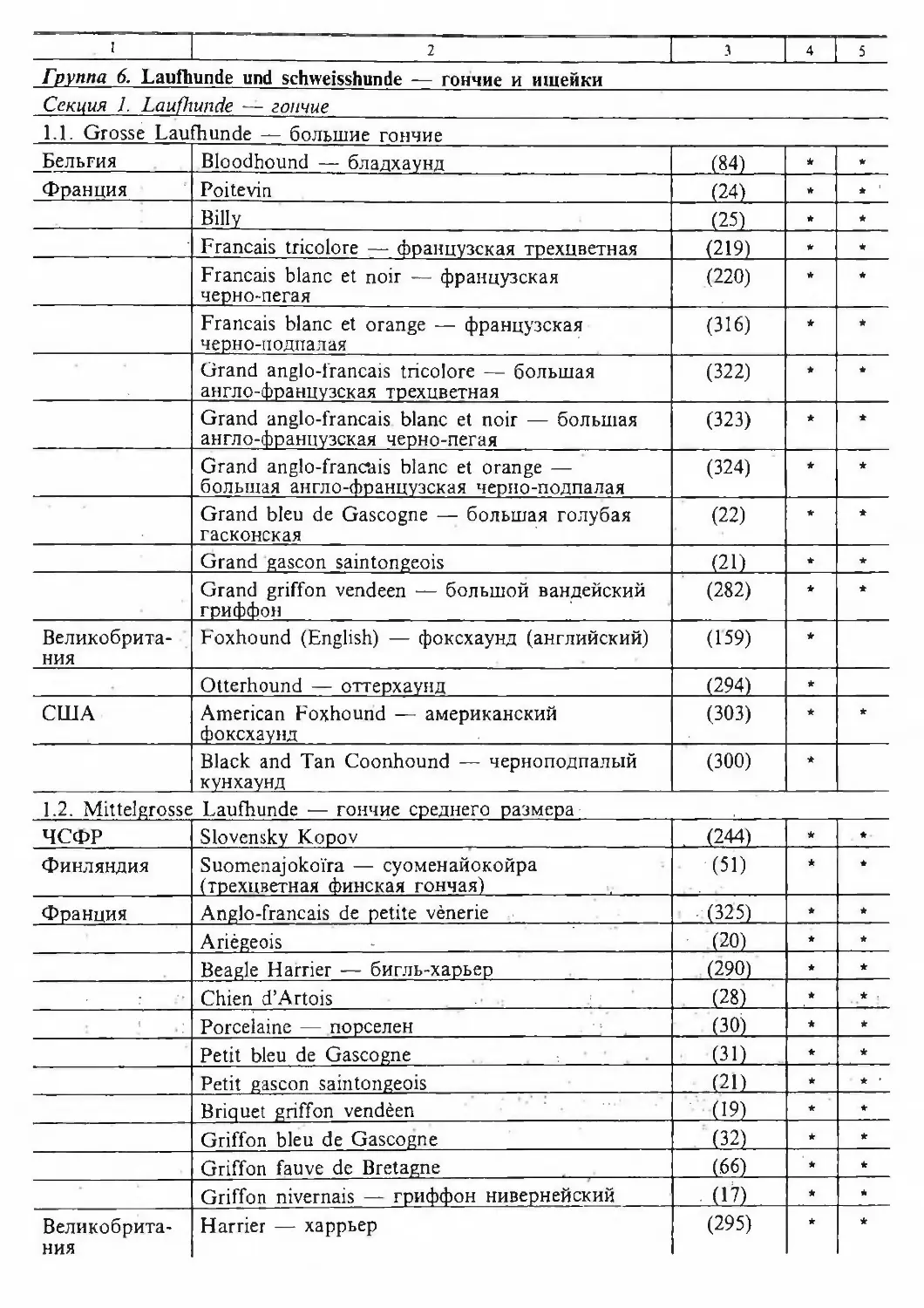
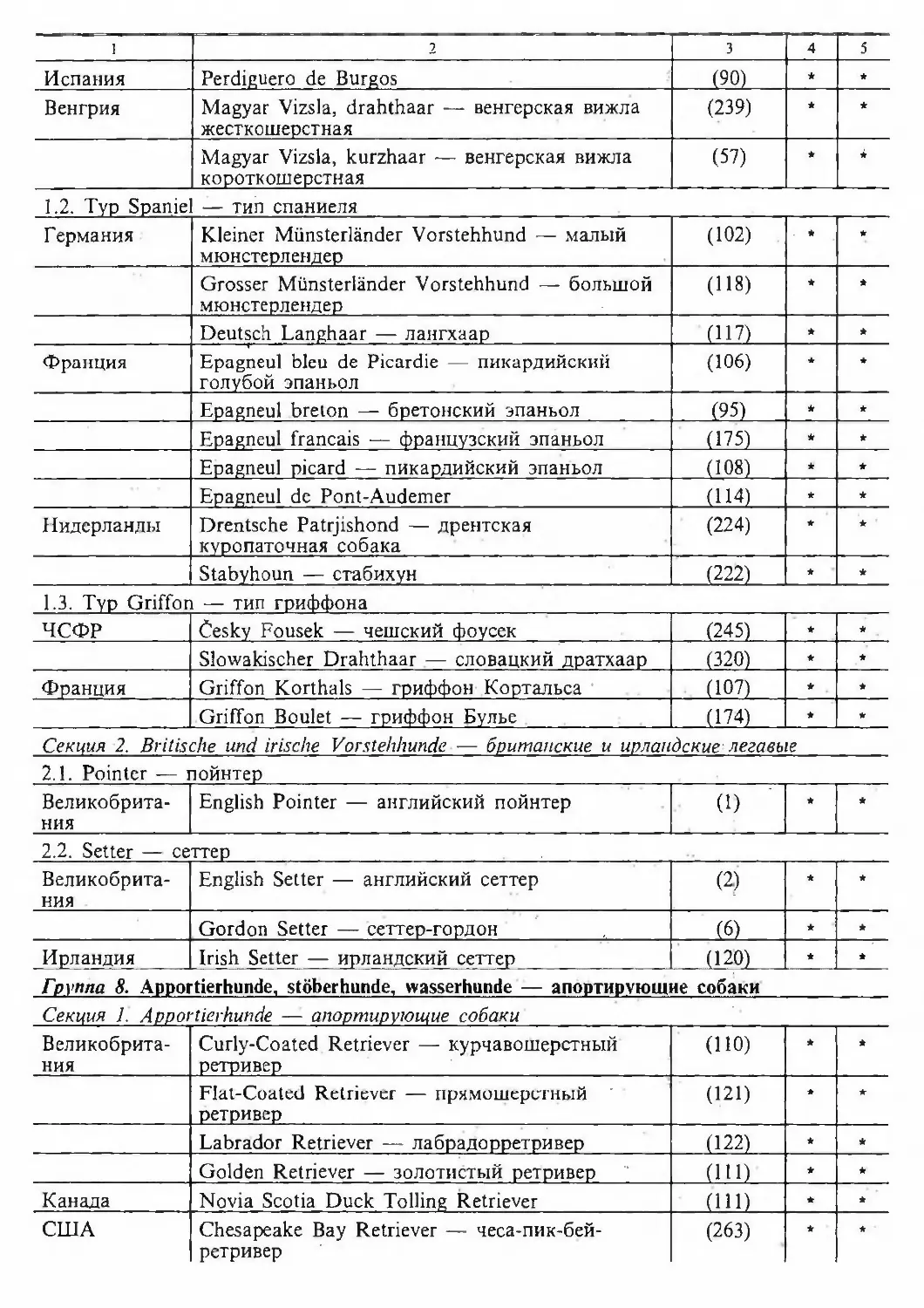
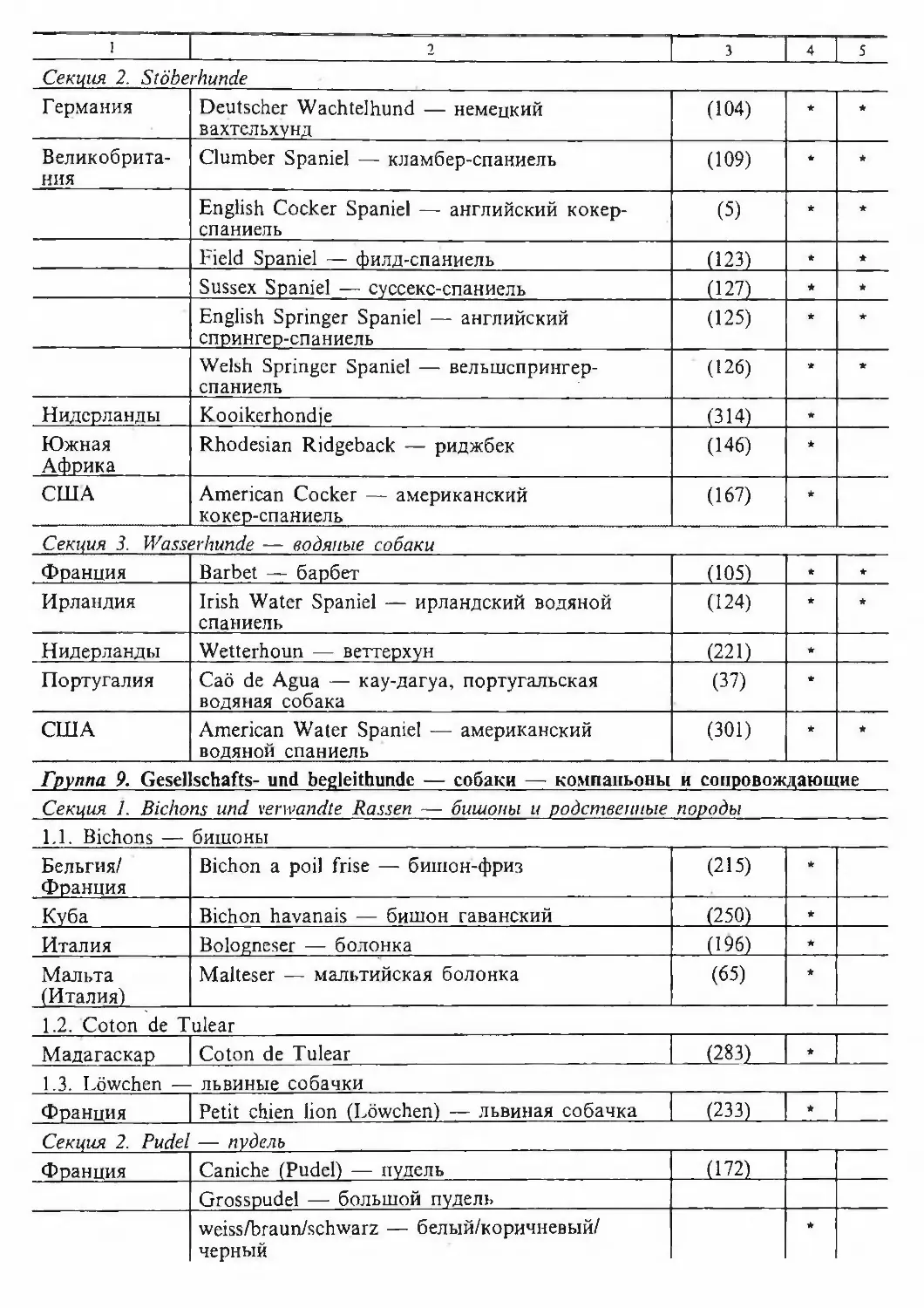
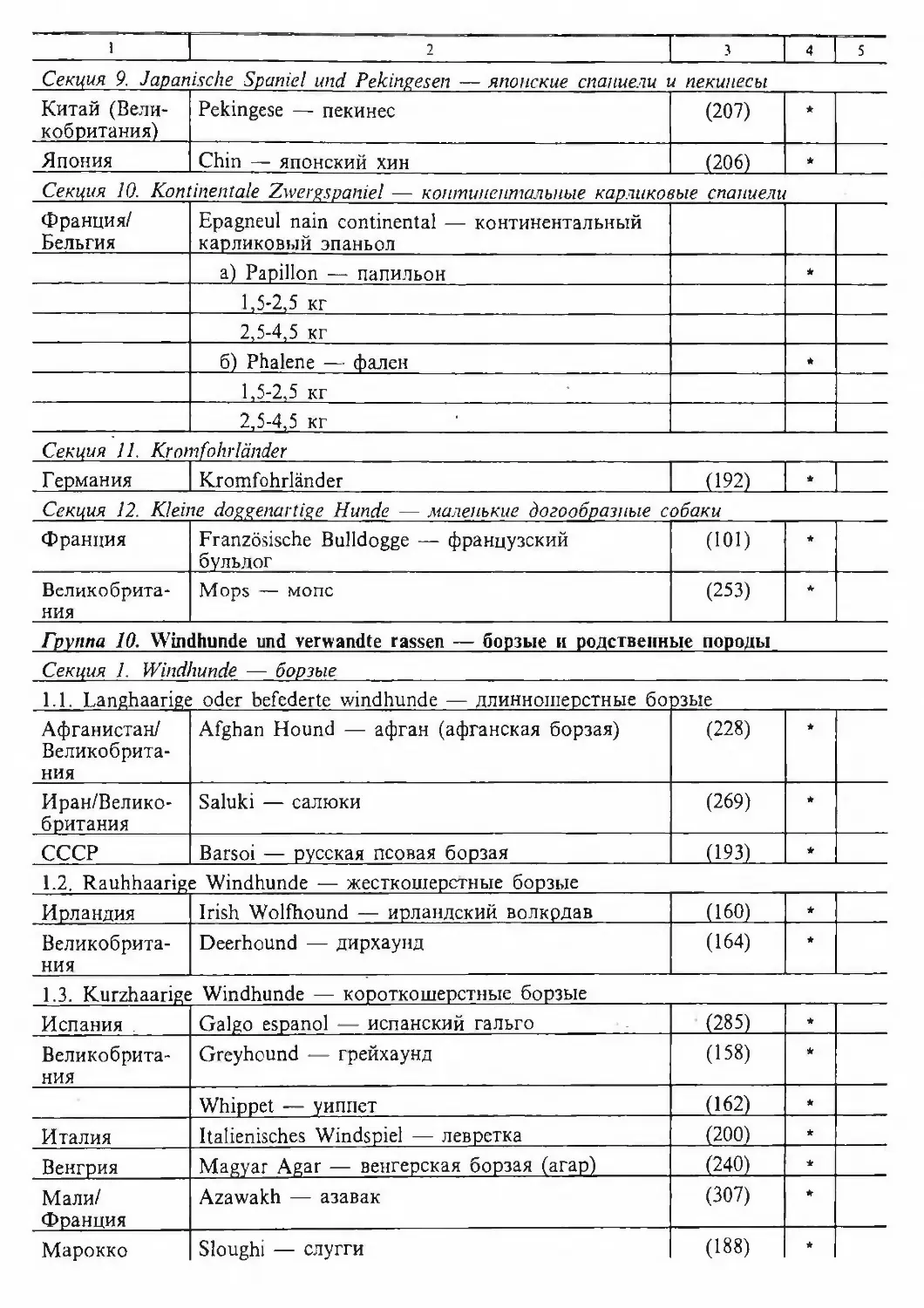
/














































































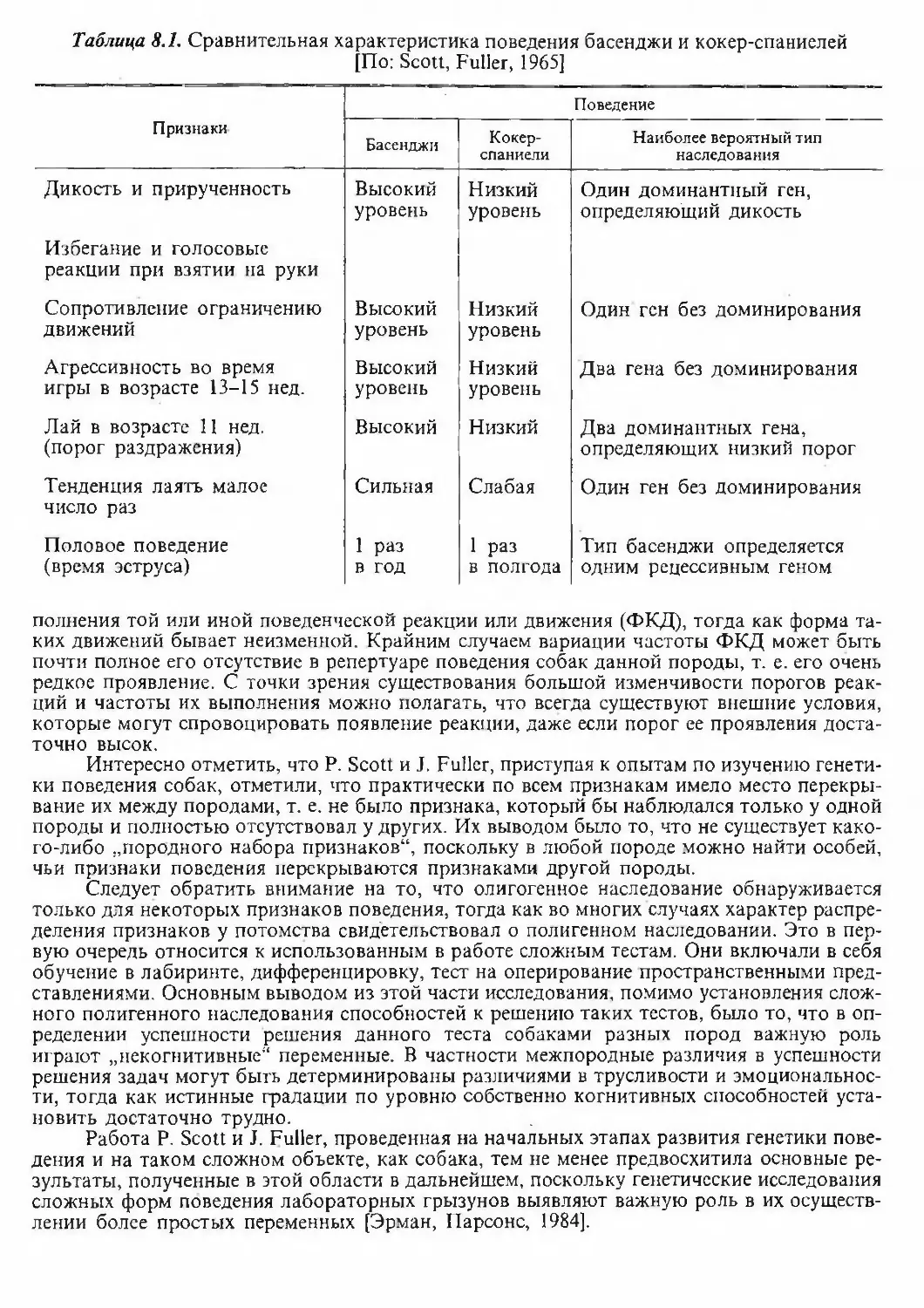











































































Текст
ГЕНЕТИКА СОБАКИ
Гпава 1
СИСТЕМАТИКА CANIDAEДомашняя собака (Canis familiaris L.) принадлежит к семейству собачьих (или волчь¬
их) (Canidae Gray) отряда хищных (Carnivora). Этот отряд включает в себя еще ряд се¬
мейств: медвежьих (Ursidae Gray), енотовых (Procyonidae Bonaparte), куницеобразных
(Mustelidae Swainson), виверровых (Viverridae Gray), гиеновых (Hyaenidae Gray) и кошачь¬
их (Felidae Gray). Спорным остается вопрос о таксономической принадлежности большой
и малой панд, Ailuropoda melanoleuca и Ailurus fulgens, своеобразие которых позволяет ря¬
ду авторов выделять их из семейства енотовых в отдельные семейства, Ailuropodidae и
Ailuridae. Несомненно, весьма близки к хищным и ластоногие (Pinnipedia), включаемые ря¬
дом авторов в отряд Carnivora в виде подотряда. Виды семейства собачьих — типичные
хищники мелких и средних размеров, приспособленные к активной, самостоятельной охо¬
те. Стройное и крепкое телосложение позволяет большинству из них добывать себе пищу
не только за счет скрадывания, но и достаточно долговременного преследования жертвы.
Длина тела от 40 см у фенека, до 160 см у некоторых подвидов волка, вес соответственно
от 1,5 до 80 кг. Волосяной покров, особенно у северных видов, густой и длинный. Окраска
чаще всего одноцветная, иногда с более светлыми пятнами. Только у песца выражено ее се¬
зонное изменение. В течение года линяют один или два раза. Череп удлиненный с узкимлицевым отделом. Зубная формула обычно 11, С |, р М =42, у кустарниковой собаки =38, у большеухой лисицы = 46-50.Распространены на всех континентах за исключением Антарктиды и некоторых ост¬
ровов. Населяют, в основном, открытые пространства. Обитают в логовах и норах, неко¬
торые хорошо роют. Держатся поодиночке, иногда парами, реже семейными стаями. Раз¬
виты слух и обоняние, значительно хуже — зрение. Активны в сумерках и ночью.
Питаются в основном животной пищей, часто падалью, ряд видов постоянно поедают рас¬
тения. Моногамны, беременность от 51 дня у фенека до 80 дней у гиеновой собаки. Дают
один помет в год, в среднем 4-7 щенков, у песца до 22. Половозрелыми становятся в возра¬
сте одного года.Семейство собачьих (Canidae) отнюдь не самое многочисленное в отряде Carnivora,
включает в себя не менее 36 видов, принадлежащих к 12 родам и 3 подсемействам [Simpson,
1945; Соколов, 1979] (табл. 1.1). Некоторые авторы предлагают сократить число таксоно¬
мических единиц в семействе, другие — увеличить, что свидетельствует о том, что система¬
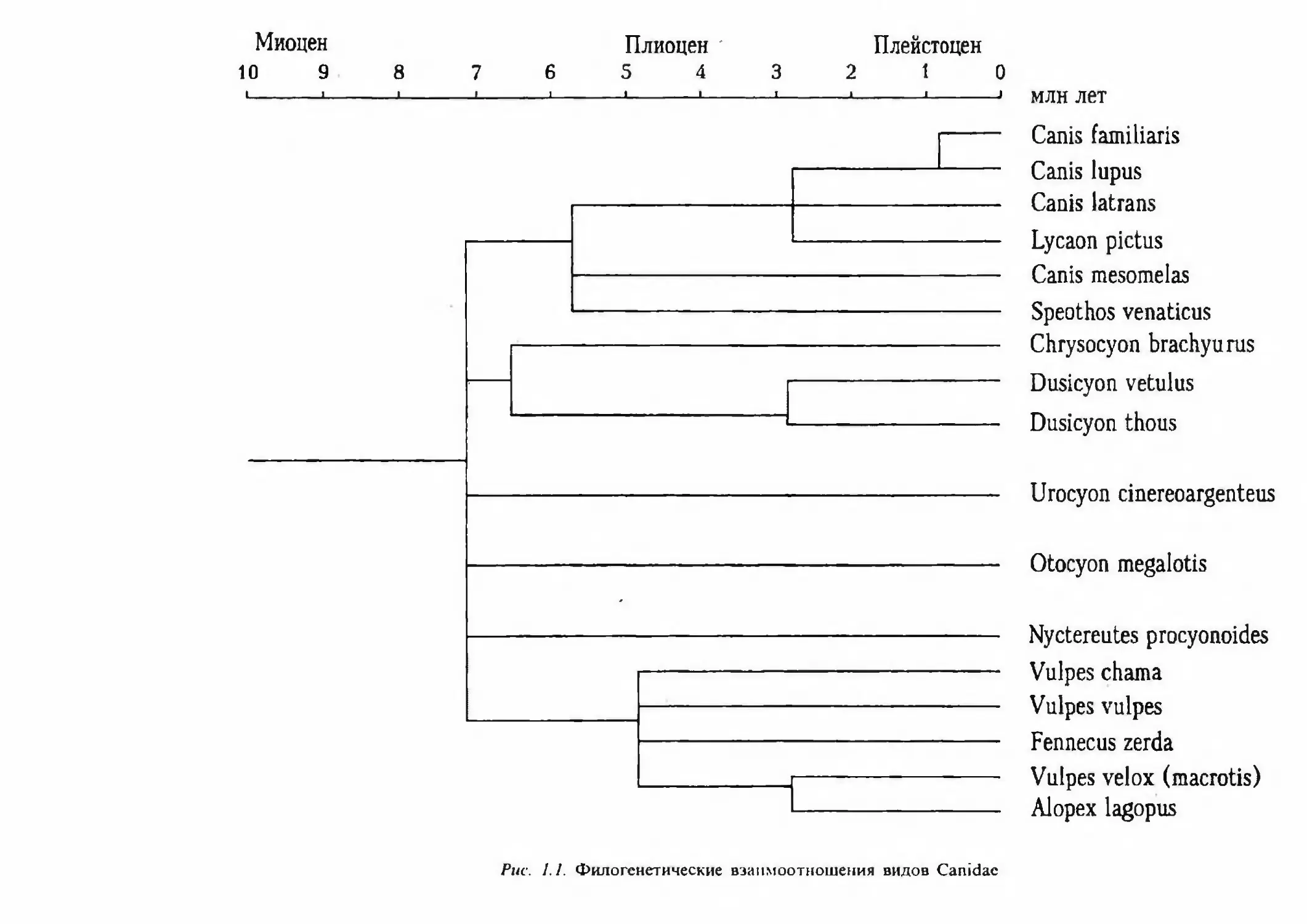
тика семейства нуждается в определенной ревизии.На рисунке 1.1 представлено филогенетическое древо семейства Canidae, основанное
на данных сравнительного изучения большой группы белков и ДНК у ряда видов [Wayne,
O’Brien, 1987]. Обращает на себя внимание то, что в целом современные Canidae представ¬
ляют собой довольно гомогенную группу видов, дивергировавших относительно недавно,
общий предок для всех современных родов существовал не более 7-8 млн лет назад. В то
же время прослеживается четкая обособленность группы волкообразных, куда входят до¬
машняя собака, волк, шакалы, гиеновая и кустарниковая собаки, от остальных групп се¬
мейства, таких, как южноамериканские лисицы и волки, большеухие лисицы, серые лиси¬
цы, енотовидная собака и Vulpes-образные лисицы. В последнюю группу входят, кроме
видов рода Vulpes, еще песец и фенек. Наиболее тесно филогенетически домашняя собака
связана с волком, Canis lupus, с которым, очевидно, она имела общего предка. По разным
Таблица 1.1. Видовой состав семейства CanidaeРодВидПодсемейство CaninaeNyctereutesN. procyonoides — енотовидная собакаCanisС.familiaris — домашняя собакаС.lupus — волкС.latrans — койотс.aureus — шакалс.mesomelas — чепрачный шакалс.audustus — полосатый шакалс.simensis — эфиопский шакалAlopexА.lagopusVulpesV.сапа — афганская лисицаV. corsac — корсакV. bengalensis — бенгальская лисицаV.vulpes — лисицаV. ferriiata — тибетская лисицаV. velox — американский корсакV.ruppeli — песчаная лисицаV.chama — южноафриканская лисицаV.pallida — африканская лисицаFennecusF.zerda — фенекUrocyonU. cinereoargenteus — серая лисица
U. littoralis — островная лисицаDusicyonD. culpaeolus — уругвайская лисица
D. culpaeus — андская лисицаD.fulvipes — чилоэсская лисицаD.griseus — южноамериканская лисицаD.gymnocercus — парагвайская лисицаD.inca — перуанская лисицаD. sechurae — эквадорская лисицаD. vetulus — бразильская лисицаD. microtis — малая лисицаD. thous — майконгChrysocyonС.brachyurus — гривистый волкПодсемейство SimocyoninaeCuonС.alpinus — красный волкLycaonL.pictus — гиеновая собакаSpeothosS.venaticus — кустарниковая собакаПодсемейство OtocyoninaeOtocyonО.megalotis — большеухая лисица
Миоцен108Плиоцен
5 4Плейстоцен
i Омлн летCanis familiaris
Canis lupus
Canis latransLycaon pictus
Canis mesomelas
Speothos venaticus
Chrysocyon brachyurusDusicyon vetulus
Dusicyon thousUrocyon cinereoargenteusOtocyon megalotisNyctereutes procyonoides
Vulpes chama
Vulpes vulpes
Fennecus zerda
Vulpes velox (macrotis)
Alopex lagopusPuc. 1.1. Филогенетические взаимоотношения видов Canidae
оценкам время дивергенции домашней собаки и волка как обособленных видов не превы¬
шает 1 млн лет, а по многим данным и того меньше, с точки зрения оценки эволюционных
событий — это почти бесконечно малая величина. Широко известны гибриды между до¬
машней собакой и волком, домашней собакой и шакалом, собакой и койотом, волком и
койотом, волком и шакалом, что подтверждает близкое филогенетическое родство между
этими видами.Особняком стоит вопрос о динго, вероятнее всего, происходящей от одомашненных
форм, завезенных предками австралийских аборигенов и одичавших по мере одичания сво¬
их хозяев. По веем генетическим характеристикам динго является самой обыкновенной до¬
машней собакой.
Гпава 2
ЦИТОГЕНЕТИКА CANIDAE•' » • •То, что собака — это собака, а не хорек или муха цеце, определяется тем, что Бог и
эволюция записали в ее ДНК, собранную у этого вида, как и у всех прочих эукариот, с по¬
мощью разнообразных белков и других сложных молекул в специальные образования, на¬
званные в конце прошлого века хромосомами. Хромосомы разнообразны по своим разме¬
рам и морфологии. У разных организмов различаются и числа хромосом, например, у
рабочих муравьев некоторых видов всего одна хромосома, у аскариды две, а у многих ви¬
дов раков их несколько сотен. Совокупность всей ядерной ДНК у вида называется гено¬
мом, совокупность всех хромосом — кариотипом.Виды млекопитающих различаются между собой по количеству ДНК в ядре незначи¬
тельно, в среднем геном содержит около 8 pg ДНК, по числу же хромосом различия весьма
существенны. Например, индийский олень мунтжак имеет в своем кариотипе 6 хромосом, а
один из видов южноамериканских грызунов — 96. Близкие размеры геномов у млекопита¬
ющих определяют то, что у видов с большим числом хромосом они, как правило, мелкие, а
у видов с небольшим числом хромосом — крупные. Естественно, что самые крупные хро¬
мосомы у мунтжака. Число хромосом у вида записывается 2п, что указывает на то, что все
хромосомы в нормальном кариотипе парные. Пол у большинства видов определяется сис¬
темой половых (непарных) хромосом. Самки имеют две абсолютно одинаковых половых
Х-хромосомы, самцы —две разных половых X- и Y-хромосомы. Таким образом, нормаль¬
ные клетки самки имеют каждую хромосому, в том числе и Х-хромосому, в двойном коли¬
честве, а в клетках самца все неполовые хромосомы (аутосомы) парные и, кроме того, не¬
парные X и Y половые хромосомы. В результате мейоза, сложного клеточного деления,
происходящего в клетках полового пути, у самки каждая яйцеклетка имеет в 2 раза мень¬
шее число хромосом (п), все аутосомы и Х-хромосома присутствуют в единственном числе.
У самцов половина сперматозоидов несет все аутосомы по одной и Х-хромосому, а вторая
половина — аутосомы и Y-хромосому. Оплодотворение яйцеклетки сперматозоидом с X-
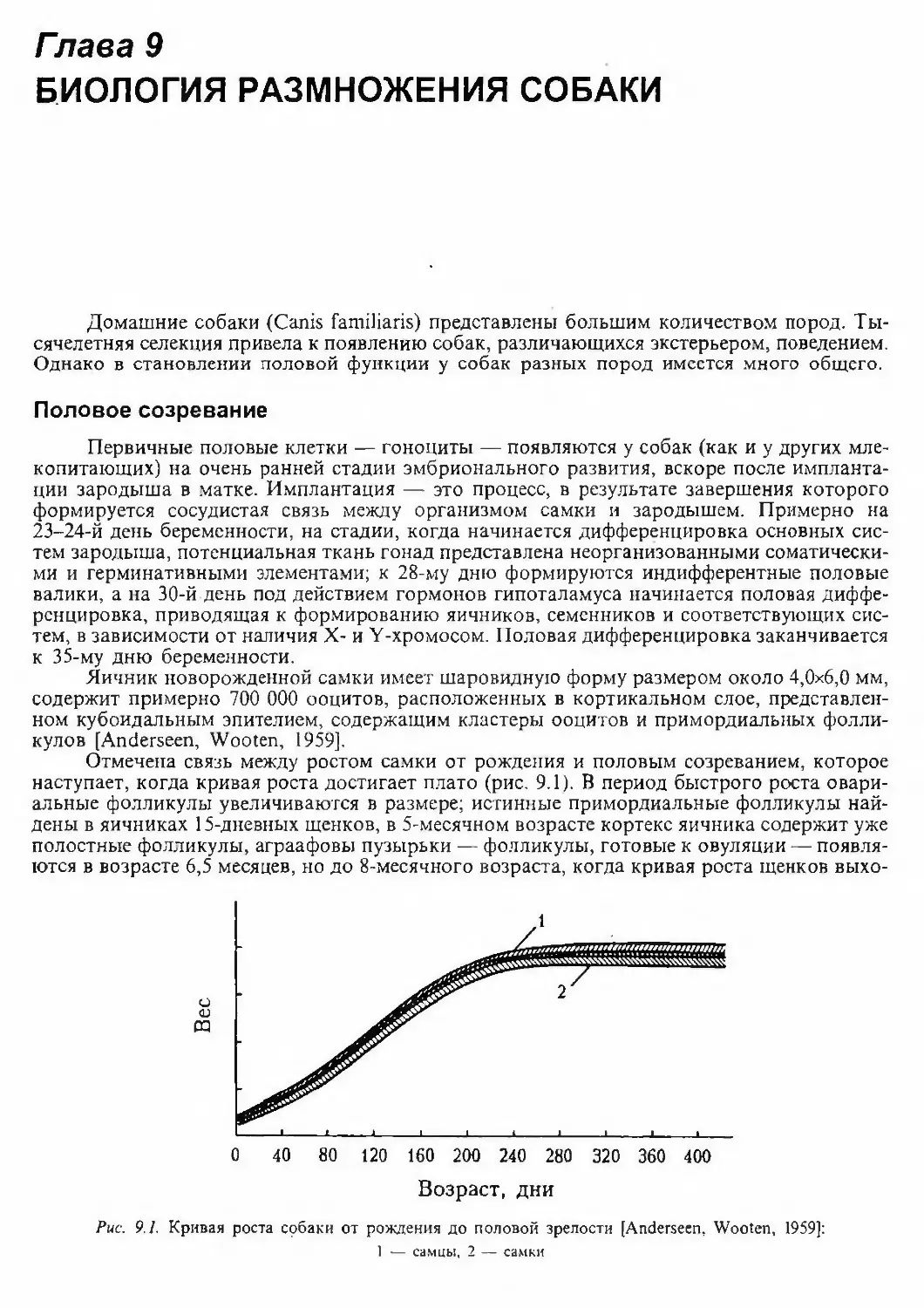
или Y-хромосомой определяет пол потомка, мужской или женский соответственно.В среднем виды млекопитающих имеют 42-48 хромосом. Из таблицы 2.1 видно, что в
кариотипах большинства видов домашних животных хромосом больше [Графодатский,
Раджабли, 1988].Наиболее высоким числом хромосом характеризуется домашняя собака. Отчасти вы¬
соким числом хромосом в кариотипе собаки, а чем оно больше, тем больше вариантов раз¬
личных сочетаний этих хромосом в клетках потомков, объясняется эволюционная пластич¬
ность собаки как вида и беспрецедентная отзывчивость его на селекцию.Хромосомы собаки. Как уже отмечалось выше, собака — самый высокохромосомный
вид среди домашних животных и, следовательно, самый неудобный для цитогенетического
анализа. Аутосомы собаки представляют собой ряд постепенно убывающих по величине
акроцентриков (одноплечих хромосом), Х-хромосома — крупный субметацентрик (двупле¬
чая хромосома), Y-хромосома — самый мелкий двуплечий элемент набора (рис. 2.1).
Структурный гетерохроматин (добавочный материал хромосом) представлен очень мелки¬
ми центромерными блоками у некоторых пар аутосом и целиком гетерохроматической
Y-хромосомой (рис. 2.2). Ядрышкообразующие районы хромосом (кластеры функциональ¬
но активных рРНК генов) расположены на теломерных концах 3 пар аутосом и
Y-хромосомы.
Таблица 2.1. Числа хромосом некоторых видов млекопитающихВидДиплоидное число
хромосом, 2nЧеловек, Homo sapiens46Домовая мышь, Mus musculus40Крыса, Rattus norvegicus42Китайский хомячок, Cricetulus griseus22Золотистый хомячок, Mesocricetus auratus44Морская свинка, Cavia porcellus64Кролик, Oryctolagus cuniculus44Корова, Bos taurus60Як, Bos grunniens60Буйвол, Bubalus bubalis48-50Коза, Capra hircus60Овца, Ovis aries54Северный олень, Rangifer tarandus70Марал, Cervus elaphus68Пятнистый олень, Cervus nippon64-68Свинья, Sus scrofa38Двугорбый верблюд, Camelus bactrianus74Одногорбый верблюд» Camelus dromedarius74Лама, Lama glama72-74Альпака, Lama pacos72-74Индийский слон, Elaphas maximus56Лошадь, Equus caballus64Осел, Equus asinus62Кошка, Felis catus ,38Домашний хорек-фуро, Mustela putorius40Собака, Canis familiaris78(I (I u It (I «»I * « » т «* <** **.' • #h v Ш-• Я »• •« II «• М'г i\'; :у *• Ч »!Г;At «А ^11- ^ «*. ' • -ЛтК -и: . '>• *И* «• 4»*т&?■Рис. 2.1. Рутинная окраска хромо¬
сом домашней собаки,
Canis familiaris, 2n=784» А*. ** ^. .• ’*". j: . **• ?.#, . 'ф-.
я* «.ft. • #f . •;AH %t .Fwc. 2.2. С-окрашенные хромо¬
сомы домашней собакиtit*и if tef fe «*» a « ; « I» иr.mШШШШ•* ш. ** :у*'.А/с. 2.5. G-окрашенные хромосомы
домашней собаки (хромосомы каж¬
дой пары представлены на разных
стадиях конденсации)
Описанные впервые в 1928 г. [Minouchi, 1928] хромосомы собаки изучались многими
авторами, в том числе и с помощью разнообразных дифференциальных окрасок хромосом,
выявляющих их гетерогенность [Selden et al., 1975; Manolache et al., 1976; Pathak,
Wurster-Hill, 1977; Larsen et al., 1978; Howard-Peebles, Pryor, 1980; Графодатский, Раджаб-
ли, 1981, 1988; Koop et al., 1982; Pathak et al., 1982; Wurster-Hill, Centerwall, 1982; Графодат¬
ский, 1983; Mayr et al., 1983; Graphodatsky, Hainan, 1989; MorenoMillan et al., 1991]. На ри¬
сунке 2.3 показаны G-окрашенные хромосомы собаки. Однако, несмотря на обилие работ
по этой проблеме, собака остается единственным видом среди домашних животных, для
которого отсутствует общепринятая номенклатура дифференциально окрашенных хромо¬
сом. Это объясняется тем, что идентификация многих пар мелких хромосом затруднена да¬
же при использовании дифференциальных окрасок высокого уровня разрешения. Однако в
ближайшие год-два потребность в номенклатуре хромосом резко возрастет. Необходи¬
мость четкой идентификации хромосомных патологий для селекционной и ветеринарной
практики, работ по картированию генома в настоящий момент стала абсолютно очевид¬
ной. С 1994 г. в рамках международного проекта ,,DogMap“ начаты работы по созданию
номенклатуры хромосом Canis familiaris.Хромосомные патологии собаки. Изменения в числе хромосом и/или их перестройки
как у человека, так и животных чаще всего ведут к проявлению множественных общих па¬
тологий, описываемых у человека как синдромы. Среди хромосомных патологий у челове¬
ка чаще всего встречается, например, синдром Дауна, обусловленный присутствием в ка¬
риотипе „лишней*1 хромосомы 21. Некоторые хромосомные перестройки, например,
центрические слияния, могут не иметь заметных морфологических проявлений у их носите¬
лей, но сказываются на их фертильности, либо обусловливают патологии у потомков. Дан¬
ные о хромосомных патологиях у собаки суммированы относительно недавно в обзоре
С. Mellink и A. Bosma [1989]. Спектр известных перестроек аутосом ограничен различными
центрическими слияниями и одним случаем присутствия в кариотипе мелкого добавочного
фрагмента. У помесного терьера [Shive et al., 1965] с множественными уродствами клетки
несли добавочную двуплечую аутосому (2п=77), образованную в результате слияния боль¬
шого и маленького акроцентриков. У кобеля малого пуделя с хондродисплазией выявлено
центрическое слияние двух акроцентриков [Hare et al., 1967]. У другого кобеля малого пу¬
деля с аномалией мочеточников и его матери обнаружено слияние хромосомы первой пары
и аутосомы средних размеров [Hare, Bovee, 1974].Центрические слияния аутосом описаны также и у фенотипически нормальных со¬
бак. У помесной между сеттером и ретривером суки обнаружено слияние (2п=77) средних
по размерам акроцентриков. Шесть особей пуделя (5 сук и 1 кобель) несли слияние первой
и мелкой аутосом в гетерозиготном состоянии и один кобель — в гомозиготном [Mayr et
al., 1986а]. R. Larsen et al. [1978; 1979] провели скрещивание между собаками со слиянием
хромосом 13 и 23. Мать, нормальный золотистый ретривер, и ее 7 потомков (3 суки и 4 ко¬
беля) несли эту транслокацию в гетерозиготном состоянии, 2n=77, XX или XY t(13q;23q).
Было проведено три скрещивания между гетерозиготами по транслокации (мать и потом¬
ки), родилось 5 нормальных гетерозиготных кобелей и 10 нормальных гетерозиготных сук.
Кроме того, 2 нормальных гомозиготных кобеля и 3 гомозиготных суки (одна нормальная
и две с паховой грыжей) имели 2n=76, XX или XY, t(13q;23q).У кобеля черного кокер-спаниеля с аномалиями в строении сердца и венозной систе¬
мы описан кариотип с 2п=79, с добавочным мелким хромосомным фрагментом [Shive et al.,
1965].У собак также описана и полиплоидия в ряде тканей, обычно не встречающаяся у
нормальных животных. Щенок большого мюнстерлендера с гидроцефалией имел хромо¬
сомный набор в клетках кожи и почек 78, XY/156, XXYY. Щенок бигля со множественны¬
ми анатомическими аномалиями имел в клетках почки кариотип 117/XXY [Bosma et al.,
1982]. Такие животные рождаются мертвыми или погибают сразу после рождения.Большинство случаев, связанных с нарушениями в балансе половых хромосом у со¬
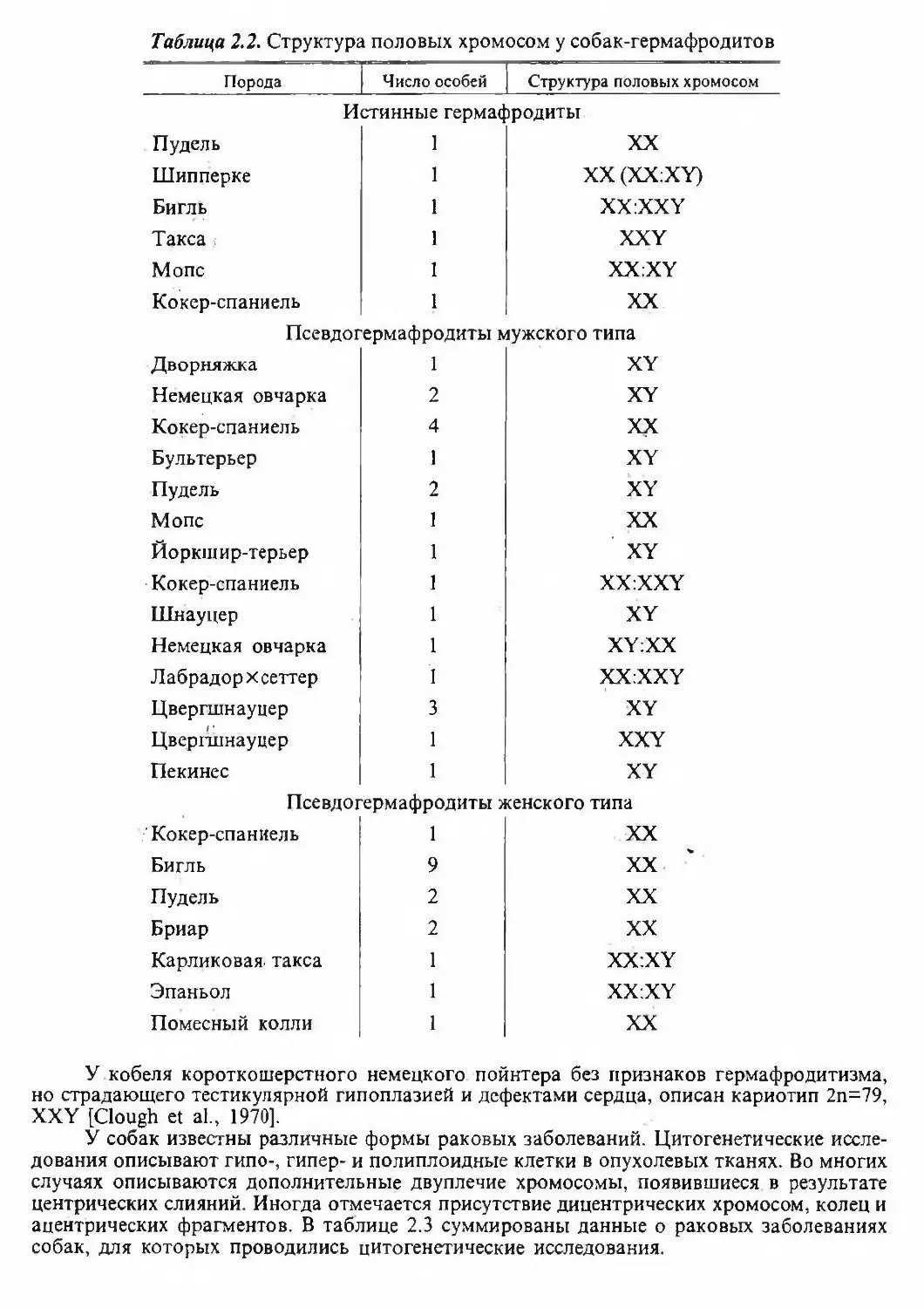
бак, были описаны на животных-гермафродитах. Их можно подразделить на 3 категории:
истинные гермафродиты, которые имеют и овариальные, и тестикулярные ткани; псевдо¬
гермафродиты мужского типа, имеющие тестикулярные ткани и дополнительные генита¬
лии женского типа; псевдогермафродиты женского типа, имеющие овариальные ткани и
дополнительные гениталии мужского типа. В таблице 2.2 суммированы данные [Hare,
1976] о собаках-гермафродитах, изученных цитогенетически.
Таблица 2.2. Структура половых хромосом у собак-гермафродитовПородаЧисло особейСтруктура половых хромосомПудельШипперкеБигльТаксаМопсКокер-спаниельИстинные гермафродиты
1
1
1
1I1XX
XX (XX:XY)
XX:XXY
XXY
XX:XY
XXПсевдогермафродиты мужского типаДворняжкаНемецкая овчаркаКокер-спаниельБультерьерПудельМопсЙоркшир-терьерКокер-спаниельШнауцерНемецкая овчарка
Лабрадор х сеттер
Цвергшнауцер
Цвергшнауцер
ПекинесXYXYXXXYXYXXXYXX:XXYXYXY:XXXX:XXYXYXXYXYПсевдогермафродиты женского типаКокер-спаниельБигльПудельБриарКарликовая такса
ЭпаньолПомесный коллиXXXXXXXXXX:XYXX:XYXXУ кобеля короткошерстного немецкого пойнтера без признаков гермафродитизма,
но страдающего тестикулярной гипоплазией и дефектами сердца, описан кариотип 2п=79,
XXY [Clough et al., 1970].У собак известны различные формы раковых заболеваний. Цитогенетические иссле¬
дования описывают гипо-, гипер- и полиплоидные клетки в опухолевых тканях. Во многих
случаях описываются дополнительные двуплечие хромосомы, появившиеся в результате
центрических слияний. Иногда отмечается присутствие дицентрических хромосом, колец и
ацентрических фрагментов. В таблице 2.3 суммированы данные о раковых заболеваниях
собак, для которых проводились цитогенетические исследования.
Таблица 2.3. Цитогенетика неоплазий у собак [Mellink, Bosma, 1989]Тип неоплазииКоличествоМодальное числоЧисло добавочныхживотныххромосомдвуплечих хромосомЛимфосаркома12780-тгттг п . .478199177,781" " 99 '1781-599 “2791 и 4" " 591813" “ 55177495 ”164-753-9• ‘ 95 14426~ 99 "156-6814-36Лимфоцитарная лейкемия2771—: - -27805-ГГ jj —17.8Есть991810Миелоцитарная лейкемия178ЕстьМоноцитарная лейкемия1 '156ЕстьМиелогенная лейкемия281,0Миелономоноцитарная лейкемия10780" ’ 99178Есть99154ЕстьХроническая миелоцитарная лейкемия185, 830Хроническая миелогенная лейкемия167-780“ >9 — -17.80Трансмиссионные венерические опухолиМного58—6015-17Фибросаркома :1(лимфатические метастазы)54-5613-25(легочные метастазы)■77, 78,0Тиреоидная аденокарцинома160-945-7Шттт190 , 93.20-3,4Оеттшркш* ■■«щвфшм .опухоль1245-5515-25Р ~?84-100Есть-- х> ~~г;194-1:05£?Т*-“ 99150-10510-30J9150-10514-30дегодаые метастазы56-10310—25К&рцидома елочной железы :дарвич&а^ с$цу*,оль178-1360-35 '112,6-167# .95 г "370-75<0•метастазы § .селе^едке;1761легочное тщщ#761
Сравнительная цитогенетика CanidaeДанные о числе хромосом и структуре кариотипов видов Canidae суммированы в ря¬
де обзоров [Wurster, Benirschke, 1968; Wurster, 1969; Орлов, Булатова, 1983]. Мы приводим
их ниже, дополнив некоторыми современными данными.1. Nyctereutes procyonoidcs procyonoides — материковая енотовидная собака, 2п=54
+2-4В. Обитающий в Китае, Корее, на Дальнем Востоке, акклиматизированный в ряде
районов России, используемый как объект пушного звероводства, этот подвид имеет в сво¬
ем наборе 5 пар двуплечих (субметацентрических) аутосом, 21 пару акроцентрических
аутосом, Х-хромосома — субметацентрик, Y-наименыний двуплечий элемент набора. Кро¬
ме хромосом основного набора для кариотипа енотовидной собаки описаны целиком гете¬
рохроматические добавочные, или В-хромосомы, варьирующие в числе, что определяет
хромосомный полиморфизм у этого подвида. Подвид достаточно подробно изучен совре¬
менными методами цитогенетического анализа [Makinen, Fredga, 1980; Wang et al., 1984;
Yosida, Shi, 1986; Ward et al., 1987; Shi et al., 1988; Wurster-Hill et al., 1988; Графодатский,
Раджабли, 1988; Graphodatsky, Hainan, 1989; Wada et a!., 1991].2. Nyctereutes procyonoides viverrinus — японская енотовидная собака, 2n=38-39 +0-4B.
В наборе у этого вида 13 пар двуплечих аутосом и 5 пар акроцентрических, X- и
Y-хромосомы, как у предыдущего вида. Отмечен случай полиморфизма (2п=39) типа цент¬
рическое слияние/разделение [Obara, Nakano, 1989]. Описана система добавочных хромо¬
сом, варьирующих от 0 до 4. От материкового подвида отличаются за счет слияния 8 пар
акроцентриков [Wurster-Hill et al., 1986; 1988; Ward et al, 1987].3. Canis familiaris — домашняя собака, 2n=78. Хромосомный набор этого вида описан
выше.4. Canis lupus — волк, 2п=78. Хромосомный набор волка идентичен таковому у соба¬
ки. Дифференциально окрашенные хромосомы описаны в ряде работ [Wurster-Hill,
Centerwall, 1982; Mayr et al., 1986a; Wayne et al., 1987a; Wurster-Hill et al., 1988].5. Canis latrans — койот, 2n=78. Хромосомный набор койота идентичен таковому у
собаки и волка. Описаны дифференциально окрашенные хромосомы, в том числе и у кой-
дога, предполагаемого гибрида между собакой и койотом [Wurster-Hill, Centerwall, 1982].6. Canis aureus — шакал, 2n=78. По числу и морфологии хромосом кариотип шакала
идентичен таковым у собаки, волка и койота. Сведений по дифференциальным окраскам
хромосом нет.7. Canis mesomelas — чепрачный шакал, 2п=78. По числу и морфологии хромосом ка¬
риотип чепрачного шакала идентичен таковым у собаки, волка, шакала и койота. Сведе¬
ний по дифференциальным окраскам хромосом нет.8. Alopex lagopus — песец, 2п=48-50. Для этого вида характерен хромосомный поли¬
морфизм по перестройке типа слияние/разделение, что приводит к вариации в числе хро¬
мосом от 48 до 50. У животных с 2п=48 все хромосомы двуплечие, у особей с 2п=50 на одну
пару двуплечих аутосом меньше, но в кариотипе присутствуют две пары акроцентриков, в
наборе песцов с 2п=49 выявляется одна непарная двуплечая хромосома и два непарных ак-
роцентрика, гомологичные плечам непарной двуплечей аутосомы. Х-хромосома песца —
субметацентрик, Y — наименьший двуплечий элемент набора. 10 пар аутосом песца несут
крупные добавочные гетерохроматические плечи. У одного самца описана добавочная (В)
целиком гетерохроматическая хромосома. Описаны дифференциально окрашенные хромо¬
сомы и у лисице-песцового гибрида [Makinen et al., 1981; Графодатский, Раджабли, 1981,
1988; Makinen, Gustavsson, 1982; Yoshida et al., 1983; Makinen, 1985b; Switonski, 1985; Mayr
et al., 1986a; Wayne et al., 1987b; Graphodatsky, Hainan, 1989].9. Vulpes corsac — корсак, 2n=36. Все хромосомы набора двуплечие, данных по какой-
-либо из дифференциальных окрасок нет.10. Vulpes bengalensis — бенгальская лисица, 2п=60. В кариотипе 5 пар двуплечих
аутосом, 24 — акроцентрических, X- и Y-хромосомы такие же, как и у других видов семей¬
ства. Данных по дифференциальным окраскам хромосом этого вида в литературе нет.11. Vulpes vulpes — лисица, 2n=34 +0-8В. Все хромосомы основного набора двупле¬
чие, мелкие добавочные (или В) хромосомы варьируют в числе от 0 до 8. Вариации в числе
В-хромосом определяют межклеточный и межиндивидуальный полиморфизм. Структур¬
ный гетерохроматин ограничен мелкими центромерными и теломерными блоками. Хромо¬
сомы лисиц, как красных диких, так и серебристо-черных клеточного содержания, изуча¬
лись многими исследователями. Описаны хромосомы лисице-песцовых гибридов [Беляев и
др., 1974; Графодатский, Раджабли, 1981, 1988; Makinen, Gustavsson, 1982; Yoshida et al.,
1983; Графодатский, 1983; Makinen, 1985a; Wayne et al., 1987b; Switonski et al., 1987;
Graphodatsky, Hainan, 1989].12. Vulpes velox (macrotis) — американский корсак, 2n=50. Хромосомный набор аме¬
риканского корсака идентичен таковому у песца с 2п=50, что подтвердилось и при сравне¬
нии G-окрашенных хромосом этих видов [Wayne et al., 1987Ь]. К сожалению, С-окрашива-
ние хромосом V. macrotis для выявления добавочных гетерохроматических плеч не
проводилось.13. Vulpes riippeli — песчаная лисица, 2п=40. Каких-либо сведений о структуре карио-
типа у этого вида, за исключением данных о числе хромосом, нет.14. Fennecus zerda — фенек, 2п=64. В кариотипе 2 пары двуплечих аутосом, двуплечие
X- и Y-хромосомы, остальные пары акроцентрические. Описаны G-дифференциально ок¬
рашенные хромосомы [Wayne et al., 1987а]. По нашим неопубликованным данным, струк¬
турный гетерохроматин фенека представлен заметными центромерными и теломерными
блоками.15. Urocyon cinereoargenteus — серая лисица, 2п=66. В наборе одна пара двуплечих
аутосом, двуплечие X- и Y-хромосомы, остальные пары акроцентрические. Имеются дан¬
ные по G-окраске хромосом [Wayne et al., 1987а].16.'Dusicyon gymnocercus — парагвайская лисица, 2п=74. Все аутосомы акроцентри¬
ческие, X- и Y-хромосомы такие же, как и у прочих видов семейства. Данных о дифферен¬
циальных окрасках хромосом нет.17. Dusicyon griseus — южноамериканская лисица, 2п=74. Все аутосомы акроцентри¬
ческие, X- и Y-хромосомы такие же, как и у прочих видов семейства. Данных о дифферен¬
циальных окрасках хромосом нет.18. Dusicyon vetulus — бразильская лисица, 2п=74. Все аутосомы акроцентрические,
X- и Y-хромосомы такие же, как и у прочих видов семейства. Данных о дифференциальных
окрасках хромосом нет.19. Dusicyon microtis — малая лисица, 2п=74+0-2В. Все аутосомы акроцентрические,
X- и Y-хромосомы такие же, как и у прочих видов семейства. Отмечено присутствие в ка¬
риотипе В-хромосом. Описаны G-дифференциально окрашенные хромосомы [Wurster-Hill,
Centerwall, 1982].20. Dusicyon thous — майконг, 2n=74. В наборе майконга 17 пар двуплечих аутосом,19 — акроцентрических, X- и Y-хромосомы такие же, как и у прочих видов семейства. Опи¬
саны G- и С-дифференциально окрашенные хромосомы, большинство двуплечих аутосом
несут небольшие добавочные гетерохроматические плечи [Wayne et al., 1987а].21. Chrysocyon brachyurus — гривистый волк, 2п=76. Все аутосомы акроцентрические,
X- и Y-хромосомы такие же, как и у прочих видов семейства. Описаны
G-дифференциально окрашенные хромосомы [Wurster-Hill, Centerwall, 1982; Wayne et al.,
1987а].22. Cuon alpinus — красный волк, 2n=78. По числу и морфологии хромосом кариотип
красного волка идентичен таковым у собаки, волка, шакала и койота. Сведений по диффе¬
ренциальным окраскам хромосом нет.23. Lycaon pictus — гиеновая собака, 2п=78. По числу и морфологии хромосом карио¬
тип гиеновой собаки идентичен таковым у собаки, волка, шакала и койота. Сведений по
дифференциальным окраскам хромосом нет.24. Speothos venaticus — кустарниковая собака, 2п=74. Все аутосомы акроцентричес¬
кие, X- и Y-хромосомы такие же, как и у прочих видов семейства. Описаны
G-дифференциально окрашенные хромосомы [Wurster-Hill, Centerwall. 1982; Wayne et al.,
1987а].25. Otocyon megalotis — большеухая лисица, 2n=72. Все аутосомы, за исключением
одной пары, акроцентрические, X- и Y-хромосомы такие же, как и у прочих видов семей¬
ства. Описаны G-дифференциально окрашенные хромосомы [Wurster-Hill, Centerwall, 1982;
Wayne et al., 1987а].В таблице 2.4 суммированы данные о размерах геномов ряда видов Canidae по срав¬
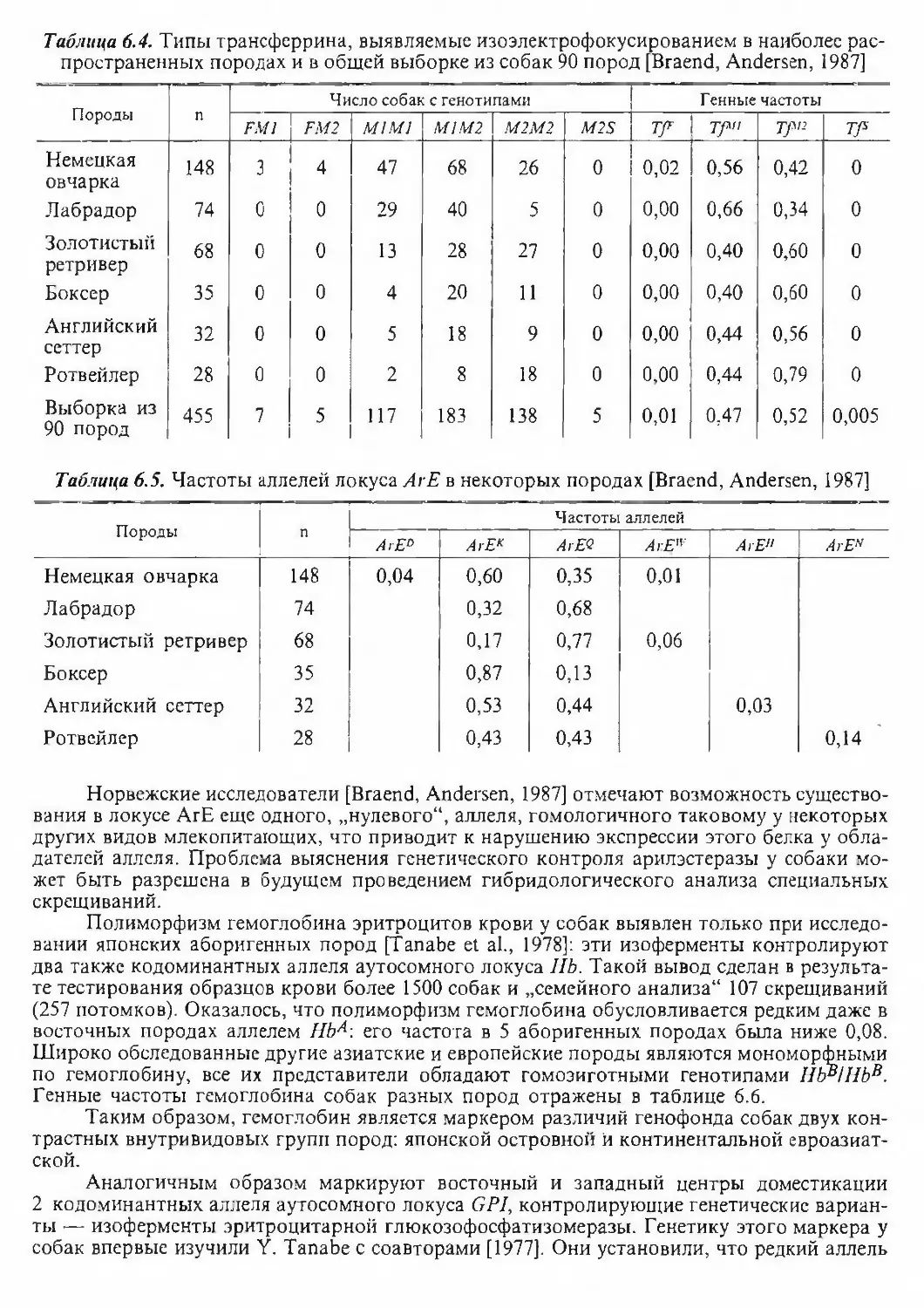
нению с количеством ядерной ДНК человека.Эти данные согласуются с результатами выявления структурного гетерохроматина у
видов семейства: виды рода Canis и красная лисица имеют минимальное для млскопитаю-
Таблица 2.4. Размеры геномов ряда видов CanidaeВидРазмеры генома,
% от Homo sapiens1. Canis lupus80,2**2. Canis latrans80,6**3. Vulpes vulpes81,4**Vulpes vulpes (fulvus)82,0*4. Alopex lagopus126,1*5. Urocyon cinereoargenteus87,6**.6. Nyctereutes procyonoides procyonoides93,4**7. Nyctereutes procyonoides viverrinus91,1*** Графодатский, Раджабли, 1981; ** Wurster-Hill et al., 1988.-щих содержание гетерохроматина в геноме, у енотовидных собак гетерохроматина заметно
больше, увеличиваются и общие размеры геномов, у песца 10 пар аутосом несут крупные
гетерохроматические плечи — размер генома самый большой в семействе. Очевидно, что
крупные размеры генома будут определены и у майконга, многие пары хромосом которого
также несут добавочные гетерохроматические плечи. Совокупно полученные данные ука¬
зывают на одну из основных тенденций в эволюции кариотипа Canidae — изменение гете¬
рохроматической компоненты геномов.Второй важной особенностью эволюции геномов Canidae является присутствие в ка-
риотипах многих видов системы добавочных или В-хромосом. Варьирующие по величине
от самых мелких у красной лисицы до довольно крупных у енотовидных собак, В-хромосо-
мы определяют межклеточный и межиндивидуальный полиморфизм. Особо следует под¬
черкнуть, что ни в одном из других семейств отряда Carnivora не выявлено видов с доба¬
вочными хромосомами.Сопоставление рисунка G-окраски хромосом видов семейства показало высокую сте¬
пень гомологии хромосом и/или их районов у видов семейства [Графодатский, Раджабли,
19,81; Makinen, Gustavssoij, 1982; Wurster-Hilj, Centerwall, 1982; Yoshida.et al., 1983; Wayne et
al., 1987a, Ь]. Показано, что кариотипы всех изученных видов построены из одних и тех же
элементов, но в различных их комбинациях. Особо важно, с нашей точки зрения, что у всех
высокохромрсомных видов, вне зависимости от их филогенетического родства, эти элемен¬
ты присутствует в неперестроенном виде, тогда как у видов с меньшим числом хромосо.м
они сливаются в различных сочетаниях, определяя кариотипическую дивергенцию в раз¬
ных таксонах. По нашему мнению, предки семейства имели кариотип .с наиболее'эддоким
числом хромосом (2п=78), таким, как у домашней собаки и волка, часть вцдов в раздэдх ро¬
дах сохранили предковый или близкий к нему кариотип, а в других группах кариотдаы из¬
менялись за счет слияния хромосом предкового кариотипа в различных сочетаниях и изме¬
нений гетерохроматической компоненты геномов. Идентичность кариогида домашней
собаки и кариотипа предковой для всего семейства формы служит косвенным подтвержде¬
нием тезиса об огромных эволюционных (а следовательно, ^ .селекционных) потенциях до¬
машней собаки как вида.Картирование генома собакиСложности в детальной идентификадии хромосомных пар собаки в значительной
степени обусловили и катастрофическое отставание работ по картированию генома у это¬
го вида по сравнению с другими видами лабораторных, домашних и п-у^цных рвотных.
До настоящего времени в плане картирования хромосом собаки в основном работала
только одна группа исследователей [Meera et al., 1993], которая использовала традицион-
нук? для этоф схему — получение панели клонов гибридных клеток собака х китайский
хомячок. Однако цитогенетически эта па,нель не анализировалась и поэтому данных о ло-
Таблица 2.5. Генетическая карта биохимических локусов у собакиХромосомаГеныU1ENOl, PGDU2PGM1изGUK1U4РЕРСU5ACPIU6MDH1U7IDH1U8ACY1, GPX1U9PGM2U10PGM3, ME], SOD2, DLA, С4U11MDH2U12АКЗU13РРАU14LDHAU15АСР2U16ESA4U17GAPD, TPI, LDHBU18ESDU19NPU20NPI, РКМ2U21PGPU22GPIU23ITPAU24SOD1U25АС02XGLA, HPRT, G6PD, F8, F9кализации того или иного гена на какой-либо из хромосом собаки (за исключением Х-хро-
мосомы) нет. Работа свелась, в основном, к определению групп синтенных (сцепленных)
генов, в таблице 2.5 обозначенных символом U.У собак так же, как у людей и сельскохозяйственных животных, огромное количество
генетических заболеваний. Диагностика таких заболеваний, выбраковка из процесса вос¬
производства генетически дефектных животных, разумное планирование селекционного
процесса — именно за .это готовы платить многие ветеринарные фармацевтические фирмы
и кинологические ассоциации. Но для этого необходима определенная предварительная
работа. В настоящее время 25 генетических лабораторий Европы и США объединились
для проведения совместных работ по картированию генома собаки (проект ”DogMap“). В
качестве стратегии для картирования выбран подход, аналогичный таковым, используе¬
мым для картирования геномов свиньи (проект PigMap) и крупного рогатого скота
(BosMap). В качестве маркеров использованы микросателлитные последовательности с
описанным к настоящему времени полиморфизмом. Источником материала стали нссколь-
ко семей (по 3 поколения) биглей и немецких овчарок, содержащихся в одном из известней¬
ших питомников в Берне (Швейцария). Хромосомное картирование осуществляется с по¬
мощью космидных и плазмидных библиотек. Основной подход в картировании — метод
флуоресцентной in situ гибридизации (FISH).Кроме непосредственных целей картирования генома собаки как одного из видов до¬
машних животных, этот проект интересен и в эволюционном плане. Ранее была проведена
работа по картированию генома лисицы и идентификации эволюционно консервативных
районов хромосом в геномах лисицы, человека, кошки и норки [Nash, О’ Brien, 1982;
Rubtsov et al., 1988; Graphodatsky, 1989] и показано, что многие консервативные для боль¬
шинства видов млекопитающих районы хромосом в ходе эволюции Canidae были сильно
перестроены, что особо выделяет это семейство среди прочих изученных групп млекопита¬
ющих. Поэтому картирование генома еще одного вида семейства, а именно домашней со¬
баки, хромосомный набор которого мы считаем „предковым“ для Canidae, представляет
огромный научный интерес.
