/
















































































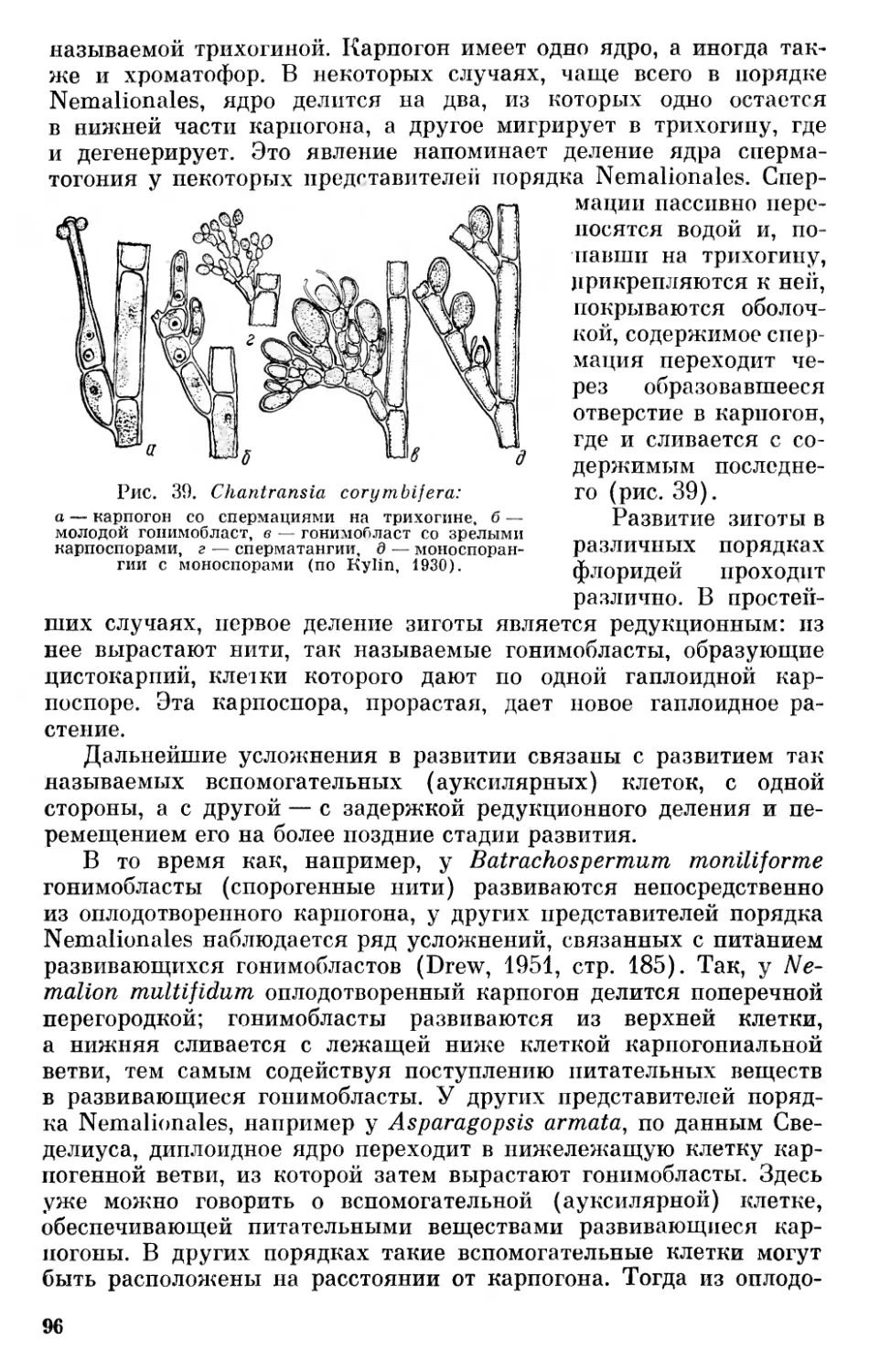








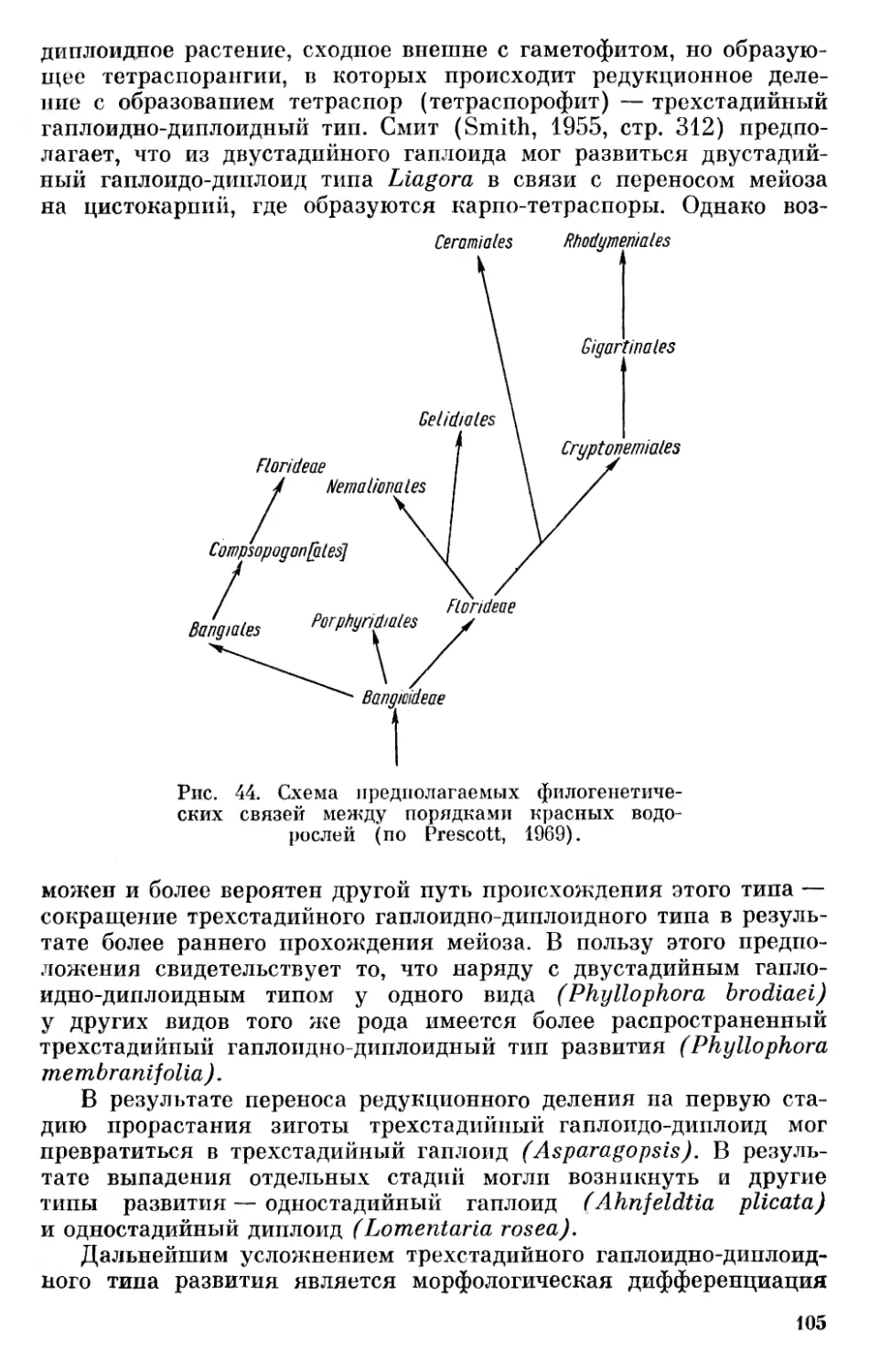







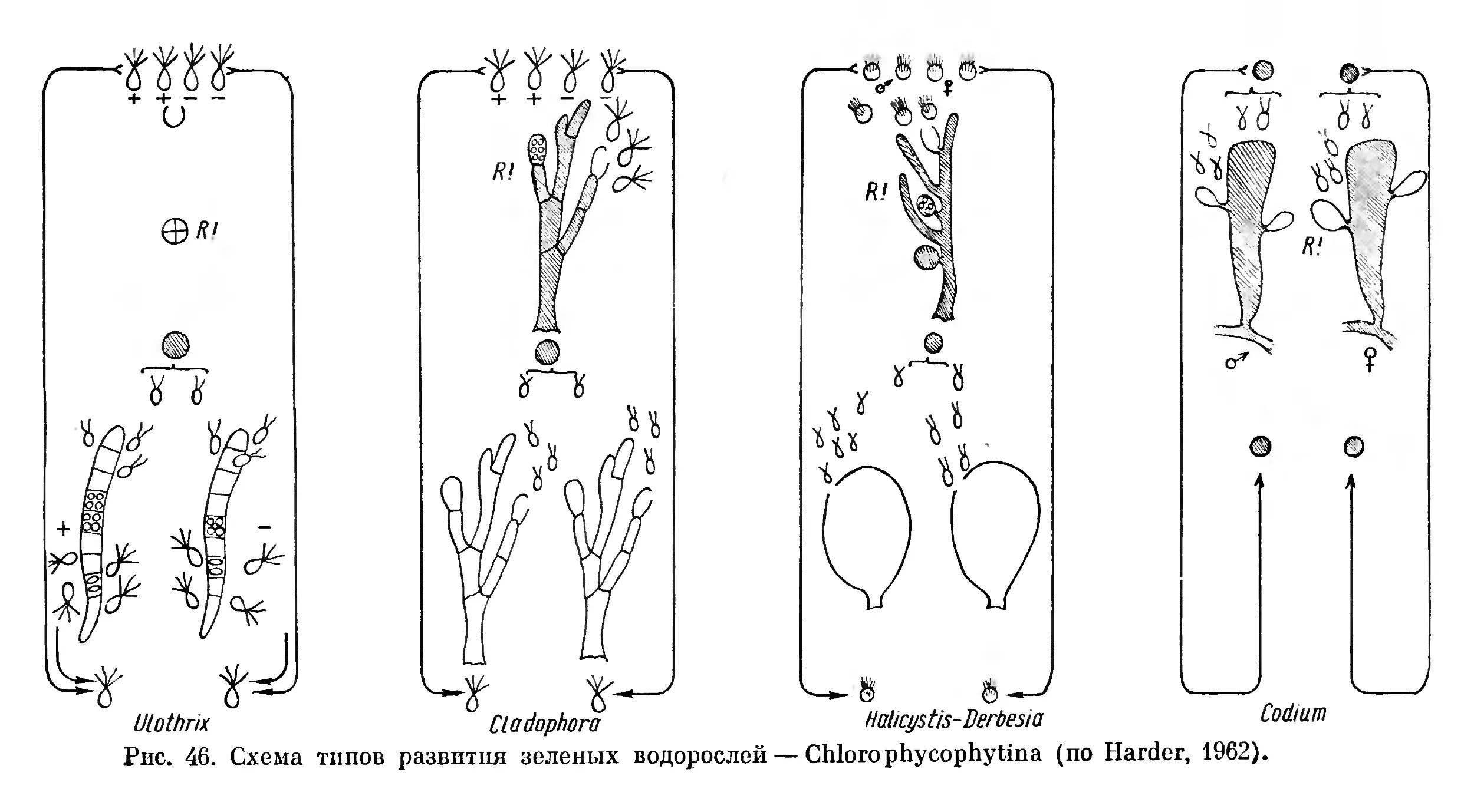











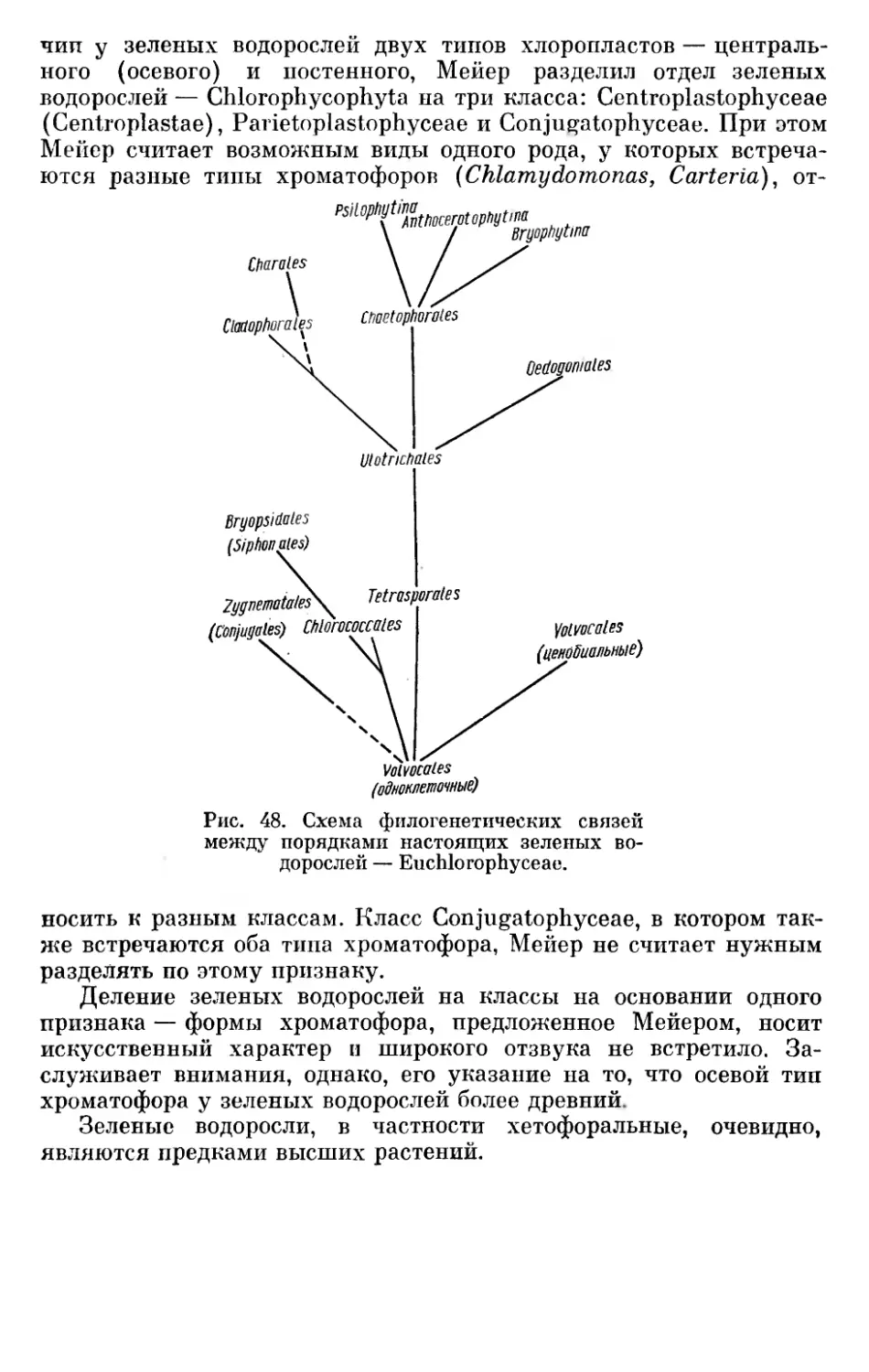




















































































































































































Текст
В монографии освещено современное состояние проблемы происхождения и развития водорослей, грибов и мохообразных. Рассмотрены основные направления развития органического мира на Земле, эволюция и основы классификации низших автотрофных организмов — водорослей, происхождение и эволюция низших бесхлорофильных растительных организмов — грибов, а также проблема происхождения и пути эволюции мохообразных. Работа снабжена резюме на английском языке. Иллюстрирована.
Рассчитана на научных работников — биологов, преподавателей, аспирантов, студентов биологических факультетов университетов и педагогических институтов.
The monograph deals with the modern state of the problem on origin and development of algae, fungi and bryophyta. Main trends in development of living world on the Earth, evolution and classification principles of lower autotrophic organisms — algae, origin and evolution of lower, chlorophyll-free plant organisms — fungi, as well as origin and evolutionary trends of bryophyta are considered. The paper is supplied with an English summary and illustrations.
It is meant for research workers engaged in biology, teachers, postgraduates, students of biological departments of universities and pedagogical institutes.
АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР ИНСТИТУТ БОТАНИКИ ИМ. н. Г. ХОЛОДНОГО
ACADEMY OF SCIENCES OF THE UKRAINIAN SSR THE N. G. KHOLODNY INSTITUTE OF BOTANY
D. K. ZEROVj
OUTLINES OF NON VASCULAR PLANT PHYLOGENY
“NAUKOVA DUMKA” KIEV — 1972
1д. К. ЗЕРОВ
ОЧЕРК ФИЛОГЕНИИ БЕССОСУ ДИСТЫХ РАСТЕНИЙ
«НАУКОВА ДУМКА» КИЕВ — 1972
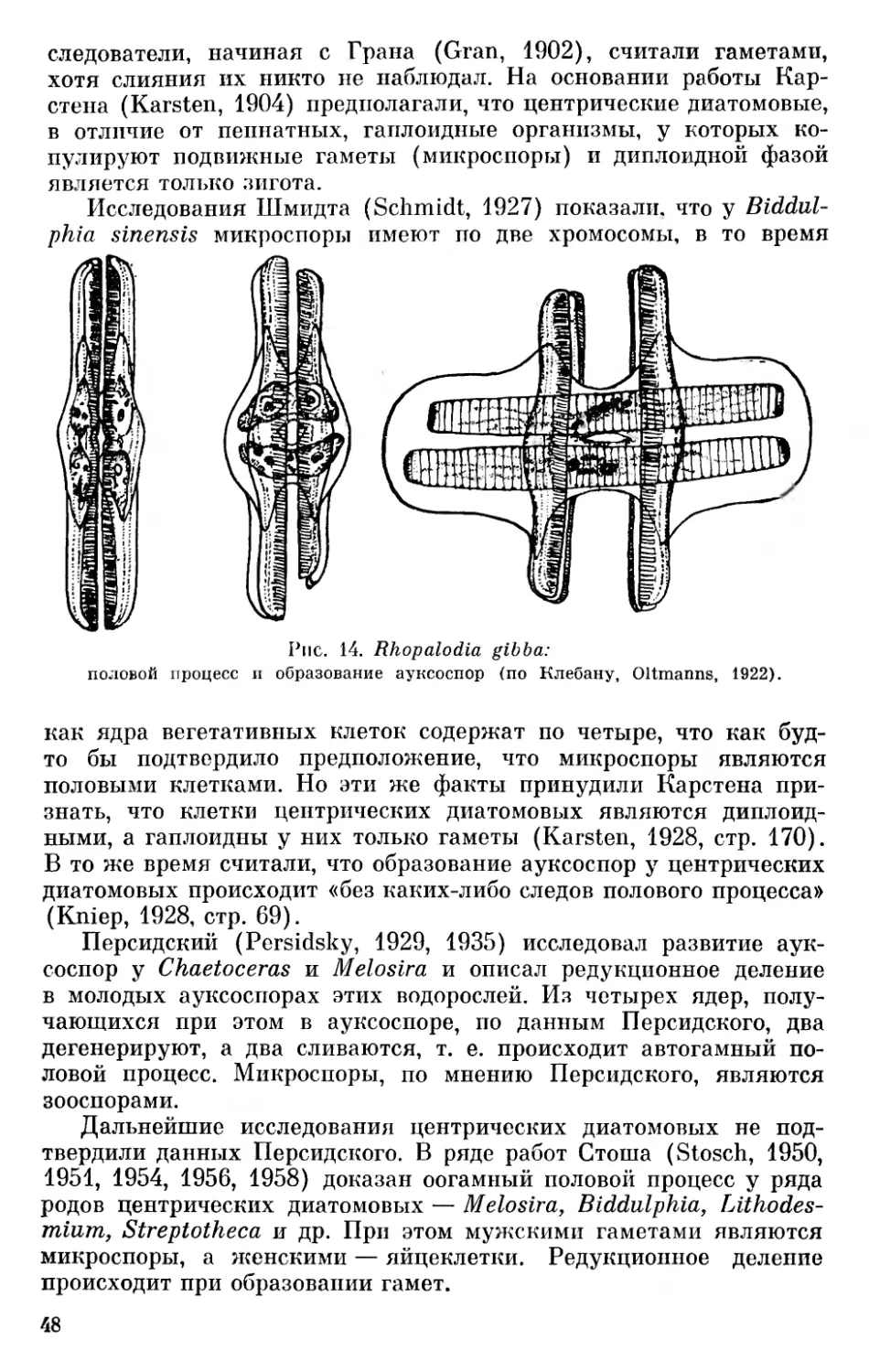
582
3-58
УДК 581: 576.12
Рецензенты:
д-р биол. наук М.Ф. Макареви ч, чл.-кор. Е. Н. Кондратюк
Редакция биологической литературы Зав. редакцией Т. Г. Кондрацкая
2—10-5 30G—72М
Киевский полиграфкомбинат
Всю область познания мы можем, согласно издавна известному способу, разделить на три больших отдела...
Ко второму классу наук принадлежат науки, msï/- чающие живые организмы. В этой области царит такое многообразие взаимоотношений и причинных связей, что не только каждый решенный вопрос поднимает огромное множество новых вопросов, но и каждый отдельный вопрос может решаться в большинстве случаев только по частям, путем ряда исследований, которые часто требуют целых столетий; при этом потребность в систематизации изучаемых связей постоянно вынуждает нас к тому, чтобы окружать окончательные истины в последней инстанции густым лесом гипотез.
Ф. Энгельс. Анти-Дюринг.
Политиздат, М., 1969, стр. 84—85.
Современный процесс эволюции живых существ в принципе представляет собою не что иное, как ряд дальнейших звеньев той непрерывной цепи превращений материи, начало которой уходит к наиболее ранним стадиям существования Земли.
А. И. О п а р и н. Возникновение жизни на Земле, 3-е изд. М., 1957, стр. 431.
ПРЕДИСЛОВИЕ
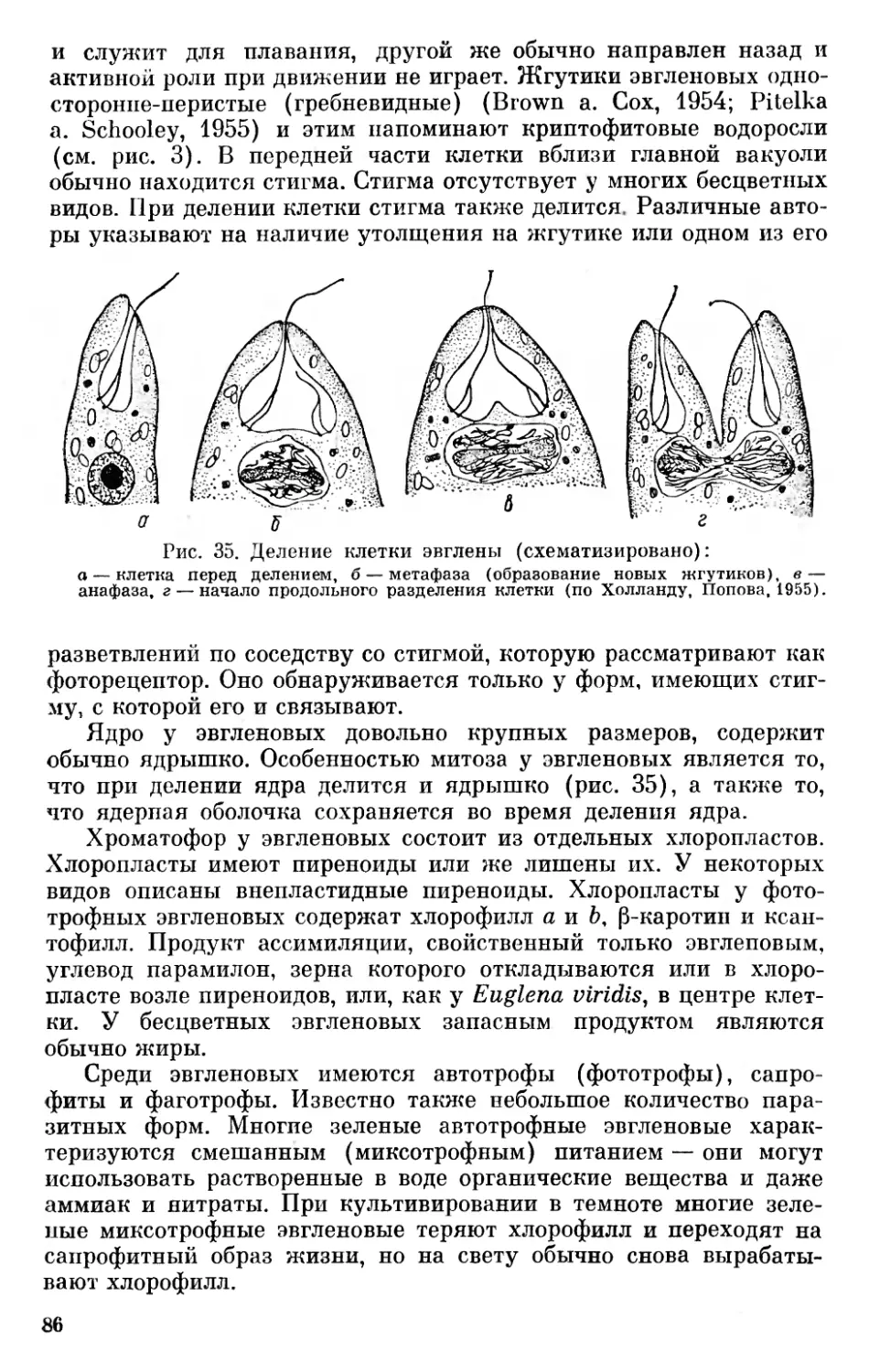
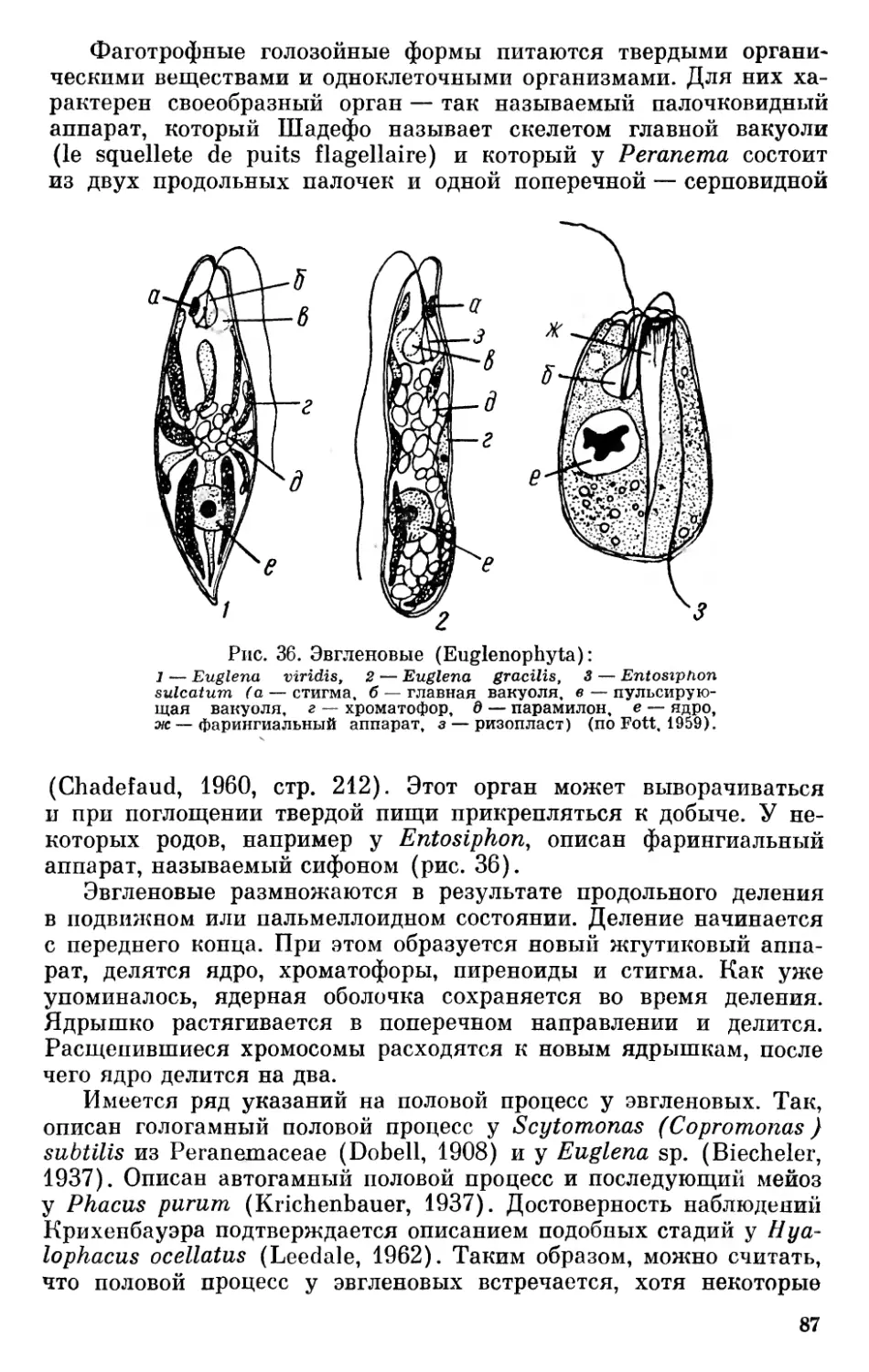
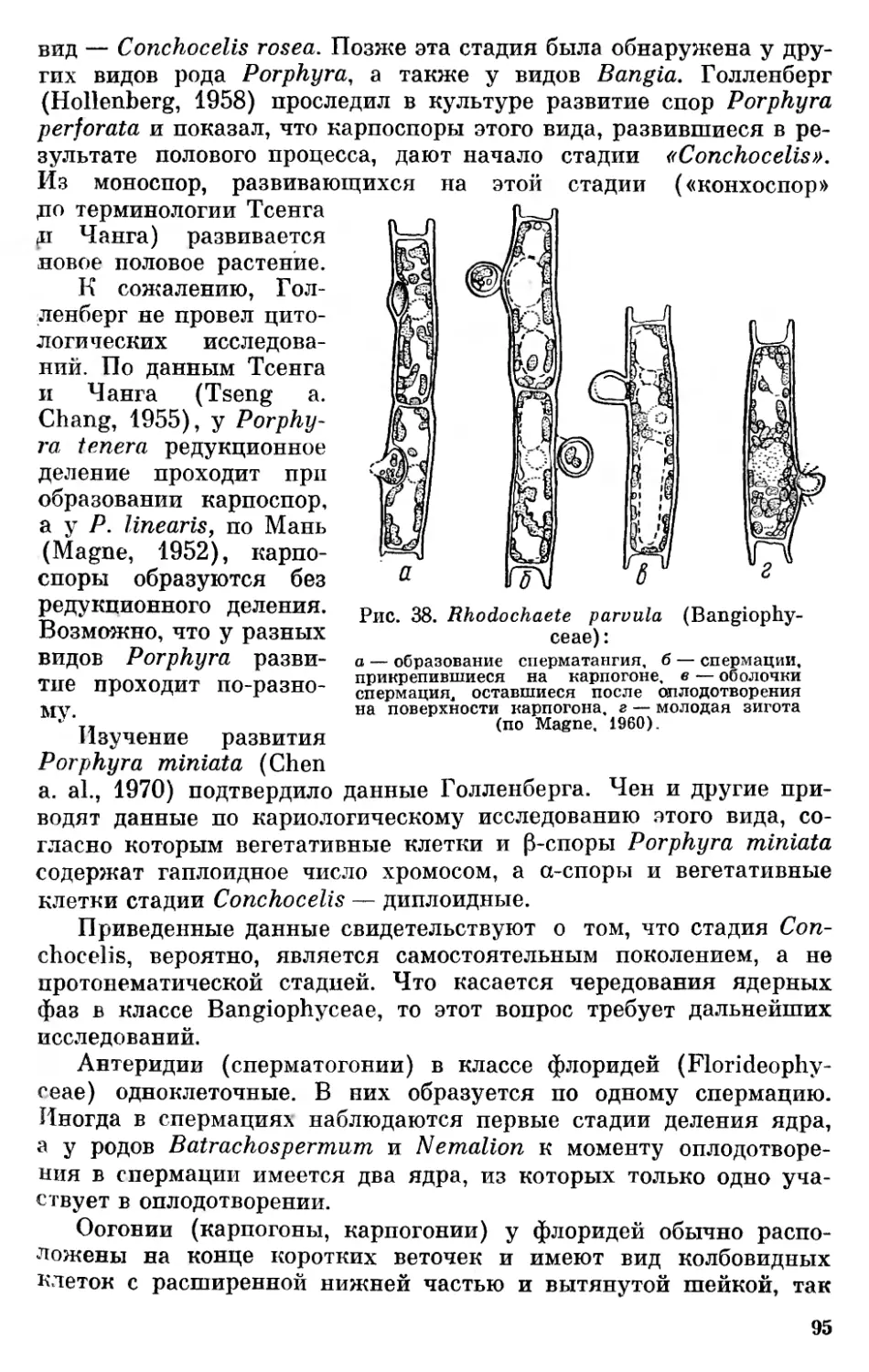
За последние годы накопилось много данных по вопросам морфологии, цитологии и онтогенетического развития низших организмов. Много нового внесло использование при их изучении электронного микроскопа и применение новых методов биохимических исследований. Эти данные дают богатый материал для освещения вопросов происхождения, эволюции и систематики низших растительных организмов.
Отсутствие в отечественной литературе последних лет обобщающих работ по вопросам филогении бессосудистых растений побудило автора сделать обобщение, которое дало бы возможность ботаникам, работающим над специальными вопросами изучения растительного мира, ознакомиться с современным состоянием общих вопросов происхождения и эволюции низших растений и мохообразных.
При освещении вопросов происхождения и эволюции отдельных фил растительного мира автор старался избегать догматизма и показать, что излагаемые взгляды являются только очередным этапом в познании путей развития органического мира.
В. И. Ленин писал: «Человеческое мышление по природе своей способно давать и дает нам абсолютную истину, которая складывается из суммы относительных истин. Каждая ступень в развитии науки прибавляет новые зерна в эту сумму абсолютной истины, но пределы истины каждого научного положения относительны, будучи то раздвигаемы, то суживаемы дальнейшим ростом знания» *. Дальнейшие исследования несомненно дадут новые данные по излагаемым вопросам, покажут в ряде случаев ошибочность современных представлений, наметят иные пути в их разрешении. Автор надеется, что настоящая работа может помочь отбросить ряд устарелых
1 В. И. Ленин. Материализм и эмпириокритицизм. Госполитиздат, М., 1937, стр. 161.
7
представлений и тем самым подготовит почву к восприятию новых данных и новых трактовок, которые дает настойчивое стремление человеческой пытливости в деле познания орга- нического мира Земли.
При написании книги была использована вся доступная автору литература, вышедшая в свет до 1971 г. Однако в связи с быстрыми темпами развития исследований и обилием новых литературных данных, возможно, что некоторые существенные работы могли ускользнуть из сферы внимания автора и не были использованы. Поэтому автор будет очень благодарен за все указания на возможные существенные пропуски в работе.
В заключение выражаю глубокую благодарность А. Н. Олешко, выполнившей кропотливую работу по подготовке всего иллюстративного материала.
Глава I
ОСНОВНЫЕ НАПРАВЛЕНИЯ РАЗВИТИЯ ОРГАНИЧЕСКОГО МИРА
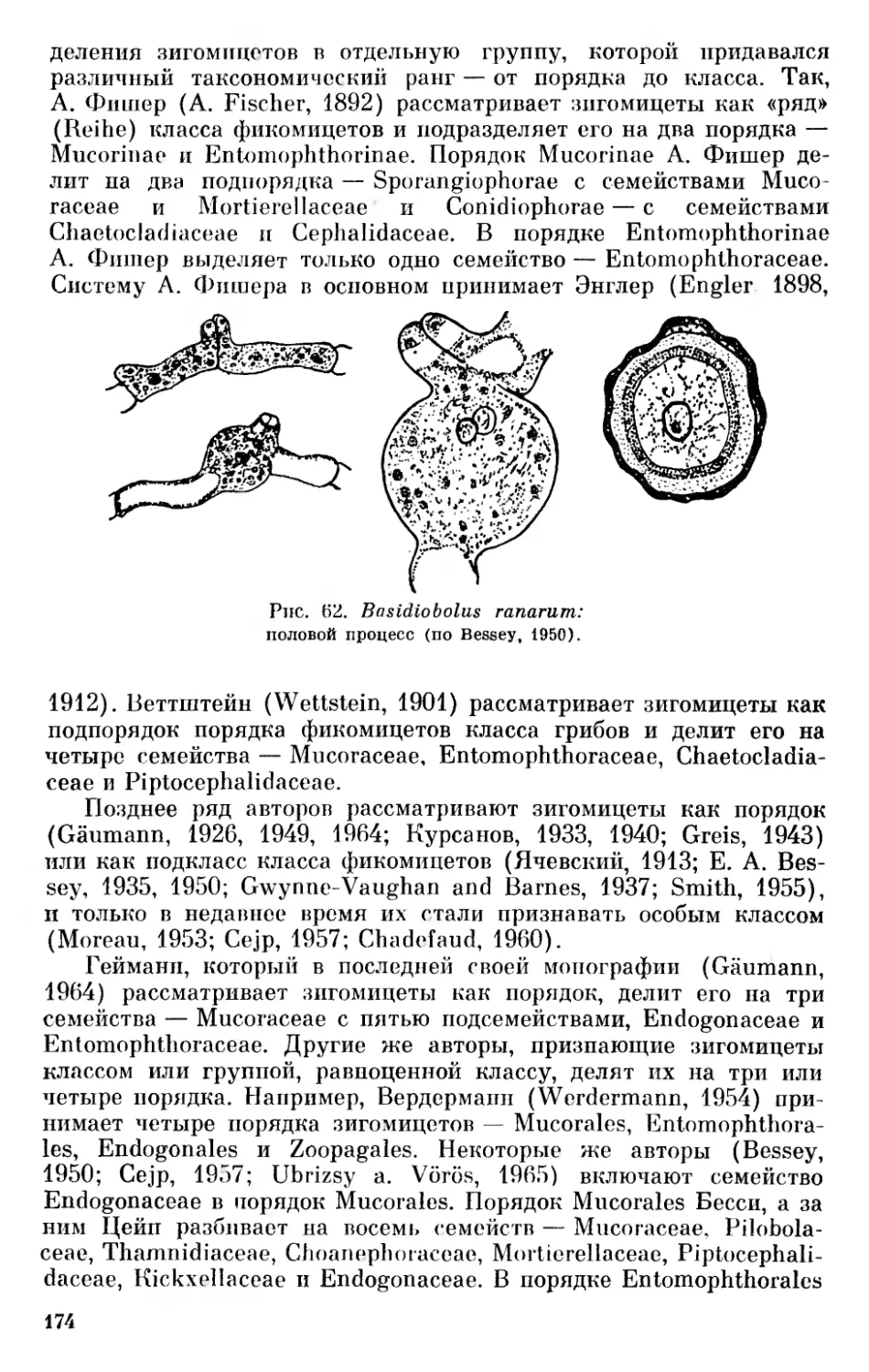
Среди широких масс населения обычно не вызывает сомнений разделение органического мира Земли на два больших отдела — «царства» — на животные и растения. Это подразделение было принято Линнеем, который различал три царства природы — неорганическое царство «камней» и два органических — растений и животных \ Однако еще Фриз в 1821 г. предложил выделять царство грибов — Regnum Mycetoideum (Fries, 1821). Это предложение, насколько нам известно, не вызвало в свое время откликов.
В прошлом столетии некоторые авторы обращали внимание на наличие группы организмов, характеризующихся одновременно рядом черт, которые, как тогда считали, свойственны или растительным, или животным организмам. Это в первую очередь различные жгутиковые, которых относили обычно к животным, несмотря на способность многих из них к фотосинтезу, низшие грибы, губки и др. В связи с этим было высказано предложение выделить в особое царство — Regne Psychodiaire — кишечнополостные и губки (Bory de Saint-Vincent, 1825, 1828).
Хогг (Hogg, 1860) предложил низшие организмы со смешанными чертами растений и животных рассматривать как особое царство примитивных организмов — Regnum Primigenum, или Protoctista. Вильсон и Кессин (Wilson a. Cassin, 1864) выделили низшие организмы в третье царство — Primalia.
Наибольшую известность имело предложение Геккеля (Haeckel, 1866, 1878, 1894) о выделении царства протистов—Protista. В это царство Геккель вначале включал и губки (1866 г.), и грибы (1878 г.), и лишь в 1894 г. он отнес к нему только одноклеточные
1 «Природными являются все тела, созданные рукою творца, образующие землю, разделенные на три царства природы, границы которых сходятся в зоофитах. Камни (lapides)—тела скученные (congesta), не живые и не чувствующие. Растения (vegetabilia) — тела организованные и живые, но не чувствующие. Животные (animalia)—тела организованные, живые, чувствующие и произвольно движущиеся» (Linné, 1767).
9
и производные от них колониальные организмы. Позже Геккель предложил органический мир делить на два царства — Protista и Histones с подразделением каждого из них на два подцарства — животных и растительных организмов (Haeckel, 1904).
Термин Геккеля «протисты» нашел широкое применение в биологии, однако различные авторы вкладывали в него различное содержание,— одни, как и Геккель в 1894 г., относили к проти- стам только одноклеточные и производные от них колониальные организмы, другие же включали в эту группу, кроме простейших и растительных флагеллат, диатомовые, красные и бурые водоросли, а также грибы (Copeland, 1938, 1947, 1956) и даже зеленые водоросли (Barkley, 1939; Rothmaler, 1948, и др.). Из этого видно, что выделение царства протистов не устраняло, а еще усугубляло трудности в разграничении основных подразделений органического мира. Если границу между протистами и многоклеточными животными — Metazoa провести легко, то автотрофные организмы совершенно невозможно разделить на «настоящие» растения и организмы, которые должны быть отнесены к протистам, особенно после того, как были установлены эволюционные филы автотрофных организмов, начинающиеся одноклеточными подвижными формами, относимыми ранее к группе флагеллат, и ведущие к «типичным» растениям (Мережковский, 1909; Pascher, 1931).
Некоторые авторы в XX ст. возвращаются к давно высказанному Фризом предложению о выделении грибов в отдельное царство. Так, Мережковский (1909) предлагал, кроме животных и растений, выделять царство микоидов^ куда он относил бактерии, синезеленые водоросли и грибы. Конард (Conard, 1939) предложил разделять органический мир на три царства — Phytalia, Animalia и Mycetalia, т. е. растения, животные и грибы. Роджерс (Rogers, 1948), сравнивая эволюционные направления этих трех групп, приходит к выводу о правильности их выделения.
Аналогичную мысль развивает и Вага (1952). Подразделение органического мира на три царства — растения, животные и грибы он основывает на особенностях питания в этих группах организмов. Вага считал, что у первичных организмов обмен веществ осуществлялся путем химических процессов между ними и соединениями, образовывавшимися в окружавшей их водной среде. Продукты распада, получавшиеся в результате отмирания первичных организмов, которые, вероятно, мало отличались от соединений, возникавших путем синтеза в окружавшей среде, могли быть использованы другими организмами. Часть организмов могла полностью перейти на сапрофитный способ питания. Такой группой, отколовшейся от общей массы первичных организмов, были первичные сапрофиты, от которых, по мнению Вага, произошли бактерии, миксомицеты и настоящие грибы.
Использование энергии солнечного света при синтезе органического вещества привело к развитию среди первичных организ¬
10
мов автотрофной ветви фотосинтезирующих организмов — первичных растений.
Вага считал, что животный способ питания, состоящий в поглощении органических тел — как живых, так и неживых — ив их переваривании внутри организма при помощи ферментов, развившийся на базе первичных хемотрофных организмов, мог привести к возникновению животных.
На базе этих гипотетических трех групп — первичных сапрофитов, первичных растений (автотрофов) и первичных животных, по мнению Вага, сформировались три царства организмов — растений, животных и грибов.
К царству растений он отнес организмы, снабженные типичным хлорофиллом, а также организмы, ныне не имеющие хлорофилла, но происшедшие от хлорофиллоносных предков, у которых вторично развился сапрофитизм, паразитизм и даже хищнический способ питания животными организмами. К царству грибов, или мицетов, Вага отнес настоящие грибы, слизевики и бактерии, относительно которых предполагается, что они произошли от первичных сапрофитов и никогда не имели предков, снабженных «типичным» хлорофиллом. Позже у представителей царства грибов могли развиться и иные способы питания — паразитизм и вторичный хемотрофизм (у бактерий). В царстве животных, характерной чертой которых, по мнению Вага, является заглатывание и переваривание органических тел, позже появляется способность к питанию живыми животными и растениями, а также паразитизм.
Морфологическим признакам, даже цитологическим особенностям различных групп организмов Вага не придавал решающего значения, в связи с чем у него бактерии и синезеленые водоросли попадают в различные царства.
В XX ст. намечается тенденция к выделению в особую группу растительных организмов дробянок — бактерий и синезеленых водорослей. Еще Кон (Cohn, 1853) считал синезеленые водоросли и бактерии родственными группами и объединял их в одном порядке (ordo) Schizosporeae в пределах класса Thallophyta (Cohn, 1872), а несколько позже назвал их Schizophyta (Cohn, 1875). Позднейшие авторы Schizophyta рассматривали обычно как подотдел (Engler, 1898) или отдел растительного мира (Wettstein, 1901; Engler, 1912, и др.).
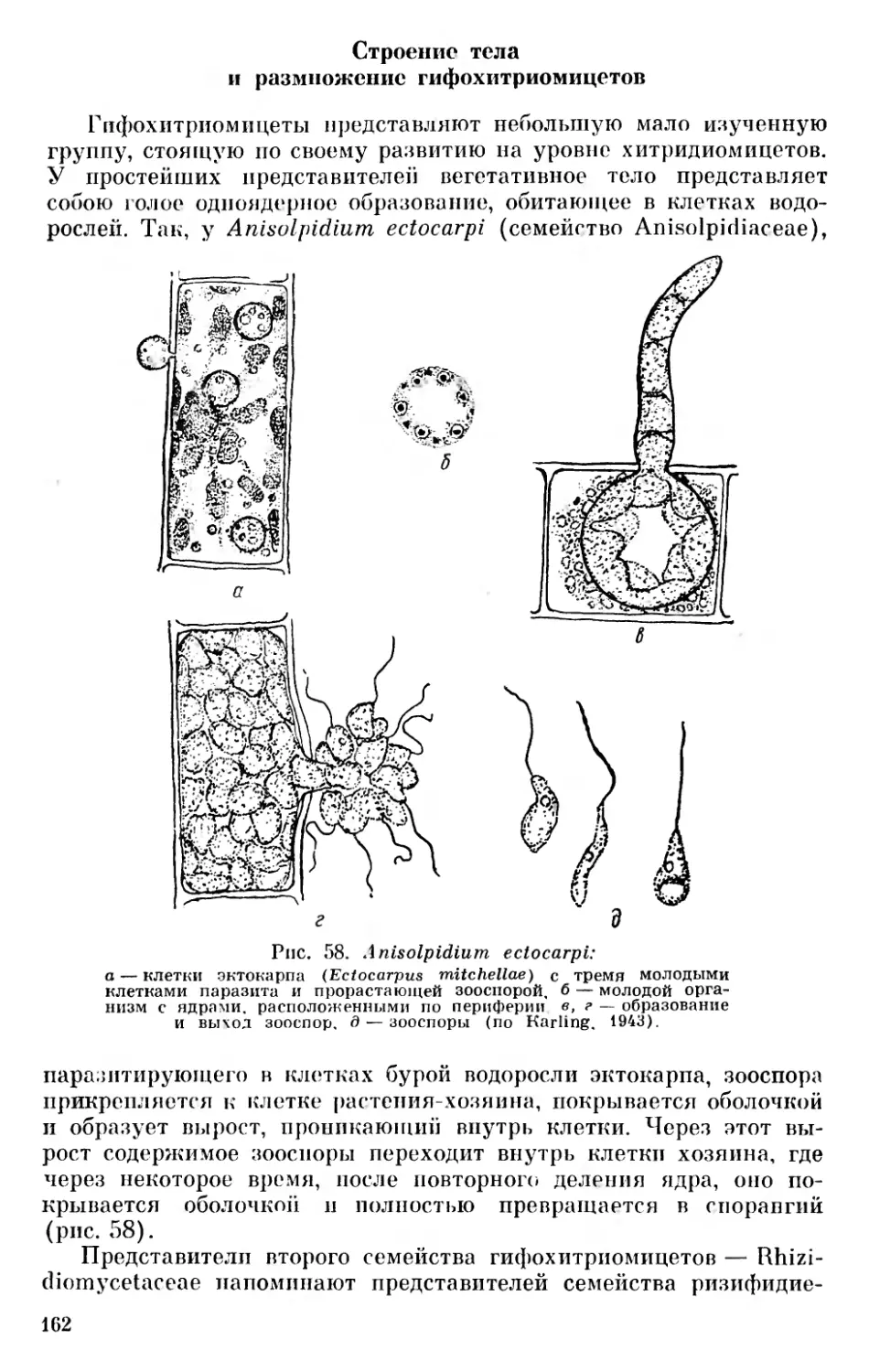
Немец (Nemee, 1929), исходя из характера строения клетки дробянок, противопоставил их как безъядерные или первичноядерные организмы (Akaryonta, Prokaryonta) всем прочим ядерным организмам (Karyonta) — животным и растениям.
Циммерманн (Zimmermann, 1930) предложил выделять среди растительных организмов три ствола (Stämme) — 1) безъядерные талломные растения (kernlose Thallophyten) — синезеленые и бактерии, 2) ядерные талломные растения (kernhaltige Thallophyten, Euthallophyten) — сборные группы — водоросли и грибы.
11
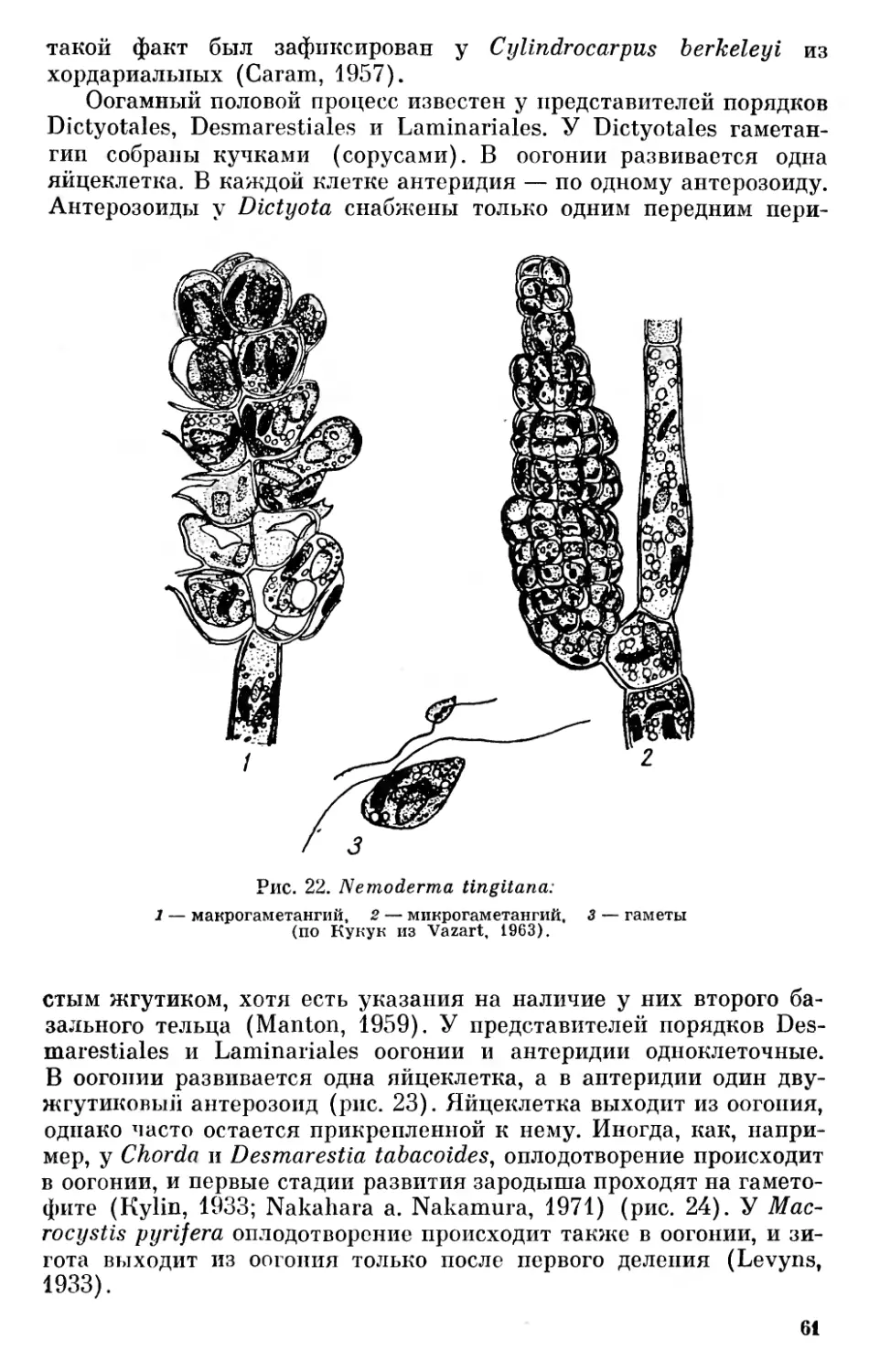
3) листостебельные растения (Kormophyten) — мхи, псилофиты, папоротникообразные и явнобрачные.
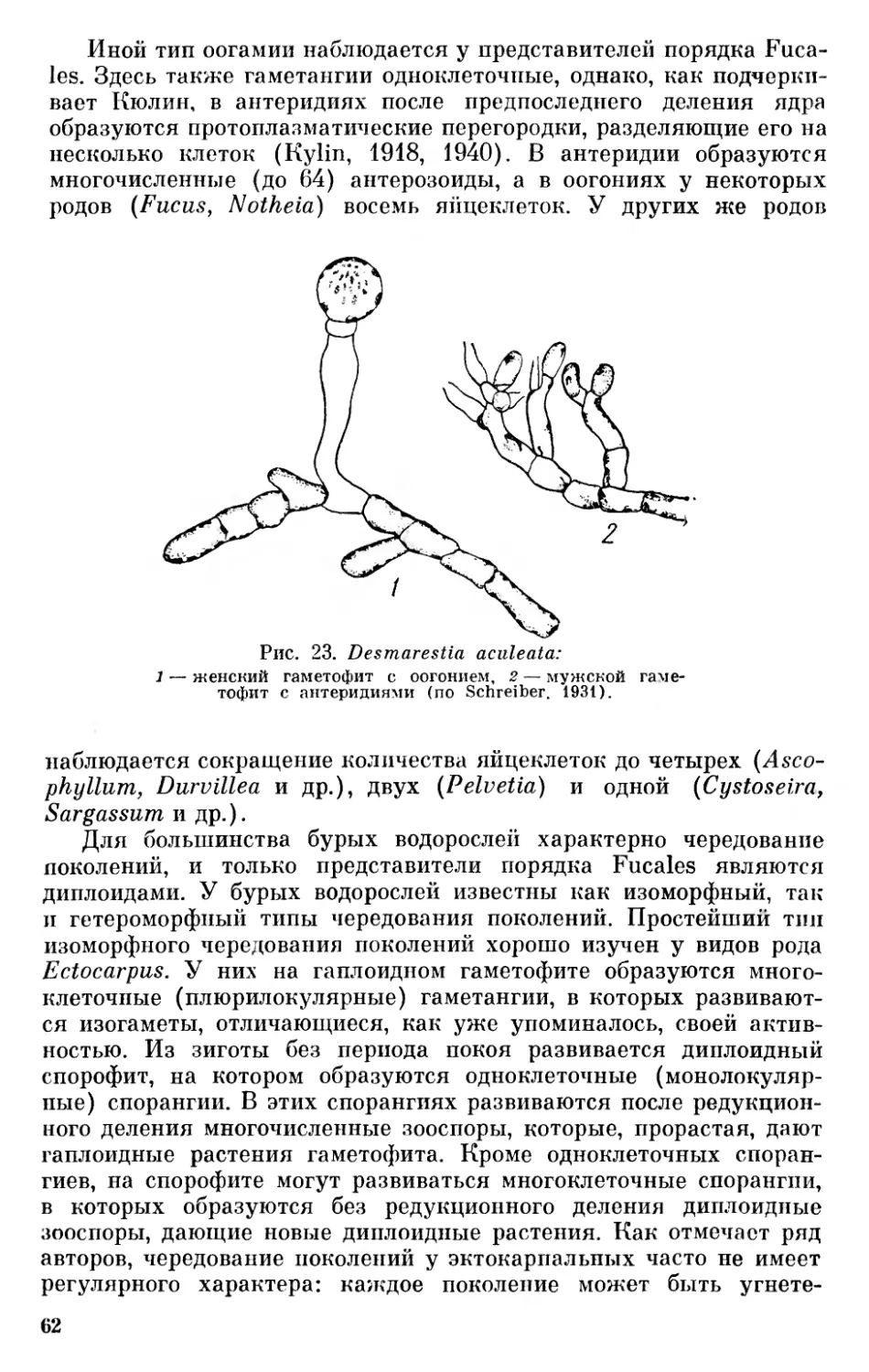
Несколько позже Пантер (Pascher, 1931) предложил царство растений разделить на два подцарства — Plantae holoplastideae, куда он включил синезеленые водоросли и бактерии, и Plantae euplastideae, куда он отнес все прочие отделы растительного мира.
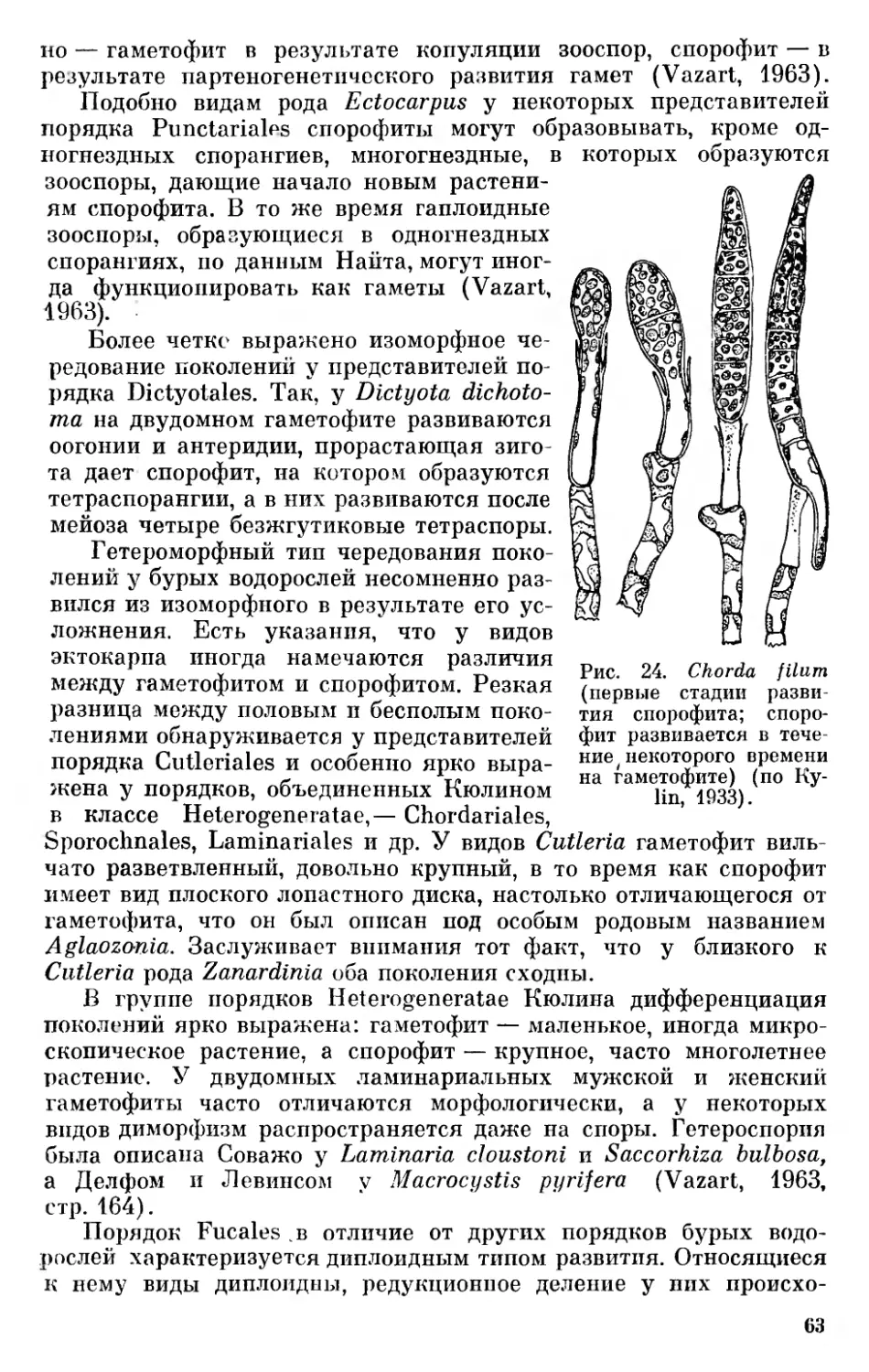
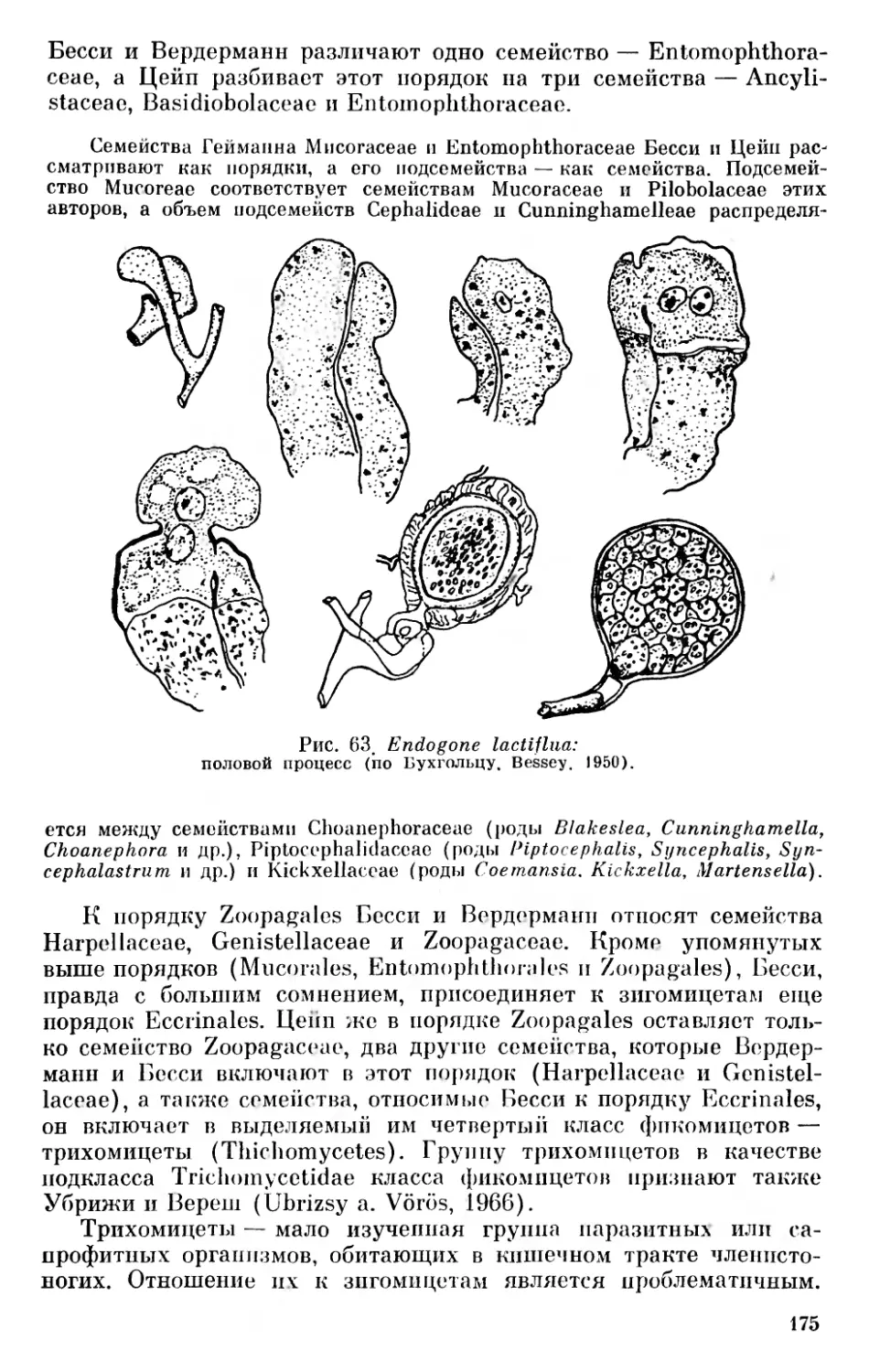
Копленд (Copeland, 1938), развивая мысль об обособленности бактерий и синезеленых водорослей, предложил делить органический мир на четыре царства — монеры, протисты, растения и животные. К царству монер (Мопега) он отнес, следуя Геккелю, безъядерные организмы, к которым причислял бактерии и сине- зеленые водоросли. Эти группы организмов Копленд считал древними, исходными формами жизни (ancestral form, the original form of life) h
К царству протистов (Protista) Копленд отнес одноклеточные и многоклеточные ядерные организмы — Flagellata (без Volvoca- les), Rhizopoda, Sporozoa, Infusoria, диатомовые, красные и бурые водоросли, а также грибы. К царству растений (Plantae) — организмы, имеющие пластиды, несущие четыре пигмента — хлорофилл а, хлорофилл Ъ, каротин и ксантофилл и образующие настоящий крахмал и целлюлозу, т. е. зеленые растения от Volvoca- les до Embryophyta. К царству животных (Animalia) Копленд отнес многоклеточные животные организмы — Metazoa «за исключением, возможно, Porifera, которые могут быть отнесены к Protista» (Copeland, 1938). Несколько позже он (Copeland, 1947) переименовал, следуя Эндерлайну (Enderlein, 1925), царство Мопега в Mychota1 2, а царство Protista — в Protoctista, приняв термин, предложенный впервые Хоггом (Hogg, 1860).
Подразделение органического мира, предложенное Коплендом, принимает Беркли (Barkley, 1939); только в царстве монер, кроме отдела и филы дробянок— Schizophyta (Photophyta), он выделяет отдел и филу Archaeophyta, к которым относит начальные, первичные организмы, вирусы и бактериофаги, а к царству протистов относит также и зеленые водоросли.
Козо-Полянский (1947) предложил царство растений разделять на три подцарства (subregna) —Schizophyta (дробянки), Мусо- phyta (грибы, включая слизевики) и Nomophyta («настоящие растения», куда он отнес все отделы автотрофных растений, кроме синезеленых водорослей). Несколько позже он же (Козо-Полянский, 1949), следуя номенклатуре Копленда (Copeland, 1947),
1 Геккель (Haeckel, 1866) безъядерные организмы, названные им Moneres, включал в царство протистов. Позже он выделял (Haeckel, 1904). в царстве Protista группы под названиями Phytomonera — синезелепые и Zoomonera — бактерии.
2 Очевидно, от греческого слова μυχός — внутренность, дубина, отдаленное помещение, в связи с предполагаемой их древностью.
12
Schizophyta переименовывает в Mychota, Mycophyta — в Protocti- sta (Leucophyta — Mycophyta, включая Myxophyta — Phytomyxi- neae), a Nomophyta — в Chlorophyta, оставляя их в ранге подцарств растительного мира. При этом он высказывает мысль, что «назрело выделение из мира собственно растений миров дробянок или «михотов» и «грибов».
Ротмалер (Rothmaler, 1948, 1950, 1951, 1955), следуя Коп- ленду и Беркли, предлагает выделять четыре царства организмов, а если считать живыми организмами вирусы и бактериофаги, то пять царств (Rothmaler, 1955):
Aphanobionta — неясные (сомнительные) организмы (Schein wesen) Вирусы и бактериофаги Akaryobionta — дробянки (Spaltwesen)
Schizophyta
Cyaiiophyta
Protobionta — первичные организмы (Urwesen)
Pyrrophyta Mycophyta
Rhodophyta Euglenophyta
Phaeophyta Chlorophyta 1
Cormobionta (Cormophyta) — растения (Pflanzen)
Psilophyta Pteridophyta
Anthocerophyta Spermatophyta Bryophyta
Gastrobionta (Metazoa) — животные (Tiere)
Включение в царство протистов. или Protobionta ряда отделов водорослей, в принадлежности которых к растительному миру нет сомнения, не может не вызывать недоумения, тем более, что общепризнанной считается теория происхождения высших растений от водорослей.
Развивая взгляды на обоснованность выделения дробянок в отдельное царство, Циммерман в позднейших работах (Zimmermann, 1949, 1959) разделил органический мир на два больших царства, существовавших уже в докембрии,— царство безъядерных организмов (Anucleobionta: бактерии и синезеленые) и царство ядерных организмов (Nucleobionta: животные и «типичные» растения). Таким образом, Циммерманн пришел к выводу, предложенному Немецом на 20 лет раньше.
Шадефо (Chadefaud, 1960) делит органический мир на две большие группы ( = царства.— Д. 3.) — Protocaryota (Protoca- ryotes) и Eucaryota (Eucaryotes) 1 2. Protocaryota он делит на Cya- noschizophyta (Gyanoschizophytes) и Racterioschizophyta (Bacte- rioschizophytes), a Eucaryota на четыре группы ( = подцарства.— Д. 3.): эвкариотические водоросли — Phycophyta (Phycophytes), грибы — Mycophyta (Mycophytes), листостебельные растения —
1 К Protobionta Ротмалер относит также Protozoa, как это он отмечал раньше (Rothmaler, 1951, стр. 264).
2 У Шадефо названия групп даны на французском языке. Мы даем их в порядке унификации в латинской транскрипции, а в скобках — оригинальные названия автора.
13
Cormophyta (Cormophytes) и животные — Animalia (Animaux или Zoaires).
Большое значение способам питания при разграничении царств живой природы придает Уиттейкер (Whittaker, 1959). По способу питания он выделяет три основные группы организмов: 1) продуценты — автотрофные организмы (producers — autotrophic organisms); 2) потребители (consumers), которые заглатывают живое или мертвое органическое вещество других организмов; 3) восстановители (reducers), также потребляющие органическое вещество, однако путем абсорбции. В результате разложения мертвых остатков других организмов восстановители возвращают в окружающую среду неорганические вещества, часть которых усваивается продуцентами. Эти три группы — продуценты, потребители и восстановители, соответствуют в общем трем основным типам питания — голофитному, голозойному и сапробному и трем или четырем большим группам организмов, а именно: продуценты — фитосинтезирующим организмам, потребители — животным, а восстановители — бактериям и грибам. Однако на схеме и в классификации фил органического мира Уиттейкер разделяет органический мир на четыре царства — протисты, грибы, растения и животные, причем, как показано на схеме самого автора, все четыре царства не имеют четких границ, они частично перекрывают одно другое. Этим четырем царствам Уиттейкер дает следующее содержание.
Царство PROTISTA
Подцарство Μ о η е г а, или Mychota
Schizomycota, или Schizomycetes Gyanophyta, или Myxophyta
Подцарство Eunucleata Chrysophyta Pyrrophyta Euglenophyta Sporozoa
Zoomastigina, или Euflagellata Sarcoflina, или Rhizopoda Ciliophora
Царство FUNGI
Подцарство Myxomycota
My corny cetes, или Mycetozoa Подцарство Eumycota
Phy corny ce tes
Ascomycètes
Basidiomycetes
Царство PZANTAE
Подцарство Bhodophyta » Phaeophyta
» Euchlorophyta, или Chlorophyta
Chlorophyta, или Phycophyta Bryophyta Tracheophyta
14
Царство ANIMALIA
Подцарство Р а г a z о а » М e s о z о а
» E и ш e t a z о а
Существуют и другие мнения о делении органического мира Земли. Так, Уолтон (Walton, 1930) признает только одно царство живых существ — Bionta, которое он делит на три подцарства — Protistodeae (протисты). Ме- taphytodeae (многоклеточные растения) и Metazoodeae (многоклеточные животные). Эта система по сути отличается от системы Геккеля только переводом его трех царств в ранг подцарств. Диллон (Dillon, 1957) признает существование одного царства — растений, дающего эволюционное древо, от которого отчленяются низшие животные и сапробы, а от них — высшие животные и грибы.
Таким образом, мы видим, что такой казалось бы простой вопрос, как разделение органического мира на основные группы, различными исследователями решается по-разному. Причиной этого, с одной стороны, является сложность филогенетических отношений между разными группами организмов, а с другой — критерии, принимаемые разными авторами при рассмотрении этого вопроса, а также цели, которые они ставят перед собой.
Если мы поставим задачу выделения основных биологических групп организмов в зависимости от характера питания, то разделение их на три группы по Уиттейкеру—автотрофные растения («производители»), животные («потребители») и грибы («восстановители») довольно точно отражает их роль в жизни природы. Но если мы хотим, чтобы наша классификация органического мира отражала происхождение и родственные связи между различными группами организмов, населяющих Землю, нам необходимо подойти к этому вопросу с исторической точки зрения и рассмотреть развитие органического мира, начиная со времени появления на Земле древнейших организмов, используя одновременно новые данные цитологии и биохимии.
* * *
Проблема происхождения жизни на Земле после первых работ Опарина (1924, 1936) и Холдейна (Haldane, 1929) привлекала внимание многих исследователей, однако в силу характера процессов и событий, протекавших более трех миллиардов лет назад в обстановке, кардинально отличавшейся от современных условий, она остается в сфере гипотез. По поводу характера первичных организмов — эобионтов, населявших нашу планету в далекие времена докембрия (архея или, возможно, даже катархея), современная наука может строить только более или менее обоснованные гипотезы.
Исходя из того, что пребиотическая атмосфера Земли не содержала или почти не содержала свободного кислорода (Haldane, 1929; Бернал, 1969, и др.), предполагают, что первые живые существа — эобионты были анаэробами и гетеротрофами, питавшимися за счет органических соединений, образовавшихся на Земле в процессе эволюции материи под действием ультрафиолетового
15
излучения солнца, не задерживаемого отсутствовавшим в то время в атмосфере озоном. В дальнейшем более перспективными могли быть организмы, выработавшие биосинтетические особенности, благодаря которым они могли использовать другие источники энергии. В процессе эволюции первичных гетеротрофных анаэробных организмов — эволюции, потребовавшей, вероятно, весьма значительное время (Klein a. Crônquist, 1967, стр. 115 и др.), развились организмы, способные к хемо-автотрофному способу питания.
В ходе дальнейшей эволюции у анаэробных хемо-автотрофов развился механизм фиксации углекислоты за счет энергии химических реакций. Примитивный фотосинтез мог не сопровождаться образованием свободного кислорода, как мы это видим у современных фотосинтезирующих бактерий (Klein a. Crônquist, 1967, стр. 116—117). Мы не будем останавливаться на возможных путях развития автотрофного питания,— этот вопрос является специальной биохимической проблемой,— отметим только, что в процессе дальнейшего развития фотосинтеза стал выделяться в атмосферу свободный кислород, а это дало возможность для развития аэробного обмена веществ, который в дальнейшем как более эффективный в энергетическом отношении стал доминирующим.
Исходя из палеоботанических данных, а также из того, что среди современных бактерий имеются анаэробные гетеро- и авто- трофы, можно предположить, что из ныне существующих групп организмов наиболее древними являются бактерии.
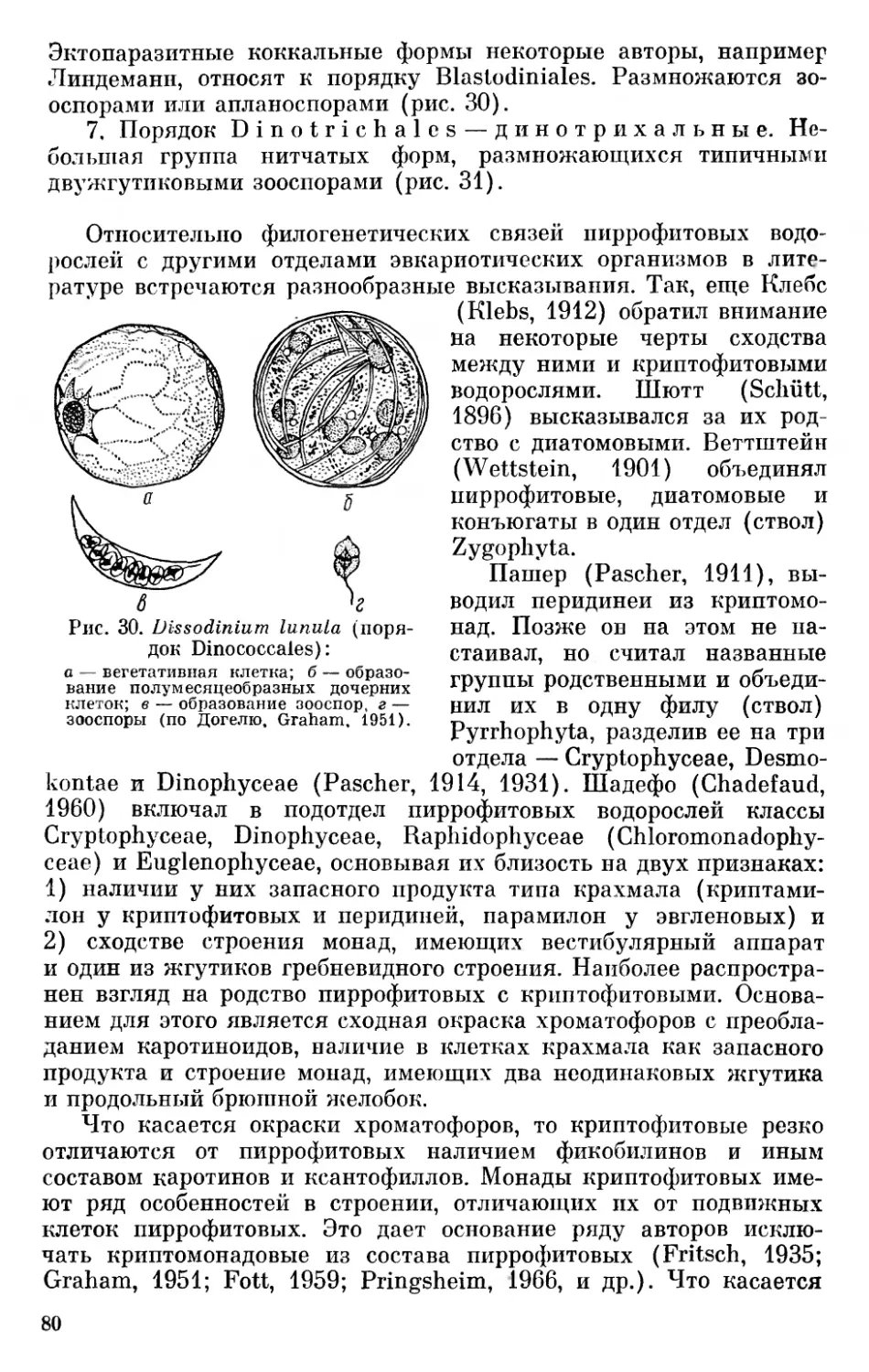
Клейн и Кронквист, резюмируя состояние вопроса о развитии органического мира, приходят к выводу, что Земля была пригодна к развитию жизни около 2,5 миллиардов лет назад. В течение приблизительно полутора миллиарда лет проходили процессы, приведшие к возникновению жизни, биогенез и эволюция первичных организмов до стадии синезеленых водорослей (Klein а. Сгоп- quist, 1967, стр. 112). Однако есть основания предполагать еще более раннее развитие жизни на Земле. Так, Баргхоорн и Тилер (Tyler a. Barghoorn, 1954; Barghoorn a. Tyler, 1965) описывают из отложений Ганфлинт на севере США, возраст которых определяют в 2 миллиарда лет и относят к нижней части среднего докембрия, остатки сферических одноклеточных и нитчатых разветвленных и неразветвленных форм неопределенного систематического положения. Несколько позже Баргхоорн и Шопф (Barghoorn а. Schopf, 1966) сообщили о находке в Южной Родезии в отложениях раннего докембрия, возраст которых определяется в 3,1 миллиарда лет, остатков бактериоподобных палочковидных организмов, названных ими Eobacterium isolatum, и нитчатых органических структур, возможно, биологического происхождения. Фишер (Fischer, 1965) сообщает, что строматолиты, возраст которых определяют в 2,7 миллиарда лет, образованы синезелеными водорослями.
Таким образом, есть достаточно данных, доказывающих, что организмы, близкие к бактериям, уже существовали в раннем
16
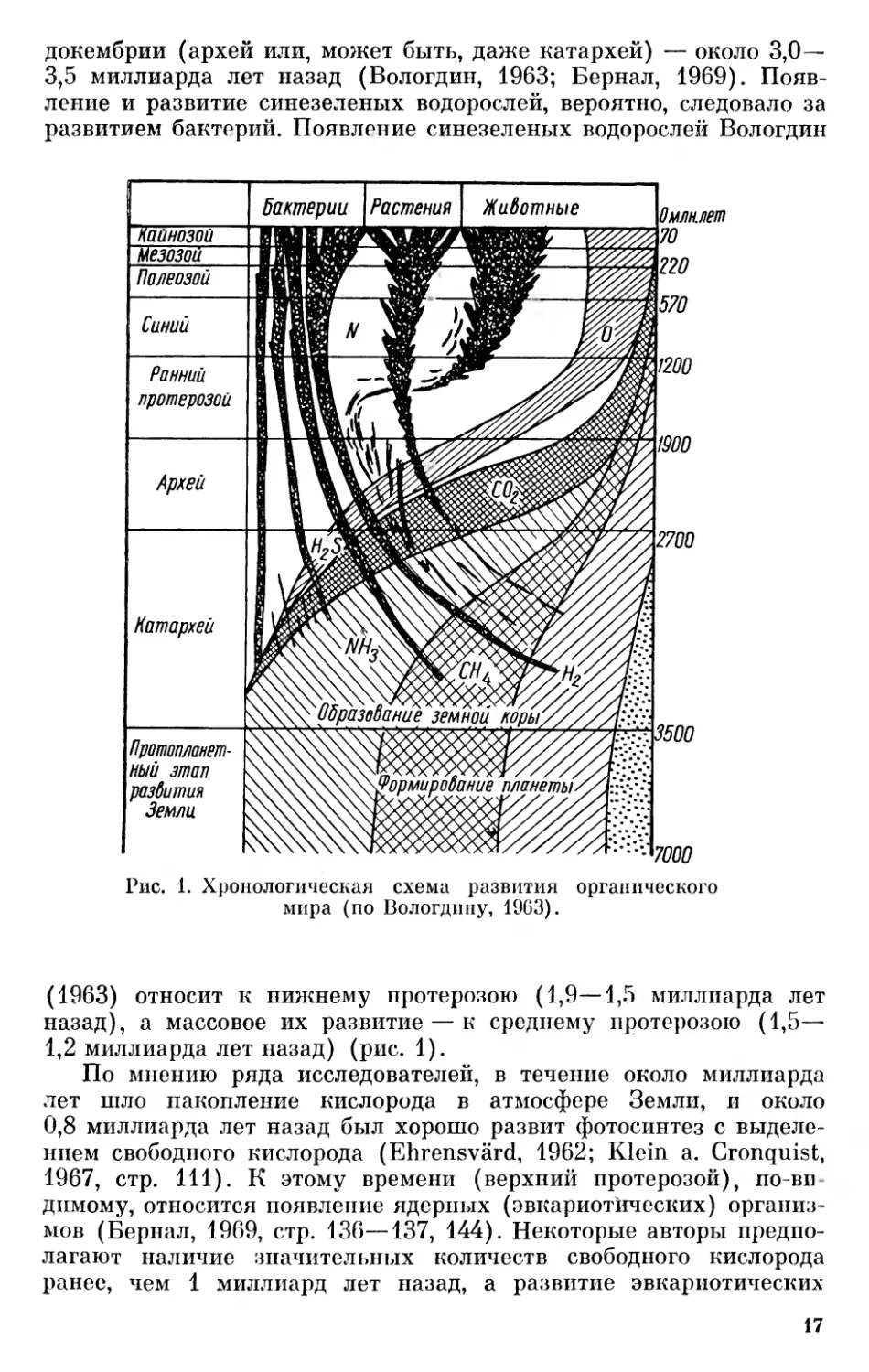
докембрии (архей или, может быть, даже катархей) — около 3,0—- 3,5 миллиарда лет назад (Вологдин, 1963; Бернал, 1969). Появление и развитие синезеленых водорослей, вероятно, следовало за развитием бактерий. Появление синезеленых водорослей Вологдин
Рис. 1. Хронологическая схема развития органического мира (по Вологдину, 1963).
(1963) относит к нижнему протерозою (1,9—1,5 миллиарда лет назад), а массовое их развитие — к среднему протерозою (1,5— 1,2 миллиарда лет назад) (рис. 1).
По мнению ряда исследователей, в течение около миллиарда лет шло накопление кислорода в атмосфере Земли, и около 0,8 миллиарда лет назад был хорошо развит фотосинтез с выделением свободного кислорода (Ehrensvärd, 1962; Klein a. Cronquist, 1967, стр. 111). К этому времени (верхний протерозой), по-видимому, относится появление ядерных (эвкариот'ических) организмов (Бернал, 1969, стр. 136—137, 144). Некоторые авторы предполагают наличие значительных количеств свободного кислорода ранее, чем 1 миллиард лет назад, а развитие эвкариотических
17
организмов — красных и зеленых водорослей и различных групп беспозвоночных животных, включая червей и кишечнополостных, относят к позднему протерозою — от 1,2 до 0,6 миллиардов лет назад (Вологдин, 1963).
Таким образом, прокариотические организмы развились значительно раньше эвкариотических. Современные бактерии и синезеленые прошли очень длительную историю и являются специализированными потомками древних примитивных докембрийских организмов.
* * *
Существенным является вопрос о филогенетических взаимоотношениях основных групп прокариотических организмов — бактерий и синезеленых водорослей между собой, с одной стороны, и с настоящими ядерными организмами — эвкариотами — с другой.
Еще Кон более ста лет назад обратил внимание на сходство бактерий и синезеленых водорослей и объединил их в одну группу (отдел) растительных организмов под названием Schizophy- ta — дробянки. Позднейшие исследователи на основании глубокого изучения этих групп подтвердили сходство цитологического строения, биохимических свойств и развития организмов, относящихся к этим группам, тем самым подчеркивая филогенетическую связь между ними.
Синезеленые водоросли и бактерии характеризуются рядом черт, отличающих их от настоящих ядерных (эвкариотических) организмов. Клетки синезеленых и бактерий характеризуются наличием лишенного ядерной мембраны нуклеоида вместо покрытого мембраной настоящего ядра, а у фотосинтезирующих форм — отсутствием покрытых мембраной пластид. У них также нет эндоплазматической сети, диктиосом (аппарата Гольджи) и типичных митохондрий. Цитоплазма у них представляет собой густой гель, стойкий к повышенной температуре, высушиванию и осмотическому воздействию. Клетки у представителей обеих групп делятся в результате врастания внутрь клетки кольцевого валика. Клеточные оболочки имеют белковую основу и характеризуются наличием муреина; типичный половой процесс отсутствует.
В то же время есть ряд существенных различий между бактериями и синезелеными водорослями. У колониальных форм сине- зеленых водорослей содержимое соседних клеток соединяется плазматическими нитями, проходящими через мелкие поры в оболочках; у бактерий такой связи между клетками нет. Споры у бактерий образуются внутри клетки из части ее содержимого, а у синезеленых водорослей в спору превращается вся клетка. Существенным различием между обеими группами Прингсгейм (Pringsheim, 1949, 1967) считает способ передвижения. Синезеле- ные водоросли передвигаются при помощи скользящего движения. Однако, как отмечает Оллсопп (Allsopp, 1969), такой способ пе¬
18
редвижения наблюдается также у миксобактерий. Фотосинтезирующие бактерии имеют специфические пигменты — бактериохло- рофилл и бактериовиридин, в то время как для синезеленых характерно наличие хлорофилла а и фикобилинов (билипротеи- дов) — фикоцианина и фикоэритрина.
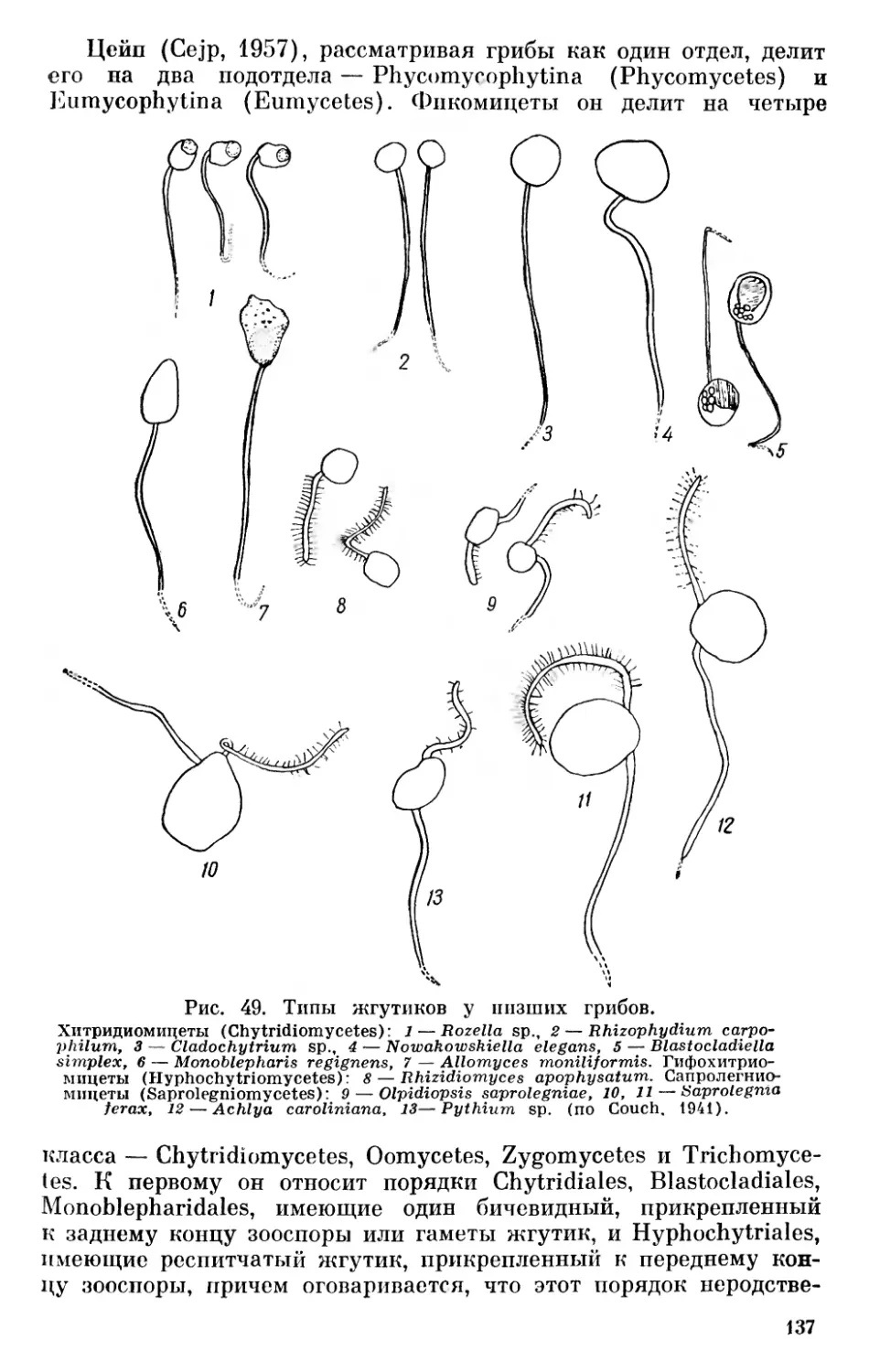
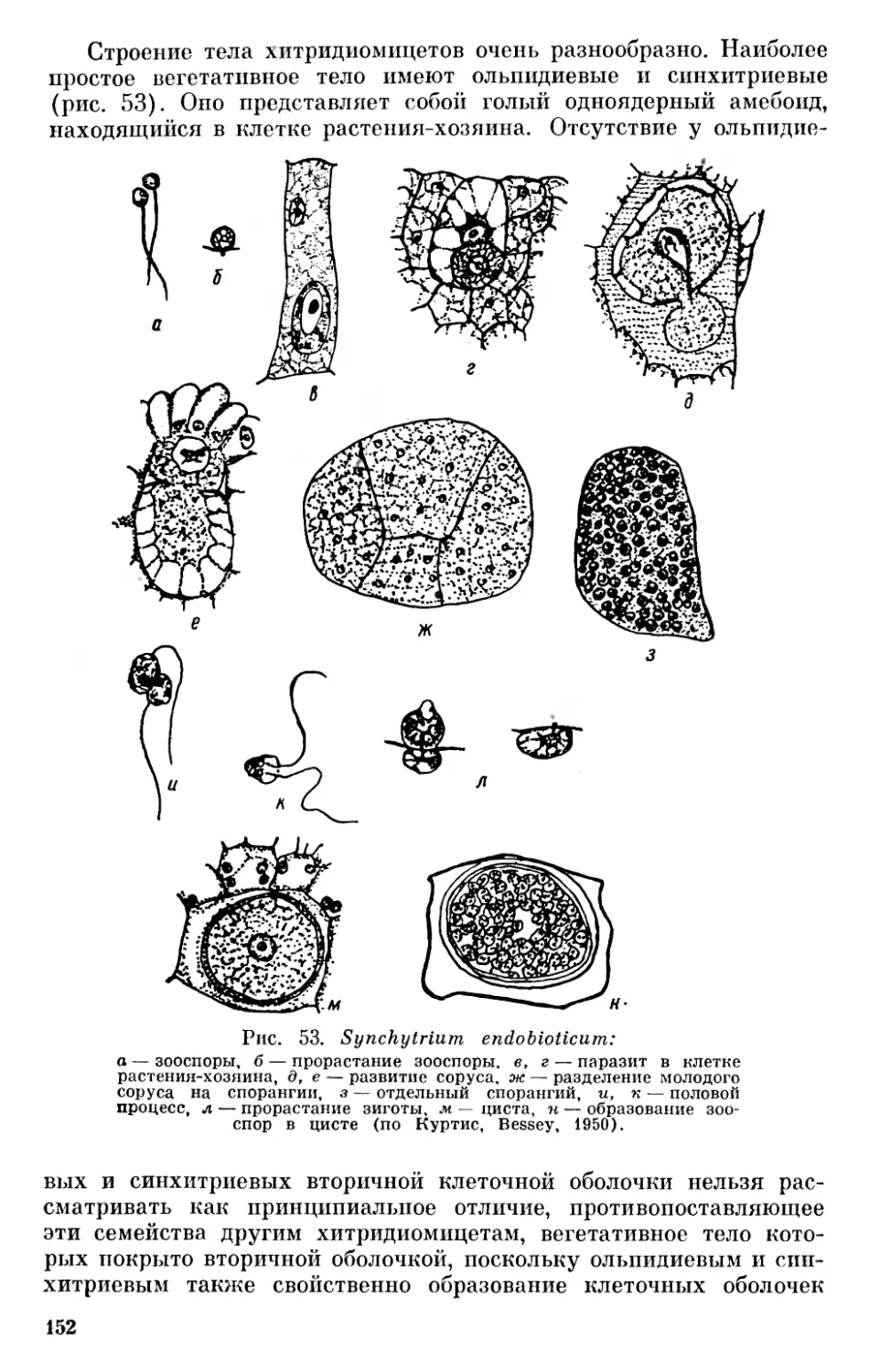
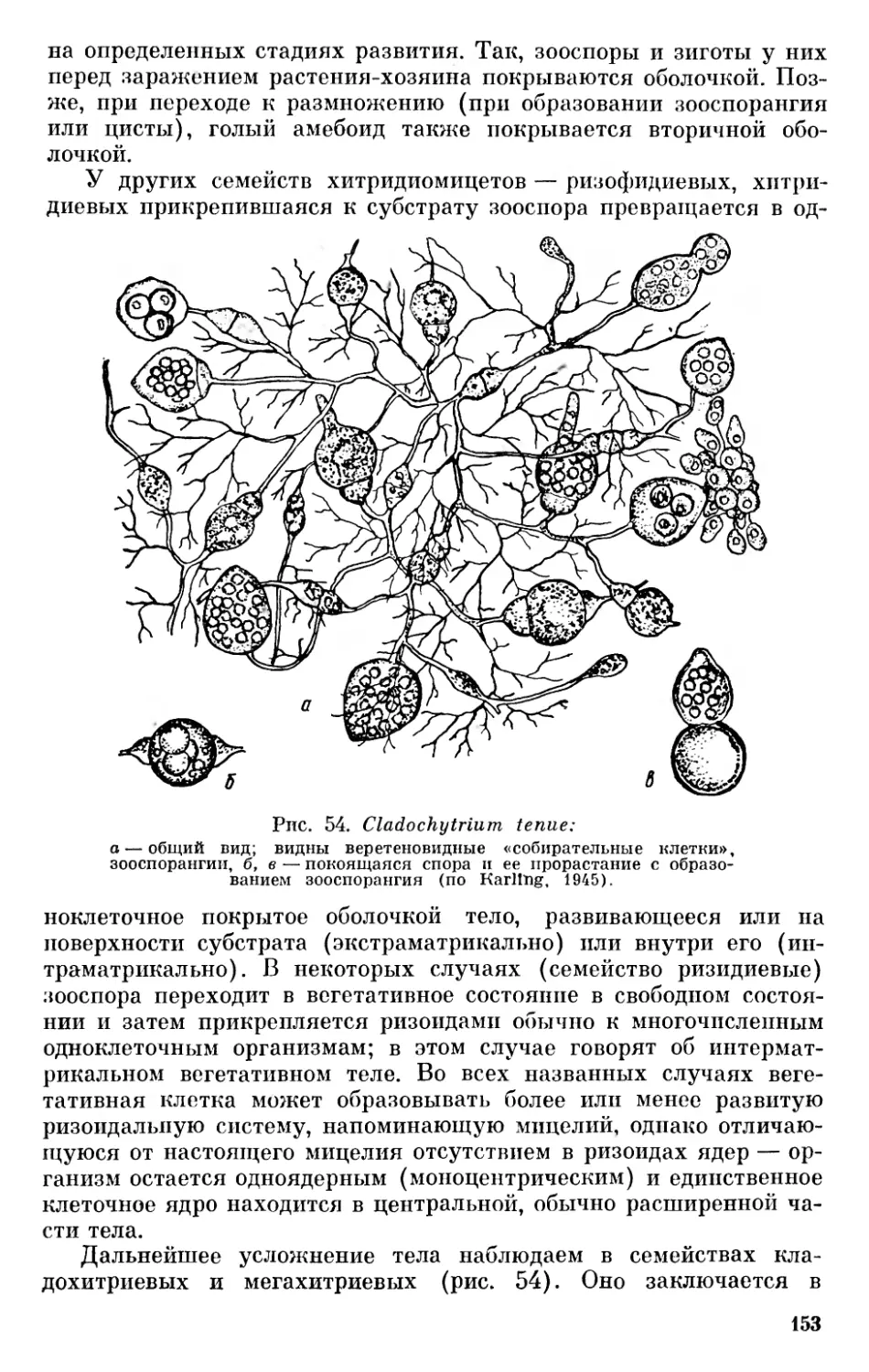
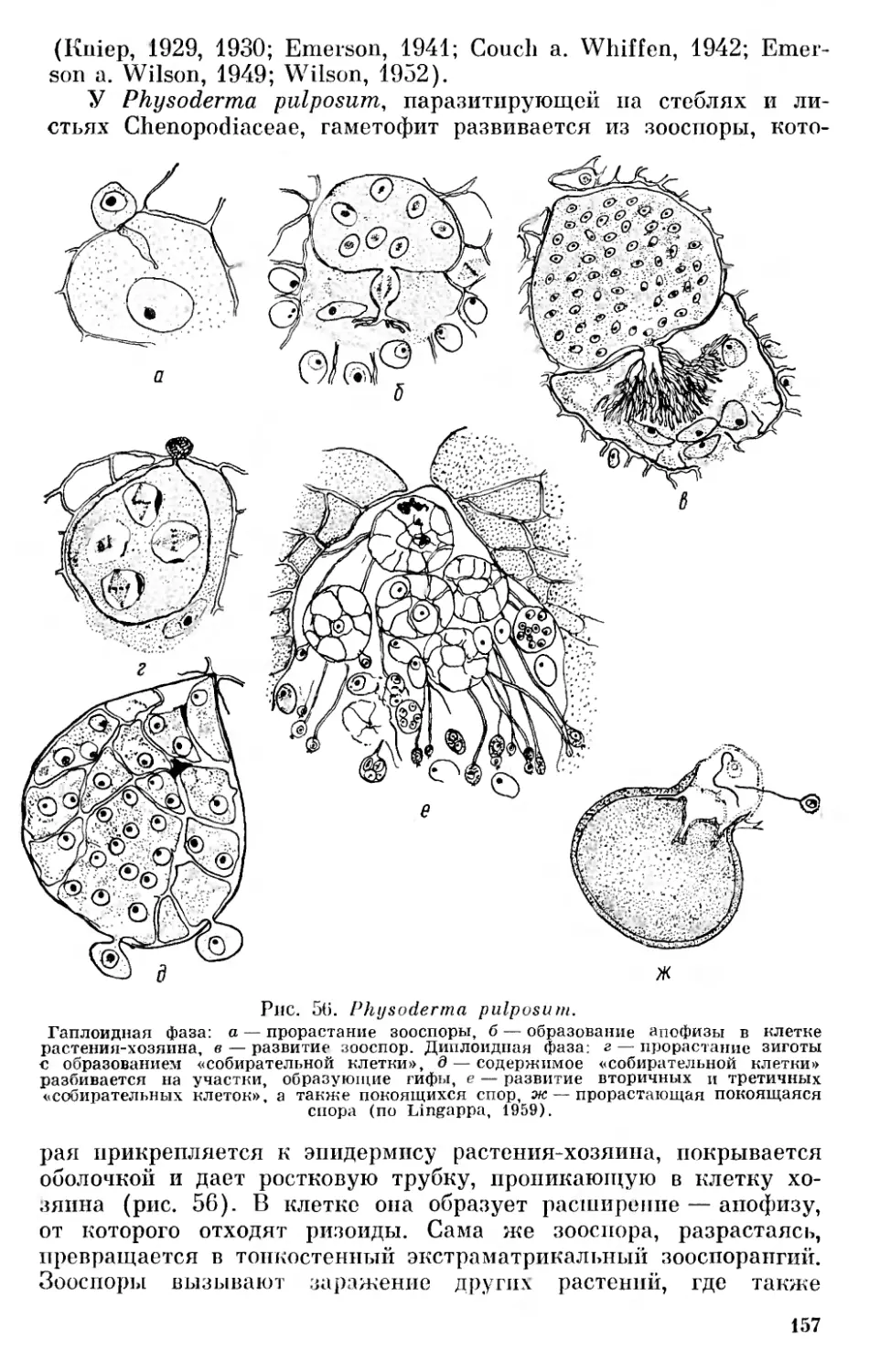
На основании сказанного выше нельзя согласиться с теми, кто отрицает всякую родственную связь между бактериями и сине- зелеными водорослями (Pringsheim, 1949, 1967), однако значительные отличия между этими группами заставляют с осторожностью относиться к распространенному мнению о происхождении синезеленых от Eubacteriales (Stanier a. van Niel, 1941, и др.). Необоснованным нам кажется предположение Шадефо (Chade- faud, 1960) о происхождении бактерий от синезеленых в результате сложного эволюционного процесса, одновременно регрессивного и прогрессивного, сопровождавшегося потерей фикобилинов, превращением хлорофилла а в специальные бактериальные пигменты или потерей его и другими изменениями. Имеющиеся в настоящее время данные свидетельствуют в пользу большей древности бактерий. Нам кажется наиболее обоснованным предположение о дивергентном происхождении бактерий и синезеленых от первичных прокариотических организмов.
В настоящее время принято рассматривать их как единую большую группу органического мира — царство Procaryota или Protocaryota, противопоставляя их настоящим ядерным организмам — царству Eucaryota К При этом бактерии и синезеленые рассматриваются или как один отдел — дробянки — Schizophyta, включающий два класса — бактерии (Bacteria, Schizomycetes) и синезеленые водоросли (Myxophyceae, Cyanophyceae, Schizophy- сеае) или же их разделяют на два отдела — бактерии (Bacteria, Bacteriophyta) и синезеленые (Cyanophyta).
До настоящего времени остается спорным вопрос о положении вирусов в системе органического мира. Начиная с Ивановского, открывшего их в 1892 г., многие авторы считают вирусы живыми организмами. Другие полагают, что вирусы являются деградированными микробами — продуктом регрессивной эволюции, связанной с паразитизмом (Green, 1935, и др.)· Высказывалось мнение, что вирусы являются ферментами, свободными генами и т. п. В последнее время проведены исследования по выяснению строения вирусных нуклеопротеидов, дающие основания для вывода, 11 Применение терминов «прокариотический» и «эвкариотический», а также названий Procaryota и Eucaryota Оллсопп (Allsopp, 1969) связывает с именами Шаттона (Chatton, 1937) и Станье и Нила (Stanier a. van Niel, 1941). Как мы уже упоминали выше, название Procaryota в несколько иной форме — Prokaryonta впервые предложил Немец (Nemee, 1929, стр. 100). От предложенных Немецом названий Acaryonta и Karyonta, а также от названий Anucleobionta и Nucleobionta, предложенных Циммерманиом, нужно отказаться в связи с тем, что и синезеленые и бактерии имеют ядерное вещество, сосредоточенное в нуклеоидах.
19
что вирусы являются самыми примитивными живыми формами на нашей планете, которые, попадая в клетки организмов, способны к репликации подобных себе форм, т. е. к размножению (Товар- ницкий, 1966). Возможно, среди более крупных форм, которые описывались как вирусы, могут встречаться и деградированные бактерии, однако сейчас имеется много данных, дающих основание считать, что «вирусы представляют собой специализированную в результате внутриклеточного паразитизма ветвь неклеточных организмов» (Сухов, 1956, стр. 273). Сухов даже допускает возможность существования в современной природе еще не обнаруженных свободных сапрофитных неклеточных организмов, однако он далее отмечает, что условия для существования сапрофитных вирусов в настоящее время могут отсутствовать, а «известные же нам патогенные вирусы представляют собой реликтовые формы, уцелевшие вследствие особо благоприятных условий, которые создал для них внутриклеточный паразитизм» (Сухов, 1956, стр. 274).
Ряд авторов считает возможным включать вирусы в систему живых организмов. Так, Беркли (Barkley, 1939) выделяет в царстве монер отдел и филу Archaeophyta, куда относит первичные организмы, вирусы и бактериофаги. Эту идею поддержал Козо- Полянский (1949, стр. 252). Ротмалер (Rothmaler, 1948, 1955) выделял вирусы и бактериофаги, правда с оговоркой, в особое царство Aphanobionta («Scheinwesen»). Сухов (1956) выделяет вирусы в особый «мир» неклеточных организмов, противопоставляя его миру клеточных организмов — растениям и животным.
Можно считать, что вирусы являются специализированными, приспособленными к внутриклеточному паразитизму потомками первичных неклеточных организмов. В таком случае их следует выделять в особое царство неклеточных организмов — Acytobionta, что мы и сделали на приведенной схеме (рис. 2).
Весьма существенным является вопрос о взаимоотношении Procaryota и Eucaryota. В результате успехов электронной микроскопии в течение последних десятилетий мы располагаем богатым фактическим материалом относительно строения клетки, который дает новые основания для решения кардинальных вопросов филогении органического мира. Исследования с применением электронного микроскопа показали удивительное однообразие строения клетки в различных группах растительного и животного мира и значительные отличия ее от клеток прокариотов — бактерий и синезеленых, что ставит под сомнение распространенный взгляд, согласно которому эвкариотические организмы возникли из прокариотических в результате прогрессивного преобразования их клеточных компонентов (Cronquist, 1960; Allsopp, 1969, стр. 607).
За последние годы на новом основании возродились старые взгляды Мережковского о происхождении ядерной клетки в результате симбиоза прокариотических одноклеточных форм.
20
Рис. 2. Схема филогенетических связей между отделами (филами) и подотделами растительного мира.
Настоящие ядерные организмы £ и сагу οΐα
Еще в прошлом столетии Шимпер (Schimper, 1883) обратил внимание на то, что хлоропласты обладают некоторыми чертами эндосимбионтов. Позже Мережковский (Mereschkowsky, 1905) высказал предположение, что они возникли из симбиотических синезеленых водорослей. Затем он (Мережковский, 1909) разработал теорию двух плазм — микоплазмы и амебоплазмы как основу симбиогенеза, поддержанную позже Козо-Полянским (1924).
По теории Мережковского известные в то время органеллы клетки — ядро и хроматофоры являются симбиотическими организмами микоидной природы, живущими в амебоидной плазме растительных и животных организмов. К микоидам он (1909) относил совокупность бактерий, грибов и синезеленых, а также симбиотически живущие с амебоплазмой хроматофоры и «некоторые ядерные элементы». Позже было высказано мнение, что митохондрии являются симбиотическими бактериями (Wallin, 1922).
Развивая взгляды Мережковского, Козо-Полянский (1924) считал, что «клетка представляет собой собрание разнородных, автономных жизненных единиц — «биотов», ведущих симбиотический образ жизни, будучи погружены в проблематическую «плазму»... (стр. 63). По мнению Козо-Полянского, «органоиды клетки не являются продуктами дифференциации, но представляют результат суммирования, присоединения и внедрения со стороны перед тем автономных и ведших самостоятельное существование жизненных единиц» (стр. 120).
В последние годы теорию симбиогенеза («а specific series of symbioses») развивает Саган-Маргулис (Sagan, 1967; Margulis, 1968). К этой теории склоняется и Бернал, который пишет, что «ядерная клетка в некоторых отношениях сильно напоминает симбиотический организм» (Бернал, 1969, стр. 124). При этом он обосновывает это мнение на известной автономности органелл и сходстве их в клетках организмов самых различных групп.
Основанием для теории симбиогенеза может быть явление внутриклеточного симбиоза среди растений и животных и особенно указания на наличие у некоторых моллюсков симбиотических хлоропластов (Kawaguti a. Yamasu, 1965; Taylor, 1967, 1968), с одной стороны, а с другой — некоторая автономность пластид и митохондрий. Явление внутриклеточного симбиоза довольно распространено, причем внутриклеточными симбионтами могут быть зеленые и синезеленые водоросли, перидинеи, бактерии; при этом обычно бывает довольно легко определить их природу. Есть указания на хлоропласты, живущие в клетках моллюсков, куда они попадают с пищей. Однако, по данным Тейлора (Taylor, 1968), они активно фотосинтезируют только в течение 24 ч после поедания водоросли. В связи с этим можно напомнить очень интересное явление, приводимое Догелем (1947, стр. 405). Некоторые брюхоногие моллюски (Aeolidae) питаются гидроидными по¬
22
липами, стрекательные капсулы которых не перевариваются и поступают в печеночные выросты моллюска, сохраняя там способность функционировать.
Приведенные факты указывают на разнообразие биологических приспособлений, но отнюдь не являются убедительным доказательством симбиотического происхождения органелл клетки. Некоторая автономность пластид и митохондрий внутри клетки, связанная с наличием в них ДНК, способность их к размножению делением является более серьезным аргументом в пользу возможности позднейшего включения их в клетку, однако строение этих органелл существенно отличается от строения клеток прокариотических организмов. Очень важными в связи с этим являются указания на ядерное происхождение промитохондрий и пропластид из инициальных частиц, отчленяющихся от ядра (Фрей-Вис- слинг и Мюлеталер, 1968), а также явления гибридизации между ДНК ядра и органелл эвглены (Richards, 1967) и между РНК хлоропластов и ядерной ДНК листьев табака (Tewari a. Wildman, 1968). Все эти данные свидетельствуют о том, что строение клетки эвкариотических организмов является результатом самостоятельной эволюции, а не результатом симбиогенеза.
