


















































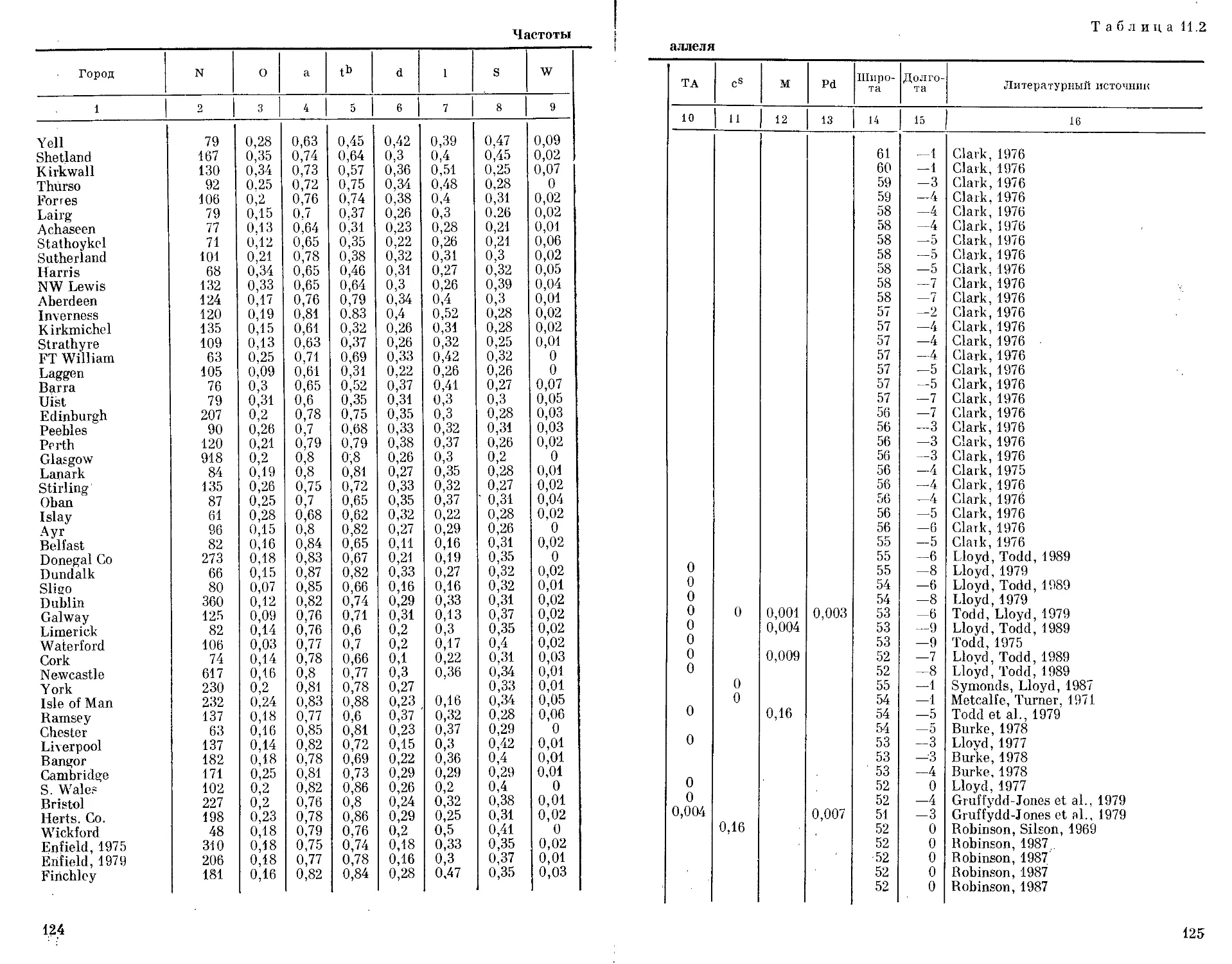
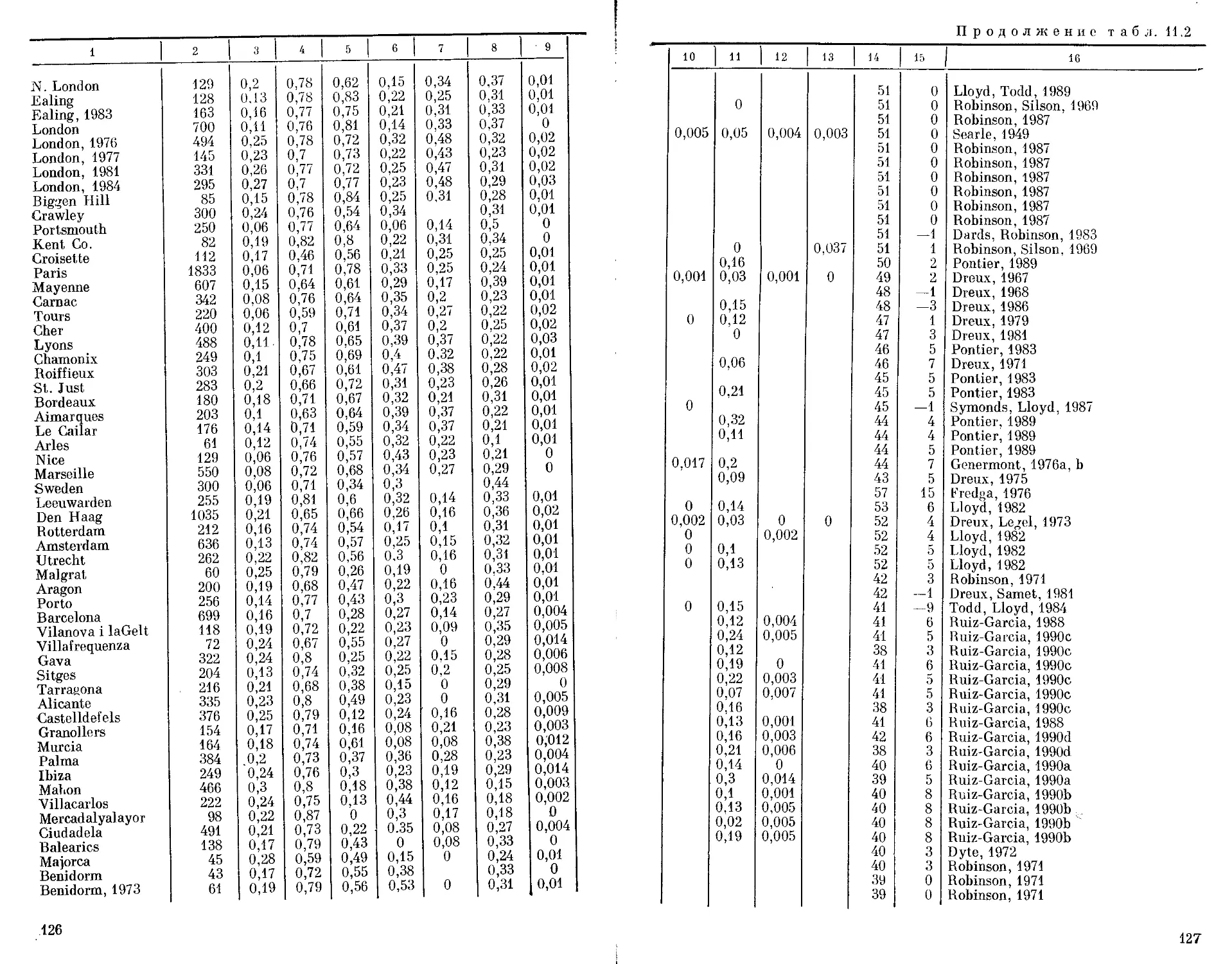
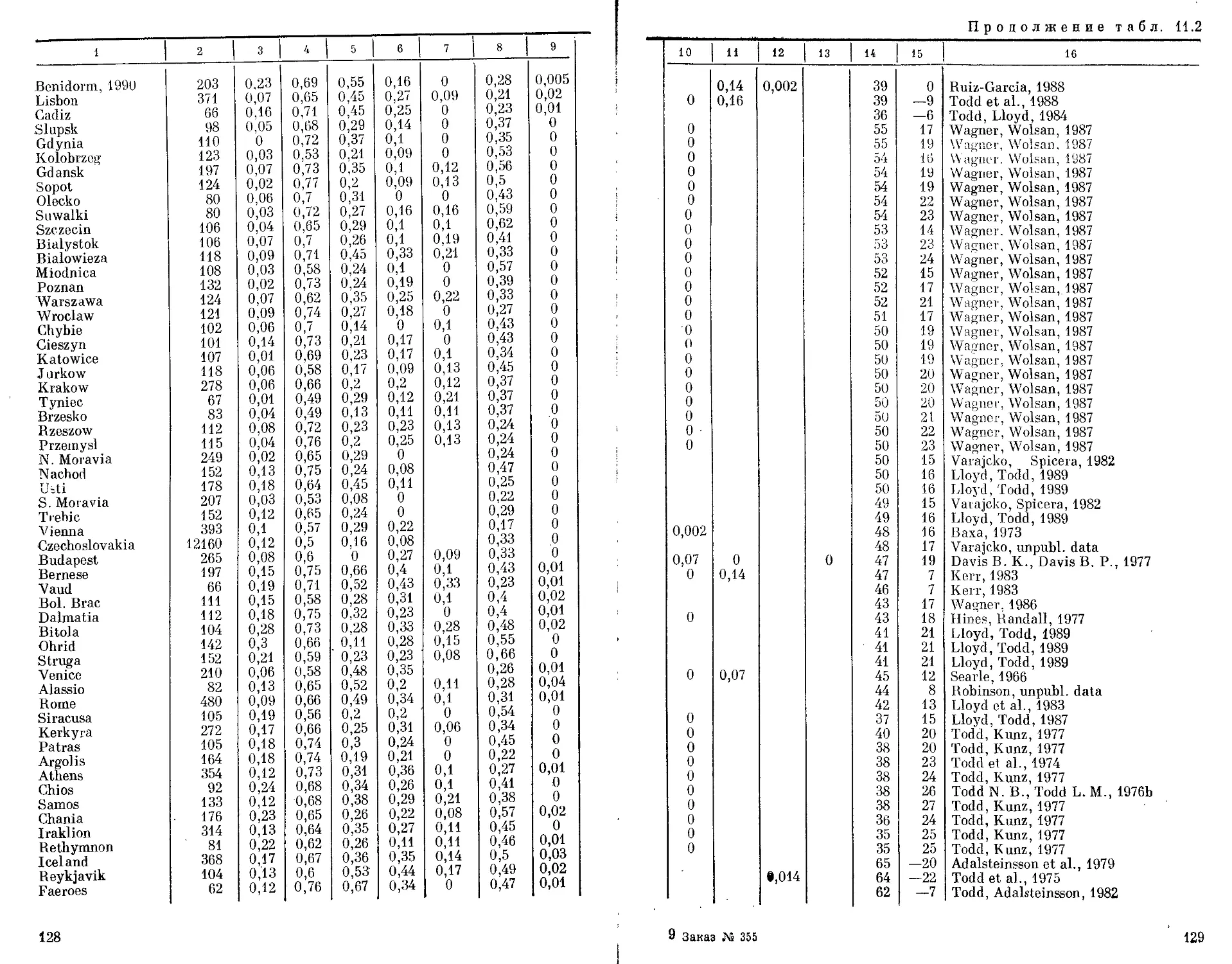
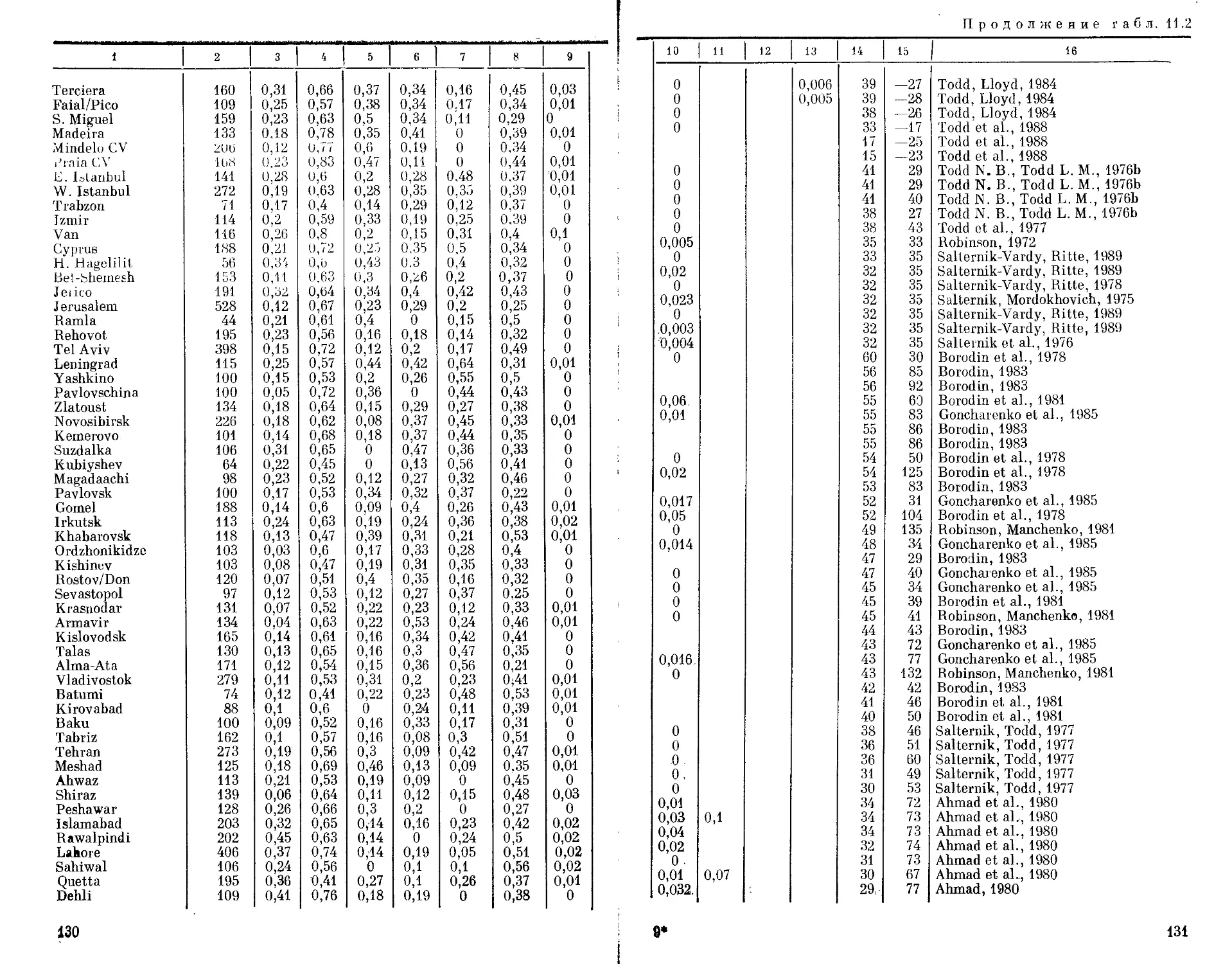
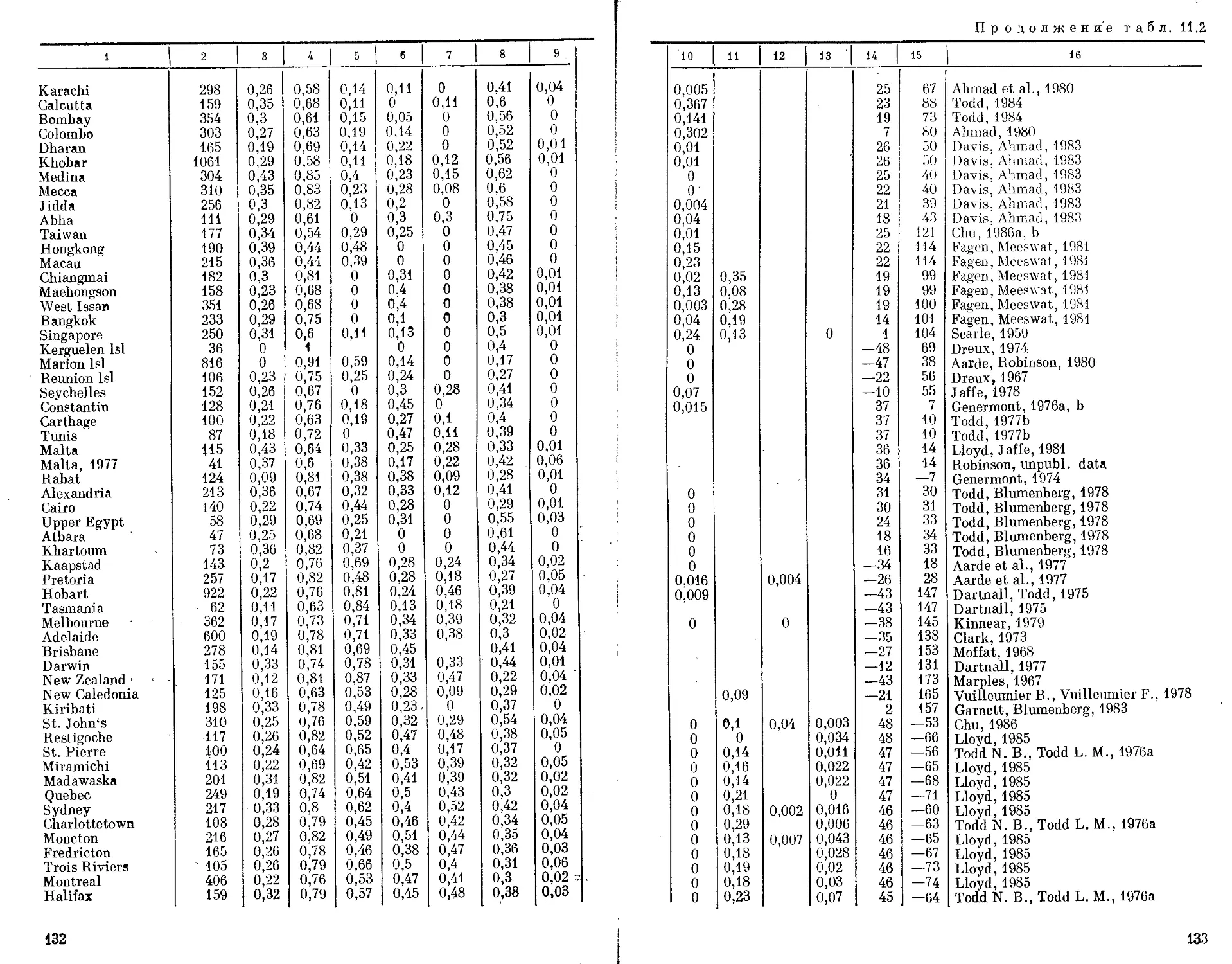
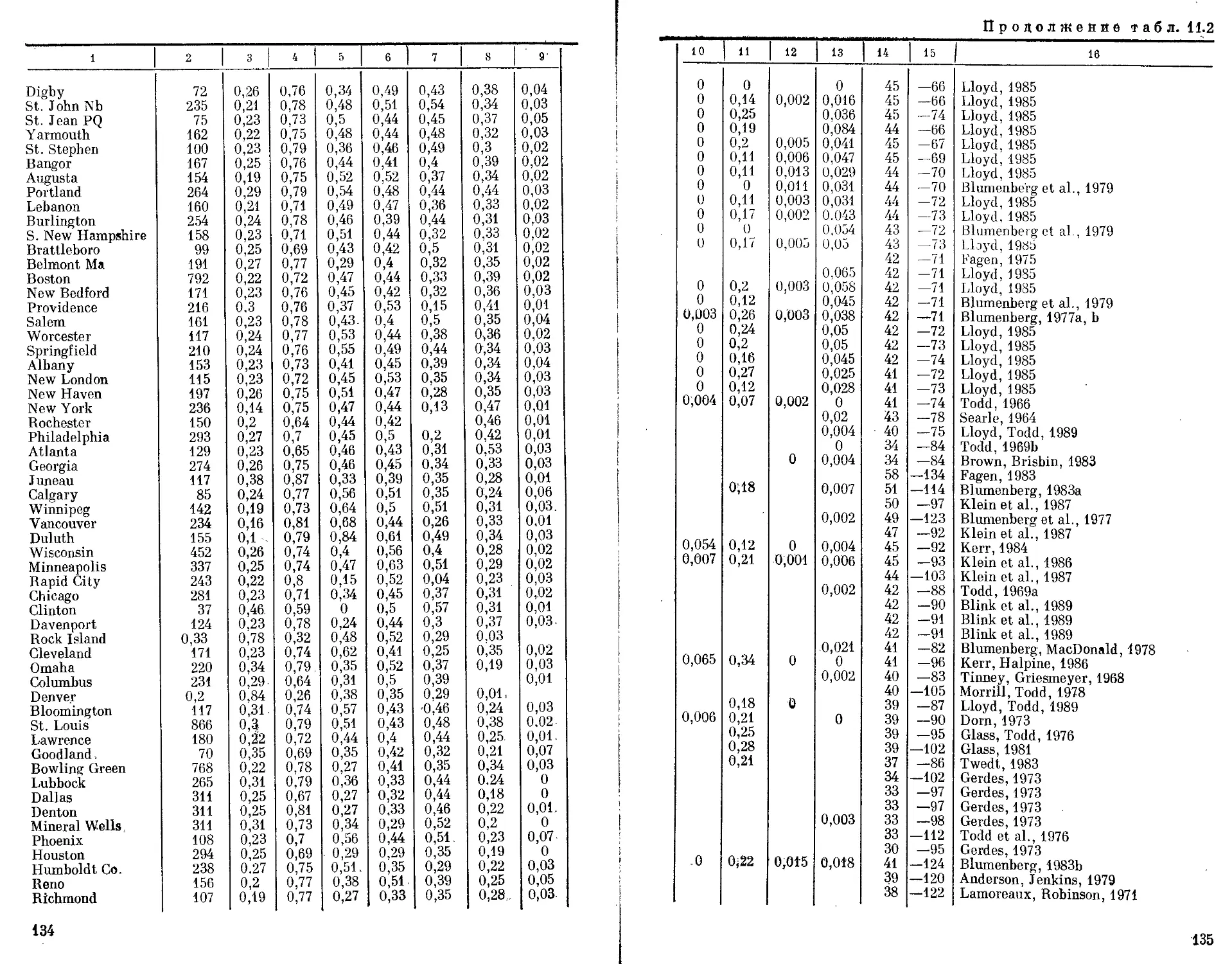






















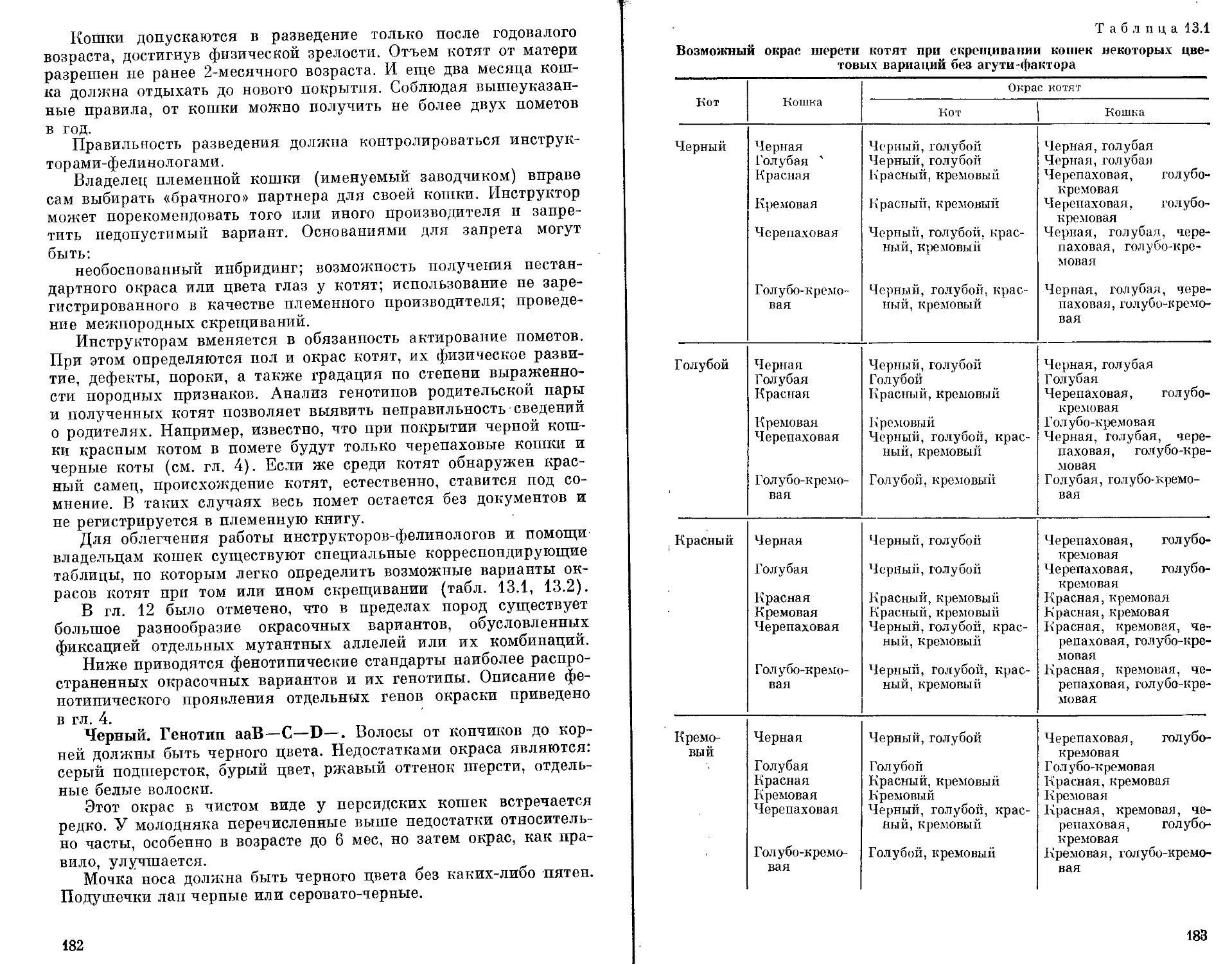














Автор: Бородин П.М. Рувинский А.О.
Теги: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез кошки биология зоология генетика
ISBN: 5-02-030567-7
Год: 1993




РОССИЙСКАЯ АКАДЕМИЯ НАУК
СИБИРСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ ЦИТОЛОГИИ И ГЕНЕТИКИ
ГЕНЕТИКА
КОШКИ
Ответственные редакторы
доктора биологических наук
П. М. Бородин, А. О. Рувинский
в
ВО «НАУКА»
НОВОСИБИРСК
1993
УДК 575.1 :636.8
Авторы
С. О’Брайен, Р. Робинсон, А. С. Графодатский, А. В. Таранин,
Т. А. Алехина, А. И. Железова, К. Клейн, О. А. Сизова
Генетика кошки/С. О’Брайен, Р. Робинсон, А. С. Графодат-
ский и др.— Новосибирск: ВО «Наука». Сибирская издательская
фирма, 1993,— 213 с.
ISBN 5-02-030567-7.
Монография представляет собой первую попытку исчерпывающего опи-
сания генетики одного вида млекопитающих — домашней кошки. Книга со-
держит сведения по эволюции и цитогенетике семейства кошачьих, полный
перечень всех известных мутаций, систематизированные данные по моле-
кулярной и биохимической генетике, генетике поведения, иммуногенетике,
популяционной генетике и биологии размножения и развития кошки. Осо-
бый интерес для любителей кошек представляют сведения об основных по-
родах кошек и принципах их разведения.
Книга будет полезна как читателям, интересующимся общими проб-
лемами генетики и эволюции животных, так и всем любителям кошек.
Табл. 9. Ил. 28. Библиогр.: 469 назв.
Рецензенты
доктор биологических наук Л. А. Васильева
кандидат биологических паук Л. В. Высоцкая
Утверждено к печати
Институтом цитологии й генетики СО РАН
1907000000 027_кб_50_191_1992
1 042(02)— 93
ISBN 5-02-030567-7
© С. О’Брайен, Р. Робинсон,
А. С. Графодатский и др., 1993
© Российская Академия наук,
1993
ПРЕДИСЛОВИЕ
«Генетика кошки» представляет собой первую книгу в заду-
манной нами серии изданий по генетике млекопитающих. Не-
смотря на постоянный интерес к генетике животных, до недав-
него времени только человек и мышь могли рассматриваться
как реальные объекты генетических исследований. Объем ин-
формации о геномах других млекопитающих, включая наиболее
распространенные виды сельскохозяйственных и домашних жи-
вотных, был невелик. Главная причина такого положения сво-
дилась к отсутствию эффективных цитогенетических и молеку-
лярно-генетических методов исследования.
Во второй половине 1970-х годов ситуация существенно из-
менилась. Появился комплекс методов, позволяющий одинаково
успешно строить генетические карты и описывать геномы даже
тех млекопитающих, которые вследствие большой протяженно-
сти их жизненного цикла, малого числа потомков, высокой стои-
мости или малой доступности объектов считались неудобными
для генетических исследований. В результате работ по сравни-
тельному картированию в последние годы сложилось представ-
ление о высокой степени гомологии геномов млекопитающих,
значительно большей, чем считали в начале 1980-х годов. Ин-
терес к сравнительно-генетическим исследованиям усилился.
Именно такой подход был взят за основу при подготовке книги
к изданию. Кроме того, мы хотели систематизировать имеющие-
ся материалы по генетике, селекции и некоторым другим свя-
занным с ними проблемам биологии кошки.
Настоящая монография является первой в отечественной
литературе попыткой детального описания генетики домашней
кошки.
Мы надеемся, что книга будет интересна для исследовате-
лей, работающих в области генетики животных, преподавателей
и студентов вузов и любителей животных.
Мы искренне признательны специалистам, откликнувшимся
на нашу просьбу принять участие в создании этой книги: Сти-
вену О’Брайену, директору лаборатории вирусного канцероге-
неза Национального института здоровья (Фредерикс, США),
Рою Робинсону, руководителю Приюта улицы Св. Стефана (Лон-
дон, Великобритания), А. С. Графодатскому, зав. лабораторией
цитогенетики животных ИЦиГ СО РАН, А. В. Таранину, зав.
лабораторией иммуногенетики ИЦиГ СО РАН, Т. А. Алехиной
и А. И. ЖелезоДой, сотрудницам лаборатории эволюционной
3
генетики ИЦиГ СО РАН (Новосибирск), Киту Клейну, про-
фессору Университета Манкато (Миннесота, США), О. А. Си-
зовой, эксперту Европейской фелинологической федерации (Мо-
сква). Большую работу по составлению и редактированию ру-
кописи провел П. М. Бородин, зав. лабораторией эволюционной
генетики ИЦиГ СО РАН. Им, а также сотрудницей этой лабора-
тории Т. Ю. Ладыгиной осуществлен перевод глав, написанных
иностранными участниками авторского коллектива. Я хотел бы
также поблагодарить коллектив ОНТИ ИЦиГ за большую ра-
боту по редактированию и подготовке рукописи к печати.
А. О. Рувинский
Глава 1
ЭВОЛЮЦИЯ СЕМЕЙСТВА КОШАЧЬИХ
И СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
ДОМАШНЕЙ КОШКИ
ВВЕДЕНИЕ
Домашняя кошка — доместицированный представитель семей-
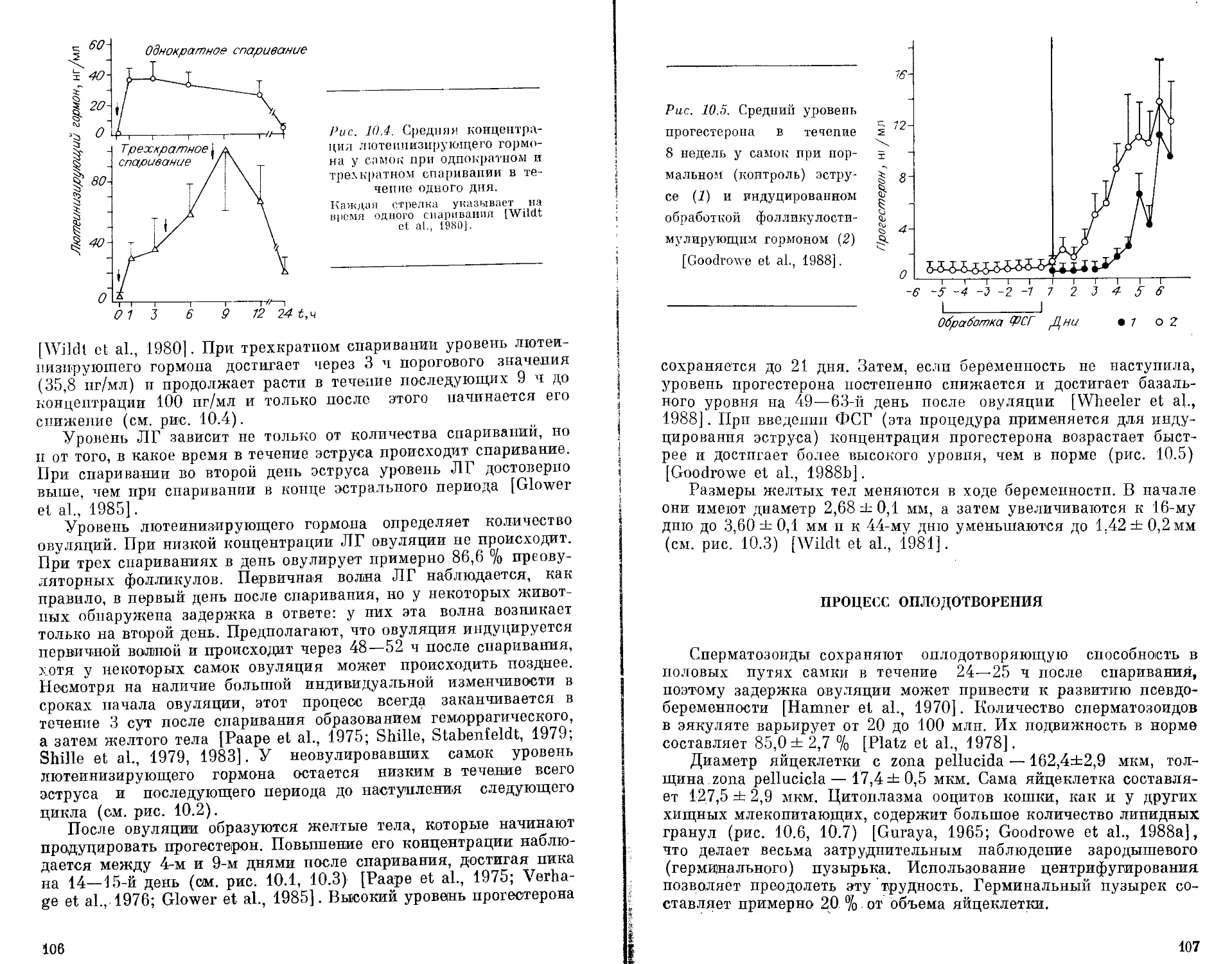
ства кошачьих. Для рассмотрения ряда вопросов генетики п фи-
зиологии вида важно ясно представлять его систематическое
положение и родственные связи с другими представителями се-
мейства. В настоящее время для решения таксономических проб-
лем активно привлекаются молекулярно-генетические подходы,
данные палеонтологии и сравнительной анатомии. Представля-
ется целесообразным дать вначале краткие сведения о суще-
стве молекулярных методов, которые в последнее время исполь-
зуются для изучения эволюционных и таксономических проблем.
МЕТОДЫ
ДНК/ДНК гибридизация. Этот метод включает в себя гибри-
дизацию радиоактивно меченой клеточной ДНК одного вида с
клеточной ДНК .другого вида и измерение стабильности фор-
мирующихся гибридных молекул ДНК. В результате этих эк-
спериментов можно получить два типа оценок: процент гибри-
дизации между видами А и Б и различия в профилях отжига
гетерологичных гибридных молекул ДНК и гомологичных ДНК.
Последняя оценка, определяемая как кТт, прямо пропорцио-
нальна степени соответствия последовательности нуклеотидов.
Оценка &Тт (или ATmR, где &Тт нормирована на процент
гибридизации) используется для составления матриц сходства
и филогенетических деревьев (дендрограмм). Использование это-
го подхода особенно эффективно для видов, дивергировавших
более 10—60 млн лет. назад. Для более поздно дивергировав-
ших видов данный метод менее пригоден [Sibley, Ahlquist, 1983;
Benveniste, 1985].
Генетические дистанции. Этот метод базируется на сравне-
нии спектров белков или изоферментов, отличающихся по элек-
трофоретической подвижности. Степень генетических различий
оценивается на основе различий в аллельных частотах для каж-
дого генного локуса. Дистанция D дает оценку числа генных
различий на локус между особями, принадлежащими к двум
популяциям. При ограничениях, налагаемых условиями самого
электрофоретического метода и темпом фиксации нуклеотидных
5
замен [Nei, 1972, 1978], дистанция между популяциями возра-
стает пропорционально времени их дивергенции от общего пред-
ка. Большинство оценок дистанций для представителей семей-
ства кошачьих было получено при анализе около 50 локусов,
однако при использовании двухмерного электрофореза анализи-
ровалось более 300 генных продуктов [Goldman et al., 1987].
Надежность использования дистанций для оценки времени
дивергенции зависит от набора локусов, включенных в анализ,
так как разные белки эволюционируют с разными скоростями.
Наиболее надежные оценки этот метод дает при времени дивер-
генции от 0,1 до 10 млн лет назад. За пределами этих времен-
ных границ связь между D и эволюционным временем стано-
вится нелинейной.
Иммунологические дистанции альбумина. Этот метод позво-
ляет оценивать иммунологические дистанции между видами. Он
базируется на учете замен аминокислот в гомологичных белках.
Замены регистрируются по смещению титрационной кривой в
реакции связывания комплемента. Несколько кроликов иммуни-
зируются очищенным сывороточным альбумином вида А. Ан-
тисыворотка, полученная от таких кроликов, титруется на об-
разцах альбумина вида Л. Для определения эволюционной ди-
станции альбумин вида Б преинкубируется с титрованной анти-
сывороткой против вида А. Связанная антисыворотка затем
вновь тестируется на альбумин гомологичного вида А. Остав-
шиеся антитела образуют комплекс с антигеном вида А и свя-
зывают комплемент в объеме, количественно пропорциональном
различиям в аминокислотах между двумя видами. Используя
антитела против разных видов, можно построить матрицу им-
мунологических дистанций и использовать ее для установления
родства.
Как и ДНК/ДНК гибридизация, данный метод полезен для
оценок далеко разошедшихся видов (5—50 млн лет назад).
Это связано с тем, что альбумин эволюционирует достаточно
медленно: одна единица иммунологической дистанции за 0,6 млн
лет. Время дивергенции 1—2 млн лет находится па нижнем
пределе разрешающей способности этого метода.
ТАКСОНОМИЯ и эволюция КОШАЧЬИХ
В гл. 3 будет подробно рассмотрена кариотипическая эволю-
ция кошачьих. Следует сказать, что кошачьи, как и представи-
тели отряда хищных, достаточно консервативны в отношении
эволюции кариотипов. Сами кошачьи, по-видимому, наиболее
близки к исходному кариотипу.
Между таксономистами нет согласия относительно родовых
подразделений в семействе кошачьих, состоящем из 37—40 ви-
дов (табл. 1.1). Одни выделяют 19 родов [Ewer, 1973], а дру-
гие — только 2 [Romer, 1968].
6
Таблица 1.1
Современные виды семейства кошачьих
Сокращенное название вида Латинское название Русское название
AJU Acinonyx jubatus Гепард
FGA Fells catus Кошка домашняя
LPA Leopardus pardalis Леопард
LSE Leptailurus serval Сервал
LCA Lynx canadensis Рысь канадская
NNE Neofelis nebulosa Леопард облачный
PLE Panthera leo Лев
PTI Panthera tigris Тигр
РТЕ Profells temmincki Кошка Темминка
РСО Puma concolor Пума
ССА Caracal caracal Каракал
FCH Fells chaus Кошка камышовая
FLI Fells libyca Кошка ливийская
FMA Fells margarita Кошка барханная
FNI Fells nigripes Кошка черноногая
FSI Fells Silvestris Кошка дикая
HYA Herpailurus yagouaroundi Ягуарунди
IPL Ictailurus plantieeps Кошка суматранская
LWE Leopardus wiedi Кошка американская
LTI Leopardus tigrina Онцилла
LGE Leopardus geoffroyi Кошка Жоффруа
LCO Lynchailurus colocolo Кошка пампасская
LLY Lynx lynx Рысь
LRU Lynx rufus Рысь красная
OGU Oncifelis guigna Кошка чилийская
ОМА Otocolobus manul Манул
PON Panthera onca Ягуар
РРА Panthera pardus Леопард
PUN Panthera uncia Снежный барс
РМА Pardofelis marmorata Кошка мраморная
РВА Pardofelis badia Кошка калимантанская
РВЕ Prionaillurus bengalensis Кошка бенгальская
PVI Prionaillurus viverrinus Кошка-рыболов
PAU Profelis aurata Кошка золотая
FBI Fells bieti Кошка китайская
LPA Lynx pardina Рысь леопардовая
MIR Mayailurus iriomotensis Кошка ириомотейская
OJA Oreailurus jacobita Кошка йокобатская
PRU Prlonailurus rubiginosa Кошка пятнисто-рыжая
Предпринята попытка решить эту проблему с использова-
нием молекулярно-генетических подходов. По альбуминовым ди-
станциям выделяются три главные ветви древа кошачьих:
1) ветвь оцелота и малых южно-американских кошек; 2) линия
7
Миоцен
Плейстоцен
Плиоцен
I I I
30
Ч-
.25
25 20 75
-Н-----'—,---L-
20 75
, ।_____ ।______|Млн лет
70 5 ИД
Г Ле1 ¥LEL РРА. PON. PUN
рм
r L Рысь(1СА)---LLY.LRU
------------HYA
— Гепард (AJU)
,____________СМ
Сервал (LSE)
,-------------1PL.PVI.PBE
г *—1— Облачный леопард (NNE)
>-------------- PAU.PBA
----- -----КошкаТемминка (РТЕ)
------ Пума (РСО)
Ветвь
пантер
---------- FMA.FN1 ’
।-------— FLI
Кошка домашняя (рСА Щ г
Ветвь домаш-
ней кошки
тт Оцелот (LPA)
l2zzzzzz
LCO.LV7I.LT1
OGU.LGE J
Ветвь оцело-
та
Гиена
Рис. 1.1. Филогенетическое древо кошачьих, построенное на основе подсче-
та реципрокных иммунологических дистанций десяти антисывороток, при-
готовленных против видов, названия которых даны полностью. Остальные
виды (обозначения см. в табл. 1.1) добавлены к древу на основании оценок
«односторонних» иммунологических дистанций.
ИД — иммунологическая дистанция.
домашней коптки и малых средиземноморских кошек; 3) ветвь
пантер, состоящая из малых и больших кошек (рис. 1.1). Вре-
мя дивергенции этих ветвей от общего ствола определяется при-
близительно в 10 млн лет.
ДНК/ДНК гибридизация позволяет выявить две ветви. Наи-
более эффективно меченая ДНК домашней кошки гибридизу-
ется с ДНК кошек той же ветви. Соответственно ДНК африкан-
ского льва — с ДНК видов ветви пантер. ,
Рассмотрим более подробно ветви древа кошачьих.
Ветвь оцелота. Эта группа малых южно-американских кошек
отделилась от общего ствола кошачьих довольно давно: около
10 млн лет назад, В то же время дивергенция видов в пределах
этой ветви произошла относительно недавно — в последние 1—
2 млн лет. Ветвь оцелота представлена 6 видами, эндемичными
для Южной Америки. По оценкам таксономистов она состоит
минимум из одного и максимум из трех родов [Hemmer, 1978;
Nowak, Paradiso, 1983]. Крупные южно-американские кошки
(ягуарунди, ягуар и пума) в нее не входят. Сравнительная
кариология также дает подтверждения такой группировке (ом.
8
гл. 3). Кошки ветви оцелота имеют 36 хромосом, тогда как все
остальные — 39. Их уникальная хромосома СЗ образовалась за
счет центрического слияния двух акроцеитриков группы F, ха-
рактерных для других кошек.
Две важные особенности этой группы: их ранняя диверген-
ция от общего ствола и поздняя дивергенция внутри ветви. Со-
гласно палеонтологическим данным, милые кошки появились в
Северной Америке 13—15 млн лет назад [Savage, Russell, 1983;
Werdelin, 1985]. Позднее в плиоцене и плейстоцене (0,5—5 млн
лет назад) в Северной Америке обитали малые кошки, похожие
на современных южно-американских [Kurten, Anderson, 1980;
Werdelin, 1985]. Видимо, именно они были предками кошек ли-
пни оцелота и не оставили других потомков в Северной Америке
и Старом Свете.
Современная радиация малых южно-американских кошек сов-
падает с образованием Панамского перешейка 2—3 млн лет
назад. До этого времени Южная Америка была изолирована
от проникновения наземных плацентарных хищников ] Patterson,
Pascual, 1972]. Поэтому быстрая дивергенция малых южно-
американских кошек была ускорена ослаблением конкуренции
с исходной североамериканской фауной. Более того, изоляция
и дивергенция могла быть ускорена ледниковой фрагментацией
фауны в плейстоцене [Gingerich, 1984]. Некоторые морфологи
считают, что основным направлением дивергенции малых кошек
этой ветви была неотеническая фиксация ювенильных признаков.
Ветвь пантер. Остальные виды кошачьих образуют гетеро-
генную группу. Они дивергировали в различное время в тече-
ние последних 5—7 млн лет (см. рис. 1.1). Наиболее поздно-
дивергировали 5 видов «рычащих» кошек рода Panthera (1—
2 млн лет назад). Довольно тесно к этой группе примыкают
рысь и мраморная кошка. Все эти виды имеют идентичные ка-
риотипы, хотя у морфологов сближение рыси с пантерами вы-
зывает возражения.
Иммунологический анализ не позволяет уточнить родствен-
ные отношения между представителями ветви пантер. Эта ветвь,
не разделимая иммунологически, включает в себя такие морфо-
логически отличные виды, как пума, золотистая кошка, гепард,
сервал, каракал и ягуарунди.
Независимые данные по дивергенции этих видов были полу-
чены при подсчете генетических дистанций, основанных на элек-
трофоретических данных (рис. 1.2) [O’Brien et al., 1987]. Этот
подход обладает более высокой разрешающей способностью.
На основе данных такого анализа можно предполагать, что
ветвь, ведущая к роду Panthera, отделилась от общего ствола
2—3 млн лет назад. До этого произошло отделение линий снеж-
ного барса (3 млн лет назад) и облачного леопарда (5—6 млн
лет назад). Внутри рода Panthera ранее всего дивергировали аф-
риканский лев и тигр, а затем ягуар и леопард. Три вида тиг-
ров разошлись друг с другом совсем недавно (0,1—0,2 млн лет
9
Миоцен
15 10
I--------'---1-------------г^~
0,5 0,4- 0,3
Плейстоцен
Плиоцен
_____Млн лет
0 г Д
г Бенгальский
004 тигр
- —I- Уссурийский тигр
„ L Суматранский
0,07 тигр
---- Лее
0,02 п
р— Ягуар
0,0^2?^^
Снежный барс
Облачный леопард
0,06
0,16
Золотая кошка
Гепард
0,17
Каракал
Пума
Гиена
Рис. 1.2. Филогенетическое древо кошачьих, построенное на основе подсче-
та неевских генетических дистанций, полученных при анализе спектров
изоферментов.
Д — генетическая дистанция.
назад). Другие четыре изученных вида (золотистая кошка, ге-
пард, каракал и пума) дивергировали от труппы около 8 млн
лет назад. Они так же сильно разошлись друг с другом, как
и с остальными кошками рода Panthera.
Морфологические и генетические данные также свидетель-
ствуют о тесной родственной связи крупных кошек. Лев, тигр,
леопард и ягуар имеют не полностью остифицированный гиоид,
что позволяет им рычать и объединяет в группу «рычащих» ко-
шек [Nowak, Paradiso, 1983]. Показано, что снежный барс,
единственный нерычащий член этого рода, наиболее сильно от-
личается от рода пантер и сохраняет ряд признаков, сходных с
пумой п ягуаром, однако его гиоид структурно похож на гиоид
пантер [Herrington, 1983]. Помещение малой южно-американ-
ской кошки ягуарунди в ветвь пантер, сделанное на основе мо-
лекулярно-генетических данных, также подтверждается морфо-
логическими данными [Werdelin, 1981].
Наиболее ранние находки представителей видов ветви пан-
тер относятся к периоду 2 млн лет назад и совпадают с моле-
кулярной оценкой времени дивергенции [Kurten, 1968; Savage,
1978; Kurten, Anderson, 1980; Savage, Russell, 1983]. Хеммер
[Hemmer, 1976] утверждает, что радиация пантер произошла
1Ь
в два этапа: сначала ягуароподобные формы распространились
в Европе, Азии, Африке и Северной Америке в раннем плейсто-
цене и затем произошла дифференциация льва, тигра, леопарда
и ягуара. Древо, построенное по молекулярным данным, согла-
суется с этой гипотезой, но с поправкой на то, что леопард
и ягуар разделились более поздно и более родственны друг дру-
гу, чем другим большим кошкам. Исторически их ареалы не
перекрываются: леопард обитает в Старом Свете, а ягуар —
в Новом Свете. Наиболее ранние находки ягуара в Южной Аме-
рике (Боливия) относятся к среднему плейстоцену (0,5 млн лет
назад) [Hemmer, 1976]. В Северной Америке ягуар обнаружен
гораздо раньше (1,5 млн лет назад) [Hemmer, 1976; Kurten,
Anderson, 1980].
Африканский лев появился в Европе и Африке около 0,5
млн лет назад [Kurten, 1968; Savage, 1978]. Его родство с
тигром довольно трудно оценить из-за большого морфологиче-
ского сходства данных таксонов [Neff, 1982]. Другие пантеры
имеют более бедную геологическую летопись. Снежный барс об-
наружен в позднем плейстоцене. Облачный леопард обнаружен
па Яве в раннем плейстоцене, хотя по молекулярным данным
время его дивергенции относится к позднему миоцену [Hem-
mer, 1976].
Ископаемые останки рыси обнаруживаются в среднем плио-
цене (3—4 млн лет назад) [Kurten, 1968]. Первые рыси мор-
фологически более сходны с ветвью домашней кошки, чем с
ветвью пантер, в которую рысь попадает по молекулярным дан-
ным. Однако рысь и ягуарунди имеют ряд сходных морфологи-
ческих признаков [Werdelin, 1981], что подтверждает их сбли-
жение на молекулярном древе.
Каракал и сервал впервые обнаружены в раннем плейсто-
цене [Savage, 1978], что согласуется с предложенным временем
дивергенции. Пума и гепард имели общего предка, как следует
из молекулярного древа. Впервые гепард обнаружен 3 млн лет
назад, но его линия может быть продлена до 4—5 млн лет
[Adams, 1979; Kurten, Anderson, 1980]. Последняя дата более
соответствует молекулярным данным.
Ветвь домашней кошки. Виды рода Felis (малые кошки) про-
исходят, по-видимому, от предков, обитавших в Средиземномор-
ском бассейне [Kurten, 1968]. Результаты анализа иммуноло-
гических дистанций позволяют группировать эти виды в кла-
стер (скорее всего, монофилетический, который мы далее будем
называть ветвью домашней кошки). В отличие от ветви оце-
лота, в данной ветви дивергенция продолжалась гораздо доль-
ше (см. рис. 1.1). Согласно иммунологическому древу, прежде
всего дивергировал манул, затем камышовая кошка, а позже —
остальные пять видов [Nowak, Paradiso, 1983].
Эти эволюционные реконструкции подкрепляются морфоло-
гическими и поведенческими данными [Hemmer, 1976, 1978;
Herrington, 1983]. По кариологии шесть видов абсолютно иден-
11
тичвы. Манул несколько отличается от них по морфологии хро-
мосомы Е4 (см. гл. 3).
Важным подтверждением монофилии рода Felis служат дан-
ные по распределению ретровирусов. Домашняя кошка имеет
по крайней мере два семейства ретровирусов FeLV (8—10 копий) и
RD-114 (около 20 копий), обнаруженных в нормальных образцах
ДНК от всех исследованных представителей. Проведен анализ эво-
люционной истории этих семейств [Benveniste, Torado, 1974;
Benveniste et al., 1975].
Исследователи тибридизовали клонированные последователь-
ности RD-114 с клеточной ДНК многих видов млекопитающих.
Хотя клеточная ДНК всех исследованных видов кошачьих по-
казывает почти 90%-ю гомологию с ДНК домашней кошки,
только у пяти видов (все они принадлежат к роду Felis): до-
машней, барханной, черноногой, камышовой и ливийской ко-
шек — обнаружены последовательности RD-114. Кроме того, по-
следовательность, родственная RD-114, обнаружена у обезьян
Старого Света. Этот результат свидетельствует, что интеграция
рет]ровируса приматов в кошачий геном произошла до диверген-
ции пяти видов рода Felis, но после отделения манула от этой
ветви. Приблизительно в то же время произошла интеграция
п FeLV, который отдаленно сходен с ретровирусом грызунов
Старого Света и обнаруживается среди фелид только у предста-
вителей ветви домашней кошки. Данные по ретровирусам хо-
рошо согласуются с гипотезой монофилетического происхожде-
ния данной ветви.
По палеонтологическим данным наиболее ранний образец ро-
да Felis в Старом Свете датируется 3 млн лет назад [Kurten,
1965], что хорошо согласуется с молекулярным древом. Но пер-
вые костные останки манула обнаружены в среднем плейстоце-
не (1 млн лет назад), или позднее, чем следует из иммунологи-
ческого древа. Европейская дикая кошка несколько старше, её
первые формы обнаруживаются в нижнем плейстоцене. Ископае-
мые останки других вадов менее изучены. ;
Таким образом, линия домашней кошки характеризуется воз-
никновением малых, морфологически сходных кошек в ходе
постепенной эволюции. Эволюция этих малых кошек происхо-
дила в Средиземноморском бассейне и в Азии, но некоторый
виды успешно расселялись по другим районам, достигая Юж-
ной Африки (черноногая кошка), холодных степей (манул) и
пустыйь (барханная кошка).
РОДСТВО КОШАЧЬИХ
С ОСТАЛЬНЫМИ ХИЩНЫМИ И ЧЕЛОВЕКОМ
Результаты ДНК/ДНК гибридизации и палеонтологические
данные показывают, что кошачьи отделились от других фелоид-
ных хищных (генеты, циветты, мангусты, гиены) около 40 млн
Рис. 1.3. Филогенетическое древо хищных, построенное на основе анали-
за термостабильпости ДНК/ДНК гибридов (/\TmIl, °C).
лет назад (рис. 1.3). Однако, как указано выше, радиация ко-
шачьих происходила гораздо позднее (около 10 млн лет назад).
Дивергенция фелоидных и каноидных (собачьи, скунсы, куни-
цеобразные, еноты, медвежьи, ластоногие) хищных имела место
в раннем эоцене (около 55 млн лет назад).
Последний общий предок кошки и человека существовал око-
ло 100 млн лет назад. Тем не менее кариотипическое сходство
кошки и человека составляет около 25 %. Некоторые хромосомы
кошки как по характеру G-бэндинга, так и по набору генов
сходны с хромосомами человека (см. гл. 3).
Сходство обнаруживается и по ряду последовательностей он-
когенов, найденных в геномах кошки и человека. Так, у чело-
века есть пять версий ras-онкогена: три функциональных и два
псевдогена. Один из псевдогенов (HRAS2) локализован в Х-хро-
мосоме. У кошки также три функциональных онкогена нахо-
дятся в синтенных группах аутосом,, один — ras-подобный сег-
мент ДНК (по-видимому, псевдоген) — в Х-хромосоме [O’Brien,
1986]. Кроме того, характерная для позвоночных версия онкоге-
на ets представляет собой два неперекрывающихся транскрип-
ционно активных гена, локализованных в разных хромосомах
у мыши, кошки и человека. Эти два гена, ETS-1 и ETS-2, на-
13
годятся в гомологичных синтечгных группах у всех трех видов.
Это свидетельствует о том, что разделение на два гена произош-
ло после дивергенции млекопитающих и птиц, но до начала ди-
вергенции самих млекопитающих [Watson et al., 1986]. Следуя
топ же логике, можно предположить, что транспозиция псевдо-
гена ras в Х-хромосому имела место у общего предка хищных
и приматов.
ЗАКЛЮЧЕНИЕ
Уже сейчас генетика кошки и ее диких сородичей достигла
весьма значительных успехов. Однако дальнейшее применение
на кошке методов, разработанных при изучении геномов мыши
и человека, открывает широкие перспективы. В нашей лабора-
тории * разрабатываются методы манипуляций с эмбрионами ко-
шек. Мы надеемся применить их для коррекции врожденных
аномалий, весьма сходных с наследственными болезнями чело-
века (ом. гл. 6). Мы планируем использовать клонированные
гены кошки и технологию культивирования и переноса эмбрио-
нов. Поскольку карта генома кошки становится все более под-
робной (см. гл. 3), цитологический анализ различных злокаче-
ственных опухолей кошек даст возможность следить за поведе-
нием онкогенов. Наконец, углубление генетического анализа се-
мейства кошачьих дает уникальную возможность вскрыть
и понять механизмы удивительных адаптаций, возникших за ве-
ка эволюционной специализации.
СПИСОК ЛИТЕРАТУРЫ
Adams D. В. The cheetah: Native American Ц Science — 1979.— Vol. 205.—
P. 1155-1158.
Benveniste R. E. The contributions of retroviruses to the study of mammalian
evolution Ц Molecular Evolutionary Genetics/Ed. R. J. MacIntyre.—
N. Y.: Plenum, 1985,— P. 359-417.
Benveniste R. E., Sherr C. J., Todaro G. J. Evolution of type C viral genes:
Origin of feline leukemia virus Ц Science.— 1975.— Vol. 190.— P. 886—
888.
Benveniste R. E., Todaro G. J. Multiple divergent copies of endogenous C-type
virogenes in mammalian cells Ц Nature.— 1974,— Vol. 252.— P. 170—
173.
Ewer R. F. The Carnivores.— Ithaca; New York: Cornell Univ. Press, 1973.
Gingerich P. G. Pleistocene extinctions in the context of origination — extinc-
tion equilibria in cenozoic mammals Ц Quaternary Extinctions/Ed. P. S.
Martin, R. G. Klein.— Tucson: Univ. Arisona Press, 1984.— P. 211—222.
Goldman D., Rathnagiri P., O'Brien S. J., Giri P. R. The molecular phylogeny
of the ursids as indicated by two-dimensional electrophoresis Ц Proc.
Natl. Acad. Sci. USA.— 1987.—Vol. 84,-P. 3307-3311. _
Hemmer H. Fossil history of the living Felidae Ц The World's Cats. Vol. 3,
N 2. Contributions to Biology, Ecology, Behavior and Evolution/Ed.
R. L. Eaton.— Seattle: Carnivore Research Institute, 1976.— P. 1—14.
* Лаборатория вирусного канцерогенеза (Национальный раковый ин-
ститут, Бетесда, США).
14
Hemmer H. The evolutionary systematics of the living Felidae. Present sta-
tus and current problems Ц Carnivore.— 1978.— Vol. 1.— P. 71—79.
Herrington S. J. Systematics of the Felidae: A quantitative analysis.— M. S.
thesis, Univ. Oklahoma, Norman, 1983.— 136 p.
Kurten B. On the evolution of the European wild cat, Felis silvestris Schre-
ber Ц Acta Zool. Fennica.— 1965.— Vol. 111.— P. 1—29.
Kurten B. Pleistocene mammals of Europe.— Chicago: Aldine, 1968.
Kurten B., Anderson E. Pleistocene Mammals of North America.— N. Y.: Co-
lumbia Univ. Press, 1980.
Neff N. A. The Big Cats.— N. Y.: Abrams Inc., 1982.
Nei M. Genetic distances between populations Ц Amor. Nat.— 1972,—
Vol. 106.— P. 283-292.
Nei M. Estimation of average heterozygosity and genetic distance from a small
number of individuals Ц Genetics.— 1978.— Vol. 89.— P. 583—590.
Nowak R. M., Paradiso J. L. Walker’s Mammals of the World.— 4th ed.—
Baltimore: Johns Hopkins Univ. Press, 1983.— Vol. 2.
O'Brien S. J. Molecular genetics in the domestic cat and its relatives Ц Trends
in Genetics.— 1986.— Vol. 2.— P. 137—142.
O'Brien S. J., Collier G. E., Benveniste R. E. et al. Setting the molecular clock
in Felidae: The great cats, P<rnthera Ц Tigers of the World/Ed. R. L. Til-
son.— Park Ridge; New York: Noyes Publications, 1987.— P. 10—27.
Patterson B., Pascual R. The fossil mammal: Fauna of South America Ц Evo-
lution, Mammals and Southern Continents/Ed. A. Keast, F. C. Erk,
B. Blass.— Albany: State Univ. New York Press, 1972.— P. 247—309.
Romer A. S. Notes and comments on vertebrate paleontology.— Chicago: Univ.
Chicago Press, 1968.
Savage D. E., Russell D. E. Mammalian paleofaunas of the world.— L.: Addi-
son-Wesley, 1983.
Savage R. J. G. Carnivora Ц Evolution of African Mammals/Ed. V. J. Maglio,
H. B. S. Cooke.— Cambridge: Harvard Univ. Press, 1978.— P. 249—267.
Sibley C. G., Ahlquist J. E. Phylogeny and classification of birds based on the
date of DNA—DNA hybridization / Current Ornithology/ Ed. R. F. John-
ston.— N. Y.: Plenum Press, 1983.— Vol. 1.— P. 245—288.
Watson D. K., Smith M. J., Kozak C. et al. Conserved chromosomal position
of dual domains of the ets protooncogene in cats, mice, and humans /
Proc. Natl. Acad. Sci. USA.— 1986,- Vol. 83,— P. 1792—1796.
Werdelin L. The evolution of lynxes / Ann. Zool. Fennici.— 1981.— Vol. 18.—
P. 37—71.
Werdelin L. Small Pleistocene felines of North America / J. Vert. Paleo.—
1985 —Vol. 5.—P. 194—210.
ПРИЛОЖЕНИЕ. МЕЖВИДОВЫЕ ГИБРИДЫ В СЕМЕЙСТВЕ КОШАЧЬИХ
(Р. Робинсон, пер. Т. Ю. Ладыгиной)
В тех случаях, когда различие в размерах тела не создает
препятствия для спаривания, многие виды семейства кошачьих
могут давать гибридное потомство. Едва ли гибридизация может
осуществляться естественным путем в природе, даже если зоны
обитания двух или большего числа видов перекрываются, по-
скольку огромное значение имеют также половое и обществен-
ное поведение, которые у разных видов значительно отличаются
друг от друга. Совсем иная ситуация в неволе, где представите-
15
ли разных пород имеют возможность привыкнуть друг к другу,
в результате чего обычное половое поведение меняется.
В 1972 г. Греем были собраны все имеющиеся литературные
данные, касающиеся гибридов семейства Felidae [Gray, 1972].
Большинство представителей крупных видов могут скрещивать-
ся друг с другом, давая жизнеспособное и часто очень сильное
потомство. Примером могут служить тигоны (гибриды тигра и
львицы), лигеры (льва и тигрицы) и леопоны (леопарда и льва).
Такие гибриды хорошо известны и появление их — не редкость.
Значительно меньше известны гибриды леопарда и ягуара, пу-
мы и тигра, льва и ягуара, ягуара и пумы. В большинстве слу-
чаев вопрос о фертильности гибридов остается открытым. Как
правило, гибридные самцы стерильны, а самки могут быть по-
луфертильными, как, например, у тигонов и лигеров [Robinson,
1976].
Подобная картина наблюдается и для более мелких видов.
Например, описаны гибриды домашней кошки и рыжей рыси;
черноногой леопардовой и тигровой кошек; камышовой и лео-
пардовой кошек [Leyhausen, Tonkin, 1966; Gray, 1972]. В этих
случаях также нет убедительных доказательств фертильности
большинства из перечисленных гибридов. Наибольший интерес
представляет гибрид домашней и тигровой кошек (онциллы),
поскольку у первой в кариотипе насчитывается 38 хромосом,
а у второй — 36 [Leyhausen, Falkena, 1966]. Несмотря на столь
серьезное кариотипическое различие, было получено семь гиб-
ридных потомков. Три из них — мертворожденные (пол не оп-
ределен), четыре оставшиеся в живых оказались кошками. Вы-
борка, конечно, очень мала, но и по ней можно сделать вывод
о том, что гибридные самцы менее жизнеспособны, чем самки.
Все гибриды оказались стерильными.
Описано несколько случаев гибридизации между домашней
и европейской кошками. Все потомки фертильны, хотя, по-ви-
димому, более чувствительны к заболеваниям, чем домашние
кошки. Гибриды домашней и степной кошек (F. libyca caudata),
как правило, также фертильны. Были получены потомки от
скрещивания европейской дикой кошки с различными подвида-
ми F. libyca, но информации о плодовитости гибридов пока не-
достаточно. Скрещивание между домашней и камышовой кош-
ками приводит к появлению жизнеспособного потомства обоих
полов, причем гибридные самки фертильны [Jackson J. М., Jack-
son J., 1967]. Плодовитость гибридов домашней и европейской
кошек или домашней кошки и африканской дикой кошки впол-
не естественна, поскольку оба эти вида рассматриваются как ди-
кие сородичи домашней кошки. Однако фертильность гибридов
домашней и камышовой кошек вызывает удивление. Вероятно,
эти две формы более тесно связаны между собой, чем предпо-
лагалось ранее (см. гл. 1).
Суммируя все вышеизложенное, есть основания думать, что
несмотря на существующие физические барьеры, все виды се-
16
мейства Felidae способны давать жизнеспособное потомство. Ка-
риотипы их, по-видимому, достаточно сходны для того, чтобы
обеспечить нормальное развитие организма (т. е. протекание
митоза), но недостаточно для того, чтобы во всех случаях мейоз
был завершен до конца.
Некоторые наблюдения за гибридными фенотипами касают-
ся формирования рисунка на шерсти [Robinson, 1976]. Полосы
на шерсти тигра значительно отличаются от рисунка, характер-
ного для других представителей семейства кошачьих, поскольку
для последних наиболее характерны пятна и разводы. Имею-
щиеся данные свидетельствуют о том, что тигровый характер
окрашивания наследуется как доминантный по отношению к
леопардовой пятнистости или отсутствию рисунка, наблюдаемо-
му у льва. У гибридных потомков тигра выявляются четкие по-
лосы, которые, правда, могут быть менее интенсивно окрашены,
не так явно выражены или переходить на отдельных частях
тела в пятна. Трудно оценить, сколько генов отвечает за фор-
мирование рисунка, поскольку до сих пор не удавалось получить
поколение F%. Исходя из того, что полосатый рисунок в поко-
лении Fi менее ярко выражен, чем у тигра, логично предполо-
жить, что в детерминацию этого признака вовлечен целый комп-
лекс генов, который, вероятно, отсутствует у леопарда и льва.
В том случае, если предложенная выше гипотеза о моногенном
наследовании, контролирующем развитие окраски у кошачьих,
верна, можно предположить, что полосы, в отличие от пятен,
возникают благодаря простому аллельному различию.
У потомков, полученных от скрещивания леопарда и льва,
наблюдается леопардовый пятнистый рисунок, хотя и в несколь-
ко ослабленном виде. Подобный характер окрашивания получен
у гибридного потомства леопарда и пумы. В обоих типах скре-
щиваний гибридные потомки как льва, так и пумы во взрослом
состоянии теряют пятнистый рисунок, характерный для них в
детском возрасте. Таким образом, вероятно, способность воспро-
изводить характер окрашивания шерсти у гибридных потомков
не теряется, а подавляется во взрослом состоянии другими ге-
нами. В геноме леопарда, по-видимому, имеются гены, которые-
способны почти полностью нейтрализовать действие этих су-
прессоров. Более того, в рассмотренном выше примере скрещи-
вания тигра со львом также наблюдается эффект подавления
супрессорных «львиных генов» генами, полученными от тигра..
СПИСОК ЛИТЕРАТУРЫ
Gray А. Р. Mammalian hybrids. — Commonwealth Agricultural Bureaux, Farn-
ham Royal, 1972.
Jackson J. M., Jackson J. The hybrid jungle cat Ц Newslett. Long Island Oce-
lot Club.— 1967,— Vol. 11,— P. 45.
Leyhausen P., Falkena M. Breeding the Brazilian ocelot cat, Leopardus tigri-
nus, in captivity Ц Int. Zoo.— 1966.— Vol. 6.— P. 176—182.
Leyhausen P., Tonkin B. Breeding the black footed cat, Felis nigripes, in can-
tivity // Int. Zoo.— 1966,— Vol. 6.— P. 176—182.
Robinson R. Homologous genetic variation in the Felidae / Genetica.— 1976.—
Vol. 46, N 1,— P. 1—31.
2 Заказ № 355
17
Глава 2
ДОМЕСТИКАЦИЯ
Из всех видов хищных животных можно выделить только
два, которые сопровождают человека всю жизнь, являясь его
постоянными спутниками,— это кошка и собака. Оба эти вида
широко распространены по всему миру, что является прямым
следствием расселения человека. Где бы ни путешествовал че-
ловек, кошка и собака всегда следуют за ним. Обычно человек
держит кошку как своего домашнего любимца, nopoii забывая
о том, что она может быть полезна на продуктовых складах
и фермах, страдающих от нашествия грызунов.
Происхождение домашней кошки достаточно хорошо изуче-
но. Она произошла от сложного вида Felis Silvestris — libyca,
представляющего собой группу небольших кошек, которые рас-
пространены преимущественно в Европе и на крупных остро-
вах — Британии, Корсике, Крите, Кипре, Сицилии, Сардинии,
на Среднем Востоке и в Северной Африке [Weigel, 1961]. Дан-
ный видовой комплекс состоит из нескольких видов, подвидов
и рас, имеющих одинаковый фенотип, идентичный кариотип и
скрещивающихся между собой, давая фертильное потомство. По-
добное явление уникально, так как межвидовая гибридизация
встречается в природе крайне редко.
Важнейшим фактором сохранения и поддержания целостно-
сти вида является географическая изоляция, которая, однако,
не всегда является прочным барьером для межвидовой гибри-
дизации. Подтверждением служит то, что разные виды кошек,
успешно вступая в гибридизацию, поддерживают существование
межвидового комплекса.
В природе можно выделить два вида, которые обнаружива-
ют близкое сходство с домашней кошкой: F. silvestris — кошка
европейская дикая и F. libyca — африканская дикая кошка. Фе-
нотипически они сильно различаются между собой. Африканская
кошка значительно крупнее, чем европейская, более гибкая, ок-
рас ее несколько светлее обыкновенного агути, а пестрый рису-
нок выражен слабее. Именно эти два вида, а также виды
F. chans — камышовая кошка и F. ornata — степная кошка, спе-
циалисты рассматривают как вероятных диких предков домаш-
ней кошки. Однако широко распространенная теория полифиле-
тического происхождения домашней кошки от четырех перечис-
ленных выше видов кажется маловероятной и подвергнута стро-
гой критике Хеммером [Hemmer, 1976а, 1978].
Согласно его взглядам, домашняя кошка произошла от F. li-
byca. Другим видом из всех перечисленных, для которого суще-
ствуют основания говорить о некотором вкладе в историю про-
исхождения домашней кошки, можно считать F. chans. Прове-
денный Моррисоном-Скоттом в 1952 г. [Morrison-Scott, 1952]
анализ огромного количества мумий кошек, обнаружен-
ие
пых в египетских пирамидах, подтвердил их принадлежность к
виду F. libyca, при этом лишь несколько экземпляров черепов
принадлежали к виду F. chaus. Учитывая такие единичные на-
ходки, едва ли можно делать вывод о том, что этот вид был
доместицирован и также играл важную роль в истории домаш-
ней кошки.
Трудно указать точное время, когда кошка начала свой дол-
гий путь рядом с человеком. Можно предположить, что доме-
стикация кошки относится ко времени расцвета цивилизации
на Среднем Востоке. Вероятно, это связано с появлением земле-
делия п строительством домов, амбаров, зернохранилищ. Это
создало новую среду обитания для мышей (Mus musculus), ко-
торые в ней быстро и успешно расселились. Об этом свидетель-
ствуют многочисленные находки костей мышей и кошек в под-
валах домов в местах археологических раскопок на Среднем
Востоке. Существует предположение о том, что, охотясь на
мышей и неотступно следуя за ними, кошка и заняла свою
нишу рядом с человеком, тем более, что он охотно поощрял
это. Сложившаяся таким образом ситуация повлекла за собой
начало доместикации кошки.
В описанный период (2 тыс. лет до н. э.) египтяне уже бы-
ли высокоразвитым народом, с прекрасными городами п сфор-
мировавшейся религией. Согласно своим религиозным убежде-
ниям они приписывали различным животным божественную си-
лу. В частности, кот ассоциировался у них с богом Ра, а кош-
ка — с богиней Бает. Считаясь «святыми» животными, кошки
содержались в египетских храмах, олицетворяя собой религиоз-
ный символ. Кроме того, кошек разводили (главным образом
для престижа) самые знатные люди Египта. Будучи очень
почитаемыми при жизни, после смерти кошки помещались »
специальные усыпальницы, размеры и великолепие которых сви-
детельствовали о степени уважения, которым кошка пользова-
лась при жизни.
В истории доместикации кошки выделяют три ключевых
этапа, связанных с отдельными событиями в истории цивили-
зации: 1) продолжительный период соседствования человека и
кошки, в течение которого кошка жила вблизи жилья человека-
и питалась мелкими грызунами (7000—2000 лет до н. э.); 2) пе-
риод зарождения процесса доместикации, в течение которого
кошка становится культовым животным (2000—1000 лет до
н. э.); 3) период перехода к полной доместикации, в течение-
которого человек стал держать кошку у себя дома (1000 лег
до н. э.— наши дни).