Гпава 3ГЕНЕТИКА ОКРАСА И ШЕРСТИОсобенности строения и окраса шерстного покрова — наиболее ярко выраженные
признаки, определяющие зачастую породность собак, их племенную и пользовательную
ценность. Кроме того, эти признаки оказались очень привлекательными и с точки зрения
изучения основ генетики вида благодаря их сильной наследственной обусловленности и
широкой изменчивости. Не случайно поэтому тип шерсти и ее окраска являются, с одной
стороны, примером генетического изучения биологических особенностей домашней соба¬
ки, а с другой — маркерами, пригодными для дальнейших углубленных исследований в
этом направлении.Шерстный (волосяной) покров собаки как типичного млекопитающего состоит из
волос нескольких типов, причем в строении каждого волоса выделяют сердцевину, корко¬
вый слой и оболочку.Покровные волосы — длинные, грубые и жесткие — являются самыми длинными и
толстыми и расположены обычно в верхней части тела собаки — на шее, спине, пояснице,
крупе. Короткошерстные собаки обычно вовсе лишены такого типа волос или имеют их в
небольшом количестве по хребту. Напротив, так называемые жесткошерстные собаки от¬
личаются сильным развитием покровных волос. У длинношерстных собак покровные во¬
лосы развиваются длинными, тонкими, шелковистыми — „украшающими", образуя „во-
ротник** или „гриву“, „очесы“, „штаны“, „подвес" — соответственно на шее, передних,
задних конечностях, на хвосте.Остевые волосы короче и тоньше покровных и там, где они растут вместе, обычно
закрываются последними. Однако у короткошерстных собак, например, шерсть зачастую
представлена только остью.Подшерсток образуют пуховые волосы, короткие, тонкие, волнистые, без сердцевин¬
ного слоя. Подшерсток имеют собаки не всех пород. Кроме того, часто в пределах одной
породы имеется изменчивость по структуре шерсти и соотношению типов волос, которое
может меняться у одного и того же животного в течение года и в разном возрасте.В окраске шерстного покрова различают собственно окрас и рисунок расцветки (ру¬
башку). Окончательное восприятие окраса обусловлено взаимодействием цвета шерсти
(покровных и остевых волос) и подшерстка, преломлением лучей света в зернах пигмента,
которые могут располагаться в разных зонах волос с разной плотностью. Если волосы не
имеют пигмента, они воспринимаются как белоснежные.Обычно выделяют два основных типа пигмента — эумеланин (в разных модификаци¬
ях обусловливает черный и коричневый цвет волос) и феумеланин (определяет рыжий
спектр окрасок).Интенсивность окраса шерсти обусловливается различной плотностью расположе¬
ния пигментных гранул в разных слоях волос. Например, черные зерна, равномерно нахо¬
дящиеся в шерстинках, формируют черный окрас, а те же зерна, расположенные только в
сердцевинной зоне, просвечивают через корковый слой осветленно и воспринимаются как
серый (голубой) окрас — генетически являющийся ослабленным черным.Возрастная изменчивость структуры волосяного покрова может вызывать и возраст¬
ное изменение окраса — как правило, его осветление.Окрас „дикого типа", характерный для диких предков и родичей собаки — „агу¬
ти* — зонарно-ссрый (волчий), обусловлен наличием по-разному окрашенных кольцевых
зон внешне „серого*1 волоса. Его кончик окрашен в черный цвет, ближе к основанию смс-
няющие друг друга желтая и вновь черная зоны. Возле основания волос опять светлый, без
пигментных гранул.Различные окрасы собак являются следствием изменения этой базовой зонарной рас¬
цветки шерстинок, когда меняется ширина и соотношение цветовых зон.Выделяют основные группы окрасов:Зонарные• агути• соболиный — при сближении черных зон и рыжей середины волос.Сплошные разной интенсивности• черный• коричневый• рыжий• белый.Пятнистые• белая пятнистость:• пегий (белые пятна в центрах депигментации — на груди, шее, концах лап,
животе, конце хвоста на основном темном фоне);• пятнистый (зоны депигментации распространяются в такой степени, что
белый цвет становится фоном, на котором расположены темные пятна
первично основного окраса);• крапчатый;• мраморный;•* желтая пятнистость:• подпалый (на темном фоне желтые отметины на морде, над глазами, на
горле, на груди, на передних и (на внутренней стороне) задних конечностях,
вокруг ануса);• чепрачный (зоны подпалин распространены по корпусу, а темный фон
остается в виде чепрака на спине и боках);• тигровый (черные поперечные полосы на рыжем фоне с разной степенью
соотношения полос и фона).Трехцветный — сочетание белой пятнистости с подпалым или чепрачным окрасами
(рыжие пятна при этом локализованы в традиционных районах расположения подпалин).Из этой классификации становится понятным, что многие любители собак генетичес¬
ки некорректно воспринимают окрасы своих питомцев, допуская путаницу в использова¬
нии терминологии и определении признака.Кроме того, нередки затруднения в понимании наследственной обусловленности ок¬
расов из-за различий в символическом обозначении генетических систем, контролирую¬
щих окрас и шерсть, введенном разными авторами.Ниже приведена генетическая символика локусов и генов, контролирующих окрас,
соответствующая системе, предложенной R. Robinson [1982], которая за минувшие годы
оказалась общепризнанной и наиболее оптимальной.Локус A (Agouti). Серия аллелей агути, контролирующих распределение пигментных
гранул как по каждому волосу, так и по всему телу собаки в целом, эти аллели детермини¬
руют несколько наиболее характерных фенотипов по окрасу.Аллель A (Agouti) — немутантный, нормальный, дикого типа, доминантный к дру¬
гим аллелям серии, обусловливает окраску „дикого типа“ — зонарно-серую, волчью, у ди¬
ких представителей Canidae и, видимо, у всех домашних, имеющих такой фенотип: у серых
норвежских лосиных собак, серых сибирских хаски и лаек, и у других северных длинно¬
шерстных. По R. Robinson [1982], видимо, серые немецкие овчарки — гомозиготы А А.Аллель As (Solid black) — мутантный доминантный по отношению к другим мутаци¬
ям данного локуса, обусловливает и „красноватые4* отметины [Little, 1957] при непол¬
ном доминировании As над АУ и а'. С. Little также считал, что гетерозиготы А5АУ имеют
красноватые отметины на голове, шее, боках и конечностях, но у собак генотипа Asa' эти
отметины располагаются только в местах, типичных для подпалин классического подпало-
го окраса гомозигот а'а'. Таким образом, неполное доминирование аллеля As вполне воз¬
можно, но R. Robinson, например, считает равномерное (не зонарное) распределение пиг¬
мента в волосе и сплошной чисто-черный интенсивный окрас по всему телу. Встречается во
многих породах, у некоторых окрас бывает не чистого тона, а коричневато-черным из-за
примеси зонарно окрашенных волосков. Одно из возможных объяснений этому — непол¬
ное доминирование аллеля As, в связи с чем генотип ASAS формирует чисто-черный фено¬
тип, а гетерозиготы ASA? или ASA могут иметь окрас коричневато-черный. При этом зо¬
нарно окрашенные волосы могут вызыватчитает необходимым иметь более достоверный
статистический материал по этому вопросу для его окончательного выяснения.Аллель АУ (Dominant yellow) — доминантный желтый — детерминирует один из ви¬
дов желтого (или рыжего) фенотипа собак, которые, имея такой ген, могут иногда быть и
чисто-желтого (рыжего) цвета, но чаще всего у них имеется и примесь некоторого количе¬
ства желтых с черными кончиками волос на голове, плечах, по хребту и хвосту. Этот ал¬
лель, перераспределяющий зону черного пигмента в кончики волос, формирует названный
окрас, известный как соболиный. Имеющие этот аллель чисто-желтые собаки должны, по
этой терминологии, называться золотисто-соболиными [Robinson, 1982]. Степень черноты
или „соболиности“ окраса изменчива и обусловливается полигенами, которые наследуют¬
ся независимо от АУ.С. Little [1957] при скрещивании соболиных собак между собой показал, что они яв¬
ляются гетерозиготами АУа1: в их потомстве наблюдалось расщепление по окрасу ры-
жие:соболиные:черно-подпалые в соотношении 1:2:1. Этот исследователь утверждал, что
таким образом получал даже черно-подпалых щенков от вязок рыжих такс „в себе“, кото¬
рые были на самом деле генотипически соболиными гетерозиготами. Впрочем, многие ав¬
торы до сих пор сомневаются в возможности такого генетического механизма названных
окрасов — по крайней мере, у такс [Robinson, 1982].Аллель asa (Saddle pattern) — рецессивная мутация чепрачного окраса, обусловлива¬
ющая очень характерный рисунок — V-образное „седло** темной пигментации на спине и
боках корпуса, как, например, у эрдельтерьера или немецкой овчарки. Собственно, суще¬
ствует большое сходство в фенотипическом проявлении мутантных аллелей asu и ано об¬
ладатели гена cisa обычно имеют большие по площади желтые отметины, чем при подпа¬
лом окрасе, особенно на морде, плечах, конечностях. Степень изменчивости размеров
чепрака, однако, весьма высока, и иногда он может быть таким распространенным, что ок¬
рас можно принять за подпалый (который варьирует не так широко).Аллель a1 (Tan pattern) — самый рецессивный в агути-серии, детерминирует очень ха¬
рактерный рисунок окраса с подпалинами, располагающимися в строго определенных мес¬
тах интенсивно рыжими отметинами (как, например, у добермана, ротвейлера и других ар-
хитипических по такому окрасу пород): темный цвет покрывает всю верхнюю сторону тела
собаки, а подпалины располагаются на внутренней поверхности конечностей, на груди, на
морде, вокруг анального отверстия. При этом характерны непременные два пятнышка
подпалин над глазами. Площадь, занимаемая подпалинами, и интенсивность (яркость)
этих отметин имеют внутрипородную изменчивость, детерминируемую комбинациями по-
лигенов-модификаторов.Резкие различия между подпалым и чепрачным окрасами проявляются в наличии или
отсутствии возрастных изменений этих расцветок. Чепрачные щенки обычно рождаются
настолько темными, что выглядят черно-подпалыми, но с возрастом светлеют, чепрак
уменьшается в размерах, и от такого псевдо“подпалого“ окраса обычно не остается и сле¬
да. Так же щенячий темный окрас может быть и при наличии желтого аллеля АУ. Это вооб¬
ще характерное явление для Canidae — даже волчата обычно гораздо темнее окрашены,
чем полновозрастные, матерые волки.Например, R. Robinson [1982] предложил два объяснения отсутствия исходного агу-
ти-аллеля А в большинстве пород (исключая эскимосских, скандинавских и северных со¬
бак). Первое — направленный отбор с самых первых этапов доместикации с целью выведе¬
ния домашних собак, резко отличающихся „рубашкой*1 от диких волков. Второе —
мутации цвета шерстного покрова, возникавшие на ранних стадиях одомашнивания вол¬
ков, использовались человеком как показатели успеха доместикации, как маркеры домаш¬
ней собаки. Другими словами, механизмом могли быть: или элиминация гена А, или изме¬
нение фенотипического проявления гена А.Еще одна интересная проблема для генетиков и кинологов: какой мутантный аллель
локуса А появился у одомашниваемых собак первым? R. Robinson [1982] полагает, что пер¬
вым мог появиться мутантный аллель asa: основа фенотипического проявления агути и
чепрачного окрасов похожа, и модификация аллеля дикого типа может достигать темного
пятна на спине как переходная форма к чепраку, а основной фон может „желтеть** из-за
увеличения желтых зон в волосах. Подтверждение тому — часто встречающийся у беспо¬
родных метисов переходный окрас — нечто между волчьим и чепрачным. Получены также
прямые доказательства, что эти два окраса могут встречаться в практически идентичных
формах [Fox, 1978]. При скрещивании койотов с биглями в потомстве выявили 4 гибрида
первого поколения и 12 — второго, у которых окрас описан как темно-соболиный в летний
период и светло-соболиный зимой. Автор использовал термин „соболиный", но окрас биг¬
ля — чепрачный, и судя по фотографиям в его статье, окрас гибридов или волчье-серый,
или светло-чепрачный.Широко известно спорное предположение С. Little [1957], что чепрачный и подпалый
окрасы детерминируются одним и тем же аллелем А-локуса, а фенотипические различия
вызваны только разными наборами генов-моди фикаторов. Однако Willis [1976] провел де¬
тальный генетический анализ и убедительно продемонстрировал существование разных
аллелей этих двух окрасов — asa и а' с доминированием первого над вторым. R. Robinson
отмечает, что и другая „спекуляция"- будто бы должно существовать несколько разных ал¬
лелей чепрачного окраса для контроля разных форм этой расцветки — также не имеет об¬
щего с действительностью. Широкий спектр от светло-чспрачного до темно-чепрачного
окрасов представляет собой плавный ряд переходов, что доказывает действие модификато¬
ров на степень яркости этого окраса.Иногда делаются попытки расширить Л-серию, добавив сюда еще „рецессивный чер¬
ный" аллель (предположительно а), так как известны изредка случаи, когда практически
черный окрас ведет себя рецессивно по отношению к соболиному, чепрачному и подпало-
му — как, например, в породе немецких овчарок. Однако, скорее всего, такой рецессивный
черный окрас чаще детерминируется геном из другого локуса (Е — extention). или же в
этих случаях играет роль модификация уже известных аллелей А.Так, профессор Н. А. Ильин [Iljin, 1941 изучил гибридизацию волка с собакой черно¬
го окраса: в первом поколении получено 7 серых и 6 черных потомков. Серые гибриды „в
себе" дали 17 серых и 3 черно-подпалых, от разведения черных „в себе" родились 12 чер¬
ных и 3 черно-подпалых. Для генетического объяснения этих фактов Ильин также постули¬
ровал черный рецессивный к гену А аллель а1, доминантный по отношению к а', и указал
генотипы родителей: серый волк Аа1, черная собака а1 а1. Однако R. Robinson считает ма¬
ловероятным, что случайно подобранные абсолютно неродственные партнеры оказались
вдруг оба носителями редкого и неизвестного до того аллеля. По его мнению, результат
опыта Ильина можно объяснить более правдоподобно: вероятнее всего, волк имел генотип
Аа', а собака — Asa'. Таким образом, расщепление волчье-собачьих гибридов согласуется с
символикой, уже известной для А-локуса.Локус В (Black —brown) имеет два аллеля: доминантный нормальный В (Black) —
аллель черного окраса (синтезирует эумеланин) и мутантный рецессивный b (brown) — ал¬
лель коричневого окраса (синтезирует феумеланин).Фенотип „дикой" окраски собачьих „агути" обусловливается наличием двух видов
пигмента — черного и желтого в виде микроскопических зерен в их волосах. Распределе¬
ние пигментных гранул в толще и в разных зонах создает окраску. Например, у черно-под-
палой собаки в местах основной — черной — окраски шерсти волосы содержат пигмент¬
ные зерна овальной формы, и каждая гранула — интенсивного черного цвета. На участках,
окрашенных подпалинами, пигментные зерна в волосах имеют значительно меньший раз¬
мер, шаровидную форму и желтоватый цвет. У собаки коричневого с подпалинами окраса
шерсть районов основного фона (шоколадного) имеет пигментные зерна более светлоко¬
ричневые, чем пигмент фона черно-подпалой собаки, а в местах, окрашенных подпалина¬
ми, пигмент обеих таких собак совершенно одинаков [Robinson, 1982]. Таким образом, му¬
тантный ген, вызывающий „шоколадный" цвет шерсти собак, действует на гранулы
черного пигмента в волосах, осветляя их цвет, что воспринимается человеческим глазом
как изменение окраски волос из черной в шоколадную. Одним из первых аллельные отно¬
шения доминирования генов В и b блестяще показал в гибридологических экспериментах с
доберманами Н. А. Ильин [1931], а чуть позже [1932] и с другими породами.Аллель b изменяет нормально (первоначально) черноокрашенные участки собаки в
интенсивно-коричневый (шоколадный) цвет, причем не только цвет шерсти, но и кожи
(особенно заметно на губах), пасти, мочки носа и когтей. Радужная оболочка глаз корич¬
невых собак также заметно светлее, чем у черных.
Первоначально коричневая мутация окраса проявилась в комбинации с геном As се¬
рии агути, при генотипе As-bb [Robinson, 1982]. Однако впоследствии возникли разные дру¬
гие генные комбинации, вызывающие разновидности шоколадной окраски.Важно отметить еще раз, что b-аллель не влияет на желтый пигмент, в связи с чем
различают два генетически особых вида рыжего окраса собак [Robinson, 1982]: „черные“
рыжие собаки генотипов АУ-В- или В-ее и „коричневые" рыжие собаки генотипов АУ-bb и
bbee. Эти два рыжих окраса, встречающиеся у домашней собаки, не полностью идентичны,
так как аллели В и b по-разному влияют на цвет мочки носа, губ, слизистой пасти, глаз.
Эти стати черного цвета у „черных" рыжих, имеющих ген В, но коричневые у „коричне¬
вых" рыжих 66-гомозигот, когда даже при генотипах соболиного или зонарного окрасов
кольца на волосе не черные, а коричневые.В свете данных о генетике 5-локуса представляется неправдоподобным, чтобы в ре¬
зультате вязок коричневых собак рождались черные щенки. Однако было одно сообщение
[Frankling, 1971] о регистрации пяти пометов черных щенков, полученных при разведении
семи „шоколадных" далматинов*. Если исключить „мезальянс" (то есть неправильности в
регистрации родительских пар), можно предположить, что коричневые фенотипы окрасов
родителей этих пометов обусловлены двумя мутациями, возникшими в разных локусах.
Такая ситуация очень редко случается у млекопитающих — известен до сих пор единствен¬
ный достоверный пример с американской норкой. В этой связи названное сообщение
Е. Frankling воспринимается как генетический нонсенс, как редчайшее исключение из пра¬
вила „от коричневых родителей не может быть черных щенков".Локус С (Albino alleles). Серия аллелей „альбино" контролирует синтез пигмента в
шерсти, интенсивность самой пигментации как таковой. Название серия получила исходя
из принципа гомологии в классе млекопитающих, так как собственно у домашней собаки
настоящий альбинизм — очень редкое явление, хотя может часто встречаться у многих
других видов млекопитающих. Более того, С-локус считают одним из наиболее мутабиль-
ных, в нем имеется целая серия разных аллелей. Интересно, что фенотипы, ими обусловли¬
ваемые, очень сходны у разных видов. Можно поэтому в силу закона гомологических ря¬
дов наследственной изменчивости [Вавилов, 1987] предполагать возможность появления
соответствующих окрасок у вида, даже если они еще у него не обнаружены, только зная
похожие формы у „дальних родственников".Что касается собаки, то у нее пока выявлены только три мутантных аллеля С-локуса.Аллель С — ген дикого типа, нормальный, доминантный, обеспечивает возможность
синтеза любого пигмента, в зависимости от генотипа по соответствующим локусам. Этим
аллелем обладают все собаки с „нормальной" окраской.Мутантный аллель cch („шиншилла") получил обозначение по гомологии с такой же
мутацией у кроликов. Он ослабляет пигментацию, причем желтый пигмент в желтом коль¬
це волоса осветляется до светло-палевого, почти белого цвета, а черный пигмент светлеет
гораздо меньше. Видимо, такая мутация закреплена в породах, где встречается палево-жел¬
тый окрас, как у норвежских лосиных собак (элкхундов). Наиболее заметно действие этого
мутантного гена у собак, имеющих в своих генотипах „желтые" гены АУ или е. Так, напри¬
мер, светло-желтые или кремовые особи в породе золотистых ретриверов обладают, скорее
всего, генотипом с(1'сс1,ее, обусловливающим некоторую вариабельность фенотипа — цвет
шерсти от кремового до почти белого — может быть, из-за модифицирующего влияния
„рыжих полигенов". Впрочем, некоторые генетики, и прежде всего С. Little [1957], считают,
что у собак возможно существование не одной шиншилловой мутации, а нескольких со
сходным действием — как у грызунов. Little даже предложил обозначение одного из таких
„других" шиншилловых аллелей — се , который якобы проявляется почти белым феноти¬
пом со светло-кремовым цветом шерсти на спине, плечах и голове. Если такой аллель дей¬
ствительно существует, он при этом окрасе может входить в генотипы: А-у-сесе или сесе ее.
Little предполагал такую генетическую сущность окраса, например, среди уэст-хайленд-у-
айт-терьеров — у них сильное осветление окраса не является альбинизмом: глаза остаются
черными. С. Little также считал, что ген пигментации С не полностью доминирует над
шиншилловым аллелем.* Далматины с коричневым крапом на белом фоне — не идеальный вариант для дискуссии о генетике
5-локуса.
Аллель cb (blue-eyed albino) — мутация, обусловливающая неполный альбинизм: го¬
мозиготные по нему собаки названы профессором Н. А. Ильиным [1932] лейцистами —
шерсть их обычно вся белоснежная из-за отсутствия в волосах пигмента, который, однако,
имеется в коже, когтях, радужной оболочке глаз — она у лейцистов голубого цвета, а не
красная, как у полных альбиносов. Такой аллель весьма редок и известен у бультерьеров,
немецких догов, борзых. Гомозиготы по нему не могут проявить окраску, детерминируе¬
мую генами других локусов и, подобно белым кошкам с голубыми глазами, почти всегда
страдают глухотой. Н. А. Ильин этот аллель обозначал по-другому — с° , но этот символ
сейчас не употребляют. В ряду доминирования аллелей серии С этот ген занимает место
над аллелем с полного альбинизма.Аллель с (albino) — самый рецессивный в локусе, в гомозиготе приводит к альбиниз¬
му, так как блокирует полностью синтез пигмента — в волосах, коже, радужке глаза, име¬
ющей красный цвет из-за просвечивающих кровеносных сосудов глазного дна. Зрение у
полных альбиносов значительно ухудшено, эта мутация встречается у собак чрезвычайно
редко, единично, и не является характерной для какой-нибудь определенной породы, за ис¬
ключением, по всей видимости, пекинесов: у этих собак чаще, чем в других популяциях, об¬
наруживалась такая мутация [Whitney, 1947; Little, 1957].Генетическая основа альбинизма — блокирование аллелем с одного из звеньев мета¬
болизма фенилаланина (превращение тирозина через 3,4-дигидроксифенилаланин в мела¬
нин [Хатт, 1969].Локус D (Dilution) определяет интенсивность пигментации, детерминируя распреде¬
ление пигментных гранул в слоях волоса.Аллель D (Dense)- доминантный, „дикого типа11, вызывает интенсивный окрас: пиг¬
мент — черный или желтый — расположен равномерно по всей толщине волоса, как в его
корковом слое, так и в сердцевине.Аллель d (dilute) ослабляет пигментацию и осветляет рыжий и коричневый окрасы до
палевого и бежевого, а также переводит черный окрас в голубой. Эти изменения фенотипа
обусловлены перераспределением пигментных гранул, которые у гомозигот dd находятся
только в сердцевине волос и тускло просвечивают сквозь непрозрачный корковый слой
[Ильин, 1931; Robinson, 1982]. Человеческим глазом окрас такой шерсти воспринимается
сильно осветленным.Четыре основных, „базовых" окраса млекопитающих — черный, голубой, шоколад¬
но-коричневый и изабелловый — обусловливаются комбинацией генов В(Ь) и D(d) при
взаимодействии с неаллельными генами As и Е:Если же в генотипе собак ген As заменяется мутацией а' в гомозиготе, то образуются
не сплошные вышеперечисленные окрасы, а соответственного основного фона с подпали¬
нами.Одновременное наличие в генотипе собаки взаимодействующих нсаллельных генов
АУ и Еь>' вызывает в комбинации с генами black и dilution разновидности тигрового окраса:ОкрасГенотипЧерныйГолубойШоколадно-коричневыйИзабелловыйA'-B-D-E-As-B-ddE-As-bbD-E-As-bbddEОкрасГенотипЖелто-тигровыйЖелто-тигровыйЧерно-тигровыйСеро-тигровыйШоколадно-тигровыйАУ—D-EK
A>'-B-ddEbr-
АУ-bbD-E1'1'-
АУ-bbddE/)|-
Ay-bbddEbr-Интересно действие гена-разбавнтеля d на цвет кожи и радужной оболочки: он очень
мало влияет на них у собак желторыжих мастей, хотя цвет шерсти осветляет до езетло-кре-
мового, а у генетически черных или коричневых ослабляет не только основной цвет шер¬
сти и осветляет кожу, но и глаза, и когти.Кроме того, при двойной гомозиготности по генам d и а1 (рецессивный аллель окраса
с подпалинами) нередко снижается жизнеспособность — например, у редких эрдельтерье¬
ров и тойтерьеров голубого окраса, изабелловых левреток, доберманов голубого с подпа¬
лами и изабеллового окрасов [Сотская, 1991].Локус Е (Extention) контролирует распространение (распределение) пигмента (феу-
меланина и эумеланина) по телу животного. В волосяном покрове „дикого типа11 представ¬
лены в каждом волосе оба этих пигмента — черный и желтый, и разные аллели £-локуса,
подобно Л-аллелям, взаимно перераспределяют их.Аллель Е (Normal extention) — нормальный ген серии, вызывает нормальное распро¬
странение черного пигмента в шерсти по всему телу — например, при чисто-черном, чеп-
рачном или черно-подпалом окрасах.Два (или три?) других аллеля локуса считаются мутациями этого нормального ге¬
на £.Аллель Ebr (Brindle) — доминантный по отношению к Е [Robinson, 1982], хотя неко¬
торые другие авторы считают, что рецессивный, — вызывает тигровый рисунок окраски
шерстного покрова: черный и желтый пигменты окрашивают определенные участки шер¬
сти, располагающиеся чаще в виде поперечных черных и желтых полос. На морде „тигро¬
вых" собак обычно сохраняется черная пигментация в виде „маски". Такой окрас весьма
типичен для собак многих пород.Аллель е (non-extention — yellow) — безусловно рецессивный в этом локусе, обуслов¬
ливает нераспространение черного пигмента в шерсти по телу и вызывает фенотип желто¬
го окраса. При этом важно подчеркнуть: черный пигмент заменяется на желтый только в
шерсти, а мочка носа, например, остается черной. Не светлеют также губы, слизистая рта,
радужная оболочка глаз.Аллельные взаимоотношения доминирования в /Г-серии окончательно не ясны до сих
пор. Например, С. Little [1957] установил, что, как показано выше, аллель тигрового окра¬
са доминантен по отношению к нормальному гену Е, хотя прежде считал его, напротив, ре¬
цессивным и даже предлагал поэтому обозначение еЬг, что привело к немалой путанице в
генетической литературе. Болес реалистично мнение R. Robinson [1982]: доминирование
аллелей в £-локусе неполное, что проявляется, например, в широкой изменчивости эксп¬
рессии тигрового окраса: от темно-тигрового с незначительным проблеском желтых поло¬
сок на черном фоне (как, например, у французских бульдогов), до светло-тигрового, когда
узкие „тигровины" пересекают занимающий основную площадь желтый фон (немецкие до¬
ги). Разумеется, такая изменчивость может обусловливаться генами-модификаторами, на¬
следующимися независимо от главного гена Е'п. Но с другой стороны, степень тигровости
собак разных генотипов по £-локусу (генотипы: Eh'Eb,\ EbrE, Ehle) вполне может быть раз¬
личной — например, при последнем генотипе собаки должны быть с менее интенсивными
полосами, чем при первом.Кроме перечисленных аллелей £-локуса, С. Little [1957] постулировал наличие доми¬
нантного аллеля, обусловливающего сохранение черной маски при желтом окрасе. Он обо¬
значил этот „масковый" ген £"' — „ген суперраспространения черного пигмента" при ос¬
ветлении пигментации шерсти тела. Однако впоследствии в литературе не опубликовано
каких-либо подтверждений наличия у собак этого аллеля. Генетика „маски" подробно об¬
суждена R. Robinson [1982], что будет отражено ниже.Неаллельное взаимодействие генов А и ЕАллелосерии эпх двух локусов контролируют образование черного и желтого пиг¬
ментов в шерсти собак большинства пород. Выше охарактеризовано действие каждого ал¬
леля из выделенных в этих локусах. Однако необходимо учитывать и их совместное влия¬
ние на окрас собак, так как существует неаллельное взаимодействие между этими генами,
обусловливающее подчас неожиданный фенотипический эффект.Нормальная экспрессия аллелей агути-локуса А, описанных выше, возможна только
при наличии в генотипе собак гена Е „дикого типа", который обеспечивает равномерное
распространение черного пигмента по всему телу собак. Если собака имеет ген Е, то ее ок¬
рас обусловливается, в частности, генотипом по Л-локусу:
ОкрасГенотипСплошной черныйА5—Е-Желтый „А"АУ—Е-ЧепрачныйasuasuE-Подпалыйа'а'Е-Однако, если у таких собак ген Е мутирует в рецессивную форму е, при гомозиготно-
сти ее все вышеперечисленные генотипы изменяются (As-ee и т. п.) и вызывают перераспре¬
деление (замену) пигмента — черного на желтый. Собаки таких генотипов оказываются
рыжими — агути-аллели не проявляются в фенотипе.Выделяют два генетически различных варианта желтого (рыжего) окраса у собак раз¬
ных пород. Они обусловлены соответственно генами АУ и е, являясь рецессивными по от¬
ношению к черному окрасу (тоже генетически неоднородному). Поэтому рыжие собаки
могут рождаться от вязок генетически различных черных генотипы черных родителей мо¬
гут быть ASA)'EE или —Ее) наряду с рождением черных щенков — в соотношении 1:3. Ры¬
жих собак, имеющих „рыжий" ген АУ, можно отличить от рыжих генотипа ее в том случае,
если от таких рыжих удастся получить, например, выщепившихся черно-подпалых потом¬
ков (генотипа а'а'ЕЕ), что иногда случается среди такс [Robinson, 1982]. Кроме того, мно¬
гие АУ — рыжие собаки имеют в разной степени еще и черный пигмент в волосах — на го¬
лове, по спине, на плечах и даже на боках. Когда таких волос с черными кончиками
оказывается достаточно много в шерсти собаки, ее окрас воспринимается как „соболи-W (Сныи .В связи с этими „следами11 черного пигмента у рыжих собак R. Robinson [1982] счита¬
ет, что „маска“ при рыжих формах окрасов в виде черной морды (иногда чернота распрос¬
траняется на голову вплоть до ушей) может быть только у Л>'-позитивных особей. В проти¬
воположность таковым, <?е-рыжие имеют иногда очень немного волос с черными
кончиками или очень „призрачную" нераспространенную маску. Впрочем, иногда по
этим признакам трудно определить, какой именно из этих генов имеет собака: некоторые
^-''-позитивные тоже проявляют лишь „следы" черного пигмента.Такие генетически разные рыжие особи могут встречаться даже в пределах одной по¬
роды [Robinson, 1982]. Это открытие сделано при регистрации черных щенков, рожден¬
ных от двух рыжих родителей. Генетически можно обозначить такое скрещивание рыжих:
АУАУЕЕхА*А*ее.Дигетерозиготные гибридные потомки генотипа AsA-vEe оказываются черными из-за
доминирования As над АУ и Е над е.Поэтому в практическом собаководстве селекционерам важно знать, с каким генети¬
ческим вариантом рыжего окраса они имеют дело, эта проблема, оказывается, весьма не¬
простая. Большинство пород, где встречаются рыжие собаки, имеют в генофонде аллель
АУ. Наличие этого гена можно доказать, если среди таких рыжих обнаружатся животные с
маской или „соболиные".Если же в какой-либо породе чепрачные или подпалые собаки произведут рыжих
щенков — значит, здесь действует „желтый" ген е, а генотипы родителей были aS(l-Ee или
а'а'Ее, соответственно. Другое доказательство наличия гена е — рождение черных щенков
от скрещивания рыжей собаки с чепрачной или черноподпалой. При этом результате ры¬
жий родитель должен иметь генотип As-ee. Выше показан еще один вариант доказатель¬
ства наличия гена е — рождение черных щенков от вязки двух рыжих партнеров.Эту проблему попытался разрешить и С. Little [1957], определив ряд пород, где ры¬
жие окрасы детерминируются только генотипом ее: английский сеттер, бигль, сеттер-гор¬
дон, далматин, золотистый ретривер, ирландский сеттер, лабрадор-ретривер, пойнтер, пу¬
дель.В дополнение к этому ряду М. Burns и М. Fraser [1966] предложили кокер-спаниелей
палевого окраса, но Little доказал, что в генофонде этой породы имеются оба рыжих ге¬
на — АУ и е, так же, как среди чау-чау, английского и филд-спаниелей.Ген ЕЬг вызывает черную пигментацию волос на рыжем фоне распространения жел¬
того пигмента в виде черных полос, также взаимодействует с аллелями А-локуса
[Robinson, 1982]:
ОкрасГенотипСплошной чисто-черныйТигровыйТемно-тигровыйAs—ЕЬг-АУ-ЕЬг-asu-Ebr-Черный с желтыми „тигровинами“ i a'a'Ebr-Гсн As эпистатичен по отношению к Еь'\ т. е. подавляет его проявление в фенотипе,
при этом собака будет чисто черного окраса, без полос. Однако в комбинации с аллелем
А У ген тигровости благополучно формирует свой „фирменный" полосатый окрас. Таким
образом, в большинстве пород тигровые собаки имеют генотип АУ-ЕЬг-. Разные другие со¬
четания гена тигровости и аллелей агути, как показано выше, обусловливают цветовые ва¬
риации тигрового окраса, весьма характерные для некоторых пород.Локус G (Progressive silvering) — прогрессирующее поседение.Аллель G — доминантный ген, вызывающий прогрессирующее с возрастом поседе¬
ние, обусловливает постепенное перераспределение пигмента из коркового слоя волоса в
его сердцевинную часть, в связи с чем черные при рождении собаки к одному-двум годам
становятся серебристо-седыми, этой мутацией обладают керри-блю-терьеры, серебристые
пудели, бедлингтонтерьеры, староанглийские овчарки-бобтейли. Кроме них, по С. Little
[1957], аллель G может быть распространен среди кэрн-, денди-динмонт-, скай- и йоркшир-
терьеров, но впоследствии получены доказательства обратного [Robinson, 1982]. При этом
считается целесообразным изучить породы австралийских терьеров и австралийских шел¬
ковистых терьеров на предмет наличия в их генофонде этой мутации.Аллель g — рецессивный ген, в гомозиготном генотипе обусловливает стойкую в те¬
чение всего онтогенеза окраску шерсти у большинства пород. Ген поседения вначале был
обозначен D. D. Whitney [1952; 1958] si (рецессивный), но затем Little [1957] установил, что
мутантный ген — неполностью доминантный, и ввел принятое сейчас его обозначе¬
ние — G. При этой неполной доминантности широко варьирует и экспрессивность гена,
поэтому необходим зачастую очень опытный глаз эксперта для правильной идентифика¬
ции всех гетерозигот. R. Robinson [1982] считает, что в разных породах или даже линиях
внутри породы могут быть как полудоминантный, так и рецессивный типы наследования
возрастного поседения шерстного покрова.Возможны разные варианты фенотипов в зависимости от соотношения седых и чер¬
ных волос в шерсти собаки. Если черных волос больше, чем седых — говорят о черной с
сединой собаке, — такой окрас и называют серебристым. Если же все наоборот — получа¬
ется белая шерсть с примесью черных волос (чалая масть). Эти термины по Robinson ха¬
рактеризуют или две различные генетические формы серебристости, или разные степени
экспрессии одного фактора. Серебристость также разделяют на постоянную — стационар¬
ную (ее степень не меняется с возрастом) и прогрессирующую (когда серебристость усили¬
вается при взрослении собаки или даже в течение всей ее жизни). Исходя из особенностей
фенотипической экспрессии гена G, вызываемую им седину считают прогрессирующей с
возрастом.Гетерозиготы Gg постепенно превращаются в собак серовато-голубого цвета, иногда
быстро, но чаще всего в течение нескольких лет. Гомозиготы GG претерпевают эти измене¬
ния в более резкой форме. При этом изменение пигментации может быть одновременным
или варьировать в разных частях тела особи (например, у бедлингтон-терьеров верхняя
часть головы .и плечи могут светлеть до палевого иди почти белого цвета).В комбинации с мутациями в других локусах гетерозиготность Gg может обусловли¬
вать незначительное ослабление окраса, тогда как гомозиготы GG вызывают сильное раз¬
бавление пигмента. Светлый окрас гомозигот GG свидетельствует об эпистатическом воз¬
действии этой мутации на другие локусы пигментации шерсти:ОкрасГенотипСеребристый
Шоколадно-серебристый
Г олубовато-серебристый
Кремово-серебристый
Светло-кремово-серебристыйAS-B-D-E-GGAs-bbD-E-GGA5-B-dciE-GGAs-B-D-eeGGAs~B-cldeeGG.
Во всех этих вариантах основной окрас может просвечивать, если серебристая собака
имеет в шерсти определенный минимум соответственно окрашенных волос. В противном
случае идентификация разновидности серебристого окраса может быть затруднена. Одна¬
ко все гомозиготы bb имеют коричневую мочку носа и светлые глаза. Их шерсть также мо¬
жет быть светло-коричневой. Подобно им, серебристые гомозиготы ее могут иметь легкий
желто-кремовый оттенок серебристого цвета шерсти. Все фенотипы вообще легко иденти¬
фицируются до поседения волос.Но и до развития поседения собака генотипа As-B-ddE-GG будет иметь голубой окрас
из-за действия гена-разбавителя d, и в этом случае по мере взросления щенка экспрессия ге¬
на G может быть почти не заметна.Локус Int (Intensification) — усиление желтого пигмента (руфизм). Интенсивность
развития желтого пигмента в шерсти собак очень сильно варьирует — от светло-кремового
оттенка до ярко-красного. Какая-то часть этой изменчивости обусловливается действием
главных генов окраски — например, гена-разбавителя d или, может быть, даже „шиншил-
лового“ гена сс1' [Robinson, 1982], но существенный вклад в это явление вносит и группа ге¬
нов-модификаторов, известных как „рыжие полигены“ — без обозначения их специальны¬
ми символами. Накопление этих действующих в „плюс-направлении" полигенов в
генотипе усиливает интенсивность окраса. Дикие родичи собаки — волки — имеют весьма
тусклый, неяркий (маскировочный) окрас, и уже первобытные собаководы стремились се¬
лекционировать собак при одомашнивании, отбирая на племя особей, наиболее отличаю¬
щихся от диких родичей яркостью окраски своей шерсти. Эта традиция сохранилась и по¬
ныне при разведении большинства пород собак.Н. А. Ильин [1932] установил в анализирующих скрещиваниях, что светло-желтый
цвет шерсти собак является доминантным признаком по отношению к ярко-рыжему, и по¬
стулировал существование локуса Int с тремя аллелями (детерминирующими разную ин¬
тенсивность оттенков желтого цвета шерсти) в следующем порядке доминирования:аллель Int — доминантный — обусловливает ослабленную пигментацию (грязно-бе-
лого цвета) перевязи (желтой зоны) на зонарном волосе;аллель int'" — неполностью доминантный — обусловливает среднюю интенсивность
пигментации желтой зоны (желто-коричневый цвет);аллель int — рецессивный — интенсивная пигментация (ярко-рыжий цвет).В той же книге Ильин постулирует еще три аллеля другого локуса Inti, меняющих
интенсивность цвета подпалин: доминантный аллель Inti („светло-белые" подпалы), рецес¬
сивный по отношению к нему Inti1” (светло-желтые подпалы) и рецессивный к двум преды¬
дущим аллелям inti (рыжие подпалы). Правда, тут же Н. А. Ильин отмечал, что эти две се¬
рии. Int и Inti могут быть аллелями одного локуса.R. Robinson, комментируя постулат Ильина о генах Int, отмечает, что идея моноген-
ного контроля главным локусом силы проявления желтого пигмента вполне реальна, одна¬
ко, по его мнению, после издания книги русского генетика в мире накоплено множество
дополнительных данных, свидетельствующих все же в пользу действия „рыжих полиге¬
нов", а не аллелей Int.В то же время экспериментальные результаты Ильина, безусловно, интересны и дос¬
товерны, так как исчезновение яркого рыжего цвета у потомков при скрещивании ярких
и светло-палевых собак наблюдали в разное время также К. Tjebbes, С. Wriedt [1927],
М. Burns, М. Fraser [1966], М. Willis [1976]. Последний отметил, что яркость этого оттенка
можно легко утерять в потомстве, а восстановить вновь — очень трудно. R. Robinson ин¬
терпретирует это обстоятельство так: „минус-рыжие полигены" доминируют над „плюс-
полигенами". Впрочем, это доминирование, справедливое в одних случаях, не проявляется
в других и в целом не является всеобщим и обязательным для всех пород.Локус М (Merle) — мраморный окрас. Временами в кинологической литературе воз¬
никает путаница с термином „мраморный окрас", но обычно этим названием обозначают
специфический рисунок „рубашки" — комбинацию белых и темноокрашенных участков
шерсти, что характерно для колли (длинношерстных и короткошерстных), шелти, пятнис¬
тых такс и немецких догов-арлекинов.От обычного пятнистого окраса мраморный отличается размерами и формой темных
пятен: они имеют характерные „рваные" границы и не сливаются в чепрак, не уменьшаясь,
однако, до размеров „тиковых" крапин.Мраморный рисунок является фенотипическим проявлением гетерозиготности по ге¬
ну М. доминантному по отношению к аллелю сплошной окраски. Светлые участки фона
обычно образованы смесью нормальных волос и волос с дефицитом пигмента (значитель¬
но больший дефицит черного/коричневого пигмента, чем желтого). Исключение — черно¬
белые доги-арлекины, у которых фон чисто-белый.Гомозиготы ММ имеют сплошной белый окрас, полностью или частично голубую
радужку глаз, уменьшенный размер самого глазного яблока, и чаще всего они полностью
или частично глухие и стерильные (бесплодные). Весь этот комплекс (синдром) аномалий
является следствием плейотропного действия гена М в гомозиготном генотипе.Различные сочетания гена М и аллелей других локусов могут проявляться фенотипи¬
чески следующим образом:ОкрасГенотипМраморно-голубой (блю-мерль)As-MmДвухцветно-мраморно-голубойа'а'МтМраморно-рыжий/соболиныйАУ-Мт.Последний вариант, например, у колли является нестандартным и появляется в ре¬
зультате неквалифицированного разведения собак — при вязках мраморно-голубых парт¬
неров с рыжими.Наиболее контрастна форма мраморно-голубого окраса — цвет шерсти собаки выг¬
лядит серым с черными неравномерно расположенными пятнами и крапинками разного
размера. Интересно, что ген М не оказывает такого ослабляющего действия на желтый
пигмент, как на черный [Robinson, 1982], но мраморный черноподпалый окрас часто оце¬
нивают как трехцветный из-за того, что этот аллель плейотропно также увеличивает пло¬
щадь белых отметин.Считается неразумным подбирать для вязок белых гомозигот ММ: более оптимально
скрещивать мраморных гетерозигот генотипа Мт с окрашенными немраморными (ими) со¬
баками: от таких вязок следует ожидать половину потомков мраморных и половину —
обычного сплошного окраса.В последние десятилетия, основываясь на межпородных различиях в фенотипах мра¬
морного окраса и данных селекции, постулировано существование не одного мраморного
аллеля, а нескольких [Schaible, Brumbangh, 1976], по крайней мере, у немецких догов-арле-
кинов современный стандарт предусматривает при мраморном окрасе не серо-голубой
фон, на котором расположены черные пятна (что бывало в этой породе прежде и характер¬
но сейчас для такого окраса у других пород: колли, такс, гончих и т. п.), а белоснежный.
Такой фенотип, считается сейчас, свойствен гетерозиготам по другому мутантному гену
М1’ (merle-harlequin). Немецкие доги-гетерозиготы по двум мутантным аллелям ММ1' фено¬
типически похожи на гомозигот ММ — с белым окрасом шерсти, частичной глухотой и
недоразвитыми глазами. Фенотип гомозиготы MhM]\ вероятно, сходен с таковым у гомо¬
зиготы ММ или сопровождается даже более серьезно выраженными аномалиями [Ro¬
binson, 1982].С учетом неаллельных взаимодействий генов окраса генотип стандартных немецких
догов-арлскинов характеризует формула: As-Mllm.Локус Ма (Mask) — детерминация маски. Под этим термином собаководы понима¬
ют распространение черного пигмента на морде, а часто и на передней части головы
вплоть до ушей. Чаще всего наблюдается у АУ — позитивных рыжих или тигровых собак,
среди которых наиболее типичны молоссоиды от мастифа до мопса. Маска может также
быть у собак чепрачных и подпалых (носителей сответствснно генов а™ и а'): распростра¬
нение желтого пигмента связано с этими генами, но может встречаться черномордость при
чепрачности или затемнение подпалин на морде в виде той же маски.Собаки, обладающие геном As, могут не иметь маску, но быть носителями ее наслед¬
ственного фактора.Степень распределения маски на голове варьирует и контролируется полигенами-мо-
дификаторами, но само наличие или отсутствие маски — моногенный признак, детермини¬
руемый одним главным геном q двумя аллелями.Аллель Ма — доминантный ген, детерминирующий образование маски, выделил
R. Robinson [1982]. С. Little [1957] провел гибридологический анализ — скрещивание ры¬
жей таксы (без маски) с мопсом палевого с маской окраса. Все восемь родившихся щенков
с первичных точек — зон депигментации на груди, нижней части головы, концах лап и
кончике хвоста. Напротив, самые стойкие к депигментации участки располагаются на вер¬
ху головы, особенно в районе ушей, и возле основания хвоста.Причина этого стойкого ряда прогрессии развития белых пятен — отсутствие пиг-
ментобразующих клеток в коже у корней волос. В процессе развития эмбриона эти клетки
перемещаются в направлении к основанию головы и вдоль спины. Если на этот процесс не¬
гативно воздействуют какие-либо помехи, щенок может родиться с белыми отметинами.
Хотя вся кожа и шерсть у него нормально развиты, эти отметины обесцвечены (человечес¬
ким глазом воспринимаются как имеющие белый цвет) из-за того, что их не достигли пиг-
ментобразующие клетки до рождения. Так как эмбриогенез имеет периодичность, то и сте¬
пень расположения зон депигментации может варьировать (см. рис. 3.1).Несмотря на эту язную тенденцию распространения депигментации, существует ши¬
рокая индивидуальная изменчивость темно-белых окрасов, особенно среди собак, имею¬
щих много белого, с темными асимметричными пятнышками на голове, плечах, конечнос¬
тях. С другой стороны, существуют породы, у которых почти без индивидуальной
изменчивости белые отметины стабильно расположены на передних ногах, плечах, груди и
шее („воротник" и „манишка") — это характерно, например, для бостон-терьеров, колли,
сенбернаров.Безусловно, крайние формы пятнистости контролируются разными мутантными ал¬
лелями, вызывающими названные выше нарушения эмбриогенеза. Но действуют также и
полигены-модификаторы, количественно варьирующие проявления главных „пегих" алле¬
лей каждого вида пегости, т. е. площади и конфигурацию депигментированных районов.С. Little [1957] постулировал три мутантных аллеля в локусе пегости, рецессивных по
отношению к доминантному гену дикого типа S (сплошной окраски):Обозначение окрасаСимволСтепеньдепигментацииСплошной без отметин50Ирландская пегость _s'1-3Пятнистость (пятнистая пегость)sP3-9Пятнистость с распространением
белого цветаs9-10„Ирландский" аллель s' образует меньшую треть ряда депигментации, в пределах от
небольших пятен на морде, груди, ногах и т. п. до ошейника на шее или холке. Типичные
гомозиготы s's‘ по С. Little — басенджи. Гомозиготы же sPsP в среднем имеют гораздо боль¬
ше белого в своих „рубашках", чем при ирландской пегости. Однако экспрессия аллеля sP
подвержена существенной изменчивости полигенами-модификаторами. Так, согласно
Little, бигли имеют генотип sl’sP, но большинство собак этой породы не очень сильно отли¬
чаются от басенджи по площади зон депигментации. С другой стороны, такие же гомози¬
готы sPsP — фокстерьеры — имеют значительно большую площадь зоны депигментации —
у их окраса уже фактически белый фон.Аллель i1*” обусловливает наибольшие площади зон депигментации, и гомозиготы по
нему воспринимаются как уже почти белые собаки, с одним или несколькими темными
пятнышками на голове или крупе. В крайнем случае пятен вообще может не быть у белой
гомозиготы j'V, но ее глаза останутся темными, с темными веками. В целом ряде пород
такой „белый" окрас закреплен стандартом. Уверенность в том, что данный фенотип обус¬
ловлен именно таким, а не другим генотипом, можно получить, если среди таких белых со¬
бак обнаружатся единичные особи с хотя бы небольшими пятнышками пигментированной
шерсти. Согласно этому критерию, гомозиготами jH'5,r являются темноглазые белые собаки
пород бультерьер, силихэм-терьер, самоедская лайка, пиренейская горная собака.Аллели пегости S'-локуса составляют по отношению друг к другу ряд неполного до¬
минирования. Однако с геном дикого типа S отношения неоднозначны: аллель S домини¬
рует над s', но не полностью доминирует над sP или sn' [Robinson, 1982]. Поэтому гетерози¬
готы S.sP и Si11' по площади белых участков на теле могут выглядеть как типичные собаки с
ирландской пятнистостью (s's1). В свою очередь, ирландский аллель s' доминантен к sP , но
не к j" , в связи с чем гетсрозиготы s's,r имеют пятнистый окрас. Не вполне ясен характер
отношений между аллелями sP и л"' из-за ограниченного объема экспериментальных мате¬
риалов [Robinson. 1982], но скорее всего, гетерозиготы, имеющие оба аллеля, имеют пятни¬
стость с большим преобладанием белого цвета. Вообще аллель sP отличается в локусс наи¬
более широкой изменчивостью экспрессии и доминирования.В проявлении депигментации очень активную роль играют и полигены-модификато¬
ры, способные сильно изменять действие главных генов локуса 5. Так, даже гомозиготы по
сплошному окрасу SS нередко имеют белые пятна в центрах депигментации. Поэтому даже
в породах, где белые отметины на сплошном окрасе считаются нежелательными и отбра¬
ковываются отбором, трудно добиться стопроцентной встречаемости сплошного, без еди¬
ного пятнышка окраса: даже элиминировав 5-аллели, собаковод в каждом поколении мо¬
жет встретить вновь особей с комбинацией полигенов, обусловливающей депигментацию
пусть даже и на самых маленьких по площади участках.Интересно, что еще L. Whitney [1947] и С. Little [1957] обнаружили различия в площа¬
ди белых отметин у черных и рыжих кокер-спаниелей: большинство собак черного окраса
имели отметины небольшие или средней величины, тогда как при рыжем окрасе отметины
обычно бывают гораздо большими. Одной из возможных причин этой связи называют
сцепление модификатора гена sP с геном е или же большую подверженность гомозигот ее
нарушениям в распределении пигментобразующих клеток при эмбриогенезе, чем собак с
£-геном. Позднее М. Bums и М. Fraser [1966] обнаружили, что шоколадного цвета кокер-с-
паниели обычно имеют меньшие отметины, чем рыжие кокеры, но большие, чем черные.
Это открытие свидетельствует, что ген коричневого пигмента b также является фактором,
влияющим на развитие белых отметин.Интересно, что для многих видов млекопитающих выявлен не один, а несколько
локусов пятнистости, в связи с чем даже F. Hutt [1979] предполагал и у собак возможность
наличия нескольких таких систем. Однако до сих пор все имеющиеся материалы генетичес¬
кого анализа однозначно свидетельствуют: у домашней собаки развитие депигментирован-
ных зон и формирование пегого/пятнистого окраса контролируют гены одного S-локуса.
Возможно, конечно, что в нем все еще не выявлены один или даже несколько аллелей от¬
дельных разновидностей такого рисунка „рубашки"; будущее покажет.Локус Sg (Slate-grey) представлен доминантным мутантным геном Sg, обусловливаю¬
щим грифельно-серый окрас, фенотипически похожий на голубой окрас гомозигот по ре¬
цессивному гену d разбавителя пигмента, но имеет в отличие от него доминантный харак¬
тер наследования. Выявлен у длинношерстных колли [Ford, 1969].Аллель sg — рецессивный, ген нормальной, не ослабленной пигментации.Локус Т (Ticking) — крапчатость.Аллель Т — доминантный, обусловливает развитие крапа — мелких темных пятны¬
шек на белом фоне отметин пегих или пятнистых собак (пойнтеров, сеттеров, немецких ле¬
гавых, спаниелей, борзых). Интересно, что крап появляется у щенков только через не¬
сколько недель после их рождения, и его экспрессия очень сильно варьирует, а у
длинношерстных собак даже имитирует чалый окрас.Аллель t — рецессивный — обусловливает отсутствие крапа. Фенотип крапчатого
окраса является результатом межгенного взаимодействия какого-либо мутантного аллеля
пегости S-локуса (чаще всего sP) и Г-аллсля. Последний не воздействует на гены пигмента¬
ции, поскольку цвет крапа соответствует цвету окраса шерсти собаки. Например, черно-бе¬
лые (с черным крапом) собаки имеют генотип As-B-D-E-sP-T-, а желто-крапчатые —
АУ-B-D-E-sP-T-. Черно-подпалые с крапом на пежинах собаки (генотип, по R. Robinson,
a'a’B-D-E-sP-T-) имеют черные крапины на белых отметинах на спине и ржаво-коричневый
крап на животе.Интерес вызывает специфический крап далматинов. В прошлом считалось, что он
обусловливается особым, независимым от Т геном, но С. Little [1957] доказал, что уникаль¬
ная рубашка далматских собак является модификацией обычной крапчатости. Так, наибо¬
лее обычные черно-белые далматины имеют генотип As-B-D-E-su's" T-, и селекционеры в
процессе отбора добились получения собак с белым фоном, по которому на всем теле рас¬
положены более крупные, чем обычно, и практически правильной круглой формы пят¬
нышки.Изменчивость степени крапчатости очень высока и наследственна: от легкого крапа
из нескольких пятнышек на ногах и животе (у легавых), до сильной испещрснности, как на-3 Закат Хе 706
пример, у гончих, гриффонов — в этих случаях даже формируется чалый окрас. Однако из¬
меняются не только степень испещренности, но и качественные особенности крапа, отли¬
чающегося наследственной „податливостью41 усилиям селекционеров. Так, С. Little
установил, что при скрещивании некрапчатых собак с далматинами у помесей первого же
поколения фирменный „далматинский" окрас превращался в обычную крапчатость.Локус R (Roan) выделен Н. А. Ильиным [1932], и позже в литературе по генетике со¬
баки этот символ не использовался.Аллель R —доминантный, вызывает у его обладателей чалый окрас (наличие беспиг-
ментных белых волос среди пигментированных), например, у немецких догов.Аллель г — полностью рецессивный по отношению к мутантному, обусловливает
слошной окрас согласно действию основных генов пигментации шерсти.R. Robinson [1982] считает, что чал ость у собак вызывается не действием особого
главного гена, а неполным доминированием в некоторых аллельных парах локусов окраса.
Однако, например, у домашней лошади действует ген чалой масти, обозначаемый также R.Локус W (White) определен Ильиным [1932], и кроме него никем позже не подтверж¬
ден.Аллель W — доминантный ген, обусловливающий, как обнаружил русский генетик,
доминирующую над другими окрасами белую расцветку шерсти у южнорусских овчарок и
некоторых лаек, причем по отношению к лайкам сам автор поставил знак вопроса: доми¬
нирует ли у них белый окрас на самом деле (соответственно, рецессивный аллель и- в гомо¬
зиготе дает возможность проявляться генам окрасов из других локусов).Однако позже L. Whitney [1947] экспериментально установил, что у сибирских лаек
белый окрас наследуется по рецессивному типу, и предположил, что у этих собак, как и
многих других, белая шерсть есть результат действия в гомозиготе аллеля s"' — гена экст¬
ремального распространения зон депигментации —• пежин. В мировой литературе неясно
до сих пор, точно ли южнорусские овчарки обладают доминантным белым окрасом, но
для всех остальных пород современная генетика идентифицирует белый окрас как резуль¬
тат действия не этого доминантного гена W, а рецессивных аллелей основных локусов пиг¬
ментации (j|r, cch чаще всего).Локус Q.Аллель Q — доминантный ген „струйчатой полосатости", также введен в генетику
собаки Н. А. Ильиным [1932], и позже символ не употреблялся (альтернативный рецессив¬
ный аллель нормы был обозначен q). Согласно автору, ген Q обусловливал у такс тигро¬
вый окрас (поперечные черные полосы на красном фоне) и сходен по действию с аллелем
тигрового окраса из £-локуса.Аллель С\ также по Ильину [1932], другой (помимо Q), ген тигрового окраса у такс,
блокирующий у них действие гена В черного пигмента. В современной генетике не под¬
твержден.Аллель R’ — доминантный мутантный ген (рецессивный аллель нормы — г) также
предложен Ильиным и употреблялся только им: обусловливает красный (рыжий) окрас у
такс. Современная генетика описывает генотипы окрасов такс без использования этого
символа.Локус Y (Eye colour, цвет глаз) выделил как диаллельный профессор Н. А. Ильин
[1932]. По его мнению, цвет глаз определяется видом пигмента в радужной оболочке и его
распределением там.У собак различают карие, коричневые, желтые, голубые, голубовато-белесые и чрез¬
вычайно редко (у альбиноидов) красные глаза. Известна разноглазость — наличие у одной
собаки разного цвета правого и левого глаз, а также мозаичность — мозаичное распреде¬
ление разных цветов в пределах одного глаза. Последние аномалии наиболее характерны
для немецких догов-арлекинов.Варьирование цвета глаз обнаруживает некоторую связь с пигментацией шерсти, хо¬
тя корреляция не сильная. Например, у доберманов коричневого с подпалинами окраса
глаза более свет лые, чем у собак черно-подпалых, и это даже оговорено стандартом. Од¬
нако и в пределах одного окраса встречаются собаки с темными и светлыми глазами. Пос¬
ледние ведут себя как менделирующий рецессивный признак, в связи с чем Н. А. Ильин
ввел обозначения соответствующих генов:Аллель Y — ген „дикого типа", доминантный аллель, обусловливающий коричневый
цвет глаз.Аллель у — рецессивный мутантный аллель голубого цвета глаз.
Таблица 3.1. Сравнительная таблица символов генов окраса собак,
использованных разными генетикамиRobinson (1982)Little (1957)1 Burns, Fraser
(1966)Dawson (1937)Winge(l 950). Ильин (1932)As .AsAEda—АУa)’аУАУA—Аaa*AСAа™—as—csu—а'a'a1afС‘Ыa1ВВВВвВВbbbbbСсСсFСCchCchcrс''f—сь—cbс*—с^Сс —сDDDDDDDdddddEbrebrePcbrePЕEEEEEЕeeeeeGG————L———к/М1'—————мMMVHHМаE"Em—cwa—РппP———PrррSS55Tss'sis'——ssPsPsPs1—s"'С/тsws"~w——ТT T5TWh--—W—работа по генетике окраса собак была издана Dawson в 1937 г., а наиболее известен труд
С. Little [1957]. В этих и некоторых других классических публикациях использованы разные
обозначения генов, да и концепции генетического контроля окраса неоднозначны.Поэтому R. Robinson составил сводную таблицу сравнительного списка символики
генов окраса собак разных авторов (табл. 3.1). которую мы несколько дополнили и пред¬
ставляем здесь вместе с таблицей 3.2, содержащей перечень всех известных мутаций окраса
собак.
Таблица 3.2. Гены, обусловливающие мутации окраса и текстуры шерстсобак,предложенные R. Robinson [1982]СимволгенаНаименование гена.,ДИКИЙ'ТИП“'А5Сплошной черный Solid blackАллель дикого типасА>'Доминантный желтый Dominant yellowА — (agouti) зонарноот*.asaЧепрачный Saddle patternа'Подпалый Tan patternЬКоричневый BrownВ — черныйCchШиншилла ChinchillaС — интенсивный» пигментсЬГолубоглазый альбинос Blue-eyed albinoсАльбино AlbinoСПCN-разбавитель CN dilutionDРазбавитель DiluteD — интенсивный шаягменпгЕь>Тигровый BrindleЕ — раепрострамеииыт жиагмент
по всему телуЕНераспространитель Non-extentionGПрогрессирующее поседение Progressive
silveringG — отсутствие поседениякКинки Kinky (курчавость)К — некурчавость.1Длинношерстность Long hairL — короткошерстностъМ1’Мраморность „арлекин" Harlequin merleМ — равномерный пигментММраморность MerleМаМаска MaskS*Ирландская пятнистость Irish spottingS — сплошной окрасSPПегость PiebaldswЭкстремально белый Extreme whiteSgГрифельно-серый Slate greyРКрасноглазый разбавитель Pink-eyed
dilution (рубиновоглазие)Р — нормальный зеленый отсвет
глазРРПуховый разбавитель Powder-puff
dilution („припудренность")РР — нормагрВолнистость RippleRepresentative — нормаТКрапчатость (тиковая испещрен ность)
TickingТ — отсутствие крапамаВолнистость шерсти WavedWa — нормаWhЖесткошерстность Whire hairwoЗавитки шерсти Hair WhorlsWo — нормауГолубоглазие [Ильин, 1932]Y — ген желто-коричневых глаз
Гпава 4ГЕНЕТИКА МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
СОБАКИПо сравнению с интенсивно развивающимися исследованиями наследственных ано¬
малий и болезней собак, а также неплохо изученной генетикой их окрасов и типов шерсти,
наследование большинства морфологических признаков до сих пор известно на уровне на¬
чального развития генетики или же вообще не изучено. Кроме того, многие морфологичес¬
кие особенности собак, определяющие степень их породной ценности, образуют так назы¬
ваемые количественные признаки, генетический контроль которых чаще всего очень
непрост и осуществляется в результате различных типов межгенных взаимодействий, без
проявления каких-либо основных, „главных" генов. Это, естественно, еще более усложняет
данный вопрос и затрудняет его изучение.Многие наследственные морфологические особенности собак, отличающие их обла¬
дателей от видовой или породной нормы, являются аномалиями, результатом нарушения
генетически обусловленного типа развития организма (см. гл. 5).Наследование типа телосложенияРост собак (определяемый промером высоты в холке) — типичный количественный
признак, детерминируемый полигенами с аддитивным взаимодействием. При скрещивании
контрастных по величине собак рост у гибридов наследуется промежуточно. Так же и фор¬
ма тела имеет промежуточный характер наследования (например, при скрещиваниях бор¬
зых с молоссоидами или доберманов с ротвейлерами [Ильин, 1932]. Интересно сформули¬
ровано Н. А. Ильиным понятие „индикаторного ядра". В него исследователь выделил
размеры нескольких основных (т. е., вероятно, жестко детерминированных наследственно¬
стью) морфометрических признаков, которые во взаимодействии со срсдовыми условиями
обусловливают размеры ряда других органов и статей.Длина конечностей, напротив, обусловливается, вероятнее всего, одним главным ге¬
ном с несколькими аллелями: аллель коротконогое™ доминирует над аллелем длинноно-
гости. Помеси бассета и боксера обладали короткими конечностями, а среди гибридов вто¬
рого поколения (от скрещивания гибридов Fi „в себе") происходило расщепление на
коротко- и длинноногих потомков [Christensen et al., 1985].Наследование формы и размера черепаХарактерная для собак ряда пород особенность пропорций черепа — короткомор-
дость — оказалась моногенно детерминируемым признаком, рецессивным по отношению
к нормальной, длинномордой форме головы. Ильин [1932] сообщал о проведенных Врид-
том в 1920-х годах скрещиваниях короткомордых пекинесов с длинномордыми шнауцера-
ми. У гибридов первого поколения проявлялись нормальные пропорции длинномордости.
Во втором поколении у части гибридов от скрещивания Fi „в себе" выщеплялся тип черепа
пекинеса.Ильин [1932] также показал менделирование при наследовании характерного видово¬
го признака, отличающего череп собаки от черепа волка, — „орбитального угла". Он оп¬
ределяется как угол между плоскостью лобной кости и плоскостью, проходящей через
глазные орбиты, и обусловливает основные особенности облика и конфигурации собачье¬
го черепа.По простому моногенному типу, видимо, осуществляется также генетический конт¬
роль ширины верхней челюсти (доминирует над узостью), ширины основания черепа, дли¬
ны теменной кости и скуловой дуги, а также угла скулового отростка верхней челюсти (до¬
минирует приближение к острому углу, характерному для волка, а рецессивный аллель
обусловливает „развернутый угол11, как у немецких овчарок [Ильин, 1932]). Этот исследо¬
ватель убедительно показал, что вообще широкая форма черепа у собак доминирует над
узкой, длинное темя рецессивно к короткой теменной кости, а длина лба наследуется по
промежуточному типу.Интересно, что проведенный нами генетический анализ наследования краниологи¬
ческих параметров при отдаленной гибридизации диких кабанов и домашних свиней [Кня¬
зев, Тихонов, 1983, 1985] свидетельствует, что среди особенностей строения черепа имеют¬
ся как промежуточно наследующиеся у гибридов (и таким образом, скорее всего,
детерминируемые полигенно. несколькими локусами), так и показывающие менделевское
наследование, с отношениями доминирования при аллельных взаимодействиях одного
главного локуса, который контролирует соответствующий признак.Наследование формы ухаСреди собак разных пород и беспородных существует огромная изменчивость по ве¬
личине ушной раковины, особенностям постава ушей (относительно других статей голо¬
вы — высокопоставленные или низкопоставленные — соответственно выше или ниже
уровня глаз и т. п.) и самому поставу (стоячие, висячие, полустоячие, „затянутые" на заты¬
лок, „разноухость" и т. п.).Профессор Н. А. Ильин [1932] выделил генетическую систему с множественным алле-
лизмом, детерминирующую постав ушей, со следующим порядком доминирования:
Ha>H>h, где Ни — пол- ностью доминантный аллель полустоячего уха „типа колли", Н —
аллель висячего уха, /г — рецессивный ген стоячего уха. При этом аллель Я неполностью
доминирует над h, поэтому отмечено следующее соответствие между генотипами и феноти¬
пами:НиНи = НаН = НаН — полустоячие уши „типа колли",НН — висячие уши,Hh — полувисячие уши,
hit — стоячие уши.Впрочем, эта схема не описывает все известные случаи наследственных особенностей
постава ушей, к тому же сам Ильин упомянул, что у некоторых собак существует и другой
тип висячих ушей, рецессивных по отношению к стоячим.Известны также модификации постава ушей: наследственно висячие уши у собак ря¬
да пород можно превратить в стоячие специальной хирургической обработкой — купиро¬
ванием (удалением) части „лопухообразной" ушной раковины. При этом известна наслед¬
ственно обусловленная предрасположенность к успеху или неуспеху при купировании из-за
разной степени твердости ушного хряща, она может подвергаться эффективной селекции
или, напротив, на протяжении многих поколений сводить на нет купирование ушей: после
операции у таких животных уши остаются висячими.
Г.пава 5НАСЛЕДСТВЕННЫЕ АНОМАЛИИАномалии — отклонения от нормы в развитии, строении и функционировании орга¬
нов и систем организма — у собак, как и других животных, могут быть различной приро¬
ды. Даже тс из них, которые являются врожденными, т. е. проявляются с момента рожде¬
ния щенков, не всегда имеют генетическую природу и не передаются поэтому от предков
потомкам в ряде поколений — не являются наследственными.Наследственны аномалии тогда, когда причиной их появления оказываются измене¬
ния наследственного аппарата. Установить, наследственна ли данная аномалия, на практи¬
ке можно, если, например, аномальные щенки обнаруживаются в пометах от производите¬
лей, имеющих родственное происхождение, или в разных пометах от какого-то
производителя. Но и это не может быть полным доказательством наследственной основы
аномалии: ведь обычно родственные животные содержатся в сходных условиях, подверга¬
ясь одинаковому воздействию окружающей среды, которая может вызывать проявление
ненаследственных аномалий — экзогенных, возникающих под влиянием тератогенов —
средовых факторов, нарушающих процесс развития эмбрионов и не затрагивающих гено¬
тип. Известны тератогены физической (ультрафиолетовое излучение), химической (многие
препараты), биологической природы (болезнетворные микроорганизмы), например, у сук,
переболевших в период щенности парвовирусным энтеритом, неоднократно рождались
щенки с разными уродствами.Различают также наследственно-средовые аномалии, проявление которых зависит
как от измененной наследственности, так и от действия внешней среды.Однако особое внимание как исследователей, так и собаководов привлекают анома¬
лии наследственные [Князев, 1989], возникающие как результат нарушения генетического
аппарата, причем нарушения, приводящие к уродствам, могут происходить на разных
уровнях организации жизни: на молекулярном (нарушения в структуре генов на уровне
точковых мутаций), на хромосомном (аберрации, перестройки хромосом) и геномном (из¬
менение числа хромосом — носителей основной наследственной информации).За рубежом известны обзоры [Robinson, 1982; Hedhammar, 1986а], содержащие весь¬
ма обширную информацию по наследственным аномалиям собак, хотя в ряде случаев (и их
число постоянно расширяется) особенности наследования изучены еще не окончательно. В
странах с наиболее развитым собаководством уже используются результаты генетических
исследований при разработке и осуществлении программ по искоренению или снижению
случаев проявления наследственных аномалий и болезней [Sundgren, 1986; Swenson. 1986].
Так, например, в Швеции Королевским университетом сельскохозяйственных наук
и Шведским кеннел-клубом такая программа с начала 1960-х годов предусматривала изу¬
чение наследования и разработку практических мероприятий по устранению в породах
дисплазии тазобедренного сустава, а с недавних пор аналогичная работа охватывает на¬
следственные заболевания глаз собак [Bjorck, 1986]. Подобные программы ветеринарно-ге¬
нетического совершенствования породистых собак разрабатываются и реализуются во
многих национальных кеннел-клубах и союзах любителей отдельных пород.Разумеется, для их успешного осуществления необходима достоверная научная ин¬
формация по генетике наследственных болезней и аномалий разных органов и систем.
АНОМАЛИИ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТАДисплазия тазобедренного сустава (ДТС) [Hip dysplasia (HDj]Эта аномалия широко известна как одна из наиболее распространенных наследствен¬
ных болезней собак [Brass, 1989], хотя диагностируется у большинства видов домашних
животных и человека; при отсутствии специальной селекции ее наблюдают у 60-70 % слу¬
жебных собак [Филиппов, Митин, 1990]. Поэтому во многих странах созданы специальные
селекционно-ветеринарные программы борьбы с ней. Единственным методом практичес¬
кой диагностики является радиография, хотя она и не лишена недостатков и не гарантиру¬
ет абсолютную точность. Например, при радиологическом обследовании 606 немецких ов¬
чарок в юго-западной части Польши у 188 (31 %) обнаружена дисплазия бедра [Siembieda,
1988]. A. Hedhammar [1986а] в числе пород, в которых обнаружена ДТС, указывает, кроме
немецкой овчарки, также швейцарских зенненхундов, бриара, чау-чау, кламбер-спаниеля,
ретриверов — золотистого и лабрадора, ньюфаундленда, бобтейля, ротвейлера, самоедс¬
кой лайки и сенбернара.Заболевание характеризуется нарушением соответствия между суставными поверхно¬
стями и ведет к возникновению в суставе вывиха или артроза. По-видимому, наследствен¬
но детерминируется недоразвитие сустава, что является пусковым механизмом сглажива¬
ния вертлужной впадины, недостаточного покрытия головки бедра ее верхним краем,
разболтанности и артроза сустава. Международная кинологическая федерация FCI приня¬
ла в 1974 г. в Утрехте пятибалльную классификацию градации аномалии: 1 балл соответ¬
ствует норме, 2 — слабой степени ДТС, 3 — средней, 4 — умеренной и 5 — сильной
[Robinson, 1982] — соответственно от HD- (frei) до HD 5. Подробно диагностика описана в
статьях Ю. Филиппова и В. Митина [1990]; а также у J. Madsen, Е. Svalastoga [1989]. При
этом освидетельствование собак на предмет ДТС проводят для мелких пород в возрасте не
менее 1 года, крупных — не менее полутора лет. Кроме того, характер рентгенограмм и
оценки степени дисплазии зависят от точности метода рентгенографии [Audell, 1986] и от
предварительной обработки собаки (внутривенное или пероральное введение успокаиваю¬
щих средств). При этом наименее стабильные результаты обусловливает применение для
анестезии ацетилпромазина [Lavellc, 1991]. У исследованных немецких овчарок ДТС заре¬
гистрирована в 69 % в возрасте 12 мес., в 83 % — в 18 мес. и в 95 % — в 24 мес. [Possi et al.,1988].Недавно предложена модификация рентгеноскопического метода для выявления
ДТС [Smith, Biery, 1991]: система количественных индексов повышает точность и снижает
вариабельность оценок с возможностью эффективного предсказания развития „синдрома
слабости суставов** и уровня их дегенерации в двухлетнем возрасте на материале тестиро¬
вания 16-недельных щенков, что очень важно для селекционеров и владельцев-собаково-
дов.Считается, что собаки обоих полов страдают ДТС с одинаковой частотой.Широкие исследования доказали наследственную природу ДТС, контролируемой по-
лигенно: гены нескольких локусов вызывают нарушение строения сустава, и при комбина¬
ции таких доминантных генов в генотипе их суммарное действие приводит к проявлению
аномалии [Robinson, 1982; Swenson, 1986]. Во многих популяциях и породах определены
значения коэффициента наследуемости ДТС: например, у ретриверов он составил 28 %, у
ротвейлеров 39 %, у немецких овчарок — 40 % [Lavellc, 1991]. В этих же пределах (40-50 %)
указана величина наследуемости в работе A. Hedhammar [1986а].Полигенное наследование ДТС не препятствует успешной работе по снижению уров¬
ня и тяжести аномалии — например, в породе немецкой овчарки Западной Германии при
20-летнем периоде специального контроля встречаемость аномалии снизилась, хотя сте¬
пень выраженности болезни сильно зависит от факторов внешней среды [Brass, 1989]. Од¬
нако стратегия отбора производителей с учетом освобождения популяции от этой анома¬
лии весьма специфична. Первоначальный „жесткий11 подход к отбраковке всех собак,
имеющих даже слабую степень дисплазии, и разведение только особей, абсолютно свобод¬
ных от ДТС, оказался неосуществимым: во-первых, от таких фенотипически здоровых пар
рождались с немалой частотой потомки с дисплазией разной степени тяжести, а во-вторых,
этот строгий отбор неизбежно сокращал племенную базу, обеднял генофонд, мог привести
к нежелательным эффектам генетико-автоматических процессов и т. д. Это оказывалось
Рис. 5.2. Схема наследования болезни Пертеса в родственной группе уэст-хайленд-уайт-терьеров:□ — кобель, О — сука; ■, • — больные собакиПитуитарная карликовость [Pituitary Dwarfism]Аллель dwСтрадающие этим заболеванием собаки рождаются нормальными, но в возрасте от 1
до 2 месяцев начинают отставать в росте и становятся карликами, сохраняя, впрочем, нор¬
мальные пропорции. Погибают в молодом возрасте. Для них характерно сохранение „ще-
нячьего“ шерстного покрова практически всю жизнь. Аномалия обусловлена пониженным
уровнем деятельности гипофиза. Доказана моногенная природа такой карликовости, ре¬
цессивный ген обозначен dw [Robinson, 1982]. Гомозиготы по мутантному аллелю имеют
дефицит гормона роста соматомсдина, тогда как гетерозиготы Dwdw характеризуются
уровнем этого гормона, промежуточным между гормональным статусом карликов и нор¬
мальных собак. Аномалия обнаружена у немецких овчарок в Дании и Австралии и карель¬
ской медвежьей собаки в Финляндии [Willeberg et al., 1975; Andresen, Willebcrg, 1976a, b;
Nicholas, 1978].Ахондроплазия [Achondroplasia]Эта аномалия, называемая также „непропорциональной карликовостью", является
следствием наиболее распространенной у животных дисплазии костей конечностей. Она
характеризуется укорочением дистальных костей конечностей; при этом голова и тулови¬
ще сохраняют „нормальные11 размеры. Среди собак наиболее типичными ахондропласти-
ческими животными являются бассеты и таксы. Конечно, нет оснований считать собак на¬
званных пород только по их экстерьеру отягощенными наследственной болезнью —
просто это самый наглядный пример явного укорочения длинных костей ног. В генетичес¬
кой литературе долго дискутировался вопрос — обусловлена ли карликовость бассетов и
такс проявлением одного главного гена „коротконогости“. С. Stockard [1941] вычленил по-
лудоминантный ген, обусловливающий укорочение конечностей путем скрещивания бассе¬
тов с немецкими овчарками. Однако Whitney [1947] при анализе беккроссов бассста и блад-
хаунда обнаружил картину полигенного наследования этого признака. При этом, однако,
укороченность ног „предоминировала“. R. Robinson [1982] считает, что эти данные не яв¬
ляются доказательством отсутствия главного гена карликовости. Он считает, что такой ген
корректнее всего можно выявить при анализирующем скрещивании внутри какой-либо по¬
роды, а не в межпородных кроссах. L. Whitney [1947] описал рецессивный аллель ас у ко-
кер-спаниелей, среди которых он наблюдал коротконогих собак, достоверно отличавших¬
ся от нормальных кокеров.Gardner [1959] сообщал об идентичном случае у пуделей.Однако К. Christensen et al. [1985] при скрещивании бассета с боксером показали, что
все гибриды первого поколения обладают типичными признаками непропорциональной
карликовости, а во втором поколении наблюдалось типичное менделевское расщепление.