Если исключить возможность симбиотического происхождения клетки эвкариотических организмов, то все же остается вопрос, можно ли считать эвкариотические организмы прямыми потомками прокариотических, как это принимают многие современные авторы (Cronquist, 1960; Klein a. Cronquist, 1967; Allsopp, 1969, и др.)· Нам это кажется мало вероятным по ряду причин. Бактерии и синезеленые являются специализированными группами, развившимися из первичных организмов — эобионтов и сформировавшимися в докембрийское время — в архейскую и раннюю часть протерозойской эры. Они отличаются рядом специфических особенностей в строении клетки, биохимическими и физиологическими свойствами. Если еще можно представить себе развитие ядра эвкариотических организмов из нуклеоида прокариотических форм, то уже труднее предположить развитие хроматофоров из фотосинтетических мембран синезеленых, являющихся выростами внутренней клеточной мембраны плазмалеммы (Пешков, 1966), так как, согласно исследованиям Фрей-Висслинга и Мюлеталера (1968), хлоропласты и митохондрии эвкариотических организмов имеют ядерное происхождение. Эквиваленты митохондрий — мезо- сомы, описанные у некоторых синезеленых и бактерий, по данным Фитц-Джемса (Fitz-James, 1960, цит. по Фрей-Висслингу и Мюлеталеру, 1968) и Кавата (Пешков, 1966, стр. 133), возникают также из складок плазмалеммы. Они имеют одну мембрану и характеризуются своеобразным строением (Фрей-Висслинг и Мюлеталер, 1968, стр. 284—285).
Характерной особенностью прокариотов является наличие клеточной оболочки, в то время как у животных и низших представителей растительного мира клетки не имеют вторичной кле¬
23
точной оболочки, а окружены только плазматической мембраной — плазмалеммой. Все филы растительного и животного мира выводят из амебоидных форм, происхождение которых от покрытых постоянной оболочкой прокариотов трудно допустить. Нужно также отметить, что жгутики всех эвкариотов состоят из одиннадцати (9 + 2) фибрилл, в то время как у бактерий они состоят из одной фибриллы, а у синезеленых жгутиков вообще нет.
Для бактерий характерен своеобразный тип передачи ядерного вещества из одной клетки (донор, «мужская» клетка) другой (реципиент, «женская» клетка) — так называемый парасексуаль- ный процесс. Правда имеются указания на «настоящий» половой процесс у некоторых бактерий (Bisset, 1955; Chadefaud, 1960), однако эти указания требуют проверки.
Что касается синезеленых водорослей, то имеющиеся данные свидетельствуют только о возможности наличия у них парасек- суального процесса, однако не дают еще сведений о его форме (Kumar, 1962; Singh a. Sinha, 1965; Bazin, 1968).
Во всяком случае описанный у бактерий тип полового процесса проходит совершенно по-иному, чем у эвкариотических организмов, где сливаются целые клетки с последующим редукционным делением.
Все сказанное выше свидетельствует о том, что Eucaryota не могли быть непосредственными потомками ни бактерий, ни сине- зеленых водорослей. Eucaryota развились из более примитивных форм, являвшихся, вероятно, общими предками прокариотических и эвкариотических организмов.
Из существующих ныне эвкариотических организмов на наиболее низкой ступени стоят амебоидные корненожки (Rhizopoda) и одноклеточные голые формы из фототрофных фил — Chryso- pliyta, Euglenophyta. Это дает основание предполагать, что исходные формы эвкариотических организмов были одноклеточными амебоидными формами, у которых на ранних стадиях эволюции сформировался ядерный и у большинства связанный с ним жгутиковый аппарат, а также способность к фототрофному питанию. Бурное развитие эвкариотов в протерозое привело к обособлению ряда стволов — фил с гетеротрофным, автотрофным, голозойным и смешанным способами питания, которые обычно принято делить на «царства» животных и растений.
Естественным результатом происхождения тех и других от общих предков явилось то, что низшие формы эвкариотов могут соединять в себе черты как «растений», так и «животных», и только в процессе дальнейшей эволюции у них выработались характерные черты животного и растительного мира. Если между Protozoa и Metazoa имеется разрыв, то между низшими одноклеточными формами растительных фил и высшими их представителями можно наблюдать постепенные переходы и усложнение. Поэтому выделение дополнительного царства протистов не только не ре¬
24
шает вопроса о разграничении животного и растительного мира, а только его усложняет.
Это видно хотя бы из того содержания, которое вкладывают различные современные авторы в понятие Protista. Так, Копленд (Copeland, 1938, 1947) к царству протистов относит Protozoa (Rhi- zopoda, Sporozoa, Infusoria), все флагеллаты — как автотрофные, так и гетеротрофные, за исключением Volvocales, диатомовые, красные и бурые водоросли, а также грибы, а к царству растений только зеленые растения — зеленые водоросли (Chlorophyceae), начиная с Volvocales, и все высшие растения. Беркли (Barkley, 1939) в царство протистов включает также зеленые водоросли. Ротмалер, как уже отмечалось, царство Protobionta, которое соответствует протистам, принимает в понимании Беркли. В результате организмы, связанные филогенетически,— зеленые водоросли и листостебельные растения (Cormophyta) попадают в различные царства природы, а ядерные организмы оказываются разделенными между тремя царствами.
Признавая общее происхождение всех фил настоящих ядерных организмов, логично рассматривать их как единую большую группу— «царство» эвкариотов (Eucaryota), противопоставляя ее другой основной группе — «царству» прокариотов (Procaryota, или Protocaryota).
Следуя традиции, филы эвкариотов с голозойным характером питания («потребители» Уиттейкера) можно рассматривать как животный мир или подцарство животных, а филы с автотрофным («продуценты») и гетеротрофным осмотическим питанием («восстановители») — как растительный мир или подцарство растений. Выделение отдельного подцарства грибов нам кажется нецелесообразным, так как клетки большинства грибных организмов, как и большинства автотрофных растений, обладают оболочками, которые обусловливают как характер их роста, так и осмотический способ питания, с одной стороны, с другой же стороны, как будет показано ниже, грибы не представляют единой филогенетической группы, а распадаются на несколько фил разного происхождения.
* * *
Не останавливаясь на разделении животного мира, который распадается на две основные группы — Protozoa и Metazoa с рядом подразделений, называемых зоологами «типами», коротко рассмотрим филы (стволы) растительного мира.
Филами мы называем основные группы органического мира, имеющие общее происхождение, развитие которых можно графически изобразить в виде разветвленного дерева. Понятие «фила» в систематике организмов целесообразно рассматривать как основную группу — то, что в ботанике принято называть «отделом», а в зоологии «типом». Мы считаем это более соответствующим принципам филогенетической классификации, чем высказанное нами недавно (Зеров, 1970) предложение различать понятия «фи¬
25
ла» и «отдел» и разбивать некоторые филы растительного мира на несколько отделов. Это, правда, несколько нарушит принятую номенклатуру основных групп зеленых растений, которые придется рассматривать как подотделы отдела (филы) зеленых растений (Chlorophyta), однако будет более последовательным.
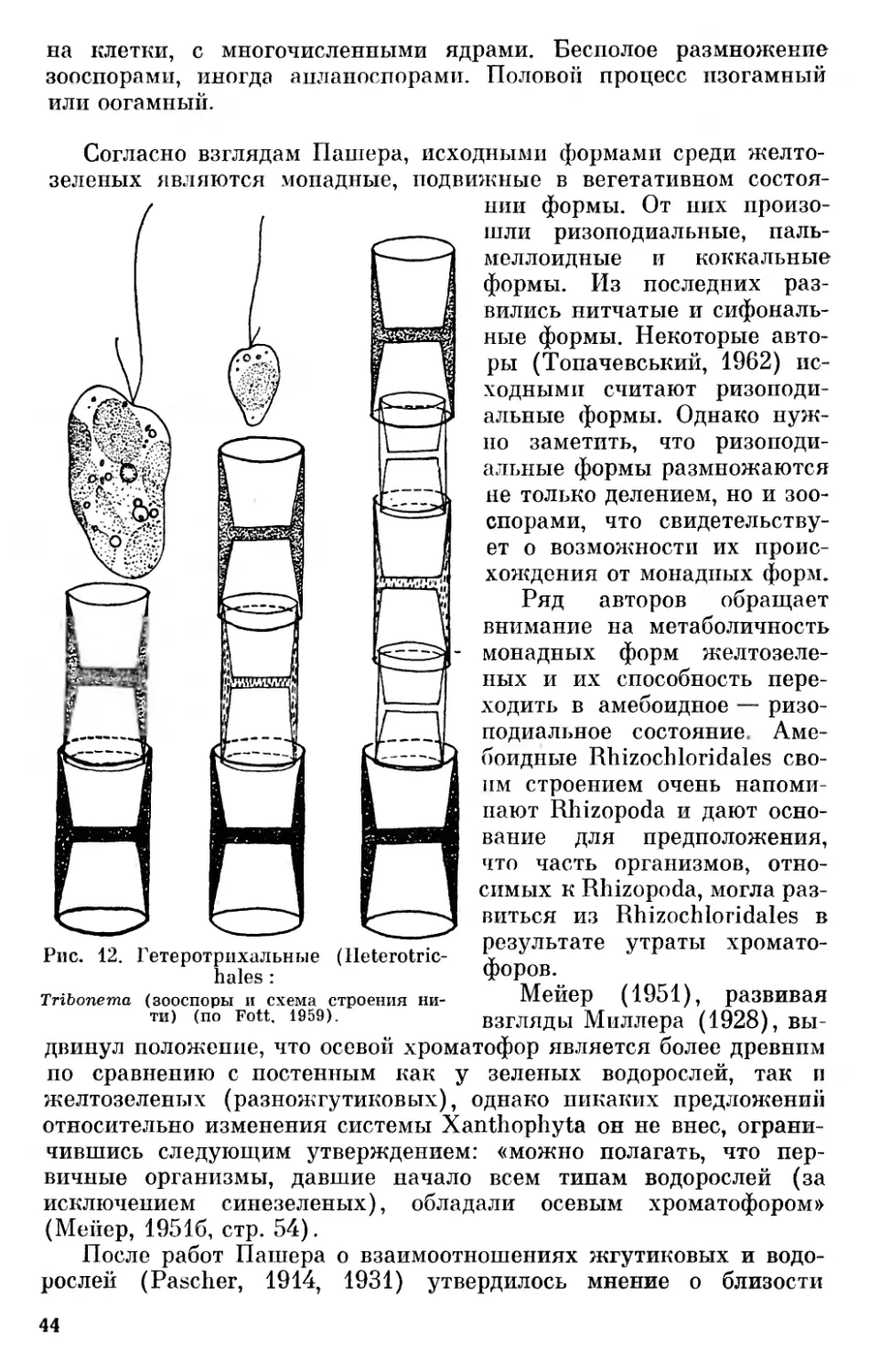
Характерной чертой всех эвкариотических фил растительного мира является то, что наиболее примитивные формы большинства из них — одноклеточные организмы, обычно лишенные вторичной оболочки, способные к амебоидному движению и снабженные в большинстве случаев жгутиками. Исключение составляют фила бурых водорослей (Phaeophyta), среди современных представителей которых нет одноклеточных форм, а также филы красных водорослей (Rhodophyta) и настоящих грибов (Ешпусо- ta), среди которых нет жгутиковых форм и одноклеточные низшие представители которых имеют вторичные клеточные оболочки. Что касается бурых водорослей, то большинство из них имеет снабженные жгутиками голые зооспоры и гаметы, что свидетельствует об их филогенетических связях со жгутиковыми формами. У красных водорослей, лишенных жгутиков, наличие голых спор и спермациев, способных к амебоидному движению, может свидетельствовать о их происхождении от голых амебоидных одноклеточных организмов. Что же касается настоящих грибов, то условия наземного существования обусловили замену голых спор спорами, снабженными вторичными оболочками, однако характер их спороношений и половой процесс указывают на происхождение от предков, общих с другими растительными филами.
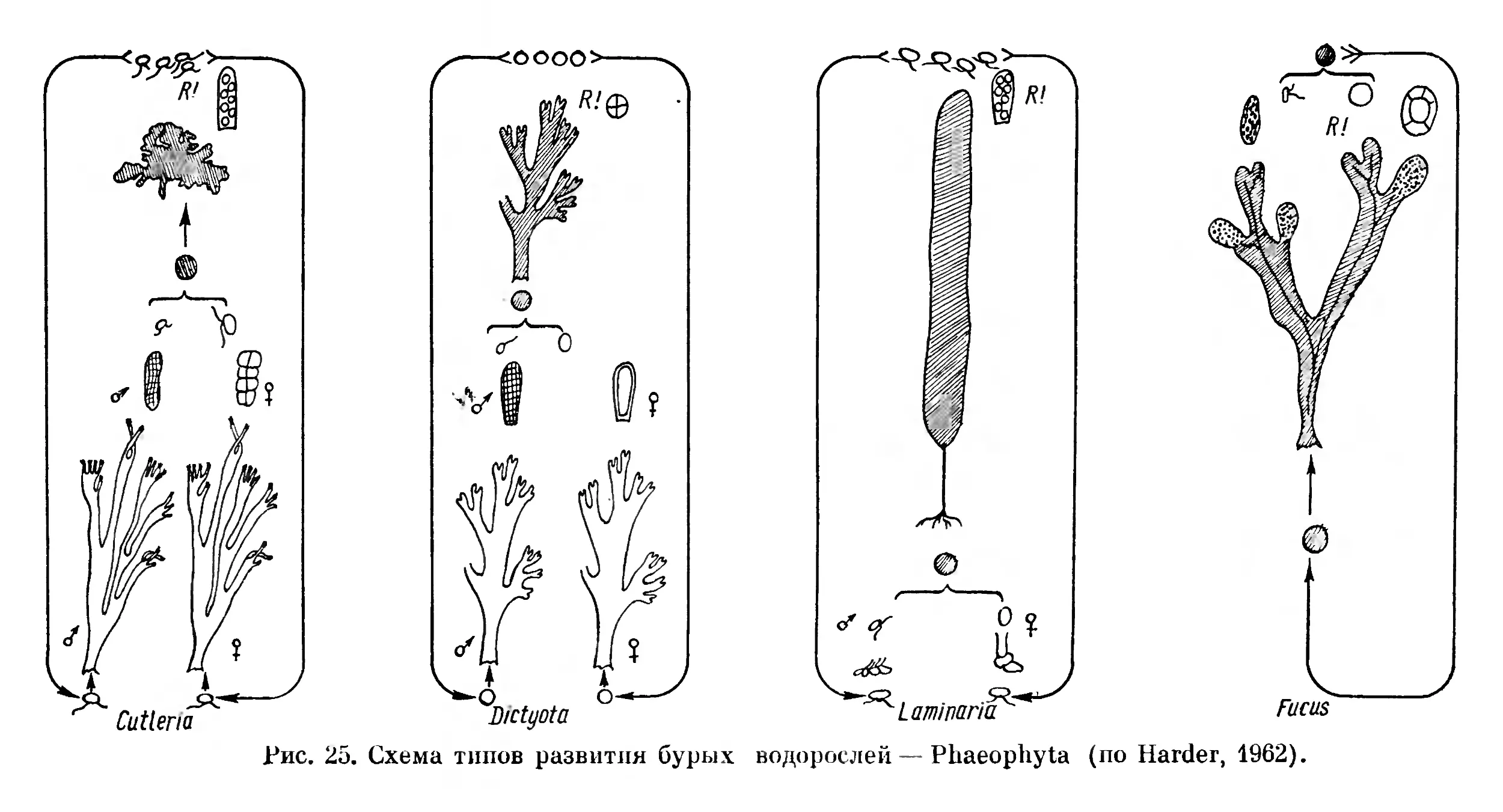
Мережковский (1909), а позже Пашер (Pascher, 1931) разработали идею полифилетического развития растительного мира из флагеллат несколькими филами (стволами). Развитие в пределах каждой филы шло от одноклеточных, обычно подвижных в вегетативном состоянии, к неподвижным многоклеточным типичным растительным формам, достигающим в пределах каждой филы разной степени усложнения. Количество фил, предлагаемых разными авторами, различно. Так, Мережковский выделял девять стволов водорослей; грибы он считал неоднородной группой гетерогенного происхождения,— низшие грибы — фикомицеты, по его мнению, представляют собой обесцветившиеся водоросли — «конечные члены разных линий эволюции», а высшие грибы — аскомицеты и базидиомицеты он выводит из бактерий через ак- тиномицеты двумя стволами.
Пашер (Pascher, 1931) подцарство Plantae holoplastideae (т. e. Procaryota) делил на две I филы—стволы — Cyanophyta и Schizomykophyta, а подцарство Plantae euplastideae, соответствующее растительным филам эвкариотических организмов,— на девять фил (стволов) автотрофных растений — Chrysophyta, Phaeophyta, Pyrrhophyta, Euglenophyta, Chlorophyta, 1 Charophyta,
1 Филу зеленых водорослей Пашер (Pacher, 1914) назвал Chlorophyta, т. е. «зеленые растения», что может привести к недоразумениям, так как
26
Rhodophyta, Bryophyta u Pterido-Anthophyta и три группы гетеротрофных — Myxophyta, Phykomykophyta и Mykophyta. Некоторые филы хлорофиллоносных организмов Пашер делит на отделы. Так, филу (ствол) Chrysophyta он делит на три отдела — Chryso- phyceae, Diatomeae и Heterocontae, филу Pyrrhophyta — тоже на три отдела — Cryptophyceae, Desmokontae и Dinophyceae, филу Chlorophyta на два отдела — Chlorophyceae и Conjugatae и филу Rhodophyta — на два отдела — Fîoridineae и Bangiineae. Группы бесхлорофильных растительных организмов Пашер считает искусственными, подлежащими расформированию, в связи с чем их нельзя сравнивать со стволами (филами) автотрофных организмов. Отделы Пашер делит на классы и порядки.
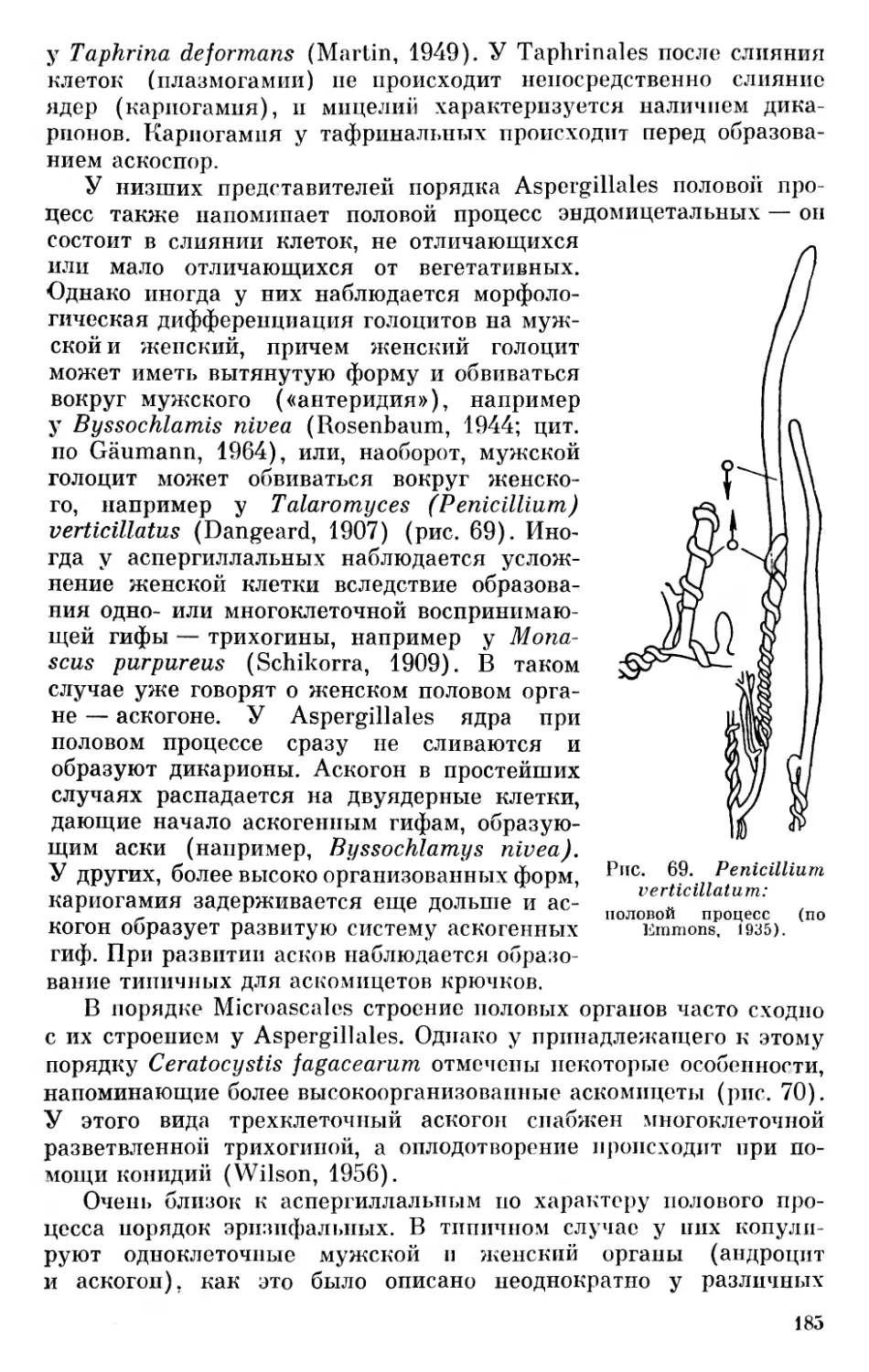
Смит (Smith, 1955, стр. 9) в отношении низших растений в основном принимает схему Пашера и делит растительный мир, включая бактерии, на шестнадцать отделов, сгруппированных в восемь стволов (фил), а именно: Chlorophyta (имеются в виду Chlorophycophyta.— Д. 3.), Euglenophyta, Pyrrophyta, Chrysophyta, Phaeophyta, Cyanophyta, Rhodophyta, Myxomycophyta, Eumy- cophyta, Bryophyta, Psilophyta, Lepidophyta, Calamophyta, Pte- rophyta, Gymnospermae и Angiospermae.
В отличие от изложенного выше взгляда на развитие эвка- риотических растительных организмов несколькими параллельными филами, некоторые авторы считают возможным выводить из бактерий синезеленые дробянки и зеленые водоросли. Из сине- зеленых они выводят красные водоросли, а из зеленых водорослей все прочие отделы растительных организмов (Cronquist, 1960; Klein a. Cronquist, 1967). Красные водоросли они увязывают с синезелеными на основании некоторых общих биохимических особенностей — наличия фикобилинов (фикоэритрина и фикоциани- на). Если основываться только на биохимических особенностях, не придавая значения цитологическим данным, то нужно выводить из синезеленых также и криптомонады — Cryptophyceae, у которых также имеются фикобилины. Однако Кронквист считает возможным криптомонады объединять вместе с перидинеями в отдел Pyrrhophyta. Что же касается выведения зеленых водорослей из синезеленых и всех других отделов растительного мира непосредственно из зеленых водорослей, то ряд структурных, цитологических и физиолого-биохимических особенностей этих отделов не позволяет считать возможным такое заключение.
зелеными растениями являются также мхи, папоротникообразные, голосеменные и покрытосеменные. Поэтому Папенфусс (Papenfuss, 1946) предложил для названий отделов водорослей окончание «pnycophyta» (Chlorophycophyta, Rhodophycophyta и т. д.). Ряд авторов, например Шадефо, Чапмен, Мейер и другие, употребляют эту форму. Применение ее не вызывается необходимостью, кроме зеленых водорослей, для которых окончание «phycophyta» нужно признать обязательным, если выделять их как отдел растительного мира, или же «phycophytina», если рассматривать их как подотдел.
27
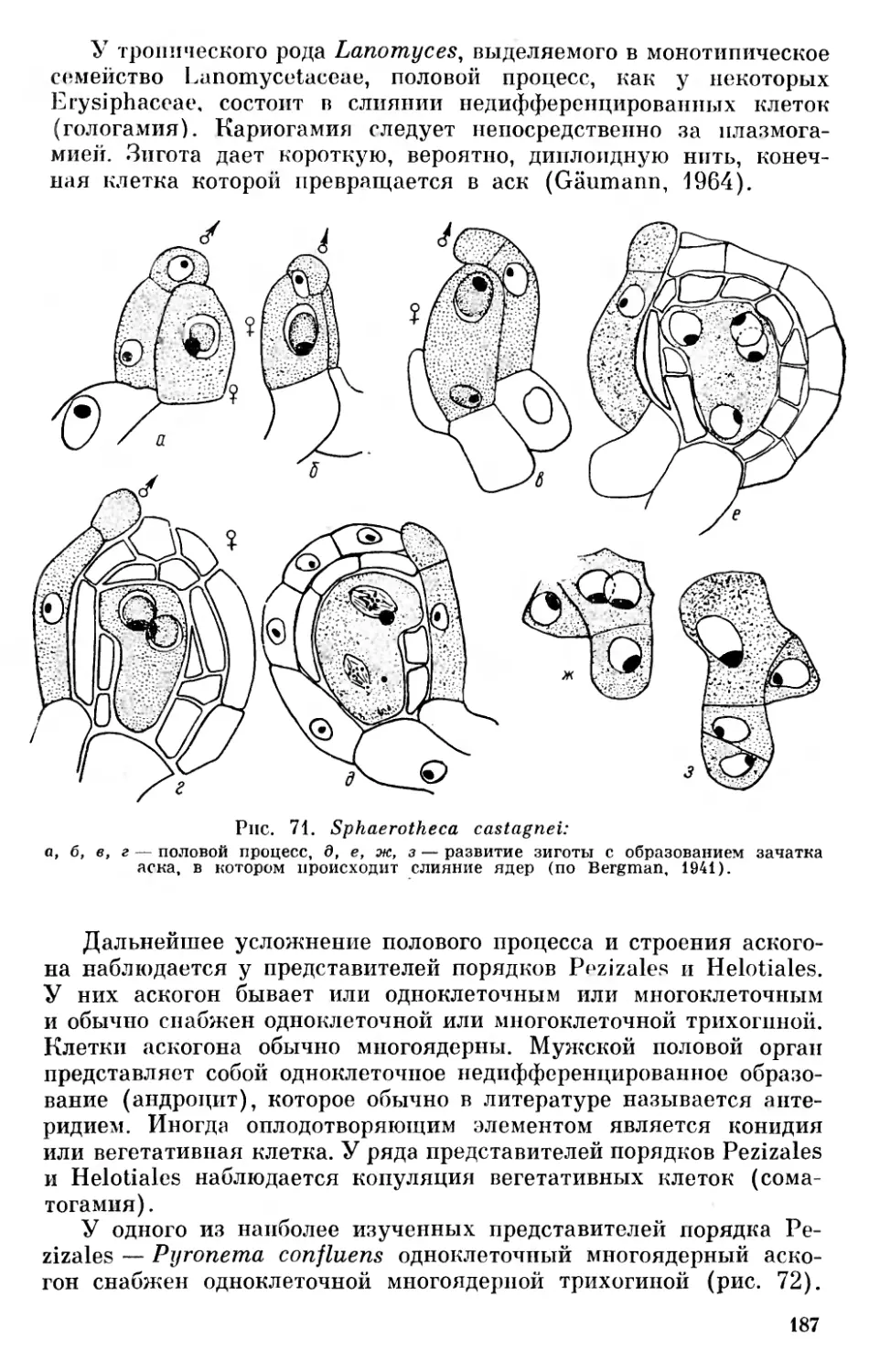
На основании современного уровня знаний о строении, разви тии и физиолого-биохимических свойствах эвкариотических организмов мы считаем наиболее обоснованным взгляды Мережковского и Пашера, согласно которым эвкариотические растительные организмы развились несколькими независимыми филами из первичных ядерных форм.
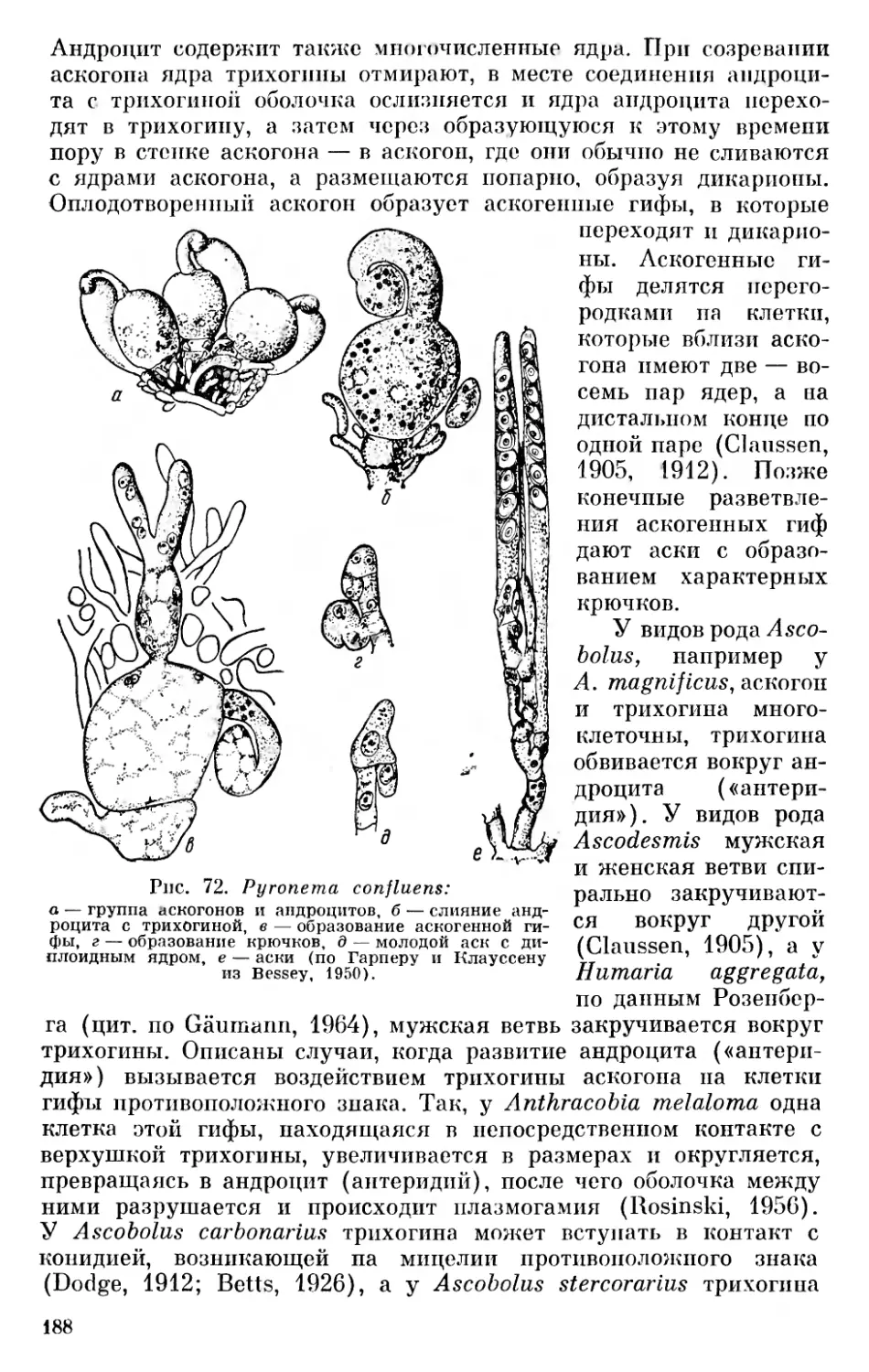
Ныне существующие организмы мы предлагаем разделять на три основные группы, «царства»: 1) неклеточные организмы — Acytobionta, к которым относятся вирусы, 2) первично-ядерные организмы — Procaryota (Protocaryota) — синезеленые водоросли и бактерии и 3) настоящие ядерные организмы — E и- caryota, которые делятся на ряд фил, относимых к животному и растительному миру. Автотрофные растительные организмы мы делим на десять фил (отделов), а именно: желтозеленые водоросли — Xanthophyta, золотистые водоросли — Chrysophyta, диатомовые водоросли — Diatomophyta, бурые водоросли — Phaeophyta, пиррофитовые водоросли — Pyrrhophyta, криптофитовые водоросли — Cryptophyta, эвгленовые водоросли — Euglenophyta, хлоромо- надовые — Chloromonadophyta, красные водоросли — Rhodophyta и зеленые растения (от зеленых водорослей до покрытосеменных) — Chlorophyta. Гетеротрофные растительные организмы (грибы) мы предлагаем делить на четыре филы (отдела): слизевики — Myxomycota, хитридиевые — Chytridiomycota, сапролегние- вые — Saprolegniomycota и настоящие грибы — Eumycota. Филу (отдел) зеленых растений мы предлагаем делить на подотделы: зеленые водоросли — Chlorophycophytina, мохообразные — Вгуо- phytina, антоцеротовые — Anthocerotophytina, псилофитовые — Psi- lophytina, плауновидные — Lepidophytina, клинолистые — Spheno- phytina (Calamophytina), папоротникообразные — Pterophytina (Filicophytina), голосеменные — Gymnospermophytina и покрытосеменные — Angiospermophytina 1 (см. рис. 2).
Обоснования для выделяемых фил (отделов) растительного мира будут более полно освещены в дальнейших главах работы.
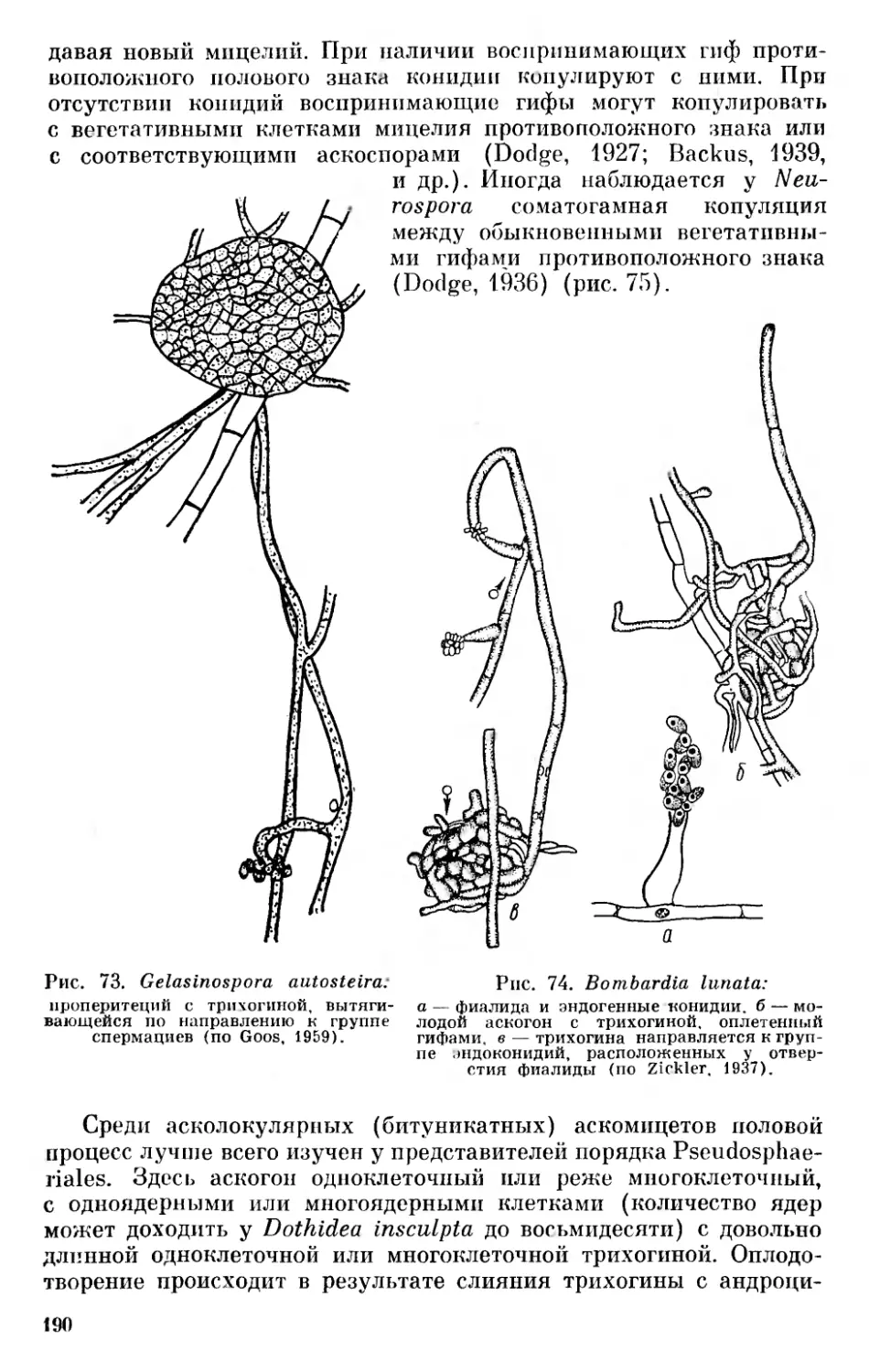
1 Возможно, количество подотделов сосудистых Chlorophyta нужно увеличить за счет выделения подотдела Tmesophytina и разделения подо.тдела Gymnospermophytina (Gymnospermae) на два — Cycadophytina и Conifero- phytina.
Глава TI
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
И ОСНОВЫ КЛАССИФИКАЦИИ НИЗШИХ АВТОТРОФНЫХ РАСТИТЕЛЬНЫХ ОРГАНИЗМОВ (ВОДОРОСЛЕЙ)
1. ОСНОВЫ КЛАССИФИКАЦИИ ВОДОРОСЛЕЙ
Термин «водоросли» (Algae) встречаем уже в работе К. Линнея (1754, 1770), однако в эту группу он включал, кроме небольшого количества родов настоящих водорослей, ряд родов печеночников и даже род Lichen. Первая трактовка указанной группы организмов в современном понимании, согласно Смиту (Smith, 1951, стр. 13), принадлежит Жюссье (de Jussieu, 1789). Позже Унгер (Unger, 1838) талломные организмы—водоросли, лишайники и грибы объединил в группу Thallophyta, которая в ранге группы или отдела растительного мира нашла себе место в ряде систем второй половины XIX ст. (Sachs, 1868, 1873; Eichler, 1883, и др.). В пределах этой группы водоросли (Algae) рассматривали как класс. Однако еще в первой половине XIX ст. среди водорослей были отмечены различные резко отличающиеся ветви — серии. По свидетельству Фотта (Fott, 1965), Ламуру, классифицируя морские Thalassiophyta, различал зеленые, бурые и красные водоросли (Lamouroux, 1813). Гарвей (Harvey, 1836) выделял среди водорослей три подкласса — зеленые (Chlorospermeae), бурые (Melanospermeae) и красные (Rhodospermeae). Синезеленые водоросли Гарвей включал в Chlorospermeae. Однако вскоре они были выделены в отдельную группу под названием Myxophyceae (Stizenberger, 1860) или Cyanophyceeae (Sachs, 1874).
Автотрофные флагеллаты вначале без колебания относили к животным организмам в связи с подвижностью в вегетативном состоянии, но уже с середины прошлого столетия была отмечена их тесная связь с водорослями. Так, Рабенгорст (Rabenhorst, 1863, 1865) отнес вольвоксовые к зеленым водорослям, а позже Энглер (Engler, 1898) включил все жгутиковые (Flagellatae) в систему растительного мира как подотдел отдела Euthallophyta.
Энглер в 1898 г. различал четыре отдела растительного мира, а именно: Myxothallophyta, Euthallophyta, Embryophyta zoidiogama (Archegoniatae) и Embryophyta siphonogama. Второй отдел — Euthallophyta он разделил на четыре подотдела: 1) Schizophyta (дробянки, т. е. бактерии и синезеле¬
29
ные водоросли), 2) Flagellatae (жгутиковые), которые разделил на три класса — Achromatoflagellatae, Chloroflagellatae и Phaeoflagellatae, 3) Eu- phyceae (настоящие водоросли) с восемью классами — Peridiniales, Bacilla- riales, Conjugatae, Chlorophyceae, Charales, Phaeophyceae, Dictyotales, Rhodophyceae, и 4) Eumycetes (Fungi — грибы). В других работах того же времени Энглер дает несколько иное подразделение отдела Euthallophyta.
Позже Энглер (Engler, 1912) распределяет водоросли между отделами — Schizophyta (класс Schizophyceae), Flagellatae, Dinof- lagellatae, Bacillariophyta, Conjugatae, Chlorophyceae, Charophyta, Phaeophyceae, Rhodophyceae и со знаком вопроса Silicoflagellatae.
Веттштейн (Wettstein, 1901) выделял восемь классов водорослей: Schizophyceae (синезеленые), которые относил к отделу
Schizophyta, Peridineae (перидинеи), Bacillarieae (диатомовые) и Conjugatae (конъюгаты), объединенные им в отдел Zygophyta, Chlorophyceae (зеленые водоросли), которые вместе с грибами он относил к отделу Euthallophyta, Phaeophyceae (бурые водоросли) — отдел Phaeophyta и, наконец, два класса красных водорослей (Rhodophyceae) — Bangiales (бангиевые) и Florideae (флори- деи). Веттштейн часть флагеллат (Peridineae) рассматривает как класс водорослей, а другие жгутиковые — хризофиты, эвглено- вые, криптомонады, подвижные в вегетативном состоянии желто- зеленые — не включает в систему растительного мира.
Борзи (Borzi, 1889, 1895) выделил из зеленых водорослей порядок Confervales. Болин (Bohlin, 1897, 1901) уточнил содержание этого порядка, а Лютер (Luther, 1899) предложил выделить их в отдельный класс под названием Heterocontae на основе их пигментов, продуктов ассимиляции и характера жгутиков.
В конце XIX ст. Клебс (Klebs, 1892, 1893) отметил гетерогенность группы флагеллат и наметил среди них ряд линий, ведущих к водорослям.
Ольтманнс (Oltmanns, 1904) совершенно ликвидирует класс флагеллат, разбивая его на четыре отдельные группы (классы) — Chrysomonadineae, Cryptomonadineae, Euglenaceae и Dinoflagellata, а конъюгаты и диатомовые объединяет в единый класс Acontae, взявши это название у Блекмана и Тенсли (Blackman a. Tansley, 1902), которые его применяли только для конъюгат. Ольтманнс различал девять групп (классов) водорослей, а именно: Chrysomonadineae, Heterocontae, Cryptomonadineae, Euglenaceae, Dinoflagellata, Acontae, Chlorophyceae с придатком Charales, Phaeophyceae и Rhodophyceae с придатком Bangiales. Включая классы флагеллат в систему водорослей, Ольтманнс подчеркивает их филогенетические связи. Мережковский (1909) также выводил все группы водорослей, которых он различал девять (Euglenaceae, Peridineae, Diatomaceae, Phaeophyceae, Chlorophyceae, Rhodophyceae, Heterocontae, Oedogoniaceae и Botrydiaceae), из флагеллат, относимых им к животному царству. Однако полную ясность в понимание отношений между флагеллатами и водорослями внес Пашер (Pascher, 1914, 1931), установивший тесную филогенети¬
30
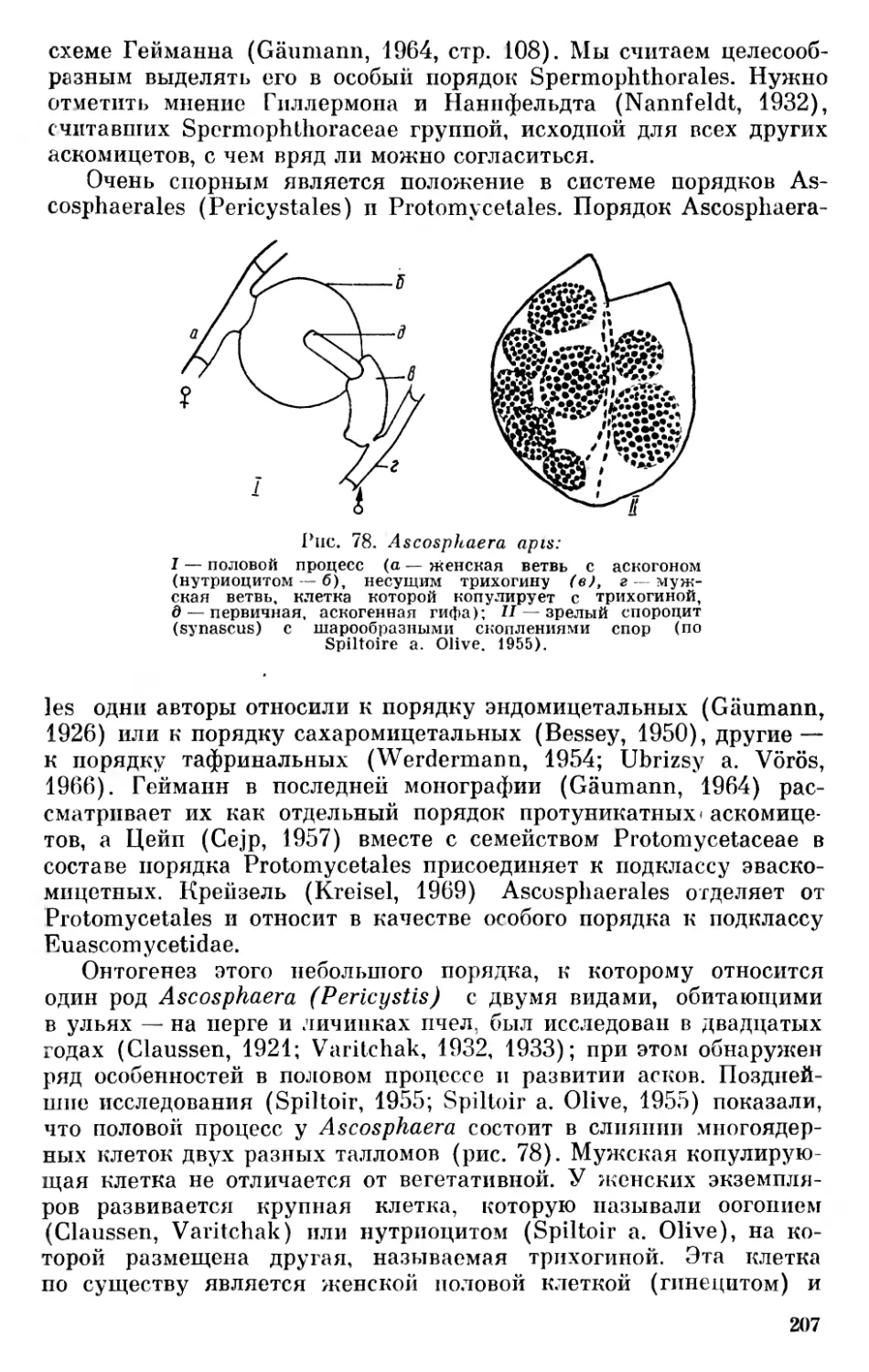
ческую связь между различными группами флагеллат и водорослей. Пашер дал систему водорослей, которая с незначительными изменениями принимается как филогенетическая до настоящего времени. На основании строения клеток, жгутиков и биохимических особенностей он выделил среди эвкариотических растительных организмов, названных им Plantae euplastideae, такие филы (Stämme) водорослей: Chrysophyta (золотистые с тремя отделами— Chrysophyceae, Diatomeae и Heterokontae), Phaeophyta (бурые), Pyrrhophyta (пиррофитовые с тремя отделами — Cryptophy- сеае, Desmocontae, Dinophyceae), Euglenophyta (эвгленовые), Chlorophyta (зеленые с двумя отделами — Chlorophyceae и Conjugatae), Charophyceae (харовые), Rhodophyta (красные). Сине- зеленые водоросли он рассматривает как филу (ствол) Plantae holoplastideae. Филы Пашера с включением Charophyta в состав отдела зеленых водорослей (Chlorophyta) принимают как отделы растительного мира Смит (Smith, 1938, 1951, 1955), Фотт (Fott, 1956, 1959) и ряд других авторов.
Несколько иные позиции в классификации водорослей занимает Фрич (Fritsch, 1935), который разделяет их на следующие одиннадцать классов: Chlorophyceae (Isocontae), Xanthophyceae (Heterocontae ), Chrysophyceae, Bacillariophyceae, Cryptophyceae, Dinophyceae (Peridinieae), Chloromonadineae, Euglenineae, Phae- ophyceae, Rhodophyceae и Myxophyceae (Cyanophyceae). Фрич исходит из позиций Пашера, только разбивает его группы Pyrrhophyta и Chrysophyta, включает харовые в состав зеленых водорослей и прибавляет класс Chloromonadineae.
В отличие от системы Пашера и Фрича, Шадефо (Chadefaud,
1960) считает возможным выделять среди эвкариотических водорослей только три крупных отдела — Rhodophycophyta, Chromo- phycophyta, включая сюда классы Cryptophyceae, Dinophyceae, Euglenophyceae, Chrysophyceae, Xanthophyceae, Bacillariophyceae, Silicophyceae, Craspedophyceae, Phaeophyceae и Chlorophycophyta с классами (подразделениями) Zygophyceae, Euchlorophyceae, Charophyceae.
Фотт, который вначале (Fott, 1956, 1959) придерживался системы Пашера, позже (Fott, 1965) разделил автотрофные эвка- риотические организмы, следуя Шадефо, на три отдела: Chromo- phyta с пятью классами — Chrysophyceae, Xanthophyceae, Bacillariophyceae, Phaeophyceae, Dinophyceae, Rhodophyta — с одним классом Rhodophyceae и Chlorophyta, куда он включил все зеленые растения от водорослей до покрытосеменных, с девятью классами, из которых три принадлежат к водорослям, а именно: Chlorophyceae, Conjugatophyceae и Charophyceae. В отличие от Шадефо, Фотт эвгленовые, криптофиты и хлоромонадовые, имеющие только монадную форму роста, выделяет в группу «остальных жгутиковых» (residual flagellates). Несколько позже он (Fott, 1967) называет их «жгутиковыми неопределенного систематического положения» (bicikovci neurcitého systematického zafazeni).
31
Чапмен (Chapman, 1964) делит водоросли на четыре отдела: Euphycophyta с классами Charophyceae, Chlorophyceae, Phaeophy- сеае, Rhodophyceae, Myxophycophyta с одним классом Myxophy-
Рис. 3. Типы жгутиков автотрофных низших организмов. Heterochrysophyceae: 1 — Ochromonas, 2 — Synura, 3 — Mallomo- nas; Isochrysophyceae: 4 — Chrysochromulina; Xanthophyceae:
5 — Vaucheria (антерозоид), 6—Vaucheria (зооспора): Phaeophy- ta: 7 — зооспоры и гаметы; Pyrrhophyta: 8 — Gyrodinium\ Cry- ptophyta: 9— Chilomonas, 10 — Cryptomonas; Euglenophyta: 11 — Euglena; Prasinophyceae: 12 — Heteromastix, 13 — Micromonas
squamata, 14 — Platymonas; организмы неопределенного систематического положения: 15 — Pedinomonas, 16 — Monomastix,
17 — Haematococcus; Euchlorophyceae: 18 — Chlamydomonas, 19 — Draparnaldia.
ceae, Chrysophycophyta с классами Chrysophyceae, Xanthophyceae, Baciliariophyceae и Pyrrophycophyta с классами Cryptophyceae и Dinophyceae.
Исходя из современного состояния знаний о строении клетки, жгутикового аппарата (рис. 3), способах размножения, биохимических особенностях растительных организмов, мы различаем среди автотрофных эвкариотических организмов десять основных стволов эволюции — фил, которые при классификации рассматриваем как отделы: Xanthophyta, Chrysophyta, Diatomophyta, Phae- ophyta, Pyrrhophyta, Cryptophyta, Euglenophyta, Chloromonado-
32
phyta, Rhodophyta и Chlorophyta. Все они, кроме последнего, включают талломные формы — то, что обычно называют водорослями, только фила (отдел) Chlorophyta включает и талломные, и сосудистые листостебельные растения. Зеленые водоросли мы выделяем как подотдел Chlorophycophytina в составе отдела зеленых растений — Chlorophyta.
2. ЗОЛОТИСТЫЕ (ХРИЗОФИТОВЫЕ) ВОДОРОСЛИ — CHRYSOPHYTA
(CHRYSOPHYCOPHYTA, CHRYSOPHYCEAE)
Золотистые (хризофитовые) водоросли являются своеобразной группой преимущественно монадных одноклеточных или колониальных организмов. Только сравнительно небольшое число форм характеризуется коккальной, ризоподи- альной (амебоидной), пальмеллевидной и нитчатой структурой.
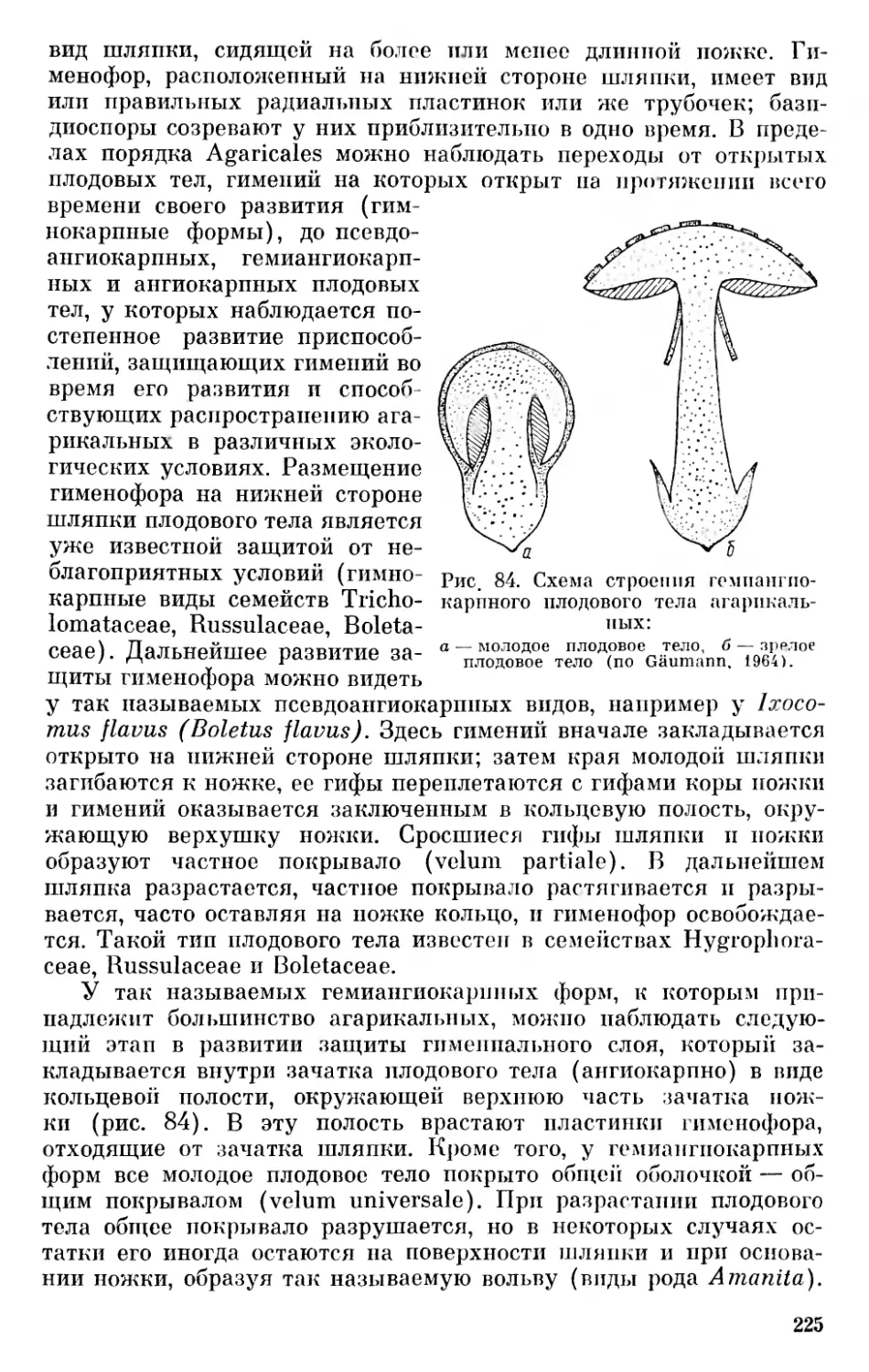
Клетки подвижных золотистых водорослей голые, покрыты только более или менее плотной пелликулой, иногда способны к метаболизму и образованию псев- доподиев. У многих золотистых водорослей клетки помещаются в своеобразных вместилищах — футлярах или, как их иногда называют, «домиках», или же покрыты панцирем. Только у неподвижных коккальных и нитчатых форм имеется вторичная клеточная оболочка.
В клетках большинства золотистых водорослей находится один или два, редко больше, постенных хроматофоров. В хро- матофорах у некоторых родов обнаружены пиреноидообразные тельца. У золотистых водорослей обнаружены такие пигменты: хлорофилл а, ß-каротин и ксантофиллы — лютеин и фукоксантин. Запасными продуктами являются углевод лейкозин и масло. Кроме автотрофных, существуют гетеротрофные формы. По-видимому, все хризофитовые одноядерные организмы.
Подвижные формы золотистых водорослей снабжены одним- двумя, редко четырьмя жгутиками равной или неравной длины. Если жгутик один, как например у хромулины, он имеет перистое строение, если два — один из них обычно перистый, другой — бичевидный (Оchromonas, Synura). У некоторых родов (Chry- sochromulina) оба жгутика бичевидные и равной длины (Bourrel- 1у, 1961). Некоторые хризофитовые из семейства Prymnesiaceae —
Рис. 4. Dinobryon borgei.
Гологамный половой процесс (по Skuja, i960).
33
Chryso ehr о mulina и другие, кроме жгутиков, имеют также так называемую гаптонему — жгутикообразное приспособление, служащее для прикрепления к субстрату.
Характерной особенностью хризофитовых является образование цист округлой или овальной формы с оболочкой, состоящей вначале из целлюлозы, а позже пропитывающейся окисью кремния. Цисты снабжены отверстием, закрытым пробкой. Однокле-
Рис. 5. Rephyriopsis cincta:
1 — вегетативная клетка, 2, 3 — копуляция и образование цисты (по Fott, 1959).
точные хризофитовые размножаются продольным делением; колониальные формы — делением колоний или же размножением клеток, освобождающихся из колонии. Неподвижные формы образуют зооспоры.
Половой процесс у хризофитовых происходит по мерогамному и гологамному типу \ Например, у Ochrosphaera neapolitana в клетке образуется четыре — шесть гамет, которые копулируют по изогамному типу (Sclnvarz, 1932). У Dinobryon sertularia клетка делится на две гаметы (Schiller, 1926), а у Dinobryon borget (рис. 4) и Rephyriopsis cincta (рис. 5) копулируют вегетативные клетки — гологамный тип (Skuja, 1950; Fott, 1959). Аналогичный процесс описан у Chrysolikos planktonicus (Mack, 1951) и у Ке- phyrion spirale (Lackey, 1938; Mack, 1953). Вероятно, половой процесс у хризофитовых нередок, на что обратил внимание еще Коршиков (1929) и другие, описавшие у ряда видов двуядерные цисты. 11 Следуя Гейманну (Gäuinann, 1926, стр. 10—11), Коршикову (1938) и Моро (Moreau, 1963), мерогамией (от греческих слов μερο — часть и
γάμος — брак, супружество), или гаметогамией, мы называем слияние клеток (гамет), возникших в результате деления содержимого материнской клетки сразу или вскоре после их освобождения из нее; а гологамией (от
греческих слов ολος — весь, целый и γάμος — брак, супружество) — слияние достигших полного развития клеток' — голоцитов (от греческих слов
ολος — весь, целый и κύτος — полость, вместилище, здесь переносно — клетка).
34
Что касается классификации золотистых водорослей, то Пашер (Pascher, 1931) рассматривал их как отдел Chrysophyceae филы (ствола) Chrysophyta наравне с диатомовыми и разножгутиковыми. Ряд последующих авторов принял это деление, низведя названные три группы до ранга класса (Krieger, 1954; Smith, 1955; Fott, 1959). Фрич (Fritsch, 1935, 1956) рассматривает золотистые водоросли как самостоятельный класс, равноценный отделу других авторов. Матвиенко (1965) рассматривает золотистые как самостоятельный отдел под названием Chrysophyta.
Пашер отдел Chrysophyceae разбил на пять классов: 1) Chry- somonadineae с тремя подклассами — Chrysomonadeae, Silicoflagei- latae и Coccolithophoreae, 2) Rhizochrysidineae, 3) Chrysocapsineae, 4) Chrysosphaerineae и 5) Chrysotrichineae.
Систему Пашера с некоторыми несущественными изменениями принимают почти все современные авторы, снижая его классы обычно до ранга порядков (Krieger, 1954; Smith, 1955; Fott, 1959, 1967) или же оставляя их в ранге классов (Fott, 1956; Матв1енко, 1954, 1965). Фрич (1935, 1956) классы Пашера Rhizochrysidineae и Chrysocapsineae включает в качестве подпорядков в порядок Chrysomonadales К
Основные подразделения хризофитовых, как и других отделов водорослей, Пашер построил на их морфологической структуре, считая основной, исходной группой Chrysomonadineae, из которых он выводил амебоидные Rhizochrysidineae, пальмеллоидные Chrysocapsineae, коккальные Chrysosphaerineae и нитчатые Chrysotrichineae. Пашер считал, что все прочие порядки развились из жгутиковых хризомонадовых, причем наиболее высокоорганизованными среди них являются нитчатые Chrysotrichineae.
Матвиенко (1954, 1965), принимая классификацию Пашера, в основании системы хризофитовых ставит как исходную группу Rhizochrysidineae (Chrysopodinophyceae).
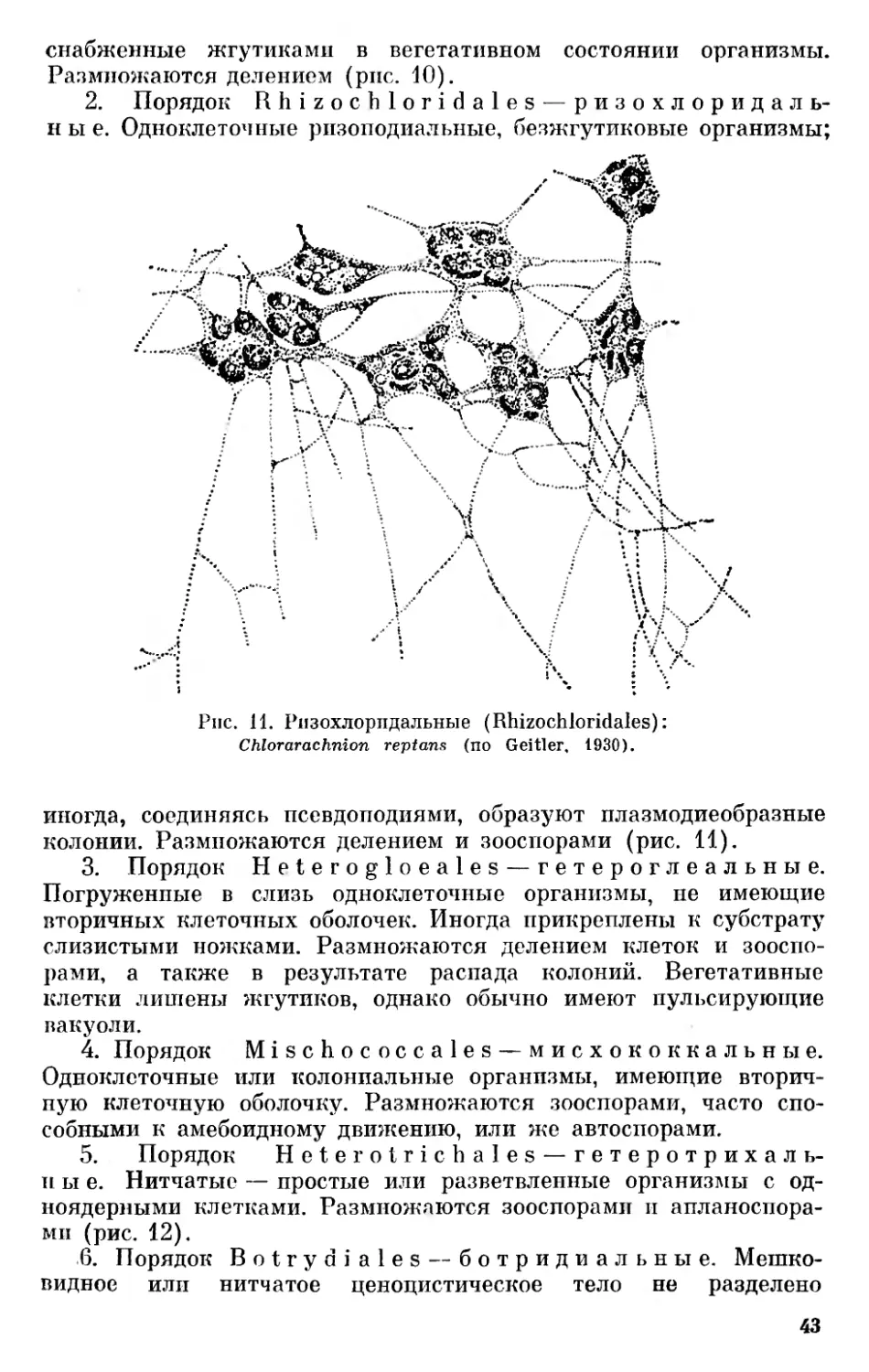
Против мнения Матвиенко можно выдвинуть следующие возражения: 1) некоторые ризохризидальные, например Myxochrysis paradoxa, образуют зооспоры, напоминающие хромулину; 2) псевдоподии ризохризидальных имеют сравнительно сложное и специализированное строение,— у них различают, кроме простых лобоподий, нитчатые ризоподии и стабильные, снабженные осевой нитью, аксоподии, что также не может свидетельствовать о примитивности этого порядка.
Наряду с классификациями золотистых водорослей, построенными на принципах Пашера, было предложено выделять хризо- фитовые водоросли с двумя бичевидными жгутиками и часто с гаптонемой между ними в особый класс (Parke, 1961), который Христенсен (Christensen, 1962, 1964) назвал Haptophyceae. 11 Фотт в последних изданиях своего руководства (Fott, 1959, 1967) делит Chrysophyceae па порядки Chrysomonadales, Rhizochrysidales, Chryso- capsales, Chrysosphaerales, Phaeotramniales.
35
Позже Бурелли (Bourrelly, 1964) дал несколько иную классификацию хризофитовых, в основу которой также положен принцип строения жгутиков. Он выделил три подкласса.
I. Acontochrysophycidae ( «провизорный под¬
класс» ) — безжгутиковые формы, без зооспор или с апланоспора- ми («зооспоры без жгутиков») с четырьмя порядками — Rhizoch- rysidales, Phaeoplacales, Stichogloeales, Chrysosaccales.
II. Heterochrysophyoidae — формы с одним жгутиком или с двумя жгутиками разной структуры, с двумя порядками — Chromulinales (с тремя подпорядками — Chromiilineae, Thallochry- sineae и Chrysosphaerineae) и Ochromonadales (с тремя подпорядками — Ochromonadineae, Phaeothamnionineae и Chrysa- pionineae).
III. I s о c h г y s о p h y c i d a e — формы с двумя жгутиками одинаковой структуры, обладающие иногда также гаптоиемой, с двумя порядками — Isochrysidales и Prymnesiales. Эту же классификацию приводит и Прескотт (Prescott, 1969).
По поводу классификации Бурелли можно заметить, что отсутствие жгутиков может быть вторичным, поэтому выделение провизорного подкласса Acontochrysophycidae нам кажется мало обоснованным. Кроме того, некоторые представители семейства Rhizochrysidaceae, отнесенного Бурелли к Acontochrysophycidae, имеют жгутики, например виды Chrysamoeba. Выделенный Бурелли подкласс Isochrysophycidae заслуживает особого внимания, так как у относящихся к нему родов, исследованных под электронным микроскопом, жгутики бичевидные, а у представителей семейства Prymnesiaceae, кроме того, имеется гаптонема. Поэтому нам кажется целесообразным разбить отдел Chrysophycophyta на два класса: 1) Heterochrysophyceae, к которому должны быть отнесены организмы с одним перистым жгутиком или с двумя жгутиками — одним перистым и другим бичевидным, и 2) Iso- chrysophyceae (Haptophyceae) — формы с двумя бичевидными жгутиками, а у некоторых (Prymnesiaceae) также и с гаптоне- мой К
В пределах класса Heterochrysophyceae, следуя Пашеру, с изменениями, предложенными Кригером (Krieger, 1954) и Фоттом (Fott, 1959), мы будем различать семь порядков — Chrysomonada- les, Coccolithales, Dictyochales (Silicoflagellatae), Rhizochrysidales, Chrysocapsales, Chrysosphaerales и Phaeothamniales (Chrysotricha- les), а в пределах класса Isochrysophyceae, согласно Бурелли (Bourrelly, 1964), два порядка — Isochrysidales и Prymnesiales. 11 Название Haptophyceae, предложенное Христенсеном, нам кажется менее удачным, так как не все Isochrysophyceae имеют гаптонему.
36
Отдел CHRYSO PH Y T A (CHRYSO PHYCO PHYT A,
CHR YSOPH YCEAE).
I. Класс HETEROCHRYSOPHYCEAE — ГЕТЕРОХРИЗОФИ- ТОВЫЕ.
Жгутиков один или два. Один жгутик перистый, другой, если он есть, бичевидный. Монадпые — одноклеточные и колониальные, ризоподиальные, пальмеллоидные, коккальные и нитчатые организмы. Семь порядков.
1. Порядок Clirysomonadales—-хризомонадаль- н ы е. Одноклеточные или колониальные организмы, снабженные
Рис. 6. Хризомоиадальные (Clirysomonadales) :
1 — Chromulina verrucosa; , 2 — Chromulina nebulosa; 3 — Chrysopsis agilis (no Fott, 1959).
одним или двумя жгутиками. Клетки или голые, часто метаболические, способные поглощать твердую пищу, или же снабженные футлярами («домиками»), или слизистыми, содержащими пектин, обвертками, покрытыми кремнистыми пластинками, снабженными различными скульптурными образованиями (рис. 6).
2. Порядок Coccolithales — к о к к о л и т а л ь п ы е, известковые хризофитовые. Одноклеточные монадные, преимущественно морские формы. Клетки покрыты слизистой обверткой, на которой и в которой откладываются своеобразные известковые тельца, так называемые кокколиты. Со временем слизистая обвертка превращается в панцирь. Клетка обычно с двумя хрома- т о фора ми и двумя жгутиками.
3. Порядок Dictyochales ( Silicoflagellatae) — д и к т и- охальные, кремнистые хризофитовые. Одноклеточные монадные морские организмы, в теле которых находится скелет,
37
состоящий из полых кремневых палочек. Клетки голые, имеют один жгутик, образуют также очень тонкие псевдоподии и снабжены мелкими хроматофорами. Диктиохальные были очень распространены в меловое и третичное время, известен ныне живущий один род Dicty- ocha с тремя видами (рис. 7).
С Dictyochales прежде увязывали небольшую группу гетеротрофных Ebriales с двумя родами, которые имеют также кремневый скелет. Их протопласт не образует псевдоподий, несет два жгутика. Скелет у них состоит из сплошных палочек, ядро по строению напоминает ядро перидиней. Структура скелета напоминает радиолярий. В то же время наличие у ископаемых Ebriales кремнистых цист дает основание выводить их из хризомонадальных.
Рис. 7. Диктиохальные (Dictyochales):
1 — Dictyocha speculum (живая клетка с жгутиком и псевдоподиями), 2 — Dictiocha fibula (кремневый скелет — вид сверху и сбоку) (по Маршаллу и Де- фландру, из Fott, 1959).
4. Порядок R h i- zochrysidales — ризохризидальные. Ризоподиальные формы, снабженные жгутиками или лишенные их, иногда снабженные футляром (Rhizaster) или образующие плазмодии (Myxochrysis).
5. Порядок Chrysocap saies — хризокаисальные. Пальмеллоидные одноклеточные или колониальные организмы, в вегетативном состоянии лишенные жгутиков.
6. Порядок Chrysosphaerales — хризосферальные. Одноклеточные или колониальные организмы, клетки которых покрыты оболочкой. Неподвижны в вегетативном состоянии, однако в клетках, особенно при размножении, иногда образуются пульсирующие вакуоли и стигма.
7. Порядок Phaeothamniales (Chrysotrichales) — фео¬
тамниальные. Нитчатые или дисковидные колониальные организмы.
И. Класс ISOCHRYSOPHYCEAE (HAPTOPHYCEAE) - ИЗО- ХРИЗОФИТОВЫЕ.
Одноклеточные или колониальные морские организмы с двумя бичевидпыми жгутиками. Клетки свободно живущие или снабженные футлярами. Автотрофные и гетеротрофные организмы. Два порядка.
38
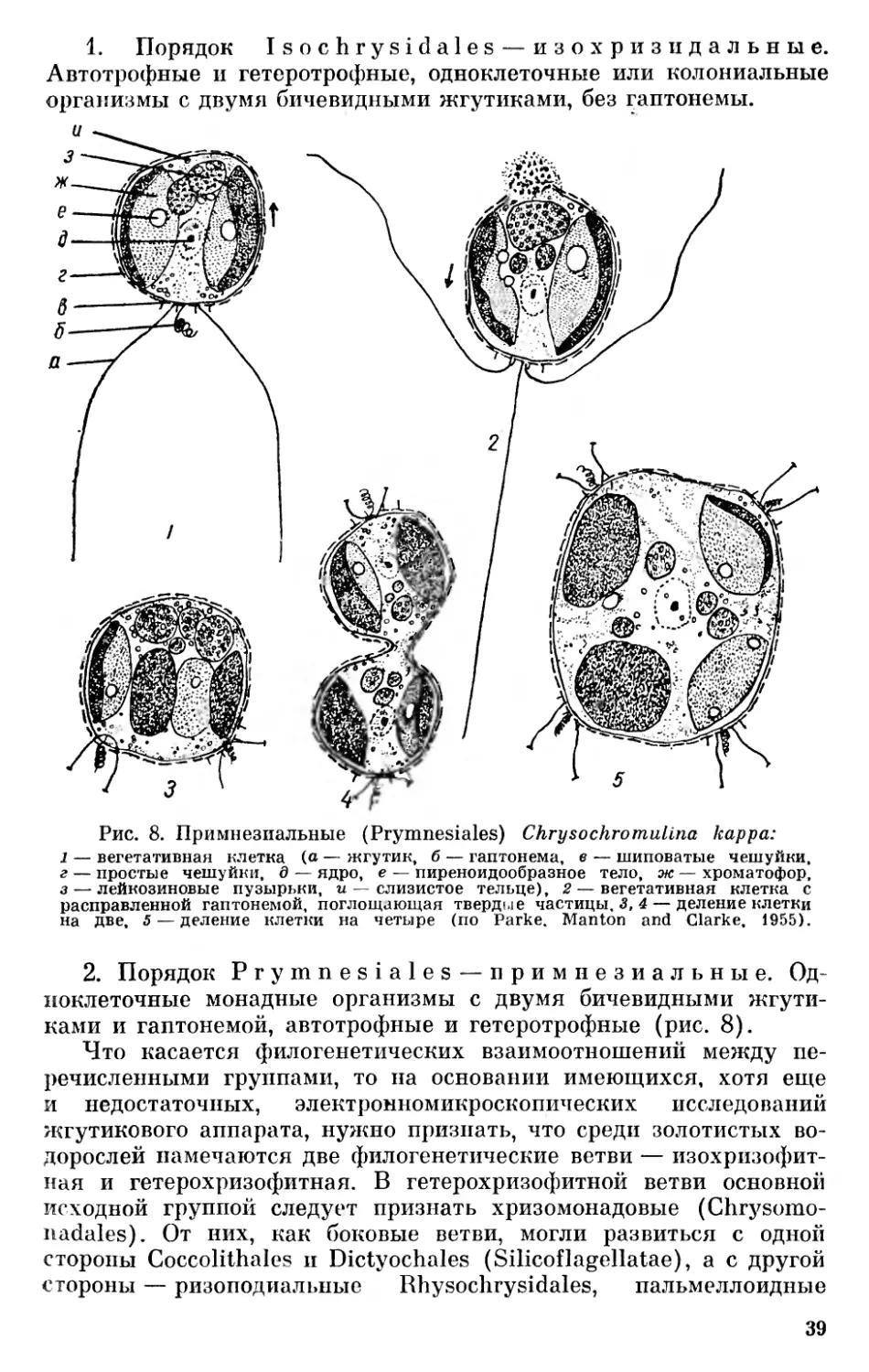
1. Порядок Isochrysidales — изохризидальные. Автотрофные и гетеротрофные, одноклеточные или колониальные организмы с двумя бичевидными жгутиками, без гаптонемы.
Рис. 8. Примнезиальные (Prymnesiales) Chrysochro mulina happa:
1 — вегетативная клетка (о — жгутик, б — гаптонема, в — шиповатые чешуйки, г — простые чешуйки, д — ядро, е — пиреноидообразное тело, ж—хроматофор, з — лейкозиновые пузырьки, и — слизистое тельце), 2 — вегетативная клетка с расправленной гаптонемой, поглощающая твердые частицы, 3,4 — деление клетки на две, 5 — деление клетки на четыре (по Parke, Manton and Clarke, 1955).
2. Порядок Prymnesiales — примнезиальные. Одноклеточные монадные организмы с двумя бичевидными жгутиками и гаптонемой, автотрофные и гетеротрофные (рис. 8).
Что касается филогенетических взаимоотношений между перечисленными группами, то на основании имеющихся, хотя еще и недостаточных, электронномикроскопических исследований жгутикового аппарата, нужно признать, что среди золотистых водорослей намечаются две филогенетические ветви — изохризофит- ная и гетерохризофитная. В гетерохризофитной ветви основной исходной группой следует признать хризомонадовые (Chrysomo- nadales). От них, как боковые ветви, могли развиться с одной стороны Coccolithales и Dictyochales (Silicoflagellatae), а с другой стороны — ризоподиальные Rhysochrysidales, пальмеллоидные
39
Chrysocapsales и коккальные Chrysosphaerales, которые в свою очередь могли дать начало нитчатым и пластинчатым Phaeotham- niales. Возможные взаимоотношения между порядками гетеро- хризофитных водорослей показаны на представленной схеме (рис. 9).
Рис. 9. Схема филогенетических связей между порядками класса Heterochrysophyceae.
Что же касается взаимоотношений классов Isochrysophyceae и Heterochrysophyceae, то они обособились, очевидно, на ранних стадиях становления отдела Chrysophyta.
О родственных связях золотистых водорослей с другими отделами сказано в разделе, посвященном желтозеленым водорослям —* Xanthophyta.
3. ЖЕЛТОЗЕЛЕНЫЕ, ИЛИ РАЗНОЖГУТИКОВЫЕ, ВОДОРОСЛИ — XANTHOPHYTA (XANTHOPHYCOPHYTA, XANTHOPHYCEAE, HETEROKONTAE)
Желтозеленые, или разножгутиковые, водоросли были выделены в отдельную группу под названием Heterokontae Лютером (Luther, 1899) из зеленых, к которым их до этого времени относили. Позже они получили название Xanthophyceae (Allorge, 1930). Это название соответствует современным правилам ботанической номенклатуры, однако оно не вполне удачно, так как вследствие преобладания в хроматофорах разножгутиковых хлорофилла они по цвету мало или совсем не отличимы от зеленых водорослей. Название «желтозеленые водоросли» (Yellow-Green Algae), которое впервые ввел, но Прингсгейму, Фрич (Fritsch, 1935, стр. 470), несмотря на свое несоответствие действительной окраске представителей этой группы, широко укоренилось, очевидно, ввиду отсутствия лучшего. Название «разножгутиковые водоросли», обоснованное как противопоставление «равножгутиковым», пока их включали в состав зеленых водорослей, потеряло свой смысл, когда их выделили в особый отдел, так как разные жгутики имеют и Chrysophyceae, и Phaeophyceae.
40
К желтозеленым водорослям относятся одноклеточные как без вторичной оболочки, так и имеющие ее, нитчатые и сифональные организмы. Среди них различают флагеллоидные, ризоподиальные, пальмеллоидные, коккоидные, нитчатые и сифональные формы.
Клетки желтозеленых водорослей обычно одноядерны, редко многоядерны; у сифональных форм (Botrydium, Vaucheria), тело которых не разделено на клетки, всегда имеется много ядер. В клетке находятся один-два или несколько постенных, обычно дисковидных хроматофоров; только у некоторых родов, например Chloromeson, хроматофоры осевые. В хроматофорах некоторых желтозеленых водорослей указано наличие пиреноидов, хотя крахмал в них не образуется (Korshikov, 1930). В хроматофорах обнаружены хлорофилл а, свойственный только этой группе организмов, хлорофилл е, ß-каротин и ксантофиллы, сведения о которых у разных авторов не совпадают. Запасным продуктом у желтозеленых водорослей является масло.
Флагеллоидные формы и зооспоры неподвижных форм снабжены двумя неравными жгутиками, из которых один обычно более длинный, перистого строения, а другой бичевидный. Иногда второй жгутик может редуцироваться. У зооспор вошерии оба жгутика каждой пары бичевидные, однако мужские гаметы вошерии имеют один перистый и один бичевидный жгутики (Bourrelly,
1961). У ряда форм обнаружены пульсирующие вакуоли.
Подвижные в вегетативном состоянии флагеллоидные формы (порядок Heterochloridales) не имеют вторичной оболочки и очень метаболичны,— они могут давать псевдоподии, которые служат как для передвижения, так и для захвата твердых частиц (голозойное питание). Близки к флагеллоидным ризоподиальные формы, в вегетативном состоянии лишенные жгутиков (порядок Rhizochlo- ridales). Некоторые ризоподиальные формы в результате объединения амебоидных клеток могут давать плазмодии, напоминающие плазмодии слизевиков (Myxochloris).
Голые одноклеточные желтозеленые могут давать пальмеллоидные формы (порядок Heterogloeales). Одноклеточные коккаль- ные формы (порядок Mischococcales) покрыты оболочкой так же, как нитчатые (Heterotrichales) и сифональные (Botrydiales).
Вторичная оболочка желтозеленых водорослей дает реакцию на пектин, и только оболочка вошерии состоит из целлюлозы. Во многих случаях, особенно в оболочках цист, обнаружен S1O2. Часто клеточная оболочка состоит из двух половинок, при этом протопласт может давать цилиндрические наперстковидные вставочные образования, благодаря которым клетка удлиняется.
Клетки делятся внутри материнской оболочки, причем каждая клетка покрывается новой оболочкой, не выходя из материнской, которая растягивается.
Флагеллоидные формы могут размножаться продольным делением в подвижном или пальмеллевидном состоянии. Коккальные, нитчатые и сифональные формы размножаются зооспорами или
М
апланоспорами. У некоторых желтозеленых водорослей образуются эндогенные цисты, оболочки которых состоят из двух неравных частей и характеризуются наличием в них кремнезема. У ботридия в конце вегетационного периода протопласт концентрируется
Рис. 10. Гетерохлоридальные (Heterochloridales).
Chloromeson agile: 1 — клетка с широкой стороны, 2 — клетка в амебоидном состоянии. 3 — деление клетки, 4 — начало образования споры, 5 — формирование споры, 6 — сформировавшаяся спора (по Pascher, 1932).
в ризоидах и распадается на многоядерные покрытые толстой оболочкой так называемые гипноспоры, при прорастании которых образуются многочисленные зооспоры.
Половой процесс у одноклеточных форм до сих пор не констатирован. Шарфель (Scherffel, 1901) описывает половой процесс у Tribonema. Он указывает, что гаметы у видов этого рода морфологически неразличимы. Однако некоторые из них («женские») теряют подвижность, округляются и оплодотворяются подвижными «мужскими». По данным Мевуса (Moewus, 1940), у Botrydium granulatum копулируют подвижные изо- или гетерогаметы. У видов Vaucheria хорошо известна типичная оогамия.
Общепринятой является система разножгутиковых, предложенная Пашером (Pascher, 1931), которую с модернизированными, согласно существующим правилам номенклатуры, названиями порядков (Pascher, 1939) приводим ниже.
Отдел XANTHOPHYTA (XANTHOPHYCOPHY- Т A, XANTHOPHYCEAE, HETERO CONTA E).
1. Порядок Heterochloridales — гетерохлорид- н ы е. Одноклеточные, лишенные вторичной клеточной оболочки,
снабженные жгутиками в вегетативном состоянии организмы. Размножаются делением (рис. 10).
2. Порядок Rhizochloridales — ризохлоридаль- н ы е. Одноклеточные ризоподиальные, безжгутиковые организмы;
Рис. И. Ризохлоридальные (Rhizochloridales): Chlorarachnion reptans (по Gei tier. 1930).
иногда, соединяясь псевдоподиями, образуют плазмодиеобразные колонии. Размножаются делением и зооспорами (рис. 11).
3. Порядок Heterogloeales — гетероглеальные. Погруженные в слизь одноклеточные организмы, не имеющие вторичных клеточных оболочек. Иногда прикреплены к субстрату слизистыми ножками. Размножаются делением клеток и зооспорами, а также в результате распада колоний. Вегетативные клетки лишены жгутиков, однако обычно имеют пульсирующие вакуоли.
4. Порядок Mischococcales — мисхококкальные. Одноклеточные или колониальные организмы, имеющие вторичную клеточную оболочку. Размножаются зооспорами, часто способными к амебоидному движению, или же автоспорами.
5. Порядок Heterotrichales — гетеротрихаль- н ы е. Нитчатые — простые или разветвленные организмы с одноядерными клетками. Размножаются зооспорами и апланоспора- ми (рис. 12).
6. Порядок Botrydiales — ботридиальньте. Мешковидное или нитчатое ценоцистическое тело не разделено
43
на клетки, с многочисленными ядрами. Бесполое размножение зооспорами, иногда апланоспорами. Половой процесс изогамный или оогамный.
Согласно взглядам Пашера, исходными формами среди желто- зеленых являются монадные, подвижные в вегетативном состоянии формы. От них произошли ризоподиальные, паль- меллоидные и коккальные формы. Из последних развились нитчатые и сифональ- ные формы. Некоторые авторы (Топачевський, 1962) исходными считают ризоподиальные формы. Однако нужно заметить, что ризоподиальные формы размножаются не только делением, но и зооспорами, что свидетельствует о возможности их происхождения от монадиых форм.
Ряд авторов обращает внимание на метаболичность монадных форм желтозеленых и их способность переходить в амебоидное — ризо- подиальное состояние. Амебоидные Rhizochloridales своим строением очень напоминают Rhizopoda и дают основание для предположения, что часть организмов, относимых к Rhizopoda, могла развиться из Rhizochloridales в результате утраты хромато- форов.
Мейер (1951), развивая взгляды Миллера (1928), выдвинул положение, что осевой хроматофор является более древним по сравнению с постенным как у зеленых водорослей, так и желтозеленых (разножгутиковых), однако никаких предложений относительно изменения системы Xanthophyta он не внес, ограничившись следующим утверждением: «можно полагать, что первичные организмы, давшие начало всем типам водорослей (за исключением синезеленых), обладали осевым хроматофором» (Мейер, 19516, стр. 54).
После работ Пашера о взаимоотношениях жгутиковых и водорослей (Pascher, 1914, 1931) утвердилось мнение о близости
Рис. 12. Гетеротрихальные (Heterotric- hales :
Tribone та (зооспоры и схема строения нити) (по Fott, 1959).
44
желтозеленых, золотистых и диатомовых водорослей. Это мнение принимается многими авторами до настоящего времени (см., например, Pringsheim, 1966). Пашер эти три группы рассматривает как родственные отделы и объединяет их в одну филу (ствол) под названием Chrysophyta. Особенное значение он придает образованию у представителей этих групп эндогенных цист, оболочки которых пропитаны кремнеземом (Pascher, 1914, 1931). Пашер обращает внимание также на то, что оболочки цист желтозеленых состоят из двух частей (створок), а цисты золотистых закрыты пробкой, которую он рассматривает как гомолог второй створки. Золотистые, желтозеленые и диатомовые водоросли не образуют крахмала; запасным продуктом у них является масло и лейкозин (хризоз, хризамилон). Для всех этих групп характерно отсутствие хлорофилла Ъ. Все это говорит в пользу предположения Пашера о родственности золотистых, желтозеленых и диатомовых водорослей, однако нельзя не отметить, что у них имеются и существенные отличия. Так, у золотистых обнаружен только хлорофилл а, у диатомовых, кроме того, имеется хлорофилл с, свойственный также бурым и пиррофитовым, а у желтозеленых обнаружен, кроме хлорофилла а, также и характерный только для этой группы хлорофилл е. Диатомовые резко отличаются от золотистых и желтозеленых циклом развития, в котором преобладает диплоидная фаза, а также своеобразным строением ядра с внешней центросомой.
Пашер, подчеркивая черты сходства между желтозелеными, золотистыми и диатомовыми водорослями, все же отмечает, что они представляют самостоятельные стволы развития (selbständige Typen), обнаруживающие между собой больше родственных связей, чем с другими отделами (Pascher, 1914, стр. 146). Поэтому мы, не отрицая общности происхождения по крайней мере желтозеленых и золотистых, считаем все же целесообразным рассматривать их как самостоятельные стволы развития — филы, а в систематическом отношении — как самостоятельные отделы автотрофных эвкариотических организмов.
4. ДИАТОМОВЫЕ ВОДОРОСЛИ - DTATOMOPHYTA (BACILLARIOPHYTA, BACILLARIOPHYCOPHYTA, BACILLARIOPHYCEAE, DIATOMEAE)
Диатомовые водоросли являются своеобразной группой одноклеточных или колониальных организмов, имеющих ряд характерных особенностей.
Клетки диатомовых одноядерны, покрыты двустворчатой кремнеземной оболочкой (панцирем), в вегетативном состоянии не имеют жгутиков, а представители класса пеннатных вообще лишены жгутиковых стадий. Хроматофоры в числе одного-двух или же многочисленны. На хроматофорах пеннатных диатомовых наблю¬
45
даются иногда ииреноидообразные тельца. Для диатомовых характерны следующие пигменты: хлорофилл а и с, ß-каротин, Е-каротин и ряд ксантофиллов — фукоксантин, неофукоксантин А и В, диатоксантин, диадиноксантин. Из них первые три являются общими с бурыми водорослями, фукоксантин обнаружен также у хризофитовых водорослей, диадиноксантин известен у пиррофито- вых и только диатоксантин специфичен для диатомовых. Запасным продуктом является масло, а также волютин и хризозы.
Клеточная оболочка диатомовых имеет характерное строение. Клетка диатомовых окружена пектиновой оболочкой, которая сверху покрыта кремнеземной, называемой панцирем, или текой (theca). Панцирь состоит из двух половинок, находящих одна на другую, как крышечка на коробку (эпитека и гипотека). Каждая половинка панциря, т. е. эпитека и гипотека, состоит из двух частей — створки и тесно с ней соединенного поискового ободка (пояскового кольца). У некоторых центрических диатомовых поисковый ободок отсутствует, и половинки панциря смыкаются краями загиба створки. У многих диатомовых наблюдается усложнение панциря — развитие между загибом створки и поисковым ободком дополнительных вставочных ободков (вставочных колец), которые могут нести на внутренней стороне более или менее развитые выступы, расположенные параллельно створке и образующие внутри клетки неполные перегородки (септы).
Створки панциря имеют сложное строение. На них различают точки и штрихи, состоящие из отдельных точек, а также так называемые ареолы, являющиеся камерами в стенках створок. По крайней мере часть точек представляет собой поры, сообщающие протопласт с внешней средой. Ареолы являются или замкнутыми полостями, или открываются наружу, или же внутрь клетки. У многих диатомовых панцирь снабжен ребрами, шипами, крыльями и другими выростами.
Для створок панциря большинства пеннатных диатомовых характерны так называемые швы и узелки. В типичном случае на створке бывает три утолщения — «узелка» — два полярных и один центральный. Шов представляет собой удлиненную щель, расположенную между центральным и одним из полярных узелков. У представителей некоторых порядков пеннатных швы имеются на обеих створках, у некоторых только на одной из них, а у некоторых — Fragilariales (Araphinales) шов вообще отсутствует и вдоль продольной оси створки проходит так называемое осевое (центральное) поле, называемое некоторыми авторами «ложным швом».
У представителей порядка Surirellales — каналошовных строение шва более сложное. Это так называемый каналовидный шов, имеющий форму канала, проходящего в створки и сообщающегося с внешней средой узкой щелью, а с полостью клетки расположенными на его внутренней стенке крупными порами (рис. 13).
46
Характерным для диатомовых является способ деления клеток, ведущий к их измельчению, и типы полового процесса, связанные с образованием ауксоспор. До сравнительно недавнего времени был хорошо изучен половой процесс только у пеннатных диатомовых, состоящий в типичных случаях в слиянии двух клеток, на основании которого авторы конца XIX и начала XX ст. сближали их с конъюгатами (рис. 14).
Перед половым процессом в клетке пеннатных диатомовых происходит редукционное деление и образуются четыре гаплоидных ядра. При этом обычно клетка делится на две; каждая клетка получает два ядра, из которых одно дегенерирует. Получившиеся таким образом гаплоидные клетки копулируют попарно с соседними клетками, в результате чего получаются две зиготы (ауксоспоры).
В некоторых случаях копулирующие клетки не делятся и тогда образуется одна зигота (ауксоспора). Однако и в этом случае ядро делится дважды, и из четырех гаплоидных ядер три отмирают. Таким образом, вегетативная клетка является диплоидной, а гаплоидны только копулирующие клетки. В некоторых случаях у пеннатных диатомовых описан автогамный половой процесс, при котором сливаются две дочерние клетки, образующиеся в результате деления вегетативной клетки (педогамия).
Сравнительно недавно описан оогамный половой процесс у пеннатной водоросли Rhabdonema adriaticum (Stosch, 19586) (рис. 15). По данным Стоша, содержимое клетки, функционирующей как оогоний, у Rhabdonema adriaticum делится на две части — богатую цитоплазмой и пластидами яйцеклетку и бедную пластидами, однако относительно крупную часть — «направляющее тело» (Richtungskörper). У другого исследованного тем же автором вида — Rhabdonema minutum вместо направляющего тела образуется вторая яйцеклетка. Сперматогонии образуются после двух — нескольких премейотических делений вегетативных клеток. В сперматогонии, как и в оогонии, происходит редукционное деление и возникают две микрогаметы, а также безъядерное остаточное тельце. Сперматогонии освобождаются из нитей, пассивно переносятся течениями и могут прикрепляться к оогонию. Микрогаметы могут передвигаться ползающими движениями к оогонию, прикрепляются к нему, и ядро микрогаметы проникает в яйцеклетку.
Половой процесс у центрических диатомовых изучен только в последние десятилетия. У них со второй половины XIX ст. известны были так называемые микроспоры, которые многие ис¬
Рис. 13. Surirella capronii:
поперечное сечение клетки с каналом-швом (а) на крыльях створки (по Лаутерборну из кн. «Диатомовый анализ», 1, 1949).
следователи, начиная с Грана (Gran, 1902), считали гаметами, хотя слияния их никто не наблюдал. На основании работы Кар- стена (Karsten, 1904) предполагали, что центрические диатомовые, в отличие от пеннатных, гаплоидные организмы, у которых копулируют подвижные гаметы ( микроспоры) и диплоидной фазой является только зигота.
Исследования Шмидта (Schmidt, 1927) показали, что у Biddul- phia sinensis микроспоры имеют но две хромосомы, в то время
Рис. 14. Rhopalodia gibba:
половой процесс и образование ауксоспор (по Клебану, Oltmanns, 1922).
как ядра вегетативных клеток содержат по четыре, что как будто бы подтвердило предположение, что микроспоры являются половыми клетками. Но эти же факты принудили Карстена признать, что клетки центрических диатомовых являются диплоидными, а гаплоидны у них только гаметы (Karsten, 1928, стр. 170). В то же время считали, что образование ауксоспор у центрических диатомовых происходит «без каких-либо следов полового процесса» (Kniep, 1928, стр. 69).
Персидский (Persidsky, 1929, 1935) исследовал развитие ауксоспор у Chaetoceras и Melosira и описал редукционное деление в молодых ауксоспорах этих водорослей. Из четырех ядер, получающихся при этом в ауксоспоре, по данным Персидского, два дегенерируют, а два сливаются, т. е. происходит автогамный половой процесс. Микроспоры, по мнению Персидского, являются зооспорами.
Дальнейшие исследования центрических диатомовых не подтвердили данных Персидского. В ряде работ Стоша (Stosch, 1950, 1951, 1954, 1956, 1958) доказан оогамный половой процесс у ряда родов центрических диатомовых — Melosira, Biddulphia, Lithodes- mium, Streptotheca и др. При этом мужскими гаметами являются микроспоры, а женскими — яйцеклетки. Редукционное деление происходит при образовании гамет.
48
Таким образом, центрические диатомовые, так же как и пен- натные, являются диплоидными организмами.
В настоящее время развитие гаметангиев, гамет и половой процесс у центрических диатомовых можно считать довольно хорошо исследованными. Клетки, превращающиеся в сперматогопии,
Рис. 15. Образование гамет и половой процес у Rhahdonema adriaticiim и видов Biddulphia:
1 — Rhabdonema adriaticum (a — яйцеклетка, б — направляющее тельце, в — сперматогонии, г — мужские гаметы), 2 — Biddulphia granulata (зрелый оогоний с двумя яйцеклетками), 3 — Biddulphia rhombus (зрелый оогоний с яйцеклеткой и направляющим тельцем), 4 — Biddulphia granulata (группы сперматогониев) (по Stosch, 1958).
могут не отличаться от вегетативных, например у рода Cyclotella, изученного Гейтлером (Geitler, 1952), или же перед формированием сперматогониев клетка проходит ряд премейотических делений и образуется группа сперматогониев, более или менее отличающихся от вегетативных клеток. Иногда все содержимое клетки — сперматогония делится на гаметы, иногда же часть содержимого не используется на образование микрогамет и остается в виде так называемого остаточного тельца. Микрогамета снабжена одним жгутиком.
49
Развитие яйцеклеток может проходить двояко. В одних случаях содержимое клетки делится на две равные части и образуется две яйцеклетки (Biddulphia granulata). В других случаях, например у Biddulphia rhombus, клетка делится на две неравные части; одна из них превращается в яйцеклетку, а другая дает дегенерирующее остаточное тельце (Stosch, 19586) (см. рис. 15). В некоторых случаях (Melosira, Cyclotella) протопласт не делится и формируется одна яйцеклетка (Steffen, 1955, стр. 68). После оплодотворения яйцеклетка увеличивается в размерах и превращается в ауксоспору (рис. 16).
* * *
На протяжении последних ста лет взгляды на положение диатомовых в системе растительного мира претерпели значительные изменения. Во второй половине XIX ст. на основании внешнего сходства полового процесса Кон (Cohn, 1872) объединил диатомовые конъюгаты в один порядок под названием Zygosporeae.
Сакс (Sachs, 1874) пошел дальше и к классу зигоспоровых (die Zygosporeen) отнес организмы, характеризующиеся конъюгацией неподвижных клеток, а именно конъюгаты, включая диатомовые, и зигомицеты.
Веттштейн (Wettstein, 1901), очевидно, под влиянием идей Кона, объединил в отделе Zygo- phyta конъюгаты, перидинеи и диатомовые. Наконец Ольтманнс (Oltmanns, 1904—1905) под общим названием Acontae (Zygophyceae) объединил конъюгаты и диатомовые. IIIютт (Schütt, 1896) считал возможным сближать диатомовые с перидинеями.
Изложенные взгляды могут иметь только историческое значение после того, как Пашер (Pascher, 1914) развил взгляды о флагеллатном происхождении различных водорослей. Однако нельзя не отметить, что Карстен в 1928 г. еще считал возможным говорить о близости диатомовых к конъюгатам (Karsten, 1928, стр. 200).
Пашер обратил внимание на сходство в строении оболочек клеток и цист диатомовых, золотистых и разножгутиковых водорослей, на наличие сходных продуктов ассимиляции и пришел к заключению, что эти группы представляют единую филу, ствол Chrysophyta, в составе которой он выделяет три отдела: золотистые водоросли — Chrysophyceae, диатомовые — Diatomeae и разножгутиковые — Heterocontae. Эти взгляды Пашера встретили широкое признание. В подтверждении взглядов Пашера Кортиков (Korshikov, 1930) приводит данные о наличии пульсирующих ва¬
Рис. 16. Μelosira varians:
1 — молодая ауксоспора со сливающимися ядрами (синка- рион), 2 — два антеридия и сперматозоиды (по Стошу из Fott, 1959).
50
куолей у некоторых диатомовых (Attheya zachariasii и Rhizosole- nia longiseta), a также указывает на наличие лейкозина у диатомовых. Несколько позже Пашер (Pascher, 1932) обнаружил сократительные вакуоли в вегетативных клетках Nitzschia.
Шадефо (Chadefaud, 1960) диатомовые рассматривает как один из классов подотдела Chrysophyceae (Lato sensu), который он разбивает на пять классов — Chrysophycineae, Xanthophycineae, Ва- cillariophycineae, Silicophycineae и Craspedophycineae (Choanofla- gellatae).
Некоторые авторы считают целесообразным рассматривать диатомовые как самостоятельный отдел, что по сути не противоречит взглядам Пашера.
В противоположность большинству отделов низших растительных организмов, благодаря кремнеземному панцирю, диатомовые хорошо сохраняются в ископаемом состоянии, что дает возможность проследить основные пути их эволюции на основании палеонтологических данных. Наиболее древние достоверные находки диатомовых относятся к нижней части юрского периода (лейас), из отложений которого описаны два вида рода Stephano pyxis (Со- scinodiscaceae) из класса центрических (Шешукова, 1949). В течение юрского и мелового периодов шло развитие центрических и только в верхнем мелу обнаружены первые представители с бисимметрическими створками, относящиеся к порядку Anaulales (Mediales) класса центрических. Переходные формы к пеннатным известны с эоцена. Возникновение пеннатных с настоящим швом относится также к палеогену — олигоцену (Жузе, 1949), а по некоторым данным, к палеоцену — олигоцену (Small, 1946; цпт. по Шешуковой, 1949, стр. 8). Широкое развитие пеннатных диатомовых происходит в неогене начиная с нижнего миоцена. Таким образом, филогенетически более древние центрические диатомовые.
Как уже упоминалось, древнейшим из описанных родов диатомовых является Stephanopyxis, виды которого имеют панцирь сферической, эллипсоидальной или цилиндрической формы, что подтверждает мысль Топачевского (1962, стр. 199) о первичности сферической формы у диатомовых. Правдоподобным является мнение, что панцири исходных форм диатомовых были без вставочных ободков (Шешукова, 1949) и характеризовались слабо развитой структурой с беспорядочным расположением пор (То- пачевський, 1962, стр. 198). В процессе дальнейшей эволюции центрических форм беспорядочное расположение пор заменяется радиальным или кольцевым, развиваются ареолы, в центре створки образуется бесструктурное центральное поле. В связи с приспособлением к планктонным условиям у центрических диатомовых вырабатываются разнообразные формы панциря и различной формы выросты.
С верхнего мела известны первые формы с билатеральной формой панциря — представители порядка Anaulales, который
51
Жузе и Прошкипа-Лавренко (1949) назвали Mediales в связи с промежуточным его положением между центрическими и пеннат- ными диатомовыми.
Характерной особенностью панциря пеннатных диатомовых является развитие на его створках шва. Среди пеннатных диатомовых различают формы бесшовные и имеющие щелевидный или каналовидный шов. При этом шов может быть только па одной или обеих створках. В зависимости от характера шва построены и принятые классификации диатомовых, из которых большое распространение имеет классификация Густедта (Hustedt, 1930), приведенная нами ниже.
A. Centrica е.
I. Discoideae.
И. Solenoideae.
III. Biddulphioideae.
IV. Rutilarioideae.
B. Pennatae.
1. Araphideae.
V. Fragilarioideae.
2. Raphidioideae.
VI. Eunotioideae. Peronieae. Eunotieae.
3. Monoraphideae.
VII. Achnanthoideae-
4. Biraphideae. VIII. Naviculoideae.
IX. Epithemioideae.
X. Nitzschioideae.
XI. Surirelloideae.
Система Густедта (Hustedt, 1930)
Створки центрического строения; структура не перистая, шов отсутствует.
Клетки дисковидные, плоские или цилиндрические, на поперечном разрезе обычно округлые, без рогов и выступов.
Клетки палочковидные, длина их значительно больше ширины, на поперечном разрезе округлые. Клетки банкообразные, створки с двумя или более полюсами, несущими большей частью рога или выступы.
Клетки с лодкообразными створками, с неправильной или радиальной структурой.
Створки зигоморфные, на поперечном разрезе обычно лодкообразные или палочковидные; структура створок перистая.
Настоящий шов отсутствует.
Имеется короткий зачаточный шов на одной или обеих створках, идущий к центру от слабо развитого полярного узелка; центральный узелок отсутствует.
Зачаточный шов только на одной створке. Зачаточный шов на обеих створках.
Одна створка с развитым швом.
Обе створки с развитым швом.
Шов расположен по парапикальной оси, без киля, а если с килем, то без Kielpunkte.
Шов расположен не по парапикальной оси, иногда на эксцентричном киле, без Kielpunkte.
Обе створки с каналовидным швом. Каждая створка с одним, часто эксцентрическим килем, расположенным в плоскости створки.
Обе створки с каналовидным швом. Каждая створка с двумя боковыми крыльями, в которых проходят каналовидные швы, таким образом развивающиеся на каждой створке в двойном числе.
Авторы «Диатомового анализа» — Жузе, Прошкина-Лавренко и Шешукова (1949—1950) дают несколько измененную схему
52
Шютта с дополнениями Карстена и Густедта, в которой они группу Rutilarioideae, как переходную между центрическими и пеннат- ными, выделяют в отдельный порядок Mediales.
Система, принятая в «Диатомовом анализе», имеет следующий вид.
Тип BACILLARIOPHYTA
Класс DIATOMEAE Порядок Centrales
Подпорядок Discineae. Семейства Coscinodiscaceae, Actinodiscaceae, Eupodiscaceae.
Подпорядок Soleniineae. Семейство Soleniaceae.
Подпорядок Biddulphiineae. Семейства Chaetoceraceae, Biddulphia- ceae.
Порядок Mediales. Семейства Anaulaceae, Rutilariaceae, Raphone- taceae.
Порядок Pennale s.
Подпорядок Araphineae. Семейства Eutopylaceae, Tabellariaceae, Fragilariaceae.
Подпорядок Raphidioineae. Семейство Eunotiaceae.
Подпорядок Monoraphineae. Семейство Achnanthaceae.
Поднорядок Biraphineae. Семейства Naviculaceae, Epithemiaceae, Nitzschiaceae, Siirirellaceae.
Классификации пеннатных диатомовых Шютта, Густедта, Карстена и авторов «Диатомового анализа» построены на принципе усложнения шва. При этом наиболее низкоорганизованными считаются бесшовные — Araphideae, более высокоорганизованными признаются примитивношовные — Raphidioideae, затем одношовные — Monoraphideae, т. е. имеющие развитой шов только па одной створке. Наконец, наиболее высокоорганизованными считаются двушовные — Biraphideae, т. е. песущие шов на обеих створках.
Признание такой прямолинейной эволюции пеннатных диатомовых вызывало ряд возражений. В первую очередь возникал вопрос о первичности бесшовных. Фрич (Fritsch, 1935, 1956) считал, что нет достаточных оснований определенно утверждать, что бесшовные являются примитивными,— некоторые из них, а возможно, и все, могут быть редуцированными. Он высказал предположение, что и примитивношовные Eunotioideae могут быть формами с редуцированным швом. Таким образом, можно «читать последовательность в противоположном направлении и рассматривать Centrales как наиболее эволюционировавшую группу». Такую попытку «прочесть последовательность в противоположном направлении» мы видим в системе диатомовых, предложенной Эструпом (Oestrup, 1910). Эструп начинает систему пеннатных с двушовных — Diraphideae и заканчивает ее бесшовными — Araphideae. Однако палеонтологические данные дают основание связывать происхождение бесшовных диатомовых с Anaulales (Mediales), как это предполагают Шентукова (1949) и Топачев- ский (1962), хотя это не исключает возможности возникновения
53
некоторых бесшовных вторичным путем в результате редукции шва (Фрич, Топачевский). Топачевский полагает, что «дальнейшие исследования в этой области выяснят историю происхождения отдельных родов и семейств его (подкласса бесшовных.— Д. 3 ). Тогда в этом подклассе останутся только первичнобесшовные» (Топачевский, 1962, стр. 205).
Дискуссионным является также вопрос о взаимосвязях одно- шовных и двушовных. В системах Густедта и других одношовные обычно ставят перед двушовными, что предполагает их бо́льшую примитивность. Топачевский (1962) приводит убедительные доказательства зависимости шва от экологии диатомовых; в частности, он объясняет развитие одношовности прикрепленным образом жизни и выдвигает предположение, что одношовные возникли из двушовных в результате перехода их из подвижного состояния к эпифитному.
Особого внимания заслуживает возникновение каналовидного шва. По мнению Топачевского, каналошовные развились параллельно с щелешовными из центрических форм. Взгляды Топачевского отражены на схеме, приведенной ниже.
Схема А. В. Топачевского (1962 г.)
Отдел (тип) BÀCILLARIOPHYTA (Diatomeae).
I класс Gentricae
II класс Mediatae
III класс Pennatae
I подкласс Araphineae
1- й порядок Araphinales. Семейства Tabellariaceae, Diatomaceae,
Fragilariaceae.
II подкласс Raphineae
2- й порядок Naviculales (Schismaraphinales)
1- й подпорядок Dischismaraphineae. Семейство Naviculaceae.
2- й подпорядок Monoraphineae. Семейство Achnanthaceae.
3- й подпорядок Brachyraphideae (Raphidioineae). Семейства Ре-
roniaceae, Eunotiaceae.
3- й порядок Aulonoraphinales.
4- й подпорядок Diaulonoraphineae. Семейства Epithemiaceae,
Rhopalodiaceae, Nitzschiaceae.
5- й подпорядок Tetraaiilonoraphineae. Семейство Surirellaceae.
В схеме Топачевского вызывает сомнение положение примитивношовных Raphidioineae, названных им Brachyraphideae. Считая одношовные редуцированными по отношению к двушовным, Топачевский пошел далее и признал мало развитой шов у представителей этой группы также редуцированным.
Резюмируя сказанное о филогении диатомовых, можно утверждать, что из двух классов диатомовых исходным был класс центрических, возникший в юрском периоде из жгутиковых форм, вероятно, близких современным золотистым — Chrysophyta. В верхнем мелу появляются двусимметричные Anaulales, давшие начало классу пеннатных диатомовых, которые характеризуются развитием шва и связанной с этим подвижностью. В основе разви¬
тия пеннатных нужно поставить первично-бесшовные, от которых пошли две ветви развития — щелешовная и каналонтовная. Одновременно с развитием класса пеннатных шла эволюция центрических диатомовых, у которых развились разнообразные приспособления к планктонному образу жизни.
В заключение мы предлагаем классификационную схему диатомовых водорослей, в основу которой положены данные Густедта,
Карстена, Топачевского, с внесением некоторых номенклатурных изменений, главным образом в соответствии с существующими правилами ботанической номенклатуры (рис. 17). Что касается названия отдела, то в соответствии с обычным названием «диатомовые водоросли» мы считаем целесообразным называть его не Bacil]ariophyta (Bacillariophycophyta), a Diatomophyta (Diatomo- phycophyta), следуя Пашеру, который называл его Diatomeae (Pascher, 1931). Названия классов и порядков согласованы с правилами номенклатуры.
Отдел DIATOMOPHYTA (В А С I L L A R I О PH YT А, BACILLARIOPHYCOPHYTA, ВАС ILLAR ЮР HY- СЕАЕ, DIATOMEAE).
I. Класс CENTRODIATOMOPHYCEAE (CENTROBACILLA- RIOPHYCEAE, CENTRICAE) - ЦЕНТРИЧЕСКИЕ ДИАТОМОВЫЕ.
1. Порядок Coscinodiscales (Discinales) — к о с ц и поди с к а л ь н ы e. Клетки короткоцилиндрические, оболочки часто с более или менее длинными шипами. Морские и пресноводные.
Семейства Coscinodiscaceae, Arachnoidiscaceae и Eupodisca- сеае.
Рис. 17. Схема филогенетических связей между порядками диатомовых водорослей (Dia- tomophyta).