Для достижения кошкой половозрелого возраста и появле-
ния у нее потомства требуется приблизительно год. Следова-
тельно, за период доместикации несколько тысяч поколений ко-
шек сменили друг друга. Это вполне достаточный период для
появления и накопления изменений в поведении. По-видимому,
кошачья популяция была генетически очень разнообразна, по-
2*
49
скольку ее члены могли адаптироваться к различным условиям:
жизни, в том числе п к неволе. Они смогли не только выжить
при близком соседстве с человеком, но и производить и воспитывать
котят в таких условиях. Наиболее критической в эволюционном
плане была вторая фаза процесса доместикации, когда кошки, став
культовыми животными, впервые были помещены в неволю, при-
чем небольшими группами. Селекция на устойчивость наиболее
эффективна в случае конкуренции между животными как внут-
ри одной группы, так и между различными группами.
Морфофпзиологические изменения, достигнутые в ходе доме-
стикации, представляют собой комбинацию трех процессов:
1) неотения, при которой такие ювенильные характеристики,
как низкая агрессивность, доверчивость, веселый нрав, сохраня-
ются у животного и во взрослом состоянии; 2) уменьшение раз-
меров органов п снижение уровня гормонов стресса; 3) умень-
шение размеров мозга, в результате чего животное теряет способ-
ность адекватно реагировать на неблагоприятные условия.
Что касается последнего процесса, то показано, что доме-
стикация проходила бы успешнее в том случае, если бы кошки
обладали мозгом незначительных размеров, что делало бы их
более податливыми к процессу доместикации [Hemmer, 1976b].
Кроме того, Хеммер установил, что из всех видов кошачьих
представители F. libyca имели мозг наименьших размеров.
В этом контексте сравнительный анализ размеров мозга у
разных видов кошек по сравнению с домашней, проведенный
Хеммером, представляет особый интерес. Среди проанализиро-
ванных видов F. silvestris имел наибольший мозг, за ним сле-
дуют F. ornata и F. libyca. Объем кошачьих черепов, обнару-
женных в египетских пирамидах, сравним с объемом черепа
F. libyca. Кошачьи черепа из коллекции городов средневековой
России имеют еще меньшие размеры. Размеры мозга некоторых
кошек на сегодняшний день колеблются от близких к F. libyca
до значительно меньших. Уменьшение объема мозга за время
доместикации составило в среднем 10—15 %. Согласно взглядам
некоторых ученых, редукция мозга возникла как ответ на от-
бор, проводимый при доместикации.
Считается, что римляне завезли кошек в Европу из завое-
ванного ими Египта. Кошка была известна в Италии в I в. н. э.,
Швейцарии во II в., в Британии в IV в., в Германии в X в.
и. э., что ясно свидетельствует о постоянной миграции живот-
ных через континент [Zenner, 1963; Robinson, 1984]. Огромная
популярность кошек во времена римлян, позднее — в средние
века — заметно снизилась. Это было связано с представлением о
кошке, как опасном дьявольском существе. Тем не менее кошки
вышли относительно невредимыми из этих событий.
Позднее появился новый способ распространения кошек по
земному шару. Он связан с тем, что стало популярным брать
с собой в путешествие на борт корабля кошек как компаньонов,
скрашивающих монотонность путешествий, и как талисман, обе-
20
регающий от опасности. Таким образом человек помогал кошке
расселяться по Старому Свету. В XVIII—XIX вв. кошка была
завезена в Северную и Южную Америку европейскими поселен-
цами, в Австралию и Новую Зеландию британскими колони-
стами.
СПИСОК ЛИТЕРАТУРЫ
Aarde R. J., Dyk A. Inheritance of the king coat colour pattern, in cheetahs Ц
J. Zool.— 1986,—Vol. 209. - P. 573—578.
Baker H. J., Lindsey J. R. Feline GM1 gangliosidosis / Amer. J. Pathol.—
1974,—Vol. 74,—P. 649—652.
Baldwin J. A. Notes and speculations on the domestication of the cat in
Egypt Ц Anthropos.— 1975.— Vol. 70.— P. 428—448.
Barnett К. C., Curtis R. Autosomal dominant progressive retinal atrophy in
Abyssinian cals // J. Hered.— 1985.— Vol. 76.— P. 168—170.
Basrur P. K., DeForest M. E. Embryologieal impact of the Manx gene / Car-
nivora Genet. Newslett.— 1979,—Vol. 3.— P. 378—384.
Bistner S. I., Aquirre G., Shively J. N. Hereditary corneal dystrophy in the
Manx cat: a preliminary report / Invest. Ophthalmol. Vis. Sci.— 1976.—
Vol. 15,—P. 15—26.
Bosher S. K., Hallpike C. S. Observations of the histological features, develop-
ment and pathonogenesis of the inner ear degeneration of deaf white
cats Ц Proc. Roy. Soc. Lond. B.— 1965.— Vol. 162.— P. 147—170.
Bosher S. K., Hallpike C. S. Observations of the histologenesis of the inner
ear degeneration of the deaf white cat/J. Laryngol.— 1966.— Vol. 80.—
P. 202—235.
Boyce J. T., Bartola S. P., Chew D. J., Gasper P. W. Familial renal amylodosis
in Abyssinian cats / Vet. Pathol.— 1984.— Vol. 21.— P. 33—38.
Centerwall W. R., Benirschke K. Male tortoiseshell and calico cats Ц J. Tie-
red.— 1973 — Vol. 64 — P. 272—278.
Centerwall W. R., Wurster-Hill D. H., Maruska E. J., Kramer L. W. Chromoso-
me identification of the rusty spotted cat Ц Amer. ,1. Vet. Res.— 1983.—
Vol. 44,— P. 856—858.
Centferwall W. R., Wurster-Hill D. H., Maruska E. J., Theobald J. A. Chromoso-
me identification of the flat-headed cat Ц Amer. J. Vet. Res.— 1977.—
Vol. 38.- 1893—1894.
Chapman V. A., Zeiner F. N. The anatomy of polydactylism in cats Ц Anat.
Res.—1961,—Vol. 141 —P. 205—217.
Chastain С. B., Guilford W. G., Schidt D. The 38, XX/39/XXY genotype in
cats // Comp. Cont. Educ. Pract. Vet.— 1988,— Vol. 10,— P. 18—22.
Chew D. J., Dibartola S. P., Boyce J. J., Gasper P. W. Renal amyloidosis in
related Abyssinian cats Ц J. Amer. Vet. Med. Assoc.— 1982.— Vol. 48L—
P. 140—142.
Clifford D. H., Soifer F. K., Wilson C. F. et al. Congenital achalasia of the
aesophagus in four cats of common ancestry Ц J. Amer. Vet. Mod. As-
soc.— 1971.— Vol. 158.— P. 1554—1560.
Cork L. C.j Munnell J. F., Lorenz M. D. et al. GM2 gangliosidosis lysomal sto-
rage disease in cats Ц Science.— 1977.— Vol. 196.— P. 1014—1017.
Cork L. C., Munnel J. F., Lorenz M. D. The pathology of feline GM2 ganglio-
sidosis // Amer. J. Pathol.— 1978,— Vol. 90,— P. 723—734.
Cotter S. M., Brenner R. M., Dodds W. J. Hemophilia A in three unrelated
cats Ц J. Amer. Vet. Med. Assoc.— 1978.— Vol. 172.— P. 166—168.
Creel D., Collier L. L., Leventhal A. G. Abnormal retinal projections in cats
with Chediak — Higashi syndrome Ц Invest. Ophthalmol. Vis. Sci.—
1962,— Vol. 23,- P. 798-801.
‘Creel D., Hendrickson A. E., Leventhal A. G. Retinal projections in tyrosinase
negative albino cats Ц J. Neurosci.— 1982.— Vol. 2.— P. 907—911.
21
Crowell W. A., Hubbell J. J., Riley J. c. Polycystic renal disease in related
cats // J. Amer. Vet. Med. Assoc.— 1979,—Vol. 175,—P. 286—288.
Danforth С. H. Morphology of the feet in polydactylv cats Ц Amer. J. Anat.—
1947a.—Vol. 80,-P. 143-171.
Danforth С. H. Heredity of polydactyly in the cat // J. Hered.— 1947b.—
Vol. 38,—P. 107-112. "
DeForest M. E., Basrur P. K. Malformations and the Manx syndrome in cats If
Canad. Vet. J.— 1979.—Vol. 20,—P. 304—314.
Desnik R. J., McGovern M. M., Schuch E. IL, Haskins M. E. Animal analogu-
es of human inherited metabolic diseases Ц Animal models of inheri-
ted metabolic disease/Ed. R. J. Desnik, D. F. Pattison, D. G. Scarpelli.—
N. Y.: Alan R. Liss Inc., 1982.
Dibartola S. P., Hill R. L., Fechheimer N. S., Powers J. D. Pedigree analysis
of Abyssinian cals with familial amyloidosis Ц Amer. J. Vet. Res.—
. 1986,—Vol. 47,—P. 2666—2668.
Dittrich L. Die vererbung des melanismus beim jaguar Ц Zool. Garten.—
1979,— Bd 49,— S. 417-428.
Dodds W. I. Second international registry of animal models of thrombosis and'
haemorrhaging diseases Ц ILAR News.— 1981.— Vol. 24.— R1—R50.
Farrell D. F., Baker H. J., Herndon R. M. ct al. Feline GM1 gangliosidosis Ц
J. Neuropathol. Exp. Neurol.— 1973 —Vol. 32,—P. 1—18.
Fraser A. S. A note on the growth of the rex and Angora coats Ц J. Genet.—
1953,— Vol. 51,— P. 237-242.
Giddens W. E., Labbe R. F., Swango L. J., Padgett G. A. Feline congenital
erythropoietic porphyria associated with severe anaemia and renal di-
sease Ц Amer J. Pathol.— 1975,— Vol. 80,— P. 367—386.
Glenn B. L., Glenn H. G., Omtvedt I. T. Congenital porphyria in the domestic
cat Ц Amer. J. Vet. Res.— 1968.— Vol. 29.— P. 1653—1657.
Gray A. P. Mammalian Hybrids.— Commonwealth Agricultural Bureaux: Farn-
ham Royal, 1972.
Green R. A., White F. Feline factor XII (Hagemann) deficiency Ц Amer. J.
Vet. Res.— 1977,— Vol. 38 — P. 893—895.
GuMlery R. W., Hickey T. L., Spear P. D. Do blue-eyed while cats have normal
or abnormal retinofugal pathways? Ц Invest. Ophthalmol. Vis. Sci.—
1981.— Vol. 21.— P. 27-33.
Haldane J. B. S. Comparative genetics of colour in rodents and carnivora /
Biol. Rev.— 1927,- VoL 2,— P. 199—212.
Haskins M. E., Aquirre G. D., Jezyk P. F., Patterson D. F. The pathology of
feline arylsulfatase В deficient mucopolysaccharidosis Ц Amer. J. Pa-
thol.— 1980,— Vol. 191,— P. 657—674.
Haskins M. E., Jezyk P. F., Desnik R. J. et al. Alpha-l-iduronidase deficiency
in a cat/Pediat. Res.— 1979a.— VoL 13.— P. 1294—1297.
Haskins M. E., Jezyk P. F., Patterson D. F. Mucopolysaccharide storage disea-
se in three families of cats with arylsulfatase В deficiency Ц Pediat.
Res— 1979b.—Vol. 13,—P. 1203—1210.
Hemmer H. Hirngrossen variation in Felis silvestris Ц Experientia.— 1972.—
Vol. 28.— P. 271—272.
Hemmer H. Zur abstammung der hausekatze: sind Siamkatzen und Perserkat-
zen polyphyletischen ursprungs? / Saugetier. Mitt.— 1976a.— Bd 24.—
S. 184-192.
Hemmer H. Man’s strategy in domestication: a synthesis of new research
trends Ц Experientia.— 1976b.— Vol. 32.— P. 663—666.
Hemmer H. Were the leopard cat and the sand cat among the ancestry of the
domesticate races? Ц Carnivore.— 1978.— Vol. 1, N 2.— P. 106—108.
Hendy-Ibbs P. M. Hairless cats in Great Britain Ц J. Hered.— 1984.— Vol. 75.—
P. 506-507.
Hendy-Ibbs P. M. Familial feline epibulbar dermoids Ц Vet. Rec — 1985.—
Vol. 116,—P. 13—14.
Henricson B., Bornstein S. Hereditary umbilical hernia in cais Ц Svensk. Vet.
Tidsk - 1965,—Vol. 17.—P. 95—97.
22
Hirsch V. M., Cunningham J. A. Hereditary anomaly of neutrophil granula-
tion in Birman cats Ц Amor. J. Vet. Res.— 1984.— Vol. 45.—P. 2170—
2174.
Iljin N. A., Iljin V. N. Temperature effects on the color of the Siamese cat Ц
J. Hered.— 1930,— Vol. 21,— P. 309—318.
Jackson 0. F. Congenital bone lesions in cats with fold-ears Ц Bull. Feline
Advis. Bur.— 1975,— Vol. 14, N 14,— P. 2—4.
James С. C., Lassman L. P., Tomlinson В. E. Congenital abnormalities of the
lower spine and spinal cord in Manx cats Ц J. Pathol.— 1969.— Vol. 97.—
P. 269—276.
Johnson К. H. Globoid leukodystrophy in the cat / J. Amer. Vet. Med. As-
soc— 1970.— Vol. 157 — P. 2051—2054.
Johnston S. O., Buoen L. C., Madl J. E. et al. X chromosome monosomy
(37, XO) in a Burmese cat with gonal dysgenesis Ц J. Amer. Vet. Med.
Assoc.- 1983,— Vol. 182,- P. 986-989.
Jones B. R., Johnstone A. C., Hancock W. S., Wallace A. Inherited hyperchy-
lomicronemia in the cat Ц Feline Pract.— 1986a.— Vol. 16.— P. 7—
12.
Jones G. R., Johnstone A. C., Cahill J. I., Hancock W. S. Peripheral neuropathy
in cats with inherited primary hyperchylomaemia Ц Vet. Rec.— 1986b.—
Vol. 119,-P. 268—272.
Jotterand M. La formule chromosomique de quatre especes de Felidae Ц Rev.
Suisse. Zool — 1971.- Vol. 78,-P. 1250-1251.
Kaas J. H., Guillery R. W. The transfer of abnormal visual field representati-
ons from the dorsal lateral geniculate nucleus to the visual cortex in
Siamese cats Ц Brain Res.— 1973.— Vol. 59.— P. 61—95.
Kier A. B., Bresnahan J. F., White F. J., Wagner J. E. The inheritance pat-
tern of factor XII (Hageman) deficiency in domestic cats Ц Canad. J.
Comp. Med.— 1980 — Vol. 44.— P. 309—314.
Kramer J. W., Davis W. C., Prier D. J. The Chediak-Higashi syndrome of
cats Ц Lab. Invest.— 1977.— Vol. 36.— P. 554—562.
Lange A. L., Berg P. B., Baker M. K. A suspected lysosomal storage disease in
Abyssinian cats / J. S. Afr. Vet. Med. Assoc.— 1977.— Vol. 48.— P. 201—
209.
Latimer H. W., Rakich P. M., Thomson D. F. Pelger-Huet anomaly in cats Ц
Vet. Pathol.— 1985,— Vol. 22 — P. 370—374.
Leipold H. W., Huston K., Blauch B., Duffy M. M. Congenital defects of the
caudal vertebral column and spinal cord in Manx cats Ц J. Amer. Vet.
Med. Assoc.— 1974.—Vol. 164.— P. 520—523.
Letard E. La constitution d’un type ethnique disparu. Sur une famille de chats
nus Ц Rec. Med. Vet.— 1938,—Vol. 114,—P. 5—13.
Leventhal A. G., Creel D. J. Projections and functional architecture of corti-
cal areas 17 and 18 in the tyrosinase negative albino cat Ц J. Neuro-
sci.— 1985,— Vol. 5,— P. 795-807.
Leventhal A. G., Vitek D. J., Creel D. J. Abnormal visual pathways in normal-
ly pigmented cats that are heterozygous for albinism Ц Science.—
1965,— Vol. 229,— P. 1395—1397.
Levick W. R., Thibos L. N., Mostyn R. Retinal ganglion cells and optic decus-
sation of white eats Ц Vision Res.— 1980.— Vol. 20.— P. 1001—1006.
Little С. C. Four ears, a recessive mutation in the cat Ц J. Hered.— 1957.—
Vol. 48,— P. 57.
Littlewood J. D. Haemophilia A (factor VIII deficiency) in the cat Ц J. Small
Anim. Pract — 1986.— Vol. 27.— P. 541—546.
Livingston M. L. A possible hereditary influence in feline urolithiasis Ц Vet.
Med. Small Anim. Clin.— 1965.— Vol. 60.— P. 705.
Loevy H. T. Cytogenetic analysis of Siamese cats with cleft palate Ц J. Dent.
Res.— 1974,— Vol. 53,— P. 453-456.
Loevy H. T., Fenyes V. L. Spontaneous cleft palate in a family of Siamese
cats / Cleft Palate J.— 1968.— Vol. 5.— P. 57—60.
. Lomax T. D., Robinson R. Tabby pattern alleles of the domestic cat Ц J. He-
red.— 1988,— Vol. 79,— P.' 21—23.
23
Mair I. W.. Elverland H. H. Hereditary deafness in the cat. An electron study
of the stria vascularis and Reisser’s membrane Ц Arch-rhino-laryngol.—•
1977,— Vol. 217,—P. 199—217.
Martin A. H. A congenital defect in the spinal cord of the Manx cat Ц Vet.
Pathol.— 1971,— Vol. 8,— P. 232—239.
Mason K. A hereditary disease in Burmese cats manifested as an episodic
weakness with head nodding and neck ventrolexion Ц J. Amer. Vet.
Med. Assoc.— 1988,— Vol. 24,— P. 147—151.
McGovern M. M., Mandell N., Haskins FL, Desnik R. J. Characterization of aryl-
fatase В mutations in homoallelic and heteroallelic (genetic compounds)
homozygotes with feline mucopolysaccharidosis 6 // Genetics.— 1985,—
Vol. 110 —P. 733—749.
McGovern M. M., Vine D. T., Haskins M. E., Desnik R. J. An improved method
for heterozygous identification in feline and human mucopolysacchari-
dosis 6 Ц Enzyme.—1931,—Vol. 26,—P. 206—210.
McGovern M. M., Vine D. T., Haskins R. J., Desnik R. J. Purification and
properties of feline and human arylsulfatase В isozymes Ц J. Biol.
Chem— 1982,—Vol. 257,-P. 12605—12610.
McKerrell R. E., Blakemore W. F., Heath M. F. et al. Primary hyperoxaluria
in the cat // Vet. Rec.— 1989 — Vol. 125,— P. 31—34.
Meyers-Wallen V. N., Wilson J. D., Griffin J. E. et al. Testicular feminization
in a cat/.T. Amer. Vet. Med. Assoc.— 1989.— Vol. 195.— P. 631—634.
Moran C., Gilles С. B., Nicholas F. W. Fertile male tortoiseshell cats J. He-
red.— 1984,— Vol. 75,— P. 397—402.
Morrison-Scott T. C. S. The mummified cats of ancient Egypt Ц Proc. Zool.
Soc. Lond— 1952 —Vol. 121,—P. 861—867.
Moutschen J. Quelques Particularities hereditaires du chat Ц Nat. Beiges.—
1950,—P. 31,-P. 20-20.3.
Narfstrom K. Heredity progressive retinal atrophy in the Abyssinian cat //
J. Hered.— 1983,— Vol. 74 — P. 273—276.
Narfstrom K. Progressive retinal atrophy in the Abyssinian cat Ц Invest. Oph-
thalmol. Vis. Sci.-- 1985,— Vol. 26,— P. 193—200.
Narfstrom K., Nilsson S. E., Andersson В. E. Progressive retinal atrophy in
the Abyssinian cat. Studies of the DC recorded electroretinogram and
the standing potential of the eye Ц Brit. J. Ophthalmol.— 1985.—
Vol. 69,—P. 618-623.
Narfstrom K., Nilsson S., Wiggert B. et al. Reduced level of interphotoreceptor
retinoid binding protein (IRBP), a possible cause for retinal degenera-
tion in the Abyssinian cat Ц Cell Tissue Res.— 1989.— Vol. 257.—
P. 631—639.
Norby D. E., Hegreberg G. A., Thuline H. C., Findley D. An XO cat Ц Cyto-
genet. Cell Genet.— 1974.— Vol. 13.— P. 448—453.
Norbv D. E., Thuline H. C. Hereditary tremor in the domestic cat // Nature.—
‘ 1970,— Vol. 227 — P. 1262—1263.
Ohno S. Sex chromosomes and Sex-linked genes — Berlin: Springer-Verlag,
1967.
Paasch L. H., Zook B. S. The pathogenesis of endocardial fibroelastosis in Bur-
mese cats Ц Lab. Invest.— 1980.— Vol. 42.— P. 197—204.
Patterson D. F. Congenital defects of the cardiovascular system of dogs Ц
Adv. Vet. Sci. Comp. Med.— 1976.— Vol. 20.— P. 1—37.
Patterson D. F., Minor R. R. Hereditary fragility and hyperextonsihility of the
skin in cats Ц Lab. Invest.— 1977.— Vol. 37.— P. 170—179.
Pearson H., Gaskell C. J., Gibbs C., Waterman A. Pyloric stenosis and oesopha-
geal disfunction in the cat / J. Small Anim. Pract.— 1974.— Vol. 15.—
P. 487—501.
Pion P. D., Kittleson M. D., Rogers Q. R., Morris J. D. Myocardial failure in
cats associated with low plasma taurine Ц Science.— 1987.— Vol. 237.—
P. 764—768.
Prieur D. J., Collier L. L. Inheritance of the Chediak-Higashi Syndrome in
cats // J. Hered.- 1981a.—Vol. 72,—P. 175—177..
Prieur D. J., Collier L. L. Morphological basis of inherited coat colour diluti-
ons of cats / J. Hered.— 1981b.— Vol. 72.— P. 178—182.
24
Rebillard M., Pujol R., Rebillard G. Variability of the hereditary deafness in
the white cat. II. Histology Ц Hear. Res.— 1981a.— Vol. 5.— P. 189—
200.
Rebillard M., Rebillard G., Pujol R. Variability of the hereditary deafness in
the white cat Ц J. Physiology. Hear. Res.— 1981b.— Vol. 6.— P. 179—
187.
Robinson R. Genetics of the domestic cat // Bibliogr. Genet.— 1959.— Vol. 18,—
P. 273-362.
Robinson R. German rex: a rexoid coat mutant in the cat Ц Genetlca.— 1968.—
Vol. 39,— P. 351—352.
Robinson R. Devon rex — a third rexoid coat mutant in the cat Ц Genetica.—
1969a.— Vol. 40 — P. 597—599.
Robinson R. The white tigers of Rewa Ц Genetica.— 1969b.— Vol. 40.—
P. 198—200.
Robinson R. Inheritance of the black form of the leopard Ц Genetica.—
1970a.— Vol. 41,— P. 190-197.
Robinson R. Homologous mutants in mammalian coat colour variation Ц Symp.
Zool. Soc. bond.— 1970b — N- 26,— P. 251—269.
Robinson R. The rex mutants of the domestic cat /[ Genetica.—1971.—
Vol. 42,— P. 466 —468.
Robinson R. Oregon rex — a fourth rexoid coat mutant in the cat Ц Geneti-
ca.— 1972a — Vol. 43,— P. 236—238.
Robinson R. The sex-linked gene tortoiseshell in the Syrian hamster Ц Gene-
tica.— 1972b.— Vol. 43 — P. 239—243.
Robinson R. The Canadian hairless of Sphinx cat Ц J. Hered.— 1973.—
Vol. 64,— P. 47—49.
Rohinson R. Genetic, aspects of umbilical hernia incidence in cats and dogs /
Vet. Rec.— 1975,—Vol. 100,—P. 9—10.
Robinson R. Cytogenetics of the Felidae / World’s Cats.— 1976a.— Vol. 3,
N 2,—P. 15-27.
Robinson R. Homologous genetic variation in the Felidae Ц Genetica.—
1976b.—Vol. 46,—P. 1—31.
Robinson R. A third hypotrichosis in the domestic cat Ц Genetica.— 1981.—
Vol. 55,— P. 39—40.
Robinson R. Dutch rex — a fifth rexoid coat mutant in the cat Ц Genetica.—
1982a.—Vol. 57,—P. 217—218.
Robinson R. Chinchilla mutant in the lion Ц Genetica.— 1982b.— Vol. 60,—
P. 61-63.
Robinson R. Cat / Evolution of domesticated animals/Ed. I. L. Mason.— L.:
Longman, 1984.
Rohinson R. Fertile male tortoiseshell cats Ц J. Hered.— 1985.— Vol. 76.—
P. 137—138.
Robinson R. Genetic defects in cats // Comp. Anim. Pract.— 1987.— Vol. 1.—
P. 10—14.
Robinson R. The American curl cat /[ J. Hered.— 1989.— Vol. 80.— P. 474—
475.
Robinson R. Genetics for cat breeders.— Oxford: Pergamon Press, 1991.
Rubin L. F. Heredity cataract in Himalayan cats Ц Feline Pract.— 1986.
Vol. 16,—P. 14—15.
Ruhin L. F., Lipton D. E. Retinal degeneration in kittens Ц J. Amer. Vet. Med.
Assoc.— 1973,—Vol. 162,—P.'467—469.
Saperstein G., Harris S., Leipold H. W. Congenital defects in domestic cats Ц
Feline Pract.— 1976,— Vol. 6, N 4,— P. 18—43.
Searle A. G. Hereditary ‘’split-hand” in the domestic cat Ц Annal. Eugen.—
1953,—Vol. 17,—P. 279-282.
Searle A. G. Comparative genetics of coat colour in mammals.— L.: Logos
Press, 1968.
Searle A. G., Jude A. C. The rex type of coat in the domestic cat Ц J. Genet.—
1956,— Vol. 54,—P. 506—512.
Severin G. A. Congenital and aquired heart disease Ц J. Amer. Vet. Med. As-
soc.—1967,-Vol. 151,—P. 1733—1736.
25
Silson M., Robinson R. Hereditary hydrocephalus in the cat // Vet. Rec.—
1969,— Vol. 84,- P. 477.
Sponenberg D. P., Graf-Webster E. Hereditary meningoencephalocele in the
cat Ц J. Hered,— 1986 — Vol. 77,— P. 60.
Suomalainen E. Hannattomyyden perytymissuhteista kissalla // Novant. Anni
Leggi Mendel.— 1956,— P. 219—234.
Thibos L. N., Levick W. R., Morstyn R. Ocular pigmentation in white and Sia-
mese cats Ц Invest. Ophthalmol. Vis. Sci.— 1980.— Vol. 19.— P. 475—
482.
Todd N. B. A pink eyed dilution in the cat Ц J. Hered.— 1961a.— Vol. 52.—
P. 207.
Todd N. B. The inheritance of taillessness in Manx cats // J. Hered.— 1961b.—
Vol. 52,— P. 228—232.
Todd N. B. The Manx factor in domestic cats Ц J. Hered.— 1964.— Vol. 55.—
P. 225—230.
Turner P., Robinson R. Melanin inhibitor: a dominant gene in the domestic
cat // J. Hered.— 1980 — Vol. 71,— P. 427—428.
Turner P., Robinson R., Dyte С. E. Blue-eyed albino: a new albino allele in
the domestic cat Ц Genelica.— 1981.— Vol. 56.— P. 71—73.
VandeVelde M., Fankhauser R., Bichsel P. et al. Hereditary neurovisceral man-
nosidosis associated with mannosidase deficiency in a family of Per-
sian cats Ц Acta Neuropathol.— 1982,— Vol. 58,— P. 64—68.
Vawer G. D. Corneal mumminification in colourpoint cats Ц Vet. Rec.— 1982.—
Vol. 109,- P. 413.
Weber S. E., Evanz D. A., Feldman B. F. Pelger-Huet anomaly of granulocytic
leukocytes in two feline litter mates Ц Feline Pract.—1981.— Vol. 11,
N 1,- P. 44-47.
Weigel I. Das fellmuster der wildelebenden katzenarten und der hausekatzo
in vergleichender und stammesgeschtliche hinsicht Ц Saugetier. Mitt-
1961.—S. 1—20.
Wenger D. A., Sattler M., Kudoh T. et al. Niemann-Pick disease: a genetic
model in Siamese cats Ц Science— 1980.—Vol. 208,—P. 1471—1473.
West C. D., Harrison J. M. Trans neuron cell atrophy in the congenitally whi-
te cat // J. Comp. Neurol.— 1973 — Vol. 151,— P. 377-398.
West-Hyde L., Buyukminci N. Photoreceptor degeneration in a family of cats /
J. Amer. Vet. Med. Assoc.— 1982.— Vol. 181.— P. 243—247.
Whiting P. W. Inheritance of white spotting and other characters in cats Ц
Amer. Nat.— 1919 — Vol. 53,— P. 473—482.
Wilier S., Wilier H. Genolte und allele mit pathologischer wirkung oder ne-
benwirkung bei der hausekatzo // Mh. Vet. Med.— 1986 —Bd 41.—
S. 781-785.
William-Jones B. G. Arrested development of the long bones of the forelimbs
in a female cat Ц Vet. Res.— 1944.— Vol. 56.— P. 449.
Woodard J. C., Collins G. H., Hessler J. R. Feline hereditary neuroaxonal dys-
trophy Ц Amer. J. Pathol.— 1974.— Vol. 74,—P. 551—566.
Wurster D. H., Benirschke K. Comparative cytogenetic studies in the order
Carnivora Ц Chromosoma.— 1968.— Vol. 24.— P. 336—382. ,
Wurster-Hill D. H., Centerwall W. R.— The interrelationships of chromosome
banding patterns in canids, mustelids, hyaena, and felids / Cytogenet.
Cell Genet— 1982 —Vol. 34,—P. 178—192.
Zeuner F. E. A history of domesticated animals.— L.: Hutchinson. 1963.
Zook В. C., Paaseh L. H., Chandra R. S., Casey H. W. The comparative patholo-
gy of primary endocardial fibroelastosis in Burmese cats Ц Virchow
Arch. Pathol. Anat.— 1981,— Vol. 390 —P. 211-227.
Zook В. C., Sostaric B. R., Draper D. J., Graf-Webster E. Encephalocele and
other congenital craniofacial anomalies in Burmese cats // Vet. Med.
Small Anim. Clin — 1983.— Vol. 78 — P. 695—701.
26
Глава 3
ЦИТОГЕНЕТИКА КОШАЧЬИХ
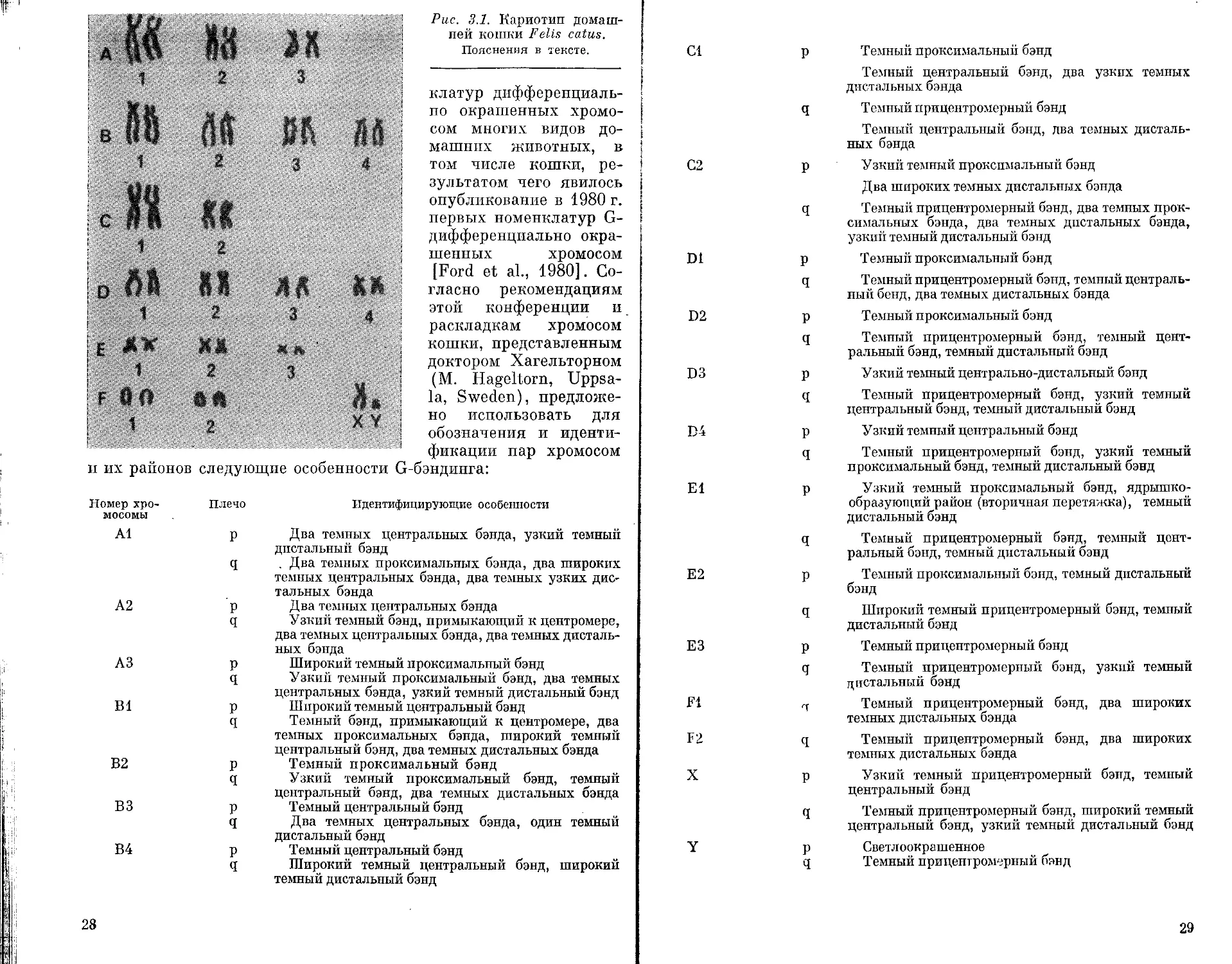
ХРОМОСОМНЫЙ НАБОР Felis catus
Цитогенетика домашней кошки проходила те же этапы раз-
вития, что п цитогенетика человека и млекопитающих в целом.
До появления методов культивирования клеток п гипотонических
обработок сложнейшей задачей было правильно определить чис-
ло хромосом. Попытки такого определения для кошки предпри-
нимались неоднократно [Minouclii, 1928а, b; Minouclii, Ohta,
1932, 1934; Matthey, 1934; Makino, Tateishi, 1952], однако толь-
ко после появления адекватных методов цитогенетического ана-
лиза стало возможным детальное описание кариотипа F. catus.
В 1965 г. в пуэрториканском городке Сан-Хуане собралась кон-
ференция из трех человек, для которых в тот момент хромо-
сомы кошки представляли хоть какой-то интерес; они решили,
что в дальнейших исследованиях будут придерживаться единого
образца в расположении каждой хромосомной пары [Jones,
1965]. Таким образом, была создана номенклатура рутинноокра-
шенных хромосом кошки, крайне неудобная, но ей продолжают
пользоваться по сей день.
В наборе домашней кошки 38 хромосом (2и=38), Х-хромосо-
ма — средних размеров субметацентрик, Y-хромосома — наимень-
ший двуплечий элемент набора. Аутосомы подразделяются на
шесть морфологических групп: группа А — 3 пары крупных
субметацентрпков; группа В — 4 пары крупных субтелоцептри-
ков; группа С — 2 пары крупных метацентриков; группа D —
4 пары мелких субмета- и субтелоцентрпков; группа Е — 3 па-
ры мелких метацентриков; группа F — две пары мелких акро-
центриков. Хромосомы первой пары группы Е (Е1) несут в ко-
ротком плече вторичные перетяжки (рис. 3.1).
Следует признать, что на уровне рутинноокрашенпых хромо-
сом сан-хуанская номенклатура была явно неработающей. Мно-
гие пары хромосом кошки близки по размерам и морфологии,
что создает непреодолимые на этом уровне трудности в иден-
тификации многих пар. Решить задачу создания реальной но-
менклатуры хромосом F. catus, как и большинства других впдов
млекопитающих, стало возможно только с появлением в прак-
тике цитогенетических исследований методов дифференциальной
•окраски хромосом. G-дифференциальная окраска хромосом кош-
ки представлена в ряде работ [Wurster-Hill, Gray, 1973; Benir-
schke et al., 1974; Ford et al., 1980; O’Brien, Nash, 1982; Wurs-
ter-Hill, Centerwall, 1982; Графодатский. Раджабли, 1988;
Graphodatsky, Hainan, 1989]. На конференции по цитогенетике
домашних животных в Рэдинге (Великобритания) в 1976 г.
было принято решение о кооперации работ по созданию номен-
27
Рис. 3.1. Кариотип домаш-
ней кошки Fells catus.
Пояснения в тексте.
клатур дифференциаль-
но окрашенных хромо-
сом многих видов до-
машних животных, в
том числе кошки, ре-
зультатом чего явилось
опубликование в 1980 г.
первых номенклатур G-
дифференциально окра-
шенных хромосом
[Ford et al., 1980]. Со-
гласно рекомендациям
этой конференции и
раскладкам хромосом
кошки, представленным
доктором Хагельторном
(М. Hageltorn, Uppsa-
la, Sweden), предложе-
но использовать для
обозначения и иденти-
фикации пар хромосом
и их районов следующие особенности G-бэндинга:
Номер хро- мосомы Плечо Идентифицирующие особенности
А1 Р Два темных центральных бэнда, узкий темный дистальный бэнд
q . Два темных проксимальных бэнда, два широких темных центральных бэнда, два темных узких дис- тальных бэнда
А2 р Два темных центральных бэнда
q Узкий темный бэнд, примыкающий к центромере, два темных центральных бэнда, два темных дисталь- ных бэнда
АЗ р Широкий темный проксимальный бэнд
q Узкий темный проксимальный бэнд, два темных центральных бэнда, узкий темный дистальный бэнд
В1 р Широкий темный центральный бэнд
q Темный бэнд, примыкающий к центромере, два темных проксимальных бэнда, широкий темный центральный бэнд, два темных дистальных бэнда
В2 р Темный проксимальный бэнд
ВЗ q Узкий темный проксимальный бэнд, темный центральный бэнд, два темных дистальных бэнда
р Темный центральный бэнд
q Два темных центральных бэнда, один темный дистальный бэнд
В4 р Темный центральный бэнд
q Широкий темный центральный бэнд, широкий темный дистальный бэнд
28
Cl
C2
DI
D2
D3
D4
El
E2
E3
Fl
F2
X
Y
p Темный проксимальный бэнд
Темный центральный бэнд, два узких темных
дистальных бэнда
q Темный прицентромерный бэнд
Темный центральный бэнд, два темных дисталь-
ных бэнда
р Узкий темный проксимальный бэнд
Два широких темных дистальных бэнда
q Темный прицентромерный бэнд, два темных прок-
симальных бэнда, два темных дистальных бэнда,
узкий темный дистальный бэнд
р Темный проксимальный бэнд
q Темный прицентромерный бэнд, темный централь-
ный бенд, два темных дистальных бэнда
р Темный проксимальный бэнд
q Темный прицентромерный бэнд, темный цент-
ральный бэнд, темный дистальный бэнд
р Узкий темный центрально-дистальный бэнд
q Темный прицентромерный бэнд, узкий темный
центральный бэнд, темный дистальный бэнд
р Узкий темный центральный бэнд
q Темный прицентромерный бэнд, узкий темный
проксимальный бэнд, темный дистальный бэнд
р Узкий темный проксимальный бэнд, ядрышко-
образующий район (вторичная перетяжка), темный
дистальный бэнд
q Темный прицентромерный бэнд, темный цент-
ральный бэнд, темный дистальный бэнд
р Темный проксимальный бэнд, темный дистальный
бэнд
q Широкий темный прицентромерный бэнд, темный
дистальный бэнд
р Темный прицентромерный бэнд
q Темный прицентромерный бэнд, узкий темный
дистальный бэнд
q Темный прицентромерный бэнд, два широких
темных дистальных бэнда
q Темный прицентромерный бэнд, два широких
темных дистальных бэнда
р Узкий темный прицентромерный бэнд, темный
центральный бэнд
q Темный прицентромерный бэнд, широкий темный
центральный бэнд, узкий темный дистальный бэнд
р Светлоокрашенное
q Темный прицентромерный бэнд
29
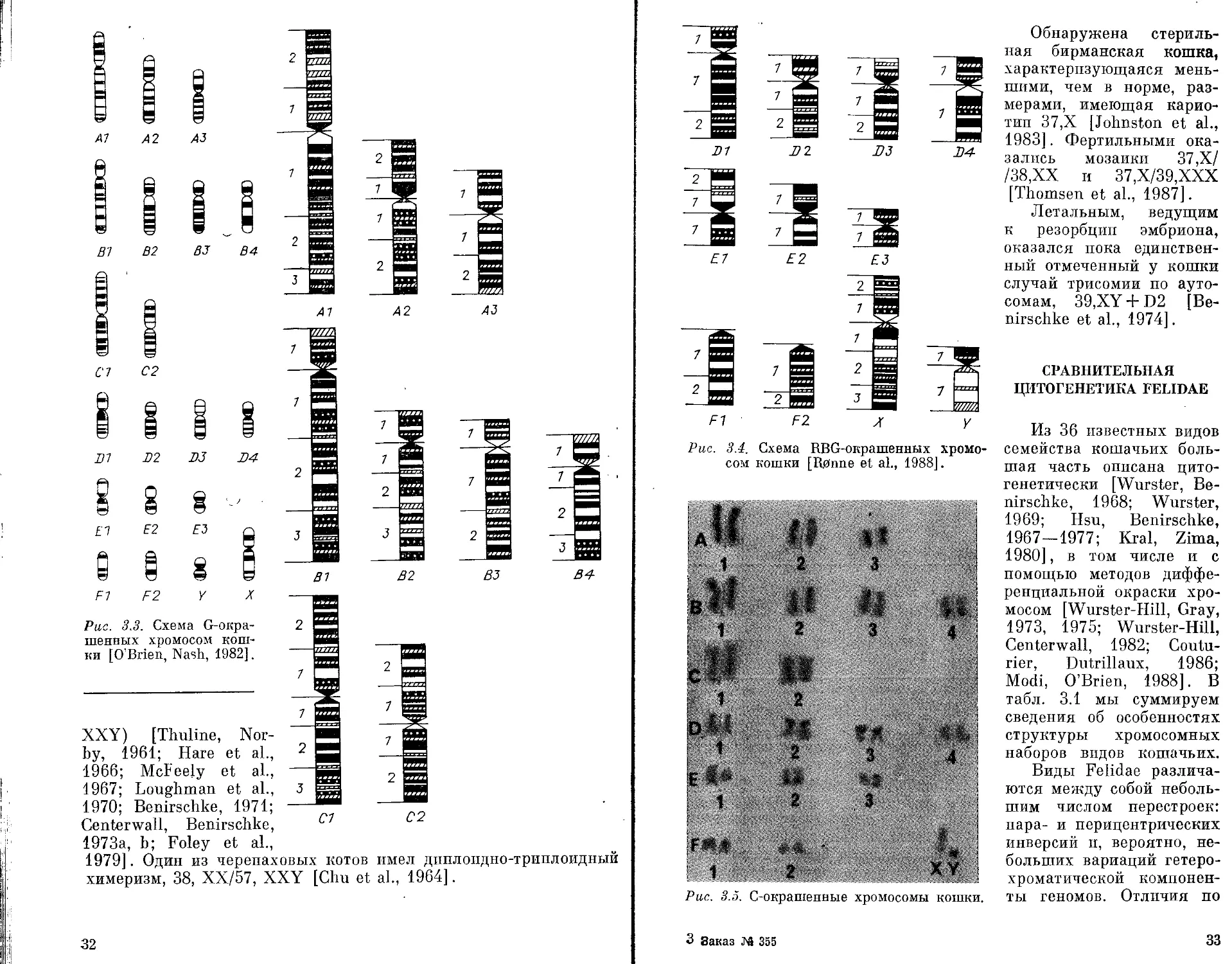
Рис. 3.2. G-окрашенные хромосомы кошки.
Анализ рисунка бэндинга G-окрашенных метафазных хро-
мосом кошки позволяет надежно идентифицировать большин-
ство крупных и средних пар хромосом кошки, однако некоторые
мелкие пары аутосом идентифицируются менее надежно. Это
обстоятельство, а также потребности сравнительно-цитогенетиче-
ских исследований, работ по генетическому картированию
хромосом и предполагаемых в ближайшем будущем работ по моле-
кулярному картированию хромосом кошки заставили исследова-
телей постепенно повышать уровень разрешения G-дифферен-
циальной окраски, для чего использовались хромосомы клеток
на ранних стадиях конденсации [O’Brien, Nash, 1982; Kiel voh
et al., 1985; Modi, O’Brien, 1988; Grapliodatsky, Hainan, 1989].
На рис. 3.2 показаны G-окрашенные хромосомы кошки на ран-
ней стадии конденсации. О’Браеном и Нэшем [O’Brien, NashT
1982] предложена для использования в цитогенетических ис-
следованиях схема G-бэндпнга хромосом кошки (рис. 3.3).
Существенно большего уровня разрешения бэндинга удалось
достигнуть при использовании RBG-дифференцпальной окраски,
в основном благодаря искусству Могенса Рённе и его сотрудни-
ков [Shibasaki et al., 1987; R^nne et al., 1988]. В последней
работе представлена номенклатура RBG-окрашенных хромосом
кошки на уровне разрешения в 580 бэндов (рпс. 3.4).
Структурный гетерохроматин хромосом кошки, выявляемый
с помощью С-метода, дифференциальной окраски, представлен
небольшими теломерными блоками аутосом группы D, неболь-
шим интеркалярным блоком длинного плеча хромосом пары Е1
и целиком С-позитивной Y-хромосомой [Pathak, Wurster-Hill,
1977; Графодатский, Раджабли, 1988; Grapliodatsky, Hainan,
1989]. С-окрашенные хромосомы кошки показаны на рпс. 3.5.
У кошки ядрышкообразующие районы (кластеры функцио-
нально активных рРНК-генов), выявляемые с помощью метода
окраски препаратов азотнокислым серебром, располагаются в зо-
нах вторичных перетяжек пары хромосом Е1 (рпс. 3.6) [Pear-
son et al., 1979; Графодатскпй, Раджабли, 1988]. Иногда яд-
рышкообразующие районы выявляются и на хромосомах пары
ЕЗ [Pearson et al., 1979].
ХРОМОСОМНЫЕ ПАТОЛОГИИ
В отличие от человека и многих других видов млекопитаю-
щих у домашней кошки не описано ни одной структурной пере-
стройки хромосом. С другой стороны, описано много случаев, на-
рушений в балансе половых хромосом [Han Qin, Hainan, 1989].
У кошек с черепаховой окраской (проявление которой невоз-
можно у самцов, так как аллельные гены черной и оранжевой
окрасок расположены на Х-хромосоме) описано много случаев
хромосомной конституции 2ra-39, XXY; либо они химерны, т. е.
имеют разные популяции клеток (38, XX/38, XY; 38, ХХ/39,
31
Al
A2
A3
i
D2
I
E2
§
D3
E3
8
F1 F2 Y X
Рис. 3.3. Схема G-окра-
шенных хромосом кош-
ки [O’Brien, Nash, 1982].
XXY) [Thuline, Nor-
by, 1961; Hare et al.,
1966; McFeely et al.,
1967; Loughman et al.,
1970; Benirschke, 1971;
Centerwall, Benirschke,
B3 B4
A2 A3
С2
1973a, b; Foley et al.,
1979]. Один из черепаховых котов
химеризм, 38, XX/57, XXY [Chu et al., 1964].
имел дпплоидно-триплоидный
32
£2
Рис. 3.4. Схема RBG-окрашенных хромо-
сом кошки [Rpnne et al., 1988].
Рис. 3.5. С-окрашенные хромосомы кошки.
3 Заказ Jti 355
Обнаружена стериль-
ная бирманская кошка,
характеризующаяся мень-
шими, чем в норме, раз-
мерами, имеющая карио-
тип 37,X [Johnston et al.,
1983]. Фертильными ока-
зались мозаики 37,X/
/38,XX и 37,Х/39,ХХХ
[Thomsen et al., 1987].
Летальным, ведущим
к резорбции эмбриона,
оказался пока единствен-
ный отмеченный у кошки
случай трисомии по ауто-
сомам, 39,XY + D2 [Be-
nirschke et al., 1974].
СРАВНИТЕЛЬНАЯ
ЦИТОГЕНЕТИКА FELIDAE
Из 36 известных видов
семейства кошачьих боль-
шая часть описана цито-
генетически [Wurster, Be-
nirschke, 1968; Wurster,
1969; Hsu, Benirschke,
1967—1977; Kral, Zinia,
1980], в том числе и с
помощью методов диффе-
ренциальной окраски хро-
мосом [Wurster-Hill, Gray,
1973, 1975; Wurster-Hill,
Centerwall, 1982; Coutu-
rier, Dutrillaux, 1986;
Modi, O’Brien, 1988]. В
табл. 3.1 мы суммируем
сведения об особенностях
структуры хромосомных
наборов видов кошачьих.
Виды Felidae различа-
ются между собой неболь-
шим числом перестроек:
пара- и перицентрических
инверсий и, вероятно, не-
больших вариаций гетеро-
хроматической компонен-
ты геномов. Отличия по
33
I
Рис. 3.6. Ag-окрашенные
хромосомы кошки.
Стрелками показаны яд-
рышкообразующие районы.
числу хромосом у оцелота, американской кошки, онциллы, кош-
ки Жоффруа и колоколо от других видов обусловлены тандем-
ным слиянием с образованием пары СЗ. Некоторые варианты
перестроек показаны на рис. 3.7, любезно предоставленном док-
тором Моди [Modi, O’Brien, 1988].
Если рассматривать вопрос об эволюции кариотипа более
широко, то следует непременно отметить, что Felidae значитель-
но отличаются от остальных групп хищных (Carnivora) мень-
шей эволюционной скоростью фиксации хромосомных перестро-
ек, т. е. характеризуются существенным хромосомным консерва-
тизмом [Wurster-Hill, Centerwall, 1982; Couturier, Dutrillaux,
1986; Grapliodatsky, 1989].
КАРТИРОВАНИЕ ХРОМОСОМ КОШКИ
Цитогенетическая карта хромосом кошки появилась в основ-
ном благодаря работам лаборатории д-ра С. Дж. О’Брайена (На-
циональный институт рака, Фредерик, США). Как и для боль-
шинства других видов млекопитающих, основой для проведения
этой работы явилось получение панели клонов гибридов сомати-
ческих клеток кошка X мышь. Полученные клоны, содержащие
хромосомы кошки в различных комбинациях, позволяют, ана-
лизируя белковые продукты гибридных клеток (если гомологич-
ные белки кошки и мыши различаются по электрофоретической
подвижности), выявить связь определенного белка и соответ-
ствующего гена с вполне определенной хромосомой набора кош-
ки. Кроме того, ДНК, выделенная из клеток различных клонов
34
Таблица 3.1
Сравнительный анализ кариотипов Felidae
Вид 2n Особенности структуры кариотипа по сравнению с F. catus
1 2 3
Felis catus — домашняя кош- ка 38
F. Silvestris — дикая европей- ская кошка 38 Хромосомный набор идентичен F. catus
F. libyca — дикая африкан- ская кошка 38 То же
F. margarita — барханная кошка 38 »
F. chaus — камышовая кош- ка 38 »
F. nigrtpes — чернояогая кошка 38 »
Prionailurus bengalensis — 38 Вероятны инверсия в коротком плече
бенгальская кошка хромосомы D2 и появление хромосомы Е4 в результате добавления/делеции (ве- роятно, гетерохроматического материала) и порицентрической инверсии Fl F. catus
Р. viverrinus — кошка-рыбо- лов 38 То же
Puma concolor — пума 38 »
Mayailurus irlomotensis — ириомотейская кошка 38 »
Lynx lynx — рысь 38 Перицентрическая инверсия В4, инвер- сия в коротком плече D2, появление F3 в результате инверсии F1 и вероятной делеции/добавления части материала
L. rufus — красная рысь 38 То же
Pardofelis marmorata — мра- морная кошка 38 »
Pantera leo — лев 38 »
Р. tigris — тигр 38 »
Р. pardus — леопард 38 5>
Р. опса — ягуар 38 »
Р. uncia — спежный барс 38 »
Leopardus pardalus — оцелот 36 Появление пары СЗ, отсутствие F1 и F2, отличие в паре D2
L. wiedi — американская кошка 36 То же
L. tigrina —• онцилла 36 Появление пары СЗ, отсутствие F1 и F2, инверсия в коротком плече D2
L. geoffroyi — кошка Жоф- фруа 36 То же
Lynchailurus colocolo (F. paj- eros) — колоколо 36 Появление пары СЗ, отсутствие F1 и F2
3*
35
Окончание т а б л. 3.1
1 2 3
Prionailurus rubiginosa — пятнисто-рыжая кошка 38 Появление хромосомы E4 в результате добавления/делеции (вероятно, гетеро- хроматического материала) и перицент- рпческой инверсии Fl F. catus, изменение рисунка F2
Ictailurus plancipes — сумат- ранская кошка 38 Перицентрическая инверсия В4, инвер- сия в коротком плече D2. Появление хро- мосомы Е4 в результате добавления/де- леции (вероятно, гетерохроматического материала) и перицентрической инвер- сии Fl F. catus
Caracal caracal — каракал 38 Инверсия в коротком плече D2
Otocolobus manul — манул 38 Появление хромосомы Е4 в резуль- тате добавления/делеции (вероятно, гете- рохроматического материала) и перицент- рической инверсии Fl F. catus
Leptailurus serial — сервал 38 Инверспя в коротком плече D2, появ- ление F3 в результате инверсии F1 и ве- роятной делецип/добавленпя части ма- териала
Projelis temmincki — кошка Темминка 38 Перестройки А1 и D1. Инверсия в ко- ротком плече D2, появление F3 в резуль- тате инверсии F1 и вероятной делеции/до- бавления части материала
Herpailurus yagouarouncli — ягуарунди 38 Инверсия в коротком плече D2. Появ- ление хромосомы Е4 и Е5 в результате добавления/делеции (вероятно, гетеро- хроматического материала) п перицент- рических инверсий F1 и F2 F. catus
Profelis aurata — золотая кошка 38 Перицентрическая инверсия В4. Инвер- сия в коротком плече D2
Neofelis nebulosa — дымчатый леопард 38 Перицентрическая инверсия В4, появ- ление F3 в результате инверсии F1 и ве- роятной делеции/добавления части мате- риала, изменение F3 по сравнению с F3 у других видов
Acinonyx jubatus — гепард 38 Перицентрическая инверсия В4, ин- версия в коротком плече D2
кошка X мышь, используется для проведения опытов по молеку-
лярной блот-гибридизации с ДНК ряда клонированных генов,
что также позволяет определить хромосомную локализацию этих
генов. Как и у всех других видов, у кошки отмечен полимор-
физм по многим из локализованных с помощью панели клонов
белков (генов). Это, в свою очередь, дает возможность опреде-
36
; ’.:e'icort*r»€
„ • 4 1ЛА<1егр % I $ *
- ,fie. .!!! --..„jlii
Рис. 3.7. Некоторые перестройки хромосом, имевшие место в эволюции Fe-
lidae [Modi, O’Brien, 1988],
Каждая хромосома обозначена первой буквой родового и двумя первыми буквами
видового латинского наименования вида (см. табл. 3.1) и буквой и номером данной
хромосомы в номенклатуре.
лять сцепление этих генов с генами, мутации которых имеют
фенотипическое проявление. Таким образом, например, у кошки
была определена хромосомная локализация гена С, мутация ко-
торого проявляется как альбинизм. По результатам работ по
картированию хромосом кошки [Nash, O’Brien, 1982; O’Brien,
Nash, 1982; O’Brien et al., 1985, 1986; O’Brien, 1990] составлена
генетическая карта кошки:
Хромосома кошки Ген
А1 А2 АЗ В1 В2 ВЗ В4 С1 С2 D1 D2 D4 ЕЗ F2 X U2 * U4 * U5 • ESD EGFR, MET, RAF1 MDH1, ACPI, ITPA, ADA, REL PEPS, IL2 MEI, PGM3, GLO, SOD2, MOS, MHC, MYB NP, MPI, PKM2, HEXA, SORD, FES, FOS, GLB1 TPI, PEPB, LDHB, GAPD, KRAS2 PGM1, PGD, IDH1, FUCA1, NRAS. PCL SOD1, ETS2, GSR HBB, HRAS, LDHA, ACP2, ETS1, GANAB, C HK1 PP, IDUA GL’SB MYC G6PD, HPRT, 0, GLA. HRASP GPI PEPA AK1
* Группы сцепления с неизвестной пока хромосомной локализацией. Краткие на-
именования локусов (генов) приведены по принятой номенклатуре [HGM 10,5, 1990].
Таким образом, для кошки получена довольно полная кар-
тина распределения групп сцепления по хромосомам. В настоя-
щее время работы по картированию хромосом ведутся примерно
для 30 видов млекопитающих: человека и 11 видов приматов,
пяти видов грызунов, четырех видов хищников, четырех видов
парнокопытных, одного вида непарнокопытных, одного вида зай-
цеобразных, пяти видов сумчатых и одного вида однопроходных.
Наиболее полная цитогенетическая карта получена для челове-
ка, на хромосомах которого локализовано около 5 тыс. генов и
анонимных последовательностей ДНК. Около 1,5 тыс. маркеров
локализовано на хромосомах домовой мыши Mus musculus — ос-
новного объекта генетических экспериментов. Во втором ряду по
изученности располагаются некоторые приматы, норка, лисица,
крыса, китайский хомячок, на хромосомах которых локализова-
но по несколько десятков генов. В эту группу входит и кошка.
Для других видов либо подробно изучено распределение генов
по группам сцепления, но чаще всего неизвестна хромосомная
локализация этих групп, как, например, для коровы, либо ра-
боты по картированию носили случайный характер, результатом
чего явилось определение локализации единичных генов или
некоторых групп сцепления. В последние годы для картирова-
ния геномов человека и млекопитающих стал широко исполь-
зоваться метод in situ гибридизации, позволяющий локализовать
38
Рис. 3.8. G-окрашенные Х-хромосомы ряда ви-
дов млекопитающих (обозначены первой буквой
родового и двумя буквами видовою латинского
наименования вида).
пробы ДНК, гены и их фрагменты либо
анонимные последовательности из геном-
ных библиотек непосредственно на теле
хромосом. Гибридизация in situ стала ос-
новным методом получения цитогенетиче-
ских карт для видов, у которых карти-
рование генома с помощью панелей кло-
нов гибридных клеток затруднено. До-
стоинство этого метода заключается преж-
де всего в том, что он позволяет прово-
дить локализацию гена на субхромосом-
ном уровне, т. е. определять место рас-
положения любой последовательности
ДНК с точностью до бэнда. К сожале-
нию, этот метод, так же как и ряд дру-
гих современных методов картирования,
для изучения генома кошки пока не ис-
пользовался.
Вместе с тем работы именно по кар-
HSA
тированию хромосом кошки стали пер-
выми в серии исследований по выявлению и анализу
закономерностей эволюции консервативных районов хромо-
сом млекопитающих. Работы по картированию хромосом разных
таксономически весьма далеких видов позволили выявить боль-
шое число консервативных групп сцепления, часто содержащих
довольно основательный набор гомологичных генов. Возникло
желание найти подтверждение такого эволюционного консерва-
тизма протяженных районов геномов млекопитающих на цито-
логическом уровне. Первыми работами этого направления, не-
сомненно, были исследования Оно, проанализировавшего генный
состав и количественные характеристики Х-хромосом млекопи-
тающих [Ohno, 1969]. Было установлено, что у многих видов
млекопитающих, в том числе у кошки, Х-хромосомы имеют
идентичный генный состав, близки по морфологии и составляют
по размерам около 5 % от суммарной длины гаплоидного набо-
ра. Анализ бэндинга позволил показать, что Х-хромосомы чело-
века, кошки, норки, собаки, лисицы, свиньи, верблюда и многих
других таксономически весьма далеких видов млекопитающих
идентичны по рисунку дифференциальной окраски [Pathak,
Stock, 1974]. Х-хромосомы такого вида обозначили как «стан-
дартный» тип, не без оснований полагая, что такую Х-хромо-
сому имели предки млекопитающих. У некоторых видов млеко-
питающих обнаружены Х-хромосомы, превышающие по разме-
рам стандартный тип. Они образовались в результате трансло-
39
Таблица 3.2
Хромосомная локализация гомологичных генов у кошки и других видов мле-
копитающих
Локус Хромосомы
FCA HS A MVI VFU CFA MMU CGR UNO оси | OAR BTA ssc
1 2 3 4 5 6 V 8 9 10 11 12 13
ESD Al 13 8 6 ul8 18 1 — 8 3 13
EGFR A2 7 — — — 11 — 14 7 —• —. —
МЕТ A2 7 —. — —. 6 — —. — — ul3 —
RAF1 A2 3 — — — 6 — 4 — — — —
MDH1 A3 2 11 16 u6 — — — u5 — — 3
ACPI A3 2 11 8 u5 12 7 6 — — —. —
ITPA A3 20 11 14 u23 2 6 4 17 ul5 ull —
ADA A3 20 11 — u23 2 6 — — ul5 ull —
REL A3 2 — — — 11 — — — —- — —
PEPS Bl 4 6 — — 5 1 —. — — 6 —
IL2 Bl 4 — — u23 3 — 2 — — u23 —
MEI B2 6 1 1 ulO 9 4 8 — u8 u2 —
PGM3 B2 6 — — ulO 9 4 — — u8 u2 —
GLO1 B2 6 1 — — — 1 20 — — 23 —
SOD2 B2 6 1 — ulO — — — — — u2 —
мне B2 6 — — ulO 17 — 20 — 20 23 7
MYB B2 6 — — — 10 — — — — — —
MOS B2 8 — — — — — 5 — — 14 —
NP B3 14 2 10 ul9 14 1 — 16 u3 u5 7
FOS B3 14 — — — 12 — 6 — — u5 —
MPI B3 15 10 15 u20 9 4 — — u9 21 7
PKM2 B3 15 — — u20 9 — 8 — u3 — 7
HEXA B3 15 — u5 —
SORD B3 15 — — — 2 6 — — — — —
FES B3 15 — — — 7 3 — — — — —
GLB1 B3 3 — — — — — 15 — — — —
TPI B4 12 9 — u21 6 8 4 4 3 5 —
LDHB B4 12 9 8 u21 6 8 4 4 — 5 —
GAPD B4 12 9 — u21 G 8 — 4 3 5 —
KRAS2 B4 12 — — — 6 — — 16 — 5 —
PEPB B4 12 9 — — 10 1 — 16 3 5 5
PGD Cl 1 2, 2 ul 4 2 5 — ul ul 6
FUCA1 Cl 1 — — — 4 2 5 — — — —
PGM1 Cl 1 2 12 u2 4 2 5 — — u6 6
NRAS Cl 1 — — — 3 — — — — — —
IDH1 Cl 2 4 16 u7 2 — — — ull 8 1
SOD1 C2 21 5 — u24 16 — — — ulO ulO 9
ETS2 C2 21 — — — 16 — — — — ulO —
GSR C2 8 6 7 — 8 1 — 19 U16 ul4 —
HBB DI 11 — — — 7 3 1 — — 15 —
40
Окончание табл. 3.2
1 2 3 4 I 5 6 7 8 9 10 11 12 13
HRAS D1 И — — 7 3 1 — — —
LDHA D1 11 7 11 и14 7 3 1 — — — —
АСР2 D1 11 7 — ul5 2 — — — — — —
ETS1 D1 И — — — 9 — — — — — —
GANAB D1 11 — — — — — — — — — —
С D1 — — — — 7 — — — — — —
НК1 D2 10 2 —. — 10 1 — — — — —
рр D4 10 2 4 1113 10 — — — — — —
IDUA D4 22 —.
GUSB ЕЗ 7 — — — 5 — — — — u8 —
G6PD X X X X X X X X X X X X
HPRT X х X X X X X X X X X X
GLA X X X X X X X X X X X X
HRASP X X — — — X X — — — —
Примечание, и — группа сцепления с неизвестной хромосомной локали-
зацией; FCA — Felis catus (домашняя кошка); HSA — Homo sapiens (человек); MVI —
Mustela vison (американская норка); VFU — Vulpes fulvus (серебристо-черная лисица);
CFA — Cdnis familiaris (собака); MMU — Mus musculus (домовая мышь); CGR — Сгг-
cetulus griseus (китайский хомячок); RNO— Rattris norvegicus (серая крыса); OCU—
Oryctolagus cuniculus (кролик); OAR — Oms aries (овца); ВТА — Bos taurus (крупный ро-
гатый скот); SSC — Sus scrofa (свинья).
каций Х-хромосом предкового типа на аутосомы либо характе-
ризовались значительным увеличением гетерохроматической ком-
поненты при неизменной эухроматической части. На рпс. 3.8
[Grapliodatsky, 1989] показаны некоторые типы Х-хромосом мле-
копитающих, в том числе группа Х-хромосом предкового типа,
к которому относится и Х-хромосома кошачьих.
Аналогичный подход, т. е. попытка определить цитологиче-
ски сходные районы хромосом на основании сведений об их
сходном генетическом содержимом, для аутосом был впервые
применен при сопоставлении цитологических и цитогенетических
карт хромосом человека и кошки [Nash, O’Brien, 1982]. В ре-
зультате этой работы, помимо известной гомологии бэндинга
Х-хромосом, у этих двух видов из разных отрядов млекопитаю-
щих было выявлено еще четыре крупных района аутосом, име-
ющих идентичное генное содержание и сходных по рисунку
G-бэндинга. В последующих работах районов гомологии хромо-
сом кошки и человека было выявлено еще больше [Kiel von et
aL, 1985]. На основании сравнительного анализа хромосом, мар-
кированных гомологичными генами, выявлены гомеологичные
районы хромосом кошки и ряда других видов млекопитающих,
для которых проводились работы по картированию [Графодат-
ский, Билтуева, 1987; Rubtsov et al., 1988; Grapliodatsky, 1989]
(табл. 3.2, рис. 3.9). Совокупно эти данные указывают на су-
ществование в эволюции млекопитающих консервативных райо-
нов хромосом, изменения (перестройки) которых, возможно, име-
41
Рис. 3.9. Некоторые гомеологпчные районы хромосом кошки, человека и ря-
да других видов млекопитающих.
Обозначения хромосом см. табл. 3.2.
ли значительные макроэволюционные последствия либо сопут-
ствовали им. В любом случае фиксация этих перестроек служит
прекрасными филогенетическими маркерами, особенно при ана-
лизе макроэволюционных событий.
СПИСОК ЛИТЕРАТУРЫ
Графодатский А. С., Бплтуева Л. С. Гомология G-окрашенных хромосом
млекопитающих // Генетика.— 1987.— Т. 23.— С. 93—103.
Графодатский А. С., Раджабли С. И. Хромосомы сельскохозяйственных и ла-
бораторных млекопитающих: Атлас Ц Новосибирск: Наука. Сиб. отд-
ние, 1988.— 127 с.
Benirschke К. Chimerism, mosaicism and hybrids Ц Proc. 4th Intern. Congr.
on Human Genetics, Paris, France.— 1971.— P. 1973.
Benirschke K., Edwards R., Low R. J. Trisomy in a feline fetus // Amer. .1.
Vet. Res.- 1974,— Vol. 35,- P. 257-259.
Centerwall W. R., Benirschke K. An animal model for the XXV Klinefelter’s
syndrome in man: tortoiseshell and calico male cats Ц Amer. J. Vet.
Res.- 1973a.— Vol. 36,- P. 1275-1280.
Centerwall W. R., Benirschke K. Male tortoiseshell and calico (T — C) cats Ц
J. Hered.— 1973b.— Vol. 64,— P. 72—78.
Chu E. H. Y., Thuline H. C., Norby D. E. Triploid-diploid chimerism in a male
tortoiseshell cat // Cytogenetics.— 1964.— Vol. 3.— P. 1—18.
Couturier J., Dutrillaux B. Chromosomal evolution in Carnivora Ц Mammalia.—
1986,— Vol. 50.— P. 124-163.
Foley C. W., Lasley J. F., Osweiler G. D. Abnormalities of companion animals.—
Ames Iowa: The Iowa Univ. Press, 1979.— P. 200—203.
Ford С. E., Pollock D. L., Gustavsson I. Proc. 1st Intern. Conf, for the standar-
dization of banded karyotypes of domestic animals. Univ, of Reading,
England Ц Hereditas.— 1980.— Vol. 92.— P. 145—162.
Graphodatsky A. S. Conserved and variable elements of mammalian chromoso-
mes Ц Cytogenetics of Animals/Ed. C. R. E. Hainan.— Oxford, UK: CAB
Intern. Press, 1989.— P. 95—123.
Graphodatsky A. S., Hainan C. R. E. Appendix 1. An atlas of selected karyoty-
pes // Ibid.- P. 321-390.
Han Qin, Hainan C. R. E. Notes on the pig, goat, sheep, hybrids and cats Ц
Ibid.— P. 267-286.
Hare W. C. D., Weber W. T., McFeely R. A., Yang T.-J. Cytogenetics in the
dog and cat Ц J. Small Anim. Practice.— 1966.— Vol. 7.— P. 575—592.
Hsu T. C., Benirschke K. An atlas of mammalian chromosomes.— N. Y.: Sprin-
ger-Verlag, 1967—1977,—Vol. 1/10.
HGM 10,5 (Human Gene Mapping 10,5). Report of the committee on compara-
tive gene mapping Ц Cytogenet. Cell Genet.— 1990.— Vol. 55.— P. 406—
433.
Johnston S. D., Buoen L., Madl J. E. et al. X chromosome monosomy (37X0)
in a Burmese cat with gonadal dysgenesis Ц JAVMA.— 1983.—
Vol. 182,- P. 986-989.
Jones T. C. San Juan Conference on karyotypes of Felidae. Special Report Ц
Mammal. Chromos. Newslett.— 1965.— Vol. 15.— P. 121—122.
Kiel K. von, Hameister H., Somsich I. E., Adalph S. Early replication banding
reveals a strongly conserved functional pattern in mammalian chromo-
somes Ц Chromosoma.— 1985.— Vol. 93.— P. 69—76.
Kral B., Zima J. Karyosystematic of the family Felidae Ц Gazella. Zoo Pra-
ha.— 1980,— Vol. 2/3,— P. 45-53.
Loughman L. S., Fraye F. L., Condon T. B. XY/XXY bone marrow mosaicism
in three tricolor cats Ц Amer. J. Vet. Res.— 1970.— Vol. 31.— P. 307—
314.
Makino S., Tateishi S. A comparison of the chromosomes in the lion, Chinese
leopard cat and house cat Ц J. Morphol.— 1952.— Vol. 90.— P. 93—102.
Matthey R. La formule chromosomiale du chat domestique Ц C. R. Soc. Biol.,
Paris.— 1934.
McFeely R. A., Hare W. C. D., Biggers J. D. Chromosome studies in 14 cases
of intersex in domestic mammals Ц Cytogenetics.— 1967,— Vol. 6.—
P. 242—253.
Minouchi O. On the chromosomes of the cat // Proc. Imp. Acad.— 1928a.—
Vol. 4,— P. 128—130.
Minouchi O. On the chromosomes of the racoon dog and the cat / Zool.
Mag.- 1928b.— Vol. 40,— P. 490-491.
Minouchi O., Ohta T. The chromosome number in the oogonium of the cat Ц
Zool. Mag.— 1932,— Vol. 44,— P. 355—362.
Minouchi O., Ohta T. On the chromosome number and sex chromosomes in
the germ-cells of male and female cats Ц Cytologia.— 1934.— Vol. 5.—
P. 355-362.
Modi W. S., O’Brien S. J. Quantitative cladistic analyses of chromosomal ban-
ding data among species in three orders of mammals: hominoid pri-
mates, felids and arvicolid rodents Ц Chromosome structure and func-
tion/Ed. J. P. Gustafsson, R. Appels.— Plenum Publ. Co., 1988.— P. 215—
242.
Nash W. G., O’Brien S. J. Conserved regions of homologous G-banded chromo-
somes between orders in mammalian evolution: carnivores and prima-
tes Ц Proc. Natl. Acad. Sci. USA.— 1982,— Vol. 79 — P. 6631—6635.
O’Brien S. J. Genetic maps: locus maps of complex genomes.—5th ed.— Cold
Spring Harbor: Cold Spring Harbor Press, 1990.— 1078 p.
O’Brien S. J., Nash W. G. Genetic mapping in mammals: Chromosome map of
the domestic cat // Science.— 1982.— Vol. 216.— P. 257—265.
O’Brien S. J., Haskins M. E., Winkler C. A. et al. Chromosomal mapping of be-
ta-globin and albino loci in the domestic cat Ц J. Hered.— 1986.—
Vol. 77,— P. 374—378.
43
О Brien S. J., Seuanez H. N., Womack J. E. On the evolution of genome or-
ganization in mammals // Molecular evolutionary genetics/Ed. R. J. Mac-
Intyre.— Plenum Publ. Co., 1985,— P. 519—589.
Ohno S. Evolution of sex chromosomes in mammals Ц Ann. Rev. Genet.—
1969,— Vol. 3,— P. 495—524.
Pathak S., Stock D. The X-chromosomes of mammals: karyolcgical homology
as revealed by banding techniques Ц Genetica.— 1974,— Vol. 78.—
P. 703-714.
Pathak S., Wurster-Hill D. H. Distribution of constitutive heterochromatin in
carnivores Ц Cytogenet. Cell Genet— 1977,—Vol. 18.—P. 245—254.
Pearson M. D., Seabright M., Maclean N. Silver staining of nucleolus organi-
zer regions in the domestic cat, Felis catus // Cytogenet. Cell Genet.—
1979.- Vol. 24,- P. 245-247.
