/








































































































































































































































Текст
Е<ч9.
Р-93.
АКАДЕМИЯ НАУК СОЮЗА ССР
ИНСТИТУТ МИКРОБИОЛОГИИ
В. Л. РЫЖКОВ
ФИТОИАТОГЕННЫЕ
ВИРУСЫ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА- 1946 -ЛЕНИНГРАД
АКАДЕМИЯ НАУК СОЮЗА ССР
ИНСТИТУТ МИКРОБИОЛОГИИ
В. Л. РЫЖКОВ
ФИТОПАТОГЕННЫЕ
ВИРУСЫ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА 1946 ЛЕНИНГРАД
Ответственный редактор
член-корреспондент АН СССР
В. Л. ИСАЧЕНКО
ПРЕДИСЛОВИЕ
Вышедшая в 1944 году наша книга «Основы учения
о вирусных болезнях растений» заключает в себе общее учение
о вирусах, тогда как настоящая работа посвящена частному
учению о вирусных болезнях растений.
Единственным руководством по вирусным болезням
растений на русском языке является наша книга, вышедшая
в 1935 г.,1 которая не только давно разошлась, но и устарела.
За 10 лет, которые прошли со дня ее выхода, учение о вирусных
болезнях растений сделало огромные успехи. Особенно
разительные успехи в этой области достигнуты советскими учеными.
10 лет тому назад было известно о существовании в СССР
нескольких вирусных болезней картофеля, мозаичной болезни
табака, мозаичного заболевания фасоли — и только. В
настоящее время, благодаря работам советских фитопатологов, список
вирусных болезней, встречающихся в СССР, можно, пови-
димому, считать в основном выявленным. По приблизительному
нашему подсчету (количества вирусных болезней растений,
зарегистрированных в различных странах) оказалось, что
на Америку приходится 82 вирусных заболевания, на Западную
Европу — 56, на СССР — 48, на Азию — 17, на Африку
и Австралию — по 12 вирусных заболеваний. Количество
зарегистрированных вирусных заболеваний растений до
известной степени является мерилом выявленностн этих
заболеваний, и мы видим, что в СССР вирусных болезней выявлена
не меньше, чем в Западной Европе. Конечно, от выявления
вирусного заболевания до строгого доказательства его вирусной
природы и подробного его изучения — еще большой путь.
В этом отношении США попрежнему находятся на первом месте.
Изучение вирусных заболеваний растений во всем мире
1 В. Л. Рыжков. Вирусные болезни растений. Москва, 1935.
развивалось в двух направлениях: с одной стороны — в
направлении описания все новых и новых заболеваний, а с другой—
в направлении установления родства — а иногда и единства —
причин заболеваний, которые сначала казались совершенно
различными. Так, нам стало известно, что желтая пятнистость
ананаса вызывается тем же вирусом, что и бронзовость
помидора, что calico картофеля вызывает вирус, известный ранее
как возбудитель мозаичной болезни люцерны.
В настоящее время все еще преобладает первое
направление, приводящее к первоначальному накоплению материала,
который дальнейшие исследования, может быть, позволят
упростить и объединить. Пока трудности, возникающие перед
каждым пытающимся систематизировать наши знания о
вирусных болезнях растений, остаются огромными. Область
систематики вирусных болезней растений все еще можно лучше
всего охарактеризовать так, как Линней в свою эпоху
характеризовал систематику простейших, — одним словом: «хаос».
Это положение вещей делает задачу составителя монографии
по вирусным болезням растений достаточно неблагодарной.
Мы имеем дело с материалом, который не всегда тщательно
описан, сведения о котором часто крайне недостаточны. Мы
сознаем все возникающие отсюда трудности для читателя,
желающего составить себе целостное представление о том
или ином заболевании, и поэтому, насколько возможно,
пытались облегчить ему эту задачу. С этой целью мы постоянно
выделяли среди огромного количества часто плохо описанных
заболеваний практически наиболее важные, наиболее часто
встречающиеся. Далее, близко стоящие друг к другу,
родственные заболевания мы объединили в одну группу. Наконец,
близкие, трудно диференцируемые болезни мы сопоставляли,
где только возможно, в отдельных таблицах, которые должны
помочь читателю уяснить себе различие между ними.
Мы не хотели исключить из нашей работы мало изученные,
плохо известные заболевания, как это сделал, например,
Holmes (1939)1, потому что хотя это облегчило бы нашу задачу,
но уменьшило бы справочную ценность книги.
В конце книги приложены таблицы для определения
важнейших вирусных заболеваний. Необходимо сказать не-
1 F. О Holmes. A handbook of phytopathojenic viruses.
Minneapolis, 1039.
4
сколько слов и о проблеме определителя вирусных болезней
растений и фитопатогенных вирусов. Первая такая попытка
была сделана Е'. Johnson (1930)г. Этот автор составил
определитель вирусных болезней табака, встречающихся в штате
Кентукки. Вторая попытка была предпринята нами, в 1935 г.
в упомянутой уже выше книге, где мы пытаемся дать в виде-
таблиц, облегчающих идентификацию вирусного заболевания,
обзор важнейших вирусных болезней по всем культурам.
J. Johnson и J. Hoggan2 в 1935 г. дали одну общую таблицу,
охватывающую важнейшие вирусы.
Здесь нет нужды оговариваться, что при современном
состоянии знаний таблицы для определения фитопатогенных
вирусов носят лишь предварительный характер, однако я
считаю их полезными: они дают в легко доступной форме обзор
важнейших вирусных заболеваний и с возможной четкостью
формулируют главные диагностические признаки их. Тот,
кто стал бы сетовать на трудности пользования этими таблицами
или на громоздкость методов идентификации вирусов (іюторая
в отдельных случаях имеет место), должен вспомнить, что
определение микробов связано с подобными же трудностями.
Само собой понятно, что определение вирусов является еще
более сложным делом.
1 Е. Johnson. Virus diseases of tobacco in Kentucky. Kentucky
Agr. Exp. Sta. Res. Bull., 306, 1930.
2 J. Johnson a. J. Hoggan. Phytopath., 25t № 3, 328—
343, 1935.
Глава I
ВИРУСНЫЕ БОЛЕЗНИ ПОРЯДКА КРАПИВОЦВЕТНЫХ
Заболевания вяза. У Ulmus в 1918 г. в штате Огайо впервые
наблюдалось заболевание, при котором пластинки листьев
¦становятся более хрупкими и закручиваются вниз; происходит
также закручивание черешков, и листья преждевременно
отмирают. Это заболевание в 1936—1937 гг. повело к массовой
гибели деревьев. Оно описано Swingle под названием
некроза флоэмы, так как при нем происходит отмирание
флоэмы и камбия как в надземных частях растений, так и в
его корне. Болезнь передается прививкой кусков корня, коры
или побегов (инкубация — от 6 месяцев до 2 лет).
Позднее Swingle, Tilford и Irish коротко описали
мозаичное заболевание вяза. На одном и том же дереве одни листья
имеют нормальные размеры, другие — мелки, морщинисты,
мозаичны и с трудом сгибаются. Дерево обнаруживает
несколько чрезмерное развитие придаточных побегов.
Заболевание передается прививкой. Из сообщения неясно отношение
описываемого заболевания к предыдущему.
Мозаичные заболевания инжира. В Калифорнии Ira Condit
(1933, 1941, 1943) описала мозаичную болезнь смоковницы,
которая по К. Smith вызывается Ficus virus l Condit and Home.
Болезнь поражает Ficus carica, F. albissima, F. Krischna,
F. tsiela и др. Она характеризуется диффузными, светло-
желтыми пятнами на листьях и плодах, причем листья
деформируются незначительно. Заболевание эгого типа также
широко распространено в СССР, например в Крыму.
Передается оно прививкой, но не соком или семенами. Возможно,
что в распространении болезни играют роль насекомые Lepi-
dosaphes fici, Eriophyes fici (в США) и Homotoma fids (в СССР).
На Кавказе, кроме этого типа мозаики, мы наблюдали
деформирующую мозаику, которая отличается тем, что
вызывает очень резкую редукцию пластинки листа, вплоть до ее
нитевидности.
На Кавказе встречается также кольцевая пятнистость
инжира, выражающаяся очень крупными кольцами бронзо-
7
вого цвета. Интересно, что этот тип кольцевой пятнистости
мы встречали также и у других растений в гербарии, собранном
в Грузии и изученном Л. Канчавели и Е. Эристави.
Возможно, что эти кольцевые пятнистости вызываются одним
и тем же вирусом, который, подобно вирусу кольцевой
пятнистости табака или бронзовости помидора, распространяется
среди представителей самых различных семейств.
Упоминается о мозаичном заболевании инжира,
культивируемого в Англии в оранжереях (Ainsworth, 1935). В
оранжерейных коллекциях инжира в СССР мозаика также не
является редкостью.
У Ficus paracelli известна мозаика ( F. Smith, 1926), ay Ficus
tsiela — болезнь, похожая на spike сандала (Lushington, 1918).
Мозаичная болезнь шелковицы. Эта болезнь подробно
изучена Endo и Kurasawa (1937—1940). Листья пораженных
растений утолщаются, происходит посветление по ходу нервов
и крапчатость. На нижней стороне листа по ходу жилок могут
развиваться листовидные выросты. Часто пластинка листа
редуцируется, и получаются нитевидные листья. Шелковичные
черви не могут питаться листьями больных деревьев.
В клетках больных растений развиваются Х-тела. Сок
больных растений обладает повышенной способностью
редуцировать метиленовую синьку. Эту особенность сохраняют
и сухие листья больных растений 9-летней давности. Болезнь
передается прививкой, но не соком.
У шелковицы, кроме того, встречается заболевание,
похожее на желтуху персиков, при котором наблюдается
курчавость листьев, хлороз и кустистость; листья переполняются
крахмалом. Инфекционный характер болезни не установлен
(Atanasoff, 1935).
Курчавость хмеля. Другие названия болезни: Kaderavost,
Krauselkrankheit, Nettlehead. По К. Smith, болезнь вызывает
Humulus virus 2, Duffield. При заболевании жилки на нижней
стороне листа становятся более выпуклыми, а вследствие
диспропорции в росте жилок и основной ткани листья делаются
морщинистыми. Верхушечная доля листа ненормально удлиняется.
В тяжелых стадиях болезни рост подавлен, побеги тонки,
слабы и многочисленны. Общий вид растения начинает
несколько напоминать крапивный куст, откуда и происходит
название nettle (крапива), head (голова). В Чехословакии
под названием Kaderavost болезнь подробно изучалась Blattny
(1930—1931). Этот автор наблюдал, в числе менее постоянных
симптомов, развитие желтых пятен и некротические явления.
В анатомическом отношении болезнь характеризовалась
гипоплазией паренхимы листа, дегенерацией хлоропластов
и образованием в клетках мелких Х-тел, интенсивно
красящихся по Гимза.
По английским данным, болезнь передается только при-
8
вивкой. Согласно Blatuiy (1931), заболевание может такжег
хотя и с трудом, передаваться соком, почвой и цикадой Chlo-
ritaflavescens. Идентичность заболевания, описанного в Англии,
с описанным в Чехословакии не может считаться прочно
установленной. Болезнь легко поражает сорт Fuggles. В Чехословакии
наблюдается курчавость хмеля, сопровождающаяся
некрозами флоэмы и мозаикой на листьях (Blaltny и Vukolov, 1935).
Болезнь, повидимому, комплексной природы, и в ней
участвует вирус курчавости.
Мозаичная болезнь хмеля. Она, как и предыдущая, описана
в Англии. Листья приобретают желто-зеленую пятнистость,
становятся морщинистыми и хрупкими. Известная хрупкость
наблюдается также и у стебля, верхушка которого утрачивает
способность виться. При тяжелых формах болезни стебель
может отмирать; происходит также частичное отмирание
корневой системы, и растение постепенно погибает. Многие сорта,
в том числе сорт Fuggles, несут вирус только скрыто. Болезнь
передается прививкой, но не соком.
По К. Smith, заболевание вызывает Humulus vifus і,
Salmon (1922). Очень сходное мозаичное заболевание было
описано также у сорта Fuggles, который, как мы видели выше,
только скрыто несет этот вирус, поэтому приходится принять,
что мозаичное заболевание этого сорта хмеля вызывается особым
вирусом, обозначаемым Humulus virus 4, Salmon и Ware (1930).
В Тасмании Мс Lennan (1931) описывает мозаику хмеля, при
которой он обнаружил в клетках своеобразные Х-тела,
принимаемые им за особый паразитический организм Leptomyxa
reticulata.
Хлороз хмеля. Болезнь вызывается Hamulus virus <?,
Salmon и Ware (1930). При ней на листьях развиваются светло-
желтые или желто-зеленые пятна, которые могут
распространяться на всю пластинку; сохранившиеся зелеными участки
вздуваются, что уродует пластинку листа. Светлые участки
листа подавлены в развитии, и количество хлорофилловых
зерен уменьшается. Болезнь передается соком и прививкой,
а также семенами. На 228 проростков от больных растений
было получено 26.8% пораженных мозаикой (Salmon и Ware,
1935).
Вирусное заболевание хмеля, встречающееся в СССР,
было подробно изучено К. С. Рахлис, которая дает следующее
описание его: «Листья бледнозеленые, очень мелкие, ломкие,
шероховатые. Некротические явления наблюдаются часто,
стебли тонкие, более светлые. Боковые ветви очень короткие,
листья на них очень мелкие. Кусты хмеля часто не доходят
до высоты шпалер».1 Автор описывает это заболевание под
названием хлороза, хотя, по вышеуказанным признакам,
1 Цит. по рукописному отчету.
'.)
оно, как нам кажется, не вполне подходит под определение
хлороза и ближе к курчавости. В некоторых случаях болезнь
удалось передать соком и прививкой, но экспериментальные
данные не особенно отчетливы. Заболевание вызывает резкое
снижение урожая и ухудшение качества сырья (уменьшение
процента содержания горьких веществ). К. С. Рахлис получила
сыворотку, специфическую к вирусу, что дает возможность
диагносцировать скрытые формы заболевания.
Меры борьбы с вирусными болезнями хмеля. Для борьбы
с вирусными болезнями хмеля рекомендуется уничтожение
больных растений, — если число их в хозяйстве не слишком
велико, — тщательный выбор здорового материала для по-
садкст, а также введение устойчивых сортов.
Вирусные болезни конопли. В Германии К. Boder (1941)
описал передающееся соком заболевание конопли,
характеризующееся хлорозом. У больных растений количество стеблей
увеличено, но они более тонки. В СССР встречается
заболевание конопли, похожее на вирусное.
Глава II
ВИРУСНЫЕ БОЛЕЗНИ СЕМЕЙСТВА CHENOPODIАСЕАЕ
Курчавость верхушки. Curly юр, или
калифорнийская болезнь, вызывается Beta virus 1, Boncquel и Har-
lung (1915). Она известна в Калифорнии с 1899 г. и получила
широкое распространение в США. В СССР и вообще в Европе
она неизвестна.
У свеклы заболевание начинается гипертрофией жилок
листа и их посветлением. С нижней стороны листа вдоль
жилок могут развиваться выросты. Наблюдается курчавость
листьев и закручивание их вверх или вниз; растение
может принять вид капустного кочана. Посветление по ходу
нервов позднее исчезает, и листья становятся темнозелеными,
толстыми, кожистыми. При тяжелых формах болезни на черешке
и: в области главной жилки развиваются коричневые
некротические участки, из которых вытекает бурая сиропообразная
жидкость. Главный корень уменьшается в размере, а боковые
развиваются в увеличенном количестве, причем усиленное
развитие боковых корней связано с их преждевременным
отмиранием. Каждый вновь возникающий корень отмирает
п заменяется новым. При заболевании наблюдаются глубокие
изменения во флоэме.
У шпината также наблюдаются посветление по ходу
нервов, курчавость и закручивание листьев. Больные растения
желтеют и часто умирают. Различные виды сорных растений,
как то: виды Polygonum, Rumex, поражаются вирусом,
вызывающим заболевания также у представителей самых
разнообразных семейств. Вызванное им заболевание помидора носит
название желтухи и приносит в Калифорнии большой
ущерб этой культуре. Вообще вредоносность описываемой
болезни очень велика. Убытки от заболевания исчисляются
в миллионах долларов.
Вирус передается цикадой Eutettix tenellus. Baker. В
Аргентине встречается заболевание, по внешним симптомам
совершенно подобное описываемому, но передающееся цикадой
Agallia stricticollis. Так как в Аргентине есть и переносчик
11
curly-top, то это говорит против идентичности обеих болезней-
Передача болезни соком обычно не удается, однако не вполне
невозможна. Согласно Bennett (1934), сок, непосредственно
вытекающий из перерезанной жилки листа, иногда вызывает
заражение при инокуляции иглой (у 14 из 124 растений). Вирус
зимует в сорняках, а также в корнях свеклы.
Вирус инактивируется при температурах 76—79°С. Он
сохраняет активность при разведении 1:20 000, сохраняется
в соке не менее 7 дней, остается активным и в сухих тканях.
Он легко проходит через свечи Шамберлена самых высоких
номеров и Беркефельда.-Вирус обнаруживает большую
устойчивость к рН среды, сохраняя активность при рН 2.9—9.1.
В абсолютном алкоголе он не инактивируется в течение 2 часов,
а в 70% алкоголе может сохранять активность в течение 56
дней (Bennett, 1935; Severin и Freitag, 1933).
Описано несколько штаммов вируса, дающих
заболевания различной силы.
Для борьбы с болезнью существенным является
своевременный посев. Семена должно высеять с таким расчетом,
чтобы ко времени появления насекомого-переносчика растения
достигли как можно более сильного развития, так как чем
выше возраст, в котором произойдет заражение, тем меньше
от заболевания страдает урожай. В настоящее время известны
сорта, высокоустойчивые к вирусу (Carsner, 1933).
Курчавость листьев свеклы. Kraupelkrankheit немецких
авторов вызывается Beta virus 3, Wille. Это заболевание свеклы
по внешним проявлениям очень похоже на предыдущее, от
которого оно отличается главным образом наличием другого
переносчика — клопа Piesma quadrata Fieb.— и географическим
распространением, так как встречается в Германии, где в
некоторых случаях бывает причиной потери 45—76% урожая.
Болезнь не передается соком. Личинки P. quadrata не
могут инфицироваться и переносить вирус, но взрослые клопы,
раз получив вирус, сохраняют его в течение всей жизни. Вирус
зимует не только в корнях свеклы, но и в насекомом.
Представители дикой флоры из семейства Chenopodiaceae
не поражаются болезнью. Удалось искусственное
инфицирование ею фасоли.
Савой свеклы. Заболевание получило название по сходству
больных растений с сортом капусты, называемой савойской.
Болезнь описана в ряде штатов США в 1937 г. Ее вызывает
Beta virus 5, Coons, Kotila и Stewarl (1937).
Признаки болезни очень сходны с признаками
предыдущей болезни; переносчиком ее является клоп Piesma cinerea.
Несомненно, что калифорнийская и аргентинская
курчавость верхушки, курчавость листьев, встречающаяся в
Германии, а также савой свеклы, известный в США, представляют
собой очень близкие по симптомам заболевания. Дальнейшие
32
исследования, может быть, покажут, что они являются как
45ы биологическими расами одного и того же вируса,
приспособившегося к разным переносчикам. Сходное заболевание
Панфилова наблюдала в Средней Азии.
[ Желтуха свеклы и шпината. Заболевание описано Roland
(1936) под названием faunisse. Его вызывает Beta virus 4,
Roland и Quanjer (1936). Первоначально заболевание неправильно
смешали с мозаичной болезнью, описание которой приводится
ниже. Желтуха вызывает уменьшение размеров, утолщение
и хрупкость листьев, а также накопление в них крахмала.
Признаки болезни особенно заметны не на самых молодых
листьях. В патолого-анатомическом отношении болезнь
характеризуется отложением гумми в ситовидных трубках. При
сухости воздуха и большой инсоляции симптомы болезни
усиливаются.
. -7
Рис. 1. Лист свеклы, больной желтухой
Болезнь не передается соком. Ее переносчиками являются
Myzns persicae Sulz. и Aphis rumicis (A. fabae). Чем
дольше насекомое находится на больном растении, тем успешнее
оно переносит вирус на здоровое.
Болезнь известна в Бельгии, Голландии и других странах
Европы, а также в США. Нами болезнь обнаружена в
Армении (по гербарным образцам — рис. 1). В Западной Европе
она является одним из главнейших заболеваний свеклы (М.
Simon, 1940).
Мозаичная болезнь свеклы. Это заболевание вызывает
Beta virus 2, Lind (1915). Оно описано впервые в 1891 г.
РгіШеаих и Delacioux.
13
У свеклы при заболевании наблюдается чрезвычайно
разнообразная по рисунку мозаичность. Пораженные участки
имеют вид пятен, колец, мелких крапинок и т. д. Иногда они
захватывают незначительную часть листа, в других же случаях
поражают его сплошь. При тяжелых формах листья могут
деформироваться. Болезнь легко маскируется при
температурах выше 21 и ниже 10°С. На плантациях широко
распространены замаскированные формы болезни, что затрудняет
отбор здоровых корней.
Хозяйственный вред от заболевания непостоянен. По
данным украинской литературы, количество сахара или совсем
не снижается, или уменьшается на 0.75—1.1%. Максимальное
уменьшение веса корня выражается в 12.9%.г
У шпината первыми симптомами заболевания являются
подавленность роста и яркожелтые лятна на молодых листьях,
которые сливаются между собой и образуют большие участки.
Вирус может поражать целый ряд сорных растений, как
то: Amarantus retroflexus, Chenopodium album, Sonchus arvensis.
Заражение соком удается с трудом, однако в последнее
время оно было осуществлено путем нежного лотирания листа
соком больных растений (Roland, 1936). Болезнь переносится
Aphis fobae, Myzus persicae, а, возможно, также и некоторыми
цикадами (Новиненко). Может быть, некоторую роль в
распространении болезни играет передача ее соком при обрезке
ботвы. При заражении свеклы первого года большую роль
играет семенная свекла, в корнях которой находится вирус.
Отсюда вытекает требование изолировать плантации свеклы,
культивируемой для семян, от посевов свеклы первого года.
Пройда- требует пространственной изоляции в 1500 м.
В пораженных участках подавлена диференцировка на
палисадную и губчатую паренхиму. Пораженные пластиды
уменьшаются в размерах и количестве и плохо окрашиваются
железистым гематоксилином. Schaffnit (1927) видел во флоэме
больных растений веретеновидные включения, которые в СССР
в дальнейшем изучались Левшиным и Сиротиной. Эти
включения были первоначально приняты за организм, являющийся
возбудителем болезни, но мы их считаем белковыми кристал-
лоидными образованиями. Последняя точка зрения была более
подробно развита К. С. Суховым. Вирус инактивируется через
10 минут при 55—60°С, устойчив к разведению до 1:1000,
сохраняется в соке 24—48 часов.
Другие вирусы у шпината и свеклы. Из других вирусов,
поражающих шпинат и свеклу, особого внимания заслуживает*
Cucumis virus 1. Этот вирус вызывает у шпината тяжелое заболе-
1 См. сб. «Мозаичные болезни сахарной свеклы». Киев, 1930.
2 Там же
14
вание, известное под названием spinach blight.1 При этой
болезни наблюдается общий хлороз листьев и отмирание ихг
причем некроз начинается с верхушки листа (Hoggan, 1933).
Нередко наблюдается также редукция пластинки листа и ее
Рис. 2. Шпинат здоровый и пораженный мозаичной болезнью (по Вовку)
уродливое развитие. Мозаика и уродование пластинки листа,
вызванные, повидимому, тем же огуречным вирусом,
наблюдались А. М. Вовком (1942) в подмосковных хозяйствах (рис. 2).
Заражение шпината вирусом обыкновенной табачной
мозаики легко удается искусственно. Наблюдается мозаич-
ность, пожелтение старых листьев, карликовость и гибель
растений через 4—6 недель после инокуляции. В естественных
условиях табачная мозаика у шпината не описана.
Вирус кольцевой пятнистости табака у свеклы и шпината
может вызывать мозаичные заболевания. Под его влиянием у
шпината развиваются крупные хлорозные кольца, причем
заболевание легко может маскироваться.
1 Blight — значит вред для растения.
Глава III
ВИРУСНЫЕ БОЛЕЗНИ КРЕСТОЦВЕТНЫХ
Первые опыты с вирусными болезнями крестоцветных
¦относятся к 1921 г. > когда Gardner и Kendrick, а также Schulz
установили инфекционную природу мозаик редьки, горчицы
и китайской капусты (Brassica chinensis). В настоящее время
известен целый ряд вирусных заболеваний крестоцветных.
Большая часть вирусов, поражающих крестоцветные,
отличается значительным сходством между собой. Это вирусы,
ішактивирующиеся при температуре 60° или ниже по С,
сохраняющиеся в соке не свыше 3 дней и передающиеся соком
я различными тлями. Большая часть этих вирусов вызывает
мозаику и некротические явления. Характерно, что эта группа
вирусов, кроме крестоцветных, может поражать пасленовые,
причем у Nicotiana tabacam обычно вызывает только местные
некрозы, а у целого ряда других видов табака, например у
N. glutinosa, общую инфекцию. Особняком стоит вирус цветной
капусты {Brassica virus 3), который отличается большей
устойчивостью к температуре и более длительным сохранением
в соке, а также тем, что не может вызывать болезненных
явлений у табака. Классификация вирусов крестоцветных еще
далеко не закончена, приводимая ниже классификация
является предварительной. Вирусные болезни крестоцветных
в СССР встречаются. Нам, совместно с Корачевским, пришлось
наблюдать мозаику и курчавость листьев у капусты,
сопровождающиеся резкой подавленностью развития растений. Там
же, на Украине, мы видели кольцевые некротические
пятнистости у капусты. Некротическую кольцевую пятнистость
у Raphanus sativus Е. М. Эристави (1941) наблюдала в Грузии
(рис. 3). Сведения о мозаичных заболеваниях крестоцветных
в СССР в литературе почти отсутствуют, так же как и их
идентификация.
Черная пятнистость капусты. Заболевание описано
Tompkins (1938) под названием cabbage black ring. Его синонимы:
wallflower mosaic и cabbage ring spot. По классификации К.
Smith, болезнь вызывает Brassica virus 1, Smith.
16
У обыкновенной капусты (В. oleracea var. capitata) болезнь
начинается хлоротическими кольцами, которые затем
становятся некротическими. Признаки болезни наблюдаются
Рас. 3. Кольце па я мозаика у редиса
(по Эристави)
только на более старых листьях, и болезнь не сопровождается
иосветлениями по ходу нервов. У цветной капусты (В. oleracea
var. botrytis) вирус вызывает общую диффузную пятнистость,
но и здесь посветления по ходу нервов не наблюдается. У старых
растений могут развиваться некротические кольца. У матиолы
(Mathiola incana и Math, incana var. annua) вирус вызывает
мозаику листьев и деформацию их, на нижних листьях могут
развиваться некрозы по жилкам. На лепестках окрашенных
цветов развиваются светлые полоски, «broken» английских
авторов. Подобные же симптомы наблюдаются у желтофиоли
{Cheiranthus cheiri).
У обыкновенного курительного табака вирус вызывает
местные некрозы, а у N. glutinosa и N. langsdorffii —
мозаичное заболевание.
Вирус передается соком и тлей Myzus persicae.
Доказательств передачи вируса семенами не имеется.
2 В. Л. Рыжков 17
Таблица 1
Отношение к внешним факторам вирусов крестоцветных
Вирус
Некротический Larson и Walker . .
Мозаики капусты Larson и Walker
Мозаики Tompkins Mathiola mild .
» » severe .
Кольцевой пятнистости Arabis hir-
Условия инактивации
вируса
температура
(в «С)
59
50
5'*
55
СО—63
75
68
58-60
58-60
60
разведение
1 :1 000
1 :500
1 :1 000
1 . 1 000
1 :3 000
1 2 000
1 : і 000 *
1 : h 000
1 : 3 000
1 : 1 000
стояние в сок&
(в сутках)
3
1
1-2
1-2
3
15
1'± *
5
7
2—3
Некротический вирус капусты. Заболевание описано Larson
и Walker (1940). Оно стоит очень близко к черной пятнистости.
При нем редко наблюдаются явления мозаичности, и совсем
не бывает посветлеиия по ходу нервов. На листьях больных
растений развиваются мелкие некротические кольца и пятна,
а на стеблях—черные и красные некротические полосы. Некрозы
наилучше развиваются при 22—25° и вполне маскируются
при 13—19° С. Кроме крестоцветных, вирус может поражать
свеклу, огурец, шпинат, Zinnia, Calendula, петунию, махорку,
N. glutinosa. К внешним факторам вирус менее устойчив,
чем вирус черной пятнистости. Дальнейшие исследования
показали, что заболевание вызывается смесью двух вирусов. Один
из них, обозначенный А, дает наиболее резкие проявления
болезни при температуре выше 25°. Он сходен с Brassica virus l.
Другой обнаруживается наиболее резкими симптомами у
пораженных растений при температуре ниже 20°. Он сходен с
Brassica virus 3 и обозначен — вирус В. (Walker, Le Beau,
Pound, 1945)
Мозаика репы. По К. Smith, болэзнь вызывает Brassica
virus 2, Hoggan и Johnson (1925). Первые симптомы болезни
у обыкновенной капусты проявляются нежной мозаикой на
молодых листьях. Мозаика состоит из крупных пятен несколько
более темнозеленого цвета, чем нормальный светлозеленый
фон, на котором они находятся. У репы (В. гара и В. nigra)
вирус вызывает тяжелые некрозы и пожелтение листьев,
которые позднее, если растение выживает, переходят в крапчатость.
От мозаичного хрена был выделен идентичный вирус. У хрена
* Не инактивируегся.
18
наблюдается посветление по нервам и крапчатость, у N. tabacum
развивается местный некроз, у N. glutinosa — общая инфекция,
так же как у шпината. Л*, langsdorffii иммунна.
У матиолы вирус вызывает мозаичность листьев, состоящую
из темнозеленых пятен. Лепестки цветов не поражаются.
Вирус передается соком, а также тлями Myzus persicae
и Brevicoryne brassicae.
Tompkins (1938) описывает мозаичную болезнь репы,
вирус которой, по его данным, несколько отличается
устойчивостью к внешним факторам.
Мозаика цветной капусты. По К. Smith, болезнь вызывает
Brassica virus 3, Tompkins (1937).
У цветной капусты первым симптомом болезни является
посветление по ходу нервов, которое затем переходит в темно-
зеленые полосы по нервам и посветление между ними. В области
хлорозных участков находятся многочисленные, неправильной
формы, темнозеленые пятна; позднее в мозаичных участках
развиваются мелкие некротические пятна. Симптомы
маскируются при 19° по С. Болезнь передается на разнообразные
сорта капусты, как то: обыкновенная капуста, брюссельская
капуста, — а также на различные другие крестоцветные,
например: Lunaria annua, Lepidium sativum., Raphanus sativus,
R. raphanistrum, Capsella Bursa pastoris, Iberis amara.
У матиолы вирус вызывает укорочение междоузлий,
розеточнып рост растения, посветление по ходу нервов и
уродование листьев. На лепестках симптомы отсутствуют.
К вирусу иммунны табак, помидор, перец, шпинат и ряд
крестоцветных, например: Cheiranthus Cheiri, Hesperis matro-
nalis, Alyssum saxatilis.
Болезнь передается целым рядом тлей: Brevicoryne
brassicae, Rhopalosiphum pseudobrassicae, Myzus persicae, Aphis
gossypii, Myzus circumflexus и др., а также может быть передана
инокуляцией сока, хотя этим способом она передается с трудом.
В Родезии описана мозаика цветной капусты,
сопровождающаяся карликовостью. Болезнь отчетливо проявляется
при низких температурах. Передается N. glutinosa и капусте
(Hopkins и Pardy, 1942).
Мозаика Brassica chinensis. Этот вирус был относительно
подробно описан Clayton в 1930 г., но с тех пор не изучен ближе.
К. Smith предполагает его тождественность с Brassica virus 27
но предварительно описывает как особый вирус — Brassica
virus 4, Clayton. Действительно, судя по описаниям Clayton,
он отличается от Brassica virus 2 хотя бы тем, что обыкновенная
капуста к нему устойчива.
Вирус вызывает наиболее тяжелую форму заболевания
у Brassica chinensis, В. alba л В. nigra. На листьях развивается
нежная пятнистость, которая с повышением температуры
переходит в общую желтую окраску, резкую подавленность
2* 1?
растений, причем развиваются некротические полоски и кольца.
Слабее проявляется болезнь у репы (В. тара) и В паро-
brassica (брюква). У этих видов некротических явлений не
наблюдается. Заражение цветной капусты удается с большим
трудом. Обыкновенная капуста не подвержена этой болезни.
Вирус передается соком, а также тлей Brevicoryne bras-
sicae. Опыты по передаче при помощи Ж", persicae дали
неопределенный результат. Последнее также отличает его от других
вирусов крестоцветных.
Мозаичные болезни матиолы. Эти болезни проявляются
.мозаикой на листьях и образованием полосок на лепестках.
Выше было сказано, что и некоторые другие вирусы
крестоцветных могут вызывать подобные явления у матиолы, но
описываемые здесь вирусы отличаются своим набором хозяев:
так, например, они не поражают обыкновенную капусту.
Tompkins (1939а) различает две мозаики у матиолы —
мягкую и тяжелую. Они отличаются между собой по силе
вызываемых ими симптомов. Вирус мягкой мозаики имеет
инкубационный период в 16—22 дня, вирус тяжелой — в 14
дней. Чувствительность этих вирусов к внешним факторам
несколько различна, как видно из выше приведенной таблицы.
Вирус мягкой мозаики поражает желтофиоль, Brassica ad-
pressa^N. glutinosaviN. langsdorffil, которые иммунны к вирусу
тяжелой мозаики. Шпинат и петуния иммунны к мягкой мозаике,
но поражаются тяжелой мозаикой.
К. Smith описывает Mathiola virus l', Tompkins, который
больше отвечает вирусу тяжелой мозаики, хотя точная их
идентификация затруднительна.
Мозаичные заболевания матиолы передаются различными
тлями и соком, но не семенами.
Мозаичная болезнь редиса. В литературе встречаются
краткие сведения о мозаичных заболеваниях у Raphanus sa-
tivus. Более подробно болезнь была описана Tompkins (1939)
в Калифорнии. Болезнь начинается маленькими округлыми
или имеющими неправильные очертания пятнами, которые
могут сливаться между собой. Некрозов не наблюдается. У В. pet-
mi вирус вызывает местные некрозы, у табака и махорки,
а также у шпината — общую инфекцию. В отличие от большей
части вирусов крестоцветных он легко передается соком без
применения карборунда, но не передается ни одной из
испытанных видов тлей (Brevicoryne brassicae, Lipaphis pseudobras-
sicae, M. persicae), Семенами вирус не передается. В литературе
имеются отдельные указания на заболевание мозаикой Lunaria
annua и различных крестоцветных Китая. Следует упомянуть,
что многие крестоцветные, например: матиола, капуста, редис,
хрен, пастушья сумка, могут поражаться вирусом Beta 1
(курчавость верхушки свеклы). Этот вирус вызывает у
крестоцветных подавленность роста, кустистость, хрупкость и скручи-
20
вание листьев. Callistephus virus 1 может поражать и Cheiran-
thus АШопіі, Malcolmia maritima, Radicula sylvestris, вызывал
здесь хлороз, ненормальную ветвистость, а у Malcolmia также1
позеленение цветов.
У Arabis hirsuta темнозеленую кольцевую пятнистость
описали К. Smith и R. Markhom (1944). Это заболевание
передается соком, хотя и нелегко. Aphis fabae и Myzus persicae
его не передают. У N. tabacum вирус вызывает хлорозные
кольца. Lycopersicum esculentum бессимптомный носитель
вируса. S. tuberosum не заболевает. Phaseolus vulgaris под
влиянием вируса развивает некрозы по жилкам и часто погибает
(сорт Canadian Wonder). У некоторых сортов Pisum sativum —
некрозы по жилкам и гибель точки роста.
Для борьбы с вирусными болезнями крестоцветных
рекомендуется уничтожение насекомых-переносчиков.
Глава IV
ВИРУСНЫЕ БОЛЕЗНИ МАЛЬВОЦВЕТНЫХ
Курчавость листьев хлопчатника (Gossypium virus 1 Far-
quharson). Это заболевание, экономически очень важное, стало
известно в Африке в 1912—1913 гг. При заболевании
наблюдается утолщение жилок листа. Утолщенные жилки имеют
более темную, чем нормально, окраску, и с нижней стороны
листа развиваются выросты, которые могут достигать 1.5 см
в поперечнике. Листья закручиваются по большей части вниз,
но иногда и вверх. К этим симптомам на листьях может
присоединиться мозаичность, развивающаяся обычно по ходу
нервов. При заболевании наблюдается укорочение черешков
листьев, а также подавляется ветвление, причем растение,
не разветвляясь в процессе роста, может оказаться выше
нормального.
В патолого-анатомическом отношении должно отметить:
1) утрату паренхимой листа диференцировки на палисадный
и губчатый слои, причем исчезает характерное для палисадных
клеток вытягивание в длину; 2) образование добавочных
цилиндров в коре стебля и черешков, которые состоят из
ксилемы, располагающейся в центре, окруженной камбием и
флоэмой.
Переполнения тканей крахмалом у больных растений не
наблюдается. Сок здоровых растений имеет красную окраску,
тогда как сок больных — зеленого цвета. Прибавление к соку
больных растений кислоты вызывает покраснение этого сока,
однако необходимо отметить, что различия в рН сока больных
и здоровых растений лишь незначительно, и разницу в окраске
сока нельзя объяснять различиями в реакции этого сока (Mas-
веу и Andrews, 1932). Болезнь очень силь'но снижает урожай.
Симптомы в описанном выше виде бывают у хлопчатника
Sakel (Gossypium peruvianurn X barbarense). У G. hirsutum
болезнь проявляется только мозаикой. Эта последняя
начинается посветлением по ходу нервов, которое переходит в
образование хлорозных пятен, часто сливающихся между собой,
причем наблюдается некоторая деформация пластинки и укоро-
22
чение междоузлий, придающее растению кустистый вид.
Болезнь от этого вида хлопчатника обратно Sakel не может быть
передана, но этот последний бессимптомно сохраняет вирус,
который снова может вызвать мозаику у G. hirsutum. Имеем
ли мы здесь дело действительно с одним вирусом или со
смешанной инфекцией — нам представляется неясным (Kirkpatrick,
1931).
В Нигерии встречаются преимущественно сорта G. реги-
•vianum и G. vitifoliam,. У них наблюдается заболевание,
выражающееся закручиванием пластинки листа вверх, причем
гипертрофия жилок листа и выросты на листьях совершенно
отсутствуют. Являются ли заболевания тождественными —
нам в данном случае представляется неясным.
Курчавость листьев хлопчатника экспериментально была
передана на целый ряд мальвоцветных, причем во всех этих
случаях наблюдаются характерные утолщения жилок листа
и закручивание пластинки листа вниз или вверх. Такие
симптомы болезни наблюдаются у кенафа (Hibiscus cannabinus),
у бамии (Hibiscus esculentus) и у Althaea rosea. С бамии вирус
не может быть возвращен на хлопчатник непосредственно,
однако, если с бамии перенести вирус на кенаф, то с этого
последнего он снова может быть передан хлопчатнику
(Kirkpatrick, 1931).
В Северной Индии у циннии описано заболевание, очень
сходное с курчавостью листьев хлопчатника и передающееся
белой мушкой — Bemisia gossypipedra,, как и курчавость
листьев (Mathur, 1932, 1933). Кроме того, в Пенджабе встречаются
заболевания картофеля, табака и огурца со сходными
признаками, однако на хлопчатнике в Индии болезнь не была найдена,
и нет доказательства тождественности всех этих заболеваний
^Husain, 1932). В Сомали у хлопчатника встречается сходное
по внешним признакам заболевание, но, по одним данным,
юно зависит от почвенных условий (Russo, 1936), а по другим, —
юно инфекционно и распространяется Empoasca faciatis (Kotone).
В Бразилии встречается два типа мозаичного заболевания
хлопчатника, отличающихся между собой следующим образом:
Мозаика жилок Обычная мозаичность
Позветление по ходу жилок Свеппгелеіьтеимоваичнкеучаеі-
ки между жилками
Морщинистость, не сопрэвожда- Морщинистость, сопровождаю-
ісщаяся пузыр іатостью; закру- щаясяпузыр атозтьк ; закручи-
чивание листьев вверч вачия вверч не набл одаегся,
Мозаикой жилок легко поражается Sakel и другие сорта
G. barbarense, и она очень легко передается прививкой, тогда
как к обычной мозаике эти сорта иммунны, и прививкой
она передается с трудом. Мозаику жилок считают (Costa и
Forster, 1932) очень похожей на курчавость листьев,
описанную в Судане.
Мы видим, таким образом, что довольно разнообразные
по симптомам заболевания ставятся в связь с описываемой
здесь болезнью хлопчатника. Обращает на себя также внимание
ее сходство с kroepoek табака. Болезнь не передается соком,
но легко передается прививкой.
Ее переносчиком является белая мушка — Bemisia gos-
sypipedra. Другие испытанные насекомые — Aphis gossypii
и Empoasca fuscialis болезни не передают (Kirkpatrick, 1937;
Golding, 1930). Для инфицирования насекомому достаточна
пребывания на больном хлопчатнике в течение 3 часов, а может
быть, и более короткого времени. Инфицированное насекомое
сохраняет вирус не менее 7 дней. Вирус не передается чере<*
яйцеклетку. Опыты по передаче вируса семенами не дали
доказательств в пользу такой передачи, но они были поставлены
в небольшом масштабе. В распространении болезни большую
роль играют зимующие на плантации отдельные экземпляры
хлопчатника, поэтому для борьбы с болезнью рекомендуется
тщательное удаление с плантаций всяких остатков хлопчатника
и такой севооборот, чтобы хлопчатник попадал снова на ту
же плантацию только раз в три года. Большую пользу
принесло введение устойчивых сортов Sakel, X 1530 и X 1730
и др. (Bailey, 1934; Lambert, 1932).
Скручивание листьев хлопчатника. В 1934 г. в Азербайджане
Д. Д. Вердеревский описал вирусное заболевание хлопчатникау
которое он считал, по всей вероятности, тождественным или
близко стоящим к описанной выше курчавости хлопчатника.
Ввиду того что тождественность данного заболевания с
суданской болезнью остается недоказанной и ввиду существенных
отличий заболевания от курчавости листьев, мы предпочитаем
назвать его скручиванием листьев. Болезнь
проявляется утолщением листьев, ломкостью листьев и стебля,,
закручиванием пластинки листа вверх, укорочением
междоузлий и общей подавленностью роста. Сильно пораженные
растения обычно своими боковыми ветвями полегают по земле.
Часто наблюдается более значительное антоцианирование
больных листьев, чем здоровых, и характерный блеск
поверхности листа. Больные растения проявляют большую
устойчивость к засухе и холоду, чем здоровые. У G. peruvianum
и его гибридов с G. barbarense листья закручиваются не вверх,
а вниз (Д. Д. Вердеревский и А. С. Барменков — рис, 4
и 5).
В патолого-анатомическом отношении болезнь
характеризуется более сильным развитием мезофилла листа, причем
палисадная паренхима чаще, чем в здоровом листе, становится
двухслойной, — переполнением пластинки листа и черешков
зернами крахмала и более слабым развитием толстостенного
24
луба, отчего зависит ломкость и полегание больных растений
(Кара-Мурза, 1938; Рыжков и Овчарова, 1941).
При заболевании повсюду в наземных частях, кроме
генеративных органов, возрастает количество крахмала и
растворимых углеводов и уменьшается содер?кание общего и белкового
азота. Таким образом, болезнь принадлежит к типу желтух.
Заметных повреждений флоэмы не обнаружено, однако
наблюдается некоторая ее гипоплазия. Причина задержки оттока
крахмала остается невыясненной. Действие пероксидазы и
амилазы у больных растений повышено. Сок здоровых растений
красный, сок больных растений часто бывает зеленым, однако
это различие, по нашим наблюдениям, не всегда выражено,
а по данным А. А. Михлиной,1 в Туркмении к концу вегетации
сок здоровых растений тоже становится зеленым. По нашим
наблюдениям, при обработке материала для определения
сахара по Бертрану, после осаждения белков уксуснокислым
свинцом сок здоровых растений приобретает окраску салатного
цвета, а сок больных становится темнозеленым.
Скручивание листьев хлопчатника весьма вредоносно:
1 Цит. по рукописному отчету.
оно снижает в целом ряде случаев вес сырца больше чем на
половину. В некоторые годы болезнь может достигать очень
значительного развития; при этом надо отметить, что с 1934
по 1938 г. наблюдалось ее нарастание (С. Н. Московец, 1940).
Рис. 5. Здоровый хлопчатник (слева) и хлопчатник того же возраста,
зараженный при помощи хлопковой тли (3 растения справа), в опытах
Барменкова
С. Н. Московец показал, что болезнь передается хлопковой
тлей {Aphis gossypii), что полностью подтверждается и в
Туркмении Д. Д. Вердеревским. Роль других тлей, например
акациевой, менее значительна. Могут ли переносить вирус, кроме
тлей, другие насекомые — точно не установлено.
Инкубационный период обычно очень длителен: он колеблется от 30 до
56 дней, однако если для эксперимента использованы
растения до начала их бутонизации, то его можно, по Д. Д. Вер-
деревскому, сократить до 18—20 дней.
Многолетние опыты С. Н. Московца говорят в пользу того,
что болезнь в небольшом проценте может передаваться семенами,
однако для точного установления этого факта, повидимому,
необходимы дальнейшие опыты. Остается невыясненным,
зимует ли вирус в каких-либо сорняках. Некоторые пасленовые
20
Азербайждана болеют несколько сходно с хлопчатником,
и в них, в первую очередь, приходится искать предполагаемых
резерваторов вируса в природе.
Для борьбы с болезнью С. Н. Московец рекомендует
удаление сорняков, уничтожение насекомых-переносчиков,
удаление остатков хлопчатника с корнями. В тех случаях, когда
количество больных растений не превышает 0.5—1%, удаление
больных растений, повидимому, может способствовать
уменьшению распространения болезни. Некоторые сорта
хлопчатника проявляют значительную устойчивость к вирусу; наиболее
перспективными в этом отношении можно считать сорта Гиза 7,
3782-1, № 1, № 2, № 75, № 7, Гиза 29—63 и др. Наиболее
сильно поражаем целый ряд египетских сортов, как, например,
Фуади (Варунцян, 1938; Московец, 1940; Вердеревский, 1938).
Вопрос о поражаемости Упландов был предметом
многолетних исследований и наблюдений. Согласно С. Н. Московцу,
они могут поражаться вирусом; по данным Д. Д. Вердерев-
ского, Упланды являются скрытыми вирусоносителями, и от них
вирус с помощью хлопковой тли может быть передан на
чувствительные сорта.
Укажем здесь важнейшие отличия курчавости листьев
от их скручивания.
Курчавость листьев
Листья закручиваются вниз или
вверх
Наблюдается разрастание жилок
и выросты с нижней стороны листа
Палисадная паренхима
подавлена в развитии
Переполнения тканей крахмалом
не наблюдается
Наблюдается образование
добавочных цилиндров
Боковое ветвление растения
подавлено
Болезнь передается Bemisia gos-
sypipedra, но не Aphis gossypii
Скручивание листьев
Листья почти всегда
закручиваются вверх
Подобі ыэ симптом л отсутствуют
Палисадная паренхима
развивается чрезмерно
Переполнение тканей крахмалом
крайне характерно
Образование добавочных
цилиндров отсутствует
Боковое ветвление хорошо
выражено
Болезнь передается Aphis gossypi
Происхождение скручивания листьев в СССР остается
невыясненным. Невозможно утверждать, чтобы болезнь не
встречалась нигде в других странах, так как картина вирусных
заболеваний хлопчатника весьма разнообразна.
Другие аномалии роста хлопчатника. На практике диферен-
циальная диагностика вирусных болезней хлопчатника
сталкивается с существенными затруднениями. Они зависят прежде
всего от того, что если симптомы курчавости или скручивания
листьев выражены не особенно резко, то они могут быть сходны
с различными физиологическими расстройствами и
возрастными явлениями. Остановимся здесь вкратце на различных
27
расстройствах у хлопчатника частью известной природы,
частью с невыясненной этиологией.
В Луизиане описана морщинистость листьев
хлопчатника (crinkle), которая не передается ни прививками,
ни соком и может быть излечена прибавкой к почве
углекислого кальция. Сходные расстройства у хлопчатника могут
быть вызваны искусственно — путем прибавления в
вегетационный сосуд солей марганца (Neal и Lovett, 1938).
Хлорирование листьев, измельчание их и
образование своеобразных морщинок перпендикулярно крупным
жилкам листа мы наблюдали в вегетационных сосудах в
экспериментальной оранжерее в Москве. Подобные же образцы
листьев нам были присланы с плантаций хлопчатника из
Туркестана.
Целый ряд аномалий развития хлопчатника описывает
Cook (1924). Даем здесь краткий перечень этих аномалий.
Crazy top, или акромания. Это заболевание
встречается в США. При нем наблюдается подавление ветвле^
ния, превр-ащение плодоносящих веток в вегетативные, а иногда
также резкое укорочение междоузлий и уродование листьев.
Б р а х и з м встречается также в США. При нем тоже
наблюдается укорочение междоузлий; цветы и листья
уменьшаются в размерах, плоды часто, недоразвившись, подсыхают,
но при этом не опадают, а остаются висеть на стебле на своих
плодоножках.
Т о н о з и с, или leaf-cut. Болезнь тоже встречается
в США. При ней наблюдается выпадение участков ткани листа;
клетки вокруг образовавшейся дырочки светлые, без
хлорофилла. Очаги поражения чаще всего начинаются возле
жировых железок.
Г и б о з и с, или leaf-curl. Это — тоже
североамериканская болезнь. Она характеризуется закручиванием
листьев вниз. Особенно сильно болезнь проявляется в холодйое
время и маскируется при наступлении летней жары.
Обращает на себя внимание ее внешнее сходство с суданской
курчавостью.
Циртозис, или club-leaf. Эта болезнь известна^
в центральной части Китая и особенно сильно выражена на
азиатском хлопчатнике. Сначала развитие идет нормально, затем
внезапно наступают болезненные симптомы. Листья стано>-
вятся мозаичными, особенно вдоль края и главных нервов;
междоузлия и черешки укорачиваются, и в то же время
наблюдается чрезмерная кустистость. Растение приобретает
своеобразный, весьма характерный облик, так как оно покрыто
густой массой мелких листьев. Сходное заболевание описано
в Индии, но там оно наблюдается редко, так как большинство
индидрких хлопчатников не подвержено этому заболеванию.
Циртозис вызывается цикадой Chlonta biguttula.
28
Вопреки Wang (1924) это, повидимому, не вирусное заболевание,
а результат энтомологического повреждения. Для борьбы
с болезнью рекомендуется опрыскивание бордосской жидкостью
<S. Н. Ои, 1934).
Стенозис, или smalling. Заболевание найдено
на Гаити. Оно сходно с только что описанным циртозисом.
Здесь также наблюдаіотся укорочение междоузлий, мозаичная
расцветка и деформация листьев. Измельчание листьев при сте-
нозисе еще более значительно выражено, чем при циртозисе.
Сорт UAB более устойчив. Сходные заболевания встречаются
у Sida acuta и S. salvaefolia (Botero, 1940). О стенозисе см. также
у Alfzal Jaggi и Singh.
На нахождение в СССР сходных с циртозисом явлений
обращает внимание Д. Д. Вердеревский.
Этиология описываемых здесь заболеваний не выяснена.
Весьма вероятно, что часть их является вирусными болезнями.
Примером генетически обусловленной карликовости,
связанной с уродованием листьев, может служить crinkled dwarf,
которую описывают Hutchinson и Ghose (1937). Эта
карликовость является мутацией, которая при скрещивании с
исходными формами то оказывается рецессивной, то дает
наследование промежуточного характера.
Dastur (1936, 1939) описывает в качестве
физиологического расстройства неизвестной этиологии заболевание
хлопчатника в Пенджабе, сопровождающееся разрушением хлоро-
пластов, переполнением тканей крахмалом и увеличением
количества танноидов. В эпидермисе возрастает количество анто-
циана и антоксаитина. Растения останавливаются в росте,
коробочки недоразвиваются.
Инфекционный хлороз мальвоцвстпых. В 1808 г. в Англии
стал известен один экземпляр Abutilon striatum с желтопсстрыми
листьями. В 1869 г. Lomoine обнаружил, что эта пестрота
путем трансплантации передается здоровым растениям. Это
удивительное для того времени явление привлекло внимание
ж садоводов-практиков, которые использовали инфекционную
пестроту в декоративных целях, и теоретиков, которые
увидали в открытом явлении новую область исследования.
В 1907 г. Lindemuth писал: «Здесь открывается широкое
поле для новых точных опытов и точных наблюдений».
К. Smith обозначает вирус инфекционного хлороза мальво-
цветных как Abutilon virus l.
У абутилона хлороз вызывает яркожелтую расцветку
листьев, симптомы становятся интенсивными при достаточном
освещении. Если растение перенести в темноту, оборвав у него
предварительно все листья, то здесь оно развивает сначала
пестрые листья, а затем исключительно зеленые. Если
выдержать растение в темноте достаточно долго, то по возвращении
на свет оно остается зеленым — оно выздоравливает. Если
2»
мозаичные листья закрыть оловом и оставить растение на
свету, то вновь развивающиеся листья не заболевают. Если:
позднее больные листья открыть, то растение все же остается
здоровым. Вирус, по выражению Hertzsch (1927), остается
как бы связанным (gebunden oder festgelegt) в тканях
больных листьев.
Инфекционный хлороз может быть передан следующим
мальвоцветным: Abutllon arboreum, Ab. africanum, Ab. atro-
sanguineum, Ab. indicum, Ab. Avicennae, Ab. Darwini, Ab. escu-
lentum, Althaea ficifolia, AL officinalis, Al. rosea, Kitaibelia
vitifolia, Lavatera arborea, Malvastrutn capense, Modiola, Sida
mollis, S. Napaea, Sidolacea Candida. Устойчивы, по этому
автору, к инфекционному хлорозу Hibiscus, Malvaviscus и Malva,
у которых инфекция или не происходит, или редко наблюдается.
Кроме того, Hibiscus и Malvaviscus плохо срастаются с абути-
лоном. Т. П. Овчарова1 произвела опыты сращивания
мозаичного абутилона с хлопчатником. Лбутилон жил на хлопчатнике
свыше месяца, после чего погибал. Инфекционный хлороз
на хлопчатник не перешел. Разные виды реагируют на вирус
различно. У Ab. indicum вирус вызывает тяжелые некрозы
и гибель растения. Отдельные побеги могут выздоравливать
(Keur, 1934).
Согласно Hertzsch (1927), можно различать два вируса
у больного инфекционным хлорозом абутилона: вирус Л
вызывает посветление по ходу нервов чаще, чем вирус В. Вирус
А может поражать Malva borealis, которая устойчива к вирусу
В; напротив, вирус В поражает Lavatera arboreum, который
устойчив к вирусу А.
Единственный известный способ передачи инфекционного
хлороза у абутилона — трансплантация. Однако на родине,
в Ост-Индии, согласно сообщению Cook (1924), заболевание
может передаваться насекомыми-переносчиками.
Мозаичная болезнь у Malva borealis. Еще в 1933 г. нами
была описана мозаика у этого встречающегося повсеместно
сорняка, которая, распространяясь по ходу нервов, вызывает
красивый сетчатый рисунок (рис. 6). Иногда хлорозные пятна
расширяются, захватывая более значительный участок листа.
По нашему поручению, Т. П. Овчарова1 произвела опыты,
которыми показала, что мозаичная болезнь мальвы очень легко
передается инокуляцией соком, а также тлями, например
Myzus persicae.
Ни соком, ни прививкой мозаика мальвы не могла быть
передана на хлопчатник, хотя срастание происходило легко
и мозаичная мальва долго жила на хлопчатнике.
Мозаичная болезнь у мальвы широко распространена
по всему Союзу. Мы встречали ее и на Кавказе и на Украине,
1 Неопубликованные данные.
М)
и в Москве. К. Smith упоминает о мозаичном заболевании
у Malva silvestris, не приводя, однако, доказательств в пользу
инфекционного характера этого заболевания.
Рис. 6. Два листа Malva borealis, обнаруживающие сетчатую
мозаику
Желтая мозаика у Hibiscous csculentus. Под этим названием
описано заболевание, сходное с leaf-curl табака. При нем
наблюдается утолщение жилок с нижней стороны листа и
подавленность роста. Болезнь не передается ни соком, ни семенами.
Ее переносчик — Bemisia gossypipedra (Uppal и др., 1940).
Глава V
ВИРУСНЫЕ БОЛЕЗНИ ВИНОГРАДА
Короткоузлис винограда. Хотя, согласно указаниям Panta-
nelli (1912), рассматриваемая здесь болезнь была известна
во Франции и ранее 1894 г., но особенно интенсивно стала
распространяться после реконструкции виноградников в
названном году, т. е. когда европейские лозы были перенесены на
корневую систему американского винограда. Viola в своей книге
о болезнях винограда, вышедшей 3-м изданием в 1903 г., назвал
заболевание roncet, что происходит от французского названия
ежевики, на которую будто бы больные кусты в известной
стадии становятся похожи. В этой книге приводятся также
и другие названия болезни, одно из которых — к о р о т к о-
узлие, или courl-noue,—было дано ей в 1900 г. В Германии
болезнь называется Reisigkrankheil, что значит
кустарниковая болезнь или кустарниковость,
а в Австрии — Krauten, что происходит от немецкого слова
Kraul — трава. Наконец, наиболее принято итальянское
название болезни arricciamento, что означает курчавость.
Многократно указывалось, что встречающаяся в Калифорнии
мелколистость (leaf-liltle) винограда представлят собой ту же
болезнь, которая известна в Европе. Так ли ото
действительно, — однако не установлено. Неизвестно также,
тождественна ли эта болезнь с очень сходным заболеванием португальских
оортов винограда в Австралии (Castella, 1934).
К. Smith описывает только одно вирусное заболевание
винограда, приписывая его Vitis virus l, Stranak (1931). Он
предполагает, что этим вирусом вызывается и мозаичная болезнь
винограда, описанная в Чехословакии, и короткоузлие.
Подобное воззрение развивает также Martinoff (1934). Однако это
совершенно невероятно, так как заболевания эти очень сильно
различаются между собой.
Приводя синонимику, мы попутно дали картину
распространения болезни. К этой картине необходимо добавить, что
¦заболевание встречается также и в СССР, причем целую кол-
32
лекцию тяжелых форм его можно было найти на Опытной
станции виноградарства и виноделия в Мэгараче.
Краткие сведения о короткоузлии в СССР можно
лайти в книге В. Л. Рыжкова (1935). Количество
иностранной литературы, особенно французской, об этой все еще
загадочной болезни огромно. Несмотря на такую обширную
литературу, до сих пор не достигнуто ни единогласия, ни
полной ясности в понимании причин болезни. Это несомненно
объясняется трудностью проблемы. Формы короткоузлия
разнообразны; с другой стороны, различные причины могут
приводить к сходным, картинам; наконец, экспериментировать
приходится с многолетним растением. К сожалению,
значительная часть выводов относительно природы короткоузлия
построена на различных наблюдениях, а не на прямом
эксперименте. Лучшими работами по короткоузлию, давшими нам
действительно известное положительное знание, остаются
немногие экспериментальные исследования. Это, прежде всего,
работы Pantanelli, давшие картину физиологии и патологической
анатомии болезни, и в особенности работы Petri (1929), который
доказал инфекционный характер короткоузлия, передачу
ого через почву. К сожалению, до сих пор не удалось провести
резкую границу между инфекционными формами
короткоузлия и различными другими аномалиями винограда, отчего
и приходится признать, что работы Petri не разрешили
проблемы полностью. Если бы мы могли с уверенностью ответить,
что, кроме инфекционных форм короткоузлия, существуют
хорошо их копирующие неинфзкционные, то и это было бы уя*е
большим достижением, но, к сожалению, пока мы не можем
сказать и этого.
Заболевание характеризуется поражениями листьев, стебля
и корневой системы. Для листьев характерны измельчание
пластинки и сближение жилок, вследствие чего лист становится
курчавым, а также резкое возрастание его изрезанности.
При тяжелых формах можно наблюдать значительную
редукцию пластинки листа, так что отдельные доли его
становятся нитевидными. У черного муската в Магараче мы
наблюдали также с нижней стороны пораженных листьев выросты,
достигающие 0.5 см в ширину. Эти выросты в том же году,
как производились наши наблюдения, описаны Gigante (1937)
в Италии. Они носят характер enation — образований, хорошо
известных при различных вирусных заболеваниях. Между
жилками листа наблюдается крапчатость, кроме того происходит
выпадение отдельных участков ткани из пластинки листа.
Болезнь характеризуется, далее, глубоким расстройством
роста, выражающимся в укорочении междоузлий, которое
заходит иногда так далеко, что очередные листья попарно
сближаются и кажутся супротивными. Наряду с укорочением
междоузлий наблюдается увеличение их числа и чрезмерная
3 В. Л. Рыжков 38
кустистость. Мы заметили, что если симптомы укорочения
междоузлий достигают максимального проявления, то листья
обычно сильно измельчены, а явления курчавости и
усиленного расчленения пластинки отступают на задний план.
Симптомы короткоузлия особенно резко выражены весной.
С наступлением летней жары они более или менее маскируются.
Наши измерения, произведенные совместно с Париевской,
показали, что летом пораженные побеги растут быстрее, чем
нормальные, так что задержка роста больных растений
приходится на более прохладный, весенний, период.
Это положение иллюстрирует табл. 2. Здесь показано,
во сколько раз за 2 месяца увеличилась длина побегов здоровых
растений, побегов, резко пораженных короткоузлием, и побегов
о выраженной курчавостью и менее выраженным
короткоузлием.
Таблица 2
Прирост побегов у виноградной лозы с 16 июня по 16 августа 1937 г.
Тип заболевания
Короткоузлие ....
Прирост
Длина (16.VIII)
Длина (16.VI)
1.714
1.524
1.152
Отличие
от
контроля
0.562
0.372
Ч^ п%
0.078
0.050
0.01
t
7.2
5.4
Чисто
намеренных
побегов
05
54
154
Представление о длине побегов, длине и числе
междоузлий у здоровой и больной виноградной лозы к концу
вегетационного периода в 1937 г. дает табл. 3.
Таблица 3
Аномалии роста при короткоузлии виноградной лозы
Средняя длина побега (в см)
Средняя длина одного междоузлия (в см)
Средняя длина одного междоузлия при
контроле, принятом за 100 . .
Среднее число междоузлий на побег .
Число исследованных побегов
Контроль
76.04
4.58
100
16
93
Короткоузлие
51.84
2.75
СО
18
(.0
Курчавость
81.52
З.()3.
79
23
47
Из приведенной таблицы видно, что к концу
вегетационного периода средняя длина побегов, пораженных курчавостью,
была не меньше, а даже несколько больше длины здоровых
побегов (последнее, однако, лежит в пределах ошибки).
34
В половине июня длина побегов, пораженных коротко-
узлием, в среднем отставала от контроля на 14.17 см, что было
статистически вполне достоверно (t=3.16). Таким образом,
больные побеги в течение лета догнали здоровые, но это
произошло не столько за счет удлинения междоузлий, сколько за
счет увеличения их количества. О распределении на побеге
коротких междоузлий см. у Маіег и Mittmann (1939).