55
2. Порядок Rhizosoleniales (Soleniales) — ризосоле- ниальные, Клетки вытянуто-цилиндрические, с многочисленными промежуточными кольцами. Большей частью морские.
Семейство Rhizosoleniaceae.
3. Порядок Biddulphiales — биддульфиа льные. Клетки банкообразные или цилиндрические, с вставочными кольцами или без них, со стороны створки эллиптические или многоугольные, па углах с выпуклостями, роговидными выростами, щетинками. Морские и пресноводные.
Семейства Chaetoceraceae и Biddulphiaceae.
4. Порядок A n a u 1 а 1 е s (Mediales) — анаулальпые. Створки двусимметричные, без правильной радиальной или перистой структуры и без осевого поля. Морские и ископаемые формы.
Семейства Anaulaceae, Rutilariaceae и Raphoneiaceae.
II. Класс PENNATODIATOMOPHYCEAE (PENNATAE) - ПЕННАТНЫЕ ДИАТОМОВЫЕ.
1. Порядок Fragilariales (Araphinales) — фрагилари- а л ь н ы е. Шов отсутствует. Имеется только осевое поле ( «ложный шов» ).
Семейства Entopylaceae, Tabellariaceae, Fragilariaceae.
2. Порядок Naviculales (Rapliinales) — н а в и к у л а л ь- н ы е. На одной или на обеих створках имеется более или менее развитый щелевидный шов.
1) . Подпорядок Eunotiineae (Raphidioidineae). С зачаточным швом на концах одной или обеих створок.
Семейство Eunotiaceae.
2) . Подпорядок Naviculineae (Biraphidineae). Хорошо развитый
шов на обеих створках.
Семейство Naviculaceae.
3) . Подпорядок Aclmanthineae (Monoraphidineae). Шов развит только на одной створке.
Семейство Achnanthaceae.
3. Порядок Surirellales (Aulonoraphinales) — сурирел- лальны е. Створки с каналовидными швами.
Семейства Epithemiaceae, Nitzschiaceae, Surirellaceae.
5. БУРЫЕ ВОДОРОСЛИ — PHAEOPHYTA (PHAEOPHYCOPHYTA, PHAEOPHYCEAE)
Строение клетки и таллома
Бурые водоросли в отличие от других отделов хлорофиллоносных талломных организмов характеризуются многоклеточным строением и своеобразным набором пигментов, отличающим их от других буроокрашенных водорослей.
56
Клетка бурых водорослей имеет обычно одно крупное ядро и хорошо заметную центросому. При митозе центросомы размещаются на поверхности ядра, которое сохраняет свою оболочку за исключением района, прилегающего к центросоме. Клеточная оболочка состоит из внутреннего — целлюлозного и наружного — пектинового слоев.
У большинства бурых водорослей главной составной частью пектинового слоя является камедеобразное вещество — альгин.
Клеточные оболочки некоторых форм содержат слизистое вещество — фукоидин.
Отложения извести в оболочках наблюдаются редко (Padina pavonia).
Хроматофоры обычно многочисленны, хотя встречаются организмы только с одним хроматофором. Они расположены чаще всего постенно, дисковидные или иногда лентовидные, без типичных пиреноидов.
В хроматофорах обнаружены хлорофилл а, хлорофилл с, ß-каротин, фукоксан- тин и ряд других ксантофиллов — виола- ксантин, неоксантин, флавоксантин, нео- фукоксантин А и неофукоксантин В. Характерно, что хлорофилл С, кроме бурых водорослей, известен также у диатомовых и перидиней, фукоксантин — у диатомовых и золотистых водорослей, а неофуко- ксантины А и Б — у диатомовых. Запасными продуктами, образующимися в процессе ассимиляции, у бурых водорослей являются различные растворимые в клеточном соке углеводы, например ламинарии, свойственный некоторым ламинариевым, и маннит (маннитоль), а также масло.
Клетки бурых водорослей имеют одну большую или несколько мелких вакуолей.
Кроме того, у них встречаются мелкие — до 4μ в поперечнике вакуоли, так называемые физоды, в которых находится фу- козан — дериват флороглицина — вещество, обладающее свойствами таннинов.
Предполагают, что фукозан является побочным продуктом обмена веществ. В клетках многих бурых водорослей накапливается йод. Например, в талломах ламинарии KJ накапливается до 0,3% к свежему весу.
Как уже упоминалось, бурые водоросли многоклеточные организмы. Наиболее простые формы имеют разветвленное нитчатое
Рис. 18. Spermatoch- nus paradoxus (гапло- стихальный тип строения таллома) (но Oltmanns, 1922).
57
строение (Pylaiella, Ectocarpus). Для эктокарпа характерно гете- ротрихальное строение (таллом состоит из двух частей — стелющихся нитей, прикрепленных к субстрату, и вертикальной разветвленной части, несущей органы размножения). Такие нитевидные формы растут интеркалярно в результате поперечного деления клеток.
Иногда боковые ветви густо покрывают клетки главной оси, образуя вокруг них как бы коровой слой (Mesoglea leveillei, Sper- matochnus paradoxus). В результате получается псевдопаренхиматическая ткань. Такой тип строения тела Куку к (Kuckuck, 1929) назвал гаплостихальным (рис. 18).
В других случаях в результате деления клеток, кроме поперечного, также и в продольном направлении развивается настоящая паренхиматическая структура (Sphacelaria и др.). Такой тип строения таллома Кукук назвал полистихальным (рис. 19).
У полистихальных бурых водорослей в меру дальнейшего продольного деления клеток проявляется тенденция к сосредоточению делений в наружных слоях таллома — образуется так называемая поверхностная меристема (меристодерма Соважо), в то время как внутренние, сердцевинные клетки не делятся и растягиваются в длину и ширину. В результате возникает два слоя — периферический, коровой слой, состоящий из мелких клеток, содержащих многочисленные хроматофоры, и внутренний, сердцевинный, состоящий из крупных клеток, бедных хро- матофорами. Иногда, вследствие натяжения тканей, связанного с быстрым ростом корового слоя, в сердцевине образуется полость. Коровая часть таллома обычно выполняет ассимиляционную и механическую функции, а сердцевинная служит для отложения запасных веществ; в ней иногда развиваются проводящие элементы, напоминающие ситовидные трубки высших растений. В коровом слое ламинариевых образуются слизевые каналы.
Рост бурых водорослей происходит или интеркалярно (например, Laminariales), или при помощи верхушечной клетки (Sphacelaria, Dictyota, Fucus), или же краевой меристемы (Padina ρανοηία). У более простых представителей эктокарпальных, в частности у многих видов рода Ectocarpus, клетки делятся без определенного порядка, и только у некоторых видов намечается ясно выраженная интеркалярная инфраапикальная меристема. Она обычно бывает расположена у основания бесцветных волосков и обусловливает так называемый трихоталлический рост, ха-
58
Рис. 19. Sphacelaria plumigera (полисти- хальный тип строения таллома) (по Olt- manns, 1922).
рактерный для многих эктокарпальных, а также десмарестиаль- ных и кутлериальных (рис. 20).
У некоторых родов порядка Chordairales, например у Acroth- rix gracilis, наблюдается типичный трихоталлический рост, а у Nemacystus flexuosus и Spermatochnus paradoxus из семейства Spermatochnaceae того же порядка — верхушечный рост при помощи полусферической апикальной клетки. Согласно данным Кукука (Kuckuck,
1929), верхушечный волосок у старых экземпляров Асго- thrix gracilis отпадает, что дает основание предполагать, что верхушечная меристема Spermatochnaceae возникла из трихоталлической. Это подтверждается тем, что у Acrothrix novae-angliae, по Тейлору (Taylor, 1928), после отпадения волоска меристема продолжает функционировать как верхушечная.
У Dictyosiphon foeniculaceus, по Совежо (Sauvageau, 1929), молодые растения имеют верхушечный волосок и трихо- таллическую меристему. Позже волосок отпадает и рост продолжается при помощи теперь уже верхушечной клеточки. Все это свидетельствует о том, что у бурых водорослей верхушечный рост возник из интеркалярного (Fritsch, 1943, стр. 66—67).
Для высших представителей бурых водорослей характерна дифференциация частей таллома. Так, у многих из них можно наблюдать в нижней части диски или крючки, служащие для прикрепления к субстрату, более или менее удлиненную стеблевидную часть — кау- лоид и плоские листовидные органы. У отдельных форм (например, Sargassum) развиваются плавательные пузыри. Некоторые бурые водоросли, особенно из порядка Laminariales, достигают очень крупных размеров — до 60 ж (род Macrocystis).
Для многих бурых водорослей характерна гетеротрихия, т. е. наличие двух стадий в развитии таллома — первоначальной, обычно нитчатой и простертой, и вторичной, обычно вертикальной.
59
Рис. 20. Mesogloea leveillei (гаплости- хальный тип строения таллома) : интеркалярная меристема (м) при основании верхушечного волоска — трихоталлический рост (по Oltmanns, 1922).
Так, у большинства хордариальных и диктиосифональных из зиготы развивается нитчатое растение, сходное с гаметофитом — плетизмоталлом (plethysmothallus), на котором вырастает как боковой вырост новое растение спорофита. В некоторых случаях у представителей диктиосифональных наблюдается размножение первичной стадии (плетизмоталлома) при помощи зооспор, образующихся в многоклеточных спорангиях.
Размножение и типы онтогенетического развития
Многие бурые водоросли размножаются вегетативно частями таллома. Виды рода Sphacelaria образуют специальные веточки, служащие для вегетативного размножения.
Бесполое размножение у бурых водорослей происходит при помощи зооспор, развивающихся в одноклеточных (унилокуляр- ных) или многоклеточных (плюрилокулярных) спорангиях. В одноклеточных спорангиях обычно образуются многочисленные зооспоры, а в многоклеточных — в каждой клетке по одной споре. В некоторых случаях (Dictyotales) бесполое размножение происходит при помощи безжгутиковых аплано- спор.
Зооспоры, как и подвижные гаметы, обычно грушевидной формы, имеют стигму, часто связанную с одним из хроматофоров, и два жгутика, из которых один — перистый, направлен вперед, а другой — бичевидный — назад.
У бурых водорослей известен изогамный, гетерогамный и оогамный половой процесс. Изогамный половой процесс в типичной форме встречается довольно редко, например, он описан у Pylaiella litoralis и состоит в копуляции двух гамет, возникших в многоклеточных спорангиях. У Ectocarpus silicalosus из эктокарпальных и Cladostephus spongiosus из сфацелариальных наблюдается дифференциация изогамет; некоторые из них («женские») малоподвижны, прикрепляются к субстрату передним жгутиком, другие же («мужские») окружают ее, и один из них сливается с неподвижной «женской» гаметой (рис. 21).
Типичная гетерогамия описана у многих бурых водорослей. Примером может быть Nemoderma tingitana из порядка эктокар- пальпых и Cutleria multifida из кутлериальных (рис. 22).
Кроме копуляции гамет, развивающихся в многоклеточных гаметангиях, у некоторых бурых водорослей описано слияние зооспор (?), развивающихся в одноклеточных спорангиях. Например,
Рис. 21. Cladostephus spongiosus (копуляция гамет) (по Schreiber, 1931).
€0
такой факт был зафиксирован у Cylindrocarpus herkeleyi из хордариальиых (Caram, 1957).
Оогамный половой процесс известен у представителей порядков Dictyotales, Desmarestiales и Laminariales. У Dictyotales гаметан- гип собраны кучками (сорусами). В оогонии развивается одна яйцеклетка. В каждой клетке антеридия — по одному антерозоиду. Антерозоиды у Dictyota снабжены только одним передним пери¬
Рис. 22. Nemoderma tingitana:
1 — макрогаметангий, 2 — микрогаметангий, 3 — гаметы (по Кукук из Vazart, 1963).
стым жгутиком, хотя есть указания на наличие у них второго базального тельца (Manton, 1959). У представителей порядков Desmarestiales и Laminariales оогонии и антеридии одноклеточные. В оогонии развивается одна яйцеклетка, а в антеридии один двужгутиковый антерозоид (рис. 23). Яйцеклетка выходит из оогония, однако часто остается прикрепленной к нему. Иногда, как, например, у Chorda и Desmarestia tabacoides, оплодотворение происходит в оогонии, и первые стадии развития зародыша проходят на гамето- фите (Kylin, 1933; Nakahara a. Nakamura, 1971) (рис. 24). У Мае- rocystis pyrifera оплодотворение происходит также в оогонии, и зигота выходит из оогония только после первого деления (Levyns, 1933).
61
Иной тип оогамии наблюдается у представителей порядка Pacales. Здесь также гаметангии одноклеточные, однако, как подчеркивает Кюлин, в антеридиях после предпоследнего деления ядра образуются протоилазматические перегородки, разделяющие его на несколько клеток (Kylin, 1918, 1940). В антеридии образуются многочисленные (до 64) антерозоиды, а в оогониях у некоторых родов (Fucus, Notheia) восемь яйцеклеток. У других же родов
Рис. 23. Desmarestia aculeata:
1 — женский гаметофит с оогонием, 2 — мужской гаме- тофит с антеридиями (по Schreiber, 1931).
наблюдается сокращение количества яйцеклеток до четырех (Asco- phyllum, Durvillea и др.), Двух (Pelvetia) и одной (Cystoseira, Sargassum и др.).
Для большинства бурых водорослей характерно чередование поколений, и только представители порядка Fucales являются диплоидами. У бурых водорослей известны как изоморфный, так и гетероморфный типы чередования поколений. Простейший тин изоморфного чередования поколений хорошо изучен у видов рода Ectocarpus. У них на гаплоидном гаметофите образуются многоклеточные (плюрилокулярные) гаметангии, в которых развиваются изогаметы, отличающиеся, как уже упоминалось, своей активностью. Из зиготы без периода покоя развивается диплоидный спорофит, на котором образуются одноклеточные (монолокуляр- ные) спорангии. В этих спорангиях развиваются после редукционного деления многочисленные зооспоры, которые, прорастая, дают гаплоидные растения гаметофита. Кроме одноклеточных спорангиев, на спорофите могут развиваться многоклеточные спорангии, в которых образуются без редукционного деления диплоидные зооспоры, дающие новые диплоидные растения. Как отмечает ряд авторов, чередование поколений у эктокарпальных часто не имеет регулярного характера: каждое поколение может быть угнете¬
62
но — гаметофит в результате копуляции зооспор, спорофит — в результате партеногенетического развития гамет (Vazart, 1963).
Подобно видам рода Ectocarpus у некоторых представителей порядка Punctariales спорофиты могут образовывать, кроме одногнездных спорангиев, многогнездные, в которых образуются зооспоры, дающие начало новым растениям спорофита. В то же время гаплоидные зооспоры, образующиеся в одногнездных спорангиях, по данным Найта, могут иногда функционировать как гаметы (Vazart,
1963).
Более четко выражено изоморфное чередование поколений у представителей порядка Dictyotales. Так, у Dictyota dichoto- та на двудомном гаметофите развиваются оогонии и антеридии, прорастающая зигота дает спорофит, на котором образуются тетраспорангии, а в них развиваются после мейоза четыре безжгутиковые тетраспоры.
Гетероморфный тип чередования поколений у бурых водорослей несомненно развился из изоморфного в результате его усложнения. Есть указания, что у видов эктокарпа иногда намечаются различия между гаметофитом и спорофитом. Резкая разница между половым и бесполым поколениями обнаруживается у представителей порядка Cutleriales и особенно ярко выражена у порядков, объединенных Кюлином в классе Heterogeneratae,— Chordariales,
Sporochnales, Laminariales и др. У видов Cutleria гаметофит виль- чато разветвленный, довольно крупный, в то время как спорофит имеет вид плоского лопастного диска, настолько отличающегося от гаметофита, что он был описан под особым родовым названием Aglaozoma. Заслуживает внимания тот факт, что у близкого к Cutleria рода Zanardinia оба поколения сходны.
В группе порядков Heterogeneratae Кюлина дифференциация поколений ярко выражена: гаметофит — маленькое, иногда микроскопическое растение, а спорофит — крупное, часто многолетнее растение. У двудомных ламинариальных мужской и женский гаметофиты часто отличаются морфологически, а у некоторых видов диморфизм распространяется даже на споры. Гетероспория была описана Соважо у Laminaria cloustoni и Saccorhiza bulhosa, а Делфом и Левинсом у Macrocystis pyrifera (Vazart, 1963, стр. 164).
Порядок Fucales в отличие от других порядков бурых водорослей характеризуется диплоидным типом развития. Относящиеся к нему виды диплоидны, редукционное деление у них происхо-
Рис. 24. Chorda filum (первые стадии развития спорофита; спорофит развивается в течение ^ некоторого времени на гаметофите) (по Ку- Ип, 1933).
63
Рис. 25. Схема типов развития бурых водорослей — Phaeophyta (по Harder, 1962).
дит при образовании гамет, а из зиготы без периода покоя развивается диплоидное растение, несущее гаметангии. Бесполого размножения у фукальных нет.
Типы развития бурых водорослей показаны на рис. 25.
Классификация и филогения
Бурые водоросли рассматривались как самостоятельная группа уже авторами первой половины XIX ст. К. Агард (С. A. Agardh, 1817) дал им название Fucoideae, а его сын различал среди них семь порядков, а именно: Ectocarpeae, Sphacelarieae, Chordarieae, Dictyoteae, Laminarieae, Sporochnoideae и Fucaceae (J. G. Agardh, 1848), которые, правда с некоторыми изменениями объема, признаются до настоящего времени. Арешуг (Areschoug, 1847) разделял Fucoideae на Episporeae и Cyclosporeae. К Cyclosporeae он относил Fucaceae, а к Episporeae все остальные Fucoideae. Тюре (Thuret, 1850) изменил название Episporeae на Phaeosporeae. По данным Кюлина (Kylin, 1937), де Бари ввел для бурых водорослей название Phaeophyceae (De Вагу, 1881), а Гаук (Hauck, 1885) разделил их на три порядка — Fucoideae, Dictyotaceae и Phaeozo- osporeae. Де Тони (De Toni, 1895) принял порядки, предложенные Гауком, давши им названия Cyclosporineae, Tetrasporinae и Phae- ozoosporinae.
Нужно отметить, что ряд авторов XIX ст. сомневался в принадлежности диктиотовых к бурым водорослям ввиду ряда особенностей в их строении и размножении. Например, Чельман (Kjelmann, 1897) рассматривал бурые водоросли — Phaeophyceae (Fucoideae) и диктиотовые — Dictyotales как независимые классы водорослей. Энгл ер в первых изданиях «Syllabus der Pflanzenfamilien» (Engler, 1898) также рассматривал бурые водоросли и диктиотовые как самостоятельные классы подотдела настоящих водорослей (Euphyceae) отдела настоящих талломных растений (Euthallophyta).
Веттштейн (Wettstein, 1901) делит отдел бурых водорослей (Phaeophyta), следуя Тюре, на два порядка — Phaeosporeae и Cyclosporeae. К Cyclosporeae он относит семейство Fucaceae, a Phaeosporeae делит на два подпорядка — Zoogonicae и Acinetae, включая в последпий семейства Tilopteridaceae и Dictyotaceae, а в первый — все остальные семейства.
Ольтманнс (Oltmanns, 1904) разделил бурые водоросли (Phaeophyceae) на три большие группы («отделы»): 1) Phaeosporeae с четырьмя семействами — Ectocarpaceae, Cutleriaceae, Sphacela- riaceae и Laminariaceae; 2) Akinetosporeae с двумя семействами — Tilopteridaceae и Choristocarpaceae и 3) Cyclosporeae с семействами Dictyotaceae и Fucaceae. Несколько позже Энглер (Engler, 1912) в отделе бурых водорослей различает три порядка — Phaeosporeae, Cyclosporeae и Dictyotales.
65
С дальнейшим изучением бурых водорослей количество выделяемых среди них порядков постепенно увеличивается. Так, Ольт- манне во втором издании своего пособия (Oltmanns, 1922) выделяет семь порядков — Ectocarpales, Sphacelariales, Cutleriales, Laminariales, Tilopteridales, Dictyotales и Fucales, а Кюлин (Kylin, 1933) дает уже двенадцать порядков. Особенностью классификации Кюлина является попытка сгруппировать порядки на основании типов смены поколений и ядерных фаз в три класса — Isoge- neratae с изоморфным типом чередования поколений (Dictyota — тип), Heterogeneratae с гетероморфным типом чередования поколений (Laminaria — тип) и Cyclosporeae с диплоидным типом развития (Fucus — тип). Тип чередования поколений с господством гаметофита (Cuttleria — тип) Кюлин во внимание не принимает, считая его «анамалией» типа Dictyota. К классу Isogene- ratae он относит порядки Ectocarpales, Sphacelariales, Cutleriales, Tilopteridales и Dictyotales. Класс Heterogeneratae Кюлин делит на два подкласса — Haplostichineae с ложным паренхимным строением, возникающим в результате склеивания разветвленных нитей, и Polystichineae с настоящим паренхимным строением, возникающим в результате продольного деления клеток. В подкласс Haplostichineae Кюлин включает порядки Chordariales, Sporochna- les и Desmarestiales, а в подкласс Polystichineae — порядки Рша- ctariales, Dictyosiphonales и Laminariales. Наконец, к классу Cyclosporeae он относит один порядок — Fucales.
Классификация Кюлина принята с теми или иными небольшими изменениями в ряде сводных работ (Smith, 1938, 1955; Рареп- fuss, 1951; Melchior, 1954; Fott, 1956, 1959, 1967). Из двенадцати предложенных им порядков обычно принимаются одиннадцать, кроме порядка Punctariales, который, по предложению Папенфус- са (Papenfuss, 1947) присоединяется к Dictyosiphonales. Фрич (Fritsch, 1945) принимает только девять порядков. Порядки Chordariales, Punctariales и Dictyosiphonales он включает в состав порядка Ectocarpales.
Что касается предложенных Кюлином трех классов, в основу выделения которых положен только один признак — характер чередования поколений, то они не могут не вызвать возражений как построенные искусственно, что было отмечено, например, Папенфуссом (Papenfuss, 1951а, стр. 7—8) и Циммерманном (Zimmermann, 1959, стр. 82).
Приводим краткую характеристику порядков бурых водорослей в трактовке Папенфусса (Papenfuss, 1951).
Отдел PHAEOPHYTA (PHAEOPHYCOPHYTA).
1. Порядок Ectocarpales — эктокарпальные. Таллом нитчатый, разветвленный или псевдопаренхиматический с ин- теркалярным ростом. Изредка только имеет место продольное деление клеток (Pylaiella). Бесполое размножение зооспорами, развивающимися на спорофите или в одногнездных спорангиях
66
с редукционным делением или в многогнездных спорангиях без редукционного деления. Половой процесс изогамный или гетеро- гамный. Чередование поколений изоморфное.
Два семейства: Ectocarpaceae и Ralfsiaceae.
2. Порядок Sphacelariaies — сфацелариальные. Таллом нитчатый, полистихальный (паренхиматический). Рост верхушечный. Имеется и интеркалярное поперечное и продольное деление. Бесполое размножение зооспорами, развивающимися в одногнездных и многогнездных спорангиях. Половой процесс изогамный или гетерогамный. Изоморфное чередование поколений.
Четыре семейства: Sphacelariaceae, Stypocaulaceae, Cladoste- phaceae, Choristocarpaceae.
3. Порядок Cutleriales — кутлериальные. Таллом паренхиматический (полистихальный). Рост интеркалярный — трихоталлический. Бесполое размножение зооспорами, развивающимися в числе 4 (Zanardina) или 4—32 (Cutleria) в одногнездных спорангиях на спорофите. Половой процесс гетерогамный; гаметангии многогнездные. Чередование поколений изоморфное (Zanardinia) или гетероморфное с преобладанием гаметофита (Cutleria).
Одно семейство — Cutleriaceae.
4. Порядок Thilopteridales — тилоптеридаль- н ы е. Таллом нитчатый, разветвленный, моностихальный в верхней части и полистихальный (паренхиматический) в нижней части. Только у Acinetospora весь таллом нитчатый моностихальный. Рост интеркалярный, трихоталлический. Бесполое размножение четырехъядерными апланоспорами, развивающимися по одной в одногнездных спорангиях. Кроме того, на половых растениях развиваются одногнездные моноспорангии, в которых образуются моноспоры, по-видимому, служащие для бесполого размножения гаметофита. Половой процесс, очевидно, оогамный. Чередование поколений изоморфного типа.
Одно семейство — Tilopteridaceae. Развитие относимых к порядку родов изучено еще недостаточно.
5. Порядок Dictyotales — диктиотальные. Таллом паренхиматический с верхушечным ростом при помощи верхушечной клетки (Dictyota) или краевой меристемы (Padina). Имеется также интеркалярное поперечное и продольное деление. Бесполое размножение безжгутиковыми тетраспорами, развивающимися в одногнездных тетраспорангиях. Половой процесс оогамный. Изоморфное чередование поколений.
Одно семейство — Dictyotaceae.
6. Порядок Chordariales — хордариал ьные. Таллом гаплостихальный, псевдопаренхиматический. Рост интеркалярный, трихоталлический. Бесполое размножение зооспорами, развивающимися в одногнездных и многогнездных спорангиях. Есть указания на то, что иногда зооспоры, развивающиеся у видов Elachista
67
в одногнездных спорангиях, очевидно, без редукционного деления, могут давать начало диплоидным растениям спорофита (Kylin, 1937). Половой процесс изогамный или реже гетерогампый (Sphae- rotrichia divaricata). Чередование поколений гетероморфное, за исключением Heterochordaria abietina, характеризующейся изоморфным чередованием поколений.
Восемь семейств: Myrionemataceae, Elachistaceae, Corynophlae- асеае, Chordariaceae, Spermatochnaceae, Acrothrichaceae, Chorda- riopsidaceae, Splachnidiaceae.
7. Порядок Sporochnales — спорохнальные. Таллом псевдопаренхиматический. Разветвления таллома заканчиваются пучком волосков. Рост интеркалярный, трихоталлический при помощи однослойной куполовидной меристемы, размещенной при основе пучка волосков. Бесполое размножение зооспорами, развивающимися в одногнездных спорангиях. Половой процесс оогам- ный. Антеридии одно-двуклеточные, оогонии одноклеточные. Чередование поколений гетероморфного типа.
Одно семейство — Sporochnaceae.
8. Порядок Desmarestiales — десмарестиальные. Таллом псевдопаренхиматический (гаплостихальный). Разветвления его заканчиваются одним волоском. Рост интеркалярный, трихоталлический. Бесполое размножение зооспорами, развивающимися в одноклеточных спорангиях. Половой процесс оогамный. Антеридии и оогонии одноклеточные. В оогонии образуется одна яйцеклетка, при созревании она выходит из оогония и часто остается прикрепленной к отверстию оогония. Антеридии расположены поодиночке или группами, каждый из них дает только один антерозоид. Чередование поколений гетероморфного типа. У некоторых видов спорофит достигает крупных размеров — до 5 м (Desmarestia).
Два семейства: Arthrocladiaceae и Desmarestiaceae.
9. Порядок Dictyosiphonales — диктиосифональ- ные (включая Punctariales Кюлина). Таллом паренхиматический (полистихальный). Рост интеркалярный или верхушечный (Dicty- osiphonaceae). Бесполое размножение зооспорами, развивающимися в одноклеточных и многоклеточных спорангиях. Половой процесс изогамный или реже гетерогамный (Soranthera ulvoidea). Гаметангии многоклеточные (плюрилокулярные). Чередование поколений гетероморфного типа.
Шесть семейств: Striariaceae, Giraudyaceae, Myriotrichiaceae, Pimctariaceae, Chnoosporaceae, Dictyosiphonaceae.
10. Порядок Laminariales — ламинариальные. Таллом паренхиматического строения. Рост при помощи интеркаляр- ной меристемы. Бесполое размножение зооспорами, развивающимися в одноклеточных (монолокулярных) спорангиях. Половой процесс оогамный. Оогонии одноклеточные; в них образуется по одной яйцеклетке, выходящей при созревании из оогония. Антеридии также одноклеточные, образуют по одному антерозоиду.
68
Чередование поколений гетероморфного типа. Спорофит иногда достигает очень крупных размеров.
Четыре семейства: Chordaceae, Laminariaceae, Lessoniaceae,
Alariaceae.
11. Порядок Fucales — фукальные. Таллом паренхима- тического строения. Растет при помощи верхушечной клетки. Бесполое размножение отсутствует. Половой процесс оогамный. В оогонии образуется от восьми до одной яйцеклетки. В антеридии образуется до 64 антерозоидов. Организм существует в диплоидной стадии. Мейоз происходит при образовании гамет. Некоторые виды достигают 10 м (Durvillea).
Семь семейств: Ascoseiraceae, Durvilleaceae, Notheiaceae, Fuca- ceae, Hemianthaliaceae, Cystoseiraceae, Sargassaceae.
Вопрос о происхождении бурых водорослей до настоящего времени мало освещен ввиду изолированности этого отдела. Достоверные ископаемые остатки бурых водорослей известны начиная с триаса. Однако есть указания, что некоторые девонские и даже верхнесилурийские ископаемые остатки, возможно, принадлежат организмам, находящимся в родстве с бурыми водорослями. Например, такими являются роды Prototaxites (Nemat оphy eus) и Ne- matothallus (Gotlian u. Weyland, 1964, стр. 61—63). Ананьев описывает из нижнедевонских отложений района Красноярска Angarolaminariopsis zinovae и Jenisseiphyton lebedevii, которые он условно относит к порядку Laminariales (Маслов, Ананьев, 1963).
Шерффель (Scherffel, 1901) связывал происхождение бурых водорослей с золотистыми (Chrysophyta). Пашер (Pascher, 1911) высказывался за филогенетическую связь бурых с криптомонадо- выми, но несколько позже (Pascher, 1914) уже не настаивал на этом, а считал, что лучше «пока» не искать предков бурых водорослей.
Нельзя не отметить, что бурые водоросли при всем их разнообразии представляют монолитную, несомненно монофилетиче- скую группу, которая характеризуется однообразными биохимическими свойствами (пигменты, продукты ассимиляции) и сходным строением подвижных стадий (Fritsch, 1945, стр. 20). Характер подвижных стадий — зооспор и гамет дает основание предполагать происхождение бурых водорослей от флагеллоидных форм с двумя разными жгутиками — одним перистым, а другим бичевидным. Такой тип строения жгутиков среди современных организмов свойственен хризофитовым и желтозеленым водорослям, что вместе с бурой окраской дало основание Шадефо (Chadefaud, I960) объединять Pyrrhophyta (Pyrrhophyceae), куда он включает, кроме перидиней, также криптофитовые и эвгленовые водоросли, Chrysophyta (Chrysophyceae), в которые он включает хризофито- вые, желтозеленые и диатомовые водоросли, и Phaeophyta (Phae- ophyceae) в один большой отдел Chromophycophyta, тем самым предполагая общность их происхождения.
69
Шадефо считает, что бурые водоросли происходят от общего корня всех Chromophycophyta, который он видит в группе Ргоруг- rophyceae. К этой группе он относит два рода — Pocillomonas и Chrysophaeum. Он обращает особое внимание на Chrysophaeum taylori, описанный из Мексиканского залива (Lewis and Bryan, 1941). Указанный вид, по его мнению, сочетает черты бурых, хризофитовых и пиррофитовых водорослей и находится у истоков (prés du carrefour à partir), из которых развились названные выше отделы растительного мира (Chadefaud, 1952, 1960). С этим родом их объединяет отсутствие крахмала, характер зооспор и гамет, пиреноиды, прикрепленные к поверхности или к краю хло- ропластов. С другой стороны они, по Шадефо, приближаются к хризофитовым строением монад, у которых мало выражен вестибулярный желобок, отсутствуют трихоцисты, строением хлоро- пластов, характером жгутиков, отсутствием крахмала и богатством пектиновых веществ в клеточных оболочках. Однако бурые водоросли не вырабатывают лейкозина, не образуют эндоцист и обладают многогнездными спорангиями и гаметангиями (Chadefaud, 1960, стр. 269).
Из всех буроокрашенных организмов по биохимическим свойствам ближе всего к бурым водорослям стоят диатомовые, что отметил еще Кюлин (Kylin, 1933, стр. 94). Действительно, диатомовые и бурые водоросли характеризуются, кроме широко распространенных у автотрофных организмов хлорофилла а и ß-каротина, такими общими пигментами, как хлорофилл с (последний свойственен также и перидинеям), фукоксантин (он встречается также у хризофитовых) и неофукоксантины А и В. Наличие фукоксантина и строение монад сближает бурые водоросли с золотистыми — хризофитами. Принимая во внимание то, что бурые водоросли, вероятно, древнее диатомовых, и большую специализацию строения клетки у последних, считать диатомовые предками бурых водорослей нельзя. С другой же стороны, ввиду наличия ряда сходных черт между диатомовыми, золотистыми и бурыми водорослями, нам кажется, можно допустить их происхождение от близких, если не общих, монадных предков.
Курсанов (1933, 1945) считал возможным происхождение бурых водорослей от окрашенных в бурый цвет жгутиковых. Он подчеркивает, что некоторые группы бурых водорослей прошли очень длительную эволюцию, в то время как другие остановились на низших ступенях развития; к числу последних он относит экто- карпальные с их простым вегетативным строением и мало дифференцированным половым процессом и чередованием поколений. В эволюции бурых водорослей Курсанов намечает две линии: одна — эктокарпальные -> кутлериальные —у ламинариальные с ин- теркалярным ростом, другая — сфацелариальные -> диктиоталь- ные —фукальные с верхушечным ростом.
Кюлин (Kylin, 1933), как мы уже упоминали, в основу классификации, а следовательно, и филогении бурых водорослей
70
положил характер смены поколений и ядерных фаз и разделил их на три основные ветви эволюции, которым он придал ранг класса. Взаимоотношения между этими классами он представил в виде воспроизводимой нами схемы (рис. 26).
Папенфусс (Papenfuss, 1951а), рассматривая схему Кюлина, отметил, что выделение этих классов исключает возможность параллельной и независимой эволюции групп с изоморфным и
Рис. 26. Обзор порядков бурых водорослей (Phaeophyta) по Ккь лину (Kylin, 1938).
гетероморфным чередованием поколений, не допускает признания Ectocarpales возможной исходной группой для других бурых водорослей и упускает из виду, что Fucales имеют паренхимное (полистихальное) строение, как и некоторые группы Isogeneratae (например, Sphacelariales, Dictyotales). Несомненно, что различные группы бурых водорослей с гетероморфным чередованием поколений произошли независимо от различных исходных форм с изоморфным чередованием поколений. Вероятно также, что различные группы с паренхимным (полистихальным) талломом развились независимо от различных форм гаплостихального строения.
Фрич (Fritsch, 1943, стр. 68), на основании всестороннего рассмотрения морфологии, размножения и чередования поколений у бурых водорослей, приходит к выводу, что эволюция в пределах этого ствола растительного мира проходила в таких направлениях.
1. Развитие организмов с плотным псевдопаренхиматозным или паренхиматозным талломом из форм с первичным гетеротрихаль- ным нитчатым строением;
2. Переход от диффузного интеркалярного роста к росту при помощи интеркалярной, обычно субапикальной меристемы (три- хоталлический рост) и к замене его верхушечным ростом;
71
3. Дифференциация паренхиматозного таллома на коровой и сердцевинный слои;
4. Образование поверхностного меристематического слоя (ме- ристодермы), обусловливающего прирост таллома в толщину;
5. Морфологическая дифференциация обоих поколений, приводящая к замене изоморфного чередования поколений гетеро- морфным;
6. Исключение дополнительного размножения спорофита при помощи спор, развивающихся в плюрилокулярных спорангиях;
7. Развитие группового (сорального — от sorus — кучка) размещения спорангиев на сапрофите;
8. Переход от изогамии к гетерогамии и оогамии.
Считая нитчатые гетеротрихальные формы типа Ectocarpales исходными для всех порядков бурых водорослей, Фрич отмечает, что специализированные порядки бурых водорослей не обнаруживают между собой родственных связей и должны рассматриваться как отдельные эволюционные линии, развившиеся из простых нитчатых форм с изоморфным жизненным циклом. У одних из них строение обоих поколений усложнилось и в то же время сохранился изоморфный тип развития (Sphacelariales, Dictyotales, род Zanardinia из Cutleriales, частично порядок Tilopteridales), у других же удержался псевдопаренхиматический тин строения, но развился гетероморфный тип развития (Chordariales, Sporoch- nales, Desmarestiales), у третьих при паренхиматическом (поли- стихальном) строении и гетероморфном цикле развития сохранился изогамный или гетерогамный половой процесс (Dictyosi- phonales, Punctariales) и только у представителей порядка Laminariales при паренхиматическом строении и гетероморфном типе развития имеется типичный оогамный половой процесс. Изолированно стоит порядок Fucales, представители которого отличаются от других бурых водорослей диплоидным типом развития. Распространено мнение, основанное на аналогии с высшими растениями и утвердившееся после открытия Соважо (Sauvageau, 1915, 1916) гетероморфного чередования поколений у Laminariales, где гаметофит представлен микроскопическим растением, что в порядке Fucales мы имеем конечное звено редукции гаметофита, где он сведен только к гаметам (Strasburger, 1906), или же совершенно исчез и гаметы фуксовых гомологичны макро- и микроспорам, принявшим на себя половую функцию (Kylin, 1916, 1918; Kniep, 1928; Zimmermann, 1959, и др.)· Этот взгляд нашел широкое признание. Однако позже Кюлин (Kylin, 1937, 1940) выдвинул возражения против возможности гомологии одноклеточных гаметангиев Fucales и спорангиев других бурых водорослей и высказал предположение, что филогенетическое развитие этого порядка проходило без чередования поколений, что предки Fucales были диплоидными организмами, образующими гаметы с редукционным делением, как это распространено в животном мире. Кюлин считает, что Fucales обособились очень рано от общего
72
ствола бурых водорослей, а не от форм, соответствующих современным Ectocarpales.
Фрич (Fritsch, 1942, 1943) предполагает, что диплоидный тип развития Fucales является результатом того, что у них закрепилась встречающаяся у некоторых других бурых водорослей копуляция спор, развивающихся после мейоза в монолокулярных спорангиях, следствием чего было исчезновение гаметофита.
Взгляд Фрича на происхождение и пути развития бурых водорослей представлен на данной в его работе схеме (Fritsch, 1943).
Рис. 27. Возможные связи порядков бурых водорослей (Phaeophy- ta) по Папенфуссу (Papenfuss, 1951).
По этой схеме развитие бурых водорослей проходило десятью независимыми стволами, выходящими из гетеротрихальных нитчатых форм. Таким образом, стволов эволюции у Фрича больше, чем принимаемых им порядков.
Более реально взаимоотношения между порядками бурых водорослей представляет схема Папенфусса (Papenfuss, 1951; см. рис. 27). Он считает наиболее простым исходным порядком бурых водорослей порядок Ectocarpales, от которого развитие бурых водорослей шло пятью ветвями. Согласно этой схеме паренхиматозное строение таллома, верхушечный рост, оогамный половой процесс и гетероморфный тип чередования поколений развивались в разных группах бурых водорослей независимо. Однако, вероятно, прав Кюлин, утверждая, что Fucales отделились от общего ствола бурых очень рано, a Ectocarpales являются потомками примитивных Phaeophyceae.
73
6. ПИРРОФИТОВЫЕ ВОДОРОСЛИ, ИЛИ ИЕРИДИНЕИ,— PYRRHOPHYTA (PYRBHOPHYCOPHYTA,
DINOPHYCEAE, PERIDINEAE)
Пиррофитовые представляют собой обособленную группу низших организмов, обитающих как в пресной, так и в морской воде. Большинство пиррофитовых — монадные одноклеточные организмы. Только небольшое количество их имеет пальмеллоидную, ри- зоподиальную (амебоидную), коккальную и нитчатую структуру.
Характерной особенностью пиррофитовых, отличающей их от других низших организмов, является своеобразное строение ядра, хроматин в котором образует четковидные нити. В ядре может быть одно или несколько ядрышек, которые обыкновенно при делении исчезают. Однако у Oxyrrhis marina ядрышко при делении сохраняется и ведет себя так, как у эвгленовых (Hall, 1925а). Ядерная оболочка при делении иногда сохраняется. Характерна связь вновь образующихся жгутиков с центросомами. Ядерное веретено у пиррофитовых не обнаружено.
Большинство пиррофитовых фототрофные организмы. В клетках автотрофных пиррофитовых имеются хроматофоры разнообразной формы; у многих видов имеются пиреноиды. В хроматофорах обнаружены хлорофилл а и с, что сближает их с диатомовыми и бурыми водорослями, ß-каротин и четыре ксантофилла, три из которых характерны только для них, а один — диадипоксаитин обнаружен также и у диатомовых. Запасными продуктами у пиррофитовых являются крахмал и масло. Кроме автотрофных, среди пиррофитовых известны также бесхлорофилльные гетеротрофные — голозойные и паразитные формы.
Клетки пиррофитовых могут не иметь вторичной оболочки, однако большинство имеет оболочку своеобразного строения, в состав которой входит целлюлоза.
Подвижные формы пиррофитовых и зооспоры неподвижных имеют два жгутика, прикрепленных или на переднем конце монады (Desmophyceae) или на брюшной ее стороне (Dinophyceae). Тонкое строение жгутиков у пиррофитовых изучено мало. Имеются указания, что у Gyrodinium из Dinophyceae поперечный жгутик имеет гребневидное строение, т. е. несет один ряд отростков (мастигонем), а продольный жгутик имеет бичевидную форму (Pitelka a. Schooley, 1955). Жгутики при основе с блефаропласта- ми и связаны с центросомой ризопластами (Hall, 1925).
У некоторых Dinophyceae обнаружены трихоцисты, сближающие их с криптофитовыми и хлоромонадовыми, а у родов Polykri- kos и Nematodinium описаны стрекательные органы (нематоци- сты), сходные с соответствующими образованиями кишечнополостных (Graham, 1951).
Характернрй особенностью клетки пиррофитовых являются своеобразные вакуоли, называемые пузулями (pusula). Для морских и некоторых пресноводных форм характерно наличие боль¬
74
шой пузули, открывающейся узким каналом у места прикрепления жгутиков. Кроме этой главной, или «собирательной», пузули, имеется мешковидная, а иногда еще несколько маленьких. Значение их до сих пор не выяснено.
Размножаются пиррофитовые делением, зооспорами или апла- носпорами. У некоторых форм описан половой процесс, состоящий в слиянии или взрослых клеток (гологамия) или одинаковых монад и апланоспор (мерогамия).
С конца XIX ст. в ботанической литературе пиррофитовые рассматривают как самостоятельную группу — класс или отдел растительного мира под названием Peridinialec (Schütt, 1896; Eng- 1er, 1898), Peridineae (Wettstein, 1901), Dinoflagellata (Oltmanns, 1904), Dinoflagellatae (Engler, 1912). Пашер (Pascher, 1914, 1927, 1931) отметил среди них пальмеллоидные, коккальные и нитчатые формы и выделил филу, или ствол, Pyrrhophyta, в которую включил также и криптофитовые, разделив ствол Pyrrhophyta на три отдела — Cryptophyceae, Desmokontae и Dinophyceae. Однако присоединение к пиррофитовым криптофитовых водорослей не было поддержано. Так, уже Линдеманн (Lindemann, 1928) исключает криптофитовые из отдела Pyrrhophyta, который он называет Peridineae (Dinoflagellatae), и делит его на три подотдела — Adi- niferae, Diniferae и Phytodiniferae, из которых первый соответствует отделу Desmokontae, а вторые два — отделу Dinophyceae системы Пашера. Позднейшие авторы рассматривают отделы Пашера Desmokontae и Dinophyceae или как подклассы единого класса Dinophyceae под названиями Desmokontae и Dinokontae (Fritsch, 1935, 1956; Graham, 1951), или же как классы Desmophyceae и Dinophyceae (Smith, 1955; Fott, 1959). Последнего деления мы будем придерживаться в дальнейшем изложении.
К классу Desmophyceae (Desmokontae) относятся в большинстве монадные формы с голыми или покрытыми оболочкой клетками. Описан только один организм — Desmocapsa gelatinosa палмеллоидного строения. У монадных форм жгутики размещены на переднем конце клетки или вблизи него; при наличии клеточной оболочки она состоит из двух половинок (створок), соединяющихся по продольной (сагиттальной) оси клетки. Особое место занимает порядок Dinophysidiales, у представителей которого оболочка по своему строению напоминает оболочку представителей класса Dinophyceae. Она состоит из двух частей — верхней — эпивальвы и нижней — гиповальвы, разделенных, как и у Dinophyceae, поперечным пояском, причем эпивальва значительно меньше гиповальвы. Оболочка у Dinophysidiales, как и у других Desmophyceae, имеет продольный шов, однако, кроме того, состоит из отдельных пластинок, что также сближает этот порядок с Dinophyceae.
Пашер (Pascher, 1931) делил Desmokontae на два класса —
1) Desmomonadineae с порядками Desmomonadales, Prorocentrales и Dinophysidiales и 2) Desmocapsineae с одним порядком Desmo-
75
capsales. Эту классификацию принимает Мельхиор (Melchior, 1954) и с небольшими изменениями Киселев (1954).
Несколько иначе подошел к классификации Desmophyceae Линдеманн (Lindemann, 1928), который разделил их на два класса — Athecatales, соответствующий порядку Desmomonadales Пантера, и Thecatales, соответствующий порядку Prorocentrales. Что
Рис. 28. Представители класса Desmophyceae:
1 — Desmomastix globosa, 2, 3 — Exuviella marina, 4, 5 — Dinophysis acuta
Cno Fott, 1959).
касается порядка Dinophysidiales Пашера, то Линдеманн относит его к Dinophyceae, которые он называет Diniferae.
Последующие авторы придерживаются классификации Пашера с теми или иными изменениями. Так, Фрич (Fritsch, 1935, 1956) ликвидирует только порядок Desmocapsales, относя единственного его представителя Desmocapsa gelatinosa к порядку Desmomonadales, и переименовывает, следуя Линдеманну, порядок Prorocentrales в Thecatales. Смит (Smith, 1955) порядок Dinophysidiales переносит в класс Dinophyceae. Фотт (Fott, 1959) делит класс Desmophyceae (Desmokontae) на два порядка— 1) Desmomasti- gales (Desmomonadales), к которому относит семейства Desmomastigaceae, Prorocentraceae и Dinopnysidaceae, 2) Desmocapsales.
Мы считаем обоснованным предложенное Пашером деление класса Desmophyceae на четыре порядка с изменением, согласно существующим правилам номенклатуры, названия Desmomonadales на Desmomastigales, а именно:
1) Desmomastigales — десмомастигальные. Подвижные формы с голыми клетками (рис. 28, 1).
2) Prorocentrales — пророцентральные. Подвижные формы, покрытые двустворчатой оболочкой (рис. 28, 2).
3) Dinophysidiales — динофизидиальные. Подвижные формы с поперечным пояском и оболочкой, состоящей из пластинок (рис. 28, 3).
4) Desmocapsales — десмокапсальные. Пальмеллоидные формы.
76
Спорное положение в системе Desmophyceae занимает порядок Dinophysidiales, который некоторые авторы переносят в класс Dinophyceae (Lindemann, 1928; Smith, 1955). Действительно, у представителей этого порядка имеются поперечный и продольный желобки (пояски), оболочка состоит из отдельных пластинок, место прикрепления жгутиков несколько сдвинуто на брюшную сторону. Однако одновременно с этими признаками, сближающими их с Dinophyceae, у Dinophysidiales имеется характерный для панцирных Desmophyceae продольный шов, разделяющий оболочку в сагиттальном направлении на две половины. Можно предположить, что развитие сходных черт у Dinophysidiales и Peridi- niales является конвергентным приспособлением к сходным условиям существования, и Dinophysidiales развились из панцирных форм типа Prorocentrales, имеющих такой же сагиттальный шов.
К классу Dinophyceae (Dinokontae) относятся голые или имеющие целлюлозную оболочку (панцирь) одноклеточные или реже колониальные формы, в большинстве случаев подвижные в вегетативном состоянии. Кроме того, встречаются пальмеллоидные, амебоидные, коккальные и нитчатые формы, принадлежность которых к Dinophyceae подтверждается строением их зооспор.
Подвижные в вегетативном состоянии Dinophyceae и зооспоры неподвижных форм характеризуются своеобразным строением. Характерным для них является поперечный желобок. Часто наблюдается также обычно менее выраженный продольный желобок. Два жгутика выходят на брюшной стороне монады в месте пересечения желобков. Один из них лежит в поперечном желобке и опоясывает клетку, другой расположен по продольной оси клет^ ки. Большинство Dinophyceae имеют оболочку, в состав которой входит целлюлоза и которая состоит из таких трех частей: верхней — эпивальвы, нижней — гиповальвы и соединяющего их кольцевидного пояска, лежащего в поперечном желобке. Каждая из этих частей образована закономерно расположенными пластинками и может иметь различного рода выросты. У многих представителей монады снабжены стигмой. Кроме монадных форм, известны амебоидные, пальмеллоидные, коккальные и нитчатые.
Большинство Dinophyceae являются фототрофными организмами, однако известны формы с голозойным питанием, а также симбионты и паразиты, обитающие в полости тела различных Copepoda.
Пашер (Pascher, 1914, 1931) различал в пределах отдела Dinophyceae на основании общего строения их тела пять классов и шесть порядков, а именно: 1) Dinoflagellatae — подвижные в вегетативном состоянии с двумя порядками — Gymnodiniales и Peri- diniales; 2) Rhizodininae — амебоидные формы с одним порядком Rhizodiniales; 3) Dinocapsineae — пальмеллоидные формы с порядком Dinocapsales; 4) Dinococcineae — коккоидпые формы с порядком Dinococcales; 5) Dinotrichineae — нитчатые формы с порядком Dinotrichales.
77
Лпндеманн (Lindemann, 1928) дает несколько иную классификацию подотдела Diniferae; он его делит на пять классов, а именно: Gymnodini- ales, Amphilothales, Kolkwitziellales, Dinophysiales и Peridiniales. K Amphilothales Лпндеманн отнес голые формы с внутренним скелетом, а к Kolkwitziellales — организмы с нерасчлененными на отдельные пластинки створками. Коккальные и пальмеллоидные формы Линдеманн выделяет в особый подотдел Phytodiniferae.
Фрич (Fritsch, 1935, 1956) Dinophyceae, которые он называет Dinokon- tae, разделяет на три порядка — Dinoflagellata, Dinococcales и Dinotrichaîes. К первому — Dinoflagellata он относит все формы, подвижные в вегетативном состоянии, а также паразитные Blastodiniaceae, коккальные и иальмеллоид- ные формы, разделяя его на три подпорядка — Peridinieae, Dinocapsineae и Rhizodinineae. Подпорядок Peridinieae Фрич делит еще на четыре группы семейств — Gymnodinioideae, Amphilothaloideae, Kolkwitzielloideae и Peridi- nioideae. Dinophysidiales Фрич оставляет в составе Desmocontae, Dinococcales и Dinotrichaîes он принимает в понимании Пашера.
Мельхиор (Melchior, 1954) принимает порядки Пашера, выделяя паразитные формы в особый порядок Blastodiniales.
Смит (Shith, 1955) принимает порядки, предложенные Пашером, и, следуя Лиидеманну (Lindemann, 1928), переносит порядок Dinophysiales из Desmophyceae в Dinophyceae.
Фотт (Fott, 1959) принимает систему Пашера, однако рассматривает Gymnodiniales, Peridiniales и Blastodiniales как подпорядки одного порядка Peridiniales и изменяет названия некоторых порядков в соответствии с действующими правилами ботанической номенклатуры. Система Dinophyceae, предложенная Фоттом, имеет следующие пять порядков: 1) Peridiniales с подпорядками Gym- nodiineae, Peridiniineae и Blastodiineae, 2) Dinamoebidiales, соответствующий порядку Rhizodiniales Пашера, 3) Gloeodiniales, соответствующий порядку Dinocapsales Пашера, 4) Dinococcales и 5) Dinotrichaîes.
В дальнейшем изложении мы будем придерживаться номенклатуры порядков, предложенной Фоттом. Что же касается их объема, то Gymnodiniales и Blastodiniales считаем целесообразным рассматривать как самостоятельные порядки. Таким образом, в пределах класса Dinophyceae мы будем различать следующие семь порядков. 11. Порядок Gymnodiniales — гимн одиниальные. Большей частью морские организмы, подвижные в вегетативном состоянии, обычно без вторичной оболочки, с довольно плотной мембраной с характерными бороздами. Размножаются делением в подвижном или неподвижном состоянии. Иногда образуются покоящиеся цисты (рис. 29).
2. Порядок Peridiniales — перид и ниальные. Преимущественно морские организмы, подвижные в вегетативном состоянии, с оболочкой, состоящей из эпивальвы, гиповальвы и пояска, образованных пластинками, расположенными в определенном порядке. Размножаются делением в подвижном или неподвижном состоянии; плоскость деления обычно расположена косо. Новая клетка получает или половину материнской оболочки,
78
или же дочерние клетки выходят из панциря и образуют новую оболочку. Описан гологамный и мерогамный половой процесс.
3. Порядок Blastodiniales — бластодиниальные. Паразитные морские организмы, обитающие или на поверхности, или внутри тела различных морских животных (ракообразных, крылоногих моллюсков, кольчатых червей, оболочников) и в клет-
Рис. 29. Представители класса Dinophyceae:
1 — Gymnodinium palustre (порядок Gymnodiniales), 2 — Polykrihos schwartzii (порядок Peridiniales), 3 — Dinamoebidium varians (порядок Dinamoebidiales): a — амебоидная стадия, б — циста, β — прорастание цисты с образованием зооспор, 4 — Gloeodinium montanum (порядок Gloeodiniales) (по Fott, 1959).
ках диатомовых водорослей. Внутри тела хозяина организм имеет вид неподвижных, иногда амебоидных клеток, дающих у некоторых форм плазмодии. Размножаются двужгутиковыми зооспорами, напоминающими своим видом гимнодиний.
4. Порядок Dinamoebidiales — динамебид и аль- н ы е. Единственный представитель порядка — Dinamoebidium varians является морским бесхлорофилльным голозойным амебоидным организмом, питающимся одноклеточными водорослями и простейшими. Образует цисты. Размножаются зооспорами типичного для Dinophyceae строения, несущими по одному жгутику.
5. Порядок Gloeodiniales—глеодиниальные. Небольшая группа пальмеллоидных форм, размножающихся типичными для Dinophyceae зооспорами.
6. Порядок Dinococcales — динококкальные. Небольшая группа прикрепленных или свободноплавающих коккаль- ных форм, не имеющих жгутиков и характерных для Dinophyceae бороздок. Автотрофные и гетеротрофные (паразитные) формы.
79
Эктопаразитные коккальные формы некоторые авторы, например Линдеманн, относят к порядку Blastodiniales. Размножаются зооспорами или апланоспорами (рис. 30).
7. Порядок Dinotrichales — динотрихальные. Небольшая группа нитчатых форм, размножающихся типичными двужгутиковыми зооспорами (рис. 31).
Относительно филогенетических связей пиррофитовых водорослей с другими отделами эвкариотических организмов в литературе встречаются разнообразные высказывания. Так, еще Клебс
(Klebs, 1912) обратил внимание на некоторые черты сходства между ними и криптофитовыми водорослями. Шютт (Schütt, 1896) высказывался за их родство с диатомовыми. Веттштейн (Wettstein, 1901) объединял пиррофитовые, диатомовые и конъюгаты в один отдел (ствол) Zygophyta.
Пашер (Pascher, 1911), выводил перидинеи из криптомонад. Позже он на этом не настаивал, но считал названные группы родственными и объединил их в одну филу (ствол) Pyrrhophyta, разделив ее на три отдела — Cryptophyceae, Desmo- kontae и Dinophyceae (Pascher, 1914, 1931). Шадефо (Chadefaud, 1960) включал в подотдел пиррофитовых водорослей классы Cryptophyceae, Dinophyceae, Raphidophyceae (Chloromonadophy- ceae) и Euglenophyceae, основывая их близость на двух признаках:
1) наличии у них запасного продукта типа крахмала (криптами- лон у криптофитовых и перидиней, парамилон у эвгленовых) и
2) сходстве строения монад, имеющих вестибулярный аппарат и один из жгутиков гребневидного строения. Наиболее распространен взгляд на родство пиррофитовых с криптофитовыми. Основанием для этого является сходная окраска хроматофоров с преобладанием каротиноидов, наличие в клетках крахмала как запасного продукта и строение монад, имеющих два неодинаковых жгутика и продольный брюшной желобок.
Что касается окраски хроматофоров, то криптофитовые резко отличаются от пиррофитовых наличием фикобилинов и иным составом каротинов и ксантофиллов. Монады криптофитовых имеют ряд особенностей в строении, отличающих их от подвижных клеток пиррофитовых. Это дает основание ряду авторов исключать криптомонадовые из состава пиррофитовых (Fritsch, 1935; Graham, 1951; Fott, 1959; Pringsheim, 1966, и др.). Что касается
Рис. 30. Dissodinium lunula (порядок Dinococcales) : а — вегетативная клетка; б — образование полумесяцеобразных дочерних клеток; в — образование зооспор, г — зооспоры (по Догелю, Graham, 1951).
80
родства пиррофитовых с диатомовыми, эвгленовыми и хлоромона- дами, то оно еще более отдаленное.
Классы пиррофитовых водорослей — Desmophyceae и Dinophy- сеае согласно принятому мнению, восходящему к Пашеру, представляют две параллельные ветви эволюции. Исходными формами для тех и других были голые монадные формы со своеобразным ядром, двумя жгутиками и наличием определенных пигментов. В классе Desmophyceae наиболее примитивный порядок — Desmo- mastigales, а в основе класса Dinophyceae должен быть поставлен порядок Gymnodiniales. Из форм, сходных с современными Gymno-
Рис. 31. Dinothrix paradoxa (порядок Dinotrichales) :
о — короткая нить, б, в — образование зооспор (по Пашеру, Fott, 1959).
Рис. 32. Схема филогенетических связей между порядками пиррофитовых водорослей (Pyrrhophyta).
diniales, развились покрытые оболочкой подвижные в вегетативном состоянии Peridiniales, амебоидные Dinamoebidiales, пальмеллоид- ные Gloeodiniales и паразитные Blastodiniales. Нитчатые Dinotri- cliales, очевидно, развились из коккоидных форм. Основанием для такого построения филогенетических отношений в классе Dinophyceae является то, что представители всех порядков этого класса, как бы они не отличались в вегетативной стадии, имеют зооспоры, сходные по строению с Gymnodinium.
Филогенетические отношения между порядками пиррофитовых водорослей представлены на схеме (рис. 32).
81
7. КРИПТОФИТОВЫЕ ВОДОРОСЛИ — CRYPTOPHYTA (CRYPTOPHYCOPHYTA, CRYPTOPIIYCEAE)
Криптофитовые представляют собой небольшую своеобразную группу организмов, в большинстве монадного строения. Только изредка встречаются пальмеллоидные формы, а существование
коккальных форм, например описанного Пашером Тetragonidium verrucosum, некоторые авторы (Fott, 1959) ставят под сомнение.
Клетки криптофитовых не имеют вторичной оболочки. Покрывающая их пелликула (плазмалемма) обычно бывает довольно плотной и часто продольно или спирально исчерчена в связи с наличием в наружном слое цитоплазмы своеобразных выделяющих слизь телец, расположенных правильными рядами. Клетка обычно имеет ас- симметричную форму — у нее можно отличить выпуклую спинную и вогнутую брюшную стороны.
От косо срезанного переднего конца по брюшной стороне проходит более или менее заметная продольная борозда, не доходящая до заднего конца клетки. На переднем конце клетки у большинства криптофитовых расположена мешковидная «глотка», или воронка. По краю и на внутренней стороне воронки часто расположены так называемые трихо- цисты в виде палочек или зернышек. При раздражении трихоцисты выбрасывают тонкие нити. На переднем конце клетки находится одна или несколько пульсирующих вакуолей, изливающих свое содержимое в воронку или борозду. На перед¬
Рис. 33. Строение клетки Cryptomonas:
1 — вид с вентральной стороны, 2 — вид сбоку, 3 — поперечный разрез, а —глотка, б — трихоцисты, в — пиреноиды, г — ядро, д — тельца Мопа́ (по Fott, 1959).
82
нем конце клетки размещены два жгутика. При основании их находятся базальные тельца (блефаропласты), от которых вглубь клетки отходит ризопласт. Данные об их строении несколько противоречивы (Bourrelly, 1961). Так, у Chilomonas, по данным Брауна и Кокса (Brown а. Сох, 1954), один жгутик имеет отростки (мастигонемы), расположенные гребневидно, другой же — бичевидный, а у Cryptomonas, по данным Пителка и Шолли (Pitelka a. Scholley, 1955), оба жгутика гребневидного строения (рис. 33). Ядро одно, расположено в задней части клетки или, реже, на ее середине, имеет ядрышко (кариосому). У ряда видов обнаружена стигма. На переднем конце клетки вблизи пульсирующей вакуоли имеются своеобразные образования — два овальные двоякопрело- мляющие свет так называемые тельца Мопа́, функция которых неизвестна.
В клетках криптофитовых один-два, редко больше хроматофо- ров, окрашенных в различные цвета: от сине-зеленого до бурого и красноватого. Состав пигментов изучен еще недостаточно. Обнаружены хлорофилл а и в небольшом количестве хлорофилл с, α-каротин, возможно Е-каротин, а также ксантофилл (Нахо a. Fork, 1959); кроме того, найдены фикоэритрин и фикоциа- нин (O’h Eocha a. Raftery, 1959). У криптофитов известны пла- стидные и внепластидные пиреноиды (амфосомы). Запасный продукт — крахмал. У гетеротрофных форм обнаружены капли жира.
Размножаются криптофитовые продольным делением в подвижном, иногда в пальмеллевидном состоянии. При размножении воронка и вакуоли делятся пополам, оба жгутика отходят к одной клетке, другая же образует их заново. Имеются указания на образование зооспор и цист. Половой процесс неизвестен.
Что касается классификации криптофитовых, то Пашер (Pascher, 1914, 1931) рассматривал их как отдел филы (ствола) Руг- rhophyta вместе с отделами Desmokontae и Dinophyceae; при этом он различал среди Cryptophyta четыре класса — 1) Cryptomonadi- пеае с порядком Cryptomonadales, 2) Rhizocryptineae (не достоверно известный), 3) Cryptocrpsineae с порядком Cryptocapsales (Phae- ocapsales) и 4) Gryptococcineae с порядком Cryptococcales. Фрич (1935, 1956) поставил под сомнение класс Cryptocapsineae (Phae- ocapsineae), так как один из относимых к нему родов (Phaeococ- eus), по-видимому, относится к хризофитовым, а другой (Phaeop- lax) представляет пальмеллоидную форму Cryptochrysis из крипто- моиадовых. Исходя из этого, Фрич в классе Cryptophyceae выделял только два порядка — Cryptomonadales и Cryptococcales. Киселев (1954) принимает оба эти порядка, относя их к соответствующим классам Патера. Шадефо (Chadefaud, 1960), кроме порядков Cryptomonadales и Cryptococcales, различает еще группу прото- зойных криптофлагеллат (Protozoaires Cryptoflagelles), к которой он относит бесхлорофилльные организмы — паразиты или фаго- трофы, питающиеся твердой пищей. Фотт (Fott, 1959) подвергает
83
сомнению самостоятельность единственного представителя порядка Cryptococcales — Тetragonidium verrucosum и оставляет в классе Cryptopliyceae только один порядок Cryptomonadales с шестью семействами.
Положение криптофитовых в системе растительного мира до настоящего времени остается спорным. Пашер высказывался за их происхождение от хризомонад в качестве боковой ветви и выводил из них перидинеи (Pascher, 1911). За родство криптомонад с перидинеями высказывался и Клебс (Klebs, 1912). Несколько позже Пашер ввел их в систему водорослей (Pascher, 1914, 1931) и включил как отдел Cryptophyceae вместе с отделами Desmo- kontae и Dinophyceae в филу Pyrrhophyta. Это объединение в одну филу предполагало происхождение отделов от общих жгутиковых предков. Ряд авторов принял предложение Пашера (Smith, 1938; Киселев, 1954; Fott, 1956; Cronquist, 1960). Мельхиор (Melchior, 1954), кроме упомянутых выше групп, отнес к Pyrrhophyta также и хлоромонады, а Шадефо (Chadefaud, 1960) присоединил к ним и эвгленовые. Однако некоторые авторы считали, что включение криптофитовых в группу ииррофитовых недостаточно обоснованно (Fritsch, 1944, стр. 270). В связи с этим Грехем (Graham, 1951) выделяет Cryptophyceae из пиррофитовых и рассматривает их как придаток — «appendix» к этому отделу. Фрич (1935, 1956) рассматривает их как самостоятельный класс, а Смит (Smith, 1955) и Фотт (Fott, 1959) выделяют Cryptophyceae, как и хлоромонад, в группу организмов «неопределенного систематического положения», причем Фотт даже ставит их вне системы растительных фил (Fott, 1959, стр. 347, 368).
В пользу близости криптофитовых водорослей к перидинеям могут быть выдвинуты следующие аргументы. В строении клеток криптофитовых намечаются некоторые черты сходства с перидинеями: гребневидное строение жгутиков и наличие брюшной бороздки, которую сравнивают с соответствующей бороздкой пери- диней. Из биохимических признаков в пользу близости криптофп- товых к перидинеям выдвигают наличие целлюлозы в оболочке цист криптофитовых и в клеточных оболочках перидиней, а также образование в клетках тех и других крахмала. С другой стороны, имеются существенные различия в строении ядра криптофитовых и перидиней, у которых хромосомы характеризуются четковидным строением. Заслуживает также большого внимания пигментный состав криптофитовых, а именно: наличие фикобили- нов, α-каротина и специфического ксантофила в отличие от свойственных перидинеям ß-каротина и перидинина.
Несмотря на упомянутые существенные различия между крип- тофитовыми и перидинеями, Клейн и Кронквист, опираясь на приведенные выше некоторые черты сходства, считают возможным связывать эти группы филогенетически и выводить их вместе с эвгленовыми из низших зеленых водорослей (Klein a. Cronquist, 1967, стр. 235). Однако ряд морфологических и биохимических
84
особенностей, отличающих криптофитовые от настоящих пирро- фитовых, свидетельствуют против их близкого филогенетического родства. Поэтому мы считаем оправданным выделение их в отдельную независимую филу.
8. ЭВГЛЕНОВЫЕ ВОДОРОСЛИ — EUGLENOPHYTA (EUGLENOPHYCOPHYTA, EUGLENOPHYCEAE, EUGLENINEAE)
К эвгленовым принадлежат почти исключительно монадные формы. Клетки эвгленовых не имеют вторичной оболочки и покрыты более или менее плотным пери- пластом (пелликулой, плазмалеммой), часто имеющим своеобразную спиральную структуру. Под перипластом обычно размещены сферические или удлиненные слизистые тельца, выделяющие слизь. У некоторых форм (Trachelomo- nas) клетки находятся в футляре («домике»), пропитанном солями железа и марганца, через отверстие которого выходит жгутик. На переднем конце тела у эвгленовых расположена полость, так называемая главная вакуоля, снабженная узким выводным каналом, открывающимся в небольшое углубление —
«воронку» (рис. 34). В резервуар главной вакуоли изливается содержимое пульсирующих вакуолей. На дне резервуара прикреплены жгутики, выходящие через канал и воронку. Жгутиков у эвгленовых один или два. Только немногие роды (Hegneria и Euglenomor- pha) имеют их больше — до шести.
Жгутики при основании снабжены базальными тельцами (блефаропластами).
Для многих эвгленовых установлена связь базальных телец с ядром при помощи тонкой нити — так называемого
ризопласта. При наличии одного жгу- 1 ^ эвглены:
тика в нижнеи части наблюдали его раз-
1 а — жгутик, о — глотка, в —
двоение, причем каждая ветвь при осно- стигма, г — фоторецептор, ь —
вании имела свое базальное тельце, а Г°РсокРа?ител?,наяЫЙ вакуоля,
внутри главной вакуоли — остаток ВТО- ж — резервуар, з — блефаро- ύ г гл пласт, и — ризопласт, к — па-
рого жгутика. dTo дает основание пред- рамилон, л — пиреноид, ж —
полагать, ЧТО исходные формы эвглено- зернышки гематохро-
вых имели два жгутика. При наличии ма, v — митохондрии, с—штри-
А хи, т — пелликула, у — слизи-
двух жгутиков ОДИН направлен вперед стые тельца (по Johnson, 1968).
85
и служит для плавания, другой же обычно направлен назад и активной роли при движении не играет. Жгутики эвгленовых односторонне-перистые (гребневидные) (Brown а. Сох, 1954; Pitelka a. Schooley, 1955) и этим напоминают криптофитовые водоросли (см. рис. 3). В передней части клетки вблизи главной вакуоли обычно находится стигма. Стигма отсутствует у многих бесцветных видов. При делении клетки стигма также делится. Различные авторы указывают на наличие утолщения на жгутике или одном из его
Рис. 35. Деление клетки эвглены (схематизировано) : о — клетка перед делением, б — метафаза (образование новых жгутиков), в — анафаза, г — начало продольного разделения клетки (по Холланду, Попова, 1955).
разветвлений по соседству со стигмой, которую рассматривают как фоторецептор. Оно обнаруживается только у форм, имеющих стигму, с которой его и связывают.
Ядро у эвгленовых довольно крупных размеров, содержит обычно ядрышко. Особенностью митоза у эвгленовых является то, что при делении ядра делится и ядрышко (рис. 35), а также то, что ядерная оболочка сохраняется во время деления ядра.
Хроматофор у эвгленовых состоит из отдельных хлоропластов. Хлоропласты имеют пиреноиды или же лишены их. У некоторых видов описаны внепластидные пиреноиды. Хлоропласты у фото- трофных эвгленовых содержат хлорофилл а и Ь, ß-каротин и ксантофилл. Продукт ассимиляции, свойственный только эвгленовым, углевод парамилон, зерна которого откладываются или в хлоропласте возле пиреноидов, или, как у Euglena viridis, в центре клетки. У бесцветных эвгленовых запасным продуктом являются обычно жиры.
Среди эвгленовых имеются автотрофы (фототрофы), сапрофиты и фаготрофы. Известно также небольшое количество паразитных форм. Многие зеленые автотрофные эвгленовые характеризуются смешанным (миксотрофным) питанием — они могут использовать растворенные в воде органические вещества и даже аммиак и нитраты. При культивировании в темноте многие зеленые миксотрофные эвгленовые теряют хлорофилл и переходят на сапрофитный образ жизни, но на свету обычно снова вырабатывают хлорофилл.
Фаготрофные голозойные формы питаются твердыми органическими веществами и одноклеточными организмами. Для них характерен своеобразный орган — так называемый палочковидный аппарат, который Шадефо называет скелетом главной вакуоли (le squellete de puits flagellaire) и который y Peranema состоит из двух продольных палочек и одной поперечной — серповидной
Рис. 36. Эвгленовые (Euglenophyta) :
I — Euglena viridis, 2 — Euglena gracilis, 3 — Entosiphon sulcatum (a — стигма, б — главная вакуоля, в — пульсирующая вакуоля, г — хроматофор, д — парамилон, е — ядро, ж — фарингиальный аппарат, з — ризопласт) (по Fott, 1959).
(Chadefaud, 1960, стр. 212). Этот орган может выворачиваться и при поглощении твердой пищи прикрепляться к добыче. У некоторых родов, например у Entosiphon, описан фарингиальный аппарат, называемый сифоном (рис. 36).
Эвгленовые размножаются в результате продольного деления в подвижном или пальмеллоидном состоянии. Деление начинается с переднего конца. При этом образуется новый жгутиковый аппарат, делятся ядро, хроматофоры, пиреноиды и стигма. Как уже упоминалось, ядерная оболочка сохраняется во время деления. Ядрышко растягивается в поперечном направлении и делится. Расщепившиеся хромосомы расходятся к новым ядрышкам, после чего ядро делится на два.
Имеется ряд указаний на половой процесс у эвгленовых. Так, описан гологамный половой процесс у Scytomonas (Copromonas ) subtilis из Peranemaceae (Dobell, 1908) и у Euglena sp. (Biecheler, 1937). Описан автогамный половой процесс и последующий мейоз у Phacus purum (Krichenbauer, 1937). Достоверность наблюдений Крихенбауэра подтверждается описанием подобных стадий у Нуа- lophacus ocellatus (Leedale, 1962). Таким образом, можно считать, что половой процесс у эвгленовых встречается, хотя некоторые
87
авторы до настоящего времени высказывали сомнения по этому вопросу (Leedale, 1968).
Что касается классификации эвгленовых, то долгое время их делили, следуя Клебсу (Klebs, 1883, 1893), по способу питания на три семейства — Euglenaceae (фототрофы), Astasiaceae (сапрофиты) и Peranemaceae (голозойные формы) (Fritsch, 1935, 1956). Пашер (Pascher, 1931) выделил эвгленовые в качестве самостоятельной филы из сборной группы жгутиковых — Flagellata и разделил их на два класса Euglenineae с одним порядком Eiiglenales и Euglenocapsineae также с одним порядком Euglenocapsales. К первому Пашер отнес формы с господствующей в жизненном цикле жгутиковой стадией, а ко второму — пальмеллоидные формы.
Деление Пашера было принято Смитом (Smith, 1938, 1955), который, не выделяя классов, разделил эвгленовые на два порядка — Euglenales и Со- laciales (Euglenocapsales), и Бегером (Beger, 1954), который признавал в отделе Euglenophyta только один порядок — Euglenales с двумя подпорядками — Eugleninales и Colaciinales. В пределах подпорядка Eugleninales Бегер различал семейства Euglenaceae, Astasiaceae, Rhynchopodaceae, Peranemaceae и Rhizaspidaceae, а к подпорядку Colaciinales относил семейство — Colaciaceae.
Попова (1951, 1955) предложила класс Euglenophyceae 1 делить на два порядка: 1) Euglenales с тремя семействами— Euglenaceae, Colaciaceae и Astasiaceae и 2) Peranematales. Разделение эвгленовых на эти порядки обоснованно в связи со значительными различиями их по строению тела и способу питания. Этого же деления придерживается и Фотт (Fott, 1956, 1959, 1967).
Не углубляясь в классификации порядков Euglenales и Peranematales, предложенные различными авторами, отметим только некоторые системы. Так, Холланд (Hollande, 1952) выделяет среди эвгленовых четыре семейства (в зоологической номенклатуре) — Euglenidae, Peranemidae, Anisone- midae и Petalomonidae. Бесцветная Astasia включена в семейство Euglenidae, которое он разделяет на три подсемейства — Eutreptiinae с двумя функционирующими жгутиками, Eugleninae — с одним функционирующим жгутиком и Euglenamorphinae — с тремя и большим числом жгутиков.
Фотт (Fott, 1959) выделяет в порядке Euglenales семейства Eutreptiaceae, Euglenaceae, Menoidiaceae, а в порядке Peranematales семейства Peranemataceae, Anisonemataceae, Petalomonada- сеае и Rhizaspidiacae. В позднейшей работе Фотт (Fott, 1967) в порядке Euglenales различает семейства Euglenaceae и Astasiaceae, а в порядке Peranematales — Peranemataceae, Anisonemataceae и Petalomonadaceae, т. е. те же семейства, что и Холланд, с прибавлением проблематического семейства Astasiaceae.
Асаул (1970) делит эвгленовые на три порядка — Euglenales, Peranematales и Euglenomophales. К Euglenales она относит орга¬
1 Вторым классом отдела («типа») Euglenophyta Попова считает хлоро^ монадовые — Chloromonadineae.
88
низмы с фототрофным и сапрофитным типами питания, которые делит на три семейства — Eutreptiaceae Euglenaceae и Menoidia- сеае. К семейству Eutreptiaceae отнесены двужгутиковые формы с фототрофным и сапрофитным способом питания и с сильно выраженной метаболией. К семейству Euglenaceae отнесены одножгутиковые организмы с разными ступенями метаболии. Семейство Astasiaceae Асаул не выделяет, а роды Distigma и Distig- mopsis как вторичные формы относит к семейству Eutreptiaceae, роды Astasia и Cyclidiopsis — к семейству Euglenaceae, а немета- боличные роды Rhabdomonas, Rhabdospira, Menoidium и Gyropaig- пе, которые не имеют близких форм среди автотрофных эвглено- вых, Асаул выделяет в особое третье семейство Menoidiaceae. В порядке Peranematales Асаул выделяет два семейства — Рега- nemataceae и Petalomonadaceae, которые она разделяет на основании наличия или отсутствия палочковидного аппарата. К порядку Euglenomorphales, выделяемому Асаул условно, отнесены эндозоофитные роды Euglenomorpha и Hegneria обитающие в пищеварительном тракте земноводных и имеющие два — шесть жгутиков.
Что касается родственных связей и эволюции в пределах порядка, то прежде всего нужно отметить, что эвгленовые водоросли в отношении морфологической дифференциации пе пошли далее пальмеллоидных форм, которые несомненно происходят от подвижных, о чем свидетельствует то, что пальмеллоидные формы (Colacium) имеют, хотя и недолговременную, жгутиковую стадию. Так как строение их близко к строению представителей семейства эвгленовых, выделение предложенного Пашером класса Euglenocapsineae или порядка Colaciales (Euglenocapsales) не нашло поддержки у последующих авторов. Сапрофитные формы, выделяемые некоторыми авторами в семейство Astasiaceae, стоят близко к Euglenaceae и, очевидно, филогенетически с ними связаны, на что указывает то, что хлорофиллоносные эвгленовые могут питаться миксотрофно, а иногда — в отсутствие света — переходят к гетеротрофному питанию и теряют хлорофилл. Для некоторых бесхлорофилльных родов можно найти окрашенные роды, из которых они, возможно, возникли, например, род Astasia из рода Euglena, Distigma из рода Eutreptia (Fott, 1959, стр. 336).
Представители порядка Peranematales значительно отличаются от эвглепальных фаготрофным способом питания и связапным с ним наличием палочковидного аппарата, а у некоторых форм — и так называемого фарингиального аппарата. В то же время о родственных связях обоих порядков свидетельствуют характер жгутикового аппарата, строение ядра, наличие слизистых телец и па- рамилона, который обнаружен у некоторых перанематальных.
Можно предположить, что эвгленальные и перанематальные произошли от общих предков, способных к смешанному питанию, которые в процессе дальнейшей специализаци дифференцировались на автотрофные и сапрофитные Euglenales и фаготрофные
89
Peranematales. С эвгленальными, очевидно, связаны и выделяемые Асаул в особый порядок Euglenomorphaceae.
Спорным является вопрос о родственных связях эвгленовых с другими филами эвкариотических организмов. Со времени выделения Пашером в 1914 г. эвгленовых водорослей в особую филу большинство авторов — ботаников рассматривает их как самостоятельную филу и отдел — Euglenophyta (Fritsch, 1935, 1956;
Smith, 1938, 1955; Jahn, 1951; Fott, 1956, 1959, 1967; Комарниц- кий, Кудряшов, Уранов, 1962, и др.).
В то же время некоторые авторы на основании наличия у эвгленовых хлорофилла а и δ, свойственного зеленым водорослям, считают возможным сближать их с зелеными водорослями (Klein a. Cronquist, 1967) или даже включать в качестве класса в отдел зеленых водорослей (Christensen, 1962). В поддержку своей мысли Клейн и Кронквист приводят такие аргументы, как отсутствие вторичной оболочки у некоторых вольвокальных, наличие у них стигмы, а у некоторых форм, относимых к вольвокальным, перистых жгутиков. Клейн и Кронквист дают филогенетическую схему, на которой выводят эвгленовые непосредственно от зеленых водорослей.
Совершенно иного мнения придерживается Шадефо (Chade- faud, 1960), который относит эвгленовые в качестве класса к подотделу Pyrrophyceae отдела Chromophycophyta вместе с крипто- фитовыми (Cryptophycineae), перидинеями (Dinophycineae) и хло- ромонадами (Raphidophycineae, Chloromonadineae). В пользу этого положения Шадефо выдвигает, с одной стороны, сходство запасных продуктов — криптамилона у Cryptophycineae и Dinophycineae и парамилона у Euglenophycineae, а с другой — сходство монад, имеющих в этих группах на переднем конце мешковидные углубления и жгутики, из которых один гребневидный. Наконец, Попова (1955) объединяет в один отдел («тип») Euglenophyta классы Euglenophyceae и Chloromonadineae.
Действительно, за связь эвгленовых и зеленых водорослей свидетельствует наличие только у этих двух групп хлорофилла а и Ь, однако уже запасный продукт — парамилон, характерный для эвгленовых, отличается от крахмала зеленых растений и стоит ближе к ламинарину бурых водорослей (Leedale, Meeuse und Pringsheim, 1965). Что же касается строения клеток эвгленовых, то они имеют ряд отличительных черт, отсутствующих у зеленых водорослей,— наличие постоянной главной вакуоли с выводным каналом, слизистых телец, характер деления ядра.
С криптофитовыми водорослями эвгленовых сближает характер строения жгутиков, наличие мешковидной «глотки» и так называемых слизистых телец, однако такие отличия, как состав пигментов (отсутствие хлорофилла b и наличие хлорофилла с и фикобилинов у криптофитовых), характер запасных продуктов (крахмал у криптофитовых и парамилон у эвгленовых), свидетельствуют против возможности непосредственной филогенетиче¬
90
ской связи между этими группами. Против объединения эвглено- вых с перидинеями говорит характер ядра последних, пигментный состав, наличие у перидиней крахмала.
Что касается связей эвгленовых с хлоромонадовыми, то пока не изучено строение жгутиков и природа пигментов у хлоромо- надовых, трудно говорить о близких родственных связях между этими группами, тем более, что отсутствие у хлоромонад крахмала или иарамилона и ряд особенностей в строении их клеток свидетельствует скорее в пользу выделения хлоромонад в самостоятельный отдел.
Резюмируя все сказанное, можно сделать заключение, что эвгленовые следует рассматривать как самостоятельную филу — отдел эвкариотических организмов — Euglenophyta, или Eugleno- phycophyta, имеющий общие корни с другими филами среди первичных эвкариотов, чем и объясняются некоторые черты сходства их клеток с монадами других фил.
9. ХЛОРОМОНАДОВЫЕ - CHLOROMONADOPHYTA (RHAPHIDOPHYCEAE, CHLOROMONADOPHYCEAE, CHLOROMONADINEAE)
Хлоромонадовые представляют собой небольшую, но довольно хорошо обособленную группу монадных организмов, насчитывающую несколько родов с небольшим количеством видов.
Клетка хлоромонадовых не имеет вторичной оболочки и покрыта только сравнительно тонким перипластом (пелликулой). На брюшной стороне клетки обычно находится заметная борозда, заканчивающаяся на переднем конце углублением, из которого выходят два жгутика, имеющие при основании блефаропласты и ризопласты. При движении один жгутик направлен вперед, другой назад. Ядро довольно крупное, с ядрышками, расположено во внутреннем слое цитоплазмы. На верхней части ядра находится широкополукруглое образование ( «супрануклеарный аппарат» ), с которым соединяются ризопласты. Во внешнем слое цитоплазмы имеются дисковидные или яйцевидные хроматофоры без пиреноидов. Запасный продукт — масло. Стигма отсутствует. В эктоплазме находятся слизистые тельца и трихоцисты, которые при раздражении выбрасывают слизистые нити. Трихоцисты часто размещены группами на переднем или заднем конце монады. В верхней части клетки под воронковидным углублением расположен вакуолярный аппарат, который состоит обычно из большой сократительной вакуоли, изливающей содержимое в углубление. В углах между большой вакуолей и супрануклеарным аппаратом находятся более мелкие вакуоли, которые, увеличиваясь, сливаются в одну большую (рис. 37).
Хлоромонады размножаются продольным делением обычно в подвижном состоянии. Половой процесс неизвестен.
91
Рис. 37. Gonyostomum ovatum (хлоромонадовые) :
а — вид с вентральной стороны, б — вид сбоку, в — поперечный разрез (по Fott, 1959).
Некоторые авторы (Melchior, 1954; Chadefaud, 1960) включают хлоромонадовые как класс в отдел Pyrrophyta, другие (Попова, 1955) — в отдел Eugleno- phyta, третьи (Smith, 1955; Fott, 1959) рассматривают их как группу невыясненного систематического положения, наконец, четвертые рассматривают их или как самостоятельный класс (Fritsch, 1935, 1956) или же как отдел — Chloromonadophyta (Prescott, 1951, стр. 8).
Наличие слизистых телец и трихоцист, а также брюшной борозды сближает хлоромонадовые с криптофитовыми; имеются некоторые черты сходства у них с пиррофитовыми и эвгленовы- ми. Однако их можно объяснять и как результат конвергенции. Во всяком случае в настоящее время, пока не изучено строение жгутиков хлоромонадовых и не исследована природа их пигментов, трудно говорить о родственных связях этой группы, а поэтому до более детального цито¬
логического и биохимического изучения хлоромонадовых их целесообразно, следуя Прескотту, рассматривать как самостоятельную небольшую филу (отдел) Chloromonadophyta.
10. КРАСНЫЕ ВОДОРОСЛИ — RHODOPHYTA (RHODOPHYCOPHYTA, RHODOPHYCEAE)
Строение клетки и таллома
Красные водоросли являются обособленной группой эвкарио- тических организмов, характеризующихся рядом свойственных только им морфологических и биохимических особенностей. К красным водорослям принадлежат как одноклеточные, так и многоклеточные — нитчатые и пластинчатые формы, у которых отсутствуют жгутиковые стадии, имеющие, кроме хлорофилла а и каротиноидов, фикоэритрин, а в некоторых случаях и фико- цианин.
92
Клетка у красных водорослей одноядерная, только иногда (порядки Gelidiales и Rhodymeniales) старые клетки могут содержать несколько ядер. Ядро обычно имеет ядрышко и хроматино- вую сеть. Клеточная оболочка состоит из внутреннего целлюлозного и внешнего пектинового слоя. Оболочки некоторых красных водорослей инкрустированы карбонатом кальция (СогаШпасеае), а иногда карбонатом магния. У простейших форм имеется один центральный хроматофор звездчатой формы с пиреноидом; у более развитых форм — несколько пластид, лишенных пиреноидов, размещенных в постенном слое цитоплазмы. В хроматофорах красных водорослей обнаружены хлорофилл а и специфический для красных водорослей хлорофилл d и α-, ß-каротины, ксантофилл лютеин и фикобилины — фикоэритрин и фикоцианин, обнаруженный не у всех красных водорослей. Продуктом ассимиляции у красных водорослей является багрянковый крахмал.
Красные водоросли делятся на две хорошо различимые группы, которые мы, как и некоторые авторы (Курсанов, 1927, 1945; Melchior, 1954; Комарницкий и др., 1962), рассматриваем как классы — бангиевые водоросли (Bangiophyceae) и флоридеи (Florideophyceae). Ряд авторов (Fritsch, 1945; Drew, 1951; Smith, 1955; Fott, 1959, 1967) рассматривают их как подклассы единого класса Rhodophyceae. Пашер (Pascher, 1931) филу Rhodophyta делит на два отдела — Floridineae и Bangiineae.
Простейшие представители красных водорослей относятся к бангиевым и представляют собой одноклеточные округлой или овальной формы или же простые, или разветвленные нитчатые, иногда пластинчатые организмы с интеркалярным ростом. Флоридеи обычно более сложного строения,— это ветвистые нитчатые организмы, которые в результате дальнейшего усложнения приобретают псевдопаренхиматическое строение и разнообразную, иногда напоминающую высшие растения форму.
Различают два типа строения таллома флоридей — одноосевой (монаксиальный) по Смиту, или «тип центральной нити» по Ольт- маннсу, и тип многоосевой (мультиаксиальный) по Смиту, или «тип фонтана», по Ольтманнсу. В первом случае (одноосевой тип) основой является одна нить, состоящая из одного ряда клеток, растущая при помощи верхушечной клетки. От этой основной, центральной нити отходят ветви, растущие каждая при помощи своей верхушечной клетки. Из этих ветвей формируются различного тина вторичные структуры. Мпогоосевой тип отличается тем, что основа таллома состоит из пучка нескольких нитей, от которых радиально отходят ветви. Рост всех ветвей верхушечный. Боковые ответвления могут, разрастаясь, тесно сплетаться и образовывать довольно плотную ткань (псевдопаренхиму), в которой отдельные ветви можно узнать только по размещению межклеточных пор. Во многих случаях можно различать в таких сложных талломах «сердцевину» и «кору», клетки которой выполняют механическую и ассимиляторную функции.
93
У некоторых флоридей, например у Batrachospermum и Lema- пеа из Nemalionales, отмечено развитие ювенильной стадии (стадия Chantransia), аналогичной протонеме мохообразных. Иногда, например при слабом освещении, стадия Chantransia задерживается на продолжительное время и организм в этой стадии размножается при помощи моноспор.
Размножение и типы онтогенетического развития
У одноклеточных красных водорослей из класса Bangiophyceae деление клеток на две дочерние, по-видимому, является обычным способом вегетативного размножения. Что же касается более высокоорганизованных многоклеточных форм, то у них изредка наблюдается вегетативное размножение частями тела.
Бесполое размножение у красных водорослей происходит при помощи безжгутиковых спор, развивающихся по одной (моноспоры) или в большем числе в клетках — спорангиях. Материнские клетки спор (спорангии) могут не отличаться от вегетативных клеток или же возникают в результате обособления при помощи косой или кривой стенки от крупной вегетативной клетки. Развитие четырех спор (тетраспор) в так называемых тетраспорангиях обычно связано с редукционным делением.
Половой процесс у красных водорослей оогамный. Оогонии здесь обычно называют карпогониями. Оплодотворяющие элементы — спермации лишены жгутиков и пассивно переносятся токами воды. Описано также амебоидное передвижение спермациев.
У представителей первого класса красных водорослей — бан- гиевых известно два способа образования спермациев. В первом случае, например у родов Porphyra и Bangia, спермации развиваются в большом количестве в результате последовательного деления материнской клетки. В другом случае развитие спермациев сходно с развитием моноспор. При этом у видов родов Erythrotri- chia и Rhodochaete сперматангий (антеридий) образуется как небольшой выступ на вегетативной клетке, от которой отделяется вогнутой перегородкой в виде чечевицеобразной клеточки, содержимое которой — спермаций выходит после ослизнения оболочки (рис. 38).
Карпогонии у бангиевых не отличаются от вегетативных клеток. На них иногда образуются небольшие выступы, к которым прикрепляются спермации. Зигота делится на четыре — восемь карпоспор, при образовании которых происходит редукционное деление (мейоз). В связи с этим принято считать, что развитие бангиевых проходит по одностадийному гаплоидному типу.
Однако имеются данные, что развитие в этом классе, по крайней мере в некоторых случаях, проходит сложнее. Так, Дрю (Drew, 1949) показала, что из спор Porphyra umbilicaris на раковинах развивается растение, описанное в свое время как особый
94
вид — Conchocelis rosea. Позже эта стадия была обнаружена у других видов рода Porphyra, а также у видов Bangia. Голленберг (Hollenberg, 1958) проследил в культуре развитие спор Porphyra perforata и показал, что карпоспоры этого вида, развившиеся в результате полового процесса, дают начало стадии «Conchocelis». Из моноспор, развивающихся на этой стадии («конхоспор» до терминологии Тсенга п Чанга) развивается новое половое растение.
К сожалению, Голленберг не провел цитологических исследований. По данным Тсенга и Чанга (Tseng а.
Chang, 1955), у Porphyra tenera редукционное деление проходит при образовании карпоспор, а у Р. linearis, по Мань (Magne, 1952), карпоспоры образуются без редукционного деления.
Возможно, что у разных видов Porphyra развитие проходит по-разному.
Изучение развития Porphyra miniata (Chen a. al., 1970) подтвердило данные Голленберга. Чен и другие приводят данные по кариологическому исследованию этого вида, согласно которым вегетативные клетки и ß-споры Porphyra miniata содержат гаплоидное число хромосом, а α-споры и вегетативные клетки стадии Conchocelis — диплоидные.
Приведенные данные свидетельствуют о том, что стадия Соп- chocelis, вероятно, является самостоятельным поколением, а не нротонематической стадией. Что касается чередования ядерных фаз в классе Bangiophyceae, то этот вопрос требует дальнейших исследований.
Антеридии (сперматогонии) в классе флоридей (Florideophy- сеае) одноклеточные. В них образуется по одному спермацию. Иногда в спермациях наблюдаются первые стадии деления ядра, а у родов Batrachospermum и Nemalion к моменту оплодотворения в спермации имеется два ядра, из которых только одно участвует в оплодотворении.
Оогонии (карпогоны, карпогонии) у флоридей обычно расположены на конце коротких веточек и имеют вид колбовидных клеток с расширенной нижней частью и вытянутой шейкой, так
Рис. 38. Rhodochaete parvula (Bangiophyceae) :
а — образование сперматангия, б — спермации, прикрепившиеся на карпогоне, в — оболочки спермация, оставшиеся после оплодотворения на поверхности карпогона, г — молодая зигота (по Magne, 1960).
95
называемой трихогиной. Карпогон имеет одно ядро, а иногда также и хроматофор. В некоторых случаях, чаще всего в порядке Nemalionales, ядро делится на два, из которых одно остается в нижней части карпогона, а другое мигрирует в трихогину, где и дегенерирует. Это явление напоминает деление ядра сперма- тогония у некоторых представителей порядка Nemalionales. Спер-
мации пассивно переносятся водой и, попавши на трихогину, прикрепляются к ней, покрываются оболочкой, содержимое спер- матдия переходит через образовавшееся отверстие в кариогон, где и сливается с содержимым последнего (рис. 39).
Развитие зиготы в различных порядках флоридей проходит различно. В простейших случаях, первое деление зиготы является редукционным: из нее вырастают нити, так называемые гонимобласты, образующие цистокарпий, клеши которого дают по одной гаплоидной кар- поспоре. Эта карпоспора, прорастая, дает новое гаплоидное растение.
Дальнейшие усложнения в развитии связаны с развитием так называемых вспомогательных (ауксилярных) клеток, с одной стороны, а с другой — с задержкой редукционного деления и перемещением его на более поздние стадии развития.
В то время как, например, у Batrachospermum moniliforme гонимобласты (спорогенные нити) развиваются непосредственно из оплодотворенного карпогона, у других представителей порядка Nemalionales наблюдается ряд усложнений, связанных с питанием развивающихся гонимобластов (Drew, 1951, стр. 185). Так, у Ne- malion multifidum оплодотворенный карпогон делится поперечной перегородкой; гонимобласты развиваются из верхней клетки, а нижняя сливается с лежащей ниже клеткой карпогопиальной ветви, тем самым содействуя поступлению питательных веществ в развивающиеся гонимобласты. У других представителей порядка Nemalionales, например у Asparagopsis armata, по данным Све- делиуса, диплоидное ядро переходит в нижележащую клетку карпогенной ветви, из которой затем вырастают гонимобласты. Здесь уже можно говорить о вспомогательной (ауксилярной) клетке, обеспечивающей питательными веществами развивающиеся кар- погоны. В других порядках такие вспомогательные клетки могут быть расположены на расстоянии от карпогона. Тогда из оплодо¬
Рис. 39. Chantransia corymbifera:
а — карпогон со спермациями на трихогине, б — молодой гонимобласт, в — гонимобласт со зрелыми карпоспорами, г — сперматангии, д — моноспорангии с моноспорами (по Kylin, 1930).
96
творенного карпогона развиваются соединительные нити и на месте соединения этой нити с вспомогательной клеткой развивается цистокарпий (рис. 40). В других случаях вспомогательные клетки могут располагаться вблизи карпогона, иногда в непосредственном соседстве с ним, причем иногда образуется так называемый прокарпий, состоящий из карпогона и двух прилегающих к нему вспомогательных (ауксилярных) клеток (рис. 41).
Как уже упоминалось, у многих немалиональных, на-
Рпс. 40. Diidresnaya coccinea:
слияние соединительной (спорогенной) нити (а) с ауксилярной (вспомогательной) клеткой (б) (по Oltmanns, 1922).
Рис. 41. Callithamnion corymbosum
прокарпий (а — карпогон, б — карпого- ниальная ветвь, в — материнские клетки ауксилярных клеток) (по Oltmanns. 1922).
пример у Scinaia jurcellata (Svedelius, 1915), Nemalion multifidam (Kylin, 1916, и др.), первое деление зиготы редукционное, клетки цистокарпия гаплоидны и дают одиночные гаплоидные карпоспоры. При этом — гаплоидном типе развития, диплоидной является только зигота.
У других флоридей при прорастании зиготы редукционного деления не происходит, клетки цистокарпия имеют диплоидные ядра. Обычно и карпоспоры диплоидны. При их прорастании развивается диплоидное растение, на котором образуются тетраспорангии, в которых происходит редукционное деление и формируются четыре карпоспоры. Этот тип развития очень распространен среди флоридей и впервые был изучен у Polysiphonia violacea (Yamanuchi, 1906). Гаплоидным здесь является гаметофит, а диплоидными — цистокарпий, который по сути представляет собой диплоидный спорофит, паразитирующий на гаметофите, в связи с чем его называют карпоспорофитом, а также диплоидное растение, несущее тетраспоры и называемое тетраспорофитом. Таким образом, в этом случае мы имеем три поколения: одно гаплоид-
97
ное — гаметофит и два диплоидных — карпоспорофит и тетраспорофит. При этом гаметофит и тетраспорофит обычно не отличаются морфологически. Однако в некоторых, правда, довольно редких случаях, они настолько отличаются, что часто описывались под особыми родовыми названиями. Такое явление давно известно у видов Galaxaura из порядка Nemalionales (Howe, 1917; Svedelius, 1942). По исследованиям Фельдманна (Feldmann, 1942),
Рис. 42. Схема типов развития красных водорослей (Rhodophyta) (по Harder, 1962).
карпоспоры Bonnemaisonia asparagoides и Asparagopsis armata из того же порядка Nemalionales, прорастая, дают микроскопические тетраспорофиты, которые описывались соответственно под названиями Hymenoclonium serpens и Falkenbergia rufolanosa. Основные типы развития красных водорослей приведены на рис. 42.
Кроме этих двух основных типов развития — гаплоидного и гаплоидно-диплоидного, описаны и другие типы, не имеющие широкого распространения. Так, известны случаи, когда редукционное деление, как у Phyllophora brodiaei, происходит на цистокарпии при образовании спор; при этом на цистокарпии развиваются тетраспорангии (Rosenvinge, 1929; Claussen, 1929; Kylin, 1930). В данном случае мы имеем чередование двух поколений — гаплоидного гаметофита и диплоидного карпотетраспорофита. По это¬
98
му типу, по-видимому, проходит развитие также Liagora tetraspo- rifera (Börgesen, 1927; Kylin, 1930) и Helminthocladia hudsonii (Feldmann, 1939). Возник этот тип, вероятно, из предыдущего в результате переноса образования тетраспор на цистокарпий. Заслуживает внимания в связи с этим тот факт, что развитие другого вида рода Phyllophora — Ph. membranifolia проходит по типу Polysiphonia (Kylin, 1928).
У Asparagopsis armata имеются все три поколения — гамето- фит, карпоспорофит и тетраспорофит, однако редукционное деление происходит при прорастании зиготы, в результате чего диплоидная фаза представлена только зиготой (Svedelius, 1933). Lo- mentaria rosea из порядка Rhodymeniales представлена только одним тетраспорофитом. Сведелиус (Svedelius, 1937) утверждает, что в тетраспорангиях у этого вида редукционное деление не происходит. У Ahnfeldtia plicata половой процесс отсутствует и организм размножается только моноспорами.
При характеристике типов развития красных водорослей мы, следуя ряду авторов (Feldmann, 1952; Drew, 1955; Chapman, 1961), различаем цитологические фазы и морфологические стадии. Мы отличаем такие цитологические фазы — гаплоид, диплоид, триплоид и т. д. в зависимости от набора хромосом в ядре. Под морфологическими стадиями мы понимаем наличие отдельных поколений — сходных или несходных морфологически, независимых или развивающихся на другом растении. В зависимости от числа морфологических стадий в процессе развития организма мы говорим об одностадийном, двустадийном, трехстадийном развитии. Отдельные типы (циклы) развития характеризуются морфологическими стадиями и цитологическими фазами.
У красных водорослей мы различаем такие основные типы развития (life history по Drew, 1951, 1955): двустадийный гаплоидный — Scinaia furcellata, Nemalion multifidum, Batrachospe- rum moniliforme; трехстадийный гаплоидно-диплоидный — Polysiphonia violacea, Ceramium и производные от них: двустадийный гаплоидно-диплоидный — Phyllophora brodiaei\ трехстадийный гаплоидный — Asparagopsis armata; одностадийный гаплоидный — Ahnfeldtia plicata; одностадийный диплоидный — Lomentaria rosea.
При двустадийном типе развития оба поколения — гаметофит и карпоспорофит — всегда различаются морфологически. При трехстадийном в типичных случаях гаметофит и тетраспорофит морфологически сходны, но в некоторых случаях (Galaxaura, Bonnemaisonia asparagoides, Asparagopsis armata) тетраспорофит резко отличается морфологически от гаметофита, и все три поколения — гаметофит, карпоспорофит и тетраслорофит различны. В данном случае, следуя Дрю (Drew, 1955), можно говорить о триморфном типе (наличие трех морфологически отличных поколений).
99
Классификация и филогения
Красные водоросли — хорошо обособленная группа, в связи с чем большинство авторов с середины XIX ст. выделяют их как особый класс или отдел растительного мира.
Шмитц (Schmitz und Hauptfleiscli, 1897) и Энглер (Engler, 1898) рассматривают их как класс и делят на два подкласса — Bangiales и Florideae. Первый из них они целят на три семейства,
а второй — на четыре порядка — Ne- malionales, Gigartinales, Rhodymeni- ales и Cryptonemiales.
Веттштейн и другие авторы начала XX ст. обычно рассматривают красные водоросли как отдел растительного мира и делят его на классы Bangiales и Florideae (Wettstein, 1901, Engler, 1912). Несколько позже Ольтманнс (Oltmanns, 1904) выделил из Rhodymeniales пятый порядок — Ceramiales.
В результате дальнейшего изучения красных водорослей намечается тенденция к увеличению числа порядков. Так, Скуйя (Skuja, 1939) делит Bangioideae на четыре порядка — Porphyri- diales, Goniotrichales, Bangiales и Compsogonales. Мельхиор (Melchior, 1954c) к ним добавляет еще пятый — Rhodochaetales с одним видом Rhodochaete pulchella.
Особое положение в системе красных водорослей занимает Cyanidium caldarium, впервые описанный из Иеллоустонского парка в конце прошлого столетия под названием Protococcus botryoides /. caldaria (Tilden, 1898). На протяжении полустолетия его неоднократно описывали под различными названиями и относили то к зеленым, то к синезеленым, то к красным водорослям. Гейтлер дал ему название Cyanidium caldarium и выделил в особое семейство Cyanidiaceae (Geitier, 1936).
Cyanidium caldarium (рис. 43) — одноклеточный организм сферической формы. В клетке имеется ядро типа красных водорослей и пластинчатый хроматофор без пиреноидов. Исследования показали наличие в хроматофоре хлорофилла а и фикоцианина. В клетке обнаружен продукт ассимиляции, близкий к багрянковому крахмалу или гликогену. Размножается Cyanidium эндоспорами. Наличие типичного ядра и хроматофора не позволяет отнести его к синезелеиым водорослям, как это сделали Гейтлер, Фрич (Fritsch, 1945) и Шмидт (Schmidt, 1954). Нельзя согласиться с мнением авторов, относивших его к зеленым водорослям (Tilden, 1898; West, 1904; Hirose, 1950; Allen, 1959) из-за наличия в его клетках фикоцианина и углевсда, близкого к багрянковому крахмалу. Очевидно, нравы авторы, относящие Cyanidium к красным водорослям (Hirose, 1958; Chapman et al., 1968). Отсутствие y Cya-
100
Pnc. 43. Cyanidium, caldarium,:
a — три взрослые клетки, б — образование четырех спор (изображены только три из них), в — молодые клетки (по Hirose, 1958).
nidium фикоэритрина, вероятно, связано с условиями существования в пресноводных водоемах, а размножение эндоспорами свойственно также Rhodospora sordida, относимой к классу Вап- giophyceae. Хирозе (Hirose, 1958) сближает Cyanidium caldariam с Porphyridium и Rhodopsora и предлагает новое название — Rhodococcus caldarius, которое не может быть принято согласно современным правилам ботанической номенклатуры. Чапмен и другие (Chapman, Cole a. Siegelman, 1968) относят Cyanidium к бангиевидным, выделяя в пределах этого класса (у авторов подкласса Bangiophycidae) два порядка — Cyanidiales и Bangiales.
Класс Florideophyceae Кюлин (Kylin, 1937b) делил на шесть порядков — Nemalionales, Gelidiales, Cryptonemiales, Gigartinales, Rhodymeniales и Ceramiales, которые нашли широкое признание. Выделение этих порядков основано, главным образом, на строении и развитии репродуктивных органов, изученных еще далеко неполно. Возможно, что дальнейшие исследования дадут новый материал, который заставит пересмотреть принятую классификацию.
Приведенная нами ниже классификация красных водорослей подана по работам Скуйя и Мельхиора (класс Bangiophyceae) и Кюлина (класс Florideophyceae).
Отдел RHODOPHYTA (R H О D О P H Y С О P H Y Т А).
I. Класс BANGIOPHYCEAE (BANGIOIDEAE, PROTOFLO- RIDEAE) - БАНГИЕВИДНЫЕ.
Одноклеточные или многоклеточные, нитчатые или же пластинчатые организмы. Хроматофор центральный, звездчатый, с пиреноидом или хроматофоры постенные, лопастные, обычно без пиреноидов. Бесполое размножение путем деления (одноклеточные формы) или моноспорами. Половой процесс оогамный, состоит в слиянии спермация с яйцеклеткой, образующейся в кар- погонии, не имеющем трихогины. Зигота делится редукционно и дает четыре — восемь гаплоидных карпоспор. По-видимому, развитие происходит по одностадийному гаплоидному типу.
1. Порядок Porphyridiales — порфиридиа льные. Организмы одноклеточные или образуют слизистые колонии. Хроматофор один, центральный, с пиреноидом или же хроматофоры в большем числе, постенные, без пиреноидов. Размножение эндогенным делением на 2 или же 4—16 клеток (так называемые автоспоры).
2. Порядок Goniotrichales — гониотрихальные. Многоклеточные нитчатые формы. Хроматофор звездчатый, центральный, с пиреноидом или хроматофоры лопастные, постенные, без пиреноидов. Размножаются делением и акинетообраз- ными гонидиями или микрогонидиями, образующимися в результате повторного поперечного и продольного деления клеток, или же моноспорами.
3. Порядок В a n g i а 1 e s— бангиальные. Нитчатые, дисковидные или пластинчатые организмы. Хроматофоры или цент¬
101
ральные, с пиреноидом или же постенные без пиреноидов. Бесполое размножение моноспорами. Известен половой процесс.
4. Порядок Compsopogonales — компсопогональ- н ы е. Разветвленные нитчатые организмы, состоящие из одного ряда центральных клеток и нескольких слоев мелких коровых клеток. Хроматофоры постенные, без пиреноидов. Размножаются моноспорами и микроспорами.
5. Порядок Rhodochaetales — родохетальные. Таллом нитчатый, разветвленный, с верхушечным ростом и порами между клетками. Размножаются моноспорами. Известен также половой процесс.
II класс FLORIDEOPHYCEAE (FLORIDEAE) — ФЛОРИДЕИ.
Таллом многоклеточный, нитчатый, обычно очень разветвленный или же дисковидный или пластинчатый. Рост верхушечный. Клетки нити соединяются порами. Бесполое размножение моноспорами или тетраспорами. Половой процесс оогамный, карпогон снабжен трихогиной. Характерной особенностью развития является образование карпоспорофита (цистокарпия) и смена поколений, которых у более развитых форм известно три — гамето- фит, карпоспорофит и тетраспорофит.
1. Порядок Nemalionales — немалиональные. Таллом многоосевой или одноосевой. Ауксилярных клеток нет совсем или же как ауксилярная функционирует одна из клеток карпо- гониальной ветви. Редукционное деление происходит в большинстве случаев сразу после оплодотворения — гаплоидный двустадийный тип; в редких случаях оно происходит при образовании карпоспор (карпотетраспор) — гаплоидно-диплоидный двустадийный тип и, по-видимому, еще реже при образовании тетраспор на тетраспорофите — гаплоидно-диплоидный трехстадийный тип.
2. Порядок Gelidiales — гелидиальные. Таллом одноосевой. Типичные ауксилярные клетки отсутствуют. Чередование трех поколений — гаплоидно-диплоидный трехстадийный тип.
3. Порядок Cryptonemiales—криптонемиаль- н ы е. Таллом одноосевой или многоосевой, часто инкрустирован известью. Ауксилярные клетки закладываются на специальных ответвлениях до оплодотворения. Обычно развитие происходит по трехстадийному гаплоидно-диплоидному типу.
4. Порядок Gigartinales — гигартинальные. Ауксилярные клетки образуются из обычных интеркалярных клеток таллома. Обычный тип развития трехстадийный гаплоидно-диплоидный. Известны случаи редукционного деления при образовании карпоспор (карпотетраспор) — гаплоидно-диплоидный двустадийный тип.
5. Порядок Rhodymeniales — родимениальные. Таллом многоосевой с короткими карпогониальными ветвями. Ауксилярная клетка образуется до оплодотворения из дочерней клетки, так называемой несущей клетки, т. е. клетки, на которой
102
образуется карпогониальная ветвь. Обычный тип развития трехстадийный гаплоидно-диплоидный.
6. Порядок Ceramiales — церамиальные. Таллом одноосевой. Ауксилярная клетка образуется после оплодотворения из несущей клетки. Обычный тип развития трехстадийный гаплоидно-диплоидный.
Красные водоросли являются одной из древних групп эвкарио- тических растительных организмов. Достоверные ископаемые остатки красных водорослей описаны из нижнесилурийских отложений (ордовик), менее достоверные указания относятся к кембрию (Маслов, 1963; Gothan-Weyland, 1964). Однако они должны были развиться гораздо раньше, так как старейшие известные ископаемые остатки относятся к высокоорганизованным формам.
Вопрос о происхождении красных водорослей и о филогенетических связях их с другими отделами растительного мира различными авторами трактуется по-разному. Одни связывают их происхождение с зелеными водорослями, другие — с синезелеными, третьи отстаивают их независимое развитие из первичных ядерных организмов.
Предположение о возможной филогенетической связи красных и зеленых водорослей основывается, с одной стороны, на сходстве строения таллома Prasiola из улотрихальных водорослей и Рог- phyra из бангиевидных. С другой стороны, некоторые авторы конца XIX ст. обращали внимание на сходство оогония Coleochae- te и его прорастающей зиготы с карпогоном и цистокарпием флоридей, считая, что это сходство может быть, как предполагали де Бари и Сакс, следствием происхождения от общих предков (Oltmanns, 1922,2, стр. 432).
Действительно, пластинчатый однослойный таллом Prasiola очень сходен с одно- или двуслойным талломом Porphyra. Сходство усиливается еще тем, что в клетках той и другой водоросли обычно находится по одному звездчатому хроматофору с пиреноидом, а также тем, что бесполое размножение Prasiola происходит при помощи апланоспор (акинет), развивающихся по одной или по несколько в апланоспорангиях и напоминающих моноспоры бангиевидных. По мнению Шмитца (Schmitz, 1897), бан- гиевидные являются слепой боковой ветвью улотрихальных (ряда Ulothrix — Ulva — Prasiola). В отличие от Шмитца, Ишикава (Ishikawa, 1921) считает, что в начале ряда бангиевидных нужно поставить одноклеточные формы (Porphyridium) с примитивным ядром. Что же касается возможного родства флоридей с Coleo- chaete, то оно основано только на внешнем сходстве.
Неодинаковый набор пигментов и характер продуктов ассимиляции у зеленых и красных водорослей свидетельствуют о различном происхождении этих групп.
В прошлом столетии Кон (Cohn, 1867) на основании наличия фикоцианина и фикоэритррша у красных и синезеленых водорос¬
103
лей и отсутствия у тех и других жгутиковых стадий высказал предположение о возможных филогенетических связях между этими группами. Этот взгляд нашел поддержку у позднейших авторов (Ishikawa, 1921; Kylin, 1930; Skuja, 1939; Klein a. Cron- quist, 1967, и др.). Однако нужно иметь в виду, что красные водоросли настолько отличаются от синезеленых строением клетки, продуктами ассимиляции, а также характером размножения, что филогенетические связи между красными водорослями и синезелеными весьма сомнительны (Fritsch, 1945). Против родства сине- зеленых и красных водорослей резко выступает Прингсгейм (Pringsheim, 1966).
Строение клетки красных водорослей, размножение, характер полового процесса свидетельствуют о принадлежности их к эвка- риотическим организмам, одним из филогенетических стволов (фил) которых их считает ряд авторов (Pascher, 1931; Zimmermann, 1959; Chadefaud, 1960, и др.). Сходство (не идентичность!) пигментного состава их и синезеленых, возможно, является следствием развития этих групп в аналогичных экологических условиях.