Rcnne M., Shibasaki Y., Strandby Paulsen B. The high resolution R-banded
karyotype of Felis catus Ц Proc. 8th Eur. Colloq. Cytogenet. Domest.
Anim. Bristol.— 1988,— P. 78—82.
Rubtsov N. B., Graphodatsky A. S., Matveeva V. G. et al. Silver fox gene map-
ping: conserved chromosome regions in the order Carnivora // Cytoge-
net. Cell Genet.— 1988,— Vol. 48,— P. 95—98.
Shibasaki Y., Flou S., Ronne M. The R-banded karyotype of Felis catus Ц
Cytobios.— 1987 — Vol. 51,— P. 35—47.
Thomsen D. P., Byskov A. G., Basse A. Fertility in two cats with X-chromoso-
me mosaicism and unilateral ovarian dysgenesis Ц J. Reprod. Fertil.—
1987,—Vol. 80,—P. 43-47.
Thuline H. C., Norby D. E. Spontaneous occurence of chromosome abnormali-
ty in cats / Science.— 1961,— Vol. 134,— P. 554—555.
Wurster D. H. Cytogenetic and phylogenetic studies in Carnivora Ц Compara-
tive mammalian cytogenetics/Ed. K. Benirschke.— N. Y.: Springer-Ver-
lag, 1969,— P. 310—329.
Wurster D. H., Benirschke K. Comparative cytogenetic studies in the order
Carnivora // Chromosoma.— 1968.—Vol. 24.— P. 336—382.
Wurster-Hill D. H., Centerwall W. R. The inter-relationships of chromosome
banding patterns in canids, mustelids, hyaena, and felids Ц Cytogenet.
Cell. Genet.— 1982,— Vol. 34,— P. 178—192.
Wurster-Hill D. H., Gray C. W. Giemsa banding patterns in the chromosomes
of twelve species of cats (Felidae) Ц Cytogenet. Cell Genet.— 1973.—
Vol.. 12,—P. 377—397.
Wurster-Hill D. H., Gray C. W. The inter-relationships of chromosome banding
patterns in procyonid, viverrids, and felids // Cytogenet. Cell Genet-
1975.— Vol. 15,— P. 306-331.
Глава 4
ГЕНЕТИКА ОКРАСОК
Окраска, унаследованная домашней кошкой от своего дикого
предка Felts libyca, представляет собой сочетание серо-коричне-
вого фона с темными разводами и вертикальными полосами.
Такая цветовая комбинация, как бы размывающая четкие кон-
туры животного, является примером защитной окраски, распро-
страненной как среди хищников, так и среди их жертв. Благо-
даря генетическому анализу удалось установить, что цвет меха
44
Рис. 4.1. Характер рас-
пределения пигмента в
просветленных остевых
волосах кошек (рисунок
Л. А, Прасоловой).
Генотип А—1: шиншилла
затушеванная. По всей
длине волоса отмечается
чередование участков с раз-
ной степенью заполнения
пигментными гранулами
коркового и сердцевинного
слоев от сильно пигменти-
рованных (1) к менее пиг-
ментированным (2, 3),
вплоть до полностью ли-
шенных пигмента. Помимо
россыпи отдельных гранул
в сердцевине иногда встре-
чаются скопления в виде
шариков (Х300).
у кошки определяется целой системой генов, отвечающих за раз-
личные окрасочные формы.
Окраска шерсти возникает в результате сочетания отдельных
волос различных цветов и типов. Типичный цвет волоса млеко-
питающих — интенсивно пигментированный (черный) на конце
и постепенно ослабевающий (до голубого) к корню. Однако в
остевых волосах (см. ниже) осветление прерывается субапи-
кальными узкими кольцами желтого пигмента (рис. 4.1). Воло-
сы на боках животного прилегают к телу, в результате чего эти
кольца становятся очень четко видны. Именно сочетание тем-
ных кончиков и желтых полос посередине волоса и приводит к
появлению серо-коричневой окраски агути.
45
Темный рисунок из полос и разводов возникает в результате
концентрации в отдельных районах равномерно окрашенных чер-
ных волос (волос теряет желтые кольца). У кошек комбинация
темных пятен на фоне агути приводит к появлению окраски,
которая называется «тэбби». Такая окраска может возникнуть
независимо от фона агути. Это объясняется наличием в геноме
кошки некоторых мутантных генов, которые могут модифициро-
вать как агути, так и тэбби-окраску шерсти. Взаимодействие
таких мутантных генов приводит к появлению всего спектра
окрасок, лежащих в основе многих пород кошек.
Локус Т (Tabby). Типичный пестрый рисунок рода F. libyca
обычно обозначается как полосатый, тигровый или макрелевый
(последний термин обычно используется специалистами, именно
он принят за стандартный или дикий фенотип). Локус, ответст-
венный за тэбби-окраску, обозначается символом Т. Он имеет
три известных аллеля, обозначаемых: абиссинский Та, полосатый
Т и мраморный tb. Характер рисунка, вызываемого аллелем Т,
считается селекционерами кошек классическим.
Аллель Та. Свое название абиссинский аллель получил в
честь породы кошек, для которой он наиболее характерен. Этот
аллель при наследовании проявляет свойства неполного доми-
нирования по отношению как к аллелю полосатой окраски Т,
так и к аллелю мраморной окраски tb. У абиссинской породы
кошек полностью отсутствует пестрый рисунок на теле, а поло-
сы сохраняются лишь на морде. Редкие отметины видны на пе-
редних лапах, бедрах и кончике хвоста. У гетерозигот Т/Та и
Ta/tb эти отметины проявляются более четко, чем у гомозигот
Та/Та. Обычно селекционеры ведут отбор на сокращение коли-
чества отметин, и лучшие выставочные представители абиссин-
ской породы не имеют полос на теле, за исключением головы и
кончика хвоста. Шерсть при этом имеет окраску осветленного
агути.
Аллель tb. Животные с мраморным рисунком в отличие от
полосатых выглядят несколько темнее. Тонкие полосы, характер-
ные для полосатых животных, вытеснены у мраморных более
темным рисунком из пятен и колец. Наиболее четко темный ри-
сунок выявляется на лапах и хвосте. Аллель, определяющий
мраморный рисунок, является рецессивным по отношению к нор-
мальному аллелю и в гетерозиготе с ним (T/tb) дает полосатую
окраску.
Локус A (Agouti). Локус агути А имеет один мутантный ал-
лель, называемый «неагути» и обозначаемый символом а. Действие
этого аллеля вызывает потерю остевыми волосами желтых ко-
лец (рис. 4.2). У гомозиготных (а/а) животных волос имеет тем-
ный кончик, а затем постепенно светлеет по направлению к кор-
ню, становясь голубым. Это объясняется тем, что пигментные
гранулы распределены у корня волоса менее плотно. Визуально
шерсть выглядит очень темной. Этот аллель обычно проявляет
эпистатичность по отношению к аллелю тэбби. Интересно, что
46
Рис. 4.2, Характер рас-
пределения пигмента в
просветленных остевых
волосах кошек (рисунок
Л. А. Прасоловой).
Генотип aal: черный дым-
чатый. Остевые волосы на-
половину пли на 1/3, начи-
ная с вершины (1, 1а), ин-
тенсивно окрашены в чер-
ный цвет за счет плотного
расположения отдельных
пигментных гранул в ко-
ре и сердцевине. Нижняя
часть волоса не содержит
пигмента (2, 2а) (хЗОО (1),
Х500 (1а, 2а)).
если разглядывать шерсть котенка в лучах яркого солнечного
света, то можно заметить на темном фоне легкий след, сохранив-
шийся от пестрого рисунка, который полностью исчезает у взрос-
лой кошки.
Локус В (Black). Локус коричневой окраски В представлен
двумя мутантными аллелями, обозначаемыми как b — коричне-
вый, и Ъ1 — светло-коричневый. Для обозначения темно-коричне-
вого цвета меха, который наблюдается у кошек, гомозиготных по
аллелю Ь, селекционерами был введен специальный термин —
шоколадный окрас. У животных, гомозиготных по аллелю Ъ1,
цвет меха значительно светлее. Гетерозиготность по двум этим
аллелям приводит к значительному уменьшению интенсивности
47
Рис. 4.3. Характер распре-
деления пигмента в про-
светленных остевых воло-
сах кошек (рисунок
Л. А, Прасоловой).
Генотип А—dd: голубой тигро-
вый. Чередование по всей дли-
не волоса участков с неравно-
мерным распределением пиг-
мента в виде отдельных гра-
нул в коре и сердцевине и ча-
сто встречающихся конгломе-
ратов, представляющих выбро-
шенные в волос безотростча-
тые меланоциты. 1 — остевой
волос, 2 — пуховый (Х300).
окраски пигментных гранул по сравнению с нормой. Светло-
коричневый окрас также называют «циннамон» (коричневый).
Локус D (Dense pigmentation). Локус D представлен един-
ственным мутантным аллелем d. Действие этого аллеля приво-
дит к склеиванию пигментных гранул. В результате их равно-
мерное поступление в растущий волос нарушается и в стволе во-
лоса возникают отдельные пустоты. Это приводит к скоплению
массы гранул в одном участке волоса и нехватке гранул в дру-
гом. В результате перераспределения гранул окраска шерсти
кажется светлее (рис. 4.3). При наличии такого аллеля-осветли-
теля темная окраска шерсти воспринимается как серо-голубая,
коричневая — как голубая с коричневатым оттенком и называ-
48
ется сиреневой, а светло-коричневая становится золотистой. Лю-
бопытно, что распределение пигментных гранул в радужной обо-
лочке глаз голубых кошек оказывается нормальным [Prieur, Col-
lier, 1981b]*.
Локус С (Color). В локусе С известно четыре мутантных
аллеля, обозначаемых как сь — бирманский альбино, cs — сиам-
ский альбино, са — голубоглазый альбино и с — альбино с розо-
выми глазами.
В гомозиготе аллель сь приводит к появлению коричневой
окраски цвета темной сепии, которая меняет свой оттенок в дор-
зально-вентральном направлении, становясь совсем светлой на
животе. Голова и лапы у таких животных значительно темнее.
В гомозиготе аллель cs вызывает у кошек появление окрасок
от светлой сепии до белоснежной с темными конечностями (нос,
лапы, хвост) и голубыми радужными оболочками глаз. Бирман-
ский и сиамский аллели являются и температурочувствительны-
ми. Это проявляется в том, что у животных, носителей данных
аллелей, при нормальной температуре тела пигментные гранулы
теряют свою окраску. Как известно, у животных температура
конечностей несколько ниже средней температуры тела и равна
критическому физиологическому уровню, что позволяет пигмент-
ным гранулам в этих районах сохранять темный цвет на фоне
светлой окраски всего тела [Iljin N., Iljin V., 1930].
Аллель са. Голубоглазые альбино имеют белую окраску
шерсти, светло-голубую радужную оболочку глаз и полупрозрач-
ные зрачки.
Аллель с. У альбино с розовыми глазами также белая
окраска шерсти, но радужная оболочка у них прозрачная, а зра-
чок розовый.
В результате можно сделать вывод, что действие аллелей
альбино заключается в ослаблении пигментации до такой степе-
ни, что не только шерсть, но и глаза практически полностью
лишены пигмента [Turner et al., 1981; Creel et al., 1982; Robin-
son, 1991].
Локус I (Melanin inhibitor). В данном локусе известен один
доминантный аллель I. Действие этого аллеля проявляется в
снижении способности волоса накапливать пигментные гранулы.
У кошек в присутствии аллеля I в верхушечной части волоса
сохраняется полный запас пигмента, но в направлении к осно-
ванию волоса наблюдается заметная его нехватка, что приводит
в итоге к полному обесцвечиванию корней волос (см. рис. 4.1,
4.2). Этот окрас иногда называют шиншилловым. Экспрессия
гена I зависит от количества пигмента, продуцируемого волосом.
Например, у кошек типа тэбби участки шерсти с окрасом агути
сильнее подвержены воздействию этого аллеля, чем темные уча-
стки. Если под действием аллеля I участки с окрасом агути
становятся почти белыми, то в области темных пятен волосы
* См. список литературы к гл. 2.
4 Заказ ^s 355
49
сохраняют способность синтезировать пигмент. Такая окраска
носит название серебряной.
С другой стороны, в комбинации с неагути присутствие ал-
леля I проявляется лишь в появлении бледно-голубого или бе-
лого подшерстка. Такой фенотип получил название «дымчатый».
Экспрессия аллеля I очень разнообразна, однако на фоне сереб-
ряного окрашивания эта вариабельность проявляется несколько
слабее по сравнению с дымчатой. Для организмов с генотипом
а/а, I/— проявление гена I настолько незначительно, что сохра-
няется обычный темный фенотип, характерный для животных аа.
У оранжевых кошек аллель I вызывает уменьшение количества
волосяного пигмента, в результате вместо кремового подшерстка
появляется белый, что приводит к общему осветлению меха.
Этот фенотип получил название «камея». В связи с сильной из-
менчивостью экспрессии аллеля I спектр оттенков окраски меха
таких животных может изменяться от едва заметного желтого
налета до темного, практически не отличимого от нормального
красного тэбби [Turner, Robinson, 1980; Prieur, Collier, 1981а].
Локус О (Orange). Хромосома X. В локусе orange известен
один доминантный мутантный аллель О. Действие этого аллеля
заключается в превращении черного/коричневого пигмента
(эумеланина) в желтый (феомеланин). Результатом является
фенотип красный тэбби, представленный желтым фоном и крас-
ными разводами на нем. По отношению к локусам агути и к ло-
кусу black аллель О проявляет эпистатичность, подавляя экс-
прессию генов в этих локусах. Поскольку локус О локализован
на Х-хромосоме, существует половой диморфизм по этой окрас-
ке, состоящий в том, что если среди самцов различаются только
две окрасочные формы: оранжевая (O/Y) и обычная (o/Y), то
среди самок встречаются также особи с черепаховой окраской
О/о. Черепаховый фенотип у гетерозиготных кошек появляется
в результате мозаицизма, возникающего при эмбриональной инак-
тивации одной из Х-хромосом. В этом случае аллель О экспрес-
сируется лишь в некоторых соматических клетках, в то время
как в других клетках экспрессируется аллель о. Подробно при-
рода этого явления обсуждалась выше (см. гл. 3).
Локус р (Pink eyed). Для животных, гомозиготных по рецес-
сивному мутантному гену р, характерно осветление окраски меха
до рыжевато-коричневой, при этом цвет глаз у них становится
красновато-розовым. О характере наследования этого признака
известно еще недостаточно, однако предварительные данные сви-
детельствуют о том, что такая окраска является рецессивной по
отношению к дикому фенотипу.
Локус Wb(Wide band)'. Пестроокрашенные кошки, произошед-
шие от гетерозиготных кошек породы шиншилла, имеют более
светлый оттенок шерсти, чем обычные тэбби. Их экстерьер на-
водит на мысль, что эти животные отличаются от обыкновенных
тэбби наличием дополнительного аллельного варианта. Известно,
что у некоторых видов млекопитающих обнаружен аллель, кото-
50
рый вызывает появление желтоватого оттенка на фоне агути в
результате расширения полосы желтого пигмента (агути бэнда).
Такой аллель назван «Wide band». Окраска, возникающая в ре-
зультате экспрессии этого аллеля, может быть названа «золотой
тэбби». Уменьшение количества черного пигмента, вызванное
данным аллелем, в сочетании с эффектом аллеля I ответственно
за формирование белого шиншиллового окраса. Существует пред-
положение о доминантном способе наследования этого признака,
который предварительно обозначается как Wb.
Локус S (Piebald spotting). Локус белой пегости представлен
полудоминантным аллелем S. Этот аллель вызывает появление
отдельных белых пятен. Степень экспрессии может варьировать
от маленьких пятнышек на носу, лапках, груди и животе до по-
явления практически полностью белой шерсти. Однако макси-
мальная форма проявления пегости, приводящая к появлению
абсолютно белой окраски у животного, встречается очень ред-
ко. Наиболее характерно наличие отдельных белых пятен на
голове и у основания хвоста. У гетерозигот по гену пегости
обычно наблюдаются на фоне окрашенного меха отдельные бе-
лые пятна на плечах и животе. Кошки со значительной долей
белого меха, как правило, имеют частично или полностью голу-
бые радужные оболочки глаз. Уменьшение уровня пигментации
у таких животных связано с нарушением синтеза меланобластов
в нейральном кресте в период эмбриогенеза. В зависимости от
степени нарушения, большая или меньшая часть меха будет
лишена меланоцитов. В очень редких случаях ген пегости вызы-
вает появление той или иной степени глухоты.
Локус W (White dominant). Локус White представлен доми-
нантным аллелем W. Кошки, носители этого аллеля, имеют аб-
солютно белую шерсть (рис. 4.4). Изредка наблюдается неболь-
шое темное пятно на голове. Это пятно появляется у всех котят,
но очень редко оно сохраняется у взрослых кошек. Любопытно,
что существует определенная корреляция между голубым цветом
глаз и нарушениями работы органов слуха: если у кошки голу-
бые глаза и абсолютно белая шерсть, то вероятность того, что
она будет страдать от глухоты, достоверно выше, чем у кошки
с желтыми глазами. Показано, что аллель W оказывает сходное,
но более серьезное влияние на пролиферацию меланобластов по
сравнению с геном пегости S. Кроме того, он имеет ряд плейо-
тропных эффектов. Максимальное воздействие он оказывает на
цвет шерсти, приводя к появлению белого окраса, несколько
меньшее — на другие признаки, вызывая частичную или полную
глухоту и появление голубого цвета глаз в результате недостатка
пигмента и полного отсутствия тапетума в радужной оболочке
[Thibos et al., 1980]. Высказывалось предположение, что аллель
White является одним из аллелей локуса пегости S [Whiting,
1919]. Однако пока экспериментальных подтверждений этому
недостаточно, а для практических целей значительно удобнее рас-
сматривать аллель White принадлежащим отдельному локусу.
4*
51
Рис. 4.4. Характер распределе-
ния пигмента в просветлен-
ных остевых волосах кошек
(рисунок Л. А. Прасоловой).
Генотип W: белый. Остевые и пу-
ховые волосы чисто-белого цвета,
не содержат пшментных гранул
по всей длине. 1 — Х300; 2 —
Х500.
| { Выше былп рассмотре-
ны окрасочные варианты,
для которых установлен
| моногенный характер на-
следования, однако суще-
ствует ряд признаков, ге-
нетическая природа кото-
• * I ' рых до К0НЧа не пзуче-
ц IfiiillS® |,а' Остановимся кратко
на них.
Имеется ряд доказа-
тельств существования ал-
леля, приводящего к по-
| f ' явлению небольших белых
’ . пятен на лапках. У ко-
шек бирманской породы,
например, выделяются бе-
лые пятнышки на лапках,
: которые селекционеры на-
зывают перчатками (glo-
ve) или чулочками (stoc-
king) в зависимости от
степени выраженности
данного признака. Ранее
появление этого признака
рассматривалось как ре-
зультат очень слабой
Д-. экспрессии аллеля S, од-
нако генетический анализ
особей бирманской поро-
ды позволил сделать заключение, что наследование этого
признака осуществляется по рецессивному типу. Таким образом,
вероятно, существует другой аллель, отличный от S, приводящий
к появлению белых пятен на лапах. Количество данных, под-
тверждающих это, невелико, но достаточно убедительно. Предва-
рительно аллелю gloving присвоен символ g [Clarke, 1983, уст-
ное сообщ.].
Яркость окраски шерсти животных находится под контролем
большого количества различных полигенов. Действие одного из
них можно обнаружить благодаря его влиянию на интенсивность
окрашивания волоса пигментом феомеланином (оранжевый, жел-
тый) . Например, дикие кошки окраса обыкновенный тэбби — се-
КО
ро-коричневые, в то же время выставочные представители тэбби
и абиссинской пород кошек имеют более яркую, красно-коричне-
вую окраску. Аналогично обыкновенный оранжевый тэбби зна-
чительно тусклее, чем выставочный красный тэбби. Только пред-
положив существование целого комплекса генов, модифицирую-
щих окраски, вызванные действием главного гена, можно объ-
яснить то разнообразие оттенков, которое можно наблюдать на
выставках кошек.
Глава 5
ГЕНЕТИКА СТРУКТУРЫ ШЕРСТИ
ВВЕДЕНИЕ
Шерсть кошки представлена тремя типами волос: первичные
направляющие, или кроющие; вторичные направляющие, или
остевые; пуховые, или подпушь.
Первичные направляющие волосы — самые длинные и прочные
из всех трех типов волос. Как правило, они прямые и лишь
иногда могут быть слегка волнистыми. По направлению от кор-
ня волоса к кончику происходит его постепенное утолщение,
достигающее максимума в субапикальном районе, верхушечная
же часть волоса имеет правильную конусовидную форму. Основ-
ная функция кроющих волос — защита кожи от повреждений.
Кроме того, они выполняют тактильную функцию, информируя
животное об опасности в случае его прикосновения к различным
предметам.
Количество вторичных направляющих волос на единицу пло-
щади в среднем значительно больше, чем кроющих. В отличие
от первичных, они сильно различаются между собой, варьируя
по толщине. Чем прочнее и ровнее вторичные волосы, тем сла-
бее проявляется волнистость шерсти в целом. На протяжении
всей длины остевые волосы имеют одинаковый диаметр, за ис-
ключением района субапикального утолщения, после которого
формируется конусовидная вершина. Главная их функция — за-
щита пуховых волос.
Пуховые волосы значительно отличаются от двух типов во-
лос, описанных выше. Они наиболее густые, очень тонкие и име-
Ьт одинаковую толщину на протяжении всей длины. Их функ-
ция — сохранение температуры тела и предохранение кожи жи-
вотного от значительных колебаний температуры окружающей
Среды. Количество изгибов и толщина пуховых волос идеальны
Для этих целей. Однако сами они нуждаются в защите кроющи-
ми волосами от повреждений.
Вибриссы представляют собой чрезвычайно толстые, длин-
ные волосы, сконцентрированные преимущественно надо ртом, так
называемые «усы». Другие расположены над глазными дугами
и на щеках. Все они выполняют тактильную функцию.
ГИПЕРТРИХОЗ
Чрезмерный рост шерсти у кошек едва ли можно рассматри-
вать как аномалию, однако, несомненно, он представляет собой
некоторое отклонение от нормы. Причина появления длинных
волос у кошек углубленно не изучалась, но, вероятно, она кро-
ется в удлинении периода нормального роста волос (фазы реге-
нерации) . Подобное явление наблюдается и у кроликов, носи-
телей родственного мутантного аллеля angora, вызывающего
обильный рост волос [Fraser, 1953]*.
ГИПОТРИХОЗ
Снижение интенсивности роста волос приводит к появлению
у животных шерсти типа «реке». Мех у кошек значительно ко-
роче и мягче, чем у нормы за счет истончения кроющих и осте-
вых волос; вибриссы короче и сильно изогнуты. Работы по изу-
чению причин, обусловливающих появление такого типа шерсти
у кошек, практически не проводились. Однако анализ роста шер-
сти у кролика, носителя гена, родственного реке-гену кошки, по-
казал, что продолжительность фазы регенерации у таких живот-
ных нормальная, но скорость роста волос заметно снижена
[Fraser, 1953].
Кошки с рекс-типом шерсти широко распространены по все-
му земному шару. Они встречаются в Австралии, Британии, Гер-
мании, Голландии, Италии, Швеции и Америке. Различают не-
сколько мутантных форм гена реке: г—корниш реке, те — девон
реке, го — орегон реке, Rd — датский реке и Se — селкирский
реке. Некоторые селекционеры выделяют аллель немецкий реке
rg, однако вероятно, что он представляет собой либо ту же му-
тацию, что и корниш реке, либо очень близкий аллель того же
самого локуса. Большинство из перечисленных аллелей являются
рецессивными по отношению к аллелю, определяющему нормаль-
ную структуру меха. Исключение составляют аллели датского и
селкирского рекса, которые наследуются как доминантные. Общей
характеристикой шерсти всех рексов является ее шелковистость
и мягкость, которые возникают в результате или полной утраты
мутантами направляющих волос или сильной их редукции п со-
хранения лишь пуховых волос. Исключение составляет девон
* См. список литературы к гл. 2.
54
реке, у которого сохраняются направляющие волосы, правда,
в сильно измененной форме. В результате мех оказывается не-
сколько грубее по сравнению с другими реке формами. Иногда
у этой породы кошек наблюдается частичное выпадение волос
[Searle, Jude, 1956; Robinson, 1968, 1971, 1972а, 1982а, 1991].
Аллель г. Мутантный аллель корниш реке (г) наследуется
как рецесспв по отношению к аллелю нормального типа шерсти.
У гомозиготных носителей данного аллеля присутствуют все три
типа волос, однако в очень укороченном и истонченном виде.
Кроме того, все они несколько закручиваются, что создает ощу-
щение мягкости и волнистости шерсти. Вибриссы у такого типа
кошек очень короткие п искривленные [Searle, Jude, 1956; Ro-
binson, 1971].
Аллель re. Аллель девон реке (ге) также наследуется по ре-
цессивному типу. Структура шерсти у животных ге/ге более
аномальна, чем у гомозигот г/г. Первичные направляющие воло-
сы практически отсутствуют, причем сохранившиеся — сильно
укорочены, изогнуты п значительно варьируют в диаметре.
Шерсть кажется очень редкой, несколько вытертой на груди,
плечах и животе. Вибриссы короткие, изогнутые, легко ломают-
ся [Robinson, 1969а].
Аллель rg. Немецкий реке (rg) внешне очень похож на кор-
нпш реке и также наследуется по рецессивному типу. Скрещи-
вания животных этих двух типов свидетельствуют в пользу того,
что либо немецкий и корниш рексы представляют собой одну
и ту же мутацию, либо представлены очень сходными аллелями;
первое предположение — наиболее вероятно [Robinson, 1968,
1971].
Аллель го. Орегонский реке (го) напоминает корниш и также
наследуется по рецессивному типу. Опыт селекционной работы
свидетельствует о том, что скрещивания орегонского рекса с
корниш рексом или девон рексом приводят к появлению котят
с нормальным типом шерсти. Это говорит о том, что мутация
орегонский реке не связана с аллелями серии реке [Robinson,
1972а].
Аллель Rd. Аллель датский реке (Rd) наследуется по доми-
нантному типу. Шерсть у таких мутантов при рождении волни-
стая, впоследствии заменяется более тонкими, но грубыми воло-
сами. На отдельных частях тела может наблюдаться полная по-
теря волосяного покрова. При этом грудная клетка, лапы и хвост
обычно остаются пушистыми. Волосы у таких кошек аномально
короткие, тонкие, неравномерно разреженные, с плохо сформи-
рованной луковицей у корня. Вибриссы сильно извиты и загнуты
[Robinson, неопубл, данные].
Аллель Se. Аллель селкирский реке (Se) является доминант-
ным по отношению к аллелю нормального типа шерсти. Обыч-
но такую шерсть описывают как густую, с характерными мягки-
ми волнами, поскольку оба типа направляющих волос на теле
отсутствуют, и лишь на хвосте сохраняются длинные тонкие
55
волосы, напоминающие направляющие. Вибриссы и шерсть над
глазами короче, чем у нормы, и извиты [Robinson, неопубл,
данные].
Другие мутантные аллели, приводящие к появлению рексопд-
ных форм шерсти, были обнаружены в Австралии, Италии и
Швеции, Британии и Америке. В отдельных случаях генетиче-
ская природа таких форм пока не установлена, в других — пред-
полагается рецессивный характер наследования [Robinson, 1991].
Связь между этими фенотипами и мутациями, описанными вы-
ше, пока не ясна. Комбинация рексоидной формы и длинных
волос приводит к появлению шерсти, длина которой больше, чем
в норме, но меньше, чем при обычной длинношерстности. При
этом очень часто шерсть становится редкой и плохого качества.
ПРОВОЛОЧНЫЙ волос
Аллель Wh. У животных с таким типом шерсти представлены
все три типа волос, правда, аномально тонкие. Остевые волосы
закручены, подпушь также волнистая. Пуховые волосы сильно
утолщены и неправильно изогнуты. На ощупь шерсть кажется
грубой и слегка упругой при поглаживании. Появление такого
типа шерсти вызывается наличием доминантного аллеля Wh
(Wire hair — волнистость шерсти) [Robinson, 1991].
РЕДКИЙ МЕХ
Аллель sf. Животные такого типа имеют тонкий, разрежен-
ный мех с отдельными участками облысения. При этом волосы
укорочены и сильно деформированы, причем вибриссы короткие
и раздвоенные. Желтовато-коричневые разводы проступают во-
круг глаз, ноздрей и рта, часто обнаруживаются на груди и жи-
воте. Веки становятся истонченными, а на глазах обнаруживают-
ся признаки сепсиса. Данные селекции свидетельствуют о рецес-
сивном характере наследования этого признака, который обозна-
чается символом sf (Sprace fur) [Robinson, 1991].
АТРИХОЗ
Существует большое количество сообщений об обнаружении в
различных уголках мира безволосых или почти безволосых ко-
шек. Их экстерьер определяется наличием мутантных аллелей,
подобных аллелям из серии реке. Безволосые котята рождаются
покрытыми подпушьподобными волосами. Как правило, впослед-
ствии они исчезают, однако изредка могут заменяться настоящи-
ми волосами.
56
Аллель h. В 1938 г. впервые установлен рецессивный харак-
тер наследования данного признака (французский вариант без-
волосости, символ h) [Letard, 1938]. Как ни странно, вибриссы
у таких животных имеют нормальную форму.
Аллель hr. Канадский вариант безволосости (hr) фенотипи-
чески подобен французскому, но вибриссы имеют загнутую фор-
му. В данном типе шерсти выделяют два типа коротких и тон-
ких волос, но волосы одного типа толще, чем другого. Оба типа
волос не имеют хорошо сформированной волосяной луковицы
[Robinson, 1973].
Аллель hd. Редкая мутация (hd) отличается от описанных
выше тем, что в этом случае шерсть у животного представлена
лишь легким налетом пуховых волос. Кожа мягкая и эластичная
при рождении, впоследствии становится очень толстой и мор-
щинистой. Вибриссы укорочены и сильно изогнуты. Коричнева-
тый оттенок проступает вокруг рта, глаз и на подбородке. Часто
обнаруживается расщепление когтей. Большинство кошек с та-
кой мутацией погибают в раннем возрасте [Robinson, 1981; Неп-
dy-Ibbs, 1984].
АСТЕНИЯ КОЖИ
Аллель Cat. У носителей данной мутации кожа имеет барха-
тистую структуру и необыкновенную растяжимость. Кроме того,
она очень восприимчива к повреждениям и даже легкие царапи-
ны после заживления оставляют белые грубые рубцы. Причина
аномалии заключается в нехватке коллагеновых волокон и фиб-
рилл в ретикулярном слое кожи. Описанный признак наследу-
ется по доминантному типу и обозначается символом Cat [Pat-
terson, Minor, 1977].
ДЛИННАЯ ШЕРСТЬ
Аллель 1. Данный локус представлен единственным мутант-
ным аллелем long hair, который приводит к появлению у кошек
длинной шерсти. Вероятно, такой эффект достигается в резуль-
тате увеличения продолжительности периода роста волос. При
скрещивании признак «длинная шерсть» наследуется как рецес-
сивный по отношению к нормальной длине волос. В зависимости
от степени увеличения длины, структуры и толщины волос в
таком признаке, как длинная шерсть, может быть выделено не-
сколько классов. Все перечисленные выше характеристики на-
следуются как полиген. Поэтому длиной и мягкостью своей шер-
сти кошки выставочных длинношерстных пород обязаны про-
должительной и кропотливой селекции.
57
Глава 6
ВРОЖДЕННЫЕ АНОМАЛИИ
И НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ
В последнее время растет поток публикаций, касающихся
различных аномалий у животных. Основные выводы на эту тему
обобщены в нескольких обзорах, где не всегда данный вопрос
рассматривается с генетической точки зрения, поскольку в не-
которых случаях врожденные дефекты, как предполагается, име-
ют негенетическую природу. Несмотря на это, изучение и такого
рода отклонений представляет большой интерес [Saperstein et al.,
1976; Desnik et, al., 1982; Wilier S., Wilier H„ 1986; Robinson,
1987, 1991]*. В настоящей главе будут рассмотрены различные
аномалии кошек, которые, как предполагается, имеют генетиче-
скую природу. Однако не во всех случаях вклад наследственных
факторов до конца ясен.
СКЕЛЕТНЫЕ АНОМАЛИИ
Брахиури
Уменьшение количества хвостовых позвонков, наследуемое
как рецессивный по отношению к норме признак, обозначается
символом br (Bhachyury) [Moutschen, 1950]. Наблюдается не-
которая вариабельность в экспрессии этого признака, но в боль-
шинстве случаев данный ген проявляет максимальную пенетрант-
ность.
Аномальные формы хвоста у мэнской породы
Аллель М. Мэнская порода характеризуется или полным от-
сутствием, или укорочением хвоста, что вызвано действием до-
минантного аллеля М (Manx). В степени проявления аномалии
обнаружено значительное разнообразие. Анализ различных фе-
нотипических форм позволил выделить 4 варианта: титру, или
истинный мэнкс,— кошки, у которых нет хвоста; rumpy-riser —
кошки, у которых сохранилось лишь несколько неподвижных
хвостовых позвонков; stumpy — количество позвонков значитель-
но больше, чем в варианте 2, хвост подвижен, но уродлив по
форме; longie — хвост укорочен, но внешне нормален.
Однако во всех описанных случаях позвоночник животных в
той или иной степени аномален, особенно в хвостовой части,
где позвонки, как правило, деформированы или слиты вместе,
а общее их количество может быть сокращено. Постоянное вы-
* См. список литературы к гл. 2.
58
деление мочи и фекальных масс — обычное явление, что, вероят-
но, является следствием нарушения иннервации соответствую-
щих сфинктеров [James et al., 1969; Martin, 1971; Leipold et al.,
1974; DeForest, Basrur, 1979].
Происхождение мэнского синдрома связано с недоразвитием
хвостовой части нейральной трубки в период эмбриогенеза. Как
правило, эта болезнь сопровождается укорочением спинномозго-
вых отростков в крестцовом отделе — районе, ответственном за
иннервацию мочевого пузыря, толстой кишки и ягодичных мышц.
Все перечисленные характеристики присущи лишь гетерозиготам
М/m, поскольку гомозиготы погибают в пренатальном периоде.
Анализ морфологии эмбрионов на пятой неделе беременности
показал, что некоторые зародыши (предполагаемые гомозиготы)
имеют шаровидную форму. У них наблюдаются нарушения в
развитии центральной нервной системы [Basrur, DeForest, 1979].
Аллель bt. Некоторые породы японского происхождения также
имеют различные аномалии хвостовой части скелета. Одна из
них получила название «бобтэйл» (Bob-tail). Их характеризует
короткий хвост, достигающий лишь 5—7 см в длину, часто за-
канчивающийся шишкой. Строгих фактов, свидетельствующих о
характере наследования такой формы хвоста, пока нет. Однако
условно селекционеры рассматривают этот признак как рецессив-
ный по отношению к нормальному хвосту.
Двойное ухо
Аллель dp. Иногда с обеих сторон головы кошки можно уви-
деть добавочные маленькие ушные раковины или разрастание
хряща. Животные с таким признаком обычно вялые, с умень-
шенным черепом и укороченными челюстями. Наследование осу-
ществляется по рецессивному типу, аллель dp (Duplicated pin-
nae). Данная аномалия носит название «четыре уха» [Little,
1957].
Эктросиндактилия
Аллель Sb. При этой болезни наблюдается нарушение разви-
тия передних лап, которое также известно как «Split-hand» (рас-
щепленная лапа). Лапа расщеплена посередине, что сопровож-
дается сращением пальцев. Степень проявления заболевания
может сильно варьировать. Иногда срастаются пястные и запя-
стные кости. Наблюдается сращение когтей с пальцами. Живот-
ные, имеющие данную аномалию, активны, могут бегать и пры-
гать, но способность к лазанью у них ограничена. Этот признак
наследуется как доминантный, обозначенный символом Sh (Split-
hand). Генетический анализ показал, что аллель Sh характери-
зуется неполной пенетрантностью [Searle, 1953].
59
Синдром плоской груди
Аллель fck. Грудная клетка котят, страдающих этим заболе-
ванием, сжата и имеет не сводчатую, как в норме, а уплощенную
форму. В зависимости от степени сжатия у котят наблюдаются
признаки затрудненного дыхания, боли, замедленный рост. При
аутопсии обнаружено нарушение положения внутренних органов,
что, вероятно, и приводит к ранней смерти. С другой стороны,
котята, у которых экспрессия гена проявляется не полностью,
имеют квазинормальную грудную клетку. Наблюдения на
бирманской породе свидетельствуют о том, что признак на-
следуется как рецессивный. Ген, его обусловливающий, обозна-
чается fck (Flat chested kitten).
Загнутое ухо
Аллель Fd. Характерной чертой данной аномалии является
загнутый вперед кончик ушной раковины. Этот признак не ха-
рактерен для новорожденных, лишь в 4-недельном возрасте на-
чинают проявляться его первые признаки. Окончательно процесс
завершается к 3-месячному возрасту. Столь необычная форма
уха вызывается действием доминантного аллеля Fd (Folded ear).
Гомозиготы Fd/Fd характеризуются не только наличием загну-
того уха, но и массивными, уродливыми костями лап и хвоста.
Лапы кошек выглядят раздутыми из-за значительного укороче-
ния и утолщения костей. Хвост, благодаря сильному укорочению
и потере подвижности, похож на обрубок. Кошки с таким гено-
типом унылые и малоактивные. Следствием такого инертного
образа жизни является неограниченный рост когтей, которые
могут врастать в подушечки лап, если их регулярно не подстри-
гать. Причиной аномалии является чрезмерный эпифизарный
рост и ухудшение окостенения [Jackson, 1975]. У гетерозигот
также наблюдается признак загнутости ушной раковины, однако
у большинства из них поражение скелета или не обнаруживает-
ся вовсе, или проявляется в небольшой степени. Исключение со-
ставляют несколько случаев гетерозигот с измененным скелетом,
обнаруженных Робинсоном [Robinson, 1991].
Гидроцефалия
Аллель hy. Несколько случаев дефектов подобного рода было
зафиксировано среди кошек сиамской породы, что связано с
выщеплением рецессивного, аутосомного аллеля, обозначаемого
символом hy (Hydrocephalus). Котята, носители этого признака,
рождались мертвыми, с гидроцефалией, и часто с раздутыми в
результате отека конечностями [Silson, Robinson, .1969].
60
Волчья пасть
Типичное проявление ранних признаков этой аномалии — не-
способность новорожденных сосать. Степень проявления этого де-
фекта очень различна, при слабой степени заболевания котят
повреждение затрагивает лишь мягкое нёбо, при более сильных
повреждениях обнаруживаются расщепление твердого нёба и
заячья губа. Характер наследования этого типа заболевания чет-
ко не определен, хотя некоторые данные свидетельствуют в поль-
зу рецессивного типа наследования [Loevy, Fenyes, 1968; Loevy,
1974].
Закрученное ухо
Аллель Си. Ушная раковина котят имеет нормальную форму
вплоть до 12—16 недель, а затем начинает загибаться по на-
правлению к спине. Степень закрученности может сильно варьи-
ровать. Ушной завиток наследуется как доминантный признак,
обозначаемый символом Си (Curl). Существуют предварительные
данные о том, что гомозиготы по этому аллелю не имеют серьез-
ных аномалий в отличие от гомозигот Fd/Fd [Robinson, 1989].
Менингоэнцефалоцеле
Аллель тс. Трещина в верхней части черепа приводит к вы-
пячиванию мозговых оболочек и тканей мозга над поверхностью
черепной коробки, формируя массивное образование на ножке.
Язык у таких животных сильно увеличен и часто бывает рас-
щеплен. Нижняя челюсть сохраняет нормальную форму, а верх-
няя — укорачивается, причем на нёбе образуется щель. Глазные
яблоки отсутствуют и видны лишь глазные впадины. Можно
наблюдать только первичные глазные ткани, включающие клетки
сетчатки глаза. Такое нарушение наследуется как рецессивный
признак me (Meningoencephalocele) и приводит к перинатальной
смерти [Zook et al., 1983; Sponenberg, Graf-Webster, 1986].
Микромелия
Существует разновидность необычных «кенгуровых» кошек,
у которых сильно укорочены кости передних лап. Несмотря на
такую серьезную аномалию, активность животных сохраняется.
Был проведен генетический анализ котят, у родителей которых
проявляется данная аномалия, но характер наследования так и
не был установлен [William-Jones, 1944].
Полидактилия
Аллель Pd. Дополнительные пальцы на лапах появляются в
результате действия доминантного аллеля Pd (Polydactyly) [Dan-
forth, 1947а, b; Chapman, Zeiner, 1961]. Количество добавочных
61
пальцев может варьировать от одного до трех. Более того, мо-
гут наблюдаться различия в количестве пальцев между всеми
лапами, или же только между передними и задними лапами
(первое — более вероятно). Предполагается, что полидактилия
появляется в результате увеличения потенциала роста преакси-
алыюго района почки конечностей [Danforth, 1947а, b; Chapman,
Zeiner, 1961].
НЕЙРОМУ ОКУЛЯРНЫЕ РАССТРОЙСТВА
Эпизодическая слабость
Аллель ew. Аномальное поведение начинает проявляться у
кошек в возрасте 4—10 недель, в среднем примерно около 7 мес.
Болезни подвержены животные обоих полов. Животные кажутся
внешне нормальными до тех пор, пока не попадают под воздей-
ствие факторов, вызывающих возбуждение или другой вид стрес-
са средней силы. Фенотипически болезнь проявляется в характер-
ном положении головы в непосредственной близости от грудной
клетки во время отдыха или ходьбы. При ходьбе голова смеща-
ется вверх-вниз, передние лапы не сгибаются, оставаясь прямы-
ми, что заставляет животное делать большие шаги. При этом
задние лапы сгибаются нормально, но двигаются более разбол-
танно. Животное может передвигаться только на короткие рас-
стояния и не в состоянии совершить обыкновенный прыжок.
Наблюдается расширение зрачка и неограниченное разрастание
когтей. Характер проявления всех этих признаков подробно изу-
чался на бирманской породе, было обнаружено, что они наследу-
ются как рецессивные [Mason, 1988]. Ген обозначен ew (Episo-
dic weakness).
Ганглиозидоз GM1
Аллель ga-1. Заболевание характеризуется прогрессивной
дегенерацией нервных ганглиев спинного и головного мозга.
У подверженных заболеванию котят не проявляются никакие
фенотипические отклонения до 2—3-месячного возраста, после
чего развивается легкий тремор головы, а .иногда и лап. К 7—
8 мес тремор усиливается и перерастает в спастическую квадри-
плегию. К годовалому возрасту у животного появляются эпилеп-
тические судороги. Со временем падает зрение. Заболевание на-
следуется как рецессивный признак и обозначается ga-1 (Gang-
liosidosis GM1).
В нервных ганглиях головного и спинного мозга обнаружи-
вается интенсивная дегенерация нейронов, проявляющаяся в их
разбухании и вакуолизации цитоплазмы. Этот процесс сопровож-
дается увеличением концентрации N-ацетилнейраминовой кисло-
ты в 2—2,5 раза в коре головного мозга. У гомозигот по ga-1
62
наблюдается достоверное снижение активности {1-галактозидазы.
У предполагаемых гетерозигот обнаруживается промежуточная
активность фермента по сравнению с гомозиготами и nopMoii.
На уровне биохимического анализа аллель ga-1 наследуется ко-
доминантно [Farrell et al., 1973; Baker, Lindsey, 1974].
Ганглиозидоз GM2
Аллель ga-2. Клинические симптомы этого заболевания по-
добны описанным выше. Легкий тремор головы становится ви-
димым к 6—10-недельному возрасту, постепенно усиливаясь и
переходя в атаксию и параплегию. Из-за постоянного тремора у
кошек возникают проблемы с едой. Череп приобретает необыч-
ную округлую форму, а роговая оболочка глаз затуманивается.
Клинически эта аномалия наследуется как рецессивный признак
(ga-2).
Клетки нервной системы, особенно те из них, которые обра-
зуют автономные ганглии, раздуваются и в них становится вид-
на пенящаяся цитоплазма. Общее содержание ганглиозного ма-
териала увеличивается в 2 — 3 раза по сравнению с нормой. Хо-
тя активность [i-галактозидазы у гомозигот ga-2 нормальна,
активность ^-гексоз амидазы значительно ниже нормы. У гетеро-
зигот уровень активности промежуточный между гомозиготами
и нормой. Биохимически аллель ga-2 наследуется кодоминантно
[Cork et al., 1977, 1978].
Глобулярная лейкодистрофия
У кошек, страдающих этим заболеванием, на 5—6-й неделях
жизни наблюдается мышечная слабость, сопровождаемая замет-
ной неустойчивостью задних лап. Состояние животного постоян-
но ухудшается, поражение распространяется и на передние ла-
пы. Появляется тремор. Обследования мозга показали, что белое
вещество мозга темнеет. Происходит значительная потеря аксо-
нов и миелина. Глобулярные клетки распределены по нехарак-
терным для них областям. Существует предположение, что недуг
наследуется как рецессивный признак, но точных доказательств
этому нет [Johnson, 1970].
Маннозидос
Аллель та. Начальными признаками заболевания являются
общая апатия, мышечная слабость и диарея. Развивается про-
грессирующий тремор, приводящий к атаксии. Желудок разду-
вается в результате отмечаемой гепатомегалии. Рассматриваемая
аномалия наследуется по рецессивному типу (mannosidiosis, ma).
Многие гомозиготы рождаются мертвыми или гибнут вскоре пос-
ле рождения.
63
В нейронах и клетках ганглиев центральной нервной системы
обнаружены четкие признаки вакуолизации. Впервые они появ-
ляются на 7-й день и развиваются к 12-му дню. Обычно вакуоли
наполнены маннозидами. Анализ активности лизосомальных фер-
ментов свидетельствует о значительном уменьшении активности
а-маннозидазы и увеличении активности семи других лизосо-
мальных ферментов [VandeVelde et al., 1982].
Мукополисахаридоз I
Аллель Mps-1. Для животных, страдающих этим заболеванием,
характерны такие морфологические аномалии, как широкий плос-
кий короткий нос, вдавленная переносица, маленькие ушные
раковины и замутненная роговица. Из поведенческих аномалий
отмечают распластанную по земле позу, с расставленными врозь
лапами. Движения в шейном и поясничном отделах причиняют
животному боль. В скелете также обнаружены многочисленные
аномалии. Шейные позвонки широкие, асимметричные и часто
слиты вместе, грудина впалая. Нередко наблюдается билатераль-
ный подвывих передних лап и бедер. У отдельных животных
обнаруживается гепатоспленомегалия, сопровождаемая утолще-
нием и побелением сердечных клапанов. Нейроны головного и
спинного мозга сильно раздуты, с вакуолизированной цитоплаз-
мой. Нейроны спинного мозга и сердечных клапанов, лейкоциты,
фибробласты глаз и селезенки содержат лизосомы, полные муко-
полисахаридов. В лейкоцитах и культивируемых фибробластах
обнаруживается сильное подавление активности а-1-идуронидазы.
Уровень активности у гетерозигот — промежуточный между го-
мозиготами и нормальными животными, что свидетельствует о
полудоминантном характере наследования активности фермента.
Аллель, вызывающий данную аномалию, обозначен Mps-1 (mu-
copolysaccharidosis I). Морфологические и поведенческие анома-
лии наследуются как рецессивный синдром [Haskins et al.,
1979а].
Мукополисахаридоз VI
Аллель Mps-6a и Mps-6b. Голова, как правило, маленькая, с
плоской мордой, широким и коротким носом' и слабовыраженной
переносицей. Верхнее веко утолщено и обвисло, в то время как
роговая оболочка непрозрачна. У некоторых особей сетчатка де-
генеративна, и зрачок сужается только при очень ярком свете.
Обычное положение кошки — прижатое к земле тело с расстав-
ленными передними лапами. К 7 мес походка таких кошек ста-
новится неуклюжей в связи с развитием легкого паралича задних
лап. Скелет аномален, особенно позвоночник, с деформирован-
ными слитыми позвонками в шейном, грудном и поясничном от-
делах. Во многих длинных и коротких костях обнаруживаются
признаки эпифизарной дисплазии.
64
Микроскопически цитоплазма фибробластов многих тканей,
таких, как сердечные клапаны, глаза, кожа, селезенка, костный
мозг, обнаруживает явные признаки вакуолизации. На уровне
электронного микроскопа видно, что клетки полны гранулярны-
ми и ламинарными включениями. Анализ шести различных фер-
ментов в культуре фибробластов показал значительное уменьше-
ние активности [3-арилсульфатазы и увеличение активности
остальных пяти ферментов. Показано, что р-арилсульфатазная
активность у гетерозпгот находится па промежуточном уровне
по сравнению с гомозиготами и нормой и частично перекрывает-
ся с нормой. Тем не менее при одновременном изучении актив-
ностей ,3- п а-арплсульфатазы отношение первого показателя ко
второму может быть использовано для определения гетерозпгот.
Биохимически уменьшение активности ферментов наследуется
по типу неполного доминирования, но клиническая картина на-
следуется по рецессивному типу.
Выявлено два различных варианта [3-арилсульфатазы. Хотя
оба они обнаружены у сиамской породы кошек, считается, что
они не связаны между собой. Межпородные гетерозпготы по
этим типам дают клинически сходные симптомы. Этот факт сви-
детельствует или об общей природе, например, повторной мута-
ции, или об аллелизме. Глубокий анализ биохимических и био-
физических характеристик этих двух вариантов [3-арилсульфа-
тазы обнаружил различия между ними. Электрофоретический
анализ показал, что ферментативные различия наследуются как
кодоминантные аллели друг к другу и к гену нормальной ак-
тивности. Эти аллелп обозначили Mps-6a и Mps-6b (mucopoly-
saccharidosis VI).
Была предпринята попытка коррекции этого заболевания.
Кошке, страдающей мукополисахаридозом, после облучения пе-
ресадили костный мозг от нормальной кошки. Последующий ка-
риотипический анализ показал устойчивый химеризм в 73 °/о
реципиентных клеток костного мозга. Клинические симптомы
свидетельствовали об улучшении состояния животного. Пол-
ностью исчезла затуманенность роговицы, форма головы норма-
лизовалась. Активность животного повысилась, походка улучши-
лась [Haskins et al., 1979b, 1980; McGovern et aL, 1981, 1982,
1985].
Нейроаксонная дистрофия
Аллель no. Кошки с такой аномалией имеют светло-серова-
тую шерсть с лиловым оттенком. В первые 5—6 недель поведение
котят не отличается от нормы, затем появляется покачивание
головой, которое, усиливаясь, переходит в дрожание всего тела.
При движении животное теряет координацию, становится не-
уклюжим, развивается атаксия. Зрачковый рефлекс затормажи-
вается, что. вероятно, свидетельствует об ухудшении зрения.
Также наблюдаются деструктивные изменения во внутреннем
5 Заказ Ю 355
65
ухе. В тканях головного мозга наблюдаются многочисленные
изменения, связанные с утолщением аксонов нейронов. Кроме
того, в нейронах обнаруживается четкая гранулярная структура
с темным образованием посередине. Вероятно, это свидетельству-
ет об интенсивном уменьшении нейронов и клеток Пуркинье и
прогрессирующей глиализации. Данная аномалия наследуется по
рецессивному типу и обозначается no (neural dystrophy) [Woo-
dard et al., 1974].
Спастический паралич
Аллель spt. Такое название получила болезнь, распростра-
ненная среди кошек породы девон реке и проявляющаяся в мы-
шечном расстройстве. Симптомы болезни начинают обнаружи-
ваться, как правило, в 4—7 недель, хотя иногда развитие болез-
ни может задерживаться до 12 — 14-й недели. Котята обыкновенно
активны, но появляются странности в движении, при этом жи-
вотное высоко поднимает лопатки, шея изгибается дугой вниз.
Во время сна кошка вытягивает тело во всю длину, наклоняя
голову на одну сторону. Процесс еды доставляет котятам много
проблем, поскольку изогнутая шея препятствует нормальному
поступлению пищи. При питье нос может быть погружен в воду.
Эти симптомы перемежаются короткими периодами, обычно из-
меряемыми несколькими днями, когда животное выглядит нор-
мально. С взрослением котят тяжесть болезни усиливается. Кош-
ка остается активной, но периоды отдыха учащаются, при этом
она либо растягивается на полу, либо опирается на вертикаль-
ные предметы. Анализ частоты рождаемости нормальных и спа-
стичных котят свидетельствует о том, что болезнь наследуется
по рецессивному типу и обозначается символом spt (Spastivity)
[Robinson, 1994].
Болезнь сфероидальных лизосом
Аллель si. Болезнь, проявляющаяся у котят в возрасте 8—12
недель, обнаруживается по легкому тремору, который переходит
в покачивание головой, а позднее — в дрожание всего тела. Жи-
вотное становится вялым, теряет способность к ориентации,
в поведении обнаруживается атаксия. Животное часто падает.
Аппетит сохраняется, однако при еде животное выглядит очень
неуклюжим. Прикосновение к котенку может вызвать у него
судорожный припадок, который, однако, вскоре проходит. Много-
численные шаровидные образования, появившиеся как резуль-
тат вздутия миелиновой оболочки, распространены по всему го-
ловному и спинному мозгу. В области лимфатических узлов и
селезенки обнаружены скопления крупных вакуолизированных
макрофагов. Это говорит о том, что подобного рода расстройства
могут возникать в результате переполнения лизосом продуктами
метаболизма. Природа этой аномалии до конца не ясна. Данная
66
дисфункция наследуется как рецессивный признак по отношению
к норме и обозначается символом si (Spheroid lysosomal disease)
[Lange et al., 1977].
Сфингомиелоноз
Аллель spl. Это нейральное расстройство развивается у ко-
шек в возрасте 4—5 мес. Первоначально обнаруживаемый лег-
кий тремор перерастает впоследствии в дисметрию, атаксию, ано-
рексию, заканчиваясь иногда летальным исходом. В тканях го-
ловного мозга таких животных обнаружен высокий уровень GM2
и GM3 ганглиозидов наряду с увеличением количества холесте-
рола и сфингомиелина в печени. Наблюдается общее отсутствие
нормальной сфингомиелиназной активности. У ожидаемых ге-
терозигот уровень активности ферментов снижен наполовину.
Клинически аномалия наследуется по рецессивному типу, но
биохимически — по полудоминант ному или кодоминантному и
обозначается символом spl (Sphingomyelinosis) [Wenger et al.,
1980].
Тремор
Аллель tr. Продолжительный тремор всего тела начинается
между 2—4 неделями после рождения. Голова и тело подерги-
ваются, совершая волнообразные раскачивания, хвост вращается
по кругу. Как правило, тремор возникает у кошки во время от-
дыха или сильной усталости. Походка у такого животного нор-
мальная, но оно начинает спотыкаться на неровностях поверх-
ности. Кроме того, теряется способность к плаванию. На электро-
энцефалограмме обнаруживается судорожная активность. Под-
верженные заболеванию котята растут значительно медленнее,
чем нормальные, они, вероятно, менее жизнеспособны. Наследо-
вание осуществляется по рецессивному типу, признак обозна-
чается символом tr (Tremor) [Norby, Thuling, 1970].
ГЕМАТОЛОГИЧЕСКИЕ РАССТРОЙСТВА
Синдром Чедиак — Хигаши
Аллель ch. При этой болезни ослабляется пигментация меха
благодаря скоплению пигментных гранул в глыбки. Шерсть ста-
новится светлее, чем при обычном фенотипе d/d (см. гл. 4). Так-
же изменяется пигментация глаз. Радужная оболочка глаз вы-
глядит красноватой вместо желтовато-зеленой в норме. Тапетум
сильно обесцвечивается или полностью теряет пигментацию.
Кроме того, выявляется судорожное подергивание глазного яб-
лока. Показано, что имеются нарушения в проводящих путях
зрительных ганглиев от сетчатки к отделам головного мозга,
5*
67
хотя в меныпей степени, чем у альбиносов. В лейкоцитах кост-
ного мозга п крови обнаружены крупные цитоплазматические
гранулы. Значительно увеличивается продолжительность крово-
течения, а в местах повреждения вен могут образовываться не-
большие гематомы, что, вероятно, связано с дисфункцией тром-
боцитов. Наследование болезни осуществляется но рецессивному
типу, признак обозначается символом ch (Chediak-Higashi syn-
drome) [Kramer et al., 1977; Prieur, Collier, 1981a, b].
Гемофилия A
Аллель Hma. Это заболевание крови возникает в связи с не-
достатком в организме фактора свертывания крови VIII. Кли-
нические симптомы заключаются в заметном удлинении времени
кровотечения, возникающего в результате незначительного ушиба
пли хирургического вмешательства. Характерно возникновение
гематом. Данное заболевание обнаруживает рецессивное,. сцеп-
ленное с полом наследование, т. е. аллель Hma (Haemophilia А)
находится на Х-хромосоме. Следовательно, как правило, этой бо-
лезни подвержены самцы, что может сокращать продолжитель-
ность их жизни при неблагоприятных условиях [Cotter et al.,
1978; Littlewood, 1986].
Гемофилия В
Аллель НшЬ. Рассматриваемая аномалия очень похожа на
описанную выше, за исключением того, что увеличение времени
кровотечения вызвано недостатком в организме фактора сверты-
вания крови IX. Локус Hmb также находится в Х-хромосоме,
и гемофилией В болеют преимущественно самцы. Самки, вероят-
но, могут заболеть лишь в том случае, если они являются потом-
ками .незначительно пораженных самцов. В целом гемофилия В
протекает несколько слабее, чем гемофилия A [Dodds. 1981].
Недостаточность фактора свертывания крови XII
Аллель hag. Нехватка фактора свертываемости крови приво-
дит к увеличению времени тромбообразовапия. Заболевание на-
следуется по рецессивному типу как клиническая аномалия (сим-
вол hag, Hageman factor XII deficiency) и как полудоминатнып
признак при гематологическом анализе. У гомозигот активность
фактора XII составляет только 1—2 %, а у гетерозигот— 64 %
от активности у нормальных гомозигот. Обычно у гетерозигот не
обнаруживается каких-либо видимых отклонений [Green, White,
1977; Kier et al., 1980] .
68
Г пперхиломпкронемпя
Аллель lice. Несмотря на то что развитие пораженных котят
проходит более или менее нормально, в крови у них обнаружива-
ется высокий уровень липидов. Приблизительно к 8—9 мес они
начинают терять способность к (поднятию век, появляются труд-
ности в принятии пищи, вытягивании пальцев. Теряется колен-
ный рефлекс. Формирование многочисленных гематом приводит к
сдавливанию периферических нервов, что вызывает потерю чув-
ствительности. Заболевание наследуется по рецессивному типу,
обозначается символом hce (Hyperchylo micronaemia) [Jones
el al., 1986a, b].
Эозинофильная зернистость нейтрофилов
Аллель ng. У здоровых кошек при окрашивании эозином ци-
топлазма нейтрофилов выглядит как светлое образование с агра-
нулярной поверхностью. Однако встречаются животные, у кото-
рых цитоплазма при окрашивании представлена множеством
мелких эозинофильных гранул. Ухудшения здоровья у таких ко-
шек не наблюдалось. Такой необычный вид цитоплазмы насле-
дуется как рецессивный признак по отношению к норме н обоз-
начается символом ng (Neutrophil granulation) [Hirsch, Cunning-
ham, 1984].
Аномалия Пелгер — Хьюта
Аллель Ph. У животных, страдающих этой болезнью, обнару-
живается уменьшение числа долей в ядрах гранулоцитов. В эри-
троцитах, лимфоцитах и тромбоцитах крови таких изменений не
наблюдается. Природа аномалии, вероятно, связана с нарушением
формирования дольчатости ядер в стволовых клетках. Этот при-
знак наследуется как доминантный, обозначается символом Ph
(Pelger — Huet anomaly) и его наличие не влечет за собой ка-
ких-либо осложнений, хотя и свидетельствует об ослаблении им-
мунной реакции организма [Weber, 1981; Latimer et al., 1985].
Порфирия
Аллель Ро. Чрезмерная продукция сложных порфиринов мо-
жет привести к их отложению в различных мягких тканях и кос-
тях. Наиболее заметными признаками, свидетельствующими об
этом процессе, являются появление коричневого оттенка зубов и
красный цвет мочи. В ультрафиолетовом свете зубы и моча при-
обретают яркий розово-красный цвет. Этот признак наследуется
как доминантный и обозначается символом Ро (Porphyria)
[Glenn et al., 1968].
Несколько иная форма порфирии была описана Гидденсом
[Giddens et al., 1975]. Симптомы заболевания были те же, что и
'69
описанные выше, .но добавлялись слабость, макроцитарная ане-
мия и почечные расстройства. Данные гибридологического анали-
за свидетельствуют о маногенном наследовании этого заболева-
ния, но более точный характер наследования пока установить не
удается.
АНОМАЛИИ ОРГАНОВ И МЯГКИХ ТКАНЕЙ
Болезни сердечно-сосудистой системы
Подобно другим животным, кошки также страдают от сер-
дечно-сосудистых заболеваний, но данные о характере их насле-
дования пока еще очень отрывочны. Однако можно остановиться
на двух аномалиях, имеющих в своей природе генетическую ком-
поненту: незаращении артериального протока [Severin, 1967] и
идиопатическом эндокардиальном фиброэластозе [Paasch, Zook,
1980; Zook et al., 1981]. Данные генетического анализа свиде-
тельствуют о том, что каждый из этих дефектов может появиться
у близкородственных кошек, однако характер наследования пока
не определен. Вполне вероятно, что генетика этих признаков
очень сложна и может быть, по-видимому, описана в терминах
пороговой концепции, выдвинутой Паттерсоном для сердечно-
сосудистых аномалий у собак [Patterson, 1976]. Согласно резуль-
татам исследований Пиона, кардиомиопатия связана с низким
уровнем таурина в плазме крови и может быть откорректирована
соответствующей диетой [Pion et al., 1987].
Гипероксалурия
Аллель ho. Кошки с такого рода заболеванием умирают от
острой почечной недостаточности в возрасте 5—9 мес. Сначала
они теряют аппетит, организм обезвоживается, затем теряют си-
лы и в течение нескольких дней после появления первых симп-
томов заболевания наступает смерть. Пальпация в брюшной об-
ласти выявляет у таких животных боли в почках. Кроме того,
наблюдаются явные признаки деиннервации мышц и накопление
филаментов в проксимальных аксонах клеток вентральных рогов
и в дорзальных корнях ганглиозных клеток спинного мозга. Почки
перестают функционировать из-за накопления в них кристаллов
оксалатов. Данная болезнь наследуется по рецессивному типу и
обозначается символом ho (Hyperoxaluria) [McKerrell et al., 1989].
Непроходимость пищевода
У больных котят возникают трудности с питанием сразу пос-
ле отъема их ог матери. Сфинктер пищевода перестает расши-
ряться, проглоченная пища остается в пищеводе и он растяги-
вается. В результате наблюдается периодическая рвота или по-
70
зывы к ней. Котенок истощается, его развитие замедляется,
иногда очень значительно. Предварительные данные говорят о
рецессивном характере наследования этого заболевания [Clifford
et al., 1971].
Поликистоз почек
Главным признаком поликистоза почек является истощение
котенка. Наблюдается разбухание брюшной полости. Смерть, как
правило, наступает в возрасте 6—7 недель. Показано, что причи-
на гибели кроется в аномальном развитии почек. Мозговой слой
становится красным, а цвет коркового слоя варьирует от белого
до желтоватого. Оба слоя пронизаны многочисленными цистами.
Печень подвержена заболеванию значительно слабее, но в ней
также обнаружено множество расширенных и склеротизирован-
ных желчных протоков. Клинические данные свидетельствуют о
семейном характере наследования этой болезни, однако генети-
ческий анализ заболевания не завершен [Crowell et al., 1979].
Стеноз привратника желудка
Основным клиническим признаком заболевания является по-
стоянная рвота. Состояние животного может облегчить операция
на привратниковой части желудка. Семейная отягощенность ио
данному заболеванию указывает на его генетическую природу,
однако характер наследования окончательно не выявлен [Pear-
son et al., 1974].
Почечный амилоидоз
Признаками болезни у молодых взрослых кошек являются
развитие грубой кожи, обезвоживание организма, потеря веса.
Наряду с этим наблюдаются недоразвитие почек, их обесцвечи-
вание и различные нарушения почечной капсулы. В почках на-
чинают развиваться многочисленные сосковидные некрозы, фиб-
розы, наблюдаются амилоидные отложения. Заболевание, как
предполагается, имеет генетическую природу, однако окончатель-
но характер наследования еще не установлен [Chew et al., 1982;
Воусе et al., 1984].
Тестикулярная феминизация
Локус tfm. До сих пор был зарегистрирован лишь один слу-
чай тестикулярной феминизации у кошек. Взрослые животные
имеют фенотип обычной кошки, однако полностью отсутствуют
признаки полового поведения. Проведенный анализ показал, что
кариотип представлен самцовым набором хромосом (XY), а по-
ловые органы развиваются по женскому типу ле полностью
[Meyers-Wallen et al., 1989]. Предполагается, что этот признак
71
наследуется как доминантный (предположительно обозначен сим-
волом tfm (Testicular feminization)) и находится на Х-хромосоме,
что и приводит к нарушению развития вторичных половых при-
знаков у самцов.
Пупочная грыжа
Накоплены данные, свидетельствующие о том, что это заболе-
вание имеет генетическую природу. Число животных, страдаю-
щих пупочной грыжей, возрастает в потомстве, (когда один, а тем
более оба родителя были больны. В одной из работ указано, что
данная болезнь представляет собой монотонный, рецессивный
признак [Henricson, Bernstein, 1965; Robinson, 1987].
Уролитиаз
Предполагается, что склонность к уролитиазу имеет генети-
ческую природу, поскольку болезнь часто обнаруживается у род-
ственных животных. Однако характер наследования пока не
определен [Livingston, 1965].
СЕНСОРНЫЕ ОРГАНЫ
Аномалии зрительных путей
У кошек-альбиносов кроме потери пигментации, которая про-
является в белой окраске меха и розовых глазах, обнаруживает-
ся также нарушение проекций ганглиозных аксонов сетчатки в
мозг. У этих животных наблюдается более высокая, чем в-норме,
концентрация нейронных путей, идущих от глаза к противопо-
ложной стороне мозга, что должно значительно ухудшать зрение.
Однако в действительности этого не происходит, так как, по-ви-
димому, мозг способен функционально компенсировать такое на-
рушение. Аллели альбинизма cs и с приводят к появлению этой
морфологической аномалии, причем особенно серьезные наруше-
ния вызывает аллель с. Несмотря на то что отсутствие пигмен-
тации наследуется как полностью рецессивный по отношению к
норме признак, у гетерозпгот C/cs и С/с также обнаруживается
-нарушение проекций зрительных нервов, правда в несколько
ослабленной по сравнению с гомозиготами форме [Kaas, Guillery,
1973; Creel et al., 1982; Leventhal, Creel, 1985].
Кошки с синдромом Чедиак — Хигаши также обнаруживают
нарушение проводящих путей, правда в ослабленной по срав-
нению с альбиносами форме. С другой стороны, ни бирманский
аллель сь, ни доминантный аллель W не приводят к нарушениям
подобного рода [Levick et al.. 1980; Guillery et aL, 1981; Creel
et al., 1982].
72
Катаракта
Билатеральная катаракта, наблюдаемая Рубином [Rubin,
1986], проявляется к 12-недельному возрасту. Несмотря на то
что наблюдается некоторая вариабельность при проявлении этого
^признака, обнаружить его достаточно просто. Незначительные ге-
нетические данные свидетельствуют в пользу рецессивного спо-
соба наследования.
Эдема роговицы
Это расстройство развивается к 4-месячном,у возрасту как
прогрессивная эдема, предшествующая эдеме роговицы. Перво-
начально появляются луковицеобразные поражения на поверх-
ности эпителия роговицы, приводящие к разрывам тканей, кото-
рые впоследствии могут инфицироваться. Существуют некото-
рые указания на генетические причины этого заболевания, одна-
ко характер его наследования пока установить не удается [Bist-
ner et al., 1976].
Мумификация роговицы
Мумификация роговицы может проявляться у кошек в воз-
расте от 10 мес до 8 лет. О существовании генетических предпо-
сылок этого заболевания можно судить по наследованию его при
возвратных скрещиваниях, проводимых в частично инбредных
линиях кошек. Однако точный характер наследования установить
до сих пор не удалось [Vawer, 1982].
Эпибульбарная дерматофиброма
Дерматома распространяется по глазному яблоку, начинаясь
у основания века в латеральном уголке каждого глаза. Как пра-
вило, эти районы пигментированы и покрыты ворсинками, что
вызывает раздражение слизистой оболочки и кератит (воспале-
ние роговой оболочки), приводящие иногда к отслаиванию рого-
вицы. Несмотря на то что это заболевание хорошо известно, ге-
нетика его не изучена [Hendy-Ibbs, 1985].
Прогрессирующая атрофия сетчатки
Клиническим симптомом прогрессирующей атрофии сетчатки
(ПАС) является сильное расширение зрачка, часто сопровождае-
мое нистагмом (судорожное подергивание глазного яблока), уве-
личением отражательной способности тапетума и прогрессирую-
щей дегенерацией кровеносных сосудов в сетчатке. Ослабление
зрения животного начинает проявляться в нерешительном пове-
дении. Кошка часто спотыкается о препятствия, легко преодоли-
мые в нормальном состоянии. Животные, страдающие этим забо-
73
леванием, проявляют симптомы ПАС уже на 3-й неделе после
рождения. В дальнейшем в течение нескольких месяцев происхо-
дит резкое утоныпение фоторецепторного слоя сетчатки.
Некоторые данные дают основание предполагать существова-
ние доминантного аллеля, обусловливающего болезнь [West-Hyde,
Buyukminci, 1982]. Результаты, представленные Рубином и Лип-
тоном [Rubin, Lipton, 1973], кажутся более убедительными и
свидетельствуют в пользу рецессивного характера наследования
(аллель обозначается символом rt (Progressive retinal atrophy)).
Клинические признаки, наблюдаемые к 12—15 неделям, четко
диагносцируются как аномальное расширение зрачка. Гистологи-
ческий анализ позволяет обнаружить утоныпение сетчатки и со-
хранение лишь остатков палочек и колбочек.
Барнет и Куртис, исследователи ПСА на абиссинской породе,
могли установить начало заболевания уже на 4—5-й неделях
[Barnett, Curtis, 1985]. Прогрессирующее изнашивание сетчатки
наступало к 12-й неделе. Данная аномалия наследуется как до-
минантный признак и обозначается символом Rdy (Retinal dystro-
phia) .
Абсолютно иные признаки протекания болезни у кошек абис-
синской породы были зафиксированы Нарфстромом. Первые при-
знаки заболевания проявляются на 16—24-м месяце после рож-
дения, а конечный этап наблюдается через 3,5—4 года. Перифе-
рические районы сетчатки поражаются, как правило, уже на ран-
них стадиях заболевания, хотя в ряде случаев нормально функ-
ционируют вплоть до поздних стадий. Электрорегинограмма
позволяет обнаружить, что аномальным изменениям подвергают-
ся прежде всего палочки в сетчатой оболочке, и лишь позднее —
колбочки. Снижение уровня интерфоторецепторного белка, связы-
вающегося с сетчаткой, также вовлечено в этиологию болезни
[Narfstrom, 1983, 1985; Narfstrom et aL, 1985, 1989]. Признак
наследуется как рецессивный и обозначается символом rdg (Re-
tinal degeneration).
Глава 7
БИОХИМИЧЕСКАЯ И МОЛЕКУЛЯРНАЯ ГЕНЕТИКА
Развитие методов биохимической и молекулярной генетики
открыло новую эру в изучении проблем наследственной изменчи-
вости. Был выявлен огромный пласт генетической вариации,
скрытой от непосредственного наблюдения. Применение этих под-
ходов в генетике домашней кошки только начинается. Однако
уже сейчас с использованием методов молекулярной и биохими-
ческой генетики получен ряд новых данных. Эти данные оказы-
ваются весьма полезными для решения проблем таксономии ко-
74
шачьих (см. гл. 1), сравнительного картирования геномов
млекопитающих (см. гл. 3) и для решения вопросов популяцион-
ной биологии (см. гл. 10).
ГЕНЫ КОШКИ, ВЫЯВЛЕННЫЕ
С ИСПОЛЬЗОВАНИЕМ МОЛЕКУЛЯРНЫХ МЕТОДОВ
Использование клонированных последовательностей ДНК, вы-
деленных из геномных библиотек человека и мыши, привело к
выявлению ряда генов в геноме домашней кошки. Позднее они
были локализованы на генетической карте. Так было обнаружено
несколько последовательностей протоонкогенов (raf, met, rel, ras,
ets) [Watson et al., 1986; Dean et al., 1987], выявлены и локали-
зованы гены, ответственные за синтез интерлейкина [Seigel et
al., 1984], рибосомальной РНК [Pearson et al., 1979] и клеточ-
ных антигенов (см. гл. 8). Особый интерес представляет серия
работ по изучению ретровирусных последовательностей, обнару-
женных в геноме кошки.
РЕТРОВИРУСНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
РНК-содержащие вирусы были впервые обнаружены в рако-
вых опухолях различных видов позвоночных. Большинство таких
вирусов содержит три сходных структурных гена: gag, pol, env.
Эти гены имеют размер, приблизительно равный 9 тыс. пар осно-
ваний. Они соединены друг с другом длинными (около 500 пар
оснований) терминальными повторами (LTR — long terminal re-
peats). Эти вирусы способны рекомбинировать с определенным
классом нормальных ядерных генов, названных онкогенами. Ре-
комбинация приводит к трансформации фибробластов в культуре
клеток и к их злокачественному росту in vivo [Bishop, 1983].
Позднее ретровирусы были обнаружены и в составе ядерной
ДНК нормальных (нераковых) клеток. Показано, что ядерная
ДНК различных видов млекопитающих содержит ряд умеренно
повторенных последовательностей, родственных геномам ретро-
вирусов. Будучи интегрированными в ядерный геном, они, как
правило, не проявляют цитопатогенной активности. Особенности
их жизненного цикла позволяют им интегрироваться в геном кле-
ток позвоночных и размножаться в составе генома хозяина. В тех
случаях, когда интеграция происходит в геном половых клеток,
ретровирусы начинают передаваться от предков к потомкам в
длинном ряду поколений, не сказываясь на приспособленности
их носителей. Число копий ретровирусных последовательностей
варьирует от одной до нескольких тысяч.