Аутосомным доминантным признаком, результатом мутации одного гена считает ахонд-
роплазию также и Т. Fjeld [1988]: гистологической причиной аномалии он отмечает преж¬
девременное, в раннем возрасте, укорачивание колонок хондроцитов в ростовой зоне
длинных трубчатых костей. Дефект образуется при энхондральном формировании кости, а
внутримембранное формирование происходит нормально. Нарушается синтез энергии в
митохондриях клеток карликов. Отмечается, что у собак-карликов ахондроплазию часто
определяют без надлежащих исследований, поэтому многие случаи костной дисплазии
классифицируются ветеринарами и собаководами неверно.Биторакальная эктромелия [Bithoracic Ectromelia]Аллель beЩенки с отсутствием конечностей изредка рождаются в разных породах, и это почти
всегда расценивается как результат нарушения эмбриогенеза, но не как наследственная
аномалия. Однако Ladrat et al. [1969] описали такое уродство как проявление в гомозиготе
рецессивного гена be. У новорожденного щенка-сучки практически полностью отсутство¬
вали передние конечности, а задние были нормальными, и щенок достаточно успешно пе¬
редвигался ползком. R. Robinson [1982] отмечает большую смертность среди таких „эктро-
мельных11 щенков.Брахидактилия [Brachydactyly]Аллель bdR. Robinson [1982] предложил обозначить символом bd гипотетический рецессивный
ген брахидактилии, основываясь на единственном сообщении Green [1957]: этот ген, види¬
мо, вызывает редукцию размеров наружных пальцев на передних, а иногда и на задних ла¬
пах собак.Подвывих запястья [Carpal Subluxation]Аллель csДефект затрагивает нижнюю часть пясти или запястных костей, из-за чего больные
животные могут передвигаться, только опираясь на прогнутую часть передней ноги от за¬
пястья до пальцев. Аномалия у нормальных с рождения щенков начинает проявляться с
3-недельного возраста, когда они становятся на ноги и пытаются ходить, и дальше стреми¬
тельно развивается вплоть до самой сильной степени, как правило, затрагивая обе пере¬
дние конечности. Для аномальных щенков характерна гиперфлексия запястья, связанная с
расслаблением сухожилия локтевого мускула. L. Vaughan [1992] диагностировал деформи¬
рующий прогиб запястья у 21 щенка в возрасте 6-12 недель, причем большинство —
17 щенков — были доберманами. Такая породная предрасположенность привела автора к
мысли о наследственном характере болезни. Еще раньше J. Pick et al. [1967] доказали на¬
следственную природу этой аномалии, сцепленной с полом: обозначили мутантный рецес¬
сивный аллель cs в локусе, расположенном в Х-хромосоме.A. Hedhammar [1986а] отметил встречаемость этой аномалии также у бостон-терье¬
ров.
Деформация шейных позвонков [Cervical Vertebral Deformity]
или нестабильность шейных позвонков [Cervical Vertebral Instability],
или цервикальная спондилопатия [Cervical Spondylopathy],
или синдром Вобблера [The Wobbler Syndrome]Причиной аномалии является сужение спинномозгового канала позвоночника, это
вызывает повышение давления в спинном мозге, а как следствие — атаксию (шаткую по¬
ходку) или паралич задних конечностей или даже всех четырех ног. Атаксия, по мнению
R. Robinson [1982], может проявляться сразу после рождения, тогда как паралич развивает¬
ся через несколько месяцев. Точный тип наследования аномалии еще не доказан.Этот синдром, обозначаемый часто CVI, достаточно подробно изучен среди добер¬
манов специалистом из Ливерпульского университета Lewis [По: Zwan, 1988. С. 11]. Оказа¬
лось, что у собак этой породы клинические проявления аномалии сильно варьируют: либо
это атаксия в виде некоординированных движений передних или задних, или всех сразу ко¬
нечностей, или „слабость ног“, или же полный паралич как экстремальная степень дефек¬
та. Могут также проявляться боли в шее, из-за чего собаки вынуждены держать головы
низко опущенными, избегать резких движений и кормления с пола. В некоторых случаях
симптомы слабости или атаксии могут недолго проявляться у больных собак, а затем по¬
вторяться вновь. Из-за такой изменчивости симптомов затруднена и диагностика CVI, ко¬
торая невозможна без рентгенографии и исключения других возможных причин.Вероятно, сложный комплекс факторов вовлечен в развитие аномалии. Lewis полага¬
ет, что высокопротеиновый рацион кормления щенков в первые 2-6 месяцев их жизни ока¬
зывается, пожалуй, самым важным таким фактором. Быстрый рост животного в этот пери¬
од также может обусловить проявление CVI в более позднее время. Другой критический
фактор конкретно для доберманов — тип их телосложения, с высокопоставленной шеей и
так называемой высокопередостью. это вызывает концентрацию всех биомеханических сил
на основание шеи (7-й шейный позвонок) для поддержания осанки и может быть причиной
развития синдрома при нарушении развития организма. Шейные позвонки у щенка до
6 месяцев не полностью окостеневают, оставаясь хрящевидными, мягкими и податливыми
к деформации. В критический период роста шейные позвонки могут деформироваться и
вызывать искривление этого отдела позвоночника, а следовательно, и проявление призна¬
ков синдрома Вобблера из-за повышения давления на спинной мозг.Хирургическое вмешательство при слабых симптомах CVI у щенков в возрасте
5-8 месяцев может быть успешным, однако Lewis, учитывая явную наследственную обус¬
ловленность аномалии, все же настоятельно рекомендует организовать скрининг CVI в по¬
пуляциях, где этот синдром обнаружен, и разводить собак, имеющих только нормальное
строение и типичный постав шеи.Расщепленное небо и заячья губа [Cleft Palate and Hare Lip]Эти две аномалии изредка встречаются в большинстве пород, и тогда их считают
следствием тератогенеза при эмбриональном развитии. Но если они необычно часто регис¬
трируются в определенной родственной группе собак — следует ожидать генетическую ос¬
нову. Наследственность этих дефектов в двух случаях зафиксировал Hutt [1979]: в семье
бульдогов наблюдалась передача в ряде поколений расщепленного неба как моногенного
рецессивного признака, тогда как у бернских зенненхундов эта аномалия наследовалась по
доминантному типу, причем у некоторых собак дефект неба сопровождался и „заячьей11 гу¬
бой. При этом R. Robinson [1982] полагает, что эти аномалии могут контролироваться по¬
лигенами с пороговым действием: „порог11 в некоторых случаях может быть принят за мо-
ногенное наследование, а в других — обусловливать другие картины наследования. Два
названных дефекта физиологически имеют много общего; и хотя один может проявляться
независимо от другого — чаще встречаются вместе, причем первый все же с более высокой
частотой.
Карликовая анемия [Dwarf-anaemia]Аллель danЭтот тип карликовости обнаружен среди аляскинских маламутов [Robinson, 1982]:
больные собаки характеризуются укороченными конечностями с увеличенными в размерах
запястными суставами. Передние ноги искривлены (вогнуты внутрь) как при рахите. При¬
чиной размягчения костей является гемолитическая анемия во время роста. Изменяется
строение эритроцитов, а клетки белой крови остаются нормальными. Рецессивный аллель
dan обусловливает эту карликовость у гомозигот; Fletch et al. [1975] с успехом выявляли
среди фенотипически нормальных родственников анемических карликов гетерозиготных
носителей мутантного аллеля dan — с помощью гематологического анализа проб крови,
что позволяет проводить эффективную селекцию против этой аномалии.Дисплазия локтя [Elbow Dysplasia]Эта аномалия локтевого сустава наиболее часто вызывает хромоту и является нару¬
шением процессов его формирования, обычно в возрасте 4-6 месяцев. Нарушения могут
затрагивать или один, или оба локтя. Хирургическая коррекция относительно несложная и
эффективная, из-за чего названный дефект кажется не настолько широко распространен¬
ным, особенно у крупных пород. Поэтому хирургическое лечение затрудняет выявление
аномальных животных кинологами и селекцию против этого дефекта. Показано, что его
наследственная природа имеет полигенный пороговый характер [Corley et al., 1968; Hutt,
1979]. R. Robinson считает, что благодаря большей изученности наследования тазобедрен¬
ной дисплазии полезно изучение данных о ней в связи с проблемой дисплазии локтя.Обширные исследования наследственной детерминации локтевой дисплазии проведе¬
ны в Швеции на бернских зенненхундах, особенно часто страдающих этим дефектом
[Klingebom, 1986]. Проанализирована статистика хирургической коррекции локтевых сус¬
тавов в связи с их дисплазией у потомков 5 кобелей-производителей, которые сами также
были прооперированы из-за этой аномалии. Например, один кобель, инбридированный на
родоначальника с оперированным локтем, от вязок с 4 суками дал потомство с наивысшим
процентом (43,8 %) оперированных собак. Интересно, что среди его потомков от других
5 сук не было никого, кому нужна была бы коррекция сустава. Другой кобель-производи-
тель имел только 2 оперированных детей среди 68 щенков от 7 партнерш. Но когда этого
кобеля повязали с сукой, у которой был прооперирован дисплазийный локтевой сустав —
получили помет из 9 щенков, причем 6 также делали операции на локте.На популяции бернских зенненхундов Швеции проведено исследование частоты
встречаемости этого дефекта: она составила 53 % по результатам рентгенографии 105 со¬
бак. Причиной остеофитных перерождений сустава является артроз, возникающий в ре¬
зультате фрагментации и/или трещины локтя [Grondalen, 1979; 1982; Klingebom, 1986]. До¬
казано, что провоцирующим механизмом развития аномалии на фоне порогового набора
полигенов в генотипе является завышенный рацион кормления быстрорастущих собак
крупных пород, в частности немецких догов [Hedhammar et al., 1974].Локтевый подвывих [Elbow Subluxation]Аллель esРецессивный аллель es аутосомного гена [Lau, 1977], по-видимому, обусловливает на¬
следственную детерминацию этого дефекта, основной симптом которого — хромота на од¬
ну или обе передние конечности, начиная обычно с возраста 3-4 месяцев [Robinson, 1982].
При передвижении очень выворачиваются локти, так что локомоция обеспечивается в ос¬
новном только работой плечевого сочленения. Локти не развиваются полноценно в про¬
цессе роста, и в этих случаях собаки испытывают и проявляют болезненность при манипу¬
ляциях. Морфологическая причина дефекта — прекращение развития дистального
локтевого эпифиза до окончания общего роста организма.Полиостотическая фиброзная дисплазия [Polyostotic Fibrous Dysplasia]Этот дефект обнаружен у доберманов и староанглийских овчарок (бобтейлей) в виде
опухоли на передних конечностях — выше запястья. Аномалия может сопровождаться хро-
Полли Николь Мелвас Муро Торнадо Фрэда—-О ГрингоОлегияштш шшаа шш шли
-1 -1 .2 -1 -1Блюстар ВальтерОклиТорквато
—□ДШШ Ш1Ш11 -1ЦевдиО-Граф ГелносГайдэШШ АДА АААААШ-1Рис. 5.5. Генеалогические схемы, иллюстрирующие рецессивный характер наследования олигодонтии по рез¬
цам в семьях доберманов:О, □ — собаки с отсутствием одного или двух резцов (указано цифрами),©, КН — гетерозиготные носители рецессивной мутации.партнершами (очевидно, свободными от рецессивной мутации) все потомки оказывались
полнозубыми, и наоборот (т. е. при скрещивании сук-носительниц с кобелями, очевидно,
разных генотипов).Всего в пометах, где обнаружены 19 аномальных щенков, было 96 потомков — рас¬
щепление составило не 3:1, а 4:1 (нормальные : неполнозубые), что говорит о неполной пе-
нетрантности гипотетического мутантного рецессивного гена отсутствия резца. Впрочем,
очень низкое значение вычисленного критерия у} равное 1,26, свидетельствует о статисти¬
чески высокодостоверном соответствии обнаруженного расщепления теоретически ожида¬
емому при моногенном рецессивном наследовании аномалии. Мутантный ген не сцеплен с
полом: среди 19 неполнозубых щенков было 9 кобелей и 10 сук.По-видимому, наследование полиодонтии (увеличения зубной формулы) осуществля¬
ется по аналогичному олигодонтии типу, но конкретных материалов, доказывающих дей¬
ствие определенных генов, пока нет.В то же время увеличение числа резцов оказалось связанным с наследственно обус¬
ловленным типом строения черепа определенных пород [Willis, 1990]: наличие лишнего
резца характерно для догообразных собак (молоссоидов) с укороченной и широкой мор¬
дой (табл. 5.4).Таблица 5.4. Встречаемость полиодонтии (7 резцов) у собак
разных пород [По: Willis, 1990]ПородаЧисло изученных собак-Частота встречаемости
собак с 7 резцами, % .Бульдог7139,4Боксер14026,0Бульмастиф368,0Мопс1014,0Пекинес741,3Мастиф230,0
В кинологической практике отношение к аномалиям зубной системы неоднозначно.
В отечественном собаководстве оно в большинстве случаев традиционно негативное: все
отклонения от ножницеобразного прикуса определяются как дисквалифицирующие. За ис¬
ключением некоторых пород с уникальным смыканием челюстей, дисквалифицируются от¬
клонения от ножницеобразного прикуса и стандартами Международной кинологической
федерации (FCI), и — в менее широких масштабах — Английского и Американского клу¬
бов собаководства. Несоответствие зубной формуле в сторону уменьшения числа зубов яв¬
ляется критерием для выбраковки при племенной работе с важнейшими служебными поро¬
дами (немецкая овчарка, доберман, ряд других) в соответствии со многими
национальными и международными их стандартами. В отношении большинства других
пород собак требования селекции против олигодонтии не так строги или даже отбор в
этом направлении не проводится, когда приоритет отдается общему уровню породности
племенных собак или степени развития наиболее характерных для породы селекционируе¬
мых признаков. В кругах, например, английских кинологов известен афоризм: если собака
хороша, ей можно простить отсутствие какого-нибудь зуба, а если плоха — то тем более.В целом генетика аномалий зубной системы собаки заслуживает гораздо более глубо¬
кого изучения.НЕРВНО-МЫШЕЧНЫЕ (НЕЙРОМУСКУЛЬНЫЕ) НАРУШЕНИЯ
[Neuromuscular maladies]В эту группу аномалий относят дефекты поведения собак и функционирования их
мускулатуры, обусловленные разного рода дегенерацией нервных центров — спинного или
головного мозга; расстройства мышечной деятельности являются вторичными, так как
движения контролируются нервной системой. Любой дефект ее сразу проявляется наруше¬
нием поведения или движений.Аморотическая идиотия [Amaurotic Idiocy]Аллель ga-2.Патология проявляется начиная с 6-месячного возраста щенков в виде усиления не¬
рвозности поведения и отсутствия способностей к дрессировке и обучению. К 9-12 меся¬
цам появляются симптомы нарушения координации движений и атаксии, слепоты и глухо¬
ты. Больные собаки обычно не живут дольше двух лет [Robinson, 1982].Дефект впервые обнаружен среди немецких курцхааров [Karbe, Schiefer, 1967]; анализ
родословных показал рецессивный характер наследования аномалии и, поскольку все
больные собаки оказались кобельками, вначале полагали, что соответствующая мутация
локализована в Х-хромосоме. Однако ныне этот ген считают аутосомным, а дефекты пове¬
дения обусловлены нарушением синтеза и неактивностью в головном мозге фермента
вследствие GM2 ганглиозидоза, из-за чего мутантный аллель и получил обозначение ga-2.Атаксия [Ataxia]Аллель atВпервые это заболевание обнаружено и изучено специалистами Королевского вете¬
ринарного колледжа в Стокгольме при поддержке и финансировании исследований Швед¬
ским кеннел-клубом начиная с 1952 г. [Bjork et al., 1957] в некоторых шведских линиях
гладкошерстного фокстерьера; правда, авторы ссылаются на литературные сведения о
врожденной атаксии у французских бульдогов, бостон-терьеров, скотч-терьеров, эрдельте¬
рьеров, но характер наследования выяснен только для фокстерьеров. Аномалия проявляет¬
ся в нарушении локомоции и вообще мышечной деятельности начиная с 4-месячного воз¬
раста, когда щенки вдруг начинают необычно лаять и выть, причем не могут правильно
ориентироваться относительно объекта облаивания; затем нарушается и затрудняется по¬
ходка и т. п. Выявлен рецессивный аллель at, обусловливающий эту аномалию [Bjork et al.,
1962]. По рецессивному типу наследуется врожденная атаксия также у крупного рогатого
скота, лошадей, кроликов, домовых мышей и куриц (у кур, правда, аномалия сцеплена с
полом); кроме того, установлено влияние среды на частоту встречаемости атаксии: наи¬
большей она оказалась в пометах щенков, рожденных в сезон с марта по июль (от 15 до
20 %), а наименьшей — с сентября по январь (от 0 до 5 %). Генеалогический анализ рас¬
пространения атаксии среди фокстерьеров в Швеции показал, что аномалия передавалась
потомству от ограниченного числа производителей, импортированных из Англии [Bjork et
al„ 1957].Несколько позднее, в 1962 г., атаксия, ассоциированная с парезами и параличами,
была обнаружена у аборигенной старинной голландской породы Kooiker, почти исчезнув¬
шей во время второй мировой войны, после которой порода была восстановлена на основе
оставшихся 15-25 собак. За период с 1942 по 1992 гг зарегистрировано 4927 животных этой
породы, из которых атаксия отмечена у 46 щенков. Подтверждено аутосомно-рецессивное
наследование аномалии [Mandigers et al., 1993], причем все больные животные родились в
результате весьма близкого инбридинга (коэффициент инбридинга в среднем равен 0,23).
Считается, что эта болезнь может рассматриваться как один из видов лейкодистрофии.Причиной заболевания являются изменения дегенеративного характера в централь¬
ной нервной системе [Robinson, 1982].Мозжечковая атаксия [Cerebellar Ataxia]Аллель сЪНаследственный характер этой атаксии выявлен среди керри-блю-терьеров: выделен
соответствующий рецессивный аутосомный аллель cb [Lahunta, Averill, 1976]. Физиологи¬
ческой причиной болезни является дегенерация коры мозжечка с разрушением клеток Пур-
кинье. Признаки проявляются у щенков в возрасте 9-16 недель: развиваются сильный тре¬
мор (дрожь) головы, несгибаемость задних ног. Затем ухудшается способность к
передвижению больных щенков, которые часто падают и не могут стоять без посторонней
помощи из-за очень сильного раскачивания головой.Лейкодистрофия Бьеркаса [Bjerkas’ Leukodystrophy]Аллель IdАномалия названа по имени исследователя, впервые изучившего ее среди далматинов
и выявившего действие рецессивного аллеля Id [Bjerkas, 1977].Дефект проявляется схожим с атаксией образом: нарушается функция вначале задних
конечностей, затем и передних. Развиваются симптомы паралича, резко ухудшается зре¬
ние. Ранние симптомы появляются в возрасте от 3 до 6 месяцев по причине дегенерации
клеток головного, спинного мозга и глазного нерва.Дегенерация мозжечка [Cerebellar Degeneration]Болезнь проявляется обычно в возрасте 4-8 недель, когда у аномальных щенков на¬
чинается нарушение координации движений задних ног. С ростом затруднение движений
усиливается, у некоторых больных собак развивается тремор головы. Аутопсия показала
прогрессирующую дегенерацию структуры мозжечка с отсутствием гранул и клеток Пур-
кинье. Хотя явный наследственный характер показан при открытии этой болезни у колли
[Hartley et al., 1978], тип наследования неясен.Эпилепсия [Epilepsy]Общепризнано, что большинство случаев проявления эпилепсии у собак имеют на¬
следственную природу [Robinson, 1982], но проблема выяснения характера наследования
этой аномалии до сих пор не решена [Hedhammar, 1986а]. Давние предположения о рецес¬
сивном типе передачи эпилепсии не подтвердились — и нет доказательств, что она обус¬
ловлена каким-либо одним простым типом наследования [Falco et al., 1974; Wallace, 1975].Отмечена относительно высокая встречаемость эпилепсии в породах ретриверов (зо¬
лотистого и лабрадора), у пуделей и кеесхонда — для последнего, как оказалось, вообще
характерна предрасположенность к этому дефекту [Robinson, 1982]. Отмечается более вы¬
сокая вероятность заболевания кобелей, чем сук.Скорее всего, наследование аномалии имеет полигенный пороговый характер, при¬
чем в качестве медиаторов проявление болезни регулируют и такие факторы, как „генотип
породы1*, половая принадлежность, возраст и влияние окружающей среды.
Лейкодистрофия глобоидных клеток [Globoid-cell Leucodystrophy]Аллель gcРецессивный ген, обусловливающий это нарушение нервной системы, выявлен у ряда
пород собак R. Hirth и S. Nielsen [1967]. Начальные симптомы заболевания в виде отсут¬
ствия координации движений появляются у аномальных щенков в возрасте нескольких не¬
дель от роду: они спотыкаются, широко расставляют задние ноги для сохранения равнове¬
сия. Нередко развиваются коллапс или тремор.Биопсия мозга больных собак выявляет множество глобоидных клеток в белом моз¬
говом веществе, которое интенсивно разрушается в ходе развития аномалии.Дефицит мышечных волокон [Muscle Fibre Deficiency]Аллель mfСтрадающие этой аномалией молодые собаки, начиная с 3-5 месяцев отличаются не¬
способностью нормально держать голову, опуская ее ниже нормального постава, это сви¬
детельство общей слабости мускулатуры всего тела, особенно у кобелей. Усугубляют де¬
фектность поведения физические нагрузки, тренинг больных собак, содержание их на
холоде, а также нервозная обстановка. При продолжительных нагрузках происходит рез¬
кое выпрямление конечностей во всех сочленениях суставов, укорочение амплитуды дви¬
жений до крошечных шагов; обе задние конечности собака перемещает одновременно, так
что может двигаться вприпрыжку. При этом передние конечности часто поражены коллап¬
сом, вызывающим падения больного животного. Правда, через несколько минут отдыха
собака вновь оказывается способной к передвижению, пока снова быстро не устанет. Эти
дефекты задерживают рост и развитие собаки, вызывая дефицит мышечной массы больно¬
го животного — за счет недостатка в мускулах волокон второго типа.Установлено, что эта аномалия наследуется как рецессивный признак под контролем
аллеля mf [Kramer et al., 1976].Миелопатический паралич [Myelopathic Paralysis]Аллель mpЭтот рецессивный ген, выявленный у афганов [Averill, Bronson, 1977], у гомозигот в
возрасте 3-8 месяцев обусловливает паралич, начинающийся слабостью задних конечнос¬
тей и затрагивающий затем также передние ноги. В итоге собака оказывается полностью
парализованной. Причиной является некроз миелиновой оболочки первичных нервных во¬
локон грудного отдела спинного мозга.Миотония [Myotonya]Дефект обусловливается чрезмерным сокращением мускулов. В возрасте около 3 ме¬
сяцев поражаются мышцы конечностей. Передние ноги при этом передвигаются совершен¬
но выпрямленными, и собака на ходу раскачивается из стороны в сторону. После несколь¬
ких таких шагов-прыжков происходит релаксация, расслабление мышц, и движения
становятся нормальными. Но спазмы могут повторяться. Дефект описан у чау-чау, причем
вначале полагали, что он передается по наследству как рецессивный признак, однако затем
оказалось, 'что не все имеющиеся данные указывают на такой простой тип наследования
этой аномалии [Wentinck et al., 1974].Нейрональная абиотрофия [Neuronal Abiotrophy]Аллель паЭтот рецессивный ген, обусловливающий в гомозиготном состоянии данную анома¬
лию, открыт при изучении дефекта у собак шведской лапландской породы [Кгоок, 1973;
Sandefeldt et al., 1973]. Аномалия нарушает функции мышц конечностей — передних или
задних, начиная с возраста 5-7 недель. В течение недели после возникновения первого сим¬
птома все конечности теряют подвижность, и собака оказывается не в состоянии даже
встать на ноги, лежа на груди с согнутыми или вытянутыми конечностями (рис. 5.6.
[Sandefeldt. Nilsson, 1986]). Мышцы ног при этом быстро атрофируются, но мышцы туло-
АНОМАЛИИ КРОВИ И СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫВрожденный лимфатический отек (водянка) [Congenital Lymphoedema]Аллель LyЭтот доминантный ген обусловливает у его носителей [Patterson et al., 1967] опухание
конечностей, которое, впрочем, оказывается не очень болезненным. Водянка может затра¬
гивать только задние ноги, либо все четыре конечности или развиваться до максимального
уровня, когда поражаются даже корпус и хвост больного животного. Аномалия заметна
уже при рождении и сопровождается высокой смертностью собак с сильным развитием де¬
фекта [Robinson, 1982]. Накопление жидкости в опухолях вызывается нарушением системы
лимфатических протоков и узлов.Циклическая нейтропения [Cyclic Neutropenia]Аллель спБолезнь известна также как синдром голубых колли, а рецессивный ген ранее называ¬
ли еще CN-разбавителем, так как он влияет и на осветление окраса [Robinson, 1982]. Ал¬
лель сп считается потенциально летальным: он вызывает периодическое изменение количе¬
ства нейтрофилов в крови с интервалами в 10-12 дней. Функции нейтрофилов связаны с
обеспечением защиты организма от патогенных бактерий, и в период снижения количества
этих защитных клеток в крови резко снижается устойчивость собаки к инфекциям.Клиническими симптомами нейтропении являются лихорадка, потеря аппетита, вос¬
паление желез и суставов, приводящее к хромоте, и развитие различных инфекционных за¬
болеваний. Диагноз может быть поставлен при анализе окраса шерсти и оормулы белой
крови (числа разных лейкоцитов). Щенки, отягощенные этой аномалией, обычно умирают
в первые же недели жизни — ни один не дожил до 6 месяцев. Правда, антибиотикотерапия
в периоды снижения числа нейтрофилов может несколько продлевать жизнь аномальных
животных, которые отстают от нормальных в размерах и чаще всего бесплодны [Ford,
1969; Lund et al., 1970].Гемолитическая анемия [Haemolytic Anaemia]Аллель ркЭта анемия, впервые обнаруженная у собак басенджи, отличается непрерывностью и
прогрессированием. Первые признаки проявляются в возрасте около года, а смерть насту¬
пает через год или два. Еще до экспрессии болезни анемичные собаки отличаются пони¬
женной активностью и противодействием тренингу. Характерны быстрая утомляемость и
осветление слизистых оболочек. Однако точный диагноз можно поставить только при ана¬
лизе эритроцитов крови, благодаря которому удается определить болезнь даже ранее 8-не¬
дельного возраста [Robinson, 1982].Дальнейшие исследования установили, что при анемии в эритроцитах значительно
снижена активность фермента пируваткиназы (РК). Количественный уровень и активность
РК регулируются геном рк. Анализ активности РК в крови фенотипически нормальных,
неанемичных собак показал, что по этому показателю такие животные разделяются на2 группы: с высоким и более низким уровнями. Соответственно собаки оказались гомо-
(РкРк) и гетерозиготами (Ркрк) по доминантному гену. Правда, между этими группами
нет резкой границы, поэтому у некоторых собак нельзя определить генотип по феноти¬
пу — активности РК. Однако достоверные различия между генотипами обнаружены по
содержанию в эритроцитах аденозинтрифосфата и пробой на фосфоэнолпируват (после¬
дний метод разделяет взрослых собак, но не щенков). Эти методы позволяют более эффек¬
тивно выявлять носителей мутации для селекции здоровых собак [Brown, Teng, 1975;
Andresen, 1977].У кэрн-терьера обнаружено, что недостаточность пируваткиназы вызывает гемоли¬
тическую анемию со вторичным гемохроматозом [Schaer et al., 1992], сопровождающимся
накоплением в различных тканях железосодержащих пигментов, циррозом печени, сахар¬
ным диабетом, дисбалансом натрия и калия в сыворотке крови, нарушением пигментации
кожи и слизистых оболочек. Первичной основой этого синдрома также признана мутация
гена пируваткиназы.
Также выявлена наследственная обусловленность дефицита пируваткиназы у собак
африканской аборигенной породы басенджи [Whitney et al., 1991]: проанализирована ак¬
тивность различных типов этого фермента у нормальных басенджи и аномальных, пора¬
женных аутосомно-рецессивным наследственным синдромом дефицита пируваткиназы
(РК). Новейшими методами иммуноблоттинга, электрофореза и ион-обменной хроматог¬
рафии установлено, что в ретикулоцитах нормальных собак выявляется матричная РНК
только R-типа пируваткиназы, а у больных басенджи — типов R и М. Сделан вывод о на¬
личии у аномальных собак мутации L-гена РК, так как именно с этого гена происходит
транскрипция R-РК- и М-РК- матричной РНК. Произвели успешное клонирование в лабо¬
ратории Университета Теннесси ДНК L-гена, ее амплификацию с помощью ПЦР (полиме¬
разной цепной реакции) и анализ для установления и картирования в дальнейшем точко-
вой мутации, вызывающей синдром дефицита пируваткиназы у собак.Гемофилия [Haemophilia], Свертываемость крови — одно из важнейших и жизненно необходимых свойств
организма, включает как минимум 13 этапов („факторов"), которые должны произойти
вплоть до полной остановки кровотечения. Нарушение какого-либо из этих этапов приво¬
дит к гемофилии — средней или сильной. На самом же деле процесс свертывания еще более
сложен [Jones, 1986].Впрочем, собаководов больше интересуют не столько физиологические тонкости
свертываемости, сколько диагностика и наследуемость разных форм гемофилии. Этим ас¬
пектам посвящены работы D. Hall [1972], W. Dodds [1974], N. Spurling [1980].Степень нарушения и генетическая природа свертывания крови могут быть различн-
ными. Однако лабораторные исследования позволяют дифференцировать наследственно и
физиологически разные болезни. Наиболее важные из них приведены в таблице 5.5.Таблица 5.5. Важнейшие виды гемофилии собак [По: Robinson, 1982]СимволгенаНазвание болезниСимптомыПримечанияDptДиспротромбинемияТяжелыеПредположительно
доминантное наследованиеheГ ипопроконвертинемия
(дефицит фактора VII)СлабыеРецессивный тип. Гетерозиго¬
тывозможно идентифицироватьНет-aГемофилия АОт среднихСцеплена с полом.(дефицит фактора VIII)до тяжелыхУ сук-гетерозигот снижена
активность фактора VIIIHcm-bГемофилия В
(дефицит фактора IX)ТяжелыеСцеплена с полом.У сук-гетерозигот активность
фактора IX снижена на 50 %HfГ ипофибриногенемия
(фактор I)ТяжелыеВероятно, доминантнаяPtaДефицит предшественникаСредниеНеполностью доминантноеStfтромбопластина плазмы
(дефицит фактора XI)Дефицит факторау гетерозигот
и тяжелые
у гомозиготнаследованиеСтюарта-Прауэра
(фактор X)»ThТромбопластинемияОт средних
до тяжелыхДоминантная. Гетерозиготы
могут быть выявленыVwdБолезнь фон ВиллебрандаОт средних
до тяжелыхДоминантная или
неполностью доминантная
Гемофилия А — наиболее характерный дефект механизма свертывания крови у со¬
бак, обнаруженный в большинстве самых распространенных пород, что свидетельствует о
высокой мутабильности гена Нет-a. Гемофилии А и В являются сцепленными с полом: оба
соответствующих локуса расположены в Х-хромосоме, поэтому большинство страдающих
этими болезнями собак — кобели, рожденные гетерозиготными матерями-носительницами
мутантных аллелей. Каждый кобель, сын такой носительницы, имеет 50 %-й риск унасле¬
довать Х-хромосому с мутантным аллелем, и в среднем половина ее сыновей неизбежно бу¬
дет поражена гемофилией, а половина дочерей окажутся носительницами гена аномалии.Собаки с тяжелыми симптомами этих форм гемофилии выявляются по гемотомасу
(подкожные опухоли, наполненные кровью, периодическая хромота, обусловленная крово¬
течением в суставах, кровотечения изо рта, носа, ануса, а также, конечно, обильные крово¬
течения в результате травм и повреждений кожи). Средней тяжести дефекты иногда могут
восприниматься как явления стресса и т. п., поэтому при подозрении на гемофилию необ¬
ходимо срочно провести диагностические анализы. Генеалогическая схема родословной
семейства шетландских овчарок из Швеции, в котором у некоторых собак диагностирова¬
ны случаи гемофилии А, приведена r работе В. Jones [1986] (рис. 5.10): типичное рецессив¬
ное наследование дефекта, ген которого локализован в Х-хромосоме. В этом исследовании
показана возможность успешного выявления случаев гемофилии А с помощью специфи¬
ческого определения коагуляции фактора F VII в шведских популяциях вельш-спрингер-
спаниеля, бордер-колли, немецкой овчарки, австралийского келпи, шелти. Встречаемость
нарушений свертываемости крови в этих популяциях была успешно сокращена путем се¬
лекции — устранения из разведения носительниц мутантных аллелей. В США при изуче¬
нии суки с гемофилией A [Murtaugh, Dodds, 1988] обсуждали возможный характер насле¬
дования этой аномалии в данном редчайшем случае: гомозиготность по рецессивному
гену, локализованному в Х-хромосоме (в этом случае один из мутантных генов должен был
передать отец-гемофилик), мутация других генов, обеспечивающих образование фактора
VIII, или мозаицизм.В Венгрии описан первый обнаруженный там случай наследственной гемофилии А у
пятилетнего кобеля породы немецкая овчарка [Horvath, Elodi, 1991]. Первичным призна¬
ком болезни обнаружена гематома на бедре, а затем клинический анализ показал дефицитАф с»446-46Ш6?-466t1 I4ii66£*666 66Рис. 5.10. Схема наследования гемофилии А у собак [Jones, 1986]:□, О — здоровые собаки, ■ — больные кобели, ф — гетерозиготные суки — носительницы гемофилии
фактора VII (уровень его активности составил только 7 % по сравнению с нормой). Боль¬
ной кобель оказался потомком интенсивно использовавшегося в породе западногерманс¬
кого производителя по кличке Канто фон Винерау, который ранее уже был определен как
носитель, распространявший гемофилию А. Больной овчарке дважды переливали по 15 мл
крови нормального донора, наступило некоторое улучшение состояния, но в третий раз
животное погибло от острого тромбоза — по-видимому, из-за иммунологической несовме¬
стимости.В. Jones [1986] рекомендовал ограничение племенного использования не только боль¬
ных особей, но и носителей гена Vwd болезни фон Виллебранда (бфВ) у скотч-терьеров. В
этой породе диагностика успешно проводилась методами коагуляции F VIII, коагуляции
комплекса антиген-F VIII, родственного антигена F VIII (восьмого фактора свертываемос¬
ти). Кстати, наследование бфВ согласно W. Dodds [1984] осуществляется по аутоеомно-ре-
цессивному типу, а не доминантному, как указано в таблице 5.5. Но в пользу наследования
этой болезни (с оговоркой — у большинства пород) по тииу неполного доминирования
склоняется J. Littlewood [1989] — автор краткого обзора по наследованию нарушений свер¬
тываемости крови у собак и кошек. БфВ менее тяжелый недуг, чем гемофилия А и В,
и характеризуется дефектом слипания тромбоцитов при кровотечениях. По-видимому, у
разных пород этот дефект действительно имеет разную наследственную природу. Так, у
10 собак породы шетландская овчарка (шелти) в США было обнаружено очень резкое
снижение в крови (< 0,1% от нормы) фактора фон Виллебранда, явившееся причиной труд-
ноостанавливаемых кровотечений [Raymond et al., 1990]. Авторы предположили, что такой
резкий дефицит обусловлен либо гомозиготностью по мутантному аллелю (в случае моно-
генного контроля аномалии), либо двойной гетерозиготностью по мутантным аллелям
двух или нескольких генов фактора фон Виллебранда. Выявленный у шелти вариант гене¬
тически детерминированного заболевания является третьим типом синдрома дефицита на¬
званного фактора (аутосомно-рецессивная форма): все больные собаки были потомками
двух животных, вероятно, гетерозиготных носителей мутации. При исследовании назван¬
ным коллективом ученых более 6000 шелти выявлено 1428 гетерозигот, у которых клини¬
чески проявлялся только первый тип болезни (мягкая форма, практически без болезненных
отклонений). В связи с такой высокой частотой распространения в этой породе вредной
мутации весьма целесообразно проведение тестирования производителей шетландских ов¬
чарок для получения здоровых потомков.По некоторым литературным сведениям [Zwan, 1989], примерно 70 % всех тестиро¬
ванных в США доберманов являются носителями бфВ (зарегистрированной у собак 54 по¬
род). Выделяют 3 типа бфВ по степени ее тяжести: если у щенка вообще отсутствует
восьмой фактор, животное нежизнеспособно, так как антиген фон Виллебранда (фак¬
тор VIII) высоко коррелирует с продолжительностью кровотечения. Среди доберманов бо¬
лезни в такой степени тяжести (тип 3) не обнаружено — все носители классифицированы
по самому слабому 1-му типу, с низким содержанием фактора VIII. Доберманы-носители
дефекта внешне нормальные и здоровые животные, но не устойчивы к действию различ¬
ных стресс-факторов, которые не могут переносить, в отличие от нормальных собак. Даже
инфекционные болезни носители переносят несравненно тяжелее (например, парвовирус-
ный энтерит). Правда, собаки с бфВ имеют пониженную вероятность развития сердечно¬
сосудистых заболеваний, особенно связанных с образованием тромбов в сосудах. Простой
тест выявления носителей бфВ — проба на продолжительность кровотечения при уколе
пальца на лапе: нормальным считается время кровотечения 4-5 минут, если же кровотече¬
ние более продолжительное (до 30 минут и дольше) — диагностируют бфВ. В США этот
тест внедрен во всех ветеринарных клиниках, особенно при подготовке собак к хирурги¬
ческим операциям (включая косметические — по купированию ушей у доберманов, напри¬
мер). БфВ выявлена и у некоторых доберманов в Европе — например, в Голландии.Тромбастения изучена впервые у выдровых гончих (оттер-хаундов), но позднее как
тромбопатия описана у бассет-хаундов [Johnstone, Lotz, 1979]. В обоих случаях дефект
свертывания наследуется как признак, детерминируемый доминантным геном, по которо¬
му гомо- и гетерозиготы могут быть дифференцированы благодаря лабораторным анали¬
зам крови. Но клинические признаки анемии, возникающие как следствие этого дефекта,
проявляются только у гомозигот по доминантному аллелю, из-за чего анемия наследуется
по „рецессивному1' типу [Robinson, 1982].
Аномалия Пелджета-Хьюта [Pelget-Huet Anomaly]Аллель PgВ норме ядра нейтрофилов, одного из классов клеток белой крови, имеют сегменто¬
образную форму, а у собак с этой аномалией нейтрофильные ядра круглые, овальные или
бобовидные. Сам по себе этот феномен вроде бы безвредный, но не исключено, что из-за
него может снижаться резистентность животного к болезням [Robinson, 1982]: уменьшается
жизнеспособность щенков и долголетие взрослых собак. Дефект обусловлен действием до¬
минантного аллеля Pg, и перечисленные симптомы характерны для гетерозигот Pgpg — го¬
мозиготы по доминантному аллелю среди собак не выявлены. Известно, что гомозигот-
ность по такому доминантному гену у крыс вызывает летальный исход незадолго до
рождения или сразу после него; но гомология у собак пока не доказана [Bowles et al., 1979].Гранулоцитопатия [Granulocytopati (швед.)]При этой болезни (дисфункции нейтрофилов) клиника проявляется в повышенной
восприимчивости к инфекциям. При изучении десяти больных щенков ирландского сетте¬
ра отмечены такие симптомы, как лихорадка, гингивит, патологические изменения кос¬
тей — нижнечелюстных, плечевых, лучевых (их деструкция и пролиферация); происходи¬
ло уменьшение фагоцитоза и способности к поглощению чужеродных элементов
лейкоцитами [Wigh, 1990]. Установлена генетическая детерминированность синдрома, на¬
следующегося по аутосомному рецессивному типу. Аномалия приводит к летальному исхо¬
ду: лечение рецессивных гомозигот антибиотиками и кортикостероидными препаратами
способствует лишь временному улучшению состояния больных щенков.Болезнь накопления гликогена типа VII
[Glycogen Storage Disease Type VII]Причиной этой аномалии, впервые обнаруженной в 1985 г. у английских кокер-спа¬
ниелей, является наследственный дефицит фосфофруктокиназы (PFK). При этом наруша¬
ется нормальная структура фермента, состоящего из 3 субъединиц L, М и С [Mhaskar et al.,
1992], в норме соотношение между ними в эритроцитах собак составляет 2:86:12, а у боль¬
ных — 31:8:61, причем у аномальных животных L- и С-субъединицы сохраняли обычную
форму и размер, а М-субъединицы были сильно укороченными — как считают, в результа¬
те рецессивной мутации. Исследователи отметили, что этот наследственный синдром у со¬
бак более сильной степени тяжести, чем у человека: из-за большего исходного поврежде¬
ния мутацией М-субъединицы PFK. У новорожденных аномальных щенков активность
фермента была высокой, но затем снижалась почти в 100 раз из-за чрезмерно сильного
процесса потери L-субъединицы белка (при росте нормальных собак также происходит по¬
теря L-субъединицы, но не так интенсивно). При этом типе болезни встречаются и другие
онтогенетические изменения состава PFK, приводящие к снижению жизнеспособности
больных собак.Врожденные пороки сердца [Congenital Heart Defects]Врожденные пороки сердца у собак являются одними из наиболее распространенных
неинфекционных болезней: в США, например, частота обращения в ветеринарные клини¬
ки в связи с ними составляет около 1 % от всех посещений [Patterson, 1989]. Названный ав¬
тор с сотрудниками в Университете Пенсильвании провел обширные исследования поро¬
ков сердца и известен компетентными обзорами на эту тему [Patterson, 1974; 1976].Чаще пороки сердца присущи чистопородным собакам, а не помесям или беспород¬
ным, которые наиболее устойчивы к этим болезням. В упомянутых публикациях указана
частота проявления разных пороков сердца среди животных определенных пород и приве¬
дены доказательства полигенной основы наследования всех этих аномалий.С. Kvart [1986] подробно обсуждает проблему распространения врожденных пороков
сердца в породах собак и считает недопустимым племенное использование производите¬
лей, от которых зарегистрированы потомки с такими дефектами. Кроме того, обращается
внимание на необходимость точной диагностики, так как нередки не врожденные, а вто¬
ричные дефекты сердечных мышц, возникающие в раннем возрасте (даже раньше 4 недель
после рождения) в результате инфекционных повреждений, например, после парвовирус-
ного энтерита.Предсердно-желудочковый створчатый эндокардит — весьма распространенное забо¬
левание среди собак. Наследственную его обусловленность доказывает сильная предраспо¬
ложенность к этому заболеванию собак породы кавалер-кинг-чарлз-спаниель в Великобри¬
тании, Швеции и Дании [Pedersen et al., 1992]. У этих спаниелей к тому же порок
развивается в более раннем возрасте, чем у других пород.Отмечены породные аспекты встречаемости субаортального стеноза, который может
быть диагностирован на основании результатов эхокардиографии [Koch et al., 1993], также
выявлена генетическая обусловленность этого заболевания.Кроме того, у бордер-колли в Австралии обнаружен артериоз открытого протока:
диагноз подтвержден некропсией и обследованием грудной клетки у 5 сук, входящих в се¬
мейство из 36 собак, происходящих от 3 сук и 2 кобелей-родоначальников, что позволяет
полагать наследственную передачу этого заболевания [Malik et al., 1991].В Германии в течение 16 лет у 17 собак были диагностирована врожденная аномалия
дуги аорты (сохранившийся артериальный проток с левым или правым шунтом) [Gerlach et
al., 1988], причем из этих 17 больных собак 10 были немецкими овчарками, что также ука¬
зывает на возможную наследственную природу врожденной аномалии.Описаны врожденные почечно-печеночные артерио-венозные (задней полой вены)
аномалии [Olson et al., 1991], но генетическая детерминация не выяснена.Врожденная повышенная активность
фермента Na, К-аденозинтрифосфатазаАномалия наследуется, по-видимому, по рецессивному типу. Пероксидирование ли¬
пидов в мембранах (оболочках) эритроцитов при этом дефекте существенно выше, чем у
нормальных собак. В целом мембраны красных клеток крови аномальных животных менее
устойчивы к окислительному воздействию бета-ацетилфенилгидразина [Ogawa et al., 1992]:
у собак, с наследственно обусловленной повышенной активностью названного фермента
происходят окислительные повреждения мембран эритроцитов, из-за чего ухудшается их
функционирование и развиваются симптомы анемии. Генетический контроль аномалии
изучен недостаточно.АНОМАЛИИ КОЖИШерсть, покрывающая кожу собак, как и почти всех млекопитающих, является по¬
рождением и неотъемлемой частью кожных покровов, и ее плотность, жесткость, длина,
интенсивность пигментации (окраса) свидетельствуют о силе и жизненности процессов
функционирования организма в онтогенезе. Многочисленные и разнообразные породы
различаются и по строению кожного (шерстного) покрова: разная длина, форма, жесткость
шерсти являются результатом мутаций и в принципе также могут рассматриваться как на¬
следственные аномалии, хотя в большинстве случаев таковыми не считаются, принадлежа
к числу важных, а нередко и главных селекционируемых породных признаков, строго рег¬
ламентируемых стандартами. Поэтому собственно аномалиями кожи и шерсти обычно
считают резкие отклонения в строении этих покровов от довольно широкого для домаш¬
ней собаки спектра „нормальной'1 изменчивости, приводящие к явному снижению жизне¬
способности и здоровья.Один из новейших обзоров по основным генетически детерминированным врожден¬
ным аномалиям кожи у собак [White, 1993] содержит клинические описания, диагностику,
лечение и данные по распространению этих дефектов в разных породах. Перечислены сле¬
дующие генетические аномалии: дерматомиозит (воспаление кожи и мышц под ней), кон-
турированный кальциноз, муциноз (микседема, болезнь шар-пеев из Китая), синдром
Элерса-Данлоса, типичный для многих видов млекопитающих и человека, а также узелко¬
вый дермофиброз, фурункулез бультерьеров, цинкзависимые дерматозы, синдром „шнау-
цер-Комедо“, воспаление сальных желез, дисплазия фолликулов черных волос, ихтиозовые
синдромы и некоторые другие. Ниже приводятся сведения по аномалиям, которые наибо¬
лее часто встречаются у собак и лучше изучены генетически.
Дисплазия фолликулов черных волос [Black Hair Follicle Dysplasia]Аллель bhОчень редкая аномалия частичного облысения шерсти на участках черного окраса.
Черно-пегие или пятнистые с белым фоном и черными пятнами (или крапом) собаки при
этом дефекте имеют нормальную шерсть без пигмента — белого цвета, а окрашенные учас¬
тки — с тусклой и редкой шерстью [Robinson, 1982], причем дефектными оказываются не
все черные участки шерсти: нормальными остаются темноокрашенные места на голове и
шее, а также пограничные с районами депигментации. Кожа на полысевших черноокра-
шенных участках темная и временами шелушащаяся. Полностью белые родичи таких де¬
фектных собак всегда оказываются с нормальной шерстью.Генетическая детерминация аномалии изучена при исследовании семьи собак, роди¬
тели которых оба были с дисплазией фолликулов: из 15 потомков 12 также страдали этой
аномалией, а остальные 3 — здоровые — оказались сплошного белого окраса. Таким обра¬
зом, аномалия проявляется у гомозигот по выявленному аллелю bh [Selmanowitz et al., 1977]
только в отсутствии гомозиготности по неаллельному гену белого окраса — по-видимому,
как следствие межлокусных взаимодействий. Отмечено, что биопсия кожных покровов
аномального потомка и предков показала, что у родителей отклонения от нормы в слое¬
нии кожи были незначительными [Post et al., 1988].Дисплазия соединительной ткани (Синдром Элерса-Данлоса)
[Connective Tissue Dysplasia (Ehlers Danlos Syndrome)]Аллель CdЭта аномалия приводит к нарушению формирования соединительных узлов и пуч¬
ков, служащих основой дермальных слоев кожи. Это приводит к ненормальной непрочнос¬
ти кожи и периферических кровеносных сосудов. Кожа становится неплотной, складчатой,
„вельветовой11, [Robinson, 1982] и легко разрывающейся, из-за чего образуются глубокие
раны. И у собак эти симптомы называют иногда синдромом Элерса-Данлоса
(Ehlers-Danlos) по аналогии (гомологии?) с такой аномалией человека.Нарушения вызывает доминантный ген Cd, и перечисленные симптомы характерны
для гетерозигот [Robinson, 1982], а гомозиготы CdCd или имеют такой же аномальный фе¬
нотип, или, чаще, погибают из-за летального действия доминантной мутации в двойной
дозе. Немногочисленные данные о скрещивании между собой редких гомозигот CdCd под¬
твердили этот феномен [Hegreberg et al., 1969].Дермоидный синус [Dermoid Sinus]Аллель dsАномалия в виде кожных синусов встречается у родезийских риджбеков впереди (на
шее) или сзади их знаменитого „гребня“ шерсти на спине, растущей в обратную сторону
(„сзади наперед11) по сравнению с остальной нормальной шерстью. Синусы представляют
собой узкие „трубки“-лакуны от кожи на спине вглубь позвоночника как результат непол¬
ного отделения кожи от позвоночника во время эмбриогенеза — маленькие отверстия, ок¬
руженные кожными валиками. У одной аномальной особи может быть один или несколько
таких синусов, которые сами по себе незлокачественные до проникновения через них
внутрь организма какой-либо инфекции [Robinson, 1982].Этот кожный синус — явно наследственная аномалия, но характер генетического
контроля долго был неясен — R. Robinson упоминал, что в литературе сообщалось о воз¬
можном доминантном, или рецессивном, или даже полигенном наследовании. Однако
скрупулезные исследования позволили установить рецессивный мутантный аллель ds
[Mann, Stratton, 1966], вызывающий в гомозиготе названный дефект кожи у родезийских
риджбеков.Бесшерстность [Hairless]Аллель HrR. Robinson считает бесшерстность весьма нередкой мутацией собак, распространен¬
ной по всему миру. Но наиболее известна она как важнейший породный признак китайс¬
кой хохлатой и мексиканской голой собачек. Бесшерстность не обязательно бывает сплош¬
ной — остатки шерстного покрова иногда сохраняются и у мутантных собак на голове,
конечностях и хвосте. Е. Letard [1930] установил, что бесшерстность обусловлена действи¬
ем доминантного гена Hr в гетерозиготном состоянии, а гомозиготы НгНг умирают при
рождении из-за нарушений в развитии головы и мозга. Гетерозиготность голых собачек
обусловливает расщепление в их потомстве по главному породному признаку. Разумеется,
это обстоятельство не значит, что и во всех других проявлениях мутации бесшерстности
гомозиготность обязательно летальна и вообще обусловлена именно этим доминантным
геном. Иногда аномалия наследуется и как рецессивный признак: так, из 14 потомков нор¬
мальных родителей 3 оказались без шерсти [Zulueta, 1949] на теле, но на морде и лапах
шерстный покров остался. Позднее описан случай рождения 2 частично бесшерстных щен¬
ков от собак с нормальной шерстью [Conroy et al., 1975]: у этих щенков при рождении
шерсть также была обычной, но в возрасте 5 недель началось прогрессирующее облысение.
Вероятно, в каждом из этих случаев наблюдали действие рецессивных генов, отличающих¬
ся от „главного" гена бесшерстности Яг.У потомков 3 бесшерстных (Hrhr) и одной нормальной (гомозиготы hrhr) сук мекси¬
канской бесшерстной породы определили содержание важнейших гормонов в крови в пе¬
риод роста от рождения до возраста 30 недель [Iwamura et al., 1992]. У щенков бесшерстных
собачек от рождения до месячного возраста оказалась выше нормального уровня концент¬
рация гидрокортизона, а начиная с двух месяцев у них проявлялись атрофия тимуса и не¬
достаточность в строении и работе иммунной системы.Пиодермия немецких овчарок [German Shepherd Dog Pyoderma]В Голландии изучен генетический контроль пиодермии немецких овчарок [Wisselink
et al., 1989] — хронической болезни кожи, развивающейся в среднем возрасте у гомозигот
по рецессивному мутантному гену, обусловливающему образование гнойников на фоне за¬
ражения больных собак коагулазапозитивными стафилококками. Кстати, именно для не¬
мецких овчарок установлено ненормально низкое по сравнению с видом в целом содержа¬
ние в крови иммуноглобулина IgA, что обсуждалось ранее как возможная причина частой
поражаемости у этой породы тонкого кишечника от разных инфекций [Whitbread et al.,
1984]. Возможно, что уникальная пиодермия, отмеченная именно у немецких овчарок, свя¬
зана с нарушениями в иммунной системе страдающих этой аномалией собак [Wisselink et
al., 1988]. Проведен анализ генеалогии 42 больных овчарок и их прямых и боковых род¬
ственников: оказалось, что в родственной группе, где обнаружены собаки с пиодермией,
встречаемость аномалии выше, чем в среднем во всей популяции; 37 из больных 42 собак
имели 3 общих предков. Частота встречаемости и тяжесть проявления пиодермии не зави¬
сят от сезона года и условий содержания собак.Это позволило сделать вывод об аутосомном рецессивном наследовании пиодермии
у немецких овчарок (хотя ранее [Muller et al., 1983] отмечалось наследование пиодермии у
собак как многофакторного признака) и о необходимости исключать из племенного разве¬
дения больных собак, а также, по возможности, их близких родственников.Цветная мутантная алопеция [Colour Mutant Alopecia]Эта аномалия вначале была обнаружена у доберманов голубого окраса, поэтому пер¬
воначально названа синдромом голубых доберманов (Blue Dobermann Syndrome [Zwan,1989]), но впоследствии зафиксирована у других „цветных11 доберманов (коричневых) и у
собак некоторых других пород (ирландские сеттеры, таксы и уиппеты голубого окраса),
поэтому названа более обобщенно.Характеризуется частичной алопецией (выпадением шерсти), сухой тусклой шер¬
стью, перхотью.Сведения о типе наследования очень скудны. Согласно исследованиям профессора
Брасса из Ветеринарной школы Ганновера [По: Zwan, 1989. С. 11], этот дефект имеет гене¬
тическую основу, будучи сцепленным с рецессивным геном d (локус £>-Dilution) — разбав¬
ления черного пигмента, который в гомозиготе dd обусловливает голубой окрас из-за рас¬
положения глыбок пигмента в глубинных слоях волоса. Большинство собак с мутантным
дефектом при рождении имеют шерсть нормального строения. Однако позже начинается
выпадение волос, и шерсть становится как бы „побитой молью“, а кожа начинает шелу¬
шиться. Волосы при этом отличаются ломкостью и сухостью, гораздо большими, чем у
нормальных однопометников. В большинстве случаев образуются папулы и прыщи. Через
несколько лет у больных собак почти вся шерсть на туловище исчезает, и места, подвер¬
женные алопеции, покрыты шелушащейся кожей. Голова, конечности и хвост при этом с
почти нормальным шерстным покровом. Болезнь неизлечима, хотя состояние кожи может
быть улучшено правильным кормлением и обработкой (тироксином, витаминами, шампу¬
нем).Этими обстоятельствами объясняется то, что в последнее время Международная ки¬
нологическая федерация по инициативе Международного доберман-клуба признала голу¬
бой окрас доберманов нежелательным и исключила его из числа стандартных.АНОМАЛИИ ОРГАНОВ ЧУВСТВАфакия [Aphakia]Эта аномалия — врожденное отсутствие в глазе хрусталика — обнаружена в семей¬
стве сенбернаров. Характер расщепления признака в потомстве нормальных родителей
свидетельствовал о моногенном рецессивном наследовании дефекта [Martin, Leipold, 1974].Катаракта [Cataract]Заболевание глаз в виде помутнения и потери прозрачности хрусталика, различимое
внешне как бельмо на зрачке. Различают катаракту первичную — если она не связана с ка¬
кими-либо другими дефектами глаз, и вторичную — если сопровождается таковыми и яв¬
ляется их следствием. Для генетиков поэтому важнее изучение первичной катаракты как
наследственной аномалии.Катаракты бывают разных видов, причем их природу можно установить только при
специальном обследовании больных животных. Обнаружены различия в возрасте проявле¬
ния аномалии при разных катарактах и у разных пород. Сводная таблица, приведенная R.
Robinson [1982], содержит совокупность сведений об этой аномалии (табл. 5.6). Оказывает¬
ся, что помутнение хрусталика вызывают многие разные гены — чуть не каждый вид ката¬
ракты является новой самостоятельной мутацией [Rubin, Flowers, 1972; Barnett, 1976; 1978;
Yakely, 1978].Кроме названных пород первичная врожденная катаракта обнаружена также у кава-
лер-кинг-чарлз-спаниелей, вельш-сприн-гер-спаниелей, больших мюнстерлендеров — в
большинстве случаев наследуется как рецессивный признак, и у доберманов, шпицев, уэст-
хайленд-уайт-терьеров, где точный тип наследования еще не установлен.Интересно, что у немецких овчарок вначале было убедительно доказано доминиро¬
вание мутантного гена катаракты над нормой [Hippel, 1939] (рис. 5.11). Однако позднее
[Barnett, 1986] в английской популяции немецких овчарок обнаружена наследственная ка¬
таракта, передающаяся как рецессивный признак (рис. 5.12): оказалось, что и в пределах
одной породы могут быть распространены несколько форм этой аномалии с различным
типом наследования из-за доминантных или рецессивных мутаций в разных локусах
[Barnett, 1986].Рис. 5.11. Доминантное наследование катаракты
у немецких овчарок в Германии [Hippel, 1939]Рис. 5.12. Рецессивное наследование катаракты у
немецких овчарок в Англии [Barnett, 1986]
Таблица 5.6. Основные виды катаракты у собак и их наследование [По: Robinson, 1982]СимволгенаПородыВозрастпроявленияПрирода катарактыНаследованиеАфган1-3 годаПрогрессирующая. Глаза
могут поражаться по-раз-
ному; дефект в передней
и задней камерахВероятно,рецессивное—Американскийкокер-спаниель6 месяцевРайон наружного ядра.
Обычно прогрессирующаяВероятно,доминантное2-4 годаП р огрессирующая.В переднем или заднем
кортикальных слояхВероятно,рецессивноеctБостон-терьер8 недельСубкапсулярная. Прогрес¬
сирующая до полного
развития к 2-4 годамРецессивноеКагНемецкая овчаркаРазныйвозрастЛамеллярная.ПрогрессирующаяДоминантноеРетриверы: золо¬
тистый, лабрадор16-12 месяцевПолярная или перифери¬
ческая. Прогрессирующая в
зрелом возрасте. Глаза мо¬
гут поражаться по-разномуВероятно,доминантноесаМиниатюрныйшнауцерС рожденияЗадний субкапсулярный
слойРецессивноеБобтейл1-2 годаОт оболочки до всей толщи
хрусталика. Связана
с нарушениями сетчаткиВероятно,рецессивноеcatСтаффордширскийбультерьерДо 1 годаСубкапсулярная. Прогрес¬
сирующая к трем годамРецессивное—Пудель большойДо 2 летСплошная.П рогрессиру ющаяВероятно,рецессивноеАномалия глаз колли [Collie Eye Anomaly (СЕА)]Аллель сеаТакой дефект строения и функционирования глаз характерен для некоторых шотлан¬
дских и шетландских овчарок — колли и шелти, и неизвестен у собак других пород. Дефект
для двух названных пород серьезен из-за высокой частоты его распространения — во мно¬
гих популяциях частота превышает 80 и даже 90 % и остается высокой даже после проведе¬
ния специальной селекции против этой аномалии: например, через 3 года интенсивной се¬
лекции частота аномалии снизилась с 97 % только до 59 % [Yakely, 1972].СЕА представляет собой результат грубого нарушения формирования глазного дна
больных собак: ненормальное строение имеют сетчатка, другие оболочки глаза, кровенос¬
ные сосуды, оптический диск, склера. В итоге минимум 8 отдельных аномалий разных час¬
тей глаза суммируются в виде синдрома СЕА [Robinson, 1982]. При этом степень тяжести
заболевания варьирует, и у разных аномальных собак проявляются различные совокупнос¬
ти клинических симптомов, которые может идентифицировать только специалист-офталь-
молог с помощью офтальмоскопа. К полной слепоте приводят только самые тяжелые ва¬
рианты СЕА.Специальный гибридологический анализ убедительно показал, что аномалия насле¬
дуется по рецессивному типу [Yakely et al., 1968] под контролем одного гена с мутантным
аллелем, обозначенным сеа. Этот ген обладает также плейотропным эффектом, обусловли¬
вая нарушения разных структур глаза.Однако F. Hutt [1979] считал, что наследственная природа СЕА иная — полиген-
ная — из-за высокой распространенности дефекта в этих двух породах и широкого варьи-
Это иллюстрирует схема, показывающая число регистраций собак с аномалией PRA в каж¬
дый год (штрихованные столбцы, шкала слева) в сравнении с общим количеством лабрадо¬
ров, зарегистрированных Клубом собаководства (верхняя линия, правая шкала); год регис¬
трации обычно является годом рождения собаки (рис. 5.15). Видно, что селекция привела к
полному исчезновению аномалии из селекционируемой популяции. В Англии также сейчас
считают полезным обязательное офтальмоскопическое обследование молодых собак при
регистрации в Клубе собаководства и выдаче племенного сертификата для участия в разве¬
дении [Bedford, 1989].Прогрессирующая ретинальная атрофия (центральная)
[Progressive Retinal Atrophy (Central) CPRA]Это вторая форма PRA, имеющая отличия от предыдущей и потому выделяющаяся в
отдельную аномалию, обозначаемую CPRA. При этом заболевании собаки лучше видят в
сумерках, чем при ярком освещении, однако зрение неострое, и больное животное в движе¬
нии часто наталкивается на неподвижные предметы и объекты. При офтальмоскопии мож¬
но обнаружить зоны с коричневым пигментом в слое сетчатки, ответственном за восприя¬
тие света глазом, эти точковые зоны не наблюдаются при периферической форме PRA, и
дегенерация сетчатки затрагивает центральную ее часть, из-за чего аномалия и носит свое
название. Развитие дефекта приводит к полной слепоте, а начинается в разном возрасте, от2 до 10 лет, но чаще в 3-6 лет. Офтальмоскопия дает возможность диагностировать анома¬
лию значительно раньше, между 18-24 месяцами от роду. А в быту, поскольку у больных
собак долго сохраняется периферическое зрение, этот вид атрофии проявляется гораздо
позже, иногда даже после 10-летнего возраста, когда владельцы обнаруживают полную
слепоту у собаки, воспринимая ее как старческое одряхление.Распространение CPRA в породах совсем не такое, как PRA: чаще всего эта анома¬
лия встречается у служебных, „рабочих11 (по английской классификации) собак, с наивыс¬
шей частотой отмечена среди колли и ретриверов. CPRA не так широко распространена у
собак, как PRA [Robinson, 1982].Тип наследования не так точно определен, как в случае с PRA, но считается, что цен¬
тральная прогрессирующая атрофия сетчатки детерминируется доминантным геном с не¬
полной пенетрантностью [Barnett, 1969] и с изменяющейся экспрессивностью.Ретинальная дисплазия [Retinal Dysplasia]Аллель rdЭто врожденная аномалия, но первые проявления заметны после прозревания щен¬
ков, в возрасте 3-6 недель. Аномальные щенки ведут себя, как слепые: менее подвижны,
чем обычно, и натыкаются на встречные предметы. Аномалия выявлена у лабрадоров и те¬
рьеров — бедлингтона, силихэм и йоркшир [Hedhammar, 1986а] и обусловлена рецессив¬
ным геном в гомозиготном состоянии [Robinson, 1982]. К. Barnett [1976] и F. Stades [1978]
путем генеалогического анализа установили в этих породах конкретных производителей,
передававших эту аномалию и оказавшихся, вероятно, обладателями спонтанно смутиро-
вавших генов.Мраморность [Merle]Аллель ММ — доминантный ген мраморного окраса, обусловливающий специфическую ру¬
башку волосяного покрова: темные пятна неправильной формы с „рваными11 краями на бо¬
лее светлом фоне, а также голубого цвета глаза — один или оба, или даже отдельный глаз
может быть частично голубого цвета, а частично обыкновенным или даже белым (если пят¬
на располагаются на белом фоне). Отсутствует пигментированный слой сетчатки, обуслов¬
ливающий восприятие глазом света в темноте. Все это характерно для гетерозигот Мт, а го¬
мозиготы ММ имеют более тяжелые проявления аномалии [Robinson, 1982]. При этом
генотипе окрас практически полностью белый, глаза только голубые, часто уменьшенного
размера (микрофтальмия) или даже иногда отсутствуют (анофтальмия). Аномальным оказы¬
вается и строение глаз. Большинство таких животных также оказываются глухими.
Глухота [Deafness]Частичная или полная глухота у собак весьма редка, но регулярно встречается, на¬
пример, у далматинов [Anderson et al., 1968], однако и у них тип наследования точно не вы¬
яснен. Если имеется моногенный контроль, то мутантный ген, видимо, обладает неполной
пенетрантностью и не сильно отличается по действию от рецессивного аллеля [Robinson,
1982]. При незначительной экспрессивности и пенетрантности мутации ее обладателей
можно и не заметить при отборе, и тогда они могут произвести на свет потомство подчас с
более тяжелыми формами дефекта.Существует и другая гипотеза наследования — полигенного, с пороговым эффектом.Причиной глухоты являются коллапс (разрушение) внутреннего уха (улитки) и деге¬
нерация кортиева органа — важнейших органов для восприятия звуков и сохранения рав¬
новесия.В Великобритании в 10 пометах доберманов выявили 21 щенка с нарушением способ¬
ности сохранять это равновесие (симптомы — плохая ориентация в пространстве, „потря¬
хивание" головой, движение по кругу — известны у некоторых доберманов в Англии и
•США). Оказалось [Wilkes, Palmer, 1992], что во всех случаях этот вестибулярный синдром
был связан с практически полной глухотой. При вскрытии погибших больных собак выяв¬
лена прогрессирующая дегенерация нейроэпителиальных клеток. Однако полностью меха¬
низм проявления этого синдрома еще неясен, так как в вестибулярном аппарате (в полу¬
кружных канальцах) не обнаружено дегенерации сенсорных клеток, в подавляющем
большинстве отолиты имелись и были нормальными. Исследователи провели генеалоги¬
ческий анализ этого врожденного синдрома в семьях доберманов, обнаружив, что он на¬
следуется по аутосомно-рецессивному типу.Цероидный липофусциноз собак [Canine Ceroid Lipofuscinosis (CLF)]Это наследственное заболевание, о типе наследования которого у собак нет еще точ¬
ных доказательств. Одно из проявлений аномалии — нарушения сетчатки глаза и потеря в
связи с этим зрения. Болезнь известна для многих пород: кокер-спаниелей, сеттеров, бал¬
канской овчарки и других. У тибетского терьера подробно исследованы сопровождающие
CLF изменения мозга (наличие аномальных дифференциально окрашенных гранул в клет¬
ках Пуркинье мозжечка и др.) и сетчатки глаза (аномальное накопление аутофлюоресцент¬
ных плотных включений, дегенерация фоторецепторных клеток) [Riis et al., 1992]. Оказа¬
лось, что у больных тибетских терьеров происходит снижение способностей к обучению и
дрессировке. У собак этой породы, больных CLF, происходит очень медленное прогресси¬
рующее поражение мозга, и функциональные отклонения в поведении проявляются лишь в
возрасте 4-6 лет, в связи с чем тибетские терьеры оказались хорошей биологической моде¬
лью для изучения CLF и других подобных болезней (например, болезни Кафа) у собак и
человека.Стойкая гиперпластическая сосудистая оболочка хрусталика/
Стойкое гиперпластическое первичное стекловидное тело
[Persistent Hyperplastic Tunica Vasculosa Lentis/
Persistent Hyperplastic Primary Vitreous (PHTVL/PHPV)]Это наследственное заболевание глаз открыл, назвал и изучил F.C. Stades [1980; 1983]
у собак породы доберман в Голландии, в связи с чем известно еще одно название анома¬
лии: „голландская болезнь14 [Zwan, 1989]. Дефект обнаружен только у доберманов: в Гол¬
ландии у 959 собак, в Швеции исследовано 6 больных доберманов [Narfstrom, 1986], но за¬
регистрирован и в других европейских странах: в Дании [Ambjerg, 1982], Бельгии,
Норвегии, Австрии, Франции, Италии, а также в США [Zwan, 1988].Причиной аномалии является нарушение эмбриогенеза глаз. В норме у собак на 20-й
день развития эмбриона происходит обособление хрусталика, и в пространстве стекловид¬
ного тела между хрусталиком и сетчаткой образуются клетки первичного стекловидного
тела (PV). От оболочки глаза начинает расти гиалоидная артерия через стекловидное тело
к хрусталику, разделяясь возле него на сеть сосудов, покрывающих хрусталик. Эта сеть и
называется Tunica Vasculosa Lentis (TVL) — сосудистая оболочка, питающая хрусталик в
период роста эмбриона до рождения щенка. На 45-й день начинается процесс отмирания
этой гиалоидной системы, и в норме на 4-6-ю недели после рождения она полностью исче¬
зает, а зрение щенка начинает развиваться. Доберманы с клиническими симптомами
PHTVL/PHPV имеют небольшие фиброваскулярные пигментные пятна на поверхности
хрусталика как остатки его неполностью исчезнувшей сосудистой оболочки. Их можно на¬
блюдать при офтальмоскопии с помощью щелевой лампы и микроскопа, эти пигментные
пятна не обязательно являются оптической преградой, но служат причиной возникновения
вторичной катаракты. Доктор F.C. Stades разработал систему классификации по степени
тяжести поражения глаз при этой аномалии, которая проявляется как результат наслед¬
ственно обусловленной дисгармонии в эмбриогенезе хрусталика и кровеносно-сосудистой
системы, вызывающей блокирование механизма регрессии сосудов перед рождением щен¬
ков.В связи с этим предложено не использовать в разведении доберманов со степенью
развития синдрома в 2 балла и выше, Доберман-клуб Нидерландов принял соответствую¬
щее положение, в результате чего в настоящее время в этой стране число собак с тяжелыми
формами заболевания снизилось и составляет от 2 до 5 % [Zwan, 1989].Разработаны методы хирургической коррекции дефекта, дающие успешные результа¬
ты, но точный тип наследования пока не выявлен.ДРУГИЕ АНОМАЛИИ
Аутоиммунная болезнь [Autoimmune Disease]R. Robinson отмечает, что распространенность этого заболевания в генеральной по¬
пуляции собаки как вида, вероятно, невелика, и в большинстве случаев истинная природа
этих дефектов остается нераспознанной. При этой аномалии механизм защиты организма
от инфекций „запускается'* против себя самого, и некоторые органы инфильтруются и под¬
вергаются атаке лимфоцитов. В результате ткани перерождаются, нарушаются их функции
и наступает хроническое воспаление.В большом питомнике биглей наблюдали две формы этой аномалии — лимфоцитар¬
ные тироидизм и орхит, при которых оказались поражены соответственно щитовидная же¬
леза и семенники (у кобелей). Названные аномалии встречались только в нескольких генеа¬
логических линиях, восходивших к общим родоначальникам, что свидетельствует о
генетической основе аутоиммунной болезни [Fritz et al., 1976]. F. Hutt [1979] считал, что эта
аномалия иммунитета имеет полигенный наследственный контроль.Собака как модель изучения патологии иммунной системы вызывает большой инте¬
рес современной науки [Lapras, Fournel, 1992].Крипторхизм [Cryptorchidism]Крипторхизм, односторонний или двусторонний (полный) — дефект развития кобе¬
лей, при котором соответственно один или оба семенника остаются в брюшной полости, не
выходя через паховый канал в мошонку. Обе формы в кинологии рассматриваются как
дисквалифицирующий порок при отборе собак на выставках и на племя. При полном
крипторхизме аномальные кобели стерильны, так как при высокой температуре внутри
брюшной полости сперматогенез блокируется, а ткани семенников перерождаются. В слу¬
чае одностороннего крипторхизма плодовитость обычно сохраняется.Аномалия относится к числу признаков, ограниченных полом (проявляется только у
самцов). Встречается у свиней, лошадей, других млекопитающих, но чаще всего у собак.
