


























































































































































































































































Автор: Саулич А.Х.
Теги: insecta hexapoda насекомые энтомология экология и география животных охрана животных биология экология мир насекомых
ISBN: 5-288-02031-0
Год: 1999




УДК 595.7
ББК 28.680
С12
Рецензенты: докт. биол. наук А.В. Селиховкин (СПбЛТА), докт. биол. наук
ГЛ Носков (СПбГУ)
Печатается по постановлению
Редакционно-издательского совета
С.-Петербургского государственного
университета
Саудия А.Х. Сезонное развитие насекомых и возможности
С12 их расселения. - СПб.: Изд-во С.-Петерб. ун-та, 1999. - 248 с.
ISBN 5-288-02031-0
В монографии обобщены новейшие сведения о сезоино-цикличес-
ких адаптациях насекомых. На основе экспериментального исследования
16 видов их трех крупнейших отрядов насекомых (Coleoptera, Heteroptera,
Lepidoptera) выявлены основные параметры фотопериодических реакций,
участвующие в синхронизации сезонного развития с ритмом климата и оп-
ределяющие возможность натурализации локальных популяций при широ-
ких внутри- и внеареальных перемещениях. Установлены основные типы
сезонных циклов; показана однотипность регуляции поливольтинного цикла
и многообразие путей осуществления моновольтинного сезонного цикла.
Предложена обобщенная схемв способов поддержания моновольтинизма.
Доказано отсутствие сезонно-циклических реакций у гомодинамных видов
и невозможность их круглогодичного обитания в умеренных широтах.
Книга предназначена для энтомологов и экологов.
ББК 28.680
Издание подготовлено при поддержке РФФИ,
грант № 98-04-62114
ISBN 5-288-02031-0
© А.Х. Саулич, 1999
© Издательство
С.-Петербургского
университета, 1999
Предисловие
В течение длительной эволюции у насекомых выработались
разнообразные жизненные стратегии, направленные на адаптацию
к периодически изменяющимся условиям внешней среды. Среди
них различные, иногда очень сложные годичные циклы, структура
которых определяется числом поколений, календарной приуро-
ченностью отдельных стадий онтогенеза, включением в цикл раз-
ных форм физиологического покоя. Исследование сезонных адап-
таций — одна из важнейших и интереснейших областей экологии
насекомых.
Изучение фенологии и сезонных циклов имеет глубокие кор-
ни, но лишь с развитием исследований фотопериодизма насеко-
мых в середине нашего века оказался возможным причинный ана-
лиз сезонного развития. Первые блестящие примеры анализа фе-
нологии на основе фотопериодических и температурных реакций
были продемонстрированы в работах А.С. Данилевского с сотруд-
никами [1961,1968]. Они отличаются убедительной корреляцией
прогноза развития с фенологическими событиями. В большин-
стве рассмотренных случаев для этого оказалось достаточно не-
большого числа параметров фотопериодической реакции, опре-
деленных в константных условиях.
Однако среди исследованных фенологических схем были и,
такие, которые не поддавались объяснению, исходя из традици-
онных представлений. При чем очевидно, что причина этого за-
ключалась не в ошибочности сложившихся в области фенологи-
ческого прогнозирования методических подходов, а в недостатке
имеющихся знаний. Так, расшифровка сложных жизненных цик-
ЙОВ некоторых жужелиц стала возможна только после обнаруже-
ния у них и исследования ступенчатых фотопериодических реак-
Ш'[Thiele, 1975; Ferenz, 1977]; при изучении сезонного развития,
3
стрекоз и некоторых сетчатокрылых была оценена чувствитель-
ность насекомых к постепенным изменениям длины дня [Corbet,
1956; Tauber, Tauber, 1973]. Последующие исследования расшири-
ли круг параметров фотопериодической реакции, характеризую-
щих ее «работу» в природе [Горышин, Гейспиц, 1975; Sauer, Spieth,
1986]. В настоящее время ни у кого не вызывает сомнения доми-
нирующая роль фотопериода и температуры в регуляции сезонно-
го развития всех живых организмов и насекомых в том числе.
С момента выхода в свет блестящего и по сей день непревзой-
денного по глубине и масштабности обобщения А.С. Данилевского
[1961], касающегося фотопериодизма насекомых, прошло более
30 лет. За эти годы все же достигнуты успехи в изучении сезонных
адаптаций и назрела необходимость их обобщения.
Настоящая монография содержит новейшие сведения по эко-
логии фотопериодизма и обобщает результаты 30-летних иссле-
дований автора, самостоятельных или в сотрудничестве с другими
специалистами, частично опубликованные в отечественных и за-
рубежных журналах. Книга, однако, не претендует на полный охват
анализируемой проблемы, в ней обсуждается следующий круг во-
просов:
— типы сезонного развития насекомых;
- роль основных параметров фотопериодической реакции в
синхронизации развития насекомых с ритмом климата;
— эколого-географическая структура вида и адаптивные воз-
можности локальных популяций разных видов при внутриареаль-
ных перемещениях;
— значение сезонно-циклических реакций адвентивных ви-
дов для формирования вторичных ареалов.
В первой части монографии излагаются современные пред-
ставления о принципах экологической регуляции сезонного раз-
вития насекомых, во второй приводятся оригинальные экспери-
ментальные данные, характеризующие реальную функцию фото-
периодической реакции в синхронизации их годичного цикла с
ритмом климата, и показана определяющая роль фотопериоди-
ческих адаптаций при расселении насекомых.
Автор пользуется случаем выразить искреннюю благодарность
своему учителю - Николаю Ивановичу Горышину и коллеге Татья-
не Анатольевне Волкович, в тесном сотрудничестве с которыми
' автору посчастливилось работать многие годы.
Часть I
ЭКОЛОГИЧЕСКАЯ РЕГУЛЯЦИЯ
СЕЗОННОГО ЦИКЛА НАСЕКОМЫХ
Глава 1. РИТМ КЛИМАТА И СЕЗОННЫЕ АДАПТАЦИИ
Среда обитания большинства живых организмов подвержена
глубоким сезонным колебаниям. Все важные для жизни физичес-
кие факторы, такие как температура, свет, количество осадков, и
связанное с ними наличие или отсутствие корма, настолько силь-
но изменяются в течение года, что от организма требуются специ-
альные адаптации, чтобы выживать в новых условиях. Такие адап-
тации чаще всего проявляются в последовательной смене физио-
логических состояний. Поскольку амплитуда сезонных измене-
ний возрастает с увеличением географической широты, то и се-
зонная ритмичность процессов жизнедеятельности ярче выраже-
на у видов, населяющих умеренные и приполярные зоны. Сход-
ные тенденции обнаруживаются и у видов горных ландшафтов, где
выражена вертикальная зональность климата. Только в тропиках
сезонные колебания значительно слабее, однако и здесь смена
сухого и влажного сезонов требует от насекомых определенных
адаптивных стратегий.
1.1. Активная жизнедеятельность и физиологический покой
Сезонные циклы насекомых представляют собой синхронное
ритму климата чередование состояний активной жизнедеятель-
ности и физиологического покоя [Данилевский, 1961].
В период активной жизнедеятельности у насекомых осуще-
ствляются важнейшие биологические функции - рост, развитие,
^змножение, расселение. Все эти проявления нормальной жиз-
недеятельности связаны с большими энергетическими затратами,
сопровождаются высоким уровнем метаболизма, что в свою оче-
редь требует интенсивного питания и комфортных условий внеш-
ней среды.
5
Жизнедеятельность насекомых как пойкилотермных организ-
мов в большой степени зависит от физических факторов, особен-
но от температуры. Она определяет саму возможность их суще-
ствования. Отклонение значения температуры от оптимального
влияет на жизнеспособность: снижает плодовитость и выживае-
мость. Скорость протекания всех процессов жизнедеятельности
также является функцией температуры.
Кроме того, температура может служить сигналом, определя-
ющим поведение насекомых, индуцирующим диапаузу и другие
сезонные феномены. В этом случае ее энергетическое действие ста-
новится второстепенным, а главнее значение приобретают такие
характеристики, как изменение действующего фактора во време-
ни (суточная и сезонная динамика) или в пространстве (темпера-
турный градиент).
Среди других климатических факторов, влияющих на разви-
тие наземных насекомых, выделяется влажность. Малые размеры
усложняют для них задачу сохранения влаги в теле и поддержания
водного баланса. Однако она успешно решается благодаря разно-
образным механизмам (Томс, 1987], ограничивающим испарение
и обеспечивающим образование метаболической воды. Основным
источником ее пополнения служит вода, получаемая с пищей, и
свободная вода. Известный физиолог В.Б. Вигглсворс [Wig-
glesworth, 1954] очень точно определил, что вся организация насе-
комых направлена на сохранение воды. Действие температуры и
влажности настолько взаимосвязаны, что говорят о комплексном
влиянии на насекомых гигротермического режима. Это влияние
многогранно.
Большое значение для жизнедеятельности насекомых имеет
световой фактор. Однако его воздействие трансформируется через
термические показатели, поэтому отметим лишь такую его сторо-
ну, как суточный ритм (смена дня и ночи). Длина светового дня,
или фотопериод — классический пример сигнального фактора.
Далее его экологическая роль будет рассмотрена подробнее в свя-
зи с регуляцией сезонных явлений и синхронизацией их проявле-
ния с ритмом климата. Здесь следует подчеркнуть, что фотопери-
од выполняет важную синхронизирующую функцию на всех уров-
нях организации живого — от клеточного до экосистемного, обес-
печивая биологическую хронометрию в широком смысле.
6
Среди факторов среды, контролирующих развитие насекомых,
особое место занимают условия питания, о чем свидетельствует
широкий круг таких адаптаций, как пищевая специализация и ре-
жимы питания, адаптации, связанные с поиском пищи, сложные
формы поведения энтомофагов и их жертв, сложившиеся в ходе
сопряженной эволюции видов, и т.д. Имеется много данных о вли-
янии качества корма на выживаемость и длительность преимаги-
нальных стадий, плодовитость имаго. Общей закономерностью
является угнетение жизнедеятельности при ухудшении условий
питания. Через главную свою функцию - поставлять энергетичес-
кие ресурсы — этот фактор во многих случаях влияет и на сезонные
циклы (Богачева, 1995].
Требования активных стадий онтогенеза насекомых к абио-
тическим компонентам среды достаточно однотипны и, как пра-
вило, соответствуют условиям вегетационного сезона. Температур-
ные различия летнего периода, определяемые географической ши-
ротой, отражаются лишь на числе поколений при поливольтин-
ном сезонном цикле, тогда как разнообразие фенологических схем,
встречающихся у насекомых, в основном создается за счет различ-
ных форм покоя, адаптированных к переживанию неблагоприят-
ных периодов года, характеризующихся низкими или высокими
температурами, засухами, отсутствием корма и т.д. В зоне умерен-
ного климата переход в состояние покоя в большей степени слу-
жит адаптацией насекомых к условиям зимы, где основными ли-
митирующими факторами выступают низкие температуры и дли-
тельный период отсутствия корма.
В настоящее время известно большое разнообразие форм фи-
зиологического покоя, различающихся интенсивностью, продол-
жительностью, стадией онтогенеза, физиологическими и биохи-
мическими особенностями. В существующих классификациях
[Muller, 1965, 1992; Mansing, 1971; Ушатинская, 1973; Меривээ,
1978] их разнообразие единодушно разделяют на два основных
типа: конзекутивный и проспективный. Покой первого типа на-
ступает под прямым давлением неблагоприятных условий среды и
заканчивается сразу после прекращения их действия. Покой вто-
рого типа формируется в качестве адаптации на повторяющиеся
^.благоприятные для активной жизнедеятельности условия внеш-
ЦСЙ среды. Ему всегда предшествует более или менее длительная
физиологическая подготовка. Возвращение в активное состояние
7
также связано со значительной физиологической перестройкой
организма.
1.2. Диапауза как наиболее совершенная форма покоя.
Краткая характеристика и значение
Типичной и наиболее изученной формой проспективного
покоя у насекомых является диапауза. Она обнаружена у предста-
вителей всех отрядов насекомых, включая первичнобескрылых
[Saunders, 1976; Danks, 1987; 1996; Leather et al., 1993; Nishizukaet
al., 1998 ].
Согласно современным представлениям диапауза насеко-
мых — это гормонально обусловленное состояние организма, ха-
рактеризующееся комплексом морфологических, физиологичес-
ких и поведенческих особенностей, известных как «синдром диа-
паузы» [Tauber et al., 1986].
При наступлении диапаузы тормозятся или приостанавлива-
ются процессы роста и морфогенеза. Резко снижается интенсив-
ность дыхания при полном или частичном замещении оксидатив-
ного (аэробного) обмена веществ аноксидативным (анаэробным).
В большинстве случаев прекращается питание. Минимальный уро-
вень жизнедеятельности поддерживается за счет накопленных ре-
зервов. Происходит значительная дегидратация организма. В со-
стоянии диапаузы повышается устойчивость к холоду, иссушаю-
щему действию среды, недостатку кислорода, увеличивается об-
щая неспецифическая устойчивость организмов к повреждающим
агентам {Lees, 1955; Ушатинская, 1957, 1958, 1980; Harvey, 1962;
Ильинская, 1963, 1965, 1966, 1971; Асахина, 1964; Лозина-Лозин-
ский, 1972; Гирфанова, 1984; Черныши др., 1986; Черныш, Симо-
ненко, 1988; Черныш, 1990; Виноградова, 1991, и др.].
У насекомых различают эмбриональную, личиночную, куко-
лочную и имагинальную диапаузу в зависимости от того, на какой
стадии онтогенеза она формируется. Несмотря на то, что диапауза
может наступать на любой его стадии, как правило, диапаузирую-
щая стадия для каждого вида специфична, и нередко состояние
покоя фиксировано даже в пределах одной стадии [Кинд, 1972;
Новак, 1972; Фалькович, 1979; Danks, 1987, и др.]. Однако встре-
чаются виды, способные формировать диапаузу не только на од-
ной стадии. Например, бархатница Pararge aegeria (Lepidoptera,
8
Satyridae) одинаково успешно зимует и диапаузирующей личин-
кой й Диапаузирующей куколкой [Lees, Tilley, 1980].
Строгое соответствие того или иного типа диапаузы опреде-
ленным систематическим группам насекомых прослеживается сла-
бо [Тыщенко, 1973; Taylor, Spalding, 1988], хотя в пределах отряда
или семейства обычно можно выделить доминирующий ее тип.
Так, у наземных полужесткокрылых (Heteroptera) чаще встречает-
ся эмбриональная и имагинальная диапауза, в пределах же семей-
ства щитников (Pentatomidae) наиболее распространена имагиналь-
ная. У исследованных в нашей лаборатории чешуекрылых обнару-
жены все типы диапаузы: от эмбриональной (Orgyia antique^ Bombyx
mori) до имагинальной (Inachis urticae, Lio и др.), но сем.
Nymphalidae, например, свойственна диапауза на стадии имаго.
Данные о встречаемости разных типов диапаузы в крупней-
ших отрядах насекомых (табл.1), представленные Х.В. Дэнксом
[Danks, 1987], уже требуют дополнений. В частности, в отряде же-
сткокрылых эмбриональная диапауза описана у листоедов Atrachya
menetriesi [Miya, 1965; Ando, 1983], Galeruca tanaceti [Siew, 1966], у
6 видов рода Diabrotica [Krysan, 1972; Krysan, 1982; Krysan et al.,
1984] и у Pyrrhalta humeralis [Nakai, Takeda, 1995]. Таким образом,
во всех отрядах отмечены все типы диапаузы. Исключение состав-
ляют Только представители Coleoptera, у которых до сих пор не
обнаружена куколочная диапауза.
Для большинства насекомых основными факторами, опреде-
ляющими индукцию диапаузы, являются температура и фотопе-
рйод, влияние которых в естественных условиях в значительной
степени взаимосвязано. Однако в настоящее время однозначно
Таблица 1. Встречаемость разных типов диапаузы в крупнейших
отрядах насекомых [по: Danks, 1987]
Отряд Кол-во видов, %, дивпаузирующих нв стадии: '4 Число ирсдедоавнных видов
яйца личинки куколки имвго
Ortho pte га 33 33 — 33 15
Homoptera 46 23 *** 31 26
Heteroptera 21 7 — 71 14
Coleoptera 0 10 0 90 39
1 Di pte га 13 32 18 37 76
>LBpldoptera 3 45 48 4 110
Hymenoptera 10 82 4 4 28
9
доказано доминирующее значение фотопериода, лишь в отдель-
ных случаях температура может выступать как самостоятельный
фактор. Так, пониженная температура оказалась основной причи-
ной, индуцирующей куколочную диапаузу у мух саркофагид
[Denlinger, 1979, 1986; Виноградова, 1991], предкуколочную диа-
паузу у видов рода Trichogramma [Сорокина, Масленникова, 1986,
1987] и имагинальную диапаузу у клопа Orius albidipennis [Chyzik
etal., 1995].
Другие факторы окружающей среды также могут участвовать в
регуляции сезонного цикла, при этом обычно прослеживается об-
щая тенденция: благоприятное для вида действие фактора спо-
собствует появлению активно развивающихся особей, а неблаго-
приятное усиливает тенденцию к формированию диапаузы.
1.3. Зимняя диапауза и холодоустойчивость
В зависимости от сезонной приуроченности и экологичес-
кой направленности различают зимнюю и летнюю диапаузу. Ги-
бернация (зимний покой) широко распространена у насекомых
умеренных и высоких широт и является основной адаптацией к
переживанию зимнего периода. Один из обязательных атрибутов
зимней диапаузы у насекомых - устойчивость к холоду [Ушатин-
ская, 1957, 1980, 1990; Baust, 1981; Lee, Denlinger, 1985; Leeetal.,
1987a; Woude, Verhoef, 1988; Denlinger, 1991; Leatheretal., 1993].
,, Среди разнообразных стратегий насекомых, направленных на
выживание в морозный период года, можно выделить две основ-
ные: холодостойкость и морозостойкость. Диапаузирующие насе-
комые чаще переживают низкие температуры в состоянии пере-
охлаждения без образования льда в клетках. При этом некоторые
виды без последствий переносят понижение температуры до -50 °C
[Somme, 1982].
Основную роль в повышении способности к переохлаждению
у зимующих насекомых играют три группы химических соедине-
ний: многоатомные спирты, моно-, ди- и полисахариды и термо-
гистерезисные белки.
Наиболее распространенным у насекомых криопротектором
из группы многоатомных спиртов является глицерин. Его концен-
трация резко повышается при понижении температуры окружаю-
щей среды от 10 до 0 °C и у некоторых зимующих насекомых мо-
10
жет составлять 25-30% массы тела [Somme, 1982]. Накопление
глицерина сопровождается повышением холодостойкости, кото-
рая достигает максимального уровня к середине декабря. Ресин-
тез глицерина в гликоген начинается с подъемом температуры до
положительных значений и, как отмечают многие авторы, может
происходить очень быстро. Например, у жужелицы Pterostichus
brevlcomis с Аляски содержание глицерина падало от 22 до 1% в
течение 36 ч [Baust, Miller, 1970].
Роль сахаров в механизме повышения холодостойкости, ви-
димо, не столь существенна. Хотя у некоторых видов, например у
короеда-типографа, глицерин обеспечивает холодозащитную фун-
кцию только во второй половине зимы, тогда как в первую полови-
ну основное значение приобретает глюкоза [Хансен и др., 1980].
В последнее время большое внимание уделяется холодоза-
щитным свойствам белков гемолимфы [Duman etal., 1982; Duman,
Horwath, 1983; Morgan, Chippendale, 1983; Horwath et al., 1996].
Присутствие термо-гистерезисных протеинов понижает темпера-
турузамерзания гемолимфы на 5-6 °C, предотвращая образова-
ние внутриклеточного льда, и способствует стабилизации состоя-
ния переохлаждения, которое в большинстве случаев очень не стой-
ко [Horwath, Duman, 1982; Zachariassen, 1985].
' Другой механизм, позволяющий выживать в условиях экстре-
мально низких температур (до -80 °C), - морозостойкость. Спо-
собность выдерживать глубокое замерзание более характерно для
насекомых, обитающих в самых суровых климатических условиях
[Лозина-Лозинский, 1972; Somme, 1985; Zachariassen, 1985; Danks,
1987; Ушатинская, 1990].
У большинства морозостойких видов обнаружены так назы-
ваемые «льдонуклеирующие агенты» (ice nucleating agents), кото-
рые синтезируются поздней осенью или ранней весной. Они слу-
жат своеобразными центрами кристаллизации, способствуя экст-
рацеллюлярному выпадению кристаллов при температурах —5...
-10 °C. До полной активации льдонуклеирующих агентов холодо-
защитные функции выполняют полиолы, повышающие способ-
ность к переохлаждению, и антифризные протеины, стабилизи-
рующие состояние переохлаждения [Duman et al., 1982].
Согласно результатам недавних исследований граница между
Устойчивыми и неустойчивыми к замерзанию видами весьма ус-
ловна: у холодостойких видов наряду с глубоким переохлаждени-
11
ем отмечается частичное образование льда в тканях [Baust et al.,
1985], а морозостойкие виды способны к неглубокому переохлаж-
дению, обеспечивающему выживание до полной мобилизации
льдонуклеирующих белков. Это положение развивается и Д.С. Бей-
лом [Bale, 1993, 1996], считающим, что по степени резистентнос-
ти к холоду насекомые образуют непрерывный ряд, на одном кон-
це которого виды с минимальной, а на другом - с максимальной
выносливостью к действию экстремально низких температур.
В связи с изложенным представляет интерес вопрос о взаи-
мосвязи холодоустойчивости и диапаузы, Есть много фактов, ука-
зывающих на то, что такая связь не обязательна. Например, высо-
кая холодостойкость обнаружена у-активных насекомых и у видов,
не имеющих диапаузы [Ильинская, 1971; Горышин, Саулич, 1982;
Horwath, Duman, 1983; Kukal et al., 1989; Chen et al., 1987;
Denlinger, 1991]. Совершенно удивительная способность повышать
устойчивость к низким температурам после кратковременного
(10 мин) выдерживания в температуре 0...—6 °C обнаружена у мухи
Sarqophaga crassipalpis [Chen etal., 1987]. При этом резистентность
к холоду повышалась не только у диапаузирующих куколок, но и у
недиапаузирующих личинок, куколок и фаратных имаго. Анало-
гичный эффект холодового шока был обнаружен у жука Xantho-
galeruca luteola и клопа Oncopeltusfasciatus [Lee et al., 1987b]. Это
дает некоторую разгадку тому, Каким образом удается насекомым
переживать кратковременные заморозки ранней весной, находясь
в состоянии активной жизнедеятельности.
Все приведенные материалы подтверждают точку зрения
Д.Л. Делингера [Denlinger, 1991] о том, что высокая холодостой-
кость может достигаться независимо от состояния диапаузы, но
она всегда является компонентом синдрома диапаузы. С этим со-
гласуются и современные представления о формировании холодо-
стойкости на основе общей неспецифической устойчивости насе-
комых к повреждающим агентам в состоянии диапаузы.
1.4. Летняя диапауза и ее экологическое значение
Состояние физиологического покоя насекомых, независимо
от его конкретной формы, имеет комплексный характер и способ-
но решать различные экологические задачи. Помимо зимовки оно
может обеспечивать переживание жарких и засушливых сезонов
12
года, сезонного недостатка пищи, синхронизировать межвидовые
и внутривидовые связи и т.д. Эстивация (летняя диапауза) насту-
пает летом и особенно часто встречается у представителей тропи-
ческого и субтропического поясов, ярко выражена у обитателей
пустынь и полупустынь, хотя наблюдается и у насекомых умерен-
ных широт [Фалькович, 1979; Masaki, 1980; Торр, 1986,1990,1991;
Ушатинская, 1987]. По своим особенностям летняя диапауза адек-
ватна зимней. Она также формируется на определенных для каж-
дого вида этапах онтогенеза, не всегда одинаковых даже у близких
видов [Masaki, Ohmachi, 1967; Ishii, Hidaka, 1982]. Подготовка к
ней происходит заранее и сопровождается значительным повы-
шением устойчивости к неблагоприятным воздействиям окружа-
ющей среды.
Основными факторами, контролирующими ее наступление,
являются длинный день и повышенная температура, а ее прекра-
щение - короткий день и пониженная температура [Данилевский,
Шельдешова, 1968]. Фактором, терминирующим летнюю диапау-
зу, может выступать и сокращающаяся длина дня [Gomi, Takeda,
1992]. Среди других факторов, регулирующих ее формирование,
можно отметить качество и количество пищи [Фалькович, 1979;
Ушатинская, 1987; Каплин, 1994,. и др.]. Особенно часто наступ-
ление диапаузы при отсутствии подходящей пищи отмечается у
хищных насекомых [Tauber, Tauber, 1973b; Hodek, 1986; Семья-
нов, 1986], У некоторых видов летняя диапауза наступает спон-
танно, независимо от внешних факторов [Thiele, 1977; Masaki, 1980;
Ishii, Hidaka, 1982].
Данные по физиологии и биохимии эстивации фрагментар-
ны, но, по мнению большинства исследователей, изменения, пред-
шествующие летней диапаузе и сопровождающие ее, отличаются
от типичных для зимней диапаузы только степенью выраженнос-
ти [Масленникова и др., 1976; Ушатинская, 1981,1987].
В некоторых случаях переживание летних неблагоприятных
условий требует более глубокой физиологической перестройки,
чем наблюдающаяся при гибернации. Так, у Coccinella septem-
pypctata brucki в Японии именно летняя диапауза по своим при-
знакам (прекращение оогенеза, снижение уровня газообмена, ре-
дукция крыловой мускулатуры, падение гормональной активное-
тц прилежащих тел) более отвечает истинной диапаузе, чем зим-
ндя [Sakurai, Takeda, 1984].
ГЛава 2. ЭКОЛОГИЧЕСКИЙ МЕХАНИЗМ РЕГУЛЯЦИИ
СЕЗОННОГО ЦИКЛА
Адаптивное значение диапаузы проявляется лишь в тех случаях,
когда наступление и окончание стадии покоя в сезонном цикле син-
хронизировано с ритмом климата. В качестве сигналов, синхронизи-
рующих раЗвитие с ритмом климата', могут выступать различные дос-
таточно Закономерно изменяющиеся факторы как абиотической, так
и биотической природы; часто их действие носит комплексный ха-
рактер. Наиболее надежным сигналом, несомненно, является длина
светового дня, изменяющаяся с астрономической точностью и не за-
висящая ни оТ каких факторов внешней среды. Физиологические ре-
акции на соотношение длины дня и ночи (фотопериод), названные
фотопериодическими реакциями (ФПР), обнаружены в самых разных
группах живых организмов и растительного и животного происхожде-
ния [Данилевский, 1961; Биологические часы, 1964; Скрипчинский,
1975; Дольник, 1976; Saunders, 1976;Хмелеваидр., 1991; Tanaka, 1991,
1992; Sounty-Grasset et al., 1994].
Согласно современным представлениям ["Лященко, 1977; Зас-
лавский, 1984] все фотопериодические эффекты у насекомых по ха-
рактеру проявления можно подразделить на два основных типа: каче-
ственные и количественные. Качественные ФПР изменяют направле-
ние морфогенетических процессов, определяя характер развития по
альтернативному принципу «да—нет» (например, диапауза — актив-
ное развитие). Количественные ФПР контролируют интенсивность
биологических процессов в пределах одного качественно однородно-
го состояния; биологический эффект при этом имеет градуальное вы-
ражение.
Видовое разнообразие реакций на длину дня можно свести к
нескольким основным типам. Наиболее распространенным у насеко-
мых умеренных широт является длиннодневный тип ФПР. Он обеспе-
14
чивает активное развитие в длиннодневных условиях и формирова-
ние диапаузы в короткодневных. Обычно длиннодневная ФПР регу-
лирует наступление зимней диапаузы при поливольтинном цикле.
Прсийвоположная зависимость развития от длины дня наблюдается
при «ороткодневвом типе ФПР. Эта реакция в основном регулирует
жизненные циклы видов тропического и субтропического происхож-
дения. У насекомых умеренных широт экологическая роль короткод-
невной ФПР связана с индукцией летнего проспективного покоя [Siew,
1966; Данилевский, Шельдешова, 1968].
У некоторых видов обнаружены ФПР промежуточного типа, ког-
да активное развитие особей или диапауза осуществляются только в
ограниченном интервале фотопериодов [Гёйспиц, 1953; Chippendale,
Reddy, 1973; Тищенко и др., 1983].
При двухступенчатых ФПР для проявления ответа необходима
смена'йсходного фотопериодического режима на альтернативный в
пределах одного онтогенетического цикла [Заславский, 1970, 1972,
1984; Thiele, 1971, и др.].
По мнению К.Ф. Гёйспиц [Гейспиц и др. ,1972], в основе всех
проявлений ФПР лежит способность формировать максимум диапау-
зы Вдйух областях фотопериодической шкалы — длиннодневной и
Длина дня, ч
Рис.1. Различные модификации фотопериодических кривых у чле-
нистоногих [по: Гейспиц и др., 1972]:
а-двухвершинная; б-длиннодневная; в-короткодневная; г—бездиапа-
узная; д - модификация с диапаузой во всех вариантах
15
короткодневной. В зависимости от степени выраженности обоих пи-
ков кривая ФПР принимает тот или иной вид. Когда тенденция к
формированию диапаузы в области обоих пиков развита одинаково
хорошо, обнаруживается двухвершинный характер кривой ФПР
(рис. 1), Этот тип ФПР служит исходным для всех остальных ее прояв-
лений и отражает адаптации к переживанию двух неблагоприятных
периодов в течение года—зимнего холодного и летнего засушливого.
2.1. Количественные фотопериодические реакции.
Их экологическая роль
Количественные фотопериодические реакции контролируют
разнообразные признаки и проявления жизнедеятельности и широ-
ко представлены у насекомых (табл. 2).
Убедительные данные фотопериодической регуляции скорости
созревания у златоглазок получила Т.А. Волкович [1987] в опытах на
Chrysoperla сатеа. Раньше всех приступали к размножению самки,
содержавшиеся при максимальной длине дня — 20 ч света в сутки. В
вариантах с меньшей продолжительностью светлой части суток сред-
ний срок начала яйцекладки в среднем возрастал и достигал максиму-
ма у самок, содержавшихся в пороговом фотопериодическом режиме
15 ч. Увеличение продолжительности созревания самок сопровожда-
лось увеличением масштаба индивидуальной изменчивости этого
показателя (рис. 2).
Очень часто количественные изменения происходят непосред-
ственно в состоянии диапаузы или предшествуют ей, обеспечивая под-
готовку к диапаузе. Так, зимующие яйца Orgyia thyellina (Lepidoptera,
Lymantriidae) [Kimura, Masaki, 1977], диапаузирующие пронимфы
лугового мотылька Loxostegesticticalis [Хомякова и др., 1986]идиапа-
узирующие куколки Mamestra brassicae (Lepidoptera, Noctuidae) [Cay-
лич, 19756] имеют большую массу, чем активные. Такая закономер-
ность проявляется и при имагинальной диапаузе у клопов Podisus
maculiventris [Горышин и др., 1988а,б] и жуков Leptinotarsa decelimeata
[Горышин и др., 19866]. При этом изменяется и длительность разви-
тия личиночных стадий. Например, у американской белой бабочки,
Hyphantria cunea Drury, в одних и тех же условиях (длина дня 14,5 ч
света в сутки, температура 25 °C) первыми окуклились гусеницы, дав-
шие активных куколок [Hidaka, 1977] (рис.З).
16
Таблица 2. Примеры количественной фотопериодической
регуляции признаков у насекомых [по: Мусолин, Саулич, 1997]
Признак Вид Источник
Продолжительность и интенсивность диапаузы Spilarctia imparil is Oedipoda miniata Teleogryllus emma Luehdorfia japonica Riptortus clavatus Calliphora vicina Chrysoperia downesi Chrysoperia carnea s.l. Kimura etal., 1982 Penner, Orshan, 1980 Masaki, 1965 Ishii, Hidaka, 1982 Numata, Hidaka, 1983 Виноградова, 1978 Tauber, Tauber, 1975 Tauber etal., 1986
Масса куколок Mamestra brassicae Acronicta rumicis Pieris brassicae Mythimna unipuncta Тыщенко, Гасанов, 1983 Саулич, 1975
Скорость созреаания самок Chrysoperia carnea sd. Вол кович, 1987
Размеры и пропорции тела или отдельных органов Euscelis incisus Stenocranus minutus Parnara guttata Muller, 1957,1979 Nakasuji et al., 1984
Длительность(скорость) личиночного развитий, иногда с появлением до- полнительных личиноч- ных врэрастоа , ь 1 > * i . - Apamea sordida Agrotis occulta A.segetum Dasychira pudibunda Pyrrharctia Isabella Pararge aegeria tircis Teleogryllus emma T.yezoemma Pteronemobius nitidus Melanoplus sanguinipes Ptprostichus oblongopunctatus Боби некая, 1966 Данилевский, 1961 Гейспиц и др., 1971 Гейспиц, 3аранкина,1963 Goettel, Philogene,1978 Nylin et al.i 1995 Масаки, 1972 Tanaka, 1983 Dean, 1982 Schaick, 1985
Интенсивность и харак- тер окраски^ » . личинок куколок имаго Plautia stall Qrgyia thyellina Riptortus clavatus •' '' i Numata, Kobayashi,1994 Kimura, Masaki, 1977 Kobayashi, Numata, 1993,1995
Плодовитость ’.'И;., Л I- 1 ' г > Plutella maculipennis Platycleis grisea Teleogryllus spp. Atwal, 1955; Harcout, Cass, 1966 Helfert, 1980 Масаки, 1972
СооГйошение’полов ’ Compoletus perdistinctus Hoelscher, Vinson, 1971
Степень развития Actolepia assectella Meudec, 1966 I
XSTwr, Drosophila melanogaster Pittendrigh, 1961 - цит. no: Saunders, 1976
Рис. 2. Влияние константных фотопериодов на продолжительность
созревания самок Chrysoperla сатеа s.l.t 25 °C [по: Волкович, 1987]:
Средние данные о сроках начала яйцекладки (с доверительным интервалом)
в опытах разных лет: 1 -1976 г., 2 —1977 г., 3 —1978 г
Аналогичное явление мы наблюдали у клопа Pyrrhocoris apterus.
В короткодневных и близких к критическому фотопериодах (см.гл.6)
число диапаузирующих особей возрастало среди имаго, окрылявших-
ся последними. Очевидно, эти различия связаны с накоплением ре-
зервных веществ, что характерно для всех диапаузирующих стадий.
Однако есть примеры, отражающие и обратное соотношение [Blau,
1981b; Tanaka, Tsubaki, 1984].
Типичные количественные реакции проявляются при регуляции
Шрк* до мушшммм, дни
Рис.З. Сроки окукливания диапаузи-
рующих (1) и иедиапаузирующих (2)
куколок ffyphantria сипеа [по: Hidaka,
1977]
продолжительности и интенсивно-
сти диапаузы. При этом для видов
умеренных широт диапауза, инду-
цированная более коротким фото-
периодом или его изменением, ха-
рактеризуется большей интенсив-
ностью. Это обнаружено, напри-
мер, у мухи Calliphoravicina [Виног-
радова, 1978], кукурузного мотыль-
ка, Ostrinia nubilalis [Beck, 1980],
клопа Riptortus clavatus [Numata,
Hidaka, 1983], златоглазки Chryso-
perla camea [Tauber, Tauber, 1970,
1972, 1973а].
18
Противоположный эффект наблюдался у златоглазки Nineta
flava'. более интенсивной была зимняя диапауза, индуцированная
длинным днем [Canard, 1983].
У Spilarctia imparilis (Lepidoptera, Arctiidae) эстивация куколок
оказывалась более продолжительной, если индуцировалась более ко-
ротким днем [Sugiki, Masaki, 1972; Kimura et al., 1982].
Нередко количественные реакции приобретают самостоятель-
ное значение, обеспечивая синхронизацию зимующей стадии с соот-
ветствующим сезоном. Классическим примером подобной функции
количественной ФПР стала регуляция темпов роста гусениц у Dasychira
pudibunda [ГЬйспиц, Заранкина, 1963].
Сходная регуляция скорости роста позже была обнаружена у не-
которых сверчков [Masaki, 1978, 1983], подгрызающих совок [ГЬры-
шин, Ахмедов, 1971; ГЬйспиц и др., 1971; Beck, 1986], медведиц
[Goettel, Philogene, 1978; Тыщенкоидр., 1985], некоторых полужест-
кокрылых [Saunders, 1983; Saulichetal., 1994; Saulich, Musolin, 1996;
Мусолин, Саулич, 1997]. При этом у одних насекомых, например у
большинства подгрызающих совок, выявлена только количественная
реакция, у других отмечены обе реакции: и качественная и количе-
ственная. В частности, как показали исследования С. Масаки [1972],
у бивольтинных популяций Pteronemobius fascipes и Р. taprobanensis ко-
роткодневные условия ускоряют развитие личинок и индуцируют эм-
бриональную диапаузу. Эти ФПР не связаны между собой, но их соче-
тание создает эффективный механизм, определяющий откладку яиц
в оптимальный период сезона. В настоящее время зарегистрировано
белее 80 видов насекомых из 9 отрядов, обладающих подобной реак-
цией [Danks, 1987]. ФПР, регулирующие скорость роста, сами по себе
йе создают повышенной устойчивости к неблагоприятным условиям,
а лишь синхронизируют определенные этапы онтогенеза с соответ-
ствующими сезонами года. Но даже эта первоначальная функция фо-
топериода как синхронизатора жизненного цикла позволяет значи-
тельно повышать приспособительные возможности насекомых и со-
здает, предпосылки для расширения их ареалов [Тыщенко, 1983/.
, , Не столь явно выражен экологический смысл в случае количе-
ственных изменений окраски у клопов [Kobayashi, Numata, 1993,
1995]. Хотя у личинок Plautia stall (Heteroptera: Pentatomidae) (рис. 4)
удалось выделить 6 четко различающихся цветовых вариаций, опре-
деляемых фотопериодическими условиями их содержания, значение
их не ясно.
19
a
$ ® s ® @ ®
1 2 3 4 5 6
6
Длина дня, ч
Рис.4. Вариации окраски личинок
V возраста Plautia stall (а) и фото-
периодические реакции (б) [по:
Numata, Kobayashi, 1994]:
ФПР, контролирующая имагиналь-
ную диапаузу, - толстые линии, детерми-
нирующая цветовые вариации личинок, -
тонкие линии. Черные символы - количе-
ство диапаузирующих самцов, %, светлые -
то же у самок. 1—6 “Цветовые градации ли-
чинок
Анализируя данные литературы, В.А. Заславский [1984] пришел
к заключению, что количественные ФПР распространены у насеко-
мыхтакже широко, каки качественные. Разница между ними вполне
ясная в резко выраженных случаях, иногда оказывается условной. Так,
самое распространенное проявление ФПР - диапауза, у некоторых
насекомых может переходить при определенных условиях в незначи-
тельные задержки развития, которые уже трудно называть диапаузой.
Например, у Dendrolimus pint диапауза гусениц, в норме обеспечиваю-
щая зимовку, при температуре 25 °C может длиться от нескольких дней
до двух месяцев [Гейспиц, 1953].
2.2. Качественные фотопериодические реакции
Качественные ФПР детерминируют общий ход морфогенетичес-
ких процессов, направляя развитие по одному из возможных путей,
предопределенных наследственностью [Тыщенко, 1973]. Согласно
имеющимся данным три группы биологических явлений контроли-
руются качественными ФПР: цикломорфозы тлей, сезонный поли-
морфизм и наступление и прекращение диапаузы.
20
л < л- л 2.2.1. Цикломорфозы тлей
Сезонный цикл большинства видов тлей усложняется чередова-
нием поколений/различающихся способом размножения. Обычно у
этих насекомых зимуют диапаузирующие яйца. Весной из них от-
рождаюгся самки-основательницы, которые дают начало ряду парте-
ногенетических поколений (virginopara). Число таких поколений не
постоянно и зависит от внешних условий. Партеногенез и живорож-
дение приводят к быстрому росту численности популяции. Появля-
ющиеся время от времени крылатые самки перелетают на новые кор-
мовые растения, при этом у многих видов наблюдается смена расте-
ний-хозяев. Например, у черемуховой тли Siphonaspispadi часть цик-
ла проходит на первичном хозяине - черемухе, затем появляющиеся
среди партеногенетических поколений крылатые митранты пересе-
ляются ни вторичных хозяев — разные виды злаков, где также прохо-
дит развитие нескольких партеногенетических поколений, с появле-
нием ремйгрантов тли возвращаются на основное растение - черему-
ху. К двудомным видам относятся также Aphis fabae, Myzus persicae,
Dysaphis plantaginea и многие другие. При отсутствии в силу разных
причин одного из растений-хозяев из нормального цикла может вы-
падать одно из звеньев, и тогда двудомные тли становятся однодом-
ными [Мргап, 1990]. Хорошо известный пример такого рода—кровя-
ная тля Eriosoma, которая на американском континенте развивается
со сменой хозяев: американский вяз—яблоня, а в Европе - только на
яблОне.
Осенью и у одно- и у двудомных видов появляются особые парте-
ногенетические особи — полоноски (ginopara), которые производят
обоеполое поколение. После спаривания самки (ovipara) откладыва-
ют диапаузирующие яйца, из которых весной выходят личинки, даю-
щие самок-основательниц. Эта типичная схема у разных видов может
усложняться или упрощаться, создавая удивительное разнообразие
жизненныхциклов.
Исследование механизмов регуляции сезонного развития тлей
начато очень даднр. Еще С. Маркович [Marcovitch, 1924] в опытах с
Aphis forbesi обнаружил, что возникновение мигрирующих форм и
смена партеногенетического размножения на обоеполое регулирует-
изменением длины дня. Эти исследования получили
Д^ййЙщее^азвитиев блестящих работах Л. Бонмезона [Bonnemaison,
1959,1964,1966], ЕХ. Шапошнико-
21
В регуляции полиморфизма тлей принимают участие генети-
ческие, гормональные и внешние экологические факторы, такие как
температура, фотопериод, состояние кормового растения, плотность
колонии. В определении пола главную роль играют генетические ме-
ханизмы, действие которых проявляется еще в начале оогенеза. Пол
зародышей зависит и от последовательности их расположения в яй-
цевых трубках: яйцеклетки, находящиеся ближе к выходу из яични-
ка, чаще дают самок, а более удаленные - самцов. В исследованиях,
выполненных на Megpura vicia Buckt., показана возможность участия
в этом процессе и экзогенных факторов [Заславский, Кац, 1987]. Диф-
ференцировка женских зародышейна девственниц и половых самок
регулируется фотопериодом и температурой. По данным А.Д. Лиса
[Lees, 1959], высокая температура и длинный день способствуют фор-
мированию партеногенетических самок, а пониженная температура
и короткий день — яйцекладущих самок обоеполого поколения. При
этом обнаружено, что основательница и первые партеногенетические
поколения не Датируют на фотопериод. Реакция на этот фактор раз-
вивается с возрастом клона. А.Д. Лис [Lees, 1966] показал, что появ-
ление половых особей в коротком дне и при пониженной температу-
ре возможно лишь в шестом партеногенетическом поколении — через
96 дней после основания колонии. Этот процесс был назван «измери-
телем интервалов» (time measurement). Интересно, что фотопериоди-
ческая чувствительность материнской особи, которая произведет яй-
цекладущих самок, начинается, когда она сама еще является эмбрио-
ном и находится в теле своей матери.
Возникновение мигрирующих форм и переселение в новые мес-
тообитания вызывается ухудшением условий питания и перенаселен-
ностью . Миграции, обеспечивая переход на более благоприятные для
питания и размножения растения, служат основным средством, под-
держивающим высокую численность тлей. Они не ограничены во вре-
мени критическими для жизни погодными условиями и не требуют
большой календарной точности, и в этом, видимо, причина того, что
фотопериодический сигнал не вовлекается в механизм, регулирую-
щий формирование мигрантов [Данилевский, 1961].
222 Сезонный полиморфизм
Сезонный полиморфизм (в зарубежной литературе — полифо-
низм) обычно проявляется в различиях окраски, размерах и пропор-
циях тела, степени развития крыльев и других структур. Сезонный
22
пояимерфизмчасто имеет выражение в тех же признаках и явлениях,
уггггуч» «играпируются количественными ФПР, но в отличие от
последних^ они регулируются по принципу «все или ничего» на каче-
ственнойоснове. Следуя предложенной классификации [Thuber et al.,
1986], нее проявления сезонного полиморфизма можно разделить на
три» типа? 1) не связанные с диапаузой; 2) так или иначе связанные с
пред- или постдиапаузным развитием либо непосредственно с самим
состоянием покоя.
Овииуый полиморфизм, не связанный с диапаузой. Самые яркие
пример» этого типа сезонного полиморфизма, безусловно, дают виды,
имеющие, гомодинамный тип развития, когда состояния проспектив-
ного покоя не бывает совсем. Так, желтушка Colias eurytheme, широко
распространенная по всему северо-американскому континенту и име-
ющая гомодинамный жизненный цикл, встречается в природе в виде
двух сезонных форм: светлой — летней и темной — весенне-осенней.
Оказалось, что регуляция этих фенотипов осуществляется длиной дня,
при которой развивались гусеницы III—ГУ возраста [Hoffmann, 1974].
Бабочки из короткодневных условий имеют небольшие размеры и
темную окраску крыльев, а из длиннодневных—более крупные разме-
рим светлую окраску крыльев.
Северо-американская белянка Nathalis iola также не имеет зим-
нейдиапаузы. Появление ее в северных регионах связано с ежегодны-
ми миграциями. Сезонный полиморфизм, проявляющийся в степе-
ни меланизации крыльев, находится под контролем фотопериода: ко-
роткий день индуцирует появление меланистических форм; В приро-
де они встречаются весной и осенью, летом доминируют светлоокра-
шенные особи [Douglas, Grula, 1978].
< Столь же отчетливо проявляется самостоятельная регуляция по-
лиморфизма у гетеродинамныхмоноцикликов. Наступление диапау-
зывкаждом поколении предопределено у них генетически и не регу-
лируется внешними факторами, а возникновение сезонных форм на-
ходите»под контролем экзогенных факторов. 1кк, у клопа слепняка
Leptoptema dolobrata (Miridae) длина крыльев определяется темпера-
турой нплотностью популяций, а диапауза наступает независимо от
вившиих условий [Braune, 1983].
^r-Мырассмотрели крайние проявления автономности сезонного
полиморфизма и диапаузы, когда последняя либо отсутствует, либо
н®<рвгуяируется факторами внешней среды. Однако у большинства
насекомых оба феномена проявляются одновременно и находятся под
23
контролем одних и тех же факторов. В этом случае самостоятельная
регуляция этихявлений выражена не столь явно, хотя большей частью
легко обнаруживается в экспериментах. Например, у водомерки
Aquariuspaludum [Harada, 1992] крыловой полиморфизм определяет'
ся длиной дня, действующей на личинок I-V возрастов, тогда как
наступление диапаузы связано с условиями содержания личинок
V возраста.
У голубянки Lycaenaphlaeas daimio ФПР определяет и наступле-
ние диапаузы, и образование сезонных форм [Sakai, Masaki, 1965; Endo
et al., 1985а]. Но если реакция, индуцирующая диапаузу, проявляется
только при 20 °C (при более высокой температуре все гусеницы раз-
вивались без диапаузы), то ФПР, контролирующая сезонный поли-
морфизм, проявляется и при повышенных температурах. В коротко-
дневных условиях развиваются светлоокрашенные особи, в длинно-
дневных - темноокрашенные. При этом установлено, что окраска ре-
гулируется длиной дня, действовавшей на гусениц ранних возрастов.
Сложная и своеобразная регуляция сезонного полиморфизма
обнаружена у толстоголовки Pamara guttata guttata [Ishii, Hidaka, 1979;
Nakasuji, Kimura, 1984]. Оказалось, что у этого вида сезонной измен-
чивости подвержена не только окраска крыльев, но и другие характе-
ристики: крыловой рисунок, размеры крыльев, степень их меланиза-
ции. Возникновение вариаций находится под контролем температу-
ры и фотопериода, соотношение которых и вызывает появление тех
или иных морфологических признаков имаго, изменяющихся неза-
висимо друг от друга. Детерминация сезонных морфотипов происхо-
дит в V гусеничном возрасте, а диапауза приурочена к IV—V возрас-
там. По мнению авторов, личиночная диапауза, индуцированная ко-
ротким днем, не связана с регуляцией сезонных форм, однако корот-
кодневные гусеницы в экспериментальных условиях давали более мел-
ких бабочек. Наблюдения за природными популяциями показали,
что среди перезимовавшего поколения были и светло- и темноокра-
шенные формы. В середине лета встречались только светлые бабочки,
в августе популяции были смешанные, а в более поздние календарные
сроки летали только темные особи. Сезонный цикл вида усложняется
ежегодными миграциями, для регуляции которых большое значение
имеет плотность популяции, поэтому в естественной природе имаго
каждой генерации морфологически значительно различаются, но
удельный вес отдельного регулирующего фактора, как оказалось, оце-
нить практически невозможно.
24
п/нуммюидмажные взаимодействия регулирующих факторов — тем-
ггертурт* мфотэпериода, в детерминации фенотипов обнаружены и у
napytttfki ibpilio xuthus [Endo, Murakami, 1985].
;; СёэОйяЫй полиморфизм, связанный с диапаузой. Анализируя раз-
тплю^ж пуимеры сезонного полиморфизма у насекомых, легко убе-
между диапаузой и этим сезонным явлением очень часто
существует самая тесная связь. Например, зимняя окраска от светло-
зеленОйДО коричневой у златоглазки Chrysoperla сатеа — внешнее
проявление репродуктивной диапаузы, индуцируемой коротким днем.
ДлиИйЫйденьвызываетпоявлениенедиапаузирующих ярко-зеленых
имаго [Md-LCOd, 1967; Honek, 1973].
У зимующих клопов Nezara viridula (Pentatomidae) преобладает
коричневая окраска, которую можно вызвать в лабораторных экспе-
риментах; воздействуя наличиноккороткимднем, начиная со II воз-
раста [Gogala, Micheli, 1966]. У клопа слепняка Notostira elongata
[Dolling; 1973] известны две сезонные морфы: длиннодневные (лет-
ние) особи зеленые, а короткодневные (осенние) коричневые. Две
формы клопа щитника Euschistus tristigmus tristigmus, ранее признава-
емыесамостоятельными видами, также оказались сезонными морфо-
типаМИ, контролируемыми фотопериодом [Mc.Pherson, 1979]. Диа-
пйУзйрующие куколки многих парусников (Lepidoptera, Papilionidae)
коричневого цвета, тогда как окраска активных куколок определяется
субстратом, на котором происходит окукливание [West et al., 1972;
Hazel/AAfest, 1983; Sims, 1983].
Изменение окраски у диапаузирующих особей может быть свя-
зано с разными процессами. В одних случаях в результате накопления
жирового тела происходит смещение цветовой гаммы в сторону свет-
лых тонов, в других усиление пигментации кутикулы приводит к по-
темнению покровов тела. Вероятно, существуют и другие причины.
У многих видов насекомых зимующие особи имеют более проч-
ные покровы, которые защищают их от повреждений в течение дли-
тельного покоя. Диапаузирующие яйца упоминавшейся выше Qrgyia
tftytttina [Kimura, Masaki, 1977] имеют более толстыйхорион, адиапа-
УЗНрующие гусеницы Euphydryas editha (Lepidoptera, Hesperiidae) [Scott,
19Wf<H- диапаузирующие куколки Heliothis armigera (Lepidoptera,
Nbdtilidae) [Roome, 1979; Hackett, Gatehouse, 1982] обладают более
ТйЯвтой и прочной кутикулой. По мнению авторов, эта особенность
®ЙСИЫсо усиливает защиту от механических повреждений,-нои регу-
Лйр$ет скорость испарения воды.
25
Для нимфалиды Апаеа andria характерен сезонный полимор-
физм, проявляющийся в форме и окраске крыльев активных и диапа-
узирующих бабочек [Riley, 1988]. Сходные эффекты описаны у Polygonia
c-aureum [Hidaka, Aida, 1963], Aclerisfimbriana [Данилевский, 1961] и
других видов. Во всех этих случаях возникновение сезонных морф свя-
зано с состоянием диапаузы и находится под контролем длины дня. В
некоторых случаях фактором, регулирующим появление сезонных
вариаций, выступает температура, причем роль ее в отдельных случа-
ях бывает доминирующей [Miyata, 1977; Shapiro, 1982].
У нимфалиды Araschnia levana-prorsa, ставшей уже классичес-
ким примером сезонного диморфизма [Данилевский ,1961], бабочки
весеннего поколения, выходящие из перезимовавших куколок, име-
ют красно-рыжую окраску с черными пятнами (форма levana)', их по-
томство, развивающееся летом, дает бабочек, имеющих черные кры-
лья с белыми пятнами (форма prorsa). Наступление зимней диапаузы
у Araschnia обусловлено продолжительностью дня в период развития
гусениц, и, таким образом, сезонная смена окраски также находится
под фотопериодическим контролем, хотя непосредственно кдиалау-
зирующей стадии (куколка) она не приурочена. Аналогичное явле-
ние обнаружено у челночницы Hylophila prasinana [Данилевский,
1961], белянок Леш лар/[Матинян, 1965] и Asciamonuste [Pease, 1962],
Eurema hecabe [Kato, Handa, 1987,1992], нимфалиды Pofygonia c-album
[Voigt, 1991] и многих других видов чешуекрылых. У американской
белой бабочки, Hyphantria сипеа, обнаружены четко выраженные се-
зонные морфотипы. Бабочки, отрождающиеся в мае-июне из диапа-
узирующих куколок, имеют темные пятна на передних крыльях, тогда
как бабочки, вылетающие в июле-августе из активных куколок, име-
ют чисто белые крылья. В экспериментальных условиях появление
меланистических форм было связано с пониженной температурой и
коротким днем [Hidaka, 1977].
С изменением длины дня связанысезонные вариации окраски и
структуры признаков и у некоторых цикадок. Так, личинки рисовой
цикадки, Nephotettix cincticeps, впадающие в диапаузу под влиянием
короткого дня, дают мелкую и относительно короткокрылую весен-
нюю форму имаго, а развивающиеся непрерывно в условиях длинного
дня — более крупную длиннокрылую летнюю форму [Kisimoto, 1959].
Под контролем фотопериода находится и возникновение сезон-
ных вариаций длины крыльев. Крыловой полиморфизм чрезвычайно
распространен в классе насекомых. Глубокое и всестороннее исследо-
26
вание этого явления связано с блестящими работами известного япон-
ского энтомолога С.Масаки. На большом числе видов сем. Gryllidae
(Orthoptera) рассмотрены разнообразные аспекты крылового полимор-
физма: распространенность, географическая изменчивость, генети-
ческая вариабельность и наследуемость соответствующих признаков,
а также вопросы видообразования и эволюции (Масаки, 1972; Masaki,
1978; 1983; Masaki, Shirado, 1987; Masaki, Senno, 1990; Masaki, Sugahara,
1992; Shimizu, Masaki, 1993].
Крыловой полиморфизм подробно исследован и у полужестко-
крылых (Heteroptera). У водомерки Gerris odontogaster [Vepsalainen,
1971,1978] встречаются короткокрылые и длиннокрылые формы. Пе-
резимовавшее поколение почти полностью состоит из длиннокры-
лых особей. Их потомство диморфно: короткокрылые отрождаются
до середины июля, длиннокрылые позднее (рис. 5). Эти длиннокры-
лые особи перелетают в места зимовки и зимуют там в состоянии реп-
Июнь Июль Август Сентябрь
Рис. 5. Крыловой диморфизм у Gerris odontogaster (no: Vepsalainen,
1971]:
a - распределение особей по длине крыльев (данные по совокупности трех
популяций, 28.07-12.08.1967 г.), На врезке - схема принятых градаций крылового
индекса: А-бескрылые, Mi-микротггера, Вг-брахиттгера, Ма~ макрошера. б-
динамика соотношения длиннокрылых (Ма) и короткокрылых (Вг и Mi) особей
в течение сезона
27
родуктивной диапаузы. В экспериментах было показано, что длин-
ный день (более 18 ч света в сутки) в совокупности с увеличением
длины дня на стадии личинки младшего возраста вызывает появле-
ние короткокрылых клопов, которые приступают к размножению.
Позднее было обнаружено, что не только длина дня, но и наслед-
ственные факторы участвуют в детерминации длины крыльев и диа-
паузы. Сходные результаты получены и в опытах с японским подви-
дом Gerris palludum insularis [Harada, Taneda, 1989]. Сложные связи
температуры, фотопериода, плотности и пищи в регуляции крылово-
го диморфизма обнаружены у другого японского клопа Microvelia
douglasi [Muraji et al., 1989].
У клопа солдатика, Pyrrhocorls'apterus, обнаружены четыре мор-
фотипа по длине крыльев: длиннокрылые (macroptera) и короткокры-
лые (brachyptera), среди которых в свою очередь выделены еще две
модификации - microptera и aptera [Honek, 1974,1986]. В естествен-
ных природных условиях преобладают формы macroptera и brachyptera.
Возникновение того или иного морфотипа, как показали исследова-
ния [Honek, 1985, 1986], определяется как наследственными факто-
рами, так и внешними условиями. Длиннокрылость - признак рецес-
сивный. Путем отбора можно получить линию с высокой частотой
встречаемости длиннокрылых особей. В экспериментальных услови-
ях в длинном дне и при высокой температуре преобладают длинно-
крылые формы, а в коротком дне и при низкой температуре — корот-
кокрылые [Honek, 1974,1981]. При этом эффект может передаваться
даже дочернему поколению [Honek, 1980]. В природе доля длинно-
крылых особей также определяется фотопериодическими и темпера-
турными условиями развития личинок. По наблюдениям А. Хонека
[Honek, 1981], в Центральной Европе личинки, окрылившиеся до
конца июля, дают в основном длиннокрылых имаго, позже домини-
руют короткокрылые особи. Интересно, что генетическая вариабель-
ность по этому признаку была идентичной у особей из Центральной
Европы, Средиземйоморья и Центральной Азии (Казахстан) [Honek,
1987].
Среди полужесткокрылых, исследованных в нашей лаборатории,
сезонный полиморфизм был обнаружен у хищного клопа Arma custos
(Pentatomidae, Asopinae) и проявился в сезонных вариациях окраски
тела. Личинки V возраста, выращенные в разных температурных и
световых условиях, различались по степени пунктированности голо-
вы, переднеспинки, щитка и зачатков крыльев. По этим признакам
28
быяияидаиены две морфологические группы. У личинок 1-го типа
укзэаниыечасти тела имели по основному желтому фону густую чер-
ную пуяктйровку которая часто переходила в сплошные черные пят-
им темно-бронзовую окраску с ярко выраженным ме-
TypnnwiwOTM блеском. Личинки, отнесенные ко 2-му типу, отлича-
лись светло-желтой окраской головы, переднее пинки, щитка и зачат-
ков крыльевс пунктировкой в виде более или менее редких черных
точек и всегда без металлического блеска [Волкович, Саулич, 1994].
При определении процентного соотношения личинок 1 - и 2-го
тиррп «^а^симпсти от температуры и длины дня (рис. 6) оказалось,
чтопри постоянной температуре 27 °C во всех фотопериодах встреча-
лись главным образам личинки 2-го типа. Появление личинок с ме-
таллическим блеском отмечено при сочетании короткодневных фо-
товериодов—12- 15ч света в сутки и терморитма 27 :13,5 ^(средне-
суточная 20,5 °C). В длиннодневных режимах, 16—18 ч, при этой тем-
пературе были личинки только 2-го типа. Однако при более низкой
средней температуре - 17,2 °C (терморитм 23,3 : 12,7 °C) личинки
Д-1» Типа появились при длине дня 16—18 ч (30 и 20% соответствен-
но); а в. короткодневных режимах 15 и 14 ч их доля возросла до SO-
SO». ' . .
\ пи Среди личинок, собранных в природе, отмечены оба типа окрас-
кипри доминировании 1-го, причем особи 2-го типа встречались толь-
ко в Июле и в начале августа. Таким образом, здесь мы также имеем
пример фотопериодически контролируемого сезонного полиморфиз-
ма» При этом диапауза и окраска—два самостоятельно регулируемых
Влияние температуры и фотопериода
на соотношение личинок двух морфологи-
’«Ярос типов у Anna custos [по: Волкович, Са-
Черное поле-изменение доли личинок 1-го типа
WhcimocTH от длины дня, заштрихованное поле - то
^Мадочннок 2-го типа. 1-Ш - температурный ре-
ЗЯКЛт.27 ‘С, II — 27:13,5" С (20,5 °C), Ш - 23:13 °C
признака, что доказывается не только их приуроченностью к разным
стадиям, но и сопоставлением температурных диапазонов их прояв-
ления. ФПР, контролирующая индукцию диапаузы, наиболее выра-
жена при высоких температурах (27-30 °C) и полностью подавляется
низкими, фотопериодический контроль окраски проявился при всех
исследованных температурах.
В целом, как свидетельствуют приведенные выше материалы, яв-
ление сезонного полиморфизма широко распространено в классе на-
секомых. В качестве факторов, контролирующих это явление, высту-
пают внешние стимулы абиотической и биотической природы, среди
которых наибольшее значение имеют фотопериод, температура и плот-
ность популяции. В некоторых случаях ответ организма определяется
сложным взаимодействием нескольких факторов, роль каждого из
которых значительно различается у разных видов. Несмотря на рас-
пространенность явления сезонного полиморфизма, адаптивный
смысл его не всегда ясен. Считается, что в период активной жизнеде-
ятельности цветовые вариации связаны с оптимизацией процессов
терморегуляции [Hoffinann, 1973; Hazel, West, 1983; Tauber et al., 1986].
Усиление меланизации весной и осенью способствует усилению по-
глощения солнечного излучения, а светлая окраска в летний период
предотвращает возможные перегревы. Показано, например, что пре-
вышение температуры тела темноокрашенного и опушенного насеко-
мого над температурой воздуха может составлять 15-30 °C [Willmer,
Unwin, 1981,Ушатинская, 1987]. Несмотря на логичность такого пред-
положения, есть примеры, прямо противоречащие ему. Так, у Lycaena
phlaeas daimio и Araschnia levana-prorsa темные фенотипы появляют-
ся, наоборот, летом, а весной и осенью бабочки имеют рыжую, более
светлую окраску. Во время зимнего покоя сезонные вариации могут
обеспечивать бблыпую автономность организма от неблагоприятно-
го воздействия внешней среды путем активизации защитных меха-
низмов, в том числе усилением критических эффектов, образовани-
ем структурных элементов, укрепляющих, например, оболочку зиму-
ющих яиц, или образованием плотных коконов и т.д. [Tauber et al.,
1986].
Интенсивные исследования последних лет пока не дают одно-
значного ответа на вопросы, связанные с механизмами регуляции се-
зонного полиморфизма. Отсутствие обязательной связи между воз-
никновением сезонных форм и диапаузой допускает известную авто-
номность обсуждаемых феноменов. Большая часть имеющихся дан-
30
ныхсв
нецдев
,ствует в пользу того, что эти явления контролируются
мн гормональными факторами [ Fukuda, Endo, 1966;
и др., 1970; Hoffmann, 1974; Awiti, Hidaka, 1982; Endo et
5,1985; Endo et al., 1992; Harada, 1992; Harada, Numata, 1993;
Kobayashi, Numata, 1993].
* 2.2.3. Индукция и терминация диапаузы
Несомненна наиболее изученным качественным проявлением
фотопериодизма является наступление и прекращение диапаузы.
Адаптивное значение ФПР определяется двумя основными па-
раметрами — критическим фотопериодом и числом свето-темновых
циклов, называемым также «пакетом фотопериодической информа-
ции». Критический фотопериод (длина дня, индуцирующая диапаузу у
50% особей) показывает, с какого срока сезона начинается индукция
диапаузы. Критическое число свето-темновых циклов определяет, че-
рез сколько дней после того, как длина дня станет меньше пороговой,
формирование диапаузы завершится у половины особей.
Восприятие и оценка длины дня осуществляется у насекомых на
специфических для каждого вида этапах онтогенеза. Чувствительность
к этому фактору может проявляться на любой стадии от яйца до има-
го, иногда она сохраняется на нескольких стадиях [Горышин, Тыщен-
ко, 1974; Saunders, 1976; Taylor, Spalding, 1988; Виноградова, 1991].
У соснового шелкопряда, Dendrolimus pint, чувствительность к
длине дня оказалась одинаковой на всем протяжении стадии гусени-
цы [Гейспиц и др.,1972]. В экспериментах независимо от того, с како-
го возраста начиналось воздействие коротким днем, диапауза у гусе-
ниц ленинградской популяции шелкопряда всегда наступала в сред-
нем через 30—39 дней (рис. 7).
Живительная способность воспринимать фотопериодические
сигналы обнаружена у эмбрионов мухи Sarcophaga crassipalpis, нахо-
дящихся в утерусе самок [Denlinger, 1971]. Существование эмбрио-
нальной чувствительности подтверждено экспериментально еще у
нескольких видов саркофагид [Saunders, 1971,1980; Виноградова, 1976],
при этом было обнаружено, что у 5. argyrostoma внутриутробная ста-
ли? развития более чувствительна к фотопериодическим воздействи-
яъ(,м^5м отродившиеся личинки.
..Чувствительность эмбрионов к фотопериодическим сигналам об-
нару<еда и у представителей других отрядов. Это, в частности, харак-
,комаров рода Aedes [Kappus, Venard, 1967; Anderson, 1970;
31
Рис, Z Фотопериодическая индукция диапаузы соснового шелко-
пряда на различных этапах развития гусениц [по: 1ЪЙспиц и др.,
1972]:
Вертикадыше линии, разделяющие поле рисунка, показывают продолжи-
тельность этапов онтогенеза в контроле, при 20 °C и непрерывном освещении; Ov -
яйцо, L1-L6-возраст гусеницы до окукливания (Р); в вариантах 1-5 заштрихован-
ные полосы и числа на них показывают начало и продолжительность воздействия
коротким днем (12 ч света в сутки) до наступления диапаузы
Pinger, Eldridge, 1977; Shroyer, Craig, 1980], сверчка Acheta commodus
[Hogan, 1960], весенней капустной мухи, Delia brassicae [Read, 1969;
ГЪльзберг и др., 1986], совки Graphiphora c-nigrum [Kowalska, 1964;
Шашенкова, 1974].
У разных видов соотношение между чувствительной и диапаузи-
рующей стадиями может быть очень разным. Встречаются насеко-
мые, у которых чувствительность к длине дня непосредственно пред-
шествует диапаузе и проявляется на этой же стадии онтогенеза. В дру-
гих случаям такая чувствительность может далеко упреждать стадию, к
которой приурочена диапауза [Горышин, 1977; Заславский, 1984;
Tauber et al., 1986; Danks, 1987].
Анализ механизма накопления фотопериодической информа-
ции у разных видов выявил наличие видовых особенностей в этом
процессе (рис. 8). Детальные исследования на совке Mdmestra brassicae
[ТЪрышин, Г. Тыщенко, 1973] позволили обнаружить, что настройка
ФПР в опытах с однократным изменением длины дня определяется
числом короткодневных (КД) импульсов, полученных гусеницами в
течение своего развития. При числе КД-импульсов меньше 14 диапа-
уза не возникает, при более 14 импульсов возникает. Длиннодневные
(ДД) импульсы не эффективны. Нечувствительность к длинному дню
32
Рис 8 Зависимость диапаузы куколок
Лсгол/сИ Л*Лй?и (1), Pieris brassicae (2) и
Mamestra brtusicae (3) от числа корот-
ких и длинных дней, воздействующих
на стадии гусеницы [по: Тыщенко, Кинд,
1983]:
Популяции - белгородские (A.rumicis,
M.brassicae) и ленинградская (Rbrassicaey, темпе-
ратурный режим—от 16 до 26 °C. Прямоугольни-
ками выделены критические пакетыкорогкоднев-
ной или длиннйдйевнЬй информации
t
Число КД Число ДД
позволяет капустной совке накапливать пакет короткодневной ин-
формации или непрерывно, или порциями в начале или в конце гусе-
ничного развития. По такому же принципу воспринимается фотопе-
риодическая информация у мухи Delia brassicae [Гользберг и др., 1986].
У большинства других видов — Acronicta rumicis [Горышин, Ты-
щенко, 1972], Pieris brassicae [Горышин, Тыщенко, 1979], Pyrrhocoris
apterus (Горышин, Волкович, 1978], Riptortus clavatus [Numata, Hidaka,
1982], Podisus maculiventris [Волкович и др., 1991], происходит сумми-
рование как короткодневных, так и длиннодневных сигналов. В силу
этого окончательный ответ зависит от того пакета фотопериодичес-
кой информации, который накапливается последним, т.е. диапауза у
эт^х видов может наступать как при избытке короткодневных, так и
при недостатке длиннодневных сигналов.
-Шма ,
Глава 3. ФАКТОРИАЛЬНАЯ ИЗМЕНЧИВОСТЬ
ФОТОПЕРИОДИЧЕСКОЙ РЕАКЦИИ
Длина дня — основной фактор, сигнализирующий о приближа-
ющихся изменениях в окружающей среде, и реакция на фотопериод
играет доминирующую роль в сезонной смене физиологических со-
стояний. Однако насекомые реагируют и на другие сопутствующие
сезонные изменения, происходящие в природе, такие как повыше-
ние или понижение температуры, изменение качества и количества
пищи и др. Их действие преломляется через факториальную измен-
чивость ФПР. Среди многообразия факторов внешней среды темпе-
ратура наиболее сильно влияет на фотопериодическую реакцию.
3.1. Температурная изменчивость основных параметров ФПР
Температура прежде всего определяет саму возможность прояв-
ления реакции. Это связано с тем, что ФПР, как и другие биологичес-
кие процессы, имеет свой температурный оптимум и пределы, за ко-
торыми световой фактор н*е эффективен. У видов с ФПР длинноднев-
ного типа высокие температуры обычно подавляют проявление реак-
ции, сцособствуя непрерывному развитию насекомого даже в услови-
ях короткого дня, а при низких температурах, наоборот, при всех све-
товых режимах наступает диапауза.
Анализ имеющихся материалов показывает, что зоны оптимума
ФПР существенно отличаются у разных видов. Например, темпера-
турный оптимум ФПР Acronicfa rumicis находится в интервале от 22
до 32 °C, у паутинного клеща Metatetranychus ulmi — от 10 до 22 °C, у
Pieris brassicae — от 12 до 28 °C [Данилевский, 1961]. Очень широкий
диапазон температурного оптимума был обнаружен у соснового шел-
копряда, Dendrolimuspini [Гейспиц, 1965]: только температура выше
27,5 °C подавляла эффект короткого дня; нижний температурный
34
ФПР установлен не был - она четко контролиро-
„оу jjifffffW развития даже при 10 °C.