В геноме домашней кошки обнаружено три различных семей-
ства ретровирусов.
75
FeLV. Первое семейство представлено группой последователь-
ностей, родственных экзогенному вирусу лейкемии кошек (FeLV).
Большинство животных, зараженных этим вирусом, вскоре поги-
бают от иммунодефицита. По симптомам это заболевание весьма
сходно со СПИДом у человека.
Эндогенная последовательность, родственная FeLV, была вы-
делена из ряда тканей нормальных (неинфицирова'нных) кошек.
Степень гомологии между экзо- и эндогенными последовательно-
стями данного семейства составляет примерно 85 % [Casey et al.,
1981]. Показано, что эндогенная последовательность отличается
от последовательности экзогенного вируса FeLV рядом последо-
вательных делеций. Можно думать, что делецип произошли в ре-
гуляторных участках ретровируса, который вследствие этого утра-
тил способность индуцировать транскрипцию соответствующих
онкогенов и вызывать злокачественную трансформацию клеток
хозяина.
Количество более или менее полных копий дайной последова-
тельности варьирует от 8 до 10 па геном. В то же время в геноме
кошки обнаруживается до 150 копий LTR данного ретровируса,
не связанных с его структурными генами gag — pol — eiiv. Проис-
хождение этих следов ретровируса остается загадкой. Они могут
маркировать сайты интеграции ретровирусных последова-
тельностей.
RD-114. Семейство ретровирусов RD-114 неродственно семей-
ству FeLV. Оно представлено вирусом, способным к репликации,
и обнаруживается приблизительно в 20 копиях на геном кошки.
Экзогенная последовательность, сходная с RD-114, выделена из
клеток рабдосаркомы человека.
Эндогенные последовательности выявляются в разных сайтах
в нескольких хромосомах. Большинегво из них не проявляет ге-
нетической активности. Анализ десяти эндогенных последова-
тельностей RD-114 показал, что все они очень сходны с экзоген-
ным вирусом, содержат нормальные гены gag и pol, связанные
LTR, но имеют ряд нуклеотидных замен в гене env. Эти замены
делают невозможным синтез белков оболочки данного вируса,
что, в свою очередь, лишает его способности к горизонтальному
переносу из клетки в клетку.
Гены gag и pol сохраняют способность к транскрипции и
трансляции. Таким образом, инактивация вирусов FeLV п RD-114
обеспечивается разными способами: в первом случае за счет
делеций в регуляторной части, во втором — благодаря заменам
в одном из структурных генов. Показательно, что в обоих случаях
инактивация происходит вскоре после интеграции вирусов в геном
хозяина до их амплификации и распространения внутри генома.
Способы распространения по геному этих двух семейств виру-
сов были разными. Последовательность RD-114 транспозировала
целиком. Что же касается FeLV, то здесь к транспозиции ока-
зались способны его отдельные фрагменты.
Среди кошек обнаружен полиморфизм по хромосоме ВЗ.
Часть особей имеют в данной хромосоме интегрированную после-
76
довательпость RD-114, способную к репликации, другие же —
только фланговые последовательности (сайты интеграции) этого
ретровируса [Reeves, O’Brien, 1984]. Данное семейство вирусов
обнаруживается не только у домашней кошки и ряда других
представителей семейства кошачьих (см. гл. 1). Сходная эндоген-
ная последовательность BaEV пайдепа в геномах обезьян Старо-
го Света.
МАС-1. В геноме домашней кошки установлено еще одно се-
мейство ретровирусных последовательностей. Его представители
сходны с эндогенным вирусом приматов МАС-1. Данное семей-
ство обнаруживается кроме кошки у всех остальных кошачьих
и у нескольких других представителей отряда хищных [O'Bri-
en, 1986].
ПОЛИМОРФИЗМ БЕЛКОВ И ИЗОФЕРМЕНТОВ
Выше описаны гены кошки, установленные с использованием
молекулярных методов и выявленные в результате изучения
ядерпой ДНК кошки. Другой подход к выявлению п локализа-
ции генов базируется на изучении генных продуктов — белков
и ферментов.
Метод электрофореза в геле позволяет выявить большое ко-
личество различных белков. Сущность метода заключается в сле-
дующем. Образцы органов и тканей животного гомогенизируют
и из гомогенатов выделяют растворимые белки. Пробы, получен-
ные таким образом, наносят на гель. Через гель пропускают в
течение нескольких часов постоянный электрический ток. Под
действием разности потенциалов белки начинают перемещаться
в геле. Скорость и направление (>к аноду или к катоду) миграции
конкретного белка зависят от его молекулярной массы и сум-
марного электрического заряда. После прекращения электрофо-
реза гель помещается в раствор, содержащий субстрат, специфи-
чески реагирующий с данным ферментом, и соединение, взаимо-
действующее с продуктами пх реакции. В результате в том месте
геля, которого исследуемый белок достиг в ходе электрофореза,
выявляется окрашенное пятно. Мутацпи, меняющие электриче-
ский заряд белка, приводят к изменению скорости его миграции
в геле. Таким образом, методом электрофореза можно выявлять
не только разные белки, но и мутантные формы отдельных бел-
ков. Формы ферментов, проявляющие одну и ту же субстратную
специфичность, но различающиеся по электрофоретической под-
вижности, называются изоферментами.
Использование методов гель-электрофореза открыло возмож-
ность для исследования изменчивости белков в природных и се-
лекционируемых популяциях разных видов.
Первая работа на домашней кошке с использованием этого
подхода выполнена О’Брайеном в 1980 г. [O’Brien, 1980] на вы-
борке, включающей в себя 56 особей из Мэриленда и Вирджи^
77
Таблица 7.
Биохимические локусы домашней кошки [по: О' Brien, 1980]
Белок Номен- клатур- ный но- мер Символ локуса Ткани, в которых белок Алле - ли
присутствует отсутствует
1 1 2 3 4 5 6
Аденин-фосфорибозил трансфераза 2.4.2.7 APRT П, Пч, Ф 3 а
Аденозин деминаза 3.5.4.4 ADA П, Ф э а, Ь, с
Аденилаткиназа 2.7.4.3 АК1 Э, П, Ф, Пч —- а
Альдолаза-А 4.1.2.13 ALDA Э. П, Пч ф а
Альфа-галактозидаза 3.2.1.22 GALA П, Пч Э,Ф а
Бета-галактозидаза 3.2.1.22 GALB П, Пч, Ф э а
Гексозаминидаза-А 3.2.1.30 HEXA П, Пч, Ф — а
Гексозаминидаза-В 3.2.1.30 HEXB П Э, Пч, Ф а
Гексокиназа-1 2.7.1.1 HK1 Э, П, Ф, Пч — а
Глиоксалаза-1 4.4.1.5 GLO1 П, Э, Ф — а
Глюкоза-дегидрогеназа-1 1.1.1.47 GDH1 П, Э Ф, Пч а
Глюкоза-дегидрогеназа-2 1.1.1.47 GDH2 п, э Ф, Пч а, Ь
Глюкоза-дегидрогеназа-3 1.1.1.47 GDII3 п, э Ф, Пч а
Глюкоза-фосфатизомераза 5.3.1.9 GPI Э, П, Ф, Пч — а, Ь
Глюкоза-6-фосфат-дегид- рогеназа 1.1.1.49 G6PD Э, П, Ф — а
Бета-глюкуронидаза 3.2.1.31 GUSB П, Пч, Ф — а
Глютамат-оксалоацетат- трансаминаза 2.6.1.1 GOT1 П, Ф э а
Глютамат-пируват-транс- аминаза 2.6.1.2 GPT э, п ф а
Г лютатион-редуктаза 1.6.4.2 GSR Э, п, Ф —. а
Диафораза-1 1.6.2.2 DIA1 П, Ф, Пч э а
Изоцитрат-дегидрогена- за-1 1.1.1.42 IDH1 П, Ф э а
Изоцитрат-дегидрогена- за-2 1.1.1.42 IDH1 П, Ф а
Кислая фосфатаза 3.1.3.2 ACPI Э, П, Пч, Ф — а
Креатинкиназа-В 2.7.3.2 CPKB Э, П, Ф, Пч — а
Ксантин-дегидрогеназа 1.2.3.2 XDHB Ф, Пч — а
Альфа-Ь-Фукозидаза 3.2.1.51 FUCA П, Ф, Пч э а, Ь
Л акт ат-дегидрогеназ а-А 1.1.1.27 LDHA Э, П, Ф, Пч — а
Лактат-дегидрогеназа-В 1.1.1.27 LDHB Э, П, Ф — а
Малат-дегидрогеназа-1 1.1.1.37 MDH1 э, п, Ф — а
М алат-дегидрогеназа-2 1.1.1.37 MDH2 Э, п, Ф — а
Малик-энзим-1 1.1.1.40 MEI П, Ф — а, Ь
Малик-знзим-2 1.1.1.40 ME2 П, Ф, э — а
М анноза-фосфат-изоме- раза 5.3.1.8 MPI Э, П, Ф . а
Пептидаза-А 3.4.11. PEPA П, Ф э а, Ь
Пептидаза-В1 3.4.11. PEPB1 П, Ф, 3 — а
Пептидаза-В2 3.4.11. PEPB2 п Ф, э а
Пептидаза-D 3.4.11. PEPD П, Ф, Пч э а
Пирофосфатаза 3.6.1.1 PP П, Ф э а
Пурин-нуклеозид-фосфо- рилаза 2.4.2.1 NP П, Ф э а
Сорбитол-дегидрогеназа 1.1.1.14 SORD Пч Э, П, Ф а
Супероксид-дисмутаза-2 1.15.1.1 SOD2 Э, П, Ф, Пч — а
Супероксид-дисмутаза-3 1.15.1.1 SOD3 э П, Ф, Пч а
Триозофосфат-изомераза 5.3.1.1 TRI Э, П, Ф — а
78
Окончание табл. 7.1
1 2 3 4 5 6
6-фосфофруктокиназа 2.7.1.11 PFK Э, Ф, Пч п а, Ь
6-фосфоглюконат дегид-
рогеназа 1.1.1.44 PGD Э, П, Ф, Пч — а, Ь
Фосфоглюкомутаза-1 2.7.5.1 PGM1 Э, П, Ф — а
Фосфоглюкомутаза-3 2.7.5.1 PGM3 П, Ф э а, Ь
Эстераза-1 3.1.1.1 ESI П, Э ф а, Ь, с
Эстераза-2 3.1.1.1 ES2 П, Пч Ф, э а
Эстераза-4 3.1.1.1 ES4 П, Пч Ф, э а
Эстераза-5 3.1.1.1 ES5 П, Ф, Э — а, b
Эстераза-6 3.1.1.1 ES6 П, Э ф а
Эстераза-7 3.1.1.1 ES7 э П, Ф а
Альбумин — ALB Плазма — а
Гемоглобин — НВ Э П, Ф, Пч а, Ь
Примечание.
П — почки, Пч — печень, Ф — фибробласты, Э — эритроциты.
нии. Было исследовано 55 биохимических локусов. Из них 12
оказались полиморфными, т. е. были представлены в популяции
двумя или более аллелями (табл. 7.1).
Ряд генов, отвечающих за синтез белков и ферментов, был
позднее локализован на генетической карте хромосом кошки с
использованием панели клонов гибридов соматических клеток
[O’Brien, Nash, 1982] (см. также гл. 3).
Выше было отмечено, что 12 локусов из 55 (22 %) были по-
лиморфными. Эта оценка уровня полиморфности хорошо согла-
суется с данными, полученными на других видах млекопитаю-
щих. Средняя доля полиморфных локусов, контролирующих
структуру белков и ферментов, для класса млекопитающих рав-
на 20,6 % [Айала, 1984].
Для большинства полиморфных систем обнаружена типичная
картина кодоминантного .наследования: у гетерозигот одновре-
менно выявляются фракции белков, характерные для обоих ро-
дителей. У кошек, гетерозиготных по локусам ES5, GRI, MEI,
PGD, наблюдается образование гибридных гетероиолимеров, что
характерно и для других млекопитающих и свидетельствует о
мультимерной природе данных ферментов.
Были проанализированы частоты аллелей полиморфных бел-
ков и изоферментов в исследованной популяции. Оказалось, что
распределение генотипов по всем локусам, за исключением ло-
куса PFK, соответствует ожидаемому при условии выполнения
соотношения Харди — Вейнберга. Эти данные указывают па на-
личие панмиксии в популяциях кошек и хорошо согласуются с
результатами, полученными при использовании окрасочного по-
лиморфизма (см. гл. 10). Средний уровень гетерозиготности по
всем локусам в дайной популяции равен 0,07. Это значение сход-
79
Таблица?.
Наследственные болезни кошки, обусловленные дефектами белков и фер-
ментов [по: O'Brien, 1986]
Болезнь Белок Литературный источник
Дпабет Гликогенов II Гемофилия А Гемофилия В Синдром Хагемана Маннозидоз Синдром Неймана — Пи- ка Мукополисахаридоз IV Мукополисахаридоз I GM1 ганглиозидоз GM2 ганглиозидоз Дерматоспоракспс Атрофия сетчатки Лейкодистрофия глобу- лярных клеток Инсулин Альфа-1,4-глюкозпдаза Фактор VIII Фактор IX Фактор XII Альфа-маннозпдаза Лизосомальная сфинго- миелиназа Арилсульфатаза В Альфа-Ь-идуроппдаза Бета-гал актозпдаза Бета-гексозампнидаза А NH2 проколлаген пепти- дазы О ртитин-дельта-амино- трансфераза Г алактоцереброзпд-бета- галактозпдаза Animal models.... 1982 Animal models..., 1982 Cotter et al., 1978 Dodds, 1978 Green, White, 1977 Animal models.... 1982 Wenger et al., 198(? Jezyk et al., 1977 Haskins et al.. 1979 Baker et al., 1971 Cork et al., 1977 Counts et al., 1980 Animal models..., 1982 Animal models..., 1982
но с оценками гетерозиготности, полученными для популяций
мыши (0,04—0,11) [Selander et al., 1969; Selander, Yang, 1969;
Berry, Peters, 1977] и человека (0,07—0,10) [Harris, Hopkinson,
1972; Nei, Roychoudhury, 1974].
Анализ спектров изоферментов у различных представителей
семейства кошачьих позволил уточнить родственные связи меж-
ду ними (см. гл. 1).
Большинство мутационных замен, приводящих к изменению
электрофоретической подвижности белков и ферментов, являют-
ся селективно нейтральными. Носители различных изоформ не
отличаются друг от друга по приспособленности. В то же время
существует ряд мутаций, приводящих к значительным измене-
ниям в функциональной активности белков и ферментов. Феноти-
пически эти мутации проявляются в виде различных наследствен-
ных заболеваний и резко снижают приспособленность их но-
сителей.
В табл. 7.2 представлены краткие сведения о таких мутациях.
Более подробные данные об их фенотипических эффектах при-
ведены в гл. 6.
Детальное изучение генома кошки только начинается. Легко
предвидеть, что в ближайшем будущем молекулярные и биохи-
мические подходы найдут свое применение в селекционно-генети-
ческой работе с породами домашней кошки, будут использованы
для моделирования на кошке наследственных болезней человека
и для разработки методов генно-инженерной коррекции этих
заболеваний.
80
СПИСОК ЛИТЕРАТУРЫ
Айала Ф. Введение в популяционную и эволюционную генетику.— М.; Мир,
1984.- 230 с.
Animal models of inherited metabolic diseases/Ed. G. Migaki, R. J. Desnick,
D. F. Patterson.— N. Y.: Alan R. Liss, 1982.
Baker II. J., Lindsey J. R., McKhann G. M., Farrell D. F. Neuronal GM1 in a
Siamese cat with beta-galactosidase deficiency Ц Science.— 1971.—
Vol. 174,— P. 838-839.
Berry R. J., Petters J. Heterogeneous heterozygosities in Mas musculus popu-
lations // Proc. Roy. Soc. Lond. B.— 1977.— Vol. 197.— P. 485—503.
Bishop J. M. Cellular oncogenes and retroviruses Ц Annu. Rev. Biochem.—
1983,— Vol. 52,— P. 301—354.
Casey J. W., Roach A., Mullins J. I. et al. The U3 portion of feline leukemia
virus DNA identities horizontally acquired proviruses in leukemic cats Ц
Proc. Natl. Acad. Sci. USA.- 1981,—Vol. 78,—P. 7778—7782.
Cork L. C., Munnel J. F., Lorenz M. D. et al. GM2 ganglioside lysosome! stora-
ge disease in cats with bela-hexosaminase deficiency Ц Science —
1977 —Vol. 196,—P. 1014—1017.
Cotter S. M., Brenner R. M., Doods W. J. Hemophilia A in three unrelated
cats Ц J. Amer. Med. Assoc.— 1978.— Vol. 172.— P. 166—168.
Counts D. F., Byers P. H., Hohnbrook K. A., Hegreberg G. A. Dermatospora-
xis in a Himalayan cat: I. Biochemical studies of dermal collagen /
J. Invest. Dermatol.— 1980.— Vol. 74.— P. 96—99.
Dean M., Kozak C. A., Robins J. et al. Chromosomal localization of the met
protooncogene in the mouse and cat genomes // Genomics.— 1987 —
Vol. 1,— P. 164—173.
Dodds W. J. Inherited bleeding disorders // Canine Pract.— 1978.— Vol. 5.—
P. 49—58.
Green R. A., White F. Feline factor XII (Hageman) deficiency // Amer. J. Vet.
Res.— 1977,— Vol. 38.— P. 893—895.
Harris H., Hopkinson D. A. Average heterozygosity per locus in man: an esti-
mate based on the incidence of enzyme polymorphisms Ц Amer. J. Hu-
man. Genet.— 1972.— Vol. 36,— P. 9—20.
Haskins M. E., Jezyk P. F., Desnick R. J. et al. Mucopolysacharidosis in a do-
mestic short-haired cat — a disease distinct from that seen in the Siame-
se cat Ц J. Amer. Vet. Assoc.— 1979,— Vol. 175,— P. 384—387.
Jezyk P. F., Haskins M. E., Patterson D. F. et al. Mucopolysacharidosis in a
cat with arylsulfatase В deficiency: a model of Maroteaux — Lamy syn-
drome Ц Science.— 1977,— Vol. 198,— P. 834—836.
Nei M., Roychoudhury A. K. Genetic variation within and between the three
major races of man, Caucasoids, Negroids and Mongoloids Ц Amer. J.
Hum. Genet.— 1974.— Vol. 26.— P. 421—443.
O’Brien S. J. The extent and character of biochemical genetic variation in the
domestic cat Ц J. Hered. — 1980.— Vol. 71.— P. 3—8.
O'Brien S. J. Molecular genetics in the domestic cat and its relatives // Trends
in Genetics.— 1986,— Vol. 2.— P. 137—142.
O’Brien S. J., Nash W. G. Genetic manping in mammals: chromosome map of
domestic cat Ц Science.— 1982.— Vol. 216.— P. 257—265.
Pearson M. D., Seabright M., Maclean N. Silver staining of nucleolar organi-
zer regions in the domestic cat Fells catus Ц Cytogenet. Cell Genet.—
1979,— Vol. 24,— P. 245-247.
Reeves R. H., O’Brien S. J. Molecular genetic characterization of the RD-114
gene family of endogenous feline retroviral sequences Ц J. Virol.—
1984,—Vol. 52,—P. 164—171.
Seigel L. J., Harper M., Wong-Staal F. et al. Gene for T-cell growth factor.
Location on human chromosome 4 and feline chromosome Bl Ц Scien-
ce.— 1984,— Vol. 223,— P. 175—178.
Selander R. K., Hunt W. G., Yang S. Y. Protein polymorphism and genetic
heterogeneity in two European subspecies of the house mouse // Evo-
lution— 1969 — Vol. 23,— P. 379—390.
6 Заказ Ns 355
81
Selander R. К., Yang S. Y. Protein polymorphism and genetic heterogeneity
in two European subspecies of the house mouse (Mas musculus) Ц Ge-
netics— 1969,— Vol. 63,— P. 653—667.
Watson D. K., McWilliams-Smith M. J., Kozak C. et al. Conserved chromoso-
mal positions of dual domains of the ets protooncogene in cats, mice
and humans // Proc. Natl. Acad. Sci. USA.— 1986.— Vol. 83.-— P. 1792—
1796.
Wenger D. A., Sattler M., Kudoh T, et al. Niemann — Pick disease: a genetic
model in Siamese cats // Science.— 1980.— Vol. 208.— P. 1471—1473.
Глава 8
ИММУНОГЕНЕТИКА
Иммуногенетику как область знания можно разделить па две
части — классическую и современную. Классическая иммуногене-
тика имеет дело с генетическим полиморфизмом сывороточных
белков и антигенов клеток крови вне зависимости от их связи
с иммунитетом. Главным основанием для использования пристав-
ки «иммуно-» в названии этой области генетики служит то, что
полиморфизм выявляется иммунохимическими методами. Совре-
менная иммуногенетика выросла из исследований генетического
контроля иммуноглобулинов и антигенов гистосовместимости и
в настоящее время рассматривается главным образом как наука,
изучающая гены, функционирующие в иммунной системе.
В отличие от человека, мыши и некоторых сельскохозяйствен-
ных видов животных исследования по иммуногенетике кошки
имеют недавнюю историю. Большая часть работ относится ко
второй половине 80-х годов. В настоящее время разрабатывается
одно основное направление — генетика главного комплекса гисто-
совместимости (МНС — major histocompatibility complex) и ге-
нетика групп крови.
ГЕНЕТИКА МНС
МНС представляет собой сложный локус, включающий несколь-
ко десятков тесносцепленных генов, разделяемых на три класса
[Klein, 1987]. Продукты генов I и II классов играют централь-
ную фоль в системе иммунной защиты млекопитающих. Их важ-
нейшая функция заключается в связывании небольших пептидов,
образующихся при внутриклеточном переваривании (процессин-
ге) различных молекул, как своего, так и постороннего происхож-
дения, и представлении этих пептидов на поверхности клеток
регуляторным и эффекторным Т-лимфоцитам, несущим антиген-
распознающие рецепторы (TCR). В частности, молекулы МНС
I класса таким образом опосредуют отторжение аллогенных
трансплантатов и цитолиз вирус-инфицированных клеток, у мы-
82
ши и человека имеется более 20 генов I класса. Все они имеют
сходную структуру, но только 2—3 гена являются «истинными»
и кодируют молекулы с перечисленными функциями. Остальные
гены — их часто называют класс 1-подобными — имеют неопреде-
ленные функции и отличаются по характеру экспрессии. Гены
III класса МНС тесно сцеплены с генами I и II классов, но ко-
дируют белки, не имеющие непосредственного отношения к анти-
генам гистосовместимости. Например, у человека к III классу
относятся гены некоторых компонентов системы комплемента:
С2, С4 и фактора В.
Характерной чертой молекул гистосовместимости I и II клас-
сов является интенсивный генетический полиморфизм. Так,
у мыши на иммунохимическом уровне описано более 50 аллелей
двух локусов молекул МНС I класса. У человека для трех ло-
кусов I класса известно около 70 аллелей. Ярко выраженный
полиморфизм длины рестрикционных фрагментов генов МНС
обнаружен также у многих других (хотя и не у всех) млекопи-
тающих при использовании в качестве зондов генов I и II клас-
сов человека и мыши. Считается, что функциональный смысл
вариабельности молекул МНС заключается в обеспечении имму-
нитета против вирусных патогенов на популяционном уровне.
Вне зависимости от того, как видоизменяется или приспосабли-
вается тот или иной вирус, в популяции всегда находится особь
или группа особей, молекулы МНС которых способны представ-
лять вирусные антигены [Klein, 1987].
Серология МНС кошки. По аналогии с МНС человека, назы-
ваемым HLA-системой, генетическая система аллоантигенов МНС
кошки получила название FLA (Feline leucocyte antigens). Пер-
вые попытки изучения FLA классическими методами с помощью
аллоиммунизаций клетками были малоуспешными. Трудности с
получением цитотоксических аллоантител, а также ряд наблюде-
ний, таких, как задержка отторжения трансплантатов и воспри-
имчивость кошек к различным вирусным инфекциям, дали осно-
вания считать, что у этого вида полиморфизм в МНС ограничен
или даже отсутствует [Tarr et al., 1979; Pollack et al., 1982, 1983;
McCarthy, Grant, 1983].
Тем не менее в последующих работах удалось продуцировать
слабые аллоантисыворотки и продемонстрировать реакции бласт-
трансформации в смешанных культурах лимфоцитов (одни из
основных тестов, позволяющих демонстрировать генетический
полиморфизм МНС) [Stif, Olsen, 1984; Wolfe et al., 1984]. С по-
мощью перекрестнореагирующих моноклональных и поликло-
нальных антител к антигенам МНС других видов удалось иден-
тифицировать молекулы I и II классов МНС кошки и изучить
их клеточное распределение и экспрессию [Neefjes et al., 1986;
Shelton, Ladiges, 1987; Aasted et al., 1988; Ladiges et al., 1988;
Pollack et al., 1988]. Нифджес и др. [Neefjes et al., 1986] исполь-
зовали иммунопреципитацию антигенов МНС из экстрактов кле-
точных мембран лейкоцитов и последующую изоэлекгрофокуси-
6*
83
ровку. Молекулы МНС кошки в этом анализе выглядели весьма
гетерогенными, что, по мнению авторов, указывало па значи-
тельный полиморфизм п множественность локусов, кодирующих
эти белки.
Позднее Винклер н др. [Winkler et al., 1989] разработали ме-
тодику индуцирования цитотоксических аллоаптител с помощью
множественных трансплантаций кожных лоскутов. В проведен-
ных экспериментах удалось получить специфичные к молекулам
МНС аллоантпсывороткп у 13 из 59 кошек, подвергнутых транс-
плантации. Из них 8 антисывороток реагировали с молекулами
I класса, 2 — с молекулами II класса и 2 содержали АТ против
обоих классов МНС. Популяционный анализ показал, что все
антисыворотки различаются по специфичности. На основании ре-
зультатов популяционного тестирования авторы сгруппировали
их в шесть кластеров, каждый из которых содержал антисыво-
ротки со статистически значимым совпадением реакций. Антиген-
ные специфичности, распознаваемые антисыворотками разных
кластеров, были обозначены FLA1 — FLAG. Семейный анализ
позволил определить 24 гаплотипа, каждый из которых сегреги-
ровал как единый мепделевский признак.
В одной из последних работ с помощью тройной пересадки
аллотрансплантата удалось получить цитотоксическую антисыво-
ротку к пепдентпфицпровапному антигену, экспрессируемому па
лимфоцитах и эритроцитах кошки. Генетический анализ показал,
что этот антиген, предварительно обозначенный F1, представляет
собой наследуемый признак и обнаруживается у 30,9 % особей
в случайной выборке [Hara et al., 1990]. Таким образом, в на-
стоящее время можно утверждать, что МНС кошки является вы-
сокополиморфным локусом. Однако для составления топкой гене-
тической карты FLA-спстемы требуется значительная работа по
детализации эпитопов, распознаваемых разными антисыворотками.
Заслуживает упоминания также С4-компонент комплемента
кошки как возможный представитель молекул III класса. У дру-
гих видов в С4-локусе обнаружен ярко выраженный полимор-
физм, функциональная роль которого неясна. Изучение С4 кошки
методом высоковольтного электрофореза в геле агарозы с после-
дующим пммуноблоттинпом и проявлением с помощью антител
к С4 человека позволило обнаружить, как минимум, четыре ва-
рианта, различающихся но подвижности [Kroon et al., 1986]. Эти
варианты были обозначены авторами С4'1, С4'2, С4'3 и С4'4.
Они наследуются кодоминантно (кодоминантное наследование
характерно для иммуногенетических систем) и, по-видимому, ко-
дируются аллелями одного гена.
В настоящее время неясно, сцеплены ли гены С4 с комплек-
сом FLA (т. е. можно ли их называть генами III класса), но
скорее всего, такое , сцепление будет продемонстрировано. Так,
генетическое картирование МНС кошки с помощью панели гиб-
ридных клеток (кошка — мышь и кошка — китайский хомячок)
показало, что гены I и II классов располагаются в хромосоме В2
84
[Yuhki, O'Brien, 1988]. Эта хромосома, как ранее было установ-
лено по результатам картирования генов некоторых изофермен-
тов, сгштенпо гомологична хромосомам, в которых находятся
МНС человека (6-я хромосома) п мыши (17-я хромосома). От-
сюда следует вывод о значительной консервативности группы
сцепления, содержащей МНС. в эволюции млекопитающих.
Молекулярная генетика МНС кошки. Результаты, получен-
ные иммунохимическими методами при изучении МНС кошки,
были во многом подтверждены методами молекулярной 'генетики.
Используя блот-аиалпз по Саузерну и гетерологичные зонды,
Юки и О'Брайен [Yuhki, O'Brien. 1988] установили, что у кошки
имеются, как минимум, 20 генов I класса и 5 генов II класса на
гаплоидный геном. Полиморфизм по длине рестрикционных фраг-
ментов генов I класса у кошки был относительно ограниченным.
В среднем разница между отдельными индивидуумами в разме-
рах рестрикционных фрагментов составляла 9 % (вычисляется
как соотношение числа различающихся между двумя индиви-
дуумами фрагментов п общего количества фрагментов обеих
особей). Эта оценка свидетельствует о существенно более низком
полиморфизме по сравнению с мышью (38 %), но близка к по-
лученной аналогичными методами оценке полиморфизма генов
МНС I класса человека (9,8%)- Однако следует учитывать, что
данным методом оценивается полиморфизм всей совокупности
гибридизирующихся генов безотносительно к их функции.
Представление о количестве функциональных генов, кодирую-
щих собственно трансплантационные антигены, можно получить,
изучая разнообразие клонов кДНК I класса. Такие исследования
провели Юки п др. [Yuhki et al., 1989]. Была получена
кДНК-овая библиотека па основе мРНК Т-клеточноп лимфомы
кошки 3201. Из библиотеки было выделено 8 клонов кДНК, ко-
дирующих молекулы МНС I класса. Определение нуклеотидной
последовательности этих клонов показало, что среди них имеются
только два варианта, которые различаются между собой по 9 %
последовательности. Клонированные гены имели высокую сте-
пень сходства с генами МНС I класса человека и мыши — соот-
ветственно 81—82% и 73—79 %- С учетом характера распреде-
ления вариабельных и постоянных участков в генах МНС I клас-
са разных видов авторы сделали вывод, что клонированные ва-
рианты представляют собой аллельные продукты одного гена.
Отсюда с высокой вероятностью следует, что у кошки имеется
только один ген МНС I класса, функционирующий как транс-
плантационный антиген. Этот результат может объяснить слож-
ности с индукцией цитотоксических аллоантител при различного
рода аллоиммунизациях. Очевидно, что чем меньше количество
полиморфных локусов, тем меньше суммарное популяционное
разнообразие по продуктам МНС.
Не исключено, что значительная часть генов I и II классов
МНС кошки является псевдогенамп. Косвенным свидетельством
этого могут служить данные, полученные другой группой иссле-
85
дователей, изучавших геномные клоны МНС кошки. Из геномной
библиотеки кошки с помощью МНС генов человека в качестве
зондов выделены два клона, содержащие последовательности ге-
нов II и I классов [Verhoeven et al., 1988; Hoof et al., 1989]. Оба
они оказались псевдогенами. Ген II класса получил обозначение
FLA-DPAi[) ввиду значительного сходства (84 %) с геном
HLA-DPa-1 человека в экзоне, кодирующем второй домен.
В этом гене имеются две мутации, нарушающие рамку считыва-
ния. Любопытно отметить, что обе мутации совпадают по поло-
жению с нарушениями в человеческом псевдогене HLA-DPA2.
По-видимому, инактивация этого гена произошла до разделения
филетических линий приматов и хищных.
Последовательность клонированного гена МНС I класса со-
держала все экзоны, кроме первого, кодирующего сигнальный
пептид (всего в этом гене 8 экзонов и отсутствие первого явля-
ется скорее всего результатом клонирования). В ней так же, как
в FLA-DPA, обнаружены несколько мутаций, нарушающих рамку
считывания, и дефектный сайт сплайсинга. Наибольшее сходство
выделенный ген имеет с HLA-A генами человека и соответствен-
но он получил обозначение FLA-Аф.
Результаты сравнения нуклеотидных последовательностей
FLA-Аф и генов I класса некоторых других млекопитающих со-
ответствуют представлениям о том, что линии приматов и хищ-
ных разделились в филогенезе млекопитающих уже после отделе-
ния линий грызунов и зайцеобразных. Более того, из этих резуль-
татов можно сделать вывод, что дупликации предкового гена
I класса, приведшие к появлению трех локусов (HLA-A, HLA-B
и HLA-C) у человека, произошли у общего предка приматов и
хищных после отделения грызунов ориентировочно около 65 млн
лет назад.
ДРУГИЕ ИММУПОГЕНЕТИЧЕСКИЕ СИСТЕМЫ
Основополагающим вкладом в классическую иммуногенетику
послужило изучение генетического полиморфизма поверхностных
антигенов эритроцитов — групп крови. В этой области особенно
много информации получено при изучении человека и сельскохо-
зяйственных животных. По сравнению с ними кошка изучена
очень слабо — имеются только фрагментарные данные.
Обнаружены, как минимум, две группы крови, обозначенные
А и В [Giger et al., 1989]. Популяционный анализ, проведенный
на случайной выборке кошек Северной Америки, показал, что
группа В встречается достаточно редко —были найдены только
две особи с таким фенотипом среди 485 кошек. В плазме крови
обеих обнаружены высокие титры изоагглютининов (естествен-
ных антител) против эритроцитов группы А. В то же время у
кошек с эритроцитами группы А изоагглютинины против груп-
пы В встречаются достаточно редко (только у 30 %) ив низком
титре [Giger et al., 1989].
86
С помощью иммунизации кроликов эритроцитами получены
антисыворотки, выявляющие еще две группы антигенов эритро-
цитов кошки, обозначенные С1 и С2. Однако каких-либо опреде-
ленных данных по наследованию С1 и С2 признаков пока не
получено [Ejima et al., 1988].
В последнее время параллельно с главным комплексом гис-
тосовместимости понемногу начинается изучение структуры и
функции ряда других генов, продукты которых принимают уча-
стие в иммунном распознавании и элиминации чужеродных анти-
генов. Полезным инструментом в этих исследованиях оказались
моноклональные антитела к различным антигенам лейкоцитов
человека, перекрестно реагирующие с антигенами лейкоцитов
кошки. Помимо молекул МНС с помощью перекрестно реагирую-
щих моноклональных антител у кошки были идентифицированы
рецепторы СЗЬ-компонента комплемента и гомолог CD8 рецептора
человека (вспомогательный рецептор, участвующий во взаимодей-
ствии Т- и В-лимфоцитов и связывании TCR с молекулами МНС
I класса) [Aasted et al., 1988]. Недавно получены моноклональ-
ные антитела и к СП4-антигену лейкоцитов кошки — другому
вспомогательному рецептору, участвующему во взаимодействии
TCR и молекул МНС II класса [Ackley et al., 1990].
В целом можно отметить, что исследования иммуногенетики
кошки имеют очевидную перспективу. Речь идет как о развитии
частной биологии этого вида, так и о решении проблем более
общего характера. Кошка может служить полезной моделью для
изучения более 20 наследуемых заболеваний, а также вирус-
индуцированных патологий клеток крови [O’Brien, 1986]. В по-
следние годы в связи с распространением AIDS большое значе-
ние приобретают модели заболеваний, приводящих к иммунодефи-
цитным состояниям. У кошки обнаружены, как минимум, два
таких заболевания. Это лимфосаркома, вызываемая экзогенным
ретровирусом FeLV [Cotter et al., 1975; McCarthy, Grant, 1983],
и синдром приобретенного иммунодефицита, вызываемый лимфо-
тропным лентивирусом [Pedersen et al., 1987; Hara et al., 1990].
He менее важное значение имеет и эволюционный аспект.
Мммуногенетика является наукой, основанной на изучении фак-
тически только двух видов — человека и мыши. Поэтому для ее
полноценного развития значительную ценность представляет воз-
можность «постороннего» взгляда на различные проблемы, от-
носящиеся к структуре и функционированию генов иммунной
системы.
СПИСОК ЛИТЕРАТУРЫ
Aasted В., Blixenkrone-Moller М., Larsen Е. В. et al. Reactivity of eleven anti-
human leucocyte monoclonal antibodies with lymphocytes from several
domestic animals // Vet. Immunol. ImmunopathoL— 1988,— Vol. 19.
Nl.—P. 31—38.
-Ackley C. D., Hoover E. A., Cooper M. D. Identification of CD4 homologue in
the cat Ц Tissue Antigens.— 1990.— Vol. 35, N 2.— P. 92—98.
87
Cotter С. M., Hardy W. D., Essex M. Association of feline leukemia virus with
lymphosarcoma and other disorders in cat Ц J. Amer. Vet. Med. As-
soc.— 1975.— Vol. 166.— P. 549—454.
Ejima II., Hara Y., Ikemoto S. Preparation of antibodies recognizing the new
group antigens of feline red blood cells Ц Jap. J. Vet. Sci — 1988.—
Vol. 50.— P. 763—768.
Giger U., Kilron C. G., Fillipson L. J., Bell K. Frequencies of feline blood gro-
ups in the USA Ц J. Amer. Vet. Med. Assoc.— 1989.— Vol. 195. N 9.—
P. 1230—1232.
Hara Y., Ejima H., Aoki S. et al. A cytotoxic antibody produced by ailo-skin
transplantation in cals Ц Jap. J. Vet. Sci.— 1990,— Vol. 52. N 3.—
P. 543—550.
Hara Y'., Ishida T., Ejima H. et al. Decrease in mutagen-induced lymphocyte
proliferative responses in cat infected with feline immunodeficiency vi-
rus Ц Ibid.— P. 573—580.
Hoof M. van, Geus J. P., Roos M. et al. The primary structure of a felloe class
I gene: striking similarity to HLA-A // Immunogenetics.— 1989.—
Vol. 30,- P. 330—334.
Klein J. The natural history of the major histocompatibility complex.— Chiche-
ster: John Wiley and Sons, 1987.
Kroon T. I. P. de, Doxiadis G., Doxiadis I., Hensen E. J. Struclure and poly-
morphism of the feline complement component C4 Ц Immunogenetics.—
1986,— Vol. 24, N 3 — P. 202—205.
Ladiges W. C., Severns E., Pesando J. M. Comparative reactivity of anti la mo-
noclonal antibodies in man and laboratory animals Ц Hybridoma.—
1988,— Vol. 7, N 5 — P. 465—470.
McCarthy J. M., Grant С. K. Cellular immune response in the blood of cats is
restricted to autochtonous feline sarcoma virus transformed cells //
Int. J. Cancer,- 1983 — Vol. 31,- P. 627—631.
Neefjes J. J., Hensen E. J., Kroon T. I. P. de, Ploegh II. L. A biochemical cha-
racterization of feline MHC products: unusually high expression of
class II antigens on periferal blood lymphocytes // Immunogenetics.—
1986,—Vol. 23,—P. 341—347.
O’Brien S. Molecular genetics in the domestic cat and its relatives Ц Trends
in Genetics.— 1986.— Vol. 2.— P. 437—448.
Pedersen N. С., Ho E. L., Brown M. L., Yamamoto J. K. Isolation of T-lympho-
tropic virus from domestic cats with immunodeficiency-like syndrome Ц
Science.— 1987,— Vol. 235 — P. 790—793.
Pollack M. S., Mastrota F., Chin-Louie J. et al. Preliminary studies of the fe-
line histocompatibility system Ц Immunogenetics.— 1982.— Vol. 16.—
P. 339—347.
Pollack M. S., Mastrota F., Chin-Louie J. et al. Additional preliminary studies
of feline major histocompatibility complex / Transplant. Proc.— 1983,—
Vol. 15,— P. 156—158.
Pollack M. S., Hayes A., Mooney S. et al. The detection of conventional
class I and class II I—E homologue major histocompatibility complex
molecules on feline cells // Vet. Immunol. Immunopathol.— 1988.—
Vol. 19, N l.-P. 79-92.
Shelton G. IL, Ladiges W. C. Murine monoclonal antibodies crossreactive
with feline peripheral blood mononuclear cells Ц Vet. Immunol. Immu-
nopathol.- 1987.— Vol. 16, N 3/4,- P. 271—276.
Stif M. I., Olsen R. G. Feline one-way mixed lymphocyte reaction Ц Vet. Im-
munol. Immunopathol.— 1984.— Vol. 7.— P. 1—9.
Tarr M., Olsen R. G., Hoover E. A. et al. The effect of melhylnitrosourea on the
immune system and hematopoietic system of the adult specific pathogen
free cats // Chem. Biol. Interact.— 1979.— Vol. 28.— P. 181—199.
Verhoeven A. J. M., Roos M. H., Ploegh H. L. A feline class II alpha gene with
striking similarity to the HLA-DPA pseudogene // Immunogenetics.—
1988,— Vol. 28,— P. 406—411.
Winkler C., Schultz A., Cevario S., O’Brien S. Genetic characterization of FLA,
cat major histocompatibility complex Ц Proc. Natl. Acad. Sci. USA,—
1989,— Vol. 86 — P. 943—947.
88
Wolfe J. L., Haskins M. E., Zmijevski С. M. Mixed lymphocyte reactivity in
cats // Transplantation.— 1984.— Vol. 37,— P. 509—513.
Vuhki N., O’Brien S. J. Molecular characterization and genetic mapping of
class J and class T1 MUG genes of the domestic cat Ц Immunogene-
tics.- 1988.— Vol. 27,— P. 414-425.
Vuhki N., Heidecker G. F., O’Brien S. Characterization of MHC cDNA clones
in the domestic cat. Diversity and evolution of class I genes Ц J. Im-
munol.— 1989.— Vol. 142, N 10,— P. 3676—3682.
Глава 9
ВРОЖДЕННОЕ ПОВЕДЕНИЕ КОШКИ
ОНТОГЕНЕЗ ПОВЕДЕНИЯ
Проследить формирование сложного поведения и оценить
вклад в него генетической компоненты можно, наблюдая онто-
генетическое развитие кошки.
Общий характер движений универсален для всех новорожден-
ных млекопитающих. Отличительные его черты — генерализован-
ность возбуждения, стереотипность движений и малая зависи-
мость от внешних факторов — характерны и для новорожденных
котят. Только что родившийся котенок, очищенный матерью от
родовых мембран, ползет вперед, загребая передними лапами и
отталкиваясь одновременно двумя задними. Мягкое прикоснове-
ние к мордочке или телу приводит к слабому повороту половы
или всего туловища с помощью чередующихся движений перед-
них пог в направлении к раздражителю. Способность ориентиро-
ваться па стимул позволяет котятам находить брюхо матери.
Котенок взбирается на пее и быстро зарывается в шерсть, там
находит выступающий сосок и начинает сосать. Таким образом,
поведение котенка сразу после рождения включает неспецифиче-
ский поисковый компопент и специфическую реакцию на сосок.
Вклад генетической компоненты в поведение новорожденных
котят оценивался в опытах по изоляции детенышей от матери.
У котят, выращенных в инкубаторе, нарушалось ориентировоч-
ное поведение на мать. Кроме того, требовалось значительное
время для того, чтобы научить пх сосать из бутылки, тогда как
«.неотлучепные» котята проявляли поведение сосания матери
сразу после рождения. Авторы эксперимента [Kovach, Kling,
1967] приходят к заключению, что поведение сосания у котят
зависит как от врожденных рефлексов, так и от поддержания
этих рефлексов контактами с матерью. Данные физиологических
экспериментов с локальным разрушением различных структур
мозга показали, что на развитие поведения сосания критически
влияет целостность промежуточного мозга и обонятельных лу-
ковиц. Однако для развития полноценного поведения с быстрой
и точной ориентировкой на мать п нахождением соска необходи-
мо сохранение связей с лимбическими и кортикальными струк-
турами [Kovach, Kling, 1967].
Дальнейшее развитие поведения вплоть до формирования
специфической для семейства кошачьих хищнической агрессии
идет по этапам, свойственным всем млекопитающим. Особенность
этого этапа онтогенеза у кошки заключается в конкретных сроках
и формах проявления поведенческих реакций. Так, например,
реакция самоочищения у кошек — груминг — отличается рядом
видоспецифических черт по сравнению с другими млекопитаю-
щими (см. раздел «Фиксированные комплексы действий» данной
главы). Для 40 котят описана достаточно жесткая последователь-
ность возникновения новых поведенческих форм с точностью до
дня [Kling et al., 1969]:
Поведенческая День появле-
ре акция ния
Открытие глаз 12
Ходьба 22
Груминг 23
Вскарабкивание 28
Игра 35
Агрессия 52
Развитие котят, лишенных материнской опеки в этот период
постнатального онтогенеза и выращенных по отдельности в инку-
баторе, существенно не влияло на формирование нормального
поведения. Было только обнаружено, что у этих котят повышена
общая возбудимость, они больше вокализируют и становление
опорно-двитательното аппарата у них происходит позже. Груминг
появляется раньше, примерно на 15-й день. Игровое и агрессив-
ное поведение, а также рост и масса котят, воспитанных матерью,
не отличаются от инкубаторских.
П. Сейтз [Seitz, 1959] изучал поведение котят, отнятых у ма-
тери в возрасте 2, 6 и 12 недель. Он обнаружил, что котята,
отделенные от матери в 2-недельном возрасте, отличаются нена-
правленной хаотической активностью. Они пугливы в новой си-
туации, их попытки достать пищу неорганизованны, при конку-
рентном кормлении они очень агрессивны, но их попытки полу-
чить корм малоуспешны. Они также медленнее обучаются на
положительном подкреплении. У кошек, отделенных от матери в
более позднем возрасте, эти же отклонения в поведении выраже-
ны слабее.
Для того чтобы узнать, сформировались ли механизмы пове-
денческих реакций до их естественного проявления, т. е. до тех
сроков, которые установлены Клингом с соавторами [Kling et al.,
1969], был проведен следующий эксперимент. Котятам имплан-
тировали электроды в миндалину и гипоталамус в возрасте от
4 ч до 50 дней. Состояние тревожности, настороженности и еома-
томоторные реакции можно было получить, раздражая обе струк-
туры во всех возрастах. Начиная с 2-недельного возраста боль-
шинство автоматических реакций вызывалось раздражением ги-
поталамуса. Это слюнотечение, уринация и дефекация. Защитные
реакции были получены раздражением гипоталамуса на 12-й
день развития и сходные ответы — раздражением миндалины в
90
Т. Соматомоторные I I I I I I I
реакции ------1-----1-----L
2. Тревожность I I I I I I I I I I I I I I
i-Принюхивание I I I I I I I I I I 1 I I I
4. Лизание __ _____________ _ ___ __ __ I I I |
5. Слюноотделение 1 I I I I I I I
6. Ури нация 1111
7. Де<рекация ___ Illi
Б.Пилозрекция I I ! I
9. Урчание ___ ____________ __ I I ! I
ТО. Шипение __ _____________||
11.Пригибания __ ___ __ __ I I I I
12. Ретропульсии ___|
73. Борьба I__I
74- Атака ___|
15. Припадки ______I I I I I I I I I
I------1 7 ill
СМ СЧ гп
2 Дни развития
Рис. 9.1. Суммарная картина поведенческих ответов котят на электрическую
стимуляцию гипоталамуса и миндалины в возрасте от рождения до 50 дней
[Kling et al., 1969].
1 — точные дни проявления поведенческих ответов; 2 — возможные дни проявления
поведенческих ответов.
возрасте трех недель. Такие реакции, как борьба и атака, появ-
лялись позже.
Результаты этого эксперимента показали, что большинство
сложных поведенческих реакций может быть вызвано до их
естественного возникновения, следовательно, они являются в сво-
ей основе врожденными. Средовые факторы, по всей видимости,
играют лишь роль пусковых механизмов включения уже готовых
адаптивных программ. Формирование такой программы происхо-
дит по мере созревания нервных структур, ответственных за ее
исполнение, и в общих чертах было исследовано Клингом и Кос-
таном [Kling et al., 1969] (рис. 9.1).
Показанные на рис. 9.1 реакции существуют уже в «готовом
виде» к 37—50-му дню с рождения кошки и составляют значи-
тельную часть поведенческого репертуара взрослого животного.
Более того, в игровом поведении кошки, характерном для этого
периода, проявляются практически все движения, необходимые
для охоты: преследование, подкрадывание, прыжки, удары лапа-
91
ми и удержание жертвы, принесенной матерью. В игре, однако,
все эти движения непоследовательны и несовершенны по срав-
нению с поведением взрослого хищника. На более поздних ста-
диях многие из этих’ реакции объединяются в так называемые
фиксированные комплексы действий (ФКД), отличающиеся
строгой последовательностью включенных в ппх реакций п отра-
жающие один из высших этапов формирования нервной систе-
мы — созревание ее координационных и интегративных центров.
ФИКСИРОВАННЫЕ КОМПЛЕКСЫ ДЕЙСТВИЙ
Наибольшей стереотипностью и низкой впдоспецифичпостыо
на всех этапах своего проявления отличается половая функция.
Половая активность кошек детерминирована достаточно жестко
п описывается в значительной степени в терминах ФКД. Пове-
денческие паттерны ухаживания, копуляции и посткопуляториой
активности (рис. 9.2) описаны Розенблаттом п Аронсоном [Ro-
senblatt, Aronson, 1958] с точностью до минут и секунд
(табл. 9.1).
Установлены также отделы нервной системы п гормональные
звенья, стимуляция которых приводит к демонстрации полового
поведения [Green et al., 1957; Porter et al., 1957; Rosenblatt,
Aronson, .1958].
Первым ФКД в онтогенезе кошки появляется груминг. Эта
форма поведения чрезвычайно важна для них и составляет зна-
чительную часть поведенческого репертуара кошки. Вылизывая
свой мех, кошка защищает себя не только от грязп и болезней,
но также и от холода и пере-
грева, витаминного дефицита,
посторонних запахов, намока-
ния кожи и даже от стресса.
Последовательность действии
такова [Morris, 1986]: 1) кош-
ка облизывает губы; 2) выли-
зывает лапу с одной стороны
до тех пор, пока она не станет
мокрой; 3) трет мокрой лапой
ухо, глаз, щеку и подбородок;
4) проделывает то же с другой
лапой; 5) вылизывает передние
лапы п плечи; 6) вылизывает
бока; 7) вылизывает генита-
лии; 8) лижет задние ноги:
9) вылизывает хвост от осно-
вания к концу (рис. 9.3).
Рис. 9.2. Позы копуляторной (7)
и посткопуляторноп (2) активно-
сти.
9 2
Эта сложная последовательность действии в очищении меха
у кошек является впдоспецпфпчной и отличает их от других
млекопитающих. Мыши и крысы, например, используют для гру-
минга одновременно обе передние лапы, а не только одну их сто-
рону поочередно.
По тину ФКД построено и охотничье поведение кошек. Все
изученные Лейхауземом [Leyhauzen, 1979] представители семей-
ства кошачьих убивают жертву, нанося ей «смертельный укус»
в области шеи, который повреждает спинной мозг и вызывает
практически мгновенную смерть жертвы. Именно наличие «смер-
тельного укуса»
отличает кошек от
отря-
других представителей
Рис. 9.4. Захват загривка против-
ником в межсамцовых столкно-
вениях.
Рис. 9.3. Последовательность
действий при груминге.
Вылизывание: 1 — передних лап,
2 — морды, з — боков, 4 — генита-
лий, 5 — задних лап.
93
Таблица 9.1
Синхронизация половой активности у самцов и самок кошки [Rosenblatt,
Aronson, 1958]
Продолжитель- ность реакции Самец Самка
10 с — 5 мин Ориентация к хвосту, обню- хивание гениталий, круговые движения, характерный клич, подход к самке Пригибается, трет нос и рот, катается, вокализиру- ет, «пританцовывает/?
П рекопуляторная активность
1—3 мин Хватает за загривок, подни- Пригибается
мает передние лапы, трется пе- редними лапами, взбирается на спину, делает тазовые толчки
Копуляция
5-Юс Эрекция, вхождение, эякуля- Спаривается, подгибает
ция зад, расширение влагали- ща, копуляторный крик, вы- тягивается вперед
Посткопуляторная активность
Менее 1 мин
Около 5 мин
Облизывает член и передние
лапы, садится рядом с самкой
Облизывает гениталии,
катается, трет нос и рот.
Наблюдает за самцом, гла-
дит лапами самца
да хищных. Одной из причин успеха в осуществлении смертель-
ного укуса Лейхаузен считает врожденные анатомические осо-
бенности строения клыков. Однако необходимым компонентом в
формировании охотничьего стереотипа является и приобретен-
ный опыт. Развитие хищнического поведения зависит от социаль-
ного опыта, полученного в игре со своими собратьями и особенно
при сопровождении матери на охоту. У котят, выросших вместе
с крысами и никогда не видевших, как мать убивает грызунов,
хищническая агрессия в полной мере не развивается. В целом же
анализ экспериментальных данных показывает, что существенных
различий в закономерностях нанесения смертельного укуса между
котятами и взрослыми кошками нет.
В опытах Рондолла [цит. по: Крушинский и др., 1983] было
продемонстрировано существование самостоятельных нейрофизио-
логических механизмов стереотипа смертельного укуса. По ре-
зультатам экспериментов основные центры этого механизма ло-
кализованы в среднем мозге.
Стереотипная последовательность реакций ориентации на
шейный отдел жертвы, используемая кошками при нанесении
смертельного укуса, включена и в поведение межсамцовых столк-
новений (рис. 9.4). Типичные этапы иоведения в межсамцовых
конфликтах следующие.
94
Доминантное животное видит соперника и приближается к
нему, приняв характерную позу —вытянувшись вверх на полно-
стью распрямленных лапах и распушив шерсть на боках. Ни на
секунду не упуская соперника из виду, атакующий кот издает
угрожающие звуки. Приблизившись на расстояние около 1 м, он
начинает делать характерные движения головой: немного подни-
мает ее и склоняет на одну сторону, затем делает небольшой шаг
вперед и склоняет голову на другую сторону. Это повторяющееся
несколько раз движение, по-видимому, обозначает намерение
атаковать.
Если коты имеют одинаковый статус, то оценивание сил про-
тивника сильно затягивается. На пути приближения друг к дру-
гу они надолго застывают в неподвижных позах.
Схватка начинается со стремительного броска вперед в по-
пытке уцепиться за загривок противника. Схваченный таким об-
разом кот тотчас начинает кружиться на месте, стараясь осво-
бодиться от соперника с помощью укусов, отбиваясь передними
лапами и лягаясь задними. Эта фаза борьбы длится недолго, по-
скольку она очень интенсивна. Клочья шерсти летят с двух ко-
тов в прямом смысле слова.
После первого столкновения противники расходятся в разные
стороны, оценивают свои потери и вновь бросаются друг на дру-
га. Борьба продолжается до тех пор, пока один из котов не обес-
силеет и не останется лежать на земле.
После этого победитель исполняет очередной характерный ри-
туал. Он обходит лежащего кота и начинает тщательно обнюхи-
вать землю, как будто она обладает необыкновенно притяга-
тельным для пето запахом. После церемонии обнюхивания побе-
дивший кот медленно удаляется, а побежденный через некоторое
время уползает в безопасное место.
Таким образом, в межсамцовых столкновениях котов прояв-
ляются видоспецифические черты поведения семейства кошачьих,
начиная с демонстрации позы угрозы и захвата загривка против-
ника и кончая ритуалом «победителя».
В целом, по-видимому, можно утверждать, что ФКД в пове-
дении кошачьих являются врожденными по своему характеру.
В индивидуальном опыте происходит лишь оптимизация навыков.
Способность к более эффективному поведению, приобретаемая в
«личном опыте», является лишь модификацией заданных типов
поведения.
КОЛИЧЕСТВЕННЫЕ ПРИЗНАКИ ПОВЕДЕНИЯ
ФКД составляют основу видоспецифического, врожденного
поведения. По представлениям Э. Майра [1968], они детермини-
руются «закрытыми» генетическими программами и для своей
реализации практически не нуждаются в поступлении информа-
ции из внешней среды. Форма и последовательность этих реакций
93
жестко, стереотипно организованы. Они в малой степени подвер-
жены модификациям в процессе развития, т. е. обладают узкой
нормой реакции. Этот тип реакций характерен для животных с
низкоорганпзованной нервной системой.
Для высших млекопитающих, к которым относится и семей-
ство Felidae, характерны реакции, детерминируемые «открыты-
ми» программами с широкой нормой реакции. Именно эти реак-
ции делают поведение высших животных пластичным, адекват-
ным изменяющимся условиям внешней среды и, следовательно,
более адаптивным. Ввиду широкой нормы реакции и различной
степени выраженности эти поведенческие реакции относят к ка-
тегории количественных признаков. Несмотря на пх очевидную
изменчивость в зависимости от внешних условий, эксперимен-
тально показано, что границы этой изменчивости наследственно
детерминированы. К количественным признакам в поведении
кошачьих можно отнести различные параметры восприятия, мо-
тивационной реактивности, обучаемости и рассудочной дея-
тельности.
Перцептивные способности. Высокоспециализировапная хищ-
ническая агрессия кошачьих определяется не только наличием в
их генетической программе ФКД по захвату и смертельному уку-
су жертвы. Точно определить, где находится жертва, им помогают
все шесть органов чувств. Главный из них —зрение.
Зрение. Кошка обладает острым зрением и способна ви-
деть в полумраке. Сужение глаз в щелочки, как и у лисиц, по-
зволяет ей точно регулировать яркость поступающего в них све-
та. Кошка, как никакое другое животное, приспособилась ви-
деть и при ослепительном свете, п в сумерках. В отличие от со-
баки, которая ориентируется в основном по запаху, кошка по
большей части использует свое острое зрение.
Слух. Однако охота по ночам делает зрение недостаточным
органом ориентировки. Уловпть, где находится жертва, ей помо-
гает слух. Кошки слышат гораздо лучше людей и не хуже, чем
собаки. По характерным звукам, которые издают грызуны, они
способны локализовать их местонахождение на расстоянии 20 м.
Известно, что гены альбинизма обладают плейотропным действи-
ем нэ слух. Голубоглазые ангорские и сиамские кошки, несущие
такие гены, в нескольких Процентах случаев оказываются
глухими.
Обоняние. Что касается роли обоняния в поведении кошек,
то здесь мнения сильно расходятся. Лейхаузеп [Leyhauzen, 1979]
считает, что это чувство играет важную роль в половом поведении
и не так существенно в агрессии, как у других хищников. Опи-
сана специфическая чувствительность кошек к запаху котовника
кошачьего (Nepeta cataria). Животные, чувствительные к этому
растению, приходят от него в сильное возбуждение и реагируют
на очень малые концентрации содержащегося в нем масла — гепе-
талактона. Реакция на котовник проявляется так: животное впа-
дает в неистовство, начинает лизать и кусать растение, трясет
96
головой, громко мурлычет, судорожно подергивается всем телом
и катается по земле. Этот приступ наркотического возбуждения
длится около 10 мип, а затем кошка возвращается в нормальное
состояние. Все виды диких кошек, даже львы, реагируют на ко-
товник кошачий, Тодд [Todd, 1962; Morill, Todd, 1978] описал
аутосомный ген, контролирующий проявление реакции кошки на
Napela cataria.
Осязание. Еше один орган позволяет кошке тонко ориен-
тироваться в окружающей обстановке. Это обонятельно-вкусовой
орган Якобсена, расположенный у нее в нёбе за передними верх-
ними зубами. Он служит для распознавания запахов, растворен-
ных в воздухе. Другие высшие млекопитающие этот орган утра-
тили, по-видимому, в связи с превалирующим развитием визуаль-
ных органов восприятия.
Вибриссы кошки представляют собой уникальный инструмент
ориентировки. Они внедрены в ткань верхней губы на глубину в
3 раза большую, чем обычные волосы, и снабжены большим чис-
лом нервных окончаний. Вибриссы могут двигаться как вперед,
так и назад, а два верхних ряда могут двигаться независимо от
двух нижних. Они жизненно необходимы кошке во время ночной
охоты, так как являются индикаторами воздушных потоков, воз-
никающих при приближении к предметам, и помогают ей бес-
шумно обходить препятствия в темноте. Кончики вибрисс кошки
определяют все мельчайшие детали в очертании добычи и подска-
зывают ей, как точно нанести смертельный укус.
Мотивационная реактивность. Под «мотивацией» понимают
«внутренний» стимул, наличие побуждения к поведению (drive).
Влияние уровня мотивационного возбуждения на проявление
пищевого и спонтанного поведения у кошек продемонстрировано
в опытах Тейтельбаума с соавторами [1982]. Снижение уровня
мотивации путем разрушения латерального гипоталамуса приво-
дило к тому, что кошки не только отказывались есть и пить, но
и теряли способность к спонтанному поведению. У них развива-
лось состояние каталепсии, при котором кошке можно было за-
вести переднюю лапу на спину, или широко раздвинуть в стороны
передние лапы и при этом она не предпринимала никаких попы-
ток привести лапы в нормальное положение. Однако если на та-
кую каталептическую кошку действовали каким-либо раздражи-
телем, например, сдавливали ей хвост, она могла легко вскараб-
каться наверх, или спрыгнуть вниз, или начать есть предлагаемую
ей пищу.
На основании этого эксперимента и по результатам аналогич-
ных опытов на других животных авторы приходят к выводу, что
все формы поведения нуждаются в наличии фонового уровня
общего тонического возбуждения. Именно этот уровень и превра-
щает разрозненные поведенческие рефлекторные акты в спонтан-
ную активность, которая часто рассматривается как антитеза не-
произвольного автоматического рефлекса. Поддержание общего
тонического возбуждения для осуществления поведенческих реак-
7 Заказ № 355
97
ций, по-видимому, обеспечивается катехоламиновой системой
мозга. Уровень катехоламинов в ЦНС, как показано на некоторых
видах животных, является чертой, наследственно детерминирован-
ной [Серова и др., 1990; Раушенбах и др., 1991].
Обучаемость и элементарная рассудочная деятельность. На
ранних этапах онтогенеза поведение кошки формируется под
ведушдм влиянием инстинктов. По мере дифференциации нервной
системы помимо инстинктов большую роль в адаптивном поведе-
нии начинают играть различные формы обучения. Дальнейшая
дифференциация конечного мозга в процессе онтогенеза приводит
к тому, что в поведении начинает играть все большую и большую
роль элементарная рассудочная деятельность. Окончательное фор-
мирование наиболее адаптивного поведения происходит под влия-
нием внешних условий в зависимости от генетически обусловлен-
ной нормы реакции обучаемости и элементарной рассудочной
деятельности. Однако надо отметить, что у кошек инстинкт пре-
валирует над приобретенными навыками. Кошки с трудом под-
даются дрессировке.
Опыты по выработке классического павловского рефлекса
зачастую приводят у домашней кошки Fells catus к формированию
условной реакции страха, из-за которой дальнейшее проведение
эксперимента оказывается невозможным [Kling et al., 1969]. По-
этому подавляющее большинство исследований по обучению кош-
ки выполнено на модели «инструментального» рефлекса, в которой
используется положительное подкрепление, или реакция-награда.
В этих экспериментах были выявлены достаточно высокие анали-
тические способности зрительного и слухового анализаторов
кошки. Оказалось, что кошки способны различать форму, размеры,
яркость, цвет, монотонность и мерцание светового сигнала [Sasaki
et al., 1971; Кратин и др., 1975; Bisli et al., 1977; Loop et al.,
1979; Кураев и др., 1990]. Что касается слуховых способностей,
то кошки способны дифференцировать звуки по частоте, интенсив-
ности, тональности, локализации источника и продолжительности
[Калмыкова, 1990].
Рассудочная деятельность отличается от любых форм обучения.
Эта форма адаптивного поведения может осуществляться при
первой встрече животного с необычной ситуацией. В том, что жи-
вотное сразу же без специального обучения может вести себя
адекватно сложившейся обстановке, и заключается уникальная
особенность рассудочной деятельности. Кошки, как было показано
Л. В. Крушинским [1977], способны к элементарной рассудочной
деятельности, но по своей способности к экстраполяции они долж-
ны быть поставлены ниже псовых.
Более трудным, чем экстраполяционная задача, для животных
является тест по оперированию с размерностью фигур. В этом
тесте тесно взаимодействуют две функции мозга: перцептивная п
рассудочная. При помощи первой осуществляется процесс опозна-
вания геометрических фигур, при помощи второй — построение
программы адаптивного поведения, основанного на оперировании
98
их размерностью. Это оперирование в отличие от перцептивного
восприятия не пассивно. Оно осуществляется па фоне чрезвычайно
высокого напряжения внимания. Кошку можно научить разли-
чать, какая из трех одновременно предъявленных форм отлича-
ется от двух других. Эта способность требует перекрестных и
круговых сравнений. Тот факт, что кошка способна решать та-
кого рода задачи, говорит о ее высоких интеллектуальных способ-
ностях.
Таким образом, по тестам на способность к экстраполяции
и к оперированию размерностью фигур кошки, несомненно, могут
быть оценены как животные, обладающие развитой элементарной
рассудочной деятельностью.
ЭВОЛЮЦИЯ
Эволюционный взгляд на поведение кошачьих позволяет выя-
вить видоспецпфические особенности этих животных и оценить
вклад неспецифических черт того филогенетического уровня орга-
низации нервной системы, к которому принадлежат кошки.
Уже в начале олигоцена кошачьи были сформировавшимися
хищниками. По сравнению с другими хищными животными их
специализация была крайне узкой. Охотничий ритуал кошки,
морфология ее скелета, черепа и зубов приспособлены для того,
чтобы тихо сидеть в засаде, неожиданно набрасываться, схваты-
вать, вонзать когти и убивать. Смертельный укус отличал кошачь-
их от других хищников на протяжении всей эволюционной
истории.
Именно высокоспециализированная способность кошки охо-
титься на крыс и мышей поддерживалась отбором со времен воз-
никновения земледелия. В летописях упоминается, что кошка
использовалась для охраны крестьянских хозяйств, амбаров, жит-
ниц и зернохранилищ от грызунов. Однако она была не единствен-
ным животным, используемым для этих целей. В Греции и Риме
держали ласок, горностаев, каменных куниц и хорьков, привезен-
ных из Северной Африки. Во Франции и Испании применяли
генету, пропорциями тела напоминающую кошку. В молодом
возрасте генета легко приручается, но домашним животным она
все-таки не стала из-за специфического запаха, присущего всем
виверровым.
Решающим фактором для одомашнивания кошек явилась их
высокая специализация, ловкость и «боеспособность» в борьбе с
грызунами, которые наряду с другими индивидуальными особен-
ностями дали им преимущество над ласками, генетами и другими
мелкими хищниками. Отбор кошек по экстерьерным свойствам
никогда не доходил до тех пределов, когда кошка становилась
неспособной выжить без помощи человека в естественных усло-
виях (кроме одной породы — рэгдолл). Это привело к поразитель-
ному разнообразию прекрасных окрасок и типов меха у кошек,
7*
99
но не дало того разнообразия типов поведения, которое мы наблю*.
даем у собак.
Домашние кошки мало различаются по размерам и форме тела.
Размах вариации по весу у них примерно в 50 раз меньше, чем
у собак. Судя по рисункам из древнеегипетских рукописей, мало
изменился и скелет кошки. Сохранение дикого хищнического сте-
реотипа — уникальная черта животного с тысячелетней историей
одомашнивания. Особенно ярко она наблюдается в популяциях
одичавших кошек. Такие кошки собираются в колонии. Любые
попытки уничтожить их оказываются безуспешными, потому что
эти колонии непрерывно пополняются. Способность кошек мгно-
венно превращаться из покорных домашних любимчиков в диких
зверей, восстанавливать свои физические силы без опеки человека
поражает. Это свойство кошачьих показывает, что, несмотря на
тысячелетнюю доместикацию, мозг и тело кошки во многом оста-
лись сходными с дикими предками.
Существенным образом в поведении кошачьих проявились не-
специфические черты организации нервной системы, характерные
и для других животных этого филогенетического уровня развития:
высокая степень индивидуализации поведения и элементарная
рассудочная деятельность. Л. В. Крушинский [1977] показал, что
чем выше уровень филогенетического развития животных, тем
большей способностью к рассудочной деятельности они обладают.
Так, по способности к экстраполяции — одному из параметров
элементарной рассудочной деятельности — кошки входят в группу
наиболее «разумных» млекопитающих, включающую обезьян,
дельфинов, волков, собак, лисиц и песцов. Крушинский расцени-
вает животных этой группы как обладающих развитой элементар-
ной рассудочной деятельностью, стоящих существенно выше зай-
цев. мышевидных грызунов и полевок.
Достаточно далеко зашла и поведенческая дивергенция пород
Felts catus. Абиссинские, норвежские лесные и американские
короткошерстные пароды отличаются независимым и свободолю-
бивым нравом. Сиамские и одна из бирманских (burmese) пород,
наоборот, не выносят одиночества. Они хорошо уживаются с дру-
гими животными и предпочитают ходить на прогулки в сопровож-
дении человека. Бирманские кошки (birman) по своей природе —
индивидуалисты, сиамские — экстраверты. Своеобразие восточных
короткошерстных и некоторых бирманских кошек (burmese) про-
является в их любознательности и привязчивости к незнакомым
людям. Но самым странным темпераментом из всех кошачьих
обладают кошки породы рэгдолл. Если такую кошку взять на
руки, она будет «свисать» с них, как тряпичная кукла (отсюда и
название породы: rag doll—тряпичная кукла). Эти животные
настолько спокойны и безмятежны, что производят впечатление
наркотизированных. Ничто не способно вызвать у них состояние
тревоги. Полное отсутствие агрессивности приводит к тому, что
они, несмотря на кошачий фенотип, по сути не имеют кошачьих
свойств поведения и не способны выжить без защиты человека.
100
Таким образом, рассмотренные в этой главе формы поведения
кошки являются по своей природе инстинктивными и наследст-
венно детерминированными. Даже спонтанное поведение удается
«разложить» на простые рефлекторные субкомпоненты. Однако
применение методов генетического анализа для изучения поведе-
ния кошки встречает трудности, общие для всех высших млеко-
питающих. Полезный материал для генетических исследований,
по-видимому, могут дать межпородные скрещивания.
СПИСОК ЛИТЕРАТУРЫ
Калмыкова И. В. Различение час тотно-моду лированных сигналов в усло-
виях свободного поведения у кошек Ц Журн. высш. нерв, деят,—
1990,—Т. 40, вып. 3,—С. 451—456.
Кратин Ю. Г., Зубкова И. А., Лавров В. В., Рассказовский С. П. Электриче-
ские реакции мозга кошки при дифференцировании световых сти-
мулов Ц Журн. высш. нерв. деят.— 1975.— Т. 25, вып. 6,— 1203 с.
Крушинский Л. В. Биологические основы рассудочной деятельности.— М.:
Изд-во МГУ, 1977,— 272 с.
Крушинский Л. В., Зорина 3. А., Полетаева И. И., Романова Л. Г. Введение
в этологию и генетику поведения.— М.: Изд-во МГУ, 1983,— 173 с.
Кураев Г. А., Соболева И. В., Сороколетова Л. Г., Пустошкин Ю. И. Особен-
ности выработки условных пищевых рефлексов на цветовые стимулы
у кошек Ц Журн. высш. нерв. деят.— 1990.— Т. 40, вып. 1,— С. 169—
173.
Майр Э. Зоологический вид и эволюция/Ред. В. Г. Гентнер.— М.: Мир, 1968.—
С. 597.
Раушенбах И. Ю., Серова Л. И., Тимохина И. С. и др. Изменение содержа-
ния биогенных аминов у двух линий Drosophila virilis и их гибридов
в онтогенезе и при тепловом стрессе // Генетика — 1991.— Т. 21,
№ 4,— С. 657—666.
Серова Л. И., Козлова О. Н., Науменко Е. В. Генетико-нейрохимический ана-
лиз участия катехоламинов головного мозга в проявлении доминант-
ного фенотипа у самцов // Генетика,— 1990,— Т. 26, № 12,— С. 2250—
2253.
Тейтельбаум Ф., Шалерт Т., де Рик М. и др. Моторные подсистемы в моти-
вированном поведении // Нейрофизиологические механизмы поведе-
ния,—М.: Наука, 1982,—С. 38—54.
Bisli S., Clement В., Maffei L. Spatial frequency and oriental turning cur-
ves of visual neurons in the cat: effects of mean luminance // Exptl.
Brain Res.— 1977,— Vol. 27, N 3,— P. 335.
Green J. D., Clemente C. D., Groot J. Rhinencephalic lesions and behavior in
cats // J. Comp. Neurol.— 1957.— N 108.— P. 505—545.
Kling A., Kovach J. K., Tucker T. J. The behaviour of cats // The behaviour
of domestic. animals/Ed. E. S. E. Hafez.— London: Bailliere, Tindall and
Cassell, 1969.— P. 482—512.
Kovach J. K., Kling A. Mechanisms of neonate sucking behaviour in the asso-
ciated structures in cats // Anim. Behav.— 1967,—N 15,—P. 91—101.
Leyhausen P. Cat behaviour. The predatory and social behavior of domestic
and wild cats.—N. Y.; L: Garland STPM Press, 1979,—340 p.
Loop M. S., Bruse L. L., Petuchovski S. Cat colour vision: the effect of stimu-
lus size and viewing distance // Vision Res.— 1979.— Vol. 19.— P. 507.
Morill R., Todd N. B. Mutant allele freouencies in domestic cats of Denver,
Colorado // J. Hered.— 1978 —Vol.'69,—P. 131—134.
Morris D. Catwatching.— Oxford: Jonathan Cape, 1986,— P. 106.
Porter R. W., Cavanauch E. B., Sawyer С. H. Localized changes in electrical
activity of the hypothalamus in oestrous cats following vaginal stimu-
lation // Amer. J. Physiol.— 1957.— N 189.— P. 145—151.
101
Rosenblatt J. S., Aronson L. R. The decline of sexual behaviour in male cats
after castration with special reference to the role of prior sexual expe-
rience / Behaviour.— 1958.— N 12.— P. 285—338.