Более высокая встречаемость этого дефекта обнаружена среди пород собак с укороченным
черепом (и крипторхизм, и укорочение черепа являются следствием нарушения гормональ¬
ного баланса): из 168 кобелей породы боксер, родившихся в 57 пометах, 23 % оказались
крипторхами [Хатт, 1969]. Полагали, что причиной такого сильного распространения
крипторхизма в этой популяции боксеров был инбридинг, усиливший концентрацию гипо¬
тетических генов этой аномалии.Наследственная природа крипторхизма не вызывает сомнений, но точный тип его на¬
следования у собак до сих пор не ясен. Ранее выдвигались гипотезы простого рецессивного
наследования [Bums, Fraser, 1966], но накопленные затем сведения не позволяют утверж¬
дать это однозначно.R. Robinson считает, что, поскольку имеются две формы — односторонний и полный
крипторхизм — значит, мы имеем дело с пороговым наследованием. При этом действуют
несколько полигенов в разных локусах, и ни один из них не может вызвать проявление
этой аномалии самостоятельно, но только во взаимодействии с подобными полигенами
при их определенной комбинации в генотипе, эти полигены распространены в породах с
низкой частотой, но могут накапливаться в некоторых линиях, семействах или у отдельных
особей. При этом встречаемость крипторхов резко возрастает.Стерильность двусторонних крипторхов, видимо, есть проявление естественного от¬
бора против этой аномалии и причина относительно низкого ее распространения. Однако
использование в племенном разведении близких родственников крипторхов, в том числе
сук, у которых генотип вообще не проявляется в фенотипе, может вызывать появление
крипторхов среди потомков следующих поколений.В связи с проблемой наследования крипторхизма представляют интерес эксперимен¬
тальные данные, полученные в Университете Айова (США) профессором М. Rothschild с
соавторами [1988] при изучении генетики свиней. Исследователи скрещивали самцов-крип-
торхов с самками, братья которых также были крипторхами. В 16 пометах получено
54 самца, из которых 8 оказались крипторхами. В ходе эксперимента показана несостоя¬
тельность гипотезы о наследовании крипторхизма под контролем рецессивного гена одно¬
го локуса, и доказано, что аномалию вызывают рецессивные аллели как минимум 2 локу-
сов.Очевидно, что для точного определения наследования крипторхизма у собак также
необходимо проведение гибридологического анализа.Диафрагмальная грыжа [Diaphragmatic Hernia]Эта врожденная аномалия заключается в эмбриональном нарушении формирования
диафрагмы — мембраны, отделяющей брюшную полость от грудной клетки. В связи с
этим некоторые внутренние органы брюшной полости располагаются в грудной клетке. В
двух пометах от одной пары фенотипически нормальных родителей получено потомство с
расщеплением 3:1 в отношении нормальные:дефектные, что свидетельствует о моногенном
рецессивном наследовании этой болезни [Feldman et al., 1968].Языко-глоточный дефект [Glossopharyngeal Defect]Аллель btРецессивный ген bt открыт F. Hutt [Hutt, Lahunta, 1971] и обусловливает сужение
языка, особенно по направлению к кончику, с краями, которые могут заворачиваться
внутрь. Эта аномалия обычно известна под названием „птичий язык" (bird tongue — отсю¬
да и символ гена bt). Родившиеся с таким дефектом рецессивные гомозиготы не могут ус¬
пешно сосать молоко из материнских сосков и погибают вскоре после рождения от исто¬
щения.Обильное выделение мочевой кислоты [High Uric Acid Excretion]Аллель uaЭта ненормальная для вида домашней собаки особенность обмена обнаружена толь¬
ко в породе далматинов, где является вполне типичным породным признаком [Trimble,
Keeler, 1938] и не связана с какими-либо иными полезными или вредными эффектами. Дал¬
матины не страдают больше других собак камнями в почках и мочевом пузыре, хотя со¬
став этих камней у них отличается [Robinson, 1982]. Поэтому у далматинов это не является
аномалией, хотя оно совершенно ненормально для других собак.Обычно у собак, как и у всех хищников, распад пуринов (одной из групп органичес¬
ких веществ, участвующих в метаболизме) приводит к образованию мочевой кислоты,
большинство которой превращается в аллантоин под действием фермента уриказы, и ал-
лантоин выводится наружу с мочой, в которой почти не содержится мочевой кислоты. У
человека совсем не так: мочевая кислота является последним звеном распада пуринов и вы¬
водится с мочой.Далматины уникальны не только своим окрасом, но и тем, что у них, как у человека,
тоже выделяется с мочой не аллантоин, а мочевая кислота. При этом фермент уриказа, рас¬
щепляющий ее до аллантоина, у них имеется, как и у других собак, но мочеточники под
действием мутантного аллеля в гомозиготе иаиа утратили способность резорбировать мо¬
чевую кислоту, свойственную другим породам.Установлено, что ген иа наследуется независимо от гена экстремально белого окра¬
са p'v и гена крапчатости Т, обусловливающих уникальный окрас далматинов.Загадкой остается происхождение феноменально высокого уровня выделения моче¬
вой кислоты в единственной породе. По-видимому, причиной может быть дрейф генов —
генетико-автоматические процессы на ранних этапах породообразования, из-за случайно¬
го наличия мутации у нескольких родственных собак, послуживших родоначальниками
всех далматинов.Атрофия поджелудочной железы [Pancreatic Atrophy]Аллель рсДоказан моногенный контроль этого дефекта и выявлен рецессивный мутантный ген
[Westermarck, 1980].Аномалия проявляется в пониженном содержании ферментов поджелудочной желе¬
зы — гидролаз — у собак в возрасте 1-3 лет. Больные собаки, гомозиготы рсрс, истоща¬
ются, несмотря на большой аппетит, хотя пытаются поедать возможно больше корма и за¬
нимаются даже копрофагией. Экскременты таких животных объемные, покрыты слизью, с
гнилостным запахом.Мутация не летальна, больные собаки остаются живыми, хотя выглядят очень изну¬
ренными.Наиболее часто аномалию диагностируют у собак 2 пород: немецкая овчарка и длин¬
ношерстная колли [Westermarck et al., 1989]. Среди колли в Финляндии встречаемость ат¬
рофии железы составляет около одного процента от численности популяции. Генеалоги¬
ческий анализ происхождения 51 колли с синдромом этой аномалии показал, что
44 животных имели родственные связи между собой. Например, одна из больных собак
имела в своей родословной 17 больных родственников из 11 разных пометов. Подтвержден
аутосомно-рецессивный тип наследования аномалии, хотя в пометах, где отмечено рожде¬
ние от нормальных родителей аномальных потомков, доля больных однопометников с ат¬
рофией поджелудочной железы оказалась в 2 раза ниже ожидаемой по менделевским зако¬
нам (0,13 вместо 0,25). Вероятно, ген рс имеет неполную пенетрантность, составляющую
около 50 %.Финские ученые отметили также возрастание частоты встречаемости аномалии среди
колли в их стране: за первые 7 лет наблюдений отмечено только 10 случаев аномалии, а за
последующие 4 года — 41. Всего диагноз атрофии поджелудочной железы авторы постави¬
ли за 10 лет 271 собаке, из них 203 были немецкие овчарки, 51 — длинношерстные колли,
15 собак относились еще к 11 разным породам и 2 были беспородными.Почечная дисфункция [Renal Dysfunction]Накопленные данные о нарушениях функций почек свидетельствуют о наследствен¬
ной их природе, хотя нет определенных сведений о типе наследования [Robinson, 1982]. Ти¬
пичные симптомы: сильная жажда (полидипсия), обильный диурез (полиурия), рвота, диа¬
рея, потеря веса больной собакой [Wilcock, Patterson, 1979]. Заболевание называют обычно
почечной кортикальной дисплазией, но R. Robinson [1982] считает более правильным на¬
звание — генерализованный гломерулонефрит. Болезнь почек отмечена у собак более 20 по¬
род, но наследственный („семейный11) ее характер зарегистрирован только для 9 пород
[Nash, 1989]: кокер-спаниель (английский), норвежская оленегонная собака, лхасский апсо,
ши-тцу, самоедская лайка, доберман, стандартный пудель, мягкошерстный пшеничный те¬
рьер, бультерьер. Сейчас точно определено наследование этой аномалии только у двух по¬
род: у кокер-спаниеля аномалия наследуется по аутосомному рецессивному типу, а у само¬
едской лайки она сцеплена с полом [Nash, 1989], т. е. ген локализован з половой
Х-хромосоме.Наследственная почечная недостаточность (гломерулопатия) самоедской лайки явля¬
ется моделью сцепленного с полом наследственного нефрита у человека [Thorner et al.,
1989]. В отличие от нормы у больных нефритом мужчин и у кобелей самоедских лаек с гло-
мерулопатией в капиллярах почечных клубочков отсутствует или изменен компонент, обо¬
значенный АГ Goodpasture (по названию синдрома Goodpaster). Этот антиген обнаружен в
составе домена NC1 молекулы коллагена типа IV, содержащегося в базальных мембранах
почечных клубочков. Молекулярно-генетический анализ показал, что мутантный белок
мономера домена NC1 вовлекается в процесс изменения молекулы коллагена типа IV, что
и является основой симптомов аномалии. При гломерулопатии происходит отложение IgA
[Miyauchi et al., 1992].Дисфункция почечных канальцев — синдром Фанкони — оказался распространенной
наследственной аномалией среди собак породы басенджи в США при апробации предло¬
женной схемы диагностики этой болезни [Noonan, Kay, 1990]. Из 959 обследованных ба¬
сенджи 96 (или 10 %) имели этот синдром, причем половина больных собак была в возрас¬
те 4-8 лет. Среди больных животных отношение суки:кобели было равно 3:1 с явным
преобладанием сук. Кроме того, среди обследованных басенджи еще у 68 имелись подозре¬
ния на синдром Фанкони, а 321 собака имела другие заболевания. Клинические признаки
синдрома у басенджи такие, как указаны выше: полидипсия, полиурия, полифагия.У собак изредка встречается врожденное отсутствие одной почки: описано несколько
случаев в семействе биглей [Fox, 1964] с видимо рецессивным наследованием. У аномаль¬
ных собак отмечено циклическое воспаление единственной почки.Болезни лизосомного накопления [Lysosomal Storage Diseases]Эти врожденные нарушения обмена веществ распространены во многих породах со¬
бак и являются специфическими, генетически детерминированными дефицитами разных
ферментов. Чаще всего каждая такая болезнь в отдельности наследуется как аутосомный
рецессивный признак [Evans, 1989]. Субстратами ферментов могут быть липиды, гликопро¬
теины или полисахариды. При недостатке ферментов они накапливаются в лизосомах, ко¬
торые набухают, причем в определенных органах и тканях.Лабораторные методы позволили, например, в Кембриджском университете успешно
выявлять [Evans, 1989] носителей этих наследственных болезней — гетерозигот. Например,
ферментный анализ дает возможность проводить выбраковку таких носителей. Рецессив¬
ные гомозиготы — больные собаки — при рождении не отличаются обычно от остальных
щенков, но в первые месяцы жизни начинают проявляться признаки аномалий. Течение бо¬
лезней лизосомного накопления имеет прогрессирующий характер, а исход — летальный.
Часто при этих болезнях наблюдаются неврологические дефекты, гепатомегалия и сплено-
мегалия, аномалии скелета.К таким болезням относится, например, фукозидоз (Fucosidosis) — наследственная
аномалия, обусловленная недостаточностью фермента альфа-Ь-фукозидазы в лизосомах.
Определяя активность этого энзима в плазме крови и лейкоцитах у спрингерспаниелей с
известными родословными, специалисты Королевского колледжа в Лондоне установили,
что фукозидоз наследуется по аутосомно-рецессивному типу [Barker et al., 1988], причем в
этой породе распространенность аномалии неоднородна и преобладает в нескольких опре¬
деленных линиях. В случае с фукозидозом ферментный тест позволяет диагностировать па¬
тологию у рецессивных гомозигот — у особей с клиническими проявлениями болезни, и у
собак, у которых еще не проявились симптомы, а также у гетерозиготных носителей мутан¬
тного гена. По результатам исследования рекомендовано включить ферментный тест опре¬
деления активности фукозидазы в селекционную программу племенного разведения сприн-
гер-спаниелей.Врожденная недостаточность кислой липазы в лизосомах у фокстерьеров является
причиной еще одной болезни — липидоза [Opitz, Loppnov, 1988]. При этом липоидном гра-
нулематозе доминируют симптомы массивной гепатоспленомегалии и кристаллогрануляр¬
ной инфильтрации в роговице. В печени накапливаются сложные эфиры холестерина. В пе¬
чени, селезенке, костном мозге, лимфатических узлах и слизистой кишечника обнаружены
скопления макрофагов, содержащих липиды. При таком липидозе обнаружена вообще не¬
характерная для собак форма цирроза.К аномалиям этой природы принадлежит и наследственная избирательная малабсорб-
ция витамина В12, которая обнаружена у собак породы ризеншнауцер при исследовании в
Университете Пенсильвании 3 сибсов (сестра и 2 брата-однопометника): у пациентов к воз¬
расту 12 недель развились постоянное отсутствие аппетита, вялость, общее истощение, за¬
держка развития. Выявили хроническую анемию, нейтропению, ацидурию и низкую кон¬
центрацию витамина В12 в сыворотке крови [Fyfe et al., 1989]. Семейный анализ
подтвердил простое моногенное аутосомное рецессивное наследование этой избиратель¬
ной малабсорции В12.К аномалиям обмена веществ у собак относится и синдром энтеропатии, связанной с
невсасыванием белка. Эта наследственная болезнь является характерной для породы лун-
дехунд — шпицеобразных собак, в течение нескольких столетий широко распространен¬
ных в Норвегии. В начале XX века порода начала исчезать, но небольшая популяция со¬
хранилась на одном из островов близ побережья Норвегии. С 1961 г. началось племенное
разведение лундехундов в Швеции, и, видимо, эффект „бутылочного горлышка" привел к
распространению энтеропатии в восстанавливаемой породе. Болезнь характеризуется диа¬
реей, рвотой, асцитом, отеками. В сыворотке крови имеется низкий уровень белка и каль¬
ция, а фракция гамма-глобулина увеличена [Ablad, 1989].
Гпава 6ГЕНЕТИКА БИОХИМИЧЕСКИХ ПРИЗНАКОВМножество генов открыто у человека и животных, в том числе у собак. Для этого ис¬
пользованы биохимические методы выявления продуктов этих генов — белков и фермен¬
тов, проявляющих полиморфизм (одновременное присутствие у особей данного вида двух
и более форм одного белка). Причем формы встречаются в таком численном соотношении,
что не могут считаться результатом повторных (обратных) мутаций. Ген, представленный
у вида более чем одним аллелем, называется полиморфным геном.Биохимический полиморфизм белков выявляют у животных на основании электро¬
фореза в геле: при постоянном электрическом токе белки из подготовленных образцов ис¬
следуемых тканей (например, крови) мигрируют в зависимости от заряда и массы молекул
к разным электродам с различной скоростью. После этого срез геля инкубируют вместе со
специфическими субстратами изучаемых ферментов и веществами, окрашивающими про¬
дукты гистохимических реакций. В итоге места расположения ферментов в геле после элек¬
трофореза окрашиваются в виде пятен и полос [Gahne et al., 1977]. Мутантные формы мо¬
лекул белков и ферментов, контролируемые полиморфными аллелями генов, при
электрофорезе располагаются в разных зонах геля — имеют, как говорят, различную элек¬
трофоретическую подвижность. Их называют изоферментами [Корочкин и др., 1977; Глаз-
ко, 1988]. Образцы электрофоретических вариантов изоферментов и полиморфных белков
эритроцитов и сыворотки крови собаки приведены на рисунке 6.1.Методы электрофоретического анализа позволили изучить наследственную изменчи¬
вость белков и ферментов в популяциях и породах собак. Эти биохимические маркеры
очень полезны для выяснения филогении — взаимоотношений между предполагаемыми
предковыми дикими и домашними формами, для анализа внутривидовых межпородных
взаимосвязей, а также внутрипородной дифференциации.У домашних собак различного происхождения установлены следующие полиморф¬
ные гены, контролирующие разные электрофоретические варианты изоферментов:
эзерин-устойчивая эстераза плазмы крови (Es),
лейцинаминопептидаза плазмы (Lap),
гемоглобин (НЬ),
эстераза-2 эритроцитов (Es-2),
эстераза-3 эритроцитов (Es-3),
кислая фосфатаза эритроцитов (Рас),
тетразолиум оксидаза эритроцитов (То),
глюкозофосфатизомераза (GPI),
ганглиозид монооксигеназа эритроцитов (Gmo),
супероксиддисмутаза (SOD),
щелочная фосфатаза плазмы (ALP),
арилэстераза (АгЕ).Полиморфными являются также гены белков плазмы крови (не обладающих фермен¬
тативной активностью и не относящихся поэтому к группе изоферментов):
альбумин (Alb),
преальбумин-1 (Ра-1),
преальбумин-4 (Ра-4),
аполипопротеин (apo-A-IV),
постальбумин (Роа),
постальбумин-3 (Роа-3),
Таблица 6.1. Генные частоты аллелей, контролирующих эзерин-устойчивые эстеразы в
плазме крови собак разных пород [Sugiura et al., 1977; Kobayashi et al., 1987; Князев и др.,1993]ПородыКоличествоЧастоты аллелейсобакEsAEsBEscЯпонские аборигенные:Сан-ин-шиба520,070,930,00Шиншу-шиба1520,070,910,02Мино-шиба710,040,910,05Акита2150,050,700,25Кишу720,020,980,00Каи1040,120,880,00Шикоку850,010,690,30Хоккайдо1110,130,860,01Аборигенные собаки-
парии Бангладеш600,150,810,04Аборигенные собаки
Центральной Евразии:Среднеазиатская овчарка270,000,930,07Кавказская овчарка270,001,000,00Западносибирская лайка340,001,000,00Породы европейского
происхождения:0,00шпиц80,060,880,06пойнтер460,340,660,00далматин120,250,750,00Английский сеттер90,390,610,00Бигль2860,220,780,00Такса110,320,680,00Мальтийская болонка330,290,710,00Английскийкокер-спаниель80,210,790,00Немецкая овчарка600,280,720,00Колли120,670,330,00Боксер200,570,430,00Доберман250,390,610,00но нарушение генетического равновесия — резко отличающееся от уравнения Харди-Вайн-
берга распределение генотипов. Австралийские кинологи сопоставили частоты генотипов
по ALP и по системе групп крови Тг (эту группу крови выявляли с помощью природного
аллоантигена QN, причем эритроцитарный Тг-антиген у собак иммунологически близок к
антигенам А человека и J крупного рогатого скота). Среди исследованных 968 домашних
собак и 31 собаки динго установлена высокодостоверная взаимосвязь: наличие
Tr-антигена сопряжено с отсутствием в плазме собак медленной изоформы щелочной фос-
фатазы ALP-S.Недавно было проведено сравнение спектров изоферментов щелочной фосфатазы,
индуцируемой кортикостероидными гормонами в печени и в клетках кишечника собаки
[Sanecki et al., 1990]. Результаты тонкого анализа показали идентичность изоферментов
ALP из разных тканей в отношении их реакций с антителами и их аминокислотного соста¬
ва, но имелись отличия по углеводному компоненту. Сделан вывод, что изученные изофер¬
менты ALP продуцируются геном одного локуса и что различия гликозилтрансферазной
активности в печени и кишечнике собак определяются разным углеводным составом изо¬
ферментов.Три фенотипа имеет и сывороточный альбумин собаки: в аутосомном локусе Alb так¬
же выявлены два кодоминантных аллеля [Day et al., 1971; Mgheni et al., 1979], кодирующие
„быструю11 (F) и „медленную11 (S) электрофоретические формы этого белка. Интересно, что
у человека и домашней лошади установлено тесное сцепление расположенных в одной хро¬
мосоме локусов Alb и белка Gc, связывающего витамин D [Sandberg, Juneja, 1978]. У соба¬
ки Gc белок до 1992 г. считался абсолютно мономорфным, поэтому такое сцепление до сих
пор и не обнаружено. Но по альбумину собак зафиксирован широкий внутривидовой по¬
лиморфизм с большими межпородными отличиями по встречаемости разных аллелей и ге¬
нотипов. Более того, датские генетики и ветеринары [Christensen et al., 1985] определили
при сравнении разных пород статистически достоверную взаимосвязь между высотой в
холке собак (т. е. длинноногостью животных) и частотой альбуминового S-аллеля. Резуль¬
таты их популяционного исследования приведены в таблице 6.2 (данные по типичным
средним размерам, характерным для пород, представлены Датским клубом собаковод¬
ства).В общей выборке частоты аллелей Albs и Altf составили соответственно 0,571 и
0,429. Оказалось, что для каждой породы в отдельности частоты генотипов по локусу Alb
находятся в соответствии с ожидаемыми, исходя из закона Харди-Вайнберга для популя¬
ций в состоянии генетического равновесия. Этого соответствия, однако, не наблюдается
для всей выборки исследованных пород в целом. Датские исследователи указали две воз¬
можные причины этого несоответствия: высокая степень инбридинга или же какая-то связь
между направлением селекции отдельных пород и типами альбумина. Но вычисленный ко¬
эффициент инбридинга оказался невысоким (F=0,33). В то же время коэффициент корреля¬
ции (взаимосвязи) между высотой, характерной для породы, и частотой встречаемости ал¬
леля Albs был достоверным, положительным и весьма высоким (г = +0,47). Немного
меньшей оказалась корреляция между живой массой собак и частотой S-аллеля (г = +0,32).Более того, К. Christensen et al. [1985] провели гибридологический анализ наследова¬
ния роста собак (их длинноногости) и типов альбумина в плазме их крови. Для этого были
получены метисы — помеси первого поколения (гибриды F]) между боксером (генотип по
локусу альбумина Albss) — представителем длинноногих собак, и коротконогим бассет-ха-
ундом с типом альбумина FF. Гибридный помет состоял из 2 кобелей и 3 сук массой от
23 до 28 кг — все они имели короткие ноги по типу бассета. При этом боксер-родитель
оказался гетерозиготным по гену тигрового окраса: среди гибридов Fi было 2 тигровых и
3 рыжих.Двух рыжих гибридов Fi повязали между собой и получили второе поколение —
3 суки и 4 кобеля. Масса их в возрасте 7 месяцев колебалась от 14 до 32 кг; шестеро из них
имели короткие конечности типа „бассет11, а один был длинноногим, как боксер. Табли¬
ца 6.3 содержит подробную информацию по гибридам F2; потомки № 6 и 7 (соответствен¬
но коротконогий и длинноногий щенки) изображены на рисунке 6.2.Среди потомков не проявилась полная конкордантность (соответствие) между длин¬
ноногостью и типом альбумина, но К. Christensen с соавторами считают, что это обуслов¬
лено рекомбинацией, кроссинговером из-за значительного генетического расстояния меж¬
ду исследованными локусами. Причиной связи между ростом собак и типом альбумина их
плазмы исследователи считают „слабое11 сцепление между геном Alb и главным геном, де¬
терминирующим длину ног собаки.Дивергенция между породами собак по частоте аллеля Albs является следствием не
инбридинга, а „дрейфа генов11 в ходе целенаправленной селекции по типу длинноногости
разных пород. Установленные в эксперименте закономерности наследования роста и длин-
Таблица 6.4. Типы трансферрина, выявляемые изоэлектрофокусированием в наиболее рас¬
пространенных породах и в общей выборке из собак 90 пород [Braend, Andersen, 1987]ПородыпЧисло собак с генотипамиГ енные частотыFM1FM2М1М1Ml М2М2 М2M2STfrТ/miТу.М2ТрНемецкаяовчарка148з47682600,020,560,420Лабрадор74002940500,000,660,340Золотистыйретривер680013282700,000,400,600Боксер35004201100,000,400,600Английскийсеттер3200518900,000,440,560Ротвейлер2800281800,000,440,790Выборка из
90 пород4557511718313850,010,470,520,005Таблица 6.5. Частоты аллелей локуса АгЕ в некоторых породах [Braend, Andersen, 1987]ПородыпЧастоты аллелейАгЕоАгЕкАгЕОАгЕ"'АгЕ"АгЕиНемецкая овчарка1480,040,600,350,01Лабрадор740,320,68Золотистый ретривер680,170,770,06Боксер350,870,13Английский сеттер320,530,440,03Ротвейлер280,430,430,14Норвежские исследователи [Braend, Andersen, 1987] отмечают возможность существо¬
вания в локусе АгЕ еще одного, „нулевого41, аллеля, гомологичного таковому у некоторых
других видов млекопитающих, что приводит к нарушению экспрессии этого белка у обла¬
дателей аллеля. Проблема выяснения генетического контроля арилэстеразы у собаки мо¬
жет быть разрешена в будущем проведением гибридологического анализа специальных
скрещиваний.Полиморфизм гемоглобина эритроцитов крови у собак выявлен только при исследо¬
вании японских аборигенных пород [Tanabe et al., 1978]: эти изоферменты контролируют
два также кодоминантных аллеля аутосомного локуса НЬ. Такой вывод сделан в результа¬
те тестирования образцов крови более 1500 собак и „семейного анализа44 107 скрещиваний
(257 потомков). Оказалось, что полиморфизм гемоглобина обусловливается редким даже в
восточных породах аллелем НЬА: его частота в 5 аборигенных породах была ниже 0,08.
Широко обследованные другие азиатские и европейские породы являются мономорфными
по гемоглобину, все их представители обладают гомозиготными генотипами Hb^lHb^.
Генные частоты гемоглобина собак разных пород отражены в таблице 6.6.Таким образом, гемоглобин является маркером различий генофонда собак двух кон¬
трастных внутривидовых групп пород: японской островной и континентальной евроазиат¬
ской.Аналогичным образом маркируют восточный и западный центры доместикации
2 кодоминантных аллеля аутосомного локуса GPI, контролирующие генетические вариан¬
ты — изоферменты эритроцитарной глюкозофосфатизомеразы. Генетику этого маркера у
собак впервые изучили Y. Tanabe с соавторами [1977]. Они установили, что редкий аллель
Таблица 6.6. Генные частоты (концентрации аллелей) эритроцитарного гемоглобина собак
разных пород [Tanabe et al., 1978; Kobayashi et al., 1987; Князев и др., 1993]Породы собакКоличество
со бак-Частоты аллелей НЬНЬЛНЬ*Японские аборигенные породы:сан-ин-шиба620,290,71шиншу-шиба1660,070,93мино-шиба990,020,98шикоку890,001,00каи1070,060,94кишу730,070,93хоккайдо1150,060,94акита2290,120,88Аборигенные собаки Бангладеш600,001,00Породы Центральной Евразии:кавказская овчарка270,001,00среднеазиатская овчарка270,001,00западносибирская лайка340,001,00Породы восточного происхождения:чау-чау50,001,00пекинес70,001,00японский хин60,001,00афган20,001,00Породы европейской селекции:мопс80,001,00терьеры (йоркшир, скоч и др.)120,001,00шпиц100,001,00пойнтер540,001,00сеттеры (английский и ирландский)160,001,00далматин140,001,00пудель50,001,00бигль2910,001,00такса140,001,00мальтийская болонка370,001,00кокер-спаниель100,001,00немецкая овчарка540,001,00колли140,001,00боксер200,001,00бобтейл40,001,00сенбернар30,001,00английский бульдог60,001,00немецкий дог50,001,00доберман150,001,00русская псовая борзая40,001,00
GPIB встречается в популяциях 5 аборигенных японских пород, а остальные исследован¬
ные ими породы восточного и западного происхождения представлены гомозиготами
GPIa/GPIa, за исключением одной особи в породе далматинов, оказавшейся гетерозиго¬
той.Среди исследованных нами аборигенных собак Центральной Евразии две кавказские
овчарки также имели гетерозиготный генотип по локусу GPI, обладая специфически „вос¬
точным" аллелем GPIB [Князев и др., 1993]. Эти различия наследственности собак Востока
и Запада иллюстрирует таблица 6.7.Эритроцитарная ганглиозид монооксигеназа у исследованных собак имеет особый
полиморфизм — в виде 2 фенотипов, так как в контролирующем ее локусе Gmo обнаруже¬
ны 2 аллеля — доминантный GmoS и рецессивный Gmou. Последний проявляется только у
гомозигот [Hashimoto et al., 1984]. Эти аллели также четко маркируют разные центры по-
родообразования и доместикации собаки: аллель GmoS найден только в азиатских породах
и популяциях и совершенно отсутствует у собак европейского происхождения.Аналогичным образом маркируют генетические взамоотношения разных форм собак
два аллеля эритроцитарной эстеразы-2, также взамодействующие по типу полного доми¬
нирования и образующие 3 генотипами лишь 2 фенотипа [Tanabe, 1980]. Доминантный ал¬
лель Es-2S назван „главным аллелем европейских пород11 [Tanabe, 1990], тогда как частота
рецессивного аллеля Es-2F намного выше в азиатских популяциях и породах.Y. Tanabe [1990] установил следы „потоков генов“ собак с материка Евразии на
Японские острова по частотам таких аллелей, как PacF, GPIB, Esc, AIbs, PoaB, Poa 3B, Tf°.
Результаты анализа показали, что в древнюю Японию собаки мигрировали двумя путями:
первый — из Юго-Восточной Азии, включая Тайвань, на все Японские острова, и вто¬
рой — с Корейского полуострова на крупнейшие острова, за исключением Хоккайдо. Эти
выводы позволили составить современное представление о происхождении нынешних за¬
водских и аборигенных собак Восточной Азии.Так, собаки с острова Хоккайдо (айну), по-видимому, являются самыми древними на
островах, мигрировав сюда 10-12 тыс. лет назад вместе с племенами джомон из Юго-Вос-
точной Азии. Другие аборигенные японские собаки рассматриваются как результат гибри¬
дизации старояпонских форм с корейскими собаками, попавшими на острова из Кореи
вместе с хозяевами — племенем яйои 1700-2300 лет назад.Используя перечисленные маркеры западного и восточного центров породообразо-
вания, мы изучили филогенетические связи аборигенных собак центра Евразии с характер¬
ными породами европейского и японского происхождения [Knyazev et al., 1992; Князев и
др., 1993]. В качестве типичных представителей центральноевразийских собак исследованы
западносибирские лайки, среднеазиатские и кавказские овчарки. Анализ генетического по¬
лиморфизма белков и ферментов крови показал: в генофонде собак этих пород имеются
как маркеры восточноазиатского центра доместикации С. familiaris — аллели Albs, Роав,
Роа-Зв, так и западного (Es-2S в гомозиготном состоянии и др.). Такой неоднородный по
происхождению аллелофонд свидетельствует о сложном характере взаимодействия разно¬
направленных потоков генов в процессе становления центральноевразийских пород собак
в их современном статусе.Для наглядного представления о филогенетических взаимоотношениях между типич¬
ными представителями разных центров доместикации и породообразования (европейского
и восточноазиатского) и собаками из Центральной Евразии мы провели специальный ана¬
лиз. Использовали метод главных компонент матрицы варианс-коварианс [Kidd et al.,
1980] на примере сравнения генных частот 15 полиморфных локусов белков и ферментов
крови у более 3 тысяч собак с построением рассеивающей диаграммы в системе координат.
Результаты анализа представлены на рисунке 6.3: в этой системе приведены относительные
филогенетические позиции 46 пород собак, определенные первой (Z1) и второй (Z2) наи¬
большими принципиальными компонентами матрицы генетических расстояний. Постро¬
енная диаграмма разделяет на несколько самостоятельных групп все изученные породы:
1) японские аборигенные собаки; 2) корейские аборигенные собаки (жиндо, чеюдо); 3) тай-
ванские аборигенные собаки; 4) китайские породы; 5) бангладешские аборигенные собаки;
6) западные породы европейского происхождения; 7) эскимосские собаки; 8) аборигенные
породы Центральной Евразии.
Таблица 6.7. Частоты аллелей глюкозофосфатизомеразы в разных породах собак [Tanabeet al, 1977; Kobayashi et al, 1987; Князев и др, 1993]Породы ссбакКоличествособакЧастоты аллелейGPHGPIBЯпонские аборигенные:сан-ин-шиба600,990,01шиншу-шиба1650,990,01мино-шиба730,900,10шикоку880,920,08каи1070,940,08кишу740,840,16акита2291,000,00Бангладешские аборигенные600,960,04Аборигенные центра Евразии:кавказская овчарка270,950,05среднеазиатская овчарка271,000,00западносибирская лайка341,000,00Восточноазиатские:чау-чау51,000,00пекинес71,000,00хин71,000,00Европейские:далматин140,960,04шпиц101,000,00пойнтер491,000,00терьеры (скоч, йоркшир, кэрн, фокс)121,000,00английский сеттер121,000,00ирландский сеттер31,000,00пудель51,000,00бигль2901,000,00такса141,000,00мальтийская болонка371,000,00кокер-спаниель101,000,00немецкая овчарка541,000,00колли141,000,00шелти61,000,00бобтейл41,000,00сенбернар31,000,00английский бульдог61,000,00боксер201,000,00немецкий дог51,000,00доберман101,000,00афган21,000,00русская псовая борзая41,000,00
Z2 (22,2 %)jPwc. 6.5. Относительное расположение 46 пород собак, определенное первой (Z1) и второй (Z2) наибольшими
принципиальными компонентами матрицы генетических расстоянии, основанными на результатах анализа ва-
рианс-коварианс генетических частот 15 полиморфных локусов. Цифры в круглых скобках показывают сте¬
пень вклада оси системы координат [Князев и др.г 1993]Обозначения пород: 1 — хоккайдо. 2 — акита. 3 — каи, 4 — кишу, 5 — шиба (Сан-ин), 6 — шиба (Шиншу), 7 — шиба (Ми-
но), 8 — шиба (Акита), 9 — сикоку, 10 — тансгашима, 11 — якушима, 12 — амамиошима, 13 — окииавахонто, 14 — ирио мо¬
те, 15 — охотничьи собаки Мис (Сима), 16 — охотничьи собаки Мне (Нанту), 17 — ики, 18 — цусима, 19 — рюкюанские со¬
баки (Янбару), 20 — рюкюанские собаки (Ишнгакп), 21 — тайванскис аборигенные собаки, 22 — жиндо, 23 — чсюдо, 24 —
бангладешские аборигенные собаки, 25 — пойнтер, 26 — мальтийская болонка, 27 — хин, 28 — боксер, 29 — немецкая ов¬
чарка, 30 — бигль, 31 — шетландская овчарка. 32 — йоркширский терьер, 33 —далматин, 34 — английский сеттер, 35 — ко¬
кер-спаниель, 36 — мопс, 37 — чау-чау, 38 —доберман, 39 — колли, 40 — померанский шпиц, 41 — такса. 42 — пудель, 43 —
эскимосская собака, 44 -г- кавказская овчарка, 45 — среднеазиатская овчарка, 46 — западносибирская лайкаПри этом между двумя группами, наиболее однородными и удаленными друг от дру¬
га генетически, — европейскими и японскими породами — промежуточное положение за¬
нимают некоторые другие группы. Так, аборигены центра Евразии — кавказские, средне¬
азиатские овчарки и западносибирские лайки — образуют на диаграмме „верхнюю"
цепочку от восточных собак к западным. Подобными „связующими11 звеньями между
крупнейшими контрастными группами пород являются в центральной части диаграммы
тайванские, бангладешские аборигенные собаки и некоторые породы китайского корня —
чау-чау, например. Таким образом, впервые изученный недавно генофонд аборигенных со¬
бак центра Евразии можно рассматривать как неизвестную до последнего времени нишу
или „переходной мост“ генного потока от восточноазиатской группы пород и популяций к
современной европейской группе.На рисунке 6.4 представлена дендрограмма, отражающая генетические дистанции
(расстояния) между 31 породой собак различного происхождения. Вычисленные по извест¬
ному методу М. Нея [Nei, 1975] также с использованием данных о частотах аллелей поли¬
морфных белков-изоферментов дистанции адекватно отражают генетические взаимоотно¬
шения разных форм собак.Изучение полиморфизма белков и ферментов крови предоставляет возможность вы¬
яснить такой важный параметр, как уровень гетерозиготности популяций. Он позволяет
оценить степень наследственной изменчивости у собак разных пород. К настоящему време¬
ни размах изменчивости средней гетерозиготности на особь в популяциях домашней соба¬
ки определен в пределах от 0,08 до 0,18 [Nozawa, 1980].Этот показатель занимает у исследованных нами среднеазиатских овчарок положе¬
ние, близкое к минимальному для вида С. familiaris (гетерозиготность 0,10); у кавказских
овчарок достигает среднего уровня (0.13) и стремится к максимальным значениям у запад-
ХоккайдоКайМино-шибаТайваньская аборигенная собакаЧау-чауТоинтерДелтиэигльТаксаМальтийская болонкаЙоркшир-терьерПудельАнглийский сеттер
Кокер-спаниель
Немецкая овчарка
Померанский шпиц
Мопс
Далматин
Доберман
БоксерКоллиАкитаШиншу-шибаКишуСан-ин-шиба
Японский хинБангладешская аборигенная собакаПекинесЩикокуЭскимосская собака
Корейская собака джиндоБ-ПJ-ЬD0.000.050.10Рис. 6.4. Дендрограмма, демонстрирующая генетические дистанции между породами собак, вычисленные по методу Нея [Kobayashi et al., 1987]
носибирских лаек (0,16). По-видимому, установленная нами изменчивость уровня гетеро-
зиготности у трех пород из центра Евразии отражает особенности микроэволюционных
процессов с разной интенсивностью селекции, различной генетической структурой популя¬
ций и влиянием на эту структуру разных факторов.Среднеазиатская овчарка как наиболее древняя порода, имеющая сравнительно кон¬
сервативный генофонд и значительно сократившиеся в последние поколения (стадия „бу¬
тылочного горлышка11) численность и ареал, характеризуется и относительно небольшим
уровнем наследственной изменчивости. Кавказская овчарка, в большей степени испытав¬
шая процесс взаимодействия разных центров доместикации при выведении и развитии по¬
роды, и во многих поколениях более широко распространенная в пределах Евразии, вклю¬
чая Западную Европу, испытывает значительные воздействия многих факторов эволюции,
в том числе селекционное давление целенаправленного искусственного отбора. Поэтому
эта порода имеет более высокий, средний для вида в целом, уровень генетической изменчи¬
вости. И, наконец, западносибирская лайка по сравнению с названными „аборигенными11
породами является в настоящее время отселекционированной, широко распространенной
заводской породой, интенсивность селекции которой отражает близкий к наивысшему для
вида уровень гетерозиготности, генетической изменчивости. Аналогичный анализ уровня
гетерозиготности в некоторых породах собак в Швеции провели Т. Aarskaug et al. [1992],
продемонстрировав возможность генетической оценки истории селекции пород и их эво¬
люционных перспектив. Авторы использовали данные по генетическому полиморфизму
7 белков (Tf, Pi-1, Ра-1, Ра-4, Prtl, A1BG, АРОА4) среди 44 норвежских лундехундов,
очень инбредной и малочисленной породы, 42 хигенхундов (норвежской кроличьей гон¬
чей), 29 английских сеттеров и 42 немецких овчарок из Швеции. Показатели гетерозигот¬
ности в этих породах приводятся в сопоставлении с их численностью (в скобках — число
ежегодно регистрируемых в стране щенков данной породы). Для немецкой овчарки сред¬
няя гетерозиготность оказалась наивысшей — 0,313 (5000 щенков ежегодно регистрируют
в Швеции), для лундехунда — наименьшей — 0,146 (59 щенков), соответственно для анг¬
лийского сеттера 0,296 (1390 щенков) и для хигенхунда 0,303 (76 щенков). Наименьшая ге¬
терозиготность популяции лундехундов, видимо, объясняется резким падением ее числен¬
ности в 1960-х годах, когда порода прошла „бутылочное горлышко11 отбора, потеряв
полиморфизм по многим локусам. В то же время у относительно редких хигенхундов гете¬
розиготность весьма высока, сопоставима с таковой у многочисленных сеттеров и немец¬
ких овчарок: по-видимому, корректная, грамотная селекция может привлекать генетичес¬
кие резервы для поддержания наследственной изменчивости и в редких, малочисленных
породах.R. Juneja et al. [1992] сообщают об успешном применении биохимических маркеров
для генетической экспертизы правильности родословных в Западной Европе. Так, за пос¬
ледние 4 года были установлены около 250 случаев неправильности племенных записей
при регистрации пометов: генетики исключали обычно одного из двух сравнивавшихся
„предполагаемых" отцов щенков. Коллектив авторов, представляющих Шведский универ¬
ситет сельскохозяйственных наук, Норвежский колледж ветеринарной медицины, универ¬
ситет Дублина (Ирландия) и Кушу Токай — университет в Японии, доказал, что возможна
объективная экспертиза подлинности родословных в 85% всех исследуемых случаев. Как
правило, с этой целью проводится типирование собак по 8 белкам плазмы (TF, PI1, РА1,
РА4, PRT1, PRT2, A1BG, АРОА4) по методу R. Juneja и Т. Shibata [1992], и одному эритро-
цитарному ферменту, SOD. Каждая из 60 исследованных этими учеными пород собак про¬
явила высокий генетический полиморфизм по 3-5 из следующих локусов: TF, PI1, A1BG,
РА1, PRT1, АРОА4.R. Juneja et al. [1992] совсем недавно обнаружили новые электрофоретические вари¬
анты:A1BG D (мигрирует несколько быстрее, чем A1BG F) — он встречается у многих по¬
род — и A1BG I (мигрирует в промежутке между A1BG F и S)- обнаружен только у курча¬
вого ретривера;PRT1 В — мигрирует впереди PRT1 D, найден у грейхаундов и уиппетов;PRT2 М мигрирует между PRT2 F и S; обнаружен только в японской породе акитаину;PI1 М (зона электрофоретической подвижности между PI1 I и S). Этот вариант обна¬
ружили [Juneja et al., 1992] только в породе сибирских хаски, а В. Morris и L. Lemon [1992]
нашли его также у турецких аборигенных собак породы акбаш с частотой, равной 0,100.
R. Juneja et al. [1992] впервые открыли генетический полиморфизм протеина Gc в
плазме собак при типировании семейства чихуахуа. Три собаки при этом имели тип Gc FS,
а все другие исследованные до этого собаки проявляли только фенотип Gc S.Таким образом, хотя в настоящее время иммуногенетика и генетика биохимических
признаков собаки и не достигли еще той высокой степени изученности, что характерна для
некоторых видов лабораторных, сельскохозяйственных животных и человека, но уже нако¬
пили немало ценных сведений, расширяющих и углубляющих наши знания о Canis
familiaris. Кроме того, эти разделы генетики предоставляют возможность использовать со¬
временные методы для анализа и контроля селекции. Генетические полиморфные системы
можно успешно использовать в следующих целях:- изучение причин и динамики генетической изменчивости, составляющей основу
эволюции;- изучение геногеографии собак;- выявление межпородной и внутрипородной дифференциации, изучение филогенеза
и аллелофонда пород, линий, семейств, а также генетических процессов в популяциях и из¬
менений генетической их структуры в процессе селекции;- контроль достоверности записей о происхождении собак;- построение генетических карт хромосом, изучение сцепления генов;- выявление их генетических взаимосвязей с селекционируемыми признаками;- использование полиморфных систем в качестве маркеров при селекции и совершен¬
ствовании пород [Петухов и др., 1989].
Гпава 7ИММУНОГЕНЕТИКА СОБАКИИммуногенетика — молодое направление в биологии, интенсивно развивается сегод¬
ня и включает в себя изучение генетического контроля антигенов сыворотки и клеток кро¬
ви, их полиморфизма (наследственно обусловленной изменчивости), а также генетики им¬
мунной системы, главным образом, антител и антигенов тканевой совместимости.Изучение иммуногенетики групп крови животных — сельскохозяйственных и некото¬
рых домашних — стало уже классикой. Под группами крови животных понимают совокуп¬
ность антигенов крови отдельных особей: антигены же — это чужеродные для организма
вещества, которые при парентеральном проникновении (минуя желудочно-кишечный
тракт) вызывают реакцию иммунного ответа, т. е. синтез антител. Разумеется, эти антитела
(специфические белки иммуноглобулины) вырабатываются организмом только против ан¬
тигенов чужеродных, отсутствующих у данного животного. При этом в его крови находит¬
ся множество антигенов собственных (контролируемых соответствующими генами) —
они-то и составляют группу крови животного. Выявить имеющиеся в крови животного ан¬
тигены, определить его группу крови можно с помощью моноспецифических реагентов —
антисывороток. Они представляют собой образцы сыворотки крови реципиентов, иммуни¬
зированных кровью доноров. Пары донор-реципиент для иммунизации подбирают с та¬
ким расчетом, чтобы донор обладал определенным антигеном, отсутствующим у реципи¬
ента, и тогда последний „вырабатывает" против чужеродного вещества антитела —
иммуноглобулины, способные соединяться только с данным антигеном. Если при иммуни¬
зации у реципиента появляются антитела к нескольким антигенам, „лишние“ из них можно
осадить адсорбцией, получив в результате специфический реагент — моновалентную анти¬
сыворотку. Набором таких реагентов тестируют образец крови изучаемого животного и
выясняют его группу крови — совокупность имеющихся у него антигенов, которые обра¬
зовали комплексы в реакциях с соответствующими антителами.Совокупность антигенов, контролируемых аллелями одного локуса, составляет гене¬
тическую систему групп крови.Антигены бывают различной природы, например, антигены белков сыворотки крови
(аллотипы), антигены, расположенные на мембранах лейкоцитов (антигены главного ком¬
плекса гистосовместимости Major Histocompatibility Complex — МНС) и эритроцитов. У
собак аллотипы пока практически не изучены: лейкоцитарные антигены МНС собаки
DLA (Dog Leucocyte Antigens) использованы для исследования генетики этого комплекса
[Rapaport, Bachvaroff, 1978] — очень сложно устроенного в виде тесно сцепленных генов
трех классов, обусловливающих активность системы иммунной защиты собаки, как и всех
млекопитающих. Не имея пока приложений к практике разведения собак, этот комплекс
весьма интенсивно исследуется методами молекулярной генетики. Например, недавно оп¬
ределена нуклеотидная последовательность некоторых клонов к ДНК — участков генов
класса I главного комплекса DLA [Sarmiento, Storb, 1990].Наиболее полно изучены у разных животных и уже достаточно широко используют¬
ся в селекции группы крови эритроцитарных антигенов [Тихонов, 1967]. Такие антигены
проявляются у животных уже с самого рождения (или даже у эмбрионов) и не изменяются в
течение всей жизни, являясь постоянной характеристикой организма — как отпечатки
пальцев у человека. Чаще всего они четко, дискретно наследуются, причем определяющие
их аллели внутри каждого локуса взаимодействуют кодоминантно; антигены, контролиру¬
емые разными аллелями, у гетерозигот проявляются одновременно, без подавления рецес¬
сивных генов доминантными. У свиней, например, открыто уже более 70 эритроцитарных
антигенов, контролируемых множеством аллелей 17 генетических систем, которые образу¬
ют у каждой особи практически уникальные сочетания — группы крови. Эти и многие дру¬
гие особенности групп крови способствовали успешному использованию их в племенной
работе в качестве генетических маркеров. Анализ генофонда, филогении разных родствен¬
ных видов, пород, популяций, частная генетика, геногеография, современные методы се¬
лекции — это не полный перечень применения иммуногенетики.В широких масштабах проводится иммуногенетическая экспертиза происхождения
племенных животных; сравнивая антигены потомков и производителей, зарегистрирован¬
ных в качестве их родителей, можно с высокой степенью достоверности обнаруживать слу¬
чаи, когда такие производители родителями молодняка быть совсем не могут (кстати, в
скотоводстве и свиноводстве ошибки в племенной регистрации весьма нередки, и торговля
племенным молодняком, особенно на экспорт, на аукционах ведется с обязательной имму-
ногенетической экспертизой его происхождения).Группы крови животных определяют [Тихонов, 1964] по результатам иммунологичес¬
ких реакций — соединению антигенов с антителами — в виде агглютинации (склеивания
эритроцитов) или гемолиза (растворения эритроцитов). Эти реакции обнаруживаются и
при переливании крови, если антигенный состав донорской крови чужероден группам кро¬
ви реципиентов.У собак впервые группы крови были обнаружены при гемотрансфузии. Например, в
1952 г. В.М. Лабунский [Западнюк и др., 1983] выявил в сыворотке крови собак два типа
антител-агглютининов, реагирующих с эритроцитами групп крови А и Вчеловека, — и тем
самым показал наличие у собак группы крови, аналогичной самой известной у людей сис¬
теме АВО. Правда, при первичных переливаниях крови у собак никаких болезненных явле¬
ний не обнаружено, и поэтому при однократной трансфузии специально подбирать донор¬
скую кровь не нужно. Однако при повторных переливаниях крови может проявляться
анафилаксия, по мнению В. Лабунского, из-за „...наличия у донора вне группового антиге¬
на, на который организм реципиента после первого переливания крови вырабатывает изо-
иммунные антитела14 [Западнюк и др., 1983. С. 102]. Этот феномен в 1950-е годы использо¬
вался для изучения изоиммунизаций у человека при сенсибилизации по резус-системе.Кстати, возможные осложнения при гемотрансфузии у собак современная иммуноло¬
гия предлагает избегать с помощью ультрафиолетового облучения донорской крови; в ре¬
зультате такой обработки у реципиентов наблюдали иммуносупрессию. Экспериментато¬
ры из японского Университета Ямагуши [Une et al., 1991] сообщили, что при этом не было
взаимодействия между лимфоцитами реципиента и донора, агглютинации эритроцитов,
образования антител и других проявлений реакций отторжения, которые неизменно разви¬
вались в контрольной группе собак после переливания необлученной крови.Кроме того, трансфузионные реакции, причем „немедленного1* типа, у собак вызыва¬
ются и антигенами тромбоцитов. С. Белоцкий [1979], применив микрометоды тромбоаг-
глютинации, лимфоцитотоксический тест и некоторые другие, выявил 4 тромбоцитарных
антигена. Они наследовались по менделевскому типу подконтролем доминантных ауто-
сомных и не сцепленных друг с другом генов. Частота встречаемости этих антигенов среди
неродственных друг другу собак варьировала от 10 до 26 %, чаще всего присутствуя в кро¬
ви животных по отдельности, хотя у небольшой части собак обнаружены двойные (попар¬
ные) и тройные сочетания антигенов.Отсутствие какой-либо положительной или отрицательной связи между встречаемос¬
тью антигенов тромбоцитов, эритроцитов и лейкоцитов собак указывает на независимость
контроля генетических систем, детерминирующих антигены разных форменных элементов
крови этих животных.По сравнению с сельскохозяйственными животными собака пока менее изучена им¬
муногенетически. При этом эритроцитарные антигены, обнаруженные разными исследова¬
телями, получали и различные обозначения. Впервые такие исследования провел L. Young
с соавторами, среди которых был и S. Swisher [Young et al., 1950]. Эти ученые впоследствии
расширили знания о группах крови домашней собаки, номенклатура, предложенная ими,
приведена в таблице 7.1.Разные системы классификации и обозначения групп крови вносили путаницу, по¬
этому в 1972 г. на 1-м Международном симпозиуме по иммуногенетике собаки (IWCI) бы-
Таблица 7.1. Первоначальные обозначения антигенов
групп крови собак, предложенные разными авторамиYoung et al., 1950Swisher, Young, 1961Swisher et al., 1973АаAlаAlaА2A2ВbВbВbСсСсСсDdDdDdЕеЕеFfFfTrtrGgHeheТаблица 7.2. Сравнение разных номенклатур антигенов
групп крови собак, предложенных на Международных
симпозиумах по иммуногенетике собаки (IWCI)Разные системы классификации и обозначения групп кровиИсходнаяIWCI, 1972IWCI, 1974(Al)CEA-1DEA 1.1(A2)CEA-2DEA 1.2(B)CEA-3DEA 3(C)CEA-4DEA 4(D)CEA-5DEA 5(E)CEA-6DEA 6(Tr)CEA-7DEA 7(He)CEA-8DEA 8ла принята числовая система обозначения антигенов с применением аббревиатуры СЕА
(Canine Erythrocyte Antigen — эритроцитарный антиген собаки [Vriesendorp et al., 1973].
Однако, поскольку оказалось, что обозначение СЕА уже применялось для системы канце¬
роэмбриональных антигенов, на 2-м симпозиуме IWCI в 1974 г. утвердили символику с аб¬
бревиатурой DEA (Dog Erythrocyte Antigen) [Vriesendorp et al., 1974; 1976] (табл. 7.2).Однако система обозначений, принятая на этих симпозиумах, не позволяла создать
классификатор для всех уже обнаруженных и еще не открытых у собаки антигенов и не со¬
ответствовала общепринятым принципам обозначения групп крови других видов живот¬
ных.Прогресс в изучении групп крови собаки обеспечили Р. Сэйзон и Д. Коллинг [1979]
из Гуэлфского университета в Канаде: предложенная ими система классификации адекват¬
но отражает взаимоотношения между антигенами, аллелями, локусами, генотипами и фе¬
нотипами и позволяет в будущем расширять спектр известных маркеров по мере их откры¬
тия и изучения (табл. 7.3).При такой номенклатуре локус гена, контролирующего генетическую систему групп
крови собак, обозначается прописной буквой (например, А), антигены обозначаются
строчными буквами следом за обозначением системы (например, Аа, Аа1), аллели, контро¬
лирующие эти антигены — в виде символа локуса с символом кодируемого антигена в „по¬
казателе степени", а генотипы составляются из пар соответствующих аллелей. Эта номенк¬
латура была принята Международным обществом по изучению групп крови животных на
Таблица 7.3. Новая номенклатура групп крови собак [Сэйзон, Коллинг, 1979]ЛокусыАнтигеныАллелиГ енотипыФенотипыАA, alAaal, Аа, А-даа1 даа1*
Аза1 да; ’
Aaal д—•
АаАа; АаА~;
А-А-A(aal),A(a),A(-)Вава, в-ВаВа; ВаВ"; B"B~B(a), B(-)СаСа; С-СаСа; СаС~; С~C(a), C(-)DаDa; D-DaDa; DaD~; D~D~D(a), D(-)FаFa; F~FaFa; FaF~; F-F~F(a), F(-)ТгТг, 0Trtr, Тг°, Тг-TrtrTrtr; TrtrTr°;
TrtrTr-; Tr°Tr°;
Tr°Tr-; TrTrTr(tr),Tr(0),Tr(-)XVI Международной конференции в Ленинграде в 1978 г. и подробно описана вместе с из¬
ложением результатов исследований иммуногенетики собак в специальном обзоре [Colling,
Saison, 1980].Так, например, эти авторы [Коллинг, Сэйзон, 1978] обнаружили еще 6 новых эритро-
цитарных антигенов: 5 из них, наследовавшиеся независимо друг от друга, отнесены к но¬
вым системам соответственно Q, К, L, М, N, а антиген О кодируется вновь открытым алле¬
лем из системы Тг. Такие выводы сделаны по результатам „семейного анализа"
расщеплений (сегрегации) потомства от скрещиваний специально подобранных родительс¬
ких пар. Авторы этого исследования полагают, что относительно скромные прежде успехи
в открытии эритроцитарных антигенов собак объяснялись применением простейших мето¬
дов регистрации иммунологических реакций „антиген-антитело“ — прямой агглютинации
или гемолиза. Оказалось, что новые антигены у собак можно обнаруживать с помощью
более сложных тестов — непрямой антиглобулиновой пробы (выявлены антигены Q, К, L,
М, N) или капиллярно-папаинового метода (антиген К). Антиген О выявлен не при обыч¬
ном температурном режиме реакции (37 °С), а при t = 4 °С.Позднее в Японии были проведены углубленные исследования генетической системы
D групп крови собак: в D-локусе обнаружены 2 аллеля, формирующие 3 фенотипа — D1,
D2 и D1/D2. Ученые Японского университета ветеринарии и животноводства [Haraet al.,
1991] использовали современный метод биотехнологии — приготовление моноклональных
антител к эритроцитарному антигену D1 из культуры гибридомных клеток мыши. Она
продуцировала в определенных условиях иммуноглобулины — антитела к D1, которые
проверяли „панелью“ из образцов крови 117 собак 5 пород (из них 78 биглей). Среди выде¬
ленных 11 клонов клеток, секретирующих моноклональные антитела, 3 клона выделяли ре¬
агенты к D1. Из 117 тестированных собак 20 животных идентифицированы как
Dl-позитивные, а остальные как D1-негативные. При использовании генноинженерного
метода Вестерн-блоттинга было установлено, что антиген D1 состоит из совокупности бел¬
ков эритроцитарных мембран с молекулярными массами 34, 53,59, 64 и 71 кД: в образцах
крови D1-негативных собак такие фракции не обнаружены. Y. Ilara с соавторами полага¬
ют, что выделенные ими моноклональные антитела перспективны для исследования как
иммуногенетического полиморфизма собак, так и биохимического анализа их
D1 -антигенного комплекса.В австралийском Университете Квинсланда М. Symons и К. Bell [1992] исследовали
иммуногенетический и биохимический полиморфизм 968 домашних собак и 31 дикой соба¬
ки динго (Canisdingo) и установили высокодостоверную корреляцию (взаимосвязь между
эритроцитарным антигеном системы Тг (который иммуногенетически близок к антигену А
человека и J-антигену крупногорогатого скота) и изоферментами щелочной фосфатазы сы¬
воротки крови (ALP). У собак имеются 2 аллеля гена ALP, контролирующие разные фор¬
мы этого фермента — „быстрый" ALP-F и „медленный" ALP-S, который, кстати, у иссле¬
дованных собак оказался очень редким и обнаружен в крови представителей всего только
6 пород. Характер обнаруженной корреляции следующий: наличие Тг-антигена было со¬
пряжено одновременно с резким дефицитомизоформы S щелочной фосфатазы. Исследова¬
тели [Symons, Bell, 1992] обсуждают возможные механизмы, обусловливающие такую взаи¬
мосвязь; один из наиболее вероятных среди них — генетическое сцепление, т. е.
расположение локусов соответствующих генов в одной хромосоме и сопряженное поэтому
наследование группы крови Тг и ALP-F.Кстати, в настоящее время биохимические маркеры в виде изоферментов крови изу¬
чены у собак более углубленно и уже достаточно широко используются не только в генети¬
ческих исследованиях, но и в селекционной практике некоторых „кинологических держав"
Западной Европы, в частности в Швеции [Juneja et al., 1992].
Гпава 8ГЕНЕТИКА ПОВЕДЕНИЯ СОБАКИО генетике поведения собак известно и мало, и много. Людям, даже мало знакомым с
этими животными, известно, что разные породы собак различаются не только размерами и
внешним обликом, но и склонностями, привычками, наконец, характером. Возбудимый,
холерический темперамент фокстерьеров и обычно уравновешенное поведение ньюфаунд¬
лендов — наиболее простые примеры таких различий. Собаководы-любители, а тем более
профессионалы могут дать подробное описание межпородных различий у собак, которые
иногда оказываются поразительными. Однако систематические исследования таких разли¬
чий проводить достаточно трудно. Вместе с тем собственно генетические исследования по¬
ведения собак имеют множество трудностей. Очевидно, что это связано и со сложностью
изучения поведения, в частности, выделения адекватных признаков для анализа (это про¬
блема, которая является общей для генетики поведения в целом), и с малой изученностью
генома собаки, и с малым числом маркеров, а также с проблемой гетерозиготности особей
породы.В связи с этим в начале главы будут рассмотрены некоторые общие проблемы гене¬
тики поведения, далее будут представлены конкретные данные по анализу поведения собак
разных пород.Общие сведения о поведении животных как объекте исследованияКак в процессе, который протекает практически непрерывно, в поведении животного
не всегда можно выделить какие-то признаки, которые могли бы стать предметом изуче¬
ния генетики количественных признаков. Такими признаками могут быть или характерные
для животных данного вида движения и позы, или степень проявления какой-либо тенден¬
ции в поведении. Относительно второй группы признаков можно сказать, что наиболее
близко к проблеме выделения признаков подходят исследования роли общей возбудимости
животного и проявления в связи с этим того или иного признака поведения. Что касается
первой категории признаков, то такими „единицами" поведения могли бы стать феномено¬
логически четкие, стереотипные и жесткие последовательности действий, которые называ¬
ются фиксированными комплексами действий (ФКД). По представлениям классической
этологии, именно ФКД и являются собственно инстинктивными движениями.Напомним, что в соответствии с этими представлениями, восходящими к У. Крэггу
[Крушинский и др., 1983], поведенческий акт можно представить схематически в следую¬
щем виде:Повышение мотивации -> поисковое поведение —> отыскание знакового стимула -»-» завершающий акт -» снижение мотивации.Начальная фаза поведенческого акта — поисковое поведение животного, в результа¬
те которого оно находит соответствующие стимулы. Поисковое поведение достаточно пла¬
стично, что обеспечивает его видоизменение как результат опыта и облегчает животному
достижение цели. В репертуар поискового поведения, наряду с общими локомоторными и
исследовательскими реакциями, могут входить и характерные для данного вида животных
ФКД. Примером могут служить специфические стереотипные движения собаки при высле¬
живании дичи, при контактах с сородичами и т. п. ФКД, которые животное обнаруживает
при поисковом поведении, бывают строго стереотипными по форме. Однако на основе ис¬
пользования накопленного опыта, с помощью механизмов научения и памяти животное
может совершенствовать ориентацию и производить корректировку таких движений. На¬
пример, некая реакция начинает проявляться только в определенной ситуации. Все это
обеспечивает пластичность поискового поведения. Оно может, таким образом, заметно ви¬
доизменяться по мере накопления индивидуального опыта. Именно во время поискового
поведения происходит процесс научения и осуществляются поведенческие акты, основан¬
ные на элементарной рассудочной деятельности.Классическим примером участия научения в формировании адаптивного поведения
на основе ФКД является описанная К. Лоренцем реакция подкатывания яйца гусем (став¬
шая хрестоматийным примером). Гнездящиеся на земле гуси возвращают выкатившееся из
гнезда яйцо с помощью стереотипного ФКД — птица вытягивает шею, загибает подклю-
вье над яйцом и катит его в гнездо, втягивая голову. Этот ФКД обнаруживается при пер¬
вой же необходимости, однако у опытной птицы появляются небольшие по амплитуде кор¬
ректирующие движения клюва в плоскости, перпендикулярной оси основного движения.
Эти движения порождаются рефлекторно, как реакция на неравномерное раздражение пра¬
вой и левой половины клгова, когда яйцо катится неровно и может откатиться вбок. Оче¬
видно, что „ловкость41 закатывания яйца в гнездо обеспечивается именно этими коррекци¬
ями, которые появляются в результате накопления индивидуального опыта.Завершающий акт — это высокоспециализированная последовательность действий,
выполнение которой обеспечивает снижение мотивации, обусловливающей возникновение
поискового поведения. Как уже упоминалось, завершающий акт — это стереотипный
ФКД, коррекции которого с помощью опыта практически не происходит. Примером сте¬
реотипно выполняемого ФКД завершающего акта может быть бросок на добычу у охотя¬
щегося животного.Эти представления изложены здесь схематично и упрощенно, однако они могут ока¬
заться полезными в качестве основы, помогающей представить себе многообразие и разно¬
плановость всех проявлений поведения.Итак, в составе поведения особей вида можно выделить четкие единицы, стереотип¬
ные „кирпичики“ поведения — ФКД, — которые могли бы служить как признаки для гене¬
тического анализа. Однако проблема оказывается значительно сложнее. Как мы знаем,
ФКД характерны для всех особей вида и, следовательно, генетическую изменчивость по
форме ФКД выявить практически не удается. В редких случаях внутривидовая изменчи¬
вость, связанная с ФКД, обнаруживается в виде наличия или отсутствия каких-либо сте¬
реотипных реакций в репертуаре поведения особей данной линии, породы или популяции.
В таких случаях проведение генетического анализа, базирующегося на конкретных ФКД,
оказывается возможным. Классический пример такого исследования представляют собой
работы Rotenbuhler по генетическому определению так называемого „гигиенического" по¬
ведения пчел [Дьюсбери, 1981]. Были исследованы 2 расы пчел, которые отличались друг
от друга по наличию и отсутствию в репертуаре ихповедения ФКД вскрывания заражен¬
ных инфекцией ячеек с личинками и стереотипных движений удаления таких личинок из
улья. Эксперименты по скрещиванию этих рас показали, что данное поведение находится
под контролем двух генов рецессивной природы. В последующих исследованиях, однако,
выявилось, что рабочие пчелы „негигиенической" расы, крайне редко, но все-таки обнару¬
живали „гигиенические" ФКД.Использование ФКД в качестве признаков для генетического анализа оказалось ус¬
пешным только для исследования поведения потомков межвидовых скрещиваний в экспе¬
риментах с птицами [Dilger, 1962; эрман, Парсонс, 1984] и с рыбами [Frank, 1974], а также с
некоторыми представителями рода Canis, скрещивания: домашняя собаках шакал [Бехтин,
Сулимов, 1985] и домашняя собака х волк [Zimen, 1969].Поскольку в пределах вида ФКД стабильны по форме, внутривидовое разнообразие
поведения (в частности, межпопуляционные и межпородные различия) может обеспечи¬
ваться различной частотой проявления отдельных реакций и разным уровнем порогов их
провокации. Иными словами, некоторые поведенческие реакции могут выглядеть как спе¬
цифические для данного генотипа, потому что они обнаруживаются чаще, а также если
внешняя стимуляция, необходимая для их провокации, оказывается слабее, чем для особей
того же вида, но имеющих другого генотипа. Генетические исследования поведения и, в ча¬
стности, селекционные эксперименты, четко показывают, что различия в поведении, конт¬
ролируемые генотипом, в значительной доле случаев можно описать как различия в часто¬
те и порогах провокации ФКД. Примером могут служить различия между инбредными
линиями крыс и мышей по уровню локомоции и ориентировочно-исследовательского по¬
ведения, выражающиеся, в частности, в большей или меньшей частоте выполнения стерео¬
типной реакции „вставания на задние лапы“ [Abeelen, 1974]. Известно, что наиболее яркие
иллюстрации упомянутых выше этологических принципов можно найти, анализируя пове¬
дение насекомых, рыб и птиц. Наблюдения за поведением млекопитающих не всегда по¬
зволяют видеть действие этих принципов. Это объясняется большой сложностью и высо¬
кой пластичностью поведения этих животных. Однако выявление в поведении
млекопитающих стереотипных движений и поз оказывается возможным даже при неболь¬
шом опыте наблюдений за ними.Частота и пороги проявления таких стереотипных реакций обнаруживают значитель¬
ную изменчивость. Общеизвестна зависимость поведения животных, в частности проявле¬
ния характерных для брачного поведения поз и движений, от гормонального статуса осо¬
би. Это в полной мере относится и к поведению собак. Вопрос заключается в том, что,
выявив достаточно мошную индивидуальную изменчивость поведения упомянутого выше
характера, следует сделать дальнейшие шаги в анализе поведенческого признака и попы¬
таться оценить генетический компонент такой изменчивости. Отметим сразу, что практи¬
чески во всех случаях выявления олигогенного наследования того или иного признака по¬
ведения у собак (примеры чему будут приведены ниже) речь может идти не о полном
наличии или отсутствии какого-либо признака, а лишь о слабой или сильной степени его
проявления, поэтому, строго говоря, в таких случаях следует констатировать детермина¬
цию признака большим или меньшим числом так называемых „главных*4 генов. Это поло¬
жение, как нам кажется, не нуждается в подробном обосновании, поскольку практически
все сложные фенотипические признаки, какими являются признаки поведения, относятся к
категории количественных.Следует также отметить, что анализируя сложное поведение такого животного, как
собака, исследователи не могут избежать искушения и не сделать попыток анализа таких
феноменов, которые в пределах поведенческого репертуара этого вида оказываются чрез¬
вычайно сложными. Речь идет о сложных феноменах типа способностей к обучению и к ре¬
шению ряда задач, именуемых в настоящее время когнитивными тестами. Межпородные
различия в таких случаях оказываются практически малоинформативными с точки зрения
выявления возможной генетической основы, так как решение подобных задач находится не
только под влиянием собственно когнитивных способностей животных, но под модулиру¬
ющим воздействием общего состояния организма, например, той или иной мотивации. Это
выявили эксперименты, проведенные P. Scott и J. Fuller [1965].Генетические различия в частотах выполнения и в порогах проявления ФКД линия¬
ми, популяциями и даже дивергирующими видами как явление, которое может вызывать
микроэволюционные события, были впервые отмечены в работе A. Manning [1967]. Однако
значительно раньше, хотя и в неявной форме подобное явление было продемонстрировано
в работах по генетике поведения собак J1.B. Крушинским [1938; 1944; 1946 а, б; 1962; 1991].
В этих исследованиях было обнаружено, что интенсивность пассивно-оборонительной ре¬
акции собак зависела от уровня общей возбудимости животного, которая, соответственно,
определяла пороги реакций, служащих у собак внешним проявлением состояния страха.Оценка генетических различий в поведении, в частности, межпородных различий в
поведении собак, с точки зрения различий в порогах проявления и частоты выполнения
ФКД, относящихся к разным формам поведения (пищевому, оборонительному, половому,
социальному), может позволить провести некоторую систематизацию обширного факти¬
ческого материала, накопленного к настоящему времени. Оценить с этих позиций данные
генетических исследований поведения собак еще только предстоит.Приведенные выше вводные замечания имеют цель помочь в систематизации разно¬
образного материала по генетике разных форм поведения собак. Нужно отметить также,
что существование разных пород собак представляет собой рекордный по длительности се¬
лекционный эксперимент, демонстрирующий, как никакой другой, фантастический раз¬
мах внутривидовой изменчивости и по морфологическим признакам, и по признакам по¬
ведения.
Подходы к описанию поведения собакиИзменчивость частоты и порогов проявления стереотипных видоспецифических пос¬
ледовательностей действий собак, разумеется, не исчерпывает феноменологии поведения
этого вида. Пластичность, и как результат, очень высокая адаптивность поведения этих
животных обеспечиваются их обучаемостью, в особенности в течение так называемых кри¬
тических периодов, и достаточно высоко развитыми когнитивными способностями к эле¬
ментарной рассудочной деятельности [Крушинский, 1986]. Ясно, что ни простые условные
рефлексы, ни более сложные интегральные реакции, обнаруживаемые при адаптивном по¬
ведении, не могут быть дискретными признаками для анализа поведения методами генети¬
ки. Это означает, что исследование подобных проявлений поведения должно опираться на
феноменологически более простые реакции или особенности строения центральной не¬
рвной системы.Немаловажную роль в исследовании генетики поведения играет возможность ис¬
пользования генетически идентичных лабораторных животных — инбредных линий мы¬
шей, крыс и других — с целью их сравнения и проведения генетического анализа. Степень
инбредности пород собак — понятие, которое редко фигурирует в литературе. Попытки
повысить инбредность обычно связаны с заботой о поддержании единообразия экстерьера
животных. Изначально при выведении некоторых пород, например, английского сеттера и
русско-европейской лайки, тесный инбридинг практиковался на протяжении ряда поколе¬
ний в период, когда порода была малочисленна (скрещивания типа I—II и II-II). Вообще же
при разведении собак чаще применяют скрещивания родственных особей по схеме II-III,
III-III и III—IV. Конечно же, ничего похожего на инбредные линии грызунов у собак нет и
быть не может. R. Robinson [1982] описал попытки выведения линейных биглей, имевшие
целью наблюдать феномен „инбредной депрессии11. Очевидно, что анализ межпородных
различий в сложных формах поведения собак может, тем не менее, дать интересный мате¬
риал для суждения о механизмах формирования поведения. Информативные данные мож¬
но также получить при сопоставлении поведения собак с таковым диких представителей
этого рода.Для выживания и сохранения вида животное должно находить пищу, укрытие, защи¬
щаться от врагов, рожать и воспитывать детенышей. Поведение членов семейства Canidae
отличается сложностью и многообразием. В стае одичавших собак или волков существует
хорошо выраженное разделение обязанностей между ее членами. В стае всегда есть и очень
пугливые особи, которые мгновенно реагируют на возможную опасность, и более реши¬
тельные, которые не стремятся скрыться при первой тревоге, а стараются определить при¬
чину ее возникновения и оценить степень опасности. Высокая степень специализации (час¬
то имеющей временный характер) ролей членов стаи обнаруживается и при охоте. Одни
волки быстро находят след добычи, другие ее окружают, третьи бывают загонщиками, чет¬
вертые убивают жертву и т. д. Некоторые животные редко участвуют в общих охотах, ос¬
таваясь в логове с детенышами.Свойственные диким представителям семейства Canidae особенности индивидуаль¬
ного поведения послужили основой для формирования специализированных пород собак в
ходе доместикации. Так, например, привязанность к определенной территории и защита ее
от членов других группировок послужили основой для формирования путем направленно¬
го отбора четко выраженной защиты дома, участка земли, хозяина и т. д. Умение отыски¬
вать добычу по следу искусственный отбор трансформировал в специфическое охотничье
поведение собак с выраженной специализацией пород. И охотничье поведение, и отыска¬
ние объекта по следу сопровождаются не только проявлением очень острого чутья, но и
специфическими позами и реакциями; например, стойка охотничьих собак. Способность
следовать за стадом травоядных животных в целях, охоты за ними послужила основой для
формирования специфических свойств овчарок — собак с „пастушеским" поведением. Та¬
ким образом, направленная селекция собак по особенностям поведения привела к появле¬
нию специализированных пород и не только к усилению некоторых особенностей поведе¬
ния у каждой из групп пород, но, соответственно, и к ослаблению некоторых тенденций.
Большинство пород охотничьих собак имеют низкий уровень агрессивности по отноше¬
нию к человеку. Отбор на высокую степень обучаемости (дрессируемость) и послушание у
немецких овчарок привел к тому, что многие представители этой породы практически не
способны к принятию самостоятельных решений и всегда ждут команды от человека.
Как отмечают P. Scott и J. Fuller [1965], установление традиций систематических выс¬
тавок собак в конце XIX века способствовало тенденции к усилению селекции на признаки
экстерьера, что иногда влекло за собой ухудшение физиологических показателей особей
какой-либо из пород. Так, например, отбор кокер-спаниелей на такой морфологический
признак, как „выпуклый лоб“ сопровождался появлением животных с признаками умерен¬
ной гидроцефалии, а отбор собак на более высокие передние ноги у ряда пород сопровож¬
дался отбором на умеренную дисплазию тазобедренного сустава. В целом поведение собак
тех пород, которые были сформированы путем отбора на „рабочие качества'4, т. е. по по¬
казателям поведения, оказывается менее изменчивым по сравнению с так называемыми
респециализированными породами, например, породами собак-компаньонов. Генетичес¬
кие механизмы, лежащие в основе таких феноменов, достаточно очевидны. Можно также
полагать, что комбинация некоторых редких рецессивных аллелей, детерминирующих
комплекс поведенческих особенностей той или иной породы, может поддерживаться в дос¬
таточно малочисленной популяции только путем специальной системы скрещиваний.Очевидно, что изучение генетики поведения собак должно начинаться с выявления и
описания различий в поведении неродственных между собой пород.Следующий этап — это проведение генетического анализа признаков поведения, об¬
наруживших контрастное проявление у выбранных пород. Классическим исследованием в
этом отношении стала работа P. Scott и J. Fuller [1965]. Краткие извлечения из богатейшего
материала, представленного в их монографии, будут даны ниже. Исследования других ав¬
торов, может быть, не столь обширные и длительные, также послужили основой для фор¬
мирования современных знаний по генетике поведения собак. Материалы большого обзо¬
ра S. Mackenzie и др. [1986] также использованы в настоящей главе.Начало исследований генетики поведения собакПервой из работ по генетике поведения собак было исследование Е. MacDowell
[1921]. Работая с таксами, он показал, что сибсы разных пометов обнаруживают индивиду¬
альные психофизиологические особенности, которые сохраняются в течение всей их жиз¬
ни. L. Whitney [1929 а, Ь] изучал характеристики поведения собак многих пород. Он отме¬
тил более высокую агрессивность по отношению к человеку у немецких овчарок по
сравнению с бладхаундами. Широко известны его работы по скрещиванию пород гончих,
работающих „с голосом11, с ищейками, которые работают по следу молча, причем у исход¬
ных пород имелись различия и в тембре голоса. Гибриды первого поколения обнаружили
доминирование признака „работа с голосом11, но при этом тембр лая был унаследован ими
от „молчаливого11 родителя. У гибридов второго поколения эти признаки распределились
независимо. Этот же автор показал доминантный характер наследования склонности к ку¬
панию в воде, характерной для собак некоторых пород (спаниели, ретриверы, лабрадоры и
др.). Его монография „How to breed dogs11 [Whitney, 1971] была и остается очень популяр¬
ной среди специалистов.Роль возбудимости нервной системы в обнаружении
генетической детерминированности некоторых реакций собакВ ранних исследованиях [Адамец, 1930] был обнаружен промежуточный тип наследо¬
вания уровня общей возбудимости животного. В качестве контрастных по этому признаку
пород собак были использованы такие породы, как возбудимые и нервозные пойнтеры и
флегматичные немецкие легавые. К высоковозбудимым собакам относили также немецких
овчарок, арабских борзых, спаниелей; к средневозбудимым — английских бульдогов, такс,
екинесов, а к маловозбудимым — французских гончих.В работе W. James [1941], проводимой в течение нескольких лет, была дана характе¬
ристика нескольких пород собак на основе классической павловской методики. Бассетов и
такс автор отнес к „летаргическому11 типу, немецких овчарок и салюк — к „активному11 ти¬
пу. При скрещивании немецких овчарок с бассетами были получены собаки с „промежу¬
точным типом высшей нервной деятельности (ВНД), которые оказались более уравнове¬
шенными, чем их родители. На основе анализа родословных собак, обладавших
аномально высокой возбудимостью, S. Hodgman [1963] показал, что это свойство имеет на¬
следственную природу и проявление его является серьезной проблемой племенного разве¬
дения собак в Великобритании.Собаки были традиционными лабораторными животными в исследованиях
И. П. Павлова. Именно в опытах на собаках была сформулирована его концепция типов
ВИД. Как известно, в соответствии с этой концепцией выделяли 4 основных типа ВИД или
темперамента — холерический, сангвинический, флегматический и меланхолический, а
также некоторое число промежуточных по характеру подтипов. Говоря схематично, каж¬
дый из этих типов ВНД предполагал у его носителя определенную комбинацию более про¬
стых свойств нервной системы, т. е. параметров так называемых основных нервных про¬
цессов — возбуждения и торможения. Под свойствами основных нервных процессов
И. П. Павлов понимал их силу, подвижность и уравновешенность.Типология ВНД, предложенная И. П. Павловым, отражала стремление подвести
строгую научную основу и дать количественную оценку индивидуальной изменчивости
признаков поведения. Следующим логическим этапом такого подхода было создание про¬
граммы изучения генетических основ индивидуальной изменчивости ВНД, точнее, генети¬
ческих основ типологии. Однако проведение генетических исследований оказалось мало¬
совместимым с длительной и очень трудоемкой процедурой определения типа ВНД,
принятой в лаборатории И. П. Павлова. В том виде, в каком они были задуманы, исследо¬
вания оказались неудачными. В то же время было проведено изучение наследования более
простых характеристик условнорефлекторной деятельности у собак, в частности, проана¬
лизировано наследование силы нервных процессов [Крушинский, 1991]. Сводку данных по
высшей нервной деятельности собак можно найти в недавно переизданном пособии по слу¬
жебному собаководству [Мазовер и др., 1994].Наиболее простым методически, но весьма перспективным с точки зрения изучения
генетико-физиологических механизмов поведения оказался анализ наследования общего
уровня возбудимости животного. Переход к оценке возбудимости собак как одного из ба¬
зовых свойств их поведения послужил далее основой для фундаментальных классических
исследований генетики поведения собак, проведенных Л. В. Крушинским [1938; 1946 а, б].