' '.Qwgfr-YFK”** интервал температурного оптимума ФПР был об-
т.арууомуKiumA.Podisus maculiventris [Горышин идр,, 1988]. Фотопе-
риодияеск^ая индукция диапаузы в самых эффективных, т.е. диапау-
зогенных фотопериодах (10, 11 и 12 ч света в сутки) максимально вы-
ражена только при температуре 19-22 °C. Как повышение температу-
ры„тМ1и ее понижение, ослабляют фотопериодический эффект
(ри₽-.?)’ . -
Эудпогическое значение температурного оптимума ФПР связа-
но с тем, что он определяет эффективность ФПР в пр иродных услови-
ях. Чем шире его диапазон, тем большее значение в регуляции сезон-
ного цикла и диапаузы приобретает длина дня. Например, у клопа
Arma custos узкий оптимум ФПР, приуроченный к области высоких
' Температура, °C
' F-< I 1
i Рис. 9. Температурный оптимум фотопериодической реакции кло-
Па Podisus maculiventris:
r 1-3-доляособей, адекватнореагирующихнадлинудняв короткодневных
фотопериодах: 12ч(1), Пч(2)и10ч(3)
35
температур, допускает активное развитие даже в условиях длинного
дня только при температуре выше 27 °C. Это, как показали проведен-
ные исследования, является одной из причин невозможности поли-
вольтинного цикла у A. custos в лесостепной здне, несмотря на то, что
наступление диапаузы у этого вида контролируется внешними усло-
виями (подробнее об этом см. гл. 6).
Наблюдения показывают, что роль температурного оптимума
ФПР в регуляции сезонного цикла насекомых значительно важнее,
чем считалось до настоящего времени. Отсутствие литературных дан-
ных в отношении этого показателя однозначно свидетельствует о том,
что значение его еще недооценено. Несомненно требуются дальней-
шие исследования в этом направлении.
Кроме сказанного выше, температура оказывает модифицирую-
щее влияние на параметры ФПР. Оно проявляется в изменении кри-
тического фотопериода и доли диапаузирующих особей в короткод-
невных и длиннодневных условиях.
В многочисленных экспериментах показано, что в случае длин-
нодневной реакции повышение температуры вызывает снижение по-
рога ФПР, и степень его температурной изменчивости сильно разли-
чается у разных видов. Например, у ивовой волнянки, Leucomasalicis,
из Армении повышение температуры на 5 °C вызывает уменьшение
критического фотопериода почти на 5 ч Данилевский и др., 1970],
тогда как у Loxostege sticticalis и северных популяций Pieris brassicae он
оставался почти постоянным в интервале температур от 18 до 25 °C.
Как обнаружено в исследованиях последних лет, термостабильность
порога не такое уж редкое свойство ФПР насекомых. Кроме назван-
ных видов она описана у листовертки Choristoneura rosaceana [Gan-
gavalli, Aliniazee, 1985], парусника Papilioxuthus [Endo, Murakani,1985],
у сухумской популяции медведицы Spilosoma menthastri [Кинд, 1985]
и многих других видов.
Относительная термостабильность порога ФПР была обнаруже-
на и у клопа Podisus maculiventris. Однако параметры ФПР в целом не
оставались постоянными при изменении температуры (рис. 10). В
наших опытах свойства ФПР подизуса исследованы при температурах
17,5; 20; 25,5 и 27,5 °C. Для 17 и 20 °C приведены усредненные кри-
вые. При повышении температуры величина критического порога ос-
тавалась относительно постоянной, тогда как количество диапаузи-
рующих особей в короткодневных условиях заметно уменьшалось. При
27,5 °C - максимальной из испытанных температур, даже в 11-часо-
36
^‘Ptic.lO. Фотопериодическая реакция клопа Podisus maculiventris при
разных постоянных температурах:
Вертикальные штрихи - предельные отклонения количества диапаузирую-
щих особей в повторностях опыта
вом режиме диапаузировал о всего 3 8 % клопов. При снижении темпе-
ратуры до 17 °C короткодневный эффект также снижался по сравне-
нию с аналогичным эффектом при более высокой температуре. При-
меры ингибирования пониженной температурой формирования диа-
паузы описаны в литературе для хлопковой совки, Chloridea obsoleta, и
восточней плодожорки, Cydia molesta Данилевский, 1961].
' Очень своеобразно проявляется влияние температуры на ФПР у
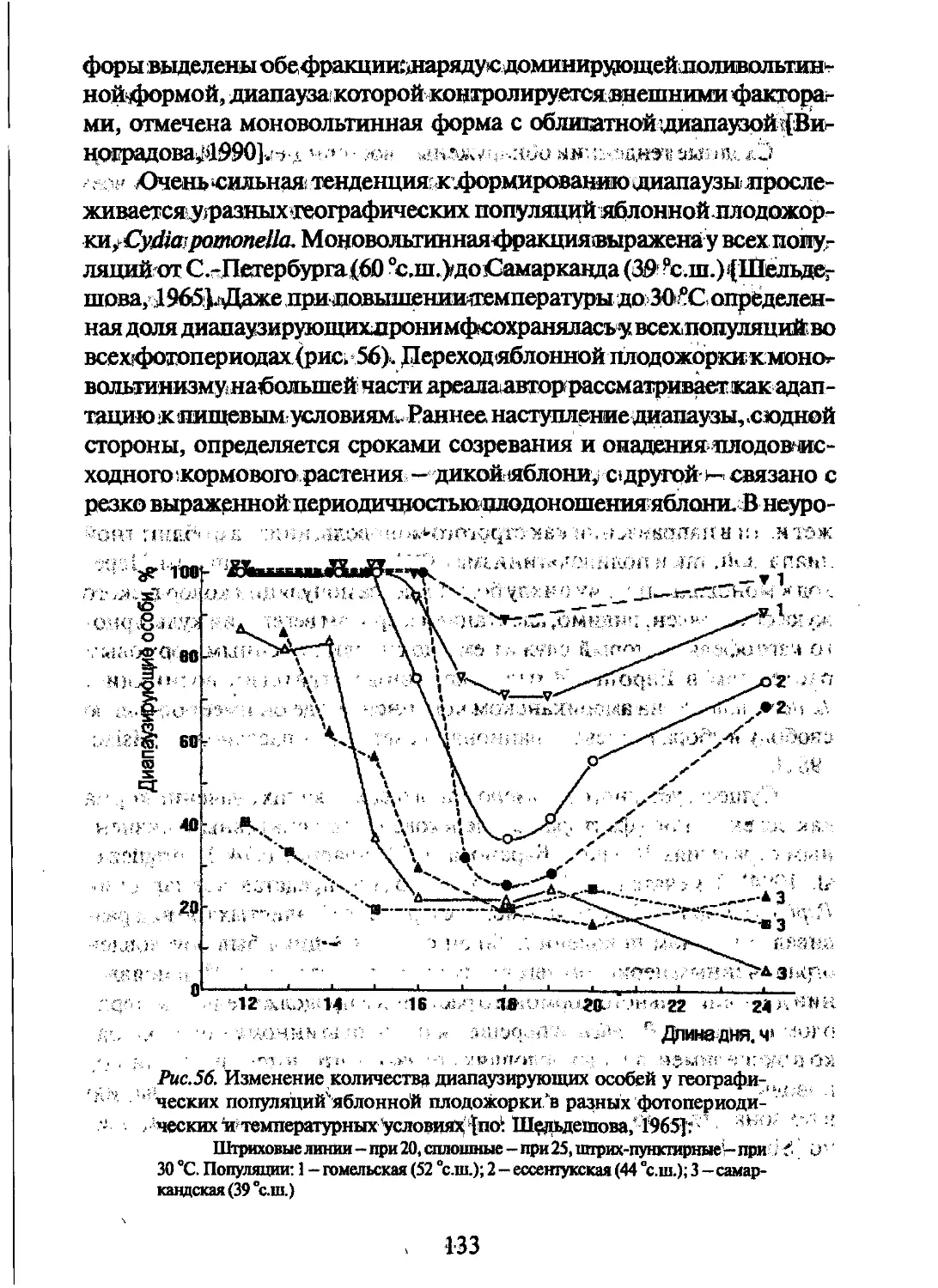
яблонной плодожорки, Cydia pomonella [Шельдешова, 1965]. У ис-
следованных популяций (от С.-Петербурга до Самарканда) незави-
симо от температуры при всех фотопериодах отмечались диапаузиру-
ющие пронимфы, число которых увеличивалось с понижением тем-
пературы. У северной гомельской популяции при 20 °C диапауза фор-
мировалась у подавляющего большинства особей независимо от фо-
топериода, а у южной самаркандской - при повышении температуры
до 30 °C эффект короткого дня подавлялся, но во всех фотопериоди-
ческих режимах отмечалось небольшое количество (от 20 до 40 %)
дйайШ/лйрующих пронимф.
У огневки Diatraea grandiosella высокая температура почти пол-
ностью подавляла индукцию диапаузы (рис. 11). При понижении тем-
перату^^бт 25 до 20 °C порог ФПР оставался неизменным, но коли-
37
Фотопериод, ч
Рис. /Л Температурная изменчивость фото-
периодической реакции Diatraea grandiosella
[по: Takeda, 1985]
чество диапаузирующих осо-
бей в короткодневных режи-
мах (от 0 до 10 ч света в сутки)
возрастало отО до 95% [Take-
da, 1985].
Необычная температур-
ная изменчивость порога
ФПР была обнаружена у со-
снового шелкопряда, Dendro-
limus pini [Гейспиц и др.,
1972]. Весенние генерации
имели термостабильный
критический порог ФПР, а
осенние — термолабильный
(рис. 12). Изменение крити-
ческого фотопериода в весен-
них опытах в интервале испытанных температур сравнимо с таковым
для северных популяций Pieris brassicae, у которых отмечается термо-
стабильная ФПР. Результаты осенних опытов, полученные при раз-
Рис. 12, Изменение критического фотопериода у соснового шелко-
пряда под влиянием температуры [по: ГЬЙспиц и др., 1972]:
а-в весеннем опыте, б-восеннемопыге. Горизонтальные стрелки показы-
вают масштабизменения!фШз1ческого фотопериода в том же диапазоне температур
У Pieris brassicae из С.-Петербурга (а) ну Acronicta ramie is из Сухуми (б)
38
ных TeBMfeparypax, нашли отражение в строгой последовательности
Размах изменчивости порога приближается к
^дж^мм.ртеггрлсгграненной норме термолабильности ФПР: повыше-
ние т^шературы на 1 °C уменьшает критический порог на 10—15 мин.
По мнению авторов, чувствительность ФПР насекомых к температур-
ным колебаниям осенью имеет адаптивное значение, поскольку бла-
годаря этому свойству осеннее понижение температуры служит до-
полштльным сигналом для ухода в диапаузу.
0’вдийриитемпературынадругой параметр ФПР-критическое
числосвето-темновых циклов—данных пока немного. Процесс фото-
периодической индукции у большинства исследованных видов не за-
висит от температуры. Это проявляется в постоянстве числа корот-
к-одиевных или длиннодневных сигналов, что необходимо для созда-
ния критической ситуации (50% диапаузирующих) при разных тем-
пературах. В качестве примера рассмотрим данные для лугового мо-
тылька(рис. 13). Очевидно, что длительность гусеничной стадии адек-
ватно изменяется с повышением температуры, тогда как критичес-
кий пакет фотопериодической информации остается Относительно
постоянным (7—8 КД- или 3—5 ДД- сигналов). Данное обстоятельство
сущесЩццоупрощает использование критического пакета инфор-
маццйДШ|^&югического анализа фенологических событий. Термо-
стабиФЙЙ®^ этого параметра обнаружена также у шелкопряда
Derufrottmus pint [Гейспиц и др., 1972], совок Acronicta rumicis и
Mamestrairassicae [Горышин, Г. Тыщенко, 19726, 1973], парусника
PapilfbxUthus [Endo et al., 1986], мухи Sarcophaga argyrostoma [Saunders,
1971, 1980], клопа Pyrrhocoris apterus [Волкович, Горышин, 1978] и
некоторых других видов.
Термолабильность критического числа свето-темновых циклов
обнаружена у скорпионовой мухи Panorpavulgaris [Sauer, Spieth, 1986]
иуклопа/Ъс?йи57иаси//уеи/га[Волковичидр., 1991]. Суть этого свой-
ства ФПР заключается в том, что для детерминации диапаузы у 50%
особей в разных температурах необходимо разное количество корот-
кодневных и длиннодневных сигналов. Так, у Р. maculiventrisllLJX-xa-
кет составил при 20 °C 10—11, апри24 °C—16 дней. ДД-пакетпритех
же температурах был соответственно 12—14 и 5—6 дней.
Соотношение двух величин — пакета фотопериодической ин-
фюмсации и длительности чувствительной стадии имеет важное зна-
-ЧЯиё при детерминации диапаузы, особенно у видов с относительно
ктостким онтогенезом, но требующих большого количества сигналов
39
a
______________С
18—*14__________ | _ ''
—i1.^- ---------l u——L—_1—_—|
20 16 12 8 4
Дни ол оирождония гдсениц go переноса
в противоположный фотопериод
Пакет КД-информации
Дни ст переноса до
образования пронимф
Рис. 1J. Критическое соотношение исходного и конечного фотопе-
риодических режимов в течение развития гусениц, индуцирующее
диапаузу у половины пронимф лугового мотылька Loxostege sticiicalis
[по: 1Ърышин и др., 1986):
а: 1-4 - у кривых указаны исходные и конечные значения фотопериодаитем-
пературы. Кружками на кривых (варианты опытов) отмечен процентдиапаузирую-
щих пронимф и число конетаых фотопериодических импульсов, полученных гусе-
ницами при переносе в условия короткого (черные точки) или длинного (белые
точки) дня.
б: гориэонтадьныепаяосы-длительность стадии гусеницы при разных тем-
пературах. Белые участки полос-дни с ДД-режимом( 18 ч света в сутки), черные-
днисКД-режимом(14чсвета в сутки). Моменгыизменениярежимовсовмещеньшо
вертикали для удобства сравнения данных
для индукции диапаузы. Так, у мухи Sarcophaga argyrostoma для этого
необходимо 13—14 коротких дней, адлительность чувствительной ста-
дии (личинка) составляет при 26 °C всего 9 дней. Все личинки пре-
40
| ууг,арим не успев набрать необходимого числа КД-сиг-
нг^^ж<ЛйЫИ5М1аузируЮ1ЦИХСРеди нихбыла очень низка [Saunders,
1^Ш^1ПЙПИП ¥аРтина наблюдалась у Р. maculiventris.
иная тенденция обнаружена у парусника Papilio
шаХк>вН(8ЬИ*44а>1985]. При нормальном метаморфозе для индук-
ции куколочкой диапаузы у 50 % особей требовалось 12 КД-сигналов,
при у^цдмл^гении продолжительности гусеничного развития в резуль-
тате адпргеугдмин прилежащих тел для создания критической ситуа-
циипрпребовалосьлишь 9,5 КД-сигналов.
3.2. Я ™яиие суточных колебаний температуры на ФПР
При анализе сезонного развития насекомых гораздо важнее знать
характер проявления фотопериодических адаптаций в условиях тем-
пдрятурнлго ритма, чем при постоянном ее уровне. Однако к настоя-
щему времени специфика действия терморитмов изучена еще недо-
статочно. Среди исследованных видов есть такие, у которых природ-
ные колебания температуры усиливают тенденцию к переходу в диа-
паузу.Например, их влияние на ФПР лугового мотылька проявилось в
увеличении критического фотопериода более чем на один час по срав-
нению,л “константными температурными условиями [Саулич и др.,
1983[ < Сводный эффект температурного ритма был отмечен в опытах
с om&BKoii Diatmea grandiosella [Chippendale et al., 1976] и бракони-
дом Afysia manducator [Зиновьева, 1976, 1991], несколько слабее он
проявилсяу листовертки Choristoneurarosaceana [Gangavalli, Aliniazee,
1985]. а.
С другой стороны, у златоглазки Chrysoperla сатеа s.l. [Волко-
вич, 1988J И хищного клопа Perillus bioculatus наблюдалось значитель-
ное снижение порога ФПР и количества диапаузирующих особей в
условиях ритмично изменяющейся температуры. В тоже время изве-
стнывиды, ФПР у которых одинаково проявляется как при изменяю-
щейся* так и константной температуре. Из исследованных нами ви-
дов таким свойством обладает колорадский жук, Leptinotarsa
decfgdineata, хищный клоп Podisus maculiventris и капустная белянка,
P^teestfcae [Горышин и др., 1985,1986,1988,1989].
□«Влияние на ФПР суточных терморитмов, несомненно, зависит
°?Л&ОДНвдитуды. Как показали исследования К.Ф. Гейспиц [1965]
с^9Ирэдпащелкопряда, тормозящий индукцию диапаузы эффект про-
яЗДЯЛЯЯдишь при терморитмах, ночная составляющая которых при-
41
ближалась к нижнему температурному порогу развития (+5 °C). Оче-
видно, более важное значение здесь имеет диапазон температурного
оптимума ФПР, нижняя граница которого, судя по результатам экспе-
риментов, приближается именно к 5 °C. Все терморитмы с амплиту-
дой в пределах оптимума ФПР по действию на критическую длину
дня были аналогичны средней постоянной температуре.
Приведенные данные свидетельствуют о том, что специфика дей-
ствия терморитмов на сезонно-циклические реакции насекомых еще
до конца не разгадана, но ее необходимо принимать во внимание при
прогнозировании сроков наступления диапаузы в природе.
3.3. Роль изменяющейся длины дня в индукции диапаузы
*
Жизнедеятельность организмов в природе происходит на фоне
сезонной динамики длины дня, поэтому не удивительно, что реак-
ции на постепенные ее изменения широко представлены у насеко-
мых [Corbet, 1956; Norris, 1965; Vepsalainen, 1971; Tauber, Tauber, 1973;
Заславский, 1975,1984].
Однако из 14видов, исследованных в нашей лаборатории [Бело-
усов, Горышин, 1982; Волкович, 1986,1996], чувствительность к по-
степенным изменениям длины дня была обнаружена только у четы-
рех. При этом роль фактора постепенности проявилась лишь в изме-
нении количества диапаузирующих особей в фотопериодах вблизи
порога ФПР.
У клопа Neacoryphus bicrucis (Lygaeidae) имагинальная диапауза
наступала только в ответ-на укорочение дня. При этом важное значе-
ние имела и абсолютная продолжительность дня: при сокращении
фотопериодас 16до 10чсветавсуткидиапаузировало95,8%клопови
только 1,3 %, если день сокращался с 18 до 14 ч [Solbreck, 1979].
Эти факты свидетельствуют о том, что сама по себе чувствитель-
ность к фактору постепенности вне связи с абсолютным значением
фотопериода, и в первую очередь с порогом ФПР, лишена смысла. Это
подтверждается и другими авторами [Ring, 1967; Tanaka, 1978].
В то же время обнаружены виды, в основе регуляции сезонного
развития которых лежит двухступенчатый контроль. Сущность двух-
ступенчатых фотопериодических реакций заключается в последова-
тельном осуществлении двух элементарных реакций — длинноднев-
ной и короткодневной. Переход длины дня через критический фото-
период в этом случае является необходимым. Разнообразие схем жиз-
42
в игтштаюитее на основе ступенчатых реакций, и соле-
мере сезонного развития жужелиц (Coleoptera,
^•1971; Ferenz, 1975; Белоусов, Горышин, 1982], ста-
' Staphylinidae) [Lipkow, 1966; Eghtedar, 1970; Topp,
ICpleoptera, Coccinellidae) [Заславский, 1965, 1984],
hKiimntera. Chrysonidae) [Tauber, Tauber, 1976; Tauber et
1988; Canard, Grimal, 1990] и др.
проблему чувствительности насекомых к изменениям
£ постепенным и резким), видимо, следует согласиться с
awyиранских энтомологов [Tauber, Tauber, 1973] о существо-
ш^^^даакразныкпугей оценки продолжительности дня. У
одюцшадФЙР работает на основе оценки абсолютной продолжи-
ypftgf у других она чувствительна к динамике длины дня, у
‘тр^урт обязательного транспорогового ее изменения.
. . г < ’ С. г . З А Друие факторы, влияющие на ФПР
У большинства насекомых фотопериод и температура играют
главною рбль вр^уляции сезонного развития. Влажность, изменение
хи^и^е&ФкЪ состава пищи для фитофагов, наличие жертвы для хищ-
нййой ЙЦЙразйТов, плотность популяции и другие внешние факторы
могут служить дополнительными источниками информации о состо-
янии Окружающей среды и принимать участие в регуляции годичного
цикла. Количественно оценить влияние этих факторов очень сложно,
но в большинстве случаев их эффект проявляется в изменении коли-
чества диапаузирукнцих особей и сказывается на числе генераций.
Кукурузный мотылек в зависимости от используемых кормовых рас-
тений и^ест два типа годичного цикла: поливольтинный на кукурузе
и моно
можн
б
тинный на конопле [Saringer, 1976; Фролов, 1984]. Воз-
перехоДа от моно- к поливольтинизму при смене местных
интродуцированными, с более длительным периодом веге-
$ описана ДЛяРарШо zelicaon [Sims, 1980].
^Особенно часто влияние пищи проявляется в условиях вблизи
- ^яе<^ой.даиныдня. Лккие данные, например, есть для колорад-
СК®Й decemlineata, хлопковой совки, Chloridea
ota(w7oiiqpjnnHH,1958,1958а], для мальвовой моли, Gelechiamalvella
плодожорки, Cydia pomonella [Jermy,
Loxostege sticticalis [Хомякова и др., 1986] и
мйй®"*ОЯидаИЧ!1*^^ случаях питание старыми и увядающи-
43
ми листьями приводило к увеличению количества диапаузирующих
особей. На примере Eurydema rugosa (Heteroptera, Pentatomidae) было
показано, что в длиннодневных условиях переход к питанию сухими
семенами Brassica napus вызывает диапаузу у 80% клопов, тогда как
при потреблении листьев этого растения все особи оставались актив-
ными [Numata, Yamamato, 1990; Ikeda-Kikue, Numata, 1994]. Для ви-
дов умеренного климата увядающий корм и переход к питанию семе-
нами, видимо так же, как пониженная температура, служит сигналом
приближения осени.
Отсутствие или недостаток корма у хищных насекомых также
способствует наступлению диапаузы. Это было обнаружено у многих
коровок [Mc.Mullen, 1967; Iperti, Hodek, 1974] ижужелиц [Penney, 1969;
Paarman, 1976]. У златоглазки Chrysoperla сатеа strain mohave корот-
кий день вызывает имагинальную диапаузу, а длинный день способ-
ствует активному развитию лишь при наличии живых тлей. Если же
тлей нет, даже в условиях длинного дня наступает диапауза [Tauber,
Tauber, 1973с].
Влияние пищевого режима на ФПР клопа P.maculiventris было
исследовано при температурах 20,23 и 25 °C (рис. 14). Наиболее чет-
ко зависимость от режима питания проявилась в пороговой зоне (и в
Длина дня, ч
Рис. 14. Влияние пищевого режима (1-6) на параметры фотоперио-
дической реакции Podisus maculiventris:
1 -гусеницы большой вощинной моли; 2 -личинки синей мясной мухи; 3 -
личинки большого мучного хрущака; 4 - личинки комнатной мухи; 5 - личинки
колорадского жука; 6 - гусеницы чешуекрылых
44
левой и в правой части фотопериодической шкалы). Доля особей, ди-
щгвузирующих в этих вариантах опыта, оказалась ниже среди клопов,
ймрЫне.нных на оптимальных видах жертвы - гусеницах большой во-
гцйнной моли, Galleria melonella, и личинках синей мясной мухи,
CdUiphora vicina, чем среди особей, выращенных на менее благопри-
йТйом корме - личинках колорадского жука, большого мучного хру-
щака, Tenebrio molitor, и комнатной мухи, Musca domestica.
При температуре 25 °C, когда тенденция к формированию диа-
паузы была слабее, чем при более низкой температуре, только в усло-
виях выращивания на личинках комнатной мухи в короткодневных
режимах диапаузировали 100% клопов. При 20 °C в короткодневных
условиях склонность к формированию диапаузы настолько велика,
что независимо от качества корма во всех вариантах опыта диапаузи-
ровали все особи. Таким образом, и в этих экспериментах проявилась
доминирующая роль фотопериода в индукции диапаузы, а пища иг-
рала лишь второстепенную роль. Подобное соотношение условий пи-
тания и фотопериодических условий свойственно большинству ис-
следованных насекомых, независимо от типа питания.
Влияние плотности популяции на индукцию диапаузы показа-
но для многих видов. В большинстве случаев увеличение скученности
особей стимулирует формирование диапаузы. В исследованиях, про-
веденных на капустной совке, Mamestra brassicae (сухумская популя-
ция), обнаружена заметная тенденция к увеличению числа диапаузи-
рующих куколок в вариантах с большей
численностью гусениц в группах [Сау-
лич, 1975]. Так, в коротком дне (10 ч
света в сутки, температура 25 °C) ми-
нимальное число диапаузирующих ку-
колок наблюдалось при наименьших
размерах группы (л = 1 и п = 2), а мак-
симальное — при наиболее скученном
содержании гусениц. Абсолютное зна-
чение полученной разницы невелико,
от 20% при л = 2 до 33% при л = 20
(рис. 15), тем не менее хорошо выра-
женная тенденция не оставляет сомне-
ния в реальности данного явления.
Сходные данные приводятся и д ля дру-
Рис. 15. Зависимость количества
диапаузирующих куколок Ma-
mestra brassicae (сухумская попу-
ляция) в короткодневном фото-
периодическом режиме (10 ч све-
та в сутки, 25 °C) от размера груп-
пы при выращивании гусениц
45
гих видов чешуекрылых [Tsuju, 1959; Iwao, 1962].
Противоположный эффект скученности был обнаружен у мухи
Sarcophaga argyrostoma. В условиях чрезмерного переуплотнения и не-
достатка пищи личинки развивались быстро и окукливались раньше,
чем набиралось необходимое число КД-сигналов, обеспечивающее,
формирование диапаузы [Saunders, 1976]. Аналогичный эффект про-
являлся при повышении температуры (см. выше).
, b ., '
Глава 4. ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
СЕЗОННЫХАДАПТАЦИЙ
" Реакции насекомых, контролирующие их сезонный цикл, под-
вержены внутривидовой дифференциации. Она неодинаково прояв-
ляйся у разных видов и затрагивает те или иные элементы жизненно-
гстфйкла. При исследовании сезонного развития насекомых наиболь-
шей Интерес представляет географическая изменчивость трех групп
реЙКЦий: по признакам вольтинизма и диапаузы, онтогенетической
приуроченности диапаузы, параметрам ФПР.
' ' 4.1. Вольтинизм и диапауза
,!Всвязи с климатической зональностью у большинства насеко-
йййгколичество генераций за вегетационный сезон увеличивается по
движения с севера на юг. При этом обычно наблюдается четкая
зйймюмерность: переход от моновольтинизма в северных частях аре-
гШ через би- и поливольтинизм к гомодинамному типу сезонного
ЦИКла на юге. Тйкая тенденция хорошо прослеживается, например, у
Pectinophora gossypiella, индийская популяция которой харакгеризу-
йТСя бездиапаузным развитием [Raina, Bell, 1974], у Papilio polyxena
(Btiu, 1981а,b], у Oncopeltu&fasciatus [Dingleet al., 1980] и многих дру-
гое видов. Однако чаще у тропических популяций диапауза не исче-
зает, а меняются только ее количественные показатели - интенсив-
ность, продолжительность, а также тенденция к формированию диа-
паузы [Jordan, 1980; Denlinger, 1986; Kurahashi, Ohtaki, 1989].
Широтный градиент тенденции к формированию диапаузы от-
ЙЙен У насекомых из разных отрядов: у мухи Drosophila auraria
Жйпига, 1983], клопа Oncopeltusfasciatus[Dingleetai., 1980], листое-
'J&Atrachya menetriesi [Ando, 1979,1983] и многих других видов.
Wflr.
47
Детальный анализ характера географической изменчивости се-
зонных адаптаций проведен Е.Б. Виноградовой [1991] в исследова-
ниях 33 географических популяций мухи Calliphora vicina с огромной
территории от Мурманска (69 °с.ш.) до Ашхабада (38 °с.ш.) и от Гам-
бурга (10 °в.д.) до Владивостока (132 °в.д.). В пределах этой террито-
рии обнаружена клинальная изменчивость как по тенденции к фор-
мированию диапаузы, так и по ее свойствам — продолжительности и
интенсивности. Самая глубокая диапауза была отмечена у северных
популяций из Мурманска и Беломорска. У южных популяций из За-
кавказья и Средней Азии непрочная диапауза формировалась только
у небольшой части особей. Для ее прекращения оказалось достаточно
нескольких дней с температурой выше 12 °C. Неожиданно высокая
прЬчность диапаузы, выпадающая*из общего характера клинальной
изменчивости данного признака, обнаружилась у сухумской популя-
ции. По мнению автора, в последнем случае диапауза выполняет дру-
гую функцию, сдерживая преждевременное развитие личинок во вре-
мя тёплой зимы, что иллюстрирует широкие возможности использо-
вания видом одной и той же адаптации - прочной личиночной диа-
паузы - для переживания столь противоположных условий как суро-
вая, холодная зима в северной части ареала и теплая мягкая зима в
зоне субтропиков.
Сходная тенденция повышения интенсивности и увеличения
продолжительности диапаузы была обнаружена у южной популяции
Acronicta rumicis [Симакова, 1971]. У белгородской популяции этого
вида при благоприятной д ля реактивации температуре диапауза про-
должалась 3—4 месяца, а у сухумской — 4,5—5 месяцев. Аналогичная
закономерность обнаружена у листоеда Diabrotica virgifera [Krysan et
al., 1977].
У Chrysopa oculata [Necholson et al., 1987] и мухи Chlorops oryzae
[Takeda, 1996] обнаружена обратная тенденция: диапауза северных
популяций характеризовалась большей глубиной.
4.2. Диапаузирующая и чувствительная
к длине дня стадии
В зависимости от географической широты у насекомых может
меняться приуроченность диапаузы к определенной стадии онтоге-
неза. У листоеда Chrysochloa basilea в суровых климатических зонах
48
.дддтт ништяки старших возрастов, в мягком климате зимовка сме-
щается на стадию имаго [Долгин, 1978].
1У комара Aedes triseriatus южные популяции обычно зимуют в
имминочном состоянии. По мере движения на север чаще наблюдает-
оаямбоиональная диапауза [Holzaphel, Bradshaw, 1981]. Сходная тен-
отмечена v японского вида A. togoffMogi, 1981].
(>аг Смещение диапаузы на стадию яйца, как у комаров, или на ста-
дИрьяичинки, как у жуков, несомненно вызвано необходимостью по-
вдвкния устойчивости к неблагоприятным условиям. Изменение он-
тогенетической приуроченности диапаузы у географических популя-
нийьииогда сопровождается смещением чувствительности к длине дня
щ^фугую стадию. Эта тенденция хорошо прослеживается, например,
умуя^Виноградова, 1973]. У популяции парусника Papilio protenor из
умеренной зоны (подвид P.p.demetrius) чувствительны к фотопериоду
гупппг11-1' двух последних возрастов, тогда как особи его субтропичес-
кагйлодвида (P.p.liukinensis) реагируют на длину дня на протяжении
всейЛусеничной стадии [Ichinose, Negishi, 1979].
Интересная особенность восприятия фотопериодических сиг-
наиввобнаружена у кукурузного мотылька. Чувствительность к фото-
ntpwiny гусеницам всех возрастов и даже эмбрионам.
ОЙЙвфиУ южных форм наибольшей чувствительностью харакгеризу-
иу>мм**Эгёеьгиць1 младших возрастов, ау северных она одинаково прояв-
ляеижиа. протяжении всего преимагинального развития: 10—16 ко-
ротких дней в любой период вызывает формирование диапаузы у всех
пронимф[Ду Чжень-Вэнь, 1961].
4.3. Параметры фотопериодической реакции
Анализируя адаптации насекомых, обеспечивающие согласова-
ние фенологии каждого вида с изменчивостью климата в пределах
ареала, А.С. Данилевский [1961] пришел к заключению, что главным
способом адаптации служат наследственные внутривидовые измене-
ния параметров ФПР. Сравнительное изучение географических попу-
ляций разных видов показало, что каждая их этих популяций имеет
свою ФПР, отличающуюся от реакции других популяций, и парамет-
РЫ ре передаются по наследству. Сравнение кривых ФПР локальных
Щфуляций одного вида показало, что у большинства из них геогра-
ЧЧНеская изменчивость носит клинальный характер, который под-
Дерхивается благодаря тому, что гибриды между особями географи-
; 49
ческих популяций наследуют ФПР с параметрами, промежуточными
по сравнению со свойствами родительских ФПР.
Обычно порог ФПР плавно возрастает у более северных популя-
ций. Благодаря этому диапауза у популяций в северных частях ареала
формируется раньше, несмотря на большую продолжительность дня
в высоких широтах. Пример клинальной географической изменчиво-
сти порога демонстрирует рисовый стеблевой мотылек, Chilo sup-
pressalis [Hidaka, 1977]. Критический фотопериод разных географи-
ческих популяций (от 32 до 44 °с.ш.) позитивно коррелирует с широ-
той и описывается уравнением линейной регрессии (рис. 16).
У яблонной плодожорки, Cydia pomonella (рис. 17), наиболее се-
верная из исследованных популяций, петербургская, слабо реагирует
на продолжительность дня: большая часть гусениц диапаузирует во
всех световых режимах. У южных, среднеазиатских, популяций, на-
оборот, только при фотопериоде 12 ч диапаузирует 100% гусениц.
Между этими кривыми располагаются кривые ФПР остальных иссле-
дованных популяций, различающиеся по значению критической дли-
ны дня [Шельдешова, 1965].
Степень изменчивости порога ФПР у локальных популяций ва-
рьирует у разных видов (рис. 18). При изменении широты местности
на 5° критический фотопериод сдвигается в среднем на 1—1,5 ч [Дани-
левский, 1961; Данилевский и др., 1970; Saunders, 1982; Tauber et
al., 1986; Danks, 1987]. Вместе с тем существует много исключений из
Рис. 16. Клинальная географическая изменчивость порога ФПР у
рисового стеблевого мотылька, Chilo suppressed [по: Hidaka, 1977]
50
Длина дня, ч
Рис. 17. Географическая изменчивость ФПР яблонной плодожорки,
Cydia pomonella. Температура 25 °C [по: Шельдешова, 1965]:
Популяции: 1 — петербургская (60 °с.ш.>; 2 - гомельская (52 °с.ш.); 3 -
полтавская (50 °с.ш.); 4 - ессентукская (44 5 - таджикская (39 °с.ш.>; 6 ~ са-
маркандская (39 °с.ш.)
этого правила. Сильно выражена географическая изменчивость поро-
га^ПР у белянки Pieris napi [Матинян, 1965], но она почти не обна-
руживается у Dendrolimuspini [Гейспиц, 1965] и Pectinophoragossypiella
[^nkersmit, Adkisson, 1967], слабо проявляется у американской белой
бабенки на Японских островах и в Европе [Hidaka, 1977; Саулич, 1994],
удугового мотылька [Горышин и др., 1986г], европейских популяций
кцдррадского жука [Арапова и Заславский, 1981; de Wilde, Hsiao, 1981;
Горышин и др., 1987], у Diatraea grandiosella [Takeda, Chippendale,
1982].
; У ряда видов обнаружен совершенно особый тип географичес-
кий изменчивости параметров ФПР. В отличие от большинства насе-
КОМых, для которых характерна клинальная изменчивость критичес-
кого порога ФПР, популяции капустной белянки, заселяющие обшир-
ную территорию Восточной Европы, распадаются на два резко обо-
ЧИэленных экотипа - северный и южный. Свойства северного экоти-
Ча исследованы на примере популяций из С.-Петербурга (60 °с.ш.),
Я|$₽лавля (56 °с.ш.), Москвы (53 °с.ш.), Бреста (52 °с.ш.), Белгорода
51
'
°с.ш. 65
Критический фстопериод, ч
Рис. 18. Географическая изменчивость критического фотопериода у
разных видов [по: Данилевский и др., 1970]:
1 Pieris brassicae (23 °C); 2 —Р. гарае (23 °C); 3~Tetranychusurticae(\% °C);
4 — Mamestra brassicae (25 °C); 5 — Р, napi(18 °C); 6—Acronicta rumicis(23 °C). Попу-
ляции из Хибин, С.-Петербурга, Витебска, Белгорода, Северного Кавказа и Черно-
морского побережья Кавказа
(50 °с.ш.) [Данилевский, Кузнецова, 1968; Горышинидр., 1986а]. При
изменении температуры от 12 до 26 °C порог ФПР разных популяций
северного экотипа остается почти неизменным (около 15 ч 40 мин).
Нельзя не согласиться с выводом А.С. Данилевского о том, что воз-
никновению клинальной изменчивости параметров ФПР у Р brassicae
в пределах Восточной и Северной Европы препятствуют свойствен-
ные ей обширные миграции, нарушающие пространственную изоля-
цию локальных популяций.
Исследование температурной и географической изменчивости у
популяций южного экотипа показало, что их ФПР проявляют кли-
нальную широтную изменчивость (рис. 19) и подвержены влиянию
температуры (рис. 20).
Кроме описанных двух резко различающихся между собой эко-
типов обнаружились переходные формы, обладающие свойствами
обоих экотипов. К ним, например, относятся популяции из Херсона
(47 °с.ш.) и Крыма (пос. Ленино, 45 °с.ш.). Критический фотопериод
52
Рис.19. Географическая изменчивость фотопериодической реакции
Pieris brassicae. Температура 25 °C:
Популяции из: 1 - Белгорода; 2 - Херсона; 3 - Крыма; 4 - Сухуми (ориг.);
5-Ленкорани [по: Мустафаева, 1974]
у'inn изменяется в соответствии с широтой, в то же время для каждой
из ййх отмечена относительная термостабильность порога ФПР, что
•£СрКМЖг. <
$, то-
-ОЖ OTG'SX
g0.
<м1нгл^<|)
:1ПФ
S во
1 П ги т&'
5 40
j_ ,, I S
, 'Rt.
'71ШАМ0- 20’
ЖХ iw -
0
6 8 10 12 14 16 18
Длина дня, ч
Температурная изменчивость фотопериодической реакции
а ' ^eris brassicae: ......
«W* , ?.? Ъ' i t Популяции из: а - Ленкорани [по: Мустафаева, 1974]; б - Сухуми
53
Рис.21. Термостабильиость фотопериодической реакции Pieris
brassicae'.
Популяции из; а - Херсона; б - Крыма [ 18 °C - по*. Масленникова и Мус-
тафаева, 1971; 25 °C—неопубликованные данные А. В. Лачининского]
Длина дня. ч
типично для популяций, обитающих на Средне-Русской равнине
(рис. 21).
Имеющиеся материалы позволяют наметить зоны распростра-
нения выявленных экотипов Р. brassicae в Восточной Европе. Ареал
северного экотипа охватывает территорию, расположенную к северу
от 47 °с.ш., область южнее 45 °с.ш. заселена популяциями южного эко-
типа. Между ними обитают формы с промежуточными свойствами.
Сходная ситуация наблюдается в Западной Европе. Большая
часть обитающих там популяций капустной белянки обладает ФПР,
зависящей от температуры и подверженной непрерывной клиналь-
ной изменчивости (величина критического порога возрастает от 12 ч
на 42,5 °с.ш. до 15 ч 50 мин на 56 °с.ш.). Форма, идентичная нашему
северному экотипу, обнаружена только на 56-й параллели [Spieth,
1985]. Можно предположить, что граница распространения южного
экотипа в Западной Европе сдвигается к северу в связи с более мягким
климатом. Сходные черты ступенчатой изменчивости критического
порога обнаружены у кукурузного мотылька, Ostrinia nubilalis [Дани-
левский, 1961], и медведицы Spilosoma menthastri [Кинд, 1985], но вы-
ражены они не столь резко.
Кукурузный мотылекв исходном европейском ареале представ-
лен двумя экотипами. На севере ареала обитает моцовольтинная, а на
54
юге—поливольтинная раса. Граница между ними в Восточной Европе
проходит по югу лесостепной зоны. Эти экотипы совпадают с отме-
ченными в литературе формами: «конопляной», развивающейся в
природе лишь в одном поколении, и «кукурузной», способной в зави-
симости от погодных условий давать одно или два поколения [Шапи-
ро и др., 1979], Для южного экотипа, заселяющего степную зону Укра-
ины, Северный Кавказ и Черноморское побережье Кавказа, характер-
на термостабильная (в пределах 18—25 °C) ФПР с порогом около 15 ч.
В экспериментах популяции из Сухуми и Днепропетровска, относя-
щиеся к этому экотипу, имели одинаковый критический фотопериод,
несмотря на значительную удаленность этих географических точек
(6° по широте). Такой же экотип заселяет юг Франции [Bonnemaison,
1974] и Венгрию [Saringer, 1976]. Северный экотип из Сумской обл.
(территориально более близкой к Днепропетровску) при той же тем-
пературе (25 °C) имел ФПР с критическим порогом около 17 ч,
зависимым от температуры [Данилевский, 1961]. У этих экотипов об-
наружена дифференциация и по длительности диапаузы, В природе
бабочки перезимовавшего поколения «кукурузной» формы вылетают
позже бабочек «конопляной» формы. Эта закономерность подтвер-
дилась и в экспериментальных условиях. Подобное временное рас-
хождение, вызывая репродуктивную обособленность особей, создает
предпосылки для разобщения экотипов, результатом которого может
быть образование новых видов [Фролов, 1984].
В Северной Америке, куда О. nubilalis был завезен в начале века,
также выделяют два экотипа, различающихся по числу поколений и
общей тенденции к формированию диапаузы [Showers et al., 1975].
Северный экотип, распространенный на юге Канады, характеризует-
ся моновольтинным циклом. Популяции южного экотипа поливоль-
тинны и имеют пониженную тенденцию к формированию диапаузы.
Однако вопрос о географической изменчивости параметров ФПР тре-
бует дальнейших исследований, поскольку граница между отмечен-
ными экотипами не обнаружена. У10 исследованных популяций ку-
к)/5рузного мотылька из Северной Америки (от 45 до 37 °с.ш.) крити-
’ШКий порог позитивно коррелировал с географической широтой
обитания популяции [Beck, Apple, 1961]. К сожалению, более север-
популяции не исследованы.
Географическая изменчивость затрагивает также параметры ФПР,
Фнтролирующей летнюю диапаузу. Впервые это явление было изуче-
ЙЪ у совки Mamestra brassicae из Японии [Masaki, 1956; Masaki, Sakai,
55
1965]. Позже аналогичные тенденции были изучены и у западно-ев-
ропейских популяций вида [Gruner, Sauer, 1988; Sauer, Gruner, 1988].
Результаты этих исследований свидетельствуют о клинальной измен-
чивости как критического порога ФПР, индуцирующей летнюю диа-
паузу (рис. 22), таки продолжительности эстивации куколок (рис. 23).
Лабораторное изучение 17 западно-европейских географических
популяций М. brassicae [Bues et al., 1991] позволило апроксимиро-
Рис.22. Клинальная изменчивость
порога фотопериодической реак-
ции, индуцирующей летнюю диа-
паузу у совки Mamestra brassicae.
Температура 25 °C [по: Sauer, Gru-
ner, 1988]*.
Белые столбики—зимняя диапауза;
заштрихованные - активное развитие; чер-
ные —летняя диапауза; п—количество ку-
колок
56
•Ш £
«I - число куколок; лр -
Длина ДНЯ
Средняя гдодопжигельндегь
Географическая
изменчивость продолжи-
тЙяЬнбсти эстивации ку-
йБок Mamestra brassicae.
ЩшЗДатура 20 °C [по:
Продолжительность эстирации, дни
ййть зависимость процента особей, уходящих в летнюю диапаузу (У),
^географической широты происхождения популяции (х) уравнение
ШГ«~0,172х+58,342.
v 'Аналогичное увеличение длительности летней имагинальной
Диапаузы у южных популяций описано у жука Catops nigricans (Са-
$Ф₽Мае, Coleoptera) (Торр, 1990].
<г 1 Изменчивость другого параметра ФПР - критического пакета
<фйТопериодической информации — исследована очень слабо. Извест-
Но, что у Acronicta runticis процесс накопления ФП-информации от-
57
л ичается у популяций из Белгорода (50 °с.ш.) и Сухуми (42 °с.ш.).
Для достижения одинакового результата (50 % диапаузирующих ку-
колок) южнойпопуляции требуется 16—18 КД-сигналов, а северной -
только 10—11 [Горышин, Г. Тыщенко*1970,19726]. У соснового шел-
копряда ленинградской популяции д ля формирования диапаузы тре-
бовалось в среднем 30-32 КД-сигнала, у более южной, гомельской, 38
[Гейспиц и др., 1972].
В исследованиях с Aedes atropalpus (Diptera, Culicidae) также было
показано, что южные популяции (14 °с.ш.) требуют большего числа
КД- сигналов для индукции диапаузы, чем северные (45 °с.ш.) [Beach,
1978] . Сходная тенденция обнаружена и у мухи Chlorops oryzae [Takeda,
1996]. У скорпионовой мухи Panorpacommunis (Mecoptera, Panorpidae)
пакет ДД-информации, индуцирующий активное развитие у север-
ных популяций (57 °с.ш.), значительно превышает пакет у более юж-
ных популяций (48 °с.ш.) и составляет соответственно 10 и 3 дня [Sauer
et al., 1986].
Несомненно географическая изменчивость величины критичес-
кого пакета ФП-информации, как и других параметров ФПР, имеет
адаптивное значение. Уменьшение количества КД, необходимых для
формирования диапаузы у северных популяций, гарантирует возмож-
ность своевременного ее наступления, соответственно уменьшение
пакета ДД-информации способствует активному развитию южных по-
пуляций.
Адаптивное значение имеет географическаяизменчивость еще
одного параметра ФПР - температурного оптимума. Основная тен-
денция его изменчивости в пределах вида, по данным А.С. Да-
нилевского [1961], проявляется в том, что у северных популяций тем-
пературный оптимум выше, чем у южных. Очевидно, что смещение
его диапазона в сторону высоких температур также направлено на уси-
ление гарантии формирования диапаузы, особенно тогда, когда по
температурным условиям возможно,развитие только одного поколе-
ния в течение вегетационного сезона,,а длинцый северный день пре-
пятствует индукции диапаузы у особей этого единственного поколе-
ния.Вэтом случае низкая температура отключает механизм фПР, и
диапауза формируется независимо от.длиныдня. . .л
В целом приведенные материалы свидетельствуют о глубокой
наследственно закрепленной дифференциации локальных рас по их
сезонно-циклическим адаптациям и строгом соответствии.сезонного
цикла каждой популяции фототермическим условиям обитания.
Часть II«
ЭКСПЕРИМЕНТАЛЬНЫЙ АНАЛИЗ
СЕЗОННОГО РАЗВИТИЯ НАСЕКОМЫХ
Глава 5. МЕТОДЫ ИССЛЕДОВАНИЯ
‘Г- “'/й ‘ф>
г. .у z'' •" У'УУГ’. У;’; ‘ r-1'o • , , , .
Основная задача экспериментальной части исследования состо-
яла в изучении сезонно-циклических реакций насекомых, отражаю-
щих их адаптацию к ритму климата и определяющих возможность
выживания «новых условиях при дальних внутри- ивнеареальных
перемещениях. '' ' • " 'Т,-: <
Объектами исследований были представители трехкрупнейших
отрядов насекомых—Coleoptera, Lepidoptera и Heteroptera. Среди них
большинство видов с имагинальной диапаузой: Leptinotarsa decern-
lineata Say (Coleoptera, Chrysame\idtt);'PyrrhocorisapterusL.
(Heteroptera, Pyrrhocoridae), Palomena prasina L. ’(Heteroptera,
Pentatomidae), Arma custos Fabr. (Heteroptera,' Pentatomidae), Рег/У/ш'
bioculatus Fabr. (Heteroptera, Pentatomidae), Podisus maculiventris Say
(Heteroptera, Pentatomidae), Grdphosoma lineatum L? (Heteroptera,
Pentatomidae), Coreus marginatas L. (Heteroptera, Coreidae). Куколоч-
ная’диапауза исследована у чешуекрылых: щавелевой совки, Acronicta
rumicis L. (Noctuidae), кайустной Совки,' Mamestra brassicae 'L.
(Noctuidae), американской белой бабочки, Hyphantria cunea Drury
(Arctiidae) и капустной белянки, Pieris brassicae LJ(Pieridae); проним-
фальная’ диапауза*— у лугового мотылька,’Xoxostege sticticalis L.
(Pyralidae). Группа исследованных видов с гомодинамным развитием
включала в себя луговую совку, Mythimna unipiincta Haw., малую на-
земную совку, SpodopteraexiguaWo. и совку гамму, Autographagamma'L. —
все из сем. Noctuidae (Lepidoptera). Всего’16 видов. ‘ ’
Экспериментальная работа проводилась в трех географических
пунктах: С.-Петербурге, 60 °с.ш. (лаб. энтомологии Биологического
института СПбГУ), в поселке Борисовка Белгородской обл., 50 °с.ш.
(заповедник «ЛеснаВорскле») и в Славянске-на-Кубани Краснодар-
ского края, 45 °с.ш. (станция защиты растений ВИЗР).
59
5.1.Фенологические наблюдения и полевые опыты
Для определения фенологических сроков появления отдельных
онтогенетическихчугадийразныхвидов.ачакжевыяснения'числа ре-
ально развивающихся поколений и сроков формирования диапаузы
насекомых ловили на свет (лампа ДРЛ-400) и собирали методом ко-
шения’в местах их естественного обитания.
В сложных случаях (каку колорадского жука или клопасолдати-
ка), когда разграничить отдельные гецррации недредставлялось воз-
можным, в природе собирали личинок последнего, V возраста, при-
надлежность которых к определенному поколению устанавливалась с
большой, степеньювероятностщи,содержали вусдовиях,, близких к
естественным.^ дальнейшемсредиокрылившихся особей определя-
лидоличестдо аетивных и диапаузирующих (см. 6.1.2, и >6.1.3).Такой
способ лозволял зафиксироватьдирамику нарастания количествадиа-
паузирующих особей в реальной обстановке и ориентировочно уста-
.новитьвозмржностьобразования,сдедующегопоколения.,,
ц Полевые рпытьвдроводились для установлениядозможности! ин-
дукции диапаузы у тестируемых объектов на,фоне динамики л рирод-
ных факторрвдразличные-календарные сроки. Они предусматривали
проведение.втечениесезона,последовательныхсерийопытов,.вкото-
рыхучитывались.сроки развития,отдельных стадийлисло акгивныхи
диацаузирующдх .особей, продолжительности, созревания и другие
показатели^ необходимые для изучеция сезонного развития. Эта ме-»
тодика, разработаннаяА.С., Данилевским иИ-А- 'Кузнецовойф1968(,
позже была апробирована на,многих, видах и оказалась перспектив-
ной для углубленного анализа закономерностей сезонного.развития
насекомых) Горышин. и др., 1980,1981МЦ98ба-в; Takeda, Chippendale;
1$8Д;Горышин,Саулич, 1983; Сауличи др* Д983; -Волкович,.! 98&К
. ..,^Црлевыерпытьввовсех.трех пео1рафических пунктах, проводи-
лись в.специально рконструированном^алюзийном домике, открьь
том с северной.стороны и защищающем насекомых от прямых сол-
нечныхлучей-Температурав домике,рщистрировадасытермометрами
(максимальный, минимальный неровный) и суточным термографом.
..•> л .* -л,,, 5*2.*11абораторныеопыты<; г'<, - .•
ч.t ;и> -‘ii.cc’ii.ofii.'H'.sB’-1 *1/’• ’йД - «ту":
,, Технинескоеоснащениелаборатории энтомологии вСтаром Пе-
тергофе ив заповедникежПес да Вррекде» обеспечивалонеобходимые
60
экспериментальные режимысавтоматиаескимжонтролем и воспро-
изведением. Колебаниятемпературы в фототермостатных камерах не
превышали^ 1°С. Для контроля температуры использовались ртут-
ные термометры. Освещенность в камерах составляла около 200 (180—
250) лк. Дляосвещенияиспользовались люминесцентные лампыДС-
20. Описания камер и автоматических программных устройств к ним
содержатся в публикациях1Горышин,’1966; Браун, Горышин,. 1978].
Влабораторныхонытахисследовалифотопсриодические(ФПР)
и термопериодические (ТПР) реакции. Схема лаб оратор ныхопыто в
по изучению ФПР была традиционной; использовались различные
константные режимы длины светового дня. Большая часть опытов
проводилась при фотопериодах, имеющих экологическое значение, —
от 12 до 1*8 ч света всутки. В отдельных случаях использовались фото-
периодыотОдо 24,ч светавсутки.Интервалы между соседними фото-
периодическими вариантами были равны 1 ч. Обычно эксперименты
выполнялись при 3—4 уровнях температуры в диапазоне от 17 до 27 "С.
Значениятемпературьгвыбирались,.исходя изтребованийвидов', но в
исследование всегда вкл ючал ись по крайнеймере три ее уровня —мак-
симальный,<минимальный1исредний,'наиболее соответствующиетем-’
пературным'условиямн природной 'обстановке^ с м на
»: При изучении^роли суточных терморитмов в'жизнедеятельнос-,
та ииндукции диапаузыбыли!применены>П-образные ритмы темпе-
ратуры, в которых ! 2-часовая< холодная компонента чередовалась ^
тепл ойикомпонентойтакойже продолжительности-Холодныйпери-!
од сочетался соскотофазойдгеплый период приходился соответствен-
но на фотофазу.Вс! 2-часовом фотопериодесинхронизациябылапол-
нойуинвариантах,с большей продолжительность^ светлого периода
пониженная температуразахватывала и часть фогофазы (рис?24)™^
Изменение температурного режима проводилос^вручнуюГут-
ромивечеромматериал переносили из одной камеры вдругую. Насе7
комые контрольных:групп содержалисьпритех жефотопериодах,но
при постоянных температурах/эквивалентных-средним и крайним
(еслиэто<>ыло'возможно}значениям терморитма. f
лоДругуювозможность'Дляизучениявлияниялеременнрйдемпе-
ратурыш а. жизнедеятельность насекомых давало-использование спе-
циальной полуавтоматической установки, с помощью которой на фоне
природноготерморитма воспроизводились пятьвариантовдлиныдня
[Браун,(Горышин, 1984]аВ сумеречноевремя принеобходимости ис-
пользовали подсветку (лампаД РЛ-400).;’" < 1 о
61':
Рис. 24. ‘Соотношение термо- я фотопериодов в разных вариантах
И1'ПОПЫТалД<:Ь-:;1Ч!«;'.‘ I »л!Л) 11 . Чр Л • ’i.T
,э <' - 4Т( U' • . * ' . '< ".
' Чувствитедьнаястадия и пакет фотопериодической информации.
Ддя> установления стадии,^воспринимающей; фотопериодическую
информацию,опыты выполняли потрадиционней схеме с одноразо-
выми изменениями альтернативных фотопериод овна разных этапах
онтогенеза (рис.25). Значение.температуры и фотопериодический
режимвыбирали с учетомособенностей.ФПРвида, При определении
пакета фотопериодической информации использовалась аналогич-
ная схема,.но альтернативныепереносыматериалаосуществлялись
; чаще (через 2-3 дня).
jar : •Плотность лопуляции. При
т изучении 'влияния этого фактора
> группы гнасекомых, состоящие из
разного, числа особей, обычно вы-
ращиваютв сосудах одинаковой ем-;
. . кости.. При>такой методике созда-
ется, опасность переуплотнения, в’
результате которого особи попада-
< ют в неравноценные условия осве-
щенности, влажности, .наконец,.
. . обеспеченности кормом. <В этом
Рис.25. Схема опытов по определе- о разграничить эффект
нию чувствительной стадии:
: Заштрихованнаяобласть— короткий ‘ « Группы,з Связанный С: ВЗаимОдеЙ-
день, светлая - длинный день; Ц-L, -личи- , цтвиеМОСОбеЙ, /ОТ Эффектов, ВЫЗВаН-'
ночный возраст
62
ных перенаселенностью замкнутого объема м качественным наруше-
нием среды обитания. Чтобы избежать этого в опытах, гусеницы по
мере-роста переводились в большие емкости. При этом -сохранялся
постоянный объем жизненногопространства, который составлял не
менее 40-50 мл на каждую особь.
Холодостойкость. Значение этого фактора оценивали термоэлек-
трическим методом по уровню предельной температуры переохлаж-
дения жидкостейтела. Измеряли температуру переохлаждения с по-
мощью тонких медно-константановых; термопар, подключенных к
зеркальному гальванометру. Его шкала позволяла считывать темпера-
туру с точностью 0,2 °C.^Надежность теплового контакта между тер-
мопарой^ телом насекомого обеспечиваласьвазелиновой прослой-
кой и: контролировалась по стабильности точки замерзания. Описа-
ние использованной термоэлектрической установки дано в книге
Н.И. Горышина [1966]. Для стандартизации и‘сравнимости измере-
ний холодостойкости в опытах использовали объекты одинаковых воз-
растных категорий: а)3-дневные яйца; б) гусеницы II—Ш и»-1У—¥воз-
растов (перед опытом гусениц выдерживали в течение 12 ч без корма
для очистки кишечника, чтобы устранить- дополнительныецентры
кристаллизации жидкости; в) 3-дневные куколки; г) имаго на 2-й.день
после вылета. ’ - ' * - .г
Устойчивость к низкой положительной температуре. Для изуче-
ния этого свойства насекомых использовали метод проб. Сущность
его заключается в том, -что разные стадии исследуемых,объектов по-
мешают на разные сроки (1,2 и 3 месяца) в условия с низкими темпе-
ратурами. Последействие холодовой выдержки оценивали по показа-
телям развития насекомого в благоприятном температурном режиме
инкубации (25 °C). -''-м -''"“т-. , -л--' - пй. к- 2 <’•.
Наступление диапаузы определяли по следующим критериям:
- имагинальная диапауза—отсутствие яйцекладки. О наступле-
нии диапаузы свидетельствовали1 также другие признаки, например
прекращение питания’и уход в почву (колорадский жук), характерные
позы покоя (полужесткокрылые).- В. сомнительных случаях через’ ме-
сяц после формирования*диапаузы- самок вскрывали и классифици-
ровали физиологическое состояние по степени развитиятонад (у по-
лужесткокрылых [Hodek, 1971с]) или по наличию в них зрелых яиц (у
колорадского жука). При Необходимости (как у P.maculiventrisy для
определения времени наступления диапаузы измеряли интенсивность
дыхания (аппарат Варбурга) й ритмы газообмена (респирометр);
63
|,ц----кукол очная диапауза-остановка развития иютсутствиев мле-
ла имаго;.' .* '>тгхуй--*.и ыЛотР .щ->н«гнб' ’лн’мщи
t от в tr- личиночная диапауза — прекращение дальнейших линек; iл
м и ; эмбриональная диапаузэъ- отсутствие отрождения личинок.
' , .£0030 СИ’/йЖЯ ЙН РМ -M.-ni* tiSHOM
/.м ^.З.Методырасчетаероковразвитияласекомых,..
жчс/'Уьрщ .м; дфчi4: ’../«ЖчК9,че»£inошвоууой моа-.'П • >•>,, пн-,'
-он Теперь необходимо остановитьсягнанекоторых методических
вопросах,связанныхсвозможностью!использованиярезультатовла-
бораторныхопытовдляанализа фенологиюнасекомых. .Главный: из
них - сопоставимостьсроков развитиявконстантных и переменных
температурных условияхвлаборатории из в природе. Имеющиеся в
распоряженииобш ирные сведения о длительности развитияразных
видов в широкомдиапазоне температур позволяют зсомпетентно-об-
СуДИТЬДИСКуССИОННЫеМОМеНТЬЬ 'Т .[сМП Ь.и::• .1 : Н
; .: х Хорошо известно,-что вопределенных (для каждого вида) грани-
цах скорость роста насекомых закономерно возрастаете повышением
температуры. Предпринималисымногочисленные'попыткидатьэтой
закономерности-математическоевыражение-лРазличные формулы,
описывающиезависимость развития •отдемпературы, были предло-
жены еще в 20—40-е годы [Pearl, Reed, 1920; Janisch, 1928,1933; Рода,
1949,<1952. Цит.пщ»Горышин,.1966)ьБольшую,известностьщолучило
«Правило .суммы^эффективных!температур»фВ1ипск, 4923? Boden-
ке1тег,1926]. 'Эта 'зависимость имеет вид таперболы и:выражается
формулой^даюгодшшг,/^! (щшэщль - ч с < «, гот-иг. .м
jirny *лг дг ичл ь*1Г,7 ’•’Г''П'’”5ч.Р Ъ.гЧ U * 1 jk > * Ч
где ЛГ— видовая константа, Y- длительность развития, / ^гемперату-
р^^.нижнийдермичеркий пррод-рдавит^я^?Ч1 3«нФч i-s. i
; j уррновноД недостаток этрйформулцтящеддекватностьотраже-
щщ]рщ^атр1ивае|мойгзависимрсти1за1пррделами1рбларти.бдадоприг-
ядных температур. Но несмотря на критику состороны ряда авторов,
вщастоящеевремя оиашироколрименяетеявэкологической практи-
кеблагодаря простотерасчетор. (Рассмотримщесколько,примеров.х
., ,р мНа1'рафике,,построенноМ|ВКОординатах«температураг7Т;скорость
и ддитедьностьразвития»<рирц26)0линиискорости,!.пересекаясь с
осью абсцисс ^отмечают общийдля дичинок и куколокпорогразви-
тия, 1-1л4 i’C. Расчетная гиперболдипостроенная на основеединий
скорр^,;соответсргеурл,среднемуд^чащк>с)ммыэффек!11двджтем-
Т I ЙЬДВКНЭЦО 1Г-4
?64
' ' ^афических!1Ю11уляций'колорадскогожука{п<к1Ьрышииидр.,>1%В7к . soи
}; q i; «а *< w 1! 1, F - линияскорости и расчетная гипербола дляличинок и куколок; 2,2' ->tl / г ; р
тожетолькодляличинок. Л—нижнийтемпературныйпорогразвития. а—скорость
развития при'постоянныхтемпературахпопуляцийжука.изБелгорода(кружки),’ '-1Э
Краснодара (квадраты) <и Владикавказа (треугольники); б - длительность развития
личинок и куколокэтихжепопуляций в условиях природногоТерморигма. Гориэон-
.... . дальние отрезки показывают амплитуду терморитмов; точки на них приурочены к
'h ‘Среднейэ<йективнойтемпераЗуретермоРигма'111’ ’Ч ' *"
-Vk’ Mqn Biting [.} Kiw-M’1!'?:.
ператур^СЭТ)'за указанный ;эга1Г<>нтагенеза^»около!320лр11-дн131ти.
данные оДНозначноТюдтверждают^чтоправияоСЭТждиапазонешем-1
пературчхНб’до130°Судовлетворительно'описываетзависимостьраз-1
вития от температуры лБлизость значений СЭТлИ нижнего темпера-
турногопорогауразных>географическихпопуляций-свидетельствует
такжео-видовом рангетемпературных норм; развития,тоделает воз-
можным^использование экспериментальных данных, характеризую-
щих;любую географическую форму, для анализа фенологии вида в
целом. >.jjiuo'X JsnoH f:>
65
. f Рис.27. Влияние температу-
; ры на развитие личинок
/ Podisus maculiventris [по: Го-
j рышин и др., 1988]:
а—скорость развития личи-
нок при константных температурах
,(1 — сигмоида, 2 - логистическая
кривая, 3 — линия регрессии для
этихвариантов); б - длительность
развития личинок в условиях при-
родныхтерморитмов; в-тожедля
лабораторныхтерморитмов. По оси
\ абсцисс символы отмечают сред-
, .. \ нюю эффективную температуру
Температура,°С -ритма
Скорость роста личинок подизуса в интервале температур от 17
до 28 °C также вполне удовлетворительно описывается уравнением
линейной регрессии 100/7= -3,564- 0,35/(рис. 27). Однако легко за-
метить, что при температуре 17-J9 °C точки скорости располагаются
несколько ниже расчетной линии, при 24 °C, наоборот, - несколько
выше, а при 27—28 °C скорость развития вновь уменьшается, что сви-
детельствует о том, что эта зависимость отличается от линейной и
для ее описания больше подходит уравнение сигмоиды
ioo/уй^Ж^5’37,^9/).
Тем не менее в среднем диапазоне использованных температур
прямая скорости (3) и сигмоида (1) близки, и лишь при 27-28 °C
линейная зависимость нарушается и нарастает торможение развития^,
что подчеркиваетнеоптимальность для вида таких температур. Таким
образом, сигмоида точнее отражает зависимостьскорости развития
от температуры. Нодля практических целей вполне правомерно ис-
пользование и формулы гиперболы, поскольку в области благоприят-
ных температур она достаточно корректно описывает обсуждаемую
закономерность. Это в настоящее время признается большинством
исследователей [Stinner etal., 1974; Маршаков, 1984; Ratte, 1985; Tauber
et al., 1987; Honek, Kocourek, 1990].
60
Применениеформулы гиперболы позволяеттакже сравнить вли-
яние постоянных и переменныхтемператур на развитиенасекомыхи
оценить возможность использования результатов лабораторных опы-
тов для анализа сезонногофазвития вшрироде-! • < н • ‘
Вопросе спецификесдействия терморитмов на скорость роста
насекомых давно привлекал внимание ученых. А.Сс Данилевский
[1947]наоснованиирезультатовсобственныхисследованийиданных
литературы пришел квыводу, чтоколебания температуры в пределах,
благоприятных для развития, не влияют на его продолжительность
по сравнению с соответствующими константными температурами; тер-
моритмы, включающие неблагоприятные повышенны&температуры,
тормозятразвитие, а включающие пониженные исубпороговые тем-
пературы, могут оказывать стимулирующее действие.»Эта закономер-
ность позжебыла подтверждена ;многими исследователями в экспе-
риментах на разныхобъектах'СМаПезоп, Decker,‘1965;Торышин,- Куз-
нецова, 1972;. Neumann, Heimbach,: 1975; Hoffmann, 1980; Rock, 1985,
и др.]. Прямое сравнение данных, полученных при коцстантныхтем-
пературахи в условияхтерморитмов, встречает известныеметодичес-
киетрудности. Возможно, с этим связаны относительная: нестабиль-
ность лабораторныхданных у разных авторов (дляразных объектов) и
противоречивость результатов, получаемых в полевыхопытах. Необ-
ходима унификация методики. Нами использован графический ме-
тод такого сравнения, уже апробированныйприменительно кдругим
объектам [Горышин, Кузнецова, 1972; Морси,1978]. - г «уч
Для колорадского жука Сравнение можно выполнить на основе
данных, приведенных на рис. 26, где гиперболические кривые (пост-
роенные по линиям скорости>могут служитьв качестве лабораторных
контролей. Любая точка гиперболы на рисункеуказываетюжидаемую
среднюю продолжительность развития изучаемойстадиипри соот-
ветствующей константной температуре. Результаты-изучения дей-
ствия терморитмов в соответствии с правилом СЭТ можно располо-
жить в избранных координатах аналогичным способом.' эксперимен-
тальная точка для данного варианта по оси ординат должна указывать
продолжительность развития изучаемой стадии, а по оси абсцисс —
среднюю эффективную температуру, при которой оно проходило. Что-
бы более полно охарактеризовать суточный терморитм, каждую экс-
периментальную точкунаграфике (см. рис.26) можно снабдитьгори-
зонтальным отрезком прямой, отмечающим его амплитуду; При та-
ком построении графика расположение экспериментальной точки
.67
вблизи пиперболыбудет означать идентинностьразвитияпри кон-
стантных температурах и терморитмелРасположениешочки нижета-
перболыбудетуказывать на ускоряющий развита е.эффекттерморит-
ма, а отклонение ее вверх — надаго тормозящее действие, >•«: <•г щ » ,<
Амплитудытерморитмоввтечениеразвитаяличиноксоставля-
ливразныхвариантах 0311,97-14,1^022,0—25,2.’С,аличинокикуко-
лок®суммеь7|ОТ;12,8г-»13,4 до 23,3—24,3'’C(cм.pиc.^26)^Bce^экcпepи-
мeнтaльныe-гонки,1 характеризующие длительность развитияличи-
нокикуколокколорадскотожукаприразныхсуточныхритмахтемпе-
ратуры,расположилисьвесьмаблизкоккривымдляконстантяыхтем-
ператур..Внекоторыхвариантахопыта,тоякилежатщагиперболе,в
друтахониотклоняются.'Вверхили внизютнее. Такие лротивополож-
ныспознакуотклоненияточекот-гиперболысвидетельствуютотом,
что они случайны. >В‘’целом,11олученные .данные позволяют считать,
что,продолжительностьразвития;личинокикуколокколорадского
жука при .константных температурахаОсуточных терморитмахприн-
одшшлыюжразштепжлщ'7'u... з пт. ч
js н < Аналогичныйвыводвытекаетииз анализа результатов-исследо-
вания клопа Л maculiventris (см.рис. j27)Он подтверждается ,и дан-
ными,шолученнымидлядругих.видов; Per.iUus.bioculatus, Amacustas,
Py/vtocorbnaptau^^ . ,•?•# тщ у.п. т {.•••кл.тх’;;
/1 ^Часто истонником ошибокири расчегах с применением правила
СЭТгявляется-неадекватный |учетдампературы. Особенно балыпая
погрешность возникает щри использовании .дляаналйзарреднедег
кадных и ,:'среднепериодныхтемператур, что щирокощрактикуется
службой>зашиты?растений. Рассмотрим.этот вопрос водробнеетаа
приме ре развития, прсимагинальных стадий^. ^их/ох п д <'в л s,; , о
j Эмбриональное развитие. Роль лемпературы вретуляпии скорос-
таразвитияяицщсследована.вщирокомдиапазонеэтого.фактора-
от 13,5 до ЗЗ^С (рис. ,28)’. Совокупность точек, характеризующих ско-
ростыэмбриональнбтразвития в разныхвариантахох1ыта,юписыва-
етсщуравнениемлинейнойрепрессии)^ткнкд<|<х».?^1из,ны:;щ,ие
N..O'OIJ •. АиИ. ;W'J , сТ.>1ИЖ'9-ТНЛ;| (.;
где/^йтельнрствразвития;/-^м^да^.., ч
) ifc ^инвяркоросздкразви^ия яиц отмщает; рдрцетно^на^риие те^р
натурного цорогд^со^аавляющ^/гр^ерцогЦ^’Ц ь,,« ч, уай
ДНатомжерцсунке прцведецысроедразвития.яиц придеремрн-
68
2S..q^.
Nt'Kig
20 ?
UXtig.
г »<&
x 32
<• t • • "" ' "•" i-iM'Ft ;•:><
Рис. 28, Влияние терморитмов rjf ,
на"сроки! развития яиц Апла'"*'
сийо5>[по-.‘ Саулич й’^Волко-ЧЗ*''
вичз J?94]:i tm>/* ч, '«''ЫЕ*
Гипербола и ее реципрока- §
зависимостьдлительностиискоро- п 20
стиразвишяйицоттемперааурыпри ; '
констаигныхусловиях. Горизонталь- ,; г| , Д®
ные линии —варианты с перемен- , £
нымитемпературами, их протяжен-1 -* iff, 12
ность — ^амплитуда тер-моритма,( п .Д
кружки итреугольникисоотэетству-,.