Sasaki H., Saito G., Bear D. M., Ervin F. R. Quantitative xariation of striate
receptive fields of the cats as a function of light-dark adaptation //
Exptl. Brain Res.- 1971,— Vol. 13, N 3,— P. 273.
Seitz P. F. D. Infantile experience and adult behavior in animal subjects Ц
Psychosom. Med.— 1959,- N 21.— P. 353-378.
Todd N. B. Behaviour and genetics of the domestic cat Ц Cornell Veterina-
rian.— 1962,— Vol. 53.— P. 99.
Глава 10
БИОЛОГИЯ РАЗМНОЖЕНИЯ
В настоящее время накоплен довольно большой материал по
биологии размножения кошки. Эти данные представляются важ-
ными для практики разведения породистых кошек. Однако особое
значение они имеют для работ по экспериментальной эмбриологии
кошки, исследований в области трансгенеза, разработки методов
криоконсервации зародышей.
СЕЗОННОСТЬ РАЗМНОЖЕНИЯ
В ходе доместикации произошло значительное изменение ряда
физиологических функций домашней кошки, в том числе и такой
важной для приспособленности функции, как размножение. Ди-
кая европейская кошка размножается только один раз в году,
причем в строго определенное время года, тогда как домашняя
кошка может приносить до трех пометов в год. Величина помета
у домашних кошек также значительно больше. Если у диких ко-
шек число котят в помете 1—2, то у домашних 5—6.
Половой зрелости самки домашней кошки достигают в воз-
расте 6—8, а самцы — 8—10 мес. После достижения половой зре-
лости самцы постоянно проявляют готовность к спариванию. У са-
мок половая активность ограничена временем течки. Как и у
многих других домашних животных, у кошки утрачена строгая
сезонность размножения. Тем не менее сезонные влияния сказыва-
ются на половой активности кошек. Пик активности приходится
на весенне-летний период. При содержании в контролируемых ус-
ловиях при стабильном световом режиме у самок формируется
постоянный 4—8-дневный цикл астральной активности.
ЭСТРАЛЬНЫЙ ЦИКЛ
При наступлении течки самки проявляют признаки полового
поведения (см. гл. 9), начинается активное выделение феромонов
специальными железами, расположенными в анальном отверстии,
102
Рис. 10.1. Среднее количество эстрадиола-17^ (I), лютеинизирующего гор-
мона (2) и прогестерона (3) во время эструса у самок, спаривающихся и
овулировавших.
Каждая стрелка указывает на время одного спаривания [Wildt et al., 19811.
Рис. 10.2. Средний уровень эстрадиола-178 (7), лютеинизирующего гормона
(3) и прогестерона (3) в течение эструса и последующих 13 дней у неспа-
ривавшихся, неовулировавших кошек.
День первый эквивалентен первому дню эструса, продолжавшегося 6,4 дня [Wildt
et al, 1981J.
происходит корнификация вагинального эпителия. Состояние теч-
ки легко диагностируется по наличию ороговевших эпителиальных
клеток во влагалищных мазках [Dawson, 1952; Michael, 1961;
Whalen, 1963; Wildt et al., 1978a].
Частота спариваний во время эструса довольно высока. Так, за
12 ч наблюдений регистрируется 14—20 спариваиий, а за 36 ч —
до 30 спариваний. Частота спаривания снижается к концу эстру-
103
Рис. 10.3. Средний уровень эстрадиола-17^, люгеиниаирующего гормона и
прогестерона в течение эструса и межэстрального интервала при овуляции,
вызванной спариванием.
День первый эквивалентен первому дню эструса и спаривания. Продолжительность
эструса, лютеальной фазы и межэстрального интервала 5,8; 38,2 и 50,3 дня соответ-
ственно [Wildt et al., 1981].
са, но возрастает его продолжительность [Glower et al., 1985;
Concanrion et al., 1989]. Продолжительность эструса не зависит от
количества спариваний и составляет в среднем 5,8 ± 0,2 дня. При
однократном спаривании она варьирует от 4,9 ± 0,9 до 5,9 ± 0,6,
а при многократном — от 4,2 ± 0,4 до 5,6 ± 0,6 дня. Продолжитель-
ность эструса не зависит также от того, произошла овуляция или
104
нет. В случае, если овуляция произошла, но беременность не
наступила, длительность лютеальной фазы составляет 38,2 ± 2,8
дня, а продолжительность межэстрального периода 50,3+2,7
дня [Wildt et al., 1981]. В том случае, если овуляции не было,
продолжительность межэстрального периода составляет 13—14
дней. Продолжительность беременности варьирует от 63 до 70
дней, составляя в среднем 65 дней.
Эндокринные механизмы астрального цикла кошек исследова-
ны достаточно подробно [Shille et al., 1979; Concannon et al.,
1980; Johnson, Gay, 1981a, b; Goodrowe, Wildt, 1987]. Развитие
фолликулов в яичниках индуцируется фолликулостимулирующим
гормоном (ФСГ). По мере созревания фолликулы начинают про-
дуцировать эстрадиол-17^, который вызывает соответствующие
изменения в половом поведении. В начале эструса его концентра-
ция возрастает от 20 до 40 пг/мл (рис. 10.1 —10.3) [Wildt et al.,
1981], а по некоторым данным и до 200 пг/мл [Schmidt, 1986].
Уровень данного гормона остается повышенным в течение всего
эструса и возвращается к базальному на 4—5-й день после ову-
ляции (см. рис. 10.1) или в течение последних 24—48 ч эструса,
если овуляция не произошла (см. рис. 10.2). В последнем случае
базальная концентрация сохраняется в течение диэструса и вновь
возрастает с наступлением эструса. Если же овуляция произошла,
снижение уровня эстрадно ла-17f> обеспечивается конкурентным
возрастанием концентрации прогестерона, продуцируемого желты-
ми телами (см. рис. 10.3).
ОВУЛЯЦИЯ
Кошки относятся к группе видов, у которых овуляция не спон-
танная, а индуцируется спариванием. Такой же тип овуляции
наблюдается у норки, хорька, кролика, белки и ряда других жи-
вотных. Спаривание приводит к выделению лютеинизирующего
гормона (ЛГ), который и индуцирует овуляцию. В отличие от
видов со спонтанной овуляцией, у кошки ооциты могут сохра-
няться в фолликулах довольно длительное время в течение
эструса. Внутрифолликулярный механизм способен поддерживать
состояние готовности ооцитов к овуляции до 6 дней и более.
В течение этого времени они остаются чувствительными к инду-
цирующему действию гонадотропных гормонов.
Интересной особенностью кошек является зависимость уровня
ЛГ и, следовательно, вероятности овуляции от продолжительности
эструса и количества спариваний [Рааре et al., 1975; Wildt et al.,
1980]. Необходимый для овуляции уровень ЛГ достигается, как
правило, после 4 копуляций н более в течение 4 ч [Concannon et
al., 1980; Wildt et al., 1981]. Единственное спаривание вызывает
через 1 ч достоверное повышение уровня ЛГ почти до порогового
значения (37 нг/мл), сохраняется на этом уровне в течение 3 ч,
а затем за 24 ч снижается до базального уровня (рис. 10.4)
105
Рис. 10.4. Средняя концентра-
ция лютеинизирующего гормо-
на у самок при однократном и
трелкратном спаривании в те-
чение одного дня.
Каждая стрелка указывает на
время одного сваривания fWildt
et al., 1980].
[Wildt et al., 1980]. При трехкратном спаривании уровень лютеи-
низирующего гормона достигает через 3 ч порогового значения
(35,8 пг/мл) и продолжает расти в течение последующих 9 ч до
концентрации 100 пг/мл и только после этого начинается его
снижение (см. рис. 10.4).
Уровень ЛГ зависит не только от количества спариваний, но
и от того, в какое время в течение эструса происходит спаривание.
При спаривании во второй день эструса уровень ЛГ достоверно
выше, чем при спаривании в конце астрального периода [Glower
et al., 1985].
Уровень лютеинизирующего гормона определяет количество
овуляций. При низкой концентрации ЛГ овуляции не происходит.
При трех спариваниях в день овулирует примерно 86,6 % преову-
ляторных фолликулов. Первичная волна ЛГ наблюдается, как
правило, в первый день после спаривания, но у некоторых живот-
ных обнаружена задержка в ответе: у них эта волна возникает
только на второй день. Предполагают, что овуляция индуцируется
первичной волной и происходит через 48—52 ч после спаривания,
хотя у некоторых самок овуляция может происходить позднее.
Несмотря на наличие большой индивидуальной изменчивости в
сроках начала овуляции, этот процесс всегда заканчивается в
течение 3 сут после спаривания образованием геморрагического,
а затем желтого тела [Рааре et al., 1975; Shille, Stabenfeldt, 1979;
Shille et al., 1979, 1983]. У неовулировавших самок уровень
лютеинизирующего гормона остается низким в течение всего
эструса и последующего периода до наступления следующего
цикла (см. рис. 10.2).
После овуляции образуются желтые тела, которые начинают
продуцировать прогестерон. Повышение его концентрации наблю-
дается между 4-м и 9-м днями после спаривания, достигая пика
на 14—15-й день (ом. рис. 10.1, 10.3) [Рааре et al., 1975; Verha-
ge et al., 1976; Glower et al., 1985]. Высокий уровень прогестерона
106
Рис. 10.5. Средний уровень
прогестерона в течение
8 недель у самок при нор-
мальном (контроль) эстру-
се (I) и индуцированном
обработкой фолликулости-
мулирующим гормоном (2)
[Goodrowe et al., 1988].
сохраняется до 21 дня. Затем, если беременность не наступила,
уровень прогестерона постепенно снижается и достигает базаль-
ного уровня на 49—63-й день после овуляции [Wheeler et al.,
1988]. При введении ФСГ (эта процедура применяется для инду-
цирования эструса) концентрация прогестерона возрастает быст-
рее и достигает более высокого уровня, чем в норме (рис. 10.5)
[Goodrowe et al., 1988b].
Размеры желтых тел меняются в ходе беременности. В начале
они имеют диаметр 2,68 ±0,1 мм, а затем увеличиваются к 16-му
дню до 3,60 ± 0,1 мм п к 44-му дню уменьшаются до 1,42 ± 0,2 мм
(см. рис. 10.3) [Wildt et al., 1981].
ПРОЦЕСС ОПЛОДОТВОРЕНИЯ
Сперматозоиды сохраняют оплодотворяющую способность в
половых путях самки в течение 24—25 ч после спаривания,
поэтому задержка овуляции может привести к развитию псевдо-
беременности [Hamner et al., 1970]. Количество сперматозоидов
в эякуляте варьирует от 20 до 100 млн. Их подвижность в норме
составляет 85,0 + 2,7 % [Platz et al., 1978].
Диаметр яйцеклетки с zona pellucida —162,4±2,9 мкм, тол-
щина zona pellucicla — 17,4 ±0,5 мкм. Сама яйцеклетка составля-
ет 127,5 ± 2,9 мкм. Цитоплазма ооцитов кошки, как и у других
хищных млекопитающих, содержит большое количество липидных
гранул (рис. 10.6, 10.7) [Guraya, 1965; Goodrowe et al., 1988a],
что делает весьма затруднительным наблюдение зародышевого
(герминального) пузырька. Использование центрифугирования
позволяет преодолеть эту трудность. Герминальный пузырек со-
ставляет примерно 20 % от объема яйцеклетки.
107
Рис. 10.6. Ооциты домашней кошки после разрушения герминального пу-
зырька на одноклеточной стадии.
а световой микроскоп; б — флюоресцентный микроскоп (Х400); в, г — после цент-
рифугирования [Goodrowe et al., 1988].
Процесс оплодотворения у кошек был исследован in vitro
(рис. 10.8) [Niwa et al., 1985]. Показано, что через 20 мин после
осеменения сперматозоиды обнаруживаются в перивителлиновом
пространстве под zona pellucida. Через 4 ч можно наблюдать
появление мужского и женского пронуклеусов. Первое деление
дробления происходит через 24—30 ч. На 6—8-й депь после спа-
ривания зародыш кошки достигает стадии морулы — бластоцисты
[Goodrowe et al., 1988а]. Имплантация бластоцист происходит
примерно на 11 — 13-й день после спаривания [Herron, 1986].
108
ЭМБРИОНАЛЬНАЯ СМЕРТНОСТЬ
Плодовитость млекопитающих определяется в основном двумя
факторами: количеством овулировавших яйцеклеток (потенциаль-
ная плодовитость) и величиной эмбриональных потерь. У кошек
число преовуляторных фолликулов варьирует от 2 до 8 и состав-
ляет в среднем 5,3 ± 0,5 на самку [Wildt et al., 1981; Schmidt et
al., 1983; Schmidt, 1986; Goodrowe et al., 1988b], Но примерно
12 — 14 % преовуляторных фолликулов не овулирует: количество
желтых тел варьирует от 4,2 до 4,9 на самку [Goodrowe et al.,
1988а].
Величину эмбриональных потерь можно оценить, сравнивая
количество овулировавших яйцеклеток (число желтых тел) с ко-
109
Рис. 10.7. Двухклеточная стадия кошачьих эмбрионов, оплодотворенных in
vitro (а), морула (б), бластоциста (в) [Goodrowe et al., 1988].
отчеством рожденных котят. Примерно 10—15 % эмбрионов гиб-
нет на разных стадиях беременности. В основном гибель прихо-
дится на первые 4 педели эмбрионального развития, т. е. на до-
имплантациопный период и первые стадии постимплантационноп>
развития. Причины гибели могут быть весьма разнообразными.
Примерно 2—3 % яйцеклеток остаются неоплодотворенными
[Schmidt, 1986: Goodrow е et al., 1988b]. Могут возникать нару-
шения в темпах дробления зародышей. Серьезным барьером в раз-
витии эмбрионов является процесс имплантации. Среди эмбрионов,
погибших после имплантации, примерно 16 % составляют носите-
110
ли хромосомных аномалий [Berepubo, 1985]. Показано, что среди
погибших эмбрионов у кошки встречаются особи с аномальным
кариотипом. Если в норме у кошек 2п = 38 хромосомам, то среди
погибших были эмбрионы с кариотипом 38,ХХ/37,ХХ или
38,ХХ/39,ХХ. Такой мозаицизм приводит к гибели зародышей на
стадии первичной дифференцировки. Сообщалось о половом мозаи-
цизме у черепаховых котов [Robinson, 1985], но в этом случае
мозаицизм связывали не с летальностью, а с бесплодием. Все
гомозиготы по мутации М, определенная часть гомозигот по алле-
лю W также гибнут после имплантации (см. гл. 3, 6).
Возраст и степень половой зрелости кошек также сказываются
на их плодовитости [Schmidt, 1986]. Первично рожающие кошки
характеризуются более высоким уровнем эмбриональной смерт-
ности и соответственно более низкой плодовитостью, чем самки,
рожающие повторно. Плодовитость кошки и жизнеспособность ее
котят постепенно возрастают к 5—7-му окотам. Затем репродук-
тивный потенциал самок снижается, хотя описаны кошки, которые
производят вполне нормальных потомков и в возрасте 8—10 лет.
Существуют межпородные различия в плодовитости кошек.
111
Рис. 10.8. Процесс оплодотворения in vitro яйцеклеток кошки, I
1 — в яйце, зафиксированном через 30 мин после осеменения, айдны головка спер- I
мия, i-e полярное тельце и хромосомы в анафазе II (х515); 2, з— сперматозоиды '
в цитоплазме яйцеклетки через 1 и 1,5 ч после осеменения (Х623); 4 — деконден-
сация головки сперматозоида (стрелка), 1-е полярное тельце и хромосомы в тело-
фазе II через 3 ч после осеменения (Х258); 5 — деконденсация головки спермато- 1
зоида (Х515); 6—трансформация головки сперматозоида в мужской пронуклеус
через 4 ч после осеменения (Х623); 7—пронуклеарные яйцеклетки, фиксированные
через 4,5 ч после осеменения; мужской (МР) и женский (FP) пронуклеусы; 8 —
яйцеклетка через 20 мин после осеменения; сперматозоид на стадии прохождения
через z. р, [Niwa et al., 1985].
'I
I
Так, средний размер помета у бурмезских кошек составляет 5,0, ?
сиамских — 4,5, персидских — 3,9, абиссинских — 3,5, шиншилле- (
вых — 2,8 [Johnstone, 1987]. >
112
ПОЛУЧЕНИЕ РАННИХ ЗАРОДЫШЕЙ И МЕТОДЫ РАБОТЫ
С НИМИ
Суперовуляция. Гормональную обработку кошек экзогенными
гормонами используют для получения индуцированного эструса,
развития фолликулов и овуляции во время анэструса. Методики,
используемые для получения астрального поведения, включают
обработку ФСГ гормоном с последующим спариванием или с обра-
боткой человеческим хориогонином для получения индуцирован-
ной овуляции, если самок осеменяют искусственно. Для искусст-
венного осеменения самок используют свежеполученную или за-
мороженную сперму. Сперму получают при электростимуляции
[Seager, 1976] или на искусственную вагину [Sojka et al., 1970].
Сразу же после сбора эякулят помещают в стерильный 0,9 %-й
раствор NaCl, доводя общий объем до 500 мкл. В том случае, если
после сбора сперму замораживают, то в раствор добавляют крио-
протектор [Platz et al., 1978]. Если же для осеменения использу-
ется свежая сперма — определяют подвижность сперматозоидов и
их морфологию. Подвижность сперматозоидов должна быть не
менее 80—85 %. Забор спермы проводится при анестезии кетамин-
хлоридом (33,0 мг/кг живого веса). Осеменение кошек проводят
на 2-й или 3-й день эструса. Овуляция у самок индуцируется спа-
риванием с вазэктомированным самцом или инъекцией 250—
500 м. е. HCG на 1-й или 2-й день эструса. При осеменении кош-
ки также слегка анестезируются кетамином (6 мг/кг живого веса).
Кошка фиксируется и раствор NaCl с содержащимися в нем:
сперматозоидами вводится пипеткой в матку. Для эффективного
осеменения необходимо вводить 50—100 млн сперматозоидов с
нормальной подвижностью.
Выход потомков при использовании замороженной и свежей
спермы одинаков, но несколько ниже, чем при естественном опло-
дотворении (1—3 и 3—6 котят на самку соответственно) [Sojka
et al., 1970; Platz et al., 1978].
Развитие работ по экспериментальной эмбриологии кошек
вызвало необходимость разработки методов для получения супер-
овуляции. Правильно подобранные дозы и сочетания гормонов
дают возможность получить большое количество яйцеклеток.
Анализ 8 различных режимов и дозировок обработки анэстраль-
ных кошек позволил Дрессеру с соавторами [Dresser et al., 1987]
рекомендовать метод, включающий последовательную обработку
фолликулостимулирующим гормоном гипофиза (ФСГ—II) в дозе
0,75 мг на самку в течение первых 5 дней и в дозе 0,25 мг на 6-й
день и введение двух доз по 375 м. е. человеческого хориогонина
на 6- и 7-й день. Такая обработка приводит к овуляции 30,9 ± 9,5
яйцеклетки, тогда как в норме овулирует около 5. Использование
сыворотки жеребой кобылы (СЖК) также приводит к суперовуля-
ции (образуется около 60 желтых тел) [Cline et al., 1980]. Однако
обработка экзогенными гормонами кроме увеличения количества
овулировавших яйцеклеток вызывает и формирование многочис-
8 заказ Х> 355
113
лениых ановуляторных фолликулов и снижение количества нор-
мально развивающихся зародышей [Goodrowe et al., 1988а]. Эти
отрицательные эффекты связывают с незрелостью, атрезией или
хромосомными аномалиями ооцитов, снижением оплодотворяе-
мости в ответ на гиперстимуляцию.
Оплодотворение in vitro. Для наблюдения за ранними ста-
диями развития зародышей кошек может быть использовано опло-
дотворение яйцеклеток in vitro. Для этой цели используют яйце-
клетки, полученные после гормональной стимуляции самок. Для
манипуляций и культивирования ооцитов in vitro чаще всего
используют модифицированный Кребс-Рингеровский раствор.
Яйцеклетки, освобожденные от клеток кумулюса, помещают в
капли под парафиновое масло и инкубируют в атмосфере, содер-
жащей 5 % СО2 при 37 °C с добавлением сперматозоидов. Для
оплодотворения in vitro может быть использована сперма, полу-
ченная от самцов методом электростимуляции или на искусствен-
ную вагину и прошедшая капацитацию в половых путях [Hamner
et al., 1970], сперма, полученная из семявыносящих канальцев
[Bowen, 1977] или придатка семенника [Niwa et al., 1985]. Сле-
дует отметить, что успешное оплодотворение оказывается возмож-
ным только в том случае, если сперматозоиды предварительно
прошли капацитацию или в половых путях самки [Hamner et al.,
1970], или в культуральной среде [Bowen, 1977; Niwa et al.,
1985].
Трансплантация эмбрионов. Первые попытки трансплантации
эмбрионов у кошки были предприняты в 1977 г. [Kramer et al.,
1979], В качестве доноров использовались самки со спонтанным
эструсом. Реципиенты спаривались с вазэктомированными сам-
цами или овуляция вызывалась у них гормональной обработкой.
В обоих случаях было продемонстрировано нормальное протека-
ние беременности, завершившееся рождением нормальных котят.
Сравнение результатов, полученных при пересадке зародышей
при естественном эструсе и гормональном стимулировании, пока-
зало, что для пересадок можно использовать и эмбрионы, получен-
ные при суперовуляции [Goodrowe et al., 1988b]. Кошки полу-
чали по 20 мг FSH (FSH — Р) в течение 5 дней — режим, эффек-
тивный для индукции эструса у анэстричных самок [Wildt et al.,
1978b]. Начиная с первого дня эструса самки-доноры спаривались
с интактными самцами по 3 раза в день с 3—4-часовым интер-
валом между спариваниями в течение 3 дней (1—3-й день эстру-
са). В экспериментах по пересадке эмбрионов кошки необходимо
добиваться строгой синхронизации сроков беременности между
донором и реципиентом. Поэтому самок-реципиентов или отбирают
по проявлению астрального поведения или используют ту же
гормональную обработку, что и для доноров, но спаривают с
вазэктомированным самцом. Для подтверждения овуляции самки
обследуются лапароскопически на 4-й или 5-й день эструса.
Сбор эмбрионов проводят на 7-й или 8-й день после фикси-
рованного спаривания. К этому сроку зародыши достигают стадии
114
поздней морулы или ранней бластулы. Для анестезии используют
кетамин-хлорид— acepromazine maleate (10:1, 20,0 мг/кг и
2,0 мг/кг). Лапаротомия проводится по белой линии живота, на
2—3 см от бифокации матки. Рога матки промываются через кате-
тер 25—30 мл теплой (37 °C) среды Ham’s —FIO. содержащей
5 % инактивированной эмбриональной сыворотки. Капли промыв-
ной жидкости просматриваются под стереомикроскопом, эмбрионы
с помощью пипетки переносятся в свежую порцию среды и оце-
ниваются по морфологии. Пересадка эмбрионов у кошки прово-
дится хирургическим методом, поэтому самки-реципиенты также
подвергаются операции лапаротомии. Пересадка зародышей осу-
ществляется в один пли оба рога матки. В стенке матки (ближе
к яичниковому концу) делается прокол, через который вводится
пипетка, в которой в капле среды находятся эмбрионы. Беремен-
ность определяется на 35-й день пальпацией живота самки или
после рождения котят.
Возможна успешная пересадка эмбрионов, полученных после
индуцированного гормонами эструса, пункции фолликулов, куль-
тивирования и осеменения in vitro в Кребс-Рингеровском растворе
с добавлением 20 % эмбриональной сыворотки фолликулярных
ооцитов [Goodrowe et al., 1988а]. В результате такой пересадки
было получено 5 пометов (4 из 18 подсаженных зародышей; 1 из
13; 3 из 8; 1 из 8 п 1 из 7). Но следует помнить, что использова-
ние суперовуляции часто ведет к образованию большого количест-
ва дефектных ооцитов и, как следствие, к высокому уровню на-
рушений в развитии эмбрионов.
Была также показана возможность успешной трансплантации
эмбрионов после их криоконсервации [Dresser et al., 1988]. После
размораживания они культивировались в среде Эрла или FIO с
добавлением 20 % эмбриональной сыворотки, а затем были транс-
плантированы в матку синхронизированных реципиентов. При-
живаемость эмбрионов составила 12,4%. К сожалению, у кошек
возможна лишь хирургическая пересадка зародышей, поэтому
оперированные самки могут быть использованы лишь один раз.
МЕТОДЫ КОНТРАЦЕПЦИИ
Наиболее эффективным и широко применяемым методом конт-
рацепции у кошек является кастрация. Однако в последнее время
разработаны фармакологические способы контрацепции. Они при-
меняются как в ветеринарной практике по просьбам владельцев
кошек, так и для контроля численности бродячих кошек.
Для прекращения эструса используется прогестерон или его
аналоги. Эффективным оказалось пероральное применение мегест-
ролацетата (Ovaban) и медроксипрогестерона ацетата (MAP, De-
po-Provera). Последний препарат прекращает половое возбужде-
ние у самок через 24 ч после приема таблетки. Делмадинон аце-
тат. снижает уровень полового возбуждения и предотвращает
8*
U5
эструс примерно па 6 мес, в зависимости от дозы. Внутримышеч-
ные инъекции препарата Rerogest в дозах от 2,2 до 4,4 мг на 1 кг
массы тела приводят к увеличению межэстрального периода. Сле-
дует соблюдать известную осторожность в применении гестагенов
па основе прогестерона. В больших дозах или при длительном
применении они могут приводить к развитию эндометрита или
диабета [Schmidt, 1986].
БИОЛОГИЯ РАЗМНОЖЕНИЯ
ДРУГИХ ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА КОШАЧЬИХ
Ранее было отмечено, что дивергенция кошачьих имела место
сравнительно недавно (см. гл. 1). Исследования показателей ре-
продуктивной функции у различных представителей этого семей-
ства выявили их значительное сходство. Как и у домашней кошки,
овуляция у пумы индуцируется спариванием или гормональной
обработкой самок. Примечательно, что характер изменений гормо-
нальных характеристик во время эструса и в ходе беременности
также оказывается сходным у пумы, тигра, ягуара, льва и домаш-
ней кошки [Rowlands, Sadleir, 1968; Seal et al., 1978; Wildt et al.,
1979; Bonney et al., 1981]. У львов описаны случаи спонтанной
овуляции. Однако если спаривание не происходит, то наблюда-
ются аповуляторпые циклы [Schmidt et al., 1979].
Продолжительность беременности у крупных кошек, принад-
лежащих к липни пантер, больше, чем у домашней кошки, и со-
ставляет 95 —110 дней [Bonney et al., 1981].
СПИСОК ЛИТЕРАТУРЫ
Berepubo N. A. A cytogenetic study of subfertility in the domestic cat IFelis
catus) Ц Canad. J. Cytol.— 1985,— Vol. 27, N 2,— P. 219-223.
Bonney R. C., Moore H. D. M., Jones D. M. Plasma concentrations of oestradi-
ol-17[l and progesterone, and laparoscopic observations of the ovary in
the puma (Felts concolor) during oestrus, pseudopregnancy and preg-
nancjr Ц J. Reprod. Fertil.— 1981.— Vol. 62, N 2.— P. 523—531.
Bowen R. A. Fertilization in vitro of feline ova by spermatozoa from the duc-
tus deferens Ц Biol. Reprod.— 1977.— Vol. 17, N 1.— P. 144—147.
Cline E. M., Jennings L. L., Sojka N. J. Breeding laboratory cats during arti-
ficially induced oestrus / Lab. Anim. Sci.— 1980.— Vol. 30.— P. 1003.
Concannon P. W., Hodgson B. G., Lein D. Reflex LH release in estrous cats fol-
lowing single and multiple copulations Ц Biol. Reprod.— 1980.— Vol. 23,
N 1,— P. 111—117.
Concannon P. W., Lein D. H., Hodgson B. G. Self-limiting reflex luteinizing
hormone release and sexual behaviour during extended periods of unre-
stricted copulatory activity in oestrous domestic cats Ц Biol. Reprod.—
1989,-Vol. 40, N 6.-P. 1179—1187.
Dawson A. B. The domestic cat Ц Care and Breeding of Laboratory Animals/
Ed. E. J. Ferris — N. Y„ 1952.— P. 202—233.
Dresser B. L., Sehlhorst C. S., Wachs К. B. et al. Hormonal stimulation and
embryo collection in the domestic cat (Fells catus) // Theriogenology.—
1987,— Vol. 28, N 6,- P. 915-927.
Dresser B. L., Gelwicks E. J., Wachs К. B., Keller G. L. Embryo cryopreserva-
tion and transfer in the domestic cat (Felis catus) Ц 11 Intern. Congr.
116
on animal reproduction and artificial insemination, University College,
Dublin, Ireland, June 26-30, 1988,— 1988,— Vol. 2, N 160,— P. 3. (Bri-
ef communications).
Glover T. E., Watson P. F., Bonney R. C. Observations on variability in LH
release and fertility during oestrus in the domestic cat {Felis catus) Ц
J. Reprod. Fertil.— 1985,— Vol. 75, N 1.- P. 145—152.
Goodrowe K. L., Wall R. J., O’Brien S. J. et al. Developmental competence of
domestic cat follicular oocytes after fertilization in vitro // Biol. Rep-
rod.- 1988a.- Vol. 39, N 2,- P. 355-372.
Goodrowe K. L., Howard J. G., Wildt D. E. Comparison of embryo recovery, em-
bryo quality, oestradiol-17^ and progesterone profiles in domestic cats
{Fells catus) at natural or induced oestrus Ц J. Reprod. Fertil.— 1988b.—
Vol. 82, N 2,— P. 553—561.
Goodrowe K. L., Wildt D. E. Ovarian response to human chorionic gonadotro-
pin or gonadotropin releasing hormone in natural and induced-oestrus
cats // Theriogenology.— 1987.— Vol. 27, N 4.— P. 811—817.
Guraya S. S. A histochemical analysis of lipid yolk deposition in the oocytes
of cat and dog Ц J. Exptl. Zool.— 1965.— Vol. 160, N 1.— P. 123—136.
Hamner С. E., Jennings L., Sojka N. J. Cat (Fells catus L.) spermatozoa re-
quire capacilation // J. Reprod. Fertil.— 1970.— Vol. 23, N 1.— P. 477—
480.
Herron M. A. Feline physiology of reproduction // Small animal reproduction
and infertility. A clinical approach to diagnosis and treatment/Ed.
T. J. Burke.— Philadelphia, USA: Lea, Febiger, 1986.— P. 13—23.
Johnson L. M., Gay V. L. Luteinizing hormone in the cat. I. Tonic secretion //
Endocrinology.— 1981a.— Vol. 109, N 1.— P. Й.0—246.
Johnson L. M., Gay V. L. Luteinizing hormone in the cat. II. Mating induced
secretion // Endocrinology.— 1981b.— Vol. 109, N 1.— P. 247—252.
Johnstone J. Reproductive patterns of pedigree cats // Austral. Vet. J.— 1987.—
Vol. 61, N 7,— P. 197—200.
Kramer D. C., Flow B. L., Schriver M. D. et al. Embryo transfer in the non-
human primate, feline and canine // Theriogenology.— 1979.— Vol. 11,
N 1,— P. 51—62.
Michael R. P. Observation upon the sexual behaviour of the domestic cat
{Fells catus L.) under laboratory conditions // Behavior.— 1961.—
Vol. 18, N 1.- P. 1-24.
Niwa K., Ohara K., Hosoi Y., Gritani A. Early events of in vitro fertilization
of the cal eggs by epidyinal spermatozoa // J. Reprod. Fertil.— 1985,—
Vol. 74, N 2,— P. 657-660.
Paapc S. R., Shille V. M., Seto П. Luteal activity in the pseudopregnant cat /
Biol. Reprod.— 1975 — Vol. 13,— P. 470—474.
Platz С. C., Wildt D. E., Seager S. W. J. Pregnancy in the domestic cat after
artificial insemination with previously frozen spermatozoa // J. Reprod.
Fertil.— 1978,— Vol. 52, N 1,— P. 279—282.
Robinson R. Fertile male tortoiseshell cats // J. Hered.— 1985.— Vol. 76, N 2.—
P. 137—138.
Rowlands G. W., Sadleir R. M. F. S. Induction of ovulation in the lion {Panthe-
ra leo) // J. Repvod. Fertil.— 1968.— Vol. 16, N 1.— P. 105—111.
Schmidt A. M., Nadal L. A., Schmidt M. J., Beamer N. B. Serum concentration
of oestradiol and progesterone during the normal oestrous cycle and
early pregnancy in the lion {Panthera leo) // J. Reprod. Fertil.— 1979.—
Vol. 57, N 1,— P. 267-272.
Schmidt P. M. Feline Breeding Management // The veterinary Clinics of North
America. Small Animal practice.— 1986.— Vol. 16, N 3.— P. 435—451.
Schmidt P. M., Chakraborty P. K., Wildt D. E. Ovarian activity, circulating hor-
mones and sexual behavior in the cat. II. Relationships during pregnan-
cy, parturition, lactation and postpartum oestrus Ц Biol. Reprod.—
1983,—Vol. 28,—P. 657—671.
Seager S. W. J. Electroejaculation of cats (domestic and captive wild felir
dae) // Applied Electronics for Veterinary Medicine and Animal Phy-
siology/Ed. W. R. Klemm.— 1976.— P. 410—418. '
147
Seal U. S,, Plotka E. D., Gray C. W. Baseline hematology, serum chemistry and
hormone data for captive tigers {Panthera tigris ssp.) and lions (Pan-
thera leo) Ц International Tiger Studbook: Congress report on 1st Intern,
symp. on the management and breeding of the tiger.— 1978.— P. 174—
192.
Shille V. M.. Lundstrom К. E., Slabenfeldt G. H. Follicular function in the do-
mestic cat as determined by estradi.ol-173 concentration in plasma: re-
lation to estrous behaviour and cornification of exfoliated vaginal epi-
thelium // Biol. Reprod.- 1979,— Vol. 21, N 5,— P. 953—963.
Shille V. M., Munro C., Farmer S. W. et al. Ovarian and endocrine responses
in. the cat after coitus // J. Reprod. Fertil.— 1983.— Vol. 68, N 1.—
P. 29—39.
Shille V. M., Slabenfeldt G. H. Luteal function in the domestic cat during
pseudopregnancy and after treatment with prostaglandin F2 Ц Biol.
Reprod.— 1979,—Vol. 21, N 6,—P. 1217—1223.
Sojka N. J., Jennings L. L., Hamner С. E. Artificial insemination in the cat
(Fells catus L.) // Lab. Anim. Care.— 1970.— Vol. 20.— P. 195—204.
Verhage H. G., Beamer N. B., Brenner R. M. Plasma levels of estradiol and
progesterone in the cat during polyestrus, pregnancy and pseudopreg-
nancy // Biol. Reprod.— 1976.— Vol. 14.— P. 579—585.
Whalen R. E. Sexual behavior of cals // Behavior.— 1963.— Vol. 20.— P. 321—
342.
Wheeler A. G.. Walker M., Lean J. Function of hormonally-induced corpora
lutea in the domestic cat // Theriogenology.— 1988.— Vol. 29, N 2.—
P. 971-978.
Wildt D. E.. Chan S. G., Seager S. W. J. Ovarian activity, circulating sexual
behavior in the cat. I. Relationships during the coitus-induced luteal
phase and the oestrus period without mating Ц Biol. Reprod.— 1981.—
Vol. 25, N l.-P. 15-28.
Wildt D. E., Guthrie S. C., Seager S. W. J. Ovarian and behavioral cyclicity
of the laboratory maintained cat Ц Horm. Behav.— 1978a.— Vol. 10.—
P. 251-257.
Wildt D. E., Kinney G. M., Seager S. W. J. Gonadotropin induced reproductive
cyclicity in the domestic cat Ц Lab. Anim. Sci.— 1978b.— Vol. 28.—
P. 301—307.
Wildt D. E., Platz С. C., Chakraborty P. K., Seager S. W. J. Oestrus and ova-
rian activity in a female jaguar (Panthera onca) Ц J. Reprod. Fertil.—
1979,— Vol. 56, N 2 — P. 555—558.
Wildt D. E., Seager S. W. J., Chakraborty P. K. Effect of copulatory stimuli
on incidence of ovulation and on serum luteinizing hormone in the cat Ц
Endocrinology.— 1980.— Vol. 107, N 4.— P. 1212—1217.
Глава 11
ГЕНЕТИКА ПОПУЛЯЦИЙ И ГЕНОГЕОГРАФИЯ
ВВЕДЕНИЕ
Изучение географического распределения различных феноти-
пов в популяциях кошек, населяющих Землю, представляет боль-
шой интерес для решения проблем эволюционной биологии. Это
животное, относительно недавно доместицированное и ставшее
надежным спутником человека, по ряду причин является идеаль-
ным объектом для такого рода исследований Популяции кошек
118
обитают в непосредственном соседстве с человеком и поэтому лег-
ко доступны для наблюдения. Генетика многих фенотипических
особенностей кошки хорошо известна. Показано, что в популяциях
кошек существует высокая степень панмиксии: ассортативный
(неслучайный) подбор пар по изучаемым фенотипическим приз-
накам не зарегистрирован. Темп размножения кошек достаточно
высок (возможно, даже слишком высок). Большие скопления ко-
шек наблюдаются практически во всех человеческих поселениях.
Популяционные исследования, базирующиеся на анализе экстерь-
ерных особенностей, сводятся к наблюдению и регистрации фено-
типов. При этом не требуется удалять животных из популяций.
Такие исследования не оказывают влияния на численность и ген-
ный состав популяций и не вызывают протеста со стороны вла-
дельцев и любителей кошек.
Экология кошки также делает этот вид удобным для популя-
ционных исследований. Большинство домашних кошек — живот-
ные, «гуляющие сами по себе», в отличие от других домашних
животных. Это дает им возможность скрещиваться без контроля
со стороны человека. Из этого, однако, не следует, что человек
вовсе не играет роли в эволюции кошки. В действительности
влияние человека как на географическое распространение, так и
па генетический состав популяций этого вида кошачьих было
весьма важным.
Кошки заселили почти всю Землю, следуя за человеком, кото-
рый тщательно документировал свои миграции. Большое число
исследований посвящено миграциям кошек и их влиянию на гене-
тическую структуру кошачьих популяций [Todd, 1977а]. Эти ис-
следования дают четкие свидетельства об историческом процессе
колонизации новых (для кошки и человека) территорий и сущест-
венно расширяют наши знания о генетических последствиях
территориальных экспансий.
В литературе до сих пор идут споры о родственных связях
домашней кошки с дикоживущими видами: Felts libyca — северо-
африканской дикой кошкой и Felis silvestris — европейской дикой
кошкой. Поскольку оба эти «вида» скрещиваются с представите-
лями домашней кошки и нет непреодолимых барьеров для мигра-
ции между Европой и Африкой, кажется вполне разумным при-
нять, что эти три группы — две дикие и одна домашняя — состав-
ляют единый вид. Фенотип дикой кошки может рассматриваться
как дикий тип.
Как представителя дикого типа следует рассматривать коротко-
шерстную кошку с окраской агути, полосатым (тигровым или
макрелевым) рисунком темных полос, не имеющую белых пятен
на шкуре. Отклонения от этого типа — это, с одной стороны, ре-
зультат доместикации, а с другой — объект исследования генети-
ков (ом. гл. 2—6).
Кошки заселяли Землю вслед за человеком и в особенности
те районы, где земледелие становилось основой выживания. Во
время великих переселений народов, связанных с образованием
119
новых поселений, кошки всегда были среди той живности, кото-
рую брал с собой человек. Как мы увидим дальше, популяция
кошек в любой географической точке содержит признаки, которые
указывают на ее происхождение как спутника соответствующей
человеческой популяции.
Гипотеза «исторических миграций» позволяет хорошо объяс-
нять закономерности распределений различных аллелей в популя-
циях кошек.
МЕТОДЫ
Сбор данных по частоте окрасочных фенотипов кошек — до-
вольно легкая задача, если есть возможность осмотреть достаточно
большое количество кошек. Методы сбора материала могут быть
разными: от поквартирного обхода владельцев кошек до разбора
кошачьих тушек в санитарных службах. Не следует регистриро-
вать одну кошку дважды и включать в анализ большое число ко-
тят, принадлежащих к одному выводку. Большинство оценок ген-
ных частот не свободны от выборочных ошибок. Чаще всего
исследователь наблюдает только небольшую часть многочислен-
ной популяции, хотя в некоторых случаях, например на островах
с ограниченной численностью кошек, удается включить в анализ
всех представителей популяции.
Как правило, в популяционных исследованиях регистрируется
стандартный набор фенотипов:
О — сцепленная с полом оранжевая окраска (альтернатива —
неоранжевая окраска);
S — доминантная белая пегость (альтернатива — отсутствие
белых пятен);
d — рецессивная ослабленная или разбавленная окраска (аль-
тернатива — яркая окраска);
а — рецессивная неагути или сплошная окраска (альтернати-
ва — агути);
1 — рецессивная длинная шерсть (альтернатива — короткая
шерсть);
W — доминантная белая окраска (альтернатива — небелая
окраска);
Т — рисунок темных полос по агути или оранжевому фону.
В популяциях встречаются три варианта: Та — доминантный абис-
синский окрас — отсутствие полос на теле, Т — дикий тип — тигро-
вый или макрелевый рисунок (рецессивен по отношению к Та и
доминантен по отношению к tb); tb — рецессивный мраморный ри-
сунок, при котором на боках животного образуются, концентри-
ческие темные круги.
Для быстрого определения генотипа кошек можно пользовать-
ся табл. 11.1, построенной по принципу определителя (тезис —
антитезис).
120
Таблица 11.1
Опр•'деление генотипов кошек
Фенотип Генотип Если «да», то
1. Кошка длинношерстная 1/1 2
Кошка имеет шерсть нормальной длины L 2
2. Кошка целиком белая W/— 3
Кошка не белая W 3
3. Кошка имеет одно или несколько белых пятен S/— 4
Кошка не имеет белых пятен 4. Кошка целиком оранжевая или кремовая (пет ни черных, ни голубых, ни агути пятен, белые э 4
пятна могут быть, могут не быть) Кошка имеет оранжевые или кремовые и черные или голубые или агути пятна (белые пятна мо о/о 6
гут быть, могут не быть) О/о о/о 5,6
Кошка не имеет оранжевых пятен 5. Кошка черная или голубая или имеет черные 5
или голубые пятна а/а 6
Кошка агути или имеет агути пятна 6. Кошка имеет осветленную окраску (кремовую, А/— 6
голубую, бледную агути) Кошка имеет окраску нормальной интенсивно- d/d 7
сти (оранжевую, черную, агути) 7. Кошка имеет мраморный рисунок, продольную полосу по спине Кошка имеет сплошные или разорванные попе- речные (тигровые'» полосы Кошка нэ гмээг рлсуякг на теле D/— tb/tb t+/— j а/ 7
Подсчет генных частот производится при допущении о рав-
новесности популяций на основе уравнения Харди — Вейнберга.
Однако при этом следует учитывать наличие эпистатических от-
ношений между некоторыми локусами окраски.
Например, белая кошка (W/—) может быть и гомозиготой
по гену О, и гетерозиготой О/о, и гомозиготой по нормальному
аллелю о/о. Но, поскольку аллель W препятствует развитию ок-
раски любого типа, вы ничего не можете сказать о том, какой из
аллелей оранжевой окраски она несет. Поэтому белых кошек
нужно исключить из выборки при подсчете частот всех остальных
аллелей окраски.
Рабочая формула для подсчета частот аллеля О будет иметь
вид
_ 2п(О/О) + п(О/о)
Р “ 2 (TV -n(W/-)) ’
В этой и во всех следующих формулах р — частота соответст-
вующего аллеля, N — общий объем выборки, п — количество ко-
шек соответствующего фенотипа.
Для аллеля 1 эта формула будет иметь вид
121
Для аллеля а —
п = 1/ " (а/а)
F дг _ n (W/—) — п (0/0) •
Мы вынуждены исключать из выборки для подсчета частоты
гена черной окраски как белых кошек (W/—), у которых никакая
окраска не проявляется, так и оранжевых (0/0 и О/Y). На фоне
оранжевой окраски невозможно установить разницу между пеагу-
ти (а/а) и агути (А/—) типом распределения пигмента в волосе.
Для аллеля ослабленной окраски
Характер рисунка не выявляется как при отсутствии окраски
(W/—), так и на фоне пеагути окраски (a/а). Поэтому формула
для подсчета частоты аллеля tb имеет вид
— 1 f_______ИдЛчО_______
> у N -n(W/-)-n(a/a) ’
Частоты доминантных мутантных генов удобнее всего рассчи-
тывать, вычитая из единицы частоты соответствующих нормаль-
ных рецессивных аллелей.
Для гена W формула будет
Для аллеля S
„ _ I /(^-»(W/-)-n(s/-))
Р&)~ 1 к ДГ—ra(W/—)
Выше были приведены мутации, которые встречаются практи-
чески во всех популяциях кошек. Есть несколько «эндемичных»
мутаций: cs—сиамский альбинизм, сь— бирманский альбинизм,
Pd — доминантная полидактилия, М — мэнская бесхвостость.
Существуют и другие мутантные аллели, влияющие на экстерь-
ер кошек, но они или крайне редки, или присутствуют только в
составе пород, и поэтому, как правило, не включаются пока в
популяционные исследования. Однако поскольку не существует
серьезных барьеров для генного потока между породами и сво-
бодно размножающимися популяциями, не исключено, что в бу-
дущем придется расширить спектр мутаций, учитываемых при
популяционно-генетических и геногеографических исследованиях.
ГИПОТЕЗЫ О РАСПРЕДЕЛЕНИИ ГЕННЫХ ЧАСТОТ
В ПОПУЛЯЦИЯХ КОШКИ
Для объяснения изменчивости кошачьих популяций были пред-
ложены следующие гипотезы:
отбор в пользу (или против) определенных признаков;
122
расселение носителей новой мутации из той популяции, где
мутационное событие произошло;
дрейф генов, случайные изменения аллельных частот в огра-
ниченной популяции;
«эффект основателя», влияние генетического состава родитель-
ской популяции на генофонд колониальной группы, заселяющей
прежде не заселенную кошками территорию.
Гипотезы, объясняющие распределение аллельных частот в
популяциях кошек действием естественного отбора, представля-
ются довольно умозрительными. Так, индустриальный меланизм,
обусловленный отбором более темных фенотипов в урбанизирован-
ных районах, не кажется мне доказанным для кошек, хотя неко-
торые авторы настаивают, что он существует.
Ниже мы обсудим возможность отбора на новизну, разновид-
ности частотно-зависимого отбора для некоторых аллелей. Такой
отбор может играть определенную роль в проникновении новых
фенотипов из одной популяции в другую и поддержании разно-
образия в популяциях [Twedt, 1983]. Плотностно-зависимый от-
бор также привлекался для объяснения различий в аллельных
частотах между городскими и сельскими популяциями, хотя при-
чины, которые должны вызывать такой отбор, остаются неясными
[Dreux, 1968, 1981, 1986; Pontier, 1983; Lloyd, 1987]. Наконец,
классический дарвиновский отбор против вредных аллелей, без-
условно, играет значительную роль в детерминации частоты хотя
бы одного аллеля из исследованных, аллеля доминантной белой
окраски (W), который характеризуется рядом вредных плейотроп-
ных эффектов для приспособленности.
РЕЗУЛЬТАТЫ ПОПУЛЯЦИОННЫХ ИССЛЕДОВАНИЙ
Систематически и целенаправленно геногеографией кошек на-
чали заниматься совсем недавно — 40 лет назад. Что же до более
древних времен, то до нас дошли только отрывочные данные.
Так, из книги Бартоломея де Гланвилля «De Proprietatibus Re-
rum» (1398 г.) [цит. по: Lloyd, 1983] мы знаем, что уже в XIII в.
в Европе были нередки оранжевые (О), черные (а) и белые (W)
кошки [Lloyd, 1983]. На японских и китайских картинах XVI —
XVIII вв. мы видим черепаховых (О/о) и пегих (S/—) кошек. Но
какова была частота этих форм — неизвестно.
Впервые частоты генов окраски были определены для лондон-
ской популяции кошек в 1949 г. [Searle, 1949]. Начаты эти ра-
боты по предложению выдающегося британского генетика
Дж. Б. С. Холдэйна. Затем они были подхвачены другими иссле-
дователями. Наибольший вклад в эту работу внесли английский
' исследователь Р. Робинсон и американский — Н. Б. Тодд.
, Результаты учета генных частот в различных популяциях ко-
шек суммированы в табл. 11.2. Размеры выборки в разных локаль-
ностях варьируют от нескольких десятков особей до нескольких
тысяч. Средний размер выборки — около 300 животных.
123
Частоты
Город N 0 a tb d 1 S W
1 1 2 3 4 5 6 7 8 9
Yell 79 0,28 0,63 0,45 0,42 0,39 0,47 0,09
Shetland 167 0,35 0,74 0,64 0,3 0,4 0,45 0,02
Kirkwall 130 0,34 0,73 0,57 0,36 0,51 0,25 0,07
Thurso 92 0.25 0,72 0,75 0,34 0,48 0,28 0
Forres 106 0,2 0,76 0,74 0,38 0,4 0,31 0,02
Lairg 79 0,15 0.7 0,37 0,26 0,3 0,26 0,02
Achaseen 77 0,13 0,64 0,31 0,23 0,28 0,21 0,01
Stathoykel 71 0,12 0,65 0,35 0,22 0,26 0,21 0,06
Sutherland 101 0,21 0,78 0,38 0,32 0,31 0,3 0,02
Harris 68 0,34 0,65 0,46 0,31 0,27 0,32 0,05
NW Lewis 132 0,33 0,65 0,64 0,3 0,26 0,39 0,04
Aberdeen 124 0,17 0,76 0,79 0,34 0,4 0,3 0,01
Inverness 120 0,19 0,81 0.83 0,4 0,52 0,28 0,02
Kirkmichel 135 0,15 0,61 0,32 0,26 0,31 0,28 0,02
Strathyre 109 0,13 0,63 0,37 0,26 0,32 0,25 0,01
FT William 63 0,25 0,71 0,69 0,33 0,42 0,32 0
Laggen 105 0,09 0,61 0,31 0,22 0,26 0,26 0
Barra 76 0,3 0,65 0,52 0,37 0,41 0,27 0,07
Uist 79 0,31 0,6 0,35 0,31 0,3 0,3 0,05
Edinburgh 207 0,2 0,78 0,75 0,35 0,3 0,28 0,03
Peebles 90 0,26 0,7 0,68 0,33 0,32 0,31 0,03
Perth 120 0,21 0,79 0.79 0,38 0,37 0,26 0,02
Glasgow 918 0,2 0,8 0;8 0,26 0,3 0,2 0
Lanark 84 0,19 0,8 0,81 0,27 0,35 0,28 0,01
Stirling' 135 0,26 0,75 0,72 0,33 0,32 0,27 0,02
Oban 87 0,25 0,7 0,65 0,35 0,37 ' 0,31 0,04
Islay 61 0,28 0,68 0,62 0,32 0,22 0,28 0,02
Ayr 96 0,15 0,8 0,82 0,27 0,29 0,26 0
Belfast 82 0,16 0,84 0,65 0,11 0,16 0,31 0,02
Donegal Co 273 0,18 0,83 0,67 0,21 0,19 0,35 0
Dundalk 66 0,15 0,87 0,82 0,33 0,27 0,32 0,02
Sligo 80 0,07 0,85 0,66 0,16 0,16 0,32 0,01
Dublin 360 0,12 0,82 0,74 0,29 0,33 0,31 0,02
Galway 125 0,09 0,76 0,71 0,31 0,13 0,37 0,02
Limerick 82 0,14 0,76 0,6 0,2 0,3 0,35 0,02
Waterford 106 0,03 0,77 0,7 0,2 0,17 0,4 0,02
Cork 74 0,14 0.78 0,66 0,1 0,22 0,31 0,03
Newcastle 617 0,16 0,8 0,77 0,3 0,36 0,34 0,01
York 230 0,2 0,81 0,78 0,27 0,33 0,01
Isle of Man 232 0,24 0,83 0,88 0,23 0,16 0,34 0,05
Ramsey 137 0,18 0,77 0,6 0,37 0,32 0,28 0,06
Chester 63 0,16 0,85 0,81 0,23 0,37 0,29 0
Liverpool 137 0,14 0,82 0,72 0,15 0,3 0,42 0,01
Bangor 182 0,18 0,78 0,69 0,22 0,36 0,4 0,01
Cambridge 171 0,25 0,81 0,73 0,29 0,29 0,29 0,01
S. Wales 102 0,2 0,82 0,86 0,26 0,2 0,4 0
Bristol 227 0,2 0,76 0,8 0,24 0,32 0,38 0,01
Herts. Co. 198 0,23 0,78 0,86 0,29 0,25 0,31 0,02
Wickford 48 0,18 0,79 0,76 0,2 0,5 0,41 0
Enfield, 1975 310 0,18 0,75 0,74 0,18 0,33 0,35 0,02
Enfield, 1979 206 0,18 0,77 0,78 0,16 0,3 0,37 0,01
Finchley 181 0,16 0,82 0,84 0,28 0,47 0,35 0,03
аллеля Таблица 11.2
TA cs M Pd Широ- та Долго- та Литературный источник
10 11 12 13 14 15 1
0 0 0 0 0 0 0 0 0 0 0 0 0,004 0 0 0 0,16 0,001 0,004 0,009 0,16 0,003 0,007 61 60 59 59 58 58 58 58 58 58 58 57 57 57 57 57 57 57 56 56 56 56 56 56 56 56 56 55 55 55 54 54 53 53 53 52 52 55 54 54 54 53 53 53 52 52 51 52 52 52 52 52 1 -1 —3 —4 —4 -4 —5 —5 —5 —7 —7 -2 —4 —4 —4 -5 -5 —7 —7 -3 —3 -3 —4 —4 -4 —5 —6 —5 -6 -8 —6 -8 —6 9 —9 —7 - 8 —1 —1 —5 —5 -3 —4 0 —4 -3 0 0 0 0 0 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark. 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1975 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Clark, 1976 Lloyd, Todd, 1989 Lloyd, 1979 Lloyd, Todd, 1989 Lloyd,1979 Todd, Lloyd, 1979 Lloyd, Todd, 1989 Todd,1975 Lloyd, Todd, 1989 Lloyd, Todd, 1989 Symonds, Lloyd, 1987 Metcalfe, Turner, 1971 Todd et al., 1979 Burke, 1978 Lloyd,1977 Burke, 1978 Burke, 1978 Lloyd, 1977 Gruffydd-Jones et al., 1979 Gruffydd-Jones et al., 1979 Robinson, Silson, 1969 Robinson, 1987 Robinson, 1987 Robinson, 1987 Robinson, 1987
124
125
1 2 * 4 * 6 8 9
N. London 129 0,2 0,78 0,62 0,15 0,34 0,37 0,01
Ealing 128 0,13 0,78 0,83 0,22 0,25 0,31 0,01
Ealing, 1983 163 0,16 0,77 0,75 0,21 0,31 0,33 0,01
London 700 0,11 0,76 0,81 0,14 0,33 0,37 0
London, 1976 494 0.25 0,78 0,72 0,32 0,48 0,32 0,02
London, 1977 145 0,23 0,7 0,73 0,22 0,43 0,23 0,02
London, 1981 331 0,26 0,77 0,72 0,25 0,47 0,31 0,02
London, 1984 295 0,27 0,7 0,77 0,23 0,48 0,29 0,03
Biggen Hill 85 0,15 0,78 0,84 0,25 0,31 0,28 0,01
Crawley 300 0,24 0,76 0,54 0,34 0,31 0,01
Portsmouth 250 0,06 0,77 0,64 0,06 0,14 0,5 0
Kent Co. 82 0,19 0,82 0,8 0,22 0,31 0,34 0
Croisette 112 0,17 0,46 0,56 0,21 0,25 0,25 0,01
Paris 1833 0,06 0,71 0,78 0,33 0,25 0,24 0,01
Mayenne 607 0,15 0,64 0,61 0,29 0,17 0,39 0,01
Carnac 342 0,08 0,76 0,64 0,35 0,2 0,23 0,01
Tours 220 0,06 0,59 0,71 0,34 0,27 0,22 0,02
Cher 400 0,12 0,7 0,61 0,37 0,2 0,25 0,02
Lyons 488 0,11 0,78 0,65 0,39 0,37 0,22 0,03
Chamonix 249 o,l 0,75 0,69 0,4 0.32 0,22 0,01
Roiffieux 303 0,21 0,67 0,61 0,47 0,38 0,28 0,02
St. Just 283 0,2 0,66 0,72 0,31 0,23 0,26 0,01
Bordeaux 180 0,18 0,71 0,67 0,32 0,21 0,31 0,01
Aimarques 203 0,1 0,63 0,64 0,39 0,37 0,22 0,01
Le Cailar 176 0,14 0,71 0,59 0,34 0,37 0,21 0,01
Arles 61 0,12 0,74 0,55 0,32 0,22 0,1 0,01
Nice 129 0,06 0,76 0,57 0,43 0,23 0,21 0
Marseille 550 0,08 0,72 0,68 0,34 0,27 0,29 0
Sweden 300 0,06 0,71 0,34 0,3 0,44
Leeuwarden 255 0,19 0,81 0,6 0,32 0,14 0,33 0,01
Den Haag 1035 0,21 0,65 0,66 0,26. 0,16 0,36 0,02
Rotterdam 212 0,16 0,74 0,54 0,17 0,1 0,31 0,01
Amsterdam 636 0,13 0,74 0,57 0,25 0,15 0,32 0,01
Utrecht 262 0,22 0,82 0,56 0,3 0,16 0,31 0,01
Malgrat 60 0,25 0,79 0,26 0,19 0 0.33 0.01
Aragon 200 0,19 0,68 0,47 0,22 0,16 0,44 0,01
Porto 256 0,14 0,77 0,43 0,3 0,23 0,29 0,01
Barcelona 699 0,16 0,7 0,28 0,27 0,14 0,27 0,004
Vilanova i laGelt 118 0,19 0,72 0,22 0,23 0,09 0,35 0,005
Villafrequenza 72 0,24 0,67 0,55 0,27 0 0,29 0,014
Gava 322 0,24 0,8 0,25 0,22 0,15 0,28 0,006
Sitges 204 0,13 0,74 0,32 0,25 0,2 0,25 0,008
Tarragona 216 0,21 0,68 0,38 0,15 0 0,29 0
Alicante 335 0,23 0,8 0,49 0,23 0 0,31 0,005
Castelldefels 376 0,25 0,79 0,12 0,24 0,16 0,28 0,009
Granollers 154 0,17 0,71 0,16 0,08 0,21 0,23 0,003
Murcia 164 0,18 0,74 0,61 0,08 0,08 0,38 0,012
Palma 384 0,2 0,73 0,37 0,36 0,28 0,23 0,004
Ibiza 249 0,24 0,76 0,3 0,23 0,19 0,29 0,014
Mahon 466 0,3 0,8 0,18 0,38 0,12 0,15 0,003
Villacarlos 222 0,24 0,75 0,13 0,44 0,16 0,18 0,002
Mercadalyalayor 98 0,22 0,87 0 0,3 0,17 0,18 0
Ciudadela 491 0,21 0,73 0,22 0.35 0,08 0,27 0,004
Balearics 138 0,17 0,79 0,43 0 0,08 0,33 0
Majorca 45 0,28 0,59 0,49 0,15 0 0,24 0,01
Benidorm 43 0,17 0,72 0,55 0,38 0,33 0
Benidorm, 1973 61 0,19 0,79 0,56 0,53 0 0,31 0,01
126
Продолжение т а б л. И .2
10 И 12 13 14 15 16
51 0 Lloyd, Todd, 1989
0 51 0 Robinson, Silson, 1969
51 0 Robinson, 1987
0,005 0,05 0,004 0,003 51 0 Searle, 1949
51 0 Robinson, 1987
51 0 Robinson, 1987
51 0 Robinson, 1987
51 0 Robinson, 1987
51 0 Robinson, 1987
51 0 Robinson, 1987
51 —1 Dards, Robinson, 1983
0 0,037 51 1 Robinson, Silson. 1969
0,16 50 2 Pontier, 1989
0,001 0,03 0,001 0 49 2 Dreux, 1967
48 1 Dreux, 1968
0,15 48 -3 Dreux, 1986
0 0,12 47 1 Dreux, 1979
0 47 3 Dreux, 1981
46 5 Pontier, 1983
0,06 46 7 Dreux,1971
45 5 Pontier, 1983
0.21 45 5 Pontier, 1983
0 45 -1 Symonds, Lloyd, 1987
0,32 44 4 Pontier, 1989
0,11 44 4 Pontier, 1989
44 5 Pontier, 1989
0,017 0,2 44 7 Genermont, 1976a, b
0,09 43 5 Dreux, 1975
57 15 Fredga, 1976
0 0,14 53 6 Lloyd, 1982
0,002 0,03 0 0 52 4 Dreux, Legel, 1973
0 0,002 52 4 Lloyd,1982
0 0,1 52 5 Lloyd, 1982
0 0,13 52 5 Lloyd, 1982
42 3 Robinson, 1971
42 -1 Dreux, Samet, 1981
0 0,15 41 -9 Todd, Lloyd, 1984
0,12 0,004 41 6 Ruiz-Garcia, 1988
0,24 0,005 41 5 Ruiz-Garcia, 1990c
0,12 38 3 Ruiz-Garcia, 1990c
0,19 0 41 6 Ruiz-Garcia, 1990c
0,22 0,003 41 5 Ruiz-Garcia, 1990c
0,07 0,007 41 5 Ruiz-Garcia, 1990c
0,16 38 3 Ruiz-Garcia, 1990c
0,13 0,001 41 6 Ruiz-Garcia, 1988
0,16 0,003 42 6 Ruiz-Garcia, 1990d
0,21 0,006 38 3 Ruiz-Garcia, 1990d
0,14 0 40 6 Ruiz-Garcia, 1990a
0,3 0,014 39 5 Ruiz-Garcia, 1990a
0,1 0,001 40 8 Ruiz-Garcia, 1990b
0,13 0,005 40 8 Ruiz-Garcia, 1990b
0,02 0,005 40 8 Ruiz-Garcia, 1990b
0,19 0,005 40 8 Ruiz-Garcia, 1990b
40 3 Dyte, 1972
40 3 Robinson, 1971
39 0 Robinson, 1971
39 0 Robinson, 1971
127
Продолжение табл. 11.2
! 1 2 1 з 1 4 5 6 1 7 8 1 9
Benidorm, 1990 203 0,23 0,69 0,55 0,16 0 0,28 0,005
Lisbon 371 0,07 0,65 0,45 0,27 0,09 0,21 0,02
Cadiz 66 0,16 0,71 0,45 0,25 0 0,23 0,01
Slupsk 98 0,05 0,68 0,29 0,14 0 0,37 0
Gdynia 110 0 0,72 0,37 0Д 0 0,35 0
Kolobrzeg 123 0,03 0,53 0,21 0,09 0 0,53 0
Gd ansk 197 0,07 0,73 0,35 0,1 0,12 0,56 0
Sopot 124 0,02 0,77 0,2 0,09 0,13 0,5 0
Olecko 80 0,06 0,7 0,31 0 0 0,43 0
Suwalki 80 0,03 0,72 0,27 0,16 0,16 0,59 0
Szczecin 106 0,04 0,65 0,29 0,1 0,1 0,62 0
Bialystok 106 0,07 0,7 0,26 0,1 0,19 0,41 0
Bialowieza 118 0,09 0,71 0,45 0,33 0,21 0,33 0
Miodnica 108 0,03 0,58 0,24 0,1 0 0,57 0
Poznan 132 0,02 0,73 0,24 0,19 0 0,39 0
Warszawa 124 0,07 0,62 0,35 0,25 0,22 0,33 0
Wroclaw 121 0,09 0,74 0,27 0,18 0 0,27 0
Chybie 102 0,06 0,7 0,14 0 0,1 0,43 0
Cieszyn 101 0,14 0,73 0,21 0,17 0 0,43 0
Katowice 107 0,01 0,69 0,23 0,17 0,1 0,34 0
J orkow 118 0,06 0,58 0,17 0,09 0,13 0,45 0
Krakow 278 0,06 0,66 0,2 0,2 0,12 0,37 0
Tyniec 67 0,01 0,49 0,29 0,12 0,21 0,37 0
Brzesko 83 0,04 0,49 0,13 0,11 0,11 0,37 0
Rzeszow 112 0,08 0,72 0,23 0,23 0,13 0,24 0
Przemysl 115 0,04 0,76 0,2 0,25 0,13 0,24 0
N. Moravia 249 0,02 0,65 0,29 0 0,24 0
Nachod 152 0,13 0,75 0,24 0,08 0,47 0
Usti 178 0,18 0,64 0,45 0,11 0,25 0
S. Moravia 207 0,03 0,53 0,08 0 0,22 0
Trebic 152 0,12 0,65 0,24 0 0,29 0
Vienna 393 0,1 0,57 0,29 0,22 0,17 0
Czechoslovakia 12160 0,12 0,5 0,16 0,08 0,33 .0
Budapest 265 0,08 0,6 0 0,27 0,09 0,33 0
Bernese 197 0,15 0,75 0,66 0,4 0,1 0,43 0,01
Vaud 66 0,19 0,71 0,52 0,43 0,33 0,23 0,01
Bol. Brae 111 0,15 0,58 0,28 0,31 0,1 0,4 0,02
Dalmatia 112 0,18 0,75 0,32 0,23 0 0,4 0,01
Bitola 104 0,28 0,73 0,28 0,33 0,28 0,48 0,02
Ohrid 142 0,3 0,66 0,11 0,28 0,15 0,55 0
Struga 152 0,21 0,59 0,23 0,23 0,08 0,66 0
Venice 210 0,06 0,58 0,48 0,35 0,26 0,01
Alassio 82 0,13 0,65 0,52 0,2 0,11 0,28 0,04
Rome 480 0,09 0,66 0,49 0,34 0,1 0,31 0,01
Siracusa 105 0,19 0,56 0,2 0,2 ' 0 0,54 0
Kerkyra 272 0,17 0,66 0,25 0,31 0,06 0,34 0
Patras 105 0,18 0,74 0,3 0,24 0 0,45 0
Argolis 164 0,18 0,74 0,19 0,21 0 0,22 0
Athens 354 0,12 0,73 0,31 0,36 0,1 0,27 0,01
Chios 92 0,24 0,68 0,34 0,26 0,1 0,41 0
Samos 133 0,12 0,68 0,38 0,29 0,21 0,38 0
Chania 176 0,23 0,65 0,26 0,22 0,08 0,57 0,02
Iraklion 314 0,13 0,64 0,35 0,27 0,11 0,45 0
Rethymnon 81 0,22 0,62 0,26 0,11 0,11 0,46 0,01
Iceland 368 0,17 0,67 0,36 0,35 0,14 0,5 0,03
Reykjavik Faeroes 104 62 0,13 0,12 0,6 0,76 0,53 0,67 0,44 0,34 0,17 0 0,49 0,47 0,02 0,01
128
10 11 12 13 14 15 16
0,14 0,002 39 0 Ruiz-Garcia, 1988
0 0,16 39 -9 Todd et al., 1988
36 -6 Todd, Lloyd, 1984
0 55 17 Wagner, Wolsan, 1987
0 55 19 Wagner, Wolsan. 1987
0 54 16 Wagner. Wolsan, 1987
0 54 19 Wagner, Wolsan, 1987
0 54 19 Wagner, Wolsan, 1987
0 54 22 Wagner, Wolsan, 1987
0 54 23 Wagner, Wolsan, 1987
0 53 14 Wagner. Wolsan, 1987
0 53 23 Wagner, Wolsan, 1987
0 53 24 Wagner, Wolsan, 1987
0 52 15 Wagner, Wolsan, 1987
0 52 17 Wagner, Wolsan, 1987
0 52 21 Wagner. Wolsan, 1987
0 51 17 Wagner, Wolsan, 1987
0 50 19 Wagner, Wolsan, 1987
0 50 19 Wagnor, Wolsan, 1987
0 50 19 Wagner. Wolsan, 1987
0 50 20 Wagner, Wolsan, 1987
0 50 20 Wagner, Wolsan, 1987
0 50 20 Wagner, Wolsan, 1987
0 50 21 Wagnor, Wolsan, 1987
0 • 50 22 Wagner, Wolsan, 1987
0 50 23 Wagner, Wolsan, 1987
50 15 Varajcko, Spicera, 1982
50 16 Lloyd, Todd, 1989
50 16 Lloyd, Todd, 1989
49 15 Varajcko, Spicera, 1982
49 16 Lloyd, Todd, 1989
0,002 48 16 Baxa, 1973
48 17 Varajcko, unpubL data
0,07 0 0 47 19 Davis В. K., Davis В. P., 1977
0 0,14 47 7 Kerr, 1983
46 7 Kerr, 1983
43 17 Wagner. 1986
0 43 18 Hines, Randall, 1977
41 21 Lloyd, Todd, 1989
41 21 Lloyd, Todd, 1989
41 21 Lloyd, Todd, 1989
0 0,07 45 12 Searle, 1966
44 8 Robinson, unpubl. data
42 13 Lloyd et al., 1983
0 37 15 Lloyd, Todd, 1987
0 40 20 Todd, Kunz, 1977
0 38 20 Todd, Kunz, 1977
0 38 23 Todd et al., 1974
0 38 24 Todd, Kunz, 1977
0 38 26 Todd N. B., Todd L. M., 1976b
0 38 27 Todd, Kunz, 1977
0 36 24 Todd, Kunz, 1977
0 35 25 Todd, Kunz, 1977
0 35 25 Todd, Kunz, 1977
65 -20 Adalsteinsson et al., 1979
•,014 64 —22 Todd et al., 1975
62 —7 Todd, Adalsteinsson, 1982
9 Заказ № 355
129
‘ 1 2 3 4 5 6 7 8 9
Terciera 160 0,31 0,66 0,37 0,34 0,16 0,45 0,03
Faial/Pico 109 0,25 0,57 0,38 0,34 0.17 0,34 0,01
S. Miguel 159 0,23 0,63 0,5 0,34 0,11 0,29 0
Madeira 133 0,18 0,78 0,35 0,41 0 0,39 0,01
Mindelo CV 20ti 0,12 0,77 0,6 0,19 0 0.34 0
draia CV IbS 0.23 0,83 0,47 0,11 0 0,44 0,01
E. Istanbul 141 0,28 0,6 0,2 0,28 0,48 0,37 0,01
W. Istanbul 272 0,19 0.63 0,28 0,35 0,35 0,39 0,01
Trabzon 71 0,17 0,4 0,14 0,29 0,12 0,37 0
Izmir 114 0,2 0,59 0,33 0,19 0,25 0.39 0
Van 116 0,26 0,8 0,2 0,15 0,31 0,4 0,1
Cyprus 188 0,21 0,72 0,25 0.35 0.5 0,34 0
H. Hagelilil 56 0,34 0,6 0,43 0.3 0,4 0,32 0
Bet-bhemesh 153 0,11 0.63 0,3 0,26 0,2 0,37 0
Jeiico 191 0,32 0,64 0,34 0,4 0,42 0,43 0
Jerusalem 528 0,12 0,67 0,23 0,29 0,2 0,25 0
Rami a 44 0,21 0,61 0,4 0 0,15 0,5 0
Rehovot 195 0,23 0,56 0,16 0,18 0,14 0,32 0
Tel Aviv 398 0,15 0,72 0,12 0,2 0,17 0,49 0
Leningrad 115 0,25 0,57 0,44 0,42 0,64 0,31 0,01
Y ashkino 100 0,15 0,53 0,2 0,26 0,55 0,5 0
Pavlovschina 100 0,05 0,72 0,36 0 0,44 0,43 0
Zlatoust 134 0,18 0,64 0,15 0,29 0,27 0,38 0
Novosibirsk 226 0,18 0,62 0,08 0,37 0,45 0,33 0,01
Kemerovo 101 0,14 0,68 0,18 0,37 0,44 0,35 0
Suzdalka 106 0,31 0,65 0 0,47 0,36 0,33 0
Kubiyshev 64 0,22 0,45 0 0,13 0,56 0,41 0
Magadaachi 98 0,23 0,52 0,12 0,27 0,32 0,46 0
Pavlovsk 100 0,17 0,53 0,34 0,32 0,37 0,22 0
Gomel 188 0,14 0,6 0,09 0,4 0,26 0,43 0,01
Irkutsk 113 0,24 0,63 0,19 0,24 0,36 0,38 0,02
Khabarovsk 118 0,13 0,47 0,39 0,31 0,21 0,53 0,01
Ordzhonikidze 103 0,03 0,6 0,17 0,33 0,28 0,4 0
Kishinev 103 0,08 0,47 0,19 0,31 0,35 0,33 0
Rostov/Don 120 0,07 0,51 0,4 0,35 0,16 0,32 0
Sevastopol 97 0,12 0,53 0,12 0,27 0,37 0,25 0
Krasnodar 131 0,07 0,52 0,22 0,23 0,12 0,33 0,01
Armavir 134 0,04 0,63 0,22 0,53 0,24 0,46 0,01
Kislovodsk 165 0,14 0,61 0,16 0,34 0,42 0,41 0
Talas 130 0,13 0,65 0,16 0,3 0,47 0,35 0
Alma-Ata 171 0,12 0,54 0,15 0,36 0,56 0,21 0
Vladivostok 279 0,11 0,53 0,31 0,2 0,23 0;41 0,01
Batumi 74 0,12 0,41 0,22 0,23 0,48 0,53 0,01
Kirovabad 88 0,1 0,6 0 0,24 0,11 0,39 0,01
Baku 100 0,09 0,52 0,16 0,33 0,17 0,31 0
Tabriz 162 0,1 0,57 0,16 0,08 0,3 0,51 0
Tehran 273 0,19 0,56 0,3 0,09 0,42 0,47 0,01
Meshad 125 0,18 0,69 0,46 0,13 0,09 0.35 0,01
Ahwaz 113 0,21 0,53 0,19 0,09 0 0,45 0
Shiraz 139 0,06 0,64 0,11 0,12 0,15 0,48 0,03
Peshawar 128 0,26 0,66 0,3 0,2 0 0,27 0
Islamabad 203 0,32 0,65 0,14 0,16 0,23 0,42 0,02
Rawalpindi 202 0,45 0,63 0,14 0 0,24 0,5 0,02
Lahore 406 0,37 0,74 0,14 0,19 0,05 0,51 0,02
Sahiwal 106 0,24 0,56 0 0,1 0,1 0,56 0,02
Quetta 195 0,36 0,41 0,27 0,1 0,26 0,37 0,01
Dehli 109 0,41 0,76 0,18 0,19 0 0,38 0
Продолжение г а б л. И .2
10 11 12 13 14 15 1 16
0 0,006 39 —27 Todd, Lloyd,1984
0 0,005 39 -28 Todd, Lloyd, 1984
0 38 -26 Todd, Lloyd, 1984
0 33 —17 Todd et al., 1988
17 —25 Todd et al., 1988
15 -23 Todd et al., 1988
0 41 29 Todd N. B., Todd L. M., 1976b
0 41 29 Todd N. B., Todd L. M., 1976b
0 41 40 Todd N. B., Todd L. M., 1976b
0 38 27 Todd N. B., Todd L. M., 1976b
0 38 43 Todd et al., 1977
0,005 35 33 Robinson, 1972
0 33 35 Salternik-Vardy, Ritte, 1989
0,02 32 35 Salternik-Vardy, Ritte, 1989
0 32 35 Salternik-Vardy, Ritte, 1978
0,023 32 35 Salternik, Mordokhovich, 1975
0 32 35 Salternik-Vardy, Ritte, 1989
.0,003 32 35 Salternik-Vardy, Ritte, 1989
0,004 32 35 Salternik et al., 1976
0 60 30 Borodin et al., 1978
56 85 Borodin, 1983
56 92 Borodin, 1983
0,06. 55 69 Borodin et al., 1981
0,01 55 83 Goncharenko et al., 1985
55 86 Borodin, 1983
55 86 Borodin, 1983
0 54 50 Borodin et al., 1978
0,02 54 125 Borodin et al., 1978
53 83 Borodin, 1983
0,017 52 31 Goncharenko et al., 1985
0,05 52 104 Borodin et al., 1978
0 49 135 Robinson, Manchenko, 1981
0,014 48 34 Goncharenko et al., 1985
47 29 Borodin, 1983
0 47 40 Goncharenko et al., 1985
0 45 34 Goncharenko et al., 1985
0 45 39 Borodin et al., 1981
0 45 41 Robinson, Manchenko, 1981
44 43 Borodin, 1983
43 72 Goncharenko et al., 1985
0,016 43 77 Goncharenko et al., 1985
0 43 132 Robinson, Manchenko, 1981
42 42 Borodin, 1983
41 46 Borodin et al., 1981
40 50 Borodin et al., 1981
0 38 46 Salternik, Todd, 1977
0 36 51 Salternik, Todd, 1977
0. 36 60 Salternik, Todd, 1977
0, 31 49 Salternik, Todd, 1977
0 30 53 Salternik, Todd, 1977
0,01 34 72 Ahmad et al., 1980
0,03 0,1 34 73 Ahmad et al,, 1980
0,04 34 73 Ahmad et al., 1980
0,02 32 74 Ahmad et al., 1980
0 . 31 73 Ahmad et al., 1980
0,01 0,07 30 67 Ahmad et al,, 1980
0,032. 29, 77 Ahmad, 1980
130
9*
131
Продолжение табл. 11.2
1 2 3 * 5 6 7 8 9 .