Результаты этих исследований, осуществленных в 1930-40-е годы, были изложены в ряде
оригинальных статей тех лет, а также в первой монографии Крушинского [1962]. В настоя¬
щее время эти работы можно найти в первом томе двухтомного издания избранных трудов
Крушинского [1991], на который мы и будем ссылаться в дальнейшем.Первым этапом этого исследования было подыскание адекватного и достаточно на¬
дежного метода оценки уровня общей возбудимости животного. В это понятие входит не
только представление о некоем фоновом уровне возбудимости ЦНС, при котором проте¬
кает вся деятельность организма и, в частности, его поведение, но и реактивность ЦНС к
внешним воздействиям. Однако по мере проведения исследований автор пришел к выводу,
что адекватным и достаточно надежным показателем общей возбудимости ЦНС собаки
может служить уровень ее двигательной активности. Этот признак оказалось довольно
просто и быстро оценивать с помощью спортивного шагомера, крепившегося на теле соба¬
ки. Показания этого устройства можно было снимать через любые промежутки времени.Основным генетическим экспериментом этой работы был следующий. Гиляцких лаек
(довоенное название одного из отродий будущей заводской породы), маловозбудимых со¬
бак, не обладавших пассивно-оборонительной реакцией (т. е. не обнаруживавших тру¬
сость), скрещивали со значительно более возбудимыми немецкими овчарками, у которых
пассивно-оборонительная реакция также отсутствовала. У потомства первого поколения
(25 особей) была резко выражена пассивно-оборонительная реакция. Подобное проявле¬
ние этой реакции в каком-то смысле не было неожиданным, поскольку в одном из более
ранних исследований такое проявление трусости у потомства Fi было обнаружено у гиб¬
ридов немецких овчарок с волками. В случае волко-собачьих гибридов их выраженная пас¬
сивно-оборонительная реакция, также отсутствовавшая у обоих родителей, была приписа¬
на специфическим особенностям генотипа волка. В случае же скрещивания упомянутых
выше двух пород дикие предки в родословной отсутствовали, и причиной повышенной
трусливости потомства был какой-то иной фактор. В то же время характерной особеннос¬
тью и волков, и гиляцких лаек была достаточно низкая общая возбудимость, проявлявшая¬
ся в невысоком уровне двигательной активности этих животных.JI. В. Крушинский предположил, что у гиляцких лаек существует породная предрас¬
положенность к проявлению пассивно-оборонительной реакции, которая не обнаружива¬
ется у них из-за низкого уровня возбудимости. Гибриды этих собак с овчарками, унаследо¬
вавшие высокую возбудимость последних, получили от другого родителя и пассивно¬
оборонительную реакцию, в результате чего это свойство поведения — повышенная трус¬
ливость — было выражено у них в полной мере. Такое объяснение было подтверждено
простым экспериментом с введением гиляцким лайкам кокаина, дофаминергического аген¬
та, который специфически активировал двигательную сферу животного. У гиляцких лаек,
но не у собак других пород, получавших инъекции кокаина, обнаруживалась четкая пас¬
сивно-оборонительная реакция. Таким образом, уровень возбудимости, который в данных
экспериментах коррелировал с уровнем двигательной активности, оказался модулятором
проявления других признаков поведения. В последующих исследованиях Л. В. Крушинс-
кий [1986] продолжил анализ проблемы возбудимости ЦНС как фактора, влияющего на
проявление и выражение генетически детерминированных особенностей поведения.Таким образом, генетически детерминированный высокий уровень признака поведе¬
ния может не обнаруживаться у животных, обладающих маловозбудимым фенотипом. В то
же время потомство, полученное от скрещивания таких собак с возбудимыми особями, об¬
наруживает эти признаки достаточно отчетливо. Классическими примерами сочетания по¬
вышенной возбудимости и пассивно-оборонительной реакции у потомков, полученных от
разных родителей, могут служить гибриды собак с дикими представителями семейства
Canidae.Изучение поведения гибридов собак с дикими формамиУже упоминались полученные Крушинским [1991] данные о сильно выраженной пас¬
сивно-оборонительной реакции у гибридов собак и волков, выращенных в Московском
зоопарке. К гибридизации собак и волков не раз прибегали как с целью изучения их пове¬
дения, так и в надежде на совершенствование рабочих качеств собак.Практически у всех таких гибридов не только первого, но и последующих поколений,
как правило, проявлялась боязнь новых предметов, ярко выраженная общая трусость, час¬
то в сочетании с повышенной агрессивностью. Контакты с такими животными нередко
представляют опасность. По данным собаководов-практиков пассивно-оборонительная
реакция таких животных выражена более резко, чем у волков. У гибридов волка и собаки
первого поколения описаны проявления пугливости и боязни новых предметов. Во втором
поколении произошло четкое расщепление по этому признаку [Iljin, 1941; Woolpy,
Ginsburg, 1967].Дрессировка волко-собачьих гибридов, по данным Е. Humphrey [1934], происходила
успешно только если животные были на привязи, но как только их отпускали, они переста¬
вали повиноваться. Склонность к одичанию, которая связана с уровнем проявления пас-
сивно-оборонитсльной реакции, отмечена у гибридов волков и овчарок [Адамец, 1930]. Эк¬
сперименты по скрещиванию пуделей с волками и с шакалами были проведены в рамках
проектов ,,Puwo“ и ,,Pusha“. Согласно результатам этих работ злобнотрусливые особи
продолжали встречаться во многих поколениях [Zimen, 1969]. На основе гибридизации со¬
бак с волками была выведена порода „чешский волчек“, и хотя волчьи предки этих собак
давно вышли за пределы родословной, эти животные характеризуются большой насторо¬
женностью и недоверчивостью по отношению к человеку [Гражданкина, 1991].Работа по скрещиванию собак (биглей) с койотами имела целью, в первую очередь,
изучить наследование звуковых сигналов у этих видов. У биглей сигнал угрозы — рыча¬
ние, тогда как койоты угрожают специфическим звуком, так называемым „зевком с шипе-
нием“. У потомства от скрещивания собак с койотами оба типа звуковых сигналов распре¬
деляются согласно менделевской схеме, причем сигнал койота оказывается рецессивным по
отношению к рычанию биглей [Smiley et al., 1977].Далее было показано, что у всех гибридов третьего поколения, имевших родителями
гомозигот по предположительно рецессивному признаку „зевок с шипением", этот тип сиг¬
нала обнаруживался [Schotte, Ginsburg, 1979].Дальнейший анализ несколько видоизменил представления о характере проявления и
наследования этих признаков. Было показано, что среди гибридов второго поколения, ко¬
торые в детском возрасте обнаруживали сигнализацию угрозы путем рычания, по достиже¬
нии половой зрелости начинал преобладать тип сигнализации койота. При этом доля „ши¬
пящих“ потомков F2 по отношению к „нешипящим11 бьша почти в 2 раза выше [Moon,
Ginsburg, 1981]. В дальнейшем были получены указания на возможное существование 3 на¬
боров генов, которые ответственны за проявление свойственного койотам сигнала угрозы
[Moon et al., 1982]. Данные наблюдений за поведением гибридов разных поколений от
скрещиваний собаки и шакала, полученные Сулимовым [Бехтин, Сулимов, 1985], подтвер¬
ждают сложность этой генетической системы.Некоторые особенности поведения собак охотничьих породГенетическому анализу поведения охотничьих собак посвящены работы М. March-
lewski [1930]. Он исследовал главным образом пойнтеров и немецких легавых. Для пойнте¬
ра характерна четкая длительная поза „стойки" перед обнаруженной затаившейся дичью.
Немецкие легавые также делают стойку, но она не имеет такой четкости и длительности.
Наследование четкой стойки пойнтеров описывается как неполное доминирование. У пой¬
нтеров обнаруживается явление так называемого „секундирования", когда собака делает
стойку не только на запах дичи, как обычно, но и при виде другой собаки, стоящей на
стойке. Очевидно, что способность к обучению такой специфической реакции, как секун-
дирование, может быть более сильно выражена у определенной породы собак. По данным
Marchlewski, эта способность четко прослеживается в отдельных семьях и линиях.М. Burns и М. Fraser [1966] цитируют более позднее исследование того же автора, в
котором он пришел к выводу, что стойка является суммарным результатом проявления не¬
скольких генетических факторов, и что между животными одной породы обнаруживается
достаточно большая изменчивость по этому сложному признаку.Интересно, что манера молодых немецких легавых преследовать добычу „с подпаи¬
ванием" оказалась рецессивным признаком по отношению к „молчаливому" способу охо¬
ты пойнтеров, что не совпадает с наблюдениями L. Whitney [1929 Ь].И Marchlewski, и Whitney указывают, что манера разыскивать птицу „нижним чуть¬
ем", т. е. двигаясь по следу с опущенной головой, есть рецессивный признак по отношению
к поиску дичи с высоко поднятой головой, с использованием „верхнего чутья", т. е. с улав¬
ливанием запаха самой птицы, а не ее следа. L. Whitney [1971] описал доминирование пове¬
дения охоты на птицу (и интереса к летающим предметам) у легавых собак.Межпородные различия в темпах созревания реакций щенков на потенциальную до¬
бычу анализировал Vauk. Таксы обнаруживали интерес к голубям уже в возрасте 22 дней,
чау-чау — в 46 дней. Пойнтеры делали стойку уже в возрасте 35 дней, тогда как у пуделей
охотничьи реакции отсутствовали.Специфические свойства поведения собак
и анализ их генетического контроляМ. Burns и М. Fraser [1966] приводят данные Kelley о том, что пастушеское поведение
колли контролируется большим числом генов аддитивного действия. Ginsburg и Zamic,
[1944. Цит. по: Mackenzie et al., 1986] исследовали в полевых условиях реакции необучен¬
ных собак на коз. Спонтанное пастушеское поведение было обнаружено у небольшой час¬
ти животных. Авторы считают, что это поведение имеет генетический контроль.Оборонительные реакции собак — их пороги и частота проявления — анализирова¬
лись многими авторами. Трусливость собак в связи с особенностями протекания процесса
индивидуального развития была впервые проанализирована в классическом исследовании
С. Н. Выржиковского и Ф. Н. Майорова [1933], проведенном в лаборатории И. П. Павло¬
ва. Они показали, что изолированное от внешнего мира воспитание щенков приводит к
развитию у них выраженного пассивно-оборонительного поведения, демонстрируя тем са¬
мым важнейшую роль внешних условий в становлении нормального поведения особи.
Павлов назвал этот феномен „неизжитым рефлексом естественной осторожности". При со¬
прикосновении щенков со всем многообразием внешнего мира происходит, по его мнению,
постепенное угасание этого рефлекса. В генетическом плане боязнь сильных раздражите¬
лей и громких звуков наследуется независимо. Павлов [1938] подчеркивал, что частота
проявления и интенсивность реакций обоих типов зависят от истории индивидуального
развития животного.
H. Trimble и С. Keeler [1939] исследовали специфическое поведение далматинов, на¬
зываемых „экипажными собаками11. Этих собак в течение многих поколений отбирали на
склонность к бегу рядом с экипажем во время его движения. Такое поведение начинает
проявляться у щенков этой породы без предварительного обучения, однако в разных семь¬
ях оно выражено неодинаково.JI. В. Крушинский [1991] изучал наследование высоких величин проявления апорти-
ровочной реакции, которая выражается в стремлении собак постоянно носить и держать
во рту разные предметы. Обычно собаки легко обучаются апортировке. Однако некоторые
животные избегают носить что-либо во рту и обучаются этому с трудом. Направленные
скрещивания и семейный анализ, проведенные Крушинским, показали доминантный ха¬
рактер наследования этой особенности поведения. Неопубликованные наблюдения авто¬
ров данной главы (М. Н. Сотская, Е. В. Мычко) подтверждают этот вывод.Наследование пассивно-оборонительной реакции у собак также исследовал Крушин¬
ский [1991]. Было проанализировано поведение 278 потомков от разных скрещиваний. По¬
казано, что высокая выраженность этой реакции наследуется как доминантный признак,
однако в определении величины признака участвует более чем один главный ген.Проявление агрессивности, а точнее, ряда агрессивных реакций собаки, и в том числе
„хватки" у собак — „активно-оборонительная реакция1' в работах Крушинского [1991] —
зависит от генотипа животного. Исследуя собак нескольких пород: эрдельтерьеров, немец¬
ких овчарок, доберманов, а также беспородных собак, воспитанных либо в условиях пи¬
томника, либо частными лицами, автор обнаружил, что хотя условия частичной изоляции,
т. е. „обедненные условия" питомника, и способствовали большей выраженности пассив-
но-оборонительной реакции, межпородные различия по этому признаку были достаточно
велики. Эксперименты по скрещиваниям показали также, что пассивно- и активно-оборо¬
нительные реакции собак наследуются независимо друг от друга. Как уже говорилось вы¬
ше, проявление и выражение этих свойств поведения собаки находятся в зависимости от
уровня общей возбудимости животного.Аналогичны данным JI. В. Крушинского результаты исследования Н. Mahut [1958],
которая исследовала собак 10 пород, выращенных в условиях обедненной среды.Опубликованные материалы F. Thorne [1944] также подтверждают полученные Кру¬
шинским результаты. Генетический анализ, проведенный в его работе на группе из 178 со¬
бак, показал, что из 82 трусливых собак 43 оказались потомками одной очень трусливой
суки бассета. Автор пришел также к выводу, что у басенджи социальная иерархия выраже¬
на более четко, чем у неагрессивных кокер-спаниелей, при этом в обеих породах суки были
менее агрессивными.Генетика поведения собак в исследованиях P. Scott и J. FullerВ середине 1940-х годов было начато наиболее полное и подробное исследование ге¬
нетики поведения собак. Результаты этой многолетней работы изложены в монографии
P. Scott и J. Fuller [1965], которая продолжает оставаться одной из наиболее авторитетных
книг на эту тему.Объектами исследования были собаки 5 пород — басенджи, бигли, американские ко¬
кер-спаниели, шетландские овчарки (шелти) и жесткошерстные фокстерьеры, а также гиб¬
ридные животные. Помимо цели сравнения поведения собак разных пород и их гибридов,
авторы были заинтересованы в изучении формирования поведения собаки в онтогенезе.
Для этого они проводили подробные систематические наблюдения щенков, начиная от
рождения и до возраста 16 недель. Всего было исследовано несколько сотен животных.Теоретической основой анализа поведения собак в этой работе было представление о
существовании достаточно жестких стереотипных поведенческих реакций, которые могут
обнаруживаться в репертуаре поведения собак той или иной породы с разной частотой. На
основе этих стереотипных реакций, которые авторы не отождествляли с ФКД, может фор¬
мироваться более сложное поведение собаки — пищевое, половое, материнское, социаль¬
ное, защитное и т. д. Авторам, в частности, импонировала классификация поведения, в
свое время предложенная JI. В. Крушинским, по которой более крупными категориями по¬
ведения следует считать так называемые „биологические формы поведения", в основном
совпадающие с категориями, перечисленными выше.
Первый этап работы включал в себя подбор пород собак и методик тестирования по¬
ведения. При исследовании развития поведения в ряде случаев щенков воспитывали „пере-
крестно“, т. е. часть помета выращивала биологическая мать, а часть — приемная, т. е. су¬
ка другой породы. Небольшое число собак всех пород было выращено не в питомнике, а в
семьях. Если поведение этих собак по отношению к человеку и было несколько иным, чем
собак питомника, то показатели большинства тестов были ни лучше, ни хуже, чем у ос¬
тальных животных. Специальное внимание исследователи уделяли условиям содержа¬
ния, выращивания и кормления животных, стремясь обеспечить их максимальное едино¬
образие.Вторым этапом исследования был генетический эксперимент, для которого были
выбраны две заведомо неродственные между собой породы собак — басенджи и американ¬
ские кокер-спаниели.Басенджи — это неспециализированная порода охотничьих собак африканских пле¬
мен. Они выраженно агрессивны и по ряду признаков напоминают диких животных. Коке¬
р-спаниели — напротив, порода с низкой агрессивностью и достаточно долгой историей
заводского разведения.По данным эксперимента относительная дикость басенджи проявляется у щенков
при попытках взять их в руки — они ведут себя подобно детенышам диких животных: из¬
дают резкий визг, пытаются вырваться, укусить и т. п. Кроме того, они резко сопротивля¬
ются попыткам обучению ходить на поводке. Гибриды первого поколения сходны по этим
признакам с басенджи. Картина расщепления признаков во втором поколении и у беккрос-
сов показала, что „дикость1* басенджи контролируется одним доминантным геном. Сопро¬
тивление ограничению свободы, повидимому, также контролируется одним геном, но без
доминирования. В этом случае нельзя также исключить и влияние материнского эффекта.P. Scott и J. Fuller изучили и такой признак, как агрессивность в игре, которая, по их
данным, может контролироваться двумя генами без доминирования, но может иметь и бо¬
лее сложное определение.Голосовые реакции авторы анализировали у щенков разного возраста в тесте на до¬
минирование, когда двух животных заставляли бороться за обладание костью в течение
10 минут. Оказалось, что голосовые реакции четко подразделяются на 2 независимых при¬
знака. Один из них — это порог провокации лая, т. е. уровень стимуляции, необходимый
для проявления реакции. Он оказался высоким для басенджи и низким для кокер-спани¬
елей. Второй признак — это длительность лая. Она была малой у басенджи и большой у
кокер-спаниелей. По данным скрещиваний низкий порог проявления лая доминирует над
высоким, а частота и длительность лая наследуются по промежуточному типу. Авторы по¬
лагают, что наследование уровня порога провокации определяется двумя независимыми
доминантными генами, тогда как длительность лая не обнаруживает доминирования ни
высоких, ни низких значений и зависит, по-видимому, от одного гена.Подобным образом были подвергнуты анализу и многие другие признаки поведения,
наследование которых также оказалось под контролем одного или двух генов. Представля¬
ет, в частности, интерес наследование характеристики эстрального цикла самок. У басенд¬
жи течка наступает один раз в год и строго приурочена к осеннему сезону. У кокер-спание-
лей течка бывает один раз в 6 месяцев в любой из сезонов года. В таблице 8.1 кратко
суммированы некоторые результаты этой работы, описание которой можно также найти в
монографии JI. Эрман и П. Парсонса [1984].P. Scott и J. Fuller [1965] оценили результаты своих исследований как довольно нео¬
жиданные, поскольку для ряда достаточно сложных признаков поведения было продемон¬
стрировано моно- и олигогенное наследование. Они отметили, что взятые для исследова¬
ния породы собак были в достаточной степени контрастны по поведению, видимо,
вследствие разных направлений отбора при их формировании. Они сделали вывод, что по¬
родное разведение собак хотя и не создает полной инбредности, но способствует появле¬
нию гомозиготности по многим аллелям. Если в скрещивании участвуют 2 неродственные
между собой породы (как было в данном случае), то во втором поколении можно получить
картину расщепления, которая свидетельствует о контроле величины признака одним или
двумя генами.Именно здесь уместно вспомнить те рассуждения, которые приводились выше. Речь
шла о том, что при внутривидовых сопоставлениях поведения и при оценке внутривидовой
изменчивости поведения практически всегда обнаруживается изменчивость по частоте вы-
Таблица 8.1. Сравнительная характеристика поведения басенджи и кокер-спаниелей[По: Scott, Fuller, 1965]ПоведениеПризнакиБасенджиКокер-спаниелиНаиболее вероятный тип
наследованияДикость и прирученностьВысокийуровеньНизкийуровеньОдин доминантный ген,
определяющий дикостьИзбегание и голосовые
реакции при взятии на рукиСопротивление ограничению
движенийВысокийуровеньНизкийуровеньОдин ген без доминированияАгрессивность во время
игры в возрасте 13-15 нед.ВысокийуровеньНизкийуровеньДва гена без доминированияЛай в возрасте 11 нед.
(порог раздражения)ВысокийНизкийДва доминантных гена,
определяющих низкий порогТенденция лаять малое
число разСильнаяСлабаяОдин ген без доминированияПоловое поведение
(время эструса)1 раз
в год1 раз
в полгодаТип басенджи определяется
одним рецессивным геномполнения той или иной поведенческой реакции или движения (ФКД), тогда как форма та¬
ких движений бывает неизменной. Крайним случаем вариации частоты ФКД может быть
почти полное его отсутствие в репертуаре поведения собак данной породы, т. е. его очень
редкое проявление. С точки зрения существования большой изменчивости порогов реак¬
ций и частоты их выполнения можно полагать, что всегда существуют внешние условия,
которые могут спровоцировать появление реакции, даже если порог ее проявления доста¬
точно высок.Интересно отметить, что P. Scott и J. Fuller, приступая к опытам по изучению генети¬
ки поведения собак, отметили, что практически по всем признакам имело место перекры¬
вание их между породами, т. е. не было признака, который бы наблюдался только у одной
породы и полностью отсутствовал у других. Их выводом было то, что не существует како¬
го-либо „породного набора признаков", поскольку в любой породе можно найти особей,
чьи признаки поведения перекрываются признаками другой породы.Следует обратить внимание на то, что олигогенное наследование обнаруживается
только для некоторых признаков поведения, тогда как во многих случаях характер распре¬
деления признаков у потомства свидетельствовал о полигенном наследовании. Это в пер¬
вую очередь относится к использованным в работе сложным тестам. Они включали в себя
обучение в лабиринте, дифференцировку, тест на оперирование пространственными пред¬
ставлениями. Основным выводом из этой части исследования, помимо установления слож¬
ного полигенного наследования способностей к решению таких тестов, было то, что в оп¬
ределении успешности решения данного теста собаками разных пород важную роль
играют „некогнитивные11 переменные. В частности межпородные различия в успешности
решения задач могут быть детерминированы различиями в трусливости и эмоциональнос¬
ти, тогда как истинные градации по уровню собственно когнитивных способностей уста¬
новить достаточно трудно.Работа P. Scott и J. Fuller, проведенная на начальных этапах развития генетики пове¬
дения и на таком сложном объекте, как собака, тем не менее предвосхитила основные ре¬
зультаты, полученные в этой области в дальнейшем, поскольку генетические исследования
сложных форм поведения лабораторных грызунов выявляют важную роль в их осуществ¬
лении более простых переменных [Эрман, Парсонс, 1984].
Корреляция между физическими и поведенческими признакамиПо мере развития генетических методов в животноводстве делались попытки исполь¬
зования их и при выведении пород собак.В диссертации М. Pfeiderer-Hogner [1968] приводятся данные Schaeller по немецким
овчаркам за период 1900-1937 гг., который провел анализ породной ценности и наследова¬
ния ряда признаков. Оказалось, что потомство от чемпионов породы не превышало по
своим качествам потомство от нечемпионов. Автор сделал вывод о том, что при разведе¬
нии собак надо использовать широкую основу, т. е. отказаться от скрещивания малого
числа выдающихся животных. В работе, проведенной на колли, было показано, что сред¬
няя оценка признака у потомства была близка к таковой у родителей [Mackenzie et al.,
1986].Оценка фенотипических, но не генотипических, корреляций между 42 физическими и
9 поведенческими признаками у немецких овчарок Швейцарии показала, что четких соот¬
ношений между ними не выявляется. Иными словами, между экстерьером собак определен¬
ной породы и особенностями их поведения не обнаруживается сколько-нибудь жесткой
связи [Humphrey, Warner, 1934]. Показано также [Ginsburg, 1958], что жесткий отбор по по¬
ведению приводит к уменьшению его изменчивости, тогда как изменчивость экстерьерных
признаков увеличивается. Аналогичное явление обнаружено в линиях английских сетте¬
ров: победители соревнований по рабочим качествам имели посредственные оценки за эк¬
стерьер и наоборот [Ginsburg, 1958]Обычно применяемый в животноводстве метод оценки производителей по потомству
был постепенно внедрен и в практику разведения собак разных пород [Pfeiderer-Hogner,
1968]. Однако если в животноводстве обычно оцениваются значительно более простые
признаки (например, молочность и т. п.), то оценка признаков поведения у собак требует
предварительной выработки строгих критериев, что не всегда оказывается просто. В то же
время применение такого метода в селекционной работе с использованием характеристик
поведения может оказаться очень удачным.В той же работе [Pfeiderer-Hogner, 1968] сообщаются данные по пойнтерам. На осно¬
ве изучения племенных книг собак этой породы и анализа данных по поведению (правда,
оценивали всего лишь 4 показателя) был сделан вывод о том, что классификация собак по
рабочим качествам осуществляется неверно. В то же время, судя по аналогичной работе,
проделанной с таксами, но на основе 12 показателей, существует нормальное распределе¬
ние признаков. В этой работе не обнаружено влияния пола на признаки поведения, но был
выявлен материнский эффект.При изучении параметров поведения собак установлено до 15 признаков поведения,
по которым можно обнаружить межпородные различия [Catell et al., 1973]. Достоверные
половые различия в ориентировочной реакции, оборонительном поведении и др. были вы¬
явлены при дрессировке немецких овчарок [Reuterwall, Ryman, 1973] и лабрадоров-рстри-
веров. Половые различия в уровне возбудимости отмечены и в исследованиях JI.B. Кру-
шинского [1991].Наряду с изучением корреляций между породными особенностями экстерьера и пове¬
дением, внимание исследователей привлекли также патологические признаки, осложняю¬
щие поддержание породы, такие, как упоминавшаяся выше дисплазия тазобедренного сус¬
тава [Castleberry et al., 1975]. При анализе наследуемости дисплазии и признаков поведения
у собак 8 пород установлена корреляция дисплазии с остротой слуха, которую определя¬
ли по громкости команды, необходимой для привлечения внимания новой в питомнике
собаки. Коэффициент корреляции составил 0,52 [Bartlett, 1976]. Достоверные величины ко¬
эффициента корреляции были найдены для таких пар признаков, как „острота слуха —
энергичность11, „тактильная чувствительность — обонятельная чувствительность", „само¬
уверенность — доверчивость", а также „острота слуха — обонятельная чувствительность".Влияние инбридинга на поведение собак — вопрос важный и в теоретическом, и в
практическом отношении. М. Marchlewski отмечал, что по его наблюдениям пойнтеры с
наиболее хорошими рабочими качествами, как правило, оказывались гетерозиготными. В
другой работе [Brown et al., 1978] сравнение высокоинбредной и неинбредной линий пойн¬
теров обнаружило, что первые проявляли враждебность к людям, тогда как вторые были
дружелюбными. Было отмечено, что враждебное отношение к человеку может контроли¬
роваться генами аддитивного действия.
Кластерный анализ признаков поведенияКорреляционный анализ в его современных модификациях использовали для оценки
породных особенностей собак в их взаимодействиях с человеком [Hart В. L., Hart L. А.,
1984]. Эта работа была проведена для выработки стандарта в рекомендациях по содержа¬
нию в домашних условиях той или иной породы собак. Для отыскания научной основы для
таких рекомендаций группе ветеринаров и экспертов-собаководов предложили провести
классификацию 56 пород собак по 13 признакам поведения. Каждому из участников нужно
было дать оценку 7 пород, выбранных случайно. 56 пород, участвовавших в общей оценке,
были отобраны по принципу участия разных групп пород: рабочие, спортивные, охотни¬
чьи, декоративные и терьеры. Анализировали возбудимость, тенденцию к голосовым реак¬
циям, чрезмерно сильный лай, сторожевой лай, общий уровень активности, злобность по
отношению к детям, стремление к доминированию над хозяином, послушание при дресси¬
ровке, легкость приучения к порядку в доме, „злонамеренность" действий, склонность к
игре.Признаки поведения оценивали по специальной Ю.-балльной шкале, что позволило
сопоставить породы по количественному выражению того или иного признака. Таким пу¬
тем были получены „профили поведения" пород. Построение подобных „профилей" облег¬
чало характеристику пород и давало шанс потенциальным владельцам собак получить
обоснованную рекомендацию.Полученные данные были также подвергнуты кластерному анализу. Предварительно
было выявлено, что взятые для исследования признаки поведения распределяются по 4 ос¬
новным факторам: реактивность, дрессируемость, агрессивность и отдельный фактор, на
который влияли показатели „злонамеренности" и склонности к игре.Для проведения кластерного анализа выраженность 3 основных факторов — реак¬
тивности, агрессивности и дрессируемости — также оценивали по 10-балльной шкале. При
этом величины факторов в интервалах 1-2,5 оценивались как очень низкая интенсивность,
2,6^4,0 — низкая, 4,1-7,0 — средняя, 7,1-8,5 — высокая, а 8,5-10 — очень высокая. Комби¬
нации определенной интенсивности каждого из факторов формировали отдельный клас¬
тер. Интересно, что кластеры, организованные таким способом, достаточно хорошо совпа¬
ли (хотя и не полностью) с традиционной классификацией пород собак по группам
использования (охотничьи, рабочие и т. п.). Однако в каждый кластер входили собаки бо¬
лее чем одной породной группы. В некоторые кластеры попали собаки со сходным типом
строения тела, но не было ни одного кластера, который включал бы в себя весь набор ти¬
пов телосложения.Эти результаты свидетельствуют о существовании каких-то сложных корреляцион¬
ных связей между телосложением и поведением, выявить которые пока не представляется
возможным. В таблице 8.2 приведено распределение проанализированных пород собак по
кластерам. Цифра в скобках обозначает генетическую дистанцию между данной породой и
центром кластера. Таблица 8.2 показывает вполне определенную тенденцию совпадения
некоторых особенностей телосложения и поведения. Из нее также видно, что, например,
терьеры, характеризующиеся сравнительно однотипным телосложением, оказались в од¬
ном кластере, заняв его почти целиком.Из данных этого исследования можно также заключить, что, по всей видимости, пра¬
вильнее не рассматривать корреляцию типа строения тела с отдельными признаками пове¬
дения, а анализировать корреляции между определенными комплексами признаков поведе¬
ния и экстерьером. Влияние пола на поведение также было выявлено в этой работе с
достаточной отчетливостью — половые различия были обнаружены в послушании при
дрессировке, общей активности, агрессивности по отношению к другим собакам, тенден¬
ции доминировать над хозяином.В поведении собаки всегда можно обнаружить некую общую схему, в соответствии с
которой оно проявляется. Этот шаблон унаследован ею от диких предков и достаточно
консервативен. Как мы видели, различия между особями или между породами собак могут
касаться чисто количественных аспектов в проявлении той или иной реакции, т. е. особен¬
ностей, которые появились как ответ на отбор на ту или иную особенность поведения со¬
баки. Такое сходство в поведении собак всех пород тем более удивительно, если иметь в
виду огромное разнообразие этих животных по физическому облику. Это обстоятельство
Таблица 8.2. Кластерный анализ пород собакКластер 1: высокая реактивность, низкая дрессируемость, средняя агрессивностьЛхасский апсо (0,93,Д)Бигль (1,83,0)Померанский шпиц (1,30,Д)Иоркшир-терьер (1,86,Т)Мальтийская болонка (1,37,Т)Веймаранер (1,94,С)Кокер-спаниель (1,57,Т)Мопс (2,02,Д)Бостон-терьер (1,63,Т)Ирландский сеттер (2,09,С)Пекинес (1,75,Д)Кластер 2: очень низкая реактивность, низкая дрессируемость, очень низкая агрессивностьАнглийский бульдог (1,21,Д)Бладхаунд (1,59,0)Староанглийская овчарка (1,23,Р)Бассет (1,77,0)Норвежская оленегонная (1,39,0)Кластер 3: низкая реактивность, низкая дрессируемость, высокая агрессивностьСамоедская лайка (0,41,Р)Боксер (1,57,Д)Маламут (0,91,Р)Далматин (1,61,Д)Сибирская лайка (0,95,Р)Дог (1,99,Р)Сенбернар (1,01,Р)Чау-чау (2,48,Р)Афганская борзая (1,20, О)Кластер 4: высокая реактивность, очень высокая дрессируемость, средняя агрессивностьКолли (1,04,Р)Стандартный пудель (2,16,Р)Ши-тцу (1,22,Д)Английский спрингер-спаниель (2,33,С)Миниатюрный (1,39,Д)Вельш-корги (2,82,Р)Той-пудель (1,74,Д).Кластер 5: низкая реактивность, высокая дрессируемость, очень высокая агрессивностьЛабрадор-ретривер (0,91,С)Чесапик-бей-ретривер (1,58,С)Выжла (1,19, С)Кеесхонд (1,59,Д)Бретонский эпаньол (1,42,0)Колли (1,60,Р)Немецкий короткошерстный пойнтер (1,48,С) Золотистый ретривер (1,83,С)Ньюфаундленд (1,50,Р)Австралийская пастушья собака (2,39,Р)Kiacmep 6: очень низкая реактивность,очень высокая дрессируемость, очень высокаяагрессивностьНемецкая овчарка (0,94,Р)Доберман (1,39,Р)Акита-ину (1,22,Р)Ротвейлер (1,40,Р)Кластер 7: высокая реактивность, средняя дрессируемость, очень высокая агрессивностьКэрн-терьер (0,79,Т) Скотч-терьер (1,70,Т)Уэст-хайленд-уайт-терьер (1,29,Т) Такса (1,73,0)Чихуахуа (1,31,Д) Цвергшнауцер (1,80,Т)Фокстерьер (1,62,Т) Силки-терьер (1,86,Т)Эрдельтерьер (2,14,Т)Примечание. Группы пород: Д — декоративные, Р — рабочие, С — спортивные, О — охотничьи, Т — терьеры.
демонстрирует, по всей видимости, проявление закономерностей, в соответствии с которы¬
ми происходит преобразование мозга животных в процессе доместикации, и которые ста¬
новятся предметом специального внимания нейрогенетики.Поведение собаки интересно также сочетанием четкого консерватизма видового по¬
ведения с необычайной пластичностью адаптивных реакций — высокой способностью к
научению и к рассудочной деятельности. Проблема генетических закономерностей в реа¬
лизации подобного сочетания уже попала в поле внимания генетики поведения, но пред¬
ставляется очень сложной.Как генетический объект собака занимает промежуточное положение между лабора¬
торными грызунами и человеком. Это крупное животное, которое трудно содержать и спе¬
циально разводить в исследовательских целях. В связи с этим большое число работ по ге¬
нетике поведения собаки базируется на обследовании животных, принадлежащих частным
владельцам. Этические соображения также ограничивают возможности экспериментов на
них.
Гпава 9
БИОЛОГИЯ РАЗМНОЖЕНИЯ СОБАКИДомашние собаки (Canis familiaris) представлены большим количеством пород. Ты¬
сячелетняя селекция привела к появлению собак, различающихся экстерьером, поведением.
Однако в становлении половой функции у собак разных пород имеется много общего.Половое созреваниеПервичные половые клетки — гоноциты — появляются у собак (как и у других мле¬
копитающих) на очень ранней стадии эмбрионального развития, вскоре после импланта¬
ции зародыша в матке. Имплантация — это процесс, в результате завершения которого
формируется сосудистая связь между организмом самки и зародышем. Примерно на
23-24-й день беременности, на стадии, когда начинается дифференцировка основных сис¬
тем зародыша, потенциальная ткань гонад представлена неорганизованными соматически¬
ми и герминативными элементами; к 28-му дню формируются индифферентные половые
валики, а на 30-й день под действием гормонов гипоталамуса начинается половая диффе¬
ренцировка, приводящая к формированию яичников, семенников и соответствующих сис¬
тем, в зависимости от наличия X- и Y-хромосом. Половая дифференцировка заканчивается
к 35-му дню беременности.Яичник новорожденной самки имеет шаровидную форму размером около 4,0x6,0 мм,
содержит примерно 700 000 ооцитов, расположенных в кортикальном слое, представлен¬
ном кубоидальным эпителием, содержащим кластеры ооцитов и примордиальных фолли¬
кулов [Anderseen, Wooten, 1959].Отмечена связь между ростом самки от рождения и половым созреванием, которое
наступает, когда кривая роста достигает плато (рис. 9.1). В период быстрого роста овари¬
альные фолликулы увеличиваются в размере; истинные примордиальные фолликулы най¬
дены в яичниках 15-дневных щенков, в 5-месячном возрасте кортекс яичника содержит уже
полостные фолликулы, аграафовы пузырьки — фолликулы, готовые к овуляции — появля¬
ются в возрасте 6,5 месяцев, но до 8-месячного возраста, когда кривая роста щенков выхо-Возраст, дниРис. 9.1. Кривая роста собаки от рождения до половой зрелости [Anderseen, Wooten, 1959]:1 — самиы, 2 — самки
дит на плато, они подвергаются резорбции. В 8-месячном возрасте созревание граафовых
пузырьков свидетельствует о наступлении половой зрелости. Однако существуют индиви¬
дуальные и породные различия в сроках наступления половой зрелости. У собак крупных
пород эструс наступает в возрасте 1,5-2 лет, но первые сигналы появляются между 10 и
14 месяцами. Определенные признаки эструса, однажды появившись, устанавливают рит¬
мичность потенциальных репродуктивных возможностей, продолжающихся в течение всей
жизни собаки [Anderseen, Wooten, 1959].У взрослой собаки размеры яичника составляют 2,5x0,7x0,5 см, вес около 3 г. До
6-9-месячного возраста поверхность яичников гладкая, позже (особенно в период течек) у
многорожавших самок становится шероховато-узловатой. Мозговое вещество (medulla
owari) богато кровеносными сосудами, нервами, соединительной тканью и содержит глад¬
кие мышечные волокна. Корковое вещество (cortex owari) состоит из плотной соединитель¬
ной ткани, в которой находятся кластеры первичных фолликулов [Miller et al., 1965; Хро¬
мов и др., 1972].Около 12-14' % собак имеют полиовуляторные фолликулы, содержащие больше од¬
ной яйцеклетки, количество ооцитов может доходить до 5 и более (до 11 [По: Anderseen,
Wooten, 1959]). Чаще такие полиовуляторные фолликулы встречаются в яичниках молодых
самок. Частота различных типов полиовуляторных фолликулов варьирует обратно про¬
порционально количеству ооцитов нафолликул. В молодых яичниках много полиовуля¬
торных фолликуловна стадии роста, но количество их уменьшается по мере созревания и
продвижения к антральной (полостной) стадии. По своим размерам они крупнее, чем од¬
нояйцевые, но ооциты в них меньше, однако они растут с увеличением массы гранулезных
клеток [Anderseen, Wooten, 1959; Tefler, Gosden, 1987].Количество фолликулов в яичниках собак уменьшается с возрастом. Так, если в яич¬
никах новорожденных сук содержится примерно 700 000 ооцитов, то ко времени наступле¬
ния половой зрелости их остается примерно половина, и только 1200-1300 достигает ста¬
дии граафова пузырька. Дегенерация, атрезия фолликулов являются нормальным
процессом, у старых самок (примерно к 10-летнему возрасту) в яичниках насчитывается не
более 500 фолликулов [Anderseen, Wooten, 1959].Размеры яйцеклетки составляют 77x90 микрон, а вместе с блестящей оболочкой, ок¬
ружающей ее, — 95x110 микрон. Яйцеклетки содержат большое количество гранул и жира,
поэтому оптически совершенно непрозрачны (рис. 9.2) [Anderseen, Wooten, 1959; Austin,
1959].Самцы достигают половой зрелости в возрасте 10-12 месяцев. Размеры семенников у
самцов коррелируют с весом собаки, и у кобеля весом в 10 кг составляют 3,0x2,1x1,9 см и
весят 8 г. Объем эякулята составляет в среднем 7,0 мл, количество сперматозоидов —
32х105, это значительно больше, чем требуется для нормального оплодотворения. Подвиж¬
ность спермы в норме составляет 89,5 %, процент аномальной спермы невелик — 5-10 %,
причем больший процент (около 10 %) составляют вторично аномальные, т. е. появляющи¬
еся после окончания сперматогенеза, чаще всего за счет быстрого прохождения через эпи-
дидимис. Сперматозоиды, находясь в половых путях самки, сохраняют оплодотворяющую
способность в течение длительного времени, по некоторым данным до 8 дней [Doak et al.,
1967; Foote et al., 1972; Concannon, 1986; Seager, 1986; England, Allen, 1989].Половое созревание собак завершается в 7-10 месяцев, в это время уже появляются
некоторые признаки полового поведения. Однако к спариванию допускаются собаки, дос¬
тигшие половой зрелости, обычно с 2-летнего возраста. Исключение делают для собак мел¬
ких пород — такс, терьеров, у них спаривание допускается уже в 15-месячном возрасте
[Anderseen, Wooten,1959; Мазовер, 1985; Слимак, Духай, 1986].Половые циклыСобаки в основном моноэстричные животные, т. е. имеют один половой цикл в тече¬
ние года, между периодами половой активности наблюдается длительный период покоя.
Весь период полового цикла может быть подразделен на несколько стадий; проэструс
(в среднем 9 дней) — стадия, предшествующая течке, эструс — стадия течки; метаэст-
рус (89-90 дней) — стадия, следующая за эструсом, анаэструс — стадия полового покоя.
Интервал между эструсами детерминируется в основном длительностью анаэструса. Про¬
должительность анаэструса может сильно изменяться в противоположность изменениям
Оптимальное время для спаривания собак может быть определено при сочетании ме¬
тодов вагинальной цитологии (появление в вагинальном мазке чешуек эпителия), морфо¬
логии, поведения собаки и ее гормонального статуса. Установлено, что наилучшие резуль¬
таты в плодовитости собак получают при спаривании, когда концентрация прогестерона
достигает уровня выше 5 ng/ml. Использование сочетания этих методов для определения
времени спаривания необходимо при искусственном осеменении собак [Шорт, 1987; Dee,
Forhhammer, 1988; Haaften et al., 1989; Jeffcoate, Lindsay, 1989; Tsutsui et al, 1989b].Обычно же наиболее целесообразно проводить вязку собак на 10-12-й день течки, в
этот период созревшие яйцеклетки в яйцеводе готовы к оплодотворению. Для оплодотво¬
рения достаточно одной вязки, уже через 15 минут сперматозоиды достигают воронки яй¬
цевода, где и происходит оплодотворение. Если же нет уверенности, что сука оплодотворе¬
на, то вязку повторяют через 12-14 часов [Мазовер, 1985; Слимак, Духай, 1986].Оплодотворяемость у собак обычно высокая, среди помесейона достигает 90 %. Пос¬
ле одного спаривания она составляет 60 %, после двух — 87-100 %, в зависимости от интер¬
вала между спариваниями. Суперплодовитость отмечается при повторном спаривании с
интервалом, не превышающим 48 часов [Anderseen, Wooten, 1959; Tsutsui, Ejima, 1988;
Haaften et al., 1989].Если не произошло оплодотворения, то у сук наступает состояние псевдобеременнос¬
ти, которое вызывается образованием желтых тел после овуляции фолликулов и их функ¬
ционированием. Для собак характерно, что продолжительность псевдобеременности почти
равнозначна беременности. В течение 5 недель, следующих за эструсом, у псевдоберемен¬
ных самок может наблюдаться увеличение матки, которое может быть определено при
пальпации [Jones et al., 1973; Хип, Флинт, 1987; Шорт, 1987].БеременностьУ беременных же самок желтые тела функционально активны до середины беремен¬
ности, под контролем гормона желтого тела — прогестерона проходят первые этапы эмб¬
рионального развития зародыша и, в частности, имплантации. Беременность самок может
быть диагностирована пальпацией на 30-й день, но с более высокой степенью достовернос¬
ти — на 40-й день. Использование метода ультразвукового сканирования позволяет опре¬
делить беременность на 20-23-й день. Наружные признаки беременности проявляются во
второй половине беременности — увеличивается объем живота, набухают молочные желе¬
зы [Anderseen, Wooten, 1959; Sokolowski, 1980].Продолжительность беременности у собак составляет от 53 до 63 дней, наиболее час¬
то щенение наблюдается через 61,41±2,14 дней после первого спаривания.Эмбриональное развитиеЭмбриональная стадия развития начинается с момента оплодотворения яйцеклетки.
У собак отмечаются значительные вариации между временем овуляции и оплодотворения,
поэтому существует широкая дивергенция в размерах и развитии между пометами одного
срока беременности, если он подсчитывается на основе времени спаривания. Оплодотворе¬
ние возможно в интервале от 3 до 4 дней, между 4-м и 8-м днем после пика лютеинизирую-
щего гормона, т. е. 2-м и 6-м днемпосле овуляции; неоплодотворенные яйца дегенерируют
на 9-й день после овуляции.Весь срок беременности может быть подразделен на 3 периода: предзародышевый —
период развития половых клеток до момента оплодотворения, зародышевый — от момен¬
та оплодотворения до формирования плода, плодный — время от конца зародышевого пе¬
риода до момента рождения.Оплодотворенные яйцеклетки в первые дни, проходя по яйцеводу, претерпевают про¬
цесс дробления, в результате которого образуется зародыш, состоящий из 16—32 клеток,
так называемая морула. Последующие деления клеток, их дифференцировка, приводят к
образованию полостного пузырька — бластоцисты (см. рис. 9.2), диаметр которого дости¬
гает 1 мм. Бластоцисты могут свободно перемещаться в рогах матки, мигрируя из одного
рога в другой. Поздние бластоцисты диаметром около 2 мм теряют блестящую оболочку
примерно на 16-й день после спаривания. На 18-20-й день на стадии примитивной полоски
отмечается имплантация, на 20-й день имплантационные места имеют диаметр около 2 см.
Имплантация у собачьих центральная, поверхностная, зародыш располагается в просвете
рогов, не погружаясь в эндометрий матки. Плацента эндотелиохориальная, зонарная, фор¬
мируется на 21-й день, образуя сосудистую связь между организмом самки и зародышем.
После формирования плаценты развитие зародыша идет быстрыми темпами, к 30-му дню
беременности у зародыша уже имеются закладки всех основных систем. В плодный период
развития (с 30-го дня беременности до щенения) идет быстрый рост плода, увеличение его
массы.В период внутриутробного развития щенков на рост и развитие очень влияют усло¬
вия кормления, и так как продолжительность беременности относительно короткая, то эм¬
брионы растут очень быстро. Это обязывает начинать усиленное кормление суки еще до
спаривания. Щенки у собачьих рождаются беспомощными, слепыми, но развиваются
очень быстро и на 20-й день у них открываются глаза [Anderseen, Wooten, 1959; Holst,
Phemister, 1971; Sokolowski, 1980; Мазовер, 1985; Burke, Badertscher, 1986; Concannon,
1986].Плодовитость собакКоличество щенков у собак во многом зависит от размеров и породы. Суки крупных
пород (например, легавые) родят 5-8, иногда до 16 щенков, некрупных 4-6, иногда 10, а
мелких, таких, как такса, терьер, — 3-6 [Слимак, Духай, 1986; Hahn, 1988].Величина помета у многоплодных животных зависит в основном от двух факторов:
количества овулировавших яйцеклеток (потенциальной плодовитости) и эмбриональной
смертности, т. е. количества эмбрионов, погибших на всех стадиях развития от овуляции
до щенения.Потенциальную плодовитость определяют на основе подсчета количества желтых
тел, образующихся после выхода яйцеклетки из фолликула. У собак количество желтых тел
может варьировать от 1 до 12, в редких случаях до 17 и зависит от размера самки. Но дале¬
ко не все яйцеклетки оплодотворяются и нормально развиваются во время беременнос¬
ти — величина эмбриональной смертности колеблется в пределах от 20 до 25 % [Doak et
al.,1967; Hahn, 1988]. Гибель щенков отмечается и после щенения, примерно в 35,5 % всех
пометов щенки гибнут в течение первых трех недель жизни, общая гибель щенков в этот
период времени составляет 10-11 %. У сук, обработанных антибиотиками во время спари¬
вания и в течение ранней беременности, отмечена тенденция к повышению оплодотворяе-
мости и увеличению величины помета по сравнению с необработанными самками [Linde
Forsberg, Forsberg, 1989].БиотехнологияИскусственное осеменение. В тех случаях, когда самки отказываются от нормального
спаривания, но в то же время от них желательно получить потомство, могут быть исполь¬
зованы методы искусственного осеменения свежей или замороженной спермой [Seager,
Fletcher, 1973]. Так, 3 группы самок (71 крупная, 70 средних и 34 мелких) были осеменены
свежей спермой, взятой от высокопродуктивных самцов, в результате чего от 60 до 70 % са¬
мок в группах принесли потомство, средняя величина помета составила 6,7, 5,4 и 3,8 щенка
на самку [Holzmann, Arbeiter, 1987].Использование замороженной спермы снижает процент оплодотворяемости до 40 %,
подвижность сперматозоидов при размораживании должна быть не меньше 60 %. И если
при искусственном осеменении свежей спермой плодовитость примерно на 20 % меньше,
чем при натуральном спаривании, то при использовании замороженной спермы она сни¬
жается еще на 20 %.Искусственное осеменение производится дважды, с интервалом в 1-2 дня. Во всех слу¬
чаях для успешного оплодотворения при искусственном осеменении необходима строгая
синхронизация стадии эструса и времени осеменения. Для уточнения стадии эстрального
цикла и используют методы вагинальной цитологии и гормонального тестирования. Стро¬
гое согласование стадии эструса и времени осеменения, повышение качества спермы увели¬
чивают оплодотворяемость до 83,8 % при осеменении свежей спермой и до 69,3 % — замо¬
роженной [Farstad, Berg, 1989].
Трансплантация эмбрионов. Операции по пересадке эмбрионов от доноров к самкам
реципиентам проводят на доимплантационных стадиях развития, на стадиях морулы и
ранней бласгоцисты до ее вылупления из блестящей оболочки. На этих стадиях зародыши
могут быть легко извлечены из матки самки реципиента промыванием рогов средой, ис¬
пользуемой для манипулирования с эмбрионами. Наилучшие результаты получают при пе¬
ресадке ранних эмбрионов и при условии синхронизации сроков беременности самок до¬
норов и реципиентов. Однако процент приживаемости эмбрионов не превышает -30 %
[Tsutsui et al., 1989а; Christiansen, 1990]. Операции по пересадке выполняются хирургичес¬
ким методом под общим наркозом.Гормональная обработка. Обработка экзогенными гормонами может быть использо¬
вана для вызывания эструса у самок, непроявляющих признаков течки. Так, ежедневная
обработка анэстричных сук в течение 10 дней PMSG (фоллигон, Intervet) дозой в 500 i.u. и
стилбэстролом дозой в 0,05-1 мг в зависимости от размера самки в течение 20 дней с интер¬
валом в 4-5 дней привела к повышению уровня гормонов до уровня, характерного для
нормально циклирующих самок [Renton et al., 1981].Особенности биологии размножения других видов семейства собачьихК семейству собачьих относятся также лисицы, песцы, енотовидная собака. Эти виды
широко используются в клеточном звероводстве и имеют много общего в физиологии раз¬
множения, прежде всего в гормональной регуляции процессов становления половой функ¬
ции, созревания и овуляции яйцеклеток, формирования желтого тела, процессов импланта¬
ции и протекания беременности [Johansson, 1940; Pearson, Enders, 1943; Venge,
1959;Stephen, Allen, 1983; Valtonen et al., 1985; Насимович,1985].Красная лисица. Самки красной лисицы становятся половозрелыми в 9-10 месяцев,
но в основном к двум годам. Самцы могут принимать участие в размножении на первом
году жизни. Лисицы моноэстричны. Сроки наступления гона зависят от географических
условий, на севере гон начинается позже, чем на юге [Вайсфельд, 1985]: с января по апрель,
а наиболее интенсивно спаривание происходит в феврале-марте. Период половой активно¬
сти может смещаться во времени подчас на 2-3 месяца. Это зависит от погоды, кормовой
базы, а также от индивидуальных особенностей отдельных лисиц.Сроки щенения также связаны с географией. Позже всего лисица приносит потом¬
ство на севере. Число лисят в потомстве подвержено географической изменчивости, на се¬
вере лисицы менее плодовиты (3-6 детенышей), в южных районах число лисят в помете мо¬
жет достигать 13.На плодовитость лисиц сильное влияние оказывает состояние кормовой базы. При
недостатке кормов более 50 % самок остаются холостыми, большой процент составляет ги¬
бель щенков.Лисица еще в конце прошлого века стала объектом клеточного разведения, в резуль¬
тате чего получено большое количество мутантных форм, прежде всего окрасочных. Неко¬
торые из окрасочных мутаций обладают плейотропным действием на плодовитость, чаще
всего снижая ее. В частности, такие мутации, как платиновая, грузинская белая при скре¬
щивании между собой приводят к снижению плодовитости на 25 % за счет эмбриональной
смертности щенков гомозиготного класса, мутация ,,star“ („звездочка") также приводит к
снижению плодовитости самок в результате увеличения гибели эмбрионов во время первой
половины беременности. В последние годы для лисиц разработаны методы искусственного
осеменения самок, в частности для получения лисо-песцовых гибридов. Разработаны и ме¬
тодики трансплантации эмбрионов у лисиц на ранних стадиях беременности [Трут, Голу¬
бица, 1979].Голубой песец (Alopex lagopus) или арктическая лисица. Голубой песец по физиологии
размножения мало чем отличается от лисицы. Сезон размножения песца начинается при¬
мерно на месяц позднее, чем у лисицы и продолжается с середины февраля до конца апре¬
ля. Пик сезона спаривания приходится на 18-19-е марта для самок 2-летнего и более стар¬
шего возраста, молодые самки в первый год часто остаются холостыми.
Продолжительность беременности у песцов, в среднем, 52 дня с очень незначительными ва¬
риациями. Песцы более плодовиты по сравнению с лисицами, плодовитость самок аляс¬
кинского песца в среднем около 10 щенков на самку, гренландского — 8,7. Детородный
возраст песцов 8-10 лет, но старые самки менее плодовиты и часто остаются пропустовав-
шими. Эмбриональная смертность у песцов составляет около 20 %.Получены межвидовые гибриды между лисицей и- песцом. Чаще всего их получают
путем искусственного осеменения самок песцов спермой лисиц, но возможно и нормальное
спаривание. Лисо-песцовые гибриды стерильны [Pearson, Enders, 1943; Venge,1959;
Fougner, 1972].Енотовидная собака. Еще одним объектом клеточного разведения в последнее время
стала енотовидная собака. В природе у енотовидной собаки половая зрелость наступает в
возрасте около 10 месяцев. Период гона приходится на март-апрель, через 2-3 недели пос¬
ле выхода зверей из нор. Зимуют енотовидные собаки в норах чаще всего парами. Длитель¬
ность периода гона составляет 2-3 недели и в значительной степени зависит от географи¬
ческих условий; как и у красной лисицы, на севере гон проходит позднее, чем в южных
районах. Продолжительность беременности 59-64 дня, в среднем 62 дня. Численность щен¬
ков в выводке до 18-19, в среднем 6-7 щенков [Насимович, 1985].
Гпава 10ГЕНЕТИЧЕСКАЯ ХАРАКТЕРИСТИКА
ПОРОД СОБАКПодробная характеристика генетики каждой породы собак — проблема актуальная
как ныне, так и в ближайшем будущем, пока что она решается лишь отчасти. Большинство
генетических сведений о породах касается наследования типа шерстного покрова и окраса
как наиболее четких породных признаков и в некоторой степени — наследственных анома¬
лий и болезней. Получены также данные о генетическом полиморфизме ряда пород и форм
собак по группам крови, белкам и ферментам (см. главы 6, 7).Наследование многих (огромного числа) других признаков и свойств, составляющих
особенности пород, до сих пор изучено недостаточно или вовсе неизвестно.Положение несколько облегчается родственными связями между некоторыми поро¬
дами, в связи с чем немало генов характерно не для какой-то одной, а для нескольких по¬
род.К сожалению, пока генетика важнейших признаков экстерьера, интерьера, поведения
собак описывается просто как детерминируемая полигенно, без вычленения главных генов,
не обозначенных никакими символами. Символика генов окраса и типов шерсти собаки
насчитывает несколько вариантов у разных авторов, наиболее удачно ее обобщил R.
Robinson [1982], по книге которого приведена она и здесь. Особенность описания геноти¬
пов пород R. Robinson заключается также в том, что он обычно перечисляет локусы с му¬
тантными генами, а немутантные гены дикого типа, если в соответствующих локусах поро¬
ды есть только такие, не приводит, чтобы не повторяться, — это, разумеется, не значит,
что такие локусы отсутствуют в геноме собак данной породы.При этом следует отметить, что многие гены по-разному проявляются у животных
различных пород. Так, например, современные генетики считают, что имеется один мутан¬
тный (рецессивный) ген длинной шерсти собак, но у длинношерстных пород длина волося¬
ного покрова сильно отличается — из-за разной экспрессивности этого гена, его модифи¬
каций, созданных в результате длительной направленной селекции. В то же время
жесткошерстность, курчавая или волнистая шерсть могут быть следствием действия неза¬
висимых мутаций: разные гены могут иметь почти идентичный фенотипический эффект (и
не только в отношении типа шерсти).Неоднозначность возможных последствий изучения таких явлений свидетельствует о
крайне желательном дальнейшем исследовании генетики пород собак: всякие новые конк¬
ретные сведения здесь будут, несомненно, полезны.Ниже приведены сведения о генетике наиболее распространенных и изученных по¬
род. Завершается эта глава Перечнем пород собак с разделением их на группы, официаль¬
но утвержденным Международной кинологической федерацией.Американский кокер-спаниель [American Cocker Spaniel]Порода выведена, видимо, на генетической базе английского кокер-спаниеля, поэто¬
му все гены последнего обусловливают и окрасы американских кокеров. В процессе микро¬
эволюции породы, естественно, возникают некоторые новые варианты, например, особо
удлиненная шерсть на конечностях, но это — следствие накопления при отборе у собак
последующих поколений генов-модификаторов, что и увеличивает длину волос на некото¬
рых статях.
R. Robinson указывает, что катаракта, проявляющаяся у американских кокеров по¬
мутнением хрусталиков уже в возрасте 6 месяцев, наследуется, видимо, по доминантно¬
му типу. В то же время еще не ясен окончательно тип наследования в этой породе другой
генетической болезни глаз — первичной глаукомы [Robinson, 1982], которая у них, и еще у
бассет-хаундов, имеет резкие отличия от глаукомы собак всех других пород: она морфоло¬
гически обусловлена закрытым иридо-корнеальным протоком, в связи с чем скапливающа¬
яся в глазном яблоке жидкость вызывает повышение внутриглазного давления и затем по¬
терю зрения.Только среди американских кокеров обнаружена еще одна наследственная болезнь
глаз — многофокусная дисплазия сетчатки — это проявление рецессивной аутосомной му¬
тации, мутантный аллель обозначен mrd [MacMillan, Lipton, 1978].Интересно, что, несмотря на филогенетическую близость американского и английско¬
го кокер-спаниелей и гомологию их генетики основных окрасов, наследственные аномалии у
этих двух пород различны (чуть не единственное исключение — прогрессирующая атрофия
сетчатки, наиболее распространенная среди представителей именно этих двух пород).Американский стаффордшир-терьер [American Staffordshire Terrier]Эта порода, как считает R. Robinson, является несколько увеличенной вариацией
стаффордширского бультерьера с идентичной генетикой основных породных признаков.Американский фоксхаунд [American Foxhound]Порода, совершенно неизвестная в России, представляет из себя увеличенную в раз¬
мерах вариацию классического фоксхаунда английского. Генетика окрасов у этих пород
одинаковая. Правда, С. Little [1957] отмечает, что в Америке чаще встречаются сейчас не¬
пятнистые собаки — со сплошным окрасом. L. Whitney [1947] сообщал, что известны и
мраморные фоксхаунды под названием „калико“ (calico).ОкрасГенотипРыжийАУ-S-Чепрачныйas“-S-Рыже-белыйАУ-sP-Чепрачный с белымasa—sP-Английский бульдог [English Bulldog]Порода ведет происхождение от древних боевых собак греческого племени молоссов,
имея родство с мастифами и прочими европейскими молоссоидами. Родоначальники буль¬
догов использовались в средневековой забаве — травле быков, запрещенной в 1835 г. [Пал¬
мер, 1988]. С конца XIX века организовано заводское разведение бульдогов как собак-ком-
паньонов.Типичный окрас короткой шерсти — сплошной рыжий разных оттенков, а также тиг¬
ровый, белый и пегий; встречается, но не обязательна черная маска на морде. Коричнева¬
то-желтый окрас является вариантом красного, разбавленного геном шиншиллы или „ми¬
нус-рыжими" полигенами. Белые отметины могут быть разной формы и величины, так как
в генофонде породы присутствуют все три аллеля пятнистости: s', sP, s"'. На белых отмети¬
нах иногда располагается крап как результат наличия гена Т.ОкрасГенотипКрасныйАУ-Е—S-СоболиныйАУ-E—S-ТигровыйАУ-Еь'-8-Красный с белымАУ-E—sP-Соболиный с белымАУ-E—sP-Тигровый с белымАУ-Eb'-sP-
Аномальное в отношении видовых параметров строение головы бульдогов вызывает
частые затруднения в дыхании этих собак [Палмер, 1988], особенно в жару, при физической
нагрузке и в стрессовых ситуациях.Установлено, что у английских бульдогов аномалия „расщепленное небо“ наследует¬
ся как моногенный рецессивный признак [Hutt, 1979] под контролем полулегального му¬
тантного гена. Для этой породы, как и для некоторых других короткомордых собак, ха¬
рактерна встречаемость, причем с рекордно высокой (около 40 %) частотой лишнего —
седьмого — резца в челюсти [Willis, 1990].Английский кокер-спаниель [English Cocker Spaniel]Английский кокер-спаниель в последние десятилетия превратился в одну из наиболее
популярных пород не только в Великобритании, но и во всем мире. Название произошло
от основного объекта охоты с этим спаниелем в период становления породы (woodcock —
вальдшнеп), хотя ее родоначальники попали в Британию из Испании, где использовались
еше в средние века на охоте с ловчими птицами [Палмер, 1988].Окрасы длинной шерсти кокера очень разнообразны и включают в себя практически
всю „собачью" гамму цветов и расцветок, с пегой пятнистостью и наличием/отсутствием
крапа — „тиковой испещренности“ по Н. Ильину [1932], не очень интенсивной и лишь
иногда формирующей чалую масть. Наиболее типичны кокеры черные, черно-подпалые,
шоколадные (или печеночные). Генетический статус собак рыжего оттенка шерсти нео¬
днозначный из-за возможного наличия в их генотипе аллелей Л? или е. По С. Little [1957],
большинство рыжих кокеров — гомозиготы ее, но некоторая часть собак такого окраса
могут иметь „рыжий“ ген АУ другого локуса, так как иногда от рыжих родителей рождают¬
ся черные щенки. Такой результат может быть при скрещивании одинаково рыжих кобеля
и суки разных генотипов: АУ- Е- х As- ее.С. Little наблюдал в питомнике Джексоновской лаборатории рождение в числе про¬
чих двух кокеров голубого окраса — это выщепление свидетельствует о наличии в гено¬
фонде породы редкого аллеля d.У черно-подпалых собак интенсивность цвета подпалин варьирует от ржаво-красно¬
го до кремового в зависимости от комбинирования в их генотипах „минус-рыжих“ полиге¬
нов [Robinson, 1982], а иногда светлые подпалы могут обусловливаться и действием редко
встречающегося аллеля cch [Little, 1957].Некоторые пегие спаниели имеют почти сплошь белый фон с цветными пятнами
только на ушах, что может быть следствием максимальной экспрессии гена sP или указы¬
вать на действие s'v, в генотипах sPsP, sPsK, j'V.Длинная шерсть кокер-спаниелей обусловлена действием гена / в гомозиготе. Отме¬
чается внутрипородная изменчивость по степени волнистости шерсти, особенно на голове
и конечностях.Установлено, что у кокеров прямая шерсть доминирует над волнистой [Whitney,
1947], но степень волнистости — признак полигенный.ОкрасГенотипЧерныйА5 В Е USЧерно-белыйAs-B-E-llsP-ttЧерно-белый с крапомA s-B-E-llSP-T-Черно-подпалыйа’а'В Е USЧерно-подпалый с белымa'a'B-E-llsP-ttЧерно-подпалый с крапома'а'В-Е UsP Т-Коричневый (печеночный)As ЬЬЕ USКоричневый с белымAs-bbE-lIsp-ttКоричневый с крапомAs-bbE~llsP-T-Коричнево-подпалыйa'a'bbE USКоричнево-подпалый с белымa!albbE-llsP-ttКоричнево-подпалый с крапомala'bbE-llsP-T-
Рыжий с черным носом
Рыжий с белыми пятнами
Рыжий с крапом
Рыжий с коричневым носом—B-eells —B-eellsP-tt
—B-eellsP-T-—BbeellS-L. Whitney [1947] сообщил об обнаружении среди английских кокеров животных с
проявлениями ахондроплазии — непропорциональной карликовости из-за резко укорочен¬
ных конечностей по сравнению со стандартом, и выделил мутацию — рецессивный ген as,
вызывающий в гомозиготном состоянии эту аномалию. Также рецессивным аллелем ss
[Pullig, 1952] в гомозиготе вызывается другая аномалия кокер-спаниелей — незарастание
швов на границах костей черепа. Дефектные щенки погибают не позже двухмесячного воз¬
раста.Породоспецифичен также для кокеров наследственный гликогеноз типа VII — дефи¬
цит фермента фосфофруктокиназы [Mhaskaret al., 1992] как следствие рецессивной мута¬
ции. Среди наследственных болезней глаз следует прежде всего выделить прогрессирую¬
щую атрофию сетчатки: у английских (и американских — редкий случай конкордантности
этих двух родственных пород) кокеров она встречается чаще, чем у собак других пород, и
наследуется также по рецессивному типу.В породе известны случаи заболевания наследственным цероидным липофусцинозом,
но тип генетического контроля этой болезни пока не установлен окончательно; но почеч¬
ная недостаточность наследуется как рецессивный признак [Nash, 1989].Английский сеттер [English Setter]Типичный представитель островных (британских) длинношерстных легавых — под¬
ружейных собак с врожденной страстью к поиску пернатой дичи и способностью к затаи¬
ванию перед сидящей птицей (стойка), выведенный в прошлом веке путем отбора сеттинг-
спаниелей и тесного инбридинга на нескольких родоначальников, главным образом в анг¬
лийском питомнике Эдварда Лаверака (1815-1877), считающегося создателем этой поро¬
ды. Среднего роста, с плавным стелющимся ходом и склонностью к „низким“ стойкам,
этот сеттер покрыт длинной шелковистой шерстью крапчатого окраса (по белому фону
крап желательно без крупных пятен черного, желтого, коричневого цветов) или трехцвет¬
ного (черный, белый, темно-рыжий). Мочка носа при этом черная или коричневая в зави¬
симости от цвета крапа.Эволюция современного окраса этих собак начиналась с типично пегих сеттеров, с
постепенным отбором на племя животных с наиболее развитыми по площади зонами де¬
пигментации вплоть до небольших пятен — на голове, ушах и крупе. Впоследствии наибо¬
лее популярными были сеттеры с типично чалой окраской, когда белые шерстинки равно¬
мерно перемежались с пигментированными, для английских сеттеров-лавераков такой
чалый рисунок имеет специальное название — бельтон (belton), причем эффект еще более
усиливает удлиненная украшающая шерсть. Вероятно, поэтому в породе одновременно
распространены аллели sP и j1,:.Трехцветный (триколор) окрас в этой породе генетически представляет собой черно-
подпалый с сильно распространенными белыми пежинами [Robinson, 1982], а согласно
С. Little [1957], желто-крапчатые собаки (окрас лемон-бельтон) — гомозиготы ее.. ОкрасГенотипЧерно-крапчатый
Коричнево-крапчатый
Черный трехцветный
Коричневый трехцветный
Желтый с черным носом
Желтый с коричневым носомAs-B-E-llsP-T-As-bbE-llsP-T-a'ci'B-E-llsP-T-a'a'bbE-llsP-T-B-eellsP-T-bbeellsP-T-
В числе наследственных аномалий, обнаруженных у английских сеттеров, нейрональ¬
ный цероидный липофусциноз. Его причина — рецессивная мутация, обозначенная li
[Robinson, 1982], что снижает уровень фермента пероксидазы уже после годовалого возрас¬
та. В результате развиваются симптомы идиотии и смерть наступает не позже двух лет.Английский фоксхаунд [Foxhound]Классическая гончая знаменитых английских парфорсных (конных псовых) охот вы¬
ведена в Британии несколько веков назад путем селекционной работы с континентальны¬
ми гончими святого Губерта и Тальбота. Типичная собака чепрачного окраса с пятнами —
пежинами. Встречаются также фоксхаунды черного, желтого и темно-рыжего окрасов с бе¬
лыми отметинами.Фоксхаунды были использованы для выведения в начале нашего столетия русских
пегих гончих в России (англо-русских) путем скрещивания их с русскими гончими и селек¬
ции помесей. Интересно, что, по сообщению Н. П. Пахомова [По: Н. Ильин, 1931], гибри¬
ды фоксхаунда с русской гончей (обладавшей чепрачным окрасом) сохраняли в первом по¬
колении „русский" чепрак — доминантный признак по отношению к пятнистости
фоксхаунда.Афган (афганская борзая) [Afghan Hound]Современная порода афганов, выведенная европейскими селекционерами из местной
афганской борзой, представлена собаками с очень длинной украшающей шерстью специ¬
фической структуры, ниспадающей на корпусе со спины, где выражен четкий „пробор".
Длинная шелковистая шерсть — маркер генотипа //.Ряд окрасов типичен для породы. Встречаемость черных и черно-подпалых афганов
свидетельствует о наличии в генофонде генов As и а'. Типичен также рыжий окрас, чаще с
черной маской и соболиным оттенком — следствие гена АУ. Эти гены обусловливают воз¬
можность рождения, например, черно-подпалых щенков от рыжих родителей [Little, 1957].