ют его среДНейтемпературе; 1,'2‘— ' I ’ fc ‘
средняя длительность развития яиц
при постояниыхтемпературах; 3,4—
o.Ms яг.и>й«ч(йгя»'
•ЧГ1 ;i’A'4
4Г
• ОСО J
W.:
.i.Wk4'/>q ХстНШрГЭС§
: ,к№ПГ;
, dk i) f •
OV M '» !
Xipki
С.ДИ,О»ЧМЖО'.
xjujqb.
~l:; w.'STjHBR
10УС‘Г-'
,ОП 'Л'.
ii. .KJ'
iujKcayuiuaHMXji»xjpaH^JHbix-irpMO-i v чл , а, ояв, , । ...........-In
ритмов; 3-тожевцдояяыхусжнё'.1Л&ц'Н1^« 21 25 29 33
виях(1,3-опыты1988г.,2,4,5- о1 «2 ^>3 «4 л5
опыты 1990г). .тзгЛ1п4- .Mi<ffcqwT®MnepsnVP07C
ритмах и соответствующих им постоянных режимах на основе линии
скорости построена'гипербола, показывающая?длительность разви-
тия яиц;при разных константныхтемпературах..Каждый варианттер-
моритмовюбозначен кружкомили треутельником. Можно видеть, что
большинство экспериментальныхТочек, отражающих развитие при
переменной температуре, расположитесь вблизи расчетнойтипербо-
лы, т.е. длительностьразвития при'терморитмахсовпадает с,длитель-
ностью развития яиц при постоянной температуре, эквивалентной
среднему значению ритма.” Заметное'различие" отмечено лишь для
вариантов, в которыхХолодная компонентабылаяиже пороговой тем-
пературы^ 13,!5°C).j •.Ой ее , v,® я-
Является ли это ускорение кажущимся или оно действительно
существует? Для того^чтобы ответить на этот вопрос, были проведены
расчеть1,, в которых исключались неэффективные для развития темпе-
ратуры. Как указано «выше, температурный порог для развития яиц
близок к 13,5 “С.Поэтому'в случае П-образного ритма, например в
69
вар. 19 (табл. 3), развитие яиц происходило только в течение полови-
ны суток (12ч термофазы), в течение второй половины суток яйца не
развивались. Внося соответствующую поправку, можно считать, что
поступательное движение в эмбриогенезе занимает не 8 дней, как за-
фиксировано в опыте, а вдвое меньше, т.е.4дня. В тех вариантах опы-
тов, где нижняя составляющая ритма равнялась 5,4 °C, истинное зна-
чение д лител ьности развития с учетом эффективных температур со-
ставило всего 4 дня. Как свидетельствуют результаты опытов при
постоянных режимах, при температуре 33,2 °C — верхней составляю-
щей ритма, эмбриогенез заканчивается через 4,дня. Точно также в
вар. 20 (табл. 3),истинное"значение длительности развития.яиц со-
ставило всего 3,1 дня.. : /. ’
Таким образом, мы еще раз получили подтверждение принято-
му положению, что терморитмы, обе составляющие которых не выхо-
дят за пределы температурных порогов, действуют аналогично» посто-
янной температуре, равной среднему значению ритма. Еслитемпера-
тура криофазы опускается ниже температурного предела1 развития, то
, Таблица 3.Сроки развития яиц Arma custos< в разных >
постоянных температурах
''Вар. hr, опыта . Температура. °C Кол-во яиц Длительность развития, дни СЭТ, гр.-ДН.
. 1990г.
. ,32J9 43 . 4,0 + 0 77,6
2 29,8 64 " 5,0±0 ,81,5
'3 ' • 29,3 ’ ' '51 1 5,2±'0,02 4 82,2
. 4':'н; 27,3 ; 73’ ’ * б,о+о 82,8
. . -uS. rJ \; г '24j0u ’•, --П 58i^> г г . 9,0 ± О 1.4 94,5
...... г-®, • , .... ,19.71 42 ; 16,4 ± 0,04 , . 101,7.
" " 7 15,5 60 24,0 + 0,20 48,0
”-”'’'+8 ? ' 14,1' ’ :. . .gQ.Sl> . Развития Нет * —
13,5 43 п ~ а . . •
» 1988г * Ьл-Л- •• -!?/'• ьч- .
32,1 20 5,0 + 0 93,0
11 29,7 38 5,0±0 ’ : 81,0
12 , 28,3 , . 63 -41 , , 6,8±0,06 100,6
13 , ., 24,2 „ 48 , 9,0 ±0 96,3
14 * 21,0 ' ' 41 11,3 + 0,09 84,7
''"м'--18,7‘- ч' " '.\Н‘"291 х 15,0+0 ’ 78,0
• ’46 .• /”й ”.16,7: . « 26,0 + 0 t 83,2
> (i .г. 17 . . .58. , , ..Развития нет
18 10,1 45 «
70
наблюдается формальное ускорение посравнению с развитием при
постоянной температуре, эквивалентной среднему значению ритма.
Но, как показано, выше, развитие в этом случае протекает только в
период термофазы, и посравнению с развитием при температуре та-
кого уровня никакого ускоряющего эффекта мы не обнаруживаем.
У насесть материалы, позволяющие сравнивать-реально необхо-
димое для развидияяиц армы значение СЭТ, с полученным в разных
экспериментальных режимах; в лабораторных (при постоянных и пе-
ременных температурах)’® полевых, Максимальна приближенных к
естественной среде.
Как свидетельствуют подученные данные (табл. 3), СЭТ, необхо-
димая для вылупления личинок в разных постоянных режимах от 33
до 17 °C, изменяется без определенной закономерности в пределах от
77,6 (вар.1) до 101,7 (вар. 6) гр.-дн. Разброс между этими крайними
значениями достигает 20%., Частично это связано с нестабильностью
величины СЭТ в математическом смысле [Кожацчикрв, 1961] , кроме
того, разброс увеличивается из-за погрешностей в регистрации тем-
пературы и непосредственно результатовопытов, хотя колебания тем-
пературы не превышали ± 1,0 °C, а наблюдения за насекомыми про-
водились через каждые 12 ч, Из всей совокупности д анных выделяет-
ся значение СЭТ, полученное для варианта с самой низкой допускают
щей развитиетемпературой15,5°С,где эта величинасоставила лишь
48 гр.-дн. Этот факт пока не поддается объяснению.
При терморитмах .(табл. 4) абсолютное значение СЭТ, рассчи-
танное с учетом нижнеготемпературного порога 13,5 °C, также колеб-
лется без определенной закономерности. Но если расчеты вести, ис-
ходя из средней температуры ритма, без учета нижнего температур-
ногопредела развития, то намечается тенденция уменыпения СЭТ с
уведичевдем амплитуды ритма. , ,3 » ,Л-‘ у: •
,, В целом СЭТ, необходимые для развития яиц в опытах с досто-
янными® переменными температурами, различались.мало;(табл.З и
4).Тоже можносказатьио результатах полевыхдпнтови за исключе-
нием случаев, когда, разница между среднесуточной, температурой и
нижним температурным порогом развития непревышала l °C.vr >_
Например, в одном из вариантов опыта температура за периоде
момента откладки яиц до,вылупления личинок (23,7 дня) колебалась
в среднем от 22,5 °Сдн₽м до 9,9 °C ночью при среднесуточном значе-
нии 14,2,°C. Если исход итьиз среднесуточной температуры запериод
наблюдений, то за 24 дня СЭТ составляет всего 16,6 гр.-дн. Наса-
71
‘Таблица 4.< Срокиоразвития: яицкАлпасс/5<оя<при‘<,терморитмах"' '-г > ‘
. W- 1 Вар. 0 C^-donw* ? I? Амплитуда ^ЬтермормтмаХ^ Средняя t, °C \К.'’ Кол-во • •’Ч1ИЦ:"''’А Длительность .. развития,удни* СЭТ,|;р,-дн. . „/ - - > ' Л
1990 г.
'19 -41 21B1H ,3 x; • 23 - 24 'V-i -< K^gg < 1 ,s 33,2-5,4 33,2-10,8 ; i33.2-T5,95 33,2>-20,7-и. 33,2-27,4 . 27,3-14,2 , ' 27,5-20,4’f -19,3 ' '‘‘•22,Ь8Н^' :o--,‘*24,6seou j,-i<‘:26»94 ,кы 30,3 20,7 . 37 ЛИ. З1.гф4 ,а. 39>мк. :-М’,а66 -4 ( 59 , 58 ^ ^50^* 8,0 ±0 ‘ б.гУо.ч’оf н <7,0 ± 0,0& а 5,2 ± 0,04 „ , 12,3+0.07,'; и Т1,Т+0,25 ! . 78,8* , 'f ‘61,0*“*' < 78,6 .193,8 [. .1 -..|87*4--;.-4 < чу 88,6 '1Т5,4
' .» 4..' . »4ui.*TV. ’• 4 • -'ll v. 4 ' 4 ggg1 ’ « I A’ ‘ • •• • .1 S'- .JU.- ... • ь'- .
5 •' -K3$H27^ ‘ 30Л-1г9,8 " S4t30,1-9(,8,;S "'•25,0 f. 00(20^*: Д ''5,7 + 0,04^ SS/ ’9,0±0, ,а «. !5 • ' 1 77,0 74,7* '
>;hix.'h<uviv;>!«?-егфйродныё терморитМы
-УаГ 129 дм ..fW: r 22,5-9,9iji 28.4-Л6.2 3 .22^9,-10,4,'г . 25^1—15,1,,. .чл । 21С1Л- :Фн.2Т,Т*'-»я - awnA6,7---;;r,t> уг'1?77<ин1 } 1|18 ^3,7‘± 0,73 ' Л1Ш,8!±Ч>,06г <!г17,0,±0,.л; ;„^8ЛР,Р4 |< !1'52,7**’”! '^йгд. ' ’ :-И- |м ^8,8
' ‘ *При Утадсчете ‘СЭ'Гиспользована температура? 'равная' верхней1 со-
ставляющей’терморитма;'- ’> чвдзоянэрущ.’! A t - > " .4 ?
„и -.- .- *^<емперд>турагучгеМ|а ио часовым: .^оказаниям термографами *й.нл
.о.кньч;-,да-до хзтэсту’оп о» влоп тхг.ф югС •«’. • • 'А
мом -делеСЭТ,рассчитаннаятючасовым'показаниям Суточного тер-
мографас Учетом 'температурногоПорога/^оставила’за'Этот период
65?9!гр.-дн.,:’ ровно столько? сколько1 йеобходимодля ^завершения
ЭМбрЙОГенеза.1 н АНН ,.ь* .''HG «МЭТ г:^гкГ 4: *»• Or 'ЛО?
1 Этотфакт/несомненно, должен бытьучтенпри'прогнозе^фено-
логических событий, который чаще всегостроитсяна’основесредне-
сугочныхилидажесреднедекадныхтемператур.Притаком учете тем-
пературпоявление’отдельных!стадийинтересующих'видовнасеко-
мыхбудетОжида'Гьсяв'бОлеепоздниесроки^емпроизойдетреалъно'.
Естестве нно,'такаяошибка'вероятнеевтепериодысезона,когд явоч-
ная температурачастоопускается нижеТемпературного'порога раз-
вития?'1 kck’W’11 v.wfii’dito вотушщдя г.ч мош'.о и дем;»?
ж *;Личийочное развитие’ Завйсимость’скорюсти ’роста' личинок
A. custos m температуры1 в целом подчиняется той же закономернос-
ти,*что?и показанная’для яиц (рис. 29),! и описывается уравнением
лийейнойре!ресс!ии'вида1'00/Т3==и-3,’6бЗ>+,01,275 f.f
72
.Линия скорости.пересекаясьсосыо-абсцисс,«отмечает.расчет-
ное значение нижнего температурного порога развитияличинокармы,
равное.13,:1°С.чгч'ч г.т!» . д-дй он? jh мбГ
"Вбольшинствевариантовопытас переменнымитемпература-
ми,где нижняятемпература ритма нежыходилазапределыэффектив-
но й,длительностьразвития личиноквгритмичес ких условиях совпа-
дала^ с продолжительностьюонто генезав констан-тныхусловиях
(pHC.29.)z; 3 ШХн .НМЬМ.Ч КМ -.д-рг»ЧСД’ПМЭТ HWlXfMi's 3 /Й
и” Особенно четкое^оответсзвие этих величин отмечено при дос-
таточновысокой среднейтемпературеритма(вар. 1—3).Чемона;ниже,
тем болыпеютклонение реальной длительности развития личинок от
расчетной(вар. 11).Особогорассмотрениязаслуживаетвар. 4, где зна-
чениемижнейсоставляющей ритма было ниже порогового. Средняя
температура вэтом вариантесоставляла 20 “G При постоянной тем-
пературе 20 ,°C клопы окрылял ись Через 56 дней, вслучае леременной
температуры - всего черет4Д дней.,Но,какуже<‘Отмечалось, ДриЛ-
образномритмеи-ночнойггемпературениже.пороговойразвитиеидет
только во время термофазы, аееуровеныв этом ва^жанте.приближает-
ся к 30 °C, Пользуясь. s' л
формулой линейной ' .-^. 72-
регрессиирполучаем,; s'gi; *
что при ‘ ПОСТОЯННОЙ м'
температуре гЗО)°С ли-.
чинки должньгокрытч Jsh56’
литьсячерез 21,5 дней. » м> ’
Близкие.результаты ’§ 48*
полуненыи;вконтроле«ё:, *
ПрН ПОСТОЯННОЙ Темк; ife i 40-^
пературе ЗО °С. Таким
образом, никакого г др 32-
ЯЗГЬГйЛ’О O’i-.M Г!- HS '
"'ТНГ.ПМВ 'SlSvi . ИКЗ? 24-
. ?*.>' ДДЫЙ ьИП Т’«',цI ' Т'^,1
Рис.29г,Влияние.,темпера- ig-
туры’на ркррость и Дли- ( / ...
тельирсть’" личиночного ' ‘
развития*'Агт»3си«Го4:!(пог’”,''!Г'' 8" J*
Саулич'И 'Волкович!,'19941: мчх
qLl_
рис. 28 !’0":
. /Д'ЕО'.' (-П •>' ,;?» да
73
корения развития личинок в этом варианте по сравнению с контро-
леммынеотмечаем. ' - -.>“?
Тем не менее давно было замечено, что развитие насекомых в
природе всеже идет быстрее, чем в лабораторных опытах, даже в слу-
чае максимал ьногоприбл ижения их условий кестественным. В опы-
тах с личинками армывовсех природных вариантах (6-1.1) окрыление
также происходило раньше, чемв контрольных лабораторных услови-
ях с близкими температурными режимами, и опережало сроки, рас-
считанные по формуле при постоянной температуре, приэтом чем
ниже была средняя температура ритма, тем больше реальные сроки
окрыления клопов отличались от расчетных значений (рис. 30). >
• Рассмотрим результаты лабораторных и палевых опытов с еще
однимхищным клопом Perillusbioculatus (рис. 31). Наибольшее уско-
рение отмечено в опытах, проведенных на фоне природного суточ-
ного хода температуры (рис. 31,.в). Опыты выполнены дважды — ле-
том11986в 1988 ТЕоМаксимальное сокращение сроков развития (на
19,5 и> 15,1 дней по сравнению со сроками развития в контрольной
группе придостоянной температуре) проявилось в вариантах 1986 г.
Анализтемпературных усло-
вий этого вегетационного се-
зона показал, что колебания
температуры втечениесуток
были очень большими и ча-
сто выходили за нижний
температурный порогразви-
" тия личинок периллюса,
расчетное значение которо-
го составило 13,9 ± 0,3 °С.:В
1988 г., несмотря на то, что
средний уровень температу-
ры лета мало отличался от
условий 1986 г., амплитуда
суточного ритма была мень-
ше, и ночная температура
только в единичных случаях
опускалась ниже 14 °C. Та-
ким образом, природные
ритмы вызывали явное уско-
рение развития по сравне-
48
<f
I 32-
5
о 24
я
х
с;
ф
m
16
8
01—J-------u.
16 17
18 19 20 21
Температура, °C :
Рис.30. Отклонения реальной длительности,
развития личинок Ата custos от теоретичес-
кой в зависимости от температуры
74
Рис.31. Влияние температурных ритмов на развитие личинок кло-
па Perillus iioculatus [по: Волкович и др., 1990]; ''
1 - линиярегрессии, отражающаязависимостьосоросги росга личинок (а) от
температуры; 2 - расчетная гипербола и фактические сроки развития личинок (б)
для вариантов при константаыхтемпературах; в—средние срокиразвигия личинок
при среднесуточной температуре в природных терморитмах; г—то же для лабора-
торныхтерморитмов.-Саризонтальные отрезки - амплитуды терморигмов
нию с таковым при постоянной температуре, причем эффект их был
тем сильнее, чем болыпе амплитуда терморитма.
Параллельно с экспериментами в природе были проведены ла-
бораторные опыты, в двух сериях которых различались как амплиту-
ды терморитма(от 16,3 до 16,8 °C водной серии иот7,9 до 10,1 °C - в
другой), так и средние значения температуры (20и 24 °C соответствен-
но) : на рис; 31 оба варианта отмечены светлыми треугольниками. Вер-
хняя составляющая в них была почти одинаковой - около 29 °C, ниж-
няя в одной серии неопускалась ниже 18,9 °C, а в другой не поднима-
75
лась выше 12,6 °C, т.е. была ниже температурного порога развития
личинок периллюса. Во всех вариантах с температурой более низкой,
чем температурный порог развития, отмечалось заметное ускорение
развития. Оно не достигало таких значений, как в природных опытах,
но в среднем’составляло 5 дней.
Результаты специальных опытов до выяснению роли амплитуды
терморитма,'выполненные также на-личинках P.bioculatus (табл. 5),
показали, что длительность их развития была достоверно меньше в
техвариантах, где амплитуда ритма был а больше. Особенно резко этот
эффект проявился на фоне менее благоприятной для периллюса сред-
ней температуры около'20 °C (вар. 1-3). Положительный эффект тер-
моритма уменьшалсяпомсре уменьшения амплитуды ритма.
Одной из биологических характеристик, используемых в форму-
ле гиперболы, является нижний температурный порог развития (био-
логический нуль). Он определяется по формуле или, как показано
выше, графически на основе данных о продолжительности развития
при разных, но-постоянных температурах. Отсюда следует, что все
факторы, влияющие на сроки развития, могут изменять и значение
нижнего температурного порога. Покажем это на конкретных при-
мерах. \
В экспериментах скорость развития дичинок клопа солдатика в
условиях терморитмов была нем но го. выше,,, чем .при постоянной
температуре. Эта разница почти не повлияла на значение СЭТ
(416 гр.-дн. в константной температуре и 429 гр.-дн. в переменной).
z и> .гит m'mwwi f • зм'мд чда
Таблица 5. Влияние температурных ритмов с разными амплитудами
и одииаковой^средней.яа продолжительност’ыличиночного развития
<огадг^..&.ет:<ь<хищнотоклопа,Рет///из,йй>си/аГиз,’эт
Вер; опыта средняя" “ f •|, р айпл «''число;, особей- *** Длительность ‘ ? ч'раэвитй^,вдни‘ ш
температура.' “С ’г'герморитма,1 •’ °C
. С za ". Л'ГрЧ? г - 20,1 • '30,0-10,2' ;.>;.72< г.; *
2 20,0 . длгбД'нлкгт’д
, нЖлЛ q |25,3—Д6»3 : т ,25,3 ±0,25* . i
20,3 22,5-18,1 115 37,1 ±0,25
। zy.»i ‘ь'АЗ1"^ ‘36,2±0,32
— ' ' ’ 1 :А Т25д.м.ад *' 16i,3i±’0l,:l2!':-!’<
iwwfoKi 3 ?;25>2м'£.. г ^,v27;&-22Jf л irilfti3[*0,22r<'
гй. JU7 гЛМ "Ш! <. '"'Г',-91 ЯШ '! 0 /Й&СН
4- !.а , '>*!Длительностъ pa38MTHnWflaWi(<M4lBapMaHTe ^ocToeep-! J л
•в кино отлимается!оттаково(бв комтрольной+руп1п^.!й » погп.с н<<:
76
Однако в первом случае ниж-
ний температурный порог
развития .оказался равным 8
12,5 °С,й во>втором - только й
9,0 °C (рис, 32>. I
х Оченьсильное влияние ё
на?длительность развития |
личинок клопа'-солдатика °
оказывает длина дня. На- ;;
пример, при. 20 °C в корот- •»
ком дне (15^ч света в сутки)
клопы развиваются почти на
месяц‘быстрее,- чем в длин-
ном дне (18^ч) [Саулич и'др.,!
1993]. Использование дан-
Рис. 32. Скорость (100/У)ки п9рог! развития
1, .личинок Pyrrhocorisapterus в условиях посто-
янных (черные точки) и переменных (свет-
'лые точки) температур. Фотопериод —15 ч
НЫХ ПО длительности онтоге- света в сутки [по: Numata et al., 1993]
неза без‘учета‘*возможного’7 X* -.л«.
влияния длиныдня можетйривдиднально изменять значение ниж-
него температуря огопорога.онределенного графическим способом,
вызывая незаслуженнуюкритикусамой'формулытиперболы'.Подоб-
ныертклонения.длительностиразвитияот обычной, диперболичес-
кой,зависимости в ррзультатерпецифичеркого, влиянияразличных
факторов среды.были ош^саныддя многихвцдовнасекомых (см.вы-
ще),С)ни отражают проявление сезонно-циклических реакций,регу-
лируюшихфенологическую приуроченность отдельных.стадий.,'.f
; к 1;В исследованияхдлительностиразвития личинок Р. maculiventris
(рцс. 33)о где варианты’отличались условиями питания.личвдрк,^е
значения представлены точками, расположенными в традиционных
ддя такого графика координатах,«температура-.скорость развитая».
Как Дыло показано, зависимость развития личинок лодизуса от тем-
пературыв.интервале17—280Снаиболееадекватноописываетсяурав,-
нением сигмоидыв. связи сзаметнымтормозящим эффектомповы-
шенной: температуры.. Однако в более узких предках температурной
шкалы, це.превыщающих 25 °C, эта зависимость.вполне корректно
опис^^етрд(И;Уравнен^)идп1нейной регр^^^ррьци мыи’^цс-
по^ьзуемся для,диализа результатов, и ,'д, PH‘hN. , ,z. 7*”,
Положение на графике экспериментальных .то^ек и линий .ско-
ррсти^рассчитанныхдрякалуюговидакорма^существенноотличает-
ся/Тонки, дляусда^и^. ^ргдадичинхи ^вду^)выращивалирь iy-
.77
Рис.33. Зависимость смертности (а) и скорости развития (б) личи- ,
,нок подизуса от температуры и условий питания:
" ’ 1—гусеницыбольшойвощинноймоли; 2—личинкисинеймясноймухи; 3— ' ; :
личинки большого мучного хрущака; 4 - личинки комнатной мухи; 5 - личинки
Колорздскогожука; б-гусеницычешуекрылых (по:1Ьрышинидр.,1988]
сеницах большой вощинной моли/ личинках синей мясной мухи и
большого мучного хрущака, смещены вверх, соответственно располо-
женыи линии скорости (1-3). Это свидетельствует обускорении раз-
вития при питании названными видами Насекомых. Клопы/ выра-
щенные на личинках комнатной мухи-и особенно на личинках коло-
радского жука, развивались медленно, что отражено в расположении
соответствующих экспериментальных точек и-линий скорости. Дан-
ные, характеризующие степень благоприятности разного корма для
питания личинок/вполне коррелируют'с влиянием качества пищи на
выживаемость личинок (см. рис. 33, а). Прямым следствием такого
влияния является йзменение наклона линий скорости. Если их про-
должить до пересечения с осью абсцисс, то они'не фокусируются в
какой-то одной точке на ней, что свидетельствовалобы о едином тем-
пературномпорогеразвития личинок. Значения графически опреде-
лённого порога’ варьируют от 6 °C у личинок/выращенных на коло-
радском жуке (линия 5), до 12 °C уличинок, питавшихся гусеницами
большой ВОЩИННОЙ МОЛИ (линия 1). '
Таким образом, следует отметить условность нижнего темпера-
турного порога, определяемого обычно графическим Методом. Осо-
78
бенно это касается развития хищных насекомых, когда температура
действует, по существу, на систему «хищник—жертва», элементы кото-
рой по-разному реагируют на изменение этого фактора.
Более точное значение нижнего термического порога развития
можно получить при прямом его определении, например по методи-
ке де Вильде [de Wilde, 1950], предложенной для определения порога
развития яиц колорадского жука. К сожалению, этот метод остался
незамеченным. Между тем он заслуживает внимания, так как позво-
ляет определить фактический порог развития при минимальном по-
вреждающем воздействии пониженных температур. В нашей лабора-
тории такой методический прием был использован для определения
реальных порогов развития даже у питающихся стадий онтогенеза(гу-
сеницы чешуекрылых), более чувствительных к неблагоприятным тем-
пературным воздействиям, чем яйца и куколки. Суть его заключается
в том, что гусеницы выдерживаются на холоде лишь часть периода
развития; остальное,время (например, 10 дней до и период после хо-
лодовой обработки до коконирования) они содержатся при оптималь-
ной температуре^ В качестве критериев оценки эффективности испы-
тываемой пониженной температуры для роста и развития гусениц ис-
пользуются: а) изменение их массы за период холодовой обработки;
б) уменьшение периода личиночного развития при оптимальной тем-
пературе (по сравнению с контролем) врезультатеего продвижения в
период холодового воздействия.. -х .. .г ;
Приведем описание экспериментов с применением указанной
методики, аде определялся нижний температурный порог развития
гусениц Acronicta rumicis. В качестве оптимальной для развития ис-
пользовали температуру 20,.оС, при,которой,длительность развития
гусениц <25 дней),служила контролем. Посхеме опыта гусеницы пер-
вые 10 дней содержались при 20 °C, а затем в течение 20 дней, -при
разных пониженных температурах (варианты опыта) и завершали раз-
витие вновь при 20 °C. , 7 ч ? г;.
и. - Результаты опытов показывают (рис. 34), чтопослрД 0 дней рдз-
витияпри 20 °Сгусеницыдостигали массы всреднем 10 мг (линия 1 -
стартовая при оценке ихростав разныхпониженных температурах).
При 14,6 °C гусеницы за время,20-дневной обработки прибавили в
массе примерно 220 мг (линия 2). Известно, что такая температура
вполне пригодна для развития гусениц щавелевой совки и они могут
успешно завершать развитие® этих условиях. При 13 °C прибавка мас-
сы задот же срок достигла 125 мг, а при 9,9 °C - всего 55 мг. Эго,
79
чжкуп яаллет > г’-е.гн.куудчс]
-un JMOHutSl'.K?1, •<»/ щгр S»n-?«!/’ И.Г.’ОИ i
;?qr>djiitfcUj£»? ?t 'WyA ч/оди,...................-;-й;-1п и,,*-,,.
wwi.Wjjfc frf.’ и К'жу FjW.Mj К экспериментальному
•ДНООгИЭИГН. • лНЁГ1:> г/нц
/ЭТ I' ЛЧТИЙ^ЛВ W-ул ХИН
Y >.* .т-Ь.Д ''’ '| . ' j
ратурного порога развития гу-
»;? •^‘‘сейицЧ:ов'Кк'^'сл>и/с/в:1 rumicis
•<'Члш 'мги-гыл' ..•« /'• ” etriiR ДпозЛЪрышин.'И др.^т1984]£
о?.? ’ <ift ГЗ’С *'- 1-средаяям»ссагусенидпе7
ред холодовым воздействием в возрас-
, рем лимчцихшм DUJMC-MViDMCM О DUJpaV-
.». . ..ЮЛ !^KiV.! ;I• л. WjhP i’г т Д .гЛ1Ы > !^10даей;2—,масса1усеницпос®е20-
-<!!U '= '•»тР1 '1 , , •- ' ", • ; г> удневногосодержания-вразныхпони-
-ипэч
-•>» пшэдп ww>s>'i «^л;-фйдачзв1^^41?*.‘яг’г«л1нажннои 'кчлт
:>г‘Л'1О0«1'^йод<л’ег.Олгкой^&п вг ииэьм лг ^йнш‘»жй (в ^.оч<4Y£nji;тк
несомненйо;*свидетельствуето том,;что'процессыфоста-®»интервале
тем пер атур'отЮдоЧЗ’Снеп сдавлены полностью, новырастить гусе-
ниц др окукливания в таких условиях не удается, При'6,2рС прибавки
массынебыло. Следовательно, 'этутемпературу следуетпризнать ниже
пороговой.'При 8‘, 5 f’C прибавка массы хотя> и была; отменена, ноока-
заласьничтожномалой(201мг:и20дией).По-видимому>этатемпера-
турафаиболее близка к'тюрогу развития.'Масса куколок (кривая З)
свидетельствует, что развитие гусениц'во всех вариантах опытатпло
нормально/^ я !*Ф* ;4энд. :‘J
:;-^Справедливость!такоговыводаподтверждаютиданные одаиг
тельности развития гусениц в этих опытах (табл. 5). В контроле1(вар. б)
при посТоянномеодержанииГусеницвусловиях20рС они завершали
развитйеаа£25 дней /Принимая' этот результат^» 100%, > проанализи-
руемстепень продвиженияразвитияввариантахопытавтеченисЗО-
дпевногохолодового<воздействия.Так,‘В'вар. Гобшаяпродолжитель-
ность развития гусениц юоставила З^дня,/в>' том-числе' 14;дней при
20'0С(10днейдохолодовоговоздействия<и4дняпосленего).Следо-
вательно.’период развития’припониженной1температуре14,6;ОС со-
ставила 44%, в’вар. 2 (приМЗ,оС)’- 32 %,’а*в?вар, 3 (при 9,9^С) этот
80
показатель снизилсядо-16%.'Ввар.4,тдепониженнаятемпература -
составила:$,-5*’С,*4в!се’развитие* гусениц3 протекало только при 2О'°С.
Его продвижение за период холодового воздействия исчезаюше мало.. >
Наконец, в вар. 5 пониженная йемпература’б,2 Х>полностыо подав-
ляла развитие гусениц.*'Не исключено ее неблагоприятное действие’/
на состояние гусениц, которое проявилось® более медленном их рази
витии при ЗО^СЛем *вконтполе. ‘'
Таблица 6 Результать/экспериментальногоопределения нижнего
температурного порога развития гусениц A. rumiciSr11 >j
М5арГ , ОЛЫ-,, та : Температура; ХОЛОДОВОЙ ' обработки,; °C » • .& Ь’СЛ .И’ •A Число ;гусениц. \ е.?. ' г' Длительностьразеития
> / общая* к ? лри20° + еХ, ’ дни и,втомчисле при 20Х .Чразвитая,
ДНИ %
14,6 - Л 47 I ч •Ш г34Г - Ч 14 ^6 /к;:44-'Д-
13,0 ч ',36; -у г -рДГ. U 17, ;,68,;
,3 , • 9,9 44, . 41 21 96 16
”'4 '8,5 ' " 33 ’ 44 - ' '24н ^ттг1’ Л ‘
-5' ‘ ;.г.18Ь« 28 too. Оч.^- i '
.Контроль • 94;:
Таким образом,обаучтенныхкритерия — рост!и продолжитель-
ностьразвития—инвариантно указываюттемпературу 8,5 “Скакочень
близкуюкреальному нижнему температурномупорогу развитиягусе-
ниц щавелевой совки.!Существенно отметить,цто!графическиопре-
деленный порог по данным' параллельных' опытов йризразных'кон-
стантныхтемпературахоказалсяравным 8,0?С(традиционно'дая этого
вида приводитсяпорогПО ’’С, чтол иш ^подчеркивает'недостаточную
изученность вопроса).
Приведенные д анные показывают, что описанный метод служит
реальным инструментом для прямого определения нижнего темпера-
турного порога развития насекомых. В эколого-физиологическом пла-
не его использование представляет несомненный интерес. С его по-
мощью, например, можно исследовать динамику реального темпера-
турного порога в онтогенезе. Для этого достаточно лишь изменить
схему опыта, чтобы холодовое воздействие приходилось на разные
(начало, середина, конец) периоды изучаемой стадии.
Таким образом, приведенные материалы свидетельствуют о том,
что правило суммы эффективных температур в пределах благоприят-
ной температурной зоны достаточно точно отражает зависимость ско-
81
ростиростанасекомых от./температуры. Отклонение расчетных зна-,
ченийдлительности.развития от реальных невелики и в большинстве .
случаев являются следствием ошибок,которыхможнолегко избежать.
Несомненно наиболеечастовозникаютпогрешности принеоб-..
ходимости прогнозировать длительность развития при переменных (
температурах как в лабораторных, так и природных условиях. Они
особенно существенны при попытках рассчитать сроки развития,на-
секомых весной и осенью с использованием среднесуточных, средне-
декадных и среднепериодных температур по показаниям стандарт-
ных метеоданных. „ж.'-<
С учетом точно определенного нижнего температурного порога
развития и при адекватной регистрации окружающей температуры в
местах, максимально приближенных к естественным местообитани-
ям объектов исследования, можно добиться очень близкого соответ-
ствия расчетных и реальных значений продолжительности развития.
Использование стандартных метеоданных требует введения до-
полнительных поправок. Положительным; примером такого подхода
могут служить поправочные коэффициенты, разработанные Л .А. Ма-
каровой и Г.М. Дорониной (1988] при прогнозировании развития
некоторых вредителей сельскохозяйственных культур. *
.Несомненно есть формулы, которые.значительцо точнее описы-
вают, обсуждаемую зависимость, и при современном состоянии вы- ,
числительцойтехникибезтрудаможнопроизводитьсложныерасче-
ты,но .в повседневной практике защиты растений где погрешность^,
пределах?—3 дней вполне,допустима, формула.гиперболы.имеет свои
преимущества, главное из которых—возможность предсказывать фе-
нологические события при. переменных температурах..,,
•’V ;’Д i-O'.'tif.1 X, <; • - SJif J i. -Uu’ U-H’I-ИД'Я’
.Г'".',:;». -< .‘Н . ’.SR/ Г Kf--. ,
f'i’R OfGbf ; л'ХДДЯа Rs’> . .
-O ' CTJ ' XKH .НОЙ.'ЛО.Д'»' ‘ 5‘1-Шй ГЛ,е':'-г, ’,ч’
.Г; '<;'Ж '' I' ЖМЬВД' . a'-'O‘ W ' •; i.' "'a'/b
- TJ'IlOsJ’’ >. dSi’Hf < :r" (V;
’ ;?Vi> i-e -.1-5 l.iOrbiB ‘Ji \ ; i - >.
t* I ' / Ь.,л’ .i b. 'HR/
.1^' -e .. -
X’MJ j\. A,; A . у „Г ?
’ Глава6. ГЕТЕРОДИНАМНОЕ РАЗВИТИЕ
Насекомые даютбогатыйматериал для исследований сезонных
адаптаций. Их жизненные стратегии отличаются большим разнооб-
разием. Но все многообразие фенологических схем легко разделяется
на два типа: гетеродинамное и гомодинамное развитие, - u k
' Гетеродинамныйтип жизненногоцикласвойствен большинству
насекомых умеренных широт, 'и в основе его лежит чередование ак-
тивных стадий истадийпокоя, каждаяизкоторых строго приурочена
к определенному времени года.Гомодинамный цикл чащевстречает-
ся ушасекомых тропического ^субтрбпическогоклймата^ характе-
ризуется ^непрерывной физиологической активностью? В-пределах
гетеродинамного типа развития можно выделить поли- И моноволь-
ТИНИЗМ. ; TI
,,'Т л??,В MS W-bl " J . IS ' A'i’itl-- МВГЛНиЫкММ'Г; ,
6.1. Поливольтинизм и его регуляция т
Основнаяособенностьполивольтинногосезонногоцикла состо-
ит в том, что перезимовавшее поколение дает начало последователь-
ным летним Поколениям, череда которых завершается1 диапаузой.
Число летних поколений зависит от тепловых ресурсов местности и
обычно увеличивается по мере движения на tor. В данной местности
число поколений будет больше у видов скороткимонтогенетическим
циклом и у видов с меньшими потребностями в сумме эффективных
температур. Поливольтинизм отличается большой экологической
пластичностью и обеспечивает наиболее полное использование веге-
тационногопериода для роста и размножения особей данного вида.'
<' < Экологическая регуляция поливольтинизма оказалась- однотип-
нойу всехи сел едо ванн ыхвидов. Она осуществляется на основе длин-
нодневной ФПР: осеннее уменьшение длины дня и понижение тем-
83
пературы индуцируют диапаузу. Надежность системы синхронизации
поливольтинного типа развития с ритмом внешних условий повыша-
ется благодаря способности воспринимать параллельное действие
разных внешних факторов — температуры, терморитмов, качествен-
ного состава корма, сезонной динамики длины дня и пр. При моди-
фицирующем влиянии внешних условий на параметры ФПР время
наступления диапаузы и число поколений могут изменяться в зави-
симости от погодных условий года. Географическая изменчивость ФПР
гарантирует- соответствие сезонного щикла -каждой географической
популяции местным особенностям климата. Рассмотрим роль от-
дельных параметров ФПР в регуляции сезонного развития насекомых
у^цдол, ДфИбодре цолнц^зученцы^р^с^еримертальцо.,
ч ' чешуекрылые'^ ' '
i.-.W.-'-inoil' г ' -’«ж -.'•'"•гипо'ч; »> ”!v . •
Лсгош'сЩ^яянск.Щавелеваясовка имеет факудьтативнуюкуко-
лочнуюдиапаузу, контролируемую ФПР длиннодневного типа. Вос-
приимчивык длине дня гусеницыД1а чувствительной стадии проис-
ходит:накоплецие какдлиннодневной,<такикороткодневнойфото-
перирдической информации,1чю.вэкспериментальных)условиях со-
здаст-усл овия для перенастройкиФПР .на альтерната вноеизменение
длины-днянаэтойстадии.Дрифотопериодическойчурствительнос-
тигусецицгвсрх возрастов результат фПРопределяетсядаиной дня*,
действующей в период перед коконированием. Для индукциидцапаг
узы у половины особей белгородской популяции А. гит/сй требуется
10-11 короткодневныхсигцалов,сухумской - 16—J8 {1Ърышин, Ты-
щенко, 19726].
?, • Величинакритического.фотопериодазависитсОТтемпературы
(табл.,7), Температура 17—18 иСу наиболее близкаякфриродной впе-
рц од .формирования, диапаузы, .находится) на. грани «оптимума ФПР
белгородской популяции. Это объясняет появлениедиапаузирующих
куколок даже при дание дня 18 ч,. 16-часовойфотрпериод всочетании
с такой низкой температурой <17—18 инвариантно-.индуцировал
диапаузуувсехкуколок.,Максимальцаяизменнивос;гьфотопериоди-
чес.ких ответов (от46,4 до92.%диапаузирующих)<выделяе;Г|Зону ,кри-
ти.ческого фотопери од а. При 20 °C одмеченскачкообразный переход
кмаксимальномудаиннодневному ответу, нозначение,критического
порогаФПРосуается.17-часовым.пВ экологическом отношении кри-
тическая длина дня представляет наиболыпийинуерес. Заметныетсо-
то*-* к й.иь ms * . и. .4' Ф щфдз*.
84
,ц-^Габяица7„ Фотопериодическая реакция Acronictemumcis yy,,-fyjc,
Д<?- .н-4>.
ФП,ч - ; i Количество диалаузйрующих куколок, %, при w /
Т7-18вС ‘ 20 °C " *.
j: у Ид , 17 16 rv- Ф ПА , : МИ'.ЙП чи 1’9,0i7,i3OJ>, чфааОа 92,0 83,0 46,6 100 100 100 1‘ М00*'л: ” ‘-',iloo->;:-’s"’*'4's01100*'' H '400.в...’.',Й-ЖХРФ':» А100^ 1 К . 3,0( у г.ПГ’ОрПС 69,0 0 । ? пР 100 . . 100 . 'юо1''’1”’*’ too1" 1 AiiopMSHC
ч V .> ч .г- 1У, 5 , '. s4tfv-’ Л J. . •' - ! Ml t ' >'. - i* s' И - Гу- Ц . ' Y! Г.’. |7. Mt sfp '??<#') ' —.
лебанияколичеетвалиапаузирующих особей при47-часовом фотопе-
риоде по годам, несомненно,;отражаю?; внутрипопуляционную не-
стабил ьность критическогофртопериода, С учетом! вероятностного
характерафеноклиматическогодрогноза,,была-рассчитанастатис1и-
ческидостовернаявеличина .критического фотопериода для.белго-
родской популяции, Arumic/s приД7-Дг8“Спосовокупностивосьми
имеющихся щ нащем<.распоряжевди»крирых<ФЦР.<бна« составила
17?ч$&мин,±Д2 мин^рис. 35).’.'- w/>; :Я , 1 о;о>юя.:>-нчьП . > а
ФПР щавелевой совкиюдинаково проявляется при константных
и изменяющи^я4ютрпериодах.[Горыц1ИН«и др,, 1979]7Ф н-’Г1“.«;,ч
Фотопериодическиерецедторы А л/дисйнувствитедьвцлкизлу-
чениювидцмойчастиспектра, втрм числесдлинрйводныДр'570 нм
[Тыщенко, 197,7].,Лучи.с длиной волны более, 600, нм исчезают,, когда
1 ‘‘ ‘ ! Рис.35. Статистическая1 норма фотопериодическойреакцииЛсголIda
! н ^гит1скХ^ МатезМ ‘Ьгазз1сае'(б) vi^Pieris Апитгс4е!(‘в)'-прй‘-,!17л“С' ‘
- >1 , > ; f‘ «Средниедлярядаповторностейзначенияпроцентадиапаузирующихкуко- Н а .>
лок (сплошные линии) и доверительный интервал (пунктирные линии) при каждом
фотопериоде. Вертикаль отмечает критический фотопериод
8;5
солнце опускается ниже линии горизонта на. 6—7° {Шаронов, 1961;
Леман, 1961]. Таким образом, гражданские сумерки оказываются почти
полностыоэффекгивными дляФПР щавелевой совки .. *
А rumicis свойственнаклинальная изменчивость критического
фотопериода у разных географических популяций [Данилевский,
1961]. • :• л ' .л*
Зная подробную экологическую характеристику A. rumicis,
рассмотрим значение разных показателей развития и параметров ФПР
для синхронизации сезонного цикла вида с климатическими услови-
ями «разных географических точках, тде сезонный цикл этой совки
различается -’числом поколений. Анализ даншых показал , что на всех
широтах отС.-Петербурга доСухуми <42 “с.ш.Эфенология щавелевой
совкиполностьюгармонируетспродолжительностъю вегетационно-
го сезона и фототермическимиусловиями обмтания. Но достигается
эта гармония’ разными путями. ФПР ленинградской популяции от-
личается высоким температурнымоптимуможг,-который, поданным
А.С. Данилевского [1961], располагается вдиапазоневысокихтемпе-
ратур - от25«до-2Ф0С;-Низкая Температура окружающей среды «от-
ключает» ФПР и , несмотря на большую продолжительность дня, в
Ленинградской обл.развиваетсяодноиоколениеЛ.гат/от.
В Белгородеимеет -место четкая регуляциясезонного цикла ме-
' стнойпоцуляциивсоответствии с фототерммческими параметрами
(рис. 36). Заштрихованное поле в нижней части рисунка отряжает
фенологию Л. rumicis в лесостепной зоне и сосэтветствуетрезультатам
учета количества бабочек на светоловушке, что совпадает с данными
А.С. Данилевского'[1961] для Полтавы. В указанной зоне развивается
два четко разграниченных поколения щавелесвой совки. Лет бабочек
перезимовавшего поколения наблюдается во второй декаде мая. Ба-
бочки первого поколения вылетают с начала иполя до середины авгус-
та. Резко выраженный пик лёта приходится на конец июля.
Экспериментально установлено, что для. раз вития одного поко-
ления необходимо 550 гр.-дн. при нижнем -температурном пороге
10 °C. Развитие двух поколений в этой зоне хорошо обеспечено тем-
пературными ресурсами (1050—1100 гр.-дн.). Основное внимание при
анализе фенологии уделялось ключевому момьенту сезонного цикла -
возникновению условий для формирования ^диапаузы. Схема поле-
вых опытовпредусматривал а проведение вс&зоне последовательной
серии опытов, в каждом из которыхпрослежжвалась судьба куколок
.. г .«а: и. <.и-..
86
Рис.МФормированиедаапаузыукуколокЛгитййвполевыхопытах '
11 ’ гВ Белгородскойоблл-' ’> КН(’ : '' Jbi'SWTO С. '
-г 1 Гориэошальные<лрезки(1-24)- варианты полевых опытов, надними -зна- ; гкрч
чениясреднейтемперазурызапериодразвигиятусениц.треугольникисщива-средний
срок вылета бабочек. Кривые: 1 - сеэонноеизменение длины дня;2- среднестати- ‘
стическоезначениепорогаФПРс доверительным интерваяомпо лабораторным!' К
даннымпри18°С(запприхованнаязона);3—нарастаниеколичествадиапаузтфую- , f,
щихкуколок,%, вполевых опытах; 4-динамика лета бабочекнасветоловушкупо
пягтвдневяымучетам.Пояснениявтексте ц ' 1 1 ' г,
; 'С' ”!' . •! ,,р>!Ц" < <<, > < , ,:у уохо?.*-S
(активно&развитие или диапауза). Опыты повторялись в течение трех
вегетационныхх:езон6»'(рис.Е^6),,.<;,,:7;-7Ч'7 .у,»п (Ьи/
Стартовые точки всехдариантов выстроены,на одной наклонной -
прямой,* чхо: позволяет сравниватьдродолжительность стадийонто-.
генеза в разных вариантахиее изменение всезоне. Как указано выше,
нормальные сроки яйцекладкибабочек перезимовавшего поколения
приходятся на вторую половину мая—начало июня., След овательно,
варианты: 1-9 наших полевых опытов соответствуют естественным ус-
ловиям развития.гусениц Ьго поколения. Продолжительность разви-
тия гусениц в этих вариантах колебалась от 33 до 38 дней.В результате
в конце июня—начале июля появились лервые коконы, а в последней
87
I
декаде июля начался вылет бабочек, вполне синхронный с лётом ба-
бочек 1-го поколения в природе (кривая 4). Развитие гусениц в этих
вариантах опыта шло на фоне длинного дня, что определило отсут-
ствие диапаузирующих куколок. Потомство этих бабочек развивалось
в августе—сентябре (вар. 19—24) и дало диапаузирующих куколок
(100%), которые и сформировали зимующий запас. Гусеницы разви-
вались при длине дня короче критической.»**’.1 ’
Таким образом,'фенология»Д>«лг/сй®.Бедгородской обл. очень
проста. Небольшие колебания погоднЫхусловий по годам могут не-
значительно сдвигать сроки развитияуотдельных стадий, но они не
вносят принципиальных изменений в схему годичного цикла вида.
Если вылет бабочек 1-го поколения-под влиянием очень теплой пого-
ды сдвинется на более ранние’,сроки,1 гусеницы следующего поколе-
ния все равно попадут в условиявкорбткого дня, вызывающие обяза-
тельное наступление диапаузы у куколок. Сумма эффективных тем-
ператур обеспечивает развитие только двух поколений'.
Теоретический интерес' представляют варианты 10—16, в кото-
рых гусеницы попадают в критическую-ситуацию. Такие сроки не ха-
рактерны для развития гусениц в природных условиях, хотя единич-
ные бабочки и отмечалисьв это время’ Рассмотрим’фотоперирдичес-
кие условцяэтого периода,сезона., Заштрихованная зона на рис. 36
(вверху) отмечает среднее значение критическорофогопериода с до-
верительным1 интервалом. Гусеницы/попадающиешосрокам разви-
тия в критическую ситуацию, 1ю-разному’«прочитывают» д лину дня.
Об этом свидетельствует разброс точек, характеризующих количество
диапаузирующих куколок,(от 50 до96%) . В этоййоне на границе длин-
нодневных и короткодневных фотопериодов приобретают реальное
экологическое значение и критический фотопериод, и пакет фотопе-
риодической (информации- ииндивидуальныеотклонения в оценке
природной длины дня. Но коллизии,•связанныес зоной критическо-
го фотоперйода, не’ возникают.1 Приуроченность пиков лета бабочек и
яйцеклаДки перезимовавшего1 и> летнего поколений «в сезоне такова,*
что чувствительные к Длине дня стадии заведомошопадают в альтер-;
нативные|фотопериодическиеусловия1<Схемасезонногоразвитияот-'
лиЧаетсяШредельной>простотой!.<т‘-'ог^отч г-..? азпи.С'
с-у Географическая.изменчивостьФПР’унасекомыхсполивольтин-
нымфазвитиемюбычно проявадется® понижении.’критической дли-
ны дня у южных форм’по сравнению ссеверными, что способствует
увеличению числа поколений на юге ареала.'Критический порогу
88
сухумской популяции щавелевой, совки составляетдпри:23°С 14 ч
2d мин. В результатедаСухуми завегетационный.сезон обычноразви-
вается Зг-4 поколения[Миляновский, 19561>.Анализфенологиивида
в этой зоне [ГЬрышин, Гейспиц, 1975] показал , что-прищаступлении.
критической длины дня вприродекуколкивпадаютв диапаузунеза-
висимо от принадлежности^ тому или, иномупоколению<рис. 37),.
Точки пересечения кривых -1. и 2 ограничивают на рисунке период
лета, когда день продол жительнеекритического.Погодные^измене-
ния температуры находят, отражениев1 форме кривой2и изменяют
сроки наступления критическойдлины дням формирования диапау?-
зы. Пакет фотопериодической информации усухумской популяции
A. rumicis составляет около<18 коротких дней. .Только по истечении
этого срока можно ожидать формирования диапаузы у 50% куколок, .
.. Приведенные материалы свидетельствуюто том, что фенология
щавелевой совки навсех широтах от С.-ПетербургаДо Сухуми строго
гармонирует с фотопериодическими и температурными условиями
.Г'”; , -НЙ » Ik-n "VlqcVIl .ГМВГ1Х TiK-RK 1 7. ТЧК-рйН , ЙиЛ* ?S>
.4»,ISO-A> 'Шьгд и. ьда.
‘ ' Рис..?7.; Схема сезоннЬго цикла 'совки Acronicta rumicis »'Сухуми (по: • <’
•д.м. Г'1Ьрышин,:.1ейспиц,-1975]} '«1?, *. '
, 1,2-эффективная длина дня (сучегом спектральной характеристики ФПР), ...
ивеличинакритическогофотопериода.привязаннаяксреднемесячнойтемперату-
ре;точки пересечениякривыхотмечают’критическуюдая^совкидаинуднявприро-» '
де;крайниевертикальныесгрелки на кривойГотмечаюгдаинудая веснойиореныо
припереходетемпературычерез 1Ь°; 3—кривая нарастания количествадиапаузиру-'
'мцих куколок В'природе;1 построенная по<фен<»1Оп<че&кх>й:схеме^«"-»(Миля-;'',= ;1>
. , >новский, 1956];«+++»-6абочки,«---»-гусенины;4-сезонноеизмене- (
ние количества диапаузирующихкуколоквсоответствиисданными лабораторных
опыпж11|>идяяиедн«14чятем11ОИ1и>е25*С' ' ' 1 г
< Г Н i-ft!
89
* - Однако, как оказалось, создавая преимущества в си нхрониза- .
ции годичного цикла различных популяций с локальными условия-1
ми,наследственнозакрепленныесвойстваФПРпрепятствуютихпе-|
ремещениюдажевпределахввдового>ареала.Так,в опытах А.С. Да-<
Милевского и И.А. Кузнецовой [1968]южные популяции A. rumicis m
могли-формировать диапаузу в условиях’Ленинградской обл. из-за!
несоответствияихкритическогофотопериодабодьшейпродолжитель-
ности дня на этой широте. В случае зимовкисеверныхформна юге и
при более раннем возобновлении развития весной, они давалитоль-.
ко одно поколение. Раносформировавшие диапаузу особи под верга-
лись длительному действию -высоких температур, это, несомненно,
снижало их жизнеспособность. Таким образом, реакции, управляют
щиедиапаузой, обеспечивают высокий уровеньприспособленности
локальных популяций к местному климату. Географическая диффе-
ренциация норм ФПР определяет эколого-географическую структуру
вида, в которой* каждая, популяция экологически ограничена в сво-
бодном перемещении даже в пределах собственного ареала. Однако,;
как выяснилось, упомянутая привязанность неодинакова у разных ви-
дов: в сильной степени она проявилась у A. rumicis, но значительно
слабее была выражена у капустной совки.
Mamestra brassicae. Капустная совка по типу температурной и
географической изменчивости очень похожа на щавелевую совку, од-
нако некоторые экологические особенности вида, обнаруженные в
экспериментальных условиях, как оказалось, приводят к заметным
отличиям в характере сезонного развития. Своеобразной .чертой в
механизме ФПР капустной совки является способность суммировать
на протяжении всей чувствительной стадии только короткодневные
сигналы {Горышин,-Тыщенко,' 1973]. В силу этого гусеницы 1-го поко-
ления, развивающиеся весной в условиях длины дня меныне крити-
ческой, начинают накапливатькороткодневную (КД) информацию
уже в младших возрастах. В случае заполнения пакета КД-информа-
ции, составляющего поданным разных.опытов10-15 сигналов, часть
гусениц 1-го поколения будет формировать диапаузу.Так происходит,
например, влесостепнойзоне.;Фенологические наблюдения и поле-
вые опыты,проведенныевБелгородской обл., подтвердили двойствен-
ное происхождение зимующего запаса:, из части' (до 40 %) куколок
1-го поколения ииз куколок 2-го поколения. Подобное явление от-
мечал А.С. Данилевский [1961] дляблизкой географической популя-
ции из Сумской обл. (51 °с.'ш.). , ' , . '
90
Другая особенность капустной совки заключается в том, что ее
южные популяции в условиях длинного дня и повышенной темпера-
туры способны.формироватьлетнююдиапаузу.В.результатеэтогопри
продвижении на юг, несмотря на увеличение тепловых ресурсовучис-
ло поколений не возрастает.Так.насевере ЯцонииДМазакцБака!,
1965] капустная совка имеет лишь зимнюю диапаузу, которая форми-
руется вусловиях короткого дня, Реактивация диапаузирующих куко-
локрозможнатолько после охлаждения, Вболее южных районахЯпог
нии у куколок наряду с зимней может, индуцироватьсяилетняядиа-
пауза, ко нтролируемая длиннымднемивысокой.температурой. Ин-
тенсивность летне й диапаузы настолько повышаетсякюгу,чтовсуб-
тропических зонах она превосходит по длительности, период зимнего
ПОКОЯ. • г и Г’зН-Г .'•> г? .
। входные, результаты получены для капустной совки в условиях
ЗападнойЕвропы[РойоиЦ Виез,1973]. Однако детальное изучение
западно-европейскихпопуляцийэтого вид a] Sane г, Gruner, 1988] пот
казало, что,только самая южная (42,5 °с.ш.) из исследованных имеет
ежегоднуюлетнюю диапаузу.Севернее (44 °с.ш.) капустная совка эст
тивирует лишь в отдельные годы, а у самой северной из тестирован-
ных популяций (48 °с.ш.) летняя диапауза,не выражена вовсе; здесь
развивается обычно двапоко-:, ?> .л-д, "
ления задето.. • -i:7-п. г • ...
; , Наконец, третьей осо-,<
бенностью капустной,совки
является отмеченная выше-
чувствительность к суточное 5
муритму температуры. Бла-,
годаря усилениютенд енции
к формированию диапаузы в
условиях терморитмов юж-
ные популяции M. brassicae
в отличие от A. rumicis спр-; *2 .
собны диапаузировать цри 0
перемещении их в северные
ШИРОТЫ (рис. 58).. , ; ;
“W
S’
; X - -
Л
с 20- * 2
ДШ'"
ЛЮ и
3
МО, о*
1
400Л?
<0
> Cp-*r9'tl ' 15 1
, -?1;'А»гует,;, ({Оняг||10К'!Т10тф. ч
_ _ Рис. 38. Формирование диапаузы (1) и изме-
.,, ч - внеООЫЧНЬГХДЛЯСухум-,,. нение массы (2) куколок у сухумской формы
СКОЙ ПОПУЛЯЦИИ УСЛОВИЯХ се- ., Mamestrahrassicae при воспитании гусениц е
вера одинаково четко про-],, даиродных условиях Ленинградскойлбл.: »
явились и пеакпия конто! ТЪчкиугазмшотсрота^
ЯВИЛИСЬ И реакция, кошро7 ..^линга-ош^кисредней^фмепгаеской 1
91
лирующая наступление'диапаузы1, и реакция, регулирующая массу
куколок. Однако формирование диапаузыв-этих опытаху южной пе-
ремещенной популяции задерживалосьпо'сравнению'с ‘местной по-
чтила месяц'. В‘этом проявилось'несоответствие сезонно-цикличес-
кихреакцийтожнойпопуляциикапустной совки’климатическим ус-
ловиям севера.- дкишжиг ши? ц’ч;
цлИнаяэкологическаяситуация былаобнаружена у видов, ФПР
которых-непроявляетобычнойтемпературной игеографической из-
менчивости. Из тесла; изученныхнамивидовтаким свойством обла-
даюткапустная-белянка и луговой мотылек. Остановимся подробнее
нарезультатахюпытовРназваннымиВидами. ’!1’ *'
и PierisAnassicae.B отличие от большинства насекомых, для кото-
рых характерна клинальная изменчивость критического порога, по-
пуляции капустной белянки/заселяющие обНшрную территорию
Восточной Европы, распадаются на' два резко1обособленных- экоти-
па "-северныйи'южный.различающихсяпосвойствам ФПР. Север-
ный экотипимеет термостабильную ФПР, критическийпорогкото-
рой не отличается у популяций, обитаюших-севернее 47 °с.ш. Юж-
ный<экотип имеет термолабильнуюФПР, подверженную клинальной
географической-изменчивости' ‘Хорошо известно,;что в природе тем-
пературные и фотопериодические условия1,'хотя и' с противополож-
ными тенденциями, но равномерно изменяются от экватора к полю-
сам. В соответствии с этим клинальная изменчивость’порога ФПР,
как это было показано экспериментально на примере Л. гит/сй, обес-
печивает согласование сезонного развития отдельных ’географичес-
ких популяцийс локальными условиямиклиматанавсех*широтах от
40 до 60 °с.ш>При этом чем южнее, тем ниже значение порога ФПР, й
тем позже наступает диапауза в природе, обеспечиваяразвитие боль-
шего числа поколений. Каким же образом годичныйцикл экотипов
Р. brassicae с резко обособленными свойствами'ФПР вписывается в
плавно меняющиеся с'-широтой условия'среды? Для этого проанали-
зируем развитие аборигенныхпопуляций Капустной белянки на раз1-
ных географическюбширотах и оценим их приспособительные воз-
можности, придальних перемещениях. J St Л /Ц;
Опытыс. капустной, белянкойв Ленинградской области были
выцолценыДС. Данилевским,и И,А. Кузнецовойх['1968]. В течение
большей «части лета .капустная белянка развивалась без дйапаузы
(рис. 39) ,Но уже с начала третьей декады августасреди’окуклившихся
резко''возрастал’цроцент дцдпауриру1рших, а После 1 сентября все ку-
92
Ионь . Ноль Аьгуст , Сентябрь . Октябрь Ноябрь
Длина1 дня, ч: и,» _ »,«
> .Рис.39. Формирование диапаузы у географических популяций"
Р. brassicae ® природных условиях Ленинградской обл. (по: Дани-
левский, Кузнецова, 1968]:
и ПопуляцииШ-ленинградская:!^^белгородская:3— ессеигукская;!4—^И‘:-»;!''
сухумская; 5—гибридаая(сухумскаясамках(ленинградский<амец}.'1рчкиуказыва- , $ц,
ют сроки окукливания ’’
' Дй: ‘.ЖлН K MPvC' ?>/5.; Я'И.,. Лйл'-J' < ’1iЖГ
колкиоставалисьзимовать. Длина дня 2(Равгуста на широте С.-Пе-
тербургас поправкой наспектрал ьную характеристику фотопериоди-
ческих рецепторов Р. brassicae [Горышин,4 Гейспиц, 1975}составляет
L5 *1 40 МИН УЧ;.?.''У 'v?*tsvО v'/i .V.1-/'<. RUCRTH30 < ?
В Белгороде, расположенном: на 10° южнее'С-Петербурга/кли-
мат теплее, а максимальная’длина дня существенно короче? Крити-
ческий’для Р. Агаадсаечротопериод отмечается уже 'в- начале* августа,
задолгодо'наступления субпороговых температур? и соответственно
на 20 дней раньше, чем в С.-ТГетербурге, создаются условия‘для ин-
дукции диапаузы уместной популяции. 'Экологическая нецелесооб-
разностьэтой тенденции очевидна: во-Нервых раннее формирование
диапаузы ограничивает использование видом тепловых ресурсов
местности,‘во-вторых,Ото неблагоприятно для диапаузирующих ку-
колок, часть которых длительное “время подвергается воздействию лет-
нихтемператур.--:'<;> ьичто:и .'л, 'п
> ' Какпоказывает анализ полученныхрезультатов, экологическая
характеристика северного экотипа'наиболее точно соответствует ус-
ловиям обитании белянок'в странах бассейна Балтийского-моря. В
лесостепной-зоне. где проводилисьнашиэксперименты, уже намеча-
ется дисгармония междупараметрами ФП Р иместнымифототерми-
ческими условиями. При неизменности порога ФПР эта тенденция к
югу должна усиливаться, все более нарушая соответствие сезонного
цикла ритму климата. Однако это предположение не подтвердилось в
93
* -too aR •
£ л
> -80 S
S' C
* Ф
60 5
r 8.
-40 t
—‘--------1-----—* * I---------------’О Рис.40. Формирование диа-
ыыюл v . ЯентЧ0₽ь> . ; v, .>> дары у капустной белянки в
М.06, г Л 3. Л, 43.03. «НС , Славянске^нагКубани
• V Г J I •' -J I. ' *Л , ‘ ! Д
опытах, проведенных вСлавянске-на-Кубани, гдеобитает популяция,
не относящаяся к[северному'Экотипу(рис.‘40).’ ! *
Критическая длина дня для местной популяции наступает в на-
чале сентября. Возможность формированиядиапаузыреализуется в
вариантах,гдегусеницыокукливались 1 иДсентября,хотячислодиа-
паузирующих ,было.невелико(.19и 23% соответственно). Но среди
окуклившихся 11 сентября все куколки были диапаузирующими, Та-
кимобразом, индукциядиапаузы уместнойпопуляциипроисходит в
соогветствиис параметрами ФПР и в оптимальные дляэтого процес-
са сроки.Рассогл асованность сезонного развитиям ритмомклимата,
наметившаяся в леростепной зоне, здесьотсутртвует.Такжев благо-
приятные сроки индуцируется диаиаузаиу более южных популяций*
Например, в Ереване среди окуклившихся 16—20сентября1963т. 45 %
куколокдиапаузировало(неопубликованныеданные В А. МаСЛеННИ-
КОВОЙ). /и».; -Г’- 11 Т;»£й., ' ЧНЬТ”: ‘ ’>'•* in.
' В опытах с перемещенными популяциями (см. с. 95) легко лри-
спосабливались только те из них. перемещение которых происходило
в пределах ареала экотипа. Так, при развитии в условиях Ленинград-
ской обл. диапауза индуцировалась у близкой по параметрам ФПР
белгородской популяции капустницы (см. рдс. 3*?), относящейся так-
же к северномуэкотипутогд а каку, сухумской белянки, принадлежа-
щей к южному экотипу, диапауза не формировалась даже у окуклив-
шихсяв октябре и началеноября, когда длина дня была гораздо мень-
ше пороговой для этойпопуляции {Данилевский,-Кузнецова, 1968].
Пр мнению авторов, индукция диапаузы у зюжной капустной белянки
.><Т -и *• Х.Ш'Л.'Ц-'ОЬ; ц; < ‘ i ГТЦ/ Н:' Wfifi J. ' ; >
94
нарушается не только большой продолжительностью дня, но и чрез-
мерно низкой температуройвпериод развития гусениц. • > и
, В наших полевых опытах, проведенных в Белгородской обл.; все
куколки,, сухумской белянки впадали в диапаузу в начале сентября.
Эти сроки более благоприятны для формированиядиапаузы, чем на-
чало августа, когда индуцируется диапауза у местной, белгородской
популяции. Способностьсухумскихкуколокформировать диапаузу в
несвойственных им условиях, вероятнеевсего, обусловлена темпера-
турной коррекцией порога ФПР, Куколки; ленинградской капустной
белянкиформировали диапаузу.одновременно с.местной. г < .,<
; Вслучае воспитания ленинградской популяции капустницы на
юге (Лазаревская* 44' с.ш.) диапауза возникала у всех куколокуже в
июне, поскольку даже максимальная длина дня на этой широте не
достигаетпорогового ддясеверного экотипа значения [Данилевский,
Кузнецова, 1968]. Этирезультаты подтверждаютнесоответствиежиз-
ненного цикла северного экотипа новым климатическим условиям.
Непривычно высокая температура на юге практически не. оказывала
влияния на сезонное развитиебелянкилвиду отмечавшейся ранее тер-
мостабильности критическогофотопериода северногоэкотипа. ; > ; > <
; Интересные материалы лолученыв опытах стформами, проме-
жуточными между северным и южнымэкотипами. Так, капустная бе-
лянкаиз Ессентуков (44°с.ш.), котораяв лабораторныхопытахпочти
не отличал ас ь,по ФПР от ленинградской, вприродев условиях С.-
Петербурга также формировала диапаузу, но в более поздниесроки,
чем местная [Данилевский, Кузнецова, 1968]. Из-за фрагментарнос-
ти данных, имеющихся в нашем распоряжении, трудно более подроб-
но обсуждать этот вопрос, однако заслуживает внимания тот факт, что
и в Белгородскойобл. промежуточные формы (из Херсона и Славян-
ска-на-Кубани) формировали зимовочное состояние в оптимальное
для этого время. - «
Таким образом, у капустной белянки внутривидовая эколого-фи-
зиологическая дифференциация; выражается, в возникновении двух
экотипов..Факториальная и географическая стабильность параметров
ФПР у северных популяций создает предпосылки для свободных пе-
ремещений их, по крайней мере, в пределах ареала северного,экоти-
па,а возникновение южного экотипа обеспечивает увеличение зоны
синхронизации фенологии видас ритмом климата. Идентичная си-
туация обнаружена у лугового мотылька, имеющего сходные черты с
капустной белянкой. .....
95
> , Тохохй^еЛййсайу.ЛуговоймотылекимееттипичнуюФПРдлин-
нодневного типа.Чувствительны к длине дня* гусеницы , диапаузиру-
ютцронимфы (завершившиепитаниегусеницы)®коконе. Величина
критическогофотопериодамалозависитоттемпературы. В интерва-
ле от 17.ДО 25’9С;он изменяется.всего от 14 ч 24>мин до 15 ч 09 мин
(pHCv.4il). . s:r.a-(w -чц.-м г: - . ui • я* у»
ч .; чСвоеобразношроявляетсяулуговогомотылькачувствительность
к постепенным изменениям'длины дня: Она'отсутствует при типич-
но длиннодневных и короткодневных фотопериодах, но максималь-
но выражена вобласти пороговых значений (Горышинидр., 1980;
Волкович,Горышин, 1982|.Врезультате факторпостепенности вы-
зываексдвиг величины критического-фотопериода примерно на
40 МИН.Н&: ЙО"«' .е ' Ний-'
» л Короткодневная индукция диапаузы у лугового мотылька про-
исходигв конце развития гусениц (чувствительная стадия). При воз-
действиибКДдиапаузирует примерно 25% пронимф, а при’9 КД -
75% „Критический пакет ФП-информации близок к7—8 дням.Опы-
ты при разныхггемпературах показали,‘что в диапазоне 17—25 ^вели-
чина критического пакета существенно не изменяется (см. рис. 1в).
Термостабильность этого параметра ФПР облегчает его использова-
ние для анализагфенологии вида.1 о ? в ; > * -и
• Критическийфотопериодоказалсясходнымупопуляцийиз Бел-
города.^ 50°с.ш.),'Хабаровска (48°с.ш.>.:Славянска-на-Кубани (45 °с.ш.)
ичНальчика.(43,ос.Ш|.')..! .. , •. чк • т <- '
''«.«'HP Mt 1 .ЛД'ЛНс'у?! .Н . ЕьГ'ЛЛ.* S/':?'
дз члнрй'П'’’;М сказ чДлинадняХ г; '<->
• Ч 1 •' > ' ’ ' г
Рис.41. Фотопериодическая реакция лугового мотылька при разных
постоянных температурах (популяция из Белгородской обл.)' ь ’
96
В отсутствии доминирующею.фактора (фотопериода)-улугового
мотылька проявляетсятермопериодическаяреакция, которая усили-
вает тенденцию к формированию диапаузы в условиях ритмично из-
меняющейся температуры. . ; , ir
Влияние природных ритмов демпературы-наиндукцию диапау-
зы было изучено с помощыоупОмянутой (см.1л„5) полуавтоматичес-
кой установки (табл. К).Опыты проводилисьдаажды: с особями 1-го
и 2-го поколений. Средняя температура’-воздуха в период развития
гусениц 1-го поколения составляла"^,“4 °C (28,7 °C днем и 15,9 °C
ночью). В период развития, гусениц 2-го поколения средняя темпера-
тура была ниже - 19,4°С (20 -14,8 °C). Контрольные группы особей
содержались при постоянноМгемпературе 20,5 °C. *
ли,! «KOT.-.-ttifti. Л ; ч ‘
‘ V»: ’ .и
Таблица в. Влияние природных ритмов температуры?;
на фотопериодическую^реакцию лугового мотылька?;
Длина ДНЯ,-ч*; Диапаузируюшие особи, % ’
1-е поколение 2-е поколение
опыт | контроль опыт | контроль
14 15 и. < и 16 ‘ ФОО 100 85,7 8,9 100 100 100 33,9, г. 22.3 0 •? MRг. - .«ншйг-мйп .я vonr-or/. • 'i
Указанные экологические свдйства,лугового модылькапозволя-
ют наметить ожидаемые сроки формирования диапаузы в разных зо-
нах ареала. Ограничим эту задачу лишь степной (Краснодарский край)
И лесЬстепной(Белгородская4обл.)’зонами(,?!где'спровОдилис^ наши
исследования/ Ч'у"• ••’"’*i
'Полевые1 опыты.1"Эти1 исследования проводились'в'Краснодар-
ском краеи в Белгородской обл. на местных й взаимно-перемещен-
ных популяциях. В- Славянске-на-Кубани за-сезон былопоставлено
25 вариантов опытов с местной, славянской иперемещенной, Белго-
род ской популяциями лугового мотылька, начатых в период сЛОиюня
по25 августа. На'графике (рис?42)'горизонтальные полосыотражают
среднююпродолжительностьразвитиягусениц в каждом-варианте. В
зависимости от условийих развития :можно'ожидать вылета бабочек
или возникновенияпронимфальной'диапаузы. -Количество' диапау-
зирующих особей (точки) календарно привязано к дню-завершения
гусеничной стадии. ПриФтом имеется‘в виду, что диапауза наступает
Рис.42. Результаты полевых опытов с луговым мотыльком в Крас-
нодарском крае (Славянск-на-Кубани) в 1980 г.:
Горизонтальные полосы—варианты полевых опытов (светлые - для славян-
ской популяции, черные - для белгородской); кружки - сроки завивки кокона.