Патологическая анатомия курчавости изучалась Panta-
nelli (1912), а затем Рыжковым и Париевской.1
Рис, 7. Схематический рисунок поперечного разреза через
лист винограда. Склеротические явления при короткоузлии
В области сближения жилок листа наблюдается
исчезновение нормального строения мезофилла. Вместо столбчатой
и губчатой паренхимы в отдельных участках мы находим здесь
клетки коленхимы. Выпадение кусочков листа легко
понять, так как под влиянием этих склеротических изменений
появляются участки листа с гораздо более прочными, твердыми
клеточными стенками, чем остальная часть листа (рис. 7).
Под влиянием ветра и других причин в этих местах особенно
легко могут происходить разрывы, что ведет к выпадению
затвердевших кусочков листа. Склеротические явления
наблюдаются, главным образом, вблизи основания листа, где имеется
главное разветвление его жилок. Клетки палисадной
паренхимы в области маленьких просвечивающих крапинок
становятся более короткими, и разница между губчатой и
столбчатой паренхимой сглаживается. Иногда на листе наблюдаются
некротические участки, вокруг них происходит усиленное
деление клеток и образуется интумесценция, ткань которой
состоит из клеток с мелкими пластидами, лишь слабо
окрашенными хлорофиллом (рис. 8 и 9).
Большое значение для диагностики болезни имеют открытые
Petri внутриклеточные палочки (cordoni endocellulari). Они
1 Неопубликованные данные.
3*
35>
представляют собой длинные нити, которые тянутся из од
ной клетки в другую, состоят из целлюлозы и похожи на
клеточные стенки. Они встречаются не только в листе (особенно
Рис. 8. Схематический рисунок поперечного среза через лист винограда,
больного короткоузлием, в области некроза и иитумесценции
в опидермисе у основания пластинки листа) и в стеблях
(особенно в поверхностных тканях), но также и в древесине и в
сердцевине, а иной раз и в корнях (рис. 10). Иногда палочки
встречаются также у внешне здоровых растений.
Рис. 9. Две клетки листа винограда с
пластидами; одна—из нормальной ткани,
имеющая крупные пластиды, и другая — из
тканей иитумесценции с мелкими
пластидами
Petri считает их в этом случае единственным признаком
заболевания. Известна форма короткоузлия без
внутриклеточных палочек, которая была названа Petri «рахитизмом».
При короткоузлии, которое встречается в Магараче, мы не
обнаружили внутриклеточных палочек, несмотря на тщательные
поиски.
В результате длительных изысканий, Petri обнаружил
в кончике молодых корней больных растений вакуолярные
тельца овальной формы, лежащие возле ядра. Gigante
сообщает, что Х-тела встречаются в клетках листьев, имеющих
описанные выше выросты на нижней стороне, у винограда,
пораженного рахитизмом.
Корневое давление у больных растений резко снижается.
Осмотическое давление сока больных растений, по Pantanelli,
36
повышено, тогда как, согласно Moreau и Venet (1938), оно резко
понижено, Транспирация на свету у здоровых растений в 10—
12 раз интенсивней, чем у больных. Количество крахмала
и нередуцирующих Сахаров в больных листьях падает5 а
количество редуцирующих
возрастает. Больные растения
содержат больше гумми, чем
здоровые. Действие амилазы,
инвертазы и пектиназы
повышено. Количество
белкового азота, так же как и
общего азота, в больных
растениях повышено. Действие
протеолитических ферментов
при заболевании возрастает*
Содержание золы в больных
растениях тоже возрастает,
причем наблюдаются и
качественные ее изменения.
Количество серной кислоты,
калия возрастает,
количество фосфорной кислоты и
кальция уменьшается, а
количество магния остается без
изменения. Дыхание больных
растений повышено, и они
Рис. 10. Поперечный разрез чере'5
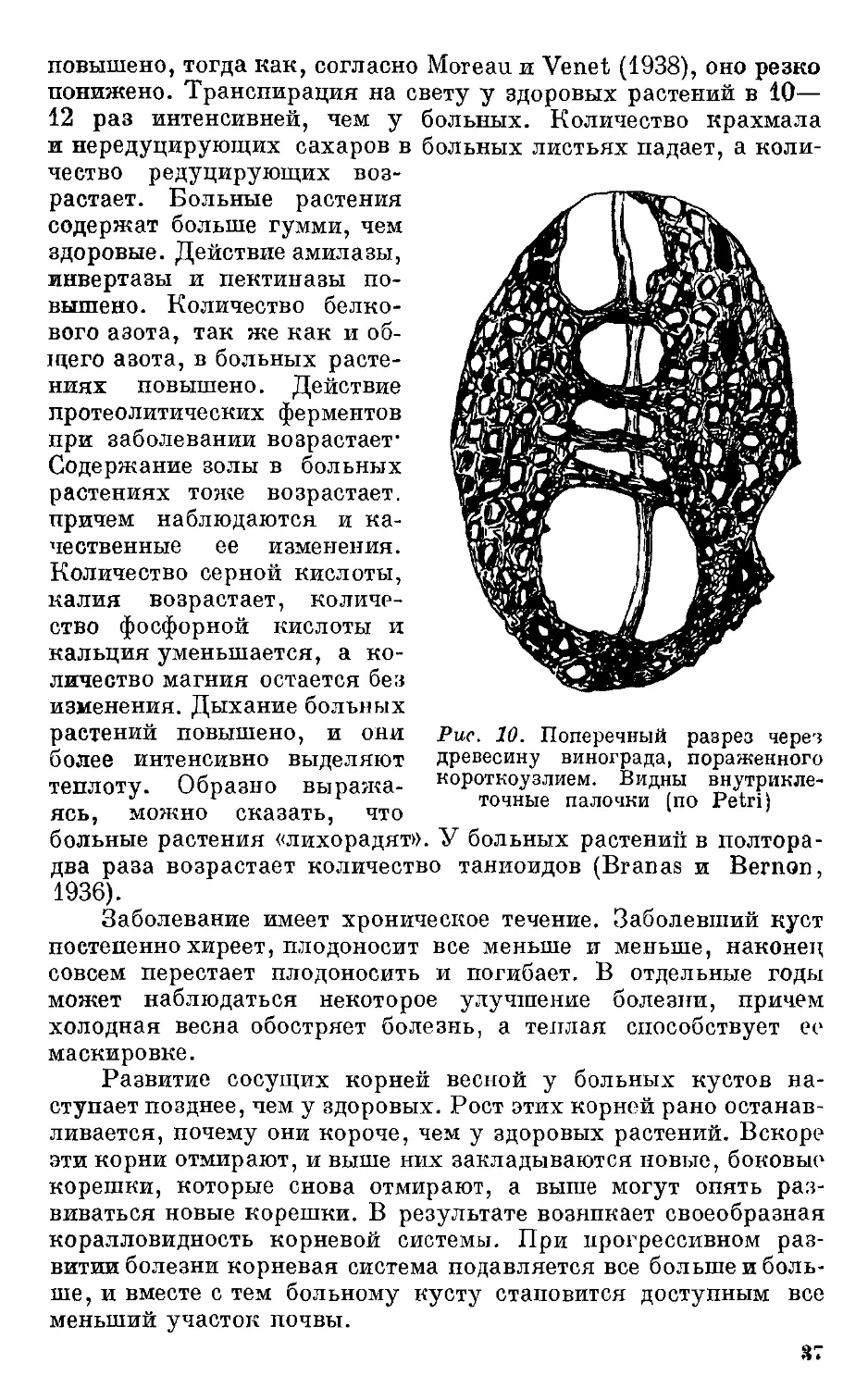
древесину винограда, пораженного
короткоузлием. Видны
внутриклеточные палочки (по Petri)
более интенсивно выделяют
теплоту. Образно
выражаясь, можно сказать, что
больные растения «лихорадят». У больных растений в полтора -
два раза возрастает количество таниоидов (Branas и Bernon,
1936).
Заболевание имеет хроническое течение. Заболевший куст
постепенно хиреет, плодоносит все меньше и меньше, наконец
совсем перестает плодоносить и погибает. В отдельные годы
может наблюдаться некоторое улучшение болезни, причем
холодная весна обостряет болезнь, а теплая способствует ее
маскировке.
Развитие сосущих корней весной у больных кустов
наступает позднее, чем у здоровых. Рост этих корней рано
останавливается, почему они короче, чем у здоровых растений. Вскоре
эти корни отмирают, и выше них закладываются новые, боковые
корешки, которые снова отмирают, а выше могут опять
развиваться новые корешки. В результате возникает своеобразная
коралловидность корневой системы. При прогрессивном
развитии болезни корневая система подавляется все больше и
больше, и вместе с тем больному кусту становится доступным все
меньший участок почвы.
37
Болезнь не передается соком. Если привить здоровое
растение на больной корень, то уже в конце перйого года
и, самое позднее, на второй год можно наблюдать первые
признаки болезни, причем, согласно Petri, прежде всего
появляются внутриклеточные палочки, а затем уже различные
внешние симптомы. Если прививка не удается, то заражения
не происходит. Согласно целому ряду авторов, если на
здоровый корень привить больной побег, то не только не
происходит заражения, но и больной побег может выздороветь;
однако Johnssen (1933) удавалось передавать болезнь *и путем
прививки больного побега здоровому растению.
Передача болезни через почву была доказана
следующими опытами. Вполне здоровые чубуки винограда были
посажены в почву из-под больных кустов, а для контроля —
в такую нее почву, но предварительно простерилизованиую,
яри температуре 120° С. На следующий год растения,
посаженные в инфицированной почве, начали проявлять корот-
коузлие, причем на третий год признаки болезни сделались
очень отчетливыми; контроль в стерилизованной почве остался
здоровым. В дальнейших опытах сажали растения в
стерилизованную почву, и только небольшую часть их корневой
системы проводили в сосуд с инфицированной почвой. И в этом
случае наблюдалось заболевание (Petri).
Инфекционность рахитизма до сих пор не установлена.
Болезнь может передаваться с чубуками от больных
растений, однако при благоприятных условиях чубуки от больных
растений могут выздоравливать. Большое значение в данном
случае имеют почвенные условия. Заболевание особенно легко
развивается на тяжелых глинистых почвах с плохой аэрацией.
Приведенные данные говорят в пользу вирусной природы
болезни. В пользу этой этиологии свидетельствуют и такие
морфологические изменения, как резкая деформация
пластинки листа, особенно выросты с нижней стороны листа,
которые вообще пока нам не известны ни при каких других
заболеваниях, кроме вирусных, и нахождение Х-тел, и опыты
зю искусственному инфицированию. Последние, однако,
пока допускают различное толкование. Неясно, почему в
подавляющем большинстве случаев растение заболевает только
на больных корнях и почему сравнительно легко происходит
выздоровление чубуков при перенесении их в новые почвенные
и климатические условия. Эти данные не могут опровергнуть
вирусной гипотезы, но могут быть использованы и против
нее. Если существует несколько форм короткоузлия, в том
числе имеющие невирусное происхождение, то проблема
становится еще более сложной. Остановимся здесь на
важнейших попытках выяснить другие этиологические факторы
короткоузлия, кроме вирусного.
Viola, который уже с конца XIX в. занимается изуче-
38
нием короткоузлия, несколько лет тому назад нашел в
сердцевине больных лоз грибок Pumilus medulla^ который, по
мнению автора, является возбудителем болезни. Возможно,
что в отдельных случаях этот грибок действительно бывает
причиной заболевания, похожего на короткоузлие, однако
достоверно известно, что типичное заболевание может
развиваться и при отсутствии этого грибка. Растения, выращенные
из семян на почве, предрасполагающей к болезни, заболевают,
хотя в их тканях и не находят грибка.
В 1932 г. Raughiano, применяя особые методы окраски,
нашел в листе здорового и пораженного короткоузлием
винограда нити неизвестного грибка. Он выдвинул сложную
гипотезу о симбиозе винограда с грибком и о нарушении
равновесия между грибком и виноградом, результатом которого
будто бы является короткоузлие. Гипотеза рухнула через
три года, когда Branas и Bernon показали, что Raughiano
отложение дубильных веществ в клетке принял за мицеллий
грибка.
Высказывалось предположение, что заболевание зависит
от неправильной и чрезмерной обрезки слабых кустов, однако
никакая обрезка не может вызвать курчавости, так же как
не может от нее предохранить.
Очень много внимания было уделено предположению,
что причиной болезни являются весенние холода. Эта
маловероятная для хронической болезни этиология была
опровергнута прямыми опытами Petri. Как уже было упомянуто выше,
большую роль в развитии болезни играют почвенные условия.
К заболеванию предрасполагает почва с мелкими частицами из
щебня и камня, с плохим стоком воды на низких местах, а также
наличность водонепроницаемой подпочвы, которая
затрудняет отток воды. Глинистые почвы часто отвечают этим
условиям, благоприятствующим развитию короткоузлия. Данные
относительно значения реакции почвы неопределенны: по
одним авторам — кислая, по другим — щелочная реакция
благоприятствует развитию болезни. Указание на роль почвы
не противоречит вирусной этиологии, так как характер почвы
может иметь значение в сохранении и распространении вируса,
а также быть условием, способствующим распространению
болезни. Branas (1937—1939) настойчиво указывает на фи-
локсеру {Phylloxera vestatrix fradicicola) как на вероятного
переносчика короткоузлия. При наличности филоксеры
болезнь может очень быстро распространяться, что,
например, автор наблюдал в Тунисе. Кроме филоксеры, болезнь
может распространяться путем вегетативного размножения
винограда.
Marsais Р. и Legol L. (1939) не наблюдали, однако, связи
между короткоузлием и филоксерой, так же как Emon (1938),
Lykiardopoulo (1940) и др.
39
В Калифорнии мелколистость винограда очень успешно-
лечат цинком, применяя для этой цели и внесение цинка
в почву вместе с сернокислым калием, и обрызгивание
растворами сернокислого цинка. Эти опыты Dufrenoy (1935),
перенес во Францию. Ему удалось достигнуть очень
хороших результатов от применения сернокислого цинка при
короткоузлии, сопровождающемся междуклеточными
палочками.
Однако не при всех формах короткоузлия сернокислый
цинк давал благоприятный эффект.
Целый ряд сортов винограда проявляет высокую степень
устойчивости к короткоузлию. К числу этих сортов
относятся Malnosio bianca, Somarella, Paga debito (см. Carpen-
tieri), а также гибриды, описанные Rives, Rous, Galles. К
сожалению, списки сортов, подверженных заболеванию, включают
несравненно больше наименований. Это относится как к
американским, так и к европейским сортам.
Цинк для борьбы с короткоузлием Dufrenoy применял
путем внесения в почву в феврале по 300 г сернокислого цинка
и сернокислого калия под каждый куст. В других случаях
он набивал кристаллическим сернокислым цинком
отверстие, сделанное в древесине лозы. Таким способом удавалось,
улучшать состояние даже таких больных кустов, которые,
казалось, уже подлежали удалению.
Для профилактики короткоузлия имеет значение выбор
места: надо избегать слишком компактных, плохо
проветриваемых почв, а также тех, в которых застаивается вода.
Большое значение имеют хорошая обработка почвы, и, в случае-
необходимости, искусный ее дренаж. Чубуки для посадки
следует выбирать здоровые, не от кустов, пораженных
короткоузлием, так как не следует возлагать слишком больших
надежд на возможное выздоровление больных чубуков, у
которых при пересадке в здоровые условия случается
наблюдать «борьбу болезненного предрасположения с
благоприятными, здоровыми условиями существования». Carpentieri
(1930), Branas и Bernon (1937) достигали улучшения в
течении болезни, применяя весной экранирование растешш
сажей: 8—10 частей сажи, 0.3 части желатина на 100 частей
воды.
Мозаичная болезнь винограда. Это заболевание, нередко»
смешиваемое с короткоузлием, в качестве самостоятельной
болезни было описано в Чехословакии. Судя по внешним
симптомам, мы встречали его и на восточном и на южном
берегу Крыма (рис. 11). При заболевании наблюдаются
различные формы мозаичности. Очень часто болезнь принимает
характер сетчатой мозаики, так как наблюдается посветление
по ходу нервов. В других случаях на листке без всякой
закономерности разбросаны светложелтые и даже белые участки.
40
причем зеленые участки могут пузыреобразно вздуваться -
Наконец, лист может полностью обесцвечиваться.
Чехословацкие авторы наблюдали при заболевании
внутриклеточные палочки, Х-тела, дезорганизацию хлоро-
пластов, некрозы во флоэме и отложение гумми в сосудах.
Рис. 11. Листья винограда с хлорозом по ходу жилок
Болезнь передается тлями и Lecanium сопьі, а также
соком и распространяется по плантации при помощи
инструментов, служащих для обрезки виноградной лозы.
Отмечается существование латентных форм болезни,
которые могут проявиться при прививке побега растения,
скрыто несущего вирус, на растения, восприимчивые к нему.
Возможно существование разных вирусов или же различных
штаммов одного и того же вируса (Blattny, 1934; Martinoff,
1934).
Для борьбы с болезнью рекомендуется уничтожение
больных кустов и стерилизация огнем инструментов, применяемых
при обрезке растений. Мозаичная болезнь, подобная
описанной в Чехословакии, встречается также в Калифорнии,
однако ее удалось передать только прививкой, но не соком
(Hewitt, 1945)
Другие вирусные заболевания винограда. Приведенными
заболеваниями, повидимому, не ограничивается список
вирусных болезней винограда. Обращают на себя внимание
заболевания, имеющие невыясненную этиологию.
В 1892 г. в Калифорнии Pierce описал болезнь
винограда, которую более подробно изучал Hewitt (1939). Эта
41
болеЭньч обычно быстро приводит к гибели пораженное ен>
растение. У- одних сортов болезнь начинается темнозеле-
нымй полосами по ходу нервов и вздутием ткани
между'нервами. На следующий год растение обычно погибает. У других
сортов Симптомы на листьях отсутствуют, но наблюдается
высыхание листьев и гибель растений в середине или в конце
лета. Также наблюдается засыхание плодов. Болезнь
передается прививкой.
Hewitt, Frazier и Houston (1939) устанавливают
передачу болезни цикадами Draeculacephala и Carneocephala и ©вязь
ее с карликовостью люцерны.
Названия этой болезни: Pieres disease, Anaheim disease,
California vine disease, Emperor disease.
Глава VI
ВИРУСНЫЕ БОЛЕЗНИ ФРУКТОВЫХ ДЕРЕВЬЕВ
ИЗ ПОРЯДКА ROSIFLORAE
Вирусные болезни фруктовых деревьев очень
многочисленны. Приступая к их изучению, необходимо учесть и
специфические особенности объекта и степень изученности
вопроса. На многолетних растениях, размножаемых
вегетативным путем, чрезвычайно сильно отражаются и возрастные
изменения и различного рода неблагоприятные почвенные
и климатические условия. Отсюда происходят многочисленные
физиологические расстройства у фруктовых деревьев, которые
то осложняют картину вирусного заболевания, то могут быть
ошибочно приняты за вирусную болезнь. Другое
обстоятельство, затрудняющее работу, состоит в том, что объект для
эксперимента в данном случае громоздкий и дорого стоящий;
кроме того, инкубационный период может быть весьма
продолжителен ¦— тянуться даже несколько лет* Поэтому самое
установление инфекционности заболеваний в данном случае
сложнее, чем при работе с однолетними растениями. Те
усложнения опытов, которые нужны для строгой идентификации
болезней, — выяснения переносчиков и т. д., — делают в
данном случае эксперименты чрезвычайно громоздкими. Этим,
невидимому, следует объяснить, что классификация вирусных
болезней фруктовых деревьев находится еще в очень ранней
стадии. Особенно затруднительно сейчас сказать, сколько
вирусов мы объединяем под общим названием «мозаичных
болезней» фруктовых деревьев.
Желтуха персиков. По классификации К. Smith, болезнь
вызывается Prunus virus l, Kunkel (1933). Она встречается
в различных штатах Северной Америки и, возможно, также
в Европе. Нередко она приобретает характер эпифитий.
Первая такая эпифития известна в 1791 г., а последняя в
1886—1888 гг.
Листья у больных растений ломкие, хрупкие,
складывающиеся вдоль жилок. В этих листьях происходит чрезмер-
43
ное накопление крахмала. Крупные листья иногда могут
быть мозаичными. К ранним симптомам относится посветление
по ходу жилок. Наблюдается усиленное ветвление вследствие
преждевременного пробуждения спящих почек. Многие ветви
тонки, проволокообразны и несут ивовидные листья. Плоды
достигают зрелости раньше, чем на здоровых деревьях, но
они ненормально велики, водянисты, безвкусны и часто имеют
красные пятна в мякоти. Больные ветви могут давать также
и здоровые плоды.
Болезнь может передаваться миндалю, абрикосу и сливе.
Встречается другой штамм вируса желтухи, который
вызывает комплекс симптомов, описанный первоначально
в Мичигане (США) как самостоятельная болезнь — мелко-
плодность персиков (little-peach). Kunkel (1936) доказал,
что вирусы обоих этих заболеваний близки друг к другу, так
как ему удалось выявить, что заражение одной из этих
болезней предохраняет от другой. Симптомы на вегетативных
органах при этом заболевании сходны с симптомами других
желтух: наблюдается усиленное ветвление и образование розеток,
состоящих из укороченных побегов, покрытых пучками
листьев. Плоды у больных растений, в отличие от плодов при
желтухе, созревают на 4—21 день позднее, чем у нормальных
растений, и наблюдается их измельчание.
Заболевание, похожее на мелкоплодность персика,
встречается в Туркмении. При нем наблюдается иволистость: листья
вдвое уже здоровых, нередко хлорозны; побеги тоньше
нормальных; междоузлия укорочены; наблюдается образование
ведьминых метел. Плоды резко уменьшены в размере.
Желтуха, как и мелкоплодность, передается не соком
и не почвой, но прививками и сохраняется при вегетативном
размножении. Переносчик болезни — цикада Macropsis Iri-
maculata. В клетках больных растений обнаружены Х-тела,
которые будто бы производят движение. Подобные же Х-тела
найдены в насекомом-переносчике (Hartzell, 1937).
Болезнь излечима при помощи высокой температуры.
Растения, помещенные в оранжерее при 34.4—36.3 С,
выздоравливают и больше не болеют. Быстрее действует теплая
ванна: погружение растений на 10 минут при 50°С
освобождает от вируса. В летнюю жару, в условиях плантации, вирус,
повидимому, сохраняется в корневой системе.
Розеточная болезнь персика. Эта болезнь вызывается,
по К. Smith, Prunus virus 2, McClintock. Она описана в 1888 г.
Е. F. Smith, известна с 1881 г. в штате Георгия (США) и
встречается, кроме США, повидимому, также и в Европе, например
в Италии. При заболевании наблюдается укорочение побегов,
каждая розетка несет сотни листочков. Болезнь нередко
приобретает острое течение, и растения могут быстро погибнуть.
Плоды, если они развиваются, морщинисты и преждевре-
44
менно опадают. Болезнь отличается от желтухи отсутствием
характерных краснопятнистых плодов и очень компактным
расположением листьев на побегах, а также более острым
течением. Заболевание розеточной болезнью не предохраняет
ни от желтухи, ни от мелкоплодия, что позволяет считать,
что ее вызывает особый вирус.
На абрикосе и сливе заболевание также обнаруяшвается
мозаикой. Болезнь передается прививкой на персик, абрикос,
сливу и вишню. При помощи повилики передается помидору
(Kunkel, J943)
Высокая температура излечивает ото заболевание, как
и предыдущее. Ferraris (1923) наблюдал розеточность в Италии,
где она излечивалась при помощи обрызгивания растворами
сернокислого железа. Вероятно, этот автор имел дело с
неинфекционным заболеванием.
Болезнь phony. Заболевание вызывается Prunus virus 3,
Hulohins (1933). Оно известно с конца XIX в. в штате Георгия
и в других штатах Северной Америки. Сходное заболевание
описано также в Италии (Curzi, 1931).
Симптомы болезни — усиленное ветвление и укорочение
междоузлий, причем листья остаются нормально зелеными
и крупными, вследствие чего больные растения приобретают
характерный облик и количество листьев на них кажется
чрезмерно большим. У больных растений наблюдается резкая
подавленность роста. Годичный прирост нормальных молодых
деревьев 30—90 см, а больных — только 5—15 см*
Для диагностики болезни служит следующая реакция.
Из корня вырезываются пластинки в. 0.5—1 мм толщиной,
и они погружаются в абсолютный метиловый алкоголь. К
каждым 25 см3 метилового алкоголя добавляют 1—5 капель
концентрированной чистой соляной кислоты. Если растение
больное, то в древесине появляются пурпурные пятнышки,
имеющие около 2 мм в диаметре.
Вирус, вызывающий болезнь, локализован в корневой
системе. Это доказывается опытами по заражению прививкой.
Прививка больных побегов на здоровые растения не
вызывает заражения. Заражение получается только при прививке
здоровых побегов на больные растения или же при прививке
кусочков корней больных растений в корни здоровых.
Прививка кусочка одной только коры корня не вызывает инфекции,
но она наступает при прививке коры с кусочком древесины.
Это говорит в пользу локализации вируса в древесине корней
(Hutchins). В опытах Hutchins (1939) молодые экземпляры
персика были заражены болезнью, а через год подвергнуты
обработке теплом. Растения, получившие теплую ванну в 48°С
в течение 40 минут, не заболели, тогда как получившие более
короткую ванну и контрольные — заболели. Вирус, подобно
вирусу желтух, легко разрушается при высоких температурах.
45
Способ распространения болезни в природе неизвестен;
возможно, что болезнь передается жуками-сверлильщиками
(Aegeria exitiosa).
Х-болезнь персика. Заболевание стало известно в 1938 г.
и с тех пор успело приобрести довольно серьезное значение
(Е. Hildebrand и D. Н. Palmiter, 1940).
Болезнь проявляется желтыми и красными пятнами на
листьях, а также продырявливанием листьев; наблюдается
преждевременное опадение листьев и нередко гибель деревьев.
Кроме персика, болезнь поражает Prunus virginiana, у которой
наблюдаются красные и желтые пятна на листьях, розеточ-
ность и гибель. Болезнь передается прививкой. При помощи
повилики передается помидору и др. травянистым растениям
(Kunkel, 1944).
Е. М. Stoddard попытался лечить заболевание путем
погружения черенков, пораженных Х-вирусом, в растворы
различных веществ. Из 10 черенков, опущенных в раствор хин-
гидрона, 8 выздоровело. Менее эффективными оказались
другие препараты, как то: 8-гидрохинолин-сульфат,
гидрохинон, пара-нитрофенол, кальциевая соль 8-гидрохинолина;
мочевина, тиосульфат натрия.
К сожалению, эти интересные результаты пока описаны
очень кратко.
Раз появившись, болезнь распространяется очень быстро.
Большое значение в распространении болезни имеет Prunus
virginiana. От этого растения она переходит на персик.
Рекомендуется уничтожение Prunus virginiana на 500 футов вокруг
садов (Beakeley, 1941).
Красный шов персика. Болезнь вызывается Prunus virus 4r
Bennett. Она известна с 1911 г. в Мичигане. Главным
симптомом заболевания является поражение плодов. Плоды больных
растений преждевременно созревают и имеют неправильные
очертания и неровную поверхность вследствие
многочисленных опухолей, Созревание плодов идет неравномерно, и раньше
всего становится сочной одна сторона плода — обычно та,
на которой находится шов. Больные плоды имеют
тёмнокрасную пигментацию, особенно интенсивную на верхушке
опухолей. На листьях наблюдается закручивание вниз их
верхушки, а также черешков листьев. Окраска листьев светлее
нормальной, а ко времени созревания плодов — желто-зеленая
или бронзовая. Нередко наблюдается также укорочение
междоузлий и розеточность.
Болезнь передается прививкой. Она излечима теплой
ванной, как и желтуха.
Инфекционное опадение листьев у персика. Заболевание
описано в 1940 г. под названием leaf casting yellow (Thomas
и др.). Листья становятся хрупкими, на них появляются свет-
лозеленые и желтые пятна, окруженные красными ободками;.
46
наблюдается закручивание листьев вверх, к концу июля они
обычно опадают. Плоды нормальны или морщинисты и
нередко опадают. Болезнь может проявляться не на всех ветвях,
а только на части их. Прививкой передается Prunus domestica,
вызывая у нее морщинистость плодов типа «оленьей кожи»
(см. ниже). Болезнь тождественна с Х-болезнью или очень
близка к ней.
Рие, /.'. Плоды вишни, пораженные buckskin (no Atanasoff)
Мозаичные заболевания косточковых. У персика, абрикосаг
сливы и различных других представителей рода Prunus широко
распространены мозаичные заболевания. Большая часть этих
заболеваний, кроме симптомов на листьях, выражающихся
в мозаичной расцветке, имеющей разнообразный рисунок,
проявляется также на плодах, давая здесь мозаику,
некротические явления, развитие гумми и уродование плодов,
связанное с ямчатостью и морщинистостью. Последнее явление
известно под названием оспы слив (рох), оленьей
кожи или замши (buckskin) вишни (рис. 12) и т. д.
В одних случаях симптомы на листьях отступают на
задний план, большую часть лета маскируются и т. д. В
Других же случаях симптомы на плодах отсутствуют совсем или
проявляются только на незначительной части плодов.
Большая часть относящихся сюда заболеваний легко
перевивается путем окулировки на различные виды рода
47
Primus, а также нередко и на других представителей
семейства розоцветных, например на розу. Все относящиеся
сюда заболевания передаются прививкой. Согласно Christoff
(1933), мозаичные заболевания также легко передаются, если
Рис. 13. Симптомы широкой полосатости
и кольцеобразной мозаики (по Ctiristoff)
под кору стебля ввести кашицу из больных растертых листьев.
Передача соком во многих случаях отрицается, однако W.
Newton (1936) описал мозаичную болезнь черешни, которая очень
легко передается соком. Классификация мозаичных
заболеваний еще далеко не закончена. К. Smith объединяет под
общим названием Prunus virus 5, Hutchins различные мозаики
вишни и слив, включая сюда также мозаику персиков. Од-
48
нако, согласно опытам (Cockran и Hulchins, 1941), между
мозаикой персика, с одной стороны, и мозаикой вишни, сливы
п абрикоса, с другой, существуют более сложные
отношения. При прививке больного мозаикой персика на один
из названных видов признаки болезни на лих не проявляются,
однако привитые растения становятся скрытыми вирусоно-
гителями и могут после такой прививки заразить здоровый
персик. Если найденный в природе мозаичный абрикос
привить персику, то у персика развиваются признаки мозаики,
которая уже может быть передана обратно на абрикос. Хотя
на персике собственная мозаика и мозаика абрикоса
проявляется совершенно одинаковым образом, однако из этих
опытов нельзя сделать заключение, что болезни тождественны,
потому что собственная мозаика персика, как мы видели,
не вызывает аналогичных симптомов у обоих названных видов
фруктовых деревьев.
Большое исследование, посвященное вирусным болезням
рода Prunus в Болгарии, произведенное ChrisLoff (1933),
позволяет различать у этого рода следующие заболевания,
встречающиеся в Болгарии, а также, по данным этого автора,
в СССР и в других страна к Европы:
Широкая п о л о с а т о с т ь и кольцевая
пятнистость (broad slreak и ring spot variegation).
Этот тип болезни характеризуется образованием колец и
полосок, имеющих по большей части в ширину 2—3 мм, но
иногда достигающих 10—15 мм в ширину (рис. 13). Полосы
имеют светлозеленый, желто-зеленый или желтый цвет.
Болезнь поражает персик, абрикос, миндаль, терн, вишню,
Pranus Pisardii, P. Maholeb и т. д. На плодах при заболевании
наблюдаются все характерные признаки оспы слив. Симптомы
на листьях особенно заметны весной, позднее они исчезают
или же проявляются только слабым хлорозом.
Болезнь может передаваться тлей Anurophis padl.
Узкая полосатость и кольцевая
пятнистость (narrow striped и ring spot variegation). При
заболевании наблюдаются узкие полосы и кольца шириной
0.5—2.0, редко 3 мм (рис. 14).
Болезнь известна у сливы, персика, черешни, миндаля,
Prunus cerasifera. Заболевание передается прививкой.
Признаки болезни могут наблюдаться с весны до осени,
но чаще всего только па первоначально развившихся листьях—
на остальных листьях болезнь не проявляется. При
заболевании можно также наблюдать мозаику и некротические
явления на плодах. Японская слива {P. salicina) является скрытым
носителем болезни.
Мозаика. Под этим названием автор объединяет
несколько типов заболеваний, из которых важнейшие два —
полигональная пятнистость и звездовидная мозаика. Звез-
4 В. Л. Рыжкоз 4?)
довидная мозаика у персика (asteroid spot) была также
описана в Америке Cockran и Smith (1938).
Мозаики, как было упомянуто выше, передаются не только
прививками, но также при инокуляции в кору дерева кашицы
инфекционных листьев. Имеются указания, что мозаика может
передаваться семенами. Она наблюдается у черешни, сливы,
персика, абрикосов, миндаля, вишни, терна и P. maholeb.
Рис. 14. Симптомы узкой полосатости и кольцеобразной мозаики
Широкую и узкую полосатость автору не удалось
передать прививкой на яблоню и грушу, тогда как подобная же
передача мозаик осуществима.
Кроме Prunus virus 5 Smith различает также другой
вирус, вызывающий мозаику у вишни, черешни и других
видов (Prunus virus 6, Thomas и Hildebrand (1937). Эта
мозаика описана названными авторами в 1936 г. Она
проявляется уменьшением размера листьев — особенно их ширины,
вследствие чего они становятся удлиненными, — хрупкостью
листьев, некоторым утолщением их пластинки,
морщинистостью краев листа и крапчатостью. Наблюдается также
укорочение междоузлий и неправильности цветка (сужение
лепестков, абортивность пестика). Больные деревья приносят
очень мало плодов, но плоды имеют нормальный вид. Prunus
virus 6 или Nanus pruni описывает также Е. W. Hildebrand
(1940, 1941).
Уже после опубликования работ Christoff (1933) в
американской литературе появилось описание целого ряда других
50
вирусных заболеваний фруктовых деревьев из рода Prunusy
точная идентификация которых весьма затруднительна.
S. Zeller и A. Evans (1941) различают Prunus virus 7У
который вызывает крапчатую мозаичность у черешни {Sweet
cherries, Pranus апат)у и Prunus virus 8. Последний вызывает
у черешни, а также у P. serrulata посветление по ходу жилок.
Края листьев станрвятоя неправильными, так как
наблюдается редукция участков листа и образуются вызубрины.
Средняя жилка может отделяться от пластинки в виде
удлиненного и тонкого выроста. На нижней стороне листа по ходу
жилок иногда происходит разрастание тканей в виде
маленьких пузырей. Поверхность листа может принимать
ненормальный, серебристый оттенок.
Междоузлия нередко подвергаются укорочению. В
середине лета больные деревья могут выглядеть увядшими.
Возможно преждевременное опадение листьев. У некоторых
сортов плоды не поражаются, у других же они заболевают.
В этом последнем случае плоды меньше нормальных и их
форма уродлива.
Оба описанные здесь вируса передаются прививкой, но
не тлей Myzus cerasi. .Вирусы 7 и 8 иногда встречаются
вместе. Ряд декоративных видов сливы, как то: P. yedoenis,
P. siebaldii, P. subhiatella, являются скрытыми носителями
вируса. Возможно, что к вирусу 8 близко стоит вирус,
вызывающий у вишни выросты на нижней стороне листа (rasp-
leaf of cherry, Bodine и Newton).
По номенклатуре Holmes, Prunus virus 7 может быть
обозначен как Мarmor cerasi, a Prunus virus 8 как Marmor
nerviclamus.
Более подробное описание мозаики, которую вызывает
Prunus virus 7, мы находим в работе Е. L. Reeves (1941). Эта
болезнь известна много лет, но только в 1935 г. доказана ее
инфекционность. При ней на листьях появляются светло-
желтые и зелено-желтые пятна. Иногда ткань по ходу нервов
остается гэленой, и тогда болезнь напоминает хлороз,
зависящий от почвенных условий. Наблюдается также морщи*
нистость между нервами. При тяжелых формах заболевания
отдельные участки листа могут выпадать, а также наблюдаются
поражения плодов. Для передачи заболевания необходимо,
чтобы при прививке произошло срастание тканей. Болезнь
поражает Prunus avium и P. emarginata. Prunus cerasus и
P. maholeb no большей части несут вирус, не проявляя никаких
симптомов этого. Вирус может перейти через ткани персика
и вызвать заболевание черешни. Нагревание черенков до 46°С
в течение 60 минут или до 49° С в течение 10 минут не инак-
тивирует вируса.
Как rusty-mottle Е. Reeves (1940) описывает в роде
Prunus (у вишни) заболевание, при котором, кроме крапча-
4* 51
тости, наблюдается быстрое развитие осенней окраски от яр-
коже лтой до красной и опадение листьев. Плоды уменьшены
в размерах, но не уродливы.
J. Л. Milbrath и S. М. Zeller (1942) описывают
заболевание декоративных видов вишни, выражающееся в том, что
в коре образуются трещины и пятна коричневого и
красноватого цвета. Дерево подавлено в росте, боковые ветки редко
развиваются, междоузлия сближены. Пластинки листьев
изгибаются вниз. Болезнь передается прививкой. Ее вирус
авторы обозначают Prunus virus 9, или Rimocortus Kwanzani.
Prunus virus 10, или Marmor pallidolimatus, у Prunus
serrulate описывают S. M. Zeller и J. Л. Milbrath. Этот вирус
вызывает заболевание, называемое banded chlorosis, которое
проявляется кольцами и полосками на листьях шириной
в 1—2 мм.
Целый ряд авторов под названием «карликовость» (dwarf)
или «желтуха» (yellow) вишни и сливы описывают тяжелые
поражения, при которых на листьях наблюдаются хлорозные
пятна, сопровождаемые опадением листьев (G. W. Keit п
С. N. Clayton, 1943)у или же общая карликовость листьев
(G. Н. Berkelej^, 1941). Штаммы этого вируса описаны у
Willison (1944).
Желтуха передается прививкой (Е. W. Hildebrand и
W. D. Milles, 1941). Она может переходить на персик, вызывая
здесь тяжелые некротические явления, хлороз и уродливость
листьев (Е. J. Rasmussen и D. Cation, 1942). Возможно, что
эта группа болезней близко стоит к Prunus virus 6.
Кольцевую пятнистость абрикоса описывают Е. W. Bodine
п W. A. Kreutzer (1942). При этом заболевании неправильные
кольца и пятна появляются как на листьях, так и на плодах.
Плоды некротизируются и часто опадают еще до созревания.
Заболевание, вероятно, близко стоит к оспе слив и других
косточковых, давно описанной в Болгарии Atanasoff и затем
Chrisioff.
Инфекционный хлоро** груш. Вызывается Pyrus virus I,
Ваш* (1907). Листья пораженных деревьев имеют желтые
верхушки, которые затем белеют; иногда весь лист может
покрыться желтыми пятнами. Пораженные участки
постепенно обесцвечиваются и затем становятся коричневыми.
Болезнь передается прививкой.
Мозаичная болезнь яблони. Болезнь вызывается Pyrus
virus 2, Bradford и Joloy (1933). Болезнь проявляется у
яблони желтыми и кремовыми пятнами и линиями
неправильных очертаний. Иногда хлорозные части отмирают.
Наблюдается также распространение хлороза по ходу нервов
(рис. 15).
Болезнь передается прививкой. Инкубационный период —
около 53 дней.
.52
Можно наблюдать передачу болезни на грушу, айву?
Cotoneaster harroviana, Eriobotrya faponica, Photinia arbuti-
foliay Sorbus pallescens и др. (Н. Thomas, 1937). Передача
соком не удалась.
Рис. 1о. Инфекционный хлороз у яблони,
листья больных растений (но Bradford и Joioy)
Пятнистость яблок. Заболевание известно также^под
другими названиями, например, «горькая ямчатость»
(bitter pit, orinkie, blotchy cork, cork, apple blister и др.).
Главным симптомом болезни являются водянистые пузырьки
на плодах. Позднее на месте этих пузырьков развивается
некроз и образуется углубление, получается пятно красного
или коричневого цвета. Возможно также образование
коричневых опробковевших участков на глубине, без выхода наружу.
Описанное поражение плодов наступает в разное время.
Иногда плоды заболевают уже вскоре после опадения
лепестков. При раннем поражении плодов они нередко осыпаются.
Иногда характерные пятна развиваются только после сбора
урожая, во время хранения плодов (Atanasoff, 1938). На
листьях пораженных деревьев иногда удается подметить
некоторую пятнистость, которая то едва выражена, то довольна
значительна. Симптомы на листьях сильно зависят от
сортовых особенностей растения и от других условий. Болезнь
передается прививкой. Увеличение питания азотом и.
кольцевание побегов повышает осмотическое давление в листьях
и чувствительность к болезни. Удаление листьев понижает
процент поражения плодов (Smock, 1941).
53
При заболевании наблюдается задержка крахмала в
пораженных клетках и образование склеренхимных клеток
вокруг поражения. Вопреки Мс Alpine, пораженные участки
не находятся в связи с сосудами (Smock и van Doren, 1938).
Каменистая пятнистость груши. Это заболевание груши
аналогично описанному выше заболеванию яблони. Первые
симптомы болезни на плодах развиваются через 10—20 дней
после опадения околоцветников. Под эпидермисом плодов
образуются темнозеленые участки, рост которых подавлен,
тогда как соседние ткани развиваются быстро. В результате
получается деформация плодов и глубокие ямки, дно которых
может некротизироваться. Вокруг каждой ямки развиваются
многочисленные каменистые клетки. Кроме симптомов на
плодах, наблюдаются на листьях светлая мозаика по ходу
нервов и нежная крапчатость. Эти симптомы легко
маскируются.
Заболеванию подвергается также кора, на которой
наблюдаются трещины и морщинистость. Под пораженными
участками коры можно видеть пятна, которые кажутся
пропитанными водой (Kienholz, 1939).
Диференциальная диагностика вирусных болезней
фруктовых деревьев. При диференциальной диагностике вирусных
болезней плодовых деревьев следует прежде всего иметь в виду
некоторые наследственные дефекты, которые могут
передаваться при вегетативном размножении, Kinman (1930) в
Калифорнии описал непродуктивные экземпляры у черешни
сорта Black Tatarian. Листья этих деревьев несколько
деформированы и имеют светлозеленые пятна. Иногда
поражено не все дерево, а только отдельные ветки. Соцветия у них
меньше нормальных, лепестки короче, и пестик сильно
редуцирован. Эти деревья обычно не приносят плодов.
Заболевание не зависит от почвенных условий; оно очень похоже
на вирусную болезнь, но прививкой не передается.
Фруктовые деревья чрезвычайно чувствительны к
различным дефектам почвы, на которые они реагируют мозаично-
стью, розеточностыо и измельчанием листьев. К числу
заболеваний с невыясненной этиологией, которое; может быть,
является вирусным, а может быть, зависит от дефицита каких-
либо солей, относится мелколистость яблони, описанная
В. Н. Клещевой1 в Куйбышевской области. Эта болезнь
встречается на сорте Анис. При ней наблюдается образование на
листьях светложелтых и белых пятен и полосок — всего
чаще у края листа, которые позднее засыхают, — а также
измельчание листьев. Измельчавшие листья встречаются,
главным образом, на верхушечных побегах. Сильно
пораженные деревья подавлены в росте, и у них наблюдается чрезмер-
1 По рукописному отчету.
54
ная кустистость и розеточность. Кроме Аниса, болезнь
встречается также на других сортах, но особенно широкого
распространения достигает у названного сорта (рис. 16, 17).
Известна мелколистность яблонь, а также косточковых,
^излечимая применением цинка. Описан некроз коры у яблонь,
%і
Рис. 16а. Яблони сорта Анис — здоровые .
сопровождающийся подавленностью роста, розеточностью
и бронзовостью листьев, при котором можно достигнуть
улучшения, применяя бор (Ark и Thomas, 1940). Были описаны
некроз на верхних листьях яблони и подавленность роста
55
в результате недостатка меди (Dunne, 1938). Дефицит магния
в почве вызывает у яблони и у других фруктовых деревьев
развитие хлорозных пятен на листьях, с последующим их
некрозом (Wallace, 1939). Для диференциальной диагностики
необходимо^ чтобы при наличии заболеваний, подозрительных
¦"
Рис. 16Ь. Яблони сорта Анис — больные (по Клещевой)
на вирусные, тотчас были заложены опыты по передаче боле.;ш>
прививкой и одновременно опыты лечения болезни солями.
Цинк можно применять путем обрызгивания деревьев
жидкостью, в которую на 4,5 килограмма сернокислого цинка
56
и 2,5 килограмма гидрата извести приходится 100 галлонов
воды. Такое обрызгивание может повредить листья, поэтому
обрызгивание следует применять после опадения листьев
или до их развития, и в таких случаях доза цинка может быть
даже увеличена. Применяется также поливка раствором
сернокислого цинка. Под четырехлетнюю сливу вносится 2,S
килограмма сернокислого цинка, под более старые деревья —
до 3,6 килограмма. Можно вносить кристаллический ZnS04
Рис. 17. Верхушки яблонь сорта Анис, пораженных мелколистостью
(по Клещевой)
в углубления, которые делаются в стволе дерева. Размер
такого углубления для старых деревьев — 3 — 4 см в
глубину и около 1 см в диаметре (Chandler, 1937).
При недостатке меди можно пользоваться обрызгиванием
бордоссяой жидкостью или же внесением в почву под дерево
1 килограмма медного купороса.
Вредоносность вирусных болезней фруктовых деревьев.
Вирусные болезни фруктовых деревьев отличаются очень
большой вредоносностью. Некоторые из этих заболеваний
имеют тенденцию быстро распространяться. Так, например,
мозаичная болезнь персика в 1934 г. в одной местности в
Колорадо была зарегистрирована на 3 тысячах деревьев, а в 1935 г.
пришлось удалить 30 тысяч деревьев, зараженных этой
болезнью.
Для борьбы с вирусными болезнями фруктовых деревьев
не известны другие меры, кроме уничтожения явно больных
57
растений и недопущения в качестве посадочного материала
черенков от больных растений. Поэтому эффективную роль
в борьбе с распространением вирусных болезней фруктовых
деревьев могут сыграть мероприятия законодательного
характера. Соответствующее законодательство уже существует
в Калифорнии. Согласно этому законодательству
организован внутренний карантин, препятствующий вывозу
посадочного материала из местностей, в которых много вирусных
болезней, а также принудительное уничтожение больных
деревьев.
Глава VII
ВИРУСНЫЕ БОЛЕЗНИ РОЗОЦВЕТНЫХ
Красная или зеленая мозаика малины. Заболевание в 1922 г.
списано Rankin и Hockey под названием красной
мозаики, однако, согласно Сооіеу (1936), его лучше называть
зеленая мозаик а по аналогии с тем, как ото принято
при других мозаичных заболеваниях. Вирус этой мозаики,
по К. Smith, называется Rubus virus I, Rankin и Hockey.
Болезнь поражает как сорта красной малины (Rubus ideus),
так и сорта ежевики или черной малины (R. Occidentalis).
Болезнь проявляется очень различно, в зависимости
от сорта. Можно различать также ее мягкую и тяжелую форму.
Мягкая форма проявляется слабой крапчатостью. При
тяжелой форме, кроме зеленой мозаики, развиваются водянистые
пятна на молодых листьях и черешках, которые приобретают
темносиний цвет. За этим следует некроз верхушечных
побегов. На другой год больные растения имеют карликовый,
розетковидный рост. Мягкая форма болезни, повидимому,
вызывается другим штаммом вируса.
Болезнь передается при вегетативном размножении
и тлёй Amphorophora rubi, а отчасти А. гиЫсоіа и A. senso-
riata. Болезнь не передается соком.
Желтая мозаика малины. Ее вирус, по классификации
К. Smith, называется Rubus virus 2, Bennett (1927). Эта
болезнь впервые была описана как одна из вариаций красной
мозаики, она часто встречается вместе с нею на одном и том
же растении. При заболевании наблюдается посветление
листьев по ходу нервов и общее резкое пожелтение. При
смешанной инфекции симптомы желтой мозаики обычно
доминируют над красной мозаикой. Болезнь поражает сорта
красной и черной малины. Болезнь передается, как и красная
мозаика малины, A. rubi.
Курчавость малины. Это заболевание описано Green в 1894 г.
По классификации К. Smith, его вызывает Rubus virus 3,
Melchers (1914) и Bennett (1927).
69
Листья пораженных растении темнозеленого цвета, но
ближе к осени могут приобретать бронзовый оттенок.
Вследствие замедленного роста жилок листа наблюдается вздутие-
ткани между нервами и закручивание листьев вниз.
Болезненные симптомы и общая подавленность растений становятся
с каждым годом все сильнее, приводя, в конце концов, растение
к гибели.
Bennett (1927) различает два вируса, вызывающих
курчавость: альфа и бета. Первый встречается только на
красных сортах малины (сорт Cuihberl). Сорта с черными
ягодами к нему иммунны (как, например, сорт Cumberland). Ко
второму вирусу оба упомянутые сорта чувствительны, но
сорта красной малины страдают от бета-вируса меньше, чем
сорта черной. Внешние проявления обоих вирусов одинаковы.
Сорт Columbian болеет от альфа-вируса очень легко, может
выздороветь и снова быть заражен утим вирусом. Бета-вирус
обычно на красной малине не встречается, но в
экспериментальных условиях красная малина может быть им заражена,
и затем он может быть возвращен на черную малину.
Экспериментально можно получить также смешанную инфекцию.
К. Smith рассматривает оба вируса как разные штаммы одного
вируса.
Болезнь передается прививкой, но не соком. Она
распространяется тлей A plus rubicolciy но не разными видами
Ampkorophora.
Полосатость малины. Другие названия болезни: eastern
blue stem, streak, severe streak, roseile.
IIo K. Smith, болезнь вызывает Rubus virus 4, Wilcox.
Можно различать тяжелую и мягкую полосатость,
причем Rankin (1931) рассматривает их как разные
заболевания. Полосатость впервые была описана Wilcox в 1922 г.
При тяжелой полосатости происходит сильное
скручивание пластинки листа вниз или закручивание ее в виде
цилиндра. Не наблюдается закручивания вниз краев листа
и резкого подавления роста жилок, которые известны при
курчавости. Листья темней нормальных, но иногда
наблюдается неопределенная пятнистость. Для заболевания
характерны темносииие полосы на стеблях, особенно сильно
развивающиеся у основания молодых побегов. Болезнь
прогрессирует из года в год. Кусты часто погибают на второй или третий
год заболевания. Болезнь поражает только сорта черной
малины (R. occidentalis). Способы распространения болезни
в природе неизвестны. Нет также данных о передаче болезни
прививкой.
Мягкая полосатость характеризуется только слабым
закручиванием листьев. Темносиние полосы здесь более узки
и отсутствуют чаще^ чем при тяжелой полосатости.
Подавленности роста почти не бывает. Пятнистость листьев здесь
60
ке встречается. Болезнь не прогрессирует л не переходит
в тяжелую полосатость. Очень своеобразно действие мягкой
полосатости на плоды. При тяжелой полосатости они
развиваются равномерно, здесь же, на одном и том же соцветии,
одни созревают раньше, другие позднее, и это очень
затрудняет сбор урожая, в чем, собственно, и состоит наиболее
чувствительный хозяйственный ущерб от этой болезни. Болезнь
встречается только у R. occidentalism наиболее часто
поражаемый сорт — Cumberland.
Распространение в природе мягкой полосатости
неизвестно. Нет также данных о передаче ее прививкой.
Карликовость малины. Вирус этой болезни, по К. Smith,
обозначается как Rubus virus 5, Zeller (1932). Болезнь
известна с 1918 г. При ней наблюдается укорочение междоузлий
и измельчание листьев. Растения кажутся чрезмерно
покрытыми маленькими листочками, листья окрашены светлее
нормального и крапчаты.
Болезнь известна на Loganberry [Rubus frnticosus), Cory,
Thornless и др. сортах. Заболевание не передается соком,
и его не удалось передать прививкой почек. Его переносит
тля Сapilophorns letrarhodus. Эта тля, взятая с дикой розы —
Rosa rubiginosa, — вызвала заболевание у малины (Zeller,
1927).
Желтая пятнистость красной малины. Е. Chamberlain (1938)
в Канаде описывает как новую вирусную болезнь, под
названием yellow-blotoh-curl, заболевание, при котором
наблюдается курчавость листьев и образование на них желтых
пятен и желтых колец. Болезнь сопровождается заметной
подавленностью роста. Она поражает различные сорта
красной малины, но черная малина к ней устойчива. Болезнь
передается прививкой. Первые симптомы ее могут проявиться
вблизи прививки уже через 45—70 дней после срастания
привоя с подвоем. Общее заболевание наступает обычно только
на другой год после прививки.
Папоротниковость листьев малины. Как и предыдущее,
заболевание это описано Е. Chamberlain (1941). Наряду
с деформацией пластинки листа (папоротниковость)
наблюдаются «жирные пятна», которые могут некротизироваться.
Количество и размер плодов уменьшаются, и плоды уродливы
п недостаточно сладки. Болезнь передается прививкой.
Вирусные заболевания малины в Европе. В СССР вирусные
заболевания малины изучались Натальиной (1938), Верто-
градовой(1938). Описанные выше заболевания изучены в США,
и нельзя с уверенностью сказать, что заболевания,
встречающиеся в СССР, дая?е внешне сходные с американскими,
с ними идентичны.
В СССР встречается желтая мозаика малины,
проявляющаяся пожелтением по ходу нервов и общим хлорозом
61
пластинки листа. Мы встречали зеленую и
коричневато-желтую мозаику на дико растущей Rubus idaeus и других видах
(рис. 18). Судя по американским данным, вирусные
заболевания дико растущей малины могут служить источником за-
Рис. 18. Маскировка мозаики у Rubus discolor
болевания культурной малины (Сооіеу, 1936). Встречается
также курчавость листьев. При этом заболевании, согласно
описаниям Вертоградовой, наблюдается подавленность роста
жилок, причем пластинки листа нередко скручиваются в
клубочек. Цвет листьев темнозеленый или светлозеленый, причем
часто встречается крапчатость, иногда почти белого цвета.
Светлозеленая окраска и крапчатость обычно появляются
весной и затем маскируются, так что в середине лета листья
имеют однообразно темнозеленую окраску. Нередко не все
побеги куста поражены заболеванием. Растения резко
подавлены в росте и обычно на второй или третий год
заболевания погибают. Болезнь может быстро распространяться
по плантации.
Вертоградова описала также новое заболевание малины
под названием ведьмины метлы. В начальной стадии
болезни высота побегов уменьшается незначительно, а коли-
02
чество их увеличивается, так что получается даже
впечатление пьшио развитого куста. Впоследствии подавленность
роста побегов и число их возрастает, так что, в конце концов,
образуются целые дерновины тесно расположенных друг подле
друга стеблей. При диаметре куста в 18 см число этих стеблей
достигает 200. Листья не скручиваются, иногда на них
наблюдаются желтые пятна диаметром до 3 см. Наблюдается также
измельчание листьев; при этом пластинки листьев сильна
уменьшаются в размерах, тогда как черешок остается длинным.
Такие листочки сильно скучены на побеге, вследствие
укорочения междоузлий, и имеют хлорозную, иногда почти белую
пластинку. Больные растения редко цветут, а если и бывает
цветение, то цветы уродливы. Наблюдается подсыхание тычинок,
удлинение цветоложа, его пролиферация, а также образование
у пестиков гинофоров, на которых они поднимаются (рис. 19).
Если развиваются ягоды, то бывают безвкусны. Болезнь
может распространяться быстро: на одном участке весной
1937 г. было 12% больных растений, а осенью 1938 г. — 80%.
В Англии Harris была произведена большая работа по
классификации типов встречающихся там мозаичных
заболеваний малины. Автор различает два заболевания:
обыкновенную мозаику и морщинистость (crinkle). При обыкновенной
мозаике наблюдается зелено-желтая пятнистость, причем
иятна обычно не резко отграничены от темнозеленых участков
и имеют в поперечнике около 2 мм. Эти пятна
сконцентрированы между главными жилками листа. Наблюдается
некоторое закручивание пластинки листа вниз, пораженные участки
слабо подавлены в росте.
У заболевания морщинистостью известны две формы —
мягкая и тяжелая. Мягкая форма проявляется хлорозными
пятнами разных размеров и неправильной формы,
распределенными^ отличие от предыдущих болезней, по всей пластинке
листа. При тяжелой форме хлорозные участки могут
приобретать яркожелтую окраску и резко выделяться на темнозе-
леном фоне. Они сильно подавлены в росте и поэтому образуют
на листе как бы углубления. Одновременно происходит
сильное закручивание листа. Обыкновенная мозаика маскируется
в летнюю жару, это же заболевание не маскируется. Мягкая
форма болезни часто внезапно переходит в тяжелую, и можно
предполагать, что болезнь вызывается смешанной инфекцией*
причем тяжелую ее форму вызывает примесь к вирусу,
имевшемуся уже в растении, другого вируса.
Американские мозаики малины передаются Amphoro-
phora гиЫ. Все попытки передачи этой тлёй мозаичных
болезней малины в Англии не увенчались успехом. Возможно,
что некоторые виды мозаики малины передают цикады (Dicker,
1940). Прививка растений, больных мозаичной болезнью,
встречающейся в Америке, к английским сортам малины дала
Рас. 19. Пролиферация цветов малины (/>, г) и «юрчальныи цветок (а)
картину болезни, резко отличную от английских мозаик.
Точно так же прививка американской курчавости листьев
на английские сорта не вызвала характерных для этой болезни
признаков, но имела своим результатом некроз побегов и смерть
растений.
У малины описана также папоротниковость листьев,
которая выражается усиленной изрезанностью пластинки
листа и ее нитевидностью. Болезнь передается тлёй Aphis
rubiphila и, по предложению Zundel (1931), описавшего ее,
вызывается огуречным вирусом.
Ксантозис клубники. Ксантозис (xanthosis) клубники
называется также пожелтением краев листа (yellow edge).
Это заболевание вызывает, по К. Smith, Fragaria virus l,Pla-
kidas. Болезнь проявляется пожелтением краев листа и
курчавостью пластинки. Черешки листьев укорочены, и у них
отсутствует красная пигментация. Наблюдается преждевре*
мснная осенняя окраска листьев.
Болезнь описана в 1926 г. Plakidas в Калифорнии.
Некоторые сорта Fragaria virginiana особенно чувствительны
к вирусу и удобны для экспериментальной работы с ним
(Hildebrand, 1941).
Вирус не передается ни соком, ни семенами.
Переносчиком болезни, по Whitehead и Wood (1941), является тля
Pentatrichopus potentillae (принятая Plakidas за Capitophorus
fragaefolii). Возможно, что и та и другая тля могут
распространять болезнь.
Морщинистость клубники* Это заболевание вызывает
Fragaria virus 2, Zeller и Vaughan (1932). Оно проявляется
неравномерным посветлением листьев, посветлением по ходу
нервов, а также образованием светлых пятен. В центре этих
пятен могут быть небольшие некротические точки. Вследствие
задержки роста нервов наблюдается морщинистость. Зубцы
листьев под влиянием заболевания часто становятся более
глубокими. Особенно резко болезнь проявляется в теплице
зимой и вообще при неблагоприятных условиях. На
плодородной почве она может маскироваться. Massee (1942)
различает две формы болезни — слабую и тяжелую, которые
сохраняют свои характерные особенности при перевивке при
помощи насекомого.
Болезнь известна в США, а также в Англии.
Заболевание не передается соком и семенами,
переносчиком его является тля Capitophorus fragaefolii.
Ведьмины метлы клубники. Заболевание вызывается
Fragaria virus 3, Zeller (1927). Больные растения имеют листья
с удлиненными, нитевидными черешками. Эти листья
развиваются в огромном количестве, их пластинки мельче
нормальных, светлозеленого цвета и несколько закручены вниз. Плети
служащие для бесполого размножения, более или менее уко-
5 В. Л. Рыжков gg
рочены, и дочерние растения, имеющие такой же вид, как
и материнское, приближены к нему. Заболевание не передается
соком, его переносчик — Capitophorus fragaefolii. Болезнь
встречается в США и, повидимому, не имеет широкого
распространения.
Низкорослость клубники. Zeller и Weaver (1941) описывают
под названием stunt disease of strawberry заболевание,
встречающееся на северо-западном побережье Северной Америки.
Оно особенно резко проявляется в холодных и влажных
условиях. Болезнь характеризуется карликовостью и
направленным вверх ростом побегов. Листья с нижней стороны
приобретают серую окраску, хлорозны, часто складываются вдоль
средней жилки, отдельные доли листа приобретают вид
чашечек; еще чаще края листочков закручиваются вниз. Плоды
уменьшаются в размере, твердые. Болезнь распространяет*
Capitophorus fragaefolii.
Окручивание листьев клубники. Болезнь характеризуется
измельчанием листьев, веретеновидностью черешков, хлороз-
ными пятнами и общим хлорозом. Часто наблюдается
морщинистость и, наконец, скручивание долек листа, причем
оно начинается у основания листочков и те принимают вид
воронкообразных трубочек (Berkeley и Plakidas, 1942).
Сомнительная (suspected) мозаика клубники. Мелкая крап-
чатость, принимавшаяся раньше за мозаичную болезнь,
согласно опытам Guba (1933), оказалась неинфекционной.
Повидимому, она является наследственным дефектом,
передающимся при вегетативном размножении. Мозаичность и
карликовость, описанные Afanasiev и Morris (1941), также,
вероятно, генетической природы.
Мозаичная болезнь розы. К. Smith различает два вируса,
вызывающих мозаичную болезнь розы: Rosa virus l, White
(1932), и Rosa virus 2, Brierley (1935). Первый из этих вирусов
вызывает светлозеленую мозаику, а также морщинистость
листьев. Мозаика может распространяться в виде посвет-
ления по ходу нервов, а также в виде более или менее крупных
пятен. Иногда можно наблюдать ненормально светлую окраску
лепестков. Болезнь передается прививкой и у разных сортов
проявляется различно.
Второй вирус вызывает желтую мозаику и тоже передается
прививкой.
Thomas и Massey (1939) различают три типа мозаики
у розы, отличающиеся между собой как симптомами, так
и неодинаковой восприимчивостью к ним различных сортов.
При первом типе наблюдается посветление по ходу нервов
и маленькие хлоротические пятна, продолговатые или
удлиненные. Второй тип характеризуется более резкой мозаикой,
причем наблюдаются хлорозные полоски и пятна. Третий тип
похож на второй, но характеризуется еще более резкой мозаикой.
66
У -некоторых еортов, устойчивых ко второму типу моааякн,
проявляется третий тип, например у Belle of Portugal. Два
последние типа иногда сопровождаются некротическими
явлениями. Мозаичная болезнь розы всего сильнее проявляется
при 15—25° С (Baker и Thomas, 1942). Мозаичные заболевания
розы передаются прививкой. Были испытаны 34 вида
насекомых, и ни одно из них не оказалось переносчиком болезни
(Вгіегіеу и Smith, 1940).
Увядание розы. Болезнь вызывается Rosa vivas 3, Grieve.
Первый симптом болезни — своеобразное закручивание
листьев молодых побегов и их хрупкость, затем начинается
опадание листьев, распространяющееся от верхушки вниз.
Листья перед опаданием становятся светло зелеными или
желтыми, затем наблюдается некроз стебля и его отмирание,
которое тоже распространяется сверху. В тканях стебля в местах
некрозов образуется пробковое вещество и откладывается
гумми, В тканях листа иногда наблюдаются возле ядра Х-тела
(В. J. Grievp, 1931).
Болезнь передается не только прививкой, но и соком.
Вирус фильтруется через фильтр Зейца. Заболевание описана
в Австралии. В Италии встречается очень сходное с ним
заболевание, также передающееся соком, переносчиком
которого являются тли из рода Macrosiphum (Gigante, 1936).
Полосатость розы. Заболевание вызывается Rosa virus 4,
Вгіегіеу (1935). При нем наблюдаются коричневые и
красноватые полоски, распространяющиеся по ходу нервов и
кольцеобразные. Подобные полосы наблюдаются не только на
листьях, но и на стеблях. Ткани в области этих полосок
подвергаются некрозу.
Болезнь передается не соком, а прививкой. Из 34
испытанных насекомых ни одно не оказалось переносчиком.
Ведьмииы метлы голодискуса. Это заболевание вызывает
Holodiscus virus 1, Zeller (1931). Болезнь проявляется у Ноіо-
discus discolor укороченными побегами и сильным развитием
придаточных побегов. Листья пораженных растений очень
уменьшаются в размерах и прижаты к стеблю; летом они
становятся красно-бронзовыми. Если весной бывает большой
холод, то эта окраска может появиться и раньше.
