pallas, 1Ы1 (СССР -Восточная Сибирь от северного Прибайкалья — Байкальского и Баргузин-ского хребтов и Верхоянского хребта до южных частей Корякского нац. окр. и Камчатки; к северу от Анадырского хребта и на Чукотке отсутствует; южная граница, проходящая, по-видимому, по Яблоновому и
Становому хребтам, не выяснена; северо-восточнее Аяна распространен, по-видимому, до Охотского побережья);
сурок Мензбира — М. menzbieri Kaschkarov, 1925 (СССР — запад Таласского и юго-запад Чаткальского хребтов, Ангренское плато и Кура-минский хребет);
альпийский сурок — М. marmota Linnaeus, 1758 (Альпы и Карпаты);
седой сурок — М. caligata Escholz, 1829 (северо-запад и запад Северной Америки-—Аляска, кроме побережья моря Бофорта, низовьев рек Юкон и Кускоквнм и западной части п-ова Аляска; Канада — провинции: Юкон, кроме крайнего севера, крайний запад Маккензи, Британская Колумбия, кроме северо-востока, узкая полоса вдоль юго-западной границы Альберта; США — полоса, проходящая от северной до южной границы через центральные районы штата Вашингтон, север Айдахо, крайний запад штата Монтана);
олимпикский сурок — М. olympus Merriam, 1898 (США — горы Олим-пикв северо-западной части штата Вашингтон);
ванкуверский сурок — М. vancouverensis Swarth, 1911 (Канада — о-в Ванкувер и провинции Британской Колумбии);
длиннохвостый сурок — М. caudata Geoffroy, 1842—1843 (от Северо-Западной Индии, Афганистана и Пакистана до Средней Азии включительно; СССР —Средняя Азия к западу от западных предгорьев Таласского и Туркестанского хребтов, к северу до предгорьев Западного Тянь-Шаня, к востоку до линии ущелье Аксу на Киргизском хребте — долина р. Джумчала — район г. Нарына — южная часть Ферганского хребта — восточные предгорья Заалайского хребта — горы Кашгарского хребта);
желтобрюхий сурок — М. flaviventris Audubon et Bachman, 1841 (Ka--нада — юг и центр провинции Британская Колумбия, крайний юг провинции Альберта; штаты США — север и восток Вашингтона, западная половина Монтаны, запад Вайоминга, Айдахо, Орегон, кроме территорий, примыкающих к тихоокеанскому побережью, северо-восток и восток Калифорнии, север Невады, Юта, кроме северо-запада и юго-востока, запад Колорадо, север Нью-Мексико; кроме того, небольшой изолированный участок ареала расположен в северо-восточном углу Вайоминга п западе Южной Дакоты);
лесной сурок — М. топах Linnaeus, 1758 (юго-восток Аляски, провинции Канады: юг Юкона, юг Маккензи, Британская Колумбия, кроме юго-запада, Альберта, кроме юга, Саскачеван, кроме юго-запада, Манитоба. кроме севера, Онтарио, юг Квебека, Нью-Брунсуик, Новая Шотландия; северо-восток США: восточная граница проходит по берегу Атлантического океана от государственной границы с Канадой на севере и до юга штата Вирджиния на юге; южная граница тянется по югу штата Вирджиния, через западные районы штатов Северная Каролина и Южная Каролина, север Джорджии, центральные районы Алабамы, огибает с севера штат Миссисипи, проходит по югу Арканзаса и переходит в западную границу, идущую по западу Арканзаса, восточной половине Канзаса, юго-востоку Небраски, северо-западу Новы, юго-западу Мин; несоты и северо-востоку Северной Дакоты; кроме того, небольшой пзо-. лированный участок ареала расположен в Канаде па востоке провинции Ньюфаундленд).
В СССР: сурки байбак, серый, тарбаган, чсрпошапочпый, Мензбира и длиннохвостый.
Б. А. Кузнецов (1965) на территории СССР насчитывает 3 вида сурков: М. caudata, М. bobac и М. menzbieri. Автор полагает, что сурки мелкосопочного Казахстана носят промежуточные признаки между байбаком и серым сурком, а сурки Хангая — между серым сурком и тарбага
ном, и все эти формы следует рассматривать как лишь подвиды единого вида — байбака. Эллермэн и Моррисон-Скотт (1966) считают гималайского сурка н тарбагана подвидом байбака, а серого и черношапочного сурка и сурка Мензбира — подвидом альпийского сурка. Дальнейшие исследования должны решить проблему системы рода сурков.
Серологические исследования сурков показали тесную связь М. baibacina и М. bobak (Баранов и Воронцов, 1973). Несколько дальше от М. baibacitia отстоят М. caudata, М. menzbieri и М. camtschatica. Дальше всех от М. baibacina удален М. camtschatica. М. menzbieri стоит несколько ближе к М. baibacina, чем М. caudata.
Сурок Мензбира включен в «Красную книгу».
Род луговых собачек
Genus Cynomys Rafinesque, 1817
Размеры средние. Длина тела 30—37 см. Длина хвоста 4—11 см. Масса до 1,4 кг. Внешним видом напоминают сурков (рис. 30). Передние конечности обладают особенно крупными и мощными когтями. Третий палец на передних лапах самый длинный. Подошвы лап покрыты шерстью. Наружное ухо короткое, широкое, умеренно опушенное. Защечные мешки невелики. Окраска спины желтовато-серая или грязно-желтая, брюшной — несколько более светлая. Хвост хорошо опушен. Сосков 4— 6 пар.
Череп массивный или широкий, угловатый. Сильно развит затылочный и сагиттальный гребни. Надглазничные отростки менее массивны, чем у сурков, относительно более длинные и несколько сильнее опушены вниз. Нижняя челюсть с сильными гребнями. Угловой отросток направлен сильно внутрь. Щечные зубы крупные, гипселодонтного типа. Верхние резцы сравнительно узкие, с уплощенной боковой поверхностью. Передняя поверхность резцов гладкая или с одной слабо выраженной продольной бороздкой.
Белохвостая и чернохвостая луговые собачки имеют в диплоидном наборе по 50 хромосом, а ютасская — 40.
Ареал охватывает в Северной Америке крайний юг провинции Саскачеван в Канаде и следующие штаты США: Северная и Южная Дакота, Небраска, Канзас, Оклахома, Техас, Монтана, Вайоминг, Колорадо, Нью-Мексико, Юта, Аризона, а также небольшие области в Северной и Центральной Мексике. Населяют пустынно-степные ландшафты равнин и предгорий. В высокой степени приспособлены к рытью нор. Живут вместе крупными сообществами, достигающими нескольких тысяч особей. Максимальная плотность достигает 8,4 особи на гектар, а средняя— 3,2. Внутри такого сообщества имеет место сложная структура отношений между отдельными особями. Норы диаметром около 15 см обычно довольно сложно устроены, в глубину могут доходить до 3—5 м. В норе имеется гнездовая камера, в которой из травы делается гнездо. Обычно перед входом есть также довольно крупная камера. На площади в 1 га в среднем насчитывается 54,1 выходных отверстий. Питаются различными травянистыми растениями, причем далеко от нор в поисках пищи нс отходят. Активны днем. Как и сурки и суслики, часто поднимаются на задние лапки, чтобы осмотреть окрестность. Если замечена какая-то опасность, луговая собачка громко кричит. Имеются несколько разных по своему характеру тревожных криков. С конца июля — августа впадают в спячку. Пробуждаются в феврале—-марте. В течение года бывает один помет. Длительность беременности 28—32 дня. В помете от 2 до
Рис. 30. Чернохвостая луговая собачка — С. ludovicianus и се череп
10 детенышей в марте, апреле или мае. На 33—37-й день у детеныша открываются глаза, примерно в 7 педель они покидают пору. Полонозрс-.тости достигают примерно в 3-летпем возрасте. Активно истребляются фермерами, так как на пастбищах являются конкурентами домашнего скота.
Род включает 5 видов:
чернохвостая луговая собачка — С. ludovicianus Ord, 1815 (Канада — крайний юг провинции Саскачеван; штаты США — Монтана, кроме крайнего запада, юго-запад Северной Каролины, Южная Каролина, кроме востока, север и восток Вайоминга, Небраска, восточная половина Коло радо, Канзас, кроме востока, западная половина Оклахомы, западная
половина Техаса, Нью-Мексико, кроме северо-запада, крайний юго-восток Аризоны; Мексика — крайний северо-восток штата Сонора и крайний север штата Чиуауа);
мексиканская луговая собачка —С. mexicanus Merriam, 1892 (Мексика— юго-запад штата Коауила и крайний север штата Сан-Луис-Потоси) ;
белохвостая луговая собачка — С. leucurus Merriam, 1890 (штаты США — крайний юг Монтаны, западная половина Вайоминга, северо-восток Юты и северо-запад Колорадо);
ютасская луговая собачка — С. parvidens J. Allen, 1905 (США — центр и юг штата Юта);
луговая собачка Гуннисона — С. gunnisoni Baird, 1855 (штаты США—центр и юго-запад Колорадо, северо-запад Нью-Мексико, крайний северо-восток Юты, центр и северо-восток Аризоны).
Мексиканская и ютасская луговые собачки включены в «Красную книгу».
СЕМЕЙСТВО ЛЕТЯГОВЫХ
FAMILIA PTEROMYIDAE BRANDT, 1855
Грызуны малых или средних размеров с длиной тела от 7 до 58 см1. Внешний облик сходен с белкой. Между передними и задними конечностями (иногда между конечностями и хвостом и передними конечностями и шеей) расположена кожная складка, служащая для планирующего полета. Передние конечности лишь немного короче задних. Передние лапы четырехпалые, задние — пятипалые. Когти относительно короткие, изогнутые, острые. Волосяной покров густой, шелковистый, обычно однотипный, охристо-серого цвета.
Череп округлый, с укороченным лицевым отделом. Скуловые дуги расставлены в стороны слабо. Орбиты крупные. Заглазничных бугров нет. Скуловая кость соприкасается со слезной. Подглазничный канал узкий и через него не проходит часть передней доли жевательной мышцы. Костные слуховые барабаны средних размеров или крупные. Зубная , . 1 0 1—2 3 IZ
формула: t — с — р —-—т~' Коронки щечных зубов низкие, корни
хорошо развиты.
К слившимся радиальной и промежуточной костям запястья при-членяются дополнительные косточки (иногда неполностью окостеневающие), поддерживающие летательную перепонку. Локтевая кость развита слабее лучевой, с небольшим локтевым отростком. Малая берцовая кость в значительной части своего дистального отдела неподвижно соединена или сращена с большой берцовой.
Распространены на северо-востоке Европы, в Северной, Восточной и Южной Азии, в Северной и Центральной Америке.
Населяют леса разных типов в умеренном и тропическом поясах. Ведут древесный образ жизни. Способны делать планирующие прыжки длиной до 130—450 м. Активны ночью. День проводят в дуплах деревьев. Питаются семенами, плодами, листьями деревьев, иногда насекомыми.
В семействе в настоящее время различают 13 родов (33 вида): гигантских летяг — Petaurisla. Link, 1795 (5 видов);
черных летяг—Aeromys Robinson et Kloss, 1915 (1 вид);
1 В пределах семейства можно выделить три группы лстяговых: мелкие—с длиной тела 7—24 см, средние —25—41, крупные — 42—58 см.
скальных летяг — Eupetaurus Thomas, 1888 (1 вид);
азиатских летяг — Pteromys G. Cuvier, 1800 (2 вида);
американских летяг—Glaucomys Thomas, 1908 (2 вида); стрелохвостых летяг — Нylopetes Thomas, 1908 (6 видов); карликовых летяг — Petinomys Thomas, 1908 (8 видов);
китайских летяг — Aeretes G. Allen, 1938 (1 вид);
сложнозубых летяг — Trogopterus Heude, 1898 (1 вид);
волосатоногих летяг — Belomys Thomas, 1908 (1 вид);
дымчатых летяг — Pteromyscus Thomas, 1908 (1 вид);
летяг пигмеев-—Petaurillus Thomas, 1908 (3 вида);
индонезийских летяг — lomys Thomas, 1908 (1 вид).
Многие ученые включают летяг в семейство белок (Эллермэн, 1940; Симпсон, 1945, и др.).
Род гигантских летяг
Genus Petaurista Link, 1795
Средних и крупных размеров летяги. Длина тела 30—58 см. Длина хвоста 36—63 см. Масса 1,1—1,3 кг. Голова укороченная, широкая. Конечности соединены широкой, покрытой шерстью летательной перепонкой, развитой относительно сильнее, чем у других представителей семейства (рис. 31). Хвост равномерно по всей его длине покрыт густой длинной шерстью. Волосяной покров мягкий, густой, особенно длинный на спинной стороне. Окраска его от желтовато-серой, ярко-коричневой до черной на спинной стороне, на брюшной она желтоватая, грязно-желтая, буроватая или белая. Хвост той же окраски, что и спина. Сосков 3 пары.
Череп характеризуется очень сильно развитыми посторбитальными отростками, отходящими от него почти под прямым углом. Костные слуховые барабаны довольно крупные. Костное небо широкое.
Распространение охватывает большую часть Южной Азии от Западных Гималаев, Северо-Восточного Китая, Японии и Тайваня на юг до Шри Ланки, Суматры, Явы, Калимантана и мелких прилегающих островов.
Населяют высокоствольные густые леса, по большей части в холмистых районах. Чаще всего встречаются на высоте 900—2000 м над уровнем моря. Ведут древесный образ жизни и по большей части держатся в кронах деревьев на высоте 15—30 м над землей. На Тайване встречаются на скалистых обрывах глубоких долин. Активность ночная. День, как правило, проводят в дуплах и активными становятся в сумерках. Держатся поодиночке, парами или семейными группами. По сучьям передвигаются так же, как и белки, но в случае необходимости делают дальние планирующие прыжки, иногда длиной до 450 м. Питаются плодами, орехами, почками, корой, листьями, а также, вероятно, насекомыми и их личинками. В помете, по-видимому, 1—2 детеныша. Продолжительность жизни в неволе до 13,5 лет.
В роде 5 видов:
изящная летяга — Р. elegans МйПсг, 1839 (о-ва Калимантан, Суматра, Ява, Натуна, п-ова Малакка. Индокитай. Юньнань в Китае; Бирма; Непал; Сикким);
гигантская летяга — Р. petaurista Pallas, 1776 (о-ва Калимантан, Ява, Суматра; п-ова Малакка, Индокитай; Таиланд; Китай — Юньнань, Фуц-зян, Сычуань, о-в Тайвань; Бирма; Непал; Индия, включая штаты Пенджаб, Кашмир; Ассам; Шри Ланка);
Рис. 31. Белорыжая летяга — Р. alborufus и череп гигантской летяги Р. petaurista
бело-рыжая летяга — Р. alborufus Milne-Edwards, 1870 (Китай — о-в Тайвань, Юньнань, Сычуань, Хубэй; Ассам; Бирма; Таиланд);
непальская летяга — Р. magnificus Hodgson, 1836 (Непал и Сикким);
японская летяга — Р. leucogenys Temminck, 1827 (Япония; Корейский п-ов; Китай — Сычуань, Ганьсу и Юньнань).
Род черных летяг
Genus Aeromys Robinson et Kloss, 1915
В роде единственный вид: черная летяга — A. tephromelas Gunther, 1873.
Размеры средние. Длина тела 25—33 см Длина хвоста 28—36 см. По внешнему виду напоминают представителей рода гигантских летяг.
Окраска волосяного покрова на спинной стороне тела от темно-бурого до черного, на брюшной — серовато-бурая. Хвост той же окраски, что и спина.
Череп сходен с черепом представителей рода гигантских летяг. Строение зубов относительно более простое, чем у последних.
Распространена на Малаккском п-ове, о-вах Пенанг, Калимантан, Северной Суматре.
Населяет леса, иногда разреженные. Активность ночная. День проводят в дуплах. Питается плодами, орехами, листьями, вероятно, насекомыми. В помете от 2 до 4 детенышей.
Род скальных летяг
Genus Eupetaurus Thomas, 1888
В роде единственный вид: скальная летяга — Е. cinereus Thomas, 1888.
Размеры крупные. Длина тела примерно 46 см. Длина хвоста 56 см. Внешний облик сходен с таковым летяги. Уши небольшие. Все туловище и особенно хвост покрыты очень длинными, густыми и мягкими волосами. Окраска волосяного покрова на спинной стороне серовато-бурая, на брюшной несколько более светлая. Конечности черновато-бурые. В отличие от представителей рода гигантских летяг скальные имеют более длинную морду, а в отличие от других представителей семейства зубы у них с более высокими коронками.
Распространена в северной части Индии, в Кашмире. Иногда этот род выделяют в отдельное семейство (Eupetauridae Schaub, 1953), на что нет серьезных оснований.
Населяет высокогорные области. Держится среди скал. Образ жизни почти не изучен. Питается, по-видимому, мхами и лишайниками.
Род азиатских летяг
Genus Pteromys G. Cuvier, 1800
Размеры мелкие. Длина тела 12—23 см. Длина хвоста 11 —13 см. Похожа на других летяг (рис. 32). Глаза очень большие, черные. Летательная перепонка между задними конечностями и хвостом не развита. Волосяной покров длинный, густой и мягкий. На спине окраска его от светло-серебристо-серого до грязно-желто-серого, на брюхе—-грязно-желто-белая. Хвост обычно серый. Волосы на хвосте имеют слабый расчес в стороны. Сосков 4 пары.
Мозговая коробка черепа уплощена в дорзо-вентральном направлении. Коренные зубы с низкими коронками. Их жевательная поверхность не имеет сложной складчатой структуры.
Распространены на обширной территории от Финляндии и северной части ГДР на восток до побережья Тихого оксана, включая Сахалин, Китай, Корейский п-ов и Японию. В горных лесах китайской провинции Ганьсу, по-видимому, имеется изолированный участок обитания летяги.
Населяют леса различного типа. Заходят в лесостепь. Ведут преимущественно древесный образ жизни. Активны в сумерки и ночью. День проводят в дуплах деревьев или гнездах, сделанных из мха и лишайников. В зимнюю спячку не впадают, но в плохую погоду подолгу не показываются из гнезда. Планирующие прыжки достигают 50 м. Легко изме-
Рис. 32. Летяга — Р volans и ее череп
няют направление полета, иногда под углом 90”, пользуясь хвостом, как рулем. Питаются почками, частично корон лиственных пород: ивы, осины, клена, различными плодами и, вероятно, насекомыми. По видимому, в году один помет не больше чем из 4 детенышей.
В роде 2 вида:
летяга —Р. volans Linnaeus, 1758 (лесная зона от Центральной и Северной Европы па восток по всей территории ( ССР до Тихоокеанского побережья; в СССР северная граница ареала тянется по Северной Карелии, берегу Белого моря до г. Мезени и по северной границе леса на восток до Пенжинской губы; на Камчатке отсутствует; южная граница ареала проходит через Центральный Китай, а в СССР через Калбипский Алтай, по правому берегу Иртыша до Омска, затем через Ялуторовск— 100
Курган —Челябинск к Южному Уралу и по рекам Белой и Каме до Казани, по Волге до Горького, по Оке до Коломны и через Смоленскую область и центральные районы Белоруссии к западной государственной границе);
малая летяга — Р. momonga Temminck, 1845 (Японские о-ва — Кюсю и Хондо).
На территории СССР встречается один вид — летяга.
Род американских летяг
Genus Glaucomys Thomas, 1908
Размеры мелкие и средние. Длина тела 21—27 см. Длина хвоста 9— 18 см. Внешний облик напоминает представителей рода летяг (рис. 33). Конечности сравнительно длинные, между ними натянута летательная перепонка. Волосяной покров густой, довольно высокий, мягкий. Спинная сторона серая с различными оттенками бурого цвета, а брюшная беловатая или кремовая. Подошвы лап покрыты шерстью. Сосков 4 пары.
Череп сверху уплощен. Лобные кости длинные и узкие. Межглазничное пространство сужено. Характерны крупные слуховые барабаны. За-глазничные отростки относительно маленькие.
Зубная формула: t — с — р ~^т~^
Ареал охватывает всю восточную часть Северной Америки, от Южной Аляски на юг до южных районов Канады и западных и восточных частей США, а также изолированные участки в Мексике и Гондурасе.
Обитают в лесных районах, преимущественно в зрелых широколиственных лесах. Активны ночью. День проводят в дуплах, где устраивают гнездо из кусочков коры, сухих листьев, мха, перьев, волос. Держатся группами, питаются и занимают одно гнездо по нескольку зверьков вместе. В спячку не впадают, ио во время холодов зимой или в дождливые дни подолгу не показываются из своего убежища. Способны делать длинные планирующие прыжки. С высоты примерно 18 м они могут планировать на расстояние до 50 м, пролетая 22 м за 12 с. Отмечена звуковая сигнализация. Питаются различными орехами, корой побегов, лишайниками. мхом, плодами, ягодами. Поедают также некоторых насекомых. Иногда делают запасы пищи. Период спаривания у южных форм растянут с февраля до конца лета, а у северных — происходит только поздней зимой. В году один помет (северная летяга) или два помета (южная летяга), в котором бывает от 2 до 6 детенышей. Беременность длится примерно 40 дней. Новорожденные голые, слепые, масса от 3 до 6 г. Глаза открываются примерно в 25-дпевпом возрасте. Уже у новорожденных детенышей летательная перепонка хорошо развита.
В роде 2 вида:
южная летяга — G. volans Linnaeus, 1758 (восток Северной Америки; северная граница ареала идет от Атлантического побережья центральных районов штата Мэйн США к юго-восточному углу провинции Онтарио, Канада, далее к восточной оконечности оз. Гурон, огибает его и оз. Мичиган и Верхнее с юга и тянется до северо-западных районов штата Миннесота, откуда поворачивает на юг, образуя западную границу ареала, которая идет вдоль границы между штатами Миннесота и Южная Дакота, к востоку от нее, пересекает северо-западный угол штата Нова, юго-восточный угол штата Небраска, идет к западу от границы между штатами Канзас и Миссури, пересекает с севера на юг середину
Рис. 33. Южная летяга — G. volans
штата Оклахома и середину Техаса и выходит к Мексиканскому заливу берег которого образует южную границу ареала-—в последний не вклю чена территория южной трети п-ва Флорида; восточная граница ареала следует по атлантическому побережью; кроме того, имеется четыре
небольших изолированных участка, три из них в Мексике: один на юго востоке штата Сонора, юго-западе Чиуауа, северо-западе Дуранго п северо-востоке Синалоа, второй — в средних частях штата Мичоакан, Гуанохуато, востоке Сан-Луис-Потоси, юге Тамаулипас, севере Идальго, средних частях Веракрус, северной половине Пуэбла и средних частях Гуерреро; третий — на севере Оахака и в центральных районах Чиапас; четвертый участок находится в центре Гондураса);
северная летяга — (7. sabrinus Shaw, 1801 (север и центр Северной Америки— юго-восток Аляски, США; провинции Канады- южная половина Юкона, запад и юг Маккензи, Британская Колумбия, Альберта, кроме юго-восточного угла, северная половина Саскачевана, Манитоба, Онтарио, южная половина Квебека, восток Ньюфаундленда, Новая Шотландия, Ныо-Брапсуик; штаты США — Мэн, Пыо-Гемпшир, Вер монт, Массачузетс, Нью-Йорк, северо-запад Пенсильвании, Западная Виргиния, крайний запад Виргинии, крайний запад Северной Каролины и восток Теннесси, Мичиган, северная половина Висконсина, северо-восток и севере запад Миннесоты, северо-восток Северной Дакоты, запал
Монтаны, Айдахо, север Вайоминга, крайний юго-запад Южной Дакоты, Вашингтон, Орегон, кроме центра и юго-востока, север Калифорнии; кроме того, имеется два небольших изолированных участка ареала: один на юге Калифорнии и второй — в штате Юта).
Род стрелохвостых летяг
Genus Hylopetes Thomas, 1908
Размеры мелкие и средние. Длина тела примерно И—33 см. Длина хвоста 10—29 см. Внешний вид не отличается от других летяг. Уши крупные, слегка заостренные. Когти на лапах короткие, тупые. Волосы на хвосте имеют ярко выраженный расчес в обе стороны. Волосяной покров мягкий, густой, средней длины. Окраска его на спине от сероватой с буроватым оттенком до желтовато-коричневой, ярко-рыже-корич-невой или черной, на брюхе беловатая, сероватая или желтоватая. Хвост обычно имеет ту же окраску, что и спина. Сосков 3 или 4 пары.
Распространены в Афганистане, Кашмире, Пенджабе, Сиккиме, Непале, Юньнане, Бирме, Таиланде, Тенассериме, п-ове Малакка, на Суматре, Яве, Калимантане и прилегающих островах, а также на Филиппинских о-вах.
Населяют изреженные джунгли, часто рядом с окультуренными пло-щадями. В горах встречаются на высоте от 150 до 3500 м над уровнем моря. Ведут древесный образ жизни. Активны ночью. День проводят в дуплах. Длина планирующего полета до 135 м. Питаются различными орехами, листьями, корой побегов, плодами, а также насекомыми, мелкими змеями.
Видовой состав стрелохвостых летяг остается не выясненным и разными исследователями трактуется по-разному. Впредь до ревизии рода в его составе можно считать 6 видов:
кашмирская летяга — Н. fimbriatus Gray, 1837 (Афганистан; штаты Северной Индии — Кашмир и Пенджаб);
бирманская летяга — Н. spadiceus Blyth, 1847 (включая Н. everetti Thomas, 1895; Н. harrisoni Stone, 1900) (Бирма, Вьетнам, п-ова Индокитай, Малакка; о-ва Натуна, Калимантан, архипелаг Риоу);
яванская летяга — Н. lepidus Horsfield, 1822 (=Н. sagitta Linnaeus, 1766) (включая Н. aurantlacus Wagner, 1841; Н. platyurus Jentink, 1890; H. belone Thomas, 1908; H. amoenus Miller, 1907; H. spadiceus Blyth, 1847) (Калимантан, Натуна, Ява, Суматра и мелкие прилегающие острова; Южная Бирма);
И. phayrei Blyth, 1859 (Бирма, Таиланд, Лаос);
пестрая летяга — Н. alboniger Hodgson, 1836 (включая Н. leonardi Thomas, 1921) (Непал; Ассам в Индии; Северная Бирма, Таиланд; п-ов Индокитай; Юньнань в Китае);
калимантанская летяга — Н. thomasi Hose, 1900 (о-в Калимантан).
Чейзен (1940) объединяет стрелохвостых летяг, населяющих Малайскую область, в 3 вида: Н. sagitta Linnaeus, 1766 (включая И. harrisoni, Н. lepidus, Н. everetti, Н. aurantlacus, Н. amoenus, Н. spadiceus, И. belone); Н. platyurus Jentink, 1890 и Н. thomasi Hose, 1900. Эллермэн (1940) выделяет в отдельные виды: II. platyurus, Н. spadiceus, Н. belone, Н. sagitta, Н. lepidus, Н. harrisoni, Н. aurantlacus, Н. amoenus, Н. phayrei, Н. everetti. Н. alboniger, Н. leonardi, Н. nigripes; Н. thomasi относит к роду Aeromys Robinson et Kloss, 1915, a H. fimbriatus выделяет в особый ^Euglaucomys Howell, 1915.
Симпсон (1945) также выделяет род Euglaucomys, а в род стрелохвостых летяг включает Petinomys Thomas, 1908.
Андерсон и Джонс (1967) насчитывают в роде 8 видов.
Предложенная в настоящем руководстве система рода базируется на данных Чейзена (1940); Харриса (1944); Эллермэна (1949) и Хилла (1960).
Род карликовых летяг
Genus Petinomys Thomas, 1908
Размеры мелкие и средние. Длина тела 13—40 см. Длина хвоста 11— 29 см. Внешне сходны с другими летягами. Голова укороченная, широкая. Волосы хвоста как бы резко расчесаны в обе стороны. Только на конце хвоста этот расчес исчезает. Волосяной покров на спине густой и довольно мягкий, на брюхе сильно изреженный. Окраска его варьирует на спине от буроватой до черной, на брюхе от сероватой до белой. Хвост обычно имеет грязно-желтую окраску. Сосков 2—3 пары.
В черепе крупные слуховые барабаны уплощены. У некоторых видов сильно развиты посторбитальные отростки и теменные гребни.
Распространены от Китая на юг до п-ова Малакка, а также в Южной Бирме, на Суматре, Калимантане, Яве, Сулавеси, Шри Ланка, в Южной Индии.
Обитают в тропических лесах, поднимаясь в горы до 1200 м над уровнем моря. Образ жизни изучен слабо. Активность ночная. День проводят в дуплах деревьев. Питаются орехами, плодами, проростками, корой деревьев, листьями и, вероятно, насекомыми.
Систематика рода окончательно не установлена. Впредь до ревизии рода в нем можно принять 8 видов:
Р. setosus Temminck, 1845 (включая Р. phipsoni Thomas, 1916, н Р. vordermannl Jentink, 1890) (Южная Бирма, п-ов Малакка, о-ва Суматра, Биллитон, Калимантан, Галанг, архипелаг Риоу);
летяга Морриса — Р. morrisi Carter, 1942 (Бирма);
Р. bartelsi Chasen, 1939 (о-в Ява);
усатая летяга — Р. genibarbis Horsfield, 1824 (п-ов Малакка, о-ва Ява, Суматра, Калимантан);
хайнаньская летяга — Р. electilis G. Allen, 1925 (о-в Хайнань у Южного Китая);
летяга Хагена — Р. hageni Jentink, 1888 (включая Р. lugens, Thomas, 1894, и Р. maerens. Miller, 1903) (о-ва Суматра, Ментавай, Северный Паги — к западу от Суматры);
филиппинская летяга — Р. crinatus Hollister, 1911 (о-в Базилан в группе Филиппинских о-в);
желтоголовая летяга — Р. juscocapillus Jerdon, 1847 (включая Р. 1ау-ardi Kelaart, 1850) (Южная Индия и о-в Шри Ланка).
Род китайских летяг
Genus Aeretes G. Allen, 1938
В роде единственный вид: китайская летяга Л. mclanoptertis Milne-Edwards, 1867.
Размеры средние. Длина тела примерно 30 см. Длина хвоста 33— 35 см. Волосяной покров низкий, но густой и мягкий. Окраска его па спине желтовато-буроватая, на брюшке сероватая до грязно-желтой, за ис
ключением белого горла. Летательная перепонка темно-коричневая. Голова обычно светлее спины. Хвост грязно-желто-серый с черным концом. Хвост покрыт более длинными волосами, чем на туловище. с расчесом на две стороны.
В черепе характерны широкие верхние резцы, имеющие вертикальную бороздку на передней поверхности.
Распространена в лесах Северо-Вос-,очного Китая.
Экология не изучена.
Род сложнозубых летяг Genus Trogopterus Heude, 1898
В роде единственный вид: сложнозу-зя летяга — Т. xanthipes Milne-Elwards, 1867.
Размеры средние. Длина тела 27— ] см, длина хвоста 27—28 см. Похожа
Рис. 34. Жевательная поверхность щечных зубов сложнозубой летяги
и других летяг. Волосяной покров густой, высокий, не очень мягкий. Ос-звпая часть волос, покрывающих спину, имеет сероватый цвет, в то вре-а как их терминальная часть бурая. У некоторых особей волосы сплошь грные. Вокруг глаз рыжеватые кольца. Хвост серый с желтым оттеп-г< Брюшная часть тела белая. У основания каждого уха расположено 1 длинному пучку черных волос. С нижней стороны хвоста волосы слег-
1 расчесаны в стороны.
Щечные зубы полугипселодонтиые, крупные, со сложноскладчатой .евательноп поверхностью (рис. 34). Костные слуховые барабаны лольшие.
Распространена в Китае—Южном Тибете, Юньнане, Сычуане, Шаньси
Населяет горные леса на высоте 1750—2150 м над уровнем моря. Эколо-ия не изучена. Ведет ночной образ жизни. Питается листьями дуба.
Род волосатоногих летяг
Genus Belomys Thomas, 1908
В роде единственный вид: волосатопогая лстйга — В. pearsoni Gray, ?842.
Размеры мелкие. Длина тела 18—20 см. Длина хвоста 10 15 см. Похожа на других летяг. Уши относительно невелики, у их основания расположены пучки длинных волос. Терминальные .части конечностей икрыты длинными волосами, почти полностью их скрывающими. Щеч-нх вибрисс нет. Волосяной покров па теле густой, мягкий, высокий. Окраска верха головы и спины рыжевато-бурая. Вершины многих волос ерные. что придает меху черноватый тон. Летательная перепонка темпо-рая с рыжеватым опенком Передние конечноегп рыжевато-бурые, •тошная сторона светло рыжая пли белая.
В черепе заглазничные отростки развиты умеренно Костные слухо-5
ад барабаны крупные. Число щечных зубов —. Зубы, особенно в ниж
ней челюсти, имеют сложную складчатую жевательную поверхность со многими выростами и углами.
Распространена в Северо-Восточной Индии — Сиккиме и Ассаме; Южном Китае; Тайване; Бирме, п-ове Индокитай.
Экология изучена слабо. Населяет густые леса. Везде малочисленна.
Род дымчатых летяг
Genus Pteroniyscus Thomas, 1908
В роде единственный вид: дымчатая летяга — Р. pulverulentus Gunther, 1873.
Размеры средние. Длина тела 25—29 см. Длина хвоста 21—23 см. Внешне сходна с волосатоногой летягой, но уши относительно меньше по размерам и не имеют у основания пучков волос. Основная окраска буровато-черная. Конечности светло-коричневые. Грудь желтоватая. Преанальная область оранжевая. Брюшная часть тела грязно-желто-белая. Хвост слегка уплощен за счет того, что растущие вбок волосы длиннее, чем растущие вверх и вниз. Окраска хвоста бледно-серовато-бурая с черноватыми волосами на вершине хвоста.
Распространена на п-ове Малакка и на о-вах Суматра и Калимантан.
Экология не изучена.
Род летяг-пигмеев
Genus Petaurillus Thomas, 1908
Размеры наименьшие в семействе. Длина тела 7—9 см. Длина хвоста 3—4 см. Внешний вид сходен с другими летягами. Окраска верхней части тела желтовато-коричневая или бледно-рыжая, нижней стороны — грязно-желто-белая. Позади ушей расположено по белому пятну. Хвост серовато-бурый с белым концом. Уши длинные, узкие, заостренные. Сосков 2 пары. В отличие от других летяг жевательная поверхность зубов не сложная. Костные слуховые барабаны крупные.
Распространены на п-ове Малакка и о-ве Калимантан.
Экология не изучена. Обитают в лесах. Ведут древесный образ жизни. Активность ночная.
В роде 3 вида:
Р. hosei Thomas, 1900 (о-в Калимантан),
Р. emiliae Thomas, 1908 (о-в Калимантан),
Р. kinlochi Rhobinson el Kloss, 1911 (п-ов Малакка).
Род индонезийских летяг
Genus lomys Thomas, 1908
•*
В роде единственный вид: индонезийская летяга - I. hnrsfiddi Water-house, 1837 (рис. 35).
Размеры мелкие. Длина тела 15—23 см. Длина хвоста 16—21 см. Внешний вид сходен с другими летягами. Уши крупные, широкие, почти безволосые. Окраска волосяного покрова рыжевато-бурая с примесью черных волос. Верхняя сторона летательной перепонки красновато-коричневая. Брюшная сторона сероватая или бледно-оранжевая. Хвост буроватый сверху и каштановый снизу. Волосы на хвосте имеют расчес в стороны; у основания волосы значительно более длинные, чем на конце.
Рис. 35. Индонезийская летяга — J. horsfieldi
Череп похож на череп волосатоногой летяги Число щечных зубов— .
3}бы квадратные в сечении с хорошо выраженными 4 вершинами на жевательной поверхности, по вершине в каждом углу
Распространена на п-ове Малакка и Калимантане, а также на мел kJ соседних островах.
Экология не изучена.
СЕМЕЙСТВО ГОФЕРОВЫХ
FAMILIA GEOMYIDAE GILL, 1872
Мелких размеров грызуны с длиной тела 13—35 см. Длина хвоста
14 см1 Самцы крупнее самок. Приспособлены к подземному образу чзни Тело утолщенное, вальковатое с коро!кимн, но сильными конечностями (рис. 36). Шея очень короткая. Глаза и уши маленькие. Сильно йцвиты слезные железы, своим секретом очищающие роговицу глаза, , э очень важно для подземпоживущего животного. Хвост короткий, ,|'«чио слабо покрытый волосами; по-видимому, это своеобразный орган кязания, конец его богато снабжен кровеносными сосудами и нервными жончаниями Конечности пятипалые. На передних конечностях трегии
-ец самый длинный, а первый и пятый наиболее короткие; все пальцы
1 В пределах семейства можно выделить 3 группы гоферовых мелкие — с длиной гла 13—19 см, средние — 20—27 см, крупные 28—35 см.
Рис 36. Равнинный гофер — G. bursarius (видны щечные мешки) и его череп
вооружены большими изогнутыми когтями. Губы могут смыкаться поза дн мощных, сильно выгнутых резцов, благодаря чему у гоферов, когда они копают резцами, земля по попадает в полость рта Но бокам головы сильно развиты два щечных мешка; они открываются наружу, покрыты изнутри волосами и служат для транспортировки пищи (рис. 36). Воло сяной покров без подпуши. Его окраска сильно варьирует от черной до почти белой.
Массивный череп уплощен в дорзо-вептралыюм направлении, угловатый, имеет широко расставленные скуловые дуги Резцовое отверстие
108
мало и находится в предчелюстной кости. Подглазничный канал длинный и узкий. Костные слуховые барабаны средних размеров. Предчелюстные кости очень большие. Носовые кости узкие. Скуловые кости короткие. Нижняя челюсть массивная с большим венечным отростком. Сагиттальный гребень развит хорошо, особенно у старых самцов. Костное небо длинное и очень узкое. Зубная формула:/— с — р -у- т —= 20. Печные зубы с постоянным ростом. Эмалевый слой на этих зубах сильно едуцирован. Предкоренные крупнее коренных. Резцы толстые. Малая берцовая кость редуцирована и сращена с большой берцовой.*
Распространены в Северной и Центральной Америке примерно от 54° с. ш. в Западной Канаде на юг до Панамы и с запада на восток ог побережья Тихого океана до Атлантического.
Населяют самые различные ландшафты от пустынных областей до верхней границы леса в горах. Ведут подземный образ жизни. Живут в сложных норах. Так, в норе Geomys bursarius имеется центральная галерея, достигающая длины 140 м, с многочисленными боковыми ответвлениями, многие из которых соединены выходными отверстиями с поверхностью, а также одна гнездовая камера примерно на глубине 60—90 см. При копании и разрыхлении земли и для перекусывания корней растений используют резцы; передними лапами отбрасывают землю под туловище п задними лапами отталкивают ее назад. Когда позади собирается достаточное количество почвы, гофер поворачивается и выталкивает ее грудью и передними конечностями через боковой ход наружу. Гоферы одинаково быстро передвигаются как головой, так и хвостом вперед. Отмечено два пика суточной активности: вечером и утром. Гоферы активны в течение круглого года и в спячку не впадают. В зимние морозы активность гоферов резко падает. Иногда они передвигаются и в снегу, устраивая туннели. В течение большей части года держатся поодиночке. Если встречаются два взрослых животных, то встреча обычно кончается дракой.
Питаются в основном подземными частями растений, но часто затаскивают в свои норы стебельки растений. Могут вредить сельскохозяйственным культурам. В южных частях ареала размножаются, по-видимо-му, круглый год, но наибольшая активность размножения приходится на позднюю зиму, весну и раннее лето. У каждой особи в течение года может быть 1 или 2 периода размножения. У самцов семенники располагаются в брюшной полости, за исключением периода размножения, когда они опускаются в мошонку. Самки большинства видов приносят от 2 до 6 детенышей, хотя в период беременности у самок находили до 13 эмбрионов. Часть эмбрионов резорбируется. Половозрслость у самок наступает в 6—9-месячном возрасте.
В семействе 8 родов и 40 видов:
восточных гоферов — Geomys Rafinc.sque, 1817 (8 видов);
западных гоферов — Thomomys Wied-Ncuwicd, 1817 (6 видов); гоферов Буллера — Pappogeomys Merriam, 1895 (2 вида);
желтых гоферов— Cratogeomys Merriam, 1895 (12 видов); гигантских гоферов — Orthogeomys Merriam, 1895 (3 вида); грубошерстных гоферов — Heterogeomys Merriam, 1895 (2 вида); центральноамериканских гоферов — Macrogeomys Merriam, 1895 (6 видов);
мичоакапских гоферов — Zygogeomys Merriam, 1895 (1 вид).
Некоторые исследователи (например, Симпсон, 1945) выделяют из рода желтых гоферов особый род Platygeomys Merriam, 1895.
Род восточных гоферов
Genus Geomys Rafinesque, 1817
Мелких и средних размеров гоферы. Длина тела 13—24 см. Длина хвоста от 5 до 12 см. Масса 226—343 г. Волосяной покров короткий. Окраска его на спине от светло-бурого до черного. Брюхо более светлое.
Верхние предкоренные не имеют по задней поверхности эмали. Верхние резцы с двумя бороздками. Диплоидный набор хромосом у тропического гофера 38, техасского — 68—72, пустынного — 70.
Распространены-в прибрежных равнинах штатов США — Джорджии, Алабамы, Флориды, а также встречаются от р. Миссисипи на востоке до Скалистых гор на западе и от канадской границы на севере до Техаса и северо-восточной Мексики на юге.
Обычно населяют легкие песчаные почвы на открытых равнинах, слабо поросших древесной растительностью. Ранней весной самцы покидают свои норы в поисках самок, но после периода размножения возвращаются к уединенному существованию. В южных частях Соединенных Штатов самки приносят детенышей с февраля по август с пиком в апреле, июне и июле. Обычно в течение года самка приносит 2 помета по 1—3 детеныша в каждом. Длительность беременности 18—19 дней. Детеныши при рождении весят от 2 до 3 г. После рождения детеныши остаются с самкой примерно 2 месяца. Лактационный период продолжается 10 дней. Половозрелости достигают примерно в 3 месяца. В сельскохозяйственных районах восточные гоферы приносят ущерб сельскохозяйственным культурам: сладкому картофелю, сахарному тростнику, бобам, фруктовым деревьям. Подземная деятельность гоферов вызывает эрозию почв на склонах холмов.
В роде, вероятно, 8 видов:
равнинный гофер — G. bursarius Shaw, 1800 (Канада —крайний юго восток провинции Манитоба; штаты США — крайний восток Северной Дакоты, крайний восток и юг Южной Дакоты, Миннесота, кроме северо-востока, запад Висконсина, Иова, север и восток Миссури, центр Иллинойса, северо-запад Индианы, Небраска, юго-восток Вайоминга, восточная половина Колорадо, Небраска, Канзас, кроме юго-востока, Оклахома, кроме северо-востока, юго-запад Арканзаса, западная половина I Луизианы, Техас, кроме довольно широкой полосы вдоль мексиканской | границы, центр Нью-Мексико);
пустынный гофер — G. arenarius Merriam, 1895 (штаты США-—юг Нью-Мексико и крайний запад Техаса; крайний север штата Чиуауа в Северной Мексике); ,
техасский гофер— G. personatus True, 1889 (семь небольших изолированных участков, пять из которых расположены на крайнем юге штата ( Техас, США и два на островах Падре и Мустанг в Мексиканском зали- ве, недалеко от берега южной части Техаса); !
тропический гофер — G. tropicalis Goldman, 1915 (крайний юго-вос- : ток штата Тамаулипас в Мексике);
юго-восточный гофер—G. pinetis Rafinesque, 1806 (штаты США— Флорида, южная половина Джорджии, юг и юго-восток Алабамы);
колониальный гофер — G. colonus Bangs, 1898 (крайний юго-восток i штата Джорджия в США па берегу Атлантического океана);
гофер Шермана— G. fontanelus Sherman, 1940 (крайний восток шта- 1 та Джорджия в США); i
кумберлендский гофер — G. cumberlandius Bangs, 1898 (о-в Кумбер- | ленд в Атлантическом океане у штата Джорджия в США).
но
Гоферы: колониальный, Шермана и кумберлендский, возможно, лишь подвиды. Холл и Келсон (1959) объединяют техасского и тропического гоферов в один вид.
Род западных гоферов
Genus Thomomys Wied-Neuwied, 1839
к
а-
Р-
Среднего и крупного размера гоферы. Длина тела 16—30 см. Длина хвоста примерно 4—9 см. Масса от 85 до 284 г. Внешний облик типичный для семейства, за исключением того, что передние конечности отличаются относительно небольшими размерами (рис. 37). Низкий, мягкий и гладкий волосяной покров варьирует по окраске на спине от почти черной, коричневой или серой до почти белой, на брюхе незначительно светлее. Число сосков от 3 до 7 пар. В отличие от представителей других родов семейства резцы не имеют на передней поверхности бороздок.
Распространение охватывает юго-западную Канаду, большую часть западной половины Соединенных Штатов и продолжается до п-ова Калифорния и Мексики.
Населяют пустыни, прерии, изреженные леса, травянистые равнины и луга от уровня моря до 4000 м над его уровнем. Норы проходят на глубине 15—91 см и в диаметре имеют 38—76 мм. В них есть особая гнездовая камера и камера для запасов пищи. Держатся поодиночке. Предпочитаемые корма — корни и луковицы. Поедают и надземные части растений, когда выходят на поверхность ночью или в пасмурные дни. Растение перегрызается на небольшие части, которые животное заталкивает передними лапами в щечные мешки. Местами причиняют вред сельскому хозяйству. В течение года дают один помет, в котором от 3 до 10 детенышей. Беременность длится примерно 19 дней (Т. bottae). Детеныш рождается совершенно беспомощный, слепой и голый, весит 2—3 г.
В роде 6 видов:
южный гофер—Т. umbrinus Richardson, 1829 (штаты США — юго-запад Орегона, Калифорния, кроме северо-востока, южная половина и восток Невады, полоса вдоль западной, южной и половины длины восточной границы Юты, юго-запад, юг и центр Колорадо, западная половина Нью-Мексико, крайний запад Техаса, примыкающий к Мексике, Аризона, кроме юго-запада; штаты Мексики — Нижняя Калифорния, северо-запад, центр и юго-запад Соноры, запад Чиуауа, Дуранго, кроме севера и северо-запада, юг Синалоа, запад Наярита, Коаула, кроме северо-запада. запад Нуэво-Леон, кроме того, имеется шестнадцать небольших изолированных участков ареала: два в штате Юта — один на севере и второй в центре, два на юге п-ва Калифорния; один на северо-западе штата Синалоа, один на юге штата Наярит, три на юге штата Закатекас, два на западе штата Сан-Луис-Потоси, один в центре штата Гуанохуато, один в центре Мичоакан, один на востоке Идальго, один в срединной части границы между штатами Веракрус и Пуэбла, один в центре и на востоке штата Мехико, на севере Морелос и северо-западе штата Пуэбла; в пределах указанного ареала Холл н Келсон, 1959 выделяют 214 подвидов);
гофер Байли •—Т. baileyi Merriam, 1901 (ареал составляют пять небольших участков: четыре в США — один на крайнем юго-востоке Аризоны и крайнем юго-западе Нью-Мехико, один на юге Нью-Мехико, два на крайнем западе Техаса и один в Мексике — в центре штата Чиуауа);
Рис. 37. Южный гофер — Т. umbrinus
гофер Таунсенда - Т townsendii Bachman, 1839 (ареал составляют пять небольших участков в США: один па юго западе штата Калифор ння, один па севере и в центре штата Невада, один на северо-западе Невады и юго-востоке Орегона, один на востоке Орегона и юго-западе Айдахо, один на юге Айдахо),
северный гофер — Т. talpoides Richardson, 1828 (провинции Кана ды — юг Британской Колумбии, центр и юго восток Альберта, южная половина Саскачевана, юго-запад Манитобы; штаты США — восточные половины Вашингтона и Орегона, Айдахо, Монтана, Северная Дакота, кроме юго-востока; крайний северо запад Миннесоты, Южная Дакота, кроме востока; крайние северо-запад и юго-запад Небраски, Вайоминг, Колорадо, кроме северо-востока и юго востока; Юта, кроме запада и юга, север и центр Невады, северо-восток Калифорнии; кроме того, имеется семь небольших изолированных участков ареала: два на западе штата Вашингтон, один на юго-западе штата Вашингтон и северо западе Орегона, один на северо западе Юты, один на юго-востоке Юты, один на севере Аризоны, один на северо-востоке Аризоны и северо-западе Нью-Мексико, один на северо-западе Нью-Мексико; Холл и Келсон, 1959, выделяют в пределах ареала 66 подвидов);
горный гофер — Т. monticola J. Allen, 1893 (штаты США — запад Орегона, север и северо восток Калифорнии, запад Невады);
Т. bulbivorus Richardson, 1829 (США — северо-запад штата Орегон).
Андерсон и Джонс (1967) выделяют в роде 9 видов.
Род гоферов Буллера
Genus Pappogeomys Merriam, 1895
Мелких размеров гоферы. Длина тела 13 17 см. Длина хвоста 5— 8 см Внешний вид типичный для представителей семейства. Волосяной покров мягкий. Окраска его на спине варьирует or оранжево-светло-коричневого до темно-бурого цвета, на брюхе несколько светлее. Характерно полное отсутствие волос на хвосте и сильное развитие когтей передних конечностей.
t В черепе сагиттальный гребень не развит. Скуловая дуга относи-fl тельно тонкая. По передней поверхности верхних резцов проходит по fl глубокой продольной борозде.
J Распространены в штатах Колима, Халиско и Наярит в Мексике.
и Экология не изучена. Обитают в горах на высоте 900—3000 м над ;| уровнем моря. Приносят существенный вред сельскохозяйственным куль-fl турам.
) В роде 2 вида:
з гофер Буллера — Р. bulleri Thomas, 1892 (пять небольших изолиро-1 ванных участков в штатах Мексики — один на юге Наярит, три в центре 1 Халиско и один на юге Халиско и западе Колима);
I гофер Алкорна — Р. alcorni Russel, 1957 (Мексика — юго-восток шта-1 та Халиско).
fl Род желтых гоферов
Я Genus Cratogeomys Merriam, 1895
Мелких и средних размеров гоферы. Длина тела 14—26 см. Длина хвоста 6—12 см. Масса 400—900 г. Внешний облик не отличается от 1 других представителей семейства. Волосяной покров мягкий. Окраска 1 его желтоватая, бурая или черноватая. У некоторых форм на туловище Я есть белые пятна. Брюхо часто бывает белым.
I Верхние резцы с одной бороздкой по передней поверхности. Сагит-я тальный гребень на черепе обычно хорошо развит.
У Распространены от южной части штата Колорадо через штаты Окла-Я хома, Нью-Мексико и Техас на юг до штата Пуэбла в Мексике. ,1 Населяют различные места: от пальмовых рощ и пустынь до лесно-fl го пояса в высоких горах; от уровня моря до 3700 м над его уровнем, fl Предпочитают песчаные почвы. Питаются различными растениями, •1 включая кактусы. Делают запасы картофеля, различных фруктов, се-I мян кукурузы. Могут приносить вред сельскому хозяйству. В году, no-fl видимому, бывает один помет — в мае или июне. Детенышей в помете Я 1-3.
1 В роде, по-видимому, 12 видов:
:i желтомордый гофер — С. castanops Baird, 1852 (штаты США — край-i ний юго-восток Колорадо, крайний восток Оклахомы, восточная полови-fl на Нью-Мексико, Техас, кроме востока и запада; штаты Мексики -— се-1 вер Тамаулипас, Нуэво-Леон, Коауила, восточная половина Чиуауа, вос-I точная половина Дуранго, восточная половина Закатекас, западная nofl ловина Сан-Луис-Потоси);
перотесский гофер — С. perotensis Merriam, 1895 (Мексика — цент-fl ральная часть штата Веракрус на границе со штатом Пуэбла);
я желтый гофер — С. fulvescens Merriam, 1895 (Мексика — централь-I ная часть штата Веракрус на границе со штатом Пуэбла и восточная I часть центрального района штата Пуэбла);
гофер Мерриама — С. merriami Thomas, 1893 (Мексика — окрест-fl нести г. Мексике и штат Морелос);
а иролосский гофер — С. irolonis Nelson et Goldman, 1934 (Мексика —
юг штата Идальго);
2 гофер Зинзера — С. zinseri Goldman, 1939 (Мексика — центр и вос-7 Я ток штата Халиско и северо-запад штата Гуанохуато);
И 1 уруапанский гофер — С. varius Goldman, 1939 (Мексика — восточная *' 1 часть центральных районов штата Мичоакан);
7 fl C.tylorhinus Merriam, 1895 (Мексика — юг штата Идальго и окрест-11 1 пости г. Мехико);
I 5-399 ИЗ
керетаросский гофер — С. neglectus Merriam, 1902 (Мексика — центр штата Керетаро);
узкомордый гофер — С. angustirostris Merriam, 1903 (Мексика — северо-восток штата Мичоакан);
дымчатый гофер — С. fumosus Merriam, 1892 (Мексика — северо-восток штата Колима);
С. gymnurus Merriam, 1892 (Мексика — центр штата Мичоакан и центр и юг штата Халиско).
Симпсон (1945) выделяет из этого рода отдельный род Platygeomys Merriam, 1895. То же делает и Эллермэн (1940), включая в этот род следующие виды: Р. gymnurus Merriam, 1892; Р. tylorhlnus Merriam, 1895; Р. neglectus Merriam, 1902; P. planiceps Merriam, 1895; P. fumosus Merriam, 1895. Правильнее всех их отнести к роду Cratogeomys.
По Андерсону и Джонсу (1967), в роде 10 видов.
Род гигантских гоферов
Genus Orthogeomys Merriam, 1895
Размеры средние и крупные. Длина тела 27—35 см. Длина хвоста 9—14 см. Внешний облик типичный для гоферов. Волосяной покров у видов тропических равнин грубый и редкий, а обитателей высокогорья— мягкий и густой. Окраска у большинства форм темно-коричневая, слегка более светлая на брюшной стороне.
Череп несколько удлинен. Межглазничный промежуток относительно широкий. Лобные кости чрезвычайно широкие и уплощенные. Скуловые дуги расставлены относительно узко. Сагиттальный гребень есть. Резцы спереди желтого или оранжевого цвета, имеют на передней поверхности по продольной бороздке.
Распространены в южной части Мексики, Гватемале, Сальвадоре, Гондурасе.
Населяют равнины, а также дубравы и сосновые леса на склонах высоких гор от уровня моря до 3000 м над его уровнем. Местами гигантские гоферы поедают на плантациях корни бананов, кофейных деревьев.
В роде 3 вида:
оахакский гофер — О. cuniculus Elliot, 1905 (Мексика — крайний восток штата Оахака);
сальвадорский гофер — О. pygacanthus Dickey, 1928 (север Сальвадора);
гигантский гофер — О. grandis Thomas, 1893 (Мексика — юг штата Мичоакан, полоса вдоль берега Тихого океана и север штата Герреро, прибрежная полоса и центральные районы штата Оахака, прибрежная полоса и центр штата Чиапас; юг Гватемалы и юг Гондураса).
Род грубошерстных гоферов
Genus Heterogeomys Merriam, 1895
Размеры средние. Длина тела 20- 25 см. Длина хвоста 7—10 см. Внешний вид типичный для гоферов. Волосяной покров может быть грубым и редким или мягким и густым. Линька происходит в июле. Окраска на спинной стороне темно-коричневая, на брюшной несколько более светлая. Конечности и хвост могут быть почти совсем лишены волос.
Череп довольно высокий и узкий. Лобные кости широкие, уплощенные. Верхние резцы имеют срединную продольную бороздку на передней поверхности.
Распространение охватывает территорию от восточной части Центральной Мексики на юг и восток до п-ова Юкатан, Гватемалы и Белиза.
Населяют прибрежные места в тропических равнинах и плато в гористых местах, поднимаясь в горы до 2000 м над уровнем моря. Представители этого рода ночью или в дождливые дни часто выходят кормиться на поверхность земли. Питаются сахарным тростником; кукурузой, бананами, папайей.
В роде 2 вида:
грубошерстный гофер — Н. hispidus Le Conte, 1852 (штаты Мексики— юг Тамаулипас, восток Сан-Луис-Потоси, север Керетаро, Веракрус, северо-восток Оахака, юг Табаско, северная половина Чиапас, восточная половина Кампече, Юкатан, Кинтана-Роо; северная и северо-восточная части Гватемалы; северная половина Белиза);
большой гофер — Н. lanitis Elliot, 1905 (Мексика — западная часть центрального района штата Веракрус).
Род центральноамериканских гоферов
Genus Macrogeomys Merriam, 1895
Размеры от мелких до крупных. Длина тела 10—30 см. Длина хвоста 5—13 см. Волосяной покров может быть мягким или грубым. Окраска его варьирует от темно-бурой до черной. У гофера Ундервуда есть белая полоска, пересекающая туловище поперек в области таза. Другие виды имеют белые отметины на голове или туловище. Верхние резцы имеют оранжевую окраску передней поверхности и глубокую борозду около внутреннего края. Лобные кости широкие и уплощенные.
Распространены от северной части Никарагуа, через Коста-Рику до Восточной Панамы.
Населяют в основном облесенные склоны гор, где предпочтительно селятся на плантациях сахарного тростника и бананов. Наибольшую активность проявляют в дождливый сезон. Предпочитаемая пища —корни бананов и сахарного тростника. Сахарный тростник животное чаще всего затаскивает целиком в нору.
В роде 6 видов:
костариканский гофер — М. heterodus Peters, 1865 (три небольших изолированных участка на юге Коста-Рики);
М. cavator Bangs, 1902 (три небольших изолированных участка — два на западе Панамы и один на юге Коста-Рики);
дариенский гофер — М. dariensis Goldman, 1912 (крайний восток Панамы) ;
гофер Ундервуда — М. underwoodi Osgood, 1931 (юг Коста-Рики);
М. cherriei J. Allen, 1893 (три небольших изолированных участка на юге Коста-Рики);
никарагуаский гофер — М. matagalpae J. Allen, 1910 (центральные районы Никарагуа).
Род мичоаканских гоферов
Genus Zygogeomys Merriam, 1895
В роде единственный вид: мичоаканский гофер—М. trichopus Merriam, 1895.
Размеры средние. Длина тела 20—25 см. Длина хвоста 9—12 см. Хвост совершенно лишен волос. Верхние поверхности конечностей по
крыты густыми волосами. На конце носа расположен участок голой кожи. Окраска спинной стороны тела варьирует от темно-серого до бурого цвета. Брюшная сторона серо-свинцовая с желтоватым оттенком.
Череп относительно длинный и узкий. Скуловые дуги расставлены нешироко. Глазничноклиновидные кости сравнительно более крупные, чем у других представителей семейства. Сагиттальный гребень может быть хорошо развит. На каждом из верхних резцов по передней поверхности проходит по две продольные бороздки.
Распространен в штате Мичоакан в Мексике.
Обитает в горах на высоте 1800—3600 м над уровнем моря в зоне сосновых лесов. Густых лесов избегает, наиболее обычен в разреженных лесах с густой травянистой растительностью. Встречается на плантациях кукурузы, пшеницы, картофеля.
СЕМЕЙСТВО МЕШОТЧАТЫХ ПРЫГУНОВ
FAMILIA HETEROMYIDAE ALLEN ET CHAPMAN, 1893
К семейству принадлежат мелкие грызуны с длиной тела 6—20 см и длиной хвоста 4,5—21,5 см1. По внешнему облику напоминают тушканчиков (Microdipodops, Dipodomys) или мышей (Liomys, Heteromys).
У некоторых представителей семейства задние конечности развиты значительно сильнее, чем передние, которые уменьшены (род Dipodomys). Хвост у всех представителей семейства длинный и у большинства хорошо оволоснен. У всех представителей, как и у гоферов, хорошо развиты открывающиеся наружу щечные мешки, которые могут сокращением особой мышцы выворачиваться наружу и таким образом освобождаться от содержимого и втягиваться опять внутрь. Внутренняя поверхность мешков покрыта волосами. Волосяной покров от мягкого, J шелковистого, до жесткого и щетинковидного, иногда напоминающего полуколючки. Окраска изменяется от серовато-черной до почти белой. Часто цвет волосяного покрова сходен с окраской субстрата местности.
Череп характеризуется очень тонкими костями. Скуловые дуги тонкие и слабо расставлены в стороны. Затылочная кость уменьшена. Нижняя челюсть небольшая и слабая. Костные слуховые барабаны крупные, слегка вздутые у родов Liomys и Heteromys и очень сильно вздутые у Dipodomys и Microdipodops. Носовые кости заходят дальше вперед, чем о . . . 1 0 1 3 оп
резцы. Зубная формула: t — с — р — т — = 2и.
Резцы тонкие, сжатые с боков. Только у рода Dipodomys щечные зубы растут в течение всей жизни. Коронки щечных зубов высокие, четырехбугорчатые. У родов Perognathus, Microdipodops и Dipodomys резцы имеют на передней поверхности продольные бороздки, а у Heteromys и Liomys их нет.
Распространение охватывает юго-западную часть Канады, западную часть США, Мексику, Центральную Америку п Южную Америку до Эквадора, Колумбии и Венесуэлы.
Населяют различные биотопы: прерии, сухие равнины, пустыни, сухие и изреженные леса (Perognathus, Microdipodops и Dipodomys), а
1 В пределах семейства можно выделить три группы: мелкие — с длиной тела 6—10 см, средние—11—15 см и крупные— 16—20 см.
также кустарниковые или густо заросшие высокой травой равнины или влажные тропические леса (Liomys и Heteromys). Обычно держатся поодиночке. Под кустами, деревьями или камнями, а иногда в песке выкапывают себе норки. Нора часто имеет несколько выходных отверстий и гнездовую камеру, а также камеру для запасов пищи. Большинство видов после возвращения в нору закрывает выход земляной пробкой. Активны ночью. Во влажную или холодную погоду зверьки не выходят наружу. Некоторые виды могут впадать в спячку. При движении представители родов Perognathus, Liomys и Heteromys употребляют все четыре конечности, а представители родов Dipodomys и Microdipodops передвигаются только на двух задних конечностях, используя хвост в качестве баланса. Питаются преимущественно семенами и другими частями растений. Поедают также и мелких животных (например, насекомых). Многие виды семейства запасают пищу, перетаскивая ее в щечных мешках. В питьевой воде не нуждаются, получая необходимую влагу вместе с пищей или в результате физиологических процессов за счет так называемой оксидазной воды. Характерен полиэстральный тип размножения. Самки приносят в гнезде от 1 до 8 детенышей (обычно от 3 до 5). Беременность длится примерно 24—33 дня. В течение года бывает один (на севере) или несколько пометов.
В семействе 5 родов и 65 видов:
мешотчатых прыгунов — Perognathus Wied-Neuwied, 1839 (26 видов); малых прыгунов — Microdipodops Merriam, 1891 (2 вида);
кенгуровых прыгунов — Dipodomys Gray, 1841 (22 вида);
колючих прыгунов — Liomys Merriam, 1902 (5 видов);
центральноамериканских прыгунов — Heteromys Desmarest, 1817 (10 видов).
Род мешотчатых прыгунов
Genus Perognathus Wied-Neuwied, 1839
Размеры мелкие и средние. Длина тела 6—12,5 см. Длина хвоста 4,5—14,5 см. Задние конечности средней длины, передние лишь незначительно короче задних (рис. 38). Хвост покрыт волосами. Некоторые виды имеют на туловище жесткий волосяной покров из щетинок или даже полуигл. Подошвы задних конечностей у них голые. У представителей других видов волосяной покров мягкий и густой и подошвы задних конечностей покрыты волосами. Окраска спинной стороны тела варьирует от бледно-желтого до темно-серого, брюшной — от грязно-желтого до белого. Волосяной покров детенышей мягкий, темно-серого цвета. В течение года бывает одна линька (в конце лета).
Череп несколько уплощен. Костные слуховые барабаны вздуты и почти соединяются между собой в передних частях. Резцовые отверстия очень малы. Костное небо широкое и достигает третьего коренного зуба. Верхние резцы с глубокими бороздами на передней поверхности.
Распространены в Юго-Западной Канаде, в западной половине Соединенных Штатов и далее па юг до п-ова Калифорния и Центральной Мексики.
Населяют низкие, засушливые равнины и пустыни; в горах встречаются редко. По земле передвигаются на всех четырех конечностях. Ведут ночной образ жизни. Держатся поодиночке. Норы обычно расположены около кустов, имеют несколько выходов, закрываемых земляными пробками. В плохую погоду впадают в спячку. Питаются в основном
семенами различных пустынных растений, в меньшей степени — зелеными частями растений и беспозвоночными животными. Запасают пищу в кормовых камерах своих нор. Размножение продолжается с апреля по сентябрь, его темп несколько снижается в июне — июле. Длительность беременности примерно 3—4 недели. В помете от 2 до 7, обычно 4 детенышей. В течение года бывает 1 или 2 помета. В неволе живут до 6 лет или немного дольше.
Систематика рода окончательно не установлена.
Ниже приводится список видов по Холлу и Колсону (1959) (26 видов):
оливковый мешотчатый прыгун — Р. fasciatus Wicd-Neuwied, 1839 (провинции Канады — крайний юго-восток Альберта, юг Саскачевана, крайний юго-запад Манитобы; штаты США — восточная половина Монтаны, Северная Дакота, кроме узкой полосы вдоль восточной границы штата, западные две трети Южной Дакоты, северо-запад Небраски, восточные две трети Вайоминга, крайний северо-восток Юты, крайний северо-запад и центр Колорадо);
желтый мешотчатый прыгун — Р. flavescens Merriam, 1899 (штаты США — юго-восток Северной Дакоты, восточная половина Южной Дакоты, юг и запад Миннесоты, север и запад Иовы, Небраска, кроме северо-запада, восточная половина Колорадо, западные две трети Канзаса, запад Оклахомы, северо-запад Техаса);
мешотчатый прыгун Мерриама — Р. merriami J. Allen, 1892 (штаты США—юго-восток Нью-Мексико, западная половина Техаса; штаты Мексики — север и северо-восток Чиуауа, северо-запад и северо-восток Коауила, северная половина Нуэво-Леон, Тамаулипас, кроме юго-запада);
шелковистый мешотчатый прыгун — Р. flavus Baird, 1855 (штаты США — юго-восток Вайоминга, западные половины Небраски, Канзаса и Оклахомы, восточная половина Колорадо, Нью-Мексико, западная половина Техаса, крайний юго-восток Юты, восточная половина Аризоны; штаты Мексики — северо-восток и центр Соноры, восточные две трети Чиуауа, восточная половина Дуранго, западная половина Коауила, За-катекас, западная половина Сан-Луис-Потоси, северо-восток Халиско, Агуаскальентес, Гуанохуато, Мексике, север Пуэбло, запад центрального района Веракрус, Тласкала, Морелос, Хидальго, кроме северо-востока) ;
Р. apache Merriam, 1889 (штаты США — юго-восток Юты, юго-запад и юг Колорадо, северо-восток Аризоны, Нью-Мексико, кроме восточной половины и юго-запада, крайний север Чиуауа);
малый мешотчатый прыгун — Р. longimembris Cones, 1875 (штаты США — крайний юго-восток Орегона, запад и южная половина Калифорнии, Невада, запад Юты, север и юго-запад Аризоны; штаты Мексики—север Нижней Калифорнии и крайний северо-запад Соноры; кроме того, два небольших изолированных участка ареала расположены: один в США на юге Юты и севере Аризоны и второй на западе Соноры в Мексике);
аризонский мешотчатый прыгун — Р. amplus Osgood, 1900 (США— запад, центр и юг штата Аризона; Мексика — северо-запад штата Сонора; кроме того, небольшой изолированный участок ареала расположен на севере Аризоны);
Р. inornatus Merriam, 1889 (США — центральная часть штата Калифорния) ;
большой мешотчатый прыгун — Р. parvus Peale, 1848 (Канада — юг провинции Британская Колумбия; штаты США — восточная половина Вашингтона, южная половина Айдахо, крайний юго-запад Вайоминга, западная половины Юты, крайний северо-запад Нью-Мексико, Невада, кроме крайнего юга, северо-восток Калифорнии);
белоухий мешотчатый прыгун — Р. alticola Rhoads, 1894 (США — два небольших изолированных участка на юге штата Калифорния);
желтоухий мешотчатый прыгун — Р. xanthonotus Grinnell, 1912 (США—южная часть центральных районов штата Калифорния);
длиннохвостый мешотчатый прыгун — Р. fortnosus Merriam, 1889 (США — запад и юго-запад Юты, Невада, кроме севера и центральных районов, крайний юг Аризоны, юго-восток Калифорнии; Мексика — восточная часть северной половины п-ва Калифорния; кроме того, изолированный небольшой участок расположен па северо-западе Аризоны) ;
мешотчатый прыгун Байли — Р. baileyi Merriam, 1894 (штаты США — юг Калифорнии, юг Аризоны; Мексика — п-ов Калифорния, западная половина штата Сонора, о-ва в Калифорнийском заливе — Мон-сургат и Тибурон);
жесткошерстный мешотчатый прыгун — Р. hispidus Baird, 1858 (штаты США — юг Северной Дакоты, Южная Дакота, кроме северо-запада и восточной половины; Небраска, крайний восток Вайоминга, кроме северо-востока; восточная половина Колорадо, Канзас, кроме востока и юго-востока; Оклахома, кроме востока и северо-востока; Нью-Мексико, кроме северо-запада; крайний юго-восток Аризоны, Техас, кроме крайнего северо-востока и юго-востока; северо-запад Луизианы; штаты Мексики— север и северо-восток Чиуауа, северо-восток Коауила, север и северо-восток Нуэво-Леон, северная половина Тамаулипас; кроме того, довольно большой изолированный участок ареала расположен в штатах Мексики — на юге Коауила, востоке и юге Закатекас, Агуаскальентес, северо-востоке Халиско, северо-востоке Гуанохуато, юге Керетаро, севере Мексике, юге Хидальго);
пустынный мешотчатый прыгун — Р. penicillatus Woodhouse, 1852 (штаты США — юг Калифорнии, крайний юг Невады, юг и юго-запад Аризоны, юг Нью-Мексико, крайний запад Техаса; штаты Мексики — север Нижней Калифорнии, север и северо-запад Соноры, Чиуауа, кроме юго-запада, Коауила, кроме севера и северо-востока, северо-восток Дуранго, север Закатекас, западная половина Сан-Луис-Потоси);
песчаный мешотчатый прыгун-—Р. arenarius Merriam, 1894 (Мексика— п-ов Калифорния, кроме севера, крайнего юга и запада центрального района);
синалоаский мешотчатый прыгун — Р. pernix J. ДПеп, 1898 (Мексика— юг штата Соноры, но без тихоокеанского побережья, штат Синалоа и крайний север штата Наярит);
скалистый мешотчатый прыгун -— Р. intermedins Merriam, 1889 (штаты США — Аризона, кроме северо-востока и северо-запада, юго-запад и центр Нью-Мексико, крайний юго-запад Техаса; Мексика — крайний северо-запад Соноры, северная половина Чиуауа);
мешотчатый прыгун Нельсона — Р. nelsoni Merriam, 1894 (США — крайний юго-восток штата Нью-Мексико и крайний юго-запад штата Техас; штаты Мексики — восточная половина Чиуауа, Коауила, восточные две трети Дуранго, Закатекас, крайний северо-восток Халиско, Агуаскальентес, западная половина Сан-Луис-Потоси, крайний юго-запад Тамаулипас, юг Нуэво-Леон);
мешотчатый прыгун Гольдемана — Р. goldmani Osgood, 1900 (Мексика— крайний юг штата Сонора и крайний север штата Синалоа);
узкочерепной мешотчатый прыгун — Р. artus Osgood, 1900 (штаты Мексики -— крайний юг Соноры, юго-восток Чиуауа, северо-восток Синалоа и северо-запад Дуранго);
полосатый мешотчатый прыгун —Р. lineatus Dalquest, 1951 (Мексика — центральные районы штата Сан-Луис-Потоси и, возможно, крайний юго-восток штата Закатекас);
мешотчатый прыгун Сан-Диего — Р. fatlax Merriam, 1889 (США — юго-запад штата Калифорния; Мексика — западная часть северной половины п-ова Калифорния);
мешотчатый прыгун Антони — Р. anthonyi Osgood, 1900 (п-в Цсррос к западу от п-ова Калифорния);
калифорнийский мешотчатый прыгун — Р. californicus Merriam, 1889 (США — западная часть южной половины штата Калифорния; Мексика— север п-ова Калифорния);
колючий мешотчатый прыгун — Р. spinatus Merriam, 1889 (США — юг штата Калифорния; Мексика — п-ов Калифорния с некоторыми мелкими соседними островами в Тихом океане и Калифорнийском заливе).
Род малых прыгунов
Genus Microdipodops Merriam, 1891
и й
?а
ч-
у-ад
;к-
ты Зи-
ек-ай-
А — во-
зрос
1889 :кси-
lA — мел-иве).
Размеры мелкие. Длина тела 6,6—7,7 см. Длина хвоста 6,4—10,3 см. Масса 10—16,9 г. Задние конечности сильно удлинены, а передние укорочены. Длинный хвост покрыт густыми короткими волосами и не име-2Т на конце кисточки. В хвосте может откладываться жир в качестве запасных питательных веществ. Подошвы задних лап покрыты густыми волосами, удлиненными по краям стопы, что увеличивает опорную поверхность конечностей. Голова сильно расширена в задней части в связи с увеличенными размерами костных слуховых барабанов. Волосяной гокров высокий, густой и мягкий. Окраска спинной стороны сероватая, черноватая, коричневая, бледно-кремовая или темно-желтая. Брюшная сторона сероватая или чисто-белая. Терминальный конец хвоста черно-гстый или хвост одноцветный по всей длине.
В черепе характерно необычайно сильное развитие костных слуховых барабанов (более сильное, чем у других представителей семейства). Они простираются далеко вперед (примерно на половину длины черепа), почти соприкасаясь друг с другом в передних отделах. Кости стопы сильно удлинены. Большая и малая берцовые кости срастаются между собой 3
на протяжении почти их длины. Шейные позвонки в большинстве случаев сращены между собой. Диплоидный набор хромосом 40 у Тем-ного и 42 у светлого малых прыгунов.
Распространены в штатах США: Орегон, Невада, Юта, Калифорния. Населяют горные районы на высоте 1200—2000 м над уровнем моря. Обитают около кустарников, растущих вокруг пересыпающихся песча-I пых барханов. Ведут ночной образ жизни. Передвигаются на задних лапах большими прыжками. Норы копают в мягком песке, вход обычно ; закрывают пробкой. Питаются исключительно семенами, иногда в качестве добавок употребляют листья растений и насекомых. На зиму, по-I видимому, впадают в спячку. Рождение детенышей происходит в мае — I июне. Число их в помете от 1 до 7, обычно от 3 до 4. В течение года I могут быть два помета.
I В роде 2 вида:
S темный малый прыгун — М. megacephalus Merriam, 1891 (штаты I США — северная половина Невады, прилегающие к Неваде районы Се-I веро-Восточной Калифорнии, северо-восток Юты, юго-восточная часть I Орегона);
| светлый малый прыгун — М. pallidas Merriam, 1901 (США — юго-I запад штата Невада и юго-восток Калифорнии; кроме того, два неболь-I ших изолированных участка ареала расположены на юго-востоке Кали-I форнии и юге Невады).
я Род кенгуровых прыгунов
л Genus Dipodomys Gray, 1841
* Размеры от средних до самых крупных в семействе. Длина тела ]0(9—20 см. Длина хвоста 10—21,5 см. Масса 35—145 г. Задние коисч- вести развиты очень сильно, передние — слабо (рис. 39). На задних ко- вечностях обычно 5 пальцев, причем первый палец, как правило, обычно очень короткий, иногда 4 пальца (первый палец отсутствует). Ступни Ж очень длинные и узкие. Подошвы задних конечностей покрыты волоса-« мп. Хвост у большинства видов густо покрыт волосами и в терминаль-
Рис. 39. Кенгуровый прыгун Мерриама — D. merriatni и череп знамехвостого кенгурового прыгуна — D. spectabilis
ном отделе имеет пучок удлиненных волос. Уши крупные. Передние конечности с очень маленьким первым пальцем. Когти длинные и острые. Волосяной покров мягкий, густой и довольно высокий. Окраска спинной стороны от бледно-желтой до темно-коричневой. Брюшная сторона, а также верхняя губа, передние лапы, пятна над глазами и позади каждого уха — белые. Хвост обычно темный сверху и снизу и белый по бокам. У большинства видов имеется белая полоса, проходящая поперек ляжек и соединяющаяся у основания хвоста. Между плечами па спине располагается специфическая кожная железа, продуцирующая жирный секрет.
В черепе очень сильно развиты костные слуховые барабаны, соединяющиеся друг с другом позади костного неба. Череп сильно расширен в мозговом отделе. Шечные зубы растут в течение всей жизни. Некоторые из шейных позвонков имеют тенденцию к срастанию.
Ареал охватывает западные части Северной Америки от Юго-Западной Канады до п-ова Калифорния и южной части Центральной Мексики; с востока распространение ограничено р. Миссури.
Населяют засушливые, пустынные и полупустынные территории.
Предпочитают места
них
лапах
длинными
с бедной растительностью. Передвигаются на зад-прыжками. Ночные животные. Питаются семе-
нами,
плодами и листьями различных растений, иногда насекомыми.
За-
пасают ются.
пищу в особых камерах в норе. Водопоем обычно не пользу-Характерно «купание» в пыли. Определенного периода
размножения снижается. В
, как правило течение года
тельность
но 2—4
беременности примерно 28-
, не отмечается, хотя зимой у одной самки может быть
35 дней.
темп размножения до 3 пометов. Дли-
мерно 6
детеныша. Молодые зверьки
В помете от 1 до 5, обыч-
остаются в
гнезде в течение при-
недель.
В роде 22 вида:
кенгуровый прыгун Орда — D. ordii Woodhouse, 1853 (Канада —
крайний
юго-
чеван; штаты
восток
США
провинции Альберта и юго-запад провинции Саска-— восточная половина Монтаны, крайний юго-запад
Северной Дакоты, юго-запад Южной Дакоты, Вайоминг, кроме запада, юг Айдахо, крайний юг Вашингтона, восточная половина Орегона, северо-восток и восток Калифорнии, Невада, кроме крайнего юга, Юта, Колорадо, кроме центральных районов, западные две трети Небраски, западные половины Канзаса и Оклахомы, западная треть и юг Техаса, Нью-Мексико, Аризона, кроме юго-запада, штаты Мексики — северо-восток Соноры, восточная половина Чиуауа, Коауила, Нуэво-Леон, юго-запад Тамаулипас, зап. половина Сан-Луис-Потоси, восток Закатекас, вост, половина Дуранго, крайний северо-восток Халиско, Хидальго, Керетаро, Гуанохуато; Холл и Келсон, 1959, считают всего 37 подвидов) ;
кенгуровый прыгун Нельсона — D. nelsoni Merriam, 1907 (штаты Мексики — Коауила, кроме северо-востока, юг Нуэво-Леон, крайний север Сан-Луис-Потоси, восток Чиуауа, восток Дуранго);
техасский кенгуровый прыгун — D. elator Merriam, 1894 (штаты США — север Техаса и юго-запад Оклахомы);
украшенный кенгуровый прыгун — D. ornatus Merriam, 1894 (штаты Мексики — юго-восток Закатекас, юго-запад Сан-Луис-Потоси и Агуаскальентес) ;
кенгуровый прыгун Филиппса — D. phillipsii Gray, 1841 (три небольших изолированных участка в Центральной Мексике — один на западе
центральной части штата Веракрус, востоке Пуэбла, и в штате Тлакска-ла, второй на севере штата Оахака, третий на востоке Мексико, юге Хидальго) ;
кенгуровый прыгун Мерриама — D. merriami Mearns, 1890 (штаты США — запад и юг Невады, юг Калифорнии, южная половина и северо-запад Аризоны, юг Нью-Мексико, юго-запад Техаса; штаты Мексики —• Нижняя Калифорния, Сонора, Чиуауа, Коауила, запад Нуэво-Леон, юго-запад Тамаулипас, западная половина Сан-Луис-Потоси, восточная половина Закатекас, северо-восток Дуранго, а также о-в Тибурон в Калифорнийском заливе);
островной кенгуровый прыгун — D. insularis Merriam, 1907 (о-в Сан-Хозе в Калифорнийском заливе к востоку от южной оконечности п-ова Калифорния);
сантамаргаритский кенгуровый прыгуи — D. margaritae Merriam,. 1907 (о-в Маргарита в Тихом океане к западу от южной оконечности п-ова Калифорния);
фресноский кенгуровый прыгун-—D. nitratoides Merriam, 1894 (Центральная Калифорния в США, в частности недалеко от Фресио);
пустынный кенгуровый прыгун — D. deserti Stephens, 1887 (штаты
США — юг и юго-запад Невады, крайний юго-запад Аризоны, юго-восток Калифорнии; штаты Мексики — крайний северо-восток Нижней Калифорнии, северо-запад Соноры):
долотозубый кенгуровый прыгун — D. microps Merriam, 1904 (штаты США — юго-восток Орегона, маленький участок на юго-западе Айдахо, Невада, кроме крайнего юга, северо-восток Калифорнии, северо-запад и юго-запад Юты, крайний северо-запад Аризоны);
панаминтский кенгуровый прыгун — D. panamintinus Merriam, 1894 (США —четыре изолированных участка: два небольших в Южной Калифорнии, один из них, в частности, включает горный хребет Панаминт; один небольшой на юго-востоке Калифорнии и крайнем юге Невады и один более крупный вытянутый с севера на юг в виде ленты от северо-запада Невады через центральные районы Южной Калифорнии);
кенгуровый прыгун Стефенса — D. stephensi Merriam, 1907 (США — юг штата Калифорния);
большеухий кенгуровый прыгун — D. etephantinus Grinnell, 1919 (США — запад южной части штата Калифорния);
узкомордый кенгуровый прыгун — D. venustus Merriam, 1904 (США — запад южной части штата Калифорния на берегу Тихого океана);
прыткий кенгуровый прыгун—D. agilis Gambel, 1848 (США — юго-запад штата Калифорния; Мексика — запад северной части п-ова Калифорния) ;
сантакатаринский кенгуровый прыгун — D. paralius Huey, 1951 (Мексика — центр п-ова Калифорния, окрестности Санта-Катарина);
полуостровной кенгуровый прыгун — D. peninsularis Merriam, 1907 (Мексика — центр п-ова Калифорния);
кенгуровый прыгун Хирманна — D. heermanni Le Conte, 1853 (штаты США — северная половина Калифорнии и небольшой участок на юге Орегоны) ;
D. gravipes Huey, 1925 (Мексика — западная часть северной трети п-ова Калифорния);
гигантский кенгуровый прыгун — D. ingens Merriam, 1904 (США — юго-запад штата Калифорния);
знамехвостый кенгуровый прыгун — D. spectabilis Merriam, 1890 (штаты США — кроме северо-востока Нью-Мексико, юго-запад Техаса, юго-восток Аризоны; штаты Мексики — север Соноры, Чиуауа, кроме юго-запада и юго-востока; а также небольшой изолированный участок расположен на юго-западе Сан-Луис-Потоси и в штате Агуаскальентес) .
По Андерсону и Джонсу (1967), род кенгуровых крыс содержит 24 вида.
Техасский и большеухий кенгуровые прыгуны включены в «Красную книгу».
Род колючих прыгунов
Genus Liomys Merriam, 1902
Размеры мелкие и средние. Длина тела 10—13,5 см. Длина хвоста 10—15,6 см. Внешний облик напоминает мешотчатых прыгунов. Задние конечности лишь незначительно длинее передних (рис. 40). Волосяной покров туловища представляет собой жесткие щетины, иногда имеющие характер полуигл; они уплощены, заострены и спереди имеют бороздки. Окраска спинной стороны варьирует от бледно-серого до рыжевато-коричневого или темно-коричневого цвета с черноватым оттенком, брюхо
IT
ю
та ие юй ще ки. ко-эхо
Рис. 40. Окрашенный колючий прыгун — L. pic-tus и его череп
видов
белое. Хвост хорошо оволоснен и обычно двухцветный; у многих по бокам проходит оранжевая или грязно-желтая продольная полоска. На втором пальце задней конечности имеется ложковидный коготь, служащий, по-видимому, для копания почвы.
Мозговой отдел черепа широкий, лицевой — узкий.
Распространение охватывает территорию от Соноры (Мексика) и крайнего юга Техаса на юг до Панамы.
Населяют пустыни и полупустыни. Активны ночью. В пищу употребляют семена, реже вегетативные части растений. Норы устраивают с выходами под камнями и у основания кустов. Вход обычно закрывают земляными пробками. В помете от 2 до 5, обычно 4 детеныша. Опрсдслси-иого периода размножения пе отмечено. По большей части самки приносят детенышей весной и ранним летом. Длительность беременности 21—28 дней.
В роде 5 видов:
мексиканский колючий прыгун—L. irroratus Gray, 1868 (США — крайний юг штата Техас; штаты Мексики — южная половина Нуэво-Леон. Тамаулипас, Сан-Луис-Потоси, кроме северо-запада, южная по-говпна Закатекас, юг Чиуауа, центральные районы Дуранго, восточная
половина Халиско, северная половина Мичоакан, Гуанохуато, Мексике, Идальго, северная половина Веракрус, Пуэбла, северо-восток Оахака, Чиапас, кроме востока и юго-запада);
окрашенный колючий прыгун — L. pictus Thomas, 1893 (штаты Мексики — центральные районы и юг Соноры, крайний юго-запад Чиуауа, Синалоа, Наярит, западная половина Халиско, Мичоакан, кроме северо-востока, Герреро, кроме северо-востока, юг Оахака, центр и юг Веракрус, запад Чиапас; запад Гватемалы) ;
халискосский колючий прыгун — L. spectabilis Genoways, 1971 (юго-восточная часть штата Халиско в Мексике);
колючий прыгун Сальвина — L. salvini Thomas, 1893 (запад Коста-Рики; юго-запад Никарагуа; юг и юго-запад Гондураса; центральные районы Гватемалы);
панамский колючий прыгун — L, adspersus Peters, 1874 (центральные районы Панамы).
Некоторые исследователи (например, Холл и Келсон, 1959) выделяют из L. irrorratus: L. bulleri Thomas, 1893 и L. guerrerensis Goldman, 1911; из L. pictus: L. annectens Merriam, 1902, L. pinetorum Goodwin, 1956; из L. salvini: L. crispus Merriam, 1902, L. anthonyi Goodwin, 1932 и L. heterothrix Merriam, 1902. Дженовейс (Genoways, 1973) показал неправомочность такого выделения.
Род центральноамериканских прыгунов
Genus Heteromys Desmarest, 1817
Размеры средние. Длина тела 12,5—16 см. Длина хвоста 13—20 см. На передних конечностях пятый палец короткий, первый палец рудиментарный; когти средних размеров. Задние конечности умеренно длинные (рис. 41) с длинными и узкими стопами; первый палец короткий. Волосяной покров, как правило, жесткий, щетинковидный и только у Н. nel-soni и Н. oresterus он более мягкий. Окраска спинной стороны варьирует от коричневой до светло-серой или черноватой. Хвост обычно коричневый сверху и белый снизу. У Н. nigricaudatus хвост весь черный. У большинства подошвы лап безволосые. Хвост покрыт редкими, короткими волосами, между которыми отчетливо видны чешуйки. Череп удлинен. Костные слуховые барабаны средних размеров.
Распространение охватывает территорию от Веракрус в Мексике до Юкатана и на юго-восток до Южной Панамы.
Населяют преимущественно влажные и тропические леса от уровня моря до 2500 м над его уровнем. Активны ночью. Норы устраивают у кустов, среди камней, под упавшими деревьями. Питаются семенами, зелеными частями растений, в основном саккулентами. Размножение, по-видимому, происходит круглогодично, с пиками весной и ранним летом. В помете от 3 до 5, обычно 4 детеныша.
В роде 10 видов:
мешотчатый прыгун Дссмарсста — Н. desmarestianus Gray, 1868 (Мексика — штаты: юг Юкатана, восток Оахака, юг Табаско и Чиапас; Гватемала, кроме севера; Белиз; юг Гондураса, север Сальвадора, центр Никарагуа, Коста-Рика; Панама; крайний северо-запад Колумбии);
мешотчатый прыгун Голдмэна — Н. goldmani Merriam, 1902 (Мексика— юг штата Чиапас; юго-запад Гватемалы);
длиннохвостый мешотчатый прыгун — Н. longicaudatus Gray, 1868 (Мексика — восток штата Табаско);
Рис. 41. Южный мешотчатый прыгун — Н. australis и череп веракрусскою мешотчатого прыгуна Н. temporalis
оахакский мешотчатый прыгун —ТУ. lepturus Merriam, 1902 (Мексика— крайний юг Веракрус и северо-восток Оахака);
веракрусский мешотчатый прыгун — Н. temporalis Goldman, 1911 (Мексика — крайний юг штата Веракрус);
мешотчатый прыгун Гаумера — Н. gaumeri J. Allen et Chapman, 1897 (штаты Мексики — север Кампече, Юкатан, север Кинтана-Роо);
южный мешотчатый прыгун — Н. australis Thomas, 1901 (крайний восток Панамы; северо-запад Эквадора; юго-запад Колумбии);
чернохвостый мешотчатый прыгун — Н. nigricaudatus Goodwin, 1956 (центр штата Оахака в Мексике);
мешотчатый прыгун Нельсона — Н. nelsoni Merriam, 1902 (юг штата Чиапас в Мексике на границе с Гватемалой);
горный мешотчатый прыгун — Н. oresterus Harris, 1932 (Коста-Рика).
Андерсон и Джонс (1967) насчитывают в роде 8 видов.
СЕМЕЙСТВО БОБРОВЫХ
FAMILIA CASTORIDAE GRAY, 1872
Крупных размеров грызуны. Длина тела до 130 см. Хвост длиной до 30 см, шириной 10—13 см. Масса до 30 кг. Хвост веслообразный, сильно уплощенный в дорзо-вентральном направлении (рис. 42). Тело приземистое, с укороченными пятипалыми конечностями; задние значительно сильнее передних. Между пальцами расположены плавательные перепонки, сильно развитые на задних конечностях и слабо — на передних. Когти сильные, уплощенные, слегка искривленные. Коготь второго пальца задних конечностей раздвоен; им зверь расчесывает свою шерсть. Волосы имеются лишь у основания хвоста. Веслообразная часть его покрыта крупными чешуйками, между которыми находятся редкие и жесткие щетинки. По средней линии верхней поверхности хвоста проходит продольный киль. Глаза небольшие, с мигательными перепонками. Уши короткие, широкие, едва выступающие над уровнем меха. Ушные отверстия и ноздри смыкаются при нырянии под воду. Выросты губ могут замыкать ротовую полость позади выступающих вперед резцов и таким образом изолировать ее от попадания воды, если бобр грызет что-либо под водой. Волосяной покров высокий, густой и мягкий, с сильно развитым пухом. Окраска его варьирует от светло- до темно-коричневой. Линька в году одна. Хвост и конечности черные. В анальной области имеются парные специфические кожные железы, продуцирующие сильно пахнущий секрет, известный под названием «бобровой струи».
Череп мощный, несколько уплощенный сверху. Лицевой отдел укорочен, мозговой сравнительно длинный и узкий. Скуловые дуги сильные, расходятся далеко в стороны. Надглазничных отростков и заглаз-ничных бугров нет. Сагиттальный и лямбдовидный гребни имеются. Предглазничные отверстия щелевидные. Костные слуховые барабаны небольшие по размерам, тонкостенные. Характерная черта — направленные вверх длинные костные трубки слухового прохода. Только у бобров среди всех грызунов на основной затылочной кости имеется довольно крупное округлое углубление. Нижняя челюсть мощная со слабо развитым угловым отростком, нижний край которого нс загнут внутрь. Зубная формула
. 1 0 1 3 оп
I — с — р — т — = 20.
1 0 1 3
Коренные зубы гипселодонтного типа с постоянным ростом. Малая берцовая кость прилегает к большой берцовой, но не срастается с ней. На
Рис. 42. Бобр — С fiber и его череп
внутренней части головки малой берцовой кости имеется длинный направленный назад клювовидный вырост. Характерны широкие, упло-енные сверху крылья подвздошных костей таза.
У бобра в диплоидном наборе 48 хромосом, а у американского боб-ц_40 (Лавров и Орлов, 1973).
Распространены в Северной Америке, Европе и Азии. В Северной ерике от Аляски и Канады на юг примерно до 30° с. ш. В Старом Снеге еще в раннеисторическое время бобры обитали па большей части Европы и Северной Азии, за исключением Сахалина и, возможно, Кам-iTKir. В настоящее время в СССР ареал охватывает лесную и отчасти •псостешпю зоны европейской части, Западную Сибирь, бассейн Епи-
гея, Хабаровский край; за пределами СССР ФРГ и ГДР, Польшу, Норвегию, северную и западную часть МНР и Северо-Западный Китай.
Предпочитают селиться на небольших, медленно текущих лесных речках или лесных озерах. Крупных водоемов избегают.
Бобры превосходно плавают и ныряют. Под водой они могут оставаться до 4—5 мин и даже несколько дольше и проплыть за это время до 750 м.
Держатся поодиночке или семьями. Полная семья состоит из двух взрослых зверей и молодых — приплода прошлого и текущего годов. Активность сумеречная и ночная. В крутых берегах делают норы сложного строения. Вход норы всегда располагается под водой. На речках с низкими берегами устраивают плотины и хатки, в которых и поселяются. Хатки строят из сучьев и скрепляют илом. Они бывают высотой до 3 м и до 10 м в диаметре. Внутри хатки имеется лаз в воду и платформа, возвышающаяся над уровнем воды.
На водоемах с изменяющимся уровнем воды бобры строят плотины. Плотину устраивают ниже по течению от своего поселения. Она представляет собой сложное сооружение из веток и стволов деревьев, скрепленных глиной, илом и дерном. Бобры могут прокапывать к местам питания каналы, по которым они сплавляют заготовленный древесный корм. Мощными резцами бобры способны перегрызать у основания даже толстые деревья. После того как дерево повалено, бобры отгрызают ветки и разделывают ствол на части. Заготовленный материал стаскивают в воду и буксируют к хатке или плотине. Питаются бобры корой и топкими ветвями деревьев, предпочитая осину, иву, а также различными водными п прибрежными травянистыми растениями (кувшинкой, кубышкой, рогозом, тростником и т. п.). Осенью бобры заготавливают большое количество корма на зиму. Чтобы корм не вмерзал в лед, бобры обычно подтапливают его ниже уровня воды под крутые нависающие берега и т. д. Таким образом, после того как водоем замерзнет, корм остается доступным для бобров подо льдом. Потомство приносят один раз в год. В помете бывает от 1 до 6 детенышей, обычно 3—5. Детеныши рождаются зрячими и покрытыми шерстью. Через 1—2 дня они уже могут плавать. В 3-недельном возрасте бобрята переходят к самостоятельному питанию, Половозрелость наступает в 2-летнем возрасте. Течка начинается в конце зимы. Длительность беременности 105—107 дней.
Бобр — ценный пушной зверь, в прошлом служил объектом интенсивной охоты. В настоящее время в некоторых странах большое количество бобров разводят на зверофермах. В СССР к началу настоящего столетия бобры были почти все истреблены. Они сохранились лишь на четырех небольших территориях, далеко отстоящих друг от друга: в бассейне р. Днепр, по рекам Березине, Сожу, Припяти и Тетереву; в бассейне Дона по притокам Воронежа (в частности по Умани), в северном Зауралье по рекам Копда и Сосьва и в верховьях Енисея по р. Азас. Общая численность бобров на этих участках не превышала 800—900 голов.
Низкая численность бобров и разобщенность участков их обитания предрешала гибель бобров как вида в случае отсутствия действенной их охраны и принятия срочных мер по увеличению их численности и реакклиматизации. Такие меры в СССР были приняты. С 1922 г. повсеместно запрещена охота на бобров. В 1923 г. в Воронежской области вдоль р. Усмань был организован охотничий заказник, который в 1927 г. преобразован в Воронежский государственный заповедник. Кроме того, в это же время созданы Березинский и Кондо-Сосьвипский государственные заповедники. Основной задачей этих заповедников была охрана
бобров и восстановление их численности. Наиболее успешно в этом плане действовал Воронежский заповедник. В 1927 г. были предприняты первые попытки расселения бобров. С 1927 по 1941 г. расселили 316 бобров в 12 областях европейской части СССР и в 2 областях Западной Сибири. Размах реакклиматизационных работ резко возрос после Вели-1 кой Отечественной войны. С 1946 по 1970 г. (включительно) были рас-» солены еще 12 071 бобр в 52 областях, краях и автономных республиках ! !!СФСР, в 3 областях БССР, в 8 областях УССР, в Литовской, Латвий-L ской и Эстонской ССР (Павлов и др., 1973). К концу 60-х годов в ре-Взультате проведенных мероприятий в СССР бобр заселил территорию, р приближающуюся по размерам к площади его ареала в XVII столетии I п общая численность этих животных достигла 85—90 тыс. особей. Возросшая численность бобров позволила организовать их промысловый отлов, который постепенно возрастал от 537 голов в промысловом сезо- не 1963—1964 гг. до 3276 — в 1969—1970 гг.
; В семействе 1 род: бобров — Castor Linnaens, 1758, и 2 вида.
Род бобров
Genus Castor Linnaeus, 1758
См. описание семейства.
В роде 2 вида:
бобр — С. fiber Linnaeus, 1758 (Норвегия, низовье Роны во Франции, бассейн Эльбы в ГДР и ФРГ, бассейн Вислы в Польше; в европейской части СССР в подзонах смешанных и широколиственных лесов и лесостепи в благоприятных для жизни бобров местах они восстановили свою численность, кроме того, в СССР обитают в Северном Зауралье (реки Конда, Сосьва, Пелым и др.) и в верховье Енисея; северная и западная части МНР — реки Урунгу и Бимен в бассейне Черного Иртыша; Гиньцзян в Северо-Восточном Китае);
канадский бобр — С. canadensis Kuhl, 1820 (большая часть Аляски, кроме восточного, северо-восточного и северного побережий; большая часть Канады, кроме крайнего севера, почти вся территория США, кроме Флориды, большей части Калифорнии и Невады; небольшая полоса территории Мексики вдоль границы с США; акклиматизирован в Финляндии и отсюда проник в Карелию).
L В СССР: оба вида бобров.
Фрейе (Freye, 1960) предполагает в роде бобров лишь один вид. Однако в опытах по гибридизации наших бобров с американскими потомства получить не удается, что свидетельствует о репродуктивной их изолированности (Лавров и Орлов, 1973) и самостоятельности этих видов.
СЕМЕЙСТВО ШИПОХВОСТОВЫХ
FAMILIA ANOMALURIDAE GILL, 1872
Мелких размеров грызуны. Длина тела 6,3—43 см. Длина хвоста 75—46 см. Масса до 2 кг. Глаза и уши крупные. Внешний вид несколько напоминает белок или летяг. Приспособлены к древесному образу жизни, За исключением представителей одного рода (Zenkerella), у всех шипохвостовых между передними и задними конечностями, а также между задними конечностями и хвостом и между передними конечностями н шеей имеется кожная летательная перепонка. От локтевого сустава в
сторону отходит своеобразный хрящевой стержень, поддерживающий эту летательную перепонку. Пальцы на конечностях хорошо развиты и снабжены острыми и сильными когтями. Хвост может иметь пучок волос в терминальном отделе. По нижней поверхности хвоста у его основания проходят два продольных ряда роговых килеобразных чешуй, черепицеобразно налегающих друг на друга. Концы чешуй вытянуты в заостренные шипы, направленные в сторону конца хвоста. Эти чешуи, возможно, служат своеобразным тормозом при посадке зверька после полета, но в основном помогают ему при лазании по гладкому стволу дерева и во время отдыха. Волосяной покров мягкий, шелковистый, обычно яркой окраски.
В черепе подглазничный канал большой и через него проходит часть жевательной мышцы. Лобные кости с очень короткими заглазничными отростками. Зубная формула: i’ -р с — т~^ = 20.
Щечные зубы имеют корни. Коронки щечных зубов уплощенные и относительно брахиодонтные с 4 или 5 гребнями. Длина коронок больше их ширины. Малая и большая берцовые кости неполностью сращены. Имеется os penis.
Распространены в Западной и Центральной Африке.
Населяют тропические и субтропические леса. Ведут древесный образ жизни. Активность ночная, а у шипохвостой белки, возможно, и дневная. День проводят, как правило, в дуплах. Держатся обычно парами, иногда небольшими группами. Делают длинные, планирующие прыжки, как летяги. Питаются плодами, семенами, орехами, листьями, корой различных деревьев, а также насекомыми. Беременные самки найдены в Камеруне в июне — июле, а в Республике Заир — феврале и июне. По-видимому, каждая самка имеет по 2 помета в год, в каждом помете от 1 до 4 детенышей.
Местное население употребляет представителей семейства в пищу.
В семействе 3 рода и 7 видов:
малых шипохвостов — Idiurus Matschie, 1894 (2 вида);
шипохвостых белок — Zenkerella Matschie, 1898 (1 вид); шипохвостых летяг — Anomalurus Waterhouse, 1843 (4 вида).
Некоторые исследователи (например, Матши, Matschie, 1917; Уокер, 1964; Андерсен и Джонс, 1967) выделяют и четвертый род — Апота-lurops Matschie, 1914. По-видимому, его более правильно включить в род Anomalurus.
Род малых шипохвостов
Genus Idiurus Matschie, 1898
Размеры наименьшие в семействе. Длина тела 6,3—9,5 см. Длина хвоста 7,5—13,3 см. Уши крупные. Длинный хвост покрыт редкими длинными волосами, сквозь которые хорошо видны чешуйки кожи. Межбедренная летательная перепонка развита слабее, чем у чешуехвостых летяг. Волосяной покров на туловище мягкий и плотный. Около кисти и стопы расположено по пучку удлиненных волос. Окраска верхней стороны тела варьирует от грязио-жслтой до коричневой; нижняя — более светлая.
Мозговая коробка черепа гладкая и округлая. Носовые кости короткие и узкие. Посторбитальпых отростков нет. Костные слуховые барабаны средних размеров. Костное небо очень узкое, продолжается назад до третьего коренного зуба. Резцовые отверстия очень малы. Резцы
э
иьно утолщены спереди назад. Щечные зубы мелкие. Особенно мал Ьетий коренной зуб.
Распространены в Сьерра-Леоне, Гане, Нигерии, Камеруне, Заире. Экология не изучена. Держатся группами до сотни особей.
В роде 2 вида:
шипохвост Ценкера — I. zenkeri Matschie, 1894 (юг и юго-запад Камеруна и Заир);
длинноухий шипохвост — I. macrotis Miller, 1898 (Сьерра-Леоне, Гана, Нигерия, юг Камеруна, Заир).
Некоторые исследователи (например Аллен, 1922, Эллермэн, Андер-н и Джонс, 1967) выделяют еще 2 вида: /. langi Alien, 1922, и I. panga »Чг,т, 1922.
Род шипохвостых белок
Genus Zenkerella Matschie, 1898
В роде единственный вид: шипохвостая белка — Z. insignis Mat-Ще, 1898.
Размеры средние. Длина тела около 20 см. Длина хвоста около 1; :м Уши крупные. В отличие от других представителей семейства ле-йтельная перепонка отсутствует (рис. 43). Передние конечности четырехпалые, задние — пятипалые. Когти на пальцах развиты относительно даее, чем у других представителей семейства. Длинный хвост густо покрыт волосами, особенно в терминальной части, где он имеет посте-шно удлиняющиеся к концу не-г,гтые волосы. По нижней основой части хвоста проходят примерно 13 килеобразных чешуи с Ьгрыми вершинками, направленной наискось в стороны. Около снования стопы расположен пу- jK удлиненных волос. В этом Ьсте располагается специфиче-гцдя кожная железа. Волосяной -«ров мягкий и густой. Верхняя асть тела пепельно-серого цвета, чжияя более светлая. Хвост чер-хый.
Череп имеет много общего с репом малых шипохвостов. бные кости относительно узкие,
д заглазничных отростков; под .эпичные отверстия крупные, твертый предкоренной и третий денной зубы сильно уменьшены (размерах. Жевательная поверх-")сть щечных зубов более про-•ая, чем у малых шипохвостов.
Распространена в Камеруне, Чю-Мунн и, возможно, в Габоне.
Населяет леса. Экология не слепа. Предполагают, что ши-лвостая белка ведет дневной раз жизни.
Рис. 43 Шипохвостая белка — Z. insignis
Род шипохвостых летяг Genus Anomalurus Waterhouse, 1843
Размеры средние и наиболее крупные в семействе. Длина тела 22—43 см. Длина хвоста 14—46 см. Масса 450—1090 г.
Вид типичный для семейства (рис. 44). Летательная перепонка развита хорошо. Хвост у некоторых видов густо покрыт довольно длинными волосами, у других — умеренно.
Концевая часть хвоста с кисточкой во-
лос. Волосяной покров высокий — до 25 мм, густой, нежно-шелковистый. Окраска его на спинной стороне тела варьирует. У некоторых животных опа сероватая, у других на сером фоне верха туловища вдоль спины располагаются желтоватые или буроватые пятна. У третьих, наконец, окраска верха тела темно-бурая, почти черная. Брюшная сторона белая, желтая, серая, бурая, золотистая или рыжеватая с ярко-оранжевыми пятнами. Через основания ушей, глаза и на нос может проходить черпая полоса. На горле и голове обычно имеются белые или светлые отметины.
Рис. 44. Шипохвост Пела — A. pell и его череп
Сосков одна пара.
Костные слуховые барабаны относительно велики в размерах. Носовые кости короткие и толстые. Скуловые кости длинные. Подглазничные и резцовые отверстия средней величины.
Распространены от Сьерра-Леоне до Уганды и Танзании и на юг до Анголы и Мозамбика, а также на о-ве Фернандо-По.
Населяют густые, влажные тропические леса, встречаются и во вторичных лесах. Экология изучена плохо. Питаются ягодами, семенами, плодами, орехами, листьями и корой (у шипохвоста Дерби основной корм — кора Strombosia schefferi). Поедают также насекомых и их личинок. Дают два помета в год, в каждом обычно 1, иногда 2 или 3 детеныша.
В роде 4 вида:
серебристый шипохвост — A. beecrofti Fraser, 1852 (от Сьерра-Леоне на восток до Заира);
шипохвост Пела — A. pelt Temmincki, 1845 (от Сьерра-Леоне на восток до Ганы);
шипохвост Дерби — A. derbianus Gray, 1842 (от Сьерра-Леоне на восток до Кении и на юг до Анголы, Замбии и Северного Мозамбика);
шипохвостая летяга — A pusillus Thomas, 1887 (Южный Камерун, Габон и Заир).
Некоторые исследователи (например, Матши, 1914; Эллермэн, 1940; Андерсон и Джонс, 1967) выделяют A. beecrofti в особый род Anomalu-rops Matschie, 1914. Иногда в роде Anomalurus (без A. beecrofti) насчитывают 5 видов (например, Уокер, 1964) или 6 (Андерсон и Джонс, 1967).
СЕМЕЙСТВО ДОЛГОНОГОВЫХ
FAMILIA PEDETIDAE OWEN, 1847
Мелкого размера грызуны. Длина тела 35—45 см. Длина хвоста 37—48 см. Масса 3—4 кг, некоторые животные достигают массы 4 кг. Внешним обликом напоминают небольших кенгуру или гигантских тушканчиков (рис. 45). Задние конечности сильно увеличены в размерах, передние сильно уменьшены. Когти уплощенные, несколько напоминают копытца. Первый палец на задних конечностях редуцирован. Пятый палеи очень короткий, но хорошо развит. Хвост длинный, густо покрыт волосами, особенно на конце. Голова укороченная, тупая спереди. Шея толстая и мускулистая. Глаза большие. Уши длинные и узкие, в высоту достигают 8 см. Они слабо покрыты шерстью в верхней половине и голые внутри, с небольшим козелком. Передние конечности пятипалые, с острыми когтями, которые используются при копании земли. Волосяной покров высокий, мягкий и густой. Спинная сторона тела имеет грязновато-желтый, буроватый или рыже-буроватый цвет или песчаный с отдельными черными или белыми волосками, брюшная сторона грязно-желто-белая или белая. Сосков одна пара — в области груди.
Череп массивный. Носовые кости широкие, лобные — чрезвычайно широкие. Подглазничный канал сильно увеличен в размерах. Лицевой отдел укороченный, широкий. Скуловые кости широкие. Заглазничные отростки сильно уменьшены в размерах или их ист вовсе. Сагиттальный и затылочный гребни отсутствуют. Костное небо широкое' и о тносительно короткое, продолжается назад только до первого или в крайнем случае второго коренного зуба. Нижняя челюсть с коротким угловым отростком посильно уменьшенным венечным отростком. Зубная формула:
. 1 0 1 3 ол
I — с — р — т — — 20.
1 0 1 3
Резцы утолщены. Щечные зубы не имеют корней. Каждый зуб подразделяется на две лопасти. Малая берцовая кость редуцирована и у взрослых животных полностью страстается с большой берцовой.
Рис. 45. Долгоног — Р. cafer и его череп
Распространение охватывает территорию Центральной и Южной Африки от Кении и Анголы до мыса Доброй Надежды.
Населяют пустынные и полупустынные равнины с бедной растительностью, иногда окультуренные площади в засушливых районах. Ведут ночной образ жизни. День проводят в норах, которые выкапывают сами. В норе живет один зверек или одна семья, иногда несколько семей вместе Как правило, несколько зверьков поселяются недалеко друг от друга. Обычно долгоноги выпрыгивают из норы сразу большим стремительным прыжком, видимо, чтобы избежать опасности нападения подстерегающего хищника. В спокойном состоянии долгоноги передвигаются не спеша на всех четырех конечностях. В случае нужды делают огромные прыжки только на задних лапах. Длина прыжков может достигать 136
2—3 м в длину, а по некоторым данным 6—9 м. Питаются надземными и сочными подземными частями растений. Могут приносить некоторый вред сельскохозяйственным культурам. Насекомые также, вероятно, употребляются ими в качестве корма. По-видимому, дают всего один помет в течение года в летний период. В помете 1, редко 2 детеныша. Новорожденные весят около 250 г, слепые (прозревают через 2 дня).
В семействе 1 род: долгоногов — Pedetes liliger, 1811, и 1 вид.
Род долгоногов
Genus Pedetes liliger, 1811
См. описание семейства.
В роде 1 вид: долгоног — Р. cafer Forster, 1778 (ареал совпадает с ареалом семейства).
Некоторые исследователи (например, Томас, Thomas, 1902; Эллермэн, 1940; Андерсон и Джонс, 1967) выделяют второй вид в роде долгоногов: Р. surdaster Thomas, 1902.
Мышеобразные
Myomorpha Brandt, 1855
В скуловой дуге скуловая кость редко заходит далеко вперед. Скуловой отросток верхнечелюстной кости сильно развит. Подглазничное отверстие увеличено в размерах и обычно щелевидно. Поверхностная часть жевательной мышцы проходит вперед перед орбитой и прикрепляется к боковому отделу рострума, в то время как глубокая часть этой мышцы входит в орбиту и проходит сквозь подглазничное отверстие. В каждой половине нижней челюсти обычно по три щечных зуба. Малая
Чтыпая берцовые кости срастаются на концах.
СЕМЕЙСТВО ХОМЯКОВЫХ
FAMILIA CRICETIDAE ROCHEBRONE, 1883
Грызуны мелких размеров. Длина тела от 5 см у карликовых хомячков, до 36 см у ондатры ’. Хвост в большинстве случаев короче тела (нередко короче его половины), обычно слабо покрыт волосами. Иногда волосы покрывают хвост довольно густо и образуют на его конце кисточку (у некоторых песчанок). Общий облик обычно мышеобразный. Задние конечности длиннее передних, но разница их в длине невелика. Лишь у некоторых видов задние конечности сильно увеличены в длину в по внешнему виду эти зверьки напоминают тушканчиков. Конечности пятипалые. Пальцы с сильными когтями. У водных форм на задних конечностях между пальцами имеются плавательные перепонки. Подземные формы имеют внешний вид, типичный для землероев; вальковатос тело с укороченным и почти неразличимым снаружи шейным перехватом. Волосяной покров у большинства видов густой и мягкий. Окраска, как правило, однотонная, на спинной стороне тела от светло- до темпо-
1 В пределах семейства можно выделить три группы хомяковых: мелкие, с длиной .5—14 см, средние — 15—25 см, крупные — 26—36 см.
коричневой. Брюхо обычно светлее спины. Иногда вдоль середины спины проходит продольная темная полоска. На спине или в передней части туловища могут быть темные и светлые пятна.
Форма черепа весьма изменчива. Скуловые дуги обычно низкие, равномерно округлые. Заглазничных отростков лобных костей нет. Сагиттальный гребень встречается редко. Подглазничное отверстие имеет округлую верхнюю часть, сквозь которую тянется жевательная мышца, и узкий нижний отдел, через который проходит нерв. Костные слуховые барабаны разной величины; часто они весьма велики по размерам. Зуб-. . 1 0 0 3 1с
ная формула: i— с — р — т — — 1ь.
Щечные зубы могут быть с низкими или высокими коронками. Жевательная поверхность щечных зубов бугорчатая или бугорчато-гребневая. Встречаются также так называемые призматические зубы, у которых бугорки превратились в спаянные между собой дентинные маленькие столбики, часть из которых имеет форму трехгранных призм. В скелете характерно срастание большой и малой берцовых костей в их нижних отделах.
Диплоидное число хромосом от 17 (орегонская полевка) и 18 (Akodon tirichi) до 66 (белогорлый рисовый хомяк) и 72 (индийская гололапая песчанка).
Распространены всесветно, за исключением Антарктики, некоторых островов — таких, как Ирландия, Исландия, некоторых арктических островов, а также Австралийско-Малайской области. В СССР представители семейства встречаются по всей территории страны.
Населяют самые разнообразные ландшафты от тундры до субнивальной зоны гор, поднимаются в Андах до высоты 5500 м, на Памире и Тибете до 4000 м над уровнем моря. Большинство ведет норный образ жизни. Некоторые перешли к чисто подземному или полуподземно му существованию. Есть виды, приспособившиеся к быстрому бегу или полуводному образу жизни. Имеются и немногочисленные лазающие виды. Питаются различными растениями, их семенами, плодами и вегетативными частями, а некоторые также и насекомыми. Отдельные представители семейства запасают пищу, причем иногда в больших количествах. У многих видов характерно периодическое изменение численности. Для некоторых видов известны дальние миграции. В южных частях ареала представители семейства размножаются в течение круглого года, а в северных, как правило, оно носит сезонный характер. Длительность беременности от 17 до 33 дней. Число детенышей в помете от 1 до 18. Половозрелость у некоторых видов наступает в возрасте 42—49 дней, иногда даже раньше. Продолжительность жизни у многих видов составляет всего 1—2 года или меньше.
Некоторые представители семейства являются носителями опасных заболеваний — туляремии, чумы, лептоспироза и др. Многие формы могут приносить значительный вред сельскому хозяйству. Отдельные виды имеют важное промысловое значение (например, ондатра). Ондатра представляет собой яркий пример успешной акклиматизации нового вида млекопитающего на огромной территории ряда стран (в частности, СССР), лежащих вне их естественного ареала.
Различные систематики не сходятся между собой во взглядах на состав семейства хомяков. Большинство авторов включает в семейство Cricetidae подсемейства Cricetinae, Nesomyinae, Lophiomyinae, Micro-tinae, Gerbillinae. Иногда ряд этих подсемейств выделяют в отдельные семейства. Некоторые ученые (Эллермэн, 1941; Б. А. Кузнецов, 1965) объединяют хомяков вместе с мышами в одно семейство Muridae.
Приводимая ниже система семейства хомяков в основном
классификации, предложенной Симпсоном В семействе 96 родов (575 видов).
(1945).
дана по
Подсемейство хомяков Cricetinae Murray, 1866
Роды:
тисовых хомяков 1 — Oryzomys Baird, 1857 (68 видов);
гигантских рисовых хомяков — Megalomys Trouessart, 1881 (3 вида);
колючих рисовых хомячков — Neacomys Thomas, 1900 (4 вида);
колючих хомячков — Scolomys Anthony, 1924 (1 вид);
плавающих хомяков — Nectomys Peters, 1861 (3 вида);
рипидомис — Rhipidomys Tschudi, 1844 (6 видов);
хомяков Томаса-—Thomasomys Coues, 1884 (25 видов);
риодежанейрских хомячков — Phaenomys Thomas, 1917 (1 вид);
колумбийских хомячков — Chilomys Thomas, 1897 (1 вид);
лазающих хомяков — Tylomys Peters, 1866 (7 видов);
большеухих хомяков — Ototylomys Merriam, 1901 (3 вида);
соневидных хомяков — Nyctomys de Saussure, 1860 (1 вид);
юкатанских хомячков — Otonyctomys Anthony, 1932 (1 вид);
бразильских древесных хомячков — Rhagomys Thomas, 1917 (1 вид); полевых хомячков — Reithrodontomys Giglioli, 1917 (17 видов);
белоногих хомячков— Peromyscus Gloger, 1841 (55 видов);
золотистых хомячков — Ochrotomys Osgood, 1909 (1 вид);
карликовых хомячков — Baiomys True, 1894 (2 вида);
кузнечиковых хомячков — Onychomys Baird, 1857 (2 вида);
южноамериканских полевых хомячков—Akodon Меуеп, 1833 (38 видов);
камышовых хомячков — Zygodontomys Allen, 1897 (6 видов);
рораимских хомячков — Podoxymys Anthony, 1929 (1 вид);
леноксусов— Lenoxus Thomas, 1909 (1 вид);
копающих хомяков — Oxymycterus Waterhouse, 1837 (8 видов);
землеройковых хомячков — Blarinomys Thomas, 1896 (1 вид);
кротовых хомячков — Notiomys Thomas, 1890 (6 видов);
водяных хомяков — Scapteromys Waterhouse, 1837 (5 видов);
бурых хомячков — Scotinomys Thomas, 1913 (4 вида);
вечерних хомячков — Calomys Waterhouse, 1837 (10 видов);
высокогорных хомячков — Eligomodontia F. Cuvier, 1837 (2 вида);
красноносых хомячков — Wiedomys Hershkovitz, 1959 (1 вид);
рато-до-мато — Pseudoryzomys Hershkovitz, 1959 (1 вид);
листоухих хомячков — Phyllotis Waterhouse, 1837 (14 видов);
чилийских хомяков — Irenomys Thomas, 1919 (1 вид);
шиншилловых хомяков — Chinchillula Thomas, 1898 (1 вид);
хомяков пуны — Punomys Osgood, 1943 (1 вид);
болотных хомяков — Neotomys Thomas, 1894 (1 вид);
кроликовых хомяков — Reithrodnn Waterhouse, 1837 (I вид);
патагонских шиншилловых хомяков — Euncoinys Cones 1874 (4 вида);
перепончатолапых хомяков — Holochilus Brandt, 1835 (2 вида);
хлопковых хомяков — Sigmodon Say ct Ord, 1825 (14 видов);
андийских хомяков — Andinomys Thomas, 1902 (1 вид);
вулкановых хомячков — Neotomodon Merriam, 1898 (1 вид);
1 Это и некоторые другие названия в буквальном переводе с английского означают с-рысы» или «мыши». Видимо, в семействе хомяков не следует пользоваться такими изваииями, а надо употреблять слова «хомяк» или «хомячок».
лесных хомяков — Neotoma Say et Ord, 1825 (22 вида);
нельсония — Nelsonia Merriam, 1897 (1 вид);
ксеномис — Xenomys Merriam, 1892 (1 вид);
рыбоядных хомяков — Ichthyomys Thomas, 1893 (2 вида);
эквадорских рыбоядных хомячков — Anotomys Thomas, 1906 (1 вид);
венесуэльских рыбоядных хомячков — Daptomys Anthony, 1929 (1 вид);
центрально американских рыбоядных хомяков — Rheomys Thomas, 1906 (5 видов);
горных рыбоядных хомячков — Neusticomys Anthony, 1921 (1 вид);
мышевидных хомячков — Calomyscus Thomas, 1905 (1 вид);
джунгарских хомячков — Phodopus Miller, 1910 (2 вида);
серых хомячков — Cricetelus Milne-Edwards, 1867 (10 видов);
обыкновенных хомяков-—Cricetus Leske, 1779 (1 вид);
средних хомяков — Mesocricetus Nehring, 1898 (4 вида);
белохвостых хомяков — Mystromys Wagner, 1841 (1 вид).
Подсемейство цокоров — Myospalacinae Lilljeborg, 1866
Род:
цокоров — Myospalax Laxmann, 1769 (2 вида).
Подсемейство мадагаскарских хомяков — Nesomyinae Major, 1897
Роды:
большеногих хомячков — Macrotarsomys Milne-Edwards et Grandi-dier, 1898 (2 вида);
зайцегубых хомяков — Nesomys Peters, 1870 (3 вида);
коротколапых хомяков — Brachytarsomys Gunther, 1875 (1 вид);
соневидных хомяков — Eliurus Milne-Edwards, 1885 (2 вида);
хомяков Роберта — Gymnuromys F. Major, 1896 (1 вид); 1
хипогеомис — Hypogeomys Grandidier, 1869 (1 вид);
короткохвостых хомяков — Brachyuromys F. Major, 1896 (2 вида).
Подсемейство косматых хомяков — Lophiomyinae Thomas, 1897
Род:
косматых хомяков — Lophiomys Milne-Edwards, 1867 (1 вид).