Кривые: 1 - сезонное изменение длины дай; 2 - среднедекадная температура; 3 -
расчетная динамика индукции диапаузывприродных условиях; 4и 5 - фактическое
"' нарастаниеколичествадиапаузирующихпронимфвопьпахсбелгородскойиславян- ‘
, скойпопуляцидатмопщцчт ; ,
вскоре после образования коконов. Кривые 4,и 5 обобщенно описы-
вают нарастание количества диапаузирующих в последовательном ряду
вариантов (соответственно для белгородской и славянской популя-
ции). Как видно, первые диапаузирующие пронимфы у обеих попу-
ляций появились в середине июля. Расчетная кривая 3 допускает по-
явление небольшого количества диапаузирующих в этот период, но в
опытах с бел гор оде кой популя циейих оказал ось заметно бол ыпеожи-
даемого (до 30%). Любопытно, что в самом конце июля наблюдалось
снижение .количества диапаузирующих особей. Таким образом по-
явился первый пик диапаузирующих пронимф при заведомо длинно-
дневдых условиях. .Слрдует заметить, что в лабораторных условиях
при длине дня. 17 ч иновда наблюдается 20—30% диапаузирующих
особей. Возможно, именно с этим связано наличие диапаузы у части
98
пронимф 1-го поколения, часто отмечаемое при полевых обследова-
ниях(Поляков и др., 1978}. Основное увеличение количества диапау-
зирующих особей наблюдалось в начале августа, а после Ю августа
диапауза формировалась'у подавляющего большинства пронимф.
‘ Сравнение полученных данныхпозволяет сделать некоторые вы-
воды. Во-первых, кривые для славянской и белгородской популяций
почти не отличаются, что подтверждает отсутствие географической
изменчивости ФПР у лугового мотылька в пределах лесостепной*»
степной зон его европейского’ареала. Во-вторых, кривые,описываю-
щие динамику нарастания количества-диапаузирующих пронимф,
хорошо согласуются с расчетной кривой 3. Разница между нею и фак-
тическими кривыми (составляющая на уровне 50 ^диапаузирующих
7—8 дней) как раз соответствует продолжительности КД-индукции,
установленной в лабораторныхопытах. - ; ! - < 4 : ;
‘Поаналогичной схеме, но в лечение 4-х лет проводились опыты
с луговым мотыльком в Белгородской обл. (рис. 43). '
п Всего с белгородской популяцией мотылька проведено 37 вари-
антов опыта, распределенных между 23 мая и 15 сентября. Кроме того,
в одном сезоне (1981) опыты с белгородской и славянской популяци-
ями проведены параллельно. Представить календарную приурочен-
< ' t ' •
Рис.43. Результаты полевых опытов с луговым мотыльком в Белго-
родской обл.: <?
” Кривые - факгическоенарастание количества диапаузирующихпронимф в;
каждом сезоне. Популяции: б - белгородская, с—славянская; остальные обозначе-
ния, как на рис. 42
99
носдьвсех этих вариантов на одном рисунке) невозможно,’Поэтому
для, каждого тода построены лишь.итоговые кривые, описывающие
изменения количества диапауз ирующих пронимф в сезоне.’ •• п д л- . ’? t
.Полученные данные далекоше идентидаы, Лишь результаты,
1981г.,(для бел городской.и-славянской’популяций) хорошо сотласу-
ются.спрогностидеской.кривой 3.,Врпытах;1976 и 1977гг.диапаузи-
рующиепронимфы(появилисьуже в>первойполови,не июляпризаве-
домо длиннодневных условиях,причемчислоихдостигалопочти80%;4
Нацрашиваетсявывод, чтораиняядиапаузавэтигодывозникалавне>
связирдлиной дня. В летние сезоны1976 иД977 гг. температурабыла
ниже средней многолетней для Бел городской обл.ъ втовремякактем-
пературные условия лета1981 г.былиблизкими кнорме.Однакотер-
мостабильность ФПР. лугового мотылька-не позволяет ссылаться на
эти особенности сезонов. СпецифическоевлИяние на формирование
диапаузыоказывают. природные терморитмы (см. табл. 8), нои с ними
надежной корреляции не обнаруживается. .Несомненно? одно:ФПР
лугового мотылька надежно«рабогает» в природе на фоне относитель-
новысоких,температур,,нонеобеснечивает’контролясезонногораз-
виргия вхолоднцегодыу . /го ыгыпо ’. 1 "’ондо з
,, . фенологические наблюдения. ВБелгородскойобл. исследования
проводились в течение 5 лет и заключались в учете лета и динамики
численности имаго лугового мотылька, что позволялоиметь представ-
ление о числе поколений и сроках развития каждого из'них. Как от-
мечалосьвыше/ддяразвития одного поколения лугового мотылька
необходима сумма эффективных температур 400—450 гр. -дн.припо-
роге*12°.В заповеднике «Лес на Ворскле» тепловые ресурсы вегетаци-
онного, сезонахоставляют около 800 гр.-дн. [Караушева, 1980],гчто
обеспечивает развитиедвух полных генераций вида. В наиболее теп-
лые годы возможен вылетбабочек 2-го поколения и появление гусе-
ниц 3-й генерации, которые едва ли могут завершить развитие.’ 1
Фенологические данные © сроках дета бабочек лугового мотыль-
ка (рис. 44)'получены путеммаршрутныхучетов вподходящих стаци-
ях и кошениясачком. Дополнительно учитывали’лет бабочек на све-
толовушку(лампа ДРЛ-400).
ЛётбабочекВ годы с их высокой численностью имел резко выра-
женный пиковый характер. При этом число пиков вылета варьирова-
ло от двух'(1976и“1978 гг.) до трех (1977 г.; такая же тенденция при
низкой.численностибабочекнаметиласьв,1979,г.). Последнее обсто-
it"- Г'1: ЗА/’ ‘ 'Н
1Q0*
ятельство заслуживает особого_внимания,.таккаклри.пика летанельзя
считдть отражением трех последовательных генераций.?’. < । ’«юи
Проеледимбол ее,детальнофенол отиюлугововомотылькав'л е-
состепной’Зонев*тенение1977г.Аналогичное.сравнениеможнопро-
вестщи паматериалам другихлет.<К. сожалению, такой возможности
нетдлятедлого19&1 г. j так как численность луфвогомотылька врай-
оне^наблюдений оказались исчезающе малойь я от? ,п>п 5 цу гяйй
h , Итак, в1977г.,первые бабочкибылиотмеченьгмежду1'5№20мая,
но, массовый лёт;перезимовавше,гопоколения"наблюдался !в конце
мая-.начале июня (рис, 44)?После.20.июня№редкавстречалисьеди-
ничные бабочки. С 10 июля начался массовый вылет бабочек летней
генерации, асначала, августа зарегистрировантретийподъемих чис-
ленности..Эти,пики, вылета, бабочек моглйибы служить отправными
точками'.даяюпределения.сроков.яйцекладки ипоявлёниягследую-
щегопоколепия.Норазвитиепреимапинальныхстадийболеегточно
характеризуют приведенные здесь жег данные* тюлевых опытов. «На
рис.(44нанесена1средняя:продолжитёльностьразвитиягусениц, про-
нимф икуколокввариантах,наиболееблизкихвсезонесоответству-
ющимщикам. Вертикальными стрелками отмечен'среднийсрок вы-
лета бабочек. Легко.виде'ЕЬ^'ЧТО интервал между первым ивторым пи-
>‘ми!!'ть'"в«ёоп0 :о5Ю:1Я!-1 а'з; *, itmtk"
• >т'ч»7#г.<’ "'’В-'’ эингргапосдГ.
.. 7;^1Н Т.П.;!Т'.''^ЧЦ№ Ч .учсбЫНС Г Ч, ' .'ЛПЮШ'!
.мц'>.11??^С-.+±^н*,«нопн »W3лк,:».?" :
^Й.,. . • . ,РИЮНЦ.-,15Ж,.,„ -р-.;,;- t ABiyCJT»‘, учу?X'i&tHfflflMЛ С
<Ш"Вис.44. Фенологические1 сроки’ вылета ‘бабочек' лугового-мотылька.' 1
1— лёт бабочек; 2—среднедекадная температура, л-число бабочек, отме- > ;
чавшихсяна50шагов. 1Ъризонтадьныелинии:сплошные-развип«ег)сениц;лунк-
4 ' ’ тарные—развигиепронимфикуколокстрелкавконце—сришийсроквылегаимаго
,приактивномразвйтйи;цифрыулиний-средняя температура за периодразвития?
гусениц и количество. % , диапаузирующихпронимф(данныедая вариаи^овполевых
'опыгов:1977г.),«+»:врамках-периоды’массовоголётабабочек J
101
ками лётабабочеквесьма точносоответствует длительности онтоге-
неза в этот период сезонами указанные пики можно считать отраже-
нием сроков вылетабабочекпоследовательныхпоколенийлугового
мотылька. Интервал между вторым и третьим пиками1 лёта слишком
мал для завершения онтогенеза;^ нормальнымбыло бы появление ба-
бочек 2-йлетнейгенерациилишь в конце августа—начале сентября.
Следует заметить, что в вариантахопыта, близких по срокам отрожде-
нця гусеницковторому пику, развитие гусениц происходилопри наи-
более высокой средней температуре и завершилось быстро, но в авгу-
сте температура стремительно снижалась, что замедлило развитие ак-
тивных пронимф, г « : ; '-л
; Аномальное положение на рисунке третьего пика подтверждает-
ся и ретроспективным анализом фенологии. Расчеты показывают, что
массовая’яйцекладка, от которой могли бы произойти бабочки тре-
тьего-вылета, должна проходить «20—25 июня, однако численность
имаго в этот период была ничтожно малой. Таким образом,‘феноло-
гия 4977 г. содержит загадочные отклонения от ожидаемой нормы,
причем подобные отклонения встречалисыи в некоторые другие годы.
Фенологически привязанныевариантыполевых опытов (рис. 44)
позволяютзаключить,чтов 1977;г зимующий запас начал формиро-
ваться уже в 1 -м поколении (от 0 до 46% диапаузирующих пронимф).
Лабораторные испытания показали,ото ранняя диапауза отличается
высокой прочностью и обеспечивает зимовку. Количество диапаузи-
рующих особей увеличивается (до 79%) среди пронимф 2-го поколе-
ния, но лишь в потомстве бабочек третьего пика лёта диапауза стано-
вится обязательной. Кажется заманчивым предположить, что третий
пик лёта бабочек формируется в результате иммиграции лугового мо-
тылька из более южных районов. Общепризнана способность этого
вида к миграциям. А.И. Мельниченко [1936], анализируя материалы
о перелетах лугового мотылька, заключил, что «случайный перелет в
системе циклонических ветров и оседание на территории выпадения
осадковявляется великолепным приспособлениемлуговогомотыль-
ка к развитию в условиях обширного ареала, в пределах которого лишь
в некоторых участках — в зоне выпавших дождей — создаются в дан-
ный период условия, экологически благоприятные для развития и
размножения». • <
В 197,8Т. мы наблюдали явление, которое свидетельствует в пользу
миграционных перелетов. Сначала весны до середины лета лугового
мотылька в зоне наблюдений почти не было, хотя предшествующий
102
год характеризовался тремя интенсивными пиками лёта. Но в период
с 25 до30 июля внезапно появилосьболыпое количество летающих
бабочек. В эти дни.шли непрерывныедожди, среднесуточная темпе-
ратура йе превышала 15—16 ?С(мин.ЛД,6 °C, макс. 22 °C).В призем-
номслое ветер был слабым, доминирующее направление от северных
румбов. Бабочки исчезли также неожиданно, как и появились, при
этом заметной их гибели не наблюдалось. Все сказанное.не. исключа-
ет двойственного состава: популяции лугового мотылька в лесостепи.
Одна часть ее, вероятно, местного происхождения, другая можетфор-
мироваться в результатемиграций.Соотношение между ними, види-
мо, непостоянно изавис ит отместныхпогодныхусловий.ичисленно-
сти вида в других зонахареала. г
Ситуация с луговым мотыльком в лесостепной зоне во многом
напоминает таковую с северным экотипом капустной белянки. Про-
слеживаются аналогиикак в экологической характеристике этих ви-
дов (термостабильность ФПР разных популяцийи одинаковый порог
ее у популяций одного экотипа); так и в результатах полевых опытов.
Отличие, заключается в том, чтошараметрьгфПР доследованных по-
пуляций дугового .мотылькащаиболее соответствуютфототермичес-
ким условиям юга Украины и Северного Кавказа, гдесинхронизация
сезонного развития видах ритмом климата оптимальна. К;северу (в
лесостепной зоне) эта синхронизация явно нарушается, и,,в отличие
от капустной белянки, у, мотылька экологический, компромисс воз-
никает в виде ранней диапаузы. бол ее сильно выраженной. чем в степ-
ной зоне. Напрашивается вывод, что именно южные зоны Европейс-
кой части России и Украины могут служить резервациейлугового мо-
тылька и источником его распространения. на север при вспышках
размножения., , ,• : , , < * . ,
6.1.2. Жесткокрылые (колорадский жук)
•История расселения колорадского жука, Leptinotarsadecemlineata
Say, хорошо известна. За какие-нибудь 70 лет этот вид освоил огром-
ную территорию в Европе, и распространениеего еще нельзя считать
завершившимся. Зонально-климатические ^условия Европейского
континента настолько разнообразны, что повсеместное распростра-
нение колорадского жука оказалосьневозможным. «Наметилась север-
ная граница ареала в северо-западной части России; свободны от жука
некоторые районы Южной Европы (в Испании?Италии, Греции). На
ЮЗ
остальнойяастиЕвропььвыделеиы,зонысразным>числомпоколений
L.ideaemlineaia. ^зависимости; отклимата изменяетсячисленность
жукалВозникаетобычнаяструктуравидовогоареала.Считается, что
неосвоенной жуком территории^расселились потомкиодной исход-
нойлопуляциивида,завезеннойв1920г.наатлантическоепобережье
Франции. Лоэтому. колорадский’жукпр ивлекаетвниманиеиссл едо-
вателейжакуникальный объектдля изученияадаптации вйдак раз-
нообразным условиям обитания,*процессоввнутривидовой диффе-
ренциациилтмикроэволюции.го > ц.ок/: a л фитисср: . u snai- ы. цС
’• нУнасекомыхФ потенциальнолюливольтинньшсезоннымцик-
лом,жкоторым10тносяти1колорадскогожука, встречаются фенологи-
ческие схемы разной сложности. Бол еешростыониувидовс четко
разграниченным и>поколениями,.как, например, <у. совки Д rumicis.
Фенологияколорад ского жука одна *из наиболеесложных, с чем со-
гласится каждый специалист, причастный кизучениюотоговида.Это
обусловлено^длительным^периодом-размноженияперезимовавших
жуковти, как следствие,шаимнымналожением вовременипоследова-
тельныхпоколений.Известно,чтомолодыхжуков легко отличить от
старых®» внешним признакам только в первые дни после окрыле-
ния, пока не завершится склеротизация покровов. Структура летней
популяции жука-еще более ’усложняется проя влением разной глуби-
ны диапаузы и-возможностью повторной зимовки имаго {Колорад-
скийкартофельныйжук... 1-981] .Вссэто-сильнозатрудняет сбордос-
товерной информации ъ полевых условиях!, стимулирует разработку
методов,1опособствукицикобъекгивньму|анайизу*фенологии'покоф-
веннымйризнаками<Р'^mow* ц л-югоЧ й.ол
’ ши Как-Известно; колорадскийжуквзависимости от условий среды
может иметь от одного до трех поколений. Принято считать, что чис-
ло поколений у него лимитируется тепловыми ресурсами местности.
Это проявляется вУвеличении числа поколений с севера на юг. Зави-
симостьразвития.этоговидаоттемпердтурыдостаточношсследована,
определены пороги развития и сумма.-эффективных температур для
каждой стадии юнтогенеза.лИ хотя;до£их порхнет единого мнения в
отношениинадежности этихпоказателейуони всочетанииюнекото-
рымипоправками-нашли,применение-для.решения конкретных фе-
нопро гностических задан'1Журавлев,1960,1964а;Сикура,1963;Ара-
по^,-*Богуш,3567^ВольвачД970, ? епг.--ш йцйрнп я»
‘ Обширный •обзоргисследований дан в^монографии «Колорад-
ский картофельный жук, Leptinotarsadecemlineata, Say» [1981] под ре-
104
дакциейР.С. Ушатинской,чтоосвобождаетнасотихдетальногорас-
смотрения. Следует заметить, однако' что’многократность попыток
разработать методику фенологическихрасчетовпоказывает существо-
вание определенныхтрудностей. Обэтом'же свидетельствуют имею-
щиеся несоответствия между расчетнойифактаческой фенологией в
зонах с разным числом поколений: реальное*число поколений часто
меньше, ’чем обеспечивают температурныересурсьТ'местности Все
это' делаетакгуальным углубленное исследован неразвитаяколорадс-
кого жука в разныхзонахетообитания.' *~ ‘ 1 X* ^виутэен
* Нижеприводятсярезулътатыфенологическихнаблюденийипо-
левыхопытов, проведенных взонедвухпоколений(Белгородскаяобл.,
где жук обитает С 1969 г.)и потенциально одного поколения'(Ленин-
градская об л.,"где жукапока нет)?ОпытывприродевоЛсех 'случаях
проводились Одновременно с необходимым лабораторным контро-
лем. Сочетание лабораторных и полевых методов исследования; как
показываетопытработырдругимиобъектами,-наиболее способствует
причинному анализу фенологии. ; - -5 w< ’; м; ? : й 1 <1
' Фенологические’наблюдения'вИБелгородской области.1 Темпера-
турные ресурсы местности для развитияколорадского жука (при по-
роге! 1,5°) по многолетним данным составляют около 800'гр.-дн. /что
согласнорасчетам обеспечивает развитие двух полных поколений. К
зоне двух’ поколений относит Лесостепь и МД'. Злотников'
Известно, что расчетпо СЭТ дает представление лишь о возможном
числе поколений, которое далеко не везде реализуется. Пределом'ак-
тивного развития и размножения поливольтанных видовчасто явля-
ется диапауза, которая уболыпинства видов умеренных'широт фор-
мируется ещевконцелетаилиранней'осеньюприз^ведомо1 благо-
приятных условиях обитания. Кроме-того?,! ‘погодные,условия-конк-
ретных лет' могут существенно отклоняться от нормы ив -холодные
годы дополнительно ограничивать число поколений.
*Фенологическиенаблюденйяза развитием колорадского’жукав
лесостепной ’зоне проводились ’нами в течение ? сезонов’(рис. 45).
Для’удобства сравнения данные по годам распОложены не в хроноло-
гическом порядке, а понисходящим-значениямсредней температу-
ры’за период развитияпреимагинальных стадий!-гопоколения. Как
видноНз рисунка, сроки появления на картофельных посадках пере-
зимовавших жуков варьируютв разные годы от конца апреля досере-
дины’мая. Точных представлений’о причинах, определяющих сроки
выхода, еще не сложилось, а данные литературы весьма противоречи-
105
вы. Одни авторы указывают,чтовыход начинается при прогредепо-
чвыв местах залегания жуков др +5...4-6 °С{Дгрп, Salinger, 1955J,apy-
гие — до,14—14,5-^(Колорадский картофельный жук... 1981].^Цопо
данным, Ц.Ф., Миндери, Э.ф., Козаржевской ,[1966], в Закарпатье ,в
ЗфЦг, выход начался,при дррщере порвьцю 1^15^]С, аХД9б2.д; -
всего до 5.—6ЛС. Вобоихслучаяхон наблюдал сядпервой декаде апре-
ля-По-видимому, противоречия здесьнет.,Цзвестно,что температур-
ный порог двигательнойаюивностижукавесьманизок-оцепенение
наступает лишь при +2 °C и ниже]ЗенякиниКарпунина, 1958J. Лемг
пература +5...+60С в.прчве, видимр,,близкакнижнему пределу, при
котором происходитвьирд жуков вприродных условиях приотсут-
ствии иных тормозящих факторов. Среди последнихследует прежде
всего назвать режим влажности почвы, при дотрром происходит вос-
становление водногобаланса.у перезимовавших, жуков, до активной
ррр^<Ущатинская,^^].)СЬ,Г1. f м >;йчоот:т -xUn м.и1.
,уг,Рассмотримданные,ларактеризующие,гигро^термический рет
жим сравниваемых годов (рис. 45). Температура концаапреля вовсех
случаях (былавыше6.°Си,вероятно, непрепятствовал аподготовке
жуков к выходу Можно заметить, что раньше в.сезоне лоявлялись
жуки в годы с, высокой влажностью и осадками в последней декад?
апреля(1981 и 1979)^В 1983 г. выход был ещеВолееускорен повышен7
цойтемпературой этого периода(+14 °С).Засушливые условия апре.г
ля задерживали выход., Этоособеннозаметнощри сравнении двух сег
зонов(1977и 1979) , сходных л о температурному режиму весеннего
выхода; -в.-1979 г. осадки,выпали на декаду раньше, соответсдвенно ус-
корился и; выходжуков.. Пониженная температуране тольковконце
апреля,, но и в первой декаде мая задержала выход жуков как при за-
сушливой (197$), так и при обеспеченной,влагой весне (.1976,1982)..
Внеожиданнослабойзависимостисрокавыходажуковоттемпе-
ратуры проявляется характерная только для имагинальной диапаузы
особенностьтвозобновление активной жизнедеятельности, не,зави-
сит от,нижнего температурного порога развития. После полного за-
вершения реактивации на дервое)месго1выходятде процессы возоб-
новления, роста и, развития, аокорее поведенческиереакции. обус-
ловленные состоянием внутреннейереды организма(особенно вод-
ного ^адарса), Д'акиефеакдии связаны, негто>кО) с вщ^1ад^до$
сигнальцымдействирмднешних.условий^,. ,. , ,f
..., . Последующиеэтапы фенологического цикла в тораздо большеД
степени определяются тепловыми ресурсами сезона:,;длительность
1Q6
-• UK*
TWV'l
-lilt
WA
-
• Л;,'И F
««К -
!.* iaUit
1Ж5тЖЭЛЦЖ4 18Ягй0.71|<ОМ7.3МСа4Рв.2hi!
>7
102 S3 10S J IM ] 167 S< ,
14.0
116
ii.tYAy
^0
103
нлк«.в. г»д| «.«m-TAW-t I «.1;1М>}АЧ H,lk« 4 t«.» |-i3.3 «А (, .u, |•.>
_____56 225 IM
№
?u
J HE1
101
oo
‘ “ . 135
10.5
103|
л ляг. i
1Гб
100
115
111
'H.HU 0
i ievv.
rjiArilHAW* iAMwi«•* Ж».ФЛАА4М
127
iST?
69
•' ;h0 de
107
135
.‘ '>кмО
; * н -..гАю-ЭЬ «.2-17.2 ДА| WA V-1 .«.*.!. «Л ЯЛ 1S.«4W\<.3 11*1 «А ®Л» 4 -ts<4 •.
190
108
Л9&
136
209
° w Х^)ЕМ0 X30^®-
Июнь Июль Август Сентябрь •
:/ЙС ,Р iXHtWx’jp •';»'''iJwt - jM' п>»'ИЦ'-v
^г3.фДлО;Д8да|^17.Д 15,006
95 ' "' ' 163
лЯ’4>-^1^^!1М.«;0г12,
240
61
..H,{— . . .
j 30 10 20
Май
JK<,
vi. ffl
• -.HLOM,
a
.№ ;
*X|W
w.;,- ^'р^с_‘4^. Феноспектры. характеризующие сезонное развитие коло-tA ^!< *'
<J радскогожукаШБелгородскоОДбл.^разНые’годы: .-ли-1<1 :o ;
, ?. 1г-перезимовашииеимаго;2—яйца;<3—личинюгЗ—жуки1-гопоколения;
’ 5 -жуки2-гопоколения; 1-среднадекаднаятемпература,”С,г-отнрщениех<?ди- , , т
..!) ‘*>1чесп1аосшхов‘ксреднем^'%.ОсталъныепоясненияТ1тексте ‘ .«ъ(...!.• л ..
•пляс; а вахкайЩ'. sei"*:.•><<).>< ->t >г
созревания самок, ‘продолжительность развития преимагинальных
стадийприналичииобилънойкормовойбазыявляютсяс известным
приближениемфункцией’температуры/Поэтомусрокинаступления
этих фаз могут* значительно 'изменяться в зависимости от погодных
усл овийконкретныхлет.Этоподтверждаютинаши наблюдения.На-
чало яйцекладкиперезимовавших самок варьируете разные 'годы от
середины'доконца мая'. Репродуктивнаяактивность самок сохраня-
етсянавысокомуровнедоконцаиюня,послечего'постепенно^атуха-
ет.Полностьюяйцекладкавприродепрекращается, примерно,к концу
107
августа, но на закл^дителвном этаперазмножения д ней участвуют не
только .перезимовавшие, но и молодыежуки летнего- поколения. В
соответствии с изменением-начала яйцекладкименяются в разные
годы наблюдений исрокипоявленияличинок.В годы с болеетеплым
летом (рис.45) личинки появлялисывпоследней декаде мая,-вхолод-
ные сезоны примернонатри недели позже. Вцелом. период, в тече-
ние котороговстречаются преимагинальные стадиихукавтоле, очень
растянут, так как отрождение личинокпроисходит по мере откладки
яиц перезимовавшими самками.Вдальнейшемсозревает часть моло-
дых самок,Дто «продлевает период яйцекладки ,п<У крайней1 мере до
серединыиликонцаавгуста.-Поэтому ддительностьиндивидуально-
го развитияютдельных стадий в полевых условиях.установить трудно.
Окрыление жуков <1 -го поколения (показатель, интегрирующий
развитие особей предшествующих.зтапов|онтогенеза) в более теплые
1979 и 1981 гг. наступало, уже в первой'декаде''июля‘(рис. 45). Лето
1983 г. было несколько'прохладнее. но очень теплый конец апреля
(среднедекадная1гемпература14,0 “С) ускорил,выход перезимовавших
жуков и1темха)иым обеспечил развитие летней Генерации. В более
холодные годыЯ982,1976,41977 июсобенно'1978 жуки1-го поколения
появились на поле лишыв’концеиюля^знанале августа, т.е. тиа месяц
позже, чемв.теплые годы, Во всех случаях выход .молодых жуков был
растянут в сезоне (в соответствии с продолжительностью яйцекладки
перезимовавших самок), что трудно отразить на феноспектрах из-за
наложения разных фенологических фаз. ' ‘ г
Описанные изменения фенологии жуков в разные годы в боль-
шой мере обусловлены прямым действием температуры, но этот фак-
тор оказывает и косвенное влияние насезонное развитие.их популя-
ций. Особенно важны последствия' изменений календарных сроков
появления жуков 1-й генерации.Какоказалось, созревают и дают по-
томство только самые ранние жуки из числа отродившихся в конце
июня-начале.июля. При более поздних сроках отрождения вероят-
ность созревания самок быстро уменьшается,.и среди окрылившихся
после 20.ИЮЛЯ приступают к яццекладке лишь единичные особи. Э,то
определяет,саму возможность появления 2-й генерации.^Молодые
яйдекл^дущие самки в годы наблюдений встречались в каждом сезо-
ну (зашсключением дета! 97,6 р.). До даже,в теплые годы, их числен-
ность и тем более вклад цобший репродуктивный потенциалшопуля-
цииневелики, ав холодные годы ничтожно малы.,Количество созре-;
вающиксямокрпределяется, фр^о^рмичестамиурдрвидми}в крто-
1ЭД
рые они попадают. В результате сроки окрыленияжуков 1-го поколе-
ния непосредственно влияют на судьбу 2.-го. Имаго 2-го поколения
появлялись лишь в наиболее теплые-годы, их окрыление в. среднем
приурочено к последней декаде августа (198Д,1979,1983 гг.). Единич-
ные особи успели'завершить развитие и вхолодном 1982 е — они по-
явились в'серединеюктя^ря.Костальные сезоны особи 2-й генерации
не достигли имагинальной стадиии погибли.....
Косвенное'влияние температуры на популяцию колорадского
жука осуществляется также^герез пищевой фактор. В районе исследо-
ваний фаза увядания ботвы картофеля наступает в среднем 12—20 ав-
густа. Жаркое исухое лето сдвигает ее на еще более ранний период. В
связи с этим окрылившиеся жуки 1-й генерации и тем более жуки,2-й
генерации оказываются необеспеченными полноценным питанием
и попадают в критические условия. Они не могут нормально подгото-
виться к диапаузе, поэтому возможность перезимовки: Этаких жуков
весьма сомнительна: значительная их часть погибаетеще осенью. В
общем балансе сезонной популяции колорадского жука численность
2-го поколения, даже в наиболее благоприятные годы, невелика, и
оно не имеет хозяйственного значения в лесостепной зоне,.
ПолевыеопытывБелгородскойобласти. Эта-исследования дол-
жны былидополнить фенологическую картину развитая колорадско-
го жука в лесостепной зонеточными данными о жизнедеятельности
каждой фракции, образующей!сезонную структуру популяции? Пред-
стояло выяснить продолжительность яйцекладам перезимовавших
жуков иихсудьбув конце сезона; способность к размножениюжуков
1-го поколениям сроки ухода на зимовку;исследоватьпотенциаль-
ную возможность развития 2-го поколениям оценить реальные усло-
вия егосуществования в природе в связи с, состоянием растений карп
ТОфеЛЯ, {<- ,, '«И::’”!' К.' чМЪ'ь < 5Т.У ’СЙ • ЙЙ Ч
« Для изучения сезонных изменений репродуктивной активности
самок (как перезимовавших, так и молодых) летом 1982г 1983 гг., да-
чиная с ! июня, проводился огбор проб из природы,(по 20,самок), в
котордгхшутем вскрытия определяли количество зрелых особей. Ее-
зультаты эт^тх учетов отражает кривая а (рис. ,46)., Кда можно видеть,
пробьцвзятые в,периоде 1 июня,цо 10июля, додержали толькозрелых
самок, причем все они принадлежали к перезимовавшей фрдкции .
Известно, что,реактивированные в ходе зимовки жуки фотопериоди-
чески нейтральны и размножаются даже в условиях короткого дня. В
дальнейшем их фотопериодическая чувствительность восстанавлива-
109
i; пИ0№>£* suou ИОЯЬ'-ХГ luH^BiycT'ьЬ'Г Сентябрь cj gmm;)?»;
г.,,- в w '.в '?(« 0Ж <„ м.ч _ . 7|JMall; '.Ю
4 Рис.46. Репродуктивная активность сформирование диапаузы, у хо-)п О1 ,
лорадского жука в Белгородской обл. (суммарные данные полевых
<' ” ’-•* 'v.-i v;. О ivr-ridt* V-нч kiuл >. j 'v jm i.L-
опытов 1>о2и lyoJiT.):
i-.OXi < >- г;в ;а*-жоличество,%,'зрелых!самок»в!пробахтаТфйроды"(даты>сборапро&^
* 'X ;1;8°™еченыт°чками);б—изменениеколичества,%,даапаузиру1ощихжуковвсеэон-;< (
ныхвариангах опыгов(точки приуроченыксреднимдатамуходаназимовкувдан-
МТ) ьлномвариа1пе;чернь[е<значки-<и1ьггы;1982,светлые-1983т);в|-смертостьжуков' ’’ 1
?;*:>.'jПВразныхвариаигах;г-сеэонж>еизм»ентедаины^мсуч^м1ражш1нских<умег,,,
рек; остальные пояснения в тексте *
ется/что делаетвозможнойиндукциюповторной диапаузыизимов-
ки {Колорадский картофельный жук.. ,«Т9И] /Однако^ольшая часть
перезимовавших’жуков позавершении периода размножения поги-
бает. В нашихюпытах 1"983т.средижуков, собранных!? и 30июня’(29
и 26 пар соответственно) и содержавшихся в полевых садках добсен-
тября, * погибли После завершения яйцекладки 81 %* самок, ушли на
повторную зимовку 19%самок.'у' -А;ь-
! •* В’пробах,1 взятых после 20-июля (рис. 46;‘кривая’а)1, количество
зрелыхсамок снижал ось/Такоеизменеиие входе кривой, несомнен-
но/связано с появлением ’примеси молодых жуков/среди которых
большаянасть’недозревала,сразуформируя диапаузу.'После20*авгу-
ста яйцекладка'в природе пре кратилас ь, В п робах встречались тблько
диапаузирующиежуки.Этоозначает,чтоперезимовавшиесамкиза-
вершиЯи размножение, погибли или *ушли на повторную зимС®ку,:а
молодые самки появлялись исходно'детерминированные надиапау-
1Ф0
зу.‘Справедливость такогозамечания подтверждается «последующих
опытах. •' •* V К. ЛМ/ЛДЛЮ ЗЛ1.-Я (S, ' х.ТМЬО у.
Оценкавозможности созревания иопределение сроковформи-
рования диапаузыулетнихжуковпроводилисьдвумя способами,,ко-
торыеотличались в деталях,:но дали сравнимые результаты, Вотпер-
вых,проводилисьопытыпопринятойунасметодикерегулярнойпро-
верки возможности индукциидиапаузы у тестируемыхобъектов(см.,
гл. 5).Дляэтого вусловиях жалюзийного домика проводилась выкор-
мкаличинокразныхсроковотрождения(варианты)ирегистрирова-
лосьфизиологическоесостояниемолодых’жуков^созревание самок
или наступление имагинальной диапаузы). Вэтих опытах насекомых
с одержал ипообщепринятой.методике{ Winning, 1951 },что.обеспе-
чивало сравнимость результатов одновременных лабораторныхипо-
левых исследований.При такойметодике создавалась возможность
точно учитывать сроки .завершения отдельныхстадийонтогенеза в
условиях, близкихклриродным.Во-вторых,регулярно.(каждые 10—
15 дней) в природе собирали взрослых личинок (за 1—2 дня до ухода в
почву)/которых докармливали в жалюзийномдомике.Благодаря та-
койметодикеусловияразвития насекомых еще болееприближались
к-естественным.'Дальнейшие наблюдения за жуками и улет данных
былитакими‘же,‘какш’предыдущйЯ5случае.л< /. л. «гл ’ з. j-' •.»< г,г:
?; в В ходе полевыхопытов получены данные о длительности разви-
тия пре имагинальных стадий колорадского жука в разные периоды
сезона/о смертности в ходе развитиям и некоторые другие.. Коснемся
лишьформированиязимующегозашса^даоявляетсяосновнымизат
ключительнымэтапомсезонногоцикла.Нарис.46вариантыопыт1ов
представлены горизонтальными отрезками, начало которых отмечает
среднюю дату отрождения имаго, аконец - средний срок завершения
преддиапаузногопитания иуход назимовку. Средние срокипоявде-
ния жуков во всех 17 вариантах для удобствасравнения выравнены по
произвольной наклонной прямой, идущей сверху вниз и направо. В
тех вариантах, где опыт начинался с момента отрождениядичинок,
штриховая часть горизонтальной прямой отмечает суммарную про-?
должительность развития личинок и куколок. В вариантах1 и 2, в
которых происходило созревание части самок, кружкамиютмечение-
риод.яйцекладки, еепрекращениеи продолжительность преддиапа-
узного питания. Средние срокиуходавпочву размножавшихсяжуков
•показаны направленными вниз стрелками на концах прямых отрез-
ков. Полученные данные показывают,.что физиологическое состоя-
ла
ниежуковвсезоне закономерно изменяется.Всамыхранних вариан-
тах опыта (1,2) часть самок (47-78%) созревала и приступала кяйце-
кладке,; которая продолжалась до 10—ЬУавгуста.Окрыление жуков в
этих вариантах.происходило при максимальной длине дня (рис. 46,
кривая г), что и определяло'возможность созревания части самок. В:
наших лабораторныхопытахпри‘17оСи18чсветавсуткиразвивалось
активнохжоло20%самок. Последующее сокращение длины дня*в;
природе, однако, вызывалорезорбциюооцитовиформированиедиа-
паузы.Другаячастьсамок(52—22%),нрприступаяк яйцекладке, ухо-
дилав почву назимовку. В более поздних.'вариантах опытавыходжу-
ковпроисходил на фонееще более‘коротко го. дня, поэтому число акг
тивных самок вних быстро уменьшалось. <Так, ® вариантах 3—5 число
их не превышало 10—17%, аввариантахб-Пдиапаузудружноформи-
ровало подавляющее.большинствоособей (кривая-6). Как показали
лабораторныеисследования, причины возникновениядиапаузы мо-
гут быть различными: участи особей диапауза индуцируется подвли-
яниемкороткого дня^у других онавозникаетспонтанно, независимо
от фотопериода. Преддиапаузное .питание в .разных вариантах поле-
выхопытовпродолжалосьв среднем 21 день.Вболеепоздниесроки
условия подготовкикзиме ухудшались, ичисло жуков, неспособных
формировать диапаузу, увеличивалось..Оченьдемонстративно;после-
дующееуменьшение количествадиапаузирующих восеннихвариан-
тах,шричемальтернативойдиапаузездесь выступаетнееостояние фи-
зиологической активности, атибельвсех особей, неспособных подго-
товиться к> зимовке (в). Лимитирующими факторами, несомненно,
были осеннее понижение температуры и, особенно, ухудшение усло-
вий питания. '< Г Г В . s «’Ski. Чйл ' j
Результаты, описанные выше,полученывусловияхжалюзийно-
го-домика, которыепредставляютсядостаточно близкими* природ-
ным. Вклиматологиивтаких условиях проводятся основные измере-
ниятемпературы ивлажностивоздуха:Различие>заключаетсялишь в
том, чтожалюзи»предохраняют от прямой солнечной радиации, спо-
собной привести кперегревусосудов с насекомыми из-за парниково-
го эффекта, Для того, чтобы дополнительно проверить сходство усло-
вий обитания насекомых в жалюзийном домике и в природе, был прог
веденэдебольшой1эксперимент ‘непосредственно ца картофельном
поле. Личинки и жуки питалисьна,растущих кустах картофеля, на-
крытыхсадками из капроновой сетки. Садки заглублялись воточву на
20 см.ЯТри снятии опыта почва была просеяна. Сравнивая результаты
112
полевыхопытоввсадкахижалюзийном домике (табл.9),можноубе-
диться в их сходстве. Число диапаузирующих среди жуков близких
сроков'отрождения отличалось мало.' Различия касались лишь про-
должительности преддиапаузной подготовки. На картофельном поле
уход жуков в почву происходил примерно на 15-й день,.что всреднем
на 6 дней раныпе, чем в условиях жалюзийного домика/ Возможно,
это связано с поведенческой реакцией поиска подходящего для зака-
пываниясубстрата, который в домике (чистый песок) отличался от
почвы на картофельном поле. Увеличение к концу сезона количества
молодыхжуков, неспособных сформировать диапаузу,также проявля-
лось сходцым образом. Как и в жалюзийном домике, оно ерпровож-
далось параллельным возрастанием смертности. Все это позволяет
считать опыты в полевых садках и в жалюзийном домике методически
сравнимыми. <1 -
Опыты с искусственно перемещенными популяциями. Сравнитель-
но-экологическое изучение географических популяций колорадско-
го жука было бы неполным без полевых опытов с популяциями, ис-
кусственноперемещенными виные условия. Параллельноегнаблюде-
ние заразвитием местной иперемещенныхпопуляцийвприродных
условиях нередко оказывается болееинформативным, чем проведен-
ное в лабораторных экспериментах. Познавательная ценность пале-
вых опытов была продемонстрирована выше. Большая чувствитель-
ность полевых опытов обусловлена, не только динамикой внешних
факторов, но и более дробным дозированием длины дня в разных ва-
риантах, чем в лаборатории. ' 1
В Белгородскойобл.(рис. 47) опыты проводилисьв1982и 1983 гг.
с популяциями из Самарской обл., Краснодарского края (у. Кропот-
кин, 45 °с.ш.) и Азербайджана (г. Хачмас, 41 °с.ш.). '
Чтобы освободить график от>второстепенной информации, на
нем оставлены только'точки, характеризующие срок ухода жуков в
почву и количество диапаузирующих самок в каждом варианте опыта.
Для оценки реакции перемещенных популяцийнаусловия лесостеп-
ной зоны в качестве фоновой на рис. 47 приведена кривая 2, перене-
сенная с предыдущего рисунка и показывающая сезонное изменение
количества диапаузирующих особей белгородской популяции. Мож-
но видеть, чтр существенных отличий в сезонном развитии местной и
перемещенных популяций нет. Сроки развития преимагинальных
стадий» сходные, что указывает на одинаковые требования всех попу-
ляций к температурным условиям^ В самых раннихсероцных вариан-
113
Таблица,?.(Формирование диапаузы-у^жуковразныхсроков окрыдениру,;
way* УМ Я Гэте-
• /4 Дата выхода z u'iji Mot.&.i’q’ ms куколок ИЗ'АД' »Л Условия л j Кол-во- Й! Количество жуков, % ЖКр
..„не.ушедших,.,,. |1; в почву др' ' ‘ " 23.09 v ;х;у
,'?'! ••••: • £4 КГ R fi." T'' •.,'" Я,1‘ ЙЯ'ГЗЭРТЛ Ч1 » ; S' 17-18.08 -«'ГкЯ.Гл'1'VJ С;Ж.ЯГГУЕ -^80^28^.0; ттотпнгсф G'f<: crif ;:х-‘3’’?31.08-02.09^ •о., г,- г,адоб-£8.О9'« -0>> 'EtjilKO йцгп -ЭН -ПХКННЙР/! О( »л\ (Tjl '1«Ж УЧ! ;К; П VTfHl >i X' гкнзу <!?• 'HMOif ЗГ ‘г! НТф.’ГЩ ЗЕ.М^ГЙМ’ 2 HRSVW' •» I- О’ я ч.ЗО /р- -з б5; ,, 1 эй^-пй. г«?Г утэе' ftpr'tgg.sx 5М2 '•"•Cgg-'J !i 29 ягаездо?* .'- МП^Сф ума? h.cn ranf?.'183.8 чу .а sq U.W’Wm R 'T* 80,7 ’°5" TB100 0i'l'JS'!”'' Я H rr-.gg^OtSQ 5ТЭ-:фФ_’ МИГ-d ПГ.'ЛГ-73^1‘Ц«)Ч 48*3 »ф?1 гявТ^Гй-^ у.!’;' *>'?•! 47Лх-'i ?< ; яб Mfc'W/.i’-';" хтжЯЬед б fcs cnS^KHHidHd г 0'>'fй!Т. тг WFGsl 19 3 .. ‘ * 1. - X 'f.Q' X -1 i \! 7 '0.01 Z М«.у7У..> ' ЙЭО!<. ЧГ‘.вСьИ ЯЗ'МЧ'Г S id ‘"2g ^<il ИТУР-.; .bSH^7M~'‘W.jp 'а v 38,а >О хэ^ёЭДАноле •'.>!; ад окга-з яФ’к <т>
тахопытов,1какяуместнойлопуляции, часть:самоксозревает^46,5%
в-1-м:и34в0% во2-м) и приступает к яйцекладке. В дальнейшем часть
из1нихпогибла,'другаяу1илавдиапаузу.Впоследующихвариайтах<3^
li’i.T- а 1»0Я
, хиа
’Ье'ЙОИ а «ЭОН
Тб CJ
• • ’. X:,-
' 34 Д
пглм <г~х;нна!.’
-гп.5ТЕ' ‘ nay? w»
яшш'шы lio-лмг
«к и нн?
'ВН?Л0гИ):м.оП .X.’irHSV'jq^TjMex.KHQ.'.' '’Г
—HUSSfiqs^rSBor-Aioq-r rr>»-,1ew
r'”WK?tvmk' ‘WArm г
^docm ‘гш.с$«
»►
, 1|Г1
ЭТ ' fti- i К £-£: н
-п л J8 НИ-
яшаЛ»
-а пчоа :)(?/
g 60-
Я .SlUlbT^'T',.
. а•«_.к s.^y*>:
.^яюэт’фг-зд.
-R-ffftifi
T-fOGHtf»
7ЯГХ
I 1 г- , л
ь'гмр.кг,
я
А .4 &OUH-..
И txp >!' '
::::V ПОТР"
MS: :
’'•Н1О.4 Ч'^ЯГОГ.
кгЛ
M>M .ВДиЭТф!ЗД йЛ$№зф /a‘f'^P№aoXN'^S<J?|^/fin^W,bHT^0F4n..ra
t; ‘;' !,'/>ис.47',ре3улЬтаты'Полевых опытов сперемещенными'популяция-1''1'
ШИЛIi ЗмйЗкалбршюкого^жука’Ъ Белгородской' обл.V! ПОД>w'• м яi fa (М эf"*Х}ап
- fЮГ :-1— сезонноеизыенениед)ииыдая;2-изменениеколтесгвадиа11аузиру- i -аю
Д14
7)-!зрелых -самок было не более 8%.Наиболееважноотметить,что
точки; характеризующиеНроцент диапаузирующих особей и срок их
уходана зимовку,'расположилисьблизко'Ккривойдлябелгородской
популяЦищполученнойвтежесезоны.Этослужитдополнительным
подтверждением-ранеесделанноговывода осл абойизменчивости
нормФПР колорадского жука 'впределахвосточно-европейского
ареала. • '-ид
^офенологические'наблюденияиопыты.вьшолненныевлесостеп-
ной зоне.позволяют глубже понятьпоследовательность'фенологи-
ческихсобытийвсезонном«цикле колорадского жукаидатьимпри-
чиннос объяснение наосновелабораторныхисследований.Получе-
но'новоеподтверждение того, что весенний выход жуков из почвы
определяетсяне толькотемпературнымиусловиями/ноижомплек-
сомфакторов, Средикоторых ведущуюфольиграютгигротермические
условияпредшествующеговыходупериода?При этом Пониженные
температуры идефицит влаги задерживаютпоявлениеЖуковипере-
ход ихкактивной жизнедеятельности. ДвигательнаяНктивностъжу-
ковимеетвесьма-низкийтемнературныЙ11редел,-близкийк2'оС|Зе-
някин и-Карпунина, 1958] . Таким образом ,выходжуков*неСвязанс
нижним температурным порогом’активногоразвития,равным 11,5 °C.1
Созревание самок,* Ио-видимому,-первое фенологическоесобытие,
непосредственно связанноес интенсивным* биосинтезомитребую-
щеевышепороговыхтемператур.Сэтого'моментавсепоследующее-
развитиежука'становится функциейтемпературы’и регламентирует-
сятепловымиресурсамиместности.Конечно,натемпературную-за-
висимость модифицирующее^влияние 'оказывают другие 'факгорй'
средыиПрймейительноккалорадскомужукутакоеНлйянйемогутока-’
зывать как'погодныеусловия.'такифакторы антропогенногопроис-1
хождения<агротехника и сортовой состав кормовыхрастений; хими-
ческая*обработкан’др'.)ц г- t-'skk.'•ц-да, •. лента ,уи.< .* .«-а-г
’Как показалирезультаты пол евыхопытов, внриродныхусловиях
существует оптимальный для подготовки к диапаузе период сезона, -
когда онауспешнозавершаетсяувсехмолодыхжуков. Влесостепной
зоне онприходитеяна'Конец июля—август.Молодые жуки,отрожда-
ющиеся позднее (независимоют-их генерации), не обеспечены на-
дежнойлшщевой базой, что всочетанииспониженнойтемпературой
приводиткихмассовойтибели осенью'илив’ходезимовки. Наиболее
ранниев<сезонежуки,которые могут появлятьсявтеплые потемпера-
зуриыму<щовиямпдаужов|Началещк)Ля,-оказыйакисянеоднород-
11$
ными по физиологическомусостоянию. Часть, из. них/обычно
большая) спонтанно уходит в,диапаузу.Как,показали исследования
Р.С,Ушатинской с сотрудниками [Колорадскийкартофельный жук,..
1981,], именноу раннихжуков диапауза наиболее глубокая, и встреча-
ются даже.случаи суперпаузы. Вместестемчастьособей,сохраняю-
щихфото периодическую чувствительность, црд влиянием длинного
дня созревают и приступают к размножению.
. Как и предполагалось на основании лабораторных исследова-
ний, фотопери одическаяреакция у белгородской популяции жукане
имеет большого экологического значения. Летний, длиннодневный
эффект проявляется в созревании части наиболее ранних жуков, при-
чем количество созревших самок при максимальной природной длин
не дня.(около18 ч) может достигать 78%. Ноуже средиокрылившихся
15-20июлятаких самокединицы.Таким образом, вивдукциирепро-.
дуктиццой активности,ФПР проявляется слабо. Тоже можно сказать в
отношении.индукции диапаузы:, даже приповышенной температуре
-28 °С ,и,оптимальной; длине дня, —18 ч, в лаборатории спонтанно
диапаузировало более половинысамрк. Влриродныхусловиях ихдоля
заметно возрастает, но сама возможность фотопериодической индук-
ции,репродуктивной активности ограничена периодом смаксималь-
ной длиной дня, в,который попадают самые ранние летние жуки. Это
накладывает отпечаток на фенологию. Районнаших исследований
относят к зоне двух генераций жука (Злотников, 1967]. По средним
многолетним метеоданным,, здесь , действительно .набирается СЭТ,
достаточнаядлядвух онтогенетических циклов. Однако спонтанное
возникновениедиапаузыуподавляющегочислажуков1-го поколе-
нияприводит к пре обладанию впопуляциимоновольтинного разви-
тия. Даже вгоды, когда темцературапревышает многолетнюю норму
тепловые ресурсы лесостепной зоны практически не используются. В
связи с этим, вероятно, заслуживают критического пересмотра сло-
жившиеся црсдставления о числе генераций жука вразныхнастях его
арЗДШ.- • .' кДЬЦГ. Я ь ИЧСТПШСП •• по сгг.-г.-г-
= Исследования выявили роль каждой фракции колорадского жука'
в динамике его численности в лесостепной зоне. В размножении уча-
ствует главным образом перезимовавшая фракция, 1 -ялетняя генера-
ция .умеренно, а.2-я генерация (когдаонаесть) не участвуетвовсе.
Жуки 1-й генерации играют ведущую роль»в формировании зимую-
щего запаса. Жуки2-й генерации едва ли влияют нанисленностьвида
в следуюшем сезоне; у них наивысшая смертн ость как восенний пе-
116
риод, так и во время зимовки. На доминирующуюрольй-й генерации
в разных климатических зонах обитания указывают и другие авторы
[Миндер, Козаржевская, 1966; Макеев, 1982]. По-видимому, это об-
щая тенденция в адаптации жука к новым условиям обитания, и ее
следует учитывать'при разработке защитных мероприятий против вре-
дителя. f ,............... ,
Полевые опыты в Ленинградской области. Как отмечалось выше,
М.Д. Злотников [19671да основании анализа СЭТ выделил на терри-
тории Европейской части России четыре зоны, различающиеся по
числу поколений/Ленинградская'обл., по его мнению,, относится к
северной зоне, гденекаждый год возможно нормальное развитие даже
одной генерации жука. В остальных зонах колорадский жук может
развиваться в Т—3-х'Поколениях. По мнению В.А.Санина [1976], Ле-
нинградская обл.относитсякзоне снеблагоприятными условиями
для развития вида.Полный цикл здесьне завершается в95% случаев,
и к следующему году увеличения численности жука1 не происходит.
Действительно, отдельные очаги размножения жука отмечаются только
в, южных районах области. В связи с этимбылоинтереснр оценить
реальные возможности обитания колорадскогожука в этих климати-
ческих условиях. Работа выполнялась по специальному разрешению
Пограничной государственной службы по карантину вредных видов
растений и животных.' . icr. ii;a >
Опытывполевых условиях Л енинградскойобл.были начаты вес-
ной 1982 г. и проводились в течение , двух календарных лет (1982 и
1983) с популяциями жука из, Самары, Белгорода, Симферополя^
Краснодара и Уральска.(рис. 48). Горизонтальными отрезками пока-
заны сроки развития отдельныхстадий жука, их пунктирное, продол-
жение означает, ,что не. все жуки,ушли на зимовку до конца октября,
Кривая отражает динамику формирования_диапаузы, точки на кри-
вой - окрыление молодыхжуковдразных вариантах опыта. Развитие
жуков .изученных попуЛяций шло, синхронно, поэтому 4 на, рисунке
обозначены сроки развития только одной (самарской) популяции.
Несмотря на то, что летние сезоны 1982 и 1983 гг.очень сильно
различались,,в развитии колорадского жука в эти годы, отмечались
общие тенденции. Оказалось, что,завегетационныйсезон;видв усло-
виях Ленинградской обл. может давать одно поколение,длязаверше-
ния которого, как указановыше,необходимооколо400.гр.-дн. Благо-
получное развитие особей этого единственного поколения оказалось
возможным.лишь в тех вариантах, где отрождение личинок происхо-
1117
O'i it* ... I I J.,
К ТЫЫТМШЯ >. ItNrtfaTKft / SrtOP Я*Й?<м*е!’’"^5Л XJH
ДОйМИХГЖ,,.., . .... , ..• ~ ' «№ г^белгародскай-’-'" <‘W
ЯГ-чЗо huhoh Я йд/л MiitUioBKpacHOflspcicenb'iij/Ht
>ф- .' . . -
£ 100г шяенси •
-«<!?© <4. t Ы!Ю
.g »go-n-Ш." "P'y ,-?«. .. .. x ,_____._чнят
CL • A \ i
IdllvV ‘Л|ИЙР'Н,'1.адО-. HYXi ад>.,.'",ч,„аос,йЗ
>fgX>,2e-’'>:'.,OHM ‘./JO Os! •.!!!.!" !!'"!..
• , , ............... .......... /\^oao. -‘ i’S!>-
>T ftp.i W MW»
хгЖ-глг <Pac.4& Результатьыюлевых опытов '«'разными теографическими'по-
7пуляциями'колорадского жука в Ленишрадскойобл--
ГШ£С- ча<У|
дило^до4,йюля<вар‘М?—3^1982*т.*/‘вар?1'-4>в*11!983'г.'):1Вболеё’п6здних-
вариантахопытамолодые*жуки,неЬ'спевали'закончитьразвитие,и
чемпозже'происходило-ихкжрыление^ем'меньшемолодыхжуков
формировалидиапаузуи!уходили!«а!зимовку.В1техвариантах,лгде
личинки'отрожд ались'после'25июля,гони*неуспевали окуклитьсяи
погибали с наступлением заморозков. .Хкппочнж н w мя ' ;
г Ул Огличителыийособенностьсезонного'развитаяжукайболеетеп-
лыхусловиях1983г.<табл. 10)\заключаласьвтом,'чтовсамыхранних
вариантах/! ’и 2) частьсамок!(до-23%);приступалакяйцекладке.-
- о Внаших лабораторныхопытах при длине дня Пч итемпературе
20 °C’диапаузировало;от 85/цо100%Ъкуков. В‘Ленинградской обл.
фотопериодуменьшается'до'тако^величиныксерединеавгустапри-
значительно более низкой температуре. Соответственно только сре-
ди жуков, окрылившихсядо;1'5августа,отмечал ис ьактивныеособи.В
болеелгозднихвариантахокрылениетфоисходилов условиях* опреде-
ляющих’диапаузу у-всех особей. В целом результаты полевых Ьйытов’
двух вегетационных'сезонов позволяют сделать вывод,'что летние ус-
ловия’в'Ленин граде кой Ъбл!. нелимитируютразвитие’колорадского
жука.Тепловые'ресурсыобеспечиваютзавершениеодногопоколения,
жукикоторогоуспевают сформироватьдиапаузу и'своевременно ухо-1
дятназимовку.Синхронное развитие особейразныхпопуляцийсви-
детельствуето виДовом'ранге температурных норм развития,'а'одина-
коваятенденциякформированиюдиапаузыподчеркиваетярковыра-
ой у Габлицаг^.О.;; Цакопдение эффективных демлерахурЛвыше 5 °С)‘ ;, , ?
Дата Средняя ...многолетняя ..... .'ТГ 1983 г. 1982 г. .’.наши I i i ЧП* s.rr Отклонение СЭТ
(f м^оголет- , ' ней „^веаг..,,
pOi'H
НЦОТ
в Vk376riu^fJ'i'}'ii:76!<4*'v4s'7<+175 а>*т М1н.200,!ад!',:''11
л;.')438пш
636
АЧйкубд^
i^goi'isr
-3X05i‘T
Г-Л>. 10.06™ -
.. . . 20.06 r
‘ " 30.06
ьр.,“. <300709'
.--да> 31.07;; 1
,..,/10.08,, ..
<ил' i- ‘
20.08
• : 31.08'
*> /. ‘410.09т’
iL«.'Bf20»09 M;
on no
лэаттЧбЗпЛ f/'I
,.:М'«290ч дд
>7 ’
' 505
#'Пс.623М^'
<*ut7S0WH£
Лчнвввцн 1
js о
. 11118'
<) ^225 "1-
ГЛ1Н302'’"-';
‘f,c-fi.1359, ц.-I
. .1397 .,
toa<£S1 ./ ?
«
382
S’; 642-
£tQ33.qv'j t>qy756ii' p
980
+ «1'077 n‘
.й-пнорда
г к 1г193ь(.
. .1177.
' *4
1282
,;;:!-1336Е!!’
. '1524 г
. 1S69
.. >ir,^148 !Ч} • 5 -Л187Г Т Hrif,
+131 +254 >
’ M',+1i46^< ':,'w+278’'
fi! -И51+' — +2501ЛСХ£>:
Ч; . +,145'>rfrt'-^?+277««rfgu
.•.Якп .. *3®9xrV > м
. +164, .......+302 .„I,
'-+t4iT "',L+289'TJ,I"''‘l
<-.a*H44F' >н*.’ИЦ®1в i; ’':1,! <
ОД11+:165оон.+цгй331>-. :с.л
Л»
* По'Агрбметеорологическому Ьюллетежо по Ленинградской обл. за 1983 г.
’И"х?нтгрмо НЫНРЯОЙ'Ч+Хй» япи ,
АЗ Нх’р. .Х'цисля ОХ .т>теиг • /
женный характер перехода к моновольтинизму, который свойствен:,
видимо, всем исследованным популяциям. — 5:
v; Следующий узловой этап сезонного цикла насекомых — зимов-
ка. Колорадский жукзимует впочве достаточно глубоко и, как прави-
ло/не подвергается'действиюлетальных отрицательных температур.
По мнению.|В. Венгорека41958}/для жуков во время зимовки губи-
тельны не отрицательные зимние, а положительные весенние темпе-
ратуры, которысйлимитируютразвитиежука: стимулируя‘их-про-
буждение, эти температуры оказываются недостаточнавысоки, что-
быжукимоглипокинутьТючву.Значительнуюгибельжуков/зймую-
щих.наглубине. до'10 см, вызывают резкиежолебаниятемпературы,:
попеременное Юттаивание’и'замерзание слоев почвы!,>гдеч>нишахо^
дятся«(Журавлев,4964б}ЛУчитываяэти^рме.ЩМ,^н6внр91рню^ние.
былоуделено-нестолькоНсследованиюсостоянияжуков'Вовремязи-
мовки,сколькооценкеихвыживаемости после зимовки и способное-
тиНфЛзмножению.^ - ;S 'Лзг.оа
а хнаОпыты Проводились ’в течение щвух зимних сезонов:1 И 1982—
1983пг.боле&подробноив1983—1984гг. Жуки, йспользованныев этих
опытах,имелидвоякое'происхождение.-Одна.часть—!это;диапаузйру-
ющиежуки из полевых опытов, другая—жуки*, собранные в природе в
разных областях'»' период преддиапаузного питания, привезенные в
119
С.-Петербургидокормленные® лабораторных условияхдо их ухода
на зимовку. -Жуки были взвешены, разделены по массе на классы и
небольшими дартиями помещены в пластиковые чашки Петри, ко-
торые были закопаны в почву на глубину 25—30 см на эксперимен-
тальном участке лаборатории энтомологии в Старом Петергофе. Ка-
рантинные ограничения не позволиливоспользоваться другими ме-
тодами'для исследования возможности зимовки,' но условия Опыта
были максимально приближены к естественным. Опытный материал
находился в почве до 10 мая, после чего жуки были перенесены в кли-
матизированную камеру с температурой 25 °C и длиной дня 18 ч света
в сутки. В этих условиях они содержались до конца августа. . После
начала яйцекладки жуков рассаживали попарно для более точного
учета их физиологического состояния^, процента яйцекладущих са-
мок, длительности яйцекладки и т.д. Полученные кладки помещали
в природные условия, где прослеживалась дальнейшая судьба отрож-
.дающихсяизнихдичинок^!
При исследовании смертности
самок колорадского жука во время
100,г₽ , зимовки была показана еедависи-
мость от; исходной,массы имаго
§ ч<(рис..49>.|Массажуков’завремязи-
<я, мовки измениласьнезначительно,
максимальноеладениеее непревы-
'.шалоКоличество погибших
I особей было больше среди более
о" .( я легких жуков и составило около
, ,, , .,48%>,vx’yл-:?;’ п ,2«<j . “.'(i-
..J^acc^caMqci» классам, мг . По литературным данным, раз-
Рис.49. 3ависимость’смертностн жу-’ ? меры смертности жуковвовремя
ков ог их .исходной массыз у.' зимовки,-варьируют»в широких преп
1 — средняя массажуков в каждом клас-
се в начале диапаузы*, 2 —то же после зимовки;
3 -процентсмертнОсии для каждогоЗсласса Г
,4 делах, Вшависимости.отклимати-
ческой зоны и погодных условий
г . количество погибших особей может
колебаться от 10 до 90 % (Венгорек,
1958; Ковальская, 1958] . В Закарпатье количествожуков,;погибших в
разные годы и в порвах разных типов, колебалось от 13до 87% [Уша-
тинская, Козаржевская, 1962; Мивдер, Козаржевская,.1966].
В наших опытах смертность жуков в различных пробах изменя-
ласьотЗдо48% всезоне:1982—1983г£*идостигала58%в 1983-1984 гг.,
120
°с.ш., за пределами еговто-
. .'x.-m-irt •61VP.’ ,Яэ1юН,iiljF
что/по-видимомусвязаносменее благоприятными* to-за большой
влажностипочвыусловиями'зимовки.Такимобразом/полученные
результаты свидетельствуютоспособности колорадскогожука пере-
живать холодныепериодыгодадаже на 60
ричного ареал ав Восточной Европе. ‘’!
‘ -Весенниййыход имаго послезимовки,'как отмечалосьвыше, яв-
ляетсяодним'изсамых'уязвимыхмоментоввгодичном'цикле^коло-
радского жука.Внаших опытахсмертность'также быланаиболыней
именно после выходажуков.изпочвы доначала питания.'За период от
Ю' до 25 мая погиблооколо’50% перезимовавших жуков. Причина
гибели не: установлена?После начала питаниясмертность1жуковне
превышалаЗ-^й.’33** -I'W' & ьнйн
* Срокисозревания самок после зимовки такжевбольшойстепе-
ниопределялисьих массой. Самки с наибольшей массой(более 160
мг) начали яйцекладку раньше других! В среднем на ! 7-й1 день после
выхода изпочвы/самкис менъшеймассой(80—Г20 мг)’— на27-й день.
Разница‘в срокахначалаяйцекладки ‘составила8(Удней(от20мая*До
9 августа}. Яйца, отложенные перезимовавшими самками/обеспечи-
лизапланированнуюсхему полевых опытов. Продолжительностьжиз-
ни перезимовавших жуков’точно не учитывалась, однакоподавляю-
щее большинство их оставались живыми до конца*сентября. Около
5% самок,неприступая,кяйцекладке,ушли на повторную зимовку.
«зд Итак.вполевыхопытахпрослеЖены все узловыеэтапыгодично-
го цикла.колорадского жука, включающие активную жизнедеятель-
ность летом, формирование1 диапаузы осенью/Зимовку/йесеннее про-
буждение, развитие'в следующем вегетационном бездне, повторную
зимовку, вновь весеннйй' выход ‘имаго и летнее развитие до ухода: на
зимовку' Экспериментально подтвержденавозможностьобитанияко-
лорадскогожукав Ленин градскойобл.,что,несомненно,следует'учи-
тыватьпри разработке карантинных мероприятий, препятствующих
дальнейшему расселению опасного вредителя;картофеля'.' ь'
- ^3. Цаяужест^крылые^слдп солдата^-j--—
ФПР’Клопа солдатика, Pyrrhocoris apterus, впервые была изучена
И. Ходеком [Hodek,1968]. В'дальнейшем были детально исследова-
ны многие вопросы,"связанные с физиологией диапаузы Р. apterus, но
такие экологическиеаспекты, как, например, сезонное развитие и его
регуляция, остались» стороне частности, обращаетна себявнима-
ние тот факт, что имея факультативную диапаузу, клоп солдатик в боль-
121
шинсдверегионов, подданным разныхавторов, развивается водном
поколении. Этоотносится кпопуляциямклопа да Украине и в Казах-
стане [Васильев,^973; Асанова, Искаков, 1977]. В Чехии (окрестнос-
ти Праги,50°с.ш.) единственна? генерация уходит в диапаузув ко нцр
лета[Нобек, 1971а; Honek, Sramkova, 1976].Популяция клопа солда-
тика из окрестностей Парижа (49 °с.ш.),также развивается пр моно-
вольтинному типу, IPouyreau, 1963]. При этом все авторы.отмечают,
что летомличинки I-V возрастов постоянно встречаются в природе,
что,затрудняетустановлениечислагенераций. . , -s ।
, Согласнорезультатамфенологическихнаблюдений,проводимых
в.заповеднике ^Лес, на.Ворскле» в течение нескольких лет, перезимо-
вавшие клопы становятся активными в последней декаде апреля. ]}
теплые солнечные дниони>спариваются,позже начинается яйце-
кладка, В дальне йшемличинки и взрослые клопы встречаются до осе-
ни. Поскольку имаго живут очень долгоиболыпе месяца сохраняют
способность откладывать яйца, разграничить генерации без исполу
зования специальных методов крайне трудно. Цугем прямых наблю-
дений можновыяснить лцшь начальные срокилоявления вприроде
молодых, имаго, которые отличаются от старых особей.бол ее яркими
краснымЦ|Пятнами и мягкими покровами,^ерез несколько дней эти
различия .исчезают.? .
г..,. Для тощунтобы разделить радельные генерации, в природе со-
бирали личинок \.возраста„принадлежность,которыхк. определен-
ному поколению устанавливаетсясбольшой вероятностью,^ содер-
жали ихгВ. условиях, близких к естественным. В дальнейшем среди
окрылившихся особей определяли долю активных и диапаузирующих.
Такой способ позволяет зафиксировать динамику,нарастания коли-
чества диапаузирующих клопов в реальной обстановкеи ориентиро-
воадо установить,возможность о^азрр^и^^зрррхрцоюрления. с ,
•MSiUW; -ТП-' '-г . ... . -
Таблица, 1 /.Количество диапаузирующих особей /= apterus , ,
' ‘ в пробах, взятых из природы летом 1991 г. *
NN вар. Дата сбора • личинок V возраста - Кол-во личинок Дата окрыления Число 'v пар имаго - ^Количество диапаузирующих самок, 96
...-hvhTt' . о ( 07.06- i 09-14.06 ' Ду -20. .i - < > :0 ;'-дс. / J
. .,2„. я рр2.О7 г. ,35.,., 04-06.07 .
" ” З5 4112.07 14-18.0Г' ^18 0
iJ ,42.07!i “ .. . v': - 21.07 ’>333 '. л:.
. ; >2Z07 • 40 - г -30-02.08 19 64,2 ч '
. . ? j ft 6 22.07 < , . .30, . <06.08 : , км5->.< , ,75,0
122
* -Наблюдения показа-'
ли (табл. 11), что в самой
раннейпробе (вар. 1)кло-
пы'начали окрыляться во
второй'декаде июня, в кон-
це июня'они приступили к-'
яйцекладке W образовали »
2-ецоколение.Яйцекладу-
щие »самки'в 'ранних про-
бахбылиобнаружены не''*
только в жарком 199 Г г., но
и в очень холодном пред-
шествующем сезоне. Начи-
наясавгуста^ впробахпре-
обладали диапаузирующие
особи.
‘Следовательно,'вус-
ловиях лесостепной зоны
Р. apterus• потенциально
может иметь 2-е поколе-
ние.Длятого,чтобывыяс-
нить реализуется ли оно в
действительности и какие
свойства Сезонно-цикли-
ческих реакций обеспечи-
вают образование'2-го по-1
коления,' были‘проведены
лабораторные и тюлевые
опыты. На основе полу-
ченных результатовбыла
составленаэкологическая'
характеристика Р. apterus.
Имагинальная диапа-
уза клопа солдатика конт-
ролируется ФПР длинно-
дневного типа. Критичес-
кий фотопериод'подверг£
жен температурной измен-
• лг./л.'.- ••‘ЧН Т И '<
Рис~50. Температурная, изменчивость, ,ФПР у
Pyrrhocoris apterus. . ;
'У"'.'! ' " s'-
123
01---------.-----—_——-
f 12 14 16 18
Длина дня, ч
Рис.51. ФПР Pyrrhocoris apterus в условиях тер-
чивости, .размах которой
отражен на рис.,50., }1Т, и,
-Порог ФПР оставался
одинаковым в условиях
ритмически-изменяющей-,
сят^мпературыиконтроле,
припостоядной темпера-,,
туре, эквивалентной сред7
нему гзнанениюп -ритма
(рисн51Х хотя-некоторая,
тенденция^ увеличению'
количествадиапаузирую-:
щихособей в пороговых и
короткодневных режимах
притерморитмах прояви-,
лась. .
Температурный- опти-
мумФПРлежитвдиапазо-
неот 20(до30°С.;Чувстви-
6 тельностью к фотопериоду,
обладаютличинки старших
возрастов ц имаго. Дляин-
дукциидиапаузы,необхо-
димр $-5 короткодневных.
моритмов:
.Тонкаялиния-ФПР при постоянной температуре,
эквивалеипюйсреднемузначению ритма.......
$—....~-.'й ( ,
ных сигналов (Волкович, Горышин, 1978].
сигналов доимагинальной
линвки,-для.индукции реп-
родуктивнойактивности--,
в дюдн₽м Д2гддщшрднев-
Клоп солдатик проявил слабую чувствительность к изменяющей7
ся длине дня только в пороговой зоне ФПР. что подчеркивает веду-
щую роль абсолютной продолжительности дня в индукци и диапаузы
[Волкович, ТЪрышин, 1979]. .игльраш.а . , W
..Количественные эффектыфотоп ер иодизма.проявил и сьв виде
задержки развития личинок в пороговой зоне.по сравнению с про-
должительностью их развития в короткодневных и длиннодневных
режимах. -Этот феноменбылвпервыеописангССаундерсом, [Saunders,
1983] у популяции клопа солдатика из Чехии (50 ?₽.ш.). У белгород-
ской его популяции подобный эффект обнаружен при всех исследо-
U4
ванных постоянных и переменных температурных режимах, разница
заключалась лишь в положении «пика» торможения личиночного раз-
вития относительно критического фотопериода (рис. 52).
Составив подробную экологическую характеристику вида, рас-
смотрим, возможно ли проанализировать фенологию клопасолдати-
ка в лесостепной зоне на основе параметров ФПР, полученных в лабо-
раторных экспериментах. На рис. 53 вертикаль показывает значение
критического порога ФПР для клопа солдатика при 22—23;°С, опре-
деленного на основе схемы (рис. 54).Т1акет короткодневной фотопе-
риодической информации по экспериментальным данным составля-
ет для белгородской популяции 4—5 сигналов. Следовательно, через
5 дней после наступления критического порога ФПР можно ожидать
формирования диапаузы уцоловины особей. Именно так происходи-
ло в полевых опытах (рис. 53). Все клопы, окрылившиеся до 10 июля
(вар. 1,2), были активными. Среди особей, окрылившихся с 10 по
20 июля, встречались обефракции — активныеи диапаузирующие, а
после 20 июля самки уже не приступали к яйцекладке. Эта законо-
мерность характерна для трехФервыхвар иантов опыта 1991 г. В более
поздних вариантах, а такжевовсех вариантах прохладного 1990 г. все
клопы были диапаузирующими. ^ '-—— —•
ЛГКЯ-.Ч -t. .,j,« f[.- £
i .I'M' 0.» r. 1 .T, . J ^-1—-—ьЦНй]------1--------------• "I
<;>. . >1 > зЖ swa1?. •
кту7 xva.»;.--
7 ! 'ft';,1 '‘""Г ' 1'0(8.».'.