Болезнь передается прививкой, переносчиком ее является
Aphis spiraella. Различные виды спиреи оказались устойчивыми
к болезни {Spiraea thunbergii, S. vanhoutei и др.)
Меры борьбы с вируспыми болезнями розоцветных.
Опасность вирусных болезней розоцветных очень велика.
Большинство культивируемых видов этого семейства размножается
вегетативно, и, следовательно, раз заболевшее растение
передает болезнь своему вегетативному потомству. Имея
хроническое течение, эти болезни розоцветных прогрессируют
и способствуют гибели растений и вырождению сортов. По-
5* 6?
давяяющее бсщьшднетво относимых сюда болезней мы еще
не умеем лечить.'Выше мы отметили, что лечению поддаются
черелки и молодые деревца персиков, пораженные некоторыми
вирусами, исчезающими после обработки теплом. По данным
Вертоградовои, орошение малины ослабляет проявление
виру сньгс заболеваний. Там, где невозможно лечение,
приходится все внимание сосредоточить на профилактике. Это
достигается путем очень тщательной браковки посадочного
материала в питомнике; просмотр этого материала должен
производиться в то время года, которое наиболее благоприятно
для проявления признаков болезни. Все больные или
подозрительные на болезнь растения, а также непосредственно
примыкающие к ним с обеих сторон, уничтожаются.
Важным мероприятием является выведение устойчивых
сортов, В Америке некоторые селекционеры малины поставили
себе задачу вывести сорта, очень слабо поражаемые насекомым-
пореносчиком, но в то же время в высокой степени
чувствительные к болезни и резко проявляющие признаки ее. Такие
растения будут редко заболевать, но, заболев, так резко
обнаружат болезнь, что их легко заметить и уничтожить. Уже
имеются сорта, в известной мере удовлетворяющие этим
требованиям (Huber и Schwartze, 1938), но, к сожалению, в СССР нет
еще сортов розоцветных, устойчивых к вирусным болезням,
й, насколько нам известно, работа по выведению таких
сортов совершенно недостаточна. Для селекционеров очень важно
знать, какие вирусы встречаются и какие из них наиболее
опасны, для того чтобы выводить сорта, обладающие
устойчивостью к этим вирусам. Поэтому следует пожелать, чтобы
была возможно скорее закончена строгая идентификация
•вирусов, встречающихся у розоцветных.
При обсуждении мер борьбы должно быть также
обращено внимание на дикие розоцветные, которые могут быть
(носителями опасных вирусов, и на борьбу с насекомыми,
переносчиками вирусных болезней.
Глава VIII
ВИРУСНЫЕ БОЛЕЗНИ БОБОВЫХ
У бобовых встречаются и мозаичные болезни и желтухи.
Мозаичные заболевания бобовых, как выяснилось в re-
чение последних лет, вызываются разнообразными вирусами.
Описание этих заболеваний представляет большие трудности,
так как они часто мало различаются между собой по внешним
симптомам болезни и по свойствам вируса. Кроме того, самая
номенклатура этих вирусов крайне запутана: многие из них
обозначаются номерами, причем разные авторы употребляют
эти обозначения в различном смысле. Попытка упорядочить
номенклатуру, сделанная К. Smith, не дала положительных
результатов, так как в новейших работах по вирусным
болезням гороха (Zaumeyer, 1940) эта номенклатура игнорируется.
Впрочем, и сам К. Smith, повидимому, без достаточного
основания попытался объединить ряд вирусов (например, Pha-
seotus virus 2 и вирус гороха, обозначаемый Zaumeyer и Wade
как Pea mosaic virus 2).
Главным критерием для диференцирования вирусов
служит разная восприимчивость к этим вирусам различных
видов и сортов. Таким образом, на практике для диференцналь-
ной диагностики необходимы эксперименты, требующие
нередко много времени. Из этого краткого вступления вытекает,
что строгая диференциация отдельных вирусов, встречающихся
у бобовых, представляет еще исследовательскую задачу, тем
более что у нас в Союзе классификация вирусов бобовых еще
не начата. При таком положении вещей для практического
работника значительно важнее общая характеристика
отдельных типичных групп вирусных заболеваний бобовых, чем
детальный разбор литературы, посвященной отдельным
вирусам.
Общая характеристика мозаичных заболеваний бобовых*
Мозаичные заболевания у бобовых различно проявляются
в зависимости от вируса, его штамма, а также вида и сорта.
Симптомы этих заболеваний следующие: мозаичная расцветка
листьев, различные степени деформации пластинки листау
подавленность роста и некротические явления.
Некротические явления характеризуют по большей части только
тяжелое течение вирусной болезни, но для некоторых вирусов
они являются правилом.
Мозаики бобовых при помощи инокуляции соком обычно
передаются с большим или меньшим трудом: в
экспериментальной работе приходится пользоваться карборундом, чтобы
облегчить искусственное заражение. Напротив, они легко
передаются тлями, причем один и тот же вирус может
передаваться различными видами тлей. Трудности, с которыми
связано искусственное заражение, несомненно будут осложнять
работу каждого систематика вирусов бобовых. Один из
вирусов, поражающих фасоль, и вирус мозаики сои очень легко
передаются семенами, что делает эти вирусы классическими
примерами передачи вирусных болезней семенами. Данные
о передаче семенами вирусных болезней гороха недостаточно
определены; некоторые из них совсем не передаются таким
путем. Большинство вирусов, поражающих бобовые, инакти-
вируются при довольно низких температурах и мало
устойчивы .
Кроме собственных вирусов, бобовые подвержены также
воздействию некоторых вирусов, широко поражающих
представителей различных семейств, как, например, огуречный
вирус, однако влияние собственных вирусов бобовых чаще
всего, ловидимому, ограничено только пределами этого
семейства. В одной работе были испытаны представители 32
семейств, 60 родов и 94 видов на чувствительность их к вирусу
мозаики гороха 3. Оказалось, что этот вирус поражает только
виды, принадлежащие к семейству бобовых. Специализация
некоторых из рассматриваемых здесь вирусов заходит еще
дальше; например, мозаика сои не передается больше ни на
одно из испытанных в этом отношении растений.
Мозаичные заболевания бобовых могут причинять
большой ущерб. Особенно опасны передающиеся семенами, так
как распространение и вредоносность их с каждым годом легко
нарастает и они могут приводить сорта к вырождению.
Меры борьбы с мозаичными заболеваниями бобовых
уводятся к введению устойчивых сортов (U. S. № 5, Refugee, сорт
красной мелкой фасоли, выведенный Murphy, и др.), сбору
семян со здоровых растений, борьбе с насекомыми и
противовирусной полке.
Приведем здесь краткую характеристику важнейших
заболеваний.
Зеленая мозаика фасоли. К. Smith обозначает вирус,
вызывающий эту болезнь, Phaseolus virus l, Pierce.
Заболевание это было известно еще Ивановскому (1899) и более полно
описано в Америке в 1908 г. (Clinton). Подробные
сведения о нем находим у Fa jar do (1928, 1930). Забодевание
те
характеризуется хлорозом листьев, причем на светлом фоне
сохраняются зеленые пятна, которые часто находятся возле
жилок. Пластинка листа может более или менее деформироваться,
закручиваясь вниз. Наблюдается задержка роста. Болезнь,
как передающаяся семенами, может проявиться уже на
семядолях. Болезнь легко маскируется, причем тенденция к такой
маскировке наблюдается при температурах выше 30° и ниже
15° С. В патолого-анатомическом отношении не наблюдается
ничего такого, что не было бы известно и для других мозаичных
заболеваний растений.
Болезнь легко передается семенами. Установлено, что
из семян, образовавшихся в течение первых 19 дней после
начала цветения, получается от 43 до 100% больных
проростков, а из позднее развившихся цветов — только 0—25%
(Harrison, 1935). Имеются также данные, что вирус может
передаваться с пыльцой больных растений. Понятно, что
практически очень важен сбор семян со здоровых растений.
Однако это затрудняется тем, что к периоду плодоношения
'болезнь легко маскируется. Болезнь распространяется при
помощи самых различных видов тлей. Как переносчики мозаики
фасоли известны следующие тли: Aphis gossypii, A, medicaginis,
A. rumicis, Л. spiraella, Brevicoryne brassicae, Hyalopterus
atriplicis, Macrosiphum ambrosiae, M. pisi, M, geiy Rhopa-
losiphum pseudobrassicae, Myzas persicae. He переносят вируса
различные другие сосущие насекомые (цикады, клопы, трипсы,
Aleyrodidae (Zaumeyer и Kearns, 1936). Почвой болезнь не
передается. Возможно заболевание здорового растения
вследствие трения его листьев об находящееся рядом больное
растение. Как видно из таблицы 4, целый ряд сортов фасоли
устойчив к вирусу фасоли 1. В качестве чувствительных
к этому вирусу видов могут быть названы: Phaseolus acuti-
fblium, var, latijolium, P. lunatus, P. calcaratus, P. aureus,
Vicia faba, V. lathyroides, Lespedeza striata и, возможно, Ме-
illotus alba, Phaseolus lunatus var. macrocarpus и P> limensis
очень трудно поддаются заражению и, возможно, устойчивы.
К вирусу устойчивы также душистый горошек и горох. Болезнь
встречается в СССР.
Желтая мозаика фасоли. Вирус, вызывающий эту мозаику,
обозначается К. Smith как Phaseolus virus 2, Pierce (1934).
Вирус этот можно считать тождественным вирусу белого
донника {White sweet clover mosaic virus), К. Smith
отождествляет его такя^е с вирусом гороха 2 (Pea mosaic virus 2,
Zaumeyer и Wade), отличающимся, однако, от него, как видно
из таблицы 4, набором видов, которые он поражает. От
предыдущего вируса он отличается: 1) характером симптомов у
фасоли, 2) тем, что не передается семенами, 3) тем, что поражает
и горох, и иммунные к вирусу 1 сорта фасоли.
Вот краткое описание симптомов, вызываемых этим ви-
71
Вирусы^ поражающие пред
Вирус
Сорта Pisutn
sativum
м
TJ1
cS
t-l
о
И
х/1
't-i
ft
и
ОТ
W
н
со
К
wS
Сорта Phaseolus
и
со
CD
J—I
о
Й
о л
ев й
Ф
d
о<2
«J
.22 й
?й
о
ни
4>
Й
о
Зеленой мозаики фасоли (1) .
Желтой » » (2) .
Вирус фасоли (4)
Мозаики сои
Деформирующей мозаики
гороха
Мозаики гороха (2)
(3)
СО
(5)
Полосатое™ гороха (1) . . .
» » (2) . . .
Мозаики люцерны (2) . . . .
Мозаики белого клевера . .
Полосатости душистого
горошка
Мозаики Trifolium hybri-
dum (1)
Мозаики Trifolium hybri-
dum (2)
Pea mottle virus
Pea wilt virus
+
4
+
+
+
+
+
+
+
+
+
-f-
+
+
+
+
-t-
- +
+
+
+
+
+
+
+
+
+
f
4
+
+
+
4
+
+
-t-
+
+
И
¦f
4-
4-
+
4-
4-
+
H
H
4-
4
+
4
+
+
4-
+
H
+
-t-
+
H
H
+
+
+
H
4-
4-
4-
+
+
г
I
T
-i
4-
4-
4-
444-
4
4
Условные Обозначения: -j-чувствительность к вирусу; — уетойчн.
72
Таблица 4
ставшпелей семействй бобовых
vulgaris
Другие виды
к
в
w
го
га
го
CD g1
wS
I«
S я
о <u
*-.ч->
•Г* re
ftS.
со
P
О
рч Ю
ДР
S «J
Р
(X CD
го
-О
гЧ
ft
"-»
о
СО
то
ев
го
¦і-Н
О
1-1
Р 3
^ чЗ
&^
ге-о
Jo
ft
*я
сд
3
ft
p
—ar
** cd
_, о
re ro
3й
re о
"-¦ i—i
'S °
о1»
,H a
Влияние внешних условии
re
ft
H
re
ft
«O
So
CD
Сохранение
Разведение
+
н —
+
+
+
4
—
4
4
—
4
f
4
4
4
:
1 г
(
і 4
• 4
4
і
г
4
4
4
4
4
4
4
4
—
—
—
—
—
Н
+
+
+
_*.
—
4
4
4
4
4
4
4
1
4
4
Н
4
4
4
—
4
4
4
~
—
4
4
¦¦' ¦'
4
Н
—
—
—
56-58
56-60
90-95
61
58
62—64
60-64
62-65
62-65
65
78—80
62-64
58
60-62
54-55
60-62
58-60
24—30 час.
24—48 »
165 час.
3—4 сут.
3 суг.
4—5 суг.
2—3 »
24—48 час.
24 час.
24 »
Свыше
41 сут.
3—4 сут.
32 час.
1—2 »
31 сут.
31 »
1 :1 000
1 :10 000
1 :500 000
1 :100 000
1 :3 000
1 :100 000
1:500
1 :10 000
1:10 000
1 :800
1 : і 000 00Q-*
1 :4 000
1:2 00Э
1:8 000
1 :8 000
1 :10 000-
1 :100 000
вость; н — местные некрозы без общей инфекции.
П
русом на фасоли. Наблюдается образование желтых пятен
на темнозеленом поле, и это пожелтение может
распространиться, давая общий хлороз листа. Закручивание листьев
вниз наблюдается только в ранних стадиях болезни, зато
характерно опускание пластинки листа вниз от места ее
прикрепления к черешку. Вначале листья становятся несколько
хрупкими, позднее приобретают характерный бдеск и
становятся слабо вогнутыми. Маскировка болезни, характерная
для поздних стадий зеленой мозаики, здесь обычно не
наблюдается. Заболевание — более тяжелое, чем зеленая мозаика,
часто сопровождается сильной подавленностью роста,
пролиферацией боковых побегов и поражением бобов (Pierce, 1934).
У гороха болезнь вызывает мозаику и посветление по
ходу нервов, а также некоторую морщинистость и уменьшение
их размеров.
У белого донника {Melilotus alba) болезнь начинается
с маленьких светложелтых пятен на листьях, которые в
дальнейшем сливаются. Наблюдается также посветление по ходу
нервов и морщинистость листьев.
Важнейшие хозяева вируса приведены в таблице.
Болезнь передается соком и тлями (Macrosiphum pisi, M. gei).
Мозаика фасоли 3. Вызывается Phaseolus virus 3, Zaumeyer
я Wade (1935), по номенклатуре К. Smith. Это заболевание
очень мало изучено. Оно описано у гибридов фасоли String-
less Green Refugee х Wells Red Kidney, у которых
выражается легким хлорозом, У сорта Stringless Green Refugee
заболевание проявляется резким хлорозом и уменьшением
размеров листьев, развивающихся непосредственно после
заражения, затем посветлением по нервам и, наконец, ярко-
желтой пятнистостью, которая не сопровождается
морщинистостью листьев.
В интересах диференциальной диагностики необходимо
упомянуть, что некоторые наследуемые дефекты хлорофилла
у фасоли могут внешне напоминать мозаичную болезнь. В
отличие от мозаики, они, разумеется, не инфекционны, и в одном
подобном случае наблюдалось расщепление в отношении 1 : 15
(Zaumeyer, 1942).
Мозаика фасоли4 (Phaseolus virus 4), Marmor laesiofaciens,
Zaumeyer и Harter (1942—1943). Это заболевание у одних
сортов ограничивается местным некрозом, который может
перейти в общий некроз, у других же выражается мозаикой.
Сорта, дающие общее заболевание, гомозиготны к
рецессивному гену; сорта, дающие местный некроз, содержат
доминантный ген. Болезнь начинается слабой мозаикой, которая
постепенно усиливается, и ткань между жилок становится
значительно светлее, чем по их ходу (veinbanding). Могут
наблюдаться морщинистость и вздутия.
Следует различать штамм А, который вызывает в ранних
74
стадиях болезни более слабые, а в поздних более сильные
симптомы, чем предыдущий.
Вирус очень устойчив к высокой температуре и другим
воздействиям, чем резко отличается от Phaseolus virus I
(табл. 3). Существуют расы фасоли, которые реагируют на
вирус только местным некрозом. Устойчивость к общей
инфекции (некротическая реакция) доминирует; во втором
поколении наблюдается расщепление 3 : 1 (Zaumeyer и Wade,
1943а).
Увядание фасоли. Это заболевание описано W. A. Jenkins
(1940). Оно начинается во время цветения, и первым
симптомом его является слабое увядание листьев. Позднее
наблюдается пожелтение нижних листьев, и увядание
распространяется все больше, причем в конце концов погибает все
растение. На стебле наблюдаются темные полосы. При
внимательном рассмотрении листьев можно обнаруяшть мозаичность.
Если растение попадает в воду или под дождь, то светлые
участки листьев инфильтрируются водой и становятся
прозрачными. На плодах можно наблюдать темные полосы. Пато-
лого-анатомические явления крайне характерны. На срезах
через стебель под микроскопом легко обнаружить потемнение
между внутренней корой и внешней ксилемой, которое может
итти глубже в ксилему и сердцевину. Это потемнение зависит
от отложения в клетках гумми.
Были сделаны многочисленные попытки выделить из
больных растений грибок или бактерию, которая могла бы
быть признана возбудителем заболевания, но эти попытки
не увенчались успехом. Искусственное заражение легко
удается путем инокуляции иглой в подсемядольное колено
или стебель. Был испытан целый ряд сосущих насекомых,
но при их помощи болезнь не удалось передать. Только в одном
¦случае наблюдалась передача, а именно — при помощи Lygus
pratensis. Заболевание еще мало изучено.
Некротическая мозаика фасоли. W. J. Virgin (1943)
описывает заболевание фасоли, при котором наблюдается
красноватая окраска узлов, распространяющаяся по жилкам листа,
а также красноватые углубления на бобах. На семенах
наблюдаются кольца и пятна. Растения подавлены в росте, бобы
мельче нормальных. Возможно, что болезнь вызывается
вирусом, близким к Phaseolus virus 2. Около 5% проростков
из семян больных растений обнаруживают заболевание.
Болезнь передается также соком.
Мозаичная болезнь сои. Заболевание впервые описано
в 1916 г. в США (Clinton). Его вызывает, по классификации
К. Smith, Soja virus 1, Gardner и Kendrick (1921). У нас в Союзе
'болезнь подробно изучена Е. Д. Якимович (1938). По внешним
симптомам болезнь похожа на мозаичную болезнь фасоли:
она сопровождается резкой, разнообразной мозаичностью
7»
и деформацией пластинки листа. Нередко наблюдается гофри-
рованность этой пластинки, сборчатость и пузырчатосты
Доли листа могут сильно редуцироваться в размерах.
Отмечено также недостаточное опушение на стебле уі бобах.
Отдельные бобы могут вовсе не иметь опушения, характерного для
Рас. 20. Листья сои морщинистой и мозаичной
(но Якимович)
данного сорта. Болезнь сильно снижает урожай. Потери
урожая колеблются, по Якимович, от 2.1% до 23.7%, но в
отдельных случаях возможны потери урожая на 84%. Симптомы
болезни могут маскироваться; значительная маскировка болезни
наблюдается, если растение развивается при коротком (10-
часовом) дне. Всхожесть семян с больных растений
пониженная.
Болезнь довольно легко передается соком. Установлена
передача мозаичной болезни сои семенами, причем, чем раньше
происходит заражение, тем больший процент семян с больных
растений дает больные проростки. Больше половины
проростков больных семян могут быть мозаичными. Вторичная
инфекция зависит от распространения болезни тлями. В
качестве переносчиков болезни в Германии указываются Doralit
frangulae, D. rhamni, D. fabae, M.acrosiphum fabae (Heinze
ж Kohler, 1940). E. Д. Якимович сообщает, что развитию
болезни на плантации предшествует массовое появление
насекомых.
Вирус вызывает местные некрозы у фасоли и у вики.
Горох иммунен. Существуют разные штаммы вируса, щ был
7в
выделен некротический штамм, который вызывает некрозы
у сои.
По Н. Heinze и Е. КбЫег, вирус, инактивируется при
61° С. Е. Д. Якимович указывает более низкую температуру
инактивации (55° С). Этот автор наблюдал искусственное за-
Рис. 21. Листья сои: слева нормальной, остальные деформированы
под влиянием мозаичного заболевания (по Якимович)
ражение при разведении сока 1 : 10, тогда как упомянутые
выше авторы наблюдали заражение даже при разведении
1 : 10 000. В соке вирус хранится 3—4 дня. . :-т,
Согласно Е. Д. Якимович, вирус обладает антигенными
свойствами. Была получена сыворотка с невысоким титром.
При помощи термической обработки семян освободить их от
вируса не удалось.
В качестве мер борьбы можно рекомендовать: сбор семйн
¦со здоровых растений, использование ранних сроков сева
и борьбу с сосущими насекомыми. Известный эффект имеет
также противовирусная полка, которая должна быть
проведена до цветения растений и которую обычно рекомендуется
применять в тех случаях, когда больных растений не больше
15>%. Необходимо выведение устойчивых сортов.
Деформирующая мозаика гороха. Другие названия
заболевания: enation pea mosaic или мозаика гороха 1. По
классификации К. Smith, болезнь вызывает Plsum virus l, Osborn.
Болезнь начинается посветлением по ходу нервов, перехо-
77
дящее затем в мбзаичн-ость. Н-аблз?даются разданные -искртв-
ления пластинки листа и черешков, а также значительная
подавленность роста. На нижней стороне листьев могут
наблюдаться выросты—enation. У некоторых сортов наблюдаются-
некротические явления (у сорта Dwarf Telephon они резко
выражены) и развивается некроз верхушки. Этот вирус
поражает сорт Perfection, что отличает его от вируса мозаики гороха
2. Заболевание, повидимому, может в небольшом проценте
передаваться семенами (0.36%). В природе оно распространяется
тлями Macrosiphum gei и М. pisi. Тля становится способной
переносить вирус после инкубационного периода в 12—24 часа
(Osborn, 1938). Тля Aphis rumicis не передает болезни.
Мозаика гороха 2 (Pisum virus 2, Osborn). По К. Smith,,
болезнь вызывается Phaseolus virus 2, однако симптомы у этого
вируса и вируса гороха несколько различны.
У гороха болезнь дроявляется посветлением по ходу
нервов, которое может сохраняться и позднее. Кроме посвет-
ления нервов наблюдается мозаичность, и может наблюдаться
легкая деформация листьев и плодов, а иногда и
некротические полосы. Общая подавленность роста обычно не резкая.
Болезнь, повидимому, передается семенами; искусственно
передается соком. В природе ее переносят тли: Macrosiphum
pisiy M. gei, Aphis rumicis. Насекомое становится
инфекционным через несколько минут питания соком больного растения,
но сохраняет вирус недолго (Osborn, 1937).
Мозаика гороха 3 (Pisum virus 3, Pierce). Эта мозаика,
по классификации К. Smith, вызывается вирусом Pisum
virus 2.1 Не следует смешивать ее с мозаикой гороха 3,
описанной Chamberlain (1939), которую мы ниже характеризуем
как полосатость гороха. При заболевании наблюдаются носвет-
ление по ходу нервов, мозаичность и очень легкая
деформация пластинки листа, выражающаяся в волнистости краев
листа.
Вирус, вызывающий эту болезнь, идентичен с вирусом
мозаики красного клевера (Tri folium pratense), обыкновенной
мозаикой гороха Osborn и, вероятно, мозаикой бобов Boning.
Описано три штамма это?о вируса, которые не передаются
Т. pratense (Stubbs, 1937). В экспериментальных условиях
болезнь передается соком, в полевых условиях ее передают
различные тли: Macrosiphum pisi, Myzus persicae, Aphis
rumicis. Семенами болезнь не передается.
Мозаика гороха 4 (Pisum virus 4, Zaumeyer и Wade). Это
заболевание описано Zaumeyer и Wade (1936). Оно
характеризуется мозаичностью, причем зеленые участки тканей со-
1 Таким образом, Pisum virus 2 в этой книге К. Smith объединяет
с Phaseolus virus 2, a Pisum virus 3 этой книги соответствует Pisum virus 2
у К. Smith.
78
храняются по большей части вблизи жилок листа. Некрозов
не наблюдается (Zaumeyer, 1940).
Мозаика гороха 5 (rea virus 8, Zaumeyer и Wade). Кроме
мозайчности, болезнь вызывает сильную подавленность роста
и укорочение междоузлий. На листьях и стеблях можно
наблюдать некротические явления. Другое название болезни
pea stunt (Zaumeyer, 1940).
Лолосатость (streak) гороха 1 (Pea virus 6, Zaumeyer
и Wade). Первоначально предполагалось, что это заболевание
вызывается вирусом люцерны, но позднее была установлена
его самостоятельность. Болезнь проявляется некротическими
полосами на стеблях, листьях и плодах. Кроме некрозов
наблюдается мозаичность (Zaumeyer, 1937).
Долосатость гороха 2 (Pea virus 7, Chamberlain). Это
заболевание описано Chamberlain (1939) в Новой Зеландии под
названием pea streak (Pisum virus 3). Болезнь проявляется
образованием пурпурных и пурпурно-коричневых полосок
на стебле, листьях и плодах. Рост растений подавлен, и многие
ив них легко погибают. Кроме бобовых вирус поражает также
тыквенные, например огурец. Как видно из сводной таблицы
вирусов бобовых, он отличается значительно большей
устойчивостью к внешним факторам, чем почти все другие вирусы
бобовых. Вирус передается соком, но не тлями.
Мозаика люцерны 1. Заболевание описано Weimer в 1931 г.
под названием обыкновенной мозаики
люцерны (common alfalfa mosaic). По К. Smith, болезнь
вызывается Medicago virus l, Weimer. Первым признаком болезни
у люцерны являются маленькие, более или менее округлые
желто-зеленые пятна; в некоторых случаях они состоят
из двух или большего количества концентрических колец.
В дальнейшем пятна разрастаются и могут сливаться, причем
из светлозеленых становятся желтыми. Наблюдаются
морщинистость листьев и уменьшение их в размере, а также общая
подавленность роста растения.
Болезнь передается не соком, а тлей Macrosiphum pisi.
На фасоль и горох заболевание не может быть передано.
Мозаика люцерны 2. По К. Smith, болезнь вызывается
Medicago virus 2, Pierce, Zaumeyer и Wade. Pierce (1934)
описывает болезнь под названием мозаика люцерны 2
(Alfalfa virus 2). Болезнь проявляется подавленностью роста,
мозаичностью и морщинистостью листьев, легко передается
соком и тлей Macrosiphum pisi на другие бобовые, а также
на пасленовые. Последнее, как мы знаем, является редким
случаем для вирусов, поражающих бобовые. Недавно
установлена чрезвычайная близость этого вируса к вирусу,
вызывающему calico у картофеля. Оба они являются родственными
штаммами (см. гл. «О вирусных болезнях картофеля»),
У фасоли вирус вызывает местные некрозы, так же как
79
и у бобов, причем у последних вслед за этим обычно
развивается некроз проводящей системы и растение погибает. У
гороха вирус вызывает некротическую полосатость, почему
полосатость гороха 1 первоначально была принята за
заболевание, вызванное этим вирусом.
Мозаичные заболевания душистого горошка (Lathyrus
odoratus). Мозаичная болезнь у душистого горошка описана
Doolittle и Jones (1925)> Она передается соком и тлей Мае-
rosiphum pisi. Из таблицы 4 видно, что различные вирусы
бобовых могут поражать душистый горошек. Следует особо
отметить полосатость (streak) душистого горошка. Эта болезнь,
описанная Ainsworth (1940), начинается посветлепием по ходу
жилок, за которым следует крапчатость на листьях и светлые
полосы на цветах. В дальнейшем развиваются некротические
полосы на стеблях и черешках, и растение часто погибает.
У гороха болезнь также начинается посветлением вдоль
нервов, а затем развиваются некрозы на вегетативных органах
и плодах. У бобов наблюдаются местные некрозы, посветление
до нервам на молодых листьях, круглые некротические пятна
на старых, а также тёмнокрасные полосы на втеблях.
Мозаика Vigna sinensis. Заболевание известно с 1921 г.
На листьях развиваются желто-зеленые пятна (D. М. Мс
Lean, 1941). Растения подавлены в росте, чрезмерно кустятся.
Семенами болезнь передается в 5% случаев, соком передается
легче при применении карборунда. Передают болезнь также
тли Macrosiphum solanifolii, M. persicae, Aphis gossypii.
Мозаика белого клевера. По К. Smith, заболевание
вызывается Trifolium virus 1, Pierce. У Trifolium repens первым
признаком болезни является слабая крапчатость: по большей
части зеленые участки сосредоточены по ходу главной жилки
и распространяются по боковым жилкам, а светлые — ближе
к краю листа. Симптомы болезни могут маскироваться (Pierce,
1936).
У Trifolium pratense и Melilotus officinalis болезнь
протекает более тяжело и вызывает морщинистость листьев и в
некоторых случаях слабые некрозы. Точно так же мозаику и
некрозы вирус вызывает у бобов и гороха. У фасоли, под влиянием
вируса, после местных некрозов развивается мозаичность,
не сопровождающаяся более тяжелыми явлениями. Было
высказано сомнение в том, что этот вирус вообще может
вызывать у фасоли общую инфекцию и что наблюдаемые
симптомы не вызваны примесью другого вируса (Zaumeyer и Wade,
1936).
Исследования F. Johnson (1942) показали, что мозаика
белого клевера действительно имеет комплексную природу
и вызывается двумя вирусами: Pea mottle и Pea wilt. Первый
из этих вирусов может передаваться Cuscuta campestris, второй
не передается. Pea wilt заражает Vigna sinensis, тогда как
$0
;>тот вид устойчив к pea mottle. По указанным признакам оба
вируса могут быть диференцированы.
Вирус pea mottle {Мarmor effeciens) вызывает у гороха
посветление по нервам, желтые пятна и подавленность роста.
Позднее симптомы становятся более слабыми. Этот вирус,
кроме бобовых, может поражать представителей различных
других семейств, например, львиный зев.
Вирус pea wilt [Marmor repens) вызывает у гороха некроз
шюкультивированных и ближайших к ним листьев, тогда
как дальше отстоящие листья не проявляют признаков
болезни. У Vigna sinensis за местным некрозом следует общая
мозаичность. Вирус заражает только представителей
семейства Legurninosae.
Мозаика Trifolium subterranum. Заболевание начинается
посветлением жилок листа и затем переходит в крапчатость;
края листа становятся волнистыми, растение — карликовым.
Передача соком удается при применении карборунда. Пови-
днмому вирус может передаваться семенами (J. Aitken и В.
Grieve, 1943).
Мозаика Trifolium hybridum 1. Заболевание описано под
названием alsike clover mosaic. У гороха она вызывает картину,
сходную с той, которую вызывает вирус гороха 3, но
симптомы менее интенсивны. Болезнь передается соком (Zaumeyer,
1936).
Мозаика Trifolium hybridum 2. Это заболевание
характеризуется гораздо более тяжелым течением: у гороха инокули-
рованные листья обычно погибают. Наряду с мозаикой
наблюдаются некротические явления. Растения резко подавлены
в росте, нередко погибает верхушка стебля, наблюдается
деформация плодов. Болезнь передается соком (Zaumeyer,
1936).
Под названием clover big-vein был описан вирус,
который, как оказалось, поражает представителей самых
разнообразных семейств и обладает исключительно интересным
действием. Он способен вызывать опухоли. На надземных
частях растения опухоли развиваются редко, и болезнь
проявляется как типичная желтуха. На корневой системе
в местах закладки боковых корней развиваются опухоли.
Вирус передается цикадой Agallia constricta и поражает
Brachycorine iberifolia. Chrysanthemum leueantheum, Ramcx
acetosa* Melilotns alba и др. (Black, 1945).
Зіозапчные заболевания Ticiafaba. Мозаика у бобов впервые
была описана Dickson в 1920 г. Подробно мозаика у бобов
изучалась Boning (1927). При заболевании наблюдаются.
согласно этому автору, мраморность листьев и посветление
но ходу жилок. Нередко наблюдается деформация листьев
и уменьшение их размеров. Заболевание может снижать
урожай примерно на одну треть. Цитируемому автору не
5 В. Л. Рыжков 81
удалось наблюдать передачу болезни соком, почвой и
семенами, другие авторы наблюдали передачу мозаики бобов
соком, однако остается невыясненным, имеем ли мы дело
с тем же самым вирусом. По Boning, болезнь может
передаваться тлями: Aphis fabae, Macrosiphum pisi, Rhopalosiphum
viciae, затем цикадами {Typhlocyba pictaW и Chlorita solani),
а также, вероятно, ее передает Thrips flavus. Заболевание,
которое наблюдал Boning (1927), вероятно, вызывается
вирусом гороха 3. Мозаика бобов изучалась также в Японии (Imai,
Muryoma, 1934) и в Италии (Gigante, 1935).
По Imai, болезнь передается соком и тлями: Aphis rumicis,
Macrosiphum pisi7 Myzus persicae. Были изучены свойства
отого вируса: он инактивируется при температуре 63° С, при
разведении 1 : 1000 и погибает в соке через 21—25 дней;
выдерживает высушивание в течение 20—25 дней; сохраняет инфек-
ционность при рН 4—8, но быстро погибает при более
щелочном или более кислом рН. Он не фильтруется через свечи Ll
и L2; очень сходен с вирусом гороха 3, но сохраняется в соке
более продолжительное время.
В Японии описана также мягкая мозаика бобов (mild
mosaic). Заболевание вызывает легкую крапчатость, посвет-
ление по ходу нервов; уродство листьев и подавленность роста
отсутствуют.
Болезнь может передаваться соком с карборундом и тлями
Macrosiphum pisi и Aphis rulicis. Тли Myzus persicae и
Macrosiphum gei болезни не переносят. Заболевание может быть
передано на люцерну, Trifolium pratense, 7\ repens, душистый,
горошек и горох.
Вирус инактивируется при 55—60° С, устойчив к
разведению 1 : 500, но инактивируется в разведении 1 : 2000. В соке
разрушается уже через 3 часа. Передачи болезни семенами
не наблюдается.
Мозаика бобовых в Италии, повидимому, вызывается
вирусом гороха 2. Она передается соком на горох, фасоль
и белый клевер.
Мозаика Dolichos biflorus. Заболевание описано Uppai
(1931) в Индии. Наблюдается посветление по ходу нервов,
хлороз и закручивание листьев вниз. Большой процент бобов
стерилен. Болезнь легко передается семенами. Она имеет
серьезное экономическое значение.
Некротическая мозаика люпинуса. Эта болезнь под
названием sore shin была описана в 1934 г. Neill, а затем
Chamberlain (1935) в Новой Зеландии; под названием Lupinusbraune
она же или очень близкая к ней по проявлениям болезнь
описана в Германии; наконец, подобное же заболевание
сотрудниками М. С. Дунина обнаружено и в СССР. По
новозеландским данным, болезнь вызывается «вирусом гороха», и К. Smith
приписывает ее Pisum virus 2 (Chamberlain, 1935), тогда какг
82
по немецким данным, заболевание вызывается вирусом, очень
близко стоящим к огуречному. При заболевании наблюдается
подавленность роста и характерное закручивание точки роста
в одну сторону. Одновременно появляются светлокоричневые
полосы по стеблю. В дальнейшем некротические явления
распространяются на листья и усиливаются на стебле. В
Германии при заболевании описывается также мозаичная
расцветка и пятнистость кольцевого характера. Согласно Kohler,
болезнь передается на табак, причем с табака на табак
передача соком удается легче, чем с табака на люпину с или с люпи-
нуса на люпинус. На табаке можно различать два штамма —
зеленый и желтый. У Lupinus angustifolius симптомы не
типичны и выражаются в общем хлорозе, подавленности роста,
развитии пазушных побегов и в отсутствии цветения.
Болезнь передается различными тлями: Myzus persicae,
Aphis rumicis, Doralis rhamni, Macrosiphum pseudosolani.
Передача болезни соком не наблюдается. Повидимому,
тем же вирусом поражаются огурец, шпинат, томаты, табак,
Zinnia elegans, Eryngium planum и др. Подобное
заболевание у разных видов я наблюдал в Башкирии. При этом
заболевании сильно некротизируется флоэма.
Огуречная мозаика у бобовых. Cucumis virus 1 может
вызывать поражения у различных бобов. У Phaseolus lunatus
macrocarpus описана мозаичная болезнь, которая вызывает
резкое уменьшение урожая и легко передается табаку и огурцу,
вызывая у них симптомы огуречной мозаики. Вирус инак-
тивируется при 70° С; сохраняется 5 дней в соке, передастся
тлями Aphis gossypii, Myzus persicae; передачи семенами не
наблюдалось. Заболевание, повидимому, вызывается штаммом
огуречного вируса. Болезнь не могла быть передана на
Phaseolus vulgaris, Lathyrus odoratus, Pisum sativum, Sofa max,
Lupinus albus, но передается на огурец, табак, помидор,
петунию, перец, синие баклажаны (Harter, 1938; Whipple
и Walker, 1938, 1941).
Согласно Ainsworth (1940), огуречный вирус у душистого
горошка вызывает некротические явления и быструю гибель.
Горох не поражается вирусом. У люпинуса К. Smith наблюдал
некротические явления под влиянием вируса и отождествляет
заболевание с тем, которое описано в Германии под названием
Lupinusbraune. О. С. Whipple и J. С. Walker описывают ряд
штаммов огуречного вируса, которые они выделили от фасоли
и гороха (1941).
Lycopersicum virus 3 у бобовых. Этот вирус, вызывающий
бронзовость листьев у помидора, может поражать различные
бобовые, вызывая у них обычно некротические явления в виде
полос и концентрических колец. Поражение этим вирусом
можно наблюдать у гороха, бобов, люпинуса и Vigna sinensis.
Фасоль им не заражается или заражается с большим трудом.
6* 88
Вирус передается Thrips tabaci, но не тлями, что может быть
использовано для идентификации вируса и диференцнро-
вания его от вирусов бобовых (Snyder и Thomas, 1938;
Whipple, 1936).
Инфекционный хлороз у Laburnum. Ото заболевание известно
с 1873 г. как сортовой признак, который, однако, передается
прививкой. При помощи пересадки болезнь может быть также
передана на Cytisus hirsutus (Baur, 1906).
Желтухи бобовых. У бобовых известен целый ряд
заболеваний типа желтух, характеризующихся, главным образом,
расстройством роста и тем, что они обычно не передаются со-
ком. Степень изученности этих заболеваний различная.
Boning (1927) описывает скручивание листьев у бобов.
Листья становятся кожистыми, толстыми, хрупкими и ложеч-
кообразно скручиваются. Иногда на листьях зіаблюдаются
немногочисленные светлые пятна. При заболевании
происходит задержка оттока крахмала. Болезнь не передается ни
почвой, ни соком, ни теми видами тлей, которые были
испытаны. Попытки передать заболевание прививкой не было
сделано. Инфекционный характер болезни остается недоказанным.
Е. Д. Якимович описывает желтуху у сои, которая
характеризуется карликовостью растения. Нижние листья таких
растений толстые, хрупкие, часто скрученные, но нормальные
по размерам, верхние же мелки и быстро желтеют. Растения
цветут нормально, но плоды дают редко, я то обычно мелкие.
Ото заболевание близко стоит к желтухе верхушки у бобовых —
болезни, которая широко распространена в восточной части
США. Встречается она, кроме сои, на фасоли, клевере.
Заболевание переносится насекомым Empoasca fabac, однако, по
Monteilh (1929), оно не инфекционно. Если части одного
растения поместить в разных ящиках, то заболевают только побеги,
подвергшиеся действию насекомого. Сок больных растении
не вызывает заражения.
Описываемые здесь опыты еще окончательно не
доказывают, что в данном случае мы не имеем дела с вирусным
заболеванием. Если это заболевание инфекционное, то, возможно,
наблюдаемые явления объясняются медленным
распространением вируса по тканям. При описываемой здесь желтухе
верхушки у люцерны, как и при инфекционных желтухах,
возрастает C/N. Количество витамина А у больных растений
значительно меньше, чем у здоровых (II. Johnson, 1939).
Остановимся здесь более детально на лучше изученных
вирусных болезнях бобовых типа желтух.
Карликовость люцерны. Заболевание известно в Южной
Калифорнии. Оно характеризуется резким укорочением
стеблей и измельчанием листьев. Цветение подавлено, хлороз
отсутствует, и листья могут казаться даже более темно
окрашенными, чем нормальные. Для заболевания характерны
84
поражения корня, которые выражаются коричневыми
полосами. Сосуды в корне переполняются гуммиподобным
веществом, и количество крахмала в нем уменьшается. Транспп-
рация у больных растений происходит значительно менее
интенсивно, чем у здоровых.
Болезнь ice передается соком, но может быть передана
прививкой кусочков больного корня с пожелтевшей от гумми
древесиной к корням здоровых растений. Распространение
болезни по полю, а такя№ то обстоятельство, что
изолированные от наземных насекомых растения не заболевают,
свидетельствует о том, что в природе существует насекомое,
являющееся переносчиком болезни (Weimer, 1936—1937).
Заболевание, по К. Smith, вызывается Mcdicago virus 3,
Weimer.
Ведьмины метлы люцерны. Ото заболевание вызывает
Medicago virus 4, Edwards. Оно распространено в Новом
Южном Уэльсе. Симптомы: карликовость растений и
усиленное развитие придаточных побегов, вследствие чего
растение приобретает характерный вид прямостоящего кустика
с множеством побегов, достигающих нескольких дюймов
в длину. Листья уменьшены в размерах, по краям часто хло-
ротичны и морщинисты. Цветение наблюдается редко. Цветы
часто подвергаются антолизу. Рано весной моя^ет наблюдаться
маскировка болезни, но в то же время заболевание легко
прогрессирует и может закончиться смертью растения.
Болезнь передается прививкой, ее переносит цикада
iHaUjmoidus acuttis (Edwards, 1935, 1936; Menzies, 1944).
Розеточная болезнь арахиса. Болезнь вызывает Arachis
virus l, Zimmermann. Заболевание характеризуется, в первую
очередь, аномалиями роста. Боковые побеги, вместо того чтобы
стлаться по земле, очень коротки, поднимаются вверх,
вследствие чего получаются характерные округлые очертания
куста. Листья хлорозны, но иногда на них наблюдаются,
особенно в ранней стадии болезни, отдельные пятна, делающие
их мозаичными. Больные растения обычно не плодоносят.
Заболевание не передается соком, семенами и почвой.
Переносчик болезни — Aphis laburni Kelt.
От заболевания особенно страдают поздние посевы. Также
более тяжело протекает п сильнее распространяется болезнь
при редком посеве, чем при густом. Главной мерой борьбы
является введение устойчивых сортов.
Метельчатость белой акации (Robinia pseudoacacia).
Болезнь вызывает Robinia virus l, Hartley и Haasis. Она
характеризуется развитием метельчатых побегов с
укороченными междоузлиями и крайне редуцированными в размерах
листьями. На стволах могут развиваться подушечки,
достигающие 18 см в диаметре п 1 см в толщину, несущие группу
почек и листьев. Эти подушечки следует рассматривать как
8*
крайнюю степень ведьминых метел. Корневая система у
больных растений развивается хуже, чем у здоровых.
Болезнь передается прививкой, другие способы ее
распространения неизвестны (Hartley и Haasis, 1929; Grandt,
1939).
Фидлодия Crotolaria juncea. Болезнь характеризуется
превращением элементов цветка в листья и явлениями
пролиферации. Заболевание может передаваться прививкой и
является, следовательно, инфекционным (Rosa и Misra, 1938).
Ведьмины метлы фасоли. Болезнь характеризуется
усиленным ветвлением. Плоды отсутствуют. Причина болезни не
выяснена (W. G. Hoyman).
Beta virus 1 у бобовых. Этот вирус, вызывающий у свеклы
курчавость верхушки, может поражать также и бобовые.
У фасоли заболевание, вызванное этим вирусом, иногда
приносит тяжелый ущерб урожаю. Наблюдается слабое
посветление по ходу жилок, закручивание листьев вниз и
карликовость, причем листья имеют более интенсивную
зеленую окраску, чем нормальные. Сходные же симптомы
наблюдаются у бобов, у белого донника и у различных видов
клевера.
В Америке получены сорта фасоли, сочетающие в себе
устойчивость к этому вирусу с устойчивостью к вирусу мозаики
фасоли (Dana, 1940).
Глава IX
ВИРУСНЫЕ БОЛЕЗНИ СЕМЕЙСТВА ЗОНТИЧНЫХ
У зонтичных растений широко распространены вирусные
-заболевания, которые по большей части вовсе не изучены.
Более детально исследованы вирусные заболевания
сельдерея, так как культура этого растения в некоторых областях
США занимает очень важное место и серьезно страдает от
вирусных заболеваний.
Западная мозаика сельдерея. Заболевание вызывает Лріит
virus 1 (Severin и Freitag, 1938). Болезнь проявляется посвет-
лением по ходу нервов и резкой мозаикой, которая может
переходить в коричневые некротические пятна. При тяжелом
заболевании наблюдается сужение долей листа и уродливость
нластинки.
Вирус может вызывать также заражение других
зонтичных — моркови, петрушки, кориандра, Carum carvi,
Anthriscus cerefolium и т. д. Представителей других семейств
он не поражает. Болезнь передается соком, а также
различного рода тлями.
Вирус инактивируется при 60° С, устойчив к разведению
1 : 4000 и сохраняется в соке не менее 7 дней; он легко
фильтруется через свечи Шамберлена.
Giganle описал в Италии мозаику сельдерея, сходную
с описываемой. Тот же автор описывает мозаику моркови.
Южная мозаика сельдерея. Заболевание было подробно
изучено Wellman (1934—1935), который считал, что
вызывающий его вирус отличается от огуречного. Однако Price
(1935) показал, что заражение Cucumis virus 1 предохраняет
от южной мозаики сельдерея, откуда можно сделать вывод,
что эта последняя вызывается одним из штаммов огуречного
вируса. Температура инактивации вируса лежит около 80° С,
следовательно, значительно выше, чем типичного штамма. Вирус
южной мозаики сельдерея, как и следовало ожидать от
огуречного, может заражать представителей различных семейств,
в том числе тыквенных и пасленовых. Южная мозаика
отличается от восточной также и симптомами. Первые симптомы
S7
обеих мозаик одинаковы и состоят в посветлении по ходу
нервов; в дальнейшем для южной мозаики характерна
желтая крапчатость листьев п желтые зигзагообразные полоски,
образующие рисунок, имеющий очертания листа дуба.
Некротические пятна на листьях здесь отсутствуют, зато
наблюдаются некротические полосы на черешках и в проводящей
системе черешка.
В опифитологии болезни большое значение имеют
сорняки, особенно однодольное Commelina undiflora, в которых
вирус может зимовать. Вирус передается соком и при помощи
тлей (DoolitLle, 1931, 1934).
Различные штаммы огуречного вируса могут вызывать
также и другие формы мозаичного заболевания у сельдерея.
Мозаика сельдерея, найденная в подмосковных хозяйствах
Л. М. Вовком, вероятно, вызывается огуречным вирусом.
Желтуха сельдерея. Это заболевание вызывает особый
штамм Callistephus virus 1 или скорее близкий к нему вид.
Первым симптомом болезни является вертикальное
положение черешков, вследствие чего листья обращены вверх.
В дальнейшем развивается общий хлороз растения,
подавленность в росте и кустистость. Болезнь передается цикадами
Cicaduladiversa, Thamnotettix montanus и Г. geminatus и
причиняет существенный ущерб культуре сельдерея в Калифорнии
и в некоторых других штатах. Этот вирус может поражать
также морковь, петрушку и пастернак (Severin, 1934).
Другие заболевания сельдерея. В литературе имеется также
краткое описание других вирусных заболеваний сельдерея
(Severin и Freitag, 1938). Calico сельдерея, вызываемое, но
классификации К. Smith, Apium virus 2, Severin и Freitag,
проявляется яркожелтыми пятнами. Иногда наблюдается
образование хлорозных колец. При дальнейшем развитии
болезни вся пластинка листа покрывается желтыми полосами,
среди которых остаются только маленькие зеленые участки.
Данные о свойствах вируса отсутствуют, он передается, по-
видимому, Aphis gossypii.
Как pseudo-calico описывается внешне сходное
заболевание, которое передается соком, но которое не удалось
передать ни одним из видов тлей, бывших в испытании.
Вирус инактивируется при 70° С, устойчив к разведению
1 : 1000 и сохраняется в соке 5 дней. Кроме зонтичных, он
может поражать представителей различных других семейств.
Морщинистость листьев (crinkle)
сельдерея начинается посветлением по ходу нервов, которое потом
переходит в резкую мозаику, причем зеленые участки пузы-
реобразно вздуваются (рис. 22). Если заражение происходит
в молодом возрасте, растения бывают резко подавлены и могут
погибнуть. Болезнь передается соком. Вирус инактивируется
при 70° С, при разведении 1 : 100 и в соке через 3 дня.
88
Желтая пятнистость (yellow spot) сельдерея
характеризуется маленькими неправильными хлорозными пятнами
на листе и желтыми полосками вдоль жилок. На черешках
часто наблюдаются белые пятна, и если содрать с черешка
Рис. 22, Кусочки листьев сельдерея. Вверху — желтая
пятнистость; посредине — морщинистость; внизу —
кольцевая пятнистость (по Severin и Freitag)
эпидермис, то можно видеть некротические полоски по ходу
жилок (рис. 22). Болезнь соком не передается, но
распространяется тлями (Severin, 1934).
Кольцевая пятнистость сельдерея
передается только зонтичными. Заражение соком удается с трудом,
легче—тлями, например RJiopalosiphum melliferum. Вирус
этой пятнистости был выделен от болиголова (Сопіит гласи-
89
latum). Он вызывает у сельдерея маленькие хлорозные кольца
или же множество хлорозных точек, которые окружают
неправильной формы зеленые участки. Это кольцо может быть
«нова окружено зеленой полоской, за которой следует светло-
желтый ободок (рис. 22).
Зонтичные могут поражаться также различными
другими вирусами. Бронзовость листьев (Lycopersicum virus 3)
вызывает у сельдерея на пластинках нижних листьев
многочисленные маленькие желтые пятна, которые скоро
становятся коричневыми, центр их отмирает. Наблюдается общая
подавленность развития растений, а также некрозы во
внутренних тканях стебля, которые обнаруживаются только при
разрезании его.
У Peucedanum sativum описана мозаика, проявляющаяся
просветлением по ходу нервов и пятнистостью (Мс Whorler
и Owens, 1940).
У Coriandrum sativum и других зонтичных встречается
позеленение цветов. Согласно нашим данным, это заболевание
вызывается тем же вирусом, что и желтуха кок-сагыза, т. е.
Calistephus virus 1 или близким к нему вирусом.
У Ligusticum scoticum в Англии описано мозаичное
заболевание с некрозами, которое не передается ни моркови, ни
сельдерею, но легко передается целому ряду пасленовых.
У курительного табака вирус вызывает местные некрозы
и затем хлорозные кольца, а у N. glutinosa — местный некроз
и затем общую слабую мозаику и некрозы (К. Smith и Mark-
hem, 1944).
У N. silvestris вирус вызывает просвечивающие пятна,
а затем мозаику и некрозы. Местные некрозы и
концентрические кольца вирус вызывает также у перца и у махорки.
У огурца под влиянием вируса развивается нежная мозаика,
а у фасоли яркожелтые пятна. Болезнь легко передается
соком у пасленовых, но только с трудом у Umbeliferae.
Персиковая тля болезни не передает.
Вирус инактивируется при 60° С, выдерживает
разведение 1 : 100, но не 1 : 1000, в соке сохраняется 7 дней.
Глава X
ВИРУСНЫЕ БОЛЕЗНИ ПАСЛЕНОВЫХ
Целый ряд представителей семейства пасленовых, как
табак, помидор, картофель, принадлежит к числу растений,
наиболее чувствительных к разнообразным вирусам. Большем
части пасленовых свойственны одни и те же вирусы;
особняком стоит картофель. Многие вирусы, поражающие это
растение, не встречаются у других пасленовых; кроме того,
картофель занимает своеобразное положение вследствие биологии
его размножения. Поэтому мы считали более целесообразным
говорить о вирусных болезнях картофеля в особой главе,
а в этой главе рассмотреть вирусные болезни пасленовых,
исключая картофель.
Обыкновенная мозаичная болезнь табака. Это заболевание,
по классификации К. Smith, вызывается Nicotiana virus l,
Mayer (Allard). Мозаичная болезнь табака описана в 1885 г.
A. Mayer и была более подробно изучена Д. Ивановским
(1892—1902), Beijerinck и др. Вирус, вызывающий эту
болезнь, и самое заболевание принадлея^ат к числу наиболее
детально изученных.
Проявления болезни могут сильно отличаться, в
зависимости от того, какой штамм вируса табачной мозаики
вызывает поражение. Ниже мы приводим прежде всего описание
наиболее обычной и распространенной картины болезни
у важнейших пасленовых.
У табака болезнь вызывает мелкие или крупные светло-
зеленые пятна (рис. 23). Эти пятна подавлены в развитии,
тогда как происходит усиленное развитие зеленых участков,
которые иногда окрашены в более темный зеленый цвет,
чем нормальный лист, и нередко вздуваются пузыреобразно.
Растения могут быть подавлены в росте. Чашечка и плоды
также часто бывают мозаичными, венчик может быть более
или менее уродливым, и у табака с розовыми цветами он несет
многочисленные светлые пятна. При тяжелом поражении
цветы могут осыпаться. Kostoff (1936) наблюдал аномалии
01
редукционного деления при созревании пыльцы у табака,
больного мозаичной болезнью.
У махорки, кроме мозаики, наблюдается общий хлороз
и некротические явления; нередко происходит отмирание
верхушки стебля.
Рас. 23. Лист табака, пораженный мозаичной болезнью
(по К. Smith)
Заболевание помидора обыкновенной табачной мозаикой
проявляется сходно с проявлением этой болезни у табака.
Можно наблюдать различные поражения, начиная от легкой
крапчатости и кончая резкой мозаикой с сильным уродованием
пластинки листа. В. Л. Рыжков и И. К. Корачевский в 1934 г.
показали, что обыкновенная табачная мозаика может
вызывать явления нитевидности и папоротниковости листьев
помидора, которые раньше были описаны у помидора как
появляющиеся под влиянием огуречной мозаики. Некоторые
штаммы Nicotiana virus 1, как и у табака, могут вызывать
у помидора некроз листьев (Doolittle и Becher).
У синего баклажана проявление болезни сильно зависит
от сорта. Некоторые сорта являются только скрытыми вирус(V
носителями, другие дают более или менее слабую, легко
маскирующуюся мозаику; наконец, у некоторых сортов могут
наблюдаться некротические явления.
Перец [Capsicum апптигі) то лишь в слабой степени
страдает от вируса, развивая легкую крапчатость, то поражается
им тяжело — очевидно, в зависимости от штамма (N. Nakata
и S. Takimo, Oliveira, 1940).
Резкой мозаикой болезнь проявляется у петунии и у видов
Pkysalis. Многие декоративные виды табака страдают от ви-
руса подобно курительному табаку: например, N. silvestris,
N. affinis, N. longiflora. N,. glutinosa и Datura реагируют на
вирус только местными некрозами (рис. 24), Atropa
Belladonna устойчива к вирусу. Кроме пасленовых, вирус табачной
мозаики может поражать
представителей различных семейств,
например шпинат, однако
практически он встречается почти
исключительно у пасленовых.
Из важнейших штаммов
должны быть отмечены прежде
всего желтые штаммы, при
поражении которыми вместо светлозе-
леной наблюдается желтая и
иногда почти белая мозаика. Один
из таких желтых штаммов описан
под названием а у к у б а - м о-
з а и к а. Он широко
распространен в Англии у помидора.
Этот штамм вызывает у N. silvestris
местный некроз, тогда как
типичный штамм вызывает общую
инфекцию. Известны также
штаммы, сильно уродующие пластинку
листа. Один из них — enation —
вызывает на нижней стороне листа
помидора развитие добавочных РиСш 24. Лист Nicotiana glu-
пластинок. Этот штамм у табака tinosa с некрозами, вызван-
сорта White Burley вызывает мест- ными Nicotiana virus l
ные некрозы, за которыми может
наступить общая инфекция. но может и не наступить
(Ainsworth, 1937). Под влиянием некоторых штаммов бывает
иногда переход мозаики в некрозы. И. П. Худына считает
вообще характерным для мозаичной болезни табака развитие
некротических пятен на нижних листьях. Один штамм вируса
табачной мозаики может вызывать на плодах помидора, вблизи
их прикрепления к стеблю, светлые полосы до 2 мм в ширину,
которые позднее некротнзируются. Под влиянием этого штамма
на листьях и стебле некрозов не наблюдается (J. К. Jones.
1940). Описаны также штаммы вируса табачной мозаики,
которые могут вызывать некротические кольца ( о
разнообразии симптомов табачной мозаики см. у F. О. Holmes,
1932).
В патолого-анатомическом отношении заболевание
характеризуется резким недоразвитием светлых участков листа. При
болезни наблюдаются Х-тела и кристаллические включения.
Последние под влиянием 0.1 нормального раствора соляной
кислоты и других кислот распадаются на веретеиовидные образования,
93
похожие на паракристаллы очищенного вируса. М. И. Гольдин
(1937, 1938—1940) показал, что для разных штаммов могут
быть характерны кристаллические включения различных
типов. Игловидные кристаллы, складывающиеся в целые нити,
которые иногда петлеобразно закручиваются в клетке,
встречаются при поражениях аукубой и близкими к ней
штаммами.
Мозаичная болезнь табака и других пасленовых резко
снижает урожаи. Особенно страдает урожай при раннем
заражении. Количество и качество табачных листьев-
с больных растений снижены. Мозаичная болезнь табака
вызывает возрастание количества белка в листьях и уменьшение
количества углеводов, что само по себе неблагоприятно
отражается на качестве сырья. При заражении табака вскоре после
посадки, по данным И. П. Худыны, урожай снижается на
40—42%; при заражении в момент первой ломки — на 17—23%.
Как показали наши опыты в Крыму, произведенные
совместно с Корачевским, средний урожай с больного куста
помидора в два раза меньше, чем со здорового.
Вирус табачной мозаики имеет чрезвычайно широкое
распространение. Он встречается повсеместно и поражает
плантации табака и помидора; по большей части он на 100%
поражает помидор в условиях крытого грунта, а также широко
распространен среди декоративных пасленовых во всех парках
и садах.
Главным источником заражения вирусом обыкновенной
табачной мозаики является сок больных растений, а главным
распространителем этой мозаики является сам человек,
переносящий инфекцию при различных хозяйственных
манипуляциях: ломке табака или общипывании помидора. В эпифито-
логии болезни играет большую роль устойчивость вируса.
Он сохраняется на парниковых рамах, корзинах, в которых
переносится рассада, и т. д., а отсюда может попасть на
рассаду, вызывая ее заражение. Вопрос о роли почвы в
распространении болезни был предметом специальных работ. В
настоящее время можно считать установленным, что при
благоприятных условиях вирус может сохраняться в почве довольно
продолжительное время и даже зимовать, служа источником
заразы (J. Johnson, 1929). Вирус сохраняется в продажном
курительном табаке, поэтому, если такой табак попадает на
руки рабочего, то и это служит источником заражения, что
неоднократно было установлено экспериментально (Valleau
и Johnson Е., 1927). Вирус табачной мозаики через семена
табака, как показывают многочисленные опыты, не передается.
Вопрос о передаче при помощи семян помидора получил в
ряде работ противоречивое разрешение. Gardner и Kendrick
(1923), наблюдая десятки тысяч проростков помидора, не могли
констатировать передачи вируса семенами; в других же слу-
94
чаях такая передача наблюдалась, особенно, если посев
производился семенами, недавно вынутыми из больных плодов
(Doolittle, 1937; Milbrath, 1939). М. И. Гольдин показал, что
вирус находится не в эмбрионах, а на поверхности семян.
При помощи 1% раствора пикриновой кислоты, 1% раствора
марганцевокислого калия или 1.5% раствора едкого натра
вирус может быть удален с поверхности семян. Особенно
удобной оказалась 5-минутная обработка 1%-марганцевокисльш
калием. Такая обработка семян резко снижает процент
инфекции. Среди растений, выращенных из протравленных семян,
0.28% больных, а среди контрольных — 12.9%. Однако при
дальнейшем развитии растений, вследствие вторичной
инфекции, это различие нивеллируется, и в конце сезона, в условиях
крытого грунта, почти все растения оказываются больными.
Хотя R. W. Samson (1940) и нашел в экстракте из семян вирусу
однако не наблюдал передачи мозаики семенами.
С табака на табак вирус мозаичной болезни тлями не
переносится. Персиковая тля, повидимому, вообще не может
переносить вируса. Некоторые другие тли, как Myzus pseudo-
solani, M. circumflexus, Macrosiphum gei, могут переносить
вирус с помидора на помидор и с помидора на табак. Передача
вируса насекомыми совершается в гораздо меньшем проценте,
чем соком, Повидимому, при распространении болезни у
помидора может играть роль также табачный трипе, поражающий
цветы (Frimmel и Lauche, 1940).
Известны сорта табака, которые только скрыто несут
вирус, не проявляя болезни, например Ambolema. М. Ф. Тер-
новский (1941) и И. П. Худына (1936, 1941) путем междувид©-
вой гибридизации (N. glutinosa X N. tabacum) получили
хорошие сорта курительного табака, которые дают только местную
реакцию на вирус и являются, таким образом, вполне
устойчивыми к нему. Сорт Ambolema и междувидовая гибридизация
были использованы для получения устойчивых сортов табака
также в США (Valleau, 1942).
В борьбе с болезнью, кроме использования устойчивых
еортов, основное значение имеют следующие санитарно-
гигиенические мероприятия, подробно разработанные для
табака И. П. Худыной.
«1. Содержание табачного рассадника в течение
всего лета в состоянии черного пара, чтобы не допустить
развития мозаики на переросшей рассаде и заражения
остатками этих растений парникового субстрата.
2. Обеззараживание парникового субстрата паром
при 100° в течение 30 минут.
3. Обеззараживание всех парников и посадочных
принадлеяшостей: а) формалином (1:25), с выдерживанием
в кучах на протяжении 3 суток, или б) паром при 100^
9*
в течение 30 минут, или в) кипячением в воде (мелкий
инвентарь) в течение 10—15 минут.1
4. Запрещение курения на рассадниках и особенно
курения «потерти», что часто имеет место в колхозах.
5. Недопущение к уходу за рассадой, выборке
рассады и посадке табака лиц, работавших в :>тот день по
обработке (по пушовке и тюковке) табака, без
предварительного трехкратного мытья рук мылом и смены рабочих
костюмов.
6. Тщательное удаление и уничтожение мозаичной
рассады специально выделенными лицами накануне
выборки рассады из парников.
7. Введение табака в севооборот, при котором
устранить посадку табака по другим пасленовым или соседство
его с ними. Лучшими предшественниками табака в борьбе
с мозаикой следует признать колосовые.
8. Удаление с корнем табачных большей на участках,
предназначенных в следующем году тоже под табак,
во всех случаях, когда к концу ломок мозаикой будет
охвачено 5% и более растений.
9. Удаление мозаичных растений с корнем на участках,
где они встречаются единично. Удаление проводится
накануне каждой культивации и последний раз перед
первой ломкой.
10. Организация выборочных ломок на участках,
где мозаикой охвачено большое количество растений.
11. Выборочное вершкование и пасынкование
мозаичных и здоровых растений (отдельными рабочими),
чтобы не допустить при проведении этих работ широкого
распространения мозаики.
12. Обязательное трехкратное мытье рук водой с
мылом при переходе от работы с мозаичными растениями
к работе со здоровыми».
Мы провели эту длинную выписку из работы И. И. Худыны
(1936), так как предлагаемые им мероприятия, с известными
видоизменениями, могут быть применены во всех случаях
борьбы с передающимися соком вирусами.
Для борьбы с мозаичной болезнью помидора можно
рекомендовать, кроме этих мероприятий, обработку семян 1%-
марганцевокислым калием в течение 5 минут, которое
производится перед обработкой протравителем, обычно применяемым
в хозяйстве (М. И. Гольдин).