Подсемейство полевок — Microtinae Miller, 1896
Роды:
копытных леммингов — Dicrostonyx Glogcr, 1841 (4 вида);
мышевидных леммингов — Synaptomys Baird, 1857 (2 вида);
лесных леммингов — Myopus Miller, 1910 (1 вид);
настоящих леммингов — Lemmas Link, 1795 (5 видов);
лесных полевок—Clethrionomys Tilesins, 1850 (7 видов);
южноазиатских полевок — Eothenomys Miller, 1896 (5 видов);
каменных полевок — Alticola Blanford, 1881 (5 видов);
кашмирских полевок — Hyperacrius Miller, 1896 (2 вида);
югославских полевок — Dolomys Nehring, 1898 (1 вид);
водяных полевок — Arvicola Lacepede, 1799 (1 вид);
ондатр — Ondatra Link, 1795 (1 вид);
флоридских ондатр — Neofiber True, 1884 (1 вид);
фенакомис — Phenacomys Merriam, 1889 (4 вида);
серых полевок — Microtus Schrank, 1798 (65 видов);
степных пеструшек — Lagurus Gloger, 1841 (3 вида); прометеевых полевок — Prometheomys Satunin, 1901 (1 вид); слепушонок — Ellobius Fischer, 1814 (3 вида).
Подсемейство песчанок—Gerbillinae Alston, 1876 Роды:
монодий — Monodia Heim de Balsac, 1943 (2 вида);
карликовых песчанок—Gerbillus Desmarest, 1804 (19 видов); гололапых песчанок — Tatera Lataste, 1882 (11 видов);
длиннолапых песчанок — Taterillus Thomas, 1910 (16 видов); короткоухих песчанок — Desmodillus Thomas, 1904 (1 вид); десмодиллискус—Desmodilliscus Wettstein, 1917 (1 вид); жирнохвостых песчанок — Pachyuromys Lataste, 1880 (1 вид); аммодил — Ammodillus Thomas, 1904 (1 вид);
малых песчанок — Meriones liliger, 1811 (13 видов);
пушистохвостых песчанок-—Sekeetamys Ellerman. 1947 (1 вид); песчанок Пржевальского — Brachiones Thomas, 1925 (1 вид); дневных песчанок — Psammomys Cretzschmar, 1828 (1 вид); больших песчанок — Rhombomys Wagner, 1841 (1 вид).
Подсемейство хомяков — Subfamilia Cricetinae Murray, 1866
Внешний вид разнообразный. Большинство имеют типичный вид хо- $
мяка. Некоторые виды приспособились к подземному (Notlomys, Blari-tiomys) или к водному (Ichthyomys) образу жизни.
Щечные зубы с корнями, бугорчатые, пластинчатые или призматические. Если на жевательной поверхности есть бугры, то они располагаются продольными рядами. У некоторых видов жевательная поверхность зубов плоская и имеет наружные и внутренние, расположенные друг против друга эмалевые складки.
Род рисовых хомяков
Genus Oryzomys Baird, 1857
Размеры от мелких до средних. Длина тела 9,3—19,7 см. Длина хвоста 10—23,5 см. Масса тела 40—85 г. По внешнему виду сходны с крысами (рис. 46). Хвост покрыт короткими и редкими волосами. Стопа узкая, три средних пальца значительно длиннее двух крайних. У некоторых видов стопа приспособлена к древесному образу жизни (панамский рисовый хомяк): опа расширена и имеет относительно длинный пятый палец. Волосяной покров грубый, по нс щетинистый. Спина серовато-коричневая или рыжевато-коричневая с примесью черного цвета. Бока светлее спины. Брюхо белое или бледпо-грязпо-желтое. Сосков 4 пары.
Череп имеет весьма разнообразное строение. У некоторых видов хорошо выражено межглазничное сужение. Лицевой отдел удлиненный или укороченный. Мозговая капсула средних размеров. Резцовые отверстия
Рис. 46. Болотный рисовый хомяк — О. palu-stris и его череп
различной длины; обычно не достигают уровня щечных зубов. Костн небо продолжается дальше рядов щечных зубов. Костные слуховые б рабаны обычно небольших размеров или очень маленькие, иногда кру ные. Верхние коренные зубы имеют по три корня, а нижние по де Верхние коренные зубы брахиодонтного типа. Число хромосом в дипл идном наборе у болотного рисового хомяка 56, О. delicatus — 60, бел горлого рисового хомяка — 66.
Распространены на большей части Южной Америки, в Централыи Америке, Мексике и на север до восточной части Канзаса, южной час Иллинойса и южной части США — Нью-Джерси, также на Галапагс ских о-вах.
Населяют, как правило, влажные места вдоль побережии, по пскот рые встречаются и в кустарниковых зарослях гор. Обычно живут гру пами. Активны круглосуточно в течение всего года. Хорошо плавают ныряют. В местах питания оставляют кормовые столики. Сооружают травы гнезда, которые располагаются выше уровня воды. В сухих n стах копают норы. Питаются сочными частями растений, а также ceiv нами, плодами, различными беспозвоночными, рыбой. В южных част ареала размножаются в течение круглого года. В дельте Миссисипи р< множение идет с февраля по ноябрь. Длительность беременности пр
мерно 25 дней. В помете бывает от 1 до 7, обычно от 3 до 4 детенышей. Масса новорожденных от 3 до 5 г. К самостоятельной жизни переходят в возрасте 11—13 дней, а размеров взрослых достигают в 4 месяца. У самок половозрелость наступает в 7 месяцев. Длительность жизни в природе составляет обычно менее года. Изменения численности наблюдаются в северных частях ареала.
В роде 68 видов.
белогорлый хомяк—О. albigularis Tomes, 1860 (Анды в Эквадоре; северо-запад Боливии, север Колумбии и хребты Центральная и Восточная Кордильеры; Перу; запад Венесуэлы);
хомяк Альфаро —О. alfaroi J. Allen, 1891 (Центральная и Северо-Западная Колумбия; северо-запад Эквадора; запад Панамы; Коста-Рика, кроме побережья Карибского моря; Никарагуа, за исключением довольно широкой полосы, примыкающей к берегу Карибского моря; Сальвадор; Гондурас, кроме его северо-восточных прибрежных районов; Гватемала; штаты Мексики: Чиапас, юг Табаско, юг и запад Веракрус, юго-запад и юг Оахака, юго-восток Герреро; северо-восток Пуэбло, север Идальго, восток Керетаро, восток Сан-Луис-Потоси, юг Тамаулипас);
андийский рисовый хомяк — О. andinus Osgood, 1914 (Северное Перу);
0. arenalis Thomas, 1913 (Северо-Восточное Перу);
0. balneator Thomas, 1900 (восток и юг Эквадора);
0. buccinatus Ollers, 1818 (Парагвай, север Аргентины);
0. capita Ollers, 1818 (северо-запад и юго-запад Эквадора, Венесуэла, Парагвай, Бразилия, юг Боливии, северо-восток и крайний север Колумбии, северо-восток Аргентины, восточная часть Анд в Перу, Гайана, о-ва Тринидад и Малые Антильские);
0. chaparensis Osgood, 1916 (восточная часть Анд в Боливии);
0. delicatus J. Allen et Chapman, 1897 (Венесуэла, Гайана, северо-восток Колумбии);
0. delticola Thomas, 1917 (Аргентина, Уругвай);
желтоватый хомяк — О. flavescens Waterhouse, 1837 (Аргентина, Уругвай);
галапагосский хомяк — О. galapagoensis Waterhouse, 1839 (Галапагосские острова);
0. intectus Thomas, 1921 (Центральная Колумбия);
длиннохвостый рисовый хомяк —О. longicaudatus Bennett, 1832 (Перу, Чили);
' хомяк Мак Коннелла — О. macconnelli Thomas, 1910 (Гайана, юг Венесуэлы; юг Колумбии и восток Эквадора);
чернобрюхий хомяк—О. melanostoma Tschudi, 1844 (Восточное Перу);
0. microtis J. Allen, 1916 (Амазонас в Бразилии);
0. munchiquensis J. Allen, 1912 (Западная Колумбия);
черный хомяк-—О. nigripes Desmarest, 1819 (Парагвай, Восточная Бразилия);
0. polius Osgood, 1913 (Северное Перу);
0. ratticeps Hensel, 1872 (Парагвай, восток и крайний юг Бразилии, северо-восток Аргентины);
0. simplex Winge, 1887 (Восточная Бразилия);
O.spodiurus Hershkovitz, 1940 (Эквадор);
0. subflavus Wagner, 1842 (Восточная Бразилия);
0. utiaritensis J. Allen, 1916 (Бразилия);
O.villosus J. Allen, 1899 (север Колумбии);
О. xantheolus Thomas, 1894 (юго-запад и запад Перу);
О. altissimus Osgood, 1933 (Анды в Перу, Эквадор);
малый рисовый хомяк—О. minutus Tomes, 1860 (Анды в Перу, хребет Западная Кордильера в Колумбии, Анды в Эквадоре);
О. caliginosus Tomes, 1860 (юго-запад, центр, крайний север Колумбии, хребет Восточная Кордильера, юго-запад, центр и северо-запад Эквадора, Панама, Коста-Рика, Никарагуа, кроме севера и юго-запада);
О. robustulus Thomas, 1914 (Эквадор);
О. zunigae Sanborn, 1949 (Перу);
двухцветный хомяк —О. bicolor Tomes, 1860 (северо-запад Боливии; Эквадор; север Перу; юг Колумбии; Бразилия, Гайана, север Венесуэлы);
О. catherinae Thomas, 1909 (юго-восток Бразилии, Гайана);
О. maculipes Pictet et Pictet, 1844 (Восточная Бразилия);
О. mamorae Thomas, 1906 (север Боливии; Эквадор; Перу);
О. marmosurus Thomas, 1899 (Гайана, Венесуэла, крайний восток Колумбии, Амазонас в Бразилии);
О. melleus Anthony, 1924 (юго-восток Эквадора);
О. mincae J. Allen, 1913 (север Колумбии);
рисовый хомяк— О. osgoodi Thomas, 1924 (Северное Перу; Эквадор);
О. phaeotis Thomas, 1901 (юго-восток Перу);
рисовый хомяк Роберта — О. roberti Thomas, 1903 (юго-запад Бразилии) ;
О. trinitatis J. Allen et Chapman, 1893 (Венесуэла; Колумбия; Панама; острова Тринидад и Малые Антильские);
рисовый хомяк Дарвина — О. darwini Osgood, 1929 (о-в Санта-Крус в группе Галапагосских о-вов);
О. indefessus Thomas, 1899 (о-в Санта-Крус в группе Галапагосских);
О. narboroughi Heller, 1904 (о-в Фернандина в группе Галапагосских) ;
О. hammondi Thomas, 1913 (Анды в Эквадоре);
О. borreroi Hernandez Camacho, 1957 (Северная Колумбия);
болотный рисовый хомяк—О. palustris Harlan, 1837 (юго-восток США — вост, треть территории штата Техас, восточная треть Оклахомы, юго-восточный угол штата Канзас, юго-западный и юго-восточный угол штата Миссури, Арканзас, кроме срединных районов на севере, крайний юг штата Иллинойс, юг и юго-запад Кентукки, штата Теннесси, кроме его крайнего северо-востока, штаты Миссисипи, Алабама, Джорджия, Флорида, Южная Каролина, крайний юго-запад и вся восточная половина Северной Каролины, восток Виргинии, восток Мэриленд, крайний юго-восток Пенсильвании, юг Нью-Джерси);
хомяк Коуеса •— О. couesi Alston, 1877 (юг США — крайний юг Техаса; Мексика, кроме северных и центральных штатов — восток Нуево-Леон, Тамаулипас, кроме крайнего запада, восточная половина Сан-Луис-Потоси, север Керетаро и Идальго, Пуэбла, Веракрус, Мексике, юг Гуанохуато, Мичоакан, Халиско, кроме северо-востока, узкая полоса вдоль берега Калифорнийского залива, проходящая в штатах Наярит, Синалоа и Сонора па север примерно до уровня о-ва Тибурон и, кроме того, все штаты, расположенные к югу от указанных выше; вся Центральная Америка на юг до северо-западных районов Коста-Рики включительно) ;
калифорнийский рисовый хомяк — О. pcninsulae Thomas, 1897 (крайний юг п-ва Калифорния в Мексике);
рисовый хомяк Томаса — О. fulgens Thomas, 1893 (Мексика, точное местонахождение не известно);
рисовый хомяк Нельсона — О. nelsoni Merriam, 1898 (о-ва Лос-Трес-Мариа у западного побережья Мексики около штата Наярит);
косумельский хомяк—О. cozumelae Merriam, 1901 (о-в Косумель у восточного побережья п-ова Юкатан);
0. azuerensis Bole, 1937 (п-ов Асуэро на юге Панамы);
гатунский хомяк — О. gatunensis Goldman, 1912 (Гатун в зоне Панамского канала в Панаме);
ямайский хомяк—О. antillarum Thomas, 1898 (о-в Ямайка);
черноухий хомяк-—О. melanotis Thomas, 1893 (юго-восток, юг и юго-запад Мексики — узкая полоса, вдоль берега Тихого океана, тянущаяся по штатам — южной части Синалоа, Наярит, Колима, Мичоакан, Герреро и Оахака, в штате Оахака пересекает его территорию на север и такой же узкой полосой следует по берегу Мексиканского залива на восток до северной оконечности п-ва Юкатан, включая штаты Табаско, Кам-печа, кроме южных районов, Юкатан и север Кинтана-Роо, другая узкая полоса тянется по побережью Мексиканского залива на север по шта-
там Веракрус, крайнему востоку Пуэбла, востоку Идальго и крайнему юго-востоку Сан-Луис-Потоси, на север ареал простирается до центральных районов штата Тамаулипас);
0. talamancae J. Allen, 1891 (крайний восток Коста-Рики, Панама);
шелковистый хомяк—О. bombycinus Goldman, 1912 (три небольших изолированных участка в центре Коста-Рики и в Панаме; в зоне Панамского канала и на крайнем востоке);
хомяк Харриса — О. aphrastus Harris, 1932 (центральные районы <та-Рики);
0. devius Bangs, 1902 (центральные районы Коста-Рики и ее берег Карибского моря недалеко от панамской границы, северо-запад Панамы) ;
пиррисский хомяк — О. pirrensis Goldman, 1913 (гора Пирри на крайнем востоке Панамы и, возможно, северо-запад Колумбии) ;
панамский рисовый хомяк — О. tectus Thomas, 1901 (два изолированных участка: один на юго-востоке Коста-Рики и крайнем юго-западе Панамы и второй в юго-восточной Панаме вдоль берега Тихого океана к юго-востоку от Панамского канала);
сентвинсентский хомяк — О. victus Thomas, 1898 (о-в Сент-Винсент из группы Малых Антильских);
карликовый рисовый хомяк —О. fulvescens Saussure, 1860 (Мексика—юг штата Наярит, западная половина штата Халиско, Колима, Мичоакан, кроме северо-востока Герреро, Оахака, Веракрус, крайний восток Пуэбло, север Идальго, восток Сан-Луис-Потоси, юг и центральные районы Тамаулипас, центральные районы востока Нуево-Леоп, а также все штаты Мексики к югу от Оахака и Веракрус; Гватемала, Белиз; Сальвадор; Гондурас, кроме крайнего северо-востока; запад и юго-запад Никарагуа, юг и юго-запад Коста-Рики; юг Панамы на восток до зоны Панамского канала);
хомяк Эндерса — О. endersi Goldman, 1933 (Панама в зоне Панамского канала);
королевский хомяк — О. trabcalus G. Allen el Barbour, 1923 (крайний юго-восток Панамы).
Систематика рода рисовых хомяков разработана недостаточно хорошо. Число видов в дальнейшем, по-видимому, будет уменьшено.
Род гигантских рисовых хомяков Genus Megalomys Trouessart, 1881
Размеры крупные. Длина тела примерно 36 см. Длина хвоста около 33 см. Уши относительно небольшие, почти полностью голые, длинный хвост покрыт редкими волосами. Стопа не специализирована. Когти довольно длинные. Волосяной покров грубый. Окраска его на спинной стороне тела черноватая или коричневая, на брюшной — белая. Секрет кожных специфических желез с сильным мускусным запахом.
Череп с узкой мозговой капсулой, слабо выраженным межглазнич ным сужением. Надглазничная область с сильными гребнями. Костные слуховые барабаны чрезвычайно малы.
Встречались на Малых Антильских о-вах: Мартинике, Барбуда и Сент-Люсия.
По-видимому, представители рода в настоящее время полностью ис чезли. Жили они среди кокосовых пальм. Убежищем служили норы. Мог ли плавать. Местные жители употребляли их в пищу. Приносили существенный вред посевам, в результате чего и были истреблены местными жителями. Вероятно, вымерли к 1900 г.
В роде 3 вида:
антильский хомяк — 44. desmarestii Fischer, 1829 (о-в Мартиника группы Малых Антильских);
барбудаский хомяк — М. andreyae Hopwood, 1926 (о-в Барбуда группы Малых Антильских);
сентлюсийский хомяк — М. luciac F. Major, 1901 (о-в Сент-Люсия группы Малых Антильских).
Эллермэн (1941) и Андерсон и Джонс (1967) различают только 2 вп да: антильского и сентлюсийского рисовых хомяков.
из
из
из
Род колючих рисовых хомячков
Genus Neacomys Thomas, 1900
Размеры мелкие. Длина тела 6,4—10 см. Длина хвоста достигает примерно длины тела. Хвост покрыт редкими волосами. Наружный палец задней конечности значительно короче трех центральных. Волосяной покров представлен смесью грубых щетин или даже игл и мягких нежных волосков. Иглы наиболее часто встречаются на спине, значительно реже на боках и почти отсутствуют на брюхе. Окраска спинной стороны тела варьирует от темно-рыжей и темно-желтоватой до оранжево-рыжей с примесью черного цвета, который имеется на вершинах игл. Боковые части тела значительно более светлые. У некоторых форм бока желтого цвета. У N. pusillus спинная часть тела отделена от брюшной оранжево-желтой продольной полосой, идущей от боков носа до бедер. Брюшная сторона тела белая, кремовая или желтоватая. Хвост буроватый сверху и более светлый снизу.
Череп небольшой, всегда с хорошо развитыми надглазничными гребнями. Костные слуховые барабаны больших размеров. Межтеменная кость хорошо развита.
Распространены в Панаме, Колумбии, Гайане, Эквадоре и Перу.
Населяют густые влажные леса от уровня моря до примерно 1100 м над его уровнем. Держатся среди скал, под камнями, упавшими деревьями, среди травы и в кустах. Образ жизни не изучен. Детенышей в помете обычно от 2 до 4.
В роде 4 вида:
украшенный хомячок — N. pictus Goldman, 1912 (восточная часть Панамы на границе с Колумбией);
гайанский хомячок — N. guiana Thomas, 1905 (Гайана, юг Венесуэлы) ;
колючий рисовый хомячок — N. spinosus Thomas, 1903 (юго-запад Бразилии, Эквадор, Колумбия, Перу);
колумбийский рисовый хомячок — N. tenuipes Thomas, 1900 (Колумбия. Эквадор);
Андерсон и Джонс (1967) насчитывают в роде 3 вида.
Род колючих хомячков
Genus Scolomys Anthony, 1924
В роде единственный вид: колючий хомячок — S. melanops Anthony, 1924.
Размеры мелкие. Длина тела примерно 9 см. Длина хвоста около 7 см. Длинные волосы на спине перемежаются с уплощенными колючками. Окраска сильно варьирует, в основном она коричневая сверху и темно-серая снизу. Конечности серые, а пальцы беловатые. Большой палец передних конечностей имеет широкий уплощенный ноготь. Сосков 3 пары.
В черепе характерен очень короткий рострум и очень маленькие щечные зубы.
Распространен в Восточном Эквадоре.
Местообитание и образ жизни не изучены. Имеющиеся в коллекции тлзсмпляры были пойманы в горах примерно на уровне 1150 м над ,ровней моря.
Род плавающих хомяков
Genus Nectomys Peters, 1861
Размеры от мелких до средних. Длина тела 11,5—25,4 см. Длина хвоста 15—25 см. Внешним обликом похож на крысу (рис. 47). Стопа крупная и широкая. На задних лапах три средних пальца длиннее двух крайних. Пальцы соединены неполной перепонкой. Длинный хвост обычно покрыт редкими короткими волосами. У N. squamipes на вентральной поверхности хвоста короткие щетиновидпые волосы образуют киль. Когти на пальцах задних конечностей удлиненные. Волосяной покров обычно мягкий, умеренной длины или длинный. Окраска спинной стороны тела рыжевато-коричневая или грязно-желтая с примесью бурой окраски. Бока более светлые. Брюшная сторона беловатая или сероватая, иногда с красновато-оранжевым оттенком.
Череп имеет хороню развитые супраорбитальные гребни. Межтеменная кость крупная. Скуловые кости короткие. Резцовые отверстия обычно широкие, продолжающиеся до зубного ряда. Костные слуховые барабаны относительно малы. Венечный отросток нижней челюсти крупный. Щечные зубы гипселодоптпого типа.
Распространены в Никарагуа, Коста-Рике, Панаме, Эквадоре, Колумбии, Перу, Гайане, Бразилии, Парагвае и о-ве Тринидад.
Населяют леса, антропогенные ландшафты, а также травянистые заросли. Поднимаются в горы до высоты примерно 2200 м над уровнем
Рис. 47. Д'. squamipes и череп N. alfari
моря. Любят селиться около болот, озер или речек. Хорошо и быстро плавают. Гнезда делают в зарослях кустарника или под упавшими стволами деревьев. Экология почти по изучена.
В роде, по-видимому, 3 вида:
N. alfari J. Allen, 1897 (юг Никарагуа, север Коста Рики, восток Па намы, север Эквадора, север Колумбии, Венесуэла);
N. dimidiatus Thomas, 1905 (юго-восток Никарагуа);
N. squamipes Brants, 1827 (Бразилия, Центральная, Восточная и Се веро-Восточная Колумбия, Эквадор, юго-восток Неру, Боливия, Гайана, Парагвай, северо-запад Венесуэлы).
Андерсон и Джонс (1967) насчитывают в роде 2 вида.
Род рипидомис
Genus Rhipidomys Tschudi, 1844
Размеры от мелких до средних. Длина тела 8—21 см. Длина хвоста обычно длиннее тела и может достигать 27 см. Хвост хорошо оволоснен, оканчивается кисточкой удлиненных волос. Кисть и стопа широкие, большие, приспособленные к лазанию, с крупными сильными когтями. На задних конечностях пятый палец удлиненный. Волосяной покров густой и мягкий, средней длины или длинный. Окраска его на спинной стороне тела серовато-грязно-желтая, рыжевато-коричневая, желтоватая, коричневая, бурая, темно-коричневая или почти черная. Брюшная сторона тела белая, кремовая, желтоватая или сероватая и резко отграничена от спинной. У многих видов по боку проходит полоса желтого цвета. Хвост обычно слегка темнее спины. Сосков 3 пары.
В черепе надглазничных гребней нет вовсе или они развиты слабо. Межтеменная кость очень крупных размеров. Костное небо короткое. Костные слуховые барабаны небольших размеров.
Распространены в Колумбии, Эквадоре, Перу, Боливии, Северной Аргентине, Восточной Бразилии, Гайане, Венесуэле и на о-ве Тринидад.
Населяют леса. Образ жизни не изучен. Хорошо лазают по деревьям , в кустам. Активны в сумерках. Самки приносят, по-видимому, двух или | трех детенышей.
В роде 6 видов:
R. scandens Goldman, 1913 (восточная часть на границе Панамы с Колумбией);
R.latimanus Tomes, 1860 (хребты Центральная и Западная Кордильеры в Колумбии, Венесуэла, Анды в Эквадоре);
белопалый хомяк — R. leucodactylus Tschudi, 1845 (крайний северо-запад Аргентины, крайний юг Боливии, Анды в Эквадоре, восток и се-i веро-запад Перу);
хомяк Макконнелла— R. macconnelll De Winton, 1900 (юго-восток Венесуэлы);
. R. mastacalis Lund, 1841 (Северо-Восточная и Центральная Брази-I дня, Восточная и Западная Венесуэла, Гайана);
хомяк Склетера — R. sclateri Thomas, 1887 (Гайана и соседние с ней I районы Бразилии, Венесуэла, о-в Тринидад).
Род хомяков Томаса
Genus Thomasomys Coues, 1884
Размеры от мелких до средних. Длина тела примерно 9—18,5 см. Длина хвоста 8,5—23 см. Хвост покрыт волосами средней густоты. Задние конечности почти нс адаптированы к лазанию по деревьям. Волосяной покров обычно густой и мягкий. Окраска спинной стороны тела варьирует от оливково-серой, оливково-желтой, желтовато-рыжей, золотисто-коричневой, красновато-коричневой, серо-коричневой до темно-коричневой или почти черной. Средняя часть спины обычно более темная. Брюшная сторона лишь незначительно светлее спинной. Сосков 3 или 4 пары.
В черепе лицевой отдел удлинен и заострен. Мозговая коробка крупных размеров, округлая. Надглазничные гребни развиты слабо. Меж-глазничное сужение выражено хорошо или плохо. Межтеменная кость
крупная. Костное небо короткое. Костные слуховые барабаны небольшие или средних размеров.
Распространены в Гайане, Венесуэле, Колумбии, Эквадоре, Перу, Боливии, Аргентине, Бразилии.
Обычно населяют леса. Поднимаются в горы до 4200 м над уровнем моря. Большинство видов ведут наземный образ жизни, некоторые — древесный. В течение года, по-видимому, бывает по два помета, в каждом из которых от 2 до 4 детенышей. Т. rhoadsi копает норы, расположенные на глубине 25—76 мм под поверхностью почвы. Экология изучена крайне слабо.
В роде 25 видов:
Т. aureus Tomes, 1860 (Анды в Эквадоре, северо-запад Перу, Колумбия) ;
Т. boeops Thomas, 1899 (западный Эквадор):
Т. bombycinus Anthony, 1925 (хребет Западная Кордильера в Колумбии) ;
Т. cinereiventer J. Allen, 1912 (хребты Восточная, Центральная и Западная Кордильеры в Колумбии, Эквадор);
Т. cinereus Tomas, 1882 (юго-восток-Эквадора, северо-восток Перу);
Т. daphne Thomas, 1917 (Центральная Боливия, юго-восток Перу);
Т. dorsalis Hensel, 1873 (Восточная и Юго-Восточная Бразилия);
Т. gracilis Thomas, 1917 (Эквадор, юго-восток Перу);
Т. hylophilus Osgood, 1912 (северо-запад Колумбии);
Т. incanus Thomas, 1894 (Анды в Перу);
Т. ischyurus Osgood, 1926 (северо-запад Перу, хребет Западная Кордильера в Эквадоре);
Т. kalinowskii Thomas, 1894 (Анды в центральных районах Перу);
Т. ladewi Anthony, 1926 (северо-запад Боливии);
Т. laniger Thomas, 1916 (запад Венесуэлы, центр и северо-восток Колумбии);
Т. lugens Thomas, 1896 (хребты Центральная и Западная Кордильеры в Колумбии, запад Венесуэлы, Анды в Эквадоре);
Т. notatus Thomas, 1917 (юго-восток Перу);
Т. oenax Thomas, 1928 (крайний юг Бразилии);
Т. areas Anthony, 1926 (Анды в Боливии);
Т. paramorum Thomas, 1898 (Анды в Эквадоре);
Т. pictipes Osgood, 1933 (северо-восток Аргентины);
Т. pyrrhonotus Thomas, 1882 (Анды на юге Эквадора, северо-запад Перу);
Т. rhoadsi Stone, 1914 (Эквадор);
Т. rosalinda Thomas et Saint Leger, 1926 (северо-запад Перу);
T. taczanowskii Thomas, 1882 (северо-запад Перу);
Т. vestitus Thomas, 1898 (запад Венесуэлы).
Некоторые ученые выделяют Т. оепах в особый род Wilfredomys Avila-Pires, 1960.
Род риодежанейрских хомячков
Genus Phaenomys Thomas, 1917
В роде единственный вид: риодежанейрский хомячок — Р. ferrugi-neus Thomas, 1894.
Размеры средние. Длина тела примерно 15 см. Длина хвоста около-19 см. Уши небольшие, густо покрытые волосами. Длинный хвост покрыт также довольно густыми волосами. Задние конечности несколько 150
адаптированы к древесному образу жизни. Густой мех на спинной стороне тела светло-красноватого или ярко-рыжего цвета. Брюхо белое пли слегка желтоватое. Сосков 4 пары.
Мозговая коробка черепа относительно узкая. Надглазничные греб-хорошо развиты.
Распространен в Восточной Бразилии.
Экология не изучена.
Род колумбийских хомячков Genus Chilomys Thomas, 1897
В роде единственный вид: колумбийский хомячок — С. instans Thomas, 1895.
Размеры мелкие. Длина тела 8,6—9,9 см. Длина хвоста 10,5—13 см. По внешнему облику сходен с мышью. Волосяной покров густой и мягкий. На спинной и брюшной стороне тела он имеет однотонный серый цвет. Морда может быть черноватая. Иногда от горла до середины брю-.а проходит белая полоса.
. В черепе мозговая коробка очень крупная и округлая. Лицевой отдел черепа узкий. Надглазничных и теменных гребней нет. Верхние резвы выступают вперед. Нижние резцы сжаты.
Распространен на северо-западе и в центральных районах Колумбии.
Населяет густые, темные леса. Поднимается в горы на высоту до 00 м над уровнем моря. Экология не изучена.
Род лазающих хомяков
Genus Tylomys Peters, 1866
Размеры средние. По внешнему облику похожи на черную крысу. Длина тела 17-—25,5 см. Длина хвоста 20—25 см. Уши крупные, голые. Длинный хвост не покрыт волосами. Задние конечности приспособлены к лазанию по деревьям: ступни расширены и укорочены. Обычно волосяной покров густой, мягкий, блестящий. Окраска его на спинной стороне тела сероватая или буроватая, на брюшной белая или желтоватая.
Череп удлинен. Мозговая капсула уплощена. Надглазничные гребни чрезвычайно сильно развиты. В затылочной области располагается низкий гребень. Резцовые отверстия широкие.
Число хромосом в диплоидном наборе 40 у мексиканского лазающего хомяка, 42 у голохвостого лазающего хомяка, 52 у панамского лазающего хомяка.
Распространены на территории от Южной Мексики на юг через Центральную Америку до Эквадора.
Населяют леса, предпочитая участки леса, окружающие скалы. Встречаются в горах на высоте 1400 м над уровнем моря. Могут лазать по деревьям, но большую часть времени, no-видимому, проводят па земле. Экология не изучена.
В роде 7 видов:
рыжебрюхий хомяк—F. fuloioenter Anthony, 1916 (Восточная Папа-тана границе с Колумбией);
мексиканский лазающий хомяк—Т. gymnurus Villa, 1941 (юго-восток Мексики— штаты: юго-восток Веракрус и север Оахапа);
голохвостый хомяк — Т. nudicaudus Peters, 1866 (Юго-Восточная Мексика: крайний восток штата Оахапа, штат Чиапас, кроме севера п
юга, Гватемала, юг Гондураса, Белиз, Сальвадор, юго-запад Никарагуа) ;
панамский лазающий хомяк — Г. panamensis Gray, 1873 (восток Панамы на границе с Колумбией);
тумбалаский хомяк — Т. tumbalensis Merriam, 1901 (Тумбала на севере штата Чиапас в Мексике);
хомяк Уотсона — Т. watsoni Thomas, 1899 (Панама, кроме части к востоку от Панамского канала);
южноамериканский хомяк—Т. mirae Thomas, 1899 (Центральная Колумбия, север Эквадора).
Род большеухих хомяков
Genus Ototylomys Merriam, 1901
Размеры от мелких до средних. Длина тела 9,5—19 см. Длина хвоста 10—19 см. Внешний вид крысовидный (рис. 48). Крупные уши и длинный хвост безволосые. Волосяной покров средней длины, мягкий, довольно густой. Спинная сторона тела буроватая или сероватая с при
Рис. 48. Короткомордый хомяк — О. brevirostrls и череп большеухого хомяка — О. phyllotis
к
я
с-н-.0-ш-
месью черных волос, брюшная — белая или сероватая. Пальцы обычно беловатые.
Череп уплощен. Межтеменная кость крупная. Костные слуховые барабаны вздутые, крупных размеров. Надглазничные гребни хорошо развиты. Венечный отросток на нижней челюсти сильно уменьшен в размерах.
Распространены в Юго-Восточной и Южной Мексике и Центральной Америке.
Обычно населяют леса от уровня моря до 1800 м над его уровнем. Хорошо лазают по деревьям, но по большей части ведут наземный образ жизни. Активность ночная. Экология не изучена.
В роде 3 вида:
гватемальский большеухий хомяк-—О. connecteus Sanborn, 1935 (север Гватемалы) ;
большеухий хомяк—О. phyllotis Merriam, 1901 (запад Коста-Рики; юго-запад Никарагуа; юго-запад Гондураса; Белиз; северная половина Гватемалы; штаты Мексики — запад Кинтано-Роо, юг Юкатана, Кампече, крайний восток Табаско, восток Чиапас);
короткомордый хомяк — О. brevirostris Laurie, 1953 (Мексика — штаты Юкатан и Кинтана-Роо; Белиз).
Род соневидных хомяков
Genus Nyctomys De Saussure, 1860
В роде единственный вид: соневидный хомяк — N. sumichrasti Saussure, 1860.
Размеры мелкие (рис. 49). Длина тела 11—13 см. Длина хвоста 8,5— 15,5 см. Уши относительно короткие, покрытые редкими, нежными волосками. Хвост покрыт редкими волосами и сквозь них хорошо видны чешуйки кожи. Только на конце хвоста волосы становятся несколько более густыми и длинными. Глаза крупные. Задние конечности приспособлены к древесной жизни — их ступни расширены и укорочены. Первый палец на них имеет коготь. Волосяной покров мягкий, короткий или длинный, на спинной стороне тела имеет грязно-желтую или рыжевато-коричневую окраску. Бока светлее спины, брюшная сторона белая. Хвост обычно коричневый, конечности белые. Вокруг глаз — темные кольца. Темные пятна расположены между глазами и в основании усов. Сосков 2 пары.
Череп с очень широкой мозговой коробкой. Межтеменная кость крупная. Лобные кости сильно расширены. Надглазничные гребни хорошо выражены и тянутся через теменные кости до затылочных. Лицевой отдел укорочен. Подглазничные отверстия большие. Костные слуховые барабаны малы. Костное небо широкое, оканчивается перед задней частью зубных рядов. Резцовые отверстия широкие, редко достигают зубных рядов. Венечный отросток нижней челюсти низкий.
Распространен от Центральной Мексики на юг до Панамы- -штаты Мексики: Колима, юг Гуанохуато, юг Герреро, юг Оахака, юг Веракрус, юг Табаско, Чиапас; южная половина Гватемалы; Сальвадор, юг Белиза, Гондурас, кроме северо-востока, Никарагуа, кроме северо-востока, Коста-Рика, запад Панамы.
Обитает в лесах. Экология изучена слабо. Ведет древесный образ жизни. Держится колониями. Активность ночная. Питается различными плодами. В помете от 2 до 4 детенышей.
Рис. 49. Соневидный хомяк—ЛЛ sumichrasti и его череп
Род юкатанских хомячков
Genus Otonyctomys Anthony, 1932
В роде единственный вид: юкатанский хомячок — О. hatti Anthony, 1932.
Размеры мелкие. Длина тепа 10,4—11,6 см. Длина хвоста примерно 12,7 см. Уши средних размеров, покрыты редкими волосками. Окраска спинной стороны тела красновато-коричневая или коричневая, брюшной—белая. Хвост буроватый, хорошо оволоснен. Сосков 2 пары.
В черепе характерны крупные костные слуховые барабаны. Щечные зубы относительно небольших размеров.
Распространен на юге Мексики в южной части штата Юкатан.
Экология не изучена. По-видимому, ведет древесный образ жизни.
Род бразильских древесных хомячков
Genus Rhagomys Thomas, 1917
В роде единственный вид: бразильский древесный хомячок: R. rufes-cens Thomas, 1886.
Размеры мелкие. Длина тела около 9,4 см. Хвост примерно такой же длины. Уши короткие, едва выдающиеся над мехом. Они покрыты густыми рыжевато-бурыми волосами. Хвост с редкими волосами. Стопа расширена и приспособлена к лазанию по деревьям. Пятый палец на ней удлинен, первый — лишен когтя. Спинная сторона тела имеет оранжево-рыжеватую окраску. Брюшная сторона несколько более светлая. Лапы желтые, хвост буроватый. Сосков 3 пары.
В черепе мозговая капсула расширена. Резцовые отверстия узкие и не достигают зубных рядов.
Распространен в Восточной Бразилии.
Экология не изучена.
Род полевых хомячков
Genus Reithrodontomys Giglioli, 1873
Размеры мелкие. Длина тела 5—14,5 см. Длина хвоста 6,5—9.5 см. Масса у взрослых особей обычно 10—20 г. Внешне сходны с мышами (рис. 50). Уши довольно крупные, почти голые. Лапы узкие, пятый палец на задних конечностях часто относительно длинный. У большинства видов волосяной покров густой и мягкий, а у некоторых более редкий и грубый. Окраска его на спине варьирует от охристо-серой до коричневой и почти черной. Бока светлее спины. Брюхо белое или сероватое. В течение года бывает одна линька. Хвост темный сверху и светлый снизу; иногда он бывает одноцветный. Сквозь довольно редкие и короткие волосы хвоста видны чешуйки кожи. Сосков 3 пары.
Мозговая коробка черепа несколько уплощена. Межглазничное сужение хорошо выражено. Надглазничные гребни не развиты. Костные слуховые барабаны небольшие. Костное небо оканчивается примерно на уровне последних щечных зубов. Резцовые отверстия узкие. Венечный отросток нижней челюсти низкий. Верхние резцы имеют по одной продольной борозде на наружной поверхности. Нижние резцы борозд не имеют.
Хромосом в диплоиднбм наборе у западного хомячка 42—49, у болотного хомячка 38, у рыжего хомячка 50.
Распространены от Юго-Западной Канады на юг через США, Мексике и Центральную Америку до Колумбии и Эквадора. Населяют также южную часть штата Огайо и долину Потомака в восточной части Соединенных Штатов.
Населяют разнообразные ландшафты от уровня моря до верхней границы леса в горах, от соляных болот до тропических лесов. Излюбленные места обитания — равнины с невысокой травой. Строят шаровидные
гнезда из травы примерно диаметром 150—175 мм. Гнезда обычно сооружаются в высокой траве, в низких кустах или даже небольших деревьях. Иногда зимние гнезда устраиваются в норах, в небольших впадинах или трещинах земли. Активны круглый год, главным образом по ночам. Могут довольно проворно лазать по деревьям. Весной и летом питаются зелеными частями растений, а поздним летом, осенью и зимой — в основном семенами и в меньшей степени вегетативными частями. Поедают также некоторых насекомых. Определенной периодичности в размножении нет, за исключением мест, где зимы бывают суровыми. В течение года, вероятно, бывает от 1 до 4 пометов. Длительность беременности примерно 21—24 дня. В помете бывает от 1 до 9, в среднем 4 детеныша. Масса новорожденного примерно 1 г. Глаза открываются на 10— 12-й день. Спустя 3 недели после рождения детеныши покидают гнездо и переходят к самостоятельной жизни. У некоторых видов самки начинают размножаться в 17-педслыюм возрасте.
В роде 17 видов:
горный хомячок — /?. monianus Baird, 1855 (западные и юго-занад-ные штаты США — юго-запад Южной Дакоты, Небраска, восточная треть Вайоминга, восточная половина Колорадо, Канзас, Нью-Мексико, кроме северо-запада, Оклахома, кроме крайнего востока, Техас, кроме
крайнего востока и юга; Мексика — штат Чиуауа, кроме юго-запада, северо-восток Соноры);
сонорский хомячок—/?. burti Benson, 1939 (Мексика — юго-запад штата Соноры и север штата Синалоа);
восточный хомячок — R. humulis Audubon et Bachman, 1841 (вост, и юго-восточные штаты США — юг Мэриленда, Западная Вирджиния, юг Огайо, Кентукки, Вирджиния, Северная Каролина, Юж. Каролина, Теннесси, северо-запад и центр Арканзаса, Миссисипи, крайний юго-восток Техаса, южная половина Луизианы, Алабама, Джорджия, Флорида);
западный хомячок — R. megalotis Baird, 1858 (юго-запад Канады — центр крайнего юга провинции Британская Колумбия, крайний юго-восток провинции Альберта и крайний юго-запад Саскачевана; запад и центр США — восточная половина штата Вашингтон, Орегон, кроме северо-запада, южная половина Айдахо, Калифорния, Невада, Юта, Аризона, Нью-Мексико, западная треть Техаса, крайний запад Оклахомы, Колорадо, кроме северо-запада, Канзас, кроме юго-востока, Миссури, крайний северо-восток Арканзаса, Иова, крайний северо-запад Иллинойса, юго-запад Висконсина, юг и юго-запад Миннесоты, Небраска, Южная Дакота, южная половина Северной Дакоты, восточная половина Монтаны, восточные две трети Вайоминга; почти вся Мексика, кроме южных двух третей п-ова Калифорния, южной половины штата Соноры, штата Синалоа, Наярит, западной половины Халиско, Колима, юго-запада н юга Мичоакан, севера и юга Герреро, юга и востока Оахака, истока и севера Веракрус, всех штатов, расположенных к востоку от Зеракрус и Оахака, восточных двух третей Тамаулипас, северной поло-шны Нуэво-Леон, восточной трети Коауила);
болотный хомячок—R. raviventris Dixon, 1908 (США — запад дентального района Калифорнии около Сан-Франциско);
вулкановый мексиканский хомячок — R. chrysopsis Merriam, 1900 (центр Мексики — крайний юго-восток штата Халиско, север Мичоакана, южная половина Мексике, Тласкала, центр западных районов Веракрус) ;
Р. sumichrasti Saussure, 1861 (три изолированных участка ареала: один на крайнем западе Панамы и в центре Коста-Рики; второй на крайнем северо-западе Никарагуа, юго-западных районах Гондураса, севере Сальвадора, центре Гватемалы и в центральных районах штата Мексики Чиапас; третий — на юге Мексики на северо-западе штата Оахака, в центре Герреро, центре западных районов Веракрус, восточной половине Пуэбла, Идальго, Мексике, северной половине Мичоакан, юге Халиско, восточной половине Кереторо);
рыжий хомячок — R. fulvescens J. Allen, 1894 (юг США — юго-восток штата Аризоны, крайний юго-восток Канзаса, крайний юго-запад Миссури, восточные две трети Оклахомы, Арканзас, кроме востока, Луизиана, .крайний юго-запад Алабамы, Техас, кроме северо-запада; почти вся Мексика, за исключением севера и центра западных районов штата Чиуауа, центра и северо-запада Соноры, юго-востока Веракрус, северо-востока Оахака; юга и севера Чиапас и штатов, расположенных к северу от Чиапас; центральные районы Гватемалы, северо-восток и восток Сальвадора, юго-запад Гондураса);
мохнатый хомячок — /?, hirsutus Merriam, 1901 (Мексика — юго-вос-I ток штата Наярит и северо-запад штата Халиско);
I изящный хомячок — R. gracilis J. Allen et Chapman, 1897 (Мексика—штаты Юкатан, Кампече, запад Кинтапо — Роо, юг Чиапас; Гватемала; Белиз, Сальвадор; юго-запад Никарагуа, северо-запад Коста-Рики);
хомячок Дариена — 7?. dariensis Pearson, 1939 (восточная половина Панамы и, вероятно, крайний северо-запад Колумбии);
мексиканский полевой хомячок — R. mexicanus Saussure, 1860 (шт. Мексики — центр южных районов Тамаулипас, крайний восток Сан-Луис-Потоси, северная половина западных районов Веракрус, северо-восток Хидальго, север Пуэбла, Оахака, кроме юга, северо-запада и северо-востока, центральные районы Герреро от северо-западной границы до юго-восточной, юго-запад Халиско, Чиапас, кроме юго-запада и северо-востока; южная половина Гватемалы, но без Тихоокеанского побережья; Сальвадор; южная половина Гондураса; западная треть Никарагуа; центральные районы Коста-Рики; запад Панамы, север и северо-запад Эквадора; запад Колумбии);
короткомордый хомячок — R. brevirostris Goodwin, 1943 (центральные районы Коста-Рики);
мелкозубый хомячок — R. microdon Merriam, 1901 (три небольших изолированных участка ареала: один в центре штата Оахака, Мексика; второй в центре штата Чиапас, Мексика и в центре Гватемалы; третий в штатах Мексики — Морелос, Мексике и северо-востоке Мичоакан);
узкомордый хомячок — R. tenuirostris Merriam, 1901 (центр Гватемалы от западной государственной границы до восточной);
хомячок Родригеса — R. rodrigtiezi Goodwin, 1943 (центральные районы Коста-Рики);
панамский хомячок — R. creper Bangs, 1902 (крайний запад Панамы, центр и юго-восток Коста-Рики).
Болотный хомячок включен в «Красную книгу».
Род белоногих хомячков
Genus Peromyscus Gloger, 1841
Размеры мелкие или средние. Длина тела 8—17 см. Длина хвоста 4—20,5 см. По внешнему виду несколько напоминают мышей или крыс (рис. 51). Уши крупные, покрыты редкими, нежными волосами. Хвост, составляющий по меньшей мере треть длины тела, покрыт хотя и короткими, но довольно густыми волосами. Часто на кончике хвоста расположен пучок удлиненных волос. Подошвы лап иногда покрыты волосами. Стопа с 5 или 6 подошвенными мозолями. Волосяной покров обычно мягкий и густой; окраска его сильно изменчива. Спинная сторона тела может быть серой, песчано-желтой, золотистой или темно-коричневой, брюшная — белая или беловатая. Некоторые виды почти целиком белые, а другие — почти черные. В течение года бывает одна линька. Сосков 2 или 3 пары.
В черепе лицевой отдел относительно длинный и узкий. Межглазничное сужение всегда имеется. Надглазничных гребней нет или они развиты слабо. Носовые кости обычно выдвинуты вперед по отношению к резцам. Межтеменная кость хорошо развита. Костное небо примерно достигает средних частей третьего коренного зуба. Резцовые отверстия длинные, щелевидные. Костные слуховые барабаны, как правило, некрупные. Венечный отросток нижней челюсти сильно уменьшен. Щеч ные зубы брахиодоптные, небольшие, с бугорчатой жевательной поверх ностыо.
Хромосом в диплоидном наборе у большинства изученных видов (хомячки калифорнийский, олений, белоногий и др.) 48 и только у скалистого хомячка 52.
Рис. 51. Белоногий хомячок — Р. leucopus и его череп
Распространение охватывает территорию от северных частей Колум-ивЮжной Америке на север до Аляски и Лабрадора.
Населяют самые разнообразные ландшафты и обычно бывают фономн видами. Активны ночью в течение круглого года. Убежищем слу-и норы, которые зверьки обычно выкапывают сами; иногда использу-норы других животных. В норах они делают гнезда из сухих расте-
1Й. Гнезда устраивают также и в дуплах упавших деревьев или во впа-нах в земле Живут обычно парами. Питаются семенами, орехами, хинами, плодами, насекомыми и другими мелкими беспозвоночными, огда падалью. Размножение происходит круглый год, за нсключепи-। холодных или очень жарких месяцев. Длительность беременности
fe-27 дней, хотя иногда может быть и несколько больше. Самка припо- от I до 9, обычно 4 детенышей. Спустя 3 6 педель после рождения теяыши переходят к самостоятельному образу жизни. Половозрелое гь кет наступать в 7-недельном возрасте. В естественных условиях лли-
тельность жизни не превышает обычно 2 лет. В лабораторных условиях могут жить до 5,5 лет.
Белоногие хомячки широко используются в качестве лабораторных животных.
В роде, по-видимому, 55 видов:
каньонный хомячок — Р. crinitus Merriam, 1891 (штаты США —юго-восток Орегона, юго-запад Айдахо, Невада, восток Калифорнии, Юта, кроме центра и северо-востока, запад Колорадо, северо-запад Нью-Мексико, север и запад Аризоны; штаты Мексики — крайний северо-запад Соноры и крайний северо-восток Нижней Калифорнии);
ложноканьонный хомячок — Р. pseudocrinitus Burt, 1932 (о-в Коро-йадос у восточного побережья п-ва Калифорния);
калифорнийский хомячок — Р. californicus Gambel, 1848 (США — юго-запад штата Калифорния; Мексика — северо-запад Нижней Калифорнии);
кактусовый хомячок — Р. eremicus Baird, 1858 (штаты США — крайний юг Невады, юг Калифорнии, кроме центра и северо-востока, юг Нью-Мексико, юго-запад Техаса; северные штаты Мексики — Нижняя Калифорния и мелкие прилежащие о-ва в Тихом океане и Калифорнийском заливе, Сонора, Чиуауа, Коауила, запад Нуэво-Леон, крайний юго-запад Тамаулипас, западные две трети Сан-Луис-Потоси, восточная половина Закатекас, северные половины Дуранго и Синалоа).
хомячок Мерриама — Р. merriami Mearns, 1896 (два небольших изолированных участка — один в центре крайнего юга штата Аризона, США и севера штата Сонора, А4ексика, второй — на крайнем юге штата Сонора);
монсерратский хомячок — Р. caniceps Burt, 1932 (о-в Монсеррат в Калифорнийском заливе; у берега п-ова Калифорния);
гуардасский хомячок — Р. guardia Townsend, 1912 (о-ва в Калифорнийском заливе: Анхель-де-ла-Гуарда, южный Сан-Лоренсо и Меджиа, у берега п-ова Калифорния);
хомячок Дикки-—Р. dickeyi Burt, 1932 (о-в Тортига в Калифорнийском заливе у берега п-ова Калифорния);
хомячок Пембертона — Р. pembertoni Burt, 1932 (о-в Сап-Педро Ноласко в Калифорнийском заливе у берега штата Сонора);
Р. stepheni Townsend, 1912 (о-в Св. Стефана в Калифорнийском заливе);
олений хомячок — Р. maniculatus Wagner, 1845 (большая часть Северной Америки; на Аляске отсутствует; северная граница ареала проходит примерно от пересечения параллели 65° с. ш. с границей провинции Канады Юкон с Аляской, далее па восток к озерам Большое Медвежье, Большое Невольничье, через северные районы провинций Саскачеван и Манитоба к середине западного побережья Гудзопова залива, огибает его с юга и через северные районы провинции Квебек выходит к побережью Атлантического океана; восточная граница ареала идет по берегу Атлантического оксана; южная граница ареала идет от штата Коннектикут к северу штата Пыо-Джерси, через северо-западные районы штатов Виргиния и Северная Каролина, север Теннесси, через юг штата Арканзас, вдается узким языком через центральные районы Техаса к берегам Мексиканского залива, огибает с северо-запада Техас и через штат Ныо-Мексико уходит в Мексику, где ареал охватывает все ее центральные районы — па юг до штата Оахака; во без прибрежных районов, на востоке к берегу Мексиканского залива ареал подходит лишь к северной части штата Веракрус, а на западе — к берегу Тихого океана только у штатов Халиско и Колима; западная граница ареала
3
>1 с e x т о ,а
«дет вдоль побережья Тихого океана примерно от 60° с. ш. до п-ва Калифорния включительно; Холл и Келсон, 1959, насчитывают в пределах ареала 66 подвидов);
сантакрусский хомячок—Р. sejugis Burt, 1932 (о-ва Санта-Крус и Сан-Диего в Калифорнийском заливе у берега п-ва Калифорния);
хомячок Слевина — Р. slevini Mailliard, 1924 (о-в Санта-Каталина в Калифорнийском заливе);
ситхасский хомячок — Р. sitkensis Merriam, 1897 (мелкие о-ва архипелага Александра в заливе Аляска);
Р. polionotus Wagner, 1843 (штаты США — западная половина Южной Каролины, Джорджия, восточная половина Алабамы, Флорида, кроме юго-запада);
черноухий хомячок — Р. melanotis J. Allen et Chapman 1897 (центральные районы Мексики — юго-запад и запад Чиуауа, Дуранго, кроме северо-востока, Закатекас, юг Нуэво-Леон, юго-запад Тамаулипас, Сан-Луис-Потоси, кроме крайнего востока, Гуанохуато, Агуаскальентес, восточная половина Халиско, северная половина Мичоакан, Мексике, Морелос, Тласкала, север Пуэбла, запад центральных районов Веракрус, Идальго, кроме северо-востока);
белоногий хомячок — Р. leucopus Rafinesque, 1818 (провинции Канады -- крайний юго-восток Саскачевана, крайний юг Манитобы, юг Новой Шотландии; восточная половина США, штаты — от южной половины Мэн, юга Вермонта и далее на запад до Монтаны от государственной границы с Канадой на юг до государственной границы с Мексикой, берега Мексиканского залива, юго-востока штата Миссисипи, центральных районов штатов Алабама и Джорджия; юго-восток Южной Каролины и западной трети Северной Каролины; западная граница ареала в США идет по центральным районам Монтаны, северо-востоку Вайоминга, юго-западу Небраски, западу Канзаса, юго-востоку Колорадо, центру и юго-востоку Аризоны и уходит на юг в Мексику; восточная Мексика, штаты— крайний северо-восток Соноры, восточная половина Чиуауа, крайний запад и северо-восток Коауила, восточная половина Дуранго, Нуэво-Леон, Тамаулипас, восточная половина Сан-Луис-Потоси, Веракрус, север Идальго, восток Пуэбло, восточная половина Оахака, Табаско, Кампече, кроме юго-востока, Юкатан, кроме северо-востока, крайний запад Кинтано-Роо);
хлопковый хомячок — Р. gossypinus Le Conte, 1853 (юго-восток США, штаты — восток Техаса, юго-восток Оклахомы, южная половина и северо-восток Арканзаса, крайний юго-восток Миссури, крайний юг Иллинойса, юг и юго-запад Кентукки, западные две трети Теннеси, Миссисипи, Алабама, Флорида с мелкими соседними о-вами, южная половина Джорджии, юго-восточная половина Южной Каролины, восток и юго-восток Северной Каролины, крайний юго-восток Виргинии);
гребенчатый хомячок — Р. boylii Baird, 1855 (юго-западные штаты США — Калифорния, кроме крайнего северо-запада и юго-востока, юг Невады, Юта, кроме севера и северо-запада, запад и юг Колорадо, Аризона, кроме юго-запада, Ныо-Мексико, Оклахома, кроме центральных районов севера, крайний юго-восток Канзаса, северо-запад Арканзаса, западная половина Техаса; почти вся Мексика, кроме южных двух третей Нижней Калифорнии, западной части Сопоры, почти всей территории Синалоа, прибрежной полосы штатов Халиско, Колима и Мичоакан; юго-восток Чиуауа, восточной трети Дуранго, северной половины Закатекас, южных двух третей Коауила, севера Нуэво-Леон и всей прибрежной зоны Мексиканского залива, включая всю территорию штатов Табаско, Кампече, Юкатан, Кинтано-Роо; в ареал входит южная половина
Гватемалы; Сальвадор, кроме узкой прибрежной полосы; центр и юго-запад Гондураса);
болотный белоногий хомячок — Р. perfulvus Osgood, 1945 (штаты Мексики — юг Халиско, юг Мичоакан, северо-запад Герреро);
оахакасский хомячок — Р. oaxacensis Merriam, 1898 (штаты Мексики — северная половина Оахака и Чиапас, кроме северо-востока и прибрежных районов);
южный хомячок — Р. hylocetes Merriam, 1898 (два изолированных участка ареала в штатах Л1ексики — один — на юге Халиско, севере Мичоакан, Мексике, севере Морелос; второй, меньших размеров, в центре Оахака);
Р. pectoralis Osgood, 1904 (юго-запад США — крайний юго-восток штата Аризона, центр и юго-запад Техаса; штаты Мексики — северо-восток Соноры, Чиуауа, кроме юго-запада и севера, Коауила, восточная половина Дуранго, Закатекас, южная половина Нуэво-Леон, западная половина Тамаулипас, Сан-Луис-Потоси, Идальго, Кереторо, Гуанохуа-то, Халиско, кроме запада и крайнего юга);
Р. polius Osgood, 1904 (Мексика — северо-запад штата Чиуауа);
Р. truei Shufeldt, 1885 (юго-западные штаты США — крайний юго-запад Орегона, Калифорния, кроме юго-востока и крайнего юго-запада, Невада, кроме севера, Юта, кроме севера и центра, Колорадо, кроме центра и северо-востока, Ныо-Мексико, кроме юго-востока, крайний северо-запад Оклахомы, Аризона, кроме юго-запада; штаты Мексики — северная треть и изолированный участок на крайнем юге Нижней Калифорнии, западная половина Чиуауа, Дуранго, крайний юго-восток Коауила, Дуранго, кроме северо-запада и северо-востока, Закатекас, Агуаскальентес, восточная половина Халиско, западная половина Сан-Луис-Потоси, Гуанохуато, северная половина Мичоакан, Керетаро, Мексика, Идальго, Тласкала, южная половина Пуэбла, север Оахака);
Р. bullatus Osgood, 1904 (Мексика — западная часть центральных районов штата Веракрус);
скалистый хомячок—Р. nasutus J. Allen, 1891 (штаты США — крайний юго-восток Юты, запад Аризоны, Колорадо, кроме северо-запада и северо-востока; Нью-Мексико, кроме юго-востока; северо-запад и запад Техаса; штаты Мексики — Чиуауа, северная половина Коауила, крайний север Дуранго);
Р. difficilis J. Allen, 1891 (центральные штаты Мексики — крайний юго-запад Чиуауа, Дуранго, кроме севера, южная треть Коауила, южная половина Нуэво-Леон, крайний юго-запад Тамаулипас, Закатекас, Сан-Луис-Потоси, Агуаскальентес, северо-восток Халиско, Гуанохуато, кроме юго-запада; Керетаро, Идальго, Мексике, восточная половина Пуэбла, Тласкала, север и центр Оахака);
Р. melanophrys Coues, 1874 (центральные и южные штаты Мексики — юго-восток Дуранго, юг Коауила, Закатекас, Агуаскальентес, северо-восточная половина Халиско, Гуанохуато, Сан-Луис-Потоси, кроме юго-востока, западная половина Идальго, Керетаро, северо-восточная половина Мичоакан, Мексике, Морелос, Тласкала, южная половина Пуэбла, северо-восточная половина Герреро, Оахака, кроме узких полос на юге и севере, юго-запад и центр Чиапас);
Р. mekisturus Merriam, 1898 (Мексика — восток штата Пуэбла и запад центра Веракрус);
тонкохвостый хомячок — Р. lepturus Merriam, 1898 (Мексика — центр штата Оахака);
гребнехвостый хомячок—Р. lophurus Osgood, 1904 (Мексика — центр и юго-восток штата Чиапас; юго-запад Гватемалы);
гондурасский хомячок — Р. hondurensis Goodwin, 1941 (юго-запад * Гондураса);
Р. simulatus Osgood, 1904 (Мексика — запад центральных районов ML.- штата Веракрус);
JT гватемальский хомячок — Р. guatemalensis Merriam, 1898 (Мекси-| ка — юго-восток штата Чиапас, юг Гватемалы, юго-запад и центр f Гондураса);
1 голоногий хомячок — Р. nudipes J. Allen, 1891 (Коста-Рика и край-( ний запад Панамы);
4 Р. altilaneus Osgood, 1904 (Гватемала);
J черноватый хомячок —Л furvus J. Allen et Chapman, 1897 (Мекси-I ка —север штата Пуэбла и запад центральных районов Веракрус);
Ж широкомордый хомячок — Р. latirostris Dalquest, 1950 (Мексика — W крайний юго-восток штата Сан-Луис-Потоси);
j краснобрюхий хомячок — Р. ochraventer Baker, 1951 (Мексика — 1 юг птата Тамаулипас);
мексиканский хомячок — Р. mexicanus Saussure, 1860 (штаты Мек-1 ыки — восточная треть Сан-Луис-Потоси, восток Пуэбла, северо-восток
Идальго, Веракрус, южная половина и крайний север Чиапас, вос-1 ток Оахака, юг Табаско; южная половина Гватемалы; север Сальвадоре ра, южная половина Гондураса; узкая полоса, проходящая с ссверо-за-пада на юго-восток через центр Коста-Рики);
I Р. allophylus Osgood, 1904 (Мексика — юг штата Чиапас);
| Р. banderanus J. Allen, 1897 (штаты Мексики — юго-запад Наярит, |Ж западная половина Халиска, Колима, Мичоакан, Герреро, юг Оахака);
юкатанский хомячок — Р. yacatanicus J. Allen et Chapman, 1897 (юг Мексики, штаты-—центр и север Кампече, Юкатан, север Киитано-в.Роо);
-V' хомячок Стиртона — Р. stirtoni Dickey, 1928 (юг Гондураса и восток Сальвадора);
fl бурый хомячок — Р. megalops Merriam, 1898 (штаты Мексики — 1 центральные районы Оахака и восток Герреро);
fl Р. sloeops Goodwin, 1955 (Мексика — крайний восток штата 1 Оахака);
fl черноногий, хомячок — Р. melanocarpus Osgood, 1904 (Мексика — fl центр штата Оахака);
fl Р. zarhynchus Merriam, 1898 (Мексика — центр и север штата Чиа-fl пас):
fl большой хомячок — Р. grandis Goodwin, 1932 (центр Гватемалы);
fl хомячок Томаса — Р. thomasi Merriam, 1898 (Мексика — центр шта-1 та Герреро);
г fl хомячок Нельсона — Р. nelsoni Merriam, 1898 (Мексика — запад s fl центральных районов штата Веракрус);
fl желтый хомячок — Р. flavidus Bangs, 1902 (крайний запад Па-а Анамы);
д пирриский хомячок — Р. pirrensis Goldman, 1912 (гора Пирри на i- fl крайнем востоке Панамы);
fl флоридский хомячок — Р. floridanus Chapman, 1889 (США — п-ов fl Флорида, кроме его юго-западной части);
fl По Андерсону и Джонсу (1967), в роде 57 видов. После ревизии чис-— fl доводов в роде, вероятно, будет значительно сокращено.
Род золотистых хомячков
Genus Ochrotomys Osgood, 1909
В роде единственный вид: золотистый хомячок — О. nuttalli Harlan, 1832.
Размеры мелкие. Длина тела 8—9,5 см. Длина хвоста 70—95 см. Внешним обликом похож на мышь (рис. 52). Волосяной покров густой и мягкий. Спина обычно золотисто-бурая, брюхо и конечности белые с оранжеватым оттенком.
Распространен в юго-восточных и южных штатах США: юг и юго-восток Миссури, крайний юг Иллинойс, Кентукки, кроме северо-запада, юг Виргинии, Северная Каролина, Южная Каролина, Джорджия,
Рис. 52. Золотистый хомячок — О nutalli и его череп
северная половина Флориды, Алабама, Миссисипи, Арканзас, Луизиана, крайний юго-восток Оклахомы, крайний восток Техаса.
Обитает главным образом в густых кустарниках. Ведет в основном ревесный образ жизни. Однако в западной части ареала, по крайней мере в Восточном Техасе, ведет наземный образ жизни. На деревьях и кустах строит гнездоподобные сооружения. Одно и то же гнездо может потребляться для нескольких поколений. Питается в основном семена-!И растений. В южных частях ареала размножение происходит, по-видимому, в холодные месяцы, начиная с сентября, в других областях— с марта по октябрь. В ряде мест размножается круглый год. Длительность беременности примерно 30 дней. В помете обычно 2— 3 детеныша.
Некоторые исследователи (например, Холл и Келсон, 1959) включают золотистого хомячка в качестве подрода в род Peromyscus.
Род карликовых хомячков
Genus Baiomys True, 1894
Размеры мелкие. Длина тела 5—8 см. Длина хвоста 3,5—5,5 см. По внешнему виду несколько напоминают мышей (рис. 53). Сходны с представителями рода белоногих хомячков, отличаясь от них меньшими размерами, относительно более короткими закругленными ушами, крупным венечным отростком нижней челюсти и некоторыми другими признаками. Хвост обычно короче % длины тела. Стопа не расширена. Подошва передней лапы с 6 мозолями. Окраска спинной стороны тела
черновато-
коричневая
или рыжевато-коричневая,
а брюшной
темпо-се-
рая, белая или грязно-желтая.
В черепе межглазничные сужения выражены слабее, чем у белоногих хомячков. Лицевой отдел черепа довольно короткий, носовые кости
по отношению к резцам не выступают вперед. Резцовые отверстия почта достигают зубных рядов. Венечный отросток нижней челюсти не уменьшен. ’ Верхние резцы крупные. Третий коренной зуб сильно реду-
цирован.
Распространены в США — штаты Аризона и Техас, Мексике, Гватемале, Сальвадоре, Гондурасе и Никарагуа.
Обычно населяют густые заросли травы, где делают многочисленные ходы. Активны преимущественно ночью. Питаются в основном зелеными частями растений, но поедают также семена и различных насекомых. Гнездо делают из сухой травы — во впадинах в земле или под упавшим стволом дерева. Размножаются круглый год. Длительность беременности примерно 20 дней. В помете от 1 до 5, обычно 3 детеныша. Масса детенышей примерно 1 г. В воспитании потомства принимают участие оба родителя (не типично для большинства грызунов). По-ловозрелость наступает примерно в 10-недельиом возрасте.
В роде 2 вида:
северный карликовый хомячок — В. taylori Thomas, 1887 (штаты США —крайний юго-восток Аризоны, юг и юго-восток Техаса; штаты 'Мексики — юг и юго-восток Соноры, западная половина Чиуауа, западные две трети Дурапго, Синалоа, Наярит, юго-запад Закатекас, Агуаскальентес, Халиско, кроме запада, восток Колима, север Мичоакан, южные две трети Гуанохуато, Керетаро, северная половина Мсксико, Идальго, север Пуэбла, север Веракрус, Тласкала, юго-восток Сан-Луис-Потоси, Тамаулипас, Нуэво-Леон, кроме юго-запада, северо-восток Коауила);
Рис. 53. Северный карликовый хомячок В taylori и его череп
южный карликовый хомячок — В. musculus Merriam, 1892 (южные штаты Мексики — центр юга Халиско, Колима, Мичоакан, кроме крайнего севера, Герреро, юг Мексике, Морелос, юг Пуэбла, Оахака, центр Веракрус, Чиапас, кроме севера; центр Гватемалы от западной государственной границы до восточной; север Сальвадора; юг Гондураса; центральная часть северо-западных районов Никарагуа).
Род кузнечиковых хомячков
Genus Onychomys Baird, 1857
Размеры мелкие. Длина гола 9 —16 см. Длина хвоста обычно 3 6 см. Масса 41—52 г. Внешне похожи па хомячков (рис. 54). Утолщенный хвост обычно меньше длины гола. Уши довольно крупные, округлые, с редкими волосами. Хвост обычно покрыт довольно густыми волосами. Подошвы лап покрыты волосами. Когти па передних лапах довольно длинные. Задние лапы несколько укорочены, первый палец на них редуцирован. Волосяной покров густой, высокий и нежный. Спинная сторона тела сероватая, коричневатая или розовато-коричие-вая, брюшная — беловатая. Хвост сверху имеет такую же окраску как и спина; кончик и низ хвоста белые. Сосков 3 пары.
В
Рис. 54. Северный кузнечиковый хомячок — О. leucogaster и его череп
черепе хорошо выражено межглазиичпое
сужение. Лицевой отдел
откий. Костные слуховые барабаны небольшие. Венечный отросток щей челюсти крупный. Резцовые отверстия большие, достигающие >зого коренного зуба.
Распространены в Западной части Северной сады на юг до Северной Мексики.
Америки or Южной
Населяют полупустыни. Обычно предпочитаю г места, покрытые ку-ipmiKOM. Активны во ночам в течение круглого года. Инг.ткиси в ос-
том насекомыми, в частности саранчовыми, их личинками, червями, также мелкими грызунами, иногда поедают семена растений. Растирная пища составляет у северного кузпечпкового хомячка 11% от
цкормов. Ведут почти исключительно наземный образ жизпн, но мо-• довольно хорошо лазать по деревьям. Гнездо сооружают в порах, ню используют норы других грызунов. Размножаются круглый год,
но пики размножения приходятся на весну и лето. В году у самки (северный кузнечиковый хомячок) бывает, по-видимому, 2 помета. Длительность беременности примерно 33 дня. В помете от 2 до б, обычно 3—4 детеныша массой 1,7—2,7 г каждый. В возрасте 15—20 дней открываются глаза. Половозрелость наступает в 3-месячном возрасте.
В роде 2 вида:
северный кузнечиковый хомячок — О. leucogaster Wied — Neuwied, 1841 (запад Северной Америки; провинции Канады — юг Альберта, южная половина Саскачевана, юго-запад Манитобы; штаты США — восточные две трети Монтаны, Вайоминг, кроме северо-запада, Северная Дакота, Южная Дакота, Небраска, кроме юго-востока, западная половина Миннесоты, северо-запад Иовы, Канзас, кроме крайнего востока, Колорадо, кроме центральных районов, Юта, кроме северо-востока, юг и юго-запад Айдахо, юго-восток Вашингтона, восточная половина Орегона, крайний северо-восток Калифорнии, северные две трети Невады, Аризона, кроме юго-запада, Нью-Мексико, Техас, кроме востока и юго-запада, западная половина Оклахомы; штаты Мексики — крайний северо-восток Соноры, крайний север Чиуауа).
южный кузнечиковый хомячок — О. torridus Coues, 1874 (юго-запад США, штаты — южная половина Калифорнии, юг Невады, Аризона, кроме северо-востока, юг и юго-запад Ныо-Мексико, юго-запад Техаса; север Мексики, штаты — крайний север Нижней Калифорнии, Сонора, крайний северо-восток Синалоа, Чиуауа, западная половина и крайний юго-восток Коауила, Дуранго, кроме юго-запада, Закатекас, кроме севера и юго-запада, юг Нуэво-Леон, крайний юго-запад Тамаулипас, Сан-Луис-Потоси, кроме юго-востока, Агуаскальентес).
Род южноамериканских полевых хомячков
Genus Akodon Meyen, 1833
Размеры мелкие. Длина тела 7,5—14 см. Длина хвоста 5—10 см. Внешним обликом сходны с хомяками (рис. 55). Плотного телосложения с короткими конечностями, относительно коротким хвостом. Уши крупные, слабо оволосненные. Хвост покрыт густыми волосами. Волосяной покров на туловище густой и мягкий. Окраска его варьириует на спине от серой до темно-коричневой, на брюхе от белой до темно-серой, иногда с желтоватым оттенком. У некоторых видов по бокам головы расположены белые отметины. Сосков 4 пары.
Межглазничная область черепа довольно широкая. Надглазничные гребни небольшие или их нет вовсе. Лицевой отдел черепа относительно укорочен. Резцовые отверстия достигают, как правило, последнего коренного зуба. Костные слуховые барабаны небольшие или средних размеров. Венечный отросток нижней челюсти средней величины или низкий. Третий верхний коренной зуб уменьшен в размерах.
Хромосом в диплоидном наборе 18 у /1. urichi, 34 у A. arviculoides, A. obscuras и A. azarae, 40 у Л. varius, A. bolivicnsis 42 43 у Л. moliiiae.
Распространены в Южной Америке - Венесуэле, Колумбии, Перу, Эквадоре, Бразилии, Боливии, Аргентине, Чили, Уругвае, Парагвае, Патагонии.
Населяют различные биотопы от относительно засушливых мест и лугов до влажных лесов на высоте от 1000 до 5000 м над уровнем моря. Некоторые виды активны только днем, другие — круглосуточно. A. arviculoides в Юго-Восточной Бразилии размножается с августа по
Рис. 55. Akodon olivaceus
j । г. Самка приносит от 3 до 7 детенышей, обычно 3—-4. Вероятно, Каждая самка имеет 2 помета в году.
В роде, по-видимому, 38 видов:
1НДИЙСКИЙ полевой хомячок — A. andinus Philippi, 1858 (центр и се-вгр Чили, Аргентина, юг Перу);
4. arviculoides Wagner, 1842 (центральная, центрально-западная и центрально-восточная Бразилия, Парагвай);
A affinis J. Allen, 1912 (Колумбия, горный хребет, Западная Кормьера);
A azarae Fischer, 1829 (восток и северо-запад Аргентины, крайний с- Боливии);
боливийский хомячок—A boliviensis Меуеп, 1833 (юг и юго-запад Uepv, северо-запад Аргентины, Анды в Боливии);
A caenosus Thomas, 1920 (горы северо-запада Аргентины, крайний or Боливии);
A. chacoensis Shamel, 1931 (север Аргентины);
A. dolores Thomas, 1916 (центр Аргентины);
A. iniscatus Thomas, 1919 (Аргентина);
A. mollis Thomas, 1894 (Эквадор, юг Колумбии, запад Перу, запад Боливии);
.4. obscuras Waterhouse, 1837 (восток Аргентины, Парагвай, юг Ботвин. Уругвай);
A olivaceus Waterhouse, 1837 (Аргентина, Чили и о-в Моча);
A. orophilus Osgood, 1913 (север и юго-восток Перу);
4. pacificus Thomas, 1902 (запад Боливии);
A. puer Thomas, 1902 (запад Боливии, юг центрального Перу);
.4. serrensis Thomas, 1902 (восток и юго восток Бразилии);
A. surdus Thomas, 1917 (Анды юго-востока Перу);
A. tapirapoanus J. Allen, 1916 (Центральная Боливия, Бразилия);
4. urichi J. Allen et Chapman, 1897 (Эквадор, Перу, Боливия, север, . о-восток и запад Венесуэлы, горные хребты Центральная и Восточная Кордильеры в Колумбии),
A. varius Thomas, 1902 (северо-запад Аргентины, центр и крайний юг • ливни, Парагвай);
A xanthorhinus Waterhouse, 1837 (восточная Аргентина, крайний юг М;
A. kempi Thomas, 1941 (о-ва в дельте Параны на границе Аргентины и Уругвая);
A. budini Thomas, 1918 (северо-восток Аргентины);
A. lasiotis Lund, 1841 (Восточная Бразилия);
черноватый полевой хомячок — A. nigrita Lichtenstein, 1829 (восток и крайний юг Бразилии);
белобрюхий полевой хомячок — A. albiventer Thomas, 1897 (крайний северо-запад Аргентины, крайний юг Боливии);
A. amoenus Thomas, 1900 (юго-восток Перу);
A. berlepschii Thomas, 1898 (запад Боливии, крайний север Чили, юго-восток Перу);
A. lactens Thomas, 1918 (северо-восток и северо-запад Аргентины, горный хребет Восточная Кордильера);
A. illuteus Thomas, 1925 (северо-запад Аргентины);
A. lanosus Thomas, 1897 (юг Аргентины, крайний восток Чили);
A. longipilis Waterhouse, 1837 (Чили, Аргентина, Огненная Земля, о-в Моча);
хомячок Санборна — A. sanborni Osgood, 1943 (Чили, Аргентина и о-в Чилоэ около Чили);
боготскин хомячок—A. bogotensis Thomas, 1895 (Восточная и Центральная Кордильеры в Колумбии, Венесуэла);
A. iheringi Thomas, 1896 (крайний юг Бразилии);
A. latebricola Anthony, 1924 (Анды в Эквадоре);
A. tnimus Thomas, 1901 (Аиды на юго-востоке Перу);
A. jelskii Thomas, 1894 (юг Боливии, центр, юго-восток, юг Перу, северо-запад Аргентины).
По Андерсону и Джонсу (1967) в роде 36 видов. Некоторые исследователи (например, Симпсон, 1945) выделяют A. affinis, A. bogotensis, A. iheringi, A. lanosus, A. latebricola в особый род Microxus Thomas, 1909.
Род камышовых хомячков
Genus Zygodontomys Allen, 1897
Размеры мелкие и средние. Длина тела 9,5—15,5 см. Длина хвоста 3,5—13 см. По внешнему виду несколько напоминают крыс. Уши относительно невелики. Хвост покрыт редкими волосами. Наружные пальцы на задних конечностях сильно уменьшены. Волосяной покров может быть густым, длинным и мягким или коротким и довольно редким. Спинная сторона тела желтовато-коричневая, рыжеватая или сероватая. Бока сероватые или желтоватые. Брюшная сторона серовато-белая или грязно-желтовато-серая. Уши и хвост коричневые. Сосков 4 пары.
В черепе, как правило, хорошо развита межтеменная кость. Надглазничные гребни обычно имеются, хотя развиты не особенно сильно. Костные слуховые барабаны небольшие. Резцовые отверстия широкие, достигают предкоренных зубов.
Распространены от западной части Коста-Рики на юг через Панаму до Перу и Восточной и Центральной Бразилии.
Населяют равнины, покрытые низким кустарником п густой травянистой растительностью. Обычно встречаются около водоемов от уровня моря до высоты примерно 600 м. По образу жизни напоминают серых полевок. Активны преимущественно ночью. В густой траве делают тропинки. Питаются семенами, зелеными частями растений, а также плодами. Сооружают гнезда из травы под землей в конце коротких нор. Длительность беременности примерно 28 дней. В помете от 2 до 8.
обычно 4 детенышей. К самостоятельной жизни детеныши переходят в возрасте 17—20 дней. Половозрелости достигают в 3—4-месячном возрасте. Камышовых хомяков используют в качестве лабораторных животных.
В роде 6 видов:
I. cherriei J. Allen, 1895 (крайний юг Коста-Рики и Панама);
сенмигельский хомячок— Z. seorsus Bangs, 1901 (остров Сан Мигель в Тихом океане у Панамы);
короткохвостый камышовый хомячок — Z. brevicauda J. Allen et Chapman, 1893 (Анды в Колумбии);
Z. lasiurus Lund, 1841 (восток и северо-восток Бразилии);
Z. microtinus Thomas, 1894 (Колумбия, Гайана, Суринам, восток и юг Венесуэлы, Бразилия);
Z. punctulatus Thomas, 1894 (северо-восток Колумбии, север Эквадора, Венесуэла).
По Андерсону и Джонсу (1967), в роде 2 вида.
Род рораимских хомячков
Genus Podoxymys Anthony, 1929
В роде единственный вид: рораимский хомячок Р. roraimae Anthony. 1929.
Размеры мелкие. Длина тела около 10 см. Длина хвоста 9,5 см. Хвост покрыт редкими волосами. Уши маленькие, почти полностью закрытые в волосяном покрове головы. Глаза маленькие. Когти на передних лапах длинные, сильно сжатые с боков, слегка искривленные. Когти на задних конечностях значительно короче. Волосяной покров высокий, на спинной стороне тела черноватый. Брюхо несколько светлее спины.
Череп с длинным и довольно тонким лицевым отделом.
Распространен на горе Рорайма в Гайане.
Обитает на склонах гор. Экология не изучена.
Род леноксусо в
Genus Lenoxus Thomas, 1909
В роде единственный вид: леноксус — L. apicalis Allen, 1900.
Размеры средние. Длина тела 15—17 см. Длина хвоста 15—19 см. Внешним видом напоминает крысу. Уши крупные. Когти не увеличены в размерах. Хвост покрыт редкими волосами. Окраска спинной стороны тела серовато-черная, по бокам она несколько светлее и серовато-бурая, иа брюхе серая или белая. Хвост коричневый, за исключением белой тер-лкпальной части.
В черепе межглазничное пространство широкое. Мозговой отдел ,’пшь незначительно шире липового. Носовые кости длинные. Межте-менная кость хорошо развита. Резцовые отверстия широки, почти достигают щечных зубов. Широкое костное небо продолжается до третьего коренного зуба. Костные слуховые барабаны маленькие. Ряды .зубоп смещены вперед.
Распространен на юго-востоке Перу и западе Боливии.
Обитает преимущественно в горных лесах на высоте 1800—2400 м вад уровнем моря. Экология не изучена.
Род копающих хомяков
Genus Oxymycterus Waterhouse, 1837
Размеры от мелких до средних. Длина тела 11,5,—17 см. Длина хвоста 7—14,5 см. Конец морды удлинен и подвижен. Хвост, как правило, покрыт редкими волосами. Когти на передних лапах крупные. Пятый палец на передних конечностях сильно уменьшен. Наружный палец на задних конечностях значительно более короткий, чем три средних пальца. Уши средних размеров. Волосяной покров, как правило, не густой. Спинная сторона тела рыжеватая, желтовато-коричневая, темно-коричневая или черноватая. Брюшная сторона грязно-желтая, серовато-коричневая или серовато-белая.
Череп длинный и узкий с незначительным межглазничным сужением. Мозговая капсула довольно крупных размеров. Лицевой отдел удлинен. Носовые кости выдвинуты вперед по отношению к верхним резцам. Надглазничных гребней нет. Костное небо продолжается до последнего коренного зуба. Резцовые отверстия очень широкие. Костные слуховые барабаны не увеличены в размерах. Нижняя челюсть с относительно низким венечным отростком. Верхние ряды щечных зубов узкие, смещены вперед. Верхний третий коренной зуб уменьшен в размерах.
Распространены в Восточной Бразилии, Уругвае, Северной Аргентине, Парагвае, Боливии и Перу.
Населяют болотистые места, равнины, поросшие кустарником, или леса. Экология изучена слабо. Ведут, по-видимому, подземный или по-луподземный образ жизни. Иногда встречаются в антропогенных ландшафтах в штате Рио-де-Жанейро в Бразилии. Период размножения у них приходится с августа по март. В течение года бывает по крайней мере 2 помета: один в октябре, другой в марте. Каждый раз самка приносит от 2 до 3, обычно 3 детенышей.
В роде, вероятно, 8 видов:
О. akodontius Thomas, 1921 (северо-запад Аргентины);
О. angularis Thomas, 1909 (восток Бразилии);
О. delator Thomas, 1903 (Парагвай);
О. hispidus Pictet, 1843 (восток и юго-восток Бразилии, северо-восток Аргентины) ;
О. inca Thomas, 1900 (запад Боливии, юго-восток и центр Перу);
О. paramensis Thomas, 1918 (крайний юг и северо-запад Боливии, север Аргентины, восток Перу);
О. roberti Thomas, 1901 (восток Бразилии);
О. rufus Desmarest, 1819 (восток и юг Бразилии, Уругвай, Аргентина, Парагвай);
По Андерсону и Джонсу (1967), в роде 9 видов.
Род землеройковых хомячков
Genus Blarinomys Thomas, 1896
В роде единственный вид: зсмлеройковый хомячок A. breuiccps Winge, 1888.
Размеры мелкие. Длина тела примерно 9,2 см. Длина хвоста около 4,5 см. Внешний вид типичен для подземного млекопитающего (рис. 56). Тело вальковатое. Конечности укорочены, голова крупная, коническая, глаза сильно редуцированы. Уши короткие, почти полностью скрытые г, волосяном покрове. Шейный перехват выражен слабо. Короткий хвост почти не имеет волос. Передние конечности с четырьмя функционирую-
Рис. 56. Землеройковый хомячок — В breviceps
щпми пальцами, пятый палец сильно уменьшен. Когти на пальцах развиты хорошо. Стопа широкая, с сильными, длинными когтями. Волостной покров густой и короткий. Окраска одноцветная темно-серая. Волосы с буроватыми вершинками. Конечности сверху коричневые.
Г Череп почти не имеет межглазничного сужения. Межтеменной кости пгт. Костное небо продолжается примерно до третьего коренного зуба. Подглазничные отверстия очень крупные. Костные слуховые барабаны передних размеров. Третий коренной зуб сильно уменьшен в размерах.
Распространен в Восточной Бразилии.
Экология почти не изучена. Обитает в холмистой местности в гус-, тых дождевых лесах, примерно на высоте 800 м над уровнем моря. Ведет подземный образ жизни. Размножающиеся самки были отмечены в [сентябре и январе.
Род кротовых хомячков
Genus Notiomys Thomas, 1890
Размеры мелкие. Длина тела 7,5—14 см. Длина хвоста 2,5—6 см. [Внешний облик зверька характерен для подземного жителя. Телосло-|Жение плотное, голова коническая, крупная, но короткая. Уши небольшие. Глаза маленькие. Конечности короткие, их подошвы голые, когти [мощные, острые и изогнутые. Пятый палец кисти сильно уменьшен в I чзмерах, но имеет коготь. Хвост относительно короткий, довольно гус-I гп покрытый небольшими волосами.
I Череп со слабо выраженным межглазпичиым сужением, широкими I юбными костями. Лицевой отдел укорочен. Мозговая капсула широкая, I <оуглая. Венечный отросток нижней челюсти относительно низкий. (Ецовые отверстия заходят между зубными рядами. Межтеменная Кутк обычно средних размеров. Третий верхний коренной зуб может I быть уменьшен в размерах.
| Распространены па большей части Аргентины и Чили.
( Населяют различные ландшафты от влажных лесов до сухих равен. Поднимаются и в горы. Большую часть жизни проводят иод зем-I лея. Экология почти нс изучена.
В роде 6 видов:
i N.angustus Thomas, 1941 (запад Аргентины);
N, delfini Cabrera, 1905 (крайний юг Южной Америки);
I 173
N edwardsi Thomas, 1890 (крайний юг Аргентины);
N. macronyx Thomas, 1894 (Аргентина, Чили);
N. megalonyx Waterhouse, 1844 (Чили);
N. valdivianus Philippi, 1858 (Чили, Аргентина, о-ва Чилоэ и Моча).
Род водяных хомяков
Genus Scapteromys Waterhouse, 1837
Размеры средние. Длина тела 16,5—22,5 см. Длина хвоста 13,5— 17 см. Внешним обликом несколько напоминают черную крысу (рис. 57). Хвост снизу довольно густо покрыт длинными волосами, образующими своеобразный киль. Конечности удлинены. На задних лапах три средних пальца удлинены, боковые пальцы укорочены; все они вооружены довольно длинными, сильными когтями. Когти на передних лапах также большие. Уши относительно небольшие. Волосяной покров густой и мягкий. Окраска спинной стороны тела серовато-черная, брюшной серовато-белая. Сосков 4 пары.
В удлиненном черепе межглазничное пространство почти не имеет сужения Гребни на нем развиты стабо. Межтеменная кость средних размеров Костное небо продолжается назад дальше рядов щечных зубов. Резцовые отверстия достигают рядов зубов.
Распространены в Северной Аргентине, Уругвае и Бразилии.
Обитают на болотах, в устьях рек или лесах. Некоторые виды ведут полуводиын образ жизни. Экология изучена слабо
В роде 5 видов
водяной хомяк — 5. aquaticus Thomas, 1920 (Аргентина);
чакосский хомяк — S. chacoensis Gyldenstolpe, 1932 (север Аргентины) ;
Рис. 57 Scapteromys tumidus
матугросский хомяк — S. gnambiquarae Ribeiro, 1914 (Мату-Гросу 5 Бразилии);
S. tomentosus Lichtenstein, 1829 (Уругвай, восток Аргентины);
S. tumidus Waterhouse, 1837 (Уругвай, восток Аргентины).
Род бурых хомячков
Genus Scotinomys Thomas, 1913
Размеры мелкие. Длина тела 8—8,5 см. Длина хвоста 4,8—7 см.
Уши относительно небольших размеров. Стопа узкая. Хвост покрыт «редкими волосами. Волосяной покров низкий и грубый, но у некоторых видов длинный и мягкий. Окраска спинной стороны тела темно-желтова-n-коричневая или черновато-коричневая. Брюшная сторона рыжевато-чичневая или сероватая. Уши, конечности и хвост обычно черноватые. В черепе надглазничные гребни развиты слабо. Межтеменная кость •Крупная. Костное небо достигает уровня задних отделов зубных рядов. «Печные зубы гипселодонтные. Верхние коренные необычно узкие.
, Распространены в Центральной Америке от Южной Мексики до Западной Панамы.
Населяют в основном скалистые участки, но встречаются также и в саваннах, и в лесных районах на высоте от 1000 до 3000 м над уровнем [Ь4оря. Экология не изучена.
В В роде 4 вида:
саванный хомячок — S. harrisi Goodwin, 1945 (центральные районы Коста-Рики);
длинноволосый хомячок — S. longipilosus Goodwin 1945 (центральные районы Коста-Рики);
панамский бурый хомячок — S. xerampelinus Bangs, 1902 (крайний запад Панамы);
центральноамериканский хомячок — S. teguina Alston, 1877 (юг Мексики—центральные районы штата Чиапас, центр Гватемалы, запад и юго-запад Гондураса, север Сальвадора, запад Никарагуа, центральные районы Коста-Рики, крайний запад Панамы).
Род вечерних хомячков
Genus Calomys Waterhouse, 1837 ( = Hesperomys Waterhouse, 1839)
Размеры мелкие. Длина тела 6—12,5 см. Длина хвоста 3—9 см. Маста взрослого С. laucha 30—38 г. Внешним видом напоминают мышей. Уши довольно крупные. Стопа и кисть узкие. Пятый палец на задних конечностях обычно сильно уменьшен. Хвост покрыт довольно густыми волосами. Волосяной покров обычно не очень густой, но мягкий. Окрас-Яка спинной стороны тела грязно-желтая, красновато-коричневая, тсмно- коричневая или сероватая, брюшной — сероватая или беловатая. У пеко-торых видов позади ушей расположено по белому пятну. Хвост обычно Жадный сверху и более светлый снизу или целиком белый. Сосков 4, 5 вли7 пар.
1 В черепе надглазничные гребни развиты слабо или их ист вовсе, .j Костное небо продолжается дальше последних щечных зубов. Мсжте-кнная кость крупная. Резцовые отверстия проникают между передними Ценными зубами. Щечные зубы с низкими коронками.
V Распространены в Перу, Боливии, Парагвае, Уругвае, Юго-Восточной «газилйи и Центральной и Северной Аргентине.
Населяют различные биотопы: леса, заросли кустарников, опушки лесов, альпийские луга, встречаются в густых зарослях травы или среди камней. Активны в сумерки и ночью. С. laucha питается в основном растительными объектами и в меньшей степени насекомыми.
В роде, по-видимому, 10 видов:
С. callosus Rengger, 1830 (Центральная Боливия, Парагвай, Северная Аргентина);
С. dubius Fischer, 1829 (Уругвай, Аргентина);
С. expulsus Lund, 1841 (Центрально-Восточная Бразилия);
С. frida Thomas, 1917 (восток и центр Перу);
С. gracilipes Waterhouse, 1837 (Аргентина);
С. laucha Desmarest, 1819 (восток и северо-запад Аргентины);
С. lepidus Thomas, 1884 (Аргентина, Боливия, Перу);
С. muriculus Thomas, 1921 (юго-восток Боливии);
С. tener Winge, 1887 (Центрально-Восточная Бразилия);
С. venustus Thomas, 1916 (Аргентина, Боливия).
По Андерсону и Джонсу (1967), в роде 4 вида.
Род высокогорных хомячков
Genus EHgmodontia F. Cuvier, 1837
Размеры от мелких до средних. Длина тела 6,5—17 см. Длина хвоста 8—14 см. По внешнему виду сходны с мышами. Уши большие, округленные. Хвост довольно густо покрыт короткими волосами. Стопа узкая с длинным пятым пальцем. Подошвы кисти и стопы покрыты волосами. Волосяной покров на туловище густой и мягкий. Окраска спинной стороны тела светло-коричневато-желтая. Брюшная сторона обычно чисто белая. Молодые зверьки более темные, чем взрослые. Сосков 4 пары.
В черепе мозговая коробка широкая. Надглазничных гребней нет. Костные слуховые барабаны относительно крупные. Венечный отросток нижней челюсти сильно уменьшен в размерах.
Распространены в Южной Америке от юга Боливии и севера Чили на востоке вдоль Анд до Южной Аргентины.
Населяют разнообразные биотопы, но обычно встречаются на сухих каменистых равнинах, в зарослях кустарников и травы, а также на каменистых склонах гор на высоте до 4500 м над уровнем моря. Очень подвижные зверьки с ночной активностью. Собственных нор, по-видимому, не роют, а занимают норы других грызунов или прячутся на день под стволами упавших деревьев или в густых кустарниках. В связи с тем, что в зимнее время их не приходилось находить, предполагают, что на зиму они впадают в спячку. Питаются предположительно в основном насекомыми.
В роде, вероятно, 2 вида:
Е. puerulus Philippi, 1896 (север и северо-восток Чили, Аргентина, Боливия, юг Перу);
Е. typus F. Cuvier, 1837 (Аргентина).
По Андерсону и Джонсу (1967), в роде один вид.
Род красноносых хомячков
Genus Wiedomys Hershkovitz, 1959
В роде единственный вид: красноносый хомячок — IF. pyrrhorhitws Wied, 1821.
Размеры мелкие. Длина тела 10—12,5 см. Длина хвоста 16—20,5 см. Внешний облик мышеобразный. Уши средних размеров. Очень длинный
хвост покрыт редкими короткими волосками. Окраска спинной стороны тела представляет смесь грязно-желтых и коричневых тонов; брюшная сторона белая. Нос, круги вокруг глаз, уши, наружные части всех четырех конечностей и крестец ярко-рыжевато-оранжевые.
Распространен в Северо-Восточной, Центральной, Восточной и Юго-Восточной Бразилии, а также, возможно, в Парагвае.
Населяет густые леса с кустарниковым подлеском. Держится преимущественно группами. Хорошо лазает по деревьям. Убежищем обычно служат гнезда птиц. Иногда строит собственные гнезда из сухих листьев и травы в дуплах деревьев или в кустарниках.
Период размножения в Северо-Восточной Бразилии, как правило, приходится на август. В помете от одного до шести, обычно пять детенышей.
Род рато-до-мато
Genus Pseudoryzomys Hershkovitz, 1959
В роде единственный вид: рато-до-мато — Р. wavrini Thomas, 1921. Размеры мелкие. Длина тела 10—12 см. Длина хвоста примерно 10,5 см. Внешний вид мышеобразный. Хвост довольно хорошо оволоснен. S Уши большие.
Окраска спинной стороны тела грязно-желто-коричневая, брюшной — Ж беловатая. Хвост коричневый сверху и белый снизу.
К Распространен в Парагвае и на севере Аргентины, fl Населяет влажные болотистые места. Экология не изучена.
I Род листоухих хомячков
Genus Phyllotis Waterhouse, 1837
Размеры от мелких до средних. Длина тела 7—15 см. Длина хвоста lfl.4,5—16,5 см. По внешнему облику несколько напоминают пищух, однако ’вотличие от последних имеют довольно длинный хвост (рис. 58). Уши «очень крупные, обычно достигают длины 20—30 мм. Пальцы нормаль-W ные. Пятый палец на задней конечности не уменьшен в размерах. По-I дошва стопы голая.
1 Волосяной покров довольно высокий, густой и мягкий. Спинная сто-рона тела сероватая, коричневатая, желтоватая, оранжевая или грязно-fl желтая. Брюшная сторона сероватая, беловатая или грязно-желтая. Сос-W ков 4 пары.
Д В черепе характерны длинные и широкие носовые кости. Межтемен-flhiafl кость крупная. Костные слуховые барабаны нс увеличены в размс-ЧРрах. Межглазничное сужение выражено хорошо. Надглазничные гребни | развиты слабо или хорошо.
« Хромосом в диплоидном наборе от 28 у Р. subfimis и Р. piclits, 38 у В.р, amicus, Р. darwini, Р. magister, Р. gerbillus и Р. haggardi, до 54 у W P.definitus, 64 у Р. andium и 70 у Р. osilae.
-Я Распространены в Южной Америке от Эквадора па юг вдоль Анд и s Тихоокеанского побережья до равнин Патагонии и Магелланова про-а лива.
I. ' Населяют обычно каменистые участки с бедной растительностью — и I от уровня моря до 5000 м над его уровнем. В лесах встречаются редко.
Рис. 58. Боливийский хомячок — Р. boliviensis
Иногда попадаются в человеческих постройках. Обычно занимают норы, выкопанные другими грызунами. Поедают семена, зеленые части растений и лишайники. Некоторые виды активны ночью, другие днем. Р. sub-limis, по-видимому, держится небольшими группами. В дождливый сезон редко появляются на поверхности земли. Возможно, впадают в летнюю спячку.
В высокогорье период размножения приходится на конец и начало года, т. е. на теплый влажный сезон, когда растительность развивается наиболее хорошо. Самка приносит от 3 до 4 детенышей.
В роде, по-видимому, 14 видов:
Р. amicus Thomas, 1900 (север и северо-запад Перу);
андийский листоухий хомячок — Р. andium Thomas, 1912 (центральные районы Эквадора и Перу);
Р. caprinus Pearson, 1958 (северо-запад Аргентины и прилегающие районы Боливии);
хомячок Дарвина — Р. darwini Waterhouse, 1837 (Чили, Перу, Боливия, Аргентина);
хомячок Хаггарда — Р. haggardi Thomas, 1908 (Лиды в Эквадоре);
Р. magister Thomas, 1912 (запад и юго-запад Перу);
Р. osilae J. Allen, 1901 (северо-запад Аргентины, юг центральных районов, юго-запад и запад Перу; Боливия);
Р. wolfsohni Thomas, 1902 (запад Боливии);
боливийский хомячок — Р. boliviensis Waterhouse, 1846 (юго-запад Боливии; Чили; юг Перу);
Р. pictus Thomas, 1884 (Анды центра Перу и запада Боливии);
Р. sublimis Thomas, 1900 (крайний северо-запад Аргентины и прилегающие районы Боливии и юг Чили; юг Перу);
Р. micropus Waterhouse, 1837 (юг Аргентины, юг Чили);
Р. domorum Thomas, 1902 (юго-запад Боливии, север и северо-запад Аргентины);
серожелтый хомячок — Р. griseoflavus Waterhouse, 1837 (восток, запад и крайний северо-запад Аргентины, крайний юг Боливии; Парагвай).
По Андерсону и Джонсу (1967), в роде 11 видов.
Рис. 59 Шиншилловый хомяк — С. sahamae
Род чилийских хомяков
Genus Irenomys Thomas, 1919
Вроде единственный вид: чилийский хомяк — /. tarsalis Philippi, |uiO.
Размеры мелкие. Длина тела 12,5—14 см. Длина хвоста 16—19 см. ли крупные. Длинный хвост покрыт довольно густыми волосами, обра-ющнми на его кончике небольшую кисточку. Стопа длинная и узкая. Вмосяной покров очень густой и мягкий. Спинная сторона тела серова-го-рыжая или серовато-светло-коричневая, брюшная светло-коричневая грязно-желтая, с розоватым оттепком. Уши черного цвета. Передние
| адние лапы беловатые. Хвост темно-коричневый.
В черепе хорошо выражено межглазничное сужение. Мозговая кап-., ia широкая. Резцовые отверстия длинные. Верхние резцы с бороздкой наружной поверхности. Верхние щечные зубы резко гипселодонтно-j типа
। Распространен в Аргентине, Чили и на о-вах Чилоэ и Гуайтекас.
Населяет влажные леса в умеренном климате. Экология изучена сла-о Ведет в основном древесный образ жизни. Отмечаются сильные изменения численности по годам, по-видимому, в зависимости от кормовых Ьювий.
Род шиншилловых хомяков
Genus Chinchillula Thomas, 1898
В роде единственный вид: шиншилловый хомяк — С. sahamae Tho-Ls. 1898.
I Размеры средние. Длина тела 15 17 см. Длина хвоста 9,5 -11 см. I : ив очень большие, длинные п широкие (рис. 59). Стона расширена и I прочена, пальцы нормальные. Хвост хороню оволосие.н. Волосяной по-I шб туловища очень густой и мягкий, шелковистый. Окраска спинной Игюпоны тела грязно-желтоватая или сероватая с черными линиями. I иршиая сторона белоснежная.
I Б черепе надглазничные гребни развиты слабо или их нет вовсе. I „обные кости относительно широкие. Межтеменная кость крупная. Кост
ное небо продолжается до последних щечных зубов. Щечные зубы резко гипселодонтного типа.
Распространен на высокогорье крайнего северо-запада Аргентины, Северного Чили, на юге Перу и юго-западе Боливии.
Населяет скалистые участки гор на высоте 4000—4600 м над уровнем моря. Активность ночная. Питается различными частями растений. Размножение приходится на октябрь — ноябрь, т. е. на конец сухого сезона, так что молодые развиваются и растут во время вегетации растений. Местами шиншилловых хомяков промышляют из-за их теплого и красивого меха.
Род хомяков пуны
Genus Punomys Osgood, 1943
В роде единственный вид: хомяк пуны — Р. lemminus Osgood, 1943. Размеры средние. Длина тела 13—15,5 см. Длина хвоста 5—7 см. Внешне похож на полевку. Конечности укорочены. Уши относительно небольшие, немного выступающие над уровнем меха. Волосяной покров высокий, густой и мягкий. Спинная сторона тела грязно-желто-коричневая или серовато-коричневая. Брюхо беловатое или сероватое.
Верхние резцы утолщенные, с гладкой передней поверхностью, нижние отделены друг от друга значительным пространством. Жевательная поверхность щечных зубов имеет сложное строение.
Распространен в Юго-Восточном Перу.
Встречается в пуне — участке высокогорья выше границы леса, на высоте 4450—5200 м над уровнем моря. Активен в течение всего дня. Населяет скалистые участки. Питается зелеными частями растений: преимущественно кустарника Senecio adenophylloides и травы Werneria digitata. Делает запасы стеблей этих растений под камнями. Самки приносят детенышей в теплый дождливый сезон с ноября по апрель. У одной самки было найдено 2 эмбриона.
Род болотных хомяков
Genus Neotomys Thomas, 1894
В роде единственный вид: болотный хомяк — N. ebriosus Thomas, 1894.
Размеры средние. Длина тела 17,6—22,5 см. Длина хвоста 7—8,8 см. Внешний облик мышевидный. Волосяной покров густой. Характерны очень длинные остевые волосы. Окраска спинной стороны тела серовато-коричневая, брюшной — беловатая. Хвост коричневый сверху и серый или желтоватый снизу. Пос ярко-рыжего цвета. Сосков 4 пары.
В черепе межглазничпое сужение резко выражено. Надглазничные гребни небольшие. Межтеменная кость крупная. Носовые кости сильно выдвинуты вперед. Резцовые отверстия длинные. Костные слуховые барабаны средних размеров. Венечный отросток нижней челюсти сильно уменьшен в размерах. Резцы очень широкие. Верхние резцы с бороздкой по наружной поверхности.
Распространен в центре и на юге Перу, юге Боливии и в Аргентине.
Населяет покрытые травой участки вдоль берегов рек и болота в горах на высоте 3400—4500 м над уровнем моря. Активен круглосуточно. Экология изучена слабо.
Род кроликовых хомяков
Genus Reithrodon Waterhouse, 1837
В роде единственный вид: кроликовый хомяк — R. physodes Olfers, 1818.
Размеры от мелких до средних. Длина тела 13—20 см, длина хвоста 8,5—10,5 см. Морда укороченная. Глаза большие. Уши довольно крупные. Наружный палец задних конечностей сильно укорочен. Между средними пальцами задних конечностей имеется перепонка. Стопа удлиненная. Ее подошва частично покрыта волосами. Хвост покрыт короткими волосами. Волосяной покров густой и мягкий. Спинная сторона тела грязно-желтая, часто с примесью сероватых и черноватых волос. Брюшная сторона беловатая или сероватая, иногда с желтоватым оттенком. Сосков 4 пары.
В черепе мозговая коробка укорочена. Лицевой отдел крупный. Надглазничные гребни развиты слабо или их нет вовсе. Межтеменная кость развита хорошо. Межглазничное сужение сдвинуто назад. Костные слуховые барабаны средних размеров. Резцовые отверстия необычайно длинные, простирающиеся от резцов до середины первого коренного зуба-. Костное небо вытянуто дальше заднего края последних коренных згбов. Верхние резцы с бороздками на наружной поверхности.
Распространен в Чили, Аргентине, Уругвае и, вероятно, также в Бра-; 1ЛИИ.
Экология изучена слабо. Населяет пампасы, включая и поля. Активен круглосуточно. Питается различными частями растений. Размножение не приурочено к какому-либо сезону. В помете от 2 до 8, обычно 3— 4 детеныша.
Род патагонских шиншилловых хомяков
Genus Euneomys Coues, 1874
I.
м )-й
ie
к> а-
10 эй
ге. о-ю.
Размеры от мелких до средних. Длина тела 10—22,5 см. Длина хвос--.1 6—10 см. Тело приземистое. Голова относительно короткая. Уши довольно крупные, голые. Хвост покрыт небольшими волосами. Когти относительно короткие. Волосяной покров необычайно густой и мягкий. Окраска его на спинной стороне тела рыжевато-коричневая или коричневатая, на брюшной грязно-желтая или сероватая. Передние и задние лапы обычно белые. Хвост коричневатый сверху и беловатый снизу. Сосков 4 пары.
В черепе мозговая капсула относительно короткая. Лицевой отдел массивный. Лобные кости сильно сжаты. Резцовые отверстия заходят между передними коренными зубами. Костное небо расширено к.задп больше, чем спереди. В задней части неба располагаются два углубления. Костные слуховые барабаны некрупные. Верхние резцы широкие, на наружной поверхности имеют продольную бороздку. Щечные зубы массивные.
Распространены в Чили и Аргентине от ее средних частей па юг до мыса Горн.
Населяют заросли кустарников и лесистые области. Экология почти ае изучена.
В роде 4 вида:
патагонский хомяк — Е. chinchitloides Waterhouse, 1839 (Чили, Огненная Земля и соседние с ней о-ва);
хомяк Петерсона — Е. petersoni J. Alien, 1903 (южные районы Аргентины и Чили);
Е. fossor Thomas, 1899 (северо-запад Аргентины);
Е. mordax Thomas, 1912 (запад Аргентины).
Некоторые ученые (например, Симпсон, 1945) выделяют Е fossor в отдельный род Chelemyscus Thomas, 1925, что, по-видимому, неправильно.
Род перепончатолапых хомяков
Genus Holochilus Brandt, 1835
Размеры мелкие и средние. Длина тела 13—22 см. Длина хвоста 13— 23 см. По внешнему облику напоминают крыс (рис. 60). Длинный хвост слабо оволоснеи. У некоторых видов по его нижнему краю удлиненные волосы образуют своеобразный киль. Между пальцами лап обычно имеется плавательная перепонка. Стона значительно крупнее кисти. На задних лапах три средних пальца значительно длиннее двух наружных. Когти на них сильные и удлиненные. Волосяной покров густой и мягкий. Окраска спинной стороны тела темпо-желтая, орапжеватая или рыжевато-коричневая с примесью черного цвета; бока светлее спины. Окраска брюшной стороны изменяется от белой до оранжевой; горло и пах всегда белые.
В черепе надглазничные гребни хорошо развиты. Имеется межтеменная кость. Лобные кости сужены. Костные слуховые барабаны средних размеров. Костное небо продолжается дальше уровня последних коренных зубов. Резцовые отверстия крупные, но уровня щечных зубов не достигают. Венечпый отросток нижней челюсти высокий. Резцы гладкие, относительно широкие. Третий верхний коренной зуб крупный.
Распространены в северо-восточной, центральной и южной частях Южной Америки.
Населяют болотистые места, поросшие густой травой берега водоемов, различные влажные участки, не покрытые лесом от уровня моря до, примерно, 2000 м над его уровнем. Ведут полуводнын образ жизни. На берегу или в водоеме в тростниках делают гнезда диаметром до
1 s
I 40 см, обычно на уровне 20—40 см над водой. Гнезда строят из стеблей & тростника и веточек. Зверьки хорошо плавают: если потревожить зверь-к ка, находящегося в гнезде, он выскакивает оттуда, прыгает в воду и W уплывает. Питаются различными болотными растениями, а также мол-I люсками. В некоторые годы происходит сильное увеличение численнос-j . ти перепончатолапых хомяков, и они начинают вредить сельскохозяй-> ственным культурам. Активность преимущественно ночная, но иногда их fl можно встретить и днем. Самки приносят детенышей обычно в сентябре. 1 В помете от 4 до 6, обычно 5 детенышей (по другим данным, от 5 до 10). 1 В роде 2 вида:
1- бразильский перепончатолапый хомяк — Н. brasiliensis Desmarest, fl 1819 (восток и северо-восток Перу, Боливия, юг и восток Бразилии, се-I веро-запад Аргентины, Гайана, Уругвай, Парагвай, восток Венесуэлы); ( большой перепончатолапый хомяк — Н. magnus Hershkovitz, 1955 fl Уругвай, Бразилия, Аргентина).
i Род хлопковых хомяков
Genus Sigmodon Say et Ord, 1825
Размеры от мелких до средних. Длина тела 12,5—20 см. Длина хвос-1 d 7,5—12,5 см. Внешне несколько напоминают крыс (рис. 61). Телосло- жение тяжелое. Уши относительно небольшие. Хвост средне или слабо } уволоснен. На кисти 6 подошвенных мозолей. На задних конечностях 1 ;i средних пальца значительно длиннее двух наружных. В большинстве fl случаев волосяной покров низкий и грубый. Его окраска на спинной сто-fl роне тела серовато-коричневая или черновато-коричневая, с примесью Я желтоватых волос, на брюшной сероватая или грязно-желтая. Хвост nep-fl иоватый сверху и светлый снизу.
$ Череп с массивным лицевым отделом, с сильно развитыми надглаз-¥ яичными гребнями. Межглазничное пространство широкое. Костные fl слуховые барабаны средних размеров. Костное небо широкое, прости-fl -дающееся несколько дальше третьего коренного зуба. Резцовые отвер-I ствия достигают щечных зубов. Венечные отростки на нижней челюсти 1 хорошо развиты. Нижний второй и третий коренные зубы имеют на же-fl нательной поверхности складку, напоминающую по форме букву S. fl Нижние резцы широкие и гладкие.
Й Хромосом в диплоидном наборе 22, 24. У колючего хлопкового хомяка а 'подвид arizonae) 28, 30 у желтобрюхого, 29, 30 у малого, 52 у желто-fl восого, белоухого хлопковых хомяков и хлопкового хомяка Аллена.
fl Распространены на обширной территории от южных районов США И на юг через Мексику и Центральную Америку до тропических областей Я северной части Южной Америки.
’ J Населяют заросли травы и кустарников. Встречаются как во влаж- пых, так и в сухих местах. Активны днем и ночью в течение круглого года. Ш Индивидуальный участок очень небольшой, радиусом примерно не бо-Я лее 30 м. По характеру питания всеядные: питаются различными рпте-Ж пнями, мелкими позвоночными животными и насекомыми. Размпожают-Я ся круглый год. Самка приносит детенышей в гнезде, которое делает Ж обычно под стволом упавшего дерева. Длительность беременности при-Ж мерно 27 дней. Число детенышей в помете от 2 до 12, обычно 5 или 6. Ж Через неделю после рождения молодые животные покидают гнездо. Поло-Ж возрелости достигают в 6-недельном возрасте. Хлопковые хомяки — иаи-W более обычные грызуны юго-восточной части Соединенных Штатов, а
1 также Мексики и Центральной Америки. Их численность подвержена
ЙГ.
значительным изменениям. В природе живут обычно менее года. Хлопковые хомяки используются в качестве лабораторных животных.
В роде, по-видимому, 14 видов:
колючий хлопковый хомяк — S. hispidus Say et Ord, 1825 (южные штаты США — крайний юго-восток Виргинии, Северная Каролина, кроме восточных районов, примыкающих к побережью Атлантического океана, Южная Каролина, кроме северо-востока, Джорджия, Флорида, Алабама, Теннесси, кроме севера, Миссисипи, Луизиана, Арканзас, юг и юго-запад Миссури, Канзас, кроме северо-запада, крайний юго-восток Вайоминга, Оклахома, Техас, кроме крайнего северо-запада, юго-восток Нью-Мексико, юго-восток, центр и юго-запад Аризоны, крайний юго-восток Калифорнии; вся Мексика, кроме северо-запада штата Соноры и Нижней Калифорнии; небольшой участок ареала проходит но границе между штатами Нижняя Калифорния и Сонора с севера па юг от государственной границы до Калифорнийского залива; вся Центральная Америка; северо-восток Колумбии, Анды в Эквадоре; север Венесуэлы; северо-запад Перу: о-в Пупа; Холл и Келсон, 1959, насчитывают в пределах ареала 33 подвида);
желтобрюхий хлопковый хомяк — S. julviventer J. Allen, 1889 (Мексика— центральные районы штата Закатекас и Дуранго);
хомяк Аллена — S. alleni Bailey, 1902 (Мексика — южная половина штата Наярит и северо-запад штата Халиско);
хомяк Мак Дугалла — S. macdougalli Goodwin, 1955 (Мексика — восточная часть штата Оахака);
вулкановый хлопковый хомяк — S. vulcani J. Allen, 1906 (Мексика — юг штата Халиско);
геррерский хомяк — S. guerrerensis Nelson et Goldman, 1933 (Мексика—центр штата Герреро);
черноухий хлопковый хомяк — S. melanotis Bailey, 1902 (Мексика — юг штата Халиско и северо-запад штата Мичоакан);
; малый хлопковый хомяк — S. minimus Mearns, 1894 (штаты США — юго-запад Нью-Мексико и юго-восток Аризоны; штаты Мексики — северо-восток Соноры, север и запад Чиуауа, центральный район севера Дуранго);
желтоносый хомяк — S. ochrognathus Bailey, 1902 (штаты США — । крайний юго-восток Аризоны, крайний юго-запад Нью-Мексико, юго-I запад Техаса; штаты Мексики — крайний северо-восток Соноры, Чиуауа, кроме юга, и крайний северо-запад Коауила);
। белоухий хомяк — S. leucotis Bailey, 1902 (штаты Мексики — запад i Дуранго и юг Закатекас);
горный хлопковый хомяк — S. alticola Bailey, 1902 (штаты Мекси-I ки—север Оахака, юго-запад Пуэбла, центр Мексике, запад Идальго, I север Керетаро);
.< большезубый хомяк — S. macrodon Goodwin, 1955 (Мексика — юго-I восток штата Оахака);
4 оахакский хлопковый хомяк — S. planifrons Nelson et Goodwin, 1933 (Мексика —юг и центр штата Оахака);
Ж хлопковый хомяк Алстона — S. alstoni Thomas, 1880 (север и восток ' Венесуэлы и Гайана).
К По Уокеру (1964), в роде 2 или 3 вида. По Андерсону и Джонсу (1967), 13. Симпсон (1945) и Эллермэн (1940) выделяют S. alstoni, S, savannarum Thomas, 1901, и S. venester Thomas, 1914, в особый род I Sigmomys Thomas, 1901. Такое выделение, видимо, неправильно, а два И последних вида следует считать синонимами первого.
| Род андийских хомяков
| Genus Andinomys Thomas, 1902
В роде единственный вид: андийский хомяк — A. edax Thomas, 1902.
Размеры от мелких до средних. Длина тела 13,5—16,5 см. Длина хвоста 11—15 см. Тело приземистое. Внешне несколько напоминает крысу. Хвост иногда частично голый. Окраска волосяного покрова на спинной стороне тела темно-желтая, на брюшной — серая. Сосков 4 пары.
В черепе хорошо выражено межглазничнос сужение. Мозговая капсула небольших размеров. Лицевой отдел массивный. Резцовые отверстия очень длинные. Носовые кости широкие, выдвинутые более вперед, чем резцы. Резцы широкие. Расстояние между зубными рядами слегка увеличивается в направлении спереди назад. Щечные зубы резко гипссло-донтного типа.
Распространен на юге Перу, в Боливии и Северо-Западной Аргентине.
Обитает в горах на высоте 1600—4800 м над уровнем моря. Населяет . каменистые берега рек; держится также в зарослях кустарников. Ведет ночной образ жизни. Устраивает гнезда на ветвях кустов или деревьев. Питается в основном вегетативными частями растений. Размножение приходится на конец сухого сезона.
Рис. 62. Вулкаиовый хомячок — N. alstoni и его череп
Род вулкановых хомячков
Genus Neotomodon Merriam, 1898
В роде единственный вид: вулкановый хомячок — N. alstoni Merriam, 1898 (рис. 62).
Размеры мелкие. Длина тела 10—13 см, длина хвоста 8—10,5 см. Масса взрослых особей 40—60 г. Внешним видом напоминает крысу. Уши средних размеров, почти голые. Подошва кисти с 6 мозолями. Длинный хвост относительно густо покрыт волосами. Волосяной покров густой и мягкий. Спинная сторона тела имеет сероватую или серовато
желтую окраску, а брюшная — беловатую. Хвост двухцветный — темный сверху и белый снизу. Сосков 3 пары.
В черепе костное небо оканчивается сразу позади последнего коренного зуба. Резцовые отверстия заходят между рядами щечных зубов. Костные слуховые барабаны небольшие. Межтеменная кость хорошо развита. Щечные зубы крупные, массивные, с корнями и плоскими коронками.
Хромосом в диплоидном наборе 48.
Распространен в Центральной Мексике: полоса, проходящая по центральным районам штата Мичоакан с запада на восток, южная половина Мексике, Морелос, юг Тласкала, север Пуэбла, запад центральных районов Веракрус.
Населяет травянистые участки лесов па склонах вулканических гор на высоте 2600—4300 м над уровнем моря, до верхней границы лесов. Ведет ночной образ жизни. День проводит в выкопанной им норе. Нора короткая, просто устроенная, расположена поверхностно, примерно 50 мм в диаметре. Иногда занимает норы других грызунов. Период размножения с начала июня по сентябрь. В течение года бывает 2 или больше пометов, в каждом из которых от 2 до 5 детенышей.
Род лесных хомяков
Genus Neotoma Say et Ord, 1825
Размеры средние. Длина тела 15—23 см. Длина хвоста 7,5—24 см. Масса тела 200—430 г. По внешнему виду несколько напоминают крысу (рис. 63). У одной группы видов хвост покрыт довольно густыми волосами, у другой —очень редкими. Пятый палец па задних конечностях относительно длинный. Волосяной покров мягкий, иногда грубый. Окраска его на спинной стороне тела варьирует от бледной и темпо-желтой до темно-серой, а на брюшной — от белой, бледно-серой или грязно-желтой > до темно-желтой. Иногда окраска меха рыжеватая. Самцы пушистохвостого хомяка имеют среднебрюшную кожную железу. Сосков 2 пары.
Череп с резко выраженным межглазничным сужением. Надглазничные гребни хорошо выражены. Лицевая часть черепа заострена. Межтеменная кость относительно крупная. Резцовые отверстия широкие, длинные, достигают рядов щечных зубов. Костные слуховые барабаны крупные. Костное небо оканчивается на уровне передней части третьего коренного зуба. Венечный отросток нижней челюсти высокий. Щечные зубы гипселодонтного типа с уплощенной коронкой.
Хромосом в диплоидном наборе 38 у сонорского, 52 у белогорлого, пустынного и мексиканского, 54 у пушистохвостого и 56 у темноногого лесных хомяков.
Распространены в западной части Северной Америки от северных районов Британской Колумбии на юг до Никарагуа и Гватемалы и на восток до Южной Дакоты, Миссури и Алабамы. Изолированный участок ареала имеется в Восточной Флориде, восточной части штата Джорджия и самой южной части штата Южная Каролина.
Населяют весьма разнообразные ландшафты. Встречаются от равнинных пустынь до влажных джунглей и каменистых склонов выше границы леса. Ведут в основном ночной образ жизни. Убежищем служат гнезда, которые лесные хомяки сооружают из веток, иногда из кактусов. Такое гнездо может достигать 2 м в диаметре. Сооружается оно чаще ! всего на земле и помещается около крупного камня, у основания дерева или куста. Иногда гнезда строят на деревьях на высоте до 6 м над зем-
Рис. 63. Пушистохвостый хомяк — W. cinerea (вверху), флоридский лесной хомяк — N flo-ridana (внизу) и череп флоридского лесного хомяка
left. Лесные хомяки, живущие на побережье, иногда устраивают гнезда на мангровых деревьях в полсотне метров от берега, но ведут они только наземный образ жизни. В случае тревоги зверек стучит задней лапкой чо земле. Питаются различными частями растений, в том числе семена-чи, а также некоторыми беспозвоночными. Иногда могут причинять вред сельскохозяйственным культурам. Обычно пьют много воды, но в сухой сезон могут обходиться без питья, употребляя в пищу части растений, содержащие большое количество влаги. Длительность беременности 27—37 дней. В помете 1—6 детенышей. В северных частях ареала в течение года бывает лишь один помет, а в южных, по-видимому, более одного, причем размножение может проходить в течение круглого года, тогда как на севере оно обычно приходится на весну (в лабораторных условиях самка приносит за год до 7 пометов). Масса новорожденных от 12
до 17 г. На 14—15 день у них открываются глаза. Половозрелость наступает в конце первого года. В неволе лесные хомяки жили до 3—4 лет.