Рис.52. Влияние фотопериода на длительность развития личинок у. ,
'' ‘ PyrrhocorisapterusB различныхТемпературных условиях (noiNumata ’ й "
et<31993>wos''i' ci" 'ослял ладшр вши л.
<,.« ^(/.ггОрел^сами^казан^и^ескиДпоро^фПРЛ ЛсТТ'.Х.’./
125
—. I I I I, l-v.: ' :r
Май Июнь Июль Август
Рис.53. Индукция диапаузы у Pyrrhocoris apterus в полевых опытах:
Горизонтальные отрезки—варианты полевых опытов. Гистограммы—коли-
чество самок и их физиологическое состояние (белые фигуры—активные, черные—
диалаузируютциесамки)ввариаитахпоежедневнымучетам. 1 —сезонное изменение
длины дня с учетом гражданских сумерек на широте Белгорода; 2—критическая
длинаднядля клопа солдатика, установленнаявлабораторныхопытахсучетюм тем-
пературы; 3,4—срзднедекадная температура воздухав 1990и 1991 ге соответственно
> s ’ с
Однаков природе (табл. 11)диапаузаформируетсяна 7 дней поз-
же, чем в нашихиолевыхопытах. Вероятно это связано с тем, что
клопы в естественных условиях выбирают наиболее прогреваемые мик-
ростации, и в этом случае критический фотопериод оказывается не-
сколько ниже„т.е. наступает несколько позже.
Полученные материалы свидетельствуют о том, что; потомство
перезимовавших особей может развиваться по одному из двух воз-
можных путей - двух- или одногенерационному пути. Два поколения
дают клопы, отрождающиеся из самых ранних кладок, отлаженных в
конце апреля-началемая. Имаго 1-гопокаления окрыляютсяв нача-
ле июля при длине дня больше критической, что определяетвозмож-
ность их размножения. Особи 2-го поколения развиваются на фоне
126
Рис.54: Влияние;фотопери- *.
ода итемпературы наиндук-f •
циюимагинальной диапау-
зы у Pyrrhocorls apterus (по:
Numata et1 al., 1993) /~1'
25
ъ?
аднй:-инм<..'лд ; чд •. rsH-^чс
!# • vH-Э S«Ff
. 7,7,1;-,₽азм|нржеми|в; чч • ч мм
'. И!/
I
Я
Диапауза
20
15
ЧН ... MTWruws.4 ' W»i-.*r!n< 'P.u
16 17 18
. ' i-< smr
Фотопериод, м
* < >< Л'УНе.::' 'Ч'Ки" •'
короткого дня, вследствие’чёго'формируют диапаузу и образуют зи-
мующийзапас. Одно поколение успевают завершить личинки,'’выхо-
дящие из поздних кладок, что'сбответствует вариантам'наших поле-
вых опытов^ поставленным после ГОиюня. Личинки старшихвозрас-
тов и имагопопадают в короткодневйые условия, и^всеклопыоказы-
ваются диапаузирующими.
Соотношение количества особей, развивающихся по двух- ‘или
одногенерационному'типу,' определяется погодой конкретного года.
В теплые по температурный условиям сезоны большая часть популя-
ции образует два поколения , в холодные увеличивается фракция кло-
пов, завершающих только одно поколение. В года, когда температура
летних месяцев особенно низкая (как в1990 г.), вся популяция P.apterus
в лесостепи'переходит к моново^ьтинйому СезоннОмуциклу.' '
J ’ При описанном сценарии развития становится ясным и эколо-
гическийсмысл'регулируемыхфотопериодомизмененийсроковраз-
вития личинок. Те особи, которые развиваются по двухгенерацион-
ному типу дважды в сезоне ^’весной и осенью - погфдгйотв условия
короткого дня, что вызывает сильное ускорение развития иповышает
шансы особей успещнб завершить цикл до наступления устойчивого
похолодания. Существенно, что чем короче день, тем быстрее растут
личинки. При одногенерационном цикле личинки развиваются на
фонедлинного дня’ когда происходитсильное торможение'роста. Бла-
годарятакой регуляции темпов развития личинок, окрыление оття-
127
гиваетсяна более поздние срокщ когда окружающие условия благо-
приятнее для формирования диапаузы.
Таким образом, фенологические наблюдения и результаты поле-
вых опытов свидетельствуют о том, что в лесостепной зоне России
клоп солдатик Р. apterus может развиваться в одном или двух поколе-
ниях, и характер его развития контролируется на основе фотоперио-
... . ...
6.2. Моповольтинизм и механизмы его поддержания
Основная стратегия сезонных адаптаций насекомых,наиравлена
на м^симальцое|цсцощ>зова1|1ие.веГ|е?|ационногосеэона,-.что достига-
ется продуцированиеммаксимальногочисла'поколенийза период
активной жизнедеятельности. Эта стратегия хорошо прослеживается
у поливольтинных видов, дающих последовательные поколения, чис-
ло которых определяется температурными условиями конкретной
местности, ц угомрдинамныхвидов,характеризующихсянепрерыв-
ной активностью..Примоновольтинномциклезасе:зон,илиза'год,
развиваетс^толькооднопоколение. Дри^ину моновольтинности ус-
матривали,!}’обязательном наступлении диапаузывкаждом онтогене-
тическом цикле независимоот внешних условий. Однако в' дальней-
шем: выяснилось,,чтосуществуют идругие способы поддержания та-
кого сезонногоцикла^аэкологические механизмы, лежащие в их ос-
нове,удается выявить тош>ко в специальных экспериментах[Дани-
левски^150^^^Ц1левср1^и др.,^970]. Рассмотрим различныепути,
обеспечивающие моновольтинноеразвитиеу насекомых умеренных
щйррт,, адаптированных к л^не-итмн®^ ^е,нргсе^зрнов.* ’*•
"В сдмыхпррртыхрлучаяхдиапауза носитласле^ственныйо^лп-
гй^/^йхаракте ринаступает независимо,отвнешних'условий,а пре-
кращает,сяобыцно цодвлиянием,пониженнойтемпературы. Приме-
ры Дидов^ дидцауза которых формируется облигатно, мы находим в
разных‘отрядах.насекомых. У^зеленой дубовой листовертки, Tortrix
у!л</оиоДДан!мевский,р5 у’непар^ого шелкопряда,
'iyniantrpa dipp^LeoD^rd,^^, ТацЬ^г et al,, ^990], диапауза форми-
руется Д?амом начале лета. Годичный цикле клад ывается следующим
;образом;зиму1от'яйр(а (у нектарного шедкопрядагусеница полностью
формируется^нутри яйца); весной выходят гусеницы, которые закон-
чив питацие^ужев июне-началеиюляокукливаются; вылетающие
бабочки в конце июля откладывают диапаузирующие яйца,' которые
'12В
остаются зимовать. Следовательно, >8—9 месяцев; выгоду .популяции:
этихвидов находятся^ состоянии диапаузы, и л ишь 2—Змесяцаиро-
должаетсяактивная жизнедеятельность.,Описанный, годичный; цикл(
сохраняется на всей территории их обширных ареалов. ? • '
ил Такую форму покоя некоторые исследователи рассматривают как
особую летне-осенне-зим нюю диапаузу., Но надежных критериев,.по-:
зволяющихразграничитьотдельныеэтапытакогопродолжительного
физиологического покоя, нет, поэтому.считать ее летнегосенне^зим-
ней пока возможно толькопо календарной приуроченности,Об,этом
жесвидетельствуютрезультатынсследования облигатной,диапаузы
непарногошелкопряда, пред принятого К. и М.Тауберами; слсотруд-
HHKaMH[Tauberiet»al.i,4990J. л '-v-
Подобнаягфеиологическая схемапрослеживаетсяуклоповсдепг
няков: Leptoptemadolobraia vttCalocoris zio5eoznacn/flto [Braune, .197.1,
1973] . Исследование этих видов вэкспериментальных условиях пр ка-
зало,>что вовсехрежимах инвариантноиндуцируетсяэмбриональная
диапауза^которая. прекращается'Только после воздействия) низких
температур. Моновольтинныйсезонный цикл е облигатной эмбрио-
нальной зимней? диапаузой описан да У листоеда.
[Nakai, Takeda, 1995].ч‘р ..
Однолетняя генерация с зимовкой на стадии яйца и ранневесен-
ним развитием гусениц может рассматриваться как стратегия, кото-
рая дает большие преимущества, позволяя избегатьконкуренцию и
ускорять онтогенез. В связи с этим заслуживают внимания порази-
тельные результаты, полученные в опытах с Epirita autumnata
(Geometridae). Оказалось, что при питании растущимидистьями гу-
сеницы удваивают свою массу при 18 °C всего за 1,2 дня, а при пита-
нии листьями, собранными всего через три дня после окончания их
роста, - за 11,2 дня [Ayres, MacLean, 1987], <
Другую группу составляют виды, облигатная диапауза которых
формируется осенью. Сюда"Относятся многие клопы, зимующие на
имагинальной стадии, в частности"Детально исследованные Palomena
prasina и Coreus marginatus. Перезимовавшие клопы весной после
непродолжительного периода питания приступают к яйцекладке,
которая может продолжаться больше месяца. Отродившиеся личин-
ки питаются, медленно растут и к середине августа окрыляются. Пос-
ле периода преддиапаузного питания клопы уходят на зимовку в со-
стоянии репродуктивной диапаузы. Никакими провоцирующими
условиями вызвать у них репродукциюне удается.Ввыравнивании
129
возрастного соетава<личиноки синхронизациидиапаузирующейста-
диисюптимальнымдля ее 'наступления'периодом сезона участвуют
количественные!фотопериодические реак11иик[!Мусолин, Саулич,
1997]. .Д.ШШЗф' :!?»<) х» «лэдньлх». <
я£>1 гкактюказали «исследования ‘{Saulich, • Musoiiny'l996],глин инки
Р.рга$7павкороткодневныхусловиях(15«светав сутки)развиваются
значительнобыстрее,чемвдлиннодневных(18ч).'Этоускорениет1ро-
являлось!во'всехтемпературных режимах (20,26ивО?С), «но максит
мальйОгоЪначениядостигалРщ>и<00^С4>(рис.г55).он;Е it -и ;;,эь
•чВпервыекатичес/лвенныередкцифрегулирующие скорость-роста
личинок'и’имеющие самостоятельное значение «поддержании моп
новольтинного цикла, были описаны у Dasychirapudibunda [Гейспиц,
Заранкйна,1963] .'Этот вид повсеместно Дает одно поколение, зимуя
диапаузирующей'куколкой.Вдлиннодневныхусловияхлетомтусе*-
ницы питаютсяимедленнорастут.С увеличениемпродолжительно-
стивегетационногосезона увсличиваетсяидлительностьразвития
гусениц.В’условиях Ленинградской обл.этазадержкасоставляетЗ—
2,5месяца,’наюге-более4 месяцев. Толькоосеньюразвитиеускорят
ется; ийаступает окукление.Какбыло-показано® эксперименталь-
ных условиях, такой цикл регулируется необычной реакцией гусениц
XHQ
Я'
'НИ
.-гг. !«ггг к пэавгjxm'a •с'ьчаг.одтэв."
пмо а
лхс'оа йнне^чнн
'Л й*
- П
nqn е ,^!4,155'^
f н.хЛ.' тг
S 50-
roan иминньснж.
ЧДдЭВМ ..ЖуА)
IJiJWiX Н'КЦГЭОЭ I
3
ч
ТЗОНТ'.<Ш1 (I >
7г’-э:*зр9И .<
:у-ОНф.;; г/ЮШТО’
О wit* «г J
yqt
I,
юг:
’ 1^88 v.
-П.КРИ гээфшпи’й
- й•. fc.j., -,чи
-<Х> ' ТНф
Т5>,
v- ' 1 . к <1 Длва;дая,»’'' >
Рис.55. Влияние, фотопериода на продолжительность личиночной
стадии Palomena prasina при постоянных температурахw‘u
130
надлинудняитемперазуру;развитиеихпривысокой;гемпературе;и
ддинномдне значительнозамедляется из-запоявления дополнитель-
ных гусеничных возрастали окукление, Hanpn»«wpi25 °С,®роисхот
дитлосдеДР-далинекз^х*'.глшвт-п-.,' т.МикГ*оц
., н <! г Очень похожиереакции быдиобнаружены поажеприизучении
сезонныхциклошнекоторых,медведиц.;Так,.у/>гйагсГ/а'45а^е//д<из
Канады зимняягусеничная диапауза наступает независимо от вне.-
шнихусловий,атемпыроста1усеницрегулируютсяжоличественной
ФПР. При длине дня более 14 ч света в сутки продолжительностьтусе-
ничнойстадии.за смет числа в03растовувеличивает£я*{О,оеНе1,
Philpgenei, J978I. м m ПЖКГ(5Р«ПОЖ>Г(ИП Я MFKHHWnoa
Особыйслучайпредставляетсобойрегуляциямоновольтинизма
наосновеЭвухстеулеячалпыхФПР,.Разнообрази9сезонныхсхем,_возг-
никающихнаосноветакихФП1^описалГ.Тиле4ТЫе1е,1977],нат1ри-
мере развития жужелиц (Carabidae). Рассмотрим сезонный циклод-
ного из исследованных им видов, относящегося по
классификации,-предложенной.автором, квидам свесеннимтипом
рЯВМИОДЕенияыг^т^дч.»^ нягЛ?> хвл w.i» .мод
;Этижужелицызимуют?встадииимагоиразмножаютсявесной.
Молодые-жукипоявляются посередине, летаюоблигатновпадаютв
состояниерепродуктивнойдиапаузы. Короткодневноестимулирова^
ниел ревителлогенеза происходитосенью крайней веснойпридлине
дняменьше 13 н; При дальнейшем увел имении длины дня.осуществ-
ляетсядлицнодневная ФИР,стимулирующаявителлогенези созрева-
ние самок. В апреле происходит яйцекладка, в маегтиюне развитие
преимагинальных стадий^ после имагинальнойлинькивновьнасту-
пасто&шгатнаярепродукжвнаядаапауза. е к н .етм
- • Регуляция моновольтинизма на основе двухступенчатых.ФПР
описана у златоглазок ^ederwe gnjcjKy(!GriniaIi,.;198j8; Canard, Grimal,
1988]® Chrysopa downesiffavber, Tauber,1976a,b]. Большую чаегьгода
онишро водят,всостояниидиапаузы,. котораянаступаетумолодых
имаго независимо от Длины дня. Экспериментально показано,!что
необходим ымусловием для быстрогосозреваниясамокияйцекладки
является носледовательцое воздействир на чувствительные-стадии
сначала короткодневных, а затем длиннодневньпсфотопериодошПри-
чемдиапаузапрсдотвращается, только в-том.случае,если(увеличение
длиныдня составляет неменее 4 ч дляСй. downesiiuni iHppH.T.gracilis.
Блаподарядакой особенности ФПР> размножение этих* видов стано-
вится возможным лишь в следующем сезоне.-Яйцекшкап&^с//»
131
начинается ;в середине лета при длине дня, №ч, [Grimal,‘1988}, араз-
множениеCh.downesi, обладающейболеенизким порогомФШЦОколо
1Фч),повсюдупроисходит в течение короткого периода ранней вес-
ной [Tauber, Tauber, 1976b]. Описанные схемыгодичныхцикловзла-
тоглазок обеспечивают существование небольших,»но относительно
стабильныхпоразмерампопуляцийв'условиях.жогдаобильныепи-
щевыересурсы —тлидоступны втечение непродолжительноговреме-
ни. В ответ на'истощение1 кормовой базы уэдатоглазокформируется
джтельнаясдиапауза. сн юп-уэя И !)<-.»>
. Ыг Приведенные примерыкасаются насекомых, у которых моно-
вольтинизм в природе определяется наступлением облигатной диа-
паузы в каждом-поколении. Однако многие видыс факультативной
диапаузой такжемогут иметьтолькооднопоколениеягоду, что обус-
ловливаетсяразнымипричинами и обеспечивается разными спосо-
бамидми и л : ;к«s- мици-чЯ/яЧ таьисьгь. шт/.ч тсига.
> >. - Существуетмножество видов, у которыхмоновольтинный цикл
свойствеилишьотдельным географическим популяциям, аневиду в це-
лом. Крайне северные формы, как правило, характеризуются усиле-
нием склонности к моновольтинизму, южные—резкимееослаблени-
ем . Сравнение отдельных видов такжевыявляетразную степеньвыра-
женноститенденциикоблигатномуформированиюдиапаузы. i о
• sчКак отмечает Е.Б. Виноградова [1990],.у мух прослеживаются
всепереходы отпочти полного моновольтинизма у изученныхпопу-
ляций до поливольтинизма с факультативной диапаузой, регулируе-
мой температурой и длиной1» дня. Так,>у-10 из12'популяций: мухи
Protophormia Zerraenovae обнаружена сильно»выраженная тенденция к
облигатному формированию диапаузьнявсе особи диапаузировали
независимо от температуры, и фотопериода. При этом установлена
значительная стабильность' проявления норм реакции на протяже-
нии нескольких лет лабораторного культивирования, которое обычно
сопровождается бессознательным отбором на'бездиапаузность. Бла-
годаря перечисленным свойствам (глубокая1, продолжительная диа-
пауза и-сильная тенденция к моновольтинности) сезонный цикл
Р. /егг^иото^хорошо^адаптирован1 к обитанию в умеренных и север-
ных широтах, где вид занимает доминирующее положение в фауне
двукрылыхихарактеризуетсяпостоянноРысокойчисленностью. '
Calliphora vicina l отличие от предыдущего видатенденция к
моновольтинизму сильно выражена лишь у самойсеверной, мурман-
ской, лопуляции и горной — из Киргизии. У ленинградской калли-
132
форы выделены обе фракции:,нарядусдоминирующей поливольтин-
ной-формой, диапауза которой контролируетсявнешними фактора-
ми, отмечена моновольтинная форма с облигатной диапаузой (Ви-
ноградова,>1990]. >
, v 'Очень сильная те нденцияк.формированию диапаузы просле-
живается уразныхтеографических популяций яблоннойплодожор-
m,Cydiapomonella. Моновольтиннаяфракциявыражена у всех попу-
ляций от С.-Петербурга (60 °с.ш.)доСамар канда (39?с.ш.)(Шельдег
шова, 1965].Даже приповышениилемпературыдо30°Сопределен-
ная доля диапаузирующихлронимфсохраняласьу всехпопуляцийво
всехфотопериодах (рис. 56). Переходяблопной плодожорки кмоно^
вольтинизму наибольшей части ареал а автор рассматривает как адап-
тацию кпищевымусловиям. Раннее наступление диапаузы, .сюдной
стороны, определяется сроками созревания и опадения плодовис-
ходного кормового растения - дикой яблони,j сдругой >- связано с
резко выраженной периодичностью плодоношения яблони. В неуро-
Рис.56. Изменение количества диапаузируклцих особей у географи-
'7 ческих популяций'яблонной плодожоркив разных фотопериоди-
- . .. ческих и температурных условиях (по: Шедьдешова,1965]: * -' '
Штриховые линии - при 20, сплошные - при 25, штрих-пунктирные—при : О'
30 °C. Популяции: 1 - гомельская (52 °с.ш.); 2 - ессентукская (44 °с.ш.); 3 - самар-
кандская (39 °с.ш.)
433
жайныетодыдиапаузирующиегусеницы 1,-гопоколенияобеспеяива-
ютсохранениепопуляцииплодожоркивлшучаегнедостаткакормадая
последующихцюколенийЗ* д .< ’ зьцднщдодоном ьи'и’што-.кы
Сходные тенденции обнаружены у восточно-европейскихпопу-
ляцийжолорадскогожукаг'моновольтиннаяфракциявыявленаувсех
9исследованных’популяцийюп Москвы.(55°с.ш.)‘доВладикавказа
(40ос.ик). Несомненно,чтоосновойдля этого служитбольшаявнут-
рипопуляционная изменчивость по; признакам, диапаузы? Исследо-
ваниеэтогосвойстваубелгородской популяции!,. decemlineatanoKi-
зало, чтовлотомстверазныхсамокколичество диапаузирующих жу-
ков'колебалосьотЗдо>96%,приэтомуровенъ1тенденциикспонтан-
ному формированию диапаузы, свойственный,исходнойсамке,!взна-
чительнойстепеникюхранялся, нетольков первомдочернем поколе^
НИИ^ЦОШШОВТСфОМС « Ш МК
он ^Генетическая вариабельность реакции на длину дня открывает
большиевозможностидля*отбора?Микроэволюциянаегоосновемо-
жет идти в направлении как строгого моновольтинизма с облигатной
диапаузой, так и поливольтинизма с ФП-крнтролем диапаузы.Пере-
ход к моноволымнному циклу большинства популяций колорадского
жука обусловлен, видимо, адаптацией к срокам вегетации культурно-
го картофеля, который служит ему почти единственным, кормовым
растением в Европе. Иные адаптивные стратегии возникли у
L. decemlineata на американском континенте, где он имеет большую
свободу выбора пищевого рациона за счет диких пасленовых [Hsiao,
1981}. '-•< л •
Существует много примеров, подтверждающих значение корма
как селективного факторапри переходе к альтернативны пожизнен-
ным стратегиям [Саакян-Баранова, 1971; Shapiro, 1974; Rodriguez et
al., 1994]. Замечательный образецтакоготипа представляет парусник
Papilio.zelicaon. B Северной Америке в районе Скалистых гор вид раз-
вивался в одном поколении. Такой сезонный цикл был обусловлен
ограниченным периодом вегетации кормовыхрастений. При появле-
нии дополнительного кормового растения с продолжительным пери-
одом вегетации. Л zelicaon перешел к поливольтинному циклу. Одна-
ко в экспериментальных условиях показано, что потенциальный мо-
новольтинизм поддерживается отборомкак адаптациях выживанию
в тех зонах’ гдепроизрастание кормрвыхрастецийвсезонеограниче-
но [Sims, 1980]
134
i Отчетливаядифференциация*посклонностиждиапаузеюбнару-
живается у всехвидовхзфакультативнойдиапаузой.Приэтомунеко-
торыхиз'них Четко намечается<ступенчатый<характер вгизменении
этогопризнака,которыйобусловливаетвозникновение*выраженных
экологических типов. Так, по данным Ходека'[Нобек,!!1960], ужоров-
киСосс/ле//л^ергетлрилс/а/а северныепопуляциихарактеризуются
моновольтинным циклом, аюжные — поливольтинным.Границамежду
нимивЗападнойЕвропе проходит примерно по 49-йпараллели. Ана-
ло1ичныетенденциипроод^живаютсяукоровки:0//(хюп«/и/ш5ф2ой«
[Заславский, 197ОД.гРезковыраженнаясклонностькмоновольтиниз-
муотмечена у северных<(ленинградской и минской)<популяций,тог-
да какразвитиеюжных(среднеазиатских)популяций контролирует-
ся обычнойФПР длиннодневноготипа.В детальных исследованиях
В^А.Заславского[1984[былобнаруженсложныйконтроль сезонного
развития - Ch. blpustslatus b природена.основе.гступенчатыхреакций
[Заславский, 1984]. УчелночницыЯу/ор/»7а ргаилала[Данилевский,
1961]на юге отСухуми(ДО Полтавыраспространена поливольтинная
форма,анасевереютБелгородадоС.-Петербурга.обитаетмоноволь-
тинная форма, у которой дажевусловияхдаинногодняжвысокой
температуры неудаетсячтолучитьбездиапаузноеразвитие-куколок.
.;':Существованиедвухэкотиповобнаруженотакже1уивовойвол-
HKHKH,Leucomasalicis. Основная<схема их годичного циклахорошо
известна.Зимуют гусеницы!! иПЬвозраста. Весеннее-развитиепро-
ходитбыстро^ивсерединелетапоявляются бабочки. Отрождающие-
сягусеницыхшедующегопоколения после2—Знедельпитаниявпада-
ют в диапаузу и уходят на зимовку. Такая сезонная стратегияитоддер-
живаетсянаобширнойтерриторииотЛенинградской'обл.доЮжно-
гсьКазахстана, только на юге ареала -«Южной Европе, в южных рай-
онах Средней Азии и в Закавказье обитает другой экотип Т. salicis,
которьгёхаракгеризуедоя поливольтиннымсезонным циклом, зсрс д
тл , Вусловиях эксцериментаЮФ.Тёйспиц [1953Ьобнаружила,что
севернойформеТ.5л//с1гсвойственналраиелсуточнаяФПРркотст)ая
допускаетбездиапаузноеразвитие при .23.-25 “Столькопридлине
'дня 20<чсветавсутки..Влриродетакое!Сочетаниетемлературыифо-
топериода'нигдене встречается,* вследствие‘этогосеверныепопуля-
лииивовоЙ1волнянкивсегдаимеютмоновальтинныйцикл.'У южных
популяций обычнаяФПРдлиннодневного типа(Данилевский, 496,1;
Хачатрян,г1984],.онимогутдавать в^зависимости'от условий?—3 по-
колениячзасезон.му;фн’Н'л*Ы*’-:ддЛ V .-‘jw. .к д 2 V .;ш
135
нтПозжеподобныереакциибылиописаныусеверных.популяций
Spilosoma теЩ/1аЦл4Кинд,19&5},летней капустноймухи, Deliafloralis
[НесинД987]лнекоторыхдрушхвидов.Диапаузаунихпотенциаль-
ноявляется'факультативной,чновреальнойприродной обстановке
возможностьактивногоразвития«ерсализуется:насевереареалаиз-
занедостаточновысокойтемпературы,анаюге —вследствиенедоста-
точнбйшродрлжительности'дня. •. ,*>>s.у.дачт'ж>м
-. ча Регуляция моновольтинизма на основе промежуточнойреакции
обнаруженаиутемнокрайнейсовки,: Axyliaputris^HO в отличиеот
предыдущих видов; ФПР которых проявляетсялишь в эксперимен-
тальных условиях, у темнокрайней'совки жизненный цикл реально
контролируетсяФПР{Ть1щенкоидр.,1983].Наосновеанализафено-
лбгишАгриПйвлесостепной зоне былюделан'вывод,что промежуточ-
ная ФПР у этого вида выполняетрегулирующую функциюв климати-
ческихзонах, температурныересурсы которых превышают потребно-
стиюдногошоколенияи втеплые годыдопускаютразвитиедаух пол-
ных поколений. Когда окукливаютсягусеницы1-го поколения, про-
межуточная ФПРоставляетв активномссостоянии некоторуючасть
популяции в качестве резерва, способного приблагоприятныхпогод-
ных условиях достичьдиапаузирующей стадии? Еслитемпературный
режимокажетсяменее благоприятным, этотпопуляционный резерв
погибает,а1зимующийзапас формируетсятолькоза счет диапаузиру-
ющих куколок1-го поколения. Таким образом, промежуточнаяФПР
может рассматриваться как специальная сезонная адаптация кпосто-
янному существованию насекомыхвзонахперехода отмоновольтин-
ного.типа.развития!кполивольтинному.т .ч >
< г Сезонный цикл с одним поколением втоду может определяться
темпершпурнылшусловиямиЖорошакзвестно, чточисло поколений,
как правило,^возрастает по мере продвижения на юг, что самымгне-
посредственным образомсвязаною суммой эффективныхтемператур
(СЭТ),необходимыхдлязавершенияонтогенеза.В техширотах, где
СЭТ достаточна только для одногоонтогенетического цикла, возни-
каетмоновольтинизм.Такихпримеровмасса.Подавляющеебольшин-
ствовидов, например, нашироте С.-Петербурга, успевает дать только
од колокол ениеза сезон.Однако и в этом: случае необходимэкологи-
ческий механизм, который синхронизировалбы появление наиболее
устойчивыхстадий с соответствующим.периодом года. - < < ;
; Эта задача очень просто и однотипнорешается многими вида-
ми. У S. menthastri, A. rumicis, М. brassicae, например, низкая темпе-
136
ратура, характерная для северных или умеренных широт, просто «от-
ключает» ФПР, которая у большинства северных популяций в отли-
чие от южных,-имеет высокийтемпературный оптимум. Вчасгности,
у ленинградской ' A. rumicis ФПР проявляется только в пределах IS-
IS °С,у5. menthastri при 18 °C влияние длины дня полностью подав-
ляется, и все куколки несмотря на большую продолжительность дня
диапаузируют. ' ’."-хх .= х..х/х.
К моновольтинному типу развития переходят популяции мно-
гих видов, обитающие в торах. Несомненно, что этов первую очередь
определяется недостатком тепла. Известно, например, что на высоте
2 000 м над уровнем моря СЭТ уменыиается почти раз. '
: Адаптация игорной зональностиияекръзняат путями. Как пока-
залА.С. Данилевский [1961], переход кмоновольтинизмууР/егюлп/»
сопровождается возникновением горного подвида P.napi bryoniae,ко-
торый по ФПР очень близок к субарктическим формам и даже при
24 °C даеттолько диапаузирующих куколокво всех фотопериодах.
В других случаях переход к моновольтинизму происходит под
прямым давлением^внешней среды. Например, у желтушки ‘Colias
alexandra[Hayes, 1982] в долинахобитают биволътинныепопуляции,
а в горах моновольтинные, что обусловлено именно недостатком
тепла. ....х . ....-х-
У гроздевой листовертки, Polychrosis botrana [Комарова, 1954],
уменьшение числа поколений в предгорной зоне также определяется
температурой, но осуществляется иначе. Низкая температура сдвига-
ет развитие чувствительной к ФП стадии на йериод с длиной дня мень-
ше критической, в результате чего все особи диапаузируют. Так же
происходит адаптация к горной зональности у ивовой волнянки в
Армении [Авакян, 1953]. ? .
Подобная регуляция моновольтинизма в Австрийских Альпах
описана у златоглазки Chrysopa perla [Gepp, 1975]. В результате 10-
летних наблюдений Дж. Гепп обнаружил, что в горах вид дает только
одно поколение, тогда как в долинах всегда два. Оказалось, что время
вылета перезимовавших имаго запаздывает на 7,5 дней на кажд ые 100 м
высоты. В результате развитие всего поколения в горах происходит
при длине дня короче критической, что однозначно определяет на-
ступлениедцапаузы. , . .. .
Факториальная регуляция моновольтинизманасекомых в горах
встречается чаще, чем обусловленная наследственностью. Для воз-
никновения наследственных различий у горных популяций, видимо,
137
требуетсяб6льшая'изоляция,пре11ятствующая постоянномуобмену
июфещйванию'междо'популяциямилалод - ••вчотол лП-о
ч <Материалы,иллюстрирующиезначенией«еш«их(0аюло!роввосу-
ществлениимоновольтинного цикла, были получены, при изучении
сезонных вдаптацийнекоторыхполужесткокрылыхв’Белгородской
обл. Клопы,.какправцло,очень.теплолюбивы.;Обычно они имеют
высокие температурные пороги и требуют большой СЭТдлязаверше-
нияонтогенезааВ силуэтого подавляющее большинство>видов дает
однопоколение>в'тоду,;развитиекоторогозавершаетсядиапаузой.на
специфическойдлявидастадии.Такая сезонная схемахарактерна для
многих паяужестк<жрылых,влесослепнойзоН|С»к < гм j Ч,'
-Однакоэкспериментальноеизучениеразныхдидоввыявило, что
вне шнеЛаднотипный; сезонный; циклэтихнасекомыхбазируется, на
разных механизмах. Если у ‘Palomenaprasinan Coreus. marginatus ana-
пауза обязательножозникаладовсехэкспериментальныхрежимах, то
у Л/тпдсш/охнаступлениедиапаузьвконтродировалось ФПР длинно-
дневного<гипа. ’Нопараметры ее таковы, что® условиях лесостепной
зоньиполивольтинныйциклникогдане, реализуется. ; „ » ,,: мtiqг
ч ни Данныедхарактеризующие.границыхпроявления ФПРA.custps
при постоянных и 'переменных лемпературах (рис. 53), доказывают,,
!fi 100
ю
- вп ади
'>Ж xfeT .T©/q№
и 40
хвпадА хсоойн
5'П * ГЖТ;
й) -
,«й«-
41 .эжг: R‘J
Ч КЦДВС'П вФ я
Мао
,ОВН
ДЦ
Щ О Г,о; ,03;
' 20
оленю । т‘-лз,*да
WW,« ОТ;'
UH ЩД.
At
" ааяжмг’ . ски.г* u i ^голкдусэбзг •'»?-ИоД
-йн снргнсондослс ,,ф > ' ••стемПер&|уреХ5а ’
Рис.57. Зависимость количества диапаузирующих особей Arma'custos* ' '4
AfcXpT; ‘от’температуры^'- j квн* у. гнци ’ пт о
гх«т итб 'лРеадлиию110ввдаиннодневш4х(18чсветавсупя)и1корогп«м1невных(14ч). = ж
„ условияхприпосгоянныхтемпературах(соп51)итерморт?4ах(ТР)сразпымиам1ии-;
’ 41' тудами(точкисоагветствуюг средним значениям ритма) 1'-'ч -та -•Л-
138
что активное развитие даже в длиннодневных условиях возможно лишь
при температуре выше 27 °C. При терморитмах диапазон температур,
допускающих проявление ФПР, несколько расширялся, но и в этом
случае при средней температуре ритма 20 °C около 60% самок диапа-
узировало в условиях длинного дня.,/ -г ,
Формирование диапаузы у всех клопов летнего поколения обус-
ловлено еще одной особенностью.вида-,чувствительностью кдлине
дня только на имагинальной стации (рис. 58). :~
' , ' . Г ' _________3
1 .2 L3 особи, %
„•‘ЮН - . г ''НДИ.Ч.
Рис.58. Чувствительность Arma custos на разных стадиях онтогенеза
к короткодневным'и длиннодневным воздействиям. 'Температура15114:
Г 25eCf?'-'сПЧЦй '- С.ДЛГВДЛОЗ ь-ч.*-.; /
• дд*. ' Г «...•ДлеВР-НО- Т рф f.ir
г В соответствии» этим виолевых .опытах, независимо откален- 4
дарнойприуроченности экспериментальных вариантов, клоп» все-;
гда формировали диапаузу. Попытки предотвратить^* наступление.,
увенчались успехом только водном случае, когда сроки развития ли-
чинок был иискусственносдвинутынаГмесяшвпередпо сравнению с
естественной фенологией. Клопы окрылились вжонце июня, и 37 % г
самок приступили к яйцекладке (рис. 59), в природе же/этого никогда
не происходит. Даже в самые жаркие годы, по.нашим наблюдениям,
первые имаго появляются не ранее конца июля, когда фотопериоди-
ческие условиязаведомонедопус каютактивногоразвития.Следова-
тельно,не облигатностьиндукции диапаузы, а особенностиФПРпод-}
держиваюпмоновольтинизмяида^в условияхлесостепи.н р о <.;. н= и
й Заслуживает в нимания тот факт, что моновольтинный сезонный •
цикл.свойствен А custos. не только* в лесостепной»зоне,.но и в Цент-'
рал ьнойЕвропе [Челнокова, 1980], наюгеУкраины,в КиргизииЩуч-
139
Вверху—сезонное изменение длины дияйЧемпературы. Горизонтальные отрезки -
сроки развитинклотюв вэкспериментальныхвариантахотвыходаличинок из яиц до
окрыления имаго; в варианте «в» в природные условия поставлены личинки IV воз-
раста. Цифры—процентдиапаузирующих клопов. 1 — имаго; 2 —яйца; 3—личинки
ков, 1961,1965), в Болгарии (Иосифов, 1981]. К сожалению, эти попу-
ляции экспериментально не исследовались, поэтому причины моно-
вольтинности вида в этих зонах пока не установлены.
У клопа солдатика Pyrrhocoris apterus регуляция моновольтиниз-
ма также внешними факторами основывается совсем на другом прин-
ципе. СамкичЕ apterus характеризуются длительным периодом яй-
цекладки, которая продолжается иногда до полугора месяцев. Этим
обусловлена разновозрастность особей в популяции (рис. 60).
Личинки из поздних кладок попадают в короткодневные усло-
вия, поэтому уже в первом поколении формируют диапаузу. Личинки
из ранних кладокразвиваются вусловиях длинного дня, поэтому де-
терминированы на активность. Сильно выраженная фотопериодичес-
кая регуляция скорости роста личинок (рис. 61) оптимизирует сроки
формирования диапаузы. и '.>
Включение втодичный цикл двух периодов проспективного покоя
(летней и зимней диапаузы) в пределах одного-онтогенетического
цикла — один из; наиболее распространенных и, надежных способов
поддержания; моновольтинизма.. Благодаря тому, что летняя диапауза
так же, как и зимняя, может формироваться на любой специфической
для вида стадии, создаются самые разнообразные комбинации летне-
140
Рис.60. Механизм образования моновольтиндой фракции;/
Pyrrhocoris apterus в лесостепной зоне:
Обозначения, как на рис. 59
го изимнегопокоя, образующие сложные «узоры» сезонные циклов.
Эти вопросы подробно представлены в сводке, С. Масаки [Masaki,
1980] по летней диапаузе насекомых. Рас- ,
смотрим некоторые наиболее яркие и хо-
рошо изученные примеры, иллюстриру-,
ющие значение летней диацаузы в подпер- ( . 1
жании моновольтинизма. s'k
Зимняя пяденица Operophtera bru- g
mate на всем протяжении ареала всегда £
дает только одно поколение в году Зимов- В ,
ка происходит на эмбриональной стадии. S
Гусеницы, выходящие весной, после пе-
риода питания в начале лета окукливают- |
ся, и куколки в состоянии летнейдиапау- ч
зы проводят все лето. Имаго появляются - 20 '
позднейрсеньюи откладывают д иапаузи-
рующие,яйца. Об? диапаузы облигатные к 8 ,5 ,6 4? 18
наступают независимо от внешних ус- " Длина дня, ч
ловий. ,.
Вероятно, большую роль в осуществ-
лении моновольтинного цикла ла всей
территории ареала у зимней пяденицы
играет географическая изменчивость лет-
ней диапаузы, которая проявляется в уве-
Рис.61. Влияние длины дня
на индукцию 'диапаузы* и дли-
тельность развития личинок
Pyrrhocoris apterus:
Сплошная линия -ФПР при
20 °C; пунктирная длительность
развития личинок
141
Рис.62. Географическая изменчивость продолжительности летней
диапаузы Operophiera'brumata в Европе [по: Masaki,11961, Ушатинс-
кая, 1987] •«*4 И0,-!Л' °я w‘
личении ее продолжительности по мере продвижения на юг (рис. 62)
[Кожанчиков,>41950^Ма5ак1, 1961]?Полмнению И.В?'к.ожанчикова,‘
сроки1 осенпеговылетаимагоопределяютсятемпературными услови-
ями. Однако более поздние исследованияйоказали' чтотерминация
летцей диапаузы не зависит от внешнихусловий?аопределяется;ге-
нетическими факторами [Bonnemaison,!4l971'; HoIIiday/lW!]: '’ i,£l’ >
. Интересную особенностьтгопуляции'зимней пяденицы изФран-
ции обнаружил Л. Бонмезон [Воппета18оп,197'1]. По его мнению, в
пределах ареала вид представленпо'крайней мере двумяйкотипами.
Один, (ИЗ Ленинградской обл.,исследованный'И.В.Кожапчиковым
[;1950],-имеет зимнюю эмбриональную диапаузуадругой —’из Сред-
ней Европы (Чехия и Франция) — неймеет. Зимовка у нихпротекает
на эмбриональной стадии без остановки развития, алишьзамедляет-
ся под влиянием низкой температуры. По наблюдениям Л . Бонмезо-
на оба экотипа встречаются вШвейцарии: в горах—'аналогичный'ле-
нинградскому, в долинах - соответствующий французскому.'Описан-
ный сезонный цикл» по-видимому,'широко!распространен'у[чешуе-
крылых. Он свойствен не только многимйяденицам, но й представи
тенямдругихремейств [Кожанчиков5,11950]'? о:''т ншо . ;
Прдрбнь^й жизненный цикл обнаружила Е.И. Глиняная [1972]у
замородковрй листовертки, Exapate congelatella. У этого видай летняя
и зимняя диапауза наступают облигатно,i нов отличие от зимней пя-
деницы у заморозковой листовертки’летняя диапауза куколок терми-
нируетрдйфотопериодическими услойийми (рис. 63)1^’* ^ ' ' ^ ’ •!
1(42
В условиях короткого дня (12 *
и 14 ч) через 90 дней вылетели все |
бабочки. С увеличением продолжи- В
тельности дня доля реактивирован-
ных куколок уменьшалась)! Повы-
шение температуры действовало §
аналогично увеличению фотопери- §
ода. При исследовании температур- ч:
ного оптимума реактивации обна-
ружена необычная смена темпера-
турных требований диапаузирую-
щих куколок E.congelatella'. нор->
мальное развитие куколок с мин и-'
мальной-смертностью отмечалось -.
лишь в вариантеопыта, когда куко- '
локподвергали последовательному
воздействию сначала высокой Рис.63..Фотопериодическая реактива-
(18 °C), азатем низкой (10°C) тем,- пня диапаузирующих куколок Exapate
пературы. Как отмечает'автор/та- congelatelia при рмиых температурах
кое явление у насекомых зарегист- j . t
рировано впервые) хотя у растений -- -" - •
оно хорошо изучено'на примере ранневесенних эфемероидов [Горы-
шина, 1969]. I' Г < . ;
Регуляция моновольтинизма на основе двух периодов покоя ха-
рактерна и для тысячедистникового листоеда, Galeruca tanaceti. Раз-
витие этого вида в природных условиях было подробно исследовано
ТА. Волкович в-Белгородскойобл. (рис?. 64, а).——-1:5
Эмбриональная'зимняя диапауза'наступает у этого’листоеда не-
зависимо от внешнихусловий/Ъесной выходятличинки, а в конце
июня - молодые жуки, летняя репродуктивная диапауза которых ре-
гулируется короткодневной ФП Реп орогомоколо; 15ч'ЗО’ мин. Попа-
дая в длиннодневныеусловия)’все'ониформирурт ^непродолжитель-
ную летнюю диапаузу, окончание которой такжеконтролируется ФП-
условиями. При уменьшении длины дня до порогового значения жуки
приступают к яйцекладке. Адаптивный смысл, летней диапаузы не
вполне’’ясен, но. исследованная
G. tanacetiiQЮжной'Ацгдиифазвигаетсяровершендрпр(трйже схе-,
ме (рис. 64, б);?Хотя сравнцваемыр популяции о^итцют нд близкой
гер^аф^тской)г^отр,чкритичесгаё(прдю1и реающй, ^^дуди^у^-
143:
.ЛИЧИНКИ»,,г t
'куколки If’-
......... .Л-
I -s.1.-.' S« з .1.
я
( Май . ,, ,Июнь . .Июль . |Август ,Сентябрь; Октябре Црябрь
' , .релродуюивная диапауза . диапауза яиц,- Г(,
: . л •• •*- -;»/
, ilPuc.64.,Схема сезонного цикла Galeruca tanacetl: . > : ; ,
, ,т а-влесосгепнойзонеРоссии;б-вЮжнойАнглии[по:51е«, 1966].Сплош-
‘ ’ ныелинии-кригическаядлинадия. Вверху-фотопериодическиереакциисоогвет-
г г* саующихпопуляцийприйО“С ' - ;,i-. >; Л •
</! J- аЖУКМг 'г ' ' ''"'V -> <Н.' j 'ЗД.' < :'<
щих ^'терминирующих диапаузу, оказались у нашей лесостепной по-
пуляции чуть выше,"примерно на 30 мин (рис. 64, вверху),' поэтому
сроки наступления и окончания летней диапаузы у нее сдвинуты к
Середине лета. Это, несомненно; связано с отличиями природных ус-
ловий в связи с континентальностью климата и отражает существова-
144
ние географическойизменчивостисезонных адаптаций-не толькопо
широте^нойтодолготе.':;’^?'':";'‘;‘‘!-,-г,':ЛГ; !/Я ?и‘-' 1 -;’-
я' У японского парусника,®1/1^^о>рй/а}орол/со;‘наоб<ч>от, летняя-^
диапаузаоблигатная, ааимняя регулируется пониженной температу-
рой. Летняя куколочная диапауза наступаете начале летанезависимо
от ‘ внешних условий, Ъднако терминируетсяподвлияниемкоротко-
го дня и пониженной температуры, что типично для летней эстива-
ции. Необычной особенностью этого вида является то, что имаги-
нальная дифференциация не завершается осенью, и летняя диапауза
куколок переходит в зимнюю диапаузу фаратного имаго. Реактивиру-
ющим фактором для имагинальной диапаузы служит воздействие по-
ниженных температур [Ishii, Hidaka, 1982]. Необходимостьхолодово-
го воздействия для реактивации у близкого вида L.puziloi не обнару-
жена. Подобным образом, видимо, контролируются сезонныециклы
насекомых в аридных и полуаридных условиях. Эстивация, переходя-
щая в зимовку, делает возможным развитие одной генерации'® усло-
виях избыточно длинного летнего сезона и синхронизирует появле-
ние питающихся стадий с относительно коротким периодом вегета-
ции их кормовых растений. Разнообразные схемы моновольтинного
сезонного цикла описаны-М.И. Фальковичем [1979] и В.Г. Каплиным
[1994] у пустынных чешуекрылых. ~ ‘ ’ Г
Наконец, чрезвычайно распространенным типом регуляции мо-
новольтинизмаявляется сочетание dejaфакультативных диапауз.
Златоглазка №ле/ара/Маимеетповсеместнов Европеодну гене-
рацию и уходит на зимовку на стадии личинок ! возраста, но зимую-
щие особи не теряют способности изредка питаться. В этом случае
под влиянием короткого осеннего дня (короче 1 $*ч)'линыайщ следу-
ющий возраст задерживается на 6—7 месяцев. Развитие личинок П и
Ш возрастов весной и'В1начале; лета, напротив, лормозится длинно-
дневными сигналами и ускоряется короткодневными. В результате
молодые имаго появлЯютсятолько в конце лета. Такойсвоеобразный
механизм осуществлениямоновольтинизма, основанный нареверсии
количественных ФПР,Затягивающих темпы роста личинок, позволя-
ет N.pallida существовать в биотопах, где численность жертвы обычно
невысока инестабйльна [Canardj’1988’, Canard, <Мта1Я988]:! *
Ущитовки Parthenolecanium comi [Шельдешова. Стекольников,
1965; Саакян-Баранова идр., 1971] также наблюдаетсядва типа'се-
зонной задержкн развития, которые авторы рассматривают хак раз-
ные формы диапаузы. Это, во-первых, относительно неглубокий пе-
145
риод.летнего покоя, сопровождающийся сильным замедлением раз-
вития личинок I стадии и прекращающийся безюхлаждения, и.во7
вторых, значительно более прочная димняя диапауздличинок 1I ста-
дии. Для возобновления развития весной здебуется .длительное, ох-
лаждение^ Цикл в целом обусловлен инверсие^ФЦР-по стадиям: ко7
роткодневцая реакция личинок! ;стадии и длиннодневная rr v ли^и?
нок II сдадии {рис.ббХр; ,
Рис. 65.’Фотопериодическая ^реакция личинок! ЯП стддиидальне-.'' ,
। . восточной популяции акациевой ложнощитовки при развитии на
,фасоли, Температура, 25 °СДпо; Саакян^рарановаидр., 197Ц,,, ,
Необычный механизм-редукции числа поколений описан у
Drosophila phalerata. Хэтоймухи, по данным К.Ф. Гейспиц и Н.П. Си-
моненко £19701,1 ^Ленинградской обл. .в июде формируется летняя
ДИапауза (рис. 66).. •, х..
f. Наиболыпееколичество диапаузирующих особей,составило 39%
д ля. ленинградской популяции .д 43% для .сухумской. Сроки наступ-
ления летней диапаузы оказались разными; дляленинградской попу-
ляцииг- начало ивдля, для сухумской,-двдуст. Длительность ее незна-
чительна, иужев конце августа-началесентября мухивозобновляют
активность. По мнению авторов, задержка развития гонад у самок 1-
го поколения возникает-в связи с нехваткой,субстрата (грибов) для
откладки яиц. Такая-фенология лоддерживается двухпиковой, ФЦР
146
Рис. 66. Формирование диапаузы у двух рас Drosophila phalerata Meig,. —
5 при воспитании вусловиях Ленинградской обл. [по: Гейспиц, Симо-
;; ненко, 1970]: ч - .z"» ! д
1-диапаузирующиесамкиленинградскойрасывприродныхпопуляциях;
;; 2-тожедлясухумскойрасыпривосшгганиивЛенинградскойоб|1.;3-даинадняв '
' , С.-Петербурге,|4-тожевСухуми ' Д'
" Ч ’ - 'if
вида: летняя диапауза регулируется длиннодневными условиями, а
зимняя — короткодневными. / у:
С На основе двухпиковой ФПР регулируется сезонный цикл ам у
коровки Coccinella novemnotata [McMullen, Д967]. ? » 5 о
i ;B заключение можноотметить, чтомоновольтинизм.'занимает
заметное место среди разнообразных жизненныхСтратегийгетероди-
намных видов. Он одинаково широко представлен во всех отрядахяа-
секомых независимо отихэволюционного возраста и свойствен-оби-
тателям всех климатических зон. Несмотря на внешнюю однотип-
ность и кажущуюся простоту этого годичногоцикла, именно на его
основе создается множество различных фенологических схем, кото-
рые осуществляются самыми разнообразными способами (рис. 67).
В простейшем случае моновольтинизм достигается благодаря
обязательному наступлению диапаузы в каждом поколении.незави-
симо от внешних условий, Такой механизм регуляции моновольти-
низма представляется более примитивным, хотя' он отмечаетеяултред-
ставителей какйрхаичных, таки более продвинутых отрядов. В других
случаях в регуляцию внешне облигатного моновольтинизма вовлека-
ются сложные экологические механизмы. При этом оказалось, что у
одних видов под контролем внешних факторов, в частности фотопе-
риода, находится только терминация диапаузы (у обладателей двух-
ступенчатой реакции); у других на первый план выдвигаются количе-
147
КОЛИЧЕСТВЕННАЯ
- РЕГУЛЯЦИЯ Й
ТЕМПОВ РОСТА
ЛИЧИНОК
;<ГК # I ?'.
ФП
Pafomana
prasfna
Tortbr
tiridana
Pterosftcfwa Д.
t^grita ^sflcls Wnrt^fs /
Laptoptarna" Dasychlra
Зо/оЬгай pudtbuhda
Pyrrhocoris Exaipata Gaferuca Utadorp^da
aptenn eongela- tanae/fk faponlca . pajffda
ДВУКСТУПЁЙЧАТЬ1Й
«фнтр|л£ ~ ;
РДЗМНО)^||4Я :
ДЕФИЦИТ
СЭТ й
Tjatferipa
gfoclBs
Chrysopf
dcwn&si
л- ; //%> I-
.'vV -! ТОрвго- АтрЫруга \1.ригПо1. Cocclneffa
\\ x 5phtera pyrtMtdea NX r novijtnno-
РЕГУЛИРУЕМАЯ
ДИАПАУЗА
йзмйй(й»м^м;играйиц:
....
$ Изменение;
норм ФПР
ФП
>
Рис.67. Способы поддержания моновольтинизма у насекомых:
- Круг-зймняйдиапауза;трбугольник-летняядиапауза
ственные ФПР, регулирующие темпы роста или продолжительность
диапаузы; у третьих обнаруживаются промежуточные или д линноднев-
ные ФПР, но проявляются они лишь при определенных температур-
ных условиях и не «работают» в природе. В последнем случае исход-
ным, вероятно, был поливольтинный сезонный цикл, а моновольтин-
ность возникла вторично при изменении условий обитания. Такой
процесс мы наблюдаем у некоторых адвентивныхвидов. Нередко мо-
новольтинизмбазируется насочетании разных типовдцапаузы (лет-
ней и зимней), разделенных периодами активности.
Адаптивный смысл моновольтинизма не всегда ясно выражен.
Переход к этой внешне упрощенно^, но более надежнойсхеме разви-
тия, диктуется, как правило, необходимостью адаптироватьсяк экст-
ремальным условиям, поэтому он чаще встречается у видов, заселяю-
щих зоны риска — высокие широты, горы, аридные и полуаридные
климаты. Он характерен для моно- и олигофагов, пищевые, ресурсы
которых ограничены во времени. Для фитофагов - это гштание эфе-
мероидами. или определенными фенофазами растений. для хищни-
ков ипаразитов— привязка к сезонномуциклу жертвы. Таким обра,-
зом,' моновольтинизмпозволяет насекомым расширить ареал.путем
использования ресурсов в течение ограниченного отрезка времени.
В цел ом, существование разнообразных сезон ных циклов, сфор-
мировавшихся в процессе конвергентной эволюции, создает предпр-
сылкидлябдоддвдгческого npoijpecpa н^де|ко>#ь1х.Оь.(.
Глава 7. ЗНАЧЕНИЕ АБИОТИЧЕСКИХ ФАКТОРОВ’
ДЛЯ ФОРМИРОВАНИЯ* ВТОРИЧНЫХ АРЕАЛОВ 1
’ АДВЕНТИВНЫХ ВИДОВ ъ '
ЙЙ. С ’'Ь ' ...Чй->Й.ОМ I; . 1 1». :
‘ Большая подвижность насекомых в ‘природе сйособствует по-
стояйному (активному илипассивному) перемещению их в простран-
стве, иногда на очень большие расстояния. Существуют поразитель-
ные примерам нашествий разных видов, приводящих к разным конеч-
ным результатам.' -=-; fv-' I- ..Щ- „ч;.. .-у,
! Исследование сезонного развития таких’адвентивных видов в их
новых’ареалах дает богатый материал для анализа приспособитель-
ных возможностей насекомых при их расселении, для понимания
процессов внутривйдовой дифференциациий микроэволюции.''v
•м '•11 С целью сравнительного анализа Этих процессов были"выбраны
колорадский жук,1 американская белаябабочкай хищный клоп поди-
зус, представляющие фазные отряды насекомых. Все три вида при-
шельцы из Северной Америки, тде они имеют широкие идентичные
ареалы и характеризуются полифагией или широкой олигофагией (как
колорадский жук) [Чураев,1962; Hsiao, 1978; McPherson, 1980]. Зиму-
ют в состоянии диапаузы, наступление которой контролируется ФПР
длиннодневного типа (Горышин, 1958а; Hidaka, 1977; Горышин идр.,
1988]. Несмотря на общность экологических черт, судьба иммигран-
тов на европейском континенте оказалась разной. Колорадский жук
акклиматизировался, освоил огромную территорию и активно рас-
ширяет свой ареал, продвигаясь дальше на восток. Американская бе-
лая бабочка акклиматизировалась, но освоенная ею территория огра-
ничена на севере 47-й параллелью, а на востоке — побережьем Кас-
пийского моря. Клоп подизус пока нигде не прижился, хотя, видимо,
существует зона его потенциальной натурализации.
Что определило судьбу интродуцентов, какие экологические осо-
бенности этих видов обусловили успех или неудачу при акклиматиза-
ции, какие факторы оказались главными при формировании вторич-
150
ных ареалов и,, наконец, каковы перспективы для ихрасширения —
вот основные вопросы, которме'МЫ раромотримщнастоящей главен
7 Д., Американская белая бабочка, Ilyphantriacunea Vrury >
у " П И)’ .1'ОТЧлШ ?Л-лД ч ' 'I.,
,, Первичный ареад американской белой бабрчки простираетсяют
юга Канады досубтропиковСША, гдерна в зависимости от климати-
веских условий может даватьртодногодоА-Фпоколенийв год [Hidaka,
1977J. «Будучи обычным элементом фауны , у себя на родине амери-
канскаязбелаябабочка не привлекала особого внимания>исследовате-;
лей. Статус ее резкр. изменился после проникновения Hai европей-
ский континент и острова Японии, где она приобрела значение
серьезнрго, вредителя .«щрво^царкрвух масавдеций/ и главнейшего
об^ектажар^нтина.; w -£; > ч-1--!,ч
В^Европу СВенгрия)-//. смиеабылдзавезенасдучайносгрузомиз
Северной Америки в 1939-1940.гг. [Warren, Tadic, 1970]. К1948 г. она
заняла почти всю территорию Венгрии,лзатем распространилась в
Чехословакию, Югославию, Болгарию, Австрию. Позже она была об-
наружена во Франции и в Греции. Цервые находки Н. сипеа в Италии
относятся лишь к началу 80-х годов, хотя появилась она там, видимо,
несколько раньше [Deseo et al., 1986].
На территорию нашей страны американская белая бабочка впер-
вые проникла в 1952 г. и заселила только равнинную часть Закарпа-
тья; барьером для дальнейшего распространения на восток оказались
горы. Позже в 1966 г. американская белая бабочка уже из Румынии
проникла в Молдавию, а впоследующие годы была обнаружении Хер-
сонской, Николаевской, Крымской, Запорожской и Донецкой облас-
тях Украины (Американская белая бабочка, 1989]. В дальнейшем она
освоила территорию Ростовской обл., Краснодарского и Ставрополь-
ского краев, Кдбардицр-Балкарии и Грузии. Распространяется аме-
риканская белая бабочка в результате активных перелетов имаго, пас-
сивного-переноса их ветром, но особенно большое значение враспро-
странении вида имеет случайный завоз диапаузирующих куколок че-
ловеком при транспортировке грузов. Так, отмечаются случаи обна-
ружения куколок при таможенном досмотреввозимых товаров - ме-
бели, швейных изделий и др. [Стригина, 1983р > • ч ла.гк -..
Экологические особенности Н. сипеа,'натурализовавшейся в Ев-
ропе и Японии, изучены лучше, чем у американской. При этом обна-
151
ружены как сходные черты, так«различия в проявлении фотоперио-
дическиходаптаций^уразныхнопуляций.r :,,v i- * < •
Куколочная диапауза Н. сипеа контролируется ФПР длинноднев-
ного типаяКритическийфотопериод/несколько’различающийся у
разных популяций, в пределах от 17 до 25 °C характеризуется относи-
тельной термостабильностью(рис. лишьповышениетемперату-
рыдо28—29°Снезначительно снижает количество диапаузирующих
куколок[Masakietal.’, 1968;Кинд,Королькова,WIJ.si - ч ж.я
Чувствительность к фотопериодическим1 сигналам*,1 по данным
японскихэнтомологов[Н1бака, 1977},’приурочена к'младшим и сред-11
ним гусеничным возрастам, такчтохарактер дальнейшего развитиях
Vвозрасту гусениц уже детерминирован (рис. 69). Д.* Ясич и ’В.’Мако'
[.Jasic, Macko, 1961} склоняются *к тому, *что чувствительны к длине*
• дня лишь гусеницы I и II возрастов. Другие авторы считают, что реах- '
ция определяется на старших гусеничных возрастах [Saringer,
- ***,'* '7 г М г адигшлА Ж-И J
»> Рйс.б-У.^Фотопериодическаяреакциягеографическихпопуляцийаме-......
риканской белой бабочки: я ?, лн т • ы >1;| '
л,; , Популяции: 1-закарпатская(25°С)[по: Кивд,Королькова, 1991];2-че-
" хословацкая (20 °C) [по: Jasic, Macko, 1961 J; 3 - японская <25 °C) [по: Hidaka, 1977];
'4-игальянская(20~24°С)[по:Ве8еоЫа1.,1986];5—BeHrepcKaa(23”C)[no:Saringer,
1974]; 6 - канадская (24 °C) [по: Hidaka, 1977]; 7,8 - краснодарская (20 и 25 °C)
152
Рис. 69. Чувствительность американской белой бабочки к длине дня.
на 'разных 'стадиях развития [пб: Hidaka, 1977]:
" Чернымобозначеикороткийдень(8ч),светлым-длинный(16ч)
• a Ъ i • , А.‘Г"I'ff' • i < ’ . ' • • ' '• ' ’
1974].Отмечается сезонная изменчивость порога ФПР и отсутствие
зависимости этого параметра от температурных ритмов [8агпц»ег,4974].
Пакет короткодневной фотопериод ической информации при 24,5 °C
составляет 15-17 сигналов [Hidaka, 1977]. ! 1 r ’ U1V '
’ "Для развитий одного поколения американской белой бабочки
требуется по сведениям разных авторов от 800 До 1000 гр.-дн. при
нижнем температурном пороге развития 10°C [Ito et al., 1968; Deseo et
al.?1986^’1 ' 1
Исследования, проведенные в Славянске-на-Кубани, показали,
что годичный цикл Я. сипеа хорошо вписывается в климатические
условия краснодарского края (рис. 70). СЭТ выше 10 °C за вегетаци-
онный сезон 2200—23QQ. гр.-дн надежно обеспечивает развитие двух
четко разграниченных поколений, а критический1 фотопериод (15 ч'
светав сутки) гарантирует активное развитие куколок 1-го поколения
и определяет обязательное наступление диапаузы у куколок 2-го: все
особи в вариантах опыта, начатых После 15 июля, Диапаузировали. По
такой же схеме американская белая бабочка развивается на юге Укра-
ины, в Ставропольском крае и Ростовской обл. <
‘ ' С продвижением на север'изменяется соотношение температур-
ных и фотопериодических условий, 'что резко влияет на характер го-
дичного цикла Н. сипеа. Опыты, проведенные в Белгородской обл.,
153'
Рис. 70. Формирование диапаузы у куколок Hyphantria сипеа в
Слав5шске-на-Кубаии:4'№'г й >«•-
1Ъризонгальные лиции - вариаигыопьпов: сплошной отрезок.-средняя дли- >
тельностъразвитиятусениц, пунктирный-тожедпякуколок-Треугольники—сред-
нийсроквылетабабочек. 1 —сезонное изменениедлины дняс 1/2гражданскихсуме-
рек; 2-значение, порога фотопериодической реакции; 3-нарастание количества, .
' диапаузируюи1йхкуквлок,1%,вполевькопыгах.Цифрынадосыоабсцисс—средне-' >- 1
оьдтэмкчьлотл ч.
Г” Н 4 , -Г ; ; -тг TV---------------------------Т“Гж
показали, что на этой широте (50 с.шп) намечается диссонанс между э
фотодермическимиусловиямцсредыи экологическими требования-
ми; вида' Hec^pptpH надо^'^тр-рЭХвь11116'10 °C длрсостеднцй зрц^т
(1600^1100,фгДН-)^ракпфур^дзвитие одного поколени^^Дютопе-^
риодические условия не'допускают формирования диапаузы у куко-^.
лок этого единственного поколения, Результате в начале августа из
них вылетают бабочки и дают начало 2-му поколецию? которое не
обеспечено тепловыми ресурсами й обречено на гибель, Для натура-
лизации американской белой .бабочки в лесостепной зоне'куколюз !
гр поколения должны формировать диапаузу Это возможно лишь при ’
условииразвитияГусениц,.ндчивдяfoil возрасталафрнедлиныдня^
меньше жритичрскойХ! 5 ч)..' длида'^цр^н;а, шцррте,Белгороду (
наступаетдолько к25 августа,тогдабольшая частьонтогенезадолжна
проходить в.сентябре, Однако, температуравоздухавсенхябре начи- {
нает резко падать, и ее среднесуточное значение .не ,превышает ! 3. °Сг ;
уже во второй декаде. Низкая^темлера'^а^не ^рзволяет^сеницам
О iqmrc>'
ii'B'AJoqj; , .! taivfi lT.rtUT •;" if ншщ
154;
Таким, образом/двеэкологическиеособенности американской
белой бабочки,акклиматизировавшейсяв>К.раснодарскомжрае, пре-
: пятствуют ее распространению далеена север-это низкийпорог ФПР
в сочетаииис болыпойсуммойтепла,необходимой для развитияюд-
•ногопоколения, .и фотопериодическая чувствительность вмладших и
среднихтусеничныхвозрастах. ! : а , хщщдс а ом® 1
t-При нал ичиитаких особенностей|никакиекратковременныеиз-
мененияпогодных условий немогутобеспечитьюинхронизациюсе-
зонного развитиявида ,с климатическимиусловиями на территории
севернее47-й параллели,жаркие годы чувствительный период мо-
жет сдвигатьсяна ещеболее-ранние календарные сроки; детермини-
руя. куколок наактивное развитие; в холодные годы гусенины обрече -
нынагибельиз-заневозможности достичьдиапаузирующейстадии.
На всей территории Восточной Европы нет зоны; где>американская
белая бабочка могла бы развиваться только® одном, поколении; чув-
ствительнаяютадиявсегда приходитсжна длинудня,больше крити-
ческойд ля завезенной®Европу ятопуляции этого вида®®'• • Gz< «
;.z ОднаковСе вернойАмерикеестьфсгионы, где//. гипса заверша-
еттолвкооднопоколенис.Например/поданнымР'Морриса[ Morris,
1Я67],>вФредериктонеК45лС.;пк)асуколки1тй генерации всегдафор-
мируютдиапаузу.Х сожалению, авторшеисследовал параметры ФПР
эдой попудяции, а лищьэкстраполировал изменениесстественной
длины дня на момент окукливания. Вычисленная величина крити-
ческоголорога составила J4,5 чсветавсутки.Совершенн ©очевидно,
чтопри таком значении лорогаиндукция диапаузы уместной попу-
ляции невозможна. Несоответствие было разрешено после исследо-
вания японскими}энтомологами‘[Н1бака, 1977|американской белой
бабочки из Онтарио (Канада, 45 °с.ш.). Оказалось, что у этой, близ-
кой к4>рсдериктонской,,лоп.уляциишоро1,ФПР составляет >16,5 ч и
рбеспечиваетлцндукцию диапаузы;улкуколок. 1-го/И единственного
поколения Т/. смлед д, указанной , зоне. Д случае, завоза* особей этой
популядиивнашустранувозможностьихакклиматизациинатерри-
'дориисевернее47-й>парадлрли<неисключена....: ь / г,-.игл,.?
. J ' Друдаивозмржцостьдкклиматизацинамериканскойгбелой баг
бочки в зоне, допускающейразвитие;только одногопоколения, мо-
жет возникнуть за счет небольшого количества особейй-го локоле-
ния. спонтанно формирующих диапаузу. Наличие единичных диапа-
узирующих куколок в 1-м летнем поколении в европейскихПопуля-
циях отмечается разными авторами [Jasic, Macko, 1961; Ушатинская,
155
Иванчик, 1974]. В наших опытах, проведенных в Белгородской обл.,
из 185куколок8былидиапаузирующими,чтосоставляет4,3%. Вопы-
тах в Краснодарском краедиапаузирующихкуколокв1-мпоколении
отмечено де было. Это позволило предположить, что диапауза в 1-м
поколении могла формироваться под влиянием внешних условий.
Однако в опытах, проведенных в Ленинградской обл., гдеможнобыло
ожидать появлениядиапаузирующих куколок, если не «большем, то
хотя бы в том же количестве, этого не наблюдалось. Температурные
условия здесь заведомо непригодны для американской белой бабоч-
ки: СЭТ выше 10 °C около 550 гр.-дн не обеспечивает развития'даже
одного поколения. Но искусственная выкормка была проведена с та-
ким расчетом стартового срока, что гусеницы успели окуклиться. Ку-
колки были получены в конце июля-начале августа, а в конце авгус-
тат-начале сентября из них вылетели бабочки. Среди Куколок не было
ни одной; диапаузирующей.
Итак,«результаты.опытов, проведенных в трех географических
точках, однозначно подтвердили вывод, чтовторичный ареал амери-
канской белой бабочки на территории Восточной'Европы сформиро-
ван, и границы.его определены экологическими особенностями заве-
зенной популяции. Можно предполагать, что* и в« Западной *Европе
ареалД силватакже будет ограничен насевере47-йпараллелью. Сход-
ный вывод .был сделан относительно возможности акклиматизации
вида,в 4exHH [Jasic, Macko, -1961],;а промелькнувшие сообщения о
натурализации* американской белой бабочкивВосточной Германии
не получили подтверждения [Braasch, 1976; Grosser, 1980]. ’
,5. .... ; ЛЭД :. rd-., .'Н ‘ ’
, 7.2. Хищный клоп ТЪЛхиглшси/п’влйгй Say
Очень ярко значениефотопериодичес ких адаптаций проявил ось
при попытках акклиматизировать американского хищного клопа
Podisus maculiventris. Этот вид был интродуцирован в Европу для био-
логического подавления! колорадского жука и американской белой
бабочки. У себя на родине подизус распространен на обширной тер-
ритории отюго-восточныхпровинцийКанады до юго-востока США
[Ижевский, Зискинд, 1981]. На основеметодаклиматических анало-
гов были намечены зоны возможнойакклиматизации хищника в За-
падной-[Couturier, 1938], а затем и в/Восточной * Европе [Власоваи
др.,1980]. - пс- - - - • ..-j-.
156
При реализации программыинтродукцииновогоэнтомофага в
нашу странубылоучтено также, чтообе потенциальные жертвы поди-
зуса (колорадский жук и американская белая бабочка), имеющиесход-
ные с ним первичные ареалы, натурализовались в Европе, поэтому
можно было ожидать, что акклиматизация подизуса на новой терри-
тории также завершится положительным результатом. Однако попыт-
ки натурализовать подизуса в естественных условиях Краснодарского
края, Черновицкой, Воронежской, Киевской и Московской областях
не достигли успеха {Колесниченко, 4980; Буракова, 1982].г-Вчем же
причина неудач? До сих пор считается, что главным условием дости-
жения успеха при акклиматизации вида (автор рассматривает только
абиотические факторы среды; безусловно,, биотические его компо-
ненты имеют не менее важное значение) является соответствие гигро-
термических условий его первичного ареала климату в предполагае-
мой зоне акклиматизации. Однако, как и следовало ожидать^'боль-
шинство неудач в этой практике связано с тем, что метод климатичес-
ких аналогов абсолютно не учитываетфотопериодического фактора,
тогда как именно он отвечает за своевременность формирования зи-
мующих'стадий и, следовательно, определяет саму возможность «оби-
тания вица на новой территорий. Проиллюстрируем* это, на примере
подизуса.' ’ •''' 4 ; •
: Многолетние исследования позволили определить основные па-
раметры" развития^ получить Четкое представление об экологичес-
кой характеристике этого вида. Вопреки литературным данным
[Couturier, 1938; Warren,-Bjegovic, 1972], было установлено, что поди-
зус имеет имагинальную диапаузу, контролируемую ФПР длинноднев-
ного типа [1орышин идр., 1988]. Чувствительностью к фотопериоди-
ческим сигналам обладают личинки, начиная с III возраста й имаго
(рис. 7Ги табл; 12). , )
’ ‘ Критическое число короткодневных сигналов, обеспечивающее
наступление диапаузы уцоловины особей, составляет 10—11 дней при
20 °C и 16 — при 24 °C. Зависимость критического пакета фотоперио-
дической информации от температуры - важное для синологическо-
го анализа свойство ФПР подизуса. Трудности длиннодневно-корот-
кодневной’перестройки Н легкость, с которой происходит возобнов-
ление яйцекладки; свидетельствуют о том, что преобладающим со-
стоянием для этого вида является репродуктивная активность. Фото-
периодическая индукция наиболее выражена при 20 °C. Диапауза у
157
Ж.ОГ
««Возраст Ъичииок”4'51
II III IV V
ННИЙЯ
-fr ! -Hi. И-., bl
Количество
' ‘даип»пи₽тюших
нГ|й р.Л /wd
.ЗТЕД 1 '
нклопиг®®"^ ь. . '
чендон гШм । г-
„ 9.1
(М,ОЦ ;.ё'П.^фЖО!-1’..р'.;
_______ 78,9__________
‘ м,
'ЖЖ
f-jqcd
”’ ?OS
-мсрр;
-па ио'
СПОЯЭС
/.wrasii
5Ж Ч ’л1
ГСЭОЛ “ '•,'•'>»•-,'Т’ V:’‘‘•ЖТП -:<ЛГ'.;•••.- .'«iji'Hb
*' Рис. 71, Чувствительность подизуса к длине дня в разные периоды
•'/?<-О:.О,\развития'^н:.1^ЯС| ЦОЬ1 Jwsiirf шимоЛр' 4лЛ1»>» 'Л<‘. >.