Оба класса красных водорослей — Bangiophyceae и Florideo- phyceae при всех их отличиях характеризуются такими общими основными чертами (набор пигментов, продуктов ассимиляции и характер полового процесса), которые, несомненно, свидетельствуют об общности их происхождения. Более простое строение тела и половых органов, прямое развитие карпоспор из зиготы дало основание ряду авторов считать возможным развитие фло- ридей из бангиевидных, в связи с чем даже было предложено для бангиевидных название Protoflorideae (Rosenvinge, 1925). Однако высказывалось и противоположное мнение — о возможном происхождении бангиевидных из флоридей как упрощенной, редуцированной группы (Pascher, 1931, стр. 328, примечание 40).
Ряд отличий между бангиевидными и флоридеями (характер роста — верхушечный у флоридей и интеркалярпый у бангиевидных, межклеточные поры у флоридей, отсутствующие у большинства бангиевидных, различия в строении половых органов, в их размещении и в развитии зиготы) дает основание для предположения, что эти два класса представляют собой дивергентные группы, происходящие от общих предков (Kylin, 1930; Fritsch, 1944, стр. 258; 1945, стр. 438).
Эволюция в пределах класса Florideophyceae проходила как по линии усложнения строения вегетативного тела, так и по линии усложнения цикла развития. Более примитивным является одноосевой тип строения, из которого развился многоосевой. Что касается типов развития, то принято считать, что более простые представители флоридей гаплоидны, и редукция хроматина у них происходит при первом делении зиготы (двустадийный гаплоид). В дальнейшем мейоз откладывается на более поздние стадии развития, в связи с чем развивается диплоидный карпоспорофит, образующий диплоидные карпоспоры, из которых развивается
104
диплоиддое растение, сходное внешне с гаметофитом, но образующее тетраспорангии, в которых происходит редукционное деление с образованием тетраспор (тетраспорофит) — трехстадийный гаплоидно-диплоидный тип. Смит (Smith, 1955, стр. 312) предполагает, что из двустадийного гаплоида мог развиться двустадийный гаплоидо-диплоид типа Liagora в связи с переносом мейоза на цистокарпий, где образуются карпо-тетраспоры. Однако воз-
Рис. 44. Схема предполагаемых филогенетических связей между порядками красных водорослей (по Prescott, 1969).
можен и более вероятен другой путь происхождения этого типа — сокращение трехстадийного гаплоидно-диплоидного типа в результате более раннего прохождения мейоза. В пользу этого предположения свидетельствует то, что наряду с двустадийным гаплоидно-диплоидным типом у одного вида (Phyllophora brodiaei) у других видов того же рода имеется более распространенный трехстадийный гаплоидно-диплоидный тип развития (Phyllophora rnembranifolia).
В результате переноса редукционного деления на первую стадию прорастания зиготы трехстадийный гаплоидо-диплоид мог превратиться в трехстадийный гаплоид (Asparagopsis). В результате выпадения отдельных стадий могли возникнуть и другие типы развития — одностадийный гаплоид (Ahnfeldtia plicata) и одностадийный диплоид (Lomentaria rosea).
Дальнейшим усложнением трехстадийного гаплоидно-диплоидного типа развития является морфологическая дифференциация
105
гаметофита и тетраспорофита, что имеет место у Galaxaura и пред-* ставителей семейства Boimemaisoniaceae.
Усложнение развития у флоридей проходило также по линии образования вспомогательных (ауксилярных) клеток, обеспечивающих снабжение питательными веществами развивающегося цистокарпия. Образование вспомогательных клеток намечается у ряда представителей порядка Nemalionales. Однако своего полного развития они достигают в порядке Cryptonemiales, Gigarti- nales, Ceramiales и Rhodymeniales. При этом у одних форм карпо- гоны и вспомогательные клетки развиваются на различных частях таллома, у других они сближены и образуют так называемый прокарпий. В некоторых случаях (порядок Ceramiales) вспомогательные клетки образуются только после оплодотворения. Указанные выше признаки развивались в различных группах флоридей не сопряженно и, вероятно, в разной последовательности, поэтому установить более тесные родственные взаимоотношения между порядками при современном знании их типов развития весьма затруднительно.
Примером предполагаемых филогенетических схем красных водорослей может быть недавно опубликованная схема Прескотта (Prescott, 1969, X, рис. 44). В ней флоридеи выводятся из бан- гиевых, причем намечается два возможных пути их происхождения — от исходных бангиевых и от форм, близких к Compsogon. Как мы уже отмечали, более вероятным является дивергентное развитие обоих классов от общих предков.
И. ЗЕЛЕНЫЕ ВОДОРОСЛИ — CHLOROPHYCOPHYTINA (CHLOROPHYCOPHYTA, CHLOROPH Y CEAE, CHLOROPHYTA)
В отличие от широко распространенного признания зеленых водорослей особым отделом растительного мира, мы, исходя из положений, высказанных рядом авторов о филогенетической связи высших растений с зелеными водорослями, представляющими единый филогенетический ствол, единую филу (Fritsch, 1945; Smith, 1933, 1938, 1955; Fott, 1959, 1965), считаем целесообразным рассматривать все зеленые растения, начиная от зеленых водорослей и кончая цветковыми растениями, как единый отдел зеленых растений (Chlorophyta) с рядом подотделов — зеленых водорослей (Chlorophycophytina), мохообразных (Bryophytina), антоцеротовых (Anthocerotophytina), псилофитовых (Psilophyti- па) и нескольких подотделов высших сосудистых растений.
Если же по традиции последних десятилетий рассматривать эти подотделы зеленых растений как отделы одной большой филы зеленых растений, то зеленые водоросли следует называть СЫо- rophycophyta согласно предложению Папенфусса ( Paperif uss, 1946; Chadefaud, 1960; Chapman, 1964; Мейер, 1961), но никак не Chlorophyta, как это делают некоторые авторы (Smith, 1938, 1955; Fott, 1959, и др.).
106
Зеленые водоросли (Chlorophycophytina) довольно обширная и разнообразная в морфологическом отношении группа, в состав которой входят как одноклеточные подвижные и неподвижные в вегетативном отношении организмы, так и ценобиальные, многоклеточные, одноядерные и многоядерные формы с расчлененным и дифференцированным телом. Общими чертами для всех зеленых растений являются: 1) наличие в хроматофорах двух форм хлорофилла — а и 6, что свойственно также, кроме отдела Chlorophyta, и эвгленовым водорослям; 2) образование в результате фотосинтеза крахмала как запасного продукта; 3) наличие целлюлозы в клеточных оболочках. Обычно принято считать характерной чертой зеленых водорослей наличие одинаковых гладких бичевидных жгутиков, которыми снабжены в числе двух — четырех, реже восьми (некоторые Polyhlepharidaceae), шестнадцати (Polychaetochloris из тетраспоровых) или же в большем числе (Oedogoniaceae, Derbesia из сифоновых) зооспоры и подвижные гаметы. Однако исследования последнего времени (Pi- telka a. Scholley, 1955; Bourrelly, 1961) обнаружили среди организмов, относимых к вольвоксовым, формы с перистыми жгутиками в числе четырех (Platymonas), двух (Bipedinomonas), одного (Pedinomonas, Micromonas squamata) или же двух неодинаковых жгутиков — одного гладкого, бичевидного, а другого перистого (Thalassomonas) (см. рис. 3). Возможно, дальнейшее изучение этих организмов покажет принадлежность их к другим разделам растительного мира, однако нужно отметить, что у Pedinomonas обнаружено наличие двух форм хлорофилла — а и b (Harder u. Koch, 1954), что свидетельствует о принадлежности этого рода к зеленым растениям. Нужно также отметить, что в роде Sphae- rella (Haematococcus), относимом часто к семейству Chlamydomo- nadaceae, описаны гребневидные жгутики с односторонне размещенными мастигонемами. Возможно предположение, что в составе зеленых флагеллат, относимых к зеленым водорослям (Chlorophycophytina), имеются представители особых еще не изученных отделов или классов растительного мира (Christensen, 1962, 1964; Manton, 1965).
Строение вегетативных органов
Строение вегетативного тела зеленых водорослей очень разнообразно — у них можно наблюдать все типы талломов, выделенные Пашером (Pascher, 1914): монадный (флагеллоидный),
пальмеллоидный, коккоидный, нитчатый и сифоновый. Нужно отметить, что флагеллоидные простые и ценобиальные формы, объединяемые в порядке Volvocales, имеют сложное строение, свидетельствующее о значительной эволюции, которая имела место в пределах этой группы после отделения от нее пальмеллоидной и коккоидной ветвей. Из монадных форм развились другие типы строения зеленых водорослей. К монадной очень близки коккоид-
107
ная и пальмеллоидная формы строения, представленные в порядках Chlorococcales и Tetrasporales. Более сложным представляется переход от коккоидных или пальмеллоидных форм к нитчатым. Фотт даже считает, что разница между теми и другими больше, чем между папоротникообразными и цветковыми растениями (Fott, 1965, стр. 121).
По предположению Пашера (Pascher, 1930), нитчатая форма возникла из коккоидной в результате образования ряда автоспор,
которые развивались при делении протопласта на две части. При этом возникающие в результате деления клетки образуют свои собственные оболочки, заключенные внутри оболочки материнской клетки, которая растет одновременно с оболочками дочерних клеток. Последующее развитие нитчатых форм привело к образованию ветвистых, а в дальнейшем гетеро- трихальиых (разнонитчатых) форм (рис. 45) и сложных форм, наблюдаемых у харо- вых водорослей. Одноклеточные коккоидные формы могли дать также начало, с одной стороны, ценобиальным коккоидным, а с другой — многоядерным сифо- нальным формам.
Кроме широко принятого мнения о жгутиковом происхождении зеленых водорослей, существуют и иные гйпотезы. Так, Ша- дефо (Chadefaud, 1952) высказал предположение ob исходных одиночных клетках (archethallus), которые дали начало жгутиковым формам, с одной стороны, и неподвижным формам водорослей — с другой. Позже Шадефо (Chadefaud, 1960, стр. 390—391) высказался за то, что вольвокальные развились из хлорококкаль- ных в результате неотении, т. е. сохранения эмбрионального строения на протяжении всего цикла развития. Топачевский (1962) для всех отделов водорослей принимает происхождение жгутиковых форм от ризоподиальных.
Рис. 45. Fritschiella tuberosa:
гетеротрихальный таллом (а — поверхность земли, б — подземная часть таллома, в — надземная его часть) (по Сингу из Fott, 1959).
Вегетативное размножение
Для ряда одноклеточных форм (Pleurococcus, десмидиевые) характерным является вегетативное размножение делением клеток. Обычный тип вегетативного размножения многоклеточных нитчатых форм зеленых водорослей состоит в их распаде, фраг- ментации. У некоторых форм наблюдается образование так называемых покоящихся спор — акинет в результате утолщения
108
оболочки вегетативных клеток. Акинеты освобождаются в результате разрушения таллома. У ряда водорослей при наступлении засухи содержимое клетки может покрываться оболочкой, превращаясь в цисту; у многоядерных сифоновых форм протопласт может распадаться на значительное число цист, иногда многоядерных (ценоцисты Protosiphon).
Бесполое размножение
Бесполое размножение зеленых водорослей происходит в результате образования зооспор или апланоспор (автоспор) внутри материнской клетки — спорангия. У одноклеточных форм, например у хламидомонады и одноклеточных хлорококкальных, в спорангий превращается вегетативная клетка. У многих просто построенных нитчатых форм (например, Ulothrix) каждая клетка может превратиться в спорангий. В других случаях образуются специальные спорангии, которые у некоторых наземных форм могут отделяться от таллома (Trentepohlia). У зеленых водорослей споры образуются по одной (Oedogonium), две, четыре, восемь (Ulothrix) или больше (Cladophora, H у dr о diet у on). Зооспоры обычно не имеют вторичной оболочки, нс зооспоры Chlamydomo- nas и споры некоторых Protococcales покрыты вторичной оболочкой.
Для зооспор различных групп зеленых водорослей характерно количество жгутиков. Так, зооспоры Chlamydomonas, Chlorococ- cales имеют два жгутика, зооспоры Ulotrichales — четыре, а зооспоры эдогониальных и Derbesia из сифоиальных снабжены многочисленными жгутиками, размещенными кольцом около переднего бесцветного кончика. На основании количества жгутиков некоторые авторы пытались построить классификацию зеленых водорослей (Blackman a. Tansley, 1902).
Половой процесс
Половой процесс у зеленых водорослей представлен в разнообразной форме, начиная от слияния двух одноклеточных организмов — типичной гологамии, до копуляции яйцеклетки с антерозоидом, т. е. типичной оогамии.
Напомним, что гологамией (от греческих слов δλος — весь, целый и γάμος — брак, супружество) мы называем слияние целых клеток — голо- цитов (от греческих слов ολος — весь, целый и χότος—полость, вместилище, здесь переносно — клетка); мерогамией (от греческих слов μέρος — часть и γάμος — брак, супружество) мы называем слияние клеток, возникших в результате деления содержимого материнской клетки сразу или вскоре после их освобождения из этой последней.
Наиболее простой тип гологамии — слияние двух одноклеточных организмов известен у Pyramimonas} Dunaliella и у описанных Коршиковым Pedinomonas, Heteromastix (Коршшов, 1938).
109
Гологамный половой процесс в типичной форме представлен также у десмидиевых и зигнемовых, где любая клетка нити может копулировать с любой клеткой другой нити, или же в некоторых случаях с клеткой той же нити. Однако у зигнемовых обычно наблюдается дифференциация сливающихся клеток — содержимое одной из них переходит в другую. Здесь уже можно говорить об андроците и гинеците, т. е. о мужской и женской клетках. Нужно отметить, что у некоторых зигнемовых (Mougeotia) не все содержимое копулирующих клеток идет на образование зиготы.
Возникновение мерогамии у зеленых водорослей можно представить в виде своеобразной неотении, когда незрелая в вегетативном отношении клетка становится способной к слиянию с другой такой же незрелой клеткой. Пример такого переходного к мерогамии процесса приводит Фотт у Polytomelia, где происходит копуляция индивидуумов меньших размеров, чем вегетативные монады (Fott, 1959, стр. 222).
Типичная мерогамия состоит в слиянии мелких монад, на которые распадается протопласт материнской клетки. В таком случае сливающиеся монады обычно называют гаметами; клетки, в которых образуются гаметы,— гаметангиями, а сам тип полового процесса гаметогамией. Различают изогамию — слияние одинаковых гамет, и анизогамию — копуляцию неодинаковых гамет с двумя вариантами — гетерогамией и оогамией. В пределах подотдела зеленых водорослей можно наблюдать различные этапы усложнения мерогамного полового процесса и формирования половых органов — гаметангиев.
В простейших случаях у одноклеточных вольвоксовых и у хлорококковых весь протопласт клетки делится на гаметы, т. е. наблюдается явление, называемое голокарпией. У многих колониальных нитчатых форм (Ulothrix, Sphaeroplea и др.) практически каждая клетка, не отличающаяся морфологически от вегетативной, может превратиться в гаметангий. У оогамных форм гаме- тангии обычно значительно отличаются от вегетативных клеток.
Среди современных зеленых водорослей наблюдаются все переходы от изогамии к оогамии. При этом нужно отметить, что иногда в пределах одного класса или семейства, а в некоторых случаях даже рода, можно наблюдать различные типы полового процесса. Так, еще Горожанкин отметил, что, кроме изогамных форм, в роде Chlamydomonas имеются гетерогамные виды — Ch. braunii (Goroschankiii, 1890) и оогамные — Ch. coccifera (Goroschankin, 1905). В порядках Volvocales, Chlorococcales, Chaetophorales наблюдаются как изогамные, так и оогамные роды.
Переход от изогамного к гетерогамному половому процессу можно видеть у Pandorina morum из вольвокальных, у видов Enteromorpha из улотрихальных, у видов Anadyomene и Micro- dictyon из сифональных. Типичная гетерогамия характерна для Pandorina charkowiensis (Thompson, 1954), Rhizoclonium из кла- дофоральных, видов Caulerpa и Codium из сифональных.
110
Интересны переходы от одного вида анизогамии к другому — от гетерогамии к оогамии. Так, принято считать, что виды родов Pandorina и Pleodorina оогамны (Корников, 1938); однако у них описаны случаи, когда женские гаметы не теряют жгутиков до оплодотворения и даже сохраняют их некоторое время после слияния гамет (Iyengar, 1937; Iyengar a. Ramanathan, 1951). У Aphanochaete repens из хетофоральных в женском гаметангии образуется одна макрогамета, очень сходная с яйцеклеткой. Однако она выходит из гаметангия и передвигается при помощи четырех жгутиков.
Интересный случай перехода от гетерогамии к оогамии описан у видов рода Sphaeroplea (Pascher, 1939). У них любая клетка простого нитчатого таллома может превратиться в гаметангий. У одних из них — антеридиях развиваются многочисленные веретеновидные, снабженные двумя жгутиками микрогаметы (антерозоиды). У других образуются до 100, а иногда и больше крупных макрогамет, обычно описываемых как яйцеклетки. Однако Пашер отметил у них наличие сократительных вакуолей и двух жгутиков, при помощи которых они медленно перемещаются внутри оогония, а через некоторое время их теряют. Есть указания, что у Sphaeroplea tenuis подвижная макрогамета оплодотворяется вне гаметангия (Fritsch, 1929).
Типичными яйцеклетками принято считать макрогаметы, лишенные жгутиков. Однако у ряда оогамных форм такая яйцеклетка все же выходит из оогония в окружающую среду, где и происходит оплодотворение. Такое явление отмечено у Chlorogonium oogamum из вольвокальных (Pascher, 1931) и у Chaetonema irregulare из хетофоровых (Meyer, 1930). У Dictyosphaerium indicum из хлорококкальных каждая клетка женской колонии превращается в оогоний и дает две яйцеклетки, которые выходят из материнской клетки в слизь, окружающую колонию. Эти яйцеклетки лишены жгутиков и стигмы, однако в них обнаружены сократительные вакуоли (Iyengar a. Ramanathan, 1940). У других хлорококкальных— Golenkiniopsis (Golenkinia) и Micractinium единственная яйцеклетка оплодотворяется в оогонии, после чего молодая зигота выходит из оогония и остается прикрепленной к его оболочке (Корников, 1937, 1953; Iyengar a. Balakrishnan, 1956). У типичных оогамных форм оплодотворение происходит внутри оогония — виды родов Volvox из вольвокальных, Coleochaete из хетофоральных, виды родов Oedogonium и Bulbochaete из эдого- ниальных, Chara и N Hella из харовых.
Смена ядерных фаз и чередование поколений
До 1925 г. господствовало мнение, что для зеленых водорослей характерен гаплоидный тип развития, при котором вегетативное тело является гаплонтом, диплоидная фаза представлена только
ill
зиготой, а редукционное деление происходит при прорастании зиготы. Шусниг (Schussnig, 1930) отмечает, что это мнение было основано на незначительном количестве достоверных фактов, из которых основным является детально исследованное редукционное деление в зиготе у Coleochaete (Allen, 1905) и косвенным указания аналогичного типа у Chara (Debski, 1896, 1898; Ernst, 1918; Lindenbein, 1927; Karling, 1926).
В 1925 г. были опубликованы результаты исследования развития сифоновой водоросли Codium tomentosum (Williams, 1925), которые показали, что редукционное деление у нее происходит при образовании гамет, а из зиготы без периода покоя развивается диплоидное растение.
Несколько позже аналогичный тип развития был описан у хло- рококкальных водорослей Apiococcus (Korschikoff, 1926) и Chio- rochytrium lemnae (Курсанов и Шемаханова, 1927), а через два года было обнаружено изоморфное чередование гаплоидного и диплоидного поколений у Ulva и Cladophora (Föyn, 1929), а также у Enteromorpha (Hartmann, 1929).
Дальнейшие исследования подтвердили наличие у зеленых водорослей трех типов развития: 1) гаплоидного, при котором растение гаплоидно, диплоидна только зигота, первое деление ядра которой является редукционным;
2) диплоидного, при котором растение диплоидно; редукционное деление происходит при образовании гамет, а зигота без периода покоя прорастает в диплоидное растение;
3) гаплоидно-диплоидного, при котором происходит чередование гаплоидного и диплоидного поколений; редукционное деление происходит при образовании спор на диплоидном поколении — спорофите; из споры образуется гаплоидное поколение — гаметофит, а зигота без периода покоя дает диплоидное поколение. Основные типы развития зеленых водорослей приведены на рис. 46.
Первый — гаплоидный тип известен у ряда зеленых водорослей — Volvocales, Ulotrichales, конъюгат, эдогониевых, харовых и др. Второй — диплоидный, кроме упомянутых выше случаев, известен у многих сифональных — Bryopsis (Zinnecker, 1935), Acetabularia (Schussnig, 1929; Hammerling, 1934), Valonia (Schussnig, 1932, 1938; Schechner-Fries, 1934), а также описан у Cladophora glomerata (Schussnig, 1928). Некоторые из этих диплоидных организмов, например Valonia, Cladophora glomerata, имеют дополнительное бесполое размножение при помощи диплоидных зооспор. Третий — гаплоидно-диплоидный тип, с чередованием гаплоидного гаметофита и диплоидного спорофита, как уже упомянуто, известен у ульвовых Ulva и Enteromoprha (Föyn, 1929, 1934; Hartmann, 1929), кладофоровых — Cladophora (Föyn, 1929, 1934; Schussnig, 1928, 1930, 1931), Chaetomorpha (Hartmann, 1929). У всех этих водорослей поколения чередуются по изоморфному типу, т. е. оба поколения по внешнему виду сходны. Несколько позже было открыто изоморфное чередование поколений у сифоновых —
112
Рис. 46. Схема типов развития зеленых водорослей — Chlorophycophytina (по Harder, 1962).
Anadyomene (Iyengar a. Ramanathan, 1940) и Microdictyon (Iyengar a. Ramanathan, 1941), а также у Draparnaldiopsis indica из хетофоральных (Singh, 1942, 1945). Вероятно чередование поколений и у Fritschiella (Fritsch, 1942).
Дальнейшее усложнение гаплоидно-диплоидного типа развития заключается в образовании морфологических различий между гаметофитом и спорофитом — в развитии гетероморфного чередования поколений. Имеются сведения о гетероморфном чередовании поколений у представителей различных зеленых водорослей. Такое явление описано у Urospora из кладофоральных, где сама Urospora представляет многоклеточную нитчастую гаплоидную водоросль, а спорофит ее — ценоцистический одноклеточный организм, сходный с хлорококковой водорослью Codiolum («Torde, 1933). Подобное чередование поколений описано у улотрихальной водоросли Monostroma, таллом которой является гаметофитом и образует двужгутиковые гаметы, а спорофит имеет вид маленького одноклеточного одноядерного растения, на котором образуются четырехжгутиковые зооспоры (Kunieda, 1934; Moewus, 1938). У хе- тофоральной водоросли Stigeoclonium subspinosum зигота без редукционного деления образует короткую диплоидную нить, в клетках которой происходит редукционное деление с образованием гаплоидных зооспор, дающих нормальное растение (Juller, 1937).
Интересный случай гетероморфного чередования поколений описан у видов рода Halicystis из сифональных. Согласно Корн- манну (Kornmann, 1938), пузыревидная водоросль Halicystis ovalis — гаплоидное растение, образующее двужгутиковые гаметы. Из зиготы развивается нитчатая водоросль, описанная как представитель другого семейства, Derbesia marina, которая образует зооспоры, снабженные венчиком жгутиков. Позже Фельдманн (Feldmann, 1950) обнаружил аналогичное явление у Halicystis parvula и Derbesia tenuissima. Описано также гетероморфное чередование поколений у Bryopsis halymeniae и Derbesia neglecta (Hustede, 1960).
Из всех типов развития зеленых водорослей — гаплоидного, диплоидного и гаплоидно-диплоидного наиболее примитивный гаплоидный, при котором диплоидной является только зигота, при прорастании которой происходит редукционное деление. Этот тип развития является основным у всех примитивных водорослей. Происхождение других типов развития, по Фричу (Fritsch, 1942), связано с мутационным перемещением редукционного деления на другие стадии. В том случае, когда зигота прорастает без периода покоя, из зиготы может развиться диплоидное растение, на котором происходит редукционное деление или при образовании зооспор или же ппи образовании гамет. В первом случае гаплоидные зооспоры дадут начало гаплоидным растениям, дающим гаметангии. В результате организм в дальнейшем будет развиваться по гаплоидно-диплоидному типу с чередованием двух поколений — гаплоидного (гаметофита) и диплоидного (спорофита).
114
В том случае, когда редукционное деление происходит при образовании гамет, прорастающая зигота дает диплоидное растение, гаплоидными будут только гаметы, а организм развивается по диплоидному типу. При этом может сохраняться и бесполое размножение зооспорами, которые образуются без редукционного деления. Фрич считает возможным происхождение диплоидного
Рис. 47. Схема возможного происхождения основных типов развития зеленых водорослей (по Fritsch, 1942).
типа из гаплоидно-диплоидного при выпадении гаплоидной фазы. Взаимоотношения между типами развития зеленых водорослей представлены на схеме (рис. 47).
Пути эволюции и классификация зеленых водорослей
Представления об объеме зеленых водорослей оформились только к началу XX ст. Авторы второй половины XIX ст. обычно рассматривали все водоросли как один класс, который разбивался на ряд подчиненных групп. Так, Сакс в первых изданиях своего пособия (Sachs, 1873) делил класс водорослей на девять групп, из которых шесть полностью или частично включали зеленые водоросли (Die Hydrodictyeen, Volvocineen, Conjugaten (включая диатомовые), Siphoneen, Oedogonieen, Coleocheten). В четвертом издании той же книги он (Sachs, 1874) разделил талломные растения (Thallophyta) по характеру полового процесса на четыре класса — Protophyten, Zygosporeen, Oosporeen и Carposporeen. При этом зеленые водоросли были распределены между всеми четырьмя классами. Очевидно, под влиянием Сакса, Веттштейн (Wettstein, 1901) разбивает зеленые водоросли между отделами Zygophyta, куда относит конъюгаты и диатомовые, и Euthallo-
115
phy ta, к которым вместе с грибами относит прочие зеленые водоросли. Такое же объединение конъюгат и диатомовых и противопоставление их зеленым водорослям видим и у Ольтманнса (Oltmanns, 1904). Ч. Э. Бесси (Ch. E. Bessey, 1907) распределял зеленые водоросли между тремя филами растительного мира — Protophyceae, Zygophyceae и Siphonophyceae, в пределах которых находим и низшие грибы, и диатомовые водоросли.
Только Энглер и Вилле (Wille, 1897; Engler, 1898) в подотделе «настоящих водорослей» — Euphyceae выделяют три класса — Conjugatae, Chlorophyceae и Charales (Characeae у Вилле), которые в основном соответствуют нашему пониманию подклассов настоящих зеленых водорослей. Позже Энглер (Engler, 1912) возводит эти классы в ранг отделов растительного мира — Conjugatae, Chlorophyceae и Charophyta. Только в XX ст. ряд авторов присоединяет конъюгаты к зеленым водорослям (Blackman а. Tansley, 1902; Lotsy, 1907; Мережковский, 1909; Pascher, 1914 и др.). Лотси связывает филогенетически с зелеными водорослями также и харовые.
До настоящего времени ряд авторов выделяет харовые в особый отдел растительного мира (Pascher, 1931; Beger, 1954; Fott, 1956), большинство же вводит их в состав зеленых водорослей, разбивая последние на три класса — настоящие зеленые водоросли — Chlorophyceae (Euchlorophyceae), конъюгаты — Conjugato- phyceae (Conjugatae, Zygophyceae) и харовые водоросли — Charo- phyceae (Курсанов и Комарницкий, 1945; Fott, 1959; Chadefaud, 1960; Комарницкий, Кудряшов и Уранов, 1962). Некоторые же авторы, не выделяя классов, разбивают зеленые водоросли непосредственно на порядки (Smith, 1955; Fritsch, 1935, 1956; Iyengar, 1951).
Одновременно с объединением в один отдел (подотдел) растительного мира различных групп зеленых водорослей в результате углубленного изучения некоторые группы были изъяты из их состава. Так, Лютер (Luther, 1899) выделил некоторые роды, до тех пор относимые к классу настоящих зеленых водорослей, в отдельный класс разножгутиковых — Heterocontae. Основанием для этого было образование у этих организмов в результате фотосинтеза масла, а не крахмала и наличие двух неравных и неодинаковых по строению жгутиков. Несколько позже Пашер (Pascher, 1914) выделил разножгутиковые в особый отдел, объединив его в филе Chrysophyta с диатомовыми и золотистыми водорослями. Несколько позже Аллорж переименовал Heterocontae в Xantho- phyceae.
Пашер (Pascher, 1931) предложил следующую систему зеленых водорослей — без харовых, которые он выделяет в особый ствол.
Ствол CHLOROPHYTA
Отдел A — Chlorophyceae 1 класс — Volvocineae
Порядки: Chlamydomonadales, Volvocales
116
2 класс — ризоподиальные организмы «не достаточно известные»
(nicht genügend bekannt)
3 класс — Tetrasporineae
Порядки: Chlorongiales, Tetrasporales
4 класс — Prutococcineae
Порядок Protococcales
5 класс — Ulotrichineae
1 подкласс — Ulotrichinae
Порядки: Ulotrichales, Chaetophorales
2 подкласс — Microsporinae
3 подкласс — Oedogoniinae
6 класс — Siphonineae
1 подкласс — Monosiphonae
Порядки: Valoniales, Caulerpales, Vaucheriales
2 подкласс — Plektenchymatae
Порядок Codiales
7 класс — Siphonocladineae
1 подкласс—Radiatae
Порядки: Siphonocladiales, Dasycladiales
2 подкласс — Cladophorinae
Порядок Cladophorales
3 подкласс —Sphaeropleinae
Порядок Sphaeropleales
Отдел В — Conjugatae
1 класс — Placodermae
Порядок Desmidiales
2 класс — Saccodermac
Порядки: Mesotaeniales, Zygnemales
Последующие авторы классы системы Пашера обычно рассматривали как порядки, частично видоизменяя их число и состав.
Так, Фрич (Fritsch, 1935, 1956) зеленые водоросли, которые он считает классом Chlorophyceae (Isocontae), делит на девять следующих порядков: Volvocales, Chlorococcales, Ulotrichales, Cladophorales, Chaetophorales, Oedo- goniales, Conjugales, Siphonales, Charales.
Смит (Smith, 1938, 1955) отдел зеленых водорослей Chlorophyta делит на два класса — Chlorophyceae и Charophyceae. Первый класс — Chlorophyceae он разбивает на двенадцать порядков — Volvocales, Tetrasporales, Ulotrichales, Ulvales, Schizogoniales (Prasiolales), Cladophorales, Oedogoniales, Zygnematales, Chlorococcales, Siphonales, Siphonocladiales, Dasycladiales; в пределах класса Charophyceae он принимает только один порядок.
Фотт (Fott, 1959) дает следующую систему зеленых водорослей.
Ствол CHLOROPHYTA
1 класс Chlorophyceae с порядками Volvocales, Tetrasporales,
Chlorococcales, Ulotrichales, Bryopsidales и Siphonocladiales.
2 класс Conjugatophyceae (Conjugatae) с порядками Mesotaeniales, Zygnematales, Gonatozygales и Desmidiales.
3 класс Charophyceae с одним порядком Charales.
В начале системы зеленых водорослей стоят одноклеточные и колониальные организмы, обычно объединяемые в порядок Volvocales, от которых, как правило, выводят тетраспоральные, хло- рококкальные и другие порядки неподвижных в вегетативном состоянии организмов.
Голые в вегетативном состоянии формы вольвокальных относят обычно к семейству полиблефаридовых — Polyblepharidaceae.
117
Еще Фрич (Fritsch, 1935, 1956, стр. 85) считал, что семейство Polyblepharidaceae представляет искусственное объединение форм. Коршиков, описавший роды Pedinomonas, Cardiomonas и Heteromastix, предложил в 1923 г. выделить эти роды, а также описанный Шерффелем род Monomastix в особый подкласс Protochlori- пеае класса Volvocineae, противопоставив его подклассу Euvolvo- cineae (Коршиков, 1923, 1938).
У представителей этой группы клетки не имеют оболочки и несут один-два, иногда три (Trichloris) жгутика. Размножаются они делением, у некоторых известен гологамный половой процесс. Коршиков считал Protochlorineae «своеобразной и безусловно гетерогенной группой, мало родственной с другими вольвоксо- выми» (Коршиков, 1938, стр. 46). Пашер (Pascher, 1927) считал, что Protochlorineae Коршикова, возможно, являются боковой ветвью криптомонад. К этой же группе Фрич (Fritsch, 1935, 1956) отнес род Monomastix. Род Pedinomonas был позже обнаружен Вишером (Vischer, 1945), который его описал под названием Chlorochytridion tuberculatum в виду того, что монады этого организма имеют один задний жгутик. Вишер считал возможным рассматривать его как представителя группы, которая дала начало хитридиевым грибам, характеризующимся также одним задним бичевидным жгутиком. Бегер (Beger, 1954), следуя Корши- кову, выделил род Pedinomonas (Chlorochytridion) в особый порядок зеленых водорослей — Chlorochytridiales (Protochlorineae). В пользу принадлежности этой группы к зеленым водорослям свидетельствует наличие пиреноидов в хроматофорах, образование крахмала, как запасного продукта, а также то, что у видов рода Pedinomonas обнаружены хлорофилл а и b (Harder u. Koch, 1954).
Иначе подошел к классификации низших зеленых водорослей Шадефо (Chadefaud, 1960, стр. 331): он предложил зеленые водоросли делить на два класса —- Prasinophyceae и Euchlorophy- сеае. Основным отличием этих классов, по Шадефо, является наличие у монад первого из них — Prasinophyceae вестибулярной ямки (la fosse vestibulaire), обычно в форме воронки, оттянутой в брюшной продольный желобок. К Prasinophyceae Шадефо отнес в основном монадные формы — Platymonas, Pyramimonas, Prasi- nochlamy do monas, Prasinocarteria, Nephroselmis, Trichloris, Dan- geardinella, некоторые формы, зачисляемые обычно к Tetrasporales — Prasinocladus, Chlorodendron, а также Halosphaera, которые, хотя и с сомнением, часто относят к Xanthophyceae.
Электронномикроскопические исследования морских зеленых флагеллат (Manton, 1959; Manton a. Parke, 1960; Parke, 1961, и др.) обнаружили своеобразное строение организмов, отнесенных Шадефо к Prasinophyceae, а именно: наличие чешуек, покрывающих их тело и жгутики, а также тонкое перистое строение жгутиков, что дало новое обоснование для выделенного Шадефо класса Prasinophyceae.
118
Христенсен (Christensen, 1962), кроме класса Prasinophyceae, выделил еще класс Loxophyceae, куда он отнес организмы, монады которых не имеют радиальной симметрии, например, Pedino- monas и Micromonas. Однако последнее предложение не встретило пока поддержки. Что же касается класса Prasinophyceae, то он принят рядом авторов (Parke a. Dixon, 1968).
Дальнейшее изучение монадных форм зеленых водорослей, особенно с применением электронного микроскопа, несомненно даст новый материал по морфологии этой еще далеко неполно изученной группы, который послужит основанием для выяснения их родственных связей и уточнения классификации. Невыясненным остается систематическое положение рода Haematococcus, относимого к вольвокальным, с его гребневидным строением жгутиков, напоминающим эвгленовые водоросли (Benesovâ, 1949).
На основании имеющихся данных необходимо признать в пределах подотдела зеленых водорослей особого класса Piasinophy- сеае, отличающегося от прочих зеленых водорослей, включаемых нами в класс Euchlorophyceae, перистым строением жгутиков, наличием чешуек, покрывающих клетку, и некоторыми биохимическими особенностями.
Систему класса Prasinophyceae мы даем по Парке и Диксону (Parke a. Dixon, 1968).
* Класс PRASINOPHYCEAE - ПРАЗИНОФИТОВЫЕ ВОДОРОСЛИ.
1. Порядок Pyramimonadales — пирамимона- д а л ь н ы е.
Семейство Nephroselmidaceae с родами Heteromastix, Micromonas, Nephroselmis.
Семейство Polyblepharidaceae с родами Asteromonas, Chloraster (?), Pyramimonas, Stephanoptera.
Семейство Platymonadaceae с родами Platymonas, Tetraselmis.
Семейство Chlorodendraceae с одним родом Prasinocladas.
2. Порядок Halosphaerales — галосферальные.
Семейство Pterospermataceae с родами Hexasterias, Pachysphae-
ra, Pterosperma, Sphaeropsis.
Семейство Halosphaeraceae с родами Halosphaera, Hyalophysa.
В основу предлагаемой ниже классификации настоящих зеленых водорослей — класса Euchlorophyceae положены системы Фри- ча, Смита и Фотта.
Класс EUCHLOROPHYCEAE - НАСТОЯЩИЕ ЗБШЕНЫЕ ВОДОРОСЛИ.
I. Подкласс Euchlorophycidae.
1. Порядок Volvocales — вольвокальные. Одноклеточные или ценобиальные организмы, подвижные в вегетативном состоянии. Клетки голые или покрытые оболочкой, снабжены
119
двумя — четырьмя жгутиками. Бесполое размножение продольным делением, зооспорами или образованием подвижных дочерних це- нобиев. Половой процесс изогамный, гетерогамный и оогамный. Гаплобионты.
Семейства: Dunaliellaceae, Phacotaceae, Sphaerellaceae, Chla- mydomonadaceae, Volvocaceae.
Объем порядка Volvocales в значительной мере условный, так как даже после выделения из него ряда форм полиблефаридовых в его составе, кроме форм с бичевидными жгутиками, еще остаются такие организмы как Haematococcus, характеризующиеся гребневидным строением жгутиков. Необходимо дальнейшее детальное изучение одноклеточных представителей этого порядка с применением электронного микроскопа.
2. Порядок Tetrasporales — тетраспора л ьные. Одноклеточные или колониальные неподвижные в вегетативном состоянии организмы, клетки или колонии которых обычно погружены в слизь и прикреплены к субстрату. Клетки покрыты оболочкой, одноядерны, кроме рода Characiosiphon, мешковидные клетки которого многоядерны. Вегетативные клетки часто имеют пульсирующие вакуоли; у некоторых форм имеются так называемые псевдоцилии, сходные с настоящими жгутиками, но лишенные дюдвижности. Бесполое размножение зооспорами или неподвижными автоспорами. Половой процесс изогамный. Монады с 2, иногда 6—16 (!) жгутиками.
Семейства (по Фотту, 1959): Hypnomonadaceae, Chlorangia- сеае, Chaetochloridaceae, Tetrasporaceae, Characiosiphonaceae.
3. Порядок Clilorococcales — хлорококкальные. Одноклеточные, многоклеточные и ценобиальные формы. Вегетативные клетки покрыты оболочкой, одноядерны или многоядерны, без жгутиков, хотя у некоторых форм наблюдали временпое образование пульсирующих вакуолей и стигмы. Хроматофор обычно чашевидный, иногда расчлененный на небольшие участки, осевой или париетальный. Бесполое размножение двужгутиковыми зооспорами или апланоспорами (автоспорами). Половой процесс обычно изогамный, реже оогамный. Гаплобионты, реже дипло- бионты.
Семейства (по Фотту, 1959): Chlorococcaceae, Chlorochytria- сеае, Characiaceae, Hydrodictyaceae, Micractiniaceae, Gloeocystida- ceae, Radiococcaceae, Botryococcaceae, Dictyosphaeriaceae, Oocysta- ceae, Scenedesmaceae, Protosiphonaceae. Фрич (1935, 1956), Кортиков (1953) и Φοττ (1967) дают иное деление порядка на семейства.
4. Порядок Ulotrichales — улотрихальные. Многоклеточные — нитчатые неразветвленные, реже паренхиматические формы с одноядерными, иногда многоядерными клетками. Хроматофор пластинчатый, постенный, иногда рассеченный или звездчатый осевой. Бесполое размножение двужгутиковыми или четырехжгутиковыми зооспорами и апланоспорами. Половой процесс
120
изогамный, гетерогамный и оогамный. Гаплобионты и дипло- гаплобионты с изоморфным чередованием поколений.
Семейства: Ulotrichaceae, Microsporaceae, Cylindrocapsaceae,
Prasiolaceae, Ulvaceae, Sphaeropleaceae.
5. Порядок Chaetop h orales — хетофоральпые. Нитчатые разветвленные формы, таллом которых часто гетеро- трихальный, т. е. расчленен на простертую, прижатую к субстрату, и вертикальную части. Многие хетофоральные снабжены волосовидными щетинками, являющимися видоизмененными клетками. Хроматофор постенный, один или несколько. Бесполое размножение зооспорами и апланоспорами. Половой процесс изогамный, гетерогамный или оогамный. Гаплобионты или дипло-гаплобион- ты с изоморфным чередованием поколений.
Семейства: Chaetophoraceae, Trentepohliaceae, Coleochaetaceae, Chaetosphaeridiaceae, Pleurococcaceae.
6. Порядок Cladophorales — кладофоральные. Нитчатые, обычно разветвленные формы с крупными многоядерными клетками. Хроматофор одиночный, сетчатый, с пиреноидами, или же хроматофоры многочисленные. Деление клетки происходит путем образования кольцевого валика. Бесполое размножение (дву-) четырехжгутиковыми зооспорами и апланоспорами. Половой процесс изогамный реже гетерогамный. Известны гапло- диплобионты с изоморфным чередованием поколений и реже дип- лобионты.
Семейство Cladophoraceae.
7. Порядок Bryopsidales (Siphonales) — бриопси- дальние, или сифональные1. Нитчатые или пузыревидные организмы, иногда сложного строения, многоядерные (цено- цистические), иногда разделенные на многоядерные участки полными или неполными перегородками. Хроматофор состоит из многочисленных дисковидных хлоропластов. Бесполое размножение зооспорами с двумя или многими (Derbesia) жгутиками или апланоспорами. Половой процесс изогамный, гетерогамный или оогамный. Диплобионты или дипло-гаплобионты с изоморфным, реже гетероморфным чередованием поколений.
Семейства: Derbesiaceae (Halicystidaceae), Bryopsidaceae, Cau- lerpaceae, Dasycladaceae, Codiaceae, Valoniaceae, Phyllosiphonaceae.
8. Порядок Oedogoniales — эдогониальные. Нитчатые неразветвленные или разветвленные формы с одноядерными клетками и своеобразным клеточным делением. Хроматофор постенный, сетчатый, с несколькими пиреноидами. Бесполое размножение многожгутиковыми зооспорами. Половой процесс оогамный. Гаплобионты.
Семейство: Oedogoniaceae.
1 Согласно существующим правилам ботанической номенклатуры названия порядков должны производиться от названий типовых семейств. Поэтому мы, следуя Фотту (Fott, 1959, стр. 281), принимаем название Bryopsidales вместо обычного прежнего Siphonales.
121
II Подкласс Zygophycidae.
9. Порядок Zygnematales (Conjugales) 1 — зигнематаль- ные. Одноклеточные, колониальные и простые нитчатые формы. Клетки одноядерные, с одним или несколькими крупными хро- матофорами. Жгутиковые стадии отсутствуют. Вегетативное размножение делением. Половой процесс — гологамия в форме так называемой конъюгации.
Семейства: Mesotaeniaceae, Zygnemataceae, Gonatozygaceae,
Desmidiaceae.
Некоторые авторы принимают 4 порядка: Mesotaeniales,. Zygnematales, Gonatozygales и Desmidiales (Fott, 1959).
III Подкласс Charophycidae.
10. Порядок Charales — харальные. Вегетативное тело сложного строения, разделено на узлы и междоузлия. Боковые ветви с ограниченным ростом. Хлоропласты многочисленные. Половой процесс оогамный, антеридии сложного строения, одноклеточные оогонии покрыты коровыми клетками. Гапло- йионты.
Семейство Characeae.
Некоторые авторы делят зеленые водоросли на большее число порядков. Так, Прескотт (Prescott, 1969) делит их на двадцать порядков за счет разделения порядков Ulotrichales, Chaetophorales и Siphonales. Например, в порядке Ulotrichales он оставляет только семейство Ulotrichaceae, а семейства Microsporaceae, Cylindrocapsaceae, Ulvaceae, Schizogonaceae (Prasiola- ceae) и Sphaeropleaceae рассматривает как самостоятельные порядки. Из порядка Cnaetopnorales он выделяет порядки Coleochaetales и Trentepohlia- les, оставляя в порядке Chaetophorales только семейства Chaetophoraceae и Protodermataceae. Порядок Bryopsidales (Siphonales) Прескотт разбивает на четыре порядка — Siphonocladales, Dasycladales, Siphonales, Derbesiales.
В начале системы настоящих зеленых водорослей — Euchlo- rophyceae стоят формы, снабженные жгутиками в вегетативном состоянии — порядок Volvocales, с которым филогенетически тесно связан порядок Tetrasporales, а также порядок Chlorococcales. Наличие сократительных вакуолей и стигмы, а также псевдоци- лий у некоторых форм Tetrasporales является убедительным доказательством этой связи. Переходный характер Tetrasporales дает основание некоторым авторам (Fritsch, 1935, 1956; Iyengar, 1951; Beger, 1954) объединять этот порядок с вольвокальными, а другим (Коршшов, 1953) сближать их с хлорококкальными. Характер размножения, образование ценобиев настолько характерны для этих трех порядков, что некоторые авторы начала двадцатого столетия (Engler, 1912) считали возможным объединять их в один класс — протококковых.
1 Согласно современным правилам ботанической номенклатуры мы для порядка конъюгат принимаем название, произведенное от названия типового семейства и предложенное Смитом (Smith, 1955) — Zygnematales вместо распространенного Conjugales (Conjugatae).
122
Происхождение нитчатых форм зеленых водорослей было значительным шагом вперед по пути эволюции этой группы. Согласно теории Пашера (Pascher, 1930) нить образовалась из одного ряда автоспор, которые остаются объединенными разрастающейся оболочкой материнской клетки. Циммерманн (Zimmermann, 1949) как прототип зачаточной нити приводит описанный им вид OU- mannsiella lineata из хламидомонадовых, клетки которого после деления остаются соединенными боковыми сторонами в однорядные колонии. Циммерманн подчеркивает, что при этом изменяется полярность клеток.
Из нитчатых форм особого внимания заслуживают сфероплей- ные (Sphaeropleaceae) и микроспоровые (Microsporaceae). Семейство сфероплейных ряд авторов в связи с многоядерностью их клеток относил к порядку Siphonocladiales (Oltmanns, 1904; Eng- ler, 1912, и др.) или к классу Siphonocladineae (Pascher, 1931), но в настоящее время многие авторы на основании характера хроматофора включают их в порядок Ulotrichales (Fritsch, 1935, 1956; Iyengar, 1951; Beger, 1954; Fott, 1959). Прескотт (Prescott, 1969) семейство Spheropleaceae выделяет в особый порядок — Sphaeropleales.
Что касается семейства Microsporaceae с одним родом Micro- spora, то его выделяет в порядке Ulotrichales своеобразное строение оболочек, напоминающее строение оболочек конфервовых и колониальных форм центрических диатомовых (Prescott, 1969).
Сифональные водоросли (Bryopsidales, Siphonales) могут быть выведены из многоядерных форм, которые известны среди СЫо- rococcales (Protosiphon) и Tetrasporales (Characiosiphon).
Много различных мнений вызывало положение в системе зеленых водорослей семейства Oedogoniaceae. Внутреннее строение клетки у представителей этого семейства типично для зеленых водорослей, однако своеобразный характер ее деления и особенно многожгутиковость зооспор и антерозоидов ставит его в обособленное положение. Одни авторы включали эдогониевые как семейство в порядок или класс нитчатых водорослей — Соп- fervales (Wille, 1897; Engler, 1898, 1912), или в порядок Ulotrichales (Oltmanns, 1904; Курсанов, Комарницкий, Флеров, 1933, 1937). Другие рассматривали эдогониевые как подкласс Oedogo- niinae класса Ulotrichineae (Pascher, 1931) или подпорядок Oedo- gonineae порядка Ulotrichales (Fott, 1959). Было высказано предложение о выделении Oeodogoniales в качестве подотдела зеленых водорослей (Chlorophyceae) под названием Stephanocontae (Bohlin, 1901) или класса, противопоставляя их настоящим зеленым водорослям — Isocontae, конъюгатам — Acontae и разножгутиковым — Heterocontae (Blackman a. Tansley, 1902).
Нужно отметить, что наличие многих жгутиков, размещенных вокруг переднего конца монады, свойственно, кроме эдогоние- вых, также и роду Derbesia из сифональных, связанному в своем развитии с другим родом из той же группы — Halicystis. Нам
123
кажется более вероятным предположение о древних связях предков эдогониевых с многожгутиковыми низшими зелеными водорослями. В пользу такого предположения можно привести тот факт, что в роде Polychaeto chlor is из тетраспоральных зооспоры обладают шестнадцатью жгутиками. Поэтому мы присоединяемся к тем авторам, которые рассматривают эдогониевые как отдельный порядок Oedogoniales настоящих зеленых водорослей (Fritsch, 1935, 1956; Smith, 1938, 1955; Iyengar, 1951; Beger, 1954; Комарницкий, Кудряшов, Уранов, 1962, и др.).
Конъюгаты (Zygnematales, Conjugales) занимают среди зеленых водорослей особое положение в связи с отсутствием у них жгутиковых форм и своеобразным половым процессом. В основе полового процесса конъюгат лежит типичная гологамия, встречающаяся у многих низших водорослей (Pyramimonas, Dunaliel- la), у которых копулируют целые клетки, правда, снабженные жгутиками. Обычно у конъюгат образуется копуляционный канал из соединяющихся выступов оболочек копулирующих клеток, однако у Spirotaenia condensata, у которой клетки окружены толстым слоем слизи, копуляционный канал не образуется (Potthoff, 1928).
Что касается отсутствия жгутиковых стадий у конъюгат, то возможны два предположения. Можно считать, что они в процессе эволюции утратили подвижные стадии или же произошли от каких-то предков зеленых водорослей, еще до возникновения мо- надной стадии морфологической дифференцировки (Топачевський, 1952, 1962; Мейер, 1961, стр. 1083).
В связи с этим вопросом Прингсгейм пишет, что многие СЫо- rococcales лишены жгутиков, а половой процесс типа конъюгации существенно не отличается от полового процесса хламидомонады. Поэтому он считает, что нет никаких оснований исключать конъюгаты из состава зеленых водорослей (Pringsheim, 1966, стр. 1Q4).
Что касается харовых водорослей, то они легко увязываются с настоящими зелеными водорослями — Euchlorophyceae, хотя многие авторы выделяют их в особый отдел растительного мира. Их нужно рассматривать как древнюю группу зеленых водорослей — ископаемые харовьте известны начиная с девона (Маслов, 1963; Gothan u. Weyland, 1964),— сильно продвинувшуюся по пути усложнения строения вегетативного тела и образования своеобразных комплексных антеридиев, однако гаплоидный тип развития и двужгутиковые антерозоиды являются бесспорным доказательством их филогенетических связей с другими зелеными водорослями.
Возможные филогенетические связи между порядками настоящих зеленых водорослей — Euchlorophyceae мы передаем на схеме (рис. 48), в основу которой положена схема Смита (Smith, 1955, стр. 27, рис. 6).
На совсем иных принципах построена филогенетическая схема Мейера (1952, 1961). Развивая взгляды Миллера (1928) о нали-
124
чип у зеленых водорослей двух типов хлоропластов — центрального (осевого) и постенного, Мейер разделил отдел зеленых водорослей — Chlorophycophyta на три класса: Centroplastophyceae (Centroplastae), Parietoplastophyceae и Conjugatophyceae. При этом Мейер считает возможным виды одного рода, у которых встречаются разные типы хроматофоров (Chlamydomonas, Carteria), от-
Рис. 48. Схема филогенетических связей между порядками настоящих зеленых водорослей — Euchlorophyceae.
носить к разным классам. Класс Conjugatophyceae, в котором также встречаются оба типа хроматофора, Мейер не считает нужным разделять по этому признаку.
Деление зеленых водорослей на классы на основании одного признака — формы хроматофора, предложенное Мейером, носит искусственный характер и широкого отзвука не встретило. Заслуживает внимания, однако, его указание на то, что осевой тип хроматофора у зеленых водорослей более древний.
Зеленые водоросли, в частности хетофоральные, очевидно, являются предками высших растений.
Глава ÏÎI
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ И ОСНОВЫ КЛАССИФИКАЦИИ НИЗШИХ ГЕТЕРОТРОФНЫХ РАСТИТЕЛЬНЫХ ОРГАНИЗМОВ (ГРИБОВ)
1. ПРОИСХОЖДЕНИЕ НИЗШИХ БЕСХЛОРОФИЛЬНЫХ РАСТИТЕЛЬНЫХ ОРГАНИЗМОВ
Со времен Линнея в систематике растений утвердилось подразделение низших растений на водоросли и грибы. В связи с тем, что в строении тела и в способах размножения у некоторых водорослей и низших грибов наблюдается ряд сходных черт, при построении естественных систем растительного мира возникли предположения о родственной близости грибов и водорослей. Было высказано даже предположение, что грибы не являются систематической группой, а только физиологической, и что водные грибы (Mycophyceae = Phycomycetes) — не естественная группа, а объединение организмов различных естественных семейств и родов на основании единственного признака — отсутствия хлорофилла (Braun, 1843, цит. по Atkinson, 1909). Кон, базируясь на «естественном» методе в систематике, предложил ликвидировать группу фикомицетов («водных грибов» — Mycophyceae) и ее роды и виды разместить между родственными родами и семействами водорослей (Cohn, 1853, стр. 108, 139—142). Следуя этому принципу, Н. Прингсгейм (Pringsheim, 1858) расположил роды сап- ролегниевых между родами сифоновых водорослей. Позже Кон (Cohn, 1872) разработал систему, в которой, отбрасывая деление талломных растений (класс Thallophyta) на водоросли, грибы и лишайники, делит их на семь порядков. В этой системе муко- ровые помещены в порядок Zygosporeae вместе с диатомовыми и конъюгатами, Chytridiaceae, Peronösporaceae и Saprolegniaceae под названием секции Leucosporeae включены в порядок Oosporeae вместе с зелеными водорослями, и только базидиомицетьт и аско- мицеты выделены в особые порядки — Basidiosporeae и Ascospo- геае.
Де Бари в предисловии к монографии, посвященной морфологии и физиологии грибов, лишайников и слизевиков (De Вагу, 1866), также связывает грибы с водорослями. Он определяет грибы как бесхлорофильные, живущие за счет органических веществ, талломные растения, тело которых состоит из гиф, «ко¬
126
ротко говоря бесхлорофильные конфервовые (сифоновые) и лишайники» (De Вагу, 1866, стр. V). Сапролегниевые, по его мнению, связывают фикомицеты непосредственно с конфервовыми (сифоновыми) и могут быть с равным правом причислены к этим водорослям, как и к грибам (De Вагу, 1866, стр. V—VII). Все же де Бари считает целесообразным в противоположность Кону оставить грибы как отдельную группу растительного мира.
В позднейшей работе де Бари (De Вагу, 1881) грибы, за исключением слизевиков и бактерий, рассматривает как единую группу, включая в нее и лишайники. От основного восходящего ряда, в начале которого он помещает пероноспоровые (Регопо- spora и Pythium) как боковые ветви отходят зигомицеты и сапролегниевые и, вероятно, хитридиевые. При этом де Бари ставит вопрос, не представляют ли хитридиевые по крайней мере две группы организмов, непосредственно между собой не связанные.
Гоби (Gobi, 1881) в основном поддерживает взгляды де Бари. Он также выводит грибы как единую группу из зеленых водорослей, однако наиболее простыми грибами считает хитридиевые и дрожжи. Из хитридиевых Гоби выводит зигомицеты, оомицеты и высшие грибы.
Несколько позже де Бари (De Вагу, 1884), связывая фикомицеты с водорослями, размещает грибы «по ходу развития» в отдельный аскомицетный ряд (1. Peronosporeen, 2. Saproleg- nieen, 3. Mucorinen oder Zygomyceeten, 4. Entomophthoreen, 5. As- comyceten, 6. Uredineen), из различных групп которого выводит хитридиевые, нротомицеты и головневые, дрожжи и базидиоми- цеты.
Аналогичные взгляды на происхождение грибов из водорослей излагает в первом издании учебника ботаники Сакс (Sachs, 1868), рассматривая грибы как класс и считая, что фикомицеты стоят ближе всего к вошериевым из сифоновых водорослей и что от сифоновых водорослей отходит ветвь, начинающаяся фикомице- тами, которая, разветвляясь, в дальнейшем приводит к различным группам грибов (Sachs, 1868, стр. 623). В последующих двух изданиях учебника Сакс (Sachs, 1870, 1873) придерживается тех же взглядов на происхождение грибов от зеленых водорослей и дает их классификацию по де Бари. Но уже в четвертом издании он (Sachs, 1874) резко меняет свои взгляды и переходит на позиции Кона, согласно которым делит талломные растения (Thallo- phyta) на основании типа полового процесса на четыре класса — протофиты, зигоспоровые, ооспоровые и карпоспоровые и в пределах каждого из этих классов размещает различные группы грибов, как ответвления бесхлорофильных организмов. Таким образом, в 1874 г. Сакс в вопросе о происхождении грибов решительно переходит на полифилетические позиции и последовательно ликвидирует грибы как самостоятельную систематическую группу, помещая дрожжи вместе с бактериями в класс протофитов, слизевики и зигомицеты — в класс зигоспоровых, сапролегниевые
127
и пероноспоровые — в класс ооспоровых, аскомицеты и базидио- мицеты — в класс кариоспоровых.
Мы не будем останавливаться на детальном разборе этой системы, построенной крайне односторонне, исключительно на типах полового процесса без учета других особенностей строения организмов и имеющей сейчас только историческое значение,— достаточно сказать, что в ней сфероплея, вошерия, эдогониевые и фукусовые попадают в один класс ооспоровых вместе с сапролегние- выми и пероноспоровыми, а колеохетовые, флоридеи и харовые — в класс карпоспоровых вместе с аскомицетами и базидиомице- тами.
Нужно подчеркнуть, что взгляды Кона и Сакса отразились на взглядах многих авторов последующих десятилетий. Так, Клементс и Шир (Clements a. Shear, 1931, стр. 18—19) считали, что грибы являются «физиологическим приспособлением» («physiological adjustment») к условиям среды и что их можно обнаружить в каждом большом отделе растительного мира; их мало среди мхов и папоротников, но они не редки среди цветковых растений, т. е. все бесхлорофильные растения, по Клементсу и Ширу, являются грибами.
Под влиянием Кона и Сакса Брефельд (Brefeld, 1889) выводил зигомицеты из зигоспоровых, а оомицеты из ооспоровых водорослей, однако считал, что высшие грибы (аскомицеты и бази- диомицеты) происходят от зигомицетов. Что касается хитридие- вых, то по мнению Брефельда, они являются редуцированными формами оомицетов (Brefeld, 1881, стр. 162). Позже Брефельд (Brefeld, 1889, примечание к таблице системы грибов) повторяет то же мнение. Его же принимает и Саккардо (Saccardo, 1888).
Таким образом, в 70—80 гг. прошлого столетия большинство исследователей придерживалось мнения о происхождении грибов от водорослей; при этом одни авторы признавали грибы группой монофилетической, а другие — полифилетической.
Изложенные выше взгляды нашли отражение в ряде сводных работ и пособий конца XIX и начала XX ст. Так, Веттштейн (Wettstein, 1901) и Энглер (Engler, 1912) считают грибы полифилетической группой, происходящей от зеленых водорослей. Веттштейн даже объединяет зеленые водоросли и грибы в один отдел Euthallophyta. Он указывал, что по морфологическому строению и характеру полового процесса низшие грибы (фико- мицеты) близки к зеленым водорослям, в частности, хитридиевые к протококковым, моноблефаридовые к эдогониевым, некоторые сапролегниевые к сифоновым и т. д. Из этого Веттштейн делает вывод, что грибы произошли полифилетически от разных групп зеленых водорослей, в частности от сифоновых, к которым он относит вошерию, и конфервовых, к которым он относит ряд семейств одноклеточных и нитчатых водорослей, начиная от тет- распоровых и плеврококковых и кончая эдогониевыми и колеохе-
128
товыми (Wettstein, I, 1901, стр. 107). Высшие грибы — аскоми- цеты и базидиомицеты Веттштейн, следуя Брефельду, выводит из низших.
Энглер считал грибы полифилетической группой, примыкающей к сифоновым и другим зеленым водорослям без более близкой связи с ныне живущими формами, с своеобразным развитием, несколько напоминающим флоридеи (Engler, 1912, стр. 37). Фи- комицеты он делит на два порядка — зигомицеты и оомицеты. В конце системы оомицетов он помещает подпорядки Chytridiineae и Ancylistineae, которые считает редуцированными.
С водорослями, правда частично, связывает грибы и Лотси (Lotsy, 1907, стр. 10). Отмечая, что среди низших грибов намечаются два ряда — одножгутиковые и двужгутиковые, он высказывает мысль, что двужгутиковые грибы происходят от равножгутиковых зеленых водорослей (Isocontae), а одножгутиковые — от гораздо более низкоорганизованных организмов («von viel niedrigeren Organismen»). Высшие грибы Лотси выводит из флоридей.
Лотси, а за ним Мережковский были сторонниками полифи- летического и неоднократного происхождения грибов от водорослей. Так, Лотси (Lotsy, 1907, стр. 111) считал, что группа грибов (die Pilzreihe) могла не только развиваться в результате дальнейшей эволюции, но и на любой ступени развития могла пополняться новыми сочленами извне, из водорослей. Мережковский признавал фикомицеты бесхлорофильными, «белыми» водорослями (Leucophyceae) и высказывал мнение, что «разные «леукофицеи» произошли от разных групп водорослей, они происходят и теперь» (Мережковский, 1909, стр. 77). Грибами (Му- cetes) Мережковский считал только высшие грибы — аскомицеты и базидиомицеты и выводил их из бактерий через актиномице- ты (!), ничем не обосновывая этого своеобразного предположения.
Полифилетических взглядов на происхождение грибов от водорослей придерживался также Бесси (Gh. E. Bessey, 1907), который в разработанной им системе растительного мира, разделенного на 15 отделов («фил»), низшие грибы распределяет между водорослями в пределах двух отделов — Protophyceae и Siphono- phyceae и только высшие грибы выделяет в отдельный чисто грибной отдел — филу Carpomycetes.
В пределах филы Protophyceae Бесси различает два класса — Proto- coccideae, к которому, кроме водорослей, относит семейство Synchytriaceae, включающее роды Olpidium и Synchytrium, и Confervoideae, который он разделяет на пять порядков; к одному из них — Chaetophorales он относит в качестве семейства моноблефаридовые, помещая их рядом с эдогониевыми. Отдел—«филу» сифоновых водорослей (Siphonophyceae) Ч# Э. Бесси делит на два класса — Vaucherioideae и Bryopsidoideae. К первому он относит два порядка водорослей — Cladophorales и Siphonales и один порядок грибов — Siphonomycetes, в который включает семейства Saprolegniaceae, С1а- dochytriaceac, Ancylistaceae, Peronosporaceae, Mucoraceae и Entomophthora- ceae, а ко второму — Bryopsidoideae он относит семейство Chytridiaceae, включая его в порядок Valoniales.
129
Из сказанного, а также из приведенной в его работе схемы родственных отношений фил растительного мира видно, что
Ч. Э. Бесси был последовательным сторонником полифилетиче- ского происхождения грибов и считал их бесхлорофильными представителями различных классов и даже отделов водорослей.
Одновременно с теорией происхождения грибов от водорослей ряд исследователей высказывал мысль о первичной бесхлорофиль- ности грибов, которые, по их мнению, произониш от бесхлоро- фильных простейших (Protozoa). Так, Гоби (1883) в отличие от прежних своих высказываний выводил грибы из амебоидных организмов (Protamoeba) через род Vampyrella. Он считал низшие хитридиевые грибы (семейство Myxochytridiaceae, по его классификации) исходной группой, из которой развились более высокоорганизованные группы грибов (Hyphomycetes). Аналогичные взгляды высказывал и Данжар (Dangeard, 1886).
По мнению А. Фишера, голокарпические немицелиальные миксо- хитридиевые являются переходными от амебоидных, образующих зооспоры Rhizopoda (von einfacheren Myzecotozoen, den Monadinen im Sinne Zopfs und specieller noch den Monadineae Zoosporeae) к голокарпическим формам микохитридиевых (A. Fischer, 1892, стр. 9). Простейшие хитридиевые он считал исходной группой для всех высших грибов (А. Fischer, 1892, стр. 2—3).
Высказанные в конце XIX ст. взгляды на происхождение грибов от бесхлорофильных животных организмов были поддержаны в первой четверти XX ст. рядом авторов. Так, из бесхлорофильных животных организмов — амебоидных и жгутиковых выводили низшие грибы также Шерффель (Scherffel, 1901, 1925) и Вил- лемен (Vuillemin, 1912). Гоби в позднейшей работе (Гоби, 1916, стр. 5, 10) повторяет свои прежние взгляды, выводя низшие грибы из Pseudopodiata, точнее из Pseudosporaceae. Эти же взгляды нашли поддержку у Генкеля (1923), Брукса (Brooks, 1923), Ячев- ского (1927).
Кук (Cook, 1928) считал исходной для низших грибов и мик- сомицетов группу Protozoa, которую он называет, вслед за Ланкастером, Prôteomyxa и относит к корненожкам Rhizopoda. Свои выводы он основывает на морфологических и цитологических данных. Аналогичные высказывания находим у Гвинни-Воган и Барнз (Gwynne-Vaughan and Barnes, 1937).
Накопление нового фактического материала по морфологии, цитологии, онтогенезу и систематике низших растений в первой половине XX ст. дало новые основания для суждений о происхождении грибных организмов.
Автор капитальной монографии по сравнительной морфологии грибов Гейманн (Gäumann, 1926) считал, что гетерогенный класс архимицетов в узком понимании (т. е. миксохитридиальные и гглазмодиофоральные) может быть выведен из группы Flagellatae — Myxomycètes, все же прочие, «настоящие» грибы монофи- летически произошли из зеленых водорослей двумя ветвями —
130
оогамной (оомицеты) и зигогамной (хитридиальные 1 — зигоми- цеты). Высшие грибы, по Гейману, произошли от зигомицетов. Этот же взгляд по сути принимает и Вердерманн (Werdermann, 1954).
В более поздней работе Гейманн (Gäumann, 1949, стр. 10) определенно высказывается за бифилетическое происхождение грибов: оомицеты он выводит из сифоновых, а все прочие грибы из жгутиковых (Flagellatae) двумя ветвями — основной, которая начинается хитридиальными и в которую входят моноблефари- дальные, бластокладиальные, зигомицеты, аскомицеты и базидио- мицеты, и боковой, включающей архимицеты.
Нужно отметить, что к архимицетам Гейман относит одноклеточные, голые в вегетативном состоянии формы (семейства Olpi- diaceae, Synchytriaceae, Plasmodiophoraceae и Olpidiopsidaceae), а к хитридиальным большей частью одноклеточные организмы, покрытые вторичной клеточной оболочкой (Rhizophidiaceae, Rhi- zidiaceae, Cladochytriaceae, Chytridiaceae, Megachytriaceae). Позднее он (Gäumann, 1964, стр. 10) дает ту же схему с той разницей, что оомицеты выведены из порядка Heterosiphonales отдела золотистых водорослей, очевидно, в связи с переносом рядом авторов вошериевых в этот отдел растительного мира, а архимицеты выведены бифилетически из флагеллат и слизевиков (Myxomycètes). Грейс (Greis, 1943) считал грибы полифилетической группой. Он полагал, что архимицеты в узком понимании, т. е. так называемые миксохитридиальные, частично являются перешедшими к паразитическому образу жизни миксомицетами, частично же паразитическими флагеллатами. Что касается самих миксоми- цетов, то по Грейсу они происходят от гетеротрофных флагеллат. Происхождение лабульбениальных Грейс увязывает с красными водорослями. Что же касается основной массы грибов — Ешпусе- tes, то по Грейсу и их происхождение, возможно, является не монофилетическим: они частично происходят от Monadophyta, частично от зеленых водорослей — Chlorophyceae. Если же учесть, что зеленые водоросли являются группой неоднородной, то картина происхождения настоящих грибов по Грейсу еще более усложняется. Во всяком случае происхождение фикомицетов представляется Грейсу также полифилетическим.
Изучение жгутикового аппарата водорослей и низших грибов дало новый материал для выяснения филогенетических связей этих групп организмов. Основываясь на строении жгутиков,
Э. А. Бесси (Bessey, 1942, 1950) считает исходной группой для всех низших грибов одноклеточные формы разножгутиковых водорослей (Heterocontae, Xanthophyceae), из которых низшие грибы развились тремя стволами. Он полагал, что у одних из них (ствол Olpidiopsidaceae — Peronosporaceae — Saprolegniaceae) жгутики сохраняют свойственное разножгутиковым строение — один
1 То есть микохитрндиевые в смысле А. Фишера (1892).
131
жгутик перистый (tinsel type), а другой бичевидный (whiplash type); у других (Hyphochytriaceae) сохраняется только перистый жгутик; у третьих (ствол Olpidiaceae — Monoblepharidaceae) сохраняется только бичевидный жгутик. Зигомицеты Бесси выводит из первого ствола; высшие грибы — из красных водорослей.
Таким образом, Э. А. Бесси дает новый вариант полифилети- ческой схемы происхождения грибов.
Смит (Smith, 1938, 1955) считает маловероятным выведение низших грибов из зеленых водорослей в связи с особенностями их метаболизма и строением жгутикового аппарата и придерживается мнения о происхождении фикомицетов от Protozoa, хотя и допускает возможность происхождения двужгутиковых фикомицетов с передним перистым жгутиком от разножгутиковых — Xanthophyceae.
Моро (Moreau, 1953) выводит грибы из зоофлагеллат, при этом переходной группой считает хитридиальные (архимицеты в смысле А. Фишера), которые на основании строения жгутикового аппарата разделяет на четыре группы: Prosomastigochytridiales с одпим передним перистым жгутиком, Opisthomastigochytridiales с одним задним бичевидным жгутиком, Dimastigochytridiales с двумя жгутиками: передним — перистым и задним — биче¬
видным и Amastigochytridiales — без жгутиков. Prosomastigochytridiales, по мнению Моро, не дали более высокоорганизованных грибов, от Opisthomastigochytridiales он выводит Blastocla- diales и Monoblepharidales, от Dimastigochytridiales — Saproleg- niales, Leptomitales, Pythiales и Peronosporales, а от Amastigochytridiales — зигомицеты, асковые и базидиальные грибы.
Спарроу в работе, посвященной филогении водных грибов (Sparrow, 1958), становится на позиции Моро, признавая независимое происхождение основных классов низших грибов — Chy- tridiomycetes, Hyphochytridiomycetes, Plasmodiophoromycetes и Phy- comycetes (т. e. ряда Leptomitales — Saprolegniales — Peronosporales). Первые три класса Спарроу выводит из «монад», а что касается Phycomycetes в узком понимании, то он считает, следуя Смиту, возможным их происхождение от разножгутиковых водорослей.
Цейп (Cejp, 1957) низшие грибы (подкласс Phycomycophytina) выводит из «сифональных водорослей или из бесцветных флагел- лат» тремя ветвями — двужгутиковой, одножгутиковой с передним жгутиком и одножгутиковой с одним задним жгутиком. Зигомицеты он выводит из двужгутиковых санролегниальных или, возможно, из одножгутиковых хитридиальных. Высшие грибы Цейп выводит из фикомицетов.
Шадефо (Chadefaud, 1960, стр. 427—428) выводит грибы из водорослей и считает, что группы грибов родственны соответствующим группам водорослей, представляя бесхлорофильные ветви этих групп. Он считает, что аскомицеты, базидиомицеты
132
и зигомицеты, возможно, связаны с красными водорослями, а фи- комицеты — с хризофитовтлми и пиррофитовыми.
Взгляды Бесси на происхождение грибов от двужгутиковых предков развивают Убрижи и Вереш (Ubrizsy and Vörös, 1966), которые выводят грибы из флагеллатных предков с двумя бичевидными жгутиками, от которых, с одной стороны, произошли слизевики, а с другой — двужгутиковые формы с одним перистым и одним бичевидным жгутиком — порядки Lagenidiales, Lepto- mitales, Saprolegniales и, возможно, Peronosporales; при этом обращенный вперед жгутик превратился в перистый. От этих форм в результате потери бичевидного жгутика образовались Hypho- chytriales, в результате потери перистого — Chytridiales. Saprolegniales, по Убрижи и Вереш, дали начало энтомофторовым и му- коральным, а от мукоральных произошли аскомицеты и базидио- мицеты.
Таким образом, вопрос о происхождении грибов до настоящего времени сводится к двум основным взглядам. Одни авторы считают, что грибы развились из водорослей в связи с потерей ими хлорофилла, при этом, по мнению некоторых, полифилетически и даже неоднократно на протяжении веков. Другие полагают, что грибы обособились на очень ранней стадии развития из низших амебоидных организмов и развивались самостоятельно как бесхлорофильный ствол (или стволы) органического мира.
Попытки выведения низших грибов из зеленых водорослей, основанные часто на внешнем сходстве вегетативных и репродуктивных органов некоторых высших фикомицетов и нитчатых водорослей, встречают серьезные возражения. Так, Смит (Smith, 1938, 1955, стр. 367) отмечает, что сапрофитные и паразитные бесхлорофильные зеленые водоросли, подобно зеленым хлорофиллоносным водорослям, отлагают в качестве запасного продукта крахмал, отсутствующий у всех грибов. Резко отличаются зеленые водоросли от фикомицетов также строением и положением жгутиков. Жгутики у зеленых водорослей всегда прикреплены к переднему концу и у двужгутиковых представителей морфологически сходны между собой, в то время как у одних фикомицетов жгутики или одиночны и прикреплены к заднему концу зооспоры или гаметы, а если в некоторых случаях прикреплены к переднему концу, то имеют перистое строение (гифохитриаль- ные), или же парные, но разного строения и направления — направленный вперед жгутик перистый, а направленный назад гладкий, бичевидный. В связи с тем, что строение жгутиков и характер обмена веществ являются существенными признаками у низших растений, Смит считает необоснованным выведение низших грибов из зеленых водорослей, с чем нельзя не согласиться.
Более обосновано предположенрге о происхождении низших грибов от одноклеточных неравножгутиковых водорослей — Hete- rocontae. Xanthophyceae (E. A. Bessey, 1942, 1950). Действительно, двужгутиковые фикомицеты (Saprolegniomycetes — Oomyce-
133
tes) имеют строение жгутикового аппарата, сходное с таковым у изученных представителей разножгутиковых водорослей. Кроме того, в их оболочке обнаружена целлюлоза: к тому же у разножгутиковых водорослей не откладывается крахмал, отсутствующий и у фикомицетов. Все это является серьезным обоснованием для предполагаемых филогенетических связей двужгутиковых фикомицетов с разножгутиковыми водорослями — не с их высшими представителями типа вошерии, а с низшими голыми одноклеточными формами, обычно объединяемыми в порядок Hetero- chloridales. Однако выведение одножгутиковых фикомицетов (класс хитридиомицеты) с прикрепленным сзади бичевидным жгутиком из тех же одноклеточных неравножгутиковых водорослей в результате утраты направленного вперед перистого жгутика маловероятно, тем более, что в отличие от двужгутиковых сапро- легниальных, в их оболочках отсутствует целлюлоза. Так же мало убедительна попытка Убрижи и Вереша (Ubrizsy a. Vörös, 1966) вывести все грибы из флагеллат с двумя бичевидными жгутиками.
Современные знания морфологии, онтогенеза и биохимических особенностей грибных организмов, нам кажется, в достаточной мере убедительно свидетельствуют в пользу высказанного еще в конце XIX ст. предположения о происхождении грибных организмов (грибов и слизевиков) от бесхлорофильных амебоидных организмов. Только относительно двужгутиковых сапролегниальных грибов и, очевидно, связанных с ними гифохитриальных, имеются веские аргументы в пользу происхождения их, если не непосредственно от низших амебоидных форм разножгутиковых водорослей (Xanthophyceae), то во всяком случае от общих с ни́ми предков.
2. ОБЩИЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ ГРИБНЫХ ОРГАНИЗМОВ И ОСНОВЫ ИХ КЛАССИФИКАЦИИ
Большинство авторов конца XIX и первой половины XX ст. при построении филогенетических классификаций грибов исходили в основном из строения вегетативного тела, органов бесполого размножения и характера полового процесса. Ряд авторов считал, что у низших грибов наблюдается постепенное усложнение тела: от одноядерной клетки голой в вегетативном состоянии к одноядерной, но покрытой оболочкой клетке, из которой в дальнейшем развился многоядерный сифоновый ценоцистический мицелий. Этот мицелий у зигомицетов проявляет тенденцию к образованию клеточных перегородок, а для большинства аскомицетов и базидиомицетов характерным является уже многоклеточный мицелий. Одновременно с усложнением вегетативного тела шло усложнение органов бесполого размножения с образованием спорангиев и конидий, а также усложнение полового процесса с пе¬
134
реходом от изогамии к оогамии и зигогамии. При этом зигогамию обыкновенно рассматривали как дальнейшее видоизменение оогамии. Грибы делили на низшие — фикомицеты и высшие — аскомицеты и базидиомицеты. В начале системы фикомицетов помещали хитридиальные, включая их в ряд (подкласс) оомице- тов (Schröter, 1897; Engler, 1898 1); или же чаще выделяя их на правах подкласса или порядка, равноценного оомицегам и зиго- мицетам (А. Fischer, 1892; Wettstein, 1901; Е. Fischer, 1912; Gwynne-Vaughan a. Barnes, 1937). Некоторые авторы считали необходимым выделять отдельный класс — архимицеты наравне с фикомицетами (Lotsy, 1907; Курсанов, 1933, 1940) или же выделяли в отдельный класс архимицетов только так называемые миксохитридиальные, а микохитридиальные включали в состав класса фикомицетов в качестве особого порядка (Ячевский, 1913; Gäumann, 1926, 1949, 1964; Werdermann, 1954). Грейс (Greis, 1943) даже счел необходимым выделить миксохитридиальные в качестве особого отдела, противопоставив его отделам Eumycetes и La- boulbeniomycetes.
Параллельно с изложенным выше взглядом на развитие низших грибов был высказан и принципиально отличный. Так, Гоби (Гоби, 1916, стр. 10) еще в 1891 г. предложил классифицировать фикомицеты в зависимости от характера жгутикового аппарата на два ряда — двужгутиковые (Biciliata) и одножгутиковые (Мопо- ciliata). Такого подразделения он придерживался и в позднейших работах, только называл их несколько иначе — Diflagelloideae и Monoflagelloideae (Гоби, 1912), Biflagellini и Uniflagellini (Гоби, 1916). Лотси, который, по-видимому, не знал о предложении Гоби, в большой группе Archimycetes — Siphonomycetes выделил два ряда — одножгутиковый (роды Olpidiopsis, Synchytrium, Rhizidi- ит, Polyphagus, Hyphochytrium, Monoblepharis и Pythium1 2), который он выводил из «более низких организмов» (von viel niedrigeren Organismen), и второй двужгутиковый (роды Pseudolpidium, Lagenidium, Myrioblepharis 3), который связывал с равножгутиковыми (Isocontae). Нужно иметь в виду, что в начале XX ст. к равножгутиковым относили также семейство вошериевых. В то же время Лотси замечает, что это предположение весьма сомнительно, так как возможно, что паразитический образ жизни привел к потере одного жгутика (Lotsy, 1907, стр. 110—111).
Несколько позже Мережковский (1909), придерживавшийся взгляда о полифилетическом и неоднократном возникновении грибов вообще, разделял фикомицеты, названные им Leucophy-
1 В более поздних изданиях «Syllabus der Pflanzenfamilien» хитридиальные рассматриваются как редуцированная группа оомицетов (Engler, 1912).
2 Отнесение Pythium к одножгутиковым основано на ошибочных наблюдениях Гессе (Gäumann, 1926, стр. 87).
3 Неисправленная еще в то время досадная ошибка (Minden, 1915; Gäumann, 1926, стр. 56; Waterhouse, 1945; Sparrow, 1960).
135
сеае, на два ряда — Monocontae, куда включал организмы, начиная с Chytridium и кончая Мonoblepharis, и Dicontae, к которым относил двужгутиковые формы, начиная с Pseudolpidium и кончая пероноспоровыми. Зигомицеты Мережковский считал производной группой от Monocontae или Dicontae, а «быть может от той и от другой группы, т. к. зигогамия встречается и у первой (Tetrachy trium, Polyphagus) и у второй группы (Lagenidium)» (Мережковский, 1909, стр. 86).
Шерффель (Scherffel, 1925), следуя Лотси, также различает среди имеющих жгутики фикомицетов, которые противопоставляются безжгутиковым зигомицетам, две главные линии развития — 1) ряд хитридиевых (die Chytridineen — Reihe) и 2) ряд сапролегниевых — пероноспоровых (die Saprolegnieen — Perono- sporeen — Reihe). Что касается мопоблефаридовых и бластокла- диевых, то их Шерффель склонен сближать с хитридиевыми.
Взгляды Гоби были приняты Ячевским (Ячевский, 1927; А. Ячевский и П. Ячевский, 1931), который выделяет в подклассе фикомицетов ряды Uniciliatae, Biciliatae, Polyciliatae и Aciliatae. За исключением основанного на недоразумении ряда Polyciliatae, эти ряды отвечают группам Мережковского. Спарроу (Sparrow, 1943, 1960) разделяет жгутиковые фикомицеты на группы Uni- flagellatae и Biflagellatae. Аналогичные ряды мы видим и у Бесси (E. A. Bessey, 1950). Только Смит (Smith, 1955) решается возвести эти группы в ранг подкласса, узаконив в системе соответствующую филогенетическую трактовку: он класс фикомицетов делит на подклассы — Uniflagellatae, Biflagellatae и Aflagellatae (Zygomycetae, Aplanatae). Гифохитриальным, названным им Ani- sochytridiales, Смит не нашел места в системе и выделил в группу «фикомицетов неясных родственных связей» (Phycomycetes of uncertain affinities). Вщсшие грибы он выводит из зигомицетов.
Моро (Moreau, 1952—1953), следуя Данжару, считает исходной группой для всех грибов хитридиальные, которых он выводит из «зоофлагеллат» (Zooflagellés). Среди хитридиальных, как мы уже отмечали, он различает в зависимости от размещения жгутиков, четыре группы: Prosomastigochytridiales, Opisthomastigochy- tridiales, Dimastigochytridiales и Amastigochytridiales. Первые, по Моро, не дали более высокоорганизованных грибов, от вторых произошла фила Opisthomastigomycetes (Blastocladiales и Мопо- blepharidales), от третьих — фила Dimastigomycetes (Saprolegnia- les, Leptomitales, Pythiales, Peronosporales) и от четвертых — филы Zygomycetes (Mucorales, Endogonales, Entomophthorales), Perias- comycetes (т. e. низшие аскомицеты — Protascales, Protomyceta- les, Ashbyales, Spermophthorales, Dipodascales, Synascales) и Dan- geardiomycetes (т. e. высшие грибы — аскомицеты и базидиоми- цеты). Несмотря на оригинальную терминологию, филы Моро очень близки классам Смита с той только разницей, что Моро хитридиальные рассматривает как единую группу, выделяя их из прочих грибов.
136
Цейп (Cejp, 1957), рассматривая грибы как один отдел, делит его на два подотдела — Phycomycophytina (Phycomycetes) и Eumycophytina (Eumycetes). Фикомицеты он делит на четыре
Рис. 49. Типы жгутиков у низших грибов.
Хитридиомицеты (Chytridiomycetes): 1— Rozella sp., 2 — Rhizophydium carpo- philum, 3 — Cladochytrium sp., 4 — Nowakowskiella elegans, 5 — Blastocladiella simplex, 6 — Monoblepharis regignens, 7 — Allomyces moniliformis. Гифохитрио- мицеты (Hyphochytriomycetes) : 8 — Rhizidiomyces apophysatum. Сапролегнио- мицеты (Saprolegniomycetes) : 9 — Olpidiopsis saprolegniae, 10, 11 — Saprolegnia ferax, 12 — Achlya caroliniana, 13—Pythium sp. (no Couch, 1941).
класса — Chytridiomycetes, Oomycetes, Zygomycetes и Trichomyce- tes. К первому он относит порядки Chytridiales, Blastocladiales, Monoblepharidales, имеющие один бичевидный, прикрепленный к заднему концу зооспоры или гаметы жгутик, и Hyphochytriales, имеющие респитчатый жгутик, прикрепленный к переднему концу зооспоры, причем оговаривается, что этот порядок неродстве¬
137
нен предыдущим и не имеет определенных родственных отношений (nejistÿch pfibuzenskÿch vstahû). Ко второму классу отнесены порядки Saprolegniales, Leptomitales, Lagenidiales и Porono- sporales; к третьему — Mucorales, Entomophthorales и Zoopagales. Четвертый класс — Trichomycetes, по словам Цейпа, неясного таксономического положения.
Стремление к построению системы низших грибов с учетом характера и строения жгутикового аппарата основано на том, что этот аппарат в условиях водной среды более устойчив, чем другие признаки, и только с переходом к наземному существованию наблюдается постепенная его утрата и замена подвижных зооспор покрытыми оболочкой и более стойкими в воздушных условиях спорами. Правда, в последнее время, особенно с развитием электронномикроскопических исследований постоянство структур жгутикового аппарата подвергается некоторым сомнениям (Bourrelly, 1961), однако характер строения жгутиков у низших грибов связан с другими признаками — строением зооспор, биохимическими особенностями клетки и, очевидно, все же является существенным признаком, имеющим важное филогенетическое значение (рис. 49).
Что касается строения вегетативного тела, то в различных группах низших грибов, как и в других отделах растительного мира, можно наблюдать его усложнение, а иногда в связи с условиями среды и упрощение. В частности, нельзя провести резкой и принципиальной границы между одноклеточными низшими грибами, голыми в вегетативном состоянии (так называемыми миксо- хитридиевыми) и покрытыми оболочкой (так называемыми мико- хитридиевыми), поскольку на известных стадиях миксохитридие- вые имеют оболочку, а микохитридиевые не имеют. Еще труднее провести такую границу у лагенидиальных, у некоторых представителей которых вегетативное плазмодиеобразное тело может продолжительное время оставаться голым, а у других очень скоро после образования покрывается оболочкой и образует гифы.
Характер полового процесса и строение половых органов являются существенным признаком для выяснения филогенетических связей и при построении системы растительного мира. Однако нужно помнить, что у низших представителей, как водорослей, так и грибов, где половой процесс состоит в слиянии неспециализированных клеток, не отличающихся от вегетативных, или от зооспор, у близких форм можно наблюдать как планетогамию, так и апланогамию, из которых у высших групп развивались такие типы полового процесса, как мерогамия (гаметогамия) и гологамия (зигогамия). Например, у Phlyctidium aenatropum из семейства ризофидиевых копулируют две свободно плавающие монады (зооиды); а у Phlyctidium apophysatum копулируют вегетативные клетки при помощи сливающихся ризоидов. У Rhizo- phydium ovatum монады перед слиянием покрываются оболочкой, а у Diplophlyctis sexualis монады (зооиды) покрываются оболоч¬
138
кой внутри спорангия и дают ризоиды, при помощи которых происходит слияние с другими клетками. В семействе хитридиевых, например у Chytridium sexuale, описаны случаи копуляции монад, которые, как у Rhizophydium ovatum, перед слиянием покрываются оболочкой, или же слияние двух неподвижных клеток при помощи популяционного канала — у Zygorhizidium willei.
Что касается типов мерогамного (гаметогамного) полового процесса, то усложнение его с переходом от изогамии к оогамии можно наблюдать в различных группах, даже в пределах одного рода* И только у высших представителей разных линий эволюции фиксируется определенный, характерный для них тип полового процесса.
Исходя из сказанного выше, нам кажется обоснованным положение, согласно которому в ходе эволюции низших грибных организмов можно выделить два основных ствола — один с одно- жгутиковьтми зооспорами, обладающими бичевидным жгутиком, прикрепленным к заднему концу зооспоры или гаметы, происхождение которого увязывается с амебоидными организмами, и второй — с двужгутиковыми зооспорами, у которых жгутики прикреплены к переднему концу грушевидной зооспоры или находятся в углублении почковидной зооспоры, причем передний жгутик имеет реснитчатое строение. Происхождение этого ствола увязывается или с амебоидными организмами или же с низшими представителями разножгутиковых водорослей (Heterocontae, Хап- thophyceae). Эти два ствола резко отличаются между собой не только строением жгутикового аппарата, но и биохимическими особенностями — двужгутиковым свойственно наличие в клеточных оболочках целлюлозы, которая отсутствует у одножгутиковых.
Согласно принципам филогенетической систематики, логично эти два ствола, развивавшиеся независимо, считать не подклассами сборной группы фикомицетов, как это делают некоторые авторы (Smith, 1955; Ubrizsy a. Vörös, 1966), или классами такого же сборного подотдела Phycomycophytina (Cejp, 1957), а отделами растительного мира (Агх, 1967; Зеров и Зерова, 1968), как это уже давно сделано при классификации водорослей,1 и называть их согласно с правилами ботанической номенклатуры, утвержденными на Седьмом международном ботаническом конгрессе в 1950 г., Chytridiomycota и Saprolegniomycota.
Нужно отметить, что некоторые современные авторы, оставляющие в системе грибных организмов класс фикомицетов, высказывали в то же время мысль о полифилетическом их происхождении (Smith, 1955, стр. 367).
Детальное изучение строения зооспор хитридиевых, проведенное с применением светового и электронного микроскопов, показало, что у некоторых хитридиальных имеется второй
1 Напомним, что Шадефо (Chadefaud, I960, стр. 427) признавал гетерогенное происхождение грибов и считал, что нет никаких оснований признавать их единой группой (une unité unique).
139
не функционирующий зачаточный блефаропласт, соединяющийся с функционирующим тонкими нитями (Koch, 1956, 1958). Наличие второго не функционирующего блефаропласта дает основание для предположения о возможном происхождении хитридиевых от двужгутиковых предков. По мнению Коха изучение блефаропласта у двужгутиковых форм может помочь решению этого вопроса. Во всяком случае, если бы даже оправдалось предположение о наличии двух жгутиков у зооспор предков хитридиевых, строение имеющегося у них одного жгутика, его расположение и биохимический состав оболочки не меняют предположения о различном происхождении и эволюции хитридиевых и сапролегниевых грибов.
Что касается гифохитриальных, обладающих одним перистым, прикрепленным к переднему концу зооспоры жгутиком, то их долгое время включали в состав порядка Chytri diales (Gäumannr 1926) и только по предложению Карлинга (Karling, 1943) стали выделять в особый порядок под названием Anisochytridiales. В ряде последующих работ гифохитриальные трактуются как порядок сборного класса фикомицетов (Bessey, 1950; Gäumann, 1964) или класса Chytridiomycetes (Cejp, 1957), или подкласса Chytridiomy- cetidae класса фикомицетов (Ubrizsy a. Vörös, 1966). Спарроу (Sparrow, 1960) относит Hyphochytriales к Uniflagellatae вместе с хитридиальными, бластокладиальными и моноблефаридальными.
Наличие, кроме хитина, также и целлюлозы в оболочках клеток гифохитриальных (Nabel, 1939; Fuller and Barshad, 1960; Fuller, 1960), а также перистое строение жгутика побудило Смита выделить их в группу «фикомицетов неопределенного родства» (Smith, 1955), а Гейманна на филогенетической схеме грибных организмов (Gäumann, 1964, стр. 10) вывести отдельной ветвыо из флагеллат. И только в последнее время высказано мнение о возможной связи гифохитриальных с сапролегниомицетами, с которыми их объединяет перистое строение жгутика и наличие целлюлозы в клеточной оболочке (Агх, 1967; Зеров и Зерова, 1968). Что касается второго — бичевидного жгутика сапролегниомицетов, то можно предположить, что он у гифохитриальных утерян в процессе эволюции. Можно напомнить в связи с этим случаи утери второго жгутика у потенциально двужгутиковых слизевиков (Ellison, 1945).
Очень спорным является происхождение третьей большой группы низших грибов — зигомицетов. По этому вопросу были высказаны различные и противоречивые мнения. Еще Сакс (Sachs, 1874) на основании внешнего сходства полового процесса объединял зигомицеты с конъюгатами. Позже Лотси (Lotsy, 1907, стр. 199—201) пытался связать с конъюгатами Basidiobolus, у которого зрелый мицелий характеризуется одноядерными клетками и половые органы имеют некоторое внешнее сходство с соответствующими органами зигнемовых. Распространено предположение о происхождении зигомицетов от оомицетов. Так, Э. А. Бесси полагает, что некоторые обитающие в почве сапролегниевые мог¬
140
ли быть исходными формами для мукоральных (Bessey, 1950, стр. 635). Смит высказывает предположение, что апланогамные изогамные фикомицеты («зигомицеты»), возможно, являются рет- рогрессивными и произошли от анланогампых оогамных форм (Smith, 1955, стр. 368). При этом считается само собой разумеющимся, что половые клетки зигомицетов — редуцированные гаме- тангии, в которых не происходит дифференциации гамет. Однако это предположение не подтверждается фактами. Гораздо больше в пользу родственных связей зигомицетов с оомицетами, нам кажется, свидетельствует характер полового процесса у наиболее простых сапролегниомицетов — лагенидиальных. Так, для рода Olpidiopsis характерен типичный гологамный половой процесс типа мукоровых. Однако этот факт, насколько нам известно, не выдвигался для подтверждения общности происхождения зигомицетов и оомицетов, так как лагенидиальные часто считают редуцированными формами оомицетов (Gäumann, 1926, стр. 54; 1964, стр. 60). Следует упомянуть, что некоторые авторы при помощи микрохимических методов доказывали наличие целлюлозы в клеточных оболочках Mucorales (Mangin, 1899; Hopkins, 1929) и En- tomophthorales (Couch, 1939). Однако другие авторы этих данных не подтвердили (Nabel, 1929). Согласно последним данным (Frey, 1950; Aronson, 1965; стр. 56—60), оболочки клеток зигомицетов целлюлозы не содержат.
Наряду с мнением о происхождении зигомицетов от оомицетов было высказано предположение, что зигомицеты происходят от хитридиальных. Еще Брефельд указывал на сходство полового процесса у этих групп (Brefeld, 1881, стр. 163). А. Фишер (A. Fischer, 1892, стр. 10) высказывал мысль, что зигомицеты и оомицеты происходят от голохитриевых, к которым он относил роды, разделенные теперь между хитридиальными (Achlyogeton), лагенидиальными (Lagenidium, Myzocytium) и энтомофторальны- ми (Ancylistes).
На филогенетических схемах Гейманна (Gäumann, 1926, стр. 590; 1949, стр. 10 и 67; 1964, стр. 10 и 84) зигомицеты выводятся из хитридиальных, хотя в первой из этих работ в другом месте (стр. 92) Геймаи высказывает мнение, что зигомицеты филогенетически связаны с хитридиальными или с примитивными оомицетами. В пользу филогенетических связей зигомицетов с хитрк- диевыми высказывались Гвинни-Воган и Барнз (1937) и Цейп (1957). Однако нужно отметить, что все упомянутые выше авторы указывают на отсутствие промежуточных форм между хитридиальными и зигомицетами.
Наконец, нужно отметить, что Шадефо выводит зигомицеты из аскомицетов, в частности из простейших форм, «предков» (ancêtres) аспергиллальных и энтомофторальных, которые, по его мнению, являются редуцированными (Chadefaud, 1960, стр. 786).
Старая гипотеза о происхождении зигомицетов от конъюгат основана на внешнем сходстве полового процесса у этих групп.
141
Конъюгаты — специализированная группа зеленых водорослей со своеобразным строением одноядерных клеток никак нельзя считать возможными предками зигомицетов с их многоядерным цено- цистическим мицелием. Появление перегородок также как и появление одноядерных клеток (Basidiobolus) — явления вторичные. Половой процесс у зигомицетов, как правило, состоит в слиянии многоядерных клеток, содержащих большое, часто огромное количество ядер (у Sporodinia больше тысячи). Слияние одноядерных клеток у базидиобола является результатом деления первично многоядерного мицелия на одноядерные участки и связывается с половым процессом мукоральных через половой процесс у видов энтомофторы и никак не может быть связан с половым процессом конъюгат.
Наличие целлюлозы в клеточных оболочках сапролегниевых грибов, отсутствующей по последним данным у зигомицетов, свидетельствует против выведения зигомицетов из сапролегниомице- тов. Хотя у низших представителей сапролегниевых грибов — в порядке Leptomitales известен гологамный половой процесс, однако он известен также и у нескольких родов хитридиевых и возник, очевидно, в различных группах низших организмов независимо. Если же принять во внимание отсутствие целлюлозы в оболочках клеток хрыридиевых, то более вероятными являются филогенетические связи зигомицетов с этим отделом грибных организмов. Однако все же нельзя не согласиться с тем, что зиго- мицеты являются группой, не имеющей определенных филогенетических связей с другими низшими грибами.
Шадефо выводит зигомицеты от предков наиболее простых аскомицетов. Будучи сторонником гипотезы о происхождении аскомицетов от красных водорослей, он в начале системы аскомицетов ставит дискомицеты, половые органы которых более, чем у других аскомицетов, напоминают половые органы флоридей. Все же остальные группы аскомицетов он считает производными и редуцированными, особенно порядки Endomycetales и Dipodas- cales, близости которых с зигомицетами он не может игнорировать. Логическим выводом из этого и было предположение о происхождении зигомицетов от наиболее простых аскомицетов. Эти вопросы мы рассмотрим детальнее позже — в связи с проблемой происхождения аскомицетов.
По вопросу происхождения так называемых высших грибов — аскомицетов и базидиомицетов, существует две основные взаимно- исключающие теории. Согласно одной, восходящей к Брефельду (Brefeld, J889), аскомицеты происходят от зигомицетов. Согласно другой, основоположником которой считают Сакса (Sachs, 1874, 1896), асковые грибы происходят от красных водорослей. Детальнее этот вопрос мы рассмотрим в главе, посвященной филогении аскомицетов. Сейчас только отметим, что более обоснована, по нашему мнению, теория происхождения аскомицетов от зигомицетов. Основанием для нее является сходство между зигомице-
142
тами и низшими аскомицетами как в строении вегетативных органов, так и в характере полового процесса и в строении органов размножения.
Филогенетическая связь базидиомицетов с аскомицетами широко признана. Поэтому пам кажется целесообразным объединять зигомицеты, аскомицеты и базидиомицеты в один отдел Eumycota, как это мы видим и у Аркса (Агх, 1967).
Что касается взаимоотношений бесхлорофильных групп растительного мира, называемых обычно грибами, и слизевиков, то нужно отметить, что «типичные» слизевики — Myxomycètes резко отличаются от хитридиевых и сапролегниевых грибов наличием плазмодия, плодовых тел, а также характером жгутикового аппарата. Как показали работы ряда авторов (Gilbert, 1927; Sino- to a. Yuasa, 1934; Yuasa, 1935; Ellison, 1945), зооспоры миксоми- цетов снабжены одним или двумя бичевидными жгутиками. Однако в тех случаях, когда у них наблюдали один жгутик, в зооспорах было по два блефаропласта, в связи с чем они должны рассматриваться как потенциально двужгутиковые (Ellison, 1945; Gottsberger, 1967 а, б).
Однако нельзя упускать из виду, что акразиевые и лабирин- туловые типичного плазмодия не имеют, как не имеют и жгутиковых стадий. У лабиринтуловых, как и у плазмодиофоромицетов, нет также плодовых тел. С другой стороны, у некоторых сапролегниевых грибов на известных стадиях образуются многоядерные плазмодии (порядок Lagenidiales). Плазмодиальные образования описывались и у некоторых водорослей, например у разножгутиковой водоросли Мyxochloris, обитающей в водоносных клетках сфагнов (Pascher, 1930). Кроме того, сходство плазмодиофоромицетов с хитридиомицетами и сапролегииомицетами проявляется также в наличии у тех и других амебоидных стадий и гологамного полового процесса. Все это свидетельствует о происхождении тех и других, если не от общих, то во всяком случае от сходных амебоидных предков. Однако дальнейшее развитие этих групп шло независимо. Поэтому мы считаем обоснованным и целесообразным рассматривать слизевики как самостоятельный отдел бесхлорофильных организмов.
Нужно иметь в виду, что некоторые авторы исключают Laby- rinthulomycetes из слизевиков (Cook, 1928; Smith, 1955), а некоторые также и Acrasiomycetes (Jahn, 1928; Martin, 1932, 1940, 1960; Alexopoulos, 1963, и др.)· Если признать аргументацию названных авторов, тогда нужно эти группы рассматривать как самостоятельные отделы растительного мира или вообще исключать их из царства растений. Не следует забывать, что многие авторы, начиная от де Бари (De Вагу, 1858, 1884) и до настоящего времени (Olive, 1970) относят слизевики к животному миру. Мы считаем этот вопрос мало существенным, исходя из признания единого царства эвкариотических организмов, низших представителей которого можно отнести как к животному, так
143
и к растительному миру ввиду отсутствия между ними принципиальных различий.
Исходя из всего сказанного выше, мы считаем вполне обоснованным выделение среди низших бесхлорофильных организмов по крайней мере четырех основных стволов эволюции, которые следует рассматривать как независимые отделы растительного мира (Агх, 1967; Зеров и Зерова, 1968), а именно:
T. Myxomycota — слизевики (Myxomycophyta Smith, 1955; Cejp, 1957; Myxomycota Whittaker, 1959; Arx, 1967; Зеров и Зерова, 1968). В этом отделе объединены организмы, у которых вегетативная стадия представлена в виде плазмодия или, реже, одноядерного или многоядерного амебоида и образующие споро- ношения в виде простого, сидящего на ножке односпорового или многоспорового спорокарпия. Амебоиды слизевиков или снабжены прикрепленными к переднему концу одним-двумя бичевидными жгутиками или же жгутиковая стадия у них может отсутствовать.
II. Chytridiomycota (Opisthomastigomycota) — хитридиевые грибы (Opisthomastigochytridiales + Opisthomastigomycetes Moreau, 1953; Uniflagellatae Smith, 1955; Chytridiomycetes Cejp, 1957; Sparrow, 1958; Chytridiomycota Arx, 1967; Зеров и Зерова, 1968). Это монолитная группа, представители которой имеют один бичевидный жгутик, прикрепленный к заднему концу монады. Название Opisthomastigomycota отражает эту характерную черту организмов, которых мы объединяем в один класс — Chytridiomycetes.
III. Saprolegniomycota (Pteromastigomycota) — сапролегниевые
грибы (Dimastigochytridiales + Dimastigomycetes + Prosomastigo- chytridiales Moreau, 1953; Biflagellatae + Anisochytridiales Smith, 1955; Oomycetes Cejp, 1953; Phycomycetes s. str.+ Hyphochytridio- mycetes Sparrow, 1958; Oomycota Arx, 1967; Saprolegniomycota Зеров и Зерова, 1968). К этому отделу мы относим формы с одним перистым, направленным вперед, и одним гладким, направленным назад, жгутиками (класс Saprolegniomycetes), а также организмы только с одним перистым жгутиком (класс Hyphochytri- diomycetes). От названия Oomycota (Oomycetes) мы вынуждены отказаться ввиду того, что псд названием Oomycetes объединяли, кроме сапролегниевых грибов, также и оогамные хитридиевые грибы (Monoblepharidales). К тому же в этот отдел входят, кроме оогамных форм, также организмы с гологамным половым процессом (Olpidiopsis из Lagenidiales). Название Pteromastigomycota отражает характерный признак отдела — наличие перистого жгутика.
IV. Eumycota (Amastigomycota) — настоящие грибы (Amasti- gochytridiales + Amastigomycetes Moreau, 1953; Zygomycetes + + Ascomycètes -f- Basidiomycctes Smith, 1955; Eumycophytina (Eumycetes) + Zygomycetes Cejp, 1957; Eumycota Arx, 1967; Зеров и Зерова, 1968). К этому отделу мы относим классы Zygo-
144
mycetes, Ascomycètes pi Basidiomycetes, которые характеризуются отсутствием жгутиковых стадий, что отражено во втогюм названии отдела — Amastigomycota.
3. СЛИЗЕВИКИ - MYXOMYCOTA (MYXOM Y СОРИ YTA, MYCETOZOA)
С конца XIX ст. в системах растительного мира принято выделять слизевики в самостоятельный отдел (Schröter, 1897; Engler, 1898; Wettstein, 1901; Lotsy, 1907; Ячевский, 1907; Werdermann, 1954; Smith, 1955; Gäumann, 1926, 1949, 1964; Cejp, 1957, и др.). Однако некоторые авторы-ботаники считали, что слизевики стоят ближе к животным, чем к растениям, и исключали их из системы растительного мира (Cohn, 1872; de Вагу, 1858, 1859, 1884). Аналогичную мысль высказывали также и Э. А. Бесси (Bessey, 1950, стр. 1—2, 22) и Олайв (Olive, 1970). С другой стороны, ряд авторов рассматривали слизевики как класс или порядок отдела грибов (Winter, 1884; van Tieghem, 1884; E. A. Bessey, 1935; Chade- faud, 1960) или как подотдел Myxomycotina отдела грибов, правда, в узком понимании, т. е. без лабиринтуловых и акразиевых (Martin, 1932, 1940, 1960; Alexopoulos, 1966), или подцарство Myxomycota царства грибов — Fungi (Whittaker, 1959), или даже включали все слизевики — Myxomycètes, Plasmodiophorales и Ас- rasiales в группу Archimycetes (Cavers, 1915).
Существует две точки зрения на объем отдела Myxomycota. Одни авторы кроме настоящих слизевиков (Myxomycètes) включали и включают в состав этого отдела и акразиевые (van Tieghem, 1884; Schröter, 1897; Engler, 1898), и лабиринтуловые (Wettstein, 1902; Lotsy, 1907; E. A. Bessey, 1950; Cejp, 1957). Другие авторы ограничивают объем отдела Myxomycota и исключают из него плазмодиофоровые, сближая их с архимицетами и хитридие- выми (Schwartz, 1914; Jahn, 1928; Gäumann, 1926, 1949, 1964; Курсанов, 1933, 1940; Greis, 1943; Schmidt и Werdermann, 1954). Некоторые авторы включают в состав Myxomycota настоящие слизевики, плазмодиофоровые и акразиевые и исключают лаби- ринтулальные, считая их родственными ризоподиальным Xantho- phyceae (Smith, 1938, 1955). Очень распространено мнение, согласно которому слизевики, как естественную группу, нужно ограничить только настоящими слизевиками (Myxomycètes), исключив из них акразиевые, лабиринтуловые и плазмодиофоровые (Lister, 1894, 1911, 1925; Jahn, 1911, 1928; Pascher, 1918; Martin, 1932, 1940, 1960; Alexopoulos, 1963, 1966). В последнее время описапа Олайвом (Olive, 1967, 1970) группа организмов, которую он относит к слизевикам и называет Protostelia.
Расхождение во взглядах па объем отдела объясняется тем, что организмы, включаемые в его состав, образуют четыре группы, которые резко различаются между собой рядом особенностей.
145
«Основной» группой среди слизевиков являются настоящие слизевики (Myxomycètes). Они характеризуются своеобразным типом развития, отличающим их от других бесхлорофильных растительных организмов. Для типа развития настоящих слизевиков характерной является вегетативная стадия в виде настоящего плазмодия. При прорастании покоящейся споры у слизевиков образуется зооспора с одним или двумя жгутиками, которая через некоторое время теряет жгутик и превращается в так называемую миксамебу. Через некоторое время миксамебы сливаются и образуют плазмодий. Перед образованием плазмодия происходит половой процесс, состоящий обычно в слиянии гаплоидных амеб, описанный впервые Яном (Jahn, 1911). Однако известны также случаи копуляции зооспор (Jahn, 1936; Ross, 1937; Me Manus, 1961, и др.). Описано даже слияние миксамебы с зооспорой (Me Manus, 1961; Benedict, 1962). Диплоидный плазмодий образует плодовые тела, в которых происходит образование спор с редукционным делением.
Следует отметить, что по данным Ульриха (Ulrich, 1943; цит. по Alexopoulos, 1963) в оболочках спор слизевиков отсутствует хитин, возможно, кроме кагшллиция Stemonitis fusca, и отмечена целлюлоза. Позже целлюлоза была обнаружена в оболочках спор, в капиллиции и пожке плодового тела трех видов Comatricha (Goodwin, 1961). Эти данные получены с применением J2KJ/H2SO4 и хлор-цинк-йода и должны быть подтверждены более совершенными методами.
Происхождение и положение в системе растительного мира другой группы, относимой к слизевикам,— плазмодиофоровых до последнего времени остается дискуссионным. Ряд авторов, например Гейманн (Gäumann, 1926, 1949, 1964), а за ним Курсанов (1933, 1940), Вердерманн (Werdermann, 1954) и другие исключают их из слизевиков и относят к низшим фикомицетам.
Некоторые авторы отмечали диморфизм ядер у плазмодио- форальных и считали, что соматическое деление ядер у них происходит по примитивному типу (промитоз), описанному у некоторых амеб (Cook, 1928). Считали, что и мейоз у плазмоди- офоральных происходит по примитивному типу, который состоит в том, что весь хроматин выбрасывается в цитоплазму, где и происходит редукционное деление по обычному типу (Cook, 1928). Однако позднейшие исследования (Р. Heim, 1955) показали, что за картины промитоза ошибочно принимали веретеновидные гаплоидные ядра. Что же касается мейоза, то и здесь имела место неточность, связанная с применением для окраски препаратов почти исключительно гематоксилина. Таким образом, в отношении клеточного деления нет существенной разницы между плазмо- диофоральными и настоящими слизевиками, у которых митоз проходит по обычному типу.
У плазмодиофоральных, как упоминалось выше, был обнаружен хитин (Ulrich, 1943). Это некоторые авторы считают аргу¬
146
ментом против признания близкого родства между слизевиками и нлазмодиофоральными. Однако нужно иметь в виду, что до применения новых методов исследования при изучении химизма клеточных оболочек спор слизевиков рискованно делать филогенетические выводы на основании современных скудных данных по этому вопросу (Alexopoulos, 1963).
Развитие Plasmodiophora brassicae проходит довольно сложно. Как показали исследования Кука и Швартца (Cook a. Schwartz, 1930), в ее развитии можно выделить две стадии: одну в корневых волосках, а другую — в клетках коры главного корня растения-хозяина. Согласно новейшим исследованиям (Kôle a. Gielink, 1962; Tommerup a. Ingram, 1971), покоящиеся споры плазмодио- форы при прорастании дают зооспоры с двумя неравными бичевидными жгутиками; зооспоры заражают клетки корневых волосков капусты, где сливаются, образуя первичный плазмодий. Первичный плазмодий через некоторое время распадается на четыре-десять зооспорангиев. По данным Келе и Гиелинка (Köle, 1959; Köle a. Gielink, 1961; цит. по Tommerup a. Ingram, 1971), зооспоры попарно копулируют без слияния ядер (плазмогамия). Предполагают, что эти двуядерные зооспоры проникают в клетки коры корня, где и формируется вторичный плазмодий. Ядра вторичного плазмодия сливаются попарно, после чего наступает редукционное деление. Есть основания полагать, что редукционное деление происходит почти непосредственно после слияния ядер, так как очень часто наблюдали в одном плазмодии ядра на поздних стадиях слияния и на ранних стадиях мейоза. Вторичный плазмодий дает гаплоидные покоящиеся споры. Аналогичное развитие, по- видимому, свойственно и Sporgospora subterranea.
Очень существенным является характер жгутикового аппарата у плазмодиофоральных. Как показали исследования Ледингема (Ledingham, 1934, 1939), для плазмодиофоральных характерно наличие двужгутиковых зооспор. Оба жгутика простые, бичевидные, неравной длины, что часто также наблюдается и у миксо- мицетов. Таким образом, строение жгутикового аппарата сближает плазмодиофоральные с настоящими слизевиками и отличает их от хитридиальных, с которыми их пытались сближать и которые имеют один направленный назад жгутик, а также от сапролегние- вых грибов, зооспоры которых имеют перистые жгутики. Между прочим в последней своей монографии Гейман отмечает, что в связи с характером жгутикового аппарата плазмодиофоровых их нельзя сближать филогенетически с архимицетами (Olpidiaceae и Synchytriaceae), а на схеме родственных отношений архимицетов он последние два семейства выводит из флагеллат, а плазмодио- форовые из настоящих слизевиков — Myxomycètes (Gäumann, 1964, стр. 14). Во всяком случае общий ход развития, строение жгутикового аппарата, характер полового процесса — все это несомненно свидетельствует о филогенетическом родстве плазмодиофоральных и настоящих слизевиков.