В роде 22 вида:
флоридский лесной хомяк — N. floridana Ord, 1818 (юго-восток США —восточные половины Техаса и Оклахомы, Канзас, кроме юго-запада, центр, центральные районы востока и северо-восток Колорадо, полоса, проходящая от южной границы штата Небраска через ее центральные районы к северной границе, небольшой участок на юго-западе Южной Дакоты, южная половина Миссури, крайний юг Иллинойса, юг
Е и юго-восток Индианы, юг Кентукки, Теннесси, Арканзас, Луизиана, Миссисипи, Алабама, Джорджия, кроме участка на северо-востоке, Флорида, кроме южной трети, южная половина и крайний северо-запад Южной Каролины, крайние юг и запад и северо-запад Северной Каролины, западная половина Виргинии, запад Мэриленда, Пенсильвания, север Нью-Джерси, крайний юго-восток Нью-Йорка);
южный лесной хомяк — N. micropys Baird, 1855 (юг США, штаты — крайний юго-восток Колорадо, юго-запад Канзаса, западная половина Оклахомы, Нью-Мексико, кроме крайнего запада и северо-запада, западная половина Техаса; северо-восток Мексики: штаты — восточная половина Коауила, Нуэво-Леон, кроме крайнего юга, Тамаулипас, кроме крайнего юго-запада, крайний север Веракрус, восточная треть Сап-
I Лунс-Потоси);
белогорлый лесной хомяк — N. albigula Hartley, 1894 (юго-запад США, штаты — юго-восток Юты, юго-запад и юго-восток Колорадо, запад Техаса, Нью-Мексико, Аризона, кроме северо-запада, крайний юго-I восток Калифорнии; штаты Мексики — крайний северо-восток Нижней I Калифорнии, Сонора, Чиуауа, Коауила, кроме северо-востока, западная I половина Нуэво-Леон, крайний юго-запад Тамаулипас, восточные две I трети Дуранго, Закатекас, Агуаскельентес, северо-восток Халиско, Гуа-I нохуато, кроме юго-запада, Керетаро, кроме северо-востока, крайний 1 север Мексики);
{ широколобый хомяк — N. latifrons Merriam, 1894 (Мексика — севе-1 по-восток штата Мичоакан);
I ’ лесной хомяк Нельсона — N. nelsoni Goldman, 1905 (Мексика — запад центральных районов штата Веракрус);
N.palatina Goldman, 1905 (Мексика — северная часть штата Халиско, граничащая со штатами Наярит, Дуранго и Закатекас);
М varia Burt, 1932 (о-в Туриер в Калифорнийском заливе);
хомяк Монтесумы — N. montezumae Goldman, 1905 (Мексика — запад штата Идальго);
пустынный хомяк — N. lepida Thomas, 1893 (юго-запад США: штаты — юго-восток Орегона, юго-запад Айдахо, Невада, Юта, кроме се-
веро-востока и юга-востока, крайний северо-запад Колорадо, западная половина Аризоны; южная половина Калифорнии; Мексика — крайний северо-запад штата Сонора, Нижняя Калифорния и большое число мелких островов, расположенных в Тихом океане и Калифорнийском заливе по соседству с п-овом Калифорния; Холл и Кел сон, 1959, выделяют в пределах ареала 30 подвидов);
хомяк Брианта — N. bryanti Merriam, 1887 (о-в Ведрос в Калифорнийском заливе);
хомяк Антони — N. anthonyi J. Allen, 1898 (о-в Тодос-Сантос в Калифорнийском заливе);
санмартинский хомяк — N. martinensis Goldman, 1905 (о-в Сан-Мартин в Калифорнийском заливе);
хомяк Бункера — N. bunkeri Burt, 1932 (о-в Коронадо в Калифорнийском заливе);
хомяк Стефенса — N. stephensi Goldman, 1903 (штаты США — центр и юго-восток Аризоны, крайний юго-восток Юты, северо-запад Нью-Мексико) ;
хомяк Голдмана — N. goldmani Merriam, 1903 (северо-запад и центр Мексики — крайний юго-восток Чиуауа, юго-запад Коауила, северо-восток Дуранго, восточная половина Закатекас, северо-запад Сан-Луис-Потоси) ;
мексиканский лесной хомяк — N. mexicana Baird, 1855 (юго-западные штаты США — юго-восточный угол Юты, юго-запад, юго-восток и центр Колорадо, Нью-Мексико, кроме юго-востока, восточная половина Аризоны, крайний юго-запад Техаса; большая часть Мексики, штаты — восточная половина Соноры, Чиуауа, Коауила, кроме северо-востока, юго-запад Нуэво-Леон, Сан-Луис-Потоси, Закатекас, Дуранго, Синалоа, кроме северо-запада, Наярит, Халиско, кроме прибрежной полосы, Гуанохуато, Керетаро, Идальго, Мексике, кроме юга, Морелос, Пуэбла, кроме севера, западная часть центральных районов Веракрус, восточная половина Герреро, Оахака, кроме северо-востока, Чиапас, кроме севера, северо-востока и юга; узкая полоса, пересекающая центр Гватемалы с запада на восток, центральная часть западных районов Гондураса на границе с Гватемалой; Холл и Келсон, 1959, выделяют в пределах ареала 25 подвидов);
никарагуасский хомяк — N. chrysomelas J. Allen, 1908 (центр и восток Никарагуа, запад Гондураса);
тамаулипасский хомяк — N. angustapalata Baker, 1951 (Мексика — юг штата Тамаулипас);
темноногий хомяк — N. fuscipes Baird, 1858 (штаты США —запад Орегона, причем только на самом юго-западе штата ареал выходит к Тихому океану, западная половина Калифорнии; Мексика — северо-запад Калифорнийского п-ова);
пушистохвостый хомяк — N. cinerea Ord, 1815 (провинции Канады — юго-восток Юкона, Британская Колумбия, кроме северо-запада и северо-востока, юг и юго-запад Альберта; штаты США — Вашингтон, Айдахо, Монтана, кроме крайнего северо-востока, юго-запад Северной и западная половина Южной Дакоты, запад Небраски, Орегон, север и северо-восток Калифорнии, Невада, кроме юга и юго-запада, Юта, северо-восток Аризоны, северо-запад Нью-Мексико, западная половина Колорадо, Вайоминг) ;
лесной хомяк Аллена — N. alleni Merriam, 1892 (штаты Мексики — крайний юго-запад Соноры, запад Наярита, юго-запад Халиско, Коли
ма, южная половина Мичоакан, юг Мексико, Морелос, южная половина Пуэбла, крайний северо-запад Оахака, Герреро, кроме юго-востока);
сонорский лесной хомяк — N. phenax Merriam, 1903 (Мексика — крайний юго-запад штата Соноры).
Род Нельсония
Genus Nelsonia Merriam, 1897
В роде единственный вид: нельсония — N. neotomodon Merriam, 1897.
Размеры мелкие. Длина тела 12—13 см. Длина хвоста 10,5—13 см. Внешним обликом несколько напоминает крысу. Уши довольно крупные, длинный хвост хорошо оволоснен и на конце имеет удлиненный пучок волос. :
Волосяной покров мягкий. На спинной стороне тела он сероватобурого цвета, а иногда темно-желтоватого. Брюхо беловатое. Конечности белые или темные.
Череп несколько уплощен. Межглазничное пространство широкое. Резцовые отверстия не достигают рядов щечных зубов. Костные слуховые барабаны довольно крупные. Резцы узкие.
Распространена в западной части Центральной Мексики — на юге штата Дуранго, юво-востоке Закатекас, крайнем севере Халиско, Агуаскальентес, северо-западе Мичоакан.
Встречается в скалах горных склонов с хвойной растительностью на высоте 1800—3000 м над уровнем моря. Ведет в основном ночной образ жизни. Питается, по-видимому, хвоей. Экология изучена слабо.
Род ксеномис
Genus Xenomys Merriam, 1892
Вроде единственный вид: ксеномис — X. nelsoni Merriam, 1892.
Размеры средние. Длина тела 15,7—16,5 см. Длина хвоста 14,3— 17 см. Внешний облик крысовидный. Длинный хвост довольно густо покрыт волосами. Крупные уши почти полностью голые. Спинная сторона тела темно-красноватого или желтоватого цвета, брюшная — кремово-'елая.
Над каждым глазом располагается по белому пятну. Верхняя ггба и щеки белые примерно до уровня половины расстояния от носа до глаза.
В черепе характерна крупная межтеменная кость и большие сильно уплощенные костные слуховые барабаны. Щечные зубы крупные с уплощенными коронками.
Третий нижний коренной зуб имеет на жевательной поверхности складку в виде буквы S.
Распространен в штатах Мексики: крайний юг Синалоа, запад Наярит, запад Халиско и Колима.
Населяет заросли кустарников и тропические леса. Держится в прибрежных районах от уровня моря до, примерно, 500 м над его уровнем. Ведет древесный образ жизни. Убежищем служат дупла деревьев. Проворно лазает по деревьям. В августе находили беременных самок. Экология почти не изучена.
Рис. 64. Хомяк Штольцмаина — /. stolzmanni
Род рыбоядных хомяков
Genus Ichthyomys Thomas, 1893
Размеры средние. Длина тела 14,5—21 см. Длина хвоста 14,5—19 см. Внешне несколько напоминает крысу (рис. 64). Адаптированы к полу-водному образу жизни. Голова несколько уплощена. Уши небольшие, голые. Глаза маленькие. Характерны длинные, жесткие вибриссы. Хвост полностью покрыт волосами. На задней конечности все пальцы хорошо развиты. Стопа задней конечности большая, широкая, с хорошо выраженным гребнем удлиненных жестких волос по краю стопы Между пальцами задней конечности расположена неполная плавательная перепонка. Передние конечности небольшие, их подошва с 5 мозолями. По нижнему краю хвоста удлиненные волосы образуют своеобразный гребень. Волосяной покров густой, но жестковатый. Окраска спинной стороны тела темно-оливково-коричневая, сероватая или черноватая, брюшная сторона беловатая. Сосков 3 пары.
Череп уплощен. Мозговая капсула длинная и широкая. Межглазничные сужения резко выражены. Межтеменная кость слабо заметна или умеренно развита. Подглазничные отверстия очень крупные. Носовые кости укорочены спереди. Костные слуховые барабаны маленькие. Костное небо, как правило, продолжается значительно дальше последних коренных зубов. Резцовые отверстия длинные, почти достигают первого коренного зуба. Верхний третий коренной зуб уменьшен. Нижние резцы узкие, верхние — широкие.
Распространены в Перу, Эквадоре, Колумбии, Венесуэле.
Населяют берега потоков и болота от 600 до 2600 м над уровнем моря. Питаются главным образом водными животными, в том числе и рыбой С этим, по-видимому, связано слабое развитие слепой кишки.
В роде 2 вида:
рыбоядный хомяк — /. hydrobates Winge, 1891 (Аиды Эквадора, запад Венесуэлы, Анды Колумбии);
хомяк Штольцмаина — /. stolzmanni Thomas, 1893 (Эквадор, Перу).
Род эквадорских рыбоядных хомячков
Genus Anotomys Thomas, 1906
Б роде единственный вид: эквадорский рыбоядный хомячок — A. leander Thomas, 1906.
Размеры мелкие. Длина тела примерно 12,8 см. Длина хвоста 12,5 см. Имеются приспособления к жизни в воде. Наружное ухо отсутствует. Ушная щель может смыкаться специальной мускулатурой во время ныряния животного. Стопа значительно крупнее кисти; все ее пальцы хорошо развиты. По краю стопы располагается гребешок жестких волос, увеличивающий гребущую поверхность при плавании. Волосяной покров очень густой и мягкий. Спинная сторона тела темно-серая, брюшная беловато-серая. Вокруг каждого ушного отверстия располагается по белому пятну.
Череп сильно уплощен. Межглазничное сужение резко выражено. Носовые кости по отношению к резцам слегка выдаются вперед. Нижние резцы узкие.
Распространен в Западном Эквадоре.
Населяет берега горных водоемов. Питается рыбами. Экология не изучена.
Род венесуэльских рыбоядных хомячков
Genus Daptomys Anthony, 1929
В роде единственный вид: венесуэльский рыбоядный хомячок — 0. venezuelae Anthony, 1929.
Размеры мелкие. Длина тела примерно 13 см. Длина хвоста около 10,5 см. Внешним обликом сходен с эквадорским рыбоядным хомячком, но адаптация к водному образу жизни выражена не так сильно. В частности, стопа не имеет столь крупных размеров. Волосяной покров содержит жесткие остевые волосы и очень мягкий и густой пух. Окраска [ спинной стороны тела черновато-бурая. Брюшная сторона сероватая. Хвост буровато-черный. Пальцы на передних конечностях беловатые, на задних буроватые.
Распространен в Северной Венесуэле.
Экология не изучена.
Род центральноамериканских рыбоядных хомяков
Genus Rheomys Thomas, 1906
Размеры от мелких до средних. Длина тела 10,5—18,8 см. Длина хвоста 9,5—15 см. По внешнему виду похожи па представителей рода рыбоядных хомячков (р. 65). Небольшие уши почти полностью скрыты в меху. На задних конечностях имеются неполные плавательные перепонки. По краю стопы проходит гребень жестких волос. Волосяной покров тела густой и мягкий. Окраска его па спинной стороне темно-бурая с примесью черных волос, на брюшной сероватая или беловатая.
Верхние резцы узкие, часто с продольной бороздкой по наружной поверхности. Нижние резцы сильно сжаты.
Распространены в Южной Мексике, Колумбии, Панаме, Коста-Рике.
Населяют мелкие речки и ручьи в джунглях. Питаются, по-видимому, водными животными — в основном моллюсками. Размножение не носит сезонного характера. Экология изучена слабо.
Рис. 65. Рыбоядный хомяк Томаса — У?, thorna-si и его череп
В роде 5 видов:
рыбоядный хомяк Томаса — R. thomasi Dickey, 1928 (Мексика — крайний юго-восток штата Чиапас; Сальвадор);
хомяк Ундервуда — R. underwoodi Thomas, 1906 (центральные районы и восток Коста-Рики и крайний запад Панамы);
хомяк Хартмаипа— R. hartmanni Enders, 1939 (крайний запад Панамы) ;
панамский рыбоядный хомяк - R. raptor Goldman, 1912 (крайний восток Панамы);
колумбийский рыбоядный хомяк — R. trichotis Thomas, 1897 (Колумбия).
Кабрера (1957—1961) считает R. underwoodi синонимом R. trichotis.
Genus Neusticomys Anthony, 1921
В роде единственный вид: горный рыбоядный хомячок —N. monticolis Anthony, 1921.
Размеры мелкие. Длина тела примерно 9,5 см. Длина хвоста около 11 см. Адаптация к водному образу жизни выражена слабее, чем у представителей рода рыбоядных хомяков. По внешнему виду представители этих двух родов сходны между собой. Характерен редуцированный первый палец на задней конечности. Ушная раковина небольших размеров. Стопа имеет по краю гребень жестких волос. Волосяной покров тела густой и мягкий. Окраска его на спинной стороне рыжевато-коричневая, на брюшной — сероватая.
Распространен в Западном Эквадоре.
Населяет берега небольших потоков. Питается, по-видимому, водными животными. Экология не изучена.
I Род мышевидных хомячков
I Genus Calomyscus Thomas, 1905
В В роде единственный вид: мышевидный хомячок — С. baiheardi Tho-Л mas, 1905.
- Размеры мелкие. Длина тела 7—8,5 см. Длина хвоста 7,5—10 см. По К внешнему виду похож на мышь (рис. 66). Уши крупные, округлые, почти голые, покрытые лишь редкими, короткими и топкими волосками. Задние К конечности значительно длиннее передних. Подошвы передних и задних лап голые. Наружный пятый палец задней конечности не укорочен. Хвост 1 довольно густо покрыт волосами и на конце имеет кисточку удлиненных 1 волос. Защечных мешков нет. Вибриссы очень длинные, обычно пре-I вышающие половину длины тела. Волосяной покров густой и мягкий. I Окраска спинной стороны тела желтовато-коричневая или серовато-ко-1 ричневая, брюшной стороны и конечностей — белая. Хвост темный сверху и белый снизу. Сосков 3 пары.
Череп с удлиненным лицевым отделом. Скуловые дуги слабо расстав-1 лены. Лобно-теменных гребней нет. Мозговая коробка несколько упло-1 щена сверху. Носовые кости выступают вперед над резцами. Резцовые I отверстия относительно короткие. Подглазничное отверстие широкое.
Костные слуховые барабаны небольшие. Венечный отросток нижней челюсти не укорочен. Третий верхний коренной зуб очень сильно уменьшен в размерах.
Хромосом в диплоидном наборе 32.
Распространен в центральных районах Передней Азии, Южной Туркмении (на Копетдаге, Больших Балхаиах и в Западном Бадхызе) и в Закавказье (на юге Нахичеванской АССР).
Населяет скалистые, засушливые горные районы, поднимаясь вверх до275Омнад уровнем моря. Держится преимущественно поодиночке плп небольшими группами. В летнее время активен ночью, а весной и осенью днем. Питается главным образом различными семенами и плодами, в меньшей степени листьями, побегами, а также и беспозвоночными жи-i вотными. Самка приносит от 3 до 5 детенышей с марта по июнь. Неко-5 торые самки имеют 2 помета в год. Детеныши рождаются голые и слепые. Глаза открываются на 13-й день после рождения. В возрасте 6—8 i месяцев они достигают размеров взрослых животных.
Рис. 66. Мышевидный хомячок — С. bailwardi и его череп
Рис. 67. Джунгарский хомячок — Р. sungarus и его череп
Род джунгарских хомячков
Genus Phodopus Miller, 1910
Размеры мелкие. Длина тела 5,3—10,2 см. Длина хвоста 0,7—1,1 см. Кю массивное, конечности короткие (рис. 67). Голова крупная, коротая. Шейный перехват выражен слабо. Уши небольшие, округлые, слабо эсрытые волосами. Глаза относительно крупные. Когти па передних лаги могут быть удлинены. На задних конечностях наружный палец не цорочен. Подошвы передних и задних лап покрыты волосами. Корот
кий хвост густо опушен волосами. Есть защечные мешки. Волосяной покров густой и мягкий. Спинная сторона тела сероватая, коричневатая или светло-палевая, брюшная — беловатая. Вдоль средней линии спины проходит темная полоска (иногда она отстутствует). Зимой мех светлеет, а в северных частях ареала полностью белеет. Сосков 4 пары.
Лицевой отдел черепа относительно короткий и высокий. Мозговая капсула не сужена. Скуловые дуги широко расставлены в стороны. Межглазничное сжатие выражено слабо. Лобно-теменные гребни не развиты. Костные слуховые барабаны несколько уплощены; их передние концы трубкообразно вытянуты. Костное небо широкое. Резцовые отверстия широкие и, как правило, не достигают уровня щечных зубов. Третий верхний коренной зуб сильно уменьшен в размерах.
Хромосом в диплоидном наборе 28 у джунгарского хомячка и 34 у хомячка Роборовского.
Распространены в МНР, восточной и западной части Северного Китая и на юге Западной Сибири (горная часть Алтая, Тувинская АССР, Забайкалье, Зайсанская котловина, Восточный Казахстан).
Обитают в степях, полупустынях и пустынях--на равнинах и в горах. В горы поднимаются на высоту до 3000 м над уровнем моря. Джунгарский хомячок выкапывает норы, достигающие глубины 1 м, слабо ветвящиеся, с несколькими отнорками и одной или двумя гнездовыми камерами. Питается различными растительными объектами (по большей части семенами). Кроме того, поедает много насекомых, их личинок и гусениц. Делает небольшие запасы пищи. Впадает в спячку, по периодически просыпается и бывает активен даже при низких температурах. Период размножения приходится на теплое время года. В течение года бывает до 3 пометов, в каждом из которых 4—8 детенышей. Характерны весьма значительные изменения численности в разные годы. Разводится в качестве лабораторного животного.
В роде 2 вида:
джунгарский хомячок — Р. sungorus Pallas, 1773 (СССР — Южное Забайкалье, южные и центральные районы Тувинской АССР, Минусинская степь, Чуйская и Агинская степи на Алтае, Восточный Казахстан и Западная Сибирь; МНР, Северный Китай);
хомячок Роборовского — Р. roborovskii Satunin, 1903 (СССР — южная часть Тувинской АССР; МНР, Северный Китай).
В СССР: оба вида.
Род серых хомячков i
Genus Cricetulus Milne-Edwards, 1867
Размеры от мелких до средних. Длина тела 8- 25 см. Длина хвоста 2,5—10,6 см. Телосложение типичное для хомяков, тяжелое, морда ту- I пая, конечности короткие (рис. 68). Уши относительно небольшие, I слегка выступающие из меха; густо покрыты топкими шелковистыми во- I лосами. Глаза средних размеров; относительно меньших, чем у джунгар- I ских хомячков. Подошвы лап голые, иногда покрытые волосами. Хвост I слабо опушен волосами или почти голый. Защечные мешки хорошо раз I виты. Волосяной покров густой, довольно длинный и мягкий. Окраска I его на спинной стороне тела обычно серая, иногда рыжеватая или темно- I желтая, на брюшной светло-серая или белая. Конечности и копчик хвое- I та белые. У даурского хомячка вдоль середины спины проходит темно-. I коричневая полоска. Сосков 4 пары. I
В черепе лицевой отдел несколько удлинен. Мозговая капсула не су-1
Рис. 68. Серый хомячок — С. migratorius и его череп
лева. Межглазничное сужение хорошо выражено. Скуловые дуги доставлены нешироко, особенно спереди. Лобно-теменных гребней обыч-юнет. Межтеменная кость широкая. Резцовые отверстия широкие, часто ютигают уровня щечных зубов. Костное небо простирается немного иыие назад уровня последних щечных зубон. Костные слуховые бара-
ины относительно крупных или средних размеров, нс уплощенные; их передний отдел не вытянут в трубку.
Хромосом в диплоидном наборе от 20 у барабипского, 24 у длиннорого и до 28—30 у крысовидного хомячка.
Распространение охватывает большую территорию от Южной и Воюй Европы, через Малую и Переднюю Азию до МНР и Китая. Сотого Приморья и Корейского п-ова включительно.
Населяют лесостепи, луговые степи, степи или полупустыни. В горы поднимаются до 4000 м над уровнем моря. Иногда заходят в леса. Один вид населяет заболоченные, поросшие кустарником долины рек. Часто селятся на окультуренных землях. Ведут преимущественно сумеречноночной образ жизни. Держатся, как правило, поодиночке. Активны круглый год. В зимнее время активность падает и в некоторых случаях может иметь место кратковременная спячка (например, у барабинского хомячка). День проводят в норах, которые устраивают довольно просто. Питаются главным образом семенами различных растений, но поедают также много насекомых. Запасают пищу, иногда в большом количестве. Например, серый хомячок запасает семян до 800 г. Размножаются, как правило, в течение всего теплого сезона. В году бывает до 3 пометов, в каждом из которых от 3 до 11 детенышей. Половозрелость наступает в возрасте 4—5 недель (барабинский хомячок). В различные годы может наблюдаться изменение численности. Иногда приносят вред сельскохозяйственным культурам, хотя этот вред относительно невелик. Могут служить промежуточными хозяевами паразитов, передающих возбудителей ряда трансмиссивных заболеваний. Некоторые виды используются в качестве лабораторных животных.
В роде 10 видов:
серый хомячок — С. migratorius Pallas, 1773 (от Восточных Балкан на западе до Алтая, западной части МНР, Северо-Восточного и Центрального Китая на востоке и от северных пределов лесостепи в СССР на севере ареала до Ирана и Афганистана, Малой Азии, Сирии, Палестины, Белуджистана и Кашмира на юге; в СССР северная граница ареала проходит примерно по линии Черновцы, Шепетовка, Житомир, Киев, Чернигов, Калуга, Рязань, Горький, Казань, Уфа, южная оконечность Урала, северное Приаралье, южный берег оз. Челкар-Тенгиз, север Бет-пак-Далы и Казахского нагорья, Семипалатинск, юго-запад Алтайского края);
длиннохвостый хомячок — С. longicaudatus Milne-Edwards, 1867 (в СССР — Тувинская автономная ССР, крайний юг Красноярского края, р. Джида в Забайкалье; МНР; Китай);
барабинский хомячок—С. barabensis Pallas, 1773 (СССР — Барабин-ская лесостепь, предалтайские степи, Тувинская котловина, Прибайкалье к северу до верховьев Лены, степи Забайкалья к северу до линии Сре-тенск—Чита—Улан-Удэ, Дальний Восток — среднее течение Амура и юг Приморья; МНР; Северный Китай);
китайский хомячок—С. griseus Milne-Edwards, 1867 (Сев. Китай);
забайкальский хомячок — С. pseudogriseus Orlov et Iskhakova, 1974 (СССР — Южное Забайкалье; восточная часть МНР);
тибетский хомячок — С. lama Вonhotc, 1917 (Тибет);
короткохвостый хомячок—С. alticola Thomas, 1917 (Тибет, Кашмир);
хомячок Эверсманна — С. eversmanni Brandt, 1859 (СССР — Заволжье, юго-восточные районы ТАССР, Южпый Урал, Волго-Уральское междуречье, Северный Казахстан; МНР; Северный Китай);
монгольский хомячок—С. curtains G. Allen, 1925 (МНР);
крысовидный хомячок — С. triton de Winton, 1899 (в СССР — юго-западная часть Приморского края; Корейский п-ов, Северо-Восточный и Центральный Китай).
В СССР: хомячки серый, барабинский, длиннохвостый, забайкальский, Эверсмаппа и крысовидпый хомяк.
Некоторые исследователи (например, Громов, 1963) выделяют хомячка Эверсманна и монгольского хомячка в отдельный род Allocrice-tulus Argyropulo, 1937.
Рис. 69 Обыкновенный хомяк — С. cricelus н его череп
Род обыкновенных хомяков Genus Cricelus Leske, 1779
Вроде единственный вид: обыкновенный хомяк—С. cricelus Linens 1758.
[ Размеры средине и крупные. Длина тела 24—34 см. Длина хвоста 3,5-5,8 см. Хвост толстый в основании, быстро утончается к концу, поган короткими и жесткими волосами. Глаза относительно небольшие. Тип средних размеров, округлые, покрыты тонкими темноватыми волос-нвш (рис. 69). Кисгь и стопа широкие. Пальцы с хорошо развитыми
когтями. Наружный палец задней конечности укорочен. Подошва ступни покрыта волосами только в задней части, в области пятки. Волосяной покров густой и мягкий. Окраска его на спине однотонная, охристо-бурая, на брюшной — черная. На боках тела спереди находятся по три больших светлых пятна, задние из которых разделены черной полосой. Позади ушей располагается по светлому пятну. Защечные мешки есть.
В черепе лицевой отдел укорочен и расширен. Лобно-теменные гребни хорошо развиты. Межтеменная кость небольших размеров. Костные слуховые барабаны маленькие, округлой формы. Лобные кости сильно сжаты. Скуловые кости очень короткие. Подглазничное отверстие в верхней своей части шире, чем в нижней. Резцовые отверстия большие, но обычно не достигают первого щечного зуба.
Хромосом в диплоидном наборе 22.
Распространен: в СССР (европейская часть от Ярославля, Горького и Кирова к югу до Крымских и Кавказских гор, Западная Сибирь к северу до южной границы тайги, на восток до Енисея, Северный и Восточный Казахстан); ГДР, ФРГ, Бельгии, Нидерландах, Северной Франции, Венгрии, Чехословакии, Румынии, Югославии.
Обычно населяет лесостепи, разнотравные и злаково-разнотравные степи. Проникает в лесную зону и в предгорья; в горы поднимается до нижней границы леса, а если лесов нет, то до горных лугов включительно. Поселяется и на окультуренных участках. Ведет сумеречный образ жизни. День проводит в норе, которую обычно делает глубокой и сложной, со многими входными отверстиями, гнездовой камерой и несколькими кладовыми. Питается различными частями растений, а также и животными. Осенью в основном поедает семена и клубни. На зиму делает большие запасы семян, иногда клубней •— до 10—16 кг. В течение года бывает 2—3 помета, в каждом из которых в среднем до 10 детенышей; наибольшее число детенышей в помете 20. Известны кормовые осенние миграции хомяков. Зимой активндсть резко снижается, обыкновенный хомяк впадает в зимний сон, который часто прерывается. Может приносить существенный вред посевам. Шкурка используется в качестве пушного сырья. Природный носитель возбудителя туляремии и одной из форм клещевых сыпнотифозных лихорадок. Продолжительность жизни около 2 лет.
Некоторые исследователи (Б. С. Виноградов и А. И. Аргиропуло. 1941; Б. А. Кузнецов, 1965) объединяют обыкновенных хомяков со средними хомяками в один род — Cricetus.
Род средних хомяков
. Genus Mesocricetus Nehring, 1898
Размеры средние и крупные. Длина тела 15—28 см. Длина хвоста примерно 2—4,2 см. По внешнему виду сходны с обыкновенным хомяком (рис. 70), отличаясь бблсе мелкими размерами, менее крупным когтем на первом пальце передней конечности, который иногда может совсем отсутствовать, и тем, что хвост короче и густо покрыт волосами. Волосяной покров густой и мягкий. Окраска спинной стороны тела буровато-коричневато-охристая или рыжевато-коричневая, брюшной — белая или кремовая. На груди или посреди брюха бывает черное пятно. На боках тела в передней части выделяется светлое шейное пятно, ограниченное спереди и сверху широкой черной полосой.
Рис. 70. Переднеазиатский хомяк — Л1. auratus и его череп
’креп похож на таковой обыкновенного хомяка.
Хромосом в диплоидном наборе 42 или 44 у предкавказского и 44 у
кднеазиатского хомяка
Распространены в Юго-Западной Европе, па Кавказе и в Закавказье, ведней и Малой Азии.
Населяют предгорные
и горные степи.
Поднимаются в юры па вы-
, до 2300—3000 м над уровнем моря. Часто поселяются вблизи перепевов. Активны преимущественно в сумерках п ночью, но в всссп-
летний периоды
— и
днем.
Живут в норах, которые делают слож-
глубокимп с несколькими выходами, гнездовой камерой, иногда /агающейся па глубине до 2 м под землей. Веспой и летом в пище ладают вегетативные части растений, осенью — семена, а также глоды. Делают запасы на зиму, иногда более 16 кг. В теплое вреда приносят 2, иногда 3 и даже 4 помета, в каждом из которых от 20 (по некоторым данным, даже до 24), в среднем около 12 дете-
нышей. На холодное время года впадают в спячку. В некоторых местах могут приносить существенный вред посевам. Природные носители возбудителя туляремии. Шкурки могут служить пушным сырьем. Используются в качестве лабораторных животных.
В роде 4 вида:
предкавказский хомяк — М. raddei Nehring, 1894 (СССР — Предкавказье на север до низовьев Дона и Маныча; северные и восточные склоны и предгорья Главного Кавказского хребта);
переднеазиатский хомяк — М. auratus Waterhouse, 1839 (Сирия, Палестина) ;
закавказский хомяк — М. brandti Nehring, 1894 (СССР -—Закавказье и Предкавказье; Западный Иран; Турция);
хомяк Ньютона — М. newtoni Nehring, 1899 (Румыния, Болгария).
В СССР: предкавказский и закавказский хомяки.
Эллермэн и Моррисон-Скотт (1966) считают предкавказского хомяка подвидом переднеазиатского хомяка. Б. С. Виноградов и А. И. Арги-ропуло (1941), Б. А. Кузнецов (1963) включают средних хомяков — Mesocricetus в род обыкновенных хомяков — Cricetus.
Род белохвостых хомяков
Genus Mystromys Wagner, 1841
В роде единственный вид: белохвостый хомяк — М. albicaudaius Smith, 1834.
Размеры от мелких до средних. Длина тела 13,6—18,4 см. Длина хвоста 5—8,2 см. Внешним видом несколько напоминают крысу. Тело приземистое. Хвост покрыт короткими жесткими волосками. Уши относительно крупные, с закругленными вершинами. Когти короткие, но острые. Защечных мешков нет. Волосяной покров густой, высокий и мягкий. На спинной стороне тела его окраска темно-желто-серая или коричневая с черноватым оттенком, на брюшной — белая. Конечности и хвост беловатые. Уши темно-коричневые. Подошвы лап частично покрыты волосами. Сосков 2 пары.
Межглазничное сужение в черепе выражено хорошо. Оно сдвинуто назад, поэтому мозговая коробка кажется укороченной. Лобно-теменные гребни появляются у старых животных. Костное небо заходит чуть дальше уровня последнего коренного зуба. Резцовые отверстия достигают зубных рядов. Венечный отросток нижней челюсти развит хорошо. Костные слуховые барабаны умеренных размеров. Третий верхний коренной зуб очень мал. Резцы светло-желтые.
Распространен в Южной Африке.
Населяет травянистые равнины и сухие песчаные районы. Активен ночью. День проводит в выкопанной им норе. Иногда занимает поры других грызунов. Питается различными растительными объектами, главным образом семенами. Размножается, по-видимому, круглый год.Дли-тельность беременности примерно 37 диен. В помете бывает 4—5 детенышей.
М. longicaudatus Noack, 1887, из Танзании, который ряд авторов (например, Эллермэн, 1941) включают в род белохвостых хомяков, по-видимому, является Praomys natalensis.
подсемейство цокоров
SUBFAMILIA MYOSPALACINAE LILLJEBORG, 1866
В подсемействе один род цокоров.
Род цокоров
Genus Myospalax Laxmann, 1769
Размеры средние. Длина тела 15,5—25 см. Длина хвоста 2,9—6,9 см. Характерны приспособления к подземному образу жизни. Тело валько-ватое (рис. 71). Шейный перехват снаружи почти незаметен. Конечности короткие. Когти сильно развиты, особенно крупных размеров достигают когти третьего и четвертого пальцев передней конечности. На передней конечности третий палец, а на задней второй и третий длиннее других. Пятый палец обеих конечностей сильно укорочен. Подошвы лап голые. Наружной ушной раковины нет. Глаза очень маленькие. Нос покрыт толстым ороговевшим слоем кожи. Хвост сильно укорочен. Волосяной покров относительно высокий, густой и очень мягкий. Окраска спинной стороны тела варьирует от почти белой, серой, серовато-коричневой до рыжей и розовато-темно-желтой. Брюшная сторона тела свет-: лее спинной. У некоторых видов хвост сероватый сверху и белый снизу. ; В области затылка иногда имеется белое пятно. У китайского цокора 1 хвост весь белый и морда спереди имеет довольно крупное белое пятно.
Сосков 3 пары.
Череп с относительно удлиненным лицевым отделом. Затылочная область высокая, почти вертикально обрывается сзади с мощным затылочным гребнем. Хорошо развитые лобно-теменные гребни располага-I ются параллельно между собой. Костные слуховые барабаны крупные. Костное небо узкое, оканчивается на уровне передней части последнего-коренного зуба. Скуловые дуги слабые.
I Хромосом в диплоидном наборе у обыкновенного цокора 44.
I Распространены в Северо-Восточном Китае, МНР, в центральных I районах Южной Сибири.
I Населяют степи, лесостепи, предгорные степи, горные луга. Предпо-I читают мягкие почвы с богатой травянистой растительностью. Живут I под землей, в сложно устроенных норах длиной до 40 м и более и глу-I биной до 1 м. Зимовочные камеры могут располагаться до 1,5 и даже до I За под землей. Кроме того, каждый зверек имеет сложную систему по-I верхностных кормовых ходов. Землю из нор выбрасывают кучками па-I ружу. Зимой роют ходы под снегом. Питаются в основном различными | подземными частями растений: корнями, клубнями, луковицами, но так-I № и зелеными растениями. На зиму делают запасы пищи. По-видимо-1 му. имеют один помет в году, в котором 2—5 детенышей. Местами могут I приносить существенный вред сельскохозяйственным культурам, особеп-v но огородным.
1 В роде 2 вида:
1 обыкновенный цокор — М. myospalax Laxmann, 1773 (СССР — Ал-Я гай с прилегающими районами Западной Сибири, приобская часть Ба-« рабпнской и Кулундинской степей, Тарбагатай, Забайкалье, по р. Амур я Приморье; северо-восточная часть МНР, Корейский п-ов и Ссверо-1 Восточный Китай);
1 китайский цокор—М. fontanieri Milne-Edwards, 1867 (Китай от 1 й.Куку-Нор, Ганьсу, Сычуань, Шэньси и Шаньси до Хэбэй).
I ВСССР один вид: обыкновенный цокор.
Рис. 71. Обыкновенный цокор — М myospalax и его череп
Большинство исследователей (например, Эллермэн и Моррисон-Скотт, 1966) отделяют от обыкновенного покора особый вид — М. psilu-rus Milne-Edwards, 1874. Б. А. Кузнецов (1965), ссылаясь па исследования А. П. Кузякина и др., утверждает, что этих покоров следует считать одним видом.
Из М. jontanieri часто выделяют в особые виды: М. rotschildi Thomas, 1911 (Ганьсу и Хэбэй) и М. stnilhi Thomas, 1911 (Ганьсу).
ПОДСЕМЕЙСТВО МАДАГАСКАРСКИХ ХОМЯКОВ
SUBFAMILIA NESOMYINAE MAJOR, 1897
Размеры мелкие, средние или крупные. По внешнему виду сходные мышами, крысами или песчанками. Щечные зубы с корнями. Их жевательная поверхность может быть уплощенной, со сложным рисунком
вдающихся в зуб эмалевых складок. Щечные зубы могут быть не уплощены и иметь низкие бугорки (Nesomys) или могут быть образованы из пастин, плотно прилегающих друг к другу (Gymnuromys).
Род большеногих хомячков
Genus Macrotarsomys Milne-Edwards et Grandidier, 1898
Размеры мелкие. Длина тела 8—10 см. Длина хвоста 10—14,5 см. Внешним видом несколько напоминают песчанок (рис. 72). Голова с вытянутым лицевым отделом. Глаза довольно крупные. Уши большие, с округлыми вершинами. Задние конечности сильно удлинены. Три сред-। них пальца на них длиннее двух крайних; пятый палец длиннее, чем первый. Длинный хвост имеет на конце кисточку удлиненных волос. Спин-I ная сторона тела коричневатого цвета, а брюхо белое.
Костные слуховые барабаны в черепе сильно увеличены.
Распространены на о-ве Мадагаскар.
। Населяют леса и равнины, поросшие травянистой растительностью и кустарниками. Убежищем служат норы длиной до 1,5 м. В некоторые рзоны в одной норе живет 4 и больше взрослых зверьков. Питаются [различными зелеными частями растений, плодами, ягодами, семенами нкорнями. В помете от 2 до 3 детенышей.
! В роде 2 вида:
I М. bastard! Milne-Edwards et Grandidier, 1898 (западный Мадагас-ар);
JM. ingens Petter, 1959 (северо-западный Мадагаскар).
Род зайцегубых хомяков
Genus Nesomys Peters, 1870
Размеры средние. Длина тела 18,6—23 см. Длина хвоста 16—19 см. Внешним обликом несколько напоминают крысу. Длинный хвост порыт довольно густыми волосами, которые иногда па конце образуют большую кисточку. Волосяной покров высокий, мягкий, блестящий, о окраска на спинной стороне тела темно-рыжевато-коричневая. Бока
Рис. 72. Macrotarsomys ingens
рыжеватые. Брюшная сторона тела и губы белые. Уши черновато-коричневые. Конец хвоста у N. andeberti белый. Мордочка удлинена. Верхняя губа раздвоена. Уши средних размеров, с круглыми вершинами. Задние конечности довольно длинные. Три средних пальца на них длиннее двух крайних. Когти на пальцах задних конечностей сильно развиты.
Череп относительно широкий. Его лицевой отдел удлинен. Межтеменная кость крупная. Широкое костное небо простирается дальше уровня третьего коренного зуба. Лобные кости широкие. Подглазничные отверстия крупные. Костные слуховые барабаны относительно крупные. Резцовые отверстия расширяются кзади, но не достигают рядов щечных зубов. Верхние резцы коричневато-оранжевые, нижние — желтоватые. Наружная поверхность резцов гладкая.
Распространены на о-ве Мадагаскар.
Населяют леса. Экология не изучена.
В роде 3 вида:
зайцегубый хомяк — N. rufus Peters, 1870 (Мадагаскар);
хомяк Ламбертона—М. lambertoni Grandidier, 1928 (восточный Мадагаскар);
хомяк Андеберта — N. andeberti Jentink, 1879 (северо-восточный Мадагаскар) .
Возможно, эти 3 вида следует объединить в один — N. rufus.
Род коротколапых хомяков
Genus Brachytarsomys Gunther, 1875
В роде единственный вид: коротколапый хомяк — В. albicauda Gunther, 1875.
Размеры средние. Длина тела 21—25 см. Длина хвоста примерно 18,5 см. Морда несколько укороченная (рис. 73). Ушные раковины пе-
Рис. 73. Коротколапый хомяк — В. albicauda
большие, округлые, почти полностью скрытые в мехе головы. Глаза маленькие. Кисть и стопа короткие и широкие. Задние конечности с хорошо развитым пятым пальцем. Наибольшей длины на них достигают три средних пальца. Пальцы с сильными когтями. Хвост покрыт редкими или средней густоты волосами. Хвост, по-видимому, хватательный. Волосяной покров мягкий, густой, серовато-бурый на спинной стороне тела, рыжеватый на боках и белый на брюшной стороне. Голова рыжевато-;оричневая. нос и губы черноватые. Проксимальная половина хвоста верная, терминальная — белая.
В черепе мозговая капсула несколько уплощена. Лобно-теменные ребни срастаются между собой в виде срединного гребня. Костные слу-товые барабаны маленькие. Резцовые отверстия средней длины. Наруж-. 1я поверхность резцов гладкая.
Распространен в восточном ^Мадагаскаре.
Экология не изучена.
Род ссневидных хомяков
Genus Eliurus Milne-Edwards, 1885
Размеры от мелких до средних. Длина тела 9,5—16 см. Длина хвоста |j'i- 18,9 см. Масса 35—103 г. Внешним видом напоминают мышей 74). Мордочка несколько вытянутая. Глаза средних размеров. Уши । них размеров и почти лишены волос. Задние конечности удлиненные, [.-.штпрованные к лазанию но деревьям. Пятый палец на них длинный. Ьюсяной покров довольно густой и мягкий. Спинная сторона тела од-устная коричневато или желтовато-серая Брюхо и конечности свет-
;< серые.
У соневидного хомяка хвост НЧфЫТ довольно длинными и иносителыю густыми темно-Мыми волосами. У малого солидного хомяка проксималь । з треть хвоста почти лишена ьлос, а остальная часть покрыта негустыми удлиненными висами. Сосков 2 пары.
Череп с округлой и широкий мозговой капсулой. Лице-pi отдел удлинен и заострен. Лйно-теменных гребней нет. Кстные слуховые барабаны I ,-ррцних размеров Подглазнич-I пне отверстия крупные. Резцо-I нне отверстия различной дли-I н; как правило, они не дости-I пт рядов щечных зубов.
Распространены на о-ве Ма-I ь .скар
I Населяют густые леса, В гша леса с густым кустар-I ыивым подлеском; встреча-I г’я по берегам рек. По-види-I , ведут древесный образ Вун лч. В то же время известно,
Рис. 74. Малый соисвиднын хомяк — Е. minor
что малый соневидный хомяк делает норы и ведет преимущественно наземный образ жизни. Экология изучена слабо. Питаются в основном семенами и плодами. Самка приносит весной 2—4 детенышей.
В роде 2 вида:
соневидный хомяк — Е. myoximus Milne-Edwards, 1885 (западное побережье о-ва Мадагаскар, а также, вероятно, реликтовые леса западных, центральных и восточных областей Мадагаскара);
малый соневидный хомяк — Е. minor F. Major, 1896 (северо-восток и восток о-ва Мадагаскар).
Род хомяков Роберта
Genus Gymnuromys Major, 1896
В роде единственный вид: хомяк Роберта — G. roberti Major, 1896. Размеры мелкие и средние. Длина тела 12,5—16,1 см. Длина хвоста 15—17,5 см. Внешним обликом несколько напоминает крысу. Стопа широкая с длинным пятым пальцем. Длинный хвост покрыт очень редкими волосами, между которыми хорошо видны кожные чешуи. Волосяной покров на спинной стороне имеет черновато-серую или серую, а на брюшной — белую или желтовато-белую окраску.
Череп с длинным лицевым отделом. Межглазничное сужение умеренное. Резцовые отверстия очень короткие, далеко впереди от рядов щечных зубов. Костные слуховые барабаны небольшие. Лобно-теменных гребней нет. Жевательная поверхность коренных зубов плоская.
Распространен на о-ве Мадагаскар.
В питании преобладают вегетативные части растений. Экология не изучена.
Род хипогеомис
Genus Hypogeomys Grandidier, 1869
В роде единственный вид: хипогеомис — Н. antimena. Grandidier, 1869. Размеры крупные. Длина тела 30—35 см. Длина хвоста 20—25 см. Глаза довольно большие (рис. 75). Уши очень крупные, достигающие в длину 5—6 см. Длинный хвост покрыт негустыми волосами, сквозь которые видны кожные чешуйки. Кисть с 5 подошвенными мозолями. Задние конечности довольно длинные, с 4 пальцевыми подушками; пальцы с мощными когтями. Наружный палец хорошо развит. Волосяной покров грубый. Спинная сторона тела серая, серовато-коричневая или рыжеватая. Голова более темная. Конечности и брюшная сторона тела белые. Хвост темный.
Крупный черен не имеет гребней. Лицевой отдел черепа вытянут. Межтеменная кость крупная. Костные слуховые барабаны очень большие. Резцовые отверстия очень крупные и длинные, простирающиеся от резцов до середины первого коренного зуба. Верхние коренные зубы с уплощенными жевательными поверхностями. Коренные зубы с высокими коронками. Нижние коренные зубы представлены серией поперечных пластин.
Рис 75. Хипогсомис —11. antiniena
Распространен на о-ве Мадагаскар.
Населяет глухие леса. В связи с тем что многие леса па Мадагаска-। вырублены, в настоящее время ареал н численность этого зверька , чво уменьшились и, но-видимому продолжают уменьшаться. Копает минные, глубокие поры, весьма сходные с норами кроликов. Питается . авным образом упавшими на землю плодами. В помете, но-видимому, । йько один детеныш.
Род короткохвостых хомяков
Genus Brachyuromys F. Major, 1896
Размеры средние. Длина тела 14,5—18 см. Длина хвоста 6—10,5 см. Пасса 85—105 г Внешним видом несколько напоминают крысу. Уши ,.1во.пы!о крупные, с округлой вершиной, опушенные. Короткий хвост ио-срыт средней густоты волосами. Волосяной покров мягкий, густой, на шиной стороне тела коричневый с примесью рыжего и черного, па тошной — рыжеватый. Сосков 3 пары.
Череп широкий и массивный. Лобно-теменные гребни развиты слаги Межтеменная коегь крупная. Носовые кости несколько расширены ссредп. Резцовые отверстия длинные, достигают первого коренного зу Костные слуховые барабаны средних размеров. Подглазничные отверстия относительно крупные. Резцы широкие- Коренные зубы боль-। ?. гнпселодонтного типа.
Распространены на о-ве Мадагаскар.
Обитают па горных плато, па влажных лугах. Живут в густой траве, кгавны круглые сутки. Экология изучена слабо.
В роде 2 вида:
B.ramirohitra F. Major, 1896 (о-в Мадагаскар);
В betsiloensis Bartlett, 1879 (о в Мадагаскар).
ПОДСЕМЕЙСТВО КОСМАТЫХ ХОМЯКОВ SUBFAMILIA LOPHIOMY1NAE THOMAS, 1897
В подсемействе один род — косматых хомяков.
1 )
Род косматых хомяков
Genus Lophiomys Milne-Edwards, 1867
I и
В роде единственный вид: косматый хомяк — L. itnftausi Milne-Edwards, 1867.
Размеры крупные. Длина тела 25,5—36 см. Длина хвоста 14—21,5см. Масса 590—920 г. Самки крупное самцов. Внешний облик очень своеобразный, с массивным неуклюжим туловищем, покрытым длинной, лохматой шерстью (рис. 76). Голова несколько напоминает голову морской свинки, а туловище немного сходно с туловищем североамериканского дикобраза. Уши небольшие, округлые, почти полностью скрыты
1-
BL ЖГ ПО ВО
I.
й,
Р-
н-
в мехе. Глаза небольшие. Конечности несколько укороченные. Стопа короткая с тремя длинными средними пальцами, укороченными боковыми и шестью подошвенными мозолями. На передних конечностях пятый палец сильно укорочен. Когти на пальцах передних конечностей крупные, сильные. Передняя конечность приспособлена к лазанию: ее первый палец частично противопоставляется остальным. Хвост покрыт густыми, длинными пушистыми волосами. От затылка вдоль всей спины проходит полоса удлиненных волос, образующих своеобразную гриву, по бокам которой растут более короткие волосы. Когда животное испу-
гано или сердится, грива волос на спине поднимается и приоткрывает железистые участки на боках туловища. Волосяной покров высокий, густой и мягкий, за исключением спинного гребня, в котором волосы довольно грубые. Окраска спинной стороны тела образована смесью двух цветов — черного и белого или коричневатого и белого с различными полосками и пятнами. Окраска брюшной стороны изменяется от серой до черной. Конечности черные. Кончик хвоста белый.
Череп весьма своеобразный. Височная ямка полностью покрыта костью, поверхность которой сильно гранулирована (грануляция ослабляется в сторону носовых костей) и образована из лобных и теменных костей, причленяющихся к середине скуловых костей. Орбиты, таким образом, очень небольшие. Затылочная область с крупным гребнем. Костные слуховые барабаны очень маленькие. Резцовые отверстия длинные, достигают первого'коренного зуба. Подглазничные отверстия в своей верхней части шире, чем в нижней. Коренные зубы увеличиваются в размерах от первого к третьему. Резцы умеренной ширины.
Распространен в Восточной Африке от Судана до Сомали, Уганды, Кении, Танзании и Эфиопии.
Населяет густые леса в горах между 1200 и 2700 м над уровнем моря. Обычно держится поодиночке. Ведет древесный образ жизни. Хорошо лазает (хотя и медленно) по деревьям. Может спускаться по стволу дерева даже вниз головой. Активен ночью. День проводит в норах или расщелинах среди камней. Питается листьями, ростками и плодами различных растений. В помете 2—3 покрытых волосами детеныша.
Эллермэн (1940) выделяет косматого хомяка в особое семейство Lophiomyidae.
ПОДСЕМЕЙСТВО ПОЛЕВОК
SUBFAMILIA MICROTINAE MILLER, 1896
Размеры мелкие, иногда средние или крупные. У большинства внешний облик полевки. Некоторые представители приспособлены к полу-водному (например, Ondatra) или подземному образу жизни (например, Ellobius).
3
I Щечных зубов др Они сложны по структуре, призматические, часто не имеют корней и в этом случае растут в течение всей жизни животного.
! * Род копытных леммингов
Genus Dicrostonyx Gloger, 1841
, Размеры мелкие и средние. Длина тела 12,5—16 см. Длина хвоста I 1—2,2 см. Масса тела 45—75 г. Особенности морфологии показывают I высокую степень адаптации к жизни в арктических условиях. Телосло-1 некие плотное, конечности короткие. Хвост очень короткий, полностью I покрыт волосами. Кисти и ступня широкие, их подошвы густо покрыты ; волосами; мозоли на подошвах отсутствуют. Глаза небольшие. Ушная
раковина в виде низкой складки кожи, скрытой в меху. Третий палец на передних и задних конечностях несколько длиннее четвертого. Первый палец передней конечности укорочен и имеет небольшой ногтеобразный коготь. Когти длинные. На передних конечностях они крупнее, чем на задних. К зимнему периоду когти третьего и четвертого пальцев передней конечности сильно увеличиваются и приобретают характерную вильчатую форму на конце. Волосяной покров высокий, густой и мягкий. В летний период его окраска варьирует на спинной стороне тела от светло-серой до темно-серой с темно-желтым или рыжевато-коричневым оттенком. Брюшная сторона тела беловатая, сероватая или темно-желтая. От головы по средней линии спины обычно проходит продольная темная полоска. В зимнее время волосяной покров приобретает белую окраску.
Череп с относительно укороченным лицевым отделом. Лобно-теменные гребни сильно развиты. Костные слуховые барабаны средних размеров. Скуловые дуги широко расставлены в стороны. Резцовые отверстия относительно широкие. Зубы гинселодонтного типа, не имеют корней и растут в течение всей жизни. Их жевательная поверхность сложная. Щечные зубы не имеют корней. Эмалевые петли на их жевательной поверхности сильно сжаты в передне-заднем направлении. На передних и задних концах М2, М3, М1 и М2 расположены дополнительные эмалевые столбики и соответствующие им петельки с обеих сторон зубов. 7И| с девятью или с десятью эмалевыми петлями.
Хромосом в диплоидном наборе 28 у лемминга Виноградова и до 60 у копытного лемминга.
Распространены в арктических областях Нового и Старого света, от Аляски до Лабрадора, включая северное побережье Гренландии и от восточного побережья Белого моря на восток до Анадырского края и Новой Земли.
Населяют равнинную и горную тундры, поднимаясь в горы до высоты 900 м над уровнем моря. Предпочитают занимать возвышенные и сухие участки. Норы обычно устраивают неглубоко в земле. Гнездовая камера довольно часто помещается в кочке. Зимой делает ходы в снегу. Питаются побегами, листьями, корой полярных кустарничков — карликовой березы, ивы, голубики, морошки. Размножение начинается в конце зимы и продолжается все лето до осени. За это время бывает 2—3 помета. В каждом помете обычно по 4—6 детенышей. Длительность беременности 20—22 дня. Численность может сильно колебаться в разные годы. Имеют большое значение как основной объект питания песца и других промысловых зверей тундры.
В роде 4 вида:
копытный лемминг — D. torquatus Pallas, 1779 (СССР — тундра и лесотундра от п-ова Канина на западе до Берингова пролива на востоке, о-ва Вайгач, Новая Земля, Новосибирские, Медвежьи; западные и северные районы Аляски, включая п-ов Аляска; север Канады — крайний север провинции Юкон, Северо-Западные территории, кроме юго-запада, Канадский Арктический архипелаг, узкая полоса но северному берегу Гренландии и по побережью Гренландского моря до Датского пролива);
лемминг Виноградова — D. vinogradovi Ognev, 1948 (о-в Врангеля); сентлоренский лемминг — D. exsul G. Allen, 1919 (о-в Сент-Лоренс в Беринговом море);
гудзонский лемминг — D. hudsonius Pallas, 1778 (северо-восток Канады •— север провинции Квебек и Ньюфаундленд, ио без о-ва Ньюфаундленда) (рис. 77).
Рис. 77. Гудзонский лемминг — D. hudsonius и его череп
В СССР: копытный лемминг и лемминг Виноградова.
Некоторые исследователи (например. Холл и Колсон, 1959) выделяют копытного лемминга, обитающего в Северной Америке, в особый вид —О- groenlandicus Traill, 1823.
Род мышевидных леммингов
Genus Synaploniys Baird, 1857
Размеры мелкие. Длина тела 10 13 см. Длина хвоста 1,6—2,7 см. Масса 19—40 г. Внешним видом несколько напоминают леммингов, хотя менее специализированы. Телосложение плотное. Конечности короток. Ушные раковины небольшие. Глаза маленькие. Подошвы лап по-I рыты относительно редкими волосами. Кисти с 6 подошвенными мозо-( ,увш. Волосяной покров не густой, довольно грубый. Окраска спинной [гороны тела серовато-коричневая, желтовато-коричневая или черная, । .ошная — беловатая или сероватая. Хвост коричневатый сверху и бе
ловатый снизу. Сосков 3 или 4 пары. Самцы имеют паховые специфические кожные железы.
Лицевой отдел черепа широкий. Лобно-теменные гребни срастаются между собой. Резцовые отверстия большие. Костные слуховые барабаны очень крупные. Верхние резцы оранжевые, необычайно широкие, с продольной бороздкой на наружной поверхности. Жевательная поверхность верхних щечных зубов с широкими петлями.
Распространены в Северной Америке от штата Виргиния и Канзас на север до Аляски, Маккензи и Лабрадора.
Населяют обычно влажные районы с каменистым грунтом. Активны круглосуточно. Подобно полевкам делают в траве тропинки. Питаются в основном вегетативными частями растений, а также различными мелкими беспозвоночными. Период размножения тянется с марта по октябрь. Длительность беременности 23 дня. В помете от 1 до 8 детенышей. За сезон бывает до трех пометов. Масса детенышей при рождении около 4 г. На 10—12 день у них открываются глаза. Самцы достигают половой зрелости на 4-й месяц. Отмечены значительные изменения численное- к ти в различные годы.
В роде 2 вида: к
южный мышевидный лемминг — S. cooperi Baird, 1858 (юго-восточ- и ная часть Канады и северо-восток США, провинции Канады — крайний юго-восток Манитобы, южная половина Онтарио, юг Квебека, Нью-Брансуик, Новая Шотландия, о-в Кейп-Бретон; штаты США — Мэн, Нью-Гэмпшир, Вермонт, Нью-Йорк; Массачузетс, Коннектикут, Род-Айленд, Нью-Джерси, Делавэр, Мэриленд, Пенсильвания, север и северо-запад Виргинии, Западная Виргиния, крайний запад Северной Каролины, крайний восток Теннесси, север и восток Кентукки, Огайо, Индиана, Мичиган, Иллинойс, Висконсин, крайний северо-восток Арканзаса, Мне- I сури, кроме юго-запада, Канзас, восточные две трети Небраски, восточ- I ная половина Южной Дакоты, Иова, Миннесота, крайний юго-восток 1 Северной Дакоты; кроме того, небольшой изолированный участок ареала I расположен на атлантическом побережье на границе между Виргинией и Северной Каролиной); I
северный мышевидный лемминг — 5. borealis Richardson, 1828 (север Ш Северной Америки — южные две трети территории Аляски, кроме п-ова Аляска; провинции Канады — Юкон, кроме северных районов, юг и f юго-запад Маккензи, в Северо-Западных территориях, Британская Ко- Д лумбия, кроме о-ва Ванкувер, Альберта, кроме юго-востока, северные S две трети Саскачевана, Манитобы, кроме крайнего севера и юго-запада, W северная половина Онтарио, северная половина Квебека, Ньюфаундленд, I но без о-ва Ньюфаундленд; небольшой изолированный участок ареала I расположен в провинциях Канады: крайнем юго-востоке Квебека, в I Нью-Брансуике и штате США Мэн).
Род лесных леммингов
Genus Myopus Miller, 1910
В роде единственный вид: лесной лемминг — М. schisticolor Lillje- Д borg, 1884. Ш
Размеры мелкие. Длина тела 9—13 см. Длина хвоста 1—2 см. По В внешнему облику сходен с лесными полевками. Морфологические приз- наки не очень специализированы. Округлые ушные раковины малы, гус- '
то покрыты волосами и скрыты в меху. Хвост короткий, покрыт густыми 1
волосами. Глаза маленькие. Когти довольно длинные, одинаково разви- I 1
ты на передних и задних конечностях. Подошва кисти не имеет волос. Пятка стопы густо покрыта волосами. Первый палец передней конечности небольших размеров, с уплощенным когтем. Волосяной покров довольно высокий, густой и мягкий. Его окраска на середине спины рыжеватая, на боках темно-серая, на брюшной стороне слегка более светлая. Конечности и хвост черные. Спинная сторона тела с характерным металлическим блеском. Сезонная изменчивость в окраске волосяного покрова не наблюдается. Зимой мех длиннее и гуще, чем летом. Сосков 4 пары.
Массивный череп с сильно выраженным межглазничным сжатием и со слившимися лобно-теменными гребнями. Скуловые дуги широко расставлены. Костные слуховые барабаны очень большие. Мозговая капсула несколько сужена, а лицевой отдел укорочен по сравнению с черепом настоящих леммингов, на который череп лесного лемминга очень похож. Характерен укороченный последний коренной зуб, особенно в нижней челюсти.
Хромосом в диплоидном наборе 32 или 34.
Распространение охватывает обширную территорию таежной зоны Евразии от Норвегии до Камчатки и Сахалина, на юг до горных районов северной части МНР и Северо-Восточного Китая. От района Нурмарк в Норвегии к северу от Осло северная граница ареала проходит по южным предгорьям Скандинавских гор до Виттанджи в Лапландии, оз Имандра, Кандалакшской губы, побережья Белого моря и Архангельска; затем поворачивает на юг к верховьям рек Северная Двина и Печора и далее на восток к пос. Питлар на Оби, низовьям Енисея и через верховья р. Анабар к северной части Верхоянского хребта, а затем проходит по низовьям рек Яна, Хрома, Индигирки, Алазея и Колымы. Восточная граница ареала неясна; известна лишь одна находка на Камчатке в Кроноцком заповеднике. Южная граница ареала начинается от побережья Японского моря близ 46° с. ш. и следует на запад через хребты Малый и Большой Хинган, обходит с севера Забайкальские степи и вдет на юг в МНР по хребту Хэнтей, затем по р. Джида — до Саян, через восточный хребет Танну-Ола — до Алтая, проходит через северную часть Новосибирской области и далее на запад через устье Иртыша, север Свердловской обл., центр Пермской обл., Костромскую обл., Вологодскую обл., ст. Нелидово в Великолукской области, северо-восточное побережье Онежского озера, следует к Южной Финляндии и подходит кМурмарке — к северу от Осло.
Населяет обычно таежные и горно-таежные леса, выбирая сырые и заболоченные места. В северо-восточной части ареала по речным долинам проникает в тундру. В Западной Сибири иногда встречается в березовых колках и островках ельника — зеленомошника среди степного ландшафта. Норы обычно устраивают в моховом покрове, под корнями деревьев, иногда поселяется в камнях. Питается в основном мхами и лишайниками. По-видимому, бывает два помета в год. В различные годы отмечены изменения численности; в годы высокой численности наблюдаются дальние миграции.
Род настоящих леммингов
Genus Lemmus Link, 1795
Размеры мелкие. Длина тела 10—15 см. Длина хвоста 1,8—2,6 см. I Высоко адаптированы к жизни в Арктике. Телосложение тяжелое. Ко-| вечности короткие. Небольшой хвост полностью скрыт в мехе. Ушные I раковины малы и не выступают из меха головы (рис. 78). Глаза неболь-
шие. Подошвы передних и задних лап покрыты гусгой шерстью. Мозоли на подошвах едва заметны. Третий палец передней конечности и четвертый палец задней длиннее других. Первый палец на передней конечности больших размеров с длинным уплощенным в боковом направлении когтем, иногда раздвоенным па конце. Когти на пальцах обеих конечностей сильно развиты, на передних они несколько длиннее, чем па задних. В зимнее время когти еще более удлиняются. Волосяной покров очень густой, высокий и мягкий. Окраска спинной стороны тела сероватая или коричневатая с темпо-желтым или гемно-коричневым оттопками, брюшной — светло-серая, темно-желтая или коричневатая. Вдоль спины может проходить продольная темная полоска. На зиму волосяной покров становится выше, гуще и светлее, иногда может стать чнето-бе-лым. Сосков 4 пары.
Лицевой отдел черепа укорочен. Скуловые дуги мощные, расставлены широко в сторону. Мозговая коробка несколько уплощена сверху.
Ш Лобно-теменные гребни хорошо развиты. Межтеменная кость крупная. • Костные слуховые барабаны небольших размеров. Резцовые отверстия не крупные. Сочленовный отросток нижней челюсти длинный и узкий. Резцы широкие, иногда с продольной бороздкой. Коренные зубы гип-селодонтного типа, не имеют корней и растут в течение всей жизни животного. Внутренние углы передних треугольных петель жевательной поверхности параллельно зубному ряду срезаны. Из всех петель слиты только задние противолежащие петли ЛР. У ЛК — пять замкнутых петель.
Хромосом в диплоидном наборе 50,
Распространены в тундре и лесотундре Европы, Азии, Северной Америки, а также в тайге Восточной Сибири.
Населяют равнинные и горные тундры. Для поселения обычно вы-j бирают увлажненные места, избегая каменистых участков. В годы повы-| шейной численности заселяют самые разнообразные биотопы, широко | распространяясь в лесной зоне. Здесь предпочитают селиться в ельни-1ках и сосняках. Активность круглосуточная. Убежищем служат норы, которые они выкапывают сами. Иногда устраивают крупные шарообразные гнезда в естественных укрытиях — в камнях, в пустотах под корнями. В толще мха и лишайников прокладывают разветвленные ходы. Зимой проделывают ходы в снегу. Питаются различными растительными объектами: мхами, ягодами, стеблями и листьями кустарничков, злаками, грибами. Зимой в основном поедают листья, побеги и кору ив и полярной березки. Размножение происходит с весны до осени, причем каждая самка в течение года дает 3—4 помета. Длительность беременности от 20 до 22 дней. В помете от 3 до 9, обычно 5—6 детенышей. В различные годы отмечаются резкие изменения численности. В годы высокой численности отмечены массовые миграции, начинающиеся еще весной, в период снеготаяния. Мигрирующих животных не останавливают даже большие водные преграды. Во время миграции основная масса их гиб* нет. Настоящие лемминги служат важным объектом питания тундровых и лесотундровых пушных зверей.
В роде 5 видов:
амурский лемминг — L. amurensis Vinogradov, 1924 (СССР — с. Пинан на р. Зее, Амурской обл., верховье р. Нельгехе в юго-восточной части Верхоянского хребта, окрестности пос. Александровский завод Читинской обл., р. Тершнак, левый приток р. Читы, 200 км к северу от Магадана) ;
сибирский лемминг — L. sibiricus Kerr, 1792 (СССР — тундра и лесотундра от Горла Белого моря до Чукотки и Камчатки, о-ва Диксон, Врангеля, Новосибирские);
чернолапый лемминг — L. nigripes True, 1894 (о-ва Прибылова в Беринговом море);
бурый лемминг — L. trimucronatus Richardson, 1825 (северо-запад Северной Америки — Аляска, кроме п-ова Аляска и юго-восточной части штата, о-в Нунивак; провинции Канады — Юкон, кроме юго-запада, северо-западные территории, кроме южных районов и Канадского Арктического архипелага, центр и центральные районы севера Британской Колумбии, крайний северо-восток Манитобы);
норвежский лемминг — L. lemmus Linnaeus, 1758 (Северная Скандинавия; в СССР — Кольский п-ов).
В СССР: амурский, сибирский и норвежский лемминги.
Сибирского, чернолапого и бурого леммингов, по-видимому, следует объединить в один вид — L. sibiricus Kerr, 1792.
Рис. 79. Рыжая полевка — Cl. glareolus и се череп
Род лесных полевок
Genus Clethrionomys Tilesius, 1850
Размеры мелкие. Длина тела 7—13 см. Длина хвоста 2,5—6 см. Ушные раковины средних размеров; едва выступают над уровнем меха (рис. 79). Глаза небольшие. Хвост покрыт средней густоты, а иногда густыми волосами. Подошвы лап голые или слабо оволоснены лишь в пяточных отделах. Имеются подошвенные мозоли. На передних конечностях третий палец длиннее четвертого, на задних эти пальцы пример
но одинаковы. Первый палец на обеих конечностях укорочен. Волосяной покров зимой густой, довольно высокий и мягкий, а летом значительно более низкий и грубый. Окраска его на спинной стороне тела в основном сероватая с рыжеватым оттенком или рыжая. Бока более серые. Брюхо серое или белое. В зимнее время волосяной покров становится более ярким, с ржаво-красными тонами. Сосков 4 пары.
Череп с укороченным лицевым отделом. Межглазничное сужение слабо выражено. Межглазничный промежуток сверху уплощен или в различной степени желобообразно углублен в продольном направлении. Лобно-теменных и затылочных гребней нет. Мозговая коробка широкая п несколько уплощенная. Костные слуховые барабаны крупные. Губчатая костная ткань в их полости не развивается. Щечные зубы имеют корни. Все петли на жевательной поверхности нижних щечных зубов, в особенности средних у М2 и М3, могут быть в различной степени слиты друг с другом, у АД число замкнутых петель варьирует от 4 до 7.
Хромосом в диплоидном наборе всех изученных видов 56.
Распространение очень обширное: в Северной Америке от северных частей этого континента (Британской Колумбии, Аляски, Юкона, Маккензи и Лабрадора) на юг до штатов США Колорадо и Северной Каро-
I липы; в Старом свете в зонах леса, лесостепи, а также в пойменных I лесах степной зоны, в горах Евразии от Пиренеев на западе до хребтов г системы Хингана на востоке, к югу до северной части Пиренейского, южной части Апеннинского п-овов, Передней Азии, Западного Закав-
I казья, МНР, Восточного Китая, Корейского п-ова и Японии, а к северу I до северных пределов лесной растительности.
ij Населяют различные биотопы, преимущественно в лиственных и I хвойных лесах. Обитают также в болотистых местах лесотундры. Под-I нимаются в горы до 3000 м над уровнем моря. Активны круглосуточно. I Убежищем служат норы, как правило, короткие и неглубокие, пустоты I и выгнивших корней или кочки. Иногда делают поверхностные норы в I толще мха или лесной подстилки. Гнезда устраивают в укрытиях на 1 поверхности почвы, реже над землей. Лесные полевки могут довольно 1 хорошо лазать по кустам и деревьям. Питаются преимущественно веге-I татнвными частями травянистых растений, в меньшей степени семенами, 1 юроп и почками кустарников и деревьев.- Поедают также различных 1 беспозвоночных животных, лишайники и мхи. Иногда делают неболь-1 шпе запасы.
1 Период размножения при благоприятных условиях начинается еще снеговом покрове и продолжается до глубокой осени. Бывает по 3— ij помета за сезон, в каждом из которых от 2 до U, обычно 5—6 детенышей. Длительность беременности 17—20 дней. Местами могут приносить вред лесным питомникам, садам, полезащитным лесонасаждениям. Относятся к природным носителям возбудителей клещевых сыпнотифозных лихорадок и лептоспирозов. Служат важным кормовым объектом пушных зверей.
В роде 7 видов:
красная полевка-—С. rutilus Pallas, 1779 (Северная Норвегия, Шведа: СССР — от Кольского п-ова на западе до Чукотки и Камчатки и о-вов Командорских, Курильских, Шантарских и Сахалина па востоке, южная граница проходит через южную Карелию, Калининскую и Горьковскую обл., Татарию, Башкирию, Южный Урал, северные области Ка-I захстана и дальше через государственную границу уходит за пределы страны; северная часть МНР; Северо-Восточный Китай; Хоккайдо; се-I веро-запад Северной Америки, кроме п-ова Аляска, Юкон, Северо-За-
паяные территории, кроме центра южных частей, крайний северо-восток ЛАанитобы, северо-запад Британской Колумбии);
рыжая полевка — С. glareolus Schreber, 1780 (Западная Европа — Англия, Франция, Бельгия, Швейцария, Италия, Норвегия, Швеция, Нидерланды, Дания, ФРГ, ГДР, Польша, Финляндия на юг до Югославии, Румынии и Малой Азии; СССР — таежная и лесотундровая зоны европейской части и Западной Сибири до Алтая на востоке, Закавказье);
тянынанская лесная полевка — С. frater Thomas, 1908 (СССР— Тянь-Шань, кроме хребтов к западу от Таласского Алатау и Ферганского хребта; восточный Тянь-Шань);
красно-серая полевка — С. rufocanus Sundevall, 1846 (Норвегия, Швеция; СССР — север европеййеой части, почти вся Сибирь и Дальний Восток, на юг до Средней Карелии, Горьковской обл., Южного Урала, Алтая, Северной части МНР; Северо-Восточный Китай);
шикотанская полевка — С. sikotanensis Tokuda, 1935 (о-в Шикотан из группы Малых Курильских о-в и Крильонский п-ов на юго-западе Сахалина) ;
полевка Гаппера — С. gapperi Vigors, 1830 (север и центр Северной Америки — северная граница ареала начинается на западе на тихооке- i анском побережье провинции Канады Британская Колумбия примерно напротив северных островов архипелага Александра и идет на восток к I юго-восточному углу провинции Юкон и далее вдоль южной границы се- I веро-западных территорий, несколько отступя к северу от нее, выходит I к западному берегу Гудзонова залива, огибает его с юга и востока, перс- I секает с запада на восток северную часть п-ова Унгава, выходит к юго- I западной части залива Унгава и далее, следуя береговой линии моря I Лабрадор и Атлантического океана, образует восточную границу ареа- I ла; о-ва Ньюфаундленд и Антикости в ареал не входят; на юг граница I ареала идет до северо-восточных районов штата Виргиния, откуда, об- I разуя южную границу ареала, сворачивает на юго-запад к юго-запад- I ному углу этого штата и далее к северо-западной оконечности Джорд- жии, поворачивает на север, доходит до центральных районов Кентукки и сворачивает на северо-восток, поднимаясь до юго-запада Пенсильва- I нии, огибает с юга озера Эри и Гурон, пересекает в средних частях Л оз. Мичиган, идет к юго-восточному углу штата Миннесота, юго-востоку Ш Южной Дакоты, юго-западному углу Северной Дакоты, центральным В районам севера Монтаны, откуда сворачивает на юго-восток к юго-вос- W точной части Вайоминга и далее на юг до центральных районов Нью- | Мексике, откуда небольшим выступом вдается в центральные районы I востока Аризоны, поворачивает к северу почти до северо-западного угла I Колорадо, делает небольшой изгиб к югу в центральные районы Юты, I поворачивает на север до срединных областей Айдахо, следует на запад I до центра Орегона, поднимается к северу до границы между Орегоном I и Вашингтоном, круто спускается на юг до северной трети штата Кали- к форния и несколько севернее Сап-Франциско выходит к Тихому океа- I ну; западная граница ареала идет по берегу Тихого океана; о-в Банку- вер в ареал не входит; небольшой изолированный участок ареала па- ходится на границе между северо-востоком штата США Вайоминг и I западом Южной Дакоты): I
западная лесная полевка — С. occidentalis Merriam, 1890 (Канада I юго-запад провинции Британской Колумбии без о-ва Ванкувер; штаты США — западные трети Вашингтона и Орегона, северо-запад Кали- I
форнии); I
В СССР: красная, шикотанская, рыжая, тянынанская лесная и крас- I
но-серая полевки.
Ряд исследователей насчитывает в роде большее число видов: Андерсон и Джонс (1967) — 10, И. М, Громов (1963) —более 20, Эллермэн (1940) — 31. Б. А. Кузнецов (1965) объединяет с рыжей полевкой тянь-шанскую лесную полевку.
Род южноазиатских полевок
Genus Eothenomys Miller, 1896
Размеры мелкие. Длина тела 9—12,6 см. Длина хвоста 3—5,5 см. Внешний вид типичный для полевок. По своим признакам занимают промежуточное положение между лесными и серыми полевками. Уши небольшие, немного выступающие над уровнем меха. Подошвы позади мозолей оволоснены. Хвост покрыт короткими, жесткими волосками; на конце короткая кисточка из удлиненных волос. Волосяной покров на туловище довольно густой, мягкий и низкий. Окраска спинной стороны темно-коричневая, рыжевато-коричневая, с характерным металлическим отблеском, брюшной голубовато-серая. Молодые имеют более темную окраску. Сосков 2 пары.
Череп слабый с относительно широким межглазничным пространством. Лобно-теменные гребни не соединяются друг с другом. Костные слуховые барабаны не содержат губчатой ткани. Щечные зубы не имеют корней.
Распространены в Китае, на севере п-ова Индокитай, в Северной Бирме, Индии — Северном Ассаме, а также на о-ве Тайвань.
Живут в горах на высоте 1800—4400 м над уровнем моря, в лесах, на горных лугах, по берегам рек. Предпочитают мягкую почву, где легко рыть норы. Активны круглосуточно. Сезонности в размножении, по-видимому, нет. В помете 1—3 детенышей.
В роде, по-видимому, 5 видов:
темнобрюхая южноазиатская полевка — Е. melanogaster Milne-Ed-v.ards, 1872 (Китай — от южного Ганьсу, Сычуаня и Юпьпана па восток до Фуцзяня и Чжэцзяна; о-в Тайвань; Северный Ассам; Северная Бир-уа: Северный Индокитай);
Е. olitor Thomas, 1911 (Китай — Юньнань);
Е. proditor Hinton, 1923 (Китай—Сычуань и Юньнань);
китайская полевка — Е. chinensis Thomas, 1891 (Китай — Сычуань и Юньнань) ;
Е. custos Thomas, 1912 (Китай — Сычуань и Юньнань).
Некоторые исследователи (Симпсон, 1945; Эллермэн, 1941) выделяют особый род Anteliomys Miller, 1896, с видами A. chinensis Thomas, 1891; .4. custos. Thomas, 1912; A. wardi Thomas, 1912. По-видимому, более правильно считать их в составе рода Eothenomys и, кроме того, Е. wardi — синонимом Е. chinensis.
По Андерсону и Джонсу (1957), в роде 8 видов.
Род каменных полевок Genus Alticola Blanford, 1881
Размеры мелкие. Длина тела 8—14 см. Длина хвоста 1,5-—4 см. Внешний вид типичный для полевок (рис. 80). Уши небольшие, слегка выступающие над мехом. Глаза средних размеров. Подошвы лап в задних отделах покрыты волосами. Хвост бывает густо, умеренно или слабо покрыт волосами. Третий палец на обеих конечностях длиннее остальных. На передней конечности первый палец может быть длинным
Рис. 80. Серебристая полевка - A. roylei и череп плоскочерепной полевки A. strelzovi
или слабо развитым с небольшим плоским коготком или даже без пего. Волосяной покров густой, мягкий, высокий. Окраска его па спинной стороне тела от серой до коричневой с различными оттопками: серебристым, желтым или рыжим. Брюшная сторона тела обычно белая или желтовато-белая. Конечности белые или серовато-белые. Хвост белый или желто-белый. В зимнее время окраска более светлая, чем летом. У одной формы мех зимой полностью белеет. Характерны очень длинные вибриссы, нередко заходящие назад за лопаточную область. Сосков 4 пары.
Череп сходен с черепом других полевок. Он несколько или сильно уплощен сверху. В области лобных костей находится своеобразная вдйв-ленность. Лобно-теменные гребни расставлены широко друг от друга.
Костные слуховые барабаны от небольших до сравнительно крупных, внутренняя полость не имеет губчатой костной ткани. В нижней челюсти сочленовный и угловой отростки относительно длинные. Щечные зубы ле имеют корней и растут в течение всей жизни животного. Треугольные петли жевательной поверхности щечных зубов небольшие, широкие, далеко отстоят друг от друга. Противолежащие петли могут частично сливаться. М3 сравнительно длинный с тенденцией к редукции наружных зубцов и образованию длинной и узкой пятки. Задний край костного неба почти доходит до линии, соединяющей передние края задних коренных зубов.
Распространены в горных районах Центральной и Северо-Восточной ,ии от Западного Тибета и Южных Гималаев до Тянь-Шаня, Памиро-:ая, Казахского Нагорья, Алтая, Саян и северо-востока Сибири.
Населяют каменистые участки от морского побережья (на Чукотке) до 6000 м над уровнем моря, а также горную тундру, придерживаясь каменистых участков. Убежищем служат пустоты среди камней, расщели-йы скал. Иногда роют норы. Гнездовую камеру часто устраивают под камнями. В моховом или лишайниковом покрове прогрызают канавки. ^Активны преимущественно днем или круглые сутки. Питаются различными растениями, причем в весеннее время преимущественно вегетативными частями, а осенью — семенами. Поедают также мхи и лишайники. Собирают запасы пищи. В течение года дают от 1 до 2—3 пометов, в каждом из которых 5—11, обычно 5—8 детенышей. Половозрелость у высокогорной сибирской полевки наступает в годовалом возрасте. Численность каменных полевок никогда не бывает высокой.
В роде, вероятно, 5 видов:
серебристая полевка—A. roylei Gray, 1842 (СССР — Памиро-Алай, Тянь-Шань, Джунгарский Алатау, Тарбагатай, Тувинская АССР; Северный Китай; МНР; Индия — Кашмир, Северный Пенджаб, Кумаоп; Афганистан) ;
гобиалтайская полевка — A. barakshin Bannikov, 1954 (горы МНР; в СССР— юго-запад Тувинской АССР);
большеухая полевка — A. macrotis Radde, 1861 (СССР — горы Южной и Восточной Сибири от низовьев Лены до Чукотки, на юг до Алтая, Саян и Амурской обл.) ;
высокогорная полевка—A. stoliczkanus Blanford, 1875 (Индия — Кашмир и, возможно, Ганьсу в Китае);
। плоскочерепная полевка — A. strelzovi Kastschenko, 1900 (СССР — [Казахский мелкосопочник, хребет Тарбагатай, Зайсанская котловина, ВАлтай, запад Тувинской АССР; северо-западная часть МНР).
И. М. Громов (1963) насчитывает в роде около 20 видов.
Род кашмирских полевок
Genus Hyperacrius Miller, 1896
В. Размеры мелкие. Длина тела 9,5—11,6 см. Длина хвоста 1,8'—4,5 см. Внешним обликом сходны с представителями рода каменных полевок. Имеются приспособления к подземному образу жизни. Вибриссы корот- кие, едва достигающие ушей. Ушные раковины сильно уменьшены. По-дошвы лап покрыты волосами лишь в задних отделах. Когти па передних конечностях длинные и сильные. Короткий хвост густо покрыт воло-сами. На стопе 5 подошвенных мозолей. Волосяной покров довольно низкий, но густой и мягкий. Спинная сторона тела рыжевато-коричневая, Идаповато-коричневая или черноватая. Брюхо охристое, сероватое или
темно-желтое. Хвост обычно двухцветный — темно-коричневый сверху и белый снизу или темно-серый сверху и светло-серый снизу. Сосков 3 пары.
Череп похож на таковой представителей рода каменных полевок. В отличие от последних форма его более угловатая, костные слуховые барабаны маленькие и лобно-теменные гребни, сливаясь между собой у взрослых животных, образуют медиальный межглазничный гребень. Лицевой отдел черепа относительно удлинен. Резцовые отверстия узкие, щелевидные.
Распространены в Кашмире и Пенджабе в Северной Индии.
Обитают в горах на высоте 2000—3500 м над уровнем моря, в сосновых лесах. Предпочитают участки, покрытые травянистой растительностью. Значительную часть времени проводят под землей. По некоторым данным, эти зверьки ведут исключительно подземный образ жизни. Экология изучена слабо.
В роде 2 вида:
Н. wynnei Blanford, 1881 (Пенджаб и Кашмир);
кашмирская полевка — Н. fertilis True, 1894 (Кашмир).
Род югославских полевок
Genus Dolomys Nehring, 1898
В роде единственный вид: югославская полевка — D. bogdanovi Martino, 1922.
Размеры мелкие. Длина тела 13—14 см. Длина хвоста 5—9 см.* Довольно крупные уши покрыты густыми волосами. Подошвы лап почти полностью голые, за исключением самых задних отделов. Стопа с 6 подошвенными мозолями. Хвост покрыт редкими волосами, за исключением удлиненных волос на копчике хвоста. Волосяной покров средней высоты, густой и мягкий. Спинная сторона тела серовато-коричневая, темно-желто-серая или голубовато-серая. Брюшная сторона обычно более светлая, часто серовато-белая. Конечности белые. Хвост обычно темно-коричневый сверху и белый по бокам и снизу. Первый палец передней конечности небольших размеров с маленьким уплощенным ногтеобразным когтем. Когти на остальных пальцах передней и задней конечностей примерно одинаковые по размерам, короткие и острые. На черепе взрослых животных лобно-теменные гребни срастаются между собой. Костные слуховые барабаны очень крупные, вздутые. Верхние резцы широкие, с бороздкой на наружной поверхности. Щечные зубы у взрослых животных имеют корни.
Распространена в Югославии.
Населяет горы на высоте 700—2100 м над уровнем моря. Активна круглый год. Убежище находит в расщелинах между скал, под камнями, где строит гнезда. Питается различными вегетативными частями растений. Запасает пищу на зиму. Даст 2 помета в году — в марте и в июне. Длительность беременности примерно 1 месяц. В помете 2—3 детеныша.
Род водяных полевок
Genus Arvicola Lacepede, 1799
В роде единственный вид: водяная полевка — A. terrestris Linnaeus, 1758.
Размеры средние. Длина тела 14—25 см. Длина хвоста составляет примерно половину длины тела. Имеются приспособления как к рытью
гмли, так и к земноводному образу жизни. Наружные ушные раковины небольших размеров Глаза небольшие (рис. 81). Внутренние выросты верхних губ позади резцов значительных размеров, густо покрыты воло-
I гами, но не срастаются между собой и не изолируют резцы полностью II от ротовой полости. Хвост покрыт редкими, жесткими и короткими воло
I сами. В поперечном сечспии хвост округлый. Первые пальцы па обеих | конечностях не укорочены. Третий палец па передней и задней конечное | тях длиннее четвертого. Когти умеренной длины. Подошвы лап без во | лес, но имеют гребневидную оторочку из жестких и густых волос по I краям. Волосяной покров довольно высокий, ।устой п мягкий с режим | разделением на остевые и пуховые волосы. Окраска спинной стороны те- ла одноцветная темно-коричневая, иногда почти черпая, брюшной — ' Н темно-желтая до серой. Сезонный диморфизм окраски слабо выражен. Как у самцов, так и у самок имеются специфические боковые кожные железы. Сосков 4 пары.
В черепе скуловые дуги широко расставлены. Лобно-теменные гребни сильно развиты. Резцовые отверстия короткие. Костные слуховые барабаны небольших размеров, тонкостенные, со слабо развитой губчатой костной тканью. У нижней челюсти угловой отросток небольшой.
Щечные зубы не имеют корней и растут в течение всей жизни животного.
Хромосом в диплоидном наборе 36.
Распространена на обширном ареале: от Атлантического побережья Европы на восток через европейскую часть СССР и Сибирь до Байкала и правобережья Лены, на юг до северного побережья Средиземного моря, Малой и Передней Азии, северных и юго-восточных частей Казахстана, Северо-Западного Китая, Алтайско-Саянской горной страны и северной части МНР, на север до кустарниковых тундр.
Держится в поймах рек, в заболоченных местах различных зон от лесотундры и южных частей тундры до степей включительно. Поднимается в горы до субальпийских лугов на высоту до 2800 м над уровнем моря. Населяет и культурные ландшафты — поля и огороды; поселяются даже в постройках. Резко выражена сезонная смена местообитаний, особенно в лесной зоне, где водяные полевки откочевывают зимой от берегов водоемов в пойменные луга или заросли кустарников. Убежищем служат норы, которые водяные полевки выкапывают неглубокими, простого строения. Гнездовая камера располагается обычно на глубине до 1 м, а кормовые ходы прокладываются на глубине 10—15 см. Осенний и зимний период жизни характеризуется активной роющей деятельностью. В теплое время года и в паводки устраивают из травы и других растительных материалов подземные или наземные гнезда. Период размножения продолжительный. В мягкие зимы он начинается уже в феврале и продолжается весь теплый период года. За это время самка приносит 4—6 пометов, в каждом из которых в среднем бывает 6—-8 детенышей. Длительность беременности около 40 дней.
В различные годы отмечены резкие изменения численности. Эти колебания особенно хорошо выражены для популяций, обитающих в поймах рек с изменяющимся паводковым режимом, а также для озер, уровень которых сильно колеблется по годам (например, в Казахстане). Во время высоких паводков численность водяных полевок резко сокращается.
Питается в основном различными растительными объектами, по большей части водными и прибрежными: тростником, рогозом, стрелолистом, осокой, кувшинками, различными луговыми травами. Поедает также и животные корма: моллюсков, насекомых, мелкую рыбу. В зимнее время питается главным образом подземными частями растений, а также корой и побегами ив, тополей, черемухи.
Зверьки некоторых популяций (на севере и востоке ареала) могут делать небольшие запасы пищи: корневища тростника, клубни карте феля.
Мех водяных полевок используется в качестве второстепенного пушного сырья.
Иногда они серьезно вредят леспым питомникам, садам, огородам, особенно расположенным в долинах рек.
Носители туляремийной инфекции и лептоспирозных заболеваний.
Некоторые исследователи (например, Андерсон и Джонс, 1967) включают водяную полевку в род серых полевок. Иногда к роду Arvicola относят Microtus richardsoni (Банфилд, 1974).
Рис. 82. Ондатра — О. zibefhicus и ее череп
Род ондатр
Genus Ondatra Link, 1795
В роде единственный вид: ондатра — О. zibethicus Linnaeus, 1758. Размеры средние и крупные. Длина тела 23—3G см. Длина хвоста Р-29,5 см. Масса 541—575 г. Ряд морфологических признаков сннде-,ствует о глубокой адаптации к водному образу жизни. Шейный пс-1т выражен слабо. Глаза небольшие, смещенные к верху головы inc. 82). Длинный хвост уплощен с боков, покрыт мелкими чешуйками едкими, короткими волосками. По его нижней стороне располагается ень удлиненных жестких волос. На задних конечностях имеются пе-Ьные плавательные перепонки. На передних конечностях самый длин-Ь палец третий, на задних— четвертый. Первый палец передних и ц-лих конечностей хорошо развит и имеет коготь. Стопа значительно ипнее кисти. Подошва стопы голая, с хорошо развитыми мозолями.
Волосяной покров высокий, густой и мягкий, резко дифференцированный на ость и пух. Окраска верхней стороны тела от коричневой до черновато-коричневой. Брюшная сторона несколько светлее спинной, иногда беловатая. Конечности и хвост темно-коричневые или черные. Сосков 3 пары. Имеются специфические перинеальные железы, продуцирующие мускусный секрет.
Череп с удлиненным лицевым отделом. Хорошо выражено сужение межглазничного промежутка. Сильно развиты лобно-теменные гребни, сливающиеся в острый срединный гребень. У взрослых животных сильно развит лямбдоидальный гребень. Резцовые отверстия узкие, но длинные. Костные слуховые барабаны относительно небольшие и не имеют губчатой костной ткани. Щечные зубы с корнями.
Хромосом в диплоидном наборе 54.
Распространена в северной и центральной части Северной Америки— северная граница ареала начинается на западе на .Аляске на побережье Чукотского моря несколько севернее залива Коцебу и идет к заливу Маккензи в море Бофорта, отсюда следует вдоль берега примерно до залива Ливерпуль и поворачивает на юго-восток к северной части Большого Медвежьего озера и далее западному берегу Гудзонова залива в месте его пересечения с 60° с. ш., затем идет по южному берегу залива до средней части восточного берега залива Джемса, поворачивает на восток, а в центральных районах п-ова Лабрадор на север и выходит к южной оконечности залива Унгава и далее следует вдоль его восточного берега до пролива Грай, откуда по берегу Атлантического океана поворачивает на юг, образуя восточную границу ареала, которая спускается к югу до южных частей штата Северная Каролина в США и сворачивает на запад, образуя южную границу ареала; она идет севернее границы между Северной и Южной Каролиной, пересекает с северо-востока на юго-запад Южную Каролину в ее западной половине, проходит по центральным районам Джорджии и тянется с востока на запад севернее границы между штатами Алабама и Флорида, затем сворачивает на северо-запад и выходит к точке, в которой сходятся границы трех штатов: Миссисипи, Арканзас и Луизиана, далее пересекает Арканзас с юго-востока на юго-запад и тянется севернее границы между Оклахомой и Техасом и далее от северо-восточного угла штата Нью-Мексико круто сворачивает на юг через крайнюю юго-западную территорию Техаса до государственной границы между США и Мексикой, затем поворачивает на северо-запад и тянется до северо-западного угла Аризоны, откуда делает поворот на восток через центральные районы Аризоны до западной части Нью-Мексико, где сворачивает вновь на запад и идет вдоль государственной границы с Мексикой, несколько к северу от нее; в юго-западном углу Аризоны выходит за пределы США, пересекает самый северо-западный участок штата Соноры в Мексике и в северо-восточной части Нижней Калифорнии сворачивает на север, образуя западную границу ареала, опять переходит в США и тянется западнее границы между Калифорнией п Аризоной, пересекает с запада на северо-восток южную оконечность Невады, доходит до центральных областей Юта. сворачивает на юго-запад, пересекает середину Невады, достигает Калифорнии и идет на север к западу от границы между Калифорнией и Невадой, поднимается к северу примерно до центральных районов Орегона, сворачивает на юго-запад, выходит к берегу Атлантического океана и далее следует к северу по береговой линии, отходя от псе к востоку на уровне о-ва Ванкувер и вновь выходя к ней примерно на уровне о-вов Королевы Шарлотты; изолированный участок ареала расположен на юго-востоке США, захватывая крайний юго-запад Алабамы, край
ний юг Миссисипи, южную половину Луизианы и крайний юго-восток Техаса.
Акклиматизирована в Европе и Азии. В Западную Европу была завезена в 1905 г. и в настоящее время обитает в Финляндии, Швеции, Норвегии, Бельгии, Нидерландах, Дании, Франции, ГДР, ФРГ, Польше, Чехословакии, Румынии, Болгарии, Югославии, Швейцарии, Австрии и Венгрии. В Англии к 1939 г. ондатра была уничтожена. В СССР в 1928— 1932 гг. 1646 ондатр было выпущено в Мурманской, Архангельской, Вологодской, Тюменской и некоторых других областях. С 1928 по 1970 г. включительно в различных областях СССР было выпущено около 300 тыс. В настоящее время ондатра заселяет водоемы севера и юга европейской части СССР, почти всю Западно-Сибирскую низменность, большую часть Восточной Сибири, ряд районов Якутии, Забайкалья и Дальнего Востока, многочисленна в Казахстане и местами в Средней Азии. Ондатра проникла в МНР, Северо-Западный и Северо-Восточный Китай. Промышляют в больших количествах, считается ценным пушным животным и к настоящему времени по ряду республик и областей СССР занимает первое место в промысле пушнины. В 1956 г. было добыто 6 млн. ондатр. В последующие годы заготовки ее снизились. Всего с 1937 го 1970 г. государство получило 99,5 млн. шкурок ондатры.
Населяет берега самых разнообразных водоемов, текучих и стоячих, пресных и солоноватоводных. В водоемах с низкими заболоченными бе-пе'ами, в болотистых местах и в плавнях устраивают гнезда — хатки диметром до 1,5 -2 м и высотой до 1 м с одной или несколькими камерами и входами, ведущими под воду. Хатки строит из различных растении. Там, где водоемы имеют высокие берега, ондатра роет сложные норы с выходами, расположенными ниже уровня воды, и гнездовой камерой—всегда выше уровня воды. Ондатра превосходно плавает и может оставаться под водой до 12 мин. Активна в течение зимы, делает отду-чшны во льду и ходы под снегом.
Питается различными прибрежными и водными растениями: тростником, рогозом, осокой, хвощом, камышом, стрелолистом. Поедает также животные корма — моллюсков, раков, лягушек, мелкую рыбу. Как и v других полевок, у ондатры имеются «кормовые столики». Иногда ондатра делает запасы. В северных частях ареала размножение начинается с весны и продолжается до осени, а у некоторых южных популяций длится круглогодично, но большинство самок приносит детенышей с ноября по апрель. В северных областях и горных районах в течение года бывает 1—2 помета, а в южных до 3 пометов. Длительность беременности составляет примерно 22—30 дней. Число детенышей в помете от 1 до 11, обычно 5—7. В первый год жизни половозрелыми становится лишь незначительное число самок. В различные годы отмечается значительное I изменение численности.
I Некоторые исследователи выделяют в роде еще один вид (например, Холл и Келсон, 1969): О. obscuras Bangs, 1894, или еще 2 вида (напри- мер, Эллермэн, 1941): О. obscuras и О. rivalicia Bangs, 1895. Более пра-I вильио считать, что в роде один вид.
к Род флоридских ондатр
pi Genus Neofiber True, 1884
В В роде единственный вид: флоридская ондатра — N. alleni True, 1884
Грис. 83).
Размеры средние. Длина тела 18,5—21,5 см. Длина хвоста 10—17 см. i jftacca 190—350 г. Адаптирована к водному образу жизни в меньшей
Рис. 83. Флоридская ондатра — N. alleru и ее череп
степени, чем ондатра. Хвост округлый в поперечном сечении с длинными, но редкими волосами, хороню развитым чешуйчатым покровом. Стопа значительно крупнее кисти. Пятый палец па передних конечностях короче трех средних, по длиннее, чем первый. Когти относительно хорошо развиты; на передних лапах они крупнее, чем на задних. Плавательные гребни волос по краям конечностей и на хвосте развиты слабо. Ушные раковины небольшие, высотой 1,5—2,2 см, и полностью прикрыты мехом головы. Волосяной покров густой, высокий и мягкий. Он резко разделяется на остевые и пуховые волосы. Общая окраска спинной стороны тела коричневая или черповато-коричпевая, брюшной — беловатая или с желтоватым оттенком. Сосков 3 пары.
Череп с сильным межглазничным сужением. Костные слуховые барабаны средних размеров. Резцы широкие. Щечные зубы не имеют корней. Третий коренной зуб на нижней челюсти небольшой.
Хромосом в диплоидном наборе 52.
Распространена в США на большей части Флориды и прилегающих островах, а также в юго-восточных районах штата Джорджия.
Населяет берега различных пресноводных водоемов, заболоченные ,а. Предпочитает участки, густо поросшие растениями. Обычно поселяется группами. Хорошо плавает. Плотность может достигать 250— 300 животных на гектар. Численность может сильно изменяться. Строит из различных растений гнезда диаметром 30—60 см, высотой 30—45 см, которые обычно имеют по два подводных входа и внутреннюю камеру диаметром примерно 10 см. Питается различными водными и прибреж-нпмп растениями. Как и для ондатр, типичны кормовые столики, расположенные недалеко от хатки. Размножение не приурочено к опреде-лсяному сезону. В течение года бывает от 4 до 6 пометов. Длительность беременности 26—29 дней. Детенышей в помете обычно 2 (от 1 до 4). Рождаются слепые и голые. Масса новорожденного 10—15 г. Полово-елость наступает в возрасте 90—100 дней.
Род фенакомис
J Genus Phenacomys Merriam, 1889
1
I'азмеры мелкие. Длина тела 10,4—10,6 см. Длина хвоста 2,6— !М. Масса 25—40 г. По внешнему облику сходны с серыми полевка-(рис. 84). Волосяной покров довольно высокий, мягкий и густой, аска его па спинной стороне тела серая или коричневая, на брюш-стороне беловатая, серая или розовато-темно-желтая. Хвост темно-1Й сверху и белый снизу. Стопа с 6 подошвенными мозолями. Сосков ры.
j черепе костные слуховые барабаны средних размеров, не содер-костной губчатой ткани. Характерно, что у щечных зубов есть ж.
фомосом в диплоидном наборе у Р. intermedins 56.
Распространены на большей части Канады и в западных районах А, вдоль Скалистых гор.
Заселяют различные биотопы: леса по берегам водоемов, участки, эытые травянистой растительностью или мхом от равнин до уровня а в горах. Экология изучена слабо. Самцы Р. longicaudus живут в ах, самки — на деревьях, где делают гнезда диаметром 20—30 см на оте 4—15 м над землей. Питаются хвоей, корой ивы, различными 1ами. Характерны неторопливые движения. Активны главным обра-ночыо — круглый год.
Р. intermedins размножается с мая по сентябрь. Длительность бере-ности 19—24 дня. За сезон бывает несколько пометов, в каждом из орых 2—8 детенышей. Масса новорожденных 2—2,7 г. На 14-й день их открываются глаза. Половозрелость наступает в 4—6 педель.
В роде 4 вида:
Р, intermedins Merriam, 1889 (провинции Канады — Ньюфаундленд, без о-ва Ньюфаундленда, Квебек, Онтарио, кроме юго-восточной око-ечности между озерами Онтарио, Эри и Гурон, Манитоба, кроме Крайли с аяаоюга, северные две трети Саскачевана, Альберта, кроме юго-востока, иная половина Северо-Западных территорий, юго-восток Юкона, Бри-
Рис. 84. Phenacomys silvicola u се череп и /’ intermedia* с 1(1-дневными детенышами
танская Колумбия, кроме узкой полосы тихоокеанского побережья; штаты США — Вашингтон, Орегон, кроме юго-востока, Айдахо, кроме юго-запада, западная половина Монтаны, северо-запад и юг Вайоминга, север и центр Юты, северо-запад, центр и центральные районы юга Колорадо, центральные районы севера Нью-Мексико; кроме того, неболь
шой изолированный участок ареала расположен на западе Невады в районе г. Карсон-Сити и востоке Калифорнии);
Р. albipes Merriam, 1901 (штаты США — запад Орегона п северо-запад Калифорнии);
Р. silvicola Howell, 1921 (США — центр крайнего запада Орегона);
Р. longicaudus True, 1890 (штаты США — крайний запад Орегона и
• северо-запад Калифорнии).
таге-та, К.о->ль-
Род серых полевок
Genus Microtus Schrank, 1798
Размеры от мелких до средних. Длина тела 11—20 см. Длина хвоста обычно меньше половины длины тела — 1,5—9,5 см. Внешний облик типичный для полевок (рис. 85). Шейный перехват выражен относительно слабо. Глаза очень небольшие у видов, ведущих преимущественно подземный образ жизни, средних размеров у слабоспециализированных или крупные у видов с преимущественно дневной активностью. Ушные раковины от очень маленьких, скрытых в шерсти головы у подземных видов, до относительно крупных. Хвост у большинства видов слабо оволоснен. У некоторых серых полевок, живущих на севере, он покрыт густыми волосами. Число подошвенных мозолей на стопе обычно 6, иногда 5. У палеарктических видов третий палец на обеих конечностях длиннее чет-чюого. Первый палец передней конечности, как правило, с тупым ног-азным когтем. У некоторых северных видов в зимнее время когти ,,,11'Овятся длиннее, чем летом. Подошвы лап, как правило, безволосые, иногда в задних отделах покрыты волосами. Волосяной покров обычно довольно высокий, густой и мягкий. Окраска его па спинной стороне тела v большинства видов серовато-коричневая, иногда черноватая или с рыжеватым или желтоватым оттенком на брюшной от сероватой или беловатой до бледно-коричневой. У видов, живущих на севере или в высокогорье, отмечен резкий сезонный диморфизм в густоте и длине волос Высокогорные с длинными вибриссами. Сосков 4, иногда 2 пары. На боках могут располагаться специфические кожные железы.
В черепе лицевой отдел укорочен. Мозговая коробка широкая. Скуловые дуги в передних отделах полого расходятся в стороны. Теменные гребни могут быть небольшие или развиты довольно сильно. Подглазничные отверстия узкие. Резцовые отверстия обычно средней длины и часто сужены. Межглазничное сужение может быть выражено хорошо ши почти полностью отсутствовать. Костные слуховые барабаны от небольших до крупных. Их внутренняя полость частично заполнена губчатой костной тканью. Щечные зубы не имеют корней. Величина и фор-~ кнель жевательной поверхности щечных зубов изменчивы. Петли от-; недалеко друг от друга и незначительно сжаты в передне-заднем явлении. На задних концах у М1 и М2 могут быть дополнительные ...пования— от небольших выступов эмали до дентинно-эмалевых нетель треугольной формы. Противолежащие петли могут сливаться, особенно у Л4з, реже у М, имеет 5, реже 6—8 замкнутых петель.
Хромосом в диплоидном наборе у серых полевок — от 17 у орегонской. 24 у горной, 32 у японской, 34 у полевки Брандта и 36 у узкоче-пеиной до 44 у полевки Томаса, 48 у М. multiplex, 50 у полевки Миддеп-дерфа и темной, 52 у большой и закаспийской, 54 у калифорнийской и афганской, 56 у снежной и длиннохвостой, 60 у скалистой.
Распространены на огромной территории умеренной субтропической а северной части тропических поясов Старого и Нового света. Населяют
Рис 85. Полевка-экоиомка— М. oeconomits и череп обыкновенной полевки — М. arvalis
большую часть Северной Америки на юг до Гватемалы, в Европе и Аиш от Атлантического до Тихою океанов па север до границы леса и южных районов тундры, а па юг до северного побережья Средиземного моря, Передней и Малой Азии, Северной Индии, центральных частей Китая; в Северной Африке — Ливию.
Населяют в Старом и Новом свете самые разнообразные ландшаф-ты, в умеренном, субтропическом и северной части тропического иоясов. Поднимаются в горы па высоту до 4500 м над уровнем моря Наиболее благоприятны для них, по-видимому, открытые ландшафты умеренною пояса. Предпочитаемые места поселения для многих видов — влажные
луга или полузаболоченные участки. Другие обитают в светлых лесах с богатым травяным покровом. Некоторые селятся в сухих степях. Ведут дневной или ночной образ жизни или активны в течение круглых суток. Обычно селятся колониями, устраивая сложные норы с несколькими выходами (от 2—3 до 300 и более), несколькими гнездовыми камерами и камерами для запасов корма. По краям колонии роют просто устро-•• енные временные норы. Выходы нор и места кормежек соединены тропиками. Норы могут быть и несложные, например у полевки-экономки; нередко у этого вида встречается наружное гнездо. Питаются преимущественно зелеными частями растений, а также корой древесных побегов, реже семенами и ягодами. Иногда в питании отдельных видов сущест-!:' венную роль играют подземные части растений. Некоторые виды запасают большое количество пищи. Размножаются преимущественно в теплый период года, но при благоприятных условиях и зимой. В течение года обычно бывает 3-—4, иногда до 7 пометов. Количество детенышей в помете обычно 5—6. В различные годы отмечены значительные изменения численности.
Могут серьезно вредить сельскохозяйственным культурам (посевам зерновых, огородным растениям, плодовым садам). Повреждают продукты, хранящиеся в складских помещениях. Некоторые виды — природные хранители возбудителей чумы, туляремии, лептоспирозных заболеваний.
В роде, по-видимому, 65 видов:
полевка Роберта—М. roberti Thomas, 1906 (Малая Азия; СССР — Западное Закавказье и западная часть Главного Кавказского хребта);
снежная полевка— М. шпа/й? Martins, 1842 (Средняя Испания; Франция; ФРГ; ГДР; Польша; Швейцария; Северная Италия; Австрия; Югославия; Румыния; Венгрия; Малая и Передняя Азия; СССР — Карпаты, Кавказ, Закавказье, Копетдаг);
I общественная полевка — М. socialis Pallas, 1773 (Малая Азия; Иран; I Сирия; Палестина; СССР — степи Крыма, юг Левобережной Украины, I Приазовье, степи Северного Кавказа, Восточное Закавказье, Калмыкия, I Казахстан, равнины Средней Азии; Китай — Джунгария);
/5 иранская полевка — М. Irani Thomas, 1921 (Иран и Ирак);
т полевка Гюнтера — М. guentheri Danford et Alston, 1880 (Греция; Ma-| тая Азия; Сирия; Палестина; Ливия);
J обыкновенная полевка—М. arvalis Pallas, 1779 (Франция; Бельгия; Нидерланды; Испания; Северная Италия; Швейцария; ФРГ; ГДР; Да-fl ния; Польша; Венгрия; Югославия; Чехословакия; Румыния; Греция; fl Малая Азия; Иран; СССР — в европейской части на север до Карелии, fl Архангельской области, Коми АССР и на юг до Кавказа и Закавказья fl включительно, Казахстан, Средняя Азия, степь и лесостепь Сибири на fl восток до верховьев Амура; МНР; Северный Китай);
fl- восточноевропейская полевка— М. subarvalis Meyer, 1972 (европей-fl ская часть СССР от Молдавии на западе и до Урала па востоке и от се-Д верных районов до Кавказа и Закавказья);
fl закаспийская полевка — Al. transcaspicus Satunin, 1905 (СССР — Копетдаг и долины рек Сырдарьи, Амударьи и Мургаба);
тянынанская серая полевка — М. ilaeus Thomas, 1912 (СССР — fl предгорья и горные долины Тянь-Шаня);
Д' монгольская полевка — М. mongolicus Radde, 1862 (северная и се-fl веро-восточная части МНР и Китая; СССР — южная и юго-восточная W части Читинской области и смежные с МНР районы Бурятии);
I японская полевка—М. montebelli Milne-Edwards, 1872 (Япония);
•s
оркнейская полевка—М. orcadensis Millais, 1904 (Оркнейские о-ва у Шотландии);
полевка Кабреры — М. cabrerae Thomas, 1906 (Испания);
югославская серая полевка — М. igmanensis Bolkay, 1929 (Югославия);
большая полевка — М. fortis Buchner, 1889 (северная часть МНР; Восточный Китай; Корейский п-ов; СССР — Западное и Восточное Забайкалье, на север до Благовещенска на Амуре и р. Селенга, а также южные районы Дальнего Востока);
восточная полевка — М. maximowiczii Schrenk, 1859 (=М. unguren-sis Kastschenko, 1912) (северная часть МНР; Северо-Восточный Китай; СССР — Забайкалье и Приамурье до низовьев Амура);
полевка Кларка — М. clarkei Hinton, 1923 (Китай —Юньнань и Северная Бирма);
тайваньская полевка — М. kikachii Kuroda, 1920 (о-в Тайвань);
темная полевка — М. agrestis Linnaeus, 1761 (Англия; Франция; Испания; Португалия; ГДР; ФРГ; Швейцария; Северная Италия; Норвегия; Швеция; Нидерланды; Дания; Польша; Венгрия; Югославия; Румыния; Финляндия.; в СССР — европейская часть от зоны тундры на юг до Северной Украины, Воронежской области и южного Урала, Западная Сибирь, бассейн Енисея, Прибайкалье и Якутия; МНР; Северный Китай);
полевка-экономка — М. oeconomus Pallas, 1776 (Норвегия; Швеция; ГДР; ФРГ; Нидерланды; Венгрия; Польша; Финляндия; СССР — зоны тундры, лесотундры, тайги и частично лесостепи к югу до Северной Украины, Воронежской области, Южного Урала, Северного Казахстана и Семиречья; МНР; Китай-д-Ганьсу и Шэньси; северо-запад Северной Америки — Аляска с соседними островами: Св. Лаврентия, Нунивак, Уналасика, Кадьяк и архипелагом Александра; провинции Канады — Юкон, кроме юго-востока, север и северо-запад Маккензи);
сахалинская полевка — М. sachcilinensis Vassin, 1955 (о-в Сахалин);
полевка Миддендорфа— М. middendorfi Poljakov, 1881 (СССР — Полярный Урал, низовья Оби, Таза и Енисея, Таймыр и Северная Якутия);
северосибирская полевка — М. hyperboreus Vinogradov, 1933 (СССР— Южный Таймыр, Верхоянский хребет, бассейны Яны, Индигирки, Колымы) ;
беренгийская полевка — М. abbreviates Miller, 1899 (о-ва Холли Святого Матвея в Беринговом море у Аляски);
сычуаньская полевка — М. millicens Thomas, 1911 (Китай — Сычуань) ;
пенсильванская полевка — М. pennsylvanicus Ord, 1815 (северная половина Северной Америки — центральные районы и юг Аляски, без п-ова Аляска; провинции Канады — Юкон, кроме северо-запада, Северо-Западные территории, кроме севера материковой части и Канадского Арктического архипелага; все остальные провинции Канады, кроме крайнего юго-запада Британской Колумбии, и о-ва Лабрадор; па о-не Ньюфаундленд в ареал входит только крайний юг; штаты США — северо-восток Вашингтона, север и восток Айдахо, центр и центральные районы севера Юты, запад, север и северо-восток Вайоминга, Моптапа, Северная Дакота, Южная Дакота, север и восток Небраски и все северо-восточные штаты на юг до центральных районов Миссури, центральных районов Иллинойса, крайнего юга Индианы, центра Кентукки, востока Теннесси, северо-востока Джорджии и крайнего юга Южной Каролины; сравнительно крупный изолированный участбк ареала расположен в
США на северо-востоке штата Нью-Мексико, крайнем северо-востоке Аризоны, в Колорадо, кроме запада и северо-запада и юго-востока, и крайнем юго-западе Небраски);
береговая полевка — М. breweri Baird, 1858 (о-в Мускегет у штата Массачузетс в США);
гуллская полевка — М. nesophilus Bailey, 1898 (о-в Большой Гулл у штата Нью-Йорк в США);
горная полевка — М. montanus Peale, 1848 (запад Северной Америки: Канада — узкая полоса, проходящая от центральных районов провинции Британская Колумбия на юг к государственной границе; штаты США—Вашингтон, кроме запада, северо-запада и крайнего северо-востока, Айдахо, кроме крайнего севера, юго-запад и центр Монтаны, Вайоминг, кроме востока и северо-востока, западная половина Колорадо, узкая полоса, пересекающая северо-западный угол Нью-Мексико с севера на юго-запад и заходящая недалеко на территорию Аризоны в ее восточной части, Юта, кроме юго-востока, Невада, кроме юга, северо-восток Калифорнии, большая часть Орегона, кроме западной прибрежной полосы; кроме того, два небольших изолированных участка ареала расположены один на юге штата Невада и второй на юге того же штата на границе с Калифорнией);
калифорнийская полевка — М. californicus Peale, 1848 (штаты США — юг и юго-запад Орегона и западная половина Калифорнии; Мексика — крайний север п-ова Калифорния; кроме того, имеется четыре небольших изолированных участка ареала: три в США Центральной и Южной Калифорнии и один в Мексике на севере п-ова Калифорния) ;
полевка Таунсенда — М. townsendii Bachman, 1839 (Канада — о-в Ванкувер; США — западные районы штатов Вашингтон и Орегон и крайний северо-запад штата Калифорния);
длиннохвостая полевка — М. longicaudus Merriam, 1888 (запад Северной Америки: крайний юго-восток Аляски; провинции Канады—южная половина Юкона, крайний юго-запад Альберта; Британская Колумбия; штаты США — Вашингтон, Орегон, север и северо-восток Калифорнии, Невада, кроме юга, Айдахо, западная половина Монтаны, Вайоминг, кроме востока, Юта, западная половина Колорадо, север и восток Аризоны и Нью-Мексико, кроме крайних востока и юга; кроме того, имеется три небольших изолированных участка ареала, расположенных: один на юге штата Калифорния, один на юго-востоке штата Аризона, один на северо-востоке Вайоминга и юго-западе Южной Дакоты);
островная полевка — М. coronarius Swarth, 1911 (о-в Коронации у Аляски);
мексиканская полевка — М. mexicunus Saussure, 1861 (юг Северной Америки, штаты США — крайний юго-восток Юты, крайний юго-запад Колорадо, восточная половина Аризоны, запад и юго-запад Ныо-Мексико; штаты Мексики — крайний запад Чиуауа, северо-восток Соноры, запад и юг Дуранго, юг Синалоа, Наярит, Халиско, Колима, Агуаскальентес, южная половина Закатекас, крайний юго-восток Коауила, южная половина Нуэво-Леон, юго-запад ТаМаулипас, Сап-Луис-Потоси, кроме запада, Идальго, Герреро, запад центрального района Веракрус, северная половина Пуэбла; кроме того, небольшой изолированный участок ареала расположен на крайнем западе Аризоны);
желтобрюхая полевка — М. fulviventer Merriam, 1898 (Мексика — центральные районы штата Оахака);
скалистая полевка-—М. chrotorrhinus Miller, 1894 (северо-восток Северной Америки, провинции Канады — южная половина Онтарио, юг и
юго-восток Квебека, Ньюфаундленд, кроме о-ва Ньюфаундленд, запад Ныо-Брунсуика; штаты США — северо-запад Мэн, север Нью-Хэмпшира, Вермонт, Массачузетс, восток Нью-Йорка, восток Пенсильвании, восток Западной Вирджинии, север Нью-Джерси, северо-запад Вирджинии, запад Северной Каролины, крайний восток Теннесси, крайний северо-восток Миннесоты);
желтощекая полевка-—М. xanthognathus Leach, 1815 (северо-запад Северной Америки — центр и центральная часть Аляски; провинции Канады — Юкон, кроме севера и юга, юг и запад Маккензи, крайний северо-восток Британской Колумбии, северная половина Альберта, север Саскачевана и север Манитобы);
афганская полевка — М. afghanus Thomas, 1912 (Афганистан; СССР — горы Большие Балканы, Копетдаг, долина Мургаба; хребты Нуратау, Гиссарский и Зеравшанский);
полевка Бедфорда — М. bedfordi Thomas, 1911 (Китай — Ганьсу);
полевка Брандта —М. brandti Radde, 1861 (МНР; Внутренняя Монголия и Северо-Восточный Китай; СССР — степи Южного Забайкалья);
китайская полевка — М. mandarinus Milne-Edwards, 1871 (Корейский п-ов; провинции Китая — Шаньси, Шэньси, Хэбэй; СССР — южная часть Бурятской АССР);
узкочерепная полевка — М. gregalis Pallas, 1779 (СССР —зоны тундры и лесотундры от Белого моря на западе до Берингова пролива на востоке, степи Северного и Восточного Казахстана: горы Средней Азии, степи Западной Сибири, Алтай, Саяны, Прибайкалье, Забайкалье, Якутия; МНР; Внутренняя Монголия и Северо-Восточный Китай);
поющая полевка—М. miurus Osgood, 1901 (юг и север Аляски, но не включая п-ов Аляска и берег моря Бофорта; провинции Канады — Юкон, кроме юго-востока и крайнего севера; крайний запад Маккензи);
тибетская полевка — М. leucurus Blyth, 1863 (Тибет, Кашмир);
сиккимская полевка — М. sikimensis Hodgson, 1849 (Сикким; Бутан); арчевая полевка — М. carruthersi Thomas, 1909 (СССР — горы Пами-ро-Алая, включая Гиссарский, Зеравшанский и Туркестанский хребты, кроме Памира, южный Бадахшан, западный Таласский Алатау; вероятно, Афганистан и Северо-Западный Китай);
памирская полевка — М. juldaschi Severtzov, 1879 (СССР — Памир и Алайская долина; Северо-Восточный Китай);
М. Irene Thomas, 1911 (Китай — Ганьсу, Сычуань, Юньнань; Восточный Тибет; Северная Бирма);
кустарниковая полевка — М. subterraneus de Selys Longchamps, 1836 (Бельгия; Нидерланды; Франция, ГДР, ФРГ; Швейцария; Северная Италия; Чехословакия; Польша; Венгрия; Югославия; Румыния; СССР — Закавказье, Северный Кавказ, Украина, Воронежская, Курская, Калининская и Вологодская области, Белоруссия);
малоазийская полевка — М. majori Thomas, 1906 (Малая Азия; СССР — Закавказье и Главный Кавказский хребет);
М. multiplex Fatio, 1905 (Швейцария и Северная Италия);
полевка Шелковникова — М. schelkovnikovi Satunin, 1907 (СССР— Талыш в Восточном Закавказье);
западноевропейская полевка—М. savil de Selys Longchamps, 1838 (Италия — Апеннинский п-ов, Сицилия; Южная Франция; Северная и Центральная Испания; Португалия);
средиземноморская полевка — М. duodecimcostatus de Selys Longchamps, 1839 (Южная Франция; Испания; Португалия);
полевка Томаса — М. thomasi Barrett-Hamilton, 1903 (Югославия, Греция);
чешская полевка — М. tatricus Kratochvil, 1965 (Чехословакия — Татры);
полевка Ричардсона — М. richardsoni Dekay, 1842 (ареал представлен двумя неравными по величине участками на западе Северной Америки: один, меньший по размерам, вытянут полосой с севера на юг от юго-западного угла провинции Британской Колумбии в Канаде, через центральные районы штатов Вашингтон, США, центр и запад Орегона, не выходя к берегу Тихого океана; второй, более крупный участок, захватывает юго-восточный угол провинции Британская Колумбия, юго-запад провинции Альберта, Канаду, крайний восток штатов Вашингтон, США, северо-восток Орегона, Айдахо, кроме юго-запада, северо-запад и запад Вайоминга, центр и центральные районы севера Юты);
орегонская полевка — М. oregoni Bachman, 1839 (Канада — крайний юг провинции Британская Колумбия; штаты США — западные полови-iH Вашингтона и Орегона и крайний северо-запад Калифорнии);
земпоалтепекская полевка — M. umbrosus Merriam, 1898 (Мексика — ора Земпоалтепек в штате Оахака).;
гватемальская полевка — М. guatemalensis Merriam, 1898 (Гвате-
>ла);
прерийная полевка — М. ochrogaster Wagner, 1842 (провинции Кана-1,1 — центр и юго-запад Альберта, юг Саскачевана, юг и юго-запад Ма-птобы; штаты США — восточные половины Монтаны и Вайоминга, Се-
рная Дакота, Южная Дакота, Небраска, Миннесота, кроме северо-остока, южная половина Висконсина, Иова, Иллинойс, Индиана, край-ий юго-запад Мичигана, Огайо, кроме северо-востока, крайний запад
Западной Виргинии, Кентукки, кроме северо-востока, крайний ссверо-за-ад Теннесси, крайний северо-восток Арканзаса, Миссури, Канзас, се-р и центральные районы Оклахомы, северо-восток Колорадо);
луизианская полевка — М. ludovicianus Bailey, 1900 (штаты США — крайний юго-запад Луизианы и крайний юго-восток Техаса);
сосновая полевка — М. pinetorum Le Conte, 1830 (восток и юго-вос-
ток США— северная граница ареала идет на восток от крайнего юга штата Мэн на берегу Атлантического океана через центральные районы Нью-Гэмпшира и Вермонта, север штата Нью-Йорк, по южному берегу Онтарио, пересекает самую южную оконечность провинции Онтарио между юго-западной оконечностью оз. Онтарио и югом оз. Гурон, огибает с юга оз. Гурон и Мичиган и несколько севернее г. Милуоки на западном берегу оз. Мичиган круто сворачивает на восток, доходит до центральных областей штата Висконсин, сворачивает на север и, не доходя до северной границы штата, поворачивает на юго-запад, превра
щаясь в западную границу ареала, которая идет через самую юго-восточную оконечность штата Миннесота, пересекает северо-запад Иовы, юго-восточный угол Колорадо, проходит с севера на юг по восточной трети Канзаса, через северо-запад Оклахомы, опускается с севера на юг по центральным районам Техаса и, дойдя примерно до 30° с. ш., сворачивает на восток, превращаясь в южную границу ареала, которая следует этой широте примерно юго-западного угла штата Джорджия, потом несколько уклоняется к северу и выходит к Атлантическому океану примерно у г. Саванна на границе штатов Джорджия и Южная Каролина; восточная граница ареала идет по берегу Атлантического океана, за исключением узкой прибрежной полосы штата Северная Каролина);
флоридская полевка — М. parvulus Howell, 1916 (США — северная часть п-ова Флорида);
центральномексиканская полевка — М. quasialer Cones, 1874 (штаты Мексики — крайний юго-восток Сан-Луис-Потоси, крайний северо-вос
ток Керетаро, северная половина Идальго, север Пуэбла, запад центральных районов Веракрус).