-<И!МОГ Т За1шрихованные4участкй —короткийдень(11,ч)>велые ~длинньгй<16'ч>. ! , (Я1
кото rt.hr (лй5№. ‘хшжкя -.' там »F лиаю id»Н..Ж
г ’ ’м)2" ''Т1-'”'••’ пт;- о-г-тг г? ;' ;-т гг-с, - •’•ч;'! -г'
всех особей формировалась при длине дня 1,0 и 11 ч светав^су^ди (см.
рис."9)?Критический фотопериод блдузд к 1£ ч 24' мин,' ]’. ” / .. ( .'
' ’’Особенно интересными и ва^ыми дляанализа сезрннрз-о^раз-
витиявидаоказалисьрезультатырпь1Тов5прд,.1.7о.С..В1опреки,ожида-.
нию короткодневная индукция диапаузыд этрм слу^ае оказалась,ме,->
нее'выраженнЬй7чрм1при^20,йС:’'цочтиУ20^шф|СВгКорртко^1евных.
режимах 8,‘1'0и 1'2 ч света в сутки'созревали и откладывали яйца.,В то
же время количестводиапаузирующих особей при 14 ч оказалось по-
чти таким’же^как дри,20 и,25 °C? Осдабление' короткоднерного эф-.
фектаи'тевдевлцщк^рми1юва1р1юдиала)^1дри#17°С^ходят;объдс7;
РИС")?}' । . г > . ) । <f ₽.?.« '!. •< и
В совркупностиполуче^ные данныр свидетельс'гвуютодом, что,
у подизуса доминирует настрой на активное рдзвихие/Дмагинальная,
диапауза возникает лишь при определенных сочетаниях длины дня и
температуры, наиболее эффективных для короткодневной индукции.
Такие’особенноста ФПР несомненно связаны с южнымдроисхожде-,
йием'популяциипод'изусаДшта/М^^и^Г^^ЗВ^р^^интроду-
ци^ованйойвнашу стра^ * ° , - Р . д
0 Проведенные эксперименту цокдзали^что критическая длина,
дня имеет одинаковое значение в условиях постоянных и перемен-1
ныхтемрератур.Но всвязи сузкимтемдердтурным’оптимумомлро-,
явления ФПР при теру орщ^мах .заметноснижается количество диа-.
паузирующихособейв короткомдне.особеннопри понижении тем-,
пёратуры ночи. Реакция на константные и изменяющиеся фотопери-
1’58’
,; :s Таблица'12. Индукцияпдиапауаыуподизуса шод;влиянием;: 7 . < g „
1.тзвДР^^,^й-к9РотюдневЧьк<йзнен^ий(/м^^^Од й, . .(т
• п '' -vr • ,cr ,;(,...
XКоличество'j; ъ: Процент << •• r'.X’N
диапау- j прекративших . v
Пирующих i яйцекладку а ХД* *
-7 ' *“ ЛЙ.Х
Л1*Шежи Югаями. переноса 2 ЪКоличестш^Й ’’ ЧмспоКТ
. чЛ h»/f^raW^q<yi2Xc.4 ‘^*a
-S I до окрыленияj •’ I
«m-mw RTHfiMOH □. шггТемпература 4 я л лиц R
Si Л Шостовы©» $2«л ч ид мюуяГцд 35аш ? i,
x;2s >.}j йс.-ргДфпм'.т? iu24r-27|.KJ(; A М энй1Р°£ • <:.. ??'«&•—’ ьэап»..-’л
ыАк- жгЗрЛгАлмцар?b'~«c^r«^Siotfsq Я“'): Л- <
4 Л-4' JD ; А) У0,0
&! IfO-lW'* р;26^т /50,0'Л1'м'*“<'4бЛ 1 ~ (-!-
•'' 6 !'Л •• Ч!№5>М Э-зНО?! г!8»-аЭ" ",8 23R>'J№ra47.8qd 58,3'Я."Н. fi’.W
• я 1 -Тп г - Y? Имаго г- -»'(> /1,с'к.сО’. .м'! .я '1<40г?г>" дон&°«r>y krхда *••
8 Постоянно в 16 ч - 33 О -.
., • ,%. Температура 24 °G ...... .<
•«’.!rSfi’Wife 5?A3h 4Э К T;
10 Л-3 , ,17-18., .23 43,5'* 0
7'H^X 3C!.R£.M.Hjj.;3.b':fi ,{₽. <.?"< ^JqHM 5 ЗОО'П ;| C И ^0'>l<^ №Г
•JF® 3Hlt?r 'Н'.1Л-4!’:ЖИи('Л74ПЗ('!’'Т.Р1ф26Л^ .йИЮОЭНЭДзОЦЗ .7,7г:-!;н:>\о
.j.fjB-&j4q')’rafqqyt8-9 кэл sn»37? v тийеАь'ф т>’> мДзжа^.’;ап
£.»;яИ|г у wn.-’fl*5 'tn цн гел^Ак?’? i и.<^;эт « ^хА?ил кгЛ
15 Имаго 0 39 0 0
‘'W’ “Имаго -Нодней1 {RBbi‘-‘ '' rwM^:o!-&q*sr
t <<дао«Ив<лоян1ю-вИвм4П й_<.еоп|цл-234;-'.;н ,‘-:qdi0»ro КГ ;,r~4'jnJ40T •;
“Г Л^гй<лг<> ₽>! OJirfEpT Г.»ТО;^А£0.з г'Г'ХЮ’?
.Ы.ч:г.ш.сгаг!*л' i'wi.Fim) ”‘°;! -шад дхжфлмэт в кпОктзс 0^
оды оказалась идентичной. Для развитияадного поколения подизуса ;
необходима сумма зффективныХ)Температур около; 400'гр.-дн...при>
nOpore'M^CwVlTH Г.:'Г*ГИЮ,1 .i-O'CHJAOT! И^шеджоп ХЭДы.'ЧГ ЖНЗШзЖГ'!
н ?1 Расчеты показывают, что подизус .мог бы нормальноразвиваться ,
набольшейчастиРоссии, давая>разноечислопоколенийвзависимо,.|
сш ютгклиматических.условий. Например^в Белгородской.обл»^теп-
ловые ресурсы д ля насекомых с таким же темпераданымгпоровом раз-.
вития, как у подизуса, £оставляют окодо;900 гр.-дн,-что,достаточно
дляззавершениявдвухпоколений хищника. Однако2-епоколение,г
котороедолжноформироватьзимующий «запасу попадаетвусловия-
длины дня.-бадьшепороговой.1Фотопериод,!допускающийиндукцию>
имагинальной диапаузы!угаоловиныюсобей, насту паетв самом конце в
сентябрялВ соответствии^ особенностямичувствительной:стадиии
величиной пакета фотопериодической информации (не менее 10—11
15#;
короткодневных сигналов при 20 °C) в условияхдлины дня ниже по-
роговой должны находиться личинки, начинаясГУ возраста (см.
табл. 12). Продолжительность развития личинок IV—V возрастов до
имагинальной линьки составляет при17—20 °C 14—16 дней Преддиа-
паузнре питание имаго до окончательного формированиядиапаузы,
как показали эксперименты, требует пребывания в течение еще
18 дней в комфортных условиях. Таким образом, с момента наступле-
ния критической длины дня'в природе до ухода клопов в диапаузу
должен пройти по крайней мере одинмесяц с температурой не ниже
15—16 °C. В районе исследований переход температуры через отметку
10—11 °C происходит ЗО сентября. Средняя температура воздуха в пер-
вой декаде октября составляет 8,7 °C; а в конце месяца — 4,4 °C. В
таких условиях успешное завершение подготовки к диапаузе у поди-
зуса невозможно. ' ?
Критический фотопериод для подизуса, равный 13 ч, с учетом
гражданских сумерек, наступает на всех широтах северного полуша-
рия примерно в одно время (23-25 сентября), приближаясь к дню
осеннего равноденствия. Этот,факт очень упрощает выделение зоны
возможной натурализации подизуса на территории нашей страны.
Для подизуса в. осенний период благоприятны природные условия
тех регионов, для которых характерна теплая продолжительная осень
с температурой октября, превышающей 15—16 °C. Такие условия в
России создаются только на крайнем юге Краснодарского края. На-
пример, в Сочи переход температуры через отметку 12 °C происходит
30 октября, а сумма температур выше 11 °C (нижний температурный
порогразвитияподизуса).завегетадионный'Сезон составляет при-
мерно 2 000 гр.-дн.ДГакая сумма эффективного-тепла обеспечиваете
появление четырех поколений подизуса, последнее из которых будет ,
развиватьсянафонефототермическихусловий/индуцирующихдиа-
паузу.Температурныетребования диапаузирующих имаготакже удов-
летворяются. Этовселяет надежду на возможность акклиматизации >
подизуса%ннэтой!Территории.?«!'''!; <-кг к-.к
о;: Следует отметить, одйако, ’что анализировались возможности^
натурализациипопуляцииподизусаиз штатаМиссури.Большие возч
можности для успешной акклиматизации, несомненно,! имеют попу- ,
ляциисеверного происхождения, тем более' что граница первичного
ареалаподизусанасевероамериканскомконтинентедостигает50 °с.ш.
Безусловно,.другие популяциилотребуют тщательного эксперимен-
I- ' : Г-Ц,- * '-4
160
тального исследования, результаты которого позволят прогнозиро-
вать зону потенциальной акклиматизации их Па новой территории.
Обобщая изложенное, можно заключить, что неудачи при по-
пытках Натурализовать подизуса на территории России связаны с ин-
тродукцией сюда популяции южногопроисхожденияХ38°с.ш.), осо-
бенности ФПР которой не соответствуют фототермическим услови-
ям большей частияерритории нашей страны. Главныеиз них:низкий
порог термостаб,ильной ФПР, узкий температурный оптимум прояв-
ления ФПР исвойства чувств ительной'Кдлине днястадии.
’ '~U~ ‘ ’ » ЧЦ’ф»
7Д?Колорадский жук,decemlineataSsy
--.ж \ ..... , .
Колорадский картофельный жук дает пример грандиозного вне-
ареального расселения, ^результате которого в Евразииформируется
вторичный ареал. Еготерритор ия, сравнимая по площади с первич-
ным а реаломжука,отличается большим разнообразием климатичес-
ких условий, адаптация к которым представляется необходимой пред-
посылкой успешного обитания. При наличии у колорадского жука
хорошо выраженного и разнообразного по глубине зимовочного со-
стояния [Ушатинская, *1966а,б) суть'такой адаптации можетзаклю-
чаться в соответствующей перестройке механизма,’ синхронизирую-
щего наступление диапаузы с сезонным ритмом климата. = »
. , По аналогиис другими поливольтиннымивидами можно было
ожидать,, что у-, колорадскогожукапо мере егораспространения на
восток и заселения новых территорий будут происходить следующие ;
адаптивные измерения параметров ФЦР: 1) критический фотопери- ‘
одбудет возрастать у северных популяций, ограничиваясезонныйникл
одной .генерацией, ,и сокращаться у южных, обеспечивая поливоль-.
тинное развитие; 2) способность к спонтан ному возникновению диа-
паузы (независимо от длины дня) у северных популяций будет усили-
ваться, д у-южных,ослабевать, также способствуя поливольтанному
развитию; 3) температурный оптимум ФПР будет повышаться с юга'
на север. Такая географическаядифференциация'по признакам ФПР
отражала бы>'фЩй*ййоВД^йацрам^иеадаптации/вида^зона5п>нр-
стидлима^а. ч ... , ,Г1 . .... .... V..7 ..Л./' .
,-;Рассмотрим, что происходит реально. Для наглядности и более
точного отражения пространственного соотношения изученных пр-
пуляций на рис. 72 каждый график территориально привязан к пунк-
ту происхождения материала, Графики построеныоднотипно. Резуль-
161
4н (Л., , ' ч . t!
.,;, iPuc.<72. -Географическая изменчивость фотопериодической .реакции t .
у. ^^ляций. колорадского жука [по: Горышин и др. , 1987):
Вертикальныелинии—максимальная длинадня втсажаойгеографической 7-
точке .BTfiKb'RH 3< 7 ЫГуИГИтЦ t>;
тать/полученыне^одновременногСлева показана ’ФПР 'берлинской
популяции,’исследованнойН'.ИТорышиным ещевТ954—1955 гг. [Го-
рышин, 1958а]. !1На ’этом графике' черным1 цветом’отмечен процент*
диапаузирующихжуковпри 26 °C ‘ прерывистой л инией — при 24 °C.
Интерполяцией'легко представить ФПР'этойнопуляции при темпе-
ратуре 25'°С,которая?была основнойв Последующихксследованиях.
Данные для' Минска'и ^Тирасполя взяты йз’работыЛ.И правовой и
В;А?^Заславского [1980]. Популяциииз Москвы, Белгорода, Симфе-
рополя, Краснодара,(Самары, Уральска исследованыв 1981—1983 гг.
изДдадикавказа - в W77-fe78 гг.АМ; чы.-’q,'••к-,.-., нм
! 1 i l При анализе1 материалов, представленных на карте, можно за-
ключить,что' у современных популяций Ькука, заселивших террито-
рию нашей страны, заметно усилилась тенденция к формированию
диапаузы. Чтобы 'более’точно'оценить степень- географической из-
менчивости ФПР колорадского жука,’обратимся к рис. 73 . Здесь дан-
ные сопоставлены’в' двух' разрезах: с ‘запаса на восток (Б’ерлин—
Минск-Белгород-Самара) и с северана юг (Москва-Белгород-Сим-
162
. :х> ' Р«с.^.-СравдениеЗфот<>периодичвской реакции, у. кодафинеских i ; ш ;
ч .популяций колорадского жукадодолготе (а) ;и.широте;(б)^<.«г >-ч;,ц u
Популяции - а: 1 - самарская, 2 - белгородская, 3-минская (по: Арапова,
1 > ‘ Заславский,1980],4ги‘5—берлинская(по:Торьш1ин, 1958а]; 6:1—московская,!— "
Н < лКЖнодарская, 3 ^белгородская! Ояновнаятемператураопигов 25 ?С >а n S . •;:! . <; ( '>
ферополь-Краснодар}.|На разрезе запад-восток (рис. 73,.а) кривые
ФПРбсрлинскойпопуляции при 24—26 °C отличаютсябольшойкру-
тизнойсчетковыраженной пороговой зоной. При длинном дне 17-
18ч(т.е.впределах экологически значимых изменений) диапаузиру-
етвсего! 5% особейрапри непрерывном освещении — почти 30%. ,,
:КривыеФПР минской, белгородской и самарской популяций
при'25 °C весьма'сближены. При экологически значимой длине дня
15,' 16 И17 чразличиямежду ними недостоверны и случайны по знаку.
Пороговая зонарастянута на 2—3 ч и более, апри непрерывном осве-
щении диапаузирует более 60% особей/По всем экологическим,цара-
метрам'ФПРэтихпопуляций оказывается менее четкой, чем у бер-
линской, которую можносчитать исходной с учетом восточного век-
тора расселенияжука.Л а разрезе «север-юг» (рис. 73,6) кривые бел-
городской и краснодарской популяций очень сходны между собой. В
обоих случаях минимальный процент диапаузирующих наблюдается
при 18—20 ч, кривые неоднократно пересекаются, что подчеркивает
недостоверностьютличиймежду ФПР этих двух популяций. Доля жу-
ков;’спонтанно формирующих диапаузу, максимальна у московской
популяции -s70-90% при длине.дня 17-24 ч. Если в эксперимен-
тальных условияхпри 25 лОи 18 ч светауровеньдиапаузирующихосо-
бей не достигает 100%, то вприродныхусловиях Московской обл. при
' U ;Т.Д ;ЛN : , ........ ,
463
средней температуре 18—19 °C развитие, несомненно,пойдет помо-
новольтинномутипу- »
Неожиданно хорошо выраженная тенденция к формированию
диапаузы была обнаружена у популяции из Ташкентской обл.
(43 °с'ш.). Только при фотопериоде 18 ч около половины жуков были
активными,тогда как при 16 и 17 ч количество диапаузирующих со-
ставляло 80 и 90%. Появление колорадского жука в Ташкентскойобл.,
несомненно, результат недавнего случайного завоза его-с картофелем
и, судя по характеру фотопериодической кривой,источником его были
либо Белоруссия, либо Московская обл» = ~ .
Приведенные материалы свидетельствуют о заметном измене-
нии параметров ФПР L. decemlineata'ROвремени по сравнению с ФПР
берлинскойпопуляции. Ойо заключаетсяв увеличениичисла особей
со спонтаннымвозникновением диапаузы. Эта тенденциянастолько
сильно выражена,чтосохраняется,ипри1ювышенной температуре,
обычно способствующей бездиапаузному'развитию. Сравнение
ФПР пяти географических
ш -у - °C
t in '-(рис. 74>показало, что даже
' при наиболее эффективном
«, • длиннодневнрмфотоперио-
ле 18ч,количестводиапаузи-
< i .|рующих’рсобей; достигает
/у»г 35-45,%.;При 16-Часовом
;//, > ..ФП/почтимаксимальная
. длинаднявСимферополе,
^Краснодаре, Владикавказе)
? „.и,- диапаузируютужебО—85%
I,, жуков-Междутемвтехжеус-
й-Доха
i J а а..а >*'*
Qi . ..I__i." I.. i——L—
/ОЬ VI;116
a . - довияхуберлинскойпопу-
•s <. >; > ляцицдиапаузировало всего
гДР%особей.(< . /hOijD
', а и Особо .следует остано-
ьр 4201 <рвидься1нагпроявлениях*ФПР
.и люл^|,. , ..ъи;кДлинвдаж1Н при1пониженных1темпераг
.Лс.^^Фотопериодаиескаяреагаимгеографи-^^ТХВ^^Ри 18 С,крт^рыебо?-
, ческих популяций калорадского;жука при 28- |Лее соответствуют УСЛОВИЯМ
"29,°,С:;'г-, ... обитания жука:влрироде.
крымскПЛХ±д^Т-^™с2Г^ Такие данные имеются не
КрЫМиКал,Ц — ЦСЛШрОДиКа>1| j"’uC|wnnUulM
164
длявсех исследованныхшопуляций(частично они представлены на
pиc^,'72^'зaцEфиxoвaнныe.yчacтки)i.i-, /> мл.? j. st, «
. Хорошовидно,-чтопонижениетемпературыдо20 °C существен-
но увеличиваетколичестводиапаузирующихжуковпридлиннодне в-
ныхфотопериодахипосугисводитк минимумуэкологическоезначе-
ние ФПР для регуляции их сезонного развития.Взонах, гдепотепло-
вымресурсамвозможноразвитиедвух-трехлокаленийзасезон,ужев
Ьйлетнейгенерациидиапаузируетбольшаячастьособей.Числен-
ность особей последующихгенераций всегдарезкопадаети неиграет
заметной роливформировании зимующего запаса; Подобная дина-
микасезонной.численностиколорадскогожукахтгмечается повсемест-
но; она проявляется и в южных районах современногоареала - в;3а-
карцатье [Колорадский картофельныйжукг Leptionotarsadecemlineata
8ау,.1981[иЗакавказье [Макеев, 1982; Ахмедов,устноесообщение].
< зРезультаты исследования географическойизменчивостиФПРко-
лорадскогожука свидетельствуют об усилениитенденциик облигат-
носхи диапаузы и моновольтинизму по мере его расселения на евра-
зийском-континенте Этатенденция проявилась не только в лабора-
торных опытах, но хорошовыраженаив природе. Полевые опытьгв
Белгородской обл.,-проведенные в течение нескольких сезонов по
традиционной методике тестирования на возможность индукции
диапаузы, ифенологическиенаблюдения, выполненныевэтомреги-
оне (см; гл. 6) ,<а также данные литературы по сезонному'развитию
жудавразныхзонах[Миндер, Козаржевская,1966^Колорадский кар-
тофельный жук,£ер//оло?агаа ,^есе/п//лед to Say, 1981;Макеев,1'982]сви-
детельствуют о том, что 1-е летнее поколение повсеместноиграет ве-
дущую роль в формировании жизнеспособного зимующего запаса,
причем у значительной части жуков диапауза возникает спонтанно.
Среди жуков последующих генераций в зоне двух-трех поколений
лишь некоторые особи-приступают кяйцекладке, небольшая их часть
успешноформирует диапаузу, а большая часть популяции погибает®
осенний период/не сформировав диапаузу, или отмирает в течение
зимовки? Они не1 имеют существенного значения для численности
вида в следующем сезоне. В связи с изложенным интересно проана-
лизировать основныетенденции, возникшие при выработке адапта-
ций к различным условиям обитания на родине колорадского жука -
в Северной Амери*®, где впрошлом векетакже шло интенсивное рас-
ширение первичного ареала жука всвязи с введением культурыкарто-
феля на континенте. 1 ?
165
Центром происхождения колорадского жукасчитаются восточ^
ные склоны южных отрогов Скалистыхтор,.откудаюнраспространилг
ся на .север и восток. К началу ХХв. колорадскийжукобосновался на
всемвосточном побережье СШАипрониквКанадудо55 °с.ш. Терри-
тория, 3SLHajMiL. decemlineata в Америке,: разнообразна похлимати-
ческимусловиям,,адаптацияккоторым, несомненно, должнабыла
вызвать изменения!ВпараметрахФПР.|Этои'было‘Показановрезуль-
татезисследования североамериканскихпопуляций 4 Hsiao,: 1981 из
четырех географическихпунктов<вл1татах Техас (26°с.ш.),Аризона
(32 |°с.ш.),1Ота'(41 °с.ш.) и;Вашингтон (4.7'₽сдл.).Самая северная по-
пуляция из штата Вашингтон <рис, 75) имееп критический фотопе-
риод 15 ч>30 мин, более южная изштата Юта -44 ч>40мин,изАризо-
.ны г-> 1 ЗчЮОмин, .наконец, у самойюжнойпопуляции из>Техаса при
всехвариантахдаиныдняколичестводиапаузирующих жуков колеба-
лосывпределах12—20%.^Представленные материалы свидетельству-
ют© внутривидовой; дивергенции по признакам ФПР, которая про-
изошла у американских популяций! колорадского жуказОтбор вел к
изменению тех признаков,хоторыеу полициклических видовтради-
ционно обеспечиваютсогласование сезонногоразвитиякаждойпо-
«пудяции о динамикой местного климата^ При этом географическая
изменчивость фотопериодического порога носит вполне определен-
ныйклинальныйхарактер. Редукция ФПР уюжнойпопуляции<Те-
xaCiX несомненно, связана» с более мягким» климатом вгэтой зоне, а
сцонтанное-формирование диапаузы участижуковнезависимоот фо-
топериодических урловийсвидетельствуетозначительнойтетероген-
ности.особей в популяции. :
100.
aS\‘ '
во
>'7бо
1^’.'
I-40
с ...
со
Д 20
14
10 12
i Рис.75. Фотопериодичес-
. «an реакция разных . гео-
графических- популяций
колорадского жука в Север-
ной Америке (по: Hsiao,
> , - Популяции: 1-Рома-Лос
16 18
Длина дня,ч
(штатТехас, 26 2—Бенсон
<штат Аризона, 3—Ло-
гаи(штатЮга,41 4 -Яки-
ма (штат Вашингтон, 47 °с.щ.).
Температура’25,’С
о
166
Позднее были! исследованы еще гдве популяции жука из (штата
Нью-Йорк [Tauber et al.,. 1988],юднамзкоторых обитает наприбреж-
ном острове в Аглантическомокеане, а другая — наматерике, вгорис--.
тоймесдности сболеежонтинентальньшклиматом. Расстояние-меж-;
ду точками сборане превышаетЗООкм.однако этих.различий оказа-
лось достаточно.чтобы возникларазница;междукривымиФПР в по-
роговой зоне (рис. 76), что определило шразную фенологию: на ост-
рове жук развивается ;в двух генерациях, а на материкел-водной.
, i Сходныеразличияв сезон номразвитии обнаруженыу двухпо-
пуляций?—! горной и равнинной, обитающие в Закарпатье [Ушатин-
ская И:Иванчик,1982].Они’отличались пофенологии, срокам весен-
него появления жуков, их плодовитости, частоте летней, зимнейш
повторнойдиапаузы/чтомоглобытьобусловленоипрямымвлияни-
ем’<климата. Но было,показано, ято особенности горной популяции,
проявляются; при переносежуковвусловия равнины. Следовательно,а
ониявляются результатом внутривидовой.дивергенции,я непрямого
влияния среды» Дальнейшие наблюдения надперемещенной горной
популяцией показали, что через 3—5 летее признаки приходят к«рав-
нинной»мнорме.. • • Ч . анпоггк, ’ Л-к'!;-?.; ,-,к
Риа76лИзменение,фотопериодической реакции колорадского.жука :
в зависимости от высоты ,и^ континентальности местности [по: р
Tauber et al., 19Й8);
Популяции:!-равнинная; 2-горная ” *’•’
167
. i Сравнение параметров ФПРколорадскогожука, заселившего За-
паднуюЕвропупоказало, чтопопуляцииего в новомареале различав
ются'значительно меныпе, чем в США; Кривые ФПР, полученные по
результатам исследования шестшпопуляций ]de Wilde, Hsiao, 1981]
из. районов, существенно различающихся1'по климату (от 37 до:
5 слп1), расположены настолькоблизко,что значения критическо- >
гофотопериодапрактнчески;одинаковы.о&. • м- j , • г i
Анализизменчивостипараметров ФПРколорадскогожука в гео-
графическомивременномаспектахвСШАивЕвропесвидетельству-
еторазныхтенденциях формирования сезонныхадаптацийу этого!
вида при заселении новыхтерриторий. У северо-американских попу-з
ляцийпри расселении сохраняласьадаптивнаястратегия,направлен- ;
ная насинхронизацию сезонною цикла с ритмом климата,, причем
фотопериод оставалсярегулирующимфактором. Этому способство-
вало наличие разных кормовых растений из семейства пасленовых, :
обеспечивающих поливольтинный сезонный: цикл. У европейских*
популяций возросла тенденция к формированию диапаузы: и моно-:
вольтинизму в результате чего ФПР потеряла,свое: доминирующее <
значение в контроле сезонного развития. В новом ареале основной:
кормовой культурой жука служит картофель, и сезонный цикл вида,
видимо, в большой степени определяется именно сроками вегетации
этого .растения. - ---- ............v
7.4. Особенности ФПР, определяющие возможность
акклиматизации разных вадов на новой территории
Адаптация насекомых к новым условиям обитания определяет-
ся различными'эколого-физиологическими особенностями вида.
Потенции разных видов в отношении акклиматизации очень специ-
фичны. Несомненно главными факторами, определяющими возмож-
ность проникновения иммигрантов на новую территорию, являются
подходящий корм и сумма эффективных температур, достаточная д ля
развития хотя бы одного поколения. В умеренном климате обязатель-
натакже диапауза или иная адаптация, обеспечивающая выживание в
зимний период. При наличии этих трех составляющих возможно пер-
вичное проникновение и выживание в новых условиях.
Рассматриваемые нами виды имеют четко, выраженную форму
покоя - диапаузу, адаптированную к достаточно суровым условиям в
первичном ареале; они многоядны и на Европейском континенте на-
168
ходят разнообразные ^климатические условия, обеспечивающиераз-
витие разного числа поколений Более того, идентичность,первич-
ных ареалов исследуемыхшидовпредполагаласходство их потенци-
альных возможностей для акклиматизации, нановой территории,
Правда, СЭТ, необходимая д ля развития одногопоколрния при близ-
ких температурных порогах развития, разная, поэтомувменее выгод-
ныеусловия попадала американская белая бабочка/поскольку для
завершения одного поколения ей требуется вдвое большетепла. чем
подизусушликолорадскому жуку. ОДндкодкак оказалось.другие мо-
менты определили судьбу иммигрантов,, и адаптациякновымуслови-
ям обитания осуществляласьунихразными путями. () lj i t
т Все европейские популяции американской белой бабочки име-
ютсходные ФПР(см. рис, 68), различия проявилисьтольковпорого-
вой зоне, В соответствии^ ее параметрами сформировался вторич-
ный ареал вида, благодарящему сезонный цикл американской белой
бабочки назанятой внастоящее времятерриториихорошо вписыва-
ется в местные условия. Вид всегда дает два поколения,1-е из них в
условиях длинного дня всегда активное, у 2-го вкороткомдне кукол-
ки всегда формируют диапаузу Никакие погодныеколебанияне из-
мени ютэтойсхемы годичного цикла.в результатечего американская
белая бабочка во вторичном ареале поддерживае'гстабильно высокую
численность. Свойства ФПР, обеспечивающие строгую синхрониза-
цию сезонного цикла с, локальными климатическими условиями, в
свою очередь являются и тормозом для расширения ареала.< Низкий
критический порог, отсутствие его температурной коррекции, чувстви-
тельность к фотопериодическим сигналам гусениц младших и сред-
них возрастов - вот главные параметры, которые определили конфи-
гурацию ареала американской белой бабочки в Восточной Европе. У
популяции, акклиматизировавшейся в России, практически нет шан-
сов для дальнейшего расширения ареала в.северном направлении.
Однако ситуацця может изменяться, что и наблюдается в насто-
ящее время, например, на Японских о-вах. Натурализовавшаяся пос-
ле случайного завоза в 1945 г. американская белая бабочка давала два
поколения на всей освоенной ею территории Японии (от 32 до
40 °с.ш.). Популяции из двух географических точек (Кобе, 35 °с.ш. и
Акита, 39°с,ш.)} исследованные более 20 лет назад [Hidaka, 1977],
имели одинаковый термостабильный в пределах 17-25 °C порог ФПР
14 ч 35 мин. Но дальнейшие 'наблюдения показали,, что параметры
ФПРяпонских популяций Н. сипеа:не остаются неизменными. У по-
169
пуляциииз Кобе(рис. 77) была обнаружена температурная изменчи-
вость критического порога: его значение при' 25 ?С оказалось на 40
мин’меньше, чём при 20 QC. В результате адаптивной изменчивости
порога ФПР эта популяция перешла от би- к тривольтинному сезон-
ному циклу’[Gomi/Takeda, 1990/1991; Gomi, 1995,4997]. Несомнен-
но за этим последует расширение ареала на юе м , > ,»г . ч
- vb Европе на всей территории вторичного1ареала американская
белаябабочкабивольтинна. Но не исключено, что и здесьпроисхоь
дит дифференциация популяций, аналогичнаятой, которая наблю-
дается у-«японских’ популяций/ Анализируяданные рис.68, можно
видеть, что кривые ФИР закарпатской и итальянской популяций хоть
и незначительно, ноЬтличаются от остальных кривых: заметно уси-
лениетенденции к формированию диапаузы вПороговом фотоперио-
де. Заслуживает’внимания и тот факт, что между исследованиями раз-
ныхавторов'существуетболыпой(более20лет)временной разрыв.
э ! Однако' нё? исключено, что здесь Имеет место1 повторный Завоз
американской белой бабочки из регионов, где обитают популяции ,
ФПР 'которых имеют более высокий критический порог. Интенсив-
ные’контакты между континентами, возросшие благодаря развитию
всех виДов транспорта, могли эффективноэтомуспособствовать. :Эту
точку зрения поддерживаюти итальянские энтомологи (Deseo<etal.,
1986). -:’!' '’><' 'Н-'/н СЛ ЧИФ /-Г'.:1?. ‘ г
' Второй из исследованных объектов—подизус поканеакклима-
тизировался в Восточной Европе, несмотря на многократные выпус-
РисЛТ. Фотопериодическая реакция; американской;белой бабочки,,
,Нур/ииЧгм сипеа.[по1. Gomi, 1^95]:.. ч ,, . ( . ... .
Пояушщии: а—из Акиты, б - из Кобе
170
ки клопов в разных регионах. Как указывалось выше, .зона его возмож-
ной аккдиматизацииопределяется осенним сезоном,когдаидет пред-
диапцузнаяподготовка. Только накрайнем юге (район £очи) склады-
ваются условия, соответствующие температурным требованиям по-
дизуса. Несомненно большие возможности дая акклиматизации ца
нашей; территории имели бы популяции северного происхождения
(например, с юга Канады), имеющие ФПРс более высоким критичес-
ким порогом. Короткодневные условия в природной обстановке на-
ступали бы для них раныле, уменьшая ч исло .поколений за вегетаци-
онный сезон, однако последнее из них развивалось была фоне более
благоприятных лляформирования.диапаузыусловий. .. ,
Наконец, колорадский жук—несомненно уникальный по своим
приспо.собитещ>цым возможностям вид. Многообразиеформ покоя,
гетерогенность особей в популяциях идругие.особенности обеспечи-
ли ему практически неограниченные возможности для расширения
аредла. Частичный отказ от фотопериодического контроля диапаузы
и переход к моновольтинизму создали еще большую независимость
колорадского жука от внешних условий. Но в отличие от американ-
ской бабочки и подизуса, у колорадского жука в выработке сезонной
стратегии большее значение приобрела зависимость от кормовых ра-
стений. Как выяснилось, в новом ареале жук оказался гораздо белее
узким олигофагом, чем в исходном, где он питался разными видами
диких пасленовых. Именно необходимость согласования сезонного
цикла с фенологией культурного картофеля вызвала переход вида к
моновольтинизму на большей части вторичного ареала. Таким обра-
зом, у колорадского жука выявилась ббльшая автономность развития
от фотопериодических и температурных условий, чем у американской
белой бабочки и подизуса, однако усилилась его зависимость от кор-
мового растения. Можно предполагать, что именно пища станет глав-
ным фактором, который определит границы распространения коло-
радского жука.
Возможность адаптации к многообразию, условий существова-
ния в пределах нового ареала посредством перехода к моновольтиниз-
му подтверждается успешной акклиматизацией амброзиевого листо-
еда, Zygogramma suturalis Е Этот вид был интродуцирован в нашу стра-
ну для биологического подавления амброзии полыннолистной (Ко-
валев, 1981; Ковалев, Медведев, 1983]. Как оказалось, Z suturalis по
своим экологическим особенностям очень близок к колорадскому жуку
и также проявил уникальные адаптивные возможности. Так же, как у
171
колорадскогожука, у него выявленошестькатегорий физиологичес-
кого покоя ибольшаяразнокачественностьособейв популяциях[Ви-
ноградова, Богданова?!988,1989]/Сходство междунимиусиливается
иболыпой склонностью кмоновольтинизму. По мнению Е.Б. Виног-
радовой; именно разнообразиепроявлений имагинальнойдиапаузы,
большая длительность! иустойчивость основных форм покоя и воз-
можность их своевременного формированияобеспечиваютамбрози-
евомулистоеду быструю^ успешную акклиматизацию в новых местах
и создают предпосылки для дальнейшего распространения на новые
территории естественным или искусственным путем.
Таким образом; колорадский жуки амброзиевый листоед дают
уникальные примеры успешнойакклиматизации на новом континен-
те.1 Нетсомнения,чтоодинаковыйположительный результат обус-
ловленотмеченным выше сходствомих физиологических реакцийна
факторы внешней'среды, пластичностью ихжизненных стратегий,- в
основе "которой лежит многообразиеформ покоя и разнокачествен-
ность особей в популяциях. 1:1 //-п /// мл мл/ ; •.' -р/
Глава 8.ГОМОДИНАМНОЕ РАЗВИТИЕ
' ' '• • ** /’г* S ' » 4 Л ' *.(
-ад ‘ .’‘Л 1; '’/„> ад ' '?. Г .‘'/•'Я ‘ ‘ ’ V ,ад !
' Главная отличительнаячерта этого типаразвития насекомых *-
непрерывная'физиологическая активность. Гомодинамные видыраз-
множаются влечение всего года, давая поколение запоколением, число
которых определяется тепловыми ресурсами местности и темпера-
турными потребностями конкретных видов; Естественна гомодинам-
ность реализуетсятолько втропикахи субтропиках'при относитель-
ном постоянстве внешних:условий. > • < ад -< > < : । <
9‘И‘ ..Я *” 'Л - ; Г v z ’ t fiu И
j ад - 8.1. Возможностьпостоянногообитанияв >ад адъ - < ад
умеренныхширотахвидовсгомодииамным развитием '-ь ч
над ч адлад-Л' ’ .><•“’ .’,,ададададададададад:;ададададад>жк ад-ъад ад-rf’
* ’ Анализ данных литературы показывает, что жизненная страте-
гия,Направленная на активность в течение !всеготода,' присуща не
только тропическим, но и широко распространенным видам. Напри-
мер,\гомодинамныйтип{газвитиясвойственшкоторым1сиНантроп-
ным мухам. Так, расселившаяся по всему земному шару комнатная
муха, Musca domestical не имея специальных адаптаций, в зоне уме-
ренного климатакруглогодичноразмножается вотапливаемых поме-
щениях, напримерживотноводческихфермах, птицефабриках ит. д.,
где при средней температуре около 20 °C она дает доЛОшоколений за
год* (Курючкин1, s 1984]. При понижений температуры ’развйгие
М. domestica затягивается, и число поколений уменьшается. Выжива-
ние комнатной мухи вне помещений в зимний период считаетсяМало
вероятным (Виноградова,1990]. Тем не менее вид существует почти
во всех климатических зонах, восстанавливая численность популя-
ции за счет-успешНо перезимовавшихособей, атакже.возможно, бла-
годаря иммиграции мух из более южных районов. Хотя Последнее пред-
ад' ад!»'- 1 . -. j'Y’ •- .’'й. ‘ ' Ыад‘ ад'Г, , ' । ад;
173
положение в специальных опытах американских исследователей
[Black, Krafsur, 1986] не подтвердилось.
С. Масаки [1972] обнаружил гомодинамное развитие у некото-
рых сверчков в Японии. Исследуя их жизненные циклы, он пришел к
заключению, что именно переход к синантропному образу жизни
обеспечивает гомодинамным насекомым возможность проникнове-
ния в области с резко выраженной сезонностью климата.
Такоеже предположение выдвинул Э. Меривээ[1971,1972] вот-
ношении капустной моли,! Pluiella jnaculipennis, считая, что этот вид
не может перезимовывать в климатических условиях Эстонии. По его
наблюдениям при открытой зимовке полностью вымерзают даже ди-
апаузирующие кукол ки Р/erw йгаи/сде. Постоянное обитание ихв этом
регионе возможнотолько при использованиив качествемест зимов-
ки различных построек.вблизигжилища людей. Подобную,защиту
создаетиснежныйпокров. • ;Г, '.
• <' Успешное существование.в умеренных широтах достигается дру-
гими гомодинамнымивидами путем использования относительно
стабильных условий почвы, где даже; на незначительной глубине се-
зонные и суточные колебания температуры сильно сглажены, а высо-
кая влажность лодцерживается круглый год. Так, тесная связь с поч-
вой на всехютадиях:развития заисключецием цмаго возродила под-
грызающим совкам распространиться далеко на север, не имея устой-
чивого физиологическогопокоя типа диапаузы [Рябов, 1956; Кожан-
чиков, 1956],. Экснериментальнысисследования Agrotts ipsilon и
Ате^ейпи.лредпринятыерядомдвторовДГорышин, Ахмедов,,. 1971;
Гейспиц и др.;,! 197 IpMatsuura, Miyashita, 1978; Beck, 1986], обнаружи-
>ЛИ, что этисовки имеют лишь количественные,фотопериодические
реакции, которые способствуют синхронизации наиболее, устойчи-
вой стадии,развития, (пронимфа, куколка) с осенне-зимним перио-
дом, чтогарантирует выживание внаиболеекритические периоду года.
1рмодинамное развитие характерно и для ряда чешуекрылых,
обитаютих на юге России. К ним «относится, например, луговая со-
вка, Mythimna.^inipuncta.yaw. Обычно в местах своего постоянного
обитания она дает 2—3 поколения за вегетационныйсезои, а небла-
ГОприятный период. проводит е состоянии оцепенения, используя
кратковременные потепления для .питания. Зимой встречаются все
стадии онтогенеза от яйцадо имаго, иуспешность зимовки, в боль-
шей степени определяетсяпогодными условиями, В силу этого круг-
логодичное существование М. unipuncta возможно лишь в зоне суб-
’ 174
тропиков или тропиковсихвечнозеленойрастительностью исравни-
тельно мягкой.зимой [Тархнишвили, 1966,1972].' К сходным выводам
пришли французские энтомологи, исследуя сезонные адаптацииме-
стных популяций этогодаида (Bues et al.,'1987]. Согласноихнаблюде-
ниям, луговая совка может перезимовывать в районах Франции между
40- и'45-й параллелями, носмертность во время зимовкибываеточень
высокой.Тем не менее уже во 2-м поколении совкздпричиняет значи-
тельный ущерб злакам, а в 3-м—ее численность настолько возрастает,
что она начинает расселяться в соседние регионы.
По сведениям'разных$авторов,'гомодинамные виды могут про-
никать далекона севериобразовыватьтам временныесезонные попу-
ляции. Так, луговая совка отмечалась в Исландии [Kaaker, Norgaard,
1970; Wolff, 1971], малая наземная совка, Spodoptera exigua — в Фин-
ляндии и других странах Скандинавии | М1кко1а, Salmensuu, 1965].
Близкий вид Spodoptera frugiperda на североамериканском континен-
те из мест постоянного обитания (юг Флориды и Техас), тоже ежегод-
но распространяется на север в континентальную часть США, где не-
редко причиняет бодыпойущерб сельскому хозяйствуДБрагкз, 1979].
7 Неопределенность сведений о зимовке названных чешуекрылых
позволяла допустить их обитание вусловиях умеренного климата на
основе неспецифической устойчивости к повреждающим факторам
зимнего периода. Однако это предположение требовало эксперимен-
тальной проверки. Не исключалось, что у таких видов обнаружатся
слабо выраженные формы покоя, способствующие зимовке.
( Объектами исследования были выбраны три вида совок (Noc-
tuidae): луговая — Mythimna unipuncta Haw. (сухумская популяция),
гамма—Autography gamma L. (белгородская и славянская популяций)
и малая наземная - Spodoptera exigua Hb. (каракалпакская популя-
ция), имеющие предположительно гомодинамный тип развития. Их
сезонные адаптации изучались д сравнен101 с адаптациями капуст-
ной совки, Mamestra Brassicae L. (сухумская популяция), характери-
зующейся.гетеродинамным типом развития (диапауза на стадии ку-
колки).....'''.С?’/' 7' 7 -.7
, Каи отмечалось выше, наиболее яркое проявление зимовочного
состояния унасекомых умеренных широт—диапауза. Однако в систе7
ме сезонно-циклических реакций имеют значение м более слабо вы-
раженныеформы покоя, способствующие зимовке. Они могут прояв-
ляться в виде задержек ростр и развития преимагинальных стадий,
прекращения питания и т.д. Такие явления описаны для Dasychira
Y15
pudibunda [Гейспици Заранкина, 1963}, для Аратеа anceps [Бобине-
кая,г1966;Ажбенов, 1995], некоторых подгрызающих совок [Гейспин
идръЛ971;Морси,<1978, ищр.].*!! некоторых случаяхпоказана роль
таких реакций урегулировании сезоннойприуроченности опреде-
ленныхфазонтогенеза.Неисключено их значение, для переживания
неблагоприятных условиймягкого- зимнего периода,, -когда лимита-
рующимфактором для выживания являетсяютсутствие корма, а не
морозы.'
8.2. Сравнитщп>ноеизучениереакцийнатемпературу и
длину дня.увидовегетеро-игомодинамным,
типомразвигия>;
. ...ДО'Щм- -.’.S&UH
штMantestra^brassicaeчр.
/ Капустная'совка принадлежит к хорошо-изученным в фотопери-
одическом отношении видам. В нашей лаборатории исследованы
популяции"изС-Петербурга,Минска,'Белгорода,Черновиц, Лаза-
ревской, Батуми Данилевский, Кузнецова, 19681], Алма-Аты [Пла-
нов,’1'979]. Вид хорошо’адаптирован к разнообразию климатических
условйй его обширного ареала. Зимовка обеспечивается куколочной
диапаузой,^ которая;контролируется фотопериодической реакцией
длиннодневного типа. / °.. _' '’’ъ
Как показали эксперименты с южной (сухумской) популяцией
капустнрй совки (рис. 78)i четкая ФПР имеет место при 20°С. В этих
условиях длиннодневные фотопериоды bf 16 до 24 ч обеспечивали
активное развитие всех куколок, апридлинедня14чикорочедиапа-
узировали !93-100% куколок. Лишь при непрерывной темноте (апе-
риодический режим) количество дйапаузирующих снижалось до 40%.
Критический фотопериод при 20 "С1 оказался1 близким к 15 ч. При
25 °С четкость й насыщенность ФП? резко ослабевали, что свидетель-
ствуетонеоптимальноститаких условий для ее проявления. Только в
зоне фотопериодов от 6 до 14ч появились диапаузирующиекуколки,
нрдажепри длине дня 12 ч доля их не превышала 50%.
Зависимость развития преимагинальных стадий капустной сов-
ки от температуры вполне удовлетворительно описывается'уравнени-
ем гиперболы и не представляет специального интереса. Однако сле-
дует отметить влияние' фотопериода на сроки развития гусениц (см.
рис. 78, внизу). При 20 °C его продолжительность в короткодневных
176
520г
S 400-
'360-
I 480 -
s
- S' ' j
Я.44С-
! ? О . 1
32ft
2Г<
Рис. 78. В лияниедлиньг^н
дня на развдетце капуст-
ной, совки: '
1—длительность раз- '
вигиятуссниц, 2—массакуко-
лок, 3- крличестводиапау-j,,,
зирующих куколокпри 20 “С;
1-Ш _тоже приЭД °C
? 38
- §
22
18
I 34-
<30-
4* -Ц.
° 26-
20 24
Длина дня, ч
зШ
, .. i
80 1
ш
60 >
-40
S,
те
-20 S
5
Jo ,
5
условиях была несколько большей (в среднем на 15%), чем вдлинно-
дневных.Аналогичнаязакономерность отмеченаи при 25 °C. Фото-
период оказываетвлияние и на массу куколок: у диапаузирующих ку-
колок из короткодневных вариаптовэтот показатель (при20°С) в сред-
нем на 38% выше, чем в длиннодневных. При 25 °C разница выражена
слабее, так'как вкороткодневных вариантах не все куколки диапа^и-
ровали.'Н'лМй г!.Г',гуо<;и^з т<'>
' Таким образом, имеющиеся данные характеризует капустную сов-
ку как насекомоес гетеродинамным типом развития. Годичный цикл
ее контролируется системой экологических адаптаций, среди кото-
рых важное место занимают фототермические реакции. • .
л * - ? 8,2.2.Луговая совка, Mythimna unipuncta
В отличие от капустной совки, у этого вида (популяция ^Суху-
ми) ни в одном опыте не были подучены диапаузирующие особи на
какой-либо стадии онтогенеза. Отсутствие диапаузы и контролирую-
177
щей ее фотопериодической реакции у луговой совки не вызывает со-
мнений, так как при проведении опытов были учтены все известные
факторы, способствующие их пррявлению. Учитывалось, что у раз-
ных видов диапауза может быть приурочена к разным стадиям онтоге-
неза, а чувствительная к длине дня и реагирующая стадии могут быть
разобщены [Горышин, Тыщенко, 19741- Принимались во внимание
описанные в литературе для некоторых видов факты влияния мате-
ринского поколения на ФПР и диапаузу дочернего поколения [Ви-
ноградова, 1.973; Гейспиц и др., 4974]. В связи с этим схема опытов
предусматривалавоспитаниевкаждомфототермическом режиме двух
полных генераций изучаемого объекта до момента отрождения из яиц
гусениц III поколения. Тем не менее развитие шло непрерывно, и ни
на одной стадии онтогенеза не возникало каких-либо признаков зи-
мовочного состояния. Г1' '
Не обнаружено определенной зависимости от длины дня и сро-
ков развития гусениц. Этот показатель при каждой из испытанных
температур проявлял’лишь случайные неболыпиеотклонения Ътсред-
х ' / "X него значения (рис?79).*06-
' ..X “ щаяпродолжительностьон-
70Г,, _____ о тогенетическогоциклалугог-
6oL°:ж”"?о** ...ь“о’Ц!*Л'войсовки— 'отяйрадовыле-
жЖ ° таимаготакженезависилаот,
х 50_ 20 °C фотопериодических усло-
нпда ж-ж » <?'• у/Жажуж'- жж: • двий^ жж'жж: ж-г; аджж
ж -вЛ’ ь>1;1''Вмест^1сдем,|,опыты:по!-
•( зволиди-выявитьслабовыраг
________________»_____д женнуюфотопериодинескую
, жж;.жи.,Ч <ЧжИ ХМЙЯ;'гию?<ре1уляцию;масадздсеницда
g 40' ° о ° ° о % ° от капустной СОВКИ, уЭТОГО
сЦ0_жтжж ь..ч;25№>жвида'->уведиче:рие.<массы на-
о^о « ’«к нмвьбдюдалрсьнрприкоротком,
отав - К-Дб'". жЛ,ж*жож ?ащр|Идлинном,дне, причем
Ш---1---1--а_—1_—1_—J—,л. масса куколок градуально
Длина дня ч возрастила с увеличением
' amir?,w ...от продолжительности дня. Не
Рис. 79. Длительность развития гусеннц луго-
вой совки (1!,2)ивсегоонтогенетического цик-'
’ла (I, II)1 в разных'фотопериодических'усло- < ’
ВИЯХ ’ ЖЖ > ’ Ж ЖЖ’ ЖузЧ i -Г,
исключено, что .это следствие
. влияния д лины дня на. дли-
‘т^ьн^твперда^ацит^и^
478
ч-м . ; , , д/.дя 'у .7/42 - ' Чв""'
* ч : • .«.>'Н .л / “Г «... г.1л. ДЛИНИПНЯ-?Ч'НО«'^ ’3*'
XI , . • jt •' ' !♦' u и -ял
( t Л/c.^ft Средняя масса куколок дутовой совки вразных фотопериог,
’s(' Одических режимах и при разных температурах *'
'HiS Vrfri^V:*- <:лфГ? К'Я'?0(Ч4'}
течение суток.Посколькупитаниелуговойсовкиприуроченоксвет-
лойчастисуток, суммарныйпериодпитаниявтечениефазвитиягусе-
ницоказываетсяпридлинномдне*больше,чем'при коротком.’ ;1 •
5 ' Отсутствие качественных проявлен ийФПРи состоян ия диапау-
зы'характеризуетлуговую'совкукаквид с гомодинамным типом раз-
ви!гия73'<ш*'|П: . ш. . ЗФ нон;. -.» \ъ><
, ‘Цм?. Т (НВ < Ли. июп е . .«циане чни
' т.; ‘ ' ,8j2:3;fCoeKa>iaMMar1Aut<^apha>gammaiи - \'i q<T \;i.
) ,и мамшназемнаякрвка;Spodoptennexigua 'шп qn , ; Л
'* Эти виды по поведению в разных фототермических условиях об-
наруживают болыпое сходство с предыдущим. Они характеризуются
полным отсутствием физиологаческих реакций на изменение длины
дня', а температура влияетдолько на продолжительность их онтогене-
тического цикла. Отличие от луговой совки заключается лишь в том,
что уэтих видов масса гусениц и куколок не зависит от фотоперцода.
Таким образом' и совку гаммуималую наземную ровкутакжеследует
рассматривать как виды с гомодинамным типом развития,.
* Описанные выше экспериментальные данные позволяют заклю-
чить, что экологические механизмы адаптации к годичному ритму
климата у четырех исследованных видов весьма различны’. V. brassicae
проявила себя'как типичный' для умеренных широт вид с факульта-
тивной диапаузой и четко выраженной системой фцзиодргических
реакций, управляющих,ее''проявлением..в резонерM^unipuncta,
A. 'gamma и 5. ех«рмадиапаузой или,иными формами, покоя, и связан-
ными с ними сезонно-циклическими реакциями не обладают.
ТТ^
Непрерывная физиологическая активность (гомодинамный тип
развития) обычнадля многих видов влажных тропиков. Она плохо
вписывается даже в аридные климаты тропического пояса и тем более
экологически неадекватна для умеренной зоны. В выполненных экс-
периментах непосредственно анализировалось действие на объекты
только двух факторов - фотопериода и температуры. Действие этих
факторов тесно взаимосвязано." Любая ФПР имеет свой температур-
ный оптимум, где она проявляется максимально. Этот оптимум спе-
цифичен для разныхвидов и даже географических популяций и опре-
деляет удельноезначенир светового фактора вэкологическом контро-
ле сезонно-циклических адаптаций. Известные данные по темпера-
турному оптимуму ФПР различных объектов позволяют утверждать,
что диапазон температур Т7—25 °C, использованный в опытах, прак-
тически гарантирует выявление ФПР любого вида, если таковая ре-
ально существует. Использованнце в опытах фотопериоды (от 0 до 24
ч) заведомо превышают пределы изменения длины дня влриродев
течение вегетационного сезцна на всех широтах,реальногораспрост-
ранения наших объектов. За пределами оптимума ФПР температура
способна оказыватьсамортоятельноевлияниеца диапаузу. При длин-
нодневной ФПР понижение температуры обычно стимулируетее воз-
никновение, а повышение - ингибирует. Диапазон испытанныхтем-
ператур 17-2S°C для многих исследованных ранее .видов наиболее
благоприятен для подготовки и формирования диапаузы. С учетом
сказанного неэффективность фототермических условий для индук-
ции зимовочного состояния у наших объектов можно считать дока-
занной^Возникает вопрос, не было ли неучтенных факторов или их
комбинаций, способных маскировать проявление диапаузы (или иной
формы покоя) и Контролирующей ее ФПР? Для некоторых видов опи-
сана причинная связь диапаузы или иных свойственных им зимовоч-
ных состояний с суточными ритмами температуры, пищевым факто-
ром, плотностью'популяций, материнским влиянием, наследствен-
но обусловленными факторами и т.д. Рассмотрим эти факторы при-
менительно кизучаемым видам. ,,
Терморитм. По ритмической'структуре этот фактор имеет сход-
ство с фотопериодом и при определенных условиях, способен само-
стоятельно индуцировать диапаузу или влиять на параметры ФПР.
ТермоперйодиЧеская реакция описана для Ndsonia vitripennis
[Saunders, 1973*],‘ Ostrinia nuliilalis^Beck, 1982, 1983, 1985], Diatraea
granJfose/Za'i'Chippendale et al.,lI976j, Musca autwnnalis [Hayes et al.,
.ISO
1983]. Не столь полные данные имеются для других видов — Acronicta
rumicisM Pieris йгалпсае [Горышин и Козлова, 1967], Lpxostege sticticalis
[Горышин и др., 1980], Perillus,bioculatus [Волкович и др., ,1991]-. Лабо-
раторные опыты при терморигмах с луговой и мал ой наземной совка-
' ми не проводились. Однако действие их на развитиеласекомых про-
верено в специальных подевых.исследованиях, результаты которых
будут представлены ниже. Они не противоречат выводу об отсутствии
специальных адаптаций к зимовке у этих видов...., ,, г .
Пищевой фактор.Известно влияние качества корма на парамет-
ры ФПР и формирование резервов эндогенного, питания, .необходи-
мых для.длительной зимовки. Использованные в одытах кормовые
растения являются обычными пищевьши рубстрадами. исследован-
ных видов и не могут препятствовать проявлению сезонно-цикличес-
ких адаптаций. ' ........
Материнское влияние. Как указано выше, схема опытов.преду-
сматривала непрерывное действиезаданныхфототермических усло-
вий в течение двух полных онтогенетических циклов, что.гарантиро-
вало возможность проявления этого фактора. , . , . , , .
Плотностьпопуляции. Ее влияние на диапаузу показанодля мно-
гих видов. Описана роль этого фактора,в возникновении миграцион-
ного состояния. Мы исследовали'влияние плотности популяции на
диапаузу и миграционное поведение луговой совки и совки гаммы
результат отрицательный [Саулич, 1975] . Таким образом, у луговой
совки,малой наземной и совки гаммы никаких признаков специаль-
ного зимовочного состояния и контролирующих его проявление се-
зонно-циклических реакций, на какой-л ибо стадии онтогенеза обна-
ружено не.было. Эго подтверждает исходный тезисогомодинамном
типе сезонного развития исследованных<видов,? . , j ;.
8.3. Холодостойкость разных стадий онтогенеза
’ ' ; ; ‘ -'i • 7 • ' a'-. i 1 ‘ ' t * ' “ . Л .'.U.'\Л Г'*. - 5 * ' . > t 1 ч - -
Исследуемые гом одинамные совки принадлежат к числу широ-
ко распространенных видов. Луговая совка—почтижосмополит, оби-
тает в тропиках и субтропикахдсех континентов,но далеко проникает
в зону умеренного климата. Повсеместно распространена в США
[Howard, 1896], Восточной Канаде [Guppy, 1961], северные находки
известны даже из Исландии [Kaaker, Norgaard, 1970; Wolff, < 1971}. В
Европе она встречается в Средиземноморье, на Кавказе ц в Крыму
[Ключко, 1964; Миляновский, 1967]. Близкийвид Mythimna separata
481
обитает на'Дальнем Востоке’[Куренцов, Груздева, 1946; Онисимова,
1949]. Северная'границаареала'остается невыясненной?!"5 N ' 1
'-Совка гамма также распространена повсеместно. Наиболее се-
верные находки отмеченывСкандинавии[8у1уеп, 1947;5Kaisila, 1962],
Восточной Европе — вАрхангельске,Сыктывкаре, Перми[Парфенть-
ев,{1923;’*Кожанчиков,Т937?Щеголев,'1964].Наюге'встречается вСре-
диземноморье/Северной Африке, Малой и Средней'Азии. На восто-
ке доходит до Приморского края, есть в Клтае, Корее, Японии [Ко-
жанчиков, 1937; ЗЬлотарейко, 1966;'Yothom^Rivnay,i1968p,‘,ai*
ы Проникновение подобных Видов в зону умеренного климата:с
достаточносуровой зимой кажетсянеожиданным, таккакне согласу-
ется с5гомодинамным типом их развития. Сведенияозимовке'иа-
званныхвидов чрезвычайно противоречивы*.*!! качестве «зимующей»
разные авторы называют любые стадии онтогенеза - от яйца до имаго
1КожанчикоВ,'1937; Breeland;i958; Pond?1960i; 'Тархнишвили, 1966,
1972]' ?Это1 лишьподчеркиваетотсутствие специализированного зи-
мовочного состояния, но не отвечает на вопрос о 'возможности и спо-
собах успешной зимовий'” 1
* м ’Ддя’зимы в умеренных широтах характерныдва'лимитирующих
фактора: отрицательнее температуры и 'значительная продолжитель-
ность'бескормного'периодаилипериода/тютемпературнымуСлови-
ям неспособствующегонормальной жизнедеятельности. Безусловно
диапауза - наиболеесовершенная адаптациякпереживаниюэтих ус-
ловий,однакоустойчивость‘кзамерзанию и голоданию проявляется
унасекомыхи всостоянии'активнойжизнедеятельности/хотянараз-
ныхстадияхонавыражена весьма различно. Не исключено.’что такая
неспецифическая устойчивость при определенных условиях окажет-
ся достаточной для зимовки видов,‘лишенных диапаузы. Для1 выясне-
ния этого вопроса была определена степень устойчивости луговой со-
вки и совки гаммы к отрицательный и низким положительным тем-
пературам; на всех стадиях онтогенеза. В сравнительно-экологичес-
ком плане одновременноисследовали’капустнуюсовку.;-
эв л . Холодостойкость оценивали термоэлектрическим (методом по
предельной температуре переохлаждения тела насекомых (Горышин,
.1966]. Способность?длительно переживать низкие положительные
йгемпературыисследоваинметодомиробного^анализад : ‘ i fc‘f.:. ‘ !
/к гВышеупоминалосьдчтодлялуговойсовкиисовкигаммы1влите-
ратурев качестве «зимующих» называются все стадии онтогенеза.. Не
вызывает сомнения, что эти стадии обнаруживаются поздней осенью
1Я9
или даже зимой при полевых обследованиях. рднакоотсутсгвуюТ|Тоя,-
ные,сведения отом,-смогли ли онипережитьдиму-умеренногоклима-
та и весной завершить развитие. Выяснение нредельных,возможное-;
тей.каждого вида кпереохлаждению наразных стадияхпредставляет в
этомцланенепосредственныйин;герес.Таккаккапустнаясовказиму-
ет всостоянии кукол очной диапаузы, другие стад ии ее,развитиямо-(
гут проявшгодицп> ^ед^ифич^^^Л'ой'щ^
, .., Согласно результатам исследования холодостой кости различных,
стадий онтогенеза капустной и луговой совок максимальная способ-
ность. к переохлаждению, у обоих видов .проявилась на ,стад ицяйца
(рис, 81 ,кривые 1) „примем у луговой совки несколько выше (среднее
значенис'гемпературыпереохлаждения-27,9 ±0,1.1оС)ьчему капуст-
ной.(-26,5 + 0,10 °С). Слабо выраженные колебания холодостойкост
ти в связи с фотопериодическими условиями развития (включая ма-г
терицское, поколение), видимо, случайны й не имекэд адаптивного
значения., Высокий уровеньпе- ,
рерхлаждения яиц не является
неожиданным иобусловлен нет ч
специфической устойчивостью
к переохлаждению, свойствен-
ной всеммелкимобъектам-гИр- ,
ве>стно,,что чемменьше разме-, ।
ры насекомого, тем легчедости-.
гается перерхлаждение,.тем.оно,.
глубже и стабильнее [Калабухов,..
1935],.Возцикновению центров,
кристаллизации у таких.объек-, ’
тов препятствуют капиллярные
силы, нацряжецие которыхвоз-
растает с.,уменьшением объема. (
Этому же, несомненно, способ-
ствует относительнаяюднород-,.
ность цитоплазмы' нйца^цана^
ле дробления.,.)О.,,, ,
Минимальной холодо-
стойкостью,отличаются гусени-,,
цы. У капустной совки средня^,
темперепурапереохлажденияту-.
л.;,..
Рис;81. Холодостойкость капустной '(а)
и луговой <б)-совок1на; разных етадиюо
> ^онтогенеза: • .--..л ' у., : , -р-и;
1 - температура переохлаждения яиц, 2А—
активных куколок, ЭД—диапаузирующих куколок,
: / 2Д‘—диапаузирующихкукояокиаЗО-йдень, 3 -
I гусениц IV-V возрастов, 4 -гусениц 11-1П-. возра-
став, 5 - фотопериодическая реакция капустной'
-Н-совкипри20”О '*з '-.я.«Д:
183
сениц младших (П-Ш) возрастов^ыла близкой к-4,0 °C (кривая 4)‘ а
для'старших(ГУ-У) возрастовсоставляла -5,9 ’С (кривая 3). У луго-
вой совки ^штриховые линии) этот показатель в среднем составлял -
4,5'°С (кривая 4) и -5,9 рС(криваяЗ) соответственно. Фотопериоди-
ческие уело виясодержаниягусениц не оказывают закономерного вли-
яния на способность их кпереохлаждению. Таким образом, по холо-
достойкости яиц икусениц оба “вида оказались сходными. 1
Наиболее интересные результаты получены по холодостойкости
куколок. Как упоминалось выше[!у капустной совки на этой стадии
происходит'зймовка. Холодостойкость активных и диапаузирующих
куколок М. brassicae из разных фотопериодическик условий на 3-й
день после окукливания оказал ась'очень ’сходной, в среднем около
-21 °C (рис. 80,2А и 2Д).Указанныйуровень холодостойкости у диа-
паузирующих куколок не изменился и после месячного их содержа-
ния при+5'°С(кривая 2Д’). ’v ’ i ь 1
s ; f Удивительное сходство температуры переохлаждения у активных
и диапаузирующих куколок капустной совкикажется неожиданным;
оно противоречит традиционному представлению о большей холодо-
стойкости насекомых в состоянии диапаузы [Ушатинская, 1957; Ло-
зина-Лозинский* ’1'972].' Это справедливо', но'неуни нереально. На-
пример, по данным Т.П. Симаковой (1971], сходную холодостойкость
имеют диапаузирующие ^активные куколки Acronictarumicis (соот-
ветственно -21,0 °C и -19,6 °C) в первый день после окукливания. Прав-
да, в дальнейшем тонка переохлаждения у диапаузирующих'понизи-
лась и достигла -29°, а у активных куколок она оставалась без измене-
ния даже после холодовой обработки (-5°).'Аналогичное сходство по-
казателей холодостойкости диапаузирующихи активныхкуколок при-
водится для капустной белянки [Данилевский, 1961] и хлопковой со-
вки{ГорышинЯ958‘]; . :
: 5 Куколки луговой совки (рис. 80, кривая 2) оказались менее холо-
достойкими'по сравнению’^ капустной совкой; йхточки переохлаж-
дения колебались в разных вариантах вокруготметки -14 °C. Для пол-
ноты картиныбыла исследована способность к переохлаждению ба-
бочек луговой совки, которые также упоминаются в качестве зимую-
щей стадии. Средний показатель их холодостойкости составил
-6,0±0,28°С. -
Опыты с совкой «гаммой (популяция из Белгородской ббл.)’вы-
полнены по земе, описанной для предыдущих видов.' Материалом-
служили живые бабочки, собранные в августе. Развитие ихпотомства
184
Л’с
-20
.Ж UI
происходило в сентябре, когда у больший-» й
стваюбитающих в лесостепной зонелоли- *
вольтинныхвидовформируется зимующий
запасжйк- я;-Л) тшшютЛ » w.
Данныепохолодостойкостивсехста-
дийсовкигаммыприведены нагистограм- .
ме (рис.82). Какшудругихисследованных.
видов;: способность к переохлаждению wr :v
оказалась ^максимальной^ яиц. {-30,4s ±- с < ш мз
±0,5б'Ю),.минимальной.у^усенициима- -;'д ,
го. Для гусениц.младшихи.старших'возра-. Рмс.&2.Температура переох-
б В
>Л '<.
5Г 'J
стовэтотпоказатель оказался одинаковым! v лажденщьсовки гаммы. на,
(всрсднем-4,4 °C). Бабочки замерзали при* 'pa?H^a^^-Pi^San-
температуре телавсего-3,4 ± 0,26 °C. Ку- пгвазрастов.в’-гусеницытч-у
колкиоказалисьнесколькоболее устойчи-. s возРастов, г^куко^л-имаго,
вымижх средняя хододостойкостьсоставь. < ‘ТТ'ТфИг: . ; ' К!.
вила -11,9'±!0Д4 °C. На всех сдадиях не проявилось существеннрго
влияниядлины, дня; наэтипоказатели, л-'-, еаохдмч
< . Таким образом, у всех трех видов обнаруженавысокаянеспеци-
фическая холодостойкость яиц, более умеренная устойчивость куког
лок и слабая — гусениц и бабочек.Полученные показатели холодов
стойкости являются предельными. При длительном воздействии от-
рицательных. температур/как-в природе) точки переохлаждения не
достигают таких значений. ? "I . /Т г <'
ч -В целом можно сделать вывод, ,что холодостойкость йгусениц и
бабочек исследованныхвидовнедостаточнадля зимовкилзонеуме-
ренногоКлимата дажеподснежнымпокровом,новполнедортаточна
дая зимовки в. субтропиках. Возможностьзимовки яиц и куколок при
отсутствии диапаузы допустима по показателям холодостойкосшэтих
стадий, но может лимитироваться продолжительностью зимнего пе^
риода. W.RQHdTO 1 'ST: ШОЮЮДТ R. .гГТШНС "Йр, '«ТЙ; : .Д>-
V Л’.Г.'ИШ! <. НИ-'- "Тй Т'Д’ЖСГ'ШХ;-i чдул Н iifth-’nSW'i .
8.4. Устойчивость к длительному действию низких’ ; / :
,т !-‘:ш:л' --о.- он о > тподожительныхтемперавд ? .teqvf кг;5:.1,..г:>
То tao?; KsSSOtiP" - о s: О W,<Tvu « К г УК) V 5, i ЯОДЬоТЯГ) f. • И-Ш -
Период,втечениекотороготемпературанепревышаетпорого-
вуюдляразвитиявеличину (длямногихвидов5—100С),продолжает-
ся в умеренных широтах втечение 4—5 месяцев и дажеболее. Онвклю-
чает позднюю осень, зиму и раннюю весну. В течение: большей .его
185
части вегетирующие растения отсутствуют^ Насекомые, попадающие
в температурные условия ниже пороговых в состоянииактивнойжиз-
недеятельности, впадают в холодовоеоцепенение.которое необлада-
ет адаптивными свойствами диапаузы. Дыхание и обмен веществ®
понижением температуры угнетаются, однако сохраняются на болев
высоком уровне, чем втехжеусловияхпридиапаузе.И.нтенсивноас7
симилирующие стадии (личинки,имаго)прекращаютлитаться—на-
ступаетвынужденное голодание.Повреждающее действие среды на-
капливаемсякумулятивно - постепенно отоприводит к истощению, а,
нередко и иссушению организма, ведущему кгибели. Таким образом;
возможность зимовки в.состоянии а ктивнойжизнедеятельности , поч
мимо морозов, можетлимитироваться еще фактором времени ^ про-
должительностью неблагоприятного периода. ; т л / *
Способностьактивных насекомых пережи вать температуры ниже
пороговых хорошо известна и широко применяется в. лабораторной
практике для кратковременного хранения живого материала. Она весь-
ма различна у разных видов и стадийразвития. По данным И.В. Ко-
жанчикова [ 1946], хранение пупариев Musca domestica в течениенеде-
ли.при температуреО, 5 и 10 °Снеоказывает существенного повреж-
дающего «эффекта. С другой стороны, яйца дубового, шелкопряда,
Antheraeapemyi,~Moiyt сохранятьЬкизнеспособность даже после 2-ме-
сячного содержанияихприО °C [Кожанников, 1949]. Аналогичные
данные можнопривести ц длядругих видов, [Кузнецова,; 1970; Зис-
кинд, 1985; Горышин, Туганова, 1989]. , ; , нгпха
ч «Всвязи® изучениемсзимовки гомодинамныхнасекомых иссле-
дованавсравнительномаспектеустойчивость капустной и луговой
совок кдлительному воздействию температур ниже пороговых. «
< « До взятияпроб в опыты материал содержался; (включая все предт
шествующее поколение) при 2Q °C вусловиях длинного (ДД) и. корот-
кого (КД)дня. Конкретная продолжительностьисходных фотопери-
одов указана при описании каждого опыта. Пробы отбирали на;ста-
дияхяйца, гусеницы и куколки и содержали в течение 1,2 и 3 месяцев
в трех температурах:>0; 5гИ 10 °C (далее приводятся точные значения
температуры в каждом опыте). Вовремягхолодового воздействия фо-
топериод равнялся 12 ч света в сутки и имитировал короткодневные
условия зимовки. После охлаждения насекомых вновь возвращали в
условия сисход ной температурой20 Ю и продолжали наблюдение за
ихразвитием вплоть до вылета имаго.. Повреждающее действие пони?