147
Нам кажется, можно согласиться с мнением ряда авторов (Cook, 1928; Pavillard, 1953), что классы настоящих слизевиков и плазмодиофоровых развивались параллельно, причем эволюция плазмодиофоровых проходила в условиях внутриклеточного па¬
Рис. 50. Labyrinthula:
1 — общий вид, 2 — клетка: о — ядро, б — вакуоля (по Young, 1943).
разитизма, в то время как настоящие слизевики приспособились к сапрофитному питанию в условиях наземного существования.
Акразиевые отличаются от настоящих слизевиков отсутствием жгутиковой стадии. Кроме того, у них вместо настоящего плазмодия развивается так называемый агрегатный, сборный плазмодий или нсевдоилазмодий, в котором миксамебы не теряют индивидуальности. Правда есть указание, что перед образованием плодовых тел у Dictyostelium миксамебы сливаются в настоящий плазмодий (Skupiencki, 1920). С другой стороны, некоторые авторы отмечают, что агрегатпый плазмодий акразиальных является фазой, связанной с плодоношением, а вегетативная стадия у них заканчивается перед образованием плазмодия (Olive, 1902; цит. по Martin, 1940). Эти особенности акразиальных послужили основанием ряду авторов для исключения их из состава слизевиков.
148
Однако общий тип развития, характер полового процесса, образование плодовых тел подтверждает то, что акразиальные и настоящие слизевики произошли если не от общих, то от близких предков. Кук (Cook, 1928) таких общих предков видит в группе Proteomyxa. Э. А. Бесси (Bessey, 1950) считает, что слизевики происходят от склонных к образованию колоний амебоидных организмов, образующих на одной из стадий своего развития инцистиро- ванные клетки. К этому взгляду склоняется и Цейп (Cejp, 1957). Заслуживает упоминания тот факт, что еще Гоби в одной из ранних работ (1883) группу Amoeboideae, куда он включал слизевики, миксохитридиевые и семейство вампирелловых, выводил из низших амебоидных организмов.
Олайв (Olive, 1970) выделяет из акразиевых как равноправную группу диктиостелиевых (Dictyostelia, Dictyosteliidae) главным образом на том основании, что у диктиостелиевых ножка спорокарпа имеет трубку, а у акразиевых ее нет.
Из организмов, относимых к слизевикам, вероятно, наименее изучена небольшая группа лабиринтуловых. Из них наиболее известен описанный еще в 1867 г. Ценковским род лабиринтула (Labyrinthula) (рис. 50). Относящиеся к нему виды в вегетативном состоянии имеют сетчатый плазмодий, состоящий из тонкой сети, образованной эластическими волокнами, по которым передвигаются веретеновидные одноядерные клетки. Эти клетки размножаются делением, иногда образуют скопления, напоминающие агрегатный мицелий акразиальных. Клетки могут инцисти- роваться или по одиночке, или образуют кучки — сорусы. При прорастании цист получаются безжгутиковые клетки, дающие новый сетчатый плазмодий. У некоторых лабиринтуловых при прорастании цист образуются монады с одним передним жгутиком (Labyrynthomyxa).
Некоторые авторы исключают лабиринтуловые из слизевиков и относят их к низшим саркодовым, в частности к группе Proteomyxa Ланкастера, вместе с некоторыми родами сомнительного положения, как Vampyrelia и Pseudospora (Cook, 1928). Смит (Smith, 1955), как уже упоминалось, также исключает Labyrin- thulomycetes из слизевиков и находит возможным сближать их с ризоподиальными водорослями, встречающимися среди разножгутиковых, например Chlamydomyxa.
Кук же относит имеющую хроматофоры Chlamydomyxa вместе с Vampyrelia и Pseudospora к Proteomyxa. Во всяком случае, Labyrinthulales стоят близко к низшим саркодовым, с которыми, очевидно, связаны филогенетически. Отсутствие жгутиковых стадий и агрегатный мицелий резко отличают акразиевые и лабиринтуловые от настоящих слизевиков и плазмодиофоровых, что свидетельствует об их филогенетической обособленности.
Недавно описанная Олайвом группа слизевиков, которую он рассматривает как один из подклассов класса Mycetozoa и называет Protostelia (Protosteliidae), характеризуется следующими
№
особенностями (рис. 51, 52). Вегетативное тело протостелиевых слизевиков представляет собой одноядерную или многоядерную амебоидную клетку или же многоядерный сетчатый плазмодий. Некоторые представители группы могут иметь жгутиковые стадии. Спороыошения имеют вид односпоровых или миогоспоровых спорокарпиев.
Олайв, кроме описанных им простых амебоидных и илазмодиальных форм, зачисляет в подкласс протостелиевых также и цератиомиксовые (Ceratiomyxidae, Ceratiomyxaceae),
относимые другими авторами к настоящим слизевикам, но отличающиеся от них одиночными спорами, сидящими на поверхности спорокарпия.
Рис. 52. Протостелиомнцеты (Protosteliomycetes): Ceratiomyxa fruticulosa (а — плодоносный выступ плазмодия. б — развитие спороношения» (по Olive, 1970).
По мнению Олайва (Olive, 1970), протостелиевые являются примитивной группой, из которой развились диктиостелиевые и настоящие слизевики.
150
Рис. 51. Протостелиомнцеты (Protosteliomycetes) : схема развития спороношения у Nematostelium ovatum (по Olive.
1970).
Мы считаем обоснованным выделение слизевиков в качестве самостоятельного отдела органического мира — Myxomycota, филогенетически связанного с низшими саркодовыми.
К этому отделу мы относим пять групп организмов, которые рассматриваем как классы, а именно:
1. Protosteliomycetes (Protostelia по Олайву) — и р о- тостелиевые слизевики с одним порядком Protosteliales.
2. Labyrinthulomycetes (Hydromycetes) — л а б и- р и н т у л о в ы е, или водные, слизевики с одним порядком Labyrinthulales.
3. Acrasiomycetes — акразиевы слизевики с одним порядком Acrasiales (Dictyosteliales).
4. Myxogastromycetes — настоящие слизевики с четырьмя порядками — Physarales, Stemonitales, Liceales и Tri- chiales.
5. Plasmo diop h oromycetes — плазмодиофоро- в ы e с л и з е в и к и с одним порядком Plasmodiophorales.
4. ХИТРИДИЕВЫЕ ГРИБЫ - CHYTRIDIOMYCOTA (OPISTHOMASTIGOMYCOTA)
Несмотря на то что еще с конца XIX ст. ряд авторов обращал внимание на необходимость при классификации низших грибов принимать во внимание характер жгутикового аппарата (Гоби, 1891, 1912, 1916; Lotsy, 1907; Мережковский, 1909), в большинстве распространенных до последнего времени систем грибов характер жгутикового аппарата не оценивался в должной степени, и низшие грибы классифицировали на основании строения вегетативного тела и характера полового процесса.
Попытки выделения одножгутиковых форм с бичевидиым жгутиком, прикрепленным к задней при движении части зооспоры (гаметы), в отдельную филогенетическую и систематическую группу видим у Гоби (1912, 1916), Мережковского (1909) и Ячев- ского (1927, 1931). Спарроу (Sparrow, 1943, 1960) разделяет водные гифомицеты на Uni flagellatae и Biflagellatae. Однако к Uniflagellatae он причисляет, кроме хитридиомицетов с задним бичевидным жгутиком, также и Hyphochytriales с одним передним перистым жгутиком. Смит (Smith, 1955) решается выделить Uniflagellatae в качестве подкласса класса фикомицетов. Позже Цейп (Cejp, 1957) возводит их в ранг класса под названием Chytridiomycetes подотдела Phycomycophytina (Phycomycetes) отдела грибов — Mycophyta. Однако он, следуя Спарроу, к хитридио- мицетам причисляет в качестве порядка также и гифохитриаль- ные, отличающиеся от них характером своего единственного жгутика. И только в последнее время (Агх, 1967; Зеров i Зерова, 1968) высказывается мнение, что группа грибов с одним задним бичевидным жгутиком представляет независимую филу и должна рассматриваться как отдел бесхлорофильных растений.
151
Строение тела хитридиомицетов очень разнообразно. Наиболее простое вегетативное тело имеют ольпидиевые и синхитриевые (рис. 53). Оно представляет собой голый одноядерный амебоид, находящийся в клетке растения-хозяина. Отсутствие у ольпидие-
Рис. 53. Sijnchytrium endobioticum: а — зооспоры, б — прорастание зооспоры, в, г — паразит в клетке растения-хозяина, д, е — развитие соруса, ж — разделение молодого соруса на спорангии, з — отдельный спорангий, и, к — половой процесс, л — прорастание зиготы, м — циста, н — образование зооспор в цисте (по Куртис, Bessey, 1950).
вых и синхитриевых вторичной клеточной оболочки нельзя рассматривать как принципиальное отличие, противопоставляющее эти семейства другим хитридиомицетам, вегетативное тело которых покрыто вторичной оболочкой, поскольку ольпидиевым и син- хитриевым также свойственно образование клеточных оболочек
152
на определенных стадиях развития. Так, зооспоры и зиготы у них перед заражением растения-хозяина покрываются оболочкой. Позже, при переходе к размножению (при образовании зооспорангия или цисты), голый амебоид также покрывается вторичной оболочкой.
У других семейств хитридиомицетов — ризофидиевых, хитри- диевых прикрепившаяся к субстрату зооспора превращается в од-
Рис. 54. Cladochytrium tenue:
а — общий вид; видны веретеновидные «собирательные клетки», зооспорангии, б, в — покоящаяся спора и ее прорастание с образованием зооспорангия (по Karllng, 1945).
ноклеточное покрытое оболочкой тело, развивающееся или на поверхности субстрата (экстраматрикально) или внутри его (ин- траматрикально). В некоторых случаях (семейство ризидиевые) зооспора переходит в вегетативное состояние в свободном состоянии и затем прикрепляется ризоидами обычно к многочисленным одноклеточным организмам; в этом случае говорят об интермат- рикальном вегетативном теле. Во всех названных случаях вегетативная клетка может образовывать более или менее развитую ризоидальную систему, напоминающую мицелий, однако отличающуюся от настоящего мицелия отсутствием в ризоидах ядер — организм остается одноядерным (моноцентрическим) и единственное клеточное ядро находится в центральной, обычно расширенной части тела.
Дальнейшее усложнение тела наблюдаем в семействах кла- дохитриевых и мегахитриевых (рис. 54). Оно заключается в
153
развитии полицентрического (многоклеточного) вегетативного тела. Так, при прорастании зооспоры у кладохитрия (Cladochytrium) образуется тонкое мицелиальное тело (ризомицелий) с веретеновидным расширением, в которое переходит ядро,— так называемая «собирательная клетка». Затем ядро делится на два, веретеновидное расширение делится перегородкой, а конец гифы растет далее, в него переходит дистальное ядро и образуется новая веретеновидная клетка. Этот процесс может повторяться, в результате чего получается довольно сложное вегетативное тело с одноядерными клетками.
В порядках бластокладиальных и моноблефаридальных наблюдаются многоядерные клетки. У более просто организованных бластокладиальных (семейства Catenariaceae и Coelomomycetaceae) вегетативное тело имеет вид более или менее разветвленного многоядерного мицелия. У видов рода Coelomomyces, паразитирующих в полости тела насекомых, вегетативный мицелий не имеет вторичной клеточной оболочки.
Вегетативное тело у родов семейства Blastocladiaceae состоит из округлой, булавовидной или разветвленной центральной (базальной) части, от которой отходят ризоиды. В роде Blastocladiella центральная часть округлой или булавовидной формы; она или непосредственно превращается в спорангий или гаметангий, или же образует его в виде выроста. В роде Blastocladia центральная часть вегетативного тела может быть более или менее разветвленной, и репродуктивные органы образуются на разветвлениях таллома. В роде Allomyces таллом состоит из цилиндрической базальной части, от которой отходят обычно дихотомически разветвленные ветви с ситовидными ненастоящими перегородками (псевдосептами). Настоящие перегородки у Blastocladiales образуются только при развитии органов размножения или повреждении таллома.
В порядке Monoblepharidales таллом представляет собой многоядерный, обычно разветвленный мицелий без перегородок. Только у видов рода Gonapodya мицелий имеет перетяжки; в суженных местах обычно образуются пробки из вещества углеводной природы, так называемого целлулина.
Параллельно с усложнением строения тела у хитридиомицетов наблюдается усложнение бесполого размножения и полового процесса. Так, у представителей Olpidiaceae и Synchytriaceae все одноклеточное тело превращается в спорангий или группу спорангиев, т. е. наблюдается голокарпия (от греческих слов όλος — весь, целый и καρπός — плод). В других семействах хитридиаль- ных наблюдается переход к эвкарпии (от греческих слов ευ— хорошо, полностью и καρπός — плод), когда органы размножения образуются только из части вегетативного тела. В дальнейшем на вегетативном теле обособляются зооспорангии и гаметангии, отделенные настоящими перегородками.
Половой процесс у хитридиевых грибов существует в разнообразной форме, и изучение его дает важный материал для филоге¬
454
нетических выводов. У низших грибов наблюдается два типа наиболее простого полового процесса, при котором сливаются две неспециализированные клетки,— мерогамия и гологамия. Мы воздерживаемся от употребления применительно к низшим грибам термина гаметогамия для первого типа — мерогамии, так как у низших представителей хитридиальных, среди которых наблю-
Рис. 55. Rhizophydium ovatum:
а, б, в — заражение клетки водоросли и развитие вегетативного тела, превращающегося в спорангий; г — зрелый спорангий с зооспорами; д — м — половой процесс
(по Couch, 1935).
дается этот тип полового процесса в виде слияния одножгутиковых подвижных клеток, ничем не отличающихся от зооспор, вероятно, функционирующих при некоторых, не всегда ясных обстоятельствах как половые клетки, нельзя еще говорить о специализированных гаметах. Эта специализация, очевидно, установилась на более поздних этапах эволюции. В типичном и наиболее простом виде мерогамия представлена, например, у Olpidium viciae, некоторых видов Synchytrium, Phlyctidium, Physoderma и других, у которых две одинаковые монады сливаются в планозиготу, сохраняющую некоторое время оба жгутика копулировавших клеток и позже, переходя в состояние покоя, покрывающуюся оболочкой.
Очень интересный половой процесс, представляющий один из возможных путей к половой дифференциации и переходу к гологамии, описан у Rhizophydium ovatum (Couch, 1935), паразитирующего на нитчатых водорослях (рис. 55). Здесь одна монада прикрепляется к субстрату и покрывается оболочкой, к ней прикрепляется другая и также покрывается оболочкой; обе клетки увеличиваются в размерах, и содержимое первой клетки, условно принимаемой за мужскую, переходит во вторую, которая превращается в диплоидную зиготу (цисту). Аналогичный процесс
155
описан у Chytridium sexuale с той только разницей, что содержимое второй клетки переливается в первую, которая, таким образом, может рассматриваться как женская (Koch, 1951).
У хитридиальных можно наблюдать переходы от мерогамии к гологамии, которые показывают, что принципиальной разницы между этими типами полового процесса у низших грибов нет. Так, например, у Rhizophydium couchii копулируют два молодые организма — клетки, образовавшиеся из зооспор. Один из них становится крупнее, в него внедряется вырост другого, через который содержимое меньшей (мужской) клетки переливается в большую (женскую), превращающуюся в зиготу (Sparrow, 1933).
У ряда представителей хитридиевых можно наблюдать разнообразные типы настоящей гологамии. Так, у Olpidium radicale, в отличие от других представителей семейства Olpidiaceae, где происходит слияние монад, голые протопласты, возникшие из двух зооспор, копулируют внутри клетки растения-хозяина (Schwartz a. Cook, 1928).
У видов рода Phlyctidium из семейства ризофидиевых наблюдается или слияние свободно плавающих гамет (Phlyctidium anat- ropum) или же копуляция происходит между двумя взрослыми особями — их ризоиды растут навстречу друг другу, набухают и копулируют, в результате чего на месте копуляции образуется зигоспора (Ph. apophysatum; Canter, 1961). У Zygorhizidium willei из семейства хитридиевых одна особь — «мужская» дает копуляционный вырост, через который ее содержимое переходит в другую — «женскую» (Loewenthal, 1905). Сходно проходит половой процесс у полифага (Polyphagus euglenae) из семейства ризидиевых. Он состоит в том, что из центральной части одного — меньшего по размерам («мужского») организма отходит длинный вырост, который прикрепляется к центральной части более крупного организма («женского»), образуя на конце вздутие. В это вздутие переходит протопласт первой клетки. Оболочка между клетками разрушается и содержимое женской клетки переходит во вздутие, превращающееся в зиготу.
Таким образом, у хитридиальных наблюдаются мерогамный и гологамный типы полового процесса в их наиболее простой форме — слиянии двух подвижных или лишенных жгутиков недифференцированных клеток. В дальнейшем происходит дифференциация половых органов и усложнение полового процесса — образуются специализированные половые клетки и в более высоко организованных порядках (Blastocladiales и Monoblepharidales) можно говорить уже о гаметогамии во всех ее типах — изогамии, гетерогамии и оогамии.
Чередование полового и бесполого поколений обнаружено в различных группах хитридиомицетов — у хитридиальных {Phy- soderma) и бластокладиальных {Blastocladiella, Allomyces), причем у физодермы описан гетероморфный тип (Lingappa, 1959), а у бластокладиальных {Blastocladiella и Allomyces) — изоморфный
156
(Kniep, 1929, 1930; Emerson, 1941; Couch a. Whiffen, 1942; Emerson a. Wilson, 1949; Wilson, 1952).
У Physoderma pulposum, паразитирующей на стеблях и листьях Chenopodiaceae, гаметофит развивается из зооспоры, кото-
Рис. 56. Physoderma pulposum.
Гаплоидная фаза: а — прорастание зооспоры, б — образование апофизы в клетке растения-хозяина, в — развитие зооспор. Диплоидная фаза: г — прорастание зиготы с образованием «собирательной клетки», д — содержимое «собирательной клетки» разбивается на участки, образующие гифы, е — развитие вторичных и третичных «собирательных клеток», а также покоящихся спор, ж — прорастающая покоящаяся спора (по Lingappa, 1959).
рая прикрепляется к эпидермису растения-хозяина, покрывается оболочкой и дает ростковую трубку, проникающую в клетку хозяина (рис. 56). В клетке она образует расширение — апофизу, от которого отходят ризоиды. Сама же зооспора, разрастаясь, превращается в тонкостенный экстраматрикальный зооспорангий. Зооспоры вызывают заражение других растений, где также
157
развиваются гаплоидные организмы. В конце вегетационного периода зооспоры, развивающиеся в перезрелых, отличающихся внешне зооспорангиях, попарно копулируют, образуя планозиго- ту, которая прикрепляется к эпидермису хозяина, втягивает жгутики и покрывается оболочкой. В ней происходит кариогамия, она дает ростковую трубку внутрь клетки, где образуется собиратель-
Рис. 57. Allomyces arbusculus:
схема чередования гаметофита (а) и спорофита (б) (по Emerson, 1938).
ная клетка. Содержимое ее делится на одноядерные или многоядерные участки, которые покрываются оболочкой и дают нежные одноядерные гифы. В это время оболочки соседних клеток растения-хозяина разрушаются, и образуется полость. Концы гиф набухают и дают новые собирательные клетки или покоящиеся споры. В покоящихся спорах после редукционного деления развиваются гаплоидные зооспоры.
В семействе бластокладиальных известно чередование поколений изоморфного и гетероморфного типов. Изоморфное чередование поколений было описано в роде Allomyces (A. javanicus, A. arbuscula) (рис. 57). Гаплоидные зооспоры у этих видов, образующиеся при прорастании толстостенных спорангиев (покоящихся цист), после редукционного деления покрываются оболочкой и развиваются в гаплоидные половые растения (гаметофит), на которых образуются макро-и микрогаметангии. Гаметы копулируют, планозйгота покрывается оболочкой и развивается в диплоидное растение (спорофит), на котором образуются тонкостенные зооспорангии и толстостенные покоящиеся цисты. В тонкостенных зооспорангиях развиваются без редукционного деления диплоид¬
158
ные зооспоры, дающие начало новым диплоидным растениям. Покоящиеся цисты прорастают обычно после периода покоя. При этом происходит редукционное деление и из гаплоидных зооспор развиваются гаплоидные растения (гаметофит).
У Allomyces cystogenus наблюдается гетероморфное чередование поколений. Гаметофит у него сильно редуцирован и представлен только цистой, развивающейся из гаплоидной зооспоры; в цисте образуется четыре гаметы. Аналогичный тип развития описан и у Blastocladiella cystogena (Couch and Whiffen, 1942).
Хитридиевые грибы мы относим к одному классу — хитридио- мицетов (Chytridiomycetes), который делим на три порядка: Chyt- ridiales, Blastocladiales и Monoblepharidales. Ниже даем их краткую характеристику.
1. Порядок Chytridiales — хитридиальные. Таллом одноклеточный (моноцентрический) или многоклеточный (поли- центрический) с одноядерными клетками. Половой процесс мерогамный (копуляция одножгутиковых монад) или гологамный. Изредка известно чередование полового и бесполого поколений (гаметофита и спорофита). Зооспоры обычно без большой ядерной шапочки (nuclear cap) или бокового тельца (side body).
По вопросу эволюции в пределах порядка Chytridiales существуют разные мнения. Так, Спарроу полагает, что эволюция в этом порядке шла двумя параллельными ветвями, различающимися по характеру раскрывания зооспорангиев: в одном ряду — Inoperculatae с зооспорами, освобождающимися через поры, возникающие в результате ослизпения оболочки папилл; в другом ряду Operculatae с зооспорангием, открывающимся в результате обособления крышечки. В обоих рядах развитие шло от моноцентрических голокарпных форм к полицентрическим эвкарпным. Он дает следующую схему классификации хитридиальных (Sparrow, 1960).
Chytridiales
Inoperculatae Operculatae
Olpidiaceae Chytridiaceae
Achlyogetonaceae Chytridioideae
Synchytriaceae Endochytrioideae
Phlyctidiaceae Megachytriaceae
Phlyctidioideae
Entophlyctoideae
Rhizidiaceae
Hhizidioideae
Polyphagoideae
Cladochytriaceae
Physodermataceae
Виффен (Whiffen, 1944) считает такое подразделение искусственным, так как способ раскрывания спорангия у хитридиевых не увязывается с другими признаками. По ее мнению, классификация хитридиевых должна базироваться на типах развития таллома. У моноцентрических эвкарпных хитридиевых она различает пять таких типов:
1. Инцпстированная зооспора непосредственно превращается в зооспорангий, как у Rhizophijdiiim и Rhizidium.
2. Инцпстированная зооспора превращается в проспорангий, из которого развивается спорангий, как у Polyphagus.
159
3. Ростковая трубка инцистированной зооспоры дает расширение — проспорангий (апофизу), а спорангий образуется из инцистированной зооспоры, как у Chytridium lagenaria — эндо-экзогенный тип развития.
4. Зооспорангий развивается из расширения ростковой трубки, как у Entophlyctis.
5. Расширение ростковой трубки дает проспорангий, на котором уже развивается зооспорангий, как у Diplophlyctis.
В связи с этим Виффен моноцентрические эвкарпные архимицеты делит на такие два семейства: Rhizidiaceae с подсемействами Phizidioideae. Polyphagoideae и Chytridioideae и Entophlyctaceae с подсемействами Entoph- lyctoideae и Diplophlyctoideae. Полицентрические хитридиевые она объединяет в одном семействе — Cladochytriaceae.
Дальнейшие исследования онтогенеза и цитологии хитридиальных, очевидно, дадут новый материал для построения филогенетической системы этого порядка. Во всяком случае исследования тонкого строения зооспор (Koch, 1956, 1958) дают материал, который свидетельствует о наличии разных типов их строения у хитридиевых грибов, что вместе с другими признаками может быть использовано для филогенетических выводов.
2. Порядок Blastocladiales — бластокладиаль- II ы е. Таллом мпогоядериый, простой или ветвистый, иногда — у высших представителей — с ненастоящими ситовидными перегородками (псевдосепты). Половой процесс изогамный или гетерогамный. У ряда видов известно чередование полового и бесполого поколений. Зооспоры с большой ядерной шапочкой и боковым тельцем.
Порядок Blastocladiales обычно разделяют на три семейства. 1. Catena- riaceae, куда принадлежат организмы, паразитирующие на червях и некоторых грибах, характеризующиеся ценоцистпческим мицелием, зооспорангиями, расположенными цепочкой и изогамным половым процессом. Один род — Catenaria. 2. Coelomomycetaceae — паразиты личинок насекомых, с це- ноцистическим мицелием, лишенным вторичной клеточной оболочки. Половой процесс неизвестен. Один род — Coelomomyces. 3. Blastocladiaceae — сапрофиты с ценоцистпческим мицелием, разделенным псевдосептами, изогамным или гетерогамным половым процессом и чередованием поколений у некоторых родов (Blastocladiella, Allomyces).
3. Порядок М onoblepharidales — моноблеф ар ιι- да льны e. Таллом в виде многоядерного разветвленного мицелия, без перегородок или иногда (род Gonapodya) с целлулиновы- ми пробками в суженных местах. Половой процесс оогамный. Чередование поколений не известно. Зооспоры на верхушке с многочисленными мелкими гранулами. Одно семейство — Monoblepha- ridaceae с тремя родами.
Спарроу (Sparrow, 1960) Monoblepharidales разделяет на два семейства — Gonapodyaceae и Monoblepharidaceae.
Что касается взаимоотношений между этими порядками, то в современной литературе принято считать наиболее примитивным порядком Chytridiales, а порядки Blastocladiales и MonobJepliari- dales производными (Bessey, 1942, 1955; Cejp, 1957). Однако были высказывания, что так называемые миксохитридиевые, т. е. семейства Olpidiaceae и Synchytriaceae, имеют вторично упрощенное строение в результате внутриклеточного паразитизма (Engler,
160
1912). Некоторые авторы признавали независимое происхождение ольпидиевых и синхитриевых и объединяли их вместе с плазмо- диофоральными в класс Archimycetes (Gäumann, 1926, 1949, 1964) или выделяли в особый отдел грибных организмов (Greis, 1943).
Мы уже отмечали, что в строении клеток, жгутикового аппарата и способах размножения нет существенной разницы между миксохитридиевыми (ольпидиевыми и синхитриевыми), с одной стороны, и микохитридиевыми — с другой. Что же касается клеточной оболочки, то и в этом нет между ними принципиального различия, так как у первых перед стадией спорообразования также образуется оболочка. Возможно, что отсутствие клеточной оболочки у ольпидиевых и синхитриевых связано с внутриклеточным их паразитизмом. Заслуживает внимания тот факт, что у некоторых родов, относящихся к другим группам низших грибов, внутриклеточный или внутриполостный паразитизм также связан с отсутствием оболочки у паразита (Coelomomyces из Blastocladiales, некоторые Lagenidiales). Однако наличие у ольпидиевых и синхитриевых наиболее простых форм полового процесса, нам кажется, свидетельствует в пользу их примитивности, а отсутствие у них клеточных оболочек в вегетативном состоянии, возможно, является первичным признаком, связанным с происхождением от голых амебоидных организмов. Внутриклеточный паразитизм только содействовал сохранению этого архаического признака.
5. САПРОЛЕГНИЕВЫЕ ГРИБЫ — SAPROLEGNIOMYCOTA (PTEROMASTIGOMYCOTA)
Организмы, включаемые нами в этот отдел, характеризуются наличием в клеточных оболочках целлюлозы, а также своеобразным строением жгутиков, из которых у саиролегниомицетов один перистый направлен вперед, а другой — бичевидный направлен назад. У гифохитриомицетов только один жгутик — перистый. Наличие целлюлозы и строение жгутиков определяют обособленное положение сапролегниевых грибов в системе бесхлорофильных растительных организмов, сближают их с разножгутиковыми водорослями и дают основание для предположения о филогенетических связях их с этим оаделом автотрофных организмов.
С другой стороны, низшие представители саиролегниомицетов и гифохитриомицеты в строении тела и в онтогенетическом развитии имеют ряд черт, сближающих их с низшими представителями хитридиевых грибов, что давало основания объединять их с хитридиомицетами.
Можно предположить, что сапролегниевые грибы представляют отдельный ствол грибных организмов, и ряд черт, сближающих низших представителей этой группы с хитридиомицетами, является результатом происхождения тех и других от примитивных организмов, давших начало различным отделам грибов и водорослей.
161
Строение тела
и размножение гифохитриомицетов
Гифохитриомицеты представляют небольшую мало изученную группу, стоящую по своему развитию на уровне хитридиомицетов. У простейших представителей вегетативное тело представляет собою голое одноядерное образование, обитающее в клетках водорослей. Так, у Anisolpidium ectocarpi (семейство Anisolpidiaceae),
Рис. 58. Anisolpidium ectocarpi: а — клетки эктокарпа (Ectocarpus mitchellae) с тремя молодыми клетками паразита и прорастающей зооспорой, б — молодой организм с ядрами, расположенными по периферии, в, ? — образование и выход зооспор, д — зооспоры (по Karling, 1943).
паразитирующего в клетках бурой водоросли эктокарпа, зооспора прикрепляется к клетке растения-хозяина, покрывается оболочкой и образует вырост, проникающий внутрь клетки. Через этот вырост содержимое зооспоры переходит внутрь клетки хозяина, где через некоторое время, после повторного деления ядра, оно покрывается оболочкой и полностью превращается в спорангий (рис. 58).
Представители второго семейства гифохитриомицетов — Rhizi- diomycetaceae напоминают представителей семейства ризифидие-
162
вых из хитридиомицетов. Например, Rhizidiomyces apophy satus, паразитирующий на оогониях сапролегниевых, образует одноклеточное тело, дающее внутри клетки растения-хозяина апофизу с ризоидными выростами.
Вегетативное тело представителей третьего семейства гифохи- триомицетов — Hyphochytriaceae напоминает представителей кла~ дохитриевых: тело у гифохитриевых полицентрическое и образует внутри субстрата довольно развитый мицелий.
Бесполое размножение гифохитриомицетов происходит при помощи зооспор. Представители семейства Anisolpidiaceae голокар- пические: у них все тело превращается в зооспорангий, образующий вырост, пробивающий стенку клетки хозяина. Через этот вырост зооспоры выходят наружу (Karling, 1943). У Rhizidiomy- cetaceae не все тело идет на образование зооспорангия (эвкар- пические формы). У Hyphochytriaceae, имеющих развитый мицелий, зооспорангии закладываются или терминально, или интерка- лярно.
Половой процесс у Anisolpidiaceae происходит или по гологамному типу, или состоит в слиянии подвижных монад. Так, у Anisolpidium ectocarpi описано слияние двух голых протопластов, возникающих из зооспор, которые прикрепляются к поверхности хозяина, покрываются оболочкой и выпускают содержимое внутрь клетки (Karling, 1943; Johnson, 1957). Ядро зиготы делится; она покрывается оболочкой и после периода покоя, прорастая, дает зооспоры. У другого рода — Reesia половой процесс состоит в слиянии двух монад.
Строение тела
и размножение сапролегниомицетов
Низшие представители сапролегниомицетов обычно выделяют в порядок лагенидиальных (Lagenidiales). Они характеризуются, с одной стороны, рядом черт, объединяющих их с высшими сапро- легниевыми (строение жгутиков, дипланетизм зооспор, наличие целлюлозы в оболочках клеток), с другой стороны, своим развитием они напоминают хитридиомицеты.
Лагенидиальные являются внутриклеточными паразитами водорослей и низших грибов. Размножаются зооспорами, которые, попадая на растение-хозяин, теряют жгутики, покрываются оболочкой и дают вырост внутрь клетки хозяина, где образуют многоядерный голый плазмодиеобразный протопласт, как, например, у паразитирующего на гифах ахлии Olpidiopsis achlyae.
В других случаях плазмодиеобразная стадия может быть недолговечной — плазмодий покрывается оболочкой и дает гифы, как, например, у паразитирующего на видах клостерия Lagenidiuni closterii.
При бесполом размножении у некоторых форм все тело превращается в один зооспорангий или в цепочку зооспорангиев
163
или же дает зооспорангии на многоядерном мицелии. Для зооспор лагенидиальных характерен дипланетизм в его различных формах, сходных с описанными у сапролегниальных.
Половой процесс у лагенидиалытых происходит по гологамному типу. Так, у видов Olpidiopsis плазмодии покрываются целлюлозной оболочкой и копулируют попарно; при этом можно различать более крупные — «женские» и более мелкие — «мужские» плазмодии. В месте соприкосновения оболочки растворяются и содержимое меньшего («мужского») плазмодия переходит в более крупный («женский»), который превращается в зиготу. Иногда с одним более крупным сливается несколько более мелких плазмодиев (Barrett, 1912; Karting, 1942). У других родов (Ру- thiella, Myzocytium, Lagenidium) наблюдается отмирание ядер копулирующих клеток, кроме одного, и, таким образом, в них копулируют одноядерные клетки.
Высшие сапролегниомицеты — сапролегниальные, лептоми- тальные и пероноспоральные характеризуются хорошо развитым ценоцистическим мицелием. Иногда (семейство Rhipidiaceae из лептомитальных) мицелий состоит из базальной, иногда древовидно разветвленной части, от которой отходят тонкие гифы. Перегородки на мицелии возникают только при образовании половых органов и обычно, но не всегда, при образовании зооспорангиев, а также при повреждении гиф или для отдаления старых отмирающих частей мицелия. У представителей порядка лептомитальных на гифах наблюдаются сужения. Суженные места гиф иногда бывают закупорены зернами целлулииа — полисахарида, близкого к клетчатке. Подобные зерна иногда встречаются и в цитоплазме.
Бесполое размножение у высших сапролегниомицетов происходит при помощи зооспор, образующихся в более или менее развитых спорангиях. У наземных форм зооспорангии могут отделяться от гиф и переноситься воздушными течениями, а в ряде случаев прорастать как конидии — ростковой гифой без образования зооспор.
Наиболее простой тип зооспорангия описан у некоторых видов рода Pythium, например у Р. adhaerens и P. dictyosporum. У них конец гифы ослизняется, протопласт выходит и образует у выхода так называемый пузырек, внутри которого формируются зооспоры. У других видов рода Pythium, как и у представителей других семейств высших сапролегниомицетов, образуются типичные спорангии, отделяющиеся перегородкой от несущих гиф. Для многих сапролегниомицетов характерен диплапетизм зооспор —вначале образуются зооспоры грушевидной формы, у которых жгутики прикреплены к переднему концу. Затем эти зооспоры теряют жгутики и покрываются оболочкой. Через некоторое время оболочка сбрасывается, и из нее выходит зооспора почковидной формы с жгутиками, прикрепленными в боковой выемке. У многих сапролегниальных, лептомитальных и лагенидиальных наблюдается
164
ослабление и выпадение стадии грушевидной зооспоры, а для перо- носпоральных характерны только почковидные зооспоры.
У наземных пероноспоралытых, как уже упоминалось, спорангии отделяются от несущей гифы и могут переноситься воздушными течениями. У некоторых родов (Phytophthora, Plasm, о para, Sclerospora, Albugo) они обычно дают зооспоры и только иногда (Phytophthora, Plasmopara, Sclerospora) могут прорастать гифой. В других случаях (Peronospora, Premia) они обычно дают ростковую гифу, как конидии.
Половой процесс в порядках Leptomitales, Saprolegniales и Ре- ronosporales описывается как типичная оогамия в виду наличия ясно выраженной дифференциации копулирующих органов, называемых оогонием и антеридием.
Антеридии сапролегниомицетов являются одноклеточными образованиями со многими ядрами. Оогонии также одпоклеточные и в начале развития многоядерпы. Дальнейшее развитие оогония проходит различно. В оогониях сапролегниальных наблюдается редукция части ядер и формирование обычно нескольких яйцеклеток; только в редких случаях описаны оогонии с одной яйцеклеткой (Aphanomyces, Leptolegnia). Подобное явление известно и в семействе Leptomitaceae, где формируется или несколько яйцеклеток (Apodachlyella completa), или только одна (Apodachlyella brachynema; Kevorkian, 1935).
Оогонии у Peronosporales и в семействе Rhipidiaceae из Leptomitales развиваются иначе. Здесь протоплазма оогония разделяется на две части — центральную, образующую яйцеклетку,— гоноплазму и периферическую — периплазму. Оогоний вначале многоядерный. В более примитивных случаях, например, у Albugo bliti и Albugo portulacae, зрелая яйцеклетка многоядерна; при оплодотворении антеридий дает вырост, который проникает в оогоний и через который проходит его протопласт со многими (до 100) ядрами. В других случаях все ядра яйцеклетки, кроме одного, гибнут (Albugo tragopogonis) или же «лишние» ядра переходят в периплазму (Albugo ipomeae-panduranae). В большинстве же случаев при разделении цитоплазмы оогония на гоноплазму и периплазму в яйцеклетке остается только одно ядро. С ядром яйцеклетки сливается только одно ядро антеридия. Зигота у ие- роноспоральных при прорастании или непосредственно дает зооспоры, или образует зооспорангий.
Вопрос о смене ядерных фаз у сапролегниомицетов до настоящего времени вызывает противоречивые суждения. С конца XIX ст. одни авторы считали, что редукционное деление у них происходит при прорастании зиготы и, таким образом, вегетативное тело имеет гаплоидные ядра, другие же полагали, что редукционное деление происходит при созревании половых ядер, зигота прорастает без редукционного деления, а вегетативное тело является диплоидным. Разноречивость мнений по этому вопросу объясняется трудностями исследования, связанными с
1G5
незначительными размерами ядер и хромосом. В последпее время изучение характера деления ядер в гаметангиях перед половым процессом (Sansome, 1961,1963, 1965; Sansome a. Harris, 1962; Barksdale, 1968), а также микроспектрофотометрические исследования количества ДНК в различных клетках сапролегнии (Bryant a. Howard, 1969) дали основания для утверждения, что редукционное деление у сапролегниевых происходит при формировании половых ядер. Цитологические исследования Фланагана (Flanagan, 1970) с подсчетом хромосом в различных клетках Saprolegnia ferax и Achlya klebsiana подтверждают это предположение.
Цитологические и микроспектрофотометрические исследования Apodachlya brachynema дали доказательства того, что и в порядке лагенидиальных редукционное деление проходит при гаметогенезе (Howard a. Bryant, 1971).
Другие исследователи последних десятилетий (Thirumalachar et al., 1949; Ziegler, 1953) утверждали, что редукционное деление у сапролегниевых происходит при прорастании зиготы. Некоторые авторы на основании генетических исследований Phytophthora capsid приходят к тому же выводу (Timmer et al., 1970).
Таким образом, цитологические и микроспектрофотометрические исследования последних лет свидетельствуют в пользу того, что редукционное деление у сапролегниевых грибов происходит при созревании половых клеток, а генетические данные — в пользу того, что мейоз происходит при прорастании зиготы. Исследования ближайших лет должны внести ясность в этот вопрос.
Эволюция и классификация сапролегниевых грибов
Как сказано выше, мы считаем возможным объединить в одном отделе растительного мира сапролегниомицеты и гифохитрио- мицеты, рассматривая их как классы отдела Saprolegniomycota (Pteromastigomycota), на основании наличия у них перистых жгутиков и целлюлозы в оболочках. Класс гифохитриомицетов включает один порядок Hyphochytriales с тремя семействами — Ani- solpidiaceae, Rhizidiomycetaceae и Hyphochytriaceae.
Что касается класса Saprolegniomycetes, то его обычно разделяют на порядки Lagenidiales, Leptomitales, Saprolegniales и Peronosporales.
Ниже даем принятую нами классификацию сапролегниевых грибов.
Отдел SAPROLEGNIOMYCOTA (PTEROMASTIGOMYCOTA).
Класс SAPROLEGNIOMYCETES - САПРОЛЕГНИОМИЦЕТЫ.
Монады с двумя жгутиками. Один, перистый, направлен вперед, а другой, бичевидный, назад.
166
1. Порядок Lagenidiales — лагенидиальные. Вегетативное тело одноклеточное, плазмодиеобразное или в виде нитей, разделенных перегородками на отдельные клетки, вначале голое, позже покрывается вторичной оболочкой. Бесполое размножение обычно дипланетическими зооспорами, половой процесс гологамный. Внутриклеточные паразиты водорослей, водных грибов, реже водных животных или же сапрофиты.
Четыре семейства: Woroniriaceae Olpidiopsidaceae, Sirolpida- сеае и Lagenidiaceae. Вероятно, к этому же порядку следует отнести семейства Ectorgellaceae и Thraustochytriaceae.
2. Порядок Leptomitales — лептомитальные. Вегетативное тело в виде ценоцистического мицелия, обычно с сужениями, иногда закупоренными зернами целлулина. У представителей семейства Rhipidiaceae таллом дифференцирован на расширенную базальную, иногда древовидно разветвленную часть и отходящие от нее гифы. Зооспоры моиоиланетические, реже ди- планетические. Половой процесс оогамный. В оогонии образуется от одной до нескольких яйцеклеток. В оогониях представителей Rhipidiaceae имеется периплазма, окружающая яйцеклетку. Водные организмы, преимущественно сапрофиты на растительных остатках и плодах.
Два семейства: Leptomitaceae и Rhipidiaceae.
3. Порядок Saprolegniales — сапролегниальные. Мицелий ценоцистический, сильно развитый. Бесполое размножение дипланетическими зооспорами. Половой процесс оогамный; в оогонии развивается большей частью несколько яйцеклеток; сапрофиты и паразиты растительных и животных организмов.
Одно семейство — Saprolegniaceae.
4. Порядок Peronosporales — пероноспоральные. Мицелий ценоцистический, сильно развитый. Бесполое размножение или зооспорами, образующимися на материнском растении, или же чаще зооспорангии отделяются от несущей гифы и переносятся воздушными течениями, причем они могут давать зооспоры или прорастать, непосредственно образуя ростковую гифу. Половой процесс оогамный. В оогонии развивается одна яйцеклетка, окруженная периплазмой.
Водные и наземные сапрофиты или же чаще паразиты высших растений.
Три семейства: Pythiaceae, Albugiriaceae и Peronosporaceae. Класс HYPHOCHYTRIOMYCETES - ГИФОХИТРИОМИ- ЦЕТЫ.
Монады с одним перистым жгутиком, прикрепленным к переднему ее концу.
Порядок Hyphochytriales — г и ф о х и т р и а л ь н ы е. Организмы моноцентрические или полицентрические, голокарпи- ческие или эвкариические представляют группу, параллельную по структуре вегетативного тела хигридиомицетам, но отличающуюся характером жгутиков и наличием в оболочках целлюлозы.
167
Карлинг (Karling, 1943) выделяет в составе порядка три семейства: Anisolpidiaceae, Rhizidiomycetaceae и Hyphochytriaceae, описание которых дано выше.
По вопросу эволюции в пределах класса сапролегниомицетов существуют различные мнения. Наряду с признанием примитивности лагенидиальных высказываются мнения, что эта группа является редуцированной в результате внутриклеточного парази¬
Рис. 59. Схема филогенетических связей между порядками сапролегниевых грибов.
тизма (Gäumann, 1926, 1949, 1964; Cejp, 1957). Так, Гейманн пишет, что лагенидиальные, возможно, происходят от низших сапролегниальных и в результате эндопаразитического образа жизни претерпели крайнюю редукцию — у конечных членов, как у Olpidiopsis, таллом вторично плазмодиообразный и голокарпи- ческий (Gäumann, 1964, стр. 81), что он и изображает, правда, со знаком вопроса на филогенетической схеме (Gäumann, 1964, рис. 47, стр. 60). Аналогичную картину видим и на схеме Цейпа (Cejp, 1957). Действительно, в случае перехода к эндопаразитизму вегетативное тело гриба может упроститься, но не нужно забывать, что и бесполое размножение, и половой процесс у лагенидиальных также наиболее примитивны в пределах класса сапролегниомицетов, и это побуждает нас считать этот порядок все же наиболее примитивным и стоящим ближе всего к исходным организмам. Переход к эндопаразитизму содействовал только сохранению у них примитивных черт, в то время как другие группы, развиваясь, ушли вперед по пути эволюции.
Наличие гологамного полового процесса у лагенидиальных дает основание предположить, что половые органы высших сапролегниомицетов образовались в результате дифференциации копулирующих голоцитов.
Эволюция в пределах класса шла, с одной стороны, в направлении приспособления к условиям водной среды — порядки Saprolegniales и Leptomitales, а с другой — к условиям паземного существования — порядок Peronosporales. Бесси (Bessy, 1950) считает возможным выводить Rhipidiaceae из семейства Leptomitaceae. Наличие в оогонии одной яйцеклетки, окруженной периплазмой у
168
представителей семейства Rhipidiaceae, с одной стороны, и порядка Peronosporales — с другой, можно рассматривать как доказательство их родства. Нужно отметить значительные изменения в развитии зооспорангиев в связи с приспособлением их к воздушной среде, что привело к превращению их в конидии.
Возможные взаимоотношения между порядками сапролегние- вых грибов показаны на поданной схеме (рис. 59).
6. ЗИГОМИЦЕТЫ - ZYGOMYCETES (MUCOROMYCETES)
Как уже упоминалось выше, мы относим зигомицеты в качестве класса к отделу высших грибов (Eumycota, Amastigomyco- ta) ввиду того, чю между ними и низшими аскомицетами можно проследить связи, свидетельствующие об их филогенетическом родстве. С другой стороны, зигомицеты довольно резко отличаются как от хитридиевых, так и особенно от сапролегниевых грибов, в связи с чем одни авторы (Smith, 1955) отмечают их обособленность, а другие (Moreau, 1953) рассматривают их как самостоятельный ствол эволюции, ведущий начало от безжгутиковых низших грибных организмов. Отсутствие целлюлозы и гологамный половой процесс, наблюдаемый у некоторых представителей хитридиевых грибов, дает некоторым авторам основание сначала с сомнением (Gäumann, 1926, стр. 93), а позже более уверенно (Gäumann, 1964) связывать происхождение зигомицетов с хит- ридиевыми грибами. Однако ряд других особенностей зигомицетов побуждает считать их группой, которая хотя и могла возникнуть от общих предков с хитридиевыми, но эти родственные связи, вероятно, относятся к ранним стадиям дифференциации амебоидных организмов.
Гораздо менее обосновано выведение зигомицетов из сапролегниевых грибов (Bessey, 1942, 1950). Против этого предположения свидетельствует наличие целлюлозы в клеточных оболочках сапролегниевых грибов, а также более примитивный тип полового процесса у низших зигомицетов, чем у сапролегниевых.
Вегетативное тело в порядке Mucorales является в большинстве случаев не септированным разветвленным ценоцистическим мицелием, иногда достигающим значительной морфологической дифференциации (Rhizopus, Absidia). В жидкой среде, содержащей сахар, мицелий некоторых мукоровых может распадаться на небольшие участки («мукоровые дрожжи»). У представителей порядка энтомофторальных молодой мицелий, обычно ценоцистиче- ский, без перегородок, но затем может делиться на клетки, в частности в семействе Entomophthoraceae прорастающая конидия дает септироваипые гифы, которые вскоре распадаются на многоядерные фрагменты — так называемые «гифальные тельца» (Hyphenkörper), которые в дальнейшем размножаются делением и почкованием.
169
Таким образом, среди зигомицетов проявляется тенденция к переходу от несептированного ценоцистического мицелия к мицелию многоклеточному, характерному для высших грибов — аско- и базидиомицетов.
Бесполое размножение у зигомицетов происходит при помощи неподвижных спорангиоспор, развивающихся в спорангиях (порядок Mucorales), или конидий, обычно активно отбрасываемых на некоторое расстояние (порядок Entomophthorales).
Наиболее простое, исходное строение спорангия можно наблюдать у представителей семейства Мисогасеае. Обычно спорангий мукоровых имеет шаровидную, головчатую форму и отделяется от несущей гифы перегородкой, которая в виде купола впячивается в полость спорангия, образуя так называемую колонку. В редких случаях спорангий имеет другую форму, например, бутылкообразную в роде Saksenaea. Содержимое спорангия или непосредственно распадается на многоядерные споры (Mucor, Rhizopus, Phycomyces и др.), или же вначале делится на одноядерные участки, так называемые протоспоры, в которых происходит многократное деление ядер, после чего они делятся на многоядерные споры (Pilot)olus). В других случаях протоспоры непосредственно превращаются в спорангиоспоры (Circinella). Споры освобождаются, обычно, в результате разрушения или расплывания оболочки. В некоторых случаях (Pilobolus) спорангий целиком с силой отбрасывается на значительное расстояние в результате разрыва верхней части спорангиеносца. Споры у зигомицетов прорастают, образуя ростковую трубку.
У одних видов рода Mucor спорангиеносец простой — несет один спорангий, у других же он разветвляется, и каждая веточка заканчивается спорангием. У видов рода Thamnidium спорангиеносец разветвлен и главная его ось заканчивается больншм многоспоровым спорангием с колонкой. Ниже спорангиеносец имеет дихотомически разветвленные веточки, несущие небольшие спорангии без колонки с немногочисленными спорами — так называемые спорангиоли, которые могут отделяться от спорангиеносца и переноситься воздушными течениями. У других представителей семейства тамнидиевых, например у Chaetostylum fresenii, конечный спорангий обычно не развивается, а у видов рода Chaetocla- dium он никогда не образуется, а количество спор в спорангиолах уменьшается до одной, в результате чего оболочка споры может срастаться со стенкой спорангиолы и прорастать как конидия (Chaetocladium brefeldii).
У представителей семейства Mortierellaceae одиночный спорангий не имеет колонки и количество спор в нем может уменьшаться до одной, а в роде Hapio sporangium количество спор, как правило, падает до двух или до одной, т. е. спорангий превращается в конидию.
В семействе Choanephoiaceae развитие спор в спорангии задерживается, а на поверхности его развиваются многочисленные
170
дочерние спорангии, называемые спорангиолами, в которых образуются немногочисленные споры (рис. 60). У родов Blakeslea и Choanephora при плохом питании образуются типичные многоспоровые спорангии с колонкой типа Mucor, а при обильном питании формирование спор задерживается и развиваются спорангио- ли, которые у Blakeslea содержат небольшое количество спор
Рис. 60. Типы спороношений зигомицетов: а, б, в — Blakeslea trispora, г — Syncephalastrum cinereum, д, e — Syncephalis pycnosperma, ж — Coemansia aciculifera (no Gäumann,
1964).
(y B. trispora — три, a у Choanephora — только одну). Syncephalastrum совсем не образует спорангиев типа Mucor, а дает только дочерние спорангии (спорангиолы), в которых образуются споры, освобождающиеся после разрушения оболочки. У рода Syncephalis споры срастаются со стенкой спорангиолы и палочковидная спо- рангиола при созревании распадается на односпоровые участки. Нужно упомянуть, что при таком переносе спорообразования в спорангиолы материнский спорангий уже не отделяется перегородкой от сиорангионосца.
У других родов (Coemansia, Piptocephalis) материнский спорангий теряет головчатую форму. У Coemansia он превращается в асимметрическое многоклеточное образование — спорокладий, на котором на коротких стеригмах сидят односпоровые спорангио- лы. У Piptocephalis материнский спорангий превращается в маленькую базальную клетку, несущую многоспоровые спорангиолы, которые распадаются на односпоровые части.
171
У Cunninghamella развивается головчатый материнский спорангий, дающий по периферии многочисленные трех — восьмиядерные спорангиолы, которые не дают спор, а, отделяясь от материнского спорангия, развиваются как конидии.
Бесполое размножение у энтомофторальных происходит при помощи конидий, обособляющихся от недифференцированной несущей гифы. Однако Basidiobolus ranarum может также образовывать интеркалярные спорангии или же его конидии могут прп прорастании образовывать не ростковую гифу, а спорангий.
Половой процесс у зигомицетов — типичная гологамия, т. е. слияние двух клеток — голоцитов, содержимое которых не распадается на гаметы. У зигомицетов сливающиеся клетки одинаковых размеров или реже различаются размерами и в зависимости от строения тела — многоядерны или реже одноядерны.
Простейший тип полового процесса представлен у родов семейства Mucoraceae. У них образуются копуляционные ветви, которые соприкасаются, и от них отчленяются перегородкой многоядерные клетки — голодиты, сидящие на обычно расширенных гифах — суспензорах. Перегородка между голоцитами растворяется, содержимое их смешивается (плазмогамия), а образовавшаяся зигоспора покрывается толстой оболочкой. У мукора ядра вскоре после копуляции сливаются попарно (кариогамия), после чего вскоре, еще до наступления периода покоя, происходит редукционное деление. У рода Phycomyces кариогамия и редукционное деление задерживаются до прорастания зигоспоры. У рода Zygorhy- nchus намечается различие в размерах между копулирующими голоцитами. В некоторых случаях (Syncephalis) зигоспора образуется в качестве выступа на одной из копулирую!цих клеток (Thaxter, 1897). Зигоспоры у мукоровых, прорастая, дают нить, несущую или спорангий, так называемый зародышевый спорангий (Mucor), или группу спорангиол (Chaetocladium).
По такому же типу происходит половой процесс у представителей порядка энтомофторальных. Так, у Conidiobolus на гифах образуются копуляционные ветви, на которых развиваются копулирующие клетки (голоциты) и суспензоры. У Conidiobolus utriculosus слившиеся клетки непосредственно превращаются в зигоспору, а у С. brefeldianus содержимое одного голоцпта переливается в другой, и этот превращается в зигоспору, т. е. здесь уже намечается половая дифференциация. У Entomophthora echi- nospora копулируют четырехъядерные гифальные тельца, а зигоспора образуется как боковой вырост. У Entomophthora fumosa копулируют также четырехъядерные гифальные тельца, но перед слиянием ядра в них делятся, после чего каждое гифальное тельце содержит по восемь ядер (рис. 61). Однако после копуляции в вырост, образующий зигоспору, заходит только одно ядро, все же остальные ядра в зиготу не проникают и погибают (Rees, 1932). У Basidiobolus ranarum, мицелий которого распадается на одноядерные участки, копулируют одноядерные клетки. Эти клетки
172
дают клювовидные выросты, куда заходит ядро и делится там на два. Одно ядро остается в выросте и дегенерирует, а другое возвращается в клетку. Затем содержимое одной клетки переходит в другую, которая в это время разрастается и превращается в зигоспору (рис. 62).
У видов рода Endogone также проявляется половая дифференциация голоцитов: одни из них — «женские» больших размеров, другие — «мужские» меньших, и содержимое меньшего голоцита переходит в больший. У одних видов, например у Endogone sphagnophila, копулируют многоядерные голоциты, а у E. lactiflua все ядра первоначально многоядерного голоцита, кроме одного, переходят в суспензор, который отделяется перегородкой, и там дегенерируют (рис. 63). Таким образом, сливающиеся голоциты являются одноядерными. Содержимое «мужского» голоцита переходит в «женский», который дает вырост, куда переходит содержимое обоих голоцитов. Вырост покрывается плотной оболочкой и превращается в зигоспору, где через некоторое время происходит слияние ядер.
У некоторых мукоровых, например у Absidia и Phy соту ces, стерильные гифы разрастаются вокруг зигоспоры, а у представителей семейств Mortierellaceae и у Endogone образуются плодовые тела, спорокарпии, напоминающие своим строением плодовые тела некоторых низших аскомицетов (семейство Gymnoascaceae из порядка Aspergillales).
Обычно считают, что копулирующие клетки зигомицетов являются редуцированными гаметангиями, в которых не происходит дифференциации гамет. Гейман подчеркивает, что при этом типе полового процесса копулируют не гаметы, а их недифференцированные многоядерные материнские клетки, ценоцистические гаме- тангии, почему он и называет этот тип полового процесса гаме- тангия (Gametangie), т. е. копуляция гаметангиев, в отличие от копуляции гамет—мерогамии (Gäumann, 1926, стр. 10—11; 1964, стр. 66 и 95). Однако ознакомление с различными типами гологамии, наблюдающимися у низших грибов, нам кажется, дает больше оснований для предположения, что половой процесс зигомицетов является дальнейшим усложнением примитивного типа гологамии — слияния двух недифференцированных вегетативных клеток.
Отсутствие подвижных зооспор и гамет, а также четко выраженный зигогамный половой процесс еще со времен де Бари и Брефельда послужили основанием многим авторам для вы-
Рис. 61. Entomophthora fumosa:
половой процесс (по Rees, 1932).
173
деления зигомицетов в отдельную группу, которой придавался различный таксономический ранг — от порядка до класса. Так, А. Фишер (А. Fischer, 1892) рассматривает зигомицеты как «ряд» (Reihe) класса фикомицетов и подразделяет его на два порядка — Mucoriiiae и Entomophthorinae. Порядок Mucoriiiae А. Фишер делит на два подпорядка — Sporangiophorae с семействами Мисо- гасеае и Mortierellaceae и Conidiophorae — с семействами ChaetocJadiaceae и Cephalidaceae. В порядке Entomophthorinae А. Фишер выделяет только одно семейство — Entomophthoraceae. Систему А. Фишера в основном принимает Энглер (Engler 1898,
Рис. 62. Basidiobolus ranarum: половой процесс (по Bessey, 1950).
1912). Веттштейн (Wettstein, 1901) рассматривает зигомицеты как подпорядок порядка фикомицетов класса грибов и делит его на четыре семейства — Miicoraceae, Entomophthoraceae, Chaetocladia- сеае и Piptocephalidaceae.
Позднее ряд авторов рассматривают зигомицеты как порядок (Gäumann, 1926, 1949, 1964; Курсанов, 1933, 1940; Greis, 1943) или как подкласс класса фикомицетов (Ячевский, 1913; Е. А. Bessey, 1935, 1950; Gwynne-Vaughan and Barnes, 1937; Smith, 1955), и только в недавнее время их стали признавать особым классом (Moreau, 1953; Cejp, 1957; Chadefaud, 1960).
Геймани, который в последней своей монографии (Gäumann,
1964) рассматривает зигомицеты как порядок, делит его на три семейства — Mucoraceae с пятью подсемействами, Endogonaceae и Entomophthoraceae. Другие же авторы, признающие зигомицеты классом или группой, равноценной классу, делят их на три или четыре порядка. Например, Вердерманн (Werdermann, 1954) принимает четыре порядка зигомицетов — Mucorales, Entomophthora- les, Endogonales и Zoopagales. Некоторые же авторы (Bessey, 1950; Cejp, 1957; Ubrizsy a. Vörös, 1965) включают семейство Endogonaceae в порядок Mucorales. Порядок Mucorales Бесси, а за ним Цейп разбивает на восемь семейств — Mucoraceae, Pilobola- сеае, Thamnidiaceae, Choanephoraceae, Mortierellaceae, Piptocephalidaceae, Kickxellaceae и Endogonaceae. В порядке Entomophthorales
174
Бесси и Вердерманн различают одно семейство — Entomophthora- сеае, а Цейп разбивает этот порядок на три семейства — Ancyli- staceae, Basidiobolaceae и Entomophthoraceae.
Семейства Гейманна Мисогасеае и Entomophthoraceae Бесси и Цейи рассматривают как порядки, а его подсемейства — как семейства. Подсемейство Мисогеае соответствует семействам Mucoraceae и Pilobolaceae этих авторов, а объем подсемейств Cephalideae и Cunninghamelleae распределя¬
ется между семействами Choanephoraceae (роды Blakeslea, Cunninghamella, Choanephora и др.), Piptocephalidaceae (роды Piptocephalis, Syncephalis, Syn- cephalastrum и др.) и Kickxellaceae (роды Coemansia. Kickxella, Martenselia).
К порядку Zoopagales Бесси и Вердерманн относят семейства Harpellaceae, Genistellaceae и Zoopagaceae. Кроме упомянутых выше порядков (Mucorales, Entomophthorales и Zoopagales), Бесси, правда с большим сомнением, присоединяет к зигомицетам еще порядок Eccrinales. Цейп же в порядке Zoopagales оставляет только семейство Zoopagaceae, два другие семейства, которые Вердерманн и Бесси включают в этот порядок (Harpellaceae и Genistellaceae), а также семейства, относимые Бесси к порядку Eccrinales, он включает в выделяемый им четвертый класс фикомицетов — трихомицеты (Thichomycetes). Группу трихомицетов в качестве подкласса Trichomycetidae класса фикомицетов признают также Убрижи и Вереш (Ubrizsy a. Vörös, 1966).
Трихомицеты — мало изученная группа паразитных или сапрофитных организмов, обитающих в кишечном тракте членистоногих. Отношение их к зигомицетам является проблематичным.
175
Рис. 63. Endogone lactifliia: половой процесс (по Бухгольцу, Bessey. 1950).
Вегетативное тело трихомицетов имеет вид ценоцистических нитей, оболочка которых дает реакцию на целлюлозу. Нижней частью таллом прикрепляется к субстрату,— чаще всего это хитиновые покровы насекомых — особенно внутри органов питания или у заднего прохода. Длина гиф достигает 100 μ, иногда мицелий имеет кустистый вид.
Вегетативное размножение у них происходит при помощи конидий (микроконидий и макроконидий), у некоторых родов семейств Harpellaceae и Genistellaceae описан половой процесс, состоящий в слиянии соседних клеток одной нити.
Дюбоск, Леже и Тюзе (Duboscq, Leger et Tuzet, 1948) считают трихомицеты растениями. Манье (Manier, 1950) высказывает мысль, что морфологические особенности трихомицетов сближают их с примитивными грибами, а развитие известных в настоящее время форм напоминает типы развития некоторых фикомицетов, у которых господствует гаплоидная стадия, а диплоидная сведена к зиготе (Tuzet et Manier, 1948). Характер полового процесса представителей семейств Herpellaceae и Genistellaceae, напоминающий половой процесс базидиобола, дает основание предполагать близость трихомицетов к порядку Entomoplithorales (Leger et Gauthier, 1935). Обычно же отношение трихомицетов к грибам многие авторы ставят под сомнение. Во всяком случае положение этой группы в системе растительного мира остается еще не ясным и требует дальнейших исследований. Цейп (Cejp, 1957) приводит пять порядков трихомицетов — Eccrinales, Amoebidiales, Palavis- ciales, Harpellales и Genistellales. Геймапп, в отличие от Бесси и Цейпа, воообще исключает из системы грибов порядок Zoopagales и группу трихомицетов.
На обзорной таблице «безжгутиковых фикомицетов» — Nonfla- gellate Phycomyceteae Бесси (Bessey, 1950, стр. 44) с большим сомнением относит к фикомицетам также порядок Protomycetales. В дальнейшем этот порядок он рассматривает вместе с Peronospo- rales. Другие же авторы (Cejp, 1957; Gäumann, 1926, 1964) роды этого порядка включают в систему аскомицетов.
Из систем зигомицетов, опубликованных в последние годы, следует отметить системы Аркса и Крейзеля. Классификация Аркса (Агх, 1967, 1968) близка к классификациям Бесси и Цейпа. Арке делит класс зигомицетов на два порядка — Mucorales с девятью семействами (к семействам Бесси и Цейпа он прибавляет еще семейство Cunninghamellaceae) и Entomophthorales с двумя семействами — Entomophthoraceae и Zoopagaceae.
Крейзель (Kreisel, 1969) подобно Вердерманну делит зиго- мицеты на четыре порядка — Mucorales, Zoopagales, Entomophthorales и Endogonales, но придает им несколько иное содержание. Порядок Mucorales он делит на шесть семейств: Mortierellaceae, Syncephalastraceae, Mucoraceae, Thamnidiaceae, Cunninghamellaceae, Choanephoraceae. Семейство Piptoccphalidaceae Крейзель вместе с семейством Zoopagaceae объединяет в порядок Zoopagales.
176
К порядку Entomophthorales on относит семейства Pilobolaceae, Basidiobolaceae и Entomophthoraceae (включая Acylistaceae). К порядку Endogonales Крейзель относит только одно семейство Endogonaceae. Трихомицеты в составе порядков Amoebidiales, Eccrinales, Harpellales (включая Genistellales) и Kickxellales Крейзель включает в 'созданный Арксом класс Endomycetes.
Резюмируя сказанное выше, мы считаем возможным принять такую систему зигомицетов.
Класс ZYGOMYCETES
1. Порядок Mu corales — мукоральные. Бесполое размножение спорангиоспорами или, реже, конидиями.
Семейства: Mucoraceae, Pilobolaceae, Thamnidiaceae, Choane- phoraceae, Mortierellaceae, Piptocephalidaceae, Kickxellaceae и Endogonaceae.
2. Порядок Entomophthorales — энтомофтораль- H ы e. Бесполое размножение только конидиями, с силой отбрасываемыми от конидиеносца.
Семейства: Ancylistaceae, Basidiobolaceae, Entomophthoraceae.
3. Порядок Zoopagales — зоопагальные. Бесполое размножение конидиями, не отбрасываемыми с силой.
Семейство Zoopagaceae.
Что касается филогенетических взаимоотношений между порядками и семействами зигомицетов, то, следуя Гейманну, мы считаем, что порядки Mucorales, Entomophthorales и Zoopagales. хотя и имеют общие корни, обособились очень рано. В основании системы мукоральных, по Гейманну, нужно поставить семейство Mucoraceae с наиболее простым типом полового процесса и многоспоровыми спорангиями. От него или близких к нему организмов развились другие семейства порядка путем усложнения спороно- шений и полового процесса.
7. АСКОМИЦЕТЫ - ASCOMYCETES Вегетативные органы
Вегетативное тело аскомицетов обычно представляет собой разветвленный мицелий с гифами, разделенными на одноядерные или многоядерные, реже двуядерные участки, обычно называемые клетками. Однако перегородки, разделяющие гифы, являются неполными. Они имеют центральное отверстие, настолько крупное, что возможен переход цитоплазмы и ядер из одного участка мицелия в другой. Иногда мицелий может распадаться на отдельные клетки — оидии. Такое состояние его является обычным для дрожжей — Saccharomycetaceae. Иногда мицелий может быть редуцированным, например у Laboulbeniales, паразитирующих на внешних покровах насекомых.
177
Бесполое размножение
Характерным способом бесполого размножения аскомицетов являются конидии. Только у немногих форм из семейства Sper- mophthoraceae известно образование интеркалярных спорангиев (см. рис. 68). Типичные конидии образуются в результате отшну- ровывания от гифы. Они могут быть одноклеточными или многоклеточными и развиваться или по одной или же образовывать цепочки. Часто конидии образуются в специальных вместилищах — никнидах. Иногда гифы, на которых образуются конидии (конидие- носцы), поднимаются над субстратом и приобретают характерную форму, например у Aspergillus или Pénicillium. Клетки конидие- носцев, от которых отделяются конидии, иногда приобретают характерную бутылковидную форму и называются фиалидами.
Особым бесполым спороношением у некоторых аскомицетов являются так называемые эндоконидии, встречающиеся у представителей порядков Aspergillales, Microascales, Sphaeriales, Laboui- beniales, Pseudosphaeriales. Образование эндоконидий изучено y Thielavia basicola (конидиалъная стадия Thielaviopsis basicola) еще в начале XX ст. (Brierley, 1915; Lehmann, 1918). У этого вида эндоконидии образуются в одноклеточных одноядерных бу- тылковидных клетках — фиалидах следующим образом. Ядро фи- алиды делится на два. Одно дочернее ядро остается в нижней части фиалиды, а другое перемещается к ее верхушке, и верхняя часть клетки отделяется перегородкой. После этого оболочка верхней клетки расщепляется на два слоя и образовавшаяся таким образом конидия приобретает собственную оболочку, внешняя же оболочка разрывается, после чего зндоконидия может выйти наружу. Ядро нижней клетки тем временем опять делится и возникает следующая конидия. В результате повторного образования конидий и удлинения шейки фиалиды может образоваться длинная цепочка конидий, которые постепенно выходят на свободу. По сути, мы имеем в данном случае акрогенные конидии, которые не отшнуровываются от образующей их клетки, а получают оболочку в результате расщепления оболочки материнской клетки. По этому типу образуются эндоконидии у представителей Microas- cales и Sphaeriales (Bombardia lunata), a также мужские половые клетки — спермации лабульбениальных.
У рода Mycosphaerella из порядка Pseudosphaeriales эндоконидии образовываются иначе. В особых или в обычных пикиида* совместно с микроконидиями развиваются материнские клетки эндоконидий (микроконидий). Ядро такой материнской клетки делится дважды, после этого протопласт делится на четыре части, которые вытягиваются бактериевидно и, покрываясь оболочкой, выходят на свободу через разрыв стенки материнской клетки. Насколько известно, они не способны прорастать вегетативно и функционируют только как мужские оплодотворяющие клетки — спермации (Higgins, 1920, 1936; Wolf, 1939).
178
Не останавливаясь на детальном описании типов конидий и способов их образования, можем сослаться на работы Хьюза (Hughes, 1953), Тубаки (Tubaki, 1958), Субраманиан (Subramanian,
1965), Дудки (1968), в которых делаются попытки использования конидиальных спороношений для классификации несовершенных стадий грибов и установления связи этих спороношений с исковыми стадиями. Наличие разнообразных конидиальных спороношений у одного вида, несомненно, является приспособлением к изменчивым метеорологическим и климатическим условиям, определяющим развитие тех или иных конидиальных спороношений (Лавров, 1951; Зерова, 1952). Разнообразные конидиальные спо- роношения обеспечивают выживание вида в крайних условиях существования.
Половой процесс
Строение половых органов у аскомицетов очень разнообразно. У некоторых представителей аскомицетов наблюдается гологамный половой процесс, при котором сливаются недифференцированные или мало дифференцированные клетки, у других же имеются одноклеточные или многоклеточные женские половые органы — аскогоны, часто снабженные воспринимающей гифой — трихогиной, а оплодотворяющие элементы представлены или одноклеточными мужскими половыми органами, обычно называемыми антеридиями, с недифференцированным содержимым, или конидиями, или же эндоконидиями, называемыми часто сперма- циями. В связи с таким разнообразием в строении половых органов, в половом процессе и, как увидим, также в последующем развитии зиготы эти особенности аскомицетов играют огромное значение при рассмотрении вопроса о происхождении и филогенетическом развитии этой крупнейшей группы грибных организмов.
Характерной чертой полового процесса аскомицетов является слияние одноядерных или многоядерных клеток, содержимое которых не образует гамет. Часто принято считать, что у низших аскомицетов, как и у зигомицетов, мы имеем дело с копуляцией гаметангиев, утративших способность давать гаметы, копуляцией «материнских половых клеток», в связи с чем этот тип полового процесса Данжар предложил называть гаметангйей, т. е. копуляцией гаметангиев, в отличие от копуляции гамет — мерогамии (Guilliermond, 1928, стр. 572; Gäumann, 1926, стр. 10—11; 1964, стр. 66, 95). Гейманн и ряд других авторов считают, что эта предполагаемая потеря способности образовывать гаметы является результатом ослабления способности к половому процессу, результатом «кризиса, нарушения сексуальности» (Gäumann, 1964, стр. 66). Эти высказывания являются отголоском взглядов Брефельда на ослабление способности к половому процессу у высших грибов.
179
Что касается половых органов аскомицетов, то в действительности нет никаких оснований считать копулирующие клетки ас- ковых грибов, как и зигомицетов, редуцированными гаметангиями- Как мы уже отмечали, ознакомление с типами полового процесса у низших грибов дает достаточные основания для того, чтобы считать, что гологамный половой процесс зигомицетов и аскомицетов является усложнением примитивного типа гологамного полового процесса — слияния двух недифференцированных вегетативных клеток, известного у хитридиомицетов. Это предположение является тем более оправданным, что у аскомицетов, а как позже увидим, и у базидиаль- ных грибов очень распространена примитивная форма полового процесса — копуляция
Рис. 64. Dipodascus uninucleatus: а, б, в — половой процесс, г, д, е — образование аска (по Biggs, 1937).
вегетативных клеток мицелия или, как часто видим у аскомицетов, копуляция женского полового органа — аскогона с клеткой вегетативного мицелия.
В процессе дальнейшего развития у аскомицетов намечается дифференциация и усложнение половых органов. В литературе их принято соответственно называть мужские — антеридиями, а женские — оогониями и аскогонами. Однако применение терминов антеридий и оогоний для гологамного полового процесса не является последовательным, так как антеридиями и оогониями называют гаметангии, внутри которых образуются гаметы. При гологамии копулируют не гаметы, а целые клетки; поэтому копулирующие клетки в этом случае правильнее называть голоцитами. В случае копуляции одинаковых голоцитов (изоцитов) можно го¬
180
ворить об изогологамии, а при копуляции голоцитов, различающихся размерами (гетероцитов),— о гетерогологамии. При наличии четких половых различий мужской голоцит можно называть андроцитом, а женский — гинецитом или, уступая традициям, ас- когоном.
У представителей порядка Endomycetales, наиболее просто организованных аскомицетов, можно наблюдать половой процесс,
Рис. 65. Myriogoniiim odontiae: развитие асков с образованием крючков (по Cain, 1948).
очень напоминающий таковой у зигомицетов и состоящий в слиянии двух клеток, не отличающихся или мало отличающихся от вегетативных. Так, у видов рода Dipodascus копулируют две соседние клетки, образуя копуляционные выросты, представляющие собою у одних видов (Dipodascus uninncleatus и D. aggregatus), обладающих одноядерными клетками мицелия, одноядерные (Biggs, 1937; Barta, 1959), а у других видов (Dipodascus albidus), с многоядерными клетками мицелия, многоядерные клетки — голо- циты, содержащие по 10—12 ядер (рис. 64). При слиянии многоядерных клеток у D. albidus часть ядер из одной — «мужской» переходит в другую — «женскую», где сливаются одно «мужское» и одно «женское» ядра; остальные ядра обеих клеток дегенериру- ют( Jnel, 1902, 1921; Dangeard, 1907). Напомним, что у Endogone lactijlua из зигомицетов наблюдается подобное явление, когда при слиянии многоядерных клеток сливаются только два ядра, а остальные в половом процессе участия не принимают и дегенерируют. При слиянии одноядерных клеток у видов Dipodascus, имеющих мицелий с одноядерными клетками (D. uninucleatus и D. aggregatus), оба ядра копулирующих клеток сливаются в диплоидное ядро зиготы. У видов рода Dipodascus зигота непосредственно превращается в аск (сумку). Однако у другого рода того же семейства — Myriogonium наблюдается повторное образование сумок при помощи специального приспособления — крючков, характерного для высших аскомицетов (рис. 65). Например, у Myriogonium
181
odontiae зигота вытягивается, крючковидно сгибается и от нее отделяется верхушечная клетка, которая затем сливается с базальной. Субтерминальная клетка увеличивается в размерах и превращается в аск. Базальная клетка, слившаяся с терминальной, дает новый крючкообразный вырост, субтерминальная клетка которого дает следующий аск. Этот процесс может повторяться и,
таким образом, в результате из одной зиготы может образоваться ряд асков (Cain, 1948).
У видов рода Eremascus — E. fertilis и E. albus копулируют две соседние одноядерные клетки; они дают выросты, которые иногда обвиваются друг около друга, как у некоторых высших аскомице- тов. Ядра этих клеток делятся на два, одно из них остается в родительской клетке, другое входит в популяционный вырост (рис. 66). У Eremascus albus, по Гарролду (Harrold, 1950), сразу после отделения копулирующей клетки — выроста или до этого ядро делится на два; одно из них переходит в верхнюю часть клетки, которая копулирует с соседней половой клеткой, после чего ядра обеих клеток сливаются и образуют диплоидное ядро. Верхние части копулирующих клеток перед слиянием ядер или после него отделяются перегородками и дают начало аску, в котором образуется восемь, а иногда большее число спор. Характер полового процесса у видов Eremascus напоминает некоторые моменты, наблюдаемые при половом процессе у Basi- diobolus из энтомофторальных, где ядра копулирующих клеток делятся на два, из которых одно дегенерирует, а другое сливается с ядром другой копулирующей клетки.
У Endomyces magnusii копулирующие клетки отличаются размерами — вначале они двух-трехъядерны; в дальнейшем одно из ядер остается у вершины клетки, а остальные уходят вниз, после чего образуется перегородка, отделяющая одноядерную копулирующую часть (Giiilliermond, 1909).
У дрожжей половой процесс состоит в слиянии вегетативных клеток (SchizoSaccharomyces octosporus, ряд рас Saccharomyces cerevisiae и S. ellipsoideus) или прорастающих аскоспор (Saccha-
Рис. 66. Eremascus albus:
половой процесс и развитие аска (по Harrold, 1950).
182
готу ces ludivigii, некоторые расы Saccharomyces ellipsoideus). У Spermophthora gossypii копулируют гаплоидные спорангиоспоры (Guilliermond, 1928). Наконец, у Nadsonia fulvescens наблюдается педогамия — копуляция материнской клетки с дочерней (Nadson and Konokotin, 1926, см. рис. 67).
Приведенные примеры свидетельствуют о большом сходстве полового процесса эндомицетальных и зигомицетов, которое указывает на их тесные филогенетические связи. Это подтверждается также сходством в дальнейшем развитии зиготы. Так, у зигомицетов (му- коральные) зигота дает начало зародышевому спорангию, в котором при образовании спор происходит редукционное деление, а у подавляющего большинства эндомицетальных из зиготы развивается аск, в котором после редукционного деления образуются гаплоидные аскоспоры. Сходство усиливается еще тем, что у некоторых родов эндомицетальных (Dipodascus, Kluyveromyces, Ashbya gossypii) количество спор в аске не является определенным, а может доходить до многих десятков.
Что касается типа развития эндомицетальных, то, кроме наиболее распространенного гаплоидного цикла, при котором, как и у мукоральных, все развитие проходит в гаплоидной фазе и только зигота имеет диплоидное ядро, у некоторых представителей наблюдается также гаплоидно-диплоидный и диплоидный типы развития. Так, у Saccharomyces cerevisiae и S. ellipsoideus прорастающая аскоспора дает сначала гаплоидные клетки. Позже они копулируют, в результате чего развивается диплоидный мицелий, на котором образуются аски. В асках после редукционного деления развиваются гаплоидные аскоспоры. Таким образом, у этих видов дрожжей мы видим чередование гаплоидного и диплоидного поколений. У некоторых рас дрожжей проявляется тенденция к сокращению гаплоидной фазы; иногда же у Saccharomyces ellipsoideus могут копулировать прорастающие аскоспоры. В этом случае все развитие гриба проходит в диплоидной фазе, а гаплоидными являются только аскоспоры. Такой цикл развития характерен для Saccharomyces ludwigii, у которого аскоспоры копулируют попарно еще в аске (Guilliermond, 1905; Winge und Laustsen, 1939; цит. но Gäumann, 1964). Иногда у некоторых рас Saccharomyces cerevisiae вместо копуляции аскоспор наблюдается автогамное слияние ядер внутри аскоспоры — ядро аскоспоры делится на два, после чего эти ядра снова сливаются (Winge a. Laustsen, 1937; цит. по Gäumann, 1964).
Рис. 67. Nadsonia fulvescens:
половой процесс и образование аска (по Guilliermond, 1928).
183
Интересный случай онтогенеза с чередованием гаплоидного и диплоидного поколений описан у Spermophthora gossypii и Ere- mothecium ashbyii — представителей своеобразного семейства Sper- mophthoraceae, причисляемого обычно к порядку эндомицеталь- ных (рис. 68). У них из гаплоидной аскоспоры развивается первичный многоядерный мицелий, не имеющий поперечных
перегородок. Затем участок гифы вблизи верхушки набухает и отделяется двумя перегородками, образуя спорангий, вначале 5—8-ядерный; после повторного деления внутри спорангия формируется до 40 спорангиоспор. При прорастании снорангиоспор образуется новый гаплоидный несептированный мицелий. Иногда же споран- гиоспоры попарно копулируют, в результате чего развивается короткий, разветвленный, обычно разделенный на одноядерные клетки диплоидный мицелий. На концах гиф этого мицелия развиваются ас- ки, в которых после редукционного деления образуется 8—12 аскоспор (Gu- illiermond, 1928, 1936).
У другого представителя семейства Spermophthora- сеае — N ematospora coryli спорангии не образуются, и аскоспоры при прорастании дают первичный мицелий. У этого вида копулируют или клетки первичного мицелия, или прорастающие споры. Зигота может некоторое время развиваться, давая гифы, образующие аски, или же она может непосредственно превращаться в аск (Guilliermond, 1928). Таким образом, у N ematospora coryli могут развиваться из зиготы гомологи аскогенных гиф, наличие которых характерно для высших аскомицетов. Аналогичный процесс описан у Nema- tospora phaseoli (Wingard, 1925).
Гологамный половой процесс присущ также представителям порядка тафринальных (Taphrinales), у которых копулируют вегетативные клетки первичного гаплоидного мицелия, как например у Taphrina (Wieben, 1927; цит. по Gäumann, 1964) или иногда
Рис. 68. Spermophthora gossypii:
a — гаплоидное растение, образующее спорангии (б, в), г — копуляция гаплоидных спор, д — диплоидный мицелий, образующий аски (е) (по Guilliermond, 1928).
184
у Taphrina deformans (Martin, 1949). У Taphrinales после слияния клеток (плазмогамии) не происходит непосредственно слияние ядер (кариогамия), и мицелий характеризуется наличием дика- рионов. Кариогамия у тафринальных происходит перед образованием аскоспор.
У низших представителей порядка Aspergillales половой процесс также напоминает половой процесс эндомицетальных — он состоит в слиянии клеток, не отличающихся или мало отличающихся от вегетативных.
Однако иногда у них наблюдается морфологическая дифференциация голоцитов на мужской и женский, причем женский голоцит может иметь вытянутую форму и обвиваться вокруг мужского («антеридия»), например у Byssochlamis nivea (Rosenbaum, 1944; цит. по Gäumann, 1964), или, наоборот, мужской голоцит может обвиваться вокруг женского, например у Talaromyces (Pénicillium) verticillatus (Dangeard, 1907) (рис. 69). Иногда у аспергиллальных наблюдается усложнение женской клетки вследствие образования одно- или многоклеточной воспринимающей гифы — трихогины, например у Мопа- scus purpureus (Schikorra, 1909). В таком случае уже говорят о женском половом органе — аскогоне. У Aspergillales ядра при половом процессе сразу не сливаются и образуют дикарионы. Аскогон в простейших случаях распадается на двуядерные клетки, дающие начало аскогенным гифам, образующим аски (например, Byssochlamys nivea).
У других, более высоко организованных форм,
кариогамия задерживается еще дольше и ас- ver.Jcl а um’
г половой процесс (по
когон образует развитую систему аскогенных Emmons, 1935). гиф. При развитии асков наблюдается образование типичных для аскомицетов крючков.
В порядке Microascales строение половых органов часто сходно с их строением у Aspergillales. Однако у принадлежащего к этому порядку Ceratocystis fagacearum отмечены некоторые особенности, напоминающие более высокоорганизованные аскомицеты (рис. 70). У этого вида трехклеточный аскогон снабжен многоклеточной разветвленной трихогиной, а оплодотворение происходит при помощи конидий (Wilson, 1956).
Очень близок к аспергиллальным по характеру полового процесса порядок эризифальных. В типичном случае у них копулируют одноклеточные мужской и женский органы (андроцит и аскогон), как это было описано неоднократно у различных
185
Рис. 69. Pénicillium verticillatum:
половой процесс (по Emmons, 1935).
186
представителей порядка, начиная с конца прошлого столетия (Harper, 1895, 1896; Blackman a. Fraser, 1905). В некоторых случаях копулирующие клетки не отличаются друг от друга (Allen, 1936). Иногда копулирующие ветви мицелия подымаются над субстратом и обвиваются одна вокруг другой, как у некоторых аспергил¬
ловых. Такое явление описано у Erysiphe graminis и E. nitida (Björling, 1946; цит. по Gäumann, 1964). В дальнейшем оплодотворенный аскогон в некоторых случаях непосредственно превращается в аск или дает зачаток аскоген- ной гифы, как это известно у некоторых рас Sphaerotheca fuliginea (Beatus, 1950; цит. но Gäumann, 1964, стр. 180). У Sphaerotheca casta- gnei дикарион в зиготе делится один раз; образовавшаяся четырехъядерная клетка делится на три клетки — две одноядерные и одну (сред нюю) двуядерную, превращающуюся в восьмисноровый аск (Bergman, 1941) (рис. 71). У других же эризифальных такая двуядерная клетка не превращается непосредственно в аск, а дает аскогенные гифы. У Phyllactinia suffulta, у которой ядро мужской клетки (андроцита) дегенерирует и образование дикариона происходит в результате деления ядра аскогона, двуядерный аскогон в результате повторного деления ядра превращается в восьмиядерный. Из него развиваются аскогенные гифы, вначале не разделенные перегородками, в которых происходит дальнейшее деление ядер. Затем аскогенные гифы делятся на клетки, из которых конечная — одноядерная, нижние — двуядерные. Предпоследняя — двуядерная клетка аскоген- ной гифы превращается в аск (Colson, 1938).
Рис. 70. Ceratiocystis fagacearum: а — популяционная ветвь, состоящая из базальной клетки и клетки — будущего аскогона, б, в — трехклеточный аскогон с трихогиной в зачатке перитеция, г — развивающийся аскогон с выростами из базальной клетки (по Wilson. 1956).
У тропического рода Lanomyces, выделяемого в монотипическое семейство Lanomycetaceae, половой процесс, как у некоторых Erysiphaceae, состоит в слиянии недифференцированных клеток (гологамия). Кариогамия следует непосредственно за плазмогамией. Зигота дает короткую, вероятно, диплоидную нить, конечная клетка которой превращается в аск (Gäumann, 1964).
Рис. 71. Sphaerotheca castagnei:
a, 6, в, г — половой процесс, д, е, ж, з — развитие зиготы с образованием зачатка аска, в котором происходит слияние ядер (по Bergman, 1941).
Дальнейшее усложнение полового процесса и строения аского- на наблюдается у представителей порядков Pezizales и Helotiales. У них аскогон бывает или одноклеточным или многоклеточным и обычно снабжен одноклеточной или многоклеточной трихогиной. Клетки аскогона обычно многоядерны. Мужской половой орган представляет собой одноклеточное недифференцированное образование (андроцит), которое обычно в литературе называется антеридием. Иногда оплодотворяющим элементом является конидия или вегетативная клетка. У ряда представителей порядков Pezizales и Helotiales наблюдается копуляция вегетативных клеток (сома- тогамия).
У одного из наиболее изученных представителей порядка Pezizales — Ругопета confluens одноклеточный многоядерный аскогон снабжен одноклеточной многоядерной трихогиной (рис. 72).
187
Андроцит содержит также многочисленные ядра. При созревании аскогона ядра трихогины отмирают, в месте соединения андроци- та с трихогиной оболочка ослизняется и ядра андроцита переходят в трихогину, а затем через образующуюся к этому времени пору в стенке аскогона — в аскогон, где они обычно не сливаются с ядрами аскогона, а размещаются попарно, образуя дикарионы. Оплодотворенный аскогон образует аскогенные гифы, в которые
переходят и дикарионы. Аскогенные гифы делятся перегородками на клетки, которые вблизи аскогона имеют две — восемь пар ядер, а на дистальном конце по одной паре (Claussen, 1905, 1912). Позже конечные разветвления аскогеппых гиф дают аски с образованием характерных крючков.
У видов рода Asco- bolus, например у A. magnificus, аскогон и трихогина многоклеточны, трихогина обвивается вокруг андроцита («антеридия»). У видов рода Ascodesmis мужская и женская ветви спирально закручиваются вокруг другой (Claussen, 1905), а у Humaria aggregata, по данным Розенберга (цит. по Gäumann, 1964), мужская ветвь закручивается вокруг трихогины. Описаны случаи, когда развитие андроцита («антеридия») вызывается воздействием трихогины аскогона на клетки гифы противоположного знака. Так, у Anthracobia melaloma одна клетка этой гифы, находящаяся в непосредственном контакте с верхушкой трихогины, увеличивается в размерах и округляется, превращаясь в андроцит (антеридий), после чего оболочка между ними разрушается и происходит плазмогамия (Rosinski, 1956). У Ascoboius carbonarius трихогина может вступать в контакт с конидией, возникающей па мицелии противоположного знака (Dodge, 1912; Betts, 1926), а у Ascoboius stercorarius трихогина
Рис. 72. Ругопета confluens: а — группа аскогонов и андроцитов, б — слияние андроцита с трихогиной, в — образование аскогенной гифы, г — образование крючков, д — молодой аск с диплоидным ядром, е — аски (по Гарперу и Клауссену из Bessey, 1950).
188
пли возникающая из нижней части аскогона воспринимающая гифа могут копулировать с прорастающей оидией, прорастающей аскоспорой или концом гифы противоположного пола (Dowding, 1931; Bistis, 1956, 1957, и др.; цит. по Gäumann, 1964, сгр. 200).
У представителей порядка Helotiales оплодотворение происходит обычно при помощи микроконидий. Например, у Blumeriella jaapii (Higginsia hiemalis), живущей на листьях видов рода Prunus, развиваются спирально закрученные аскогоны, состоящие из 8—12 одноядерных клеток и многоклеточной трихогины, поднимающейся над субстратом. Оплодотворение происходит при помощи одноклеточных микроконидий, образующихся на разветвленных конидиеносцах (Backus, 1934).
Такое же строение половых органов характерно для дискоми- цетных лишайников (Lecanorales), например для рода Collema, где они были описаны еще в конце прошлого столетия (Stahl, 1877; Baur, 1898, и др.). Аскогон и трихогина у видов этого рода многоклеточные, оплодотворение происходит при помощи конидий, возникающих в пикнидах. У некоторых родов лишайников обнаружено, что такие конидии способны прорастать и давать нормальные талломы (Möller, 1887, 1888). У Collemodes bachman- nianum конидии образуются на гифах, не заключенных в пик- ниды.
В порядке Sphaeriales половой процесс хорошо исследован у представителей семейства Sordariaceae. Аскогон и трихогина, если они имеются, многоклеточные. Оплодотворение осуществляется или при помощи мало дифференцированной мужской клетки — андроцита (антеридия) или спорами экзогенного типа — макро- II микроконидиями или же эндогенными конидиями, называемыми часто спермациями. Иногда аскогон сливается с недифференцированной вегетативной клеткой. Так, у Sordaria fimicola и некоторых других видов описано слияние трихогины с мужской клеткой— андроцитом (антеридием), а при его отсутствии — с вегетативной гифой (Gäumann, 1964, стр. 257—258).
У Gelasinospora autosteira (G. calospora var. autosteira) оплодотворение происходит в результате копуляции трихогины с конидией (микроконидией) экзогенного типа (Tylutki, 1958; Sloan a. Wilson, 1958; Goos, 1959) (рис. 73), а у Bombardia lunata — с эндогенными конидиями, образующимися в фиалидах (Zickler, 1934, 1937) (рис. 74).
Детально изучен половой процесс у N e urospora sitophila. Аскогон у этого вида спирально закручен и состоит из многоядерных клеток. Настоящая трихогина отсутствует; вместо нее верхние клетки аскогона дают одну или несколько септированных воспринимающих гиф. N eurospora sitophila — вид гетероталлический, дающий две различающиеся в генетическом отношении линии, которые могут образовывать аскогоны, одноядерные эндогенные ми- кроконидпи и многоядерные макроконидии. Конидии не являются специализированными мужскими клетками и могут прорастать,
189
давая новый мицелий. При наличии воспринимающих гиф противоположного полового знака конидии копулируют с ними. При отсутствии конидий воспринимающие гифы могут копулировать с вегетативными клетками мицелия противоположного знака или с соответствующими аскоспорами (Dodge, 1927; Backus, 1939,
и др.)· Иногда наблюдается у Neu- rospora соматогамная копуляция между обыкновенными вегетативными гифами противоположного знака (Dodge, 1936) (рис. 75).
Рис. 73. Gelasinospora autosteira:
проперитеций с трихогиной, вытягивающейся по направлению к группе спермациев (по Goos, 1959).
Среди асколокулярных (битуникатных) аскомицетов половой процесс лучше всего изучен у представителей порядка Pseudosphae- riales. Здесь аскогон одноклеточный или реже многоклеточный, с одноядерными или многоядерными клетками (количество ядер может доходить у Dothidea insculpta до восьмидесяти) с довольно длинной одноклеточной или многоклеточной трихогиной. Оплодотворение происходит в результате слияния трихогины с андроци-
190
Рис. 74. Bombardia lunata: а — фиалида и эндогенные конидии, 6 — молодой аскогон с трихогиной. оплетенный гифами, в — трихогина направляется к группе эндоконидий, расположенных у отверстия фиалиды (по Zickler, 1937).
том (антеридием), материнской клеткой конидии, эндогенной микроконидией (спермацием) или вегетативной клеткой. У гомо- таллической Leptosphaerulina gaeumannii (Pleospora gaeumannii) описан одноклеточный аскогон с трихогиной, копулирующей с одноклеточным андроцитом — антеридием (Müller, 1951). У гетеро- таллического Cochliobolus sativus аскогон одноклеточный, много- ядерный, с длинной нитчатой трихогиной. Оплодотворение происходит при помощи эндокоиидий
(спермациев), образующихся в фиалидах, расположенных в пикнидах ( сперхмогониях ). Эндоконидии у этого вида являются специализированными оплодотворяющими клетками, не способными прорастать на питающих средах (Shoemaker, 1955).
У видов рода Mycosphae- rella аскогон одноклеточный, одноядерный с длинной трихогиной, выступающей над стромой и эпидермисом растения-хозяина. В особых пикнидах (спермогониях) или в обычных пикнидах, вместе с макроконидиями, возникают эндогенные микроконидии (спермации). Образуются они следующим образом: ядро материнской клетки микроконидии
дважды делится, протопласт распадается на четыре микроконидии, которые выходят из материнской клетки, покрываются оболочкой и могут копулировать с трихогиной (Higgins, 1920, 1936; Wolf, 1939) (рис. 76). Микроконидии у видов Mycosphaerella не могут прорастать на питательной среде.
Особый интерес представляет половой процесс у представителей порядка Laboiilbeniales в связи с наличием у некоторых представителей голых эндоконидий (спермациев), гомологизируемых некоторыми авторами со спермациями красных водорослей. Женский половой орган (архикарп) лабульбениальных обычно состоит из трех клеток — аскогона, трихофорной клетки и три- хогины. Оплодотворяющими элементами являются или экзогенные конидии (семейство Ceratomycetaceae), или эндоконидии — спермации, возникающие в фиалидах, подобно тому, как это происходит у Bombardia из порядка Sphaeriales (семейство Laboulbenia- сеае). Иногда фиалиды собраны группами во вместилищах и эндоконидии из них выходят сначала в это общее вместилище (семейство Peyritschiellaceae). В случае отсутствия спермациев оплодотворение происходит в результате слияния аскогона с трихофорной клеткой. Леки развиваются из двуядерных аскогенных клеток следующим образом. Оба ядра аскогенной клетки делятся.
Рис. 75. Neurospora tetrasperma:
схематический рисунок соматогамной копуляции (по Dodge, 1936).
101
Одна пара ядер остается в аскогенной клетке, другая переходит в вырост клетки — будущий аск, где оба ядра сливаются. Затем происходит новое деление ядер аскогенной клетки и образование нового аска и т. д. В результате получается пучок асков.
Таким образом, в основе полового процесса у аскомицетов, как и у зигомицетов, лежит гологамный половой процесс — слияние двух недифференцированных клеток. В различных порядках аскомицетов можно наблюдать усложнение строения, главным образом аскогона, который может быть одноклеточным и многоклеточным; на нем развиваются специальные органы, воспринимающие мужские оплодотворяющие элементы,— трихогина или воспринимающие гифы. Мужская же клетка — андроцит (антеридий) остается почти неизменной. Характерным для аскомицетов является то, что функции андро- цита часто берут на себя вегетативные клетки и конидии экзогенного и эндогенного происхождения. У представителей одного семейства или даже рода могут быть представлены различные типы оплодотворяющих элементов.
Тот факт, что оплодотворение у аскомицетов может происходить при помощи вегетативных клеток и клеток бесполого размножения — конидий, свидетельствует о том, что вегетативный мицелий у грибов сохраняет всю необходимую наследственную информацию, результатом чего и являются его потенциальные половые возможности. Это явление никоим образом нельзя рассматривать как ослабление сексуальности.
В цикле развития у асковых грибов наблюдаем усложнение строения аска, развитие аскогенных гиф, плодовых тел и разнообразных органов бесполого размножения.
Аски являются характерным органом размножения аскомицетов. У низших представителей класса количество аскоспор иногда бывает непостоянным в результате повторного деления ядра, однако для подавляющего большинства аскомицетов это число обычно стабильное и равняется восьми или в результате недоразвития части спор — четырем, двум и реже одной. Количество спор иногда увеличивается в результате их повторного почкования в аске, но это явление вторичное. Характерным для асков является наличие двух ядер, их слияние — кариогамия и последующее редукционное деление, приводящее к образованию гаплоидных аскоспор, а также способ их формирования, так называемое свободное образование, состоящее в том, что после деления ядра вокруг
192
Рис. 76. Mycosphae- rella fraxinicola:
a — карпогон, б, в — образование эндоконидий (по Wolf. 1939).
каждого из них обособляется часть цитоплазмы и покрывается оболочкой, образуя аскоспоры, которые окружены не использованной на образование спор цитоплазмой, так называемой периплазмой или эпиплазмой, образующей иногда дополнительный слой оболочки спор — эписпорий.
Оболочка аска у нескольких порядков, объединяемых иногда в группу протуникатных аскомицетов, отличается простым строе-
Рис. 77. Типы строения асков: а—Uncinula (Erysiphales), б — Humaria (Pezizales), в — Sordaria (Xylariales), г — Cainia (Xylariales), д — Microglossum (Helotiales), e — Cordiccps (Hypocreales), ж — Caloplaca (Lecanorales), з — Cladonia (Lecanorales) (no Kreisel, 1969).
нием, и споры из них освобождаются в результате разрушения или ослизнения оболочки. Более сложно построены оболочки асков у так называемых эвтуникатных аскомицетов, у которых в процессе эволюции образовались специальные приспособления для активного рассеивания спор. По строению оболочек асков различают две группы эвтуникатных аскомицетов — с одной оболочкой — однооболочковые (унитуникатные) и с двумя обособленными оболочками — двуоболочковые (битуникатные). Однооболочковые аски открываются на верхнем конце в результате обособления крышечки или образования щели, причем аскоспоры активно выбрасываются из аска. Представление об аппарате раскрывания асков у однооболочковых дают примеры, приведенные на прилагаемом рисунке (рис. 77).
У двуоболочковых аскомицетов стенка аска двойная и состоит из наружного слоя — тонкого и жесткого, не растяжимого и внут¬
193
реннего — толстого и эластичного.
При созревании аска внешняя оболочка лопается на вершине и внутренняя выходит через разрыв внешней в виде длинного цилиндрического мешка и затем выбрасывает споры по одной через верхушечную пору.
Одним из характерных признаков высших грибов, в том числе и аскомицетов, является образование плодовых тел — вместилищ, несущих основные органы спороношения. Единичные примеры таких образований можно наблюдать и у низших грибов. Так, у Absidia и Phycomyces из порядка Mucorales из подвеска выроста- ют темные плотные гифы, окружающие зиготу, а у Mortierella такие кроющие гифы дают плотную обвертку, обволакивающую копулирующие ветви мицелия. Начиная с Aspergillales, у аскомицетов, как правило, наблюдаем плодовые тела различного типа, образованные тифами гаплоидного мицелия. Образование плодовых тел у одних аскомицетов происходит вследствие того, что гифы мицелия обрастают группы половых органов или молодых сумок, которые в большинстве случаев дают так называемый ги- мениальный слой, и образуют плотную покровную ткань. Такой тип образования плодовых тел называют за Наннфельдтом (Nann- feld't, 1932) аскогимениальным. По этому типу образуются полностью закрытые плодовые тела — клейстотеции или клейстокар- пии, характерные для эризифальных и большинства аспергиллаль- ных, закрытые плодовые тела, открывающиеся при созревании отверстием,— перитеции, свойственные семейству Melanosporaceae из аспергиллальных, порядкам Microascales, Laboulbeniales, Sphae- riales, Xylariales, Diaporthales, Clavicipitales, совершенно открытые в зрелом состоянии блюдцевидные плодовые тела — апотеции, в наиболее типичном виде наблюдаемые в порядках Pezizales и Helotiales, а также их различные производные, свойственные родам семейства Helvellaceae или порядка Tuberales. Если плодовые тела — перитеции или апотеции возникают по аскогимениальному типу на плотных образованиях мицелия — стромах, то каждое плодовое тело имеет свою собственную стенку, отграничивающую его от ткани стромы.
В некоторых порядках аскомицетов плодовые тела образуются по иному типу. У них вначале развивается строма, а затем внутри стромы закладываются половые органы и развиваются аскоген- ные гифы и сумки. Вокруг каждой сумки или группы сумок образуется полость (loculus), в результате чего в типичном случае каждая сумка или их группа помещается в особой камере. Такой тип развития плодовых тел называется асколокулярным, а самое плодоношение — аскострома или аскома. По асколокулярному типу развиваются плодовые тела в порядках Phacidiales, Myriangia- les, Pseudospliaeriales и Dothiorales. Иногда локулы сливаются, верхняя часть ткани стромы разрушается и плодовое тело приобретает вид апотеция (порядок Dothiorales). Если ткань между полостями (локулами) растворяется, ее остатки могут образовы¬
19/.
вать интертециальные волокна — парафизоиды или псевдопара- физы, отличающиеся по своему происхождению от настоящих па- рафиз, свойственных аскогимениальным грибам. В отличие от плодовых тел аскогимениального типа локулы не имеют собственной стенки и представляют собой полости в ткани стромы. Развитие спороиошений по асколокулярному типу является дальнейшим приспособлением асковых грибов к наземным условиям существования, так как развитие половых органов в локулах предохраняет их от неблагоприятных условий внешней среды.
Происхождение и основные направления эволюции аскомицетов
Относительно происхождения аскомицетов с 80-х годов прошлого столетия и до настоящего времени существуют две основные теории. По одной, восходящей к де Барн (De Вагу, 1884) и Бре- фельду (1889), аскомицеты происходят от зигомицетов, причем промежуточным звеном между ними является порядок эндо- мицетальных. Эту теорию поддерживали Аткинсон, Данжар, Ячевский, Гиллермон; в последнее время ее развивали многие исследователи, в частности Вердерманн (Werdermann, 1954), Цим- мерманн (Zimmermann, 1959) и Геймаин (Gäumann, 1904). Другие авторы, следуя Саксу (Sachs, 1874), выводят асковые грибы из красных водорослей — флоридей и наиболее близкими к фло- ридеям, а следовательно исходными для аскомицетов, считают порядки Laboulbeniales, Lecanorales, Sphaeriales и некоторые Pezizales, у которых часто наблюдается оплодотворение при помощи микроконидий (спермациев), копулирующих с трихогиной. Эту теорию поддерживали Харпер, Лотси, Додж, Джексон и ряд других авторов. Из современных сторонников этой теории можно назвать Э. А. Бесси (Bessey, 1950) и Шадефо (1960). Были и другие высказывания по этому вопросу, не нашедшие отклика, например предположение Мережковского (1909) о происхождении высших грибов от бактерий через актиномицеты.
Существует мнение, по которому аскомицеты бифилетического происхождения — основная масса их происходит от зигомицетов, а лабульбениальные — от флоридей (Nannfeldt, 1932; Greis, 1943). Исходя из этой теории, Грейс выделяет лабульбениальные в особый отдел растительного мира — Laboiilbeniomycetes (Greis, 1943, стр. 314).
Сторонники происхождения аскомицетов от флоридей считают основным аргументом в пользу этой теории сходство в строении половых органов и в характере полового процесса между флори- деями и некоторыми группами аскомицетов, заключающееся в следующем:
1) наличие на карпогоне у флоридей и аскогоне многих сумчатых грибов воспринимающих выростов или нитей — так называемой трихогины;
195
2) оплодотворение при помощи безжгутиковых клеток, обычно называемых спермациями, образующихся эндогенно в специальных одноклеточных органах (антеридиях) ;
3) образование у флоридей из оплодотворенного карногона спорогениых нитей или гонимобластов и сходных образований у высших аскомицетов в виде аскогенных гиф;
4) образование у некоторых флоридей и у многих аскомицетов вместилищ, заключающих в себе у флоридей — карпоспорангии, а у аскомицетов — конидии и аски.
Кроме того, аргументом в пользу филогенетических связей аскомицетов и флоридей считают также наличие пор в поперечных перегородках талломов флоридей и асковых грибов.
Действительно, в строении половых органов высших аскомицетов и флоридей наблюдаются на первый взгляд поразительные черты внешнего сходства, заслуживающие внимательного рассмотрения. Так, одноклеточные карпогоны флоридей снабжены выростом — трихогиной. Есть указания на наличие особого ядра в трихогине некоторых флоридей (Kniep, 1928, стр. 210—211; Fritsch, 1945, стр. 592). Все это вместе с наличием у некоторых флоридей сужения при основании трихогины дает основание предполагать, что у их предков трихогина была самостоятельной клеткой (Bessey, 1950). Однако среди ныне живущих флоридей нет форм с независимой трихогиной. У более простых красных водорослей — бангиевидных иногда наблюдается образование небольших выпячиваний карпогона. Все это дает основания предполагать, что трихогина флоридей является только выростом клетки карпогона. Напомним, что оогоний Coleochaete из зеленых водорослей также имеет колбовидную форму с вытянутой шейкой. Что же касается трихогины аскомицетов, то она представляет собой одноклеточное или часто многоклеточное образование, причем клетки ее всегда содержат одно или несколько ядер. Можно допустить, что образование трихогины у аскомицетов связано с дифференциацией аскогенной ветви и усложнением ее строения, причем в аскогон превращается не конечная клетка ветви. Необходимо также отметить, что карпогон у флоридей обычно одноядерный, а аскогон сумчатых грибов может состоять, как и трихогина, из одноядерных или многоядерных клеток.
Вторым основным аргументом в пользу выведения аскомицетов из флоридей является сходство оплодотворяющих элементов флоридей — так называемых спермациев с оплодотворяющими элементами пекоторых аскомицетов из порядков Laboulbeniales, Sphaeriales, Pseudosphaeriales, Pezizales и некоторых лишайников из порядка Lecanorales, у которых оплодотворение происходит при помощи эндоконидий, называемых по аналогии с оплодотворяющими элементами флоридей спермациями. Однако развитие спермациев флоридей и эндоконидий аскомицетов происходит по- разному. У всех красных водорослей — у бангиевидных и у флоридей — спермации образуются по одному в антеридии подобно
196
характерным для красных водорослей моноспорам, гомологами которых они являются. Возникновение же микроконидий (спер- мациев) у сумчатых грибов происходит, как это детально изучено у Thielavia basicola из Eurotiales (Aspergillales), в так называемых фиалидах последовательно и является видоизменением образования обычных конидий (эктоконидий). По этому же типу образуются микроконидии (спермации) у Microascales, Sphaeriales и Laboulbeniales. Несколько иначе образуются микроконидии у Му- cosphaerella из порядка Pseudosphaeriales, где ядро материнской клетки микроконидий делится повторно на четыре, после чего формируются четыре микроконидии, которые покрываются оболочкой и последовательно выходят из материнской клетки. Этот тип образования микроконидий (настоящих эндоконидий) можно рассматривать как дальнейшее развитие эндогенного образования конидий типа Thielavia. Он резко отличается от развития сперма- циев красных водорослей, у которых спермации, как уже упоминалось, всегда образуются в материнских клетках по одному.
У некоторых флоридей (большинство Nemalionales) из оплодотворенного карпогона развиваются спорогенные нити, дающие более или менее компактное образование (карпоспорофит) ; конечные клетки этих нитей дают по одной карпоспоре. Редукция хроматина при этом происходит при первых делениях ядра зиготы. У других же флоридей (например, у Cryptonemiales и Ceramia- les) из оплодотворенного карпогона вырастают спорогенные нити (гонимобласты), которые после слияния с вспомогательными (авксилярными) клетками образуют карпоспорофиты, дающие карпоспоры. При этом, обычно, редукционного деления не происходит и карпоспоры являются диплоидными. Из карпоспор развивается особое растение — тетраспорофит, образующее тетраспорангии, в которых происходит редукционное деление ядра и образование четырех тетраспор. Некоторые авторы высказывали предположение, что аскогенные гифы, образующиеся из оплодотворенного аскогона сумчатых грибов, являются гомологами го- нимобластов. Однако нужно отметить, что ядра клеток гонимобла- стов, как и карпоспор, являются диплоидными; карпоспоры возникают по одной в клетках карпоспорофита; редукционное деление происходит у высших флоридей на другом растении — тетраспорофите, гонимобласты сливаются с вспомогательными клетками. Все это свидетельствует против предположения о гомологии гонимобластов и аскогенных гиф. У некоторых флоридей (Liagora tetrasporifera, H elminthocladia, Phyllophora brodiaei) редукционное деление происходит при образовании спор на кар- поспорофите, в клетках которого формируются по четыре карпо- тетраспоры, что напоминает развитие аскоспор в сумках. Однако этот способ образования тетраспор является редким и, безусловно, вторичным.
Иногда указывают на сходство вместилищ (нематециев), в которых у некоторых флоридей образуются спермагонии и карноспо-
197
рангии с перитециями и пикнидами сумчатых грибов. Однако вместилища, в которых размещены органы размножения, известны и у бурых водорослей, и развитие их в различных отделах растительного мира могло происходить независимо.
Как доказательство родственных связей аскомицетов и флори- дей приводят наличие у тех и других пор в межклеточных перегородках, но нужно помнить, что перегородки у флоридей разграничивают обычно одноядерные клетки, а у аскомицетов клетки часто бывают многоядерными. По сути, аскомицеты являются це- ноцистическими организмами, таллом которых, подобно многим сифоновым водорослям, разделен неполными перегородками на отдельные участки.
Таким образом, у высших аскомицетов и высших красных водорослей — флоридей имеется ряд сходных черт в строении таллома и органов размножения, которые последователи Сакса рассматривают как доказательство филогенетических связей между этими группами организмов. Однако эти взгляды встречают, как уже отмечалось, ряд возражений. Очень серьезным аргументом против филогенетических связей сумчатых грибов и флоридей являются биохимические данные. Так, клеточная оболочка флоридей состоит из целлюлозы с примесью пектиновых веществ, а у исковых грибов она имеет хитин; у флоридей запасным углеводом является багрянковый крахмал, а у аскомицетов, как и у других грибов,— гликоген. Против этого может быть выдвинуто возражение, что своеобразный химизм грибов связан с условиями гетеротрофного питания, однако нужно помнить, что для некоторых групп грибов также характерно при гетеротрофном питании наличие целлюлозы (сапролегниомицеты).
Если стать на позиции последователей Сакса, то необходимо признать, что наиболее близки филогенетически к флоридеям высшие аскомицеты, которые обнаруживают наибольшие черты сходства с ними. Все более простые группы аскомицетов — эндо- мицетальные, эвротиальные (аспергиллальные), эризифальные нужно признать вторичными, упрощенными формами. Так и поступают последовательные сторонники этой теории Э. А. Бесси и Шадефо. Последний, ввиду сходства в строении и развитии зи- гомицетов и эндомицетальных, рассматривает и зигомицеты как наиболее упрощенную и наиболее удаленную от флоридей группу настоящих грибов (Mycomycophyta).
Главным основанием для теории происхождения аскомицетов от зигомицетов является сходство в строении таллома, половых органов и спороношений между низшими аскомицетами и зи- гомицетами, с одной стороны, и низшими аскомицетами и высшими представителями этого класса — с другой. Наличие промежуточных форм часто облегчает установление филогенетических связей.
Таллом у зигомицетов обычно многоядерный, ценоцистический. Однако у энтомофторальных мицелий может быть разделен на отдельные клетки.
198
Половой процесс зигомицетов и низших аскомицетов сходен не только в общих чертах (гологамия), но и в деталях. В то время как у некоторых форм (Mucor и др.) при слиянии многоядерных клеток в зиготе происходит попарное слияние многих ядер, у ряда высших форм зигомицетов (например, Endogone lactiflua, Entomophthora fumosa) при копуляции многоядерных клеток сливается только по одному ядру каждой копулирующей клетки (голоцита); остальные ядра дегенерируют. Аналогичный процесс встречаем у Dipodascus albidus из эндомицетальных.
У представителей порядка мукоральных ядра копулирующих клеток обычно сливаются сразу после слияния клеток, но уже у Phycomyces и у Endogone lactiflua слияние ядер задерживается, и в зиготе можно наблюдать типичный дикарион, что характерно для аскомицетов. У зигомицетов, как и у низших аскомицетов, можно иногда наблюдать морфологическую дифференциацию сливающихся клеток, в связи с чем можно уже говорить о мужских и женских клетках (Zygorhynchus, Endogone). Наконец, количество аскоспор у низших аскомицетов (род Dipodascus), а иногда как вторичное явление и у высших (роды Thecotheus и Ryparo- bius из Pezizales), может быть неопределенным, как и в зародышевых спорангиях зигомицетов. Все это дает основание проводить гомологию между зародышевым спорангием зигомицетов и сумкой (аском) аскомицетов. Сходно также образование зачаточных плодовых тел у некоторых зигомицетов (Mortierella, Endogonaceae) и у низших аскомицетов (Gymnoascaceae из эвротиальных), имеющих вид закрытых плодовых тел — клейстотециев.