В СССР: полевки Роберта, снежная, общественная, обыкновенная, восточноевропейская, монгольская, закаспийская, тяньшанская серая, большая, восточная, темная, экономка, Миддендорфа, сахалинская,.северосибирская, афганская, Брандта, китайская, узкочерепная, арчевая; памирская, кустарниковая и малоазийская и полевка Шелковый-кова.
Систематика рода серых полевок остается до конца не установленной и трактуется различными учеными по-разному. Так, иногда в этот род включают водяных полевок (Андерсон и Джонс, 1967); из М. nivalis выделяют М. gud Satunin, 1909 (Эллермэн и Моррисон-Скотт, 1966); М. transcaspicus и М. mongolicus включают в М. arvalis (Б. А. Кузнецов, 1965, Эллермэн и Моррисон-Скотт, 1966); к этому же виду относят М. maximowiczii и М. fortis (Б. А. Кузнецов, 1965); М. orcadensis считают подвидом М. arvalis (Циммерман, 1959); М. curruthersi Thomas, 1909, объединяют с М. juldaschi (Б. А. Кузнецов, 1969), Е. Н. Ляпунова и Т. Б. Фишер (1969) подчеркивают разницу в хромосомных наборах у этих двух форм, однако В. Н. Большаков и др. (1969), В. Н. Большаков и А. В. Покровский (1969), по-видимому, ошибочно указывают на отсутствие у них репродуктивной изоляции; М. leucurus, М. kimensis, М. juldaschi, М. Irene, М. subterraneus, М. savii и М. duodecimcostatus выделяют в род Pitymys Murtrie, 1831 (Эллермэн и Моррисон-Скотт, 1966), что, вероятно, неправильно (справедливо, видимо, сомнение, высказываемое Б. А. Кузнецовым, 1965, по поводу отнесения европейских кустарниковых полевок к американскому роду Pitymysy, М. schelkovnikovi включают в М. subterraneus (Эллермэн и Моррисон-Скотт, 1966); из М. maximowiezi выделяют М. sachalinensis Vassin, 1955 (Б. А. Кузнецов, 1965); М. afga-nus выделяют в особый род Blanfordimus Argyropulo, 1933 (Эллермэн и Моррисон-Скотт, 1966); имеется и много других высказываний по поводу состава и структуры рода Microtus.
Microtus pennsylvanicus provectus и M. р. breweri включены в Красную книгу.
Род степных пеструшек
Genus Lagurus Gloger, 1841
Размеры от мелких до средних. Длина тела 12—21 см. Длина хвоста 4,5—6,5 см. По внешнему виду несколько сходны с леммингами (рис. 86). Телосложение тяжелое; туловище приземистое, конечности короткие, глаза маленькие, уши небольшие, едва выступающие над мехом головы. Короткий хвост покрыт довольно редкими волосами. На передних и на задних конечностях третий палец несколько длиннее четвертого. Первый палец передних конечностей сильно укорочен и имеет небольшой тупой коготь. Когти на остальных пальцах умеренной длины, в зимнее время немного длиннее, чем летом. Подошвы лап покрыты волосами. У основания пальцев расположены подошвенные мозоли. Выросты губ, загибающиеся в ротовую полость позади резцов, невелики. Волосяной покров относительно высокий, густой и мягкий. Окраска спинной стороны тела коричневато-охристая, иногда с продольной черной полоской вдоль середины спины. Бока светлее спины, брюхо серебристое или беловатое, иногда темно-желтоватое. У желтой пеструшки в зимнее время волосяной покров значительно выше и несколько бледнее летнего. Сосков 4 пары.
Рис 86 Степная пеструшка — L lagurus и ес череп
Череп угловатый, уплощенный сверху. Лобно-теменные гребни разлиты сильно, но не сливаются между собой в межглазничном промежут-х. Скуловые дуги широко расставлены. Межглазничное сужение хорошо сражено и у старых зверьков имеет продольные желобообразные углубления. Костные слуховые барабаны от небольших до крупных. Значительная часть их внутренней полости содержит губчатую костную ткань. Резцовые отверстия короткие. Щечные зубы не имеют корней и растут в течение всей жизни. Треугольные петли жевательной поверхности зубов сильно сжаты в передне-заднем направлении. Противолежащие петли не сливаются между собой. На передней стенке средних входящих углов М1 и М2 есть небольшой выступ.
Хромосом в диплоидном: наборе 54 у степной и 56 у желтой пеструшек.
Распространены в Старом и Новом свете. В Евразии па западе от Полтавской области до Присписсйскпх степей, .западной и южной частей МНР и Северного Китая па востоке, па север заходят в лесостепь. В Новом свете населяют запад CILIA н крайний юго-запад Канады.
Характерно обитание в засушливых местах, сухих степях, полупустынях и пустынях. Встречаются в южных частях лесостепи. Поселяются также и на пашнях, залежах, выгонах, по краям дорог. Активны в течение круглого года главным образом ночью, по часто деятельны и в тсче-
ние дня. Из нор выходят лишь на короткое время. Копают сложные норы с многочисленными выходами и несколькими гнездовыми камерами. У желтой пеструшки подземные ходы достигают в длину несколько десятков метров и занимают площадь до 200—400 м2. Кроме постоянных нор, у нее есть и временные с 1—4 входами, соединенными подземными ходами на глубине 20—40 см. Постоянные норы могут иметь 6—10 входов и подземные ходы на глубине до 50—60 см с 3—4 гнездовыми камерами овальной формы на глубине 40—80 см. В камерах устраивается подстилка из листьев злаков, реже полыни. Уборные находятся в от-норках недалеко от гнезд. Питаются различными растительными объектами, предпочитая узколистные злаки и белую полынь. Поедают также и некоторых беспозвоночных. Делают запасы пищи в норах.
Лемминговая пеструшка размножается в течение круглого года, а степная пеструшка — в летний период, в течение которого дает до 6 пометов. В помете в среднем бывает по 5—6 детенышей, иногда до 12. Половая зрелость наступает у нее в 1,5-месячном возрасте. Длительность беременности примерно 24—26 дней. Детеныши рождаются слепыми, с закрытыми ушами, почти голые. В различные годы отмечаются сильные изменения численности. В годы массового размножения совершают кочевки. Местами вредят сельскохозяйственным культурам, особенно пшенице, просу, овсу. Существенный вред во время массового размножения приносят пастбищам. Природные носители возбудителя туляремии.
В роде 3 вида:
степная пеструшка — L. lagurus Pallas, 1773 (СССР — европейская часть к востоку от Днепра и к югу от Рязанской и Горьковской областей, Татарская АССР, Западная Сибирь на восток до Енисея, большая часть Казахстана; Китай-—Джунгария; западная часть МНР);
желтая пеструшка — L. luteus Eversmann, 1840 (прежде была широко распространена в степях Казахстана, Китае — Джунгарии и МНР; в последние десятилетия в Казахстане почти вымерла; сохранилась лишь в Зайсанской котловине);
лемминговая пеструшка — L. curtatus Соре, 1868 (южные районы провинции Канады — юго-восток Альберта и юго-запад Саскачевана; штаты США — Монтана, кроме северо-запада и юго-востока, восток Северной Дакоты, Вайоминг, кроме северо-востока, северо-запад Колорадо, Юта, кроме юга и юго-востока, Невада, кроме крайнего юга, крайний северо-восток Калифорнии, восточная половина Орегона, центральные районы и центр юга штата Вашингтон).
В СССР: степная и желтая пеструшки.
А. А. Громов (1963) полагает заслуживающим выделение лемминго-вой пеструшки в особый род Lemmiscus Thomas, 1912.
Род прометеевых полевок
Genus Prometheomys Satunin, 1901
В роде единственный вид: прометеева полевка — Р. schaposchnikovi Satunin, 1901.
Размеры средние. Длина тела 13,5 17 см. Длина хвоста 3,1 6,5 см. Обладает рядом приспособлений к подземному образу жизни. Тело валь-коватое, с незаметным шейным перехватом (рис. 87). Голов,а крупная, укороченная. Глаза маленькие. Небольшие наружные уши скрыты в шерсти головы. Конечности короткие. Третий палец на передних конечностях значительно, а на задних немного длиннее остальных. Первые пальцы на передних и задних конечностях сильно укорочены, с исболь-
Рис. 87. Череп прометеевой полевки — Р. schaposclinikovi
шими когтями. Когти на остальных пальцах сильно развиты, особенно иа пальцах передних конечностей. Подошвы лап почти голые; волосы имеются только в их задних отделах. Хвост покрыт густыми волосами, образующими иногда на его конце слабо выраженную кисточку. Волосяной покров относительно низкий, густой и мягкий. Окраска спинной стороны тела коричневато-охристая, брюшной — серая с желтоватым оттенком.
Череп несколько укорочен, с широким лицевым отделом, тонкими скуловыми дугами. Межглазничное сужение хорошо выражено. Крупные лобно-теменные гребни сливаются в сагиттальный гребень. Межтеменная кость очень маленькая. Небольшие костные слуховые барабаны не имеют губчатой костной ткани. Резцовые отверстия удлиненные, крупные. Щечные зубы с корнями. Треугольные петли жевательной поверхности щечных зубов сильно заострены. Большая часть петель чередуется и слита только их средняя пара у Задние коренные укорочены. У М| на жевательной поверхности 3—5 петель. Верхние резцы со слабо выраженной продольной бороздкой па передней поверхности.
Хромосом в диплоидном наборе 56.
Распространена в СССР на Главном Кавказском хребте от западных окраин субальпийского пояса на восток до Военно-Грузинской дороги.
Населяет субальпийские и альпийские луга, а также высокотравные луга верхней части лесного пояса, гор на высоте примерно 1500— 1800 м над уровнем моря. Ведет почти исключительно подземный образ жизни. Норы, как правило, располагаются неглубоко под поверхностью земли и имеют сложную систему кормовых ходов. Гнездовая камера находится обычно на глубине 50—60 см от поверхности. В норе может жить 2—3 семьи, состоящие из значительного числа зверьков разного возраста. Питается различными растениями. В летпее время поедают, по-видимому, в основном надземные части растений, за которыми выходят на поверхность земли, но затем утаскивают их в норы и
там поедают. В зимнее время питаются подземными частями растений. Зимой в спячку не впадают. По-видимому, в течение года дают 2 помета, в каждом из которых от 2 до 5, в среднем 3 детеныша.
Род слепушонок
Genus Ellobius Fischer, 1814
Размеры мелкие. Длина тела 10—15 см. Длина хвоста 0,5—2,2 см. Хорошо выражены приспособления к подземному образу жизни. Тело вальковатое с незаметным шейным перехватом, несколько укороченной, клинообразной головой (рис. 88)..Гл аза очень небольшие, наружная ушная раковина редуцирована; вокруг наружного слухового отверстия есть лишь небольшая складка кожи. Короткие конечности с голыми подошвами. Третий палец на обеих конечностях немного длиннее четвертого. Первый палец на передних и задних конечностях хорошо развит. Когти на пальцах небольшие. По краю подошвы задней лапы волосы образуют своеобразную щетку. Внутренние выросты губ полностью изолируют сильно выступающие вперед резцы — от ротовой полости. Хвост покрыт негустыми волосами, на конце образующими небольшую кисточку. Волосяной покров довольно высокий, густой и мягкий. Окраска спинной стороны тела от светлой, палевой до темной, коричневатосерой. Голова обычно темнее спины. Брюшная сторона тела сероватая, коричневатая или беловатая.
Череп с удлиненным лицевым отделом (рис. 88). Скуловые дуги широко расставлены в стороны. Межглазничное сужение выражено слабо. Лобно-теменные гребни имеются. Костные слуховые барабаны уплощенные; в них развита крупноячеистая костная ткань. Резцовые отверстия укороченные. Щечные зубы с корнями. Противолежащие петли жевательной поверхности щечных зубов слиты между собой и часто соединяются с соседними петлями. .
Хромосом в диплоидном наборе 32, 52, 54 у обыкновенной, 36 у афганской и 17 у закавказской слепушонок.
Распространены в Евразии: от Южной Украины, горных степей Закавказья и Передней Азии до Приалтайских и Тувинских степей, МНР и Северо-Западного Китая; на север проникают в лесостепь.
Населяют лесостепи, степи и полупустыни равнин, а также высокогорные луга и степи. Поднимаются в горы до 4000 м над уровнем моря. Ведут подземный образ жизни, очень редко показываются на поверхности земли. Сложную систему кормовых ходов прокладывают близко к поверхности, а гнездовые камеры располагают на глубине 1—4 м. В одной норе, как правило, живут до десятка особей, принадлежащих к одной семье. Питаются в основном различными подземными, а также надземными частями растений. Активны круглый год. Во время летней жары и засухи активность резко понижается. Делают запасы пищи. Биология размножения изучена недостаточно. По-видимому, в некоторых участках ареала слепушонки могут размножаться круглый год. За теплый период времени года, вероятно, бывает 3—4 помета, в каждом из которых от 2 до 5, чаще 3 детеныша. Своей роющей деятельностью слепушонки оказывают существенное механическое и химическое воздействие на почву, перераспределяя значительные объемы почвы. Повреждают различные корнеплоды, огородные и бахчевые культуры, подгрызают кору и корни у саженцев плодовых культур, портят стенки оросительных каналов.
Рис. 88. Обыкновенная слепушонка — £. talpinus и ее череп
В роде 3 вида:
обыкновенная слепушонка — Е. talpinus Pallas, 1770 (СССР — степи Крыма, Южной Украины, Предкавказья, Калмыкии, Нижнего Поволжья, Татарии, Южного Урала, Зауралья, предал гайских районов, почти всего Казахстана, а также горы и равнины Средней Азии; МНР; Северный Китай);
афганская слепушонка — Е. fuscocapillus Blyth, 1841 (СССР — горы Копетдага и их предгорья, Бадхыз и Карабиль; Белуджистан; Афганистан; Иран);
закавказская слепушонка — Е. lutesccns Thonias, 1397 (СССР Талыш; Западный Иран; Турция).
В СССР: все три вида слепушонок.
Некоторые исследователи (например, Б. А. Кузнецов, 1905) объедн |яют афганскую и закавказскую слепушонок в одни вид — Е. fuscoca fillus
ПОДСЕМЕЙСТВО ПЕСЧАНОК
SUBFAMILIA GERB1LL1NAE ALSTON, 1876
Размеры от мелких до средних. Приспособлены к наземному существованию в аридной зоне.
Щечные зубы с высокой коронкой, у большой песчанки с постоянным ростом. У специализированных форм они состоят из поперечных пластин, разделенных внутренними и наружными глубокими складками. У примитивных форм зубы, как правило, не разделяются на пластины; их складки выражены слабо, на жевательной поверхности заметны следы бугорков.
Род монодия
Genus Monodia Heim de Balsac, 1943
Размеры мелкие. Длина тела примерно 5,5 см. Длина хвоста около 7,5 см. Хвост покрыт короткими, довольно густыми волосами, удлиняющимися к концу. Волосяной покров высокий и мягкий. Окраска спинной стороны тела песчаного цвета, брюшной — белая. Хвост желтовато-серый.
Распространены в Мавритании и Сомали.
В роде 2 вида:
М. mauritaniae Heim de Balsac, 1943 (Мавритания);
M. juliani Saint Leger, 1935 (Северное Сомали).
Род карликовых песчанок
Genus Gerbillus Desmarest, 1804
Размеры мелкие. Длина тела 8—13 см. Длина хвоста 7,5—12 см. Внешним видом напоминают тушканчиков (рис. 89). Задние конечности удлинены. Подошвы их голые (за исключением карликовой песчанки). Уши крупные. Глаза большие. Подошвы, стопы и кисти покрыты волосами (у ряда видов голые). Первый и пятый пальцы задней конечности относительно длинные. Когти пальцев передней конечности хорошо развиты. Хвост покрыт довольно густыми, по короткими волосами, удлиняющимися к концу его в небольшую кисточку. У многих видов по верхней стороне терминальной половины хвоста проходит невысокий гребень из удлиненных волос. Волосяной покров относительно высокий и мягкий. Окраска спинной стороны тела от бледно-желтовато-серой или песчано-темно-желтой до песчано-рыжеватой или серой, темножелтой или ярко-рыжевато-коричневой. Вдоль середины спины обычно проходит более темная полоска. У некоторых видов поперек туловища в области плеч располагается белая полоса. Бока светлее спины. Брюхо белого или кремового цвета, иногда светло-серое. У многих видов над глазом или позади ушей или в обоих этих местах имеются светлые пятна. Сосков 3 или 4 пары.
Череп с широкой мозговой капсулой и узким относительно длинным лицевым отделом. Лобно-теменные гребни обычно хорошо развиты, но иногда их нет вовсе. Скуловые дуги расставлены нешироко. Подглазничные отверстия узкие. Костные слуховые барабаны вздуты, их размеры сильно варьируют у разных видов. Резцовые отверстия длинные. Хорошо развитая вторая пара резцовых отверстий располагается между рядами щечных зубов. Они не такие длинные, как отверстия первой пары. Верхние резцы сжаты и имеют на наружной поверхности по одной бороздке.
Рис. 89. Карликовая пес шика — G gerb tins
Хромосом в диплоидном наборе 36 у южноафриканской песчанки, 40 v палестинской, 40, 52, 66 у египетской, 43 у карликовой, 52 у белуджистанской, 56 у североафриканской.
Распространены на обширной территории от Южной Африки па се-вер до Марокко и АРЕ и на восток через Сипай, Палестину, Сирию, северную часть п-ова Аравия, Ирак до Пакистана.
Населяют открытые сухие песчаные равнины. Активны преимущественно ночью. День проводят в порах, которые выкапывают короткими, простого строения. Часто поселяются сообществами, вырывая поры поблизости одна от другой. Питаются различными надземными и подземными частями растений, поедают также насекомых. Длительность беременности 20—21 день. В году дают от 2 до 4 пометов, в каждом из которых от 1 до 7, обычно 4 детеныша. В неволе жили до 5 лет.
В роде, по-видимому, 19 видов:
североафрпканская песчанка — G. campestris Levaillant, 1857 (от Марокко, Туниса, Ливии на восток до АРЕ и Сомали включительно, па юг до Судана);
аденская песчанка — G. poccilops Yerbury cl Thomas, 1895 (окрестности Адена);
чернохвостая песчанка — G. jamulus Yerbury et Thomas, 1895 (Южная Саудовская Аравия);
белуджистанская песчанка — G. nanus Blanford, 1875 (вся Сахара, Судан, Сомали и на восток до Южного Ирана п Белуджистана);
G. garamantis Lataste, 1881 (Тунис, Алжир);
G. dasyurus Wagner, 1842 (Западная Индия, Ирак, Саудовская Аравия, АРЕ, Ливия, Алжир);
G. henleyt de Winton, 1903 (АРЕ, Синай и Алжир);
карликовая песчанка — G. gerbillus Olivier, 1800 (от Западной Сахары по Палестины, Ирака и Восточного Ирана, па юг до Судана, Северной Нигерии, Уганды; возможно, также Южная Африка);
индийская карликовая песчанка — G. glcadowi Murray, 1886 (Северо-Западная Индия);
египетская песчанка — G. pyramidum 1. Geoffroy, 1825 (вся Сахара на восток до Сипая и Палестины);
песчанка Чизмапа — G. cheesmani Thomas, 1919 (Прак, Саудовская Аравия);
G. bottai Lataste, 1882 (Судан);
палестинская песчанка — G. allenbyi Thomas, 1918 (Палестина); сомалийская песчанка — G. dunni Thomas, 1904 (Сомали);
суданская песчанка — G. nancillus Thomas et Hinton, 1923 (Судан);
нигерийская песчанка — G. nigeriae Thomas, 1920 (от Нигерии до Судана);
южноафриканская песчанка — G. paeba Smith, 1836 (Южная Африка) ;
G. vallinus Thomas, 1918 (Ангола, Южно-Африканская Республика);
G. peeli de Winton, 1898 (Сомали);
G. simoni Lataste, 1881 (Алжир);
По Андерсону и Джонсу (1967), в роде 32, а по Уокеру (1964) — 54 вида.
Иногда G. paeba и G. vallinus выделяют в особый род G. erbillurus Shortridge, 1942, включая сюда же известный лишь по черепам из совиных погадок G. tytonis Bauer et Niethammer, 1959; G. peeli — в род Microdillus Thomas, 1910, a G. simoni — в род Dipodillus Lataste, 1881.
Род гололапых песчанок
Genus Tatera Lataste, 1882
Размеры от мелких до средних. Длина тела 9—19 см. Длина хвоста 12—24,5 см. Масса 30—100 г. Глаза крупные. Уши относительно длинные, округлые. Задние конечности значительно длиннее передних. Подошвы лап голые. Стопа узкая. Пятый палец па задних конечностях значительно короче трех центральных. Когти на пальцах передних конечностей часто удлиненные. Хвост полностью покрыт волосами, на конце может иметь небольшую кисточку волос. Волосяной покров мягкий или грубоватый. Окраска спинной стороны тела варьирует от светло-песчано-серой или темно-желто-серой до темно-желтой, черновато-коричневой, песчано-коричневой или темно-серой. Брюшная сторона тела белая или беловатая. Конечности светлые. Хвост у большинства видов слегка темнее, чем спина. Его конечная часть беловатая, а кисточка темная. Позади глаз и над ними и ушами, а также по бокам носа обычно расположены светлые или белые пятна. Сосков 3 или 4 пары.
Мозговая капсула черепа, как правило, не столь увеличена и расширена в задней части, как у большинства других родов песчанок. Обычно имеются лобно-теменные гребни, но, как правило, небольшие. Лицевой отдел узкий, заостренный. Костные слуховые барабаны крупные. Подглазничные отверстия сужены, особенно в нижней части. Передние резцовые отверстия длинные, обычно достигают зубных рядов. Задние резцовые отверстия различных размеров, иногда очень небольшие. Верхние резцы с продольной бороздкой.
Хромосом в диплоидном наборе 36 у песчанки Кемпа, 40 у чернохвостой гололапой песчанки, 42 у белобрюхой, 44 у песчанки Браптса и южноафриканской гололапой песчанки, 46 у гребнехвостой, 48 у гвинейской, 52 у саванной.
Распространены па о-ве Шри Лапка, в Индии, Пакистане, Турции, Иране, Ираке, Сирии, Саудовской Аравии и большей части Африки, за исключением области экваториальных лесов.
Населяют равнины, саванны и леспые участки. Активность, как правило, ночная. День проводят в норах, которые выкапывают часто сложными, уходящими на глубину до 1 м, со многими выходами и гнездовой
камерой — обычно в центре системы ходов. Выходы часто закрывают пробками земли. Как правило, эти зверьки передвигаются не спеша по поверхности на всех четырех конечностях, спасаясь же бегством, делают огромные прыжки до 1,5 м в высоту. Индийская гололапая песчанка . способна делать прыжки до 3,5 м в длину. Питаются побегами расте-j ний, луковицами, корневищами, а также семенами и насекомыми. Индийская гололапая песчанка, кроме того, поедает мелких млекопитающих, а также яйца и птенцов птиц. У некоторых видов (песчанка Брантса) размножение продолжается в течение всего года, у других, к как южноафриканская гололапая песчанка, размножение происходит Ж в определенный сезон. Длительность беременности в среднем 22 дня. В Обычно в помете бывает от 4 до 8 детенышей. Они остаются в гнезде К до 3—4-недельного возраста. Некоторые виды — природные хранители В возбудителей чумы.
W В роде, по-видимому, 11 видов:
J индийская гололапая песчанка — Т. indica Hardnicke, 1807 (Шри
Ланка, Индия, Пакистан, Турция, Иран, Ирак, Сирия, северная часть Д Аравийского п-ова);
Д. песчанка Боема — Т. boehmi Noack, 1887 (Африка — от экватора на Ж юг до 14° ю. ш.; Юго-Восточная Уганда, Юго-Западная Кения, Руанда, В Бурунди, Западная Танзания, Заир, Малави, Замбия, Ангола);
В белобрюхая песчанка — Т. leucogaster Peters, 1852 (от нижнего те-Ж чения Оранжевой реки на юго-западе и Зулуленда на юго-востоке да-В лее на север до Анголы, юга Заира и юго-запада Танзании);
В чернохвостая гололапая песчанка — Т. nigricauda Peters, 1878 (се-В', веро-восток Уганды, Кения, юго-запад Эфиопии и северо-восток Танза-Т‘ нии);
< гребнехвостая песчанка — Т. robusta Crcter, 1826 (от Западной Аф-1 рики до Эфиопии и Сомали и на юг до северных районов Центральной 1 Танзании);
южноафриканская гололапая песчанка — Т. afra Gray, 1830 (Южно-Lj Африканская Республика);
саванная песчанка — Т. valida Bocage, 1890 (от Западной Африки £ до Судана к югу от 15ю с. ш.);
? песчанка Брантса — Т. brantsii Smith, 1834 (Южно-Африканская 4 Республика, юг Анголы, юго-запад Родезии, северо-восток Ботсваны); ; Т. inclusa Thomas et Wroughton, 1908 (от Родезии на север через 1 .Мозамбик до Северо-Восточной Танзании);
.j песчанка Кемпа — Т. kempii Wroughton, 1906 (Южная Нигерия);
| гвинейская песчанка — Т. guineae Thomas, 1910 (Гвинея-Бисау, Га-( на, Гамбия);
; Система рода точно не установлена. По Андерсону и Джонсу (1967), Е в роде 25 видов.
I Род длиннолапых песчанок
I Genus Taterillus Thomas, 1910
Размеры мелкие. Длина тела 10—14 см. Длина хвоста 14—17 см. Т Глаза очень крупные. Задние конечности длинные, обычно с голой no-fl дошвой, иногда покрытой волосами в пяточной области. Хвост покрыт .1 довольно густыми и короткими волосами. На конце хвоста удлиненные 1 волосы образуют кисточку (рис. 90). Окраска спинной стороны тела от 1 бледно-желтой и светло-рыжевато-коричневой до темно-коричневой. 1 251
Рис. 90 Нигерийская длиннолапая песчанка — Т. nigeriae
Бока более светлые. Брюшная сторона белая или почти белая. По бокам морды выше глаз и на задних частях ушей часто бывают белые пятна. У некоторых видов на морде видны темные отметины. Хвост примерно такой же окраски, как и спина.
Череп напоминает таковой гололапых песчанок. Верхние резцы имеют по бороздке на наружной поверхности. Лобно-теменные гребни имеются, иногда очень небольших размеров. Костные слуховые барабаны относительно невелики.
Хромосом в диплоидном наборе 30 у нигерийской длппполапоп песчанки, 36 у стройной, 44 у Т. emini и 54 у конголезской песчанки.
Распространены в Африке: в Сенегале, Нигерии, Центрально-Африканской Республике, Судане, Заире, Уганде, Кеппи, Эфиопии.
Населяют безлесные равнины, покрытые густыми кустарниками. Живут сообществами. Роют довольно сложно устроенные норы. Активны ночью. Экология изучена плохо.
В роде, по-видимому, 16 видов:
стройная песчанка — Т. gracilis Thomas, 1892 (Гамбия и Северная Нигерия);
нигерийская длиннолапая песчанка — Т. nigeriae Thomas, 1911 (Северная Нигерия);
Т. lacustris Thomas et Wroughton, 1907 (окрестности оз. Чад); конголезская песчанка — Т. congicus Thomas, 1915 (Заир);
Т. emini Thomas, 1892 (Уганда, Эфиопия, Судан);
песчанка Осгуда — Т. osgoodi Wroughton, 1910 (Кения);
Т. nubilis Dollman, 1911 (Кения);
Т. lowei Dollman, 1914 (Уганда);
Т. melanops G. Allen, 1912 (Кения);
Т. tenebricus Dollman, 1911 (Кения);
песчанка Харрингтона — Т. harringioni Thomas, 1906 (восточное побережье оз. Рудольфа);
рыжая песчанка — Т. rufus Wettstcin, 1916 (Судан);
Т. gyas Thomas, 1918 (Судан);
Т. clivosus Thomas et Hinton, 1923 (Судан);
T. perluteiis Thomas cl Hinton, 1923 (Судан);
песчанка Бутлера — Г. butleri Wroughton, 1910 (Судан).
По Андерсону и Джойсу (1967), в роде 11 видов. Эллермэн (1941) предполагает возможность объединения всех перечисленных выше видов в один вид.
Ж?
Рис. 91. Короткоухая песчанка — D. auricularis
Род короткоухих песчанок Genus Desmodillus Thomas et Schwann, 1904
В роде единственный вид: короткоухая песчанка — D. auricularis К. Smith, 1834.
Размеры мелкие. Длина тела 9—12,5 см. Хвост несколько короче. Уши относительно короткие, округлые, глаза крупные (рис. 91). Подошва покрыта волосами. Волосяной покров высокий и мягкий. Окраска спинной стороны тела варьирует от орапжеватого до темно-коричневого. Позади ушей и на затылке обычно расположены белые пятна. Брюхо и конечности белые. Сосков 4 пары.
В черепе костные слуховые барабаны сильно вздуты. Продольная бороздка на наружной поверхности верхних резцов выражена слабо.
Хромосом в диплоидном наборе 52.
Распространена в Юго-Западной и Южной Африке.
Населяет открытые песчаные равнины, иногда окультуренные земли. Убежищем служит выкопанная обычно на глубину не более 2 м нора простого строения. В конце норы располагается гнездовая камера. Живут поодиночке, парами или семейными группами. Активна в основном ночью. Питается различными семенами, а также насекомыми. Во время засухи, в бескормицу, могут мигрировать.
Род десмодиллискус
Genus Desmodilliscus Wettstein, 1917
В роде единственный вид: десмодиллискус — D. braueri Wettstein, 1917.
Размеры мелкие. Длина тела 5—7 см. Длина хвоста составляет примерно 75% длины тела. Подошвы лап голые. Хвосг покрыт средней густоты волосами; кисточки на его конце нет. Волосяной покров мягкий. Спинная сторона тела желто-песчапая, бока светлее, брюхо белое. Есть защечные мешки.
Череп напоминает таковой короткоухой песчанки. Костные слуховые барабаны сильно вздуты. Костное небо широкое. Передняя пара резцовых отверстий широкая, вторая — широкая и длинная, почти такой же
Рис. 92. Жирнохвостая песчанка — Р duprasi
длины, как ряды зубов. Венечный отросток нижней челюсти очень мал.
Характерно, что в нижней челюсти всего два коренных зуба.
Распространение охватывает Северную Нигерию и Судан.
Экология не изучена.
Возможно, этот вид следует включить в род короткоухих песчанок.
Род жирнохвостых песчанок
Genus Pachyuromys Lataste, 1880
В роде единственный вид: жирнохвостая песчанка — Р. duprasi La-taste, 1880.
Размеры мелкие. Длина тела 10,5—13,5 см. Длина хвоста до 4,5— 6,0 см. Телосложение довольно тяжелое (рис. 92). Глаза большие. Уши короткие, с округлыми вершинами. Короткий хвост утолщен па конце и покрыт довольно густыми волосами. Задние конечности незначительно длиннее передних. Первый палец на них короткий. Задняя часть подошвы передних и задних конечностей голая. Когти на передних конечностях сильно развиты. Волосяной покров мягкий. Окраска его па спинной стороне тела светло-желто-серая или темно-коричневая с белыми пятнами позади ушей. Брюшная сторона и конечности белые. Хвост двухцветный. Окраска дорзальной его части совпадает с окраской спины, вентральной та же, что и на брюхе.
В черепе костные слуховые барабаны очень сильно увеличены и сдвинуты далеко назад по отношению к большому затылочному отверстию. Лобно-теменные гребни имеются. Верхние резцы несут на наружной поверхности по одной бороздке.
Хромосом в диплоидном наборе 54.
Распространена в Северной Африке от Алжирской Сахары до юго-западной части АРЕ.
Населяет песчаные районы с бедной растительностью. В помете от 3 до 6 детенышей. Беременность длится 19—22 дня.
2S4
Род аммодил
Genus Ammodillus Thomas, 1904
В роде единственный вид: аммодил-—A. imbellis de Winton, 1898. Размеры мелкие. Длина тела 11 см. Длина хвоста около 14,4 см. Задние конечности длиннее передних. Длинный хвост не очень густо покрыт волосами и имеет на конце кисточку. Стопа узкая с голой подошвой. Когти на передних пальцах хорошо развиты. Окраска спинной стороны тела рыжевато-желто-коричневая с черноватым оттенком. Брюхо и конечности белые. У основания ушей и над глазами располагаются белые пятна. Хвост по верхней стороне более темный.
Череп расширен в задней части. Коренные зубы гипселодонтного типа. Носовые кости выступают далеко вперед над резцами. Костное небо расширено спереди; ряды щечных зубов сходятся в задних отделах. В нижней челюсти нет венечного отростка. Вторая пара резцовых отверстий почти незаметна.
Распространен в Сомали.
Населяет песчаные равнины. Экология не изучена.
Род малых песчанок
Genus Meriones liliger, 1811
Размеры от мелких до средних. Длина тела у мелких форм до 15 см, у крупных — до 18,5 см. Внешним видом несколько напоминают крысу (рис. 93). Уши округлые, средних размеров или крупные, у индийской песчанки небольшие. Глаза крупные. Хвост опушен довольно густыми волосами и обычно имеет на конце небольшую метелку. Первый палец на передних конечностях довольно хорошо развит, всегда с крупным когтем. Стопа узкая, ее подошва обычно покрыта волосами, иногда голая. Волосяной покров у большинства видов высокий и мягкий, но у некоторых видов (королевская и индийская песчанки) очень короткий и грубый. Окраска спинной стороны однотонная, охристо-песчаная, иногда сероватая или коричневатая. Бока обычно светлее спины. Брюхо и конечности белые, бледно-желтоватые, темно-желтые или светло-серые. На морде иногда бывают светлые участки.
Лицевой отдел черепа удлинен и сжат с боков. Мозговая коробка сверху уплощена. Лобно-теменных гребней нет. Межглазничное сужение сильно выражено. Костные слуховые барабаны очень велики. Щечные зубы с корнями, но с высокой коронкой. Верхние резцы с продольной бороздкой по наружной поверхности.
Хромосом в диплоидном наборе 38 у пышнохвостой песчанки, 40 у индийской песчанки и гребенчуковой, 44 у песчанки Виноградова, когтистой, африканской и краснохвостой, 50 у полуденной, 60 у песчанки Сундевалла.
Распространение охватывает степи, полупустыни и пустыни Северной Африки, Передней и Малой Азии, Кавказа, северо-восточного Предкавказья, Средней Азии, Казахстана, Ирана, Забайкалья, МНР и Китая.
Обычно населяют сухие степи, полупустыни и пустыни на равнинах, в предгорьях, иногда в горах. Местами поднимаются в горы на высоту до 3000 м над уровнем моря. Активны преимущественно ночью. День проводят в норах, которые выкапывают сами. Норы часто бывают сложно устроены и имеют много отнорков. Зимовальные норы более
Рис. 93. Полуденная песчанка — М. meridianus и ее череп
глубокие. У полуденной песчанки длина их достигает 2 м с общей длиной ходов до 4 м. У некоторых песчанок поры более простые (гребен-чуковая песчанка). Норы иногда располагаются группами, образуя своеобразные городки (краснохвостая песчанка). Питаются семенами, зелеными и подземными частями растений, а также, насекомыми и некоторыми другими животными. Многие виды запасают корм па зиму. В течение года бывает 2—3 помета, в каждом из которых от 1 до II (в среднем 4—6) детенышей. Длительность беременности примерно 25—29 диен. Численность малых песчанок может сильно изменяться по годам. Местами они приносят вред посевам зерновых, хлопку. Имеют важное эпидемиологическое значение и качестве природных носителей возбудителен чумы, клещевого возвратного тифа, кожного лейшманиоза и некоторых других заболеваний.
В роде 13 видов:
персидская песчанка — М. persicus Blanford, 1875 (Иран, Афганистан, Белуджистан, Турция; СССР — горы Копетдаг и Большие Балхаша, южная Армения, Нахичеванская АССР, Карабах, Талыш);
королевская песчанка — М. rex Yerbury et Thomas, 1895 (южная часть Аравийского п-ова);
индийская песчанка — М. hurrianae Jerdon, 1867 (Северная и Западная Индия, Афганистан, Иран);
песчанка Виноградова — М. vinogradovi Heptner, 1931 (СССР — Нахичеванская АССР и юго-восточная Армения; северо-западный Иран, северо-восточная Турция); '
гребенчуковая песчанка—М. tamariscinus Pallas, 1773 (СССР — на север до Элисты, Енотаевска, Актюбинска, оз. Аккуль, нижнего течения Сарысу, центральной части пустыни Бетпак-Дала, северного Прибалхашья и оз. Алаколь, на юг до песков Каракум, Самаркандского и Бухарского оазисов и Ферганской долины, в долинах озер Зайсан и Иссык-Куль; Северо-Западный Китай);
малоазиатская песчанка — М. blackleri Thomas, 1903 (Турция; Иран; Сирия; СССР — южная Армения, Нахичеванская АССР, по р. Куре от Тбилиси до Евлаха, Карабах, Мильская и Мургабская степи, предгорья Талыша);
когтистая песчанка — М. unguiculatus Milne-Edwards, 1867 (СССР — южное Забайкалье и юг Тувинской АССР; МНР; Северный и Северо-Восточный Китай);
полуденная песчанка — М. meridianus Pallas, 1773 (СССР — Восточное Предкавказье, Нижнее Поволжье, Казахстан, равнины Средней Азии, степи Тувинской АССР; Северный Афганистан; Северо-Восточный Иран; МНР; Северный Китай);
африканская песчанка — М. shawl Duvernoy, 1842 (Марокко; Алжир; Тунис; Ливия; АРЕ; Палестина);
аравийская песчанка—М. arimalius Cheesman et Hinton 1924 (центральная часть Аравийского п-ова);
краснохвостая песчанка — М. libycus Lichtenstein 1823 (СССР — равнины и предгорья Средней Азии и Южного Казахстана к северу до низовьев Урала, Устюрта, Аральских Каракумов, среднего течения Сарысу, Прибалхашья; Китайский Туркестан; Белуджистан; Афганистан; Иран; Ирак; Палестина; Сирия; Саудовская Аравия; АРЕ; Ливия; Алжир и до Рио-де-Оро);
песчанка Сундевалла — М. crassus Sundevall, 1842 (Алжир; Ливия; АРЕ; Саудовская Аравия; Синай; Палестина; Иран; Ирак; Афганистан; северо-западная часть Индии);
песчанка Зарудного — М. zarudnyi Heptner, 1927 (Иран; Афганистан; СССР —юг Туркмении).
В СССР: песчанки персидская, гребенчуковая, малоазиатская, когтистая, полуденная, краснохвостая и Зарудного.
Род пушистохвостых песчанок
Genus Sekeetaniys Ellerman, 1947
В роде единственный вид: пушистохвостая песчанка —S. calurtis Thomas, 1892.
Размеры мелкие. Длина тела 10—12,5 см. Длина хвоста И—16 см. Общий вид, как у других песчанок (рис. 94). Вибриссы очень длинные. Характерно, что хвост густо покрыт довольно длинными волосами, об-
Рис. 94. Пушистохвостая песчанка — S. calurus
разующими на конце кисточку. Подошвы задних лап голые. Окраска спинной стороны тела желтоватая или рыжеватая с черным оттенком. Брюшная сторона тела и конечности белые. Хвост коричневатый, кисточка его белая.
Распространена в Палестине и на Сипайском п-ове.
Населяет пустыни и каменистые склоны холмов. Характерно обитание на твердом грунте. Способны быстро лазать ио камням. Норы часто роют под камнями. Размножение идет круглый год. В помете бывает в среднем 3 детеныша, максимум до 6.
По Андерсону и Джонсу (1967), в роде 2 вида. Эллермэн (1966) включает пушистохвостую песчанку в род Meriones в качестве подрода.
Род песчанок Пржевальского
Genus Brachiones Thomas, 1925
В роде единственный вид: песчанка Пржевальского — В. przewalskii Biichner, 1889.
Размеры мелкие. Длина тела 8—9,5 см. Длина хвоста 7—8 см. Телосложение тяжелое. Конечности несколько укорочены. Хвост покрыт довольно густыми волосами. Уши маленькие. Концевой кисточки иа хвосте нет. Когти на передних конечностях длинные. Подошва стопы полностью покрыта волосами, подошва кисти голая. Окраска на спинной стороне тела бледно-серовато-желтая, бледпо-жел говатая или песчаносерая. Конечности и брюшная часть тела белые. Хвост темно-желтый или беловатый.
Распространена в Синьцзяне и в МНР (в пустыне Гоби).
Населяет пустыни. Экология не изучена.
Род дневных песчанок
Genus Psammomys Cretzschniar, 1828
В роде единственный пид: дневная песчанка — Р obesus Cretzsch-шаг, 1828.
Размеры мелкие и средние. Длина тела 13—18,5 см. Длина хвоста 11—15 см. Телосложение тяжелое. Задние конечности незначительно 258
длиннее передних. Хвост полностью покрыт волосами и имеет на конце небольшую кисточку. Подошвы лап покрыты волосами. Уши небольшие, округлые и толстые. Окраска спинной стороны тела рыжеватая с черноватым оттенком, желтоватая, рыжевато-коричневая или охристая. Брюхо и конечности беловатые.
Череп сходен с черепом представителей рода малых песчанок, но межглазничная область сжата сильнее, а лобно-теменные гребни более крупные. Костные слуховые барабаны вздуты. Верхние резцы не имеют продольной борозды. Щечные зубы с корнями, но с высокой коронкой.
Хромосом в диплоидном наборе 48.
Распространена в Алжире; Тунисе; Ливии; АРЕ; Судане; Израиле; Саудовской Аравии.
Населяет песчаные равнины с бедной растительностью. Активна в основном днем. Живет колониями. Строит сложные норы с многими выходными отверстиями. Питается различными растениями, запасая их в своих норах. По-видимому, самки дают по одному помету в год с марта по апрель; от 3 до 5 детенышей.
Род больших песчанок
Genus Rhombomys Wagner, 1841
В роде единственный вид: большая песчанка — R. opimus Lichtenstein, 1823.
Размеры от мелких до средних. Длина тела 13,5—20 см. Длина хвоста 12,6—16 см. Общий облик несколько напоминает крысу (рис. 95). Морда притупленная. Ушные раковины небольшие. Задние конечности незначительно длиннее передних. Первый палец задней конечности укорочен. Когти на пальцах лап довольно длинные. Подошвы передних в задних лап покрыты волосами. Хвост опушен довольно густыми волосами, образующими на конце кисточку. Волосяной покров густой и мягкий. Для него характерен сезонный диморфизм: в зимнее время он гуще и выше, чем летом. Окраска спинной стороны тела песочно-желтая, оранжеватая, темно-желтая или темно-серовато-желтая. Брюшная поверхность беловатая. Имеется среднебрюшная кожная железа.
В черепе лицевой отдел укорочен. Мозговая коробка уплощена сверху. Резцовые отверстия короткие. Нижняя челюсть со вздутой наружной поверхностью и отклоненным наружу венечным отростком. Щечные зубы гипселодонтного типа, без корней. Верхние резцы на наружной поверхности с двумя продольными бороздками.
Хромосом в диплоидном наборе 40.
Распространена в равнинных и предгорных полупустынях и пустынях от восточного Прикаспия и центральной части Передней Азии до южного Прибалхашья, МНР, Северо-Западного и Северного Китая; в СССР северная граница ареала тянется от правого берега р. Урал к северу от г. Гурьева, пересекает р. Эмбу несколько севернее ее большой излучины, севернее г. Челкара, к югу от г. Иргиза огибает с востока Приаральские Каракумы, проходит через низовья Сарысу по пустыне Бетпак-Дала, достигает западного берега оз. Балхаш, следует по его южному побережью до р. Лепсы и по восточным предгорьям Заилийско-го Алатау доходит до государственной границы СССР; к югу от этой северной границы большие песчанки встречаются во всех подходящих для ее жизни местах вплоть до государственной границы СССР.
Населяет бугристые и грядово-бугристые песчаные пустыни, а также глинистые и лессово-песчаные пустыни предгорных районов. Обычно ио-
Рис. 95. Большая песчанка — /?. opimus и ее череп
селяется в межгрядовых понижениях, в нижней части склонов песчаных бугров. Селится колониями, сооружая сложные подземные городки. Норы располагаются в несколько этажей, нижний из которых может находиться на глубине более 2 м. В зависимости от возрасту колонии она может иметь более сотни н даже несколько сотой выходных отверстий, много камер для запасов пищи н гнездовую камеру. Камера для запасов иногда достигает до 1 м в длину и до 25 см в высоту. Жилая камера находится обычно на значительной глубине иногда до 3 м. Активны в основном днем. В жаркое летнее время активность смещается на ранние утренние и сумеречные вечерние часы. Зимой во время сильных морозов или снегопадов по нескольку дней не выходит па поверхность. Характерен тревожный крик или топанье задней ногой по почве при виде врага.
1 Питается растительной пищей, в том числе ветвями пустынных кус-I тарников. Лазает по кустарникам и обгрызает веточки. При запасании ? пищи существует разделение труда, когда одна песчанка сгрызает ветки | с кустарников и сбрасывает на землю, а другие затаскивают эти веточ-' ки в нору. Размножение начинается весной, оканчивается с наступлением жаркого лета. Иногда размножение идет и осенью. За весну бывает i 2, иногда 3 помета, в среднем по 5-—6 детёнышей в каждом.
1 Характерно изменение численности в различные годы. Местами при-1 нссит вред посевам зерновых. При рытье нор иногда разрушают берега L арыков, железнодорожное полотно. Имеет важное эпидемиологическое К значение в качестве природного носителя возбудителей чумы, сыпноти-К фозных лихорадок, клещевого возвратного тифа, лейшманиоза и неко-И торых других. В Средней Азии и Казахстане проводятся широкие меро-S приятия по уничтожению больших песчанок в районах, имеющих эпи-Т демиологическое значение.
1 СЕМЕЙСТВО СЛЕПЫШОВЫХ
| FAMILIA SPALACIDAE GRAY, 1821
I Грызуны мелких размеров. Длина тела 23—25 см. Длина хвоста до I 3,6 см. В высокой степени приспособлены к подземному образу жизни. А Телосложение тяжелое, туловище вальковатое. Шейный перехват сиа-Л: ружи незаметен (рис. 96). Голова короткая, тупая, сверху сильно упло-S щенная. По бокам головы располагаются два жестких голых кожистых В канта. Небольшие глазные яблоки находятся под кожей. Глазные мышцы и глазной нерв развиты слабо или отсутствуют. Наружное ухо имеет вид небольшого кожного валика. Резцы используются в качестве органа рытья. Внутренние выросты губ изолируют резцы от ротовой полости и при копании земля в ротовую полость не попадает. Конечности пятипалые, сильно укороченные. Когти на всех пальцах хорошо развиты. Когти тупые, с закругленными концами. Волосяной покров довольно низкий, но густой и очень мягкий. Окраска его варьирует от темносерого до светло-песчаного, иногда охристо-коричневая.
Череп характерен клиновидными очертаниями Затылочная область черепа сильно увеличена в размерах и наклонена вперед. Скуловые дуги относительно слабые, широко расставлены. Надглазничных отростков у лобных костей нет. Сагиттальный гребень крупный. Ламбдовидный гребень имеется. Характерно, что скуловая кость не соприкасается с слезной. Подглазничные отверстия крупные. Костные слуховые барабаны небольших размеров, уплощенные. Нижняя челюсть с небольшим угловым отростком. Венечный отросток нижней челюсти широкий, длинный, а сочленованный сильно укорочен. Характерно, что па осевом черепе имеется две сочленовные поверхности для нижней челюсти; на передней из них нижняя челюсть располагается при рытье и грызс-нии, на задней — при жеванйи и в состоянии покоя. Зубная формула: i—- с — р — т —= 16. Коронки коренных зубов средней высоты, полу-гипселодонтные, с корнями, хотя и слабо развитыми. Жевательная поверхность складчатая. Резцы, особенно нижние, очень мощные. В скелете туловища характерны длинные и тонкие ключицы, узкая лопатка, сильно редуцированная лобковая часть в тазе. Кости конечностей укорочены.
Хромосом в диплоидном наборе от 48, 50, 54 и 56 у малого до 60 у обыкновенного и 62 у гигантского слепышей.
Рис. 96. Палестинский слепыш — S ehrenbergi и череп S. microphtalmus
Распространены в восточной частя Средиземноморья и Юго-Восточной Европе — Австрии, Венгрии, Румынии, Югославии, Греции, Турции, степи и лесостепи европейской части СССР па восток до р. Эмбы, в Иране, Ираке, Сирии, Израиле, Иордании, северных областях ЛРЕ и Ливии.
Населяют лесостепные, стенные, полупустынные и пустынные районы. Поднимаются в горы до 2400 м над уровнем моря. Ведут подземный образ жизни, за исключением периода расселения молодых животных. Кормовые ходы располагаются неглубоко иод поверхностью почвы п отличаются значительной длиной и сложными разветвлениями. Откры тых выходов из поры пет. Гнездовая часть располагается па глубине до 3,5 м. Здесь же кроме гнездовой камеры помещаются и кладовые, иногда их до 10. Во время копания слепыш разрыхляет почву своими сильными резцами, рыхлые частицы откидывает лапами назад, затем
поворачивается в норе и уплощенной сверху мордой выталкивает землю на поверхность через отнорок.
Питаются в основном различными подземными частями растений, но также и зелеными надземными, втаскивая их в нору. На зиму делают большие запасы пищи. Активны круглый год. Длительность беременности, по-видимому, около месяца. В течение года бывает один помет из 2—4 детенышей. В области Средиземного моря самки приносят /етенышей с января по март. Новорожденные безволосые, беспомощные, весят в среднем примерно 5 г, в длину достигают 5 см. Примерно через 2 недели после рождения они покрываются длинными серыми волосами и достигают примерно 8 см. В возрасте 4—6 недель покидают гнездо.
Местами слепыши могут приносить вред огородным культурам, корнеплодам на полях; в лесных питомниках уничтожают прорастающие желуди, корни дубовых сеянцев.
В семействе 1 род слепышей — Spalax Giildenstaedt, 1770, и 4 вида.
Род слепышей
Genus Spalax Giildenstaedt, 1770
См. описание семейства.
В роде 4 вида:
малый слепыш — S. leucodon Nordmann, 1840 (Югославия, Венгрия, Румыния, Болгария, Греция, Турция, Малая Азия; СССР — Молдавия и Южная Украина к востоку до р. Буг, па север до Балты и Первомайска, кроме того, Северо-Западная Армения и Юго-Западная Грузия) ;
обыкновенный слепыш — S. microphtalmus Giildenstaedt, 1770 (Румыния, Венгрия, Болгария, Польша, в европейской части СССР на север примерно до линии, проходящей через Ровно — Киев—-Чернигов — Тела — Пенза — Ульяновск, на восток до Волги и на юг до подножий Большого Кавказа и Перекопского перешейка).
гигантский слепыш — S. giganteus Nehring, 1898 (северо-восточное Предкавказье, между низовьями рек Кума и Терек, к югу примерно до линии Махачкала-Гудермес, а к северу до южных районов Астраханской обл.; изолированные колонии встречаются к востоку от р. Урал по рекам Уилу, Темиру, верхнему и среднему течению Эмбы);
палестинский слепыш — S. ehrenbergi Nehring, 1898 (Сирия, Палестина, АРЕ и Ливия).
Некоторые исследователи (например, Эллермэн и Моррисон-Скотт, 1966) рассматривают гигантского слепыша в качестве подвида обыкновенного слепыша.
В СССР: слепыши малый, обыкновенный и гигантский.
СЕМЕЙСТВО БАМБУКОВЫХ КРЫС
FAMILIA RHIZOMYIDAE MILLER ET GIDLEY, 1918
Мелких и средних размеров грызуны с. длиной тела 16,5—48 см. ЬДлина хвоста 5—15 см. Приспособлены к подземному образу жизни. Кло массивное, шейный перехват снаружи почти незаметен (рис. 97). Голова укороченная, тупая, крупная. Ушные раковины очень малы и Ггкрыты в волосах головы (Тachyor у etc s) или выступают над волосяным
покровом (Rhizomys). Глаза небольшие. Конечности короткие. Когти на пальцах укороченные и уплощенные, сходные с ногтями. Хвост без чешуй, покрыт редкими волосами. Волосяной покров высокий, густой и мягкий или короткий и грубый (большая бамбуковая крыса). Окраска спинной стороны тела серая (например, седая бамбуковая крыса), коричневато-серая (например, китайская бамбуковая крыса) или каштаново-коричневая, брюшной — пепельно-серая (например, малая бамбуковая крыса). Сосков 1 или 2 пары грудных 2 или 3 пары паховых.
На структуре черепа сказалась подземная жизнь этих грызунов. Он несколько уплощен в дорзо-вентральном направлении. Скуловые дуги сильные, широко расставленные в стороны. Подглазничное отверстие сильно уменьшено. Его нижний край почти прямой и расположен на верхней границе скуловой пластины. Невральная часть подглазничного отверстия уменьшена или полностью отсутствует. Сагиттальный и ламбдоидальные гребни сильно развиты. Зубная формула
. 1 0 0 3 ,п.л. .1 0 I 2
t — с — р — т—— 16(Эллермэн, 1940) или i — с — р — т—= 16
(Грассе, 1955). По-видимому, правильнее считать все три щечных зуба коренными. Коренные зубы имеют корни и высокие, плоские сверху коронки. У Tachyoryctes коренные зубы с постоянным ростом в течение всей жизни. Резцы крупные, белые или оранжевые.
Толстый отдел кишечника может быть длиннее тонкого. В слепой кишке есть спиральная складка.
Хромосом в диплоидном наборе от 50 у большой бамбуковой крысы до 60 у малой бамбуковой крысы.
Распространение охватывает территорию Китая к югу от р. Янцзы, Индию, Непал, Бирму, Таиланд, Вьетнам, Лаос, Камбоджу и п-ов Мал-лакку до Суматры, а также тропическую Восточную Африку (Эфиопию, Сомали, Кению, Уганду, Танзанию, Заир).
Населяют равнинные, песчаные или поросшие травянистой растительностью участки, леса, заросли бамбука. В горы поднимаются на высоту до 4000 м над уровнем моря. Строят сложные подземные туннели. Значительную часть времени проводят в норах. Активны в основном ранним утром или вечером, когда животные появляются на поверхности земли в поисках пищи. Питаются различными подземными частями растений, в частности бамбука, а также семенами и плодами. Превосходно копают землю.
Представители родов Rhizomys и Cannomys роют землю при помощи зубов и когтей. Представители рода Tachyoryctes употребляют при копании когти; зубы используются ими только при разгрызании корней. Размельченные частицы земли они подгребают передними лапами под брюхо и затем откидывают задними лапами назад. Когда позади животного скапливается небольшая кучка земли, опо поворачивается к ней мордой и проталкивает по поре грудью. В помете от 1 до 3—5 голых, слепых детенышей. Длительность беременности, по-видимому, 21—49 дней. Местные жители в некоторых районах употребляют бамбуковых крыс в пищу.
В семействе 3 рода (6 видов):
африканских бамбуковых крыс Tachyoryctes Riippell, 1835 (2 вида);
бамбуковых крыс — Rhizomys Gray, 1831 (3 вида);
малых бамбуковых крыс—Cannomys Thomas, 1915 (1 вид).
Род африканских бамбуковых крыс
Genus Tachyoryctes Riippell, 1835
Рис. 97. Африканская бамбуковая крыса — 7. splendeus
Размеры средние. Длина тела 18 25 см. Длина хвоста Гт- 9 см. Масса 160—280 г. Внешним видом несколько напоминают кротсш (рис. 97). Уши и глаза очень маленькие. Короткий хвост обычно покрыт довольно густыми волосами. На передних конечностях третий палец самый длинный. Первый палец рудиментарный. На задних конечностях самый длинный второй палец. Пятый палец уменьшен. Когти не очень крупные. Волосяной покров относительно высокий, густой и мягкий. Окраска его варьирует от почти черного до коричневого, рыжевато-коричневого, светло-серого цвета. Часто встречаются альбиносы. Молодые животные, как правило, черные. Брюшная сторона несколько светлее спинной.
В черепе лобные кости сильно сжаты. Костные слуховые барабаны относительно крупные. Лицевой отдел черепа утолщен. Очень толстые резцы оранжевого цвета. Щечные зубы гипселодонтного типа. Верхние коренные зубы несколько наклонены назад и наружу.
Распространены в Восточной Африке: Эфиопии, Сомали, Кении, Уганде, восточной части Заира и на севере Танзании.
Населяют различные биотопы от песчаных равнин до бамбуковых лесов, предпочитая степи и саванны. В горы поднимаются на высоту до 3700 м над уровнем моря. Держатся поодиночке. Ведут почти исключительно подземный образ жизни, лишь в ночное время иногда выходят на поверхность земли. Норы устраивают обычно па глубине 15- 30 см, а в сухой сезон — до 1 м. Местами африканские бамбуковые крысы очень многочисленны. Питаются корнями различных растений, а в меньшей степени их зелеными частями.
У африканской бамбуковой крысы самка приносит от 1 до 4 детенышей (как правило, 1 или 2) в дождливый сезон. Продолжительность беременности 46—49 дней.
В роде, по-видимому, 2 вида:
эфиопская бамбуковая крыса — Т. macrocephalus Riippell, 1842 (Эфиопия);
африканская бамбуковая крыса — Т. splendeus Riippell, 1835 (восточ-
пая часть Заира, Уганда; Бурунди; Кения; Северная Танзания; Эфиопия; Сомали).
Эллермэн (1940) относит род бамбуковых крыс вместе с родом Brachyuromys в подсемейство Tachyoryctinae семейства Muridae, отдельно от родов бамбуковых крыс и малых бамбуковых крыс В роде он насчитывает 14 видов: Т. macrocephalus Rijppell , 1842 (Эфиопия); Т. splendens Rijppell, 1835 (Эфиопия, Сомали, Кения); Т. pontifex Neumann et Rilmmler, 1928 (Эфиопия); T. cheesmam Thomas, 1928 (Эфиопия); T. annectens Thomas, 1891 (Кения); T. storeyi Thomas, 1909 (Кения); T. naivashae Thomas, 1909 (Кения); T. spalacinus Thomas, 1909 (Кения); T. audax Thomas, 1910 (Кения); T. rex Heller, 1910 (Кения); T. ruddi Thomas, 1909 (Кения); T. ankoliae Thomas, 1909 (Уганда); T. daemon Thomas, 1909 (гора Килиманджаро); Т. ruandae Lonnberg et Gyldenstolpe, 1925 (Заир).
Род бамбуковых крыс
Genus Rhizomys Gray, 1831
Размеры наибольшие в семействе. Длина тела 23—48 см. Длина хвоста 5—15 см В большей пли меньшей степени приспособлены к подземному образу жизни. Хвост слабо оволосиен. Уши небольшие, слегка выступающие над уровнем меха (рис. 98). Глаза маленькие. Когти па лапах крупные, сильные. На передних конечностях третий палец наиболее длинный, второй немного короче четвертого. Пятый самый короткий. В северных частях ареала волосяной покров густой и мягкий, у тропических форм — редкий и грубый. Окраска его на спинной стороне тела от серой до розовато-серой или коричнсвато-серой, на брюшной — обычно несколько более светлая Сосков 4, иногда 5 пар.
В черепе лобные кости сильно сжаты. Сагиттальный гребень может тянуться до ламбдоидального. Костные слуховые барабаны крупные. Скуловые дуги широко расставлены. Костное небо узкое. Резцовые ог-
Рис. 98. Большая бамбуковая крыса — R sumatrensis
верстия небольшие. Резцы очень широкие, сильные, оранжевого цвета. Верхние резцы направлены почти вертикально вниз. Щечные зубы полу-пшселодонтного типа.
Распространены в Южном Китае, Бирме, на п-ове Индокитай, в Таиланде. на п-ове Малакка, Суматре.
Населяют бамбуковые заросли. Большую часть времени проводят в норах, которые выкапывают при помощи резцов и когтей. Обычно каждая бамбуковая крыса имеет по нескольку нор. Питаются в основном корнями бамбука, а также зелеными частями растений, их семенами и различными плодами. В помете бывает от 3 до 5 детенышей. Длительность беременности примерно 21 день.
В роде 3 вида:
китайская бамбуковая крыса — /?. sinensis Gray, 1831 (Южный Кичи'! и Северная Бирма);
седая бамбуковая крыса — A*. pruinosus Blyth, 1851 (Юньнань и Гуандун в Южном Китае; Ассам; Восточная Бирма; п-ов Индокитай; Таиланд; п-ов Малакка);
большая бамбуковая крыса — R. sumatrensis Raffles, 1821 (о-в Суматра; п-ов Малакка; Таиланд; п-ов Индокитай; Бирма).
Род малых бамбуковых крыс
Genus Cannomys Thomas, 1915
В роде единственный вид: мал^я бамбуковая крыса С. badius Hodgson, 1841.
Размеры от наименьших в семействе до средних. Длина тела 16,5— 25,5 см. Длина хвоста 6—7 см. По внешнему облику сходна с представителем рода бамбуковых крыс. Хвост покрыт редкими волосами. Волосяной покров густой и мягкий. Одноцветная по всему туловищу окраска изменяется от коричневой до серой, иногда по верху головы проходит продольная белая полоска. Такая же белая полоса тяпется от подбородка к горлу. Сосков 4 пары.
Череп сходен с черепом бамбуковых крыс, но резцы не такие мощные. Подглазничное отверстие очень мало. Щечные зубы уменьшаются спереди назад.
Распространена в Непале; Индии — Ассаме; Бирме и Таиланде.
Населяет травянистые равнины, леса, иногда встречается в садах. ЬЗемлю роет при помощи резцов и когтей, устраивая глубокие и сложные ►ходы. Может копать очень твердую почву. Вечером иногда показываются на поверхности земли. Питается различными растениями. Экология изучена слабо.
СЕМЕЙСТВО МЫШИНЫХ
FAMILIA MURIDAE GRAY, 1821
К семейству принадлежат различные по размерам грызуны от мелких до средних. Длина тела от 5 до 48,5 см1. Телосложение у болыипп-I ства относительно стройное. Шейный перехват хорошо выражен. Коиеч-I цепи обычно средней длины, причем задние немного длиннее передних,
1 В пределах семейства можно выделить три группы: мелкие с длиной тела 5—19 см, средние — 20—34 см, крупные — 35—48 см.
реже значительно превышают их или длина передних и задних конечностей примерно одинаковая. Длинный хвост, как правило, лишен волос, иногда покрыт редкими волосами. На хвосте расположены хорошо выраженные кожные чешуи. У некоторых видов хвост покрыт довольно густыми волосами или имеет на конце кисточку. У некоторых лазающих форм хвост может быть полухватательным. Конечности пятипалые с различной .степенью редукции крайних пальцев. У некоторых древесных видов первый палец на передних конечностях или на задних противопоставляется остальным пальцам и имеет вместо когтя ноготь. Подошвы конечностей безволосые. У полуводных видов на задних конечностях между пальцами могут быть плавательные перепонки. Представители трех родов (Beamys, Saccostomus и Cricetomys) имеют защечные мешки. Волосяной покров разнообразный. Он может быть однородный, шелковистый и мягкий или резко подразделяться на удлиненную, грубую ость и тонкий мягкий пух или превращаться в короткие, нетолстые иглы. Окраска спинной поверхности и тела обычно бурая или серая с рыжим или черным оттенками. Половой диморфизм в окраске отсутствует.
Череп удлиненный. Мозговая капсула выпуклая или несколько уплощенная. Лицевой отдел иногда укорочен. Скуловые дуги, как правило, слабые. Подглазничное отверстие несколько вытянуто в высоту, с расширенным верхним отделом, через который проходит часть передней доли жевательной мышцы. Изредка подглазничное отверстие больших размеров, округлое. Сагиттального гребня, как правило, нет. Ламбдоидаль-ного тоже нет или оп слабо развит. Лобно-теменные гребни обычно имеются. Костные слуховые барабаны различных размеров, у большинства небольшие и тонкостенные. В нижней челюсти венечный отросток часто слабо развит, а сочленовный отросток довольно крупный. Типичная зубная формула: i 1 с р т— 16. Число коренных зубов мо-
2 1
жет быть уменьшено до — или до — (род Mayermys).
Щечные зубы имеют корни или не имеют их. Жевательная поверхность щечных зубов, как правило, имеет выступы или поперечные гребни, причем выступы обычно располагаются тремя продольными рядами. Коронки щечных зубов низкие или средней высоты, лишь редко высокие. Размеры зубов почти всегда уменьшаются в направлении спереди назад. Os penis имеется.
Плацента хориоаллантоидная, дискоидальная.
Хромосом в диплоидном наборе от 32 у Oenomys, 40 у Mils до 50 у Thamnomys и Aethomys и 60 у Micromys.
Распространены почти по всему земному шару, за исключением самых высоких широт. Наибольшее число из 399 видов семейства встречается в Юго-Восточной Азии. Некоторые виды, распространяясь за человеком, стали космополитами.
Представители семейства населяют самые разнообразные ландшаф ты. Ведут наземный или нолудревесный (большинство видов), редко по луводный образ жизни. Некоторые виды могут передвигаться прыжками на задних конечностях. Почти все представители приспособлены к рытью нор, хотя адаптаций к исключительно подземному существованию нет. Убежищем служат поры, вырытые самими грызунами, или пустоты под камнями, упавшими стволами деревьев, иногда дупла, птичьи гнезда, постройка человека. Активны днем или ночью, причем древесные формы обычно активны ночью. В спячку не впадают. Некоторые виды держатся поодиночке, другие парами или семейпыми группами, третьи образуют крупные группы или колонии.
Большинство видов питается различными растительными объектами и беспозвоночными. Некоторые виды поедают и мелких позвоночных — амфибий, пресмыкающихся, птиц, их яйца, мелких грызунов, иногда рыб. Есть всеядные формы. В помете от 1 до 22 детенышей. Длительность беременности от 18 до 42 дней. Половозрелость может наступать в 35-дневном возрасте (домовая мышь), или в возрасте нескольких месяцев (большинство видов). В южных частях ареала, как правило, размножаются круглый год, часто с несколькими пиками в активности размножения. Продолжительность жизни в естественных условиях 1—3 года. Численность некоторых видов может сильно изменяться по годам. Некоторые виды приносят существенный вред посевам и запасам продуктов. Есть виды, имеющие важное эпидемиологическое значение.
В семействе, по-видимому, 100 родов (400 видов).
Ь Подсемейство мышей — Murinae Murray, 1866
! Роды:
I длиннохвостых мышей — Hapalomys Blyth, 1859 (1 вид);
I . лазающих мышей — Vernaya Anthony, 1941 (1 вид);
f пальмовых мышей— Vandeleuria Gray, 1842 (1 вид);
I колючих мышей — Tokudaia Kuroda, 1943 (1 вид);
I мышей-малюток — Micromys Dehne, 1841 (1вид);
I лесных и полевых мышей — Apodemus Каир, 1829 (5 видов);
I африканских мышей — Thamnomys Thomas, 1907 (4 вида);
} кустарниковых крыс — Grammomys Thomas, 1915 (6 видов);
к лузонских крыс — Carpomys Thomas, 1895 (2 вида);
к минданаосских крыс — Mindanaomys Sanborn, 1953 (I вид);
К лесных крыс — Batomys Thomas, 1895 (2 вида);
К красных крыс — Pithecheir F. Cuvier, 1838 (1 вид);
г гигантских крыс — Hyomys Thomas, 1903 (1 вид);
кроличьих крыс — Conilurus Ogilby, 1838 (2 вида);
толстохвостых крыс — Zyzomys Thomas, 1909 (3 вида);
мезембриомис — Mesembriomys Palmer, 1906 (2 вида); рыженосых крыс-—Oenomys Thomas, 1904 (1 вид);
бороздчатозубых крыс-—Mylomys Thomas, 1906 (1 вид);
лохматоволосых крыс — Dasymys Peters, 1875 (1 вид);
травяных мышей-—Arvicanthis Lesson, 1842 (4 вида); манипурских'кустарниковых крыс — Hadromys Thomas, 1911 (1 вид); индийских крыс— Golunda Gray,, 1837 (1 вид);
бороздчатозубых болотных крыс — Pelom'ys Peters, 1852 (8 видов); полосатых мышей — Lemniscomys Trouessart, 1881 (4 вида);
полосатых полевых мышей — Rhabdomys Thomas, 1916 (1 вид); однополосых мышей — Hybomys Thomas, 1910 (2 вида);
мягкошерстных крыс — Millardia Thomas, 1911 (3 вида); большезубых крыс — Dacnomys Thomas, 1916 (1 вид);
сулавесских мягкошерстных крыс — Eropeplus Miller et Hollister, 1921(1 вид);
узкоголовых крыс — Stenocephalemys Frick, 1914 (1 вид);
чрыс—Rattus Fischer, 1803 (97 видов);
акациевых крыс — Aethomys Thomas, 1915 (10 видов);
праомис — Praomys Thomas, 1915 (22 вида);
темных крыс — Thyphomys Miller, 1910 (1 вид);
австралийских полевых мышей — Gyomys Thomas, 1910 (8 видов);
. австралийских крыс — Leporillus Thomas, 1906 (3 вида);
южных мышей — Pseudomys Gray, 1832 (11 видов);
юзаичнохвостых крыс — Pogonomelomys Rummler. 1936 (4 вида);
банановых крыс — Melomys Thomas, 1922 (17 видов);
голохвостых крыс — Solomys Thomas, 1922 (3 вида);
белохвостых крыс-—Xenuromys Tate et Archbold, 1941 (1 вид); чешуехвостых крыс — Uromys Peters, 1867 (6 видов);
большеухих крыс — Malacomys Milne-Edwards, 1877 (1 вид);
малых мышей •— Heteromys Thomas, 1911 (2 вида) ;
азиатских древесных мышей — Chiromyscus Thomas, 1925 (1 вид); широкоголовых мышей — Zelotomys Osgood, 1910 (2 вида);
манипурских мышей — Diomys Thomas, 1917 (1 вид);
полосатоспинных мышей — Muriculus Thomas, 1902 (1 вид);
мышей — Mus Linnaeus, 1758 (25 видов);
австралийских мышей — Leggadina Thomas, 1910 (7 видов); африканских крыс — Colomys Thomas et Wroughton, 1907 (1 вид); керамских крыс — Nesoromys Thomas, 1922 (1 вид);
филиппинских болотных мышей — Crunomys Thomas, 1897 (2 вида); новогвинейских крыс — Macruromys Stein, 1933 (2 вида);
кенгуровых мышей — Lorentzimys Tentink, 1911 (1 вид); жестковолосых мышей — Lophutomys Peters, 1874 (6 видов); бороздчатозубых мышей — Leimacomys Matschie, 1893 (1 вид); тушканчиковых мышей — Notomys Lesson, 1842 (10 видов); широкозубых крыс — Mastacomys Thomas, 1882 (2 вида);
колючих крыс — Echiothrix Gray, 1867 (1 вид);
длиннокоготных мышей — Melasmothrix Miller et Hollister, 1921 (1 вид);
иглистых мышей — Acomys I. Geoffroy, 1838 (5 видов);
крупнозубых мышей — Uranomys Dollman, 1909 (1 вид);
бандикот — Bandicota Gray, 1873 (2 вида);
пластинчатозубых крыс — Nesokia Gray, 1842 (1 вид); треугольнохвостых крыс — Beamys Thomas, 1909 (2 вида); мешотчатых крыс — Saccostomus Peters, 1846 (1 вид);
хомяковидных крыс — Cricetomys Waterhouse, 1840 (2 вида); анизомис — Anisomys Thomas, 1903 (1 вид).
Подсемейство лазающих мышей — Dendromurinae Allen, 1939
Роды:
африканских лазающих мышей — Dendromus A. Smith, 1829 (5 видов);
дендроприономис — Dertdroprionomys Petter, 1966 (1 вид); длинноухих мышей — Malacothrix Wagner, 1843 (1 вид);
древесных мышей — Prionomys Dollman, 1910 (1 вид);
скалистых мышей — Petromyscus Thomas, 1926 (2 вида);
болотных мышей-—Delanymys Hayman, 1962 (1 вид);
толстых мышей — Steatomys Peters, 1846 (11 видов);
конголезских мышей—Deomys Thomas, 1888 (1 вид);
Подсемейство болотных крыс Otoinyiiiae Thomas, 1897
Роды:
болотных крыс— Otomys F. Cuvier, 1823 (10 видов);
паротомисов — Parotomys Thomas, 1918 (2 вида).
Подсемейство тонкохвостых крыс — Phloeomyinae Alston, 1876
Роды:
сулавесских крыс — Lenomys Thomas, 1898 (1 вид);
цепкохвостых крыс — Pogonomys Milne-Edwards, Г877 (9 видов);
соневидных мышей — Chiropodomys Peters, 1868 (2 вида);
древесных крыс — Mallomys Thomas, 1898 (1 вид);
флоресских крыс — Papagomys Sody, 1941 (1 вид);
тонкохвостых крыс — Phloeomys Waterhouse, 1839 (2 вида);
пышнохвостых крыс — Craterotnys Thomas, 1895 (1 вид).
Подсемейство хоботковых крыс — Rhynchomyinae Thomas, 1897
Роды:
хоботковых крыс — Rhynchomys Thomas, 1895 (1 вид);
Подсемейство водяных крыс — Hydromyinae Alston, 1876
Роды:
полосатых крыс — Chrotomys Thomas, 1895 (1 вид);
лузонских водяных крыс — Celaenomys Thomas, 1898 (1 вид);
безухих крыс — Crossomys Thomas, 1907 (1 вид);
ложных водяных крыс — Xeromys Thomas, 1889 (1 вид);
австралийских водяных крыс — Hydromys Е. Geoffrey, 1805 (3 вида);
горных водяных крыс — Parahydromys Poche, 1906 (1 вид);
неохидромис — Neohydromys Laurie, 1962 (1 вид);
очковых мышей — Leptomys Thomas, 1897 (1 вид);
микрохидромис — Microhydromys Tate et Archbold, 1941 (1 вид);
паралептомис — Paraleptomys Tate et Archbold, 1941 (2 вида);
байянкасских крыс — Baiyankamys Hinton, 1943 (1 вид);
новогвинейских мышей — Pseudohydromys Rmnmler, 1934 (2 вида);
мышей Майера — Mayermys Laurie et Hill, 1954 (1 вид).
Некоторые систематики (например, Эллермэн, 1941; Кузнецов, 1965) объединяют Muridae вместе с Cricetidae в одно общее семейство Muridae. Здесь, по Симпсону (1945) и многим другим исследователям, эти семейства разделены.
ПОДСЕМЕЙСТВО МЫШЕЙ
SUBFAMILIA MURINAE MURRAY, 1866
Размеры от мелких до средних и крупных. Внешний вид разнообразный, но крайние формы адаптаций к водному или подземному образу жизни отсутствуют. Имеются формы, в разной степени приспособленные к жизни на деревьях.
В черепе подглазничное отверстие крупное. Сквозь него проходит о , .10 0 3
порция жевательных мышц. Зубная формула: i— с р —т — .
10 0 3
Коренные зубы с корнями. Верхние коренные пластинчатые или бугорчатые, причем бугорки располагаются тремя продольными рядами.
Род длиннохвостых мышей Genus Hapalomys Blyth,
В роде единственный вид: длиннохвостая мышь — Н. longicaudatus Blyth, 1859 (рис. 99).
Размеры мелкие. Длина тела 12—13,5 см. Длина хвоста 14—17 см. Хвост покрыт средней густоты волосами. На кончике хвоста волосы иногда удлинены. Задние конечности приспособлены к древесному образу жизни: подошвенные мозоли крупные, первый палец противопоставляется остальным и не имеет когтя, четвертый палец наиболее длинный; пятый палец примерно равен ио длине второму. Волосяной покров густой и мягкий. Окраска его па спинной стороне темпо-желтая, рыжевато-серая или буровато-серая.
Рис. 99. Длиннохвостая мышь — II longicaudatus
Бока несколько светлее спины. Брюхо и конечности ниже локтевых н коленного сочленений белые. Вибриссы хорошо развиты и несколько длиннее головы. Сосков 4 пары
Мозговая коробка черепа очень широкая. Лицевой отдел укорочен. Лобно-теменные гребни хорошо выражены. Костное небо относительно узкое. Костные слуховые барабаны крупные. Подглазничное отверстие узкое. Резцовое отверстие средней длины. Резцы сильно расширены, мощные.
Длиннохвостая мышь — единственный представитель семейства мышей, имеющий на жевательной поверхности нижних щечных зубов три ряда бугорков. Это, по-видимому, очень примитивный признак.
Распространена на п-ове Малакка, в Южной Бирме, Таиланде, п-ове Индокитай, и о-ве Хайнань.
Ведет полудревесный образ жизни Экология по изучена.
Род лазающих мышей
Genus Vernaya Anthony,1941
В роде единственный вид: лазающая мышь — V. fuloa G. Allen, 1927.
Размеры мелкие. Длина тела примерно 9 см. Длина хвоста около 11,5 см. На конце хвоста кисточки пет. Первый палец па передних конечностях развит слабо и имеет очень маленький уплощенный ноготь. Остальные пальцы передних конечностей длинные, с заостренными когтями. Окраска волосяного покрова рыжеватая.
Резцы нс имеют бороздок.
Распространена в Северной Бирме и Юго-Западном Китае.
Населяет горные склоны,ттокрытые зарослями кустарников, со скальными выходами на высоте примерно 2250 м над уровнем моря. Экология не изучена.
Род пальмовых мышей
Genus Vandeleuria Gray, 1842
В роде единственный вид: пальмовая мышь — V. oleracea Bennet, 1832.
Размеры мелкие. Длина тела 5,5—8,5 см. Длина хвоста 9—13 см. Хвост довольно густо покрыт короткими волосами, не удлиненными на его конце. Пятый палец передней конечности уменьшен, коготь его напоминает по форме ноготь. На остальных пальцах когти небольшие. Второй палец также сильно уменьшен и значительно короче двух средних. На задней конечности пятый палец не имеет когтя; первый палец полностью противопоставляется остальным. Волосяной покров густой, очень мягкий. Окраска спинной стороны от светло-желтой до темно-желто-бу-рой и темно-рыжевато-бурой. Брюшная сторона белая или кремовая. Сосков 4 пары.
В черепе лицевой отдел не укорочен. Лобные кости сильно сжаты. Лобно-теменных гребней обычно нет. Жевательная поверхность щечных зубов с бугорками.
Хромосом в диплоидном наборе, по данным различных исследователей, 29 или 46.
Распространена в южной части Индии, а также в Кумаоие и Ассаме; Непале; на о-ве Шри Ланка; Бирме, на п-ове Индокитай в Таиланде, а по Аллену, 1938, 1940, также в Юньнане-— в Китае.
Ведет, по-видимому, древесный образ жизни. Активна ночью. День проводит в гнезде, которое строит из травы или сухих листьев на кустах, и деревьях или в дуплах. Иногда зверек цепляется хвостом за ветку. На землю спускается редко. В помете обычно 3—4, иногда до 6 детенышей. Питается плодами, почками, ростками кустарников и деревьев.
Род колючих мышей
Genus Tokudaia Kuroda, 1943
В роде единственный вид: колючая мышь — Т. osimensis Abe, 1934. Размеры мелкие. Длина тела 12,5—17,5 см. Длина хвоста 10—12,5 см. По внешнему облику несколько напоминает крупную полевку. Шерстный покров состоит из относительно редких, мягких и топких волосков и грубых, довольно длинных, толстых иголок, имеющих па своей поверхности бороздки. Иглы встречаются по всему телу, за исключением областей вокруг рта, ушей, конечностей и хвоста. Иглы на спине черные, на брюхе белые с рыжеватой вершиной. Окраска спины смешанная —-черная с оранжево-желтым, брюха — серовато-белая с оранжевым оттенком. Хвост тоже двухцветный. Сосков 4 пары.
Распространена на о-вах Амамиосима и Окинава из группы Рюкю.
Населяют густые заросли подлеска. Экология не изучена.
Популяция, обитающая на о-ве Окинава, нуждается в охране и включена в «Красную книгу».
Род мышей-малюток
Genus Microniys Dehne, 1841
В роде единственный вид: мышь-малютка —М. minutus Pallas, 1771.
Размеры мелкие. Длина тела до 7 см. Длина хвоста тоже до 7 см. Голова укороченная, притупленная спереди Глаза маленькие. Уши небольшие, округлые (рис. 100). Слуховое отверстие может закрываться специальной кожистой складкой, расположенной в основании заднего края уха. Хвост полухватательный, его терминальная часть безволосая. Стопа удлинена и расширена. Третий палец на обеих конечностях несколько длиннее остальных. Пятый палец передних и первый палец задних конечностей укорочены. Первый палец задней конечности имеет коготь и неполностью противопоставляется остальным. Волосяной покров густой и мягкий. Окраска спины буроватая с желтоватым оттенком, брюшной — белая или темножелтая. В зимнее время волосы длиннее, чем летом. Сосков, 4 пары.
В черепе лицевой отдел сильно укорочен, а мозговая коробка крупная и вздутая. Лобно-теменных гребней пет. Скуловые дуги расставлены нешироко. Резцовые отверстия относительно длинные и обычно достигают уровня переднего коренного зуба. Костное небо широкое. Костные слуховые барабаны крупные. Щечные зубы имеют относительно высокие коронки.
Хромосом в диплоидном наборе 68.
Распространена в Евразии от Англии и Пиренеев на западе и до берегов Тихого океана на востоке. В СССР обитает почти по всей европейской части страны, в Северном Казахстане, Южной Сибири, па Дальнем Востоке; северная граница ареала проходит от Центральной Карелии через южные районы Архангельской обл., бассейн р. Конди, Нарым-ский край и центральную Якутию.
Населяет лесостепную зону, поднимается в горы выше верхней гра ницы леса в горные луга до 2200 м над уровнем моря. Предпочитает высокотравные луга в речных поймах, по которым проникает в тайгу и степь, где населяет заросли бурьяна на пустошах, залежах, межах. Осенью и зимой многочисленна в скирдах. Хорошо лазает но высокой траве. Гнездо делается из травы шарообразное, диаметром 60—130 мм. Подвешивает на высоких стеблях травянистых растений или на ветках 274
кустов на высоте не выше 1—1,3 м над землей. В гнезде выводит детенышей, а в холодное время года живет в просто устроенных норах. Активна обычно днем. Питается преимущественно различными семенами, а ;акже насекомыми. Делает небольшие запасы пищи на зиму. За лето бывает 3—4 помета, в каждом из которых 5—8, иногда до 12 детенышей. Длительность беременности примерно 21 день. Отмечены изменения численности по годам. В годы с повышенной численностью мыши-малютки могут приносить существенный вред посевам.
Род лесных и полевых мышей
Genus Apodemus Каир, 1829
Размеры мелкие. Длина тела 6—15 см. Хвост несколько короче длины тела, равен ей или немного длиннее. Масса взрослых 15—60 г. Голова с удлиненной лицевой частью, крупными глазами и относительно большими ушами (рис. 101). Хвост не хватательный, покрыт волосами средней густоты. Стопа длинная и узкая. Наружные пальцы немного укороченные. Волосяной покров обычно мягкий. Остевые волосы в большинстве случаев редкие, сравнительно тонкие. Окраска спины серовато-темно-желтая, серовато-коричневая или коричневая. У полевой мыши вдоль спины проходит темная полоска. Брюхо белое или серое, иногда с желтым оттенком. Конечности белые. У некоторых видов на груди расположено рыжевато-желтое пятно. Сосков 3 или 4 пары.
В черепе лицевой отдел удлинен. Мозговая коробка относительно чтдутая, крупная. Носовые кости обычно выдаются над резцами. Лобно-сменные гребни либо отсутствуют, либо хорошо развиты. Резцовые отверстия короткие и широкие; обычно достигают передних щечных зубов. Скуловые дуги расставлены в стороны нешироко. Костное небо относительно широкое. Костные слуховые барабаны средних размеров. Жевательная поверхность верхних щечных зубов сложной конфигурации.
Хромосом в диплоидном наборе 48 у лесной и горной мышей, 46— 31 у восточноазиатской и 48 или 50 у полевой мыши.
Распространены на обширной территории, охватывающей лесную, лесостепную и степную зоны и северные полупустыни Евразии от Швеции л Англии до Японии, Сахалина, Восточного и Северо-Восточного Китая, Малой и Передней Азии и Северной Индии, а также Северной Африки.
Населяют широколиственные, смешанные леса, а также лесостепь, луговые степи на равнинах и в горах до высоты 3500 м над уровнем мо-. Активны обычно ночью. День проводят в просто устроенных норах ;„ш в дуплах деревьев на разной высоте над землей. Иногда в качестве убежища используют пустоты под корнями, расщелины между камней. Некоторые виды хорошо лазают по кустам и деревьям. Питаются различными частями растений: листьями, семенами, ягодами. Существенное значение в летнем питании имеют насекомые. На зиму некоторые виды запасают пищу (обычно различные семена). У желтогорлоп мыши запасы могут достигать 4 кг.
Размножаются в теплое время года и за этот период дают 3—4, иногда до 5 пометов, в каждом из которых обычно 5—7 (до 9) детенышей. Длительность беременности 27—29 дней. Половозрелость наступает в возрасте 3—3,5 месяца. В различные годы происходит значительное изменение численности. Некоторые виды наносят существенный вред сельскому хозяйству (полевая мышь). Являются природными носителями возбудителей туляремии, лептоспироза и некоторых других заболеваний.
В роде, по-видимому, 5 видов:
Рис. 101 Полспая мышь—И agrarius п се череп
полевая мышь — A agrartus Pallas, 1771 (ГДР, ФРГ, Нидерланды, Дания, Австрия, Венгрия, Польша, Югославия, Румыния; СССР — боль шинство районов европейской части, кроме Крыма и северных районов, Северный и Восточный Казахстан, Северная Киргизия, южные районы Западной и Средней Сибири на восток до Байкала, в Забайкалье нет, далее по Амуру и Приморье);
восточноазиатская мышь — A. speciosus Temminck, 1845 (СССР — Восточная Сибирь, Прибайкалье, Саяны, Алтай, Дальний Восток; Япония, Китай, МНР);
лесная мышь — A. syloaticus Linnaeus, 1758 (Англия, Гебридские о-ва, Ирландия, Исландия, Франция, Бельгия, Нидерланды, Испания, Португалия, Италия, Сицилия, Корсика, Швейцария, Норвегия, Шве-пия, ГДР, ФРГ, Венгрия, Чехословакия, Югославия, Румыния, Греция, о-в Крит; СССР — европейская часть на север до Центральной Карелии, южных районов Архангельской обл. и Коми АССР, Кавказ, почти вся Средняя Азия и Казахстан, исключая некоторые пустынные районы, южная часть Западной Сибири и на восток до Алтая включительно; Синьцзян-Уйгурский автономный р-он в Китае, северный Ассам, Бирма, Пенджаб; Малая Азия, Иран, Палестина, Марокко, Алжир, Тунис);
желтогорлая мышь — A. flavicollis Melchior, 1834 (Англия, Шетландские о-ва, часть Гебридских, Франция, Швейцария, Норвегия, Швеция, ГДР, ФРГ, Дания, Нидерланды, Венгрия, Австрия, Югославия, Румы-' ния, Греция, Финляндия; СССР — западные, центральные и южные, районы европейской части и Закавказья;
северная граница ареала идет по балтийскому побережью Литовской, Латвийской и Эстонской ССР, Карельскому перешейку, через Ленинградскую, Калининскую, Смоленскую, Тульскую, Рязанскую и Горь-ковскую области, Татарскую, южную часть Удмуртской и Башкирскую АССР к Южному Уралу, откуда сворачивает на юго-запад, образуя южную границу ареала, которая проходит по Оренбургской и Уральской областям и по западным окраинам прикаспийских степей выходит к предгорьям Кавказа, в ареал включены Черноморское побережье Кавказа, Украины и Молдавии, западное и большая часть центрального Закавказья, западная часть Главного Кавказского хребта, горный' Крым);
горная мышь — A. mystacinus Danford et Alston, 1877 (Югославия, Греция, Малая Азия, Палестина; СССР — Западное Закавказье).
В СССР: все перечисленные в роде виды.
Уокер (1964) и Андерсон и Джонс (1967) насчитывают в роде 7 видов, А. А. Громов (1963) — 10.
Род африканских мышей
Genus Thamnomys Thomas, 1907
Размеры мелкие (рис. 102). Длина тела 10—14,5 см. Длина хвоста 14—20 см. Хвост покрыт относительно густыми волосами; на конце хвоста они несколько удлинены. Стопа короткая и широкая, приспособлена к древесному образу жизни. Пятый палец примерно такой же длины, как и второй. Пальцы на задней конечности короткие; первый палец относительно длинный с когтем и, по-видимому, по противопоставляется ос-; тальному. Волосяной покров высокий и мягкий у одних видов (особенно высокогорных) и более грубый у других. Окраска варьирует от темно-коричневой до светло-рыжевато-коричневой или оранжево-рыжей. Брюхо белое. Сосков 2—3 пары.
Череп с относительно длинным лицевым отделом и широкой мозговой коробкой. Лобно-теменные гребни крупные. Резцовые отверстия длинные. Костные слуховые барабаны средних размеров. Подглазничные отверстия относительно большие.
Хромосом в диплоидном наборе у Т. rutilans 50.
Рис. 102. Thamnomys venustus и череп Т. ги-titans
Распространена в Африке от Ганы на восток до Западной Уганды.
Населяют леса и кустарниковые заросли. Предпочитают вторичные леса и сельскохозяйственные угодья. В горы поднимаются на высоту до 4000 м над уровнем моря. Ведут, по-видимому, почти исключительно древесный образ жизни. Активны ночью. Строят гнезда из листьев и сухой травы в развилках ветвей или в дуплах деревьев. По характеру питания растительноядные. Беременность около 25 дней.
В роде 4 вида:
Т. kuru Thomas et Wroughton, 1907 (Заир);
Т. rutilans Peters, 1876 (Камерун);
Т. venustus Thomas, 1907 (Уганда, Заир);
Т. kempi Dollmann, 1911 (Заир);
По Андерсону и Джонсу (1967), в роде 3 вида, а по Мизонну (1968),— 4: Т. venustus Thomas, 1907 (включая Т. kempi)-, Т. rutilans Peters, 1876; Т. cometes Thomas et Wroughton, 1908; T. dolichurus Smuts, 1832.
Род кустарниковых крыс
Genus Grammomys Thomas, 1915
Размеры мелкие. Длина тела 10—16 см. Длина хвоста 13—20,5 см. Внешним видом напоминает африканских лесных мышей, но конечности менее приспособлены к древесному образу жизни (рис. 103). Хвост покрыт редкими короткими волосами, за исключением концевой трети, где волосы более густые и несколько более удлиненные. Волосяной покров на туловище средней длины, мягкий и довольно густой. Окраска спины варьирует от темно-коричневой или рыжевато-коричневой до коричйе-вато-серой. Брюхо белое, кремово-белое, серое или темно-желтое. Окраска спинной и брюшной сторон тела резко отделена одна от другой. Сосков 2 или 3 пары.
Череп имеет расширенную мозговую коробку. Лобно-теменных гребней нет или они развиты слабо. Костные слуховые барабаны обычно ^небольшие.
Распространены от Судана на юг через Кению, Уганду, Заир, Малави и Анголу до Южно-Африканской Республики.
Населяют леса, выбирая более сухие места, чем африканские лесные мыши. Ведут древесный образ жизни. Активны ночью. День проводят в гнездах, которые делают на деревьях из сухих листьев. Питаются листьями и семенами. Экология изучена слабо.
В роде, по-видимому, 6 видов:
крыса Макмиллана — G. macmillani Wroughton, 1907 (Эфиопия, Судан, Кения, Мавритания);
G. ibeanus Osgood, 1910 (Кения);
G. dryas Thomas, 1907 (Уганда);
либерийская крыса — G. buntingi Thomas, 1911 (Либерия);
G. dolichurus Roberts, 1931 (Южно-Африканская Республика, Мозамбик, Южная Родезия, Замбия, Малави, Танзания, Кения, Заир, Эфиопия, Судан, Либерия);
мозамбикская крыса—G. ruddi Thomas et Wroughton, 1908 (Мозамбик) .
Уокер (1964) насчитывает в роде 8 видов. Некоторые исследователи (например, Робертс, 1954; Мизонн, 1968) включают виды этого рода'в состав рода Thamnomys.
Род лузонских крыс
Genus Carpomys Thomas, 1895
Размеры мелкие. Длина тела 17,5—20 см. Длина хвоста 16 -21 см. Стопа широкая, приспособленная к древесному образу жизни, по се первый палец имеет коготь и не противопоставляется остальным; пятый валец удлинен. Первый палец передних конечностей с крупным ногтем. Волосяной покров густой, мягкий. Спинная сторона тела желтая, брюшная— желтовато-белая. У С. melanurus хвост блестящего черного цвета, основание хвоста на расстоянии 25-—50 мм покрыто удлиненными волосами. У С. phaeurus хвост темно-бурого или черноватого цвета н основание хвоста на протяжении пе более 25 мм покрыто удлиненными волосами. Сосков 2 пары.
Лицевой отдел черепа несколько укорочен. Мозговая коробка округлая. Лобно-теменных гребней нет или они развиты слабо. Лобные кости сильно сжаты. Костные слуховые барабаны средних размеров. Костное небо широкое, не продолжается назад за окончания рядов щечных зубов. Резцовые отверстия длинные.
Распространены в северной части о-ва Лузон из группы Филиппинских.
Населяют горные леса на высоте 2100—2400 м над уровнем моря. Экология не изучена.
В роде 2 вида:
чернохвостая лузонская крыса — С. melanurus Thomas, 1895 (северная часть о-ва Лузон);
лузонская крыса — С. phaeurus Thomas, 1895 (северная часть о-ва Лузон);
Возможно, оба вида следует объединить в один.
Род минданаосских крыс
Genus Mindanaomys Sanborn, 1953
В роде единственный вид: минданаосская крыса — М. sa.lomon.seni Sanborn, 1953.
Размеры мелкие. Длина тела 17,5 см. Длина хвоста 14 см. Волосяной покров мягкий и густой. Спина темно-бурая, коричневая. Бока несколько более светлые, брюхо серое с желтоватым оттенком. Хвост черновато-коричневый сверху и снизу, опушен слабо.
Череп с резко выраженным межглазничиым сужением и широкой и короткой мозговой коробкой. Подглазничные отверстия узкие. Резцовые отверстия широкие. Костные слуховые барабаны небольшие. Щечные зубы гипселодонтного типа.
Распространена на о-ве Минданао из группы Филиппинских.
Встречается в горах на высоте примерно 1600 м над уровнем моря. Экология не изучена.
Род лесных крыс
Genus Batomys Thomas, 1895
Размеры средние. Длина тела около 20 см. Длина хвоста 12—18 см. Внешним видом напоминает лузонских крыс. Окраска спины может быть желтоватая, черная или светло-коричневая, брюха — серовато-желтая или охристо-желтая. У В. granti глаза окружены слабо покрытым волосами участком кожи. Хвост покрыт значительно более густыми волоса ми, чем у В. dentatus.
Череп напоминает череп минданаосских крыс; характерны сильное межглазничное сужение и широкий, укороченный мозговой отдел. Лицевая часть черепа вытянутая. Лобно-теменные гребни имеются. Подглаз ничные отверстия сверху и снизу сильно сужены. Костные слуховые ба рабаны относительно невелики. Костное небо широкое. Резцовые отверстия широкие, средней длины. Щечные зубы гипселодонтного типа. Резцы тонкие.
Распространены в северной части о-ва Лузон из группы Филиппинских.
Населяют горные дубовые и сосновые леса с густым подростом на высоте 2100—2400 м над уровнем моря. Экология не изучена.
В роде 2 вида:
лесная крыса — В. granti Thomas, 1895 (о-в Лузон);
зубатая крыса —В. dentatus Miller, 1911 (о-в Лузон),
Род красных крыс
Genus Pithecheir F. Cuvier, 1838
В роде единственный вид: красная крыса — Р. melanurus F. Cuvier, 1838.
Размеры средние. Длина тела 20,9—22,7 см. Длина хвоста 14— 18,6 см. Хвост хватательный, покрыт очень редкими волосами. Стопа адаптирована к древесному образу жизни: первый палец с небольшим когтем, по-видимому, полностью противопоставляется остальным пальцам; пятый палец длинный. Первый палец передней конечности лишь незначительно уменьшен в размерах. Волосяной покров высокий, густой. Окраска его рыжеватая на спине, темно-желтая на боках, чистобелая на брюхе. Хвост, уши и лапы рыжеватого цвета. У некоторых особей конечности белые.
В черепе лобно-теменные гребни у взрослых животных развиты сильно. Костные слуховые барабаны очень крупные, сильно вздутые. Костное небо широкое. Резцовые отверстия широкие и длинные. Резцы средних размеров.
Распространена в южной части п-ова Малакка и на о-вах Суматра и Ява.
Населяет густые леса в горах на высоте 1300—1600 м над уровнем моря. Ведут древесный образ жизни. Двигается медленно, помогая себе цепляющимся за ветви хвостом. Убежищем служат гнезда, которые зверьки сооружают на деревьях, иногда дупла. Когда детеныши еще малы, самка покидает гнездо, унося их с собой. При этом детеныши захватывают соски резцами. Питаются зелеными растениями.
Род гигантских крыс
Genus Hyomys Thomas, 1903
. В роде единственный вид: гигантская крыса — Н. goliath Milne-Edwards, 1900.
Размеры средние и крупные. Длина тела 29,5—39 см. Длина хвоста 25,5—38 см. Уши небольшие. Чешуйчатый хвост почти полностью лишен волос. Чешуи на хвосте очень грубые, направлены остриями назад и, возможно, служат для опоры при лазании по деревьям. Первый палец передних конечностей имеет ноготь. Волосяной покров грубый и низкий. Окраска его на спине серая или темпо-серая, па брюхе сероватая, желто-серая или темпо-желто-бслая. Сосков 2 пары.
В черепе лицевой отдел расширен, мозговой несколько сужсп. Лобнотеменные гребни мощные. Костное небо узкое и короткое. Резцовые отверстия очень короткие и узкие. Костные слуховые барабаны небольшие. Резцы очень широкие и мощные. Верхние щечные зубы гипсслодонтного типа.
Распространена на Новой Гвинее.
Экология не изучена.
Род кроличьих,крыс
Genus Conilurus Ogilby, 1838
Размеры мелкие. Длина тела 16,5—20,0 см. Длина хвоста 18—21,5 см. Уши относительно большие. Волосы равномерно покрывают хвост и лишь на конце хвоста его удлиняются, образуя небольшую кисточку. Задние конечности несколько удлинены. Волосяной покров у С. albipes мягкий, плотно прилегающий, а у С. penicillatus очень грубый, напоминающий колючки. Окраска спины варьирует от черновато-песчаной. Брюхо белое или темно-желтое. Хвост может быть полностью черный или с черным терминальным отделом, или бурый сверху и белый снизу.
В черепе хорошо выражено межглазничное сужение. Лобно-теменные гребни небольшие или их нет вовсе. Костное небо широкое. Резцовые отверстия длинные, достигают первого коренного зуба. Костные слуховые барабаны средних размеров. Венечный отросток нижней челюсти почти не заметен.
Распространены в Южной и Северной Австралии и на юге Новой Гвинеи.
Населяют различные биотопы, берега водоемов, болота, травянистые равнины, леса. Ведут ночной образ жизни. Убежищем служат дупла. Имеется указание, что самка таскает детенышей до перехода их к самостоятельному образу жизни на себе; детеныши висят на сосках, захватив их резцами.
В роде, вероятно, 2 вида:
С. albipes Lichtenstein, 1829 (Южная Австралия);
С. penicillatus Gould, 1842 (Северная Австралия и южная часть Новой Гвинеи).
По Андерсону и Джонсу (1967), в роде 3 вида, а по Эллермэну (1941), — 4: С. albipes, С. penicillatus, С. melibius Thomas, 1921, С. hemi-leucurus Gray, 1858.
Род толстохвостых'крыс
Genus Zyzomys Thomas, 1909
Размеры мелкие. Длина тела 8,5—17,7 см. Длина хвоста 9,5—13,5 см. Хвост довольно хорошо оволоснен и утолщен в проксимальной части. У старых особей утолщение хвоста развито сильнее. Это утолщение не связано с отложением жира. Волосяной покров сильно изреженный, грубый, превратившийся почти в колючки. Окраска на спине буроватая, сероватая, темно-желтая или рыжевато-песчаная, на брюхе беловатая. Конечности белые. Хвост может быть двухцветным— коричневым сверху и белым снизу или целиком белым. В последнем случае па его конце имеется кисточка. Конец хвоста легко отрывается.
Череп с относительно удлиненным лицевым отделом и широкой мозговой коробкой. Костные слуховые барабаны небольшие. Лобно-теменных гребней нет. Костное небо широкое. Резцовые отверстия достигают уровня щечных зубов.
Распространены в Северной и Западной Австралии и Квинсленде. Населяют скалистые холмы и скалистые обрывы. Активность ночная. Экология не изучена.
В роде 3 вида:
Z. argurus Thomas, 1889 (Южная и Северо-Западная Австралия);
Z. pedunculatus Waite, 1896 (Центральная Австралия);
Z. woodwardi Thomas, 1909 (Северо-Западная Австралия).
Некоторые исследователи (например, Эллермэн, 1941; Симпсон, 1945) выделяют два последних вида в род Laomys Thomas, 1909.
Род мсзембриомис
Genus Mesembriomys Palmer, 1906
Размеры средние. Длина тела 24—35 см. Длина хвоста 27—39 см. Уши средних размеров. Хвост довольно густо покрыт волосами, за исключением проксимального отдела, где сквозь волосы видны кожные чешуи. На конце хвоста — пучок удлиненных волос. Когти на пальцах передних и задних конечностей крупные. Стопа относительно широкая; первый и пятый палец не уменьшены. Волосяной покров грубый. Спина ярко-рыжая, темно-желто-коричневая, желтовато-серая или желтовато-коричневая. Брюхо беловатое, кремовое, серое или кремово-белое. Конечности белые или черные. Кисточка на хвосте белая. Сосков 2 пары.
В крупном, массивном черепе лицевой отдел утолщен. Мозговая коробка невелика. Лобно-теменные гребни небольшие. Лобные кости вздуты сразу позади их сочленения с носовыми. Резцовые отверстия крупные, однако уровня щечных зубов не достигают. Костное небо широкое. Костные слуховые барабаны большие. Венечный отросток нижней челюсти низкий.
Распространены в Северной Австралии на восток до Квинсленда, а также на о-ве Мелвилл.
Населяют светлые, чаще эвкалиптовые леса. Ведут древесный образ жизни. Активность ночная. День обычно проводят в гнездах, которые они сооружают в дуплах из листьев и коры. Питаются в основном плодами пальмы (Pandanus). Самки приносят одного или двух детенышей.Местные жители употребляют мезембриомис в пищу.
Род содержит 2 вида:
крыса Гулда — М. gouldi Gray, 1843 (Австралия — Северная территория и Квинсленд);
М. macrurus Peters, 1876 (северная часть Западной Австралии).
Род рыженосых крыс
Genus Oenomys Thomas, 1904
В роде единственный вид: рыженосая крыса — О. hypoxanthus Puche-ran, 1855.
Размеры мелкие. Длина тела 13,5—18 см. Длина хвоста 17—20,5 см. Уши большие, округлые, покрытые волосами (рис. 104). Хвост почти лишен волос. Кожные чешуи, покрывающие его поверхность, крупные. Подошва стопы голая с крупными мозолями; пятый палец немного длиннее первого. Волосяной покров густой и мягкий. Окраска спины варьирует от рыжеватой или оливковой до серой с черноватым оттенком. Брюхо белое, иногда с желтым оттенком. Конечности белые, корнчнсвато-се-рые или рыжие. Носовая часть и бока морды рыжие. Сосков 2—4 пары.
Лицевой отдел черепа массивный, удлиненный. Межглазничное сужение выражено слабо. Лобно-теменные гребни крупные. Костное небо относительно узкое. Резцовые отверстия большие и удлиненные. Костные слуховые барабаны крупные. Верхние щечные зубы широкие со сложной структурой жевательной поверхности, имеющей очень высокие бугры и глубокие продольные углубления.
Рис. 104 Рыжспосая крыса — О. hypoxanlhus
Хромосом в диплоидном наборе 32.
Распространена в Африке от Сьерра-Леоне и Ганы до Северной Анголы; изолированные участки ареала имеются в Южной Катанге, Южном Судане и Кении.
Населяет разреженные леса, в горных районах на высоте от 300 до 3000 м над уровнем моря. Держатся обычно поодиночке. Ведет полудре-весный образ жизни. Делает поверхностно расположенные подземные ходы с гнездовой камерой. Активна в основном ночью. Питается зелеными частями растений, а также насекомыми. Период размножения приходится, по-видимому, на период с октября по апрель. В Западной Уганде и Восточном Заире сезонности в размножении, возможно, нет. Самка приносит от 3 до 4, иногда до 6 детенышей. Местные жители употребляют рыженосых крыс в пищу.
Эллермэн (1941) выделяет в роде второй вид: О. ornatus Thomas, 1911.
Род бороздча гозубых крыс
Genus Mylomys Thomas, 1906
В роде единственный вид: бороздчатозубая крыса—М. dybowskyi Pousargues, 1893 (=М. cuninghamci Thomas, 1906).
Размеры мелкие. Длина тела 13,5—19 см. Длина хвоста 10—18 см. Хвост покрыт довольно длинными, блестящими, средней густоты волосами. Первый и пятый пальцы на задних и особенно на передних конечностях сильно укорочены. Спина темно-желтая или коричневая с при
месью черных волос. Брюхо обычно белое. Хвост темный сверху и более-светлый снизу.
В черепе лицевой отдел массивный. Лобно-теменные гребни сильно развиты. Межглазничное сужение выражено умеренно. Межтеменная кость крупная. Костное небо узкое. Длинные резцовые отверстия сужаются кзади, достигая уровня передних щечных зубов. Костные слуховые барабаны довольно крупные. На наружной поверхности верхних резцов по продольной бороздке. Нижние резцы бороздок не имеют.
Распространена в Африке: Берег Слоновой Кости, Гана, юг Камеруна, север и восток Заира, Уганда, Кения, Руанда.
Экология изучена слабо. Населяет равнины, луга, заросли кустарников. Поднимается в горы до 2400 м над уровнем моря. Питается зелеными частями растений. Активна днем. В Западной Уганде размножение приходится на март — июнь и сентябрь — декабрь.
Род лохматоволосых крыс
Genus Dasymys Peters, 1875
В роде единственный вид: лохматоволосая крыса —D. incomtus Sun-devall, 1847 (рис. 105).
Размеры мелкие. Длина тела 13,5—19 см. Длина хвоста 10,5—18,5 см. Телосложение плотное. Уши средних размеров, округлые. Стопа несколько расширена, с хорошо развитыми пальцами. Пятый палец передних конечностей укорочен. Волосяной покров может быть длинным, мягким, шелковистым или более редким и грубым. Хвост и проксимальные отделы конечностей почти лишены волос. Спина оливково-коричневая, желтовато-бурая, с примесью черного цвета или серо-черная. Брюхо сероватобелое, белое или оливково-желтое. Кисть и стопа обычно светло-коричневые. Хвост весь темно-коричневый или слегка более светлый снизу. Сосков 3 пары.
Лицевой отдел черепа массивный, несколько укороченный. Межглазничное сужение очень сильно выражено. Лобно-теменные гребни обычно большие. Носовые кости широкие. Костное небо сильно сужено. Резцовые отверстия узкие, длинные, достигают уровня щечных зубов. Костные слуховые барабаны средних размеров. Верхние и нижние резцы очень широкие. Борозд на них нет.
Хромосом в диплоидном наборе 38.
Распространена в Африке; от Сьерра-Леоне до Судана и Эфиопии и далее на юг до Южно-Африканской Республики; в Юго-Западной Африке найдена только на крайнем северо-востоке.
Населяет равнинные и горные саванны и леса. Поднимается в горы до 4 тыс. м над уровнем моря. Предпочитает берега водоемов, болотистые места, хотя встречается и в относительно сухих районах. Активна круглосуточно. Ведет полуводный образ жизни; хорошо плавает и ныряет. Питается различными прибрежными и водными растениями. Собственных нор не делает; убежище находит в различных углублениях в берегах выше уровня воды, где строит гнездо из травы. В помете бывает от 2 до 4 детенышей. В Юго-Западной Африке массовое размножение приходится на период с июня по октябрь.
Некоторые исследователи (например, Эллермэн, 1941; Андерсон и Джонс, 1967) выделяют в роде еще один вид: D. orthos Heller, 1911.
Род травяных мышей
Genus Arvicanthis Lesson, 1842
Размеры мелкие. Длина тела 12—19 см. Длина хвоста 9—16 см. Уши довольно крупные, округлые (рис. 106). Хвост хорошо оволоспен, Первый и пятый пальцы задних конечностей сильно уменьшены; три средних пальца длинные. Пятый палец передних конечностей сильно укорочен, однако имеет коготь. Волосяной покров низкий, грубый, у некоторых видов волосы превратились в полуколючкп. Окраска спинной стороны тела варьирует от оливковой, серовато-оливковой до светлосерой и черновато-коричиевой. У некоторых видов вдоль середины спины проходит темная полоска. Брюшная сторона тела несколько светлее спинной. Уши обычно красноватые. Стопа и кисть серые, желтоватые пли темные, как спина.
Лицевой отдел черепа широкий и укороченный. Мозговая коробка не расширена. Межглазничное сужение выражено не резко. Лобпо-темен-
Рис. 106. Травяная мышь — A niloticus и ее череп
ные гребни сильно развиты. Костное небо несколько сужено. Костные слуховые барабаны крупные или очень крупные. Резцовое отверстие длинное, сужается в задней части, обычно достигая уровня щечных зубов. Резцы без борозд, утолщенные. Щечные зубы широкие, несколько укороченные.
Хромосом в диплоидном наборе у Л. niloticus 5G и у A. abyssinicus62.
Распространены в Саудовской Аравии и на большей части Африки от дельты Нила вверх по его долине через АРЕ, Судан и Эфиопию до Сенегала, на севере и востоке Заира; в Кении, Уганде, Танзании, Замбии.
Населяют различные биотопы: саванны, кустарниковые заросли, леса от уровня моря и до 2000 м над его уровнем. Ведут наземный образ жизни. Убежищем служат норы. Гнезда делают как в норах, так (иногда) и на поверхности земли. В густой траве прокладывают тропинки. Часто травяные мыши живут колониями. Активны в основном днем. Питаются листьями, проростками, семенами различных трав, а также сладким картофелем. Самка за сезон дождей приносит несколько пометов, в каждом из которых бывает по 4—6 детенышей.
В роде 4 вида:
травяная мышь — A. niloticus Desmarest, 1822 (АРЕ, Судан, южная часть Саудовской Аравии, Западная Сахара, Сьерра-Леоне, Гана, Гви-нея-Бисау);
A. lacematus Riippell, 1842 (Эфиопия);
эфиопская мышь — A. abyssinicus Riippell, 1842 (Эфиопия, Кения, Уганда, Танзания, восточная часть Заира);
сомалийская мышь — A. somalicus Thomas, 1902 (Сомали, Кения);
Возможно, все эти виды следует объединить в один — A. niloticus Desmarest, 1822.
Род манипурских кустарниковых крыс
Genus Hadromys Thomas, 1911
В роде единственный вид: манипурская кустарниковая крыса — Н. humei Thomas, 1886.
Размеры мелкие. Длина тела 9,5—12,5 см. Длина хвоста 10,5—14 см. Хвост покрыт густыми волосами. Пятый палец передней конечности-очень короткий, но имеет коготь. Стопа узкая; первый и пятый пальцы сильно укорочены. Волосяной покров густой и мягкий. Спинная сторона тела черновато-серая, причем область посредине спины более темная, чем голова и плечи. Брюшная сторона тела желтоватая или оранжевая. Стопа и кисть желтовато-белые. Хвост двухцветный: черный сверху и беловатый снизу. Сосков 4 пары.
В черепе лицевой отдел расширен и укорочен. Лобно-теменные гребни хорошо выражены. Резцовые отверстия длинные, узкие. Резцы относительно мощно развитые и широкие, бороздок на передней поверхности нет.
Распространена в Индии — Манипуре и Ассаме.
Населяет высокие травянистые заросли парковых ландшафтов с редкими высокими дубами и с густой кустарниковой растительностью. Обитатель склонов холмов на высоте примерно 1200 м над уровнем моря. Питается листьями травянистых растений.
Род индийских крыс
Genus Golunda Gray, 1837
В роде единственный вид: индийская крыса—G. ellioti Gray, 1837.
Размеры мелкие. Длина тела 11 —15 см. Длина хвоста 9—13 см. Сложение тяжелое. Внешне похожа па полевку (рис. 107). Голова несколько укороченная. Уши округлые, большие, покрытые волосами. Хвост относительно хороню оволоснсп. Первый и пятый пальцы задней конечности очень короткие, три средних — длинные. Пятый палец передних конечностей сильно уменьшен. Характер волосяного покрова значительно варьирует: у некоторых особей он мягкий, лишь с отдельными щетинкоподобными волосами, у других — грубый, с длинными, уплощен-
Рис 107. Индийская крыса—G ellioti
ними колючками, имеющими продольные бороздки. Спинная сторона тела сероватая, желтовато-коричневая, рыжевато-коричневая, иногда темно-коричневая, брюшная — светло-серая, голубовато-серая или бслова тая. Сосков 4 пары.
В черепе резко выражено межглазничпое сужение. Лицевой отдел сильно укорочен. Лобно-тсмеппые гребни крупные. Костные слуховые барабаны большие Верхние резцы широкие, с одной продольной бороздкой на наружной поверхности. Нижние резцы спереди гладкие. Костное небо относительно суженное. Резцовые отверстия длинные, сужающиеся кзади. Щечные зубы сильно расширены.
Хромосом в диплоидном наборе 52.
Распространена на о-ве Шри Ланка, в Пенджабе (Индия), Пакистане, Непале и Бутане.
Населяет различные биотопы, болотистые места, травянистые равнины, бурьяны по краям полей, джунглей. Живет поодиночке или семейными группами. Наибольшая активность в раннеутрепине часы и вечером. Ведет в основном наземный образ жизни, хотя хорошо лазает по де ревьям. Питается главным образом различными растительными объектами. Когда на Шри Ланке в больших масштабах возделывали кофейные плантации, численность индийских кустарниковых крыс резко возросла. Они питались почками и цветками кофейных деревьев. В связи с сокращенем площадей кофейных плантаций в последние годы их численность значительно уменьшилась. Норы копает редко. Гнездо диаметром 15—23 см строит из растительных волокон; чаще на земле или низко над землей в густой траве. В помете бывает от 3 до 4 детенышей.
Род бороздчатозубых болотных крыс
Genus Pelomys Peters, 1852
Размеры от мелких до средних. Длина тела 12,5—21,5 см. Длина хвоста 10—18 см. Хвост довольно хорошо оволоспеп (рис. 108). На задних конечностях первый и пятый пальцы сильно укорочены У некоторых видов коготь на пятом пальце передних конечностей можег отсутствовать, Волосяной покров довольно грубый. Спинная сторона тела темно-жел-
Й53ЙЖЧ
Рис 108. Pelomys сатрапае тая, желтая или черная, иногда с зеленовато-коричневым или оливковым блестящим металлическим отблеском. Брюшная сторона серая, темно-желтая или беловатая. У некоторых видов вдоль середины спины проходит темная полоска. Хвост темный сверху и светлый снизу. Уши покрыты редкими, тонкими, рыжеватыми волосами. Сосков 4 пары.
Верхние резцы имеют продольную бороздку на наружной поверхности. Передняя поверхность нижних резцов гладкая. Щечные зубы широкие.
Хромосом в диплоидном наборе у Р. сатрапае 48.
Распространены в Африке от Мозамбика и Юго-Западной Африки на север до Заира, Уганды, а также на о-вах Комэ и Буссу озера Виктория.
Обычно держатся по берегам водоемов, во влажных местах, иногда по опушкам лесов, в зарослях травы. Активны днем. Хорошо плавают, ныряют. Питаются различными прибрежными и болотными растениями. Как правило, нор не делают. Ведут наземный образ жизни. В помете от 2 до 3 детенышей. В Замбии сезонности в размножении, не видимо му, нет.
В роде, по-видимому, 8 видов:
Р. isseli de Beaux, 1924 (о-ва Комэ и Буссу оз. Виктория в Уганде);
крыса Гопкинса — Р. hopkinsi Ilaym.ni, 1955 (Уганда);
Р. rex Thomas, 1906 (Эфиопия);
крыса Харрингтона — Р. harringtoni Thomas, 1902 (Эфиопия);
Р. dembeensis Ruppell, 1845 (Эфиопия);
Р. minor Cabrera et Ruxton, 1926 (Катанга, Северо-Восточная Ангола и,возможно, Северо-Западная Замбия);
Р. fallax Peters, 1852 (Заир, Руанда, Юго-Западная Уганда, Танзания, Южная Кения, Замбия, Родезия, Малави, Северный Мозамбик);
Р. сатрапае Huet, 1888 (низовье р Конго, запад Анголы).
Рис. 109. Полосатая мышь — L. striatus
Андерсон и Джонс (1967) насчитывают в роде 9, а Эллермэн (1941) — 10 видов. Некоторые исследователи (например, Грассе и Декейзер, 1955) выделяют Р. rex, Р. harringtoni и Р. dembeensis в особый род Desmomys Thomas, 1910, а Р. minor и Р isseli — в род Kotnemys de Beaux, 1924.
Род полосатых мышей
Genus Limniscomys Trouessart, 1881
Размеры мелкие Длина тела 10—14 см Длина хвоста 10,2—15,5 см. Хвост довольно густо покрыт короткими волосами. Стопа узкая; первый и пятый пальцы сильно укорочены На передних конечностях три средних пальца развиты сильно, два боковых укорочены, причем пятый палец не имеет когтя. Волосяной покров, как правило, редкий и очень грубый. Спина от темно-коричневой до почти черной с продольными, светлыми пятнами, образующими вытянутые вдоль спины ряды, иногда эти пятна сливаются в сплошные полосы (рис. 109). Число полос (или рядов пятен) по 5—6 на каждом боку и одна срединная, более длинная полоса проходит от лобной части головы до основания хвоста Светлые полосы могут быть развиты в различной степени; у некоторых видов они почти отсутствуют, за исключением лишь срединной. Окраска брюха варьирует от желтовато-белой до желтовато-серой. Сосков 4 пары — 2 грудных и 2 паховых.
Череп напоминает таковой травяных мышей.
Хромосом в диплоидном наборе у I. barbarus 54, а у L. striatus 48.
Распространены па большей части Африки.
Населяют различные биотопы, заросли травы около болот и по берегам рек, тростниковые заросли, кустарники, леса. В горы поднимаются на высоту до 2100 м над уровнем моря. Ведут наземный образ жизни. Активны преимущественно днем. Делают наземные гнезда из травы, иногда занимают норы других грызунов. Питаются различными частями растений. В помете обычно от 2 до 5, но иногда до 12 детенышей. Некоторые виды размножаются круглый год, другие, по-видимому, в оп-
Рис. ПО. Полосатая полевая мышь — R. ршпШо
ределенные периоды. Для L. striatus установлена беременность в 28 дней и способность менее чем в 4 месяца принести 4 помета.
В роде, по-видимому, 4 вида:
L. griselda Thomas, 1904 (от Сенегала, Заира, Руанды и Кеппи па юг до Южно-Африканской Республики);
L. barbarus Linnaeus, 1758 (Танзания, Кения, Заир, Марокко, Судан);
полосатая мышь —A. striatus Linnaeus, 1758 (от Сьерра-Леоне до северной Танзании, север Заира и северо-запад Анголы; изолированные участки в Замбии и Малави; Дарфур в Судане; Эфиопия);
L. macculus Thomas et Wroughton, 1910 (север и восток Заира; Уганда; юг Судана; Кения).
Эллермэн (1941) кроме этих видов выделяет еще L. olga Thomas et Hinton, 1921; L. lynesi Thomas et Minton, 1923. По Андерсону и Джонсу (1967), в роде 5 видов.
Род полосатых полевых мышей
Genus Rhabdomys Thomas, 1916
В роде единственный вид: полосатая полевая мышь — R. pumilio Sparrmann, 1784 (рис. НО).
Размеры мелкие. Длина тела 9—13 см. Длина хвоста 8—13,5 см. Хвост покрыт довольно густыми, короткими волосами. Стопа узкая, три средних пальца удлинены, а два наружных сильно укорочены. Па передних конечностях удлинены также три средних пальца. Пятый палец не имеет когтя. Волосяной покров грубый. На спине проходят полосы. Окраска их значительно варьирует у разных особей: обычно вдоль средней линии спины от шеи до основания хвоста проходит широкая светлая полоса от бледно-желтовато-серого до темпо-желтого цвета, ее окаймляют темные полосы от светло-золотисто-коричпевого до темпо-шоколадного или почти черного. Обычно темных полос бывает четыре, иногда две. Остальная часть спинной поверхности туловища имеет окраску
желтовато-серую, бледно-желтовато-коричневую или светло-серо-корич-невую. Брюшная сторона светлее спинной. Сосков 4 пары.
Череп сходен с таковым травяных мышей. Лицевой отдел укорочен. Лобно-теменные гребни обычно имеются. Костные слуховые барабаны относительно крупные. Резцовые отверстия длинные. Щечные зубы массивные.
Хромосом в диплоидном наборе 48.