женныхтемператур оценивали по показателям смертности в ходе экс-
186
позиции и жизнеспособностиподопытных насекомых после их воз-
вращениям исходные благоприятные условия' (смертность, способ-
ностью завершению ЬнтогенезаиТс размножению).^ -Ч-Щ 'Л
1 Яйца,отложенныебабочкамикапустнойХ:овкиизальтернатив-
ныхфотопериодов(12и’18чсветавсутки),были помещены вусловия
10;6ш1 °C. Уже через месяц во всехтемпературныхвариантах опыта
наблюдалась большая гибелыматериала: при WX) погиблоют ЗФдо
48%. Примечательно,‘чтовзтих близких к пороговым условиях через
20 дней из части яиц отродились гусеницы. В дальнейшем все они
погибли. При 6 °C смертностьза первый месяц опыта была еще выше —
более70%,а после 2-месячногоюхлаждения погибли все яйца.'Разви-
тие яиц, ‘перенесших месячное пребывание при 6’°С, в дальнейшем
(при 20 0С)было'медленнее,чемв контроле.<ТемпературуГ°С яйцане
выдержали втечение одного месяца. Смертностьв исходныхусловиях
(контроль) варьировала между 11 и 34%,-^развитие яиц инвариантно
завершал ось забдней. 1 алж гйгж
Яйца луговой совки выдерживались при 11; 5и0°С. В целом они
проявили несколько меньшую устойчивость к пониженным темпера-
турам, чем яйца капустной совки. При 11 °C в разных,вариантах их
погибло от 43 до 60%, из остальных через 24.дня появились Гусеницы.
Оставленные в этих условиях, они питались, медленно росли и посте-
пенно отмирали. К концу 3-го месяца в двух вариантах,.(из исходных
КД-фотопериодов 10 и 13 ч) еще оставались в живых 8—9% гусениц,
которые достигли V возраста. Эти гусеницы были мелкими, лишь
ширина головной капсулы соответствовала возрастной норме. При
более низких температурах яйца не развивались, а смертность почти
во всех вариантах резко возросла. Д вухмесячногоиспытания при 0 и
5 °C яйца луговой совки не выдержали. Приведенные данные надеж-
но свидетельствуют, что Несмотря на Высокую холодостойкость яиц
(температура переохлаждения в среднем -27,9 ±0,11 °C), зимовкалу-
говой совки на этой стадии невозможна. ж ‘
При зимовке настадии гусеницы лимитирующим фактором, по-
мимо морозов; может быть отсутствие корма или нарушение функ-
ции питания. У насекомых с полным превращением на личиночной
стадии происходит интенсивный рост. Полноценное питание в соче-
тании с оптимальным уровнем,физических факторов среды — норма
существования этой стадии. Л ишь хищники и паразиты способны к
длительному голоданию. По данным’А.С. Данилевского'[’1935], при
оптимальной дляразвития температуре последствия вынужденного
187
голодания гусеницХохоу/е^еЦ/сПса/и и,других исследованных видов
всегданеблагоприятны,ноповреждающеедействиезависитотпро-
должительности голодания ивозраста гусениц. При голоданиив мл ад-
шихдозрастах задержка в ростеможет компенсироватьсяпосле возоб-
новлениядитания.Голодание встаршихвозрастахприводиткпояв-
лению «карликовых; особей игибели. В связи сосказанным свежий
корм был достоянным фоном «аших оцытов. Голодание моглогбыть
лишь результатом нарушения функции питания пониженными темг-
ц₽ратурами1<ч-,и Н го ирдт; 'п
>. Гусеницмободх видов оказались сцособнымипитаться изавер-
шить развитие лри Л °C (рис. .83). Вопытах,начатыхс П-Ш возраста,
окукливаниенаблюдалось примерно через 2 месяца (средний срок
окукливания отмеченчерным кружком насоответствующих кривых),
а вопытахс1У—У возраста—через,месяц.Из-куколок после переноса
их в благоприятные температурные условиявыл стели способные к
размножению бабочки. Гибель гусениц в течение экспозиции при 11 Ч?
-'У I fit, : УСВВНА.ЙШ-'Г ЧА<'?3
' '‘Рис.83. Выживаемость; %, гусениц-П—1П (а,б) ^и' IV-V (в,г)'возрас- u f
.~товкапустной;(а,в):илуговой(б,г)совокпри1длительном'содержа- ч\!
нии в низких температурах (J0,.,6 и 1 °C):, ’ г : «г /
* К-момент окукливания, ДЦ-длинный день, КД—короткий день. Для
з луговойсовкиприведеныданные.суммированные'потремисходнымфотопериодам' 11'
18»
былаоченьболыпойв опыте.начатомс младших возрастов. При6°С
гибель былавысокрй-среди всех возрастных групп.Около50% гусе*
ниц,.погибло через.месяц,а через2-3месяцапогибдивсе гусеницы
обои хвидов, (рис. 83),Интересно, чтопри.этой температурепроисхо-
дилопитание и.даже динькигусениц. Так,вопытах„начатыхс JI—III
возраста, насекомые достигли даже IV возраста.!! редел ьнымсроком
выживаниягусениц капустной илуговой совокпри,6,°Сможно при-
нять2месяца(при большой гибели).3авершениеметаморфоза при
переносе гусениц из.условий с 6 °C в исходные 20 °С:оказалось воз-
можным лишь после содержаниявхолодевтечениенеболееЗмесяца.
При температуре ! ?С все.гусеницы капустнойсовки погибалшчерез
10—12дней,агусеницылуговой совки —дерез! месяц. В заключение
следуетотметить, что длина днядо содержания, в холодели на его. про-
тяжении не влияла достовернонавыживаемостьгусениц.,^
, я Куколкиобоихвидов,.как11оказаноранее,независимоотихфи-
з иол огическогоюостоянияммеют холодостойкость,достаточную для
зимовки в.почвеч(под<снегом1;взрне умеренного климата.. Ку колки
существуютзасчетрезервов, накопленных настадцигусеницы, ине
зависят от наличия корма. У капуст ной совки куколки зимуют в со-
стоянии диапаузы. Все это подчеркивает особенную важность выяс-
нения способности куколок переживать длительное воздействие низ-
ких положительных температур.
У капустнойсовки предшествующие фотопериодическиеусло-
вия (длина дня, 10 и 18 ч) индуцировали соответственно диапаузуили
активное развитие,куколок.-,При всех испытанных температурах( 1;6;
10°С)какактивные, так и диапаузирующие куколки проявили высо-
кую,жизнеспособность.;При10°С погибла неболъшая.часты(до 12%)
активных 1куколок,изостальныхчерез1,5месяца,вылетели'бабочки.
ПрибгХ-погибло 5%,активных куколок. Остальныекуколки:во»всех
вариантах,!и том числе и после З^месячногр содержания на холоде,
остались живыми. Вскрытие длиннод невных куколок, , проведенное
через 4,5месяца показало, что онипогиблинастадииполностью сфор-
мировавшихся бабочек.Та ким образом,температура 6 “Снеюстанав-
ливалащолностью развития куколок, но имагинальнаялинькавотих
усдрвияхрказаласьневозможной- г - -льнгг ’•>» ятямиклл хк..
Разница между активными и диапаузирующими куколками ка-
пустной совкишроявилась вскорости потери,массы в ходелолодовой
“экспозиции”.Массадиапаузирующихкуколок,заЗмесяца экспери-
мента существенно не изменилась. ]Исходносредняя масса,ихсостав-
189
ляла 463,3 мг, а к концу эксперимента 426,9 мг.Таким«образом,
потерямассысоставилавсего26,4мг (5,7%)..Активныеже‘куколки
потеряли 77,'8мг,‘что!составило 22,4%. Это свидетельствует обол ее
интенсивной тратерезервных веществ активными куколками лгусле-
довательно,ограниченной-их способности' Существовать* при *пони-
женныхтемпературах. мя.йгмх и .«>.; чячг. ьт.-- -го-.
Куколки луговой совки независимо отнсходной длины*дня ( 10,
>13 и 16чсветансутки)развивались без диапаузы. При11 “С их смерт-
ностьчерезмесяц достигла40—43%, а через2месяца—78—100%. При
5 °С смертность>была почти,такой же. При 1 "С к концу первогбмеся-
цасмертностьдостигла75-100,%,ачерез2 месяца погибли все кукол-
ки. Трехмесячного-содержания на холоде‘куколки луговой совки не
перенесли при всех испытанных температурах. Таким образом, не-
специфическая устойчивостькуколок луговой совкикпониженным
температурамоказаласьниже, чем у «капустной совки, и совершенно
недостаточной Для переживаниязимнегопериода даже вюжныхрай-
онахзоныумеренногоклимата. Рассматриватьих вкачестве «зимую-
щей» стадии-не представляется возможным,' несмотря на относитель-
но® ысокуюхолодостойкость. . ; <
ч..е, «з ус и, 8.5.Сезошюеразвитие я
Представленные материалы демонстрируют глубокие различия
между насекомымис'гетеро- и гомодинамным развитием. Первые
(капустная совка) характеризуются наличием диапаузы в годичном
цикле и системойжонтроля за своевременностью бе формирования.
Возникновение диапаузы связано с накоплением энергетических ре-
зервов для длительного существования в отсутствие корма, резким
снижениемюбмена! веществ, повышенной устойчивостью к высыха-
нию и «повреждающим «факторам внешней среды. Специфическая
потребность диапаузирующих насекомых умеренных широт в холо-
довой‘реактивации превращаетпониженные температуры осенне-зим-
негопериодаи31неблагоприятныхвнеобходимые условияихсуще-
ствованияд Все этообеспечиваетюбитание гетеродинамныхшасеко-
мых в климатических зонах с резко Выраженной сезонностью , в{том
числен продолжительным зимнимшериодом. * < - * и
i ч 'Насекомые с гомодинамным развитием «на всех стадиях своего
жизненногоциклаинвариантнонаходятсявфизиологически актив-
ном состоянии. В условиях сезонности климата их выживание воз-
190
можно лишь на основе неспецифической устойчивости к неблаго-
приятным условиям. При этом неспецифическая устойчивость к эк-
стремальным факторам зимы (морозы), достаточно высокая на неко-
торых стадиях онтогенеза (яйцо, куколка), оказалась важным, но не
решающим условием зимовки, Лимитиррватьсе могут ограниченная
способность к длительному голоданиюприпониженных температу-
рах и отсутствие специальных энергетических ресурсов. Эксперимен-
тальная проверка таких потенций у луговой совки заставляет сделать
вывод о неспособностиее и, очевидно, других подобных видов к дли-
тельной зимовке в условиях умеренного климата. Окончательной про-
веркой этоговывода служили испытаниягомодинам ных видов в поле-
вых опытах.
Такие Опыты былипроведены в трех географических точках, зна-
чительно удаленных друг от друга: С.-Петербург, 60 °с.ш.; Белгород,
50 °с.ш. и Славянск-на-Кубани, 45 °с.ш. Этизоны существенно раз-
личаются по тепловым ресурсам. Так, сумма температур выше 10 °C в
них соответственносоставляет 570,1030® 2100 гр.-дн. Разная степень
континентальности климата сильно влияет на продолжительность
безморозноропериода,Осенние заморозкив.С.-Петербургенаступа-
ют в среднем в начале октября, в.Белгороде в середине октября, а на
Черноморском побережье Крарнодарского край в ноябре (Агрокли-
матическийатласмира, 1972). Параллельное изучение гомодинам-
ных и гетеродинамных видов в полевых условиях предоставляет наи-
бодееде^д™сд)дащ1тель1«>-экологи5[ес1а1йматериац5!г5
8.5.1. Капустная совка,'Mamestrdbrassicae '* Н
же.-- _ -тол <<
О Результаты полевых опытов, выполненных в Ленинградской обл.
с.разными нопуляциями капустной совки, показали, чтоосновные
различия».между популяциями проявляются-в сроках формирования
диапаузы .(рис.84)ч.^ЛестнаяпопуляциякапустнойсовкИ'Даетв
С.-Пея»рбурге толькоюднугенерацию.Даже прикирождениигусе-»
ниц в наиболее, ранние изинаблюдаемых фенологические: сроки .(на
рис»84штриховкойпоказанынормальныесрокияйцекладки) диапа-
уза имеетместо.у,всехкуколок(вар.'5, Л), Следовательно, несмотряна
большую продолжительность дня (в июле более 20-я с учетом граж-
данских сумерек)^ ФПР нафоне весьма умеренныхлгриродных темпе-
ратур о казываетсяне эффективной. Этому способствует'высокое зна-
чение критическогофотопериодауленинградской популяции (даже'
> - <- ' ‘.и; " - ’ • -.ЧД
1Я
Май Июнь ‘ Июль Август Сентябрь Октябрь
' •> " «.UH.!..--.и.»
Рис.84. Формирование диапаузы у географических популяций .
rMarnestra brassicae.B условиях С.-Петербурга:.. , , , , . ч
1-10 - сезонная привязка вариантов опытов с сухумской популяцией (С);
сплошныеагрезки-стадиятусеницы.ихиггриховоепродалжеиие'-стадия'куколки;' "
начало и конец стадии отмечены вертикальными рисками. Л —данные для ленин-
градской,.Б—длябелгородскойпопуляцийЩанилевский.Кузнецова, 1968].Числа .
' возле полос- количество диапаузирующих куколок, %. <+>"—гибель отзамОроз-
’хов.Штриховкойклмеченыпредельныефенологическнесрокиийцекладки местной1
,популяции,.Кривая-^среднедекэдтдотемперазураддяС^ . .... , >1;у]
при 25 °C он близок к‘18ч), 'скрытый образ жизни йвыраженйая тен-'
денция к формированию диапаузы.
Белгородская популяция совки, имеющая более короткий кри-
тический фотопериод (15 ч 30 мин при 25 °С, >17 ч 30 мин при 18 °C),
оказаласьспособнойразвиватьсяв условиях С.^Петербургабездиа-
паузы.Однакодажешварианте 3,Б (одноммзнаиболеераннихХ где
отрождениезгусеницотмечалосьзначительно ’раньше «нормальных
фенолопическихсроковипроисходиловпериод«белыхночей»,все
жедиапаузировало>56%жуколок.Приотрождениигусеницв*начале
фенологическогосрока(вар.5,Б)долядиапаузирующихкуколоквоз-,
росладо£^,аафиотрождении.в;концеш»госрока<вари7!/Б>диапа-
узироваливсе куколки. о< .о я ai е
. :> Болееюжная1сухумская популяция, исследованная нами наибо-
лее подробно (критическийфотопериод’при 20 °С близокк15 ч), фор-
мирует диапаузув ещеболеепоздние сроки..Первые диапаузирую-
щие куколки (4%) этой популяции появились от гусениц, отродив-
192
шихся Юиюля (вар. 6), авследующемвариантеопыта диапаузирова-
ли уже’все куколки;{Интересно отметить,что особи южных популя-
цийкапустнойсовки проявили способность формировать диапаузу
приотрождении гусеницв нормальныедляС.-Петербур гасрокираз-
вития этотовида.Этообеспечивает даже-для сухумской!'популяции
возможность в первом жевегетационномсезоне сформировать зиму-
ющий запасна севере ареала. Такая способностьобусловленане толь-
котермолабильностьюФПР,но'вещебольшейстепенивлияниемсу-
точных терморитмов 'На.тенденцию к формированию диапаузы [Да-
нилевский, Кузнецова, 1968]. Она предоставляет значительнуюсво-
боду дая внутриареальных перемещений разных' популяций, Свой-
ственную ДаЛекоНевсемисследованнымвидам/ £ кы •:н:.?;п
1 а( Легко представить дальнейшую судьбу 'особей южных популя-
ций М. brassicae на севере ареала. Так как холодостойкость и темпера-
турные нормы реактивации—признаки видовогоранга.весенний вылет
южных бабочек произойдет синхронно свылетом бабочек Уместной
популяции и в нормальные фенологические сроки. Отбор будет дей-
ствовать в направлении оптимизации параметровФПР, в первую оче-
редьв сторонуувеличениякритического фотопериода.'Ещеболее силь-
ное воздействиенаперемещеннуюпопуляциюокажетскрещиваниеС
местными особями, при1котором "будет’иметь место промежуточное
(сне которым1 преобладаниемпризнаковсамки)наследованиетакого
признака,'как критический фотопериод . В результате'экологические
реакции впотомствеперемещенной популяциипридутв соответствие
с фототермическими условиями ленинградского климата. На Приме-
ре развития капустнойсбвки видно, что завершающим сезонное ргй-
витие этапом является формирование диапаузы. Сказанное в Полной
мере касается и других гетеродинамных видов с поливольтинным цик-
ЛОМ раЗДИТИЯ. /'Т/ -ХОП ЙкГ ОН.ПТ
* ‘ .Специальный интерес представляют данные о жизнедеятельно-
сти активных стадий гетеродинамных’ видов в природных условиях
(особенновконце вегетационного сезона), так как эти стадии пофи-
зиологическомусостояниювполнеподобны таковым гомодинамных
видов. Исследованные географические популяциикапустной совки в
период активного роста и развития Идентично реагируют на сезон-
ные изменения условий (рис. 84). Небольшиеразличия впродолжи-
тельности’развития1 (личиночного и куколочногорвблизкихпосро-
кам опытах обусЛовленыНогодными отличиями разных сезонов1. -!
•,-ЧН Я ’ й</ :: rs-jObMS'riiliHXSH J .-I <
193
ш Опыты с сухумской популяцией М. .brassicae выполнены по наи-?
более полнойсхеме, пр^тому остановимсяна приученных результатах
более.подробно Всего в сезоне с этой популяциейбылолроведено 10
вариантов опыта, различающихся сроками отрождения гусениц: пер-
ваяихцартия; был а перенесена из лаборатории в> природныеусловия,
21мая, последняя—Д5августа. Продолжительность развития гусениц
в каждом вариантеопытаобнаруживает тесную корреляцию^ сезон-;
ной динамикой температуры. Наиболее! медленно гусеницы развива-,
лись ввесеннийи осенний периоды, когдасреднесуточная темпера-
тура не превышала 12 °С (температурный дорог развития по лабора-
торным данным близок, к 6 °C). Минимальная продолжительность
развития гусениц (28 дней), наблюдалась в варианте 6 при среднесу-
точной температуре Д 8,5°. Аналогичную, зависимость сроков разви-
тияот демпературы .можно проследить идля куколок. Суммарная про-,
должительность личиночного и кукол очного развития в вариантах 1 -
б оказалась почти стабильной. о н:
;Наблюдения заразвитием капустной ссвки в условияхЛенин-
градскойсбл.. позволило заметить наличие предельных сроков отрож-,
дения гусенищ нозжекоторыхони не успевают завершить ростиокук-
литься. Такимкритическимсрокомв наших опытах оказалась 1 -я де-
када августа- Гусеницы, отродившиеся в середине августа (вар. 10), уже
нремогли окуклиться ипогиблид началеноября-отзаморозков, Не-
сомненно, что критический срокотрождения гусениц зависит от осо-
бенностей сезона и варьирует по годам, по меньше, чем ожидалось,
Так,вопытах А.С. Данилевского и И.А. Кузнецовой [1968], выпол-
ненные зд есь в 1962 т,у критическим сроком для М..brassicae оказа-
лось 5 августа. Существенно отметить,<что он оказался одинаковым у
разных географическихпопуляций. Реальность предельных сроков для
начала личиночного развития показана для ряда видов ипопуляций
чешуекрылых с куколочной диапаузой. Раньще других такой рубеж
наступает у Poliaoleracpa (конециюля); к 1-й декаде августа онпри-
уронен yMCRBeawwSpilosoma.menthaslrin щавелевойсовки^Укапуст-
нойбелянкинесмогдизавершитьразвитиеиокуклитьсягусеницы,
отродившиеся 20 августа. Последовательность, видов в приведенном
перечне неслучайна. Она позволяет заключить, чтр предельный срок
в,сезоне.зависитрт температурных норм.развития (дорога и суммы
эффективных температур). У капустной белянки продолжительность
стадии гусеницы {при равных условиях) короче, чем у других назван-
ных видов, а нижний температурный порог развития ниже, что и по-
194
зволяет ей достигнуть стадии куколки до наступленияэкстремальных
условий при более позднем отрождении гусениц./Наступление суб-
пороговых температур делает невозможными линьку на куколку и
последующее формирование диапаузы, а низкая холодостойкость ак-
тивно развивающихся гусениц приводит их к гибели при заморозках.
Можно предполагать, что при личиночной диапаузе критические сро-
ки отрождения гусениц окажутся-более поздними.
Многочисленные полевые опыты сгетеродинамными видами по
идентичной схеме, проведенные нами в Белгородской обл. с местны-
ми популяциями Л. rumicis, Р. brassicae, Leptinotarsadecemlineata, Ата
custos и других видов, показали, что достижение стадии, к которой
приурочена диапауза, является непременным условием существова-
ния гетеродцнамныхнасекомых в зоне умеренного климата.
-8.5.2. Полевыеопытысгомодинамнымивидами
‘ «Результаты лабораторных исследований с луговой, малой назем-
ной совками и совкой гаммой показали отсутствие у них выраженного
зимовочного состояния Неспособность всех стадий онтогенеза дли-
тельно переживать не только отрицательные, но и низкие цолржи-
тельные температуры. Полевыеопыты сназванными, объектами име-
ли'задачей окончательно Проверить вывод о гомодинамном тице их
развития и проследить, какие этапы онтогенеза могут, выступать в ка-
чествезавершающихИегетационный'сезон. . (.7?'
Малая наземнаясовка, Spodoptera exigua. Полевые опытысэтим
видом проводились по той же схеме, что ирхйяты с капустной совкой.
На 'основании полученных данных (рис. 85) можно заключить, что
малая наземная совка - более теплолюбивый'вид, чем капустная совг
ка. Развитие гусениц до окукления оказалось возможным лишь впер,-
выхтрехъариантах опыта, начатых соответственно’Зиюня,1 и 7 июля.
В вар. 4 от 28 июля гусеницы достигли стадии пронимфы, но не рмог-
ли окуклиться и погибли от заморозков в середине октября. Такая же
судьба постигла гусениц из вар.% а сроки последующие вариантов
оказались' еще’более неблагоприятными: гусеницыпогиблив октяб-
ре, не закончивпитания. ' ’
Гусеницы капустной совки (см. рис. '83)успешно завершилг^раз-
витие даже при отрождении из яиц 11 августа с окукливанием в цачале
октября. Несомненно, что отмеченные различия между видами обус-
ловлены разными температурными порогами развития(6°С у капуст-
195
j a ^v/аемиой <й>Вки8«*Ленинград-
' + ской обл.:
io aXik 20 frwta ' \1-^г«риш™«»п<»(а-
™ «.м . .V tu .u XV uv гусеницы,б-пронимфы,в-кукол-
Июнь Июль Август Сентябрь Октябрь ки):-<-+>—гибельотзаморозков
4.SH fee,.ЧбЖЖ!.-, xbJt,;-. sv/L,-..j...! <'< -г-.;’i О-/ы-tu : Г .,tn ..
s . .7 , f " ", 1' ' '! ' x'-.-- ЙОП
ной совкии11 °C у малой наземной (результаты лабораторных опытов
и расчетов)При'близкой сумме эффективных температур(сордвет-
ственно346и'296гр.-дн.) указанная разница-в порогахрезкоограни-
чивает'возможности развития ф^р^над^
СКОЙОбл'.'0’'^''''^ .
<х В ‘пределах сезона, допускающего формальное развитие малой
наземной совки в Ленинградской обл', изменение скорости развития
гусени^ и куколок'обнаружило близкую к гиперболической зависи-
мость от'температуры.В соответствии с динам икойтемпературыпро-
должительность развития гусениц уменьшалась от 40 дней в вар. 1 до
32и26днейввариантах2иЗ.Длительностьразвитиякуколок,наобот
рот, была:Минимальной в вар.1 (всего!*! дней), но сильноувеличива;
ласьв двухследующих вариантах (соответственно 25 и,45;дней)^,
• с?! Как и в лабораторных исследованиях, ни в одном варианте поле-
вых'опытовнебыл и получены диапаузирующие особи, хотя опыты
проводились на фоне удлиняющегося дня, в период самых.длинных
дней и при сокращающемся дне,1 при суточных терморитмах и других
факторах, образующих сезонную динамику природных условий. Это
окончательно доказывает гомодинамный характер развития, малой
наземнойсбвки. < - , ........
J Луговая совка, Mythimna unipuncta. С этим видом в Ленинград-
ской обл. удалось провести лишь наиболее важные осенние варианты
196
опыта, так как материал в нужном количестве был получен только к
началу августа. Как и у малой наземной совки, гусеницы, отродивши-
еся после 1 августа, Не заканчивают развитий. К концу сентября на
50—55-й день гусеничногоразвитияони достигли массы 500—600 мг
(в лабораторных опытахпри достижении такой массы было возможно
окукливание), а в дальнейшем гусеницы теряли в массе и в начале
ноября при резком заморозке погибли. ' \ ;й ,t
Совка гамма, Autographagamma.B С.-Петербурге долевые экспе-
рименты с совкой гаммой были проведены на материале, доставлен-
ном из Краснодарского края. Развитие гусени цпрои сходил о с конца
июня досерединыавгуста. Из всех куколок вылетели бабочки, кото-
рые отложили яйца. В вариантах опыта, начатых в августе, гусеницы
погибли. Наиболее’важно отметить, что в условиях Ленинградской
обл., где.больший ствогетеродинамных полицикликовможет давать
только одно поколение, у совки гаммы вылетающие самки откладыва-
ли яйц^ и давали дач^лр следующемупоколению.Этобылортмечено
не тольков экспериментах: очень«свежие»,хотяи редкие!, бабочки
этого вида, встречающиеся в природе в августе/свидетельствуют, что
так происходит и в естественной обстановке.
В Белгородскойобл . совка гамма—вид весьмарбычный. Вовсех
вариантах полевых опытов гусеницы завершали развитие, окуклива-
лись, из куколок появлялись бабочки,' которые.приступали, к дйце-
кладке (рис. 85). Куколки'из последнего варианта. Ю'сентября были
доставлены в С.-Петербург, где их развитие также завершилось выле-
том бабочек (на рис. фигурной скобкой отмечен расчетный срок за-
вершений их развития в падевых1 условиях с учетом сезонного хода
температуры). Лимитированные сроки экспедиционной работы гге
позволилилровестиболеепоздниеварианты'опытаиррпосредс^вен-
нонаблюдатьзавершающийэтап сезонногоразвитияэтого дмда^Од7
накоегоможно пропгозировать'наосновании динамики температу-
ры‘и экспериментально установленных параметров развития. ,
' u В Белгородской обл. у исследованных нами гетеродицамныхви-
дов уже *в июле’возникает возможность формирования драпгузр
(рис. ‘86, П, 1П). Совка гамма в значительно более поздних вариантах
(в том числе начатом'15 августа) развивалась без задержек,"и flta,всех
стадияхонтогенезанаходиласьв физиологически активномсостоя-
нии.’Возможности ее нормальной жизнедеятельности ужев октябре
будут ограничен ыв связи с сезоннымпонижениемтемпературы(сред-
немеся^нси? значенирпр многолетнимдацдьш .близко к 6,3*.С).,Ср--
J97
• л :а. ь : Рис*86. Развитие;совки гаммы; капустной я щавелевой совок (Мест-
. цыешрпуляции);^ ,условиях Белгородской обл.: *
1—5-варианты опытов с совкой гаммой (1—гусеницы, 2 — куколки.З — ...
бабочки);а—расчетныйсрок вылета бабочек впоследнем варианте (куколки были ’ '
р ; г < доставления С.-Петербург Юсеитября). Кривые: Г—среднедекаднаятемпература/';
. , .воздуха, П-сроки формированиядиапаузыу^апустной cobjch, Ш-тоже у щаве- ,
‘левой'совки’, ‘ ’*"лл>л- Jji - ... - 1 ——
й.--..-,, ,т. . < ,'1ДЛ.,Г.
гласно экспериментальным данным возможность длительно пережи-
вать субпороговые температуры в состоянии активной жизнедеятель-
ности у всех исследованных видов ограничена. Совокупность всех фак-
тов'позволяет сделать вывод, что у совки гаммы нет предпосылок для
успешной зимовки в условиях белгородской обл. ' ’., ’
'4 ‘ Опыты в Славянске-на- Кубани продолжались с 10 июня до сере-
дины сентября ивключали 8 вариантов; “Высокие значения среднесу-
точнойтемпературы, характерныездесьдлязначительнойчастивеге-
тационногосезонаХдажесредняя температура сентября П^С),обес-
печили высокие темпы развития совки гаммы. При этом средняя дди-
тельностьразвития гусениц составляла около 20 дней (15—24), куко-
лок - не более 8—11 дней. Лишь в последнем варианте отмечена зна-
чительная' задержка развития (гусеницы окуклились к 21 августа, а
бабочки вылетели в среднем'через 20'Дней). Но сам’вылет бабочек
свидетельствует, что задержка развития гусенцц не имеет экологичес-
когосмысла:'ихпотомствоужепопадаетвкритическиепотемперату-
ре условиями едва ли Достигнет стадии куколки Ддительный вегета-
ционный период (средняятемператураоктября! 1,5°С и'лишь в но-
ябре наступает пора заморозков), к сожалению, не позволил в экспе-
диционных условиях проследить завершающий этап сезонного цик-
198
ласовки гаммы в Краснодарском крае. Однако в.рпытахс местными,,
популяциями гетеродинамных видов (JLoxostege sticlfcalls, Mamestra
brasslc/ae, ffyphantiiafcunea) !obuio доказано^дтр у^цк 15-20^£усхау,
90-100% особей индуцируется диапауза.., ’
. 4; Результаты лабораторных и полевых исследований позволяют '
бол ее лол но охарактеризовать основные особенностисезонного цик-
ла гетеродидамных, и гомодинамных насекомых Полное.сходство
между ними наблюдаетсялишь в течецие весенне-летнего сезона, koi^
у тех и других доминируют рроцессы активнойжизнедеятельности—.
роста, развития и размножения. Принципиальные отлидия появля-
ются в перирд, когдау гетеродинамных полициюгикрв начинав? фор-
мироваться диапауза. Какдоказано на,примере дапустноцровки, ус-
ловия, дляфотопериодической индукции диапаузы (даже у .южных
популяций, испытанных в С.-Петербурге) возникают в сезоце,еще
летом, авконце июля-августеуже появляются диапаузируюшие ку-
колки.®, дальнейшем ихдоля сред иску кл ивающихсяособейбыстро
растет и достигает 100%. С этого момента прекращается .пополнение
активной фракции популяции, начидаетсянакопледиезинующего
запаса. Сезонная заблаговременность указанных событий позволяет,
как правило„рсновной,массе гусениц достичькукол очной стадии др
наступления субпорогорыхдрмцератур ивстретить зимувсрстоядид
диапаузы,, Г
, .. Црдобная схема.сезонцрто развития капуртнойсовки достаточ-
но универсальна длягетеродинамныхполицикликовОнаподтвррж-,
дается данными полевых, исследований, выполненных на, разных
объектах. Видовые отличия у совместно,обитающих насекомых лро-
являлись лишь в сроках фррмирования диапаузы вприродныхусло-
виях, что зависит от параметров физиологических реакций, контро-
лирующих дидпаузу. и погодных особенностей конкретдого ррзона.
Достижение стадии, щкоторой цриуроченадиапауза, -завершающий,
этапсезоннопо развития любогодетеродинамного вида-Дсеособщне^
достигшиеэтойртадиидокодцавегетационногосезона, обреченына
гибель.. Таким образом» диапауза выступает в качестве наиболее со-
вершенной, адаптации к переживанию зимнего периода.
Экологическое значение диапаузы не ограничивае?сяре?истен-
тностью насекомых ^неблагоприятным условиям зимы. Формирова-
ние зимующего запаса на характерной дая вида диапаузируюшейста-
дии онтогенеза приводитк выравниванию физиологического рортоя-
ния данной популяции в предзимний период, следовательно, спо-
199
собствует более дружномувозобновлению‘развития в следующемсе- -
зоне.'Существенно,чтоу большинствавидов умеренныхшироте ди-1
апаузой связано адаптивное’ изменение экологических требований^
среде. В зоогеографическом аспекте температурные Нормы холодо-
вой’реактивации могут определять южныеграницы ареалов Многих
вид овумеренных широт[Данилевский,1961].Всесказанноесвиде-1
тельствует о том' чтодиапауза - уникальное свойство'гетеродинам-
ныхнасекомых, определяющеемхадаптацию ксезонности и зональ-
ности климата. Она способна решать комплекс экологических задач,
в‘томЧислёпроблему’зимовки.!Л!*’! ‘ вд-‘м<<н f'i ?> «йтк, .« , .
Основное свойствотомодинамныхвидов— непрерывное состоя-
ние физиологическойактивности — ярко проявилосьв полевыхопы-
тахс совкой'гаммой/луговой'ималой наземнойсовками.Как и в ис-
следованияхс капустнойсовкой,схема обеспечивала одновременное?
наличие; вбпытах всех фаз онтогенеза. Приотсутствии диапаузы есте-
ственный пред елразвитияэтих’видоввсезонеобусловлентолькона-
ступлениемсубпороговыхтемператур.' ; з
Полученные результаты'подтверждают’сложившееся прение о
том,1чтосовка‘гаммаймалаяназемнаясовка«зазимовывают»на всех
стадиях ра!звития[Кожанчиков,1937;Брянцевй|ДоброзраковаД1960;’
Космачевский,' 1966,'идр.]'. Вопрос заключается втом, какиеименно
стадии и насколько успешно могут пережить зиму. Лабораторные ис-
следованияне дают оснований для оптимистических прогнозов в от-
ношении‘зимовкийсследованныхтомодинамных ИидовдПри^отно-
сительно высокой холодостойкости отдельных стадий (яйцо, кукол-
ка) лимитировать зимовку может ихНеспособностьдлительно пере-
носитьдаже низкие положительныетемпературы. ' * - 1 * ’ w а>;
-'Эти данные согласуются с Мнением большинствазарубежных ис-
следователей1, что совка"'гамма, например, Не может зимовать в Цент-
ральной Европе fMell,’ 1953; ’Koch,11965;'*VojnitS, 1966!,]', а В-Англии
перезимовываеттольковтодыС'наиболеемягкой‘зимой*[Р15Ьег,1938;
Cooper’ 1946; Huggins,’Т958]1.кОни подкрепляют 'гипотезу о миграци-
онном Проникновении1 гомодинам ных видов1 в умеренные широты
[Kanervo, 1947;!8у1уеп,!1947;Ка1811а,!'19б2].|Одншсотрудно;предста-
вить какой-либо реальный механизм миграций/способный обеспе-
чить ежегодное возобновление популяций за пределами их основных
ареалов'. Но широкое распространение той же совки’гаммы в Пале-
арктике, ее регулярное появление в’Средней полосе Россиимежду 52
и 58 °с.ш. [Щеголев, 1964], наши собственные наблюдения за лётом
200
бабочек этого вида В Белгородской обл. заставляют с'вниманием от-
нестись к имеющимсяФлитературесведениям о перезимовке совки
гаммы в умереиной зоне. Так, нарядус признанием:того,что?на зи-
мовкууходятже стадии развития,длякаждого вида называют наибо-
лее зимостойкие:тусенйцуйгкуколку ’для совкигаммыЧДрахов-
с кая,1962; Novak,19ОД,'куколку длжмалой наземной совки[Брянцев
иДоброзракова, 1960] .Несмотря'на противоречивостьданныхЛите-
ратуры, нельзяПсеже исключить, чтов конкретнойприродной обста-
новке могутвоаникать какие-то локальные условия,‘обеспечивающие
зимовку отдельныхособейгомодинамнЫхвидовна основе неспеци-
фическойустойчивости.Постоянно’низкаячисленностьсовки гам-
мы в средних широтах при ее высокой потенции кразмножениюпо-
зволяет предположить, чтотакиеусловияскладываются крайнередко
и основная масса «зазимовавших» особей влечение 'зимы гибнет. as г
Интересные данные о'всйышке массового размножения малой
наземнойсовкив Воронежской обл. Приводит В.Ф. Палий[1956] . При
обслеДовании почвы в период с' 15 сентября по Ю октября^было обна-
руже но мн ого кукол о к/авесь октябрь продолжался вылет бабочек.*®
результате кайме куколок не осталось. абабочки не перенесли зимов-
ки . На следующий 'год не было найдено Ни одной бабочки или гусе-
НИЦы5. ex/g«a. ' 1 ./"г г< Л Ур i
'= 4 На основании многолетних>наблюденИй( Дж.' Кайсиля {Kaisila,
‘1962] выдвинулпредположение, чтопопуляциясовкигаммыв Фин-
ляндии состоит из особей двух типов.<3постоянных£обитателей',’мало-
численных,никогда не дающих вспышек массового размножения, но
успешно акклиматизировавшихся в новой местности, и мигрантов,
бабочек и их потомства/имеющих южное происхождение,-неспособ-
ных зимоватьвсеверных широтахи снаступлениемхолодов либовоз-
вращающихся:обратнб,либопогибающих.Численностьустойчивой
^местным условиям популяции'порой-бывает ничтожно малой, но
всеже достаточнойдля ее’поддержания.Подъемчисленностипроис-
ходитблагодаря массовому притокуособей из центральной-части аре-
ала. Подтверждением и развитием этого предположения служат на-
блюдения И. Новака [Novak, 1968] над местной (Чехия) популяцией
совки гаммы. Собранные осенью в природе гусеницы резко различа-
лись по характеру развития: одни из них развивались аналогично лет-
ним и в октябре дали имаго; другие гусеницы росли очень медленно и
только в декабре из них вылетели бабочки; оставшиеся прекратили
метаморфоз и в III возрасте зазимовали. При экспериментальной оцен-
201
ке способности этих гусениц противостоять охлаждению обнаружи-
лось, что температура переохлаждения у них достигает -22 °C.
> f В.наших опытах сбелгородской и славянской популяциями сов-
ки гаммыподобная разнокачественностьособей в природных выбор-
ках не была обнаружена, а температура переохлаждения гусениц млад-
ших возрастов, вое питанных-вразных фототерм и ческих режимах с
учетом условий содерхфния материнского поколения, инвариантно
не опускаласьниже -4...-6 °C. Остается предположить, что среди осо-
бей,: использованных в наших опытах, были только иммигранты или
их потом ки. Это вполне, вероятно, если, согласиться с тем, что основ-
ная.часть популяций совки гаммы в умеренной зоне.состоит именно
из|них»|аж&’!!мска.’н«
. гп: В связи сэтим заслуживают внимания и наблюдения Л.П. Бер-
гер [1985] дальневосдочной луговой совки, Муthimna.separata. Она
предполагает, чтоМ separata, также имеющаягомодинамный тип раз-
!вития, может перезимовывать на Дальнем Востоке в условиях хрлми-
стого-рельефа на стадии куколки в состоянии оцепенения ,в техмик-
ростациях/гдедлительноевремя сохраняется температура, близкая к
ft’jC-йЭто предположение подтверждаемся анализом фенологии вида в
весенний,период. Нермотря на разнокачественный сослав популяции
перед уходом на зимовку, весной луговая совка обнаруживается толь-
ковоиремялёта бабочек. Успешно перезимовывают куколки, кото-
рые;сформировались вболеепоздниесроки, непосредственно перед
устойчивымлонижением температуры, Весной,когда набирается,СЭТ
порядка<120--140 гр.-дн., необходимая длязавершения развития ку-
колок,происходит одновременный вылет бабочек из благополучно
церезимовавшихжуколок. Однако увеличение численности луговой
сов ки,на Дальнем Востокеопределяются заносами.бабочекиих миг-
рациямиизочагов цостоянногоразмножения [Бергер,1985;Макаро-
вд, Доронина,. 1988,1994],. Сходным образом, возможно, формируют-
ся популяции у совки Agrotis ypsilonnневерных частях ареала на севе-
роамерцканскрм континентеJKaste^-SJwwe^l^]^ в Евррце^оуак,
Spitzer»'1972]...
Глава 9. МИГРАЦИИНАСЕКОМЫХИ ИХ МЕСТО В СИСТЕМЕ
........СЕЗОННО-ЦИКЛИЧЕСКИХ АДАПТАЦИЙ
‘. Большая подвижность насекомых создает исходные; предпосыл-
кик миграциям как средству избегания неблагоприятных условий.
Они проявляются в самых широких градациях - от микромиграций
до дальних перемещений, вполне сравнимых,с перелетами птиц., ,
Простейшей формой миграций в любое время сезона могут^слу-
жить1 перемещения и поисгах пищи, мест для откладки яищу^рд из
некомфортныхусловий поградиенту экстремального фактора —тем-
пературы/влажности, освещенности и ад. Следующий этап развития
миграций связан с( запрограммированными в онтогенезе изменения-
ми поведенческих реакций. 'Благодаря .этому у ряда видов насекомых
происходит изменение среды обитания — уход в понву личинок для
окукливания. Аналогичное миграционное поредение (мржет ,проис-
ходить и дри формировании диапаузы, что существенно; повышаетее
протекторные функции, так как обеспечивает защипу от прямогр дей-
ствия экстремальных условий. я,
На осноре микромиграций, по-видимому, моглиразвиться и дру-
гиеболее сложные формы, миграционногощоведения, Однаизних—
сезодные миграции,.которые имедхг строго закономерный характери
являются. как правил о,неотъемлемой частью жизненного цикла,вида.
Оциприурочены крпределенным,этапам индивидуальногоразвития,
происходят в. определенные резоны рода и приводят к адаптивным
сменам местообитаний. Такие миграции рассматридаются.как прояв-
ление особого поведенческого и физиологического синдрома, назы-
ваемого миграционным состоянием [Johnson, 1909]. . . ..,
Мигрирующие особи характеризуются повышенной дви^тель-
ной активностью и торможением, других функций, особенно размно-
жения и питания. Исследования показали, что у большинства видов
миграции совершаются в состоянии репродуктивной диапаузы, на-
203’
ступление которой регулируется внешними факторами [Kennedy, 1961;
Johnson, 1969; Dingle, 1978; Rankin, Rankin, 1980].
Сезонные миграции свойственны насекомым из разных отря-
дов. Например, некоторые жуки осенью перелетают в места зимовок.
Так, у коровок Semiadalia undecimnotata, Coccinella septempunctata,
Hippodamia convergenus, Coleomegilla maculata [Hodek, 1973, 1984; Ce-
мьянов, 1978,1979; Obrycki, Tauber, 1979] диапаузирующие имаго миг-
рируют, как правило, в горные ландшафты и зимуют там, образуяболь-
шие скопления. Вравнинныхусловиях зимовка происходит на опуш-
ках лесов, в лесополосах, на склонаховрагов.' Весной реактивирован-
ные жуки возвращаются в места размножения. Такие же миграции
свойственны полужесткокрылым. Например, хорошо известнысезон-
ныеперемещениявместа зимовки черепашек’рода Eurygaster [Ар-
нольди,!955;Андреев,Молчанова,1964;Викторов, 1967].Кми гра-
циям такогожехарактера относятся осейнйе миграции тлей',’ связан-
ные с откл адкой'зимующих яиц.Летние Перемещен иякрылатыхтл ей
на'новыерастения-хОзяеватакженосят'закономерный характер, рд-
нако'неимеют'строгой календарной'приуроченности',илосновным
фактором'Ьпределяющимихначало,‘выступаетскученность![П1апош-
iгиков, 19591, Lees,z 1966]. Миграции1 к!местам эстивации' описаны у
coBOK*(Lepidoptera) [Pruessl,l'1967f'Okui, Kobayashi, l:978';lOku,?li983]?i£
*' * У многих полужесткокрылых формирование миграционного со-
стоянияусложняетсякрыловым полиморфизмом.Вэтом случаевоз-
никаютсложные взаимодействия трех сопряженных явлений - Диа-
паузы, сезонногополиморфизма и MnrpannftfVepsalainen,1974, ‘1978;
Dingle^WTB; Zera', m5;^uisa^'1989/1993;? Harada, 1992; Harada;
Numata, 1993]. •"4”
t Наиболее подробно регуляция этих'явлений изучена' у'клопа
Oncopel^fasciatu^^yg^idaey^hi^ei^l^ieS’, 1;974Ь;'11978;!'1980;о198!5,
1991 и;др!|. 'Этот вид 'замечателен тем,чтосовершает дальние‘ежегод-
ные *миграции,вкоторыхучаствуютособиразных1генераций. Вумег
ренной зоне Северной Америки' мигранты появляются в конце вес-
ны-началелета/Здесьюни размножаются/питаясъ’наразныхрасте-
нйях семейства’ молочайных‘(ЕирЬогЫасеае). Осенью в ответ на ко-'
роткийдень’уклоповИндуцируетсяимагинальнаяДиапауза, которая
сопровождается формированием Миграционного состояния, что при-
водит=к!массовЬймитрацийклойовнаЩг.''н
' Более южные популщции (Флорида)‘ размножаются‘ большую
частьТода.' Толькой зимние месяцы в результате наступления йепро-
>.Н . I il. ‘Г7 Т' t'o'-Xi. :И-; МГЭ(1 • >л 1 "Л Л'и № л , '
204
должительной лабильной диапаузы, под влиянием короткого дня.,и
низкой температуры клопы нанекотороевремяпрекращают;актив-
ную жизнедеятельность. Эти. популяции обладаю^ меньд}ей мигра-
|щрцнрй.ак1иен9стьк’7чемсеверцыеJPingleetjal.,Д93О].Г, тонщ
, . Тропические популяции активны круглый год, у них нет состоя-
нияпокоя .в виде диапаузы июни не.склоцны к дальним перемещени-
ям^, Дрзнапительные .цррелеты^у, ццх связаны' лирь^цоисКрт^опртг
мальныхгу«ловий{питанця {J)iij^lerB31din^l^3]M^ ?s m? j > w v
. p Экспериментальнымиисследованиями .цроведеннымис особя-
ми-митр ирующихпопуляций изАйовь! ,(42 °сдп.) и, немигрирующих
из Пуэрто-Рико(18 Ос.ш.),обнаружены достоверные различияв мор-
фологических показателях, характеризующих миграционное состоя-
ние.. Мит-ран.тыотличались. большими размерами тела,большей дли-
ной крыльев и более длительным ,периодомсозревания.самок [Р1гщ1е,
1991], Испытаннырна «каруседи* мигрирующие осо(бипроявилизна-
чительное превосходство над немитрирующим^в способностикдли-
тельцому'цолсгу ,(рис.. ^7)л.В9^да-ДРадесса,
формирование миграционногосостоянияи активная репродукция
регулируются, нейроэндокринной системой.Высокийуровень,юве-
нильного гормона,определяет размножение, средний - миграцион-
ную готовность, анизкий вызывает диапаузу (Капк1п,1978].участие
ювенильного гормона в регуляции миграционного состояния дбнарут
'«* .вхгщпщч • IW4V7-Л'-чА
1 . ' А : дз-даос.
мок Oncopeltus fasciatus [по: Dingle, 1978]
205 ;
7ггПозжемодобные,реакциибыли1Описаныусеверных.популяций
Spilosoma rnenthastri>\KMHp, 1985},«летней капустноймухи, Deliafloralis
[Несин,, 1987[инекоторыхдрушхвидов.Диапаузаунихпотенциаль-
но.являетсяфакультативной^новреальнойприродной обстановке
возможность активногоразвития негреализуетсягнасевереареала из-
занедостаточновысокойтемпературы,анаюге-вследствиенедоста-
точнЬйшродалжительностидня. w--. т_
- 1А Регуляция моновольтинизма на основе промежуточной'реакции
обнаружёна,и> у темнокрайней' Совки^ Лху/м />«/ги,|НО вотличие »от
предыдущих видов; ФПР которых проявляетсялишь в эксперимен-
тальных условиях; у темнокрайней'совки жизненный цикл реально
контролируетсяФПР{Тыщенкоидр.,1983].На основеанализа фено-
логии:Л.ди/швлесостепной зоне былсделанвывод, что промежуточ-
ная ФПР у этого вида выполниетрегулирующую функцию в климати-
ческихзонах,температурныересурсы которыхпревышают потребно-
стиодноптпоколенияи втеплые годыдопускаютразвитие-двух пол-
ныхпоколений. Когда окукливаются гусеницы Тго поколения, про-
межуточнаяФПР;оставляетв^ активномссостоянии некоторуючасть
популяции’вкачестве резерва, способного приблагоприятныхпогод-
ных условияхдостичьдиапаузирукяцей стадии; Если температурный
режим окажется менее благоприятным, этот популяционный резерв
погибает,азимующий запас формируется только за счет диапаузиру-
ющих куколок1-го поколения. Таким образом,промежуточная ФПР
может рассматриваться как специальная сезонная адаптация ^посто-
янному существованию насекомыхвзонахперехода отмоновольтин-
ного.типаразвитиямполивальтинному;г . ?и * л.: г;
> ' г Сезонный цикл с одним поколением втоду может определяться
температурнымиусловиями. Хорошоизвестно, что число поколений,
как правило,эвозрастаетло мере продвижения на юг, что самым не-
посредственным образом связаноссуммойэффективныхтемператур
(СЭТ),'необходимыхдлязавершенияонтогенеза.В тех широтах, где
СЭТ достаточна толькодляодногоонтогенетического цикла, возни-
каетмоновольтинизм.Такихпримеровмасса.Подавляющеебольшин-
ствовидов, например, нашироте С.-Петербурга,успеваетдать только
однопоколениеза сезон. Однако и в этом случае необходим эколоди-
ческий механизм, который синхронизировал.бы появлениенаибол ее
устойчивыхетадийссоответствующимпериодомгода.' г;
; Эта задача очень просто и однотипно решается многими вида-
ми. У S. menthastri, A. rumicis, М. brassicae, например, низкая темпе-
136
ратура, характерная для северных или умеренных широт, просто «от-
ключает» ФПР, которая у большинства северных популяций в'отли-
чие от южных,-имеет высокийтемпературный оптимум. В частности,
у ленинградской'^, rumicis ФПР проявляется только в пределах 25-
28 °С, у 5. menthastrinpn 18 °C влияние длины дня полностьюподав-
ляется, и все куколки несмотря на большую продолжительность дня
ДИапауЗИруЮТ. ' 1
К моновольтинному типу развития переходят лопуляции мно-
гихвидов, обитающие вторах. Несомненно, что этов первую очередь
определяется недостатком тепла. Известно, например, что на высоте
2 000 м над уровнем моря СЭТ уменьшается почти в> 10 раз.
i Адаптация игорной зональности идет разными путями. Как пока-
зал А.С. Данилевский [1961],переходкмоновольтинизмууР/еп₽лар/
сопровождается возникновениемгорного подвида P.napi b/yoniae,ко-
торый по ФПР очень близок к субарктическим формам и: даже при
24 °C даеттолько диапаузирующих куколок во всех фотопериодах.
В других случаях переход к моновольтинизму происходит под
прямым давлением^внешней среды. Например, у желтушки Colias
a/exanrfra [Hayes, 1982] в дол инах обитают бивольтинные популяции,
а в торах моновольтинные, что обусловлено именно недостатком
тепла. ....с .. ...,,
У гроздевой листовертки, Polychrosis botrana [Комарова, 1954],
уменьшение числа поколений в предгорной зоне также определяется
температурой, но осуществляется иначе. Низкая температура сдвига-
ет развитие чувствительной к ФП стадии на период с длиной дня мень-
ше критической, в результате чего все особи диапаузируют. Так же
происходит адаптация к горной зональности у ивовой волнянки в
Армении [Авакян, 1953]. ? .
Подобная регуляция моновольтинизма в Австрийских Альпах
описана у златоглазки Chrysopa perla [Gepp, 1975]. В результате 10-
летних наблюдений Дж. Гепп обнаружил, что в горах вид дает только
одно поколение, тогда как в долинах всегда два Оказалось, что время
вылетаперезимовавшихимагозапазцываетна7,5днейнакаждые 100 м
высоты. В результате развитие всего поколения в горах происходит
при длине дня короче критической, что однозначно определяет на-
ступление,дцапаузы. , ,
Факториальная регуляция моновольтинизманасекомых в горах
встречается чаще, чем обусловленная наследственностью. Для воз-
никновения наследствецных различий у горных популяций, видимо,
!жено ‘гакже'у коровок (Coccinellidae) и листоедов’(Chrysomelidae)
[Rankin,^Rankin, 1980;Coates fetal;, 1987]. w
’ •iHУникальный пример сезонных’миграцийсредичешуекрылых
дают бабочкиймонархи,;|2)ала«л plexippus. В течение летних месяцев
они встречаются на болыней'части территорииСШАи южной ‘Кана-
ды,¥де дают несколькопоследовательных генераций. Осеньюбни'со-
вершаютзимовочныемиграции двумяотдельнымитруппамисосвои-
ми местами зимовки кзападу и востоку от Скалистых гор,'тде’образу-
ютогромные скопления/Летомбабочкивновьвозвращаютсяв север-
ные районы,гдеприступаюткразмножению[Urquhart, Urquhart; 1'978;
Schmidt-Koenig,1985; Brower, Malcolm,’1991]’. Монархи/ совершаю-
щие миграциинакк,находятся в соетоянии репродуктивной диапау-
зы,’ которая"индуцируется вириродныхусловияхи поддерживается
короткимдаем и низкойтемпературой; Длинный д ень и высокие тем-
пературы’вызываютреактивацию и стимулируютоогенез.‘Интересно,
чтовэкспериментальныхусловиях'реактивациядиапаузирующихба-
бочек достигалась не только действием обычных’экологических фак-
торов, но и'искусственнойЦнъекцией аналогов ювенильногогормона
[Herman,*1973,1981]:, *тогдакакиндуцировать диапаузу'истимулиро-
вать полет в южном направлении привыпуске бабочек в природу ока-
залосьвозможным покатолько с помощью гормональных препаратов
[Htennaniit99'3J,'c,*<' ’ • ^йом,д.--. .‘юна' rwv
Примечательно, что популяции монарха, недавно интродуци-
рованные в Австралию, по-видимому, утратили способность к даль-
ним миграциям. Скопления особей зимующих популяций были об-
наружены возле Сиднея и Аделаиды, недалеко от мест размножения.
Более того, исследования показали, что и состояние покоя у австра-
лийских монархов значительно отличается по глубине от диапаузы
американских популяций [James, 1982,1983].
Все перечисленные примеры относятся к видам, у которых миг-
рации связаны с диапаузой и являются обязательным элементом го-
дового цикла. Они изученыу немногих видов, но имеющиеся матери-
алы позволяют довольно четко понять суть этого явления. В боль-
шинстве случаев миграции регулируются внешними условиями (тем-
пература, длина дня, плотность популяции)-и характеризуются осо-
бым миграционным состоянием, альтернативным репродуктивному.
Расстояние, которое преодолевают мигрирующие особи, колеблется
у разных вид рв,не.превшая нескольких кидометровутодних ид ости-
’’'•У'' ’Г- - и ; : У
206
гая сот ен километров удругих.Однако все эта перемещения происхо-
дят внутри ареалов. Их адаптивноезначение не требует доказательств.
, , л Но существуют миграции, до сих пор представляющие собой, за-
гадочное явление. Кним,относятся,’например, дальние сезонные пе-
рем ещениянасекомых за пределы их видового ареала, имеющие эпи-
зодический характер. Такие миграции позволяют лишь временноис-
пользодать дополнительное жизненное пространство путем создания
сезонных популяций, однако они вносят большие изменения встра-
тегию-сезрнного развития видов, обладающих такой способностью.. ”
По мнению разных авторов, список насекомых, для котор ых ха
рактерен такой тип миграций, насчитывает 200—300 видов [Johnson,
19,69] . К их чисду из европейских чешуекрылых относятся, нацример,
Autographagamma, Mythimnaunipuncta,Agrotisypsilon (NoctuidaeVP/w/e/fc
maculipennis (Plutellidae)', Pyrameis cardui (Nymphalidae),1 Coliascroceus
(Pieridae) и многие другие. Они представляютраЗные семейства,что
свидетельствует ораспространенности этого явления вотряде.’ ,
Принадлежность вида к мигрантам устанавливается разными
способами. Наряду р пря^иыми наблюдениями за массовыми пере-
движениями насекомых в рдном направлениик их мечением ^Walker,
Wineriter.l 98 i; Drake, $9Т; Walkey,1991] существуют косвенные дока-
зательства — внезапноедгоявление или исчезновение большого числа
особей одного вида.беззаметной’смертности;' появление некоторых
видов в областях, где,климатические условия' не позволяют им круг-
логодично существовать' высокая'синхронность развития отдельных
вцдов на значительной’территории и T.fl‘/[Ni'elsen,! 1964; Taylor et
aj.,1980]. Метод ы изученця Миграций постоянно совершенствуются’,
предлагаются новые, иногда неожиданные решения.'Так, миграции
лугового мотылька исследовали на основе анализа содержания хими
цеских элементов в организме бабочек, которые, как оказалось, раз-
личались ,у разных^ографических популяций. Сравнение элемент-
ного, состава позволидоустановитьрайонпроисхождения мигрантов
[Наумоваидр., 1994]/Ранееэтотметодиспользовался "А.В. Жулидо-
BbiM.cfcoaBTopaMH.[1982] для изучения перемещений’четырех в^дов
совок на Северном Кавказе.' 1 3'i?‘
„С наступление^ неблагоприятных условий, не имея специаль-
ных адаптаций для переживания суровых зим, потомки мигрантов
должны,откочевывать обратно,'в места свдего постоянного обитания.
Несмотря на загадочность возвратного механизма, существование та-
койвозможностиц!оказанодля некоторыхвидов [Novak, Spitzer,1972;
ж
Poitout et al., 1975; Shapiro, 1980;Kaster,'Showers, 1982; Showers et all,
Г993]‘.'В большинстве случаев массовый перелет насекомых происхо-
дит'вовполнеопределенномнаправлении, ’часто, совпадающем с на-
правлением господствующих ветров, ифриурочен к периоду актива-
ции циклонической деятельности. Напримере 'многих видов доказа--
на возможность дальнего их перемещения при определенной синоп-
тичрской ситуации исопутствующейейсистеме ветров[Мельничен-
ко, T936;’l^ielsen,1964;11ау1ог ef^
Scott, АсЬ1ете1егД987;Макарова,ДоронинаД988Д994; Drake, ’Farrow,
1988j L'“' *,4!' «nwm лизшк
так, бабочки совки SpodopteraexiguaYSb.в результате действия
силъныхдрозовыхфронтов,двигающихсявсеверо-западномнаправ-
лении с территории СреднейАзии , систематически заносятся в Скан-
ди,цавские страны. При этом они преодолеваютрколо3—3,5тыс.!км
от их основного местообитания [Mikkola, Г967, 1'970; Aarvik,'>1981;];
Кацустная моль, Plutella /ласи/фелли,вспьппки!размножеииякото-
рой происходили в Центральных районах зоны 'Балтийского ’моря,
пролетев!,5-2,'5тыс.кмоседалав1массев‘Англии;иИсландии‘[¥ап-1-
Chu, 1986] . Дальние миграции с помощью атмосферных течений опи-
саны и’у cowm'Agiytis ipsilon [Showers et al.,11993].'w,‘ c u {’*
'* Тщательный анализ роли атмосферных явлений в миграциях во-
сточной луговой совки,' Mythinina separata,проведенный Л.А:'Ма-
каровой и Г.М. Дорониной [1988] / показал,' что'время и Интенсив-
ностьлёта лугрвой совки в условияхПриамурья не зависит от местных
атмосферных циркуляций, а связан с характером перемещений'воз-
духа впределахвсего Дальневосточного региона. Массовый лет бабо-
чек наблюдается там обычно1 в годы с активной цйклонической дея-
тельностью при преобладании'воздушных потоков1 южного направле-
ния.^Число%пиков»’ лёта бабочек хорошо совпадает с количеством
проходящих циклонов, а его инте нсивность закономерно возрастает
по мере снижения атмосферного давления в центральной’зоне цик-
лона1. Максимальное количество бабочек (700-1000 и'болеёУ отлав-
ливается после перемещения глубоких циклонов’с давлением' в цент-
ре ниже 985 гПа. •. ( ;.......... (.
, - .. .Как свидетельствуют имеющиеся данные , дальние сезонные миг-
рации наблюдаются и у гомо-и у гетеродинамных видов. Попытаемся
оценить биол огическийсмыслтакихмиграцийприменительно к двум
типам развития! Тетеродинамный цикл,’выражающийся в чередова-
нии активных и диапаузирующих стадий, характеризуется 'глубокой
208
адаптацией^ ритму климата. Специальный экологический механизм
синхронизирует этот цикл е1 ритмикой внешнихусловий.Реакции,
управляющие диапаузой, характеризуются разнообразием и лабИльг
ностыо.Именно ониобеспечивают высокую приспособленность ло-
кальных популяций к местному климату,создавая сложную’эколого-
географическуюструктурувида/котораяограничиваетвозможности
свободного Перемещения даже в пределах собственного ареал a. sz
'! 'Миграции'за пределы ареалапочти неизбежно должны приво-
дить кдиссонансу между системой’экологических реакций вида и рит-
мом климата в районе залета, что вызывает элиминацию всех особей с
неадаптивными реакциями. Однако у экологически лабильных видов
индиЬидуальная изменчивость НормФПР создаетПредпосылки для
адаптации'к 'новым условиям1 путем отбора'. По-видимому, именно
отбор играет решающую роль в натурализации вида и его последую-
щей зонально-климатической дифференциации в новом районе оби-
тания. Результаты наблюдений над интродуцированными в Америку
видами показывают, что такая географическая дифференциация уже
произошла не только у Cydia pomonella [Шельдешова, 1967; Riedl, Croft,
1978; Riedl, 1983], завезенной из Европы более 200 лет назад, но так-
же у Ostrinia nubilalis и Pectinophoragossypiella, расселившихся на этом
континенте уже в нашем столетии [Beck, Apple, 1962; Ankersmit,
Adkisson, 1967]. Подобный процесс осуществляется и у колорадского
жука в Европе. Диффузные миграции являются одним из механизмов
поддержания клинальной изменчивости норм таких экологических
реакций, как ФПР. Благодаря промежуточному характеру ее наследо-
вания [Данилевский, 1961], гибриды от скрещивания мигрантов и
местных особей оказываются более приспособленными к новым фо-
тотермическим условиям, чем их залетные родители. Таким образом,
мигранты поглощаются местной популяцией. Наконец, диффузные
миграции позволяют насекомым восстанавливать популяции в тех
регионах, где они по каким-то случайным причинам частично или
полностью погибают. Как отмечалось выше, даже у таких обычных
для фауны Европы видов, как Р. brassicae, зимующий запас в отдель-
ные годы полностью вымерзает во многих северных областях, и вос-
становление популяции происходит за счет бабочек иммигрантов
с юга.
Гомодинамные виды лишены адаптаций к ритму климата. Их
устойчивость к повреждающим воздействиям среды носит неспеци-
фический характер. Следовательно, для таких видов внутриареаль-
209
ные перемещения не ограничены сезонно-циклическими реакция-
ми. В тожевремя натурализация ихза пределами ареала при дальних
миграциях или. завозе представляется мало вероятной. Исключение
могут составить некоторые синантропные .виды, (Обитатели убежищ и
паразитытеплокровныхживртных, которые существуют при относи-
тельно постоянных у словиях-Обитаниеми гр антов в районахс летне-
зимней сменой сезонов рграничено вегетационным периодом, когда
условиявыравниваютсявщиротном направлении. Димовка таких ви-
довокологически не обеспеченней их эпизодическое появление в уме-
ренных широтах следует связывать., с.высокой миграционной актив;
НрСТЬЮ.!1,Л{,.К«'<ГГ 8'0 'ГО' !;Н ХСЗДГЧЧМ
.1 s Вщелом дальниесезонные миграциинасекомыхлредставляют-
ся пока западрчными. Однако новейшие техническиевозможности,
современные методы и научные подходы,к их изучению,, позволяют
надеяться наразгадкуэтого явления.[Dicksonet д1., 1983; Wellington,
УКАЗАТЕЛЬ ЛИТЕРАТУРЫ ;
! •-ov
1 АвакянГД. Новые'данные о биологии ивового шелкопряда // Изв. АД
Арм’ ссрЛШ!т/б,‘выл.‘io. ,0.83-87.'..,' '/у
Агроклиматический атлас мира'. М'.-Л., 1972?Н7 с. ~ v
1‘Агроклиматические ресурсы’Белгородской области. Л., 1972. 9Г'с..
Ажбенов В.К. Серая зерновая совка {Apamea anceps Schiff.'Npctuidae,
Lepidopteia)1 В Казахстане,' (биология, 'экология, ^система прогнозов): Авуореф.
дис... д-ра биол. наук. Алматы, 1995, 43 с. л'\‘ь * "
, , 1Е .! Американскаябелая'бабочка// Защита'раст. 1980.’Вып. 1. С. 36—40. ,,
‘ ' Андреев С.В., Молчанова В.А. Радиомаркировка при изучении насекомых
вредителей // Вести, с.-х. науки. 1964. Вып. 2. С. 122—128. „ Л
“ ' ' Арапова Л.И., Богуш ‘ЯЯ Биология колорадского жука ^ условиях юго-
'запада* Белорусской ССР //Труды ВИЗРа.г 1'967.'Т. *27/ 0. 33-45? '?/ ( '
Арапова Л,И., Заславский В.А. Фотопериодические реакции двух попу-
ляций колорадского жука (Leptinotarsa1 decemlineata SayY // ЗоолДжурн. ,1980.
*т>9;>п! У?d 1035-1059.^''д
' 1 Арнольду К.В. Зимовка вредной "черепашки (Eurygaster integriceps Tfat )'Д
горах Кубари цЬ исследованиямТ949—1953"гг. // Вредная ререпашка. Т.У. М.,
1955. С. 1'36-269 ' V'* /“Л '
‘ 'Асанова Р.Б.',' Искаков Б.В. Вредные и 'полезные полужесткокрылые Ка-
'захстана.'Ал^р-Ата,;^??'.^.1 124—126; 1 ‘ " £ ; л.
Асахина1 С. Принципы 'механизма морозоустойчивости у растительных' и
животных1 клеток //Хлетка’и температура среды'. М.-Л;, 1964. С.'Тб—19.
' ' ' ^Белоусов иЛ^Торышин' Я.Я.ЧЭ' функцйональной автономности'Элемен-
тарных* реакций в двухступенчатой фотопериодической реакции жужелицы
Agonum 'assimile Payk1. (Coleoptera, Carabidae).// Вести. 'Ленингр.'ун-Та '1982.
;Вып.82|1.!С.|'5-12:;'''‘=-^1'л''(Г''' "1 д.«‘у
'£" ! ’Бергер Л.Я. Прогноз динамики'популяций луговой совки на Дальнем
Востоке // Труды ВИЗРа. Пути автоматизации фитосанитарной диагностики.
Л’ г 1985 С 68“7*5'if ' 1 14 ;i'’ ‘*лЛг“' 1 *''-'S ' д'»;.
'Биологические'19б4/сб94‘с; * ' * s ' п ’ ' ’
Бабинская С.Г. Влияние экологических условий на жизненный цикл'се-
poft3epHOBoii coBkH!(Hadenasordida) //Зоол.'жури. 1966. Т. 45, Вып'.Т1.'С. 1650-
’U'^59^' 'r'i ” ‘ Л А.; г i Д ‘ 'Л Ш'С; d. И
’ *'*' БогачеЧа И.А. 'Ос'оббнности адаптивных стратегий листогрызущйх насе-
комых Субарктики//Зоол. жури. 1995. Т. 74,'вып/8. С. 83—93?' ‘
:211
Браун В.А., Горышин Н.П. Климатизированные камеры с программирова-
нием фотопериода и температурных ритмов для экологических исследований
// Вести. Ленингр. ун.-та. 1978. Вып. 3. С. 26-34.
Браун В.А., Горышин Н.П. Установка для фотопериодических исследова-
ний в условиях ритма температуры // Зоол. журн. 1984. Т. 63, вып. 2. С. 272—
276.
Брянцев Б.А., Доброзракова Т.Л. Защита растений от вредителей и болез-
ней. М.-Л., 1960. 480 с.
Буракова В.И. Энтомофаг колорадского жука — клоп подизус — в усло-
виях Воронежской области // Биологические и химические методы защиты
растений. Воронеж, 19,82.-С. .28—29. -
Васильев В.П. Вредители сельскохозяйственных культур и лесных на-
саждений. Киев, 1973. С. 340—341.
Венгорек В. Влияниепродолжительности дня и качества корма на био-
логию'колорадского жука Leptinotarsa decemlineata Say // Колорадский жук.и
меры борьбы с ним. Т. 2. М-, 1958. С. 129-135. * , , ’ , ,
Викторов ГА. Проблемы динамики численности насекомых (на примере
вредной черепашки). М., 1967. 271 с. 1 ' ‘ J
' Виноградова Е.Б. Материнское влияние на диапаузу потомства у насеко-
мых // Чтения памяти Й.А. Холодковркого. Л., 1973. С. 39—66.
(Виноградова Е.Б.) Vinogradova "Ге. В.. The pattern of reactivation ofdiapausing
larvae in the flowfly, Calliphora'vicina II J, Insect Physiol. 1974. Vol. 20, N 12.
P. 2487-2496.’ \
(Виноградова E.E.)"Vinogradova Ye.B, Embryonic photoperiodic sensitivity in
two species of flesh flies,Tarasar.cophaga similis and Boettcherisca septentrionalis
// J. Insect Physiol. 1976? Vol. 22; N 6. P. 819-821. \ ‘ ' , *
7 Виноградова E.Б.‘ Зависимость гдубины диапаузы личинок Calliphora vicina
R.-D. (Diptera, Calliphoridae) от фототермических условий при их развитии и
при содержании самок родительского поколения // Фотопериодические реак-
ции насекомых. Л.,' 1978. С. 95—10-1 (Труды ЗИН АН 'СССР. Т. 69). ( V
Виноградова’Е.Б. Диапауза мух и ее регуляция.- СПб., 199-1- £55 с.,
Виноградова Е.Б., Богданова Т.П. Особенности репродукции и формиро-
вания' диапаузы у полосатого амброзиевого'листоеда Zygogramma suturalis F. в
Ставрополье // Энтомол. обозр. 1988. Т. 67, вып. 3. С. 468—478.
Виноградова Е.Б,, Богданова Т.П, Особенности сезонного развития амб-
розиевого полосатого листоеда Ц Труды’ Зоол. ин-та. 1989? Т. 189. С. 62—75.
, / Власова В.А., Зискинд Л.А,, Грлубева Н.П.,. Ижевский С-С- О возможнос-
ти акклиматизации цодизуса // Защита растений. 1980* Вып. 4'..,С. 46—47..
Волкович Т.Л. Экологическое значение постепенных* изменений^ длины
дня в фотопериодизме ндрекомых: Ащоррф, дис.... канд. биод.^наук. Л,, 1983.
J ' '-0 ' >. ’Л --- .1
Волкович Т.А. Экологическая роль постепенных изменений длины дня в
регуляции развития и диапаузы насекомых // Труды ВЭО. 1986. Т. 68.С. 99—
102, 7,\''' 7Х.<
Волкович Т.А. Роль световых и температурных условий в контроле актив-
ного развития и диапаузы златоглазки обыкновенной Chrysopa earned Steph.
(Neuroptera, Chrysopidae).,!. Фотопериодическая, реакция при константной и
постепенно изменяющейся'длине дня//Энтомол. обозр. 1987. Т. 66, выр,.!.