Если принять точку зрения Сакса — Бесси, то, исходя из того, что наибольшие черты сходства наблюдаются между высшими флоридеями и высшими аскомицетами из порядков Pseudosphae- riales, Spliaeriales и Laboulbeniales, необходимо признать, что эти порядки должны стоять ближе всего к исходным формам аскомицетов и из них или близких к ним форм нужно выводить все остальные порядки, считая их производными и упрощенными. Это можно видеть на схеме эволюционных связей аскомицетов, данной Э. А. Бесси (E. A. Bessey, 1942, 1950). На этой схеме исходными порядками приняты, с одной стороны, Pezizales и Lecano- rales, с другой — Sphaeriales и Hypocreales, от которых Бесси производит все остальные порядки аскомицетов, кроме Laboulbeniales, которые он выводит как отдельную ветвь, наиболее близкую к красным водорослям. Основанием для такого построения Бесси считает то, что у представителей упомянутых порядков встречается «много форм со свободными спермиями и трихогинами». От дискомицетной ветви (Pezizales и Lecanorales) Бесси выводит, хотя и с сомнением, базидиальные грибы. При построении этой схемы он не придает особого значения строению клеток, наличию разнообразных типов полового процесса, строению сумок и плодовых тел. Недочетом гипотезы Бесси, по нашему мнению, является и то, что он не видит никакой связи между зигомицетами, с
199
одной стороны, и эндомицетальными, с другой. Первые он выводит из сапролегниевых, а вторые считает упрощенной группой, происходящей от сфериальных через Eurotiales (Aspergillales).
Более последователен Шадефо, который, как уже упоминалось, зигомицеты выводит из аскомицетов в порядке их дальнейшего упрощения.
Слабым местом теории Сакса — Бесси является также то, что авторы ее без всякого объяснения считают основным направлением эволюции грибов редукцию и упрощение как вегетативного тела, так и органов размножения. Эта теория не объясняет, почему в процессе развития аскомицетов, если принять как основное направление их эволюции редукцию, должно происходить упрощение строения сумок и плодовых тел, почему должны исчезнуть аскогенные гифы и разнообразные конидиальные спороношения, т. е. ряд черт строения, связанных с приспособлением к суровым условиям наземного существования.
Если же мы станем на позиции де Бари — Геймана и поставим в основе системы аскомицетов более простые порядки — эндомицетальные и эвротиальные (аспергиллальньте), то увидим, что все развитие сумчатых грибов, а также усложнение их строения и биологии тесно связано с приспособлением к разнообразным условиям наземного существования. Эти приспособления касаются главным образом органов, связанных с размножением. Необходимо отметить при этом, что в строении вегетативного тела в процессе эволюции аскомицетов не наблюдается значительных изменений; у них сохраняется цитологическая и генетическая примитивность, что проявляется в том, что вегетативные клетки сохраняют способность функционировать в качестве половых элементов. Этим и объясняется тот факт, что видоизмененные вегетативные клетки — конидии в условиях наземного существования смогли выполнять функцию оплодотворяющих элементов.
При переходе от более примитивных форм к более сложным в пределах класса аскомицетов мы наблюдаем усложнение строения сумки — переход от примитивной сумки с недифференцированной стенкой, споры из которой освобождаются в результате ее разрушения (протуникатный тип), к сумкам, у которых наблюдается полярность и вырабатываются различные структуры в оболочке, способствующие активному выбрасыванию спор (эвтуни- катный тип). Параллельно с усложнением строения сумки наблюдается образование и усложнение плодовых тел, усложнение строения архикарпа, развитие аскогенных гиф, происхождение многочисленных типов конидий, обеспечивающих массовое размножение вида. Особенный интерес представляет обеспечение защиты половых органов от неблагоприятных условий. В то время как у так называемых аскогимениальных форм развитие плодового тела происходит после закладки половых органов, у аско- локулярных форм раньше образуется строма (аскострома, аскома), внутри которой формируются полости — локули, в которых уже
200
развиваются половые органы, аскогенные гифы и сумки. Параллельно с этим идет и развитие оплодотворения при помощи конидий — спермациев, переносимых воздушными токами, и образование разветвленных воспринимающих гиф. Если у более примитивных форм (Ascobolus carbonarius из порядка Pezizales) оплодотворение при помощи конидий является случайным замещением слияния трихогины с вегетативной клеткой, то у некоторых порядков (Sphaeriales, Pseudosphaeriales) конидии, как правило, принимают на себя функции оплодотворяющих элементов, а в дальнейшем даже могут терять свою основную функцию — органов бесполого размножения.
Э. А. Бесси считает трудно объяснимым появление трихогины у аскомицетов, если принимать их происхождение от зигомицетов. Однако развитие аскогоиа у некоторых низших грибов, например Eurotiaceae (Aspergillaceae), где Monascus purpureus имеет одноклеточную трихогину, или даже у таких представителей порядка пезизовых, как Ругопета, позволяет предположить, что трихогина у них возникла в результате того, что в аскогон превращается не конечная клетка аскогенной ветви, а вторая или даже, как это видим у рода Ascobolus и других родов, одна из нижних клеток.
Мы считаем, что при построении системы аскомицетов следует принимать за основу усложнение строения их тела, полового процесса и строения органов бесполого размножения, и, особенно, асков, развитие которых проходило в условиях приспособления к рассеиванию спор в условиях наземного существования. Некоторые группы аскомицетов, приспособившиеся к своеобразным условиям существования, например Laboulbeniales, естественно, обнаруживают и ряд особенностей, отличающих их от других групп аскомицетов.
Теоретические взгляды на происхождение и развитие аскомицетов отражались и на попытках построения филогенетических систем этого класса. Так, сторонники происхождения аскомицетов от флоридей (Бесси, Шадефо) в начале системы аскомицетов ставят сравнительно сложные формы — лабульбениевые и пезизо- вые, а такие порядки, как эвротиальные (аспергиллальные), эн- домицетальные и эризифальные, считают группами редуцированными. Большинство же авторов связывают аскомицеты с низшими грибами и принимают как основу их развития усложнение структуры в связи с приспособлением к наземным условиям.
Еще в конце XIX ст. Шретер (Schröter, 1897) разделял класс аскомицетов на два подкласса: 1. Hemiasceae с одним порядком Hemiascineae и 2. Euasceae, который он разбивал на девять порядков — Protoascineae, Protodiscineae, Helvellineae, Pezi- zineae, Phacidiineae, Hysteriineae, Tuberineae, Plectascineae и Pyrenomycetineae. Последний порядок Линдау в том же издании разделяет на четыре подпорядка — Perisporiales, Hypocreales, Do- thideales и Sphaeriales.
201
Большое распространение имела система аскомицетов Э. Фишера (Е. Fischer, 1912), который разделял аскомицеты тоже на два подкласса: A. Protoascineae с двумя порядками — Endomy- ceteae и Saccharomyceteae и В. Ascomyceteae typicae с восемью порядками — Plectascineae, Perisporineae, Pyrenomyce- teae, Hysterineae, Discomyceteae, Tuberineae, Exoasceae и Laboul- beniaceae.
По мере углубления изучения грибов количество порядков, на которые разбиваются аскомицеты, увеличивается. В ранней системе Гейманна (Gäumann, 1926) аскомицеты также разделены на два подкласса — Н e m i a s с i (Protascineen) и E и a s с о my- се t e s.
К первому подклассу он относит два порядка — Endomycetales и Exoascales, а ко второму — двенадцать: Plectascales, Perisporia- les, Myriangiales, Hypocreales, Sphaeriales, Dothideales, Hysteriales, Hemisphaeriales, Phacidiales, Pezizales, Tuberales и Laboulbeniales. Эту же систему с незначительными изменениями принимает и Курсанов (1933, 1940). Порядки второго подкласса — Euascomy- cetes он объединяет в три группы: Cleistomycetes с вполне закрытыми плодовыми телами (клейстокарпиями), Pyrenomycetes с плодовыми телами, снабженными отверстием на вершине (перитециями), и Discomycetes с плодовыми телами, в зрелом состоянии открытыми (апотеции). Однако нужно отметить, что сам автор относительно группы Pyrenomycetes отмечает, что это понятие употребляется «не в смысле определенной таксономической категории, а лишь как объединение нескольких порядков плодо- сумчатых аскомицетов, характеризующихся тем, что плодовые тела их построены по типу перитециев» (Курсанов, 1940, стр. 265). Для «удобства обзора» в своем учебнике Курсанов (1933, 1937, 1945) рассматривает как порядки укрупненные группы пиреноми- цетов и дискомицетов, разбивая их на подпорядки.
Иное направление в вопросе филогении и классификации аскомицетов связано с работой Наннфельдта (Nannfeldt, 1932). Развивая взгляды Генеля и других исследователей, которые обратили внимание на различный характер строения плодовых тел и процессы развития в них асков у высших аскомицетов, он предложил разделять аскомицеты на три группы, каждая из которых представляет восходящий ряд развития, а именно: Plecta- scales, Ascoloculares и Ascohymeniales. Первая группа — Plecta- scales, по Наннфельдту, характеризуется тем, что аскогоны и антеридии у них закладываются открыто и только аскогенные гифы покрываются оболочкой из стерильных гиф. У низших плекта- екальных (Gymnoascaceae) плодовое тело представляет собой клубок гиф, в котором разбросаны аски. У высших — наблюдается дифференциация на плотный перидий и центральную рыхлую ткань, в которой размещены аски; строматические формы отсутствуют. Для второй группы — Ascoloculares, к которой Наннфельдт относит порядки Myriangiales, Pseudosphaeriales, Hemisphaeriales
202
и Trichothyriales, характерным является развитие аскогонов в строматической ткани, которая позже — во время роста аскоген- ных гиф и асков, частично вытесняется, частично резорбируется. В третьей группе — Ascohymeniales, куда относятся дискомицеты и «настоящие» пиреномицеты, аскогон и антеридий размещены или открыто, или в строме. После образования аскогенных гиф, но перед развитием асков образуется стенка плодового тела и на внутренней ее стороне формируется гимений из размещенных параллельно гиф — будущих парафиз, между которыми образуются аски. У асколокулярных аски толстостенные, с двуслойной стенкой, а у аскогимениальных они тонкостенные и утолщены только на верхушке.
Ряд авторов в последние десятилетия обратил внимание на разнообразие строения асков и различные приспособления для рассеивания аскоспор. Это дало основание Латреллу (Luttrell, 1951) делить высшие аскомицеты на однооболочковые (Unitunica- tae) и двуоболочковые (Bitunicatae). Первые в основном отвечают аскогимениальным, а вторые — асколокулярным Наннфельдта, хотя и наблюдаются некоторые расхождения. Так, Гейман отмечает, что, например, у Phacidiales развитие асков идет по аско- локулярному типу, а сами аски однооболочковые. У псевдосфе- риального рода Cochliobolus и, вероятно, у Ophiobolus плодовое тело развивается, как у аскогимениальных грибов, а аски хотя и битуникатные, но оба слоя оболочки являются тонкими, а споры не выбрасываются, а освобождаются после разрушения стенки аска, который отделяется целиком.
Позже Латтрелл (Luttrell, 1955), считая асколокулярные пиреномицеты единой филогенетической группой, предложил делить аскомицеты на три подкласса — Hemiascomycetes, Euascomycetes и Loculoascomycetes (Ascoloculares, Bitunicatae). К первому подклассу — Hemiascomycetes он отнес порядки Dipodascales, Endomycetales, Taphrinales и Pericystales. Ко второму — Euascomycetes — порядки Eurotiales, Tuberales, Ophiostomatales, Erysiphales, Laboulbeniales, Hypocreales, Diaporthales, Coryneliales, Coronophorales, Xylariales, Ostropales, Pezizales, Helotiales, Phacidiales и Lecanorales. К третьему подклассу — Loculoascomycetes Латтрелл отнес шесть порядков — Myriangiales, Dothi- deales (Pseudosphaeriales, Capnodiales, Dothiorales), Trichothyriales, Pleosporales, Hysteriales и Mycrothyriales (Hemisphaeriales, Asterinales).
В последнем издании своей монографии Гейманн (Gäumann, 1964), отбрасывая деление на первичносумчатые и настоящие аскомицеты, в основу классификации аскомицетов кладет строение аска и делит их на два подкласса — протуникатные (die protunicaten Ascomyceten) с недифференцированной стенкой аска и эвтуникатные (die eutunicaten Ascomyceten) со сложным строением стенки аска, обусловливающим активное рассеивание спор. К первому подклассу — протуникатным он относит порядки
203
Endomycetales, Aspergillales, Microascales, Coronophorales, Onyge- nales, Meliolales, Laboulbeniales, Ascosphaerales и Protomycetales. Второй подкласс — эвтуникатыые Гейманн, следуя Латтреллу, делит на две группы порядков — унитуникатные (Unitunicatae) и битуникатные (Bitunicatae). Нужно отметить, что Гейманн, проводя деление аскомицетов по строению асков, отмечает, что его подкласс — протуникатные не представляет естественной филогенетической группы, а только ступень развития, объединяющую несколько не родственных между собой ветвей развития (Gäumann, 1964, стр. 106), а по поводу групп унитуникатных и би- туникатных пишет, что они являются также гетерогенными. По его мнению, возможно, некоторые унитуникатные и некоторые битуникатные находятся между собой в бо́льшем родстве, чем с другими представителями своей группы, например унитуникатные Sordariaceae и битуникатные Sporormiaceae. Далее он говорит, что битуникатные не представляют естественной группы родственных форм, а только конечную ступень дифференциации стенки аска, которой могли достигнуть независимо представители нескольких линий эволюции. В то же время на филогенетической схеме Гейманн (Gäumann, 1964, стр. 10) выводит все порядки битуникатных (асколокулярных) грибов одной ветвью из ас- пергилловых, тем самым подчеркивая их филогенетическое родство.
По поводу предложенного деления на асколокулярные и ас- когимениальные были выдвинуты возражения, которые в основном сводятся к тому, что эти группы не однородны по своему происхождению. Так, Окснер (1956, стр. 23) отмечает, что деление на асколокулярные и аскогимениальные иногда проходит через группы, филогенетическое единство которых не вызывает сомнений.
Нельзя все же не признать, что строение аска и развитие плодовых тел являются весьма существенными признаками, которые могут пролить свет на ряд вопросов эволюции аскомицетов и дать основания для разработки филогенетической системы этого класса грибов. Нужно отметить, что если аскогимениальные (унитуникатные) несомненно являются группой гетерогенной и заключают в себе, вероятно, несколько линий эволюции, то группа асколокулярных, характеризующаяся своеобразным развитием плодового тела — аскостромы (аскомы) и свойственным только им сложным строением аска, по всей вероятности, является единой ветвью эволюции, что и отображено на схеме Гейманна.
В системах аскомицетов, предложенных в течение последних двух десятилетий, в отличие от последней системы Гейманна (1964) сохраняется деление на подклассы Protascomycetidae и Еи- ascomycetidae, а деление на асколокулярные и аскогимениальные имеет вторичное значение (Gäumann, 1949) ; сохраняется также деление на пиреномицеты и дискомицеты (Красильников, Курса- нов и Наумов, 1954), иногда же как вспомогательное применяется
204
деление и на асколокулярные и аскогимеииальные наравне с выделением среди аскогимениальных пиреномицетов и дискомицетов (Werdermann, 1954; Ubrizsy and Vörös, 1966).
В системе Цейпа (Cejp, 1957) подкласс Euascomycetes (Carpo- ascomycetes) разделен на пять групп — Synascomycetes (порядок Protomycetales), Plectomycetes [порядки Eurotiales (Plectascales), Myriangiales и Erysiphales], Pyrenomycetes [порядки Dothideales, Pseudosphaeriales, Microthyriales (Hemisphaeriales), Hysteriales, Hypocreales и Xylariales], Discomycetes [a) Inoperculatae — Ostro- pales, Helotiales, Cittariales, b) Operculatae — Pezizales, c) Hypogaei — Tuberales] и как «дополнение к аскомицетам» — Laboulbe- niomycetes. Деления на асколокулярные и аскогимеииальные Цейп в систему не вводит.
Из классификаций последних лет упомянем еще классификацию Аркса (Агх, 1967), который выделяет класс Endomycetes с такими порядками: Endomycetales, Torulopsidales, Sporobolomy- cetales, Taphrinales (включая Protomycetaceae), Exobasidiales (!) и Ustilaginales (!). Класс Ascomycètes в узком понимании, т. е. без указанных выше порядков, Арке делит на два подкласса — Ascohymenomycetidae с порядками Pezizales, Tuberales, Eurotiales, Erysiphales, Sphaeriales, Phacidiales, Helotiales и Ascoloculomyce- tidae с порядками Pseudosphaeriales, Dothiorales и Myriangiales. Порядок Laboulbeniales дан как добавление к классу аскоми- цетов.
В опубликованной недавно обзорной работе Крейзель (Kreisel, 1969), следуя Арксу, выделяет класс Endomycetes (Hemiascomyce- tes), куда он, кроме первичноасковых Spermophthorales и Endomycetales, относит, как и Арке, Taphrinales, включая в их состав Photomycetales, и Ustilaginales, но без семейства Tilletiaceae, а также трихомицеты и порядок Kickxellales, т. е. создает еще менее объяснимую сборную группу, чем та, которую дает Арке. Аскомицеты в узком понимании, т. е. без первичносумчатых и Taphrinales, Крейзель делит на два подкласса — Euascomycetidae (Ascohymeniales) с двенадцатью порядками и Loculoascomycetidae (Ascoloculares) с девятью порядками. Порядки эти большей частью совпадают с порядками Гейманна. Заслуживает внимания тот факт, что Крейзель включил в систему Ascomycètes семейства асковых лишайников («лихенизированных грибов»), распределив их между соответствующими порядками.
Краткий обзор главнейших систем аскомицетов показывает, что большинство авторов (Gäumann, 1926, 1949; Greis, 1943; Красильщиков, Курсанов и Наумов, 1954; Smith, 1955; Werdermann, 1954; Cejp, 1957; Ubrizsy and Vörös, 1966; Alexopoulos, 1966) придерживается деления аскомицетов на два подкласса — первичносумчатые и настоящие сумчатые, которые согласно современным правилам ботанической номенклатуры должны называться Prot- ascomycetidae (Hemiascomycetidae) и Euascomycetidae, как их и называют Вердерманн, Алексопулос, Убрижи и Вереш.
205
Большинство упомянутых авторов к первому подклассу — Protascomycetidae относят порядки Endomycetales и Taphrinales (Exoascales). Некоторые из них в состав порядка Taphrinales включают также семейство Protomycetaceae (Gäumann, 1926; Красильщиков и др., 1954), а некоторые также и семейство Ascosphae- raceae (Pericystaceae) (Werdermann, 1954); другие же выделяют семейства Protomycetaceae и Ascospliaeraceae (Pericystaceae) в особый — третий порядок (Ubrizsy a. Vörös, 1966).
Включение порядка Taphrinales в подкласс Protascomycetidae является очень спорным, так как исследования последних лет (Schneider, 1956) показали, что оболочка аска у видов рода Taphrina состоит из двух слоев — более плотного внешнего и более нежного внутреннего, разделенных слоем слизи. На верхушке аска намечается линия отделения крышечки. После созревания спор шапочковидная часть внешнего слоя оболочки отпадает, внутренний слой оболочки под давлением содержимого клетки разрывается и аскоспоры выбрасываются на расстояние до 1,2 см (Wolf, 1958). Другой особенностью Taphrinales, отличающей его от типичных Protascomycetidae, является развитие дикариофитного мицелия, который в типичных случаях, например у Taphrina epi- phylla, образуется в результате плазмогамии, происходящей при слиянии гаплоидных ростковых клеток, или же в других случаях, например у Taphrina deformans, в результате деления ядра, не сопровождаемого образованием клеточной перегородки. Кроме того, у Taphrinales аски расположены плотным слоем, напоминающим гимений дискомицетных грибов. Все это дает основание Гей- манну (1964) относить порядок Taphrinales к высшим аскоми- цетам, помещая их в подкласс эвтуникатных, и сближать с некоторыми Helotiales (Ascocorticium), для которых характерно простое строение аска. Напомним, что еще Шретер (Schröter, 1897) выделял Taphrinaceae (Exoascaceae) в отдельный порядок Proto- discineae, к которому относил и семейство Ascocorticiaceae (!). В последнее время к порядку Taphrinales относит Ascocorticiaceae Цейп (Cejp, 1957). С другой стороны, также трудно согласиться с мнением Бесси (Bessey, 1950), который считает Taphrinales группой редуцированной, происходящей от высших дискомицетных форм, например, от Pezizales. Эту же мысль отражает и схема Гейманна (Gäumann, 1964, стр. 10). Сравнительно простое строение аска Taphrinales не позволяет выводить их из высших диско- мицетов. На основании сказанного мы считаем обоснованным отнесение порядка Taphrinales к подклассу Euascomycetidae, где они должны быть помещены среди более примитивных порядков.
К порядку Endomycetales обычно относят описанное Гиллер- моном (Guilliermond, 1928) семейство Spermophthoraceae. Это семейство характеризуется рядом особенностей в развитии (образованием спорангия, называемого Гиллермоном гаметангием, копуляцией спор, чередованием поколений), дающих основание предполагать филогенетическую его обособленность, что показано на
206
схеме Гейманна (Gäumann, 1964, стр. 108). Мы считаем целесообразным выделять его в особый порядок Spermophthorales. Нужно отметить мнение Гиллермона и Наннфельдта (Nannfeldt, 1932), считавших Spermophthoraceae группой, исходной для всех других аскомицетов, с чем вряд ли можно согласиться.
Очень спорным является положение в системе порядков As- cosphaerales (Pericystales) и Protomycetales. Порядок Ascosphaera-
Puc. 78. Ascosphaera apis:
I — половой процесс (а — женская ветвь с аскогоном (нутриоцитом — б), несущим трихогину (в), г — мужская ветвь, клетка которой копулирует с трихогиной, д — первичная, аскогенная гифа); II— зрелый епороцит (synascus) с шарообразными скоплениями спор (по Spiltoire a. Olive. 1955).
les одни авторы относили к порядку эндомицетальных (Gäumann, 1926) или к порядку сахаромицетальных (Bessey, 1950), другие — к порядку тафринальных (Werdermann, 1954; Ubrizsy a. Vörös, 1966). Гейманн в последней монографии (Gäumann, 1964) рассматривает их как отдельный порядок протуникатных! аскомицетов, а Цейп (Cejp, 1957) вместе с семейством Protomycetaceae в составе порядка Protomycetales присоединяет к подклассу эваско- мицетных. Крейзель (Kreisel, 1969) Ascosphaerales отделяет от Protomycetales и относит в качестве особого порядка к подклассу Euascomycetidae.
Онтогенез этого небольшого порядка, к которому относится один род Ascosphaera (Pericystis) с двумя видами, обитающими в ульях — на перге и личинках пчел, был исследован в двадцатых годах (Claussen, 1921; Varitchak, 1932, 1933); при этом обнаружен ряд особенностей в половом процессе и развитии асков. Позднейшие исследования (Spiltoir, 1955; Spiltoir a. Olive, 1955) показали, что половой процесс у Ascosphaera состоит в слиянии многоядерных клеток двух разных талломов (рис. 78). Мужская копулирующая клетка не отличается от вегетативной. У женских экземпляров развивается крупная клетка, которую называли оогонием (Claussen, Varitchak) или нутриоцитом (Spiltoir a. Olive), на которой размещена другая, называемая трихогиной. Эта клетка по существу является женской половой клеткой (гинецитом) и
207
копулирует с мужской клеткой (андроцитом). После слияния клеток зигота дает внутрь нутриоцита первичную аскогенную гифу, которая в дальнейшем внутри нутриоцита дает начало аскогенной системе, состоящей из аскогенных гиф с крючками и асками. Аски, содержащие, вероятно, по восемь аскоспор, собраны в шары, заключенные в оболочку разросшегося нутриоцита (synascus). Нутриоцит является своеобразным образованием, свойственным только порядку Ascosphaerales. Развитие аскогенных гиф с крючками и асков свидетельствует о принадлежности порядка аскосфе- ральных к аскомицетам.
Что касается порядка Protomycetales в узком понимании, т. е. семейства Protomycetaceae, то его положение в системе еще менее ясно. У представителей этого порядка мицелий разделен на многоядерные участки. На мицелии образуются многоядерные спорангии («хламидоспоры»), в которых после повторного (редукционного?) деления ядер образуются многочисленные споры. Для ряда видов имеются указания на попарную копуляцию спор. Клеточные оболочки у них с хлор-цинк-йодом дают реакцию на целлюлозу. Все это послужило основанием Фитцпатрику (Fitzpatrick, 1930), а за ним и Бесси (Bessey, 1950) сближать порядок Protomycetales с пероноспоровыми, т. е. с сапролегниомицетами. Другие же авторы (Gäumann, 1926; 1964; Cejp, 1957; Ubrizsy a. Vörös, 1966) относят их к аскомицетам — к подклассу Protascomycetidae (Ubrizsy а. Vörös, 1966) или к подклассу Euascomycetidae (Cejp, 1957). При этом Цейп, Убрижи и Вереш объединяют их с семейством Asco- sphaeraceae (Pericystaceae) в один порядок Protomycetales, хотя между этими семействами нет никаких общих черт, кроме образования многоспоровых органов, которые имеют разное происхождение.
Принимая деление класса аскомицетов на два подкласса — Protascomycetidae (Hemiascomycetidae) и Euascomycetidae, мы считаем, что к подклассу Protascomycetidae должны быть отнесены организмы с простым строением асков, развивающихся из зиготы без аскогенных гиф, и не образующие плодовых тел. К этому подклассу мы относим только порядки Endomycetales и Spermo- phthorales, объединяющие организмы, несомненно близкие к исходным формам аскомицетов, а также, хотя и с сомнением, порядок Protomycetales, оставляя в нем только семейство Protomy- cetaceae. Что же касается семейства Ascosphaeraceae (Pericystaceae), то на основании того, что у его представителей имеются аскогенные гифы, образующие крючки и аски, мы считаем возможным отнести его к эваскомицетам, выделяя в особый порядок Ascosphaerales (Pericystales).
Подкласс Euascomycetidae отличается большим разнообразием. В настоящее время в его составе отмечают до двадцати порядков, характеризующихся разнообразными формами полового процесса, различным строением плодовых тел и сумок. Если к тому же принять во внимание далеко не достаточную изученность
208
их строения и онтогенеза, то станет ясным, что говорить о построении детальной филогенетической системы подкласса еще невозможно, и те классификационные схемы, которые мы упоминали, являются, очевидно, очень недолговечными и будут кардинально перестраиваться по мере накопления нового фактического материала.
Среди большого количества порядков, семейств и родов, относящихся к этому подклассу, в зависимости от характера полового процесса, строения асков и характера плодовых тел намечаются группы более примитивные, которые могут быть поставлены в начале системы, и более сложные, более прогрессивные по своему строению и развитию.
К более примитивным порядкам Euascomycetidae обычно относят Eurotiales (Aspergillales), Microascales, Coronophorales, Ony- genaies, Ascosphaerales, Taphrinales, Erysiphales. Среди них намечается несколько направлений эволюции, часть из которых заканчиваются тупо, а другие, вероятно, дали начало более высокоорганизованным группам. Среди этих порядков у некоторых форм появляются уже признаки, характерные для более высокоорганизованных форм — усложнение строения аска (Taphrinales, Erysiphales), развитие плодовых тел типа перитециев (Meliolates, Microascales), асколокулярное заложение асков (Melio- lales, Phaeotrichum hystricinum из Eurotiales), многоклеточная и даже разветвленная трихогина (Ceratocystis jagacearum из Microascales), оплодотворение при помощи конидий — спермаци- ев (тот же Ceratocystis fagacearum; см.: Wilson, 1956).
От некоторых примитивных порядков или скорее от групп, близких к ним, произошли, вероятно, более высоко огранизован- ные порядки подкласса Euascomycetidae. Особенно резко выделяется среди них группа асколокулярных — порядки Myriangiales, Dothiorales, Pseudosphaeriales, которые отличаются от других как способом развития плодовых тел, так и своеобразным строением асков.
Что касается группы аскогимениальных, которую Наннфельдт считает монофилетической, то по мнению других исследователей (Munk, 1953; Окснер, 1956; Gäumann, 1964, и др.) она является гетерогенной и, нужно полагать, дальнейшие исследования прольют свет на пути эволюции в пределах этой группы.
Изолированное положение среди аскомицетов занимает порядок Laboulbeniales. Некоторые авторы рассматривают ее как «придаток» к этому классу (Cejp, 1957; стр. 458; Агх, 1967), другие же считают их особым отделом грибных организмов (Greis, 1943). В отличие от этих авторов Гейманн (Gäumann, 1964) находит возможным связывать происхождение лабульбениальных с порядками Eurotiales (Aspergillales) и Microascales, исходя из следующих положений. Некоторые эндопаразитические формы из лабульбениальных, как например Trenomyces histophthoriis, паразитирующие на куриных клещах, обладают мицелием, разви¬
209
вающимся в жировой ткани хозяина. У других форм мицелий образуется только вблизи места заражения. У большинства же эктопаразитических форм мицелий редуцирован и вегетативное тело состоит из одной клетки, которой гриб прикреплен к покровным тканям хозяина. Оплодотворение у лабульбениальных происходит при помощи конидий — спермациев, развивающихся или экзогенно (семейство Ceratomycetaceae) или эндогенно — в фиа- лидах, напоминающих фиалиды Thielavia и Sordariaceae. Аски у них образуются не на аскогенных гифах, а непосредственно из аскогенной клетки в результате повторного ее деления. Аски простого строения (протуникатный тип) и сидят пучком, как у Thie- laviopsis или Melanospora из порядка Eurotiales (Aspergillales).
Резюмируя сказанное, мы даем ниже краткий обзор системы аскомицетов. Порядки, объем которых мы принимаем в основном по Гейманну (Gäumann, 1964), распределены между двумя подклассами: Protascomycetidae и Eiiascomycetidae.
Класс ASCOMYCETES.
1. Подкласс Protascomycetidae (Hemiascomycetidae) — первичные аскомицеты.
Половой процесс гологамный — копулируют вегетативный клетки, аскоспоры или спорангиоспоры. Аски образуются или непосредственно из зиготы или на диплоидном мицелии (порядок Spermophthorales). У представителей относимого с большим сомнением к аскомицетам порядка Protomycetales образуется многоядерный сборный аск (synascus), протопласт которого распадается на материнские клетки спор, дающие при делении тетрады эндоспор.
Порядки: Endomycetales, Spermophthorales, Protomycetales.
2. Подкласс Eiiascomycetidae — настоящие а с к о - м и ц е т ы.
Половой процесс гологамный — состоит в слиянии одинаковых или отличающихся морфологически клеток (изоцитов или гете- роцитов) или же при дальнейшем усложнении — в слиянии аско- гона (гинецита) с андроцитом или же конидией. Аски обычно образуются на аскогенных гифах; стенка аска у высших представителей имеет сложное строение. Большинству порядков свойственно развитие плодовых тел.
Порядки: Eurotiales (Aspergillales), Microascales, Coronophora- les, Onygenales, Meliolales, Laboulbeniales, Ascosphaerales (Peri- cystales), Erysiphales, Taphrinales, Pezizales, Tuberales, Helotiales, Pliacidiales, Sphaeriales, Xylariales, Diaporthales, Clavicipitales, Myriangiales, Dothiorales, Pseudosphaeriales.
В качестве примера возможных путей развития аскомицетов приведем филогенетические схемы Гейманиа (Gäumann, 1964) и Латтрелла (Luttrell, 1955) (рис. 79 и 80). Обе эти схемы, безусловно, не отражают всех сложных взаимоотношений между многочисленными порядками, на которые разбит в настоящее
210
Рис. 79. Схема филогенетических связей порядков аскомицетов по Гейманну (Gäumann, 1964).
Рис. 80. Схема филогенетических связей порядков аскомицетов по Латтреллу (Lut- trell, 1955).
время класс аскомицетов (дело будущих исследователей уточнить эти вопросы), однако в обеих схемах показан переход от более простых, примитивных форм, близких к зигомицетам, к формам с более сложным строением асков, плодовых тел и конидиальных спороношений, приспособленных к разнообразным условиям наземного существования.
Трудно согласиться с Гейманном по вопросу происхождения Taphrinales, которые он считает редуцированными формами Helotiales. Характер строения сумки у этих порядков скорее свидетельствует о том, что Helotiales через формы типа Ascocorticium связаны с предками Taphrinales и являются более прогрессивной ветвью с более сложным строением плодовых тел и асков. Еще менее правдоподобным нам кажется выведение Erysiphales из Pezizales. Вряд ли можно все порядки высших аскомицетов выводить из Aspergillales — корни некоторых их ветвей, вероятно, нужно искать глубже — в подклассе Protascomycetidae.
Что касается схемы Латтрелла, то мало убедительно в ней выведение дискомицетных порядков из Xylariales. Эти порядки (Helotiales, Pezizales, Phacidiales), вероятно, представляют самостоятельную ветвь эволюции, идущую от низших аскомицетов.
8. БАЗИДИОМИЦЕТЫ — BASIDIOMYCETES Вегетативные органы базидиомицетов
Вегетативное тело базидиомицетов имеет вид многоклеточного разветвленного мицелия. Из прорастающей гаплоидной базидио- споры обычно развивается многоядерный первичный ценоцисти- ческий мицелий с гаплоидными ядрами, который вскоре делится перегородками на одноядерные клетки. Только изредка, например у Stereum hirsutum или Coniophora cerebella, клетки мицелия бывают многоядерными. Первичный гаплоидный мицелий у базидиомицетов обычно недолговечен — в результате гологамного слияния клеток (цитогамии или плазмогамии), при котором ядра не сливаются, он превращается во вторичный мицелий, в клетках которого неопределенно долгое время могут существовать дикарио- ны, представленные неслившимися ядрами копулировавших клеток. Слияние ядер — кариогамия, т. е. завершение полового процесса, происходит в молодой базидии перед образованием бази- диоспор.
Изредка первичный мицелий хможет совершенно отсутствовать. Это происходит в тех случаях, когда базидиоспора бывает двуядер- ной и непосредственно дает дикариофитный мицелий, например у Hypochnus terrestris (Kniep, 1913, 1928), или в случае копуляции базидиоспор или клеток базидий (промицелия), как это описано у многих головневых.
Иногда же плазмогамия может задерживаться и происходить непосредственно перед образованием базидий; в таком случае вто¬
212
ричный (дикариофитный) мицелий или совершенно исключается или же существование его является кратковременным как, например, у Ustilago zeae, Doassansia sagittariae из головневых, у ржавчинных грибов типов «эндо» (О, I) и «микро» (О, III). Эти случаи безусловно являются вторичными и могут наблюдаться в систематических группах, где другие виды характеризуются нормальным развитием.
Вторичный, диплоидный, дикариофитный мицелий, кроме ди- карионов, часто, но далеко не всегда, имеет так называемые пряжки. Образование пряжек детально описал Книп (Kniep, 1915)
(рис. 81).
Этот процесс проходит следующим образом. Оба ядра верхушечной клетки гифы располагаются по продольной оси клетки. Между ними на боковой стенке образуется мешковидный вырост. Оба ядра делятся одновременно (конъюгированное деление) и нижнее дочернее ядро верхней пары заходит в вырост, после чего вырост, содержащий это ядро, отделяется перегородкой. Одновременно верхнее дочернее ядро нижней пары, которое находится в это время у верхнего конца выроста, отделяется перегородкой от сестринского ядра, в результате чего новая верхушечная клетка становится двуядерной, а нижняя клетка — временно одноядерной. Затем мешковидный вырост соединяется со стенкой нижней клетки, перегородка между ними ослизняется, и ядро из выроста переходит в нижнюю клетку, которая, в свою очередь, становится двуядерной. Таким образом, в каждой новой клетке имеется по два ядра, происходящих от разных ядер материнской клетки.
У некоторых базидиальных грибов с многоядерными клетками (Coniophora cerebella, Stereum hirsutum) описано мутовчатое образование пряжек. Книп и ряд других авторов считают образование пряжек гомологичным образованию крючков на аскогенных гифах и делают соответствующие выводы о происхождении бази- диомицетов от аскомицетов, имеющих развитые аскогенные гифы; другие же авторы, например Савиль (Savile, 1955), образование пряжек считают биологическим приспособлением, обеспечивающим правильное деление ядер дикариона и возникшим независимо в разных группах базидиомицетов. Свой вывод эти авторы основывают на том, что пряжки обычны на диплоидном мицелии, имеющем узкие гифы, и часто отсутствуют на мицелии с широкими гифами (Martens, 1932; Nable, 1937). Есть указания на то, что в пределах рода Coprinus одни виды не имеют пряжек, дру¬
Рис. 81. Образование пряжек на мицелии базидиальных грибов (по Kniep, 1929).
213
гие же близко родственные виды имеют мицелий, обильно их образующий (Brunswik, 1924). Такие факты во всяком случае свидетельствуют о том, что образование пряжек в процессе эволюции может выпадать в различных группах базидиальных грибов.
Бесполое размножение
Бесполое размножение у базидиальных грибов, кроме ржавчинных, характеризующихся наличием разнообразных спороноше- ний, происходит при помощи конидий и оидий, образующихся часто группами или даже сидящих на конидиеносцах типа Botrytis. Оидии могут образовываться как на первичном мицелии так и на вторичном, например у Tremella mesenterica, где согласно Данжару (Dangeard, 1895) они могут возникать на плодовом теле. У представителей семейства Polyporaceae описано образование конидий и хламидоспор на плодовых телах (E. A. Bessey, 1950, стр. 487). Обильное образование хламидоспор описано у Nyctalis asterophora из Tricholomataceae. Есть указания, что вообще у гименомицетов нет настоящих конидий, а их следует рассматривать как различные формы хламидоспор (Oehm, 1937). Все же нужно отметить, что в отличие от аскомицетов базидиаль- ные грибы, за исключением ржавчинных, характеризуются значительно более слабым развитием конидиальных спороношений.
Половой процесс и развитие базидий
Половой процесс у базидиомицетов представлен типичной изогологамией — слиянием двух недифференцированных клеток; этот процесс называют еще соматогамией (Gäumann, 1964). Описаны случаи копуляции гаплоидного мицелия с прорастающей оидией (Gäumann, 1964, стр. 342). У головневых грибов давно известен случай копуляции базидиоспор (Tilletia tritici). Аналогичное явление описано у Ustilago bromivora (Bauch, 1925) и других головневых (Kniep, 1928). У ряда видов рода Ustilago давно известны случаи копуляции клеток базидии («промицелия») между собой и с прорастающими базидиоспорами. Ряд усложнений полового процесса известен также у ржавчинных грибов. Они сопровождаются развитием дополнительных спороношений — пикнид («спермогониев»), эцидий, уредо- и телейто-спороношений.
Впервые половой процесс у ржавчинных грибов, протекающий по типу изогологамии, был описан Кристменом на Phragmidium speciosum (Christman, 1905), а позже другими исследователями (Moreau, 1914, и др.) у ряда иных видов. Происходит он в молодых эцидиальных спороношениях — цеомах, где закладывается слой вытянутых прилегающих друг к другу клеток. Эти клетки делятся на меньшую верхнюю стерильную и нижнюю базальную клетки. Оболочки двух соседних базальных клеток ослизняются сверху вниз и их протопласты сливаются, образуя двуядерную
214
клетку, отчленяющую последовательно материнские клетки эци- диосиор, которые, в свою очередь, дают эцидиоспоры и промежуточные клетки. Блекман (Blackman, 1904) описал проникновение другого ядра в базальную клетку из клетки, расположенной под ней.
В половом процессе ржавчинных грибов принимают участие также споры и в первую очередь пикноконидии. Пикноконидии ржавчинных грибов долгое время были загадкой для исследователей, которые не наблюдали их дальнейшего развития. Им приписывали функции оплодотворяющих элементов, в связи с чем называли спермациями, а пикниды соответственно спермогониями, но только Крейджи (Graigie, 1927, 1931) удалось доказать их участие в половом процессе. В случае попадания пикноконидии, переносимой насекомыми, привлекаемыми сладким соком, который выделяют пикниды, на воспринимающие гифы, развивающиеся в пикнидах другой половой расы, она сливается с ней (рис. 82). Если же вблизи нет воспринимающей гифы, то пикноконидия дает ростковую трубку, проникающую прямо в пикниду или же через устьице непосредственно в ткани растения-хозяина и там сливается с клеткой гаплоидной гифы противоположной расы (Hanna, 1929; Allen, 1934). Ядро пикноконидии («спермация»), попавши в гифу другой половой расы, переходит из клетки в клетку, пока не попадет в зачаток эцидия, где проникает в базальную клетку (Savile, 1939). Иногда образование дикариона может происходить ранее — в вегетативной клетке (Wang et Martens, 1939).
Описаны случаи, когда двуядерные уредоспоры Puccinia helian- thi и Phragmidium speciosum прорастали вблизи гаплоидного мицелия и копулировали с его клетками, причем в эту клетку переходило одно ядро прорастающей уредоспоры (Brown, 1932, 1940; Oliveira, 1939; цит. по Gäumann, 1964, стр. 445). В других случаях прорастающие эцидио- или уредоспоры копулировали у Puccinia graminis с клетками воспринимающей гифы пикниды, причем одно из ядер двуядерной ростковой трубки переходило в воспринимающую гифу (Cotter, 1960).
Таким образом, мы видим, что у ржавчинных грибов, кроме обычной для базидиомицетов изогологамии, наблюдается слияние с клетками гаплоидного мицелия или специализированной воспринимающей гифы пикиоконидий (спермациев) или же переход в них
Рис. 82. Puccinia graminis:
спермогонии, образующие спермации, и выходящие из спермогониев воспринимающие гифы (по Buller, 1938).
215
одного из ядер дикариофитной ростковой гифы уредо- или эци- диоспоры.
Особенностью полового процесса базидиомицетов, как и большинства асковых грибов, является разделение неопределенным промежутком времени двух стадий полового процесеа — слияния клеток — цитогамии (плазмогамии) и слияния ядер копулировавших клеток — кариогамии. В результате этого в цикл развития базидиомицетов вклинивается своеобразное диплоидное поколение — дикариофит или так называемый вторичный мицелий, клетки которого содержат пары ядер различного происхождения.
Характерной чертой базидиомицетов является развитие у них тетраполярного гетероталлизма. В этом случае, в отличие от обычного — биполярного гетероталлизма, представленного, например, у Coprinus comatus, наблюдается четыре половых группы, причем особи первой группы реагируют только с представителями второй групп, а третьей группы только с представителями четвертой, например, у Aleurodicus polygonius, Schizophyllum commune и др. (Kniep, 1928). Подобное явление не встречается ни в какой другой группе растительного мира и может быть объяснено, согласно Книпу (Kniep, 1922, 1928), наличием двух пар половых факторов (генов).
Все сказанное выше о половом процессе у базидиомицетов явно противоречит распространенному мнению, что в этой группе наблюдается угасание сексуальности, завершающееся полной редукцией половых органов. С этим положением никак нельзя согласиться, поскольку весь онтогенез базидиомицетов, характерной чертой которого является образование половым путем диплоидного (дикариофитного) мицелия, распространенность в этой группе грибов биполярного и тетраполярного гетероталлизма, развитие своеобразных форм полового процесса ржавчинных грибов свидетельствует о существенной роли полового процесса в жизни бази- диальных грибов. У базидиомицетов половой процесс является неотъемлемой частью нормального онтогенеза, однако он сохранился в весьма примитивной форме изогологамии, унаследованной от далеких зигомицетообразных предков, в отличие от аскомице- тов, где изогологамия дала начало гетерогологамии, усложнение которой привело к вторичному образованию половых органов сложного строения (Zimmermann, 1959, стр. 93).
Завершающим этапом онтогенеза базидиальных грибов является образование базидий с базидиоспорами. Базидии образуются или непосредственно на дикариофитном мицелии (папример, у Exobasidiales) или на плодовых телах, являющихся особой формой существования этого мицелия, или же на покоящихся клетках (хламидоспорах, склеробазидиях, телейтоспорах), возникающих на дикариофитном мицелии (Uredinales, Ustilaginales). В молодой базидии завершается половой процесс — происходит слияние клеток дикариона (кариогамия) и редукционное деление с образованием четырех — восьми гаплоидных ядер. Обычно на ба-
216
зидии образуются четыре, реже две — восемь или еще реже большее число одноядерных базидиоспор. При количестве ядер, превышающем число образующихся базидиоспор, часть ядер отмирает, иногда же образуются двуядерные базидиоспоры, при прорастании которых развивается дикариофитный мицелий.
Юэль (Juel, 1898) обратил внимание на расположение ядерного веретена при делении диплоидного ядра базидиоспоры. У некоторых форм, например у Auricularia, Dacryomyces и других, оно расположено по продольной оси базидии; их Юэль назвал стихо- базидиальными; у других же ядерное веретено расположено перпендикулярно к продольной оси базидии (например, Tremellales, большинство Agaricaceae), такие формы он назвал хиастобазиди- альными. Юэль придавал этому признаку филогенетическое значение, однако нужно отметить, что у близких форм могут встречаться различные типы расположения ядерных веретен (например, род Exobasidium, семейство Clavariaceae), и поэтому этот признак может играть только ограниченную роль при классификации базидиомицетов.
Происхождение и основные направления эволюции базидиомицетов
По вопросу происхождения базидиомицетов со времен де Бари господствует мнение о тесных филогенетических связях их с ас- комицетами, хотя нужно отметить, что некоторые авторы конца XIX и первой половины XX ст. считали, что аскомицеты и ба- зидиомицеты развились как две параллельные группы или от фикомицетов (Brefeld, 1889; Курсанов, 1933, 1940), или от красных водорослей (Lotsy, 1907), или даже от различных групп низших грибов, а именно: аскомицеты от двужгутиковых, а ба- зидиомицеты — от одножгутиковых (Гоби, 1912).
Родственные связи асковых и базидиальных грибов обосновывают гомологией аска и базидии. Действительно, оба эти органа возникают в результате полового процесса, в обоих присутствует дикарион, в обоих происходит кариогамия и редукционное деление, в результате которого образуется после повторного деления четыре или восемь гаплоидных ядер и соответствующее число ас- коспор и базидиоспор. Ряд авторов проводит гомологию также между крючками на аскогенных гифах аскомицетов и пряжками на гифах вторичного мицелия базидиомицетов (Kniep, 1915; Greis, 1938, и др.)· Исходя из этого, некоторые авторы выводят бази- диомицеты от аскомицетов с развитыми аскогенными гифами, т. е. от высокоорганизованных асковых грибов. Так, Бесси (E. A. Bessey, 1950) выводит базидиомицеты от форм типа дис- комицетов, хотя и отмечает, что ржавчинные и головневые грибы произошли из каких-то Ascomycètes, возможно, до того, как дифференцировались Pezizales, Sphaeriales и Dothideales («possibly before the Pezizales, Sphaeriales and Dothideales had become strongly
217
distinguished from one another», стр. 645). Грейс (Greis. 1938) выводил базидиомицеты от форм, близких к Sordaria (Sphaeriales) и Tuber, Роджерс (Rogers, 1932, 1934) и в более поздней работе Грейс (Greis, 1943) — от форм типа Ascocorticium (Helotiales). Линдер (Linder, 1940) искал предков базидиальных грибов среди организмов типа Pyrenomycetes (Dothideales). На схемах Гейма- нна (Gäumann, 1926, 1964) базидиомицеты выводятся от порядка Pezizales. В отличие от этих авторов, другие связывают происхождение базидиальных грибов с низшими аскомицетами типа Taphrinales — гипотетическая «Prototaphrina» (Savile, 1955).
Выведение базидиомицетов от высокоорганизованных аскоми- цетов со сложными, характерными для высших представителей этого класса плодовыми телами и высокоспециализированными половыми органами нам кажется мало обоснованным и противоречит одной из основных закономерностей эволюции — развитию новых групп от неспециализированных предков. Более правдоподобным является мнение, согласно которому обе эти группы высших грибов обособились на ранних стадиях их становления, вероятно, более ранних, чем Taphrinales, в результате дифференциации зародышевого спорангия или примитивного аска. Напомним, что у некоторых мукоральных, например у Choanephora, спорангии могут давать экзогенные односпоровые спорангиоли, а по некоторым указаниям (Sadebeck, 1884) в роде Taphrina аски иногда могут давать почковидные выросты, из которых образуются споры на подобных стеригмам ножках.
По вопросу, какая группа базидиомицетов стоит ближе всего к исходным формам этого класса, существуют разные мнения. Так, Роджерс (Rogers, 1932) считал исходною базидию типа Ти- lasnella (Tremellales). Мартин (Martin, 1945) указывал, что для Tremellales характерным является наибольшее количество примитивных черт; по его мнению, в этой группе есть формы, от которых можно выводить, с одной стороны, Uredinales, а с другой — Homobasidiomycetes. Р. Эйм (R. Heim, 1957) также считал формы типа Tulasnella первичными и связующими базидиомицеты с аскомицетами. Грейс (Greis, 1943) исходным для базидиомицетов считал семейство Thelephoraceae, от которого выводил Holobasi- diomycetes и Phragmobasidiomycetes. На схеме Э. А. Бесси (Bes- sey, 1950) базидиомицеты выведены из дискомицетов двумя ветвями — фрагмобазидиальной, где основной группой являются Uredinales, a Ustilaginales и Auriculariales — производными, и голоба- зидиальной, в основании которой стоят Thelephoraceae.
Ряд авторов (Linder, 1940; Jackson, 1944; Chadefaud, 1960) считают исходной группой класса базидиомицетов ржавчинные грибы на основании наличия у них, как и у высших аскомицетов, от которых они выводят базидиомицеты, воспринимающих гиф и пикиид («спермогониев»). Савиль (Savile, 1955) также считает первичной фрагмобазидию; в то же время он выдвигает предположение, что первичные базидиомицеты были паразитными орга¬
218
низмами и выводит из гипотетической «Prototaphrina», с одной стороны, ржавчинные грибы, а с другой — паразитические Auricu- lariales, которые считает исходной группой для всех прочих ба- зидиомицетов.
Наконец, Гейманн (Gäumann, 1964) первичной группой для всех базидиомицетов ставит Aphyllophorales, но склоняется к мысли, что развитие базидиомицетов шло не монофилетически, а по- лифилетически, что базидиомицеты возникали из аскомицетов неоднократно, а различные типы базидий развились из асков в результате задержки спорообразования и перемещения спор в выросты материнской клетки.
Таким образом, рассмотрение эволюции внутри класса базидиомицетов упирается в разрешение двух вопросов, с одной стороны, являются ли они группой монофилетической или полифи- летической, а с другой — какой тип базидии является первичным, и в связи с этим, какая из существующих групп базидиомицетов стоит ближе всего к исходным формам класса.
Идея полифилетического происхождения базидиомицетов не имеет широкого распространения. Даже Гейманн, высказывая ее, не намечает среди базидиальных грибов групп, происхождение которых было бы независимым. Однотипность онтогенеза, характер развития базидии, даже характер рассеивания спор — все это свидетельствует об общности происхождения базидиомицетов. Бесси (Bessey, 1950, стр. 648) особенно подчеркивает как доказательство единства базидиомицетов способ рассеивания спор. Как показали исследования Вуллера (Buller, 1909), базидиоспоры у большинства базидиомицетов, кроме ангиокарпных форм, асимметричны и косо прикреплены к стеригме. Перед отделением споры сбоку от места прикрепления появляется капля жидкости, образование которой происходит в течение 5—20 секунд. После сформирования капли спора отбрасывается на расстояние, превышающее ее размеры в 10—20 раз. По мнению Бесси, базидиомицеты с симметрически прикрепленными и не отбрасываемыми базидиоспорами, встречающиеся у закрытоплодных Agaricales и Gasteromycetales, являются вторичными, и эти их черты связаны со строением плодовых тел. Следует также иметь в виду, что два типа базидии — одноклеточная и многоклеточная связаны переходами, что также подтверждает мысль об общем происхождении фрагмобазидиальных и голобазидиальных базидиомицетов.
Что касается вопроса какой тип базидии является первичным, то исходя из положения, что базидия является гомологом аска аскомицетов и зародышевого спорангия зигомицетов, а эти органы имеют одноклеточное строение, есть все основания предполагать, что исходная базидия также была одноклеточной. Роджерс (Rogers, 1932) считал такой базидию типа Tulasnella. Как уже упоминалось, Мартин отмечал, что для Tremellales характерен ряд примитивных черт и что в этом порядке имеются формы, от которых можно вывести как ржавчинные — через Auriculariales,
219
так и гомобазидиальные грибы — от Tulasnellaceae. В пределах двух более примитивных порядков базидиомицетов — Tremellales и Thelephorales базидии характеризуются большим разнообразием и неустойчивостью строения и развития. Этим же порядкам свойственно примитивное строение плодовых тел или даже их отсутствие, а также простые конидиальные спороношения. В порядке Tremellales встречаются как одноклеточные базидии (Tulasnella, Dacryomyces, Ceratobasidium), так и снабженные обычно продольными, иногда неполными перегородками (Tremella); иногда же перегородки базидий могут быть вертикальными, косыми и даже поперечными (Tremella steidleri). У других представителей порядка перегородки на базидиях также могут быть косыми (Si- robasidium) и поперечными (Galzinia).
Развитие базидии и образование базидиоспор в порядке Tremellales проходит различно. Так, в роде Tulasnella описаны дву- ядерные базидиоспоры, возникающие в результате послемейоти- ческого деления ядер; иногда же ядра делятся в базидии и тогда образуется до семи базидиоспор на одной базидии. У ряда представителей порядка Tremellales прорастающая базидиоспора может давать конидию — так называемую «вторичную базидиоспору», которая, в свою очередь, может давать вместо мицелия конидию. Для ряда представителей порядков Thelephorales и Tremellales характерно образование пробазидий или гипобазидий. Все это дает основание считать, что порядки Tremellales и Thelephorales стоят ближе всех к исходным формам класса базидиомицетов.
Авторы, которые считают начальной группой базидиомицетов ржавчинные грибы (Linder, 1940; Jackson, 1944; Chadefaud, 1960), исходят из того, что ржавчинные грибы, характеризующиеся наличием пикнид («спермогониев») и воспринимающих гиф, стоят ближе всего к высшим аскомицетам, непосредственными потомками которых, по их мнению, они являются.
Для обоснования происхождения ржавчинных грибов от высших аскомицетов Линдер разработал гипотезу возникновения базидий ржавчинников из битуникатных асков высших аскомицетов. Согласно этой гипотезе, аск гомологизируется с телейтоспорой, а молодая базидия (промицелий), развивающаяся из телейтоспо- ры,— с содержимым аска, покрытым внутренней эластической оболочкой, четыре клетки фрагмобазидии — с аскоспорами, а базидиоспоры— с конидиями (вторичными спорами). Гомологи Линдера совершенно не обоснованы. Линдер ничего не говорит о возникновении своеобразных типов спороношений ржавчинных грибов и на том основании, что сокращенные циклы развития у них являются производными от полного цикла, считает, что в результате дальнейшего сокращения и выпадения уредо- и те- лейтоспороношений произошли головневые и аврикуляриальные, а после упрощения базидии — Tremellales и Holobasidiomycetes.
Как уже было сказано выше, нельзя допустить, чтобы такая специализированная группа облигатных паразитов, как ржавчип-
220
ники, могла быть исходной для разнообразных групп базидиоми- цетов, среди которых мы видим ряд сапрофитных, паразитных и симбиотических организмов и наряду с высокоорганизованными формами порядки Tremellales, Thelephorales и Exobasidiales, включающие ряд неспециализированных организмов сравнительно простого строения.
Савиль (Savile, 1955) также ставит ржавчинные грибы в начале филогенетического дерева базидиомицетов, однако считает их слепой ветвью. Он полагает, что исходной группой для прочих базидиомицетов являются паразитические аврикуляриальные, которые он ставит вместе с ржавчинными в начале системы базидиомицетов, выводя их из гипотетической нрототафрины.
В отличие от Линдера Савиль много внимания уделяет развитию различных спороношений ржавчинных грибов и дает обоснованную картину эволюции в пределах этого порядка. Однако его концепция первичности ржавчинных и аврикуляриальных совсем не обоснована.
Исходя из всего сказанного выше, мы считаем, что в основании филогенетического дерева базидиальных грибов должны быть поставлены организмы с простым типом развития и одноклеточной неспециализированной базидией. Из современных базидиомицетов к первичным базидиомицетам стоят ближе всего, как мы уже упоминали, порядок Tremellales и низшие формы порядка Thelephorales. От подобных примитивных форм произошли другие группы базидиомицетов двумя основными ветвями — фрагмобази- диальной и голобазидиальной.
От форм типа Tremellales с базидиями, разделенными косыми и поперечными перегородками, могли произойти Auriculariales с их базидиями, разделенными поперечными перегородками, и с простыми плодовыми телами.
У некоторых представителей порядка Thelephorales, например у Brachybasidium и Vuilleminia \ между кариогамией и мейозом проходит некоторый срок, что связано с тем, что у них вклинивается стадия пробазидии, в которой происходит кариогамия и которая в процессе развития дает уже базидию, где происходит мейоз и образование базидиоспор. Аналогичное явление описано также в семействе Cystobasidiaceae (Platygloea nigricans) из порядка Auriculariales. У других представителей этого семейства, например у Cystobasidium lasioboli, пробазидия имеет плотную оболочку, здесь мы уже наблюдаем переход к склеробазидии — органу, приспособленному к переживанию неблагоприятных условий. В семействе Septobasidiaceae из того же порядка, наряду с развитием базидий по типу Auricularia, т. е. без стадии пробазидии, часто можно видеть хорошо развитые склеробазидии (Septo- basidium pseudopedicellatum (рис. 83), UredineUa coccidiophaga). 11 Гейманн (Gäumann, 1964) Brachybasidium относит к семейству Ехо- basidiaceae.
221
P»tc. 83. Septobasidium pseudo ре dic ellat um:
развитие базидии из пробази- дии (склеробазидии) (по Couch, 1935).
Заслуживают внимания дополнительные спороношения у представителей семейства Septobasidiaceae. Так, у Goplana dioscoreae описано образование на нижней стороне листьев ацервулярных спороношений, которые очень напоминают уредоспороношения
ржавчинных грибов (Cummins, 1935; Viennot-Bourgin, 1953; цит. по Gäumann, 1964, стр. 438).
У Uredinella coccidiophaga (Couch, 1937), кроме склеробазидий, напоминающих телейтоспоры, формируются толстостенные двуядерньте образования, которые, прорастая, образуют тонкостенные споры; их сравнивают с материнскими клетками уредоспор. Склеробази- дии аврикуляриальных можно считать прототипом телейтоспор ржавчинных и хламидоспор головневых грибов.
Наиболее примитивным из порядка Uredinales является семейство Puccini- astraceae (Savile, 1955; Gäumann, 1964), представители которого паразитируют da папоротниках, покрытосеменных (уредо- и телейтоспоры) и на хвойных (эцидии и пикниды) и характеризуются наиболее простым строением пикнид и телейтоспор. Их пикниды имеют вид простых ацервулярных спороношений. Телейтоспоры у них тонкостенные и бесцветные, развиваются у Uredinopsis и Pucciniastrum в межклетниках, а у Milesia и Hyalo- spora внутри клеток растения-хозяина. Эцидии и уредоспороношения также имеют примитивное строение. Если принять во внимание данные Артура и других (Arthur et al., 1929) о том, что эцидии увязываются генетически с уредоспороношениями, а морфологические и цитологические данные ясно свидетельствуют о родственности уредо- и телейтоспороношений (Savile, 1955), а также наличие в семействах Cystobasidiaceae и Septobasidiaceae порядка аврикуляриальных склеробазидий, ничем принципиально не отличающихся от телейтоспор, а также спороношений типа «уредо» у родов Goplana и Uredinella (сем. Septobasidiaceae), можно представить себе развитие ржавчинников от форм, близких к современным Auriculariales (сем. Septobasidiaceae), в результате дифференциации и усложнения спороношений, идущих параллельно со специализацией их паразитизма. Наивысшего развития порядок Uredinales достигает в семействе Pucciniaceae.
Головневые грибы (Ustilaginales) сходны с Uredinales наличием у них хламидоспор типа телейтоспор (склеробазидий), однако отличаются наличием пряжек и отсутствием характерного для Uredinales разнообразия спороношений. Кроме того, развитие баз ид ии у головневых проходит несколько иначе, а именно: гаплоидное ядро, обычно, не переходит в молодую базидиоспору («спо-
222
ридий»), а делится, причем одно дочернее ядро переходит в ба- зидиоспору, а другое остается в клетке и после ряда повторных делений может дать ряд базидиоспор («споридий»), число которых может доходить до нескольких десятков и даже до 150 (Neovossia indica из Tilletiaceae). Характерным для головневых является наличие, кроме четырехклеточной, также и одноклеточной бази- дии (Tilletiaceae), а также неустановившийся тип полового процесса. Половой процесс у Ustilaginales может происходить путем слияния базидиоспор («споридий»), клеток гаплоидного мицелия, а также клеток базидии между собой или с базидиоспорой. Все эти черты являются признаками более примитивной организации, а не резулыатом упрощения, как полагает Гейманн (Gäumann, 1964) или даже вырождения, как считает Бесси (1950). Головневые несомненно ближе к исходным формам бизидиомице- тов, чем ржавчинные. Они могли развиться, как и аврикуляриаль- ные, от начальных базидиомицетов с примитивной базидией не- установпвшегося строения, развивающейся из пробазидии.
Нельзя согласиться с мнением Линдера (Linder, 1940) о происхождении головневых от «эндо» форм ржавчинников, у которых выпадают стадии «уредо» и «телейто», а функции телейтоспор — образование базидий выполняют эцидиоспоры. Против этой гипотезы свидетельствует наличие у головневых пряжек, отсутствующих у ржавчинников, неустановившееся строение базидий и более примитивный характер полового процесса. Наличие многоклеточной базидии в семействе Ustilaginaceae и одноклеточной — в семействе Tilletiaceae, является также свидетельством близости Ustilaginales к примитивным базидиомицетам с их разнообразным строением базидий.
Что касается происхождения одноклеточной базидии Tilletiaceae, то заслуживает внимания гипотеза Линдера (Linder, 1940) о возможности ее развития из многоклеточной базидии семейства Ustilaginaceae через базидии таких организмов, как Ustilago domestica и Tuburcinia trientalis. У Ustilago domestica базидиоспоры образуются не последовательно по одной на клетке базидии, а в большом количестве и почти одновременно, формируя мутовки базидиоспор. У Tuburcinia trientalis из Tilletiaceae содержимое базидиоспоры с ядрами перемещается по мере роста базидии к ее верхушке, а нижняя часть клетки по мере вытягивания базидии отделяется перегородками. Базидиоспоры образуются на верхушке перегородчатой базидии. Следующий этап—Tilletia tritici, у которой базидия уже не делится перегородками.
С организмами типа Tremellales легко увязываются низшие представители порядков гомобазидиомицетов — Thelephorales и Exobasidiales. В этих порядках встречаются организмы с неуста- новившимся строением базидии, примитивными плодовыми телами или первичным их отсутствием. Так, у относимого к семейству Corticiaceae, рода Vuilleminia (Cejp, 1958; Gäumann, 1964) и у рода Brachybasidium из порядка Exobasidiales (Gäumann, 1964)
223
наблюдается образование пробазидии, что давало основание выделять эти роды вместе с семейством Tulasnellaceae в отдельный порядок — Tulasnellales (Gäumann, 1926). В роде Tomentella (Thelephorales) нет сформированного гимениального слоя. У рода Kordyana (Exobasidiales) базидии размещаются одиночно среди пучков парафиз, выходящих из устьиц питающего растения.
В дальнейшем эволюция гомобазидиомицетов шла в направлении образования плодовых тел, защиты гимениального слоя от неблагоприятных условий и обеспечения рассеивания спор. Давно установилось мнение, что это развитие шло двумя путями — через образование открытых и закрытых плодовых тел. В связи с этим гомобазидиомицеты (голобазрщиомицеты, автобазидиомицеты) делили на две группы, обычно рассматриваемые как порядки — Hymenomycetes и Gasteromycetes (Gastromycetes). Неоднократно высказывалось мнение, что эти группы неоднородны и объединяют организмы сходные по внешнему виду, но отличающиеся по внутреннему строению и происхождению. Эти теоретические соображения, а также разнообразие типов плодовых тел, дали основание разбивать гименомицеты и гастеромицеты сначала на ряд нодпорядков (Engler, 1912; Е. Fischer, 1933; Werdermann, 1954), а затем и порядков (Gäumann, 1926; Konrad et Maublanc, 1948; Cejp, 1958). Необходимо отметить, что в последнее время проявляются тенденции к укрупнению порядков базидиомицетов (Gäumann, 1964; Ubrizsy a. Vörös, 1966).
Как мы уже отмечали, на основании сравнительного изучения строения плодовых тел гомобазидиомицетов обычно принято считать, что усложнение плодовых тел в пределах этой группы шло двумя путями — по линии гименомицетов и по линии гастероми- цетов. Эта теория подытожена в последней монографии Гейманна (Gäumann, 1964). Ее придерживается ряд авторов, из которых упомянем Маленсона, Эйма, Мартина, Пилата, Э. А. Бесси и др.
У низших представителей гомобазидиомицетов (Tomentella- сеае из Thelephorales, род Kardyana из Exobasidiales) базидии размещены одиночно или образуют пучки. В семействе Corticia- сеае они смыкаются, образуя гимениальный слой на гладкой или морщинистой поверхности чешуйчатого плодового тела. В более высоко организованных группах можно наблюдать тенденцию к усложнению гименофора — образование ямок, зубцов, выступов, складок и поднятие гименофора над поверхностью субстрата (Ро- lyporaceae, Hydnaceae, Clavariaceae) с образованием расширений в верхней части плодового тела типа примитивной шляпки, на складках нижней стороны которой размещен гимений (Cantharel- laceae). Все эти семейства иногда объединяют в одну группу (порядок) Aphyllophorales, характеризующуюся простыми гимнокарп- ными плодовыми телами, гимений которых не защищен от влияния неблагоприятных метеорологических условий.
Дальнейшее усложнение плодового тела наблюдается у Aga- ricales. Оно заключается в том, что зрелое плодовое тело имеет
224
вид шляпки, сидящей на более или менее длинной ложке. Ги- менофор, расположенный на нижней стороне шляпки, имеет вид или правильных радиальных пластинок или же трубочек; бази- диоспоры созревают у них приблизительно в одно время. В пределах порядка Agaricales можно наблюдать переходы от открытых плодовых тел, гимений на которых открыт на протяжении всего времени своего развития (гим- покарпиые формы), до псевдо- ангиокарпных, гемиангиокарп- ных и ангиокарпных плодовых тел, у которых наблюдается постепенное развитие приспособлений, защищающих гимений во время его развития и способствующих распространению ага- рикальных в различных экологических условиях. Размещение гименофора на нижней стороне шляпки плодового тела является уже известной защитой от неблагоприятных условий (гимно- карпные виды семейств Tricho- lomataceae, Russulaceae, Boleta- ceae). Дальнейшее развитие защиты гименофора можно видеть у так называемых псевдоангиокарпных видов, например у Ixoco- mus flavus (Boletus flavus). Здесь гимений вначале закладывается открыто на нижней стороне шляпки; затем края молодой шляпки загибаются к ножке, ее гифы переплетаются с гифами коры ножки и гимений оказывается заключенным в кольцевую полость, окружающую верхушку ножки. Сросшиеся гифы шляпки и ножки образуют частное покрывало (velum partiale). В дальнейшем шляпка разрастается, частное покрывало растягивается и разрывается, часто оставляя на ножке кольцо, и гименофор освобождается. Такой тип плодового тела известен в семействах Hygropliora- сеае, Russulaceae и Boletaceae.
У так называемых гемиангиокарпных форм, к которым принадлежит большинство агарикальных, можно наблюдать следующий этап в развитии защиты гимениального слоя, который закладывается внутри зачатка плодового тела (ангиокарпно) в виде кольцевой полости, окружающей верхнюю часть зачатка ножки (рис. 84). В эту полость врастают пластинки гименофора, отходящие от зачатка шляпки. Кроме того, у гемиангиокарпных форм все молодое плодовое тело покрыто общей оболочкой — общим покрывалом (velum universale). При разрастании плодового тела общее покрывало разрушается, но в некоторых случаях остатки его иногда остаются на поверхности шляпки и при основании ножки, образуя так называемую вольву (виды рода Amanita).
225
Рис. 84. Схема строения гемиангио- карпного плодового тела агарикаль- ных:
а — молодое плодовое тело, б — зрелое плодовое тело (по Gäumann, 1964).
Затем разрывается ткань между краем шляпки и ножкой (частное покрывало), остатки которой могут оставаться на ножке, образуя кольцо. Иногда верхние концы пластинок гименофора срастаются с тканью ножки; в таких случаях при освобождении гименофора верхняя его часть, сросшаяся с ножкой, остается в виде так называемого манжета (arvilla). В некоторых случаях, например в роде Amanita, пластинки гименофора закладываются одновременно с образованием кольцевой полости, которая в таких случаях не является сплошной, а разделена пластинками гименофора на многочисленные узкие радиальные участки. Споры у гемиангио- карпных форм созревают после раскрывания плодового тела. Геми- ангиокарпные плодовые тела встречаются в семействах Tricholoma- taceae, Amanitaceae, Agaricaceae, Coprinaceae и др.
По мнению Гейманна, с различными семействами порядка Aga- ricales филогенетически связан ряд семейств, обычно относимых к гастеромицетам — к порядкам Podaxales (Secotiaceae и Podaxa- сеае) и Hymenogastrales (Rhizopogonaceae и Hydrangiaceae). Эйм, а за ним Гейманн (Gäumann, 1964) объединяют их в особый порядок Agaricogastrales и рассматривают как регрессивные серии (ein Bündel von Rückbildungsserien), происхождение которых связано с переходом от гемиангиокарпии к ангиокарпии и от наземного к подземному образованию плодовых тел. Происхождение других порядков и семейств гастеромицетов Гейманн увязывает с низшими гомобазидиомицетами — «еще неизвестными Aphyllo- phorales» и считает их группой полифилетической, которая образует ряд параллельных ветвей эволюции. Он отмечает усложнение строения гимения, плодовых тел и способов рассеивания спор.
Э. Фишер (Е. Fischer, 1912) высказывал сомнение в том, можно ли считать гастеромицеты филогенетически единой группой, но в более поздней работе (Е. Fischer, 1933) считает их единой группой, которая ведет начало от форм с базидиями, однообразно разбросанными в ткани плодового тела или размещенными в его лакунах. Фишер считает возможным выводить гастеромицеты из примитивных гомобазидиальных грибов с недифференцированным рыхлым плодовым телом, не имеющим еще гимения, типа Нуро- chnaceae (порядок Thelephorales). Лаваг (Lohwag, 1926) связывает гастеромицеты с Corticiaceae (порядок Thelephorales). Э. Фишер выделяет связанные между собой переходами несколько типов строения плодовых тел гастеромицетов, а именно: 1) однообразный тип, у представителей которого базидии равномерно разбросаны в ткани плодового тела (Tulostoma); 2) лакунарный тип, при котором базидии размещены беспорядочно или образуют гимений в многочисленных полостях (Melanogastraceae, Sclerodermataceae, Nidulariaceae) ; 3) кораллоидный тип, при котором от компактной базальной или осевой плотной ткани в радиальных направлениях расходятся складки, коралловидно разветвляющиеся, а между складками размещаются камеры глебы (Hymenogastraceae, Ly- coperdaceae, Hysterangiaceae) ; 4) многошляпочный тип, при ко¬
226
тором радиально расходящиеся коралловидные ветки на концах — у перидия образуют щитовидные расширения, на которых и между которыми размещены пластинки стромы (Hysterangiaceae, Clathraceae); 5) одношляпочный тип, при котором имеется один
Рис. 85. Схемы строения типов ангиокариных плодовых тел гастеромицетов: а — лакунарный, б — коралловидный, в — многошляпочный, г — одношляпочный (по Ed. Fischer, 1933).
тяж плотной ткани, заканчивающийся шляпообразным или щитовидным расширением (Phallaceae) (рис. 85). Фишер считает, что в пределах этих типов можно наблюдать переходы от более примитивных форм к более сложным.
В отличие от Гейманна и Фишера, Цейп (Cejp, 1958) считает возможным выводить порядки гастеромицетов от порядков гимено- мицетов, в основании которых он ставит семейство Corticiaceae.
Ряд авторов высказывает совершенно противоположный взгляд на происхождение агарикальных грибов. Так, по Зингеру (Singer, 1962), еще Брефельд высказывался за происхождение агарикальных грибов от гастеромицетов. Позже эти взгляды были развиты Бухгольцем и Ловагом. В последнее время их настойчиво развивает Зингер (Singer, 1936, 1939, 1949, 1958, 1962). Основаны они на отмеченном выше сходстве представителей порядков Podaxales и Hymenogastrales с агарикальными. Зингер приводит ряд примеров сходства родов из разных семейств Agaricales с родами указанных порядков гастеромицетов (Singer, 1962,
227
стр. 124). Он особо подчеркивает анатомическое сходство — наличие сфероцист у представителей семейства Russulaceae и семейства Hydnangiaceae. Однако это сходство с равным успехом можно объяснить и происхождением Hydnangiaceae от Russulaceae (Tricholomataceae). Бесспорным в данном случае является то, что эти семейства занимают особое положение в системе гомобазидио- мицетов, в связи с чем их может быть следовало бы выделить в отдельную или отдельные таксономические группы.
С другой стороны, нужно помнить, что строение и отбрасывание базидиоспор, характерное для гимнокарпных и гемиангиокарпных базидиомицетов, отличает их от гастеромицетов, и признание происхождения агарикальных от гастеромицетов предполагает повторное развитие этого характерного для открытоплодных базидиомицетов признака, что маловероятно (E. A. Bessey, 1950).
Эволюция наземных грибов связана с двумя характерными биологическими приспособлениями, обеспечивающими борьбу за существование,— защитой гимения от неблагоприятных метеорологических условий, с одной стороны, и обеспечением рассеивания спор — с другой. Это в наиболее совершенной форме, хотя и различными путями, осуществляется у высших представителей гименомицетов и гастеромицетов. В результате приспособления к одинаковым условиям существования в этих группах могли возникнуть сходные морфологические признаки — плодовые тела с закрытым гимением, ножка, поднимающая гименофор или глебу над субстратом и т. п.
Порядки Podaxales и Hymenogastrales с закрытыми до созревания спор плодовыми телами, обнаруживающие ряд общих черт с порядками Agaricales и Boletales, являются специализированными группами, развившимися в ксеротических условиях, и выводить из них агарикальные и другие гименомицеты нам кажется маловероятным.
На основании всего сказанного выше мы считаем, что класс базидиомицетов целесообразно разделить на три подкласса:
1. Protobasidiomycetidae — протобаз и диаль- н ы е гриб ы, характеризующиеся рядом примитивных черт: не- установившимся строением базидий, первичным отсутствием плодовых тел или их примитивным строением. Один порядок Tremellaies, в который мы включаем также выделяемые некоторыми авторами порядки Tulasnellales и Dacryomycetales;
2. Sclerobasidiomycetidae — склероба зидиаль- н ы е грибы с базидией в большинстве случаев разделенной поперечными перегородками, развивающейся обычно на специальной споре (склеробазидии, телейтоспоре, хламидоспоре). Мы считаем целесообразным к этому подклассу отнести как исходный порядок Auriculariales, в котором происходит становление склеробазидии и который связывает типичные склеробазидиальные — ржавчинные и головневые — с протобазидиальными;
228
3. Homobasidiomycetidae — гомобазидиальные грибы, характеризующиеся одноклеточной базидией и постепенным развитием плодовых тел. К этому подклассу относятся порядки как с открытыми (гимнокарпные) и полузакрытыми (ге- миангиокарпные), так и с закрытыми (ангиокарпные) плодовыми телами, а именно: Exobasidiales, Thelephorales, Clavariales, Hyd- nales, Polyporales, Cantharellales, Agaricales, Boletales, Podaxales,
Corticiaceae
Рис. 86. Схема филогенетических связей гомобази- диомицетов (по Cejp, 1958).
Hydnangiales, Protogastrales, Hymenogastrales, Sclerodermatales, Nidulariales, Lycoperdales, Hysterangiales, Phallales.
От выделения укрупненных единиц — подклассов или групп порядков — Aphyllophorales, Hymenomycetes и Gasteromycetes мы считаем целесообразным отказаться, так как в этих группах искусственно объединяются группы грибов различного филогенетического происхождения.
Как пример филогенетической схемы гомобазидиомицетов даем схему Цейпа (Cejp, 1958), который выводит все порядки бази- диомицетов из семейства Corticiaceae (порядок Thelephorales) (рис. 86).
9. ЛИШАЙНИКИ - LICHENES И ИХ МЕСТО В СИСТЕМЕ РАСТИТЕЛЬНЫХ ОРГАНИЗМОВ
Своеобразную группу грибных организмов представляют лишайники. Линней, которому было известно около девяноста видов лишайников, включал их вместе с печеночниками в состав наземных водорослей (Linné, 1753, 1770). Однако уже Ахариус в начале XIX ст. выделяет их в самостоятельную группу (Acharius, 1803, 1814). Ряд авторов XIX ст. рассматривает лишайники как группу талломных организмов наравне с водорослями и грибами (Endlicher, 1836—1840). В то же время были тенденции относить их к грибам (Adanson, Sachs, 1868, 1873).
После того, как исследования Фаминцына и Баранецкого (1867) и Швенденера (Schwendener, 1867) разъяснили природ} лишайников многие авторы продолжали и продолжают рассматривать их как самостоятельную группу — класс или отдел расти¬
229
тельного мира. Так, Маттик (Mattick, 1954) выделяет лишайники в особый отдел растительного мира, Комарницкий и др. (1962) рассматривают их также как отдел — «тип» лишайники (Lichenes).
Однако с конца XIX ст. ряд авторов настойчиво высказывается против признания лишайников самостоятельной группой растительного мира. Например, Вайнио (Wainio, 1890) считал, что лишайники тесно связаны с грибами и не являются особой систематической группой. В пособии Веттштейна (Wettstein, 1901) лишайники названы «грибами, приспособившимися к симбиозу с водорослями»; Энглер (Engler, 1898, 1912, и др.) хотя и выделяет лишайники в особую группу, но не дает им ранга самостоятельной систематической единицы, а называет «дополнительным классом» (Nebenklasse).
Особенно резко выступает против признания лишайников самостоятельной группой растительного мира Наннфельдт (Nannfeldt, 1932), взгляды которого вызвали широкий отклик.
Широкое применение в связи с этим нашел термин «лихенизированные грибы».
В конце XIX и в начале XX ст. была распространена классификация лишайников Вайнио (Wainio, 1890):
ASCOPHYTA TL. FR.
I. Gymnocarpeae
1. Discolichenes
A. Cyclocarpeae Y a i n.
B. Graphideae E s c h w.
G. Coniocarpeae (Meyer) Vain.
II. Pyrenocarpeae Vain.
1. Pyrenolichenes.
Appendix.
Lichenes imperfecti, куда внесены и базидиальные лишайники.
Широкое распространение приобрела классификация Цальбрукнера (Zahlbruckner, 1926), основу которой приводим ниже.
LICHENES (Лишайники)
1. Подкласс Ascolichenes
1. Порядок Pyrenocarpeae
2. Порядок Gymnocarpeae
1. Подпорядок Coniocarpineae
2. Подпорядок Graphidineae
3. Подпорядок Cyclocarpineae
2. Подкласс Hymenolichenes.
По сути классификация Цальбрукнера является «перевернутой» классификацией Вайнио.
Из позднейших систем, являющихся дальнейшим усложнением системы Цальбрукнера, укажем систему Маттика (Mattick, 1954).
Отдел LICHENES
1. Класс Phycolichenes. Сомнительная группа, к которой относят одноклеточный гриб (фикомицет), в клетках которого живет водоросль.
Один порядок Geosiphonales.
230
2. Класс Ascolichenes
1. Подкласс Pyrenocarpeae
Порядки: Verrucariales, Pyrenulales, Pyrenidiales, Dermatinales.
2. Подкласс Gymnocarpeae
1- я группа порядков Goniocarpiidae
Порядок Caliciales.
2- я группа порядков Graphidiidae
Порядки: Graphidales, Roccellales.
3- я группа порядков Cyclocarpiidae
Порядки: Thelotrematales, Cyanophilales, Lecideales, Lecanora¬
les, Caloplacales.
3. Класс Basidiolichenes
Порядок Corales.
Эта система отличается от системы Цальбрукнера в основном тем, что все систематические группы подняты в ранге — Pyrenocarpeae и Gymnocarpeae из порядков переведены в подклассы, а подпорядки Coniocarpineae, Graphidineae и Cyclocarpineae рассматриваются как группы порядков и разбиты на несколько порядков.
Авторы, отрицающие самостоятельное систематическое положение лишайников, включают их в систему грибов. Так, Клементс и Шир (Clements a. Shear, 1931) распределяют семейства лишайниковых грибов между порядками Sphaeriales, Dothideales, Phacidiales и Pezizales. Э. А. Бесси (E. A. Bassey, 1950) асковые лишайники относит к аскомицетам, распределяя их между двумя порядками — Lecanorales, куда он включает дискомицетные лишайники (Gymnocarpeae, Discolichenes) и Pyrenulales — перите- циальные лишайники (Pyrenocarpeae, Pyrenolichenes).
В связи с различным строением сумок и наличием различных типов развития плодовых тел у асковых лишайников, аналогичным строению сумок и плодовых тел у асковых грибов, все более утверждается мысль о полифилетическом развитии лишайникбв.
Окснер (1956) делает попытку связать происхождение отдельных систематических групп лишайников — порядков и семейств с соответствующими группами грибов и дает схему филогенетических их связей. Однако нужно отметить, что порядки, приводимые Окснером позже — во «Флоре лишайников Украины» (1956,
1962), не всегда соответствуют группам, поданным на этой схеме.
Сантессон (Santesson, 1953) категорически отбрасывает старые классификации лишайников и, следуя Наннфельдту, делит асковые лишайники на две группы — асколокулярные (Ascoloculare) и аскогимениальные (Ascohymeniales). В пределах асколокуляр- ных он выделяет четыре семейства, которые относит к порядку Pseudosphaeriales. Среди Ascohymeniales Сантессон различает три порядка, к которым относятся «лихенизированные грибы»: Sphaeriales, Lecanorales и Calicales — последний с вопросом.
Шадефо (Chadefaud, 1960) считает, что на основании изучения строения асков лишайников, выполненного им, а также Ма- нем, Галену и Кульберсоном, асковые лишайники могут быть распределены, хотя и с оговоркой на недостаточность исследований строения асков и аскокарпов, между различными группами
231
дискомицетных и пиреномицетных нелихенизированных грибов, а именно: Discolichenes между порядками Lecanorales (большинство видов), и Helotiales (?), Graphidales, Leotiales и Caliciales, a Pyrenolichenes — между порядками Dothideales (большинство« видов) и Sphaeriales.
Окснер (1956, 1968) делит асковые лишайники на 15 порядков: Sphaeriales, Dothideales, Graphidales, Arthoniales, Pilocarpales,, Chrysotrichales, Caliciales, Thelotremales, Coenogoniales, Collema- les, Pannariales, Stictales, Gyalectales, Lecideales, Lecanorales.
Крейзель (Kreisel, 1969) асковые лишайники размещает по филогенетическим связям между шестью порядками своей системы аскомицетов:
Порядок Helotiales — семейство Baeomycetaceae.
Порядок Sphaeriales — семейства: Verrucariaceae, Stauro- theleaceae, Pyrenulaceae, Stringulaceae.
Порядок Dothideales — семейства: Arthopyreniaceae, Try- petheliaceae, Dermatinaceae.
Порядок Lecanorales (со включением порядков Buelliales, Caloplacales, Cyanophilales, Lecideales и Thelotremales) с семействами: Lecanoraceae, Acarosporaceae, Pertusariaceae, Umbilicaria- ceae, Lecideaceae, Cladoniaceae, Teloschistaceae, Buelliaceae, Gyal- ectaceae, Collemataceae, Lichinaceae, Peltigeraceae.
Порядок Hysteria les — семейства: Graphidaceae, Arthonia- ceae, Roccellaceae.
Порядок Caliciales — семейства: Cypheliceae, Sphaero-
phoraceae, Caliciaceae.
Из всех шести порядков только один — Lecanorales содержит почти исключительно лишайниковые грибы.
Количество порядков в системе Крейзеля уменьшено по сравнению с системой Окснера за счет уплотнения их объема. Так, порядки Graphidales и Arthoniales у Крейзеля объединены в порядке Hysteriales, несколько последних порядков от Thelotremales до Lecanorales системы Окснера объединены в порядке Lecanorales и только один порядок — Helotiales выделен Крейзелем из объема порядка Lecideales системы Окснера.
Таким образом, исходя из принципов филогенетической систематики мы должны признать лишайники группой гетерогенного происхождения и размещать их роды в пределах порядков грибов, с которыми они генетически связаны. Небольшая группа базидиальных лишайников, тесная связь которой с семейством Thelephoraceae доказана, должна быть отнесена к этому семейству. Заслуживает внимания спорадическое нахождение водорослей в плодовых телах некоторых других базидиальных грибов — Clavaria, Polyporus, виды которых не связаны с водорослями.
Все это является подтверждением того, что лишайники представляют собой пе систематическую группу, а один из видов симбиоза гетеротрофных организмов с автотрофными, так широко распространенного в растительном мире.
Глава IV
ПРОИСХОЖДЕНИЕ И ПУТИ эволюции МОХООБРАЗНЫХ
1. ПРОИСХОЖДЕНИЕ МОХООБРАЗНЫХ Предки архегониалъных растений
Фила зеленых растений — Chlorophyta отличается от других фил автотрофных эвкариотических организмов тем, что высшие ее представители являются типичными наземными растениями. Особое место среди них занимают мохообразные, которые многими авторами рассматриваются как особый отдел растительного мира — Bryophyta. Считая целесообразным филогенетические стволы — филы при классификации отождествлять с отделами растительного мира, мы мохообразные рассматриваем как подотдел Bryophytina отдела зеленых растений — Chlorophyta, выделяя из них как особый равноценный подотдел Anthocerotophytina. Ряд авторов связывает происхождение мохообразных с зелеными водорослями или непосредственно или же через сосудистые архего- ниальные растения. Фрич (Fritsch, 1945), а за ним и другие авторы предками архегониат считают хетофоральные или организмы, близкие к хетофоральным, с гетеротрихальным строением таллома. При этом как прототип архегониат рассматривается наземная водоросль Fritschiella.
Фрай и Кларк (Frye a. Clark, 1937—1947, стр. 10, 169) выводят из нитчатых разветвленных водорослей, с одной стороны, харо- вые водоросли, а с другой — первичные печеночники. Они также допускают общее происхождение печеночников и папоротникообразных, причем общие их предки, вероятно, были ближе к печеночникам, чем к папоротникообразным, так как при таком предположении не нужно принимать дегенерацию корней, сосудистой системы и листоносных побегов у спорофита.
Были и другие высказывания относительно происхождения мохообразных. Так, Шенк (Schenk, 1909) считал возможным выводить мхи и папоротникообразные из бурых водорослей. Аналогичные взгляды высказывали Курсанов (1909) и Мейер (Meyer, 1912). Позже Мейер (1946, стр. 114) высказывается более неопределенно, считая возможными предками мохообразных как зеленые, так и бурые водоросли. Из бурых водорослей, а именно из
233
Fucaceae (!) выводил архегониальные растения Гейнтце (Hein- tze, 1927).
Ван Тигем (Van Tieghem, 1884) считал красные водоросли — Florideae непосредственными предками мохообразных на том основании, что только у них среди талломных растений яйцеклетка развивается на материнском растении в новый организм, дающий споры — un embryon sporifere (т. е. карпоспорофит.— Д. 3.). Козо- Полянский (1927) также считал, что «наиболее близкими к древнейшей наземной флоре кажутся красные водоросли». Он, как и ван Тигем, полагал, что цистокарпий является как бы прототипом сиорогона мохообразных; прокарпий высших флоридей, по мнению Козо-Полянского, близок к архегонию, а гаплоидно-диплоидный тин чередования поколений, наблюдаемый у многих флоридей «как бы в исходном виде для высших растений,— род обобщающего явления для смены и типа мхов, и типа папоротникообразных зара́з» (Козо-Полянский, 1927, стр. 67). Ввиду больших отличий красных водорослей в биохимическом отношении Козо-Полянский не считал возможным выводить архегониаты непосредственно из флоридей, а выводил их из гипотетических Chloroflorideae. Но и в этом случае нельзя согласиться с предположением о происхождении архегония из прокарпия. К тому же многоклеточные антеридии архегониат никак нельзя увязать с одноклеточными сперматогониями флоридей. Нужно также иметь в виду полное отсутствие жгутиковых стадий у красных водорослей. Поэтому гипотеза о происхождении архегониат из красных водорослей может иметь только исторический интерес. Что касается бурых водорослей, как возможных предков архегониальных растений, то аргументом в пользу этого предположения является наличие у них типичных многоклеточных гаметангиев, которые можно рассматривать как возможный прототип половых органов архегониат. Однако, как отметил Фрич (Fritsch, 1916), биохимические данные — различный набор пигментов и характер продуктов ассимиляции, а также различное строение жгутиков бурых водорослей и архегониат являются кардинальным препятствием для признания бурых водорослей возможными предками мохообразных и папоротникообразных.
В настоящее время широко принятым считается происхождение архегониальных растений от зеленых водорослей. Основанием для этого являются как биохимические, так и морфологические данные — пигментный состав, крахмал как запасный продукт, характер строения жгутиков. Основным затруднением в вопросе признания зеленых водорослей предками архегониальных растений является обычно одноклеточное строение их гаметангиев *. 11 Фрич (Fritsch, 1916) для объяснения отсутствия среди зеленых водорослей форм с многоклеточными гаметангиями высказал даже предположение, что более прогрессивные зеленые водоросли погибли в тяжелых условиях приспособления к наземному существованию.
234
Однако у Chaetonema irregulare Мейер (Meyer, 1930) описал многоклеточные гаметангии, что укрепляет предположение Девиса (Davis, 1903), Веттштейна (Wettstein, 1903 )и ряда других авторов о филогенетических связях архегониальных с зелеными водорослями.
Что касается происхождения половых органов архегониат, то наиболее правдоподобна теория Девиса (Davis, 1903), согласно которой архегонии и антеридии мохообразных и папоротникообразных развились из многоклеточных гаметангиев, сходных с га- метангиями бурых водорослей. В связи с выходом на сушу внешний слой клеток таких гаметангиев стерилизовался и превратился в защитную стенку. В дальнейшем, в связи с усилением половой дифференциации, мужские гаметы превратились в типичные сперматозоиды. Что же касается женского гаметангия, то макрогаметы уменьшились в количестве, потеряли подвижность, а затем количество их сократилось до одного ряда, причем плодущей осталась только нижняя из них, а все остальные превратились в канальцевые клетки. Развитие уродливых половых органов, например архегониев, в которых канальцевые клетки превращались в материнские клетки сперматозоидов или в дополнительные яйцеклетки, или антеридиев, в которых развивались, кроме сперматозоидов, также и яйцеклетки (Holferty, 1904; Meyer, 1912, и др.), подтверждает обоснованность гипотезы Девиса.
* * *
Для выяснения происхождения мохообразных важное значение имеет вопрос о чередовании поколений в растительном мире. Основы изучения чередования поколений у растений были заложены классической работой Гофмейстера (Hofmeister, 1851), который исследовал процессы оплодотворения и развития зародыша в архегониях, смену поколений — гаметофита и спорофита, установил гомологии между спорофитами и гаметофитами в различных группах высших растений. Устанавливая эти гомологии, Гофмейстер имел в виду только выявление общего плана в строении и развитии растений Гофмейстер разместил высшие растепия по степени развития спорофита и редукции гаметофита в линейный ряд (мохообразные ->- папоротникообразные -> голосеменные ->
покрытосеменные) и считал, исходя из принципа редукции гаметофита и развития спорофита, что «только один шаг» отделяет мхи от харовых.
В отличие от Гофмейстера Прингсгейм (Pringsheim, 1855) видел зачаток спорофита мохообразных в зиготе Oedogonium и «плоде» — прорастающей зиготе Coleochaete.
После победы эволюционного учения установленные Гофмейстером гомологии стали трактовать как результат общего хода развития растительного мира, а морфологический ряд как филогенетический.
235
На основании имевшихся во второй половине XIX ст. данных Челаковский предложил так называемую антитетическую (интер- каляриационную) теорию смены поколений, согласно которой спорофит развился из зиготы водорослей как новообразование, вклинившееся между двумя нормальными поколениями (Celakov- sky, 1874). Дальнейшее развитие взгляды Челаковского нашли в работах Боуэра (Bower, 1890, 1908), который считал, что спорофит возник как приспособление к наземному существованию. При переходе на сушу, где благоприятные условия для оплодотворения подвижными антерозоидами могли быть только при наличии достаточной влаги, должны были выработаться новые средства размножения — споры, приспособленные к рассеиванию в воздушной среде. Увеличение количества спор дает больше шансов для выживания и распространения. Но увеличение количества спор требует большего количества питательных веществ. В результате у спорофита вырабатывается способность к ассимиляции и он развивается морфологически. По Боуэру, процесс развития спорофита проходил такие этапы: 1) частичная стерилизация спорогенных клеток, 2) разделение спорообразующей ткани на отдельные спорангии и 3) образование на спорофите дополнительных органов — листьев, корней и т. п., что привело в конце концов к полной его независимости. Частичная стерилизация спорогенных клеток, по Боуэру, осуществляется уже у мхов, а последующие этапы — у папоротникообразных. Веттштейн во втором томе своего учебника (Wettstein, 1903) указывает, что чередование поколений у мохообразных является следствием приспособления к жизни в двух различных в отношении влажности средах, а папоротникообразные рассматривает как промежуточную группу между мохообразными и цветковыми растениями. Наглядно это положение отражено на предложенной им схеме (Wettstein, 1903, стр. 14—15). В позднейшей монографии Боуэр придерживается тех же взглядов (Bower, 1935), несмотря на многочисленные новые данные о наличии чередования поколений в различных группах водорослей. Он только отмечает, следуя Ольтманнсу, что подобно тому как половой процесс развивался в различных группах растительного мира повторно и независимо, так и чередование поколений вырабатывалось независимо в различных группах низших организмов. Земноводные условия, но Боуэру, упрочили, «стандартизировали» правильное чередование поколений в цикле развития архегониат в противоположность, как он считал, относительной нестойкости его в жизненном цикле водорослей.
Взгляды Боуэра и Веттштейна встретили широкое признание и на некоторое время оставались господствующими. Однако открытие чередования поколений в различных группах водорослей опровергло взгляд Боуэра на его возникновение как приспособления к наземным условиям существования. Характер чередования поколений у бурых и зеленых водорослей, наличие разных его типов, в том числе широкое распространение изоморфного типа,
236
дало поддержку теории происхождения чередования поколений путем дифференциации организмов, несущих половые органы и органы бесполого размножения — так называемой гомологичной теории, восходящей к Прингсгейму (Pringsheim, 1876, 1878). В связи с этим ряд авторов (Мейер, 1922; Zimmermann, 1930; Smith, 1938, 1955; Chadefaud, 1950; Christensen, 1954, 1957; Schuster, 1966) высказались за происхождение архегониальных растений от водорослей с изоморфным чередованием поколений. По сути на этих же позициях стоит и Лазаренко, который считает, что происхождение спорофита у предков мохообразных шло «путем расчленения индивида, совмещающего способность к образованию гамет со способностью производить споры» (Лазаренко, 1961, стр. 380) h
Однако нужно помнить, что кроме изоморфного и происходящего от него гетероморфного чередования поколений у водорослей имеется еще иной тип чередования поколений, одним из членов которого является спорофит, возникающий на гаметофите, с ним связанный и с самого начала отличающийся морфологически от других поколений, а именно карпоспорофит красных водорослей. Подобно тому, как карпоспорофит (цистокарпий) у более низкоорганизованных флоридей является гаплоидным, а у более прогрессивных становится диплоидным, так же можно предположить задержку мейоза в многоклеточном образовании типа «плода» Caleochaete и образование диплоидного спорофита, давшего начало спорофиту мохообразных. Что касается происхождения чередования поколений папоротникообразных, то его, действительно, легче вывести, согласно гомологической теории Принсгейма, из изоморфного чередования поколений типа зеленых водорослей. Эта мысль впервые высказана Фричем (Fritsch, 1916), позже поддержана нами (Зеров, 1945).
Своеобразные взгляды на происхождение наземных растений высказал Черч (Church, 1919). По его мнению, исходными формами наземных растений были морские зеленые водоросли па- ренхиматического строения с изоморфным чередованием поколений. Ко времени выхода на сушу эти поколения уже несколько отличались между собой и спорофит был прикреплен к гаметофиту. Водоросли — предки высших растений должны были, по Черчу, обладать метаболизмом зеленых водорослей, строением бурых и типом развития более высокостоящих, чем тип Dictyota. Эти гипотетические организмы должны были иметь оли- ственные гаметофит и спорофит. С выходом на сушу связаны 11 На схеме возникновения чередования поколений Лазаренко'изобразил не предков мохообразных, а лиственные мхи и хотя оговаривается, что схематические рисунки мхов он предлагает рассматривать как символ, все же через две строки отмечает, что олиственные формы считает исходными для Bryophyta. Процесс развития спорофита, по Лазаренко, шел «путем редукции сексуальной потенции, потери филлоидов и изменения полителомного спорофита в монотеломный...» (стр. 381). Взгляды эти близки к взглядам Веттштейна, Черча и Эванса.
237
процессы редукции гаметофита, а у мохообразных и спорофита. Гипотеза Черча встретила ряд возражений (Fritsch, 1921).
Сходные со взглядами Черча соображения высказаны были недавно Джеффри (Jeffrey, 1962) с той только разницей, что исходными формами для архегониальных растений он считает пресноводные водоросли.
Филогенетические связи мохообразных и папоротникообразных
Если проблема происхождения архегониальных и вообще высших растений от зеленых водорослей в настоящее время может считаться в основном решенной положительно, то вопрос о непосредственных предках мохообразных и о взаимоотношении между этой группой растительного мира и папоротникообразными до сих пор остается спорным. Авторы конца XIX и начала XX ст., как мы уже упоминали, трактовали морфологические гомологии Гофмейстера филогенетически и, считая, что развитие чередования поколений связано с выходом на сушу, строили систему высших растений на основе редукции гаметофита и прогрессивного развития спорофита. В связи с этим Боуэр считал первичными формами наземных растений организмы с наименее развитым спорофитом типа риччии.
Ундервуд (Underwood, 1894) из Anthocerotales с их более или менее независимым спорофитом выводил папоротникообразные и другие высшие растения. Такого же взгляда придерживался и Веттштейн (Wettstein, 1903).
Основным и по нашему мнению непреодолимым препятствием для выведения спорофита папоротникообразных из спорогона мохообразных является его крайняя специализация.
Лотси (Lotsy, 1909) внес некоторые поправки в теорию Боуэра. Ввиду того, что гаметофит риччии характеризуется значительной сложностью строения и несомненно является результатом длительной эволюции, он предложил в качестве исходной формы архего- ниат гипотетическую сферориччию (Sphaeroriccia) с простым га- метофитом типа Sphaerocarpus и спорофитом типа Riccia. Кроме того, Лотси высказался за дивергентное происхождение мохообразных и папоротникообразных 1.
Позднейшие исследования чередования поколений в различных группах водорослей, как мы уже отмечали, опровергли тезис 11 Нужно отметить, что гораздо раньше Сакс писал о происхождении мохообразных так: «Если по различным основаниям сомнительно, что моховые имеют общее происхождение с определенными типами водорослей, то несомненно, что в дальнейшем своем развитии они все больше удалялись не только от водорослей и харовых, но также от сосудистых тайнобрачных; моховые не являются низшей ступенью последних, но представляют самостоятельную и своеобразную ветвь в родословном дереве тайнобрачных» (Sachs, 1868, стр. 273).
238
Боуэра о связи возникновения чередования поколений с переходом к наземным условиям и открыли иные возможные пути развития архегониальных растений. Тем не менее некоторые авторы все же выводили папоротникообразные из мохообразных. Так, Кемпбелл (Cambell, 1940), развивая гипотезу Ундервуда о происхождении папоротникообразных от антоцеротальных, выводил из спорогона Anthoceros спорогон псилофитов в результате перемещения (shifting) меристемы из основания спорогона на его верхушку, дихотомического разветвления точки роста и сосредоточения спорообразования только на верхушках разветвлений спорофита. Взгляды Ундервуда — Кемпбелла были поддержаны Смитом (Smith, 1938, 1955).
На первый взгляд гипотеза Ундервуда — Кемпбелла может показаться убедительной, однако при ближайшем рассмотрении она оказывается маловероятной. Трудно представить себе причины и пути перемещения меристемы па верхушку спорогона. Трудно также допустить, что строение спорогона антоцероса, как исходной формы папоротникообразных, сохранилось без существенных изменений со времени раннего палеозоя.
Изучение чередования поколений у разных групп водорослей укрепило взгляд, высказанный Лотси, о дивергентном происхождении мохообразных и папоротникообразных. Фрич (Fritsh, 1916) считал предками высших растений зеленые водоросли с гетеро- трихальным строением таллома, т. е. с талломом, расчлененным на простертую (стелющуюся) и прямостоящую части. Такими водорослями Фрич считает организмы, сходные с современными хетофоральными (Chaetophorales). По его мнению, мохообразные и папоротникообразные возникли дивергентно. Папоротникообразные, по Фричу, возникли в результате дифференциации двух поколений из форм с изоморфным чередованием поколений (по гомологическому типу). При этом в дифференцировавшихся поколениях гаметофит развился из простертой части гетеротрихаль- ного таллома, а спорофит — из вертикальной. Предками мохообразных, по Фричу, были водоросли типа Coleochaete с простертым талломом и спорофитом, возникшим путем дифференциации зиготы (по антитетическому типу). Фрич полагал, что гаметофит печеночников развился из простертой части гетеротрихалыюго таллома водорослей, а мхов — их прямостоячей.
В противоположность Фричу, Мейер (1922) и Циммерманн (Zimmermann, 1930), исходя из чередования поколений, свойственного бурым и зеленым водорослям, у которых наблюдается смена поколений изоморфного и гетероморфного типа, предложили гипотезу о происхождении архегониат от водорослей с изоморфным чередованием поколений, обладающих многоклеточными гаметан- гиями. В процессе дифференциации обоих поколений развился гетероморфный тип чередования поколений с господством гамето- фита у мохообразных и спорофита у папоротникообразных. Возникает вопрос, почему в одном случае (папоротникообразные)
239
редуцировался гаметофит, а в другом (мохообразные) в условиях того же наземного существования — спорофит, казалось бы более приспособленный к наземным условиям. Однако ни Мейер, ни Циммерманн на этот вопрос ответа не дают.
Были и иные попытки установления предков мохообразных. Как отмечает Хаскелл (Haskell, 1949), в свое время Скотт (Scott, 1911) высказал мысль, что, возможно, мохообразные произошли как редуцированная группа от неизвестных высших групп, связанных с сосудистыми криптогамами. Скотт обосновывал это предположение наличием устьиц на спорогонах мхов.
Кашьяп (Kashyap, 1919) на основании сходства заростка хвощей с гаметофитом Anthoceros высказал мысль о возможности происхождения Anthocerotales от папоротникообразных. Происхождение мохообразных от папоротникообразных отстаивает Штейнбек (Steinböck, 1954). Исходя из имеющихся в то время данных о возрасте мохообразных, а именно из того, что самые старые печеночники известны были из верхнего карбона, а мхи из еще более поздних отложений, Штейнбек приходит к выводу, что мохообразные являются относительно молодыми, во всяком случае значительно моложе папоротников. На основании того, что мхи более родственны папоротникам, чем водоросли, а протонема мхов и заростки папоротников сходны, Штейнбек приходит к выводу, что мхи происходят от папоротникообразных. При этом гаметофит мха — протонема и гаметофор — гомологичен заростку. Спорогон мхов, по Штейнбеку,— редуцированный спорофит папоротникообразных. Для подтверждения этого он приводит такие доводы: а) наличие гаусториев у спорогонов некоторых мохообразных (Anthocerotales, Diphyscium, Eriopus), которые обеспечивают почти независимое их развитие; б) наличие хлорофилла в спорогонах; в) наличие у мхов устьиц, часто редуцированных; г) сходство перистома мхов с кольцом (annulus) многих лепто- снорангиальпых папоротников. Штейнбек считает, что Hymeno- phyllaceae с их слабо развитым спорофитом стоят близко к стадии «мохообразования» (die Mooswerdung). Что же касается споро- гона юнгерманниевых и маршанциевых то, по мнению Штейн- бека, он является новообразованием. С целью доказательства родства мохообразных и папоротникообразных Штейнбек приводит данные о непостоянстве числа жгутиков у мохообразных. В заключение он высказывает мысль, что мохообразные это придаток к папоротникам («ein Anhängsel der Farne»).
Если принять во внимание новейшие данные об ископаемых мохообразных (Нейбург, 1956, 1960; Hueber, 1961), то отпадает основной аргумент Штейнбека о сравнительной молодости мохообразных. Вряд ли можно считать приемлемым и утверждение его о различном происхождении спорогонов мхов и печеночников.
Изучение псилофитов и близких к ним организмов дало новый материал для решения вопроса о происхождении папоротникообразных. Правда, гаметофит у псилофитов до сих пор неизвестен,
240
однако исследование спорофита представителей этой группы помогло выяснить пути развития основного поколения папоротникообразных — спорофита. Изучение псилофитов побудило ряд авторов выводить из них также и мохообразные. Так, Козо-Полян- ский (1927) высказал предположение о связи мохообразных с псилофитами и считал, что мхи представляют собой «ветвь от оснований ствола плауновых». Более определенно связывают мохообразные с псилофитами Гейнтце (Heintze, 1927), Дженнингс (Jennings, 1928) и Скотт (Scott, 1929). Этот взгляд был поддержан и развит рядом последующих авторов (Козо-Полянский, 1947, 1949; Haskell, 1949; Тахтаджян, 1950, 1953, 1956; Schuster, 1966, II др.)·
По Гейнтце (Heintze, 1927, цит. по Jennings, 1928), фукусовые дали начало гипотетическому сапрофитному «Protocormophyton» силурийских морей, от которого произошли опять-таки гипотетические «Arcliipsilophytaceae» и ископаемые Rhyniaceae и Astero- xylaceae. Archipsilophytaceae, по Гейнтце, дали начало мхам и через антоцерос — печеночникам. Из печеночников через риччиевые Гейнтце выводит харовые (!).
Дженнингс (Jennings, 1928) считал, что псилофиты в результате дифференциации дали начало таким группам растительного мира как Sphenopsida, Pteropsida, Lycopsida и Bryopsida К Он полагал, что следствием утраты спорогонами мохообразных фото- синтетической функции является редукция устьиц.
Козо-Полянский (1947, 1949) считал исходной группой для высших растений, в том числе и мохообразных, псилофиты, которые, в свою очередь, произошли от водорослей — Phycophyta (Pro-Rhodophyceae?). От псилофитов он выводит антоцеротовые, объединяемые вместе с псилофитами в отдел Procormophyta, и мохообразные — Bryophyta, включаюнще сфагновые мхи — Sphagnobrya, к которым он отнес также и андреевые, настоящие мхи— Eubrya и печеночники (без антоцеротовых).
Хаскелл (Haskell, 1949) пытается дать следующее обоснование для гипотезы происхождения мохообразных от псилофитов. Он указывает на сходство ископаемых спорангиев Sporogonites и IIornea со спорогонами мхов и повторяет высказывание Черча (Church, 1919), что Нornea почти «слегка разветвленный и свободно растущий антоцерос» (Hornea is little more than a slightly ramified and free-growing Anthoceros). Хаскелл отмечает более или менее выраженную независимость спорофита Anthoceros как связующего звена между мохообразными и псилофитами и отмечает, что спорогенная ткань нижней части коробочки у Anthoceros может стерилизоваться, а колонка играть роль проводящего пучка. Хаскелл также ссылается на сходство проводящих элемен-
1 Шустер (Schuster, 1966, стр. 275) утверждает, что Дженнингсу принадлежит приоритет в применении термина Bryopsida. Позже этот термин предложили Тахтаджян (1943) и Лам (Lam, 1948).
241
тов у псилофитов и у гаметофитов мхов. Особое значение он придает наличию устьиц, часто зачаточных, на спорогонах мхов. Хаскелл считает, что у предков мохообразных спорофит был более развит и что они родственны псилотовым, а через них, возможно, и псилофитовым. По мнению Хаскелла, мхи являются редуцированной группой, причем их редукция связана со вторичным приспособлением к водной среде, куда они были вытеснены папоротникообразными.
Тахтаджян (1950, 1953) считает мохообразные прямыми потомками псилофитов. Свои выводы он основывает, вслед за Хаскеллом, на сходстве спорангиев Нornea и Sporogonites со споро- гонами сфагновых и андреевых мхов (наличие колонки, не достигающей верхушки спорангия и покрытой сверху спорогенной тканью), на наличии у антоцеротовых и многих мхов устьиц типичного для высших растений строения, а также на наблюдаемом у мхов тератологическом явлении — дихотомическом разветвлении коробочек спорогона. Отдел высших растений (Cormophyta, или Telomophyta) Тахтаджян (1956) делил на следующие шесть подотделов («типов»): Psilopsida, Bryopsida, Lycopsida, Tmeso- psida, Sphenopsida и Pteropsida. При этом Тахтаджян подчеркивает, что мохообразные «являются, вероятно, прямыми потомками пси- лофитовых» (1956, стр. 43).
Более решительно поступил при построении системы высших растений Шварц (Schwarz, 1955), который листостебельные растения (Kormobionta) разделил на четыре ствола (Stämme) — Lepidophyta, Stachyophyta, Psilophyta и Phyllophyta. Ствол Psi- lophyta он делит па два «подствола» (Unterstämme) — Psilophy- tina и Bryophytina. При этом Bryophytina он выводит из Psilo- phytina через Horneophyton и Anthocerotales и делит их на пять отделов — Anthoceropsida, Hepaticopsida, Andreaeopsida, Bryopsida и Spagnopsida.
Шадефо (Chadefaud, 1950), основываясь на предположении, что предки архегониат характеризовались изоморфным чередованием поколений, считает, что наиболее архаичными среди известных архегониат являются те, у которых спорофит и гаметофит наиболее сходны между собой, например, псилотовые (Psilotum, Tmesipteris). Шадефо допускает, что гаметофит у предков псило- товых, подобно спорофиту, состоял из корневищной части и оли- ственного побега. В дальнейшем, по Шадефо, олиственная гипотетическая часть гаметофита редуцировалась, возможно, в связи с микотрофным питанием гаметофита. На основании этих допущений и смелых аналогий Шадефо считает возможным сделать вывод, что мохообразные и папоротниковые развились из форм, у которых гаметофит и спорофит были сходны и облиственны, т. е. повторяет высказывания Черча (Church, 1919).
Христенсен (Christensen, 1954, 1957) более осторожно высказывается в пользу того, что общий предок мохообразных, папоротникообразных и семенных растений должен был быть примитивным
242
папоротникообразным (must be classified as a primitive pteri- dophyte).
Mexpa и Ханду (Mehra a. Handoo, 1953) считают, что Antho- cerotales и Rhyniaceae произошли в додевонское время от общего корня — Antho-rhyniaceae, которые, по их мнению, произошли от еще более древних предпеченочников (pro-liverworts), которые дали начало также маршанциальным, юнгерманниальным и сфе- рокарпальным. Предпеченочники произошли от вышедших на сушу зеленых водорослей.
Шустер (Schuster, 1966) склонен выводить мохообразные и высшие псилофиты (Psilophy tales) из первичных сосудистых растений — Prototracheophyte-Antho-rhyniaceae (по Мехра и Ханду), которые, в свою очередь, произошли от зеленых водорослей с изоморфным чередованием поколений и изогамным половым процессом. Шустер считает характерной чертой эволюции мохообразных редукцию спорофита, которую он объясняет возраставшим паразитическим образом жизни его на гаметофите. В связи с редукцией спорофита в размерах и сложностью строения, по Шустеру, стоит и редукция вентиляционной системы и, в частности, устьиц. Сам же гаметофит не мог достигнуть больших размеров и сложности, так как «его эволюционный потенциал на суше ограничен» в связи с потребностью в воде как среде для оплодотворения.
Оставляя в стороне такие трудно приемлемые предположения, как происхождение архегониальных от фукусовых (Гейнтце) или от гипотетических Pro-Rhodophyceae (Козо-Полянский) или же происхождение харовых от риччиевых, основные положения сторонников происхождения мохообразных из псилофитов путем редукции сводятся к: а) сходству строения спорангиев Нornea и Sporogonites и коробочек спорогона мохообразных, б) наличию на спорогонах мхов устьиц, часто редуцированных, в) таким тератологическим явлениям, как ветвление спорогонов мохообразных, г) наличию проводящих элементов в спорогонах антоцерото- вых и д) относительной его независимости у антоцерота и некоторых мхов.