Афганы голубого окраса встречаются, но редко. С. Little [1957] также отмечал, что гибри¬
дологическим анализом, проведенным в знаменитой Джексоновской лаборатории (США),
установлено, что некоторые рыжие афганы могут иметь и генотип ее.Кроме того, известны афганы палевые, кремовые и почти белые — удивительно кра¬
сивые животные, особенно с черной маской на морде. Little полагал, что оттенки палевого
окраса обусловливает ген c(,t во взаимодействии с АУ и е. Аллель сс1’ работает как разбави¬
тель желтого, но не черного пигмента. Впрочем, равновероятно, что „рыжие" полигены де¬
терминируют рыжий цвет шерсти, а „минусовые" полигены обусловливают желтый и кре¬
мовые окрасы. Длинная шерсть вносит дополнительный эффект в образование
специфических окрасов — благодаря палевому подшерстку, модифицирующему цвет по
сравнению с палевым окрасом короткошерстных пород. При этом некоторые палевые аф¬
ганы могут иметь в генотипе ген-разбавитель (dilution) d, особенно собаки без темной мас¬
ки. При наличии к тому же гена ссЛ или „минус-рыжих" полигенов такие особи будут
иметь особо светлый окрас — светло-кремовый, почти белый.Окрас ГенотипЧерный As-sP-
Желтый АУ-sP-
Чепрачный asa-sP-ОкрасГенотипЧерныйЧерно-подпалыйРыжий/желтыйГолубойКремовыйAS-D-Ua’a'-D-lIАУ-D-llAsAsddllA>—ddll
Из наследственных аномалий афганов следует упомянуть катаракту (прогрессирую¬
щее помутнение хрусталика) у собак в возрасте 1-3 лет: видимо, она имеет рецессивный ха¬
рактер [Robinson, 1982]. В породе выявлен также рецессивный аутосомный ген тр, вызыва¬
ющий у гомозигот в 3-8 месяцев миелопатический паралич [Averill, Bronson, 1977].Аффен-пинчер [Affenpincher]В самостоятельную породу, отделенную от общей породы пинчеров, эти маленькие
собаки попали в 1896 г. на Берлинской выставке по главному признаку, отличающему их
от гладкошерстных пинчеров — по специфическому шерстному покрову. Шерсть аффен-
пинчеров „должна быть короткой и густой в одних местах и длинной и лохматой — в дру¬
гих, особенно у глаз, на морде и на подбородке, что придает собаке сходство с обезьяной"
[Палмер, 1988. С. 23], „аффен“ в переводе с немецкого — „обезьяний".Обычно это собаки черного окраса с длинной жесткой шерстью. Впрочем, иногда
встречаются также черно-подпалые и соболиные различных оттенков. При этом у некото¬
рых особей бывает черная маска на морде [Little, 1957].ОкрасГ енотипЧерныйAs-llWh-Черно-подпалыйa'a'llWh-СоболиныйАУ-llWh-Басенджи [Basenji]Своеобразный облик этих собак, называемых также собаками Занде, лесными соба¬
ками из Конго, конго-терьерами или ньямньям-терьерами, запечатлен на гробницах древ¬
неегипетских фараонов, но в новейшей истории их обнаружили только в середине прошло¬
го века в Конго и Южном Судане [Палмер, 1988], откуда басенджи попали к собаководам
Европы и Америки.Среди басенджи впервые открыта наследственная аномалия собак — гемолитическая
анемия. Рецессивная мутация рк [Robinson, 1982] снижает при этом активность фермента
пируваткиназы, вызывая проявление признаков анемии к годовалому возрасту и гибель
гомозигот к двум годам. В американской популяции басенджи (в США) с высокой часто¬
той распространен наследственный синдром Фанкони — дисфункция почечных канальцев.Басенджи — подвижные собаки, не умеющие лаять (хотя голосовые связки у них в
норме), имеют обычно светло-рыжий окрас — либо сплошной, либо со светлыми пятнами
и черной спиной. Возможен также черно-подпалый окрас, а в прошлом известны и сплошь
черные особи.ОкрасГ енотипЧерныйA(l-s'-Черно-подпалыйa'a’s'-РыжийАУ-s'-Бассет-хаунд [Basset Hound]Низкорослые гончие для охоты на зайца выведены в Великобритании путем скрещи¬
вания артезиано-нормандских бассетов французского происхождения с бладхаундами. Эти
приземистые коротконогие собаки с проявлением ахондроплазии обладают, видимо, гена¬
ми коротконогости. Дж. Палмер [1988] ссылается на французских кинологов (бассет по-
французски — коротконожка): первые бассеты появились в помете нормальных длинноно¬
гих гончих, откуда и повелась порода. Если это так, то ген, выщепившийся у коротконогих
гомозигот, — рецессивный. Однако гибридологический анализ скрещивания бассетов с
боксерами свидетельствует о доминировании коротконогости [Christensen et al., 1985]. По¬
зднее также получены подтверждения того, что такая коротконогость — следствие доми¬
нантной точковой мутации одного гена [Fjeld, 1988].
Так же доминантной мутацией обусловливается наследственная болезнь свертывания
крови бассетов — тромбастения. Однако клинические признаки проявляются только у го¬
мозигот по доминантному аллелю, из-за чего тип наследования аномалии приобретает „ре¬
цессивный" характер [Robinson, 1982].Типичный окрас — рыжий или черно-подпалый, но чаще с чепраком и белыми пежи¬
нами. Все собаки имеют белые отметины и разной степени пегость, бывает крап, особенно
вокруг морды. R. Robinson считает, что, видимо, это следы проявления гена крапчатости Т
с неполной экспрессивностью.ОкрасГ енотипЖелтый с белымАУ-sP-Черно-подпалый с белымasa-sP-Для бассет-хаундов, как и для американских кокер-спаниелей, характерна уникаль¬
ная форма наследственной первичной глаукомы — с закрытым иридо-корнеальным прото¬
ком, через который в норме удаляется излишняя жидкость из глазного яблока, но тип на¬
следования окончательно еще не выяснен.Бедпингтон-терьер [Bedlington Terrier]Первый клуб любителей бедлингтон-терьеров был организован в Англии в 1875 г.,
но уже за 50 лет до этого стали известны первые типичные их представители, разводившие¬
ся в графстве Нортумберленд, в городке, по названию которого порода и получила свое
наименование. В числе возможных предков, очевидно, английские борзые грейхаунд и уип-
пет, а также, несомненно, денди-динмонт-терьер [Палмер, 1988].Согласно С. Little [1957], палевая гамма, типичная для окрасов этой породы, обуслов¬
лена доминантным геном G „посерения“ — поседения. В случае голубого окраса щенки
рождаются черными и с возрастом изменяют цвет шерсти до светло-голубого. Считалось
при этом, что у особей всех окрасов присутствует ген G.Однако R. Robinson считает эту классификацию ошибочной. По его мнению, в дис¬
куссии о генетике окрасов бедлингтона Little допустил несколько ошибочных постулатов.
Предположи тельно, аллель А5 детерминирует сплошные окрасы; однако, поскольку иног¬
да осветленные участки расположены в местах, традиционно занимаемых подпалинами, в
генофонде должен быть и ген а1. Видимо, имеется и ген cch — на это указывает факт, что
иногда собаки голубого окраса имеют очень светлый оттенок, а некоторые „песочные"
становятся кремовыми при возрастном становлении расцветки, этот аллель мало влияет на
голубой окрас, и если необходимо вовлекать второй ген-разбавитель, то им более вероятно
может быть d. С. Little рассматривал песочных бедлингтон-терьеров как печеночных с раз¬
бавителем cch (или d). Это предположение вполне может быть истинным, но более вероятно
действие АУ или е [Robinson, 1982].ОкрасГенотипГолубойAs-B-D-G-Голубой с подпаломa'a'B-D-G-ПеченочныйAs-bbD-G-Печеночный с подпаломtfa'bbD-G-ПесочныйАУ D GКремовыйАУ ddGУ бедлингтонов совершенно особенной структуры шерсть, и до сих пор нет точных
данных о типе ее наследования: предполагается, что это модификация жесткошерстного
покрова, детерминируемого геном Wh, но это пока только предварительная гипотеза
[Robinson, 1982].Рецессивный ген rd [Robinson, 1982] в гомозиготном состоянии обусловливает у бед¬
лингтонов, как и у некоторых других терьеров, ретинальную дисплазию (аномалию сетчат¬
ки глаза).
Бельгийские овчарки [Belgian Shepherd Dogs]Основные генетические особенности бельгийских овчарок рассматривают как гомо¬
логичные ряды родственной им немецкой овчарки, в том числе и по генофонду локусов,
детерминирующих окрас и структуру шерстного покрова [Robinson, 1982]. В общую груп¬
пу под названием „бельгийские овчарки" объединяют фактически четыре породы, считаю¬
щиеся сейчас разновидностями породы: грюнендаль, малинуа, тервюрен, лакенуа.Бернская пастушья собака (бернский зенненхунд)
[Bernese Mountain Dog]Под этим названием также известна группа швейцарских альпийских пород пастушь¬
их собак, имеющих обшее происхождение с центральноевропейскими догообразными по¬
родами и относящихся поэтому по классификации FCI ко второй группе пород (см. При¬
ложение). Бернские пастушьи собаки имеют унифицированный и очень характерный
именно для них окрас длинной шерсти: черно-подпалый, с белыми отметинами, в идеале
расположенными на груди, лапах и лбу. Поэтому генотип по локусам шерсти: afa'lls'-.Кроме бернского зенненхунда (рост 58-70 см), выделяют большого зенненхунда (до
71 см, гладкошерстный — гомозигота LL), аппенцеллер-зенненхунда (рост до 58 см) также
гладкошерстного и с закрученным на спину хвостом и невысокого коренастого, сложением
и подпалым окрасом (но с белыми отметинами) напоминающего ротвейлера энтлебух-зен-
ненхунда (до 46 см). Все они составляют гомологический ряд близкородственных и облада¬
ющих сходными экстерьерными и поведенческими особенностями форм.Среди зенненхундов относительно широко распространены наследственные анома¬
лии сочленений конечностей. A. Hedhammar [1986b] отмечал среди них проявления диспла¬
зии тазобедренных суставов. Однако наиболее весомым генетическим грузом в популяциях
этих тяжелых, массивных собак является наследственная дисплазия локтевого сустава —
частота ее встречаемости, например, среди бернских зенненхундов в Швеции превышает
50 % [Klingebom, 1986]. Генетическая основа этой аномалии имеет полигенный пороговый
характер [Corley et al., 1968; Hutt, 1979].Знаменитый F. Hutt [1979] установил, что у бернских зенненхундов по доминантному
типу наследуется аномалия „расщепленное небо“, причем у части дефектных животных от¬
мечена также „заячья губа“.Бигль [Beagle]Низкорослые (высота в холке до 38 см) гончие, известные в Англии по крайней мере
с 1475 г. [Палмер, 1988]. Для породы характерен чепрачный окрас особого типа. Рисунок
расцветки весьма стабилен, хотя имеется вариация степени распространения чепрака и ин¬
тенсивности его пигментации. Яркость рыжего цвета вне чепрака также варьирует, эта из¬
менчивость, скорее всего, обусловливается разными наборами полигенов в генотипах раз¬
ных особей. Для породы обычен белого цвета „ошейник" (или пежина на шее). У
некоторых особей может встречаться также ген Т, обусловливающий крап („тиковую ис-
пещренность"). Генотип окраса: asa-sl- или asa-sP-.Среди 160 пометов биглей, полученных в Джексоновской лаборатории, родилось не¬
сколько щенков редких, необычных окрасов [Little, 1957]. Печеночный, голубой, желтый и
особенно тускло-желтый цвет на нормальном рыжем фоне (без чепрака, вероятно, типа
„шиншилла") — такие экзотические окрасы зарегистрированы в единичных случаях при
чистопородном разведении. Эти щенки обладали, видимо, генами b, d, е, с*-'1’ соответствен¬
но. Однако такие совершенно нетипичные и очень редкие цветовые вариации рассматрива¬
ются как латентный след проявления межпородного скрещивания в отдаленном прошлом
или следствие мутации. Считается поэтому, что у биглей предоминирует один стандарт¬
ный окрас.Изучена наследственная природа у биглей первичной глаукомы (с открытым иридо¬
корнеальным протоком): тип наследования — моногенный рецессивный [Gelatt, 1981].
Необычно рано (в возрасте 6-18 месяцев) по сравнению с другими породами проявляются
у биглей симптомы этой болезни.Рецессивная мутация вызывает иногда у биглей врожденное отсутствие одной из двух
почек [Fox, 1964].
Бладхаунд [Bloodhound]Эта ищейка, одна из старейших пород собак, сохранившихся в чистоте, сформирова¬
лась в Англии, куда попала, как считают, в XI веке вместе с Вильгельмом Завоевателем.
Самые типичные окрасы ее густой недлинной шерсти — красный (красно-коричневый) и
так называемый черно-подпалый, который у данной породы генетически является окрасом
чепрачным, с распространенным за обычные границы чепраком. Некоторые рыжие блад-
хаунды имеют и черную маску на морде. Известны также изредка рождающиеся собаки пе¬
ченочного окраса и чепрачные с голубым чепраком.Бобтейл (староанглийская овчарка) [Old English Sheepdog]Крупная собака с древней историей: полагают, что выведена в Великобритании пу¬
тем скрещивания бриара с центральноевропейскими пастушьими собаками типа венгерс¬
ких [Палмер, 1988]. Название („куцый хвост“) отражает породную особенность — гомози-
готность по рецессивному гену, обусловливающему рождение щенков с сильно
укороченным хвостом.Густая длинная шерсть может иметь любой оттенок серого, голубого или голубова¬
то-мраморного, с белыми отметинами или без них. Дисквалифицируются оттенки бурого
или коричневого.С. Little [1957] считал, что бобтейли имеют ген G поседения шерсти, но при этом
часть собак могут быть и первично серыми из-за гомозиготности по рецессивному гену
разбавления пигмента d Он также полагал вероятным наличие в генофонде породы гена
мраморного окраса М.Генотип типичной староанглийской овчарки серого оттенка шерсти: As-G-llsP -.Эти мощные, массивного сложения собаки предрасположены к дисплазии тазобед¬
ренных суставов [Hedhammar, 1986b], наследующейся полигенно. Кроме того, у них обна¬
ружен еще один вид дисплазии — полиостатическая фиброзная — в виде опухолей на пере¬
дних конечностях. Конкретный тип ее наследования пока не определен. В то же время
катаракта, связанная у бобтейлов также с нарушениями строения сетчатки глаза, наследу¬
ется в этой породе, скорее всего, как рецессивный признак [Robinson, 1982].Боксер [Boxer]Порода сформировалась в начале нашего века в результате селекции помесей потом¬
ков древних молоссоидов — английского бульдога и брабантских травильных собак —
булленбейсеров, с последующей целенаправленной племенной работой. В результате со¬
зданы гармонично сложенные собаки с отличными служебными качествами и характерной
формой головы (короткая морда). Специфично строение зубной системы и смыкание че¬
люстей боксеров, с выходящей вперед нижней челюстью.Стандарт допускает только два окраса — рыжий и тигровый, хотя раньше характер¬
ными были и пятнистый и даже белый. Стандартные рыжие и тигровые собаки различаются
по интенсивности пигментации и могут иметь белые отметины — пежины, как следы „ир¬
ландской пятнистости". Максимальное развитие центров депигментации при генотипе s'V
вызывает белый окрас, который ныне считается дисквалифицирующим пороком в породе.ОкрасГ енотипКрасныйЧепрачный (,,черно-подпалый“)
Чепрачный (печеночный)
Чепрачный (голубой)АУ—D-
asa-B-D-
asa-bbD-
as“-B-ddОкрасГ енотипТигровыйРыжийТигровый с белым
Рыжий с белымAy-Ebr-S-
АУ-Е—S-
АУ-E^-s'-
АУ-Е—s'-
Кроме специфического прикуса, аномального для большинства собак, боксеру свой¬
ственна полидонтия — нередко встречаются особи с увеличенным числом зубов, особенно
резцов, с частотой более 25 % [Willis, 1990]. В породе открыта рецессивная мутация ра
[Griffiths et al., 1980], обусловливающая у гомозигот развитие прогрессирующей аксонопа-
тии — нейромускулярной аномалии. Благодаря знаменитой книге Ф. Хатта [1969] известен
факт необычно высокой (с частотой 23 %) встречаемости крипторхизма среди изученных
боксеров американской популяции.Бордер-колли [Border Collie]Порода представляет собой относительно нестарую линию потомков южной популя¬
ции колли из пограничных с Англией районов Шотландии, разводившуюся с отбором пре¬
имущественно по служебным качествам этих популярных пастушьих собак. Из-за такой се¬
лекции экстерьерные характеристики бордер-колли необычно широко варьируют, а,
например, по строению головы, весьма широкой в черепе, с выраженным переходом ото
лба к морде современная пастушья собака довольно резко отличается от своей родона¬
чальницы — шотландской овчарки колли. Окрас бордер-колли очень разнообразный и так
же не напоминает строгие окрасы колли-родоначальников. Наиболее часто встречаются
бордер-колли черного окраса длинной шерсти с белыми отметинами (черно-пегие), их ге¬
нотип As-s‘-.У бордер-колли Австралии обнаружили [Malik et al., 1991] в родственной группе,
происходящей от 2 кобелей-родоначальников и 3 сук-родоначальниц, 36 их потомков,
страдающих артериозом открытого протока: видимо, эта аномалия сердечно-сосудистой
системы наследуется по рецессивному типу.Рецессивный ген рта в гомозиготе вызывает развитие прогрессирующей атрофии сет¬
чатки — аномалии, частота которой является среди бордер-колли одной из самых высоких
среди всех обследованных пород.Бостон-терьер [Boston Terrier]Название порода получила в 1893 г. в честь города, где ее вывели в результате скре¬
щиваний привезенного из Англии в США метиса бульдога и терьера с английскими и
стаффордширскими бультерьерами (первоначально такие собаки и назывались американс¬
кими бультерьерами). Стандарт очень строго регламентирует окрас бостон-терьеров: тиг¬
ровый, равномерно распределенный по всему телу с непременно симметричными белыми
отметинами на морде, лбу, шее (ошейник), груди и конечностях. Разумеется, при такой
трактовке немало рождающихся чистопородных щенков оказываются нестандартными,
так как наряду с тигровым в породе превалирует и черный окрас, а из-за комбинации мо¬
дификаторов отметины не всегда располагаются симметрично. С. Little [1957] указывал,
что в этом случае наиболее перспективна селекция окраса „ирландский пятнистый11 на мак¬
симальную его экспрессию, что и дает натуральный симметричный рисунок отметин, тогда
как обычный пятнистый окрас так себя не ведет. Рождающиеся в породе белые щенки, по
Little, — результат сегрегации гена $>*■', так как типичные бостон-терьеры с белыми отмети¬
нами должны иметь генотип Ss'v. Нестандартные черные бостон-терьеры имеют окрас, ли¬
бо обусловленный действием черного гена As, либо у них очень темные тигровины слива¬
ются в сплошную рубашку без разделения полос волосами с желтым пигментом.ОкрасГенотипТигровыйAy-Ebr-si-ЧерныйAs s‘-Широко известны проблемы со здоровьем бостон-терьеров: в частности, часто воз¬
никающие у них затруднения с дыханием [Палмер, 1988], которыми „наградили14 их пред¬
ки — английские бульдоги.Кроме того, A. Hedhammar [1986а] обнаружил среди бостон-терьеров встречаемость
наследственного прогибания запястья. Рецессивный ген cs, вызывающий этот сцепленный
с полом дефект, локализован в половой Х-хромосоме [Pick et al., 1967]. В породе также
встречали животных с проявлениями наследственной атаксии [Bjork et al., 1957]. Судя по
анализу этой аномалии у фокстерьеров, она наследуется по аутосомно-рецессивному типу.Аналогично наследуется у бостон-терьеров первичная катаракта — мутантный ген,
выявленный именно в этой породе, обозначенный ct [Robinson, 1982].Бульмастиф [Bullmastiff]Эти сильные и свирепого вида собаки выведены, вероятно, два-три столетия назад в
Великобритании в результате скрещивания бульдогов с мастифами. Генетика окраса ко¬
роткой шерсти бульмастифов гомологична генетике окраса боксеров, за исключением то¬
го, что в данном случае стандарт не допускает белых отметин, кроме небольшого пятна на
груди (нежелательного). Типичен окрас, обусловленный желтым пигментом — от красного
до палевого, в зависимости от набора „рыжих“ полигенов. При этом обязательна черная
маска на морде. Однако С. Little [1957] отметил, что в 1950-е годы в США лишь некоторые
бульмастифы имели маску.ОкрасГенотипТигровыйАУ-ЕЬг-Красный, рыжийАУ-Е-Для бульмастифов, как и для некоторых „короткомордых“ пород, весьма характерна
встречаемость лишних резцов в челюсти с частотой около 8 % [Willis, 1990].Бультерьер [Bull Terrier]Современная история породы имеет начало на границе прошлого и нынешнего ве¬
ков, после того как в Бирмингеме Дж. Хинкс [Палмер, 1988] скрестил белого английского
терьера с английским бульдогом и далматином, и впервые выставил в 1862 г. на выставке в
Лондоне результат своей селекции — белую суку Пусс [Хорнер, 1992], которой новая поро¬
да обязана ростом популярности. Сейчас порода разделяется на две цветовые вариации —
белых и цветных собак, и дискутируется вопрос о возможном в будущем разделении вариа¬
ций на две разные породы [Robinson, 1982]. Обычные окрасы цветных бультерьеров — чер¬
ный, тигровый и рыжий (или желтый, в зависимости от набора рыжих полигенов). У ры¬
жих и желтых может быть разной интенсивности маска на морде, и у многих цветных
собак бывает отметина — белый ошейник — следствие гомозиготности s's‘. Прежде
встречались черно-подпалые бультерьеры (генотип а!аг) и голубые (As-dd). Белые бульте¬
рьеры имеют генотип Л'1' (иногда у белых собак бывают черные пятна на голове). Впро¬
чем, белые собаки могут иметь в своем генотипе и названные выше гены цветных окрасов,
но они маскируются супрессорами — генами белого окраса.ОкрасГенотипЧерныйAs-E—s'-РыжийАУ-Е—s'-ТигровыйAy-Ehr-s‘-Уникальной наследственной болезнью является аномалия кожи — так называемый
фурункулез бультерьеров [White, 1993]. Наследственный характер определен для бультерье¬
ров (наряду с еще восемью породами) и относительно почечной недостаточности [Nash,
1989]. Ген Сь обусловливает иногда неполный альбинизм и проблемы со слухом [Ильин,
1932].Веймаранер (веймарская легавая) [Weimaraner]Превосходная порода рабочих собак, не только подружейных охотничьих, но и слу¬
жебно-розыскных, выведена в замке герцога Веймарского в Германии в XVII веке на осно¬
ве старонемецких легавых и гончих святого Губерта [Палмер, 1988]. Уникален благород¬
ный окрас веймаранера, называемый серебристо-серым, с металлическим блеском. R.
Robinson объясняет этот феномен необычной для собак комбинацией генов bud при дей¬
ствии черного аллеля As, вероятно, имеющегося у этих собак в гомозиготном состоянии.
Известны как традиционно короткошерстные веймаранеры (L-), так и длинношерстные
(И). Генотип по генам окраса: ASAS bbdd.Вельш-терьер [Welsh Terrier]Создан в Уэльсе путем целенаправленной селекции на генетической базе старинного
кельтского жесткошерстного черноподпалого терьера. Порода официально признана в
1887 г.Воспринимается как уменьшенная копия эрдельтерьера — по крайней мере, во всем,
что касается типа шерсти и окраса, хотя чепрак не настолько густо-черного цвета. Генотип
генов шерсти включает в гомозиготном состоянии аллели asu и Wh.Среди вельш-терьеров известны случаи неполнозубости (олигодонтии), причем прак¬
тически всегда недостает вторых премоляров в нижней челюсти Гибридологический ана¬
лиз указывает на возможный рецессивный характер наследования этого дефекта.Грейхаунд [Greyhound]Грейхаунды, английские борзые, являются одной из самых древних пород: на основа¬
нии сходства их облика с собаками на древнеегипетских барельефах выводят прямое про¬
исхождение от ловчих собак из долины Нила (IV тысячелетие до нашей эры) [Палмер,
1988]. Рекордсмен по резвости среди собак — рекорд грейхаунда на беговой дорожке кино¬
дрома в соревнованиях за искусственным „зайцем“ более 60 км/ч.Окрасы короткой гладкой шерсти — черный, белый, рыжий, голубой, палевый, тиг¬
ровый — сплошные или пегие этих же цветов. Нетипичны только черно-подпалый и пече¬
ночный окрасы. Варьирование желтого пигмента от рыжего до палевого и кремового
обусловлено действием гена d в сочетании с модификаторами — „рыжими“ полигенами
[Robinson, 1982]. С. Little [1957] полагал, что пегие окрасы грейхаундов могут контролиро¬
ваться любым из всех трех аллелей пятнистости S'-локуса, но более всего вероятно действие
аллеля sP. Все грейхаунды являются, кроме того, гомозиготами по гену В [Ильин, 1931], по¬
этому среди них не встречаются особи коричневых окрасов.ОкрасГенотипЧерныйAS-D-E—S-Г олубойAs-ddE—S-Черно-тигровыйA.v-D-Ehr-S-Серо-тигровыйAy-ddEb'-S-Рыжий и желтыйАУ-D-E—S-Палевый и кремовыйАУ-ddE—S-Черно-пегийAs-D-E—sP-Серо-пегийAs-ddE—sP-Тигрово-пегийAy-D-Ebr-sP-Рыже-пегийАУ-D-E—sP-Следует отметить идентичность генетики окраса грейхаунда и его „уменьшенных11копий — уиппета и левретки (у которой, правда, отсутствует тигровый Окрас)., , . * 1 \ . . .Гриффон (брюссельский и брабантский) [Griffon Bruxellois]Бельгийская порода маленьких собак (от 2,5 до 5 кг), выведенная для охраны, глав¬
ным образом, конюшен и истребления там же крыс. Две разновидности породы различа¬
ются только шерстным покровом [Палмер, 1988]: брюссельский имеет жесткую шерсть (ге-
нотип WhWh), а брабантский гриффон — гладкую (генотип L-). Впервые показаны на
Брюссельской выставке в 1880 г. и ведут происхождение от аффен-пинчеров (брабантский,
видимо, и от мопсов).Окрасы — черный, черно-подпалый и рыжий (желательно с черной маской и черны¬
ми ушами).ОкрасГенотипБрабантский:ЧерныйAs-whwhЧерно-подпалыйatatwhwhРыжийАУ-whwhБрюссельский:ЧерныйAs-Wh-Черно-подпалыйa'a'Wh-РыжийАУ-Wh-Далматин [Dalmatian]Судя по названию, порода происходит из Далмации, области на Балканах, но ее за¬
водское разведение и совершенствование в основном начались в Великобритании —
с XVII века. Уникальный окрас этих собак (чисто-белый фон с разбросанными по нему
равномерно четкими и круглыми 1-2-сантиметровыми пятнами черного или коричневого
цвета) — результат труда собаководов над бесчисленным рядом селекционируемых поко¬
лений животных. По С. Little [1957], генотип окраса далматинов sn'swT-. Таким образом,
далматины — белые собаки с геном крапчатости Т, но крап в виде круглых пятнышек .
сформировался под действием уникальной комбинации модификаторов в ходе селекции.
Little указал на встречаемость собак также с пятнами желтого цвета, и такие далматины
могут быть с мочкой носа черной или коричневой.Окрас (цвет пятен)ГенотипЧерныйAs-B-E-swswT-КоричневыйAs-bbE-swswT-Желтый (черный нос)—B-eeswswT-Желтый (коричневый нос)—bbees'vsn'T-Лейкодистрофия Бьеркаса — наследственная нейромускулярная аномалия, впервые
открытая среди далматинов. Нарушение движений, паралич из-за дегенерации клеток моз¬
га обусловливаются действием в гомозиготе рецессивного аллеля Id.Не вполне ясна генетическая природа регулярно встречающейся среди далматинов
глухоты [Anderson et al., 1968]: видимо, мутантный ген обладает неполной пенетрантнос-
тью.Но самая поразительная генетическая особенность далматинов — повышенный уро¬
вень (по сравнению с собаками других пород) выделения мочевой кислоты: аномальное
для всех остальных собак, это явление типично для чистопородных далматинов, не отлича¬
ющихся при этом повышенной частотой почечно-каменной болезни или какой-либо иной
возможной патологией мочевыделения. Установлено, что эта особенность детерминирует¬
ся мутантным рецессивным геном иа, по которому далматины гомозиготны. Он не сцеплен
с генами рп' и Т, обусловливающими „фирменный11 окрас этих собак. Генетиков и селекци¬
онеров интригует механизм приобретения всеми далматинами такой наследственной осо¬
бенности обмена веществ — по-видимому, причина в дрейфе генов на ранних этапах созда¬
ния породы, когда возникшая впервые у родоначальника мутация закрепилась в его
потомстве путем инбридинга и гомогенных подборов.
Доберман [Dobermann]Во второй половине 19-го века в германской Тюрингии (город Апольда) сборщик на¬
логов Карл Льюис Фридрих Доберман занимался разведением злобных собак, пригодных
для надежной охраны хозяина при его работе по взиманию податей — скрещивая с этой
целью „самых кусачих1' местных собак — чаще всего старонемецких пинчеров, босеронов
и ротвейлеров. Впоследствии оказалось, что эти „собаки Добермана" пригодны для разно¬
сторонней работы, в том числе в армии и полиции, и с конца века началось заводское раз-
ведение доберман-пинчеров, как тогда называлась порода. На первом этапе ее совершен¬
ствования, может быть, были использованы скрещивания также с грейхаундом,
манчестерским терьером [Zwan, 1989], а по некоторым сведениям, и с немецкими догом, ов¬
чаркой и веймарской легавой [Мазовер, 1960]. В начале породообразования доберманы
имели рост 50-60 см, а современный международный стандарт FCI ограничивает высоту в
холке от 63 до 68 см (для сук) и от 66 до 72 см (для кобелей). В США, где действует особый,
американский стандарт, собаки этой же породы значительно крупнее: большинство кобе-
лей-чемпионов Америки имели высоту в холке около 75 см и даже больше [Stepansky,
1974].В настоящее время доберман — классическая служебная собака, популярная у дрес¬
сировщиков и любителей во всем мире. Головные организации по селекции породы: До¬
берман-союз (Dobermann-Verein, Германия) и Международный доберман-клуб (IDC,
International Dobermann Club) — большое внимание уделяют комплексной селекции, со¬
вершенствуя экстерьер, породность, красоту сложения собак наравне с сохранением и раз¬
витием их рабочих качеств, уравновешенности характера, крепости конституции, здоровья
[Wiblishauser, 1990].Доберман — одна из классических „архитипических“ пород подпалого окраса корот¬
кой гладкой жесткой шерсти (в первые десятилетия разведения породы собаки имели гус¬
тую относительно удлиненную шерсть с развитым подшерстком, который теперь не допус¬
кается стандартом). Ржаво-красные подпалины желательно как можно более интенсивного
оттенка располагаются на строго оговоренных стандартом местах и четко отграничены от
основного фона, который может быть черного, коричневого, голубого или изабеллового
цвета (современный международный стандарт допускает только черный и коричневый ок¬
расы, так как последние два — голубой и изабелловый — связаны с целым рядом общекон¬
ституциональных явлений ослабления крепости собак, что обусловлено, в частности, гомо-
зиготностью таких доберманов по рецессивным генам ряда локусов и другими
причинами).В генетику добермана большой вклад внес своими исследованиями в 1920-е годы
русский ученый профессор Н. А. Ильин. В опубликованной в 1931 г. статье он представил
обширные данные и результаты гибридологического анализа, создав убедительную схему
наследования окраски доберман-пинчеров. Кроме генетики основных типов окраса,
Н. Ильин [1931] установил также существование „начальных точек депигментации'1 как
точно локализованных участков (на груди, шее и лапах), на которых у некоторых собак
бывают белые отметины.ОкрасГенотипЧерно-подпалыйa'a'B-D-LLS-Коричневыйa'a'bbD-LLS-Голубойa'a'B-ddLLS-Изабелловыйa'a’bbddLLS-Окрас с белыми отметинами s‘s‘При всех перечисленных окрасах цвет подпалин доберманов варьирует весьма ши¬
роко — от наиболее желательного ржавокрасного оттенка до желтовато-белесого через
промежуточные формы. Полагают обычно, что эта изменчивость (в некоторой степени мо¬
дифицируемая средовыми факторами) детерминируется действием полигенов-модификато¬
ров. Однако К. Stepansky [1974] высказал мнение о действии на оттенок подпалин доберма¬
на аллельной пары генов локуса С: якобы собаки с яркими красными отметинами
являются доминантными гомозиготами СС, обладатели подпалин средненасыщенного цве¬
та — гетерозиготы Ccch, а доберманы с „осветленным" подпалом оказываются гомозиго¬
тами по рецессивному „шиншилловому" аллелю-осветлителю {<fh) пигментации.Но существование более чем трех вариаций оттенков подпалин в породе, на наш
взгляд, свидетельствует о более сложной природе генетико-средового контроля интенсив¬
ности пигментации подпалин: имеет место, видимо, влияние как полигенов-модификато¬
ров, так и аллелей одного локуса (С), а также действие паратипических факторов среды.Короткий хвост добермана — результат купирования в младенчестве (оставляют 2 по¬
звонка); кроме того, с самого начала разведения породы купируют и уши добермана -^ на¬
следственно висячие „лопухообразные" (генотип „вислоухости" Н — по Н. А. Ильину
[1932]), которым в результате операции и тренировки придают стоячее положение. Созда¬
тели породы справедливо считали, что тем самым собака приобретает более завершенный
облик и более „строгое" поведение. Правда, современное движение „зеленых" привело в
ряде европейских стран; в том числе на родине породы, к запрету на купирование (пока
только ушей) — и сразу проявились негативные последствия отсутствия модифицирующе¬
го влияния купирования — вислоухие доберманы не только потеряли во внешней привле¬
кательности, но и их „рабочее" поведение формируется гораздо труднее — вислоухая соба¬
ка стала менее резкой, потеряв в значительной мере знаменитый свой „напор“.В целом доберман — весьма здоровая порода. Однако селекционеры, пристально от¬
слеживающие параметры жизнеспособности при разведении доберманов, обнаружили
встречаемость, обычно невысокую, нескольких наследственных болезней. Как и у многих
крупных собак, у доберманов наблюдали дисплазию тазобедренного сустава. Большин¬
ство европейских и международных доберман-клубов и союзов проводят обязательное
рентгенографическое тестирование всех племенных производителей и производительниц и
допускают к разведению только животных, свободных от ДТС, или с невысокой степенью
ее проявления [Zwan, 1988].У доберманов зарегистрирован так же, как у некоторых других пород, деформирую¬
щий прогиб запястья. Его вызывает рецессивный аллель cs, локализованный в Х-хромосо-
ме. Полиостатическая фиброзная дисплазия обнаружена только у доберманов и бобтейлов
в виде опухолей на передних конечностях. Но определенного типа наследования пока не
установлено.Другая аномалия — Вобблер-синдром — видимо, специфичен только для доберма¬
нов. Этот дефект называют также цервикальной (шейной) спондилопатией. Он представля¬
ет наследственно обусловленную деформацию шейных позвонков, провоцируемую высо¬
копротеиновым рационом кормления щенков в период наиболее интенсивного их роста.
Сейчас дискутируется вопрос о целесообразности тестирования (рентгенографического)
доберманов перед разведением с целью выбраковки из племенного состава собак, облада¬
ющих этой аномалией даже со слабой степенью проявления.Наследственно обусловленные дефекты зубной системы — неправильный прикус и
неполнозубость, а также крипторхизм -^ предмет заботы селекционеров: современный
стандарт требует дисквалификации доберманов с проявлениями отклонений от нормы по
этим параметрам.Из аномалий крови у доберманов встречается болезнь фон Виллебранда — наслед¬
ственное нарушение свертывания крови из-за дефицита фактора 8: обычно дефект у добер¬
манов выражен слабо (дефицит фактора неполный) и не вызывает серьезного снижения
жизнеспособности от кровотечений, хотя и сопровождается большей чувствительностью в
разных стрессовых ситуациях.Так называемая цветная мутантная алопеция так же, как и Вобблер-синдром, впер¬
вые обнаружена у доберманов, причем голубого (а затем и коричневого) окраса. Характе¬
ризуется частичным облысением и наследственной связью с генами осветления пигмента
шерсти.В породе обнаружена первичная врожденная катаракта, но тип наследования окон¬
чательно не ясен, так же, как и для породоспецифической стойкой гиперплазии первичного
стекловидного тела/стойкой гиперплазии сосудистой оболочки хрусталика
(PHTVL/PHPV). Эта аномалия, называемая по региону, где обнаружена впервые, „голлан¬
дской болезнью", является объектом внимания доберманистов Голландии, где, благодаря
селекционному контролю, частота сильного проявления дефекта в последнее время значи¬
тельно снижена. Но она известна и в других популяциях породы.
Энтропион (заворот век) и почечная недостаточность также изредка встречаются у до¬
берманов как наследственные болезни, хотя пока точный тип их наследования не известен.По аутосомно-рецессивному типу у доберманов наследуется вестибулярный синдром,
проявляющийся глухотой и нарушением способности сохранять равновесие гомозиготами.
Они пока обнаружены только в Англии и США [Wilkes, Palmer, 1992].Золотистый ретривер [Golden Retriver]Замечательные своим характером и нарядным обликом эти превосходные подружей¬
ные собаки стали в последние годы одной из самых популярных пород во всем мире как
идеальные друзья дома и семьи хозяина. Происхождение породы не документировано, но,
скорее всего, это продукт скрещивания лабрадор-ретривера со спаниелем. Известны в Анг¬
лии со второй половины прошлого века.Стандартный окрас — сплошной любых оттенков золотистого и кремового цвета, но
не красного и рыжего. Допускаются небольшие пятна на груди, но не на голове, шее или
конечностях.Гомозиготы по гену длинношерстности /, золотистые ретриверы имеют гомозигот¬
ный генотип и по гену е, а изменчивость интенсивности цвета шерсти контролируется ры¬
жими полигенами.Правда, встречаемость палевых ретриверов в породе вызывает предположения о на¬
личии в генофонде шиншиллового аллеля сс'г.Дисплазия тазобедренного сустава (ДТС) так же наследуется полигенно в этой поро¬
де, как и в остальных. Относительно высока в этой породе встречаемость эпилепсии, но
тип наследования ее не изучен окончательно.Предположительно доминантна у ретриверов первичная катаракта, развивающаяся у
них к 6-12 месяцам от роду и прогрессирующая с возрастом. Другой дефект зрения ретри¬
веров — прогрессирующая ретинальная атрофия — встречается у них чаще, чем у многих
других собак, имея рецессивный характер. Наиболее типичная для этой породы еще одна
форма патологии сетчатки глаза — центральная прогрессирующая ретинальная атро¬
фия — напротив, контролируется, вероятно, доминантным геном с неполной ггенетрант-
ностыо и с изменяющейся экспрессивностью.Ирландский волкодав [Irish Wolfhound]Громадного роста (бывают и выше метра), эти собаки отличаются от других борзых
более крепким телосложением, хотя и не таким массивным, как у мастифов. Попав на ост¬
рова вместе с кельтами еще до нашей эры [Палмер, 1988], уже многие века волкодав обла¬
дает характерными и отличительными признаками породы, в том числе жесткой шерстью,
своеобразными движениями и окрасом: наиболее типичны тигровый, серый, рыжий, а так¬
же черный, белый, палевый. При „фирменном" тигровом рисунке волкодавы, по мнению
R. Robinson, имеют генотип а5а-Е°1 -, так как часто полосы-тигровины располагаются на
серо-черном, а не желтом фоне. Однако С. Little [1957] считает, что тигровость обусловли¬
вает и взаимодействие аллелей АУ и ЕЬг — при желточерном оттенке окраса. Точность оп¬
ределения истины в этих спорных случаях затруднена цветовыми эффектами жесткой гру¬
бой шерсти.Относительно генетического статуса белых волкодавов Little полагал, что этот ок¬
рас — осветленный палевый в результате неаллельного взаимодействия генов АУ и сс1\ или
ослабления „минус-рыжими“ полигенами желтого пигмента, синтезируемого в шерсти со¬
бак, обладающих геном 'А.Не вполне ясен генотип волкодавов по локусам длины и текстуры шерсти: проблема¬
тично, имеют ли они ген длинношерстности I или же только аллель L [Robinson, 1982].ОкрасГ енотипЧерныйAs-E-ТигровыйАУ-ЕЫ-Рыжий, палевыйАУ-Е-
Ирландский сеттер [Irish Setter]Прекрасная легавая с огневым темпераментом и уникальным „огненным11 окрасом.
Порода признана в 1876 г. и считается самой „молодой11 из островных пород легавых, вы¬
веденной в результате скрещивания ирландских водяных спаниелей, спрингер-спаниелей,
испанских легавых и сеттеров — английских и шотландских [Палмер, 1988].Насыщенный темно-рыжий цвет „красного дерева11 без примеси черного, но с допус¬
тимыми белыми отметинами — „визитная карточка11 породы. С. Little [1957] показал, что
отсутствие черного пигмента обусловлено контролем окраса ирландцев со стороны гена е.
Относительная длинношерстность обусловлена гомозиготностью II. Таким образом, гено¬
тип „шерстных11 локусов: As-eell. Богатство насыщенного цвета создается максимальной
концентрацией „плюс-рыжих“ полигенов-модификаторов пигментации.Правда, в прошлом от рыжих родителей рождались иногда черные щенки ирландско¬
го сеттера [Winge, 1950], а С. Little [1957] сообщил: среди 1197 щенков этой породы, полу¬
ченных в Джексоновской лаборатории, родился один черный. М. Burns и М. Fraser [1966]
собрали информацию от заводчиков, у которых очень редко рождались в этой породе со¬
баки черно-подпалые, черные и соболиные. Эти данные говорят о том, что большинство
рыжих ирландских сеттеров, вероятно, имеют генотип As-eeSS, а некоторые — АУ-EESS.
С белой проточиной ирландцы — гомозиготы s's‘, а редкие рыже-пегие — гомозиготы sPsP.Среди ирландских сеттеров профессор Н. А. Ильин [1932] наблюдал проявление
иногда гена р (ра) — „рубиновоглазие“ — из-за ослабленной пигментации радужной обо¬
лочки у гомозигот при освещении под определенным углом глаза сверкают красным отсве¬
том, а не зеленым, как обычно. Зрение у таких собак нормальное.Цветная мутантная алопеция характеризуется частичным выпадением шерсти у свет¬
лоокрашенных сеттеров.В породе впервые обнаружен мутантный рецессивный аллель qu [Palmer et al., 1973],
обусловливающий в гомозиготном генотипе квадриплегию — „плывущие движения11 не
встающих на ноги щенков с рядом других нейромускулярных отклонений.По аутосомному рецессивному типу у ирландских сеттеров наследуется также специ¬
фичная только для этой породы гранулоцитопатия — аномалия активности нейтрофилов,
приводящая к повышенной чувствительности гомозигот к инфекциям [Wigh, 1990]. Мута¬
ция летальная. Напротив, мутантный и тоже рецессивный ген рга обнаружен не только у
ирландских сеттеров, но и у многих других собак. Он вызывает у гомозигот прогрессирую¬
щую атрофию сетчатки. Но именно у ирландцев [Aquirre et al., 1978] установлено, что эта
аномалия глаза связана со снижением активности ключевого для жизнеобеспечения и мета¬
болизма в клетках сетчатки фермента — циклической GMP-фосфодиэстеразы. Таким обра¬
зом, при исследовании именно этой породы выяснена причина дегенерации зрения, вызы¬
ваемая геном рга.Ирландский терьер [Irish Terrier]В современной кинологии считается, что эта порода в нынешнем виде представляет
собой потомков английских черно-подпалых жесткошерстных терьеров и старинных терь¬
еров Ирландии, из-за чего вплоть до 1879 г. ирландские терьеры представляли множество
локальных географических форм разного экстерьера, размера и окраса. И только с конца
прошлого века поголовье было стандартизировано и приобрело знакомые нам черты. Же¬
сткая шерсть имеет сплошной ярко-рыжий окрас, у некоторых собак осветляющийся до
пшеничного и золотистого оттенков под действием „рыжих11 полигенов. В генетической
формуле ирландских терьеров по локусам шерсти выделяются аллели АУ и Wh.Среди ирландских терьеров обнаружена уникальная для них сцепленная с полом ми-
опатия, детерминируемая рецессивным аллелем ту, который локализован в Х-хромосоме
[Wentinck et al., 1974], — она проявляет свое действие практически всегда только у гемизи-
готных кобелей (т. е. имеющих только один аллель в единственной Х-хромосоме).Йоркшир-терьер [Yorkshire Terrier]Относительно молодая порода миниатюрных чрезвычайно популярных ныне собак,
созданная около ста лет назад от скрещивания скай-терьера со староанглийскими черно-
подпалыми терьерами [Палмер, 1988]. Английский кеннел-клуб официально признал со¬
временное название этой породы в 1886 г., а до того собаки были известны, как ни стран¬
но, под именем шотландских терьеров [Fletcher, 1977]. Возможно, именно эта „путаница"
была причиной разных взглядов на происхождение „йорки“: считалось даже, что порода
выведена от скотч-терьеров и мальтийских болонок [Senglaub, 1978].Масса взрослых „йорки“ до 3 кг, длинная шерсть ниспадает с боков прямо и ровно с
четко выраженным пробором от носа до конца хвоста. Щенки рождаются полностью чер¬
ными, и только с возрастом формируется настоящий окрас: голубовато-стальной цвет на
спине и золотисто-палевый — на груди и голове. С генетической точки зрения это чепрач¬
ный окрас, только необычного оттенка и рисунка, следствие действия гена asa. Little дока¬
зал, что в детерминировании окраса участвует ген поседения G, хотя R. Robinson считает
это мнение несколько спекулятивным: возможно, что окрас йоркшир-терьера есть резуль¬
тат селекции на удлинение прямой шерсти с максимальным проявлением грифельно-голу-
бого подшерстка.Среди „йорки“, как и некоторых других миниатюрных собак, встречается аномалия
строения эпифизов бедренных костей — болезнь Пертеса, причем именно для этой породы
недавно [Robinson, 1992] определен генетический контроль мутантным рецессивным алле¬
лем pd. У гомозигот по другому рецессивному гену rd проявляется дисплазия сетчатки.Кавалер-кинг-чарлз-спаниель [Cavalier King Charles Spaniel]Добродушная и гармонично сложенная собака, ведущая происхождение от собак-
компаньонов короля Карла (Чарлза) Первого, изображенных на полотнах Ван Дейка, но
окончательно в современном облике сформированная в начале нынешнего века английски¬
ми заводчиками. Стандарт допускает 4 варианта окраса длинной шелковистой шерсти:
черный с рыжими подпалинами классического рисунка (черно-подпалый), трехцветный (на
белом фоне черные крупные пятна с рыжими отметинами — подпалинами), рубиновый
(сплошной ярко-рыжий) и бленхейм — „фирменный11 окрас этой породы — рыжие пятна
на белом фоне с уникальным и особо ценимым белым „ромбом1* на почти плоской голове
между ушами. Генетическая детерминация этих окрасов очень трудноустановима. Еще
С. Little [1957] дал генетическую формулу рыжего оттенка окрасов кавалер-кинг-чарлзов
как ее, но признал, что в породе эти же оттенки могут обусловливаться и геном АУ. Труд¬
ность дифференцировки этих альтернативных типов генетического контроля заключается
в том, что при генотипе ее рыжие разводящиеся „в себе“ должны давать только рыжих по¬
томков, но при наличии аллеля АУ родители могут давать также черно-подпалых щенков.
При этом оба названных окраса у потомков в действительности могут проявляться в ком¬
бинированном с пегостью виде — в окрасах трехцветном и бленхейм.ОкрасГенотипЧерно-подпалыйa'a'B-llS-Трехцветныйa'a'E-llsP-Рубиновый eellS-Бленхейм eellsP-Из наследственных болезней в породе встречается катаракта — помутнение хруста¬
лика глаза (наследуемая как рецессивный признак), и предсердно-желудочковый эндокар¬
дит. Наследственная обусловленность этого порока сердца была доказана благодаря силь¬
ной предрасположенности к нему именно собак данной породы — в Англии, Швеции,
Дании [Pedersen et al., 1992]. Тип наследования точно еще не определен, но эта аномалия у
кавалер-кинг-чарлз-спаниелей развивается в более раннем возрасте, чем у собак других по¬
род.Кардиган-вельш-корги [Welsh Corgi, Cardigan]Одна из любимейших национальных британских пород пастушьих (скотогонных) со¬
бак, имеющая многовековую историю: в Южном Уэльсе она была известна уже во времена
Вильгельма Завоевателя (XI век).
Наиболее типичны окрасы соболиный, тигровый, черно-подпалый и мраморно-голу-
бой. Большинство собак имеют при этом на морде, груди и животе белые отметины, детер¬
минируемые аллелем ирландской пятнистости (У). Однако Little [1957] наблюдал в породе
и собак с более распространенными белыми зонами, видимо, из-за аллеля sP. Мраморные
кардиганы имеют на сероватом фоне черные и голубоватые пятнышки.ОкрасГенотипСоболиныйАУ-Е-—ттТигровыйАУ-Еь,'-ттЧерно-подпалыйа'а'Е—ттМраморныйа'а'Е—МтКерри-блю-терьер [Kerry Blue Terrier]Выведен в ирландском графстве Керри с использованием генофонда ирландских те¬
рьеров, а также бедлингтонов и бультерьеров. Первоначально эти голубые терьеры ис¬
пользовались по прямому назначению — как охотничьи собаки, но теперь в основном
они — компаньоны и прекрасные друзья семьи.Чрезвычайно знаменит шерстный покров этих собак — пышная, волнистая и длин¬
ная шерсть при рождении щенков густого черного цвета, но с возрастом (после года) свет¬
леет до сероголубого оттенка разной интенсивности от slate — грифельно-шиферной — до
светло-серебристой. Более темные тона на морде, ногах и хвосте; маска может быть не у
всех собак. Такая возрастная трансформация окраса обусловлена С-геном поседения во¬
лос, причем, вероятно, аллель G не проявляет эффекта разбавления пигмента в названных
зонах максимальной окраски. Генотип по шерстному покрову, очевидно, таков: As -G-ll.Из наследственных аномалий в этой породе известна встречающаяся иногда олиго-
донтия. Выявлен уникальный для керри мутантный рецессивный ген cb [Lahunta, Averill,
1976], обусловливающий у гомозиготных щенков в возрасте от 9 до 16 недель развитие
спинномозговой атаксии. Ее признаки — вначале тремор (дрожь) головы и слабость зад¬
них ног, затем — нарушение способности щенков самостоятельно подниматься и стоять
из-за дегенеративных процессов в спинном мозге и почти полного отсутствия в нем клеток
Пуркинье.Китайская хохлатая собачка [Chinese Crested Dog]Ее еще называют китайской голой собачкой, известность в Америке и Европе она по¬
лучила только после 1966 г., хотя, несомненно, имеет древнюю историю. Один из типич¬
ных примеров мутации бесшерстности (ген Hr) среди собак. При этом китайские собачки
сохраняют остатки шерстного покрова в виде хохолка на голове, а также на нижней части
конечностей и на хвосте — в отличие от фактически полностью голой мексиканской собач¬
ки. Интересно, что Н.А. Ильин [1932] сообщал о плейотропном действии гена бесшерстно¬
сти, обусловливающего также часто дефектное развитие зубной системы голых собачек: у
некоторых из них вырастает только по одному боковому зубу в каждой челюсти (премоля-
ру или моляру)! Голая кожа животных этой породы может быть сплошь пигментирован¬
ной, или же с зонами депигментации в виде пятнистости или пегости. При этом часто бе¬
лые пежины соседствуют с черными пятнами на основном „сером11 фоне. R. Robinson
отмечает трудность предположений („спекуляций") о наличии каких-либо генов окраса
шерсти в генофонде породы — из-за отсутствия самой шерсти! — и предложил следующие
формулы:ОкрасГ енотипСплошнойHr-llS-ПятнистыйHr-llsP-T-
Кламбер-спаниель [Cljjmber Spaniel]Самый тяжелый из современных спаниелей, с рыхлой конституцией, кламбер, види¬
мо, ведет происхождение от французских бассетов и исчезнувших ныне альпийских спание¬
лей [Палмер, 1988], но сформировался в британском поместье герцога Ньюкастлского, ку¬
да родоначальники попали из Франции во времена Великой французской революции.Генетической особенностью породы является наличие в генофонде аллеля вместо
обычного для спаниелей sP, так как стандартный окрас — белый с небольшими лимонны¬
ми (но допускается и с оранжевыми) пятнами, и белый тон очень распространен по телу. С.
Little [1957] полагает, что цвет отметин детерминируется геном в. Мочка носа кламбера
обычно коричневого или телесного цвета, что подтверждает его гомозиготность по Ь. Та¬
ким образом, генотип по окрасу и шерсти кламбера: bbeells'vsw.Кламбер — относительно невысокая собака, но очень массивная и весьма тяжелая, а
потому в этой породе, как и среди наиболее крупных собак, обнаружена полигенно насле¬
дуемая дисплазия тазобедренного сустава.Колли длинношерстная (шотландская овчарка) [Rough Collie] и
Шелти (шетландская овчарка) [Schetland sheep dog]Их предки — пастушьи овчарки — были завезены в Шотландию 400 лет назад из Ис¬
ландии [Палмер, 1988] для пастьбы и охраны овец особой породы — с черными мордами и
ногами, называвшихся „коллиз“ (отсюда произошло и название собак-пастухов). Ныне по¬
пулярная, в основном, декоративная собака — друг дома, хотя прежде успешно использо¬
валась как служебная. Впервые колли появились на выставке собак (в Бирмингеме) в
1860 г., несколько позже началось их заводское разведение.Стандарт допускает три типа окраса: соболиный с белым (рыже-пегий), трехцветный
(черно-подпалый с белыми отметинами) и мраморно-голубой (серебристый фон с неболь¬
шими черными пятнами).По-видимому, у соболиных колли в окрасе проявляется ген АУ, а у трехцветных —
классический аллель подпалин ас. Мраморные колли являются гетерозиготами по „мра¬
морному" гену — Мт. Белые отметины, часто в виде „воротника" на шее, пятен на груди,
конечностях и конце хвоста (типичный пегий окрас) обусловлены действием скорее всего
аллеля s‘, а не sP, который вызывает пятнистость. Little [1957] отметил, что в породе иногда
встречается ген-разбавитель пигмента (d), вызывая ослабление окраса, — проявляется у
светло-палевых, голубых и голубых с подпалинами колли.Только длинношерстным колли свойствен особый ген Sg (Slate-grey), вызывающий
так называемый грифельно-серый или голубой окрас (ранее входил в число стандартных
окрасов этой породы). Фенотипически он напоминает голубой окрас гомозигот по рецес¬
сивному гену-разбавителю пигмента d, но наследуется доминантно.Родственная форма — короткошерстная колли — имеет такую же давнюю генеало¬
гию и отличается от вышеописанной колли только короткой гладкой шерстью (ген L).На Шетландских островах, вблизи от Великобритании, сформировался ряд типично
островных форм домашних животных: маленькие лошадки (шетландские пони), мини¬
атюрные овцы. И пастушьи собаки — шетландские овчарки — являются миниатюрными
„копиями11 колли.Шелти филогенетически близка колли, и генетика окраса и длины их шерсти абсо¬
лютно идентична [Robinson, 1982].У этих родственных пород и наследственные аномалии сходны. Только среди них
распространена называемая по имени породы аномалия глаз колли. Этот дефект глазного
дна собак — гомозигот по мутантному рецессивному гену сеа. Интересно, что мутантный
аллель сеа имеет плейотропный эффект, обусловливая дефекты в разных тканях глаза.ОкрасГенотипСоболиныйЧерно-подпалыйМраморно-голубойАУ-D-llmms
aWD-llmms1-
dafddllMms1-
Центральная прогрессирующая атрофия сетчатки глаза как самостоятельная наслед¬
ственная аномалия наиболее широко распространена среди колли (и еще среди ретриве¬
ров) — результат действия мутантного доминантного гена с неполной пенетрантностью.
Так называемая церебеллярная дегенерация тоже обнаружена только у колли, но тип на¬
следования еще не ясен.Не установлен пока и характер генетического контроля встречающейся среди колли
эпилепсии.Специфична только для колли циклическая нейтропения — ее еще называют синдро¬
мом голубых колли. Аномалию обусловливает рецессивная мутация „разбавления пигмен¬
та" — аллель сп. Он плейотропно вызывает периодическое снижение количества нейтрофи¬
лов в крови и тем самым устойчивости к инфекции. Плейотропно этот ген приводит к
летальному исходу болезни.У шелти в Швеции обнаружены случаи гемофилии А, наследуемой как типичная
сцепленная с полом патология свертывания крови (из-за дефекта фактора VIII) — ее рецес¬
сивный ген локализован в Х-хромосоме. А в США у некоторых шелти в крови оказалось
резко сниженным содержание фактора фон Виллебранда, приводящее к другой аномалии
свертывания крови.Мутантный рецессивный ген рс (с неполной пенетрантностью, равной примерно
50 %) вызывает у гомозиготных колли (кроме них — еще у немецких овчарок) атрофию
поджелудочной железы. В Скандинавии, например, где наиболее хорошо изучены популя¬
ции колли в этом отношении, частота этой аномалии весьма высока и в последние годы
имеет тенденцию к росту. Это вызывает тревогу ветеринаров и селекционеров, считающих
необходимым учитывать носительство гена рс при отборе производителей.Одна из характерных породных особенностей этих собак — „фирменный" постав
ушей, так называемые „уши типа колли": у настороженной собаки стоячие в основании,
верхняя треть ушной раковины направлена вперед — параллельно земле. Этот признак, по
Н. А. Ильину [1932], детерминируется доминантным аллелем На, доминирующим над алле¬
лями Я (висячее ухо) и h (стоячее). По-видимому, в генофонде породы встречается и рецес¬
сивный аллель h : подчас (прежде гораздо чаще) встречаются колли и с полностью стоячи¬
ми ушами (а бывает и „разноухость").Кэрн-терьер [Cairn Terrier]Выведен в Шотландии как типичный охотничий терьер для охоты на зверей в норах,
в современном облике известен более 150 лет, но до 1924 г. вполне обычным методом раз¬
ведения породы было скрещивание кэрнов с „уэсти" — уэст-хайленд-уайт-терьерами [Пал¬
мер, 1988]. Характерна взъерошенная, довольно длинная и густая шерсть. Интересно, что
на выставках кэрны оцениваются без тримминга, в „натуральном виде". Первоначальным
окрасом считается тигровый [Robinson, 1982], причем полосы бывают как четкими, так и
расплывчатого рисунка. Цвет шерсти варьирует от черноватого до красноватого, со мно¬
гими промежуточными оттенками, в зависимости от соотношения количества черно- и
желтоокрашенных волос. Интенсивность желтого цвета также варьирует, потому что неко¬
торые кэрны имеют желторыжую „подцветку", тогда как другие особи бывают кремовыми
или почти белыми, и многие цветовые варианты обусловливаются комбинацией „рыжих"
полигенов. Контраст между темной головой и часто гораздо более светлым телом обуслов¬
ливается удлиненной шерстью на туловище (на голове шерсть короче) и черной маской на
морде. Красный, желтый и рыжий окрасы обусловлены, вероятно, действием гена АУ, с
разной экспрессивностью из-за взаимодействия с „рыжими" полигенами [Robinson, 1982].
При этом весьма возможно, что некоторые цветовые вариации, определяемые как тигро¬
вый окрас, на самом деле есть проявление соболиного (sable) окраса.ОкрасГенотипТигровыйАУ-ЕЬг-11Красный, рыжийАУ-Е—ИДля породы характерна наследственная гемолитическая анемия, вызываемая у гомо¬
зигот рецессивной мутацией (аллель назван рк, так как он снижает активность в крови фер¬
мента пируваткиназы). Именно у кэрн-терьеров эта мутация является первопричиной ком¬
плекса дефектов — синдрома анемии со вторичным гемохроматозом [Schaer et al., 1992].
Лабрадор-ретривер [Labrador Retriever]Одна из популярнейших сейчас пород собак, предки которой были завезены в начале
XIX века в Великобританию с острова Ньюфаундленда. На своей далекой родине первона¬
чально они использовались рыбаками для вытаскивания сетей из моря. Превосходные
пловцы, они работают на охоте как ретриверы — подают битую дичь из воды, а чаще со¬
держатся как любимцы семьи хозяина. Видимо, имеют родство с ньюфаундлендами. Ко¬
роткая плотно прилегающая шерсть лабрадоров (с густым подшерстком) имеет сплошной
окрас: черный или желтый, хотя допустим и любой другой оттенок без белых отметин — за
исключением возможного небольшого пятна на груди [Палмер, 1988]. По R. Robinson ин¬
тенсивность цвета шерсти определяется комбинацией полигенов.Проведено глубокое клиническое, морфологическое и генетическое изучение наслед¬
ственной болезни сетчатки глаз лабрадоров — ретинальной дисплазии, приводящей к по¬
тере зрения и слепоте [Коек, 1974]. Оказалось, что, например, в Швеции популяция лабра¬
доров содержит в весьма высокой концентрации мутантный ген, обусловливающий у
гомозигот аномальное развитие: глаза с дефектной сетчаткой растут в размерах быстрее,
чем нормальные глаза здоровых щенков-ретриверов. Генеалогический анализ 19 пометов
лабрадоров, в которых зарегистрирован хотя бы один щенок с дисплазией сетчатки (в них
рождены 33 дефектных — 13 кобельков и 20 сучек — и 124 нормальных щенка), а также
специальные скрещивания в разных комбинациях показали, что аномалия контролируется
аутосомным рецессивным аллелем rd, как и у других пород, где она выявлена. В 8 специ¬
ально полученных пометах от обоих родителей-лабрадоров с дисплазией сетчатки роди¬
лось 25 щенков, из которых не было ни одного здорового — все с этой аномалией (11 кобе¬
лей и 14 сук). Доктор Е. Коек отмечает, что все лабрадоры в Швеции, больные
ретинальной дисплазией, и носители мутации происходят от племенного кобеля-произво-
дителя, рожденного в Англии в 1934 г. и распространившего аномалию в шведской попу¬
ляции.У этих ретриверов также встречается дисплазия тазобедренного сустава с полиген-
ным наследованием. Сложно наследуется у них и эпилепсия — нет доказательств ее моно-
генного контроля.У лабрадоров, в соответствии с законом гомологических рядов Н.И. Вавилова [1987],
много генетических особенностей, сходных с таковыми у золотистых ретриверов. Так, и
катаракта у них встречается одинаковая — прогрессирующая и наследующаяся, вероятно,
доминантно.Лапландский шпиц (лапхунд) [The Lapland Dog]Это аборигенная скандинавская порода, выведенная в Швеции, где разводится в ста¬
тусе редкой исчезающей национальной породы. Является примитивной шпицеобразной
формой, по размерам уступая среднему шпицу (высота кобелей 45-50 см и сук — 40-45 см).
Голова широкая между ушами и куполообразная, в противоположность другим шпицам.
Стоячие широкие уши заметно расставленного постава. Средней длины хвост завернут в
кольцо.Превосходная пастушья собака, она издавна разводилась в регионах северного оле¬
неводства Скандинавии, но в современных условиях численность породы снижается:
Шведским кеннел-клубом зарегистрировано в 1950-1970 гг. только 2069 лапландских со¬
бак — всего 0,38 % от общего числа всех зарегистрированных собак в стране. В шведской
армии в последнее время стали использовать этих собак в защитно-караульной службе
[Палмер, 1988].ОкрасГенотипЧерныйКоричневыйЖелтый с черным носом
Желтый с коричневым носомAs-B-ee
A5-hhee
—В-ее
—bbee
Порода известна в кинологии и генетике как популяция с широко распространенной
уникальной наследственной нейрональной абиотрофией. Клинически эта аномалия харак¬
теризуется тетраплегией (параличом всех конечностей) и мышечной атрофией у молодых
лапхундов. Установлена гомология в подобных нейромускулярных нарушениях лапхунда
и человека.При исследовании 12 пометов, в которых рождено 19 щенков с этой аномалией, не
установлено достоверных различий во встречаемости болезни у кобелей и сук и доказан
аутосомный рецессивный характер наследования болезни [Sandefeldt et al., 1973]. Мутант¬
ный ген характеризуется 100 %-й пенетрантностью и независимостью в проявлении от фак¬
торов окружающей среды.Левретка (маленькая итальянская борзая) [Italian Greyhound]Эта старинная порода „кроличьих" борзых, как видно из ее англоязычного названия,
многими воспринимается как миниатюрная (масса 2,5-4 кг) копия грейхаунда. По крайней
мерс, R. Robinson полагает, что генофонд левретки по генам окраса ее гладкой короткой
шерсти идентичен генофонду грейхаунда, за исключением отсутствия у итальянских бор¬
зых гена тигровости Е°г, так как среди левреток нет собак такой расцветки.Маламут [Alaskan Malamute]Основной окрас этих собак — волчий (черый) с вариациями оттенков от светло-серо¬
го до почти черного. Некоторые собаки при этом имеют черную маску на морде, а у дру¬
гих морда светло-серая. R. Robinson [1982] полагает, что генотип такого окраса, видимо,
Asa- или а-, если оба гена имеют идентичный эффект.Дж. Палмер [1988] отмечает, что стандартным является также сплошной белый окрас.
Видимо, при этом генотип может быть swsw.Среди маламутов встречается иногда наследственная гемералопатия — „дневная сле¬
пота", обусловленная действием у гомозигот рецессивной мутации гена he [Rubin et al.,
1967].Мальтийская болонка [Maltese]Одна из древнейших европейских пород комнатных собачек, но не ясно, имеет ли она
чисто западные корни или предки ее были завезены в средние века (на Мальту?) из Восточ¬
ной Азии.Чрезвычайно густая длинная шерсть имеет чисто-белый цвет, хотя иногда встречают¬
ся болонки с допускаемыми стандартом светло-палевыми отметинами. С. Little [1957] пока¬
зал, что белизна болонок обусловлена действием „экстремально-белого" аллеля j11', а не
комплементарным взаимодействием желтого гена (АУ или е) с шиншилловым аллелем cch
и/или „минус-рыжими“ полигенами. Он установил: желтые отметины белых болонок, рас¬
полагающиеся на голове или туловище, детерминируются гомозиготностью sPsP. Однако,
если отметины на корпусе отличаются большей площадью пигментации — например, на
всей спине — более вероятно, что в этих случаях собаки имеют генотипы АУ-ссЬ — или
сс,1-ее.Мопс [Pug]Эта порода маленьких со своеобразной головой собак имеет историю, уходящую в
глубь веков, где корни ее филогенетического древа четко не просматриваются. В Европу
(во Францию) мопс попал вместе с турецким флотом в 1553 г. [Палмер, 1988], затем был по¬
пулярен в Нидерландах, откуда завезен в Англию — там и сформировался современный
облик этих собак.Окрас короткой (LL) шерсти однотонный, четко выраженный, в контрасте с черной
маской на морде и ремнем от затылка вдоль спины до хвоста. Цвета — серебристый, абри¬
косовый, бежевый. Но бывают и черные мопсы. Осветленные тона окраса обусловливают¬
ся комбинацией „минус-рыжих“ полигенов.
ОкрасГенотипЧерныйAs-C-БежевыйАУ-С-СеребристыйАУ-ссН-Среди мопсов обнаружены собаки с болезнью Пертеса — наследственной аномалией,
проявляющейся в некрозе бедренной кости. Судя по гибридологическому анализу в семьях
миниатюрных терьеров, этот дефект детерминирует у собак рецессивный мутантный ал¬
лель pd [Robinson. 1982]. Встречаются у мопсов и лишние резцы в нижней челюсти — как и
у большинства короткомордых собак [Willis, 1990].Немецкая овчарка [German Shepherd Dog]Одна из самых распространенных пород собак в мире, немецкая овчарка, по праву
знаменита своими прекрасными способностями к разнообразной дрессировке и несению
службы, высоким адаптационным потенциалом в любых условиях жизни и работы. Перво¬
начально типичные овчарки — пастушьи собаки (известные на территории современной
Германии как минимум с XVII века) как заводская порода — претерпели за последние сто
лет огромные преобразования в породном облике, телосложении, экстерьере и характере.
Впервые представленные как порода на выставке в Ганновере в 1882 г., эти собаки были
вовлечены в интенсивный селекционный процесс под руководством талантливого киноло¬
га Макса фон Штефаница, в результате чего сформировалась популяция животных, при¬
годных практически ко всем видам служб в армии, полиции и т. п.Стремительная эволюция породы отмечена созданием ряда внутрипородных генеа¬
логических и конструктивных типов, наиболее современный и перспективный из кото¬
рых — „овальный" — завоевал сейчас практически повсеместное лидерство на выставках
красоты и соревнованиях/испытаниях рабочих качеств. Головные организации— Немец¬
кий и Всемирный союзы владельцев немецкой овчарки — проводят комплексную селекцию
с целью совершенствования всех наиболее ценных породных качеств этих собак: гармонич¬
ного сложения, крепкого характера, работоспособности, здоровья и адаптационного по¬
тенциала. Эта работа ведется весьма успешно благодаря верной стратегии и высокой ин¬
тенсивности отбора: объем племенного ядра, одного из крупнейших по сравнению с
другими породами собак (почти во всех странах немецкая овчарка входит в „первую десят¬
ку" по числу регистраций чистопородных щенков), дает возможность повышать селекци¬
онный дифференциал и, следовательно, эффективность селекции.В породе сейчас встречаются несколько четко различающихся окрасов (при сильной
степени индивидуальной изменчивости). Рыжий окрас может варьировать от желтого до
сильно затемненного, а чепрачный — от светлоокрашенного чепрака на почти белом фоне
до почти черного. Наименее изменчив черно-подпалый окрас, но и здесь возможно умень¬
шение распространения подпалин — например, на морде, где может быть черная маска,
типичная для большинства овчарок за исключением черных, у которых морда вся сплошь
черная. Интенсивность желтого пигмента колеблется от золотого до палево-кремового
(почти белого оттенка). При этом немецкие кинологи — законодатели мод в оценке эксте¬
рьера этой породы — справедливо предпочитают в качестве более породных овчарок с ин¬
тенсивно пигментированным шерстным покровом: еще в эмбриогенезе пигментобразую-
щие клетки и клетки нервной системы закладываются из одного зародышевого листка,
неврального ствола, и интенсивность окраса положительно коррелирует с силой и прочно¬
стью нервной системы собаки.В генофонде породы распространены гены As, A v, asa, а1. Изменчивость окраса, обус¬
ловливаемая аллелем А>', не отличается от таковой у других пород собак, но заслуживает
точного изучения вариация активности аллеля asa в его взаимодействиях с рыжими полиге¬
нами [Robinson, 1982]. В итоге V-образный чепрак может быть узким или широким, вплоть
до чепрака от плеч на все бока. Он может быть интенсивно черным или светлым, с приме¬
сью желтых волос, и т. д. Поэтому чепрачный окрас немецких овчарок — сложное явление
и рассматривается генетиками как распространение желтой пигментации от области ста¬
тей „верха" собаки в нижние части ее тела. При этом, как считает R. Robinson, наличие
крапчатости — тиковой испещренности — может маскировать этот окрас вплоть до ти-
и потому такая форма стандартом не допускается как желательная. Эти гены (dd) обуслов¬
ливают и превращение черной маски у тигровых и палевых догов в грязно-серую, что так¬
же не допускается стандартом.Согласно данным С. Little [1957], мраморный окрас формируется гетерозиготным ге¬
нотипом по мраморному гену М в комбинации с одним из аллелей черно-белой пятнистос¬
ти локуса 5 — вероятно, аллеля sP. Бывают доги и пятнистые (генотип mmsP -), но они так¬
же нестандартны — их „рубашка11 с черной спиной и белым животом отличается от
мраморного рисунка, и граница черного и белого цветов не такая „рваная", как должно
быть у арлекинов. При гомозиготности по аллелю d мраморные доги имеют серые пятна
на белом фоне, что современный стандарт не допускает. Стандартный окрас догов-арлеки-
нов отличается от окраса мраморных колли и такс, у которых М-ген вызывает расцветку
смешения нормальных черных и более светлых — голубоватых — пятен. Черные пятна у
догов мраморного окраса располагаются по всему телу, что свидетельствует об отсутствии
в их генотипе экстремально-белого аллеля j11', в качестве гена, контролирующего белый
фон. R. Robinson поэтому полагает, что арлекиновый окрас немецких догов обусловлен ге¬
ном, подобным гену, обозначенному R. Schaible и J. Brumbaugh [1976] как ген мраморнос-
ти „арлекин11 'Н1'. У собак Hhh мраморность может быть не черно-серая, а черно-белая, как
и бывает у немецких догов. Поэтому такая гипотеза (в пользу наличия в генофонде немец¬
кого дога не традиционно приписываемого ему гена М, а гена другого локуса Hh или, по
крайней мере, особого черно-белого аллеля Mh) представляется альтернативной, если не
более приемлемой и самой корректной. В пользу этой гипотезы служат и встречавшиеся
нам немецкие доги нестандартного мраморного окраса — черные пятна на сером фоне, и
то, чт© не все почти чисто-белые доги оказываются с дефектами зрения и слуха, как долж¬
но быть в случае действия на мраморный окрас только гена М (известно, что у гомозигот
ММ окрас из мраморного становится белым с плейотропно вызываемой при этом глухо¬
той). Таким образом, вероятно, требуется признать, что генетический контроль мраморно¬
го окраса осуществляется в этой породе не одной парой аллелей М-т, как это считалось
ранее и что отражает классическая схема генетики окрасов догов (по R. Robinson), где чер¬
но-белый арлекин признается гетерозиготой по особому гену М1’.ОкрасГенотипЧерныйAs-D-E—mmS-Палевый (черная маска)АУ-D-E—mmS-ТигровыйAy-D-Ebf-mmS-ГолубойAs-ddE—mmS-Разбавленный палевый с серой маской (нестандарт)АУ-ddE—mmS-Серо-тигровый (нестандарт)As-ddEbi—mmS-Арлекин черно-белыйA S-D-E—M,lmsP-Арлекин серо-белый (нестандарт)As-ddE—MhmsP-Арлекин черно-серый(нестандарт — черные пятна на серо-голубом фоне)As-D-E—MmsP-Дисплазия, в частности, локтевых суставов [Hedhammar et al., 1974], — обычный на¬
следственный дефект крупных и тяжелых собак. Типичен он и для немецких догов. Генети¬
чески эта аномалия контролируется полигенами с пороговым действием, а фенотипически
проявляется в результате взаимодействия генотипа и условий среды. Особенно резко про¬
воцирует этот дефект излишне калорийный рацион кормления молодых догов в период их
интенсивного роста.Видимо, как рецессив наследуются случаи олигодонтии — недостатка зубов, причем
у немецких догов наиболее часто недостает первых или четвертых премоляров в нижней
челюсти.Явно наследственную природу у немецких догов имеет паралич Стокарда, но генети¬
ческий контроль аномалии осуществляет более чем один локус: видимо, дефект наследует¬
ся по пороговому типу.