Karachi 298 0,26 0,58 0,14 0,11 0 0,41 0,04
Calcutta 159 0,35 0,68 0,11 0 0,11 0,6 0
Bombay 354 0,3 0,61 0,15 0,05 0 0,56 0
Colombo 303 0,27 0,63 0,19 0,14 0 0,52 0
Dharan 165 0,19 0,69 0,14 0,22 0 0,52 0,01
Khobar 1061 0,29 0,58 0,11 0,18 0,12 0,56 0,01
Medina 304 0,43 0,85 0,4 0,23 0,15 0,62 0
Mecca 310 0,35 0,83 0,23 0,28 0,08 0,6 0
Jidda 256 0,3 0,82 0,13 0,2 0 0,58 0
Abha 111 0,29 0,61 0 0,3 0,3 0,75 0
Taiwan 177 0,34 0,54 0,29 0,25 0 0,47 0
Hongkong 190 0,39 0,44 0,48 0 0 0,45 0
Macau 215 0,36 0,44 0,39 0 0 0,46 0
Chiangmai 182 0,3 0,81 0 0,31 0 0,42 0,01
Maehongson 158 0,23 0,68 0 0,4 0 0,38 0,01
West Issan 351 0,26 0,68 0 0,4 0 0,38 0,01
Bangkok 233 0,29 0,75 0 0,1 0 0,3 0,01
Singapore 250 0,31 0,6 0,11 0,13 0 0,5 0,01
Kerguelen Isl 36 0 1 0 0 0,4 0
Marion Isl 816 0 0,91 0,59 0,14 0 0,17 0
Reunion Isl 106 0,23 0,75 0,25 0,24 0 0,27 0
Seychelles 152 0,26 0,67 0 0,3 0,28 0,41 0
Constantin 128 0,21 0,76 0,18 0,45 0 0,34 0
Carthage 100 0,22 0,63 0,19 0,27 0,1 0,4 0
Tunis 87 0,18 0,72 0 0,47 0,11 0,39 0
Malta 115 0,43 0,64 0,33 0,25 0,28 0,33 0,01
Malta, 1977 41 0,37 0,6 0,38 0,17 0,22 0,42 0,06
Rabat 124 0,09 0,81 0,38 0,38 0,09 0,28 0,01
Alexandria 213 0,36 0,67 0,32 0,33 0,12 0,41 0
Cairo 140 0,22 0,74 0,44 0,28 0 0,29 0,01
Upper Egypt 58 0,29 0,69 0,25 0,31 0 0,55 0,03
Atbara 47 0,25 0,68 0,21 0 0 0,61 0
Khartoum 73 0,36 0,82 0,37 0 0 0,44 0
Kaapstad 143 0,2 0,76 0,69 0,28 0,24 0,34 0,02
Pretoria 257 0,17 0,82 0,48 0,28 0,18 0,27 0,05
Hobart 922 0,22 0,76 0,81 0,24 0,46 0,39 0,04
Tasmania 62 0,11 0,63 0,84 0,13 0,18 0,21 0
Melbourne 362 0,17 0,73 0,71 0,34 0,39 0,32 0,04
Adelaide 600 0,19 0,78 0,71 0,33 0,38 0,3 0,02
Brisbane 278 0,14 0,81 0,69 0,45 0,41 0,04
Darwin 155 0,33 0,74 0,78 0,31 0,33 • 0,44 0,01
New Zealand 171 0,12 0,81 0,87 0,33 0,47 0,22 0,04
New Caledonia 125 0,16 0,63 0,53 0,28 0,09 0,29 0,02
Kiribati 198 0,33 0,78 0,49 0,23, 0 0,37 0
St. John's 310 0,25 0,76 0,59 0,32 0,29 0,54 0,04
Restigoche 117 0,26 0,82 0,52 0,47 0,48 0,38 0,05
St. Pierre 100 0,24 0,64 0,65 0,4 0,17 0,37 0
Miramichi 113 0,22 0,69 0,42 0,53 0,39 0,32 0,05
Madawaska 201 0,31 0,82 0,51 0,41 0,39 0,32 0,02
Quebec 249 0,19 0,74 0,64 0,5 0,43 0,3 0,02 -
Sydney 217 0,33 0,8 0,62 0,4 0,52 0,42 0,04
Charlottetown 108 0,28 0,79 0,45 0,46 0,42 0,34 0,05
Moncton 216 0,27 0,82 0,49 0,51 0,44 0,35 0,04
Fredricton 165 0,26 0,78 0,46 0,38 0,47 0,36 0,03
Trois Riviers 105 0,26 0,79 0,66 0,5 0,4 0,31 0,06
Montreal 406 0,22 0,76 0,53 0,47 0,41 0,3 0,02
Halifax 159 0,32 0,79 0,57 0,45 0,48 0,38 0,03
To 11 12 13 14 15 1 16
0,005 25 67 Ahmad et al., 1980
0,367 23 88 Todd, 1984
0,141 19 73 Todd,1984
0,302 7 80 Ahmad, 1980
0,01 26 50 Davis, Ahmad, 1983
0,01 26 50 Davis. Ahmad, 1983
0 25 40 Davis, Ahmad, 1983
0 22 40 Davis, Ahmad, 1983
0,004 21 39 Davis, Ahmad, 1983
0,04 18 43 Davis, Ahmad, 1983
0,01 25 121 Chu, 1986 a, b
0,15 22 114 Fagen, Meeswat, 1981
0,23 22 114 Fagen, Meeswat, 1981
0,02 0,35 19 99 Fagen, Meeswat, 1981
0,13 0,08 19 99 Fagen, Meeswat, 1981
0,003 0,28 19 100 Fagen, Meeswat, 1981
0,04 0,19 14 101 Fagen, Meeswat, 1981
0,24 0,13 0 1 104 Searle, 1959
0 -48 69 Dreux, 1974
0 -47 38 Aarde, Robinson, 1980
0 —22 56 Dreux, 1967
0,07 -10 55 Jaffe, 1978
0,015 37 7 Genermont, 1976a, b
37 10 Todd,1977b
37 10 Todd,1977b
36 14 Lloyd, Jaffe, 1981
36 14 Robinson, unpubl. data
34 —7 Genermont, 1974
0 31 30 Todd, Blumenberg, 1978
0 30 31 Todd, Blumenberg, 1978
0 24 33 Todd, Blumenberg, 1978
0 18 34 Todd, Blumenberg, 1978
0 16 33 Todd, Blumenberg, 1978
0 —34 18 Aarde et al., 1977
0,016 0,004 —26 28 Aarde et al., 1977
0,009 —43 147 Dartnail, Todd, 1975
-43 147 Dartnail, 1975
0 0 —38 145 Kinnear, 1979
-35 138 Clark, 1973
—27 153 Moffat, 1968
—12 131 Dartnall, 1977
-43 173 Marples, 1967
0,09 —21 165 Vuilleumier B., Vuilleumier F., 1978
2 157 Garnett, Blumenberg, 1983
0 0,1 0,04 0,003 48 —53 Chu,1986
0 0 0,034 48 —66 Lloyd, 1985
0 0,14 0,011 47 -56 Todd N. B., Todd L. M., 1976a
0 0,16 0,022 47 —65 Lloyd,1985
0 0,14 0,022 47 -68 Lloyd, 1985
0 0,21 0 47 —71 Lloyd, 1985
0 0,18 0,002 0,016 46 -60 Lloyd,1985
0 0,29 0,006 46 -63 Todd N. B., Todd L. M., 1976a
0 0,13 0,007 0,043 46 -65 Lloyd, 1985
0 0,18 0,028 46 —67 Lloyd, 1985
0 0,19 0,02 46 —73 Lloyd,1985
0 0,18 0,03 46 -74 Lloyd, 1985
0 0,23 0,07 45 —64 Todd N. B., Todd L. M., 1976a
132
133
1 2 3 3 8 9
Digby 72 0,26 0,76 0,34 0,49 0,43 0,38 0,04
St. John Nb 235 0,21 0,78 0,48 0,51 0,54 0,34 0,03
St. Jean PQ 75 0,23 0,73 0,5 0,44 0,45 0,37 0,05
Y armouth 162 0,22 0,75 0,48 0,44 0,48 0,32 0,03
St. Stephen 100 0,23 0,79 0,36 0,46 0,49 0,3 0,02
Bangor 167 0,25 0,76 0,44 0,41 0,4 0,39 0,02
Augusta 154 0,19 0,75 0,52 0,52 0,37 0,34 0,02
Portland 264 0,29 0,79 0,54 0,48 0,44 0,44 0,03
Lebanon 160 0,21 0,71 0,49 0,47 0,36 0,33 0,02
Burlington 254 0,24 0,78 0,46 0,39 0,44 0,31 0,03
S. New Hampshire 158 0,23 0,71 0,51 0,44 0,32 0,33 0,02
Brattleboro 99 0,25 0,69 0,43 0,42 0,5 0,31 0,02
Belmont Ma 191 0,27 0,77 0,29 0,4 0,32 0,35 0,02
Boston 792 0,22 0,72 0,47 0,44 0,33 0,39 0,02
New Bedford 171 0,23 0,76 0,45 0,42 0,32 0,36 0,03
Providence 216 0,3 0,76 0,37 0,53 0,15 0,41 0,01
Salem 161 0,23 0,78 0,43. 0,4 0,5 0,35 0,04
Worcester 117 0,24 0,77 0,53 0,44 0,38 0,36 0,02
Springfield 210 0,24 0,76 0,55 0,49 0,44 0,34 0,03
Albany 153 0,23 0,73 0,41 0,45 0,39 0,34 0,04
New London 115 0,23 0,72 0,45 0,53 0,35 0,34 0,03
New Haven 197 0,26 0,75 0,51 0,47 0,28 0,35 0,03
New York 236 0,14 0,75 0,47 0,44 0,13 0,47 0,01
Rochester 150 0,2 0,64 0,44 0,42 0,46 0,01
Philadelphia 293 0,27 0,7 0,45 0,5 0,2 0,42 0,01
Atlanta 129 0,23 0,65 0,46 0,43 0,31 0,53 0,03
Georgia 274 0,26 0,75 0,46 0,45 0,34 0,33 0,03
Juneau 117 0,38 0,87 0,33 0,39 0,35 0,28 0,01
Calgary 85 0,24 0,77 0,56 0,51 0,35 0,24 0,06
Winnipeg 142 0,19 0,73 0,64 0,5 0,51 0,31 0,03.
Vancouver 234 0,16 0,81 0,68 0,44 0,26 0,33 0,01
Duluth 155 0,1 . 0,79 0,84 0,61 0,49 0,34 0,03
Wisconsin 452 0,26 0,74 0,4 0,56 0,4 0,28 0,02
Minneapolis 337 0,25 0,74 0,47 0,63 0,51 0,29 0,02
Rapid City 243 0,22 0,8 0,15 0,52 0,04 0,23 0,03
Chicago 281 0,23 0,71 0,34 0,45 0,37 0,31 0,02
Clinton 37 0,46 0,59 0 0,5 0,57 0,31 0,01
Davenport 124 0,23 0,78 0,24 0,44 0,3 0,37 0,03.
Rock Island 0,33 0,78 0,32 0,48 0,52 0,29 0.03
Cleveland 171 0,23 0,74 0,62 0,41 0,25 0,35 0,02
Omaha 220 0,34 0,79. 0,35 0,52 0,37 0,19 0,03
Columbus 231 0,29 0,64 0,31 0,5 0,39 0,01
Denver 0,2 0,84 0,26 0,38 0,35 0,29 0,01,
Bloomington 117 0,31 0,74 0,57 0,43 0,46 0,24 0,03
St. Louis 866 0,3 0,79 0,51 0,43 0,48 0,38 0.02
Lawrence 180 0,22 0,72 0,44 0,4 0,44 0,25. 0,01.
Goodland, 70 0,35 0,69 0,35 0,42 0,32 0,21 0,07
Bowling Green 768 0,22 0,78 0,27 0,41 0,35 0,34 0,03
Lubbock 265 0,31 0,79 0,36 0,33 0,44 0.24 0
Dallas 311 0,25 0,67 0,27 0,32 0,44 0,18 0
Denton 311 0,25 0,81 0,27 0,33 0,46 0,22 0,01.
Mineral Wells 311 0,31 0,73 0,34 0,29 0,52 0,2 0
Phoenix 108 0,23 0,7 0,56 0,44 0,51. 0,23 0,07
Houston 294 0,25 0,69 0,29 0,29 0,35 0,19 0
Humboldt Co. 238 0.27 0,75 0,51. 0,35 0,29 0,22 0,03
Reno 156 0,2 0,77 0,38 0,51 0,39 0,25 0,05
Richmond 107 0,19 0,77 0,27 0,33 0,35 0,28. 0,03-
134
Продолжение табл. 11.2
10 и 12 1 13 14 15 16
0 0 0 45 -66 Lloyd, 1985
0 0,14 0,002 0,016 45 -66 Lloyd, 1985
0 0,25 0,036 45 -74 Lloyd, 1985
0 0,19 0,084 44 -66 Lloyd, 1985
0 0,2 0,005 0,041 45 —67 Lloyd, 1985
0 0,11 0,006 0,047 45 -69 Lloyd, 1985
0 0,11 0,013 0,029 44 -70 Lloyd, 1985
0 0 0,011 0,031 44 —70 Blumenberg et al., 1979
0 0,11 0,003 0,031 44 —72 Lloyd, 1985
0 0,17 0,002 0.043 44 —73 Lloyd, 1985
0 0 0,054 43 —72 Blumenberg et al., 1979
0 0,17 0,005 0,05 43 —73 Lloyd, 1985
42 —71 Lagen, 1975
0,065 42 —71 Lloyd, 1985
0 0,2 0,003 0,058 42 —71 Lloyd, 1985
0 0,12 0,045 42 —71 Blumenberg et al., 1979
0,003 0,26 0,003 0,038 42 —71 Blumenberg, 1977a, b
0 0,24 0,05 42 —72 Lloyd, 1985
0 0,2 0,05 42 —73 Lloyd, 1985
0 0,16 0,045 42 —74 Lloyd, 1985
0 0,27 0,025 41 —72 Lloyd, 1985
0 0,12 0,028 41 -73 Lloyd, 1985
0,004 0,07 0,002 0 41 —74 Todd,1966
0,02 43 -78 Searle, 1964
0,004 40 -75 Lloyd, Todd, 1989
0 34 -84 Todd,1969b
0 0,004 34 —84 Brown, Brisbin, 1983
58 —134 Fagen, 1983
0,18 0,007 51 —114 Blumenberg, 1983a
50 -97 Klein et al., 1987
0,002 49 —123 Blumenberg et al., 1977
47 —92 Klein et al., 1987
0,054 0,12 0 0,004 45 —92 Kerr, 1984
0,007 0,21 0,001 0,006 45 -93 Klein et al., 1986
44 —103 Klein et al., 1987
0,002 42 —88 Todd,1969a
42 -90 Blink et al., 1989
42 -91 Blink et al., 1989
42 -91 Blink et al., 1989
0,021 41 -82 Blumenberg, MacDonald, 1978
0,065 0,34 0 0 41 -96 Kerr, Halpine, 1986
0,002 40 -83 Tinney, Griesmeyer, 1968
40 -105 Morrill, Todd, 1978
0,18 0 39 —87 Lloyd, Todd, 1989
0,006 0,21 0 39 —90 Dorn, 1973
0,25 39 -95 Glass, Todd, 1976
0,28 39 —102 Glass, 1981
0,21 37 —86 Twedt, 1983
34 —102 Gerdes, 1973
33 —97 Gerdes, 1973
33 —97 Gerdes, 1973
0,003 33 -98 Gerdes, 1973
33 —112 Todd et al., 1976
30 -95 Gerdes, 1973
.0 0,22 0,015 0,018 41 —124 Blumenberg, 1983b
39 —120 Anderson, Jenkins, 1979
38 —122 Lamoreaux, Robinson, 1971
135
1 2 3 4 5 6 8 9
San Francisco 120 0,32 0,7 0,5 0,13 0,35 0,02
San Francisco 75 0,31 0,94 0,32 0,49 0,38 0,29 0,03
Los Mocliis 141 0,31 0,71 0,39 0,24 0,31 0,32 0,01
Veracruz 55 0,21 0,69 0,44 0,19 0,47 0,35 0,01
Mexico City 170 0,16 0,62 0,23 0,29 0,57 0,29 0,02
J amaica 96 0,15 0,81 0,65 0,34 0,32 0,32 0,04
Willemstad 151 0,14 0,84 0,19 0,19 0,28 0,34 0,02
Caracas 164 0,13 0,79 0,32 0,1 0,33 0,33 0,01
Porto Alegre 489 0,16 0,68 0,26 0,28 0,27 0,31 0,02
Curitiba 327 0,17 0,71 0,25 0,21 0,27 0,35 0,02
Rio de Janeiro 1545 0,24 0,74 0,32 0,27 0,19 0,38 0,04
Sao Paolo 1164 0.22 0,71 0,26 0,18 0,35 0,42 0,03
Belo Horizonte 859 0.18 0,71 0,26 0,25 0,22 0,36 0,02
Brasilia 492 0,26 0,63 0,28 0,19 0,18 0,31 0,01
Campo Grande 512 0,19 0,64 0,27 0,2 0,24 0,29 0,02
Cuiaba 302 0,16 0,66 0,2 0,12 0,1 0,29 0,04
Salvador 959 0,2 0,57 0,42 0,14 0,16 0,47 0,02
Rio Branco 235 0,25 0,49 0,29 0,06 0,06 0,34 0
Juazeiro do Norte 503 0,33 0,6 0,24 0,21 0,05 0,39 0
Teresina 994 0,18 0,69 0,33 0,26 0,01 0,35 0,03
Fortaleza 1254 0,2 0,66 0,27 0,29 0,08 0,5 0,02
Sao Luis 1323 0,24 0,66 0,19 0,24 0 0,53 0,01
Manaus 993 0,18 0,68 0,32 0,06 0,06' 0,47 0,03
Belem 909 0,22 0,65 0,25 0,14 0,05 0,42 0,01
Buenos Aires 295 0,27 0,83 0,31 0,45 0,4 0,28 0,02
Набор мутантных аллелей практически одинаков почти во всех
популяциях. Исключение составляет аллель 1 (длинная шерсть),
гомозиготы по которому не были обнаружены во многих по-
пуляциях.
Данные были собраны по популяциям, населяющим практи-
чески весь земной шар, за исключением нескольких неисследо-
ванных областей (наиболее интересные из них — Южная и Цент-
ральная Америка и Китай). Совокупность этих данных представ-
ляет собой крупнейшую популяционную выборку для одного вида.
Более изучен в этом отношении только человек.
РАСПРЕДЕЛЕНИЕ МУТАНТНЫХ АЛЛЕЛЕЙ
При популяционных исследованиях обычно предполагают а
priori, что центр видового ареала должен характеризоваться наи-
более значительным генетическим разнообразием. Что касается
разнообразия по окраске и длине шерсти, то кошка является
исключением из этого правила.
Есть свидетельства того, что кошка была впервые доместици-
рована на Среднем Востоке, однако разнообразие по исследуемым
признакам в этом районе ниже, чем где бы то ни было. Среди
объяснений, используемых для этого феномена, наиболее правдо-
136
Окончание табл. 11.2
10 И 12 13 14 15 16
0 38 —122 Searle, 1964
38 —122 Blumenberg В., Blumenberg G., 1976
26 —109 Todd et al., 1976a
19 -96 Todd et al., 1976a
19 -99 Todd et al., 1976a
18 —77 Lloyd et al., 1981
12 -69 Todd et al., 1976b
10 —67 Todd et al., 1976b
-30 -51 Watanabe, 1983
-25 -49 Watanabe, 1983
-23 -43 Watanabe, 1983
-23 —47 Watanabe, 1981
-20 -44 Watanabe, 1983
-16 —48 Watanabe, 1983
—20 —55 Watanabe, 1983
-16 —56 Watanabe, 1983
—13 -38 Watanabe, 1983
-10 -68 Watanabe, 1983
— / -39 Watanabe, 1983
—5 -43 Watanabe, 1983
—4 -39 Watanabe, 1983
—3 -44 Watanabe, 1983
—3 —60 Watanabe, 1983
—1 —48 Watanabe, 1983
0,01 -35 —39 Kajon et al., in press
подобным кажется представление о том, что люди предпочитают
редких кошек и тем самым способствуют сохранению новых вари-
антов, где бы они ни возникали [Lloyd, Todd, 1989]. Такой
частотно-зависимый отбор на новизну сыграл важную роль в эво-
люции популяций кошек во всем мире и остается существенным
фактором, определяющим генные частоты.
Происхождение домашней кошки затеряно во мгле времен.
Однако у нас есть некоторые надежды, что можно установить
происхождение некоторых мутантных аллелей и понять их совре-
менное распределение. Домашнюю кошку как вид в исторической
перспективе можно представить как большую группу изолирован-
ных субпопуляций, каждая из которых прошла свой собственный
путь более или менее независимой эволюции. По этому сценарию,
мутация, возникнув в некотором месте, может стать превалирую-
щей в этой локальной субпопуляции. По мере размножения попу-
ляции и, в особенности, если есть тенденция к экспансии этой
популяции, новый аллель может проникнуть в соседние популя-
ции. В таком случае на карте генных частот мы должны наблю-
дать что-то вроде кругов по воде, когда изоклины равных аллель-
ных частот расходятся вокруг точки возникновения мутации в
виде концентрических окружностей. Мутация мраморного рисунка
V является, по-видимому, лучшим примером именно такого рас-
пределения генных частот у млекопитающих [Todd, 1977а].
10 Заказ № 355
137
Концентрация аллеля мраморной окраски tb максимальна на
Британских островах, чуть ниже во Франции и далее падает
постепенно по мере удаления от центра распространения на вос-
ток, север п юг. Второй центр распространения аллеля tb находит-
ся в Иране.
Кошки, однако, мигрируют не только по суше, но и по морю.
Более того, морской путь миграции часто оказывается более эф-
фективным, чем сухопутный. Мы легко можем увидеть, что у ко-
шек в портовых городах частота аллеля tb выше, чем на более
удаленных от моря территориях. В Хабаровск и Владивосток мра-
морные кошки проникли явно с моря, а не по Транссибирской
магистрали.
Большинство исследователей сходятся во мнении, что аллель
мраморной окраски возник сравнительно недавно. Центром его
возникновения и всемирного распространения были Британские
острова. Там частота его наиболее высока и, по-видимому, возрас-
тает постоянно от поколения к поколению. На чем базируется
последний вывод? На сравнении частот этого аллеля в Британии
и ее бывших колониях.
Заселение новых территорий любым видом часто приводит к
тому, что в эволюционной генетике называется «эффектом осно-
вателя». В генетическом смысле популяция колонистов может
рассматриваться как выборка генотипов из родительской популя-
ции. В ходе колонизации небольшое количество основателей вно-
сит непропорционально значительный вклад в генетический состав
популяций следующих поколений. Заселение Америки, Австралии,
Новой Зеландии, Сибири и Дальнего Востока эмигрантами из
Европы .(кошками и их компаньонами — людьми) ясно показы-
вает действие эффектов основателя.
Данные, полученные при анализе колониальных популяций,
подтверждают представления о том, что немедленно после коло-
низации начинался быстрый рост численности популяций. Коло-
ниальные популяции Северной и Южной Америки существенно
отличаются от европейских, от которых они произошли, по часто-
там ряда аллелей. Особенно яркий пример — изменение частоты
аллеля tb.
Британская колониальная экспансия началась в XVII в. В хо-
де колонизации происходило мощное вселение кошек в новые
районы Земли.
Попробуем сопоставить частоту аллеля tb в популяциях, ныне
обитающих на территории того или иного из бывших британских
владений, со временем основания этой колонии.
В Нью-Йорке, Филадельфии и Бостоне, основанных в середи-
не XVII в., частота аллеля tb — 0,45. В Галифаксе и Сент-Джон-
се, основанных столетием позже, —0,55—0,60. В австралийских
городах Брисбене и Аделаиде, возникших в середине XIX в.,—
0,70. И, наконец, в Южной Англии сейчас частота этого аллеля
превышает 0,80.
Такцм образом, вырисовывается четкая, почти линейная зави-
симость между временем основания колонии и частотой аллеля
138
tb: чем раньше основана колония, тем ниже в ней частота кошек
с мраморным окрасом.
Наиболее простое объяснение этой зависимости таково. Ал-
лель tb возник в Британии. Отбор на этой территории способст-
вовал постепенному и неуклонному возрастанию частоты дан-
ного аллеля примерно на 10 % за столетие' или на 0,1 % за по-
коление. Те кошки, которые эмигрировали из Великобритании,
освобождались от действия этого отбора и сохраняли па новых
территориях ту концентрацию аллеля, которая была в материн-
ской популяции в момент образования дочерней. Аналогичную
картину мы наблюдаем, сравнивая частоты аллеля tb в метропо-
лии и Колониях Франции, Испании и Португалии [Todd, 1977а].
Американские популяции существенно отличаются от попу-
ляций Старого Света по частотам аллелей неагути а, ослаблен-
ной окраски d и оранжевой окраски О, которые в Северной и
Южной Америке выше, чем в Европе [Lloyd, Todd, 1989].
Отдельные популяции содержат в высокой концентрации ал-
лели, редкие для всех других популяций. Можно думать, что это
именно те популяции, где данные аллели возникли.
Так, о-в Мэн был местом возникновения мутации мэнской
бесхвостости М, что следует хотя бы из названия этой мутации.
Частота ее на острове равна 0,16, тогда как практически во всех
остальных популяциях она не обнаруживается, а в тех немно-
гих; где она есть, ее частота не превышает 0,01. Показательно,
что эти территории или были британскими колониями (Новая
Англия), или имели устойчивые коммерческпе связи с Брита-
нией (Голландия, Исландия).
Аллель полидактилии Pd также встречается довольно редко.
Он наиболее част в Новой Англии. В смежных с ней областях он
также встречается, но с меньшей частотой. Все неамериканские
популяции, где данный аллель обнаруживается, населяют или
порты (Бристоль) или острова (Азоры). Из этого следует, что
данный аллель возник в Новой Англии, п его частота может
быть использована для оценки уровня миграции новоанглий-
ских кошек на другие территории.
Аллель абиссинский тэбби Та наиболее част в популяциях
Калькутты и Коломбо. Его частота ниже, но все же значитель-
на в Бомбее, Гонконге, Мако, Монгсоне и Сингапуре. Во всем
остальном мире он или отсутствует вовсе, или крайне редок. Ис-
ходя пз этого, можно предположить, что оп возник в районе Бен-
гальского залива и отсюда распространился в другие популя-
ции. Как и многие другие редкие аллели, он встречается за пре-
делами своего «естественного» распространения только в разви-
тых странах, где практикуется разведение экзотических пород
кошек. Это отражает влияние импорта редких производителей
на генный пул локальных популяций.
Гораздо более важным для популяционных генетиков явля-
ется широкое распространение по всему миру аллеля сиамского
альбинизма cs. Именно фенотип, контролируемый данным ал-
10*
139
лелем, ассоциируется у большинства людей с образом породи-
стой кошки. Хотя частоты его в популяциях Юго-Восточной
Азии, ло-видимому, отражают реальный статус аллеля с5 в этих
популяциях (он, скорее всего возник именно там), но частота
его, скажем, в Омахе (США) или в Бордо (Франция) так вы-
сока, что довольно трудно отвергнуть гипотезу, что данные рай-
оны являются центрами возникновения этой мутации. Наиболее
вероятно, что высокие частоты, которые исследователи обнаружи-
вают в Юго-Восточной Азии, обусловлены включением в выбор-
ку породистых кошек, а не значительной концентрацией данного
аллеля в свободно размножающихся популяциях.
Сходные эффекты можно, по-видимому, обнаружить и для дру-
гих аллелей, поддерживаемых в генофондах пород. Объектами для
будущих исследований являются редкие аллели бирманского аль-
бинизма с*’, шиншилловой окраски I, коричневой окраски b и рек-
соидпой шерсти Re. С возрастанием популярности этих мутаций
среди любителей породистых кошек следует ожидать увеличения
частот этих мутаций в популяциях.
ЧИСЛЕННЫЙ АНАЛИЗ ПОПУЛЯЦИОННЫХ ДАННЫХ
Одно из преимуществ работы с популяциями кошек заключа-
ется в том, что используя распределение фенотипов, контроли-
руемых сцепленным с полом локусом оранжевой краски О, можно
оценить равновесность по Харди — Вейнбергу и, следовательно,
наличие панмиксии в локальных популяциях. В огромном боль-
шинстве популяций, где такой анализ был проведен, не было
обнаружено существенных отклонений от равновесного состояния.
Это указывает на то, что в отношении аллелей данного локуса
(и, по-видимому, других локусов тоже) популяции кошек явля-
ются папмиксическими' Этот факт имеет особое значение для город-
ских популяций, где количество породистых кошек и их вклад
в генный пул неизвестны. Если чистопородные кошки вносят
вклад в генный пул (а данные по аллею с8 свидетельствуют об
этом), то они это делают случайно, не искажая общую картину
панмиксии. Менее надежные, по также свидетельствующие в
пользу гипотезы о панмиксии данные получены при анализе фе-
нотипов, контролируемых локусом белой пятнистости S. В боль-
шинстве исследований оценивалась доля белого в окраске пегих
кошек с градацией от 0 (нет белых пятен) до 10 (вся шкура
белая). Градация 0 соответствует отсутствию белых пятен (ре-
цессивная гомозигота), градации 1—5 относят к гетерозиготам,
носителей градаций 6—9 рассматривают как доминантных гомо-
зигот, а градацию 10 приписывают носителям гена доминантной
белой окраски W, эпистатичной над S. Анализ данных по рас-
пределению частот разных градаций в популяциях дает хорошее
соответствие наблюдаемых частот с ожидаемыми при выполнении
условий соотношения Харди — Вейнберга.
,140
РАСПРЕДЕЛЕНИЕ ЧАСТОТ МЕЖДУ ПОПУЛЯЦИЯМИ
Если взять данные для каждого локуса отдельно и оценить
распределение частот по локальным популяциям, то полученную
кривую распределения можно анализировать, используя метод
Райта [Wright, 1965] для популяционной структуры. Нормальное
распределение аллельных частот, которое было найдено для ло-
кусов оранжевой О и ослабленной d окрасок [Lloyd, Todd, 1989],
характерно для популяций со слабой степенью структурирован-
ности. Причины наблюдаемого распределения частот кроются
в особенностях или исторической миграции (выборочная вариация
при общем происхождении), или современной миграции, которая
проявляется как поток генов, или во взаимодействии этих двух
факторов. В исследованных популяциях оба фактора имеют оди-
наковое значение. В Северной Америке эффект генного потока
проявляется в существенном выравнивании генных частот, ведя
к образованию очень большой, свободно скрещивающейся попу-
ляции [Blink et al., 1989]. В Европе и Азии распределение частот
до сих пор следует нормальному распределению, что, скорее всего,
следует приписать исторической миграции. Достоверные разрывы
до сих пор существуют на государственных границах и вдоль
естественных барьеров миграции. Таким образом, политика может
играть роль также и в жизни кошек.
Распределения в частотах некоторых аллелей оказываются
существенно ограниченными на краях. Например, не существует
достаточно крупных популяций с частотой аллеля а выше, чем
0,87. Для аллеля О максимальная частота равна 0,5. Если бы
стохастика вносила существенный вклад в распределение частот,
то частоты должны были варьировать от 0,0 до 1,0. Конечно,
сам характер распределения определяется структурой популяции,
но его границы должны быть именно такими: от нуля до едини-
цы. Настоящие причины этих ограничений остаются непонятными,
но в качестве объяснения можно принять гипотезу об отборе
новых или редких фенотипов (частотно-зависимом отборе). Вла-
делец кошки предпочитает держать такую кошку, которая отли-
чается от той, что держит его сосед. Это предполагает, что попу-
ляция кошек в данном районе достаточно велика, чтобы обеспе-
чить возможность выбора. Интересные примеры дают островные
популяции, такие, как популяции островов Кергелен и Марион.
На о-ве Кергелен пять из семи обычно исследуемых аллелей
фиксированы, включая неагути а. Единственным полиморфным
локусом был локус белой пегости (S). Его частота равна 0,4.
Таким образом, только два фенотипа поддерживались в этой
очень ограниченной популяции: черный и черно-белый [Dreux,
1974]. На о-ве Марион, где популяция кошек была больше, чем
на Кергелене, фиксировались только два аллеля из обычного
набора: о и L [Aarde, Robinson, 1980].
Некоторые аллели проявляют бимодальное распределение.
Наиболее интересно распределение аллеля длинной шерсти 1.
Этот аллель отсутствует в большинстве популяций, а в тех, где
141
он есть, обнаруживается зависимость его частоты от климата.
Для популяций Северо-Восточной Америки и для популяций
Британских островов были обнаружены достоверные корреляции
между среднегодовой температурой и частотой аллеля 1, хотя
объединение этих двух групп популяций ведет к тому, что кор-
реляция исчезает [Lloyd, 1987]. Бразильские популяции также
демонстрируют высокую корреляцию длинной шерсти с клима-
том (г = 0,95, А =16) [Watanabe, 1983]. Однако если мы рас-
сматриваем все популяции земного шара вместе, то не обнару-
живаем такой корреляции.
Индексы генетического сходства были использованы для оцен-
ки сходства между различными популяциями кошек [Klein et al.,
1986]. Преимущество этого подхода заключается в том, что можно
свести данные, характеризующие сходство популяций по частотам
нескольких аллелей, к одному индексу. Недостатком же является
то, что при таком подходе невозможно сказать, какой из локусов
вносит наибольший вклад в различия между популяциями. Это
означает, что индексы генетического сходства не коммутативны,
и выводы, полученные на одной группе популяций, не могут быть
перенесены на другую группу. Тем не менее эти индексы могут
быть вычислены для всех возможных комбинаций (более 50000).
Было показано, что индексы генетического сходства варьиру-
ют от 1,0 для тесно связанных популяций (например, Миннеапо-
лис и Стивен Канти, Миннесота) до 0,65 для очень отдаленных
популяций (Британские острова и Бангкок) [Klein, неопубл,
данные]. Такие различия в индексах вполне естественны при
допущении, что основную роль в дивергенции генетических про-
филей популяций играет географическая изоляция. Обнаружено
несколько случаев, когда пространственно близкие популяции
менее сходны друг с другом, чем пространственно далекие. Как
правило, это наблюдается в популяциях, разделенных естест-
венными барьерами, такими, как Гималайский хребет.
Попытки использовать киевские индексы сходства для по-
строения дендрограмм не привели к большим успехам [Ruiz-
Garcia, 1991b]. В общем географически соседствующие популяции
находятся рядом друг с другом и на дендрограмме. Точность
дендрограмм сильно зависит от количества локусов, использован-
ных для их построения. Поэтому данный метод не может быть
использован для популяций настолько далеких, что поток генов
между ними ограничен. Главное, что вызывает сомнение при
интерпретации любых дендрограмм, это уровень значимости каж-
дого конкретного индекса, полученного от сравнения двух попу-
ляций. Анализ индексов генетического сходства дает, таким обра-
зом. очень грубую картину дивергенции кошачьих популяций.
Более ясную картину структуры размножения популяции
кошек можно получить, вычисляя F-статистики (уровни инбри-
динга) на основе кумулятивных данных по частотам каждого
мутантного аллеля [Weir, Cockerham, 1984]. Эта оценка вклада
популяционной структурированности в инбридинг важна потому,
142
что она позволяет оцепить генный поток {Slatkin, 1987]. Анализ
региональных популяций продемонстрировал наличие относитель-
но сильного потока генов между популяциями на весьма большой
территории [Blink et al., 1989]. Исследования, в которых для
выделения репродуктивных сообществ использовались этологиче-
ские критерии, показали, что уровень инбридинга на микропопу-
ляционном уровне довольно высок [Ruiz-Garcia, 1990с]. Таким
образом, популяция большой территории (государства, острова
и т. д.) может быть в равновесии по аллельным частотам, при-
ближаясь к глобальной норме, но локальные репродуктивные со-
общества могут быть довольно далеки от нормы. Только последо-
вательные локальные вымирания и заместительная колонизация
могут поддерживать генетическое единство популяции кошек как
целого. Долговременные исследования демографии популяций ко-
шек свидетельствуют, что эти популяции локально вариабельны,
но глобально стабильны [Pontier, 1989].
ЗАКЛЮЧЕНИЕ
Исследование популяций кошек важно для понимания эволю-
ционного процесса. По-видимому, ни один из связанных с чело-
веком видов не имеет такого широкого распространения и не
пользуется такой свободой скрещивания, как кошка. Значение
кошки как модели для исследования эволюции природных попу-
ляций обусловлено этими особенностями ее поведения. Также
полезным является наличие в большинстве популяций полимор-
физма по моногенным признакам. Никакой другой вид не может
соперничать с кошкой в смысле доступности для популяционно-
генетического анализа.
Что же мы можем извлечь из этого анализа? Наиболее инте-
ресен тот эффект, который вызвала доместикация этого широко
расселенного вида. Ослабление давления естественного отбора
привело к формированию такой фенотипической пластичности, ко-
торая не наблюдается у диких видов. В некоторой степени естест-
венный отбор замещен отбором со стороны человека. Этот отбор
является по своей природе частотно-зависимым и способствует
накоплению редких фенотипов и генотипов.
Важными представляются и отношения между разными попу-
ляциями. Данные показывают, что обмен генами между ними
весьма значителен. Нет популяций настолько далеких, чтобы
можно было с уверенностью сказать, что обмен генами между
ними полностью отсутствует, хотя, конечно, географическая уда-
ленность влияет на интенсивность обмена. В то же время кошки
образуют глобально структурированную популяцию. Микрогеогра-
фические различия могут быть весьма значительными. Их обычно
приписывают стохастическому процессу — эффекту основателя.
Последствия эффекта основателя могут быть выявлены макрогео-
графически при сравнении популяций кошек, возникших в то
время, когда человек заселял земной шар.
143
Картина эволюции кошачьих популяций — это картина, харак-
терная для весьма преуспевающего, интенсивно расселяющегося
вида. Кошки распространили свой видовой ареал на весь земной
шар, заселив прежде пустые ниши во множестве экологических
систем. В генетическом смысле это было связано с ослаблением
давления отбора на целостный фенотип, что привело к возникно-
вению огромного фенотипического разнообразия. Для популяций
кошек характерна фрагментация па дискретные локальные груп-
пы, которые могут существенно отличаться по частотам аллелей,
контролирующих различные части фенотипа котики. Каждая из
этих микропопуляций может исчезнуть и ее место могут занять
мигранты из общей популяции. Это ведет к интенсивному потоку
генов и к общей стандартизации глобальной популяции. Колони-
альные популяции дивергируют от популяций метрополии благо-
даря частотно-зависимому отбору, сохраняя при этом следы своего
происхождения за счет эффекта основателя.
Кошка являет собой модель для исследования факторов, кото-
рые формируют судьбу всех преуспевающих видов, включая наш
собственный вид.
Автор хотел бы выразить благодарность людям, которые под-
держали его в этом исследовании или предоставили свои данные:
более всего Эндрю Ллойду из Тринити колледжа в Дублине, ко-
торый составил оригинальный каталог генных частот для различ-
ных популяций кошек всего мира и тем самым, избавил всех
популяционистов от бесчисленных часов библиографической ра-
боты, и который любезно предоставляет этот каталог всем, кто
в нем нуждается. Я также признателен Мануэлю Руис-Гарсии из
Барселонского университета за новые данные и комментарии к
методам их анализа. Наконец, я хотел бы поблагодарить Павла
Бородина из Института цитологии и генетики в Новосибирске,
давшего мне возможность синтезировать все эти исследования па
кошках при написании данной главы.
СПИСОК ЛИТЕРАТУРЫ
Aarde R. J. van, Erasmus В. И., Blumenberg В. Mutant allele frequencies for
the Pretoria and Cape Town cat populations / South African J. Sci.—
1977- P. 168—171.
Aarde R. J. van, Robinson T. J. Gene frequencies in feral cats on Marion Is-
land // ,1. Hered.— 1980,-Vol. 71,—P. 366-368/
Adalsteinsson S., Sigurjonsson T., Jonsson G. Variation in colour gene fre-
quencies among Icelandic cats Ц Carnivore Genet. Newslett.— 1979.—
Vol. 3. N 10 — P. 359—372.
Ahmad M., Blumenberg B., Chaudhary M. F. Mutant allele frequencies and ge-
netic distance in cat populations of Pakistan and Asia Ц J. Hered.—
1980 — Vol. 71,— P. 323-330.
Andersen M. M., Jenkins S. H. Gene frequencies in the domestic cats of Reno,
Nevada: Confirmation of a recent hypothesis Ц J. Hered.— 1979.—
Vol. 70,— P. 267—269.
Ваха H. Gene frequencies in stray cats from Vienna Ц Genetica.— 1973.—
Vol. 44,— P. 25-30.
Blink T. Y., Klein K., Kerr S. J. Mutant allele frequencies in Eastern Iowa and
Western Illinois cats / J. Hered.— 1989.— Vol. 80.— P. 332—335.
144
Blumenberg В. Mutant allele frequencies in the domestic cats of California,
Texas and New Spain Ц Carnivore Genet. Newslett.— 1977a.— Vol. 3.—
P. 2-9.
Blumenberg B. Mutant allele frequencies in the domestic cats of Eastern Mas-
sachusetts Ц Genetica.— 1977b.— Vol. 46.— P. 9—14.
Blumenberg B. The genetic profile of the cats of Calgary, Alberta: A prelimi-
nary assessment // Carnivore Genet. Newslett.— 1983a.— Vol. 4.—
P. 215—217.
Blumenberg B. Mutant allele frequencies in the domestic cals of Humboldt
county, California // Ibid.— 1983b.— P. 218—223.
Blumenberg B., Blumenberg G. Cat gene frequencies in the San Francisco Bay
region, California Ц Genetica.— 1976.— Vol. 46.— P. 385—389.
Blumenberg B., Fish C.. MacDonald B. Mutant allele frequencies in three do-
mestic cat populations of New England Ц Carnivore Genet. Newslett.—
1979,— Vol. 4,— P. 1—12.
Blumenberg B., Lowry R. B., Blumenberg G. Mutant allele frequencies in the
domestic cats of Vancouver, В. С. Ц J. Hered.— 1977.— Vol. 68.—
P. 333—336.
Blumenberg B., MacDonald B. A population genetic analysis of the domestic
cals of Cleveland, Ohio, and the Midwestern United Slates: Proc. 1st
Intern, conf, domestic cat popul. genet, ecol. Ц Carnivore Genet. News-
letl - 1978,— Vol. 3,— P. 227—240.
Borodin P. M., Bochkarev M. N., Smirnova I. S., Manchenko G. P. Mutant al-
lele frequencies in domestic cat populations of six Soviet cities Ц J.
Hered.- 1978,-Vol. 69,-P. 169—174.
Borodin P. M., Manchenko G. P., Robinson R., Todd N. B. New studies in mu-
tant allele frequencies on USSR cats: Proc. 2nd Intern, conf, domes-
tic cat popul. genet, ecol. Ц Carnivore Genet. Newslett.— 1981.— Vol. 4.—
P. 153—167.
Brown C. J., Brisbin I. L. Genetic analysis of Pariah cat populations from the
Southeastern United States // J. Hered.— 1983.— Vol. 74.— P. 153—167.
Chu К. C. A study of mutant allele frequencies in the domestic cats of St.
John's, Newfoundland Ц Carnivore Genet. NewsletL— 1986a.— Vol. 4.—
P. 210—212.
Chu К. C. Mutant allele frequencies in the domestic cals of Taiwan // J. He-
red.—1986b.—Vol. 77.—P. 277—278.
Clark J. M. Gene frequencies in the domestic cats of Adelaide // Austr. J. Bi-
ol. Sci.— 1973,—Vol. 26,—P. 1215—1219.
Clark J. M. The effects of selection and human preference on coat color gene
freauencies in urban cals Ц Heredity.— 1975.— Vol. 35.— P. 195—210.
Clark J. M. Variations in coat color gene frequencies and selection in cats of
Scotland I/ Genetica.— 1976.— Vol. 46.— P. 401—412.
Dards J. L., Robinson R. Gene frequencies in a population of feral cats in Port-
smouth naval dockyard Ц Theor. Appl. Genet.— 1983.— Vol. 64.—
P. 197—204.
Dartnail J. A. Gene frequencies in the feral domestic cats of Tasmania // Car-
nivore Genet. Newslett.— 1975,— Vol. 2.— P. 248—250.
Dartnail J. A. Domestic cat gene frequencies in Parwin, North Australia // Car-
nivore Genet. Newslett.— 1977.—Vol. 3.— P. 137—139.
Dartnall J. A., Todd N. B. A study of the population genetics of Hobart Ц
Austr. J. Zool.— 1975.— Vol. 23.— P. 405—409.
Davis В. K., Ahmad M. Allele frequencies in Saudi Arabian cat populations /[
Carnivore Genet. Newslett — 1983.— Vol. 4.— P. 224—229.
Davis В. K., Davis В. P. Allele freauencies in a cat population in Budapest Ц
J. Hered.— 1977,— Vol. 68 — P. 31—34.
Dorn C, R. Cat gene frequency in St. Louis Ц Carnivore Genet. Newslett.—
1973,—Vol. 2.-P. 106—111.
Dreux Ph. Gene frequencies in the cat population of Paris Ц J. Hered.—
1967,— Vol. 59,— P. 89—92.
Dreux Ph. Gene freauencies in the cat population in a French Rural district Ц
J. Hered.- 1968,- Vol. 59,- P. 37-39.
145
Dreux Ph. Genetique des populations de chats domestiques dans le Vallee de
Chamonix (Haute — Sovaie) Ц Ann. Genet. Sei. anim.— 1971.— Vol. 3.—
P. 145—151.
Dreux Ph. The cat population of the Peninsule Corbet, Isles de Kergulen: an
example of the Founder effect / Polar Records.— 1974.— Vol. 17.—
P. 53-54.
Dreux Ph. Genetique des population des chats domestique de Marseille (Bou-
che-de-Rhone, France) Ц Ann. Genet. Sei. anim.— 1975.— Vol. 7.—
P. 23—33.
Dreux Ph. Frequences de genes a effets visible dans les populations de chats
de la Ville de Tours (Indre et Loire, France) Ц Ann. Genet. Sei. anim.—
1979.—Vol. 11,—P. 391—396.
Dreux Ph. Genetique des populations de chat domestique dans un District
Rural du Cher (France) // An. Genet. Sei. anim.— 1981.— Vol. 13,—
P. 363—370.
Dreux Ph. Genetique des populations de chats domestique dans un District de
Bretagne (Carnac, Mor bihan, France) // Coll. Nat. CNRS Biologie des
Populations, Septembre 1986.— P. 312—316.
Dreux Ph., Legel J. C. Gene frequencies in the cat population of the Haque Ц
J. Hered.- 1973,— Vol. 64,— P. 337-339.
Dreux Ph., Samet D. Frequences des genes dans populations Rurales Espag-
noles de chats domestiques // Carnivore Genet. Newslett.—1981.—
Vol. 4,—P. 127—129.
Fagen R. M. Coat color gene frequencies in the cats of Watertown and Bel-
mont, Massachusetts Ц Carnivore Genet. Newslett.— 1975.— Vol. 2.—
P. 251-255.
Fagen R. M. Domestic cat demography and population genetics in a Midwes-
tern USA Metropolitan Area Ц Carnivore.— 1978.— Vol. 1.— P. 60—67.
Fagen R. M., Meeswat K. Population genetics and demography of the domes-
tic cat (F. ca-t'As L.: Carnivora, Felidae) in Thailand Ц Nat. Hist. Bull.
Siam Soc.—1981 —Vol. 29 —P. 137—150.
Garnett M., Blumenberg B. Mutant allele frequencies in feral and domestic
cat populations of Christmas Island, Kiribati, Central Pacific Ц Carni-
vore Genet. Newslett.— 1983.— Vol. 4.— P. 230—234.
Genermont J. Estimation des frequences de quelque genes determinant la cou-
leur du pelage dans le population de chats domestiques de Rabat (Ма-
тос) /7 Carnivore Genet. Newslett.— 1974.— Vol. 2.— P. 181—182.
Genermont J. Gene frequencies in the cat population of Constantine with ge-
neral considerations of the breeding systems of cats Ц Carnivore Genet.
Newslett.— 1976a.— Vol. 3.— P. 83—87.
Genermont J. Gene frequencies in the cat population of Nice // Carnivore Ge-
net. Newslett.— 1976b.— Vol. 3.— P. 356—357.
Gerdes R. A. Cat gene frequencies in five Texas communities / J. Hered.—
1973 — Vol. 64 — P. 129—132.
Glass G. A preliminary survey of domestic cat gene frequencies in Goodland,
Kansas: Proc. 2nd Intern, conf, domestic cat popul. genet, ecol. // Car-
nivore Genet. Newslett.— 1981.— Vol. 4.— P. 130—132.
Glass G., Todd N. B. Gene frequencies of the domestic .cats of Lawrence, Kan-
sas Ц Carnivore Genet. Newslett.— 1976.— Vol. 3.— P. 28—31.
Goncharenko G. G., Lopatin О. E., Manchenko G. P. Coat color mutant genes
in domestic cat populations of Middle Asia and European part of the
USSR (in Russian) Ц Genetika.— 1985.— Vol. 21.— P. 1151—1158.
Gruffydd-Jones T., Jaffe P., Lloyd A. T. et al. Mutant allele frequencies in do-
mestic cat populations of Bristol and South Wales Ц Carnivore Genet.
Newslett.— 1979.— Vol. 4,— P. 13—17.
Hines W, Randall J. Estimates of mutant allele frequencies in domestic cats
of the Dalmatin coast Ц Carnivore Genet. Newslett.— 1977.— Vol. 3.—
P. 140—142.
Jaffe P. Cat gene frequencies in the Seychelles: Proc. 1st Intern, conf, do-
mestic cat popul. genet, ecol. // Carnivore Genet. Newslett.— 1978.—
Vol. 3,— P. 339—345.
146
Kajon A., Garcia D. C., Ruiz-Garcia M. Gene frequencies in the cat populati-
on of Buenos Aires (Argentina) and the possible origin of this popu-
lation (in press).
Kerr S. J. Mutant allele frequencies in Swiss Rural cat populations Ц J. He-
red.— 1983,- Vol. 74,— P. 349-352.
Kerr S. J. Mutant allele frequencies in Rural Wisconsin cats Ц J. Hered.—
1984.—Vol. 75,—P. 203—206; Revised J. Hered.—Vol. 76,—P. 216.
Kerr S. J., Halpine T. Mutant allele frequencies in the cat population of Oma-
ha, Nebraska // J. Hered — 1986,— Vol. 77.— P. 460—462.
Kinnear J. F. Mutant allele frequencies in domestic cats of Melbourne: Proc.
1st Intern, conf, domestic cat popul. genet, ecol. Ц Carnivore Genet.
Newslett.— 1979,— Vol. 3,— P. 48—417.
Klein K., Smith S., Schmidt J. et al. Mutant allele frequencies in Upper Mid-
western cat populations Ц J. Hered.— 1987.— Vol. 79.— P. 389—393.
Klein K., Wong L., Olson J., Amundson S. Mutant allele frequencies in cats of
Minneapolis/St. Paul Ц J. Hered.— 1986.— Vol. 77.— P. 132—134.
Lamoreaux L.. Robinson R. Cat gene frequencies in Richmond, California Ц
Genetica.— 1971,— Vol. 42 —P. 61—64.
Lloyd A. T, Surveys of gene frequencies in England Ц Carnivore Genet. News-
lett.— 1977,— Vol. 3,— P. 168.
Lloyd A. T. The population genetics of cats in Northwest Ireland: Proc. 1st In-
tern. conf, domestic cat popul. genet, ecol. / Carnivore Genet. News-
lett.— 1979 — Vol. 3 — P. 373—377.
Lloyd A. T. Mutant allele frequencies in the domestic cat populations of the
Netherlands // Genetica.— 1982 — Vol. 58 — P. 223—228.
Lloyd A. T. Cats of the thirteenth centry: evidence of mutant phenotypes in
Bartholomew de Glanville’s De Proprietatibus Rerum Ц Carnivore Ge-
net. Newslett.— 1983,— P. 265—267.
Lloyd A. T. Geographic distribution of mutant alleles in domestic cat popu-
lations of New England and the Canadian maritimes Ц J. Biogeogr.—
1985,— Vol. 12 — P. 315—322.
Lloyd A. T. Cats from history and history from cats // Endeavour.— 1987.—
Vol. 11, N 3.-P. 112-115.
Lloyd A. T., Jal'fe P. Mutant allele frequencies in the domestic cats of Malta Ц
Carnivore Genet. Newslett.— 1981.— Vol. 4.— P. 206—209.
Lloyd A. T., Todd N. B. Domestic cat gene frequencies, a Catalog and Biblio-
graphy.— Newcastle-Upon-Tyne, UK: Tetrahedrom Publ., 1989.
Lloyd A. T., Todd N. B., Baldwin J. A. Mutant allele frequencies in the cats
of Kingston, Jamaica: Proc. 2nd Intern, conf, domestic cat popul. genet,
ecol. Ц Carnivore Genet. Newslett.— 1981.— Vol. 4.— P. 203—205.
Lloyd A. Jq Todd N. B., Dyte С. E. et al. Towards a comprehensive picture
of the Mediterranean: population genetics of the cats of Rome, Italy Ц
Carnivore Genet. Newslett.— 1983.— Vol. 4.— P. 235—241.
Marples B. J. Notes on the phenotypes of cats observed in New Zealand and
Thailand Ц Carnivore Genet. Newslett.— 1967.— Vol. 1.— P. 43—44.
Metcalfe J. A., Turner J. R. G. Gene frequencies in the domestic cats of New
York: evidence for Selection Ц Heredity.— 1971.— Vol. 26.— P. 259—
268.
Moffat B. W. Cat gene frequencies in two Australian cities Ц J. Hered.—
1968,—Vol. 59 —P. 209—211.
Morrill R. B., Todd N. B. Mutant allele frequencies in the domestic cats of
Denver, Colorado Ц J. Hered.— 1978.— Vol. 71.— P. 2—8.
Mutant allele frequencies in domestic cats of Siracusa, Sicily: Proc. 1st In-
tern. conf, domestic cat popul. genet, ecol. // Carnivore Genet. News-
_ lett.— 1979,— Vol. 3,— P. 418—420.
Pontier D. Frequences genetiques chez le chat domestique, Felis catus, dans
quatre communes rurales francais Ц Genetica.— 1983.— Vol. 62.—
. P. 69—73.
Pontier D. Annual variations of coat genes frequencies in the domestic cat
Felis catus in France/Ed. A. T. Lloyd, N. B. Todd.— 1989.— P. 17—20.
Robinson R. On gene frequencies in cats of Benidorm, Spain Ц Carnivore Ge-
net. Newslett.— 1971.— Vol. 2.— P. 32—33.
147
Robinson R. Mutant allele frequencies in the cats of Cyprus Ц Theor. Appl.
Genet.— 1972,— Vol. 42,— P. 293—296.
Robinson R. Mutant gene frequencies in cats of the greater London area //
Theor. Appl. Genet. — 1987,— Vol. 74 — P. 579—583.
Robinson R., Manchenko G. P. Cat gene frequencies in the USSR Ц Geneti-
ca.— 1981,— Vol. 55.—P. 42—46.
Robinson R., Silson M. Mutant allele frequencies in cats of Southern England Ц
Theor. Appl. Genet.— 1969.— Vol. 39,— P. 326—329.
Ruiz-Garcia M. Frequencias alelicas mutantes en una poblacion de gatos
domesticos urbanos (Barcelona) у en una poblacion de gatos rurales
(Castelldefels Rural) en Cataluna, Espana Ц Genet. Iber.— 1988.—
Vol. 40,— P. 157—187.
Ruiz-Garcia M. Frequencias alelicas en la poblacion de gatos domesticos de
Palma de Mallorca e ibiza у relaeiones geneticas con poblaciones de
gatos europeos у Norteafricanos Ц Evol. Biol.— 1990a.— Vol. 4.—
P. 189—216.
Ruiz-Garcia M. Frequencias alelicas en la poblacion de gatos domesticos de la
Isla Menorca (Baleares): differentes modelos de evolution colonizado-
ra / Evol. Biol.— 1990b.— Vol. 4.— P. 307—342.
Ruiz-Garcia M. Mutant allele frequencies in domestic cat populations on the
Spanish Mediterranean Coast and genetic distances from other Europe-
an and North African cat populations Ц Genetica.— 1990c.— Vol. 82.—
P. 215-221.
Ruiz-Garcia M. Mutant allele frequencies in domestic cat populations in Ca-
talonia, Spain, and genetic relationships between Spanish and English
colonial cat populations Ц Genetica.— 1990d.— Vol. 82.— P. 209—215.
Ruiz-Garcia M. Mas sobre la genetica de poblaciones de Felis catus en la Cos-
ta Mediterranea Espanola: un analisis de la estructura genetica de las
poblaciones naturales de gatos // Evol. Biol.— 1991a (in press).
Ruiz-Garcia M. Genetic profiles of Balearic cat populations: II. An analysis
of the genetic diversity within and between Balearic and Spanish cat
populations.— 1991b (in preparation).
Salternik R., Mordokhovich D. Gene frequencies of coat color and length in do-
mestic eats of Jerusalem I/ Carnivore Genet. Newslett.— 1975.— Vol. 2.—
P. 259—262.
Salternik-Vardy R., Ritte U. Coat color gene frequencies in the domestic cat
population of Jerico: Proc. 1st. Intern, conf, domestic cat popul. genet,
ecol. Ц Carnivore Genet. Newslett.— 1978.— Vol. 3.— P. 332—337.
Salternik-Vardy R., Ritte U. Cats of Israel/Ed. A. T. Lloyd, N. B. Todd.—
1989,—P. 14—15.
Salternik R., Shoenbaum M., Ritte U. Frequencies of mutant alleles for coat
color and length in cats of Tel Aviv, Israel / Carnivore Genet. News-
lett.—1976,—Vol. 3,—P. 35-42.
Salternik R.. Todd N. B. Population genetics of domestic cats in Iran Ц Car-
nivore Genet. Newslett.— 1977.— Vol. 3.— P. 148—154.
Searle A. G. Gene frequencies in London’s cats Ц J. Genet.— 1949.— Vol. 49.—
P. 214—220.
Searle A. G. A study in the variation in Singapore cats Ц J. Genet.— 1959.—
Vol. 56,—P. 1—16.
Searle A. G. The gene geography of cats Ц J. Cat. Genet.— 1964.— Vol. 5.—
P. 18—24.
Searle A. G. Coat colour gene frequencies in Venetian cats Ц Carnivore Ge-
net. Newslett.- 1966,-Vol. l.-P. 6—7.
Slatkin M. Gene flow and the structure of populations Ц Science.— 1987.—
Vol. 236,— P. 787—792.
Symonds J. E., Lloyd A. T. Gene frequencies of the domestic cats of Urban
and Rural Bordeaux Ц Genetica.— 1987.— Vol. 72.— P. 133—136.
Tinney L. M., Griesmeyer R. A. Gene frequencies in the cats of Columbus,
Ohio, USA, and a comparison of Northeastern populations / Carnivore
Genet. Newslett.— 1968.— Vol. 1.— P. 96—99.
Todd N. B. Gene frequencies in the cat population of New York city Ц J. He-
red.— 1966 — Vol. 57,— P. 185—187.
148
Todd N. В. Cat gene frequencies in Chicago and other populations of the Uni-
ted States Ц J. Hered.— 1969a.— Vol. 60.— P. 273—277.
Todd N. B. The cats of Atlanta, Georgia Ц Carnivore Genet. Newslett. -
1969b — Vol. 1,—P. 171—199.
Todd N. B. Mutant allele frequencies in domestic cats of Dublin and Lime-
rick Ц Proc. Royal Irish Acad.— 1975.— Vol. 77(B).— P. 201—205.
Todd N. B. Cats and commerce Ц Sci. Amer.— 1977a.— Vol. 237.— P. 100—
107.
Todd N. B. Mutant allele frequencies in the cats of Tunis and Carthage Ц
Carnivore Genet. Newslett.— 1977b.— Vol. 3.— P. 155—160.
Todd N. B., Adalsteinsson S. Mutant allele frequency estimate for some Faeroe-
se cats Ц Carnivore Genet. Newslett.— 1982.— Vol. 4.— P. 22—23.
Todd N. B., Blumenberg B. Mutant allele frequency and genetic distance rela-
tionships in domestic cat populations of Lower Egypt and the Eastern
Mediterranean Ц Theor. Appl. Genet.— 1978.— Vol. 52.— P. 257—262.
Todd N. B., Fagen R. M., Fagen K. Gene frequencies in Icelandic cats // He-
redity.— 1975,— Vol. 35,— P. 172—183.
Todd N. B., Garrad L. S., Blumenberg B. Mutant allele frequencies in domes-
tic cats of the Isle of Man: Proc. 1st Intern, conf, domestic cat popul.
genet, ecol. Ц Carnivore Genet. Newslett.— 1979.— VoL 3.— P. 257—262.
Todd N. B., Glass G. E., Creel D. Cat population genetics in the US Southwest
and Mexico Ц Carnivore Genet. Newslett.— 1976a.— Vol. 3.— P. 43—54.
Todd N. B., Glass G. E., McLure I. Gene frequencies in some cats of South
America: Caracas, Venezuela and Willemstad, Curacao Ц Carnivore Ge-
net. Newslett.— 1976b.— Vol. 2.— P. 230—235.
Todd N. B., Kunz T, H. Mutant allele frequencies in domestic cat populations
of Greece Ц Biologia Gallo-Hellenica.— 1977.— Vol. 6.— P. 289—310.
Todd N. B., Lloyd A. T. Mutant allele frequencies in the cats of Dublin and
Vicinity // Carnivore Genet. Newslett.— 1979.— Vol. 4.— P. 24—30.
Todd N. B., Lloyd A. T. Mutant allele frequencies in domestic cats of Portu-
gal and the Azores Ц J. Hered.— 1984.— Vol. 75.— P. 495—497.
Todd N. B., Robinson R., Clark J. M. Gene frequencies in domestic cats of
Greece // J. Hered — 1974,— Vol. 65,— P. 227—231.
Todd N. B., Sawyer L., Todd L. M. Mutant allele frequencies in the cats of
Van, Turkey Ц Carnivore Genet. Newslett.— 1977.— Vol. 3.— P. 161—
167.
Todd N. B., Lloyd A. T., Todd A. C. Population genetics of the cats of Madeira
and Cabo Verde: the Portuguese Atlantic Empire Explored Ц J. Hered.—
1988,- Vol. 80.
Todd N. B., Todd L. M. Mutant allele frequencies among domestic cats in so-
me Eastern areas of Canada Ц J. Hered.— 1976a.— Vol. 67.— P. 368—
372.
Todd N. B., Todd L. M. Mutant allele frequencies in domestic cats of Turkey
and Greece Ц Genetica.— 1976b.— Vol. 46.— P. 183—192.
Twedt D. J. Influence of survey methods and sample sizes on estimated gene
frequencies in a domestic cat population Ц J. Hered.— 1983.— Vol. 74.—•
P. 121—123.
Varajcko J., Spicera J. Allele frequencies in a domestic cat population in a
country area of Northern Moravia Ц Genetica.— 1982.— Vol. 58.—
P. 137-140.
Vuilleumier B., Vuilleumier F. Mutant allele frequencies in the domestic cats
of New Caledonia: Proc. 1st Intern, conf, domestic cat popul. genet,
ecol. Ц Carnivore Genet. Newslett.— 1978.— Vol. 3.— P. 350—355.
Wagner A. Polimorfizm genetyczny barwy siersci w populacji koti domowego
г miejscwosci Bol na wyspie Brae, Jugoslavia / Zesz. Nauk UJ. Prace
Zoologiczne.— 1986.— Vol. 31.— P. 7—14.
Wagner A., Wolsan M. Pelage mutant allele frequencies in domestic cat po-
pulations of Poland Ц J. Hered.— 1987.— Vol. 78.— P. 197—200.
Watanabe M. A. Mutant allele frequencies in the domestic cats of San Pau-
lo, Brasil: Proc. 2nd Intern, conf, domestic cat popul. genet, ecol. Ц
Carnivore Genet. Newslett.— 1981,— Vol. 4,— P. 168—177.
149
Weir В. S., Cockerham C. G. Estimating E statistics in the analysis of popu-
lation structure / Evolution.— 1984.— Vol. 38.— P. 1358—1370.
Wright S. The interpretation of population structure by F-statistics with spe-
cial regard to systems of Mating // Evolution.— 1965.— Vol. 19.—
P. 395—420.
Г л а в a 12
ПОРОДЫ КОШЕК,
ИХ ПРОИСХОЖДЕНИЕ И СТАНДАРТЫ
ВВЕДЕНИЕ
Породы существуют у всех одомашненных видов животных.
Порода с биологической точки зрения — это достаточно боль-
шая для разведения без родственных скрещиваний группа живот-
ных одного вида, имеющих одинаковые биологические и морфоло-
гические признаки, которые передаются по наследству.
У разных видов домашних животных различия между поро-
дами могут быть незначительными (по качеству шерсти, неболь-
шим изменениям экстерьера), а могут и очень широко варьиро-
вать, как, например, у собак (от крошечной чихуа-хуа до массив-
ного сенбернара). Кошки в меньшей степени, чем собаки, подвер-
жены влиянию доместикационных изменений. Большинство из
них до сих пор сохраняют неизменными признаки, характерные
для их диких предков.
Каждая из существующих пород кошек имеет описательный
стандарт и балльную шкалу (по .100-балльной системе) оценок,
градированную в зависимости от приоритета отдельных экстерь-
ерных характеристик у каждой конкретной породы.
Существует несколько крупных международных организаций,
занимающихся разведением кошек. Каждая из них имеет своп
стандарты пород, но, как правило, они едины в своих требованиях
к экстерьеру.
В нашей стране разведением кошек начали заниматься только
с 1987 г., после создания первого любительского клуба «Фауна»
в Москве. Сегодня этот и ряд других клубов' различных городов
объединены в организацию, получившую название «Ассолюкс».
Эта крупная организация является с 1 января 1990 г. полноправ-
ным членом Международной федерации любителей кошек —
ФИФе (FIFe — Federation Internationale Feline). В данной главе
приведен перечень пород и различных цветовых вариаций, при-
знанных ФИФе, с указанием индексов, применяемых этой органи-
зацией для идентификации пород. „
Описание наиболее известных пород приводится в алфавитном
порядке. Не все из них сегодня признаны ФИФе, но познако-
миться с ними тем не менее интересно.
150
Стандарты окрасов у разных пород практически одинаковы.
Эти требования с указанием генотипов цветовых вариаций приве-
дены в гл. 13.
В этой главе использованы литературные данные из следую-
щих публикаций [Austin, 1980; Leigh, 1984; Muller, 1984; Muller-
Girard, 1988; Konarska-Szubska, 1989; Pintera, 1989; Непомнящий,
1991; Стандарты ФИФе; Регистрационный журнал ВО КЛК
«Ассолтокс»].
АБИССИНСКАЯ КОШКА
Происхождение точно не установлено. Предполагают, что эта
порода происходит от кошки по кличке Зула, которая была до-
ставлена в 1868 г. в Англию из Эфиопии. Она напоминала древ-
неегипетские изображения священных кошек. Вероятно, эта кош-
ка (или ей подобные) действительно была родоначальником по-
роды. Достоверно известно, что селекция велась с использованием
обыкновенных короткошерстных домашних кошек, сиамских и
бурманских. Селекционеры добивались отсутствия всякого рисун-
ка на теле животного, и их труды увенчались успехом. В 1929 г.
был утвержден стандарт, вполне соответствующий настоящему.
Кошки абиссинской породы, пожалуй, самые гармоничные из
всех домашних кошек и справедливо пользуются большой попу-
лярностью во всем мире.
Это средних размеров кошка с крепким, гибким и мускулис-
тым телом. Ноги длинные, стройные с узкими овальной формы
лапками. Голова среднего размера, слегка клиновидной формы,
с мягким овальным контуром, подбородок сильный. В профиль
выражен переход ото лба к носу плавной линией с небольшим
изгибом. Уши относительно большие, широкие у основания и ши-
роко расставленные. На кончиках ушей желательно наличие кис-
точек. Большие глаза имеют миндалевидную форму и интенсивно
окрашены в янтарный или зеленый цвет. Темный ободок вокруг
широко расставленных глаз придает им выразительность. Шерсть
короткая, плотно прилегающая к телу, густая, тонкая и мягкая
на ощупь. Хвост длинный.
К недостаткам экстерьера относят маленькие и заостренные
уши, сиамский тип, круглую голову, отсутствие ободка на глазах,
круглую форму глаз и их неопределенный смешанный цвет, рез-
кий переход ото лба к носу.
Первоначально, добившись «копии» нубийской кошки, сел-ек-
ционеры занимались особями дикого окраса. Еще в 80-х годах
прошлого столетия было сообщено о первой красной (соррель)
абиссинской кошке. Появлялись они и в более позднее время, но
тогда носители окраса соррель выбраковывались. Лишь в 1963 г.
красные абиссинские кошки получили признание в качестве вто-
рой цветовой вариации породы.
В 1982 г. были официально признаны голубой и бежевый
(цвета косули) абиссинские окрасы.
151
В 1983 г. к ним добавилось несколько вариаций окрасок сереб-
ристых абиссинских кошек. Появились эти разновидности в ре-
зультате скрещиваний с серебристыми британскими короткошер-
стными кошками.
В настоящее время ведутся работы по созданию других окра-
сов: шоколадного, лилового, красного (на основе мутаций, в отли-
чие от цвета соррель, который обусловлен аутосомными генами),
кремового, камео.
В нашу страну до настоящего времени привозились единич-
ные экземпляры абиссинских кошек, по их разведение до сих пор
не ведется.
Балльная шкала оценки
Тело, конечности, хвост 20
Голова, уши 15
Глаза 10
Окрас 25
Типинг 15
Текстура шерсти 10
Кондиция 5
АМЕРИ КАНСКАЯ ЖЕСТКОШЕРСТНАЯ
В американском штате Нью-Йорк близ г. Вернон в 1966 г.
в помете американской короткошерстной кошки появился коте-
нок, отличавшийся от своих братьев и сестер жестковатой волнис-
той шерстью. Он получил кличку Адам. Генетический анализ по-
казал, что он является носителем мутации Wh (Wire hair — про-
волочный мех) (см. гл. 5).
Для закрепления нового признака (красно-белый окрас) Ада-
ма скрестили с сестрой черепахового окраса. В 1967 г. родились
четверо котят, из них две кошки имели жесткую шерсть, выжила
только одна из них — Ами, которая за свою жизнь принесла мно-
го пометов с жесткошерстными котятами.
Селекционеры добились стабильности породы и ее признания
в 1977 г.