Спорангии Нornea и Sporogonites действительно напоминают спорогоны родов Sphagnum и Andreaea, а также молодые споро- гоны видов рода Anthoceros наличием в них не доходящей до верху колонки, покрытой сверху спорогенной тканью. Однако бесплодная колонка имеется далеко не у всех мохообразных: у представителей класса Hepaticopsida она отсутствует, как отсутствует и у большинства известных родов псилофитов. Кроме того, нужно отметить, что археспорий у мохообразных образуется то из внутреннего слоя амфитеция (Sphagnales, Anthoceros), то из эндотеция (Bryales, Andreaeales, Jungermanniales, Notothylas из антоцеро- тальных). Все это свидетельствует о правильности предположения Ланга (Lang, 1907), что эндотеций является потенциально спорогенной тканью. Частичная стерилизация спорогенной ткани у не¬
243
которых мохообразных и псилофитов могла независимо привести к образованию бесплодной колонки (Zimmermann, 1932, стр. 459; Зеров, 1951). Поэтому наличие ее в спорогонах мхов и антоцеро- товых и в спорангиях некоторых псилофитов, нам кажется, нельзя считать убедительным доказательством происхождения мохообразных из псилофитов в результате редукции спорофита.
Устьица, наличие которых на спорогонах антоцеротовых и многих мхов ряд авторов, начиная со Скотта (Scott, 1911), считают доказательством происхождения их от организмов с более развитым спорофитом — псилофитов или папоротникообразных, также могли развиться независимо в различных группах растительного мира. Если же принять мнение ТахтадЖяна, что мхи (Bryopsida) произошли от вымерших юнгермапниальных, что нам кажется весьма правдоподобным, то придется признать, что и колонка в спорогонах и устьица у них развились вторично, так как у всех известных юнгерманниальных спорогоны не имеют ни колонки, ни устьиц.
Что касается таких тератологических явлений как раздвоение коробочек у некоторых мхов и печеночников, принимаемых как доказательство происхождения спорогонов мхов от дихотомически разветвленных плодущих теломов псилофитов, то нужно иметь в виду, что разветвленные коробочки образуются в результате повреждения зачатка спорогона. Разветвленные спорогоны, так же как и другие тератологические явления в их строении, свидетельствуют только о ненормальностях их развития, но никак не являются атавистическими. Что касается тератологических явлений, наблюдаемых в спорогонах мохообразных, то бо́льшего внимания заслуживают указания на то, что у видов рода Porella из юнгерманниальных наблюдалось превращение тканей ножки и стопы спорогона в спорогенную ткань, что можно трактовать как подтверждение теории Боуэра о том, что бесплодная часть спорогона развилась из плодущей, спорогенной ткани (Smith, 1938, стр. 5).
Известную независимость спорогона Anthoceros, обладающего неограниченным ростом и способностью к автотрофному питанию и могущего, по свидетельству Кемпбелла (Campbell, 1924), в известных случаях самостоятельно существовать, что он склонен трактовать как залог дальнейшего прогрессивного его развития в спорофит псилофитов и папоротникообразных, сторонники регрессивного развития мохообразных рассматривают как свидетельство обратного явления — редукции спорофита, которая завершается у мхов и печеночников. В различных группах мохообразных, кроме антоцеротовых, наблюдаются случаи более или менее независимого существования спорогона (например, Buxbaumia), однако они возникают независимо в результате приспособления к экологическим условиям и не могут свидетельствовать ни о происхождении мохообразных от псилофитов или папоротникообразных, ни о развитии этих групп из мохообразных.
244
Если принять происхождение мохообразных от псилофитов или папоротникообразных в результате редукции спорофита, то прежде всего возникает вопрос, почему в условиях наземного существования, в условиях, благоприятствующих развитию спорофита и рассеиванию покрытых оболочкой спор, в одном случае (папоротникообразные — цветковые растения) спорофит пышно разветвляется при одновременной редукции гаметофита, требующего для оплодотворения водной среды, а в другом случае (мохообразные) он регрессирует.
Хаскелл пытается объяснить упрощение спорофита у мохообразных вторичным переселением их в водную среду. Однако нужно иметь в виду, что большинство мохообразных обитает не в водной среде, а часть их — типичные ксерофиты.
Шустер считает, что упрощение строения спорофита мохообразных связано с переходом его к паразитическому существованию, возникшему в результате того, что яйцеклетка после оплодотворения у них остается и развивается в брюшке архегония. Однако и у папоротникообразных зигота вначале также развивается на гаметофите, а у семенных растений спорофит развивается паразитически до стадии зародыша — у голосеменных за счет гаметофита (первичного эндосперма), а у покрытосеменных — за счет вторичного эндосперма, и это отнюдь не способствует его деградации. Да и можно ли считать редуцированным органом спорогон мохообразных с его сложным строением, приспособленным к существованию в разнообразных наземных условиях?!
Считая мало вероятной гипотезу о происхождении мохообразных регрессивным путем от папоротникообразных или псилофитов, а еще менее обоснованным и устарелым предположение о развитии папоротникообразных из мохообразных, мы склоняемся к теории о происхождении тех и других от общих водорослеобразных предков с многоклеточными гаметангиями и разными типами чередования поколений. Образование архегониев и анте- ридиев с защитной стенкой из стерильных клеток связано, вероятно, как предполагал Девис, с переходом к наземному существованию. Водоросли с гомологической сменой поколений, у которых гаметофит и спорофит вначале были независимыми растениями, дали начало папоротникообразным. Оплодотворение яйцеклетки внутри женского гаметангия и развитие в нем зародыша могли происходить еще в водной среде, как это намечается у некоторых ламинариевых и обычно у красных водорослей. Эта связь спорофита с гаметофитом у папоротникообразных закрепилась, вероятно, уже в наземных условиях в связи с завершением формирования архегония, в котором оплодотворение яйцеклетки и прорастание зиготы происходит на месте.
Что касается происхождения мохообразных, то нам кажется более вероятным предположение о их происхождении от водорослей с антитетическим чередованием поколений, при котором
245
спорофит вклинился между двумя половыми поколениями в результате развития зиготы на материнском растении, как это мы видим у Coleochaete, с задержкой мейоза до спорообразования (Fritsch, 1916; Зеров, 1945). Происхождение мохообразных от водорослей с гомологической сменой поколений с преобладанием гаметофита над спорофитом, например у кутлерии (Cutleria), нам кажется менее вероятным, так как аналогичное явление наблюдается очень редко. Предположение о выходе из воды развитого облиственного спорофита, сидящего на вертикальном гаме- тофите, как это предполагают Черч (Church, 1919) и Эванс (Evans, 1939), недопустимо с экологической точки зрения (Fritsch, 1921).
В то время как псилофиты и папоротникообразные вышли на сушу с автономным спорофитом, обладавшим развитой вегетативной системой, которая в условиях наземного существования могла развиться в мощное растение, судьба мохообразных была иной. Спорофит первичных мохообразных не был настолько сложным органом, как спорогоны Buxbaumia или Anthoceros, которые, хотя и обладают некоторой автономностью, все же остаются только более или менее долгоживущими спорогонами. Однако при выходе на сушу он был уже настолько специализированным спороносным органом, что не мог развиться в самостоятельное растение, как предполагал Боуэр, хотя и обладал способностью к фото· трофному питанию.
Развитие гаметофита, который у мохообразных является основным вегетативным телом, лимитировалось с одной стороны необходимостью сохранения хотя бы временной связи с водной средой, необходимой для оплодотворения, а с другой — выполнением функций фотосинтеза. В силу взаимодействия этих двух противоречивых обстоятельств гаметофит мохообразных не мог развиться в мощное растение, но и редукция его ограничивалась необходимостью обеспечения развития спорогона. Таким образом, гаметофит мохообразных сохранил при своих сравнительно незначительных размерах способность к самостоятельному существованию и способность обеспечивать процесс оплодотворения и развитие спорогона, служащего для образования и рассеяния спор.
Нельзя думать, что эволюция мохообразных проходила исключительно под знаком прогрессивного развития. Во время длительного процесса их становления и развития в силу тех или иных экологических условий имели место и явления редукции, регресса — достаточно вспомнить, например, редукцию перистома у некоторых мхов, сокращение гаметофора почти до его выпадения, например у Ephemeropsis tjibodensis, Protocephalozia ephemeroi- des, видов Buxbaumia, редукцию листьев у некоторых печеночников, например у некоторых видов Zoopsis. Однако общее развитие мохообразных шло прогрессивно в направлении наибольшего приспособления к разнообразным условиям наземного существования.
246
2. ПУТИ ЭВОЛЮЦИИ МОХООБРАЗНЫХ И ИХ КЛАССИФИКАЦИЯ
Основные подразделения мохообразных
Еще в конце XVIII ст. установлены были группы мхов (Musci) и печеночников (Hepaticae). Группу мхов (Musci) встречаем уже у Линнея (Linné, 1753, 1770, 1780) *; печеночники же, за свидетельством Ундервуда (Underwood, 1894), выделены Адансо- ном. Жюсье (Jussieu, 1789) эти группы называет семействами. В системах растительного мира XIX ст. обычно находим отдел Bryophyta с двумя классами — Hepaticae и Musci (Endlicher, 1836—18401 2; Sachs, 1873; Eichler, 1883; Engler, 1898; Wettstein, 1903, и др.).
Однако наряду с делением мохообразных на два класса — печеночники (Hepaticae) и мхи (Musci) с последней четверти XIX ст. было выдвинуто предложение выделить из состава печеночников третий класс — антоцеротовые. Шустер (Schuster, 1966, стр. 369, сноска 2-я), ссылаясь на Хи (Ну, 1884), приводит цитату из работы Янчевского (Janczewski, 1872), который предложил выделить семейство антоцеротовых в третий класс мохообразных, равноценный печеночникам и мхам. Позже Хоу (Howe, 1899) также выделил антоцеротовые в особый класс. Однако это предложение широкого отклика долгое время не находило. В лучшем случае антоцеротовые выделяли в особый подкласс в пределах класса Hepaticae, противопоставляя их подклассу Hepatî- cales (Verdoorn, 1932). Только в последнее время проявляется снова тенденция выделять их в особый класс — Anthocerotae или Anthocerotopsida (Smith, 1938, 1955; Тахтаджян, 1950, 1953; Schuster, 1953, 1958, 1966; Parihar, 1959, 1965; Зеров, 1964, и др.), равноценный классам Hepaticopsida (Hepaticae) и Bryopsida (Musci).
Мехра и Ханду (Mehra, a. Handoo, 1953; Mehra, 1964) считают, что имеется ряд оснований рассматривать Anthocerotales как самостоятельную группу того же ранга, что и печеночники. Многочисленные хлоропласты, поверхностное положение половых органов, способ развития спорофита, эндотециальное происхождение спорогенной ткани у печеночников они противопоставляют единичным хлоропластам, погруженным половым органам, способу развития спорофита и амфитециальному происхождению споро¬
1 У Линнея двадцать четвертый класс — Cryptogamia имеет такие подразделения: Filices, Musci, Algae и Fungi. Ко мхам отнесены: кроме настоящих мхов (роды Sphagnum, Phascum, Splachnum, Polytrichum, Mnium, Bry- um, Hypnum, Fontinalis, Byxbaumia), также роды Lycopodium и Porella. Роды же печеночников Jungermannia, Targionia, Marchantia, Blasia, Riccia, Anthoceros отнесены к водорослям (Algae).
2 У Эндлихера вместо Bryophyta находим не нашедшее признания название Acrobrya anophyta.
247
генной ткани у антоцеротальных. Отличает их также и основное число хромосом. Мехра и Ханду, как уже упоминалось, считают вероятным, что Anthocerotales и Rhyniaceae произошли в пред- девонское время от общего корня — Antho-rhyniaceae. Эта группа в свою очередь произошла от предпеченочников (pro-liverworts), которые дали начало также маршапциальным, юнгерманниальным и сферокарпальным.
Ротмалер (Rothmaler, 1951, 1955) считает необходимым выделять антоцеротальные в особый отдел царства Cormobionta (растений) под названием Anthocerophyta, равноценный отделам Bryophyta, Psilophyta, Pteridophyta и Spermatophyta. Лазаренко (1961, стр. 381), очевидно, независимо от Ротмалера, высказался за выделение антоцеротовых в особый отдел («тип»).
Нужно отметить, что имелись тенденции не делить мохообразные на классы. Так, Кейверс (Cavers, 1910—1911) отдел Bryophyta делил на десять следующих порядков: Sphaerocarpales,
Marchantiales, Jimgermaimiales, Anthocerotales, Sphagnales, And- reaeales, Tetraphidales, Polytrichales, Buxbaumiales и Eu-Bryales.
Своеобразное строение спорофита антоцеротовых и особенности строения их клетки, а именно наличие хлоропластов с пиреноидами и отличный от других мохообразных набор хромосом, дают основание для выделения их в особый класс Anthoceroto- psida. Если же принять во внимание высказывания некоторых авторов об иных путях происхождения этой группы (Mehra а. Handoo, 1953), будет обоснованным выделение их в особый подотдел зеленых растений (Chlorophyta) под названием Anthocero- tophytina.
Ввиду того, что все три группы мохообразных обособились очень рано, очевидно еще в начале палеозоя, и развитие их проходило независимо, процесс эволюции печеночников и мхов рассмотрим раздельно.
Пути эволюции печеночников и вопросы их классификации
Во второй половине XIX ст. был распространен взгляд, что печеночники с их в основном более простым строением гамето- фита и спорофита являются более примитивной группой. На основании взглядов Гофмейстера об усложнении спорофита при переходе от низших архегониат к высшим, в начале системы мохообразных ставили печеночники, в частности маршанциальные с их более простым спорофитом.
Веттштейн (Wettstein, 1903), основываясь на том, что эволюция растительного мира в наземных условиях шла по линии редукции гаметофита, считал, что более простой гаметофит печеночников вторичный, упрощенный, и что основной исходной группой мохообразных являются лиственные мхи. На схеме Веттштейн (Wettstein, 1903, стр. 26) изображает два независимых ствола —
248
печеночников и мхов, происходящих от «гипотетических исходных форм мохообразных». Ряд других авторов начала XIX ст. высказывается также за параллельное развитие печеночников и мхов (Velenovsky, 1905; Lotsy, 1909).
Авторы первой половины XIX ст. в работах, вышедших в свет до появления труда Гофмейстера (Hofmeister, 1851) уже различали основные группы печеночников и при построении системы размещали их по степени усложнения строения либо в восходящем (Endlicher, 1836—1840), либо в нисходящем порядке (Gotische, Lindenberg et Nees von Esenbeck, 1844—1847). В капитальной работе Готтше, Линденберга и Нееса печеночники разделяются на таких пять «триб»: Jungermannieae с подтрибами — Foliosae и Frondosae, Monocleae, Marchantiae, Anthocerotae и Riccieae.
Не останавливаясь на ряде систем, не имевших широкого распространения и не повлиявших на развитие взглядов на эволюцию печеночников (Dumortier, 1874; Trevisan, 1877; Spruce, 1884—1855, и др.), отметим работы Линдберга (Lindberg, 1869, 1875), который разделил печеночники на такие три основные группы: Jungermanniaceae, Marchantiaceae и Anthocerotaceae. Эти группы в том или ином значении до сего времени фигурируют в системах печеночников.
После выхода в свет работы Дарвина «Происхождение видов путем естественного отбора» (1859), когда гомологии, установленные Гофмейстером, стали трактовать как отражение хода эволюции, и мохообразные начали рассматривать как промежуточную ступень между водорослями и папоротникообразными, системы печеночников стали строить главным образом по степени усложнения спорофита, который считался наиболее приспособленным к условиям наземного существования. При этом в начале системы ставили талломные маршаициальные, затем юнгермапниальные — вначале талломные анакрогинные, затем листостебельные акро- гинные, а в конце системы — антоцеротальные. Теоретическое обоснование усложнения спорофита было детально разработано Боуэром (Bower, 1890, 1908), который рассматривал спорофит как приспособление к наземным условиям существования.
Веттштейн (Wettstein, 1903), который стоял на тех же позициях антитетической смены поколений, что и Боуэр, и который тоже считал, что возникновение и развитие спорофита у высших растений связано с переходом к наземным условиям, свои выводы о происхождении мохообразных и сосудистых растений построил на редукции гаметофита в условиях наземного существования. Ввиду того, что у печеночников гаметофит имеет более простое строение, чем у мхов, Веттштейн считал печеночники производной группой (starker abgeleitet). По его мнению, выведение печеночников из типа мхов не представляет трудностей; кажущееся простое строение их гаметофита является вторичным, среди печеночников имеются формы, которые обнаруживают четкие связи с папоротникообразными, тогда как таких форм среди мхов нет.
249
По мнению Веттштейна, лиственные формы печеночников, в частности акрогинные, еще близки во многих отношениях к лиственным мхам. У акрогинных печеночников наблюдается переход к дорзивентральному строению, с которым Веттштейн связывает редукцию листьев на гаметофите, в результате чего возникает таллом, наблюдаемый у антоцеротовых, который очень походит на талломный гаметофит папоротникообразных.
В дальнейшем, как мы уже отмечали, оформились два основных направления в вопросе происхождения мохообразных. Одни авторы выводили мохообразные непосредственно от водорослевых предков, другие же считали их боковой ветвью основного ствола псилофитов — папоротникообразных.
В зависимости от взглядов авторов на происхождение мохообразных формировались и их взгляды на направления эволюции в пределах отдельных классов этой группы. В частности, по поводу эволюции в пределах класса печеночников, включая и антоце- ротовые, наметилось два крайних мнения. Согласно одному, восходящему к Лейтгебу (Leitgeb, 1881) и позже поддержанному и развитому рядом авторов (Underwood, 1894; Schiiïner, 1893— 1895, 1917; Bower, 1908, 1935; Cavers, 1910-1911; Campbell, 1905, 1925, 1940; Мейер, 1922, 1947; Zimmerman, 1930, 1959; Frye a. Clark, 1937-1947; Smith, 1938, 1955; Müller, 1906-1916, 1951 — 1958; Reimers, 1954; Кудряшов, 1962; Зеров, 1966, и др.), эволюция печеночников шла в основном от простых талломных форм, с одной стороны, к сложным талломным, а с другой — к листостебельным. Согласно другому мнению, впервые высказанному Веттштейном (Wettstein, 1903), процесс эволюции печеночников шел регрессивным путем — от более сложных форм к простым. Этого мнения придерживался ряд исследователей (Church, 1919; Kashyap, 1919; Verdoorn, 1932; Evans, 1939; Haskell, 1949; Тахта- джян, 1950, 1953, 1956; Chadefaud, 1960; Soô, 1961; Schuster, 1953, 1958, 1966, и др.)· Эти теоретические взгляды разных авторов отразились и на размещении материала в предлагаемых ими системах.
Как мы уже отмечали, в основе систем печеночников, построенных по принципу прогрессивной эволюции, лежит система Лейтгеба (Leitgeb, 1881), который разделил класс печеночников (Hepaticae) на три порядка — Marchantiales, Jungermanniales (с двумя подпорядками — Jungermanniales anakrogynae и Jungermanniales akrogynae) и Anthocerotales.
В пределах этих порядков материал размещен по восходящей линии — от более простых форм к более сложным. Система Лейтгеба легла в основу сводных работ Шиффнера (Schiffner, 1893— 1895) и К. Мюллера (К. Müller, 1906—1916).
Сторонником восходящего развития печеночников был также Кейверс (Caves, 1910—1911). По его мнению, мохообразные представляют восходящий ряд с прогрессивным развитием спорофита. Следуя Тенсли, он выводит из «архегониальных водорослей»
250
независимыми ветвями и первичные мохообразные (Protobryophy- ta) и первичные папоротникообразные (Protopteridophyta). От первичных мохообразных через сферориччию он выводит две ветви — маршанциальную и сферокарпальную. От сферокарпальной ветви Кейверс выводит два ответвления — юнгерманниальные и анто- церотальные, а от последних — мхи. Кейверс делит печеночники на четыре порядка — Sphaerocarpales, Marchantiales, Jungerman- niales и Anthocerotales, причем семейства в них размещены по степени усложнения строения. Обсуждая взгляды Гебеля по во- просу развития маршанциальных, Кейверс считает возможным регрессивное развитие некоторых рядов этого порядка в связи с приспособлением к влажным и затененным условиям, однако он возражает против предположения, что весь ряд маршанциальных является редукционным тем более, что большинство представителей этого порядка в большей или меньшей степени ксеро- фильны.
Особого внимания заслуживает классификация Мюллера, поданная в последнем издании его монографии по печеночникам Европы (К. Müller, 1951—1958). Мюллер на основании сравнительно морфологических, систематических и географических данных считает талломные формы печеночников более древними, чем лиственные. Антоцеротальные Мюллер считает очень примитивными. Относительно сферокарпальных он нерешительно говорит, что этот порядок является очень примитивным или же сильно редуцированным. Однако в системе он ставит его в начале порядка Marchantiales. Семейство риччиевых Мюллер считает редуцированным и допускает редукционное развитие некоторых семейств маршанциальных. Мюллер считает, что современные юнгерманниальные (акрогинные) представляют собой конечные члены различных эволюционных рядов. В целом же Мюллер признает прогрессивное развитие печеночников, что видно из его системы приведенной ниже.
Система печёночников К. Мюллера (1951 г.)
Порядок I. Anthocerotales
Семейства: Anthocerotaceae, Notothylaceae.
Порядок II. Marchantiales Подпорядок A. Sphaerocarpineae
Семейства: Sphaerocarpaceae, Riellaceae.
Подпорядок В. Marchantiineae a) Caudiciformes
Семейства: Targioniaceae, Grimaldiaceae, Conocephalaceae, Lunula- riaceae, Cyathodiaceae. в) Quadranticae
Семейства: Cleveaceae, Marchantiaceae, Monocleaceae, Exormotheca- ceae, Corsiniaceae.
Поднорядок: C. Ricciineae
Семейства: Oxymitraceae, Ricciaceae.
Порядок III. Jungermanniales
Подпорядок A. Anakrogynineae (Metzgerineae)
Семейства: Metzgeriaceae, Aneuraceae, Pelliaceae, Dilaenaceae, Blasi- aceae, Treubiaceae, Codoniaceae.
251
Подпорядок В. Calobryineae (Haplomitrineae)
Семейство Haplomitriaceae.
Подпорядок С. Akrogynineae (Jungermanineae) a) Jungermanieae
Семейства: Herbertaceae, Ptilidiaceae, Trichocoleaceae, Blepharosto- maceae, Hygrobiellaceae, Lophocoleaceae, Lophoziaceae, Marsupella- ceae, Jungermaniaceae, Southbyaceae, Plagiochilaceae, Schistochila- ceae, Scapaniaceae, Cephaloziellaceae, Harpanthaceae, Cephaloziaceae, Odontoschismaceae, Lepidoziaceae, Calypogeiaceae, Radulaceae, Goe* beliellaceae, Pleuroziaceae, Madothecaceae. в) Jubuleae
Семейства: Frullaniaceae, Lejeuneaceae.
Очень близка к системе Мюллера система печеночников Рей- мерса (Reimers, 1954). Отличается она только тем, что сферокарповые выделены в особый порядок Sphaerocarpales и тем, что порядок Jungermanniales поставлен после порядка Anthocerotales перед порядками Sphaerocarpales и Marchantiales.
Больше отличается от системы Мюллера система, принятая нами во «Флоре» печеночных и сфагновых мхов Украины (Зеров, 1964).
Мы разделили мохообразные (Bryophyta) на три класса — антоцеротовидные ( Anthocerotopsida), печеночники (Hepatico-
psida, Hepaticae) и мхи (Bryopsida, Musci).
Для класса печеночников мы приняли следующую классификацию.
Класс ПЕЧЕНОЧНИКИ — HEPATICOPSIDA (HEPATICAE)
Подкласс I. Marchantiidae
Порядок I. Сферокарпальные — Sphaerocarpales Семейства: Sphaerocarpaceae, Riellaceae.
Порядок II. Маршанциальные — Marchantiales Подпорядок I. Ricciineae
Семейства: Ricciaceae, Oxymitraceae.
Подпорядок II. Marchantiineae
Семейства: Targioniaceae, Grimaldiaceae, Cleveaceae, Conocepha- laceae, Lunulariaceae, Cyathodiaceae, Marchantiaceae, Monoclea- ceae, Exormothecaceae, Corsiniaceae.
Подкласс II. Jungermanniidae Порядок I. Метцгер и альн ы e — Metzgeriales
Семейства: Aneuraceae, Metzgeriaceae, Pelliaceae, Blasiaceae, Di- laenaceae, Treubiaceae, Codoniaceae.
Порядок II. Гапломи три альные — Haplomitriales Семейства: Haplomitriaceae, Calobryaceae.
Порядок III. 10 н герма нн и альные — Jungermanniales Подпорядок I. Jungermanniineae
Семейства: Hygrobiellaceae, Herbertaceae, Lophoziaceae, Junger- manniaceae, Plagiochilaceae, Southbyaceae, Schistochilaceae, Lophocoleaceae, Harpanthaceae, Cephaloziaceae, Odontoschismata- ceae, Ptilidiaceae, Trichocoleaceae, Blepharostomataceae, Lepidoziaceae, Calypogeiaceae, Cephaloziellaceae, Marsupellaceae, Scapaniaceae, Radulaceae, Goebeliellaceae, Pleuroziaceae, Porellaceae. Подпорядок II. Frullaniineae
Семейства: Lejeuneaceae, Frullaniaceae.
В основе систем авторов, рассматривающих печеночники как регрессивную группу, лежит система Веттштейна, который в
252
начале системы класса печеночников поставил листостебельные юнгерманниальные, от которых выводил талломные анакрогинные юнгерманниальные и маршанциальные.
Веттштейн, принимая подразделения Линдберга и Шиффнера, дает такую классификацию класса печеночников — Hepaticae (Wettstein, 1903).
1 порядок Junge г mann i а 1 e s
1 сем. Haplomitriaceae
2 » Acrogynaceae
3 » Anacrogynaceae.
2 порядок Marchantiales
1 сем. Marchantiaceae
2 » Ricciaceae.
3 порядок Anthocerotales
1 сем. Anthocerotaceae.
Веттштейн, считая лиственные мхи исходной группой архе- гониальных растений, развившейся из водорослей, ничего не говорит о происхождении облиственного гаметофита мхов. Попытку ответа на этот вопрос находим в работе Черча (Church, 1919), который считал, что вышедшие на сушу предки архегониальных растений, в том числе и печеночников, имели прямостоящие облиственные гаметофит и спорофит. Дальнейшее развитие печеночников шло по линии упрощения этого строения. Редукция гаметофита печеночников, по Черчу, проявлялась в переходе от вертикального побега к простертому, в развитии дорзивентральности, анизофиллии, исчезновении амфигастриев, уплощении стебля и потере боковых листьев. По мнению Черча, все талломные печеночники развились из облиственных форм.
Редукция спорофита, по Черчу, выражалась в потере листьев в результате «интенсивной инсоляции и высыхания», в исчезновении аэрационной системы, редукции фотосинтезирующей ткани и устьиц. Одновременно с этим развивались структуры, связанные с распространением спор.
Горячим сторонником взглядов Черча был Эванс (Evans, 1939). Он развил мысль Черча о регрессивной эволюции в пределах юнгерманниальных и маршанциальных. Что касается антоцеро- тальных, то Эванс считал спорофит антоцерота также редуцированным, однако в меньшей степени, чем спорофит других печеночников, а гаметофит — очень упрощенным по сравнению с предполагаемыми облиственными предками.
Особенно окрепла теория регрессивного развития печеночников после изучения псилофитов, которых стали считать возможными предками мохообразных.
Взгляды Веттштейпа, Черча и Эванса нашли отражение в позднейших классификациях печеночников. В тридцатых годах XIX ст. было опубликовано несколько классификаций, построенных на принципах Веттштейна — Черча, а именно: классификации Фер- доорна (Verdoorn, 1932), Буха (Buch, 1936), Буха, Эванса и Фер- доорна (Buch, Evans a. Verdoorn, 1938) и Эванса (Evans, 1939).
Фердоорп делит печеночники — класс Hepaticae на два подкласса — Hepaticales и Anthocerotales. В подклассе Hepaticales он различает четыре порядка: Jungermanniales acrogynae (с че¬
253
тырьмя подпорядками — Haplomitrieae, Macvicariaea \ Jungerman- nieae и Jubuleae), Jungermanniales anacrogynae, Sphaerocarpales и Marchantiales; в подклассе Anthocerotales — один порядок, под тем же названием. Фердоорн акрогинные печеночники поставил в начале системы, начинает их подпорядком Haplomitreae и закапчивает подпорядком Jubuleae, т. е. размещает подпорядки в восходящем порядке, что же касается анакрогинных и маршан- циальных, то семейства в них размещены в нисходящем порядке. При этом, анакрогинные выводятся, очевидно, из листостебельных акрогинных, что же касается происхождения таких сложных организмов, как представители семейства Marchantiaceae, поставленных в начале порядка Marchantiales, то по этому вопросу автор никаких соображений не высказывает.
Ярким отражением идей Черча в системе печеночников является система Эванса (Evans, 1939), которую приводим ниже без перечисления родов.
Порядок 1. Jungermanniales
Подпорядок 1. Haplomitrineae Семейство Haplomitriaceae.
Подпорядок 2. Jungermannineae (Jungermanniales Acrogynae)
Семейства: Ptilidiaceae, Lepidoziaceae, Calypogeiaceae, Cephaloziaceae, Cephaloziellaceae, Harpanthaceae, Jungermanniaceae, Marsupellaceae, Plagiochilaceae, Scapaniaceae, Schistochilaceae, Porellaceae, Goebeliella- ceae, Radulaceae, Pleuroziaceae, Frullaniaceae, Lejeuneaceae.
Подпорядок 3. Metzgeriineae (Jungermanniales Anacrogynae)
Семейства: Treubiaceae, Fossombroniaceae, Pelliaceae, Blasiaceae, Pal- laviciniaceae, Metzgeriaceae, Riccardiaceae, Monocleaceae.
Порядок 2. Marchantiales
Семейства: Marchantiaceae, Rebouliaceae, Sauteriaceae, Targioniaceae, Corsiniaceae, Ricciaceae.
Порядок 3. Sphaerocarpales
Семейства: Sphaerocarpaceae, Riellaceae.
Порядок 4. Anthocerotales Семейство Anthocerotaceae.
По тому же принципу построены и системы Шустера (Schuster, 1953, 1958, 1966). Он придерживается деления Bryophyta на три класса — Anthocerotae, Hepaticae и Musci, а в пределах порядка Jungermanniales выделяет подпорядки. В отличие от Эванса Шустер в начале порядка Jungermanniales ставит подпорядок Jungermannieae, который начинает семейством Lophoziaceae.
Приведем последнюю систему Шустера (Schuster, 1966).
Класс I. HEPATICAE
Подкласс A. Jungermanniae Порядок 1. Takakiales
Семейство Takakiaceae.
Прядок 2. Calobryales
Семейство Haplomitriaceae.
1 Выделение подпорядка Macvicarieae и семейства Macvicariaceae основано на неправильном понимании рода Macvicaria, близкого к роду Morelia или даже идентичного с ним.
254
Порядок 3. Jungermanniales Подпорядок A. Herbertinae
Семейства: Herbertaceae, Vetaformaceae, Blepharostomaceae,
Chaetocoleaceae, Isotachidaceae.
Подпорядок B. Ptilidiinae
Семейства: Ptilidiaceae, Lepicoleaceae, Chaetophyllopsidaceae,
Trichocoleaceae, Lepidolaenaceae.
Подпорядок D. Jungermanniinae
Семейства: Lepidoziaceae, Calypogeiaceae.
Подпорядок D. Jungermanniinae
Семейства: Lophoziaceae, Jungermanniaceae, Gyrothyraceae, Mar- supellaceae, Scapaniaceae, Delavayellaceae, Balantiopsidaceae, Schistochilaceae, Lophocoleaceae, Chonecoleaceae, Plagiochilaceae, Acrobolbaceae, Southbyaceae, Antheliaceae, Cephaloziaceae, Ce- phaloziellaceae, Adelanthaceae.
Подпорядок E. Perssoniellinae Семейство Perssoniellaceae.
Подпорядок F. Radulinae Семейство Radulaceae.
Подпорядок G. Porellinae
Семейства: Porellaceae, Frullaniaceae, Goebeliellaceae, Lejeunea- ceae.
Подпорядок H. Pleuroziinae Семейство Pleuroziaceae.
Порядок 4. Metzgeriales Подпорядок Treubiinae Семейство Treubiaceae.
Подпорядок Codoniinae
Семейство Fossombroniaceae.
Подпорядок Aneurinae
Семейства: Blasiaceae, Dilaenaceae, Hymenophytaceae, Aneura- ceae, Metzgeriaceae.
Подпорядок Phyllothalliinae Семейство Phyllothalliaceae.
Подкласс B. Marchantiae
Порядок 5. Sphaerocarpales Подпорядок A. Naiaditinae Семейство Naiaditaceae.
Подпорядок B. Riellinae Семейство Riellaceae.
Подпорядок C. Sphaerocarpinae Семейство Spnaerocarpaceae.
Порядок 6. Monocleales Семейство Monocleaceae.
Порядок 7. A. Marchantiales Подпорядок A. Corsiniinae
Семейства: Corsiniaceae, Monocarpaceae.
Подпорядок В. Targioniinae.
Семейство Targioniaceae.
Подпорядок C. Marchantiinae
Семейства: Grimaldiaceae, Conocephalaceae, Lunulariaceae, Clevea- ceae, Marchantiaceae, Exormothecaceae.
Подпорядок D. Ricciinae
Семейства: Oxymitraceae, Ricciaceae.
Класс II. ANTHOCEROTAE
Порядок 8. Anthocerotales Семейство Anthocerotaceae.
Особенностью этой системы является введение порядка Така- kiales и разделение порядка Jungermanniales на восемь, порядка
255
Metzgeriales на четыре, порядка Sphaerocarpales на три, а порядка Marchantiales на четыре подпорядка. Выделение такого числа подпорядков, например в порядке Sphaerocarpales, где каждый из трех подпорядков включает по одному семейству, нам кажется нецелесообразным.
* * *
Ввиду того, что в современной литературе существуют два противоположных взгляда на происхождение и эволюцию печеночников, рассмотрим фактический материал, который может помочь объективно разрешить этот вопрос.
Начнем наше рассмотрение с палеоботанических данных. Принято считать, что палеоботанические данные по мохообразным очень скудны и дают очень мало для восстановления их истории.
До некоторой степени это справедливо, однако в течение последних десятилетий накоплен материал, который в значительной мере помогает выяснить геологическую историю мохообразных. Имеющиеся палеоботанические данные дают основание для вывода о древности мохообразных, возраст которых восходит по крайней мере до первой половины палеозоя. Савич-Любицкая и Абрамов (1958, 1959) предполагают, что основные классы мохообразных обособились, по крайней мере, в силуре, а может быть еще раньше. В пользу такого допущения свидетельствуют данные Наумовой (1949, 1950, 1953), которая в нижнекембрийских и нижнесилурийских отложениях Прибалтики и в девонских отложениях Центральной платформы европейской части СССР обнаружила споры типа мохообразных и папоротникообразных, а также исследования Нокс (Knox, 1949, 1950), обнаружившей в отложениях продуктивного карбона Великобритании споры мохообразных, в частности антоцеротальиых и маршан- циевых. Однако нужно отметить, что по мнению Проскауэра (Proskauer, 1960) споры, описанные Нокс как принадлежащие антоцеротальным, может быть, являются спорами горнеофитона. Козловский и Грегуш (Kozlowski a. Greguss, 1959; Greguss, 1961) описывают остатки примитивных талломных растений из ордович- ских известковых валунов, занесенных в Польшу из Прибалтики во время четвертичных оледенений, принимаемых ими за предшественников мохообразных. Однако нужно отметить, что эти
Рис. 87. Sciadophytopsis tirassi- сит:
а — общий вид, б — спороношение (?) (по Ищенко, 1968).
256
находки вызывают сомнения в правильности определения (Schuster, 1966; Jovet-Àst, 1967),
В настоящее время самой древней несомненной находкой ископаемого мохообразного является Hepaticites devonicus, описанный Хюбером (Hueber, 1961) и переименованный Шустером в Palla- vicinites devonicus (Schuster, 1966, стр. 352). Как показывают рисунки Хюбера, это несомненный представитель порядка Metzge- riales.
Заслуживает большого внимания в связи с происхождением мохообразных описанный Ищенко (1968) из нижнего девона Приднестровья род Sciadophytopsis (рис. 87) и отнесенный нею к пси- лофитам (семейство Sciadophytaceae). Виды этого рода имеют розетковидные, лопастные на краях талломы 0,4—3,5 см в диаметре. На розетках, кроме лентовидных лопастей, имеются сидящие на коротких ножках округлые образования 0,5—2 мм в диаметре, принимаемые автором за спороношения. Поверхность этих телец слегка бугорчатая. Розетковидные талломы внешним видом напоминают гаметофиты Anthoceros. Для окончательного решения о систематической принадлежности трех описанных Ищенко видов Sciadophytopsis необходимо изучить взаимоотношение спороносных выростов и розетковидных талломов; к сожалению, автор обнаружил только отпечатки.
Несомненные остатки мохообразных известны из карбона (Renault et Zeiler, 1888; Lignier, 1914; Walton, 1925, 1928). При этом остатки печеночников, описанные под названием Hepaticites, обнаруживают тесные связи с современными метцгериальными (Harris, 1939), a Muscites polytrichaceus и М. bertrandi — с современными Bryopsida. Остатки маршанциального — Marchantites lorea Залесский (1937) приводит для среднепермских отложений Урала. Из пермских отложений Кузнецкого и Тунгусского бассейнов и Печорского края Нейбург (1956, 1960) приводит ряд настоящих мхов. Особенно интересны описанные ею роды Jiin- jagia, V or cut annular ia и P roto sphagnum, которые она объединяет в порядок Protosphagnales и которые являются возможными предшественниками рода Sphagnum. Эти роды характеризуются своеобразным строением листьев, в котором можно видеть образующие сеть хлорофиллоносные клетки; в петлях сети размещены, как у сфагнов, водоносные клетки. Отличает их от рода Sphagnum наличие средней жилки в листьях.
Для мезозоя известна находка Hepaticites cyathodoides триасовых отложениях Южной Африки (Townrow, 1959). В верхнетриасовых отложениях Англии (рет) в середине прошлого столетия был обнаружен организм, описанный Букменом (Buck- man, 1950) иод названием Naiadita lanceolata. Харрис (Harris, 1938, 1939) дал детальное его описание и отнес его к риелловым (Riellaceae), а Шустер (Schuster, 1966) выделил в особый иод- порядок Naiaditinae порядка сферокарпальных. Для мезозоя (триас — юра) Харрис указывает четыре вида Hepaticites, близких
257
к современным метцгериальным (Harris, 1939). Из ретско-лейас- ских отложений Швеции Лундблад (Lundblad, 1954) описала Ricciopsis florini, R. scanica и Marchantiolites porosus. В этой же работе по спорам был описан Ricciosporites tuberculatus. Одпако позже Лундблад (Lundblad, 1959) высказала сомнение в правильности отнесения этих спор к риччиевым. Для юрских отложений Донецкого бассейна (бат — келловей) Станиславский (1957) указывает Thallites toretziensis, Th. plicatus и Thallites sp. Первый из них, по словам автора, сходен с Ricciopsis florini, а второй и третий, несом ненно, относятся к метцгериальным. Автор сравни вает Thallites plicatus с триасовой (ретекой) Palae- ohepatica rostafinskii(Thallites rostafinskii). Споры сфагновых мхов ( Sphagnum-sp or ites ) известны из верхнетриасовых отложений (рет) Тюрингии, юрских отложений Австралии, отложений мелового периода Канады, Австралии и Сибири. Листья и споры сфагновых мхов обнаружены в юрских отложениях Баварии и СССР и в верхнемеловых отложениях Гренландии (Са~ вич-Любицкая и Абрамов,
1958; Jovet-Ast, 1967). Из верхнемеловых отложений Рис. 88. Хронологическая таблица развития США описаны остатки основных групп мохообразных по палеон- Mescites lesquereuxi (Вег- алогическим Данным (по Jovet-Ast, 1967). гу, 1928; Steere, 1946). В отложениях мелового периода Европы и Северной Америки обнаружены остатки, которые отнесены к сборному роду Jungermannites; это первые достоверные сведения об остатках юнгерманниальных (Jovet-Ast, 1967).
Гораздо богаче и разнообразнее мохообразные из третичных отложений. По данным Савич-Любицкой и Абрамова (1958), известно для третичного периода 13 родов и 34 вида печеночников и 63 рода с 105 видами мхов. Большинство третичных родов мохообразных существует до настоящего времени. В третичной флоре хорошо представлены Jungermanniales, Bryidae, Spliagnidae.
258
Рис. 88. Хронологическая таблица развития основных групп мохообразных по палеонтологическим данным (по Jovet-Ast, 1967).
Резюмируя сведения об ископаемых мохообразных (см. рис. 88), мы можем сказать, что в настоящее время наиболее древние достоверные остатки мохообразных относятся к порядку метцге- риальных (девон). Остатки маршанциальных известны начиная с карбона, причем представители риччиевых встречаются уже с юрского периода. С юрского периода известны сферокарпальные. Что касается юнгерманниальных (акрогинных), то они появляются только в верхнемеловых отложениях. Достоверные остатки Bryidae известны начиная с верхнего карбона, близкие к сфагновым мхам Protosphagnales известны из перми, а настоящие сфагновые — с юры. Остатки антоцеротальных известны только начиная с верхнетретичного времени, а андреевые в ископаемом состоянии неизвестны. Савич-Любицкая и Абрамов (1958) делают вывод, что «судя по имеющимся палеонтологическим данным, ... печеночники являются более древней группой по сравнению с листостебельными мхами, причем слоевищные формы возникли ранее листостебельных...» (стр. 1416). Таким образом, имеющиеся палеоботанические данные дают основание считать, что просто построенные талломные формы печеночников являются первичными, а не результатом упрощения организмов с вегетативным телом сложного листостебельного строения.
Исходя из данных сравнительной морфологии вегетативных органов печеночников, казалось бы на первый взгляд естественным предположить, что эволюция в этой группе растительного мира шла от форм с простым талломом к более сложным талломным и листостебельным формам. Однако нельзя забывать, что вегетативные органы, наиболее связанные со средой, быстрее всего реагируют на ее изменения; редукция вегетативных органов — явление очень распространенное. Поэтому часто бывает очень трудно решить, является ли простота строения организма первичной или результатом вторичного его упрощения.
Сторонники концепции регрессивного развития печеночников в подтверждение своих взглядов приводят следующие доводы. Веттштейн (Wettstein, 1903) считал основной причиной регрессивного развития в классе печеночников переход листостебельных форм юнгерманниальных к плагиотроиному росту, результатом чего, по его мнению, явилась редукция листьев и превращение стебля в талломное образование. Доказательством происхождения талломных маршанциальных от листостебельных форм, по Ветт- штейну, является наличие брюшных чешуек, которые он считает рудиментами листьев. Эванс (Evans, 1939), кроме того, ссылается на наличие листообразных чешуек на верхней поверхности таллома у Pallavicinia и Symphyogyna, а также на существование в порядке листостебельных юнгерманниальных форм с редуцированными листьями (Zoopsis, Pteropsiella, Schiffneria).
Гебель (Goebel, 1930) в результате детального изучения брюшных чешуек у маршанциальных и метцгериальных пришел к выводу, что эти чешуйки в своем происхождении тесно связаны
259
со слизистыми железками (Schleimpapillen) и никоим образом не могут считаться редуцированными листьями. Что же касается листообразных чешуек па верхней поверхности таллома у представителей семейства Dilaenaceae, то они являются выростами средней жилки таллома и по происхождению напоминают пе- рихеции (обвертки), окружающие гаметангии (Müller, 1951, стр. 86—87).
Следуя Веттштейну и Эвансу, некоторые авторы при попытках доказать редукционное развитие печеночников от листостебельных форм до талломных особое значение придают переходу юнгерманниальных от ортотропного роста к плагиотропному и как доказательство приводят формы с редуцированными листьями (Zoopsis, Pteropsiella, Schiffneria) или же с господствующей про- тонематической стадией (Protocephalozia, Мetzgeriopsis). Действительно, переход от ортотропного роста к плагиотропному является причиной анизофиллии — различия в форме боковых и брюшных листьев; этот переход мог вызывать иногда даже полную редукцию брюшных листьев (амфигастриев). Однако полное исчезновение листьев и превращение листостебельного побега в таллом никак не может быть непосредственным следствием перехода к плагиотропному росту, так как в порядке юнгерманниаль ных имеется много родов и семейств с плагиотроиным ростом, у представителей которых не только не наблюдается исчезновение листьев, а наоборот — усложнение их строения (Scapania- сеае, Porellaceae, Radulaceae, Frullaniaceae, Lejeuneaceae и др.). Правда, иногда в процессе эволюции и приспособления к определенным условиям существования у некоторых юнгерманниальных действительно наблюдается редукция листьев на вегетативных побегах или же у них господствующей является иротонематиче- ская стадия. Однако у этих форм побеги, несущие половые органы, всегда имеют листостебельное строение. Нужно также отметить, что другие роды семейств (Gephaloziaceae и Lejeuneaceae), к которым относятся эти редуцированные роды, и даже виды того же рода (Zoopsis) имеют нормальное листостебельное строение. Гебель, склонный к объяснению редукцией многих морфологических особенностей печеночников, после рассмотрения всех этих случаев пришел к заключению, что нет никаких убедительных оснований считать, что все талломиые формы печеночников произошли от листостебельных. Он даже идет дальше и заключает, что «все это говорит скорее за то, что лиственные формы развились из талломных» (Goebel, 1930, стр. 738). Гебель обращает внимание на тот факт, что у многих лпстостебельных форм при прорастании споры на первых стадиях развития проростка обычно наблюдается талломное строение. Правда, у талломной Blasia pusilla на ростках из выводковых телец развиваются листовые лопасти, более заметные, чем па взрослом растении.
В свое время Лейтгеб (Leitgeb, 1881) считал, что у маршан- циальных размещение половых органов на талломе постепенно
т
усложнилось: у более простых форм гаметангии разбросаны на поверхности таллома, затем они концентрируются в группы на верхней его поверхности, затем образуются особые подставки, которые являются видоизмененными ветвями таллома и наибольшей сложности достигают у маршанции. Гебель (Goebel, 1910, 1930, стр. 831—841) проводил противоположную идею, что развитие шло в обратном направлении — от высокоразвитых подставок, какие мы находим у маршанции, путем упрощения к формам с наиболее простым размещением гаметангиев на верхней поверхности таллома. Кашъяп (Kashyap, 1919) также указывал на наличие редукционных рядов у маршанциальных. Гебель ничем не обосновывает упрощения размещения гаметангиев на талломах маршанциевых и не дает никаких соображений по поводу происхождения сложных форм подставок, которые должны были выработаться в процессе длительной эволюции как приспособление для защиты гаметангиев от неблагоприятных условий и облегчения рассеивания спор. Самое строение подставок, их онтогенез, наличие реликтовых форм, у которых как бы зафиксированы промежуточные стадии развития (например, Neohodgsonia mirabilis; см.: Mehra, 1969) — все это свидетельствует о прогрессивном развитии маршанциальных, достигшем у представителей семейства маршанциевых своей наивысшей формы, как и строение вегетативной части таллома. Что же касается упрощения строения таллома у некоторых маршанциальных (Cyathodium, Dumortiera, Monocleae), проявляющегося в редукции воздушных камер или основной ткани, то Герцог (Herzog, 1925, стр. 25) и Гебель (Goebel, 1930, стр. 760—762) объясняют его условиями избыточной влажности, с чем нельзя не согласиться.
Никак нельзя рассматривать как доказательство регрессивного развития изменение формы антеридиальных кроющих листьев и амфигастриев (Fulford, 1965, стр. 15). Тут мы видим обратный процесс — усложнение и специализацию листьев в связи с изменением их функции, но никак не редукцию, ведущую в конечном счете к превращению листостебельиого побега в талломное образование, как это принимают для метцгериальных и маршанциальных сторонники регрессивного развития печеночников.
На протяжении многих миллионов лет в процессе эволюции у печеночников, как и в других группах растительного мира, были возможны и имели место случаи регрессивного развития, связанные с приспособлением к определенным условиям среды. Однако эти случаи ne свидетельствуют о том, что все талломпые формы печеночников произошли от листостебельных.
Кроме простоты строения вегетативного тела простых таллом- пых форм метцгериальных в пользу признания первичности их талломного строения свидетельствует также наличие в роде Ric- cardia внутриклеточных выводковых телец, образование которых сходно с образованием зооспор у водорослей. Аналогичным с развитием зооспор водорослей является также образование «прида¬
261
точных побегов» у видов метцгерии, возникающих в результате деления содержимого материнской клетки (Goebel, 1930, стр. 807— 809).
Строение спорогона печеночников, основной функцией которого является продукция спор и который в отношении питания гораздо менее, чем гаметофит, зависит от условий среды, может дать более надежные данные для выяснения вопроса о примитивности или вторичности той или иной группы.
Наиболее простое строение спорогона характерно для сферокарповых и риччиевых. Слабая их морфологическая дифференциация, однослойная стенка, клетки которой лишены утолщений, примитивный способ освобождения спор, отсутствие элатер, которым у сферокарповых соответствуют стерильные археспориаль- ные, так называемые питающие клетки,— все это является определенным свидетельством в пользу примитивности их спорогона.
В других порядках печеночников наблюдаем спорогоны более сложного строения, имеющие ряд приспособлений для распространения спор в воздушной среде. Так, у маршанциальных, хотя коробочка спорогона имеет однослойную стенку и только верхняя ее часть бывает многослойной, у большинства родов имеются утолщения стенок коробочки; почти у всех родов, кроме примитивного рода Corsinia, имеются хороню развитые элатеры; разнообразны способы освобождения спор, причем у некоторых форм верхняя часть коробочки обособляется в форме крышечки. У маршанциальных мало развита ножка спорогона, что компенсируется развитием подставки — карпофора.
У гапломитриальных спорогон имеет хорошо развитую ножку, а его коробочка, кроме верхней части, имеет однослойную стенку с утолщениями оболочек клеток, раскрывается двумя-тремя, иногда четырьмя створками; кроме спор, имеются элатеры — свободные и прикрепленные ко дну коробочки и к концам створок.
Наиболее сложным строением коробочки характеризуются метц- гериалыше и юнгерманниальные. У представителей этих порядков стенка коробочки многослойная, с утолщениями (кроме рода Blasia из метцгериальных), обычно раскрывается четырьмя створками (только в семействе Codoniaceae из метцгериальных она распадается на неправильные куски); кроме спор образуются элатеры — свободные и иногда прикрепленные (Aneuraceae, Pel- Насеае, Metzgeriaceae из метцгериальных и Frullaniaceae из юнгер- манниальных).
Не останавливаясь на оценке особенностей спорогона ангоце- ротальных, которые мы рассматриваем как отдельпый подотдел зеленых растений, мы можем констатировать, что наиболее простые спорогоны характерны для порядка сферокарнальных и семейства риччиевых из маршанциальных; много примитивных черт в спорогонах маршанциальных, гапломитриальных и некоторых метцгериальных (Codoniaceae); наиболее сложны спорогоны у прочих метцгериальных и юнгерманниальных.
262
Большой интерес для филогении представляют данные кариологии. Правда, в настоящее время они сводятся в основном к изучению чисел хромосом и их морфологии, однако накопленный в результате исследований Гейтца, Лорбеера, Мехра, Проскауэра, Татуно и др. авторов материал по кариологической характеристике печеночников позволяет делать ряд выводов и обобщений по их филогении.
По данным Татуно (Tatuno, 1959), наименьшее количество хромосом среди печеночников имеет Takakia lepidozioides, для которой характерным является п = 4; две хромосомы из четырех имеют коленчатую форму и две палочковидную (кариотип К = = 2v + 2ι). Татуно подчеркивает, что такое же количество хромосом характерно для некоторых улотрихальных водорослей, например Ulothrix zonata и Draparnaldiopsis indica.
В порядке маршанциальных основными числами хромосом являются п = 8 (Riccia) и /2 = 9 (большинство родов порядка Marchantiales). Кроме того, в этом порядке известны случаи удвоения и утроения этих чисел. По мнению Татуно, набор хромосом п = 8 возник в результате удвоения примитивного числа п = 4. Так, для Riccia fluitans кариотип характеризуется четырьмя коленчатыми и четырьмя палочковидными хромосомами (К = = 4v + 4ι). Что же касается набора /2 = 9, то он, по мнению Татуно, возник в результате образования еще одной палочковидной хромосомы (К = 4γ + 5ι). Обычпо наименьшая из палочковидных хромосом, а иногда и одна коленчатая являются гетерохромосомами.
Для порядка сферокарпальных характерным является гг = 8 (Sphaerocarpus) и п = 9 (Riella, у видов которого известна девятая маленькая хромосома, которая у R. affinis, по Проскауэру, является гетерохромосомой). По мнению Татуно, набор хромосом у видов Riella является более примитивным, чем у сферокар- па, и образовался в результате удвоения примитивного числа п = 4 и образования одной дополнительной хромосомы, а набор хромосом сферокарна (п — 8) возник из набора Riella в результате выпадения этой дополнительной хромосомы. Мне кажется более естественным признать, что набор хромосом сферокарпа возник в результате удвоения первоначального числа п = 4, как это признает Татуно для рода Riccia, а набор Riella является результатом усложнения этого пабора по типу, описанному Татуно для маршанциальных.
Для гапломитральных (Haplomitriales) характерен набор хромосом п = 9 (К = 4v + 5ι), причем у Calobryum rotundifolium одна коленчатая и одна палочковидная хромосомы являются гетерохромосомами.
В порядке метцгериальных основное число хромосом обычно равняется девяти, иногда оно бывает равным восьми (Pallavici- nia) или десяти (Riccardia pinguis, R. latifrons, R. palmata).
Наблюдается также удвоение или утроение основного числа.
263
Так, у Pellia borealis п = 18, Riccardia multifida η = 20, R. sinuata η = 30.
В порядке юнгерманниальных основное число хромосом п = 9, изредка оно равно 8 (Radula lindbergiana, многие виды рода Porella, мужские экземпляры Frullania dilatata); часты случаи удвоения, утроения и даже учетверения числа хромосом (п = 18у п = 27, п = 36). Набор хромосом усложняется в порядках метц- гериальных и юнгерманниальных наличием гетерохромосом.
Образование восьмихромосомных кариотипов у метцгериаль- ных и юнгерманниальных Татуно объясняет выпадением одной палочковидной хромосомы. Кариотип, равный 5ν + 5ι, свойственный некоторым видам Riccardia, происходит, по его мнению, в результате увеличения числа палочковидных хромосом.
Антоцеротальным свойственно число хромосом п = 6, реже п = 5. Интересно, что у одного и того же вида, например у Antho- ceros laevis, были обнаружены шестихромосомная и пятихромосомная расы.
У авторов первой трети XX ст. (1899—1930 гг.) имеются указания на четыре хромосомы у видов антоцерота. Мехра и Ханду (Mehra a. Handoo, 1953) высказывают предположение, что эти сведения являются результатом того, что исследователи пропускали одну хромосому. Татуно считает возможным выводить кариотип 3ν + 3ι антоцеротальных из четыреххромосомного кариотипа Takakia в результате удвоения числа и выпадения двух гетерохромосом — коленчатой и палочковидной. Что же касается набора хромосом п = 5, то он, по мнению Татуно, является следствием выпадения одной хромосомы из шестихромосомного набора.
Попытка Татуно свести кариотип антоцеротальных к кариотипу Takakia является мало убедительной. Проскауэр (Proskauer, 1957) дает иное объяснение его происхождения. По Проскауэру кариотип Anthoceros характеризуется пятью хромосомами, из которых одна гетерохроматическая микрохромосома (К = 4 + 1). Между прочим, Проскауэру удалось наблюдать расу Anthoceros punctatus var. punctatus с набором η = 8 + 1, который он считает диплоидным с выпадением одной гетерохромосомы. Этот набор не отличается от обычного набора хромосом у большинства печеночников. На основании этого Проскауэр высказывает предположение об общности происхождения антоцеротальных и печеночников.
Мехра и Ханду (1953) считают наличие микрохромосом характерной чертой печеночников и антоцеротовидных, на основании чего они высказывают предположение об общности происхождения этих групп. Различия в основном числе хромосом в этих двух классах дает им основание считать, что антоцеротальные обособились от печеночников на очень ранней стадии развития.
Имеющиеся кариологические данные свидетельствуют о возможности происхождения мохообразных и в том числе печеноч¬
264
ников от зеленых (улотрихальных) водорослей; печеночники и антоцеротовидные обособились, очевидно, очень рано. Наиболее простым кариотипом среди печеночников обладает явно реликтовый род Takakia, который сохранил, очевидно, кариотип древнейших печеночников. Просто построенные виды рода Sphaerocar- pus обладают простым кариотипом так же, как виды рода Riccia среди маршанциальных. В порядках маршанциальных, метцгери- альных и юнгерманниальных наблюдается усложнение основного набора хромосом (п = 9) путем полиплоидизации и дифференциации хромосом, а также изредка упрощения кариотипа вследствие выпадения одной из хромосом. Данные кариологии свидетельствуют в основном о прогрессивном направлении развития в классе печеночников.
Изучение систематического состава свидетельствует о большой древности классов печеночников и антоцеротовидных. Такие порядки, как антоцеротальные, а также недавно открытые така- киальные, являются типичными олиготипными реликтовыми группами. Их олиготипность находится в созвучии с их примитивным строением. Несколько более богаты родами и видами порядки Metzgeriales и Marchantiales, однако и в этих группах преобладают монотипные и олиготипные семейства и роды. Из всех Metzgeriales только два рода — Metzgeria и Riccardia содержат более чем по сто видов, а в порядке Marchantiales более или менее богаты видами роды Riccia — около 100 видов и Fimbriaria — около 90 видов.
Несравненно богаче в родовом и видовом отношении порядок юнгерманниальных. В его составе сейчас насчитывают 25 семейств, включающих до 180 родов и более 8000 видов, что составляет более 80% всех родов и видов печеночников. В этом порядке также имеется ряд олиготипных семейств и родов, однако порядок Jungermanniales содержит наряду с этим ряд семейств и родов, представленных многочисленными видами. Заслуживает упоминания семейство Lejeuneaceae, в составе которого в настоящее время различают около семидесяти родов с многочисленными видами. Очень богаты видами роды Frullania, Drepanolejeiinea, Microlejeunea, Cololejeunea, Porella, Radula, Bazzania, Plagiochila, Lophocolea, Chiloscyphus и ряд других. Все это свидетельствует, что в составе порядка Jungermanniales, кроме древних реликтовых систематических единиц, вероятно, мезозойского возраста, имеется ряд развивающихся в настоящее время семейств и родов. Интересно то, что многие процветающие роды характеризуются довольно сложным морфологическим расчленением тела при характерном для этого порядка плагиотропном росте, который, как упоминалось выше, некоторыми авторами считается одной из причин, якобы обусловивших у юнгерманниальных редукцию листьев и переход к талломному строению.
Географическое распространение печеночников свидетельствует о большой древности этой группы растительного мира. Еще
265
Кемпбелл (Campbell, 1907) обратил внимание на тот факт, что многие роды печеночников, особенно относящиеся к талломным порядкам, характеризуются широким распространением, а ряд родов и значительное количество их видов почти космополитны. Фулфорд (1951) на основании изучения географического распространения родов юнгерманниальных в южном полушарии пришла к заключению, что в юрское время уже существовали основные филогенетические линии юнгерманниальных печеночников и ряд сохранившихся до настоящего времени родов. Основанием для такого вывода является дизъюнктивное распространение некоторых родов в южном полушарии (Южной Америке, Австралии, Новой Зеландии, Южной Африке и Северо-Восточной Индии (Сикким)), свидетельствующее о флористических связях, возможных, по крайней мере, в среднем мезозое.
Проведение широкого сравнительного изучения географического распространения печеночников затрудняется неудовлетворительным состоянием изученности видового состава флоры печеночников тропических областей в результате крайне некритического описания видов в прошлом. Древность основных разделов печеночников, в результате чего по сути во всех группах наблюдается очень широкое, часто космополитное распространение многих родов, наличие явления сравнительно молодого видообразования в пределах некоторых древних родов,— все это затрудняет возможность широких филогенетических выводов на основании современного распространения печеночников.
Таким образом, и на основании имеющихся данных можно отметить наибольшее развитие иеоэндемизма в порядке юнгерманниальных, который, по-видимому, является и наиболее молодым.
Приведенные выше данные палеоботаники, сравнительной морфологии, цитологии, систематики и географического распространения печеночников, несмотря на свою неполноту, все же дают основания для выводов о направлениях эволюции в пределах этой группы. Древнейшие палеоботанические остатки печеночников (девон) относятся к типичным талломным формам типа метц- гериальных, позже появляются формы, близкие к маршанциаль- ыым (пермь), в триасе известны риелловые (Naiadita lanceolata), с начала юры — типичные маршанциальные типа риччиевых и маршанциальных, с конца мезозоя развиваются юнгерманниаль- ные. Это дает основание считать талломные метцгериальные, если не самой древней, то во всяком случае одной из древнейших групп печеночников, достигшей значительного развития еще в палеозое. Поэтому мы считаем совершенно необоснованным предположение о происхождении метцгериальных из листостебельных юнгерманниальных.
Особенности морфологического строения и набор хромосом (п = 4), сходный с набором хромосом некоторых зеленых водорослей у Takakia lepidozoides свидетельствует о древности этого
26G
рода, в связи с чем Мехра (Mehra, 1964) высказал предположение, что род Takakia близок к предкам печеночников. Нельзя не согласиться с мнением Мехра, что Takakiaceae и, по-видимому, родственные им гапломитриальные близки к тому исходному ядру, из которого путем усложнения листостебельного побега развились юнгерманниальные.
Простота строения гаметофита и спорофита сферокарпа, а также тот факт, что из ретских слоев Англии известен относимый к риелловым довольно высоко развитый печеночник Naiadita, дает основание предполагать древний возраст и порядка сферо- карпальных.
Что касается происхождения маршанциальных, то указание Залесского на их наличие в пермских отложениях и несомненные находки представителей риччиевых и маршанциевых в нижнеюрских отложениях свидетельствуют также о сравнительной древности порядка маршанциальных. Возможно, что развитие аридных условий в конце палеозоя и в начале мезозоя на значительной части земного шара привело к возникновению сложно устроенных талломных форм маршанциальных (Lundblad, 1954, стр. 384).
Сторонники регрессивного развития маршанциальных, как уже упоминалось, считают исходными для всего этого порядка наиболее сложные формы, принадлежащие к семейству маршанциальных. При этом они не пытаются объяснить пути возникновения этих сложных форм, которые несомненно должны были развиться путем длительной эволюции из более простых талломных форм.
Существуют различные взгляды на происхождение строения таллома маршанциальных. По Лейтгебу (Leitgeb, 1881), воздушные камеры маршанциальных образуются как углубления верхней поверхности таллома в результате задержки роста в определенных точках. Позднейшие исследования (Pietsch, 1911; Orth, 1929; цит. по Goebel, 1930, стр. 747—752) показали, что камеры возникают как межклетники в результате расщепления многослойного, вначале плотного тела. По данным Орта (Orth, 1929), воздушные камеры у Marchantia и Preissia возникают под верхним слоем клеток, после чего образуется канал вследствие расщепления клеток. У других же маршанциальных они образуются как углубления на поверхности. У риччии также происходит расщепление клеток. Об этом же свидетельствуют и наблюдения Проскауэра над Carrpos (Proskauer, 1962).
Мехра, оставляя в стороне данные онтогенеза, пытается обосновать происхождение сложного таллома маршанциальных от лиственных печеночников (Mehra a. Vashisht, 1950; Mehra, 1957). Согласно разработанной Мехра «конденсационной теории» первым этапом в развитии маршанциальных было срастание боковых листьев облиственных печеночников с образованием поперечных вертикальных пластинок, как у видов Petalo phy Пит, с последую¬
267
щим заворачиванием и срастанием их, результатом чего было образование воздушных камер.
Конденсационная теория Мехра вызывает ряд возражений. Прежде всего нужно иметь в виду, что по характеру и размещению масляных телец в талломе маршанциальные отличаются от метцгериальных и юнгерманниальных, и сходны со сферокарпаль- ньтми. Строение спорогона у маршанциальных проще и разнообразнее, чем у метцгериальных и юнгерманниальных и характеризуется рядом архаических черт. Данные онтогенеза таллома маршанциальных тоже не дают подтверждений конденсационной теории Мехра.
Учитывая характер размещения масляных телец в талломе, тип прикрепления жгутиков у антерозоидов, особенности развития архегония и антеридия, способ раскрывания антеридия, строение спорогона, форму материнских клеток спор, нам кажется более обоснованным выведение маршанциальных из простых таллом- ных форм типа сферокарпальных.
Многие авторы считают, что риччиевые являются редуцированной группой, возникшей из более развитых маршанциальных, на том основании, что спорогон риччиевых имеет простое строение — не расчленен на стопу, ножку и коробочку. Однако своеобразное сложное строение таллома риччиевых и палеоботанические данные свидетельствуют о том, что Ricciineae и Marcha- ntiineae обособились рано — не позже начала мезозоя и с того времени развивались независимо. Некоторые черты строения спорогона риччиевых — однослойная стенка, отсутствие элатер, наличие питающих клеток — являются весьма архаичными и могут быть объяснены тем, что спорогон в этой группе сохранил ряд особенностей древних предков.
Происхождение порядка Jungermanniales, как мы уже отмечали, увязывается с такими примитивными реликтовыми группами как такакиальные и гапломитриальные. Как отмечал еще Кайверс (Cavers, 1910—1911, стр. 298), примитивными признаками у представителей порядка Jnngermanniales нужно считать симметричные, двураздельные, плоские или слегка желобчатые листья, наличие амфигастриев, размещение архегониев на главной оси, свободный цилиндрический или трехгранньтй периантий, нитчатую протонему, ортотропный рост. Вторичными признаками Кейверс
Рис. 89. Изофильные формы юнгерманниальных :
1 — Pachyglossa tenacifolia, 2 — Her- zogiaria teres (no Fulford, 1963).
268
считает листья цельнокрайиие или расщепленные на большее, чем две, число лопастей или же на две неравные лопасти, отсутствие амфигастриев, размещение архегониев на специальных коротких веточках, срастание периантия с кроющими листьями, уплощение периантия или образование на нем крыльев, дисковидную протонему, образование специальных мешков на листьях, парафиллий, марсупиев. Сомнение вызывает признание вторичным признаком цельных листьев, так как у гапломитриальных они более или менее цельнокрайние.
Имеются все основания предполагать, что первичные юнгер- манниальные были прямостоячими растениями с трехрядным размещением двураздельных или цельных листьев, с верхушечным размещением архегониев, скрытых в простом периантии, образованном в результате срастания трех одинаковых кроющих листьев, и антеридиями, размещенными в пазухах не только боковых листьев, но и амфигастриев. В настоящее время известен главным образом из южного полушария ряд родов, характеризующихся рядом этих признаков, или близких к таковым (Herzog und Grolle, 1958; Fulford, 1963, 1965). Особого внимания заслуживают роды Herzogiaria с многослойными двураздельными листьями, со сходными с вегетативными листьями антеридиальными кроющими листьями и амфигастриями, Pachyglossa с цельными многослойными трехрядными листьями (рис. 89), Triandrophyllum с двураздельными листьями и амфигастриями и с антеридиями в пазухах как листьев, так и не отличающихся от них амфигастриев. От таких примитивных форм развилось разными ветвями все многообразие юнгерманниальных в результате перехода к плагио- тропному росту и усложнению формы вегетативных и кроющих листьев, периантия, строения стебля, размещения на нем половых органов, строения спорогона.
Пути эволюции мхов и вопросы их классификации
Начало классификации мхов восходит к работам Гедвига (Hedwig, 1804), который сделал попытку разделить мхи по характеру перистома на три класса: 1) Peristomio milio, 2) Peristomio nudo и 3) Peristomio effigurato. Последний класс разделен на две группы — с простым перистомом (ordine simplici) и с двойным перистомом (ordine duplici). Первая группа — с простым перистомом разделена на две подгруппы — с зубчатым и реснитчатым перистомом, а вторая — с двойным перистомом на три группы — с зубчато-реснитчатым; с зубчато-перепончатым и зубчато-сетчатым перистомом.
Эндлихер (Endlicher, 1836—1840) делил уже класс мхов (Musci) на три порядка (ordo) — Andreaeceae, Sphagnaceae и Вгуасеае, однако это подразделение не нашло отклика у современников. Например, Мюллер (С. Müller, 1849—1851) разделил
269
мхи на три класса— 1) Schistocarpi, куда отнесены Andreaeaceae с коробочкой, открывающейся щелями; 2) Cleistocarpi — клейсто- карпные формы, не имеющие крышечек и 3) Stegocarpi — крышкоплодные, разделенные на два подкласса — Acrocarpi и Pleuro- carpi. Сфагновые мхи отнесены к акрокарпным.
Один из авторов монографии «Bryologia europaea» — Шимпер (Schimper, 1860) дал систему, за основу которой принято размещение архегониев и спорогонов. Шимпер делит мхи на три секции: 1) Musci acrocarpi (верхоплодные, акрокарпные мхи),
2) Musci pleurocarpi (бокоплодные, плеврокарпные мхи) и 3) Musci schizocarpi (щелеплодные мхи), куда отнесены андреевые (Andreaeaceae). Акрокарпные мхи разделены на два порядка (ordines): клейстокарпные — Cleistocarpi и крышкоплодные — Stegocarpi. Сфагновые мхи даны без всякого ранга под названием Sphagna.
Очень широко была распространена система Лимприхта, положенная в основу его работы по флоре мхов Средней Европы (Limpricht, 1890—1904). Лимприхт делил мхи (Laubmoose) на четыре порядка — Sphagnaceae, Andreaeaceae, Archidiaceae и Bry- ineae. Последние разделены на две трибы — Cleistocarpae и Ste- gocarpae, a Stegocarpae на две подтрибы — Асгосаграе и Pleuro- саграе.
Эта система была принята в первом издании справочника Энглера и Прантля (Ruhland, Warnstorf, Brotherus, 1893—1909) с той разницей, что порядки были переименованы в подклассы — Sphagnales, Andreaeales и Bryales. Archidiaceae включены в подкласс Bryales, который был разделен на две группы семейств — Acrocarpi — верхоплодные и Pleurocarpi — бокоплодные. Группа клейстокарпных мхов, как искусственная, была ликвидирована, а клейстокарпные формы размещены в соответствующих семействах.
Радикальные изменения в систему мхов внес Флейшер (Fleischer, 1906—1908, 1920). Основные черты системы Флейшера сводятся к следующему. Ликвидируется деление настоящих мхов на акрокарпные и плеврокарпные; вместо этого в основу классификации положен характер перистома.
Система Флейшера была принята с незначительными изменениями Бротерусом во втором издании справочника Энглера и Прантля (Brotherus, 1924—1925), где она представлена в таком виде.
Класс MUSCI (LAUBMOOSE)
1 подкласс — Sphagnales
2 » — Andreaeales
3 » — Bryales
1. Группа порядков Eubryinales с порядками: Fissidentales, Dicranales, Pottiales, Grimmiales, Funariales, Schistostegales, Tetraphidales, Eubry- ales, Isobryales, Hookeriales, Hypnobryales.
2. Группа порядков Buxbaumiinales с одним порядком Bux bau niai es.
3. Группа порядков Polytrichinales с двумя порядками: Polytrichina- les, Dawsoniales.
270
Почти без изменений эта же система положена Бротерусом в основу его сводки по флоре мхов Финляндии (Brotherus, 1923) и работы Менкемейера по флоре мхов Европы (Mönkemeyer, 1927). В несколько упрощенном виде — без групп порядков, с некоторым изменением размещения их и с модернизацией названий согласно правилам современной ботанической номенклатуры эта система принята Абрамовой, Савич-Любицкой и Смирновой (1961) и другими авторами флористических пособий.
Диксон разработал систему мхов, в которой в основу классификации настоящих мхов (Bryidae) положено строение перистома (Dixon, 1924, 1932). Даем позднейший вариант этой системы (Dixon, 1932).
Класс MUSCI
Подкласс I. Sphagnales
Подкласс II. AncJreaeales
Подкласс III. В г у а I e s
Диксон разделил Bryales на две группы, названные им «кланами», а именно:
1. Клан Nematodonteae.
Перистом с плотными зубцами без поперечных полосок, образован несколькими концентрическими слоями клеток.
Порядки: Tetrapnidales, Calomniales, Schistostegales, Buxbauminales,
Polytrichales.
2. Клан Arthro donte ae.
Зубцы перистома тонкие, перепончатые, образованы одним слоем клеток, с поперечными полосками, членистые, или перистом отсутствует. Этот клан Диксон, в свою очередь, делит на три подклана.
1) Подклан Haplolepideae
Зубцы перистома часто вверху вильчатые, при основании образованы двумя слоями пластинок; с наружной стороны один вертикальный ряд пластинок, с внутренней — два ряда. Почти исключительно акрокарпные мхи.
Порядки: Fissidentales, Grimmiales, DicranaJes, Syrrhopodontales, Pot- tiales.
2) Подклан Heterolepideae
Зубцы перистома очень разнообразны. Перистом простой или двойной, гаплолепидейный или диплолепидейный или же отсутствует.
Порядок Encalyptales.
3) Подклан Diplolepideae.
Перистом обычно двойной. Каждый зубец внешнего перистома состоит из двух слоев пластинок, с наружной стороны из двух вертикальных рядов пластинок, с внутренней — из одного. Внутренний перистом, если имеется, обычно более нежный, состоит из основного цилиндра с шестнадцатью выростами, противостоящими или чаще чередующимися с зубцами наружного перистома. Часто между зубцами внутреннего перистома имеются нитчатые реснички. Плеврокарпные или акрокарпные мхи.
Порядки: Orthotrichales, Funariales, Eubryales, Isobryales, Hookeriales, Hypnobryales.
Реймерс в двенадцатом издании Энглеровского «Syllabus der Pflanzenfamilien» (Reimers, 1954) класс мхов делит на пять подклассов.
271
Класс MUSCI
1- й подкласс Sphagnidae с порядком Sphagnales.
2- й подкласс Andreaeidae с порядком Andreaeales.
3- й подкласс В г у i d а е с порядками: Archidiales, Dicranales, Fissidenta-
les, Pottiales, Grimmiales, Funariales, Schistostegales, Tetraphidales, Eubry-
ales, Isobryales, Hookeriales, Hypnobryales.
4- й подкласс Buxbaumiidae с порядком Buxbaumiales.
5- й подкласс Polytrichidae с двумя порядками Polytrichales, Daw-
soniales.
Система Реймерса, по существу, отличается от системы Бро- теруса только тем, что три группы порядков, на которые Бротерус делит подкласс Bryales, у Реймерса рассматриваются как самостоятельные подклассы.
Таким образом, мы видим, что новейшие системы Бротеруса, Диксона и Реймерса совершенно отбрасывают принятое Лимприх- том деление настоящих мхов на клейстокарпные и стегокарпные и на акрокарпные и плеврокарпные ввиду того, что эти группы не являются филогенетически оправданными. Нужно также от-
Рис. 90. Схема филогенетических связей между порядками мохообразных.
метить, что основные подразделения (подклассы) у новейших авторов более или менее совпадают, что объясняется хорошей их обособленностью, восходящей, очевидно, еще к палеозою.
Как уже упоминалось, из отложений карбона Рено, Цейлером и Линье описаны Muscites polytrichaceus и М. bertrandi, напоминающие современные Bryidae, а из пермских отложений Печорского края и Кузнецкого бассейна Нейбург описала ряд родов, несомненно относящихся к мхам, и среди них роды, напоминающие строением клеток виды рода Sphagnum. Споры сфагновых мхов указаны для верхнетриасовых отложений Германии и юрских отложений разных стран, листья и споры типичных сфаг-
272
новых мхов описаны из юрских отложений Баварии и СССР. Все это говорит о древности мхов.
Мхи, несомненно, имеют общие корни с печеночниками, однако эти группы обособились, очевидно, еще на ранних стадиях становления мохообразных. Такое предположение подтверждается тем, что при общем типе развития мхи отличаются от печеночников рядом характерных черт. Так, у мхов нет столь характерных для печеночников масляных телец. Листья у мхов цельные, в то время как для печеночников характерны листья, разделенные на лопасти. В спорогоне большинства мхов имеется колонка и перистом, отсутствующие у печеночников \ и в то же время у мхов нет ни элатер, ни питающих клеток.
На прилагаемой схеме филогенетических связей мохообразных (рис. 90) они выведены из зеленых водорослей параллельно с антоцеротовыми и псилофитами. Общий ствол мохообразных очень рано разделился на две крупные ветви — печеночники и мхи. Развитие печеночников, по нашему мнению, пошло тремя ветвями — сферокарпальной, которая разделилась на два разветвления — сферокарпальные и маршанциальные, метцгериальной и гапломитриально-юнгерманниальной. Среди мхов очень рано обособились три основные группы — андреевые, сфагновые и настоящие мхи.
Не отрицая возможностей отдельных случаев упрощения строения организмов, редукции их в связи с приспособлением к некоторым специфическим условиям существования, например переход из наземных условий в водную среду, мы на основании изложенного материала все же считаем, что эволюция в пределах мохообразных в основном проходила прогрессивно — от простых талломных форм с одной стороны к более сложным талломным, а с другой — к листостебельным. Наибольшего родового и видового разнообразия достигли листостебельные юнгерманниальные и настоящие мхи.
1 Колонка есть у антоцеротовидных, которые мы выделяем в особый подотдел зеленых растений.
ЗАКЛЮЧЕНИЕ
В настоящей работе мы попытались осветить современное состояние проблемы происхождения и основных направлений развития бессосудистых растений, т. е. низших автотрофных и гетеротрофных организмов, а также мохообразных. Нужно отметить, что использованное в заглавии работы выражение «бессосудис- тые растения» имеет только описательное значение, но никак ие систематическое, так как оно охватывает разнородные организмы, общей чертой подавляющего большинства которых является отсутствие проводящей системы, столь характерной для высших растений, Tracheophyta.
Мы принимаем деление органического мира нашей планеты на три основные отдела — «царства» — неклеточные организмы (Acytobionta), к которым относятся вирусы и бактерии, прокариотические организмы (Procaryota, Protocaryota), включающие филы бактерий и синезеленых водорослей, и эвкариотические организмы (Eucaryota), объединяющие филы животного и растительного мира, за исключением дробянок — бактерий и синезеленых.
Принимаемые нами три основных отдела («царства») органического мира нельзя рассматривать как последовательные ступени эволюции, выводя их одно из другого. Каждое царство является своеобразной основной группой, прошедшей независимую эволюцию, в процессе которой оно дало ряд больших подразделений — растительных и животных фил. Это отражено па нашей схеме, согласно которой первичные организмы дали три основные отдела — «царства», у представителей которых на общей исходной основе по разному в условиях реальной среды успешно решается вопрос развития жизни.
Автотрофные растительные организмы, на основании принципов Мережковского и Пашера и с учетом данных других авторов, мы делим на десять фил, которые при классификации рассматриваем как отделы растительного мира.
Мы считаем целесообразным предложение о включении в филу (отдел) зеленых растений (Chlorophyta) всех зеленых организ¬
274
мов, имеющих хлорофилл а и b и образующих крахмал, начиная с зеленых водорослей и кончая покрытосеменными.
Отдел золотистых (хризофитовых) водорослей, следуя Бурел- ли, мы делим на основании строения жгутикового аппарата на два класса: Heterochrysophyceae, имеющие один перистый и один бичевидный жгутики или же один перистый, и Isochrysophyceae с двумя бичевидными жгутиками и часто с гаптонемой.
Отдел желтозеленых (ксантофитовых) водорослей мы рассматриваем как один класс, который, следуя Пашеру, делим на шесть порядков с некоторыми номенклатурными изменениями.
Диатомовые водоросли разделены на два класса — Centrodia- tomophyceae (Centricae) и Pennatodiatomophyceae (Pennatae), из которых исходным является первый, развившийся в юрском периоде из жгутиковых форм, возможно, близких к современным золотистым водорослям. В верхнем мелу появляются двусимметричные Anaulales, давшие начало пеннатным диатомовым. В основании класса пеннатных диатомовых нужно поставить первичнобесшовные, от которых пошли две линии развития — щелешовная и каналошовная.
Бурые водоросли представляют собой монолитную монофиле- тическую группу, развившуюся из флагеллоидных форм с двумя разными жгутиками — одним перистым, другим — бичевидным, с одним классом Phaeophyceae, который мы, следуя Паиенфуссу, делим на одиннадцать порядков.
Нами принято предположение Пашера о развитии пиррофито- вых двумя ветвями, представляемыми классами Dinophyceae и Desmophyceae. Исходными для тех и других были монадные формы, близкие к предкам криптофитовых.
После рассмотрения взглядов различных авторов на возможные родственные связи криптофитовых, на основании ряда морфологических и биохимических особенностей, мы считаем целесообразным выделение криптофитовых в самостоятельный отдел.
Точно также эвгленовые водоросли представляют самостоятельную филу и должны рассматриваться в систематике как особый отдел, отличный от зеленых и пиррофитовых водорослей.
Для более детального изучения строения жгутикового аппарата и природы пигментов хлоромонад целесообразно предложение Прескотта рассматривать их как самостоятельную филу (отдел).
Строение клетки красных водорослей и их органов размножения и характер полового процесса свидетельствуют о принадлежности их к эвкариотическим организмам. Что же касается сходства (но не идентичности) состава пигментов красных и синезеленых водорослей, то оно, вероятно, является следствием развития этих групп в аналогичных экологических условиях, а не результатом общности происхождения. Классы Bangiophyceae и Florideophyceae при всех отличиях имеют ряд общих основных черт, что свидетельствует об общности их прорхсхождения.
Зеленые водоросли рассматриваются как подотдел зеленых
275
растений. Исследования последних лет подтвердили мнение Кор- шикова о неоднородности группы монадных форм, относимых к порядку вольвокальных, и дали обоснование предложению Шадефо о выделении класса Prasinophyceae. В предлагаемой классификации зеленые водоросли разделены на классы Prasinophyceae и Euchlorophyceae. Класс Euchlorophyceae разделен на подклассы Euchlorophycidae, Zygophyeidae и Charophycidae.
В отличие от обычно принятого разделения гетеротрофных растительных фил на слизевики (Myxophyta) и грибы (Fungi), мы считаем необходимым, исходя из полифилетичности грибных организмов, выделять среди них четыре филы (отдела) — Мухо- mycota, Chytridiomycota, Saprolegniomycota и Eumycota. Основанием для этого является строение монад, в частности жгутикового аппарата, биохимические особенности, характер полового процесса и органов размножения. Из них Myxomycota, Chytridio- mycota и Eumycota являются первичными гетеротрофами, в то время как Saprolegniomycota по характеру жгутикового аппарата и наличию целлюлозы связываются с золотистыми водорослями.
Принимая во внимание исследования Олайва, выделившего нротостелиомицеты, мы считаем обоснованным деление слизевиков на пять классов, а именно Protosteliomycetes, Labyrinthulomy- cetes, Acrasiomycetes, Myxogastromycetes и Plasmodiophoromycetes.
В отделе хитридиевых грибов мы принимаем один класс — Chytridiomycetes, который делим на три порядка — Chytridiales, Blastocladiales и Monoblepharidales.
К отделу сапролегниевых грибов отнесены организмы как с двумя жгутиками — одним перистым и другим бичевидным (класс Saprolegniomycetes), так и организмы с одним перистым жгутиком (класс Hyphochytriomycetes).
Отдел настоящих грибов (Eumycota) мы делим на три класса: Zygomycetes, Ascomycètes и Basidiomycetes. Филогенетические связи аскомицетов с зигомицетами подтверждаются наличием переходных форм. Гомология аска и базидии, сходство полового процесса у базидиальных грибов и аскомицетов свидетельствует о их происхождении от общих предков. Что же касается происхождения зигомицетов, то связи их с сапролегниевыми и хитри- диевыми грибами являются очень проблематичными. Более вероятным является предположение о развитии зигомицетов от предков, близких к хитридиевым. Однако эти связи, по-видимому, относятся к ранним стадиям дифференциации низших эвкариоти- ческих организмов.
Класс аскомицетов мы делим на два подкласса — Protoascomy- cetidae и Euascomycetidae. К первому отнесены организмы с аска- ми простого строения, развивающимися из зиготы без образования аскогенных гиф и плодовых тел, а ко второму — организмы, характеризующиеся разнообразными формами полового процесса, сложным развитием и строением сумок и в большинстве случаев наличием плодовых тел.
276
Что касается класса базидиомицетов, то его происхождение связывается с исходными формами аскомицетов. В основании класса должны быть поставлены организмы с простым типом развития и одноклеточной неспециализированной базидией. Из современных базидиомицетов ближе всего к исходным формам стоят порядок Tremellales и простейшие роды порядка Theleophorales. Из подобных примитивных форм развились прочие базидиомице- ты двумя основными ветвями — склеробазидиальной и гомобази- диальной. В связи с этим предлагается деление класса Basidiomy- cetes на три подкласса: Protobasidiomycetidae, Sclerobasidiomyceti- dae и Homobasidiomycetidae.
Лишайники являются гетерогенной группой. Поэтому мы считаем правильным распределение семейств лишайниковых грибов между соответствующими порядками классов Ascomycètes и Ва- sidiomycetes.
Мохообразные и антоцеротовые рассматриваются нами как подотделы отдела зеленых растений — Chlorophyta. Мы считаем наиболее обоснованной теорию их происхождения от зеленых водорослей. Из зеленых водорослей развились отдельными стволами антоцеротовые, мохообразные (печеночники и мхи) и псилофи- товые, которые рассматриваются как подотделы отдела Chlorophyta. От псилофитовых произошли субфилы сосудистых растений.
На основании данных сравнительной морфологии, систематики, цитологии, палеоботаники и географического распространения наиболее древними в классе печеночников нужно считать простые талломные формы, известные уже из отложений девонского времени. От простых талломных форм развились с одной стороны сложные талломные формы (подкласс Marchantiidae), а с другой — листостебельные (подкласс Jungermanniidae).
В пределах класса настоящих мхов рано обособились три ветви, рассматриваемые нами как подклассы — Andreaeidae, Sphagni- dae и Bryidae.
Так в основных чертах представляем мы развитие основных подразделений бессосудистых растений. Интенсивное развитие исследований в течение последних десятилетий дало новые материалы, которые побудили к пересмотру многих казалось бы установившихся представлений. Несомненно, дальнейшие исследования с применением новой методики, новых приборов позволят глубже познать строение и развитие низших растительных организмов, дадут новые данные для изучения их эволюции и построения 9истемы, покажут в некоторых случаях ошибочность наших современных представлений. Однако это не должно нас смущать в дальнейших исканиях на путях к познанию органического мира, его происхождения и путей развития. В связи с этим нельзя не напомнить высказывание В. Гете, который писал, что «заблуждения нас никогда не оставляют; однако высшая потребность всегда влечет пытливый ум медленно ввысь к истине» (В. Гете. Четыре времени года. Осень, 58).
277
ЛИТЕРАТУРА
Абрамов И. И. и С а в и ч - Л ю б и ц к а я Л. И. Тип Bryopsida — мохообразные.— В кн.: Основы палеонтологии. Изд-во АН СССР, М., 1963, 344—414.
Абрамова А. Л., С а в и ч - Л ю б и ц к а я Л. И., Смирнова 3. Н. Определитель листостебельных мхов Арктики СССР. Изд-во АН СССР, М.- Л., 1961.
А с а у л 3. I. Основш принципи класифшацп евгленових водоростей.— Укр. бот. жури.,27, 1970, 545—556
Бернал Д ж. Возникновение жизни. «Мир», М., 1969.
Вага А. Я. Филема органического мира.— Бот. жури., 37, 1952, 639—647.
Вологдин А. Г. Земля и жизнь. Эволюция среды и жизни на Земле. М.— Л., 1963.
Г е и к е л ь А. Г. Новые взгляды на систему низших споровых в связи с изменением номенклатуры.— Изв. Биол. н.-и. ип-та и биол. станции . при Пермском ун-те, 2, 5, 1923.
Гоби X. Низшие споровые или глеофиты (Gloeophytae). СПб, 1883.
Гоби X. Общий курс ботаники. Лекции, читанные в С.-Петербургском университете в 1911—12 уч. году. Споровые растения, СПб, 1912.
Г о б и X. Обозрение системы растений. Птр., 1916.
Голлербах М. М. и Полянский В. И. Пресноводные водоросли и их изучение. Определитель пресноводных водорослей СССР, 1. «Советская наука», М., 1951.
Дедусенко-Щеголева H. М., Матвиенко А. М., Шкорба- т о в Л. А. Зеленые водоросли. Класс вольвоксовьте. Определитель пресноводных водорослей СССР, 8. Изд-во АН СССР, М.—Л., 1959.
Догель В. А. Зоология беспозвоночных, изд. 4. «Советская наука», М., 1947.
Дудка I. О. Нова система порядку Hyphomycetales (Mondiales).— Укр. бот. журн., 25, 2, 1968, 3—16.
Ж у з е А. П. и П р о ш к и и а - Л а в р е π к о А. И. Порядок Mediales.— В кн.: Диатомовый анализ. Госгеолиздат, Л., 2, 1949, 210—224.
Ж у з е А. П. и Шешукова В. С. Часть палеоботаническая.— В кн.: Диатомовый анализ. Госгеолиздат, Л., 1,1949, 103—184.
Забелина М. М., К и с е л е в И. А., Прошкин а-Лавренко А. И., Шешукова В. С. Диатомовые водоросли. Определитель пресноводных водорослей СССР, 4. «Советская наука», М., 1951.
Залесский М. Д. Описание некоторых ископаемых растений из бардин- ского яруса Урала.— Пробл. палеонтол., 2—3, 1937, 37—101.
3 е р о в Д. К. Проблема происхождения архегониатных растений.— Сов. бот., 13, 1945, 3—13.
278
Зеро в Д. К. Фшогешя мохопод1бш1Х ί ϊχ М1сце в систем! рослинного cbî- ту.— Бот. жури. АН УРСР, 8, 4, 1951, 3—14.
3 е р о в Д. К. Флора печшочних i сфагнових moxîb Украши. «Наукова думка», К., 1964.
Зеров Д. К. Вопросы филогении печеночников (Hepaticopsida).— Бот. журн., 51, 1966, 3—14.
Зеров Д. К. Сучасний стан проблемп походження рослинного свИу.— Укр. бот. журн., 27, 1970, 144—157.
Зеров Д. К. i Зеров а М. Я. Основш наирямки еволюци грибних орга- ΗΪ3ΜΐΒ.— Укр. бот. журн.,25, 5, 1968, 3—17.
3 е р о в а М. Я. Грибш хвороби вид1в клена на Правобережж1 Украш- cbKoï PGP.- Бот. журн. АН УРСР, 9, 1, 1952, 42-44.
3 е р о в а М. Я. Питания класифшацп аскових rpii6ÎB.— Укр. бот. журн., 21, 1, 1964, 3-17.
Зингер Р. О. О новых данных для систематики и филогении Agaricales как потомков Gastromycetes.— Сов. бот., 1, 1939, 95—98.
И ид е н к о Т. А. Флора верхов нижнего — низов верхнего девона Подольского Приднестровья.— В кн.: Палеонтология и стратиграфия нижнего палеозоя Волыно-Подолшь «Наукова думка», К., 1968, 80—113.
Киселев И. А. Пиррофитовые водоросли. Определитель пресноводных водорослей СССР, 6. «Советская наука», М., 1954.
Козо-Полянский Б. М. Новый принцип биологии. Очерк теории сим- биогенеза. «Пучина», М.— Л., 1924.
К о з о - П о л я н с к и й Б. М. Происхождение высших растений. 1. Первенцы сухопутной флоры. Вологда, 1927.
К о з о П о л я н с к и й Б. М. О новой системе растений — ДАН СССР, 56.
3, 1947, 309-311.
Козо-По л янский Б. М. Способы изображения эволюционной системы растений.— Бот. журн. АН СССР, 34, 1949, 245—252.
К о з о - П о л я н с к и й Б. М. К модернизации системы растительного мира.— Тр. Воронежск. ун-та, 15, 1949, 76—129.
К о м а р и и ц к и й Н. А., Кудряшов Л. В. и У ран о в А. А. Систематика растений. Учпедгиз, М., 1962.
К о р in и к о в A. A. Protococcineae, новая группа зеленых Flagellata.—Архив Русск. Протистол. Общ., 2, 1923, 148—169.
К о р ш i к о в О. А. Про статевнй ироцес у Micractinieae.— Уч. зап. Харшв- ського ун-ту, 10, 1937, 109—126.
К о р ш i к о в О. A. Volvocineae. Визпачннк щйсноводиих водоростей УРСР,
4. В-во АН УРСР, К., 1938.
ΚορυιίκοΒ О. А. Шдклас προτοκοκοΒΐ (Protococcineae). Визначник npic- иоводних водоростей Украшсько'1 PCP, 5. В-во АН УРСР, К., 1953.
Кудряшов Л. В. Тип Мохообразные (Bryophyta) — В кн.: Комарниц- кий Н. А., Кудряшов Л. В., Уранов А. А. Систематика растений. «Учпедгиз», М., 1962.
К у р с а и о в Л. И. Бурые и красные водоросли. Изд-во МГУ, М., 1927.
К у р с a π о в Л. И. Микология. «Учпедгиз». 1-е изд., М., 1933; 2-е изд., 1940.
Курса нов Л. И., Комарницкий Н. А. и Флеров Б. К. Курс низших растений, 1-е изд. «Медгиз», М.—Л., 1933; 2-е изд., «Биомедгиз», М.— Л., 1937.
К у р с а и о в Л. И. и Комарницкий Н. А. Курс низших растений, 3-е изд. «Советская наука», М., 1945.
К у р с а и о в Л. 11., 11 а умов Н. А., Красильников Н. А., Г о р- л е и к о М. В. Определитель низших растений, 3 — Грибы. «Советская наука», М., 1954.
К у р с a π о в Л. И. и Шемаханова H. М. К вопросу о смене ядерных фаз у зеленых водорослей 1. Цикл развития Chlor о chy trium lemnae Cohn.—Русск. apx. протистол., 6, 1—4, 1927, 131—146.
Лавров Н. И. Флора грибов и слизевиков Сиби́ри, 5. Очерк микофлоры злаков Сибири, 1951,
279
Лазаренко А. С. Вопросы генезиса чередования поколений у моховидных.— Журн. общ. биологии, 22, 1961, 372—382.
Маслов В. П. Тип Charophyta. Лучшщы или харовые водоросли.—В кн.: Основы палеонтологии. Изд-во АН СССР. М., 1963, 224—242.
Маслов В. П. Тип Rhodophyta. Красные или багряные водоросли.— В кн.: Основы палеонтологии. Изд-во АН СССР, М., 1963, 243—260.
Маслов В. П. и Ананьев А. Р. Тин Phaeophyta. Бурые водоросли.— В кн.: Основы палеонтологии. Изд-во АН СССР, М, 1963, 188—192.
Матвиенко А. М. Золотистые водоросли. Определитель пресноводных водорослей СССР, 3. «Советская наука». М., 1954.
Мат big и ко О. М. Золотист) водоросы — Chrysophyta. Визначник npicHO- водних водоростей Украшсько!’ PCP, 3, 1, «Наукова думка», К., 1965.
Мейер К. И. Происхождение наземной растительности. М., 1-е пзд., 1922; 4-е пзд. «Советская наука», 1946.
Мейер К. И. Морфология и систематика высших растений. 1. Архегони- альные растения. «Советская наука», М., 1947.
Мейер К. И. К филогении зеленых водорослей.— Бюл. Моек, о-ва йен. прир., отд. биол., 56, 1, 1951а, 53—65.
Мейер К. И. К филогении разножгутиковых водорослей,— Бюл. Моек, о-ва исп. прир., отд. биол., 56, 4, 19516, 50—54.
Мейер К. И. Опыт филогенетической системы зеленых водорослей.— Бюл. Моек, о-ва исп. прир., отд. биол., 57, 5, 1952, 56—64.
Мейер К. И. О филогении зеленых водорослей (ChloiOphycophyta).—Бот. журн, 46, 1961, 1073-1086.
Мережковский К. С. Конспективный курс споровых растений, ч. 1 и II. Казань, 1909а.
Мережковский К. С. Теория двух плазм, как основа симбпогенезиса, нового учения о происхождении организмов. Казань, 19096.
Миллер В. В. К филогенетической систематике зеленых водорослей.— Дневник Всесоюзного съезда ботаников в Ленинграде в 1928 г. Л.. 1928, 1955-1956.
Наумова С. Н. Споры нижнего кембрия.— Изв. АН СССР, сер. геол, 4, 1949.
Наумова С. Н. Споры нижнего силура —Тр. конф. по спорово-пыльцевому анализу, 1948 г, 1950, 165—190.
Наумова С. Н. Спорово-пыльцевые комплексы верхнего девона Русской платформы и их значение для стратиграфии.— Тр. Ин-та геол. наук АН СССР, 143, сер. геол, 60, 1953.
Нейбург Μ. Ф. Открытие листостебельных мхов в пермских отложениях СССР.- ДАН СССР, 107, 2, 1956, 321-324.
Нейбург Μ. Ф. Листостебельные мхи из пермских отложений Ангариды.— Тр. Геол. ин-та АП СССР, 19, М, 1960.
О кс не р А. М. Систематичне положения лишайниюв та основш фшоге- нетичш лшп ïx розвитку.—Укр. бот. журн, 13, 1, 1956, 15—30.
Окснер А. М. Флора лишайниюв Украши. К, 1, 1956; 2, вып. 1, К, 1968.
Опарин А. И. Происхождение жизни. М, 1924.
Опарин А. И. Возникновение жизни на Земле. 1-е пзд, М, 1936; 2-е пзд, М.— Л, 1941; 3-е изд, М, 1957.
11 е ш к о в М. А. Сравнительная цитология сине-зеленых водорослей, бактерий и актиномицетов. «Наука», М, 1966.
Попова Т. Г. Эвгленовые (Ëuglenineae) Европейского Севера СССР.— Тр. Бот. ин-та АН СССР, сер. II, 7, 1951.
Попова Т. Г. Эвгленовые водоросли. Определитель пресноводных водорослей СССР. 7. «Советская наука», М, 1955.
С а в и ч - Л ю б и ц к а я Л. И. и Абрамов И. И. Геологическая летопись мохообразных.— Бот. журн, 43, 1958, 1409—1417.
Стан и с л а в с к и й Ф. А. Ископаемая флора батско-келловитскпх отложений Донецкого бассейна и Днепровско-Донецкой впадины. К, 1957.
Сухов К. С. Вирусы. Изд-во АН СССР, М, 1956.
280
T а X т а д ж я и А. Л. Соотношения онтогенеза и филогенеза у высших растений.— Т|». Ерев. Гос. унив., 22, 1943, 71 —176.
ï а X т а д ж я и А. Л. Филогенетические, основы системы высших растений.— Бот. журн., 35, 1950, ИЗ—139.
T а X т а д ж я л А. Л. Высшие растения. 1. От псилофитовых до хвойных.
М.— Л., 1956.
Т о в а р н и ц к π и В. И. Основные этапы развития молекулярной биологии вирусов.— В кн.: Молекулярные основы биологии вирусов. М., 1966, 379—410.
Топачевськнй О. В. Походження кон’югат i типи статевого процесу у них.— Бот. журн. АН УРСР, 9, 2, 1952, 32—35.
Топа ч е веки й А. В. Вопросы цитологии, морфологии, биологии и филогении водорослей. Изд-во АН УССР, К., 1962.
Ф а м и н ц ы и А. и Баранец кий И. К истории развития гонидий и образования зооспор у лишайников.— Изв. АН, сер. 7, И, 9, 1867.
Фрей-Висслинг А., Мюлет а л ер К. Ультраструктура растительной клетки. М., 1968.
III о ш у к о в а В. С. Введение в систематику диатомовых водорослей.— В кн.: Диатомовый анализ. Госгеолиздат, М., 2, 1949, 5—13.
Я ч е в с к и й А. А. Микологическая флора Европейской н Азиатской России. т. 2. Слизевики. М., 1907.
Я невский А. А. Определитель грибов, 2-е изд., 1. Совершенные грибы. СПб, 1913.
Ячевский А. А. К филогенетике грибов.—В кн.: Юбилейный сборник, посвященный И. И. Бородину. Л., 1927.
Ячевский А. А. и Я ч е в с к и й П. А. Определитель грибов. Совершенные грибы (диплоидные стадии), 1. Фикомицеты, М.— Л., 1931.
А с h а г i и s E. Methodus Lichenum. Stockholm, 1803.
А с h а г i и s E. Synopsis methodica Lichenum. Lund, 1814.
A g а г d h С. A. Synopsis algarum Scandinaviae. Lund, 1817.
A g a r d h J. G. Species, genera et ordines algarum. 1, Lund. 1848.
Alexopoulos C. J. The Myxomycètes IL— Bot.Rev., 29, 1963, 1—78.
A 1 e X о p о и 1 о s C. J. Einführung in die Mycologie. Jena, 1966.
Allen C. E. Die Keimung der Zygote bei Coleochaete.— Ber. Deutsch, bot. Ges., 23, 1905, 285—292.
Allen M. В. Studies with Cyanidium caldarium, an anomalously pigmented Chlorophyte.— Arch. Mikrobiol., 32, 1959, 270—277.
Allen R. F. A cytological study of heterothallism in Puccinia sorghi J. Agr. Res., 49, 1934, 1047—1068.
Allen R. F. A cytological study of Erysiphe polygoni on Delphinium.— J. Agr. Res., 53, 1936, 801-818.
Alio r go P. Iléterocontes ou Xanthophycées? — Rev. Algol., 5, 1930, 230.
A11 s о p p A. Phylogenetic relationships of the Procaryota and the origin of the eucaryotic cell.— New Phytol., 68, 1969, 591—612.
Areschoug J. E. Phyceae scandinavicae marineae.— Nova Acta reg. soc. sc. Upsal., 13, 1847 (цит. no: Kylin, 1933).
Aronson J. M. The cell wall.—In: G. C. Ainsworth and A. S. Sus- sman (ed.). The Fungi. An advanced treatise. N. Y. and London, I, 1965, 49—76.
Arthur et al. The plant rusts (Uredinales). N. Y., 1929.
A rx J. Λ., von. Pilzkunde, Lehre, 1967; 2-te Aufl., 1968.
Atkinson G. F. Some problems in the evolution of the Lower Fungi.— Ann. MycoL 7, 1909, 441—472.
Atkinson G. F. Phylogeny and relationships in the Ascomycetes.—Ann. Missouri Bot, Garden, 2, 1915, 315—376.
Backus M. P. Initiation of the ascocarp and associated phenomena in
Coccomyces hiemalis.— Contr. Boyce Thompson <Inst., 6, 1934, 339—379; (цит. no: Gäu rn a n n, 1964).
Backus M. P. The mechanics of conidial fertilization in Neurospora sito~ phila.— Bull. Torrey Bot. Club, 66, 1939, 63—76.
281
B a r g h о о г η E. S. and Tyler S. Л. Microorganisms from the Gunflint Chert.— Science, 147, 1965, 568—577.
Barghoorn E. S. and S c h о p f J. W. Microorganisms three billion years old from the Precainbrian of South Africa.— Science, 152, 1966, 758—763.
Barkley F. A. Keys to the phyla of organisms. Missoula, Montana, 1939.
Barksdale A. W. Meiosis in the antheridium of Achlya ambisexualis E 87.—J. E. Mitchell Sei. Soc,, 84, 1968, 187—194.
Barrett J. T. Development and sexuality of some species of Olpidiopsis (Cornu) Fische r.— Ann. Bot., 26, 1912, 209—288.
В a г y A., d e. Ueber die Myxomyceten.— Bot, Zeitung, 16, 1858, 357—358; 361-364; 365-369.
Вагу A., de. Die Mycetozoen. Ein Beitrag zur Kenntnis der niedersten Thiere.— Zeitschr. Wiss. Zool., 10, 1859, 88—175.
В а г у A., de. Morphologie und Physiologie der Pilze, Flechten und Myxomyceten. Leipzig, 1866.
Вагу A., de. Zur Systematik der Thallophyten.—Bot. Zeitung, 39, 1881, 1, 1-17, 2, 33-36.
В а г у A., de. Vergleichende Morphologie und Biologie der Pilze, Mycetozoen und Bactérien. Leipzig, 1884.
B a t r a L. R. A comparative morphological and physiological study of the species of Dipodascus.— Mycologia, 51, 1959, 329—355.
Bauch R. Untersuchungen über die Entwicklungsgeschichte und Sexualphysiologie der Ustilago bromivora und Ustilago grandis.— Zeitschr. Bot., 17, 1925, 129-177.
B a u r E. Zur Frage der Sexualität der Collemaceen.— Ber. Deutsch, bot. Ges., 16, 1898, 363-367.
Bazin M. J. Sexuality in a blue-green alga: genetic recombination in Ana- cystis nidulans.— Nature, 218, 5138, 1968, 282—283.
B e g e r H. Euglenophyta.— In A. En gler’s Syllabus der Pflanzenfamilien, 12-te Aufl. Berlin — Nikolassee, 1954, 26—64.
B e g e r H. Chlorophyta — Grünalgen.— In: A. E n g 1 e r’ s Syllabus der Pflanzenfamilien, 12-te Aufl. Berlin — Nikolassee, 1954, 86—107.
Benedict W. G. Haplophase activity in Stemonitis fusca R о t h.— Canad. J. Bot., 40, 1962, 71-76.
Benesovâ VI. Fouets pleuronématés et disposition ampliconte chez VHaematococcus pluvialis F 1 о t о w.— C. R. Acad. Sc., Paris, 228, 1949, 1883-5.
Bergman B. Zytologische Studien über die Befruchtung und Askokarpbil- dung bei Sphaerotheca caslagnei L c v.— Svensk Bot. Tidskr., 35, 1941, 194-210.
Berry E. W. Contributions to the mesozoic flora of the Atlantic coastal plain, XV.- Bull. Torr. Bot. CL, 55, 1928, 441-448.
Berthold. Zur Kenntnis der Siphoneen and Bangiaceen.— Mitt. Zool. Stat. Neapel, 2, 1880, 72.
Bessey Ch. E. A synopsis of plant phyla.—Univ. Studies of the University of Nebraska, 7, 4, Linkoln, 1907, 275—358.
Bessey E. A. A text-book of mycology. Philadelphia, 1935.
Bessey E. A. Some problems in fungus phylogeny.—Mycologia, 34,4, 1942.
Bessey E. A. Morphology and taxonomy of Fungi. Philadelphia — Toronto, 1950.
Betts E. M. Heterothallism in Ascobolus carbonarius.— Amer. J. Bot., 13, 1926, 427-432.
Biecheler B. Sur l’existence d’une copulation chez une Euglène verte et sur les conditions globales qui la déterminent.— Compt. Rend. Soc. Biol., 124, 1937, 1264-1266.
B i g g s R. Dipodascus uninucleatus.— Mycologia, 29, 1937, 34—44.
Bisset K. A. The cytology and life-history of bacteria. 2-nd ed. London, 1955.
282
Bi st i s G. Sexuality in Ascobolus stercorarius. I. Morphology of the ascogo- nium; plasmogamy; evidence for a sexual hormonal mechanism.— Amer. J. Bot., 43, 1956. 389—394.
Bistis G. Sexuality in Ascobolus stercorarius II. Preliminary experiments on various aspects of the sexual process.— Amer. J. Dot., 44, 1937, 436—443.
Blackman V. H. On the fertilization, alternation of generations, and general cytology of the Uredineae.—Ann. Bot., 18, 1904, 323—373.
Blackman V. II. a. F r a s e г H. C. J. Fertilization in Sphaerotheca.— Ann. Bot., 19, 1905, 567—569.
Blackman F. F. a. T a n s 1 e y A. G. A revision of the classification of the green Algae.— New Phytol., I, 1902, 17—24, 47—48, 67—72.
В о h 1 i n К. Studier öfver nägra slägten af Alggruppen Confervales В о r z i.— Bih. Svensk. Vet.-Akad. Hand!.. 23, Afd. 3, 3, 1897, 3-56.
Bo hl in K. Utkast tili de gröna Algernas och Arkegoniaternas fylogeni. Uppsala, 1901.
Bonner J. T. The cellular slime molds. Princeton Univ. Press. Princeton, 1959.
Bonnet J. Reproduction sexuée et alternance des générations chez les algues.— Progressus rei botanicae, 5, 1914, 1—126.
Börgesen F. Marine Algae from the Canary Islands. III. Rhodophyceae.— Kgl. Danske Vidensk. Selsk., Biol. Meddel., 6, 1927.
В о r z i A. Botrydiopsis, nuovo genre di alghe verde.— Boll. Soc. Ital. Micro- scopisti, I, 1889, 66—70.
В о r z i A. Studi algologici, 2, 1895.
В о r y de Saint-Vincent. Histoire naturelle.— Dictionnaire classique d’Histoire naturelle, 8, Paris, 1825.
В о r y de S a i n t - V i n c e n t. Psychodiaire.— Dictionnaire classique d’Histoire naturelle, 14, Paris, 1928.
Bourrelly P. La structure fine des flagelles et la classification des algues.— Rev. génér. de Botanique, 68, 804, 1961, 332—336.
В о u r r e 11 y P. La classification des Chrysophycées: ses piOblems.— Tenth intern, bot. congress. Abstracts of papers. Edinburgh. 1964, 109.
Bourrelly P. Les Algues d’eau douce, I. Les Algues vertes. Paris, 1966.
Bower F. O. An antithetic as distinct from homologous alternation of generations in plants.—Ann. Bot,, 4, 1890, 347—370.
Bower F. O. The origin of a land flora. London, 1908.
Bower F. O. Primitive land plants also known as the Archegoniatae, 1935.
Bref eld O. Botanische Untersuchungen über Schimmelpilze. Ill: Basidio- myceten I. Leipzig, 1877.
Brefeld O. Botanische Untersuchungen über Schimmelpilze. IV. Leipzig. 1881.
Brefeld. O. Untersuchungen aus dem Gesammtgebiete der Mycologie. VIII: Basidiomyceten Ш, Leipzig, 1889.
Brier ley W. В. The endoconidia of Tkielavia basicola.— Ann. Bot., 29. 1915, 483-493.
Brooks F. T. Some present day aspects of mycology.—Trans. Brit. Mycol. Soc., 9, 1923.
Brother us V. F. Die Laubmoose Fennoskandias. Helsingfors, 1923.
Brother us V. F. Unterklasse Bryales. Spezieller Teil.—In: A. E n g 1 e r, (ed.) Die natürlichen Pflanzenfamilien, 2te Aufl. Leipzig, 10, 1924, II, 1925.
Brown A. M. Diploidization of haploid by diploid mycelium of Puccinia
helianthi S c h w.— Nature, 130, 1932, 111.
Brown A. M. The sexual behaviour of several plant rusts.—Canad. J. Res., Sect., G, 18, 1940, 18—25.
Brown II. F. a. Cox A. An electron microscope study of protozoan flagella.-Amer. Midi. Natur., 52, 1954, 106-117.
Brunswik U. Untersuchungen über die Geschlechts- and Kern Verhältnisse der Hvmenomycetengattung Coprinus.— Bot. Abhandl. herrausgeg. von Dr. K/Goebel, I, 5, 1924, 1—152.
283
Bryant T. R. a. II о w a r d K. L. Meiosis in the Oomycetes. T. A microspe- ctrophotometric analysis of nuclear deoxyribonucleic acid in Saprolegnia terrestris.— Amer. J. Bot., 56, 1969, 1075—1083.
Buch H., Evans A. a. Verdoorn Fr. A preliminary check list of the Hepaticae of Europe and America (north of Mexico).—Ann. Bryol., 10, 1938, 3-8.
Buckman J. On some fossil plants from the lower Lias.— Quat. J. Geol. Soc., 6, 1850, 413—418.
Buller A. H. R. Researches on Fungi. London, I, 1909; 2, 1909; 3, 1924; 4, 1931; 5, 1933; 6, 1934.
Gain R. F. Myriogonium, a new genus among simplified Ascomycetes.— Mycologie, 40, 1948, 158-167.
Campbell D. H. The structure and development of mosses and ferns. 2nd ed., 1905.
Campbell D. H. On the distribution of the Hepaticae and its significance.— New Phytol., 6, 1907.
Campbell D. H. A remarcable development of the sporophyte in Antho- ceras fusiformis A u s t.— Ann. Bot., 38, 1924, 473—483.
Campbell D. H. The relationship of the Anthocerotae.—Flora, N. F. 18— 19, 1925, 52-74.
Campbell D. H. The evolution of land plants. Stanford. Calif., 1940.