Е. В. Шатова (1941) для борьбы с мозаичной болезнью
помидора рекомендует также применять бордосскую жидкость
1 Для дезинфекции, эффективной по отношению к вирусу табачной
мозаики, предложены также щелок (Kohler, 1941), Na3P04 (Nei, 1940),
экстракты из растений, содержащие дубильные вещества, и т. д. (van
Schreven, 1941).
96
и другие химикалии, инактивирующие вирус. Е. В. Шатова
перед пикировкой рассады обрызгивала ее 1%-бордосской
жидкостью или 2%-известково - серным отваром или 1%
раствором кремнефтористого бария. Таким способом ей удалось
снизить заболеваемость мозаикой помидора и табака.
Огуречная мозаика пасленовых. Заболевание вызывается,
по классификации К. Smith, Cucumis virus 1, который более
детально описан в главе о вирусных заболеваниях тыквенных.
Внешняя картина болезни у табака и помидора мало
отличается от поражений вирусом табачной мозаики. Желтые
штаммы вируса вызывают желтую мозаику. Явления редукции
пластинки листа (нитевидность и папоротниковость листьев)
встречаются чаще, чем при табачной мозаике.
Кристаллические включения, характерные для табачной мозаики,
при заболевании отсутствуют. Некоторые штаммы могут
вызывать у N. tabacum (сорт Burley) местные некрозы кольцевого
характера (Phillips, 1939). Заболевание у пасленовых
встречается гораздо реже, чем обыкновенная табачная мозаика.
У помидора оно было подробно изучено Mogendorff (1930).
У этого вида огуречный вирус вызывает резко выраженную
нитевидность и папоротниковость листьев. При поражении
перца этим вирусом наблюдаются общий хлороз (причем
кончики листьев часто более темного зеленого цвета, чем их
основания), измельчание листьев, кустистость и
подавленность роста. У петунии, кроме мозаики и деформации пластинки
листа, вцрус может вызывать некротические явления. У N. glu-
tinosa вирус вызывает мозаику и редукцию пластинки листа.
Вирус с трудом передается соком, чем он резко отличается
от вируса табачной мозаики, и легко — тлями. Для борьбы
с вирусом главное значение имеет борьба с
насекомыми-переносчиками.
Пестрица табака. Вирус, вызывающий это заболевание,
по данным И. П. Худыны (1941), по серологическим
особенностям и другим признакам близко стоит к огуречному вирусу.
Наиболее ярким симптомом болезни являются мелкие
белые некротические пятна, округлые, угловатые или имеющие
вид полосок, прямых или дугообразно изогнутых. На нижних
листьях сначала появляются хлоротические пятна, которые
затем некротизируются по краю, так что получается
некротическое кольцо. В дальнейшем вся ткань внутри кольца
отмирает, и получается пятно. На верхних листьях появляются
многочисленные мелкие пятна, которые позднее
некротизируются. Листья приобретают более светлую окраску, и
нормально-зеленая ткань сохраняется только по ходу жилок.
При анатомическом исследовании можно видеть, что
некрозу подвергается, в первую очередь, палисадная паренхима,
причем клетки ее заполняются содержимым, которое
интенсивно окрашивается анилиновыми красками и совершенно
7 В. Л. Рыжков **7
закрывает нормально присущие клетке органоиды. Сходная
картина заболевания может быть получена у махорки (В.
Рыжков).
Вирус только с большим трудом передается соком;
искусственное заражение удалось только при натирании листа
вирусным соком с наждачной пылью или инъекцией этого сока
в пазуху листа. В естественных условиях болезнь передается
персиковой тлей, а также очень легко — семенами.
Вирус вызывает заражение при разведении 1:1000; инакти-
вируется при 60° С в течение 10 минут.
Для борьбы с заболеванием имеет большое значение
термическая обработка семян (см. В. Рыжков, «Основы учения
о вирусных болезнях растений», 1944, стр. 179 или у И. П. Ху-
дыны).
Кольцевые пятнистости пасленовых. Различные вирусы
могут вызывать у табака и других пасленовых хлоротические
или некротические кольца, которые часто бывают
концентрическими. Из этих заболеваний наиболее подробно изучена
широко распространенная кольцевая пятнистость табака,
описанная Fromme и др. в 1927 г.
Вирус, вызывающий это заболевание, описывается К. Smith
под названием Nicotiana virus 12, Fromme.
Первым признаком заболевания является развитие
некротического кольца, вокруг которого через несколько дней может
развиться второе кольцо, а затем третье. Ткань внутри кольца
остается зеленой или некротизируется.Кроме колец, на листьях
могут наблюдаться зигзагообразные хлоротические и
некротические линии. Иногда после периода резкого проявления
симптомов может наступить более или менее полное выздоровление,
причем растения, не проявляющие болезни, являются скрытыми
носителями вируса (рис. 25).
Вирус способен поражать представителей различных
семейств. У помидора заражение соком не удается, но болезнь
мояшо передать на него прививкой. После прививки на листьях
наблюдаются хлоротические кольца и полосы, которые могут
некротизироваться.
При болезни встречаются Х-тела, имеющие удлиненную
форму, зернистое содержимое и вакуоли. Кроме того, в клетках
обнаружены мелкие ромбические кристаллы.
Вирус передается соком лишь с трудом и не постоянно.
Способ распространения вируса в природе, несмотря на явную
большую инфекционность болезни, не выяснен. В целом ряде-
случаев установлена передача вируса семенами табака и
петунии.
Valleau (1932) описал желтую и зеленую кольцевую
пятнистость табака; вирусы, вызывающие эти пятнистости, являются,
повидимому, только штаммами описываемого здесь вируса.
Они вызывают общее посветление листа у табака, причем
98
зеленый штамм вызывает его светлозеленую окраску, а желтый—¦
желтую, а также некротические кольца. Все три вируса
способны взаимно предохранять друг от друга, почему и могут
рассматриваться как штаммы одного вируса.
Рис. 26. Различные проявления кольцевой пятнистости у табака
(по Price)
Вирус кольцевой пятнистости получен в очищенном виде.
Под названием Nicotiana virus 13, Price К. Smith
описывает вирус кольцевой пятнистости, очень близкий к
предыдущему. Впервые он был описан в 1936 г. Price. Nicotiana
virus 14, Smith также вызывает у табака кольцевую
пятнистость и близок к предыдущему, однако эти два вируса не
обладают взаимно-предохраняющим действием и не
предохраняют от Nicotiana virus 12- ттоптому их следует рассматривать
как особые вирусы
В оранжерейных условиях К. Smith и R. Markham (1944)
7* 89
нашли кольцевую пятнистость таоака, при которой кольца
часто носят прерывистый характер (tobacco broken ring spot
virus). Вирус часто вызывает местную реакцию в виде маленьких
светлых пятен и колец у N. tabacum, сорт Bnrley, Phaseolus
vulgaris, Cucumis, За местной реакцией следует общее заболе-
вание.
Перечисленные здесь вирусы отличаются между собой
и симптомами заболевания, которые они вызывают у разных
растений, и разным отношением к внешним воздействиям
(табл. 5 и 6).
У помидора также известны заболевания типа кольцевой
пятнистости. Одно из них описано Imle и Samson в 1937 г. При
отом заболевании наблюдаются коричневые некротические
кольца на листьях, некротические полосы на черешках и
стеблях, а также некротические кольца на плодах. Помидор может
выздоравливать, оставаясь скрытым носителем вируса.
Способность давать инфекционный иммунитет характерна для всех
вирусов рассматриваемой здесь группы. Вирус может заражать
различных представителей семейства пасленовых. Соком вирус
передается только с трудом, прививкой легче. Он может быть
передан дурману, картофелю, табаку, синим баклажанам,
Capsicum, Hyoscyamus, Nicandra physaloides, Solarium pseudo-
capsicum, S. carolense, Amarantus retroflexas, A. tricolor, Marty-
nia luisiana. Вирус очень близок к Lycopersicum virus 4, но
несколько более устойчив и не дает бронзовости листьев у
помидора. Вирус инактивируется при 56 — 58° С, выдерживает
разведение 1:500, в соке инактивируется за 27 часов.
Другая кольцевая мозаика помидора описана в 1930 г.
Valleau и Johnson. Ее вызывает Lycopersicum virus 2. Ее
симптомы: круглые водянистые и некротические пятна до 0.6 см
в диаметре на месте проникновения вируса; зараженный лист
постепенно отмирает; в дальнейшем наступает общее
заболевание, при котором также наблюдаются концентрические
желтые кольца и пятна, имеющие очертания листьев дуба.
Подобные же пятна наблюдаются на плодах. У табака
под влиянием болезни также наблюдаются некротические
кольца.
Вирус инактивируется при 80° С; он может сохраняться
годами в сухой ткани и проходит через свечу Ьг— L3.
Под названием редкая вирусная болезнь
т а б а к а в 1940 г. J. Johnson и Fulton описали еще один тип
кольцевой пятнистости табака, который характеризуется
более широкими кольцами, чем обычная кольцевая пятнистость,
и не предохраняет от обычной кольцевой пятнистости табака.
По мнению автора, этот тип кольцевой пятнистости близок
к описанному раньше у помидора. Вирус поражает горох,
огурец, фасоль и многие другие виды, принадлежащие к разным
семействам (J. Johnson и R. W. Fulton, 1942).
100
Таблица 5
Влияние внешних воздействий на вирусы кольцевых пятнистостей
В іі р у с
1° С
Nicotiana virus 12
Nicotiana virus 13
Nicotiana virus 14
Прерывистая кольцевая пятнистость
табака '. . .
Кольцевая пятнистость помидора . . .
Широкая кольцевая пятнистость
табака
Lycopersicum virus 2, YalleaunE.
Johnson
GO
60
80
GO
56-58
52
80
Условия инактивации
разведение
1 : 10 000
1 : 1 000
1 : 1 000
1 :100
1 :500
1 :750
сохранение
в соке
4 сут.
4 »
12 »
6 »
27 час.
36—48 час.
Поражение Х-вирусом пасленовых. Х-впрус более подробна
описан в главе о вирусных болезнях картофеля. Он является
весьма вирулентным для пасленовых, причем различные штаммы
его вызывают у табака заболевания различной силы, начиная от
маскированных форм и кончая формами, дающими очень
тяжелые некрозы. Впервые картины болезни у табака были
описаны J. Johnson (1925) под названием крапчатость
(mottle) и некротическая пятнистость (spot
necrosis), которые были вызваны.у табака соком внешне
здорового, но скрыто несущего вирус картофеля.
Заболевание у табака проявляется крапчатостью, которая
иногда светлозеленая, но чаще желтая. Пятна нередко носят
характер колец и могут некротизироваться. Болезнь обычно
проявляется не на самых молодых листьях, а по мере старения
листа.
У помидора заболевание проявляется желтой мозаикой,
обнаруживающейся опять-таки не у самых молодых листьев.
Согласно Kohler (1935), следует различать два типа X-
вирусов. Первый вызывает крапчатость у табака (X), а
второй— кольцевую пятнистость (Х2), которая, в отличие от
описанных выше кольцевых пятнистостей, может быть названа
картофельной кольцевой пятнистостью.
Эти две группы не могут взаимно предохранять друг от друга*
Они были подробно изучены Koch в 1933 г.
Х-вирус вызывает мозаику у дурмана и у N. glutinosa.
Последний вид тяжело поражается вирусом, и, кроме мозаики,
у него наблюдаются морщинистость листьев и некрозы.
Некротические иолосатости. Различные вирусы, как мы
видели уже выше, могут вызывать некротические полосы у
пасленовых, но для некоторых вирусов этот симптом особенно
характерен.
101
Таблица 6
Реакция разных растений на вирусы типа кольцевых пятнистостеи
Вирусы
Nicotiana virus
VI
Nicotiana virus
13
JV icotiana virus
l'i
Lycopersicum
virus 2, Val-
leau и
Johnson
N. glutinosa
Общая инфекция,
хлорозные
кольца
Не заболевает
Местные некрозы,
посветление по
нервам
Только местные
некрозы
Дурман
Общий некроз,
гибель
Некротические
пятна
Общая инфекция,
мозаика
Местные некрозы,
редко — общая
инфекция
Помидор
Общая крапча-
тость
Общая инфекция,
некротические
кольца
Нежная
пятнистость, сетчатый
некроз
Хлорозные,
некротические
кольца и
полоски
Некротические полосатости, обозначаемые обычно
американскими и английскими авторами как streak, наблюдаются
прежде всего под влиянием смешанной инфекции. При
смешанной инфекции Х-вируса и обыкновенного табачного вируса
у табака (рис. 26), а также у помидора развиваются тяжелые
некротические явления. Некрозы у помидора наблюдаются не
только на листьях и стебле, но и на плодах. Поражения плодов
делают болезнь особенно вредоносной. Valleau и Е. Johnson
изучали влияние на помидор различных комбинаций вирусов.
X-вирус вызывает некрозы не только при соединении с табачным
вирусом, но также и с огуречным, с вирусом гравировки и
вирусом кольцевой мозаики.
Кроме двойной некротической мозаики, как можно
назвать полосатость, вызванную соединением двух вирусов, можно
различать полосатости, которые вызваны одним вирусом.
Boning в 1928 г. описал вирус, обозначаемый К. Smith
как Nicotiana virus 5. Он вызывает мозаичность верхних листьев,
за которой следует некроз молодых листьев и стебля. Этот
вирус передается на махорку, но его не удалось передать на
картофель и томат. Он описан только в Баварии.
Более подробно описана полосатость табака в 1936 г.
J. Johnson. Вирус, ее вызывающий, обозначается Nicotiana
virus 8. Болезнь проявляется посветлением по ходу нервов
и некротическими симптомами, причем мозаики обычно не
наблюдается. Некрозы имеют вид пятен, полосок и колец,
причем они сосредоточены, главным образом, в нижней части
листа.
102
Болезнь может поражать также помидор, картофель,
синий баклажан, перец и дурман. Она передается соком, но
не насекомыми. Вирус инактивируется при 53° С, не выдер-
Рис. 26. Streak табака (по Quanjer)
живает разведения 1:30 и разрушается в соке через 24—36
часов.
Большое значение^ для помидора крытого грунта имеет
волосатость, вызванная особым вирусом, который описан у
К. Smith под названием Lycopersicum virus l, Bewley и
подробно изучен Ainsworth (1933). Этот вирус вызывает у
помидора крапчатость, уродование листьев, некоторую
подавленность роста, некротические полосы на стеблях и листьях, а
также некротические пятна на плодах.
У N. glutinosa, Datura stramonium N. tabacum вирус
вызывает только местные некрозы.
Вирус инактивируется при 90° С, может годами сохраняться
в соке и проходит через фильтр Шамберлена Lx — L8; по своим
свойствам очень близко стоит к Nicotiana virus l, Allard и,
вероятнее всего, является одним из его штаммов.
Е. А. Осницкая (1936) подробно изучала полосатость
у помидора в подмосковных хозяйствах. Она нашла, что это
юа
заболевание, иногда сильно распространяющееся и приносящее
большой убыток, может передаваться соком, а также семенами.
Вирус инактивировался при 70—75° С и реагировал с
антисывороткой к Y-вирусу. Согласно нашим наблюдениям, в
подмосковных хозяйствах встречается полосатость, вызванная
вирусом, сходным с Lycopersicum virus 1. Этот вирус вызывал
местные некрозы на листьях N. tabacum и TV", glutinosa и
инактивировался только при температуре выше 80° С.
Присутствия Х-вируса при этой полосатости нами не обнаружено,
и она вызывается, очевидно, одним вирусом. Вероятно,
встречаются также другие типы некротических мозаик.
Некротические полосатости оранжерейного помидора очень
сильно зависят от внешних условий развития. В последнее
время для их контроля рекомендуются понижение влажности
оранжерейного воздуха, хорошая вентиляция и хорошее
удобрение растений, в особенности калийное (Selman, 1943).
Вронзовость помидора. По классификации К. Smith,
заболевание вызывает Lycopersicum virus 3, Brittlebank. Этот
вирус первоначально был описан в Австралии и в настоящее
время известен в различных странах, в том числе в СССР
(Е. М. Эристави, 1941). Английское название заболевания
spotted wilt (см. подробное описание болезни у Samuel, Bald
и Pittman, 1930).
Болезнь обнаруживает себя бронзовостью листьев, которая
поражает уже молодые листочки и развивается обычно на
верхней стороне; однако, если- лист закручен вверх, то она
развивается на морфологически нижней стороне. Вронзовость
связана с действием солнечного света, окраска колеблется от
едва заметной бронзовости до вполне черного окрашивания.
При очень слабом проявлении болезни бронзовый налет на
листьях не появляется, а имеются только слабая морщинистость
листьев и светлые кольцеобразные пятна. Рост больных растений
резко подавлен, главная точка роста часто погибает, и тогда
обычно развиваются боковые побеги. Только сравнительно
редко болезнь совсем убивает растение.
У табака вирус вызывает концентрические кольца более
или менее крупные или мелкие. Нередко болезнь вызывает
общий некроз и быструю гибель растения. У дурмана также
наблюдаются концентрические кольца под влиянием
заболевания, а также пятна, напоминающие очертания дубового листа.
У петунии вирус вызывает через 48—72 часа местные
кольцеобразные некротические пятна. Общая инфекция у этого
вида наблюдается редко. N. glutinosa очень чувствительна
к вирусу и дает местные некрозы уже на четвертый день; они
постепенно увеличиваются в размерах, образуя концентрические
кольца.
Вирус может поражать многочисленных представителей
различных семейств, как, например, Delphinium Sp., Papaver
104
nadicaule, Plantago major, Tropoeolum mafus, Begonia, Phaseolus
vulgaris, Pisum sativum, Callistephus chinensis, Cineraria,
Dahlia, Zinnia elegans, Primula, Campanula, Gloxinia, Hip-
peastrum.
Этот вирус очень широко распространен у декоративных
растений, которые могут быть источником для заражения
помидоров. В Харьковском ботаническом саду мы наблюдали
однажды развитие поражения помидора бронзовостыо листьев.
Эти помидоры стояли в глиняных вазонах недалеко от далий,
которые были поражены мозаикой. Бронзовость листьев
зависит от некроза, которому подвергается эпидермис листа.
Из эпидермиса некроз может распространяться вглубь.
Заболевание, вызываемое рассматриваемым здесь вирусом,
было описано под разными названиями. Сейчас можно считать,
что diaback помидора, описанная К. Smith (1931), кольцевая
пятнистость Solanum capsicastrum, а также африканская коль-
певая пятнистость табака (kromnek) идентичны с
бронзовостыо помидора. Желтая пятнистость (yellow spot) ананаса
также вызывается этим вирусом.
Переносчиком болезни являются различные виды трипса,
как то: Frankliniella insularis, F. occidentalis, F. moultoni,
Trips taeaci.
Болезнь может передаваться только насекомыми,
питавшимися на больных растениях уже в личиночной стадии.
Инкубационный период в насекомом длится не менее 5—6 дней.
Данные по эпифитологии болезни см. у Gardner, Tompkins
и Thomas (1937); об устойчивых видах помидора — у Wein-
holz (1940). Для борьбы с насекомыми-переносчиками
рекомендуется рвотный камень или парижская зелень (С. Mage и
W. Morgan, 1942).
Инокуляция соком — дело капризное и не всегда удается;
сильно увеличивает успех этой операции прибавка к соку
карборунда. У Lycopersicum pimpinellifolia встречаются
устойчивые к вирусу линии (P. G. Smith, 1944).
Вирус бронзовости принадлежит к числу самых
малостойких. Он не выдерживает высушивания, при комнатной
температуре в соке он обычно утрачивает активность в
несколько часов; быстро инактивируется окислителями. Из всех
фитопатогенных вирусов он самый нестойкий к действию
высокой температуры, подвергаясь инактивации при 42° С, однако
выдерживает разведение до 1:100 000.
Под названием tomato top-blight в США описано
заболевание томата, при котором наблюдаются некрозы на листьях,,
стеблях и плодах, а также отмирание верхушки. У табака
чаше всего вирус вызывает только местные некрозы, иногда
и на молодых листьях. У дурмана вирус вызывает
некротические пятна и отмирание листьев. Это заболевание очень
похоже на бронзовость. Вирус, вызывающий его, с трудом
104
лередается соком, перепосчиком его является табачный трилс.
Вирус инактивируется при 40—41.5° С, утрачивает инфек-
дионность в соке менее чем в течение одного часа и становится
неинфекционным уже при разведении 1:50. Вирус, однако,
не тождествен с вирусом бронзовости, так как не предохраняет
от этого последнего (J. Milbrath, 1939).
Гравировка табака. По классификации К. Smith, эту
болезнь вызывает Nicotiana virus 7, Е. Johnson (1930). Болезнь
описана под названием etch, что значит гравировка. На
молодых листьях заболевание обнаруживает себя маленькими
хлоротическими пятнами, появляющимися между нервами. В
дальнейшем пятна некротизируются. При развитии болезни
листья несут многочисленные мелкие некротические кольца и
некротические узоры разной формы; иногда некроз
распространяется по ходу нервов.
Сходная картина болезни наблюдается у N. rustica, N.
alata, N. langsdorfii. Datura stramonium,, Hyoscyamus niger.
У N. glutinosa, Hyoscyamus — хлороз и уродливое развитие
верхних листьев, а также некрозы по жилкам. На листьях
ломидора наблюдается мозаика, а на плодах желтые
концентрические кольца. В полевых условиях симптомы лучше
проявляются ранней весной и осенью, чем летом. У Physalis peruviana
вирус вызывает местные некрозы (Holmes, 1942).
Solanurn nodiflorum и S. melongena иммунньт к вирусу.
При заболевании в ядрах развиваются кристаллические
включения. Описаны гранулярные Х-тела в цитоплазме, которые
распадаются на игловидные кристаллы от 2 цдо 10 и в длину.
Согласно Kassanis (1941), вирус переносит тля Myzus
persicae. По данным F. Bawclen и В. Kassanis (J941), вирус
инактивируется при температуре в 54° С (слабый штамм) и
температуре 58° С (суровый штамм), выдерживает разведение 1:5000
В соке слабый штамм инактивируется через 8 дней, суровый —
через 13 дней. Заражение суровым штаммом гравировки
предохраняет от Y-вируса и Ну-3 вируса.
Е. Johnson (1930) различает также грубую гравировку
(coarse etch), при которой наблюдаются крупные некротические
кольца, однако мельче тех, которые наблюдаются при
Nicotiana virus 12.
Некроз табака. Сюда относятся поражения, вызываемые
Nicotiana virus 11, Smith (1935 а, 1937 а). Этот вирус
занимает совершенно исключительное положение. В естественных
условиях он лишь в очень редких случаях вызывает заражение
табака. Обычно он накопляется в корнях различных видов,
что внешне никак не отражается на растении. Однако, если
натереть листья соком корней, в которых находится вирус,
то на них развиваются многочисленные некрозы; с этих листьев
вирус может быть передан на новые и т. д. У табака при
естественной инфекции наблюдаются некрозы по ходу жилок ни-
106
жних листьев. Молодые растения могут быть убиты некрозами,
тогда как у старых дело обычно ограничивается отмиранием
нижних листьев и растение в дальнейшем выглядит нормально.
Вирус вызывает местные некрозы у N. taeacum, N. gluti-
nosa, Phaseolus vulgaris, Raphanas sp., Spinacia oleracea,
Cardans sp., Sonckus oleraceus.
У Polygonum, кроме местных некрозов, изредка возникают
отдельные некротические очаги и на вновь развившихся листьях,
что зависит от случайного распространения отдельных частичек
вируса по растению.
Вирус обнаружен в корнях: Pelargonium zonale, Lycoper-
ъісит esculentum, Primula овсопіса и др.
Вирус обычно передается почвой, в которую он попадает
или с пылью из воздуха или же с водой из чанов, в которых
хранится вода для поливки оранжерейных растений.
Вирус получен в чистом виде. Он образует пластинчатые
кристаллы. Устойчив к высушиванию, сохраняется в соке
не менее 12 дней, инактивируется в соке при 72° С, а в сухом
виде — только при нагревании до 100°. Bawden и Рігіе (1942)
выделили в чистом виде шесть разных вирусов, вызывающих
некроз, из которых только два серологически идентичны.
Отношения родства между этими вирусами не выяснены.
Едва ли этот вирус имеет большое экономическое значение,
однако он чрезвычайно интересен в теоретическом отношении.
Курчавость листьев табака. Болезнь известна также под
названиями: kroepoek, Faltenzweig, curl, gila, crinkle-disease,
rrinkle-dwarf. Ее вызывает Nicotiana virus 10, Storey (1931,
1932, 1935). При заболевании происходит утолщение жилок
листа, которые разнообразно искривляются, придавая всей
пластинке уродливый, измятый вид. Очень часто на нижней
•стороне листа вдоль жилок развиваются вторичные пластинки,
достигающие 1.5 см в длину и имеющие нередко форму асцидий.
Болезнь может быть передана махорке, TV* glauca, а также
зі помидору. Кроме того, заболевание это наблюдается у целого
ряда представителей не-пасленовых, например: у Zinnia ele-
gans, Vernoca sp., Althala, Cueumis sativa, Ageratum conyzoides
и др. (Pruthi, 1939). В патолого-анатомическом отношении
заболевание характеризуется гипертрофией флоэмы и пери-
цикла, причем в перицикле может закладываться новый
цилиндр, имеющий свою флоэму и древесину.
Болезнь не передается соком, но легко передается
прививкой, а также белой мушкой Bemisia gossypiperda из
семейства Aleyrodidae. Заболевание встречается в Индии, Южной
Африке, на Яве, Мадагаскаре, в Занзибаре, в Румынии, а
также в небольшом количестве в СССР.
Matsumoto (1938) описывает на о. Формоза заболевание,
-очень похожее на курчавость листьев; при нем наблюдаются
карликовость, уменьшение размера листьев, хрупкость их
107
и закручивание вниз, а также утолщение жилок листа. Пови-
димому, болезнь передается паутинным клещиком.
Совместно с Михайловой и Пивоваровой (1936) нами было
описано своеобразное заболевание помидора, которое, может
быть, близко стоит к курчавости листьев табака. При этом-
Рис. 27. Курчавость листьев помидора; слева —
пораженный побег, справа — здоровый
заболевании пластинка листа закручивается вниз таким образом;
что край пластинки плотно прижимается к нижней стороне
листа. Все растение выглядит очень ненормально: толстые,
сильно одеревеневшие стебли сплошь покрыты прилегающими
к ним, из-за закручивания черешков, листьями (рис. 27).
При заболевании наблюдаются гипертрофия флоэмы и развитие
в ней древесины. Болезнь передается прививкой.
Желтухи. Beta virus 1, вызывающий курчавъсть верхушки
у свеклы, поражает также пасленовые. У помидора при
заболевании наблюдаются пожелтение листьев, их огрубение и
закручивание вверх. Растения антоцианируют, останавливаются
в росте и накопляют в вегетативных частях крахмал (Shapova-
fov, 1930).
Внешне сходное заболевание описывает Ивахненко близ
Харькова. У заболевших растений развиваются общий хлороз,
хрупкость листьев и подавленность роста. Причины заболевания
остаются невыясненными.
Под названием psyllid yellow описано заболевание
помидора, при котором наблюдаются хлороз, антоцианирование
108
и скручивание листьев. Болезнь вызывается пснллидами и
после их удаления проходит. Повидимому, она не является
инфекционным заболеванием (Blood, Richards, Wann, 1933).
Столбур. Заболевание также называется у табака мокрый
м о н т а р ь. Различные английские названия заболевания —
big bud, tomato fruit "woodiness.
По классификации К. Smith, его вызывает Lycopersicum
virus 5, Samuel, Bald и Eardley.
Болезнь давно известна в Австралии и в СССР. Первое
описание заболевания как вирусного было одновременно
сделано в СССР Рыжковым, Корачевским и Михайловой и в
Австралии — Samuel, Bald и Eardley (1933). Следует отметить,
что в Крыму Горбань правильно распознал столбур как
вирусное заболевание еще в 1931 г., хотя и не привел
экспериментальных доказательств в пользу этого. К сожалению, его подробные
и для своего времени чрезвычайно интересные исследования
остались неопубликованными.
Болезнь обычно появляется на плантации в конце июля.
У пораженного помидора происходит посветлеиие
развивающихся после заражения листьев. Эти листья хлорозны,
антоцианируют, развертываются медленнее, чем нормальные,
п их доли сильно измельчены (рис. 28). Пораженные листья
утолщены и хрупки. Очень характерны изменения цветов —
они подробно описаны в общей части этого руководства. У
больных растений можно видеть различные формы разрастания
чашечки (которая часто остается сростнолистой и пузыре-
образно раздувается), редукции венчика и тычинок и распада
на листочки пестика, а также пролиферации оси цветка.
Плоды, развившиеся после заболевания, ребристы даже у тех
сортов, которым в норме это несвойственно, деревенеют и
лишаются вкуса.
У табака болезнь мало заметна на вегетативных органах.
Поражение столбуром у этого растения мы впервые узнаем
по характерным изменениям соцветия, которое обнаруживает
кустистость и недоразвитие цветов (рис. 29). Вместо нормальных
цветов табака имеются мелкие цветочки, внутренние круги
которых совсем не видны. При внимательном изучении мы
обнаруживаем в этих цветах различные стадии позеленения и
пролиферации. Листья больного табака только с трудом высыхают.
В тюки они обычно попадают не до конца высохшие, легко
плесневеют и загнивают, способствуя порче находящихся
вместе с ними и нормальных листьев; в этом-то и состоит
главный вред столбура для табачной промышленности. У табака
в Австралии болезнь описана А. V. Hill (1937).
Синий баклажан также серьезно поражается столбуром,
причем наблюдаются недоразвитие плодов, трещины на них,
а также позеленение цветов и бесплодие.
Корачевскому в 1941 г. не удалось заразить картофель
1?'*>
Рис. 28.. Столбурный помидор
столбуром. Однако, согласно В. Э. Понер (1941), можно
произвести это заражение. Возможно, что одни сорта устойчивы
к столбуру, а другие им болеют. Сухов и Вовк (1945)
описывают в Краснодарском крае увядание картофеля столбур-
ного происхождения.
Рис. 29. Соцветие табака: нормальное —слева и столбурное—справа
Из других пасленовых столбур широко распространен
у дурмана и у белладонны. Симптомы столбура и здесь
состоят в измельчании листьев, переполнении их крахмалом
и характерном поражении цветов.
Судя по симптомам, столбур широко распространен у
обычного сорняка помидорных плантаций в Крыму — вьюнка.
Больной вьюнок имеет измельчавшие листья, укороченные
побеги и редко цветет. Если у такого вьюнка развиваются
цветы, то они характерным образом изменены: чашечка
разрастается, внутренние круги подавлены и наблюдается
пролиферация. Экспериментально передача столбура на вьюнок
удалась И. П. Худыне.х
При столбу ре наблюдается гипертрофия флоэмы.
Сосудисто-волокнистые пучки плодов, пораженных столбуром.
содержат увеличенное количество механических элементов.
1 Сообщение в письме.
Ш
Гипертрофия п одеревенение этих пучков в плодах легко
видны даже невооруженным глазом.
Столбур ¦— типичная желтуха: при нем наблюдаются
переполнение зеленых частей растения углеводами и уменьшение
количества общего п белкового азота. Количество алкалоидов
у больной белладонны уменьшается.
Столбур легко передается прививкой. Инкубационный
период длится 18—45 дней, в зависимости от вида растения
и различных других условий. На плантации очень часто
встречается смешанная инфекция столбура с мозаичной болезнью
табака. Соком столбур не передается, не удалось установить
также передачи его почвой или семенами. Насекомое —
переносчик столбура в Краснодарском крае—цикада Hyalesthes
obsaletus (Сухов и Вовк, 1946).
Согласно А. V. Hill (1943), в Австралии big bud
передается цикадой Thamatettix argentaia и может, кроме
пасленовых, поражать виды, принадлежащие к различным другим
семействам, что для Convolvulus arvensis впервые установлено
нами. Передача болезни цикадой, которая является также
переносчиком желтухи сельдерея, а также широкое
распространение столбура среди разных семейств сближают вирус
столбура с Calistephus virus 1. Как далеко идет эта близость,
покажут дальнейшие исследования! Повидимому, картину
столбура в разных случаях вызывают различные вирусы, может
быть, иногда более или менее родственные между собой. Об
идентичности столбура с желтухой астр, вопреки Сухову и Вовку
(1945), говорить невозможно, т. к. вирус желтухи астр не
вызывает у помидора столбура.
Столбур развивается во второй половине лета. В Крыму
впервые стол бурные растения чаще всего можно наблюдать
в конце июня и начале июля. Вслед за появлением столбура
обычно вскоре появляется большое количество больных
растений, позднее болезнь распространяется гораздо медленнее.
На одной плантации помидора под Симферополем с 20 июля
по 10 августа заболело 5369 кустов помидора, а с 10 августа
по 20 сентября — только 2429 кустов.
Несомненна связь развития болезни с экологическими
условиями. Она распространена в южной части нашей страны.
Харьковская область, повидимому, является уже северной
ее границей, и здесь заболевание встречается редко и в
небольшом проценте. В различных частях Крыма столбур
распространен не одинаково: он сильно свирепствует в степной
части Крыма и на восточном побережья, а на южном берегу
встречается лишь изредка.
Существуют указания на то, что плантации помидора
в -фруктовых садах почти не поражаются столбуром, однако
О. Н. Вертоградова наблюдала сильное его развитие и в этих
условиях.
112
Существует мнение, что столбур связан с засухой —
с ксерофитными условиями существования. Это не совсем
верно. По данным И. К. Корачевского, усиленная поливка
помидора не предохраняет его от столбура и даже, наоборот,
способствует более сильному его развитию. О. Н. Вертоградова
(1941) считает, что в условиях искусственного орошения
столбур, как правило, появляется прежде всего в наиболее
орошаемых участках. Наблюдения за целый ряд лет показывают,
что столбур в Крыму особенно сильно развивается в годы
наиболее резкого перехода от влажной первой половины лета
к большим засухам в начале июля.
По нашему мнению, экологические условия в развитии
столбура играют роль главным образом вследствие влияния
их на насекомое-переносчика. Наша гипотеза такова: столбур
переносится насекомым, которое первую часть лета развивается
в степи на диких растениях; вместе с наступлением засухи
и выгоранием диких трав насекомые переселяются на помидор,
перенося с собой столбур. Чем резче контраст между
влажностью первой части лета и засушливостью второй, тем резче
контраст между выгорающей травой и прекрасным развитием
листьев у культурных растений, тем сильнее привлекательность
этих последних для насекомых. В условиях более влажной
второй части лета стимул для миграции насекомых слабее,—и
меньше заболеваний столбуром. Миграция должна происходить
в самом начале июля; насекомые, заселяющие плантацию
помидора, повидимому, скоро вымирают. Если помидор
высадить в грунт в июле, то обычно он почти не заболевает
столбуром.
Вопрос о борьбе со столбуром был предметом
многочисленных исследований И. К. Корачевского, А. И. Серебрякова,
П. И. Сербинова, Ю. М. Райдмана и др.1
Так как столбур появляется во второй половине лета,
то все меры, направленные к более раннему получению урожая,
содействуют ограждению значительной его части от столбура.
Следует поэтому рекомендовать возможно более ранние и
сжатые сроки высадки, а также введение самых ранних сортов
помидора. В этом отношении в Крыму хорошие результаты
дал сорт «Бизон».
Установлено, что мульчирование плантаций соломой сильно
снижает процент столбура. Мера эта, к сожалению, по
различным причинам мало применима в хозяйственных условиях.
Применялись также различные приемы затенения помидора.
Экранирование помидора бордосской жидкостью может
способствовать снижению процента заболеваний столбуром, но
особенно эффективно загущение посадок. В одном опыте в Крыму
с помидором сорта «Чудо рынка» было найдено, при площади
1 См. «Вирусные болезни растений и меры борьбы с ними», М., 1941.
^ В. Л. Рыжков 11&
питания 100X60 см, 46% столбурных растений, и валовой
урожай, при пересчете на гектары, равнялся 37.5 т, а при
площади питания 70X^0 см — 24% столбурных растений, и
урожай на гектар равнялся 47.3 т. Подобный же эффект дает
загущение посадок синего баклажана.
Относительно устойчивы к столбуру некоторые сорта
помидора из группы Vallidum, например «Стоит». Синий
баклажан «Болгарский № 14» очень устойчив к столбуру, тогда
как его селекционный сорт
7/14, устойчивый к
увяданию, болеет стол буром
наряду с помидором. По
данным Шмидта, тетраплоидныіі
синий баклажан болеет
столбу ром в меньшей мере, чем
диплоидный. По выведению
сортов, устойчивых к
столбуру, еще необходима
большая работа.
В Калифорнии описано
заболевание помидора,
чрезвычайно похожее на стол-
бур. Подобные же симптомы
наблюдаются в Калифорнии
у целого ряда других
культурных растений: сои,
люцерны, тыквы и моркови
(Dana, 1940). В настоящее
время неизвестно, имеем ли
мы в этих случаях дело со
столбуром. В Индии
наблюдается мелколистость синего
баклажана,
сопровождающаяся позеленением цветов.
Эта болезнь прививкой
передается на дурман, табак и
помидор. Судя по
фотографиям, приводимым Thomas
и Krishnaswarii (1939), она
тождественна со столбуром
или, по крайней мере, стоит
Рис. 30. Часть листа помидора с
поражениями, вызванными Ly coper-
sicum virus 4 (по Ainsworth)
к ней близко. Заболевание с непостоянным успехом удалось
передавать при помощи цикад Empoasca devastans, Eutettir
phycotis. -
Кустистая карликовость (bushy stunt). Возбудитель болезни
Lyсо per si cum virus 4, Smith.
Болезнь описана впервые в 1935 г. У помидора после
заражения наблюдается местная некротическая кольцевая
114
пятнистость. Зараженный лист обычно вскоре желтеет и
отмирает; в дальнейшем развивается общее заболевание, которое
проявляется желтыми кольцевыми пятнами. Эти пятна могут
Рис. 31. .Задержка роста у помидора,
которая вызвана вирусом bushy - stunt (no
Ains worth)
приобретать красную окраску и некротизироваться/: Иногда
светлые кольца наблюдаются также на плодах. Нижние листья
постепенно отмирают. Нередко погибает точка роста, вслед
за чем обычно развиваются боковые побеги, придающие
растению кустистый розеточный вид (рис. 30, 31). При заражении
очень молодых растений некротические явления могут быть
выражены еще сильнее; иногда наблюдаются некроз стебля
вблизи его основания и гибель всего растения.
У N. tabacum и N. glatinosa вирус обычно вызывает только
местные симптомы в виде некротических концентрических
колец. Заболевание описано в Англии, а в других странах
пока неизвестно. Болезнь передается соком. Насекомые —
переносчики болезни, а также возможность передачи ее
семенами неизвестны.
8*
115
Вирус, вызывающий заболевание, легко выделяется в
чистом виде и образует настоящие кристаллы. Этот вирус
ішактивируется при нагревании до 80° С, становится неинфек-
циониым в соке через 33 дня, причем ослабление его инфек-
цнонности происходит значительно раньше. При тщательном
высушивании вирус теряет активность.
Кустистость верхушки помидора. Болезнь, названная
по-английски bunchy top, вызывается Lycopersicum virus 6,
Мс.Clean (1935).Болезнь описана вТрансваале в1926г.(Мс.С1еап,
1935). У помидора она выражается тем, что в молодых,
нарастающих частях растения задерживается развитие междоузлий,
вследствие чего листья оказываются на верхушке растения
сближенными, прикрывающими друг друга. Пластинки листьев
имеют сильно измельчавшие доли. Нередко наблюдаются
некрозы на стебле и листьях. После периода подавленности
в росте стебель опять приобретает способность удлиняться
более или менее нормально, но первоначально укороченная
часть так и остается короткой, и новый стебель значительно
тоньше нормального. Придаточные побеги усиленно
развиваются. Наблюдается переполнение тканей крахмалом.
К вирусу чувствительны различные пасленовые, в том
числе: перец, синий баклажан, табак, петуния, физалис и ни-
кандра.
Вирус передается соком. Он инактивируется при 70° С,
а иногда и при более низкой температуре; инактивация вируса
в соке происходит в 12—24 часа.
О. Н. Вертоградова наблюдала в Саратовской области
заболевание, внешне очень похожее на эту болезнь, или же
на bushy stunt, описанную выше.1
Мало изученные вирусные болезни пасленовых. У
пасленовых описано большое количество вирусных и подозрительных
на вирусные заболеваний, которые либо до сих пор точно не
идентифицированы, либо были встречены только один раз.
Приводим здесь перечень этих заболеваний.
Прежде всего надо остановиться на заболеваниях, которые
были очень коротко описаны J. Johnson в 1927 г. в его
классификации вирусов табака. Это целый ряд болезней табака,
свойства вирусов которых автор описал довольно подробно,
но симптомы которых были им описаны только очень бегло.
Большая часть этих заболеваний в дальнейшем не встречалась.
Они таковы:
Пятнистая мозаика (speckled mosaic.) По K.Smith, ее
вызывает Nicotiana virus 2, Johnson. При болезни наблюдается
пятнистость листьев табака; у петунии вирус вызывает, кроме
того, подавленность роста, уродование листьев и некрозы;
у перца Physalis никаких симптомов не наблюдается. Вирус
1 Сообщение в письме.
11G
инактивируется при 90° С, сохраняется в соке несколько
месяцев, к азотистой кислоте менее устойчив, чем Nlcotiana virus l.
Мягкая мозаика табака (mild mosaic). Это
заболевание вызывает Nicotiana virus 3 (Johnson, 1937). У табака
наблюдается слабая крапчатостъ, так же как и у махорки.
У N. glutinosa наблюдаются слабый хлороз и подавленность
роста. Физалис и N. glauca не поражаются вирусом. Вирус
разрушается при 60° С и утрачивает инфекционность через
6 дней хранения в соке.
Белая мозаика (bleaching mosaic). По К. Smith, ее
вызывает Nicotiana virus 4 (Johnson, 1937). Этот вирус был
выделен от перца, у которого на листьях наблюдались белые и
кремовые участки: подобные же явления наблюдались у табака.
Пораженные участки могут позднее некротизироваться, и
старые листья отмирают. Вирус инактивируется при 75° С и
после 3 дней хранения в соке.
Мс. Kinney (1929) во время своего путешествия на
Канарские острова изолировал Nicotiana virus 6 из табака,
пораженного зеленой мозаикой. Вирус легко передается соком и
устойчив к высушиванию. Он получен в очищенном виде, и его пара-
кристаллы мало отличаются от паракристаллов обыкновенного
вируса табачной мозаики, с которым он чрезвычайно сходен.
Различие между этими вирусами состоит в том, что Nicotiana
virus 6, Мс. Kinney не иммунизирует табак против желтого
штамма Nicotiana virus 1, а также в тем, что вирус,
происходящий с Канарских островов, по имеющимся данным, не передается
на помидор. Нам представляется не вполне установленным
существование этого вируса. Возможно, что он является только
одним из штаммов вируса табачной мозаики, так как разные
штаммы не всегда взаимно предохраняют друг от друга.
Как средняя мозаика (medium mosaic) табака
описано заболевание, по силе симптомов промежуточное между
обыкновенной табачной мозаикой и мягкой мозаикой.
Отношение к внешним факторам у вируса такое же, как у вируса
обыкновенной табачной мозаики. J. Johnson (1927) описал
отот вирус как Tobacco virus 7, однако, вероятно, он
является только штаммом вируса обыкновенной табачной мозаики.
Некроз стебля томата (tomato stem necrosis, J.
Johnson, 1927), вирус которого у томатов вызывает отмирание
тканей стебля, а у табака слабую пятнистость. Он устойчив
к нагреванию, хранению в соке, как вирус обыкновенной
табачной мозаики, но менее устойчив к азотистой кислоте.
Возможно, что целый ряд других, описанных в качестве
самостоятельных, вирусов являются только штаммами вируса
табачной мозаики 1. Сюда следует отнести Nicotiana virus 9,
Jochems (по классификации К. Smilh), вызывающий
Роттердамскую болезнь у табака, которая проявляется светлозелеными
и желтыми участками, быстро отмирающими. Наблюдаются
И"
также некрозы в стебле. В отличие от вируса обыкновенной
табачной мозаики вирус этой болезни лишь с трудом проходит
через фильтр Беркефельда (Jochems, 1928).
Под названием желтая карликовость (yellow
dwarf) в Австралии описано заболевание табака, при котором
наблюдаются карликовость и пожелтение растений. Болезнь
передается прививкой, но не соком. Переносчиком болезни
является Thamnotettix argentata (Hill, 1941). Биология этого
переносчика изучена J. Johnson (1939).
Helson (1942) описывает коричневую корневую
гниль табака. При этом заболевании наблюдаются
некрозы периферических, а иногда и глубже лея?ащих тканей
корня. Болезнь передается почвой, причем заразное начало
нельзя из нее экстрагировать. Оно инактивируется при 45° С
в 24 часа. При медленном высушивании оно разрушается,
а при быстром не инактивируется. Видимых под микроскопом
возбудителей не обнаружено. Причины болезни неизвестны.
В Родезии описана розеточнаяболезнь табака,
при которой также наблюдается карликовость; вследствие
укорочения главного стебля растение принимает вид розетки.
Болезнь соком не передается, ее переносчиком, повидимому,
является персиковая тля (Wickens, 1938). Datura virus 1,
Smith и d'Oliveria вызывает у дурмана при инокуляции местные
некрозы, за которыми иногда следует общая инфекция,
выражающаяся в желтой пятнистости. У табака за местным
некрозом следуют посветление по ходу жилок, развитие
многочисленных мелких некрозов и, наконец, некротических колец,
похожих на те, которые вызываются картофельным Х-вирусом.
У помидора тоже наблюдаются местные некрозы. Общая
инфекция или не наступает, или же проявляется мелкими
некротическими пятнами Картофель устойчив к вирусу.
Вирус инактивируется при 50° С, при разведении 1:10 000;
сок остается инфекционным 9 дней. Инфекционность
утрачивается, если сок стоит 12 дней. Кроме сока, болезнь, повидимому,
передается также персиковой тлей.
Генетик Blakeslee (1921) описал у дурмана заболевание,
которое назвал quercina, оттого что листья больного дурмана
становятся похожими на листья дуба. При этой болезни
наблюдается также подавление развития игл на коробочке,
вследствие чего на ней появляются характерные плеши. Болезнь
легко передается семенами, пыльцой, прививкой, но не
инокуляцией соком.
У белены был описан целый ряд вирусных болезней, но
позднее, после идентификации, остался только один вирус,
неизвестный ранее, название которого, по Hamilton,—Ну III,
а по К. Smilh,— Hyoscyamus virus 1, Hamilton. Этот вирус
вызывает у белены сначала посветление по ходу сосудов, а
затем образование темнозеленой каемки по их ходу и желтую
118
мозаику. Сходные симптомы развиваются также у табака,
причем здесь к ним могут присоединиться некротические пятна.
Нередко табак несет вирус скрыто. У N. glutinosa, кроме
характерной темнозеленой окраски по ходу жилок, которая может
маскироваться у старых листьев, наблюдаются белые полоски
на венчике.
Вирус передается соком, а также персиковой тлей. Он
пнактивируется при температуре 50° С и сохраняет инфекцион-
ность в соке не более 24 часов. Он проходит через свечу L1?
но не L3- В Румынии описано мозаичное заболевание у синего
баклажана; при этом заболевании листья покрываются
желтыми пятнами, размер которых постепенно увеличивается,
и затем больные листья погибают. Мозаика проявляется только
на тех листьях, которые достигли известного возраста. Болезнь
передается соком на картофель, разные виды табака, черный
паслен и дурман. Повидимому, в природе болезнь
распространяет персиковая тля.
Вирус инактивируется при температуре 60° С, но остается
инфекционным при разведении 1:1000 (Alexander, 1939).
S. S. Ivanofl в Техасе описывает желтую пятнистость
¦синего баклажана, которая передается на огурец и вызывается,
вероятно, Cucumis virus 1. S. E. Jones (1942) предлагает
опыление серой, пиретрумом и т. д. для борьбы с этой болезнью.
К. М. Smith (1943) описывает мозаичное заболевание
Atropa Belladonna, при котором, кроме мозаики, наблюдаются
некрозы на листьях и стебле. Вирус вызывает у Hyoscyamus
niger местный некроз и общую мозаику, также поражается
N. tabacum и N. glutinosa. Под влиянием вируса развивается
местный и общий некроз. У Datura Stramonium и S, nigrum
вирус вызывает только слабую мозаику.
Вирус инактивируется при температуре между 75—80° С,
выдерживает разведение 1:10 000, в соке сохраняется 6 дней;
через 11 дней в этих условиях вирус становится инактивным.
Заболевания пасленовых, внешне похожие на вирусные.
Здесь следует отметить целый ряд заболеваний, имеющих
большое внешнее сходство с вирусными, инфекционность
которых, однако, крайне сомнительна; или же такие
заболевания, относительно которых мы достоверно знаем, что они
неинфекционны. К числу таких болезней относится
скручивание листьев помидора, которое подробно описано М. А. Ря-
ховским. При этой болезни наблюдается пожелтение листьев;
пораженные листья становятся хрупкими, более толстыми,
чем здоровые, и закручиваются вверх вдоль средней жилки,
принимая форму трубочки или лодочки. Болезнь очень легко
передается семенами, но не передается соком. В течение ряда
лет мы ставили опыты по передаче заболевания прививкой,
но оказалось, что не только при прививке пораженных побегов
.к здоровым растениям, но и при прививке здоровых побегов
119
к растениям со скручиванием листьев передачи заболевания
не наблюдается. Можно было бы думать, что происходит
инфицирование эмбрионов, однако семена, собранные с плодовг
развившихся на здоровых побегах, привитых к больным
растениям, дали совершенно здоровое потомство.
Под названием ш у р г а л у нас описано заболевание
табака, при котором листья становятся светлозелеными и до
неузнаваемости узкими. Повидимому, оно соответствует болезни,
описанной в США как frenching табака, и на Суматре как
Daon lidah. Это — загадочное заболевание. Попытки передать
его соком или трансплантацией обычно остаются
безуспешными. Причины его ищут в почвенных условиях, однако — от
каких именно почвенных условий зависит болезнь, не выяснено.
Возможно, что дело — в целом комплексе этих условий. По
Karraker и Borlner (1934), болезнь развивается при рН почвы
5.8 и выше. Кроме благоприятного для развития болезни рН,
нужен недостаток азота или фосфора, или калия, или всех
этих элементов, вместе взятых. При прибавлении указанных
элементов болезнь исчезает. Отравление солями таллия может
вызвать картину болезни, очень похожую на frenching.
Гипотеза о том, что и в естественных условиях болезнь зависит
от интоксикации таллием, не подтверждается спектральным
анализом золы больных растений: в ней этот элемент не удается
обнаружить (Shear и Ussery, 1940; Spencer и Lavin, 1939).
К остов описывает заболевание, очень похожее на frenching,
которое, однако, было инфекционным. Оно передавалось
прививкой, но заражение наступило лишь после длительного
периода инкубации.
По поводу всех этих случаев хотелось бы заметить, что
непередача болезни прививкой довольно убедительно
свидетельствует против ее инфекционности, а следовательно, против ее
вирусного происхождения; однако, пока достоверно не
установлена ее причина, преждевременно делать окончательные выводы
о ее невирусном происхождении, так как условия заражения
при трансплантации могут быть очень сложными.
Целый ряд расстройств у пасленовых, зависящих от де
фектов в минеральном питании, могут вызывать мозаичную
расцветку и некрозы. Недостаток в почве фосфора вызывает,
например, у табака карликовость, темнозеленую окраску и
многочисленные мелкие круглые некрозы. Ионы хлора
вызывают у помидора мозаику. Следует остерегаться принимать
эти и подобные им физиологические расстройства за вирусные
заболевания. Целый ряд работ специально посвящен симптомам
физиологических расстройств у табака, зависящих от
недостатка питательных веществ (М. Mes, 1930, и др.).
Заключение. Очень большое количество вирусов,
поражающих пасленовые, а также большое сходство симптомов,
вызываемых разными заболеваниями, побуждают нас в заключение
120
сделать краткий обзор важнейших заболеваний пасленовых,
с которыми практически наиболее часто приходится
сталкиваться.
Прежде всего необходимо считаться с мозаичной болезнью
табака, вызываемой Nicotiana virus 1. Несмотря на большое
количество штаммов, свойственных этому вирусу, чаще всего
наблюдается типичная форма болезни, выражающаяся в светло-
зеленой мозаике и деформации листьев. Болезнь широко
распространена у табака, помидора и декоративных пасленовых.
Диагносцировать ее легко по гексагональным кристаллам
в клетках, серологически или по некротической реакции у
N. glutinosa. Далее идут различные некротические мозаики,
среди которых надо различать две группы: кольцевые
пятнистости и полосатости. Большая часть этих болезней нелегко
передается соком, часто они передаются семенами. Вызывающие
их вирусы обнаруживают разную устойчивость к внешним
воздействиям. Немногие из них устойчивы почти так же, как
вирус обыкновенной табачной мозаики; наименее устойчивым
среди них является вирус бронзовости помидора. Кроме этих
мозаичных заболеваний, большое значение имеет одна
желтуха. Это с т о л б у р, поражающий табак, помидор и сити ^
баклажаны и др. пасленовые.
Глава XI
ВИРУСНЫЕ БОЛЕЗНИ ПАСЛЕНОВЫХ
(Картофель)
Изучение вирусных болезней картофеля связано с целым
рядом специфических трудностей. Вследствие вегетативного
размножения картофеля клубнями у этого растения широко
распространено вирусоносительство. Клубни являются как
бы тем резервуаром, который из поколения в поколение может
накоплять различные вирусы; поэтому у картофеля также
широко распространены смешанные инфекции, что затрудняет
анализ и идентификацию вирусов. Длительный симбиоз вируса
и растения, вероятно, является причиной того, что вирусы,
поражающие картофель, имеют многочисленные штаммы. Это
обстоятельство снова затрудняет идентификацию вирусов .
J Пир око распространено у картофеля скрытое
вирусоносительство, вследствие чего внешне здоровые растения могут в
действительности иметь вирус (о сортах-вирусоносителях — см.
Whitehead, 1937). Реакция разных сортов на один и тот же
вирус чрезвычайно различна. Изучение вирусных болезней
у картофеля параллельно развивалось в Европе, где начало
отим исследованиям было положено Quanjer (1913), и в Америке.
В настоящее время производится работа по сличению и
идентификации вирусов, встречающихся у картофеля и в Европе
и в Америке (Koch и Johnson, 1936; Dykstra, 1929).
Вирусные болезни картофеля часто сильно зависят от
экологических условий. Может быть, это обстоятельство,
наряду с тем, что доказательства инфекционности вирусного
заболевания картофеля иногда связаны о известными
специальными условиями, которые не соблюдаются не-специалистами,
было причиной того, что существование целого ряда вирусных
болезней картофеля отрицалось. Merkenschlager (1932) отрицает
вирусную природу скручивания листьев у картофеля, считая
его вырождением под влиянием экологических условий.
Подобного же взгляда деряштся Schweizer (1930). Рожалин (1940)
считает морщинистую мозаику картофеля результатом вырож-
і'22
деныя. Названные авторы иногда правильно подмечают влияние
тех или других экологических факторов на развитие болезни,
однако странным образом игнорируют совершенную
достоверность того, что заболевания, выдаваемые ими за вырождение,
в действительности являются инфекционными болезнями. Ро-
жалин считает, что передача прививкой не доказывает
инфекционное™. Он отказывается также видеть доказательство инфек-
ционности в заражении соком, который вводится капиллярной
трубочкой, рассматривая такое введение как своеобразную
¦((подкормку», от которой может меняться природа растения.
Не решаясь, очевидно, утверждать, что и передача инфекции
при помощи насекомого неубедительна, он, вопреки строгой
доказанности того, что морщинистая мозаика передается тлями,
утверждает, что такая передача не доказана. При этом он
ссылается на таблицу в книге В. Л. Рыжкова,1 где приводятся
данные о насекомых — переносчиках скручивания листьев
у картофеля, игнорируя тот факт, что если роль отдельных
насекомых в происхождении этого явления спорна, то
значение других насекомых как переносчиков скручивания
листьев картофеля совершенно достоверно. Передача
скручивания листьев персиковой тлей, как видно из названной
таблицы, была констатирована шестью авторами, и никто из
лих не получил отрицательного результата.
Было бы грубой ошибкой недооценивать роль
экологических факторов в развитии вирусных болезней и
возможность борьбы с этими болезнями путем вмешательства в условия
жизни растений, однако не только неверно, но и практически
опасно также игнорирование того факта, что мы имеем дело
в данном случае с заболеваниями, в высокой степени
инфекционными и, при благоприятных условиях, быстро
распространяющимися путем заражения здоровых растений от больных.
Некротические мозаики картофеля. Сюда относятся
заболевания, вызываемые различными вирусами, а еще чаще смесью
их, которые могут разнообразно проявляться, в зависимости
от характера вируса и еще в большей мере от особенностей
сорта. Мозаичные заболевания картофеля впервые описал
Orion (1914). Эти заболевания могут быть латентны, могут
проявляться нежной крапчатостью и носят тогда название
мягкой м'озаики (mild mosaic). Они могут также
вызывать, наряду с мозаичностью, более или менее значительную
морщинистость листьев и тогда обозначаются как различные
типы crinkle (складчатость). Наконец, эти заболевания могут
вызывать некрозы, нередко приводящие растения к гибели.
Из некротических мозаик наиболее известны
морщинистая мозаика (rugose) и п о"л осатая мозаика
(streak). Морщинистая мозаика характеризуется некрозами,
1 В. Л. Рыжков. Вирусные болезни растений. М., 1935.
123
... і
резче выражающимися на нижних листьях (рис. 32). Часта
они распространяются по ходу нервов, но встречаются и вне
нервов; имеются также на черешках и в стебле. Иногда они
находятся на самой поверхности стебля и черешков в виде темно-
коричневых и черных полосок, иногда же залегают под
эпидермисом, образуя темные пятна,
просвечивающие из глубины ткани
черешка.
Анатомическое исследование
показывает, что некрозы начинаются
чаще всего в коленхиме и оттуда
распространяются глубже внутрь. С
нижней стороны листа часто
выделяются капли жидкости и наблюдается
преждевременное отмирание нижних
листьев. Стебель может вовсе
оголиться и отмереть. Болезнь особенно
резко проявляется на второй и
третий год заболевания. Существование
подлинных отличий между полосатой
и морщинистой мозаикой довольно
сомнительно. Schultz и Folsom (1923)
пишут, что полосатая мозаика редко
встречается отдельно от
морщинистой. Эти авторы указывают, что
первая отличается от второй тем,
что при ней посветление наступает
незадолго до некроза и мозаичности, собственно, не
наблюдается. Многие авторы (например, Schultz и Folsom) отрицают
существование двух резко отграниченных типов мозаики —
полосатой и морщинистой. При обоих типах отмирание идет
снизу вверх, и Quanjer (1931) описывает это заболевание
как акропетальный некроз, в отличие от некроза верхушки,
при котором отмирает верхняя часть растения и некрозы
закладываются прежде всего в области флоэмы, а оттуда
распространяются. Quanjer (1931) считал, что акропетальный некроз
и некроз верхушки вызываются разными вирусами, однако
большое значение здесь имеют также сортовые особенности.
Вирус, вызывающий у одного сорта некроз верхушки, у
другого может его не вызвать. Первый анализ некротических
мозаик связан с именем К. Smith, который в 1928 г. обнаружил,
как он писал тогда, курьезный факт: что заражение табака
от больного морщинистой мозаикой картофеля дает совершенно
разную картину болезни в зависимости от того, передана ли
болезнь инокуляцией сока или при помощи персиковой тли. Этот
автор назвал Х-вируссм тот заразный фактор, который тлями
не передается, а передается только уколом иглы, и Y-вирусом,
— фактор, передаваемый и уколом иглы и тлями.
124
Рис. 32. Морщинистая
мозаика картофеля,
вызванная Y-вирусом (по
Rosendaal)
Мозаики рассматриваемой здесь группы могут вызываться
различными вирусами и смесями их. К рассмотрению этих
вирусов мы и переходим, причем наряду с другими названиями
вируса везде будем приводить наименование вируса по
классификации К. Smilh.
X вирус. Другие названия этого вируса—в и р у с
латентной мозаики, вирус здорового
картофеля и, наконец, Solanum virus 1, Orion. Этот вирус в очень
многих сортах картофеля находится в скрытом виде. Будучи
привит на другие сорта, он вызывает у них заболевание. Он ла-
тентея в сорте Up-lo-Date, при прививке на сорт President
вызывает мозаичность, а при прививке на Epicure—некроз
верхушки, или, как его иначе называет Quanjer (1931), — а к р о-
н е к р о з. У табака этот вирус вызывает мозаику и кольцевую
пятнистость, которые описаны в предыдущей главе.
При заболевании в клетках часто встречаются Х-тела,
имеющие более или менее округлую или удлиненную форму,
6—50 а в поперечнике. Эти Х-тела гомогенны или зернисты
и лежат возле ядра. Вирус передается соком. Насекомые —
переносчики вируса — неизвестны.
Фитопатологическое значение Х-вируса состоит главным
образом в том, что он обладает способностью усиливать
действие других вирусов. Редко встречаются сорта картофеля,
не несущие латентно этого вируса. Согласно Bald и Norris,
даже латентное носительство вируса отражается на урожае
(1940).
Х-вирус получен путем химической очистки в виде
высокомолекулярного аморфного нуклеопротеида. Вирус инактиви-
руется при 66—70° С, редко сохраняет инфекционность при
разведении 1:10 000. В соке он сохраняет инфекционность в
течение 2—3 месяцев; при высушивании он в одних случаях инак-
тивируется, а в других сохраняет инфекционность (Burnett,
1934). Инактивация вируса при высушивании зависит от
неконтролируемых условий; он проходит через свечу L5.
Описано большое количество штаммов рассматриваемого
здесь вируса. Данные об его изменчивости см. В. Л. Рыжков —
«Основы учения о вирусных болезнях растений».
Y-вирус. Другие названия этого вируса: вирус Ну II,
вирус акропетального некроза и Solanum
virus 2, Orton.
Этот вирус у разных сортов картофеля вызывает
заболевание различной силы. Резкие симптомы он дает у картофеля
President, Up-lo-Date и др. Через 15—20 дней после заражения
развивается посветление по ходу нервов, мозаика,
морщинистость, некрозы и отмирание нижних листьев (рис. 33).
У табака вирус вызывает сначала посветление по ходу
нервов, а затем образование темнозеленой каемки,
обозначаемое американскими авторами как veinbanding. Сходные симп-
125
томы вирус вызывает у белены и петунии. У помидора и черного
паслена наблюдаются первичные симптомы в виде лосветления
с последующим потемнением по ходу жилок; позднее симптомы
исчезают, хотя растение остается носителем вируса. Вирус
поражает также Solarium nodiflorinn. Дурман иммунен к вирусу.
У Lyciurn eareatum на десятый
день после инокуляции
наблюдаются местные некрозы
в виде коричневых колец, а
общая инфекция не наступает
(Dennis, 1938).
Y-вирус является одним
из самых главных
компонентов, вызывающих у
картофеля очень тяжелые
некротические мозаики.
Присутствие Х-вируса усиливает
его действие даже в том
случае, если сам по себе X-
вирус остается в данном сорте
картофеля латентным.
Существует большое
количество штаммов Y-вируса,
которые подробно изучал
КбЫег (1935—1941). Из
многочисленных сортов
картофеля в разных странах
выделен целый ряд штаммов этого
вируса. Описанный в
Америке вирус veinbanding (окай-
мленность жилок) следует
рассматривать как один из
штаммов Y-вируса. Один из
вариантов Y-вируса описан
Smith и Dennis (1940).
Вирус передается при
помощи инокуляции соком, а также тлями, в особенности
персиковой тлей.
Есть указания, что при помощи персиковой тли Y-вирус
может быть перенесен на турнепс, капусту, красный клевер,
горох, Convolvulus arvensis, причем они становятся скрытыми
вирусоносителями (Salaman и Wortley, 1939).
Вирус инактивируетея при температуре 52° С; при
разведении 1:1000 он дает заражение только изредка. При
комнатной температуре он инактивируетея в соке через 24-—36
часов; при более низких температурах, а также при прибавлении
редуцирующих веществ может сохраняться более
продолжительное время.
126
Рис. 33. Вторичные симптомы • у
картофеля, вызванные У-вирусом
(но Kosendaal)
При высушивании происходит инактивация вируса.