Размер животного от среднего до крупного. Тело коренастое,
сильное, мускулистое. Ноги средней длины, сильные, с круг-
лыми лапами. Овальная, пропорциональная телу голова с
развитыми скулами и сильным подбородком. Выражен небольшой
переход ото лба к носу. Уши средних размеров, широко расстав-
лены и имеют закругленные кончики. Большие глаза окрашены
в золотистый, зеленый, оранжевый или голубой цвет, в зависи-
мости от окраса шерсти. Хвост средней длины, сильный, закруг-
ленный на конце. Шерсть густая, жесткая, кудрявая, «проволоко-
образная». Тип шерстного покрова является основной характе-
ристикой этой породы.
В ФИФе порода не признана.
152
АМЕРИКАНСКАЯ КОРОТКОШЕРСТНАЯ
С первыми поселенцами в Северную Америку прибыли и кош-
ки. Разведение начали с британских короткошерстных кошек, ко-
торые скрещивались с акклиматизировавшимися в местных усло-
виях другими кошками. Кличка первой зарегистрированной в
Племенной книге щипки была Красавица Бредфорда. К концу
1950-х годов в Племенной книге числилось 50 кошек данной по-
роды, а в 1965 г. они получили признание под названием «амери-
канская короткошерстная порода». В улучшении типа использо-
вались персидские кошки, и это привело к формированию типа,
очень близкого к британскому.
Стандарт предусматривает кошек от среднего до крупного раз-
мера, с коренастым, сильным телом, хорошо развитой грудной
клеткой. Ноги средней длины (но длиннее, чем у британских),
сильные, с круглыми лапами. Хвост средней длины с закруглен-
ным кончиком. Голова крупная, как и у европейских, длина го-
ловы больше, чем ширина, но ненамного. Хорошо развиты щеки.
Заметен переход ото лба к носу. Уши средних размеров с закруг-
ленными кончиками. Глаза круглые и широко расставленные.
Цвет глаз зависит от окраса. Шерсть короткая, мягкая по тек-
стуре.
В ФИФе порода не признана.
АНГОРСКАЯ
Кошки этой породы были получены в Турции и уже несколь-
ко столетий тому назад элегантные животные вывозились в дру-
гие страны. Кстати, в литературе иногда упоминается, что в
прошлом наиболее достойными внимания для импорта из Турции
считались турецкие ковры и ангорские кошки.
Попав в Европу, ангорские кошки послужили основой для
создания персидской породы, а их прежний фенотип был утерян.
Лишь в 60-х годах XX в. американские любители кошек снова
заинтересовались ангорской и восстановили исходный фенотип.
В 1973 г. в Америке признали белую ангорскую кошку трех ва-
риаций: с голубыми глазами, с желтыми и с различно окрашенны-
ми (один глаз голубой, другой — желтый). К концу 70-х годов
были признаны ангорские с цветной шерстью.
В ФИФе ангорская порода принята совсем недавно (1987 г.)
и только классического белого цвета. В Европе ангорских кошек
пока мало. Вероятно, это связано с тем, что Турция очень неохот-
но расстается со своими племенными кошками. Приобретение,
а тем более вывоз из страны кошки ангорской породы с родослов-
ной сопряжены для иностранца с огромными трудностями и прак-
тически невозможны, особенно это касается производителей.
В нашей стране встречаются кошки, отвечающие всем требова-
ниям стандарта ангорской породы. Они включены в эксперимен-
11 Заказ Ns 355
153
талыюе разведение. Некоторые из них на международных выстав-
ках получали высокие оценки. Титул «чемпион» впервые был
присвоен в 1990 г. кошке Гайдэ.
Требования стандарта следующие: среднего размера удлинен-
ное тело на высоких тонких ногах. Лапы маленькие, округлые.
Задние ноги немного выше передних. Голова несколько удлинен-
ная, клиновидной формы. Нос средней длины. Легкий изгиб ли-
нии профиля. Большие и широкие уши высоко поставлены на го-
лове, кончики ушей заостренные. Глаза большие, миндалевидной
формы и слегка раскосые (не путать с косоглазием!). Хвост длин-
ный, обильно поросший шерстью. У ангор слабо развит подшер-
сток, а остевая шерсть тонкая, шелковистая и как бы стекающая
по телу.
В некоторых описаниях отмечается, что при беге ангорские
кошки закидывают хвост на спину, подобно белкам.
БАЛИНЕЗИЙСКАЯ
Появились балинезийские кошки в результате спонтанной
мутации в локусе 1 у сиамских кошек. Первоначально котята
с удлиненной шерстью выбраковывались как несоответствующие
стандарту. В 1950-х годах некоторые владельцы начали целе-
направленно разводить таких кошек и в 1961 г. впервые про-
демонстрировали их на выставке в Нью-Йорке под названием
«длинношерстные сиамские». Позже, в 1963 г., новая порода
получила в США признание и стала именоваться балинезийской.
Экстерьер балинезийских кошек полностью идентичен внеш-
нему облику сиамских кошек. Они имеют тонкий костяк, сильно
вытянутое тело на высоких ногах, причем задние ноги должны
быть немного длиннее передних. Лапы маленькие, овальные.
Хвост длинный и тонкий, заостренный на конце. Шея длинная
и стройная. Голова также вытянутая, клиновидной формы. Нос
длинный, прямой, продолжает линию лба. Подбородок сильный.
Глаза средней величины, миндалевидной формы, раскосые. Цвет
глаз должен быть интенсивным голубым. Уши большие, широко
расставленные, с заостренными кончиками. Как и у сиамских,
анфас должен иметь форму равностороннего треугольника, одна
из граней которого соединяет кончики ушей, а две другие про-
ходят по сторонам морды и включают в себя наружные края
ушей. Шерсть средней длины, шелковистая. Подшерсток очень
слабо развит.
В 1970 г. в Америке были зарегистрированы дополнительные
окрасы: с голубыми, шоколадными и лиловыми отметинами, до-
бавившиеся к уже утвержденному сил-пойнту.
В ФИФе породу признали в 1983 г. со всеми окрасами, за-
регистрированными у сиамских кошек.
В нашей стране среди обычных домашних кошек с неизве-
стным происхождением выявляются обладатели стандартного
экстерьера, которые успешно конкурируют с зарубежными пред-
154
ставителями на международных выставках. С 1990 г. появились
и первые чемпионы: с темно-коричневыми отметинами кот Эве-
рест-Эсти и с красными отметинами кот Бадрис Кристалл.
БИРМАНСКАЯ
Порода была выведена во Франции целенаправленным скре-
щиванием сиамских и персидских кошек. В 1930 г. появился
первый французский стандарт породы. Кошки уже успели за-
воевать во Франции популярность, когда вторая мировая война
привела к гибели почти всех племенных животных. К счастью,
сохранились кот Орлофф и кошка Ксения из питомника де Ка-
аба. Они и составили племенную основу современных кошек
бирманской породы.
Презентация породы в Англии состоялась в 1960 г. ФИФе
признала бирманскую породу в 1964 г. в двух цветовых вариа-
циях: сил-пойнт и блю-пойнт. В США стандарт утвердили поз-
же, чем в Европе,— в 1967 г.
В настоящее время разводятся бирманские кошки несколь-
ких окрасов, как и персидские колор-пойнты.
Требования стандарта: слегка вытянутое туловище с отно-
сительно короткими и сильными ногами. Лапы круглые. Хвост
средней длины, обильно опушенный. Голова широкая, мощная,
круглая. Нос короткий и не должен быть прямым (как у си-
амских), не должен иметь выраженную «седелку» (как у пер-
сов). Хорошо развиты щеки и сильный подбородок. Уши ма-
ленькие, широко расставленные. Глаза круглые или слегка
овальные, темно-голубого цвета. Шерсть средней длины, густая,
шелковистая.
Балльная шкала оценки
Тело 20
Г'олова п глаза 20
Окрас 15
Симметричность «носочков» на перед-
них ногах 5
На задних ногах 5
Равномерность окраса «носочков» 5
Форма задних «носочков» 5
Хвост 10
Шерсть 10
Кондиция 5
БИРМАНСКАЯ КОРОТКОШЕРСТНАЯ (БУРМАНСКАЯ) *
Первая коричневая кошка, похожая на очень темную сиам-
скую, была привезена в Америку из Бирмы доктором Дж. Томп-
соном в 1930 г. Кошка имела кличку Вонг Мау. В 1932 г. она
была показана на выставке в Сан-Франциско.
* При переводе названий пород на русский язык возникают проблемы
правильной транскрипции. Слово «Бирма» в английском языке имеет два
написания: Birma и Burma. Во избежание путаницы мы предпочли транс-
крипцию переводу и обозначили Birman breed как бирманскую, a Burmese
breed как бурманскую породы.
11* 155
Генетики предположили, что окрас у Вонг Мау вызван ал-
лелем альбинизма, отличным от сиамского. На первых этапах
бурманскпх кошек скрещивали с сиамскими производителями.
Селекционная работа п исследования дали желаемые результа-
ты. В 1936 г. коричневая бурманская порода получила офици-
альное признание.
В 1950 г. порода признана ФИФе.
Постепенно бурманская короткошерстная порода пополнилась
новыми окрасами и сегодня разводится в нескольких цветовых
вариациях.
Тело у бурманских кошек средней длины, мускулистое,
с сильной грудной клеткой, овальной в профиль. Линия спины
прямая. Ноги относительно тонкие, стройные, с изящными ла-
пами овальной формы. Средней длины прямой хвост не должен
быть толстым у основания. Кончик хвоста заострен. Голова кли-
новидной формы, вверху слегка овальная. Скулы широкие. Че-
люсти п подбородок массивные. Средних размеров уши широко
расставлены, копчики с легким закруглением. В профиль уши
немного наклонены вперед. Отчетливо выражен переход ото лба
к носу. Глаза большие, широко и слегка раскосо посаженные,
причем верхняя линия глаз заметно наклонена, а нижняя —
овальная. Цвет глаз может варьировать от светло-желтого до
янтарного, но предпочтение отдается золотисто-желтому. Бле-
стящая, тонкая, бархатистая, короткая шерсть плотно прилегает
к телу. К недостаткам относят сиамский, а также британский
или европейский типы, круглые глаза, их зеленый цвет и впалые
щеки.
Балльная шкала оценки
Тело (включая ноги и хвост)
Голова, уши
Глаза
Текстура шерсти
Окрас
Кондиция
20
25
20
15
15
5
БОМБЕЙСКАЯ
Кошки этой породы возникли путем скрещивания коричне-
вой бурманской и американской черного окраса короткошерстных
кошек в конце 1950-х годов.
В США стандарт утвержден в 1976 г. и до настоящего вре-
мени бомбейские кошки признаны только в Америке.
По требованиям стандарта тело у кошек среднего размера,
мускулистое, не должно быть ни приземистым, ни вытянутым.
Длина ног и хвоста пропорциональна туловищу. Голова округ-
лая, с хорошо развитыми щеками и сильным подбородком. Уши
156
средних размеров, широко расставленные, с широким основани-
ем и слегка направленные вперед. Нос широкий. Сильно выра-
жен переход ото лба к носу. Глаза круглые, широко расставлен-
ные, желтого или темно-медного цвета. Шерсть ярко-черная,
гладкая, короткая, шелковистой текстуры, плотно прилегающая
к телу.
БРИТАНСКАЯ КОРОТКОШЕРСТНАЯ
В Англии под названием британской породы начали разво-
дить обыкновенных короткошерстных кошек и долгое время они
были идентичны европейским короткошерстным. С 1980 г. их
начали выставлять в Англии.
Для улучшения типа британской породы стали использовать
завезенных в Англию персидских кошек. Это придало британ-
ским кошкам округлость головы и крепость костяка, у них уко-
ротился нос. Но наряду с положительным эффектом улучшения
типа проявляется и недостаток такого вводного скрещивания:
в пометах британских кошек иногда выщепляются пушистые
потомки.
Официальное признание порода получила только в 1984 г.
Представители породы имеют размеры от средних до достаточ-
но крупных. Тело приземистое, мускулистое, несколько массив-
ное, с мощной грудной клеткой. Ноги относительно короткие,
сильные. Лапы крупные, круглой формы. Хвост короткий и тол-
стый, закругленный на конце. Череп массивный, широкий и
круглый. Шея короткая, сильная. Глаза большие, круглые, ши-
роко расставленные. Цвет глаз зависит от окраса: у белых бри-
танских кошек радужка голубая, оранжевая или на одном гла-
зу — голубая, на другом — оранжевая; у серебристых — зеленая,
но допускаются желтовато-зеленая и даже желтая; у шиншилл
и затушеванных шиншилл — зеленая или зеленовато-голубая;
у остальных окрасов — от темно-оранжевого до медного. Пред-
почтение отдается наиболее интенсивному п чистому цвету глаз.
Уши у британских кошек короткие, широко расставленные.
Шерсть короткая, густая, очень мягкая, неприлегающая, с хоро-
шо развитым подшерстком, который почти равен по длине осте-
вой шерсти.
В нашей стране чистопородные британские животные соста-
вили основу для селекции породы. Первые их представители
были импортированы из Чехо-Словакии и Германии. Первым
чемпионом стал в 1991 г. черный британский кот чехо-словац-
кого разведения.
Балльная шкала оценки
Телосложение, ноги и хвост 20
Г олова (включая уши и глаза) 30
Цвет глаз 10
Окрас 25
Текстура шерсти 10
Кондиция 5
157
ВОСТОЧНАЯ КОРОТКОШЕРСТНАЯ (ОРИЕНТАЛЬНАЯ)
Перед второй мировой войной Ф. Швангарт, известный не-
мецкий зоолог из Дрездена, занимался разведением одноцвет-
ных кошек сиамского типа.
Самое раннее упоминание о них относится к 1888 г., когда
была описана одноцветная шоколадная сиамская кошка. А в
1894 г. в Англии на выставке демонстрировалась коричневая
кошка под названием «швейцарская горная».
В 50-х годах нашего столетия выведением ориентальных ко-
шек занимались как в Англии, так и в Европе. В разведении
использовали сиамские и европейские породы кошек. Постепен-
но добились соответствия экстерьера восточных короткошерстных
и сиамских кошек, за исключением полной пигментации тела
и зеленого (кроме кошек белого окраса) цвета глаз.
В 1972 г. в ФИФе признали ориентальных кошек новой по-
родой. А некоторые окрасы получили специфические названия.
Так, восточные короткошерстные коричневые кошки были на-
званы «Гавана». У этих кошек окрас шерсти насыщенного ко-
ричневого цвета, что вызывает ассоциацию с цветом гаванских
сигар. Черные, как смоль, ориентальные получили название
«эбони», что означает «черное дерево». За белыми кошками за-
крепилось название «иностранная белая» (хотя в Англии до сих
пор «иностранными» именуются все одноцветные вариации ори-
ентальных кошек).
Десять лет спустя были признаны дополнительные окрасы:
варианты черепаховых и тэбби.
Экстерьер и оценочная шкала аналогичны сиамской породе.
Когда восточных короткошерстных кошек начали разводить
в чистоте, без прилития сиамских кровей, было обнаружено, что
интенсивный зеленый цвет глаз легче получить у гетерозиготных
СС8 ориенталов, в то время как гомозиготные особи СС часто
имеют глаза нестандартного цвета.
Следует отметить также, что процесс изменения ювенильного
голубого цвета глаз в зеленый может продолжаться до двух-
летнего возраста.
Племенная работа с ориентальными кошками базируется на
импортных кошках. В последнее время их количество значи-
тельно увеличилось благодаря усилиям наших фелинологов.
В 1990 г. появились первые чемпионы: лиловая черепаховая
кошка и коты эбони и гавана.
ДЕВОН РЕКС
В английском графстве Девоншир в 1960 г. родился голубой
кудрявый котенок, сходный по типу шерсти с носителями му-
тантов кошек, родоначальников корниш реке. Кота назвали Кир-
158
ли и неоднократно скрещивали с самкой породы корниш реке.
Появлявшиеся от таких спариваний котята имели нормальную
шерсть. Таким образом стало известно, что одинаковый тип шер-
сти был сформирован благодаря действию совершенно различ-
ных генов, не аллельных друг другу.
В 1959 г., основываясь на экстерьерных данных кота Кирлп,
был разработан стандарт породы, получившей название девон
реке. Порода признана в 1967 г. При. селекции был поставлен
запрет на скрещивание с породой корниш реке.
В США породу признали в 1979 г.
Тело девон реке среднего размера, мускулистое. Horn очень
высокие и тонкие. Лапы маленькие, овальные. Относительно
широкая грудная клетка и длинная, тонкая шея. Хвост тонкий,
заостренный. Клиновидная голова короткая, с сильно развитыми
скулами и щеками. Хорошо выражен переход ото лба к носу.
Череп имеет плоскую форму. Большие миндалевидные глаза
широко расставлены. Цвет глаз должен гармонировать с основ-
ным окрасом, но при этом быть чистым, равномерным. Уши у
девон рексов очень большие и широкие, с закругленными кон-
чиками и складками кожи на наружной стороне. Шерсть очень
короткая, тонкая, волнистая и мягкая благодаря отсутствию
остевых волосков. Вибриссы загибаются. Окрасы для этой поро-
ды не ограничены. Очень оригинально выглядят девопы с си-
амскими отметинами.
Иногда у животных наблюдаются участки тела без шерсти.
Если у молодняка это расценивается как недостаток, то у взрос-
лых служит поводом для дисквалификации.
Балльная шкала оценки
Тело (включая ноги, хвост и
шею} 20
Голова и глаза 20
Уши 10
Шерсть 40
Вибриссы 5
Кондиция 5
ДОМАШНЯЯ КОШКА
Это видовое название закрепилось за простыми кошками, не
претендующими на какую-либо породу.
С 1982 г. на выставках, организуемых ФИФе, домашние
кошки не только участвуют в показе, но и проходят оценочный
конкурс в самостоятельном классе домашних кошек. Критерия-
ми оценки при этом служат: пропорциональность и гармонич-
ность телосложения, отсутствие физических недостатков, чистота
окраса, ухоженность животного и, что очень важно, спокойный,
добрый, непугливый характер.
159
ЕВРОПЕЙСКАЯ КОРОТКОШЕРСТНАЯ
Основу породы п ее стандарта составили обыкновенные ко-
роткошерстные домашние кошки. Селекция была направлена на
стабилизацию типа и укрепление костяка. Европейские корот-
кошерстные кошки с 1982 г. разводятся по следующему стан-
дарту: тело от среднего до крупного размера, плотное, мускули-
стое. Грудная клетка хорошо развита. Сильные, средней длины
ноги, равномерно сужаясь, переходят в круглые лапы. Хвост
средней длины, широкий у основания, кончик хвоста закруглен-
ный. Голова относительно крупная. Длина головы несколько
больше, чем ее ширина. Хорошо развитые щеки и сильный под-
бородок. Голова слегка округлой формы. Нос средней длины,
выражен переход ото лба к носу. Уши средних размеров с за-
кругленными кончиками, прямо поставленные. Глаза большие,
широко расставленные. Признанные окрасы и цвет глаз абсо-
лютно идентичны таковым у британских короткошерстных
кошек.
В нашей стране начальное поголовье для разведения соста-
вили домашние кошки, экстерьером и окрасом отвечавшие тре-
бованиям стандарта. Титул «чемпион» первыми получили в
1991 г. белый с различно окрашенными глазами кот по кличке
Белый и красный тигровый кот Томас.
Балльная шкала оценки
Тип 25
Голова (включая уши и гла-
за) 25
Цвет глаз 10
Качество шерстя 10
Окрас 25
Кондиция 5
ЕГИПЕТСКИЙ МАУ
Об их происхождении нет достоверных сведений. Некоторые
авторы утверждают, что пара кошек (Гера и Лудол) была им-
портирована в 1953 г. в США из Египта и уже тогда кошки
имели черты породы, названной впоследствии «египетский мау».
Другие считают, что появление египетских мау — результат
скрещиваний сиамских кошек тэбби-пойнт с домашними или
бурманскими кошками. Селекционеры изначально поставили пе-
ред собой цель добиться восстановления породы, существовав-
шей в Египте задолго до распространения кошек в Европе.
В 1968 г. порода была признана в США.
Стандарт предусматривает средней длины тело с пропор-
циональными ему конечностями. Задние ноги немного длиннее
передних; создается впечатление, что кошка стоит как бы на
кончиках пальцев передних ног. Голова имеет слегка округлую
клиновидную форму. Линия носа прямая. Уши относительно
160
большие, с заостренными концами, широким основанием и ши-
роко расставленные. Глаза слегка миндалевидной формы. До
18-месячпого возраста допускается янтарный оттенок в цвете
глаз, но в более позднем возрасте радужная оболочка должна
принять светло-зеленый цвет. Хвост средней длины, толстый у
основания и заостренный на конце. Шерсть шелковистая. Осо-
бое внимание уделяется рисунку на теле: пятна могут быть раз-
личного размера, но ни в коем случае не допускается слияние
их в полосы.
В ФИФе египетские мау не признаны, но любители породы
есть и в Европе.
В литературе отмечается биологическая особенность египет-
ских мау — беременность в норме длится от 63 до 73 дней.
КАРТЕЗИАНСКАЯ (ШАРТРЕЗ)
Корин происхождения у шартрезов уходят в глубину веков.
Более 500 лет тому назад они были привезены из Африки во
Францию, в картезианский монастырь «La Grande Chartreuse».
Эту же породу завезли в Англию, где использовали при соз-
дании британской короткошерстной породы. Затем был долгий
период смешивания двух пород между собой и с персидскими:
кошками. Разделили породы в 1966 г., утвердив самостоятель-
ные стандарты. Все же картезианские и британские кошки на-
столько похожи, что даже опытнейший специалист затруднится
определить породу по внешнему виду.
Картезианские кошки имеют средние размеры, тело мускули-
стое, скорее даже массивное. Ноги относительно короткие, силь-
ные, заканчиваются круглыми, мощными лапами. Хвост средней
длины с закругленным кончиком. Голова широкая, формы об-
ратной (перевернутой) трапеции. Хорошо развиты щеки. Нос
широкий, прямой. Уши средних размеров, не должны быть по-
ставлены прямо и высоко на круглом черепе. Глаза большие,
круглые, окрашены в темно-оранжевый или медный цвет. Шерсть
густая, мягкая, шелковистая, не прилегающая к телу, подшер-
сток равен по длине покровной шерсти.
Балльная шкала оценки
Тело и ноги 25
Голова 25
Глаза 10
Текстура шерсти и окрас 25
Кондиция 5
КИМРИК
В 60-х годах в США в породе мэнских бесхвостых кошек
возникла мутация 1 (long hair)—длинная шерсть. В результате
планомерной работы с носителем этой мутации селекционеры
161
получили новую породу под названием «кимрик». Полное соот-
ветствие экстерьера мэнским кошкам, такой же способ передви-
жения (скачками, по-заячьи), но удлиненная шерсть, формиру-
ющая густой воротник и пушистые «штаны»,— вот облик этих
кошек. Название породе дано в честь кельтских племен — древ-
него населения п-ова Уэльс (Великобритания)— киммерийцев.
Окрас допускается любой. Цвет глаз должен соответствовать
окрасу шерсти.
КОРАТ
Родиной коратов является одноименная провинция (Корат)
Таиланда. Эти кошки играли большую роль в обычаях и куль-
туре страны. До сих пор в Таиланде бытует поверье, что эле-
гантные кораты приносят счастье. Об этом свидетельствует вто-
рое (местное) название породы—«си-сават» (si-sawat — счастье),
сават — это и название тропического плода, цвет которого на-
поминает цвет шерсти кората.
В 1959 г. первая пара коратов была привезена в Америку.
Стандарт утвердили в 1965 г.
В Англии с представителем породы корат познакомились
на выставке значительно раньше — в 1896 г., но дальнейшая
его судьба неизвестна.
В ФИФе коратов признали только в 1982 г.
Кошки породы корат средней величины, сильные, муску-
листые. Ноги умеренной длины, пропорциональные телу,
с овальными лапами. Хвост средней длины, толстый у основа-
ния, с закругленным кончиком. Спина слегка вытянутая. Если
на голову кошки смотреть спереди или сверху, она имеет серд-
цевидную форму. Верхнюю часть «сердца» образуют сильные
надбровные дуги и широкий лоб. Нос средней длины. Мочка
носа резко, как у льва, идет вниз и окрашена в темно-голубой
или лавандовый цвет. Недостатком является как слишком длин-
ный, так и слишком короткий нос. Переход ото лба к носу дол-
жен быть резко выражен. Полные щеки и сильный подбородок
также вносят вклад в создание общей формы «сердца». Уши
большие, широкие у основания, с закруглёнными кончиками,
высоко посаженные. Глаза большие, круглые, широко расстав-
ленные. Стандарт предусматривает зеленый цвет глаз, но допу-
скается янтарный и янтарно-зеленый, особенно до двух лет,
когда радужная оболочка приобретает окончательный цвет.
Шерсть густая, прилегающая, тонкая по текстуре. Окрас сереб-
ристо-голубой, кончики волос серебристые. Предпочтение имеет
как можно более интенсивная серебристость, в образовании лег-
кого серебристого тикинга у коратов ген I (ингибитор мелани-
на) участия не принимает. Подушечки лап темно-голубого или
лилового цвета, допускается розовый оттенок.
162
Балльная шкала оценки
Телосложение и хвост 25
Голова п уши 20
Форма и расположение глаз 15
Цвет глаз 5
Окрас 20
Текстура шерсти 10
Кондиция 5
КОРНИШ РЕКС
Рексовая мутация возникла в 1950 г. в помете кошки на
п-ове Корнуэлл (Англия). Один из котов этого помета имел
волнистую шерсть кремового цвета. Тип шерсти вызвал у хо-
зяйки ассоциацию с мехом кроликов породы реке и это назва-
ние перешло к созданной в дальнейшем породе кошек. С мо-
мента появления первых мутантов и до утверждения породы
корншп реке (1967 г.) прошло 17 лет упорной селекционной
работы. Использовали и тесный инбридинг, и скрещивания с
кошками различных пород. Включали в разведение обнаружен-
ных вновь рексовых мутантов и проводили возвратные скрещи-
вания.. В результате были получены две породы: корниш реке
и девон реке (см. выше).
В ФИФе порода корниш реке была признана в 1968 г.
Представители этой породы имеют средние размеры, тело
стройное, худое, мускулистое. Ноги длинные, тонкие, с малень-
кими овальными лапками. Кошки производят впечатление высо-
коногих животных. Голова клиновидной формы, длина головы
примерно на 1/3 больше ее максимальной ширины. Подбородок
сильный. В профиль плоский череп продолжается прямой лини-
ей от середины лба до кончика носа. Уши большие, широкие
у основания, высоко поставленные, с закругленными кончиками,
покрытые тонким волосяным покровом. Глаза средней величи-
ны, миндалевидной формы, их цвет должен гармонировать с
окрасом и сохранять чистоту и ясность. Характерным призна-
ком, как и у всех рексов, является короткая волнистая шерсть,
лишенная остевых волосков. Вибриссы должны завиваться по
всей длине. Хвост тонкий, длинный, покрыт плотной волнистой
шерстью.
Балльная шкала Оценки
Телосложение и ноги 25
Голова и уши 20
Форма и цвет глаз 5
Шерсть 35
Хвост и его шерстный покров 5
Вибриссы 5
Кондиция 5
МЭЙН-КУН
Дословный перевод названия породы означает «майская ено-
товая». Считают, что название дано из-за схожести окраски голо-
вы у мэйн-кунов и енотов или из-за богато опушенного хвоста,
163
подобного хвосту енота. Название прижилось не только в амери-
канском штате Мэн (Maine — мэн — принятая, хотя и неверная
транскрипция), но и во всем мире.
О происхождении этой породы бытует много легенд. Одна из
них рассказывает, что в Новый Свет предки этих животных были
привезены викингами. И действительно, между породами мэйн-кун
и норвежской лесной очень много общего.
Суровые условия Северной Америки оказали влияние на ко-
шек. ведущих вольный образ жизни. Появились очень крупные
(массой до 15 кг) и выносливые животные, покрытые густой
шерстью с водоотталкивающим свойством.
Впервые на всеобщее обозрение мэйн-куны были представ-
лены в 1860 г., задолго до кошачьих выставок, на сельскохозяй-
ственном празднике. Но при всей любви американцев к этим
кошкам, как порода они не представляли интереса до 50-х годов
нашего века, лишь с 1953 г. началось планомерное разведение
этих животных. Окончательно в США породу мэйн-кун признали
в 1976 г., хотя стандарт был принят большинством американских
клубов в 1967 г.
В Европе первая демонстрация этих кошек состоялась в 70-х
годах на выставке в ФРГ. Огромные размеры мэйн-куна произвели
сильное впечатление на любителей кошек, и европейцы долгое
время считали, что было представлено нетипичное для породы
животное. В 1982 г. мэйн-куны признаны в ФИФе, и теперь
общеизвестно, что «гиганты»— вовсе не редкость для этой породы.
Мэйн-куны характеризуются размерами от среднего до круп-
ного, крепким, мускулистым телосложением. Грудь сильная и
широкая. Шея относительно длинная, особенно у кошек. Тело
удлиненное, высокие ноги пропорциональны туловищу. Большие
круглые лапы имеют длинные (более 2 см) и густые пучки
шерсти между пальцев. Хвост длинный, широкий у основания,
с очень длинной остевой шерстью, сохраняющейся и в летнее
время. Голова средняя по ширине, у котов с возрастом череп
становится шире. Нос средней длины, прямой. Подбородок силь-
ный, в профиль как бы соединен одной линией с мочкой носа
и верхней губой. Благодаря развитым подушечкам вибрисс морда
не выглядит острой. Уши большие, широкие у основания, высоко
и прямо поставленные. Кончики ушей заостренные, предпочти-
тельно с кисточками. Глаза большие, круглые, слегка раскосо
посаженные. Допускается любой цвет глаз. Шерсть средней дли-
ны, короче на голове и плечах. Покровная шерсть слегка жирная,
что предохраняет кошек от намокания. По текстуре шерсть до-
статочно мягкая, шелковистая. Окрас допускается любой, но при
индексации подразделяется на 4 группы: агути, агути с белым,
неагути, неагути с белым.
Балльная шкала оценки
Тело 35
Голова 30
Текстура шерсти 20
Окрас 10
Кондиция 5
464
МЭНСКАЯ БЕСХВОСТАЯ (МЭНКС)
Популяция бесхвостых кошек была обнаружена на о-ве Мэн,
расположенном в Ирландском море. Описание гена мэнской бес-
хвостости дано в гл. 6. Гомозиготные особи погибают в эмбрио-
нальный период. В гетерозиготном состоянии ген М такще влияет
на жизнеспособность кошек. Кроме отсутствия или укорочения
хвоста достаточно часты сращения позвонков, деформация костей
таза и крестца, а как следствие — затруднения уринации и дефе-
кации; в результате нарушения развития спинного мозга возмо-
жен паралич задних конечностей. Это должно бы заставить
любителей кошек задуматься о гуманности и целесообразности
разведения животных с физическими дефектами. Тем не менее
порода существует и признана в крупнейших фелинологических
организациях. Основным аргументом в пользу культивирования
мэнских бесхвостых кошек является их достаточно стабильное
существование на острове в течение нескольких столетий без
видимого вреда для популяции.
Стандарт мэнксов: кошки средней величины с короткой спи-
ной и широкой грудной клеткой. Бока несколько впалые. Задние
конечности длиннее передних. Таз должен быть поднят высоко,
короткая спина и глубокий прогиб в спине. Особенностью по-
роды является передвижение кошек скачками, подобно кролику.
На конце позвоночника, где у нормальных кошек других пород
начинается хвост, у мэнских — ямка, куда, согласно стандарту,
должны помещаться два пальца. Голова крупная. Щеки полные.
Средней длины нос соединен со лбом по изогнутой линии, но без
седелки. Уши средних размеров, с заостренными кончиками.
Шерсть короткая, тонкая, с плотным подшерстком. Окрас допус-
кается любой. Большие круглые глаза должны иметь цвет, соот-
ветствующий окрасу, аналогично британским короткошерстным
кошкам.
Балльная шкала оценки
Тело 25
Голова и уши 15
Спина 10
Отсутствие хвоста 25
Шерсть 15
Глаза 5
Кондиция 5
НЕМЕЦКИЙ РЕКС
Родоначальница немецких рексов — кошка но кличке Лэм-
хен — была обнаружена в клинике Бух в Берлине в 1951 г.
Сначала скрещивания Лэмхен велись с обыкновенными котами,
затем с ее собственными сыновьями. Позже получали потомков
от полусибсов, а в дальнейшем применялись вводные и возврат-
ные скрещивания с кошками пород девон реке и корниш реке.
165
На международной выставке в Праге в 1970 г. впервые была
показана кошка породы немецкий реке по кличке Жаннет фон
Грунд.
ФИФе признала новую породу в 1982 г.
Размеры этих кошек средние. Тело сильное, мускулистое, но
не массивное. Округлая в профиль грудь сильно развита. Спина
прямая. Ноги относительно тонкие, средней длины. Лапы имеют
овальную форму. Хвост средней длины, сильный у основания,
сужающийся к концу, равномерно покрытый шерстью. Голова
округлая, с сильным подбородком и хорошо развитыми щеками.
Уши большие, широко расставленные, кончики их слегка закруг-
лены. Переход ото лба к носу отмечен легким изгибом. Глаза
средней величины, открытые, их цвет должен гармонировать
с окрасом. Остевые волоски отсутствуют в шерстном покрове,
благодаря чему шерсть очень мягкая, «плюшевая», напоминаю-
щая шкурку крота. Шерсть слегка волнистая. Котята рождаются
с волнистой шерстью, затем волоски распрямляются и в период
смены зубов шерсть снова становится волнистой. Иногда жи-
вотные вырастают с прямой шерстью, но они не исключаются
из разведения при условии соответствия всем остальным тре-
бованиям стандарта. Правильный выбор партнеров в разведении
позволяет получить от них хорошее потомство. Окрас допускается
любой.
Балльная шкала оценки
Тело 15
Голова 10
Уши 5
Глаза 10
Хвост 5
Шерсть 50
Кондиция 5
НОРВЕЖСКАЯ ЛЕСНАЯ
Существуют две основные версии появления кошек с удлинен-
ной шерстью на территории Норвегии. Одна из них предполагает
доставку в XVI в. из Италии ангорских кошек, которые затем
скрещивались с местными, и теплоизоляционные свойства шерсти
ангорских, привнесенные после гибридизации, оказались полез-
ными для выведения породы. По другой — мутация long hair (1)
могла возникнуть спонтанно и сыграть свою роль в естественном
отборе.
В скандинавских мифах и сказках упоминаются длинношерст-
ные кошки. А богиня Фрея часто изображалась в колеснице,
запряженной кошками, очень похожими на современных норвеж-
ских лесных.
Полувольные и одичавшие пушистые и грациозные кошки
встречаются в лесных окрестностях норвежских ферм. Первым
166
описание этих животных сделал в 1599 г. священник-натуралист
К. Фринс, правда, отнеся их к самостоятельному виду дикой
кошки.
Программа разведения породы была составлена в 1930 г.,
и вскоре представители норвежских лесных были высоко оце-
нены на выставке в г. Осло. Вторая мировая война нарушила
планомерность разведения. Лишь в начале 70-х годов было отоб-
рано около 200 особей, которые и составили основу генофонда
породы. На родине, в Норвегии, порода признана в 1972 г.
В ФИФе норвежские лесные заняли полноправное место среди
признанных пород кошек с 1976 г.
Это грациозные кошки с сильным, длинным телом на высоких
ногах. Голова треугольной формы, неширокая. Подбородок силь-
ный. Нос относительно длинный и прямой. Глаза большие, слегка
раскосо посаженные. Цвет допускается любой. Уши большие, вы-
соко поставленные. Заостренные кончики ушей должны иметь
кисточки, как у рыси. Длинный и пушистый хвост дополняет
вытянутые формы животного. Покровная шерсть обладает водоот-
талкивающими свойствами благодаря длинным, слегка масля-
нистым остевым волоскам. Подшерсток густой, в зимнее время
формируется длинный воротник на шее. Летняя шерсть значи-
тельно короче зимней, обычно сохраняются пушистыми лишь
«штаны» и хвост. Окрас допускается любой, кроме сиамского;
на выставках выделяются 4 группы: агути, агути с белым, не-
агути, неагути с белым.
У котят ость вырастает в возрасте от 3 до 5 мес. Качество
шерсти — один из основных критериев оценки норвежских лес-
ных кошек.
Балльйая шкала оценки
Тело 25
Голова 20
Глаза 5
Уши 10
Хвост 10
Шерсть 25
Кондиция 5
оцикэт
Американские селекционеры скрещиванием сиамских и абис-
синских кошек добивались получения пойнтового окраса с отме-
тинами абиссинского тэбби. В одном из пометов появился коте-
нок с шерстью цвета слоновой кости, испещренной пятнышками
цвета корицы. Котенок был назван Тонга. Так как он своим
окрасом очень напоминал котенка оцелота, комбинация двух
слов — оцелот и кэт( cat—кошка), образовала название породы.
При создании пород в скрещивания вовлечены американские
короткошерстные кошки.
Оцикэты достаточно крупные (масса 5—7 кг), что генетики
объясняют стойким эффектом гетерозиса.
167
В Америке признали породу в 1966 г., но в Европе оцикэты
до сих пор не зарегистрированы в качестве официально признан-
ной породы из-за большой схожести с египетскими мау.
ПЕРСИДСКАЯ
Происходят персидские кошки от ангорских. Первые были
привезены в Европу итальянским путешественником Пиетро
делла Валле в 1526 г. из персидской провинции Хорассаи. Эти
животные были далеки от признанного в наше время экстерьер-
ного типа, но пушистая шерсть привлекала к ним особое вни-
мание.
Направленное разведение началось в Англии в 70-х годах
прошлого столетия, и в течение почти 30 лет порода включала
в себя только два окраса: белый и голубой. Эти классические
цвета до сих пор самые распространенные у персидских кошек.
В результате тщательной селекции голубые персидские сегодня
имеют наиболее выраженный тип породы и используются для
улучшения экстерьера других окрасов.
Первые белые персидские кошки, как и их ангорские пред-
ки. были голубоглазыми и глухими. Среди них и сегодня до
40 % — глухие животные. Кроме того, белые голубоглазые кошки
часто имеют более слабое телосложение.
Скрещиванием белых кошек с голубыми и черными персид-
скими получили белых с оранжевыми и с различно окрашенными
глазами. Признали их в 1938 г.
Одна из старейших цветовых вариаций — черный дымчатый
окрас. Впервые описан в 1860 г., а первый чемпион был заре-
гистрирован в 1890 г.
В 1888 г. на выставке в Хрустальном Дворце в Лондоне
произвел сенсацию кот по кличке Сильвер Ламбкин. Он произо-
шел от скрещивания животных черного серебристого тэбби и чер-
ного дымчатого окрасов и оказался первой в истории персидской
шиншиллой. Правда, его шерсть была более темной, чем у совре-
менных шиншилл, да и на лапах отчетливо проступали полосы.
Тем не менее после 17 лет жизни кота его чучело было помещено
в Национальный музей естествознания.
С 1880 г. под названием «оранжевая персидская» были извест-
ны красные и красные мраморные окрасы, которые считали одной
цветовой вариацией. В 1890 г. появились кремовые персидские
кошки. В этом же году были впервые представлены и черепаховые
персидские. Голубо-кремовые окрасы стали признавать значи-
тельно позднее — около 1930 г.
В 1934 г. были получены кошки окраса камео. Их системати-
ческое разведение началось в Америке с 1950 г. Признаны
в 1954 г., ФИФе зарегистрировала окрас только в 1974 г.
Биколоры выставлялись и раньше, но без собственного на-
звания. Они экспонировались как «персидские, другие окрасы».
Утверждены в Англии (GCCF) в 1966 г., в Европе (FIFe)
168
в 1969 г., в США (CFA) в 1971 г. В 1986 г. ФИФе признала
арлекиновые окрасы.
Шоколадные персидские кошки были получены скрещиванием
с гаваной. Впервые описаны в 1971 г. Официально признаны
в 1984 г., одновременно с лиловыми.
В конце прошлого столетия популярностью пользовались чер-
ные мраморные персидские. В 1962 г. в Америке признали
голубых мраморных, а 'в 1980 г. с большим запозданием они
были зарегистрированы и в ФИФе.
Следует отдельно остановиться на окрасах колор-нойнт, ко-
торые иногда выделяются в самостоятельную породу длинношер-
стных кошек.
Первые опытные скрещивания сиамских и персидских кошек
с целью получения длинношерстных животных сиамского окраса
были проведены в 1924 г. К. Тьеббесом в Швеции. Затем в 30-х
годах подобные эксперименты велись американским исследовате-
лем К. Килером. Целенаправленное разведение началось около
1940 г. в Англии и Америке. На раннем этапе колор-пойнты имели
шерсть более короткую и менее густую, чем персидские, на хвосте
часто появлялись изломы и узлы. Далеки от идеала голова
и облегченный тип телосложения. И по сей день большим не-
достатком колор-пойнтов является тенденция возврата экстерьера
к сиамскому типу.
В Европе колор-пойнты получили официальное признание
в 1955 г. в качестве цветовой вариации персидских кошек. Перво-
начально эти удивительно красивые животные носили название
кхмерских кошек. В Америке употребляется и другое назва-
ние — гималайские (это название происходит из-за схожести по
окрасу колор-пойнтов и гималайской породы кроликов).
Сегодня наибольшей популярностью у любителей кошек поль-
зуются шиншиллы и колор-пойнты.
Стандарт породы одинаков для всех цветовых вариаций пер-
сидских кошек.
Тело от среднего размера до крупного, приземистое, мас-
сивного сложения. Ноги сильные, короткие. Лапы большие, круг-
лые, с густыми пучками шерсти между пальцев. Грудь, плечи
и спина широкие, мощные, мускулистые. Хвост короткий, пушис-
тый, с закругленным кончиком. Круглая голова на короткой
и толстой шее. Череп широкий с выпуклым лбом и полными
щеками. Одной из типичных характеристик породы является
ярко выраженная глубокая седелка (или «стоп») — вмятина
в месте перехода от носа ко лбу. Нос короткий и широкий. Очень
мощный подбородок. Профиль считается идеальным, когда прямая
линия соединяет крайнюю точку лба, мочку носа и подбородок.
Мочка носа не должна быть выше нижнего века. Уши маленькие,
со слегка закругленными кончиками, широко расставленные. Хо-
рошо развиты ушные пучки шерсти (не путать с кисточками).
Большие круглые глаза широко расставлены, блестящие и вы-
разительные. Глаза не должны быть ни слишком глубоко поса-
12 Заказ № 355
169
жены, ви излишне выдаваться из орбит. У большинства вариаций
глаза имеют цвет от оранжевого до медного. Белые персидские
могут иметь голубые глаза, а колор-пойнты — только голубогла-
зые. Все варианты шиншилл, а также персидские серебристых
окрасов обладают зеленой радужной оболочкой глаз. Во всех
случаях цвет глаз должен быть чистым и предпочтение отдается
наиболее интенсивным оттенкам. Важнейшим признаком породы
является длинная, густая и очень пушистая шерсть. Покровные
волоски настолько тонкие, что часто не отличаются от подшерстка.
Балльная шкала оценки
Тип 20
Голова 30
Цвет глаз 15
Шерсть 30
Кондиция 5
В системе бывшего Всесоюзного объединения любителей ко-
шек титул «чемпион» впервые был присвоен в 1990 г. черному
дымчатому коту из Чехо-Словакии и колор-пойнту с темно-
коричневыми отметинами американского разведения. Первым
чемпионом отечественной селекции стал белый с оранжевыми
глазами кот Август Грифон. В 1991 г. зарегистрированы первые
международные чемпионы — голубой с белым и черный дымча-
тый коты.
ПИК-ФЭЙС
Дальнейшая селекция персидских кошек на укорачивание
носа привела к созданию новой самостоятельной разновидности
кошек. Стремились достигнуть как можно большей схожести
с собаками пекинесами (отсюда и название). Селекционеры доби-
лись желаемого результата, и кошки этой породы имеют настоль-
ко короткий нос, что он угадывается по двум отверстиям нозд-
рей между глаз. Седелка носа поднята к уровню верхних век.
Эта форма приводит к очень сильному сужению слезных про-
токов, вплоть до их полной непроходимости. Отсюда постоянные
слезные истечения из глаз, что совсем не способствует нормаль-
ному здоровью животных.
Прочие экстерьерные черты идентичны персидским кошкам.
Признание в США пик-фэйсы получили в 1971 г., в ФИФе не
признаны.
РУССКАЯ ГОЛУБАЯ
В происхождении породы много неясного, но ее родиной счи-
тают окрестности Архангельска, где, предположительно, обита-
ли во множестве кошки голубого цвета. Русские голубые привлек-
ли внимание не цветом, а короткой, густой, мягкой, как бы
плюшевой шерстью.
Есть легенда, что несколько голубых архангельских кошек
были подарены английской королеве. Правда это или вымысел —
170
остается неизвестным. Реально одно, что разведение породы на-
чалось в Англии, а основу селекции составили импортированные
из России кошки. Изначально экстерьер у этих животных был
весьма разнообразен, а глаза окрашены в желтый цвет. Этих
кошек называли русскими голубыми, архангельскими голубыми,
иностранными голубыми и даже мальтийскими голубыми.
В 1939 г. сделаны первые записи в английскую племенную книгу
кошек под названием «русская голубая». После второй мировой
войны в скрещивание были введены сиамские и восточные ко-
роткошерстные кошки. Русская голубая приобрела зеленый цвет
глаз п ярко выраженный восточный тип телосложения. В 1965 г.
английские селекционеры разработали новый стандарт породы,
в котором уделялось особое внимание качеству шерсти, а сиамский
тип был признан нежелательным. В Америку русские голубые
попали около 1900 г., а признаны с 1949 г. Австралийские се-
лекционеры в 1971 г. составили программу по выведению русской
белой вариации при условии сохранения качества шерсти и стан-
дартного экстерьера. Сегодня известна и русская черная вариация,
признанная в США.
Организации ФИФе допускают для селекции только класси-
ческую голубую форму этой породы.
В бывшем СССР встречаются иногда особи, отвечающие тре-
бованиям стандарта, но основу селекции составили все же им-
портированные из Чехо-Словакии представители русской голубой
породы.
К стандарту предъявляются следующие требования. Тело вы-
тянутое, крепкое, грациозное. Шея длинная и прямая. Высокие
и тонкие ноги оканчиваются узкими овальными лапками; хвост
длинный, неширокий у основания, заканчивающийся заостренным
кончиком. Голова относительно короткая, клиновидная с длинным
и плоским в профиль черепом. Точка разделения носа со лбом
находится на уровне бровей. Четко обозначены подушечки виб-
рисс. Уши большие, заостренные, широко расставленные и вер-
тикально посаженные. Кожа ушей тонкая и прозрачная. С внут-
ренней стороны уши практически лишены шерсти. Большие гла-
за имеют миндалевидную форму, широко расставлены и окрашены
в зеленый цвет. Шерсть густая, очень тонкая, шелковистая и,
как у плюшевого медвежонка, прямостоящая. Подшерсток оди-
наковой длины с покровной шерстью. Голубой окрас должен быть
с отчетливым серебристым отливом.
Недостатками экстерьера считаются: приземистый илп тяже-
лый и, напротив, облегченный сиамский типы, круглые глаза,
желтый цвет глаз, прилегающая шерсть, короткий и широкий
хвост.
Балльная шкала оценки
Тело и хвост 30
Голова и уши 25
Глаза 10
Текстура шерсти 20
Окрас 10
Кондиция 5
12*
171
РЭГДОЛЛ
Порода получена скрещиванием персидских колор-пойнтов и
биколоров. Описывая рэгдоллов, многие авторы определяют их
экстерьер как смесь бурманской породы, колор-пойнта и би-
колора.
Иногда в публикациях можно встретить несколько иную ис-
торию происхождения рэгдоллов. Хозяйка ангорской кошки Жо-
зефины заметила, что эта кошка очень легко переносила болевые
ощущения. Владелица Жозефины решила закрепить у потомков
эту особенность в совокупности с ласковым характером. Она
скрестила свою кошку с бирманским котом и обнаружила, что
полученные котята унаследовали от матери все желаемые ка-
чества. Она же предложила название породы — рэгдолл, что в пе-
реводе означает «тряпичная кукла». Дальнейшее разведение стро-
го контролировалось. Рэгдоллы отличаются от других кошек
флегматичностью и неспособностью к самозащите.
Другие владельцы серьезно занялись разведением приобре-
тенных рэгдоллов, формируя из них породу с новым, отличным
от других кошек, экстерьерным типом. В 1979 г. породу признали
в Америке.
С января 1992 г. рэгдоллы признаны в качестве породы в ор-
ганизациях ФИФе.
Рэгдоллы — это крупные кошки, достигающие массы 10 кг.
Голова широкая, средней длины, клиновид,ной формы. Подбородок
круглый, сильный. Уши широко расставлены. Щеки полные
и придают голове некоторую сердцеобразную округлость. Тело
вытянутое. Ноги средней длины, крепкие, с круглыми и мощными
лапками. Хвост пропорционален телу, очень хорошо опушенный.
Шерсть шелковистая, густая, с хорошо развитым подшерстком.
Окрас можно охарактеризовать как сиамский с белой пятнис-
тостью, причем белые пятна могут располагаться в любых частях
тела и их размеры не ограничиваются.
СИАМСКАЯ
Некоторые биологические особенности сиамских кошек, как,
например, 68-дневная беременность, позволяют предположить,
что они происходят от бенгальской кошки (Felis bengalensis).
Считается, что на Востоке был самостоятельный очаг одомаш-
нивания этих животных.
Мутация, известная сегодня под названием «сиамский аль-
бинизм», возникла у кошек Таиланда (бывшего Сиама), а жи-
вотные — ее обладательницы — стали специально культивиро-
ваться.
Появившиеся в Европе первые сиамские кошки обладали
хвостами с изломами, узлами, их глаза сильно косили. К со-
172
жалению, эти физические недостатки долгое время считались
непременным признаком породы.
Кроме того, прежний тип значительно отличался от сегод-
няшнего стандарта. Кошки имели круглые головы с высоко по-
ставленными ушами, несколько массивный костяк и напоминали
бурманских кошек.
Необычных кошек в 1884 г. привез в Англию английский
консул в Бангкоке. Это были Пхо и Миа, пара сиамских кошек.
На выставке в Хрустальном Дворце в Лондоне кошки демонстри-
ровались уже со своим потомством, вызывая всеобщий интерес.
Порода была описана под названием «королевские кошки Сиама».
В 1892 г. был утвержден первый стандарт породы для сил-пойнто-
вого окраса.
В 1901 г. в Англии образовался «Клуб сиамских кошек»,
выработавший в 1902 г. новый стандарт породы. В 1936 г. признан
голубой окрас (блю-пойнт), в 1950 г.— шоколадный. Сиамские
кошки с лиловыми отметинами зарегистрированы в 1960 г., еще
через 6 лет признали рэд-пойнтов (с красными отметинами).
Популярность сиамских кошек в Англии была очень высокой:
в 1961 г. из всех зарегистрированных кошек 75 % составили
сиамские.
В 1958 г. был пересмотрен стандарт породы и утвержден
новый, соответствующий современному. Сиамские и восточные ко-
роткошерстные кошки имеют одинаковый экстерьер, отличаясь
лишь цветом шерсти и глаз.
Это стройные и грациозные животные с вытянутым телом.
Описание стандарта приведено в разделе, посвященном восточным
короткошерстным. Шкала оценки используется та же.
В нашей стране большинство претендентов на породу своим
экстерьерным типом не вписываются в рамки стандарта. Они
часто имеют изломы на хвосте, порой не один, а несколько, или
отличаются короткими, как обрубки, хвостами. Импортированные
чистопородные сиамские кошки и полученные от них потомки
теперь наглядно демонстрируют, какой должна быть порода.
СИБИРСКАЯ
Трудно сказать, когда кошки впервые появились в Сибири.
Активное освоение этого таежного края началось в XVI в. Пере-
селенцы, естественно, везли с собой и кошек. Неизвестно, были ли
среди этих кошек особи с длинной шерстью. Вполне вероятно,
что эта мутация появилась спонтанно, а затем была подхвачена
естественным отбором.
Упоминания о сибирских кошках можно встретить в трудах
А. Брема и некоторых других авторов. Эти животные не только
у нас в стране, но и за рубежом снискали славу очень выносливых
кошек и непревзойденных охотников на грызунов.
173
В настоящее время найти типичных сибирских кошек ока-
залось большой проблемой. Сибирь продолжала заселяться, си-
бирские кошки скрещивались с завезенными животными, выво-
зились в европейскую часть страны, где также не разводились
в чистоте. В итоге кошки, которых относят к сибирским, обладаю^
широким спектром экстерьерных типов.
Стандартный сибирский тип имеет следующие характеристики:
тело крепкое, сильное, мускулистое, несколько массивное. В це-
лом животные довольно крупные. Ноги средней длины, толстые,
с крупными круглыми лапками и густыми пучками) шерсти между
пальцев. Голова относительно большая, широкая в лобной части,
трапециевидной формы. Нос средней длины, широкий. Хорошо
развиты подушечки вибрисс. Вибриссы сильные. Уши средней
величины, широкие у основания, формы равностороннего тре-
угольника, широко расставленные. Кончики ушей слегка закруг-
ленные, удлиненные волоски формируют кисточки. Глаза круг-
лые, широко расставленные. Хвост длинный. Шерсть средней
длины, особенно удлиненная на шее (образует пышный воротник)
и на хвосте. Подшерсток (в домашних условиях) развит умеренно.
Остевые волоски жесткие. Окрас допускается любой, но пред-
почтителен с агути-фактором. Цвет глаз должен гармонировать
с окрасом шерсти.
Стандарт принят и утвержден в 1987 г. вместе с селекционной
программой по созданию и стабилизации отечественной породы.
В этом же году началась экспериментальная племенная работа
с животными сибирского типа. Интересно, что особи, обладающие
отличным сибирским экстерьером, встречаются практически во
всех городах нашей страны. Они были представлены на выстав-
ках в Москве, Санкт-Петербурге, Челябинске, Минске, Красно-
ярске, Киеве, Ташкенте, Риге и в других городах, но повсюду
единичными экземплярами. Кошки, причисленные к сибирским
первого поколения, далеко не всегда дают потомство желательного
типа. Предстоит долгая и кропотливая работа, чтобы сибирские
кошки устойчиво передавали породные черты из поколения в по-
коление.
В последнее время за рубежом возрос интерес к сибирским
кошкам. В большом количестве вывозят из страны животных,
приобретенных без родословных и выдаваемых за сибирских.
Экстерьер этих кошек далек от желательного для сибирской по-
роды. Известно, что в некоторых странах разводят сибирских
кошек п используют стандарт породы, основанный на животных,
не имеющих с настоящими сибирскими ничего общего. К со-
жалению, и в нашей стране многие клубы рекламируют и про-
дают за рубеж нетипичных для сибирской породы кошек.
ВО КЛК «Ассолюкс» придает большое значение престижу
отечественной породы, и ввиду сложности селекционной ра-
боты в настоящее время прекращен экспорт племенных сибир-
ских кошек до полной стабилизации поголовья и признания
породы.
174
Балльная шкала оценки
Тело и конечности 25
Голова 25
Глаза 5
Уши 10
Шерсть 20
Хвост 10
Кондиция 5
СИНГАПУРСКАЯ
Американские селекционеры обратили внимание на популяцию
кошек в Сингапуре, которые отличались небольшой массой
(1,8—2,7 кг в зависимости от пола) и изящным телосложением.
«Дети сточных канав Сингапура»—называют иногда этих кошек,
предполагая, что среда обитания повлияла на их облик. С 1976 г.
этих кошек можно видеть на выставках в США. Стандарт, вы-
работанный по образцу первых импортированных кошек, похож
на стандарт абиссинских. В 1981 г. порода получила официальное
признание в Америке. В ФИФе она не признана.
Сингапурские кошки, по-видимому, происходят из Юго-Вос-
точной Азии (Малайзии или Индии).
Отмечается, что кошки этой породы после рождения разви-
ваются медленно и отнимать от матери их можно не ранее
5-недельного возраста. Более позднее у них и физиологическое
созревание.
Кроме мелких размеров стандарт предусматривает средней
длины, крепкое, мускулистое, слегка приземистое тело. Спина
немного выпуклая. Средней длины ноги с маленькими и узкими
лапами. Хвост средней длины и толщины, заканчивается тупым
кончиком. Голова круглая, тупая, с коротким и широким за-
тылком. Подбородок сильный. Переход ото лба к носу отмечен
небольшим изгибом. Уши большие, слегка заостренные на кон-
чиках, широкие и выпуклые у основания. Глаза большие, мин-
далевидной формы, раскосо посаженные. Они могут быть зеле-
ными, золотистыми или цвета лесного ореха. Очень тонкая, шел-
ковистая, плотно прилегающая к туловищу шерсть должна об-
ладать двойным темно-коричневым тикингом на светлом, цвета
•слоновой кости, фоне. Кончики шерстинок темные, допускается
темная полоса на спине. Требования к окрасу такие же, как и у
Абиссинских кошек.
СКОТТИШ ФОЛД
Шотландская вислоухая — таково название породы в пере-
воде с английского языка (Scottish Fold) и этим в полной мере
определяется оригинальность облика скоттиш фолдов.
Из литературы известно, что вислоухие кошки уже в древ-
ности разводились в Китае. Подобные кошки описаны и в книгах
А. Брема.
175
Впервые в плане разведения кошкой с висящими ушами за-
интересовался шотландский овцевод, случайно обнаруживший
в 1961 г. на хуторе соседа маленькую белую кошку по кличке
Зузи. Провели анализирующие скрещивания для того, чтобы
определить, как наследуется вислоухость. Было показано доми-
нирование данного признака. Скрещивания таких кошек между
собой показали, что в гомозиготном состоянии ген Fb приводит
к возникновению физических аномалий: деформации скелета, не-
подвижности хвоста, параличу задних ног (см. гл. 6). Этот факт
в 1973 г. послужил поводом для запрещения разведения скоттиш
фолдов в Англии. Но селекция породы была продолжена в Аме-
рике. Чтобы избежать появления в пометах котят-уродов, висло-
ухих кошек стали скрещивать только с особями, имеющими нор-
мальную форму ушей. В разведение были вовлечены представи-
тели американской короткошерстной, экзотической короткошерст-
ной и даже персидской пород. В итоге был сформирован экстерьер-
ный тип скоттиш фолдов, очень схожий с британскими коротко-
шерстными кошками.
Размеры вислоухих кошек средние. Тело коренастое, как
у животных британского типа. В отличие от британских кошек
допускается небольшая седелка на переносице. Отличительный
признак — маленькие уши наклонены вперед так, чтобы закруг-
ленными кончиками касаться шерсти перед ушной раковиной.
Кошка дисквалифицируется, если одно ухо из двух прямое.
Сразу после рождения нельзя точно установить, у каких
котят уши нормальные, а у каких—«породные». Только после
трех недель жизни у вислоухих особей кончики ушей наклоня-
ются. У некоторых котят сразу появляется желаемая форма ушей,
а затем уши постепенно выпрямляются или одно из них стано-
вится прямым. Возможны случаи, когда уши выпрямляются
у беременных кошек. Все это, конечно, создает дополнительные
сложности при разведении породы.
В ФИФе шотландские вислоухие кошки пока признания не
получили, хотя в Европе занимаются их разведением.
СОМАЛИЙСКАЯ
Спонтанные мутации возникали периодически в пометах
абиссинских кошек, но животные с удлиненной шерстью, счи-
таясь нестандартными, выбраковывались. Однако в 1965 г., во-
преки общепринятым традициям, мутантная длинношерстная
абиссинская кошка была выставлена в Австралии под названием
«сомалийская кошка». В 1972 г. американцы основали «Клуб лю-
бителей сомалийских кошек» и начали добиваться признания
этой породы. Зарегистрировали сомалийскую породу в 1979 г.
В Европе порода признана в 1982 г.
Сегодня спонтанное возникновение мутации, зафиксированной
в породе, оспаривается. Было обнаружено, что котята с длинной
176
шерстью стали появляться после завоза из Англии кошки Раби
Чуффа оф Селене, родословная которой ведется от неизвест-
ного кота в IV поколении по материнской линии. Предполагается,
что этот кот имел длинную шерсть или был гетерозиготным по
гену длинношерстности.
Сомалийские кошки имеют тело средней длины, мускулистое
и грациозное. В целом они крупнее и сильнее абиссинских. Грудь
округлая, спина слегка выпуклая, что создает впечатление', гото-
вящейся к прыжку кошки. Ноги и хвост пропорциональны телу.
Лапы овальные. Пушистый хвост у основания утолщен, а к концу
сужается. Голова не имеет плоских линий, форма слегка закруг-
ленная клиновидная. Переход ото лба к носу легкий, плавный.
Затылок выпуклый. Морда не должна быть заостренной. Подбо-
родок хорошо развит. Уши большие, широкие, чашеобразной фор-
мы у основания, широко расставленные и немного сдвинуты
назад. Кончики ушей слегка заостренные. Большие, миндалевид-
ной формы и широко расставленные глаза окрашены в янтарно-
желтый или зеленый цвет. Темный ободок окаймляет глаза и при-
дает им особую выразительность. Средней длины шерсть очень
тонкая, мягкая и густая. Хвост обильно опушен. Предпочтение
отдается кошкам с хорошо развитыми воротником и «штанами».
В отличие от абиссинских кошек, сомалийские имеют тикинг, со-
стоящий из 10—12 зон. Окончательно окрас формируется до
2-летнего возраста, поэтому на выставках к молодым животным
с недостаточно выраженным тикингоМ относятся снисходительно.
Балльная шкала оценки
Тело 20
Голова и уши 15
Глаза 10
Общий окрас 15
Тикинг 10
Длина шерсти 15
Текстура шерсти 10
Кондиция 5
СФИНКС
Мутации, приводящие к отсутствию шерсти, известны у раз-
ных видов животных. Что касается кошек, то безволосые предста-
вители их были знакомы уже ацтекам. В начале нашего столетия
безволосые кошки разводились в Мексике, но от них сохранилось
только название — мексиканская голая.
Появление кошек, лишенных шерстного покрова, отмечалось
во Франции, Марокко, Индии, Парагвае — среди животных раз-
личных пород.
Название «сфинкс» получили кошки, разведение которых на-
чалось в 1966 г. в штате Онтарио (Канада).
Наряду с отсутствием шерсти на теле эти кошки имеют за-
медленный рост и утолщенную кожу. Из-за нарушения термо-
177
регуляции сфинксы нуждаются в постоянном тепле, кроме того,
они в большей степени, чем другие кошки, подвержены всевоз-
можным повреждениям. Одно из достоинств кошек породы
сфинкс — возможность тесных контактов с ними людей, страдаю-
щих аллергической реакцией на шерсть.
Еще одна особенность бесшерстных кошек: гомозиготные осо-
би, скрещиваясь между собой, остаются бесплодными.
Проблематичные во многих отношениях и к тому же мало-
эстетичные животные признаны лишь в некоторых фелинологи-
ческих организациях.
ТОНКИНСКАЯ
Как таковая эта группа кошек породой не является, потому
что при разведении «в себе» дает в потомстве расщепление фе-
нотипов (пример промежуточного наследования). Поэтому дол-
гое время тонкинским кошкам отказывалось в признании. Тем
не менее, в мае 1984 г. порода официально зарегистрирована
в американских клубах.
Эти кошки впервые появились перед 1930 г., после того как
провели экспериментальные скрещивания для получения кошек
с бурманским телосложением и сиамским окрасом. В первом по-
колении рождались котята ровного кремового цвета, а затем они
вырастали в Тонкинских кошек, демонстрируя классический при-
мер кодоминирования генов cs и сь.
Название породе было дано по расположенной в северной части
Вьетнама провинции Бак-бо, ранее именовавшейся Тонкинг.
И хотя происхождение современных тонкинских кошек не под-
вергается сомнению, можно предположить, что среди первых
сиамских кошек, завезенных в Англию в конце прошлого века,
находились и Тонкинские.
Происходя, с одной стороны, от голубоглазых сиамских пред-
ков и от желтоглазых бирманских короткошерстных — с другой,
тонкипские кошки имеют голубовато-зеленый цвет глаз.
ТУРЕЦКИЙ ВАН
Это представитель кошек, которые обитали свободной попу-
ляцией без систематического разведения в турецкой провинции
около оз. Ван.
Они не только охотно плавают, но и свободно ловят рыбу.
Второе название —«плавающие кошки»— отражает эту их осо-
бенность. Первую пару ванов завезли в Англию в 1955 г. Затем
через несколько лет были завезены еще три экземпляра. Эти
кошки стали родоначальниками новой породы. В 1969 г. порода
была официально зарегистрирована в Англии и в 1971 г,—
в ФИФе.
178
Турецкие ваны имеют мускулистое вытянутое тело средних
размеров. В целом у них более грубая конституция, чем у ан-
горских кошек, а тело короче. Ноги; средней длины с изящными,
круглой формы лапками. Хвост средней длины, покрыт остевой
шерстью. Шея сильная, короткая голова имеет форму тупоуголь-
ного треугольника. Подбородок сильный. Нос средней длины, пря-
мой. Глаза большие, миндалевидные, слегка раскосо посаженные,
светло-янтарного цвета. Подушечки лап, мочка носа и окаймляю-
щая линия век розового цвета. Уши большие, широкие у осно-
вания, поставлены прямо и имеют слегка закругленные кончики.
По сравнению с ангорскими кошками у турецких ванов более
широкая и короткая голова, а уши имеют меньшие размеры.
Шерсть ванов мягкая и шелковистая, лишенная подшерстка. Окрас
является своеобразной «визитной карточкой» породы: строго обя-
зательны два каштаново-красных пятна на голове, разделенных
на лбу белым, и такого же цвета хвост, окрашенный от крупа
до кончика, допускаются легкие темные разводы или полосы по
хвосту. Остальные части туловища чисто-белого цвета.
С 1984 г. стали разводить особей с голубыми и различно окра-
шенными глазами.
Балльная шкала оценки
Телосложение, включая ноги и хвост 25
Голова, включая форму глаз 25
Цвет глаз 10
Уши 10
Текстура и окрас шерсти 25
Кондиция 5
ЭКЗОТИЧЕСКАЯ КОРОТКОШЕРСТНАЯ
Для простоты экзотических кошек можно охарактеризовать
как персидских кошек с короткой шерстью.
В начале 60-х годов нашего столетия американские селекцио-
неры улучшали тип своих американских короткошерстных кошек,
скрещивая их с персидскими. Особи, получавшиеся от такого раз-
ведения, имели высокую конкурентоспособность на выставках.
Тогда в 1966 г. одна из американских фелинологических органи-
заций приняла решение регистрировать скрещивания между
персидскими и короткошерстными американскими кошками.
С 1967 г. утвержден стандарт новой породы, получившей назва-
ние экзотической короткошерстной, и началось ее планомерное
разведение. Полностью экзотические кошки были признаны
в Америке в 1977 г., с тех пор селекционное развитие породы
идет параллельно с развитием породы персидских кошек с пере-
крестными скрещиваниями. Отмечено, что разведение экзоти-
ческой короткошерстной породы в чистоте, лишает животных ха-
рактерного для породы мягкого, очень густого и не прилегающего
к телу шерстного покрова.
179
Отличаясь от персидских качеством и длиной шерсти, по, всем
остальным экстерьерным чертам экзотические кошки имеют аб-
солютно идентичный стандарт. Цветовые вариации также соот-
ветствуют персидским.
Первые представители этой породы были импортированы из
Канады в московский питомник «Сад Марош».
ЯВАНСКАЯ
Порода выведена в начале 1980-х годов в результате скре-
щивания балинезпйских кошек с восточными короткошерстными.
В результате облик яванских можно кратко описать как ориен-
тальные с удлиненной шерстью или как балинезпйские с полным
окрасом. Цорода имеет еще одно название—«мандарин», но
в классификации ФИФе употребляется вынесенное в заглавие
наименование. Зато в Америке название «яванские» применяется
к балинезийским кошкам нетрадиционных цветов (т. е. кроме
окрасов с темно-коричневыми, голубыми, шоколадными и лило-
выми отметинами).
ЯПОНСКИЙ БОБТЭЙЛ
Несколько столетий короткохвостые кошки были традиционны
в Японии. Особенно почитались трехцветные (черепаховые с бе-
лым), получившие название Ми-Ke, и, по поверью, приносящие
счастье. Японцы любят дарить друг другу фигурки кошек Ми-Ке
с поднятой для приветствия лапкой. В Токио на фасаде одного
из храмов запечатлена такая же кошка.
В 1968 г. пара японских короткохвостых кошек попала в Аме-
рику. Через год число животных значительно увеличилось, и уже
спустя два года порода японский бобтэйл получила официальное
признание.
Кошки имеют средние размеры. Это мускулистые животные на
длинных ногах с овальными лапками, задние ноги длиннее пе-
редних. Голова треугольная с легким пинчем. Нос длинный.
Щеки округлые и хорошо развитые. Большие уши широко рас-
ставлены. Глаза большие, миндалевидные, их раскосое располо-
жение создает яркой впечатление восточного типа. Шерсть у боб-
тэйлов короткая, мягкая, шелковистая. Главный породный приз-
нак — хвост, вернее, короткий обрубок длиной от 3 до 8 см.
Часто хвост искривлен и неподвижно сращен с телом. Шерсть
на хвосте длиннее, чем на теле, и растет, топорщась во все сто-
роны, подобно кухонному ершику.
В противоположность бесхвостым мэнским кошкам японские
бобтэйлы имеют полигенное наследование характерного признака.
Кроме того, возникновение короткого хвоста не сопровождает-
ся какими-либо деформациями в развитии животных. Напротив,
180
поклонники этой породы свидетельствуют о необыкновенной жиз-
нестойкости и выносливости своих любимцев.
При разведении японских бобтэйлов в себе все потомки имеют
короткие хвосты, п никогда не появлялись бесхвостые животные
или кошки с хвостами нормальной длины.
В Европе эта интересная порода пока не получила распро-
странения и соответственно признания.
СПИСОК ЛИТЕРАТУРЫ
Непомнящий И. И. Кошки.-- М.: Агропромпздат, 1991.— 207 с.
Регистрационный журнал ВО КЛК "Ассолюкс”.
Стандарты ФИФе.
Austin М. Polygenes and breeding for tvpe Ц Cats Australian style.— Februa-
ry, 1980,— P. 13—16.
Konarska-Szubska A. Koty rasowc.—Warszawa: Panstwowe Wydawhictwo Rol-
nicze i Lesne, 1989.— 228 s.
Leigh D. Elementary genetics // Catnips.— May, 1984.
Muller U. Das Nene Katzenbuch.— Munchen: Grate und Unzer GmbH, 1984.—
144 S. '
Muller-Girard C. Bi-Lexikon Rassekatzen.— Leipzig: VEB Bibliographisches
Institut, 1988.— 496 S.
Pintera A. Kocky, kocouri a kot’ata.— Praha: Prace, 1989.— 336 s.
Глава 13
ЦВЕТОВЫЕ ВАРИАЦИИ ПОРОД КОШЕК
И ИХ СТАНДАРТЫ
ПЛЕМЕННАЯ РАБОТА С КОШКАМИ
Племенная работа в основном строится на разведении чисто-
породных кошек. Чтобы животное было поставлено на племенной
учет, необходимо получить па выставке оценку в открытом клас-
се (в возрасте старше 10 мес). Для кошек обязательна оценка
пе ниже, чем «очень хорошо», для котов — не ниже, чем «отлич-
но» (при отсутствии дисквалифицирующих пороков). Оценка
действительна 2 года, затем для племенного использования соот-
ветствие животных стандарту должно быть подтверждено па оче-
редной выставке.
Если кошка имеет неполную родословную или ее происхож-
дение неизвестно, она должна получить подтверждение соответст-
вия стандарту породы в классе новичков (с 6-месячного возраста)
у двух разных экспертов. И лишь затем кошка заявляется в от-
крытый класс и, получив высокую оценку у третьего эксперта,
может занять свое место среди племенных животных. Беремен-
ные (сроком более месяца) п кормящие кошки в выставках не
участвуют. * ;
181
Кошки допускаются в разведение только после годовалого
возраста, достигнув физической зрелости. Отъем котят от матери
разрешен не ранее 2-месячного возраста. И еще два месяца кош-
ка должна отдыхать до нового покрытия. Соблюдая вышеуказан-
ные правила, от кошки можно получить не более двух пометов
в год.
Правильность разведения должна контролироваться инструк-
торами-фелинологами.
Владелец племенной кошки (именуемый заводчиком) вправе
сам выбирать «брачного» партнера для своей кошки. Инструктор
может порекомендовать того или иного производителя и запре-
тить недопустимый вариант. Основаниями для запрета могут
быть:
необоснованный инбридинг; возможность получения нестан-
дартного окраса или цвета глаз у котят; использование не заре-
гистрированного в качестве племенного производителя; проведе-
ние межпородных скрещиваний.
Инструкторам вменяется в обязанность актирование пометов.
При этом определяются пол и окрас котят, их физическое разви-
тие, дефекты, пороки, а также градация по степени выраженно-
сти породных признаков. Анализ генотипов родительской пары
и полученных котят позволяет выявить неправильность сведений
о родителях. Например, известно, что при покрытии черной кош-
ки красным котом в помете будут только черепаховые кошки и
черные коты (см. гл. 4). Если же среди котят обнаружен крас-
ный самец, происхождение котят, естественно, ставится под со-
мнение. В таких случаях весь помет остается без документов и
не регистрируется в племенную книгу.
Для облегчения работы инструкторов-фелинологов и помощи
владельцам кошек существуют специальные корреспондирующие
таблицы, по которым легко определить возможные варианты ок-
расов котят при том или ином скрещивании (табл. 13.1, 13.2).
В гл. 12 было отмечено, что в пределах пород существует
большое разнообразие окрасочных вариантов, обусловленных
фиксацией отдельных мутантных аллелей или их комбинаций.
Ниже приводятся фенотипические стандарты наиболее распро-
страненных окрасочных вариантов и их генотипы. Описание фе-
нотипического проявления отдельных генов окраски приведено
в гл. 4.