Ньюфаундленд [Newfoundland]Происхождение знаменитых „водолазов", впервые ставших известными на северо-во¬
сточном побережье Канады, точно не выяснено: видимо, они выведены в результате скре¬
щивания крупных „корабельных" догообразных собак, попадавших в этот регион на ры¬
бацких судах со всех уголков Атлантики, с местными собаками. Одно из первых
достоверных описаний ньюфаундлендских собак было сделано Бюффоном [По: Goerttler,
1978], который в своей монографии по этой породе привел также и схему филогенеза нью¬
фаундленда в контексте развития молоссоидов начиная с тибетского волка (рис. 10.3).Заводское разведение ньюфаундлендов началось в Германии, а затем в Англии, отку¬
да новая порода распространилась по всему миру — в виде двух вариаций окраса. Первая
вариация: наиболее характерный сплошной окрас — черный, но встречаются также пече¬
ночные (шоколадные) ньюфаундленды, хотя L. Whitney [1947] сообщал о редких выщепле-
ниях в породе красного и голубого окрасов. Вторая вариация: пегие, черно-белые ньюфа¬
ундленды стали известны под названием лендзиров по имени английского художника,
изображавшего их па своих картинах. Сейчас разделяют эти вариации па ДБе самостоя¬
тельные породы, причем ньюфаундленд зарегистрирован в реестре этой организации как
канадская порода, а лендзир — немецко-швейцарская, в честь стран, где они сформирова¬
лись. Известны случаи, когда при получении пегих собак от черных родителей последние
могут передавать им не проявляющийся, но имеющийся в генофонде у ньюфаундлендов
ген Т, что обнаружено по черному крапу на участках депигментации таких лендзиров.
Длинная шерсть — следствие гомозиготности по гену I.ОкрасГенотипЧерныйAs-B-D-S-КоричневыйAs-bbD-S-ГолубойAs-B-ddS-ЛендзирAs-B-D-sP-Среди ньюфаундлендов нередко встречаются больные собаки с дисплазией тазобед¬
ренного сустава (дефект наследуется полигенно).Сенбернарf Ньюфаундленд
Античный молосс ^ ^t IДревний ассирийский iДОГ ,' t !Тибетский дог |С. moIossus tibetanustДикая форма: Canis niger
(черный тибетский волк)tЦентр образования
догообразных (молоссоидов) - ТибетРис. 10.3. Схема эволюции догообразных собак ,и
выведения ньюфаундлендов по Келлеру [Goerttler, 1978]
Пекинес [Pekingese]Предки современных пекинесов разводились многие века в столичном Дворце китай¬
ских императоров в качестве талисмана членов императорской фамилии. Наиболее ста¬
ринные источники, описывающие „львиных дворцовых собачек в Пекине11, датированы
VIII—X столетиями [Wittenberger Н., Wittenberger G., 1981], а в 1860 г., после захвата Двор¬
ца английскими войсками, они попали в Европу, где с тех пор разводятся в чистоте заводс¬
кими методами.Типичная масса взрослых собачек 3-5,5 кг, но нередко появление „карманных11 осо¬
бей в обычных пометах (весом не более 267 г) — следствие выщепления гомозигот по ре¬
цессивному гену карликовости.Длинная шерсть может быть разнообразных окрасов, среди которых наиболее попу¬
лярны рыжие и соболиные. У многих пекинесов имеется черная маска на морде. Встреча¬
ются светлые особи — от палево-кремовых до почти белых (но не альбиносы, которые,
впрочем, периодически иногда рождаются и дисквалифицируются), их генотип может быть
таким же, как у „желтых шиншилл" {АУ -cch -) или у рыжих, но с концентрацией „минус-ры¬
жих" полигенов, осветляющих цвет. С. Little [1957] получал черных пекинесов от рыжих
родителей и установил наличие в породе гена е, так же, как и аллеля АУ у рыжих. Белые от¬
метины вполне характерны для этих собак, которые могут иметь в генотипах любой из
трех аллелей пятнистости. Little также встретил одного пекинеса голубого окраса (гомози¬
готу dd).ОкрасГенотипЧерныйAS-IIS-Красно-рыжий, соболиныйАУ-llS-Черный с белымAs-llsP-Рыжий с белымАУ-llsP-В 1920-е годы в породе обнаружен рецессивный ген, вызывающий расщепление мяг¬
кого неба [Ильин, 1931] — полулегальный, так как дефектные щенки погибают вскоре пос¬
ле рождения от голода: молоко при сосании вытекает через щель.Пемброк-вельш-корги [Welsh Corgi, Pembroke]Имеющая общее с кардиганами (и такое же древнее) происхождение, эта форма уэль¬
ских скотогонных собак выделена в самостоятельную породу только в 1934 г., а до этого
пемброки и кардиганы даже выставлялись на выставках в одном общем ринге.Отличается от кардиганов меньшим разнообразием расцветки шерсти. Обычны собо¬
линый разной интенсивности и черно-подпалый окрасы. Допускаются небольшие отмети¬
ны на морде, и вообще белые пятна уменьшенного размера бывают на животе, груди, пере¬
дних ногах.ОкрасГенотипСоболиныйАУ-S1'-Черно-подпалыйa!als'-Пинчер [Pinscher]Старинная немецкая порода, филогенетически занимающая одно из центральных по¬
ложений в группе родственных друг другу пинчеров и шнауцеров. Гладкая короткая
шерсть (генотип LL) бывает черного, черно-подпалого, коричневого, голубого окрасов.
Известны очень редкие и эффектные пинчеры мраморного окраса — следовательно, ген М
имеется в генофонде породы, правда, с очень невысокой частотой.
ОкрасГенотипЧерныйA'-B-D-ШоколадныйAs-bbD-ГолубойAs-B-ddЧерно-подпалыйa'a'B-D-Коричнево-подпалыйa'a'bbD-Голубой с подпаломalalB-ddРыжий с черным носомАУ-B-D-Рыжий с коричневым носомАУ-bbD-Пойнтер [Pointer]Широко распространенная короткошерстная „островная” легавая, знаменитая ост¬
рым чутьем, огневым стилем работы по перу и „скульптурной11 стойкой. Порода сформи¬
ровалась в Великобритании на генетической базе староиспанских легавых с участием в по¬
родообразовательном процессе бладхаундов, фоксхаундов и грейхаундов.Наиболее типичен двухцветный окрас пойнтера: на белом фоне пятна и крап по все¬
му телу черного, желтого, красного, кофейного цветов. Обычно считают, что желтый пиг¬
мент в шерсти пойнтеров детерминируется геном е [Little, 1957]:ОкрасГенотипЧерно-пегийAs-B-E-sPsPКофейно-пегийAs-bbE-sPsPЖелто-пегий с черным носомAs-B-eesPsPЖелто-пегий с коричневым носомAs-bbeesPsPОднако R. Robinson сообщал, что некоторые пойнтеры желтого цвета имеют „сле¬
ды" маски и „мазаные“ уши, свидетельствующие, что их желтый пигмент обусловливается
желтым аллелем другого локуса — АУ, а не геном е. Если это так, в генотипах желтых пой¬
нтеров следует заменить гены As и ее на аллели АУ- и Е'.Иногда собаки этой породы встречаются не пегих, а сплошных окрасов — они имеют
в генотипе вместо аллеля sP доминантный ген S.Пегость и белые отметины пойнтеров являются проявлениями действия гена sP. Од¬
нако С. Little [1957] подметил, что в США пегие пойнтеры имеют „рубашки" с большей
площадью белых отметин, чем в Великобритании и вообще в Европе. Соглашаясь, что пе¬
гие европейские пойнтеры являются гомозиготами по пегому аллелю sPsP, он полагал, что
в американской популяции пегий окрас этих собак детерминируется генотипом Л"' — го¬
мозиготным по „экстремальному" аллелю распространения зон депигментации.У большинства пегих собак этой породы на белых отметинах располагаются окрашен¬
ные в тон пятен крапины (тиковая испещренность), результат действия гена крапчатости Т.Померанский (карликовый) шпиц [Pomeranian]Представитель шпицеобразных, ведущих происхождение от семейства больших ездовых
собак из околополярных районов Евразии, этот карликовый шпиц выведен в прибалтийской
Померании. Очень распространенная в Европе в XVIII веке порода имела иной облик (масса
до 14 кг) и несколько утратила популярность после увлечения любителей декоративных соба¬
чек пекинесами. С конца прошлого века начался ренессанс померанцев в Англии, где их дер¬
жала королева Виктория: за несколько поколений селекционеры уменьшили размеры собак
чуть не в 10 раз, ныне их масса только 1,4-3 кг [Fletcher, 1977]. Исключительно длинная, пыш¬
ная шелковистая шерсть однородного и насыщенного цвета. Типичны белый, черный, темно-
и светло-коричневый, голубой, оранжевый и соболиный окрасы — результат действия почти
всех главных генов пигментации. Как случайность встречается пегий окрас (ген sP), в комбина¬
ции белых отметин с пятнами любого из перечисленных цветов.
ОкрасГенотипЧерныйAs-B-D-llКоричневыйAs-bbD-llГолубойAs-B-ddllКрасный (черный нос)АУ-B-D-llКрасный (коричневый нос)АУ-bbD-llСоболиный (черный нос)АУ-B-D-llСоболиный (коричневый нос)АУ-bbD-llКремовый (темный нос)АУ-B-ddllКремовый (светлый нос)АУ-bbddllПудель [Poodle]Ныне одни из популярнейших собак, пудели разводятся в виде нескольких вариаций
размеров (большой, малый, карликовый и той-пудель), считаясь французской породой.
При этом чисто французским происхождение этих собак большинство кинологов не счита¬
ют [Палмер, 1988], признавая паритет на авторство и германских собаководов. Имея в
предках пастушьих собак и ретриверов, эти собаки именно в Германии использовались для
апортирования стреляной дичи из воды (немецкое ,,pudel-nass“ — промокший до нитки).Современных пуделей как одну породу подразделяют по их размерам на стандарт¬
ных (больших, или королевских — самых крупных), малых, карликовых и той-пуделей (са¬
мых крошечных, ростом менее 28 см). В настоящее время разводят каждую из этих разно¬
видностей „в себе“- как изолированные популяции.С длинной пышной шерстью, подвергаемой специальным стрижкам, пудели выделя¬
ются разнообразием окрасов, среди которых европейские стандарты признают сплошные
(генотип SS) следующих цветов: белый, кремовый (черные губы и нос), коричневый, абри¬
косовый, черный, серебристый, голубой. Имеющийся в генофонде породы ген раннего по¬
седения шерсти G может модифицировать названные окрасы в слабой или сильной степе¬
ни: при этом G-позитивные собаки рождаются интенсивно окрашенными, но в процессе
роста к полутора годам сильно светлеют. Это наиболее характерно для серебристого (серо¬
го окраса). Правда, по мнению московского кинолога JI. И. Московкина, возрастной „пе-
рецвет“, например, коричневых пуделей, возможно, и не имеет отношения к генетической
основе такого менделирующего посветления серебристых собак и может просто свидетель¬
ствовать о генетической нестабильности организма. С. Little полагал, что красные и кре¬
мовые пудели обладают геном е, а не А-1’, но R. Robinson отмечал, что подтверждений этой
точки зрения не получено.ОкрасГенотипЧерныйAs-B-D-E-ggГолубойAs-B~ddE-ggКоричневыйAs-bbD-E-ggСеребристыйAs-B-D-E-G-Светло-красный—bbd-eeggКремовый bbddeeggАбрикосовый -D-eeG-Белый— ddeeG-Болезнь Пертеса (некроз головки бедра) — характерная наследственная аномалия
миниатюрных собак, обнаружена у карликовых и той-пуделей. Скорее всего, она детерми¬
нируется рецессивным геном pd.
Непропорциональная карликовость — ахондроплазия — наблюдалась у пуделей
[Gardner, 1959] как следствие действия рецессивного аллеля ас. В то же время характер на¬
следования эпилепсии в этой породе, как и у других, еще не выяснен.По моногенному рецессивному типу наследуется обнаруженная у малых пуделей
(развивается после 8-летнего возраста) первичная глаукома с открытым иридо-корнеаль-
ным протоком, а у карликовых — гемералопатия („дневная слепота"), которую контроли¬
рует мутантный аллель he.Рецессивное наследование характерно у пуделей (малого и той-пуделя) и для прогрес¬
сирующей атрофии сетчатки.Несомненно наследственна у большого пуделя почечная недостаточность, но точный
характер наследования этого дефекта пока не определен.Риджбек [Rhodesian Ridgeback]Название породы („гребень на спине“) отражает уникальную и не имеющуюся ни у
одной другой породы особенность этих прекрасных сторожевых собак африканского про¬
исхождения. Гребень из волос, растущих на спине против шерсти, тянется от холки (где об¬
разует два завитка) до крупа. Правда, такие завитки и небольшие гребни, в соответствии с
законом гомологических рядов Н. И. Вавилова [1987], вполне могут проявиться как итог
гомологических мутаций и у других видов — известно нечто подобное у лабораторных ми¬
ниатюрных свиней.В современном виде эти могучие гармоничные собаки являются результатом двухсот¬
летнего разведения при скрещивании готтентотских аборигенных охотничьих собак, об¬
ладавших названным шерстным гребнем, с привезенными колонистами крупными собака¬
ми европейских служебных и охотничьих пород.Знаменитый спинной гребень из шерсти — признак наследственный, ведет себя как
рецессивный. Установлено, что он контролируется аутосомным рецессивным геном ds, ко¬
торый иногда плейотропно вызывает аномалию кожи — дермоидный синус, узкие трубоч¬
ки-каналы от поверхности в глубь кожи. Трубки как следствие неполного отделения кожи
от позвоночника в эмбриогенезе могут располагаться впереди или сзади „гребня" риджбе¬
ков, причем у гомозиготы их может быть одна или несколько. Дермоидный синус сам по
себе не имеет злокачественной природы, но открывает пути для проникновения инфекций
внутрь тела животного, что рано или поздно приводит к другим серьезным заболеваниям
гомозиготных по аллелю ds собак. К счастью, аллель ds имеет неполную пенетрантность
при контроле кожных синусов. Поэтому встречаемость больных риджбеков относительно
невысокая.Шерсть короткая (генотип LL). Окрас желтовато-красный разной интенсивности (от
светло-пшеничного до рыже-пшеничного) с черной маской на морде — в генофонде ридж¬
беков имеется аллель АУ.Ротвейлер [Rottweiler]Порода сформировалась в южном немецком городе, по имени которого и названа, в
земле Вюртемберг, неподалеку от границы со Швейцарией, Италией, Францией. Так назы¬
ваемые ротвейльские мясницкие собаки, помогавшие охранять и перегонять гурты скота с
альпийских пастбищ в центр Европы, ведут происхождение от средневековых догообраз¬
ных травильных собак и от античных молоссов. Сейчас широко распространенная порода
рабочих собак, применяющихся во многих службах.Одна из ярко архитипичных пород, с закрепленным единственным окрасом — чер¬
ным с подпалинами. Современные ротвейлеры — гомозиготы а‘а‘ВВ, хотя раньше в попу¬
ляции встречался и рецессивный аллель коричневого пигмента Ь. Подпалины имеют оттен¬
ки от светло-рыжего до тона красного дерева, хотя R. Robinson считает, что изменчивость
интенсивности пигментации этих отметин не такая широкая, как у некоторых других по¬
род черно-подпалых собак, где, как, например, у сеттера-гордона, она может быть след¬
ствием не просто разной комбинации полигенов (как это бывает, видимо, у ротвейлеров), а
в результате наличия в генотипах мутаций главных генов окраса.Интересно, что для породы ротвейлер, особенно несколько десятилетий назад, была
характерна врожденная куцехвостость: всем щенкам при рождении хвосты купируют, но в
некоторых сочетаниях выщепляются гомозиготы по рецессивно укороченному хвосту свое¬
образной „грушевидной" формы [Ильин, 1932]. Интересно, что этот признак ротвейлеры
передали выведенным при их участии доберманам, у которых врожденная куцехвостость
сочеталась с другим маркером ротвейльских собак — массивным и грубым сложением
[Мазовер, 1947]. Будучи весьма характерной чертой доберманов на ранних этапах породо-
образования, эта конституциональная грубость была выбракована среди доберманов в
процессе селекции, и одновременно почти перестали рождаться щенки с укороченными
хвостами.Тяжелые и массивные, ротвейлеры весьма нередко страдают дисплазией тазобедрен¬
ного и локтевого [Grondalen, 1982] суставов в результате взаимодействия определенной
комбинации полигенов этих аномалий со средовыми факторами — условиями кормления и
выращивания молодняка.Русская псовая борзая [Barzoi]Порода в современном виде сформировалась в последней четверти XIX века на гене¬
тической основе старинных русских борзых — густопсовой, псовой, чистопсовой, с участи¬
ем в породообразовательном процессе борзых восточноазиатского и западного происхож¬
дения, а также русских остроухих охотничьих собак — лаек [Казанский, 1984; Сабанеев,
1987]. Такое многообразие генетических ресурсов, использованных в породообразователь¬
ном процессе, и уникальные требования, предъявляемые охотниками-собаководами к псо¬
вым борзым, сформировали комплекс феноменальных породных черт: кроме изумитель¬
ной красоты и характерного экстерьера, этим собакам свойственны также высокий рост,
сила, резвость, особенно „накоротке" (способность к „броску" при поимке зверя) и злоб¬
ность, позволявшая успешно охотиться даже на волков.Уникальны форма и постав ушей псовой борзой, отличающие ее от других длинно¬
шерстных борзых: в спокойном состоянии собака держит свои тонкие, недлинные уши „за¬
тянутыми" на затылке (где они иногда даже перекрещиваются кончиками), а настороже
поднимает их в положение „ухо типа колли", при этом обычно верхняя треть уха направле¬
на вперед, параллельно земле. Иногда русские псовые в возбуждении ставят уши „конем",
полностью стоячими. Видимо, эта породная особенность обусловлена модифицированным
взаимодействием главного гена (скорее всего, На) с полигенами-модификаторами.Чрезвычайно характерен шерстный покров этих знаменитых собак: шерсть русской
псовой длинная, шелковистая, волнистая или в крупном завитке, с развитыми подвесом на
хвосте и очесами на конечностях. Окрас разнообразный, от чисто-белого до редкого чер¬
ного, который, впрочем, в кругах охотников не популярен. Наиболее типичен пегий окрас
красно-рыжих оттенков разной интенсивности, определяемой, видимо, „красными полиге¬
нами", с белыми отметинами на голове, шее, груди, конечностях и хвосте, причем при яр¬
ких окрасах нередка маска на морде и зонарная пигментация остевых волос. Генотип чер¬
ных борзых по R. Robinson: As-llsP-; а борзых красно-желтых окрасов (красного, полового,
муругого и др.): АУ-llsP-. Кроме того, русские псовые нередко имеют и черно-подпалый (с
белым) окрас, что свидетельствует о наличии в генофонде породы аллеля ас, а возможно и
сс/] (при сочетании его с АУ образуется полово-подпалый окрас). Серо-пегие борзые, види¬
мо, имеют гены В и в гомозиготе d, а иногда встречающийся нежелательный крап на белых
отметинах — свидетельство действия гена Т.Самоедская лайка (самоедский шпиц) [Samoyed]Шпицеобразные собаки выведены как ездовые и оленегонные ненцами, по старому
названию которых (самоеды) и названы.Густая пышная шерсть чаще всего белого окраса. Самоедские лайки при белой шерсти
имеют черные глаза, поэтому отсутствие пигментации волос обусловливается гомозиготностью
по гену sw. Изредка (по С. Little, примерно с частотой 9 %) у собак этой породы бывают неболь¬
шие пятна пигментации кремового цвета, занимающие не более 10 % поверхности тела. Эти
данные R. Robinson также считает подтверждением названного генотипа самоедской лайки.К. Tjebbes, С. Wriedt [1927] вывели более полную формулу окраса собак этой породы:
As-eesnsw. Они провели скрещивание самоедов с волчье-серой эльзасской овчаркой, гетеро¬
зиготной по гену а1, что дало пять черных с белыми отметинами щенков. В другом скрещи¬
вании с тигровой собакой, гетерозиготной по аллелю е, получены один тигровый щенок и
два потомка, масть которых охарактеризована как палево-замшевая. Этот генетический
анализ свидетельствует, что самоедские лайки имеют в своих генотипах „минус-рыжие“
полигены, модифицирующие осветление синтезируемого в их шерсти желтого пигмента.Встречающиеся самоеды кремового окраса должны иметь ослабляющий цвет аллель
альбиноидной серии, скорее всего, ген „шиншилла14, и их генотип может быть сс1,сс1,ее или
ее с „рыжими" полигенами-модификаторами, ослабляющими окрас, как это имеет место в
генофонде золотистых ретриверов. Структура шерстного покрова свидетельствует о гомо-
зиготности по гену длинношерстности /.Из наследственных болезней этой породы отмечены дисплазия тазобедренного суста¬
ва, наследующаяся полигенно, а также почечная недостаточность, которая имеет у самоед¬
ских лаек уникальный тип наследования, сцепленного с полом: мутантный рецессивный
ген, обусловливающий развитие дефекта, локализован в их Х-хромосомах, тогда как у дру¬
гих изученных пород эта аномалия контролируется, скорее всего, аутосомным геном.Сенбернар [St. Bernard]Порода выведена в одноименном монастыре в Швейцарских Альпах (для поиска и
спасения заблудившихся в снегах путников) двести лет назад на генетической базе потом¬
ков античных молоссоидов. Заводское разведение и большая популярность у любителей
начались с начала нынешнего столетия, главным образом в Великобритании, хотя порода
традиционно считается швейцарской.Окрас традиционно красно-пегий с различной степенью затемнения пигмента
(ген АУ). Чаще собаки имеют черную маску на морде. Обязательный белый „ошейник" сви¬
детельствует о гомозиготности s's1. Однако встречаются сенбернары и пятнистого окраса,
имеющие аллель sP.Шерсть может быть или гладкой жесткой короткой (ген L) или длинной (гомозиго¬
ты II).Обладая огромной массой и в молодости большой скоростью роста, сенбернары не¬
редко страдают дисплазией тазобедренных суставов. Известен у них (как и у немецких до¬
гов) так называемый паралич Стокарда, наследуемый также полигенно, — видимо, с по¬
роговым эффектом. Как моногенный рецессивный признак наследуется у сенбернаров афа-
кия — врожденное отсутствие хрусталика [Martin, Leipold, 1974].Сеттер-гордон (шотландский сеттер) [Gordon Setter]Знаменитая островная порода легавых, отличающихся от двух других сеттеров более
крепкой конституцией, тяжелым телосложением, более уравновешенным характером. Пер¬
воначально собаки назывались „сеттеры из замка Гордон" в Банффшайре, владелец кото¬
рого герцог Ричмондский и Гордонский в 70-х годах XVIII века начал заводское разведе¬
ние этих легавых с использованием прилития крови колли и бладхаунда [Палмер, 1988].Единственный стандартный окрас средней длины с украшающим волосом шерсти —
черный с красными подпалинами (генотип а'а'В-Щ, хотя встречаются в некоторых линиях
собаки с осветленным подпалом, по предположению С. Little [1957] — из-за действия шин¬
шиллового гена cch. В случае справедливости утверждений Little, в породе встречаются ге¬
нетически различные виды подпалого окраса, хотя, видимо, нельзя исключить и переход¬
ных форм по яркости подпала из-за модификаторов — „рыжих" полигенов. Интересно,
что Little [1957] и Winge [1950] сообщали об установленных ими случаях рождения сетте-
ров-гордон рыжего окраса. Если это были и вправду чистопородные шотландские сеттеры,
то их генотип a'a'eell по R. Robinson [1982].Кроме того, еще в 1920-е годы профессор Н.А. Ильин [1931] обнаружил у части сетте-
ров-гордон рубиновоглазие — красный, а не зеленый, как в норме, отсвет при особом осве¬
щении их глаз — действие рецессивного гена р (ра).Скай-терьер [Skye Terrier]Красивая небольшая собака удлиненного формата (длина ее корпуса в два раза боль¬
ше высоты в холке) выведена на маленьком одноименном острове у берегов Англии, но
филогения точно не ясна. В противоположность подавляющему большинству ныне суще¬
ствующих пород других терьеров, выведенных не раньше прошлого века, современные
длинношерстные скаи полностью соответствуют описанию, данному этой породе Джоном
Кайусом четыре столетия тому назад [Fletcher, 1977]. Происходя от мелких терьеров Шот¬
ландии, скай, видимо, имеет родство с кэрн-терьером. По крайней мере, С. Little [1957] по¬
лагал, что генетика окраса этих двух пород сходная. Он наблюдал у некоторых скай-терье-
ров светлый хохолок шерсти на голове, как у бедлингтонов. R. Robinson считает, что
серый окрас скаев обусловливается действием гена седины G.Скотч-терьер (шотландский терьер) [Scottish Terrier]Скотч-терьер, „скотти“, раньше назывался абердинским терьером и многие века раз¬
водился как аборигенная порода в Шотландии в разных формах. В современном виде по¬
рода стандартизирована впервые в 1892 г. при создании первого клуба ее любителей.Жесткая шерсть, подвергающаяся специальному триммингу, формирует характер¬
ный облик этих собак, которые могут быть черного, тигрового или пшеничного окраса.
Тигровый окрас скотти, вероятно, подобен таковому у кэрн-терьера, обусловленный геном
ЕЬг, в виде смеси волос черного и желтого цветов с оптической „модификацией1' из-за
длинной жесткой шерсти. Насыщенность цвета волос с желтым пигментом варьирует от
желтого до светло-кремового под влиянием „минус-рыжих“ полигенов. Пшеничные „скот-
ти“ с геном АУ имеют вариабельную интенсивность из-за тех же полигенов. Полосы тигро¬
вых собак варьируют по цвету от черного до светлого независимо от цвета фона, соответ¬
ственно пропорции смешения черных и желтых волос. Самые темные тигровые легко
могут быть приняты за черных собак, не обязательно имея при этом ген As [Robinson,
1982].ОкрасГенотипТигровыйAy-Ebr-llWh-ПшеничныйАУ-E—llWh-Из генетических особенностей породы наиболее известна наследственная аномалия,
названная по имени породы — скотти крамп (судороги скотча). Вызывает нейромускуляр-
ную болезнь рецессивный аллель sc. Обнаружена и наследственная атаксия под контролем
рецессивного мутантного гена at. Недостаток фактора F VIII свертывания крови, вызыва¬
ющий болезнь фон Виллебранда, также является рецессивной аномалией.Стаффордширский бультерьер [Staffordshire Bull Terrier]Выведен несколько веков назад для популярной тогда в Англии травли быков и мед¬
ведей, а затем собачьих боев, запрещенных в прошлом веке.Окрасы короткой шерсти (ген L) — рыжий, палевый, белый, черный, голубой и тиг¬
ровый — сплошные или пегие, с белыми отметинами.Интересно, что генетика окраса этой породы идентична таковой американского
стаффордшир-терьера, имеющего несколько иное происхождение [Robinson, 1982].ОкрасГенотипЧерныйAs-D-E-РыжийАУ-D-E-ТигровыйAy-D-Ebr-ГолубойAs~ddE-ПалевыйАУ-ddE-Тигрово-голубойAy-ddEbl-В этой породе выявлен рецессивный ген cat, вызывающий в гомозиготном генотипе
возникновение к годовалому возрасту первичной катаракты хрусталика — субкапсуляр-
ной; прогрессирующей до достижения больными собаками трех лет.
Такса [Dachshund]Такса, несомненно, — одна из старейших немецких охотничьих собак, известна из
глубины веков. Таксообразные коротконогие собаки запечатлены на барельефах Древнего
Египта, но нынешние таксы, вероятнее всего, ведут происхождение от средневековых гон¬
чих, у которых известны мутантные формы, отличающиеся коротконогостью (как, напри¬
мер, бассеты), — переходные формы от гончих к таксам имеют название „таксообразные
бракки“. Такая непропорциональная коротконогость — ахондроплазия — наследственная
аномалия для обычных собак, у такс не считается уродством, став важнейшим породным
признаком, обусловливающим наряду с качествами бойцовского характера их высокую
рабочую продуктивность как норных охотников.В настоящее время существует большое разнообразие среди такс по их шерстному
покрову, окрасу и даже размерам (от обычных до карликовых), но первичной, самой древ¬
ней формой является стандартного размера короткошерстная разновидность (происходя¬
щая от старинной немецкой бобровой собаки). От скрещивания короткошерстных такс с
денди-динмонт-терьерами и другими терьерами произошла жесткошерстная такса, а длин-
' ношерстная — продукт скрещивания короткошерстных со спаниелем (и старонемецкой ле¬
гавой штеберхунд) [Палмер, 1988]. Этот породообразовательный процесс начался после со¬
здания в 1888 г. в Германии Клуба любителей такс — тогда еще только короткошерстных,
со складками кожи на ногах (эти складки сейчас у такс уже не встречаются). Явная корот¬
коногость и удлиненный формат — типичные признаки этих отличных охотничьих (нор¬
ных) собак.Большинство рыжих такс обладают аллелем АУ, о чем свидетельствует встречаемость
слабой маски на морде у собак соболиного цвета. Черно-подпалые и коричнево-подпалые
таксы имеют классический генотип для этих расцветок. Необычные прежде пятнистые
(мраморные) таксы являются гетерозиготами по гену М. Также уникальный волчий окрас
(его еще называют медвежьим и барсучьим) жесткошерстных такс может быть следствием
действия гена дикого типа А или же светло-чепрачного asa. Короткая, длинная или жесткая
шерсть детерминируются соответственно генами L, I и Wh.ОкрасГенотипЧерно-подпалый
Коричнево-подпалый
Рыжий с черным носом
Рыжий с коричневым носом
Пятнистый (черный, голубой)
Пятнистый (коричневый, сизый)
Пятнистый (красный, желтый)aWB-L-mmwhwh
aWbbL-nimwhwh
АУ—B-L-mmwhwh
A s—bbL-mmwhwh
aWB-L-Mm wh и 7/
dafbbL-Mmwhwh
АУ—L-MmwhwhУ длинношерстных такс соответствующие генотипы отличаются отсутствием гена L
и гомозиготностью по аллелю /, а у жесткошерстных — наличием аллелей L и Wh.Таксы — весьма здоровые животные, хотя, как и у некоторых других чрезмерно „уд-
линенных“ собак, у них с возрастом возникают иногда признаки слабости задних конечно¬
стей. Из наследственных болезней у такс (длинношерстных) обнаружена прогрессирующая
атрофия сетчатки глаз, передающаяся в поколениях по рецессивному типу.Уэст-хайленд-уайт-терьер [West Highland White Terrier]. . , . v- ... •В 1905 г. в Англии были организованы первые клубы любителей этих терьеров, со¬
зданных на основе линии белых скотчтерьеров селекции полковника Малколма из Полтал-
лоха (полталлох-терьеры, [Палмер, 1988]).Длинная жесткая шерсть этих белых терьеров имеет окрас, обусловленный, по
С. Little, межлокусным взаимодействием неаллельных генов А У и c°h.Это взаимодействие (генотип АУ-ссЬ cfh) образует светло-кремовый цвет или даже бе¬
лый при направленной селекции на отбор самых светлых особей. Генотип по гену длинной
шерсти гомозиготен (И).
Рецессивная мутация (ген pd) обусловливает у гомозиготных терьеров развитие бо¬
лезни Пертеса — некроз бедренной кости. При исследовании этих собак в Швеции уста¬
новлено, что болезнь Пертеса распространена в шведской популяции „уэсти“ с частотой
около 4 % [Wallin, 1986].В Германии недавно выяснена наследственная природа черепно-челюстной остеопа¬
тии уэст-хайленд-уайт-терьеров [Stur et al., 1991]: установлен рецессивный ген, вызываю¬
щий эту аномалию, и рекомендовано проводить рентгенологическое обследование при от¬
боре собак для разведения.Характер наследования в этой породе особой формы катаракты к настоящему време¬
ни еще не установлен.Фокстерьер [Fox Terrier]Фокстерьеров сейчас разделяют на две породы — жесткошерстных и гладкошерст¬
ных — в зависимости от структуры их шерстного покрова (соответственно гены Wh и L),
причем гладкошерстная разновидность — более древняя и более популярная раньше, а же¬
сткошерстные, появившись на рингах лишь в конце прошлого века, стали в настоящее вре¬
мя гораздо более многочисленными. Впрочем, фокстерьеры и сейчас во многих странах
мира не только эффектные выставочные „бойцы“ с абсолютно квадратным форматом, но
и отважные охотники-„норники“, какими их и выводили англичане более ста лет назад
[Marx, Mohrke, 1983].У обеих разновидностей окрас с преобладанием белого цвета, чаще черно-пегий и
чепрачный, при котором у собаки V-образный чепрак чисто-черного цвета и отметины
(подпалы) от красного до золотистого цвета. Желто-пегие фокстерьеры имеют в генотипе
аллели А у или е, или asa, в последнем случае чепрак встречается только вместе с желтыми
подпалинами [Robinson, 1982].ОкрасГенотипЧерно-пегийAs-sP-Чепрачныйasa-sP“Олигодонтия у фокстерьеров, подробно изученная чешским исследователем Ф. Гора-
ком, наследуется, видимо, как рецессивная аномалия, причем чаще всего в этой породе от¬
мечался недостаток третьих моляров.У гладкошерстных фокстерьеров всесторонне изучено наследование нейромускуляр-
ных аномалий — атаксии, контролируемой рецессивным аутосомным аллелем at [Bjork et
al., 1962], и миастении (рецессивный аллель mg [Miller et al., 1984]).Французский бульдог [French Bulldog]Порода этих „очаровательных, с чувством юмора“ [Палмер, 1988] собак, как явствует
из названия, выведена во Франции в XIX веке, но до начала эпохи заводского собаковод¬
ства, методами „народной селекции", по-видимому, с использованием мелких английских
бульдогов в скрещиваниях с местными французскими комнатными собачками. Видимо,
благодаря прилитию „местной" крови французские бульдоги гораздо здоровее английских
предков: не страдают, например, затруднением дыхания.У этих собак три стандартных окраса: тигровый (частые мелкие черные полосы
на бронзовом, рыжем или палевом фоне) с тенденцией к более темному оттенку; бронзо¬
вый (с черной маской и черной окантовкой век) и пятнистый (белый с весьма крупными
черно-рыже-бронзовыми пятнами при черной окантовке век). У тигровых и бронзовых
бульдогов могут быть небольшие белые отметины.Генетической основой бронзового окраса является аллель АУ. Тигровый окрас обус¬
ловлен геном ЕЬг. „Нестандартные" черные бульдоги, нередко рождающиеся у породистых
родителей, являются генетически тигровыми с непроявляющимися или очень небольшими
желтыми отметинами. Генетика белых отметин у тигровых и бронзовых с одной стороны и
пятнистых с другой — одна и та же: площадь зон депигментации сильно варьирует от на¬
бора полигенов, но сами белые пежины обусловлены, очевидно, одним аллелем sP, хотя
R. Robinson [1982] высказал предположение, что в генофонде породы имеются все три ал¬
леля пятнистости.ОкрасГенотипБронзовыйАУ-Е—S-ТигровыйAy-Ebr-S-Бронзовый с белымАУ-E—sP-Тигровый с белымA>-Ebr-sP-Атаксия — нейромускулярная наследственная аномалия — обнаружена в числе пер¬
вых у французских бульдогов, а позднее выявлен рецессивный ген at, обусловливающий
эту болезнь.Чау-чау [Chow Chow]По-видимому, эта порода является прямым потомком древнекитайских собак, тибет¬
ских мастифов [Палмер, 1988], и в то же время самой древней породой шпицев. Распрост¬
раненная по всему миру порода в современном варианте сформирована в Великобритании,
где заводское разведение ее организовано в 1895 г. Уникальная особенность, отличающая
чау-чау от всех других пород домашней собаки, — черный (темно-фиолетовый) язык. Ок¬
рас: сплошной черный, красно-рыжий, голубой, кремовый разных оттенков (иногда удли¬
ненная шерсть на внутренней стороне задних ног светлее основного тона). Черный окрас
обусловлен геном As, тогда как голубой — гомозиготностью по d. Согласно С. Little [1957],
рыжие чау могут иметь ген АУ или быть гомозиготами по другому локусу — е. Светлоокра¬
шенные — палевые или кремовые — могут иметь аллель d или набор полигенов „минус-
рыжей“ группы. Шерсть „классического" чау-чау длинная, равномерно покрывающая те¬
ло, с выраженным украшающим волосом (генотип //), но бывают и короткошерстные чау-
чау (ген L).ОкрасГенотипЧерныйAs-D-ГолубойAs-ddКрасныйАУ-D-КремовыйАУ-ddУ чау-чау выявлены случаи дисплазии тазобедренного сустава (полигенное наследо¬
вание). Впервые в этой породе описана миотония — спазматическое сокращение мускула¬
туры. G. Wentinck et al. [1974] сообщили: считалось, что у чау-чау этот дефект наследуется
под контролем одного рецессивного аллеля, но затем накопленные данные не позволили
принять гипотезу о простом моногенном наследовании миотонии. Она, скорее всего, де¬
терминируется взаимодействием генов нескольких локусов.Чихуахуа [Chihuahua]Очень маленькие собаки прославили эту породу, чрезвычайно популярную во мно¬
гих странах мира. Названная по имени одного из штатов Мексики, своей родины, чихуа¬
хуа имеет древнюю историю, ее предки были некогда священными собаками инков. Впро¬
чем, вопрос филогении породы этих самых маленьких собак нередко дискутируется и не
вполне ясен: некоторые авторы утверждают, что порода происходит из Египта или других
стран Средиземноморья, проникнув на запад через порты Мальты [Fletcher, 1977]. Стан¬
дарт разрешает любой окрас для этих собак двух разновидностей — короткошерстной и
длинношерстной. Встречаются собачки черные, черно-подпалые, рыжие, печеночного ок¬
раса, печеночного с подпалинами, и печеночно-соболиные. С. Little [1957] сообщал, что из¬
вестны черного окраса чихуахуа, полученные от скрещивания обоих рыжих родителей, при
этом в популяции (породе) может быть распространен ген е желтого пигмента, неизвестно,
однако, насколько широко. Встречаются различные оттенки рыжего и соболиного окра¬
сов — это может быть обусловлено или действием гена сс,\ или „минус-рыжих“ полигенов,
комбинация которых может создавать даже почти белый цвет шерсти. Голубой окрас в по¬
роде редок. Очень типичен, кроме одноцветных окрасов, также пятнистый обоих видов:
„ирландский" и „пегий“ При этом может быть и максимальное распространение белых пе¬
жин из центров депигментации, поэтому могут встречаться разные аллели из 5-серии: s', sP,
s'v, S.Длинношерстные чихуахуа — рецессивные гомозиготы //, а у короткошерстных мо¬
жет быть генотип LL или Ы.ОкрасГенотипЧерныйAs-B-D-S-Черно-подпалыйaWB-D-S-ПеченочныйAs-bbD-S-Печеночный с подпалинамиa'a'bbD-S-Г олубойAs-B~ddS-Голубой с подпалинамиa’a'B-ddS-СоболиныйАУ-B-D-SПеченочно-рыжийАУ-bbD-S-Красный с черным носомАУ-B-D-S-Красный с коричневым носомАУ-bbD-S-Знатоки породы отмечают [Fletcher, 1977], что, несмотря на миниатюрные размеры
(максимальный вес 3 кг), эти собаки отличаются крепким здоровьем, редко болеют и жи¬
вут много дольше, чем большинство других пород. Правда, для них характерны аномалии
зубной системы.Шнауцеры [Schnauzer]Под этим наименованием группы близкородственных собак известны три самостоя¬
тельных породы — ближайших родичей группы пинчеров: миттельшнауцер (средний шна-
уцер), карликовый шнауцер (цвергшнауцер) и гигантский шнауцер (ризеншнауцер). Цент¬
ральная фигура в филогении шнауцеров — средний, или, как иногда его называют,
стандартный шнауцер — древняя немецкая порода: статуя городского стража с собакой,
типичным шнауцером, в южном немецком городе Штутгарте (земля Вюртемберг) датиро¬
вана 1620 годом. Типично деревенские скотогонные собаки и крысоловы, они лишь в нача¬
ле нынешнего века привлекли внимание городских собаководов, и ныне весьма популяр¬
ная порода во многих странах. По-видимому, карликовый шнауцер — продукт скре¬
щивания в прошлом веке миттеля с аффен-пинчером [Палмер, 1988], а ризеншнауцер, стан¬
дартизированный лишь в 1925 г., принял современый вид после скрещиваний разнообраз¬
ных немецких пастушьих собак, в том числе, видимо, овчарок, со средними шнауцерами.Все три породы объединяет сходство многих черт облика, в том числе типа шерст¬
ного покрова — жесткой шерсти с „бородой“ и „усами“ — и окраса двух видов: густо-чер¬
ного и так называемого „перец с солью“. Этот последний, по словам R. Robinson, сильно
заинтриговал в свое время генетиков. Однако ныне считается, что „перец с солью" — уни¬
кальная для шнауцеров вариация чепрачного окраса, его видоизменение. При этом желтый
пигмент фона, на котором расположен черный чепрак у других пород, у шнауцеров такой
расцветки сильно осветлен за счет действия „минус-рыжих“ полигенов. Длинная и жесткая
(генотип Wh-lt) шерсть еще более скрывает истинную „чепрачную“ природу собак окраса
„перец с солью“. Очевидно, черные шнауцеры несут доминантный ген Л5, а „перец с со¬
лью” обусловлен рецессивным аллелем asa [Robinson, 1982]. Редкие наблюдаемые нами слу¬
чаи рождения черно-подпалых шнауцеров могут быть или следствием сочетания „рыжих“
полигенов, отделяющих „распространенный*1 чепрак и желтый фон в местах расположения
традиционных подпалин, или выщепления гомозигот по имеющемуся, видимо, у некото¬
рых шнауцеров рецессивному аллелю подпалого окраса апопавшего когда-то в генофонд
этих популяций от давних родственников — пинчеров. У ризеншнауцеров отмечали случаи
дисплазии тазобедренного сустава, характерные для крупных собак вообще, и уникальную
только для данной породы избирательную малабсорбцию витамина В12, наследующуюся
по простому аутосомно-рецессивному типу [Fyve et al., 1989].Характерной для миттелыпнауцеров является специфическая наследственная анома¬
лия кожи, называемая синдромом ШнауцерКомедо [White, 1993]. А у цвергшнауцеров вы¬
явлен также специфический только для них рецессивный ген са, вызывающий у гомозигот
врожденную, т. е. проявляющуюся сразу при прозревании, первичную катаракту — помут¬
нение заднего субкапсулярного слоя хрусталика.Эрдельтерьер [Airdale Terrier]Один из самых крупных терьеров выведен на генетической базе старинных терьеров
графства Йоркшир в долине реки Эйр, с прилитием крови оттерхаунда. Окончательно
сформировался как порода во второй половине прошлого века. Изначально рабочие соба¬
ки эрдельтерьеры успешно использовались во многих службах, даже в обеих мировых вой¬
нах.Порода состоит из собак исключительно только чепрачного окраса жесткой шерсти,
с четко выраженными V-образными черными пятнами чепрака на каждом боку. Спина то¬
же черная. Ярко-рыжие подпалы на конечностях и морде. На спине могут встречаться в не¬
значительном количестве белые шерстинки. Генотип шерсти и окраса: asa-Wh-.Японский хин [Japanese Chin]Древняя восточная порода, многие века разводившаяся во Дворце японских импера¬
торов, а в Европе заводским разведением хинов занялись после того, как в 1853 г. пара со¬
бачек была подарена британской королеве Виктории. Кинологи отмечают возможное род¬
ство хина с пекинесом и кинг-чарлз-спаниелем, в связи с чем и его иногда рассматривают
как настоящего спаниеля [Палмер, 1988].Обильная шелковистая шерсть имеет специфический пегий окрас — белый с черными
или рыжими разных оттенков пятнами, которые на голове и ушах располагаются симмет¬
рично, а на туловище площадь пигментированных участков должна быть больше зоны де¬
пигментации.ОкрасГенотипЧерный с белымAs-llsP-Рыжий с белымАУ-llsP-
Приложение
ОФИЦИАЛЬНАЯ КЛАССИФИКАЦИЯ СОБАК
ПО ГРУППАМ ПОРОД**В последнее время FCI введены некоторые изменения в эту классификацию, но в дос¬
тупной литературе они пока не опубликованы. Изменения сводятся к следующему:— во вторую группу впервые помещена среднеазиатская овчарка под номером 326;— во вторую группу из третьей переведен черный (русский) терьер;— из девятой группы в шестую переведен далматин.Обозначения в таблице:А — возможность присуждения сертификата CACIВ (кандидат в международные
чемпионы красоты).Б — необходимость прохождения испытаний рабочих качеств для получения серти¬
фиката международного чемпиона.Страна выведения
породыНазвание породы№ стандарта
породыАБ12345Группа 1. Hiitehunde und treibhunde (ausgenommen Schweizer Sennenhunde) — пастушьи и
скотогонные собаки (за исключением швейцарских зенненхундов)Секция 1. Schaferhunde — овчаркиАвстралияKelpie — келпи(293)**БельгияChien de berger beige (Belgische Schaferhunde) —
бельгийские овчарки(15)- Groenendael — грюнендаль- Laekenois — лакенуа**- Malinois — малинуа**- Tervuercn — тервюрен**Schipperke — шипперке(83)★ЧехословакияSlovensky Cuvac — словацкий чувач(142)*Г ерманияDeutscher Schaferhund — немецкая овчарка(166)**ФранцияBerger de Beauce — босерон Гберже де Бос1(44)**Berger de Brie — бриар Гберже де Бри1(113)*Г*Berger de Picardie — пикардийская овчарка
[берже де Пикарди](176)★*Принята Международной кинологической федерацией (Federation cynologique internaticnale, FCI) в1988 г. („Der Hund“, 1990, № 4, S. 23-26).
12345[Berger des Pyrenees a poil long] пиренейская
горная овчарка(141)**[Berger des Pyrenees a face rase](138)**Bearded Collie — бородатая колли\2>11) .*Border Collie — бордер-колли(297)★*Collie Rough — колли длинношерстная(156)★Collie Smooth — колли гладкошерстная(296)*Bobtail — бобтейл(16)*Sheltie — шелти(88)*Welsh Corgi Cardigan — велып-корги-кардиган(38)*Welsh Corgi Pembroke — вельш-корги-пемброк...09) *ИталияCane da Pastore Bergamasco — бергамская
пастушья собака(194)*Cane da Pastore Maremmano Abruzzese —
мареммоабруцкая овчарка(201)ЮгославияHrvatski OvcSar — хорватская овчарка(277)*Krazski Ovcar — краинская овчарка(278)Sarplaninac(41)*НидерландыHollandse Herdershond — голландская овчарка
хердер(223)*a) kurzhaar — короткошерстная*6) langhaar — длинношерстная*в) rauhhaar — жесткошерстная★Saarloos Wolfhond — голландский волчий
шпиц(311)itSchapendoes(313)*ПольшаPolski Owczarek Nizinny — польская низинная
овчарка(251)*Polski Owczarek Podhalanski — польская
подгалянская овчарка(252)*ПортугалияCao da Serra de Aires — айрская овчарка(93)ИспанияPerro de Pastor Mallorquin — маллоркуинская
овчарка(321)*Gos d’Atura — атурийская овчарка(87)ВенгрияKomondor — комондор(53)Kuvasz — кувас(54)Mudi — муди(238)Puli — пули(55)a) weiss — белый6) sonstige Farben — прочие окрасыPumi — пуми(56)СССРSiidrussischer Schaferhund — южнорусская
овчарка(326)*
Секция 2. Treibhunde (ausgenommen Schweizer Sennenhunde) —
собаки скотогонные (исключая швейцарских зенненхундов)АвстралияAustralian Cattledog — австралийская коровья
собака(287)**БельгияBouvier des Ardennes — бувье де Арденн(171)**Бельгия/ФранцияBouvier des Flandres — бувье де Фландр(191)**Группа 2. Pinscher und schnauzer, molosser und schweizer sennenhunde — пинчеры и
шнауцеры, молоссы и швейцарские зенненхунды [альпийские пастушьи собаки!Секиия 1. Pinscher und Schnauzer — пинчеры и шнауцеры1.1. Pinscher — пинчерыГерманияDobermann — доберман(132)★*Pinscher — пинчер(184)★Zwergpinscher — цвергпинчер [карликовый
пинчер1(185)*Affenpinscher — аффенпинчер(186)*АвстрияOsterreichischer kurzhaariger Pinscher —
австрийский короткошерстный пинчер(64)★1.2. Schnauzer — шнауцерыГерманияRiesenschnauzer — ризеншнауцер(181)a) pfeffersalz — окрас „перец с солью“**б) schwarz — черный**Mittelshnauzer — миттельшнауцер(182)a) pfeffersalz — окрас „перец с солью“*б) schwarz — черный*Zwergschnauzer — цвергшнауцер(183)a) pfeffersalz — „перец с солью“*б) schwarz — черный★в) schwarz-silber — черно-серебристый*г) weiss — белый.13. Smoushond — смосхондНидерландыHollandse Smoushond — голландский смосхонд(308)★Секция 2. Molosser — молоссы2.1. Doggenartige Hunde — догообразные собакиАргентинаDogo Argentino — аргентинский дог(292)*БразилияFila Brasileiro — бразильская фила(255)*Китай/Велико¬британияShar Pei — шар-пей(309)*ДанияBroholmer — броголмер(315)*ГерманияDeutsher Boxer — боксер(144)**Deutsche Dogge — немецкий дог(235)a) gelb — палевыйб) gestromt — тигровый★
12345в) schwarz — черныйг) gefleckt — мраморный★д) blau — голубой*Rottweiler — ротвейлер(147)**ФранцияBordeauxdogge — бордосский дог(116)*Великобрита¬нияEnglish Bulldog — английский бульдог(149)*Bullmastiff — бульмастиф(157)*Mastiff — мастиф(264)*ИталияMastino Napoletano — неаполитанский мастиф(197)*ЯпонияTosa — тоза(260)*2.2. Berghunde — горные собакиКанадаNeufundlander — ньюфаундленд(50)*ГерманияHovawart — ховаварт(190)**Leonberger — леонбергер(145)*Германия/ШвейцарияLandseer — ландсир(226)*ФранцияChien des Montagne des Pyrenees —
пиренейская горная собака(137)*МароккоAidi — аиди(247)*Португалиябао da Serra da Estrela — эштрельская
овчарка(173)a) langhaar — длинношерстная*6) kurzhaar — короткошерстная*Cao de Castro Laboreiro(170): *Rafeiro do Alentejo(96)*ШвейцарияBernhardiner — сенбернар(61)*a) kurzhaar — короткошерстный*6) langhaar — длинношерстный*ИспанияMastin Espanol — испанский мастифin .*Mastin de los Pirineos — пиренейский мастиф(92)★Perro de Presa Mallorquin тю★Тибет (FCI)Tibetdogge — тибетский дог(230)*СССРKaukasischer Schaferhund — кавказская
овчарка(328)*Секция 3. Schweizer Sennenhunde — швейцарские зеппенхупдыШвейцарияAppenzeller Sennenhund — аппенцеллер
зенненхунд(46)*Berner Sennenhund — бернский зенненхунд(45)*Entlebucher Sennenhund — энтлебух зенненхунд(47)*Grosser Schweizer Sennenhund — большой
швейцарский зенненхунд(58)*
Группа 3. Terrier — терьерыСекция 1. Hochldufige Terrier — высоконогие терьерыГ ерманияDeutscher Jagdterrier — немецкий ягдтерьер(103)**Великобрита¬нияAiredale Terrier — эрдельтерьер(7)*(*)Bedlington Terrier — бедлингтон-терьер(9)*Border Terrier — бордер-терьер(10)*Fox Terrier glatth. — Фокстерьер
гладкошерстный(12)*(*>Fox Terrier drahth. — Фокстерьер
жесткошерстный(169)*(*)Lakeland Terrier — лейкленд-терьер(70)*Manchester Terrier — манчестерский терьер(71)*Welsh Terrier — вельш-терьер(78)*ИрландияGlen of Imaal Terrier — глен-овимаал-терьер(302)*Irish Terrier — ирландский терьер(139)*Kerry Blue Terrier — керри-блютерьер(3)*Soft-coated Wheaten Terrier — мягкошерстный
пшеничный терьер(40)*ЯпонияTerrier iaponais — японский терьер(259)*СССРSchwarzer Terrier — черный терьерti 1*Секция 2. Niederlaufige Terrier — низконогие терьерыАвстралияAustralian Terrier — австралийский терьер(8)*ЧССРCesky Terrier — чешский терьер(246)*Великобрита¬нияCairn Terrier — кэрн-терьер(4)*Dandie Dinmont Terrier — денди-динмонт- терьер(168)*Norfolk Terrier — норфолк-терьер(272)*Norwich Terrier — норвич-терьер(72)*Scottish Terrier — скотч-терьер(73)*Sealyham Terrier — силихэм-терьер(74)*Skye Terrier — скай-терьер(75)*West Highland White Terrier — уэст-хайленд-
уайт-терьер(85)*Секция 3. Bullartige Terrier — буллеобразные терьерыВеликобрита¬нияBull Terrier (english) — бультерьер
(английский)(И)a) normal — нормальный*6) miniature — миниатюрный*Staffordshire Bull Terrier — стаффордширский
бультерьер(76)*СШАAmerican Staffordshire Terrier — американский
стаффсрдшир-терьер(286)*Boston Terrier — бостон-терьер(140)*
'2345Секция 4. Zwersterrier — карликовые терьерыАвстралияSilkv Terrier — силки-терьер(236)*Великобрита¬нияToy Terrier (english, black and tan) —
той-терьер (английский, черно-подпалый)(13)*Yorkshire Terrier — йоркшир-терьер(86)*Группа 4. Dachshunde — таксыГ ерманияDachshund — такса(148)a) Normalgrosse — нормального размера —
kurzhaar короткошерстная**- langhaar — длинношерстная**- rauhhaar — жесткошерстная**б) Zwerg — карликовая- kurzhaar — короткошерстная★*- langhaar — длинношерстная**- rauhhaar — жесткошерстная**в) Kaninchen — кроличья- kurzhaar — короткошерстная**- langhaar — длинношерстная*★- rauhhaar — жесткошерстная*★Гvynna 5. Spitze und urtyp — шпицы и примитивный типСекция 1. Nordische Schlittenhunde — северные ездовые собакиСкандинавияGronlandhund — гренландская собака(274)*СССР (Скан¬
динавия)Samojede — самоедская лайка(212)*СШАAlaskan Malamute — маламут(243)*Siberian Husky — сибирский хаски(270)*Секция 2. Nordische Jagdhunde — северные охотничьи собакиНорвегияNorsk Elghund Gra — норвежская лосиная
собака Грэ(242)**Norsk Elghund Sort — норвежская лосиная
собака Сорт(268)**Norsk Lundehund — норвежская лундехунд(265)**СССРLaika, russisch-europaisch — русско-европейская
лайка(304)*★Laika, ostsibirisch — восточно-сибирская лайка(305)★*Laika, westsibirisch — западно-сибирская лайка(306)**ШвецияJamthund(42)**Nordbottenspets — норботтен-шпиц(276)★*ФинляндияKarelischer Barenhund — карельская медвежья
собака(48)★Suomenpystykorva — финский шпиц(49)*Секция 3. Nordische Wach — und Hiitehunde — северные караульные и сторожевыесобакиИсландияIceland Dog — исландская собака(289)*
12345НорвегияNorsk Buhund — норвежский бухунд(237)*ШвецияLapphund — лапхунд(135)*Vastgotaspets — вестготская собака04)* .ФинляндияLapinkoi'ra(189)*Lapinporokoira(284)*■Секция 4. Europaische Spitze — европейские шпицыГ ерманияDeutscher Spitz — немецкий шпиц(97)a) Wolfsspitz — вольфшпиц*б) Grossspitz — большой шпиц- weiss — белый*- braun/schwarz — коричневый/черный*в) Mittelspitz — средний шпиц- weiss — белыйг1*- braun/schwarz — коричневый/черный*- orange — оранжевый*- wolfsgrau (graugewolkt) — волчье-серый*г) Kleinspitz — малый шпиц- weiss — белый*- braun/schwarz — коричневый/черный★- orange — оранжевый*- wolfsgrau (graugewolkt) — волчье-серый★д) Zwergspitz — карликовый шпиц- weiss/braun/schwarz/orange/wolfsgrau
(graugewolkt) — белый/коричневый/
черный/оранжевый/волчье-серый* * :ИталияVolpino italiano(195)*Секция 5. Asiatische Spitze und verwandte Rassen — азиатские шпицы и родственные
породы Китай (Вели¬
кобритания)Chow Chow (rot/schwarz, blau) — чау-чау
(красный/черный, голубой)(205)*Г ерманияEurasier — евразиер(291)*ЯпонияAkita — акита(255)*Hokkaido — хоккайдо(261)**Kai — кай(317)**Kishu — кишу(318)**Japanischer Spitz — японский шпиц(262)★Shiba — шиба(257)**Shikoku — шикоку(319)** .Секция 6. Urtvp— первобытный типИзраильCanaan Dog — ханаанская собака(273)*Заир (Велико¬
британия)Basenji — басенджи(43)*
Группа 6. Laufhunde und schweisshunde — гончие и ищейкиСекция 1. Laufhunde —■ гончие1.1. Grosse Laufhunde — большие гончиеБельгияBloodhound — бладхаунд(84)*★ФранцияPoitevin(24)★*Billy(25)**•Francais tricolore — французская трехцветная(219)★★Francais blanc et noir — французская
черно-пегая(220)**Francais blanc et orange — французская
черно-подпалая(316)**Grand anglo-francais tricolore — большая
англо-французская трехцветная(322)**Grand anglo-francais blanc et noir — большая
англо-французская черно-пегая(323)**Grand anglo-francais blanc et orange —
большая англо-французская черно-подпалая(324)**Grand bleu de Gascogne — большая голубая
гасконская(22)★*Grand gascon saintongeois(21)★*Grand griffon vendeen — большой вандейский
гриффон(282)**Великобрита¬нияFoxhound (English) — фоксхаунд (английский)(159)*Otterhound — оттерхаунд(294)*СШАAmerican Foxhound — американский
фоксхаунд(303)*★Black and Tan Coonhound — черноподпалый
кунхаунд(300)★1.2. Mittelgrosse Laufhunde — гончие среднего размераЧСФРSlovensky Kopov(244)**ФинляндияSuomenajokoira — суоменайокойра
(трехцветная финская гончая)(51)**ФранцияAnglo-francais de petite venerie(325)**Ariegeois. (20)**Beagle Harrier — бигль-харьер(290)**ГChien d’Artois(28)**Porcelaine — порселен. (30)**Petit bleu de Gascogne(31)**Petit gascon saintongeois(21)*★ 'Briquet griffon vendeen(19) **Griffon bleu de Gascogne .(32.)...*★Griffon fauve de Bretagne(,66)**Griffon nivernais — гриффон нивернейский(17)**Великобрита¬нияHarrier — харрьер(295)**
12345ГрецияHellinikos Ichnilatis(214)**ИталияSegugio Italiano — итальянская гончая(198)a) a pelo raso — гладкошерстная**б) a pelo forte — грубошерстная**ЮгославияBalkanski Gonic — балканская гончаяп®*★Bosanski Ostrodlaki Gonic-Barak(155)**Istarski Kratkodlaki Gonic (a poil court)(151)**Istarski Ostrodlaki Gonic (a poil dur)(152)**ftJugoslovenski Planinski Gonic(279)**Jugoslovenski Trobojni Gonic(229)**Posavski Gonic(154)**НорвегияDunker — дункер(203)*★Haldenstovare — халденстёваре(267)★★Hygenhund — хигенхаунд(266)**АвстрияBrandlbracke(63)**Steirische rauhhaarigeHochgebirgsbracke (Peintinger Bracke)(52)**Osterreichische glatthaarige Bracke (Tiroler
Bracke) — тирольский бракк(68)*★ПольшаOgar Polski — польский orap(52)*★ИспанияSabueso Espanol(204)**ШвейцарияSchweizerische Laufhunde — швейцарская
гончая(59)a) Berner Laufhund — бернская гончая**6) Jura Laufhund Тур Bruno und Typ Sankt
Hubert*★в) Luzerner Laufhund**r) Schweizer Laufhund**ШвецияHamilton Stovare — гамильтонстёваре (гончая
Г амильтона)(132)**Smalandsstovare — смоландстёваре(129)**Schiller Stovare — шиллерстёваре(131)★*ВенгрияErdelyi Коро..(241).. .★*1.3. Kleine Laufhunde — маленькие гончиеГерманияDeutsche Bracke — немецкий бракк(299)★*Westfalische Dachsbracke — вестфальский
таксообразный бракк(ЮО)**ФранцияBasset artesien normand —
артезиано-нормандский бассет(34)★*Basset bleu de Gascogne — голубой гасконский
бассет(35)**Basset fauve de Bretagne — рыжий бретанский
бассет(36)**Grand Basset griffon vendeen — большой
вандейский бассет-гриффон(33)**
12345Petit Basset griffon vendeen — малый
вандейский бассет-гриффон(67)**Великобрита¬нияBasset Hound — бассет-хаунд(163)**Beagle — бигль(161)**АвстрияAlpenlandische Dachsbracke(254)**ШвейцарияSchweizerische Niederlaufhunde — швейцарская
низинная гончая(60)a) Berner Niederlaufhund — бернская
низинная гончая**6) Jura-Niederlaufhund — юрская низинная
гончая**в) Luzerner Niederlaufhund — люцернская
низинная гончая**г) Schweizer Niederlaufhund — швейцарская
низинная гончая**ШвецияDrever — древер.. Ш'О*★Секция 2. Schweisshunde — ищейкиГерманияBayrischer Gebirgsschweisshund — баварская
горная ищейка(217)**Hannoverscher Schweisshund — ганноверская
ищейка(213)**Группа 7. Vorstenhunde — легавыеСекция 1. Kontinentale Vorstehhunde — континентальные легавыеДанияGammel Dansk H0nsehund(281)★*ГерманияDeutsch Kurzhaar — курцхаар(119)**Deutsch Drahthaar — дратхаар(98)★*Pudelpointer — пудель-пойнтер(216)**Deutsch Stichelhaar — немецкая иглошерстная
легавая(213)**Weimaraner — веймаранер(99)a) kurzhaarig — короткошерстный«г*6) langhaarig — длинношерстный**ФранцияBraque de l’Ariege(177)**Braque d’Auvergne(180)**Braque du Bourbonnais(179)**Braque Dupuy(178)**Braque francais, type Gascogne (grande taille) —
французский брак к. гасконский тип(133)**Braque francais, type Pyrenees (petite taille) —
французский бракк, пиренейский тип(134)**Braque Saint-German — сен-жерменский бракк(115)**ИталияBracco italiano — итальянский бракк(202)**Spinone italiano — спиноне(165)**ПортугалияPerdigueiro Portugues(187)*★
12345ИспанияPerdiguero de Burgos(90)**ВенгрияMagyar Vizsla, drahthaar — венгерская вижла
жесткошерстная(239)**Magyar Vizsla, kurzhaar — венгерская вижла
короткошерстная(57)**1.2. Тур Spanie— тип спаниеляГ ерманияKleiner Miinsterlander Vorstehhund — малый
мюнстерлендер(102)**Grosser Miinsterlander Vorstehhund — большой
мюнстерлендер(118)**Deutsch Langhaar — лангхаар (П7)**ФранцияEpagneul bleu de Picardie пикардийский
голубой эпаньол(106)★*Epagneul breton — бретонский эпаньол(95)*kEpagneul francais — французский эпаньол(175)*kEpagneul picard — пикардийский эпаньол(108)*kEpagneul de Pont-Audemer(114)*kНидерландыDrentsche Patrjishond — дрентская
куропаточная собака(224)*kStabyhoun — стабихун(222)*k1.3. Тур Griffon — тип гриффонаЧСФРCesky Fousek — чешский фоусек(245)**Slowakischer Drahthaar — словацкий дратхаар(320)**ФранцияGriffon Korthals — гриффон Кортальса(Н§**Griffon Boulet — гриффон Булье. 1.174) _**Секция 2. Britische und irische Vorstehhunde — британские и ирландские легавые2.1. Pointer — пойнтерВеликобрита¬English Pointer — английский пойнтер(1)★*ния2.2. Setter — сеттерВеликобрита¬нияEnglish Setter — английский сеттер(2)**Gordon Setter — сеттер-гордон(6)**ИрландияIrish Setter — ирландский сеттер(120)**Группа 8. Apportierhunde, stoberhunde, wasserhunde — апортирующие собакиСекция 1. Apportierhunde — апортирующие собакиВеликобрита¬нияCurly-Coated Retriever — курчавошерстный
ретривер(110)**Flat-Coated Retriever — прямошерстный
ретривер(121)**Labrador Retriever — лабрадорретривер(122)**Golden Retriever — золотистый ретривер(HD**КанадаNovia Scotia Duck Tolling Retrievernil)**СШАChesapeake Bay Retriever — чеса-пик-бей-
ретривер(263)* '*
I 23 4 5Секция 2. StoberhundeГерманияDeutscher Wachtelhund — немецкий
вахтельхунд(104)**Великобрита¬нияClumber Spaniel — кламбер-спаниель(109)**English Cocker Spaniel — английский кокер-
спаниель(5)*★Field Spaniel — филд-спаниель(123)**Sussex Spaniel — суссекс-спаниель(127)**English Springer Spaniel — английский
спрингер-спаниель(125)**Welsh Springer Spaniel — вельшспрингер-
спаниель(126)**НидерландыKooikerhondje(314)★ЮжнаяАфрикаRhodesian Ridgeback — риджбек(146)*СШАAmerican Cocker — американский
кокер-спаниель(167)*Секция 3. Wasserhunde — водяные собакиФранцияBarbet — барбет(105)**ИрландияIrish Water Spaniel — ирландский водяной
спаниель(124)**НидерландыWetterhoun — веттерхун(221)*ПортугалияCad de Agua — кау-дагуа, португальская
водяная собака(37)*СШАAmerican Water Spaniel — американский
водяной спаниель(301)**Группа 9. Gesellschafts- und begleithunde — собаки — компаньоны и сопровождающиеСекция 1. Bichons und verwandte Rassen — бишоны и родственные породы1.1. Bichons — бишоныБельгия/ФранцияBichon a poil frise — бишон-фриз(215)*КубаBichon havanais — бишон гаванский(250)*ИталияBologneser — болонка(196)*Мальта(Италия)Malteser — мальтийская болонка(65)★1.2. Coton de TulearМадагаскарСо ton de Tulear(283)*1.3. Lowchen — львиные собачкиФранцияPetit chien lion (Lowchen) — львиная собачка(233)*Секция 2. Pudel — пудельФранцияCaniche (Pudel) — пудель(172)Grosspudel — большой пудельweiss/braun/schwarz — белый/коричневый/
черный*
]2345silber/apricot — серебристый/абрикосовый*Kleinpudel — малый пудельweiss/braun/schwarz — белый/коричневый/
черный*silber/apricot — серебристый/абрикосовыйHrZwergpudel — карликовый пудель*weiss/braun/schwarz — белый/коричневый/
черный*silber/apricot — серебристый/абрикосовый★Toy-Pudel — той-пудель★Секция 3. Kleine belsische Hunderassen — малые бельгийские породы3.1. Griffons — гриффоныБельгияGriffon beige (81)/bruxellois (80) — гриффон
бельгийский/брюссельский★3.2. Petit brabancon — малый брабансонБельгияPetit brabancon — малый брабансон(82)*Секция 4. Haarlose Hunde — бесшерстные собакиКитай (Вели¬
кобритания)Chinese crested dog — китайская хохлатая
собачка (китайская голая собачка)(288)★МексикаXoloitzcuintle — ксолоитцкуинтли
(мексиканская голая собачка)(288)*a) gross — большая*б) miniature — миниатюрная*ПеруPerro sin pelo del Peru(310)a) gross★6) mittel★•в) klein*Секция 5. Tibetanische Hunderassen — тибетские породыТибет (Вели¬
кобритания)Lhasa Apso — лхасский anco(277)*Shih Tzu — ши-тцу(208)*Tibetan Spaniel — тибетский спаниель(231)*Tibetan Terrier — тибетский терьер(209)*Секция 6. Chihuahua — чихуахуаМексикаChihuahua — чихуахуа(218)a) kurzhaar — короткошерстная*б) langhaar — длинношерстная*Секция 7. Dalmatiner — далматинЮгославияDalmatiner — далматинЛ153)*Секция 8. Englische Gesellschaftsspaniel — английские спаниели-компаньоныВеликобрита¬нияCavalier King Charles Spaniel — кавалер-кинг-
чарльз-спаниель(136)★King Charles Spaniel — кинг-чарлз-спаниель(128)*
Секция 9. Japanische Spaniel und Pekingesen — японские спаниели и пекинесыКитай (Вели¬
кобритания)Pekingese — пекинес(207)*ЯпонияChin — японский хин(206)*Секция 10. Kontinentale Zwergspaniel — континентальные карликовые спаниелиФранция/БельгияEpagneul nain continental — континентальный
карликовый эпаньолa) Papillon — папильон*1,5-2,5 кг2,5-4,5 кгб) Phalene — фален*1,5-2,5 кг2,5-4,5 кгСекция 11. КгomfohrlanderГерманияKromfohrlanderД921Секция 12. Kleine doggenartige Hunde — маленькие догообразные собакиФранцияFranzosische Bulldogge — французский
бульдог(101)*Великобрита¬нияMops — мопс(253)*Группа 10. Windhunde und verwandte rassen — борзые и родственные породыСекция 1. Windhunde — борзые1.1. Langhaarige oder befederte windhunde — длинношерстные борзыеАфганистан/Великобрита¬нияAfghan Hound — афган (афганская борзая)(228)*Иран/Велико¬британияSaluki — салюки(269)★СССРBarsoi — русская псовая борзая(193)*1.2. Rauhhaarige Windhunde — жесткошерстные борзыеИрландияIrish Wolfhound — ирландский волкодав(160)*Великобрита¬нияDeerhound — дирхаунд(164)*ИспанияGalgo espanol — испанский гальгог ......*Великобрита¬нияGreyhound — грейхаунд(158)*Whippet — уиппет(162)*ИталияItalienisches Windspiel — левретка(200)*ВенгрияMagyar Agar — венгерская борзая (агар)*Мали/ФранцияAzawakh — азавак(307)★МароккоSloughi — слугги(188)★
Секция 2. Verwandte Rassen (Hunde mil Stehohr fur die Hetzjagd) — родственные
породы (собаки, пригодные для псовой охоты) ИспанияPodenco Canario — поденгу-канариу(329)Podenco Ibicenco — поденгу-иби-сенку(89), ■ИталияCimeco dell’Etna(199)* ‘Мальта (Вели¬
кобритания)Pharao Hound — фараонова собака(248)*ПортугалияPodengo Portugues — поденгу-португеш(94)a) kurzhaar — короткошерстный- gross — большой★- mittel — средний*- klein — малый★6) langhaar — длинношерстный- gross — большой*- mittel — среднийsklein — малый*
ЛитератураАдамец J1. Общая зоотехния. — М.; Л.: Сельхозгиз, 1930. — 674 с. — Цит. по: Крушине-
кий Л.В. Избр. тр. — М.: Наука, 1991.Белоцкий С.М. Тромбоцитарные антигены собак: особенности специфичности // Материалы
XVI Междунар. конф. по группам крови и биохимическому полиморфизму животных. — Л.: ВИР,1979. -г- Т. III. — С. 235-237.Беляев Д.К., Волобуев В.Т., Раджабли С.И., Трут Л.Н. Полиморфизм и мозаицизм по доба¬
вочным хромосомам у серебристо черных лисиц // Генетика. — 1974. — Т. 10. — С. 58-67.Бехтин И.Н., Сулимов К.Т. Скрещивается собака с шакалом // Природа. — 1985. — № 3. —
С. 63-71.Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости [Избр. тр.]. — Л.:
Наука, 1987. — 260 с.Вайсфельд М.А. Красная лисица // Промысловые животные СССР и среда их обитания. Пе¬
сец, лисица, енотовидная собака. — М.: Наука, 1985. — С. 73-115.Войлочников А/Г., Войлочникова С.Д. Охотничьи лайки. — М.: Лесн. пром-сть, 1982. — 256 с.Выржиковский С.Н., Майоров Ф.Н. Материалы к вопросу о влиянии воспитания на склад
высшей нервной деятельности у собак // Тр. физиол. лаб. академика И.П. Павлова. — 1933. —
Т. 5. — С. 171-192.Гейц А.В. О прикусах собак //Охота и охотничье хозяйство. — 1972. — № 7. — С. 23.Глазко В.И. Генетика изоферментов сельскохозяйственных животных // Итоги науки и техни¬
ки. — М.: ВИНИТИ. Сер. Общая генетика. — 1988. — Т. 10. — С. 4-212.Гражданкина А.Н. Интерканис-89 // О собаке. — М. — Ташкент, 1991. — С. 115-120.Графодатский А.С. Сравнительная цитогенетика трех видов собачьих (Carnivora, Canidae).
Сообщение III // Генетика. — 1983. — Т. 19. — С. 778-783.Графодатский А.С., Раджабли С.И. Сравнительная цитогенетика 3 видов собачьих (Carnivora,
Canidae) // Генетика. — 1981. — Т. 17. — С. 1500-1507.Графодатский А.С., Раджабли С.И. Хромосомы сельскохозяйственных и лабораторных мле¬
копитающих: Атлас. — Новосибирск: Наука, 1988. — 128 с.Дьюсбери Д. Поведение животных. Сравнительные аспекты. — М.: Мир, 1981. — 479 с.Западнюк И .П., Западнюк В.И., Захария Е.А., Западнюк Б.В. Лабораторные животные. Разве¬
дение, содержание, использование в эксперименте. — 3-е изд. — Киев: Вища шк. Головное изд-во,1983. — 383 с.Ильин Н.А. О наследовании окраски у доберман-пинчера // Труды по динамике развития
(Продолжение „Трудов Лаборатории экспериментальной биологии Московского зоопарка4') / Под
ред. проф. М. Завадовского. — М.: Госмедгиз, 1931. — Т. VI. — С. 241-251.Ильин Н.А. Генетика и разведение собак. Генетическое введение в кюнологию. — М.-Л.: Гос.
изд-во с.-х. и колх.-кооп. лит., 1932. — 164 с.Казанский В.И. Борзые. — М.: Лесн. пром-сть, 1984. — 199 с.Князев С.П. Селекция против аномалий в экстерьере / Клуб служебного собаководства. — М.:
Изд-во „Патриот", 1989. — С. 88-102.Князев С.П., Тихонов В.Н. Гибридологический анализ и значение краниологических особен¬
ностей диких и домашних форм свиней // Докл. АН СССР. — 1983. — Т. 270, № 4. — С. 976-978.Князев С.П., Тихонов В.Н. Краниологические особенности европейских и азиатских диких
кабанов и их гибридов с домашними свиньями // Морфология и генетика кабана. — М.: Наука,1985. — С. 33-49.