Canter H. M. Studies on British chytrids. XIX. On Phlyctidium apophysatum emend. C a n t e r.—Trans, Brit. Mycol. Soc., 44, 1961. 522—528.
Caram B. Sur la sexualité et le développement d’une Phèophycée: Cylindro- carpus berkeleyi (Grev.) Crouan.—C. R. Acad. Sc. Paris, 245, 1957, 440—443.
Cavers F. The interrelationships of the Brvophyta.—New Phytol. 9, 1910. 81-112, 157-186, 193-234, 269-304, 341-353; 10, 1911, 1-46, 84-86.
Cavers F. Interrelationships of Protista and primitive fungi.— New Phytol., 14, 1915, 94-104, 164-168, 223—227, 275-280, 302-304.
Cejp K. Houby. Praha, I, 1957; 2, 1958.
Celakovsky L. Über die verschiedenen Formen und die Bedeutung des Generationswechsels der Pflanzen.— Sitzungsber. k. böhm. Gesellsch. Wiss., Sitzg. d. math.-nat. Klasse, 1874, 21.
Chadefaud M. Les Psilotinées et l’évolution des Archégoniates.— Bull. Soc. Bot. France, 97, 1950, 99—100.
Chadefaud M. La leçon des Algues.—Ann. bioL, 28, 1952, 19—25.
Chadefaud M. Une Algue unicellulairo marine des îles Tortugas et la notion de Chromophycées.— Bull. Soc. Bot. France, 99, 1952, 84—86.
Chadefaud M. Les végétaux non vasculaires (Cryptogamie).—In: M. Chadefaud etL. Emberger, Traité de Botanique Systématique, I, Paris, 1960.
Chapman D. J. a. C h a p m a n V. J. Life histories in the Algae.— Ann. Bot., N. S. 25, 1961, 547-561.
Chapman D. J., Cole W. J. a. S i e g e 1 m a n H. W. Phylogenetic implications of phycocyanobilin and c-phycocvanin.— Amer. J. Bot., 55, 1968, 314-316.
Chapman V. J. The Algae. London, 1964.
Chatton E. Titres et Travaux scientifiques. Sottano (цит. но: Stanier and van N i e 1, 1962).
C h e n L. C — M., Edelstein T., O g a t a E., and McLachlan L. The life history of Porphyra miniata.— Canad. J. Bot., 48, 1970, 385—389.
Christensen T. Some considerations on the phytogeny of the Bryo- phyta.— Bot. Tidskr., 51, 1954, 53—58.
Christensen T. Remark on the phylogeny of the Bryophyta.— Bot. Tidskr., 53, 1957, 317.
Christensen T. Botanik. Copenhague, 2, 1962 (цит. no: Bourre 11 у, 1966).
Christensen Т. The gross classification of Algae.—Algae and Man. Plenum Press. N. Y., 1964, 59—64.
284
Christman A. H. Sexual reproduction of the rusts.— Bot. Gaz., 39, 1905, 267—275.
Church A. H. Thalassiophyta and the subaerial transmigration. Oxford. 1919.
Claussen H. Zur Entwicklungsgeschichte von Phyllophora brodiaei.— Ber. Deutsch. Bot. Ges., 47, 1929, 544—547.
Claussen P. Zur Entwicklungsgeschichte der Ascomyceten. Boudiera.— Bot. Zeitung, 63, 1905, 1—28.
Claussen P. Zur Entwicklungsgeschichte der Ascomyceten. Pyronema confluens.— Zeitschr. f. Botanik, 4, 1912, 1—64.
Claussen P. Entwicklungsgeschichtliche Untersuchungen über den Erreger der als „Kalkbrut“ bezeicnneten Krankheit der Bienen.— Arb. biol. Reich- sanst, Land-u. Forstw., 10, 1921, 467—521.
Clements F. E. a. Shear C. L. The genera of Fungi. N. Y., 1931.
Cohn F. Untersuchungen über die Entwicklungsgeschichte der mikroskopischen Algen und Pilze.— Nova Acta Acad. Caesar. Leop. Carol., 24, 1853, 103-256.
Cohn F. Beiträge zur Physiologie der Phycochromaceen und Florideen.— Arch. Mikrosk. Anat. Entw. Mech., 3, 1867.
Cohn F. Conspectus familiarum cryptogamarum secundam methodum naturalem dispositarum.— Hedwigia, 9, 2, 1872, 17—20.
Cohn F. Untersuchungen über Bakterien II.—Beitr. Biol. Pfl., I, 3. 1875. 141—207.
Colson В. The cytology and development of Phyllactinia corylea. L é v.— Ann. Bot., N. S. 2, 1938, 381-402.
Conard H. S. Plants of Iowa, being a fifth edition of the Grinnell Flora, revised and enlarged.— Iowa Acad. Sei., Biol. Surv. Publ., 2, 1939. 1-95.
Cook W. R. I. The interrelationships of the Archimycetes.—New Phytol.. 27, 1928, 230-260; 298-320.
Cook W. R. I. a. Schwartz E. J. The life — history, cytology and method of infection of Plasmodiphora brassicae Woron., the cause of finger and toe disease of cabbage and other Crucifers.— Phil. Trans. R. Soc., London, ser. B, 218, 1930, 283-314.
Copeland IL F. The kingdoms of organisms.—Quart. Rev. Biol., 13, 1938, 383-420.
Copeland H. F. Progress report on basic classification.—Amer. Natur., 81. 800, 1947, 340-361.
Copeland H. F. The classification of lower organisms.—Pacific Books. Palo Alto, 1956.
Cotter R. U. Fertilization of pycnia with urediospores and aeciospores in Puccinia graminis.— Phytopath., 50, 1960, 567—568.
Couch J. N. New or little known Chytridiales.— Mycologia, 27, 1935, 160— 175; 376-387.
Couch J. N. A new fungus intermediate between the rusts and Septobasi- dium.— Mycologia, 29, 1937, 665—673.
Couch J. N. The structure and action of the cilia in some aquatic Phycomy- cetes.— Amer. J. Bot., 28, 1941, 704—713.
С о u c h J. N. a. WhiffenA. J. Observations on the genus Blastocladiella.— Amer. J. Bot., 29, 1942, 582—591.
Craigie J. H. Discovery of the function of the pycnia of the rust fungi.— Nature, 120. 1927, 765-767.
Craigie J. H. An experimental investigation of sex in the rust fungi.— Phytopathol., 21, 1931, 1001—1040.
Cronquist A. The divisions and classes of plants.—Bot, Rev., 26, 1960. 425-482.
Cummins G. B. Notes on some species of the Uredinales.—Mycologia. 27, 1935, 605-614.
D a n g e a r d P. A. Recherches sur les organismes inférieurs.— Ann. Sc. Nat., Bot., ser. 7, 4, 1886.
283
D a n g e a r d P. A. Mémoire sur la reproduction sexuelle des Basidiomyce- tes.— Botaniste, 4, 1895, 119—181 (цит. по: Gäumann. 1964; С e j p, 1958).
Dangeard P. A. Recherches sur le développement du perithèce chez les Ascomycètes.— Botaniste, 10, 1907, 1—385.
Davis B.' M. The origin of the archegonium.— Ann. Bot., 17, 1903, 477—492.
D e b s к i B. Beobachtungen über die Kerntheilung bei Chara fragilis.— Pringsh. Jahrb. wiss. Bot., 30, 1896.
Debski B. Weitere Beobachtungen an Chara fragilis.—Pringsh. Jahrb. wiss. Bot., 32, 1898.
Dillon L. S. Progressive evolution. A new approach to the development of life.— Inpubl. abstr. of paper given at A. A. A. S. meetings in Indianapolis, dec. 27, 1957.
Dixon H. N. Classification of Mosses.— In: Fr. У e г d о о r n (ed.), Manual of Bryology, Hague, 1932, 397—412.
Dobell С. C. The structure and life-history of Copromonas subtilis n. g., n. sp.— Quart. J. Microscop. Sei., 52, 1908, 75—120.
Dodge B. O. Methods of culture and the morphology of the archicarp in certain species of the Ascobolaceae.— Bull. Torrey Bot. Club, 39, 1912, 139-197.
Dodge B. O. Nuclear phenomena associated with heterothallism and homo- thallism in the Ascomycete Neurospora.— J. Agr. Res., 35, 1927, 289— 305.
Dodge B. O. Facultative and obligate heterothallism in Ascomycetes.— Mycologia, 28, 1936, 399-409.
Dowd in g E. S. The sexuality of Ascobolus stercorarius and the transportation of the oidia by mites and flies.— Ann. Bot., 49, 1931, 621—637.
Drew K. M. Conchocelis-Phase in the life-history of Porphyra umbilicaris (L.) Kütz.— Nature, 164, 748, 1949.
Drew K. M. Rhodophyta.—In: G. M. Smith (ed.). Manual of Phycology. Waltham, 1951, 167—191.
Drew K. M. Life histories in the Algae with special reference to the Chlorophyta, Paeophyta and Rhodophyta.— Biol. Rev., 30, 1955, 343.
Drew K. M. Reproduction in the Bangiophycidae.— Bot. Rev., 22, 1956, 553-611.
Duboscq O., Léger L. et Tuzet O. Contribution à la connaissance des Eccrinides. Les Trichomycètes.—Arch. Zool. Exp., 86, 2, 1948.
Dumortier B. C. Hepaticae Europeae, Jungermannideae Europae past se* miseculum recensitae, adjunctis Hepaticis,— Bull. Soc. Bot. Belg., 13, 1874, 1-202.
Ehrensvard G. Life, origin and development. Chicago, 1962.
E i c h 1 e r A. W. Syllabus der Vorlesungen über specielle und medicinisch- pharmaceutische Botanik. 3 Aufl. Berlin, 1883.
Elliot E. W. The swarm cells of Myxomycètes.— Mvcologia, 41, 1949, 141— 170.
Ellison B. R. Flagellar studies on zoospores of some members of the Mycetozoa, Plasmodiophorales, and Chytridiales.— Mycologia, 37, 1945, 444-459.
Emerson R. An experimental study of the life cyclesand taxonomy of AUomyces — Lloydia, 4, 1941, 77—144.
Emerson R. a. Wilson C. M. The significance of meiosis in Allomy- ccs.— Science, 110, 1949, 86—88.
Endlicher S. Genera plantarum secundum ordines naturales disposita. Vindobonae, 1836—1840.
E n gl er A. Syllabus der Pflanzenfamilien, 2te Aufl., Berlin, 1898; 7te Aufl., Berlin, 1912.
Ernst A. Bastardierung als Ursache der Apogamie in Pflanzenreich. Jena, 1917 (цит. no: S c h u s s n i g, 1930).
Evans A. W. The classification of the Hepaticae.— Bot. Rev., 3, 1939, 49-96.
286
Feldmann J. Une Némalional à carpotetraspores Helminthocladia Hudsont (C. A g.) J. A g r.— Bull. Soc. Hist. Nat. Afrique du Nord., 30, 1939.
F e 1 d m a η n J. et G. Recherches sur les Bonnemaisoniacées et leur alternance de générations.— Ann. Sei. Nat., Η-e serie, Bot., 3, 1942, 75— 175.
Feldmann J. Sur l’existence d’une alternance de générations entre VHali- cystis parvula Schmitz et le Derbesia tenuissima (De Not.) Grn.— Comptes rend. Acad. Sc. Paris, 230, 1950, 322—323.
Feldmann J. Les cycles de reproduction des Algues et leurs rapports avec la phylogénie.— Rev. Cytol., 13, 1952, 1.
Fischer A. Phycomycetes.— In: L. Babenb arst’s Kryptogamen-Flora von Deutschland, Oesterreich u. d. Schweiz, 2-te Aufl., IV Abt. Leipzig, 1892.
Fischer E. Pilze.— In: Handwörterbuch der Naturwissenschaften. Jena, 7, 1912, 880—929.
Fischer E. Reihe Gastromyceteae.— In: A. E n g 1 e r und K. P г a n 11, Die natürlichen Pflanzenfamilien, 2-te Aufl. Leipzig, 7a, 1933, 1—122.
Fischer A. G. Fossils, early life, and atmospheric history.— Proc. Nation. Acad. Sci. U. S. A., 53, 1965, 1205-1215.
Fitzpatrick H. M. The Lower Fungi — Phycomycetes. N. Y. and London, 1930.
Flanagan P. W. Meiosis and mitosis in Saprolegniaceae.— Canad. J. Bot., 48, 1970, 2069—2076.
Fleischer M. Die Musci der Flora von Buitenzorg, I—IV, Leiden, 1900— 1922.
F о 11 B. Sinice a fasy. Praha, 1956; 2-е изд., 1967.
Fott B. Algenkunde. Jena, 1959.
F о 11 B. Evolutionary tendencies among Algae and their position in the plant kingdom.—Preslia, 37, 1965, 117—126.
F ö y n В. Untersuchungen über die Sexualität und Entwicklung von Algen. IV. Vorläufige Mitteilung über die Sexualität und den Generationswechsel von Cladophora und Ulva.— Ber. Deutsch. Bot. Ges., 47, 1925, 495— 506.
Föyn B. Lebenszyklus, Cytologie und Sexualität der Chlorophycee Cladophora suhriana K ü t z i n g.— Arch. Protistenk., 83, 1934a, 1—56.
Föyn B. Lebenszyklus und Sexualität der Chlorophycee Ulva lactuca L.— Arch. Protistenk., 83, 1934b, 154—177.
Föyn B. Über die Sexualität und den Generationswechsel von Ulva mutabilis (n. sp.).—Arch. Protistenk., 102, 1958, 473—480.
Frey R. Chitin und Zellulose in Pilzzellwänden.— Ber. Schweiz. Bot. Ges.. 60, 1950, 199-230.
Fries E. Systema mycologicum, sistens Fungorum ordines, genera et species, usque cognitas. Lund, 1, 1821, 1—520.
Fritsch F. E. The algal ancestry of the higher plants.— New Phytol., 15, 1916, 233—250.
Fritsch F. E. Thalassiophyta and the algal ancestry of the higher plants.— New Phytol., 20, 1921, 165-178.
Fritsch F. E. The genus Sphaeroplea.— Ann. Bot., 43, 1929, 1—26.
Fritsch F. E. The structure and reproduction of the Algae. Cambridge, 1, 1935, 1956; 2, 1945.
Fritsch F. E. Studies in the comparative morphology of the Algae. I. Heterotrichy and juvenile stages.— Ann. Bot., N. S. 6, 1942, 317, 11. The algal life-cycle.—Ann. Bot., N. S. 6, 1942, 533—563; III. Evolutionary tendencies and affinities among Phaeophyceae.— Ann. Bot., N. S. 7, 1943, 63—87. IV. Algae and Archegoniate Plants.— Ann. Bot., N. S. 9. 1945, 1—29.
Fritsch F. E. Present-day classification of Algae.— Bot, Rev., 10, 1944, 233—277.
Frye T. C. a. Clark L. Hepaticae of North America. Seattle, 1937—1947.
F u 1 f о r d M. Recent interpretations of the relationships of the Hepaticae.— Bot, Rev., 14, 1948, 127—173.
287
Fnï ford Μ. II. Manual of the leafy Hepaticae of Latin America.—Mem. N. Y. Bot. Gard., 11, p. 1, 1963; p. II, 1966; p. Ill, 1968.
F u 1 f о r d M. Evolutionary trends and convergence in the Hepaticae.— Bryo- logist, 68, 1965, 1—31. '
Fuller M. S. Biochemical and microchemical study of the cell walls of Rhizidiomyces sp.— Amer. J. Bot., 47, 1960, 838—842.
Fuller M. S. a. Bars had I. Chitin and cellulose in the cell walls of Rhizidiomyces sp.— Amer. J. Bot., 47, 1960, 105—109.
Gäumann E. Über die Entwicklungsgeschichte von Lanomyces etc.— Ann. Jard. Bot. Buitenzorg, 32, 1922, 43—63.
Gäumann E. Vergleichende Morphologie der Pilze. Jena, 1926.
Gäumann E. Die Pilze. Grundzüge ihrer Entwicklungsgeschichte und Morphologie. Basel, 1949, 2-te Aufl., 1964.
G ei tier L. Oogamie, Mitose, Meiose und metagame Teilung bei der zentrischen Diatomee Cyclotella.— Oesterr. bot. Z., 99, 1952, 506—520.
Gibbs S. P., Lewin R. A. a. Philpott D. E. The fine structure of the flagellar apparatus of Chlamy do monas moewusii.— Experim. Cell. Res., 15, 1958, 619-622.
Gilbert F. A. On the occurence of biflagellate swarm cells in certain Myxomycètes.— Mycologia, 19, 1927, 244—283.
Gobi C h. Grudzüge einer systematischen Eintheilung der Gloeophyten (Thallophyten Endl.).— Bot. Zeitung. 29, 1881, 489—501; 505-568.
Goebel K. Organograpnie der Pflanzen. 3-te Aufl. Jena, 1, 1930; 2, 1930.
Goodwin D. C. Morphogenesis of the sporongium of Comatricha.— Amer. J. Bot., 48, 1961, 148—154.
G о о s R. D. Spermatium — trichogyne relationship in Gelasinospora calospora var. antosteira.— Mycologia, 51, 1959, 416—428.
Goroschankin J. Beiträge zur Kenntnis der Morphologie und Systematik der Chlamydomonaden I. Chlamydomonas braunii.— Bull. Soc. Nat. Moscou, N. S. 4, 1890, 498—520.
Goroschankin J. Beiträge zur Kenntnis der Morphologie und Systematik der Chlamydomonaden. III. Chlamydomonas coccifera mihi.— Flora, 94, 1905, 420—423.
G о t h a n W. u. W e y 1 a n d H. Lehrbuch der Paläobotanik, 2-te Aufl. Berlin,
1964.
Gottsberger G. Geissein bei Myxomyceten (Elektronenoptische Studie).— Nova Hedwigia, 13, 1—2, 1967a, 235—243.
Gottsberger G, Geissein bei Myxomyceten.— Nova Hedwigia, 14, 4, 1967b, 415-420.
Gottsche C. M., Lindenberg J. B. G. et N e e s von Esenbeck C. G. Synopsis Hepaticarum. Hamburgi, 1844—1847.
Graham H. W. Pyrrophyta.— In: G. M. Smith (ed.) Manual of Phyco- logy, Waltham, 1951, 105—118.
Gran H. II. Das Plankton des Norwegischen Nordmeeres.— Rep. on Norw. Fischery- and Marine-Investigations, 2, 5, 1902 (цит. no: Kniep, 1928).
Green R. On the naturn of filterable viruses.—Science, 82, 1935, 443— 445.
Greguss P. Die Entdeckung von Urkormophyten aus dem Ordovizium (2).— Acta Biol. Univ. Szeged., n. ser., 7, 1—2, 1961, 3—30.
Greis H. Die Entstehung der Basidiomycetenschnallen aus den Ascomy- cetenhaken.—Jahrb. wiss. Botan., 86, 1938, 81—106.
Greis H. Eumycetes: Allgemeiner Teil. Bau., Entwicklung und Lebensweise der Pilze.— In: A. E n g 1 e r u. K. P r a n 11. Die natürl. Pflanzenfamilien, 2-te Aufl., 5a, I. Berlin, 1943.
Guilliermond A. Recherches sur la germination des levures.— Rev. Gêner. Bot., 17, 1905, 337—376.
Guilliermond A. Recherches cytologiques et taxonomiques sur les Endo- mycétées.— Rev. Gêner. Bot., 21, 1909, 353—391, 401—419.
Guilliermond A. Recherches sur quelques Ascomycètes inferieurs isolés de la stigmatomycose des graines de catonnier. Essai sur la phylogénie
288
des Ascomycètes.— Rev. Gêner. Bot., 40, 1928, 328—342, 397—414, 474— 485, 555—574; 606-624, 690—697.
Guilliermond A. L' Eremothecium ashbyii, nouveau champignon, parasite des capsules du cotonnier.— Revue de Mycol., I, 1936, 117—156.
Gwynne-Vaughan H. С. I. a. Barnes B. The structure and development of the Fungi, 2-d ed. Cambridge, 1937.
Haeckel E. Generelle Morphologie derOrganismen.il. Allgemeine Entwicklungsgeschichte der Organismen. Berlin, 1866.
Haeckel E. Das Protistenreich. Leipzig, 1878.
Haeckel E. Systematische Phylogenie. Entwurf eines natürlichen Systems der Organismen auf Grund ihrer Stammesgeschichte. Teil I. Systematischen Phylogenie der Protisten und Pflanzen. Berlin, 1894.
Haeckel E. Die Lebenswunder. Gemeinverständliche Studien über biologische Phylosophie. Stuttgart, 1904.
Haldane J. B. S. The origin of life.— Rationalist Annual, 1929 (цит. по русскому переводу. См: Бернал, 1969).
Hall R. Р. Binary fission in Oxyrrhis marina D u j a r d i n.— Univ. Calif. Publ. Zool., 26, 1925a, 281—324.
Hall R. P. Mitosis in Ceratium hirundinella O. F. M., with notes on nuclear phenomena in encysted forms and the question of sexual reproduction.— Univ. Calif. Publ. Zool., 28, 1925b, 29-64.
Hammerling J. Über die Geschlechtsverhältnisse von Acetabularia mediterranea und Acetabularia wettsteinii.— Arch. Protistenk., 83, 1934, 57—97.
Hanna W. F. Nuclear association in the aecium of Puccinia graminis — Nature, 124, 3120, 1929, 267.
Harder R. Phycophyta, Algen.—In: E. Strasburger, F. Noll,
H. Schenk und A. F. W. S c h i m p e r. Lehrbuch der Botanik, 28 Aufl., Jena, 1962.
Harder R. u. Koch W. Über die Plastidenfarbstoffe von Pedinomonas (Protochloridales).—Arch. Mikrob., 21, 1954, 1—3.
Harper R. A. Die Entwickelung des Peritheciums bei Sphaerotheca casta- gnei — Ber. Deutsch. Bot. Ges., 13, 1895, 475—481.
Harper R. A. Ueber das Verhalten der Kerne bei der Fruchtentwicklung der Ascomyceten.— Jahrb. wiss. Bot., 29, 1896, 655—685.
Harris T. M. The British rhaetic flora.— British Museum Nat. Hist. London, 1938.
Harris T. M. Naiadita, a fossil Bryophyte with reproductive organs.— Ann. Bryol., 12, 1939, 57—70.
Harrold С. E. Studies in the genus Eremascus. 1. The rediscovery of Eremascus albus Eidam and some new observations concerning its life- history and cytology.— Ann. Bot., N. S. 14, 1950, 127—148.
Hartmann M. Untersuchungen über die Sexualität und Entwicklung von Algen. III. Über die Sexualität und Generationswechsel von Chaetomorpha und Enteromorpha.— Ber. Deutsch. Bot. Ges., 47, 1929, 485—494.
Harvey W. H. Algae.—In: J. T. M a c k a y, Flora Hibernica, Dublin, 1836.
Haskell G. Some evolutionary problems concerning the Bryophvta.— Bryolo- gist, 52, 1949, 49—57.
Hauck F. Die Meeresalgen Deutschlands und Oesterreichs.— ln: L. Rabenhorst, Kryptogamen-Flora, 2-te Aufl. Leipzig, 2, 1885.
Il a x о F. T. a. Fork D. C. Photosynthetically active accessory pigments of cryptomonads.— Nature, 184, 4692, 1959, 1051—1052.
H aye A. Über den Exkretionsapparat bei den Protisten etc.— Arch. Protistenk., 70, 1930, 1-86.
H e d w i g J. Species muscorum frondosorum descriptae et tabulis aeneis coloratis illustratae, 1801.
Heim P. Le noyau dans le cycle évolutif de Plasmodiophora brassicae Wo r о n.— Revue de Mycologie, 20, 2, 1955.
Heim R. Les champignons d’Europe, 1, 2. Paris, 1957.
H e i n t z e A. Cormofytermas Fylogeni. Lund, 1927.
289
II е г z о g T h. Anatomie der Lebermoose. In K. L i n s b a u e r (ed.), Handbuch der Pflanzenanatomie, IL Abt., 2. Teil. Bryophyten, Berlin, 1925.
Herzog Th. u. Grolle R. Was ist Pachyglossa? — Revue bryol. et liche- nol., 27, 1958, 147-165.
Higgins В. В. Morphology and life-history of some Ascomycetes with special reference to the presence of spermatia.— Amer. J. Bot., 7, 1920, 435—444.
Higgins В. B. Morphology and life-history of some Ascomycetes with special reference to the presence and function of spermatia. III.— Amer. J. Bot., 23, 1936, 598-602.
H i г о s e H. Rearrangement of the systematic position of a thermal alga, Cyanidium caldarium.— Bot. Mag., Tokyo, 71, 1958, 347—352.
Hofmeister W. Vergleichende Untersuchungen über Keimung. Entfaltung und Fruchtbildung bei höheren Kryptogamen. Leipzig. 1851.
Η о g g J. On the distinctions of a plant and an animal, and on a fourth kingdom of nature.— Edinb. new Phil. J., N. S. 12, 1860, 216—225.
Holferty G. M. Development of the archegonium of Mnium cuspidatum.— Bot., Gaz., 37, 1904, 106-126.
Hollenberg G. J. Culture studies of marine Algae. III. Porphyra perforata.— Amer. J. Bot., 45, 1958, 653—656.
Hopkins E. W. Microchemical tests on the cell walls of certain fungi. Cellulose and chitin.— Trans. Wisconsin Acad. Sei., 24, 1929, 187—196.
Howard K. L. a. Bryant T. R. Meiosis in the Oomycetes: II. A micro- spectrophotometric analysis of DNA in Apodachlya brachynema.— Myco- logia, 63, 1971, 58-68.
Howe M. A. The Hepaticae and Anthocerotes of California.— Mem. Torrey Bot. Club, 7, 1899, 1-208.
H owe M. A. A note on the structural dimorphism of sexual and tetrasporic plants of Galaxaura obtusata.— Bull. Torrey Bot. Club, 43, 1917, 621—624.
Hueber F. M. Hepaticites devonicus — a new fossil liverwort from the Devonian of New York.—Ann. Missouri Bot. Garden, 48, 1961, 125—131.
Hughes S. J. Conidiophores, conidia and classification.— Canad. J. Bot., 31, 1953, 577-659.
Hustede H. Über den Generationswechsel zwischen Derbesia neglecta Berth, und Bryopsis halymeniae Berth.— Naturwiss., 47, 1960, 19.
Hustedt F. Die Kieselalgen Deutschlands, Österreichs und der Schweiz, Teil I. — In:L. Rabenhorst’s Kryptogamenflora von Deutschland, Österreich und der Schweiz. Leipzig, 7, 1930.
Hy M. Recherches sur l’archégone et le développement du fruit des Musciné- es.— Ann. Sei. Nat, Sér 6, 18, 1884, 105—206 (цит. no: Schuster, 1966).
I s h i к a w a M. Cytological studies on Porphyra tenera K j e 11 m.— Bot. Mag. Tokyo, 35, 1921, 206-218.
Iyengar M. O. P. Fertilization in Eudorina elegans Ehrenberg.— J. Ind. Bot. Soc., 16, 1937, 111—118.
I e y n g a r M. O. P. Chlorophyta.— In: G. M. Smith (ed.), Manual of Phy- cology. Waltham, 1951, 21—67.
Iyengar M. O. P. a. Balakrishnan M. S. On sexual reproduction in a new species of Golenkinia.— J. Ind. Bot. Soc., 35, 1956, 371—373.
Iyengar M. O. P. a. R a m a n a t h a n K. R. On sexual reproduction in a Dictyosphaerium.— J. Ind. Bot. Soc., 18, 1940, 195—200.
Iyengar Μ. O. P. a. R a m a n a t h a n K. R. On the life-history and cytology of Microdictyon tenuis (A g.) Decsne (Preliminary note).—J. Ind. Bot. Soc., 20, 1941, 157—159.
Iyengar M. O. P. a. Ramanathan K. R. On the structure and reproduction of Pleodorina sphaerica Iyengar.— Phytomorphology, I, 1951, 215—224.
Jackson II. S. Life cycles and phylogeny in the higher fungi.— Trans. Roy. Soc. Can., V, 38, 1944, 1—32.
J a h η E. Myxomycetenstudien. 8 Der Sexualakt.— Ber. Deutsch. Bot. Ges., 29, 1911, 231-247.
290
Jahn E. Myxomycetenstudien, 12. Das System der Myxomyceten.— Ber. Deutsch. Bot. Ges., 46, 1928, 8—17.
Jahn E. Myxomycetenstudien. 16. Die Kernphasen und die Zahl der Chromosomen.— Ber. Deutsch. Bot. Ges., 54, 1936, 517—528.
Jahn E. Myxomycètes.—In: A. Eng 1er u. K. Prantl, Die natürl. Pflan- zenfam., 2-te Aufl., 2, 1928, 304—339.
Jahn T. L. Euglenophyta.—In: G. M. Smith (ed.), Manual of Phycology. Waltham, 1951, 69—81.
Janczewski E. Vergleichende Untersuchungen über die Entwicklungsgeschichte des Archegoniums.— Bot. Zeit., 30, 1872, 377—394, 401—420, 440—443.
Jeffrey C. The origin and differentiation of the Archegoniate land-plants.— Botaniska Notiser, 115, 1962, 446—454.
Jennings O. E. The ancestry of the mosses.— Bryologist, 31, 1928, 10—15.
Johnson L. P. The taxonomy, phylogeny, and evolution of the genus Euglena — In: D. E. Buetow (ed.), The biology of Euglena. N. Y. and London, 1, 1968, 1—25.
Johnson T. W. Resting spore development in the marine Phycomycete Anisolpidium ectocarpi.— Amer. J. Bot., 44, 1957, 875—878.
Jorde I. Untersuchungen über den Lebenszyklus von Urospora A re sch und Codiolum A. Brau n.— Nyt Mag. Naturv., 73, 1933, 1—20.
Jovet-Ast S. Bryophyta.—In: E. Boureau. Traité de Paléobotanique, II, Paris, 1967, 17-186.
J u e 1 H. O. Die Kernteilungen in den Basidien und die Phylogenie der Basi- diomyceten.— Jahrb. wiss. Bot., 32, 1898, 361—388.
J u e 1 H. O. Ueber Zellinhalt, Befruchtung und Sporenbildung bei Dipodas- cus.— Flora, 91, 1902, 47—55.
J u e 1 H. O. Zytologische Pilzstudien. II. Zur Kenntnis einiger Hemiasceen.— Nova Acta R. Soc. Sei. Upsal., ser. 4, 5, 1921, 1—43.
J u 11 e r E. Der Generations- and Phasenwechsel bei Stigeoclonium subspino- sum.— Arch. Protistenk., 89, 1937, 55—93.
Jussieu A. L., de. Genera plantarum secundum ordines naturales disposita. Paris, 1789.
Just T h. The proper designation of the vascular plants.—Bot. Rev., 11, 1945, 299-309.
K a r 1 i n g J. S. Nuclear and cell division in Nitella and Chara.— Bull. Torrey Bot. Club, 53, 1926, 319—379.
K a r 1 i n g J. S. A new fungus with anteriorly uniciliate zoospores: Hypho- chytrium catenoides.— Amer. J. Bot., 26, 1939, 512—519.
К а г 1 i n g J. S. The simple holocarpic biflagellate Phycomycetes. N. Y., 1942.
Karling J. S. The life history of Anisolpidium ectocarpi gen. nov. et sp. nov., and a synopsis and classification of other fungi with anteriorly uniflagellate zoospores.— Amer. J. Bot., 30, 1943, 637—648.
Karsten G. Die sogenannten „Mikrosporen“ der Plankton-diatomeen und ihre weitere Entwicklung, beobachtet an Corethron valdiviae n. sp.— Ber. Deutsch. Bot. Ges., 22, 1904, 544—554.
Karsten G. Abteilung Bacillariophyta (Diatomeae).— In: A. E n g 1 e r und K. Prantl. Die natürlichen Pflanzenfamilien, 2-te Aufl., Leipzig, 2, 1928.
K a s h y a p S. R. The relationships of liverworts, especially in light of some recently discovered Himalayan forms.— Proc. Asiat. Soc. Bengal, N. S. 15, 4, 1919, 152—166 (цит. no: M e h r a a. H a n d о o, 1953).
Kawaguti S. a. Yamasu T. Electron microscopy on the symbiosis between an elysioid gastropod and the chloroplasts of a green algae.— Biol. J.Okayama Univ., 3, 1965 (цит. no: A 11s op p, 1969).
Kevorkian A. G. Studies in the Leptomitaceae. II. Cytology of Apodachlya brachynema and Sapromyces reinschii.— Mycologia, 27, 1935, 274—285.
K j e 11 m a n F. R. Phaeophyceae (Fucoideae) .— In: A. E n g 1 c r
und K. P r a n 11, Die natürlichen Pflanzefamilien. Leipzig, 1, 2, 1897, 176-290.
291
К jell ma n F. R. Dictyotales.—In: A. Γη gier und K. P г a n 11. Die natürlichen Pflanzenfamilien, Leipzig, 1, 2, 1897, 291—297.
Klebs G. Über die Organisation einiger Flagellatengruppen und ihre Beziehungen zu Aglen und Infusorien.— Untersuch. Bot. Inst. Tübingen, 1, 1883, 233—360.
Klebs G. Flagellatenstudien, I. II.—Zeitschr. wiss. Zool., 55, 1893. 265—351; 353—445.
Klebs G. Über flagellaten- und algenähnliche Peridineen.— Verh. nat-, med. Ver. Heidelberg. 11, 1912, 369—451.
Klein R. M. a. Cronquist A. A consideration of the evolutionary and taxonomic significance of some biochemical, micromorphological. and physiological characters in the Thallophytes.— Quart. Rev. Biol., 42, 1967, 105-296.
К n i e p H. Beiträge zur Kenntnis der Hymenomyceten. I. Die Entwicklungsgeschichte von Hypochnus terrestris nov. spec.— Zeitschr. Bot., 5, 1913, 593-609.
Kniep H. Beiträge zur Kenntnis der Hymenomyceten. III. Über die konjugierten Teilungen und die phylogenetische Bedeutung der Schnallenbildungen.— Zeitschr. Bot., 7, 1915, 369—398.
Kniep H. Über Geschlechtsbestimmung und Reduktionsteilung.— Verh. Phy- sik.-mediz. Ges. Würzburg, 47, 1922, 1—28.
Kniep H. Die Sexualität der niederen Pflanzen. Jena, 1928.
Kniep H. Allomyces javanicus n. sp., ein anisogamer Phycomycet mit Planogameten.— Ber. Deutsch. Bot. Ges., 47.1929, 199—212.
Kniep H. Ueber den Generationswechsel von Allomyces.— Zeitschr. Bot., 22, 1930, 433—441.
K n о X E. M. Microspores and their significance in biological problems.— Trans. Proc. Bot. Soc. Edinb., 35, 2, 1949, 109—119.
Knox E. M. The spores of Lycopodium, Phyloglossum, Selaginella and lsoëtes, and their value in the study of microfossils of Palaeozoic age.— Trans. Proc. Bot. Soc. Edinb., 35, 1950, 3.
Koch W. J. Studies in the genus Chytridium with observations on a sexually reproducing species.—J. Elisha Mitchell Sc. Soc., 67, 1951, 267—278.
Koch W. J. Studies of the motile cells of Chytrids. I. Electron microscope observations of the flagellum, blepharoplast and rhizoplast.— Amer. J. Bot., 43, 1956, 811-819.
Koch W. J. Studies of the motile cells of Chytrids. II. Internal structure of the body observed with light microscopy.— Amer. J. Bot., 45, 1958, 59—72.
Ko le A. P. Plasmodiophora brassicae and Spongospora subterranea, points of resemblance and difference.— Tijd. Plantenziekt, 65, 1959, 47—55.
Ko le A. P. a. Gielink A. J. Electron microscope observations on the flagella of the zoosporangial zoospores of Plasmodiophora brassicae and Spongospora subterranea.— Proc. К. nederl. Acad. Wetensch., ser. C, 64, 1961, 157—161.
Ko le A. P. a. Gielink A. J. Electron microscope observations on the zoospores arising from the resting spores of Plasmodiophora brassicae.— Proc. К. Nederl. Acad. Wetensch., ser. C, 65, 1962, 117—121.
К о 1 e A. P. a. Gielink A. J. The significance of the zoosporangial stage in the life-cycle of the Plasmodiophorales.— Neth. J. PI. Path., 69,1963, 258.
Konrad P. et Maublanc A. Les Agaricales — Agaricaceae. Paris, 1948.
Kornmann P. Zur Entwicklungsgeschichte von Derbesia und Halicystis.— Planta, 28, 1938, 464-470.
Korschikoff A. On some new organisms from the groups Volvocales and Protococcales, and on the genetic relations of these groups.— Arch. Pro- tistenk., 55, 1926, 439-503.
Korschiko V A. Studies on the Chrysomonads. I.— Arch. Protistenk., 67, 1929, 253-290.
Korshikov A. A. On the origin of the diatoms.—Beih. Bot. CentralbL, 46, Abt. I, 1930a, 460-469.
292
Ко г sh i к ον A. A. On the occurence of pyrenoids in Heterocontae.— Beih. Bot. Centrait)!.. 46, Abt. 1, 1930b, 470—478.
Kozlowski B. a. G г e g u s s P. Discovery of Ordovician land plants (preliminary communication).— Acta Palaeontol. Polon., 4, 1, 1959,
1—9.
Kreisel И. Grundzüge eines natürlichen Systems der Pilze. Jena, 1969.
К richenbauer H. Beitrag zur Kenntnis der Morphologie und Entwicklungsgeschichte der Gottungen Euglena und Phacus.— Arch. Protistenk., 90, 1937, 88-123.
Krieger W. Abteilung Chrysophyta.— ln: A. E n g 1 e r’ s Sullabus der Pflanzenfamilien, 12-te Aufl., Berlin — Nikolassee, 1, 1954, 73—85. Kuckuck P. Fragmente einer Monographie der Phaeosporeen (edit, by W. Nienburg).—Wiss. Meeresunters, Abt. Helgoland, N. F. 17, 1929 (цит. по: Fritsch, 1943).
Kumar H. D. Apparent genetic recombination in a blue-green algae.— Nature, 196, 4859, 1962, 1121—1122.
Kunieda H. On the life-history of Monostroma.— Proc. imp. Acad. Japan.
10, 1934, 103—106 (цит. no: Vazart, 1963).
К у 1 i η Η. über die Befruchtung und Reduktionsteilung bei Nemalion multifidum— Вег. Deutsch. Bot. Ges., 34, 1916a, 257—271.
Kylin II. Über den Generationswechsel bei Laminaria digitata.— Svensk. Bot. Tidskr., 10, 1916b, 551-561.
Kylin H. Studien zur Entwicklungsgeschichte der Phaeophyceen.— Svensk Bot. Tidskr., 12, 1918, 1—64.
К у 1 i η Η. Entwicklungsgeschichtliche Florideenstudien.— Lunds Univers.
Àrsskr., N. F., Avd. 2, 24, 4, 1928, 1-127.
Kylin H. Über die Entwicklungsgeschichte der Florideen.— Lunds Univ.
Àrsskr. N. F., Avd. 2, 26, 6, 1930a. 1-104.
Kylin H. Some physiological remarks on the relationship of the Bangiales.— Bot. Notiser,1930b, 417—420.
Kylin H. Über die Entwicklungsgeschichte der Phaeophyceen.— Lunds Univ.
Àrsskr. N. F., Avd. 2, 29, 7, 1933, 1-102.
Kylin H. Anatomie der Rhodophycen. Berlin, 1937a.
Kylin H. Bemerkungen über die Entwicklungsgeschichte einiger Phaeophyceen.— Lunds Univ. Àrsskr., N. F., Avd. 2, 33, 1, 1937b, 1—34.
Kylin H. Zur Entwicklungsgeschichte der Ordnung Fucales.— Svensk Bot. Tidskr., 34, 1940, 301—314.
Lackey J. B. Scioto River Valley forms of Chrysococcus.— Amer. Midi. Natur., 20, 1938, 619-623.
Lam H. J. Classification and the new morphology.— Acta biotheor., 8, 1948, 107-154.
Lam о u roux J. V. F. Essai sur les genres de la famille des thaîassiophytes non articulées.— Mem. Mus. Nat. Hist. Paris, 20, 1813, 21—47; 115—139; 267-293.
L e d i n g h a m G. A. Zoospore ciliation in the Plasmodiophorales.— Nature. 133, 1934, 534.
L e d i n g h a m G. A. Zoospore ciliation in the Plasmodiophorales.— Canad. J. Res. G., 17, 1939, 38.
Lee dale G. F. The evidence for a meiotic process in the Euglenineae.— Arch. Mikrobiol., 42, 1962, 237—245.
Lee da le G. F. The nucleus in Euglena.— In: D. E. Buetow (ed.), The biology of Euglena. N. Y. and London, I, 1968, 185—242.
L e e d a 1 e G. F., M e e u s e B. J. D. a. P r i n g s h e i m E. G. Structure and phvsiology of Euglena spirogyra. I, II.— Arch. Mikrobiol., 50, 1965, 68—102!
Leger L. et G a u t h i e r M. La spore des Harpellacées (Leger et Dubos cq), Champignons parasites des insects.—Comptes rend. Acad. Sei.. Paris, 200, 1935, 1458—1460.
293
Lehman S. G. Conidial formation in Sphaeronema fimbriatum.— Mycologia, 10, 1918, 155—163.
Leitgeb H. Untersuchungen über die Lebermoose. VI. Die Marchantieen und allgemeine Bemerkungen über die Lebermoose. Jena, 1881.
Levy ns M. R. Sexual reproduction in Macrocystis pyrifera A g.— Ann. Bot, 47, 1933, 349—353.
Lewis I. F. a. Bryan H. F. A new Protophyte from the Dry Tortugas.— Amer. J. Bot., 28, 1941, 343—347.
L i g n i e r O. Sur une Mousse houillère à structure conservée.— Bull. Soc. Linn. Normandie, Sér. 6, 7, 1914, 128—131.
Limpricht G. Die Laubmoose Deutschlands, Oesterreichs und der Schweiz.— In: L. Rabenhorst’ s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz, IV. Leipzig, 1890—1904 (Abt. I—1890; Abt. 11-1895; Abt. III—1904).
Lindberg S. O. Musci novi scandinavici. Notis. Sällsk. Fauna et Flora Fennica Förhandl., 9, 1868, 255—299.
Lindberg S. O. Hepaticae in Hibernia mense Julii lectae.— Acta Soc. Sc. Fennica, 10, 1875, 467—559.
Lindemann E. Abteilung Peridineae (Dinoflagellatae).— In: A. E n g 1 e r, Die natürlichen Pflanzenfamilien, 2-te Aufl., 2. Leipzig, 1928, 3—104.
Lindenbein W. Beitrag zur Cytologie der Charales.— Planta, 4, 1927.
Linder D. H. Evolution of the Basidiomycetes and its relation to the terminology of the basidium.— Mycologia, 32, 1940, 419—447.
L i n g a p p a I. The development and cytology of the epibiotic phase of Physoderma pulposum.— Amer. J. Bot., 46, 1959, 145—150, 233—240.
Linné C. Species Plantarum, 1753.
Linné C. Systema Naturae, ed. 13, Vindobonae, 1, 1767; 2, 1770.
Linné C. Systema Plantarum, IV, Francofurti ad Moenum, 1780.
Lister A. A monograph of the Mycetozoa. London, 1894; ed. 2, 1911; ed. 3, 1925.
L о h w a g H. Zur Entwickelungsgeschichte und Morphologie der Gastromy- ceten.— Beih. Bot. Centralbl., Abt. II, 42, 1926, 177—334.
Lotsy J. P. Vorträge über botanische Stammesgeschichte, Jena, I: Algen und Pilze, 1907; II: Cormophyta zoidogamia, 1909.
Lundblad B. Contributions to the geological history of the Hepaticae. Fossil Marchantiales from the Rhaetic — Liassic coalmines of Skromberga (prov. of Scania), Sweden.— Svensk Bot. Tidskr., 48, 1954, 381—417.
Lundblad B. On Ricciisporites tuberculatus and its occurence in certain strata of the “Hollviken II” boring in S. W. Scania.— Grana Palinolog., N. S. 2, 1959, 1—10.
Luther A. Über Chlorosaccus, eine neue Gattung der Süsswasseralgen, nebst einigen Bemerkungen zur Systematik verwandter Algen.— Bih. Kgl. Svensk Vetensk.-Akad. Handl., 24, Afd. 3, 13, 1899, 1-22.
L u 11 r e 11 E. S. Taxonomy of the Pyrenomycetes.— Univ. Missouri Stud., 24, 3, 1951, 1-20.
Lu tt re 11 E. S. The ascostromatic Ascomycetes.— Mycologia, 47, 1955, 511— 532.
Mack B. Morphologische und entwicklungsgeschichtliche Untersuchungen an Chrysophyceen.— Oesterr. Bot. Zeitschr., 98, 1951, 249—279.
Mack B. Untersuchungen an Chrysophyceen I—III.—Oesterr. Bot. Zeitschr., 100. 1953, 147—152.
M c Manus M. A. Culture of Stemonitis fusca on glass,— Am. J. Bot., 48, 1961, 582-588.
Me Manus M. A. Laboratory cultivation of Clastoderma debaryanum,.— Am. J. Bot., 48, 1961, 884—888.
Magne F. La structure du noyau et le cycle nucléaire chez le Porphyra linearis G r e v i 11 e.— Compt. Rend. Acad. Sc. Paris, 234, 1952, 986—988.
Magne F. Le Rhodochaete parvula Thure t (Bangioidée) et la reproduction sexuée.— Cahiers Biol, шаг., 1, 1960, 407—420.
294
Manier J. F. Recherches sur les Trichomycétes.— Ann. Sc. Nat., Bot,, 11. Ser., 11, 1950, 53—162 (цит. no: G e j p, 1957—1958).
M a n t ο η I. Observations on the internal structure of the spermatozoid of Dictyota — J. Exper. Bot., 10, 1959, 448—461.
Man ton I. Electron microscopical observation on a very small flagellate: the problem of Chromulina pusilla Butcher.— J. mar. biol. Ass. U, K., 38, 1959, 319-333.
M ant on I. Electron microscopical observations on a clone of Monomastix Scherffel in culture.— Nova Hedwigia, 14, 1967, 1—11.
M a n t ο n I. a. Parke M. Further observations on small green flagellates with special reference to possible leratives of Chromulina pusilla Butcher.- J. mar. biol. Ass. U. K., 39, 1960, 275—298.
Margulis L. Evolutionary criteria in Thallophytes: a radical alternative.— Science, 161, 1968, 1020—1022.
Martens P. L’origine du «crochet» et de l’«anse d’anastomose» chez les Champignons supérieurs.— Bull. Soc. Мус. France, 48, 1932, 259—281.
Martin E. M. The morphology and cytology of Taphrina deformans — Amer. J. Bot., 27, 1940, 743—751.
M a г t i n G. W. Systematic position of the slime molds and its hearing on the classification of the fungi.— Bot. Gaz., 93, 1932, 421—435.
Martin G. W. The Myxomycètes.— Bot. Rev., 6, 1940, 356—388.
Martin G. W. The classification of the Tremellales.— Mycologia, 37, 1945, 527—542.
Martin G. W. Are Fungi plants? — Mycologia, 47, 1955, 779—792.
Martin G. W. The systematic position of the Myxomycètes.— Mycologia, 52, 1960, 119-129.
Mattick F. Abteilung: Lichenes. Flechten.— In: A. E n g 1 e r’s Syllabus der Pflanzenfamilien, 12-te Aufl. Berlin — Nikolassee, 1954, 204—218.
M e h r a P. N. A new suggestion on the origin of thallus in the Marchanti- ales. I. The thallus structure.— Amer. J. Bot., 44, 1957, 505—512,— II. The theory.— Amer. J. Bot., 44, 1957, 573—581.
M e h r a P. N. Classification and phyletic relationships in the Hepaticae.— Tenth intern, bot. congress. Abstracts of papers. Edinburgh, 1964, HOHL
M e h r a P. N. Evolutionary trends in Hepaticae with particular reference to the Marchantiales.— Phytomorphology, 19, 1969, 203—218.
Me hr a P. N. a. H a n d о о О. N. Morphology of Anthoceros erectus and A. himalayensis and the phytogeny of the Anthocerotales.— Bot. Gaz., 114, 1953, 271-381.
Me hr a P. N. a. V a s h i s h t B. R. Embryology of Petalophyllum indicum K a s h., and a new suggestion of the evolution of thalloid habit from foliose forms.— Bryologist, 53, 1950.
Melchior H. Abteilung Pyrrophyta.— In: A. E n g 1 e r’s Syllabus der Pflanzenfamilien, 12-te Aufl., Berlin, 1, 1954a, 64—72.
Melchior H. Abteilung: Phaeophyta. Braunalgen (Melanophyceae).— In: A. Engle r’s Syllabus der Pflanzenfamilien, 12-te Aufl., Berlin, 1954b, 109—123.
Melchior H. Abteilung: Rhodophyta, Rotalgen, Sea — mosses.— In: A. E n g - le r’s Syllabus der Pflanzenfamilien, 12-te Aufl., Berlin, 1954c, 123—138.
M e r e s c h к о w s к y C. Über Natur und Ursprung der Chromatophoren im Pflanzenreiche.— Biol. Zentralbl., 25, 1905.
Meyer K. Zur Frage von der Homologie der Geschlechtsorgane und der Phylogenie der Archigoniums.—Биол. журн., II, 1912.
M ey er К. I. Über den Befruchtungsvorgang bei Chaetonema irreguläre Nowak.—Arch. Protistenk., 72, 1930, 147—157.
Minden N. Oomyceten.— In: Kryptogamenflora der Mark Brandenburg, 5, 1915, 209-630.
Moewus F. Die Sexualität und der Generationswechsel der Ulvaceen und Untersuchungen über die Parthenogenese der Gameten.— Arch. Protis- tenk., 91, 1938, 357—441.
295
Моеwus F. Uber die Sexualität von Botrydium granulatum.— Biol. Zentralbl., 60, 1940, 484—498.
Möller Л. Ueber die Kultur flechtenbildender Ascomyceten.— Unters. Bot. Inst. Münster, 1887 (цит. по: K n i e p, 1928).
Möller A. Ueber die sogenannten Spermatien der Ascomyceten.— Bot. Zeitung, 48, 1888, 421—425 (цпт. по: К n i e p. 1928).
M ö n к e m e у e г W. Die Laubmoose Europas.— In: L. R a b n e h о r s t s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz. Leipzig, 1927.
Moreau F. Les phémomènes de la sexualité chez les urcdinées.— Botaniste. 13, 1914, 145—284.
Moreau F. Les Champignons. Physiologie, morphologie, développement et systématique, Paris, I, 1952; II, 1953.
M u n к A. The system of the Pyrenomycetes,— Dansk Botanisk Arkiv. 15, 2, 1953, 1-163.
Müller C. Synopsis muscorum frondosorum omnium hucusque cognitorum. Berolini, I, 1849, 2, 1851.
Müller E. Die schweizerischen Arten der Gattungen Clathrospora, Pleospora, Pseudoplea and Pyrenophora.— Sydowia, 5, 1951, 248—310.
Müller K. Die Lebermoose Europas. Leipzig, I, 1954 (1951—1954); II, 1957 (1956-1958).
Nabel K. Uber die Membran niederer Pilze, besonders von Prizidiomyces bivellatus nov. spec.— Arch. Mikrobiol., 10, 1939, 515—541.
Nadson G. u. Konokotin A. Étude cytologique sur les levures à copulation hétérogamique du genre «Nadsonia» Svd.— Ann. Sci. nat. bot., sér. 10, 8, 1926, 165-182.
N a k a h a r a H. a. N a k a m u r a J. The life history of Desmarestia tabacoides O k am u r a.— Bot. Mag. Tokyo, 84, 1971, 69—75.
Nannfeldt J. A. Studien über die Morphology und Systematik der nicht- lichenisierten Inoperculaten Discomyceten.— Nova Acta Reg. Soc. Scient. Upsal., ser. IV, 8, 2, 1932, 1—368.
Nèmec B. Üvod do vseobeené biologie. Praha, 1929.
Noble M. The morphology and cytology of Typhula trifolii Rost r.— Ann. Bot., N. S., 1, 1937, 67—98.
Oehm G. Kritische Bemerkungen zur Nebensporenfrage bei höheren Pilzen mit besonderer Berücksichtigung der Gattung Ceriomyces.— Beih. Bot. Centralbl., Abt. A, 57, 1937, 252—263.
Oestrup E. Danske Diatoméer. Kopenhagen, 1910 (цит. no: Hustedt, 1930).
O’ h Eocha C. a. Raftery M. Phycoerythrins and phycocyanins of Cryptomonads.— Nature, 184, 4692, 1959, 1049—1051.
Olive E. W. Monograph of the Acrasieae.— Proc. Bost. Soc. Nat. Hist., 30, 1902, 451—513.
Olive L. S. The Protostelida — a new order of the Mycetozoa.— Mycologia, 59, 1967, 1—29.
Olive L. S. The Mycetozoa: a revised classification.— Bot. Rev., 36, 1970, 59—89.
Oltmanns F. Morphologie und Biologie der Algen. Jena, I, 1904; 2, 1905; 2te Aufl., 1, 1922, 2, 1922.
Orth R. Vergleichende Untersuchungen über die Luftkammerentwicklung der Marchantiaceen.— Flora, 1929, 124.
Papenfuss G. F. Proposed names for the phyla of Algae.— Bull. Torrey Bot. Club, 73, 1946, 217-218.
Papenfuss G. F. Extension of the brown algal order Dictyosiphonales to include the Punctariales.— Bull. Torrey Bot. Club, 74, 1947, 398—402.
Papenfuss G. F. Problems in the classification of the marine Algae.— Svensk Bot. Tidskr., 45, 1951a, 4—11.
Papenfuss G. F. Phaeophyta.— In: G. M. Smith (ed.), Manual of Phy- cology. Waltham, 1951b, 119—158.
Parihar N. S. An introduction to Embryophyta. I. Bryophyta, Allahabad, 3d ed., 1959; 5th ed., 1965.
296
Parke M. Some remarks concerning the class Ghrysophyceae.—British Phycol. Bull., 2, 1961, 47—55.
Parke M. Electron microscope observations on scale-bearing Chrysophyseae.— Recent Advances in Botany, Sect. 3. University of Toronto Press, 1961. 226—229.
Parke M. a. D i x ο n P. S. Check-list of British marine Algae — second revision.— J. mar. biol. Ass. U. K., 48, 1968, 783—832.
Pascher A. Über die Beziehungen der Cryptomonaden zu den Algen.— Ber. Deutsch. Bot. Ges., 29, 1911, 193-203.
Pascher A. Über Flagellaten und Algen.— Ber. Deutsch. Bot. Ges., 32, 1914, 136—160.
Pascher A. Über die Myxomyceten.— Ber. Deutsch. Bot. Ges., 36, 1918, 359-380.
Pascher A. Über die Übereinstimmungen zwischen den Diatomeen. Heterokonten und Chrysomonaden.— Ber. Deutsch. Bot. Ges., 39, 1921, 236.
Pascher A. Die Süsswasser-Flora Deutschlands, Oesterreichs und der Schweiz, 4. Volvocales-Phytomonadineae. Jena, 1927.
Pascher A. Über einen grünen, assimilationsfähigen plasmodialen Organismus in den Blättern von Sphagnum.— Archiv Protistenk., 72, 1930.
Pascher A. Systematische Übersicht über die mit Flagellaten in Zusammenhang stehenden Algenreihen und Versuch einer Einreihung dieser Algenstämme in die Stämme des Pflanzenreiches.— Beih. Bot. Centralbl., 48, II Abt., 1931a, 317-332.
Pascher A. Über einem neuen einzelligen Organismus mit Eibefruchtung.— Beih. Bot. Centralbl., 48, Abt. A, 1931b, 466-480.
Pascher A. Über die Verbreitung endogener bzw. endoplasmatisch gebildeter Sporen bei den Algen.— Beih. Bot. Centralbl., 49, Abt. I, 1932a. 293-308.
Pascher A. Über das Vorkommen von kontraktilen Vakuolen bei pennaten Diatomeen.— Beih. Bot. Centralbl., 49, Abt.I, 1932b, 703—709.
Pascher A. Über geisselbewegliche Eier, mehrköpfige Schwärmer und voll- ständingen Schwärmervelust bei Sphaeroplea — Beih. Bot. Centralbl., 59, Abt. A, 1939, 188—213.
P a V i 11 a r d J. Sur les affinités des Plasmodiophorales.— Bull. Soc. Bot, France, 100, 1953, 81.
Persidsky В. M. The development of the auxospores in the group of the Centricae (Bacillariaceae). Moskau. 1929 [Рефераты: P. Schmidt in Zeitschr. f. Bot., 22, 1929, 459—461; Kolbe in Bot. Centralbl., N. F. 17 (159), 1930, 235-236].
Persidsky В. M. The sexual process in Melosira varians.— Beih. Bot. Centralbl., 53, Abt. A, 1935, 122-132.
Pietsch W. Entwicklungsgeschichte des vegetativen Thallus, insbesondere der Luftkammern der Riccien.— Flora, 103, 1911, 347.
P i t e 1 к a D. R. a. S c h о 11 e y C. N. Comparative morphology of some pro- tistan flagella.—Univ. Calif. Publ. Zool., 61, 2, 1955 (цит. no: Bourrel- ly, 1961).
Potthoff H. Zur Phylogénie und Entwicklungsgeschichte der Conjugaten.— Ber. Deutsch. Bot. Ges., 46, 1928, 667—673.
Prescott G. W. History of Phycology.—In: G. M. Smith (ed.). Manual of Phycology, Waltham, 1951, 1—11.
Prescott G. W. The Algae: a review. Frome and London, 1969.
Pringsheim E. G. The relationships between Bacteria and Myxophyceae.— Bacteriol. Rew., 13, 1949, 43—98.
Pringsheim E. G. Die Grundlagen eines taxonomischen Systems der Algen.— Z. Pflanzenphysiol., 54, 1966, 99—105.
Pringsheim E. G. Bactérien und Cyanophyceen. Übereinstimmungen und Unterschieden.— österr. Bot. Zeitschr., 114, 1967, 324—340.
Pringsheim N. Über die Befruchtung und Keimung der Algen etc.— Monatsber. Akad. Wissensch., Berlin, 1855.
297
Pringsheim N. Beiträge zur Morphology und Systematik der Algen. II Die Saprolegnieen.—Jahrb. Wiss. Botanik, I, 1858.
Pringsheim N. Über den Generationswechsel der Thallophyten und seinen Anschluss an den Generationswechsel der Moose.— Monatsber. K. Akad. Wiss., Berlin, 1876, 869—911.
Pringsheim N. Über Sprossung der Moosfrüchte und den Generationswechsel der Thallophyten.—Jahrb. wiss. Bot., 11, 1878, 1—46.
Proskauer J. Studies on Anthocerotales V.— Phytomorphology, 7, 1957, 113-135.
Proskauer J. Studies on Anthocerotales. VI.— Phytomorphology, 10, 1960, 1-19.
Proskauer J. On Carrpos 1.—Phytomorphology, 11, 1962, 359—378.
Ra ben borst L. Kryptogamen-Flora von Sachsen, der Ober-Lausitz, Thüringen und Nordböhmen etc. Abt. I. Leipzig, 1863.
Raben borst L. Flora europaea Algarum aguae dulcis et submarinae. Lip- siae, I, 1864; II, 1865; III, 1868.
R a w i t s c h e r F. Beiträge zur Kenntnis der Ustilagineen. L— Zeitschr. Bot., 4, 1912, 673—706.
Rees O. L. The morphology and development of Enteromorpha fumosa.— Amer. J. Bot., 19, 205-217, 1932.
Reimers H. Abteilung: Bryophyta. Moose.— In: E n g 1 e r’ s Syllabus der Pflanzenfamilien, 12-te Aufl., I. Berlin — Nikolassee, 1954, 218—268.
Renault B. et Zeiller R. Flore fossile de Commentry.— Bull. Soc. In- dustr. Min., 41, 1888, 34.
Richards O. C. Hybridization of Euglena gracilis chloroplast and nuclear DNA.— Proc. Nat. Acad. Sc. USA, 57, 1967.
Rogers D. P. A cytological study of Tulasnella.— Bot. Gaz., 94, 1932, 86— 105.
Rogers D. P. The basidium.— Univ. Iowa Stud, in Nat. Hist., 16, 1934, 160-183.
Rogers D. P. A comparison of evolutionary tendencies in plants, fungi, and animals.— Bull. Torrey Bot. Club, 75, 1948, 442—443.
R о s e n V i n g e L. K. Remarques sur les Protoflorideae.— Nuov. Notarisia, 36, 1925, 189-190.
Rosenvinge L. K. Phyllophora brodiaei and Actinococcus subcutaneus.— Kgl. Danske Vidensk. Selsk. Biol. Meddel., 8, 4, 1929, 1—40.
Rosin ski M. A. Development of the ascocarp of Anthracobia melaloma.— Mycologia, 48, 1956, 506-533.
Ross I. K. Syngamy and plasmodium formation in the Myxogastres.— Amer. J. Bot., 44, 1957, 843—850.
Rothmaler W. Über das natürliche System der Organismen.— Biol. Zent- ralbl., 67, 1948, 242-250.
Rothmaler W. Die Abteilungen und Klassen der Pflanzen.—Feddes Re- pert. sp. nov. regni veget., 54, 1951, 256—266.
Rothmaler W. Allgemeine Taxonomie und Chorologie der Pflanzen. Jena, 1 Aufl., 1950; 2 Aufl., 1955.
Ruhland W., Warnstorf C., Brotherus V. F. Klasse Musci (Musci frondosi), Laubmaose.—In: En gl er and Prantl, Die natürlichen Pflanzenfamilien, I, 3. Leipzig, 1893—1909.
Saccardo P. A. Sylloge fungorum, VII, 1888.
Sachs J. Lehrbuch der Botanik. Leipzig, 1868; 2te Aufl., 1870; 3te Aufl., 1873; 4te Aufl., 1874.
Sadebeck R. Untersuchungen über die Pilzgattung Exoascus — Jahrb: Hamburg. wiss. Anstalt, 1, 1884, 93—124 (цит. no: S a v i 1 e, 1955).
Sagan L. On the origin of mitosing cell.— J. Theoret. Biol., 14, 1967, 225.
Sager R. a. Pa la de G. E. Structure and development of the chloroplast in Chlamy do monas. I. The normal green cell.— J. Biophys., Biochem., Cytol., 3, 1957, 463-488.
Sansome E. Meiosis in oogonium and antheridium of Pythium debaryanum Hess e.— Nature, 191, 1961, 827—828.
298
S an so me E. Meiosis in Pythium debaryanum Hesse and its significance in the life history of the biflagellatae.— Trans. Brit Mycol. Soc., 46, 1963, 63—72.
Sa n some E. Meiosis in diploid and polyploid sex organs of Phytophthora and Achlya — Cytologia, 30, 1965, 103—117.
Sansome E. and Harris B. J. Use of camphor-induced polyploidy to determine the place of meiosis in fungi.— Nature, 196, 1962, 291—292.
Santesson R. The new systematics of Lichenized Fungi.—Proc. 7th Intern. Bot. Congr. Stockholm. July, 12—20, 1950. Stockholm, 1953, 809—810.
Savicz-Ljubitzkaja L. I. a. Abramov I. I. The geological annals of Bryophyta.— Rev. bryol. et lichenol., 28, 1959, 330—342.
Sauvageau C. Sur la sexualité hétérogamique d’une Laminaire (Saccor- hiza bulbosa).— Comptes rend. Acad. Sci. Paris, 160, 1915, 557—559.
Sauvageau C. Sur les gemétophytes de deux Laminaires (L. flexicaulis et L. saccharina).— Comptes rend. Acad. Sei. Paris, 162, 1916, 601—604.
Sauvageau C. Sur le développement de quelques Phéosporées.— Bull. Stat. Biol. Arcachon, 26, 1929, 253—420 (цит. no: Fritsch, 1943, 67).
S a V i 1 e D. B. 0. Nuclear structure and behavior in species of the Uredi- nales.—Am. J. Bot., 26, 1939, 585—609.
S a vile D. B. O. A phylogeny of the Basidiomycetes.—Canad. J. Bot., 33, 1955, 60-104.
Schechner-Fries M. Der Phasenwechsel von Valonia utricularis (Roth) A g.— Oesterr. Bot Zeitschr., 83, 1934, 241—254.
Schenk H. Über die Phylogénie der Archegoniaten und der Characeen.— Engler’s Bot. Jahrbücher, 1909, 42.
Scherffel A. Kleiner Beitrag zur Phylogenie einiger Gruppen niederer Organismen.— Bot Ztg., 59, Abt. I, 1901, 143—158.
Scherffel A. Endophytische Phycomyceten-Parasiten der Bacillariaceen und einige neue Monadinen. Ein Beitrag zur Phylogenie der Oomyceten.— Arch. Pratistenk., 52, 1925, 1—141.
Schiffner V. Klasse Hepaticae (Lebermoose).— In: Engle r und Prantl (ed.), Die natürlichen Pflanzenfamilien. Leipzig, Teil, Abt. 3, 1893—1909, 3-141.
Schiffner V. Die systematisch-phylogenetische Forschung in der Hepa- ticologie seit dem Erscheinen der Synopsis Hepaticarum und über die Abstammung der Bryophyten und Pteridophyten.— Progr. rei botanicae. 5, 1917, 387—520.
Schikorra W. Ueber die Entwicklungsgeschichte von Monascus.— Zeitschr. Bot, I, 1909, 379-410.
S c h i 1 1 e r J. Über die Fortpflanzung, geissellose Gattungen und die Nomenklatur der Coccolithophoraceen nebst Mitteilung über Copulation bei Di- nobryon.— Arch. Protistenk., 53, 1926, 326—342.
S c h i m p e r A. F. W. Über die Entwickelung der Chlorophyllkörner und Farbkörner.— Bot. Zeitung, 41, 1883, 105.
Sch im per W. P h. Synopsis muscorum europaeorum. Stuttgartiae, 1860.
Schmidt E. W. Abteilung Cyanophyta. Blaualgen, Spaltalgen.— In: E n g- ler’ s Syllabus der Pflanzenfamilien, 12te AufL, I. Berlin — Nikolassee, 1954a, 50-55.
Schmidt E. W. Abteilung Myxophyta. Schleimpilze.—In: Engler’s Syllabus der Pflanzenfamilien, 12te Aufl., I. Berlin — Nikolassee, 1954b. 57-62.
Schmidt P. Ist die scharfe Trennung zwischen zentrischen und pennaten Diatomeen haltbar? — Int. Rev. d. Hydrobiol. u. Hydrogr., 17, 1927. 274—288.
Schmitz F. u. Hauptfleisch P. Rhodophyceae.—ln: Engle г und Prantl, Die natürlichen Pflanzenfamilien, 1, 2. Leipzig, 1897, 298—544.
Schreiber E. Über die geschlechtliche Fortpflanzung der Sphacelariales.— Ber. Deutsch. Bot. Ges., 49, 1931, 235—240.
Schröter J. Myxomycètes (Mycetozoa, Schleimpilze, Pilzticre).—In: Entier und Prantl, Die natürl. Pflanzenfam., I, 1. Leipzig, 1897, 1—41.
299
Schröter J. Fungi (Pilze).—- In: E n gl e r und Prantl, Die natürl. Pflan- zenfam. I, 1. Leipzig, 1897.
Schussnig B. Die Reduktionsteilung bei Cladophora glomerata,— Oesterr. Bot. Zeitschr., 77, 1928a, 62—67.
Schussnig B. Zur Entwicklungsgeschite der Siphoneen.— Ber. Deutsch. Bot, Ges., 46, 1928b, 481—490.
Schussnig B. Zur Entwicklungsgeschite der Siphoneen. II. Ber. Deutsch. Bot. Ges., 47, 1929, 266-274.
S c h u s s n i g В. Der Generations- und Phasenwechsel bei den Chlorophyce- en (Ein historischer Bückblick).— Oesterr. Bot. Zeitschr., 79, 1930,
58—77.
Schussnig B. Die somatische und heterotype Kernteilung bei Cladophora suhriana К ü t z.— Planta, 13, 1931, 474—528.
S c h u s s n i g В. Der Generations- und Phasenwechsel bei den Chlorophyceen.
III.— Oesterr. Bot. Zeitschr., 81, 1932, 296—298.
Schussnig B. Der Kernphasenwechsel von Valonia utricularis (Roth) A g.- Planta, 28, 1938, 43—59.
Schuster R. M. Boreal Hepaticae. A manual of the liverworts of Minnesota and adjacent regions—Amer. Midi. Naturalist, 49, 1953, 257—684.
Schuster R. M. Keys to the orders, families and genera of Hepaticae of America (north of Mexico).— Bryologist, 61, 1958, 1—66.
Schuster R. M. The Hepaticae and Anthocerotae of North America, I. N. Y. and London, 1966.
Schütt F. Peridiniales — In: E n g 1 e r u. Prantl, Die natürlichen Pflanzenfamilien, I, 1. Leipzig, 1896, 1—30.
Schütt F. Bacillariales (Diatomeae).— In: Eng le г und Prantl, Die natürlichen Pflanzenfamilien, I, lb. Leipzig, 1896, 31—150.
Schwartz E. J. The Plasmodiophoraceae and their relationship to the Mycetozoa and the Chytrideae.— Ann. Bot., 28, 1914, 227—240.
Schwartz E. J. a. Cook W. R. I. The life history and cytology of a new species of Olpidium: Olpidium radicale, sp. nov.— Trans. Brit. Mycol. Soc., 13, 1928, 205-221.
Schwarz E. Der Formwechsel von Ochrosphaera neapolitana.— Arch. Pro- tistenk., 77, 1932, 434—462.
Schwarz O. Die Entstehung der Blüte und die systematische Stellung der Phanerogamen.— Mitteil. Thiiring. Bot. Ges., 1, 1955, 292—302.
Schwendener S. Über die Natur der Flechten.— Verhandl. Schweiz. Na- turf. Ges. 51, 1867.
Sco tt D. H. The evolution of land plants. London, 1911.
Scott D. H. Aspects of fossil botany. Ferns and seed-ferns.—Nature, 123, 1929, 350-352.
Shoemaker R. A. Biology, cytology, and taxonomy of Cochliobolus sativus.— Ganad. J. Bot., 33, 1955, 562—576.
Singer R. Das System der Agaricales.— Ann. Mycol., 34, 1936a, 286—278.
Singer R. Studien zur Systematik der Basidiomyceten.— Beih. Bot. Centralbl., 56, АЫ. B., 1936b, 137—174.
Singer R. The Agaricales (Mushrooms) in modern taxonomy.—Lilloa, 22, 1949, 5—832.
Singer R. The meaning of the affinity of the Secotiaceae with the Agaricales.— Sydowia, 12, 1958, 1—43.
Singer R. The Agaricales in modern taxonomy, 2d ed. Weinheim, 1962.
S i n g h R. N. Reproduction in Drap arnaldiop sis indica Bharadwaj a.— New Phytol., 41, 1942, 262-273.
Singh R. N. Nuclear phases and alternation of generations in Drapamaldi- opsis indica Bharadwaj a.— New Phytol., 44, 1945, 118—129.
S i ng h R. N. a. S i n h a R. Genetic recombination in a blue-green alga, Cylindrospermum majus К u e t z.— Nature, 207, 4998, 1965, 782—783.
Skupienski F. X. Récherches sur le cycle évolutif de certains Myxomycètes. Paris, 1920 (цит. no: Martin, 1940).
S к u j a H. Versuch einer systematischen Einteilung der Bangioideen oder
300
Protoflorideen.— Acta Horti Bot. Univ. Latviensis, 11—12, 1939, 23—38.
Skuja И. Körperbau und Reproduktion bei Dinobryon borget Lemm.— Svensk Bot. Tiskr., 44, 1950, 96—107.
S i η о t о Y. a. Y u a s a A. Studies in the cytology of reproductive cells. I. On the planocytes in five forms of Myxomycètes.— Bot. Mag. (Tokyo), 48, 1934. 420-729.
Sloan B. J. a. Wilson G. В. The functions of the microspores of Gela- sinospora calospora var. autosteira.— Mycologia, 50, 1958, 111—116.
Small J. Quantitative evolution. VIII. Numerical analysis of tables to illustrate the geological history of species-number in Diatoms; an introductory summary.— Proc. Roy. Irish Acad., 51, Sect. B., 4, 1946.
S m i t h A. L. Lichenes. Cambridge, 1921.
Smith G. M. The fresh-water algae of the United States. N. Y., London, 1933; 2nd ed., N. Y., Toronto, London, 1950.
Smith G. M. (ed.), Manual of Phycology. Waltham, 1951.
Smit h G. M. Cryptogamie Botany, I. Algae and Fungi. N. Y. and London, 1938; 2nd ed. N. Y.— Toronto — London, 1955.
S ο ό R. The present aspect of the evolutionary history of Telomphyta.— Ann. Univ. Sc. Budapest de R. Eötvos nominatae, sect, biol., 4, 1961.
Sparrow F. K. Inoperculate chytridiaceous organisms collected in the vicinity of Ithaca, N. Y., with notes on other aquatic fungi.— Mycologia, 25, 1933, 513-535.
Sparrow F. K. Interrelationships and phylogeny of the aquatic Phycomyce- tes.— Mycologia, 50, 1958, 797—813.
Sparrow F. K. Aquatic Phycomycetes. Ann Arbor, 1943; 2nd ed., 1960.
Spiltoir Ch. F. Life cycle of Ascosphaera spis (Pericystis apis).— Amer. J. Bot, 42, 1955, 501-508.
Spiltoir C h. F. a. Olive L. S. A reclassification of the genus Pericystis Bett s.— Mycologia, 47, 1955, 238—244.
Spruce R. Hepaticae amazonicae et andinae.— Trans. Proc. Bot. Soc. Edinburgh, 15, 1884-1885, I—XI, 1-590.
S t a h 1 E. Beiträge zur Entwicklungsgeschichte der Flechten. I. Leipzig, 1877.
S t a n i e r R. Y. a. Niel C. B., van The main outlines of bacterial classification.— J. Bacter., 42, 1941, 437—466.
S t a n i e r R. Y. a. N i e 1 C. B., van. The concept of a bacterium.— Arch. Mikrobiol., 42, 1962, 17-35.
S t e e r e W. C. Cenozoic and mesozoic Bryophytes of North America.— Amer. Midi. Nat., 36, 1946, 298-324.
Steffen K. Entwicklungsgeschichte und Fortpflanzung.—Fortschr. Bot., 17, 1955, 61-106.
S t e i n b ö c k H. Gedanken zur Phylogénie der Moose. Eine entwicklungsgeschichtliche Studie.— Agronomia lusitana, 16, 1954, 115—149.
Stizenberger E. Dr. L. Rabenhorst’s Algen Sachsens, resp. Mittol-Euro- pa’s. Decaden I—G. Systematische geordnet mit Zugrundelegung eines neuen Systems. Dresden, I860 (цит. по: Smith, 1951).
S t о s c h H. A. Oogamie in a centric diatom.— Nature, 165, 1950, 531—533.
S t о s c h H. A. Entwicklungsgeschichtliche Untersuchungen an zentrischen
Diatomeen. I. Die Auxosporenbildung von Melosira varians.— Arch. Mikrobiol., 16, 1951, 101—135.
Stosch H. A. Die Oogamie von Biddulphia mobiliensis und die bisher bekannte Auxosporenbildung bei den Centrales.— 8me Congr. int. Bot., Paris. Rep. Com. Sect. 17, 1954, 58—66.
Stosch II. A. Entwicklungsgeschichtliche Untersuchungen an zentrischen
Diatomeen. II. Geschlechtszellenreifung. Befruchtung und Auxosporenbil¬
dung einiger grundbewohnender Diatomeen der Nordsee.— Arch. Mikrobiol, 23, 1956, 227—265.
Stosch II. A. Entwicklungsgeschichtliche Untersuchungen an zentrischen
Diatomeen. III. Die Spermatogenese von Melosira moniliformis A g a r d h.— Arch. Mikrobiol., 31, 1958a. 274—282.
Stosch II. A. Kann die oogame Araphidee Rhabdonema adriaticum als Bin-
301
desglied zwischen den beiden grossen Diatomeengruppen angesehen werden?— Ber. Deutsch. Bot. Ges., 71, 1958b, 241—249.
Strasburger E. Zur Frage eines Generationswechsels bei den Phaeophyce- en.— Bot. Zeitg., 64, 2 Abt., 1906, 1—7.
Subramanian С. V. Spore type in the classification of the Hyphomyce- tes.— Mycopath. et mycol. appl., 26, 1965.
Svedelius N. E. Zytologisch-Entwicklungsgeschichtliche Studien über Scinaia furcellata.— Nova Acta Reg. Soc. Sc. Upsal., Ser. 4, 4, 1915, 1—55.
Svedelius N. Einige Bemerkungen über Generationswechzel und Reduktionsteilung.— Ber. Deutsch. Bot. Ges., 39, 1921, 178—187.
Svedelius N. Alternation of generations in relation to reduction division.— Bot. Gaz., 83; 1927, 362-384.
Svedelius N. E. On the development of Asparagopsis armata Harv. and Bonnemaisonia asparagoides (Woodw.) A g.— Nova Acta Reg. Soc. Sc. Upsaliensis, Ser. 4, 9, 1933, 1—61.
Svedelius N. E. The apomeiotic tetrad division in Lomentaria rosea in comparison with the normal development in Lomentaria clavellosa.— Symbolae Bot. Upsaliensis, 2, 1937, 1—54.
Svedelius N. E. Zytologisch-entwicklungsgeschitliche Studien über Gala- xaura, eine diplobiontische Nemalionales-Gattung.— Nova Acta Reg. Soc. Sei. Uposaliensis, Ser. 4, 13, 1942, 1—54.
T a к h t a j a n A. L. Phylogenetic principles of the system of higher plants.— Bot. Rev., 19, 1953, 1-45.
T a t u η о S. Chromosomen von Takakia lepidozioides und eine Studie zur Evolution der Chromosomen der Bryophyten.— Cytologia, 24, 1959, 138— 147.
Taylor D. L. The occurence and significance of endosymbiotic chloroplasts in the digestive glands of herbivorous opisthobranchs.— J. Phycol., 3, 1967 (цит. по: А 11 so pp, 1969).
Taylor D. L. Chloroplasts as symbiotic organelles in the digestive gland of Elysia viridis (Gastropoda: Opisthobranchia).—J. mar. biol. Ass. U. K., 48, 1968, 1-15.
Taylor W. R. A species of Acrothrix on the Massachusetts Coast.— Amer. J. Bot., 15, 1928, 577—583.
Taylor W. R. Phaeophycean life-histories in relation to classification.— Bot. Rev., 2, 1936, 554-563.
Thaxter R. New and peculiar Zygomycetes. 2. Syncephalastrum and Syn- cephalis.— Bot. Gaz., 24, 1897, 1—15.
Thirumalachar M. J., Whitehead M. D. a. Boyle J. S. Gameto- genesis and oospore formation in Cystopus (Albugo) evolvuli — Bot. Gaz., 110, 1949, 487-491.
Thompson R. H. Studies in the Volvocales. I. Sexual reproduction of Pandorina charkowiensis and observations on Volvulina steinii.— Amer. J. Bot., 41, 1954, 142—145.
T h u r e t G. Recherches sur les zoospores des algues.— Ann. des. sc. nat, Botanique, S. 3, 14, 1850.
T i e g h e m P h., v a n. Traité de Botanique. Paris, 1884.
T i m m e r L. W., Castro J., Erwin D. С., В e 1 s e r W. L. and Zentmy- e r G. A. Genetic evidences for zygotic meiosis in Phytophthora capsici.— Amer. J. Bot, 57, 1970, 1211-1218.
T i w a r i K. K. a. W i 1 d m a n S. G. Function of chloroplast DNA. I. Hybridization studies involving nuclear and chloroplast DNA with RNA from cytoplasmic (80 S) and chloroplast (70 S) ribosoms.— Proc. Nat. Acad. Sei., 59, 1968 (цит. по: A 11 s о p p, 1969).
T о mine ru p I. C. a. Ingram D. S. The life-cycle of Plasmodiophora brassicae Wo го n. in Brassica tissue cultures and in intact roots.— New Phytol., 70, 1971, 327—332.
T ο n i J. B., de. Sylloge algarum. Patavii, 3, Fucoideae, 1895.
Townrow J. A. Two triassic Bryophytes from South Africa.— J. South Afric. Bot, 25, 1959, 1-22.
302
Trevisan V. Schema di una nuova classificazione delle Epatiche.— Mem. Reale 1st. Lomb., ser. 3, 4, 1877, 383—451 (цит. no: Schuster, 1966).
Tseng G. K. a. Chang T. J. Studies on Porphyra. II. On Conchocelis-phase of Porphyra tenera К j e 11 m. and its conchospores.— Acta. Bot. Sinica, 4, 195b a, 27-46.
Tseng С. K. a. Chang T. J. Studies on Porphyra. III. Sexual reproduction of Porphyra.— Acta Bot. Sinica, 4, 1955b, 153—166.
T u b a k i K. Studies on the Japanese Hyphomycetes. V. Leaf and stem group with a discussion of the classification of Hyphomycetes and their perfect stages.— J. Ilattori Bot Lab., 20, 1958.
T u z e t O. et Manier J. F. La sexualité et les spores chez Palaviscia philosciae T u z e t et Manier et chez P. sphaeromae nouvelle espèce de Palavisciées parasite de Sphaeroma serratum F.— Comptes Rend. Acad. Sei. Paris, 226, 1948.
T y 1 e r S. A. a. B a r g h о о r η E. S. Occurence of structurally preserved plants in precambrian rocks of the Canadian Shield.— Science, 119, 1954, 606— 608.
T y 1 u t k i E. E. Some aspects of morphology, genetics, and cultural behaviour of Gelasinospora calospora var. autosteira.— Mycologia, 50, 1958, 333—356.
U b r i z s y G. a. V ö r ö s J. A new conception in the review of the phylogeny and system of fungi.— Acta Bot. Acad. Scient. Hungariae, 12, 1966, 199— 219.
Ulrich R. Les constituents de la membrane chez les champignons.— Revue Mycol., Mém. Hors-Sér., N 3, 1943, 1—44.
U n d e r w о о d L. M. The evolution of the Hepaticae.— Bot. Gaz., 19, 1894, 347—361.
Unger F. Aphorismen zur Anatomie und Physiologie der Pflanzen. Wien, 1838 (цит. no: Jus t, 1945).
Varitchak B. L’évolution nucléaire chez le Pericystis apis M a a s s e n.— С о m p t. Rend. Acad. Sc. Paris, 194, 1932, 300—302.
Varitchak B. Deuxième contribution à l’étude du développement des Ascomycètes: Evolution nucléaire dans le sac sporifére de Pericystis apis Maassen et sa signification pour la phylogénie des Ascomycètes.— Botaniste, 25, 1933, 343—390.
Vazart B. Différenciation des cellules sexuelles et fécondation chez les Cryptogames. Wien, 1963.
Verdoorn F. Classification of Hepatics.—In: F. Verdoorn (ed), Manual of Bryology. Hague, 1932, 413—432.
V i s c h e r V. Uber einen pilzähnlichen Mikroorganismus Chlorochytridion,
einige neue Protococcalen und die systematische Bedeutung der Chloro- plasten.— Verh. Naturf. Ges. Basel, 56, 1945, 41—59.
Y u i 1 1 e m i n P. Les champignons. Essai de classification, Paris, 1912.
W a i n i о E. A. Lichens du Brézil I, IL— Acta Soc. Fauna et Flora Fenn., 7, 1890.
W a 11 i n I. E. On the nature of mitochondria. — Amer. J. Anat., 30, 1922. 203 (цит. no: Allsopp, 1969).
Walton J. Carboniferous Bryophyta, I. Hepaticae.—Ann. Bot., 39, 1925, 563—572. II. Hepaticae and Musci.—Ann. Bot., 42, 1928, 707—716.
Walton L. B. Studies concerning organisms occuring in water supplies, with particular reference to those found in Ohio.— Bull. Ohio Biol. Surv., 24, 5, 1, 1930, 1—86.
Wang Y.-C h. et Martens P. Sur l’origine de la dicaryophase chez quelques Urédinées.— Cellule, 48, 1939, 213—245.
Waterhouse G. M. The true nature of Myrioblepharis T h a x t e r.— Trans. Brit. Mycol. Soc., 28, 1945, 94—100.
Werdermann E. Pilze.—In: E n g 1 e r’s Syllabus der Pflanzenfamilien, 12 Aufl. Berlin, 1954.
Wettstein R. Handbuch der systematischen Botanik. Leipzig und Wien, 1, 1901; 2, 1903—1908.
303
VV h i f f e n A. J. A discussion of taxonomie criteria in the Chytridiales.— Farlowia, I, 1944, 583—597.
Whittaker R. H. On the broad classification of organisms.— Quart, Rev. Biology, 34, 1959, 210-226.
Wie be r M. Die Infektion, die Myzelüberwinterung und die Kopulation bei Exoasceen.— Forschungen a. d. Geb. d. Pflanzenkrankh. herausgeg. v. Schaffnit, Jena, 3, 1927, 139—176 (цит. по: Gäumann, 1964).
Wille N. Conjugatae. Chlorophyceae. Characeae.— In: E n g 1 e г und P r a n 11, Die natürlichen Pflanzenfamilien, 1, 2. Leipzig, 1897, 1 —175.
Williams M. The cytology of the gametangia of Codium tomentosum.— Proc, Linn. Soc. New South Wales, 50, 1925, 98—111.
Wilson G. M. Meiosis in Allomyces.— Bull. Torrey Bot. Club, 79, 1952, 139—160.
Wilson Ch. L. Development of the ascogonium and perithecium of Endo- conidiophora fagacearum.— Phytopathology, 46, 1956, 625—632.
Wilson T. B. a. C a s s i n J. On a third kingdom of organized beings.— Proc. Acad. Nat. Sei. Philadelphia, 15, (1863) 1864, 113-121.
W i n g a r d S. A. Studies on the pathogenicity, morphology and cytology of Nematospora phaseoliBull. Torrey Bot. Club, 52, 1925, 249—290.
Winter G. Pilze.—In: L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz, 2te Aufl., I, 1. Leipzig, 1884.
Wolf F. A. Leafspot of ash and Phyllosticta viridis.— Mycologia, 31, 1939, 258-266.
Wolf F. A. Spore discharge in Taphrina.— J. E. Mitchell Sc. Soc., 74, 1958, 161—162.
Y amanouchi S. The life-history of Polysiphonia violacea, I—IL—Bot. Gaz., 41, 1906, 425—433; 42, 1906, 401—449.
Yu as a A. Studies in the cytology of reproductive cells. III. The genesis of the flagellum in the planocytes of Fuligo septica G m e 1 i n.— Bot. Mag. (Tokyo), 49, 1935, 538-545.
Zahlbruckner A. Lichenes (Flechten). Speziell er Teil. In: Engle r, Die natürlichen Pflanzenfamilien, 2te Aufl, 8. Leipzig, 1926.
Z i с к 1 e r H. Genetische Untersuchungen an einem heterothallischen Ascomy- zeten (Bombardia lunata nov. spec.).— Planta, 22, 1934, 573—613.
Zick 1er H. Die Spermatienbefruchtung bei Bombardia lunata.— Ber. Deutsch. Bot. Ges., 55, 1937, 114—119.
Ziegler A. W. Meiosis in the Saprolegniaceae.— Amer. 40, 1953, 60—66.
Zimmermann W. Phylogenie.— In: Fr. Verdoorn (ed.), Manual of Bryology. Hague, 1932, 433—464.
Zimmermann W. Geschichte der Pflanzen. Stuttgart, 1949.
Zimmermann W. Die Phylogenie der Pflanzen, Jena, 1930; 2te Aufl., Stuttgart, 1959.
Zinnecker E. Reduktionsteilung, Kernphasen Wechsel und Geschlechtsber- timmung bei Bryopsis plumosa (Huds.) A g.—Oesterr. Bot. Zeitschr., 84, 1935, 53-72.
OUTLINES OF NONVASCULAR PLANT PHYLOGENY
Summary
The present paper is aimed at generalizing modern data on origin and principal trends in development of phyla of lower autotrophic and heterotro- phic plant organisms and bryophyta.
The paper consists of four chapters. Chapter i deals with the problem on main trends in development of living world on the Earth and on classification principles of it. Chapters II, III and IV consider the problem on evolutionary trends and classification principles of lower autotrophic organisms (algae), lower heterotrophic plant organisms (fungi) and bryophyta, respectively.
I. Main Trends in Development of Living World
In the second half of the 18th century C. Linnaeus distinguished three kingdoms of nature — mineral kingdom of “stones” — lapides and two organic ones — plants (vegetabilia) and animals (animalia). This division corresponded to the usual ideas of that time and resulted from the fact that lower organisms were completely unstudied then. In connection with a successful study of lower organisms in the 19th century which showed the presence among them of a series of groups with mixed features of plants and animals, a number of suggestions was advanced on separating lower organisms into a special kingdom (Bory de Saint-Vincent, Hogg, Haeckel). Haeckel’s suggestion on separation of the kingdom of Protista was particularly popular, though it should be mentioned that Haeckel himself during 1866—1894 more than once changed the composition of this kingdom. The later authors (Copeland, Barkley, Kozo-Poliansky, Rothmaler) complicated still more conception of the Protista (Protobionta) kingdom volume. Assumptions were also advanced on separating fungi into a special kingdom (Fries, Meresch- kowsky, Kozo-Poliansky, Vaga). Divergences in opinions of many authors in conception of the Protista kingdom volume evidence for its artificial character. “Typical” animals and plant organisms originate from common ancestors anl therefore it is natural that lower representatives of animals and
305
plant phyla have similar, “mixed” characters. The same concerns the attempt to separate fungi into a special kingdom.
A study of the cell structure showed, on the one hand, a single-type character in principle of a plant cell and an animal one, on the other — a profound difference of them from the cell structure of Schizophyta — bacteria and blue-green algae. Nemec (1929) was the first to make a suggestion of dividing the living world according to the cell structure into two main parts, kingdoms — Akaryonta, or Prokaryonta, on the one hand, and Karyon- ta — on the other. However Nëmec’s suggestion did not find a response, and only much later the division of the living world into two kingdoms based on the cell structure proposed by Némec was widely recognized (Chatton, 1937; Stanier a. van Niel, 1941, and others). The names of the kingdoms were some what changed — Procaryota (Protocaryota) and Eucaryota which are more precise in reflecting peculiarities in the cell structure of the representatives of these kingdoms. Following Rothmaler’s suggestion (1948), the third kingdom containing non-cellular organisms — viruses and bacteriophages, which we suggest to call Acytobionta, should be added to the mentioned two. Division of the living world based on the nutrition character and suggested by Whittaker (1959) reflects the role of some groups of organisms in nature life, but it cannot in any way be considered as phylogenetic. This is evident from the fact that Whittacker’s kingdoms have no clear boundaries and often overlap each other.
As far as recent representatives of the kingdom Acytobionta — viruses are concerned, they may be considered as specialized, adapted to intracellular parasitism descendants of primary non-cellular organisms. Procaryota — Cy- anophyta and Bacteriophyta might also originate from primary non-cellular organisms (Archaebionta, Eobionta).
As to interrelations between Procaryota and Eucaryota, an opinion is spread according to which the latter developed from the former as a result of progressive transformation of their cell components (Cronquist, 1960; All- sopp, 1969), or as a result of symbiogenesis (Mereschkowsky, 1909; Kozo- Poliansky, 1924; Sagan-Margulis, 1967, 1968). Researches of the cell structure in Procaryota and Eucaryota give no convincing evidence in favour of the theory of symbiogenesis. Direct development of Eucaryota from those close to recent Schizophyta is also hardly probable. Schizophyta — bacteria and blue-green algae are specialized groups developed from primary organisms — Eobionta and formed in the Pre-Cambrian period. They are notable for a series of specific peculiarities in the cell structure, biochemical and physiological properties differing them from Eucaryota. This indicates that Eucaryota can be direct descendants neither of bacteria, nor of blue-green algae. Eucaryota developed from more primitive forms which might be common ancestors of Procaryota and Eucaryota.
Initial forms of Eucaryota were simple unicellular amoeboid organisms in which the nuclear apparatus and flagellar one connected with it as well as ability to phototrophic nutrition formed at early stages of evolution. Rapid development of Eucaryota in the Proterozoic era resulted in isolation of a number of phyla with heterotrophic, autotrophic, holozoic (phagotrophic) and mixed modes of nutrition. Development of secondary cellular wall in
306
representatives of a number of phyla stimulated an osmotic mode of nutrition and formation of characteristic features of plant ogranisms.
We distinguish ten phyla of autotrophic plant organisms and four phyla of heterotrophic ones which are suggested to be considered in classification as divisions of the plant kingdom, namely: autotrophic — Xanthophyta, Chry- sophyta, Diatomophyta, Phaeophyta, Pyrrhophyta, Cryptophyta, Euglenophyta, Ghloromonadophyta, Rhodophyta, Chlorophyta (all the green plants from green algae to angiosperms), heterotrophic — Myxomycota, Chytridiomycota, Sapro- legniomycota, Eumycota. The phylum (division) Chlorophyta we suggest to divide into subphyla (subdivisions) Chlorophycophytina, Bryophytina, Antho- corophytina, Psilophytina, Lepidophytina, Sphenophytina, Pterophytina, Gym- nospermophytina, Angiospermophytina. Perhaps, the number of subdivisions of higher Chlorophyta should be increased by isolating the subdivision.
Tmesophytina and dividing the subdivision Gymnospermophytina into two — Cycadophytina and Coniferophytina.
Interrelations between the kingdoms of organisms and phyla (divisions) of plant organisms are presented in Fig. 2.
II. Evolutionary Trends and Classification Principles of Lower Autotrophic Plant Organisms (Algae)
On the basis of the data available on the cell structure, reproduction and biochemical peculiarities, we distinguish among autotrophic Eucaryota the above-mentioned nine phyla and the subphylum Chlorophycophytina which are usually referred to a biological group of algae.
Chrysophyta (Chrysophycophyta, Chrysophyceae). Pascher (1931), proceeding from the vegetative body structure, divided Chrysophyta into five classes — Chrysomonadineae, Rhizochrysidineae, Chrysocapsineae, Chrysosphaerineae and Chrysotrichineae. However further studies detected in Chrysophyta a difference in the type of the flagellar apparatus. On the basis of its structure, two groups are outlined among them: 1) heteroflagellate organisms, with one pennate (tinsellike) and the other whiplashlike flagella and 2) organisms with two flagella and often with a haptonema between them (Parke, 1961). This induces to consider them as two subphyla, and hence two classes, which we, according to Bourrelly (1964), call Heterochrysophyceae and Isochryso- phyceae (Haptophyceae). The name Haptophyceae suggested by Christensen (1962) we consider to be less suitable as not all Isochrysophyceae have a haptonema. Possible interrelations between the Heterochrysophyceae orders are shown in Fig. 9.
Xanthophyta (Xanthophycophyta, Xanthophyceae, Heterocontae). Classification of Xanthophyta proposed by Pascher (1931) is generally adopted. We follow it with some nomenclature changes in conformity with modern rules of botanical nomenclature. Some morphological and biochemical peculiarities make us to consider Xanthophyta as a special phylum which might have common ancestors with Chrysophyta and Diatomophyta.
Diatomophyta (Bacillariophyta, Bacillariophycophyta, Bacillariophyceae, Diatomeae). Taking into account views of different authors (Hustedt, 1930;
307
Zhiize, Proshkina — Lavrenko and Sheshukova, 1940—1950; Topachevsky, 1962) on origin and evolution of Diatomophyta as well as paleobotanical and comparative-morphological data, the author comes to a conclusion that from two classes of Diatomophyta Centrodiatomophyceae (Centricae) is an initial one which originated in the Jurassic period from the flagelloid forms, probably close to recent Chrysophyta. During the Upper Cretaceous period there appear bisymmetrical Anaulales which gave rise to the class Pennatodiatomophyceae (Pennatae). Primitive araphinate forms should be assumed as a basis for the above class. They generated two branches of development — with slit-raphe and with canal-raphe. The author gives classification of Diatomophyta based on systems of Hustedt, Karsten and Topachevsky with some nomenclature changes. Supposed interralations between the Diatomophyta orders are shown in Fig. 17.
Phaeophyta (Phaeophycophyta, Phaeophyceae). On considering the data on comparative morphology of vegetative organs, cytology, types of reproduction and development, the author makes a conclusion that Phaeophyta represent an undoubtedly monophyletic group characterized by uniform biochemical properties and similar structure of motile reproductive cells. The character of motile stages — zoospores and gametes gives reason to suppose origin of brown algae from flagellate forms with two different flagella, one being tinsel-like and the other — whiplashlike. The author suggests the Phaeophyta system as interpreted by Papenfuss (1951) and diagrams of Kylin (1933) and Papenfuss (1951) of possible interrelations between this division orders (Fig. 26 and 27).
Pyrrhophyta (Pyrrhophycophyta, Dinophyceae, Peridineae). The author agrees with Pascher’s suggestion on Pyrrhophyta development in two branches represented by the classes Dinophyceae (Dinokontae) and Desmophy- ceae. Naked monad forms with a peculiar nucleus structure, two flagella and presence of specific set of pigments close to the Cryptophyta ancestors were initial for the both branches. The system proposed by Pascher (1931) is adopted in the paper with some changes. Fig. 32 presents a diagram of possible interrelations between the division orders.
Cryptophyta (Cryptophycophyta, Cryptophyceae). After considering the views of different authors as to possible related ties of Cryptophyta, the author draws a conclusion on the basis of some morphological and biochemical peculiarities on expediency of considering Cryptophyta as an independent phylum.
Euglenophyta (Euglenophycophyta, Euglenophyceae, Euglenineae). Examination of suggestions made by some authors concerning related ties and taxonomic position of Euglenophyta enabled the author to make a conclusion that this group differs from green algae and Pyrrhophyta by a number of peculiarities and should be considered as a special phylum (division) of plant organisms. Within the division two groups are outlined which the author considers, according to Popova (1951, 1955), as orders — autotrophic and saprophytic Euglenales and phagotrophic Peranematales differing from Eugle- nales in a nutrition character and in the presence of a rod-like apparatus, in some forms —of pharyngial apparatus as well, connected with the nutrition character. The flagellar apparatus type, nucleus structure, presence of
308
mucous bodies and paramylon which was detected in some Peranematales evidence for related ties of the Euglenales and Peranematales.
Chloromonadophyta (Rhaphidiophyceae, Chloromonadophyceae, Chloromo- nadineae) represent a small group which is insufficiently studied till now. The presence of mucous bodies, triehocysts and ventral furrow make them closer to Cryptophyta; they have some common features with Pyrrhophyta and Euglenophyta. However, until the Chloromonadophyta flagellar structure and nature of their pigments are studied in detail, it is difficult to speak on related ties of this group and it is expedient according to Prescott (1951), to consider them as an independent phylum (division.)
Rhodophyta (Rhodophycophyta, Rhodophyceae). The problem on origin of Rhodophyta and on their phylogenetic relations with other phyla of the plant kingdom is interpreted differently. A view is widely accepted on their connection with blue-green algae based on similarity in their pigment sets. But the cell structure of red algae, their reproductive organ structure, character of the sexual process indicate their belonging to Eucaryota. Similarity, but not identity, of the pigment set in them and blue-green algae might be a result of these phyla development under analogous ecological conditions. The classes Bangiophyceae and Florideophyceae, with all their differences, are characterized by such common features as a pigment set, assimilation products and the character of the sexual process. Therefore their common origin cannot raise any doubts. The both classes are most likely to represent groups which have common ancestors and were isolated at early stages of development. The recently published diagram of Prescott (1969) is presented as an example of the supposed phylogenetic relations of Rhodophyta. As seen from the diagram (Fig. 44), phylogenetic relations between Bangiophyceae and Florideophyceae are far from being clear to the author of the diagram.
Chlorophycophytina (Chlorophycophyta, Chlorophyceae) are considered by the author subdivisions of the Chlorophyta phylum.
As to the volume of the subdivision, the author thinks the involvement of both conjugates and charophytas into its composition to be quite resonable. Recent researches (Parke, Manton, and others) with application of electron- microscopic technics supported Korshikov’s opinion on heterogeneity of the group of monad forms, with are usually referred to the order Volvocoles, and substantiated Chadefaud’s suggestion (1960) on separation of the class Prasi- nophyceae which differ by a peculiar structure of flagella and by the presence of scales covering the cell and flagella. Necessity of isolating the class Loxophyceae (Christensen, 1962) requires an additional control as well as elucidation of taxonomic position of some other forms, for instance the genus Haematococcus characterized by a pectinate structure of flagella. The author gives the class Prasinophyceae system according to Parke and Dixon (1968), and the presented system of Euchlorophyceae bases on the systems of Fritsch, Smith and Fott. Possible phylogenetic relations between the class Euchlorophyceae orders are presented in Fig. 48.
309
III. Evolutionary Trends and Classification Principles of Lower Heterotropliic Plant Organisms (Fungi)
As it was already mentioned in Chapter I, the author distinguishes among the lower heterotrophic plant organisms four phyla which are considered by him as independent divisions — Myxomycota, Chytridiomycota, Saprolegniomycota, and Eumycota. The structure of monads, in particular of flagellar apparatus, biochemical peculiarities, character of the sexual process and reproductive organs were assumed as a basis for distinguishing such divisions.
Myxomycota (Myxomycophyta, Mycetozoa). The author thinks separation of myxomycètes into an independent division of Eucaryota to be substantiated and refers to it five groups considered by him as classes, namely: Protostelio- mycetes, Labyrinthulomycetes, Acrasiomycetes, Myxogastromycetes and Plas- modiophoromycetes.
Chytridiomycota (Opisthomastigomycota). Attempts to isolate uniflagellate forms with a posteriorly attached whiplashlike flagellum into a separate phylogenetic and taxonomic group were made by Gobi, Mereschkowsky and Jaczevsky. However, only in recent years it was suggested to consider this group of organisms as an independent division (Arx, 1967; Zerov and Zerova, 1968). The author points to the complication in the structure of a vegetative body, reproductive organs and sexual process within the limits of the orders Chytridiales, Blastocladiales and Monoblepharidales.
Saprolegniomycota (Pteromastigomycota). A peculiar structure of the flagellar apparatus in representatives of Saprolegniomycota and the presence of cellulose in their cell wall composition give ground to suppose that this division is most likely to originate from the forms close to Chrysophyta. Besides heteroflagellate Saprolegnimycetes, the author refers to Saprolegniomycota Hyphochytriomycetes as a separate class characterized by the presence of one tinsel-like flagellum. Possible interrelations between the Saprolegnio- mycetes orders are presented in Fig. 59.
Eumycota. The division Eumycota is divided by the author into three classes — Zygomycetes (Mucoromycetes), Ascomycetes and Basidiomycetes. Phylogenetic relations of Zygomycetes with the class Ascomycetes are confirmed by the presence of some intermediate forms. Homology of ascus and basidium, similarity of the sexual process in Basidiomycetes and Ascomycetes testify to their origin from common ancestors. As far as Zygomycetes origin is concerned, their relations with Saprolegniomycota and Chytridiomycota are rather problematic because of the absence of intermediate forms. The supposition on the development of Zygomycetes and so the whole phylum Eumycota from the ancestors close to Chytridiomycota is more probable, however these relations might be confined to early stages of differentiation of lower Eucaryota.
As to the phylogenetic interrelations between the orders and families of Zygomycetes, the author considers that the orders Mucorales, Entomophthora- les and Zoopagales were isolated very early. The family Mucoraceae should be taken, following Gäumann, with the simplest type of the sexual process and multisporous sporangia as a basis for the system of Mucorales.
After detailed consideration of the existing theories on the origin of the class Ascomycetes the author makes a conclusion on their connection
310
with Zygomycetes. Accepting the division of Ascomycetes into two subclasses — Protoascomycetidae and Euascomycetidae, the author to the first subclass refers organisms with a simple structure of asci developing from a zygote without forming ascogenous hyphae and fruit bodies, i. e. the orders Endomycetales and Spermophthorales. The much greater subclass Euascomycetidae involves up to 20 orders characterized by various forms of the sexual process and different structure of fruit bodies and asci. Eurotiales (Aspergillales), Microascales, Coronophorales, Onygenales, Ascosphaerales, Ta- phrinales, Erysiphyles are usually referred to more primitive orders of Euascomycetidae.
Among them several evolutionary trends are outlined, some of them were likely to give rise to more high-developed groups. Among more high- organized Euascomycetidae the group of ascolocular ones — the orders My- riangiales, Dothiorales and Pseudosphaeriales — is especially pronounced. As far as the group of ascohymenal orders is concerned it might be heterogeneous. The schemes of Gäumann (Fig. 71) and Luttrell (Fig. 80) are presented as an example of possible phylogenetic relations and evolutionary trends of Ascomycetes.
As to the class Basidiomycetes, the author considers that the organisms with a simple type of development and unicellular non-specialized basidium should be placed for the basis of this class phylogenetic tree. Among recent Basidiomycetes the order Tremellales and lower genera of the order Thelepho- rales are most close to the initial forms. From the similar primitive forms the other groups of Basidiomycetes developed in two branches — heterobasidial and homobasidial. In this connection the author suggests to divide the class Basidiomycetes into three subclasses — Protobasidiomycetidae, Sclerobasidio- mycetidae and Homobasidiomycetidae. To the subclass Protobasidiomycetidae only one order Tremellales is referred involving the orders Tulasnellales and Dacryomycetales. In the subclass Sclerobasidiomycetidae the order Auri- culariales is the most primitive in which the formation of sclerobasidia takes place and which connects the typical sclerobasidial Uredinales and Ustilagina- les with Protobasidiomycetidae. The subclass Homobasidiomycetidae is characterized by a unicellular basidium and gradual development of fruit bodies. This subclass includes the orders with opened (gymnocarpous), semiclosed (he- miangiocarpous) and closed (angiocarpous) fruit bodies. The author believes the separation of the enlarged groups — Aphyllophorales, Hymenomycetes and Gasteromycetes to be unreasonable as they artificially unite fungi of different phylogenetic origin.
Because of different phylogenetic origin of lichens the author considers their distribution between the respective orders of the classes Ascomycetes and Basidiomycetes to be correct.
IV. Origin and Evolutionary Trends of Bryophyta
Bryophyta are considered as a subdivision of green plants’ division — Chlorophyta. On analyzing the main theories of Bryophyta origin, the author considers as the most substantiated one the theory of their origin from green
311
algae, which gave the rise to separate branches that are considered as subdivisions, namely — Anthocerotophytina, Bryophytina and Psilophytina. Within the limits of the subdivision Bryophytina two groups considered by the author as classes — Hepaticopsida and Bryopsida were isolated early.
As to the evolution of the class Hepaticopsida, the author, proceeding from the data of comparative morphology, taxonomy, cytology, paleobotany and geographic distribution, arrives at a conclusion that simple thalloid forms known already from the Devonian deposits are the most ancient within the limits of this class. The simple thalloid forms gave rise to the complex thalloid forms, on the one hand, and to foliose ones, on the other, among which orthotropic Takakiales and Ilaplomitriales are most primitive. Within the limits of the youngest order Jungermanniales a transition can be observed from orthotropic isophyllous forms to plagiotropic heterophyllous ones.
Within the class Bryopsida three branches were isolated at early stage of development considered as the subclasses — Andreaeidae, Sphagnidae and Bryidae.
Possible phyllogenetic connections within the limits of Bryophytina and their relation to other subphyla of green plants are presented in the scheme (Fig. 90.)
СОДЕРЖАНИЕ
Предисловие 7
Глава I. Основные направления развития органического мира 9
Глава II. Направления эволюции и основы классификации низших автотрофных растительных организмов (водорослей) 29
1. Основы классификации водорослей 29
2. Золотистые (хризофитовые) водоросли — Chrysophyta
(Chrysophycophyta, Chrysophyceae) 33
3. Желтозеленые, или разножгутиковые, водоросли — Xan¬
thophyta (Xanthophycophyta, Xanthophyceae, Heterocon¬
tae) 40
4. Диатомовые водоросли — Diatomophyta (Bacillariophyta,
Bacillariophycophyta, Bacillariophyceae, Diatomeae) 45
5. Бурые водоросли — Phaeophyta (Phaeophycophyta, Phaeo¬
phyceae) 56
6. Пиррофитовые водоросли, или перидинеи,— Pyrrhophyta
(Pyrrhophycophyta, Dinophyceae, Peridineae) 74
7. Криптофитовые водоросли — Cryptophyta (Cryptophycophyta,
Cryptophyceae) 82
8. Эвгленовые водоросли — Euglenophyta (Euglenophycophyta,
Euglenophyceae, Euglenineae) 85
9. Хлоромонадовые — Chloromonadophyta (Rhaphidophyceae,
Chloromonadophyceae, Chloromonadineae) 91
10. Красные водоросли — Rhodophyta (Rhodophycophyta, Rho¬
dophyceae) 92
11. Зеленые водоросли — Chlorophycophytina (Chlorophycophyta,
Chlorophyceae, Chlorophyta) 106
Глава III. Направления эволюции и основы классификации низших гетеротрофных растительных организмов (грибов) 126
1. Происхождение низших бесхлорофильных растительных организмов 126
2. Общие направления эволюции грибных организмов и основы
их классификации 134
3. Слизевики — Myxomycota (Myxomycophyta, Mycetozoa) 145
4. Хитридиевые грибы — Chytridiomycota (Opisthomastigomy¬
cota) 151
5. Сапролегниевые грибы — Saprolegniomycota (Pteromastigomy¬
cota) 161
6. Зигомицеты — Zygomycetes (Mucoromycetes) 169
313
7. Аскомицеты — Ascomycetes 177
8. Базидиомицеты — Basidiomycetes 212
9. Лишайники — Lichenes и их место в системе растительных
организмов 229
Глава IV. Происхождение и пути эволюции мохообразных
1. Происхождение мохообразных 233
2. Пути эволюции мохообразных и их классификация 247
Заключение 274
Литература 278
Резюме (на английском языке) 305
CONTENTS
Preface 7
Chapter I. Main Trends in Development of Living World 9
Chapter II. Evolutionary Trends and Classification Principles of
Lower Autotrophic Plant Organisms (Algae) 29
1. Bases of Alga Classification 29
2. Chrysophyta (Chrysophycophyta, Chrysophyceae) 33
3. Xanthophyta (Xanthophvcophyta, Xanthophyceae, Heterocon¬
tae) 40
4. Diatomophyta (Bacillariophyta, Bacillariophycophyta, Bacilla¬
riophyceae, Diatomeae) 45
5. Phaeophyta (Phaeophycophyta, Phaeophyceae) 56
6. Pyrrhophyta (Pyrrhophycophyta, Dinophyceae, Peridineae) 74
7. Cryptophyta (Cryptophycophyta, Cryptophyceae) 82
8. Euglenophyta (Euglenophycophyta, Euglenophyceae, Eugleni¬
neae) 85
9. Chloromonadophyta (Rhaphidophyceae, Chloromonadophyceae,
Chloromonadineae) 91
10. Rhodophyta (Rhodophycophyta, Rhodophyceae) 92
11. Chlorophycophytina (Chlorophycophyta, Chlorophyceae, Chlorophyta) 106
Chapter III. Evolutionary Trends and Classification Principles of
Lower Heterotrophic Plant Organisms (Fungi) 126
1. Origin of Lower Achlorophyllous Vegetative Organisms 126
2. General Trends of Fungus Organisms Evolution and Principles
of Their Classification 134
3. Myxomycota (Myxomycophyta, Mycetozoa) 145
4. Chytridiomycota (Opisthomastigomycota) 151
5. Saprolegniomycota (Pteromastigomycota) 161
6. Zygomycetes (Mucoromycetes) 169
7. Ascomycetes 177
8. Basidiomycetes 212
9. Lichenes and Their Place in the System of Vegetative Organisms 229
Chapter IV. Origin and Evolutionary Trends of Bryophyta
1. Origin of Bryophyta 233
2. Evolutionary Trends of Bryophyta and Their Classification 247
Conclusion 274
References 278
Summary 305
315
Дмитрий Константинович Зеров
ОЧЕРК ФИЛОГЕНИИ БЕССОСУДИСТЫХ РАСТЕНИЙ
Печатается по постановлению ученого совета Института ботаники АН УССР им. Н. Г. Холодного
Редактор Г. И. Дружина Художественный редактор Р. И. Калыш Оформление художника Козюренко Технический редактор Б. М. Кричевская Корректор И. А. Каплан
Сдано в набор 24. 11. 1971 г. Подписано к печати 16. 6. 1972 г. БФ 00552. Заказ
№ 1—2355. Изд. Кя 133. Тираж 1500. Бумага № 1, 60 X 90‘/i6. Усл.-печ. листов 19,75. Учетно-издат. листов 22,31. Цена 2 руб. 53 коп.
Издательство «Наукова думка». Киев, Репина, 3.
Киевский полиграфический комбинат Комитета по печати при Совете Министров УССР, ул. Довженко, 3.