Черный. Генотип ааВ—С—D—. Волосы от кончиков до кор-
ней должны быть черного цвета. Недостатками окраса являются:
серый подшерсток, бурый цвет, ржавый оттенок шерсти, отдель-
ные белые волоски.
Этот окрас в чистом виде у персидских кошек встречается
редко. У молодняка перечисленные выше недостатки относитель-
но часты, особенно в возрасте до 6 мес, но затем окрас, как пра-
вило, улучшается.
Мочка носа должна быть черного цвета без каких-либо пятен.
Подушечки лап черные или серовато-черные.
182
Таблица 13.1
Возможный окрас шерсти котят при скрещивании кошек некоторых цве-
товых вариаций без агути-фактора
Кот Кошка Окрас котят
Кот Кошка
Черный Черная Голубая 1 Красная Кремовая Черепаховая Голубо-кремо- вая Черный, голубой Черный, голубой Красный, кремовый Красный, кремовый Черный, голубой, крас- ный, кремовый Черный, голубой, крас- ный, кремовый Черная, голубая Черная, голубая Черепаховая, голубо- кремовая Черепаховая, голубо- кремовая Черная, голубая, чере- паховая, голубо-кре- мовая Черная, голубая, чере- паховая, голубо-кремо- вая
Голубой Черная Голубая Красная Кремовая Черепаховая Голубо-кремо- вая Черный, голубой Голубой Красный, кремовый Кремовый Черный, голубой, крас- ный, кремовый Голубой, кремовый Черная, голубая Голубая Черепаховая, голубо- кремовая Голубо-кремовая Черная, голубая, чере- паховая, голубо-кре- мовая Голубая, голубо-кремо- вая
. Красный Черная Голубая Красная Кремовая Черепаховая Голубо-кремо- вая Черный, голубой Черный, голубой Красный, кремовый Красный, кремовым Черный, голубой, крас- ный, кремовый Черный, голубой, крас- ный, кремовый Черепаховая, голубо- кремовая Черепаховая, голубо- кремовая Красная, кремовая Красная, кремовая Красная, кремовая, че- репаховая, голубо-кре- мовая Красная, кремовая, че- репаховая, голубо-кре- мовая
Кремо- вый Черная Голубая Красная Кремовая Черепаховая Голубо-кремо- вая Черный, голубой Голубой Красный, кремовый Кремовый Черный, голубой, крас- ный, кремовый Голубой, кремовый Черепаховая, голубо- кремовая Г олубо-кремовая Красная, кремовая Кремовая Красная, кремовая, че- репаховая, голубо- кремовая Кремовая, голубо-кремо- вая
183
Цвет шерсти котят при скрещивании
Кот Кошка
WW белая АА агути аа черная aadd голубая aacbcb коричневая бур- ма
WW белый Белые Белые Белые Белые Белые
АА агути Белые Агути Агути Агути Агути
аа Белые Агути Черные Черные Черные
черный
aadd голубой Белые Агути Черные Голубые Черные
aacb cb Коричневые
коричневая бурма aabb Белые Агути Черные Черные бурманские
Белые Агути Черные Черные Черные
коричневый aabbdd Белые Агути Черные Голубые Черные
лиловый
о Белые ц агути, <? черные, черные, д' черные,
красный $торби $ черепа- $ черепа- $ черепахо-
ховые ховые вые
ОО — dd Белые агути, $ торби с? черные, 9 черепа- & голубые, $ голубо- с? черные,
кремовый ховые $ черепахо-
кремовые вые
aaSS Белые Агути с бе- Черные с Черные с Черные с бе-
черный арлекин лым белым белым ЛЫМ
Голубой. Генотип ааВ—С—dd. Окрас должен быть однотон-
ным и равномерным по всему телу. Допускаются все тона голу-
бого цвета, но предпочтение отдается наиболее светлому окрасу.
Недостатками являются: контраст между цветом подшерстка и
покровной шерсти, бурые или ржавые оттенки, наличие рисунка,
белые волоски.
Мочка носа и подушечки лап серовато-голубые.
Шоколадный. Генотип ааЬЬС—D—. Допускаются все тона
коричневого цвета. Окрас должен быть ровным. Недостатками
являются: любой, отличный от коричневого, оттенок, наличие
рисунка, серый подшерсток, белые волоски.
Мочка носа молочно-шоколадного цвета.
184
Таблица 13.2
гомозиготных кошек некоторых окрасов
aabb коричневая aabbdd лиловая 00 красная OOdd кремовая aaSS черный арлекин
Белые Белые Белые Белые Белые
Агути Агути красные $ торби красные 9 торби Агути с белым
Черные Черные красные 9 черепаховые красные 9 черепаховые Черные с бе- лым
Черные Голубые красные, 9 черепаховые кремовые, 9 голубо-кре- мовые Черные с бе- лым
Черные Черные красные, 2 черепаховые с? красные, 2 черепаховые Черные с бе- лым
Коричневые Коричневые с? красные, ? черепаховые с? красные, 9 черепаховые Черные с бе- лым
Коричневые Лиловые красные, 9 черепаховые кремовые, 9 голубо-кре- мовые Черные с бе- лым
<У черные, § черепа- ховые черные, 2 черепаховые Красные Красные черные с бе- лым, 9 черепахо- вые с белым
черные, $ черепа- ховые <? голубые, $ голубо-кре- мовые Красные Кремовые <? черные с бе- лым, 9 черепаховые с белым
Черные с белым Черные с бе- лым красные с белым, О черепаховые с белым с? красные с белым, 9 черепаховые с белым Черные арле- кины
Подушечки лап должны быть от молочно-шоколадного до шо-
коладного цвета.
Лиловый. Генотип ааЬЬС—dd. Цвет шерсти — ровный светло-
лиловый с легким розовым оттенком. Недостатками являются:
любой оттенок другого тона, рисунок, серый подшерсток, белые
волоски.
Мочка носа и подушечки лап — лавандово-розового цвета.
Белый. Генотипы W/—, саса; сс. Окрас должен быть чисто-бе-
лым без примеси какого-либо другого цвета или оттенка (W—).
У гетерозиготных котят на голове могут быть черные, голу-
бые, красные или кремовые пятна, что не снижает оценку молод-
няка. С возрастом эти пятна обычно исчезают.
13 Заказ М5 355
185
Мочка носа и подушечки лап розовые.
Красный. Генотип С—D—0(0). В идеале шерсть должна быть
как можно более интенсивного красного цвета, ровно окрашенная
по всей длине шерстинок. Типичными недостатками являются:
наличие рисунка, неравномерность окраса, осветленный кончик
хвоста. Абсолютно однородного цвета добиться трудно, особенно
у короткошерстных пород. Поэтому у животных допускаются
легкий рисунок на лбу и незамкнутые кольца на ногах.
Мочка носа и подушечки лап должны быть окрашены в кир-
пично-красный цвет без пигментных пятен.
Кремовый. Генотип С—ddO(O). Шерсть окрашена в ровный
кремовый цвет. Предпочтение отдается более светлым пастель-
ным тонам. Недостатки: наличие рисунка, светлый подшерсток,
неравномерный окрас. Как и в случае с красными, у кремовых
кошек допускается легкий рисунок на лбу и ногах. Но в целом
у кремовых животных тенденция к выраженности рисунка ниже,
чем у красных.
Мочка носа и подушечки лап розовые, без пигментных пятен.
Черепаховый. Генотип ааВ—С—D—iiOoss. Окрас выражен со-
четанием черных и красных пятен. Цвета могут быть четко раз-
граничены или перемешаны. Желательно равномерное и гармо-
ничное распределение цветов по всему телу. Красный может
быть различных оттенков. Предпочтительно наличие так назы-
ваемого «огонька» —оранжевого пятна на лбу. Недостатками ок-
раса являются серый подшерсток и полосатый рисунок. У молод-
няка дымчатый подшерсток — довольно обычное явление, но с
возрастом окрас должен стабилизироваться. Тигровый или мра-
морный рисунок чаще проявляется у животных в виде крупных
пятен красного цвета, а в случае смешанного окраса вероятность
такого недостатка практически исключена.
Мочка носа и подушечки лап могут быть окрашены в розо-
вый, черный цвета или иметь черные пятпа на розовом фоне и
наоборот.
Шоколадный черепаховый. Генотип ааЬЬС—D—iiOoss. В дан-
ном окрасе сочетаются пятпа молочно-шоколадпого и кремового
цветов различной интенсивности. Как и у животных с черепахо-
вым окрасом, желательны равномерное распределение цветов и
наличие кремового «огонька». Встречаются те же, что и у чере-
паховых, недостатки.
Мочка носа и подушечки лап шоколадного цвета различных
оттенков, розовые или пятнистые.
Голубой черепаховый (голубо-кремовый). Генотип ааВ—С—
—ddiiOoss. Голубой и кремовый цвета в окрасе, предпочтительно
светлых тонов, у персидских кошек обычно перемешаны, у ко-
роткошерстных пород чаще располагаются пятнами. Требования
к окрасу те же, что и у черепаховых кошек.
Мочка поса и подушечки лап серо-голубого, розового цвета
илй пятнистые.
Лиловый черепаховый. Генотип ааЬЬС—ddiiOoss. Сочетание
светло-лилового и светло-кремового цветов, а также выполнение
186
всех условий черепахового окраса — необходимые требования к
лиловым черепаховым кошкам.
Мочка носа и подушечки лап лавандовые, розовые, лавандово-
розовые или пятнистые.
Черный дымчатый. Генотип ааВ—D—I—. Очень контрастный
окрас. Остевые волоски на 1/3 длины и более окрашены в чер-
ный цвет. Подшерсток пепельно-белого цвета. При этом у длин-
ношерстных кошек наиболее интенсивно окрашены голова и ко-
нечности, а воротник и ушные пучки шерсти почти полностью
светлые. В спокойном состоянии дымчатые кошки, особенно ко-
роткошерстные, выглядят как черные, и лишь в движении ярко
выражается эффективный окрас. Недостатками являются: нали-
чие рисунка, пятен или оттенков других цветов, обратный конт-
раст в окрасе. Молодняк часто имеет эти недостатки, но с возра-
стом они обычно исчезают. У взрослых животных допускается
легкий осветленный рисунок на голове.
Мочка носа и подушечки лап должны быть окрашены в чер-
ный или коричнево-черный цвет.
Голубой дымчатый. Генотип ааВ—ddl—. Окрас аналогичен
черному дымчатому, но остевая шерсть окрашена в голубой цвет.
Мочка носа и подушечки лап серо-голубого цвета.
Шоколадный дымчатый. Генотип aabbD—I—. В отличие от
предыдущих дымчатых окрасов, у кошек покровная шерсть шо-
коладного цвета. Мочка носа коричневого, подушечки лап розо-
вато-коричневого цвета.
Лиловый дымчатый. Генотип aabbddl—. Окрашенная шерсть
лилового цвета с легким розоватым оттенком. Мочка носа и по-
душечки лап лавандово-розовые. Недостатки такие же, как и у
прочих дымчатых окрасов.
Красный камео, красный затушеванный камео и красный дым-
чатый. Генотип D—I—0(0). Эти три окраса имеют одинаковый
генотип, а различия внешнего проявления вызываются действием
генов-модификаторов. Впрочем, отличаются эти окрасы друг от
друга лишь длиной типинга, т. е. длиной окрашенной части осте-
вой- шерсти.
У камео около 1/8 части длины шерстинок красного цвета, у
затушеванных камео — примерно 1/3 часть, а у дымчатых ости
окрашены на большую длину, вплоть до самых корней. Подшер-
сток в любом случае чисто-белого цвета. Типинг должен быть
равномерно распределенным. Недостатки этих окрасов: полоса-
тый рисунок, возможность выраженности которого возрастает с
увеличением длины типинга, замкнутые кольца на ногах, оттенок
коричневого цвета, а у красных дымчатых следует обращать вни-
мание на те же недостатки, что и у других дымчатых окрасов.
Мочка носа и подушечки лап окрашены в розовато-красный
цвет.
Кремовый камео, кремовый затушеванный камео, кремовый
дымчатый. Генотип ddl—0(0). Окрасы полностью аналогичны
предыдущим, исключая кремовый цвет типинга: те же требования
13*
187
и те же недостатки. Общий той окрасов значительно светлее со-
ответствующих им красных и, как в случае однотонных кремовых
окрасов, проявление полос заметно меньше.
Мочка поса и подушечки лап розовые.
Черепаховый дымчатый, черепаховый затушеванный, черепа-
ховый завуалированный. Генотип В—D—I Оо. Как и все чере-
паховые, такие окрасы встречаются только у самок. Стандарты
те же, что и для описанных выше пород, имеющих геи I.
Мочка поса и подушечки лап могут быть однотонными — чер-
ного, розового цвета или иметь черные и розовые пятна.
При гомозиготности по аллелю d на фоне данного генотипа
образуются осветленные цветовые вариации: голубой чере-
паховый дымчатый, голубой черепаховый затушеванный, го-
лубой черепаховый завуалированный. Генотип В—ddl—Оо
Аналогично: шоколадные, генотип bbD—I—Оо и лиловые, ге-
нотип bbddl—Оо
Шиншилла, шиншилла затушеванная (серебристый с тенью).
Генотип А—В—D—I—
Голубая шиншилла, голубая затушеванная шиншилла. Гено-
тип А—В—ddl—
Шоколадная шиншилла, шоколадная затушеванная шиншилла.
Генотип А—bbD—I—
Лиловая шиншилла, лиловая затушеванная шиншилла. Гено-
тип А—bbddl—
Перечисленные группы окрасов, как и дымчатые, обусловлены
действием гена ингибитора меланина. Шиншиллы имеют типинг
на длину 1/8 части остей, затушеванные вариации — на длину
1/3 части.
Шиншиллы, особенно персидские, на первый взгляд кажутся
белыми, но они как бы покрыты легкой темной вуалью. Чисто
белыми должны быть подбородок, ушные пучки шерсти, грудь,
живот, внутренняя сторона ног и конец хвоста. Эффектное допол-
нение окраса — зеленые глаза и интенсивная пигментация век.
Недостатками являются: наличие тигрового рисунка, пятна
любых оттенков, отличный от белого цвет подшерстка, желтый
или ореховый цвет глаз. Затушеванные шиншиллы внешне вы-
глядят темнее классических шиншилл, по требования к живот-
ным этих окрасов предъявляются такие же, допускается лишь
наличие незамкнутых колец на ногах. Кроме того, у кошек зату-
шеванных вариаций подушечки лап темные, равномерно окра-
шенные.
Мочка носа должна быть кирпично-красного цвета, окантова-
на черным, голубым, коричневым или лавандово-розовым обод-
ком, в зависимости от окраса.
Подушечки лап соответственно имеют цвета: черный или ко-
ричневато-черный, серовато-голубой, от цвета корицы до шоко-
ладного, лавандово-розовый.
Черный серебристый тигровый/мраморный. Генотип А—В—
С D I-T /t’t”
188
Голубой серебристый тигровый/мраморный. Генотип А—В—
С—<MI—T-/tbtb
Шоколадный серебристый тигровый/мраморный. Генотип
AbbCDIT /Ль
Лиловый серебристый тигровый/мраморный. Генотип А—ЬЬС—
ddl—Т—/tbtb
Серебристые окрасы отличаются наличием тикинга, т. е. че-
редованием темных и светлых (в данном случае белых) зон по
всей длине шерстинок, а также четко выраженным темным ри-
сунком. Окрас определяется цветом темных зон тикинга и фор-
мой рисунка. Рисунок во всех случаях должен быть отчетливым,
хорошо разграниченным основным серебристым фоном.
Мраморный рисунок образует букву «М» па лбу, две-три спи-
рали на щеках, две полосы, подобные ожерелью, на шее и так
называемую «бабочку» на плечах. От плеч по спине проходит
темная полоса, оканчивающаяся у самого кончика хвоста. Ещё
две продольные полосы проходят по бокам, от верхней полосы
они отделяются серебристой шерстью. На боках широкие изви-
листые разводы, на ногах поперечные полосы. По всей длине
хвоста равномерно расположены кольца, заканчивается он тем-
ным кончиком. По груди и животу проходит двойной ряд точек.
Наиболее эффектны короткошерстные черные серебристые мра-
морные кошки. У персидских же кошек рисунок имеет слабую
контрастность из-за длинной шерсти.
Тигровый рисунок—па лбу также выделяется буква «М»,
щеки украшены двумя-тремя спиралями, а шея окольцована дву-
мя полосами, не прерывающимися на груди. Вдоль по спине до
кончика хвоста проходит широкая полоса, от которой отходят,
спускаясь к животу, тонкие вертикальные полосы, разделенные
основным серебристым фоном. Ноги и хвост покрыты кольцами. Кон-
чик хвоста черный. От груди по животу проходят два ряда точек.
Пятнистый рисунок является модификацией тигрового. При
этом тело должно быть равномерно покрыто пятнами круглой
или овальной формы. Пятна обязательно должны быть четко от-
делены друг от друга. Этот рисунок введен в стандарты только
короткошерстных кошек, вероятно, из-за невозможности иденти-
фикации его у кошек с удлиненной, а тем более с длинной
шерстью.
Мочка носа соответственно окрасам должна быть кирпично-
красная с черным, голубым или шоколадным ободком и розовая
с лавандово-розовым ободком.
Подушечки лап черные или коричневато-черные, голубовато-
серые, коричневые различных тонов, лавандово-розовые.
Черный тигровый/мраморный. Генотип А—В—С—D—ИТ—/tbtb.
Черный тигровый — это самый естественный для кошек окрас,
так называемый дикий тин.
Черные мраморные, в отличие от тигровых, несут в гомозигот-
ном состоянии рецессивный ген мраморного рисунка.
В любом варианте шерсть кошек имеет зональное распределе-
ние пигментов: чередование черных и рыжих зон по всей длине
189
шерстинок. Выраженность рисунка должна быть аналогичной
окрасам серебристых тэбби интенсивного черного цвета. Недостат-
ками считаются: серый подшерсток, белые остевые волоски,
слишком светлый или белый подбородок, белесый кончик хвоста.
Мочка носа должна быть окрашена в кирпично-красный цвет
с черным ободком.
Подушечки лап черные или коричневато-черные.
Голубой тигровый/мраморный. Генотип А—В—С—ddiiT—/tbtb.
В отличие от предыдущих окрасов тикинг образован чередова-
нием голубых и цвета слоновой кости зон. Рисунок темно-голубо-
го цвета. Остальные требования идентичны требованиям, предъ-
являемым к предыдущему окрасу.
Мочка носа бледно-розового цвета с темно-голубой окантовкой.
Подушечки лап серовато-голубые.
Шоколадный тигровый/мраморный. Генотип А—ЬЬС—D—
iiT—/tbtb. Отличаются от голубых: чередованием на шерсти зон
цвета слоновой кости с зонами шоколадного цвета, темно-шоко-
ладным рисунком, коричневым ободком вокруг бледно-розовой
мочки носа и шоколадными подушечками лап.
Лиловый тигровый/мраморный. Генотип А—ЬЬС—ddiiT—/tbtb.
Особенность и отличие от приведенных выше: лиловый цвет зон
и контрастный лиловый рисунок, лавандово-розовый ободок во-
круг мочки носа и розовые или лавандово-розовые подушечки лап.
Золотая (завуалированная) шиншилла, золотая затушеванная
шиншилла. Генотип А—В—D—ii. Эти окрасы зарегистрированы
только у персидских кошек.
Подшерсток может быть окрашен в тона, варьирующие от
светло-кремового до коричневатого, но предпочтение отдается на-
сыщенному кремовому цвету. У классических золотых шиншилл
типинг занимает 1/8 части остей, а у затушеванных — 1/3 длины
покровной шерсти. При этом типинг должен равномерно распро-
страняться по спине, бокам, голове, верхней части хвоста и на-
ружным сторонам ног без какого-либо рисунка (у завуалирован-
ных допускается лишь легкая полосатость на ногах). Все осталь-
ные части тела окрашены в пастельные тона цвета подшерстка.
Кроме явно выраженного рисунка недостатками считаются
замкнутые кольца на ногах, коричневый подшерсток, белый или
почти белый подбородок, слишком светлые грудь, живот и уши.
Мочка носа темно-розового цвета с коричневой, или коричнева-
то-черной окантовкой. Подушечки лап должны иметь коричнева-
то-черпый цвет.
ДВУХЦВЕТНЫЕ КОШКИ (БИКОЛОРЫ)
Черные с белым. Генотип ааВ—D—iiS—
Голубые с белым. Генотип ааВ—ddiiS—
Шоколадные с белым. Генотип aabbD—iiS—
Лиловые с белым. Генотип aabbddiiS—
190
Красные с белым. Генотип D—iiO/0/S—
Кремовые с белым. Генотип ddiiO/0/S—
Идеальным вариантом биколоров считают кошек, у которых
белый рисунок занимает от 1/3 до 1/2 поверхности тела и рас-
положен симметрично, причем верхняя часть туловища должна
быть окрашена, а нижняя — белая. На морде желателен белый
рисунок в виде печатной буквы «Л», глаза же должны разме-
щаться в окрашенной зоне. Требования по расположению белой
пятнистости особенно строго соблюдаются у короткошерстных
кошек.
Но для всех пород, включающих биколорпые окрасы, обяза-
тельны чистота и равномерность основного цвета, четкость отгра-
ничения от белого. Не допускается тигровый или мраморный
рисунок. И лишь у полудлинношерстных пород (норвежская лес-
ная, мэйн-кун, сибирская) не придается никакого значения ва-
риантам сочетания белой пятнистости с любым другим окрасом.
Существует другой стандарт для двухцветных кошек: если
белый цвет значительно преобладает, то такие окрасы носят на-
звание «арлекинов» (черный, арлекин, голубой арлекин, красный
арлекин и т. д.). В этих случаях по белому фону разбросаны от-
дельные цветные пятна, которые должны быть чистыми и четко
отделенными от осиовпого белого фона. Особой популярностью
пользуются короткошерстные кошки, у которых по белоснежному
телу разбросаны пятна идеальной круглой формы размером с мо-
нету серебряного доллара. Они чрезвычайно редки и появляются
случайно.
В подавляющем большинстве случаев биколорпые кошки гете-
розиготны по гену белой пятнистости, а арлекины гомозиготны.
Мочка носа и подушечки лап у двухцветных кошек окрашены
соответственно цвету окраса или в розовый цвет.
ПОЙНТОВЫЕ ОКРАСЫ С СИАМСКИМИ ОТМЕТИНАМИ
Ген сиамского альбинизма в сочетании с другими генами,
влияющими на цвет шерсти, дает целую гамму окрасов, признан-
ных у кошек ряда пород: персидских (колор-пойнты), балине-
зийских, сиамских. Морда, уши, ноги и хвост имеют цвет, соответ-
ственно которому и называется каждый окрас. Остальное тело
должно быть светлым.
. Отметины (интенсивно окрашенные части тела) должны быть
равномерными по тону, четко ограниченными. Часто встречаю-
щимся недостатком является наличие темных пятен па спине и
животе. Дисквалифицирующим дефектом считаются белые пятна
и белые пальцы.
С темно-коричневыми отметинами (Seal-point). Генотип ааВ—
cscsD—. Отметины очень интенсивного темно-коричневого цвета,
тело с легким бежевым оттенком.
.Мочка носа и подушечки лап коричневато-черного цвета.
191
С голубыми отметинами (Blue-point). Генотип ааВ—c8c"dd.
Отметины голубовато-серого цвета, тело слегка голубоватого от-
тенка.
Мочка носа и подушечки лап голубовато-серого цвета.
С шоколадными отметинами (Chocolate-point). Генотип
aabbc3csD—. Отметины цвета молочного шоколада, тело цвета
слоновой кости.
Окраска мочки носа и подушечек лап варьирует от цвета ко-
рицы до цвета молочного шоколада.
С лиловыми отметинами (Lilac-point). Генотип aabbcsc3dd. От-
метины лиловые, а тело практически белого цвета с легким ро-
зоватым оттенком.
Мочка поса и подушечки лап лавандово-розового цвета.
С красными отметинами (Red-point). Генотип aacsc3D—0(0).
Отметины оранжевого цвета, тело практически белое, лишь с лег-
ким кремовым оттенком.
Мочка носа и подушечки лап розовые.
С кремовыми отметинами (Cream-point). Генотип aac3c3dd—О (О).
Отметины пастельного тона кремового цвета, тело белое с очень
легким кремовым оттенком.
Мочка носа и подушечки лап розовые.
С темно-коричневыми черепаховыми отметинами (Seal-tortie-
point). Генотип ааВ—csc3D—Оо. Отметины в виде темно-коричне-
вых п красных пятен, расположение которых желательно как
можно более равномерное, пятна должны быть четко отделены
друг от друга. Красный цвет может быть различных оттенков.
Как и у всех черепаховых окрасов, предпочтительно наличие
«огонька» на лбу. Тело имеет светлый бежевый пвет.
Мочка носа и подушечки лап темпо-коричневые, розовые или
пятнистые.
Черепаховые пойптовые вариации с генами ослабленных ок-
расов аналогичны. Пигментация мочки носа и подушечек лап
идентична прочим черепаховым окрасам. Поэтому здесь мы пе-
речислим лишь их наименования с указанием генотипов.
С голубыми черепаховыми отметинами (Blue-tortie-point). Ге-
нотип ааВ—c3c3ddOo
С шоколадными черепаховыми отметинами (Chocolate-tortie-
point). Генотип aabbcscsD—Оо
С лиловыми черепаховыми отметинами (Lilac-tortie-point). Ге-
нотип aabbcscsddOo
Следующая группа пойнтовых окрасов — тэбби-пойнт, или с по-
лосатыми отметинами.
С темно-коричневыми полосатыми отметинами (Seal-tabby-
point). Генотип А—В—cscsD—
С голубыми полосатыми отметинами (Blue-tabby-point). Гено-
тип А—В—csc3dd
С шоколадными полосатыми отметинами (Chocolate-tabby-
point). Генотип А—bbcscsD—
С лиловыми полосатыми отметинами (Lilac-tabhy-point). Гено-
тип А—bbcscsdd
192
С красными полосатыми отметинами (Red-tabby-point). Гено-
тип А—cscED—0(0)
С кремовыми полосатыми отметинами (Cream-tabby-point). Ге-
нотип A—csc3ddO(O)
С темно-коричневыми черепаховыми полосатыми отметинами
(Seal-tortie-tabby-point). Генотип А—В—cscsD—Оо. Прочие tortie-
tabby подобны всем черепаховым.
Непременным условием окрасов тэбби-пойпт является нали-
чие четких полос в районе отметин и даже на вибриссах. Лишь
уши должны быть свободными от полос. Полосатый рисунок
везде одного типа, соответственно окрасу.
Бирманские кошки и кошки породы рэгдолл также имеют
Окрасы, полученные на основе гена сиамского альбинизма, но в
отличие от всех остальных пойнтовых пород они имеют белые
пятна. И если у рэгдоллов белая пятнистость занимает достаточ-
но обширные участки поверхности тела, то у бирманских она
не распространяется дальше конечностей, так как кошки этой по-
роды обладают аллелем g (Gloving), ограничивающим распростра-
нение белого цвета, например:
Бирманские с темно-коричневыми отметинами. Генотип
aacsc8D—gg
Бирманские с голубыми отметинами. Генотип aacscsddgg
Таким же образом, включая, разумеется, рецессивные гены
SbSb, выписываются генотипы и остальных окрасов бирманских
кошек.
В связи с наличием белых «носочков» к бирманским предъ-
являются дополнительные требования по окрасу: белый цвет дол-
жен быть чистым, без примеси цветных оттенков или пятен, на
передних ногах «носочки» не должны заходить дальше запястья,
а на задних ногах они заканчиваются не выше скакательного су-
става, и, конечно, отдается предпочтение симметричности.
БУРМАНСКИЕ ОКРАСЫ
Общие требования к окрасу; равномерность распределения
цвета по телу с легким контрастом более темных отметин, на жи-
воте шерсть может быть несколько светлее, чем на спине, у мо-
лодняка возможны следы тигрового рисунка, недопустимы белые
пятна или выраженная тигровость.
Коричневая бурма. Генотип ааВ—cbcbD—
Голубая бурма. Генотип ааВ—cbcbdd
Шоколадная бурма. Генотип aabbcbcbD—
Лиловая бурма. .Генотип aabbcbcbdd
А .также другие окрасы (красный, кремовый, черепаховый и
т. д.)..
. Цв,ет мочки носа и подушечек лап идейтичен соответствую-
щим пойнтовым окрасам. < .
В американских.стандартах цветовые вариации .бурманских ко-
шек носят поэтические , названия: соболь- (темно-коричневая),
шампань (шоколадная), платина’ (лиловая)’.
193
АБИССИНСКИЕ ОКРАСЫ
Зарегистрированы для абиссинских и сомалийских кошек в не-
скольких вариациях, из которых мы рассмотрим основные.
Абиссинский дикий окрас. Генотип А—В—С—D—ПТа—г
Характерен равномерный по всему телу двойной или тройной
тикинг, не формирующий какого-либо рисунка. Чередующиеся
участки шерсти имеют красно-коричневый и черный цвета. Кончи-
ки остей окрашены в черный цвет. Вдоль позвоночника допуска-
ется более темный тон окраса. Подшерсток окрашен в оранжевый
цвет без зональности, поэтому живот, внутренняя сторона ног,
а у сомалийских также грудь и «штаны», имеют ровный оранже-
во-абрикосовый цвет различной интенсивности. Пучки шерсти
между пальцами и подошвы до скакательного сустава черные.
Мочка носа кирпично-красная с черной окантовкой.
Подушечки лап черные, у сомалийских кошек могут иметь
коричневый оттенок.
Основные недостатки окраса: недостаточно выраженная зональ-
ность; серый, осветленный или, напротив, коричневый и черный
подшерсток; наличие рисунка на теле, а также полос на ногах,
колец на хвосте, ожерелья на шее.
Следует отметить, что у сомалийских кошек окрас окончатель-
но формируется к 2-летнему возрасту, что учитывается при оцен-
ке молодняка.
Красно-коричневый абиссинский (соррель). Генотип
A bb’C D iiT1—
Окрас кошек варьирует от рыжевато-коричневого до медно-
красного цвета. В данном случае красный цвет не сцеплен с по-
лом, а вызван действием гена светло-коричневого разбавления.
Корни остей и подшерсток окрашены в темно-абрикосовый цвет.
Тикинг образован зонами интенсивного красно-коричневого цве-
та, что особенно ярко выражено на кончике хвоста и задних но-
гах от подушечек до скакательного сустава.
Мочка носа и подушечки лап розового цвета, но мочка носа,
кроме того, обведена красно-коричневым ободком.
Голубой абиссинский. Генотип А—В—С—ddiiTa—
Кошки имеют общий серо-голубой тон окраса. Зональность
сформирована голубым со стальным оттенком, цветом, переме-
жающимся со светло-кремовыми или бежевыми участками шерсти-
нок. Ступни выделяются более темным голубым цветом.
Мочка носа окрашена в кирпично-красный цвет с голубой
окантовкой. Подушечки лап серо-голубого цвет'а.
Цвет косули (бежевый). Генотип А—ЫЫС—ddiiTa—
Как свидетельствует название, окрас, особенно у абиссинских,
напоминает цвет тела косули, т. е. матово-бежевый вперемежку
с неярким кремовым. Тикинг из темных и светлых кремовых збн.
Мочка носа розовая с грязновато-розовым ободком. Подушеч-
ки лап грязновато-розового или выцветшего розового цвета,
, У всех перечисленных окрасов недостатками считаются те же
признаки, что и у классических абиссинских кошек. 11
194
Приложение
СПИСОК ПОРОД КОШЕК
Индекс Название
1 2
1 1 ь 1 с 2 2а 2Ь 3 4 6 6а 6Ь 6 с 6 d SL 6 dSD 6 d Sm 6 dd SL 6 dd SD 6 dd Sm 6 e 6? 6h 6j 6 e SL 6 e SD 6 g SL 6 g SD 6 h SL 6 h SD 6 jSL 6jSD 7 7 a 7 b Длинношерстные кошки Персидские кошки Черная персидская кошка Шоколадная персидская кошка Лиловая персидская кошка Белая голубоглазая персидская кошка Белая персидская кошка с оранжевыми глазами Белая персидская кошка с различно окрашенными глазами Голубая персидская кошка Красная персидская кошка Кремовая персидская кошка Дымчатая персидская кошка Дымчато-голубая персидская кошка Дымчато-шоколадная персидская кошка Дымчато-лиловая персидская кошка Персидская кошка красного камео Персидская кошка красного затушеванного камео Красно-дымчатая персидская кошка Персидская кошка кремового камео Персидская кошка кремового затушеванного камео Кремово-дымчатая персидская кошка Черепахово-дымчатая персидская кошка Кремово-голубая дымчатая персидская кошка Шоколадная черепахово-дымчатая персидская кошка Лиловая черепахово-дымчатая персидская кошка Завуалированная черепаховая персидская кошка (shell) Затушеванная черепаховая персидская кошка (shaded) Завуалированная голубая черепаховая персидская кошка Затушеванная голубая черепаховая персидская кошка Завуалированная шоколадная черепаховая персид- ская кошка Затушеванная шоколадная черепаховая персидская кошка Завуалированная лиловая черепаховая: персидская кошка Затушеванйая лиловая черепаховая персидская кошка Серебристо-черйая мраморная персидская кошка Сёрёбристо-чголубая мраморная персидская кошка Серебристо-шоколадная Мраморная персидская кошка
195
Продолжение приложения
1 2
7 с Серебристо-лиловая мраморная персидская кошка
8 Черная мраморная персидская кошка
8 а Голубая мраморная персидская кошка
8b Шоколадная мраморная персидская кошка
8 с Лиловая мраморная персидская кошка
8 GSL Завуалированная золотая персидская кошка
8 GSD Затушеванная золотая персидская кошка
9 Красная мраморная персидская кошка
9f Кремовая мраморная персидская кошка
10 Персидская кошка шиншилла
10 а Персидская кошка голубая шиншилла
10 b Персидская кошка шоколадная шиншилла
- 10 с Персидская кошка лиловая шиншилла
10SS Серебристая затушеванная персидская кошка
10SS а Серебристо-голубая затушеванная персидская кошка
10 SS ь Серебристо-шоколадная затушеванная персидская кошка
10SS с Серебристо-лиловая затушеванная персидская кошка
11 Черепаховая персидская кошка
11 ь Шоколадно-черепаховая персидская кошка
И с Лилово-черепаховая персидская кошка
12 Персидская кошка черепаховая с белым
12 b Персидская кошка голубая черепаховая с белым
12 ch Персидская кошка шоколадная черепаховая с белым
121 Персидская кошка лиловая черепаховая с белым
12 а Черно-белая персидская кошка
12 а Ы Голубая с белым персидская кошка
12 a ch Шоколадная с белым персидская кошка
12 al Лиловая с белым персидская кошка
12 аг Красная с белым иерсидская кошка
12 а сг Кремовая с белым персидская кошка
13 Кремово-голубая персидская кошка
Персидская кошка с отметинами — colour point
. 13bSP
13 b BP
13 b ChP
13 b LP
13bRP
. 13 b GrP
13 b STP
13 b BTP
13 b ChTP
1 13 b LTP
13 b Tb SP
, 13 b Tb BP
. 13bTbChP
, 13 b Tb LP
Персидская кошка с темно-коричневыми отметинами
Персидская кошка с голубыми отметинами
Персидская кошка с шоколадными отметинами
Персидская кошка с лиловыми отметинами
Персидская кошка с красными отметинами
Персидская кошка с кремовыми отметинами
Персидская кошка с темно-коричневыми черепахо-
выми отметинами
Персидская кошка с гояубймй черепаховыми отме-
тинами
Персидская кошка с шоколадными черепаховыми от-
метинами ,
Персидская кошка с лиловыми черепаховыми отме-
тинами
Персидская кошка с темно-коричневыми полосатыми
отметинами
Персидская кошка с голубыми полосатыми отмети-
нами
Персидская кошка с шоколадными полосатыми от-
метинами
Персидская кошка с лиловыми полосатыми отмети-
нами
|96
Продолжение приложения
1 9
13bTb RP Персидская кошка с красными полосатыми отмети- нами
13 b ТЬ СгР Персидская кошка с кремовыми полосатыми отме- тинами
13 b Tb STP Персидская кошка с темно-коричневыми черепахо- выми полосатыми отметинами
13 b Tb BTP Персидская кошка с голубыми черепаховыми поло- сатыми отметинами
13 b Tb ChTP Персидская кошка с шоколадными черепаховыми по- лосатыми отметинами
13 b Tb LTP Персидская кошка с лиловыми черепаховыми поло- сатыми отметинами^
13 x Пока неизвестная вариация персидских кошек
Полудлинношерстные кошки
13cSP
13 с ВР
13 с СИР
13 с LP
13 d
13 NF А
13 NF’Aw
13 NF В
13 NF Bw
13 SO
13 SOI
13 SO bl
13 SO fa
13 SO Sv
13 SO R Sv
13 SO bl Sv
13 SO fa Sv
13 MCA
13 MC Aw
13 MG В
13MCBw
13 BA SP
13 BA BP
13 BA ChP
13 BA LP
13 BA RP
13 BA CrP
13 BA STP
13 BA BTP
13BAChTP
13 BA LTP
13 BA Tb SP
13 BA Tb BP
Бирманская кошка с темно-коричневыми отметинами
Бирманская кошка с голубыми отметинами
Бирманская кошка с шоколадными отметинами
Бирманская кошка с лиловыми отметинами
Турецкая кошка
Норвежская лесная кошка с агути-фактором
Норвежская лесная кошка с агути-фактором с белой
окраской
Норвежская лесная кошка без агути-фактора
Норвежская лесная кошка без агути-фактора с белой
окраской
Сомалийская коптка дикого окраса
Красная сомалийская кошка
Голубая сомалийская кошка
Сомалийская кошка цвета косули (beige fawn)
Черная серебристая сомалийская кошка
Красная серебристая сомалийская кошка
Голубая серебристая сомалийская кошка
Цвета косули серебристая сомалийская кошка
Манская енотовая кошка с агути-фактором
Манская енотовая кошка с агути-фактором с белым
окрасом
Манская енотовая кошка без агути-фактора
Манская енотовая кошка без агути-фактора
с белым окрасом
Балинезийская с темно-коричневыми отметинами
Балинезийская с голубыми отметинами
Балинезийская с шоколадными отметинами
Балинезийская с лиловыми отметинами
Балинезийская с красными отметинами
Балинезийская с кремовыми отметинами
Балинезийская с черепаховыми отметинами
Балинезийская с голубыми черепаховыми отметинами
Балинезийская с шоколадными черепаховыми отме-
тинами
Балинезийская с лиловыми черепаховыми отметинами
Балинезийская с темно-коричневыми полосатыми от-
метинами
Балинезийская с голубыми полосатыми отметинами
197
Продолжение приложения
1 2
13 В A Tb ChP Балинезийская с шоколадными полосатыми отмети- нами
13 BA Tb LP Балинезийская с лиловыми полосатыми отметинами
13 В A Tb STP Балинезийская с темно-коричневыми черепаховыми полосатыми отметинами
13 В A Tb ВТР Балинезийская с голубыми черепаховыми пятнистыми отметинами
13 BA Th ChTP Балинезийская с шоколадными черепаховыми поло- сатыми отметинами
13 BA Tb LTP Балинезийская с лиловыми черепаховыми полосаты- ми отметинами ,
13 ТА Ангорская кошка (турецкая ангора)
13 а Пока неизвестная порода длинношерстных кошек Короткошерстные кошки
Экзотические короткошерстные ксшки
1 ЕХ Черная экзотическая короткошерстная кошка
1 EX b Шоколадная экзотическая короткошерстная кошка
1 ЕХ с Лиловая экзотическая короткошерстная кошка
2ЕХ Белая голубоглазая экзотическая короткошерстная кошка
2ЕХ а Белая экзотическая короткошерстная кошка с оран- жевыми глазами
2ЕХ b Белая экзотическая короткошерстная кошка с раз- лично окрашенными глазами
ЗЕХ Голубая экзотическая короткошерстная кошка
4ЕХ Красная экзотическая короткошерстная кошка
5ЕХ Кремовая экзотическая короткошерстная кошка
6 ЕХ Черная дымчатая экзотическая короткошерстная кош- ка
6ЕХ а Голубая дымчатая экзотическая короткошерстная кошка
6 EX b Шоколадная дымчатая экзотическая короткошерстная кошка
6 ЕХ с Лиловая дымчатая экзотическая короткошерстная кошка
6 EX d Sin Красная дымчатая экзотическая короткошерстная кошка
6 EX dd Sin Кремовая дымчатая экзотическая короткошерстная кошка
6 EX d SL Экзотическая короткошерстная кошка красного камео
6 EX d SD Экзотическая короткошерстная кошка красного за- тушеванного камео
6 EX dd SL Экзотическая короткошерстная кошка кремового ка- мео
6 EX dd SD Экзотическая короткошерстная кошка кремового затушеванного камео
6 EX e Черепаховая дымчатая экзотическая короткошерст- ная кошка
6 EX g Кремово-голубая дымчатая экзотическая короткошер- стная кошка
6 EX b Шоколадная черепаховая дымчатая экзотическая ко-
6 EX j роткошерстная кошка
Лиловая черепаховая дымчатая экзотическая корот-
. , - • • • кошерстная кошка
1«Я.
П р о д о л же ии е приложения
1 1 2
6 EX е SL Завуалированная черепаховая экзотическая коротко-
6 EXе SD шерстная кошка Затушеванная черепаховая экзотическая короткошер- стная кошка
6 EX g SL Завуалированная голубая черепаховая экзотическая
6 EX gSD короткошерстная кошка Затушеванная голубая черепаховая экзотическая ко-
6 EX h SL роткошерстная кошка Завуалированная шоколадная черепаховая экзотиче-
6 EX h SD ская короткошерстная кошка Затушеванная шоколадная черепаховая экзотическая
6 EX j SL короткошерстная кошка Завуалированная лиловая черепаховая экзотическая
6 EX j SD короткошерстная кошка Затушеванная лиловая черепаховая экзотическая ко-
7 EX роткошерстная кошка Серебристая черная мраморная экзотическая корот-
7 EX a кошерстная кошка Серебристая юл у бая мраморная экзотическая корот-
7 EX b кошерстная кошка Серебристая шоколадная мраморная экзотическая ко-
7 EX c роткошерстная кошка Серебристая лиловая мраморная экзотическая корот-
8 EX кошерстная кошка Черная мраморная экзотическая короткошерстная кошка
8 EX a Голубая мраморная экзотическая короткошерстная кошка
8 EX b Шоколадная мраморная экзотическая короткошерст-
8 EX c ная кошка Лиловая мраморная экзотическая короткошерстная кошка
8 EX G SL Завуалированная золотая экзотическая короткошерст-
8 EX C SD ная кошка Затушеванная золотая экзотическая короткошерстная кошка
9 EX Красная мраморная экзотическая короткошерстная
9 EX f кошка Кремовая мраморная экзотическая короткошерстная кошка
10 EX Экзотическая короткошерстная кошка черная шин- шилла
10 EX a Экзотическая короткошерстная кошка голубая шин- шилла
10 EX b Экзотическая короткошерстная кошка шоколадная шиншилла
10 EX c Экзотическая короткошерстная кошка лиловая шин- шилла
10 EX SS Серебристая черная затушеванная экзотическая ко-
10 EX SS a роткошерстная кошка Серебристая голубая затушеванная экзотическая ко-
10 EX SS b роткошерстная кошка Серебристая шоколадная затушеванная экзотическая
10 EX SS c короткошерстная кошка Серебристая лиловая затушеванная экзотическая ко-
11 EX роткошерстная кошка Черепаховая экзотическая короткошерстная кошка
199
Продолжение приложения
2
11 EX b
И EX с
12 EX
12 EX b
12 EX ch
12 EX 1
12 EX a
12 EX a bl
12 EX a ch
12 EX al
12 EX ar
12 EX a cr
13 EX
13EXbSP
13 EX b BP
13 EX b ChP
13 EX b LP
13EXbRP
13 EX b CrP
13 EX b STP
13 EX BTP
13EXbChTP
13 EX b LTP
13 EX b Tb SP
13 EX b Tb BP
13 EX b Tb ChP
13 EX b Tb LP
13EXbTb RP
13 EX b Tb CrP
Шоколадная черепаховая экзотическая короткошерст-
ная кошка
Лиловая черепаховая экзотическая короткошерстная
кошка
Экзотическая короткошерстная кошка черепаховая
с белым
Экзотическая короткошерстная кошка голубая чере-
паховая с белым
Экзотическая короткошерстная кошка шоколадная
черепаховая с белым
Экзотическая короткошерстная кошка лиловая чере-
паховая с белым
Черная с белым экзотическая короткошерстная кошка
Голубая с белым экзотическая короткошерстная кошка
Шоколадная с белым экзотическая короткошерстная
кошка
Лиловая с белым экзотическая короткошерстная кош-
ка
Красная с белым экзотическая короткошерстная кош-
ка
Кремовая с белым экзотическая короткошерстная
кошка
Кремово-голубая экзотическая короткошерстная кош-
ка
Экзотическая короткошерстная кошка с темно-корич-
невыми отметинами
Экзотическая короткошерстная кошка с голубыми
отметинами
Экзотическая короткошерстная кошка с шоколадными
отметинами
Экзотическая короткошерстная кошка с лиловыми
отметинами
Экзотическая короткошерстная кошка с красными
отметинами
Экзотическая короткошерстная кошка с кремовыми
отметинами
Экзотическая короткошерстная кошка с черепаховы-
ми отметинами
Экзотическая короткошерстная кошка с голубыми
черепаховыми отметинами
Экзотическая короткошерстная кошка с шоколадными
черепаховыми отметинами
Экзотическая короткошерстная кошка с лиловыми
черепаховыми отметинами
Экзотическая короткошерстная кошка с темно-корич-
невыми полосатыми отметинами
Экзотическая короткошерстная кошка с голубыми
полосатыми отметинами
Экзотическая короткошерстная кошка с шоколадными
полосатыми отметинами
Экзотическая короткошерстная кошка с лиловыми
полосатыми отметинами
Экзотическая короткошерстная кошка с красными
полосатыми отметинами
Экзотическая короткошерстная кошка с кремовыми
полосатыми отметинами
200
Продолжение приложения
1 2
13 EX b Tb STP 13 EX b Tb ВТР 13 EX b ТЬ СЬТР 13 EX b Tb LTP Экзотическая короткошерстная кошка с темно-корич- невыми черепаховыми полосатыми отметинами Экзотическая короткошерстная кошка с голубыми черепаховыми полосатыми отметинами Экзотическая короткошерстная кошка с шоколадными черепаховыми полосатыми отметинами Экзотическая короткошерстная кошка с лиловыми черепаховыми полосатыми отметинами
Британские короткошерстные кошки
14
14 а
14 b
14 х
15
15 b
15 с
16
17
17 d
18 btch
18 tig
18 a btch
18 a tig
18 b btch
18 b tig
18 c btch
18 c tig
19 btch
19 tig
19 f btch
19 f tig
20 btch
20 tig
20 a btch
20 a tig
20 b btch
Белая голубоглазая британская короткошерстная кош-
ка
Белая британская короткошерстная кошка с оран-
жевыми глазами
Белая британская короткошерстная кошка с различно
окрашенными глазами
Британские короткошерстные кошки неизвестных по-
ка вариаций
Черная британская короткошерстная кошка
Шоколадная британская короткошерстная кошка
Лиловая британская короткошерстная кошка
Голубая британская короткошерстная кошка
Кремовая британская короткошерстная кошка
Красная британская короткошерстная кошка
Серебристо-черная мраморная британская коротко-
шерстная кошка
Серебристо-черная тигровая британская короткошер-
стная кошка
Серебристо-голубая мраморная британская коротко-
шерстная кошка
Серебристо-голубая тигровая британская короткошер-
стная кошка
Серебристо-шоколадная мраморная британская корот-
кошерстная кошка
Серебристо-шоколадная тигровая британская корот-
кошерстная кошка
Серебристо-лиловая мраморная британская коротко-
шерстная кошка
Серебристо-лиловая тигровая британская коротко-
шерстная кошка
Красная мраморная британская короткошерстная
кошка
Красная тигровая британская короткошерстная кошка
Кремовая мраморная британская короткошерстная
кошка
Кремовая тигровая британская короткошерстная кош-
ка
Черная мраморная британская короткошерстная кош-
ка
Черная тигровая британская короткошерстная кошка
Голубая мраморная британская короткошерстная кош-
ка
Голубая тигровая британская короткошерстная кошка
Шоколадная мраморная британская короткошерстная
кошка
14 Заказ Л» 355
201
Продолжение приложения
1 2
20 b tig Шоколадная тигровая британская короткошерстная кошка
20 с btch Лиловая мраморная британская короткошерстная кошка
20 с tig- 21 21 Ь Лиловая тигровая британская короткошерстная кошка Черепаховая британская короткошерстная кошка Шоколадная черепаховая британская короткошерст- ная кошка
21 с Лиловая черепаховая британская короткошерстная кошка
22 Британская короткошерстная кошка черепаховая с
22 Ы Британская короткошерстная кошка голубая черепа- ховая с белым
22 ch Британская короткошерстная кошка шоколадная че-
22 1 репаховая с белым Британская короткошерстная кошка лиловая черепа- ховая с белым
28 30 Кремово-голубая британская короткошерстная кошка Темно-коричневая пятнистая британская коротко- шерстная кошка
30 а Голубая пятнистая британская короткошерстная кош-
30 Ь Шоколадная пятнистая британская короткошерстная кошка
30 с Лиловая пятнистая британская короткошерстная кош-
30 О Красная пятнистая британская короткошерстная кош-
30 f Кремовая пятнистая британская короткошерстная кошка
30 S-Sv Серебристо-черная пятнистая британская коротко- шерстная кошка
30 В-Sv Серебристо-голубая пятнистая британская коротко- шерстная кошка
30 Ch-Sv Серебристо-шоколадная пятнистая британская корот- кою третная коьша
30 I-Sv Серебристо-лиловая пятнистая британская коротко- шерстная кошка
31 31 Ы 31 ch Черно-белая британская короткошерстная кошка Голубая с белым британская короткошерстная кошка Шоколадная с белым британская короткошерстная кошка
31 1 31 r 31 ci- 36 36 a 36 b Лиловая с белым британская короткошерстная кошка Красная с белым британская короткошерстная кошка Кремовая с белым британская короткошерстная кошка Дымчатая британская короткошерстная кошка Дымчато-голубая британская короткошерстная кошка Дымчато-шоколадная британская короткошерстная кошка
36 c 37 37 a Дымчато-лиловая британская короткошерстная кошка Британская короткошерстная кошка шиншилла Британская короткошерстная кошка голубая шин- шилла
37 b Британская короткошерстная кошка шоколадная шин- шилла
202
Продолжение приложения
2
37 с Британская короткошерстная кошка лиловая шин-
шилла
37 SS Серебристая затушеванная британская короткошерст- ная кошка
37SS а Серебристо-голубая затушеванная британская корот- кошерстная кошка
37 SS'b Серебристо-шоколадная затушеванная британская ко- роткошерстная кошка
37 SS с Серебристо-лиловая затушеванная британская корот- кошерстная кошка
Европейские короткошерстные кошки.
14 Е
14 Е а
14 ЕЬ
14 Е х
15 Е
15 Е Ь
15Е с
16 Е
17 Е
17 Ed
18 Е btch
18 Е tig
18 Е a btch
18 Е a tig
18 Е b btch
18 Е b tig
18 Е с btch
18 Е с tig
19 Е btch
19 Е tig
19 Е f btch
19 Е f tig
20 Е btch
20 Е tig
20 Ё a btch
Белая^ голубоглазая европейская короткошерстная
кошка
Белая европейская короткошерстная кошка с оран-
жевыми глазами
Белая европейская короткошерстная кошка с раз-
лично окрашенными глазами
Европейские короткошерстные кошки еще неизвест-
ных цветовых вариаций
Черная европейская короткошерстная кошка
Шоколадная европейская короткошерстная кошка
Лиловая европейская короткошерстная кошка
Голубая европейская короткошерстная кошка
Кремовая европейская короткошерстная кошка
Красная европейская короткошерстная кошка
Серебристо-мраморная европейская короткошерстная
кошка
Серебристо-тигровая европейская короткошерстная
кошка
Серебристо-голубая мраморная европейская коротко-
шерстная кошка
Серебристо-голубая тигровая европейская коротко-
шерстная кошка
Серебристо-шоколадная мраморная европейская ко-
роткошерстная кошка
Серебристо-шоколадная европейская короткошерстная
кошка
Серебристо-лиловая мраморная европейская коротко-
шерстная кошка
Серебристо-лиловая тигровая европейская коротко-
шерстная кошка
Красная мраморная европейская короткошерстная
кошка
Красная тигровая европейская короткошерстная кош-
ка
Кремовая мраморная европейская короткошерстная
кошка
Кремовая тигровая европейская короткошерстная
кошка
Черная мраморная европейская короткошерстная Кош-
ка
Черная тигровая европейская короткошерстная кошка
Голубая мраморная европейская короткошерстная
кошка
14*
203
Продолжение приложения
1
20 Е a tig Голубая тигровая европейская короткошерстная крш-
20 Е Ь btch Шоколадная мраморная европейская короткошерст- ная кошка
20 Е b tig Шоколадная тигровая европейская короткошерстная кошка
20 Е с btch Лиловая мраморная европейская короткошерстная кошка
20 Е с tig Лиловая тигровая европейская короткошерстная кош-
21 Е 21 Е b Черепаховая европейская короткошерстная кошка Шоколадная черепаховая европейская короткошерст- ная кошка
21 Е с Лиловая черепаховая европейская короткошерстная кошка
22 Е Европейская короткошерстная кошка черепаховая с
22 Е Ы Европейская короткошерстная кошка голубая чере- паховая с белым
22 Е ch Европейская короткошерстная кошка шоколадная че-
22 Е 1 репаховая с белым Европейская короткошерстная кошка лиловая чере- паховая с белым
28 Е ЗОЕ 30 Е а Кремово-голубая европейская короткошерстная кошка Темно-коричневая пятнистая короткошерстная кошка Голубая пятнистая европейская короткошерстная кош-
ЗОЕ b Шоколадная пятнистая европейская короткошерстная кошка
30 Е с Лиловая пятнистая европейская короткошерстная кошка
ЗОЕ d Красная пятнистая европейская короткошерстная кошка
30 Е f Кремовая пятнистая европейская короткошерстная кошка
30 Е S-Sv Серебристо-черная пятнистая европейская коротко- шерстная кошка
30 Е B-Sv Серебристо-голубая пятнистая европейская коротко-
30 Е Ch-Sv шерстная кошка Серебристо-шоколадная пятнистая европейская ко-
30 Е L-Sv роткошерстная кошка Серебристо-лиловая пятнистая европейская коротко-
31 Е 31 Е Ы 31 Е ch шерстная кошка Черная с белым европейская короткошерстная кошка Голубая с белым европейская короткошерстная кошка Шоколадная с белым европейская короткошерстная кошка
31 Е 1 31 Е г 31 Е сг Лиловая с белым европейская короткошерстная кошка Красная с белым европейская короткошерстная кошка Кремовая с белым европейская короткошерстная кош-
36 Е 36 Е а Дымчатая европейская короткошерстная кошка Дымчато-голубая европейская короткошерстная кош- ка '
36 Е b Дымчато-шоколадная европейская короткошерстная кошка - ’
304
Продолжение приложения
1 2
36 Е с Дымчато-лиловая евроиейская короткошерстная кош-
ка
Остальные короткошерстные кошки
16 а Русская голубая кошка
16 F Картезианская кошка
23 Абиссинская кошка дикого окраса
23 а Красно-коричневая абиссинская кошка
23 Ы Голубая абиссинская кошка
23 fa Абиссинская кошка цвета косули
23 Sv Рыжевато-коричневая серебристая абиссинская кошка
23 a Sv Красная серебристая абиссинская кошка
23 Ы Sv Голубая серебристая абиссинская кошка
23 fa Sv Абиссинская кошка цвета косули серебристая
25 Манская кошка
25 ST М.шская кошка с коротким хвостом (manx stumpy)
" 27 Коричневая бирманская кошка
27 a Голубая бирманская кошка
27 b Шоколадная бирманская кошка
27 c Лиловая бирманская кошка
27 cl Красная бирманская кошка
27 о Черепаховая бирманская кошка
27 f Кремовая бирманская кошка
27 <? Голубая черепаховая бирманская кошка
27 h Шоколадная черепаховая бирманская кошка
- 27 j Лиловая черепаховая бирманская кошка
33 Корниш реке
33 a Девон реке
33 GK Немецкий реке
34 Корат
26 Остальные, пока неизвестные короткошерстные кошки
Сиамские и восточные короткошерстные кошки
Сиамские кошки
24 Сиамская кошка с коричневыми отметинами
24 a Сиамская кошка с голубыми отметинами
24 b Сиамская кошка с шоколадными отметинами
24 c Сиамская кошка с лиловыми отметинами
32 a Сиамская кошка с красными отметинами
32 c Сиамская кошка с кремовыми отметинами
32 b SP Сиамская кошка с темно-коричневыми черепаховыми
отметинами
32 b BP Сиамская кошка с голубыми черепаховыми отмети-
нами
32 b ChP Сиамская кошка с шоколадными черепаховыми от-
32 b LP метинами
Сиамская кошка с лиловыми черепаховыми отметина-
ми
' 32 SP Сиамская кошка с темно-коричневыми пятнистыми
отметинами
- 32 BP Сиамская кошка с голубыми пятнистыми отметинами
32 ChP Сиамская кошка с шоколадными пятнистыми отме-
тинами
32 LP Сиамская кошка с лиловыми пятнистыми отметинами
- '32 STP Сиамская кошка с темно-коричневыми черепаховыми
пятнистыми отметинами
205
Продолжение приложения
1 | 2
32 ВТР Сиамская кошка с голубыми пятнистыми отметинами
32 ChTP Сиамская кошка с шоколадными черепаховыми пят- нистыми отметинами
32 LTP Сиамская кошка с лиловыми черепаховыми пятнисты- ми отметинами
Восточные короткошерстные кошки
29
29 sb
29 а
29 с
29 d
29 е
29 f
29 %
29 h
29 j
35
29 S Tb btch
29 S Tb tig
29 В Tb btch
29 В Tb tig
29 Ch Tb btch
29 Ch Tb tig
29 L Tb btch
29 L Tb tig
29 R Tb btch
29 R Tb tig
29 Cr Tb btch
29 Cr Tb tig
29 S Tb sp
29 В Tb sp
29 Ch Tb sp
29 L Tb sp
29 R Tb sp
29 Cr Tb sp
29 S-Sv Tb btch
29 S-Sv Tb tig
29 В-Sv Tb btch
29 В-Sv Tb tig
Коричневая восточная короткошерстная кошка «Га-
вана»
Черная восточная короткошерстная кошка эбеновая
Голубая восточная короткошерстная кошка
Лиловая восточная короткошерстная кошка
Красная восточная короткошерстная кошка
Черепаховая восточная короткошерстная кошка
Кремовая восточная короткошерстная кошка
Голубая черепаховая восточная короткошерстная
кошка
Шоколадная черепаховая восточная короткошерстная
кошка
Лиловая черепаховая восточная короткошерстная
кошка
Белая восточная короткошерстная кошка
Черная мраморная восточная короткошерстная кошка
Черная тигровая восточная короткошерстная кошка
Голубая мраморная восточная короткошерстная кош-
ка
Голубая тигровая восточная короткошерстная кошка
Шоколадная мраморная восточная короткошерстная
кошка
Шоколадная тигровая восточная короткошерстная
кошка
Лиловая мраморная восточная короткошерстная кош-
ка
Лиловая тигровая восточная короткошерстная кошка
Красная мраморная восточная короткошерстная кош-
ка
Красная тигровая восточная короткошерстная кошка
Кремовая мраморная восточная короткошерстная кош-
ка
Кремовая тигровая восточная короткошерстная кошка
Черная пятнистая восточная короткошерстная кошка
Голубая пятнистая восточная короткошерстная кошка
Шоколадная пятнистая восточная короткошерстная
кошка
Лиловая пятнистая восточная короткошерстная кошка
Красная пятнистая восточная короткошерстная кошка
Кремовая пятнистая восточная короткошерстная кош-
ка
Серебристо-черная мраморная восточная коротко-
шерстная кошка
Серебристо-черная тигровая восточная короткошерст-
ная кошка
Серебристо-голубая мраморная восточная коротко-
шерстная кошка
Серебристо-голубая тигровая восточная короткошерст-
ная кошка .
206
Окончание приложения
1 2
29 Ch-Sv Tb btch Серебристо-шоколадная мраморная восточная корот- кошерстная кошка
29 Ch-Sv Tb tig Серебристо-шоколадная тигровая восточная коротко- шерстная кошка
29 L-Sv Tb btch Серебристо-лиловая мраморная восточная коротко- шерстная кошка
29 L-Sv Tb tig Серебристо-лиловая тигровая восточная короткошер- стная кошка
29;S-Sv Tb sp Серебристо-черная пятнистая восточная короткошерст- ная кошка
29 В-Sv Tb sp Серебристо-голубая пятнистая восточная коротко- шерстная кошка
29 Ch-Sv Tb sp Серебристо-шоколадная пятнистая восточная коротко-
29 L-Sv Tb sp шерстная кошка Серебристо-лиловая пятнистая восточная коротко-
29 x шерстная кошка Сиамские и восточные короткошерстные кошки пока неизвестной цветовой вариации
ПОСЛЕСЛОВИЕ
Дорожите вашими исключениями.
У. Бетсон
Уже на заре генетики выделился довольно узкий круг излюб-
ленных объектов исследований: горох, кукуруза, дрозофила,
мышь. Львиная доля всех генетических работ была выполнена и
выполняется до сих пор на этих объектах. Генетики руководство-
вались простым соображением: что верно для дрозофилы, то вер-
но и для слона, зачем тогда мучиться со слоном. С этим утверж-
дением можно согласиться, когда речь идет о законах Менделя.
Они действительно одинаково справедливы и для дрозофилы,
и для слона. Хотя даже для того чтобы в этом убедиться, пришлось
анализировать не только дрозофилу. Однако чем сильнее генети-
ки углублялись в изучение наследственности^ тем больше обнару-
живалось различий в устройстве генетического материала у раз-
ных видов. Становилось все более ясно — несмотря на удиви-
тельное сходство фундаментальных принципов организации гене-
тического материала у разных видов живых организмов, каждый
из них имеет совершенно уникальный геном, каждый из них
прошел CBoii путь эволюции. Да и сами общие принципы выяви-
лись только тогда, когда составилась достаточно полная картина
генетики отдельных видов.
Медленно, но неуклонно расширялся круг генетических объек-
тов. Это было на первых порах трудной и неблагодарной работой:
получать потомство от медленно размножающихся видов, оцени-
вать расщепления у малоплодных животных. Нужно было про-
анализировать мутации альбинизма у мыши, кролика, крысы и
кошки, чтобы прийти к заключению об их гомологии. Генетиче-
ская карта мыши, безусловно, интересна сама по себе. Но гораздо
важнее те выводы, которые следуют из сравнения ее с картами че-
ловека и кошки. То, что у кошки 38 хромосом — само по себе не
более, чем констатация факта. Нам нужно знать, сколько и каких
хромосом у пумы, тигра и ягуарунди, чтобы понять, почему у нее
38 хромосом.
Первые генетические исследования на кошке были выполнены
в 20-е годы нашего века, хотя некоторые данные о наследовании
ее экстерьерных признаков можно обнаружить и в более ранних
работах. Например, еще Ч. Дарвин отмечал плейотропные эф-
фекты гена доминантной белой окраски. Однако наиболее серьез-
ный прогресс в накоплении и систематизации генетических зна-
ний о данном виде был достигнут благодаря работам одного из
авторов этой книги, английского генетика Роя Робинсона. Он ус-
208
тановил характер наследования некоторых экстерьерных призна-
ков и опубликовал ряд превосходных сводок по генетике кошки.
Примечательно, что все это он сделал, будучи самостоятельным
п самофинанспруемым исследователем. Рой Робинсон — один из
последних представителей редкого ныне племени натуралистов,
которые занимаются изучением природы вещей из чистого любо-
пытства и за собственный счет. Так работал Ч. Дарвин в XIX в.,
так работает Рой Робинсон в наши дни. Организация, в которой
он работает, имеет в штате одного человека — его самого — и на-
зывается Приют улицы Св. Стефана. Его дом стоит на этой лон-
донской улице и во дворе дома он разводит кошек, крыс и хомя-
ков. Зачем? Он их продает? Нет, ему интересно, как наследуется
окраска их шерсти. Ему платят за эти исследования? Нет, ему
просто интересно этим заниматься. Редактор этой книги чувство-
вал себя очень неловко, когда, придя в Приют улицы Св. Стефа-
на, просил его владельца написать четыре главы для «Генетики
кошки». Он честно признался, что ему нечем платить за эту ог-
ромную работу. «Не беспокойтесь,— сказал Рой Робинсон.— Мне
будет интересно это сделать. Не хотите ли еще чаю?».
Из чистого любопытства была сделана и та гигантская рабо-
та по геногеографии популяций кошек, результаты которой при-
ведены в гл. 11. Почти все участники этого уникального между-
народного проекта — натуралисты-любители. Они проводили эти
исследования в свободное от основной работы время. Роль коор-
динатора проекта играл американский генетик Нил Б. Тодд, ру-
ководитель Центра исследований генетики хищных (кроме него
в центре работают его жена и дочь). Этой группой натуралистов
проанализировано более 300 популяций. И ныне с геногеографи-
ческой картой кошки может соперничать лишь геногеографиче-
ская карта человека.
Как возник этот международный проект? Как все начина-
лось? Вот что об этом пишет пионер геногеографических иссле-
дований сотрудник отдела радиобиологии Медицинского исследо-
вательского совета (Соединенное Королевство) Антони Серл.
«Я думаю, что все это началось в гениальной голове профес-
сора Дж. Б. С. Холдейна, члена Королевского общества, генетика
экстраординарного и выдающегося во многих отношениях, в том
числе и в отношении его телосложения. Он держал в доме ко-
шек (белую глухую кошку, один глаз которой был желтым,
а другой — голубым, ее способности он часто демонстрировал
своим студентам; и орехового абиссинского тэбби, очень изящно
сложенного)^и знал все о генетике кошек. В частности, он знал
о наследовании сцепленного с полом гена рыжей окраски и по-
нимал, что этот ген очень удобен для проверки соблюдения соот-
ношения Харди — Вейнберга в популяциях кошек. Нет сомне-
ний, что он держал эту мысль в голове, когда в 1947 г. я на-
брался, наконец, смелости постучать в его дверь в Университет-
ском колледже Лондона. Я пришел к нему просить совета, ка-
кую тему мне выбрать для курсовой работы. Он немедленно
209
предложил мне исследовать лондонских кошек. Я учился тогда
(в 1947 г.) на втором курсе, по так как я, вернувшись с войны,
чувствовал себя древним старцем — мне очень хотелось опубли-
коваться.
Холдейн отнесся ко мне с симпатией (хотя в первый момент
я привел его в ярость, усевшись по ошибке в его любимое кресло)..
Он объяснил в нескольких словах, почему считает этот проект стоя-
щим — регистрацию разнообразия по сегрегирующим признакам
окраски шерсти в большой выборке лондонских кошек. Несмотря
па мое полное незнание генетики кошек, я быстро согласился.
„Проф“ п его замечательная жена и бывшая ученица доктор
Элеи Спаруэй немедленно приобщили меня к этой науке. Они
взяли меня на своего рода вечернее сафари по страшно разбомб-
ленным районам вокруг Университетского колледжа Лондона.
Именно там Холдейны провели со мной семинарские занятия,
там же и проэкзаменовали по распознаванию генотипов и фено-
типов кошек. За всем этим последовал ужин в их доме, где я
познакомился с их „коллекцией11 кошек, которые служили объ-
ектом этологических исследований для Элеи.
Было не так уж трудно найти подходящее место для учета
кошек, поскольку в то время довольно много „организаций по
спасению животных11 занимались отловом и уничтожением бро-
дячих кошек. Я не стану утверждать, что внимательное исследо-
вание мусорных баков, заполненных кошачьими трупами, было
очень приятным занятием, но по крайней мере такой подход поз-
волил мне набрать большой материал за короткое время. Хол-
дейн постоянно следил за моей работой и очень помог мне в ана-
лизе результатов и подготовке статьи „Генные частоты у лондон-
ских кошек11 для „Journal of Genetics11, который он тогда редак-
тировал. Я помню, что на определенном этапе работы ему в го-
лову пришла идея, что я должен представить мои результаты на
заседании Королевского общества, потому что, как он считал,
„вещи такого рода должны понравиться этим простодушным пар-
иям11, но к моему облегчению, он не стал настаивать на этом.
Статья появилась в 1949 г. В ней было показано, что кошки
(и, видимо, только они, единственные из всех доместицирован-
пых видов) следуют уравнению Харди — Вейнберга и ведут себя
в этом отношении как природная популяция. Я писал в этой
статье: „...биологическое значение генных частот не может быть
оценено, если мы не имеем данных по генным частотам в не-
скольких районах11.
Тремя годами позже Таку Комаи описал генные частоты для
популяции кошек из Мишины, Япония. (Показательно, что его
внимание к моей статье привлек сэр Сьюэл Райт.) Через 10 лет я
сам провел такую же работу в Сингапуре. Я ясно представляю
теперь, что если бы эти три исследования дали одинаковые ре-
зультаты, на этом бы все дело и кончилось. Вместо этого они
выявили очень значительные различия в генных частотах по оп-
ределенным генам, что свидетельствовало о наличии глобальных
210
клинов генных частот. Необходимость продолжения исследований
стала действительно очевидной...»
И оно не замедлило последовать. Одиако, несмотря на боль-
шое количество исследований, геногеографическая карта попу-
ляций кошек все еще далеко не полна. Усилий редактора — ав-
тора послесловия — и нескольких его друзей и коллег было недо-
статочно, чтобы сделать точную геногеографическую карту быв-
шего Советского Союза. Попытки использовать данные, собран-
ные зрителями его телевизионных передач, окончились неудачей.
Обнаружилось, что для проведения исследований, даже таких,
казалось бы, простых, как подсчет количества кошек разных ок-
расов, недостаточно одного энтузиазма, требуются серьезные ге-
нетические знания. Внимательный читатель этой книги их уже
приобрел и вполне подготовлен к такого рода исследованию.
Современная наука становится все более дорогим удовольст-
вием, уделом профессионалов. Натуралисты-любители, трудами
которых развивалась биология в прошлом, сейчас остались не
у дел. Их средств не хватит для серьезных научных исследова-
ний. Это в основном верно, но остались области, которые могут
развиваться усилиями любителей. По-видимому, именно в их ру-
ки и попадет большинство экземпляров этой книги, потому что
профессионалов в генетике кошек во всем мире едва ли больше
двух десятков. И вот они-то, любители кошек, и могут сущест-
венно способствовать развитию той области науки, которой посвя-
щена эта книга. Они могут участвовать в составлении геногео-
графической карты популяций кошек. Для этого им требуется
всего четыре вещи: пара крепких ботинок, записная книжка,
карандаш и знание генетики окраски кошек. Любители, и толь-
ко они, могут внести вклад в расширение реестра мутантных
аллелей кошек. Один из отцов-основателей генетики Уильям
Бетсон советовал: «Дорожите вашими исключениями». Вся ге-
нетика возникла на основе анализа исключений — носителей му-
тантных аллелей. Если ваша кошка принесла необычного котен-
ка — дорожите им, проведите его генетический анализ. Помните,
что мутации возникают редко и каждой из них нет цены. Вы
можете внести свой вклад в генетику кошки. Сделайте это.
II. М. Бородин
ОГЛАВЛЕНИЕ
Предисловие (.4. О. Рувинский)............................ 3
Глава 1. Эволюция семейства кошачьих и систематическое положе-
ние домашней кошки (С. О’Брайен, пер. П. М. Бородина) 5
Глава 2. Доместикация (Р. Робинсон, пер. Т. Ю. Ладыгиной) . . 18
Глава 3. Цитогенетика кошачьих (Л. С. Графодатский) ... 27
Глава 4. Генетика окрасок (Р. Робинсон, пер. Т. Ю. Ладыгиной) . 44
Глава 5. Генетика структуры шерсти (Р. Робинсон, пер. Т. Ю. Лады-
гиной) .................................................. 53
Глава 6. Врожденные аномалии и наследственные болезни (Р. Ро-
бинсон, пер. Т. Ю. Ладыгиной)..............................58
Глава 7. Биохимическая и молекулярная генетика (С. О’Брайен, пер.
П. М. Бородина) ............ 74
Глава 8. Иммуногенетика (Л. В. Таранин) ....... 82
Глава 9. Врожденное поведение кошки (Т. А. Алехина) ... 89
Глава 10. Биология размножения (А. И. Железова)..........102
Глава И. Генетика популяций и геногео.графия (Я. Блейн, пер.
П. М. Бородина) .................................118
Глава 12. Породы кошек, их происхождение и стандарты (О. А. Си-
зова) ................................................... 150
Глава 13. Цветовые вариации пород кошек и их стандарты (О. А. Си-
зова) ................................................. 181
Приложение. Список пород кошек (О. А. Сизова)............195
Послесловие (77. М. Бородин) . . 208
Научное издание
О’Брайен Стивен
Робинсон Рой
Графодатский Александр Сергеевич и др.
ГЕНЕТИКА
КОШКИ
Редактор Н. Л. Шварцкопп
Художественный редактор В. А. Реймхе
Художник А. И. Смирнов
Технический редактор Г. Я. Герасимчук
Корректор В. В. Игнатьева
ИБ № 51050
Сдано в набор 24.08.92. Подписано к печати 27.11.92. Формат 60Х90‘/:б Бумага
тип. № 2. Высокая печать. Усл. иеч. л. 13,5. Усл. кр.-отт. 13,8. Уч.-изд. л. 15,5.
Тираж 1350 экз. Заказ № 355. С027.
Ордена Трудового Красного Знамени ВО «Наука», Сибирская издательская фирма.
630099 Новосибирск, ул. Советская, 18.
Новосибирская типография № 4 ВО «Наука». 630077 Новосибирск, ул. Станислав-
ского, 25.