Князев С.П., Тихонов В.Н., Танабе Ю. Филогенетические связи аборигенных собак Кавказа,
Средней Азии и Сибири с породами Европы и Японии, выявляемые анализом полиморфизма бел¬
ков и ферментов крови // Генетика. — 1993. — Т. 297, № 3. — С. 449-459.Коллинг Д., Сэйзоп Р. Сообщение о 5 новых эритроцитарных группах у собак и новой аллели
эритроцитов в Тг-системе // XVI Муждунар. конф. по группам крови и биохимическому полимор¬
физму животных: Тезисы. — Л.: ВИР, 1978. — С. 175.Корочкин Л.И., Серов О.Л., Пудовкин А.И. Генетика изоферментов. — М.: Наука, 1977. —280 с.Крушинский Л.В. Исследование по феногенетике признаков поведения у собак // Биол.
журн. — 1938. — Т. 7, № 4. — С. 869-892. То же // Избр. тр. — М.: Наука, 1991. — Т. 1. — С. 7-31.Крушинский Л.В. Наследственное „фиксирование “ индивидуально приобретенного поведе¬
ния животных и происхождение инстинктов II Журн. общ. биологии. — 1944. — Т. 5, № 5. —
С. 261-283. То же // Избр. труды. — М.: Наука, 1991. — Т. 1. — С. 32-52.Крушинский Л.В. Роль наследственности и условий воспитания в проявлении и выражении
признаков поведения у собак // Изв. АН СССР. Сер. биол. наук. — 1946а. — 1. — С. 69-82. То же II
Избр. труды. — М.: Наука, 1991. — Т. 1. — С. 96-108.Крушинский Л.В. Корреляция между конституциональным строением тела и поведением со¬
бак // Докл. АН СССР. — 19466. — Т. 52, N 7. — С. 639 642. То же // Избр. труды. — М.: Наука,1991. — Т. 1. — С. 109-113.Крушинский Л.В. Формирование поведения животных в норме и патологии. — М.: Изд-во
МГУ, 1962. — 264 с.Крушинский Л.В. Биологические основы рассудочной деятельности. •— М.: Изд-во МГУ,1986. — 2-е изд. — 270 с.Крушинский Л.В. Избр. труды. — М.: Наука, 1991. — Т. 1. Эволюционно-генетические аспек¬
ты поведения. — С. 7-52; 96-114.Крушинский Л.В., Зорина З.А., Полетаева И.И. и др. Введение в этологию и генетику поведе¬
ния. Пособие для студентов биологических факультетов. — М.: Изд-во МГУ, 1983. — 183 с.Мазовер А.П. Экстерьер и породы служебных собак. — М.: Изд-во ЦС Осоавиахима, 1947. —246 с.Мазовер А.П. Племенное дело в служебном собаководстве. — М.: Изд-во ДОСААФ, 1960. —296 с.Мазовер А.П. Охотничьи собаки. — М.: Агропромиздат, 1985. — 239 с.Мазовер А.П., Крушинский Л.В., Израилевич И.Е. и др. Служебная собака: Руководство по
подготовке и содержанию служебных собак. — Д., ВАП, 1994. — 576 с.Меркурьева Е. О прикусах собак // Охота и охотничье хозяйство. — 1968. — №9. — С. 24-26.
Насимович А.А. Енотовидная собака II Промысловые животные СССР и среда их обитания.
Песец, лисица, енотовидная собака. — М.: Наука, 1985. — С. 116-158.Орлов В.Н., Булатова Н.Ш. Сравнительная цитогенетика и кариосистематика млекопитаю¬
щих. — М.: Наука, 1983. — 405 с.Павлов И.П. Двадцатилетний опыт объективного изучения высшей нервной деятельности
(поведения) животных. — М.; Л.: Биомедгиз. — 1938. — 6-е изд. — 771 с.Палмер Дж. Ваша собака. — М.: Мир, 1988. — 247 с.Петухов В.Л., Князев С.П., Христенко В.Т., Гудилин И.И. Генетика свиньи. — Новосибирск:
Новосибирский СХИ, 1989. — 80 с.Сабанеев Л.П. Собаки охотничьи... Борзые и гончие / Сост. Е.А. Калганов. — М.: Физ-ра и
спорт, 1987. — 571 с.Слимак К., Духай И. Охотничьи собаки /Ред. B.C. Лобачев. — М.: Лесн. пром-сть, 1986. —335 с.Соколов В.Е. Систематика млекопитающих. — М.: Высш. шк., 1979. — 528 с.Сотская М.Н. Окраска собак и основные принципы ее наследования II О собаке. — М., Таш¬
кент: Изд-во „Улей“, 1991. — С. 91-105.Сэйзон Р., Коллинг Д. Предлагаемая номенклатура для эритроцитарных антигенов собак //
Материалы XVI Междунар. конф. по группам крови и биохимическому полиморфизму живот¬
ных. — Л.: ВИР, 1979. — Т. 111. — С. 231-233.Тихонов В.Н. Изучение групп крови животных. — Новосибирск: Ред.-изд. отдел СО АН
СССР, 1964. — 63 с.Тихонов В.Н. Использование групп крови при селекции животных (с основами иммуногене¬
тики). — М.: Колос, 1967. — 391 с.
Трут JI.H., Голубица А.Н. Трансплантация бластоцист у серебристо-черной лисицы // Докл.
АН СССР. — 1979. — Т. 249. — С. 471^73.Филиппов Ю.И., Митин В.Н. Дисплазия тазобедренных суставов у собак // Ветеринария. —
1990. — № 4. — С. 66-69.Хатт Ф.Б. Генетика животных. — М.: Колос, 1969. — 439 с.Хип Р., Флинт А. Беременность // Гормональная регуляция размножения у млекопитающих /
Под ред. К. Остина, Р. Шорта. — М.: Мир, 1987. — С. 193-244.Хорнер Т. Все о бультерьерах. — М.: Изд-во фирмы „Скиф“, 1992. — 102 с.Хромов Б.М., Короткевич Р.С., Павлова А.Ф. и др. Мочеполовая система // Анатомия собаки /
Под ред. Б.М. Хромова. — Л.: Наука, 1972. — С. 153-169.Шерешевский Э. Еще раз о неправильных прикусах // Охота и охотничье хозяйство. —
1972. — № 1. — С. 32-33.Шорт Р.В. Эстральный и менструальные циклы // Гормональная регуляция размножения у
млекопитающих /Под ред. К. Остина, Р. Шорта. — М.: Мир, 1987. — С. 145-191.Эрман Л., Парсонс П. Генетика поведения и эволюция. — М.: Мир, 1984. — 566 с.Aarskaug Т., Lingaas F., Grondalen J., Juneja U.K. Genetic variation in seven protein systems in dog
breeds with different population size // XXIII Intern. Conf. on Animal Genetics. — Berne, 1992. — P. 25.Abeelen J.H.F. van Genotype and the cholinergic control of exploratory behaviour in mice // The
genetics of behaviour /Ed. J.H.F. van Abeelen. — Amsterdam: North-Holland Publ. Co., 1974. —
P. 347-374.Ablad B. Proteinlosing enteropati hos lunnehund // Svensk Veterinartidn. — 1989. — Vol. 41,
№ 14. — P. 857-863.Aguirre G.D., Rubin L.F. Progressive retinal atrophy in the Miniature Poodle // J. Amer. Vet. Med.
Assoc. — 1972. — Vol. 160. — P. 191-201.Aguirre G., Farber D., Lollcy R. et al. Red-cone dysplasia in Irish Setters: a defect in cyclic GMP
metabolism in visual cells II Science. — 1978. — Vol. 201. — P. 1113-1134.Anderseen A.G., Wooten E. The estrous cycle of the dog // Reproduction in domestic animals / Eds.
H.H. Cole, P.T. Cupps. — N.Y.; London: Acad. Press, 1959. — Vol. 1. — P. 359-397.Anderson М., Henricson B., Lundequist P.G. et al. Genetic hearing impairment in the Dalmatian
dog // Acta Otolaryngol. — 1968. — Suppl. — Vol. 232. — P. 1-34.Andersson B., Andersson M. Skottekramp och splay: — sjukdomsbild, sannolik nedarvning och
orsak // Svensk Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 166-171.Andrescn E. Haemolytic anaemia in Basenji dogs // Anim. Blood Grps Biochem. Genet. — 1977. —
Vol. 8. — P. 149-156.Andresen E., Willeberg P. Pituitary dwarfism in Carelian Bear dogs // Hereditas. — 1976a. —
Vol. 84. — P. 232-234.Andresen E., Willeberg P. Pituitary dwarfism in German Shepherd dogs // Nord. Vet. Med. —
1976b. — 28. — 481-486.Ambjerg J. Persistent hyperplastic Tunica Vasculosa Lentis (PHTVL) in dogs II Nord. Vet. Med. —
1982. — 34. — 394.Audell L. Hoftledsdysplasi en utvecklingsrubbning 11 Svensk Veterinartidn. — 1986. — Vol. 38,
№ 15. — Suppl. 11. — P. 60-62.Austin C.R. Fertilization and development of the egg // Reproduction in domestic animals /Eds.
H.H. Cole, P.T. Cupps. — N.Y.; London: Acad. Press, 1959. — Vol 1. — P. 399-432.Averill D.R., Bronson R.T. Inherited necrotizing myelopathy of Afghan Hounds II J. Neuropath.
Exp. Neurol. — 1977. — Vol. 36. — P. 734-737.Barker C.G., Herrtage M.E., Shanaham F., Winchester B.G. Fucosidosis in English springer spaniels:
results of a trial screening programme // J. Small Anim. Pract. — 1988. — Vol. 29, № 10. — P. 623-630.Barnett K.C. Primarv retinal dystrophies in the dog // J. Amer. Vet. Med. Assoc. — 1969. —
Vol. 154. — P. 804-808.Barnett K.C. Comparative aspects of canine hereditary eye disease // Adv. Vet. Sci. Comp. Med. —1976. — Vol. 20. — P. 39-67.Barnett K.C. Hereditary cataract in the dog // J. Small Anim. Pract. — 1978. — Vol. 19. —
P. 109-120.Barnett K.C. Hereditary cataract in the German Shepherd Dog II J. Small Anim. Pract. — 1986. —
Vol. 27. — P. 387-395.Bartlett C.R. Heritabilities and genetic correlations between hip dysplasia and temperament traits of
seeing-eye dogs // Masters Thesis, Rutgers Univ., New Brunswick, NJ., 1976.Bedford P.G. The aetiology of canine glaucoma II Vet. Rec. — 1980. — Vol. 107. — P. 76-82.
Bedford P.G.C. Control of inherited retinal degeneration in dogs and cats in the United Kingdom //
J. Small Anim. Pract. — 1989. — Vol. 30, № 3. — P. 172-177.Bjerkas L Hereditary ,,cavitating“ leukodystrophy in Dalmatian dogs // Acta Neuropathol. —
1977. — Vol. 40. — P. 163-169.Bjorck G. N£gra tidiga erfarenheter av utredning och bekampning av arftligt betingade sjukdomar //
Svensk Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 20-25.Bjork G.S., Dyrendahl S.E., Oisson S.-E. Hereditary ataxia in smooth-haired Fox Terriers // Vet.
Res. — 1957. — Vol. 69. — P. 871-876.Bjork G., Mar W., Oisson S.G., Sourander P. Hereditary ataxia in Fox Terriers // Acta Neuro¬
pathol. — 1962. — Suppl. 1. — P. 45-58.Boom H.P.A. Anomalous animals // S. Afr. J. Sci. — 1965. — Vol. 61. — P. 159-171.Bosma A.A., Baumans V., Buwalda G., de Haan N.A. Triploidy in a newborn dog // Proc. 5th Eur.
Coll. Cytogenet. Dorn. Anim. — 1982. — P. 392-398.Bowles C.A., Alsaker R.D., Wolfe T.L. Studies of the Pelget-Huet anomaly in Foxhounds // Amer. J.
Pathol. — 1979. — Vol. 96. — P. 237-248.Bracnd М., Andersen E. Variation of transferrin and esterase in sera of dogs // Acta Vet. Scand.1987. — 28. — 435-444.Brass W. Hip dysplasia in dogs // J. Small Anim. Pract. — 1989. — Vol. 30, № 3. — P. 166-170.Brown C.J., Murphree O.D., Newton J.E.O. The effect of inbreeding on human aversion in Pointer
dogs II J. Hered. — 1978. — Vol. 69. — P. 362-365.Brown R.V., Teng T.S. Studies of inherited pyruvate kinase in the Basenji // J. Amer. Anim. Hosp.
Assoc. — 1975. — Vol. 11. — P. 362-365.Burke T.J., Badertscher R.R. Pregnancy diagnosis II Small animal reproduction and infertility. A
clinical approach to diagnosis and treatment / Eds. T.J. Burke. — Philadelphia, USA, 1986. — P. 85-92.Burns М., Fraser M. Genetics of the dog. — Edinburgh, London: Oliver and Boyd, 1966. — 564 p.Carrig C.B., Seawright A.A. Familial canine polyostotic fibrous dysplasia with subperiosteal cortical
defects // J. Small Anim. Pract. — 1969. — Vol. 10. — P. 397-405.Carver E.A. Coat color genetics of the German Shepherd dog // J. Hered. — 1984. — Vol. 75,
№ 4. — P. 247-252.Castleberry М., Linn J.M., Nyland T.G., Leighton E.A. Development and evaluation of improved
biological sensory systems. Research in biological and medical sciences, including biochemistry, commu¬
nicable diseases and immunology, internal medicine, nuclear medicine, physiology, psychiatry, surgery and
veterinary medicine // Annu. Progr. Rep.-Water Reed Army Institute of Research: Washington, DC.,
1975. — Vol. 1. — P. 1589-1595.CateLI R.B., Bolz C.R., Korth B. Behavioral types in purebred dogs objectively determined by
taxonomy // Behav. Genet. — 1973. — Vol. 3, № 3. — P. 205-216.Christensen K., Arnbjerg J., Andresen E. Polymorphism of serum albumin in dog breeds and its
relation to weight and leg length // Hereditas. — 1985. — Vol. 102. — P. 219-223.Christiansen I.J. Reproduction in small domestic animals. Use of biotechnology for dogs and cat //
Dansk. Veterinaertids. — 1990. — 73, № 14. — 745-749.Christie D.W., Bell E.T., Horth C.E., Palmer R.F. Peripheral plasma progesterone levels during the
canine oestrous cycle // Acta Endocrinol. — 1971. — Vol. 68. — P. 543-550.Clough E., Pyle R.L., Hare W.C.D. et al. An XXY sex-chromosome constitution in a dog with
testicular hypoplasia and congenital heart disease // Cytogenetics. — 1970. — Vol. 9 — P. 71-77.Colling D.T., Saison R. Canine blood groups II Anim. Blood Groups and Biochem. Genet. —
1980. — Vol. 11. — P. 1-12.Concannon P.W. Canine physiology of reproduction // Small animal reproduction and infertility. A
clinical approach to diagnosis and treatment /Ed. T.J. Burke. — Philadel phia, USA, 1986. — P. 23-77.Concannon P.W., Hansel W., Visek W.J. The ovarian cycle in the bitch: plasma oestrogen, LH and
progesterone // Biol. Reprod. — 1975. — Vol. 8. — P. 74-82.Conroy J.D., Rasmusen B.A., Small B. Hypotrichosis in Miniature Poodle siblings // J. Amer. Vet.
Med. Assoc. — 1975. — Vol. 166. — P. 697-699.Cork L.C., Griffin J.W., Munnell J.F. et al. Hereditary canine spinal muscular atrophy // J.
Neuropath. Exp. Neurol. — 1979. — Vol. 38. — P. 209-221.Cork L.C., Struble R.G., Gold B.G. et al. Changes in size of motor axons in hereditary canine spinal
muscular atrophy // Lab. Invest. — 1989. — Vol. 61, № 3. — P. 333-342.Corley E.A., Sutherland T.M., Carlson W.D. Genetic aspects of canine elbow dysplasia // J. Amer.
Vet. Med. Assoc. — 1968. Vol. 153. — P. 543-547.
Curtis R.L., English D., Kim Y.J. Spina bifida in a „stub" dog stock, selectively bred for short tails //
Anat. Rec. — 1964. — Vol. 148. — P. 365.Day M.E., Kraay G.J., Stevens R.W.C. Polymorphism of canine serum albumin II Anim. Blood
Grps Biochem. Genet. — 1971. — Vol. 2. — P. 195-199.Dee J., Forchhammer S. Measurement of serum progesterone in dogs. Determination of time of
mating II Dansk Veterinartids. — 1988. — 71, № 8. — 425-430.Dilger W. The behavior of lovebirds II Sci. Am. — 1962. — Vol. 206. — P. 88-98.Doak R.R., Holl A., Dale H.E. Longevity of spermatozoa in the reproductive tract of the bitch II
J. Reprod. Fert. — 1967. Vol. 13, № 1. — P. 51-58.Dodds W.J. Hereditary and acquired hemorrhagic disorders in animals // Progress in Hemostasis
and Thrombosis /Ed. Т.Н. Spaet. — Grune and Stratton, 1974.Dodds W.J. von Willebrand’s disease in dogs II Mod. Vet. Pract. — 1984. — Vol. 65, № 9. —
P. 681-686.Eisner E. Malokklusionen bei Hund und Katze // Vet. — 1990. — Vol. 5, № 2. — P. 10-17.England G.C.V., Allen W.E. Seminal characteristics and fertility in dogs II Vet. Records. — 1989. —
Vol. 25, № 15. — P. 399.Evans R.I. Lysosomal storage diseases in dogs and cats II J. Small Anim. Pract. — 1989. — Vol. 30,
№ 3. — P. 144-150.Falco M.J., Barker J., Wallace M.E. Genetics of epilepsy in the British Alsatian // J. Small Anim.
Pract. — 1974. — Vol. 15. — P. 685-692.Farstad W., Berg K.A. Factors influencing the success rate of artificial insemination with frozen
semen in the dog // J. Reprod. Fert. — 1989. — Suppl. 39. — P. 289-292.Feldman D.B., Bree M.M., Cohen B.J. Congenital diaphragmatic hernia in neonatal dogs II J. Amer.
Vet. Med. Assoc. — 1968. — Vol. 153. — P. 942-944.Ferwandes P.A., Bowen R.A., Sawyer H.R. et al. Concentration of receptors for estradiol and
progesterone in canine endometrium during estrus and diestrus // Amer. J. of Vet. Res. — 1989. — Vol. 50,'
№ i. _ p. 64-67.Fjeld T. Achondroplastisk dvergvekst // Norsk Veterinaertidsskr. — 1988. — 100. № 10. —
P. 711-714.Fletch S.M., Pinkerton P.H, Brueckner P.J. The Alaskan Malamute chrondrodysplasia
(dwarfism-anemia syndrome) II J. Amer. Anim. Hosp. Assoc. — 1975. — Vol. 11. — P. 353-361.Fletcher R. Dogs of the world. — Toronto; a.o.: A Ridge Press Book, 1977. — 159 p.Foote R.H., Swierstra E.E., Hunt W.L. Spermatogenesis in the dog II Anat. Rec. — 1972. — Vol. 73,
№ 3/4. — P. 341-351.Ford L. Hereditary aspects of human and canine cyclic neutropenia // J. Hered. — 1969. —
Vol. 60. — P. 293-299.Fougner J.A. Embryonic death in the blue fox II Nordisk Vet. Med. — 1972. — 24. — 25-31.Fox M.W. Inherited polycystic mononephrosis in the dog II J. Hered. — 1964. — Vol. 55. —
P. 29-30.Fox M.W. The Dog: Its Domestication and Behaviour. — Garland STPM Press, 1978.Frank D. The genetic basis of evolutionary changes in behavioural patterns // The Genetics of
Behaviour /Ed. J.H.F. van Abeelen. — Amsterdam: North-Holland Publ. Co., 1974. — P. 119-140.Frankling E. The Dalmatian II Popular Dogs, 1971.Fritz T.E., Lombard L.S., Tyler S.A:, Norris W.P. Pathology and familial incidence of orchitis and
its relation to thyroiditis in a closed Beagle colony // Exper. Molec. Pathol. — 1976. — Vol. 24. — P.
142-158.Fyfe J.C., Jezyk P.F., Giger U., Patterson D.F. Inherited selective malabsorption of vitamin B12 in
Giant Schnauzers // J. Amer. Anim. Hosp. Assoc. — 1989. — Vol. 25, № 5. — P. 533-539.Gahne B., Juneja R.K., Gromus J. Horizontal polyacrylamide gradient gel electrophoresis for the
simultaneous phenotyping of transferrin, post-transferrin, albumin and postalbumin in the blood plasma of
cattle // Anim. Blood Grps Biochem. Genet. — 1977. — Vol. 8. — P. 127-137.Gardner D.L. Familial canine chondrodystrophia foetalis II J. Path. Bact. — 1959. — Vol. 77. —
P. 243-247.Garmer L. Linsluxation hos Tibetansk Terrier // Svensk Veterinartidn. — 1986a. — Vol. 38,
№ 15. — Suppl. 11. — P. 132-133.Garmer L. Progressiv retinal atrofi (PRA) hos Labrador Retriver // Svensk Veterinartidn. —
1986b. — Vol. 35, № 15. — P. 120-123.Gelatt K.N. Familial glaucoma in the Beagle dog // J. Amer. Anim. Hosp. Assoc. — 1972. —
Vol. 8. — P. 23-28.
Gelatt K.N. Inheritance of primary glaucoma in the Beagle // Amer. J. Vet. Res. — 1981. —
Vol. 42. — P. 1691—1693.Gerlach K., Skrodzki М., Trautvetter E. Kongenitale Anomalien des Aortenbogens beim Hund //
Kleintierpraxis. — 1988. — Bd. 33, № 9. — S. 355-356, 358-363.Ginsburg B.E. Genetics as a tool in the study of behavior II Persp. Biol. Med. — 1958. — Vol. 1. —
P. 397-424.Goerttler V. Neufundlender. — Wittenberg Lutherstadt: A. Ziemsen Verlag, 1978. — 160 s.Graf K.J. Serum oestrogen, progesterone and prolactin concentration in cyclic, pregnant and
lactating Beagle dogs II J. Reprod. Fert. — 1978. — Vol. 52, № 1. — P. 9-14.Graphodatsky A.S. A conservative and variable elements of mammalian chromosomes II Cyto¬
genetics of Animals / Ed. C.R.E. Hainan. — Oxon: CAB Intern. Press, 1989. — P. 95-123.Graphodatsky A.S., Hainan C.R.E. Appendix 1. An atlas of selected karyotypes II Cytogenetics of
Animals IEd. C.R.E. Hainan. Oxon: CAB Intern. Press, 1989. — P. 321-390.Green E.L. Mutant stocks of cats and dogs offered for research // J. Hered. — 1957. — Vol. 48. —
P. 56-57.Griffiths I., Duncan I., Barker J. Progressive axonopathy of Boxer dogs // J. Small Anim. Pract. —
1980. — Vol. 21. — P. 29-43.Grondalcn J. Arthrosis with special reference to the elbow joint of young rapidly growing dogs. II.
Occurence, clinical and radiographical findings II Nord. Vet. Med. — 1979. — 31. — P. 69-75.Grondalen J. Arthrosis in the elbow joint of young rapidly growing dogs. VII. Occurence in the
Rottweiler breed // Ibid. — 1982. — 34. — 76-82.Gruneberg H., Lea A.J. An inherited jaw anomaly in long haired Dachshunds // J. Genet. —
1940. — Vol. 39. — P. 285-296.Gustavsson E., Swenson L. Glaukom hos Springer Spaniel // Svensk Veterinartidn. — 1986. —
Vol. 38, № 15. — Suppl. 11. — P. 149-151.Haaften B. van, Dieletman S.J., Okkens A.C., Willemse A.H. Timing the mating of dogs on the basis
of blood progesterone concentration II Vet. Rec. — 1989. — Vol. 125, № 21. — P. 524-526.Hahn S. Variation of skull traits and reproduction on breeds of small dogs. — Thesis. —
Tierarztliche Hochschule Hannover, G.F.R. — 1988. — S. 130.Hall D.E. Blood Coagulation and its Disorders in the Dog. — Balliere Tindall, 1972.Hansen H.-J. Konstitutiones roll for uppkomsten av sjukdomar och skador II Svensk Veterinar¬
tidn. — 1986. — Vol. 38, N 15. — Suppl. 11. — P. 206-211.Нага Y., Ejima H., Aoki S. et al. Preparation of monoclonal antibodies against dog erythrocyte
antigen D1 (DEA-3) // J. Vet. Med. Sci. — 1991. — Vol. 53, N° 6. — P. 1105-1107.Hare W.C.D. Intersexuality in the dog // Can. Vet. J. — 1976. — Vol. 17. — P. 745.Hare W.C.D., Bovce K. Chromosomal translocation in Miniature Poodles // Vet. Rec. — 1974. —
Vol. 95. — P. 218-219.Hare W.C.D., Wilkinson J.S., McFeely R.A., Riser W.H. Bone chondroplasia and a chromosome
abnormality in the same dog // Am. J. Vet. Res. — 1967. — Vol. 28. — P. 583-587.Hart B.L., Hart L.A. Selecting the best companion animal: breed and gender specific behavioural
profiles. The Pet Connection. — Censfare, Minneapolis, Minnesota, 1984. — P. 180-192.Hartley W.J., Barker J.S.F., Wanner R.A., Farrow B.R.H. Inherited cerebellar degeneration in the
Rough Coated Collie // Aust. Vet. Pract., 1978. — Vol. 8. — P. 79-85.Hashimoto Y., Yamakawa Т., Tanabe Y. Further studies on the red cell glycolipids of various breeds
of dogs. A possible assumption about the origin of Japanese dogs II J. Biochem. 1984. — Vol. 96. —
P. 1777-1782.Hedhammar A. Arftlight betingade sjukdomar hos hund — en gemensam angelagenhet II Svensk
Veterinartidn. — 1986a. — Vol. 38, № 15. — Suppl. 11. — P. 10-19.Hedhammar A. Dagslaget avseende bekampning av hoftledsdysplasi i Sverige // Svensk
Veterinartidn. — 1986b. — Vol. 38, № 15. — Suppl. 11. — P. 68-7.Hedhammar A., Wu F., Krook L. et al. Overnutrition and skeletal disease. An experimental study in
growing Great Dane dogs II Cornell Vet. — 1974. — Vol. 64. — Suppl. 5.Hegreberg G.A., Padgett G.A., Gorham J.R., Henson J.B. A connective tissue disease of dogs //
J. Hered. — 1969. — Vol. 60. — P. 249-254.Hippel E. von. Embriologische untersuchungen uber Vererbung angeborener Katarakt, uber
Schichtstar des Hundes sowie uber eine besondere form von kapselkatarakt // Graefes Archiv fur
Ophthalmologic. — 1939. — 124. — 300.Hirth R.S., Nielsen S.W. Familial canine globoid cell leukodystrophy // J. Small. Anim. Pract. —1967. — Vol. 8. — P. 569-575.
Hodgman S.F.J. Abnormalities and defects in pedigree dogs. I. An investigation into the existence of
abnormalities in pedigree dogs in the British Isles // J. Small Anim. Prac. — 1963. — Vol. 4. — P. 447-456.Holst R.A., Phemister R.D. The prenatal development of the dog: preimplantation events // Biol.
Reprod. — 1971. Vol. 5. — P. 194-206.Holzmann A., Arbeiter K. Die Samenubertragung bei der Hundin — eine Analyse klinischer Fall //
Deutsche Tierarztliche Wochenschrift. — 1987. — Bd. 94, № 10. — S. 562-563.Hopkins A.L. Canine myasthenia gravis II J. Small Anim. Pract. — 1992. — Vol. 33, № 10. —
P. 477-484.Horvath L., Elodi Z. Oroklodo verzekenyseg (haemophilia-A) esete egy nemet juhaszkutyaban //
Magy. allatorv. lap. — 1991. — 46, N° 1. — P. 29-32.Howard-Peebles P.N., Pryor J.C. The R-banding pattern of the canine karyotype // J. Hered. —1980. — Vol. 71. — P. 361-362.Humphrey E.S. „Mental tests“ for Shepherd Dogs. An attempted classification and evaluation of the
various traits that go to make „temperament41 in the German Shepherd Dog // J. Hered. — 1934. —
P. 129-136. — Цит. no: Mackenzie S.A., Oltenacu E.A.B., Houpt K.A., 1986.Humphrey E.S., Warner L. Working dog — an attempt to produce a strain of German Shepherd
which combines working ability and beauty 01 conformation. — Baltimore, MD.: John Hopkins Press,
1934. — Цит. no: Mackenzie S.A., Oltenacu E.A.B., Houpt K.A., 1986.Hutt F.B. Genetics for Dog Breeders. — Freeman, 1979.Hutt F.B., Lahunta A. A lethal glossopharyngeal defect in the dog // J. Hered. — 1971. — Vol. 62. —
P. 291-293.Iljin N.A. Wolf-dog genetics // J. Genet. — 1941. — Vol. 42. — P. 359-414.Iwamura S., Koizumi N., Matsubara Y. Changes in the level of plasma cortisol, progesterone and
total testosterone in developing hairless dogs Hi. Vet. Med. Sci. — 1992. — Vol. 54, N 6. — P. 1217-1218.James W.T. Morphologic form and its relation to behavior. The genetic and endocrinic basis for
differences in form and behavior /Ed. C.R. Stockard. — Phyladelphia, PA: The Wistar Institute of
Anatomy and Biology, Sect. VI, 1941.Jeffcoate I.A., Lindsay F.F. Ovulation detection and timing of insemination based on hormone
concentrations, vaginal cytology and endoscopic appearance of the vagina in domestic bitches I! J. Reprod.
Fert. — 1989. — Suppl. 39. — P. 277-287.Johansson I. Oestrus and mating in silver fox II Ann. Agric. Coll. Sweden. — 1940. — № 9. —
239-263.Johnstone I.B., Lotz F. An inherited platelet function defect in Basset hounds II Canad. Vet. —1979. — Vol. 20. — P. 211-215.Jones B. Arftlight betingade hemostas-rubbningar hos hundar i Sverige // Svensk Veterinartidn. —1986. — Vol. 38, N 15. — Suppl. 11. — P. 172-176.Jones G.E., Boyns A.R., Cameron E.H.D. et al. Plasma oestradiol, luteinizing hormone and pro¬
gesterone during pregnancy in the beagle bitch Hi. Reprod. Fert. — 1973. — Vol. 35, № 1. — P. 187-189.Jones G.E., Boyns A.R., Cameron T.H.D. et al. The ovarian cycle of the bitch: plasma estrogen and
progesterone // Biol. Reprod. — 1975. — Vol. 13..— P. 112-121.Juneja R.K., Christensen K., Andresen E., Gahnc B. Frequencies of transferrin types of various
breeds of domestic dogs // Anim. Blood Grps Biochem. Genet. — 1981. — Vol. 12. — P. 79-88.Juneja R.K., Gahne B. Lukka М., Ehnholm C. A previously reported polymorphic plasma protein of
dogs and horses, identified as apolipoprotein A-IV // Anim. Genet. 1989. — Vol. 20, № 1. — P. 59-63.Juneja R.K., Lingaas S.F., Shibata T. et al. Paternity testing in dogs by typing of plasma proteins:
further data and some new variants // XXIII Intern. Conf. on Animal Genetics. — Berne, 1992. — P. 26.Juneja R.K., Shibata T. Genetic polymorphism and close linkage of two plasma protein loci in
dogs II Anim. Genet. — 1992. — Vol. 23, N° 2. — P. 143-150.Karbe E., Schiefer B. Familial amaurotic idiocy in male German short-haired pointers II Pathol.
Vet. — 1967. — Vol. 4. — P. 223^233.Kidd K.K., Stone W.H., Crimella C. Immunogenetic and population genetic analysis of Iberian
cattle // Anim. Blood Grps Biochem. Genet. — 1980. — Vol. 11. — P. 21-38.Klingeborn B. Indikationer pa en arftlig bakgrund till arm bags ledsforandringar hos Berner
Sennenhund // Svensk Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 1. — P. 102-107.Knyazev S., Tikhonov V., Tanabe V. Phylogenetic study of native dog breeds from Middle Asian,
Caucasian and Siberian regions by blood protein polymorphisms // XXIII Intern. Conf. on Animal
Genetics. — Berne, 1992. — P. 24-25.Kobayashi R., Miyakawa Т., Tanabe Y. Blood protein polymorphism in Bangladesh native dogs //
Genetic studies on breed differentiation of the native domestic animals in Bangladesh, Pt 2. Hiroshima
Univ., 1987. — P. 93-103.
Коек Е. Retinal dysplasia. A comparative study in human beings and dogs. — Karolinska Instituted
Stockholm, 1974. — 70 p.Koch J., Pedersen H., Flagstad A. Subaortal-stenose hos hund // Dan. Veterinaertidsskr. — 1993. —
76, № 6. — 231-234.Koop E., Mayr B., Schleger W. Nucleolus organizer regions on chromosomes of the domestic dog //
J. Hered. — 1982. — Vol. 73. — P. 73-74.Koppang N. Neuronal ceroid-lipofuscinosis in English setters // J. Small Anim. Pract. — 1970. —
Vol. 10. — P. 639-644.Kramer J.W., Hegreberg G.A., Bryan G.C. et al. A muscle disorder of Labrador Retriever
characterized by a deficiency of tupe II muscle fibres // J. Amer. Vet. Med. Assoc. — 1976. — Vol. 169. —
P. 817-820.Krook L. Hereditary neuronal abiotrophy in the Swedish Lappland dog // The Cornell Veterina¬
rian. — 1973. — Vol. 63, Suppl. 3. — P. 1-41.Kvart C. Kongenitala hjartfel hos hund // Svensk Veterinartidn. — 1986. — Vol. 38, № 15. —
Suppl. 11. — P. 184-188.Ladrat J., Blin P.C., Lauvergne J.J. Ectromelie bithoracique hereditaire chez le chien // Ann. Genet.
Select. Anim. — 1969. — Vol. 1. — P. 119-130.Lahunta A., Averill D.R. Hereditary cerebellar cortical and extrapyramidal nuclear abiotrophy in
Kerry Blue Terriers // J. Amer. Vet. Med. Assoc. — 1976. — Vol. 168. — P. 1119-1124.Lapras М., Fournel C. Recherches a l’ecole veterinaire de lyon sur les maladies auto-immunes et
Pimmunopathologie chez le chien // Bull. Acad. Vet. Fr. — 1992. — 65, 1. — 63-76.Larsen R.E., Dias E., Cervenka J. Centric fusion of autosomal chromosomes in a bitch and
offspring // Amer. J. Vet. Res. — 1978. — Vol. 39. — P. 861-864.Larsen R.E., Dias E., Flores G., Seldcn J.R. Breeding studies reveal segregation of a canine
Robertsonian translocation along Mendelian proportion II Cytogenet. Cell Genet. — 1979. — Vol. 24. —
P. 95-101.Lau R. Inherited premature closure of the distal ulnar physis // Amer. Anim. Hosp. Assoc. —1977. _ Vol. 13. — P. 609-612.Lavelle R.B. Radiographic techniques and radiological interpretation. The important environmental
components in hip dysplasia II 9th Meet. Int. Vet. Radiol. Assoc. (IVRA): Proc. S.l. — 1991. — P. 30.Lefflcr M. CEA-Diagnostisk tillforlitlighen — symtombild hos foraldrar och avkomma // Svensk
Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 144-148.Letard E. Experiences sur heredite mendelienne du charactere peau nue dans Г espece chien // Rev.
Vet. J. Med. Vet. — 1930. — 82. — 553-570.Linde-Forsberg G.C., Forsberg M. Fertility in dogs in relation to semen quality and the time and site
of insemination with fresh and frozen semen // J. Reprod. Fert. — 1989. — Suppl. 39. — P. 299-310.Lingaas F., Klemetsdal G. Breeding values and genetic trend for hip dysplasia in the Norwegian
Golden Retriever population // Z. Tierzucht. und Ziichturgsbiol. r— 1990. — Bd. 107, № 6. — S. 437-443.Little C.C. The inheritance of coat colour in dogs. — Cornell Univ. Press, 1957.Littlewood J.D. Inherited bleeding disorders of dogs and cats // J. Small Anim. Pract. — 1989. —
Vol. 30, № 3. — P. 40-143.Lund J.E., Padgett GA., Gorham J.R. Inheritance of cyclic neutropenia in the dog II J. Hered. —
1970. — Vol. 61. — P. 46-49.MacDowell E.C. Heredity of behavior in dogs /Ed. C.B. Davenport. — Dept, of Genetics 101-56,
Yearbook of the Carnegie Institution, 1921. — N 20. — P. 128-133. — Цит. no: Mackenzie S. A., Oltena-
cu E.A.B., Houpt K.A., 1986.Mackenzie S.A., Oltenacu E.A.B., Houpt K.A. Canine behavioral genetics. A review // Appl. Anim.
Behav. Sci. — 1986. — Vol. 15. — P. 365-393.MacMillan A.D., Lipton D.E. Heritability of multifocal retina dysplasia in American Cocker
Spaniels II J. Amer. Vet. Med. Assoc. — 1978. — Vol. 172. — P. 568-572.Madsen J., Svalastoga E. Hofteledsdysplasi hos hund. En oversight II Dan. Veterinaertidsskr. —1989. — 72, № 4. — 178-188.Mahut H. Breed differences in the dog’s emotional behaviour // Can. J. Psychol. — 1958. —
Vol. 12. — P. 35-44.Makinen A. The standard karyotype of the silver fox chromosomes (Vulpes fulvus Desm.) II
Hereditas. — 1985a. — Vol. 103. — P. 171-176.Makinen A. The standard karyotype of the blue fox (Alopex lagopus L.) II Ibid. — 1985b. —
Vol. 103. — P. 33-38.
Makinen A., Fredga K. Banding analysis of the somatic chromosomes of raccoon dogs; Nyctereutes
procyonoides, from Finland // Proc. 4th Eur. Col). Cytogenet. Dom. Anim., 1980. — P. 392-398.Makinen A., Gustavsson I. A comparative chromosome banding study in the silver fox, the blue fox,
and their hybrids // Hereditas. — 1982. — Vol. 97. — P. 289-297.Makinen A., Lohi O., Juvonen M. Supernumerary chromosomes in the chromosomally polymorphic
blue fox (Alopex lagopus) II Ibid. — 1981. — Vol. 94. — P. 277-279.Malik R., Turnbull G.R., Black A.P. Patent ductus arteriosus in five related female Border Collies II
Austral. Vet. Pract. — 1991. — Vol. 21, № 1. — P. 2-4.Mandigers P.J.J., Van Nes J.J., Knol B.W. et al. Hereditary Kocker dog ataxia // Tijdschr.
Diergeneesk. — 1993. — 118, Suppl. №1.-65.Mann G.E., Stratton J. Dermoid sinus in the Rhodesian Ridgeback II J. Small Anim. Pract. —
1966. — Vol. 7. — P. 631-642.Manning A. Genes and evolution of insect behaviour // Behaviour genetic analysis /Ed. J. Hirsch. —
McGraw Hill Book Co. — 1967. — P. 44-60.Manolache М., Rose W.M., Schmid M. Banding analysis of the somatic chromosomes of the
domestic dog (Canis familiaris) // Canad. J. Genet. Cytol. — 1976. — Vol. 28. — P. 513-518Marchlewski M.T. Genetic studies on the domestic dog II Bull. Int. Acad. Pol. Sci. Lett., Cl. Sci.
Math. Nat. Ser. B. — 1930. — P. 117-145.Martin C.L., Leipold H.W. Aphakia and multiple ocular defects in St. Bernard puppies II Vet. Med.
Small Anim., 1974. — Vol. 69. — P. 448-453.Martin C.L., Wyman M. Glaucoma in the Basset Hound // J. Amer. Vet. Med. Assoc. — 1968. —
Vol. 153. — P. 1320.Marx A., Mohrke A. Foxterrier. — Hamburg; Berlin: Verlag Paul Parey, 1983. — 70 s.Mayr B., Geber G., Auer H. et al. Heterochromatin composition and nucleolus organizer activity in
four canid species II Can. J. Genet. Cytol. — 1986a. — Vol. 28. — P. 744-753.Mayr B., Krutzler J., Schleger W., Auer H. A new type of Robertsonian translocation in the
domestic dog Canis familiaris II J. Hered. — 1986b. — Vol. 77. — P. 127.Mayr B., Schweizer D., Schleger W. Characterization of the canine karyotype by
counterstain-enhanced chromosome banding // Canad. J. Genet. Cytol. — 1983. — Vol. 35. — P. 616-621.Meera K., Brahe C., Wijnen L.M.M. Domestic dog (Canis familiaris) (2n=78) II Genetic Maps. 6th
edition /Ed. SJ. Brain..— Cold Spring Harbor Laboratory Press, 1993. — P. 4254-4255.Mellink C.N.M., Bosnia A.A. The karyotype of the domestic dog (Canis familiaris L.) // Cytogenetics
of Animals /Ed. C.R.E. Hainan. — Oxon, UK: CAB Intern. Press, 1989. — P. 151—157.Meyers K.M., Padgett G.A., Dickson W.M. The genetic basis of a kinetic disorder of Scottish Terrier
dogs II J. Hered. — 1970. — Vol. 61. — P. 189-192.Mgheni М., Christensen K., Andersen E. Albumin polymorphism in domestic dog breeds II
Hereditas. — 1979. — Vol. 91. — P. 307-308.Mhaskar Y., Harvey J.W., Dunaway G.A. Developmental changes of 6-phosphofructo-l-kinase
subunit levels in erythrocytes from normal dogs and dogs affected by glycogen storage disease type VII II
Compar. Biochem. Physiol. B. — 1992. — Vol. 101, № 3. — P. 303-307.Miller M.E., Christensen G.C., Evans H. Anatomy of the dog. Philadelphia, London, 1965.Miller L.M., Hegreberg G.A., Prieur D.J., Hamilton M.J. Inheritance of congenital myasthenia
gravis in smooth Fox Terrier dogs II J. Hered. — 1984. — Vol. 75, № 3. — P. 163-166.Millichamp N.J., Arden G.B. Transretinal mass receptor potentials recorded from the canine retina
in vitro // Amer. J. Vet Res. — 1989. — Vol. 50, № 10. — P. 1710-1714.Minouchi O. On the chromosomes of the dog // Jap. J. Zool. — 1928. — Vol. 1. — P. 255-268.Miyauchi Y., Nakayama H., Uchida K., Uetsuka K. Glomerulopathy With IgA deposition in the
dog // J. Vet. Med. Sci. — 1992. — Vol. 54, № 5. — P. 969-975.Mohr E. Ungarische Hirtenhunde. — Wittenberg Lutherstadt: A. Ziemsen Verlag, 1969. — 146 s.Moon A.A., Ginsburg B.E. Double trouble in F2 canine hybrids // Paper presented to the 11th
Annual Meeting of the Behavior Genetics Association, Purchase, NY, Abstr. in Behav. Genet. — 1981. —
Vol. 11.Moon A.A., Ginsburg B.E., Fine M.D. Thresholds of gene expression in coyote x beagle hybrids II
Paper presented at the 12th Annual Meeting of the Behavior Genetics Association, Ft. Collins, Co. Abstr.
in Behav. Genet. — 1982. — Vol. 12.Moreno-Millan М., Rodero A., Alonso F.J., Falcon C. Contribution to the establishment of the
R-banded karyotype in dogs // Genet. Sel. Evol. — 1991. — Vol. 23, Suppl. 1. — P. 179-182.Mori M. Thesis for Master of Science. — Gifu University: Faculty of Agriculture, 1982.
Morris B.G., Lemon L. A new allele in the 1-Protease Inhibitor (Pi-1) system in dogs I IXXIII Intern.
Conf. on Animal Genetics. — Berne, 1992. — P. 25.Muller G.H., Kirk R.W., Scott D.W. Small animal dermatology. 3rd ed. — Philadelphia: W.B. Saun¬
ders Co., 1983. — 890 p.Murtaugh R.J., Dodds W.J. Haemophilia A in a female dog // J. Amer. Vet. Med. Assoc. — 1988. —
Vol. 193, № 3. — P. 351—352.Muto N. Thesis for Bachelor of Science. — Faculty of Agriculture, Gifu University, 1983..Narfstrom K. PHTVL/PHPV : Beskrivning av en ogonsjukdom hos Dobermann // Svensk
Veterinartidn. — 1986. — Vol. 38, N 15. — Suppl. 11. — P. 134-139.Nash A.S. Familial renal disease in dogs II J. Small Anim. Pract. — 1989. — Vol. 30, № 3. —
P. 178-183.Nash W.G., O’Brien S. J. Conserved regions of homologous G-banded chromosomes between orders
in mammalian evolution: carnivores and primates // Proc. Natl. Acad. Sci. USA. — 1982. — Vol. 79. —
P. 6631-6635.Nei M. Molecular population genetics and evolution. — Amsterdam: North Holland Publ. Comp.,
1975. — 281 p.Nicholas F. Pituitary dwarfism in German Shepherd dogs II J. Small Anim. Pract. — 1978. —
Vol. 19. — P. 167-174.Noonan C.H.B., Kay J.M. Prevalence and geographic distribution of Fanconi syndrome in Basenjis
in the United States II J. Amer. Vet. Med. Assoc. — 1990. — Vol. 197, N 3. — P. 345-349.Nozawa K. Phylogenetic studies on the native domestic animals in east and southeast Asia. Proc. of
SABRAO Workshop on Animals // Genet. Res. Asia and Oceania. — 1980. — P. 23-43.Obara Y., Nakano T. Robertsonian fission polymorphism in the nothern Honshu population of the
Japanese raccoon dog, Nyctereutes procyonoides viverrinus // J. Mammal. Soc. Jap. — 1989. — Vol. 14. —
P. 19-25.Ogawa E., Kawakami A., Yagi T. et al. Oxidative damage to the membrane of canine erythrocytes
with inherited high Na-K-ATPase activity // J. Vet. Med. Sci. — 1992. — Vol. 54, № 1. — P. 57-62.Olson L.C., Walker D.M., Hall W.C. Portal-caudal vena cavalrenal-hepatic arteriovenous
malformation in a dog // Lab. Anim. Sci. — 1991. — Vol. 41, № 6. — P. 635-638.Opitz М., Loppnow H. Ein weiterer Fall von Cholesterinester — Speicherkrankheit bei einem
Foxterrier // Kleintierpraxis. — 1988. — Bd. 33, № 9. — S. 369-376.Palmer A.C., Payne J.E., Wallace M.E. Hereditary quadriplegia and amblyopia in the Irish Setter //
J. Small Anim. Pract. — 1973. — Vol. 14. — P. 343-352.Patel V., Koppang N., Patel B., Zeman W. Phenylenediamine-mediated peroxidase deficiency in
English Setters with neuronal ceroid-lipofuscinosis // Lab. Invest. — 1974. — Vol. 30. — P. 366-368.Pathak S., Van Tuinen P., Merry D.E. Heterochromatin synaptonemal complex, and NOR activity
in the somatic and germ cells of male domestic dog, Canis familiaris (Mammalia, Canidae) II Cytogenet.
Cell Genet. — 1982. — Vol. 34. — P. 112-118.Pathak S., Wurster-Hill D.H. Distribution of constitutive heterochromatin in carnivores //
Cytogenet. Cell Genet. — 1977. — Vol. 18. — P. 245-254.Patterson D.F. Pathologic and genetic studies of congenital heart disease in the dog II Advance.
Cardiol. — 1974. — Vol. 13. — P. 210-249.Patterson D.F. Congenital defects of the cardiovascular system of dogs // Adv. Vet. Sci. Comp.
Med. — 1976. — Vol. 20. — P. 1-37.Patterson D.F. Hereditary congenital heart defects in dogs II J. Small Anim. Pract. — 1989. —
Vol. 30, № 3. — P. 153-165.Patterson D.F., Medway W., Luginbuhl H., Chacko S. Congenital hereditary limphoedema in the
dog II J. Med. Genet. — 1967. — Vol. 4. — P. 145-152.Pearson J.P., Endcrs R.K. Ovulation, maturation and fertilization in the fox // Anat. Rec. —
1943. — Vol. 85. — P. 69-83.Pearson K., Usher C.H. Albinism in dogs // Biometrika. — 1929 — Vol. 21. — P. 144-163.Pedersen H.D., Koch J., Flagstad A. Endokardiose hos Cavalier King Charles Spaniel // Dan.
Veterinaertidsskr. — 1992. — 75, N 5. — 165-166, 168.Pfeiderer-Hogner M. Moglichkeiten der Zuchwertschatung beim Deutchen Schaferhund anhand der
Schutzliundpriifung I: Doctoral Thesis. — Munich, W. Germany: Ludwig-Maximilians Universitat,1968. — S. 187.Phillips J.M. Pig jaw in the Cocker Spaniel // J. Hered. — 1945. — Vol. 36. — P. 177-181.Pick J.R., Goyer R.A., Graham J.B., Rcnwick J.H. Subluxation of the carpus in dogs // Lab. In¬
vest. — 1967. — Vol. 17. — P. 243-248.
Pidduck H., Webbon P. The genetic control of Perthes’ disease in Toy Poodles — a working
hypothesis // J. Small Anim. Pract. — 1978. — Vol. 19. — P. 7-12.Possi L., Корег H., Mucha W. Esame radiologico delle giunture sacroiliache del cane pastore tedesco
(PT), affetto da displasia bilaterale congenita dell,anca (DA) // Boll. Assoc. Ital. Vet. Piccoli Anim. —1988. — 27, № 1. — P. 39-46.Post K., Dignean М., Clark E. Hair follicle dysplasia in a Siberian Husky II J. Amer. Anim. Hosp.
Assoc. — 1988. — Vol. 24, № 6. — P. 659-662.Priester W.A. Canine progressive retinal atrophy: occurrence by age, breed and sex II Amer. J. Vet.
Res. — 1974. — Vol. 35. — P. 571-574.Pullig T. Inheritance of a scull defect in Cocker Spaniels II J. Hered. — 1952. — Vol. 43. — P. 97-99.Pullig T. Anury in Cocker Spaniels // J. Hered. — 1953. — Vol. 44. — P. 105-107Pullig T. Brachyury in Cocker Spaniels // J. Hered. — 1957. — Vol. 48. — P. 75-76.Rapaport F.T., Bachvaroff R.J. Experimental transplantation and histocompatibility systems in the
canine species II Adv. Vet. Sci. Comp. Med. — 1978. — Vol. 22. — P. 195-219.Raymond S.L., Jones D.W., Brooks M.B., Dodds D.J. Clinical and laboratory features of a severe
form of von Willebrand disease in Shetland sheepdogs // J. Amer. Vet. Med. Assoc. — 1990. — Vol. 197,
№ 10. — P. 1342-1346.Renton J.P., Munzo C.D., Heathcote R.H., Carmichael S. Some aspects of the aetiology, diagnosis
and treatment of infertility in the bitch II J. Reprod. Fert. — 1981. — Vol. 61, № 2. — P. 289-294.Reuterwall C., Ryman N. An estimate of magnitude of additive genetic variation of some mental
characters in Alzatian dogs II Hereditas. — 1973. — Vol. 73. — P. 277-283.Riis R.C., Cummings J.F., Loew E.R., De Lahunta A. Tibetan terrier model of canine ceroid
lipofuscinosis // Amer. J. Med. Genet. — 1992. — Vol. 42, № 4. — P. 615-621.Robinson R. Genetics for dog breeders. — Oxford a.o.: Pergamon Press Ltd, 1982. — 264 p.Robinson R. Legg-calve-Perthes disease in dogs: genetic aetiology //J. Small Anim. Pract. — 1992. —
Vol. 33, № 6. — P. 275-276.Rothschild M.F., Christian L.L., Blanchard W. Evidence for multigene control of cryptorchidism in
♦swine // J. Hered. — 1988. — Vol. 79, № 4. — P. 313-314.Rubin L.F., Bourns T.K.R, Lord L.H. Hereditary of hemeralopia in Alaskan Malamutes // Amer.
J. Vet. Res. — 1967. — Vol. 28. — P. 355-357.Rnbin L.F., Flowers R.D. Inherited cataract in a family of Standard Poodles // J. Amer. Vet. Med.
Assoc. — 1972. — Vol. 161. — P. 207-208.Rubtsov N.B., Graphodatsky A.S., Matveeva V.G. et al. Silver fox gene mapping: conserved
chromosome regions in the order Carnivora // Cytogenet. Cell Genet. — 1988. — Vol. 48. — P. 95-98.Sandberg K., Juneja R.K. Close linkage between albumin and Gc loci in the horse // Anim. Blood
Grps Biochem. Genet. — 1978. — Vol. 9. — P. 169-173.Sandefeldt E., Cummings J., Lahunta A. de, et al. Hereditary neuronal abiotrophy in the Swedish
Lappland dog // The Cornell Veterinarian. — 1973. — Vol. 63. — Suppl. № 3. — P. 1-41.Sandefeldt E., Nilsson L. Hereditar juvenil neuronal muskelatrofi hos lapphund — nulaget // Svensk
Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 162-164.Sanecki R.K. Hoffman W., Dorner E., Kuhlenschmidt M. Purification and comparison of
corticosteroid-induced and intestinal isoenzymes of alkaline phosphatase in dogs II Amer. J. Vet. Res. —
1990. — Vol. 51, № 12. — P. 1964-1968.Sarkany P., Ocsag I. Hungarian dog breeds. — Budapest: Corvina Kiado, 1986. — 216 p.Sarmiento U., Storb R. Nucleotide sequence of a dog class I cDNA clone II Immunogenetics. —1990. — Vol. 31, № 5/6. — P. 400-404.Schaer М., Harvey J.W., Calderwood-Mays М., Giger U. Pyruvate kinase deficiency causing
hemolytic anemia with secondary hemochromatosis in a Cairn Terrier // J. Amer. Anim. Hosp. Assoc. —1992. — Vol. 28, № 3. — P. 233-239.Schaible R.H., Brumbangh J.A. Electron microscopy of pigment cells in variegated and
nonvariegated piebald spotted dogs // Pigment Cell. — 1976. — Vol. 3. — P. 191-220.Schotte C.S., Ginsburg B.E. Evidence for mendelian inheritance of gape-hiss in coyote-beagle
hybrids. — Draft of paper presented at the Portland Wolf Symposium. — Portland, OR, 1979.Scott P.J., Fuller J.L. Genetics and the social behaviour of the dog. — Univ. of Chicago Press,
1965. — 349 p.Seager S.W.J. Artificial insemination in dogs II Small animal reproduction and infertility. A clinical
approach to diagnosis and treatment /Ed. T.J. Burke. — Philadelphia, USA, 1986. — P. 23-77.Seager S.W.J., Fletcher W.S. Collection, storage and insemination of canine semen II Lab. Anim.
Sci. — 1973. — Vol. 22. — P. 177-182.
Sdden J.R., Moorhead P.S., Oehlert M.L., Patterson D.F. The Giemsa banding pattern of the canine
karyotype II Cytogenet. Cell Genet. — 1975. — Vol. 15. — P. 380-387.Selmanowitz V.J., Markofsky J., Orentreich N. Black hair follicular dysplasia in dogs // J. Amer.
Vet. Med. Assoc. — 1977. — Vol. 171. — P. 1079-1081.Senglaub K. Wildhunde — Haushunde. — Leipzig-Jena-Berlin: Urania-Verlag, 1978. — 240 s.Shi L., Tang L., Ма К., Ма C. Synaptonemal complex formation among supernumerary В
chromosomes: an electron microscopic study on spermatocytes of Chinese raccoon dogs II Chromo¬
soma. — 1988. — Vol. 97. — P. 178-183. “Shive R.J., Hare W.C.D., Patterson D.F. Chromosome studies in dogs with congenital cardiac
defects II Cytogenetics. — 1965. — Vol. 4. — P. 340-348.Siembieda J. Wystepowanie dysplazji biodrowej u owczarkow niemeckich w poludniowo-zachodniej
Polsce II Med. Vet. — 1988. — Vol. 44, № 4. — P. 221-226.Simpson G.G. The principles of classification and a classification of mammals // Bull. Amer. Mus.
Nat. Hist. — 1945. — Vol. 85.Smiley L.E., Schotte C.S., Ginsburg B.E. An analysis of vocalizations of coyote-dog hybrids: a
preliminary report // Paper presented at the Animal Behavior Soc. Meetings, Univ. Park, PA, 1977.Smith G.K., Biery D.N. Canine hip dysplasia diagnosis: a comparative study // 9th Meet. Int. Vet.
Radiol. Assoc. (IVRA). — Proc. Suppl. 1. — 1991. — P. 31.Sokolowski J.H. Normal events of gestation in the bitch and methods of pregnancy diagnosis II
Current therapy in theriogenology /Ed. D.A. Morrow. — Toronto: Saunders Co. 1980. — P. 590-595.Spurting N.W. Hereditary disorders of haemostasis in dogs // Vet. Bull. — 1980. — Vol. 50. —
P. 151-173.Stades F.C. Retinal dysplasia in the dog // Tijdschr. Diergeneesk. — 1978. — 103. — 1087-1090.Stades F.C. Persistent hyperplastic Tunica vasculosa lentis and persistent hyperplastic primary
vitreous (PHTVL/PHPV) in 90 closely related Dobermann Pinshers: Clinical aspects // JAAHA. —1980. — Vol. 16. — P. 739.Stades F.C. Persistent hyperplastic Tunica vasculosa lentis and persistent hyperplastic primary
vitreous in Dobermann Pinshers: Genetic aspects // Ibid. — 1983. — Vol. 19. —- P. 957.Stepansky K. Sluzebni a pracovni plemena psu // Statni zemedelska nakladatelstri. — Praha,1974. — 296 s.Stephen H. A., Allen H. Comparison of red fox litter sizes determined from counts of embryos and
placental scars // J. Wildl. Manage. — 1983. — Vol. 473. — P. 860-863.Stockard C.R. An heredity lethal for localized motor and preganglionic neurons with a resulting
paralysis in the dog II Amer. J. Anat. — 1936. — Vol. 59. — P. 1-53. — Цит. no: Hatt, 1969.Stockard C.R. The genetic and endocrine basis for differences in form and behavior, as elucidated by
studies of contrasted pure-line dog breeds and their hybrids II Am. Anat. Mem. 19 Wistar Ints. Anat. Biol.,
Philadelphia, 1941.Stur I., Mayrhofer E., Kopf N., Muller S. Craniomandibulare Osteopathie beim West Highland
White Terrier II Kleintierpraxis. — 1991. — Bd. 36, № 9. — S. 491-492; 494; 499-500.Sugiura S., Tanabe Y., Ota K. Genetic polymorphism of eserine resistant esterases in canine
plasma II Anim. Blood Grps Biochem. Genet. — 1977. — Vol. 8. — P. 121-126.Sundgren P.-E. Risker forknippade med avelsarbete i sma populationer // Svensk Veterinartidn. —1986. — Vol. 38, 15. — Suppl. 11. — P. 28-31.Swenson L. Betydelsen av foraldradjurens egenstatus for uppkomsten av hoftledsdysplasi hos
avkomman II Svensk Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 64-67.Swisher S.N., Bull R., Bowdler J. Canine erythrocyte antigens. Joint Rep. of 1st Intern. Workshop
on Canine Immunogenetics // Tissue Antigens. — 1973. — Vol. 3. — P. 164-165.Swisher S.N., Young L.E. The blood group systems of dogs // Physiol. Rev. — 1961. — Vol. 41. —
P. 495-520.Switonski M. Heterochromatin heteromorphism (С-band) in the karyotype of the blue fox (Alopex
lagopus) // Rosz. Nauk. Zoot. — 1985. — Vol. 12. — P. 29-35.Switonski М., Gustavsson I., Hojer K., Ploen L. Synaptonemal complex analysis of the
B-chromosomes in spermatocytes of the silver fox (Vulpes fulvus Desm.) II Cytogenet. Cell Genet. —1987. — Vol. 45. — P. 84-92.Symons М., Bell K. Canine plasma alkaline phosphatase polymorphism and its relationship with the
canine Tr blood group system // Anim. Genet. — 1992. — Vol. 23, № 4. — P. 315-324.Tanabe Y. Phylogenetic relationships of dog breeds especially of Japanese native dog breeds
determined by the blood protein polymorphism II Rep. Soc. Res. Native livestock. — 1980. — Vol. 9. —
P. 162-227.
Tanabe Y. Genetic relationships among dog breeds with special reference to Asian dog breeds
studied by biochemical polymorphisms of blood proteins // Isozymes: structure, function and use in
Biology and Medicine. Wiley-Liss, Inc. 1990. — P. 619-637.Tanabe Y., Omi Т., Ota K. Genetic variants of glucose phosphate isomerase (E.C.5.3.1.9) in canine
erythrocytes // Anim. Blood Grps Biochem. Genet. — 1977. — Vol. 8. — P. 191-195.Tanabe Y. et al. Genetic variants of hemoglobin in canine erythrocytes // Ibid. — 1978. — Vol. 9. —
P. 79-83.Tanabe Y., Sugiura S., Asamona М., Ota K. Genetic polymorphism of leucine aminopeptidase in
canine plasma II Anim. Blood Grps Biochem. Genet. — 1974. — Vol. 5. — P. 225-230.Tefler E., Gosden R.G. A quantitative cytological study of polyovular follicles in mammalian ovaries
with particular reference to the domestic bitch (Canis familiaris) // J. Reprod. Fert. — 1987. — Vol. 81,
№ 1. — P. 137-147.Tesch E. Der Pudel. — Wittenberg Lutherstadt: A. Ziemsen Verlag: 1980. — 95 s.The inheritance of shyness in dogs // J. Genet. Psychol. — 1944. — Vol. 65. — P. 275-279.Thorner P., Baumal R., Valli V.E.O. et al. Abnormalities in the NCI domain of collagen type IV in
GBM in canine hereditary nephritis // Kidney Int. — 1989. — Vol. 35, № 3. — P. 843-850.Tjebbes K., Wriedt C. The albino factor in the Samojed dog // Hereditas. — 1927. — Vol. 10. —
P. 165-168.Trimble H.D., Keeler C.E. The inheritance of high uric acid excretion in dogs // J. Hered — 1938. —
Vol. 19. — P. 280-289.Trimble H.D., Keeler C.E. Preference of Dalmatian dogs for particular position in coach running,
and inheritance of this character // Nature. — 1939. — Vol. 144, № 3650. — P. 671-672. — Цит. по: Кру¬
шинский, 1991.Tsutsui T, Gamete physiology and timing of ovulation and fertilization in dogs II J. Reprod. Fert. —1989. — Suppl. 39. — P. 269-275.Tsutsui Т., Ejima H. Experimental induction of superfecundation in the dog // Jap. J. Vet. Sci. —1988. — Vol. 50, № 2. — P. 581-583.Tsutsui Т., Shimada K., Nishi M. et al. An experimental trial on embryo transfer in the dog II Jap.
J. of Vet. Sci. — 1989a. — Vol. 51, № 4. — P. 797-800.Tsutsui Т., Shimizu Т., Ohara N. et al. Relationship between the number of sperms and the rate of
implantation in bitches inseminated into unilateral uterine horn II Ibid. — 1989b. — Vol. 51, № 2. —
P. 257-263.Une S., Taura Y., Nakama S., Ejima H. Immunosuppression in dogs by pretransfusions with
ultraviolet (UV)-irradiated whole blood // J. Vet. Med. Sci. — 1991. — Vol. 53, № 5. — P. 811-817.Valtonen М., King W.A., Gustavsson I., Makinen A. Embryonic development in the blue fox //
Nordisk Veterinaermedicin. — 1985. — 37. — 243-249.Vaughan L. Flexural deformity of the carpus in puppies // J. Small Anim. Pract. — 1992. — Vol. 33,
No 8. — P. 381-384.Venge O. Reproduction in the fox and mink // Anim. Breed. Abstr. — 1959. — Vol. 27, № 2. —
P. 129-145.Vriesendorp H.M. et al. Joint Rep. of 1st Intern. Workshop on Canine Immunogenetics II Tissue
Antigens — 1973. — Vol. 3. — P. 147-172. — Цит. по: Материалы 16 Междунар. конф. по группам
крови и биохимическому полиморфизму животных. — J1.: ВИР, 1979. — Т. 3. — С. 233.Vriesendorp H.M. Hartog В., Smid-Mered B.M.J., Weslbrock D.L. Immunogenetic markers in
canine paternity cases II J. Small Anim. Pract. — 1974. — Vol. 15. — P. 693-699.Vriesendorp H.M. et al. Joint Rep. of the 2nd Workshop on Canine Immunogenetics //
Transplantation Proc. — Vol. 8. — 1976. — P. 289-314. — Цит по: Материалы 16 Междунар. конф. по
группам крови и биохимическому полиморфизму животных. — Л.: ВИР, 1979. — Т. 3. — С. 233.Wada M.Y., Lim Y., Wurster-Hill D.H. Banded karyotype of a wild-caught male Korean raccoon
dog, Nyctereutes procyonoides koreensis // Genome. — 1991. — Vol. 34. — P. 302-306.Wallace M.E. Keeshonds: a genetic study of epilepsy and EEG readings // J. Small Anim. Pract. —1975. _ Vol. 16. — P. 1-10.Wallin B. Perthessjukdom hos west highland White Terrier, en genetisk studie // Svensk Veteri¬
nartidn. — 1986. — Vol. 38, № 15. — Suppl. 1. — P. 114-118.Wang Z., Quan G., Yie Z., Wang S. Karyotypes of three species of carnivora // Acta Zool. Sinica. —1984. — Vol. 30. — P. 188-195.Ward O.G., Wurster-Hill D.H., Ratty E.J., Song Y. Comparative cytogenetics of Chinese and
Japanese raccoon dogs, Nyctereutes procyonoides // Cytogenet. Cell Genet. — 1987. — Vol. 45. —
P. 177-186.
Warren D, Dingwall J. Legg-Perthes disease in the dog — a review 11 Can. Vet. Journ. — 1972. —
Vol. 13. — P. 135-137.Watson A.D.J., Dixon R.T. Cyctic bone lesions in related old English Sheepdogs // J. Small Anim.
Pract. — 1977. — Vol. 18. — P. 561-571.Wayne R.K., O’Brien S.J. Allozyme divergence within the Canidae // Syst. Zool. — 1987. —
Vol. 36. — P. 339-355.Wayne R.K., Nash W.G., O’Brien S.J. Chromosomal evolution of the Canidae. I. Species with high
diploid numbers // Cytogenet. Cell Genet. — 1987a. — Vol. 44. — P. 123-133.Wayne R.K., Nash W.G., O’Brien S.J. Chromosomal evolution of the Canidae. II. Divergence from
the primitive carnivore karyotype // Ibid. — 1987b. — Vol. 44. — P. 134-141.Wcntinck G.H., Hartman W., Doeman J.P. Three cases of myotonia in a family of chows // Tijdschr.
Diergeneesk. — 1974. — Bd. 99. — S. 729-731.Westermarck E. Hereditary nature of canine pancreatic degenerative atrophy in the German
Shepherd dog // Acta Vet. Scand. — 1980. — Vol. 21. — P. 389-394.Westermarck E., Pamilo P., Wiberg M. Pancreatic Degenerative Atrophy in the Colli Breed: a
hereditary disease // J. Vet. Med. — 1989. — Vol. 36. — P. 549-554.Whitbread T.J., Batt R.M., Garthwaite G. Relative deficiency of serum IgA in the German Shepherd
dog: A breed abnormality II Res. Vet. Sci. — 1984. — Vol. 37. — P. 350-352.White S.D. Congenital and genetic skin disease II Tijdschr. Diergeneesk. — 1993. — 118,
Suppl. 1. — 53-57.Whitney D.D. Silver Poodles II Popular Dogs. — 1952. — August.Whitney D.D. Black and Silver Poodles // Popular Dogs. — 1958. — August.Whitney L.F. Inherited mental aptitudes in dogs // Eugenics. 1929a. — Vol. 2, № 4. — P. 8-16. —
Цит. по: Крушинский, 1991.Whitney L.F. Heredity of the trail-barking propensity in dogs // J. Hered. — 1929b. — Vol. 20. —
P. 561-562. — Цит. по: Крушинский, 1991).Whitney L.F. How to Breed Dogs // Orange Judd Publishing Co., 1947.Whitney L.F. How to Breed Dogs. — N. Y.; Howell, 1971. — 528 p.p.Whitney K.M., Goodman S.A., Bailey E.M. et al. Canine pyruvate kinase deficiency due to L gene
defect // Blood. — 1991. — Vol. 78, № 10. — Suppl. 1. — P. 406.Wiblishauser H. Der Dobermann. — Hamburg; Berlin: Verlag Paul Parey, 1990. — 96 s.Wigh G.T. Granulocytopati hos Irlandsk Setter II Svensk Veterinartidn. — 1990. — Vol. 42, № 5. —
P. 221-222.Wikstrom B. Collie eye anomaly Redovisning av ogonspeglingar av Collie i Mellansverige
1974-1984 // Svensk Veterinartidn. — 1986. — Vol. 38, № 15. — Suppl. 11. — P. 140-143.Wilcock B.P., Patterson J.M. Familial glomerulonephritis in Doberman Pincher dogs II Canad.
Vet. J. — 1979. — Vol. 20. — P. 244-249.Wildt D.E., Panko W.B., Chakraborty P.K., Seager S.W.J. Relationship of serum estrone,
estradiol-17 and progesterone to LH, sexual behaviour and time of ovulation in the bitch // Biol. Re-
prod. — 1979. — Vol. 20. — P. 648-658.Wilkes M.K., Palmer A.C. Congenital deafness and vestibular deficit in the Dobermann II J. Small
Anim. Pract. — 1992. — Vol. 33, №5. — P. 218-224.Willeberg P.K.W., Kastrap W., Andresen E. Pituitary dwarfism in German Shepherd dogs: studies on
somatomedin activity // Nord Vet. Med. — 1975. — 27. — 448-454.Willis M. Zur Genetic der Schoheit // Unser Dobermann. — 1990. — № 7. — S. 2-5.Willis T.B. The German Shepherd Dog. — К & R Books, 1976 .Willis M.B., Barnett K.C., Tempest W.M. Genetic aspects of lens luxation in the Tibetan Terrier //
Vet. Rec. — 1979. — Vol. 104. — P. 409-412.Winge O. Inheritance in Dogs. — Constable, 1950.Wisselink M.A., Bernadina W.E., Willemse A., Noordzij A. Immunologic aspects of German
Shepherd dog Pyoderma // Vet. Immunol. Immunopathol. — 1988. — Vol. 19. — P. 67-77.Wisselink M.A., Bouw J., der Weduwen S.A., Willemse A. German Shepherd dog pyoderma: a
genetic disorder // Vet. Quart. — 1989. — Vol. 11, № 3. — P. 161-166.Wittenberger H.? Wittenberger G. Der Pekinese. — Hanau: Verlag Werner Dausien, 1981. — 104 s.Woolpv J.H., Ginsburg B.E. Wolf socialization: a study of temperament in a social species // Amer.
Zool. — 1967. — Vol. 7. — P. 357-363.Wurster D.H. Cytogenetic and phylogenetic studies in Carnivora // Comparative mammalian
cytogenetics /Ed. K. Benirschke. —.N.Y.: Springer-Verlag, 1969. — P. 310-329.
Wurster D.H., Bcnirschke К. Comparative cytogenetic studies in the order Carnivora //
Chromosoma. — 1968. — Vol. 24. — P. 336-382.Wurster-Hill D.H., Centerwall W.R. The inter-relationships of chromosome banding patterns in
canids, mustelids, hyaena and felids // Cytogenet. Cell Genet. — 1982. — Vol. 34. — P. 178-192.Wurster-Hill D.H., Ward O.G., Kada H., Whittemore S. Banded chromosome studies and В
chromosomes in wild-caught raccoon dogs, Nyctereutes procyonoides veverrinus // Cytogenet. Cell Ge¬
net. — 1986. — Vol. 42. — P. 85-93.Wurster-Hill D.H., Ward O.G., Davis B.H. et al. Fragile sites, telomeric DNA sequences, В
chromosomes, and DNA content in raccoon dogs, Nyctereutes procyonoides, with comparative notes on
fox, coyote, wolf, and raccoon // Cytogenet. Cell Genet. — 1988. — Vol. 49. — P. 278-281.Yakely W.L. Collie eye anomaly: decreased prevalence through selected breeding // J. Amer. Vet.
Med. Assoc. — 1972. — Vol. 161. — P. 1103-1107.Yakely W.L. A study of heritability of cataracts in the American Cocker Spaniel // Ibid. — 1978. —
Vol. 172. — P. 814-817.Yakely W.L., Wyman М., Donovan E.F., Fechheimer N.S. Genetic transmission of an ocular fundus
anomaly in Collies // Ibid. — 1968. — Vol. 152. — P. 457-461.Yoshida M.A., Takagi N., Sasaki M. Karyotypic kinship between the blue fox (Alopex lagopus
Linn.) and the silver fox (Vulpes fulva Desm.) // Cytogenet. Cell Genet. — 1983. — Vol. 35. — P. 190-194.Yosida Т.Н., Shi L.M. Cytogenetic studies on the Japanese raccoon dog. XIII. A preliminary note
on the karyotype of a Chinese specimen with 57 chromosomes collected in Yunnan // Proc. Jap. Acad. —1986. — Vol. 8. — P. 49-52.Young L.E., Christian R.M., Ervin D.M. Erythrocyte — isoantibody reactions in dogs // Proc.
Intern. Soc. Hematol. — 1950. — P. 226-233.Zimen E. Von wolfen, hunden und kreuzungen // J. Internat. Hunde Rev. — 1969. — № 2. —
S. 16-18.Zulueta A. The hairless dogs of Madrid II Proc. VIII Congr. Genet. — 1949. — P. 687-688.Zwan J.M. van der. A hundred Dobermann dogs. — Meppel: RACMO, 1988. — 142 p.Zwan J.M. van der. A hundred Dobermann-bitches. — Meppel: RACMO, 1989. — 136 p.