Распространена в Африке от Танзании, Кении и Уганды до южной оконечности материка.
Населяет сухие участки долин рек, покрытые высокой травой, кустарниковые заросли, опушки лесов, подножья холмов, окультуренные земли. Встречается рядом с поселениями человека. Ведет наземный образ жизни. Активна днем. Выкапывает просто устроенные, с одним входом норы, иногда делает наземные гнезда из травы, листьев, мха и растительных волокон. Хорошо лазает по ветвям кустов. Питается различными растительными объектами (травой, корой, корнями, семенами, ягодами и т. д.), а также насекомыми, моллюсками. Период размножения длится с сентября по апрель. В течение года имеет, по-видимому, до. 4 пометов, в каждом из которых от 4 до 12 детенышей.
Род однополосых мышей
Genus Hybomys Thomas, 1910
Размеры мелкие. Длина тела 10,8—13 см, длина хвоста 9,5—11,5. Хвост покрыт редкими волосами (рис. 111). Стопа узкая, с сильно редуцированным пятым пальцем. Пятый палец передней конечности средних размеров. Подошва стопы имеет пять, а подошва кисти шесть хорошо развитых подошвенных мозолей. Волосяной покров обычно мягкий, лишь иногда грубый. Окраска спинной стороны тела от светло-желтокоричневой до темно-коричневой, почти черной. По середине спины обычно проходит продольная черная полоса, у некоторых особей, кроме того, еще две боковые темные полосы. Брюшная сторона тела темно-желтая, охристая или серовато-белая. Сосков 2 или 3 пары.
Череп укороченный, широкий. Межглазничное сужение выражено слабо. Лицевой отдел широкий и относительно длинный. Лобно-теменные гребни развиты хорошо. Костные слуховые барабаны средних размеров. Подглазничное отверстие относительно крупное. Резцовые отверстия средней длины или короткие. Щечные зубы довольно широкие.
Хромосом в диплоидном наборе у Н. univittatus 48.
Распространены в Западной, Центральной и Восточной Африке.
Населяют леса. Предпочитают держаться около водоемов. Ведут наземный образ жизни. Хорошо плавают. Активны преимущественно днем. Питаются различными плодами. При достаточном количестве пищи сезонности в размножении ист. Беременность 30 дней. В помете 1—3 детеныша. Самка достигает половозрслости в возрасте 77— 126 дней.
В роде 2 вида:
Н. univittatus Peters, 1876 (юг Камеруна, о-в Ферпапдо-По, Центрально-Африканская Республика, Габон, Заир, Конго, Уганда, Берег Слоновой Кости, Либерия, Сьерра-Леоне, Гвинея, Гапа);
Н. trivirgatus Temminck, 1853 (запад Нигерии, Гана, Берег Слоновой Кости, Либерия, Гвинея).
Рис Hi. Ilybnmys trivirgatus и cc череп
Род мягкошерстных крыс
Genus Millardia Thomas, 1911
Размеры мелкие. Длина тела 9,7—16 см. Длина хвоста 7,5—16 см. Внешне напоминают серую крысу. Уши крупные, округлые. Хвост хорошо оволоснеп Подошва стопы имеет 4 или 5 мозолен. Пятый пален задних конечностей сильно укорочен. Пятый палец передних конечностей средних размеров. Волосяной покров мягкий. Спинная поверхность тела имеет песчано-серую, серовато-коричневую или светло-серую окрас
ку, брюшная — белую или сероватую. Хвост светло-коричневый сверху и белый снизу. Сосков от 2 до 4 пар.
Лицевой отдел черепа не укорочен. Межглазничное сужение хорошо выражено. Лобно-теменные гребни имеются. Резцовые отверстия очень, длинные, достигают уровня первых щечных зубов. Костные слуховые барабаны крупные. Щечные зубы мощные.
Хромосом в диплоидном наборе у М. meltada 50, а у Л1. gleadowi 40.
Распространены на о-ве Шри Ланка, в Индии — на север до Пенджаба, Пакистане и Бирме.
Населяют болотистые места, поля, скалистые участки. Живут группами от 2 до 6 особей. Убежищем служат короткие норы, которые они выкапывают сами, пустоты под камнями или норы других грызунов. Питаются болотными растениями, а также зерном на полях. Самка приносит в помете от 6 до 8 детенышей. В некоторые годы отмечается значительное увеличение численности этих зверьков.
В роде 3 вида:
мягкошерстная крыса — М. meltada Gray, 1837 (о-в Шри Ланка, п-ов Индостан на север до Паланпура, Кача, Синда, Катхиавара, Пенджаба, Непала);
бирманская крыса — М. kathleenae Thomas, 1914 (Центральная Бирма);
желтая крыса — М. gleadowi Murray, 1885 (п-ов Индостан от Западного Синда до Катхиавара и западного Пенджаба).
Род большезубых крыс ।
Genus Dacnomys Thomas, 1916
В роде единственный вид: большезубая крыса — D. millardi Thomas, 1916.
Размеры средние. Длина тела 22,8—29 см. Длина хвоста 30,8— 33,5 см. Внешним обликом напоминает серую крысу. Хвост покрыт тонкими, короткими и редкими волосами. Подошвенные мозоли на ступне большие, округлые. Пятый палец задней конечности без когтя. Волосяной покров довольно грубый, низкий, редкий. Основная окраска спинной стороны тела от оливково-коричневой до серой или дымчато-коричневой, брюшной — обычно светло-буроватая. Горло, паховая и подмышечная области белые. Сосков 4 пары.
В черепе лобно-теменные гребни хорошо развиты. Резцовые отверстия очень широкие, сужающиеся спереди и сзади. Они достигают уровня щечных зубов. Костные слуховые барабаны очень маленькие. Щечные зубы массивные; их ряд удлинен.
Распространена в Восточных Гималаях, Сиккиме, Ассаме, Лаосе.
Встречается в горах на высоте 1000—1800 м над уровнем моря. Экология не изучена.
Род сулавесских мягкошерстных крыс
Genus Eropeplus Miller et Hollister, 1921
В роде единственный вид: сулавесская мягкошерстная крыса — Е. canus Miller et Hollister, 1921.
Размеры средние. Длина тела 19,5—23 см. Длина хвоста 26,5—29см. Хвост покрыт средней густоты волосами. Волосяной покров очень мягкий. Спинная сторона тела буровато-серая, брюшная — светло-серая.
Терминальная треть или половина хвоста белая. Конечности едва покрыты редкими черными волосами.
Череп с массивным, удлиненным лицевым отделом и значительным межглазничным сужением. Лобно-теменные гребни хорошо развиты. Костное небо сильно сужено. Резцовые отверстия длинные. Костные слуховые барабаны средних размеров. Щечные зубы крупные.
Распространена на о-ве Сулавеси.
Встречается в горах на высоте около 1800 м над уровнем моря.
Сулавесская мягкошерстная крыса близка к роду крыс и, возможно, должна быть включена в этот род в ранге подрода.
Род узкоголовых крыс
Genus Stenocephalemys Frick, 1914
В роде единственный вид: узкоголовая крыса — S. albocaudata Frick, 1914.
Размеры мелкие и средние. Длина тела 14,5—19 см. Длина хвоста 14,5—16,5 см. Хвост покрыт довольно густыми волосами. Волосяной покров мягкий, длинный, очень густой. Окраска спины темно-коричневая с черноватым оттенком, брюхо серое. Хвост белый.
Лицевой отдел черепа массивный, длинный. Мозговая капсула укороченная. Межглазничное сужение выражено очень сильно за счет сильно сжатых лобных костей. Резцовые отверстия длинные, достигают уровня щечных зубов. Подглазничные отверстия относительно большие.
Распространена в Южной Эфиопии.
Населяет горы на высоте примерно 3000 м над уровнем моря. Эко-i логия не изучена.
По своим признакам близка к роду крыс и, возможно, должна быть включена в этот род в ранге подрода.
Род крыс
Genus Rattus Fischer, 1803
Размеры от мелких до средних. Длина тела 8—30 см. Длина хвоста | может быть короче или несколько длиннее тела. Внешний облик также । значительно варьирует. Телосложение может быть тяжелое или легкое. | Морда удлиненная, глаза крупные (рис. 112). Средних размеров округ-I лые уши безволосые или с тонкими редкими волосками. Хвост обычно покрыт редкими, короткими волосами. Стопа удлиненная, относительно : узкая, обычно с шестью подошвенными мозолями. У некоторых видов она значительно модифицирована за счет приспособления к древесному образу жизни; крайние пальцы могут в различной степени противопоставляться средним. Пятый палец передних конечностей, за исключением некоторых лазающих форм, укорочен. Волосяной покров у некоторых видов мягкий, у других грубый. Волосы могут быть удлинены или превращены в своеобразные щетинки или даже иглы. Спинная сторона тела черная, сероватая, коричневая, желтоватая или рыжая. Брюхо обычно серое или беловатое. Сосков от 2 до 6 пар.
। В черепе лицевой отдел относительно удлинен. Мозговая коробка » крупная. Лобно-теменные гребни, как правило, хорошо развиты. Резцо-| вне отверстия длинные. Костные слуховые барабаны средних размеров I нли крупные.
| Хромосом в диплоидном наборе от 36 у крысы Бланфорда и R. rajah,
Рис. 112. Серая крыса — 2? norvegicus и ее череп
38 и 42 у черной крысы, до 42 у R. lutreolus, малой и серой, 42—44 у туркестанской и 46 у темпохвостой, R. niviventer и каштановой крыс.
Наибольшее число видов распространено в тропической и Юго-Восточной Азии и Африке. За человеком крысы расселились почти по всему земному шару, за исключением Лшаркгпды и некоторых океанических островов. В Европе серая крыса была, по-видимому, неизвестна до 1553г. В Северную Америку она была завезена человеком в 1775 г.
Населяют самые разнообразные ландшафты. Предпочитают лиственные леса гор, речных долин тропического, субтропическою и южной части умеренного пояса. В северной части умеренного пояса, в высокогорье, пустыне и к северу от лесной зоны ворочаются обычно лишь в жилищах человека. Среди представителей рода есть виды, хорошо лазающие по деревьям, плавающие в воде. Убежищем служат норы, кото-
1 рые они выкапывают сами, или норы других грызунов, пустоты под кам-| ними и под упавшими деревьями, иногда гнезда на деревьях. Активность может быть дневная или ночная.
Питаются различными растениями и животными. Некоторые виды всеядны. Например, серая крыса в природных условиях питается раз-; личными животными кормами — рыбой, моллюсками, мелкими мышевидными грызунами, а также различными растениями. Черная крыса, напротив, в основном питается плодами, побегами и корой растений и в меньшей степени животными. У туркестанской крысы преобладают растительные корма. В природных условиях она делает большие запасы (до 14 кг орехов, 7 кг яблок).
Размножаются большую часть года. Пик размножения у серой крысы приходится на весенне-летний период. У крыс, обитающих в жилищах человека, размножение продолжается круглый год, однако в зимний период интенсивность размножения снижается. Черная крыса приносит 2—3 помета в год, серая — до 3, а туркестанская крыса — до 4. Число детенышей в помете в среднем бывает у серой крысы 7 (от 1 до 17), черной —6, туркестанской — 8 (от 2 до 13). Длительность беременности составляет 21—22 дня. Половозрелость наступает в 3—4-месяч-i ном возрасте.
Численность может изменяться в разные годы. Хозяйственное значе- ние некоторых видов крыс очень велико, так как они сильно вредят сельскому хозяйству и портят продукты питания человека и корма для домашних животных в складах. Велико эпидемиологическое значение крыс. Они — природные носители возбудителей чумы, туляремии, клещевых сыпнотифозных лихорадок, тифа, риккетсиозов, лептоспирозной желтухи, бешенства и других заболеваний. Альбиносов серых крыс используют в качестве лабораторных животных.
Род крыс — один из наиболее многочисленных по числу видов среди млекопитающих. Его систематика разработана недостаточно. Разные исследователи насчитывают в роде различное число видов: Андерсон и Джонс (1967) — 120, Эллермэн (1940)—276—279. Исходя из работ Чэйзена (1940), Эллермэна (1941, 1949), Эллермэна, Моррисон-Скотта и Хеймана (1953), Лори и Хилла (1954), Хилла (1960), Эллермэна и Моррисон-Скотта (1966), Муссера (1969—1971) и некоторых других в роде можно выделить 97 видов (приведенный ниже перечень в ряде случаев не отражает филогенетическую близость видов):
крыса Бланфорда — R. blanfordi Thomas, 1881 (о-в Шри Ланка и Ин-дия к северу до Центральной провинции и провинции Бихар);
черная крыса — R. rattus Linnaeus, 1758 (включая R. lalolis Tate et ’Archbold, 1935; R. atridorsum Miller, 1903, R. flebilis Miller, 1902) (pac-, пространена как дикое животное на большей части п-ова Индостан, о-ве Шри Ланка, в Бирме, Южном Китае, п-овах Индокитай, Малакка, ; о-вах Ява, Суматра, Калимантан, Сулавеси, Филиппинских; в качестве комменсала человека распространилась почти по всему земному шару, в ’ частности, в СССР встречается в ряде областей европейской части стра-। ны от Архангельска до южных районов Закавказья, а также на Даль-i нем Востоке);
крыса Гоффмана — R. hofftnani Matschie, 1901 (включая R. tatei Ellerman, 1941) (о-в Сулавеси);
крыса Даммермаиа — R. dammcrmani Thomas, 1921 (северный Су-j лавеси);
1 R. fuscipes Waterhouse, 1839 (штаты Австралии — Южная и Юго-За- падная Австралия, Северная территория и Квинсленд);
| R. glauerti Thomas, 1926 (о-в Альброхос и у Западной Австралии);
’ 299
•а
/?. lutreolus Gray, 1841 (Тасмания и Южная Австралия);
туркестанская крыса — A rattoides Hodgson, 1845 (включая R. сап-па Swinhoe, 1870) (в СССР — Таджикистан, Узбекистан, Южная и Западная Киргизия и Южный Казахстан, Афганистан; Кашмир, Пенджаб; Непал; Южный и Юго-Восточный Китай и, возможно, Тайвань);
горная крыса — R. montanus Phillips, 1932 (о-в Шри Ланка);
гималайская крыса — R. nitidus Hodyson, 1845 (Индия — Кумаон; Ассам; Непал, Северная Бирма; Таиланд; п-ов Индокитай; Китай — от Фуцзяна до Юньнаня, Сычуаня, Ганьсу и Хайнаня);
малайзийская крыса — R. annandalei Bonhote, 1903 (Малайзия, о-в Суматра);
сантбарбская крыса — R. fulmineus Miller, 1913 (о-в Сант-Барб в Южно-Китайском море);
пальмовая крыса — R. palmarum Zeleboz, 1869 (включая R. stoicus Miller, 1902, и R. taciturnus Miller, 1902) (Никобарские о-ва);
R. zamboangae Mearns, 1905 (о-в Минданао из группы Филиппинских) ;
крыса Келлера — R. kellerl Mearns, 1905 (о-в Минданао из группы* Филиппинских);
большемордая крыса — R. magnirostris Mearns, 1905 (о-в Минданао из группы Филиппинских);
базиланская крыса — R. coloratus Hollister, 1913 (Базилам, Филиппинские о-ва);
кагайанская крыса — R. robiginosus Hollister, 1913 (Кагайан, Филиппинские о-ва);
добосская крыса — R. doboensis Beanfort, 1911 (о-в Добо из группы Ару);
тикаосская крыса — R. tyrannus Miller, 1910 (Тикао, Филиппинские о-ва);
тринаутская крыса — R. burrus Miller, 1902 (о-в Тринаут из группы Никобарских);
Никобарская крыса — R. burrulus Miller, 1902 (о-в Никобар, Никобарские о-ва);
R. burrescens Miller, 1902 (о-в Большой Никобар из группы Никобарских);
R. pulliventer Miller, 1902 (о-в Большой Никобар из группы Никобарских) ;
малая крыса—R. exulans Peale, 1848 (Бирма, п-ов Индокитай, Таиланд, п-ов Малакка, о-ва Суматра, Ява, Калимантан, Сулавеси, Филиппины, Новая Гвинея, Соломоновы, Фиджи, Бисмарка, Молуккские);
R. xanthurus Gray, 1867 (о-в Сулавеси);
R. everetti Gunther, 1879 (Филиппинские о-ва);
R. celebensis Gray, 1867 (о-ва Сулавеси, Лузон, Минданао);
квинслендская крыса — R. leucopus Gray, 1867 (Квинсленд в Австралии) ;
R. facetus Miller et Hollister, 1921 (центральные районы о-ва Сулавеси);
R. hamatus Miller et Hollister, 1921 (центральные районы о-ва Сулавеси);
R. taerae Sody, 1932 (северный Сулавеси);
R. marmosurus Thomas, 1921 (включая R. tondanus Sody, 1932) (северный Сулавеси);
R. arcuatus Tate et Archbold, 1935 (южный Сулавеси);
R. salocco Tate et Archbold, 1935 (южный Сулавеси);
R. punicans Miller et Hollister, 1921 (центральный Сулавеси);
серая крыса — R. norvegicus Berkentheut, 1769 (включая R. humilia-lus Milne-Edwards, 1868) (почти весь земной шар, кроме полярных областей и пустынь; СССР — большая часть территории, кроме Крайнего Севера, где, однако, встречается в некоторых портах, таежных районах Сибири и Дальнего Востока, пустынь, где, однако, обитает в Ташкентском оазисе, и полупустынь Казахстана и Средней Азии);
R. adspersus Miller et Hollister, 1921 (центральный Сулавеси);
R. andrewsi Allen, 1911 (о-в Бутон к югу от Сулавеси);
R. coelestis Thomas, 1896 (южный и юго-восточный Сулавеси);
R. sordidus Gould, 1858 (включая R. vanheurni Sody, 1933) (Квинсленд, Северная, Южная и Северо-Восточная Австралия, Новый Южный Уэльс, Новая Гвинея);
R. villosissisimus Waite, 1898 (Центральная Австралия и Новый Южный Уэльс);
R. rennelli Troughton, 1945 (Соломоновы о-ва);
R. paraxanthus Sody, 1941 (северный Сулавеси);
крыса Симпсона — R. simpsoni Ellerman, 1941 (северный Сулавеси);
крыса Маклеара — R. macleari Thomas, 1887 (о-в Рождества к югу от Явы);
R. nativitatis Thomas, 1888 (о-в Рождества);
крыса Мюллера — R. miilleri Jentink, 1879 (о-ва Суматра, Калимантан, Никобарские, Сиантан, Банка, Бату, Рио, Натуна, п-ов Малакка и на север до Тенассерима);
маратуаская крыса — R. тага Miller, 1913 (о-в Маратуа у Калимантана) ;
энганосская крыса — R. enganus Miller, 1906 (о-в Энгапо к югу от Суматры);
крыса Роджерса — R. rogersi Thomas, 1907 (Южные Андаманские о-ва);
R. ruber Jentink, 1879 (включая R. ringens Peters et Doria, 1881) (Новая Гвинея);
R. verecundus Thomas, 1904 (Новая Гвинея);
R. callitrichus Jentink, 1878 (северный Сулавеси);
калимантанская крыса — R. infraluteus Thomas, 1888 (о-ва Калимантан и Ява);
крыса Боверса — R. bowersi Anderson, 1879 (включая R. kennethi Kloss, 1918) (Ассам; Бирма; Южный Китай; п-ов Индокитай; Таиланд; п-ов Малакка);
R. dominator Thomas, 1921 (включая R. frosti Ellerman, 1941) (северный и центральный Сулавеси);
R. callitrichus Jentink, 1879 (юго-восточный Сулавеси);
моротаисская крыса—R. morotaiensis Kellogg, 1945 (о-в Моротаи из группы Молуккских);
овисская крыса — R. owiensis Troughton, 1946 (о-в Ови у Новой Гвинеи);
R. chrysocomus Hoffman, 1887 (центральный и северный Сулавеси);
R. beccarii Jentink, 1879 (северный Сулавеси);
R. shawmayeri Hinton, 1943 (Новая Гвинея);
R. coxingi Swinhoe, 1864 (о-в Тайвань, п-ов Индокитай; Китай — Юньнань, Сычуань, Шэньси; Северная Бирма);
R. niviventer Hodgson, 1836 (Востбчпый и Южный Китай; Индия — восточный Пенджаб; Ассам; Непал; Бирма; п-ов Индокитай; Таиланд; п-ов Малакка; о-ва Суматра, Ява, Бали, Тайвань и, возможно, Калимантан);
R. huang Bontohe, 1905 (Китай — Фуцзян, Гуандун. Ганьсу, Хайнань; п-ов Индокитай);
каштановая крыса — R. fulvescens Gray, 1847 (Индия— Кумаон; Ассам; Непал; Бирма; Юго-Восточный Тибет; п-ова Индокитай, Малакка; о-ва Суматра, Ява);
R. brahma Thomas, 1914 (Северная Бирма);
темнохвостая крыса—R. cremoriventer Miller, 1900 (о-ва Суматра, Ява, вероятно, Калимантан, п-ов Малакка, юг Бирмы, Ассам, Таиланд, п-ов Индокитай);
дымчатобрюхая крыса — R. eha Wroughton, 1916 (Непал; Сикким; Северная Бирма; Китай — Юньнань);
крыса Бартелса — R. bartelsi Jentink, 1910 (о-в Ява);
R. inflatus Robinson et Kloss, 1916 (западная Суматра);
R. baeodon Thomas, 1894 (о-в Калимантан);
охиясская крыса — R. ohietisis Phillips, 1929 (о-в Шри Ланка);
суматранская крыса —R. hylomyoides Robinson et Kloss 1916 (западная Суматра);
R. alticola Thomas, 1888 (о-в Калимантан);
крыса Мусшенброка-—R. musschenbroeki Jentink, 1879 (включая R. sakeratensis Gyldenstolpe, 1916) (северный Сулавеси, северный Калимантан, Таиланд, п-ов Малакка);
R. rajah Thomas, 1894 (включая R. surifer Miller, 1900) (о-ва Калимантан, Палаван, Ява, Суматра, п-ов Малакка, юг Бирмы, Таиланд, п-ов Индокитай);
крыса Хеллвалда — R. hellwaldi Jentink, 1878 (о-в Сулавеси);
крыса Доллмана — R. dollmani Ellerman, 1941 (о-в Сулавеси);
R. niobe Thomas, 1906 (Новая Гвинея);
филиппинская крыса — R. datae Mayer, 1899 (о-ва Лузон и Минданао из группы Филиппинских о-вов);
R. hylocoetes Mearns, 1905 (о-ва Минданао из группы Филиппинских);
R. itisignis Mearns, 1905 (о-в Минданао);
R. petraeus Mearns, 1905 (о-в Минданао);
R. major Aliller, 1911 (о-в Лузон);
R. musculus Miller, 1911 (о-в Лузон);
R. hollisteri Ellerman, 1949 (Филиппинские острова);
крыса Эдвардса — R. edwardsi Thomas, 1882 (Сикким; Ассам; Северная Бирма; Южный Китай; п-ова Индокитай, Малакка; о-ва Суматра и Сипора);
R. sabanus Thomas, 1887 .(о-ва Калимантан, Ява, Суматра, п-ов Малакка с прилегающими островами; юг Бирмы; Таиланд; п-ов Индокитай; Ассам);
манипурская крыса — R. manipulus Thomas, 1916 (Западная Бирма и Ассам);
R. berdmorei Blyth, 1851 (Таиланд);
качская крыса — R. cutchicus Wroughton, 1912 (Кач, п-ов Катхиавар, Раджастан, Бихар и южная часть п-ва Индостан);
индонезийская крыса — R. canus Miller, 1903 (о-ва Суматра, Калимантан, Ява и п-ов Малакка);
R. legatius Thomas, 1906 (о-в Рюкю);
R. faberi Jentink, 1883 (северный Сулавеси);
таиландская крыса — R. bocourti Milne-Edwards, 1876 (Таиланд).
После ревизии рода число видов в нем будет, по-видимому, значительно сокращено.
В СССР: черная, серая и туркестанская.
Эллермэн (1941), Симпсон (1945) и некоторые другие исследователи выделяют виды /?. datae, R. hylocoetes, R. insignis, R. petraeus, R. major, R. musculus, R. microdon, в род Apomys Mearns, 1905.
Род акациевых крыс
Genus Aethomys Thomas, 1915
Размеры мелкие. Длина тела 12,8—15,8 см. Длина хвоста 13—21 см.
Уши средних размеров, округлые, покрыты редкими волосами. Хвост
опушен довольно густыми волосами. Конечности относительно короткие.
На
задних
конечностях наружный палец
сильно
редуцирован. Волося-
ной покров высокий, мягкий или довольно грубый. Спинная сторона тела темная, бока и морда серые, брюхо белое. Хвост коричневый или черный. Сосков 3 пары.
Череп с массивным лицевым отделом. Носовые кости широкие. Межглазничное сужение хорошо выражено. Есть лобно-теменные гребни. Резцовые отверстия длинные, проникают между рядами щечных зубов. Костные слуховые барабаны крупные или очень крупные. Щечные зубы относительно широкие со сложным рельефом жевательной поверхности.
Распространены в Западной, Центральной, Восточной и Южной Аф-
рике.
A. chrysophilus населяет саванны; держится небольшими группами. A. hindei предпочитает степи, а также поля. A. longicaudatus — лесной житель, встречается в основном по берегам водоемов, в заболоченных местах, иногда на плантациях. Активны преимущественно ночью. Пита-
ются упавшими на землю плодами, семенами, листьями, проростками. Беременность 21—23 дня. У A. chrysophilus и A. longicaudatus сезонности в размножении нет.
В роде 10 видов:
западноафриканская крыса — A. defua Miller, 1900 (Западная Африка от Сьерра-Леоне до Ганы);
длиннохвостая крыса — A. longicaudatus Tullberg, 1893 (Центральная Африка от Юго-Восточной Нигерии до восточных и южных областей Заира);
крыса Гранта—A. granti Wroughton, 1908 (Южная Африка);
намакуасская крыса — A. namaquensis A. Smith, 1834 (Южная Африка к югу от Замбези);
золотистая крыса — A. chrysophilus de Winton, 1897 (Южно-Африканская Республика, Юго-Западная Африка и на север до Анголы, Заира и Кении);
крыса Томаса — A. thomasi de Winton, 1897 (Западная и Центральная Ангола);
малавийская крыса — A. nyikae Thomas, 1897 (Малави и восточная часть Южной Родезии);
крыса Кайзера—A. kaiseri Noack, 1887 (юг п восток Заира; Замбия; Восточная Ангола; Танзания);
A. hindei Thomas, 1902 (север и северо-восток Заира, Уганда; Кения; северо-восток Танзании; юг Судана па восток до Северной Лиге-, рии);
ангольская крыса — A. bocagei Thomas, 1904 (Центральная и Запад-* ная Ангола).
Некоторые исследователи (например, Эллермэн и др., 1953 включают этот род в род Rattus. Иногда A. defua и A. longicaudatus выделяют в особый род Stochonomys Thomas, 1926.
Род праомис
Genus Praotnys Thomas, 1915
Размеры мелкие. Длина тела около 10—10,5 см. Длина хвоста примерно 12—13 см. Голова удлиненная с вытянутой мордой. Уши крупные, округлые, почти голые. Хвост покрыт редкими, короткими и тонкими волосками. На передней конечности первый палец рудиментарен, вместо когтя имеет плоский ноготь. Третий и четвертый пальцы самые длинные. Стопа удлинена; три средних пальца значительно длиннее крайних. Волосяной покров мягкий. Его окраска на спинной стороне тела варьирует от серой до ярко-рыжей. Брюшная сторона белая или сероватая. Молодые животные темно-серые. Сосков 3 пары.
В черепе лицевой отдел и мозговая коробка умеренно вытянуты. Межглазничное сужение хорошо выражено. Имеются лобно-теменные гребни. Резцовые отверстия длинные, почти достигают рядов щечных зубов или заходят между ними. Костные слуховые барабаны средних размеров.
Распространены в Западной, Центральной, Восточной и Южной Африке.
Населяют в основном дождевые леса; предпочитают вторичные насаждения около водоемов. Р. natalensis приспособилась к жизни в деревнях. Активны ночью и в сумерках. Хорошо лазают по деревьям. Питаются плодами пальм, зелеными частями растений, семенами, насекомыми. Делают гнезда на ветвях деревьев, иногда в дуплах. Размножаются в течение круглого года. В году дают несколько пометов, в каждом из которых от 2 до 6, обычно 4—5 детенышей. Длительность беременности 26—27 дней. Детеныши рождаются голыми и слепыми. Глаза открываются на 15-—19-й день после рождения. Половозрелость наступает примерно в возрасте 2,5 месяцев. Р. natalensis имеет важное значение в распространении чумы.
В роде, по-видимому, 22 вида:
Р. pernanus Kershaw, 1921 (Кения и Северная Танзания);
Р. natalensis Smith, 1834 (Наталь; в деревнях по всей Африке к югу от Сахары, а также Марокко);
Р. erythroleucus Temminck, 1853 (Африка к югу от Сахары и Южное Марокко);
Р. angolensis Bocade, 1890 (Ангола и Заир);
Р. shortridgei St. Leger, 1933 (Юго-Западная Африка);
Р. albipes Riippell, 1842 (Эфиопия, Судан);
Р. fumatus Peters, 1878 (Танзания, Кения, Сомали, Эфиопия, Южный Судан, Уганда);
Р. verreauxi Smith, 1934 (Южно-Африканская Республика);
Р. butleri Wroughton, 1907 (юго-запад Судана);
Р. daltoni Thomas, 1892 (Западная Африка от Сенегала до Нигерии);
Р. tullbergi Thomas, 1894 (Западная Африка от Сьерра-Леоне и Гвинеи до Ганы);
Р. morio Trouessart, 1881 (Камерун, Центрально-Африканская Республика; Нигерия, Габон; Конго; Заир);
Р. jacksoni de Winton, 1897 (от Камеруна па запад до Южного Судана, Уганды, Кении, Танзания и Малави);
Р. delectorum Thomas, 1910 (Малави, Танзания, Кения);
Р. baeri Heim de Baisac et Aellen, 1965 (Берег Слоновой Кости);
P. aeta Thomas. 1911 (Камерун, о-в Ферпапдо-По);
P. carillus Thomas, 1904 (Камерун; Габон; Конго; Заир; Западная Уганда; Северная Ангола; о-в Фернандо-По);
P. denniae Thomas, 1906 (Уганда; Замбия; Танзания; Кения);
Р. parvus Brosset, Dubost et Heim de Balsac, 1965 (Габон и Конго);
P. fumosus Brosset, Dubost et Heim de Balsac, 1965 (Габон; возможно, Центральноафриканская Республика);
Р. alleni Waterhouse, 1837 (о-в Фернандо-По);
Р. stella Thomas, 1911 (от Гвинеи до Заира, Уганда, Кения).
Некоторые исследователи (например, Уокер, 1964) включают мягкошерстных крыс в род Rattus. Р. parvus и Р. fumosus, возможно, представляют один вид; Р. alleni и Р. stella в настоящее время, по-видимому, вымерли.
Род темных крыс
Genus Tryphomys Miller, 1910
В роде единственный вид: темная крыса — Т. adustus Miller, 1910.
Размеры мелкие. Длина тела 17,4 см. Длина хвоста 15 см. Внешним обликом напоминает серую крысу. Хвост почти не покрыт волосами; чешуи на нем крупные, грубые. Когти на пальцах задних конечностей удлинены; пятый палец сильно уменьшен. Ноготь на первом пальце передней конечности небольшой. Подошвы конечностей голые. Волосяной покров грубый. Вершинки коротких волос имеют тенденцию загибаться в сторону головы, придавая меху своеобразный ворс. Окраска спинной стороны тела коричневая или черная, брюшной—желтовато-белая. Хвост одноцветный, темно-коричневый. Сосков 5 пар.
Распространена на о-ве Лузон из группы Филиппинских.
Обитает в горах на высоте примерно 2500 м над уровнем моря. Экология не изучена.
Сходна с представителями рода крыс и, возможно, ее следует включить в этот род в качестве подрода, как это, например, делают Андерсон и Джонс (1967).
Род австралийских полевых мышей
Genus Gyomys Thomas, 1910
Размеры мелкие. Длина тела 6,8—13,2 см. Длина хвоста 6,1—13,7 см. Характерна относительно крупная голова с небольшими или среднего размера ушами (рис. ИЗ). Хвост покрыт средней густоты волосами, на конце несколько более удлиненными. У большинства видов волосяной покров на туловище мягкий, у G. deserter — грубый. Окраска его на спинной стороне тела варьирует от бледно-серой до песчано-коричневой и коричневато-оливковой. Брюшная сторона и конечности белые. Хвост темный сверху и белый или темно-желтый снизу. Сосков 2 пары.
В черепе мозговая капсула широкая. Межглдзничпая перетяжка резко выражена. Лобно-теменных гребней нет. Костные слуховые барабаны небольшие. Резцовые отверстия средних размеров. Венечный отросток нижней челюсти сильно редуцирован. Распространены почти по всей Австралии, за исключением части Северной территории.
Населяют песчаные равнины, каменистые гребни, леса п участки с зарослями высокой травы. Экология изучена слабо. G. albocinereus выкапывает в песке норы до 1 м глубиной с несколькими выходами. По-ви-димому, эврифаги: питаются различными насекомыми, семенами и другими частями растений.
Рис. 113. Западноавстралийская мышь G. albocinereus
В роде 8 видов:
G. glaucus Thomas, 1910 (южный Квинсленд);
G. berneyi Trough ton, 1936 (Квинсленд);
G. pumilus Troughton, 1936 (побережье Квинсленда);
G. fumeus Brazenor, 1934 (Виктория);
G. novaehollandiae Waterhouse, 1843 (Новый Южный Уэльс);
южноавстралийская мышь— G. apodemoides Finlayson, 1932 (Южная Австралия);
центральноавстралийская мышь— G. desertor Troughton, 1932 (Центральная Австралия);
западноавстралийская мышь — G. albocinereus Gould, 1845 (Западная Австралия).
Андерсон и Джонс (1967) включают австралийских полевых мышей в род австралийских ложных мышей — Pseudomys Gray, 1832.
Род австралийских крыс
Genus Leporillus Thomas, 1906
Размеры от мелких до средних. Длина тела 14—20 см. Хвост нссколь ко короче или немного длиннее тела. Уши большие, округлые (рис. 114). Хвост хорошо оволоснен. В его герминальном отделе волосы удлинены. Стона умеренной длины. Волосяной покров густой, мягкий Окраска его на спинной стороне тела светло-коричневая, с сероватыми или желтоватыми оттенками, па брюшной — обычно сероватая или белая. Сосков 2 пары.
В черепе лобно-теменных гребней пег. Резцовые отверстия крупные, достигают уровня щечных зубов. Костные слуховые барабаны большие. Щечные зубы массивные.
Рис. 114. Lepor Ulus conditor
Распространены в южной части Центральной Австралии, в Новом Южном Уэльсе, а также па о-вах Франклина и Нуит у побережья Южной Австралии.
L. conditor и L. jonesi строят крупные «дома» до 6 м в диаметре и 125 см в высоту из веток, до 1 м длиной и 25 мм толщиной. Часто такие «дома» сооружают вокруг небольшого куста или около камня, скалы Иногда на верх «дома» или на его степы затаскивают небольшие камни, предотвращающие разрушение ветром В «доме» устраивают несколько травяных гнезд. Заселяют его несколько пар. Активны в основном ночью. Питаются преимущественно растениями.
В роле 3 вида:
L. apicalis Gould, 1851 (Южная Австралия);
L. jonesi Thomas, 1921 (о-в Франклин, архипелаг Нуит, Южная Австралия) ;
L. conditor Sturt, 1848 (Новый Южный Уэльс).
Род ложных мышей
Genus Pseudomys Gray, 1832
Размеры мелкие. Длина тела 7,6—16 см Длина хвоста 7—18 см Уши довольно большие (рис. 115). Хвост хорошо опушен. Стопа несколько удлинена. Волосяной покров мягкий. Спинная сторона тела коричневая или желтая, брюшная — желговато-белая, желтая или серовато-белая. Хвост у некоторых видов темный сверху и белый снизу. Сосков 2 пары.
Лобные кости в черепе сильно ежа гы. Резцовые отверстия крупные. Подглазничные отверстия суженные. Костные слуховые барабаны средних размеров.
Хромосом в диплоидном наборе у Р. higginsi 48
Распространены в Юго-Западной, Центральной, Восточной Австралии, на Тасмании
Населяют разнообразные биотопы: травянистые равнины, песчаные холмы, покрытые лесом; болотистые участки, поросшие кустарником; берега мелких солоноводпых лагун и т. д. Убежищем служат поры, обычно
Рис. 115. Pseudomys ferculinus
просто устроенные, мелкие, с одним выходом. Активны в основном ночью. Питаются главным образом различными частями растений. В помете от 2 до 4 детенышей.
В роде 11 видов:
Р. australis Gray, 1832 (Новый Южный Уэльс);
Р. minnie Troughton, 1932 (Южная Австралия и Квинсленд);
Р. rawlinnae Troughton, 1932 (Западная Австралия);
Р. auritus Thomas, 1910 (Южная Австралия);
Р. shortridgei Thomas, 1907 (Юго-Западная Австралия);
Р. higginsi Trouessart, 1897 (Тасмания);
Р. gracilicaudatus Gould, 1845 (юг Квинсленда);
Р. gouldi Waterhouse, 1839 (Новый Южный Уэльс);
Р. nanus Gould, 1858 (Новый Южный Уэльс);
Р. praeconis Thomas, 1910 (Западная Австралия);
Р. ferculinus Ihomas, 1902 (Западная Австралия).
Андерсон и Джонс (1967) включают сюда род Gyomys Thomas, 1910, поэтому насчитывают в роде Pseudomys 22 вида.
Род мозаичнохвостых крыс
Genus Pogonornelomys Riimmler, 1936
Размеры мелкие. Длина тела 13—19 см. Длина хвоста 14—21 см. Характерен хватательный хвост, поверхность которого покрыта шестиугольными кожными чешуями, причем на каждую чешую приходится обычно I—3 волоса (иногда больше). Стопа с сильными копями. Волосяной покров довольно грубый. Спинная сторона тела коричневатая, рыжеваю-коричневая или темно-коричневая, брюшная — беловатая, сероватая или
темно-желтая. Конечности желтовато-коричневые или беловатые. Сосков 2 пары.
Распространены на Новой Гвинее.
Населяют равнинные и горные леса. Ведут древесный образ жизни. Экология не изучена.
В роде 4 вида:
Р. mayeri Rotschild et Dollman, 1932 (запад Новой Гвинеи);
Р. bruijni Peters et Doria, 1876 (запад Новой Гвинеи);
Р. sevia Tate et Archbold, 1935 (северо-восток Новой Гвинеи);
Р. ruemmleri Tate et Archbold, 1941 (запад Новой Гвинеи).
Некоторые исследователи (например, Эллермэн, 1940) включают мозаичнохвостых крыс в род банановых крыс. Систематическое положение этих двух родов должно быть уточнено.
Род банановых крыс
Genus Melomys Thomas, 1922
Размеры мелкие. Длина тела 9—17,5 см. Длина хвоста 11—17 см. Полухватательный хвост почти полностью лишен волос. Чешуя на нем не налегают друг на друга, а в виде мозаики прилегают одна к другой. Широкие и несколько укороченные конечности с крупными мозолями на подошвах приспособлены к древесному образу жизни. Когти на пальцах длинные. Пятый палец задних конечностей не укорочен. Волосяной покров обычно мягкий, густой и довольно высокий. Окраска спипной стороны тела от коричневатой до красновато-коричневой. Брюшная сторона белая, кремовая или светло-серая.
Лицевой отдел черепа массивный. Мозговая коробка относительно широкая. Лобно-теменные гребни небольшие или их нет вовсе. Костные слуховые барабаны средних размеров или очень малы.
Хромосом в диплоидном наборе у М. cervitiipes 52.
Распространены на Новой Гвинее, о-вах Керам и Мелвилл, на Северной территории Австралии, в Квинсленде, Новом Южном Уэльсе, на архипелаге Бисмарка и Соломоновых о-вах.
Населяют леса, покрытые травой равнины, иногда плантации сахарного тростника. Обычно держатся около воды. Ведут в основном древесный образ жизни. Убежищем служат гнезда, которые они делают на деревьях из травы и листьев. Иногда гнезда сооружаются в дуплах или у основания дерева, среди его корней или под камнями. Питаются в основном плодами и ягодами. Период размножения приходится на дождливый сезон — с ноября по март. Примерно в течение первых двух недель после рождения детеныши удерживаются на сосках матери своими острыми резцами, а самка лазает по деревьям вместе с ними. Полово-зрелость у самок наступает в 7 месяцев.
В роде 17 видов:
М. albidens Tate, 1951 (запад Новой Гвинеи);
М. fellowsi Hinton, 1943 (северо-восток Новой Гвинеи);
М. levipes Thomas, 1897 (юго-восток Папуа, северо-восток и запад Новой Гвинеи);
М. aerosus Thomas, 1920 (о-в Керам);
М. rubicola Thomas, 1924 (Папуа);
М. moncktoni Thomas, 1904 (Новая Гвинея);
М. obiensis Thomas, 1911 (о-в Оби из группы Халмахера);
М. platyops Thomas, 1906 (Новая Гвинея);
М. rubex Thomas, 1922 (Новая Гвинея);
M. lutillus Thomas, 1913 (Новая Гвинея);
М. rufescens Alston, 1877 (о-в Герцога Йоркского, Новая Ирландия и Новая Британия, Новая Гвинея, Соломоновы о-ва);
М. leucogaster Jentink, 1908 (Новая Гвинея);
М. fulgens Thomas, 1920 (о-в Керам и Талауд);
М. arcium Thomas, 1913 (о-в Россел у восточной оконечности Новой Гвинеи);
М. porculus Thomas, 1904 (Соломоновы о-ва);
М. fraterculus Thomas, 1920 (о-в Керам);
М. cervinipes Gould, 1852 (Квинсленд; Северная Австралия).
Некоторые исследователи (например, Эллермэн, 1941) включают в род Melomys роды Pogonomelomys и Solomys.
Род голохвостых крыс
Genus Solomys Thomas, 1922
Размеры средние. Длина тела 23—33 см. Длина хвоста 21,5—34 см. Хвост хватательный, лишенный волос почти по всей длине. Подошвенные мозоли хорошо развиты. Когти на пальцах крупные. Волосяной покров мягкий или довольно грубый. Окраска тела одноцветная — коричневато-черная. Сосков 2 пары.
Характерны сильные и широкие резцы.
Распространены на Соломоновых о-вах.
Населяют густые леса. Ведут древесный образ жизни. Экология не изучена.
В роде 3 вида:
сантаисабельская крыса — S. sapientis Thomas, 1902 (о-в Санта-Исабель) ;
бугенвильская крыса-—S. salebrosus Troughton, 1936 (о-в Бугенвиль) ;
голохвостая крыса — S. ponceleti Troughton, 1935 (о-в Бугенвиль).
Некоторые исследователи (например, Симпсон, 1945; Эллермэн, 1941) включают голохвостых крыс в качестве подрода в род Melomys.
Род белохвостых крыс
Genus Xenuromys Tate et Archbold, 1941
В роде единственный вид: белохвостая крыса — X. barbatus Milne-Edwards, 1900.
Размеры средние. Длина тела 27,5—31 см. Длина хвоста 22—28,1 см. Хвост почти лишен волос; чешуи, располагаясь кольцевидно, налегают одна на другую, как у представителей рода крыс. Конечности почти голые. Окраска волосяного покрова спинной стороны тела рыжевато-коричневая или серая, брюшной-—темпо-желтая или белая. Хвост белый по крайней мере па половине своей длины. Кисть п стопа почти лишены волос.
Распространены на Новой Гвинее.
Населяет густые леса.
Эллермэн (1940) включил белохвостую крысу в род Uromys в качестве подвида IJ. caudimaculatus barbatus. Лори и Хилл (1954) в своей монография вообще не упоминают о роде Xenuromys и виде X. barbatus.
Рис. 116. Uromys caudimaculatus
Род чешусхвостых крыс
Genus Uromys Peters, 1867
Размеры от средних до крупных. Длина тела 29—35,5 см. Длина хвоста 25,5—35 см. Уши крупные (рис. 116). Укороченные и широкие кисти и стопа приспособлены к древесному образу жизни. Пальцы с сильными когтями. На хвосте кожные чешуи в виде мозаики плотно прилегают друг к другу. У каждой чешуи — один волос. Пятый палец задних конечностей относительно длинный. Четвертый палец немного длиннее третьего. Волосяной покров низкий и грубый, но густой. У U. caudimaculatus волосы сильно изрежены. Окраска спинной стороны тела от коричневатой до серо-черной и сероватой, брюшная — серая или белая. У некоторых видов хвост полностью черный, у других терминальная часть белого или желтоватого цвета. Сосков 2 пары.
В черепе мозговой отдел относительно узкий. Лицевой отдел средних размеров. Лобно-теменные гребни развиты слабо. Костные слуховые барабаны очень малы. Задняя часть костного неба широкая, закапчивается сразу позади третьего коренного зуба. Резцовые отверстия сильно укорочены и располагаются далеко впереди щечных зубов. Резцы массивные.
Хромосом в диплоидном наборе у U. caudimaculatus 54.
Распространены па Новой Гвинее, о-вах Ару и Кен, архипелаге Бисмарка, о-вах Соломоновых, Новой Британии и в Квинсленде в Австралии.
Населяют леса. Ведут древесный образ жизни. Превосходно лазают по деревьям. Убежищем служат дупла. Питаются в основном плодами кокосовых пальм.
В роде 6 видов:
U. caudimaculatus Krefft, 1866 (северный Квинсленд в Австралии, Новая Гвинея, о-ва Ару и Кей);
U. anak Thomas, 1907 (северо-восток Новой Гвинеи);
U. neobritannicus Tate et Archbold, 1935 (о-в Новая Британия);
V. rex Thomas, 1888 (о-в Гвадалканар из группы Соломоновых);
U. imperator Thomas, 1888 (о-в Гвадалканар);
U. salamonis Ramsay, 1883 (о-в Флорида из группы Соломоновых).
Эллермэн (1941) включает в этот род Xenuromys barbatus в качестве подвида U. caudimaculatus.
Род большеухих крыс
Genus Malacomys Milne-Edwards, 1877
В роде единственный вид: большеухая крыса — М. longipes Milne-Edwards, 1877. ।
Размеры мелкие. Длина тела 13—18 см. Длина хвоста несколько превышает длину тела. Уши большие. Хвост покрыт редкими волосами, иногда почти голый. Стопа узкая, длинная; пятый палец относительно удлинен. Волосяной покров густой и мягкий, почти как у землероек. Окраска спинной стороны тела темно-коричневая, с сероватым оттенком, брюшной — белая. Бока охристые. На лбу обычно находится сероватобелое пятно. Вокруг каждого глаза может быть по черному пятну. Сосков 3 пары.
Череп удлиненный (особенно лицевая часть), узкий. Мозговая коробка ненамного шире лицевого отдела. Межглазпичпое сужение почти не выражено. Лобно-теменных гребней, как правило, нет. Резцовые отверстия короткие, обычно не достигают зубных рядов. Костные слуховые барабаны небольшие. Ряды щечных зубов сильно укорочены.
Хромосом в диплоидном наборе 48.
Распространена в Восточной, Центральной и Западной Африке: Гвинее, Либерии, на Берегу Слоновой Кости, в Гане, Южной Нигерии, Камеруне, Центральноафриканской Республике, на о-ве Фернандо-По, в Габоне, Заире, Конго, Уганде, Северо-Западной Замбии.
Экология изучена слабо. Населяет лесные болотистые места или участки с густой травянистой растительностью около водоемов. Из травы и листьев делает гнезда в углублениях почвы, между корнями деревьев. Держится поодиночке. Активна в основном ночью. Питается растениями и животными. В Центральной Африке размножение, по-видимо-му, приходится на период с марта по июнь.
Эллермэн (1940) выделяет в роде еще один вид: М. edwardsi Ro-chebrune, 1885.
Род малых мышей
Genus Haeromys Thomas, 1911
Размеры мелкие. Длина тела примерно 7,6 см. Длина хвоста 11— 14,4 см. Хвост покрыт короткими, редкими волосами. Конечности приспособлены к древесному образу жизни. Па задних конечностях пятый палец примерно такой же длины, как и второй; первый палец противопоставляется другим и имеет очень маленький коготь. На передних конечностях первый палец имеет крупный ноготь, остальные пальцы с короткими, изогнутыми, острыми когтями. Морда укороченная. Уши средних размеров, овальные, покрыты редкими волосами. Волосяной покров
относительно высокий, мягкий. Окраска спинной стороны тела рыжеватая или коричневато-рыжеватая, брюшной — белая. Хвост коричневый или зеленовато-серый. Сосков 3 пары.
Мозговая коробка черепа очень широкая, округлая. Лицевой отдел относительно короткий. Костные слуховые барабаны средних размеров. Ряды щечных зубов относительно короткие. Резцовые отверстия короткие.
Распространены на о-вах Калимантан и Сулавеси.
Ведут древесный образ жизни. Экология не изучена.
В роде, вероятно, 2 вида:
Н. margarettae Thomas, 1893 (о-в Калимантан);
Н. minahassae Thomas, 1896 (северная часть о-ва Сулавеси).
Андерсон и Джонс (1967) выделяют в роде 3 вида. j
Род азиатских древесных мышей
Genus Chiromyscus Thomas, 1925
В роде единственный вид: азиатская древесная мышь — С. chiropus Thomas, 1891.
Размеры мелкие. Длина тела 12,7—16 см. Длина хвоста 20,4—24 см. Красноватые уши покрыты короткими волосами. Волосы на хвосте тонкие, короткие, редкие. Стопа приспособлена к лазанию: она расширена, имеет большие пальцевые мозоли, первый палец противопоставляется другим и не имеет когтя. На остальных пальцах когти короткие. Волосяной покров довольно грубый. Окраска спинной стороны тела темножелтая с черноватым оттенком, брюшной — кремовая. Каждый глаз окружен темным пятном. Сосков 4 пары.
В черепе мозговая коробка широкая. Лобно-теменные гребни хорошо развиты. Костные барабаны небольшие. Межтеменная кость увеличена в размерах. Резцовые отверстия длинные.
Распространена в Бирме, Таиланде, на п-ове Индокитай.
Встречается в лесах. Ведет, по-видимому, древесный образ жизни. Экология не изучена.
Род широкоголовых мышей
Genus Zelotomys Osgood, 1910
Размеры мелкие. Длина тела 11,5—13,7 см. Длина хвоста 8,8—11,5 см. Голова несколько расширена. Уши средних размеров, округлые. Хвост покрыт средней густоты волосами. Волосяной покров на спинной стороне тела имеет окраску от темно-серой и желтовато-коричневой до розовато-темно-желтой. Брюшная сторона тела светло-серая, беловатая или розовато-темно-желтая. Сосков 5 пар.
В черепе мозговая коробка укорочена. Межглазничное сужение резко выражено. Лобно-теменных гребней нет. Межтеменная кость относительно небольшая. Костные слуховые барабаны средних размеров. Резцовые отверстия очень длинные, заходят между передними коренными зубами. Верхние резцы длинные, сильно выдаются вперед. Щечные зубы с крупными выступами на жевательной поверхности.
Распространены в Африке от Кении на юг через Заир до Южной Родезии и на запад до Анголы.
Экология не изучена. Населяют влажные травянистые саванны. Питаются насекомыми. В помете 5—7 детенышей.
В роде, по-видимому, 2 вида:
Z. hildegardeae Thomas, 1902 (Ангола, Замбия, Малави, Танзания, Центральная и Южная Кения, Западная Уганда, Руанда, Бурунди, Заир);
Z. woosnami Schwann, 1906 (Юго-Западная Африка).
Уокер (1964) считает в роде один вид: Z. hildegardeae.
Эллермэн (1941) выделяет 3 вида: Z. hildegardeae, Z. instans Thomas, 1915 и Z. shortridgei Hinton, 1920.
Род манипурских мышеи
Genus Diomys Thomas, 1917
В роде единственный вид: манипурская мышь — D. crumpi Thomas, 1917.
Размеры мелкие. Длина тела 10—13,5 см. Длина хвоста 10,5—13,5 см. Хвост покрыт редкими волосами. Волосяной покров густой, мягкий. Спинная сторона тела черновато-серая, брюшная беловато-серая. Хвост черный сверху и белый снизу.
Верхние резцы почти не выдаются вперед, а направлены вертикально вниз. Распространена в Бихаре (Индия).
Встречается в горах на высоте примерно 1300 м над уровнем моря.
Род полосатоспинных мышей
Genus Muriculus Thomas, 1902
В роде единственный вид: полосатоспинная мышь — М. imberbis Ruppell, 1845.
Размеры мелкие. Длина тела 7—9,5 см. Длина хвоста 4,5—6 см. Голова относительно укороченная. Уши средних размеров, округлые. Когти на пальцах короткие. Волосяной покров мягкий или довольно грубый. Волосы на спинной стороне тела имеют голубовато-серые основания и темно-желтовато-бурые вершины. Вдоль середины задней половины спины (иногда от лопаток) до основания хвоста обычно проходит темная полоса. Брюшная сторона тела кремовая или почти белая. Хвост обычно темно-коричневый сверху и желтовато-серый снизу. Иногда спереди у основания ушей располагается по пучку удлиненных волос.
В черепе мозговой отдел широкий. Верхние резцы сильно направлены вперед, узкие, с гладкой передней поверхностью.
Распространена в Эфиопии.
Населяет открытые травяные или каменистые склоны гор, поднимаясь до верхней границы леса примерно на высоту до 3200 м над уровнем моря. Экология не изучена.
Род мышей
Genus Mus Linnaeus, 1758
Размеры мелкие. Длина тела до 12,5 см, по обычно меньше 10 см — чаще всего 6,5—9,5 см. Длина хвоста 6—10,5 см. Морда несколько укороченная (рис. 117). Уши довольно большие, округлые. Хвост покрыт короткими, тонкими, редкими волосами. Стопа, как правило, узкая; наружные пальцы имеют тенденцию к укорочению. Пятый палец на задней
Рис. 117. Домовая мышь — М. musculus
конечности несколько длиннее, чем первый. Волосяной покров может быть мягким или грубым, иногда даже иглистым. Окраска тела почти одноцветная; спинная сторона от светло-темпо-желтой или светло-серой до темно-серо-коричневой или темно-желтовато-коричневой. Брюшная сторона тела обычно несколько светлее спинной. Сосков 5 пли (> пар.
Череп несколько уплощен. Лобно-теменные гребни развиты слабо пли их нет вовсе. Резцовые отверстия длинные п относительно узкие. Костные слуховые барабаны средних размеров. Резцы топкие. Щечные зубы имеют высокие коронки и хорошо обособленные бугорки па жевательной поверхности.
Хромосом в диплоидном наборе от 18—36 у карликовой, 26—40 у домовой, 20—22 у М. triton, 22—26 у индийской колючей до 32—34 у М. haussa, 40 — желто-коричневой и 48 у сиккимской мышей.
Распространение охватывает практически почти весь земной шар; во всяком случае, за малым исключением, все места, где обитает человек.
Населяют самые разнообразные биотопы, однако избегают пустынь, тундр, сплошных лесов таежного типа и высокогорья. Являясь комменсалами человека, распространились вслед за ним по всем ландшафтам. В природе представители рода активны главным образом ночью Домовая мышь активна круглосуточно. Многие виды хорошо лазают. Домовая мышь хорошо плавает. В природе роют поры пли используют в качестве убежищ различного рода укрытия. Поры Moryi быть простого устройства. Иногда же сложного строения; например, курганчпковые мыши насыпают очень большую кучу земли в виде кургана над сложно устроенными ходами и кладовыми, которые вмещают большие запасы корма (до 10 кг зерна). Домовая мышь всеядна. В природе поедает в основном семена, листья п стебли растений, а также насекомых.
Размножение в южных частях ареала и в постройках человека по всему ареалу продолжается круглый год. В течение года бывает до 5, иногда даже больше пометов Длительность беременности — 17—21 день. Число детенышей в помете колеблется от 3 до 12, обычно 4—7. Полово-зрелость наступает в возрасте около 2 месяцев. Продолжительность жизни в неволе иногда достигает 6 лет. Для популяции домовой мыши,
населяющей естественные биотопы, характерны резкие изменения численности в различные годы. Некоторые виды (в частности, домовая мышь) наносят значительный вред сельскохозяйственным и огородным культурам, а также продуктам, хранящимся в складах. Имеют большое эпидемиологическое значение, являясь природными носителями возбудителей чумной и туляремийной инфекций и др. Альбиносы домовой мыши широко используются как лабораторные животные.
В роде, по-видимому, 25 видов:
домовая мышь — Mus musculus Linnaeus, 1758 (ареал совпадает с ареалом рода; СССР — почти по всей стране, кроме Крайнего Севера, некоторых глухих таежных и ряда высокогорных районов);
индийская мышь — М. booduga Gray, 1837 (Бирма, Индия);
желтокоричневая мышь — М. cervicolor Hodgson, 1845 (о-в Шри Ланка, Индия, Непал, Бирма, п-ов Индокитай, Таиланд, о-ва Рюкю);
М. famulus Bonhote, 1898 (Северо-Восточная Индия, Бирма; Китай — Юньнань и, вероятно, Таиланд);
колючая мышь — М. platythrix Bennett, 1832 (п-ов Индостан и Центральная Бирма);
шриланкская мышь — М. fernandoni Phillips, 1932 (о-в Шри Ланка);
сиккимская мышь — М. pahari Thomas, 1916 (Сикким; Ассам; Бирма; Таиланд; п-ов Индокитай и, вероятно, Ява);
мышь Майора — М. mayori Thomas, 1915 (Шри Ланка);
карликовая мышь —-М. minutoides A. Smith, 1834 (Южно-Африканская Республика, Мозамбик, Южная Родезия, Юго-Западная Африка, Малави, Замбия, Ангола и иа север до Заира, Северной Нигерии и Эфиопии);
М. triton Thomas, 1909 (Ангола, Замбия, Заир, Уганда, Танзания, Кения);
мышь Каллеваерта — М. callewaerti Thomas, 1925 (Ангола, юг Заира);
М. bufo Thomas, 1906 (Кения, Заир);
кенийская мышь — М. gratus Thomas et Wroughton, 1910 (Кения);
Al. musculoides Temminck, 1853 (Гана, Уганда);
камерунская мышь — Al. paulina Thomas, 1918 (Камерун);
Al. mahomet Rhoad, 1896 (Сомали);
Al. proconodon Rhoad, 1896 (Сомали, Заир);
Al. haussa Thomas et Hinton, 1920 (Нигерия);
Al. gerbillus G. Allen et Loveridge, 1933 (Танзания);
Al. tenellus Thomas, 1903 (Судан, Танзания);
M. wamae Heller, 1911 (Кения);
Al. matschieWettstein, 1916 (Судан);
Al. birungensis Lonnberg et Gyldenstolpc, 1925 (Заир);
филиппинская мышь — M. castaneus Waterhouse, 1843 (Филиппинские о-ва);
Al. crociduroides Robinson et Kloss, 1916 (о-ва Суматра и Ява).
Число видов в роде в будущем, очевидно, будет уменьшено. Уокер (1964) насчитывает в роде 14 видов. Некоторые исследователи рассматривают Al. platythrix priestleyi Thomas, 1911, в качестве представителя рода Pyromys Thomas, 1911 (Р. priestleyi); М. mayori — в качестве представителя рода Coelomus Thomas, 1915; М. crociduroides — в качестве представителя рода Mycteromys Robinson et Kloss, 1916; Al. callewacr-ti — в качестве представителя рода Hyelenomys Thomas. 1925 (Эллермэн, 1941; Симпсон, 1945; Грассе и Декейзер, 1955). По-видимому, правильнее включать всех их в состав рода мышей.
В СССР: один вид — домовая мышь.
Рис. 118. Leggadina delicatula
Род австралийских мышей
Genus Leggadina Thomas, 1910
Размеры мелкие. Длина тела 5—9,7 см. Длина хвоста 5—9 см. Голова широкая, притупленная спереди (рис. 118). Уши средних размеров. Хвост покрыт относительно густыми волосами. Пятый палец задней конечности не уменьшен. Волосяной покров у большинства видов мягкий, у L. forresti — грубый. Спинная сторона тела бледно-серая, желтовато-коричневая или светло-коричневая, брюшная — беловатая или сероватобелая. Сосков 2 пары.
Череп не имеет лобно-теменных гребней. Костные слуховые барабаны небольшие.
Хромосом в диплоидном наборе у L. hermannsburgensis 48.
Распространены в Австралии.
Населяют сухие травянистые равнины, леса на песчаных холмах и прибрежные районы. Ведут наземный образ жизни. Убежищем служат норы, иногда различные углубления в земле. Норы обычно просто устроенные, имеют гнездовую камеру с гнездом из сухой травы. Питаются семенами, корнями и другими частями растений. Размножаются в течение всего года, однако пик размножения приходится на период после сильных дождей. В помете обычно 3—4 детеныша.
В роде, вероятно, 7 видов:
L. forresti Thomas, 1906 (Северная территория Австралии);
L. messorius Thomas, 1925 (Южная Австралия);
L. delicatula Gould, 1842 (Северная Австралия);
L. hermannsburgensis Waite, 1896 (Центральная Австралия);
L. patrius Thomas et Dollman, 1909 (Квинсленд);
L. fieldi Waite, 1896 (Центральная Австралия);
L. waitei Trough ton, 1932 (Центральная Австралия).
Андерсон и Джонс (1967) насчитывают в роде 6 видов.
Род африканских крыс
Genus Coloinys Thomas et Wroughton, 1907
В роде единственный вид: африканская крыса — С. goslingi Thomas et Wroughton, 1907.
Размеры мелкие. Длина тела 14—14,8 см. Длина хвоста 14.5—18 см. Уши крупные, почти без волос. Хвост покрыт редкими волосами. Стопа
длинная, очень узкая; первый и пятый пальцы короче средних, но относительно длиннее, чем пальцы большинства других видов. Удлинение стопы, по-видимому, связано с жизнью на топких болотистых почвах. Передняя конечность с четырьмя длинными пальцами. Волосяной покров густой, короткий, очень мягкий. Окраска спинной стороны тела от светло-коричневой до темно-коричневой и черновато-серой; брюшная — белая. Уши сероватые. Позади ушей расположено по белому пятну. Окраска хвоста сверху от светло-коричневой до почти черной; снизу слегка более светлая. Сосков 4 пары.
В черепе мозговая коробка широкая, округлая. Лобные кости значительно сжаты. Лобно-теменных гребней нет. Костное небо широкое. Резцовые отверстия длинные, достигают щечных зубов. Костно-слуховые барабаны маленькие. Ряды щечных зубов смещены вперед.
Распространена в Африке: Камеруне, Центральноафриканской Республике, Заире, Габоне, Анголе, Кении, Северо-Западной Замбии, Эфиопии.
Поднимается в горы на высоту до 2400 м над уровнем моря. Живет на берегах небольших водоемов. Ведет полуводный образ жизни. Экология не изучена.
Подвид африканской крысы С. goslingi plumbeus Osgood, 1928 (возможно, он является отдельным видом — С. plumbeus Osgood, 1928) некоторые исследователи (например, Эллермэн, 1941; Симпсон, 1945; Уокер, 1964) выделяют в особый род Nilopegamys Osgood, 1928, с одним видом N. plumbeus Osgood, 1928. Такое выделение, по-видимому, неверно.
Род керамских крыс
Genus Nesoromys Thomas, 1922
В роде единственный вид: керамская крыса — N. ceramicus Thomas, 1920.
Размеры мелкие. Длина тела около 13,5 см. Длина хвоста примерно 14 см. Морда несколько вытянутая. Стопа узкая. Хвост почти лишен волос. Волосяной покров мягкий, густой. Спинная сторона тела оливковобурого цвета, брюшная несколько светлее. Короткие уши черные. Кисть, стопа и хвост темно-коричневые.
В черепе лицевой отдел удлинен. Лобно-теменные гребни слабо развиты. Костные слуховые барабаны средних размеров. Костное небо очень широкое, простирается далеко назад позади рядов щечных зубов. Относительно короткие, суженные спереди резцовые отверстия располагаются впереди рядов щечных зубов. Подглазничные отверстия большие, более узкие в нижних отделах.
Распространена на о-ве Керам — к западу от Новой Гвинеи.
Населяет густые джунгли в горах на высоте примерно 1800 м над уровнем моря. Экология не изучена.
Род филиппинских болотных мышей
Genus Crunomys Thomas, 1897
Размеры мелкие (рис. 119). Длина тела 9,8—12,2 см. Длина хвоста 6,8—7,9 см. Хвост покрыт средней густоты волосами. Стопа узкая. Воло-, сяной покров низкий, но густой с примесью уплощенных игл. Окраска спинной стороны тела у С. fallax светло-сероватая с примесью желтого.
Рис. 119. Crunomys fallax
Иглы имеют белое основание и черные вершины. Брюшпая сторона серовато-белая. Хвост черный сверху и несколько более светлый снизу. Окраска спинной стороны тела С. melanius черновато-коричневая. Брюшная сторона лишь немного светлее спинной. Хвост одпоцвстпо-чсрпый. Сосков 3 пары.
Мозговой отдел черепа крупный. Межглазпичпый промежуток очень широкий. Лобно-теменных гребней пет. Резцовые отверстия короткие. Костные слуховые барабаны средних размеров. Подглазничные отверстия узкие. Ряды щечных зубов смещены вперед. Третий верхний щечный зуб очень мал.
Распространены на о-вах Лузон и Минданао в группе Филиппинских.
Населяют леса. Ведут, по-видимому, нолуводиын образ жизни. Экология не изучена.
В роде 2 вида:
С. fallax Thomas, 1897 (о-в Лузон);
С. melanius Thomas, 1907 (о-в ^Минданао).
Род новогвинейских крыс
Genus Macruromys Stein, 1933
Размеры мелкие и средине. Длина тела 15—25 см. Длина хвоста 20,5—34 см. Уши короткие. Хвост покрыт крупными чешуямп п редкими, короткими волосами. Стопа сильно сужена. Когти крупные. Волосяной покров довольно грубый. Окраска спинной стороны тела Серова о-корпч-невая или желтовато-черная, брюшной — сероватая пли белов,i гая. У М. elegans хвост темный сверху и белый снизу. У А4. major терминальные две трети хвоста белые.
В черепе мозговой отдел массивный, лицевой относительно удлиненный Костные слуховые барабаны очень маленькие. Подглазничные отверстия средних размеров. Резцовые отверстия широкие, располагаются впереди щечпых зубов. Костное псбо широкое, не продолжается позади рядов зубов. Щечные зубы сильно уменьшены и сдвину 1Ы довольно далеко вперед.
Распространены на Новой Гвинее.
Рис. 120. Кенгуровая мышь — L. nouhuysi
Населяют горные склоны на высоте 1200—1800 м над уровнем моря. Экология не изучена.
В роде 2 вида:
М. elegans Stein, 1933 (западная часть Новой Гвинеи);
М. major Riimmler, 1935 (северо-восточная часть Новой Гвинеи).
Род кенгуровых мышей
Genus Lorentzimys Jentink, 1911
В роде единственный вид: кенгуровая мышь — L. nouhuysi Jentink, 1911.
Размеры мелкие. Длина тела 7,5—8,5 см. Длина хвоста 10,3—11 см. Уши довольно крупные, узкие, с закругленной вершиной. На конце хвоста волосы удлинены. Задние конечности несколько удлиненные, с пятью пальцами. 4 внутренних пальца имеют острые когти, а пятый очень мал и сохраняет лишь небольшой уплощенный ногтеобразный коготь. Волосяной покров высокий и мягкий или более короткий и грубый. Окраска спинной стороны тела рыжевато-коричневая, брюшной — темно-желтая или рыжевато-коричневая. Терминальные части конечностей беловатые. Сосков 3 пары. Череп короткий и широкий. Резцовые отверстия короткие.
Распространена на Новой Гвинее.
Населяет склоны гор на высоте 600—3000 м над уровнем моря. Экология не изучена. По мнению одних исследователей, ведет древесный образ жизни, других — это наземный зверек, передвигающийся прыжками.
Род жестковолосых мышей
Genus Lophuromys Peters, 1874
Размеры мелкие (рис. 121). Длина тела 10 14,5 см. Длина хвоста 5—11,1 см. Хвост у одних видов довольно густо покрыт волосами, у других он почти голый и имеет только редкие, короткие и тонкие волоски. Стопа короткая; три средних пальца относительно длинные и хорошо развиты, а два боковых очепь маленькие. Копи на пальцах передних лап крупные, пятый палец сильно уменьшен. Волосяной покров довольно высокий, густой и грубый. Спинная сторона тела темпо-коричпевая пли оливково-серая, иногда песчаио-темпо-желтая, оливково-коричневая пли темно-коричневая с различными штрихами и пятнышками белого или
Рис. 121. Lophuromys sikapusi и ее череп
оранжеватого цвета, изредка без этих пятен. Брюшная сторона тела оранжевая, коричневатая или кремовая. У некоторых видов основная часть волос на туловище имеет оранжевую окраску. Верхняя поверхность хвоста обычно темная, нижняя — светлая.
Череп широкий, несколько укороченный. Лицевой отдел варьирует по длине, но относительно короткий. Межглазпнчпое сужение почти отсутствует. Подглазничные отверстия очень большие, округлые. Резцовые отверстия длинные, заходят между рядами щечных зубов. Лобно-теменные гребни небольших размеров или их пет вовсе. Костное небо относительно широкое. Костные слуховые барабаны небольшие. Ряды щечных зубов сильно сдвинуты вперед. Резцы тош не, слегка направлены вперед. Щечные зубы с крупными выступами па жевательной поверхности.
Хромосом в диплоидном наборе у L. sikapusi GO.
Распространены в Африке: в Эфиопии, Кении, Уганде, Танзании, Замбии. Анголе, Заире, Камеруне, Габоне и Гапе.
Населяют травянистые долины, болотистые участки, леса, заросли кустарников, окраины полей. Поднимаются в горы до верхней границы леса на высоту до 3700 м над уровнем моря. Активность примерно одинакова в течение суток. Питаются корнями некоторых растений, а также
червями, личинками насекомых, иногда земноводными. Гнезда из сухой травы и листьев устраивают под стволами упавших деревьев или под камнями. Могут также копать длинные и глубокие норы. В помете от 1 до 4 детенышей.
В роде, по-видимому, шесть видов:
желтобрюхая мышь — L. luteogaster Hatt, 1934 (Заир);
мышь Вуснама — L. woosnami Thomas, 1906 (Уганда);
L. ra/wz Verheyen, 1964 (западный берег озера Киву);
желтоточечная мышь— L. flavopunctatus Thomas, 1888 (Эфиопия, Кения и юг Заира, Северо-Восточная Ангола, Северо-Восточная Замбия, Малави, Мозамбик, Руанда);
L. sikapusi Temminck, 1853 (от Сьерра-Леоне до крайнего юга Судана; север и восток Заира, Руанда, Уганда, Западная Кения, Северная Танзания, Северо-Западная Ангола, Габон, Конго, о-в Фернандо-По, Малави);
габонская мышь — L. naso Thomas, 1911 (Габон).
Андерсон и Джонс (1967) насчитывают в роде 8 видов, а Эллермэн (1941) — 11.
Род бороздчатозубых мышей
Genus Leimacomys Matschie, 1893
В роде единственный вид: бороздчатозубая мышь — L. bilttneri Matschie, 1893.
Размеры мелкие. Длина тела примерно 11,8 см. Длина хвоста около 3,5 см. Уши маленькие, покрыты довольно густыми волосами. Когти на передних пальцах передних конечностей длинные, а на задних еще длиннее. Окраска спинной стороны тела изменяется от темно-коричневой до серовато-коричневой. Конечности покрыты короткими коричневыми волосами.
Верхние резцы имеют по глубокой бороздке на наружной поверхности.
Распространена в Западной Африке.
Экология не изучена.
Род тушканчиковых мышей
Genus Notomys Lesson, 1842
Размеры мелкие. Длина тела 9,1—17,7 см. Длина хвоста 12,5— 22,5 см. Внешним видом напоминают тушканчиков (рис. 122). Уши крупные, глаза большие. Длинный хвост обычно покрыт густыми волосами, на конце образующими кисточку. Кисть небольшая, пальцы нормальные. Задняя конечность очень сильно удлинена, стопа очень узкая, приспособленная к движению скачками. Третий палец на стопе самый длинный, четвертый и второй слегка укороченные, пятый палец чрезвычайно уменьшен, хотя все же он несколько более длинный, чем первый. Подошва стопы лишена волос; число мозолей на ней уменьшено до 4 или до 3. Волосяной покров густой и мягкий. Окраска спинной стороны тела от бледно-песчапо-коричневой до желтовато-коричневой или сероватой. Брюшная сторона белая, иногда с примесью серого цвета. Вибриссы очень длинные. Сосков 2 пары. На горле расположена специфическая кожная железа.
Мозговая коробка черепа округлая. Лицевой отдел относительно
Рис. 122. Тушкапчиковая мышь -Nntomys sp.
длинный. Межглазничное сужение выражено слабо. Лобио теменных гребней нет. Костные слуховые барабаны обычно очень большие. Костное небо довольно широкое, не доходит назад до уровня последних щечных зубов. Резцовые отверстия длинные, широкие.
Распространены на большей части Австралии.
Населяют равнины, разреженные леса, степные участки, песчаные дюны. Активность наступает в сумерках. При медленном движении передвигаются на четырех конечностях, а при быстром прыгают на задних ногах. День проводят в норах, которые выкапывают сложными и уходящими вглубь до 1,5 м. В норе имеется гнездовая камера с гнездом из травы и листьев. Питаются ягодами, листьями, семенами и т. п. В помете от 2 до 5 детенышей. Продолжительность жизни до 3 лет.
В роде, по-видимому, 10 видов:
мышь Митчелла — N. mitchelli Ogilby, 1838 (Новый Южный Уэльс и Южная Австралия);
N. mordax Thomas, 1922 (Южный Квинсленд);
N. aquilo Thomas, 1922 (Северный Квинсленд) ;
N. megalotis Iredale et Troughton, 1934 (Западная Австралия);
мышь Ричардсона — A’, richardsoni Gould, 1851 (Западная Австралия);
N. alexis Thomas, 1922 (Южная Австралия);
N. cervinus Gould, 1851 (Южная Австралия);
длиннохвостая мышь — N. longicaudatus Gould, 1844 (Западная Австралия) ;
N. sturti Thomas, 1821 (Новый Южный Уэльс);
мышь Алстона — N. alstoni Brazenor, 1934 (Южная Австралия)
Андерсон и Джонс (1967) насчитывают в роде 14 видов.
Род широкозубых крыс
Genus Mastacomys Thomas, 1882
Размеры мелкие. Длина тела 14,5—18,5 см. Длина хвоста 9,5— 11,5 см. По внешнему виду напоминают полевок (рис. 123). Относительно короткий хвост хорошо оволоснен. Волосяной покров высокий, густой, мягкий. Окраска его одноцветная, от черновато-бурой до серо-бурой. Сосков 2 пары.
Мозговая коробка и лицевой отдел черепа несколько укорочены. Межглазничное сужение резко выражено. Лобно-теменные гребни небольшие. Костное небо сильно сужено. Резцовые отверстия длинные, очень узкие, особенно в задней части. Костные слуховые барабаны относительно небольшие. Резцы широкие. Щечные зубы очень широкие, массивные.
Распространены в Южной Австралии и Тасмании.
Обитают в густых зарослях высокой травы примерно на высоте 900 м над уровнем моря. В траве делают тропинки. Населяют также густые, влажные леса. Экология не изучена.
В роде 2 вида:
М. fuscus Thomas, 1882 (Тасмания);
М. mordicus Thomas, 1922 (Южная Австралия).
Род колючих крыс
Genus Echiothrix Gray, 1867
В роде единственный вид: колючая крыса — Е. leucura Gray, 1867.
Размеры средние. Длина тела 20—25 см. Хвост несколько короче тела. Голова, особенно морда, сильно удлинены. Хвост почти лишен волос и покрыт крупными чешуями. Стопа узкая. Волосяной покров представлен комбинацией мягких и очень жестких щетинковидных волос,
Рис. 123. Mastacomys fuscus
лногда напоминающих колючки. Спинная сторона тела сероватая или гемно-серовато-коричневая. Брюхо беловатое, желтоватое или рыжевато-коричневое. Сосков 3 пары.
ААозговой отдел черепа широкий, массивный, лицевой сильно вытянутый (до половины длины черепа), узкий. Резцовые отверстия длинные, хотя и располагаются впереди рядов щечных зубов. Подглазничные отверстия большие, слегка сужающиеся книзу. Межглазничное сужение хорошо выражено. Костные слуховые барабаны средних размеров. Носовые кости спереди несколько загнуты вверх. Нижняя челюсть со слабо развитым венечным отростком. Короткие верхние резцы имеют по две продольные бороздки на наружной поверхности. Нижние резцы крупные, выгнутые и настолько широко расставлены друг от друга, что, по-видимому, не соприкасаются с верхними резцами.
Распространена на о-ве Сулавеси.
Экология не изучена.
Род длиниокоготных мышей
Genus Melasmothrix Miller et Hollister, 1921
В роде единственный вид: длиннокоготная мышь — М. naso Miller et Hollister, 1921.
Размеры мелкие. Длина тела примерно 12,4 см. Длина хвоста 9 см. Морда удлиненная. Уши короткие. Кольцевидно расположенные чешуйки на хвосте почти полностью скрыты волосами. На пальцах передней конечности когти длинные, за исключением очень небольшого первого пальца, имеющего уплощенный иогтеобразпый коготь. Волосяной покров густой, мягкий. Окраска его на спинной стороне тела одноцветная, черноватая; брюшная сторона немного светлее спинной.
Распространена в центральных районах о-ва Сулавеси.
Экология не изучена.
Род иглистых мышей
Genus Acomys I. Geoffroy, 1838
Размеры мелкие. Длина тела 7—12,5 см. Длина хвоста 6—12,5 см. Уши крупные, округлые (рис. 124). Хвост почти без волос. Стопа широкая. Пальцы ее не уменьшены. Спина покрыта многочисленными иглами бледно-желтоватого, рыжевато-коричневого, рыжеватого или темносерого цвета, брюшная сторона — мягкими белыми волосами. Сосков 2 или 3 пары.
Мозговая коробка черепа широкая. Лицевой отдел средней длины. Межтеменная кость очень крупная. Лобно-теменные гребни хорошо выражены. Костные слуховые барабаны средних размеров. Подглазничные отверстия крупные, слегка сужающиеся книзу. Резцовые отверстия длинные, продолжающиеся между рядами зубов. Нижняя челюсть с очень небольшим венечным отростком.
Хромосом в диплоидном наборе у A. cahirinus 3G, 38; у A. russatus 66, у A. subspitiosus 64.
Распространены на Крите, Кипре, в Передней Азии, Саудовской Аравии и на большей части Африки.
Населяют скальные выходы. Иногда обитают на песчаных участках. Убежищем служат норы; часто используют норы других грызунов. Ак-
Рис. 124. Acomys cahirinus п cc череп
тивны преимущественно рано угром и поздно вечером. Питаются в основном растительными кормами. Самки приносят детенышей в основном в период с февраля по сентябрь. Длительность беременности около 42 дней В помете бывает от 1 до 5, в среднем 3 детеныша. Половозре-лость у самцов наступает примерно в возрасте семи недель.
В роле, по-видимому, 5 видов:
A. cahiritius Е. Geoffrey, 1803 (АРЕ, Ливия, Мавритания, Нигерия, Судан, Эфиопия, Кения; Индия — Западный Синд; Южный Иран, Палестина, Саудовская Аравия, Кипр и Крит);
A. dimidiatus Crestzschmar, 1826 (АРЕ, Судан, Нигер, Сомали, Кения и Юго-Западная Азия);
A. russatus Wagner, 1840 (восток АРЕ, Синай, Палестина, Саудовская Аравия);
A. spinosissimus Peters, 1852 (Африка от северной части Мозамбика на запад через Южную Родезию);
A. subspinosus Waterhouse, 1838 (вероятно, вся Южная и Восточная Африка, но достоверно известна из Южно-Африканской Республики, Кении, Сомали, Уганды и Судана).
Эллермэн (1941) насчитывает в роде 26 видов, а Андерсон и Джонс (1967) — 18.
Род крупнозубых мышей
Genus Uranomys Dollman, 1909
В роде единственный вид: крупнозубая мышь—U. ruddi Dollman, 1909.
Размеры мелкие (рис. 125). Длина тела 9—13,4 см, длина хвоста 5,3—7,7 см. Хвост покрыт редкими волосами; темный сверху и слегка более светлый снизу. Стопа широкая, укороченная; первый и пятый пальцы короткие. Тело покрыто длинными щетипкоподобными волосами. Окраска одноцветная, серая, иногда с желтоватым или коричневатым оттенками. Брюхо у некоторых видов белое или сероватое. Сосков 6 пар.
В черепе мозговая коробка расширена; лицевой отдел относительно укорочен. Межглазничное сужение хорошо выражено. Лобно-теменные гребни обычно имеются. Костные слуховые барабаны средних размеров. Костное небо широкое. Резцовые отверстия длинные, широкие, продолжаются назад примерно до второго коренного зуба. Верхние резцы сильно направлены вперед. Нижние резцы длинные.
Хромосом в диплоидном наборе 52.
Распространена в Малави, Кении, Уганде, на северо-востоке Заира, в Северной Нигерии, Того, Гвинее и на Берегу Слоновой Кости.
Экология не изучена. Населяет травянистую саванну и вторичную саванну. Держится поодиночке или парами. Встречается редко. Питается преимущественно насекомыми. Активна ночью.
Эллермэн (1941) выделяет в роде 7, а Андерсон и Джонс (1967) — 5 видов.
Род бандикот
Genus Bandicota Gray, 1873
Размеры от мелких до крупных. Длина тела 16—36 см. Длина хвоста 14—26 см. Масса от 685 до 1132 г. Морда укорочена и расширена (рис. 126). Хвост покрыт редкими волосами. Когти па передних конечностях крупные. На стопе 6 подошвенных мозолей. Волосяной покров высокий, мягкий, довольно густой или грубый и изреженпый. Окраска спинной стороны тела от светло-серой до коричневой или почти черной. Брюшная сторона тела грязно-белая. Сосков 6—9 пар.
Череп относительно узкий. Костные слуховые барабаны крупные. Костное небо немного сужено. Резцовые отверстия узкие, достигают
Рис. 125. Крупнозубая мышь—U. г и deli и ее череп
щечных зубов. Венечный отросток нижней челюсти высокий. Щечные зубы представлены серией поперечных пластин. Резцы желтого или оранжевого цвета.
Распространены на о-ве Шри Ланка, в Индии — Ассаме, Бангладеш, Непале, Бирме, Южном Китае, Таиланде, п-ове Индокитай, о-вах Суматра, Пенанг, Ява, Тайвань (на Тайване представители рода были акклиматизированы) .
Населяют джунгли или заросли кустарников, а индийская бандпко-товая крыса стала комменсалом человека (во всяком случае, в некото-
Рис. 126. Индийская бандикота— В. indica (вверху) и бенгальская бандикота — В. bengalensis
рых областях). Выкапывают глубокие сложно устроенные норы с несколькими кладовыми, в которых запасают зерна, плоды, орехи. Актив ны в сумерках. Могут хорошо плавать. В помете 10—12 детенышей. Размножение происходит, по-видимому, круглый год. В культурных шндшафгах этот грызун приносит значительный вред посевам.
Хромосом в диплоидном наборе у бенгальской бандикоты 41—42, а у индийской — 42.
В роде 2 вида:
бенгальская бандикота—В. bengalensis Gray, 1835 (Ассам; Бангла деш; Шри Ланка; Непал; Бирма; о-ва Пенанг; Суматра; Ява);
индийская бандикота — В. indica Bechstcin, 1800 (Ассам; Бангладеш, Шри Ланка; Непал; Бирма; Китай — Юньнань; п-ов Индокитай; Таиланд; о-ва Суматра, Яза, Тайвань).
Род пластинчатозубых крыс
Genus Ncsokia Gray, 1842
В роде единственный вид: пластинчатозубая крыса — N. indica Gray, 1830.
Размеры мелкие и средине. Длина тела 14—22 см. Длина хвоста по солее 2/з длины тела Морда тупая и широкая. Глаза маленькие. Уши относительно короткие (рис. 127). Толстый в основании хвост, быстро опчающийся к концу, покрыт редкими волосами. Ступни широкие, с
короткими пальцами. Первый палец передней конечности сильно укорочен и имеет вместо когтя ноготь. Волосяной покров может быть относительно короткий и грубый. Особенно длинные волосы располагаются на крестце. Окраска спинной стороны тела от желтовато-коричневой до серовато-коричневой с рыжеватым оттенком. Брюшная сторона тела сероватая или беловатая. Сосков 4 пары.
Мозговая коробка черепа относительно невелика. Лицевой отдел короткий. Лобно-теменные гребни хорошо развиты. Носовые кости укорочены. Скуловые дуги широко расставлены- Резцовые отверстия короткие и узкие. Костные слуховые барабаны крупные, сильно вздутые. Костное небо относительно узкое. Нижняя челюсть с высоким венечным отростком. Жевательная поверхность щечных зубов плоская; бугорки па ней заметны только у совсем молодых животных. Верхние резцы широкие, направлены вперед.
Хромосом в диплоидном наборе 42.
Распространена в СССР в бассейнах рек Средней Азии — Зеравшаиа, Амударьи, Мургаба, Теджепа н в горах Коиетдага; Афганистане, западных районах Китая, северной Индии, Пакистане, Северной Африке, Малой и Передней Азии.
Населяет влажные участки в различных биотопах or пустынь до лесов и культурных ландшафтов. Поднимается в горы до 1500 м над уровнем моря. Большую часть времени проводит в порах под землей. Норы представляют собой очень сложные сооружения с многочисленными, поверх-
Рпс. 128. Bcamys major
o-
11- a o-.a-
СОТО-;ne
I hoctho расположенными кормовыми ходами и более глубокой гнездовой частью. Ведет полуколониальный образ жизни. Активна круглый год, преимущественно в сумерки и ночью. Из норы появляется редко и отходит от нее недалеко. Питается в основном подземными частями растений, в летнее время поедает и зеленые, в меньшей степени использует животные корма (насекомых). В значительных количествах запасает пищу, в основном различные семена. Отмечены небольшие кормовые миграции. В течение года бывает не менее 3 пометов но 3—5 детенышей в каждом. ’ Местами наносит серьезный вред посевам и бахчевым культурам, а также плодовым деревьям. Поселяясь в домах, может портить продукты, фуражные корма.
Род треугольнохвостых крыс
1ое ст-
Genus Beamys Thomas, 1909
{ей Размеры мелкие. Длины тела 11—16 см. Длина хвоста 10,5—14,5 см.
ие,
на, ад-Ма-
|*сов нем ред-,ерх-
I Хвост почти полностью голый (рис. 128); он несколько расширен вдоль I нижней поверхности и сужен вдоль верхней, так что в поперечнике имеет t форму треугольника. Копен хвоста белый. Стопа широкая, прпспособ-I денная к лазанию по деревьям. Пятый палеи ошосительпо длинный, пер-j вын палец средних размеров. Имеются защечные мешки. Волосяной покров мягкий. Его окраска на спинной стороне тела сероватая или серова-I то-корпчневая, па брюшной — белая.
В черепе лицевой отдел относительно длинный. Лобпо-темеппые греб-’ ни развиты слабо. Резцовые отверстия очень короткие, расположены * далеко впереди от рядов щечных зубов. Подглазничные отверстия средних размеров. Костные слуховые барабаны маленькие. Костное небо ши-
рокое. Бугорки на жевательной поверхности щечных зубов хорошо выражены, острые.
Распространены в Африке: Кении, Малави и Замбии.
Экология изучена слабо. Населяют леса от уровня моря до 2100 м над его уровнем. Предпочитают места с песчаными и легкими почвами, в которых выкапывают норы Выбирают участки рядом с водоемами. В норах длиной до 9 м и глубиной до 60 см устраивают кладовые и гнездовые камеры. Гнездо делают из сухих листьев. Питаются преимущественно семенами и плодами. У В. hindei в Малави период размножения совпадает с сезоном дождей; самка приносит в помете 4—7 детенышей, которые остаются с матерью, пока не достигнут длины 100—120 мм.
В роде 2 вида:
В hindei Thomas, 1909 (юг Кении);
В. major Dollman, 1914 (Малави и Замбия).
Род мешотчатых крыс
Genus Saccostomus Peters, 1846
В роде единственный вид: мешотчатая крыса — S. campestris Peters, 1846.
Размеры мелкие. Длина тела 12—19 см. Длина хвоста 3—8 см. Масса 40—85 г. Тело неуклюжее, приземистое. Голова массивная, широкая. Уши короткие, округлые (рис. 129). Глаза маленькие. Короткий хвост утолщен у основания и покрыт редкими волосками. Конечности сильные, укороченные. Стопа широкая, сильно укороченная. Большие защечные мешки открываются с внутренней стороны рта около губ и продолжаются назад почти до плеч. Волосяной покров довольно высокий, густой, мягкий. Окраска спинной стороны и боков тела серая или коричневато-серая, несколько более темная по средней линии спины. Брюшная чистобелая или желтовато-белая, резко отграниченная от боков. Хвост двухцветный, темный и белый снизу. Сосков 5—6 пар.
Череп относительно узкий со значительным межглазничным сужением. Лицевой отдел не укорочен. Лобно-теменные гребни развиты слабо или их нет вовсе. Резцовые отверстия длинные, обычно достигают рядов щечных зубов. Костное небо несколько расширено. Костные слуховые
Рис. 129. Мешотчатая крыса — S. campestris
барабаны относительно крупные и вздутые. Нижняя челюсть с хорошо развитым венечным отростком. Бороздок на наружной поверхности резцов нет.
Хромосом в диплоидном наборе 44, 46.
Распространена в Африке от крайнего юга Сомали. Южной Кении, Восточной и Северо-Восточной Уганды и Восточной Танзании на юг до Южно-Африканской Республики, кроме ее крайнего юга и западного берега.
Населяет кустарниковые заросли в долинах, травянистые поляны в разреженных лесах, антропогенные ландшафты, песчаные холмы. Предпочитает селиться около водоемов. Ведет ночной образ жизни. Выкапывает простые норы с гнездовой камерой и кладовыми. Иногда занимает чужие норы. Держатся поодиночке. Передвигается обычно медленно. Питается различными частями растений, в частности семенами; реже ловит насекомых. Семена запасает летом и использует их в течение зимы. Периодичности в размножении нет. Самка приносит в помете до 8 детенышей. Новорожденные покрыты волосами.
Иногда в роде выделяют 4 вида:
S. campestris Peters, 1846; S. isiolae Heller, 1912; S. mearnsi Heller, 1910; S. cricetulus Allen et Lawrence, 1936.
Род хомяковидных крыс
Genus Cricetomys Waterhouse, 1840
Размеры средние и крупные. Длина тела 24—45 см. Длина хвоста 36,5—46 см. Масса 1—1,5 кг. Большая голова удлинена и несколько сужена. Уши средней длины, округлые, почти голые. Хвост покрыт редко расположенными волосами, чешуйчатый. Стопа широкая с шестью подошвенными мозолями; крайние пальцы не уменьшены в размерах, средние не удлинены. Кисть нормального строения. Имеются защечные мешки объемом до 100 мл. Волосяной покров довольно редкий, у С. gambianus высокий, грубый, у С. emini низкий, относительно мягкий. Окраска его на спинной стороне тела темно-серо-коричневая, сероватая с коричневатым оттенком или рыжевато-коричневая. Бока чуть светлее спины. Брюхо белое или кремовое. Вокруг глаз могут быть темные кольца. Проксимальный конец хвоста темно-серый, дистальный —белый. Передняя часть спины может быть покрыта пятнами. Сосков 4 или 5 пар.
Узкий череп удлинен в основном за счет лицевого отдела. Носовые кости расширены несколько спереди. Лобно-теменные гребни хорошо развиты и проходят назад до затылочной области. Костные слуховые барабаны маленькие. Резцовые отверстия короткие, расположены главным образом в предчелюстных костях. Подглазничные отверстия очень широкие, округлые, часто немного сужены книзу. Нижняя челюсть с широким венечным отростком. Резцы спереди бороздок не имеют.
Хромосом в диплоидном наборе у С. gambianus 78.
Распространены на большей части Африканского континента от Гамбии на восток до Судана и Кении и па юг до Северного Трансвааля в Южно-Африканской Республике.
Населяют леса и заросли кустарников. Засушливых открытых мест обычно избегают. В горы поднимаются до 3500 м над уровнем моря. Заселили некоторые города Западной Африки. Активны ночью, иногда и днем. Обычно держатся поодиночке. Убежищем служат естественные пустоты в земле, трещины, дупла упавших деревьев, термитники. Могут также выкапывать норы, обычно располагаются у основания дерева, в
густых зарослях и имеют от 2 до 6 выходных отверстий, гнездовую камеру и кладовые. Могут хорошо плавать. При передвижении по земле хвост не волочится, а поднят кверху и выполняет роль своеобразного баланса. В поисках пищи неплохо лазают по кустам и деревьям. Питаются различными растительными кормами, в частности плодами, семенами, а-иногда и насекомыми. Могут делать запасы корма. Отмечена копрофагия. Период размножения не приурочен к определенному сезону года. Длительность беременности около 32 дней. В помете 1—4,’обычно 2— 3 детеныша. Новорожденные голые с закрытыми глазами и ушами. Волосы появляются на 5-й день и детеныши покрываются волосами на третью неделю. На 4-й день открываются глаза. Продолжительность жизни примерно 4,5 года. Хомяковидных крыс местные жители используют в пищу. Зверьки могут хорошо уживаться в неволе и быстро приручаются.
В роде, вероятно, 2 вида:
С. gambianus Waterhouse, 1840 (от Сенегала на восток до озера Чад и Центрального Судана и на юг до Анголы и Южно-Африканской Республики) ;
С. emini Wroughton, 1910 (от Сьерра-Леоне и о-ва Фернандо-По на восток до озера Танганьика).
Некоторые исследователи (Эллермэн, 1941) выделяют в роде один вид — С. gambianus.
Род анизомис
Genus Anisomys Thomas, 1903
В роде единственный вид: анизомис — A. imitator Thomas, 1904.
Размеры средние. Длина тела 24,4—30 см. Длина хвоста 28,5—33 см. Уши короткие. Почти голый хвост покрыт крупными шестиугольными чешуями. Задние конечности с укороченной стопой, приспособлены к лазанию по деревьям; пятый палец относительно хорошо развит, четвертый— самый длинный, первый палец имеет уплощенный ногтевидный коготь. На передней конечности самый короткий палец пятый, второй палец тоже сильно укорочен. Волосяной покров низкий, грубый. Спинная сторона тела черновато-коричневая, брюшная-—тускло-темно-желтая или беловатая. Голова почти черная. Сосков 3 пары.
Череп с расширенным и удлиненным лицевым отделом, слабо выраженным межглазничным сужением, небольшими лобио-теменными гребнями. Костные слуховые барабаны чрезвычайно малы. Резцовые отверстия очень укороченные, располагаются далеко впереди рядов щечных зубов. Верхние резцы нормальной ширины, нижние очень сильно сужены. Щечные зубы небольших размеров и у взрослых особей состоят из вертикальных пластин.
Распространена в Новой Гвинее.
Населяет горные леса на высоте от 900 до 2700 м над уровнем моря. Экология не изучена.
ПОДСЕМЕЙСТВО ЛАЗАЮЩИХ МЫШЕЙ
SUBFAMILIA DENDROMURINAE ALLEN, 1939
Размеры от мелких до средних. Сходны с представителями подсемейства Murinae. Внутренний ряд бугорков жевательной поверхности верхних щечных рядов сильно редуцирован, так что функционируют только 334
внутренние бугорки на первом и втором коренных зубах. Третьи нижний и верхний коренные зубы сильно уменьшены
Род африканских лазающих мышей Genus Dendromus A. Smith, 1829
Размеры мелкие. Длина тела 5—10 см. Длина хвоста 6,5—11,3 см.
Масса 7—21 г. Длинный, почти лишенный волос хвост полухватательного
типа (рис. 130). Передние конечности имеют всего по 3 хорошо развитых пальца. Задние конечности с очень укороченным I пальцем, пятый палец хотя и длинный, но часто не имеет когтя. Волосяной покров густой и мягкий. Спинная сторона тела сероватая или коричневатая с одной, иногда тремя темными срединными полосками вдоль спины. Брюхо беловатое или желтоватое. Вокруг глаз темные «очки». Нижняя поверхность хвоста иногда светлее верхней. Сосков 4 пары.
Череп с округлой средних размеров мозговой коробкой. 1Межглазничное сужение резко выражено. Лобно-теменные гребни слабо развиты. Резцовые отверстия длинные, широкие, обычно достигают уровня середины первого коренного зуба. Костное небо широкое. Подглазничное отверстие очень крупное. Костные слуховые барабаны относительно крупные. Верхние резцы сжаты и имеют по одной бороздке, проходящей по наружной поверхности. Щечные зубы сильно сужены. Третий коренной зуб небольших размеров. На нижней челюсти последний щечный зуб также мал. Нижние резцы с гладкой наружной поверх-
Рис. 130. Dendromus mcsotnelas и череп D. melanotis
ностью
Хромосом в диплоидном наборе у D. melanotis 36, у D. mystacalis—38. Распространены в Африке к югу от Сахары до южной оконечности материка.
Населяют прибрежные биотопы, болотистые участки, места, покрытые густой травянистой растительностью или кустарником. Встречаются от уровня моря до 4300 м над его уровнем. Избегают густых лесов и засушливых местностей. Активны ночью. D. mesamelas активна также и днем. Хорошо лазают по высокой траве, кустарникам и деревьям. Однако большую часть времени проводят на земле. Убежищем служат округлые гнезда из травы, расположенные в зарослях кустарников или травы, иногда на высоте до 3 м над землей, изредка в норах. Питаются различными ягодами, семенами, а также насекомыми, мелкими ящерицами и яйцами птиц. В помете 3—5 детенышей. У D. mesomelas размножение носит сезонный характер.
В роде, по-видимому, 5 видов:
D. mesomelas Brants, 1827 (от Южно-Африканской Республики на север до Танзании, Кении, Эфиопии, Заира и Южной Нигерии);
D. melanotis Smith, 1834 (Южно-Африканская Республика, север Юго-Западной Африки, Ангола, Малави, Нигерия, Танзания, Эфиопия и Северная Нигерия, Гвинея);
• D. mystacalis Heuglin, 1863 (от Южно-Африканской Республики до Эфиопии и Нигерии);
D. lovati de Winton, 1899 (высокогорье Эфиопии);
D. nyikae Wroughton, 1909 (Южно-Африканская Республика, Родезия, Восточная Замбия, Центральная Ангола, Северная и Центральная Малави);
Уокер (1964) насчитывает в роде 3 вида.
Род дендроприономис
Genus Dendroprionomys Petter, 1966
В роде единственный вид: дендроприономис — D. rousseloti Petter, 1966.
Размеры мелкие. Хвост длиннее тела. Имеются промежуточные признаки между Dendrom.us и Prionomys. Уши большие. Хвост чешуйчатый. Передние конечности четырехпалые. Мозоли на подошвах развиты относительно слабо. Задние конечности пятипалые. Пятый палец сильно развит, находится сзади и ниже других пальцев и сгибается латерально, как и у Dendromus. Волосяной покров мягкий, густой. Окраска его рыжая, более темная на спинной стороне тела, чем на боках. Брюхо белое. Вокруг глаз — зона черноватых волос. Уши серые.
Череп с крупной мозговой коробкой и укороченным лицевым отделом. Межглазничное сужение выражено умеренно. Подглазничные отверстия спереди имеют бобовидную форму. Резцовые отверстия крупные, по не достигают уровня первого щечного зуба. Верхние резцы с продольной бороздкой на наружной поверхности.
Распространен в Конго.
Экология не изучена. По-видимому, ведет древесный образ жизни Питается, вероятно, насекомыми.
Род длинноухих мышей
Genus Malacothrix Wagner, 1843
В роде единственный вид: длинноухая мышь — М. typica Smith, 1834. Размеры мелкие. Длина тела 6,5—9,5 см. Длина хвоста 2,8—4,2 см. Уши большие, округлые. Короткий хвост покрыт редкими волосами. Стопа очень узкая, относительно длинная, имеет только 4 пальца — первый палец отсутствует (исключительный случай в семействе). Подошва покрыта волосами. Волосяной покров высокий, густой, мягкий. Окраска его на спинной стороне тела от бледно-коричневой до рыжевато-коричневой. На спине обычны более или менее заметные темные полоски. Верх головы темный. Брюшная сторона тела, конечности и хвост белые. Сосков 4 пары.
В черепе лицевой отдел очень узкий. Мозговая коробка укороченная, округлая, несколько скошена книзу в задней части. Межглазничное сужение выражено чрезвычайно сильно. Скуловые дуги широко расставлены. Ряды щечных зубов смещены в черепе относительно далеко вперед. Костное небо очень широкое. Костные слуховые барабаны средних размеров. Резцовые отверстия крупные, длинные; достигают уровня середины первого коренного зуба, вперед доходят до резцов. Верхние резцы сжаты, на наружной поверхности имеют по одной борозде. Нижние резцы гладкие. Верхние щечные зубы чрезвычайно узкие, со сложной структурой жевательной поверхности.
Хромосом в диплоидном наборе 38.
Распространена в южной части Африки от Южной Анголы через Юго-Западную Африку, Малави и Трансвааль до провинции Кейп в Южно-Африканской Республике.
Обитает на песчаных равнинах. Ведет ночной образ жизни. Активный грызун, способный за ночь уходить за 4 км от своей норы. Сложную нору с гнездовой камерой выкапывает на глубине 60—120 см от поверхности. Гнездо делает из травы, перьев птиц. Питается в основном семенами различных растений. Бывает 3 помета в год, в каждом из которых от 3 до 7 детенышей. Половозрелости достигает в 3-месячном возрасте.
Род древесных мышей
Genus Prionomys Dollman, 1910
В роде единственный вид: древесная мышь — Р. batesi Dollman, 1910.
Размеры мелкие. Длина тела около 6 см. Длина хвоста примерно 10 см. Уши небольшие, округлые. Хвост слабо оволоснен; возможно, хватательный. Передние конечности четырехпалые, первый палец отсутствует, второй и четвертый пальцы умеренно удлинены и имеют короткие когти. Наружный палец достигает всего половины длины двух средних и имеет небольшой ноготь. Пятипалая задняя конечность приспособлена к лазанию по деревьям, укорочена и имеет короткую стопу. Все пальцы с довольно сильными когтями. Первый и пятый довольно длинные, причем первый палец, по-видимому, противопоставляется остальным. Волосяной покров мягкий, густой и низкий. Окраска его па спинной стороне тела розовато-шоколадная, брюшной — серовато-розовато-желтая. Вока тела немного светлее спины. Глаза окружены узкой черноватой полоской. Хвост темно-коричневый, почти черный.
В черепе мозговой отдел массивный, лицевая часть относительно короткая. Лобно-теменные гребни крупные. Скуловые дуги широко расстав
лены. Резцовые отверстия относительно короткие. Подглазничные отверстия крупные. Костное небо очень широкое. Костные слуховые барабаны некрупные. Нижняя челюсть с низким венечным отростком. Верхние резцы направлены вперед, несколько укорочены, с гладкой наружной поверхностью. Нижние резцы небольшие.
Распространена в Африке — Южном Камеруне и Центральноафриканской Республике.
Экология не изучена.
Род скалистых мышей
Genus Petromyscus Thomas, 1926
Размеры мелкие. Длина тела 7—9 см. Длина хвоста 8—10 см. Уши длинные, округлые. Хвост довольно густо покрыт короткими волосами, не прикрывающими кожные чешуи. Конечности укорочены. Стопа короткая, широкая с относительно длинными первым и пятым пальцами. Волосяной покров мягкий, несколько изреженный. Окраска спинной стороны тела от темно-желтой до коричневатой или желтовато-серой. Брюхо сероватое. Хвост и конечности одноцветные — белые или темно-желтые. Иногда хвост сверху серый, снизу белый. Сосков обычно 3, иногда 2 пары.
Череп с широкой, уплощенной мозговой коробкой. Межглазничное сужение выражено не очень сильно. Костное небо широкое. Резцовые отверстия длинные. Верхние резцы сжаты; их наружная поверхность гладкая. Верхний третий щечный зуб сильно уменьшен.
Распространены от Юго-Западной Анголы, через Юго-Западную Африку до провинции Кейп в Южно-Африканской Республике.
Населяют аридные горные районы: предпочитают держаться в местах скальных выходов. Активны ночью. День проводят в пустотах под камнями, расщелинах между скал. Всеядные.
В роде 2 вида:
Р. monticularis Thomas et Hinton, 1925 (Берсеба в Большом Намак-валенде в Юго-Западной Африке);
Р. collinus Thomas et Hinton, 1925 (юго-западная и западная части провинции Кейп в Южно-Африканской Республике, Южно-Западная Африка и Южная Ангола).
Род болотных мышей
Genus Delanymys Hayman, 1962
В роде единственный вид: болотная мышь — D. brooksi Hayman, 1962.
Размеры мелкие. Длина тела 5—6 см. Длина хвоста 8,7—10,5 см. Масса около 5 г. Хвост пс-лухватательный. Задние лапы с 4 подвижными пальцами и слабо развитым первым пальцем. Волосяной покров густой и мягкий. Окраска спинной стороны тела красновато-коричневая, брюшной— темно-желтая. С каждой стороны морды между глазом и носом находится по черному пятну.
В черепе мозговая коробка крупная, округлой формы. Лицевой отдел укорочен, узкий. Межглазничное сужение выражено не очень резко. Костно-слуховые барабаны небольшие.
Распространена на крайнем юго-западе Уганды, а также в горах к западу от озера Киву.
Рис. 131. Steatomys pratensis и череп S. caurinus
Населяют заросли камыша в болотистых местах, в горах на высоте примерно между 1700 и 2600 м над уровнем моря. Экология не изучена. Хорошо лазает по камышам, траве. Убежищем служат небольшие травяные гнезда, которые располагаются на высоте около 50 см над землей на ветвях кустов или на стеблях травы. Питается в основном семенами трав, а также ягодами. В помете 3—4 детенышей.
Род толстых мышей
Genus Steatomys Peters, 1846
Размеры мелкие. Длина тела 5—14 см. Длина хвоста 3,3—7,2 см. Сложение несколько тяжелое, туловище относительно утолщенное (рис. 131). Уши большие, округлые. Толстый хвост покрыт редкими волосами. На передней конечности только четыре функционирующих пальца, на задней—пять. Пальцы передних конечностей длинные. Волосяной покров густой, низкий, мягкий. Окраска верхней стороны тела от коричневой до темно-желтой, часто с примесью черных тонов. Брюшная сто
рона белая. Хвост обычно двухцветный; сверху темный, снизу — белый, иногда весь хвост белый. Сосков 4—7 пар.
Череп с заостренным лицевым отделом. Межглазничное сужение выражено умеренно. Лобно-теменных гребней нет. Подглазничные отверстия очень крупные. Костные слуховые барабаны крупные. Верхние резцы имеют на наружной поверхности по бороздке. Нижние резцы снаружи гладкие.
Хромосом в диплоидном наборе у S. pratensis 68.
Распространены по большей части Африки от Судана на юг до провинции Кейп в Южно-Африканской Республике.
Населяют засушливые области: песчаные равнины, саванны, скальные участки, а также леса и заросли кустарников. Активны преимущественно ночью. Держатся поодиночке или парами. Выкапывают норы длиной 1,5—1,7 м, обычно с крупной гнездовой камерой на глубине до 90— 120 см. К осени зверьки становятся очень жирными. На зиму впадают в спячку (или сон), которая продолжается примерно с апреля по октябрь. Питаются в основном семенами, луковицами растений, насекомыми. Может запасать корм. В помете 4-—6 детенышей.
В роде, по-видимому, 11 видов:
S. pratensis Peters, 1846 (Южно-Африканская Республика, Мозамбик, Родезия, Юго-Западная Африка, Ангола, Малави, Замбия, Нигерия, Заир, Судан, Танзания, Кения);
S. muanzae Kershaw, 1923 (Танзания);
S. parvus Rhoads, 1896 (Эфиопия);
S. aquilo Thomas et Hinton, 1923 (Судан);
S. cuppedius Thomas et Hinton, 1920 (Нигерия);
S. bocagei Thomas, 1892 (Ангола);
S. jacksoni Hayman, 1935 (Гана);
S. caurinus Thomas, 1912 (Нигерия);
S. gazellae Thomas et Hinton, 1923 (Судан);
S. opimus Pousargues, 1894 (Заир);
S. athi Heller, 1910 (Кения).
Андерсон и Джонс (1967) насчитывают в роде 13 видов, Уокер (1964) —2, Кингдом (1974) —3; S. pratensis, S. opimus и S. minutus.
Род конголезских мышей
Genus Deomys Thomas, 1888
В роде единственный вид: конголезская мышь — D. ferruginous Thomas, 1888.
Размеры мелкие. Длина тела 12—16 см. Длина хвоста 15—21,5 см. Масса 40—70 г. Морда вытянутая, заостренная. Уши крупные. Хвост почти голый в проксимальной части, в дистальной покрыт более густыми волосами; возможно, хватательный. Конечности длинные. Стопа узкая, длинная, с пятью подошвенными мозолями; наружные пальцы сильно уменьшены. Волосяной покров высокий, очень грубый. Окраска его на спинной стороне тела бледно-рыжая или рыжевато-коричневая, смешанная с черным. Часть остевых волос на спине черного цвета. Брюшная сторона белая. Вокруг глаз черноватые круги. Хвост двухцветный: серый сверху и белый снизу, иногда конец хвоста весь белый. Сосков 2 или 3 пары.
Череп с длинным лицевым отделом и гладким, округлым мозговым. Лобные кости сильно сжаты, хорошо развиты. Лобно-теменные гребни хорошо развиты. Костное небо расширено спереди и сужено сзади. Рез
цовые отверстия широкие, средней длины, не достигают уровня первого коренного зуба. Костные слуховые барабаны маленькие. Резцы суженные; верхние имеют на наружной поверхности по две мелкие бороздки, нижние снаружи гладкие. Верхние резцы оранжевого цвета, нижние — желтого. В верхней челюсти третий коренной зуб небольших размеров.
Распространена в восточной части Заира, Уганде, южном Камеруне, юго-западной части Центральноафриканской Республики, Габоне, юго-западном Конго, о-ва Фернандо-По.
Населяют леса. В горы поднимаются до 1800 м над уровнем моря. Экология изучена слабо. Ведет ночной образ жизни. Может лазать по ветвям кустов, по земле передвигается прыжками. Питается в основном насекомыми, а также водными ракообразными плодами. Сезонность в размножении ие выражена. В помете 1—3, обычно 2 детеныша.
ПОДСЕМЕЙСТВО БОЛОТНЫХ КРЫС
SUBFAMILIA OTOMYINAE THOMAS, 1897
Размеры от мелких до средних. Внешне похожи на крыс или полевок. „ г- 3
Коренные зубы —крупные, образованы сериями поперечных пластинок.
Верхний третий коренной зуб больше второго и почти всегда крупнее первого. Резцы толстые, с бороздками на передней поверхности.
Род болотных крыс
Genus Otomys F. Cuvier, 1923
Размеры от мелких до средних. Длина тела 12,4—21,7 см. Длина хвоста 5,5—17 см. Масса 100-—200 г. Внешним видом несколько напоминают полевок (рис. 132). Голова несколько укороченная и округлая. Уши относительно малы. Туловище утолщенное, приземистое. Хвост довольно густо покрыт волосами. Волосяной покров обычно высокий, густой, мягкий. Окраска верхней части тела от темно-желтой до темно-коричневой. Брюшная сторона белая, кремовая, темно-желтая, коричневатая или темно-серая. Хвост сверху темнее, чем снизу.
Череп с резко выраженным межглазничным сужением и крупными лобно-теменными гребнями. Носовые кости спереди сильно расширены, костное небо очень узкое. Резцовые отверстия узкие и длинные, почти достигают первого коренного зуба. Костные слуховые барабаны средних размеров, несколько вздуты. Угловой отросток нижней челюсти слегка повернут наружу. Верхние резцы сильно загнуты назад и имеют по одной глубокой бороздке и иногда второй, более мелкой на наружной поверхности. Нижние резцы тоже с двумя бороздками. Иногда бороздок на резцах нет.
Хромосом в диплоидном наборе у О. irroralus 28, а у О. angonien-sis 56.
Распространены по большей части Африки от Судана па юг до южной оконечности континента.
Населяют влажные места, болота, берега различных водоемов. Некоторые виды встречаются иа лесистых склонах гор, другие — в сухих областях с песчаной почвой, в зарослях кустарников. По данным разных авторов, держатся поодиночке или колониями. Большинство видов строят гнезда из растительного материала на поверхности почвы. Иногда живут в норах, которые выкапывают сами. Активны круглосуточно, но главным
Рис. 132. Otomys irroratus и ее череп
образом в сумерках утром и вечером. Могут плавать, даже нырять. Питаются листьями различных трав, семенами, ягодами, корнями, корой, иногда муравьями. Размножаются круглый год. В течение года бывает до 5 пометов — от 2 до 4, обычно 3 детеныша в каждом. Рождаются с открытыми глазами, покрытые шерстью, способны убегать сразу после рождения. Масса новорожденного 12 г. В 2 недели неотличимы от взрослых. Половозрел ость наступает в возрасте около 3 месяцев. В Южной Африке О. irroratus вредит иосадкам в лесных питомниках.
В роде, по-видимому, 10 видов:
О. tijpus Heuglin, 1877 (горы Килиманджаро, Кения, Абердер, Рувензори, Элгон в Уганде, горы Эфиопии, ила го Нинка в Замбии; южный Судан);
О. laminatus Thomas et Schwann, 1905 (юг Африки на север до 23° ю. ш.);
О. anchietae Bocage, 1882 (Ангола, кроме юга, Южная Танзания и, вероятно, гора Элгон в Уганде);
О. angoniensis Wroughton, 1906 (от Капской провинции в Южно-Африканской Республике до Анголы на западе и Кении на севере);
О. sautidersiae Roberts, 1929 (Южно-Африканская Республика);
О. tropicalis Thomas, 1902 (Заир, Камерун, Уганда, Кения);
О. irroratus Brants, 1827 (Южно-Африканская Республика, Родезия);
О. denti Thomas, 1906 (горные районы в Западной Руанде, Уганде, хМалави, Замбии, Танзании);
О. sloggetti Thomas, 1902 (Южно-Африканская Республика к югу от 27° ю. ш.);
О. uriisulcatus Cuvier, 1829 (Южно-Африканская Республика, Капская провинция).
Андерсон и Джонс (1967) насчитывают в роде 8 видов, Уокер (1964)—24. Некоторые исследователи (например, Уокер, 1964) выделяют О. unisulcatus в особый род Myotomys Thomas, 1918.
Род паротомисов
Genus Parotomys Thomas, 1918
Размеры мелкие. Длина тела 13,5—17 см. Длина хвоста 7,5—12 см.
Внешним видом несколько напоминают болотных крыс. Уши небольшие. Волосяной покров мягкий. Окраска спинной стороны тела ржавожелтая, с черноватыми или коричневатыми оттенками, рыжевато-коричневая или темно-желтая. Брюхо серовато-белое. Хвост может быть черным или рыжевато-оранжевым в основании и коричневато-рыжим па конце. Сосков 2 пары.
Череп очень крупный, вздутый. Лобно-теменная область сужена. Однако межглазничное сужение не очень сильно выражено. Носовые кости несколько расширены спереди. Костные слуховые барабаны крупные. Верхние резцы обычно с двумя бороздками на наружной поверхности (иногда бороздок нет), нижние резцы снаружи гладкие.
Распространены в Южно-Африканской Республике и Юго-Западной Африке.
Обитают на песчаных почвах с бедной растительностью. Активны днем. Держатся группами. Убежищем служат норы, которые зверьки выкапывают сами. Над норой устраивают гнездо высотой до 60 см из ветвей и травы. Питаются различными частями растений-саккулентов, в частности солянок. По-видимому, дают 4 помета в год, в каждом из которых от 2 до 4 детенышей. Половозрелость наступает в 3-месячном возрасте.
В роде 2 вида:
Р. brantsi Smith, 1840 (центр, север и запад Капской провинции в Южно-Африканской Республике и юг Юго-Западной Африки);
Р. littledalei Thomas, 1918 (запад и север Капской провинции в Южно-Африканской Республике и Юго-Западная Африка).
ПОДСЕМЕЙСТВО ТОПКОХВОСТЫХ КРЫС SUBFAMILIA PHLOEOMYINAE ALSTON, 1876
Размеры мелкие, средние или крупные. В большей или меньшей степени приспособлены к лазанию по деревьям. Коренные зубы со сложной структурой жевательной поверхности, состоящей из трех рядов крупных бугорков; средний ряд, как правило, развит особенно сильно. Верхний первый коренной зуб имеет 8, 9 или 10 бугорков. Иногда (Phloeomys) коренные зубы образованы серией поперечных пластин.
Род сулавесских крыс
Genus Lenomys Thomas, 1898
В роде единственный вид: сулавесская крыса — L. meyeri Jentink, 1879.
Размеры средние. Длина тела 23,5—29 см. Длина хвоста 21—28,5 см; он почти лишен волос. Волосяной покров густой, мягкий. Окраска спинной стороны тела оливково-черная с беловатыми отметинами. Середина спины более темная. Брюхо темно-желто-белое. Кочености коричневатосерые. Дистальная половина хвоста (иногда три пятых длины хвоста) розового цвета. Сосков 2 пары.
В черепе лобно-теменные гребни сильно развиты. Подглазничные отверстия узкие. Резцовые отверстия короткие. Костные слуховые барабаны большие и вздутые. Костное небо сильно сужено, особенно спереди. Жевательная поверхность коренных зубов сложная.
Распространена в центральных и северных районах о-ва Сулавеси. Экология не изучена.
Род цепкохвостых крыс
Genus Pogonomys Milne-Edwards, 1877
Размеры мелкие. Длина тела 11,5—17 см. Длина хвоста 16,5—25 см. Внешним видом напоминают представителей рода крыс. Хвост покрыт редкими волосами, причем дистальная его часть совсем безволосая и, по-видимому, хватательная. Кисти и стопы ног широкие и укороченные, приспособленные к лазанию по деревьям. Пятый палец задней конечности удлинен, но первый палец не противопоставляется другим пальцам и имеет коготь. Волосяной покров густой, мягкий. Окраска спинной стороны тела от сероватой, рыжеватой и рыжевато-коричневой до темно-коричневой и почти черной. Брюшная сторона белая, темно-желтая, желтоватая или рыжеватая. Сосков 3 пары.
Мозговая коробка черепа округлая. Лицевой отдел длинный или короткий. Межглазничное сужение выражено хорошо. Лобно-теменные гребни хорошо развиты. Скуловые дуги широко расставлены. Костные слуховые барабаны очень маленькие. Костное небо широкое. Резцовые отверстия укороченные и расположены далеко впереди щечных зубов Резцы обычно широкие и мощные. Верхние коренные зубы сложного строения.
Распространены на Новой Гвинее и прилегающих островах.
Ведут древесный образ жизни. Поднимаются в горы на высоту до 2700 м над уровнем моря, хотя встречаются также и в долинах на уровне моря. Экология не изучена.
В роде 9 видов:
Р. macrourus Milne-Edwards, 1877 (о-ва Новая Гвинея, Япсп, Новая Британия);
Р. sylvestris Thomas, 1920 (юго-восток Новой Гвинеи);
Р. mollipilosus Peters et Doria, 1881 (Новая Гвинея);
Р. fergussoniensis Lauric, 1952 (о-в Фергюссоп);
Р. shawmayeri Laurie, 1952 (о-в Фергюссоп);
Р. lamia Thomas, 1897 (юг Папуа);
Р. kagiPate, 1951 (Папуа);
Р. vates Thomas, 1908 (Папуа);
Р. forbesi Thomas, 1888 (Папуа, о-ва Гудипар и Фергюссоп).
Рис. 133. Сопевидиая мышь — С. gliroides
Род соневидиых мышей
Genus Chiropodomys Peters, 1868
Размеры мелкие. Длина тела 7,6—13,3 см. Длина хвоста 8—15,5 см. Уши средних размеров, тонкие, почти без волос (рис. 133). Хвост покрыт редкими волосами, удлиненными на конце, не хватательный. Стопа с крупными мозолями на пальцах приспособлена к лазанию по деревьям; пятый палец имеет коготь и достигает такой же длины, как и второй, первый палец короткий, противопоставляется остальным и не имеет когтя. На передней конечности пятый палец имеет коготь, первый палец сильно уменьшен. Волосяной покров густой, мягкий. Окраска спинной стороны тела темно-серовато-коричневая или темно-желто-коричневая. Брюшная сторона белая или бледно-кремовая. Хвост темпо-сероватый. Вибриссы многочисленные и длинные. Сосков 2 пары.
В черепе характерны массивная округлая мозговая коробка и относительно короткий лицевой отдел. Профиль черепа снижается к затылочной области. Межглазиичное сужение выражено не очень резко. Лобнотеменные гребни имеются. Скуловые дуги расставлены узко. Костное небо широкое. Резцовые отверстия широкие и короткие, не достигают зубных рядов. Костные слуховые барабаны относительно небольшие. Верхние щечные зубы суженные. Жевательная поверхность пнжпих и верхних щечных зубов сложная.