Вследствие большой способности абсорбироваться, он не проходит
через бактериальные фильтры.
Y-вирус был получен путем ультрацентрифугирования
в относительно чистом виде. Несмотря на серологическое
родство с Cucumis virus 1, он существенно отличается от этого
последнего в отношении видов, которые он поражает; например,
дурман, чувствительный к огуречному вирусу, устойчив
к Y-вирусу. Огуречный вирус не предохраняет от Y-вируса.
А-вирус. Этот вирус обозначается иначе Solarium virus 3
Murphy и Мс. Kay, 1932, super-mild mosaic virus, Quanjer.
Подобно Х-вирусу этот вирус может вызывать у известных
сортов картофеля некроз верхушки; поэтому первоначально
он считался родственным первому, хотя, по КбЫег, он очень
близко стоит к Y-вирусу и, согласно КбЫег (1937), соединен
с этим последним рядом штаммов, являющихся как бы
переходными. Он идентичен с описанным в Германии Р-вирусом.
В отличие от Y-вируса, он не может поражать Nicotiana gluti-
nosa. У табака симптомы, развивающиеся под влиянием этого
вируса, сходны с теми, которые вызывает Y-вирус. А-вирус
не защищает от Y-вируса и, по F. С. Bawden и F. Sheffield,
не имеет отношения ни к нему, ни к Х-вирусу.
У картофеля самостоятельно или в смеси с другими
вирусами этот вирус вызывает различные мозаичные заболевания, —¦
в частности, мягкую мозаику и складчатую мозаику (Crinkle).
При пересадке на особо чувствительные сорта, как выше было
упомянуто, он вызывает некроз верхушки.
Соком вирус передается с большим трудом, особенно-
на картофель; облегчает передачу примесь к соку карборунда.
Передается вирус персиковой тлей. Согласно КбЫег (1937),
он несколько устойчивей к температуре, чем Y-вирус, и не инак-
тпвируется при 58° С.
При разведении инактивация вируса происходит, пови-
дпмому. в пределах 1:50—1:100. Инактивация в соке наступает
через 24 часа.
В-впрус. Другие названия: Solarium virus 4, Murphy,
Up-to-Date virus. Этот вирус находится в латентном состоянии
в сорте картофеля Up-to-Date. При прививке на President
вызывает у последнего некроз верхушки, тогда как при
передаче иглой не вызывает никаких явлений или ведет к легкой
крапчатости.Повидимому,этот вирус обычно встречается в смеси
с Х-вирусом. У дурмана и табака он вызывает симптомы,,
подобные тем, которые развиваются под влиянием Х-вируса.
Вообще В-вирус сходен с этим последним и не может заражать,
сорт картофеля US 41956 (Bawden F., 1936). Clinch (1942)
считает В-вирус только штаммом Х-вируса. К тому же
мнению приходят F. Bawden и F. Sheffield (1944).
Вирус инактивируется при 70° С, сохраняет инфекцион-
127
ность в соке табака при разведении 1:100 000 и сохраняется
до шести недель. Вирус фильтруется через свечу L3 (Dennis,
1939).
С-ви рус. Другие его названия: Solarium virusb, Salaman,
Di Vernon Streak. Этот вирус, латентный в сорте Di Vernon.
при прививке на President вызывает некроз верхушки;
встречается обычно в смеси с Х-вирусом. С-вирус отличается от
других главным образом по реакции на него различных сортов.
По F. Bawden и F. Sheffield (1944) этот вирус предохраняет
табак от заражения Y-вирусом и близко к нему стоит.
D-вирус. Другие его названия: Solarium virus 6, МигрЬл ;
potato foliar necrosis virus, Bawden (1936).
Он стоит близко к X - вирусу и реагирует с его антп-
сывороткой. Он дает на дурмане и табаке более слабые
симптомы, а на многих вариациях картофеля более сильные, чем
типичный Х-вирус.
Вирус инэктивируется при 67° С; его устойчивость к
разведению сильно вариирует в зависимости от разных условий.
Сок больного, но молодого, быстро растущего табака сохраняет
инфекционность при разведении 1: 5000, а сок старого —
только при разведении 1:1500. Сок картофеля, в зависимости
от возраста растения, в разведении 1:750—1:2000. При
комнатной температуре вирус инактивируется в соке через 6—7
дней, на холоде его активность сохраняется значительно
дольше. Вирус проходит через свечу L5.
Е-вирус. Его другие названия: Solarium virus!, Salaman n
Le Pelley (1930); paracrinkle virus. Существование этого вируса
в латентном виде у сорта King Edward устанавливается по
симптомам, развивающимся у других сортов картофеля, например
Arran Victory, после прививки. Другие способы передачи вируса
неизвестны, и заболевание, им вызванное, наблюдалось только
в экспериментальных условиях. Оно характеризуется посвет-
лением по ходу нервов, мозаичной пятнистостью и развитием
некротических пятен. Мозаика у дурмана наблюдается только
после прививки, но не при инокуляции соком. Табак иммунен
к вирусу.
Мы привели здесь список большого количества вирусов,
которые оказалось необходимым различать для того, чтобы
объяснить разное отношение различных сортов картофеля
к перекрестным прививкам. Ниже мы приводим в виде таблицы
сравнение отношения к этим вирусам некоторых сортов. Все
эти вирусы, повидимому, принадлежат к двум группам.
Наиболее изученным представителем одной из них является X-
вирус, а другой — Y-вирус. Наличие многочисленных штаммов
этих вирусов, то обстоятельство, что они встречаются в
разнообразных смесях и что разные сорта реагируют на них различно,
дает необычайное разнообразие мозаичных заболеваний
картофеля, вызываемых этими группами вирусов.
128
Таблица 7
Реакция разных сортов картофеля на вирусы некротических мозаик
картофеля
Название
вируса
А
В
<:
D
Е
JL-J
X
У
Передача
инокуляцией
С трудом
—
—
+
+
4
Up-to-Date
Некроз
верхушки
—
Epicure
—
President
—
Аггатх "Victory
—
Некроз верхушки
Некроз верхушки
Некроз
верхушки
—•
Акропе-
тальный
некроз
Некроз
верхушки
Некроз
листьев
—
Акропе-
тальный
некроз
—
—
—
Мозаичные заболевания картофеля, подавляющее
большинство которых вызвано комбинацией перечисленных здесь
вирусов, приносят очень большой ущерб во всех странах
Европы и Америки. В США в 1918 г. потеря урожая от этих
мозаик исчислялась в 36 000 т, т. е. в количестве, которое могло
бы прокормить в течение года 170 000 человек.
Мозаики, сопровождающиеся некрозами клубней. Сюда
относится своеобразная группа заболеваний, вирологически
проанализированная лишь недавно. При этих заболеваниях
наблюдается некроз в паренхиме клубня, располагающийся
вблизи внешней и внутренней флоэмы. Этот некроз обычно
развивается только при лежании клубней. Он был описан
под названием псевдонекроза Quanjer (1931) в
качестве самостоятельного вирусного заболевания клубней.
Позднее оказалось, однако, что существует целая группа очень
близко стоящих друг к другу вирусов, которые вызывают
в клубнях описанный некроз и в то же время могут вызывать
на листьях характерные яркожелтые пятна. Этот последний
симптом то отсутствует, то резко выражен, то сопровождается
некротическими явлениями на листьях и стеблях. Форма
заболевания с резкой выраженностью мозаичных симптомов
на листьях давно известна под названием а у к у б а-м о-
заики картофеля. Симптомы, вызываемые этой
группой вирусов на листьях, становятся особенно резкими
при смешанной инфекции с А-вирусом (рис. 34).
Отправной точкой к изучению этой группы вирусов была
маленькая заметка Loughnane и Clinch (1935), опубликованная
еще в 1930 г. В этой работе было показано, что так называемая
мозаика между нервов (interveinal) картофеля
вызывается двумя вирусами, из которых один принадлежит
9 В. Л. Рыжкоз I29
к группе Х-вируса и не передается тлями, а другой может
ими передаваться. Последний был освобожден от
сопутствующего Х-вируса проведением через сорт картофеля Arran Crest.
На сорте President этот вирус вызывает слабую крапчатость
Рис. Si. Лист картофеля, пораженный вирусом
аукуба-мозаики
и в то же время рассеянные некротические пятна на коре
и мякоти клубней. По этому характерному признаку вирус
получил название tuber blotch virus, что значит виру с
пятнистости клубней. Из картофеля сорта Mono-
crat был выделен вирус, не дающий у этого сорта симптомов,
ноу других сортов вызывающий желтую крапчатость нижних
листьев, а также некротические явления в клубнях (Murphy
и Loughnane, 1936, 1937). Согласно Dykstra (1939), очень
близко к этим вирусам стоит вирус канадской полосатости
(Canadian streak), который, наряду с яркой пятнистостью
нижних листьев, у ряда сортов может вызывать некротические
полосы на черешках и стеблях. Серологическое родство этого
вируса с другими вирусами рассматриваемой группы установить
130
пока не удалось. Один очень вирулентный штамм LuLer blotch
вируса вызывает некроз верхушки у картофеля сорта Irish
Chieftoin (Clinch, 1944).
Относящиеся сюда вирусы К. Smith описывает как два
разных Solarium virus 8 и 9, однако, собственно говоря, они
принадлежат к одной и той же группе, обозначаемой Murphy
и др. как F-вирус.
В области некроза паренхиматозные клетки клубня
переполняются зернистым коричневым содержимым и отмирают;
вокруг них можно наблюдать зону гипертрофированных клеток,
не содержащих крахмала или содержащих его очень мало.
Относящаяся сюда группа вирусов у табака, а также
N. glutinosa, N\ silvestris (Dykstra, 1939) может вызывать
неправильные желтые пятна, то сосредоточенные в верхней
части листа, то равномерно распределенные по всему листу.
В других случаях наблюдается скрытое вирусоносительство.
У перца на месте инокуляции появляются коричневые
или пурпурные кольца, а затем развивается общее заболевание,
начинающееся посветлением вдоль нервов и переходящее
в яркожелтую мозаику, и отмирание листьев.
Приводим здесь в табл. 8 данные об отношении вирусов
группы F к различным внешним воздействиям.
Таблица 8
Отношение к внешним факторам вирусов группы F
В к р у с
Aucuba (potato virus 8)
Canadian streak . . . .
Pseudo net necrosis . .
Tuber blotch
Вирулентный штамм
tuber blotch
Температура
инактивации
1000
1000
500
500
200
Разведение
68
68
65
65
62
Устойчивость
в соке
(часов)
96
У6
96
72
"2-96
Кроме данных, приведенных в таблице, имеются также
сведения об устойчивости этих вирусов к различным рН.
Большая часть их сохраняет активность при рН 3.8—8.55.
Неинфекционные некрозы клубней. Следует отметить, что
наряду с некрозами в клубнях картофеля, которые являются
одним из симптомов вирусного заболевания, широко распро1-
странены заболевания клубней неинфекционного характера,
при которых также наблюдаются некротические явлений.
Работы Рожалина устанавливают зависимость
концентрического некроза картофеля от почвенных условий. Van Schreven
(1934) также указывает на возможность развития некрозов
в клубнях под влиянием неблагоприятных почвенных условий.
9* Ш
Wenzel пытался соком и прививкой передать покоричневе-
ние мякоти клубней картофеля и не мог установить инфек-
ционности этого явления. Имеются также данные о разной
устойчивости различных сортов картофеля к некротическим
явлениям в клубнях (Philipp, 1939).
Calico. Это заболевание, по классификации К. Smith,
называется Potato virus 10, Hungerford (1920). Оно проявляется
яркожелтыми или желто-белыми, очень крупными пятнами
на листьях картофеля. Иногда светлый участок занимает
большую часть листа. Самостоятельное существование вируса,
вызывающего описываемую картину болезни, очень
сомнительно. Black и Price обнаружили близкое родство вируса
с Medicago virus 2. Этот вирус передается соком и Macrosipkum
pisi. Перенесенный с бобовых на картофель, он вызывает
симптомы calico, а перенесенный с картофеля на бобовые, он дает
у соответствующих видов картину, характерную для мозаики
люцерны.
КбЫег (1940) выделил в трех случаях у картофеля,
обнаруживавшего симптомы calico, вирус кольцевой пятнистости
табака {Nicotiana virus 12).
Веретеновидность клубней картофеля. Другое название
болезни: potato marginal leaf-roll. По К. Smith, болезнь
вызывается Solarium virus 12, Schultz и Folsom (1923). Это
заболевание впервые описано в США. При нем наблюдается
уменьшение размеров листьев, причем листья обращены вверх.
Наиболее характерным признаком являются изменения в
клубнях, состоящие в том, что клубни приобретают удлиненную
форму даже у т^х сортов, у которых нормально они более
или менее шарообразны. В антоцианосодержащих клубнях
при заболевании могут наблюдаться пятна, лишенные анто-
циана. У некоторых сортов при заболевании наблюдаются
своеобразно выдающиеся «брови».
Болезнь очень легко передается соком, а также тлями
и различными другими сосущими насекомыми — цикадами,
клопами, личинками колорадского жука и т. п.
Вирус инактивируется при 60—65° С, сохраняет инфек-
, ционность при разведении 1:1000, утрачивает ее при 1:10 000.
При высушивании, листья некоторое время сохраняют инфек -
ционность, но затем ее утрачивают.
Сходное с веретеновидностью клубней заболевание было
описано на Украине А. И. Терещенко в 1941 г. под названием
готика. Это название было дано вследствие своеобразного
облика больных кустов картофеля, у которых все листья
обращены вверх. Больные растения подавлены в развитии, и у них
появляется характерная веретеновидность клубней (рис. 35,
36). Болезнь легко передается соком, а также наземными
насекомыми, так что изоляция картофеля от этих последних
предохраняет от готики. Готика чрезвычайно распространенное,
132
очень вредоносное заболевание. Балашев (1941) наблюдал
готику в Узбекистане.
Закручивание листьев. Заболевание, называемое
американцами leaf-rolling, не следует смешивать со скручиванием
листьев, т. е. leaf-roll. Его вызывает Solarium virus 11 (Schulu
Рис, 35. Кусты картофеля: слева — здоровый, справа —
пораженный готикой (но Терещенко)
и Folsom, 1923). При этой болезни наблюдаются слабая
диффузная пятнистость листьев и закручивание верхних листьев,
похожее на то, которое можно наблюдать в различных случаях
увядания картофельного куста, зависящего от поражений
корневой системы. От скручивания листьев болезнь отличается тем,
что при ней не наблюдается ни общего хлороза, ни хрупкости
листьев.
Болезнь передается соком только с трудом; ее
переносчиком является персиковая тля, а также другие виды тлей.
Возможно, что заболевание родственно paracrinkle, известному
в Европе и вызываемому упомянутым выше Е-вирусом (Dyk-
stra, 1939). Возможно, что близко сюда примыкает вирус К,
описанный КбЫег (1941). Он латентен в полевых условиях,
но проявляется в оранжерее.
Желтая карликовость картофеля. Эта болезнь впервые
наблюдалась в штате Нью-Порк в США в 1917 г. Болезнь вызы-
138
вается Solanum virus 16, Barrus и Сішрр (1922). Болезнь
проявляется у картофеля резким хлорозом и карликовостью;
стебель и листья часто некротизируются, причем некрозу
подвергаются, главным образом, верхние части растения.
Очень характерным признаком болезни является раетрескп-
Рис. 36. Клубни картофеля: вверху — здорового растения, внизу —
пораженного готикой (по Терещенко)
вание клубней, описанное более подробно в общей части
книги «Вирусные болезни растений».
Вирус может быть передан на клевер (Trifolium pratensr,
Medicago lupulina, а также на виды табака: например: N. ги-
stica, N. glutinosa. Он поражает, кроме того, Kalanc/iae diag-
remontiana, Barharea vulgaris, Capsella bursa pastoris.
Chrysanthemum leucanthemum (Younkin, Hausing, 1942).
Первоначально за переносчика вируса ошибочно были приняты
некоторые тли. Black (1934) нашел подлинного переносчика
болезни — цикаду Aceratogallia sanguinolenta, обитающую
на клевере. Переселяясь с клевера на картофель, эта цикада
приносит с собой инфекцию на картофельные плантации.
]Зольными оказываются растения, зараженные лишь от цикады.
Передача инфекции от одного куста картофеля другому почти
ire играет роли, так как цикады после переселения с клевера
вскоре погибают.
Соком вирус передается с трудом. Легче он передается
при смеси сока с карборундом, причем легче на виды табака,
чем на картофель (Black, 1937).
В соке N. ghitinosa вирус разрушается при 50° С; инак-
тпвируется при разведении 1:1000—1:1000 000. В соке
corn
храняется 2.5—12 часов. В сухих листьях вирус инактиви-
руется (Black, 1938).
Для борьбы с болезнью рекомендуются опыление
пиретрумом, серой, применение бордосской жидкости, удаление
больных растений (Hausing, 1942).
Непятнистая курчавая карликовость. Заболевание
называется Solanum virus 13, Schultz и Folsom (1923).
Оно описано Orton в 1914 г. Болезнь характеризуется резкой
подавленностью роста; листья зеленого цвета обнаруживают
слабую хрупкость и часто становятся морщинистыми;
наблюдаются веретеновидность и растрескивание клубней.
Болезнь передается очень легко соком. Переносчики
ое те же, что л в веретеновидности клубней.
Вирус инактивируется при 75—85° С, сохраняет инфек-
ционность при разведении 1:1000, повидимому, очень быстро
утрачивает инфекционность в соке. Schultz и Folsom считали
заболевание смешанной инфекцией результатом соединения
вируса закручивания листьев и веретеновидности клубней.
Пятнистая курчавая карликовость бесспорно является
смешанным заболеванием.
Ведьмины метлы картофеля. Заболевание вызывается
Solanum virus 15, Hungerford и Dana (1924). Впервые оно
описано в 1915 г. Ранние симптомы болезни состоят в пожелтении
верхних листьев и карликовости. Позднее главный стебель
может отмирать; появляется множество пазушных побегов,
наблюдается усиленное развитие мелких клубней; иногда клубни
развиваются над землей. Один куст может нести до 200 таких
маленьких клубней, которые нередко, не переходя в период
покоя, развивают побеги.
Болезнь передается прививкой, насекомое-переносчик
неизвестно. Заболевание может быть передано на помидор и на
табак, у которых оно также проявляется в резкой задержке
роста и кустистости. При помощи повилики болезнь
передается Vinca major и здесь излечивается теплой ванной
(Kunkel, 1943).
Верхушечное скручивание листьев картофеля. Болезнь
вызывается Solanum virus 17, Whipple (1919). Она известна
также под названием желтухи верхушки.
Начинается болезнь закручиванием верхушечных листьев, как это
бывает при первичных симптомах настоящего скручивания
листьев. В отличие от этого последнего, и на следующий год
наблюдаются скручивание только верхушечных листьев. Часто
обнаруяшвается известная карликовость растений и усиленное
развитие • мелких клубней. Болезнь передается прививкой
кусочков клубней (core grafts).
Скручивание листьев картофеля. По классификации
К. Smith, заболевание вызывается Solanum virus 14, Appel
и Quanjer. Под названием скручивание листьев
135
оно было впервые описано в 1905 г. Арреі, который, однако,
приписывал его грибку Fusarium. Впервые Quanjer описал
характерные для болезни анатомические признаки и
квалифицировал ее как инфекционное
вирусное заболевание. (История
вопроса — у Pethybridge, 1939).
Болезнь начинается
первичными симптомами, которые
состоят в закручивании верхних
листьев, причем в этих листьях
наблюдается задержка оттока
крахмала. Эти симптомы
появляются в год заражения, но
могут и совсем отсутствовать.
На другой год развиваются
вторичные симптомы. Листья
становятся кожистыми,
хрупкими, хлорозными, иногда
также красноватыми, фиолетовыми
и даже бронзовыми, вследствие
развития антоциана. Доли
листа скручиваются вдоль главной
жилки, причем цоражение
листьев распространяется
начиная от более нижних, старых*
(рис. 37). Материнский клубень
листа остается часто
нетронутым. У многих сортов вирус
скручивания листьев вызывает
сетчатый некроз клубня,
распространяющийся по ходу
сосудисто-волокнистых пучков¦
(Atanasoff, 1926; Huber, 1940). ,
Болезнь легко передается клуб-*
нями и из года в год
прогрессирует.
Вирус скручивания листьев,
может быть передан целому
ряду пасленовых. У томата он
вызывает очень слабые
симптомы в виде легкой хрупкости и слабого закручивания
листьев; у дурмана наблюдаются хлороз и слабое закручивание
листьев. Подобные же симптомы наблюдаются у перца. По
Salaman u Wortley (1939), капуста, турнепс и Campanula могут-
. быть скрытыми вирусоносителями. Это не подтверждается
в работе Helson и Norris (1943). Болезнь легко передается
прививкой, различного рода тлями, а также, повидимому,
некоторыми другими насекомыми; главным переносчиком яв-
136
Рис. 37. Скручивание листьев у
картофеля
Таблица 9
Передача leaf-roll насекомыми
-f- передается; — не передается; ? — неясный результат
Насекомое
Aphis fabae
» »
» gossypii
» »
» rhamnl
» »
» abbreviata ....
Calocoris norvegicus . . .
» >> ...
Chlorita viridula ....
Nobis alterinidis ....
Epitrix cucumens . . .
» subcTinata . . .
Eupteryx auratus ....
» » ....
Empoasca filamenta . . .
Leptinotarsa decemlineata
Philaenus leucophtkalonius
Lygus Sp.
» pabulinus ....
» pratensis
Macrosiphum gei ....
» » . . . .
» » . . . .
» pseudosolanl
» solanifolii .
Mysus circumflexus .
» pelargoni . .
» persicae . . .
» ornatus . .
Psyilliodes affinis
» »_
Typhlocyba ilmi
» solatii .
Tipula paludosa (личинки)
Передача
+
+
+
+
+
+
+
?
+
+
+
+
Автор
Elze (1931)
К. Smith (1931)
Elze (1927, 1931)
K. Smith (1931)
Elze (1927, 1931)
K. Smith (1931)
D. Folsom (1942)
Murphy (1923), Murphy u Mc. Kay
(1926)
K. Smith (1929)
K. Smith (1929)
Dykstra (1938)
Schultz u Folsom (1929); D. Folsom
(1942)
Dvkstra (1938)
Elze (1927)
K. Smith (1929)
Dykstra (1938)
Schultz u Folsom (1923); D. Folsom
(1942)
Dykstra (1938)
Elze (1927)
К Smith (1929)
Dykstra (1938)
Murphy u Mc Kay (1929); Mc. Kay
u Dykstra (1932)
K. Smith, Cottier (1931)
Whithead (1931)
Elze (1931); K. Smith (1931); Murphy
u Mc. Kay (1929;
Elze (1931); Murphy u Mc. Kay
(1929); Dykstra (1938); D. Folsom
(1942)
Whithead (1931); Smith (1931)
Mc. Kay u Dykstra (1932)
K. Smith (1929); Elze (1927, 1931);
Murphy u Mc. Kay (1929);
Dykstra (1938)
Loughnane (1939)
Elze (1927)
K. Smith (1929)
Murphy u Mc. Kay (1929)
Elze (1927)
Elze (1931)
137
ляется персиковая тля (табл. 9); соком не передается. Данные
о передаче болезни при помощи семян не вполне определенны.
Quanjer наблюдал передачу 7 растениям на 42. Согласно
Elze (1931), проростки из больных семян имели внешне
здоровый вид, но при пересадке на здоровые растения могли
инфицировать эти последние, т. е. были носителями вируса.
При болезни наблюдается некроз флоэмы, который обычно
не распространяется на соседние ткани. Описаны также при
заболеваний различные Х-тела. Nelson описывает их как трипа-
нозомы. Он видел у них даже ядра и блефаробласты. Schaffnil
(1933) во флоэме больных растений видел спиральные тельца
без ядер. По Clinch (1932), Х-тела яря скручивании листьев
у картофеля отсутствуют.
Характерным признаком болезни является задержка
оттока крахмала. Она наблюдается в качестве первичного
симптома у верхушечных листьев; при развитии вторичных
«симптомов наиболее сильно она выражена у нижних и средних
листьев.
Целый ряд немецких авторов отрицал инфекционный
характер скручивания листьев у картофеля, рассматривая его
как вырождение. Эти авторы отрицали инфекционный характер
скручивания листьев, встречающегося в Германии, однако
и для Германии было доказано, что скручивание листьев может
передаваться и прививкой и насекомыми (Schaffnit, J933;
Kohler, 1933).
Наряду с некротическими мозаиками скручивание листьев
картофеля принадлежит к числу наиболее распространенных
и вредоносных вирусных заболеваний этого растения. В США
в 1926 г. от этой болезни погибло 381 628 т картофеля.
Другие вирусные и подобные им заболевания картофеля.
Мы дадим здесь краткий перечень мало изученных и не
получивших широкого распространения болезней картофеля, а также
ряд заболеваний с невыясненной этиологией, обнаруживших
известное сходство с вирусными.
Псиллидная желтуха картофеля вызывается,
по К. Smith, Solatium virus 18, Richards (1928).
Однако существуют большие сомнения, является ли это
заболевание, встречающееся также у помидора, вирусным,
или же оно — результат действия токсинов насекомого
(цикады Paratrloza cockerelli).
Под названием purple top will описано заболевание
картофеля, при котором наблюдается измельчание листьев,
становящихся пурпурными по ходу жилок. Столоны несут
многочисленные мелкие клубни, а также многочисленные нитевидны^
корни. Нередко столоны выходят вверх, появляясь над землей
в виде коротких зеленых побегов. Многие клубнп у места
прикрепления стебля имеют некрозы; встречаются также некрозы
в области проводящей системы в стебле. Часто клубни мягки,
J3S
а также морщинисты, с очень маленькими поверхностными
глазками. Болезнь переносит Macrosteles (Cicadula) diversa, н
она вызывается вирусом желтухи сезідерея (S. G. Younkin,
1942, 1943).
Как круглолистость картофеля может быть
обозначено заболевание, описанное Сладкомедовой (1941) в
Харьковской области. Главным признаком болезни являются
измельчание листьев и хлорозность их краев. Эти симптомы
во вторую часть лета могут совершенно маскироваться, однако
пораженные кусты или дают очень мало клубней, или совсем
их не дают. По характеру распространения болезни можно
предполагать, что она является инфекционной.
Мелколистость картофеля описана в Узбекистане.
При этом заболевании, кроме уменьшения пластинки листа,
наблюдается хлороз листьев; их края становятся волнистыми.
Подобное заболевание встречается в Киргизии. Оно резко
снижает урожай (Балашевг 1941).
Гигантизмом (giant hil) у картофеля называется
заболевание, при котором наблюдается пышный рост
надземных побегов, тогда как число клубней часто уменьшается.
Нередко клубни больных растений, без периода покоя,
прорастают в землю. Наблюдалось, будто бы, что при близком
соседстве больных кустов со здоровыми болезнь постепенно
распространяется по плантации; указывалось также на
передачу ее при помощи core grafts. По другим данным, болезнь
неинфекционна и пересадкой не передается (Мс. Кау и Dyk-
stra, 1932).
Brehmer (1930) наблюдал в эспериментальных условиях
у растений, выращенных в изоляции на стерилизованном
торфе, заболевание, которое он назвал Knotchenkrank-
h е і t. Оно проявляется после того, как растение достигает
15 см в высоту. Листья становятся морщинистыми и желтеют,
междоузлия укорачиваются.
Болезнь передается трансплантацией. Возбудителем этой
болезни, как и различных вирусных заболеваний, автор считает
плазмодия со сложным циклам развития.
Под названием блестящая болезнь (Glanzkrank-
heit) Kohler (1938) описывает заболевание картофеля. При этой
болезни листья с нижней стороны приобретают характерный
блеск, наряду с этим наблюдается их закручивание вверх;
иногда раньше отмирают верхние листья и вся верхушка,
в других случаях средние и нижние листья. Подобное же
заболевание, сопровождающееся блеском листьев с нижней
¦стороны, наблюдалось у перца и дурмана. Все эти явления
наблюдались у экспериментальных растений, произраставших
в оранжерее, и были связаны с появлением белой мушки
^Aleyrodidae), которую автор считает переносчиком болезни.
Purple dwarf — заболевание, описанное в Канаде, при
139
котором рост побегов останавливается, листья закручиваются
вверх и антоцианируют по краю. Корни засыхают. Флоэма
некротизируется. Болезнь передается прививкой (Sanford
п Clay, 1941).
Эпифитология вирусных болезней картофеля. В
распространении вирусных болезней картофеля огромную роль играет
вегетативное размножение: обычно вирусы зимуют в клубнях
и вместе с этими клубнями передаются. Распространение
в природе важнейших вирусных болезней картофеля происходит
довольно быстро; способ его ясен в тех случаях, когда имеется
насекомое-переносчик. Как мы видели выше, большинство'
вирусных болезней картофеля действительно распространяется
насекомыми. Не совсем ясно, как распространяется в природе
Х-вирус. Насекомое-переносчик этого вируса неизвестно,
по он лег;ко распространяется соком, и, вероятно, главную
роль играет соприкосновение с больными растениями.
Заражение может происходить не только в поле, но и во время
зимнего покоя клубней, особенно если они начинают
прорастать до посадки. Тли, попадая на молодые побеги,
способствуют заражению клубней (Y-вирус).
Различные авторы ставили опыты с целью выяснить, как
быстро распространяется та или иная вирусная болезнь
картофеля в зависимости от расстояния здоровых растений от
первоначального очага болезни (Murphy и Loughnane, 1937; Clinch)
1938). А. И. Терещенко (1941) ставил подобные опыты с
готикой и показал, что здоровый картофель, высаженный рядом
с больным, дал до 100% больных кустов, а высаженный на
расстоянии 200 м дал лишь 4% больных кустов. Автор считает,
что пространственная изоляция в 500 м почти гарантировала
бы от распространения болезни.
Не подлежит сомнению, что распространение вирусных
болезней очень сильно зависит от экологических условий.
В пользу этого свидетельствует уже то обстоятельство, что*
многие вирусные болезни приурочены главным образом к
областям с более теплым и сухим климатом. У нас в Союзе
морщинистая мозаика и готика наибольшего распространения
достигают в южных областях. Точно так же в США
морщинистая мозаика и веретеновидиость клубней в северных штатах
распространены гораздо меньше, чем в южных. В 1928 г. Quan-
jer, Elze и Blattny изучали распространение вирусных болезней
картофеля в зависимости от географических условий (см.
Quanjer и Gaumann, 1935). В Чехословакии были одновременно
сделаны посадки здорового картофеля в долине и в горах, э
к нему был подсажен картофель, больной морщинистостью
и скручиванием листьев. В горах обе болезни
распространялись гораздо медленнее, чем в долине. Одновременно были
сделаны посадки в Голландии. Здесь указанные заболевания
распространялись так же медленно, как и в горах. Авторы
140
считают, что более медленное распространение вирусных
болезней зависит от меньшего количества персиковой и других
тлей, являющихся переносчиками этих болезней. Давно
известно, что горный климат благоприятствует получению
здорового картофеля. Согласно Constantin (1935) и его школе, в
горных условиях наблюдается оздоровление картофеля, больного
скручиванием и мозаикой. Этот автор находит в корнях
картофеля, произрастающего в горах, микоризу, которой он и
приписывает значение в его оздоровлении. Quanjer (1935) произвел
посадку картофеля в Альпах на высоте 1680 м над уровнем
моря и одновременно в долине на высоте 455 м над уровнем
моря и не наблюдал выздоровления больного картофеля. В
горах могут наблюдаться очень тяжелые формы мозаики,
приводящие растение к гибели, однако распространение вирусных
заболеваний в горах и в этих опытах шло более медленно,
чем в долине. Меньшее распространение вирусных болезней
в горах объясняется обычно меньшим количеством там тлей,
однако первичное заражение вирусными болезнями не
исключено и в горах. Chouard и Duffrenoy (1938) в горных условиях
высадили свободный от скручивания листьев и Y^Bnpyca
картофель, который был отделен от возможных источников
инфекции сосновым питомником. Несмотря на это, наблюдалось
заболевание картофеля. Живая персиковая тля найдена на
высоте 2200 м над уровнем моря. Урожай, собранный на высоте
1850 м над уровнем моря, был значительно лучше, чем урожай
в условиях равнины. Указать точно, на какой высоте над
уровнем моря может быть получен наиболее здоровый урожай
картофеля, невозможно, так как это зависит от разных, трудно
учитываемых условий (С. Сгёріп, 1938, и др.). Merkenschlager
(1931, 1932), рассматривающий вирусные болезни картофеля
как результат вырождения, видит причину его в
неблагоприятных тяжелых глинистых почвах, которые для картофеля
затрудняют извлечение воды; напротив, почвы песчаные и вообще
легкие являются в этом отношении благоприятными. Quanjer
сообщает, что картофель, изолированный от больного, много
лет сохранялся здоровым на тяжелых глинистых почвах,
а наоборот, на легких песчаных почвах его труднее сохранить
здоровым, так как ветер вместе с песком легко переносит тлей,
а это способствует распространению по полю заразного
начала.
Кіарр (1933) указывает на то, что вырождение картофеля
зависит от несоответствия экологического комплекса
нормальному циклу развития картофеля; при этом большое значение
в развитии депрессии картофеля имеют удлинение периода
вегетации и укорочение периода покоя, создающие
перенапряжение, в деятельности растения.
Т. Д. Лысенко (1939) считает, что вырождение картофеля
на юге зависит от ускоренного старения картофеля. Картофельу
Ш
выращенный на юге, подвергаясь действию летней жары,
стадийно является старым.
Устанавливается значение дождей для здоровья картофеля.
Для его развития наилучшими являются те области, где
прохладное и влажное лето и много дождей; по мнению Davidson
(1938), при дождях главный переносчик вирусных болезней —
персиковая тля — мало активен, так же как и во влажных
и ветреных условиях, а в безветреные и солнечные дни она
становится весьма активной. Blattny (1935) установил, что<
поливка картофеля вызывает маскировку скручивания и
некротических мозаик. Согласно этому автору, при поливке
уменьшается также количество тлей, а вместе с тем и скорость
распространения болезни- Очень важные данные о зависимости
развития вирусных болезней картофеля от распространения
тлей имеются в отчете о дискуссии по поводу вирусных болезней
картофеля,1
Меры борьбы с вирусными болезнями картофеля. Мы очень
мало знаем о сорняках, в которых резервировались бы вирусы,
поражающие картофель. Сохранение вируса в сорняках в
данном случае не играет сколько-нибудь существенной роли,
так как вирусы зимуют в клубнях и таким способом
сохраняются. Получение здорового посадочного материала является
одной из самых существенных мер в борьбе с вирусными
болезнями картофеля. Эта задача может быть разрешена
различными способами.
В целом ряде стран стали на путь получения здорового
посадочного материала в специальных условиях.
Французскими авторами широко рекомендуется выращивание
картофеля для посева в горах, где легче удается сохранить это
растение здоровым. В Ирландии, под руководством Murphy,
с 1928 г. была предпринята работа по отбору, изоляции и
размножению картофеля, свободного от вирусов, который
специально служит посадочным материалом (Murphy и Loughnane,
1937).
Очень успешно задача получения здорового посадочного
материала была разрешена Т. Д. Лысенко (1939) для юга нашего
Союза. Т. Д. Лысенко исходит из положения, что главной
причиной вырождения картофеля в южных, жарких районах
является высокая температура почвы во время развития
клубней и особенно действие ее на пробужденные глазки клубней.
Последнее часто имеет место во вторую половину лета, во время
лежки клубней под кустами, а также во время осенне-зимнего
хранения семенного картофеля. 2 С целью избежать
неблагоприятного влияния высоких температур был разработан метод
летних посадок картофеля (в условиях южных районов в конце
1 См. Annals of Appl. Biol. 1943. 30, № 1.
2 Лысенко иФаворов. Летние посадки картофеля, стр. 6,1930,
142
июня или в начале июля). При такой посадке большая часть
клубней развивается в сентябре, т. е. при более низких
температурах. Посевный материал от летних посадок и при весенней
посадке на следующий год дает значительно лучший урожай,
чем материал от весенних посадок. Так, на полях Всесоюзного
селекционно-генетического института при посадке сорта
«Ранняя роза» материалом Горьковской области (семеноводческие
колхозы Арзамасского района) урожай получен в 80 ц с гектара;
при посадке материалом Московской области (Институт
картофельного хозяйства) — 91 ц с гектара, а при посадке материалом
летней репродукции Всесоюзного селекционно-генетического
института — 120 ц с гектара.1 Картофель, полученный от
летних посадок, при соблюдении соответствующих
агротехнических правил, можно собрать в достаточно большом
количестве; следует также отметить, что клубни от летних посадок
крупнее и имеют больше крахмала, чем клубни от весенних
посадок.
Опыты, произведенные за границей, также говорят в
пользу летних посадок. Wartenberg и др. в продолжение
целого ряда лет проводили опыты, получившие название Tages-
parzellenversuch. В этих опытах в течение всего лета каждый
день производилась псоідка картофеля, причем оказалось,
что поздние посадки дают меньший процент больных вирусными
болезнями растений, чем ранние. Согласно Berkner (1936),
в апрельском посеве было 45% больных растений, в майском —
36%, в июньском — 66%, в июльском — 1.5%. Урожай по
;>тим срокам посадки относился как 100:110:35:115. Сходные
опыты, но в меньшем масштабе, были поставлены Blattny
(1935), который мог констатировать, что при поздних посадках
вирусные болезни картофеля распространяются медленнее,
чем при ранних. По данным этого автора, на поздних посадках
на 38% меньше вирусных заболеваний. На этих посадках
также встречается меньше тлей. На картофеле, посаженном
весной, приходилось 289 тлей на куст, а на летних посадках
только 56 тлей. Согласно данным Davies (1934), вирусные
болезни картофеля в Англии наблюдаются в тех местностях,
где много персиковой тли, и нет их там, где персиковой тли
мало или она появляется очень поздно. Вопрос о роли тлей
в экологических условиях в развитии вирусных болезней
картофеля снова обсуждался в дискуссии, которая имела место
в Английском обществе прикладной биологии в 1943 г.2 K5hler
(1935а) подверг весенне-летние посадки картофеля вирологи-
ческому анализу. Он нашел, что весенние посадки почти сплошь
имеют Y-вирус, в июльских посадках он имеется только у 5%
растений, а августовские посадки от него совершенно свободны.
1 Лысенко и Фаворов. Летние посадки картофеля, стр. 7, 1939.
2 Отчет об этой дискуссии см. Ann. of Appl. Biol. 1943, 30, № i.
Можно считать вполне прочно установленным, что летние
посадки картофеля практически сводят на-нет некоторые
вирусные заболевания. Механизм действия летних посадок
в данном случае не вполне выяснен, В первую очередь следует
иметь в виду уменьшение количества тлей, а затем и более
быстрые сроки развития картофеля. Чем позднее произойдет
заражение, чем время его ближе ко времени сбора урожая,
тем меньше вероятность проникновения вируса в клубень
и, следовательно, больше шансов получить здоровый урожай.
Следует отметить, что летние посадки представляют
неодинаковую ценность для борьбы с различными вирусными
болезнями. Сами по себе летние посадки не могут, например,
значительно ограничить развитие готики картофеля.
Неоднократно рекомендовался для борьбы с вирусными
болезнями сбор недозрелых клубней картофеля. Так как такие
клубни плохо переносят зиму, то с целью сделать их несколько
более устойчивыми можно обрезать ботву, оставив их до осени
в земле. Эта мера, не привившаяся в хозяйстве, очевидно,
основана на том, что при применении ее уменьшаются шансы
проникновения вируса в клубни. А. И. Терещенко (1941)
указывает, что ранние сроки уборки картофеля способствуют
ограничению распространения готики. Чем раньше производится
уборка картофеля,тем меньший процент растений заболевает готикой.
Из вышеприведенного видно, что здоровье посадочного
материала очень важно; поэтому понятно, что был предложен
целый ряд методов, которые должны помочь диагносцировать
вирусные заболевания уже в клубнях.
Наиболее надежным, хотя и очень громоздким методом
является так называемый tuber-index, который состоит в том,
что еще до посадки клубней рано весной от каждого
испытуемого клубня отрезают кусок, нумеруют и кусок и клубень,
а затем кусок проращивают в теплице на песке. Получают
проростки, несли проросток болен, то бракуется весь клубень.
Kohler (1935) несколько упростил этот метод, используя
для такого предварительного проращивания совсем маленькие
кусочки клубня с глазками.
Несмотря на громоздкость описанного метода, он не вполне
гарантирует отбор здоровых клубней, потому что вирус моя^ет
быть распределен в клубне неравномерно и здоровые глазки
могут встретиться и у больных клубней; следует также учесть
случаи маскировки болезни. Еще менее надежны другие методы.
Не без успеха может быть применен серологический метод,
так как присутствие вируса в клубнях можно обнаружить
при помощи специфической сыворотки.
Далее был предложен целый ряд проб и реакций, которые
более подробно описаны в нашей книге «Вирусные болезни
растений» и на которых мы не считаем нужным здесь
останавливаться ввиду того, что ни одна из них не оказалась достаточно
144
Таблица 10
Методы диагностики вирусных заболеваний по клубням
Метод
Окисление хингидрона
фенола
бензидина
Восстановление селенита
натрия
Восстановление пермаита-
ната
Обесцвечивание метилено-
вой синыш псрмангаиа-
тои
Разложение клубня при
брожении
Потемнение клубня от
медной пластинки
Окрашивание сока
растворами NaOH и CuS04
Изменение разности
потенциал эв
Флуоресценция в
ультрафиолетовых лучах
Развитие проростков в
различных концентрациях
сахара
Отдача воды клубнем в 96°
алкоголе
Проницаемость для
электролитов
Буферность сока клубней
Окрашивание сока
клубней иодом
Здоровый
быстрое
быстрое
быстрое
быстрое
быстрое
слабое
медленное
значительное
желтое
не выше 150
милливольт
зеленая
быстрое
больше
меньшая
меньшая
сохраняется
синим
Больной
медленное
медленное
медленное
медленное
медленное
сильное
быстрое
слабое
фиолетовое
выше 190
милливолы
грязно-
молочно-
коричнево-
зеленая
медленное
меньше
большая
большая
обесцвечивается
Литература
¦
1 Магх и Мег-
Г kenschlager (1932)
)
Рузинов, г 1930
Boas, 1931
Bechold и Erbe,
1932
Friedrich, 1937
Ней, 19322
Stary, 1937
Магх и Мег-
Kenschlager, 1932
Kosmat, 1932
Hathsackn Hur-
wiis, 1932
Kaho, 1935
Warienberg, 1937
Wartenberg и
Kinkowski, 1937
практичной. Мы ограничимся здесь только краткой сводкой
предложенных методов в виде таблицы 10, воспроизводимой
с изменениями п дополнениями, из упомянутой выше книги.
В борьбе о вирусными болезнями картофеля некоторое
значение может иметь также противовирусная прополка, под
которой понимается удаление больных растений. Само собой
разумеется, что эту меру можно применять только при
отсутствии массового поражения. Обычным сроком прочистки
является июль, однако А. И. Терещенко (1941) рекомендует
1 Отрицательные результаты получены Пивоваровой и Горелик
(1936).
2 Этот метод, на который возлагались большие надежды, не
оправдал себя (О. П. Чернышева, Friedrich, 1937).
J0 В. Л, Рыжкоз 145
переместить прочистку на более ранние сроки, с последующей
тщательной проверкой и в более поздние сроки.
Можно возлагать большие надежды в деле борьбы с
вирусными болезнями картофеля на устойчивые сорта. Имеется
целый ряд работ в этом направлении. Можно различать сорта?
имеющие различную степень устойчивости. КоЫег (1940)
обращает внимание на то, что разные сорта картофеля
отличаются между собой также в отношении скорости, с которой
вирус проникает в клубни.
При одновременном заражении Y-вирусом у одних сортов
вирус проникает в большую часть клубней, у других же только
в незначительное их число.
Выведен абсолютно иммунный к Х-вирусу сорт US 41956.
Сорт Ка tab din весьма устойчив к Х-вирусу в полевых условиях,
но легко заболевает при прививке. Мягкая мозаика картофеля
зависит от сочетания Х-вируса и А-вкруса. Получен сорт,
устойчивый к обоим вирусам (Schultz, Clark и Stevenson, 1940).
Вполне устойчивые к скручиванию листьев сорта картофеля
еще неизвестны (I. F. Stevenson, D. Folsom и Т. Dykstra, 1943).
Необходимо обратить внимание также на приобретенный
иммунитет к вирусным болезням у картофеля. Известен целый
ряд случаев, когда при первичном заражении картофель дает
тяжелые симптомы, которые в дальнейшем сильно ослабляются,
и растения, хотя и продолжают болеть, но страдают от
заболевания уже в меньшей степени. Следует считать перспективной
разработку метода искусственной вакцинации картофеля от
различных вирусных болезней. Ослабленные штаммы вируса,
размножаясь в растении и передаваясь вегетативным
поколениям, могли бы в отдельных случаях дать сорта картофеля,
устойчивые к более сильным штаммам того же самого вируса.
Рекомендуется также борьба с тлями — переносчиками
вирусных болезней. Хорошие результаты дало опрыскивание
фруктовых деревьев во время их покоя, так как на них зимуюг
тли, и опрыскивание самого картофеля никотином или бордос-
ской жидкостью.
Ефейкин (1940) указывает на то, что «вырожденный))
картофель с юга оздоровляется под влиянием культуры в Чувашии.
Интересно было бы изучить это явление более подробно и с-
вирологической точки зрения.
Глава XII
ВИРУСНЫЕ БОЛЕЗНИ ТЫКВЕННЫХ
Обыкновенная огуречная мозаика. Заболевание ото описано
под различными названиями, важнейшими из которых
являются: cucumber mosaic и cucumber white pickle.
Заболевание вызывает Cucumis virus 1, Doolittle, по классификации
К. Smith.
Мозаичная болезнь огурца впервые была описана в 1903 г.
в штате Огайо в США Selby. В 1910 г. тот же автор доказал
инфекционыость болезни. Более подробно заболевание изучено
Doolittle (1920). На листьях огурца болезнь проявляется
светлозеленой и желтой мозаикой, причем окрашенные в темно-
зеленый цвет участки нередко вздуваются и вся пластинка
листа становится морщинистой и неправильной. У более
старых листьев болезнь иногда проявляется в хлорозе и в
отмирании участков листа. Цветы больных растений могут быть
несколько подавлены в развитии, и число женских цветов
уменьшено по сравнению с числом их у здоровых растений.
Заболевание резко отражается на плодах, которые у больных
растений окрашены в светлозеленый или почти белый цвет
и несут на этом белом фоне многочисленные темнозеленые
вздутия — бородавки.
Проявления болезни у тыквы и дыни, а также у других
тыквенных, например, Bryonia alba, В. dioica, Micrampelis
lobata, сходны с описанными признаками болезни у Огурца.
Арбуз или не подвержен заболеванию, или дает только очень
слабые признаки мозаики.
Заболевание огурца мозаикой иногда распространяется
очень быстро, причиняя большие убытки. В 1922 г. в некоторых
местностях США убытки от мозаики огурца достигали 75 тысяч
долларов.
Обыкновенная огуречная мозаика встречается также
в Европе, в том числе и в СССР.
Заболевание передается соком и различными насекомыми-
переносчиками. Передача болезни соком не всегда удается
легко. В распространении ее в природных условиях, во всяком
Ю* 147
случае, главную роль играют насекомые. Болезнь могут
передавать различные виды тлей, например: Myzus persicae, M. pseu-
dosolani, М. circumflexus, М. gei. Aphis gossypii, а также
питающиеся листьями жуки {Diabrotica sp.). Насекомое является
механическим переносчиком Cucumis virus 1. В некоторых
случаях болезнь может передаваться семенами. Такая передача
наблюдалась у дыни и дикого тыквенного Micrampelis lobatay
хотя огурец, повидимому, семенами ее не передает.
Вирус обыкновенной огуречной мозаики фильтруется через
свечу Беркефельда, но не Шамберлена. В последней он
задерживается вследствие адсорбции. При высушивании вирус
инактивируется, его инактивация также происходит при 60—¦
70° С и при разведении 1:10 000. В соке он инактивируется
в 72—96 часов.
Вирус обыкновенной огуречной мозаики является одним
из самых мало специализированных, он поражает
представителей разнообразных семейств, вызывая такие хорошо
известные болезни, как огуречная мозаика табака, огуречная
мозаика сельдерея и мозаичные заболевания различных
однодольных. Эти заболевания описываются в соответствующих
главах. В зимовке вируса большую роль играют сорняки,
например, виды физалиса. Известны различные штаммы вируса,
желтый и др.
Для борьбы с обыкновенной огуречной мозаикой можно
рекомендовать уничтожение сорняков, больных растений, если
число их на плантации невелико, и усиленную борьбу с
насекомыми-переносчиками. Известны устойчивые к вирусу сорта
огурца, например Китайский длинный (Chinese long).
Беттендорфская мозаика огурца. Заболевание описано
Porter (1930) под названием огуречная мозаика 2.
К. Smith рассматривает вирус, вызывающий эту мозаику,
как штамм D. Cucumis virus 1, не приводя к этому, однако,
достаточных оснований и считая свое мнение предварительным.
Беттендорфская мозаика не вызывает такой резкой
подавленности роста, как обыкновенная огуречная мозаика; симптомы
на плодах при ней отсутствуют. Она поражает арбуз и сорт
огурца Chinese long, которые устойчивы к обыкновенной
огуречной мозаике. По Chester, вирус Doolittle и Porter не
обладают серологическим родством, однако, по Price, они взаимно
предохраняют друг от друга.1
При скрещивании сортов Chinese long, устойчивого к
обычной мозаике, с White spine, устойчивого к Беттендорфской
мозаике, получается первое поколение — неустойчивое к обоим
вирусам, а во втором наблюдается сложное расщепление;
признак устойчивости рецессивен.
1 См. В. Л. Р ы ж к о в. Основы учения о вирусных болезнях
растений. Мм 19'*4, стр. 115.
148
Мозаичная болезнь арбуза. Walker (1933) описывает мозаику
на арбузах, которая проявляется в мозаичности и в искривлении
пластинки листа, и эта пластинка может редуцироваться вплоть
до того, что сводится к одной средней жилке. Иногда
наблюдается деформация цветов, и цветы часто увядают до
созревания. Плоды мозаичные и нередко имеют уродливую форму.
При заболевании наблюдаются подавленность роста,
укорочение междоузлий, мозаичность и искривление пластинки
листа. Каким именно вирусом вызывается заболевание —
неизвестно.
Английская огуречная мозаика. Первое сообщение об этой
болезни было сделано в 1921 г. W. Bewley (см. W. Bewley,
1923). Более подробно она изучена A ins worth (1935). По
классификации К. Smith, болезнь вызывает Cucumis virus 2. Другие
названия этого вируса: Cucumber mosaic virus 3 (зеленый)
и 4 (желтый). Известны два штамма этого вируса — зеленый
и желтый; первый вызывает светлозеленую мозаику, второй —
желтую и почти белую.
У огурца болезнь проявляется крапчатостью, которая
развивается на молодых, возникших после заражения, листьях.
Растения подавлены в росте, пластинка листа может
уродоваться вследствие выпячивания его зеленых участков.
Плоды или не страдают, или на них наблюдаются светло-
зеленые или желтые полосы. Сходны проявления болезни
у дыни и арбуза; тыква не болеет. Вирус, в отличие от вируса
обыкновенной огуречной мозаики, не поражает ни одного
представителя других семейств, кроме тыквенных. Болезнь
не передается насекомыми, но чрезвычайно легко передается
соком.
Вирус очень устойчив: он инактивируется при 90-—95° С,
может сохранять активность в соке свыше года, не
инактивируется при высушивании и проходит через свечу Шамберлена
ht — L7.
Этот вирус методом химической очистки получен в чистом
виде и по серологическим свойствам, а также по некоторым
другим признакам близок к вирусу обыкновенной табачной
мозаики.
Кроме Англии, болезнь известна в СССР. А. М. Вовк
(1942) нашел ее у огурца в крытом грунте.
Наиболее распространенными вирусными заболеваниями
тыквенных являются заболевания, вызываемые огуречным
вирусом 1 и огуречным вирусом 2. Эпифитология этих болезней
совершенно различна, как и свойства вирусов, их вызывающих.
Первый является вирусом, широко распространенным у
различных семейств ж передаваемым преимущественно тлями;
второй встречается только у тыквенных и распространяется
исключительно при соприкосновении растений или же при
заражении их соком. Внешне оба вируса вызывают чрезвы-
140
чайно сходное заболевание. Удобный диагностический признак
описан А. М. Вовком. При погружении среза листа,
пораженного огуречным вирусом 2, в 0.1-нормальный раствор соляной
кислоты в клетках появляются многочисленные веретено-
видные паракристаллы. Этого не наблюдается ни в клетках
здорового огурца, ни при поражении огуречным вирусом 1.
В последнее время кратко описано нескодько вирусов,
поражающих тыквенные. Одни из них передаются тлями и
Diabrotica trivittata, другие, как кольцевая пятнистость тыквы
и мозаика Echinocystis, — только Diabrotica. Вирус мозаики,
передаваемый Diabrotica, инактивируется при 75° С,
сохраняется 6 недель in vitro и выдеряшвает разведение 1:1 000 000
(Freitag, 1941).
Глава XIII
ВИРУСНЫЕ БОЛЕЗНИ СЕМЕЙСТВА СЛОЖНОЦВЕТНЫХ
Вирусные заболевания сложноцветных мало изучены.
К числу более детально исследованных принадлежат только
вирусные болезни хризантемы, далии и салата.
Желтуха астры (Aster yellow). Заболевание вызывает
Callistephus virus 1, Kunkel (1924, 1926). Оно проявляется
у китайской астры [Callistephus chinensis) или хризантемы
общим хлорозом листьев, которому предшествует посветление
по ходу нервов. Наблюдаются подавленность роста и
усиленная кустистость. Соцветия подавлены в развитии, причем
можно наблюдать явления позеленения.
Вирус, вызывающий это заболевание, может поражать
различные сложноцветные, а также представителей разных
других семейств. У салата заболевание известно под названиями:
lettuce white-heart, lettuce «rabbit-ear», lettuce Rio ¦
Grande disease.
Пораженный салат хлорозен и страдает карликовостью.
Нормальное закручивание листьев в головку отсутствует, но
зато развиваются многочисленные боковые побеги. По краю
больных листьев часто образуются коричневые пустулы с
засохшим млечным соком. Вирус поражает одуванчик, циннию,
ноготки, Tagetes, различных представителей сем. Scrophu-
lariaceae (Kunkel, 1931).
Экономическое значение имеют заболевания, вызываемые
вирусом у зонтичных. Описание этих заболеваний дается
в соответствующей главе.
Вирус не передается соком, его переносчиком является
Cicadula (Macrosteles) diversa Uhl., ранее неправильно
определенная как С. sexnotata Калифорнийский штамм этого
вируса, который поражает преимущественно сельдерей,
передается двумя другими видами цикады: Thamnotettix montanus
Van D., Т. geminatus Van D. Первая из них может переносить
вирус и на хризантему, а вторая переносит его на сельдерей,
но не на хризантему (Severin, 1934). Вирус имеет в
насекомом-переносчике длительный инкубационный период. За-
151
болевание известно в СШАТ
Канаде, Японии (Fukushi)
и, вероятно, встречается
также в Западной Европе.
В СССР встречается
заболевание, которое В. Л.
Рыжков описал как желтуху
кок-сагыза. Это заболевание
вызывает подавленность
роста, хлороз, задержку
оттока углеводов и другие
характерные для желтухи
симптомы. Цветы под
влиянием заболевания
подвергаются антолизу. Заболевание
поражает Taroxacum, Cicho-
rium (рис. 38), Cirsiumr
Chrysanthemum, виды Plan-
tago, а также представителей
других семейств, например:
Valerianaceae, Solanaceae.
По нашему поручению
был выявлен переносчик
болезни, который оказался
цикадой Macro steles quadri
punctata. По всей вероятности,
заболевание вызывает один
из штаммов Callistephus virus
1 (Сухов и Вовк).
Для борьбы с болезнью
рекомендуется огораживать
грядки, на которых
культивируется Callistephus или
салат, сеткой из проволоки
или из легкой ткани, высотой
в 1.5 м. Этого барьера
достаточно для прекращения
доступа цикад.
Vinca rosea, пораженная
Callistephus virus 1,
излечивается водной баней в 32 —
42°С (Кшікеі, 1941, 1941а).
Борьба с насекомыми при
помощи пиретрума и других
способов уменьшает поражение салата (Linn, 1940).
Мозаичная болезнь далии (георгины). Наиболее
характерным признаком болезни является хлороз, который
распространяется диффузно по ходу жилок, но обычно не
152
Рис. 38. Соцветие цикория, больного
желтухой
образует правильной сети. В зависимости от сорта могут
наблюдаться также различные другие симптомы, как-то: светло-
зеленая и желтая пятнистость, — причем остающиеся
нормальными участки листьев вздуваются, — а также некротические
полосы- Нередко болезнь вызывает подавленность в росте
и усиленную кустистость. На венчике могут появляться
светлые полосы.
В клетках больных растений обнаружены Х-тела, которые
чаще прилегают к ядру и имеют круглую, овальную или
удлиненную форму.
Болезнь широко распространена в США и в Европе,
встречается также в СССР. Переносчик болезни — персиковая
тля; соком болезнь не передается. Возбудитель ее, по
классификации К. Smith, Dahlia virus 1, Brandenburg (1928).
Для борьбы с мозаикой далии рекомендуются отбор
здоровых клубней и систематическая борьба с тлями.
Кольцевая пятнистость далии (георгины). Болезнь
вызывает Dahlia virus 2, Brier ley (1933).
При заболевании наблюдаются зигзаги и
концентрические кольца, нередко образующие очень тонкий и красивый
рисунок на листе. Они имеют светлозеленый или желтый цвет
и позднее некротизируются. Встречается также заболевание,,
при котором рисунок отличается яркожелтой окраской. Вирусг
вызывающий его, повидимому, является одним из штаммов
Dahlia virus 2.
Болезнь не передается соком, и ее не удалось передать
персиковой тлей; способы ее распространения в природе
неизвестны.
Oakleaf далии. Заболевание вызывает Dahlia virus 3,
Brier ley (1933). Болезнь проявляется хлорозными полосками
неправильной формы, которые дают очертания, напоминающие
очертания листа дуба, откуда и происходит английское
название болезни (дуболистость). Иногда также наблюдаются
крупные хлоротические кольца.
Мозаичная болезнь салата. Заболевание известно в США,
Германии и Англии.Хотя строгой идентификации форм болезни,
встречающихся в разных странах, и не было произведено,
но можно думать, что болезнь вызывается везде одним и тем
же вирусом Lactuca virus l, Jagger (1921), который более
подробно был исследован Ainsworlh и Ogilvie, 1939.
У салата наблюдается посветление по ходу молодых
листочков, которое затем переходит в мозаичность или общий
хлороз. Листья больных растений более морщинисты, чем
это нормально свойственно растению. Наблюдается
подавленность роста, и иногда растение образует только розетку листьев.
Болезнь может быть передана целому ряду сложноцветных,
например, Senecic vulgaris, Sonchus asper.
Из сложноцветных к ней устойчив Sonchus oleraceus. BnpyG
153
вызывает тяжелые поражения у гороха и душистого горошка,
причем у первого заболевание ограничивается мозаикой,
а у второго развиваются и некротические явления. Передать
болезнь помидору, дурману, огурцу и капусте не удалось.
Заболевание легко передается семенами (Newhall, 1923),
соком, а также тлями. В качестве тлей-переносчиков в Америке
указаны Myzus persicae Sulz, в Англии, кроме того, Масго-
siphum gei, хотя последняя вызвала заражение только в одном
случае из 23. Целый ряд бывших в испытании тлей не
передавал болезни.
Вирус инактивируется при 60° С, сохраняет инфекцион-
ность в соке не более 48 часов и при разведении до 1:100. Он
не переходит через фильтры Шамберлена, а также через тонкий
слой инфузорной земли.
Для борьбы с болезнью следует рекомендовать отбор
семян от здоровых растений, усиленную борьбу с сорняками
и насекомыми, выбор более устойчивых сортов. На
возвышенных и открытых местах наблюдается меньше тлей и меньше
мозаичных растений, чем на низких местах и под деревьями.
Мозаика у салата встречается также в СССР. Другой вид
мозаичной болезни, переходящей к салату от одуванчика,
описан в Англии. Болезнь переносят Myzus ornatus и
Ж. pseudosolani, но не М. persicae (Kassanis, 1944).
Коричневая болезнь салата. Это заболевание описано Jag-
ger (1940) под названием brown blight. Очень близко к нему
стоит более тяжелое заболевание big vein (Jagger и Chandler,
1934). Болезнь выражается подавленностью роста и развитием
-желтых пятен; позднее пятна некротизируются, и листья
засыхают. Наблюдаются также подавленность в развитии
корней и отмирание. При big vein происходит припухание
ткилок листа, а также описанные выше некротические явления.
Болезнь передается почвой. На инфицированной почве
получается 100% больных растений. Этиология болезни неизвестна;
овоею связью с почвой она похожа на мозаичную болезнь
пшеницы, но неизвестно, является ли она вирусным
заболеванием. В эпидермисе корней больных растений часто
встречается грибок Asterocystis radicis, а у здоровых растений только
изредка.
Другие вирусные болезни сложноцветных. Сложноцветные
часто поражаются различными другими вирусами. Особого
упоминания заслуживает бронзовость листьев (Lycopersicum
virus 3), часто встречающаяся у далии и салата. Этот вирус
вызывает кольцевую пятнистость, некрозы и бронзовость.
Далия может быть резерватором этого вируса, и от нее он
может переходить на другие культурные растения, в частности
на помидор (Gardner, 1935; Harris, 1939; Pittmann, 1934).
У подсолнечника мне приходилось наблюдать мозаичное
заболевание, сопровождающееся резкой деформацией листьев.
154
Мозаика у подсолнечника описана также в Аргентине (Soriano $
1932). В. П. Ягодкина (1941) описывает у этого растения целый
ряд подозрительных на вирусные заболеваний: обыкновенную
зеленую мозаику, сопровождающуюся
деформацией листьев; желтую мозаику,
при которой деформации листьев не
наблюдается; резкую деформацию всего
растения, сопровождающуюся
скручиванием листьев в виде лодочки,
узловатостью стебля, ветвлением пучками из
одного узла и деформацией корзинок;
редукцию генеративных органов и,
наконец, некроз семян.
Данные об инфекционности
большей части этих болезней отсутствуют.
Опыты, которые должны
свидетельствовать в пользу вирусной природы
некроза семян, очень мало убедительны.
У подсолнечника иногда
наблюдается массовое превращение цветов в
короткие вегетативные побеги, в пазухе
листьев которых могут сохраняться
семяпочки. При этом заболевании
наблюдается некроз проводящей системы,
и, возможно, оно имеет вирусное
происхождение (Рыжков, 1939).
У топинамбура (Heliantus tuberosus)
B. П. Ягодкина (1941) наблюдала
мозаичное заболевание, которое соком
может быть передано на подсолнечник.
Мозаичные заболевания
встречаются также Gaizotia, Erigeron, y Cirsium
arvcnse и у лопуха. Мозаика лопуха
чрезвычайно широко распространена.
Иногда при ней наблюдается (рис. 39)
резкая деформация листьев, которые
становятся перистыми и нитевидными.
В анатомическом и морфологическом
отношении эта болезнь была описана
C. М. Шапиро. Из диких
сложноцветных мозаика встречена была у Doronicum на высоте 1300 м
над уровнем моря и автором этой книги — у одного из видов
Eapatoriam вблизи Карадагской биостанции.
Emilia scabra на Цейлоне страдает мозаикой,
распространяющейся по ходу нервов. Эта мозаика передается
прививкой (Loos., 1942).
Рис. 39. Лист лопуха,
пораженного мозаичной
болезнью
Глава XIV
ВИРУСНЫЕ БОЛЕЗНИ ЗЛАКОВ
Из многочисленных описанных в литературе заболеваний
злаков в настоящее время в СССР известны только закукли-
вание и мозаичная болезнь яровой и озимей пшеницы. С
описания этих заболеваний мы и начнем.