Распространены в Ассаме, па п-ове Малакка, па Суматре, Яве, Калимантане с мелкими прилегающими островами.
Населяют лесные районы. Ведут древесный образ жизни. Активны в основном ночью. Питаются различными растительными кормами.
В роде 2 вида:
соневидная мышь — С. gliroides Blyth, 1856 (Ассам; Бирма; Тепассе-рим; Южный Китай; п-ов Индокитай, Таиланд, п-ва Малакка; о-ва Суматра, Натунас, Ява и Калимантан);
ниасск-ая мышь — С. niadis Miller, 1903 (о-в Ниас у Суматры).
Чейзен (1940) выделяет, кроме С. gliroid.es, в особые виды С. major Thomas, 1893; С. legatus Thomas, 1911; С. pictor Thomas, 1911, и C. pusil-lus Thomas, 1893, обитающих на о-ве Калимантан, а также С. anna Thomas et Wroughton, 1909, с о-ва Ява. По Андерсону и Джонсу (1967), в роде 6 видов.
С. fulvus G. Allen, 1927, включенный Эллермэном (1941) в этот род, большинство систематиков выделяют в особый род Vernaya Anthony, 1941.
Род древесных крыс
Genus Mallomys Thomas, 1898
В роде единственный вид: древесная крыса — Al. rotschildi Thomas, 1898.
Размеры крупные. Длина тела 34,5—44 см. Длина хвоста 36,5—42 см. Морда несколько укороченная. Хвост почти лишен волос и покрыт крупными, грубыми чешуями. Стопа приспособлена к лазанию по деревьям, относительно короткая, массивная. Пальцы с крупными когтями. Первый палец передних конечностей имеет короткий ноготь. Волосяной покров высокий, густой, мягкий. Окраска его на спинной стороне тела темно-коричневая или сероватая с белым оттенком (остевые волосы имеют белые вершинки). Брюшная сторона кремневая или тускло-белая. Конечности и вибриссы черные. Проксимальная часть хвоста коричневатая, дистальная — белая.
Череп очень массивный. Лобно-теменные гребни крупные. В лобном отделе имеется широкая вогнутая площадка. Межглазничное сужение выражено не очень резко. На чешуйчатых костях есть небольшие гребни. Передние отделы лобных костей сильно вздуты. Резцовые отверстия очень широкие. Костное небо сильно сужено. Костные слуховые барабаны очень маленькие. Щечные зубы очень широкие, с массивными выступами на жевательной поверхности. Наружная поверхность резцов гладкая.
Распространена на большей части Новой Гвинеи.
Обитает среди скал в горах на высоте от 1300 до 2700 м над уровнем моря. Возможно, ведет древесный образ жизни. Экология не изучена.
Род флоресских крыс
Genus Papagomys Sody, 1941
В роде единственный вид: флоресская крыса — Р. armandvillei Jentink, 1892.
Размеры крупные. Длина тела 41—45 см. Длина хвоста 33—37 см. Хвост покрыт толстыми, удлиненными чешуями, каждая из которых имеет по 3 коротких грубых волоска. В волосяном покрове туловища встречаются иглы.
Череп сходен с таковым гигантских древесных крыс, однако широкой вогнутой площадки в лобном отделе нет. Лобно-теменные гребни сильно развиты.
Распространена на о-ве Флорес к востоку от Явы.
Экология не изучена.
Род тонкохвостых крыс
Genus Phloeomys Waterhouse, 1839
Размеры наибольшие в семействе. Длина тела 37,5—48,5 см. Длина хвоста 20—35 см. Морда притуплена. Уши короткие, округлые. Хвост покрыт длинными, густыми волосами (рис. 134). Конечности приспособлены к лазанию по деревьям. Стопа и кисть широкие. Все пальцы развиты хорошо. Когти на передних конечностях крупные. Волосяной покров обычно густой и мягкий, иногда грубый. Окраска его на спинной стороне тела черновато-коричневая с желтоватым или рыжеватым оттенком или серовато-коричневая. Брюхо светлее спины. Уши могут быть черные. Щеки серые. Хвост черный или коричиевато-черный. На спине иногда выделяется неправильной формы рыжевато-коричневое пятно. Сосков 2 пары.
Череп с укороченным широким лицевым отделом. Мощные лобно-теменные гребни проходят далеко назад. Межглазничное сужение выражено слабо и сдвинуто назад. Имеются заглазничные отростки, между которыми располагается впадина. Костные слуховые барабаны очень небольшие. Костное небо довольно широкое. Резцовые отверстия укороченные и не достигают рядов щечных зубов. Скуловые дуги массивные. Резцы очень широкие. Щечные зубы представлены серией поперечных пластин.
Распространены на Филиппинских о-вах.
Встречаются от уровня моря до высокогорья Ведут древесный образ жизни. Экология не изучена.
В роде 2 вида:
Р. camingi Waterhouse, 1839 (Филиппинские о-ва);
Р. pallidas Nehring, 1890 (о-в Лузон из группы Филиппинских островов) .
Рис. 134. Phloeomys cumingi
Рис. 135. Пышнохвостая крыса — С. schadenbergi и жевательная поверхность ее верхних (Л) и нижних (Б) щечных зубов
Род пышнохвостых крыс
Genus Crateromys Thomas, 1895
В роде единственный вид: пышпохвостая крыса — С. schadenbergi Meyer, 1895.
Размеры средние и крупные. Длина тела 32,5—36 см. Длина хвоста 35,5—40,7 см. Голова укороченная. Глаза средних размеров. Уши небольшие (рис. 135). Хвост покрыт густыми, длинными волосами и напоминает хвост белки. Конечности приспособлены к лазанию по деревьям. Стопа укороченная; пятый палец примерно такой же длины, как и три средних, первый палец также длинный, ио, по-видимому, не противопоставляется остальным. Пальцы как на передних, так и па задних конечностях с крупными когтями. Только первый палец передней конечности имеет уплошенпып ноготь. У основания каждого когтя растет пучок удлиненных волос. Волосяной покров очень высокий и мягкий, с сильно развитым пухом. Его окраска значительно варьирует: обычно опа бывает темно-коричневая или черная па спинной стороне тела и темпо-серая или серая на боках и брюшной стороне У некоторых особей передняя часть тела беловатая или коричневатая.
Череп сильно сжат в лобной области, поэтому его мозговая коробка кажется укороченной. Скуловые дуги широко расставлены в стороны. Костное небо относительно узкое. Резцовые отверстия средней длины, не достигают рядов щечных зубов. Костные слуховые барабаны небольшие, уплощенные.
Распространена в северной части о-ва Лузон из группы Филиппинских островов.
Населяет горные леса. Ведет древесный образ жизни. Активна в основном в сумерках и ночью. Убежищем служат дупла или впадины между корнями деревьев. Питается почками и корой молодых сосен, а также различными плодами. Экология изучена слабо.
ПОДСЕМЕЙСТВО ХОБОТКОВЫХ КРЫС SUBFAMILIA RHYNCHOMYINAE THOMAS, 1897
В подсемействе единственный род — хоботковых крыс.
Род хоботковых крыс / Genus Rhynchomys Thomas, 1895
В роде единственный вид: хоботковая крыса — Д. soricoid.es Thomas, 1895.
Размеры средние. Длина тела 18,8—21,5 см. Длина хвоста 14,2— 14,6 см. Морда сильно удлинена, что придает зверьку сходство с землеройкой. Уши средних размеров, округлые. Глаза небольшие. Хвост покрыт средней густоты волосами. На задней конечности первый и пятый пальцы значительно короче трех средних, первый палец с широким ногтем. Волосяной покров густой и мягкий, но низкий, плотно прилегающий к телу. Одноцветная темно-оливково-серая окраска спинной стороны тела постепенно переходит в грязно-серую на брюшной стороне. На горле иногда расположено белое пятно. Хвост черноватый сверху и слегка более светлый снизу.
В черепе очень сильно вытянут тонкий лицевой отдел. Межглазничное сужение выражено слабо. Лобно-теменных гребней нет. Передний конец носовых костей повернут вверх. Костные слуховые барабаны относительно небольшие. Скуловые дуги расставлены нешироко. Нижняя челюсть с хорошо выраженным венечным отростком. Резцовые отверстия 2
длинные. Щечных зубов у, их размеры чрезвычайно малы. Верхние резцы очень маленькие. Нижние резцы в меньшей степени уменьшены, острые, но очень тонкие. Верхние резцы белые, нижние светло-желтые.
Распространена на о-ве Лузон в группе Филиппинских островов.
Населяет густые кустарники и овраги в лесах на склонах гор на высоте 2000—2500 м над уровнем моря. Экология не изучена. Возможно, питается насекомыми и червями.
ПОДСЕМЕЙСТВО ВОДЯНЫХ КРЫС SUBFAMILIA HYDROMYINAE ALSTON, 1876
Размеры мелкие, средние или крупные. В большей или меньшей степени адаптированы к водному образу жизни. Жевательная поверхность коренных зубов несложная, лункообразиая. Третий верхний и третий нижний коренной зубы отсутствуют или очень малы.
Род полосатых крыс
Genus Chrotomys Thomas, 1895
В роде единственный вид: полосатая крыса — С. whiteheadi Thomas, 1895.
Размеры средние. Длина тела около 20 см. Длина хвоста примерно 11 см. Морда удлиненная. Уши средних размеров, округлые. Глаза небольшие. Хвост покрыт относительно редкими волосами. Стопа узкая; пятый палец короткий. Первый палец передней конечности имеет округлый ноготь. Все остальные пальцы обеих конечностей со слегка искривленными когтями. Волосяной покров густой, мягкий. Окраска его на спинной стороне тела серовато-коричневая с довольно широкой оранжевой продольной срединной полосой, окаймленной по бокам почти черными полосками, проходящими от середины морды до основания хвоста. Брюшная сторона серая.
Череп с удлиненным лицевым отделом и широкой мозговой коробкой. Резцовые отверстия очень короткие, расположенные примерно посредине между резцами и щечными зубами. Скуловые дуги слабо расставлены в з стороны. Костные слуховые барабаны маленькие. Щечных зубов-g-. Третий щечный зуб очень маленький. Верхние резцы широкие, направлены несколько вперед, нижние сильно удлинены. Все резцы бледно-желтого цвета.
Распространена в северной части о-ва Лузоп из группы Филиппинских островов.
Населяет горные районы на высоте примерно 2,5 тыс. метров над уровнем моря. Питается сладким картофелем и различными травами. Экология не изучена.
Род лузонских водяных крыс
Genus Celaenomys Thomas, 1898
В роде единственный вид: лузонская водяная крыса — С. si.la.ceus Thomas, 1895.
Размеры средние. Длина тела примерно 19,5 см. Длина хвоста около 11 см. По внешнему виду сходна с землеройками. Уши средних размеров, округлые. Глаза маленькие. Хвост покрыт редкими волосами. Волосяной покров мягкий, густой, но низкий. Спинная сторона тела серая, брюшная светло-серая. Боковые части морды почти черные. Уши сероватые.
Череп лузонской водяной крысы сходен с черепом полосатой крысы.
2 Щечных зубов —.
Населяет овраги в горных лесах на высоте 2—2,5 тыс. метров пад уровнем моря.
Распространена на о-ве Лузон из группы Филиппинских островов.
Экология не изучена.
Род безухих крыс
Genus Crossomys Thomas, 1907
В роде единственный вид: безухая крыса — С. moncktoni Thomas, 1907.
Размеры средние. Длина тела около 20,5 см. Длина хвоста примерно 22 см. В высокой степени приспособлена к полуводному образу жизни. Уши сильно уменьшены. У пятипалых задних конечностей основания
пальцев соединены плавательной перепонкой. Передние конечности очень малы и имеют только по четыре функционирующих пальца. По нижней стороне хвоста располагаются удлиненные жесткие волосы, образующие плавательный гребень, который у основания хвоста разделяется на два. Волосяной покров очень густой, мягкий, высокий. Окраска спинной стороны тела коричневато-серая с желтовато-оливковым оттенком. Брюхо белое. Хвост светло-серый сверху и белый снизу. Волосяной гребень на его нижней стороне белый. Сосков 2 пары.
Череп с очень крупным мозговым отделом округлой формы. Костные слуховые барабаны очень маленькие. Резцовые отверстия средней длины. Подглазничные отверстия очень большие. Ряды щечных зубов сдви-2
нуты вперед. Коренных зубов-g-. Размеры их относительно малы. Резцы узкие.
Распространена на Новой Гвинее.
Встречается на берегах рек и мелких потоков в горах на высоте от 600 до 3000 м над уровнем моря. Экология не изучена. Питается, по-видимому, различными мелкими водными животными.
Род ложных водяных крыс
Genus Xeromys Thomas, 1889
В роде единственный вид: ложная водяная крыса — X. myoides Thomas, 1889.
Размеры мелкие. Длина тела 11—11,4 см. Длина хвоста примерно 8,8 см. Внешним видом напоминают домовую мышь. Уши средних размеров, широкие. Глаза маленькие. Хвост почти не имеет волос. Стопа нормального строения, но пятый палец значительно укорочен. Пятый палец передней конечности также очень мал. Волосяной покров довольно высокий, густой, мягкий. Окраска его на спинной стороне тела темносерая, на брюшной несколько более светлая. Сосков 2 пары.
В черепе межглазничное сужение выражено слабо. Носовые кости спереди несколько укорочены. Лобно-теменных гребней нет. Костные слуховые барабаны маленькие. Резцовые отверстия средней длины. Щечных 2
зубов—. Резцы толстые, слегка направлены вперед. Верхние резцы
оранжевого цвета, нижние — белые.
Распространена в Квинсленде в Австралии.
Населяет болотистые районы с густыми зарослями высокой травы и кустов. Экология изучена слабо. Питается, вероятно, водными животными и растениями.
Род австралийских водяных крыс
Genus Hydroniys Е. Geoffгоу, 1805
Размеры средние и крупные. Длина тела 20,5—35 см. Длина хвоста 20—35 см. Адаптированы к водной жизни, но в гораздо мсныней степени, чем безухая крыса. Наружные ушные раковины небольшие. Голова длинная, уплощенная. Глаза поставлены высоко (рис. 136) Хвост густо покрыт короткими, жесткими волосами. Передние конечности с четырьмя функционирующими пальцами. Задние конечности значительно больше передних. Стопа очень широкая, крупная с пятью пальцами, частично соединенными плавательной перепонкой; три средних пальца наиболее длинные. Волосяной покров образован многочисленными грубыми, высокими остевыми волосами и густым, очень мягким пухом Окраска спин-
Рис. 136. Златобрюхая крыса — Н. chrysogaster
ной стороны тела от темно-коричневой и черной до золотисто-коричневой и темно-серой, брюшной — от коричневатой до желтовато-белой или ярко-оранжевой. Хвост с белым кончиком.
В черепе мозговая коробка узкая. Лобные кости сильно сжаты. Лобно-теменные гребни развиты слабо или их нет вовсе. Дорзальный профиль черепа уплощен. Костные слуховые барабаны очень маленькие. Резцовые отверстия короткие, расположены далеко впереди от рядов щечных зубов. Ряды щечных зубов несколько сближены спереди; число 2
щечных зубов—. Они крупные, причем первый коренной значительно
больше второго. Резцы довольно мощные.
Хромосом в диплоидном наборе у Н. chrysogaster 48.
Распространены на Новой Гвинее, в Тасмании, Австралии, на о-вах Мелвилла, Новая Британия, Ару, Кей и в некоторых других местах.
Населяют берега различных водоемов и болота в горах. Ведут полу-водный образ жизни. Активность сумеречная и ночная. День проводят в убежищах различного типа, иногда в норах, сделанных в берегах. Питаются моллюсками, ракообразными, улитками, а также рыбами, лягушками, водными птицами, их яйцами и различными растениями. За водяными крысами активно охотятся из-за хорошего меха. В некоторых областях вследствие малочисленности зверьков необходимо наладить нх защиту.
В роде 3 вида:
златобрюхая крыса — Н. chrysogaster Е. Geoffrey, 1804 (о-ва Бруни у Тасмании; Ару и Кей; Южная и Западная Австралия, Квинсленд, Северная территория);
новобританская крыса — Н. neobritannicus Tate et Archbold, 1935 (о-ва Новая Британия из группы Соломоновых);
Н. habbema Tate et Archbold, 1941 (западная часть Новой Гвинеи).
Род горных водяных крыс
Genus Parahydromys Poche, 1906
В роде единственный вид: горная водяная крыса — Р. asper Thomas, 1906.
Размеры средние. Длина тела около 24 см. Длина хвоста примерно 26 см. Уши средних размеров, округлые. Задние конечности значительно длиннее передних. Между тремя средними пальцами задних конечностей имеется неполная плавательная перепонка. Подошва стопы гладкая. Волосяной покров низкий, сильно изреженный, короткий. Окраска спинной стороны тела коричневато-серая, брюшной — белая. Хвост коричневато-черный с белым концом. Сосков 2 пары.
Череп сходен с таковым австралийской водяной крысы. Мозговые барабаны предельно маленькие. Лобно-теменных гребней нет. Резцовые отверстия относительно короткие. Резцы довольно слабые, узкие.
Распространена на Новой Гвинее.
Населяет горные склоны, предпочитая скалистые участки в лесах на высоте 600—2700 м над уровнем моря. Несмотря на то .что горные водяные крысы имеют некоторые адаптации к водной жизни, с водой они, по-видимому, не связаны и ведут чисто наземный образ жизни. Экология не изучена.
Род неохидромис
Genus Neohydromys Laurie, 1952
В роде единственный вид: неохидромис — N. fuscus Laurie, 1952.
Размеры мелкие. Длина тела около 9,2 см. Длина хвоста примерно 7,8 см. По внешнему виду похож на мышь. Морда несколько вытянутая. Волосяной покров низкий, довольно густой, мягкий. Спинная сторона тела серая, брюшная слегка более светлая. Хвост коричневатый с белым концом.
Щечные зубы маленькие.
Распространение охватывает северо-восточную часть Новой Гвинеи. Населяет леса на горных склонах на высоте 2400—3000 м над уровнем моря. Экология не изучена.
Род очковых мышей
Genus Leptomys Thomas, 1897
В роде единственный вид: очковая мышь — L. elegans Thomas, 1897. Размеры мелкие. Длина тела 14,4—16,2 см. Длина хвоста 15—16 см. По внешнему виду похожи на мышей. Уши средних размеров, почти полностью лишенные волос. Глаза маленькие. Чешуйчатый хвост голый. Стопа удлиненная, узкая; три средних пальца значительно длиннее пятого, который немного короче первого. На подошве стопы 6 мозолей, на кисти — 5. Волосяной покров густой, мягкий, довольно низкий. Спина рыжевато-желто-коричневая. Глаза окружены черным кольцом. Плечи, бока и крестец ярко-рыжего цвета. Верх морды черный. Щеки, внутренние стороны передних конечностей и брюхо кремово-белые. Брюхо может быть и серовато-коричневым. Хвост коричневый сверху и белый снизу; его кончик весь белый.. Сосков 2 пары.
В черепе межглазничное сужение выражено слабо. Лобно-теменных гребней нет. Резцовые отверстия короткие, не достигают коренных зубов.
Скуловые дуги расставлены узко. Число щечных зубов—. Третий щечный зуб очень мал. Резцы широкие, уплощенные, имеют желтую окраску.
Распространена на Новой Гвинее.
Населяет горы от уровня моря до высоты примерно 3000 м над его уровнем. Экология не изучена.
Род микрохидромис
Genus Microhydromys Tate et Archbold, 1941
В роде единственный вид: микрохидромис — М richardsoni Tate et Archbold, 1941.
Размеры мелкие. Длина тела 8 см. Длина хвоста 9,2 см. Морда укорочена. Конечности не приспособлены для плавания. Спинная сторона тела серовато-черная, брюшная немного светлее. Конец хвоста белый.
В черепе характерны относительно крупные костно-слуховые барабаны. Верхние резцы с продольными бороздками на наружной поверхности.
2
Число щечных зубов — .
Распространение охватывает северо-восток Новой Гвинеи.
Населяет горные склоны. Экология не изучена.
Род паралептомис
Genus Paraleptomys Tate et Archbold, 1941
Размеры мелкие. Длина тела 12—14 см. Длина хвоста 13—14 см. По внешнему виду сходны с представителями рода очковых мышей. Задние конечности не удлинены. Окраска спинной стороны тела серовато-коричневая, брюшной грязно-белая. Хвост серый сверху и белый снизу; его кончик весь белый. Р. rufilatus отличается от Р. Wilhelmina белым горлом, широкой, рыжевато-бурой боковой полосой и более рыжими головой и задними конечностями.
2
Число щечных зубов — .
Распространены в западной части Новой Гвинеи.
Встречаются в горах на высоте 1400—2700 м над уровнем моря. Экология не изучена.
В роде 2 вида:
Р. Wilhelmina Tate et Archbold, 1941 (центральный район западной части Новой Гвинеи);
Р. rufilatus Osgood, 1945 (северо-восток западной части Новой Гвинеи).
Род байянкасских крыс
Genus Baiyankamys Hinton, 1943
В роде единственный вид: байянкасская крыса — В. shawmayeri Hinton, 1943.
Размеры мелкие. Длина тела примерно 14,6 см Длина хвоста около 17,6 см. Имеет морфологические приспособления к плаванию Уши средних размеров. Стопа крупная, средние пальцы соединены небольшими плавательными перепонками. Волосяной покров густой, мягкий Окраска
его темно-серая на спинной стороне тела и светло-серая на брюшной. Хвост покрыт довольно густыми волосами; его конец белый.
В черепе мозговая коробка крупная с умеренно выраженным меж-
2
глазничным сужением. Число щечных зубов — .
Распространена на северо-востоке Новой Гвинеи.
Встречается на берегах горных водоемов. Ведет водный образ жизни.
Род новогвинейских мышей
Genus Pseudohydromys Rummler, 1934
Размеры мелкие. Длина тела 8,5—11,5 см. Длина хвоста 9—9,5 см. Внешним обликом сходны с мышами. Конечности имеют подошвенные мозоли, как у наземных форм. Морфологические приспособления к водному образу жизни отсутствуют. Уши средних размеров, округлые, почти лишенные волос. Окраска спинной стороны тела темно-серая, брюшной— несколько более светлая. Число щечных зубов .
Распространены в Новой Гвинее.
Населяют горные склоны на высоте 2100—3600 м над уровнем моря. Ведут наземный образ жизни. Экология не изучена.
В роде 2 вида:
Р. murinus Rummler, 1934 (северо-восток Новой Гвинеи);
Р. occidentalis Tate, 1951 (западная часть Новой Гвинеи).
Род мышей Майера
Genus Mayermys Laurie et Hill, 1954
В роде единственный вид: мышь Майера — М. ellertnani Laurie et Hill, 1954.
Размеры мелкие. Длина тела 9—10,5 см. Длина хвоста 10—И см. Хвост покрыт короткими, довольно редкими волосами. Пальцы соединены слабо развитой перепонкой. Окраска волосяного покрова спинной стороны тела серая с одним или двумя беловатыми пятнами, брюшной немного светлее. На груди может быть белое пятно. Хвост буроватый с белым концом.
Мышь Майера от всех других грызунов (и всех млекопитающих) отличается тем, что в верхней и нижней челюсти имеется только по одному коренному зубу. Резцы хорошо развиты, бороздок снаружи не имеют. Резцовые отверстия маленькие. Костные слуховые барабаны небольшие, уплощенные.
Распространена на северо-востоке Новой Гвинеи.
Населяют лесные горные склоны на высоте 1800—2700 м над уровнем моря. Экология не изучена.
СЕМЕЙСТВО СОНЕВЫХ
FAMILIA GLIRIDAE THOMAS. 1897
Грызуны мелких размеров. Длина тела 8—19 см. Длина хвоста 5,5— 16 см. Внешний вид мышеобразный (у наземных форм) или белкообразный (у древесных форм). Уши средних размеров, округлые. Глаза крупные. Длинный хвост слегка уплощенный, по крайней мере в концевом
•отделе. У наземных форм хвост покрыт редкими, а у древесных длинными, густыми волосами. Передние конечности четырехпалые, задние пятипалые. Наружные пальцы могут сильно отодвигаться в сторону, в известной степени противопоставляясь остальным. Первый палец задней конеч-
ности иногда лишен когтя, на остальных пальцах когти хорошо развиты; они небольшие, но очень острые. За исключением мышевидных сопь у всех остальных представителей семейства сильно развиты подошвенные мозоли. Волосяной покров густой и мягкий, но низкий. Окраска однотонная от серой до коричневой. Сосков от 4 до 6 пар.
Мозговой отдел черепа округлый. Сагиттального гребня нет. Ламб-доидальный гребень очень небольшой или его нет вовсе. Надглазничные отростки лобных костей и заглазничные бугры отсутствуют. Межглазничное сужение выражено обычно слабо. Сверху эта часть черепа плоская или имеет незначительные продольные углубления. Скуловые дуги тонкие, нешироко расставленные в стороны. Костные слуховые барабаны
большие, тонкостенные. В нижней челюсти угловой отросток, если он имеется, загнут несколько наружу. Сочленованный отросток хорошо развит. Зубная формула: г— с — р — т—=20. Щечные зубы брахиодонт-10 13
«ого типа с хорошо развитыми корнями. Их жевательная поверхность у примитивных форм бугорчато-гребенчатая, а у более специализированных представлена поперечными низкими валиками эмали. Верхние резцы крупнее тонких нижних. Кости предплечья и голени удлиненные. Малая и большая берцовые кости в дистальных отделах срослись.
Распространены в Африке в изолированном участке на северо-западе и к югу от Сахары, в лесной и лесостепной зонах Европы от Южной Скандинавии до Средиземноморья и на восток до Урала, Малой Передней и Средней Азии, на Алтае, в Северо-Западном Китае, Японии.
Населяют лесную и лесостепную зоны. В горы поднимаются до высоты 3500 м над уровнем моря. Предпочитают широколиственные и смешанные леса. Активны, как правило, ночью. В различной степени приспособлены к древесному образу жизни. Некоторые (соня-полчок) почти •совсем не спускаются на землю, другие (мышевидная соня) ведут в основном наземный образ жизни. Питаются в основном плодами, орехами, семенами, в больших количествах поедают насекомых, а также яйца птиц и мелких птиц. К осени становятся очень жирными и на холодный период времени года впадают в спячку. В течение года, как правило, бывает один (иногда 2) помет, в каждом из которых от 2 до 9 детенышей. Длительность беременности 21 — 28 дней. Самка приносит детенышей в гнезде, которое устраивает в дуплах деревьев или среди ветвей, иногда в какой-нибудь впадине или расщелине земли. Длительность жизни в естественных условиях от 2 до 5,5 лет. Местами могут приносить известный вред плодовым насаждениям. Шкурки крупных сонь используются в качестве второстепенной пушнины.
В семействе 7 родов (11 видов):
сонь-полчков— Glis Brisson, 1762 (1 вид);
орешниковых сонь — Muscardinus Капр, 1829 (1 вид);
садовых сонь — Eliomys Wagner, 1843 (1 вид);
лесных сонь — Dryomys Thomas, 1906 (1 вид);
японских сонь—Glirulus Thomas, 1905 (1 вид);
мышевидных сонь — Myomimus Ognev, 1924 (1 вид);
африканских сонь— Graphiurus Smuts, 1832 (5 видов)
Род сонь-полчков
Genus Glis Brisson, 1762
В роде единственный вид: соня-полчок — Clis glis Linnaeus, 1766 (рис. 137).
Наиболее крупный представитель семейства. Длина тела до 19 см. Длина хвоста до 16,5 см. Масса около 170 г. Внешним обликом напоминает белку. Хвост покрыт длинными, густыми волосами с расчесом па две стороны. Задние конечности лишь ненамного длиннее передних. Кисть и стопа широкие с жесткими подошвенными мозолями. Четвертый палец задней конечности самый длинный, третий палец немного короче его. На передних конечностях наиболее длинный третий и четвертый пальцы. Небольшие, округлые уши гусю покрыты короткими волосами. Волосяной покров относительно низкий, густой, мягкий. Спина серебристо-серая
или буровато-серая, бока более светлые, брюхо белое или желтоватое. Сосков 6 пар.
Мозговая капсула черепа несколько уплощенная. Лицевой отдел средней длины. Лобно-теменные гребни хорошо выражены и у взрослых особей могут соприкасаться с передней частью мозговой капсулы. В области межглазничного сужения имеется продольное углубление. Скуловые дуги относительно широко расставлены. Костные слуховые барабаны несколько вытянуты.
Хромосом в диплоидном наборе 62.
Распространена в Северной Испании, Франции, Швейцарии, ФРГ и ГДР, Голландии, Италии, Сицилии, Сардинии, Австрии, Югославии, Венгрии, Румынии, Чехословакии, Болгарии, Польше, Малой и Передней Азии; в СССР на Кавказе, в Закавказье, западных и центральных районах европейской части страны, к северу до Прибалтики, Тульской и южных районов Горьковской обл., правобережье Татарии, на восток до Волги, на юг до линии Саратов — Воронеж — Чернигов — Киев — Кишинев, возможно, встречается в горах Копетдага. Акклиматизирована в Англии.
Населяет широколиственные леса на равнинах и в горах. Встречается также в смешанных лесах, фруктовых садах с примесью лещины и диких фруктовых деревьев. Активна в сумерки и ночью. День проводит, как правило, в дуплах. Иногда селится в просто устроенных норах между корнями деревьев или делает наружные гнезда. На зиму впадает в спячку, продолжающуюся обычно с сентября по апрель. Зимует чаще всего в дуплах, иногда собираясь вместе по нескольку особей. Питается желудями, орехами, различными плодами и их косточками, семенами, ягодами. Поедает также насекомых, яйца птиц и мелких птиц. В течение года бывает один помет с июня по август. Число детенышей в помете от 2 до 8. Местами причиняет значительный вред садам.
Род орешниковых сонь
Genus Muscardinus Каир, 1829
В роде единственный вид: орешниковая соня — М. avellatiarius Linnaeus, 1758.
Размеры мелкие: длина тела до 9 см. Длина хвоста до 7,7 см. Хвост по всей длине равномерно и густо покрыт короткими волосами без расчеса снизу (рис. 138). Пальцы относительно длинные; на задних конечностях четыре пальца длинные, а первый укорочен и лишен когтя (изредка коготь имеется). Уши невысокие, глаза крупные. Морда притупленная. Окраска спинной стороны тела желтовато-бурая до желтовато-рыжей. Горло и грудь кремово-белые. Сосков 4 пары.
Мозговой отдел черепа округлый, вздутый. Лицевая часть черепа укороченная. Лобно-теменных гребней нет. Резцовые отверстия довольно длинные. Костное небо очень короткое. Костные слуховые барабаны умеренно вздутые. Характерен сложный желудок.
Хромосом в диплоидном наборе 48.
Распространена в Западной Европе до Великобритании на западе и Швеции на севере; в европейской части СССР на север до северной Белоруссии, Калининской, Ивановской, Горьковской областей и Татарии, на восток до Предуралья, на юг до Куйбышевской, Харьковской, Полтавской, Днепропетровской и Одесской областей; в Крыму и на Кавказе отсутствует. Встречается в Малой Азии..
Обитает в широколиственных (особенно дубовых), а также смешанных
Рис. 138. Орешниковая соня — М avellanarius и ее череп
лесах, преимущественно там, где есть подлесок из лещины, липы, клена. Ведет древесный образ жизни. Активность сумеречная и ночная Убежищем служат дупла или шаровидные гнезда, которые устраивает на ветвях кустарников из листьев, растительного пуха, измочаленных полосок коры. Иногда заселяет искусственные гнездовья птиц Зимнюю спячку проводит в подземных норах. В Англии спячка продолжается с конца октября до начала апреля. Питается различными семенами древесных и кустарниковых пород, желудями, орехами, а также насекомыми, япцами птии и птенцами. Местами может приносить незначительный вред лесному хозяйству Дает два помета в год: первый в мае—июне, второй в июле—августе. Детенышей в помете обычно 4—5 (от 3 до 7) Длительность беременности примерно 21—28 дней В различные годы численность может значительно изменяться.
Род садовых сонь
Genus Eliomys Wagner, 1843
В роде единственный вид: садовая соня — Е. quercinus Linnaeus, 1766.
Размеры средние. Длина тела до 14 см, длина хвоста до 12,1 см. Волосы равномерно покрывают весь хвост, постепенно удлиняясь от его основания к концу, где образуют широкую уплощенную кисточку с расчесом в обе стороны снизу. Задние конечности длиннее передних (рис. 139). На задних конечностях первый палец укорочен, пятый примерно такой же длины, как и три средних. Передние конечности с четырьмя хорошо развитыми пальцами. Ступни сравнительно узкие и длинные. Уши довольно длинные, покрыты редкими, тонкими и короткими волосками. Ок-
Рис. 139 Садовая соня — Е quercinus и ее череп
раска волосяного покрова на спиннои стороне от сероватой до коричневой, на брюшной кремовая или белая. От носа через глаз к уху, огибая его сверху и особенно широко снизу, проходит черная полоса. Терминальная половина хвоста сверху черная. Сосков 4 пары.
В черепе мозговой отдел высокий, но несколько уплощенный сверху. Лицевая часть относительно длинная. Череп сильно сужен в области лобных костей. Лобно-теменные гребни развиты слабо или их нет вовсе. Углубления на межглазничном пространстве, как правило, нет. Скуловые кости относительно длинные. Костное небо широкое. Костные слуховые барабаны крупные, вздутые. Подгланизничые отверстия узкие.
Хромосом в диплоидном наборе 52.
Распространена в Европе от Пиренейского п-ова и островов Средиземного моря на восток до Урала; в СССР в западных и центральных районах европейской части страны к северу до Южной Карелии, Калининской, Горьковской и Пермской областей, на юг до Оренбургской, Куйбышевской, Ульяновской, Орловской областей, Украины и Молдавии включительно. Изолированные участки ареала имеются в Северной Африке (Марокко, Алжир, Тунис, Ливия) и на западе Малой Азии.
Населяет смешанные и широколиственные леса. Поднймается в горы на высоту до 2,5 тыс. м над уровнем моря. День проводит в дуплах деревьев, иногда строит гнезда на ветвях на высоте 0,8—3 м над землей. Нередко поселяется в строениях человека в лесу. В некоторых областях ареала на холодное время года впадает в спячку. Питается различными семенами древесных пород, а также орехами, плодами, насекомыми, мелкими грызунами, птенцами. Местами может значительно снижать численность мелких птиц. Период размножения приходится на май — октябрь. В течение года бывает 2 помета, в каждом из которых от 2 до 7 детенышей. Длительность беременности составляет 22—28 дней. Глаза открываются примерно на 21-й день после рождения. Продолжительность жизни около 5,5 лет.
Иногда в роде выделяют второй вид — Е. melanurus Wagner, 1840.
Род лесных сонь
Genus Dryomys Thomas, 1906
В роде единственный вид: лесная соня — D. nitedula Pallas, 1779.
Размеры мелкие: длина тела до И см. Длина хвоста до 11,3 см. Опушение хвоста такое же, как у сони-полчка. Задние конечности заметно длиннее передних (рис. 140). Стопа несколько удлинена. Уши средних размеров, округлые, покрыты редкими, короткими волосками. Спина и бока серовато-коричневые или желтовато-коричневые; брюхо грязно-темно-желтое или белое. Сосков 4 пары.
В черепе лицевой отдел укорочен. Мозговая коробка округлая. Лобно-теменных гребней нет. В области межглазпичного сужения находится небольшое продольное углубление.
Хромосом в диплоидном наборе 48.
Распространена в Западной Евразии от Швеции до Северной Италии на западе и Алтая и Тянь-Шаня па востоке, в СССР в западных, Центральных и Южных районах европейской части страны; к северу до Литовской ССР, Калининской, Рязанской, Горьковской обл. и Татарии, и на восток до Волги, а также на Кавказе, Закавказье, в горных лесах Средней Азии, Восточного Казахстана и Южного Алтая. Встречается в Малой Азии, Иране, Афганистане.
Населяет широколиственные леса и лесостепь. В горы поднимается до 3500 метров над уровнем моря. Активна преимущественно в сумерки и ночью, а в южных частях ареала и днем. Па зиму (в центральных районах европейской части СССР с сентября — половины октября по апрель) впадает в спячку, которая в южных частях ареала может прерываться. Убежищем служат дупла деревьев, иногда гнезда, устроенные на ветвях деревьев, кустарников. На зиму делает поры среди корней деревьев, уходящие глубоко под поверхность почвы. Поедает различные ягоды, плоды, орехи, желуди, семена, почки, кору молодых побегов, а также насекомых, яйца птиц, птенцов и мелких грызунов. Самка приносит 1, а в некоторых районах 2 помета в год. Число детенышей в помете 3—7. Местами вредит плодовым насаждениям.
Рис. 141. Японская сопя—G. japotucus
Род японских сонь
Genus Glirulus Thomas, 1905
В роде единственный вид: японская соня— G. japonicus Schinz, 1845.
Размеры мелкие, длина тела до 8 см. Длина хвоста до 5,5 см. В общем сходна с садовой соней (рис. 141). Морда укороченная. Уши относительно небольшие. .Хвост уплощен ог основания до копчика. Волосяной покров высокий, густой, мягкий. Окраска его бледно-оливково-коричневая с темно-коричневой или черной продольной полосой по середине спины. Перед ушами располагается по пучку удлиненных волос. Сосков 4 пары.
В черепе характерны маленькие уплощенные костные слуховые барабаны.
Распространена в Японии на о-вах Хонсю, Сикоку и Кюсю.
Населяет горные леса на высоте 400—1800 м над уровнем моря. Иногда поднимается почти до 3000 м над уровнем моря. Ведет древесный образ жизни. Активна ночью. День проводит в дуплах деревьев. Иногда устраивает гнезда на ветвях кустов и деревьев. На холодное время года впадает в спячку. Питается плодами, в течение года бывает один, иногда два помета. Обычно самка приносит 3—4 (редко до 7) детеныша в июне или в июле (второй помет возможен в октябре). Длительность беременности примерно равна месяцу.
Род мышевидных сонь
Genus Myominius Ognev, 1924
В роде единственный вид: мышевидная соня — М. personatus Ognev, 1924.
Размеры мелкие. Длина тела до 10,2 см. Длина хвоста до 7,8 см. Хвост в отличие от других представителей семейства покрыт редкими
короткими волосками, не прикрывающими чешуйки кожи. Задние конечности лишь не намного длиннее передних. Стопа узкая и длинная. Мозоли в основании пальцев передних конечностей развиты относительно слабо. На обеих конечностях самые длинные пальцы — третий и четвертый. Уши короткие, округлые, густо покрытые короткими, темными волосками. Окраска верхней стороны тела серая или охристо-серая. По средней линии ее проходит широкая размытая темная полоса. Брюхо белое. Хвост неясно двухцветный, сверху той же окраски, что и спина, снизу беловатый.
Череп с округлой, несколько уплощенной сверху мозговой коробкой. В межглазничной области имеется небольшое продольное углубление. Костные слуховые барабаны крупные. Резцовые отверстия удлиненные. Подглазничное отверстие щелевидное, вытянутое вертикально.
Распространена в Юго-Восточной Болгарии, а также в СССР—в Южной Туркмении.
Обитает в каменистой кустарниковой горной степи, где обнаружена примерно на высоте 900 м над уровнем моря. Ведет наземный образ жизни. Экология не изучена.
Род африканских сонь
Genus Graphiurus Smuts, 1832
Размеры от мелких до крупных. Длина тела 7,5—16,5 см. Длина хвоста 5,8—13,5 см. Масса около 30 г. Хвост довольно густо покрыт волосами, несколько удлиняющимися в его терминальной части (рис. 142). Задние конечности с пятью пальцами, причем пятый относительно длинный, а первый короткий. Стопа приспособлена к лазанию по деревьям, укороченная, широкая. Волосяной покров у одних видов очень мягкий, у других немного грубее, у третьих довольно грубый. Окраска спинной стороны тела от серой до темно-серой или темно-желтой и рыжевато-коричневой. Брюшная сторона белая или сероватая, часто с желтоватым или рыжеватым оттенком. На морде есть черные и белые отметины. Конец хвоста обычно черный или темно-бурый. Сосков 3—4 пары.
Мозговой отдел черепа гладкий и округлый. В области лобных костей череп сужен. Костное небо относительно широкое. Костные слуховые барабаны крупные, вздутые.
Распространение охватывает большую часть Африки от Сахары и Судана до южной оконечности материка.
Населяют леса, саванны, скалистые районы, сухие плато — обычно вдоль водоемов, антропогенные ландшафты. Поселяются и в строениях человека. Активны ночью, но в густых лесах часто и днем. Убежищем служат дупла деревьев, гнезда среди ветвей, иногда расщелины скал, пустоты под камнями. Питаются различными семенами, орехами, плодами, насекомыми, яйцами змей и молодыми змеями, ящерицами и птицами. Устраивают запасы корма (обычно орехов). В некоторых частях ареала на зиму впадают в спячку. Самки приносят детенышей в период с октября по январь. Длительность беременности около 24 диен. В помете обычно от 2 до 5 детенышей.
В роде, по-видимому, 5 видов:
южноафриканская соня—G. ocularis Smith, 1829 (провинции Южно-Африканской Республики — Трансвааль, Оранжевая, Капская);
G. monardi St. Leger, 1936 (Северная Ангола и Северо-Западная Замбия, возможно, также низовье р. Конго и Западная Катанга в Заире);
Рис. 142. Craphiurus murinus
G. hueti Rochebrune, 1883 (от Сенегала до Центрально-Африканской Республики);
G. platyops Thomas, 1897 (Южно-Африканская Республика, Замбия, Южная Малави, Восточная Ангола, юг Заира);
G. murinus Desmarest, 1822 (Африка к югу от Сахары)
Некоторые исследователи насчитывают в роде большее число видов; например, Эллермэн (1940) —39, Уокер (1964) —20, Андерсон и Джонс (1967) — 17.
СЕМЕЙСТВО КОЛЮЧИХ СОНЬ
FAMILIA PLATACANTHOMYIDAE MILLER ET GIDLEY, 1918
По внешнему виду напоминают сонь (рис. 143). Размеры мелкие. Длина тела 7—21,2 см. Длина хвоста 7,5—13,5 см. Базальная часть
хвоста покрыта короткими волосами, постепенно удлиняющимися в терминальной половине, где они образуют пышную кисточку. Задние конечности длиннее передних. Первый палец на задних конечностях силь-
но укорочен. Когти сжатые с боков, небольшие. Первый палеи передних конечностей лишен волос. У китайской сони волосяной покров мягкий, а у колючей сони на спинной стороне много твердых уплощенных колючек. Окраска спинной стороны серая пли рыжевато-коричневая, брюшной — белая или бело-серая.
Лицевой отдел черепа несколько укорочен. Мозговая капсула округлая. Лобно-теменные гребни имеются. Скуловые дуги слабо расставлены в стороны. Костные слуховые барабаны небольшие Зубная форму-. 1 0 0 3 ,*,,
ла: i — с — р — т — = 16.Коренпые зубы с высокими коронками, на
жевательной поверхности имеют поперечно расположенные эмалевые гребни. Первый коренной зуб крупнее прочих. Слепой кишки пет или опа развита слабо.
Рис. 143. Колючая соня — Р. lasiurus
Распространение охватывает Южную Индию, Юго-Восточный Китай и северную часть п-ова Индокитай.
Населяет скалистые, поросшие лесом долины или горные леса. Убежищем служат дупла деревьев или расщелины скал. Питаются различными растительными объектами. Экология изучена слабо.
В семействе 2 рода (2 вида):
колючих сонь — Plata-canthomys Blyth, 1859 (1 вид);
китайских сонь — Ту-phlomys Milne - Edwards, 1877 (I вид).
Некоторые ученые (например, Эллермэн, 1940) включают колючих сонь в качестве подсемейства в семейство сонь, а другие (например, Вуд, 1955) — в качестве подсемейства в семейство Cricetidae. По-видимому, правильнее считать колючих сонь отдельным семейством.
Род колючих сонь
Genus Platacanthomys Blyth, 1859
В роде единственный вид: колючая сопя — Р. lasiurus Blyth, 1859.
Размеры крупные. Длина тела 13—21,2 см. Длина хвоста 7,5—10 см. Стопа очень широкая. Ее первый палец сильно редуцирован и не имеет когтя. Морда заостренная, глаза небольшие (рис. 143). Уши относительно крупные, лишены волос. Волосяной покров спины и верхних частей боков имеет многочисленные, острые, уплощенные колючки, между которыми находятся тонкие и мягкие волосы. Спинная сторона тела светло-рыжевато-коричневая, брюшная — грязно-белая. Конечности беловатые. Хвост сверху несколько темный, и только самый кончик хвоста белый.
В черепе мозговой отдел широкий, с очень крупной межтеменной костью. Лобные кости широкие. Лобпо-темепные гребни крупные. Костное небо широкое. Резцовые отверстия смещены далеко вперед. Костные слуховые барабаны очень маленькие. Нижняя челюсть с низким венечным отростком. Угловой отросток повернут несколько внутрь.
Распространена в Южной Индии.
Населяет горные долины на высоте 600—900 м над уровнем моря. Экология изучена слабо. Ведет в основном древесный образ жизни. Делает гнезда из листьев и мха в дуплах деревьев, среди ветвей и в расщелинах скал. Питается плодами, семенами, корнями растений.
Род китайских сонь
Genus Typhlomys Milne-Edwards, 1877
В роде единственный вид: китайская соня — Т. cinereus Milne-Edwards, 1877.
Размеры мелкие. Длина тела 7—9,8 см. Длина хвоста 9,6—13,5 см. Внешне сходна с мышами. Хвост покрыт чешуйками и в основной части лишен волос. В терминальной части хвоста волосы образуют небольшую кисточку. Стопа не очень сильно расширена; пятый палец относительно длинный. Задние конечности удлинены. Когти на всех пальцах небольшие, сжатые с боков. Уши вытянутые, слабо оволосненные. Глаза очень маленькие. Волосяной покров плотный, густой, мягкий, без колючек. Окраска на спинной стороне тела серая, на брюшной светло-серая. Конечности беловатые. Конец хвоста белый.
В черепе мозговой отдел округлый. Межглазничного сужения почти нет. Костные слуховые барабаны маленькие. Венечный отросток нижней челюсти низкий. Угловой отросток сильно уменьшен и повернут внутрь. Коренные зубы узкие. Имеется небольшая слепая кишка.
Распространена в Юго-Восточном Китае и северной части п-ова Индокитай.
Населяет леса на горных склонах, встречаясь на высоте 1200—2100 м над уровнем моря. Экология не изучена.
СЕМЕЙСТВО СЕЛЕВИНИЙ
FAMILIA SELEVINIIDAE ARGYROPULO ET VINOGRADOV, 1939
Размеры мелкие. Длина тела до 95 мм. Длина хвоста до 75 мм. Хвост тонкий, покрыт густыми короткими волосами и мелкими кольцеобразными чешуйками. Передние четырехпалые и задние пятипалые, конечности примерно одинаковы по длине (рис. 144). Конечности по строению сходны с таковыми представителей семейства сонь, но подошвенные мозоли развиты слабее и пальцы относительно более короткие, особенно пятый палец задних ног. Пальцы вооружены небольшими когтями. Уши довольно крупные, подвижные, способные складываться в трубку. Волосяной покров очень густой, мягкий. Окраска спинной стороны сероватая, брюшной — беловатая. Подошвы передних и задних конечностей голые.
Мозговая коробка в черепе округлая. Лицевой отдел несколько укорочен. Скуловые дуги слабо расставлены в стороны. Надглазничных отростков и заглазничных бугров нет. Межглазничная область имеет небольшое продольное углубление. Лобно-теменные и сигиттальный гребни отсутствуют. Ламбдоидальный гребень иногда встречается, ио развит слабо. Подглазничные отверстия небольшие. Часть передней доли жевательной мышцы проходит через короткий подглазничный капал. Костные слуховые барабаны очень крупные, тонкостенные. Венечный отросток нижней челюсти хорошо развит. Зубная формула:
1 1 0 — 2 3 , с
— с — р-------m — = 16 — 20.
0 0 3
Рис. 144. Сслевиния — S. betpakdalensis и ее череп
Щечные зубы очень маленькие, с низкими коронками; имеют корни. Коренные зубы обычно выпадают и у взрослых зверьков частично или полностью отсутствуют. Второй коренной зуб верхней и нижней челюстей самый крупный. Жевательная поверхность зубов гладкая, несколько углубленная в средней части. Верхние резцы большие, с глубоким желобо-образным углублением, проходящим вдоль зуба по его наружной поверхности и с углообразпым вырезом на нижнем крае. Нижние резцы тонкие.
Распространена только в пределах СССР, в центральной и восточной частях Казахстана: в пустыне Бетпак-Дала, северном Прибалхашье и в котловине озера Алакуль.
Населяет щебнистые, глинистые, солончаковые пустыни с преобладанием полыней и солянок, особенно боялыча (Salsola larictfolia). На зиму впадает в спячку обычно с начала октября по февраль включительно. Активна вечером и утром. Подвижный зверек, передвигающийся
прыжками. Может хорошо лазать по кустам. Убежищем служат, по-видимому, норы различных грызунов. Питается в основном насекомыми, в особенности саранчовыми. Беременные и лактирующие самки встречались с конца мая до конца июля. Детенышей в помете до 8. Редко встречающийся и мало изученный грызун.
В семействе 1 род: селевиний — Selevinia Belosludov et Bashanov, 1938, и 1 вид.
Род селевиний
Genus Selevinia Belosludov et Bashanov, 1938
См. описание семейства.
В роде 1 вид: селевиния — S. betpakdalensis Belosludov et Bashanov, 1938 (ареал совпадаете ареалом семейства).
СЕМЕЙСТВО МЫШОВОК
FAMILIA ZAPODIDAE COUES, 1875
Мелкие грызуны. Длина тела 5—10 см. Длина хвоста 6,5—16 см. Масса от-6 до 28 г. Внешним видом сходны с мышами или несколько напоминают тушканчиков. Мордочка удлинена. Уши небольшие с округлыми вершинами; обычно покрыты тонкими и редкими волосами. Глаза маленькие. Задние конечности удлинены у Eozapus, N apaeozapus и Zapus и слабо удлинены у Sicista. Передние и задние конечности пятипалые. Пальцы хорошо развиты, с когтями. Хвост покрыт короткими и редкими волосами. У Sicista хвост полухватательный. Имеются внутренние защечные мешки. Волосяной покров низкий, грубоватый, окраска на спине серая или коричневая, на брюхе белая или желто-белая. Часто по средней линии спины проходит продольная темная полоса. Сосков 4 пары.
В черепе мозговая коробка овальная. Межглазничное сужение выражено слабо. Имеется крупная межтеменная кость. Слезные кости маленькие. Костные слуховые барабаны небольшие, невздутые. Нижняя челюсть слабая, в ней хорошо развит венечный отросток. Подглазничное отверстие большое. Сквозь него проходят жевательная мышца и нерв. Зубная формула: i— с — Р 16—18.Щечные зубы брахио-
донтного (Sicista) или полугипселодонтного (Zapus) типа. Шейные позвонки и метатарзальные кости не срастаются между собой.
Распространение охватывает Северную и Восточную Европу, Центральную и Восточную Азию, а также Северную Америку от арктической Канады на юг примерно до 35° с. ш.
Населяют лесную, лесостепную и степную зоны; в горах поднимаются до альпийского пояса включительно — до 3000 м над уровнем моря. При движении некоторые представители семейства способны делать большие прыжки до 2—4 м длиной. Длинный хвост служит балансиром. Представители родов Sicista и Napaeozapus могут хорошо лазать по низкорослым растениям, а представители рода Zapus — хорошие пловцы. Активны в основном ночью. Держатся обычно поодиночке или парами. Убежищем служат норы, которые они выкапывают сами, или занимают норы других грызунов. Иногда поселяются под упавшими стволами деревьев. Осенью становятся жирными и па зиму впадают в спячку, длящуюся 6—8 месяцев. Питаются различными растительными объектами, а также мелкими беспозвоночными, главным образом насекомыми.
В течение года самка приносит 1—2 помета; в каждом от 1 до 8 детенышей. Длительность беременности от 18 до 35 дней.
В семействе 4 рода (11 видов):
мышовок — Sicista Gray, 1827 (6 видов);
полутушканчиков — Zapus Cones, 1875 (Звида);
китайских мышовок — Eozapus Preble, 1899 (1 вид);
полумышей — Napaeozapus Preble, 1899 (1 вид).
В СССР представители рода мышовок —Sicista.
Некоторые исследователи (например, Эллермэн, 1940, Виноградов и Громов. 1952, Громов и др., 1963) включают мышовок в качестве подсемейства в семейство тушканчиков. По-видимому, более правильно рассматривать мышовок в качестве самостоятельного семейства.
Род мышовок
Genus Sicista Gray, 1827
Длина тела 5—9 см. Длина хвоста 6,5—11,6 см. Внешний вид мышеобразный (рис. 145). Задние конечности не более чем в два раза пре вышают длину передних. Первый палец на задней конечности очень мал Стопа укороченная.
Окраска спины от светло- до темно-коричневой или коричневато желтой Степная и лесная мышовки имеют резко выраженную темную почти черную полосу, проходящую вдоль тела по средней линии спины Форма черепа напоминает таковую мышей. Наибольшая скуловая ширина приходится на середину или передний отдел скуловых дуг. Ниж ний и верхний края подглазничного отверстия находятся в одной верти калькой плоскости. Верхние предкорепные зубы имеются.
Хромосом в диплоидном наборе у степной и лесной мышовок по 32, а у рыжей — 42.
Распространение охватывает лесную, лесостепную и степную зоны, а также горы Евразии от Норвегии, Швеции и ФРГ до Тихого океана, на юг до Кавказа, Тянь-Шаня и Гималаев.
Населяют лесную, лесостепную и степную зоны, а в горах — лесной, субальпийский и альпийский пояса. Активны в сумерках. Убежищем служат норы мышевидных грызунов, пустоты между корнями деревьев и кустарников, трухлявые пни. При понижении температуры впадают в оцепенение, зиму проводят в спячке. Питаются семенами, ягодами, сочными надземными и подземными частями растений, а также насекомыми и другими беспозвоночными. В течение года, по-видимому, быващ один помет. Число детенышей 3— 7. Стенная мышовка размножается в конце мая — начале июня, кавказская — в конце июля — начале августа, тяньшанская — в первой половине июля.
В роде, по-видимому, 6 видов:
степная мышовка — S. subtilis Pallas, 1773 (Чехословакия, Румыния, Польша; СССР — северная граница проходит по Северной Украине, Воронежской области, Татарии, Южному Уралу, Омской, Новосибирской и Иркутской обл.; южная граница — по побережью Черного и Азовского морей, предгорьям Крыма, Предкавказью, северному побережью Каспия, Северному Казахстану от р. Эмбы через Тургай, район г. Караганды, котловину Алакуля; также в кузнецких, минусинских и прибайкальских степях);
лесная мышовка-—S. betulina Pallas, 1779 (Норвегия, Швеция, Дания, ГДР, Венгрия, Чехословакия; в СССР — к северу до Карелии, Архангельской обл., низовьев Печоры, среднего течения Таз, на юг до Северной Украины, Северного Кавказа, Северного Казахстана; на восток —-до Приморья включительно);
серая мышовка — S. pseudonapaea Strautman, 1949 (СССР — Южный Алтай);
рыжая мышовка — S. параеа Hollister, 1912 (СССР — горы и предгорья Алтая);
одноцветная мышовка — S. concolor Buchner, 1892 (СССР — о-в Сахалин, Приморский край, Тянь-Шань; Китай — Сычуань, Ганьсу, Синьцзян, Индия — Кашмир);
кавказская мышовка — S. caucasica Vinogradov, 1925 (СССР — главный Кавказский хребет и Армения).
В СССР: все указанные виды.
Некоторые исследователи выделяют из вида одноцветная мышовка в качестве особых видов S. caudata Thomas, 1907 (например, Громов и др„ 1963; Эллермэн и Моррисон-Скотт, 1966), и S. tianschanica Salen-sky, 1903 (Громов и др., 1963). А- А. Громов и другие включают серую мышовку в качестве подвида лесной мышовки. А. П. Кузякин (см. Бобринский и др., 1965) предполагает возможность объединения рыжей, серой, кавказской и одноцветной мышовок в один вид.
Род полутушканчиков
Genus Zapus Coues, 1875
Длина тела 7,6—11 см. Длина хвоста 11—16,5 см. Масса 11—32 г. Задние конечности значительно длиннее и сильнее передних (рис. 146). Стопа узкая; три средних пальца длиннее; пятый палец достигает основания четвертого, а передний палец сильно укорочен. Кисть также узкая; третий и четвертый пальцы слегка длиннее второго и пятого, первый палец рудиментарен. Длинный хвост покрыт редкими или средней густоты волосами, сквозь которые видны кожные чешуйки. На конце хвоста волосы несколько удлинены. Имеются защечные мешки. Волосяной покров низкий, густой. Окраска его серовато-коричневая па спине, желтовато-коричневая на боках и белая на брюхе. Хвост темно-коричневый сверху и желтовато-белый снизу. Сосков 4 нары.
Резцовые отверстия черепа крупные. Верхние резцы узкие, имеют по продольной бороздке на наружной поверхности. Верхние предкоренпые небольшие. Нижние резцы гладкие. Третий коренной меньше других коренных.
Хромосом в диплоидном наборе у лугового и западного полутушканчиков по 72.
Распространение охватывает лесную зону Северной Америки, Аляски и от Канады на юг до северной Каролины, Миссури, Ныо-Мексико и Калифорнии.
Рис. 146. Луговой полутушканчик и его череп
Населяют влажные районы лесов пли безлесные травянистые равни ны, предпочитая берега водоемов. Активны в сумерки и ночью. Передвигаются прыжками до 2—2,5 м, иногда до 4 м длиной. Хвост используется как балансир. Некоторые виды (луговой полутушканчик) хорошо плавают и даже ныряют. Питаются в основном семенами, а также плодами, ягодами и насекомыми, иногда делают запасы в гнезде. Летом зверьки строят наземное гнездо округлой формы из различных растений. В остальное время года сооружают гнезда под упавшими стволами деревьев или под землей в норах, иногда почти на 2-метровоп глубине. Осенью становятся очень жирными и па холодный период года впадают в спячку (луговой полутушканчпк — с конца сентября — октября по май) Даже в летнее время при похолодании впадают в оцепенение. Обычно в году один помет, иногда 2, даже 3. Дли тельноегь беременное тп 13— 19 дней. В помете от 2 до 8 детенышей — обычно с мая по сентябрь. Глаза у детенышей открываются на 22—25-й день. К самостоятельной жизни переходят в возрасте 26—33 дней. Самки в возрасте 2 месяцев становятся половозрелыми.
В роде 3 вида:
луговой полутушканчик — Z. hudsonius Zimmermann, 1780 (южная половина Аляски, включая п-ов Аляска; провинции Канады — южная по-372
ловина Юкона, юг Маккензи, Британская Колумбия, кроме Тихоокеанского побережья, крайнего юга и юго-востока, северная половина Альберта, северная половина и крайний юго-восток Саскачевана, Манитоба, кроме крайнего северо-востока, Онтарио, Квебек, кроме севера и северо-запада, Ньюфаундленд, на о-ве Ньюфаундленд отсутствует, Нью-Брансуик, Новая Шотландия; США — восточную границу ареала образует атлантическое побережье на юг до штата Северная Каролина, южная граница ареала, пересекая юго-восточную часть этого штата, идет на юго-запад через центральные районы Южной Каролины, середину штата Джорджия, доходит до центра Алабамы и круто сворачивает к северу до центра Теннесси, откуда вновь поворачивает на запад, отсекает юго-восток этого штата, проходит по границе между штатами Арканзас и Миссури, захватывает крайний северо-восток Оклахомы, сворачивая на северо-запад, пересекает центральные районы Канзаса и юго-запад Небраски и от северо-западного угла этого штата круто поворачивает на юг и вдается языком в центр штата Колорадо, откуда сворачивает на север, образуя западную границу ареала, которая проходит с юга на север по середине штата Вайоминг, отсекает юго-восточный угол Монтаны и уходит за государственную границу);
западный полутушканчик — Z. princeps J. Allen, 1893 (провинции Канады — центральные районы юга Юкона, Британская Колумбия, кроме крайних северо-востока и юго-запада, южная половина Альберта, южная треть Саскачевана, крайний юго-запад Манитобы; штаты США — Северная Дакота, кроме юго-запада, крайний северо-восток Южной Дакоты, Монтана, кроме юго-востока, Вайоминг, кроме северо-востока, центральные районы Колорадо от северной границы штата до южной, узкая изогнутая полоса, проходящая от середины северной границы штата Нью-Мексико до ее центральных районов, поворачивающая на запад и заходящая небольшим выступом в центральные районы востока Ари-зоны, центральные районы и север Юты и Невады, Айдахо, крайние северо-восток и юго-восток штата Вашингтон; восточная половина Ориго-на, северо-восток Калифорнии);
тихоокеанский полутушканчик — Z. trinotatus Rhoads, 1895 (Канада— крайний юго-запад провинции Британская Колумбия; США — западная половина штата Вашингтон, узкая полоса вдоль Тихоокеанского побережья штата Орегон с небольшим выступом с севера в центральные районы этого штата, узкая полоса вдоль Тихоокеанского побережья штата Калифорния от северной границы штата на юг до Сан-Франциско) .
Эллермэн (1940) и Уокер (1964) насчитывают в роде 11 видов.
Род китайских мышовок
Genus Eozapus Preble, 1899
В роде единственный вид: китайская мышовка — Е. sctchaanus Pou-sargues, 1896.
Длина тела 8—10 см. Длина хвоста 10—15 см. Задние конечности значительно длиннее передних. Внешним обликом напоминают полутушкан-чиков, отличаясь от них окраской. Верхняя сторона тела рыжевато-коричневая, нижняя белая с медиальной темно-желтой полосой. Хвост темный сверху и белый снизу; копчик его белый.
Череп также сходен с таковым полутушканчиков. Отличия имеются в характере жевательной поверхности щечных зубов. Имеются предко-репные зубы. Верхний четвертый коренной зуб сильно уменьшен.
Рис 147. Полумышь — N. insignis
Распространена в провинциях Сычуань и Ганьсу в Китае.
Населяет высокогорные леса, встречаясь по берегам водоемов. Экология не изучена.
Некоторые исследователи (например, Симпсон, 1945) включают китайскую мышовку в состав рода полутушканчиков.
Род полумышей
Genus Napaeozapus Preble, 1899
В роде единственный вид: полумышь — N. insignis Miller, 1891 (рис.147).
Длина тела 8—10 см. Длина хвоста 11,5—16 см. Масса 17—31 г. Самки немного крупнее самцов. Внешним обликом сходна с полутушканчиками. Задние конечности значительно длиннее передних. Волосяной покров грубый. Спинная сторона тела желтовато-коричневая. Бока золотисто-желтые, рыжевато-желтые или темно-желтые. Брюшная сторона белая. Хвост двухцветный: серовато-коричневый сверху и белый снизу, обычно с белым кончиком.
По характеру жевательной поверхности щечных зубов отличаются от полутушканчиков, хотя большинство специфических черт в черепе сильнее выражено, чем у полутушканчиков. Подглазничное отверстие крупное, овальной формы. Зубная формула: i с — р — т -у- = 16.Резцы
оранжевые или желтые. Верхние резцы с продольной бороздкой на наружной поверхности.
Хромосом в диплоидном наборе 70 —72.
Распространена в провинциях Канады — па краппом юго востоке Манитобы, юге Онтарио, юге Квебека, юго востоке Ньюфаундленда (на о-ве Ньюфаундленд отсутствует), Ныо-Брапсупкс, Новой Шотландии; в штатах США — Мэн, Ныо-Гэмпшир, Вермонт, Нью-Йорк, Массачузетс, северо-западе Коннектикута, севере Нью-Джерси, Пенсильвании, кроме юго-востока, северо-востоке Огайо, восточной половине Западной Виргинии, крайнем западе Мэриленда, западе Виргинии, крайнем юго-востоке Кентукки, крайнем востоке Теннесси, западе Северной Каролины, крайнем северо-западе Южной Каролины, крайнем северо-востоке
Джорджии, северо-восточной половине Мичигана, северной половине Висконсина, на северо-востоке Миннесоты.
Населяет леса, предпочитает опушки полян около водоемов. Держатся группами. Образ жизни ночной. Убежищем служат норы, которые выкапывают сами или используют чужие норы других грызунов или насекомоядных. Иногда устраивают гнезда из травы под упавшими стволами деревьев. Питается семенами, плодами, мелкими беспозвоночными. Пищу не запасает. Хорошо плавает. Может лазать по ветвям кустов. Осенью сильно жиреет и впадает в спячку: обычно с сентября — ноября по конец апреля или начало мая. В течение года дают один или два помета; первый в июне или июле, второй в августе. Длительность беременности 23—29 дней. В помете от 2 до 7, обычно 4 детеныша. Новорожденные массой около 1 г, голые, слепые, без вибрисс. Глаза открываются на 26-й день.
СЕМЕЙСТВО ТУШКАНЧИКОВЫХ
FAMILIA DIPODIDAE WATERHOUSE, 1842
Размеры мелкие. Длина тела 4—26 см. Длина хвоста 7 — 30,8 см *. Внешность весьма характерная с удлиненными, сильными задними конечностями (рис. 148). Тушканчики передвигаются при медленном движении на всех четырех ногах, а при быстром — только на двух задних. Внутренние и наружные метатарзальные кости уменьшены или их нет вовсе, а три средние метатарзальные кости сливаются между собой в так называемую «цевку» (за исключением представителей родов Саг-diocranins и Salpingotus). Задние конечности пятипалые. Когти па пальцах хорошо развиты, причем на пальцах задних конечностей они обычно длиннее, чем на передних. Длинный хвост часто имеет на конце уплощенную кисточку из удлиненных волос — так называемое «знамя». Хвост служит балансиром при прыжках. Уши, как правило, большие, покрыты редкими волосами, иногда в основании срослись в трубку. Глаза большие. Волосяной покров довольно густой и мягкий. Окраска спинной стороны тела обычно однотонная, коричневатая или охристопесчаная. У некоторых тушканчиков темные продольные полосы проходят по спине или есть светлые пятна на бедрах. У видов, живущих в песчаных районах, жесткие и упругие волосы на стопе могут образовывать своеобразную «щетку», увеличивающую площадь опоры при движении. Сильно развитые вибриссы могут достигать длины тела.
В черепе, как правило, лицевой отдел несколько укорочен. Подглазничный канал крупный: сквозь него проходит жевательная мышца. Костные слуховые барабаны большие. Нижняя челюсть относительно слабая. Зубная формула: t с р °— 1 т = 16 — 18. Щечные зубы имеют корни; их коронки средней высоты. Верхний четвертый предкорепной зуб небольших размеров или его нет совсем. Шейные позвонки могут срастаться между собой. Малая и большая берцовые кости срастаются в дистальных отделах.
Распространение охватывает Северную Африку, южную часть Восточной Европы, Малую Азию, Северный Китай, МНР.
Населяют лесостепи, степи, пустыни и полупустыни. При быстром передвижении тушканчики могут делать большие прыжки длиной до 3 м.
1 В пределах семейства можно выделить три группы тушкапчиковых: мелкие с длиной тела 4—11 см, средние—12—19 см, крупные — 20—26 см.
Рис 148 Четырехпалый тушканчик — Л tetradactyla
Активны ночью. День проводят в выкопанных ими порах. На зиму большинство видов впадают в спячку. Питаются подземными и надземными частями различных растений. Поедают также насекомых. В некоторых районах могут причинять вред бахчевым культурам. У многих видов бывает 2 помета в год. Длительность беременности примерно 25—42 дня. Детенышей в помете от 2 до 6.
В семействе 10 родов (27 видов)-
земляных зайцев — Allactaga F. Cuvier, 183G (включая Scarturus Gloger, 1841) (10 видов);
земляных зайчиков—• Alactagulus Nehring, 1897 (1 вид), толстохвостых тушканчиков — Pygerettnus Gloger, 1841 (3 вида); емуранчиков — Stylodipus G. Allen, 1925 (1 вид);
мохноногих тушканчиков — Dipus Zimmermann, 1780 (1 вид);
гребнепалых тушканчиков — Paradipus Vinogradov, 1930 (1 вид);
песчаных тушканчиков — Jacidus Erxleben, 1777 (включая Eremodi-pus Vinogradov, 1930) (5 видов);
пятипалых карликовых тушканчиков — Cardiocranius Satunin, 1902 (1 вид);
трехпалых карликовых тушканчиков — Salpingotus Vinogradov, 1924 (3 вида);
длинноухих тушканчиков — Euchoreules Sclatcr, 1891 (1 вид).
В СССР: представители всех родов, за исключением длинноухих тушканчиков.
Род земляных зайцев
Genus Allactaga F. Cuvier, 1836
Размеры от мелких до наибольших в семействе. Длина тела 9,5— 26 см. Длина хвоста 16—30,8 см. Лицевая часть головы несколько укорочена. Нос спереди приплюснут в виде «пятачка» Глаза крупные Уши
длинные, не образуют в основании трубку. Передние и задние конечности пятипалые (только четырехпалый тушканчик имеет на задних конечностях по 4 пальца). Первый палец на передних конечностях и боковые пальцы на задних укорочены. Нижняя поверхность ступни и пальцев задней конечности, как правило, не покрыта волосами. «Знамя» на хвосте развито хорошо. Жировых отложений в хвосте не бывает.
Волосяной покров средней,высоты, мягкий, густой. Окраска спинной с-ороны тела от песчаной или серовато-темно-желтой до красновато-коричневой или черноватой. Брюшная сторона беловатая.
В черепе лицевой отдел укорочен. Мозговая коробка очень широкая, крупная. Лобные кости широкие. Слезные кости большие. Подглазничные отверстия очень велики. Костные слуховые барабаны относительно тт 4
невелики. Число щечных зубов -5-, они полугипселодонтного типа, со О
сложной жевательной поверхностью. Нижняя челюсть сравнительно длинная,низкая.
Хромосом в диплоидном наборе у всех изученных видов по 48.
Распространение охватывает Северный Египет, северную часть Аравийского п-ова, Малую Азию, Кавказ, юг европейской части СССР, Северный Иран, Афганистан, Среднюю Азию, Казахстан и территорию далее на восток до Алтая, МНР и Северного Китая.
Населяют степи, степные участки лесостепи, полупустыни и пустыни. Поднимаются в горы до 3—3,5 тыс. метров над уровнем моря. Обычно роют два типа нор: простые временные и постоянные — летние и зимовочные, значительно более глубокие и сложные. В постоянных норах на глубине 40—60 см устроены одна или несколько гнездовых камер. Продолжительность зимней спячки 4—7 месяцев. Питаются клубнями, луковицами, вегетативными частями растений и их семенами. Некоторые виды поедают также насекомых. Как правило, приносят один помет в году, а некоторые виды —два. Детенышей в помете от 2 до 8. Период размножения растянут. Местами причиняют вред сельскому хозяйству. Носители возбудителей чумы.
В роде 10 видов:
тушканчик-прыгун — A. sibirica Forster, 1778 (СССР — от восточного побережья Каспийского моря и Приуралья до южного Забайкалья; МНР; Северного и Северо-Восточного Китая; изолированный участок ареала имеется в горах восточной Киргизии);
малый тушканчик —A. elater Lichtenstein, 1825 (полупустыни Азербайджана, горные степи Армении, Восточное Предкавказье, Калмыкия, Южное Заволжье, Казахстан, равнины Средней Азии, а также Джунгария, юго-запад МНР и северо-восток Синьцзян-Уйгурского автономного района Китая, Северный Иран, Афганистан, Пакистан, Белуджистан);
евфратский тушканчик — A. euphratica Thomas, 1881 (Ирак, Трансиордания, Сирия, северо-восток Аравийского п-ова);
горный тушканчик — A. williamsi Thomas, 1897 (СССР — Закавказье; Малая Азия, Иран, Афганистан);
тушканчик Хотсона — A. hotsoni Thomas, 1920 (юго-восток Ирана); монгольский тушканчик — A. bullata G. Allen, 1925 (МНР);
тушканчик Бобринского — A. bobrinskii Kolesnikov, 1937 (СССР — пустыни Средней Азии);
тушкачик Северцова — A. severtzovi Vinogradov, 1925 (СССР — равнинные пустыни Южного Казахстана и Средней Азии к северу до северного Устюрта, Приаральских Каракумов, низовьев р. Чу, озера Балхаш, на юг до юго-западной Туркмении, Сарыкамышских озер, низовьев Амударьи);
большой тушканчик — A. major Kerr, 1792 (СССР — от Николаева, Кривого Рога, Умани, правобережья Днепра в районе Черкасс, Киева и Новозыбкова на западе до Алтая и Джунгарского Алатау на востоке, на север до Чернигова, Брянска, Калуги, Горького, Казани, нижней Камы, р. Белой, Южного Урала, Свердловска, южных районов Тюменской и Омской обл., Новосибирска и южного Алтая; на юг до северного побережья Черного моря, северного предгорья Крымских гор, побережья Азовского моря, Кавказского хребта, побережья Каспийского моря, Устюрта, юго-восточного берега Аральского моря, р. Сырдарьи, северного предгорья Тянь-Шаня);
четырехпалый тушканчик — A. tetradactyla Lichtenstein, 1823 (APE, Ливия).
В СССР: тушканчик-прыгун, малый, горный, Бобринского, Северцова и большой тушканчик.
Род земляных зайчиков
Cenus Alactagulus Nehring, 1897
В роде единственный вид: земляной зайчик — A. pygmaeus Pallas, -1779.
Размеры мелкие. Длина тела до 12 см. Длина хвоста до 13 см. Внешне сходен с мелкими представителями рода земляных зайцев. В отличие от последних уши у земляного зайчика несколько короче, голова более широкая и укороченная. «Знамя» на хвосте выражено слабее. Передние и задние конечности пятипалые. Боковые пальцы задних конечностей укорочены. Подошва стопы без волос, но окаймлена ими по краю. Окраска спинной стороны тела песчано-желтая с черноватым оттенком. Брюхо белое.
Череп сходен с таковым мелких представителей рода земляных зайцев, однако в отличие от последних в верхней челюсти всего три коренных зуба.
Хромосом в диплоидном наборе 48.
Распространен в Восточном Предкавказье, Нижнем Поволжье, Казахстане (на север до Чапаевска, Актюбинска, Тарбагатая), Средней Азии (кроме гор и Ферганской долины), Л1НР, Северном Китае.
Населяет пустыни и полупустыни, предпочитая солончаки и такыры. Роет норы двух типов: временные и постоянные. Постоянные норы могут быть значительной длины — до 6 м, с гнездовой камерой. В зимовочных норах обычно находится несколько «камер спячки» па разной глубине, не имеющих подстилки в отличие от гнездовой камеры. В весеннелетнее время питается в основном стеблями различных злаков и луковицами, осенью — семенами. В течение года дают два помета. Детенышей в помете 2—6. На зиму впадает в спячку, которая в теплые дни может прерываться.
Род толстохвостых тушканчиков
Genus Pygerethmus Gloger, 1841
Размеры мелкие. Длина тела 8—12,6 см. Длина хвоста 9,5—13,4 см. Внешне сходны с земляным зайчиком, однако отличаются от него несколько укороченной ступней, утолщением средней части хвоста (за счет жировой клетчатки) и отсутствием «знамени» па хвосте (рис. 149). В хвосте происходит отложение жира, и в осеннее время он сильно утол-
Рис. 149. Череп толстохвостого тушканчика — Р. platyurus
щается. Передние и задние конечности пятипалые. Окраска волосяного покрова песчано-бурая на спинной стороне тела и белая на брюшной.
Череп по строению сходен с таковым земляных зайчиков (рис. 149).
Хромосом в диплоидном наборе у всех трех видов рода по 48.
Распространены от низовьев р. Урал до южного Устюрта и Восточного Казахстана.
Населяют полупустыни и пустыни, предпочитая глинисто-щебнистые пустыни и солончаки. Экология изучена слабо. Тушканчик Житкова роет короткие и неглубокие временные и тоже неглубокие, постоянные норы длиной до 5—6 м. Питаются в основном зелеными частями растений. Период размножения растянут. Пометов в течение года, по-видимому, не менее двух; 2—4 детеныша в каждом. На зиму впадают в спячку.
В роде 3 вида:
тушканчик Житкова — Р. zhitkovi Kuznetzov, 1930 (Восточный Казахстан) ;
толстохвостый тушканчик — Р. platyurus Lichtenstein, 1823 (пустыни по нижнему течению р. Урал и по Эмбе, Устюрт, северное и северо-восточное побережье Аральского моря, степи по р. Иргизу и Тургаю);
тушканчик Виноградова — Р. vinogradovi Vorontzov, 1958 (Зайсан-ская котловина в Восточно-Казахстанской обл.).
Род емуранчиков
Genus Stylodipus G. Allen, 1925
В роде единственный вид: емуранчик обыкновенный — S. telum Lichtenstein, 1823.
Размеры мелкие. Длина тела до 12,5 см. Длина хвоста до 16,3 см. Уши средней длины, в основании трубчатые. Морда несколько вытянутая. Задние конечности трехпалые. Средний палец относительно длиннее, чем у других трехпалых тушканчиков. Жесткие, короткие, темные волосы образуют на стопе щетку. Па конце хвоста волосы образуют лишь широкую негустую кисточку без белого окончания. Окраска спинной стороны тела песчаная или темно-желтая.
В черепе крупные костные слуховые барабаны соприкасаются между собой в передних отделах. Верхний предкорепной имеется или отсутствует. На наружной поверхности верхних резцов бороздки нет.
Рис. 150. Мохноногий тушканчик — D sagitta
Хромосом в диплоидиом наборе 58
Распространен в СССР — Восточном Предкавказье, Нижнем Поволжье, Казахстане, а также в Алешкпнских песках на левом берегу Нижнего Днепра и в песках в низовьях Дона, а также в Китае —Джунгарии; МНР.
Населяет степи, песчаные, глинистые и щебнистые пустыни, полупустыни. Число выходных отверстий постоянных нор бывает от 1 до 6. Временные норы обычно просто устроены и не имеют гнездовой камеры. В летне-осенний период питается, по-видимому, преимущественно семенами. В году дает один помет — 3—4 детеныша. Период размножения сильно растянут, и беременные самки встречаются с середины марта до середины августа (большинство в апреле). Осенью перед спячкой сильно жиреют, причем жир откладывается и в хвосте. Начало спячки совпадает с первыми заморозками, а пробуждение — с началом вегетации растений. В некоторых районах могут причинять вред бахчевым культурам.
Род мохноногих тушканчиков
Genus Dipus Zimmermann, 1780
В роде единственный вид: мохноногий тушканчик — D. sagitta Pallas, 1773.
Размеры средние Длина тела до 14 см. Длина хвоста до 18,5 см.Морда несколько удлиненная (рис. 150). Уши средней длины, в основании
•сложены в трубку. Задние конечности трехпалые. Подошва стопы и нижняя поверхность пальцев несут щетку коротких, жестких волос. Длинный хвост имеет хорошо развитое «знамя». Жировых отложении в хвосте не бывает. Волосяной покров средней высоты, густой, мягкий. Окраска спинной стороны тела от бледной до охристо-коричневой, брюшной — белая.
В черепе скуловые дуги относительно слабо расставлены в стороны. Межглазничный промежуток широкий. Костные слуховые барабаны
4 большие, вздутые. Число щечных, полугипселодонтных зубов-^". В скелете задних конечностей средние метатарзальные кости срастаются в цевку. Фаланги боковых пальцев полностью редуцируются. Шейные по-1 звонки, за исключением первого, срастаются между собой.
Хромосом в диплоидном наборе 48.
Распространение охватывает пустыни и полупустыни Восточного Предкавказья, Нижнего Поволжья, Волго-Уральского междуречья, Казахстана, равнинных областей Средней Азии, Центральной Азии к востоку до Северного Китая.
Населяет, как правило, пески различного типа — от барханных до закрепленных. Строит норы с 1—3 запасными выходами. Зимовочные норы сложные, достигают в глубину 3 м. Питается травами, подземными частями растений, зелеными веточками кустарников, семенами. Дополнительным объектом питания служат насекомые. Хорошо лазает по веткам кустов. Длительность периода размножения географически варьирует. Обычно бывает один помет из 2—5 детенышей. В северных частях ареала на зиму впадает в спячку, в южных активен круглый год, за исключением особо суровых зим. Местами вредит посадкам саксаула.
Род гребнепалых тушканчиков
Genus Paradipus Vinogradov, 1930
В роде единственный вид: гребнепалый тушканчик — Р. ctenodactylus Vinogradov, 1929.
Размеры средние. Длина тела до 15 см. Длина хвоста до 22 см. По внешнему виду сходен с мохноногим тушканчиком (рис. 151). Уши относительно длинные. Средний палец задней конечности сильно удлинен. Жесткие короткие и длинные волосы на подошвах и пальцах задней конечности образуют двухъярусную щетку. Окраска спинной стороны тела желтовато-песчаная. В задних частях щек расположено по неяркому рыжеватому пятну. Брюхо белое.
В черепе скуловые дуги расставлены относительно нешироко. Кост-з
ные слуховые барабаны крупные, вздутые. Число щечных зубов—. Верхнего предкоренного нет.
Хромосом в диплоидном наборе 48.
Распространен в Каракумах и Кызылкумах.
Населяет различного типа пески от незакрепленных барханов до закрепленно-бугристых. Имеет временные и постоянные поры. Постоянные зимовочные норы достигают длины 4,5 м, а в глубину идут до 3 м, где оканчиваются гнездовой камерой. Питается в основном вегетативными частями растений. Животную пищу, как правило, не употребляет. Может лазать по ветвям кустарников, обгрызая и поедая ветки.
Рис. 151. Тушканчики А — гребнепалый, Р. ctcnodactylus, Б — пятипалый карликовый — Cardiocranius paradoxus,
Род песчаных тушканчиков
Genus Jaculus Erxleben, 1777
Размеры от мелких до средних. Длина тела 10—15 см. Длина хвоста 15—25 см. Уши довольно крупные. Передние конечности пятипалые, задние трехпалые со щетками жестких волос на подошвах. «Знамя» на хвосте развито относительно слабо. Волосяной покров густой, мягкий
Рис. 151. Продолжение. В — жирнохвостый тушканчик Salpingotus crassicauda
Окраска спинной стороны тела бледно пли темно песчаная Брюхо беловатое.
В черепе характерны очень широкие лобные кости, узкая лицевая часть и крупные слезные кости. Костные слуховые барабаны сильно вздуты. Верхние резцы имеют на наружной поверхности ио одной бо-роздке. Число щечных зубов—; коронки у них высокие. Бедренная кость н «цевка» относительно длинные.
Хромосом в диплоидном наборе у египетского и туркменского тушканчиков по 48.
Распространение охватывает Северную Африку, Аравийский п-ов, Переднюю и Среднюю Азию.
Населяют песчаные и глинисто-песчаные пустыни. Убежищем служат норы обычно длиной в 1,5—2 м, в глубину уходят па 50 60 см. Питаются преимущественно семенами. На зиму впадают в спячку. В период размножения дают, по-видимому, 2 помета. Детенышей в помете 2—3.
В роде 5 видов:
тушканчик Лихтенштейна — J. Uchtensteini Vinogradov, 1927 (СССР — пустыни Каракумов, Кызылкумов, Северного Прпаралья, Юж ного Прибалхашья);
египетский тушканчик — J. jaculus Linnaeus, 1758 (Сахара от Средиземного моря до южной Мавритании, Тимбукту, оз. Чад, Судана и С о мали; Палестина; Сирия; Ирак; Аравийский н ов);
туркменский тушканчик — J. turkmenicus Vinogradov et Bondar, 1949 (СССР — песчаные пустыни Туркмении);
тушканчик Бланфорда — J. blanfordi Murray, 1884 (Праи; Белуджистан в Пакистане; запад Афганистана);
восточный тушканчик — J. orientalis F.rxleben. 1777 (Средиземноморское побережье Африки от Танжера до Суэца);
Некоторые исследователи (например, Виши радов и I ромов, 1952: Громов и др., 1963) выделяют тушканчика Лихтенштейна в особый род
Eremodipus Vinogradov, 1930. В. Г. Гептнер (1975) включает в этот род египетского тушканчика, а туркменского тушканчика считает подвидом тушканчика Бланфорда.
В СССР 2 вида: туркменский тушканчик и тушканчик Лихтенштейна.
Род пятипалых карликовых тушканчиков
Genus Cardiocranius Satunin, 1903
В роде единственный вид: пятипалый карликовый тушканчик — С. paradoxus Satunin, 1903.
Размеры мелкие. Длина тела до 7,5 см. Длина хвоста до 7,5 см. Хвост
утончен в основании, расширен в средней части и постепенно сужается
к концу, где имеется небольшая кисточка волос (рис. 151). Уши короткие. Передние и задние лапы пятипалые. Пятый и первый пальцы на задних конечностях укорочены. На подошве стопы имеется пучок жестких волос. Спинная сторона тела песочного цвета. Брюхо белое. Хвост
светло-бурый сверху и белый снизу.
В черепе крупные костные слуховые барабаны соприкасаются передними частями между собой. Число щечных зубов—. Верхние резцы име-
3
ют бороздки на наружной поверхности.
Хромосом в диплоидном наборе 48.
Распространен в МНР и в СССР — в южной части Тувинской автономной республики.
Населяет глинистые и песчаные пустыни. Питается различными растительными объектами, в основном семенами, а также насекомыми. Экология изучена слабо.
Род трехпалых карликовых тушканчиков Genus Salpingotus Vinogradov, 1924
Размеры мелкие. Длина тела 4,1—5,7 см. Длина хвоста 9,3—12,6 см. Голова относительно крупная (рис. 151). Ее лицевая часть укорочена и несколько притуплена. Наружные ушные раковины небольшие, трубкообразные. Характерны очень длинные вибриссы. Передние конечности пятипалые, задние — трехпалые. На подошве и нижней поверхности пальцев задних конечностей сильно развита щетка жестких удлиненных волос. Хвост не имеет «знамени» и покрыт редкими, короткими волоса-
ми, перемежающимися с длинными волосами, более многочисленными на его конце. Проксимальная треть или половина хвоста утолщена из-за жировых отложений. Волосяной покров высокий, густой, мягкий. Окраска спинной стороны тела светло-песчаная, иногда темпо-жслтая, брюшной — беловатая или светло-желтоватая. Сосков 4 пары.
В черепе лицевой отдел укороченный, узкий. Мозговая капсула трапециевидной формы, сильно расширенная, сужающаяся кпереди. Кост-
ные слуховые барабаны крупные, вздутые, соприкасающиеся между собой передними отделами. Нижняя челюсть высокая, короткая. Число
4
щечных зубов— . Верхние резцы па наружной поверхности не имеют бо-3
роздок.
Хромосом в диплоидном наборе у жирнохвостого тушканчика 46.
Распространены в Центральной Азии и на юго-востоке Казахстана.
Экология изучена слабо. Населяют пустыни и степи. Жирнохвостый
тушканчик обитает в ковылы-ю-полынных песчаных степях, а также в закрепленных бугристых песках. Выкапывает временные простые и постоянные, более сложные, длиной более 3 м норы. Питается различными растительными объектами и насекомыми.
В роде 3 вида:
тушканчик Козлова — S. kozlovi Vinogradov, 1922 (Гоби в МНР);
жирнохвостый тушканчик — S. crassicauda Vinogradov 1924 (степи Джунгарии в МНР и Китае; в СССР — Зайсанская котловина);
тушканчик Томаса — S. thomasi Vinogradov, 1928 (Афганистан).
В СССР: жирнохвостый тушканчик.
Род длинноухих тушканчиков Genus Euchoreutes Sclater, 1841
В роде единственный вид: длинноухий тушканчик — Е. naso Sclater, 1891.
Размеры мелкие. Длина тела до 9 см. Длина хвоста примерно 15 см. Лицевой отдел головы относительно удлиненный. Уши очень крупные (рис. 152). Их длина равна примерно половине длины тела. Хвост покрыт густыми короткими волосами, на конце имеет «знамя» из черных и белых волос. Передние и задние конечности пятипалые. Задние конечности узкие и очень сильно удлинены, боковые пальцы на них сильно укорочены. Волосяной покров высокий, густой, мягкий. Спинная сторона тела рыжевато-желтая или бледно-красновато-коричневая. Брюхо белое. Пальцы задних конечностей окаймлены гребешками жестких удлиненных волос. Сосков 4 пары.
В черепе характерно удлинение лицевого отдела. Слезные кости небольшие. Костное небо широкое, продолжающееся позади уровня третьих коренных зубов. Костные слуховые барабаны вздутые, их вершины
Рис. 152. Длинноухий тушканчик — Е. naso
соприкасаются между собой. Резцовое отверстие крупное. Число щечных зубов —. Они гипселодонтного типа. Резцы белого цвета, верхние не 3
имеют бороздки по наружной поверхности.
Распространен в Китае — северо-восточной части и Внутренней Монголии, и на юге МНР.
Населяет песчаные пустыни. Экология изучена слабо.
Дикобразные
Hystricomorpha Brandt, 1855
Череп с массивной скуловой дугой, в которой скуловая кость не поддерживается скуловым отростком верхнечелюстной кости. Подглазничное отверстие чрезвычайно крупное, обычно больше затылочного. Через подглазничное отверстие проходит значительная часть передней доли жевательной мышцы. Зубная формула одинакова для всех семейств, входящих в Hystricomorpha: i-p с -—р -1-m -|-=20.Большая и малая берцовые кости не срастаются между собой.
СЕМЕЙСТВО ДИКОБРАЗОВЫХ
FAMILIA HYSTRICIDAE BURNETT, 1830
Размеры от мелких до средних. Длина тела 38—90 см. Масса от 2—3 до 18—27 кг. Телосложение тяжелое, довольно неуклюжее (рис. 153). Конечности короткие, сильные. Голова крупная с укороченной притупленной мордой. Шейный перехват выражен слабо. Хвост короткий или длинный, но меньше длины тела. Большая часть поверхности тела покрыта иглами, часть из которых достигает значительной длины (до 35 см у представителей рода дикобразов). Волосы сохраняются на брюшной поверхности тела, нижних частях конечностей и морде. На хвосте имеются своеобразные измененные иглы бокаловидной формы или щетины. Передние конечности трех- или четырехпалые, задние пятипалые. Первый палец задних конечностей сильно уменьшен. Пальцы с сильными когтями. Конечности стопоходящие. Подошвы безволосые, гладкие. Окраска тела однотонная, буроватая или черноватая. Длинные иглы имеют чередующиеся черные и белые кольца. Сосков 2 или 3 пары.
В черепе лицевой отдел удлинен, мозговой укорочен. Лобные кости у некоторых видов имеют тенденцию к пневматизации. Скуловые дуги слабо расставлены в стороны. Надглазничных отростков и заглазнич-ных бугров нет. Межглазничная область почти не имеет сужения, сверху выпуклая. Теменные гребни образуют сагиттальный. Ламбдоидаль-ный гребень хорошо выражен. Подглазничное отверстие крупное. Костные слуховые барабаны небольших размеров. В пижпей челюсти венечный отросток небольшой, а сочленованный — крупный. Щечные зубы имеют корни. Высота коронок обычно значительная. Щечные зубы складчатые, внутренние края складок часто усложнены вторичной складчатостью. Ключица хорошо развита.
Распространение охватывает большую часть Африки. Южную Европу, Южную Азию на север до Кавказа, гор Средней Азии и Гималаев.
Населяют, как правило, лесные и лесостепные районы субтропиков
и тропиков. Встречаются в предгорьях, горах, иногда в пустынях. Поднимаются в горы до 3900 м над уровнем моря. Активны в сумерках и ночью. Убежищем служат или естественные пещеры и углубления, или сложно устроенные, длинные, глубокие норы, обычно с гнездовой камерой. Временные норы простые, короткие и неглубокие. О г пор на места кормежек могут уходить па 10—15 км. Питаются надземными и подземными частями различных растений. В спячку не впадают, но в холодную погоду мало активны. В течение года, по-видимому, самки имеют один помет из 1—4 (обычно 1—2) детенышей. Длительность беременности от 42 до 110 дней. Детеныши рождаются с мягкими иглами и открытыми глазами. За первую неделю жизни иглы становятся жесткими. Продолжительность жизни 12—15 лет в естественных условиях и до 20 лет — в неволе. Местами могут причинять вред сельскому хозяйству. Мясо дикобразов можно использовать в пищу.
В семействе 4 рода (13 видов):
индонезийских дикобразов — Thecurus Lyon, 1907 (2 вида);
дикобразов — Hystrix Linnaeus, 1758 (6 видов);
кистехвостых дикобразов — Atherurus F. Cuvier, 1876 (4 вида); длиннохвостых дикобразов — Trichys Gunther, 1876 (1 вид).
Симпсон (1945) выделяет из рода дикобразов род малайских дикобразов— Acanthion Cuvier, 1822. По-видимому, более правильно объединить их вместе.
В СССР: род Hystrix.
Род индонезийских дикобразов
Genus Thecurus Lyon, 1907
Мелких и средних размеров дикобразы. Длина тела 45—54 см. Внешним обликом сходны с представителями рода дикобразов. Все тело густо покрыто уплощенными иглами, особенно длинными на спине и боках (рис. 153). Иглы имеют продольные борозды. На хвосте кроме обычных расположены бокаловидные, укороченные иглы, которые при движении хвоста производят специфический шум. Между иглами находятся тонкие, короткие волоски. На брюхе также имеются большие бороздчатые иглы. Конечности покрыты щетинковидпыми грубыми волосами. Окраска передней части спины темно-бурая, задней — почти черная. Бока тела более светлые, чем спина, испещрены белыми пятнами. Брюшная сторона беловато-желтая. Морда и конечности бурые. Некоторые иглы имеют светлое основание и светлую вершину.
В черепе ростральный отдел сужен. Череп несколько уплощен. Лобные кости длиннее носовых. Носовые кости узкие. Имеется незначительное межглазничное сужение. Небные отверстия обычно очень маленькие. Костные слуховые барабаны относительно небольшие. Щечные зубы резко гипселодонтного типа.
Распространены на Суматре, Калимантане и Филиппинских о-вах.
Населяют леса. Экология не изучена.
В роде 2 вида:
филиппинский дикобраз — Т. pumilis Gunther, 1879 (о-ва Палаван, Балабок и Каламиан из группы Филиппинских);
жесткоиглый дикобраз — Т. crassispinis Gunther, 1876 (восточная часть Суматры и Калимантан).
Эллермэн (1940) считает жестконглого дикобраза с о-ва Суматра особым видом — Т. sumatrae Lyon, 1907. По Чейзену (1940), здесь жестко-иглые дикобразы с о-вов Суматра и Калимантан объединены в один вид. Чейзен (1940) относит жестконглого дикобраза к роду Hystrix, что, по-видимому, неправильно.
Род дикобразов
Genus Hystrix Linnaeus, 1758
Наиболее крупные представители семейства. Длина тела 60—90 см. Длина хвоста 12—15 см. Иглы достигают большой длины. На загривке имеется грива из длинных, упругих, тонких, неострых игл (рис. 154). Задняя часть спины и бока покрыты длинными острыми цилиндрическими иглами черного, темно-бурого, белого, желтовато-белого цвета, часто с черными и белыми кольцами. Брюшная сторона тела обычно черноватая. Волосяной покров редуцирован в значительно большей степени, чем у других представителей семейства. Брюшная сторона покрыта гру-
II. indica
•быми щетинковидными волосами или небольшими иглами. Па хвосте расположены бокаловидные пустотелые нглы-hoi ремушки. Передние конечности четырехпалые, задние—пятипалые. Пальцы с сильными когтями. Первый палец задней конечности развит относигельно хорошо. Сосков 3 пары.
В черепе лицевой отдел удлинен. Лобные кости длинные, широкие. Затылочный отдел укорочен и песет крупные гребни. Костные слуховые барабаны относительно маленькие. Широкое костное небо ищется назад до уровня последних щечных зубов. Венечный отросток низкий Резцы широкие.
Хромосом в диплоидном наборе у индийского дикобраза 66, у гребенчатого 60, 66.
Распространение охватывает большую часть Африки (без Мадагаскара), Италию, включая о-в Сицилию, Закавказье, Палестину, Сирию, Среднюю Азию, Южный Казахстан, Белуджистан, Индию, Шри Ланка, Непал, Сикким, Бутан, Ассам, Бирму, Южный Китай, п-ов Малакку, о-ва Суматру, Яву, Калимантан, Сумбаву и Флорес.
Населяют преимущественно горные районы, поднимаясь в горы до 3900 м над уровнем моря. Обитают также в предгорьях, на равнинах, иногда в пустынях. Часто встречаются около населенных пунктов. Ведут сумеречный и ночной образ жизни. Убежищем служат естественные пещеры или сложные норы, в которых дикобразы могут обитать в течение многих лет. Б поисках корма уходят нередко за 10—16 км от нор, но к утру возвращаются обратно в свое убежище. Питаются различными частями растений. Посещают посевы, бахчи, плодовые сады, где поедают опавшие фрукты. В спячку не впадают, но в холодное время активность снижена. В течение года дают один помет 2—5 детенышей. Длительность беременности примерно НО дней. Продолжительность жизни в неволе около 20 лет. Местами могут причинять вред сельскому хозяйству. Мясо можно использовать в пищу.
В роде, по-видимому, 6 видов:
яванский дикобраз — И. javanica Cuvier, 1822 (включая Н. sumbawae Schwarz, 1911) (о-ва Ява и Флорес);
малайский дикобраз — Н. brachyura Linnaeus, 1758 (включая Н. yunnanensis Anderson, 1878) (п-ов Малакка; Юньнань в Китае; о-ва Калимантан и Суматра);
гималайский дикобраз — Н. hodgsoni Gray, 1847 (включая Н. sub-cristata Swinhoe, 1870 = 77. klossi Thomas, 1916) (Непал; Ассам; Бирма; Таиланд; п-ов Индокитай; Юго-Восточный Китай на север до Шэньси);
индийский дикобраз — 77. indica Kerr, 1792 (=77. leucurus Sykes, 1831 (о-в Шри Ланка; Индия; Непал; Иран; Ирак; Палестина; Сирия; Малая Азия; южная часть Аравийского п-ова; СССР — Азербайджан, большая часть Средней Азии на север до Мангышлака, Центральных Каракумов, Ташкентского оазиса, Киргизского хребта и Заилийского Алатау);
гребенчатый дикобраз — 77. cristata Linnaeus, 1758 (включая 77. ga-leata Thomas, 1893) (Центральная и Южная Италия, включая Сицилию; северная половина Африки на юг до Заира, Уганды, Кении и Танзании);
южноафриканский дикобраз—77. ajricaeaustralis Peters, 1852 (включая Н. stegmanni Muller, 1910) (Южная Африка к югу от линии, проходящей от устья р. Конго до Руанды и Южной и Западной Танзании).
В СССР: индийский дикобраз.
Юньнаньский дикобраз — (77. junnanensis), возможно, является подвидом не малайского, а яванского дикобраза (Эллермэн и Моррисон-Скотт, 1968). Чейзен (1940) считает яванского дикобраза подвидом малайского. По-видимому, его следует выделить в особый вид (Эллермэн, 1949). Эллермэн и Моррисон-Скотт (1966) предполагают возможность принадлежности дикобраза из Азербайджана к II. cristata. Симпсон (1945) выделяет яванского, гималайского и малайского дикобразов в особый род Acanthion Cuvier, 1823.
Род кистехвостых дикобразов
Genus Atherurus F. Cuvier, 1829
Мелких размеров дикобразы. Длина тела 40—55 см. Длина хвоста 15—25 см. Масса 1,5—4 кг. Большая часть спины и боков покрыта иг-
Рис. 155. Большехвостый дикобраз — Л. та-crourus и череп дикобраза Турнера — Л. tur-neri
ламп, которые наибольшей длины достигают по средней линии спины (рис. 155). Большинство игл уплощенные с бороздками. Хвост относительно длинный с пучком длинных, грубых, белых щетин на конце. Щетины имеют своеобразные четковпдные перетяжки. Голова, конечности и брюшная часть тела покрыты мягкими иголками. Основная окраска спинной стороны тела серовато-коричневая, отдельные иглы имеют белые вершины. Бока светлее спинной стороны тела, брюхо белое.
В черепе носовые кости короткие, лобные — длинные, иногда арковидно выпуклые. Обычно посторбитальных отростков нет. Мсжглазпич-ное сужение выражено слабо. У взрослых особен имеется крупный сагиттальный гребень. Костные слуховые барабаны относи I ел ыю небольшие. Нижняя челюсть с низким венечным н сочленованным отростками.
Распространены в Южном Китае, Яссами, Юго-Восточной Азии, Суматре, а также в Центральной Африке от Кеппи на запад через Заир до Сьерра-Леоне.
Населяют леса обычно около водоемов. Поднимаются в горы до 2300 м над уровнем моря. Активны ночью. Лень проводят в различных естественных пещерах, углублениях, расщелинах между скал. Иногда в одном месте собирается до 6—8 особей вместе. Довольно хорошо плавают, могут лазать по деревьям. Питаются в основном плодами, корнями,
клубнями, а также насекомыми. Иногда несколько животных собираются вместе в поисках корма. Длительность беременности 100—110 дней. В помете бывает 1—4 (обычно 2) детенышей. Местное население употребляет кистехвостых дикобразов в пищу.
В роде 4 вида:
большехвостый дикобраз — A. macrourus Linnaeus, 1758 (Суматра; п-ова Малакка, Индокитай; Китай — Хайнань, Сычуань; Лесам);
конголезский дикобраз — A. centralis Thomas, 1895 (Заир);
дикобраз Турнера — A. turnerl St. Leger, 1932 (Западная Кения);
африканский дикобраз — A. africanus Gray, 1842 (Сьерра-Леоне).
Род длиннохвостых дикобразов
Genus Trichys Gunther, 1876
В роде единственный вид: длиннохвостый дикобраз — Т. Прага Gunther, 1876 (рис. 156).
Размеры мелкие. Длина тела 38—46 см. Длина хвоста 17—23 см. Туловище покрыто уплощенными, гибкими иглами средней длины с про-392
дольными бороздками. Они чередуются с грубыми щетинковидными волосами. Основание хвоста покрыто колючками, а средняя часть — чешуя-ми и лишена волос или игл; на конце хвоста расположен пучок длинных грубых щетин, не имеющих четковидных перетяжек. Голова и брюшная сторона тела покрыты грубыми волосами. Кисти широкие, четырехпалые, с хорошо развитыми толстыми когтями; первый палец редуцирован. Задние конечности пятипалые, первый палец развит слабо. Окраска спинной стороны тела коричневатая. Бока более светлые. Брюхо белое.
Череп сильно удлинен и сужен. У взрослых особей имеется хорошо выраженный сагиттальный гребень. Посторбитальные отростки крупные. Носовые кости короткие, узкие, короче лобных. Затылочная область черепа высокая, сильная. Костные слуховые барабаны относительно маленькие. Венечный отросток нижней челюсти высокий.
Распространен на п-ose Малакка и о-вах Калимантан и Суматра.
Населяет лесные районы, обычно около водоемов. Экология изучена слабо.
Некоторые исследователи (например, Уокер, 1964) выделяют длиннохвостого дикобраза с Суматры в особый вид: Т. macrotis Miller, 1903.
СЕМЕЙСТВО АМЕРИКАНСКИХ ДИКОБРАЗОВ
FAMILIA ERETHIZONTIDAE THOMAS, 1897
Мелких, средних и крупных размеров грызуны. Длина тела 30— 103 см. Длина хвоста 7.5—45 см1. Масса тела от 7 до 18 кг. Самцы крупнее самок. Телосложение тяжелое и неуклюжее. Конечности относительно короткие. Хвост разной длины; у цепкохвостого дикобраза он хватательный. На туловище и хвосте имеются короткие острые иглы, вершины которых покрыты зазубринами, направленными острием назад. На спине обычно расположены длинные, волнистые щетины. Конечности приспособлены к лазанию по деревьям. Стопа и кисть расширены. На передних и задних конечностях по 4 функционирующих пальца. Первый палец задней конечности имеет широкую, подвижную подушку. Когти длинные, искривленные. Окраска черноватая или коричневая с желтыми или белыми полосами на иглах и волосах. Сосков 2 пары.
В черепе костные слуховые барабаны крупные. На лобных костях имеются заглазничные отростки. У тонкоиглого дикобраза в отличие от большинства грызунов орбита почти полностью окружена костным мостиком.
Щечные зубы с корнями, имеют довольно сложные эмалевые складки на жевательной поверхности. Os penis имеется.
Распространены в Северной (кроме тундр), Центральной п Южной Америке.
Населяют, как правило, лесные районы умеренной и тропической зон. Ведут наземный и древесный образ жизни. Лазают по деревьям хорошо, хотя и медленно. Прыгать по деревьям не могут. Довольно хорошо плавают. Активность преимущественно сумеречная и ночная, иногда дневная. Убежищем служат различные естественные углубления, пещеры, расщелины, дупла. Во время холодной погоды малоактивны. Зимой индивидуальный участок у представителей рода североамериканских
1 В пределах семейства можно выделить три группы: мелкие с длиной тела 38— 53 см, средние —54—77 см, крупные-—78—103 см.
дикобразов достигает 5 га. Питаются листьями, иглами хвойных деревьев, корой, проростками, плодами, цветками, корнями. За кормом могут взбираться на деревья на высоту до 18 м над землей. Из органов чувств лучше всего развиты слух и обоняние. Длительность беременности от 60—70 дней у Coendou до 7 месяцев у Erethizon. В помете от 1 до 4 детенышей; рождаются с открытыми глазами. Местами американские дикобразы могут приносить серьезный вред лесным посадкам.
В семействе 4 рода (11 видов):
североамериканских дикобразов—Erethizon F. Cuvier, 1822 (1 вид);, цепкохвостых дикобразов — Coendou Lacepede, 1799 (8 видов);
амазонских дикобразов — Echinoprocta Gray, 1865 (1 вид); тонкоиглых дикобразов — Chaetomys Gray, 1843 (1 вид).
Род североамериканских дикобразов
Genus Erethizon F. Cuvier, 1822
В роде единственный вид: североамериканский дикобраз — Е. dorsa-tum Linnaeus, 1758.
Размеры средние и крупные. Длина тела 64—103 см. Длина хвоста 14—30 см. Взрослые самцы могут достигать массы 18 кг. Голова небольшая, конечности укороченные (рис. 157). Окраска спинной стороны тела обычно темно-коричневая или черноватая. Спинная часть тела покрыта толстыми (около 2 мм в диаметре), острыми и относительно короткими (до 7,5 см) иглами с многочисленными, обращенными назад зазубринами на вершине. Иглы располагаются вперемежку с густыми, длинными, грубыми остевыми волосами и густым подшерстком. На брюхе игл нет. Короткий, толстый хвост почти по всей длине покрыт иглами вперемежку с волосами. Кисти широкие, с четырьмя функционирующими пальцами. Пятипалые задние конечности обладают хорошо развитым первым пальцем, который, однако, короче остальных. Подошвы передних и задних лап голые. Когти изогнутые, мощные. Сосков 2 пары.
В черепе ростральный отдел широкий. Носовые и лобные кости широкие. Подглазничное отверстие крупное. Сагиттальный гребень хорошо развит. Костное небо короткое, узкое спереди и очень широкое сзади. Костные слуховые барабаны крупные. Нижняя челюсть с низкими венечным и угловым отростками. Резцы относительно тонкие, желтые или оранжевые.
Хромосом в диплоидном наборе 42.
Распространение охватывает лесные области: Аляски, кроме западной половины п-ова Аляска и прибрежной полосы морей Берингова, Чукотского и Бофорта; Канады, кроме северо-восточной и восточной частей Северо-Западных территорий, севера Квебека и о-ва Ньюфаундленда; США, кроме крайнего юго-запада, включающего Тихоокеанское побережье и юг Калифорнии, крайний юго-запад Аризоны и юго-восток Нью-Мексико и юго-востока страны, где граница проходит через крайний юго-запад Техаса, восток Нью-Мексико, север Техаса, центральные районы Оклахомы, крайний юго-восток Канзаса, центральные районы Иовы, середину Иллинойса, юго-запад Индианы, середину Кентукки, центр Теннесси и Северной Каролины и в юго-восточной части Вирджинии выходит к Атлантическому океану; Северную Мексику — север штатов Сонора и Чиуауа.
Населяет лесные районы, главным образом хвойные леса. Ведет ночной образ жизни, проводя день в различных укрытиях, дуплах, пещерах..
Как правило, держатся поодиночке, ио иногда собираются небольшими группами, особенно зимой. В спячку нс впадает, однако в холодную погоду (ниже —18°С) активность резко снижается. Американский дикоб раз — мирное животное и по собственной инициативе по нападает. Обо роняясь, поворачивается к врагу задом и быстро двигает хвостом. Колючек не теряет и не «выстреливает» ими, однако они настолько слабо держатся в кожном покрове дикобраза, что, проникнув даже неглубоко в кожу врага, там и остаются благодаря своим зазубренным вершинам. Иглы, попавшие в мышцы, могут проникать в тело со скоростью 1 мм в час и даже быстрее и часто вызывают смерть у животного. Утерянные
иглы вырастают вновь через 10—42 дня. Хорошо лазает по деревьям и плавает. Зимой питается в основном камбиальным слоем и корой деревьев, а также хвоей; в весенне-летнее время предпочитает почки, проростки,, листья, траву, ягоды, цветки, семена, орехи. Спаривание происходит осенью или ранней зимой (ноябрь — декабрь). Длительность беременности 120—217 дней. Самка приносит за сезон, как правило, одного детеныша с открытыми глазами и мягкими колючками, которые становятся твердыми через час. Масса новорожденного от 340 до 640 г. Через два дня после рождения уже может лазать по деревьям. Половозре-лость наступает в возрасте 2,5 года.
Местами североамериканский дикобраз может приносить небольшой вред лесным посадкам. В зимнее время, когда снег очень глубокий, может жить на одном и том же дереве, пока не объест с него всю кору. Длительность жизни в неволе до 10 лет.
Род цепкохвостых дикобразов
Genus Coendou Lacepede, 1799
Размеры от мелких до средних. Длина тела 30—60 см. Длина хвоста-33—45 см. Масса 0,9—4,3 кг. Спинная сторона тела и бока покрыты короткими, толстыми иглами (рис. 158). У некоторых видов иглы скрыты в длинном довольно густом волосяном покрове. Волосы на груди и брюхе обычно мягкие, иногда щетинистые. Длинный хвост хватательный и лишен игл. На конце его имеются своеобразные мозоли. На передних и задних конечностях всего по 4 функционирующих пальца, вооруженных длинными искривленными когтями. Подошва стопы имеет сильно развитую мозоль, внутри которой находится костное образование — своеобразное приспособление к лазанию. Окраска спинной стороны тела от светло-желтой до бурой, почти черной. Иногда на спине имеются пятна. Брюшная сторона обычно серая.
Череп широкий, сильно выпуклый в области лобных костей. Лицевой отдел укорочен. Носовые кости обычно короткие. Костное небо широкое. Небные отверстия короткие.
Распространение охватывает территорию от Мексики на юг через Центральную Америку до Колумбии, Венесуэлы, Бразилии, Боливии, Перу и Эквадора.
Населяют лесные районы. Ведут древесный образ жизни. Активны ночью. Передвигаются медленно. День проводят в сне среди густых ветвей деревьев, иногда в дуплах. Питаются листьями, почками, плодами. С февраля по май самки приносят по одному детенышу. Масса его от 280 до 426 г.
В роде 8 видов:
мексиканский дикобраз— С. mexicanus Kerr, 1792 (юг Мексики, штаты: крайний юго-восток Сан-Луис-Потоси, восток Идальго, северо-восток Пуэбло, запад и юг Веракрус, Оахака, кроме севера и юга, центральные районы восточной части Герреро, Чиапас, кроме юга и юго-запада, восточная половина Кампече, Юкатан, кроме полосы вдоль берега Мексиканского залива, западная половина Кинтано-Роо; Гватемала, кроме северо-востока и полосы вдоль берега Тихого океана; север Сальвадора, южная половина Гондураса, центральные районы Никарагуа и Коста-Рцки, крайний северо-запад Панамы);
светлый дикобраз — С. pallidas Waterhouse, 1848 (Вест-Индия; точное место находки неизвестно);
дикобраз Ротшильда — С. rotschildi Thomas, 1902 (Панама);
Рис. 158. Цепкохвостый дикобраз —С. preliensillis и его череп
двухцветный дикобраз —С. bicolor Tschudi, 1844 (Неру, Анды в Эквадоре, Боливия, север Колумбии);
цепкохвостый дикобраз — С. prchensillis Linnaeus, 1758 (Боливия, Центральная и Восточная Бразилия, Гайана и восток Венесуэлы);
бразильский дикобраз —С. insidiosus Kuhl, 1820 (Бразилия);
колючий дикобраз—С. spinosus F. Cuvier, 1822 (Бразилия, Параг вай, северо-восток Аргентины);
венесуэльский дикобраз —С. vestitus Thomas, 1899 (Венесуэла, Колумбия).
Андерсон и Джонс (1967) насчитывают в роде 5 видов.
Род амазонских дикобразов
Genus Echinoprocta Gray, 1856
В роде единственный вид: амазонский дикобраз — Е. rufescens Gray, 1865.
Размеры мелкие. Длина тела и длина хвоста вместе составляют около 50 см. Хвост короткий •— немного длиннее, чем стопа; покрыт волосами и нехватательный. Задние конечности и мозоль на подошве такие же, как у цепкохвостых дикобразов. Иглы в передней части туловища щетинковидные, но по мере приближения к крестцу становятся длиннее, толще и жестче. Голова покрыта короткими, острыми иглами. Окраска спины и боков светло-коричневая или черноватая. Брюшная сторона светло-коричневая. Конечности и хвост темно-серые или черные. Голова беловатая с бурыми или черными пятнами.
Череп походит на таковой мелких видов цепкохвостых дикобразов. В межглазничной области — небольшое сужение. Резцы тонкие, желтого цвета.
Распространен в Колумбии в Южной Америке.
Экология не изучена. По-видимому, ведет древесный образ жизни, поднимаясь в горы до 1200 м над уровнем моря.
Род тонкоиглых дикобразов
Genus Chaetomys Gray, 1843
В роде единственный вид: тонкоиглый дикобраз — С. subspinosus Kuhl, 1820.
Размеры мелкие. Длина тела 43—45 см. Длина хвоста 25—28 см (рис. 159). Спина покрыта в основном жесткими волнистыми щетинками. На голове, шее и передних конечностях густо расположены короткие и острые иглы. Поверхность длинного хвоста чешуйчатая, и только по его нижней стороне растут короткие грубые волосы. Хотя конец хвоста обнажен и подобен хвосту цепкохвостых дикобразов, по-видимому, он не хватательный. Передние и задние конечности имеют по 4 пальца с длинными, искривленными когтями; первые пальцы очень небольшие. Основная окраска спинной стороны тела тускло-буроватая, иногда серовато-белая, брюшной — рыжевато-коричневая. Хвост и конечности буровато-черные.
В черепе лицевой отдел укороченный, широкий. Лобные кости очень широкие, но перед посторбитальными отростками есть сужение. Костное небо узкое и короткое. Небные отверстия очень короткие. Костные слуховые барабаны относительно крупные. Посторбитальные отростки хорошо выражены и вместе со средними частями скуловой дуги образуют почти замкнутую костную орбиту. Нижняя челюсть с низким венечным и небольшим угловым отростками. Резцы узкие.
Распространен в Восточной, Центральной и Северо-Восточной Бразилии.
Населяет кустарники по окраинам открытых саванн. Очень редок. Экология не изучена. Хорошо лазает по деревьям. Медлительное животное. Питание растительное.
Включен в «Красную книгу».
Рис. 159. Тонкопглый дикобраз — С subspinosus и его череп
СЕМЕЙСТВО СВИНКОВЫХ
FAMILIA CAVIIDAE WATERHOUSE, 1839
Грызуны от мелких до средних размеров. Длина тела 22—75 см. Масса 0,5—16 кг. Телосложение тяжелое, конечности короткие. Или телосло жение довольно легкое, конечности длинные и тонкие. Шея укороченная. Голова крупная. Уши короткие или довольно длинные. Глаза крупные. Хвост короткий (до 5 см), иногда едва заметный. Верхняя губа цельная. Когти короткие и острые (Cavia) или тупые (Kerodon.) или даже копытоподобные (Dolichotis). Передние конечности четырехпалые, .задние трехпалые. Подошвы лап голые (Cavia, Kcrodon) или покрыты волосами (Dolichotis}. Волосяной покров, как правило, грубый. Окраска спины сероватая, оливковая, светло-коричневая, иногда с черноватым оттенком; брюхо белое или темно-желтое. Сосков одна пара.
В черепе костное небо короткое. Угловой отросток нижней челюсти отогнут назад и не выгнут наружу. На нижней челюсти под щечными зу-
бами располагается борозда, к которой прикрепляется глубокая порция жевательной мышцы. Слезные кости относительно крупные.
Желудок простой. Слепая кишка крупная и сложная. Щечные зубы без корней, гипселодонтного типа. Резцы короткие. Малая и большая берцовые кости частично срастаются между собой. Грудина состоит из 6 частей. Ключица развита слабо.
Распространены в Южной Америке на юг до Южной Аргентины, за исключением большей части Чили и, вероятно, части Восточной Бразилии.
Населяют различные биотопы от болотистых участков тропических лесов до сухих каменистых лугов в горах на высоте до 4000 м над уровнем моря, а также пампасы и саванны, кустарниковые и скалистые участки. Густых джунглей обычно избегают. В густой траве проделывают тропинки. Питаются различными растительными кормами. Некоторые виды роют норы в земле или среди камней. Могут использовать и норы других животных. Быстрые зверьки, бегающие на всех четырех конечностях. Представители рода Dolichotis прыгают наподобие зайцев, Kerodon может вскарабкаться на деревья в поисках пищи. Активны в сумерках и ночью (Cavia, Kerodon, Microcavia) или днем (Dolichotis, Galea). В спячку не впадают
В помете 1—2 (Kerodon) или от 2 до 5 (Dolichotis) детенышей. Беременность длится около двух месяцев. В течение года обычно бывает 2 помета (в неволе число пометов возрастает). Часто держатся парами или более крупными группами от 8 до 30 особей (Dolichotis). Galea обычно живут колониями. Местное население употребляет морских свинок в пищу. Cavia porcellus была одомашнена и употреблялась индейцами в пищу еще до того, как в Перу впервые появились конквистадоры. Морских свинок широко используют в качестве лабораторных животных.
В семействе 5 родов (15 видов):
морских свинок — Cavia Pallas, 1766 (6 видов);
моко — Kerodon F. Cuvier, 1825 (1 вид);
куи — Galea Меуеп, 1831 (3 вида);
горных свинок — Microcavia Gervais et Ameghino, 1880 (3 вида);
мара — Dolichotis Desmarest, 1820 (2 вида).
Некоторые исследователи (например, Уокер, 1964) считают Galea и Microcavia подродами рода Cavia. Более правильно выделять их в особые роды (Симпсон, 1945). Кабрера (1960) выделяет из рода Dolichotis род Pediolagus Alarelli, 1927, что, по-видимому, неправильно.
Род морских свинок
Genus Cavia Pallas, 1766
Размеры мелкие. Длина тела до 35 см. Телосложение тяжелое (рис. 160). Конечности укороченные. Голова крупная. Шейный перехват выражен слабо. Уши относительно короткие, округлые. Хвост снаружи незаметен. Задние конечности длиннее передних, трехпалые, третий палец самый длинный. Передние конечности четырехпалые, третий палец самый длинный, пятый наиболее короткий, четвертый немного длиннее второго. Волосяной покров грубый и длинный. Окраска его у диких форм обычно сероватая или буроватая. Одомашненные морские свинки имеют разнообразный по мягкости, густоте и длине волосяной покров самых различных расцветок.
В черепе хорошо выражено межглазничное сужение. У взрослых особей имеется развитый сагиттальный гребень. Костные слуховые барабаны 400
Рис. 160. Свинка Чуди — С. tschudi
относительно крупные. Короткое костное небо заканчивается па уровне передней части третьего коренного зуба. Небные отверстия короткие и узкие. Подглазничное отверстие снизу шире, чем в верхней части. Резцы белые.
Хромосом в диплоидном наборе у морской свинки 64.
Распространены в Южной Америке от Бразилии, Гайаны, Венесуэлы и Колумбии к югу до Северной Аргентины.
Населяют самые различные биотопы, саванны, опушки лесов, оолота, скальные участки в горах. Норы обычно роют сами, по могут занимать и норы других животных. Иногда объединяются в небольшие группы до 5—10 особей. Активны в сумерках и ночью. Питаются различными растениями. Размножаются в течение всего года. Длительность беременности 60—70 дней. Самка приносит от 1 до 4 детенышей. Одомашненные формы рождают большее число детенышей. Через несколько часов после рождения детеныши способны самостоятельно передвигаться. Лактационный период продолжается около 3 недель. Половозрелое™ достигают в возрасте 55—70 дней. Продолжительность жизни около 8 лет. Одомашненные морские свинки (С. porcellus porcellus) широко применяются в качестве лабораторных животных.
В роде, по-видимому, 6 видов:
С. aperea Erxleben, 1777 (восток и юго-восток Бразилии. Парагвай, север Аргентины);
С. fulgida Wagler, 1831 (Восточная Бразилия);
малая свинка — С. папа Thomas, 1917 (Западная Боливия); пампасская свинка — С. pamparum Thomas, 1901 (Аргентина); морская свинка — С porcellus Linnaeus, 1758 (Колумбия, Гайана, Южная Венесуэла и, возможно, север Бразилии);
свинка Чуди — С. tschudi Fitzingcr, 1867 (Перу, крайний юг Боливии, северо-запад Аргентины).
Андерсон и Джонс (1967) насчитывают в роде 3 вида, а Эллермэн (1940) — 10.
Род моко
Genus Kerodon F. Cuvier, 1825
В роде единственный вид: моко — К. rupestris Wied, 1820.
Внешне сходны с морскими свинками, однако конечности относительно более высокие. Размеры примерно такие же или несколько более крупные, чем у морских свинок. Масса тела 0,9—1 кг. Хвоста нет или он едва заметен. Окраска спинной стороны тела сероватая с белыми или черными пятнами. Горло беловатое. Брюшная часть тела желтоватобурая. Характерны тупые ногти.
Череп сходен с таковым морских свинок. Сагиттального гребня нет. Лицевой отдел относительно узкий. Костные слуховые барабаны средних размеров. Небные отверстия очень узкие.
Распространен в Северо-Восточной Бразилии.
Населяет засушливые склоны гор, холмов. Убежищем служат пустоты под камнями или расщелины скал; иногда делают норы под камнями. Активность сумеречио-ночная. В поисках пищи хорошо лазает по деревьям. Голос — характерный громкий свист. В течение года у самки бывает, по-видимому, два помета по 1—2 детенышей. Местное население охотится за моко из-за мяса. Моко легко одомашнивается.
Род куи
Genus Galea Meyen, 1831
По внешнему виду сходны с морскими свинками. Строение щечных зубов более простое. Резцы пигментированные.
Распространены в Боливии, Северной Аргентине, Чили и Бразилии.
В роде, вероятно, 3 вида:
G. jlavidens Brandt, 1835 (Бразилия);
G. musteloides Meyen, 1833 (Боливия, Аргентина, Перу);
G. spixii Wagler, 1831 (Боливия, Бразилия)
Эллермэн (1940) выделяет, кроме перечисленных выше трех видов, еще 3: G. minimus Molina, 1782; G. wellsi Osgood, 1915; G. palustris Thomas, 1911.
Некоторые исследователи (например, Уокер, 1964) включают куи в род морских свинок.
Род горных свинок
Genus Microcavia Gervais et Ameghino, 1880
По внешнему виду сходны с морскими свинками.
В черепе у взрослых имеется сагиттальный гребень. Костные слуховые барабаны относительно крупные. Небные отверстия относительно большие. Резцы не пигментированы. Щечные зубы сходны с щечными зубами куи.
Распространены в Боливии и Аргентине.
В роде 3 вида:
М. australis I. Geoffrey et D’Orbigny, 1833 (Аргентина);
M. shiptoni Thomas, 1925 (Северо-Западная Аргентина);
М. niata Thomas, 1898 (Анды в Боливии).
Рис. 161. Патагонская мара — D. patagona
Род мара
Genus Dolichotis Desmarest, 1820
Размеры средние и крупные. Длина тела 45—75 см. Длина хвоста не превышает 5 см. Масса 9—16 кг. Внешне эти звери несколько напоминают длинноногих кроликов или зайцев (рис. 161). Сложение довольно легкое. Уши длинные. Конечности высокие; задние конечности трехпалые, каждый из пальцев имеет копытовидный коготь. Передние конечности четырехпалые, пальцы несут острые когти. Волосяной покров густой, мягкий. Окраска его на спинной стороне тела сероватая, брюшной — беловатая.
В черепе носовые и особенно лобные кости расширены. Костные слуховые барабаны средних размеров. Костное небо очень короткое, продолжается назад только до уровня второго коренного зуба. Небные отверстия длинные, узкие. Скуловые кости широкие, короткие. Слезные кости очень крупные. Щечные зубы с постоянным ростом, гипселодонтного типа.
Хромосом в диплоидном наборе у патагонской мары 64.
Распространены в Аргентине, Парагвае и Боливии.
Населяют засушливые места со скудной растительностью, низкой травой, редкими кустами. Норы выкапывают сами, по иногда занимают и поры других животных. Активны днем. Любят принимать солнечные ванны. Обычно держатся небольшими группами по 3—4 особи. Иногда собираются в более крупные стада до 40 особен. Хотя мара прыгают наподобие зайцев, их бег не очень быстрый. Питаются различными рас
тениями. Самка приносит от 2 до 5 детенышей в гнездовой камере норы.
В роде 2 вида:
патагонская мара — D. patagona Zimmermann, 1780 (Аргентина — Патагония);
D. salinicola Burmeister, 1875 (Северо-Западная Аргентина; крайний юг Боливии; Парагвай).
Некоторые ученые, например Кабрера (1960), выделяют D. salinicola в особый род Pediolagus Marelli, 1927.
СЕМЕЙСТВО ВОДОСВИНКОВЫХ
FAMILIA HYDROCHOERIDAE GILL, 1872
Размеры наиболее крупные среди современных грызунов. Длина тела достигает 100—130 см. Хвост очень короткий. Высота в плечах около/ 50 см. Масса И. hydrochaeris может несколько превышать 50 кг, а Н. isth-mius — 27 кг. Сложение тяжелое. Внешним видом напоминают гигантских морских свинок (рис. 162). Конечности относительно короткие. Голова крупная, массивная. Морда тупая и широкая. Верхняя губа толстая. Уши короткие, округлые. Глаза маленькие, помещаются высоко на голове и отнесены несколько назад. Передние конечности четырехпалые (третий палец наиболее длинный), задние — трехпалые. Пальцы соединены неполной плавательной перпонкой и имеют короткие сильные когти. Волосяной покров образован длинными (до 10 см) грубыми волосами, настолько редко расположенными, что сквозь них видна кожа. Спинная сторона тела рыжевато-бурая или сероватая, брюшная — желтовато-бурая. У половозрелых самцов на верхней части морды расположен участок кожи с многочисленными крупными сальными железами.
Череп массивный. Носовые кости широкие, лобные — широкие, длинные. Затылочный отдел относительно узкий, без сагиттального гребня. Слезная кость крупная. Костные слуховые барабаны относительно небольшие. Скуловые дуги широкие, массивные. Подглазничное отверстие не имеет канала для прохождения нерва. Костное небо сужено спереди. Щечные зубы без корней, растут в течение всей жизни животного. Правые и левые ряды щечных зубов сближены в передних отделах. Третьи коренные зубы верхней и нижней челюсти крупнее всех остальных коренных зубов и образованы поперечными пластинками, соединенными необычайно большим количеством цемента. Резцы широкие, белые. Верхние резцы имеют продольную бороздку на наружной поверхности. Малая и большая берцовые кости частично срастаются между собой. Ключицы нет.
Хромосом в диплоидном наборе у водосвинки 66.
Распространены в Южной Америке к востоку от Аид, на юг до Уругвая и Южной Бразилии, на запад до Атлантического побережья.
Населяют густо заросшие берега различных водоемов, болота. Держатся в травянистых зарослях, в лесах. Обычно встречаются семейными группами или стаями по 10—20 особей.
Активны преимущественно днем; в местах, где их часто тревожат, переходят на ночной образ жизни. Устраивают лежки на земле. Поры или специальные гнезда не устраивают. Хорошо плавают и ныряют. При плавании над водой возвышаются только ноздри, уши и глаза. От врагов часто спасаются в воду. Любят лежать подолгу в грязевых ваннах. Питаются в основном различными водными и прибрежными растениями. Дают один помет в год. Беременность длится 104—111 дней. Детенышей в помете от 2 до 8. Новорожденные хорошо развиты. Масса 5-дневных
Рис. 162. Водосвинка — //. hydrochaeris и ее череп
900—1350 г. С матерью остаются е течение долгого времени. Продолжительность жизни 8—10 лет. Главный враг водосвинок (кроме человека)— ягуар, а для молодых животных — и кайман. Мясо употребляется местным населением в пищу. Водосвинки хороню приручаются.
Некоторые исследователи (например, Эллермэн, 1910) рассматривает семейство водосвинок в качестве подсемейства семейства морских свинок Caviidae. По-видимому, более правильно считать водосвинок отдельным семейством.
В семействе 1 род: водосвинок — Hydtochoerus Brisson, 1762, и 2 вида.
Род водосвинок
'Genus Hydrochoerus Brisson, 1762
Описание рода приведено в описании семейства.
В роде 2 вида:
водосвинка — И. hydrochaeris Linnaeus, 1766 (от Венесуэлы и Гайаны на юг до Уругвая и Восточной Аргентины и от Колумбии и Перу на восток до атлантического побережья);
малая водосвинка — Н. isthmius Goldman, 1912 (Панама, север Колумбии, запад Венесуэлы).
Кабрера (1960) объединяет оба вида в один — И. hydrochaeris.
СЕМЕЙСТВО ПАКАРАНОВЫХ
FAMILIA DINOMYIDAE ALSTON, 1876
Грызуны средних размеров. Длина тела 73—79 см. Длина хвоста около 20 см. Масса 10—15 кг. Телосложение тяжелое. Внешне несколько напоминает морских свинок (рис. 163). Голова широкая. Уши небольшие, округлые. Конечности короткие, четырехпалые. Стопа широкая. Когти длинные, слабо изогнутые и мощные. Хвост по всей длине покрыт волосами. Характерны длинные вибриссы. Волосяной покров грубый, низкий, редкий. Окраска тела черная или бурая. По обе стороны от средней линии спины, вдоль нее, проходят две широкие белые полосы, иногда превращающиеся в ряд белых пятен. По бокам тела располагаются два коротких ряда белых пятен. Брюшная сторона несколько более светлая, чем спина, и не имеет пятен.
Череп массивный, широкий с длинными, расширенными лобными костями. Сагиттального гребня нет. Костные барабаны средних размеров. Костное небо продолжается назад чуть дальше уровня последних щечных зубов. Резцовые отверстия небольшие. Слезные кости крупные. Щечные зубы резко гипселодонтные, с постоянным ростом в течение жизни животного. Резцы широкие, массивные. Каждый щечный зуб образован серией поперечных пластин.
Встречается на западе Южной Америки: в горах от Колумбии до юго-востока Перу и запада Боливии.
Населяет долины и нижние части склонов Анд. Держится в лесах, на скалистых склонах. Встречается редко и, по-видимому, близка к полному исчезновению. Ведет наземный образ жизни. Активна днем. Двигается медленно, «флегматично». Питается преимущественно плодами, листьями, нежными проростками. Самка приносит, по-видимому, двух детенышей.
В семействе один род:
пакаран — Dinomys Peters, 1873 (1 вид).
Род пакаран
Genus Dinomys Peters, 1873
См. описание семейства.
В роде 1 вид:
пакарана — D. branickii Peters, 1873 (ареал совпадает с ареалом семейства).
Рис. 163. Пакарана—D. branickii и ее череп
СЕМЕЙСТВО СЕМИЗУБОВЫХ
FAMILIA HEPTAXODONTIDAE MILLER ET GIDLEY, 1918
Грызуны средних размеров, тяжелого сложения, теперь, по-видимому, полностью вымершие. Известны только но скелетным остаткам, обнаруженным на Больших и Малых Антильских о-вах. Представители семейства, вероятно, жили в историческое время. Найденные остатки костей свидетельствуют о том, что люди использовали этих грызунов в пишу. Период их исчезновения, по-видимому, приходится па то время, когда здесь был Колумб.
Череп массивный. Характерно строение щечных зубов, образованных 4—7 поперечно расположенными пластинами. Каждая пластина имеет эмаль только на передней поверхности. Щечные зубы гипселодонтные с постоянным ростом в течение жизни.
Распространение охватывало Большие и Малые Антильские о-ва.
Экология неизвестна. Все виды, вероятно, вели наземный образ жизни.
В семействе 2 рода (2 вида), представители которых жили в историческое время:
эласмодонтомис — Elasmodontomys Anthony, 1916 ( = Heptaxodon Anthony, 1917) (1 вид);
куэмизия — Quemisia Miller, 1929 (1 вид)
Род эласмодонтомис
Genus Elasmodontomys Anthony, 1916
В роде единственный вид: эласмодонтомис — Е. obliquus Anthony, 1916.
Череп эласмодонтомис напоминает череп нутрии. Общая длина черепа примерно 12,5 см. Верхние резцы со слабо выраженными продольными 4
бороздками. Число щечных зубов: — . Коронка их образована пятью эма-4
левыми пластинами, соединенными дентином. По-видимому, этот грызун имел внешний вид, сходный с пака. Тяжелое телосложение, короткие пальцы свидетельствуют о том, что он вел наземный образ жизни.
Был распространен на о-ве Пуэрто-Рико.
Род куэмизия
Genus Quemisia Miller, 1929
В роде единственный вид: куэмизия — Q. gravis Miller, 1929.
Размеры, по-видимому, были сходны с пака. Характерны короткие нижние резцы, длинное сочленение между половинками нижней челюсти, продолжающееся назад до уровня первого коренного зуба, и необычные извилины эмали на жевательной поверхности щечных зубов. Куэмизия, по-видимому, еще не вымерла, когда испанцы впервые появились в Центральной Америке. Она исчезла, вероятно, в первой половине XVI столетия.
Была распространена на о-ве Гаити.
СЕМЕЙСТВО АГУТИЕВЫХ
FAMILIA DASYPROCTIDAE SMITH, 1842
Размеры мелкие или средние. Длина тела 32—80 см. Длина хвоста 1—7 см. Масса взрослой особи достигает 1 —10 кг. Самцы (по крайней мере, у пака) немного крупнее самок. Приспособлены к быстрому бегу. Телосложение легкое. Конечности относительно высокие, особенно задние. Передние конечности четырехпалые, задние трех- или пятипалые. Пальцы с толстыми когтями, иногда копытовидпыми. Волосяной покров грубый, средней высоты, довольно густой. Окраска его черноватая, коричневатая, рыжеватая или желтоватая. У пака рыжевато-коричневая окраска спины испещрена белыми пятнами и линиями. Брюхо более светлое, чем спина. Имеются своеобразные специфические анальные кожные железы, продуцирующие секрет с сильным запахом. Характерны защечные мешки. Сосков 2 или 4 пары.
Слезная кость в черепе относительно крупная. Резцы сравнительно тонкие. Ключица недоразвита.
Распространены в тропической Северной Америке, Центральной и Южной Америке, к востоку от Анд, к югу до Боливии, Парагвая и Аргентины, а также на Малых Антильских о-вах.
Населяют тропические леса. Часто держатся около водоемов. Ведут, как правило, одиночный образ жизни. Активны ночью (пака) или днем’ (агути). Убежищем служат норы, которые они самостоятельно выкапывают. Пака, спасаясь от врагов, бросается в воду — она не только хорошо плавает, но и ныряет. Агути — более наземное животное. Он также хорошо плавает, но не ныряет. Питаются различными растительными кормами (листьями, плодами, корнями, корой). Агути может запасать пищу. Сезонности в размножении нет. Самки приносят, по-видимому, 2 помета в году. Число детенышей в помете 1—6. Длительность беременности 64—104 дня. Детеныши рождаются хорошо развитыми, массой около 210 г. Продолжительность жизни в неволе составляет 15—20 лет. Местное население охотится за всеми представителями семейства и употребляет их в пищу.
В семействе 4 рода (16 видов):
пака — Cuniculus Brisson, 1762 (= Agouti Lacepede, 1799) (1 вид); горных пака—Stictomys Thomas, 1924 (1 вид);
агути — Dasyprocta liliger, 1811 (12 видов);
акуши — Myoprocta Thomas, 1903 (2 вида). ।
Некоторые исследователи (например, Кабрера, 1960) относят пака й горных пака к отдельному семейству Agoutidae, что, по-видимому, неправильно.
Род пака
Genus Cuniculus Brisson, 1762
В роде единственный вид: пака — С. раса Linnaeus, 1756.
Размеры крупные. Длина тела 60—80 см. Длина хвоста 20—30 см. Масса взрослой особи до 10 кг. Туловище массивное (рис. 164). Голова большая. Уши средних размеров, округлые. Передние конечности четырехпалые, задние пятипалые; первый и пятый пальцы развиты очень слабо. Волосяной покров образован грубыми остевыми волосами без подшерстка. Окраска спины коричневатая или черноватая. По бокам тела расположены четыре ряда белых пятен, иногда образующие продольные полосы. Брюхо беловатое или темно-желтое. Сосков 2 пары.
Череп весьма своеобразен. Большая часть верхнечелюстных и скуловых костей расширена и образует крупные костные щечные пластины с сильно складчатой поверхностью. Эти пластины направлены вниз й прикрывают большую часть нижней челюсти. Носовые кости широкие, относительно короткие. Лобные кости широкие, очень длинные. Теменные кости сжаты. У взрослых особен есть сагиттальный гребень. Хорошо выражены заглазничные отростки. Костные слуховые барабаны относительно небольшие. Костное небо широкое, продолжается назад до второго коренного зуба. Резцы тонкие, сжатые. Щечные зубы сильно гипсело-донтные.
Хромосом в диплоидном наборе 74.
Распространена в северной части Центральной Мексики, в штатах: крайнем востоке Сан-Луис-Потоси, Веракрус, северной половине Чиапас, Табаско, южной половине Кампече, крайнем юге Кинтано-Роо; Белизе; северной половине Гватемалы; Гондурасе, кроме полосы, проходящей вдоль границы с Сальвадором; Никарагуа, кроме области вдоль Тихоокеанского берега; Коста-Рике, кроме Тихоокеанского побережья; Пана-
Рис. 164. Пака — С. раса и ее череп
ме; северной и центральной частях Южной Америки от Венесуэлы па севере до Парагвая на юге.
Населяет различные биотопы, по обычно встречается в лесных районах около воды. Активна ночью. День проводит в норе, которую выкапывает в берегах водоемов, на склонах холмов, среди корней деревьев или под камнями. Обычно делает несколько выходов. Держатся поодиночке. Ведет наземный образ жизни, хотя охотно входит в воду, хорошо плавает и ныряет. От врагов часто спасается в воде. Питается листьями, стеблями, корнями, плодами. В течение года, по-видимому, дает 2 помета, по одному, редко 2 детеныша. В неволе живет примерно до 16 лет.
Некоторые исследователи (например. Эллермэн, 1940; Уокер, 1964) включают в род Cuniculus род Stictomys. По-видимому, более правильно считать их отдельными родами, как это делают Симпсон (1945), Андерсон и Джонс (1967) и др.
Род горных пака
Genus Stictomys Thomas, 1924
В роде единственный вид: горная пака—S. taczanowskii Stolzmann, 1865.
Внешне сходна с пака. Волосяной покров более густой, когти сильнее сжаты с боков, подошвы лап более гранулированы.
В черепе носовые кости относительно длинные, орбиты — небольшие.
Распространена в Эквадоре, на северо-востоке Венесуэлы, на севере и юге Колумбии.
Встречается в Андах на высоте около 3000 м над уровнем моря. Держится около групп деревьев рядом с водоемами. Экология не изучена.
Род агути
Genus Dasyprocta liliger, 1811
Размеры средние. Длина тела 41—62 см. Длина хвоста 1—3,5 см. Масса взрослой особи 1,3—4 кг. Телосложение легкое, приспособленное к быстрому бегу (рис. 165). Конечности, особенно задние, удлинены. Кисть узкая. Голова массивная, уши короткие, округлые. Передние конечности четырехпалые, задние — трехпалые с острыми копытовидными когтями. Средний палец стопы наиболее длинный, четвертый немного короче второго, подошва голая. Волосяной покров обычно довольно грубый, глянцевитый; в задней части туловища он удлинен и более густой. Окраска спинной стороны тела варьирует от бледно-оранжевой до почти черной. У некоторых видов на спине проходят слабо выраженные светлые полосы. Брюхо беловатое или желтоватое. Сосков 4 пары.
Череп менее массивный, чем у пака. Лобные кости широкие, длиннее носовых костей. У взрослых особей есть небольшой сагиттальный гребень. Костные слуховые барабаны относительно крупные. Резцовые отверстия короткие, сдвинуты вперед. Слезные кости сильно увеличены. Угловой отросток нижней челюсти повернут наружу.
Хромосом в диплоидном наборе у бразильского агути 64.
Распространены от Южной Мексики на юг через Центральную Америку до Южной Бразилии. Встречаются также на Малых Антильских о-вах.
Населяют прохладные, влажные места, низменные леса, берега водоемов, поросшие травянистой растительностью, густые кустарники, а также высокие сухие склоны холмов, саванны, иногда антропогенные ландшафты. Ведут дневной образ жизни, но в тех местах, где их часто беспокоят, переходят к ночному существованию. Норы выкапывают сами. Бегают с большой быстротой. Питаются плодами, различными вегетативными частями растений, в том числе сельскохозяйственных культур. По-видимому, дают 2 помета в году. Длительность беременности около 3 месяцев. Детенышей в помете от 2 до 4 (обычно 2). Продолжительность жизни в неволе 13—20 лет. Хорошо приручаются. Местное население охотится за агути из-за их вкусного мяса.
В роде, вероятно, 12 видов:
бразильский агути — D. aguti Linnaeus, 1766 (Бразилия, Гайана, Гвиана, Венесуэла, о-в Тринидад);
D. azarae Lichtenstein, 1923 (юг Бразилии, Парагвай и северо-восток Аргентины);
гребенчатый агути— D. cristata Dcsmarcst, 1822 (Суринам);
D. fuliginosa Wagler, 1832 (Западная Колумбия, Бразилия, Южная Венесуэла, Перу);
агути Калиновского — D. kalinowskii Thomas, 1897 (юго-восток Перу);
D. prymnolopha Wagler, 1831 (северо-восток Бразилии, Гайана);
агути — D. punctata Gray, 1842 (Южная Мексика, штаты — юг Чиапас, восток Табаско, Кампече, Юкатан, Кинтано-Роо; Белиз; Гватемала,
кроме северо-запада; Сальвадор; Гондурас; Никарагуа; Коста-Рика; Панама; юг и северо-запад Колумбии, крайний северо-запад Венесуэлы; юго-запад Бразилии; Перу; Эквадор; Боливия);
сентвинсентский агути — D. albida Gray, 1842 (о-в Сент-Винсент в группе Малых Антильских островов);
антильский агути — D. antillensis Sclater, 1875 (о-в Сент-Люсия в группе Малых Антильских островов);
гваделупский агути — D. noblei G. Allen, 1914 (о-в Гваделупа в группе Малых Антильских островов);
мексиканский агути — D. mexicana Saussure, 1860 (Мексика — южная половина штата Веракрус, западная половина штата Табаско);
руатанский агути — D. ruatanica Thomas, 1901 (о-в Руатан около Гондураса).
Число видов в роде, вероятно, должно быть уменьшено. Андерсон и Джонс (1967) насчитывают в роде 7 видов.
Род акуши
Genus Myoprocta Thomas, 1903
Размеры мелкие. Длина тела 32—38 см. Длина хвоста 4,5—7 см. Внешне напоминают агути. Окраска спинной стороны тела рыжеватая или черноватая, иногда темно-зеленоватая. У некоторых особей морда и бока головы ярко-желтого, оранжевого или рыжего цвета. Брюшная -сторона рыжеватая, желтоватая, коричневая или беловатая.
Строение черепа сходно со строением черепа агути, но зубы меньше.
Хромосом в диплоидном наборе у акуши 62.
Распространены в Гвиане, Бразилии, Колумбии, Эквадоре, Перу, Венесуэле.
Населяют влажные леса. Экология изучена слабо.
В роде 2 вида;
акуши — AL acouchy Erxleben, 1777 (Гвиана, Бразилия, Эквадор);
М. pratti Pocock, 1913 (Бразилия, Венесуэла, юг Колумбии, северо-восток Эквадора, север Перу).
СЕМЕЙСТВО ШИНШИЛЛОВЫХ
FAMILIA CHINCHILLIDAE THOMAS, 1896
Телосложение довольно изящное. Размеры мелкие или средние. Длина тела 22—66 см. Длина хвоста 7.5—32 см. Масса взрослой особи варьирует от 0,5 до 7 кг. У Lagostomus самцы крупнее самок, а у Chinchilla, наоборот, самка крупнее самцов. Голова остносительно крупная, несколько уплощенная. Морда широкая. Глаза большие. Уши округлые, маленькие у Lagostomus и большие у Lagidium и Chinchilla. Хвост по всей длине густо покрыт волосами. Задние конечности длинные, сильные. Стопа удлинена и имеет 3 {Lagostomus) или 4 пальца {Lagidium, Chinchilla). Передние конечности короткие. Когти на задних лапах крупные, острые у Lagostomus и слабые, тупые у Lagidium и Chinchilla. Подошвы голые. Волосяной покров довольно высокий, густой, очень мягкий. Окраска спинной стороны тела варьирует от желтоватой до темно-серой или сероватой с белыми полосами, проходящими по бокам морды от носа до глаз. При испуге могут терять часть или весь волосяной покров (линька испуга). Вибриссы обычно длинные. Сосков одна пара — по бокам тела в области груди.
В черепе скуловые кости обычно соприкасаются с большими слезны ми костями. Костные слуховые барабаны у Lagostomus небольшие, у Lagidium средних размеров и у Chinchilla крупные. Скуловая дуга без: щечных пластин, утолщенная спереди. Костное небо сужено в переднем отделе. Резцовые отверстия длинные, узкие. Щечные зубы не имеют корней и растут в течение всей жизни животного; они образованы поперечными пластинами без цемента. Резцы относительно узкие. Малая берцовая кость небольшая и не сращена с большой берцовой. Ключица хорошо развита.
Распространены в Южной Америке: на равнинах Аргентины, в высокогорьях Перу, Боливии и Аргентины, на юг примерно до 52° ю. ш.
Населяют склоны гор от 800 до 6000 м над уровнем моря или пампасы. Активны днем или ночью. Держатся группами от нескольких особей до ста. Быстрые зверьки, передвигающиеся на всех четырех конечностях. Норы выкапывают сами. В одной норе обычно живет семейная группа из 2—5 животных. Иногда поселяются в расщелинах скал, в щелях под камнями. Питаются различными растительными кормами. Длительность беременности примерно 3—5 месяцев. В течение года бывает 1—3 помета от 1 до 6 детенышей в каждом. Детеныши рождаются хорошо развитыми. Лактационный период короткий; вскоре после рождения детеныши начинают питаться растениями. Животных промышляют из-за ценного меха. Разводят на зверофермах. Мясо употребляют в пищу. В результате охраны численность в последние годы увеличилась.
В семействе 3 рода (6 видов):
равнинных вискаш — Lagostomus Brooks, 1828 (1 вид);
горных вискаш — Lagidium Meyen, 1833 (3 вида);
шиншилл — Chinchilla Bennett, 1829 (2 вида).
Род равнинных вискаш
Genus Lagostomus Brooks, 1828
В роде единственный вид: равнинная вискаша — L. maximus Des-marest, 1817.
Размеры наибольшие в семействе. Длина тела 47—66 см. Длина хвоста 15—20 см. Масса до 7 кг. Голова большая, с короткой мордой, уши относительно короткие (рис. 166). Передние конечности четырехпалые, с тупыми когтями. Задние конечности длинные, трехпалые, с чрезвычайно’ толстыми острыми когтями. На третьем пальце растут длинные щетинковидные волосы. Волосяной покров сравнительно менее мягкий, чем у других представителей семейства. Окраска его темно-серая на спинной стороне, белая на брюшной. На морде имеются продольные белые и черные полосы.
Череп уплощен. Лобные кости широкие. Заглазпичпые отростки хорошо выражены. Носовые кости относительно длинные и узкие. Сагиттальный гребень крупный. Костные слуховые барабаны небольших размеров. Подглазничные отверстия имеют канал для нерва. Костное небо-сильно сужено в передней части. Скуловые кости резко подняты в передней части и соприкасаются со слезными костями. Нижняя челюсть имеет довольно крупный гребень рядом с сочленовапной головкой. Венечный-отросток низкий. Угловой отросток слегка повернут наружу. Резцы средней ширины, их поверхность покрыта мелкими продольными бероздками.
Хромосом в диплоидном наборе 56.
Распространена в Аргентине и на крайнем юге Парагвая.
Населяет пампасы и заросли кустарника на равнинах. Иногда под-
яимается в горы до 2000 м над уровнем моря. Живут колониями. В колонии многочисленные норы связаны подземными переходами. Некоторые норы используются равнинными вискатами в течение столетни. В колонии живет обычно 15—30 особей, возглавляемых взрослым самцом. Зверьки очень чистоплотны и вытаскивают из нор на поверхность все ненужные остатки. Норы имеют округлые камеры, расположенные примерно на глубине 1,5 м от поверхности. Временных пор обычно нс делают. Могут передвигаться со скоростью до 40 км в час, делать прыжки до 3 м длиной. Во время бега делают резкие повороты в стороны. Активны >ночыо. Питаются надземными и подземными частями различных расте-
Рис. 167. Горная вискаша — L. viscaccia: Л — кисть, Б — стопа
ний. Продолжительность беременности около 5 месяцев. Самки приносят детенышей преимущественно в июле — сентябре. В течение года бывает один помет, обычно из 2 детенышей. Лактационный период короткий. Половозрелое™ достигают примерно в 8 месяцев.
L. crassus, обитавшая в Неру вымерла, по-видимому, в недалеком прошлом.
Род горных вискаш
Genus Lagidium Meyen, 1833
Размеры мелкие и средние. Длина тела 32—40 см. Длина хвоста 23— 32 см. Масса взрослых особей 0,9—1,6 кг.По внешнему виду несколько напоминают кроликов, но с длинным хвостом (рис. 167). Уши крупные. Стопа удлиненная, узкая. Передние и задние конечности четырехпалые с тупыми слабыми когтями. Волосяной покров очень густой, мягкий, за исключением хвоста, где он довольно грубый. Окраска спинной стороны тела темно-желтая или темпо-серая. Брюшная сторона тела обычно беловатая, желтоватая или светло-серая. У некоторых форм вдоль середины спины проходит черная полоса. Кончик хвоста черный или рыжевато-коричневый. Сосков одна пара.
Череп узкий, с длинным лицевым отделом. Мозговая коробка уплощенная, округлая. Сагиттального гребня нет. Костные слуховые барабаны очень большие. Резцовые отверстия длинные и узкие. Костное небо сильно сужено спереди. Подглазничные отверстия не имеют канала для нерва. Скуловые кости очень широкие и спереди соприкасаются со слезными костями. В нижней челюсти рядом с сочленованной головкой расположен небольшой гребень.
Распространены в Перу, Боливии, Чили, Аргентине, Патагонии. Южная граница ареала проходит примерно на уровне 52° ю. ш.
Населяют сухие скалистые участки с очень бедной растительностью. Встречаются на высоте от 900 до 5000 м над уровнем моря. Часто держатся около водоемов. Активны днем. Живут колониями от нескольких особей до 80. Убежищем служат расщелины скал, пустоты под камнями. Собственные норы выкапывают редко. Проворно лазают по скалам и между камнями. В одном и том же убежище обычно держится небольшая, по-видимому, семейная группа из 2—5 особей. Питаются травами, мхами, лишайниками. Период спаривания приходится на октябрь — ноябрь. Беременность длится около 3 месяцев. В течение года может быть 2 или 3 помета, обычно по одному детенышу. Половозрелости достигают в возрасте около 7 месяцев. Длительность жизни 2—3 года.
В роде 3 вида:
перуанская вискаша — L. peruanum Meyen, 1833 (Перу);
горная вискаша — L. viscaccia Molina, 1782 (Северо-Западная, Западная и Юго-Западная Аргентина, Южная и Западная Боливия, север и центр Чили, Перу);
вискаша Вольффсона— L. wolffsohni Thomas, 1907 (Юго-Западная Аргентина и прилегающие районы Чили).
Эллермэн (1949) насчитывает в роде 4 вида.
Род шиншилл
Genus Chinchilla Bennett, 1829
Размеры наименьшие в семействе. Длина тела 22—38 см. Длина хвоста 7,5—15 см. Масса взрослой особи от 0,5 до 1 кг. Глаза большие, черные. Уши крупные (рис. 168). Есть рудиментарные защечные мешки. Стопа удлиненная, узкая. На внутреннем пальце имеется пучок длинных жестких волос. Передние конечности четырехпалые, задние — также четырехпалые, но с сильно укороченным пятым пальцем. Пальцы вооружены слабыми когтями. Волосяной покров необычайно густой, очень мягкий. Окраска спинной стороны тела голубовато-серая или коричневатосерая с черноватым оттенком. Брюшная сторона желтовато-белая. Хвост покрыт удлиненными грубыми волосами темно-серого или коричневого цвета. Многочисленные вибриссы белые или черные, очень длинные.
В черепе хорошо выражено межглазничное сужение. Костное небо узкое, сильно суженное спереди. Костные слуховые барабаны сильно вздуты. Скуловые кости обычно не достигают слезной кости. В подглазничном отверстии канала для нерва нет. Резцовые отверстия очень длинные. Нижняя челюсть с узким угловым отростком, резко направленным вверх. Расположенный на ней гребень рядом с сочленовным отростком слабо развит.
Хромосом в диплоидном наборе у шиншиллы 64.
Распространены в Андах, Аргентины, Перу, Чили и Боливии.
Держатся на скалистых участках высокогорья па высоте 3— 6 тыс. метров над уровнем моря. Живут колониями. Убежищем служат расщелины скал, пустоты под камнями. Раньше встречались крупными колониями до сотни и более особей. Активны ночью. Питаются различными растениями высокогорья. По-видимому, моногамы. Дают от 1 до 3 пометов в течение года, в каждом от 1 до 6 детенышей. Длительность беременности составляет примерно 112 дней. Детеныши способны быстро передвигаться уже спустя несколько часов после рождения, а через несколько дней начинают питаться самостоятельно. Половозрелости дости-
Рис. 168. Шиншилла — С. lanlger и ее череп
гают в 5—8 месяцев. Продолжительность жизни около 10 лет; в неволе, однако, животные могут жить до 20 лет. Шиншиллы усиленно промышляются из-за ценного меха и были почти уничтожены в результате неумеренного промысла. В настоящее время благодаря охране их численность несколько возросла. Шиншилл разводят па зверофермах. В последние годы сделаны успешные попытки разведения их па фермах в СССР. В 1964—1970 гг. пробовали акклиматизировать шиншиллу на Западном и Восточном Памире. Было выпущено 211 зверьков; успеха добиться не удалось.
В роде, по видимому, 2 вида:
короткохвостая шиншилла — С. brevicauda Waterhouse, 1848 (Анды Южной Боливии, Северо-Западной Аргентины и Южного Перу);
шиншилла — С. laniger Molina, 1828 (север Чили).
Некоторые исследователи (например, Эллермэн, 1940; Уокер, 1967) объединяют оба вида в один — С. laniger.
Шиншилла включена в Красную книгу.
СЕМЕЙСТВО ХУТИЙ
FAMILIA CAPROMYIDAE SMITH, 1842
Грызуны мелких и средних размеров. Длина тела 30—50 см. Длина хвоста 3,5—30 см. Масса взрослой особи может достигать 7 кг. Сложение тяжелое. Голова крупная с притупленной широкой мордой. Глаза маленькие. Уши небольшие, мясистые. Хвост голый или покрыт волосами, иногда хватательный. Конечности относительно короткие. Кисть и стопа широкие. Пальцы с сильными когтями. Первый палец передней конечности уменьшен. Подошвы голые, без мозолей. Волосяной покров грубый. Окраска его серая или коричневая, иногда с красноватым оттенком. Сосков 2 пары. Семенники располагаются под кожей в паховых областях.
Гипселодонтные щечные зубы растут в течение всей жизни животного. Складки на их жевательной поверхности заполнены цементом. Резцы узкие. Os penis имеется.
Распространены на Больших Антильских и Багамских о-вах.
Населяют лесные, иногда холмистые районы, антропогенные ландшафты. Ведут наземный или древесный (Capromys) образ жизни. Питаются травянистыми растениями, листьями, плодами, корой деревьев. Самка приносит до 4 детенышей, коГорые рождаются полностью покрытые волосами, с открытыми глазами.
В семействе 3 рода (И видов):
кубинских хутий — Capromys Desmarest, 1822 (4 вида); короткохвостых хутий — Geocapromys Chapman, 1901 (3 вида); гаитянских хутий — Plagiodontia F. Cuvier. 1836 (4 вида).
Некоторые исследователи (например, Эллермэн, 1940; Симпсон, 1945) включают в семейство хутий нутрий.
Иногда (Грассе и Декейзер, 1955) Plagiodontia переносят в семейство Myocastoridae.
Семейство, по-видимому, включает также 3 рода: Isotobodon J. Allen, 1916; Aphaetreus Miller, 1922, и Hexolobodon Miller, 1929, представители которых вымерли в недалеком прошлом (но еще до того времени, когда Колумб достиг Америки).
Род кубинских хутий
Genus Capromys Desmarest, 1822
Длина тела 30—50 см. Длина хвоста 15—30 см. Масса взрослой особи 4—7 кг. Внешним обликом несколько напоминает больших крыс (рис. 169). Длинный хвост слабо оволоснен; может быть хватательным (С. prehensilis). Пальцы широких лап с длинными когтями. Первый палец передних конечностей мал, задних — средних размеров. Грубый волосяной покров состоит из длинных, грубых остевых волос и довольно редкого пуха. Окраска спинной стороны тела темпо-желтая, сероватая, коричневатая, красноватая или почти черная. Брюшная сторона обычно более светлая. Одноцветный хвост темно-желтого или темно-красновато-коричневого цвета. Характерно подразделение желудка на 3 камеры; это один из наиболее сложных желудков в отряде грызунов.
Череп удлинен и уплощен. Теменные кости могут иметь сильно развитой гребень. Скуловые дуги сильные с крупным, направленным назад отростком. Подглазничные отверстия не имеют канала для нервов. Костные слуховые барабаны крупные. Костное небо слегка сужено в переднем
Рис. 169. Кубинская хутия — С. pilorid.es
отделе. Резцовые отверстия средних размеров. Нижняя челюсть с угловым отростком, наклоненным назад и наружу. Венечный отросток низкий. Резцы узкие.
Хромосом в диплоидном наборе у С. pilorides 40.
Распространены на о-ве Куба.
Населяют лесные угодья. Живут обычно парами. Активны днем. Обитают на деревьях. Питаются плодами, листьями, мелкими животными, в том числе пресмыкающимися. Беременность длится 17—18 недель. В помете 1—3 детеныша. В возрасте 10 дней детеныши начинают питаться твердой пищей. Местное население употребляло кубинских хутий в пищу. В настоящее время охраняются государством.
В роде 4 вида:
кубинская хутия — С. pilorides Say, 1822 (о-ва Куба и Пинос);
цепкохвостая хутия — С. prehensilis Poeppig, 1824 (о-ва Куба и Пи-нос) ;
кистехвостая хутия — С. melanurus Peters, 1864 (о-в Куба);
крошечная хутия — С. папа G. Alien, 1917 (о-в Куба).
Procapromys gleayi Pousargues, 1899, описанная по одному экземпляру из Венесуэлы, по-видимому, молодая С. pilorides, завезенная туда человеком.
Кистехвостая и крошечная хутии как редкие звери внесены в Красную книгу.
Род короткохвостых хутий
Genus Geocapromys Chapman, 1901
Длина тела 33—45 см. Длина хвоста 3,5—5,7 см. Сходны с кубинскими хутиями, но хвост значительно более короткий и когти менее длинные. Первый палец передних конечностей редуцирован сильнее. Небольшие, округлые уши покрыты короткими волосами. Низкий, плотный мех имеет на спинной стороне тела желтовато-серую, темно-коричневую или черноватую окраску, на брюшной — темио-желто-серую или темно-коричневую.
Рис. 170. Череп хутии Брауна — G. brownii
Череп и зубная система сходны с черепом и зубами кубинских хутий (рис. 170). У взрослых животных может быть развит сагиттальный гребень.
Хромосом в диплоидном наборе у хутии Брауна 88.
Распространены на о-вах Ямайке и Багамских.
Населяют скалистые районы. Ведут наземный образ жизни. Убежища расположены под камнями, в дуплах упавших деревьев, среди древесных корней. Держатся, по-видмому, поодиночке. Пища в основном состоит из травянистых растений, листьев кустов и, вероятно, также плодов и грибов. В свое время многочисленные короткохвостые хутии служили важным источником пищи для местного населения. В настоящее время они стали редки вследствие не только неумеренной охоты, но и уничтожения их мангустами, которые были ввезены на о-в Ямайку в 1872 г. С 1922 г. охраняются государством.
В роде 3 вида:
хутия Брауна — G. brownii Fischer, 1829 (о-в Ямайка);
хутия Инграхама—G. ingrcthctmi J. Allen, 1891 (Багамские о-ва);
короткохвостая хутия — G. columbictnus Chapman, 1892 (о-ва Куба и Пинос).
Хутия Инграхама и 2 подвида хутии Брауна — G. brownii brownii Fischer, 1830, и G. brownii thoracatus True, 1889, включены в Красную книгу.
Род гаитянских хутий
Genus Plagiodontia F. Cuvier, 1836
Длина тела 31—40 см. Длина хвоста 12—15 см. Напоминают других представителей семейства. Кисть и стопа крупные. Хвост лишен волос и покрыт чешуйками. Когти па лапах сильно развиты. Пятый палец задних конечностей относительно длинный. Первый палец передних конечностей имеет короткий, тупой ноготь. Волосяной покров относительно низкий, плотный. Окраска его на спинной стороне тела коричневатая или сероватая, на брюшной — темно-желтая.
В черепе скуловые кости не имеют отростков. Межглазпичное сужение выражено слабо. Щечные зубы с постоянным ростом. Верхние корен-
иые зубы только с одной очень длинной и глубокой складкой, проникающей глубоко в зуб и заполненной цементом. Нижние коренные зубы с двумя длинными и глубокими внутренними и одной мелкой наружной складками.
Распространены на о-ве Гаити.
Экология изучена слабо. По-видимому, могут лазать по деревьям. День проводят в дуплах. Активность в основном ночная. Питаются главным образом корнями и плодами. В помете один детеныш. Численность невелика, в частности, вследствие уничтожения их мангустами, ввезенными на о-в Гаити.
В роде 4 вида:
Р. aedium F. Cuvier, 1836 (восток о-ва Гаити);
Р. ipnaeum Johnson, 1948 (восток о-ва Гаити);
доминиканская хутия — Р. hylaeum Miller, 1927 (восток о-ва Гаити); малая хутия — Р. spelaeum Miller, 1929 (о-в Гаити).
Р. ipnaeum и Р. spelaeum вымерли.
Доминиканская хутия и Р. aediurn включены в Красную книгу.
Грассе и Декайзер (1955) относят гаитянских хутий к семейству нутрий. По-видимому, более правильно считать этот род. в пределах семейства хутий.
СЕМЕЙСТВО НУТРИЕВЫХ
FAMILIA MYOCASTORIDAE MILLER ET GIDLEY, 1918
Грызуны средних размеров. Длина тела до 60 см. Длина хвоста до 45 см. Максимальная масса самца до 8,2 кг, самки — 8,0 кг. Ряд морфологических признаков обусловлен приспособлением к водному образу жизни. Телосложение тяжелое, довольно неуклюжее (рис. 171). Голова треугольной формы. Глаза и уши маленькие. Конечности относительно короткие. Задние длиннее и массивнее передних. Стопа значительно крупнее кисти. На передних конечностях 4 пальца развиты хорошо, а первый палец рудиментарный. Задние конечности пятипалые, причем 4 внутренних пальца соединены между собой сильно развитой плавательной перепонкой. Когти острые и высокие. Хвост округлый в сечении, почти лишен волос и покрыт чешуйками. Вибриссы длинные. Волосяной покров густой, мягкий, высокий. Окраска спинной стороны тела темно-желтовато-коричневая или красновато-коричневая. Бока и брюхо несколько более светлые. Характерно расположение на боках туловища 4— 5 пар сосков. У самцов и самок имеется крупная анальная железа.
Череп уплощен сверху. Лицевой отдел укороченный, с сильно развитыми гребнями. Мощные скуловые дуги расширены в задней части. Надглазничные отростки небольшие. Костное небо спереди сильно сужено. Костные слуховые барабаны небольшие, толстостенные. В нижней челюсти угловой отдел имеет загнутый наружу нижний край и длинный отогнутый внутрь угловой отросток. Венечный отросток сильно редуцирован, сочленованнын—короткий и массивный.
Гипселодонтные щечные зубы уменьшаются в размерах сзади наперед. Корни их нечетко выражены. Коронки расширенные, с крупными наружными и внутренними складками. Резцы мощные, широкие.
Хромосом в диплоидном наборе 42.
Распространение охватывает южную часть Южной Америки от Парагвая и Южной Бразилии на юг до Магелланова пролива. Акклиматизирована во многих местах Европы и Северной Америки.
Рис. 171. Нутрия и ее череп
Ведут полуводный образ жизни. Населяют берега медленно текущих рек, озер, пресноводных болот. Встречаются также и около солоноватых водоемов. Устраивают поры в берегах или гнезда пал водой в густых зарослях водных растений. На берегу в густой траве протаптывают тропы. Обычно не уходят от норы дальше 200 м. Хорошо плавают п пыряют. Питаются различными водными растениями. Животные корма поедают редко; в основном при недостатке растительных. Для самок характерен полиэстральнын тип размножения. Эстральный цикл продолжается от 24 до 29 дней. Длительность эструса от 2 до 4 дней. Беременность длится примерно 4 месяца. В течение года бывает 2 помета: в каждом от 2 до 8, в среднем 5 детенышей. Половозрелости достигают в 3—8 месяцев. Молодые рождаются хорошо развитые, зрячие, покрытые
шерстью. Живут парами. Нутрия акклиматизирована в ряде стран мира, в том числе и в СССР. В 1930—1932 гг. в нашу страну было завезено 2656 нутрий, в том числе 676 из Аргентины и 1980 со звероводческих ферм Англии и Германии. С 1930 по 1963 г. в разных участках южных районов Советского Союза было выпущено 6270 зверьков. Акклиматизация удалась в Закавказье в районе Ленкорани, в бассейне Нижней Куры в Грузии, а также в южном Таджикистане. Известный успех имеет и клеточное разведение нутрий.
В семействе один род: нутрий — Myocastor Kerr, 1792, и 1 вид.
Симпсон (1954) включает нутрий в семейство хутий, даже без выделения их в отдельное подсемейство. Некоторые другие исследователи, например Холл и Келсон (1959), выделяют нутрий в качестве особого подсемейства в семействе хутий. Эллермэн (1940) рассматривает нутрий в качестве подсемейства семейства Echimyidae Miller et Gidley, 1918. По-видимому, более правильно выделить нутрий в особое семейство, как это делают, например, А. А. Громов (1963), Б. А. Кузнецов (1965), Андерсон и Джонс (1967) и др. Грассе и Декейзер (1955) относят род Р1а-giodontia к семейству нутрий, что, по-видимому, неправильно. Целесообразнее отнести этот род к семейству хутий.
Род нутрий
Genus Myocastor Kerr, 1792
См. описание семейства.
В роде 1 вид: М. coypus Molina, 1782 (ареал совпадает с ареалом семейства).
СЕМЕЙСТВО ВОСЬМИЗУБОВЫХ
FAMILIA OCTODONTIDAE WATERHOUSE, 1839
Грызуны мелких размеров. Длина тела 12,5—19,5 см. Длина хвоста 4—18 см. Телосложение довольно тяжелое. Внешним обликом несколько напоминают крыс. Некоторые представители имеют приспособления к подземному существованию. Голова относительно крупная. Уши средних размеров, округлые. Вибриссы длинные. Хвост покрыт грубыми короткими волосами, которые удлиняются к его концу, и могут образовывать кисть. Когти средних размеров искривленные и острые. Щетки грубых волос на пальцах задних лап выдаются за пределы когтей. Первый палец на передних конечностях редуцирован. Волосяной покров на туловище обычно высокий, густой, мягкий. Его окраска варьирует от сероватой до красноватой. Окраска может быть одноцветная или брюшная сторона тела резко контрастирует со спиной своей белой окраской. Сосков 4 пары. Из них 3 передние пары располагаются по бокам тела, а задняя — в паховой области.
В черепе костные слуховые барабаны сильно или умеренно вздуты. Костное небо закапчивается между рядами щечных зубов. Жевательная поверхность щечных зубов имеет эмалевые петли в виде цифры 8, иногда бобовидной формы (рис. 172). Резцовые отверстия небольшие. Вертикальный диаметр подглазничного отверстия больше горизонтального диаметра.
Распространение охватывает Южную Америку от Южного Перу до Северо-Западной Аргентины и Чили.
Населяют различные биотопы, включая антропогенные ландшафты и
скалистые склоны гор. В горы поднимаются до 3,5 тыс. метров над уровнем моря. Убежищем служат пустоты под камнями, расщелины скал. Могут выкапывать собственные норы со сложной системой ходов и несколькими выходами. Устраивают гнезда в норах или у корней в густых кустах. Иногда поселяются группами. Питаются в основном луковицами, клубнями, побегами и корой. Могут запасать пищу в подземных камерах. Активны как днем, так и ночью. Некоторые виды ведут преимущественно подземный образ жизни (Spalacopus). Могут лазать по ветвям кустарников. В течение года дают 2 помета, в каждом, по-видимому, по 2 детеныша. Лактационный период продолжается около 2 месяцев.
В семействе 5 родов (8 видов):
восьмизубов—Octodon Bennett, 1832 (Звида); верхтость^ер^ни^мГй соневидных восьмизубов — Octodontomys нижних (Б) щечных зу-
Palmer, 1903 (1вид); бов вискашевой крысы
слепышовых восьмизубов — Spalacopus Wag-
ler, 1832 (1 вид);
крысиных восьмизубов — Aconaemys Ameghino, 1891 (1 вид);
вискашевых крыс — Octomys Thomas, 1920 (2 вида).
Род восьмизубов
Genus Octodon Bennett, 1832
Длина тела 12,5—19,5 см. Длина хвоста 10,5—16,5 см. Уши крупные (рис. 173). Передние конечности с четырьмя сильно развитыми пальцами и рудиментарным первым. Задние конечности с укороченным первым пальцем и пятым более коротким, чем три средних. Когти на пальцах небольшие. Кисточки удлиненных волос на конце хвоста нет. Волосяной покров грубее, чем у других представителей семейства. Окраска спинной стороны тела сероватая или коричневая, часто с оранжевым оттенком, брюшной — кремово-желтая.
Рис. 173. Дсгу — О. degus
Череп не имеет сужения в межглазничной области. Костные слуховые барабаны относительно небольшие. 8-образная форма эмалевых петель жевательной поверхности щечных зубов выражена нечетко. Третий коренной зуб самый маленький.
Хромосом в диплоидном наборе у дегу 58.
Распространены в Чили и Перу.
Населяют прибрежные холмистые районы. Поднимаются в горы до 1200 м над уровнем моря. Экология изучена слабо. Ведут наземный, образ жизни. Активность дневная.
В роде 3 вида:
О. bridgesi Waterhouse, 1844 (Анды в Чили);
дегу — О. degas Molina, 1782 (Чили);
О. lunatus Osgood, 1843 (Центральные районы Чили).
Род соневидных восьмизубов
Genus Octodontomys Palmer, 1903
В роде единственный вид: соневидный восьмизуб — О. gliroides Gervais et D’Orbigny, 1844.
Длина тела примерно 18 см. Длина хвоста около 15 см. На конце хвоста хорошо развитая кисточка из удлиненных волос. Волосяной покров густой, мягкий. Окраска его на спинной стороне тела темно-серая, смешанная с черным, на брюшной — снежно-белая. Конечности снаружи имеют ту же окраску, как и спина, а внутренняя часть их белая.
В черепе костные слуховые барабаны относительно невелики, однако более крупные, чем у рода восьмизубов. Подглазничные отверстия имеют канал для нерва. Наружные и внутренние выемки жевательной поверхности щечных зубов не соединяются друг с другом посредине этих зубов, образуя незамкнутую посредине восьмерку.
Хромосом в диплоидном наборе 38.
Распространен в Андах, Юго-Западной Боливии и Северо-Западной Аргентине.
Населяет высокогорные районы. Экология изучена слабо. Держится среди скал и кактусов. Активен в основном ночью.
Род слепышовых восьмизубов
Genus Spalacopus Wagler, 1832
В роде единственный вид: слепышовый восьмизуб — S. cyanus Molina, 1782.
Длина тела 14—16 см. Длина хвоста 4—6 см. Внешний облик имеет ряд признаков приспособления к подземному образу жизни. Сложение тяжелое. Голова крупная. Уши небольшие (рис. 174). Крупные, длинные, сильно расширенные верхние резцы выдаются изо рта вперед. Конечности укороченные. Когти не очень сильно увеличены. Хвост короткий, покрыт волосами. Волосяной покров мягкий, блестящий. Окраска его коричневато-черная или черная, одноцветная по всему туловищу.
В черепе лобные кости сужены. Костные слуховые барабаны меньших размеров по сравнению с другими представителями семейства. Резцовые отверстия небольшие. Подглазничные отверстия не имеют канала для нерва. Жевательная поверхность щечных зубов имеет эмалевые петли 8-образной формы, но выемки в середине зуба не соединяются.
Рис. 174. Слепышовый восьмизуб — 5. cyanus и его череп
Распространен в Чили.
Населяет холмистые и горные районы, а также прибрежные области. В горы поднимается примерно до высоты 3000 м над уровнем моря. Экология изучена слабо. Ведет преимущественно подземный образ жизни. Строит длинные п сложные норы. Питается в основном подземными частями растений. На зиму запасает луковицы и клубни. В течение года, по-видимому, бывает 2 помета.
Род крысиных восьмизубов
Genus Aconaemys Ameghino, 1891
В роде единственный вид: крысиный восьмизуб — A. fuscus Waterhouse, 1841.
Длина тела 15—18 см. Длина хвоста 5,5—7,5 см. Имеет ряд приспособлений к подземному образу жизни. Телосложение тяжелое. Конечнос
ти укороченные. Когти крупные. Уши средних размеров. Хвост покрыт редкими волосами. Волосяной покров довольно мягкий. Окраска его одноцветная, темно-коричневая. Хвост может быть двухцветный: черный сверху, белый снизу.
Лицевой отдел черепа укорочен. Костные слуховые барабаны небольшие или средних размеров. Резцы сильно расширены. Эмалевые петли на жевательной поверхности щечных зубов имеют четко выраженную форму восьмерки, причем внутренние и наружные выемки встречаются посредине зуба.
Распространен в Чили и, возможно, в прилегающих районах Аргентины.
Населяет прибрежные области или склоны гор. Экология изучена слабо. Копает сложно устроенные, поверхностно расположенные подземные ходы. Зимой делает ходы под снегом.
Род вискашевых крыс
Genus Octornys Thomas, 1920
Длина тела 16—17 см. Длина хвоста 17—18 см. На конце хвоста кисточка удлиненных волос. Волосяной покров очень мягкий. Окраска его на спинной стороне тела темно-желтая, на брюшной и на конечностях белая.
Череп с заостренным, но не укороченным лицевым отделом. Сужения в межглазничной области нет. Костные слуховые барабаны относительно наиболее крупные в семействе, сильно вздутые. Резцовые отверстия короткие, широкие. Подглазничные отверстия с каналом для нерва. Эмалевые петли жевательной поверхности щечных зубов имеют отчетливо выраженную форму восьмерки. Наружные и внутренние выемки соприкасаются посредине зуба.
Распространены в Аргентине.
Обитают в скалистых областях гор на высоте 1000—1200 м над уровнем моря. Численность невелика, встречаются редко. Экология не изучена. Ведут ночной образ жизни. День проводят в норах, которые выкапывают сами. В течение года бывает несколько пометов.
В роде 2 вида:
вискашевая крыса — О. тутах Thomas, 1920 (горы Западной Аргентины) ;
О. barrerae Lawrence, 1941 (центральные районы Западной Аргентины) .
Некоторые исследователи (например, Кабрера, 1960) выделяют О. barrerae в особый род Tympanoctomys Yepes, 1940.
СЕМЕЙСТВО ТУКОТУКОВЫХ
FAMILIA CTENOMYIDAE ТАТЕ, 1935
Размеры мелкие. Длина тела 17—25 см. Длина хвоста 6—11 см. Масса взрослого животного 200—700 г. Морфологические признаки показывают высокую степень приспособленности к подземному образу жизни. Телосложение тяжелое. Голова крупная, шея толстая и короткая. Небольшие глаза расположены высоко на голове (рис. 175). Ушные раковины сильно редуцированы. Морда несколько уплощенная. Туловище цилиндрическое. Конечности укороченные, мускулистые. Хвост корот-
Рис. 175. Ту ко ту ко — Ctenomys sp и череп С. tuconax
кий, округлый в сечении. Передние конечности несколько короче задних. Кисть и стопа широкие, пятипалые. Все пальцы с длинными, мощными когтями, развитыми сильнее на передних конечностях. Стопа окаймлена щеткой жестких щетинковидных волос. Хвост покрыт редкими короткими волосами. Волосяной покров разной высоты и мягкост. Окраска его темно- или светло-коричневая, темно серовато-желтая пли темпо-желтая. Сосков 3 пары.
Череп с широким лицевым отделом. Заглазппчные отростки обычно имеются. Теменные кости с хорошо развитыми гребнями. Костные слуховые барабаны крупные. Подглазничные отворены не имеют канала для нерва. Скуловые кости с очень крупным, поднимающимся вверх отростком. Резцы мощные Веркине резцы несколько загнуты концами назад. Корни верхних резцов продолжаются далеко назад. Щечные зубы сверху уплощены, внутренняя складка их эмалевой стоики отсутствует. Последний коренной зуб верхней и нижней челюстей небольших размеров.
Хромосом в диплоидном наборе о г 26 у С. opimus, 36 у С. magella-nicus, до 48 у С. taiarum и С. porteousi и 61 у С. tuconax.
Распространены в Южной Америке от Южного Перу и Мату-Гросу в Бразилии на юг до Огненной Земли.
Обитают обычно в умеренной и субтропической зонах. Поднимаются в горы на высоту до 5000 м над уровнем моря, предпочитая высокогорные неокультуренные участки в различных биотопах. Ведут преимущественно подземный образ жизни. Выбирают, как правило, рыхлые или песчаные почвы, хотя встречаются и в самых разнообразных почвах с различной влажностью. Водяной туко-туко строит норы по берегам потоков, и, по-видимому, ведет полуводный образ жизни. Активны в вечерние и раннеутренние часы. На поверхность земли выходят редко. Роют сложную систему подземных нор, сообщающихся с центральной гнездовой камерой. Есть камеры и для запасов пищи. Землю из нор выталкивают задними конечностями. Питаются в основном подземными, сочными частями растений. Характерен громкий крик: как бы «туку-туку-туко» или «тлок-ток-тлок». В течение года обычно бывает один помет из 1—5 детенышей. Беременность длится 103—107 дней. В Уругвае период спаривания приходится на июль — октябрь. Новорожденные хорошо развиты. У С. peruanus детеныши почти сразу после рождения способны покинуть гнездо и питаться зелеными частями растений. Продолжительность жизни составляет примерно 3 года. В последнее время численность туко-туко резко упала.
В семействе 1 род: туко-туко — Ctenomys de Blainville, 1826, и 27 видов.
Род туко-туко
Genus Ctenomys de Blainville, 1826
См. описание семейства.
В роде 27 видов:
боливийский туко-туко — С. boliviensis Waterhouse, 1848 (Центральная и Южная Боливия);
бразильский туко-туко—С. brasiliensis Blainville, 1826 (Восточная Бразилия);
С. colburni J. Allen, 1903 (Юго-Западная Аргентина);
С. dorsalis Thomas, 1900 (Парагвай);
С. emilianus Thomas et Saint Leger, 1926 (Аргентина);
C. frater Thomas, 1902 (Северо-Западная Аргентина, Южная Боливия);
желтый туко-туко — С. fulvus Philippi, 1860 (Западная Аргентина, Северное Чили);
С. khighti Thomas, 1919 (Западная Аргентина);
С. leucodon Waterhouse, 1848 (Юго-Западная Боливия и Юго-Восточное Перу);
водяной туко-туко — С. lewisi Thomas, 1926 (Южная Боливия);
магелланский туко-туко — С. magellanicus Bennett, 1835 (Чили, Аргентина, Огненная Земля);
С. maulinus Philippi, 1872 (Чили);
С. mendocinus Philippi, 1869 (Аргентина);
малый туко-туко — С. minutus Nehring, 1887 (Бразилия);
туко-туко Наттерера — С. nattereri Wagner, 1848 (Бразилия);
С. opimus Wagner, 1848 (Северо-Западная Аргентина, Южное Перу, Юго-Западная Боливия и прилегающие районы Северного Чили);
С. perrensis Thomas, 1898 (Северо-Западная Аргентина);
перуанский туко-туко — С. peruanus Sanborn et Pearson, 1947 (крайний юг Перу);
С. pontifex Thomas, 1918 (Западная Аргентина);
С. porteousi Thomas, 1916 (Восточная Аргентина);
С. robustus Philippi, 1896 (Северное Чили);
С. saltarius Thomas, 1912 (Северная Аргентина);
С. sericeus J. Allen, 1903 (Юго-Западная Аргентина);
туко-туко Стейнбаха — С. steinbachi Thomas, 1907 (Боливия);
С. taiarum Thomas, 1898 (Восточная Аргентина);
С. tuconax Thomas, 1925 (Северо-Восточная Аргентина);
С. conoveri Osgood, 1946 (Парагвай).
Эллермэн (1940) относит Ctenomys к подсемейству Octodontinae семейства Echimyidae. По-видимому, более правильно выделять туко-туко в особое семейство.
СЕМЕЙСТВО ШИНШИЛЛОКРЫСОВЫХ
FAMILIA ABROCOMIDAE MILLER ET GIDLEY, 1918
Размеры мелкие. Длина тела 15—25 см. Длина хвоста 6—18 см. Внешним видом несколько напоминают крыс (рис. 176). Голова крупная, заостренная к носу. Конечности укороченные. Подошвы голые, покрыты мелкими бугорками. Хвост довольно густо покрыт короткими волосами. Уши округлые, относительно крупные. Глаза большие. Передние конечности короче задних и имеют по четыре пальца; когти на них маленькие. Задние конечности с небольшим первым пальцем, пятый палец короче трех средних. Когти как на задних, так и на передних конечностях слабые. Три средних пальца задних конечностей несут пучки удлиненных щетинкообразных волос, выдающихся за концы когтей. Волосяной покров мягкий,густой.
Окраска спинной стороны тела серебристо-серая или коричневатосерая, брюшной — белая, желтоватая или коричневатая. Под беловатым или желтоватым пятном на груди расположена крупная кожная специфическая железа. Сосков 2 пары.
В черепе лицевой отдел узкий, длинный. Мозговая капсула округлая, без граней. Лобные кости сильно сжаты. Костное небо узкое, короткое. Скуловые дуги простые. Подглазничные отверстия не имеют канала для нерва. Резцовые отверстия длинные, узкие. Костные слуховые барабаны очень крупные. Угловой отросток нижней челюсти длинный, слабый, венечный отросток низкий. Резцы узкие. Щечные зубы с открытыми корнями, растут в течение всей жизни. Верхние коренные зубы имеют одну внутреннюю и одну наружную выемки, нижние —две резко выраженные острые внутренние выемки и одну наружную. У A. bennetti 17 пар ребер — больше, чем у каких-либо других грызунов.
Распространены в Южной Америке: Перу, Боливии, Северо-Западной Аргентине, Чили.
Населяют скалистые области в горах па высоте от 1800 до 5000 м над уровнем моря. Часто встречаются в очень холодных местах, малопригодных для жизни других млекопитающих. Убежища находят под камнями и в расщелинах скал. Могут выкапывать собственные норы. Хорошо лазают по кустам и деревьям. По-видимому, ведут колониальный
Рис. 176. Шиншилловая крыса Беннетта — A. bennetti и ее череп
образ жизни. Встречаются редко; экология слабо изучена. Питаются различными растительными кормами. В помете, по-видимому, 2 детеныша.
В семействе 1 род: шиншилловых крыс — Abrocoma Waterhouse, 1837, и 2 вида.
Род шиншилловых крыс
Genus Abrocoma Waterhouse, 1837
См. описание семейства.
В роде 2 вида:
шиншилловая крыса Беннетта — A. bennetti Waterhouse, 1837 (Чили);
серая шиншилловая крыса —A. cinerea Thomas, 1919 (Южное и Юго-Восточное Перу, Южная Боливия, Северо-Западная и Западная Аргентина) .
Эллермэн (1940) считает шиншилловых крыс подсемейством семейства Echimyidae, а Ландри (1957)—подсемейством семейства Octodon-tidae.
СЕМЕЙСТВО колючих шиншилл
FAMILIA ECHIMYIDAE MILLER ET GIDLEY, 1918
Размеры от мелких до средних. Длина тела 8—48 см. Длина хвоста 4,5—43 см 1. Внешний облик колючих шиншилл напоминает таковой крыс (рис. 177). Морда заострена к носу или иногда притуплена. Уши и глаза средних размеров. Ушные раковины округлые, немного выдаются над мехом. Конечности средней длины. Первый палец на передних конечностях рудиментарный, остальные пальцы короткие или средней длины (Echimys, Diplomys, Isothrix) или сильно удлинены; иногда имеет место синдактилия. Хвост довольно густо покрыт короткими волосами или почти голый. Волосяной покров от мягкого (Cercomys) до грубого и иглистого. В последнем случае он представлен уплощенными острыми щетинковидными волосами, почти иглами. Окраска спинной стороны тела красновато-коричневая, темно-коричневая или черноватая. Брюхо сероватое или беловатое. Сосков 3 пары: две грудных и одна паховая.
В черепе лобные кости расширены. Костные слуховые барабаны довольно крупные. Коренные зубы с корнями и уплощенными коронками.
Распространение охватывает Центральную Америку (на север до Никарагуа) и Южную Америку на юг до Парагвая и Южной Бразилии, о-в Мартинику в группе Малых Антильских, а также о-в Пуэрто-Рико. На нем обитали представители вымерших в историческое время родов Iso-lobodon Allen, 1916; Heteropsomys Anthony, 1916, и Homopsomys Anthony, 1917.
Населяют лесные районы или разреженные леса, обычно около водоемов. Ведут в основном наземный или древесный (Euryzygomatomys) образ жизни. Хорошо плавают. Активны в сумерки или ночью. Питаются травой, плодами, орехами. Пьют много воды. Роют норы или используют в качестве убежищ пустоты под камнями и стволами деревьев. Некоторые живут в дуплах деревьев, иногда поселяются в постройках человека (Proechimys). Иногда держатся небольшими группами. В течение года дают, по-видимому, 2 помета, в каждом 1—6, обычно I—3 детеныша. Детеныши рождаются покрытые волосами, с открытыми глазами. Примерно через 11 дней они переходят на самостоятельное питание. Легко теряют кожу с хвоста, если хищник схватит за хвост. Численность в некоторых местах весьма велика. Некоторые виды колючих шиншилл местное население употребляет в пищу.
В семействе 14 родов (44 вида):
щетинистых крыс — Proechimys J. Allen, 1889 (12 видов); толстоиглых крыс — Hoplomys J. Allen, 1908 (1 вид);
гиар — Euryzygomatomys Goeldi, 1901 (1 вид);
клиомис-—Clyomys Thomas, 1916 (1 вид);
картеродон — Carterodon Waterhouse, 1848 (1 вид);
пунарес— Cercomys F. Cuvier, 1829 (I вид);
мезомис — Mesomys Wagner, 1845 (Звида);
лоихотрикс — Lonchothrix Thomas, 1920 (1 вид);
1 В пределах семейства можно выделить три группы колючих шиншилл: мелкие с длиной тела 8—20 см, средние — 21—34 см и крупные — 35—48 см.
Род щетинистых крыс
Genus Proechimys J. Allen, 1889
Размеры мелкие и средние. Длина тела 16—32 см. Длина хвоста 12— 32 см. Хвост безволосый или покрыт волосами средней .густоты (рис. 177). Стопа длинная, узкая. Первый палец на передней конечности рудиментарен. В волосяном покрове всегда имеются иглы. Окраска спинной стороны тела оранжево-коричневая или светло-желтая, смешанная ( черным, брюшная обычно беловатая. Хвост бурый сверху и белый снизу.
В черепе лицевой отдел относительно узкий, заостренный. Супраорбитальные гребни обычно хорошо развиты. Гребни есть и на теменных костях. В подглазничных отверстиях канал для нерва развит слабо или его нет вовсе. Резцовые отверстия довольно крупные. Ряды щечных зубов укорочены и сдвинуты вперед. Костные слуховые барабаны крупные. Нижняя челюсть с сильными гребнями. Ее угловой отросток повернут наружу; венечный отросток низкий.
Хромосом в диплоидном наборе у Р. guyannensis 46.
Распространены в Центральной Америке, южной и центральной части Южной Америки примерно от широты 12° с. ш. до 22° ю. ш., а также на о-ве Тринидад.
Населяют прибрежные области, леса около водоемов, у подножий холмов; поднимаются в горы на высоту до 1200 м над уровнем моря. Часто встречаются в антропогенных ландшафтах, поселяются в постройках. Ведут ночной образ жизни. Питаются растениями, плодами. Обычно бывает 2 помета в год, каждый из 2—3, иногда 6 детенышей.
В роде, вероятно, 12 видов:
Р. canicollis J. Allen, 1899 (Северная Колумбия, Южная Гайана); щетинистая крыса Гелди — Р. goeldii Thomas, 1905 (Бразилия); гайанская щетинистая крыса — Р. gujannensis Е. Geoffroy, 1803 (Бразилия, Боливия, Колумбия, Гайана, Эквадор. Венесуэла, Перу);
Р. hendeei Thomas, 1926 (Северо-Восточное Перу, Эквадор);
длиннохвостая щетинистая крыса — Р. longicaudatus Rengger, 1830 (Бразилия, Центральное, Восточное и Юго-Восточное Перу, Северный Парагвай, Западная Боливия);
Р. quadruplicatus Hershkovitz, 1948 (о-в Ллунги на р. Напо в Эквадоре);
Р. semispinosus Tomes, 1858 (Южная Венесуэла, Бразилия; Эквадор, северо-восток Перу, Панама, Никарагуа, Коста-Рика);
белоиглая щетинистая крыса — Р. albispinus I. Geoffroy, 1838 (Бразилия) ;
Р. dimidiatus Gunther, 1876 (Бразилия);
Р. iheringi Thomas, 1911 (Бразилия);
Р. myosurus Lichtenstein, 1818 (Бразилия);
Р. setosus Desmarest, 1917 (Бразилия).
После ревизии рода число видов в нем, по-видимому, будет уменьшено. Эллермэн (1940) насчитывает в роде 21 вид.
Род толстоиглых крыс
Genus HoplomysJ. Allen, 1908
В роде единственный вид: толстоиглая крыса —II. gymnurus Thomas, 1897.
Размеры мелкие и средние. Длина тела 22—32 см. Длина хвоста 150—250 мм. Внешним видом напоминают щетинистых крыс. В волося-
пом покрове (особенно по средней линии спины) много игл, более многочисленных, чем у других представителей семейства. Длина игл до 33 мм, толщина до 2 мм. В нижней части они белые, на вершине — темные. Между иглами расположены мягкие волосы, обычно оранжевой или желтоватой окраски Брюшная сторона белая, почти лишенная волос. Хвост покрыт чешуями; имеет коричневатую окраску сверху и беловатую снизу. Почти безволосые уши темно-коричневые или черные. Стопа задних лап длинная, узкая.
Череп толстоиглых крыс сходен с черепом представителей рода щетинистых крыс Костные слуховые барабаны несколько меньших размеров Лобные и теменные кости имеют сильно развитые гребни.
Распространена от Никарагуа в Центральной Америке на юг через Коста-Рику и Панаму до северо-запада Колумбии, северо-запада Эквадора и юго-востока Венесуэлы.
Населяет дождевые леса, заросли кустарников, вырубки с густым травянистым покровом, где обычно встречается около гниющих стволов деревьев. Экология изучена слабо Ведет, по видимому, наземный образ жизни.
Род тиар
Genus Euryzygomatomys Goeldi, 1901
В роде единственный вид: гиара— Е. spinosus Fischer, 1814.
Размеры мелкие и средние. Длина тела 17—27 см. Длина хвоста 5— 5.5 см (рис. 178) Волосяной покров щетинистый, на спине имеются иглы, развитые немного слабее, чем у щетинистых крыс. Стопа узкая. Когти на передних конечностях несколько удлиненные. Хвост короткий, по-
крыт редкими волосами. Окраска волосяного покрова темно-коричневая на спинной стороне тела и беловатая — на брюшной.
Череп со слабо развитыми надглазничными гребнями, относительно широким лицевым отделом, довольно крупными костными слуховыми барабанами. Костное небо узкое и довольно короткое. Резцовые отверстия широкие, короткие. Подглазничные отверстия имеют канал для нерва. Скуловая дуга длинная, значительно утолщенная спереди.
Распространена в Юго-Восточной, Восточной и Южной Бразилии, Парагвае и Северо-Восточной Аргентине.
Населяет открытые пространства, покрытые травянистой растительностью и кустарником. Экология не изучена. По-видимому, ведет подземный образ жизни.
Род клиомис
Genus Clyomys Thomas, 1916
В роде единственный вид: клиомис — С. laticeps Thomas, 1909.
Размеры мелкие. Длина тела 10—20 см. Длина хвоста 5,5—8 см. Внешне сходна с гиара. Волосяной покров щетинистый. Окраска спинной стороны тела серовато-коричневая или черная с рыжеватым, брюшной— беловатая или темно-желтая. Передние конечности с крупными когтями.
В черепе сильно развиты костные слуховые барабаны.
Населяет открытые пространства. Живут, по-видимому, колониями. Копает сложно устроенные норы.
Распространен в Восточной Бразилии и Парагвае.
Экология не изучена.
Род картеродон
Genus Carterodon Waterhouse, 1848
В роде единственный вид: картеродон — С. sulcidens Lund, 1841.
Размеры мелкие. Длина тела 15,5—20 см. Длина хвоста 6,8—8 см (рис. 179). Стопа узкая, длинная. Когти средних размеров. Волосяной покров содержит многочисленные иглы. Хвост густо покрыт волосами. Спинная сторона тела желтовато-коричневая с черноватым оттенком. Бока сероватые. Брюхо посредине белое, по бокам желтовато-красное. Горло красноватое. Хвост черный сверху, беловато-желтый снизу.
В черепе скуловые дуги утолщены в передних частях. Надглазничные гребни хорошо развиты. В межглазпичной области иногда есть небольшое сужение. Верхние резцы на наружной поверхности несут по про-
Рис. 179. Картеродон — С. sulcidens
дольной бороздке. Наружная сторона верхних резцов желтая, внутренняя — белая.
Распространен в Восточной Бразилии.
Населяет открытые пространства типа саванн. Экология не .изучена. Встречается в засушливых районах. Выкапывает норы с гнездовой камерой. Питается различными растительными кормами.
Род пунарес
Genus Cercomys F. Cuvier, 1829
В роде единственный вид: пунарес — С. cunicularis Cuvier, 1829.
Размеры средние. Длина тела 20—29 см, длина хвоста 18—22 см. Внешним обликом несколько напоминает крысу. Волосяной покров мягкий, щетин или игл нет. Хвост густо покрыт волосами. Окраска спинной стороны тела темно-коричневая, брюшной — сероватая или беловатая.
Череп широкий, с менее выраженными гребнями, чем у щетинистых и толстоиглых крыс. Мозговая капсула относительно более широкая. Скуловые дуги не утолщены в передних отделах. Костные слуховые барабаны крупные. Подглазничные отверстия имеют канал для .нерва. Резцовые отверстия очень широкие.
Распространен в Восточной и Северо-Восточной Бразилии и Парагвае.
Населяет различные биотопы. Встречается даже в болотистых местах, однако предпочитает густо поросшие скалистые участки. Строит гнезда в дуплах упавших деревьев или под камнями. Поедает хлопковые семена, во время сухого сезона питается сочными частями некоторых кактусов. Спаривание происходит в августе и сентябре. В помете от 1 до 4, обычно 2 детенышей.
Эллермэн (1940) и Уокер (1964) насчитывают в роде 2 вида (дополнительно еще С. inermis Pictet, 1841).
Род мезомис
Genus Mesotnys Wagner, 1845
Размеры мелкие. Длина тела 15—20 см. Длина хвоста 12—22 см. В волосяном покрове имеются многочисленные иглы. Стопа широкая и, по-видимому, приспособлена к древесному образу жизни; пятый палец длинный, когти длинные. Окраска спинной стороны тела коричневая, а брюшной — оранжевая, темно-желтая или беловатая. Хвост покрыт довольно редкими волосами, которые на конце образуют небольшую кисточку.
В черепе лицевой отдел короткий, узкий. Надглазничные гребни хорошо развиты. Лобные кости очень широкие. Костные слуховые барабаны относительно крупные. Подглазничное отверстие не имеет канала для нерва. Резцовые отверстия короткие. Ряды щечных зубов сдвинуты вперед.
Распространены в Бразилии (за исключением ее южных областей), Восточном Перу и Эквадоре.
Иногда встречаются в горах па высоте до 2000 м над уровнем моря. Экология не изучена.
В роде, вероятно, 3 вида:
М. didelphoides Desmarest, 1817 (Бразилия);
М. hispidus Desmarest, 1817 (Северное Перу, Бразилия, Эквадор, крайний юг Венесуэлы);
M. obscurus Wagner. 1840 (Бразилия);
Эллермэн (1940) и Уокер (1965) насчитывают в роде 6 видов (дополнительно еще М. ferrugineus Gunther, 1876, М. leniceps Thomas, 1926, и М. stimulax Thomas, 1911).
Род лонхотрикс
Genus Lonchothrix Thomas, 1920
В роде единственный вид: лонхотрикс — L. emiliae Thomas, 1920.
Размеры мелкие. Длина тела около 20 см. Длина хвоста 19 см. Все тело густо покрыто иглами; даже на брюшной стороне встречаются полуиглы. Стопа и кисть широкие, по-видимому, приспособлены к древесному образу жизни. Хвост покрыт редкими волосами, за исключением концевой части, где удлиненные более густые волосы образуют кисточку. Окраска спинной стороны тела темно-коричневая, брюшной — темножелтая.
Череп лонхотрикс сходен с черепом представителей рода мезомис.
Распространен в Бразилии.
Экология не изучена. Судя по строению конечностей, вероятно, ведет древесный образ жизни. Населяет леса.
• Род коноконо
Genus Isothrix Wagner, 1845
Размеры средние. Длина тела 22—29 см. Длина хвоста около 25 см. Стопа широкая, приспособленная к лазанию по деревьям. Волосяной покров мягкий; не имеет ни колючек, ни щетин. Хвост густо покрыт длинными волосами. Окраска меха темно-коричневая или черная, иногда с белыми пятнами по бокам туловища, на шее и по бокам головы.
В черепе лобные кости широкие. Костные слуховые барабаны большие. Костное небо узкое. Резцовые отверстия короткие. Щечные зубы сложные.
Распространены в Венесуэле, Колумбии, Бразилии и Перу.
Экология изучена слабо. Держатся около водоемов. Убежищем служат норы между корнями деревьев. Активность, по-видимому, сумеречная или ночная.
В роде 3 вида:
/. bistriata Wagner, 1845 (Бразилия, Южная Венесуэла, Колумбия);
I. picta Pictet, 1841 (Восточная Бразилия);
/. villosa Deville, 1852 (Северо-Восточное Перу).
Эллермэн (1940) насчитывает в роде 5 видов (дополнительно еще 1. pachyura Wagner, 1845, и /. pagurus Wagner, 1845).
Род дипломис
Genus Diplomys Thomas, 1916
Размеры средние и крупные. Длина тела 25—48 см. Длина хвоста 20—28 см. Хвост покрыт довольно редкими волосами, сквозь которые видны кожные чешуи. На конце хвоста волосы несколько более удлинены. Стопа широкая, приспособленная к лазанию по деревьям. Когти короткие, но широкие и сильные. Ушные раковины широкие, округлые. Волосяной покров грубый, без колючек. Окраска его на спинной стороне
тела темно-коричневая, красноватая или рыжая, смешанная с черным, брюшной — темно-желтая, рыжеватая или рыжевато-белая.
Череп дипломис сходен с черепом представителей рода Echimys.
Распространены в Панаме, Колумбии, Эквадоре и на о-ве Сан-Мигель (около Панамы).
Населяют лесные участки. Экология изучена слабо. Хорошо лазают по деревьям. Активны ночью.
В роде 3 вида:
D. labilis Bangs, 1901 (Панама, о-в Сан-Мигель);
D. caniceps Gunther, 1876 (Западная Колумбия и Северный Эквадор);
D. rufodorsalis J. Allen, 1899 (Северо-Восточная Колумбия).
Эллермэн (1940) насчитывает в роде 4 вида (дополнительно еще D. darlingin Goldman, 1912).
Род коро
Genus Echimys F. Cuvier, 1809
Размеры от мелких до крупных. Длина тела 17—35 см. Длина хвоста 15—30 см (рис. 180). Стопа широкая, приспособленная к лазанию по деревьям; ее пятый палец относительно длинный. Когти на задних конечностях сильные. У некоторых видов хвост густо покрыт волосами. У других почти голый. Кисточки волос на конце хвоста нет. Волосяной покров грубый, щетинистый или иглистый. Спинная сторона тела рыжеватая, светло-темно-желто- или золотисто-коричневая или черная, брюшная — обычно белая или темно-желтая.
Череп с широкими лобными костями и обычно с хорошо выраженными надглазничными гребнями. У крупных видов теменные кости с крупными гребнями. Подглазничные отверстия не имеют канала для нерва. Костные слуховые барабаны крупные, костное небо обычно узкое. Резцовые отверстия короткие. Скуловая дуга относительно широкая. Скуловая кость, как правило, имеет верхние и нижние отростки. Нижняя челюсть с крупными гребнями. Щечные зубы сложные. У каждого верхнего зуба обычно имеется по 2 наружные и 2 внутренние складки, а у нижнего — по одной наружной и 2 внутренние складки, за исключением предкоренных зубов, у которых 3 или иногда 4 внутренние складки.
Распространены в северной и центральной части Южной Америки. На о-в Мартиника, по-видимому, завезены человеком.
Населяют леса, предпочитая места рядом с водоемами. Экология изучена слабо. Ведут древесный образ жизни. Строят в дуплах гнезда из сухих листьев. Живут небольшими группами. Активны ночью. Характерен громкий крик, напоминающий звуки «кро-кро» или «тро-тро». Самки обычно приносят 1—2 детенышей.
В роде И видов:
Е. armatus I. Geoffrey, 1838 (Гвиана, Южная и Северная Венесуэла, Северный Эквадор, Западная Колумбия и о-в Тринидад);
коро Блайнвилля — Е. blainvillei Cuvier, 1837 (Юго-Вост. Бразилия);
бразильский коро- Е. braziliensis Waterhouse. 1848 (Восточная Бразилия) ;
Е. chrysurus Zimmerman, 1780 (Суринам,- Северо-Восточная Бразилия) ;
Е. dasythrix Hensel. 1872 (Юго-Восточная и Восточная Бразилия);
Е. grandis Wagner, 1845 (Бразилия и Северо-Восточное Перу);
Е. macrurus Wagner, 1842 (Бразилия);
Е. nigrispinus Wagner, 1842 (Восточная Бразилия);
Рис. 180. Echimys sp. и череп Е. arrnaius
Е. saturnus Thomas, 1928 (Эквадор);
Е. semivillosus I. Geoffrey, 1838 (Венесуэла и Северная Колумбия); одноцветный коро — Е unicolor Wagner, 1842 (Бразилия).
Эллермэн (1940) насчитывает в роде 21 вид.
Род дактиломис
Genus Dactylomys I. Geoffroy, 1838
Размеры средние. Длина тела 25 ДО см. Длина хвоста 32—43 см. Волосяной покров мягкий, без щетин или игл. Спинная сторона тела желтовато-коричневая, оливково-серая или серая, часто с ржавым оттенком, брюшная — беловатая. Длинный хвост лишен волос, за исключением проксимального отдела. Третий и четвертый пальцы передних и задних конечностей удлинены и расширены. Когти слабые.
Череп со слабо выраженными посторбитальпыми гребнями, но с крупным сагиттальным гребнем. Относительно широкие скуловые кости не
утолщены в передних отделах. На верхних и нижних краях их имеются небольшие отростки. Костные слуховые барабаны средней величины. Подглазничное отверстие не имеет канала для нерва. Костное небо сужено спереди, так что предкоренные зубы правого и левого рядов почти касаются друг друга. Щечные зубы брахиодонтного типа, широкие, массивные.
Распространены в Эквадоре, Перу, Боливии и Бразилии.
Населяют лесные районы. Экология изучена слабо. Хорошо лазают по деревьям. Активны ночью.
В роде 3 вида:
боливийский дактиломис — D. boliviensis Anthony, 1920 (Центральная Боливия и Юго-Восточное Перу);
бразильский дактиломис — D. dactylinus Desmarest, 1817 (Бразилия, Эквадор и, вероятно, Колумбия);
перуанский дактиломис — D. peruanus J. Allen, 1900 (Юго-Восточное Перу);
Некоторые исследователи (например, Уокер, 1964) выделяют D. peruanus в особый род Lachnomys Thomas, 1916, или (например, Симпсон, 1945) включают D. peruanus в род Kannabateomys.
Род каннабатеомис
Genus Kannabateomys Jentink, 1891
В роде единственный вид: каннабатеомис — К. amblyonyx Wagner, 1845.
Размеры средние. Длина тела примерно 25 см. Длина хвоста около 32 см. Стопа широкая. Третий и четвертый пальцы на передних и задних конечностях удлинены и расширены. Волосяной покров мягкий, густой, лишен щетин и игл. Окраска спинной стороны тела темно-желтая, брюшной — более светлая или белая. Хвост относительно хорошо оволоснен, особенно в проксимальной части.
Череп каннабатеомис сходен с черепом представителей рода дактиломис. Костное небо, однако, лишь слегка сужено спереди. Сагиттального гребня обычно нет.
Распространен в Парагвае, Северо-Восточной Аргентине и Восточной Бразилии.
Обитает в густых зарослях кустарников и травянистой растительности, обычно вблизи водоемов. Поднимается в горы на высоту около 2000 м над уровнем моря. Экология не изучена. Хорошо лазает по кустам, захватывая ветви между третьим и четвертым пальцами. Активен, по-видимому, ночью.
Род тринакодус
Genus Thrinacodus Gunther, 1879
Размеры от мелких до средних. Длина тела 8—24 см. Длина хвоста до 35 см. Когти на передних конечностях более острые, чем у других представителей подсемейства. Первый палец стопы сильно уменьшен. Волосяной покров густой и мягкий, не имеет щетин или игл. Окраска его на спинной стороне тела рыжевато-коричневая или желтовато-коричневая, на брюшной желтовато-белая или белая. Хвост почти лишен волос или довольно густо покрыт в терминальной половине.
Череп и зубы сходны с таковыми представителей рода Dactylomys.
Распространены в Колумбии и Венесуэле.
Населяют склоны гор на высоте 2000—2800 м над уровнем моря. Экология не изучена. Держатся в зарослях кустарников. Голос напоминает громкий свист.
В роде 2 вида:
белохвостый тринакодус—Т. albicauda Gunther, 1879 (Северо-Западная и Центральная Колумбия);
Т. edax Thomas, 1916 (Западная Венесуэла).
Эллермэн (1940) насчитывает в роде 3 вида (кроме упомянутых двух, еще Т. apolinari Allen, 1914).
СЕМЕЙСТВО ТРОСТНИКОВЫХ КРЫС
FAMILIA THRYONOMYIDAE РОСОСК, 1922
Размеры мелкие и средние. Длина тела 35—61 см. . Длина хвоста 6,5—26 см. Масса взрослых тростниковых крыс около 4—7 кг, а отдельные самцы достигают даже 9 кг. Телосложение тяжелое. Внешний вид несколько напоминает нутрию (рис. 181). Ушные раковины короткие, округлые, едва возвышающиеся над мехом. Подошвы передних и задних лап голые. На передних конечностях 3 средних пальца хорошо развиты. Первый палец редуцирован, пятый очень небольшой, не функционирует. На задних конечностях первого пальца также нет. Когти сильные, толстые. Волосяной покров жесткий, щетинистый. Щетипковидпыс волосы несколько уплощены и имеют продольные борозды па наружной’поверхности. Подшерстка нет. Окраска спинной стороны тела желтовато- или серовато-коричневая, брюшной — сероватая или беловатая. Хвост покрыт редкими, короткими жесткими волосами, между которыми видны чешуи. Окраска его коричневая сверху и темно-желто-белая снизу. Сосков две или три пары — по бокам тела.
Череп массивный, обычно с очень крупными гребпями. Лицевой отдел черепа высокий, широкий. Скуловая кость почти соприкасается со слезной. Лобные кости широкие. Подглазничное отверстие очень большое, с хорошо выраженным каналом для нерва. Затылочная область черепа очень сильно развита. Костные слуховые барабаны средних размеров. Широкое костное небо продолжается назад немного дальше уровня третьего коренного зуба. Резцовые отверстия очень широкие, большие. Нижняя челюсть с венечным отростком средних размеров, низким угловым отростком. Резцы на передней поверхности оранжевого цвета, очень большие — относительно более крупные, чем у других грызунов. Верхние резцы имеют на наружной поверхности по 3 продольные бороздки. Щечные зубы с корнями.
Хромосом в диплоидном наборе у большой тростниковой крысы 44.
Распространение охватывает территорию Африки к югу от Сахары.
Большая тростниковая крыса населяет густые заросли по берегам различных водоемов, а также болотистые места, малая тростниковая крыса — сухие участки влажных саванн, кустарники среди скал на склонах гор на высоте до 2600 м над уровнем моря. Оба вида придерживаются густых зарослей травянистых растений, где проделывают тропинки. Большая тростниковая крыса ведет полуводный образ жизни, хорошо плавает и ныряет. Малая тростниковая крыса также может плавать; представители этого вида держатся семьями. Большая тростниковая крыса часть года встречается группами, смешанными по возрасту и полу. В сухой сезон самки объединяются отдельно большими группами, а сам-
Рис. 181. Большая тростниковая крыса — Т. swinderianus и череп малой тростниковой крысы Т. gregorianus
цы живут поодиночке. Активны ночью, а малая тростниковая крыса иногда и днем. День проводят обычно среди густых зарослей пли в неглубоких норах. Могут занимать поры других животных. Питаются различными сочными частями растений, а также семенами, орехами, плодами, малая тростниковая крыса — и корой деревьев. В году обычно бывает 2 помета; у большой тростниковой крысы в Южной Африке с июня по август, а в Западной — с октября по январь. Детенышей в помете от 2 до 6, обычно 4. У новорожденных глаза открыты тело покрыто волосами. Половозрелость у большой тростниковой крысы наступает в годовалом возрасте. Продолжительность жизни в природе, по-видимому, не превышает 3 года. В некоторых местах приносят вред плантациям сахарного тростника. Местное население употребляет тростниковых крыс в пищу.
В семействе 1 род: тростниковых крыс -Thryunomys Fit7ingcr, 1867, и 2 вида.
Род тростниковых крыс
Genus Thryonomys Fitzinger, 1867
См. описание семейства.
В роде, по-видимому, 2 вида:
большая тростниковая крыса — Т. swinderianus Temminck, 1827 (от Гамбии до Камеруна, северной части Заира и Южного Судана и далее на юг до северной части Юго-Западной Африки и Южно-Африканской Республики включительно);
малая тростниковая крыса — Т. gregorianus Thomas, 1894 (Южная Родезия. Мозамбик, Малави, Замбия, Танзания, Южная и Западная Кения, Уганда, Заир, Южный Судан, Центральноафриканская Республика, Камерун).
Эллермэн (1940), Андерсон и Джонс (1967) насчитывают в роде 6 видов.
Эллермэн (1940) рассматривает тростниковых крыс в качестве подсемейства Echimyidae. По-видимому, более правильно считать тростниковых крыс отдельным семейством. Некоторые исследователи (например, Грассе и Декейзер, 1955) выделяют из рода тростниковых крыс род Choeromys Thomas, 1922, с одним видом — малой тростниковой крысой.
СЕМЕЙСТВО СКАЛЬНЫХ КРЫС
FAMILIA PETROMYIDAE TULLBERG, 1899
Размеры мелкие. Длина тела 14,5—19 см. Длина хвоста 13—18 см. Внешний вид несколько напоминает белку (рис. 182). Хвост покрыт довольно длинными волосами, особенно на конце. Уши короткие. Передние конечности четырехпалые, задние пятипалые. Первый палец задних конечностей укорочен. Стопа узкая. Подошвы голые. Когти короткие, тупые. На концах пальцев имеются пучки щетинковидных волос. Волосяной покров туловища мягкий, шелковистый, но пуха нет. Спинная сторона тела спереди серая или желтоватая, сзади рыжевато-коричневая, оранжевая или желтоватая. Брюшная сторона тела сероватая или желтоватая. Сосков 3 пары — расположены высоко на боках тела.
Череп широкий, уплощенный. Сужения в межглазничной области нет. Костные слуховые барабаны сильно вздуты. Подглазничные отверстия с каналом для нерва. Резцовые отверстия глубокие и длинные; достигают уровня щечных зубов. Относительно узкое костное небо продолжается чуть дальше назад рядов щечных зубов. Угловой отросток нижней челюсти несколько повернут наружу. Резцы узкие, с гладкой наружной поверхностью. Щечные зубы имеют корни и высокие коронки. Характерно очень подвижное сочленение ребер, так что скальную крысу можно сильно сжать в дорзо-вептралыюм направлении без вреда для нее. Это помогает ей проскальзывать в самые узкие щели среди скал. Os penis имеется.
Распространена в Юго-Западной Африке и Южно-Африканской Республике.
Населяет скалистые участки склонов холмов и гор. Ведет наземный образ жизни. Активна днем. Подвижный зверек, быстро бегает между камней. Держатся поодиночке или парами. Питается различными растительными кормами, ягодами, семенами. В Южном полушарии период
Рис. 182. Скальная крыса — Р. typicus и ее череп
спаривания приходится на ранние летние месяцы — с ноября по декабрь. В течение года бывает один помет из I—2 детенышей. Новорожденные покрыты короткими волосами.
Эллермэн (1940) рассматривает скальных крыс в качестве подсемейства Petromyinae семейства Echimyidae. По-видимому, более правильно выделять их в качестве отдельного семейства. Симпсон (1945) в написании рода употребляет «Petromys» вместо «Petromus», предполагая, что при первой публикации была внесена ошибка.
В семействе 1 род: скальных крыс — Petromus A. Smith, 1831, и 1 вид.
Род скальных крыс
Genus Petromus A. Smith, 1831
См. описание семейства.
В роде 1 вид: скальная крыса — Р. typicus A. Smith, 1831 (ареал совпадает с ареалом семейства).
Hystricomorpha incertae sedis
СЕМЕЙСТВО ЗЕМЛЕКОПОВЫХ
FAMILIA BATHYERGIDAE WATERHOUSE, 1841
Размеры мелкие. Длина тела 8—33 см. Длина хвоста 1—7 см. Многие признаки показывают высокую степень приспособленности к подземному существованию. Сложение тяжелое. Голова относительно крупная. Шея сильная, укороченная. Хвост и конечности короткие. Глаза маленькие. Наружные ушные раковины представлены небольшим валиком кожи, окружающим слуховое отверстие. Кисть и стопа крупные, подошвы голые. Конечности пятипалые. Пальцы с короткими или длинными когтями. Обычно когти на задних конечностях короче, чем на передних. Волосяной покров густой или сильно изрежеп, а у Heterocephalus представлен отдельными волосками. Окраска серая, коричневая или черная, иногда имеются белые пятна. Сосков 3 или 4 пары.
Череп массивный. Костные слуховые барабаны крупные, но уплощенные. Характерны малые размеры подглазничного отверстия. Сквозь подглазничное отверстие проходит лишь небольшая порция жевательной мышцы или она не проходит совсем. Скуловой отросток верхнечелюстной кости не расширен. В сочетании эти два признака говорят о примитивности черепа. В нижней части угловой отросток повернут наружу. Зуб-] о 2______________________3 Q___3
ная формула: i — с — р -—- т -—- =12 — 28. Щечные зубы с высокими коронками, но не имеют постоянного роста, с корнями. Резцы крупные. У некоторых видов они имеют бороздку на наружной поверхности. Корни верхних резцов очень велики и продолжаются до крыло-видных костей. Малая берцовая кость уменьшена и срастается с большой.
Распространены в Африке, от Судана, Эфиопии и Сомали на востоке и от Ганы на западе на юг до мыса Доброй Надежды.
Населяют открытые пространства. Обитают во влажных и засушливых местах вплоть до пустынь. В горы поднимаются до 1500 м над уровнем моря. Ведут в основном подземный образ жизни. Копают поверхностно расположенные подземные туннели при помощи когтей передних лап {Bathyergus) или нижних резцов {Cryptomys). Питаются подземными сочными частями растений. Корпи и луковицы запасают в подземных кладовых. Превосходно ориентируются под землей. Если туннель разрушить, способны вырыть другой, точно параллельный. По-видимому, размножение носит сезонный характер. В помете 1—5 детенышей.
В семействе 5 родов (14 видов):
капских землекопов — Georychus liliger, 1811 (1 вид);
пескороев — Cryptomys Gray, 1864 (8 видов);
тенелюбов — Heliophobius Peters, 1846 (3 вида);
капских пескороев — Bathyergus liliger, 1811 (1 вид);
голых землекопов — Heterocephalus Riippell, 1842 (1 вид).
Род капских землекопов
Genus Georychus liliger, 1811
В роде единственный вид: капский землекоп — G. capensis Pallas, 1779.
Длина тела 15—20 см. Длина хвоста 1,5—4 см. Хвост покрыт волосами, более длинными на его боках. Когти средних размеров, относительно слабые. Волосяной покров густой, высокий, мягкий. Спинная сторона тела темно-желтая или темно-рыжая, часто с буроватыми пестринками, брюшная светлее спины. Конечности и хвост белые. Голова имеет черную или темно-коричневую окраску с белыми пятнами. Сосков 3— 4 пары.
Резцы выдаются изо рта. Снаружи они белые и не имею борозд.
Хромосом в диплоидном наборе 54.
Распространен в Южно-Африканской Республике.
Населяет, как правило, местности с песчаными почвами. Строит поверхностно расположенные подземные ходы, которые ведут в центральную Гнездовую камеру. При рытье использует в основном резцы. Запасает в кладовых корни, луковицы и клубни растений. Местами может приносить некоторый вред сельскому хозяйству. Разможение не изучено.
Род пескороев
Genus Cryptomys Gray, 1864
Длина тела 10—21 см. Длина хвоста 1—3 см. Масса около 200 г
(рис. 183). Сильно редуцированы глаза, видимо, не несут зрительной функции. Когти на пальцах небольшие. Волосяной покров низкий, густой, мягкий. Окраска его изменчива: беловатая, желтоватая, желтовато-ко
ричневая, серовато-коричневая, черноватая. На голове могут быть белые
пятна. Сосков обычно 2 или 3 пары.
В черепе верхние резцы гладкие. Число щечных зубов:
4
4
Распространены в Африке к югу от Сахары.
Населяют, как правило, песчаные почвы. Иногда встречаются в за
рослях кустарников, в изреженных лесах или на скалистых участках. Поднимаются в горы на высоту до 2200 м над уровнем моря. Пустынь и сплошных лесов избегают. Держатся группами до 10—12 особей. Норы роют при помощи резцов и передних конечностей. В холодные месяцы в Южной Африке, по-видимому, впадают в спячку или, во всяком случае, снижают свою активность. Питаются в основном луковицами, корнями, корневищами и листьями, иногда и почвенными беспозвоночными. Большинство видов запасает пищу в больших подземных кладовых. Местами наносят значительный вред сельскому хозяйству. В течение года, по-видимому, бывает 1 помет в дождливый сезон. В каждом от 1 до 5 детенышей. В Анголе самки приносят одного-двух детенышей в январе и феврале.
Систематика рода разработана недостаточно. По-видимому, в роде 8 видов:
готтентотский пескорой — С. hottentotus Lesson, 1826 (Южно-Африканская Республика, Южная Родезия, Мозамбик, Малави, Танзания, Уганда, Заир, Судан, Нигерия, Того);
Рис. 183. Суданский пескорой — С. ochraceocinereus и его череп
большой пескорой — С. holosericeus Wagner, 1843 (Южно-Африканская Республика, Мозамбик, Ботсвана);
ангольский пескорой — С. mechowi Peters, 1881 (Ангола, Замбия, Заир);
конголезский пескорой — С. lechei Thomas, 1895 (Заир);
С. kummi Thomas, 1911 (Центральноафриканская Республика);
нигерийский пескорой — С. foxi Thomas, 1911 (Северная Нигерия);
тоголезский пескорой — С. zechi Matschie, 1900 (Того);
суданский пескорой — С. ochraceocinereus Hcuglin, 18G4 (Судан).
Андерсон и Джонс (1967) насчитывают в роде 15 видов, а Эллермэн (1940) —45.
Род тенелюбов
Genus Heliophobius Peters, 1846
Длина тела 10—20 см. Длина хвоста 1,5—4 см. Ушных раковин нет. На задних конечностях второй палец наиболее сильно развит. Когти не очень крупные. Волосяной покров густой, высокий, мягкий. Окраска его светло-песчаная, рыжеватая или сероватая на спине и несколько более светлая на брюхе. У некоторых особей имеются белые пятна на лбу и брюхе.
Характерно очень узкое костное небо. Полное число щечных зубов: —. Однако все зубы редко функционируют одновременно. Обычно перед
ние предкоренные теряются прежде, чем прорежутся коренные. Поэтому число зубов обычно— или — . Костное небо чрезвычайно узкое и в от
личие от родов капских землекопов, пескороев и капских пескороев не продолжается кзади дальше окончания ряда щечных зубов. Подглазничное отверстие чрезвычайно мало.
Распространены в Восточной и Центральной тропической Африке: Заире, Кении, Танзании, Южной Родезии, Мозамбике, Замбии.
Населяют в основном песчаные почвы на высоте 750—1500 м над уровнем моря. Обычно под землей вырывают основной главный туннель с многочисленными боковыми отнорками в среднем длиной до 47 м. Выкапывают также большие подземные кладовые, где запасают луковицы, корни и корневища. Питаются в основном сочными подземными частями растений. В году, по-видимому, бывает 1 помет. Самка приносит 1—4 детенышей в начале дождливого сезона. Местами вредят сельскому хозяйству. Местное население употребляет тенелюбов в пищу.
В роде 3 вида:
слепышовый тенелюб — И. spalax Thomas, 1910 (Танзания);
серебристосерый тенелюб — И. argenteocinereus Peters, 1852 (Южная Родезия, Мозамбик, Заир, Танзания, Кения, Малави, Замбия);
конголезский тенелюб — Н. mottoulei Schuteden, 1913 (Заир).
Кингдом (1974) считает слепышового тенелюба подвидом серебристого тенелюба.
Род капских пескороев
Genus Bathyergus liliger, 1811
В роде единственный вид: капский пескорой — В. suillus Schreber, 1782.
Длина тела 17,5—33 см. Длина хвоста 4—7,3 см. Ушных раковин нет. Хвост толстый, несколько уплощенный, с длинными волосами по бокам, придающий ему вид пера. На передних конечностях сильно развиты когти, особенно на втором, третьем и четвертом пальцах. Второй палец длиннее третьего. Задняя конечность с наиболее сильно развитым средним пальцем и вторым, слегка более длинным, чем четвертый. Когти па задних конечностях средних размеров. Волосяной покров густой, мягкий. Окраска спины светло-коричневая, темно-серая или серебристо-желтоватая, а у большинства видов такая и на брюхе; у одного вида брюхо черное.
В черепе затылочная область несет очень сильные гребни, особенно у старых зверьков. Лобные кости сильно сжаты. Верхние резцы на наружной поверхности имеют по одной глубокой продольной борозде, ниж
ние резцы гладкие. Корни верхних резцов не продолжаются до задних щечных зубов.
Распространен в Южно-Африканской Республике.
Населяет песчаные почвы на высоте 1200—1500 м над уровнем моря. Копает почву в основном передними конечностями. Питается луковицами и сочными корнями. Запасает пищу в подземных кладовых. Может причинять существенный вред сельскому хозяйству.
Некоторые исследователи (например, Эллермэн, 1940; Андерсон и Джонс, 1967) выделяют в роде второй вид: В. janetta Thomas et Schwann, 1904.
Род голых землекопов
Genus Heterocephalus Riippel, 1842
Рис. 184. Голый землекоп — II glabcr и его череп
В роде единственный вид: голый землекоп — И. glaber Riippel, 1842. Размеры небольшие. Длина тела 8—9,2 см. Длина хвоста 3—4,4 см. Масса взрослой особи 30—80 г. Характерно отсутствие сплошного воло-
сяного покрова.
Все тело покрыто редко расположенными отдельными вибриссовидными, относительно длинными волосами. Окраска кожи по всему телу светло-рыжеватая или желтоватая. Кожа на туловище очень подвижна и образует многочисленные большие и мелкие складки. Глаза и ушные отверстия чрезвычайно малы. Ушной раковины нет совсем (рис. 184). Передние конечности имеют пять широких уплощенных пальцев, каждый
из которых с небольшим коническим когтем. Третий палец на передней конечности достигает наибольшей длины. По краям стопы и кисти рас-
тут гребешки жестких волос.
В черепе лобные кости слегка сужены, а носовые кости относительно более широкие, чем у других родов семейства. Костное небо короче по сравнению с другими родами семейства, за исключением рода тенелю-3 2
бов. Нормальное число щечных зубов —, иногда бывает — .
3 2
Распространен в Эфиопии, Сомали и Кении.
Встречается в различных биотопах, но предпочитает жить в песчаных почвах засушливых районов на высоте от 600 до 1800 м над уровнем моря. Роет различные по глубине (иногда до 1 м) подземные туннели. Туннели могут располагаться на нескольких уровнях и расходиться от центральной гнездовой камеры в радиусе 30—40 м. Держатся группами до 30 особей (иногда до 100 — в Сомали). Наиболее активен утром и вечером. Почву копает резцами, помогая конечностями. На поверхность почву выталкивает сначала головой, потом задними конечностями. Характерна относительно низкая температура тела, обычно около 30—32°С. Питается корнями, луковицами, корневищами. По-видимому, поедает также почвенных беспозвоночных. Отмечена копрофагия. Размножение не изучено. В помете может быть до 5 детенышей. Масса новорожденного около 2 г.
Hystricomorpha или Myomorpha incertae sedis
СЕМЕЙСТВО ГУНДЯЕВЫХ
FAMILIA CTENODACTYLIDAE ZITTEL, 1893
Сложение тяжелое. Тело компактное. Внешне несколько напоминает пищух (рис. 185). Размеры мелкие. Длина тела 16—24 см. Длина хвоста 1—5 см; он полностью покрыт волосами. Глаза относительно крупные. Ушные раковиы небольшие, округлые. Передние и задние конечности четырехпалые. Стопа относительно узкая. Когти па пальцах небольшие. Два внутренних пальца на задних конечностях имеют гребневидные пучки жестких волос. Волосяной покров высокий, густой, мягкий. Окраска его на спинной стороне тела темно-желтая, пепельно-серая, коричневая или желтовато-рыжая, на брюшной — рыжеватая, серовато-белая или белая.
Череп уплощен в дорзо-вентральном направлении, сужен в лицевом отделе и расширен в задней части. Лобные кости широкие. Подглазничный канал увеличен. Скуловой отросток верхнечелюстной кости узкий.
Рис. 185. Гуиди — С. gundi и его череп
Слезная кость крупная. Костные слуховые барабаны сильно вздутые. Надглазничные гребни относительно хорошо развиты. Резцовые отверстия большие. Нижняя челюсть не имеет венечного отростка. Ее угловой о , .1 о 1—2 3 ол Ол
отросток удлинен. Зубная формула i — с—р_______т—= 20— 24.
Щечные зубы растут в течение всей жизни. Наружная поверхность резцов гладкая и лишь иногда имеет слабо выраженные бороздки. Малая и большая берцовые кости нс срастаются между собой.
Распространены в Северной Африке от Сенегала и Марокко до Сомали.
Населяют полуаридпые и аридные области. Обычны в скалистых и каменистых участках. Экология изучена слабо, за исключенном представителей рода гуиди. Активны днем. Подобно опоссуму, могут имитировать смерть, что служит защитной реакцией от врагов. Характерны крики, напоминающие писк и чириканье птиц. Живут, по-видимому, семейными группами. Детенышей приносят в январе и апреле. Как правило, в
помете бывает 1—2 детеныша. Новорожденные покрыты волосами, с открытыми глазами; способны сразу к самостоятельному передвижению. Убежищем служат пустоты под камнями, расщелины в скалах. Растительноядны.
В семействе 4 рода (5 видов):
гунди — Ctenodactylus Gray, 1830 (2 вида) ;
кистехвостых гунди — Pectinator Blyth, 1856 (1 вид);
массутьер — Massoutiera Lataste, 1885 (1 вид);
феловиа — Felovia Lataste, 1886 (1 вид).
Род гунди
Genus Ctenodactylus Gray, 1830
Длина тела 16—20 см. Длина хвоста 1—2 см (рис. 185). Окраска волосяного покрова на спинной стороне тела темно-желтая, иногда розовато-желтая, на брюшной — более светлая, обычно беловатая или сероватая.
В черепе костное небо относительно длинное. Костные слуховые ба-4
рабаны сильно вздуты. Полное число щечных зубов j-, но, как правило,
. з
их бывает —
3
(предкоренной зуб рано выпадает).
Хромосом в диплоидном наборе у гунди 40.
Распространены в Ливии, Тунисе и Алжире.
Населяют скалистые склоны холмов и гор. Превосходно лазают, иногда почти по отвесным скалам. Активны преимущественно в сумерки. Питаются в основном растительными кормами.
В роде, по-видимому, 2 вида:
гунди — С. gundi Rothmann, 1776 (Тунис, Восточный Алжир, Северо-Западная Ливия и горы Высокий Атлас в Марокко);
С. vali Thomas, 1902 (Алжир, Ливия к югу и юго-востоку от Триполи и Южное Марокко).
Некоторые исследователи насчитывают в роде один вид С. gundi (например, Уокер, 1964) или 3 вида: С. gundi Rothmann, 1776; С. vali Thomas, 1902, и С. joleaudi Heim de Balsac, 1936 (например, Эллермэн, 1940).
Род кистехвостых гунди
Genus Pectinator Blyth, 1856
В роде единственный вид: кистехьостый гунди — Р. spekei Blyth, 1855.
Длина тела примерно 17 см. Длина хвоста 4—5 см. Хвост покрыт густыми волосами. Стопа относительно узкая. Волосяной покров очень мягкий. Окраска спинной стороны тела пепельно-серая с черным или коричневым оттенками. Бока сероватые. Брюшная сторона тела серовато-белая.
В черепе костное небо более короткое, чем у представителей рода гунди, и не продолжается дальше рядов щечных зубов. Костные слуховые барабаны относительно вздуты. Полное число щечных зубов —
5 (интересно отметить, что кистехвостый гунди, а также Heliophobius из семейства Bathyergidae — единственные современные грызуны, у которых число щечных зубов превышает . Третьи коренные зубы проре
заются поздно. Как правило, предкоренные зубы выпадают до того, как прорезывается задний коренной зуб. Нижние коренные зубы более сложные по структуре, чем у других представителей семейства. Они имеют две острые, хорошо выраженные внутренние складки и по одной наружной.
Распространен в Эфиопии и Сомали.
Населяет скалистые участки. Экология изучена слабо.
Род массутьер
Genus Massoutiera Lataste, 1885
В роде единственный вид: массутьера — М. mzabi. Lataste, 1881.
Длина тела 17—24 см. Длина хвоста около 3,5 см. Хвост покрыт густыми, довольно длинными волосами (рис. 186). Волосяной покров высокий, густой, мягкий. Окраска желтоватая или коричневая.
Череп массутьеры сходен с черепом представителей рода гунди. Щечные зубы более сложные. Костные слуховые барабаны наиболее вздуты по сравнению с таковыми других представителей семейства. По внутреннему краю ушей растут гребневидные волоски, направленные вершинами внутрь, как у многих пустынных грызунов.
Распространена в западных и центральных частях Сахары, юго-восточной части Алжирской Сахары, на плато Аир в Нигере, нагорье Тибес-ти в Чаде и. вероятно, также в юго-западной Ливии.
Экология не изучена.
Некоторые исследователи (например, Эллермэн, 1940; Андерсон и Джонс, 1967) насчитывают в роде 3 вида: Л1. mzabi Lataste, 188I; Al. roth-schildi Thomas et Hinton, 1921; M. harterli Thomas, 1913.
Рис. 186. Массутьера — M. mzabi
Род феловиа
Genus Felovia Lataste, 1886
В роде единственный вид: феловиа-—F. vae Lataste, 1886.
Длина тела 17—23 см. Длина хвоста примерно 3 см. Хвост покрыт удлиненными, густыми волосами. Волосяной покров мягкий, густой, высокий. Окраска его темно-желтовато-рыжая на спинной стороне тела и рыжеватая на брюшной.
Череп такой же, как у представителей рода гунди. Костные слуховые барабаны вздуты умеренно. Верхние резцы имеют на наружной поверхности слабо намеченную продольную бороздку.
Распространена в Сенегале, Мавритании, Мали.
Экология не изучена.
ЛИТЕРАТУРА
Баранов О. К., Воронцов Н. Н. Серологическая дифференциация пяти палеарктических видов — Marmota (Rodentia, Sciuridae). Зоол. журн., 1973, т. 52, вып. 3, с, 577—583.
Бобринский Н. А., Кузнецов Б. А., К у з я к и н А. П. Определитель млекопитающих СССР. Изд. 2-е. М., «Просвещение», 1965.
Большаков В. Н., Покровский А. В. О степени репродуктивной изоляции между памирской (Microtus juldaschi Severtzov) и арчевой (Al. carruthersi Thomas) полевками. ДАН СССР, 1969, т. 188, № 4, с. 940—941.
Большаков В. Н., Россолимо О. Л., Покровский А. В. Систематический статус памиро-алтайских горных полевок группы Microtus juldaschi (Mammalia, Cricetidae). Зоол. журн., 1969, т. 48, вып. 7, с. 1079—1089.
Виноградов Б С., Громов И. М. Грызуны фауны СССР. М.—Л., Изд-во АН СССР, 1963.
Воронцов Н. Н., Ляпунова Е. А. Хромосомные числа и видообразование у наземных беличьих (Sciuridae: Xerinae ct Marmotinae) Голарктики. Бюлл. МОИП, 1970, отд. бнол., т. 75, вып. 3, с. 112—126.
Воронцов Н. Н., Л я п у н о в а Е. А., 3 а г о р у й к о Н. Г. Сравнительная кариология и становление изолирующих механизмов в роде Marmota. Зоол. журн., 1969, т. 48, вып. 3, с. 317—334.
Гептнер В. Г. Материалы по морфологии и систематике трехпалых тушканчиков рода Jaculus Erxl., 1777, и близких к нему форм (Mammalia, Dipodidae). Бюлл. МОИП, 1975, т. 80, вып. 3, с. 5—15.
Громов И. М., Бибиков Д. И., Калабухов Н. И., Мейер М. Н. Наземные беличьи (Marmotinae). Фауна СССР. Млекопитающие. М.—Л., «Наука», 1965, т. III, вып. 2. ,
Громов И. М., Гуреев А. А., Н о в и к о в Г. А., С о к о л о в И. И., С т р е л-к о в П. П., Ч а п с к и й К. К. Млекопитающие фауны СССР. М.—Л., Изд-во АН СССР, 1963.
Кузнецов Б. А. Отряд грызунов. — В ки.. Бобринский Н. А., Кузнецов Б. А., К у з я к и н А. П. Определитель млекопитающих СССР. Изд. 2-е. М., «Просвещение», 1965.
Лавров Л. С., Орлов В. И. Кариотипы и таксономия современных бобров (Castoridae, Mammalia). Зоол. жури., 1973, т. 52, вып. 5. с. 734—742.
Ляпунова Е. Н., Фишер Т. Б. Хромосомные наборы и систематическое поло жение Phaiomys juldaschi Severtzov и Phaiomys carruthersi Thomas. Млекопитающие. Тезисы к II Всес. совещ. по млекоп. Новосибирск, с. 131—133.
Огнев С. И. Звери СССР и прилежащих стран. М—Л., Изд-во АН СССР, т. IV. 1940, т. V, 1947, т. VI, 1948, т. VII, 1950.
Павлов М. П., Корсакова И. Б., Тимофеев В. В., Сафонов В. Г. Акклиматизация охотничье-промысловых зверей и птиц в СССР. Ч. I. Киров, Волго-Вятское книжное издательство, 1973.
Соколов В. Е. Систематика млекопитающих. Отряды: однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылов, приматов, неполнозубых, ящеров. М., «Высшая школа», 1973.
А п germa п п R. Beitrag zur Kenntnis der Gattung Lepus (Lagomorpha Lepori-dae). Mitteilungen aus dem Zoologischen Museum in Berlin, 1966, 42 В. H. 1. S. 127-—144.
В a n f i e 1 d A. W. F. The Mammals of Canada. University of Toronto Press. Toronto and Buffalo, 1974.
В r a n d t J. B. Betrage zur nahern Kenntnis der Saugethiere Russlands. Mem. Acad. Imp. Sci. St-Peterbourg, 1855, ser. 6, v. 9, p. 375.
Cabrera A. Catalogo de los mamiferos de Americo del Sur. Revista del Museo Argentine de ciencias naturales «Bernardino Rivadavia» e Institute nacional de investigation de las ciencias naturales. Buenas Aires, 1960, t. IV, N 2.
Chasen F. N. A handlist of Malaysian Mammals. Bull. Raffles Museum. Singapore. 1940, N 15.
Ellerman J. R. The families and genera of living rodents. V. I, II, III. British Museum (Natural History), 1940—1941—1949.
Ellerman J. R. and Morrison-Scott T. C. S. Checklist of palaearctic and indian mammals. 1758 to 1946. Second Edition. British Museum (Natural History). London, 1966.
Ellerman J. R., Morrison-Scott T. C. S. and Hayman R. W. Southern african mammals. 1758 to 1951: a reclassification. British Museum (Natural History). London, 1953,-
Fraye H. A. Zur Systematik der Castoridae (Rodentia, Mammalia). Mitt. Zool. Mus. Berlin, 1960, 36, L. s. 105—122.
Ge noway s H. H. Systematics and evolutionary relationships ol spiny pocket mice, Genus Liomys. Special publications the museum Texas Tech. University. 1973, N 5, p. 1—268. Texas Tech. Press. Lublock Texas.
Grasse P. P. et D e Keyser P. L. Ordre des Ronguers. In.: Grasse P. P. Ed. Traite de Zoologie. T. XVII. 2 Fasc. p. 1320—1525. Masson K- R- et Cie Paris.
Hall E. R. and Kelson К R. The mammals of North America. V. I. The Ronald Press Co. New York, 1959.
Harris W. P. Additions and corrections to the section on Sciuridae in Ellermah’s families and genera of living rodents. Occasional papers of the Museum of Zoology University of Michigan, 1944, 484, p. 1—21.
H i 11 J. E. The Robinson collection of malaysian mammals. Bulletin of the Raffles Museum, Singapore, 1960, N 29, p. 1—109.
Kingdon J. East African Mammals. VII. B. 1974. Academic Press. London a. New York, 1—704.
Landry S. O. The interelationships of the New and Old World histricomorph rodents. Univ. Calif., 1957, Press Zool. 56, p. 1—118.
Laurie E. M. O. and Hill J. E. List of land mammals of New Guinea, Celebes and adjacent islands. London, 1954.
Medway L. The wild mammals of Malaya and offshore islands including Singapore. Kuala Lumpur. Singapore. Oxford University Press. London, 1969.
Miller G. S. and G i d 1 e у J. W. Synopsis of the supergeneric groups of Rodents. Jour. Wash. Acad. Sci., 1918, p. 431—448.
M i s о n n e X. Rodentia. Smithsonian Institution, 1968.
Moore J. C. Relationships among the living squirrels of the Sciurinae. Bull, of the American Museum of Natural History, 1959, v. 118, 4 p. 159—206.
Petter F. Elements d’une revision des lievres africains du sous-genre Lepus. Mammalia, 1959, v. 23, n. 1, p. 41—67.
Petter F. Nouveaux elements d’une revision des lievercs africains. Mammalia 1963, v. 27, N 2, p. 238—255.
Pocock R. J. The classification of the Sciuridae. Proc. Zool. Soc. London, 1923, p. 209—246.
Roberts A. The mammals of South Africa. Second Ed. Published by the trustees of the «Mammals of South Africa» Book Fund, 1954.
Rosevear D. R The rodents of West Africa. British Museum (Natural History). London, 1969.
Schaub T. S. Remarks on the distribution and classification of the «Hystricomorpha». Verb. Naturf. Ges. Basel, 1953 a, 64.
Schaub S, La trigonodonties des rongeurs sirnplicidentes Ann. Paleont., 1953 b, 39, p. 29—57.
St ehl in H. G. und Schaub. Die Trigonodontie der simplicedentaten Nager. Schweiz. Pal. Abh, 1951, v. 67, S. 1—385.
Simpson G. G. 1945. The principles of classification and a classification of mammals. Bull, of the American Museum of Natural History, 1945, v. 85.
Taylor E. H. Phillipine land mammals. Bureau of science. Manila. Monograph. 1934, 30, p. 1—548.
T h e n i u s E. und Hofer H. Stammesgeschichte der Saugetiere. Spring-Verlag. Berlin—Gottingen—Heidelberg, 1960.