Закукливание. Первые сведения в литературе о закук-
ливании относятся к 1922 г., однако есть основания
предполагать, что болезнь встречалась и раньше на территории
Западной и Восточной Сибири и ДВК. Первое подробное
описание болезни принадлежит Донченко (1929). О причинах
этой болезни существовали различные предположения. Многие
рассматривали ее как физиологическое расстройство,
вызванное неблагоприятными экологическими условиями. Мураш-
кинский допускал тождественность заболевания с розеточнои
болезнью пшеницы, описанной в Америке. Работами Сухова
и Вовка (1938—1939) было установлено, что закукливание —
вирусная болезнь, передающаяся темной цикадой. Дальнейшее
описание заболевания дается преимущественно на основе работ
этих авторов.
Болезнь наиболее сильно и характерно поражает овес.
Первым признаком заболевания является мозаика, состоящая из
светлозеленых полосок (рис. 40). При очень слабых формах
болезнь проявляется только мозаикой, при более тяжелых
наблюдаются задержка роста, чрезмерная кустистость и порая?ение
цветов (рис. 41). При очень тяжелых поражениях больные
растения представляют собой карликов с огромным количеством
побегов; корневая система у таких карликов развита очень
слабо. Тяжело больные растения могут совсем не давать
метелки, в других же случаях метелка развивается ненормально
и цветы находятся в различных стадиях превращения в
вегетативные побеги. Нередко встречается развитие мелких корешков
в соцветии. Даже в тех случаях, когда метелка выглядит
более или менее нормальной, больные растения все же по
большей части стерильны.
156
У Setaria viridis при закукливании также наблюдается
пролиферация цветов. У других злаков, страдающих закукли-
ванием, болезнь проявляется мозаикой, карликовостью и
кустистостью, но соцветия не подвергаются пролиферации,
а только слабее развиты и стерильны.
Рас. 40. Листья овса: внизу — здоровый, вверху — пораженный
закукливанием (по Сухову)
В патолого-анатомическом отношении для болезни
характерен некроз флоэмы, который очень слабо выражен у
нормально развитых растений и достигает максимального
выражения у карликов.
В клетках больных растений встречаются крупные
белковые включения, имеющие вид колец, восьмерок, перевитых
жгутов'и более сложных фигур. Часто можно наблюдать рас-
лад этих включений на множество тонких кристаллов (рис. 42).
Описанные включения являются хорошим диагностическим
признаком закукливания. При закукливании встречаются
также описанные при розеточной болезни пшеницы Х-тела.
Согласно Рыя^кову, Громыко и Воробьевой, при
закукливании происходит переполнение надземных и вегетативных
органов крахмалом и растворимыми углеводами. Количество
общего, и особенно белкового, азота у больных растений по
большей части резко падает.
Закукливание не передается соком и почвой. Опыты по
выяснению роли семян неизменно давали отрицательный
результат — передачи семенами не наблюдалось. Переносчик
болезни — темная цикада: Delphax striatella Fall.
Закукливание, кроме овса, поражает пшеницу, ячмень,
рожь, кукурузу, просо, рис, а также целый ряд диких злаков,
например, Calamagrostis epigeios. Закукливанием страдают
157
самые различные виды и сорта овса и пшеницы. Рожь очень
мало чувствительна к заболеванию и поражается им в
небольшом проценте.
Развитие закукливания находится в тесной связи с
развитием темной цикады и миграцией ее на посевы овса.
Эта цикада зимует в стадии личинки третьего возраста,
причем, повидимому, главное значение в зимовке вируса имеют
личинки, в теле которых вирус может
сохраняться. Цикада располагается на
зимовку вблизи поля, где находился
овес. На расстоянии 1 м от поля овса
было найдено от 700 до 800 личинок
на 1 м% тогда как на расстоянии 2 м
их было только 20 на ту же самую
площадь. Переселение цикад на
посевы овса в Омской области обычно
начинается во второй половине мая.
Закукливанием наиболее
поражаются посевы овса, которые к этому
времени достигают ранних стадий
развития. Таблица 11 указывает на
связь, которая существует между
сроком посева овса, заселением ее
цикадами и процентом
закукливания.
У Изреженные посевы овса
страдают закукливанием значительно
сильней, чем густые. В соответствии с
этим установлено, что число
насекомых, приходящихся на одно
растение на густых посевах,
значительно меньше, чем на изреженных.
На посевах, имевших 800 растений
на 1 м2, на одно растение
приходилось от 0.09 до 0.19 цикад, а на
посевах, имевших 300 растений на
1 м2, на одно растение приходилось
от 0.67 до 1.50 цикад.
Овес поражается тем сильней, чем раньше он был заражен,,
поэтому повторные генерации цикад не имеют такого большого
значения в распространении болезни, как первая их генерация.
Поражение озимой пшеницы закукливанием, как показали
наблюдения Вовка, также находится в корреляции с
миграцией цикады. Наиболее ранние посевы озимой пшеницы
поражаются наименее. Именно эта пшеница успевает ко времени
появления новой генерации цикад достаточно подрасти, чтобы
не страдать сильно от болезни.
Радикальные меры борьбы с болезнью в настоящее время
158
Рис. 41. Овес, больной
закукливанием
Таблица 11
Поражение ранних и поздних посевов овса темной цикадой
и закукливанием (по Сухову и Петлюку)
Срок посева
8 мая
11 »
20 »
1 июня
Срок учета
насекомых
10 июня
10 »
10 »
10 »
Число насекомых
на 1 ма
570
200
немного окз.
70
Срок учета
болезни
24 июня
29 »
29 »
24 »
% закуклива-
ния
89*7
41.0
3.8
10 0
еще не разработаны. Следует всячески избегать возможного
разрежения посевов овса. Вопрос о сроках сева очень сложен
и должен решаться конкретно в применении к различным
Рис. 42. Кристаллы вакукливания в клетках пшеницы
районам (Хайрулин, Сухов, Вовк, Брызгалова1 (1941). Наи~
большая поражаемость овса наблюдается на апрельских
посевах, а наименьшая — на посевах последней декады мая.
Выбор сроков сева овса, конечно, решается не только распро-
1 См. в Сб. «Вирусные болезни растений и меры борьбы с ними»-,
., 1941.
159
етранением закукливания, но и, в первую очередь, всей
совокупностью хозяйственно-экономических соображений.
Все меры, направленные к уничтожению темной цикады,
особенно во время ее зимовок, должны дать существенный
эффект в деле борьбы с закукливанием. Следует рекомендовать
после сбора урожая стерню лущить и как можно скорее
запахивать, избегать посевов овса по овсу как предшественнику
и вести борьбу с сорняками, особенно из семейства тех злаков,
которые являются основными резерваторами вируса и цикад
в поле.
До сих пор неизвестны сорта, устойчивые к закуклива-
нию, — выведение таких сортов является первоочередной
задачей. Следует обратить внимание на то, что пырей в
высокой степени устойчив к закукливанию и некоторые из пше-
нично-пырейных гибридов Цицина наследуют эту
устойчивость.
Закукливание злаков широко распространено в
азиатской части нашего Союза; оно переходит также Урал и
проникает в европейскую часть Союза. По указаниям Петруковича,
оно встречается в Кировской области. Дальше на запад
закукливание сменяется мозаичной болезнью озимой и яровой пшеницы,
а если и встречается в Центральной части нашего Союза, то в
совершенно ничтожном количестве.
Мозаичная болезнь озимой и яровой пшеницы. Мозаичная
болезнь озимой и яровой пшеницы, так же как и закукливание,
первоначально была принята за розеточную болезнь.
Заболевание было изучено Зажурило и Ситниковой (1939, 1940, 1941),
которые нашли и его переносчика.
Первые симптомы мозаичной болезни на озимой пшенице
появляются осенью, на 15—20-й день после появления всходов.
Мозаичные полосы бывают сначала светлозеленого цвета,
но позднее принимают интенсивную желтую окраску. У боль-
пых растений наблюдается карликовость и чрезмерная
кустистость. У озимой пшеницы болезнь наиболее выражена
на верхних листьях (рис. 43), тогда как нижние могут совсем
не обнаруживать мозаики. У яровой пшеницы, наоборот,
наиболее резко мозаика выражена на нижних листьях. В
зависимости от сортов и других условий пораженные участки
бывают в большей или меньшей степени лишены хлорофилла.
У овса и ячменя мозаика выражена желто-зелеными полосами.
При мозаичной болезни пшеницы, как и при закукливании,
наблюдаются некрозы флоэмы и внутриклеточные вакуолярные
тельца. Кроме того, отмечается гипертрофия ядер,
сопровождающаяся увеличением числа ядрышек и недоразвитостью
пластид. Кристаллические включения в клетках больных
растений при мозаичной болезни не обнаружены. Сухов показал,
что если к соку, выжатому из больных мозаикой растений,
прибавить раствор какой-либо кислоты до рН 4—5, то можно
160
наблюдать выпадение в этом соке игловидных кристаллов
и тонких нитей, что происходит также в слабых растворах
кислоты. В дальнейшем Сухову при применении кислых
Рис. 43. Пшеница: слева — здоровая,
справа—пораженная мозаичной
болезнью (по Запурилло)
—г
фиксаторов удалось обнаружить кристаллические включения
и в клетках пораженных мозаикой растений. В живых клетках
ігри мозаичной болезни пшеницы кристаллы отсутствуют;
Л В. Л. Рыжков 161
не бывает их также при фиксации материала спиртом, тогда
как при закукливании овса спирт хорошо сохраняет кристаллы.
При мозаичной болезни кристаллы обнаружены только в
клетках, окаймляющих сосудистоволокнистый пучок, а при
закукливании они находятся в различных тканях мезофилла.
При мозаичной болезни, как и при закукливании,
количество растворимых углеводов и крахмала в листьях пораженных
растений возрастает (Ряховский, 1941).
Мозаичная болезнь, изученная в Воронежской области,
не передается ни соком, ни почвой, ни семенами. Из большого
количества испытанных насекомых ее не передают цикады
Cicadula sexnotata^ Delphax striatella, трипе — Haplothrips
tritici, виды черепашки Eurygaster integriceps, E. maurus,
E. austricus и клоп Aelia acuminata.
Единственный переносчик болезни — полосатая цикада
(Deltocephalus striatus). Это насекомое инфицируется только
в личиночном возрасте, через яйцо вирус не передается. Так как
цикада зимует в виде яиц, то резерватором вируса на зиму
она служить не может. Вирус сохраняется в озимой пшенице
и, вероятно, также в многолетних сорняках.
Мозаикой озимой пшеницы могут болеть различные виды
пшеницы, ячмень, рожь, овес, просо, могар, рис. Последние
три рода мало подвержены заболеванию. Не поражаются
воронежской мозаикой кукуруза, суданка, ползучий пырей,
безостый костер, зеленый мышей.
Густые посевы поражаются заболеванием меньше, чем
изреженные. Для борьбы с болезнью рекомендуются лущение
и запашка жнивья, что ведет к уменьшению количества
цикад.
Мозаичные заболевания пшеницы, описанные в США.
Весной 1919 г. в штате Иллинойс была обнаружена на озимой
пшенице розетковидность и было доказано, что она отличается
от австралийской болезни take-all, которая вызывается ОрМ-
obolus granitis (Мс. Kinney, 1923). В дальнейшем Мс. Кіпшу
(1930) показал, что эта болезнь может быть отнесена к числу
вирусных и встречается в двух формах — зеленой и желтой
мозаики. Впоследствии тот же автор подверг сравнительному
изучению целый ряд мозаичных заболеваний пшеницы.
Учитывая, что симптомы болезни сильно варьируют в зависимости
от внешних условий и от сорта, все опыты были поставлены
на одном стандартном сорте и при относительно константных
условиях: в камерах при 15.5° С и при продолжительности
освещения — 8 часов ежедневно. Результатом работы было
описание семи разных вирусов или, точнее, разных картин
болезни, потому что, как признает сам автор, некоторые из
вирусов представляют только штаммы одного и того же вируса.
Можно различать два типа описываемых ниже вирусных
заболеваний; одни из них передаются почвой, другие не пегреда-
162
ются. Первые преимущественно встречаются в западной части
США, вторые — в восточной.
Остановимся, прежде всего, на розеточной болезни. Кроме
США, она, повидимому, встречается в Японии (Е. Wada и
Н. Tukado, 1937). Болезнь поражает озимые сорта пшеницы.
Известны две формы этой мозаики — зеленая и желтая.
Главное различие между ними состоит в том, что при первой
пораженные участки светлозеленые, а при второй — желтые. При
зеленой мозаике мозаичность выражена очень слабо и часто
сменяется расстройствами роста. Карликовые и кустистые
Рас. 44. Пшеница: здоровый колос (А) и
пораженные ролегочной болезнью (В) (по Мс. Kinney)
растения вовсе не несут мозаики, — напротив, они окрашены
в зеленый цвет более интенсивно, чем нормальные растения.
При тяжелых формах болезни сильно страдает колос: он несет
меньше нормальных зерен, и размер этих зерен уменьшается
(рис. 44). Возможна более или менее полная маскировка болезни,
особенно к концу сезона. На отдельных растениях озимой
пшеницы болезнь появляется уже поздно осенью.
Необходимо иметь в виду возможность смешать розеточную
болезнь с некоторыми другими поражениями.
Карликовость, зависящая от неблагоприятных почвенных
условий, появляется на поле, так же как и розеточная болезнь,
в вице группы низкорослых растений, однако при розеточной
И* 10&
болезни, переход от нормального участка поля к пораженному
бывает обычно резким, а при карликовости пшеницы, зависящей
от дефектов почвы, он носит менее резкий характер.
Поражение злаков гессенской мухой дает такую же темно-
зеленую окраску, как и розеточная болезнь, однако насекомое
не вызывает усиленного развития придаточных побегов.
Труднее диагностики диференциальная диагностика
мозаичной болезни от мозаики, возникающей на листьях злаков
вследствие расстройства минерального питания. Решающую
роль в диференциальном диагнозе здесь должен играть
эксперимент, доказывающий инфекционность болезни.
В природных условиях розеточная болезнь
распространяется через почву. Заразное начало в известных почвах моя^ет
храниться не меньше шести лет. Были поставлены опыты,
показавшие, что заражение пшеницы происходит и тогда,
з^огда инфекционная почва расположена ниже зерна
(28—55% заражения), и тогда, когда она находится выше
зерна (22—39% заражения). Если пораженная почва
находится и сверху и снизу зерна, то процент зарая^ения будет
особенно велик (70—95%). Вирус не может быть извлечен
из почвы водой, и мы до сих пор не знаем, в каком состоянии
он находится в почве и каков механизм заражения.
Дезинфекция почвы формалином в разведении 1:49 уничтожает
инфекцию. Заражение в инфицированной почве наилучше
удается при посеве растений в эту почву с осени. Болезнь
передается также инокуляцией сока, причем успех такого
заражения довольно непостоянен: он колеблется от 0 до 45%
для зеленой мозаики и от 16 до 100%—для желтой. Интересно,
что при перевивке вируса зеленой мозаики этот последний
постепенно утрачивает вирулентность и перестает давать
заражение. Успех искусственного заражения сильно зависит от
температурных условий. Оптимальной для развития болезни
является температура 15° С.
Вирус розеточной болезни у К. Smith назван Triticum
virus 1, Мс. Kinney, а соответственный вирус желтой мозаики
рассматривается как желтый штамм этого вируса. У Мс. Kinney
(1937) первый вирус обозначается № 1, а желтый — № 3.
Приводим ниже краткое описание остальных вирусов
из работы Мс. Kinney
Вирус пшеницы 2 не передается почвой. Он обладает
способностью заражать ползучий пырей {Triticum repens),
который устойчив ко всем остальным известным вирусам
пшеницы. Соком он передается в 60—87% случаев. Он вызывает
очень слабую светлозеленую крапчатость, легко
маскирующуюся после выхода листа из трубки. Образования розеток
этот вирус не вызывает.
Вирус пшеницы 3, вызывающий желтую мозаику,
передающуюся почвой, описан выше.
1в4
Вирус пшеницы 4 вызывает светлозеленую мозаику.
Большая часть листьев пораженного растения имеет светлозеленые
или желто-зеленые полосы, а иногда эту же окраску принимают
и более крупные участки листа. Вирус вызывает подавленность
роста у пшеницы, но не приводит к образованию розеток. Почвой
он не передается, успех искусственного заражения соком
выражается в 0—100%. Вирус обнаружен в штате Канзас.
Вирус пшеницы 5 вызывает симптомы, очень сходные
с симптомами поражения вирусом 3, однако желтая окраска
пораженных участков здесь выражена сильнее. Растения очень
подавлены и часто погибают. При инокуляции соком
получается 5—85% удачного заражения. Вирус обнаружен также
в штате Канзас.
Вирус пшеницы 6 вызывает па листьях пшеницы светло-
зеленые или желто-зеленые полосы, которые тянутся вдоль
всего листа. Растения подавлены в росте, но не образуют
розеток. Удачное заражение выражается в 20—100%. Вирус
обнаружен в штате Канзас.
Вирус пшеницы 7 также вызывает полосы, которые часто
тянутся по всей длине листа, но они желтого цвета, кроме
того наблюдается яркожелтая пятнистость. В пораженных
участках хлорофилл редуцирован меньше, чем это бывает
при мозаике, вызванной вирусами 3 и 5, но в большей степени,
чем при мозаике от вирусов 4 и 6. Вирус обнаружен также
в штате Канзас и, как все вирусы этой группы, не передается
на пырей, — нет также данных о передаче его почвой.
Приведенный перечень показывает, в каком
неразработанном состоянии находится область изучения вирусных болезней
злаков в США. Наилучше описана розеточиая болезнь, однако
здесь связь инфекции с почвой осталась до сих пор
неразъясненной. Что касается группы мозаик из восточных штатов
США, то мы о них почти ничего не знаем.
Карликовость риса. Первые симптомы болезни,
вызываемой, согласно номенклатуре К. Smith, Oryza virus 1, Fukushi,
состоят в желто-белых полосках вдоль жилок листа на вновь
развившихся листьях. Симптомы болезни особенно резко вы-
ражены'в середине июля. Развитие растений резко подавлено,
наблюдаются укорочение междоузлий и кустистость
пораженных экземпляров. Особенно тяжелые симптомы наблюдаются
у рано заболевших растений.
В пораженных клетках наблюдаются Х-тела, имеющие
овальную или амёбовидную форму и снабженные вакуолями.
Они достигают 10 у. в длину и 8.5 \і в ширину.
Болезнь известна в Японии. Пи соком, >и почвой, ни
семенами она не передается; ее переносчиком является цикада
Nephotettlx apicalis (Fukushi, 1934).
От карликовости риса отличается заболевание, описанное
в Японии в 1931 г. под названием slripe-clisea&e и вызываемое
1«Г>
Oryza virus 2, Kuribayashi (1931). Это заболевание, как и
предыдущее, проявляется карликовостью растений, и, кроме того
при нем наблюдаются желтые полосы вдоль листа. Болезнь,
передается цикадой Delphax stiratella, которая является также
переносчиком закукливания злаков.
Выше было упомянуто, что рис болеет закукливанием.
Интересно было бы выяснить, не является ли описываемая
здесь болезнь, о которой мы располагаем лишь краткими
сведениями, тождественной с закукливанием или близко к нему
стоящей. При этой болезни, однако, кристаллов в клетках
не обнаружено, но имеются в мезофилле и в замыкающих
клетках маленькие Х-тела (1.5 ^ — 4.5 \і) — см. J. Kawai.
Необходимо также отметить, что у риса встречаются
разнообразные заболевания невыясненной этиологии, например
запал, часть которых может оказаться известными или еще
неизвестными вирусными заболеваниями.
Вирусные болезни кукурузы. Описан целый ряд вирусных
заболеваний кукурузы, известных главным образом в
тропических странах. Внешне они обладают очень сходными
признаками, но если принять во внимание, что симптомы каждой
болезни, в связи с сортовыми отличиями п условиями
существования, сильно варьируют, то мы должны будем сказать,
что хотя по внешним симптомам эти болезни трудно отличимы,
но различаются между собой довольно хорошо, потому что
по большей части имеют разного переносчика. Выше мы
видели, что кукуруза может болеть закукливанием. Мы
располагаем данными о том, что в СССР у кукурузы встречаются
заболевания, похожие на вирусные, но ближе мы о них ни
чего не знаем (см., например, Е. М. Эристави, 1941).
Дадим здесь краткий перечень известных в литературе
заболеваний кукурузы.
Заболевание кукурузы вирусом мозаики
сахарного тростника встречается в странах, где
распространена культура сахарного тростника. Оно проявляется
светлыми полосами неправильной формы. Пораженные участки
имеют светлозеленый или желтый цвет. Болезнь распространяет
тля Aphis rnaidis Fitch.
Трансваальская мозаика трав поражает кукурузу и
различные виды сорго. Она, как показывает название, встречается
в Трансваале. Симптомы, вызываемые этой болезнью, не
отличимы от симптомов, которые вызывает у кукурузы Saccharum
virus l.
Переносчик этой болезни, как и предыдущей, — Aphis
rnaidis. Главное отличие ее от мозаики сахарного тростника
состоит в том, что она не передается сахарному тростнику.
К. Smith считает вирус, вызывающий ее, штаммом вируса
мозаики сахарного тростника, хотя экспериментальные
доказательства этого отсутствуют (Storey, 1929).
1W
Мозаичная болезнь кукурузы вызывается, по К. Smith,
Zea Virus 1, Kunkel (1927). Другое ее название — stripe disease.
Первые симптомы болезни обнаруживаются в виде маленьких
белых полосок у основания молодых листьев. Эти полосы
удлиняются и нередко сливаются между собой: в отдельных
случаях получаются довольно широкие ленты желтого цвета.
У некоторых сортов на пораженных листьях позднее
развиваются тёмнокрасные полосы, которые затем, вследствие
оттирания ткани, коричневеют. В пораженных клетках можно
наблюдать распад пластид. Иногда во флоэме образуются
некрозы. Переносчиком заболевания является Peregrinus
maidis Ashm. Инокуляцией иглой болезнь не передается.
Болезнь не передается на сахарный тростник, но в Тринидаде
встречается в естественных условиях на сорго (Kunkel, 1927).
Заболевание известно на Кубе, Гавайских островах, в
Тринидаде. При заболевании наблюдается некроз флоэмы (М. Cook,
1936).
Полосатость маиса или streak, по классификации К. Smith,
вызывается Zea virus 2, Storey (1925). Болезнь проявляется
-светлыми и часто белыми полосами, которые несколько
отличаются от полос, наблюдаемых при мозаичных заболеваниях,
тем, что они почти никогда не бывают, непрерывными.
Переносчики болезни — цикады: Cicadulina mbila Naudi, С. zeae,
С. nickolsi (Storey, 1931), — причем первая из этих цикад
является важнейшей. Соком болезнь не передается. Вирус
лишь с трудом фильтруется'через свечу Шамберлена и не
фильтруется через диски Зейца (Storey, 1932). Болезнь встречается
в различных частях Африки.
Крапчатость маиса, возбудителем которой является Zea
virus 3, Storey (1937), переносится теми же насекомыми, что
и полосатость. Болезнь эта проявляется только на молодых
листьях диффузными крапинками нормально зеленой ткани
на более светлом фоне. У взрослых листьев всякие признаки
болезни исчезают, и она снова проявляется на молодьіх листьях.
Болезнь известна в Танганайке (Вост. Африка). Под названием
crazy-top у маиса описано заболевание неизвестной этиологии,
при котором наблюдаются кустистость соцветия и превращение
его в вегетативный побег (КбЫег). В Калифорнии описана
мозаика, сопровождающаяся уродованием соцветий и, повиди-
мому, передающаяся цикадой Baldulus maidis (Frazier, 1945).
Мозаичная болезнь сахарного тростника. По К. Smith,
ее вызывает Saccharum virus l, Brandes (1920). Болезнь впервые
стала известна в 1892 г. на Яве. В 1908 г. она становится
известной на Гавайских островах, в 1919 г. — на Филиппинах,
а в 1921 г. обнаружена в Индии. В настоящее время заболевание
встречается в Аргентине, Перу, Индии, на Формозе, на Фиджи,
в Австралии, в Новой Гвинее, на Кубе, в Порто-Рико,
в Египте и т. д.
167
Первичные симптомы болезни выражаются в появлении
на листе полосок светлозеленого или желтого цвета Ткани
пораженных участков иногда отмирают, и тогда полоски
приобретают коричневую или красную окраску. При развитии
вторичных симптомов наблюдается более или менее
значительная задержка роста. Растения достигают только трети
нормального роста, стебель значительно тоньше нормального,
и на нем появляются полосы, которые могут некротизироваться.
Иногда на стебле наблюдаются трещины, и стебель может
искривляться, что является следствием тяжелых повреждений
с одной стороны стебля. К основному заболеванию могут
присоединиться повреждения грибками.
Болезнь переносит Aphis rnaidis, а также Corolinaia
cyperi, Hysteroneura setariae (Ingram и Summers, 1938), хотя
эти последние, как переносчики, имеют меньшее значение
(Tate Н. D. и Vandenberg, 1939). Кукурузная тля живет на
различных культурных и диких злаках; на кукурузе, Рапісит
colonum, Paspolum sanguinale, Eleosine indica, Setaria
saleata.
Во время засухи насекомые обычно эмигрируют на
сахарный тростник (Chardon и Veve, 1923). Инкубационный
период — около 18 дней. Соком болезнь почти не передается^
хотя в отдельных случаях ее передача удавалась.
Из культурных злаков описываемый здесь вирус поражает
кукурузу, просо и сорго. К нему устойчивы рожь, пшеница,
ячмень, овес и рис.
Вирус в соке при комнатной температуре быстро
утрачивает активность (в течение 2 часов), но при — 6° Сможет
сохранять активность в течение 27 дней. При разведении 1:100
вирус становится неактивным, так же как и после нагревания
до 53—54° С. Вирус не проходит через фильтр Шамберлена
и задерживается даже фильтровальной бумагой (Rafay).
Он обладает антигенными свойствами, и была получена
специфическая к нему сыворотка (Desai, 1935).
Мозаичная болезнь сахарного тростника приносит
огромный ущерб, как это можно видеть из следующих цифр,
заимствованных у Lyon, 1921 (Гавай):
Урожай с 1 акра здорового и больного сахарного тростника
(в т)
Тростника Сахара
От здоровых растений 101,13 14.98
От больных растений 56.24 8.4-*
Разница 44.89 6.55
В борьбе с мозаичной болезнью сахарного тростника
решающую роль сыграло введение устойчивых сортов. Эти
сорта получены путем скрещивания разных видов сахарного
тростника с Saccharum officinarum. К числу устойчивых видов
US
относятся: Saccharum spontaneum, S. robustum. Из устойчивых
сортов сахарного тростника можно привести Р. О. J. 2714,
2725, 2727, Р. О. J. 36, 213 и т. д. Введение устойчивых сортов
лишило болезнь того грозного значения, которое она нередко
приобретала.
Полосатость сахарного тростника. Болезнь известна в
Южной Африке, где она нередко дает до 100% поражения. Болезнь
проявляется неправильными хлорозными полосами, которые
позднее некротизируются и коричневеют. Заболевание не
передается соком, и его переносчиком являются те же цикады,
которые передают полосатость кукурузы.
Заболеванию подвержены некоторые сорта сахарного
тростника, которые иммунны к Saccharum virus 1, например
Uba. Заболевание, повидимому, вызывается штаммом того
же вируса, который вызывает полосатость у кукурузы. Вирус
полосатости сахарного тростника вызывает у кукурузы лишь
очень слабое заболевание, точно так же полосатость кукурузы
при переносе на сахарный тростник дает только у немногих
сортов слабое заболевание, причем больные растения легко
выздоравливают, совсем утрачивая вирус. Полосатость от маиса
может быть передана сорному злаку Digitaria horizontals,
а от этого злака она легко передается сортам сахарного
тростника, которым непосредственно от маиса не может быть
передана. Очевидно, мы имеем дело с большой лабильностью
вируса, свойства которого меняются при пассаже через разные
организмы (Storey и Мс. Lean, 1930).
Не будем здесь останавливаться сколько-нибудь подробно
на заболевании, вызываемом Saccharum virus 4, Storey, очень
похожем на полосатость кукурузы, но которое не передает
кукурузе или сахарному тростнику, ни Cicadulina, ни Percgrimis
maidis. Это заболевание встречается только у одного сорта
сахарного тростника—RP 8. Также не останавливаемся на
полосатости, описанной в 1931 г. (Cuban streak) у отдельных
экземпляров сорта P. O.J. — 2725, переносчик которого также
неизвестен (Proide, 1933).
Болезнь Sereh. Это заболевание известно с 80-х годов
прошлого века, когда оно внезапно появилось в восточной
части Явы. В настоящее время оно встречается на островах
Малайского архипелага, в Индии и Австралии. Свое название
болезнь получила вследствие того, что пораженные растения
приобретают внешнее сходство с явайской лимонной травой
(Andropogon Schoenanthus L.), носящей туземное название
sereh. Возбудитель этого заболевания — Saccharum virus
3, Kunkel.
Болезнь характеризуется резким сближением узлов
вследствие сильного укорочения междоузлий. Получаются кусты,
несущие на коротком стебле многочисленные листья. Очень
часто у больных растении развиваются придаточные корни
из узлов у основания лиственных влагалищ. В анатомическом
отношении болезнь характеризуется отложением красного
гуммиподобного вещества в сосудах, особенно в области узлов.
Сердцевина нормального сахарного тростника наполнена
сочными клетками. При заболевании они отмирают и
становятся сухими.
Неизвестен другой способ распространения болезни, кроме
передачи ее дри вегетативном размножении черенками.
Имеется указание Wilbrink (1923), что нагревание черенков
до 50° С вызывает их выздоровление. Надежной мерой борьбы
с заболеванием может служить введение устойчивых сортов,
к числу которых принадлежит Р. О. J. 2878. Введение
этого сорта на Яве сделало болезнь незаслуживающей там
внимания.
Болезнь Фиджи. Заболевание вызывает Saccharum virus 2,
Kunkel. Впервые оно стало известно на о. Фиджи в 1910 г.
на культурном и диком сахарном тростнике. В настоящее
время мы его знаем на Яве, Филиппинах, Новой Гвинее и
в Австралии. Это вирусная болезнь, при которой
развиваются опухоли в виде удлиненных галлов по ходу
нервов с нижней стороны листа. В течение нескольких
месяцев галлы могут быть единственным симптомом
заболевания. Позднее внезапно возникают тяжелые поражения молодых
листьев, разворачивание которых затрудняется, и эти листья
имеют уродливый, обожженный вид. Вскоре после этого
растение погибает. Болезнь не передается соком. В Квинсленде
болезнь передает цикада Perkinsiella satcharicida Kirk. (Mun-
gomery и Bell, 1933), а на Филиппинах Perkinsiella vastatrix
(Ocfemia, 1934).
Борьба с болезнью ведется путем введения устойчивых
сортов. К сожалению, неизвестно ни одного сорта, абсолютно
иммунного к болезни.
Карликовость сахарного тростника. Заболевание вызывает
Saccharum virus 5, Bell. Оно описано в Квинсленде Bell в 1933 г.
Его симптомы — карликовость и своеобразное уродование
листьев, зависящее от того, что сосудисто-волокнистые пучки
листа гипертрофируются, приобретают неправильное
расположение и сливаются между собой. Хлорофиллоносные
влагалища пучков редуцируются до немногих клеток, сильно
гипертрофируется древесинная часть пучка; флоэма либо
отсутствует, либо разбита на ряд пучков одеревеневшей ткани.
Близко к этой болезни стоит multiple bud disease, описанная
в Луизиане (Tims, 1939).
Огуречный вирус (Cucumis virus 1) у злаков. Огуречный
вирус может поражать рожь, кукурузу, а также Euchlaena
mexicana, Sorghum. Эти поражения, однако, наблюдались
при искусственном заражении. У кукурузы этот вирус вызывает
на месте питания вирофорной тли маленькое первичное нора-
170
жение в виде светлой полоски, за которым развивается общая
мозаика, состоящая из многочисленных эллиптических пятен
различной длины и ширины. Иногда присоединяются
некротические явления, и растения могут быть резко подавлены
в росте (Wellman, 1934).
Вирусная болезнь сорго, Chertan и Kulasam в 1937 г. описали
мозаичное заболевание сорго. При тяжелой форме болезни
(frecklead yellow) наблюдается сильная деформация листьев.
Болезнь передает Peregrinus maidis. Возможно, болезнь
вызывает Zea virus l.
Глава XV
ВИРУСНЫЕ БОЛЕЗНИ ОТРЯДА ЛИЛЕЙНЫХ
Мозаичная болезнь лука. Заболевание было описано в 1937 г-
В. Л. Рыжковым и А. М. В обком.
Болезнь начинается светложелтыми или белыми полосами
па листьях, в дальнейшем наступают общий хлороз листьев
и их полегание. Соцветие у больных растений часто резко
изменяется, цветоножки неравномерно удлиняются (рис. 45),
венчик становится сростнолепестным, тычинки некротизируют-
ся, а пестик нередко распадается на листочки. Луковицы
больных растений имеют удлиненную форму и часто не
переходят в период покоя, легко прорастая зимой (рис. 46). Они
нередко используются в хозяйствах для зимней выгонки лука.
При заболевания наблюдается уродливое развитие устьич-
ного аппарата (рис. 47). В пораженных клетках пластиды
резко измельчены, в клетках возле ядра нередко наблюдаются
крупные Х-тела.
Болезнь легко передается соком, а также различными
видами тлей, например Myzus persicae, Aulacorthum pelar-
gonii. В распространении болезни, вероятно, главную роль
играет посев лука рядом с плантациями семенного лука,
который резервирует в себе вирус. По невыясненным причинам
плантации семенного лука, засаженные более мелкими
луковицами, страдают от болезни меньше, чем засаженные более
крупными. В большом опыте, охватывавшем свыше 30 тысяч
растений, у лука, выросшего из луковиц, имевших 3 см в
диаметре, было только 0.54—2.43% больных растений, а у лука
с луковицами диаметром 5—7 см — от 14 до 32%.
Инактивация вируса происходит при температуре около
70° С. Вирус не фильтруется через свечи Шамберлена.
Искусственное заражение легко удается кашицей больных
листьев, но соком, процеженным через фильтровальную бумагу,
оно уже не удается.
Н. И. Андреев (1937) и Brenner (1937) смешали ото
заболевание с описанной ниже желтой карликовостью. От этой
последней мозаичная болезнь лука отличается порая^ением
172
луковиц, цветов, а также тем, что вирус мозаичной болезни
не фильтруется через бактериальный фильтр.
Желтая карликовость лука. Болезнь вызывается Allium
virus l, Melhus (1929). Начинается болезнь узкими желтыми
полосками, которые позднее увеличиваются в размерах.
Листья закручиваются вниз, и растение
подавлено в развитии. Нередко
наблюдается маскировка болезни:
соцветие нормально, но уменьшено в
размерах, луковицы также имеют
нормальные очертания. Вирус может
поражать чеснок, виды лилий и
нарциссы.
Заболевание известно в США и
было указано для СССР и Германии,
где его смешали с мозаичной
болезнью. Возможно, однако, что, наряду
с мозаичной болезнью, в этих странах
встречается и желтая карликовость.
В окрестностях Лиссабона болезнь,
кроме лука, поражает Muscari ca-
mosum, Narcissus tazelta и чеснок
(d'Olviera, 1941).
Болезнь передается соком и
самыми различными видами тлей.
Было испытано более 50 видов тлей,
и все они оказались способными
переносить болезнь (Drake. 1933, и др.)-
В то же время другие сосущие
насекомые — трипсы, червецы и
цикады—не могут быть ее переносчиками.
Инактивация вируса (Tate, 1940)
происходит при 75—80° С; он
утрачивает инфекционность при разведении
1:10 000. В соке вирус утратил
активность через 100 часов.
Для борьбы с вирусными
болезнями лука можно рекомендовать
пространственную изоляцию лука
первого года от лука, культивируемого
для семян, и усиленную борьбу с
тлями.
Полосатость тюльпана. Это заболевание известно под
названием breaking. При нем наблюдаются тёмнокрасные и
лишенные антоциана полосы на лепестках. Этот признак ранее
считался сортовым, и он, по сообщению Celusius, известен
в турецких садах с 1555 г. Celusius упоминает, что луковицы
передают окраску венчика, характерную для этого заболевания,
173
Рис. 45. Цветы лука:
слева — нормальный,
справа — уродливый
тогда как при разведении тюльпана семенами получить
ее нельзя.1 В 1928 г. было установлено, что мы имеем дело
в данном случае с вирусным заболеванием, которое может
передаваться и соком и тлями (Сауіеу, 1932; Мс. Kinney, 1931).
Рис. 46. Луковицы: вверху — здорового лука, внизу — больного
мозаичной болезнью
По классификации К. Smith, болезнь вызывается Т и 1 і р а
virus 1, Cayloy, однако Mc.Whorter (1932, 1938, 1939)
различает два вируса: один из них вызывает утрату эпидермисом
лепестков антоциана, а другой, напротив, усиление красной
окраски. Эти вирусы довольно трудно различимы. Температура
инактивации обоих лежит между 65—70° С. Оба они
выдерживают разведение до 1:100 000 и устойчивы к высушиванию.
Отбирая растения, у которых почти отсутствуют более темные
полосы на лепестках, удается отделить в чистом виде вирус,
вызывающий утрату антоциана. То же может быть сделано
для вируса, вызывающего усиление антоциановой окраски,
но с гораздо большим трудом. В чистом виде его удалось
изолировать только от одного растения. Вирус, вызывающий белые
полосы на лепестках, также вызывает хлорозные крапинки
на листьях, второй вирус этим действием не обладает. Путем
смешивания сока растений, из которых каждое поражено только
одним из этих вирусов, и инокуляции его здоровым тюльпанам
1 Цит. по Jones.
174
можно снова соединить оба вируса, но это не всегда удается,
так как вирус, удаляющий антоциан, часто берет верх и
проявляется один. Смесь вируса, удаляющего антоциан, с вирусом*
усиливающим его образование, в отношении 1:10 в опытах
Ри< 47 Уродливые устьица мозаичного лука
Мс. Whorler (1938) наиболее часто давала синтез типичного
breaking.
При заболевании в клетках образуются шарообразные
и эллипсоидные Х-тела, чаще лежащие возле ядра. Они
нередко имеют вакуоли. Сухов и Вовк (1945) нашли в больных
растениях белковые кристаллы. В кислых фиксаторах Х-тела
распадаются на игловидные кристаллы.
При иску ссі венном заражении тюльпана заболевание
по большей части проявляется только на другой год. У
некоторых видов лилии, например L. longiflorum, болезнь
проявляется уже на 14-й день характерной крапчатостью листьев
(Вгіегіеу и Doolittly, 1940). Различные штаммы вируса,
поражающие лилии, описаны у Вгіегіеу и Smith (1944). У многих
3 75
ізидов лилий вирус не вызывает признаков болезни и сохраняется
латентно. Согласно Atanasoff, 1928, вирус может поражать
гиацинт и нарцисс*. Главный переносчик болезни — Myzus
persieae, меньше значения имеет Macrosiph. solanifolii (Brierley
н Мс. Kay, 1938).
Огуречная мозаика лилейных. Cucumis virus 1 был
неоднократно выделен при мозаичных заболеваниях лилейных.
Он встречается здесь по большей части в форме штамма, который
у табака и у огурца вызывает только местную инфекцию в виде
некротической реакции (Price, 1937; Brierley, 1939; Ainsworlh,
1938). Судить о том, что мы здесь имеем дело действительно
з огуречным вирусом, можно по опыту перекрестной
иммунизации.
Price показал, например, что если Zinnia elegans заразить
Cacumis virus 1, то она становится невосприимчивой к вирусу,
выделяемому от лилий.
Ainsworth (1938) у Ыііит longiflorum var. insulare
(L. formosum) под влиянием огуречного вируса наблюдал
крапчатость, некрозы на листьях и тяжелые уродства цветов.
brierley (1940) описывает у лилии латентную инфекцию
различными штаммами огуречного вируса. При заражении
вирусом тюльпана растений, скрыто несущих огуречный вирус,
развивается, кроме мозаики, характерной для этого
последнего, также некротическая пятнистость. Согласно Gadd и Loos
(1940), огуречный вирус может вызвать у лилии светлозеленые
или желтые полосы на листьях, закручивание листьев и
отмирание их.
Вирусные заболевания лилии. Кроме перечисленных выше
вирусов, виды лилии могут поражаться вирусами,
вызывающими другие различные болезни. Эти последние мало изучены,
и мы ограничимся их кратким перечислением.
Розеточная болезнь, или yellow-flot, описанная
Ogilvie в 1928 г., проявляется общим хлорозом листьев, их
закручиванием вниз и карликовостью всего растения, которое
образует прижатую к земле розетку. Болезнь не передается
соком, ее переносчиком является Aphis gossypii Glover. Вирус
:>той болезни обозначается: Lilium virus 1, Ogilvie. Болезнь
известна в Европе, в США и в Азии. О мозаичном заболевании
лилии в Англии см. Green и Timker (1940).
Кроме мозаичных заболеваний и желтой карликовости,
в Японии у лилии описаны следующие заболевания:
Crook neck (свернутая шея). Заболевание
наблюдалось у L, auratum, тогда как L. longiflorum и L. speciosum
им не порая^аются. Наблюдаются пожелтение листьев и их усы-
хание. Процесс этот начинается сверху и заканчивается полной
утратой листьев.
Pimple-leaf (pimple — прыщик). Растения резко
подавлены^ на листьях развиваются бугорки, зависящие
176
от гипертрофии палисадных субэпидермальных клеток (Kawa-
mura, 1938).
Вирусные заболевания нарциса. В роде Narcissus
встречается по крайней мере два различных вирусных заболевания.
Первое из них может быть названо мозаичной
болезнью нарциса. При нем появляются светлозеленые
и желтые прерывистые полосы на листьях, а также
характерные поперечные ребристые возвышения. Последние зависят
от гипертрофии и тангентального деления клеток палисадной
паренхимы. Клетки эпидермиса либо вовлекаются в этот
процесс, либо, под давлением пролиферирующих клеток
палисадной паренхимы, отмирают, и последние выходят к
поверхности (Caldwell и James, 1938; Caldwell и Prentice, 1943).
Инактивация вируса происходит при 70—75° С. При
разведении 1:100 он сохраняет активность и сохраняется в соке 72 часа,
через свечи Шамберлена не проходит (llaasis, 1939). Болезнь
передают Aphis rumicis, Macrosiph. solanifol. {gel), M. rosae,
M. convolvuli, Anuraphis roseus, Macrosiph. pisi (Blonthon
и Haasis); соком болезнь передается с трудом.
Второе заболевание, обозначаемое как белая пол о-
с а т о с т ь (while streak), или же серая болезнь
(gray disease), отличается от предыдущего белыми полосами,
а также отсутствием пролиферации в тканях; напротив, белые
полосы подавлены в развитии.
Оба описываемые здесь заболевания вызываются вирусами,
которые и серологически отличны между собой. Сыворотка,
полученная к вирусу белой полосатости, не реагирует с соком
растений, больных мозаичной болезнью (Мс. Whorlher, 1939,
1940 и более ранние работы того же автора; Bijl и v. Slogteren,
1939).
Вирусные болезни других лилейных. Мозаичные
заболевания известны также у других представителей отряда лилейных.
Мозаичное заболевание ириса проявляется хлорозными
полосами на листьях, подавлением роста и полосами на
лепестках. Эти полосы чаще всего имеют более темную окраску,
чем нормально окрашенный фон, но изредка и более светлую.
Путем впрыскивания сока болезнь передается с трудом. Тли
Macrosiphum get, Myzus persicae ее легко переносят (Brierley
и Мс. Whorter, 1936).
Мозаичное заболевание фрезии (Freesia) вызывает на
листьях водянистые пятна, которые после высыхания остаются
белыми. При тяжелых формах болезни эти пятна сливаются
и могут убить растение. В лепестках иногда развивается
хлорофилл, и часто происходит опадение цветов. Болезнь описана
в Америке, Англии и Италии. Повидимому ее переносчиком
является Myzus convolvuli Ко!I (Woodward1).
1 См. по К. Smith Textbook of Plant virus diseases. 1937.
12 В. Л. Рыжков 1" 7
Мозаичная болезнь гладиолуса {Gladiolus) описана Dos-
dall, 1928. Листья мозаичны и достигают только половины
длины нормальных и тоньше этих последних. Происходит
также деформация листьев и развитие некротических участков
в эпидермисе; последнее вызывает бронзовую окраску листа.
На лепестках могут развиваться светлые полосы. В пораженных
клубнях наблюдается своеобразная бугристость.
Atanasoff (1928), который посвящает большую статью
мозаичным болезням лилейных, упоминает также о мозаике
у Hyacinthus, Crocus, Muscari сото sum, М. сот-pactum, М. botry-
oides, Nerine sarmiensis, Allium neapolitanum, Al. moly.
Blattny сообщает о мозаичной болезни у Convallaria mafalis
в Чехословакии, a Ogilvie — у Convallaria lineata на
Бермудских островах. Мне пришлось видеть под Москвой заросли
ландыша с мозаичной расцветкой листьев.
Мозаичное заболевание у Ornithogalum thyrsioides описано
F. F. Smith и Ph. Brierley (1944). Болезнь передается
различными видами тлей (Myzus persicae. Aphis gossypii и др.).
Она поражает также Galionia, Hyacinthus. Вирус обозначается
как Marmor scillearum.
У ананаса {Ananas sativa) описана желтая пятнистость,
которая проявляется желтыми полосами на листьях, позднее
некротизирующимися и принимающими бронзовую окраску.
Переносчиком болезни является табачный трипе. Из сорняков
болезнь широко распространена у Emilia sagittata (Linford,
1932). Любопытно, что при большой поражаемости сорняка
и сильном распространении трипса плантации ананаса могут
оставаться здоровыми, но стоит только приступить к полке
сорняков или же сорнякам пострадать от засухи, как трипе
переселяется с сорняков и вызывает массовое поражение
ананаса. В настоящее время установлено, что заболевание
вызывает штамм Lycopersicum virus 3, (Paris, 1940; Sakimura, 1940).
У агавы {Agave rigida v. sisalana) Stoner (1929) описал
в Бельгийском Конго заболевание, характеризующееся светло-
зелеными просвечивающими пятнами, которые позднее
становятся желтыми, с коричнево-красным краем. Предполагается,
что болезнь вирусная. Есть также упоминание о мозаичном
заболевании Agave cantala и A. sisalana.1
Мозаичные заболевания описаны также у Anthurium sp.
(Verplancke, 1930) и Hippeastrum equestre. У последнего при
мозаичной болезни наблюдаются в клетках Х-тела, которые
были предметом специального исследования (Holmes, 1928;
Kunkel, 1922).
1 Annual Report of the Bureau of Agric. Philippine Islands, .V 25„
1926.
Глава XVI
ВИРУСНЫЕ БОЛЕЗНИ ПРЕДСТАВИТЕЛЕЙ РАЗЛИЧНЫХ
СЕМЕЙСТВ
В предыдущих главах были рассмотрены семейства и роды,
вирусные болезни которых принадлежат к числу
наиболее изученных. Теперь нам остается познакомиться с
различными вирусами и подозрительными на вирусные
заболевания, большая часть которых изучена лишь очень слабо.
В эту главу мы включили также некоторые хорошо изученные
вирусные заболевания, которые, однако, поражают растения,
не культивируемые в СССР и потому не представляющие для
нас актуального интереса, как, например, spike санадала.
Рассмотрим прежде всего вирусные заболевания двудольных.
Семейства будут расположены в порядке, принятом в
систематике Е. Warming.
Порядок Querciflorae. В статье Atanasoff (1935) имеются
указания на мозаику у Corylis, выражающуюся посвет-
лением по ходу нервов. У того же автора мы находим
упоминание о крапчатости у бука, о мозаике у каштана и граба.
Семейство Juglandaceae. Американский орех Hicoria pecan
страдает заболеванием, называемым bunch disease, которое
проявляется хлорозом, утолщением листьев и
преждевременным отмиранием. Происходит образование ведьминых метел.
Болезнь встречается в США, передается прививкой (Cole,
1937).
Семейство Salicaceae. У тополя (Populas balsamifera) описана
мозаика, переходящая у старых листьев в некроз; деревья
подавлены в росте. Летом болезнь маскируется. Инфекционный
характер заболевания не доказан (Atanasoff, 1935).
Семейство Polygonaceae. У щавеля (Rumex lanceolatus
и R. sanguineus) J. Grainger и G. Cockerham (1930) описали
мозаичное заболевание. Листья пораженных растений имеют
желто-зеленые пятна; растения подавлены в росте. Вирус,
вызывающий заболевание, обозначается Rumex virus l,
Grainger. Этот вирус инактивируется при 30° С при разведении
1:100; он выдерживает высушивание и двухнедельное пребы-
12* 179
вапие в соке. Вирус не проходит через фильтр Беркефельда,
передается при помощи трения, но не инокуляции иглой.
У К. Smith в его «Textbook» имеется упоминание о мозаичном
заболевании ревеня. Polygonum hydropiper поражается
кольцевой пятнистостью табака.
Семейство Рірегасеае. У Piperomia описана кольцевая пят
нистость вирусного происхождения (Creager, 1941).
Семейство Caryophyllaceae. Известна желтуха у садовых
рас Dianthaus, которая передается прививкой (Peltier, 1916).
По исследовании Jones (1945), желтуха гвоздики
вызывается смесью двух вирусов—мозаики и полосэтости.
Первый из них вызывает светлозеленую мозаику и слабую по-
лосатость на лепестках. Этот вирус передается механически
и илактивируется при температуре 60° и через 42 часа in
vUro. Вирус полосатости вызывает желто-красные полосы
и пятна на листьях. Он передается только при помощи
персиковой тли.
Семейство Cactaceae. Мозаичное заболевание известно у
целого ряда кактусов, например Epiphyllum, Hvllocactus, Rhyp
salts (Pape, 1932; Blattny и Vukulov, 1939).
Согласно Blattny и Vukulov, мозаичная болезнь ЕpiphyHum
выражается появлением светлозеленых неправильных пятен,
укорочением и искривлением междоузлий. Соком болезнь
удалось передать в 3.3%. Инкубационный период длился
7 месяцев. Переносчик болезни — Orthiezia insignia.
Удается в 13% произвести заражение также растертым
соком насекомого-переносчика.
Порядок Poly carp icae. В семействе лютиковых описан целый
ряд вирусных заболеваний. Многочисленные интересные
исследования Klebahn (1926, 1928) посвятил аллоиофилии анемона
{Anemone nemorosa, A. ranunculoides, A. trifolia). Болезнь
проявляется утолщением листьев и стебля, листья уродливы,
цветы или не развиваются вовсе, или деформированы. Вирус
легко передается соком. Растение также заболевает, если
инфицировать почву остатками больных растений. Этот вирус
обозначается как Anemone virus l, Klebahn. Delphinium
поражается различными вирусами. Delphinium virus 1, Burnett
(1934) вызывает ведьмины метлы или bushy stunt этого растения.
Пораженные растения резко подавлены в росте, листья имеют
мозаичный рисунок и некрозы. Характерны поражения цветов:
соцветие имеет вид ведьминой метлы, наблюдается
пролиферация и позеленение цветов. Болезнь может быть передана
представителям различных семейств. У табака вирус
вызывает посветление по ходу нервов, а также кольца и
неправильные полоски, которые могут некротизироваться. У
помидора наблюдается карликовость и посветление по ходу нервов,
у огурца — симптомы, близкие к огуречной мозаике.
Delphinium virus 2, Valleau (1932) вызывает хлорозные
180
кольца; вскоре после цветения листья больных растений обычно
отмирают. У табака вирус вызывает некротические кольца
и рисунок, похожий на тот, который бывает при грубой
гравировке. В дальнейшем развиваются симптомы, похожие на
характерные для огуречной мозаики. Оба описываемые здесь
вируса Delphinium, согласно К. Smith, близко стоят к Cucumis
virus 1, а согласно Valleau (1932) — к вирусу кольцевой
пятнистости.
Delphinium при помощи цикад Thamnotettix montanus
и Г. geminatus может быть от сельдерея заражен Callistepkus
virus 1.
У пиона во Франции (Duffrenoy, 1934) и в Англии (Green,
1935) встречается желтая мозаика; желтые пятна могут иметь
очертания колец. Кроме того, на листьях иногда развиваются
мелкие некротические пятна. Болезнь может быть передана
соком на табак. Описываемую здесь кольцевую пятнистость
пиона вызывает Paeonia virus 1, Duffrenoy, Мозаика
встречается также у Aconitum (Blattny, 1938).
В семействе лавровых известно вирусное заболевание
у авокадо (Persea gratissima). Эта болезнь проявляется па листях.
молодых стеблях и плодах желтыми пятнами и полосками,
Первоначально заболевание было приписано ожогу под
влиянием солнца, почему и получило название sun blottch. Болезнь
передается прививкой (Ноте и Parker, 1931). У Berberis
vulgaris встречается мозаика по ходу нервов (Blattny, 1933).
Вирусные заболевания у Papaveraceae почти неизвестны.
У Papaver nudicaule встречается Lycopersicum virus 3 (броизо-
вость). Этот мак также подвержен Beta virus 1, которым
поражается также Eschscholzia californica.
Семейство Violaceae. Различные виды фиалок могут
поражаться следующими вирусами: Beta virus I, Beta virus 3,
Cucumis virus 1, Nicotiana virus 12,
Семейство Tern&troemiaceae. У камелии под названием
желтая пятнистость описана крапчатость листьев,
сопровождаемая образованием бугорков и опробковением
эпидермиса; могут наблюдаться пятна в виде колец, пожелтение
по ходу нервов и по краям. Бугорки иногда некротизируются.
Болезнь передается прививкой (Milbralh и Мс. Worter, 1940;
Band, 1944). У чая встречаются заболевания с невыясненной
этиологией, подозрительной на вирусную. Одно из них —
курчавость листьев, сопровождающаяся некрозом
флоэмы. Соком заболевание не передается, но может быть
передано прививкой кусочков корня. Болезнь встречается
на Цейлоне на высоте 3400 футов и выше (С. Н. Gadd, 1939;
Bond, 1944). Другое получило название ведьминых
метел. В золе больных растений обнаружен хром, что дает
повод предполагать, что болезнь является результатом
интоксикации хромом (Keiler, 1939).
181
Семейство Sterculiaceae. Серьезным заболеванием
какаового дерева является поражение побегов, выражающееся
в развитии вздутий (swollen-shoot). Эта болезнь передается
прививкой и семенами, соком не передается (Poshette, 1941).
Тли и псиллиды не переносят болезнь. Разные штаммы
вируса переносятся различными червецами: Ferrisiona virgata,
Pseudococcus exitiabilis, Pseud, vitri (Box, 1945). Тяжелые формы
болезни вызывают общий некроз и гибель растения, а легкие—¦
слабый хлороз, частичную утрату листьев и утолщение
стеблей. У какаового дерева известны также мозаичные
заболевания: красная пятнистость и посветление по жилкам. Обе
болезни передаются прививкой (Posnette, 1944).
Семейство Euphorbiaceae. Pape (1931) описал скручивание
листьев у декоративного молочая Euphorbia pulcherrima.
Большой экономический ущерб в тропических странах приносит
мозаичная болезнь кассавы. Ее вызывает Manihot virus l,
Dammer (1885). На пораженных листьях развиваются я^елтые
пятна, зеленые участки листа развиваются значительно лучше
желтых и вздуваются. Пластинка листа может быть
изуродована и резко редуцирована в размерах, растения подавлены
в росте, междоузлия укорочены; иногда также происходит
усиленное развитие боковых побегов (Н. Muller, 1931; Golding,
1935; Silberschmidt, 1931; Williams, 1940).
Болезнь известна в Африке, на Яве и Мадагаскаре. В
некоторых случаях наблюдалась передача болезни соком, хотя,
по другим авторам, соком она не передается. Легче происходит
передача болезни, если кусочки порайонной ткани ввести
в стебель здорового растения. Переносчик вируса — Bemisia
gossypipedra, no Muller, 1931, также Lecanium sp. и термиты.
Интересны работы, в которых изучалась зависимость
скорости распространения болезни от сезона. Н. Storey и R. F.
Nikolas (1938) в течение двух лет производили на протяжении
всего года посадку здоровых растений среди больных и
наблюдали затем за ходом их заражения. Оказалось, что при посадке
в июне растения наиболее долго сохраняют здоровье, почти
так же благоприятны май, июль и август; наиболее быстро
инфекция распространяется на посадках с декабря по апрель.
Мозаика кассавы может уносить свыше 40% урожая.
В борьбе с ней главную роль играют устойчивые сорта, для
получения которых применяется междувидовая гибридизация
(Williams, 1940 и др.).
Известны разные штаммы вируса, вызывающего мозаику
кассавы. У кассавы Storey (1936) кратко описано заболевание,
поражающее стебель. Оно вызывается Manihot virus 2, Slorey.
На листьях появляются желтые пятна, а на стебле коричневые
полосы. Стебель становится хрупким и часто обламывается.
От этих заболеваний, по Costa (1941), отличается обыкновенная
мозаика и мозаика по нервам манигота. Вирус первой пере-
182
дается соком. Он устойчив in vitro 48 часов, инактивируется
при 65—75° С и выдерживает разведение 1:5000. Вирус второй
передается прививкой, но не соком. Пути распространения
этих заболеваний в природе остаются невыясненными.
У тунга (виды Aleurites Sp.) описан целый ряд
заболеваний, этиология которых не выяснена. Важнейшие из них
приводятся у Plakidas (1937). Знакомясь с насаждениями тунга
на Кавказе, в связи с гибелью отдельных деревьев тунга (Иванов
и Иванова), мы наблюдали на его листьях ряд пятнистостей,
подобных тем, которые описаны в Луизиане. Важнейшие из
них — покоричневение между жилками. Заболевание
начинается диффузным хлорозом между жилками; позднее
наметившиеся хлорозные пятна приобретают бронзовую и коричневую
окраску вследствие некроза, который начинается с эпидермиса
и распространяется вглубь листа. Другая пятнистость,
описанная под названием translucent spot, т. е. просвечивающие
пятна, которую наблюдали также и мы, проявляется
неправильно разбросанными просвечивающими пятнами с
некротической точкой в середине. Анатомическая картина этого
заболевания совсем другая. Некроз здесь начинается чаще всего
с нижней стороны листа, и по периферии некроза происходит
резкое разрастание клеток губчатой паренхимы. Мы видели
также гербарные экземпляры тунга, обнаруживающие мозаику
или кольцевую пятнистость. Весьма вероятно, что некоторые из
этих заболеваний — вирусного происхождения. Внешнее
сходство с вирусными заболеваниями имеет бронзовость тунга,
зависящая от недостатка цинка (Newell, 1935).
Семейство Geraniaceae. У пеларгонии описана курчавость
листьев, вызываемая Pelargonium virus 1, Раре. Этот вирус
поражает Pelargonium zonale и P. hederaceum. Болезнь резко
проявляется весной, на черенках наблюдаются многочисленные
округлые или звездовидные светлые пятна, которые могут
некротизироваться (рис. 48). Иногда пятна просвечивают
и имеют маслянистый вид, могут также быть кольцеобразны.
Листья морщинисты. Часто болезнь при дальнейшем развитии
растений маскируется. Вирус не передается соком, но
прививкой передается легко. Можно предполагать, что переносчиком
его является тля Macrosiph. pelargonii. Для борьбы с болезнью
рекомендуется не брать черенков от больных растений,
обрызгивать растения содержащими серу смесями; пересадка растений
из оранжереи под открытое небо способствует ослаблению
признаков болезни (Раре, 1927; Рігопе, 1938, 1940). Явления
курчавости уступают также действию FeS04 (Jones, 1938,
1940).
Кроме курчавости листьев у пеларгонии встречаются,
повидимому, и другие заболевания, которые коротко описаны
Blattny (1933). Мозаика между жилками резко выражена
у молодых листьев и маскируется у более старых. Болезнь
183
передается прививкой. Кольцевая пятнистость, которая не
передается соком, но, возможно, распространяется трипсом,
описана в Бразилии (Silveira, 1941).
Семейство Rutaceae. У цитрусовых известно большое
количество заболеваний, имеющих невыясненную этиологию.
Часть их является функциональными расстройствами, а часть,
может быть, принадлежит к вирусным болезням. Atanasoff
(1935а) посвятил большую статью этим заболеваниям,
рассматривая их преимущественно в качестве вирусных. Важней-
шим из них являются инфекционный хлороз, который
характеризуется появлением светлозеленых и желтых крапинок,
а также деформацией пластинок листа. Заболевание, по
классификации Н. S. Fawcett, вызывает Citrivir italicum. Пластинки
листьев могут опадать, а черешки остаются на дереве (Petri,
1931; Fawcett и Klotz, 1938, 1939 и др.). Передача болезни
путем прививки больной почки была осуществлена Tralrut
(1933). Возмоя^но, что в природе болезнь распространяет тля
Toxoptera auranti. Petri (1931) передача прививкой или каким-
нибудь другим способом не удалась. Несомненно, что
существует внешне очень сходное заболевание, излечимое цинком
и носящее неинфекционный характер (Batcheler и др., 1934),
а также хлороз, зависящий от недостатка магния (Oberhol-
7or, 1941).
II с о р о з и с цитрусовых представляет собой
заболевание, которое может проявляться на листьях слабой мозаич-
ностью (Wager, 1941), однако главным ого симптомом является
тяжелое поражение коры деревьев. По Fawcett, вирус,
вызывающий эту болезнь, должен называться Citrivir psorosis.
Болезнь никогда не начинается у деревьев ранее пяти-шести-
летнего возраста, чаще развитие ее начинается в более позднем
возрасте. Заболевание начинается небольшой пустулой на
коре, которая постепенно углубляется и расширяется. На
дне такой язвы, которая может достигать древесины, отлагается
гумми. Работами Fawcett Н. S. (1938) было установлено, что
болезнь может передаваться прививкой. Этот автор указывает
на то, что во Флориде сходные симптомы наблюдаются также
у одного вида дуба Quercus larifolia, растущего близко к
плантациям цитрусовых (Fawcett и Rhoades, 1939). Fawcett (1941)
различает целый ряд штаммов вируса, вызывающего псорозис.
Для борьбы с болезнью применяется уничтожение сильно
пораженных растений, посадка исключительно здорового
материала, строгая регистрация всех плантаций, где имеются
больные растения (Hewitt, 1939). Паллиативом может служить
удаление кусков пораженной коры. О псорозисе см. также
Doidge, 1939. Bitancourt (1941) считает заболевание излечимым
путем опрыскивания известково-серным отваром.
При диференциальном диагнозе вирусных болезней
цитрусовых должны быть учтены: mal secco — болезнь, вызываемая
184
грибком, который, поражая древесину, окрашивает ее в красно-
оранжевый цвет, и Gelbnervigkeit — заболевание,
развивающееся при неправильной посадке деревьев (Gassner, 1940).
У Ptelea Sp. известен инфекционный хлороз, имеющий
декоративное значение, передача которого прививкой известна
с 1870 г. (см. по Ваш, 1908).
Рис. 48. Курчавость листьев пеларгонии (по К. Smith).
Семейство Anacardiaceae. У фисташкового дерева (Pistacia
vera) В. Э. Крейцберг в 1940 г. описал розеточную болезнь.
У больных деревьев листья уменьшаются в размерах, жилки
листьев становятся выпуклыми, особенно с верхней стороны,
пластинки листа в большей или меньшей степени
деформируются. Наблюдается усиленное ветвление и образование
розеток, несущих суженные листья, скрученные вдоль краев.
Соцветия, и женские и мужские, подвергаются позеленению и
пролиферации (рис. 50). Ветвление этих соцветий увеличено,
они очень плотны и уменьшены в размерах. Плоды уродуются,
число их уменьшено, при тяжелых формах болезни образование
плодов вовсе не происходит. В анатомическом отношении
болезнь характеризуется подавленностью в развитии палисадной
паренхимы, клетки которой становятся очень различных
размеров; число хлорофилловых зерен в клетках уменьшается,
чрезмерно развивается ксилема, от чего и зависит гипертрофия
жилок листа; плодоножки покрыты деревянистыми опухолями,
185
и на них появляются добавочные цилиндры. Отток ассимилятов
из растений задерживается.
Болезнь приносит очень значительный ущерб. Она не
передается соком, но легко передается прививкой.
Переносчиком болезни является фисташковый трипе Liothrips pistaciae.
Семенами болезнь передается легко. Болезнь проявляется
уже на всходах. Тщательное удаление пораженных веток
может дать оздоровляющий эффект.
Семейство Aceraceae. Atanasoff (1935) упоминает о
мозаичных заболеваниях Acer Negundo, A. pseudoplatanus,
передаваемых пересадкой. Не следует их смешивать с широко
распространенными пестролистными формами различных видов клена.
Семейство Sapindaceae. У Atanasoff имеется упоминание
о мозаичном заболевании конского каштана (Aesculus hippo-
castanum).
Порядок Rhamnales. Должны быть отмечены: мозаика
у Rharnnus, известная с 1887 г. (см. Atanasoff, 1935),
инфекционный хлороз у Ilex aqui folium^ передача которого
пересадкой наблюдалась еще в 1719 г. (см. Atanasoff, 1935),
а также инфекционный хлороз у Evonymus /аропіса. Этот
хлороз проявляется посветлением по ходу нервов, слабой
крапчатостью и интумесценциями. У этого декоративного
кустарника встречаются ташке многочисленные пестролистные
расы. Пестролистность может маскировать существование
инфекционной мозаичности, так как последняя выражается
гораздо менее резко, чем первая (Baur, 1908; Rischkov, 1927).
Порядок Thymclaeinae. У декоративных видов
Daphne в Новой Зеландии описана мозаичная болезнь. На
листьях развиваются крупные, неправильной формы пятна.
Пластинка листа часто деформирована. Соком болезнь
передается с трудом, легко —¦ прививкой (Е. Е. Chamberlain, 1941).
Порядок Passiflorales. У пассифлоры Noble (1928)
описал одеревенение плодов— заболевание, приносящее
существенный экономический ущерб в Австралии. Его вызывает
Passiflora virus l, Noble. На листьях болезнь обнаруживается
резкой мозаикой и деформацией пластинки. Плоды
пораженных растений имеют неправильную форму, поверхность их
часто морщиниста. Наиболее характерной является
деревянистость плода, которая зависит от усиленного развития склерен-
химных элементов в перикарпе. Если смочить кусочек ваты
в соке больного растения и ввести такой ваточный томпончик
в проводящую систему здорового растения, то таким способом
можно вызвать заражение. Болезнь передается персиковой
тлей и другими афидами (Noble, 1939).
В Восточной Африке, в Кении Natbrass (1939, 1940) описал
одеревенение плодов пассифлоры, отличающееся тем, что
болезнь почти совершенно не обнаруживает себя на пластинке
листа, а также тем, что не может быть передана инокуляцией сока.
186
Представитель другого семейства (Сагісасеае) того
же порядка, дынное дерево (Сатіса Papaya) болеет целым
рядом вирусных заболеваний. Одно из них, описанное под
названием leaf crinkle, проявляется морщинистостью листьев,
закручиванием их вниз и утолщением их пластинки. Болезнь
передается прививкой (Thomas и Krishnasawami, 1939).
Заболевание этого типа встречается в Порто-Рико. В Тринидаде
описана мозаичная болезнь, проявляющаяся маслянистыми
пятнами на листьях и плодах. Могут наблюдаться также общий
хлороз, некрозы и гибель растения. Болезнь передается
прививкой (Baker, 1939). На Гавайях описано заболевание,
проявляющееся морщинистостью листьев, мозаикой и некрозами
Растения подавлены в развитии, листья преждевременно
опадают. Болезнь удается передать соком с карборундом
(Parris, 1939).
Семейство Santalaceae. У сандала давно известно и очень
детально изучено заболевание, носящее название spike. Его
вызывает Santalum virus l, Coleman (1918). Листья больного
растения уменьшаются в размерах — не столько в длину,
сколько в ширину, вследствие чего становятся очень узкими.
В то же время наблюдается утолщение пластинки, междоузлия
укорочены, и сближенные вследствие этого листья торчат
на стебельке, придавая ему вид колоса, откуда и происходит
название болезни (spike — колос). Наблюдается также
образование «ведьминых метел». В клетках пораженных растений
встречаются Х-тела овальной формы, часто близко лежащие
к ядру; цветы часто подвергаются позеленению.
Болезнь может передаваться прививкой (Sreenivasaya и
Naid, Rdo, 1933; Jyengar, 1938, 1940). Переносчиком ее, как
видно, является насекомое, которое до сих пор точно не
установлено. Возможно, что такие переносчики — некоторые
активные ночью насекомые (Jyengar и Griffith, 1940). В
физиологическом отношении болезнь принадлежит к желтухам, и при
ней происходит переполнение тканей крахмалом. Благодаря
работам Jyengar (1938) это заболевание является одним из
наилучше изученных в патофизиологическом отношении.
Целый ряд видов, у которых паразитирует сандал, могут болеть
spike (Rangaswami, 1939). Другая болезнь сандала носит
название курчавость листьев. Ее вызывает
Santalum virus 2, Rao (1933). В отличие от предыдущей болезни,
при этом заболевании листья уменьшаются в размерах
одинаково, как в длину, так и в ширину. Пластинка их утолщена,
хрупка, и нередко они преждевременно опадают. В ранних
стадиях заболевания наблюдается мозаичность, нередко края
листьев приобретают красно-коричневый цвет. Болезнь
передается прививкой, но не соком.
Семейство Ribesiaceae. У черной смородины встречается
заболевание, называемое махровостью или р е в е р-
187
спей. Она приписывается Ribes virus 1, Amos и Hatton (1926).
Главным симптомом болезни являются превращения, которые
испытывают цветы. В зависимости от силы поражения они
различны. В случае полной махровости цветка все наружные
круги его превращаются в узкие чешуйки, завязь становится
верхней и распадается на листочки. Пораженные соцветия
могут засыхать с неразвитыми бутонами. Иногда встречается
пролиферация оси цветка. Количество махровых кистей
на одном и том же кусте черной смородины в разные годы
бывает различным. Наблюдается также изменение пластинки
листа, причем листья махровых кустов становятся длиннее
и уже. Число боковых жилок у главной доли листа у больных
растений бывает обычно меньше пяти, тогда как у здоровых
пять и более (Lees, 1922). Изменения пластинки листа,
однако, не являются надежным признаком болезни, так как
могут отсутствовать.
Болезнь инфекционна; указания на то, что она
передается прививкой (Amos и Hatton, 1926), получили полное
подтверждение в работах нашего аспиранта Алексеевой;
махровость передает клещик Eriophyes ribis, паразитирующий в
почках смородины.
Болезнь обладает значительной вредоносностью и
распространением. В некоторых садах в СССР процент пораженных
кустов может доходить до 92, тогда как в других садах
болезнь вовсе отсутствует или встречается в ничтожных
размерах.
Для борьбы с болезнью особое значение имеет здоровый
посадочный материал. Большую пользу при апробации
посадочного материала может принести метод, предложенный
Т. С. Алексеевой (1945). Оказывается, что, просматривая
заложившиеся с осени почки с помощью бинокулярной лупы,
легко отличить нормальные почки от почек с махровыми цветами,
так как в последних зачатки листочков обособлены и завязь
верхняя, тогда как в нормальных почках уже на этой стадии
можно видеть нормально организованные элементы цветка.
Кроме махровости, у черной смородины встречается мозаичное
заболевание. Больные растения подавлены в росте, и
продуктивность их уменьшена. Е. М. Hildebrand (1939) удалось
передать болезнь при помощи прививки. Мы часто встречали
у различных видов Ribes мозаику, распространяющуюся
по ходу жилок и дающую листу красивый сетчатый рисунок.
Этиология ее нам не известна. Е. М. Hildebrand (1941—1942)
наблюдал у черной смородины кольцевую пятнистость, которая
моя^ет быть передана на табак, вызывая у последнего симптомы,
характерные для кольцевой пятнистости томата. Заражение
табака этим вирусом предохраняет от кольцевой пятнистости
томата, описанной Imle и Samson (см. «Вирусные болезни
пасленовых»). Обратно передать заболевание от табака сморо-
188
дине не удается. Посветление по ходу нервов и морщинистость
листьев у красной смородины описал Winter (1940).
Семейство Pittosporaceae. Pittosporum variegatum страдает
инфекционным хлорозом, который в 1887 г. был передан
пересадкой (см, Atanasoff, 1935).
Семейство Согпасеае. У кизиля мозаичное заболевание
наблюдал ALanasoff (1935).
Семейство Rhodoraceae. У рододендрона Раре (1931)
описывает мозаичную болезнь, при которой светлые участки резко
подавлены в росте, а зеленые вздуваются.
Семейство Уассіпіасеае. У Vaccinium macrocarpon описано
заболевание, известное под названием false-blossom. Его вирус
называется Vaccinium virus l, Dobroscky (1931).
Оно передается Euscelis striatulus. Главным симптомом
болезни является позеленение цветов, вследствие чего они
становятся бесплодными; наблюдается карликовость и усиленная
кустистость растений. К вирусу чувствителен также V. оху-
coccus (Beckwith, 1933; Wilcox и Beckwith, 1931). Vaccinium
australe страдает подавленностью роста, кустистостью и
хлорозом, которые, возможно, имеют вирусное происхождение
(R. Wilcox, 1931). При помощи повилики болезнь
передается помидору и здесь вызывает типичные для столбура
симптомы (Kunkel, 1945 .
Семейство Primulaceae. У Primula sinensis мы часто
встречали мозаичное заболевание. Возможно, оно тождественно
с описанным Раре в 1924 г. Виды примулы, по литературным
данным, могут поражаться огуречным вирусом 1 (К. Smith),
а также Lycopersicum virus 3.
С. М. Tompkins (1941) описывает заболевание Primula
оЪсопіса, которое проявляется резкой мозаичностью и
деформацией листа; последний может стать нитевидным. Размеры
венчика уменьшаются и появляется его пятнистость. Болезнь
передается сохлом на различные виды примулы. Nicotiana glutin-
osa не поражаются. При помощи тлей Myzus persicae и
М. circumflexus болезнь не удалось передать. Болезнь не
передается и семенами. От больных растений было получено 20
тысяч здоровых проростков.
В СССР часто встречается не изученное ближе мозаичное
заболевание Primula sinensis.
Семейство Convolvulaceae. Convolvulus arvensis страдает
заболеванием, выражающимся в хлорозе, измельчании листьев
и укорочении междоузлий. Больные растения редко цветут,
а если и цветут, то цветы обычно имеют столбуриый тип
поражений. Многолетние наблюдения свидетельствуют с полной
определенностью, что мы имеем дело с поражением вьюнка
столбуром. По наблюдениям Корачевского1, столбуром также
1 Устное сообщение
189
болеет Ipomea Batatus. У этого вида, кроме того, описано
мозаичное заболевание, при котором наблюдается гипертрофия
средней жилки листа, морщинистость, укорочение
междоузлий. Растения нередко имеют розеточный облик. Если
привить к здоровым растениям побеги больных, то на другой год
из клубней растений, служивших подвоем, получаются больные
растения (Ноте и Parker, 1931; Weber, 1933). Другое
заболевание, желтая пятнистость, также передается только
прививкой, его вызывает вирус, который Doolittle и Hartor
назвали Flavimacula іротеае.
Рядом работ установлено, что виды Cuscuta могут
переносить различные вирусы (F. Johnson, 1941). Вирус табачноймоза-
ики иногда передается при помощи Cuscuta campestris, но не
может репродуцироваться в этом растении. Cucumis virus lnVac-
cinum virus 1 сохраняются в Cuscuta campestris и после
прохождения через несколько экземпляров иммунных к этим вирусам
растений и могут, очевидно, в повилике репродуцироваться
(A. Costa, 1941, 1944). Целый ряд видов Cuscuta, например
С. californica, являются латентными носителями вируса,
который вызывает мозаику и некроз у сахарной свеклы, дыни-кан-
толупы, томата, сельдерея и других растений. Brassica incana
является скрытым носителем вируса. Вирус передается с
семенами Cuscuta campestris, но не дыни. Инактивация вируса —
56°—60°; он сохраняет инфекционность в соке Phytolacca decandra
при разведении 1:3000 и не инактивируется через 48 часов.
Высушивание в соке выдерживает около 48 часов, но не в целом
растении. Соком передается легко Phytolacca, у которой
вызывает местные некрозы; труднее — Chenopodium murale, свекле
и Cuscuta campestris. Другим растениям соком не передается
(С. W. Bennett, 1944).
Порядок Personatae. Вирусные болезни поражают
норичниковых. У львиного зева (Antirrhinum ma/us) может,
например, встречаться Lycopersicum virus 3 и Nicotiana virus 12.
У губоцветных в целом ряде случаев были описаны мозаичные
заболевания. Liro (1930) в окрестностях Гельсинки нашел
мозаичную Prunella vulgaris. Мозаика передавалась соком. Blattny
(1933) описывает у Lamium galeobdolon мозаику типа аукуба.
Растения произрастали на высоте 1200 м над уровнем моря.
В Боржомском парке мы наблюдали в большом количестве
Salvia glutinosa с чрезвычайно типичными симптомами
мозаики. Желтые участки листа были подавлены в росте, а зеленые
пузыреобразно вздуты. Имеются указания на то, что Nepeta
cataria болеет огуречной мозаикой и кольцевой пятнистостью
табака.
У кунжута (Sesamum indicum), под названием филлодия,
описано заболевание, при котором главным симптомом является
позеленение и пролиферация цветов. Болезнь эта приносит
очень большой ущерб и была впервые описана в Индии, но встре-
190
чается также и у нас. Она не передается соком, но может быть
передана прививкой (Pall и Nath, 1935). Насекомое —
переносчик болезни — остается невыясненным.
Декоративные виды Gloxinia поражаются Callistephus
virus 1, Lycopersicum virus 3.
ТаЬеЬиіа pallida из семейства Bignoniaceae страдает ведь-
миными метлами. Болезнь передается введением в стебель
кусочков ткани (Cook, 1937).
Из представителей Verbenaceae у Stachytarpheta jamaiceay
растущей дико на Цейлоне, описано 3 вирусных заболевания:
желтая пятнистость, мозаика по жилкам и розеточность. Они
могут передаваться прививкой (Loas, 1942).
Семейство Rubiaceae. У Coffea агаЫса описано
заболевание, связанное с гипертрофией и некрозом флоэмы, причем
ткани переполняются крахмалом, а проводящую систему
наполняет гумми. Деревья увядают и высыхают. Заболевание
это по своим признакам очень похоже на вирусное, но Stahel
(1931—1933) описывает в качестве его возбудителя Phytomonas
leptovasorum — жгутиконосца. Болезнь передается пересадкой
кусков корня, но не стебля. Это заболевание нам кажется весьма
подозрительным на вирусное. К. Silberschmidl (1941) описывает
в Бразилии болезнь кофе и других Rubiaceae (Borreria, Ri-
chardsonia), похожую на кольцевую пятнистость табака,
вызывающую также язвы в коре. Эта болезнь не передается соком
и семенами, но легко передается прививкой.
Семейство Оіеасеа. У ясеня встречается целый ряд
вирусных заболеваний. Инфекционный хлороз проявляется
красивой яркожелтой пятнистостью типа аисиЪа (Baur, 1907)
и имеет декоративное значение (Fraxinus pubescens и Fr. anculifo-
lia). Кроме того, Atanasoff (1935) в Болгарии наблюдал мелко-
листость ясеня и хлорозные пятна мея^ду жилками. Болезнь
удалось передать прививкой. В СССР В. Ф. Купревич (1940)
описывает розеточную болезнь Fraxinus americana, при которой
деревья подавлены в развитии и стволы покрыты большим
количеством веток, образуются многочисленные розетки, у которых
сближены листья. Листья могут иметь мозаичную расцветку у
флоэма подвергается некрозу.
У сирени встречается целый ряд вирусных заболеваний.
Atanasoff (1935) описывает кольцевую пятнистость сирени.
Светлозеленого цвета кольца рассеяны по всему листу, что
нередко ведет к преждевременной его гибели. При скручивании
листьев у сирени пластинки листа становятся хрупкими,
толстыми и ложечкообразно закручиваются. Ткани листа
переполняются крахмалом (Atanasoff, 1935).
Давно известны инфекционные хлорозы, передающиеся
прививкой, у Ligustrum vulgare и Jasminium officinale
(Atanasoff, 1935). У маслины Pesante (1938) описал заболевание,
при котором наблюдается бронзовость листьев и их опада-
191
ние. В листах накопляется крахмал. Флоэма некротизи-^
руется.
Семейство Caprifoliaceae. У бузины (Sambucus) Blattny
(1938) описал мозаичную болезнь, при которой хлороз
распространяется но ходу нервов, растения подавлены в росте
и большая часть цветов стерильна.
Семейство Valerhmaceae. У валерьяны (Valeriana
officinalis) имеется два заболевания, похожих на вирусные.
Одно из них мы наблюдали в Симферополе. Оно проявляется
хлорозными пятнами, которые позднее приобретают
бронзовую окраску. Другое принадлежит к типу желтух. При этой
болезни пластинки листа утолщаются, становятся хрупкими
и ложечкообразно закручиваются (рис. 49). В соцветии
наблюдаются антолиз и пролиферация. Изменения цветов были
подробно описаны В. Л. Рыжковым (1930). Заболевание это,
судя но нашим наблюдениям, вызывается тем же вирусом, что
и желтуха кок-сагыза.
Обратимся теперь к однодольным. У банана (Musa)
известен ряд вирусных заболеваний. Кустистость верхушки
(bunchy 1ор)гвызывается Musa virus I, Magee (1940). Эта болезнь
поражает Musa cavendishii,
М. sapientum, M. texlilis.
Первым симптомом ее
являются темнозеленые
полосы по ходу главной и
боковых жилок,
развертывание листьев более или
менее подавлено. Первые
листья, развивающиеся
после заболевания,
приподняты вверх, позднее
листья вообще не
развертываются, и получается
нечто вроде капустного
кочана. Характерным па-
тол ого-анатомическим
симптомом является развитие
богатых хлорофиллом
паренхиматозных клеток
в сосудисто-волокнистых
пучках, которые более или
менее вытесняют
механические элементы и
ситовидные трубки. Передача
болезни соком весьма сомнительна. Переносчиком ее является
тля Pentalonia nigronervosa. Для приемки вируса этой тле
необходимо 17—24 часа. Тли сохраняют инфекционность не менее
13 дней и являются, повидимому, «механическими» переносчи-
192
Рис. 49. Лист валерианы, больной
желтухой.
ками болезни. В листьях, развившихся до заражения, вирус
отсутствует (Magee, 1940 а). Кроме поражения Musa virus 1, Musa
і
Вверху
Рис. 50. Ветки женских экземпляров фисташки.
здорового; внизу (Зольного розеточной болезнью (по Крейцбергу)
textilis поражается еще другим вирусом, переносчиком которого
является также тля P. nigronervosa. Заболевание, вызываемое
этим вирусом, очень похоже на предыдущее. Однако от тек-
J3 Ь. R. Рыжков *"**
стильного банана к упомянутым выше видам банана болезнь
не может быть передана, почему и приписывается особому
Musa virus 2, Ocfemia (1937).
У Musa textilis также описано мозаичное заболевание г
которое не передается Pentalonia nigronervosa. «Механическим»
переносчиком этого заболевания являются тли (Aphis gossypiiy
Aphis maidis, Rhopalosiphum nympheae) и другие виды этого
рода (Celino, 1940; Celino и Ocfemia, 1941). Musa cavendishii
страдает инфекционным хлорозом (другие названия:
мозаичная болезнь, сердцевинная гниль). Это-
заболевание выражается яркожелтой пятнистостью, а также
некротическими явлениями, в которые вовлекается точка роста,
что приводит растение к гибели. Некрозы обычно развиваются
только в течение зимних месяцев. Болезнь описана в
Австралии, но встречается также и в Бразилии.
К. Smith приписывает ее Musa virus 3. Magee, однако,
недавно доказал, что вирус тождествен Cucumis virus l (Mageer
1940). Он передается теми же видами тлей, что и этот последнийг
и может быть перенесен на огурец, помидор и табак. Еще раньше
было известно, что огуречный вирус от сельдерея может быть
перенесен на банан.
Возможно, что мозаичная болезнь, передаваемая разным
видам Musa textilis и кукурузе, также вызывается огуречным
вирусом.
Известное декоративное растение Richardia africana из
семейства Агассае может страдать от Lycopersicum virus 3.
На листьях, стебле и цветочных почках развиваются желтые
пятна. Болезнь переносится табачным трипсом (Ainsworth,
1935).
Семейство Commelinaceae. Виды Соттеііпа поражаются
мозаичными заболеваниями. Эти заболевания предохраняют
от заражения Cucumis virus 1 и вызываются, очевидно,
штаммами этого последнего (Price, 1941).
ПРИЛОЖЕНИЯ
Таблица I
Вирусы^ вызывающие местные некрозы у растений - индикаторов
Р аст сние-инд ик а то р
Nicotiana tabacum
Nicotiana rusUca
Nicotiana glutmosa
Nicotiana silvestris
Petunia kybrida
Capsicum frutescens
(некоторые сорта)
Capsicum annuurn
Solanum nodiflorum
Solanum lycopersicum
Solanum tuberosum
(Arran Victory,
President)
Phaseolus vulgaris
Вирус
Brassica virus 1
Brassica virus 2
Nicotiana virus 11
Nicotiana virus 12-
Ligusticum virus 1
Lycopersicum virus
Lycopersicum virus
Lycopersicum virus
Lycopersicum virus
Datura virus 1
Peonia virus 1
Nicotiana virus 1
Nicotiana virus 1
Nicotiana virus 11
-14
1
2
3
4
Cucumis virus 1 Штамм 5
(Price)
Nicotiana virus 14
Ligusticum virus 1
Nicotiana virus 1
Штамм аукуба
Lycopersicum virus
Nicotiana virus 1
Solanum virus 8
Solanum virus 9
Solanum virus 8
Solanum virus 9
Lycopersicum virus
Lycopersicum virus
Datura virus 1
Solanum virus в
Trifolium virus 1
3
2
4
Наступает ли общая
инфекция
Нет
»
»
Да
»
Нет
Да
»
Нет
Да
»
»
Нет
»
Да
» '
»
»
Нет
»
Да
»
»
»
»
»
»
»
Да (местные
некрозы непостоянно^
*
195
Таблица I (продолжение)
Растение-индикатор
Vigna sinensis
Zinnia elegans
Phytolacca decandra
Вирус
Medicago virus 2
Nicotiana virus 1
Nicotiana virus 11
Nicotiana virus 12-
Datura virus 1
Cucumis virus 1
Nicotiana virus 12-
Lycopersicum virus
Datura virus 1
Pea wilt virus 1
Cucumis virus 1
Nicotiana virus 1
—14
-14
4
Латентный вирус Cuscuta
Наступает ли общая
инфекция
Нет
Нет(местную
реакцию дают
некоторые сорта,
напр Early Golden,
GlustST тя. Robust)
Нет
Да
»
Нет (кроме
некоторых)
Общий некроз
Нет
Да
»
»
Нет
Да
Таблица II
Вирусы, поражающие представителей многих семейств
1. Мозаичные заболевания, могущие сопровождаться некрозами—2.
Заболевания, характеризующиеся расстройствами роста, хлорозом.
Пластинка листа часто становится ненормально хрупкой, нередко
наблюдаются уродливые изменения цветов — 4.
2. Мозаичное заболевание, редко сопровождающееся некрозами;
рисунок обычно не имеет вида колец. У Vigna sinensis, но не у Phaseo-
lus vulgaris — местные некрозы, у N. tabacum — общее заболевание
Вирус легче передается тлями, чем иглой — Cucumis virus l.
Заболевание сопровождается некрозами, часто встречается
кольцевой рисунок — 3.
3. Вирус вызывает местные некрозы у Petunia hybrida, листья
больного помидора принимают бронзовый оттенок, N. glutinosa очень
чувствительна к вирусу, который инактивируется при 42°С — Lyco-
persicum virus 3.
— У Petunia hybrida — общее заражение, у Phaseolus vulgaris —
местные некрозы в виде концентрических колец.. Помидор мало
подвержен заболеванию. Температура инактивации — 60° по G — Nicotiana
virus И2.
4. Морщинистость и закручивание листьев отсутствуют, в природе
чаще встречается у сложноцветных и сельдерея. Листья хлорозны,
растения подавлены в росте, развиваются многочисленные придаточные
побеги, часто наблюдается антолиз. Переносчики вируса — цикады: Сіса-
dula diversa, Thamnotettix montanusy Т. gemintus и, возможно, другие
виды. Соком болезнь не передается — Callistephus virus 1 и
родственные виды.
196
— Морщинистость, закручиваниа листьеа отсутствуют. . шшсда
выражена мозаика. Болезнь передается соком (некоторые случай
поражения Cucumis 1, когда мозаика не выражена или слабо выражена
и на первый план выступают карликовость и кустистость) — 2
Морщинистость и закручивание листьев резко выражены — 5.
5. Часто встречаются выросты на нижней стороне листа. Листья
закручиваются главным образом вниз. Во флоэме развивается
древесина. Переносчики болезни — Aleyrodidae — Nicotiana virus 10.
Выростов на нижней стороне листа не бывает, однако жилки могут
припухать. Древесина во флоэме не встречается, но флоэма нередко
гипертрофирована. Болезнь чаще встречается у свеклы и помидоров.
Переносчик болезни — Eutettix tenellus — Beta virus 1.
Таблица III
Вирусные болезни хмеля
1. Мозаичная окраска отсутствует, листья закручиваются вверх
и вниз, побеги слабые, многочисленные—Humulus virus 1.
— Мозаичность или хлороз ¦— 2.
2. Мозаика, сопровождающаяся повреждением корневой системы,
которая частично погибает. Листья хрупкие, закручивающиеся — 3.
— Корневая система не поражается, зеленые участки часто
развиваются лучше светлых и вздуваются — Humulus virus 3.
3. Мозаика проявляется в начале лета, сорт Fuggles не поражается—
Humulus virus l.
— Мозаика проявляется к концу сезона, -сорт Fuggles
поражается — Humulus virus 4.
Таблица IV
Вирусные болезненные семейства Chenopodiacexe
1. Желтухи — 2.
— Мозаики — 3.
2. Листья закручиваются вверх, жилки с нижней стороны опухают
и выделяют сок.
Болезнь вызывается одним из трех вирусов: a) Beta virus 1
(переносчик — Eutettix tenellus), b) Beta virus 3 (переносчик — Piesma qua-
drata Fieb.), c) Beta virus 5 (переносчик — Piesma cinerea).
— Хлороз и измельчение листьев — Beta virus 4.
3. Резко выраженная мозаичность, часто посветление по ходу
нервов; табак не заражается — Beta virus 2.
— Мозаичность выражена слабее, иногда рисунок в виде
зигзагов и колец; табак заражается — 4.
4. У свеклы рисунок в виде зигзагов, у табака — хлорозные
некротические кольца — Nicotiana virus 12.
— Обычно резко выраженная редукция пластинки листа (ните-
видность, уродства и т. д.), часто выделяется из шпината, легко
передается тлями, у табака вызывает мозаичное заболевание — Cucumis
virus 1.
Таблица V
Вирусные болезни крестоцветных
1. На листьях капусты хлорозные некротические кольца, каемка
по ходу жилок отсутствует — 2.
— Кольцевой характер рисунка отсутствует, часто каемка по
ходу жилок — 3.
2. Вирус инактивируется при 59° С — Brassica virus l.
197
— Вирус инактивируется при 50° С — некротический вирус
капусты (Larson и Walker) —два близко стоящих вируса.
3. Вирусы, не поражающие капусты, —¦ 4.
— Вирусы, поражающие капусту, — 5.
4. Вирус, выделенный от редьки, вызывает, общее заболевание
,у табака — вирус мозаики редьки.
— Вирус выделен от Brassica chmensis — Brassica virus 4.
— Вирус выделяется от Malthwla — 5.
5. Вирус заражает N. glutmosa. Петуния иммунна.—Мягкая мо
заика Matthwla.
— N. glutmosa иммунна, петуния поражается.—Тяжелая мозаика
Matthwla (2 близких вируса; Matthwla, кроме того, поражается
brassica virus 1 и Brassica virus 3, из которых первый, как и вирус Matthwla^
вызывает полоски на лепестках, а второй — не вызывает).
Таблица VI
Вирусные болезни мальвоцветных
1. Заболевания хлопчатника — 2
— Заболевания других мальвоцветных — 4.
2. Скручивание и морщинистость листьев при слабой мозаике или
отсутствии ее — 3.
— Мозаичное заболевание (мало изученное) — Бразильская
мозаика хлопчатника.
3. Листья закручиваются чаще вниз, с нижней стороны листа по
ходу жилок нередко выросты, задержка крахмала в черешках не
наблюдается — Gossypium virus 1.
— Листья закручены чаще вверх, выростов не бывает, черешки
переполнены крахмалом — Gossypium virus 2.
4. Мраморно-желтая мозаика абутилона и других мальвоцветных —
Abutdon virus I,
— Мозаика по ходу нервов у различных видов мальвы — Malva
virus 1.
Таблица VII
Вирусные болезни семейства миндальных
1. Желтухи — 2.
— Мозаичное заболевание — 6.
2. Наблюдаются поражения коры (трещины коричнево-красной
окраски) — Prunus virus 9.
— Кора не поражается — 3.
3. Листья нормального размера, нормально окрашенные. Болезнь
проявляется только розеточностью и чрезмерным ветвлением — Prunus
virus 3.
— Листья более или менее хлорозны, иногда мозаичны.
Междоузлия укорочены — 4.
4. Листья расположены компактными розетками. Плоды
морщинисты, быстро опадают или отсутствуют. Течение острое — Prunus
virus 2.
— Несмотря на розеточность, компактность не выражена. Плоды
не морщинисты — 5.
6. Плоды крупные, водянистые, безвкусные, с красными пятнами
внутри — Prunus virus l (yellow).
— Плоды мельче. Созревают позднее нормальных — Prunus virus
1 A (Little peach).
6. Листья приобретают красно-желтую осеннюю окраску и
преждевременно опадают — Х-болезнь персика и rusty-mottle вишни.
— Мозаичность носит другой характер — 1.
198
7. Мозаика по ходу нервов, резко выражена деформация пластинки
листа — Prunus virus 8.
— Другие типы мозаики — 8.
8. Кольца и полосы на листьях, а иногда и на плодах [Prunus
virus 4 и 5) — 9.
— Другие типы мозаики (Вирусы группы Prunus virus 6 и 7) —10.
9. Полосы шириной от 2 до 15 мм — Broad streak.
— Полосы шириной от 0.5 до 2 мм, редко в 5 мм—Narrow striped
variegation.
10. Пятна в форме звезд — Asteroid mosaic.
Крапчатость между нервами, морщинистость — Prunus virus 7.
Таблица VI I
Вирусные болезни малины
1. Мозаика или общее пожелтение. Иногда следы некроза и
слабая подавленность роста — 2.
— На первый план выступают другие симптомы — 6.
2. Мозаика в виде колец, подавленность рода значительна — Yel-
lowbloth
— Мозаика не в виде колец — 3.
3. Заболевания, передаваемые видами Amphorophora (главным
образом rubi), известны в США на сортах Cumberland и King — 4.
Заболевания, не передаваемые Amphorophora rubi, описанные
е Англии на сортах Mitchel и Baumforth, — 5.
4. Зеленая мозаика, которая резче проявляется весной, — Rubus
virus 1.
— Общая желтизна пластинки, пожелтение по ходу нервов.
Заболевание не маскируется летом. Часто встречаются смешанные формы
(Rubus virus 1 -f- Rubus virus 2) — Rubus virus 2.
5. Светлозеленые и желтые пятна разного размера, граница между
желтой и зеленой тканью не резкая, слабое закручивание листа.—
Болезнь № 1 (тип В).
— Светлозеленые и желтые пятна, более резко ограниченные
пораженные участки, значительно тоньше нормальных. Деформация
пластинки значительная. — Болезнь № 2 (симптомы слабее выражены
у типа А и сильнее у типа С).
6. На стебле синие и некротические полосы, морщинистость листа
слабо выражена — Rubus virus 4.
— Синие полосы на стебле отсутствуют — 7.
7. Пластинки листьев нитевидны и папоротниковидны — Сие и-
mis virus l.
— Другие признаки — 8.
$. Резко выражена морщинистость листьев, имеющих темнозе-
леный цвет и закручивающихся вниз, — Rubus virus 3.
— Другие признаки — 9.
9. Растения карликовые, с укороченными междоузлиями. Листья
значительно мельче нормальных. Вдоль жилок листа иногда некрозы —
Rubus virus 5.
— Чрезмерная кустистость, бесплодие, пролиферация цветов —
Ведьмины метлы.
Таблица IX
Вирусные болезни клубники
1. Черешки листьев сильно удлинены, количество листьев
увеличено, растение кустистое — Fragaria virus 3.
— Совокупность признаков другая — 2.
2. Поражаются главным образом молодые листья. Черешки
укорочены. Края листьев хлорозные. Х-тела в клетках — Fragaria virus l.
199
— Хлорозные пятна на листьях, иногда с некротическими точками
в центре. Морщинистость листьев, иногда пооветление по ходу нервов —
Fragana virus 2 (Часто встречаются смешанные формы Fragaria virus 3~f-
-\-Fragaria virus 2,. которые отличаются особо тяжелым течением).
Таблица X
Вирусные болеши фасоли
1. Мозаичные заболевания — 2
— Мозаика отсутствует или слабо выражена — 6.
2. Мозаика передается тыквенным, например, огурцу — 3.
— Мозаика не передается тыквенным — 4.
3. Мозаика, кроме тыквенных, передается пасленовым — Cucumis
virus l.
— Мозаика пасленовым не передается — Pea mottle virus.
4. Общей инфекции предшествует местный некроз; вирус инакти-
вируется только при 90° С и поражает сорта, которые иммунны к Pha-
seolus virus 1% например Idago Hefugi, Robust — Phaseolus virus 4.
— Мозаика развивается без местного некроза, инактивация
происходит при г° ниже 70°С — 5.
б. Зеленая мозаика. Пластинка листа часто закручивается вниз,
пороху болезнь «е передается — Phaseolus virus l.
•— Желтая мозаика. Закручивание вниз пластинки листа
наблюдается только в самых ранних стадиях болезни Характерно опускание
пластинки листа вниз от места ее прикрепления к черешку. Болезнь
гороху передается —• Phaseolus virus 2.
6*. Пораженные растения увядают На стебле темные полосы, некроз
флоэмы — увядание фасоли.
— Окраска листьев интенсивно-зеленая, по ходу нервов может
наблюдаться посветление, листья закручиваются вниз — beta virus l.
Таблица XI
Вирусные болезни гороха
1. У проростков погибают первые 2—3 листа. Общие симптомы
отсутствуют или выражены очень слабо — Pea wilt.
— Картина болезни другая — 2.
2. Кроме бобовых, вирус вызывает местные или общие поражения
у тыквенных, например, огурца — 7.
— Заболевание не передается тыквенным — 3.
3. Заболевание не передается фасоли — 5.
— Болезнь передается фасоли, например сорту Robust — 4.
4. Болезнь не передается Trifohum repens и Mehlotus albus —
Pisum virus 2
— Болезнь передается Trifohum repens и Mehlotus albus — Pisum
virus 4.
б. Некротические полосы и стебли на листьях, мозаичность,
Trifohum repens поражается — Streak 1.
— Мозаика без некрозов, а если некрозы есть, то Trifohum repens
не заболевает — 6.
6, Более или менее резкая деформация пластинки листьев, иногда
выросты на нижней стороне листа и некрозы Trifohum repens не
заболевает — Pisum virus l
— Мозаика без деф рмации, болезнь может передаваться
Trifohum repens — Pisum virus 3.
7, У огурца только местная реакция, выражающаяся желтыми
пятнами — вирус Pea mottle.
— У огурца общее заболевание — 8.
200
8. Некротические полосы, температура инактивации вируса 78—
80ГС. Тлями не передается — Streak 2.
— t° инактивации — 70°-. Легко передается тлями — Cucumis
virus 1.
Таблица XII
Вирусные болезни пасленовых кроме картофеля
1. Мозаичные заболевания или скрытое вирусоносительство — 2,
— Расстройство роста и хлорозы без мозаики — 14.
2. Местные некрозы у N. glutmosa — 3.
— Местные некрозы у N. glutmosa отсутствуют — 9.
3. Местные некрозы у N. tabacum — 4.
—- Местные некрозы у N. tabacum отсутствуют. Вирус не погибает
при высушивании — Nicotiana virus ll.
4. Вирус выделяется из корней внешне здоровых растений —
Nicotiana virus 11.
— Вирус выделяется не только из корней, но и из других частей,
растения — 5.
5. Общая инфекция у N. tabacum не наступает — 6.
— За местной реакцией следует общая инфекция — 7.
6. У помидора желтые пягна, часто отмирание точки роста- Lyco-
persicum virus 4.
¦— У помидора мозаика, некрозы, некротические полосы на стебле —
Lycopersicum virus 1.
7. Помидор сильно поражается — 8.
— Помидор не поражается или поражается очень слабо, без
некрозов — 9.
8. Вирус инактивируется при 70° С, вызывает у табака и помидора
кольцевую мозаику и некрозы без бронзовой окраски — Lycopersicum
virus 2.
— Вирус инактивируется при t° ниже 50° С, кроме колец, вызывает
бронзовую окраску — Lycopersicum virus 3.
9. Кольцевой рисунок не характерен, болезнь легко передается
тлями ¦— 10.
—• Характерные кольца и некротические пятна — 11.
10. Рисунок в виде сетки по ходу нервов. Тыквенные и N. gluti-
nosa иммунны — Solarium virus 2.
— Рисунок в виде мозаики, тыквенные и N. glutmosa поражаются —
Cucumis virus l.
11. Вирус легко передается соком, некрозы вызывает обычно только
й смеси с N. virus 1. Помидор очень чувствителен к вирусу,
температура инактивации — 70° по С — Solarium virus l.
Температура инактивации не выше 60°, помидор и N. glutmosa
мало чувствительны к вирусу и инокулируются с трудом — 12.
12. Вирус передается тлями, вызывает у табака мелкие
некротические пятна или сложный некротический рисунок — 13.
— Насекомое-иереиссчик не известно; резко выражен кольцевой
характер рисунка — Nicotiana virus 12 и другие родственные вирусы
13. Рисунок простой, состоящий в некротических пятнах и
некрозах по ходу жилок на старых листьях — Иестрица
— Рисунок в виде сложных зигзагов, колец и т. д. В цитоплазме
и ядрах пораженных клеток — кристаллы—Nicotiana virus 7.
1 У некоторых сортов табака отдельные штаммы вируса дают
некрозы, за которыми следует общее заражение. Вирус легко распознается
по гексогональным кристаллам в клетках и высокой устойчивости к
внешним воздействиям.
201
14. Болезнь легко передается соком. Задержка роста мзждоузлий,
измельчание листьев, после ряда коротких междоузлий дальнейшие
могут быгь нормальны—Lycopersicum virus 6.
— Болезнь соком не передается, совокупность признаков
другая — 15.
16. Резкая морщинистость листьев, часто на нижней стороне листа
выросты, во флоэме появляется древесина — Nicotiana virus 10.
— Совокупность признаков другая — 16.
16. Уродства цветов, флоэма гипертрофирована, листья утолщены
,и плохо высыхают —- Lycopersicum virus 5.
—• Хлороз и хрупкость листьев, посветление по нервам,
карликовость, некрозы флоэмы — Beta virus 1.
Таблица XIII
Вирусные заболевания картофеля
1. Мозаичные заболевания, а также вирусы, латентно находящиеся
у внешне здорового картофеля — 2.
— Желтухи и другие заболевания, при которых мозаичная
расцветка не наблюдается, — 12.
2 Яркожелтые пятна на листьях, которые могут сливаться и
занимать большую часть пластинки, клубни нередко с некрозами в области
сосудисто-волокнистого пучка. Иногда симптомы только на листьях
или только в клубнях — 3.
— Мозаичные пятна светлозеленые или желтоватые — 5
3. Яркожелтые пятна, сливающиеся между собой. Вирус передается
люцерне N. tabacum и N. glutmosa — Medwago virus 2, Solarium virus
10, Calico
— Мелкие, круглые пятна на листьях, иногда единичные, клубни
часто с некрозами. Симптомы могут быть только на листьях или только
в клубнях. У перца на месте инокуляции — желтые пятна (вирусы
группы F) — 4.
4. Вирус вызывает желтые пятна у помидора и S. dulcamarum —
Solarium virus 9.
— Вирус не вызывает пятен у этих видов — Solarium virus 8.
5. Молодые листья слабо закручиваются вверх, мозаика диффузная
и легко маскирующаяся — Solatium virus 11.
— Крапчатость, морщинистость, часто некроз верхушки или
нижних листьев. Картина болезни меняется в зависимости от сорта
картофеля, вирусов и штаммов, ее вызывающих. Часто смешанная инфекция
— 6.
6. У N. tabacum при инокуляции соком развиваются хлорозные
некротические пятна, часто имеющие вид колец. N. glutmosa легко
поражается вирусом (главным образом вирусы группы X) — 7.
—¦ N. tabacum — не поражается или у этого вида наблюдается
слабый рисунок по ходу жилок — 10.
/. Вирус соком не передается, обычно встречается в смеси с Х-ви-
русом, вызывает тяжелый некроз у картофеля сортов Аггап Coasal,
Up-to-Date, Duke oi York — Solarium virus 5 (вирус С).
— Легко передается соком на N. tabacum—8.
8. Выдерживает разведение 1 : 1000 и хранение в соке 4 суток — 9.
—• Не выдерживает этих условий и может быть отделен от смеси
с типичным Х-вирусом при помощи заражения сорта картофеля US
41956, иммунного к этому вирусу—¦ Solanum virus 4 (Вирус Bj
9. У дурмана вызывает мозаичное заболевание — Solarium virus l.
— Дурман является скрытым носителем вируса или проявляет
заболевание очень слабой, маскирующейся мозаикой —- Solarium virus 6
(Вирус D).
10. Вирус передается соком на N. txbacum (обычно в смеси с
Х-вирусом) и тлями (этим путем может быть отделен от X-вируса), вызывая
202
у N. tabacum иосветление по ходу нервов, а затем более интенсивную
зеленую окраску на тех же местах. N. glutmosa и дурман вирус не
поражает (преимущественно вирусы группы Y — И).
N. tabacum вовсе не поражает, на дурман и картофель передается
прививкой — Solanum virus 1 (вирус 1l).
11. У картофеля сорт President через 3—4 недели после
инокуляции вызывает тяжелый некроз, у N. tabacum —¦ посветление по ходу
нервов — Solanum virus 2
— У картофеля сорт President вызывает только слабую мозаику,
N. tabacum иммунен — Solanum virus 3 (вирус А).
12. Пластинка молодых или более старых листьев закручивается
вверх, резко выраженных некрозов стебля или верхушки растения не
бывает — 13.
— Картина болезни другая. Если закручивание листьев и
наблюдается, то верхушка растения некротизируется и погибает —15.
13. Закручиваются только доли листьев у основания черешка,
поражаются преимущественно молодые листья, закрученные участки
-хлорозны и антоцианируют, сильное развитие боковых побегов с
розетками листьев и воздушными клубнями — Solanum virus 18 [Psylhd
yellow).
— Картина болезни другая — 14
14. Закручиваются пластинки только верхних листьев, иногда
наблюдается образование клубочков, состоящих из многочисленных
мелких клубней, но этот признак менее выражен, нем при ведьминых
метлах — Solanum virus \1.
—¦ При первичном заражении закручиваются верхние листья,
а на следующий год, главным образом,—¦ нижн-ие, листья хлорозны и
хрупки, клубочков клубней не наблюдается, во флоэме мелкие некрозы —¦
Solanum virus 14.
15. Сильное развитие мелких наземных и подземных побегов
Многочисленные мелкие подземные клубни, собранные клубочком, а также
воздушные клубни, на второй год болезни — отмирание главного стебля—
Solanum virus 15.
— Совокупность признаков другая — 16.
16. Карликовость, о.мирание главного стебля, некроз стебля,
особенно сердцевины и коры узлов; вирус передается махорке и Trifohum
pratense при помощи сока и цикады Aceratogallia mngumolenta—Solanum
virus 16.
— Совокупность признаков другая—17.
17. Стебель и листья резко обращены вверх, листья несколько
хрупки, клубни удлинены, что особенно ваметно у сортов с
шарообразными клубнями, трещин клубней не наблюдается — Solanum virus 12
— Наряду с веретеновидностью клубней резко выражены
продольные трещины на клубнях, тогда как веретеновидяость мо-кет и
отсутствовать — Solanum virus 13.
Таблица XiV
Вирусные болезни далии
1. Мозаика без колец, зеленые участки вздуваются — Dahlia
virus 1.
'— Мозаика кольцами и зигзагами — 2
2. Болезнь передается соком и трипсами, заражаются различные
виды разных семейств, в том числе N. glutmosa — Lycopersicum virus 3.
— Болезнь не передается ни соком, ни трипсами — 3
3. Рисунок в виде тонких полосок, образующих кольца и зигзаги —
Dahlia virus 2.
— Более широкие хлорозные линии, образующие очертания,
которые напоминают дубовые листья — Dahlia virus 3.
ЛИТЕРАТУРА
Крапивоцветные
К анчавели Л А иЕ М Эристави 1945, Сообщение Академии
Наук Груз ССР 6, № 5, 369—376
Эристави Е М 1941 Сб «Вирусные болезни растений и меры
борьбы с ними» М , 294—309
Ainsworth G 1935 Jour Royal Hort Soc , 60, JY° 12, 533
Atanasoff D 1935 Phytopath Ztschr , 8, № 2, 197—223
В 1 a 11 n у С 1930 Sbor vyzkumnych ustavu zemedelsk RCS, 56,
1—44
В 1 a 11 n у С 1931 Rec d Trav Inst Ann Appl Biol , 8, № 2—3,
51—56
Blattny С a Vukolov V 1935 Rec Inst Rech Agr Rep
Teshac 137 3 18
Conditlia'a Home W T 1933 Phytopath , 23, № 5, 887.
Conditl I a HorneWT 1941 Phytopatn , 31, JV> b
Conditl I a HorneW T 1943 Phytopath , 33, № 8, 719—
723
Duiheld С A 1925 \nn of Appl Biol , 12, № 4, 536—543
En do Y a Kuia&ava T 1937 Bull Ser Silk Ind Uyeda,
9, Ко 2, 115—132
Endo Y a KurasavaT 1938 Japan Jour Bot 9, № i 34
En do Y. 1939 Bull Sera Silk Ind Uyeda, 11, № 4, 203—218
Endo Y 1940 Japan Joui Bot, 9, JY° 1
Lu&hmgton P M 1918 Ind Forester, 44, 114—117, 439—460
McLennan E T 1931 Aubtr Jour Exp Biol Med Sc , 5, № 1,
9—44
Roder К 1941 Faserforsch , IS, 77—81
balmon С S 1922 Jour Mm Agr , 29, № 10, 927—934
Salmon E S a WareW M 1925 Gard Chron , 77, 320—322.
Salmon E E a Ware, W M 1930 Ann of App] Biol , 22 № 4.
Smith F F 1926 Ann Mi^sour Bot Gard , 13, JV 1, 425—484
Swingle R U 1938 Phytopath, 19, Л'« Ю, 757—759
SwingJe R U 1940 Phytopath , 30, № 1, 23
Swingle R U, Tilford P. S, Irish С F. 1941
Phytopath , 31, № 1, 22
S w l n g I e R U 1942 Circ U S Dept Agr , 640, 8
Chenopodiaceae
В об к A M 1942 Микробиология, 11, № 4, 169—177
Левшин 4. H 1930 Сб «Мозаичные болезни сахарной свеклы>>
Киев 177—179
«Мозаичные болезни сахарной свеклы) 1930. Сч под ред Муравьева
Киев Работы Новиненко и др
Сиротина М 1932 Наук зап цукер промисл 9, № 24, 195—216.
Сухов К С 1940 Микробиология, 9, JV» 2, 188—195
Bennett С W 1934 Jour of Agr Res , 48, 665— 701
204
Bennett G. W. 1935. Jour, of Agr. Res. 50, 211—241.
Bonoquet P. a. Hartung W. 1915. Phytopath., 5, № 6, 348—
349.
Coons G. H., К о til a J. E., Stewart D. 1937. Phytopath.,
27 125.
CarYnerE. 1933. U. S. Dept. Agr. Techn. Bull., 360.
Hogg on J. 1933. Phytopatn., 2j3, 446.
Lind J. 1915. Tidssknft for Planteave, 22, 444—457.
Quanjer H. M. 1936. Tijdschr. for Plantenziekt., 46, 46—60.
Prillieaux et Delacioux. 1891. Bull. Soc. Mycol. France,
7 22 25.
Roland G. 1936. Tijdschr. Plantenziekt., 46, 54—70.
Schaffnit u. Weber. 1927. Forsch. auf d. Geb. d. Pflanzenkr.
u. d. Immun. in Pflanzenreich, 1, № 4, 23—42.
Sever in H. H. P. a. F r e i t a g. 1933. Hilgardia, 8, 1— 48.
Simon M. 1940. Publ. Inst. Belg. Amelion Better., 8, 1, 20—22.
Wille J. 1928. Arb. biol. Reichsanstalt, 16, 115—167.
Walker I. C, de Blau F. I,. Paund G. S., 1945, J. agr. гез. 7J,
373—404.
Крестоцветные
Dana B. a. Mc.Whorter F. 1932. Phytopath., 22, № 12, 1000—
1001.
Clayton M. E. 1930. Jour, of Agr. Res., 40, № 3, 263—270.
Gardner M. a. Kendrick J. 1921. Jour, of Agr. Res., 22, № 2,
111—114.
Hoggan I. a. Johnson J. 1925. Phytopath., 25, 6.
Hopk'ins a. P a r d у М. 1942. Rhod. agr. Jour., 30, JY° 5, 376—383.
Larson R. H. 1939. Jour, of Agr. Res., 59, № 5, 367—392.
Larson R. H. a. Walker, J. G., 1940. Phytopath., 30, № 11,
15.
Ling L. a. Yong J. Y. 1940. Phytopath., 30, JV 4, 338—342.
S с u 1 t z E. 1921. Jour, of Agr. Res., 22, JY» 3, 173—179.
Smith K. a. Markham R. 1944. Phytopath. <34, № 3, 324—329.
Tompkins C. M. 1934. Phytopath. 24, Лг° 10, 1136.
Tompkins С. М. 1934. Pnytopath., 24, J4» 10, 1137.
Tompkins C. M. 1937. Jour, ot Agr. Re». 55, № 1, 33—45.
Tompkins С M. Gardner M. W. a. Thomas H. R.
1937. Phytopath. 27, № 9, 955—956.
Tompkins С. М. 1938. Jour, of Agr. Res. 57, .TV» 8, 589—602.
Tompkins С M. Gardner H. W. a. Thomas H. R.
1938. Jour. As?r. Res. 5Г, № 12, 929—943.
Tompkins С. М. 1939 Jour, of Agr. Res. 58, № 2, 119—130.
Tompkins C. M 1939a. Jour, of Agr. Res. 58, № 1, 63—77.
Мальвоцпетные
В а рун ц ян И. 1938. Сов. хлопок, № б, 76—81.
Вердеревский Д. Д. 1938. «Вирусные болезни ростений». М.
29—55.
Кара-Мурза Л. X. 1938. Сб. «Вирусные болезни растений».
Вып. 2. М. 56—72.
Кара-Мурза Л. X. 1941. Сб. «XX лет Азербайджанского
института». Кировабад, 18—26.
Кара-Мурза Л. X. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М. 197—203.
«Курчавость листьев — вирусная болезнь хлопчатника». 1937. Сб. под
ред. Запрометова. Ташкент.
Московец С. Н. 1938, Сов. хлопок, № 6, 74—76.
Московец С. Н. 1940. Вирусное заболевание хлопчатника и меры
борьбы с ним. Ташкент.
205
Московец С. Н. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М. 173—191.
Рыжков В. Л. и Овчаров Т. П. 1941. Сб. «Вирусные болезни.
растений и меры борьбы с ними». М. 191—197.
Afzal Н. М. Jaggi S. S. a. Singh В. 1935. Ind. Jour. Agr.
Sc. 5, № 1, 624—631.
Pailey M. A. 1934. Emp. Cott. Grow. Rev. 11, № 4, 280—288.
В о t e г о К. О. 1940. Publ. Min. Econ. Colombia.
Cook О. Е. 1924. Jour, of Agr. Res. 28, № 8, 803.
Costa A. tf. a. Forster R. 1938. Rev. Agr. S. Paulo, 13, № 3/4„
187—191.
Cotone. 1931. Rev. Appl. Ent., 731.
U a s t ц r R. H. 1936. Ind. Jour. Agr. Sc. 6, 377—395.
h>astur R. H. 1939. Ind. Jour. Agr. Sc. 9, № 9.
IJavis E. F. 1929 Ann. Miss. BoL Gard. 16, № 2, 385.
folding F. D. 1930. Emp. Cott. Grow. Rev., 7, № 2, 120—126.
*arquh arson. 1913. Report of the MycoJogist Nigeria.
Hertzsch W. 1927. Ztscnr. f. Bot., 20.
Husain M. 1932. Nature, ISO, № 3278, 312.
Hutchinson a. Ghose. 1937. Jour, of Genet.
?eur J. Y. 1934. PhytopaLh., 24, № 1, 12—13.
Kirkpatrick T. W. 1931. Bull, of Ent. Res., 22, 3.
Lambert A. R. 1938. Emp. Cott. Grow. Rev., 15, № 1.
Ь e m о i n e M. 1869. Jour, de la Soc. Imp. Centr. Hort. d. France, 554.
Lindsmuth H. 1907. Landw. Jahrb., 36, 807—861.
Masse у R. E. a. Andrews F. W. 1932. Emp. Cott. Grow. Rev.,
9, 1, 32—45.
Mathur R. N. 1932. Nature, 129, № 3265, 797.
M a t h u r R. N., 1933. Ind. Jour. Agr. Sc., 3, № 1, 89—96.
Neal D. C. a. Lovett H. C. 1938. Phytopath., 28, № 8, 582—
587.
Ou S. H. 1934. Sinensia, 5, № 5—6.
Russo G. 1935. Agr. Colon., 29, № 2, 78—95.
Silberschmidt K. 1943. Arquiv. do Inst. Biolog. Sao Paulo,
14, № 9, 105—156.
Teng S. C. 1936. Sinensia, 7, № 1, 63—79.
U p p a 1 B. N.. Varma'P. M., С о р о о r S. P. 1940. Current
Sc. 9 № 5 227- ^28
Wong s! C". 1924. Res. Lab. Wat. South: East. Univ., 28.
Виноград
Рыжков В. Л. 1935. «Вирусные болезни растений). М.
Blattny С. 1934. Ochr. Rostl., 13, 104—105.
Branas J. et Bernon G. 1935. Ann. Ec. Nat. Agr. Montpellier,
23, № 3.
Branas J. et Bernon G. 1936. Ann. Ec. Nat. Agr., Montpellier,
24, № 1.
Branas J. et Bernon G. 1937. Ann. Ec. Nat. Agr., Montpellier,
24, № 3, 249—252.
Branas J., Bernon С et Levadoux.' 1937. Rev. Vit., 87,
№ 2258, 263—268.
Branas J. 1938. Progr. Agr. Vit., 110, № 2, 25—31.
Branas J. 1938. Progr. Agr. Vit., 110, № 44, 373—379.
Branas J. 1939. Progr. Agr. Vit., 112, № 37, 202—204.
Carpentieri. 1930. Trattato di Vitic6ltura moderna, Roma.
С a s t e 1 1 a F. 1934, Jour. Agr. Victoria, 32.
Duffrenoy J. 1935. Progr. Agr. Vit., 102, № 42, 43, 44, pp. 177—
182, 405—411, 429—432.
Em on J. 1938. Rev. Vit., 87, № 2287, 303—305.
206
G a 1 1 e s P. 1937 Progr Agr Vit , 107, JY° 15, 346—347,
G l g a n t e 1937 Boll R Staz Path Veg , Roma, 17, № 2, 164—192.
d. H. 1936. Progi Agr Vit , 104, № 40, 328—329
Hewitt W В , Jacob H E, Proebstiry E. L, La-
miman J E 1939 Phytopath, 29, 110
Hewitt W. B, Frazier B. N, vV Houston. 1942
Phytopath , 32, 1
H e w l t t W. B. 1945 Phytopath 35, № 11, 94C- 942.
Johnbsen A 1933 Dtscn Wembau, № 17—20,
Lykiardopulo T L. 1940 Progr Agr. Vit , 93, 12—13.
Maier W u Mittmann-Maier G. 1939, Wem u Rehe, 21r
251—272
MarsaibP a Segal L 1939 Rev Vit , 91, 257—264
MartinoffS G 1934 Agriculture, Fofia, 33, № 2, 6
MoreauL etVmetE 1938 G R. Ac Agr. Fr , 24, № 2, 709—
714
P a n t a n e 111 . 1912—1913 Zeit. f. Pflanzenkrankheiten.
Petri L 1911 С R. Ac Lincei, 25, № 2, 155—160.
Petri L 1934 Boll. R. Staz Path Veg Roma, 14, 273—277
Ranphiano 1932 Rev de Viticulture
Rives L 1935 Progr Agr Vit, 104, JV* 33, 159—162.
Rous L 1937 Progr Agr Vit, 107, № 12, 285—287.
Stranak F 1931 II me Congr. Int Pathol, comp , 1, 367
Vial a P 1903 Maladies de la vigne. Pans.
Viala P et Marsais. 1934. Ann. Inst. Agr.
Розоцветные. I
Ark P. A aThomasH E 1940 Phytopath , 30, № 2, 148—154.
Atanasofl D 1933 Yearbook Umv Sofia, 13, 31—67.
В a ur E 1907 Ber d. D Bot. Ges , 25, № 7, 410—413
Berkeley G H 1941 Publ. Div Bot PI Path Ottava, 679, p 2
BodineE W. a. KreutzerW. A. 1942 Phytopath , 39, J\« 2.
В о d 1 n e E. W. a. Newton J. H. 1942. Phytopath , 32, № 4,
333—334
Bradford F. С a JolayL. 1933 Jour of Agr Res , 46, JV« 10,
901—908
Dunne T С 1938 Joui Dep Agr W, Austr , 16, 120—126.
Cation D. 1941 Phytopath, 31, № ц, 1004—1010
Cation D 1942 Phytopath, 32, JV> 1.
Chandler. 1937. Bot. Gaz , 98, № 4.
С h r 1 s t о f f A 1938 Phytopath. Ztg , 11, JY° 4, 360—422.
Cochran L. C. a Smith С О. 1933 Phytopath , 28, № 9, 278—
281.
Cochran L. С a Hutchins L. M. 1938. Phytopath., 28,
№ 12, 890—892
Cochran L. С a Hutchins L. M. 1941. Phytopath.» 31t
JV» 9, 860
С u r z 1 M. 1931. Boll. R. Staz. Path. Veg , Roma, № 3, 11, № 3f 221—
243.
Ferraris J. 1928. Guriamo le Piante, Torino, 6, 101—114.
H a r t z e 1 1 A. Phytopath , 27, JY> 2, 130.
Hartzell A. 1937 Contrib. Boyce Thompson Inst , 8, JY° 5, 375—388,
HildebrandE M. a. Palmiter 1940. Phytopath , 30, JY° 1, 10.
Hildebrand E. M. a. Mi lies W. D. 1941. Phytopath., 31y
№ k, 355—356
Hutchins L. M. 1933, Off. St. Ent. Georgia Bull., 78, 1—55.
Hutchins L, M. 1939 Phytopath., 29, JY° 1, 12
Keit G. W. a. Clayton С N. 1941. Phytopath., 30, 14.
Keit G. W. a. Clayton C. N. 1943. Phytopath., 33, JV- 6, 449—
461.
207
Kienholz I. R. 1939. Phytopath., 29, JY° 3, 260—267.
К i n m a n C. F. 1939. Jour, oi Agr. Res., 41 ... 327—355.
Kunkel L. O. 1933. Contrib. Boyce Тдолірзол Inst., 5, 19—20.
Kunkel L. O. 1935. Phytopath., 25, № 1, 24.
Kunkel L. 0. 1936. Phytopath., 26, № 2, 201—219.
KunkelLO. 1943. Offprint fron Virus dislaie New York, 63—32.
Kunkel Ti. 0. 1944. Phythopath, 34, 12, 1006.
Milbrath I. A. a. Zeller S. W. 1942. Phytopath., 32, JY° 5,
423—430.
Newton W. 1936. Better Fruit. 31, JY° 3, 7—14.
Rankin W., Hockey Ja. M с Gurry. 1922. Phytopath.,
12, 58.
Reeves E. L. 1940. Phytopath., 30, JY» 9, 729.
Reeves E. L. 1941. Jour, of Agr. Res., 62, № 9.
Rasmussen E. I. a. Cation D. 1942. Phytopath., 32, JY° 1,
15.
Siraonds A. O. a. Bodine E. W. 1943. Science, 97, 587—588.
Smith E. I. 1888. U. Sf D. A. Div. of BoL Bull., 9, 254.
Smock R. M. 1941. Proc. Amer. Soc. Hort. Sc, 32, 7.
Smock a. van Doren. 1938. Proc. Amer. Soc. Hort. Sc, 3*,
176—179.
Stoddard Е- М., 1940. Phytopath., 32, 2, 17.
Thomas a. Hildebrand. 1936. Phytopath., 26, JV» 12, 1145—
1148.
Thomas H. E. 1937. Hilgardia, 10, № 14, 581.
Thomas H. E., Rawlins T. E., Parker K. G. 1940.
Phytopath., 30, JY> 3, 322—328.
Wallace T. 1939. Jour. PomoL, 17, № 2, 150—166.
Wi Hiss on A. S. 1944. Phytopath. 34, 12, 1037—3 049.
Zeller S. M. a. Evans A. W. 1941, Phytopath., 31, № 5, 463—
467.
Zeller S. M. a. Milbrath J. A. 1942. Phytopath., 32, JV» 7,
Розоцветные. II
Вертоградова О. Н,. 1938. Сб. «Вирусные болезни растений
и меры борьбы с ними». Вып. 2. М., 232—237.
Натальина О. Б. 1938. Сб. «Вирусные болезни растений». Вып. 2.
М., 181—196.
ТетерниковаД. Н, иКечекН.А. 1943. Болезни ягодных культур.
Арм. ССР, Ереван, 25 стр. .
A f a n a s і е v М. М. а. М о о г г і S. 1941. Phytopath., 31, W 1, 1.
Baker К. F. а. Т h о m .a s Н. S. 1942. Phytopath., 32, JV 4.
Bennett С. W. 1927. Agr. Exp. St. Techn. Bull., 80, 1—38.
Berkeley G. H. a. Plakidas A. G. 1942. Pnytopath., 32t
№ 7.
Brierley Ph. 1935. Phytopath., 25, № i, 7.
Brierley Ph. 1935a. Pnytopath., 25, № i, 8.
Brierley P. a. Smith F. F. 1940. Jour, of Agr.. Res,, 61, № 9,
625—666.
Chamberlain E. E. 1938. Can. Jour. Res. Sect. G., 16, № 3,
118—124.
Chamberlain E. E. 1941. Sc. Agr., 22, JY° 2, 119—124.
С о о 1 e у L. M. 1936. Phytopath., 26, № і, 44.
€ooley L. M. 1936. N. I. Stat. Techn. Bull., JY° 665.
Dicker G. H. L. 1940, Jour. PomoL, 18, JY° 3, 275—286.
G i g a n t e R. 1936. Bull. R. Staz. Path. Veg., Roma, N. S., .10, № 1,
76—94.
Ore en» S. B. 1894. Minnesota Agr. Expt. Stat. Rpt., 230.
208
Grive В. J. 1931. Auslr. Jour. Exp. Biol. a. Med. Sc., 11, JY« 1, 56—
76.
Orive B. J. 1942. Proc. Rov. Soc Victoria, 229—238.
Ouba E. F. 1933. Phytopath., 23, № 8, 654—661.
Harris R. V. 1933. Jour, of Pom. a. Hort. Sc, 11. № 1, 56—76.
Harris R. V. 1933. Jour, of Pom. a. Hort. Sc, 11, Л'? 3, 237—255.
Harris R. V. 1937. Ann. Rep. East. Mall. Res. St. 1936.
Harris R. V. 1940. Jour, of Pom. a. Hort. Sc, 17, № 4, 318—343.