Tullberg T. Uber das System der Nagethiere: eine phylogenetische Studies. Nova Acta Reg. Soc. Scient. Upsala, 1899. ser. 3, 18, S. 1—514.
W a 1 к e r E. P. Mammals of the world. The Johns Hopkins Press Baltimore, 1964.
Weber M. Die Saugetiere IL Verlag von Gustav Fischer. Jena, 1928.
W i n g e H. Rodentia, Carnivora, Primates. Pattedyr—Slaegter. V. 2. Copenhagen, H. Hagerups Forlag, 1924.
Wood A. E. A revised classification of the rodents. Journal of Mammalogy, 1955, v. 36, p. 165—187.
Wood A. E. Eocene radiation and phylogeny of the rodents. Evolution, 1959, v. 13, p. 354—361.
Wood A. E. Grades and classes among rodents. Evolution, v. 19, p. 115—130.
Zimmermann K- Uber die Kreuzung von Unterartcn der Feldmaus, Microtus arvalis. Zool. Jahrb., 1959, Abt. 3, 87, 1/2, S. 1—12.
УКАЗАТЕЛИ
Указатель русских названий животных 1
Агути 409, 411, 412
— антильский 412
— бразильский 411, 412
— гваделупский 412
— гребенчатый 412
— Калиновского 412
— мексиканский 412
— руатанский 412
— сентвинсентский 412
Агутиевые 408
Акушн 409, 413
Аммодил 141, 255
Аиизомис 270, 334
Аплодонт 41
Аплодонтиа 40, 41
Аплодонтовые 39
Байбак 91, 92, 93, 94
Бандикота 270, 327
— бенгальская 329
— индийская 329
Беличьи 41, 44
Белка 32, 42, 44, 97, 131
— Аберта 48
— Александра 62
— Аллена 48
— Алстона 65
— аризонская 49
— африканская 59
— балабакская 66
— белохвостая 66
— бокуетская 50
— бороздчаторезцовая 42, 49, 50
— Винсента 62
— гамбийская 63
— гигантская 42, 57, 58
— гималайская 67
— Двухцветная 58
— длинноносая 42, 68
— Дугласа 54, 55
— желтобрюхая 51
— желтолапая 65
1 Полужирным шрифтом даются ссылки на описание рода.
Белка зеленая 62
— земляная 42, 72
— — красная 73
— золотистобрюхая 47
— золотоспинная 65
— индийская 58
— ирравадийская 65
— кавказская 47, 49
— кайбабская 48
— калималтанская 42, 70
— карликовая 42, 50
— — сулавесская 42, 52
— каролинская 47
— кистсухая 42, 53
— конголезская 61
— красная 42, 53, 55
— — земляная 73
— краснобрюхая 65
— красноногая 63
— красномордая 51
— краснощекая 70
— кремовая 58
— крошечная 42, 71
— Купера 62
— кустарниковая 41, 61, 62, 63
— Лайарда 57
— Лови 66
— магрибская 42, 72
— малайская 42, 69
— масличная 42, 58, 59
— минданаосская 66
— мпогополосая 42, 67, 68
— Моллендорфа 66
— мышиная 41, 42 63, 64
— папосская 51
— наяритская 49
— Нельсона 47
— Общественная 48
— обыкновенная 30, 44, 45, 46, 49, 53
— оранжевобрюхая 70
— оранжевоголовая 61
— палаваиская 66
— пальмовая 42, 56, 57
— — африканская 42
— — северная 56, 57
— Перни 70
— перуанская 50
Белка пестрая 48
— пигмей 42, 51, 52
— полосатая 42, 60, 63
— — кустарниковая 62
— поэзская 50
— Превоста 64
— прекрасная 42, 64, 65, 67
— Ричмонда 49
— рубиновая 42, 52
— рувензорская 63
— рыжая 62
— сабаниллская 51
— самарская 66
— серая 46, 48
— — мексиканская 47
— серожелтая 48
— синалоасская 47
— солнечная 42, 62
— сонорская 47
— стройная 66
— сулавесская 42, 65
— тонкохвостая 59
— трехполосая 57
— филиппинская 66
— Финлайсона 64, 65
— флоренциасская 51
— черная 48
— чернобрюхая 66
— чернорыжая 62
— черноухая 42, 71
— — самарская 71
— чирикаусская 49
— шипохвостая 132, 133
— юкатанская 48
Белкообразные 39
Беляк 6, 16, 17, 18, 19, 20
Бобр 129, 130, 131
— американский 129
— канадский 131
Бобровые 128
Бурундук 42, 75, 80, 82
— азиатский 76, 77, 79
— альпийский 78
— беличий 42, 80
— Буллера 79
— восточный 77
— Давида 80
— длинноухий 77, 79
— колорадский 79
— малый 75, 78
— Мерриама 78
— Палмера 79
— панамннтский 79
— рыжехвостый 79
— сероворотничковый 79
— скалистый 77, 78
— сономский 78
— сосновый 75, 78
— Таунсенда 77, 78
— уинтасский 77, 79
— Форреста 80
Вискаша 416
— Вольффсона 417
— горная 414, 416, 417
— перуанская 417
— равнинная 414,4415
Водосвинка 405, 406
— малая 406
Водосвинковые 404
Восьмизубовые 424
Восьмизуб 425
— крысиный 425, 427
— слепышовый 425, 427
— соневидный 425, 426
Гиар 433, 436
Гофер 109
— Алкорна 113
— Байли 111
— большой 115
— Буллера 109, 112, 113
— восточный 109, 110
— гигантский 109, 114
— горный 112
— грубошерстный 109, 114, 115
— дариенский 115
— дымчатый 114
— желтый 109, ИЗ
— желтомордый 113
— западный 109, 111
— Зинзера 113
— иролосский 113
— керетаросский 114
— колониальный НО, 111
— костариканский 115
— кумберлендский ПО, III
— Мерриама 113
— мичоаканский 109, 115
— пикарагуасский 115
— оахакский 114
— перотесский 113
— пустынный 110
— равнинный 108, 110
— сальвадорский 114
— северный 112
— Таунсенда 112
— техасский 110, 111
— тропический 110, 111
— узкомордый 114
— Ундервуда 115
— уруапанский 113
— центральноамериканский 109, 115
— Шермана ПО, 111
— юго-восточный 110
— южный 111, 112
Гоферовые 107
Грызуны 7, 29, 31, 32, 393
Гунди 453, 454, 456
— кистехвостый 454
Гундневые 452
Дактиломис 434
— боливийский 442
— бразильский 442
— перуанский 442
Дсгу 425, 426
Депдроприономис 270, 336
Десмодиллнскус 141, 253
Дикобраз 388, 390, 395
— амазонский 394, 398
— американский 393
— африканский 392
— большехвостый 391, 392
— бразильский 397
— венесуэльский 397
— гималайский 390
— гребенчатый 390
— двухцветный 397
Дикобраз длиннохвостый 388, 392, 393
— жесткоиглый 387, 388
— индийский 389, 390
— индонезийский 388 '
— кистехвостый 388, 390, 392
— колючий 397
— конголезский 392
— малайский 388, 389, 390
— мексиканский 396
— Ротшильда 396
— светлый 396
— североамериканский 393, 394, 395, 396
— тонкоиглый 393, 394, 398,
— Турнера 391, 392
— филиппинский 388
— цепкохвостый 393, 394, 396, 397, 398
— южноафриканский 390
— юньнаньский 390
— яванский 390
Дикобразные 386
Дипломис 434, 439
Долгоног 136, 137
Долгоноговые 135
Дремомис 42, 69
Емуранчик 376, 379
— обыкновенный 379
Зайцевые 7, 11
Зайцеобразные 5, 7
Зайчик земляной 378, 379
Заяц 6, 7, 9, 12, 13, 15, 16, 18
— американский 16, 17, 18, 19
— антилоповый 17, 19
— африканский 13, 16
— белобокий 20
— белохвостый 17
— бесхвостый 19
— бирманский 18
— бушменовый 13, 15
— Гейлларда 20
— желтоватый 20
— жесткошерстный 16
— земляной 376
— калифорнийский 17, 20
— капский 17, 18
— китайский 17, 19
— курчавый 19
— кустарниковый 18
— лазающий 13
— маньчжурский 19, 20
— полосатый 13, 14
— рыжебокий 18
— сиамский 18
— темношейный 17, 18
— Уайта 18
— чернобурый 20
• — щетинистый 15
— эфиопский 18
— яркендский 19
Землекоп голый 447, 451, 452
— капский 447, 448, 450
Землекоповые 447
Иктидомис 42, 83, 89
Каннабатеомис 434, 442
Капибара 29
Картеродон 433, 437
Клиомис 433, 437
Коноконо 434, 439
Коро 434, 440
— Блайнвилля 440
— бразильский 440
— одноцветный 441
Кролик 6, 7, 12, 13, 27
— айдахский 23, 25
— американский 13, 22
— амильтемский 25
— белохвостый 20
— бесхвостый 11, 13, 21, 22
— болотный 24
— бразильский 23
— водяной 22, 25
— дикий 27, 28
— калифорнийский 24, 25
— мексиканский 25
— новоанглийский 22, 24
— Нуталла 22, 24
— островной 25
— степной 22, 25
— флоридский 22, 23, 24
— южноафриканский 13, 25, 26
Крыса 31, 141, 269, 297
— австралийская 269, 306
— акациевая 269, 303
— ангольская 303
— африканская 270, 317, 318
— базиланская 300
— байяикасская 271 354
— бамбуковая 263, 264, 266
— — африканская 264, 265
— — большая 264, 266, 267
— — китайская 264, 267
— — малая 264, 266, 267
— — седая 264, 267
— банановая 270, 309
— Бартелса 302
— безухая 350, 351
— белохвостая 270, 310
— бирманская 296
— Бланфорда 297, 299
— Боверса 301
— болотная 270, 341
— — бороздчатозубая 269, 290
— большезубая 269, 296
— большемордая 300
— большеухая 270, 312
— бороздчатозубая 269, 285
— бугенвильская 310
— вискашевая 425, 428
— водяная 271, 349
— — австралийская 271, 351
— — горная 271, 353
— — ложная 271, 351
— — лузонская 271, 350
— гигантская 269, 282
— гималайская 300
— голохвостая 270, 310
— Гопкинса 291
— горная 300
— Гранта 303
— Гулда 284
— Гоффиана 299
— Даммермана 299
— Длиннохвостая 303
Крыса добосская 300
— Доллимана 302
— древесная 271, 346
— дымчатобрюхая 302
— желтая 296
— западноафриканская 303
— златобрюхая 352
— золотистая 303
— зубатая 282
— индийская 269, 289, 290
- - индонезийская 302
— кагайаиская 300
— Кайзера 303
— калимантанская 301
— качская 302
— каштановая 298, 302
— квинслендская 300
— Келлера 300
— керамская 270, 318
— колючая 270, 324
— красная 269, 282
— кроличья 269, 283
— кустарниковая 269, 279
---индийская 290
— — манипурская 269, 289
— лесная 269, 281, 282
— либерийская 279
— лохматоволосая 269, 286, 287
— лузонская 269, 280, 281
--- чернохвостая 281
— Маклеара 301
— Макмиллана 279
— малавийская 303
— малайзийская 300
— малая 298, 300
— манипурская 302
— маратуаская 301
— мешотчатая 270, 332
— миндаиаосская 269, 281
— мозанчнохвостая 269, 308, 309
— мозамбикская 279
— моротансская 301
— Мусшенброка 302
— Мюллера 301
— мягкошерстная 269
сулавесская 269, 296, 297
— намакуасская 303
— Никобарская 300
— новобританская 352
— новогвинейская 270, 319
— овисская 301
— охиясская 302
— пальмовая 300
— пластинчатозубые 270, 329, 330
— полосатая 271, 350
— пышиохвостая 348
— Роджерса 301
— рыженосая 269, 284, 285
— сантбарбская 300
— сантаизабельская 310
— серая 32, 295, 298, 299, 301. 302
— Симпсона 301
— скальная 445, 446, 447
— сулавесская 271, 344
— суматранская 302
— таиландская 302
— темная 305
— темнохвостая 298, 302
— тикаосская 300
— толстоиглая 433, 435
— толстохвостая 269, 283
— тонкохвостая 271, 343, 347
— Томаса 303
— треугольнохвостая 270, 331
— тринаутекая 300
— тростниковая 443, 444, 445
----большая 443, 444, 445
----малая 443, 444, 445
— туркестанская 298, 299, 302
— узкоголовая 269, 297
— филиппинская 302
— флоресская 271, 346
— Харрингтона 291
— Хеллвалда 302
— хоботковая 271, 349
— хомяковидная 270, 333, 334
— цепкохвостая 271, 344
— черная 174, 298, 299, 302
— чешуехвостая 270, 311
— шиншилловая 432
----Беннетта 431, 432
---- серая 432
—• широкозубая 270, 324
— щетинистая 433, 435, 436
----белоиглая 435
---- гайанская 435
----Гел ди 435
— — длиннохвостая 435
— Эдвардса 302
— эпгапосская 301
Ксепомис 140, 191
Куп 400, 402
Куэмизия 408
Лемминг 32
— амурский 218, 219
— бурый 218, 219
— Виноградова 215
— гудзонский 214, 215
— копытный 140, 213, 214, 215
— лесной 140, 216
— мышевидный 140, 215
----северный 216
----южный 216
— настоящий 140, 217
— норвежский 219
— сентлоренский 214
— сибирский 219
— черполапый 219
Леноксус 139, 171
Летяга 44, 97, 100, 101, 131
— азиатская 97, 99
— американская 97, 101
— белорыжая 98
— бирманская 10.3
— волосатоногая 97, 105, 106
— гигантская 96, 97, 98, 99
— дымчатая 97, 106
— желтоголовая 104
— изящная 97
— индонезийская 97, 106, 107
— калимантанская 103
карликовая 97, 104
кашмирская 103
— - китайская 97, 104
— малая 101
— Морриса 104
Летяга непальская 98
— пестрая 103
— пигмей 97, 106
— северная 101, 102
— скальная 97, 99
— сложнозубая 97, 105
— стрелохвостая 97, 103, 104
— усатая 104
— филиппинская 104
— Хагена 104
— хайнаньская 104
— черная 96, 98
— шипохвостая 132, 134, 135
— южная 101, 102
— яванская 103
—’японская 98
Летяговые 96
Лонхотрикс 433, 439
Мара 400, 403
— патагонская 403, 404
Массутьера 454, 455
Мезембриомис 269, 284
Мезомис 433, 438
Микрохидромис 271, 354
Мышеобразные 137
Мышиные 267
Мышовка 369, 370
— кавказская 370, 371
— китайская 370, 373, 374
— лесная 370, 371
— одноцветная 371
— рыжая 371
— серая 371
— степная 370, 371
— тяньшанская 370
Мышь 138, 165, 175, 269, 270, 271, 314, 316
— австралийская 270. 317
— — ложная 306
— — полевая 306
— Алстона 323
— африканская 269, 277
— болотная 270, 338
— — филиппинская 318
— бороздчатозубая 270, 322
— восточноазиатская 275, 277
— Вуснама 322
— габонская 322
— горная 275, 277
— длиннокоготная 270, 325
— длинноухая 270, 337
— длиннохвостая 269, 272, 323
— домовая 32, 269, 315, 316
— древесная 270, 337
— — азиатская 270, 313
— желтобрюхая 322
— желтогорлая 275, 277
— желто-коричневая 315, 316
— желтоточечная 322
— жестковолосая 270, 320
— западноавстралийская 306
— иглистая 270, 325
— индийская 316
— — колючая 315
— Каллеваерта 316
— камерунская 316
Мышь карликовая 315, 316
— кенгуровая 270, 320
— кенийская 316
— колючая 269, 273, 316
— конголезская 270, 340
— крупнозубая 270, 327, 328
— курганчиковая 315
— лазающая 269, 272, 334
— — африканская 270, 335
— лесная 269, 275, 275, 277
— ложная 269, 307
— Майера 271, 355
— Майора 316
— малая 270, 312
— малютка 269, 274
— манипурская 314
— Митчелла 323
— ниасская 345
— новогвинейская 271, 355
— однополосая 269, 294
— очковая 271, 353, 354
— пальмовая 269, 273
— полевая 269, 275, 275, 276
— — австралийская 269, 305
— — полосатая 269, 293
— полосатая 269, 292, 293
— Ричардсона 323
— сиккимская 315, 316
— скалистая 270, 338
— сомалийская 289
— соневидиая 271, 345
— толстая 270, 339
— травяная 269, 287, 288, 289, 294
— тушканчиковая 270, 322, 323
— филиппинская 316
— центральноавстралийская 306
— широкоголовая 270, 313
— шрилаикская 316
— эфиопская 289
— южноавстралийская 306
Нельсония 140, 191
Неохидромис 271, 353
Нутриевые 422
Нутрия 419, 422, 423, 424
Ондатра 32, 137, 138, 140, 229 — флоридская 140, 231, 232
Пака 408, 409, 410, 411
— горная 409, 411
Пакарана 406
Пакараиопые 406
Паралептомис 271, 354
Наротомис 270, 343
Песец 32
Пескорой 447, 448
— ангольский 449
— большой 449
— готтентотский 448
— капский 447, 450
— конголезский 449
— нигерийский 449
— суданский 449
— тоголезский 449
Пеструшка желтая 32, 242, 243, 244
— леммингов ая 244
— степная 141, 242, 243, 244
Песчанка 141, 248
— аденская 249
— аравийская 257
— африканская 255
— белобрюхая 250
— белуджистанская 249
— Боема 251
— большая 141, 248, 259, 260, 261
— Брантса 250, 251
— Бутлера 252
— Виноградова 255, 257
— гвинейская 250, 251
— гололапая 141, 250, 252
---индийская 138, 251
— — чернохвостая 250, 251
— — южноафриканская 250, 251
— гребенчуковая 255, 256, 257
— гребнехвостая 250, 251
— длиннолапая 141, 251
— — нигерийская 252
— дневная 141, 258
— египетская 249
— жирнохвостая 141, 254
— Зарудного 257
— индийская 255, 257
— карликовая 141, 248, 249
— — индийская 249
— Кемпа 250, 251
— когтистая 255, 257
— конголезская 252
— королевская 255, 257
— короткоухая 141, 253, 254
— красиохвостая 255, 256, 257
— малая 141, 255
— малоазиатская 257
— нигерийская 250
— Осгуда 252
— палестинская 249, 250
— персидская 257
— полуденная 255, 256, 257
— Пржевальского 141, 258
— пушистохвостая 141, 257, 258
— пышнохвостая 255, 257
— рыжая 252
— саванная 250, 251
— североафриканская 249
— сомалийская 250
— стройная 252
— суданская 250
— Сундевалла 255, 257
— Харрингтона 252
— Чизмана 249
— южноафриканская 249, 250
Пика 8, 9, 10, 11
Пищуха 6, 7, 9, 10, 12
— алтайская 9, 10, 11
— большеухая 10, II
— воротничковая 9, 11
— даурская 9, 10, 11
— камская 10
— китайская 10
— Козлова 11
— красная 9, 10, 11
— ладакская 11
— монгольская 9
— непальская 10
Пищуха палласова 9, 10, 11
— рыжеватая 9, 11
~~ северная 9, 10, 11
— степная 9, 10, 11
— тибетская 11
— Томаса 11
— черногубая 11
Пищуховые 7, 9
Полевка 140, 213
— арчевая 240, 242
— афганская 235, 240, 242
— Бедфорда 240
— береговая 239
— беренгийская 238
— большая 235, 238, 242
— Брандта 235, 240, 242
— водяная 140, 226, 227, 228, 242
— восточная 238, 242
— восточноевропейская 237, 242
— высокогорная 225
— Таппера 222
— гватемальская 241
— гобиалтайская 225
— горная 235, 239
— гуллская 239
— Гюнтера 237
— длиннохвостая 235, 239
— желтобрюхая 239
— желтощекая 240
— закаспийская 235, 237, 242
— западноевропейская 240
— земпоалтепекская 241
— иранская 237
— Кабреры 238
— калифорнийская 235, 239
— каменная 140, 223
— кашмирская 140, 225, 226
— китайская 223, 240, 242
— Кларка 238
— красная 221, 222
— красно-серая 222
— кустарниковая 240, 242
— лесная 32, 140, 220, 221
— — западная 222
— — тяньшанская 222, 223
— луизианская 241
— малоазийская 240, 242
— мексиканская 239
— Миддендорфа 235, 238, 242
— монгольская 242
— общественная 237, 242
— обыкновенная 237, 242
— орегонская 138, 235, 241
— оркнейская 238
— островная 239
— памирская 240, 242
— пенсильванская 238
— нлоскочсрепиая 224
— поющая 240
— прернйиая 241
— прометеева 141, 244, 245
— Ричардсона 241
— Роберта 237, 242
— рыжая 220, 222, 223
— сахалинская 238, 242
— северосибирская 238, 242
— серая 32, 141, 228, 235, 242
— — тяньшанская 237, 242
— — югославская 238
Полевка серебристая 224, 225
— сибирская высокогорная 225
— сиккимская 240
— скалистая 235, 239
— снежная 235, 237, 242
— сосновая 241
— средиземноморская 240
— сычуаньская 238
— тайваньская 238
— Таунсенда 239
— темная 235, 238, 242
— тибетская 240
— Томаса 235, 240
— узкочерепная 32, 235, 240, 242
— флоридская 241
— центральномексиканская 241
— чешская 241
— Шелковникова 240, 242
— шикотанская 222
— экономка 32, 238, 242
— югославская 140, 226
— южноазиатская 140, 223
— — темнобрюхая 223
— японская 235, 237
Полумышь 374
Полутушканчик 370, 371, 374
— западный 371, 373
— луговой 371, 372
— тихоокеанский 373
Праомис 269, 304
Прыгун кенгуровый 117, 121
— — большеухий 124
— — гигантский 124
— — долотозубый 124
— — знамехвостый 122, 124
— — малый темный 121
— — Мерриама 122, 123
— — Нельсона 123
— — Орда 123
--- островной 123
— — панаминтский 124
— — полуостровной 124
— — прыткий 124
— — пустынный 123
---сантакатаринский 124
---сантамаргаритский 123
— — Стефанса 124
— — техасский 123, 124
— — узкомордый 124
— — украшенный 123
— — Филиппса 123
— — фресноский 123
— — Хирманна 124
— колючий 117, 124
— — мексиканский 125
— — окрашенный 125, 126
— — панамский 126
— — Сальвина 126
— — халискосскин 126
— малый 117, 121
— — светлый 121
— — темный 121
— мешотчатый 116, 117, 124
— — Антони 120
— — аризонский 119
— — Байли 119
— — белоухий 119
— — большой 119
— — веракрусский 127, 128
Прыгун мешотчатный Гаумера 128
— — Голдмэна 126
--- Гольдемана 120
— — горный 128
— — Десмареста 126
— — Длиннохвостый 119
— — желтый 119
— — жесткошерстный 120
— — калифорнийский 120
— — колючий 120
— — малый 119
— — Мерриама 119
— — Нельсона 120, 128
— — оахакский 128
— — оливковый 118
— — песчаный 120
— — пустынный 118, 120
— — Сан-Диего 120
— — синалоаский 120
— — узкочерепной 120
— — чернохвостый 128
— — шелковистый 119
— — южный 127, 128
— центральноамериканский 117, 126
Пунарес 433, 438
Рато-до-мато 139, 177
Рипидомис 139, 149
Русак 17, 18, 20
Свинка горная 400, 402
— малая 401
— морская 400, 401, 402, 404, 406
— пампасская 401
— Чуди 401
Свинковые 399
Селевиния 367, 368, 369
Слепушонка 141, 246
— афганская 246, 247
— закавказская 246, 247
— обыкновенная 246, 247
Слепыш 263
— гигантский 261, 263
— малый 261, 263
— палестинский 262, 263
— обыкновенный 261, 263
Слепышовые 261
Собачка луговая 43, 94
— — белохвостая 94, 96
— — Гуннисона 96
— — мексиканская 96
— — чернохвостая 94, 95
— — ютасская 94, 96
Соболь 32
Сониевые 355
Соня 31, 366
— африканская 356 , 364
— китайская 366, 367
— колючая 366, 36G
— мышевидная 356, 363
— лесная 356, 361, 362
— орешниковая 356, 358, 359
— полчок 356, 357
— садовая 356, 360
— южноафриканская 356, 364
— японская 356, 363
Сундасциурус 42, 66
Сурок 41, 42, 89, 91, 92, 93, 94
— альпийский 91, 93, 94
Сурок ванкуверский 91, 93
— гималайский 92, 94
— длиннохвостый 91, 93
— желтобрюхий 91, 93
— лесной 91, 92, 93
— Мензбира 91, 93, 94
— олнмпикский 91, 93
— седой 91, 93
— серый 91, 92, 94, 94
— черношапочный 90, 92, 93, 94
Суслик 42, 82, 83, 85, 87, 94
— айдахский 88
— алашанский 88
— американский 86, 87, 89
— антилоповый 42, 83, 89
— Белдинга 86, 88
— белохвостый 83
— большой 89
— Таунсенда 88
— голохвостый 82
— горный 88, 89
— даурский 86, 88, 89
— Длиннохвостый 86, 87, 89
— европейский 89
— желтый 86, 89
— золотистый 82
— калифорнийский 80, 81
— Каскадных гор 82
— колумбийский 86, 87
— кольцехвостый 82
— крапчатый 86, 87, 89
— краснощекий 86, 89
— малый 85, 86, 87, 88, 89
— малоазиатский 88, 89
— мексиканский 84
— мохавесский 82
— Нельсона 83
— островной 83
— перотесский 84
— пустынный 42, 82, 89
— пятнистый 84
— реликтовый 86, 87, 89
— Ричардсона 86, 87
— скалистый 42, 80, 81, 82, 89
— сьеррамадресский 82
— техасский 83
— тоиколапый 42, 74
— тринадцатиполосый 84
— тропический 82
— уинтский 86, 88
— Франклина 84, 86
— Харриса 83
— черноголовый 81
Тамиопс 42, 66, 67
Тарбаган 32, 92, 93, 94
Тенелюб 447, 450
— конголезский 450
— серебристо-серый 450
— слепышовый 450
Толай 17, 18, 20
Тринакодус 434, 442
— белохвостый 443
Туко-туко 429, 430, 431
— боливийский 430
— бразильский 430
— водяной 430
— желтый 430
— магелланский 430
Туко-туко малый 430
— Наттерера 430
— перуанский 431
— Стейнбаха 431
Тукотуковые 428
Тушканчик 137, 370
— Бобринского 377, 378
— Бланфорда 383, 384
— большой 378
— Виноградова 379
— восточный 383
— горный 377, 378
— гребиепалый 376, 381, 382
— длинноухий 376, 385
— евфратский 377
— египетский 383, 384
— жирнохвостый 383, 384, 385
— Житкова 379
— карликовый пятипалый 376, 382, 384
— — трехпалый 376, 384
— Козлова 385
— Лихтенштейна 383, 384
— малый 377, 378
— монгольский 377
— мохноногий 376, 380
— песчаный 376, 382
— прыгун 377, 378
— Север нова 377, 378
— толстохвостый 376, 378, 379
— Томаса 385
— туркменский 383, 384
— Хотеоиа 377
— четырехпалый 376, 378
Тушканчиковые 375
Феловиа 454, 456
Фенакомис 140, 233
Хипогеомис 140, 210, 211
Хомяк 138, 139, 141, 168
— Альфаро 143
— Андеберта 208
— андийский 139, 185
— антильский 146
— Антони 190
— барбудаский 146
— белогорлый 143, 187
— белопалый 149
— белоухнй 183, 185
— белохвостый 140, 204
— болотный 139, 180
— большезубый 185
— большеухий 152, 153
— — гватемальский 153
— Брианта 190
— Бункера 190
— водяной 139, 174
— галапагосский 143
— гатупский 145
— геррерский 185
— Голдмана 190
— голохвостый 151
— двухцветный 144
— древесный бразильский 139, 155
— желтоватый 143
— желтоносый 183, 185
Хомяк зайцегубый 140, 207, 208
— закавказский 204
— копающий 139, 172
— королевский 145
— коротколапый 140, 208
— короткомордый 152, 153
— короткохвостый 140, 211
— косматый 140, 212, 213
— косумельский 145
— Коуеса 144
— кроликовый 139, 181
— крысовидный 200
— лазающий 151
— — мексиканский 151
— — панамский 151, 152
— Ламбертона 208
— лесной 140, 187
— — Аллена 190
— — белогорлый 187, 189
— — голохвостый 151
— — мексиканский 187, 190
— — Нельсона 189
— — пушистохвостый 187, 188, 189
— — сонорский 187, 191
— — флоридский 189
— — южный 189
— мадагаскарский 140, 206
— Мак Дугалла 185
— Мак Коннелла 143, 149
— малый рисовый 144
— матугросский 175
— Монтесумы 189
— никарагуасский 190
— Ньютона 204
—.обыкновенный 140, 201, 202, 203, 204
— патагонский 181
— — шиншилловый 139, 181
— Петерсона 182
— переднеазиатский 203, 204
— перепончатолапый 139, 182
— — большой 183
— — бразильский 182, 183
— пирисский 145
— плавающий 139, 147
— предкавказский 203, 204
— пуны 139, 180
— пустынный 187, 189
— пушисто хвостый 187, 188, 190
— рисовый 141, 144, 145
--- андийский 143
— — белогорлый 138, 142
— — болотный 142, 144
— — гигантский 139, 146
— — Дарвина 144
— — длиннохвостый 143
— — калифорнийский 144
— — карликовый 145
---малый 144
— — Нельсона 145
— — панамский 141, 145
— — Роберта 144
— — Томаса 144
— Роберта 140, 210
— рыбоядный 140, 192
— — колумбийский 194
— — панамский 194
— — Томаса 194
— — центральноамериканский 140, 193
Хомяк рыбоядный Хартманна 194
— рыжебрюхий 151
— санмаринский 190
— сентвинсентский 145
— сентлюсийский 146
— Склетера 149
— соневидный 139, 140, 153, 154, 209, 210
— — малый 209, 210
— средний 140, 202, 204
— Стефенса 190
— тамаулипасский 190
— темноногий 187, 190
— Томаса 139, 149
— тумбаласский 152
---Харриса 145
— хлопковый 139, 183, 184
— — Аллена 183, 184
— — Алстона 185
--- белоухий 183
— — вулкановый 185
— — горный 185
— — желтобрюхий 183, 184
— — желтоносый 183, 185
— — колючий 183, 184
— — малый 183 185
— — оахакский 185
— — Харриса 145
— — черноухий 185
— Ундервуда 194
— Уотсона 152
— чакосскнй 174
— чернобрюхий 143
— черноухий 145
— черный 143
— чилийский 139, 179
— шелковистый 145
— шиншилловый 139, 179, 181 .
— — патагонский 139, 181
— широколобый 189
— Штольцманна 192
— Эндерса 145
— южноамериканский 152
— ямайский 145
Хомяковые 137
Хомячок 166
— барабинский 199, 200
— белоногий 139, 158, 159, 160 161. 165
— — болотный 162
— боготский 170
— боливийский 169, 178
— болотный 155, 157, 158
— большеногий 140, 207
— большеухий 139, 152
— большой 163
— бурый 139, 163, 175
— — панамский 175
— вечерний 139, 175
— восточный 157
- — вулканный мексиканский 157
— вулкановый 139, 186
— высокогорный 139, 176
— гайанский 147
— гватемальский 163
— голоногий 163
— гондурасский 163
— горный 156
— гребенчатый 16<
Хомячок гребнехвостый 162
— гуардасский 160
— Дарвина 178
— Дариена 158
— Дикки 160
— джунгарский 140, 197, 198
— длинноволосый 175
— длиннохвостый 199, 200
— древесный бразильский 155
— желтый 163
— забайкальский 200
— западный 155, 157
— землеройковый 139, 172, 173
— золотистый 164
— изящный 157
— кактусовый 160
— калифорнийский 158, 160
— каньонный 160
— камышовый 130, 170
— — короткохвостый 171
— карликовый 137, 139, 165
— — северный 165, 166
— — южный 166
— китайский 200
— колумбийский 139, 151
— Колючий 139, 147
— короткомордый 158
— короткохвостый 200
— краснобрюхий 163
— красноносый 139, 176
— кротовый 139, 173
— крысовидный 200
— кузиечиковый 139, 166
— — северный 167, 168
— — южный 168
— лазающий 139, 151
— листоухий 139, 177
— — андийский 178
— ложноканьонный 160
— мексиканский 163
— мелкозубый 158
— Мерриама 160
— монгольский 200
— монсерратский 160
— мохнатый 157
— мышевидный 140, 195, 196
— Нельсона 163
— оахакасский 162
— олений 158, 160
— панамский 158
— Пембертона 160
— пирриский 163
— полевой 139, 155
— — андийский 169
— — белобрюхий 170
— — мексиканский 158
— — черноватый 170
— — южноамериканский 139, 168
— риодежанейрский 139, 150, 156
— рисовый колумбийский 147
— — колючий 139, 146, 147
— Роборовского 198
Хомячок Родригеса 158
— рораимский 139, 171
— рыбоядный венесуэльский 140, 193
— — горный 140, 195
— — эквадорский 140, 193
— рыжий 155, 156, 157
саванный 175
— Санборна 170
— сантакрусский 161
— сенмигельский 171
— серожелтый 178
— серый 140, 198, 199, 200
— ситхасский 161
— скалистый 158, 162
— Слевина 161
— сонорский 157
— Стиртона 163
— тибетский 200
— тоикохвостый 162
— Томаса 163
— узкомордый 158
— украшенный 147
— флоридский 163
— Хаггарда 178
— хлопковый 161
— центральноамериканский 175
— черноватый 163, 175
— черпоногий 163
— черноухий 161
— широкомордый 163
— Эверсманна 200
— южный 162
— юкатанский 139, 154, 163
Хутия 419, 422, 424
— Брауна 421
— гаитянская 419, 421, 422
— доминиканская 422
— Инграхама 421
— кистехвостая 420
— короткохвостая 419, 420, 421
— крошечная 420
— кубинская 419, 420
— малая 422
— цепкохвостая 420
Цокор 140, 205
— китайский 205
— обыкновенный 205, 206
Шиншилла 414, 417, 418
— колючая 433
— короткохвостая 418
Шиншилловые 413
Шипохвост Дерби 135
— длинноухий 133
— малый 132
— Пела 134, 135
— серебристый 135
— Ценкера 133
Шинохвостоные 131
Эласмодонтомис 408
Указатель латинских названий животных
Abrocoma 432 — bennetti 431, 432 — cinerea 432 Abrocomidae 431 Abrocomyinae 434 Acanthion 388, 390 Acomys 270, 325 — pahirinus 325, 326, 327 — dimidiatus 327 — r uss at us 325, 327 — spi nosissi mus 327 — subspinosus 325, 327 Aconaemys 425, 427 — fuscus 427 Aeretes 97, 104 Aeromys 96, 98, 103 — tephromelas 98 Aethomys 268, 269, 303 — bocagei 303 — chrysophilus 303 — defua 303 — granti 303 — hindei 303 — kaiseri 303 — longicaudatus 303 — namaquensis 303 — nyikae 303 — thomasi 303 Agouti 409 Agoutidae 409 Akodon 139, 168 — affinis 169, 170 — albivente 170 — amoenus 170 — arvicuioides 168, 169 — azarae 168, 169 .— berlepschii 170 — bogotensis 170 — boliviensis 168, 169 — budini 170 — caenosus 169 — chacoensis 169 — dolores 169 — iheringi 170 — iiiuteus 170 — iniscatus 169 — jeiskii 170 — kempi 170 — lactens 170 — lanosus 170 — lasiotis 170
Akodon latebricoia 170
— longipilis 170
— molinae 168
— mollis 169
— mimus 170
— nigrita 170
— obscurus 168, 169
• — olivaceus 169
— orophilus 169
— pacificus 169
— puer 169
— sanborni 170
— serrensis 169
— surdus 169
— tapirapoanus 169
— urichi 138, 168, 169
— varius 168, 169
— xanthorhinus 169
Alactagulus 376, 378
— pygmaeus 378
Ailactaga 376
— bobrinskii 377
— bullata 377
— euphratica 377
— elater 377
— hotsoni 377
— major 378
— severtzovi 377
— sibirica 377
— tetradactyla 376, S78
— williamsi 377
Aliocricetulus 200
Allolagus 20
Alticola 140, 223
— barakshin 225
— macrotis 225
— roylei 224, 225
— stoliczkanus 225
— strelzovi 224, 225
Ampbilaginae 7
Ainmodillus 141, 255
— imbellis 255
Ammospermophilus 42, 83
• — harrisii 83
— insularis 83
— interpres 83
— leucurus 83
— nelson i 83
Anotomys 140, 193
— ieander 193
Andinomys 139, 185
Anisomys 270, 334 — imitator 334 Anomaluridae 131 Anomaiurops 132, 135
Anomalurus 132, 134, 135
— beecrofti 135
— derbianus 135
— peli 134, 135
— pusillus 135
Aphaetreus 419
Aplodontia 41 — rufa 40, 41 Aplodontidae 39 Apodemus 269, 275 — agrarius 276 — flavicollis 277 — mystacinus 277 — speciosus 277 — sylvaticus 277
Apomys 303
Arvicanthis 269, 287
— abyssinicus 288, 289
— iacernatus 289
— niloticus 288, 289
— somalicus 289
Ar'vicola 140, 226, 228
— terrestris 226, 227
Atherurus 388, 390
— africanus 392
— centralis 392
— macrourus 392
— turneri 391, 392
Atiantoxerus 42, 72
— getulus 72
Baiomys 139, 165
— musculus 166
— taylori 165, 166
Baiyankamys 271, 354
— schawmayeri 354
Bandicota 270, 327
— bengalensis 329
— indica 329
Bathyergidae 447, 454
Bathyergomorphi 32
Bathyergus 447, 450
— janetta 451
— suillus 450
Batomys 269, 281
— dentatus 281, 282
— granti 281, 282
Beamys 268, 270, 331
— hindei 332
— major 331, 332
Belomys 97, 105
— pearsoni 105
Bunoiagus 13, 15
— monticularis 15
Blanlordimys 242
Blarinomys 139, 141, 172
— breviceps 172, 173
Brachiones 141, 258
— przewalskii 258
Brachytarsomys 140, 208
— albicauda 208
Brachyuromys 140, 211, 266
— betsiloensis 211
Brachyuromys ramirochitra 211
Brachylagus 13
Caiomys 139, 175
— caliosus 176
— dubius 176
— expulsus 176
— frida 176
— gracilipes 176
— laucha 175, 176
— iepidus 176
— muriculus 176
— tener 176
— venustus 176
Calomyscus 140, 195
— bailwardi 195, 196
Callosciurus 42, 52, 64
— albescens 65
— alstoni 65
— caniceps 65
— elbertae 65
— finlaysoni 65
— llavimanus 65
— erythraeus 64, 65
— jentinki 65
— leucomus 65
— mowewensis 65
— nigrovittatus 65
— notatus 65
— pygerythrus 65
— quinquestriatus 65
— prevosti 64
— rubriventer 65
— sarasi norum 65
Cannomys 264, 267
— badius 267
Caprolagus 13, 15, 20
— aileni 16
— brachyurus 16
— dayanus 15
— hainanus 16
— hispidus 15
— marjorita 16
— nigricollis 15
— peguinsis 15
— ruficaudatus 15
— siamensis 15
— sinensis 15
— yarcandendensis 16
Capromyidae 419, 434
• Capromyinae 434
Capromys 419
— melanurus 420
— nana 420
— pilorides 420
— prehensilis 419, 420
Cardiocranius 375, 376, 384
— paradoxus 382, 384
Carpoinys 269, 280
— melanurus 280, 281
— phaeurus 280, 281
Carterodon 433, 437
— sulcidens 437
Castor 131
— canadensis 131
— fiber 129, 131
Castoridae 128
Cavia 399, 400
— aperea 401
Cavia fulgida 401
— папа 401
— porcellus 400, 401
— tschudi 401
Caviidae 399, 405
Choeromys 445
Chaetomys 394, 398
— subspinosus 398, 399
Celaenomys 271, 350
— silaceus 350
Cercomys 433, 438
— cunicularis 438
— inermis 438
Chelemyscus 182
Chilomys 139, 151
— instans 151
Chinchilla 413, 414, 417
— brevicauda 418
— ianiger 418
Chinchillidae 413
Chinchiiiula 139, 179
— sahamae 179
Chiromyscus 270, 313
• — chiropus 313
chiropodomys 271, 345
anna 346
— fulvus 346
— giiroides 345, 346
— iegatus 346
— major 346
— niadis 345
— pictor 346
— pusillus 346
Chrotomys 271, 350
— whiteheadi 350
Citellus 42, 82, 83, 84, 85, 89
— aiaschanicus 88
— armatus 88
— beidingi 88
— citellus 89
— columbianus 87
— dauricus 88
— erythrogenys 89
— fulvus 89
— major 89
— musicus 88
— parryi 87
— pygmaeus 85, 88, 89
— relictus 87
— richardsoni 87
— suslicus 88
— undulatus 87, 89
— townsen di 88
— xanthoprymnus 88
Clethrionomys 140, 220
— gapperi 222
— giareolus 220, 222
— fra ter 222
— occidentalis 222
— rufocanus 222
— rutilus 221
— sicotanensis 222
Clyomys 433, 437
— iaticeps 437
Coeiomys 316
Coendou 394, 396
— bicolor 397
— insidiosus 397
— mexicanus 396
Coendou pallidus 396
— prehensiliis 397
— rotschildi 396
— spinosus 397
— vestitus 397
Colomys 270, 317
— goslingi 317, 318
— plumbeus 318
---plumbeus 318
Conilurus 269, 283
— albipes 283
— hemiieucurus 283
— melibiis 283
* — penicillatus 283
Cunicuius 409, 411
— paca 409, 410
Crateromys 271, 348
— schandenbergi 348
Cratogeomys 109, 113, 114
— angustirostris 114
— castanops 113
— fulvescens 113
— fumosus 114
— gymnurus 114
— irolonis 113
— merriami 113
— neglectus 114
— perotensis 113
— tylorhinus 113
— varius 113
— zinseri 113
Cricetidae 137, 138, 366
Cricetinae 138, 139
Cricetomys 268, 270, 333
— emini 333, 334
— gambianus 333, 334
Cricetulus 140, 198
— aiticoia 200
— barabensis 200
— curtatus 200
— eversmanni 200
— griseus 200
— lama 200
— iongicaudatus 200
— migratorius 199, 200
— pseudogriseus 200
— triton 200
Cricetus 140, 201, 202, 204
— cricetus 201
Crossomys 271, 350
— moncktoni 350
Crunomys 270, 318
— fallax 318, 319
— melanius 319
Cryptomys 447, 448
— foxi 449
— holosericciis 449
— hotlenlotiis 448
— kuinnii 449
— lechei 449
— mechowi 449
— ochraceocinereus 449
— zechi 449
Ctenodactilidae 452
Ctenodactylus 454
— gundi 453, 454
— joleaudi 454
— vaii 454
Ctenomyidae 428
Ctenomys 429, 430 — boliviensis 430 — brasiiiensis 430 — conoveri 431 — colburni 430 — dorsalis 430 — emilianus 430 — frater 430 — fulvus 430 — magelianicus 429 — maulinus 430 — knighti 430 — mendocinus 430 — minutus 430 — leucodon 430 — iewisi 430 — nattereri 430 — opimus 429, 430 — perrensis 430 — peruanus 430 — pontifex 431 — porteousi 429, 431 — robust us 431 — saltarius 431 — sericeus 431 — steinbachi 431 — taiarum 429, 431 — tuconax 429, 431 Cynomys 43, 94 — gunnisoni 96 — leucurus 96 — ludovicianus 95 — mexicanus 96 — par videos 96 Dendromus melanotis 335, 336 — mesomelas 335, 336 — mystacalis 336 — nyikae 336 Dendroprionomys 270, 336 — rousseloti 336 Deomys 270, 340 — ferrugineus 340 Desmodilliscus 141, 253 — braueri 253 Desmodillus 141, 253 — auricularis 253 Desmomys 292 Dicrostonyx 213 — exsul 214 — groenlandicus 215 — hudsonius 214, 215 — torquatus 214 — vinogradovi 214 Dinomyidae 406 Dinomys 406 — branickii 406 Diomys 270, 314 — crumpi 314 Diplomys 433, 434, 439 — caniceps 440 — darlingin 440 — iabiiis 440 — rufodorsalis 440 Dipodidae 375 Dipodomys 117, 121 — elator 123 — neisoni 123 Dipodiiius 250 Dipus 376, 380 — sagitta 380
Dacnomys 269, 296 — millardi 296 Dactyiomyinae 434 Dactyiomys 441, 443 — boliviensis 442 — dactylinus 442 — peruanus 442 Daptomys 140, 193 — venezueliae 193 Dasymys 269, 286 — incomtus 286, 287 — orthos 286 Dasyprocta 409, 411 — aguti 412 — azarae 412 — albida 413 — antillensis 413 — cristata 412 — fuliginosa 412 — kalinowskii 412 — mexicana 413 — noblei 413 — prymnolopha 412 — punctata 412 — ruatanica 413 Dasyproctidae 408 Delanymys 270, 338 — brooksi 338 Delichotis patagona 404 Dendromurinae 270, 334 Dendromus 270, 335, 336 — lowati 336 Dipodomys 116, 117, 121 — agiiis 124 — deserti 123 — elator 123 — heermanni 124 — gravipes 124 — elephantinus 124 — ingens 124 — insularis 123 — margaritae 123 — merriami 123 — microps 124 — neisoni 123 — nitratoides 123 — ordi 123 — ornatus 123 — panamintinus 124 — paralius 124 — peninsularis 124 — phillipsii 123 — spectabilis 122, 124 — stephensi 124 — venustus 124 Dolichotis 399, 400, 403 — patagona 404 " — salinicoia 404 Dolomys 140, 226 — bogdanovi 226 Dremomys 42, 69 — everetti 70 — lokriah 70 — pernyi 70 rufigenis 69, 70
Dryomys 356, 361
— nitedula 361, 362
Duplicidentata 7
Echimyidae 424, 431, 432, 433, 434, 445,
446
Ecliimyinae 434
Echimys 433, 434, 440, 441
— armatus 440 , 441
— blainvillei 440
— traziliensis 440
— chrysurus 440
— dasythrix 440
— grandis 440
— macrurus 440
— nigrispinus 440
— saturnus 441
— semivillosus 441
— unicolor 441
Echinoprocta 394, 398
— rufiscens 398
Echiothrix 270, 324
— leucura 324
Elasmodontomys 408
— obliquus 408
Eligomodontia 139, 176
— puerulus 176
— typus 176
Eliomys 356, 360
— melanurus 361
— quercinus 360
Eliurus 140, 209
— minor 209, 210
— myoximus 210
Ellobius 141, 213, 246
— fuscocapiilus 247
— lutescens 247
— talpinus 247
Eothenomys 140, 223
— chinensis 223
— custos 223
— melanogaster 223
— oiitor 223
— proditor 223
— wardi 223
Eozapus 369, 373
— setchuanus 373
Epixerus 42, 59
— ebii 59
Eremodipus 384
Eretizontidae 393
Erethizon 394
— dorsatum 394, 395
Eropeplus 269, 296
— canus 296
Euchoreutes 385
— naso 385
Euglaucomys 103, 104
Euneomys 139, 181
— chinchilioides 181
— lessor 182
— mordax 182
— petersoni 182
Eupetauridae 99
Eupetaurus 97
— cinereus 99
Eurymyiidae 7
Eurymylus 7
Euryzygomatomys 433, 436
Euryzygomatomys spinosus 436
Eutamias 80
Epixerus 59
— ebii 59
— wilsonl 60
Exiiisciurus 42, 71
— exiiis 72
— whitheadi 72
Felovia 454, 456
— vae 456
fimbriatus 103
Euglaucomys 103, 104
Funambulus 42, 56
— layardi 57
— palmarum 57
— pennanti 56
— sublineatus 57
— tristriatus 57
Funisci urus 42, 60
— anerythrus 61
— bayonii 61
— congicus 61
— ieucogenys 61
— pyrrhopus 60, 61
— substriatus 61
Galea 400, 402
— flavidens 402
— minimus 402
— musteloides 402
— palustris 402
— spixii 402
— wellsi 402
Geocapromys 419, 420
— brownii 421
— — brownii 421
— — thoracatus 421
— columbianus 421
— ingrahami 421
Geomyidae 107
Geomys 109, 110
— arenarius 110
— bursarius 108, 109, 110
— coionus 110
— cumberlandius 110
— fontanelus 110
— personatus 110
— pinetis 110
— tropicaiis 110
Georychus 447, 448
— capensis 448
Geosciurus 73
Gerbillinae 138, 141, 248
Gerbiilus 141, 248
— alieyi 250
— bottai 249
— campestris 249
— cheesniani 249
— dasyurus 249
— dunni 250
— erbillurus 250
— gararnaiitis 249
— gerbiilus 249
— gleadowi 249
— henleyi 249
— nancillus 250
— nigeriae 250
— paeba 250
— peeii 250
Gerbillus poecilops 249 — pyramidum 249 — simoni 250 — tytonis 250 — vallfnus 250 Glaucomys 97, 101 — volans 101, 102 Glires 7 Gliridae 355 Glirulus 356, 363 — japonicus 363 Glis 356, 357 — glis 357 Glyphotes 42, 70 — simus 70 Golunda 269, 289 — eliioti 289, 290 Grammomys 269, 279 — hunting! 279, 280 — dolichurus 279 — dryas 279 — ibeanus 279 — macmillani 279 — ruddi 279, 280 Graphiurus 356, 364 —' hueti 365 — ocularis 364 monardi 364 — murinus 365 — platyops 365 Guerlinguetus 49 Gymnuromys 140, 207, 210 — roberti 210 Gyomys 269, 305 — apodemoides 306 — berneyi 306 — deserter 305, 306 — fumeus 306 — giaucus "306 — novaehollandiae 306 — pumilus 306 Heteromys goldmani 126 — lepturus 128 — longicaudatus 126 — neisoni 126, i28 — nigricaudatus 126, 128 — oresterus 126, 128 — temporalis 127, 128 Heterogeomys 109, 114 — hispidus 115 — lanius 115 Heteropsomys 433 Hexolobodon 419 Histricomorphi 32 Holochilus 139, 182 — brasiiiensis 183 — magnus 183 Hopiomys 433, 435 — gymnurus 435 Homopsomys 433 Hybomys 269, 294 — trivirgatus 294, 295 — univittatus 294 Hydromyinae 271, 349 Hydrochoeridae 404 Hydrochoerus 405, 406 — hydrochaeris 404, 405, 406 — isthmius 406 Hydromys 271, 351 — chrysogaster 352 — habbema 352 — neobritannicus 352 Hyeienomys 316 Hylopetes 97, 103 — alboniger 103 — amoenus 103 — aurantiacus 103 — belone 103 — everetti 103 — fimbriatus 103 — harrisoni 103 — leonardi 103
Hadromys 269, 289 — humei 289 Haeromys 312 — margaettae 313 — minahassae 313 Hapalomys 269, 272 — longicaudatus 272 Heliophobius 447, 450, 454 — argenteocinereus 450 — mottoulei 450 — spalax 450 Helisciurus 42, 43, 62 — bayonii 63 — gambianus 63 — lucifer 63 — poensis 63 — rufobrachium 63 — ruwenzorii 63 — vexiliarius 63 Heptaxodon 408 Heterocephalus 447, 451 — glaber 451, 452 Heteromyidae 116 Heteromys 116, 117, 126 — australis 127, 128 —- desmarestianus 126 — gaumeri 128 — lepidus 103 — nigripes 103 — phayrei 103 — platyurus 103 — sagitta 103 — spadiceus 103 — thomasi 103 Hyperacrius 140, 225 — fertilis 226 — wynnei 226 Hypogeomys 140, 210 — antimena 210, 211 Hyosciurus 42, 43, 65 — heinrichi 65 Hystricidae 386 Hystricognathi 32 Hystricomorpha 31, 32, 386, 447, 452 Hystrix 388 — africaeaustralis 390 — brachyura 389, 390 — cristata 390 — galeata 390 — hodgsoni 390 — indica 389, 390 — javanica 390 — junnanensis 390 — klossi 390 — leucurus 390
Histrix stegmanni 390 — subcristata 390 — sumbawae 390 Leimacomys bflttneri 322 Lemmus 140, 217 — amurensis 218, 219 — lemmus 219
Ichthyomys 140, 141, 192 — hydrobates 192 — stolzmanni 192 Ictidomys 42, 83 — frankiinii 84 — mexicanus 84 — perotensis 84 — spilosoma 84 — tridecemlineatus 84 Idiurus 132 — langi 133 — macrotis 133 — panga 133 — zenkeri 133 lomys 97, 106 — horsfieldi 106, 107 Irenomys 139, 179 — tarsalis 179 Isolobodon 419, 433 Isothrix 433, 434, 439 — bistriata 439 — pachyura 439 — pagurus 439 — picta 439 — viliosa 439 — nigripes 219 — sibiricus 219 — trimucronatus 218, 219 Lemniscus 244 Lemniscomys 269, 292 — barbarus 292, 293 — griselda 293 — lynesi 293 — macculus 293 — olga 293 — striatus 292, 293 Lenomys 271, 344 — meyeri 344 Lenoxus 139, 171 — apicalis 171 Leporidae 11 Leporilius 269, 306 — apicalis 307 — conditor 307 — jonesi 307 Leptomys 271, 353 — elegans 353 Lepus 13, 16 — aegyptius 20 — alleni 19, 20 — americanus 19
Jaculus 376, 382 — blanfordi 383 — jaculus 383 — lichtensteini 383 — orientalis 383 — turkmenicus 383 — arabicus 18, 20 — arcticus 19, 20 — atlanticus 18, 20 — brachyurus 19, 20 — caiifornicus 20 — capensis 18, 20 — chadensis 20
Kannabateomys 434, 442 — amblyonyx 442 Kerodon 399, 400, 402 — rupestris 402 Komemys 292 — crawshay 18 — dayanus 18, 20 — europaens 18 — flavigularis 20 — gaillardi 20
Lachnomys 442 Lagidium 413, 414, 416 — peruanum 417 — viscaccia 416, 417 — wolffsohni 417 Lagomorpha 5, 7 Lagostomus 413, 414 — crassus 416 — maximus 414, 415 Lagurus 141, 242 — curtatus 244 — lagurus 243, 244 — luteus 244 Laomys 284 Lariscus 42, 69 — hosei 69 — insignis 69 Leggadina 270, 317 — delicatula 317 — fieldi 317 — forresti 317 — hermannsburgensis 317 — patrius 317 — messorius 317 — waitei 317 Leimacomys 270, 322 — habessinicus 18 — hainanus 18, 20 — kakumegae 20 — mexicanus 20 — microtis 20 — nigricollis 18, 20 — oiostolus 19 — orachyurus 20 — othus 19, 20 — peguensis 18, 20 — ruficandatus 20 — saiae 20 — siamensis 20 — sinensis 20 — tlbetanus 20 — timidus 20 — tolai 20 — victoriae 20 — zechi 20 — yarcandensis 19, 2 Liomys 116, 117, 12' — adspersus 126 — annectens 126 — anthonyi 126 — buiieri 126 — crispus 126 — guerrerensis 1
Liomys heterothrix 126
— irroratus 125, 126
— pictus 125, 126
— pinerotum 126
— salvini 126
— spectabilis 125
Lonchothrix 433, 439
— emiiiqae 439
Lophiomyidae 213
Lophiomyinae 138, 140, 212
Lophiomys 140, 212
— imhausi 212
Lophuromys 270, 320
— flavopunctatus 322
— luteogaster 322
— naso 322
— rahmi 322
— sikapusi 321, 322
— woosnami 322
Lorentzimys 270, 320
— nouhuysi 320
Macrogeomys 109, 115
— cavator 115
— cherriei 115
— dariensis 115
— heterodus 115
— matagalpae 115
— underwood! 115
Macrotarsomys 140, 207
— bastardi 207
— ingens 207
Macruromys 270, 319
— elegans 319, 320
— major 319, 320
Maiacomys 270, 312
— edwardsi 312
— longipes 312
Malacothrix 270, 337
— typica 337
Mailomys 271, 346
— rotschiidi 346
Marmota 43, 89
— baibacina 92, 94
— bobac 92, 93, 94
— caiigata 93
— — brewer i 91
— camtschatica 90, 92, 94
— caudata 93, 94
— flaviventris 93
— himalayana 92
— marmota 93
— menzbieri 93, 94
— monax 93
— Olympus 93
— sibirica 92
— vancouverensis 93
Massoutiera 454, 455
— harterti 455
— mzabi 455
— rotschiidi 455
Mastacomys 270, 324
— fuscus 324
— mordicus 324
Mayermys 268, 271, 355
— eiiermani 355
Megaiomys 139, 146
— audreyae 146
— desmarestii 146
Megaiomys luciae 146
Melasmothrix 270, 325
— naso 325
Melomys 270, 309, 310
— aerosus 309
— aibidens 309
— arcium 310
— cervinipes 310
— feiiowsi 309
— fraterculus 310
— fulgens 310
— monctoni 309
— leucogaster 310
— levipes 309
— lutillus 310
— obiensis 309
— platyops 309
— porcuius 310
— rubex 309
— rubicoia 309
— rufescens 310
Menetes 42, 67
— berdmorei 67
Meriones 141, 255, 258
— arimalius 257
— biackieri 257
— crassus 257
— hurrianae 257
— iibycus 257
— meridiamus 256, 257
— persicus 257
— rex 257
— shawi 257
— tamariscinus 257
— unguicuiatus 257
— vinogradovi 257
— zarudnyi 257
Mesembriomys 269, 284
— gouidi 284
— macrurus 284
Mesocricetus 140, 202, 204
— auratus 203, 204
— brandti 204
— newtoni 204
— raddei 204
Mesomys 433, 438
— didelphoides 438
— ferrugineus 439
— hispidus 438
— leniceps 439
— obscurus 439
— stimuiax 439
Microdipodops 116
Miliardia 269, 295
— gieadovi 296
— kathlcenac 296
— meltada 296
Mindanaornys 269, 281
— saloiiionseni 281
Microcavia 400, 402
— australis 402
— niata 402
— shiptoni 402
Microdill us 250
Microdipodops 116, 117, 121
— megacephaius 121
— paiiidus 121
Microhydromys 271, 354
— richardsoni 354
Microlagus 13, 25
Micromys 268, 269, 274
— minutus 274
Microsciurus 42, 50
— alfari 50
— avunculus 51
— boquetensis 50
— flaviventer 51
— florencial 51
— isthimius 50
— manarius 51
— mimulus 50
— napi 51
— otinus 50
— paimeri 50
— peruanus 50
— rubrirostris 51
— sabanillae 51
— septentrionalis 51
— similis 50
— simonsi 50
Microtinae 138, 140, 213
Microtus 141, 235
— abbreviatus 238
— afghanus 240, 242
— agrestis 238
— arvalis 237, 242
— bedfordi 240
— brandti 240
— breweri 239
— cabrerae 238
— californicus 239
— carruthersi 240, 242
— chrotorrhinus 239
— clarkei 238
— coronarius 239
— duodecimcostatus 240, 242
— fortis 238, 242
— fulviventer 239
— gregalis 240
— guatemalensis 241
— gud 242
— guentheri 237
— hyperboreus 238
— igmanensis 238
— irani 237
— irene 240, 242
— iiaeus 237
— juldaschi 240
— kikachii 238
— kimensis 242
— leucurus 240, 242
— longicaudus 239
— ludovicianus 241
— rnajori 240
— mandarinus 240
— maximowiczii 238, 242
— mexicanus 239
— middendorfi 238
— millicens 238
— miurus 240
— mongolicus 237, 242
— montanus 239
— montebell i 237
— multiplex 235, 240
— nivalis 237, 242
— nesofilus 239
— ochrogaster 241
— oeconomus 238
Microtus orcadensis 238, 242
— oregoni 241
— parvuius 241
— pennsyivanicus 238
— — breweri 242
— — provectus 242
— quasiater 241
— pinetorum 241
— richardsoni 228, 241
— roberti 237
— sachalinensis 238, 242
— savii 240, 242
— schelkovnikovi 240, 242
— sikimensis 240
— social is 237
— subarvaiis 237
— subterraneus 240, 242
— tatricus 241
— thomasi 240
— townsendii 239
— transcaspicus 237, 242
— umbrosus 241
— xanthognathus 240
Microxus 170
Micteromys 316
Monodia 141, 248
— juiiani 248
— mauritaniae 248
Muridae 138, 266, 267
Murinac 269, 271
Muriculus 270, 314
— imberbis 314
Mus 268, 270, 314
— birungensis 316
— booduga 316
— bufo 316
— callewaerti 316
— castaneus 316
— cervicolor 316
— crociduroides 316
— famulus 316
— fernandoni 316
— gerbiilus 316
— gratus 316
— haussa 315, 316
— mahomet 316
— matschie 316
— mayori 316
— minutoides 316
— musculoides 316
— muscuius 315, 316
— pahari 316
— pauiina 316
— platythrix 316
— — priesieyi 316
— proconodon 316
— tenellus 316
— triton 315, 316
— wamae 316
Muscardinus 356, 358
— avellanarius 358, 359
Mylomys 269, 285
— cuninghamei 285
— dybowskyi 285
Myocastor 424
— coy pus 424
Myocastoridae 419, 422, 434
Myocastorinae 434
Myomimus 356, 363
Myomimus personatus 363
Myomorpha 31, 32, 137, 452
Myomorphi 32
Myopus 140, 216
— ’ schisticolor 216
Myoprocta 409, 413
— acouchy 413
— pratti 413
Myosciurus 42, 64
— pumilio 63, 64
Myospaiacinae 140, 205
Myospalax 140, 205
— fontanieri 205, 206
— myospalax 205, 206
— psilurus 206
— rotschildi 206
— smithi 206
Myotomys 343
Mystromys 140, 204
— albicaudatus 204
— longicaudatus 204
Nannosciurus 42, 71
— concinnus 71
— iuncefordi 71
— melanotis 71
— samaricus 71
— surittilus 71
Napaeozapus 369, 370, 374
— insignis 374
Neacomys 139, 146
— guiana 147
— pictus 147
— pusillus 146
— spinosus 147
— tenuipes 147
Nectomys 139, 147
— alfari 148
— dimidiatus 148
— squamipes 147, 148
Nelsonia 140, 191
— neotomodon 191
Neofiber 140, 231
— alleni 231, 232
Neohydromys 271, 353
— fuscus 353
Neotoma 140, 187
— aibigula 189
— alleni 190
— angustapalata 190
— anthonyi 190
— bunkeri 190
— bryanti 190
— chrysomeias 190
— cinerea 188, 190
— floridiana 188, 189
— fuscipes 190
— goidmani 190
— latifrons 189
— iepida 189
— martinensis 190
— mexicana 190
— micropys 189
— montezumae 189
— neisoni 189
— paiatina 189
— phenax 191
— stephensi 190
— varia 189
Neotomodon 139, 186
— alstoni 186
Neotomys 139, 180
— ebriosus 180
Nesokia 270, 329
— indica 329, 330
Nesolagus 13, 14
— netscheri 14
Nesomyinae 138, 140, 206
Nesomys 140, 207
— andeberti 208
— iambertoni 208
— rufus 208
Nesoromys 270, 318
— ceramicus 318
Neusticomys 140
— monticoiis 195
Nilopegamys 318
— plumbeus 318
Notiomys 139, 141, 173
— angustus 173
— delfini 173
— edwardsi 174
— macronyx 174
— megalonyx 174
— valdivianus 174
Notomys 270, 322, 323
— alexis 323
- — alstoni 323
— aquilo 323
— cervinus 323
— longicaudatus 323
- — mcgalotis 323
— mitchelli 323
— mordax 323
— richardsoni 323
— sturni 323
Nyctomys 139, 153
— sumichrasti 153, 154
Ochotona 10
— alpina 11
— camensis 10
— collaris 11
— curoniae 11
— daurica 10, 11
— erythrotis 10, 11
— hyperborea 11
— kamensis 10
— • koslovi 11
— ladacensis 11
— macrotis 10, 11
— nepaiensis 10, 11
— pallasi 11
— princeps 8, 11
— pusiila 11
— roylei 10. 11
— rufescens 11
— rulila 10, 11
— thomasi 11
— thibetana 11
Ochotonidae 7
Ochrotomys 139, 164
— nuttalii 164
Octodon 425
— bridgesi 425, 426
— degus 426
— lunatus 426
Octodontidae 424, 432
Octodontinae 431, 434
Octodontomys 425, 426
— gliroides 426
Octomys 425, 428
— barrerae 428
— my max 428
Oenomys 268, 269, 284
— hypoxanthus 284, 285
— ornatus 285
Ondatra 140, 213, 229
— obscurus 231
— rivalicia 231
— zibethicus 229
Onychomys 139, 166
— leucogaster 167, 168
— torridus 168
Orthogeomys 109, 114
— cuniculus 114
— grandis 114
— pygacanthus 114
Oryctolagus 13, 27
— cuniculus 27
Oryzomys 139, 141
— albigularis 143
— alfaroi 143
— altissimus 144
— andinus 143
— aphrastus 145
— arenalis 143
— antillarum 145
— azuerensis 145
— b al neater 143
— bicolor 144
— bombycinus 145
— borreroi 144
— buccinatus 143
— cal igi nosus 144
— capito 143
— catherinae 144
— chaparensis 143
— couesi 144
— cozumel ae 145
— darwini 144
— delicatus 143
— delticola 143
— devius 145
— endersi 145
— flavescens 143
— fulgens 144
— fulvescens 145
— galapagoensis 143
— gatunensis 145
— hammondi 144
— indefessus 144
— intectus 143
— longicaudatus 143
— macconnelli 143
— maculipes 144
— mamorae 144
— marmosurus 144
— mel anostoma 143
— melanotis 145
— melleus 144
— microtis 143
— mincae 144
— minutus 144
— munchiquensis 143
— narboroughi 144
— nelsoni 145
Oryzomys nigripes 143 — osgoodi 144 — palustris 142, 144 — phaeotis 144 — peninsulae 144 — pirrensis 145 — polius 143 — ratticeps 143 — roberti 144 — robustulus 144 — simplex 143 — spodiurus 143 — subflavus 143 — talamancae 145 — tectus 145 — trabeatus 145 — trinitatus 144 — utiaritensis 143 — victus 145 , — villosus 143 — xantheolus 144 — zunigae 144 Otomyinae 270 Otomys 270, 341 — anchietae 342 — angoniensis 341, 343 — denti 343 — irroratus 341, 342, 343 — laminatus 342 — saundersiae 343 — sloggetti 343 — tropicalis 343 — typus 342 — unisulcatus 343 Otonyctomys 139, 154 — hatti 154 Otospermophilus 42, 80 — akocetus 82 — annulatus 82 — atricapillus 81 — beecheyl 81 — lateralis 82 — madrensis 82 — saturatus 82 — variegatus 81 Ototylomys 139, 152 — brevirostris 152 153 — connecteus 153 — phyllotis 152, 153 Oxymycterus 139, 172 — axodontius 172 — angularis 172 — delator 172 — hispidus 172 — inca 172 — paramensis 172 — roberti 172 — rufus 172
Pachyuromys 141, 254
— duprasi 254 Palaeolagidae 7 Palaeolaginae 7 Papagoinys 271, 346 — armandvillei 346 Pappogeomys 109, 112 — alcorni 113 — bulleri 113
Paradipus 376, 381
Paradipus ctenodactylus 381, 382
Parahydromys 271, 353
— asper 353
Paraleptomys 271, 354
— rufilatus 354
— Wilhelmina 354
Paramyidae 32
Paraxerus 42, 61
— alexandri 62
— antoniae 62
— boehmi 62
— bridgemani 62
— byatti 62
— cepapi 62
— cooper! 62
— emini 62
— fluvivittis 62
— lucifer 62
— ochraceus 62
— palliatus 62
— poensis 62
— sponsus 62
— vexillarius 62
— vincenti 62
— vulcanorum 62
Parotomys 270, 343
— bransti 343
— littledalei 343
Pectinator 454
— spekei 454
Pedetes 137
— cafer 136, 137
— surdaster 137
Pedetidae 135
Pediolagus 400, 404
Pelomys 269, 290
— campanae 291
— dembeensis 291, 292
— fallax 291
— harringtoni 291, 292
— hopkinsi 291
— minor 291, 292
— isseli 291, 292
— rex 291, 292
Pentalagus 13
— furnessi 13
Perognathus 116, 117
— alticoia 119
— amplus 119
— anthonyi 120
— apache 119
— arenarius 120
— artus 120
— baileyi 119
— californicus 120
— fallax 120
— fasciatus 118
— flavescens 119
— fiavus 119
— formosus 119
— goldmani 120
— hispidus 120
— inornatus 119
— intermedins 120
— longimembris 119
— merriami 119
— nelsoni 120
— parvus 119
— pernix 120
Perognathus penicillatus 118, 120
— spinatus 120
— xanthonotus 119
Petromyinae 434 ["•
Peromyscus 139, 158, 165
— allophylus 163
— altilaneus 163
— banderanus 163
— boylii 161
— buiiatus 162
— californicus 160
— caniceps 160
— crinitus 160
— dickeyi 160
— difficilis 162
— eremicus 160
— flavidus 163
— floridanus 163
— furvus 163
— gossypinus 161
— grandis 163
— guardia 160
— guatemalensis 163
— hondurensis 163
— hylocetes 162
— latirostris 163
— lepturus 162
— leucopus 159, 161
— iophurus 162
— maniculatus 160
— megalops 163
— mekisturus 162
— melanocaprus 163
— melanophrys 162
— melanotis 161
— merriami 160
— mexicanus 163
— nasutus 162
— nelsoni 163
— nudipes 163
— ochraventer 163
— oaxacensis 162
— pectoralis 162
— pembertoni 160
— perfulvus 162
— pirrensis 163
— polionotus 161
— polius 162
— pseudocrinitus 160
— truei 162
— sejugis 161
— simulatus 163
— sitkensis 161
— sievini 161
— slolops 163
— steplieni 160
— stirtoni 163
— thomasi 163
— yacatanicus 163
— zarliynchus 163
Petaurillus 97, 106
— emiliae 106
— hosei 106
— kiniochi 106
Petaurista 96, 97
— alborufus 98
— elegans 97
— leucogenys 98
— magnificus 98
Petaurista petaurista 97, 98
Petinomys 37, 104
— bartelsi 104
— crinatus 104
— electilis 104
— fuscocapillus 104
— genibarbis 104
— hageni 104
— layardi 104
— lugens 104
— maerens 104
— morrisi 104
— phipsoni 104
— setosus 104
— vordermanni 104
Petromus 446, 447
— typicus 446, 447
Petromyidae 434, 445
Petromyinae 434, 446
Petromyscus 270, 338
— collinus 338
— monticularis 338
Phaenomys 139, 150
— ferrugineus 150
Phenacomys 140, 233
— albipes 235
— intermedins 233, 234
— longicaudus 233, 235
— silvicola 234, 235
Phodopus 140, 197
— roborovski 198
— sungorus 197, 198
Phloeomyinae 271, 343
Phloeomys 271, 347
— cumingi 347
— pallidus 347
Phyllotis 139, 177
— amicus 178
— andium 177, 178
— boliviensis 178
— caprinus 178
— darwini 177, 178
— definitus 177
— domorum 178
— gerbillus 177
— griseoflavus 178
— haggard! 177, 178
— magister 177, 178
— micropus 178
— osilae 177, 178
— pictus 177, 178
— sublimis 177, 178
— wolfsohni 178
Pithecheir 269, 282
— melanurus 282
Pitymys 242
Plagiodontia 419, 421, 424, 434
— codium 422
— hylaeum 422
— ipnaeum 422
— spelaeum 422
Plagiodontinae 434
Platacanthomyidae 365
Plata canthomys 366
— lasiurus 366
Platygeomys 109, 114
— fumosus 114
— gymnurus 114
— neglectus 114
Platygeomys planiceps 114
— tylorhinus 114
Podoxymys 139, 171
— roraimae 171
Poelagus 13, 16
— marjorita 16
Pogonomelomys 269, 308, 310
— bruijni 309
— mayeri 309
— ruemmleri 309
— sevia 309
Pogonomys 271, 344
— fergussoniensis 344
— forbesi 344
— kagi 344
— lamia 344
— macrourus 344
— mollipilosus 344
— shawmayeri 344
— sylvestris 344
— vates 344
Praomys 269, 304
— albipes 304
— alta 304
— alleni 305
— angolensis 304
— baeri 304
— butleri 304
— carillus 304
— daltoni 304
— delcctorum 304
— denniae 305
— erythroleucus 304
— fumatus 304
— fumosus 305
— jacsoni 304
— morio 304
— natalensis 304
— parvus 305
— pernanus 304
— short ridgei 304
— Stella 305
— tullbergi 304
— verreauxi 304
Prionomys 270, 336, 337
— batesi 337
Procapromys 420
— gleayi 420
Proechimys 433, 435
— albispinus 435
— cancollis 435
— dimidiatus 435
— goeldii 435
— gujannensis 435
— hendeei 435
— iheringi 435
— longicaudatus 435
— myosiirus 435
— quadruplicate 435
— scmispinosus 435
— setosus 435
Prometheoinys 141, 244
— schaposchnikovi 244, 245
Pronolagus 13, 25
— barretti 26
— caucinus 26
— crassicaudatus 25, 26
— curryi 26
— kobosensis 26
Pronolagus melanurus 26
— randensis 26
— rupestris 26
— whitei 26
Prosciurillus 42, 43, 52
— abstrurus 52
— evidens 52
— leucomus 52
— murinus 52
Protoxerus 42, 58
— aubinii 59
— stangeri 59
Psammomys 141, 258
— obesus 258
Pseudohydromys 271, 355
— murinus 355
— Occidentatus 355
Pseudomys 269, 306, 307, 308
— australis 308
— auritus 308
— ferculinus 308
— gouldi 308
— gracilicaudatus 308
— higginsi 307, 308
— minnie 308
— nanus 308
— praeconis 308
— rawlinnae 308
— shortridgei 308
Pseudoryzomys 139, 177
— wavrini 177
Pseudosciuridae 32
Pteromys 97, 99
— momonga 101
— volans 100
Pteromyscus 97, 106
— pulverulentus 106
Punomys 139, 180
— iemminus 180
Pygerethmus 376, 378
— platyurus 379
— vinogradovi 379
— zhitkovi 379
Pyro my s 316
— priestleyi 316
Quemisia 408
— gravis 408
Rattus 269, 297, 303, 305
— adspersus 301
— alticola 302
— andrewsi 301
— annandalei 300
— arcuatus 300
— atri dorsum 299
— baeodon 302
— bartelsi 302
— beccarii 301
— berdmorei 302
— blanfordi 299
— bocourti 302
— bowersi 301
— brahma 302
— burrescens 300
— burrulus 300
— callitrichus 301
Rattus canus 302
— celebens 300
— chrysocomus 301
— coelestis 301
— coloratus 300
— coxingi 301
— cremoriventer 302
— cutchicus 302
— datae 302, 303
— dammermani 299
— doboensis 300
— dollmani 302
— dominator 301
— enganus 301
— edwardsi 302
— eha 302
— everetti 300
— exulans 300
— faberi 302
— flebilis 299
— frosti 301
— fulmineus 300
— fulvescens 302
— fuscipes 299
— glauerti 299
— hamatus 300
— hellwaldi 302
— hoffmani 299
— hollisteri 302
— huang 302
— humiliattis 301
— hylocoetes 302, 303
— hylomyoides 302
— inflatus 302
— infraluteus 301
— insignis 302, 303
— kennethi 301
— kelleri 300
— lalolis 299
— legatius 302
— leucop us 300
— lutreolus 298, 300
— macleari 301
— manipulus 302
— magnirostris 300
— major 302, 303
— mara 301
— marmosurus 300
— microdon 303
— montanus 300
— morotaiensis 301
— miilleri 301
— muscuius 302, 303
— musschenbroeki 302
— nativitatus 301
— niobe 302
— nitidns 300
— niviventcr 298, 301
— norvegicus 298, 301
— ohiensis 302
— owiensis 301
— palinarum 300
— paraxanthus 301
— petraeus 302, 303
— pulli venter 300
— punicans 300
— rajah 297, 302
— rattoides 300
— rattus 299
Rattus rennelli 301
— ringens 301
— robiginosus 300
— rogersi 301
— ruber 301
— sabanus 302
— sakeratens 302
— shawmayeri 301
— salocco 300
— simpsoni 301
— sordidus 301
— stoicus 300
— surifer 302
— taciturnus 300
— taerae 300
— tatei 299
— tyrannus 300
— tondanus 300
— vanheurni 301
— verecundus 301
— villosissisimus 301
— xanthurus 300
— zamboangae 300
Ratufa 42, 52, 57
— affinis 58
— bicolor 58
— indica 58
— macroura 57, 58 Rhabdomys 269, 293 — pumilio 293 Rhagomys 139, 155 — rufescens 155 Reithrodon 139, 181 — physodes 181 Reithrodontomys 139, 155 — burti 157 — brevirostris 158 — chrysopsis 157 — creper 158 — dariensis 158 — fulvescens 156, 157 — gracilis 157 — hirsutus 157 — humulus 157 — megalotis 157 — mexicanus 158 — microdon 158 — montanus 156 — raviventris 157 — rodriguezi 158 — sumichrasti 157 — tenuirostris 158 Rheithrosciurus 42, 52, 53 — macrotis 53 Rheomys 140, 193 — hartmanni 194 — raptor 194 — thomasi 194 — trichotis 194 — underwoodi 194 Rhinosciurus 42, 43, 68 — laticaudatus 68 Rhynchomyinae 271, 349 Rhynchomys 271, 349 — soricoides 349 Rhipidomys 139, 149 — latimanus 149 — leucodactylus 149 — macconnelli 149
Rhipidomys mastacalis 149
— scandens 149
Rhizomyidae 263
Rhizomys 264, 266
— pruinosus 267
— sinensis 267
— sumatrensis 266, 267
Rhodopus 140, 198
— roborovskii 198
— sungorus 197
Rhombomys 141, 259
— opimus 259, 260
Rodentia 29
Romerolagus 11, 13, 20
— diazi 20, 21
Rubrisciurus 42, 43, 52
— rubriventer 52
Saccostomus 268, 270, 332
— campestris 332, 333
— cricetulus 333
— isiolae 333
— mearnsi 333
Salpingotus 375, 376, 384
— crassicauda 383, 385
— kozlovi 385
— thomasi 385
Scapteromys 139, 174
— aquaticus 174
— chacoensis 174
— gnambiquarae 175
— tomentosus 175
— tumidus 174, 175
Scarturus 376
Sciuravidae 32
Sciuridae 41, 43
Sciurillus 42, 50, 52
— evidens 52
— murinus 52
• — pusillus 52
Sciurognathi 32
Sciuromorpha 31, 32, 3$
Sciuromorphi 32
Sciurotamias 42, 80
— davi dianus 80
— forresti 80
Sciurus 42, 44, 49, 64, 65
— aberti 44, 48
— alleni 48
— alphonsei 49
— anomalus 47
— apache 49
— argentinus 49
— arisonensis 49
— aureogaster 47
— boliviensis 49
— candelensis 49
— carolinensis 47
— chapmani 49
— cliiricahuae 49
— colliaci 47
— dcppci 48
— duida 49
— durangi 49
— ferminae 49
— flammifer 49
— gerrardi 49
— granatensis 49
Sciurus griseimembra 49
— griseoflavus 48
— griseogena 49
— griseus 48
— hoffmani 49
— ignitus 49
— igniventris 49
— ingrami 49
— kaibabensis 48, 49
— langsdorffi 49
— miravallensis 49
— nayaritensis 49
— • negligens 49
— neisoni 47
— nesaeus 49
— niger 48
— — cinereus 49
— — vulpinus 49
— nigratus 49
— oculatus 48
— poasensis 50
— poliopus 47
— pucheranii 49
— pusillus 51
— pyrrhinus 49
— pyrrhonotus 49
— rhoadsi 49
— richmondi 49
— sanborni 49
— sinaloensis 47
— social is 48
— splendidus 49
— stramineus 49
— tricolor 49
— truei 47
— variegatoides 48
— vulgaris 30, 44, 45, 46
— yucatanensis 48
Scolomys 139, 147
— mel anops 147
Scotinomys 139, 175
— harrisi 175
— longipilosus 175
— teguina 175
— xerampelinus 175
Sekeetamys 141, 257
— calurus 257, 258
Selevinia 369
— betpakdalensis 368, 369
Selevinidae 367
Sicista 369, 370
— betulina 370, 371
— caucasica 371
— caudata 371
— concolor 371
— napaea 371
— pseudanapaea 371
— subtilis 371
— tianschanica 371
Sigmodon 139, 183
— alleni 184
— alstoni 185
— aiticola 185
— fulviventer 184
— guerrerensis 185
— hispidus 184
— — arizonae 183
— leucotis 185
— macdougalli 185
Sigmodon macrodon 185 — melanotis 185 — minimus 185 — ochrognathus 185 — planifrons 185 — savannarum 185 — venester 185 — vulcani 185 Sigmomys 185 Simplicidentata 32 Spalacidae 261 Spalacopus 425, 426 — cyanus 426, 427 Spalax 263 — ehrenbergi 262, 263 — giganteus 263 — leucodon 263 — microphtalmus 262, 263 Solomys 270, 310 — ponceleti 310 — salebrosus 310 — sapientis 310 Spermophilopsis 42, 74 — leptodactylus 74 Spermophilus 85 Steatomys 270, 339 — aquillo 340 — athi 340 — bocagei 340 — caurinus 339, 340 — cuppedius 340 — gaze Пае 340 — jacksoni 340 — minutus 340 — muanzae 340 — opimus 340 — parvus 340 — pratensis 339, 340 Stenocephalemys 269, 297 Stictomys 409, 411 — taczanowskii 411 Stochonomys 303 Stylodipus 376, 379 — telum 379 Sundasciurus 42, 66 — albicauda 66 — brookei 66 — hippurus 66 — juvencus 66 — lowi 66 — melanogaster 66 — mindanensis 66 — moliendorffi 66 — philippinensis 66 — samarensis 66 — steeri 66 — tenius 66 Sylvilagus 12, 13, 22 — aquaticus 12, 25 — audoboni 25 — bachmani 24 — brasiliensis 23, 25 — cunicularis 25 — floridaiius 23, 24 — gabbi 25 — graisoni 25 — i dachoensis 23 — insonus 25 — mansuetus 25
Sylvilagus nuttalli 24
— palustris 24
— transitionalis 24
Synaptomys 140, 215
— borealis 216
— cooperi 216
Syntheosciurus 42, 49
— brochus 49, 50
— poasensis 50
Tachyoryctes 263, 264, 265
— ankoliae 266
— annectens 266
— audax 266
— cheesmani 266
— daemon 266
— macrocephalus 265, 266
— naivashae 266
— pontifex 266
— rex 266
— ruandae 266
— ruddi 266
— spalacinus 266
— splendens 265, 266
— storeyi 266
Tachyoryctinae 266
Takudaia 269, 273
— osimensis 273
Tamias 42, 75, 79
— alpinus 78
— amoenus 78
— bulleri 79
— cinereicollis 79
— dorsalis 78
— merriami 78
— minimus 78
— palmeri 79
— panamintinus 79
— quadrimaculatus 79
— quadrivittatus 79
— ruficaudus 79 *
— sibiricus 76, 79
— sonomae 78
— speciosus 77, 79
— striatus 77, 80
— townsendi 78
— umbrinus 79
Tamiasciurus 42, 53
— douglasi 55
— fremonti 56
— hudsonicus 55
— regalis 56
Tamiops 42, 65, 66
— maclellandi 67
— swinhoe 67
Tatera 141, 250
— afra 251
— boehmi 251
— brantsii 251
— guineae 251
— inclusa 251
— indica 251
— kempii 251, 255
— leucogaster 251
— nigricauda 251
— robusta 251
— valida 251
Taterillus 141, 251
Taterillus butleri 252
— clivorus 252
— emini 252
— gracilis 252
— gyas 252
— harringtoni 252
— lacustris 252
— leucogaster 251
— lowei 252
— melanops 252
— nigeriae 252
— osgoodi 252
— perluteus 252
— rufus 252
— tenebricus 252
Thamnomys 268, 269, 277, 279
— cometes 279
— dolichurus 279
— kempi 279
— kuru 279
— rutilans 277, 278, 279
— venustus 278, 279
Thecurus 388
— crassispinis 387, 388
— pumilis 388
— sumatrae 388
Theriomyidae 32
Thomasomys 139, 149
— aureus 150
— boeops 150
— bombycinus 150
— cinereiventer 150
— cinereus 150
— daphne 150
— dorsalis 150
— gracilis 150
— hylophilus 150
— incanus 150
— ischyurus 150
— kalinowskii 150
— ladewi 150
— laniger 150
— lugens 150
• — notatus 150
— oenax 150
— oreas 150
— paramorum 150
— pictipes 150
— pyrrhonotus 150
— rhoadsi 150
— rosalinda 150
— taczanowkii 150
— vestitus 150
Thomomys 109, 111
— baileyi 111
— bottae 111
— bulbivorus 112
— monticola 112
— talpoide# 112
— townsen di i 112
— umbrinus 111, 112
Thrinacodus 434, 442
— albicauda 443
— apolinari 443
— edax 443
Thryonomycidae 434
Thryonomydae 434
Thryonomyinae 434
Thryonomys 444, 445
Thryonomys gregorianus 445
— swinderianus 445
Tokudaia 269, 273
— osimensis 273
Trichys 388, 392
— lipura 392
— macrotis 393
Trogopterus 97, 105
— xanthipes 105
Thyphomys 269, 305
— adustus 305
Tylomys 139, 151
— fulviventer 151
— gymnurus 151
— mirae 152
— nudicaudus 151
— panamensis 152
— tumbalensis 152
— watsoni 152
Tympanoctomys 428
Typhlomys 366, 367- -— cinereus 367
Uranomys 270, 327
— ruddi 327, 328
Uromys 310, 311
— anak 312
— caudimaculatus 311, 312
— — barbatus 310
— fumosus 305
— imperator 312
— neobritanicus 312
— rex 312
— salamonis 312
Vanderleuria £69, 273 <
— oleracea 273
Vernaya 269, 272, 346
— fulva 272
Wiedomys 139, 176
— pyrrhorhinos 176
Wilfredomys 150
Xenomys 140, 191
— neisoni 191
Xenuromys 310
— barbatus 310, 312
Xeromys 271, 351
— myoides 351
Xerospermophilus 42, 82
— mohavensis 82
— tereticaudus 82
Xerus 42, 72
— erythropus 73
— inauris 73
— princeps 73
— rutilus 73
Zapodidae 369 Zapus 369, 370. 371 — hudsonius 372 — princeps 373 — trinotatus 373 Zelotomys 270, 313 — hildegardeae 314 — instans 314 — shortridgei 314 — woosnami 314 Zenkerella 131, 132, 133 — insignis 133 Zygodontomys 139, 170 — brevicauda 171 — cherriei 171 — lasiurus 171 *
— microtinus 171 — punctulatus 171 — seorsus 171 Zygogeomys 109, 115 — trichopus 115 Zyzomys 269, 283 — argurus 283 — pedunculatus 283 — woodwardi 284
ОГЛАВЛЕНИЕ
От автора ................................................................... 3
Отряд зайцеобразных, Lagomorpha Brandt, 1855 . . ......... 5
Семейство пищуховых, Ochotonidae Thomas, 1897 ......... 7
Род пищух, Ochotona Link, 1795 ................................. 10
Семейство Зайцевых, Leporidae Gray, 1821 ...........................11
Род лазающих зайцев, Pentalagus Lyon, 1903 . ......................... 13
Род полосатых зайцев, Nesolagus Major, 1899 .......................... 14
Род бушмевовых зайцев, Bunolagus Thomas, 1929 ........................ 15
Род щетинистых зайцев, Caprolagus Blyth, 1845 ........................ 15
Род африканских зайцев, Poelagus St. Leger, 1932 ..................... 16
Род зайцев, Lepus Linnaeus, 1758 ................................. 16
Род бесхвостых кроликов, Romerolagus Merriam, 1896 ................... 20
Род американских кроликов, Sylvilagus Gray, 1867 .............. 22
Род южноафриканских кроликов, Pronolagus Lyon, 1903 ........... 25
Род кроликов, Oryctolagus Lillijeborg, 1874 ...................... 27
Отряд грызунов, Rodentia Bowdich, 1821 29
Белкообразные, Sciuromorpha Brandt, 1855 .............................. 39
Семейство аплодонтовых, Aplodontidae Trouessart, 1897 .............. 39
Род аплодонтов, Aplodontia Richardson, 1829 ...................... 41
Семейство беличьих, Sciuridae Gray, 1821 ...........................41
Род белок, Sciurus Linnaeus, 1758 .................................... 44
Род бороздчаторезцовых белок, Syntheosciurus Bangs, 1902 ............. 49
Род карликовых белок, Microsciurus J. Allen, 1895 .................... 50
Род белок-пигмеев, Sciurillus Thomas, 1914 ......................52
Род карликовых сулавесских белок, Prosciurillus Ellerman, 1949 ... 52
Род рубиновых белок, Rubrisciurus Ellerman, 1954 ................. 52
Род кистеухих белок, Rheithrosciurus Gray, 1867 .................. 53
Род красных белок, Tamiasciurus Trouessart, 1880 ..................... 53
Род пальмовых белок, Funambulus Lesson, 1832 ..................... 56
Род гигантских белок, Ratufa Gray, 1867 .......................... 57
Род масличных белок, Protoxerus Major, 1893 ..................... 58
Род африканских белок, Epixerus Thomas, 1909 ......................... 59
Род полосатых белок, Funisciurus Trouessart, 1880 .................... 60
Род кустарниковых белок, Paraxerus Major, 1893 ................. 61
Род солнечных белок, Heliosciurus Trouessart, 1880 .............. 62
Род мышиных белок, Myosciurus Thomas, 1909 ........................... 64
Род прекрасных белок, Callosciurus Gray, 1867 ........................ 64
Род сулавесских белок, Hyosciurus Tate et Archbold, 1935 ............. 65
Род сундасциурус, Sundasciurus Moore, 1958 ........................... 66
Род тамиопс, Tamiops J. Allen, 190C>...................................66
Род многополосых белок, Menetes Thomas, 1908 ......................... 67
Род длинноносых белок, Rhinosciurus Gray, 1843 ....................... 68
Род малайских белок, Lariscus Thomas et Wroughton, 1909 ...............69
Род дремомис, Dremomys Heude, 1898 ................................... 69
Род калимантанских белок, Glyphotes Thomas, 1898 ..................... 70
Род черноухих белок, Nannosciurus Trouessart, 1880 ................... 71
Род крошечных белок, Exilisciurus Moore, 1958 ........................ 71
Род магрибских белок, Atlantoxerus Major, 1893 ...................... 72
Род земляных белок, Xerus Hemprich et Ehrenberg, 1832 ............... 72
Род тонкопалых сусликов, Spermophilopsis Blasius, 1884 .............. 74
Род бурундуков, Tamias liliger, 1811..................................75
Род беличьих бурундуков, Sciurotamias Miller, 1901.................. 80
Род скалистых сусликов, Otospermophilus Brandt, 1814..................80
Род пустынных сусликов, Xerospermophilus Merriam, 1892 .............. 82
Род антилоповых сусликов, Ammospermophilus Merriam, 1892 .... 83
Род иктидомисов, Ictidomys Allen, 1877 .............................. 83
Род сусликов, Citellus Oken, 1816 ( = Spermophilus F. Cuvier, 1825) . . 85
Род сурков, Marmota Frisch, 1775 .................................... 89
Род луговых собачек, Cynomys Rafinesque, 1817.........................94
Семейство летяговых, Pteromyidae Brandt, 1855 ........................... 96
Род гигантских летяг, Petaurista Link, 1795 ......................... 97
Род черных летяг, Aeromys Robinson et Kloss, 1915.....................98
Род скальных летяг, Eupetaarus Thomas, 1888 ......................... 99
Род азиатских летяг, Pteromys G. Cuvier, 1800 ....................... 99
Род американских летяг, Glaucomys Thomas, 1908 ..................... 101
Род стрелохвостых летяг, Hylopetes Thomas, 1908 .................... 103
Род карликовых летяг, Petirut/nys Thomas, 1908 .................. 104
Род китайских летяг, Aeretes G. Allen, 1938 ........................ 104
Род сложнозубых летяг, Trogopterus Heude, 1898 .................. 105
Род волосатоногих летяг, Eelomys Thomas, 1908 .................. 105
Род дымчатых летяг, Pteromyscus Thomas, 1908 ...................106
Род летяг-пигмеев, Petaurillus Thomas, 1908 ........................ 106
Род индонезийских летяг, lomys Thomas, 1908 ........................ 106
Семейство гоферовых, Geomyidae Gill, 1872 ............................ . 107
Род восточных гоферов, Geomys Rafinesque, 1817 ..... . . . ПО
Род западных гоферов, Thomomys Wied-Neuwied, 1839 ...................Ill
Род гоферов Буллера, Pappogeomys Merriam, 1895 ..................... 112
Род желтых гоферов, Cratogeomys Merriam, 1895 ...................... 113
Род гигантских гоферов, Orthogeomys Merriam, 1895 .................. 114
Род грубошерстных гоферов, Heterogeomys Merriam, 1895 .............. 114
Род центральноамериканских гоферов, Macrogeomys Merriam, 1895 . . . 115
Род мичоаканских гоферов, Zygogeomys Merriam, 1895 ................. 115
Семейство мешотчатых прыгунов, Heteromyidae Allen et Chapman, 1893 . . 116
Род мешотчатых прыгунов, Perognathus Wied-Neuwied, 1839 .... 117
Род малых прыгунов, Microdipodops Merriam, 1891......................121
Род кенгуровых прыгунов, Dipodomys Gray, 1841........................121
Род колючих прыгунов, Liomys Merriam, 1902 ......................... 124
Род центральноамериканских прыгунов, Heteromys Desmarest, 1817 . . 126
Семейство бобровых, Castoridae Gray, 1872 .............................. 128
Род бобров, Castor Linpaeus, 1758 .................................. 131
Семейство шипохвостовых, Anomaluridae Gill, 1872 ....................... 131
Род малых шипохвостов, Idiurus Matschie, 1898 ...................... 132
Род шипохвостых белок, Zenkerella Matschie, 1898 ................... 133
Род шипохвостых летяг, Anomalurus Waterhouse, 1843 ................. 134
Семейство долгоноговых, Pedetidae Owen, 1847 ........................... 135
Род долгоногов, Pedetes liliger, 1811................................137
Мышеобразные, Myomorpha Brandt, 1855 ................................. 137
Семейство хомяковых, Cricetidae Rochebrune, 1883 ....................... 137
Род рисовых хомяков, Oryzomys Baird, 1857 .......................... 141
Род гигантских рисовых хомяков, Megalomys Trouessart, 1881 . . . 146
Род колючих рисовых хомячков, Neacomys Thomas, 1900 ........... 146
Род колючих хомячков, Scolomys Anthony, 1924 .................147
Род плавающих хомяков, Nectomys Peters, 1861.........................147
Род рипидомис, Rhipidomys Tschudi, 1844 ........................ . 149
Род хомяков Томаса, Thotnasotnys Cones, 1884 ....................... 149
Род риодежанейрских хомячков, Phaenomys Thomas, 1917 . . . 150
Род колумбийских хомячков, Chilomys Thomas, 1897 .... 151
Род лазающих хомяков, Tylomys Peters, 1866 . . . ............. 151
Род большеухих хомяков, Ototylomys Merriam, 1901 ..... 152
Род соневидных хомяков, Nyctomys De Saussure, 1860 ..................153
Род юкатанских хомячков, Otonyctomys Anthony, 1932 ................. 154
Род бразильских древесных хомячков, Rhagomys Thomas, 1917 . . . 155
Род полевых хомячков, Reithrodontomys Gigiioli, 1873 ........... 155
Род белоногих хомячков, Peroinyscus Gloger, 1841.....................158
Род золотистых хомячков, Ochrotomys Osgood, 1909 .... . . 164
Род карликовых хомячков, Baiomys True, 1894 ...................... . 165
Род кузнечиковых хомячков, Onychomus Baird, 1857 ................. 166
Род южноамериканских полевых хомячков, Akodon Меуеп, 1833 .... 168
Род камышовых хомячков, Zygodontomys Allen, 1897 ................. 170
Род рораимских хомячков, Podoxymys Anthony, 1929 ................. 171
Род леноксусов, Lenoxus Thomas, 1909 ............................. 171
Род копающих хомяков, Oxymycterus Waterhouse, 1837 ............... 172
Род землеройковых хомячков, Blarinomys Thomas, 1896 .............. 172
Род кротовых хомячков, Notiomys Thomas, 1890 ..................... 173
Род водяных хомяков, Scapteromys Waterhouse, 1837 ................ 174
Род бурых хомячков, Scotinomys Thomas, 1913........................175
Род вечерних хомячков, Calomys Waterhouse, 1837 (=Hesperomys Waterhouse, 1839)...................................................... 175
Род высокогорных хомячков, Eligmodontia F. Cuvier, 1837 ............ 176
Род красноносых хомячков, Wiedomys Hershkovitz, 1959 ................ 176
Род рато-до-мато, Pseudoryzomys Hershkovitz, 1959 ................... 177
Род листоухих хомячков, Phyllotis Waterhouse, 1837 .................. 177
Род чилийских хомяков, Irenomys Thomas, 1919..........................179
Род шиншилловых хомяков, Chinchillula Thomas, 1898 ..................<179
Род хомяков пуны, Punomys Osgood, 1943 ............................. 180
Род болотных хомяков, Neotomys Thomas, 1894 ....................... 180
Род кроликовых хомяков, Reithrodon Waterhouse, 1837 ............ ... 181
Род патагонских шиншилловых хомяков, Etineornys Coues, 1874 .... 181
Род перепончатолапых хомяков, Holochilus Brandt, 1835 ....... 182
Род хлопковых хомяков, Sigmodon Say et Ord, 1825 ................... 183
Род андийских хомяков, Andinomys Thomas, 1902 ...................... 185
Род вулкаиовых хомяков, Neotomodon Merriam, 1898 ................... 186
Род лесных хомяков, Neotoma Say et Ord, 1825 ...................... 187
Род нельсония, Nelsonia Merriam, 1897 .............................. 191
Род ксеномис, Xenomys Merriam, 1892 .................................191
Род рыбоядных хомяков, Ichthyomys Thomas, 1893 ...................... 192
Род эквадорских рыбоядных хомячков, Anotoniys Thomas, 1906 ......... 193
Род венесуэльских рыбоядных хомячков, Dapiomys Anthony, 1929 .... 193
Род центральноамериканских рыбоядных хомяков, Rheomys Thomas, 1906 . 193
Род горных рыбоядных хомячков, Neusticomys Anthony, 192]..............195
Род мышевидных хомячков, Calomyscus Thomas, 1905 .................... 195
Род джунгарских хомячков, Phodopus Miller, 1910.......................197
Род серых хомячков, Cricetulus Milne-Edwards, 1867 ................. 198
Род обыкновенных хомяков, Cricetus Leske, 1779 ..................... 201
Род средних хомяков, Mesocricetus Nehring, 1898 ..................... 202
Род белохвостых хомяков, Mystromys Wagner, 1841 .................... 204
Подсемейство цокоров, Myospalacinae Lilljeborg, 1866 ................... 205
Род цокоров, Myospalax Laxmann, 1769 ................................ 205
Подсемейство мадагаскарских хомяков, Nesomyinae Major, 1897 ............ 206
Род большеногих хомячков, Macrotarsomys Milne-Edwards et Grandidier, 1898 207
Род зайцегубых хомяков, Nesomys Peters, 1870 ........................ 207
Род коротколапых хомяков, Br achy tarsomys Gunther, 1875 ............ 208
Род соневидных хомяков, Eliurus Milne-Edwards, 1885 ................. 209
Род хомяков Роберта, Gymnuromys Major, 1896 ......................... 210
Род хипогеомис, Hypogeomys Grandidier, 1869 ......................... 210
Род короткохвостых хомяков, Brachyuromys F. Major, 1896 ............. 211
Подсемейство косматых хомяков, Lophiomyinae Thomas, 1897 ............... 212
Род косматых хомяков, Lophiomys Milne-Edwards, 1867 ................. 212
Подсемейство полевок, Microtinae Miller, 1896 .......................... 213
Род копытных леммингов, Dicrostonyx Gloger, 1841......................213
Род мышевидных леммингов, Synaptomys Baird, 1857 .................... 215
Род лесных леммингов, Myopus Miller, 1910.............................216
Род настоящих леммингов, Lemmus Link, 1795 .......................... 217
Род лесных полевок, Clethrionomys Tilcsius, 1850 .................... 220
Род южноазиатских полевок, Eothenomys Miller, 1896 . 223
Род каменных полевок, Allicola Blanford, 1881...................... 223
Род кашмирских полевок, Ilyperacrius Miller, 1896 ................... 225
Род югославских полевок, Dolomys Nehring, 1898 ...................... 226
Род водяных полевок, Arvicola Lacepede, 1799 ........................ 226
Род ондатр, Ondatra Link, 1795 ...................................... 229
Род флоридских ондатр, Neofiber True, 1884 .......................... 231
Род фенакомис, Phenacomys Merriam, 1889 ............................. 233
Род серых полевок, Microtus Schrank, 1798 ........................... 235
Род степных пеструшек, Lagurus Gloger, 1841 ......................... 242
Род прометеевых полевок, Prometheomys Satunin, 1901 ................. 244
Род слепушонок, Ellobius Fischer, 1814 .........................246
Подсемейство песчанок, Gerbillinae Alston, 1876 .................... 248
Род монодия, Monodia Heim de Balsac, 1943 ....................... 248
Род карликовых песчанок, Gerbillus Desmarest, 1804 .............. 248
Род гололапых песчанок, Tatera Lataste, 1882 250
Род длиннолапых песчанок, Taterillus 'Thomas, 1910................251
Род короткоухих песчанок, Desmodillus Thomas et Schwann, 1904 . . 253
Род десмодиллискус, Desnwdilliscus Wettstein, 1917................253
Род жирнохвостых песчанок, Pachyuromys Lataste, 1880 ............ 254
Род аммодил, Ammodillus Thomas, 1904 ............................ 255
Род малых песчанок, Meriones liliger, 1811........................255
Род пушистохвостых песчанок, Sekeetamys Ellerman, 1947 .... 257
Род песчанок Пржевальского, Brachiones Thomas, 1925 .............. 258
Род дневных песчанок, Psammomys Cretzschmar, 1828 ............... 258
Род больших песчанок, Rhombomys Wagner, 1841 .................... 259
Семейство слепышовых, Spalacidae Gray, 1821 .......................... 261
Род слепышей, Spalax Giildenstaedt, 1770 ......................... 263
Семейство бамбуковых крыс, Rhizomyidae Miller et Gidley, 1918 .... 263
Род африканских бамбуковых крыс, Tachyoryctes Rfippell, 1835 .... 265
Род бамбуковых крыс, Rhizomys Gray, 1831 ......................... 266
Род малых бамбуковых крыс, Cannomys Thomas, 1915...................267
Семейство мышиных, Muridae Gray, 1821 ................................ 267
Подсемейство мышей, Murinae Murray, 1866 ........................... 271
Род длиннохвостых мышей, Hapalomys Blyth; 1859 ................... 272
Род лазающих мышей, Vernaya Anthony, 1941 ........................ 272
Род пальмовых мышей, Vandeleuria Gray, 1842 ................. 273
Род колючих мышей, Tokudaia Kuroda, 1943 ...................... 273
Род мышей-малюток, Micromys Dehne, 1841 ...................... 274
Род лесных и полевых мышей, Apodemus Каир, 1829 ...... 275
Род африканских мышей, Thamnotnys Thomas, 1907 . . . . 277
Род кустарниковых крыс, Grammomys Thomas, 1915 .... . . 279
Род лузонских крыс, Carpomys Thomas, 1895 ...................... 280
Род минданаосских крыс, Mindanaomys Sanborn, 1953 ................ 281
Род лесных крыс, Batomys Thomas, 1895 ............................ 281
Род красных крыс, Pithecheir F. Cuvier, 1838 ..................... 282
Род гигантских крыс, Hyomys Thomas, 1903 ......................... 282
Род кроличьих крыс, Conilurus Ogilby, 1838 .................... ... 283
Род толстохвостых крыс, Zyzomys Thomas, 1909 ..................... 283
Род мезембриомис, Mesembriomys Palmer, 1906 ...................... 284
Род рыженосых крыс, Oenomys Thomas, 1904 ......................... 284
Род бороздчатозубых крыс, Mylotnys Thomas, 1906 .............. 285
Род лохматоволосых крыс, Dasymys Peters, 1875 .................... 286
Род травяных мышей, Arvicanthis Lesson, 1842 .................... 287
Род манипурских кустарниковых крыс, Hadromys Thomas, 1911 . . . 289
Род индийских крыс, Golunda Gray, 1837 ...................... 289
Род бороздчатозубых болотных крыс, Pelotnys Peters, 1852 .... 290
Род полосатых мышей, Limniscomys Trouessart, 1881 ................ 292
Род полосатых полевых мышей, Rhabdomys Thomas, 1916................293
Род однополосых мышей, Hybomys Thomas, 1910........................294
Род мягкошерстных крыс, Millardia Thomas, 1911.....................295
Род большезубых крыс, Dacnomys Thomas, 1916........................296
Род сулавесских мягкошерстных крыс, Eropeplus Miller et Hollister, 1921 ......................................................... 296
Род узкоголовых крыс, Stenocephalemys Frick, 1914..................297
Род крыс, Rattus Fischer, 1803 ................................... 297
Род акациевых крыс, Aethomys Thomas, 1915..........................303
Род праомис, Praomys Thomas, 1915..................................304
Род темных крыс, Tryphotnys Miller, 1910...........................305
Род австралийских полевых мышей, Gyotnys Thomas, 1910.............305
Род австралийских крыс, Leporilltis Thomas, 1906 .............. 306
Род ложных мышей, Pseudomys Gray, 1832 ........................... 307
Род мозаичнохвостых крыс, Pogonomelomys Riimmler, 1936 .... 308
Род банановых крыс, Melotnys Thomas, 1922 ......................... 309
Род голохвостых крыс, Solomys Thomas, 1922 ....................... 310
Род белохвостых крыс, Xenuromys Tateet Archbold, 1941..............310
Род чешуехвостых крыс, Uromys Peters, 1867 .................... 311
Род большеухих крыс, Malacomy$ Milne-Edwards, 1877 ............... 312
Род малых мышей, Haeromys Thomas, 1911.............................312
Род азиатских древесных мышей, Chiromyscus Thomas, 1925 .......... 313
Род широкоголовых мышей, Zelotomys Osgood, 1910.....................313
Род манипурских мышей, Diomys Thomas, 1917..........................314
Род полосатоспиниых мышей, Muriculus Thomas, 1902 ................. 314
Род мышей, Mus Linnaeus, 1758 ..................................... 314
Род австралийских мышей, Leggadina Thomas, 1910.....................317
Род африканских крыс, Colomys Thomas et Wroughton, 1907 .... 317
Род керамских крыс, Nesoromys Thomas, 1922 ........................ 318
Род филиппинских болотных мышей, Crunomys Thomas, 1897 .... 318
Род новогвинейских крыс, Macruromys Stein, 1933 ................... 319
Род кенгуровых мышей, Lorentzimys Jentink, 1911.....................320
Род жестковолосых мышей, Lophuromys Peters, 1874 .................. 320
Род бороздчатозубых Вышей, Leimacomys Matschie, 1893 .............. 322
Род тушканчиковых мышей, Notomys Lesson, 1842 ..................... 322
Род широкозубых крыс, Mastacomys Thomas, 1882 ..................... 324
Род колючих крыс, Echlothrix Gray, 1867 ........................... 324
Род длиннокоготных мышей, Melasmothrix Miller et Hollister, 1921 . . 325
Род иглистых мышей, Acomys I. Geoffroy, 1838 ...................... 325
Род крупнозубых мышей, Uranomys Dollman, 1909 ..................... 327
Род бандикот, Bandicota Gray, 1873 ............................. 327
Род пластинчатозубых крыс, Nesokia Gray, 1842 ..................... 329
Род треугольнохвостых крыс, Beamys Thomas, 1909 .............. 331
Род мешотчатых крыс, Saccostomus Peters, 1846 ..................... 332
Род хомяковидных крыс, Cricetomys Waterhouse, 1840 .............. 333
Род анизомис, Anisomys Thomas, 1903 ............................... 334
Подсемейство лазающих мышей, Dendromurtnae Allen, 1939 .............. 334
Род африканских лазающих мышей, Dendromus A. Smith, 1829 . . . 335
Род дендроприономис, Dendroprionomys Petter, 1966 ................. 336
Род длинноухих мышей, Malacothrix Wagner, 1843 .................... 337
Род древесных мышей, Prionotnys Dollman, 1910......................337
Род скалистых мышей, Petromyscus Thomas, 1926 .................... 338
Род болотных мышей, Delanymys Hayman, 1962 .................... 338
Род толстых мышей, Steatomys Peters, 1846 ......................... 339
Род конголезских мышей, Deomys Thomas, 1888 .................... 340
Подсемейство болотных крыс, Otomyinae Thomas, 1897 .................. 341
Род болотных крыс, Otomys F. Cuvier, 1923 341
Род паротомисов, Parototnys Thomas, 1918............................343
Подсемейство тонкохвостых крыс, Phloeomyinae Alston, 1876 ........... 343
Род сулавесских крыс, Lenomys Thomas, 1898 344
Род цепкохвостых крыс, Pogonomys Milne-Edwards, 1877 ............ 344
Род соневидных мышей, Chiropodomys Peters, 1868 ............... 345
Род древесных крыс, Mallomys Thomas, 1898 ......................... 346
Род флоресских крыс, Papagomys Sody, 1941 ......................... 346
Род тонкохвостых крыс, Phloeomys Waterhouse, 1839 ................. 347
Род пышнохвостых крыс, Crateromys Thomas, 1895 ............... 348
Подсемейство хоботковых крыс, Rhynchomyinae Thomas, 1897 .... 349
Род хоботковых крыс, Rhynchomys Thomas, 1895 ...................... 349
Подсемейство водяных крыс, Hydromyinae Alston, 1876 ................. 349
Род полосатых крыс, Chrotomys Thomas, 1895 ........................ 350
Род лузонских водяных крыс, Celaenomys Thomas, 1898 ............... 350
Род безухих крыс, Crossomys Thomas, 1907 .....................' . 350
Род ложных водяных крыс, Xeromys Thomas, 1889 ..................... 351
Род австралийских водяных крыс,Hydromys Е. Geoffroy, 1805 .... 351
Род горных водяных крыс, Parahydromys Poche, 1906 .............. 353
Род неохидромис, Neohydromys Laurie, 1952 ........................ 353
Род очковых мышей, J.eptomys Thomas, 1897 ........................ 353
Род микрохидромис, Microhydromys Tate et Archbold, 1941 .... 354
Род паралептомис, Paraleptomys Tate et Archbold, 1941 ... . 354
Род байянкасских крыс, Baiyankamys Hinton, 1943 .................. 354
Род новогвинейских мышей, Pseudohydromys Riiinmler, 1934 .......... 355
Род мышей Майера, Mayertnys Laurie et Hill, 1954 .................. 355
Семейство соневых, Gliridae Thomas, 1897 ............................. 355
Род сонь-полчков, Glis Brisson, 1762 ............................ 357
Род орешниковых сонь, Muscardinus Каир, 1829 ................. 358
Род садовых сонь, Eliomys Wagner, 1843 ........................ 360
Род лесных сонь, Dryomys Thomas, 1906 ............................ 361
Род японских сонь, Glirulus Thomas, 1905 .......................... 363
Род мышевидных сонь, Myomimus Ognev, 1924 ......................... 363
Род африканских сонь, Graphiurus Smuts, 1832 ...................... 364
Семейство колючих сонь, Platacanthomyidae Miller et Gidley, 1918 .... 365
Род колючих сонь, Platacanthomys Blyth, 1859 .................... 366
Род китайских сонь, Typhlomys Milne-Edwards, 1877 ................... 367
Семейство селевиний, Selevinidae Argyropulo et Vinogradov, 1939 . . . 367
Род селевиний, Selevinia Belosludov et Bashanov, 1938 ............... 369
Семейство мышовок, Zapodidae Cones, 1875 ............................ 369
Род мышовок, Sicista Gray, 1827 ..................................... 370
Род полутушканчиков, Zapus Cones, 1875 ....................... 371
Род китайских мышовок, Eozapus Preble, 1899 .................... 373
Род полумышей, Napaeozapus Preble, 1899 ....................... 374
Семейство тушканчиковых, Dipodidae Waterhouse, 1842 .............. 375
Род земляных зайцев, Allactaga F. Cuvier, 1836 ...................... 376
Род земляных зайчиков, Alactagulus Nehring, 1897 .............. 378
Род толстохвостых тушканчиков, Pygerethmus Gloger, 1841 ............. 378
Род емуранчиков, Stylodipus G. Allen, 1925 .......................... 379
Род мохноногих тушканчиков, Dipus Zimmermanp, 1780 .... 380
Род гребнепалых тушканчиков, Paradipus Vinogradov, 1930 .... 381
Род песчаных тушканчиков, Jaculus Erxleben, 1777 .............. 382
Род пятипалых карликовых тушканчиков, Cardiocranius Satunin, 1903 . 384
Род трехпалых карликовых тушканчиков, Salpingotus Vinogradov, 1924 384
Род длинноухих тушканчиков, Euchoreutes Sclater, 1841 ............... 385
Дикобразные, Hystricomorpha Brandt, 1855 ............................ 386
Семейство дикобразовые, Hysiricidae Burnett, 1830 ....................... 386
Род индонезийских дикобразов, Trecurus Lyon, 1907 ................... 388
Род дикобразов, Hystrix Linnaeus, 1758 ............................ 388
Род кистехвостых дикобразов, Atherurus F. Cuvier, 1829 .... 390
Род длиннохвостых дикобразов, Trichys Gunther, 1876 ................. 392
Семейство американских дикобразов, Erethizontidae Thomas, 1897 . . . 393
Род североамериканских дикобразов, Erethizon F. Cuvier, 1822 . . 394
Род цепкохвостых дикобразов, Coendou Lacepede, 1799 ................. 396
Род амазонских дикобразов, Echinoprocta Gray, 1856 .................. 398
Род тонкоиглых дикобразов, Chaetomys Gray, 1843 ..................... 398
Семейство свинковых, Caviidae Waterhouse, 1839 .................... 399
Род морских свинок; Cavia Pallas, 1766 .............................. 400
Род моко, Ktrodon F. Cuvier, 1825 ................................... 402
Род куи, Calea Меуеп, 1831 .......................................... 402
Род горных свинок, Microcavia Gervais et Ameghino, 1880 . . . 402
Род мара, Dolichotis Desmarest, 1820 ................................ 403
Семейство водосвинковых, I/ydrochoeridae Gill, 1872 ..................... 404
Род водосвинок, Hydrochoerus Brisson, 1762 .......................... 406
Семейство пакарановых, Dinomyidae Alston, 1876 .......................... 406
Род пакаран, Dinotnys Peters, 1873 ............................ 406
Семейство семизубовых, Heptaxodontidae Miller et Gidley, 1918 .... 407
Род эласмодонтомис, Elasmodontomys Anthony, 1916......................408
Род куэмизия, Quemisia Miller, 1929 ................'................ 408
Семейство агутиевых, Dasyproctidae Smith, 1842 .................... 408
Род пака, Cuniculus Brisson, 1762 ................................... 409
Род горных пака, Stictomys Thomas, 1924 .......................... . 411
Род агути, Dasyprocta liliger, 1811...................................411
Род акуши, Myoprocta Thomas, 1903 ................................... 413
Семейство шиншилловых, Chinchillidae Thomas, 1896 ....................... 413
Род равнинных вискаш, Lagostomus Brooks, 1828 ....................... 414
Род горных вискаш, Lagidium Меуеп, 1833 ............................. 416
Род шиншилл, Chinchilla Bennett, 1829 ............................... 417
Семейство хутий, Capromyidae Smith, 1842 ................................ 419
Род кубинских хутий, Capromys Desmarest, 1822 ....................... 419
Род короткохвостых хутий, Geocaprotnys Chapman, 1901 ................ 420
Род гаитянских хутий, Plagiodontia F. Cuvier, 1836 .................. 421
Семейство нутриевых, Myocastoridac Miller et Gidley, 1918.................422
Род нутрий, Myocastor Kerr, 1792 .................................... 424
Семейство восьмизубовых, Octodontidae Waterhouse, 1839 .................. 424
Род восьмизубов, Octodon Bennett, 1832 .............................. 425
Род соневидных восьмизубов, Octodoniomys Palmer, 1903 ............... 426
Род слепышовых восьмизубов, Spalacopus Wagler, 1832 ................. 426
Род крысиных восьмизубов, Aconaemys Ameghino, 1891 .................. 427
Род вискашевых крыс, Octomys Thomas, 1920 .................... 428
Семейство тукотуковых, Ctenomyidae Tate, 1935 ........................... 428
Род туко-туко, Ctenomys de Blainville, 1826 ......................... 430
Семейство шиншиллокрысовых, Abrocomidae Miller et Gidley, 1918 . . . 431
Род шиншилловых крыс, Abrocoma Waterhouse, 1837 .................... 432
Семейство колючих шиншилл, Echimyidae Miller et Gidley, 1918 .... 433
Род щетинистых крыс, Proechimys J. Allen, 1889 ...................... 435
Род толстоиглых крыс, Hoplomys J. Allen, 1908 ....................... 435
Род гиар, Euryzygomatomys Goeldi, 1901 .............................. 436
Род клиомис, Clyomys Thomas, 1916.....................................437
Род картеродон, Carterodon Waterhouse, 1848 ......................... 437
Род пунарес, Cercomys F. Cuvier, 1829 ............................... 438
Род мезомис, Mesotnys Wagner, 1845 .................................. 438
Род лонхотрикс, Lonchothrix Thomas, 1920 ...................... 439
Род коноконо, Isothrix W'agner, 1845 ................................ 439
Род дипломис, Diplomys Thomas, 1916...................................439
Род коро, Echimys F. Cuvier, 1809 ................................... 440
Род дактиломис, Dactylomys I. Geoffroy, 1838 ........................ 441
Род каннабатеомис, Kannabateomys Jentink, 1891 ................ 442
Род тринакодус, Thrinacodus Giinther, 1879 .......................... 442
Семейство тростниковых крыс, Thryonomyidae Pocock, 1922 ................. 443
Род тростниковых крыс, Thryonomys Fitzinger, 1867 .............. 445
Семейство скальных крыс, Petromyidae Tullberg, 1899 ..................... 445
Род скальных крыс, Petromus A. Smith, 1831 .......................... 447
Hystricomorpha incerttie sedis........................................447
Семейство землекоповых, Bathyergidae Waterhouse, 1841 ................... 447
Род капских землекопов, Georychus liliger, 1811.......................448
Род пескороев, Cryptomys Gray, 1864 ................................. 448
. Род тенелюбов, Heliophobius Peters, 1846 ........................... 450
Род капских пескороев, Bathyergus liliger, 1811.......................450
род голых землекопов, Heterocephalus Riippel, 1842 .................. 451
Hystricomorpha или Myomorpha incertae sedis..........................452
Семейство гундиевых, Ctenodactylidae Zittel, 1893 ....................... 452
Род гунди, Ctenodactylus Gray, 1830 ................................. 454
Род кистехвостых гунди, Pectinator Blyth, 1856 ...................... 454
Род массутьер, Massouiiera Lataste, 1885 455
Род феловиа, Felovia Lataste, 1886 .............................. 456
Литература 457
Предметный указатель ........................................................460
Соколов В. Е.
С59 Систематика млекопитающих (Отряды: зайцеобразных, грызунов). Учеб, пособие для ун-тов. М., «Высш, школа», 1977.
494 с. с ил. Библиогр.: 457—459.
Первая часть «Систематики млекопитающих» вышла в 1973 г. Вторая часть посвящена систематике, морфологии, экологии зайцеобразных и грызунов. Приводятся подробные характеристики отрядов, семейств, кратко — родов. Дается полный перечень видов и их ареалы. Указаны виды, вошедшие в «Красную книгу».
Книга хорошо иллюстрирована.
Предназначается для студентов университетом. Может быть использовано преподавателями биологии средней школы, преподавателями вузов и научными сотрудниками.
60000—059
001(01)—77
69—77
596.5
ВЛАДИМИР ЕВГЕНЬЕВИЧ СОКОЛОВ
СИСТЕМАТИКА МЛЕКОПИТАЮЩИХ
И. Б. № 958
Редакторы: Л. П. Петровская, Т. А. Рыкова Художник А. В. Алексеев Художественный редактор Т. А. Коленкова Технический редактор А. К. Нестерова Корректор С. К. Марченко
Сдано в набор 23/IV-76 г. Подп. к печати 24/XI-76 г. Формат 70X100716- Бум. тип. № 2. Объем 31 печ. л. Усл. п. л. 40,3. Уч.-изд. л. 41,87. Изд. № Е-285. Тираж 20 000 экз. Зак. 399. Цена 1 р. 82 к.
План выпуска литературы для вузов и техникумов издательства «Высшая школа» на 1977 г.
Позиция № 69
Издательство «Высшая школа», Москва, К-51, Пеглипиая ул., д. 29/14
Ярославский нолиграфкомбииат Союзполиграф-прома при Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли. 150014. Ярославль, ул. Свободы, 97.