Hildebrand A. 1941. Canad. Jour. Res. Sect. C, 19, № 7, 225—233.
IF liber G. A. a. Schwartz E. D. 1938. Jour, of Agr. Res., 57,
JV« 8, 623—633.
Mass ее. 1942. Jour, of Pom., 20, Л» 1—2, 42—47.
Mel с hers L. 1914. Ohio Naturalist, li, 281—288.
JMakidas A. G. 1927. Jour, of Agr. Res., 35, № 12, 1057—109o.
Rankin W., Hockey J. a. M с. С u r r y. 1922. Phyiopath.,
12, 58.
Hankin W. H. 1931. N. У. St. Agr. Exp. Stat. Techn. Bull., Л?> 175,
24.
Thomas H. E. a. Massev L. M. 1939. Hilgardia, VI, ,V« 10,
647—663.
White R. P. 1932. Phytopath., 22, № 1, 53—69.
Whitehead a. Wood G. 1941. Nature, 148, 597—598.
Wilcox R. B. 1922. United Stat. Dept. Agr. Giro. Л? 227, 3—12.
Я e 1 1 e r S. M. 1927. Phytopath., 17, Л'ь 9, 629 -648.
teller S. M. a. V а у g h a n E. K. 1932. Phytopath., 23, JV 8,
709—713.
/eller S. M. 1931. Phytopath, 21, 923—925.
teller S. M. a. Weaver L. E. 1941. Phytopath., 31, № 9,
849—851.
7j n n d e 1 G. L. 1931. Phytopath., 21, 755—757.
Бобовые
Д и t m a p. 1936. Болезни бобовых. М.
Я к и м о в и ч Е Д. 1938. Сб. Тр. НИ. Сев. аерн. хоз. Вып. 3, 79—112.
A it ken Y. а. В. 1. G rive. 1943. I. Aibtr. Insr. Agric Sc 9, 2, 81—82.
Ains worth G. C. 1940. Ann. of Appl. Biol., 27, Л* 2, 218—226.
Baur E. 1906. Ber. d. D. Bot. Ges., 24, 416—428.
BlacV L. M. 1945. Amer. J. Bot. 32, № 7.
В б n j n g. 1927. Forsch. auf d. Geb. d. Pflanzenkv. u. d. Immuii. im
Pflanzenreich, № 4, 43—111.
Chamberlain E. E. 1936. N. Seel. Jour, of Agr., 51, № 2, 86—92.
Chamberlain E. E. 1935. N. Zool. Jour. Sc Techn., 20, № 6,
365—371.
Clinton G. P. 1909. Connec State Agr. Exp. Stat. Ann. 1908. Rpt.,
742—743.
Clinton G. P. 1916. Connec State. Agr. Exp. Stat. Ann. 1915. Rpt.,
446—447.
Dana B. F. 1940. Phytopath., 30, № 9, 781.
Doolittle S., Parones F. 1925. Phytopath., 15, № 2, 763—
771.
Dickson. 1922. Mc. Donal College Techn. Bui., A1- 2, p. 123.
Edwards. 1935. Journ. Ausrt. Inst. Agr. Sc, 1, № 1, 31—32.
Edwards. 1936. Sc Bull. Dept. Agr. New. Soath Wales. № 52, p. 3J.
Faijardo T. G. 1928. Phytopath., 18, № 1, 155.
F a i j a r d о Т. G. 1930. Phytopath., 20, № 6, 469—494.
Faijardo T. G. 1930. Pnytopath., 20, № ц, 883—888.
Gardner M. W. a. Kendrick J. 1921. Jour, of Agr. Re$M
22, № 2, 111—114.
¦Gardner M. W. 1925. Jour, of Agr. Res., 30, Ле 9, 871—888.
3 4 в. л. Рыжков 200
Grant T. J. 1939. Phytopath., 29, № 1, 8.
Gigaute K. 1935. Boll. K. Staz. Pat. Veg. 15, Al> 4, 533—547.
Harrison. 1935. Teciin. Bull. Stat. Agr. Exp. Stat. № 4, 236.
Harter J:. L. 1938. Jour, ol' Agr. Res., 56, № 12, 825—906.
Hartley G. a. Haasis F. M. 1929. Phytopath., 19, № 2, 162—
166.
Heinze K. u. Kohler E. 1940. Phytopath. Zbxmr., 7<3, H. 3,
207—242.
Hoy man \V- G. 1944. Phytopath., 34, № 5, 505—506.
Imai J. 1934. Trans. Sapporo Nat. Hist. Soc, 13, K° 3, 241—245.
Jenkins W. A. 1940. Jour, of Agr. Res., 60, A<> 4, 279—288.
Johnson H. W. 1938. Phytopatn., 28, AT» 1, 10.
Johnson H. W. a. Lefebvre G. L. 1938. Phytopath., 28,
№ 1, 10—11.
Johnson IT. VV. 1938. Phytopath., 28, A* 4, 273—277.
Johnson F. 1942. Pnytopath., 3^, As 2, 103—116.
Ivanowski D. 1899. Ceafcr. f. Bakterioiog., 5, 250—254.
Kohler B. 1935. Augew. Bot., -77, JVs 5, 276.
Meiuies I. D. 19k4. P.iyiopalu., 34, in, 936.
McLean D. M. 1941. Pnytopath., 31, A* 5, 420—430.
M о n 11 e i t h a. Hollowell. 1929. Jour, of Agr. Res., 38, № 12,
649—677.
Murayama D. 1938. Trans. Fac. Agr. Sapporo, A* 44, 1—32.
Murpnv D. M. Ш0. Phytopath., 30, A? 9, 779—784.
Neill J. C, Br ion R. M. a. Chamber Jain E. К 1934.
N. Z. J(Mi\ Agr., </0, Ac 3, 139—146.
О s b о r n H. T. 1937. Phytopath., 27, A* 5, 589 -603.
Osborn H. T. 1938. Pnytopath., 28, A"» 10, 749—754.
О s born IT. T. 1938a. P.iylopath., 28, A? 12, 923—936.
Pierce W. II. a. II u n g e r f о г d, 1929. Agr. exp. Stat. lies. BuL
Idago, Af> 7, 37.
Pierce \V. H. 1934. Phytopath., ,?/, A* 2, 87 — 115.
P i с rr e AV. II. 19:i,'). Jour, of Agr. Res., 57, A? 11, 1017—1039.
Pinrce W. II. 1937. Phytopath., 27, A<> 8, 836—842.
Rosa H. I), a. Misra S. I). 1938. lnd. Joir. of Agr. Sc, 8, № 4r
417-418.
Snyder W. G. a. T h о m a s. 1938. Hilgardia, 10, A« 8, 257—262.
Snyder W. G. 1942. Phytopath., 32, № 6.
St abbs M. W. 1937. Phytopatn., 27, № 5, 242—266.
U p p a 1 A. 193». Inter. Bull. Plant. Prot., 5, A? 9, 163.
Virgin W. J. 1943. Jour, of Phytopath., 33, A° 8.
Weimer J. L. 1936. Jour, of Asfr. Кеч., 53, 333—347.
Weimer J. L. 1937. Jour. Agr. Res., 55, № 2, 87—104.
Weimer J. J,. 1937. Phytopath., 27, № 6, 697—702.
Whipple O. G. 1936. Pnytopath., 26, № 9, 918—920.
Whipple O. C. a. Walker J. G. 1938. Phytopath., 28, № 1, 22.
Whipple O. G. a. Walker J. G. 1941. Jour, of Agr. Res., 62y
A? 1, 27—60.
Zaumeyer W. J. а. К earns G. W. 1936. Phytopath., 26, № 1,
614—629.
Zaumeyer W. J. 1937. Phytopath., 27, A? 2t 144.
Zaumeyer W. J. 1938. Jour, of Agr. Res., 56, № 10, 747—772.
Zaumeyer W. J. 1940. Jour, of Agr. Res., 60, Ai> 7, 433—452.
Zaumeyer W. J. Jour, of Agr. Res., 64, № 2, 119—127.
Zaumeyer W. J. a. Harter L. L. 1942. Phytopath., 32, A1? 5;
438—439.
Zaumeyer W. J. a. Harter L. L. 1942. Phytopath., 32, № 5,
428—439.
Zaumeyer W. J. a. Harter L. L. 1943. Jour, of Agr. Res.,
67, № 8, 297—320.
Zaumeyer W. J. a. H a r t e r L. L. 1943. Jour, of Agr. Res.
67, № 7, 287—292.
Zaumeyer W, J. a. Harter L. L. 1944. Phytopath., 34, № 5,
510—512.
Zaumeyer W. J. a. Wade B. L. 1935. Jour, of Agr. Res., 51,
№ 8, 715—749.
Zaumeyer W. J. a. Wade B. I. 1936. Jour, of Agr. Res., 53,
№ 3, 161—185.
Зонтичные
В 1 a tt n у С. 1933. Ochr. Rostl., 13, № 3—4, 145—146.
Doolittle S. P. 1931. Phylopath., 21, JN° 1, 114.
D о о I i t t 1 e S. P. a. Wellnian F. L. 1934. Phytopath., 24,
№ 1, 48—61.
Gigante R. 1936. Boll. R. Staz. Path. Veg. Roma. N. S., 21, № 3, 2.
Mc. Whorther a. Owens. 1940.-Plant Diss. Rep., 24, № 9. 176.
Price W. 1935. Phytopath., 25, № ю, 947—954.
Sever in H. H. P. 1934. Hilgardia, 8, № 10, 334—361.
Sever in H. H. P. a. Freitag Т. Н. 1938. Hilgardia, 11, № 9,
* 439—558.
Smith К. М. a. Markham R. 1944. Phytopath., 34, № 3, 335—
350.
Well man F. L. 1934. Phytopath., 24, № 7, 695—725.
Well man F. L. 1935. Phytopath., 25, № 4, 377—404.
Пасленовые (кроме картофеля)
Вертоградов О. И. 1941. Сб. «Вирусные болезни растений и
меры борьбы с ними». М., 278—286.
Гольдин М. И. 1937. ДАН СССР, 15, Л* 9, 559—561.
Гольдин М. И. 1938. Микробиология, 8, Л° 5, 613—619.
Гольдин М. И. 1940. Микробиология, 9, JV» 7—8, 713—723.
Ивановский. 1902. Мозаичная болезнь табака. Варшава.
И в а х н е н к о А. И. 1938. Зап. Харьк. с.-х. ин-та, 1, Л° 2, 179—282.
Корачевский И. К. 1941. Сб. «Вирусные болезни растений
и меры борьбы с ними». М., 255—264.
К о с т о в Д. 1936. Труды конференции по ультрамикробам,
фильтрующимся вирусам и бактериофагу. М., 391—417.
Осницкая Е. А. 1936. Плодоовощное хозяйство, 1, JV« 2, 23—25.
П о н е р В. О. 1941. Сб. «Вирусные болезни растений и меры борьбы
с ними». М., 227—245.
Рыжков В. Л. и Корачевский И. К. 1934. Сб. «Вирусные
болезни растений в Крыму и на Украине». Симферополь, 1-—30.
Рыжков В., Михайлова TL, Пивоварова Р. 1936.
Сб. «Вирусные болезни растений»'. Вып. 1, М.
Р я х о в с к и й Н. А. 1936. Сб. «Вирусные болезни растений». 1, М.
-Су х о в К. С. и В о в к А. М. 1946. ДАН СССР, 53, № 2, 153.
Терновский М. Ф. 1941. Сб. «Вирусные болезни растений и
меры борьбы с ними». М., 219—227.
Худы на И. П. 1937. Вирусные болезни табака. Краснодар.
Худына И. П. 19*1. Сб. «Вирусные болезни растений и меры
борьбы с ними». М., 203—219.
Шатова Е. В. 1941. Сб. «Вирусные болезни растений и меры борьбы
с ними». М., 286—294.
пристава Е. М. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М., 294—309.
Ainsworth G. С. 1933. Ann. of Appl. Biol., 2Q, № 3, 421—428.
Ainsworth G. C. 1934. Ann. of Appl. Biol., 21, № 4, 566—580.
Ainsworth G. С 1937. Ann. of Appl. Biol., 24, № 3, 545—556.
Alexandri A. V. 1938. Rev. Appl. Mycol., 17, № 9, 571.
В. Л. Рыжков
2ii
Bawden F. С. a. Kassanis B. 1941. Ann. of Appl. Biol., 28*
№ 2, 107—118.
Bawden F. G. a. Pirie N. 1942. Brith. Jour. Exp. Path., 2t3>
314—328.
Bei jerinck M. W. 1898. Verhandl. K. Akad. Wetensch. Amsterdam
Sect. 2, 6, № 5, 1—22.
В lake si ее A. F. 1921. Jour. Gen., 11, № 1, 17—36.
Blood H. L., Richards B. L. a. Wann F. R. 1933.
Phytopath., 23, № 11, 930.
Boning K. 1928. Arb. aus d. Bayer Lands. Pflanzenb. Pflanzansch.,.
4, 36—40.
Boning K. 1931. Zeitschr. Parasitenkund., , 103—144.
Dana B. F. 1940. Phytopath., 30, № 9, 785.
Doolittle S. a. Beecher F. 1937. Phytopath., 27t 800—801,
Doolittle S. a. Beecher F. 1943. Exp. tot. Rec, 88, 3.
Frimmel F., Lauche K. 1940. Obst. u. Gemuseb., 86, № 1, 2—3.
From me F. D., Win gar d S. A., Priode G. N. 1927.
Phytopath., 17, № 5, 321—328.
Gardner M. W. a. Kendrick J. B. 1923. Phytopath., ty,
№ 8, 372—375.
Gardner M. W., Tompkins a. Thomas. 1937. Phytopath.„
27, № 1.
Hamilton M. A. 1932. Ann. of Appl. Biol., 19, № 4, 550—567.
H e 1 s о n G. A. H., 1942. Jour. Coun. for Sc. a. Industr. Res. Austr.
bull., 15, № 2, 175—184.
Hill A. 1937. Jour. Coun. for Sc. a. Industr. Res., 10, № 4, 309—312.
Hill A. 1941. Jour, for Sc. Industr. Rec, 14, 181—186.
Hill A. 1943. Jour. Coun. for Sc. Industr. Res., 16, № 2, 85—90.
Hoggan J. a. Johnson J. 1936. Jour. Agr. Res., 52, 271—294.
Holmes F. O. 1932. Contr. f. Boyce Thompson Inst., 4, № 3, 323—357.
Holmes F. O. 1941. Phytopath., 30, № 1, 12.
Holmes F. O. 1942. Phytopath., 32, № 12, 1058—1067.
Imle E. F. a. Samson R. W. 1937. Phytopath., 27, № 2, 132.
I van off S. S. 1942. Phytopath., 42, JV* 1 (Abstr.).
Jochems S. 1928. Deli Proefst. Med. Sumata Bull., 26, 5—26.
Johnson E. M. 1930. Kent. Agr. Exp. Stat. Bull., 306, 289—415.
Johnson J. 1925. Wisconsin Agr. Exp. Stat. Res. Bull. JY° 3, p. 12.
Johnson J. 1927. Wisconsin Agr. Exp. Stat. Res. Bull., № 76, p. 15.
Johnson J. 1929. Wisconsin Agr. Exp. Stat. Res. Bull., № 95, 25.
Johnson J. 1936. Phytopath., 26, 285—292.
Johnson J. 1937. Jour, of Agr. Res., 54, 239—273.
Johnson J. 1939. Jour. Agr. Res., 58, № 11, 843—863.
Johnson J. a. Fulton R. W. 1940. Phytopath., 30, № 4 13.
Johnson J. a. Fulton R. W. 1942. Phytopath., 32, № 7.
Jones L. K. 1940. Phytopath., 30, № 6.
Jones S. E. 1942. Bull. Tex. Agr. Exp. Stat. № 623, 17.
Kassanis B. 1941. Ann. of Appl. Biol., 28, № 3, 238—243.
Karraker a. Bortner. 1934. Kent. Agr. Exp. Stat. Bull., № 349.
Koch K. L. 1933. Phytopath., 23, № 4, 319—342.
Kohler E. 1935. Naturwiss., 23, 828—830.
Kohler E. 1941. Zeit. Bakter., Parasit. u. Infektionskrankh.,
103, № 18/20, 325—334.
К os toff D. 1933. Genetica, 15, 301—314.
Mage С a. Morgan W. 1942. Jour, of Austr. Indust. Agric. Sc.,.
8, № 3, 115.
M c. Kinney. 1929. Jour. Agric. Res., 39, № 8, 557—578.
Matsumoto J. 1938. Trans. Nat. Hist. Soc. Formosa, 28, № 176„
123—137.
Mayer A. 1886. Landw. Versuchst., 32, 451—467.
Mc. Clean. 1935. Sc. Bull. Dept. Agr. South. Afr., 139, 5—34.
212
Mes M. 1930. Phytopath. Ztschr., 2, 594.
Milbrath J. A. 1937. Phytopath., 27, № 8, 868—869.
Milbrath J. A. 1939. Phytopath., 29, № 2, 156—168.
Mogendorff N. 1930. Phytopath.» 20, № 1, 25—46.
Nakata K. a. Takimo S. 1940. Bull. Sc. Fakult. Kunshu Univ.
Fukuoka, 9, JY° 2, 167—189.
Price W, G. 1936. Phytopath., 26, № 7, 655—675.
Prut hi. 1939. Ind. Jour, of Agr. Sc., 9, 223—275.
Rischkow, Koratschewsky, Mic. hailowa. 1933.
Zeitschr. f. Pflanzenkr. und Pflanzenschutz, 43, 496—498.
Samson R. W. 1940. Phytopath., 30, № 1, 21.
Samson R. W. a. Imle N. P. 1942. Phytopath., 32, № 12, 1037—
1047.
Samuel G., Bald J. a. P i t t m a n H. 1930. Jour. Count, f,
Sc. a. Industr. Res. Austr. Bull. № 44, 64.
Samuel G., Bald J. a. Eardley E. 1933. Phytopath., 23
№ 8, 641—653.
Schreven D. A. 1941. Naturwetensch. Tijdschr. Nederl. Indie 101.
113—114.
Shapovalov. 1928. U. S. Dept. Agr. Misc. Publ., № 13.
Shapovalov M. a. Beecher F. 1930. U. S. Dept. Agr, Techn.
Ball., № 189.
Shear G. M. a. Ussery H. D. 1940. Jour, of Agr. Res., 60, № 2
129—139.
Smith K. 1931. Ann. of Appl. Biol., 18, № 1, 1—15.
Smith K. 1935. J. Roy. Hort. Soc, 60, 448—451.
Smith K. 1935 a. Parasitology, 27, № 2, 231—245.
Smith K. 1937. Parasitology, 29, № 1, 70—84.
Smith K. 1943. Parasitology, 35, № 3, 159—160.
Smith K. a. Markham R. 1944. Phytopath., 34, № 3, 324—329.
Smith P. G. 1944. Phytopath., 34, № 5, 504—505.
Spenser E. L. a. Lavin G. J. 1939. Phytopath., 29, № 6, 502—
503.
Storey H. H. 1931. Nature, 128, 187—188.
Storey H. H. 1932. Rhod. Agr., 29, № 3, 186—192.
Storey H. H., 1935. East Afr. Agr. Jour., 1, № 2, 148—153.
Thomas K. M. a. Krishna. 1939. Proc. Ind. Acad. Sc. Sect. B,
10, 201—212.
Thomas W. a. Mack W. B. 1941. Proc. Am. Soc. Hort. Sc, 39y
319—328.
d'O 1 i v e i r a Maria. 1940. Agron. lusit,, 11, № 3, 209—223.
Valleau W. a. Johnson E. 1927. Phytopath., 17, № 8, 523—528.
Valleau W. a. Johnson E. 1930. Kent. Agr. Exp. Stat. Res.
Bull., JV* 309, 475—507.
Valleau W. a. Johnson E. 1930. Phytopath., 20, № ю, 831—839.
Valleau W. 1932. Kent. Agr. Exp. Stat. Bull., № 327.
Valleau W. 1942. Jour. Bacter., 43, № 2, 272.
W e 1 i van der. 1940. Meded. Deli Praef. Medan, 3, № 6, 22.
Wenholz. 1940. Rev. Appl. Мус, 19, № 3, 169.
Wickens G. M. 1938. Rod. Agr. Jour., 35, 842—849.
Картофель
Б а л а ш e в. 1941. Докл. С.-х. акад. им. Ленина. М.
Ефейкин А. К. 1940. Сов. бот., 6, № і, 22—27; 5—6, 242—251.
Купрэвіч В. Ф. 1938. 36. Прац. інст. біял. Ак. нав. БССР, 7,
31—72.
Лысенко Т. Д. и Фаворов А. М. 1939. Летние посадки кар*
тофеля. Сельхозгиз. М.
21&
Пивоварова Р. М. и Горелик И, Г. 1936. Сб. «Вирусные
болезни растений». М.
Рожалин Л. В. 1936. Сб. «Вирусные болезни растений». М. 69—74.
Рожалин Л. В. 1940. Вестн. с.-х. наук, Овощ, и карт. № 1.
Рузинов П. Г. 1930. Болезни растений, 19, № 3—4, 148—159.
Сладкомедова А. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М., 309—316.
Терещенко А. И. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М., 321—326.
Чернышева О. П. 1936. Сб. «Вирусные болезни растений». М.,
42—51.
Ар pel О. 1906. Mitt. Kais. Biol. Anst. f. Land. u. Forstw., 2, 10.
Atanasoff D. 1926. Phytopath., 16, № 12, 929—940.
Black L. M. a. Price W. 1940. Phytopath., 30, № 5,444—447
Barrus M. a. Chupp Ch. 1922. Phytopath., 12, № 3, 123—132.
Black L. M. 1934. Amer. Potato Jour., 11, 148—152.
Black L. M. 1937. Cornell Univ. Agr. Exp. Stat. № 209, 23.
Black L. M. 1938. Phytopath., 38, № 12, 863—874.
В a 1 d J. G. a. N о г г і s D. O. 1940. Jour. Count, for Sc a. Ind. Res.
Austr., 13, 252—254.
В a w d e n F. С 1936. Ann. of Appl. Biol., 23, JY° 3, 487—497.
В a w d e n F. G. a. S с h e f f i e 1 d F. M. L., 1944. Ann. of Appl.
Biol. 31, № 1, 33—40.
Bechholdu. Erbe. 1932. Vorl. Mitt. Arb. Biol. Reichsanstalt
Land- u. Forstwirt., 20, 111—139.
Berkner F. 1936. Landw. Jahrb., 78, 791—831.
Black L. M. 1938. Phytopath., 28, № 12, 863—874.
Black L. M. 1938. Phytopath., 28, № i, 3.
Black L. M. a. P r i с e W. С 1940, Phytopath., 30, № 5, 444—447.
Blattny a. Vulkanov. 1935. Stud. Path, chmele a bramboru.
Praga, 48 p.
Boas F. 1931. Nachrichtenbl. Pflanzenschutz, 11, 66.
Brehmer W., von u. Barner J. 1930. Arb. Biol. Reichsanstalt
Land- u. Forstwirt, 18, № 1, 1—54.
Breutzal W. E. 1938. Plant Dis. Rept., 22, № 2, 44—51.
Burnett. 1934. Phytopath., 24, № 3, 215—227,
Chouard P. et Duffreaoy J. 1938. Bull. Soc. Acclim., 85,
№ 1—20, 40—42.
Clinch Ph. 1932. Sc. Proc. Roy. Dublin Soc, 20, № 15, 143—172.
Clinch Ph.,Loughnane J. B. a. Murphy P. A. 1938. Sc.
Proc. Roy, Dubl. Soc, 22, № 2, 17—31.
Clinch Ph. 1942. Sc. Proc Roy. Dubl. Soc, 23 (4), 18—34.
Clinch Ph. 1944. Sc. Proc Roy. Dubl. Soc, 23, № 28, 273—299.
Constantin J. et Marou J. 1935. Rev. Pat. Veg. Entom. Agr.,
22, № 1, 60—62.
CrepinCMBustarret J., Chevalier A. R. 1938. Ann. Epi-
phit., 4, № 3, 449—479.
Davidson W. D. 1938. Jour. Dept. Agr. Eire, 35, № 1, 201—224.
D a v i e s ' W. H. 1934. Ann. of Appl. Biol., 21, № 2, 283—299.
Dennis R. W. G. 1938. Nature, 142, № 3586, 154.
Dennis R. W. G. 1939. Phytopath., 29, № 2, 168—177.
Dykstra T. P. 1930. Phytopath., 20, № 10, 853.
DykstraT. P. a. Whitaker W. С 1938. Jour, of Agr. Res., 51,
№ 5, 319—333.
Dykstra T. P. 1939. Phytopath., 29, № 11, 917—935.
E 1 z ? D. L. 1927. Meded. Lanbbouwhoog. Wageningen, 31, № 1, 1—90.
Elze D. L. 1931. Phytopath. Ztschr., 3, № 4, 449—460.
Folsom' D. 1942. Bull. Meogrie Exp. Sta. № 410, 215—250.
Friedrich H. 1938. Phytopath. Ztschr., 11, № 2, 202—207.
Friedrich H. 1937. Phytopath. Ztschr., 10, № 6, 559—577.
314
Hausing E. D. 1942. Phytopath., 32, № 1, 7.
Hey. 1932. Vorl. Mitt. Arb. Biol. Reichsanstalt Land-und Forstw., 20r
79—96.
Huber G. A. 1940. Phytopath., 30, № 9, 787.
Hungerford Ch. W. 1920. Idaho Agr. Exp. Stat. Bui. № 122.
H u n g e r f о г d G h. W. a. D a n a B. F. 1924. Phytopath., 14, № 8,
372—383.
К a ho H. 1935. Phytopath. Ztschr., 8, № 2, 157—164.
К 1 a p p E, L. 1933. PfJanzenbau, Pflanzenschutz PflanzenzuchV
10, № 5, 161—197.
KochK.a. Johnson.J. 1935. Ann. of Appl. Biol., 22, № 1, 37—54.
KohlerE. 1933. Phytopath. Zeitschr., 6, № 4, 359—369.
Kohler E. 1935. Zuchter, 9, № i, 62—65.
К 6 h 1 e г E. 1935 a. Angew. Bot., 17, № ц, 60—74.
KohlerE. 1937. Phytopath. Zeitschr., 10, № 1....
Kohler E. 1938. Angew. Bot., 20, № 5, 373—380.
Kohler E. 1939. Phytopath. Ztschr., 12, № 5, 480—489.
Kohler E. 1940. Zuchter, H. 11, 273—275.
Kohler E. 1940. Angew. Bot., 22, H. 6, 385—399.
Kohler E. 1941. Naturwissensch., 29, № 26, 390.
Kosmat H. 1932. Fortschr. Landw., 7, № 15, 395—397.
Leach J. G. 1939. Phytopath., 29, № 1, 14.
Loughnane J. B. a. Clinch P. 1935. Nature, 135, № 3420, 8S3>
Loughnane J. B. 1939. Nature, 144, № 3653, 785—786.
Mc. Kay a. Dykstra. 1932. Oregon Agr. Exp. Stat. Bull. № 294.
McKay, Dykstra a. all. 1933. U. S. D. A. Circ. № 271.
Marx u. Merkenschlager. 1932. Landw. Jahrb., 76, 733—744.
Marx u. Merkenschlager. 1932. Arb. Biol. Reichsanst.
Merkenschlager F. 1932. Forschungen u. Fortschritte, 8, 58.
Merkenschlager F. et all. 1931. Arb. Biol. Reichsanst., 19, № 2»
199 210.
Murphy P. A. 1923. Sc. Proc. Roy. Dublin. Soc, 17, № 5, 163—184.
Murphy P. A. a. M с Kay R. 1932. Sc. Proc. Roy. Dubl. Soc,
20, № 30, 224—277.
Murphy P. A. a. Loughnane J. B. 1936. Sc. Proc. Roy. DubL
Soc, 19, № 20/28, 343—353.
Murphy P. A. a. Loughnane J. B. 1936. Sc. Proc Roy. DubL
Soc, 21, № 40, 419—430.
Murphy P. A. a. Loughnane. 1937. Sc. Proc. Roy. Dubl. Soc,
21, J\& 50, 567—579.
О r t о n W. A. 1914. Bull, of U. S. Dep. of Agr. JY° 62, 48.
Pethybridge G. H. 1939. Phytopath., 12, NO 3, 283—291.
Philipp W. 1939. Kranke Pflanze, 16, H. 4, 69—71.
Quanjer H. M. 1913. Meded. von de Rijks Hoogere, Land, Tuinen
Boschbouwsool, 6, 41.
Quanjer H. M. 1931. Phytopath., 21, № 6, 577—613.
Quanjer H. M. a. Gaumann E. 1935. Phytopath. Ztschr., 8r
№ 4, 307—321.
Rathsack u. Hurwitz. 1932. Fortschr. Landw., 7, № 22, 553—558.
Richards. 1928. Phytopath., 18, № 1, 140—141.
Salaman a. le Pelley. 1930. Proc. Roy. Soc. Bot., Ser. В., 106,
№ 13, 140—175.
galaman R. N. 1932. Proc. Roy. Soc. Bot., Ser. B, 110, 186—224.
Salaman R. N. 1938. Phil. Trans. Roy. Soc, London, 229, № 559r
137—217.
Salaman R. N. a. W or tie у W. R. S. 1939. Nature, 144t
№ 3660.
Sandford G. B. a. Clay S. B. 1941. Canad. Jour, of Res., 19;
№ 3, 68—74.
215
Schaffnit u. Johnssen. 1933, Phytopath. Ztschr., 5, № 6,
603—612.
Schr'even D. A. 1934. Tijdsch. Plantenzickt., 40, 225—266.
Schultz E. E.a.Folsom D. 1923. Jour, of Agr. Res., 25, № 2,
43—117.
Schultz E. S., Clark G. F. a. Stevenson F. G. 1940.
Phytopath. Ztschr, 12.
Schweizer G. 1930. Phytopath. Zeitschr., 2, № 6, 557—591.
Smith K. 1928. Nature, 127, № 3214, 852—853.
Smith K. 1929. Ann. Appl. Biol., 16, № 2, 209—229.
Smith K. 1931. Proc. Roy. Soc. Bot., 109, 251—267.
Smith K. a. D e n n i s R. W. G., 1940. Ann. of Appl. Biol., 21.
Soevenson 1. Т., Folsom D. a. Dykstra T. P. 1943.
Amer. Potato Jour., 20, 1—10,
*Stary 1937. Ochr. Rost., 14, 53—55.
Wartenberg H. и др. 1935. Angewand. Botan., 17, № 1, 74—93.
Wartenberg H. 1937. Phytopath. Ztschr., 9, № 1.
Wartenb-erg H. u. Klinkowski. 1937. Phytopath. Ztschr.,
10, ДЬ 1, 107—109.
Wenzel H. 1939. Phytopath. Ztschr., 12, H. 4, 351—359.
Whipple О. В. 1919. Montana Agr. Exp. Stat. Bull., ISO, 1—29.
Wh i t e h e-a d. 1931. Ann. of Appl. BioL, 18, 299—304.
Whithead T. 1937. Ann. of Appl. BioL, 24, № 2, 323—341.
Younkin S. G. 1942. AmeF. Potato Jour., 19, 1.
You n kin S. G. 1943. Phytopath., 3$, 1, 16.
Younkin S. G. 1943. Amer. Potato Jour., 20, 7, 177—183.
Тыквенные
Вовк A. M. 1942. Микробиология, 11, № 4, 169—177.
Bewley W. F. 1923. Diseases of Glasshouse Plants. London.
Ainsworth G. G. 1935. Ann. of Appl. BioL, 22, № 1, 55—67.
D о о 1 i 11 1 e S. P. 1920. U. S. Dept. Agr. Bull. № 879.
Doolittle S. P. a. W a 1 k e r M. N. 1926. U. S. Dept. Agr. Bull.
№ 1461, 1—14.
Doolittle S. P. a. Walker M. N. 1925. Jour, of Agr. Res.,
SI, № 1, 1—58.
Freitag Т. Ы. 1941. Phytopath., SO, № 1, 8.
Porter R. H. 1930. Phytopath., 22, № 1, 113.
Selby A. D. 1903. Ann. Rep. Ohio Stat. Hort. Soc. 109. 1910. Ohio
Agr. Exp. Stat. Bull. JY° 214, 394.
Walker M. N. 1933. Phytopath., 23, № 9, 741—744.
Сложноцветные
Рыжков В. Л. 1939. Сб. «Президенту АН GCCP академику В. Л.
Комарову^, 696—706.
Рыжков В. Л. 1943. ДАН, 41, № 2, 90—92.
Сухов К. С. и В о в к А. М. 1945. ДАН. 48, 387—390.
Ягодкина В. П. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М. 333—338.
Ainsworth G. С. 1936. Nature, 137, 868.
Ainsworth G. С. a. Ogilvie L. 1939. Ann. of Appl. BioL, 26,
№ 2, 279—296.
Brandenburg E. 1928. Forsch. Geb. d. Pflanzenkrankh. u. d.
Immun. im Pflanzenreich, 5, № 1, 39—72.
Brier ley P. 1933. Contrib. Boyce Thompson Inst. Plant. Res., 5,
№ 2, 235—288.
Campbell A. H. 1934. The Dahlia Yearbook. 14—23.
frukushi T. 1930. Jap. Jour. Bot., 2, 31.
216
О а г d n е г М. W., Tompkins С. М., Whipple О. С. 1935.
Phytopath, 25, № 1. 17.
Harris М. R. 1939. Bull. Dept. Agr. Calif., 28, № 3, 201—213,
J agger J. G. 1921. I. Agr. Res., 20, № 10( 737—740.
Jagger J. С a. Chandler N. 1934. Phytopath., 24, № 11, 1253—
1256.
Jagger J. C. 1940. Phytopath., 30t № 1, 53.
Kassanig A. 1944. Nature 154, №3896.
Kunkel L. O. 1924. Phytopath., 14, № 1, 524.
К u n к e 1 L. O. 1926. Amer. Jour. Bot., 13, № 10, 646—705.
Kunkel L. O. 1931. Contrib. Boyce Thompson Inst., 3, № 1, 85—123.
Kunkel L. O. 1932. Contrib. Boyce Thompson Inst., 4, № 3, 405—414.
Kunkel L. O. 1941. Phytopath., 34), № 1, 14.
Kunkel L. O. 1941a. Amer. Jour, of Bot., 28, № 9, 761—769.
Linn M. B. 1940. Bull. Cornell. Agr. Exp. Stat. № 742, 33.
Loos C. A. 1942. Trop. Agr., 97, № l, 18—21.
NewhalL A. G. 1923. Phytopath., 13? № 2, 104—106.
Pittmann H. A. 1934. Jour. Dep. Agr. West. Austr., J. 2, 11, № 1,
123—140.
Sever in H. 1934. Hilgardia 8, № 10, 305—306.
Soriano. 1932. Physis (Rev. Soc. Agr. Gen. Nat. Buenos Aires), 11,
38, 87—90.
Злаки
Донченко. 1929. Новая болезнь овса. Омск.
Зажурило В. К. и Шварц П. 1939. ДАН СССР, 24, № 2,
186—189.
Зажурило В. К. и Сйтникова Г. М. 1939. ДАН СССР,
26, № 9, 796—799.
Зажурило В. К. и Сйтникова Г. М. 1940. ДАН СССР,
29, № 5—6, 480—484.
Зажурило В. К. и Сйтникова Г. М. 1941. ДАН СССР,
30, № 7, 659—661.
Ряховскйй Н. А. и Федулаев А. Л. 1941. ДАН СССР,
30, № 7, 662—663.
Сухов К. С. и В о в к А. М. 1940. Закукливание культурных
злаков и пути его распространения в природе. М. — Л.
Сухов К. С. и Петлюк П. Т. 1940. ДАН СССР, 26, № 5, 489—
492.
Эристави Е. М. 1941. Сб. «Вирусные болезни растений и меры
борьбы с ними». М., 294—309.
Brandes Е. W. 1920. Jour. Agr. Res. 19, № 3, 134—138.
С h а г d о п С. Е. а. V е v e R. А. 1923. Phytopath., 13, № 1, 24—29.
•Cherian a. Kulasam. 1937. Proc. Ass. Econ. Biol. Commatore,
4, 57—63.
Cook M. T. 1936. Phytopath., 26, № 2, 90.
D e s a i S. V. 1935. Curr. tic, 3, № 7, 28.
Frazier N. W. 1945. PL Dis. Rep. 29, 212—233.
F u k u s h i F. 1934. Jour. Fac. Agr. Hokkaido Imp. Univ. Fukusbi. 37,
41—164.
Ingram J. W. a. Summers E. M. 1938. Jour, of Agr. Res.,
52, № 11, 879—887.
Kawai J. 1940. Ann. Phytopath. Soc. Japan, 9, № 2, 97—100.
Koehler B. 1939. Phytopath., 29, № 9.
Kunkel L. O. 1921. Bull. Exp. Stat. Hawaian Sug. Planters Assoc,
3, № 1—14.
Kunkel L. O. 1927. Phytopath., 17, № 1, 41.
Kuribayashi K. 1931. Nagano Agr. Exp. Stat. Bull., 2, 45—69.
Lyon H. L. 1921. Plant. Ass. Exp. Stat. Bull., Bot. S«r., 3, № 1, 1—43.
Mc.Kinney H. H, 1923. Jour, of Agr. Res., 23, № 10, 778—810.
217
Mc Kinney Н Н 1930 Jour of Agr Res , 40, № 6, 547—556
Mc Kinney H H 1937 U S Dept Agr Girc , №442, 1—22
MungomeryR W a Pell A F 1933 Queensland Ber Sug
Exp Stat Bull , № 4, 28 p
Ocfemia G О 1934 Amer Jour Bot , 21, 113—120
Pnode G N 1933 Phytopath , 23, JY° 8, 674—676
Rafay S A 1935 Ind Jour Agr «c , 5, 663—670
Storey H H 1925 Ann of Appl Biol , 12, № 4, 422—439
Storey H H 1929 Ann of Appl Biol , 16, № 4, 525—532
Storey H H a Mc Lean A 1930 Ann ol Appl Biol , 17f
№ 4, 691—719
Storey H H 1931 II Congr Inter de Path Gomp , 2, 470—479
Storey H H 1932 Ann Appl Biol, 14, № l, l-—15
Storey H H 1937 Ann of Appl Biol, 12, 422—439
Tate H D a Vandenberg 1939 Jour of Agr Res , 59, JY* 1,
73—79
Tims E G 1939 Proc 6 Gongr Int ^oc Sug Cane Techn , 467—471.
WadaE a FukanoH 1937 Jour Imp Agr Exp Stat Tokyo,
3, JY° 1
Wellman F. L. 1934 Phytopath, 24, № 9, 1035—1057
Wilbrink G 1923 Med ed Profestat Java buikermd, 1, 1—15
Лилейные
Андреев H И. 1937 Сб научно исслед работ Аз Черн с х
ин та, Ростов/Д , 5, 125
Вовк А М 1944 Микробиология, 13, № 4, 180—184
РыжковВ Л иВовкА М 1937 ДАН СССР, 16, № 1, 69—72
Сухов К С и Вовк А М 19*6 ДАН СССР (в печати)
AmsworthG С 1938 Ann of Appl Biol , 25, JV» 4, 867—869
Ainsworth D 1928 Pull de la Societe bot de Bulgane, 2, 51—60
BijiJ P a van blogteren 1939 Ztbl Bakt , Abt 1, 144y
№ 1—5, 109—110
BlonthonF S a HaasisF 1940 Jour Econ Ent , 33, 6, 942
Brenner, 1937 Phytopath Ztschr , 10, H 1, 79—105.
Brierley Ph. a Mc Whorter F P 1936 Jour Agr Res „
63, 621—635
Brierley Ph. Mc Kay 1938 Phytopath, 28, JY° 2
Brierley Ph 1939 Phytopath, 29, JY° 1, 3
Brierley Ph a DoolittleS P 1940 Phytopath , 30, № 2„
171—174
Brierley Ph 1940 Phytopath , 30, JY° 3, 250—257
Brierley Ph a. DoolittleS P 1940 Phytopath , 30 JV« 2,
171—174
Caldwell J a JamesA R 1938 Ann of Appl Biol , 25, № 2„
244—253
Caldwell J a Prentice J 1943 Ann of Appl Biol , 30, 1,
27—32
Gayley Dorothy M 1928 Ann of Appl Biol , 15, № 4, 529—539
Gayley Dorothy M 1932 Ann of Appl Biol , 19, № 2, 153—172
Dosdall Louise 1928 Phytopath, 18, 3Y° 2, 215—217
Drake С J, Tate H D a Harris H M 1933 Jour Econ
Ent ( 26, JY» 4, 841—846.
GaddC H a Loos С A 1940 Trop Agr , 94, N> 3, 160—167
Gigante H 1938 Boll R Staz Path Veg Roma N S , 18, JV 4,
429—454
Green D E a TimkerM 1940, Lily Jearbook, 9, 28—33
H a a s 1 s F A 1939 Mem Corn Agr Exp btat , 224, 22
Haasis F A 1939 Phytopath, 29, № 10, 890—895
Holmes F. О 1928 Bot Gaz , 86, № 1, 50—58
218
Henderson W. J. 1932. Phytopath., 22, № 1, 11.
Jones L. 1934. Science, 75, № 2052, 385.
Kawamura. 1938. Ann. Phytopath. Soc. Japan., 7, № 3—4.
Kunkel L. 1922. Science, o5, 73.
Linfo'rd M. B. 1931. Phytopath., 21, № 10, 999.
McKinney H. a. Hughes A. W. 1931. Ann. of Appl. BioL
18, № 1, 16—29.
Mc. Whorter У. P. 1932. Phytopath., 22, № 12, 998.
Mc. Whort er F. P. 1938. Ann. of Appl. Biol., 25, № 2, 254—270.
M с Wh о r t e r F. P. 1939. Phytopath., 29, № 9, 826.
Различные семейства
Алексеева Т. С. 1946. Махровость смородины. Известия АН СССР.
Сер. биолог, наук (в печати).
Иванов С. М. и Иванова В. И. 1940. Сов. субтроп., № 10„
26—32.
Крейцберг В. Э. 1940. ДАН СССР, 27, № 6, 615—618.
Купревич. 1940. Совет, бот. № 3, 110—125.
Рыжков В. Л. 1939. Сб. «Президенту АН СССР академику В. Л.
Комарову», 696—706.
Ainsworth G. С. 1935. Gard. Chron., 97, № 2507, 31.
Amos J. a. Hatton R. 1927. Jour. Pom. a. Hort. Sc, 6, № 3,
167—183, 1928, № 4, 282—295.
Atanasoi'f D. 1935. Phytopath. Ztschr., 8, № 2, 197—223.
Atanasoff D. 1935 а. Годишник на Софийский унив., Агрон. фак.,
13, 1, 14.
Batcheler et al. 1934. Citrographic, 19, № 5.
В a u r E. 1908. Ber. d. Bot. Ges., 26, № 18, 711—713; Ber. d. D. Bot.
Ges., 25, № 7, 410—413.
Beckwith C. S. 1933. New Jersey Agr. Exp. Stat. Giro., № 275,
4 p.
В e k e r R. E. D. 1939. Trop. Agr. 16, № 7, 159—163.
Bennett С W. 1944. Phytopath., 34, № 1, 77—91.
В 1 a 11 n у С. 1933. Ochr. Rostl., 13, № 3—4, 145.
Blattny С a. Vukulov V. 1932. Gartenbauwiss., 6, № 4, 425—
432.
Blattny С 1938. Ochr. Rostl., 14, № 2, 80—81.
Bitancourt A. A. 1941. Biologico, 7, №6, 149—152.
Bond Т. Е. 1944. Ann, ol Appl. Biol., 31, № 1, 40—46.
Box H. E. 1945. Nature 155, бой—608.
Burnett G. 1934. Phytopath., 24, № 5, 467—481.
G a 1 i n i s e n M. R. 1939. Philipp. Jour. Agr., 10, № 2, 121—130.
Celino M. S. 1940. Philipp. Jour. Agr., 29, 379—403.
Celino M. S. a. Ocfemia G. O. 1941. Philipp. Jour. Agr., 30,
№ 1, 70—78.
Chamberlain E. E. a. Mathewes R. E. 1941. N. Z. Jour.
Sc. Techn. A, 23, 254—256.
Cole J. R. 1937, Phytopath., 27, № 5, 607—612.
Coleman L. C. 1918. Mysore Stat. Dept. Agr. Bui. Mycol. ser. Bull
№ 3, 58.
Cook M. 1937. Phytopath., 27, № 1, 125.
Costa A. iS. 1941. Jour. Agron. (Sz. о Paulo), 3\ ... 3, 239—248,
Casta A. S. 1944. Phytopath., 34, 2, 151—162.
Creager D. B. 1941. florists Rev., 87.
Greager D. B. 1943. Pnytopath., 33, № 9, 823—27.
Dade H. A. 1931. Gold Coast Dept. Agr. Yearbook, 1930, Bull. 23r
245—247.
D о Ь г о s с k у J. D. 1931. Contrib. Boyce Thompson Inst. Plant Res.,
3, № 1, 59—83.
219
Doidge E. M. a. Turner F. A. S. 1939. Farm, of South. Afr.,
14, № 161, 363—364.
Doidge E. M. a. Turner F. A. 1939. Farm, of South Afr., 14,
№ 162, 363,
Do oli ttle S. P. a, Harter L. L. 1945. Phytopath. 35, 695— ?04.
Duffrenoy J. 1934. G. R. Soc. Biol. Paris. 1171 346—348.
Fawcett H. S. a. Atherton H. 1926. Citrus Diseases and their
Control.
Fawcett H. S. 1934. Phytopath., 24, № 6, 659—668.
Fawcett H. S. 1938. Phytopath., 28, № 9, 669.
Fawcett H. S. a. Klotz. 1938. Phytopath., 28, № 9, 670.
Fawcett H. S. a. Rhoads A. S. 1939. Phytopath., 29, № 10,
907—908.
JFawcett H. S. a. Klotz. 1939. Phytopath., 29, № 10, 911— 912.
Fawcett H. S. 1941. Phytopath., 31, 4, 356—357.
Oadd С. Н. 1926, Trop. Agr. (Ceylon), 66, № 1, 3—9.
Gadd G. H. 1939. Tea Quart., 12, № 3, 110—116.
Gassner G. 1940. Phytopath. Ztschr., 13, 2, 27—125; 1, 1—90.
•G о 1 d i n g F. D. 1935. Trop. Agr. (Trinidad), 12, № 8, 215.
Grainger J. a. Cockerham G. 1930. Proc. Leeds Phil. Soc,
2, № 120—124.
Green D. 1935. Gard. Chron., 98, № 2543, 213.
Harter L. L. a. W e i m e r. 1929. U. S. Dept. Agr. Techn. Bull.
№ 99,
Hewitt J. L. 1939. Phytopath., 29, № 9, 825.
Hildebrandt E. M. 1939. Phytopath., 29, № 4, 369—371.
Hildebrandt E. M. 1941. Phytopath., 30, JV* 1, 11.
Hildebrandt E. M. 1942. Amer. Jour. BotM 29, № 5, 362—366.
Horns T. a. Parker E. R. 1931. Phytopath., 21, 235.
Jones L. 1938. Phytopath., 28, № 1.
Jones L. 1940. Wash. Agr. Exp. Stat. Bull., 390, 1—19.
Jones L. K. 1945. Phytopath. 35, 37-46.
Jyenger A. V. 1938. Jour. Ind. Inst. So., 21, № 8, 89—161.
Jyenger A. V. a. Griffith. 1940. Ind. For. Rec. N. S. Ent.,
6, 86—196.
К e i 1 e r P. A. 1939. Rev. Appl. Mycol., 18, № 11, 713.
Klebahn H. 1926. Planta, 1, № 4, 419—440.
Klebahn H. 1928. Angew. Bot., 10, № 5, 470.
Lees A. H. 1922. Ann. of Appl. Biol., 9, № i, 49—68.
L i г о J. I. 1930. Ann. Soc. Zool. Bot. Fenn. Vanamo, 11, N0 2, 143—149.
Loos С A. 1942. Trop. Agr., 98, № 1, 8—12.
Ma gel С J. P. 1940. Jour. Austr. Inst. Agr. Sc, 6, № 1, 44—47.
Magel C. J. P. 1940a. Jour. Austr. Inst. Agr. Sc. 6, № 2, 109—110.
Mc. Whorter F. P. 1940. Phytopath., 30, № 9, 788.
JM e 1 h u s J. H., R e d d у С. G., Henderson W. J.,
Vestal E. F. 1929. Phytopath., 19, № 1, 73—77.
JMilbrath I. A. a. Mc. Whorter F. P. 1940. Phytopath., 30,
№ 9, 788.
M tiller H. R. A. 1931. Inst. Plantenziekten Bull., 24, 1—17.
Nattrass R. M. 1939. East Afr. Agr. Jour., 2, 130—133.
Nettrass R. M. 1940. East. Afr. Agr. J., 6, JY° i, 54.
Newell W., Nlowrey H., Barnett'e R. M. 1935. Agr.
Exp. Stat., Gaiesville Florida Bull. № 280.
Noble R. J. 1928. Jour. Proc. Roy. Soc. N. S. W., ?2, 78—79.
Noble R. J. a. Noble N. S. 1939. Jour. Roy. Soc. N. S. W.,
72 293 317
Oberholzer.' 1941. Farm, of South Afr., 14, 184, 235—236.
Ocfemia G. O. 1934. Philipp. Agr., 23, № 3, 174—186.
Ocfemia G. O. 1937. Philipp. Agr., 26, Л* 4, 338—440.
Ogilvie L. 1928. Ann. of Appl. Biol., 21, 113—120.
?20
Pal В. P. a. Nath P. 1935. lad. Jour. Agr. Sc, 5, № 4, 517—522.
Рар? Н. 1924. Angew. Bot., 6, 255—275.
Раре Н. 1927. Gartenwelt, 31, № 22, 320—331.
P a p e H. 1931. Gartenwelt, 35, № 45, 621.
P a p e H. 1932. Gartenwelt, 36, 707—708, 731—732.
Peltier. 1916. Proc. Amer. Garn. Soc, 25, 29-35.
P arris S. G. K. 1939. Ргоз. Amer. Soc. Hort. Sc. 36,. 263—268.
Parr is S. G K. 1940. Phytopath., 30, 4, 299—312.
PesonteA 1933. Ball. R. Staz. Path. Veg. Roma. N. S., 28, 401—428.
Petri L. 1931. Boll. R. Staz. Path. Veg., Roma. N. S.,11, №2, 105—114.
P i г о n e P. P. 1938. Plant Diss. Rep. 21, № 9, 146.
Pirone P. P. 1940. Plant. Diss. Rep., 24, № 6.
P 1 а к i d a s A. G. 1937. Louisiana Bull., JY° 282.
Poshette A. F. 1941. Trop. Agr. 18, № 5, 87—90.
Posnette A. F. Trop. Agric. Trin. 21, № 105—106.
Price W. G. 1937. Phytopath. 27, № 2.
Price W. G 941. tpath., 35, 8, 756—758.
Rangaswami P. a. Griffitz A. L. 1939. Ind. For., 65, № 6,
335—345.
Rao Venkata. 1933. Mysore Sandal Spike Invest. Comm. Bui. № 3.
Rischkow V. 1927. Biol. Zentr., 47, № 12, 752—764.
Sakimura K. 1940. Phytopath., 30, № 4, 281—299.
Schmidt A. 1932; Gartenflora, 81, № 2, 40.
Sever in H. H. P. a. Oliver S. J. 1939. Phytopath., 29, № 9,
826.
.Silberschmidt K. 1936. Biologico, 4, JY» 6, 177—181.
Silberschmidt K. 1941. Biologico, ?, 4, 93—99.
Silveira. 1941. BoL Soc. Brasil. Agr., 4, № 1, 120.
Smith F. E. 1932. Jour. Jamaic. Soc, 36.
Smith K. 1935. Ann. of Appl. Biol., 22, 231.
Smith T. S. a. Brierley Ph. 1944. Phytopath., 34, № 5.
S t a h e 1 G. 1931—1933. Phytopath. Ztschr., 4, № 1, 65; 4, № 5, 539;
6, № 4, 335.
Staner P. 1929. Int. Bull. Plant. Protect., 3, № 12, 183.
Stevins H. E. 1939. Phytopath., 29, № 6, 537—541.
Storey H. H. 1936. Ann. Rep. East Afr. Agr. Res. Stat., 8, 11.
Storey H. H. a. Nichols R. F. W. 1938. Ann. of appl. Biol.,
25, № 4, 790—806.
Storey H. H. a. Nichols R. F. W. 1938. -East Afr. Agr. Jour.,
3, № 6, 446—449.
Tate H. D. 1940. Jowa St. Coll. Jour. Sc, 14, 3, 267—294.
Thomas K. M. a. Krishnaswani L. S. 1939. Gurr. Sc, 8,
№ 7, 316.
Tompkins G. M. 1941. Jour, of Agr. Res., 6в, № 11, 671—679.
Trabut. 1913. Gompt. Rend. Acad. Sc, Pans, 156, 243—244.
Valleau W. D. 1932. Kent. Agr. Exp. Stat. Bull. Щ327, 43—80.
Verplancke G. 1930. Сотр. Rend. Soc. de Biol., 103, № 7, 524—526.
Verplancke G. 1932. Bull. Sc. Acad. Roy. Beig. ScL, 5, 18, № 3,
Wager V." A. 1941. Fr. South Afr. 178, 29—30.
Weber G. 1923. Phytopath., 13, JY° 1, 42—43.
Wilcox R. a. Beck with S. S. 1931. Jour, of Agr. Res., 47,
№ 8.
Wil 1 i am s T. L. 1940. Pop. Third. North Afr. Agr. Conf., 1, 45—60.
Winter G. 1940. Zeitr Pfianzenkrank., 60, 512—520.
О ГЛАВЛ ЕНИЕ
Стр.
Предисловие 3
Глава I. Вирусные болезни порядка крапивоцветных
Заболевания вяза 7
Мозаичные заболевания инжира 7
Мозаичная болезнь шелковицы 8
Курчавость хмеля 8
Мозаичная болезнь хмеля д
Хлороз хмеля 9
Меры борьбы с вирусными болезнями хмеля 10
Вирусные болезни конопли 10
Глава II. Вирусные болезни семейства Chenopodiaceae
Курчавость верхушки И
Курчавость листьев свеклы 12
Савой свеклы 12
Желтуха свеклы и шпината .' IS
Мозаичная болезнь свеклы 13-
Другие вирусы у шпината и свеклы 14
Глава III. Вирусные болезни крестоцветных
Черная пятнистость капусты 16
Некротический вирус капусты 16
К рестоцветных • 1 &
Мозаика репы 18
Мозаика цветной капусты 19
Мозаика Brassia ekijiensis 19
Мозаичные болезни матиолы 20
Мозаичная болезнь редиса 20
Глава IV. Вирусные болезни мальвоцветных
Курчавость листьев хлопчатника 22
Скручивание листьев хлопчатника 24
Другие аномалии роста хлхшлатшша 27
Морщинистость листьев 28
Хлорирование листьев 28'
Crazy top или акромания 28
Брахизм 28
Тонозис или leaf-cut 28
Гибозис или leaf-curl 28
Циртозис или club-leaf 28
Стенозис или smaling 29
Инфекционный хлороз мальвоцветных 29
222
Стр.
Мозаичная болезнь у Malva borealis 30
Желтая мозаика у Hibisous esculentus 31
Глава V. Вирусные болезни винограда
Короткоузлие винограда 33
Мозаичная болезнь винограда 40
Другие вирусные заболевания винограда 41
Глава VI, Вирусные болезни фруктовых деревьев из порядка Rosiflorae
Желтуха персиков « 43
Розеточная болезнь персика 44
Болезнь phony 45
Х-болезнь персика 46
Красный шов персика 46
Инфекционное опадание листьев у персика 47
Мозаичные заболевания косточковых 49
Широкая полосатость и кольцевая пятнистость .... 49
Узкая полосатость и кольцевая пятнистость 49
Мозаика (полигональная пятнистость и звездовидная) . 50
Инфекционный хлороз груш 52
Мозаичная болезнь яблони 52
Пятнистость яблок 53
Каменистая пятнистость груши 54
Диференциальная диагностика вирусных болезней фруктовых
деревьев 54
Вредоносность вирусных болезней фруктовых деревьев ... 57
Глава VII. Вирусные болезни розоцветных
Кратная или зеленая мозаика малины 59
Желтая мозаика малины 59
Курчавость малины 59
Полосатость малины 60
Карликовость малины 61
Желтая пятнистость красной малины 61
Папоротниковость листьев малины 61
Вирусные заболевания малины в Европе 61
Ксантозис клубники 65
Морщинистость клубники 65
Ведьмины метлы клубники 65
Низкорослость клубники 66
Скручивание листьев клубники 66
Сомнительная мозаика клубники 66
Мозаичная болезнь розы 66
Увядание розы 67
Полосатость розы 67
Ведьмины метлы голодискуса 67
Меры борьбы с вирусными болезнями розоцветных .... 67
Глава VIII, Вирусные болезни бобовых
Общая характеристика мозаичных заболеваний бобовых . . 69
Зеленая мозаика фасоли 70
Желтая мозаика фасоли 71
Мозаика фасоли 3 74
Мозаика фасоли 4 74
Увядание фасоли 75
Некротическая мозаика фасоли 75
Мозаичная болезнь сои ' 75
228
Стр.
Деформирующая мозаика гороха ¦ 77
Мозаика гороха 2 7&
Мозаика гороха 3 7&
Мозаика гороха 4 78
Мозаика гороха 5 79
Полосатость гороха 1 79
Полосатость гороха 2 79
Мозаика люцерны 1 78
Мозаика люцерны 2 79
Мозаичные заболевания душистого горошка 80
Мозаика Vigna sinensis 80
Мозаика белого клевера 80
Мозаика Trifolium subterranum 81
Мозаика Trifolium hybridum 1 81
Мозаика Trifolium hybridum 2 81
Мозаичные заболевания Vicia faba 8J
Мозаика Dolichos bifiorus 82
Некротическая мозаика лонинуса 82
Огуречная мозаика у бобовых 83
Lycopersicum virus 3 у бобовых 83'
Инфекционный хлороз у Laburnusn 84
Желтухи бобовых 84
Карликовость люцерны 84
Ведьмины метлы люцерны 85
Розеточная болезнь арахиса 85
Метельчатость белой акации 85
Филлодия Crotolaria juncea 86-
Ведьмины метлы фасоли 88
Beta virus 1 у бобовых 89
Глава IX. Вирусные болезни семейства зонтичных
Западная мозаика сельдерея 87
Южная мозаика сельдерея 87
Желтуха сельдерея 88
Другие заболевания сельдерея 88
Pseudo-calico 88
Морщинистость листьев сельдерея 88
Желтая пятнистость сельдерея 89
Кольцевая пятнистость сельдерея 89
Глава X. Вирусные болезни пасленовых
Обыкновенная мозаичная болезнь табака 91
Огуречная мозаика пасленовых 97
Пестрица табака 97
Кольцевые пятнистости пасленовых 98
Поражение Х-вирусом пасленовых 101
Некротические полосатости 101
Бронзовость помидора 104
Гравировка табака 106
Некроз табака 107
Курчавость листьев табака 108
Желтухи 109-
Столбур 111
Кустистая карликовость 116'
Кустистость верхушки помидора 116
Мало изученные вирусные болезни пасленовых 116-
Пятнистая мозаика ¦ 117
Мягкая мозаика табака 117
224
Cmpr
Белая мозаика 117
Средняя мозаика ЦТ
Некроз стебля томата 117
Желтая карликовость 118-
Коричневая корневая гниль табака 118
Розеточная болезнь 118-
Заболевания пасленовых, внешне похожие на вирусные 1191
Шургал табака 120
Заключение 120
Глава XI Вирусные болезни пасленовых (картофель)
Некротические мозаики картофеля 123
Х-вирус . . . . 125
У-вирус . . ... . . . 125
А-вирус ... . . 127
В-вирус ..... . . 127
С-вирус . . . .... 128
Д-вирус . 128
Е-вирус 128
Мозаики, сопровождающиеся некрозами клубней 129
Неинфекционные некрозы клубней 129
Calico 131
Веретеновидность клубней картофеля 132
Закручивание листьев 132
Желтая карликовость картофеля 133
Непятнистая курчавая карликоЕОСть 135
Ведьмины метлы картофеля 135
Верхуіьечное скручивание листьев картофеля 135
Скручивание листьев картофеля 138
Другие вирусные и подобные им заболевания картофеля 138
Псиллидная желтуха 139
Круглолистость картофеля 139
Мелколистосіь карюфеля .... . 139
Гигандиум . . . .139
Knotchenkrankheit . . 139
Блестящая болезнь . . 139
Purple dworf . . . 139
Эпифитология вирусных болезней картофеля 140
Меры борьбы с вирусными болезнями картофеля 142
Глава XII. Вирусные болезни тыквенных
Обыкновенная огуречная мозаика 147
Беттендорфская мозаика огурца 148
Мозаичная болезнь арбуза . 148
Английская огуречная мозаика - 149
Глава XIII Вирусные болезни семейства сложноцветных
Желтуха астры 151
Мозаичная болезнь далии 152
Кольцевая пятнистость далии 153
Oakaleaf далии 153
Мозаичная болезнь салата . 153
Коричневая болезнь салата . 154
Другие вирусные болезни сложноцветных 154
Глава XIV. Вирусные болезни злаков
Закукливание . . . 156
Мозаичная болезнь озимой и яровой пшеницы . , 160
Мозаичные заболевания пшеницы, описанные в США 162
225
Стр.
-Карликовость риса 162
Вирусные болезни кукурузы 166
Трансваальская мозаика трав 166
Мозаичная болезнь кукурузы 166
Полосатость макса 167
Крапчатость маиса 167
Мозаичная болезнь сахарного тростника 167
Полозатость сахарного тростника 169
Болезнь Sereh 170
Болезнь Фиджи 170
Карликовость сахарного тростника 170
Огуречный вирус у злаков 171
Вирусная болезнь сорго 171
Глаз а XV. Вирусные болезни отряда лилейных
Мозаичная болезнь лука 172
Желтая карликовость лука 173
Полосатость тюльпана J 73
Огуречная мозаика лилейных 176
Вирусные заболевания лилии 176
Розеточная болезнь 177
Сгозк neck (свернутая шея) 177
Pimple leaf (прыщик ) 177
Вирусные заболевания иарциса 177
Вирусные болезни других лилейных 177
Глава XVI. Вирусные болезни представителей различных семейств
Порядок Quercijlorae 179
Семейство Juglandaceae 179
Семейство Salicaceae 179
Семейство Polygonaceae, . t 179
Семейство Piperaceae. . 179
Семейство Caclaceae 180
Порядок Polycarpicae 180
Семейство Lauraceae . . 180
Семейство Vwlaceae # 181
Семейство Temstroemiaceae 181
Семейство Sterculiaccae . 181
Семейство Тіііасеае . . 191
Семейство Euphorbiaceae . . - 182
Семейство Geraniaceae. . . 183
Семейство Rutaceae я 183
Семейство Anacardiaceae t 185
Семейство.4сетсеае . . 186
Семейство Sapindaceae 186
Порнд ж Rhamnales , . 187
Порядок Thymelaeinae 187
Порядок Passifiorales 187
Семейство Сагісасеае 187
Семейство Saatalaceae 188
Семейство Ribesiaceae 188
Семейство Pittosporaceae 189
Семейство Согпасеае .,..-. 189
Семейство Rhodoraceae 189
Семейство Vacciniaceae 189
Семейство Primulaceae ' 190
Семейство Convolvulaeeae 190
226
Стр.
Порядок Personate 191
Семейство Rubiaceae 191
Семейство Oleaceae 1Ь.2
Семейство Gaprifoliaceae 1S2
Семейство Valerianaceae 192
Однодольные 193
Семейство Соттеііпасеае 195
Приложения:
Таблицы I—XIV ; 195
Литература 204
Печатается по постановлению
Редапционно-издателъского совета
Академии Наук СССР
*
РИСО АН СС^Р .№ 2291. А- 05858 Изд. № 651
Тип. заказ №358. Лодп. к печ. 2/1X1918 г. Формат
бум. 60Х92/ів. Печ. л. 14,25. Уч.-пзд. л. 15,75
Тираж 2500
2-я типография Издательства Академии Наук СССР-
Москва, Шубинский пер., 10.