212
С. 3-12. II. Сезонное развитие ^.Белгородской области// Там ркек 1988. Т. 67,
ВЫП. Д. <2, ,3—10. - ч-:г- . ;• twW-«г Л> фн s '•!>(•
Волкович Т.А., Горышин Н.И. Оценкам накоплениефотопермодической
информации у клопа солдатика Pyrrhocoris apterus (Byrrhocoridae, Hemiptera)
при.ицдукции, яйцекладкщ//, 3рол. журн.'.1978. .T.,-57^ рып. IjG.*46-55. » .
Волкович Т.А., Горышин Н.И. Влияние ^константных и постепенно увели-
чивающихся-фотопериодов на. индукциюрепродуктивдцй активности у^ клопа
солдатцкд .(.Pyrrh^toris apterus L.) // Зорл,;журй. ,1979,,Т. 5$, вы^9.иС.~ 132Ztt
1332., ' \ J " .... А - , . I, Г -А ЛА
ВолковичТ.А..Горышин Н.И. Индукция пронимфальиой диапаузы у луго-
вого мотылька* Loxostege sticticplis L. {Lepidoptera, РугаШдаеД.под влиянием
сокращающейся длины дня //.Докл. AHjCCCB^SS.^. ,276,- вып.1. С„240г-
242. - , -i.,- е a.,.{
i. Волкович Г..А., Колесниченко Л.И, Саулич А.Х. Роль терморитмовв .раз-
витии хищного клопа PerillusMQCulatus Fabr. (Hemiptera^Pentatomidae) //Зоол.
журн. 1990. Т, 69, вып. 6. С. 70—81. ч
- Волкович Г.А., Саулич Л.Х Хищный клоп Arma custos'. фотопериодичес-
кий и температурный контроль диапаузы и окраски?// Зорл. журн. 1994. T. 7J3,
ВЫП, 10. С. 26-37. ,'M.rC ; ,
Волкович Т.А., Саулич. А.Х., Горыши» Я.Я..Чувствительная'К-длине дня
стадия м накопление фотопериодической информации ухищногоклрпа Podisus,
maculiventris Say., (Heteroptera, Pentatornjdae)// Энтомол- 'о.бозр. 1991.. T. 70,
вып. 1. С. 14-22. ...-ч • ...Г-
Вольвач В.В. Оценка точности метода сумм эффективных температур-при
расчетах продолжительности, от дельных стадий .колорадского жука Leptinotarsa
decemlineata.SuyJf *Ц>УШ иц-та эксдер. метеорол.: 197О.Т. 11, Gi *68—75, •’ *«, <
Вольвач В.В. Моделирование влияния агрометеорологических условий
на.развитие колорадского жука. Л., 1987. 239 с. -• -. ,: u i л
Гасанов О.Г. Влияние температуры на качественнее и количественные
фотопериодинеские-реакции у совр,К|//,ЗоолЛ1Жури. 1979.:<. ;58,,выц,6.«С. 926—
928. *.• . - , ; л -а- .. Аг " АГ.
Гейспиц К.Ф. Реакция моновольтинных чешуекрылых (Lepidoptera) на
продолжительность дня // Энтомол. юбозр. 1953. ,Т. 33. С. 17,—31.. .,л >
Гейспиц К®- Фотопериодические и температурные реакции,-определя-
ющие сезонное развитие хвойных шелкопрядов Dendrolimus pintL. u D.sibiricus
Tschetw^ (Lepidoptera,,tLasiocajppi|dae),// Энтомол, обозр. 1965.:T.,.44i вып. 3.
С..5Д8-553. ... , . a a,,,. , a , ,a ro a г
Гейспиц К.Ф. Генетические аспекты изменчивости фотопериодических
адаптаций //, фотопериодические адаптации .у насекомых ижлещей,. Л;, 1968.
С,}52г—791. A'V' г " ‘ ' А . ' t - А * ; - . • , г г1. ‘ 7 Ап- г А
Гейспиц К.Ф., Глиняная Е.Н, Сапожникова Ф.Д.^закыосвязыразлячных
типов, фотопериодической реакции насекомых //Проблемы фотопериодизма и
диапаузы насекомых. Л, 1972. С. 74-8i8, чгг* .. г. »; а'--."ал,
Гейспиц К. Ф. Г Гяиняная> Е.И, Сапожникова Ф.Д., - Симоненко, Н. П. Соот-
ношение, эндогенных,-и экзогенных факторов в регуляции{сезонных* измене-
ний,фотопериодической реакции членистоногих //Энтомол. обозр. 1974.’ Т. 53,
вып. 3. С. 523-534.
213
’’ ТЪйспицХ.Ф., Заранкина’А.И. Особенности’фотопериОдической реакции
шерстолапки плодовой Dasychira pudibunda L. (Lepidoptera, Orgyidae) // Эн-
томол«. обозр'.1'1963i, TJ 42лВып.-1.'С.'29— 38. ' 5 »'
- "^'рЛйслиф-ХФ./Пенязъ М.И., Шашенкова Д.Х/Фотопериод и «температура
как факторы в развитии совки 4grorir5egetu/n(Lepidoptera;Noctuidae)//3oofl.
журн'.4971?Т 50/вЫп41.‘С14«74-1685. ' ' ««"•-V- !• \
,п>- ^ГейспицК.Ф.^Сапожникова^Ф.ДХ'Симоненко Я.Л :©кол<иические’прин-
ципы1 регуляции чередования активных ’и диапаузирующих фаз'развития чле-
нистоногих // Фотопериодизм животных и растений. Л., 1976. С. 26—46.
' «\1’ ГейспицгК.Ф.,!Симоненко Я.Л? Экспериментальный 'анализ1 сезонных из-
менений'-фотопериодической 'реакции Drosophila1 phalerata' Meig.'-'fDiptera?
Drosophilidae)11// Энтомол/обозр.' 1970'. T. 49, вып. 1. С.!83—96?' !i' ' л •'
Гирфанова Ф.К. Изменение устойчивости Calliphora vicinae (Diptera,
Calliphoridae)<K«повреждающим факторам -В связи с формированием и 'прекра-
щением диапаузы'// Зоол/журн. 1984. Т. 63-,’1выП/’31.*С.-406—410.°' ' f' «
Глиняная Е.И. Роль фотопериодических'условий В процессах'реактивации
при1 зимнейи летней-диапаузе // Проблемы фотопериодизма и диапаузы насе-
комых'.'Л'Д 1972. С. 88—102. 1 *'« ' • " 1 • н.ч.
Гользберг К.Л., Акимов А.Г., Анисимов А.И. Особенности фотоперио-
дической" индукции дйапйузыу весенней капустной'мухи — Delia brassicae
(Diptera/Anthomylidae) //‘ЭоОл'.',журн,.'-1986.,Т>65,'11вЫп!,'12. <!’. 1858—1863. '
•5' 1 Горышин -Н.И. Влияние длины -дня нй формирование диапаузы' у - 'коло-
радского жука (Leptinotarsa decemlineata Say) // Колорадский "жук. Т.2. М.’,‘
1958а. •СЛвв-'149;1,-^>--9‘*Ф‘- «'•.»><' ""/wн
? <1 - ГорышинН.И. Экологический анализ сезонного"циклй 'развития хлопко-
вой cGevM^Chloridedlobsoleta F.’)b севериых-’районах'ее распространения // Уч.
заИ. ЛГУ/ 19586/Т.'240. С.'3-20. нг,.-...л з ./ ' 9, «. .««
Горышин НИ. Техническое оснащение экологическихИсследований В эн-
тоМологиИ.'Ш., 4966. 235 c/ 'S''"«/ -R't/’w*" owhI'T’-I! tev,.',' ,
•Горышин Н.И. Свойства<чувствительной к длине' дня СтадиИИ ее роль в
механизме фотопериодической реакции насекомых // Журн. общей биол. 1977.
Т*38,1вып/4;.аС? 559-572/*/-мйляо:- <: -i».»
Горышин Н.И., Ахмедов' Р:М. Фотопериод и температура Как факторы в
разантии coBkwAgrotis ypsilon (Lepidoptera, Noctuidae) // Зоол. журн. 1971.
T 50- вып51.'С.‘56-66.‘ О.'-л:- UW'-лг хлмгэ-ч с
' ^ Горышин И.И., Волкович Т.А! 'Оценка м' накопление фотопериодической
информации у клопа-солдатика (Pyrrhocoris apterus) при индукции яйцекладки
Ц Зоол.'жури. 1978i Т.,57,<|Вып.гТ.!СЛ46—54. ' >• -
’ ‘ Горышин Н.'И1, Волкович Т.А ;'СауличА.Х1 Сравнительная эффективность
константных и постепенно изменяющихся фотопериодов для трех видов че-
шуекрылых //-3оол. журн. 1979. Т. 58,'вып. 1. С. 44—53. . *’4 4 > ч j?
^' г Горышин ДИ.,''Волкович Т.А:, Саулич А:Х."Некоторые итоги исследований
фотопериодизма насекомых в заповеднике «Лес на Ворскле» // 'Комплексные
исследования «биогеоценозов лесостепных дубрав."Л .,4986а? С. 74—92.'
Горышин! Н.И., Волкович Т.А.;тСаулйч АХ Новые Исследования по*эколо-
гии колорадского жука.1 «Рукопись депонирована В«®ИНИТИ. Т986б.-3997-В86.
91 с. Ь"’ «• i
214
, Горышин Н.И.,.Волкович .Tut,,..Саулич(А.Х. и др,..Роль температуры и фо-
топериода, в контроле развития и диапаузы хищного Kjiona^opfc^ maculiyeHtris
(Hemiptera, Pentatomidae)h,//)3oon. жури. 1988a. Т.67, ври. 8' С. 1148—1161.
,,г)1 Горышин Н.И., Волкович T.А., Саулич A.X, и др/Экспериментальный ана-
лиз, сезонного .развития колорадского, жука в лесрстепно,й зрне. ^Особенно-
сти фотопериодической,реакции // Зоол. журн.’ ДЙЗ. Т. 64, иып.,.^.С. 1349—
1359,-Л.ДДолрдые опыты, # ^нр^<^ // 4TaMvixe.t 1986в.
Т^5^1ВЫЦ. A* I-'* 52^7-535. , Л,-
, ГаррииинЛ^И., BoeKoeu4i T.A.,. fiiay^u4 А.Х. зл др. Сравнительно-;эколоп1-ч
ческое изучение популяций колорадского xyKuLeptinotarsadecemlineojaSzy
(Colppptera» Chryspinelidae) из Европейской „части рССР Эш^ирд^рбозр.
1987.,Т. 66,-вып. ДДЗ" ^5-^236,‘7 •>> ..v Л"--.,-,
ГорышинЛ.И.,Гейсниц Л".Ф. Некоторые актуальные вопросы фенологи-
ческого анализа р энтомологии // Зоол..журн. 1975..Д. 54, вып. 6„ С. 895—912.
, . ГорышинН.И., Козлова.Р,Н. Термопериодизм как факторвразвитии,^?-,
комых //Журн. общей биол. 196?. Д’. 28, выи. 3. ,С. 278—288. ,
Горышин Л-И.j ,Кузнецова И,А. .Продолжительность развитид насекомых
при постоянных и пе^м^дат^^атур^ // Двддр „ВДЗ^а,. 1,9,72г .Т^З?.
г 14 » • <• ’ !.• ' w . , " t " V -
Горышин НИ., Саулич А.Х. Изучение .сезонных адаптаций у гомодинам-
ных чешуекрылых. I, Реакции нд температуру и длину-дня // Зоол. журр. 1981.
Т. 60, вып.12. С. 1784—1791» -Холодостойкость, и выживаемость прц,низких
положительных температурах,//.Тамже. 1982. Т(61, выц. 1. С, ^6-46/JII.
Развитие в полевых условиях умеренного климата //Там же. 1983. Т. 62, выц. 2.,
С- 222—232»-.<. и ,П ')!«; п гз. г.; ,, "Г it . •> Т» ‘1
Горышин Л.И., Саулич А.Х., Волкович Т.А. Исследование экологической
роли.фотопериодической реакции у трех видов чешуекрылых // Фотоперио-
дическая регуляция ссзондых. ЧРЯерий У чл,енистрдогих и, растений., 'JL, 1981..
С. 60-81. ' ..... ....... ................. '
. ^ Горышин Саулич А.Х., Волкович Т.А. Фототер,миче|С|кие1 адаптации
луговргогмотылькаиРукописьгдерониррвана|д.,В|ИрЦ4ТИ,:2929-В. 1986я 56.С.,
Горышин Н.И., Саулич А.Х., Вол^орич Т.А. цдр.,Роль температуры <и фотог
периода д регудяции развитиями диапаузы у лугового yt^^bK^Loxostege
sffcticplisi ;//=s3ooa. зяурн,|1980. Д^ 59, вьцц 4., /С.,^33-5.45., , ,
Горышин Л И., Саулич А.Х., Волкович Т.А. и др. Влияние .пищевого факто-
ра ,на развитие и;фртодерирдическу10 реа;кцию,:хищного ^слопа Д’оЛтит
maculiventris (jgeniipteya;;Pe^tatooijdaei);l//Зорд1журв^ Д38^1!Т‘>^77«ВДП'.9.
С. 1324-1332. . .' "-у '
ю Гррышин^Н.И.цСаулиу А.Х., Морей; М А. ^(методике, экспериментального
определения-,нил^едолТ^пературно^ррро.гд развития, насекомдгх //^Зррл.
журн. 1984. Т. 63, вып. 1. С. 126— 129v-c !Vi’ ,' i; •! г .г-и- • -- ...,-
Горышин НИ., Туганова^И-А. гРдкимидадия условий кратковременного
Зфанения; яищ хищнрго клопа PoBfsus ^ac^Uvgntris .i^y^tefiiL, JPepjatp^aidae}
//Зоол. журн. 1989. Т. 68, вып. 6. С 111—119. . ,
:л... Горышин НИ., Тыщенко B.7E 3BeHo.naMa^4.Hrer9MeqTp в механизме фото-
периодической реакции насекомых // Журн. общей бнол, 1974. "Г. 35, вып. 4.
С> 518—538. ,' , ч --
215
Горышин НИ., Тыщенко Г.Ф. Отермостабильности процесса восприятия
фотопериодическойинформации у совки Acronyctarumicis'L. (Lepidoptera,
Noctuidae)// Докл. АН СССР. 1970. Т.Ч931. С. 458-4ЙГ.* ; ’ •
ГорышинДИ., Тыщенко Г.Ф. Экспериментальный ’анализ процессов фо-
топериодической индукции'диапаузы у насекомых '// Механизмы регулятор-
ных процессов.‘Л;, 1972а. С. 68—89 (Труды ПетерГофск. ’биол. 'ин-та. Т,21);
Горышин'Н.И.,'Тыщенко Г.Ф. Влияние-температуры и продолжительности
свето-темнового цикла на критическое число фотопериодов, индуцирующее
диапаузу у совки ЛсголусГа rumicis (Lepidoptera, Noctuidae) //'Проблемы1 фото-
периодизма'П диапаузы'насекомых.1 Л.,49726. С. 1’58—17-4;*' ’
'Горышин Н.И., Тыщенко ГФ. О'ткотшети фотопериодической-инфор-
мации у капустной совки Barathra brassicae (Lepidoptera, Noctuidae) При ин-
дукции диапаузы 7/ Энтомол/обозр1-Т973'. Т/52', вып.‘2. С. 249—255. ‘ ‘
Горышин Н:И., Тыщенко Г.Ф. Влияние фототермических условий на-ин-
дукцию "и прочность диапаузы у'белянкй!'Р/еги"brassicae (Lepidoptera, 'Pieridae)
//Зоол. журн. 1979. Т. 58, вып. 9; С. 1327-1333. ( ’ ‘ *'' ’ ‘ ‘ 1
-'* ’Горышина Т.К. Ранйевесеннйе эфемероидьГлесОстепных дубрав (иссле-
дования по'экологии, физиологии и1 фитоценологии). Л., 1969. 165‘с. '
Данилевский А. С. Роль питающих растений в биологии лугового мотыль-
ка// Энтомол/обозр; 1935? Т.!'26'. С.й9Т-1О0. ' ' - А * ьим.,
’ Данилевский А. С. О действиипостоянной’и’перёменной температуры' 'на
развитиеяицайлантового Philosamia cynthia Dr.’ и дубового Antheraea pernyi
G.-M. шелкрпрядоЬ //’Труды Ленингр/о-ва естествоиспыт. 1946. Т. ОД'вып.Ч1.1
С. 46—69 ' • ' ‘ -i-v.
Данилевский А.С. Фотопериодизм и сезонное развитие насекомых? Л.,
I961'.:243’lc/'4S '’ = '» -А ,л.л >>»w. .; » л ,.аа
°' • ДанилевскийА.С.Сисге.ма экологических'адаптаций насекомыхк'сезон-
ности климата // ПроблеМыфотопериодизмаН’диапаузы насекомых; Л., 1972'.
С. 15-25.
' ‘ Данилевский А'С.,1 Бёй-Биенко' Я.'/?‘Дубовая- листовёртка/ Tortrix’ viridaпа
L.) и вопрос об устойчивости форм дубак ‘повреждениям // Уч; зап; ЛГУ. Сер.
биол. 1958; N W, нов 'выйМб.С; 61-7O.u’' • ! ‘ ;-
<Г1 Данилевский А. С., Горышин Н.И.,ТыщенкоВ:П. 'Механизмы’'экологичес-
ких ритмов у наземных членистоногих //ТрудыГ1еТергофск;биол.ин-та.
1970. Т. 2O’.ICl.l!4O—S8.‘; ’ ь- ’ > ' -->.А 1 ’ -•
Данилевский’ДС, Кузнецова Я.А.’ВнутривидовЫе’адаптацииНасекомых
к климатической' зональности //Фотопериодические адаптации у Насекомых
и клещей. Л., 1968. С. 5—51. :
" ki 1 Данилевский А. С. .'Шельдешова^Г. Г.Приспособителъное • значение фото-
периодической’и холодовой' реактиваций // Фотопериодические ’адаптации у
насекомых и клещей. Л., 1968. С. 80—99. •’ ' 1 г ' ; 'Г;.'
" - Долгинf ^биолог^^СБг^лоёМоА bdsilea Get»!.1 (Coleoptera,
Chrysomelidae)‘Ha Алтае'
С. 154-161.
' ' ’ ДольНик ВД: Фотопериодизм'у ‘птиц //Фотопериодизм животных и рас-
тений Л., 1976; С. 47-81;' « "л-»
Драховская М. А. Прогноз в защите растений. М., 1962. 352 с.
// Насекомые 1-Воёточной‘Якутии. ''Иркутск’, ‘-'WW.
216
h Дружелюбова. T.C. Специфичность реакций озимой совкина среду и их
учет при лрогнозечиследности // Методы прогноза развития вредителей и
болездей сельском», культур.. М., 1979. С. 84—11)5. X' . ,'Х
. Ду, Чжень-ВэньДРоль .сезонных адаптаций в фенологии и распростране-
нии кукурузного дотыды^а (Ду/auf ta пиМ^/и НЬ.^ АвТрреф '^дис,... гацд. биол.
наук. Л., 1961. 20 с. 4 - v;
, . Жулидов А.В., Полтавский А.Ц., Емец В.М. Изучение миграций совок в
горных районах Северного Кавказа. Рукопись деп.в ВИНИТИ. 5309-82,1982.
15с. к ..........* ” ................. '
Журавлев,В.Е,Сумма эффективных температур и возможность ее приме-
ненияпри определении сроков развития крлорадского жука,// .Краткие .итоги
научных исследований по защите растений в.Прибалтийскойдоне СССР^Рига,
1960. С. 45—48.., . . .. ....... , ., ..... ... . . ........
, .Журавлев, В.Я. Зависимость сроков выхода перезимовавшего колорадско-
го.жука и продолжительность развитияотдельныхего фазрт внешних услрвий
// .Прорноз,.в защитерастений о^вредитрдей, и;болезне^,,(Ригд^,Д96|а^!4С, 65—
7Д-t .Л...;.. - ...
Журавлев В.Н. Биология колорадского жука Leptinotarsa decemlineata Say
и его зндчрнир.как вредителя ^артофмя в KariHjHM^ipaAciwii,,области,: Авто-
реф. дис... кацд. биол. наук. Л.»,19646.-23 с. ' ,,
. " Заславский В,А. Географические расы Chilocorus bipustulatusX. j(Coleoptera,
Coccinellidae). 1. Два типа фотопериодической реакции, контролирующей
имдгинадьнуюдц^цдузу„у северной расы ,/ДЗоол,, „журн. 197'0. Т. 49,-вып. 9.
1954^1956. . ‘ ' /’ ’? ‘-77 .
Засравский В.А., Двухступенчатые фотопериодические реакции , как ос-
нова для, разработки, модели фотопериодического контроля рдзвц^ия ’Гленцс-
тоногих // Энтомол. обозр. 1972.,Т. 51,, вып. 2, С..2Д7—239.', . > . .
,. Заславский В.А. Распространенность ступенчатых фотопериодических
реакций средд насекомых д клещей ^/Энтомол.’обозр. ,1975. ‘,Т. 54, выд. 2.
С..291-304. X Х'5"*'4"..' ; 'X "" X" ’ ; 'Х""к
. • Заславский,В.А.,Взаимосвязь;количественных качественных реакций в
фотопериодизме насекомых //'Ьоол. жури. 1975/Т. 54, ’вып. 'б. С/913—921.
. . . ,3пслпвскгш В.А. Фртрперирдичесуий^и.температдуньн! контроль разви-
тия насекомых. Л.,1984. 184 с. ‘ ‘4 4 \ л '* "'У.
Заславский В,А,, Кац Т-Сг Фотопериодическое определение пола у тли
Megoura vipia BuckL (Homopteray^Ai^iidida^y/ Днтоирл^ о^озр. 1987. Т, 6$,
вып. 4- С. 681—688. ”*-* ’ ’
( , Заславский В. А., Семья нов В. П. Миграционное состояние у семцточеч-
fHpjif коровки;(^о1сс/^//4-^р^^дпсЩД1Х//- ^ррлл журн,. l^Sk.XX^J *ВЫЛ-, &
С 8^8—891-. т. * - ’
ЗенякинЛ.А., Карпунина Н.Н. Изучение путей расселения и полетов ко-
лорадского жука Leptinotarsa decemlineata р зависимости от метеорологических
факторов й’физиологического состояния’// 'Колорадский жук и меры борьбы
с ним. М., I95&. Т. 1. С. 60—72. г . ,
! Зиновьева К.Б. Роль фото- и терморитмов в индукции диапаузы у н'аезд-
/дика’ Alysia manducator Panz. (IJymenppfera, Bracon^daey// Энтомол,' обозр.
1976". T."35, выд. 3.' С.' 517—524.." ’.............. '* . '
. ’ ' .'-с ' 1.'». л/ : - s.
217
Зиновьева'КБ. Влияние*суточных ритмовтемпературы при разных фото-
периодах'на’индукцию и ^реактивацию Alysiamanducator‘Pa.nz. (Hymenoptera,
Braconidae) // Энтомол. обозр. 1991. Т. 70, вып. 2. С. 265—280. ’
Зискинд Л.А.Хищный клоп Podisus maculiventris Say и его совместное
использование^ пестицидами против колорадского жука: "Автореф.’дис1..; канд.
биол. наук. М., 1985. 15 с. 1 1
Злоп1ников‘'М:Д. 'Возможный* ареал распространения и'сроки1' развития
колорадско'го жука'в'Европейской части СССР* // Труды ВИЗРа. Т^б/.1 Т.27.
С. 68—74.
1 Золотаренко Г.С. Совкй-металловидки рода Plusia Ъ. (Lepidoptera,
Phytometrinae) Лесостепной зоны Западной Сибири // ‘Фауна иэкологияЧле-
нистоногих Сибири. Новосибирск,у'1'9б6. С. 174—177.'
Ижевский С.С. Интродукция и применение энтомофагов. М, 1990'. 222 с.
‘ ‘‘Ижевский '’С/С.’ "Зискинд Л.Л. Перспективы использования’интродуци-
рованных хищных'Клопов* Perillus bioculatus (Fabr.),‘ Podisus maculiventris’ (Say),
Oplomusnigripennisvdr.pu/cAerDull.'(Pentatomidae:*Hemiptera)npoTHB£epn'notor3a
decemlineata Say (Chrysomelidae: Coleoptera) // Биологическое подавление ка-
рантинных вредителей'и сорняков. М., 1981. С. 20-37. f
‘Ильинская Н.Б.'РеакДДя ‘мышечной ткани активных и диапаузирующих
гусениц соснового шелкопряда на действие гипер- и гипотонических раство-
ров хлористого натрия**// Реакция клеток на экстремальные воздействия. М,-
Л„ il,963.tC.i8b-96.’ ; : •'-*
Ильинская ~Н.Б. Солеустойчивость мышц активных и диапаузируюГцих
пронимф яблонной плодожорки // Цитология. 1965. Т. 7, вып. А. С. 5’38—546.
Ильинская И Б. ’Устойчивость клеток диапаузирующих насекомых к по-
вреждающимвоздействиям // Реакция Клеток' и их белковых компонентов на
экстремальные воздействия. М.-Л./1966. С. 59—72: ’* ’ ’ ’ .
'' ’ЯльинскаяЯ.КСоотношение холодоустойчивости и неспецифической
устойчивости мышц насекомых во время диапаузы’// Холодостойкость ‘насе-
комых и клещей. Материал. Симп. 19—21 апреля 1971. Тарту, 1971'.*С. 29—3L
\’;"^осифод ^.В.‘Фауна ^а'*Бъл^ария. HeterpPtera'.'Pentatomidae.Cd^HH, 1981.
' 'Калабухов И.И ^Анабиоз» у животных притемпецдтур^х ниже 0“ // Зоол.
журн. 1935. Т. 14, вып. 15. С. 97-111. ’ ;; У :
* ‘ Каплйн'И.Т. 'Сезонные' ритмы развития открытоживущих Насекомых се-
менных растений1 Каракумов // Журн.общейбиоУ 1994. Т‘ 55, вып. 6. С. 716—
736. i-
Караушева А.И. Особенности климата'и характер погоды'на территории
леростепной дубравы «Лес на Ворскле»'Белгородской области. *1. Особенности
климатообразующих факторов /^ Вести. Ленингр. ун-та. 1980. Т? 9? вып. 2.
У 'Кинд Т’В‘. Эндокринная детерминация эмбриональной диапаузь! у ан-
тичной волнянки Orgyia antiqua L. (Lepidoptera, Lytnanthriidap) '// Цроблемы
фотопериодизма и диапаузы насекомых. Л., 19/2. С. 2Г0—228. ’. **' *
' ' Кинд ‘Т.В. Влияние длины дня и температуры на индукцик/диапаузы у
белгородской и‘ сухумской популяции* медведицы .Spitosomd menthastri
(Lepidoptera, Arctiidae) // Зоол. журн. 1985. Т.”б4, вып. 3. С. 377—383.
Кинд Т.В., КорольковаЕ.Д. Индукция и прекращение'диапаузы у.закар-
патской популяции американской белой бабочки Hyphantria cunea Drury
(Lepidoptera.Arctiidae) // Энтомол. обозр, 1991.-Т. 70, вып. 1. С. 28—35.
. КлючкоД. Ф. Луговая>совка на Украине // Защитарастений от вредителей
и бодезней,.'1964,. Выц. JO.,r.C. .44... ,
Ковалев О.В. Интродукция и акклиматизация фитофагов амброзий
{Ambrosia L., Asteraceae) в СССР// Вопросы общей энтомол. Л., 198L..C. 9-11.
Ковалев О.В., Медведев Л.Н. Теоретические основы, интродукции амбро-
зиевых листоедов pppnZygogramma Clevz. (Coleoptera, Chrysomelidae) в СССР
для биологической борьбы с амброзией // Энтомол, обозр. ,Д9|83,.Т. 62, выл.1.
С.; 17-32. ... , . . - • ,! . -л v. ... „м..л
Ковальская Т. Влияние физиологического состояния уходящего на зи-
мовку колорадского жука Leptinotarsa decemlineatarSayna течение диапаузы и
смертность во время зимовки // Колорадский, жук м меры борьбы с hhm. T..2i.
М„ 1958. С. 66-73. ... . .-.--.о,
Кожанчиков И.В. Некоторые данные, по влияниютемпературы ивлаж-
ности на развитие совки .гаммы // Защита растений- 1937>. Вып. 14, С. 49-61.
Кожанчиков И. В. О нижнем термическом пороге развития’насекомых//
Докл, АН СССР. 1946,»Т. 51, вып. 3. С. 237-240.
Кожанчиков-И.В. Черты влияния отрицательной температуры на эмбрион
нальное развитие насекомых // Журн. общей биол. 1949. Т. 10, вып. 11. С. 50—
67.. , '• ' -.„V ' ' г'. ' '
Кожанчиков И.В. Цикл развития и географическоераспространениезим-;
ней пяденицы (Operophtera brumata L.) // Энтомол. обозр, 1950.. Т. ЗД»,вып.; l-p
2..С 178-192. ' / - , Лг
Кожанчиков И Ji. Особенности зимовки и холодоустойчивости карадри-
ны (Laphygma exiguajfb., Lepidoptera, Insepta)// Дркд- АЦ СССР. 1955.-Т.ДОЗ,
вып. 3. С. 517—519. •••• - <v
Кожанчиков И.В. Видовой-состав, особенности биологии и географичес-
кое распространение вредных , подгрызающих совок в, СССР /ДСб. работ. Ин-;
та прикладной зоол.ги фитопатод. 1956. Вып. 4. С. 5—51. , < i <
Кожанчиков И.В. Экспериментально-экологические методы14исследова-
ния в энтомологии. Л., 1961. 260 с.-, ... s : ,и • -
Колесниченко Л.И. Сравнительная фенология подизуса и колорадского
жука в условиях Черновицкой области // Исслед. по энтомол. и акарол. на
Украине. Тез докл. -2-го съезда УЭО. Ужгород, 1980, Киев, 1980. С. 104—105.
Колорадский картофельный жук, Leptir^ptqrsa decemlineata^py / Иод ред.
Р.С..,УшатинскойиМ., 1981. 375, с. - > < .
, Комарова0. С. Жизненный цикл и условия развития гроздевой листовертки
(Polycjtrosis botrana Schifl) // Зоол. журн. 1954. Т. 33, вып. 1, С. 102—113, ' *•>
Космачевский А.С. Борьба с вредителями полевых; и овощных кулыур.
Краснодар, 1966}- 156 с. ; > ; , ' . А
Кузнецова ИА. Факторы, вызывающие наступление диапаузы, у, мальво-
вой моли Pectinophpra majvella Hb. (Lepidoptera:,,GBlechiidae) // Энтомол,,обозр.
1962. ,Т. 41, вып.. 3. С. 510-515., ' .... . ;
Кузнецова Ю.И. Исследование возможностей хранения'яиц златоглазки
Chrysopa carnea,Steph, (hleuroptera, СЬту5ор1бае) припрниженных темцерату-
pax //Дрод.^урц.,б^дцп.ДО, С.Д505-1614. . , . ns
219
ЗиновьеваК.Б. Влияние'1 суточных ритмов температуры при разных фото-
периодах'^ индукцию 'и ‘реактивацию Alysid'mariducatoP Banz; (Hymenoptera,
Braconidae)//Энтомол. обозр. 1991. Т. 70, вып. 2. ‘С, 265—280. '
Зискинд Л.А; Хищный клоп Podisus maculiventris’ Sky и его совместное
использованием пестицидами против колорадского жука’: Автореф.’дис1..; канд.
биол. наук. М., 1985. 15 с. '*
Злотников МД.'Возможный’ ареал распространения и сроки развития
колорадского жука'в' Европейской части СССР* // Труды ВИЗРа. г19б7/ Т.;27.
С. 68—74.
*' ‘ЗолотаренХоГ.^. Совки-металловидки рода Plusia О. (Lepidoptera,
Phytometrihae) лесостепной зоны Западной Сибири //’Фауна и"'экологияЧле-
нистоногих Сибири. Новосибирск,**1966. С. 174-177.' - « *.
Ижевский С.С. Интродукция и применение энтомофагов. М, 1'990’. 222 с.
ЯзкевскийС.С.,Зискин<)#.Л.Перспективыиспользования‘интродуци-
рованныххищных'клоповРел7/и5Й/оси/аШ5 (РаЪг!),1 Podisus maculiventris’(Say),
Oplomusntgripennis Vdr.pulcher Dull?(Pentatomidaet*Hemiptera) Против’ Leptinotarsa
decemlineata Say (Chrysomelidae: Coleoptera) // Биологическое подавление ка-
рантинных '"вредителей*и сорняков. М.,' 1981.*С. 20—37. *
’ " Ильинская1 Н.Б.'Реакция ‘мышечной ткани активных’и диапаузирующих
гусениц соснового шелкопряда на действие гипер- И гипотонических раство-
ров хлористого натрий // Реакция'клеток на экстремальные воздействия. М,-
Л.,-!Г963.^?8Ь-96;’':/'ч';л’; ‘ ‘ -
’ Ильинская Я.Л.Солеустойчивость мышц активных и диапаузирующих
пронимф яблонной плодожорки // Цитология. 1965. Т. 7, вып. 4. С. 5’38—546.
Ильинская Я.Я/Устойчивостьклетокдиапаузирующих насекомых к по-
вреждающим воздействиям // Реакция Клеток'и их белковых компонентов на
экстремальные воздействия. М.-Л./ 1966. С. 59-72. '*
Ильинская Н.Б./ Соотношение холодоустойчивости и неспецифической
устойчивости мышц насекомых во время диапаузы ’// Холодостойкость насе-
комых и клещей. Материал. Симп. 19—21 апреля 1971. Тарту, 1971/С. 29— 31.
* ‘ 'Иосифов M-В. Фауна даБългария. Heteroptera.Pentatomidae. София, 1981.
т. 12'205 с.' ?;л ' .. "
1 Калабухов НИ, «Анабиоз» у животных при'температурах ниже 0" // Зоол.
журн. 1935. Т. 14, вып, 15. С. 97-111. ’ ; ; .
‘ ' Каплин'В.Т. Сезонные ритмы развития открытоживущих насекомых се-
менных растений1 Каракумов // Журн. общей биол. 1994. Т. 55, вып. 6. С. 716—
736.,, , , t, , . .
1 ’КараушеваА.ИОсобенности климата'и характер погоды на территории
лесостепной дубравы «Лес на Ворскле» Белгородской области. *1. Особенности
климатообразующих факторов /J Вести. Лерингр. ун-та. 1980. Т.‘ 9, вып. 2.
Кинд Т!В. Эндокринная детерминация эмбриональной диапаузы у ан-
тичной волнянки* Orgyia'antiqua L. (Lepidoptera, Lymanthriidap) '// Проблемы
фотопериодизма и диапаузы насекомых. Л„ 1972, С, 210—228. " .
* ’ ’ Кин<Гт!В. Влияние длины дня и температуры на индукцию*диапаузы у
белгородской и‘ сухумской популяции' медведицы .Spilosoma menthdstri
(Lepidoptera, Arctiidae) // Зоол. журн. 1985. Т.”б4, вып. 3. С. 377—383.
218
Кинд. Т.В., Королькова. ЕЛ. Индукция и прекращение'диапаузы у. закар-
патской популяции американской белой бабочди ffyphaittria сипеа Drury
(Lepidoptera,-Arctiidae) // Энтомол. обозр. 1991.тТ. 70, вып. Г. С. 28—35.
. Ключко^З.Ф. Луговая.совка на Украине // Зашита .растений от ^вредителей
н-бцлезней,.- 1964,, Вып. 10. С. 44.. , ........- ; •
Ковалев О.В. Интродукция и акклиматизация фитофагов амброзий
(Ambrosia L., Asteraceae) в СССР // Вопросы общей энтомол. Л., 198L С. 9—11.
Ковалев О.В., Медведев Л.Н. Теоретические основы интродукции- амбро-
зиевых листоедов рода Zygqgnamma Clevz. (Coleoptera, Chrysomelidae) в СССР
ддя биологической борьбы о амброзией // Энтомол, обозр. ,1983.,Т. 62,вып.1.
С.17-32. - ,
Ковальская Т. Влияние физиологического,состояния уходящего на зи-
мовку колорадского жука Leptinotarsa decemlineata.SayHa течение диапаузы и
смертности вовремя зимовии,// Колорадский, жук.и меры борьбы с ним.-ТЛ
М„ 1958. С. 66-73. - й!
Кожанчиков И.В. Некоторые данные по, влиянию температуры ^влаж-
ности на развитие совки гаммы//Защита растений. 1937.. Вып.14, С. 49-61.
Кожанчиков И.В. О нижнем термическом пороге развития -насекомых .//
Докл, АН СССР. 1946. Т<51., вып.-З. С. 237-240. ь,
Кожанчиков-И.В. Черты влияния отрицательной температуры на эмбрио-,
нальное развитие насекомых // Журн. общей биол. 1949. Т. 10, вып. 11. С. 50—
67. -.-V ' WS-, ч.ч '?»
Кожанчиков И.В. Цикл развития и географическое распрортраление.зим-;
ней пяденицы (Operophtera brumata L.) // Энтомол. обозр, 1950, Т. ЗД^<вып.,1-г
2..С, 178—197,- - v * . . . . лг
Кожанчиков ИЛ- Особенности -зимовки и холодоустойчивости карадри-
ны (Laphygma exigua Hb., Lepidoptera, Insecta) // Двд, AH СССР. 1955. T.,103,
вып. З.С. 517-519. ; ...
Кожанчиков И.В. Видовой, состав, особенности биологии и географичес-
кое распространение вредных, подгрызающих совок в.СССР/ДСб.работ Ин-;
та прикладной зоол. и фитопатол. 1956. Выц. 4. С. 5—51. i... , ;
Кожанчиков И.В. Экспериментально-экологические методы,исследова-
ния вэнтомологии.Л., 1961. 260с. S- , !
Колесниченко Л.И. Сравнительная фенология -подизуса и колорадского,
жука в условиях Черновицкой;области // Исслед. по энтомол. и акарол. на
Украине. Тез докл; 2-го съезда УЭО», Ужгород,; 1980. Киев, 1980. С. .104—105,
Колорадскийкартофельный жук, LeptiqptifrsftJ. Под ред.
Р.С-Хшатинской^КД., 1981. 37-5-ст 1 1 , - ;
, Комарова. О. С, Жизненный цикл и условия-разаития гроздеврй листовертки
(Polychrosis botrana Schiff) // Зоол. журн. 1954. Т.,33, вып. 1, С. 102-113. , .
Космачевский А.С. Борьба с вредителями.полевых; и овощных культур.;
Краснодар, 196(>. 15$ с. / • , . ..’
Кузнецова /М..Факторы, вызывающие наступление-диапаузы-у.мальво-
вой моли Pectinophpra ma/ve//a Hb. (LepidopterajXlelechiidae) // Эитомол,,обозр.
1962. Т-41, вып.; З. С. 510-515., \ ; -.« л;
Кузнецова Ю И. Исследование возможностей хранения'яиц златоглазки
Chrysopa carnep .Steph. (Neuroptera, Chxysopidae). при пониженных температу-
рах //ДрОДчХТУРН., 1Ж 7, 69иЖД. ,10» С.;Д505,—1^14. :l i:.
219
’Куренцов А.И.,Груздева-Н.П. ЛуговаясовкавПриморскомкрае // Труды
Далъневост. Таежнд'станции. 1946. Т? 5.'С.<235^-256и
Курючкин В.А.Некоторыеособенности размножения? комнатной мухи я
осенней’жигалки.в условиях: жнвотноводческихпомещений // Морфология и
экология двукрылых — потенциальных переносчиков заразных заболеваний.
Иваново,». 1984. С, 43^-54* <- .-• ч/-иг.'-до-р sb Ай; w.v'.a
I •‘Леман'-'ВДЛ'.Курс’светокультуры' растеняК/М?, 4964? 206 с? ’
• г‘Лозина-ЛозиМки^Л.К.1 Очерки по.криобиологии. "Л’., 1972.-288 С?
i > " Макарова Л.А., ДоронинаГ.М.'Агрометеорологические предикторы про-
гноза>размножения вредителей сельскохозяйственныхкультур. Л., 1988. 213 с.
Макарова Л.А., Доронина Г.М. Синоптический метод прогноза -дальних
миграций вредных насекомых.'СПб.,.1994.‘Л98;с.г? с J 1 . . .. г г А
» «'• *: Макеев^ГИ:Экологическое обоснование Прогноза* численности/вредо^
носкости колорадского: жука и меры ’борьбысним'наСеверномКавказегАв-
тореф. дис... канд. биол. наук. Л., 1982. 25 с. о'г scv1. .л:.
- si р Маршаков .В.ЛВлияниепостоянных непеременных температур ;на дли-
тельность искоростьразвития’насекомых //> Энтомол'? ббoзp;.r’1984’., Т.'бЗ*,
вып.4.С. 3-423 ик.ои! '? ньл-'-'А
Масаки С. (Masaki S). Фотопериодизм в сезонном жизненном'цикле
сверчков // Проблем ыфотопериодизмаи’диапаузы насекомых?Л-.,1972.<3. 25—
50/. /' I лТ Г .У//; .«0 й исуЖ • \ г/гтш: ... яиш/.
J \/ Масленникова В.А., Абдель-Наби А.А., Черныш С.И. Титр экдизона при
индукции’летией'н*зймней‘д|иапаузы 'капустной coBKu'<Barathra'brassicae L
(Lepidoptera. Noctuidae): // Энтомол. обозр. 1976.‘-Т.55,'вып. 4. С.!768—776.!Н
Масленникова В.А., Мустафаева Т.М. Анализ фотопериодическйх’адап-
„ тацийу географических Популяций 4/WH/e/e5^/o/nera/ur(Hynienoptera,
Вгасотбае)‘и 'Pieris drass<cae(Lepidoptera, Р1епбае) //:Энтомол.!обозр.‘1971.
Т. 50, вып. 3. С. 497-505. - ! i f
‘’^Матиняй Т;£>Адаптапииисезонн&х циклов?Кгеографическнм1изменени-
ям 'длины дня и температуры у крестоцветиых &enHHOK (Pieris napi L.rst P.rapae
L. // Изв. АН Арм. ССР. Биол. науки. 1965. Т-48/-вып.'Н).1С?'81'-97|.«м«'’'' -
е> ’1-Мелъниченко А.И. Закономерности массовых перелетов лугового мотылька
и проблема построения прогноза его залетов// Труды’по^зашите^растеиий1
1986;вып.Ч«.»С.’‘1г5б.-|л<'’,1--'^Ф ’«:/ • V.t.
<*' Меривэз Э.Э.' Эко-физиологическая'характеристика холодоустойчивости
насекомых: Автореф. дис... канд. биол.' наук-. Тарту, 1972.''39 с. ‘‘ ! /, w ’ : '
- я ‘Меривээ^Э.Э. Холодоустойчивость-насекомых;'Таллин/1978; IWc.
Меривэз Э.Э., Хансен Г.Э. О прогнозировании *на основе°точекПереох-
лаждения Зимней-смертности насекомых. 'зимующих выше линии сНежного
покрова //'Холодостойкость насекомых' иклещей. МатериалМ'симпМ^-гТап-
реля 1971/Тарту,<1|971.Ю.»90-931 н 1 «их;:-.у-шзп'А
Миляновский Е. С. К фауне чешуекрылых (Macrolepidoptera) Абхазии’ //
Труды' ин-та^ЗоологиИ'АИ.ЧрузССР. Ii956. Т.1‘5. С. 5'1'-100/ : ' i
/г, Миляно1вский E.CJ JlymvaaicoBKa^^rphis^unipancta На^.)///'МатериалЫ'
сессии Закавк. совета по координации н.-и. работ'по защите растений/Бреван,
1967V/C. 523—527’Я!::-Ч,?./' nj.j/.repu ".
! s-ккМдер^И.'Ф. Холодоустойчивость >кблорадскогОгжука:‘|1е1р//и1о/аг5в
decemlineata Say // Периодичность индивидуальното развития насекомых. М.,
1969. С. 99-130.
220
> ч Миндер ,И.ф., ХозаржевскаяЭ.Ф.лЭкологнязимпей диапаузы колорадс-
кого жука <// Экология и «физиология диапаузы «колорадского жука. <<А966ц
С. 45-68. i й- ’ ? -чч
:< • \iq iMopcu A JJj Термопериодизм и»его роль В1ситеме1сезонно-цикдических
реакцийчнасекомых: Автореф.дис. канд.биол. иаук.: Л., 1978.22 с» . ; m
Мусолин Д.Л., Саулич А.Х. Фотопериодическая регулйцияфоста. личи-
нок у иастоящих полужесткокрылых (Heteroptera) //Зоол. жури»* 1997. Т. 76,
_вып.<2.' С. 530—542; \ ч . адп'эоч лчгэй--- »У гу
Мустафаева Т.М. Экспериментальный анализ сезонно-циклических адап-
таций некоторых энтомофагов: Автореф.'дис... канд. биол. наук. Баку, 1974.
25 с1. '‘Ч^- _ ;•» fyyj- ,(fW ’(Т! ТГ'Ч-Ь.Ч-Д 't V Ч-,,-чп'» I. !< Ь'»Л». ?
’1 ' Наумова Е.Н., ГорбунатБ.З., .КнорПЕДи др.Изучениемассовых миграций
бабочек лугового мотылька Loxostege sticticalis L, (Lepidoptera, Pyralidae) JHa
основании* анализа-их элементного! состава?1//- Экология1.'? '1994;' ®ын. >4. С. 75—
810,;Ч-;п . ' •.I.s f.jSИ -Я* ’• V Л'ЙчЛ' ' si£W Я
' '? < Несин АЛ. Фотопериодическая'реакция, регулирующая диапаузу улет-
ней капустной мухи Delia floralis Fall. (Diptera, Anthomylidae) // Вестн. Ле-
нингр. 'ун-та. 1987."Вып.'2.“С: 122—123. v .чМ А\.а .аул'ч-' У
н Яовак В.Л. Тормональные основы диапузы у насекомых // Проблемы
фотопериодизма и диапаузы насекомых. Л., 1-972. SC.. 193— 209. '•" яьиогг.си'
в ч .Онисимова 3. Г.'Луговая совка^вредитель-зерновых; культур Приморья и
меры борьбы>С'4ней. Владивосток, 1949л95с.' " 1 ' .is;1 -w лзчж
Палий В.Ф. О вспышке массового размножения совки карадрины
Laphygma exigua Hb. (Lepidoptera, ЬкхИшбае/вВоронежской области //Энто-
мол. обозр. 1956. T. 35, вып. 4. С. 799-802. •>'< А / г ; .
ПарфентъевИ.А.Обзорновейшей?литературы по Phytometra gamma L.
1914-19227/'Изв. Моск.'энтомол<.'*о-|ва."1923?Т.;*7,''вып.12лС.1в8—44;. 1 *'
Поляков И.Я., Гейспиц К.Ф., Горышип Н.П. и др. Совершенствование-Me-;
тодов прогноза распространения лугового мотылька и снгнализации сроков
борьбы с-ним //'Методы прогноза развития1 вредителей и болезней сельскохо-
зяйственных'культур.?'М.,'19<78‘. С. 5—59.-1' га'1 'й мячАчАп
Пучков В.Г. Фауна Украины. Щитники. Киев, 1961. Т. 21, вып. 1. 367 '0?
чПучков В./’АЩитМйкимСредней 'Азии. Фрунзе, 1965 . 329 сА*7" '
<'?‘ Агбдв'ЛУ.Л1|*Типыг годичных- циклоВ'зеМлянЫх'"подгрызающих совок
(Lepidoptera, Noctuidae) //!Энтомол» обозр. 1956. Т?:31, !вып/ 1. С?-69—79. ‘','-
-вай<1Саакян-‘£аранова А.А., 'Сугоняев Е.С., Шельдешова Г.'Е. 'Акациевая лож-
нощитовка иеепаразиты.,Л.,'1971.165'е.г'''' .“.sot»'i'f.A'.?-.! V
Сонин В.А Колорадский жук.'М.,197б. 108 с. ’ : ч «и? ' <
.д .пчсаулич А.4. Влияние фотопериода и'величииы «группы на развитие луго-
вой (Cirphis unipuncta Haw.) и малой наземной (Laphygma exigua Hb.) совок
(Lepidoptera. Noctuidae) //‘Энтомол. обозр. 1975а. Т. 54‘, :вып. 2."С.*314—325.
он ^ 'Саулич^А.Х. Эффект* группыгпрй фотопериодической индукции диапаузы
у капустной совки Barathea brassicae // Зоол. Журн'. 19756.гТ.‘65!, вып?91'С.г1335—
1329> 111 ,i! i!‘ 'Щ'.'-'А!.' ’£: , iiL'.t'JsO Я ‘ : 4’. 1
... л» Саулич 4.Х?Сравнительно-экологическое?изучение гомодинамных и ге-
теродинам ных чешуекрылых в связи с проблемой -миграций: Автореф'. дис...
' канд.4биол> наук.'1 Л., 1977.’22"с. - '<'
....и \\ s:.-f тА
221
Т'.г ~Саулин;А.Х. Значениеабиотических факторов при формировании вто-
ричных ареалов адвентивныхлвидов насекомых //(Энтомол. обозр. 1994./Г. 73,
вып. 3. С. 591-605. -Ч
. ' Саулич А.Х.:, Горышин: И.И.,Волкович Т.А. Фотопериодический контроль
развития лугового мотылька в природных условиях//Зоол.журн..1983.Т.62,
ВЫпи2. С. Т663-1.675. ГЧ
.. ’ Саулич А,Хи, Мусолин AJI.r(Saulich A.H., Musolin D.L.). Univoltinism and
its regulation in some temperate true bugs (Heteroptera) // European J. Entomol..
1996лVol. 93,?N 3. P. 507—518. s t. »< ,: ; ? - . < «
>' vi Саулич A.X., Волкович Т.А., Нумата Х. .(Saulich A.H., Volkovich Т.А.,
Numata H.). Control of seasonal development by photoperiod and temperature in
the*linden bug,;.Pyrrhocoris apterus in Belgorod, Russia,//Zool. sci. 1994.,Afol. 11.
t -r ' Ч-. „ 7С.Л .
>J Семаков В.В. Поведение .некоторых вредителей крестоцветныхрастений
в связи с изменением трофических цепей // Поведение насекомых как основа
для. разработки мерборьбы с вредителямисельскогоилесного хозяйства.
Киев,|1975,. С. 136-141.,?;;., " с •' ,, " -•
Семьянов В.П. Миграции и миграционное состояние У семиточечцой. бо-
жьей коровки (Coccinelja septempunctata L) // Морфология, систематика и
эволюция животных/ Л1., 1978. С. 75—76. ; » >
Семьянов, В.П, Места зимовки и миграции семиточечной коронки на
южных склонах Гиссарского хребта // Изв. АН Тадж. ССР. Отд. биол., наук.
1979. Вып. Л.;С.: 32-36. . > .. ,г. м. . . *
Семьянов-В.П. Эстивация у кокцинеллид //.Труды Всесоюз. энтомол.
о-ва. 1986. Т 68. С. 111-114. Т , !й : , -- « - ,
, Сикура Л.В. О метода^ прогнозирования сроков: развития отдельных ста-
дий колорадского жука Leptinotarsa decemlineata Say //,3оол. журн. 1963. Т..42,
ВЫН. 7. С. .W41.-1044. ' 1 ч >
Симакова Т.П., Географическая, изменчивость реакций на условия зимов-
ки уловки Acronycta rumicis // Зоол. журн. 1971 . Т. 50, вып. <3. С. 373—382.
Скрипчинский В.В. Фотопериодизм, его происхождение и эволюция., Л>,
1975. 299 си-. . л
Сорокина А.П., Л/асленидковлВ.Л.Особенности.фототермических реак-
ций .некоторых видов рода Trichagramma (Hymenoptera, Trichogrammatidae) //
Вести. Ленингр. ун-та, 1986. Сер. 3. Вып. ,1. С. 9—14. . • , > > .• .•;..
, . Сорокина А.П., Мррленникова В.А. Температурный оптимум формирова-
ния диапаузы у видов рода Trichogramma Westw. /Hymenoptera. Twhogramn
matidae)//Энтомол. обозр. 1987. Тлбб.хвып. 4..С.: 689-Л>97. >
- Стрыгина С.П. Американская,-белая бабочка,// Защита раст, 1983, Вып. 6.
С. 52— 53... . , Ч/ >,*, , • ,, ,- .;V. 'Ч. '
Тархнишвили А-А. Некоторые моменты, биологии, Pteuda/e/m unipuncta
(Haw.) в Грузии,и меры борьбы с ней// Материалы сессии Закавказ. совета по
координации н-и.^работно защите растений Баку,.1966,. С..366—370.. ,
Тархнишвили А.А. Особенности биологии, экологии и динамики числен-
ности лутовой совки (Psepdaletia.urtipupcta),д условияхДрузии; Артод^Ф^ дне...
кацд. бцол. цдук. Тбилиси, 1972. 24.С. . .. , , : Г",-
Томс С.В. Эколого-физиологические адаптации полужесткокрылых
(Heteroptera) к обитанию в аридных условиях // Вопросы экологической фи-
зиологии насекомых. М., 1987. С. 93-128.
222
Тыщенко В.П. О происхождении* и эволюцииофотопериодической реак-
ции насекомых //Журн. 'общейбиол. 1973. Т. ‘34, вып?'4'50'539—558. к-т .
, Тыщенко В.П. Физиология5 фотопериодизма насекомых // ТрудыВсесо-
юз. энтомол. о-ва. 1977ЛГ/59. 155 сП' ' '• -.......*v «t,«. v.-sWl
*' Тыщенко В.П. Сигнальное и витальное действие экологических факторов
//Журн. общей биол. 1980. “Г. 41; вып. 5.'С!. *655^6671 m <•
‘Тыщенко В.П. Эволюциясезонныхадаптацийунасекомых // Журн. об-
щей биол. 198J.'T. 44,вып. «1*. С. 10-22. • <*» ’* ’• панк <t < u., »><т
Тыщенко В.П., Волкович Т.А., ЛаневичВП. Промежуточная фотоперио-
дическая реакция и ее роль в регуляции сезонного; развития темнокрайней
совки Axyliaputris L. (Lepidoptera, Noctuidae) // Энтомол. обозр-.? 1983. Т. 62,
вып. 2. С. 225—234. 5 в <
Тыщенко В.П.', Гасанов О.Г. Сравнительное-изучениефотопериодической
регуляции диапаузы ивеса куколок у нескольких видов чешуекрылых
(Lepidoptera) //"Зоол. журн.1983. Т. 62, вып. 1. 063—68. ’ ‘ - < < ?
Тыщенко В.П., Кинд Т.В. Нейроэндокринные механизмы регуляции'се-
зонных циклов // Гормональная1 регуляция развитиянасекомых.Л-., 1983.
С. 82-117. (Труды Всесоюз. энтомол. о-ва. Т-64.).т -«ь >-? !< >< г,
" Тыщенко В.П., Ланевич В.П., Квитко Н.В. Сезонные адаптации 'медведи-
цы ' Callimorpha dominula L. (Lepidoptera, -Arctiidae)- //•-ВестнС Ленингр.- ун.-та.
1985. N 24.0 104-106. О- О"' Л адхч.,,.- - ' <?,
Тыщенко В.П., Тыщенко Г.Ф. Сезонные адаПтации совки-с-черное
GrapMphora C-nigrum L.’(Lepidoptera, Noctuidae)1 //Эитомолюбозр. 1988. T. 67,
вып. 1. С. 11—20. ;> ’ ! !
Ушатинская Р.С. Основы холодостойкости насекомых. М.;,’495/. 314 с.
Ушатинская Р.С. Некоторые физиологические и биохимические особен-
ности диапаузы колорадского жука Leptinotarsa decemlineata Say// Колорад-
ский;жук’и'меры1 борьбы с ним. Т.’2.;М./1958. С.450—185? -
1 Ушатинская Р.С. Многообразие форм'физио логического-покоя у чсоло-
радского жука Leptinotarsa decemlineata Say как одна Из причин расширения
ареала // Экология и физиология диапаузы колорадского жука. М., 4966а.
С. 5-22. " О . \ Р
Ушатинская Р. С. Физиологическая разнокачественность особей в'зиму-
ющих Популяциях 'колорадского жука1 (Leptinotarsadecemlineata) // Экология
и физиология диапаузы1колорадского1 жука/М.1, 1966б.'’С.-144и.179О .- < «><
Ушатинская Р.С.Диапауза насекомыхчгее модификации //’Журн. общей
биол. 1973, Т. 34, вытг.25. С. 194-215. '
' Ушатинская Р;С. Эколого-физиологическая 'приспособленность насе-
комых к холодному сезону года // Адаптации животных к зимним условиям.
М„ 1980. С. 117-125. 1 1,1
Ушатинская"' PiC. Терморегуляция в классе насекомых // Вопросы эко-
логической физиологии насекомых. М1., 1987-.'С. 5—46/! "* > к
Ушатинская Р. С. Летний покой (эстивация) у насекомых'//Там 1же. С. 140—
" г"< , - -- - -к -if--- 1 л , i - ’
Ушатинская Р.С. Скрытая жизнь и анабиоз.‘*М.,гГ990. 182 с.
Ушатинская Р.С,, Иванчик Е П. Обратимостьэколого-физиолОгических
адаптаций'у колорадского жука Leptinotarsa decemlineata ^ay // ЗОол. журн.
1982. Т. 62, вып. 3. С. 358-369.
223
Ушатинская PC., Козаржевская Э.Ф. Диапауза и зимовка колорадского
жука Leptinotarsa decemlineataпервой генерации при зимовке в почвах разного
типа>1//Зоол. журн. 1962. Т. 41, вышЛЬС.41166—1174. ,. (> ,, > . Л '
Ушатинская Р.С., Чеснек С.И. Переохлаждение изамерзание,в-онтоге-
незе колорадскогозжука 1ед«ло/дгга^есе/и/г>|е4/а|5ау//1Вопросыэкологичес-
кой физиологии беспозвоночных.'М,, 4974, С.129—143, Wi и; , s ‘
о .Фалькович ЗУ.Лц.Сезонноеразвитие.пустынныхчешуекрылых (Lepi-
doptera) Средней Азии и его историко^фаунистмческий:анализ •// Энтомол.
обозр. 1979. Т.^в^выгыЗ. С. 26(0-281. \ ; ; н ". - ??
'л.<Фролов-<АШ. :Биотаксономический> анализ вредных: видов:,рода.fistrifUa
Hbn. // Этология насекомых, Л.» 1984..С, .4—l.WMJpiyflM Всеррюз. энтомол.
о-ва. Т. 66). ,, ? : . ?,
I. .. . Хансен Т’.Э.Исследование содержания многоатомных спиртов в зимую-
щих- стадиях насекомых в связке их холодостойкостью // Холодостойкости
насекомых и клещей. Материалы симп. 19-21яапредя 197LТарту, 1971.С. 137-
142.: ц‘>. -. г , .-р я •» •, .< >- ,
*’ Хансен Т.Э.;11ВийкМ.1О.,<ЛуйК!А. Биохимические изменения и холодо-
стойкость у зимующих жуков короеда типографа, Ips typographus-L. (Coleoptera,
Ipidae) /А Энтомол .зобозр: 1980. Т. 69, вып. 2. С. ,249—253. , -
Хачатрян А.Г.-Изучение фотопериодической реакции армянских попу-
ляций ивовой волнянки Leucoma salicis L., Lepidoptera», Lymantriidac // Мате-
риалы УПзсъездаВЭО.Киев, 1984.‘С. 219—220.
Хмелева. Н.Н^, Рощина Н.НрФилюковаТ.А, Фотопериодизм водц^х бес-
позвоночных. Мн., l^l. 109 с. . ' ;: г . ,
> lit Хомякова В.О., Дыкова Е.Л^ Узихина В.С, Влияние корма,и^фотоперио-
дцческих условий' иа разаитие лутовогомотылька Loxostege s/icficalis L.
(Lepidoptera, Т^гаН4ае)н///Энтомлобозр. 1.986..Хг,65>1рыц,2. С.,255—260.
Челнокова Т.А-Экологическая характеристика, и биотодическое разме-
щение хищных полужесткокрылых (Hemiptera-Heteroptera) семейства Pentato-
либае;лесной»зоны Среднего Поволжья,//.ТРУДЫ Куйбыди дед, ин-тд.(.1,980.
Т. 243. С. 82—86. :• л о,'..| и: :!}«, .(«' • i л; v
Черныш С.И. Явление неспецифической резистентности насекомых: Ав-
тореф. дис... д-ралбиол.;наук.!оЛ., 1990^,35:0. , .я -ч л. , v
,ч ч ч - Черныш , С.И., <Симоненко Н.Л. Взаимосвязь устойчивости к повреждени-
ям, уровня'метаболизма'-и прочности диапаузы у,1ли.чинок,Са(/г7>Аага угсгла
s1(Diptera,f,Calliphpridae>l// Зоол. журн. 1988, Тм 67, вып. 8. С. 1186-1193. 4
Черныш С.И., Гирфанова Ф.К., Тыщенко В. JL Состояние неспецифичес-
кой1резистентности!при диапаузе насекомых // ,Труды ВЭ[0.1986. ХА8.. с- 92-
..95*д1о, 1,-vhv> » йп-.«8Н.к их: .-о?, но: vs. ин- -'.из ц .
Чураев И.А. Американская белая бабочка. М., 61 с.< _
i Шапиро И.Д., Переверзев Д.С., Хроменко А.С. О характере,пищевых свя-
зей стеблевого мотылька в условиях цеитральной., лесортепи Хкраины // Эко-
логия, ОДЯлВшМнС. Z5r-7|9.j
Шапошников Г.Х. Становление смены хозяев и диапаузы у тлей (Aphididae)
в процессе приспособлеция к годичным циклам, их, кормовых, растений // Эн-
томрд, Обозр1959.- Т. 38, выц. .3f С.483-504.
; ; л 1СГ1а№шликов>ХХ4Эволюцця1тлей, всвязи, со специализацией и сменой
хозяев: Автореферат дис... д-ра биол. наук. Л., 1967. 41 с.
224
i Шаронов B.B. Таблицы для- расчетов’ природной’освещенностии видимо-
сти. M.-JIlvd945.T98a№j<’:c: ?««nuА " ,' :-awiH'asi »».':< к, ?ciissisf»
Шаронов В.В. Свет и цвет. М., 1961. 311 с. <*'' 'i
i' ,Шашенкова'Д.Х-.хЭкспериментальное изучение’ особенностей сезонных
адаптаций совки с-черное // Физиологические и биологические основы защи-
ты растений. Материалы'научно-произв. конф.Вороиеж,' 1974с С. 54—59.
Шельдешова Г. Г. Географическая изменчивость фотопериодической''ре-
акции.и.сезонного развития яблоиной плодожорки Laspeyresiapomonella L.
(Lepidoptera, Tortricidae) // Экология вредных насекомых^иэнтомофагов.Л.,
1.965uC.<6.-25.:v<:; •ф,; ss.'Vip) . ’’ :> . .Д V :
глг ШельдешоваГ.Г. Экологические факторы,•определяющиеареал’яблон-
ной плодожорки, Laspeyresia’pomonella L. в северном и ‘южном''полушариях ’//
Энтомоллобозр.Ы967«Т.?46',>вып.13>:С'..'583—60бфЯ Я р'Л ,.В ’.‘.ewS .
Шельдешова Г.Г. Стекольников А.А. О?смене тйпов фотолериодической
реакцииспри развитии акациевой ^ожнощитювки ^ 'Рдг/Лело^есдади'т corn!
Boucher (Homoptera, Coccidae) // Экология ’вредныхнасекомых’иихЬнтомо-
фагов.М.-Л.,; 1965.С..26—ЗО(ТрудыЗИН’AHCCCP;.T./36‘). i " ,
" : Щееолев В.Н. Насекомые, вредящие полевым 'культурам,. М.ИИ., 1964.
464 с. - ггь ^ц..л .i а н тл*
Штейнберг Д.М., Каменский С.А. !(Steinberg D)M., Kamenskii‘lS.A.) Les
premisses oecologiques de la diapause de Loxostege sticticalis L. // Bui. Biol, Paris.
1936. Volc70J N'-2..S.-445-4«3^'/'w..- <> .к/ >И|1Я1 -ЛЬ ^дат:;’Т ->
AarvikL. The migrant moth ‘Spodopteraexigua (Hubner) (Lepidoptera,
Noctuidae)‘'recorded in 'Norway'//' Fauna,now. 4984WBd.>287W2i P.'9O—92.
Adedokun T.A., Denlinger D.L- Cold-hardiness: a component of the diapause
syndrome in pupae 'of the’flesh flies, Sarcophaga'crassipalpis andl’5.Z>ri//ata//Physiol.
Entomol? 1984. i-VoL’9.'P.’ 361-364. «-да ч'ИШ n m-W-1»
unii.p' Anderson‘ J.\F. Induction and ’termination of embryonic diapause in the salt
marsh'"mosquito, 4eder TO//icrra«i (Diptera:r<Culicidae) '// ConnJ Agr. Exp. Sta. Bul.
1970. Vol. 711. P. 1-22. Й 5
.iAndo< Ko Geographic; variationrin the incidence of non-djapause eggs of the
falsehnelon beetle, Atrachya, menetriesi:Falderman (Coleoptera. Chrysomelidae) //
Appl. Ent. Zool. 1979. Vol. 14, N 2. P. 193-202, i ! W i г. г
’ uAndo KcDiapause and- geographicrvariation in a ieafsbeetle .//Brawe V-1L-,
Hodek I. (Eds).'Diapause and life cyclestrategies in insects. Junk, The Hague; 1983.
(Seiies’Entomological 1Уо1<.'23).'Р,'Ф27—147.;,: 1 ^«4
Ankersmit G.W., Adkisson P.L. Photoperiodic ^responses of certain’geographical
strains<iof.Pecti«o/>Aor«'gess)'/>feWof://JyTnseCtiiPhysiobii967,’VoL 13!. .P. '553—564.
' i'it; ie AsaMna'E. ^Freezing' and. fiostansisttace in'insects';//'Cryobiology.» London^
New-York, 1966. P. 45:1-^86. ! hvr’-y.-.H v. ,g<. Л.)
у; Influence 'ofitemperature,iphotoperiod andTfood on the speed of
development, longevity, fecundity. and'*other qualities’ of the»diamond-back moth
PZ<<re//atffiocuhpewus>(Curtis)'(Tineidae^lxpidoptera)7/Ausf. J. Zook 1955. Vol. 3..
P.185— 221Vi S’jiOJ*-. -X’:.!??. ><'!’- ,:А A1'.
Awiti L.R.S., Hidaka T. Neuroendocrine mechanism involved in: pupal’colour
dimorphism in^wdlowtail P«p#/o-xu//w.rL. //TnsecfcScLiAppl. 1982. Vok’B^lP. 181—
19L . ‘ .' !'; Ar'i ' .cj, <<A ' ma W .i,.1 ’ «•>. , ..
225
Ay res I M.P. ,Mac LeanS. F. Development of birch .leaves and the1'growth
energetics of Eppihta autumnata (Geometridae) // Ecology., 1987. Vol.. 68, N. 3.
P. 558-568. u ' , / ...<.VvV'.
- < t Bale /wSvClasses pf insecLcold hardiness //. Funci4Ecol.U993. Afol.,7. P. 751-
753^ •(•>. ’ Ч' 'П .< r’ *’ ' /' !,1,i ’l> •' ' ч Uf’’1-' ‘ 1 "
• -Bale J.S. Insectcoldhardiness/Amatter of life.anddeath //Eur. Ji. Entomol.
199& Vol. 93v Nr;3 P, 369-382., — >. 4 ' J
-Baust J.G. Biochemical correlates to cold hardening in insects //- Cryobiology.
1981. Vol. 18,f-N,2.'P.-l&6^198.‘ -v = v!. ,<>>.. hi/-.
Baust J. G., Millerr L.K. Variations in glycerol content and its influence on cold
hardiness in the Alaskan carabidBeetle, Ptetostichus brevicomis //J.InsectPhysiol.
1985.рШ;16,1Р..!979-99(Ь • ,.G , н/г-л. г л.-
Baust J.G., Rojas R.R. Review-insect cold hardiness: factand rfancy // I.-Insect
PtoBhrf4tlMSaVU.H31,<N-10. Р-Й55-759-. > •.», ;i , t
Baust J.G., Rojas R.R., HamilionM.D. Life at low temperatures, representative
insect adaptations,//iCRYO-Lett.rsrf985. Vob6, N 3. P. .199—210. > ’ . . .
Beach R. The-required day number and .timely-induction of diapause in
geographic J strains of the mosquito, Aedesatropalpus// J. Insect-Physiol. 1978.
Vol. 24, N 5. P. 449-455.
Beck\S.Insect photoperiodism. Second, edn. Academic P.ress,New-York,
198ft. 38,7'pp.r," • Д* • <; •<'; >,i! ., ед -</ ,
Beck S.D. Thermoperiodic induction of larval diapause in the* European corn
borer,(Ostrinla nubflalis //>3- Insect Physiol,- 1982- Ablu29,.N I. P.. 107.-1 IX
Beck^S.]), Insect ^hermopeiaodismi//An; Rew Entomol. 1983. Vol, 28, P.91—
.108. '-'.„Ъ, ' Л',\ Д П5Д/ Ъ'.АЛ
, , Beck S.D.-i Effects of thermoperiodnon photoperiodic determination of larval
diapause in Ostrinia itubilalis // J. Insect Physiol.-,1985. Vol. ЗЕ- N liPi 41-46,/
. Recfc &D. Effect of photoperiod and thermoperiod ongrowthof^rotw ipsilon
(Lepidoptera, Noctuidae) // An./Entomol. Soc. Amer. 1986. Vol. 79, N?5. P. 821—
828.
Beck S.D.,Apple J.W. Effect of temperature sand photpperiod on voltinism of
geographical populations of the Europeancorn borer Pyrausta nubilaltsf/3. Econ.
Entomol. 1961. Vol. 54, N 3. P. 550—559. *. .. 51 h •' : >
Black JF.G, -Krafsur E.S. Population biology and genetics of winter.house fly
populations//»An, iEntomol. sSoc. Amer. 1986, Vol. 79,».bb 4. P. 636>-644.1 ,*
Blau H'S. Life history variation in, the black wallowtail butterfly//!Oecologia.
1981a. Vol. 48v.N l. P- H6-42X - - л/ ’
f i, Blau W.S. Latitudinal variation in theslife(histories of insects occupying disturbed
habitats: ac^sestudy ЦОанпа, Dingle,(lEds-X-lnsect Life-History .Patters: Habitat and
Geographic variation. New-York, Heidelberg, Berlin, 1981b. P. 75—95. о * :
n Blunck .ff. DietEntwicklung von;Dydsc«j^marginatus.L. yma Ei «bis,sur imago
// Zeitschr. viss. Zoologie. 1923.;Bd. 121. S. 279^321. ,«
<i Bodenheimer F.S: Lieber die Voraussage der Generationenzahl von Insekten.
Die Bedeutung des Klimas fur die landwirtschaftliche Entomologie // Z. Angew.
Entomol, 1926. Bd»> 12. S^9L—122;?. . . v.. * >
Bonnemaison L. Contribution aT6tude des facteurs provoquant 1’apparition
des formes ailhes et sexudchez les Aphidinae // An. Epiphyt. 1951. Vbl. 2. P. 1-380.
226
Bonnemaison L.’Action de la photoperiode sur la production des gynopares
ailees"de DysapMs planlaginea Pass'1//' An’.-ZooFEcoL Anim? 1970? Vol.*;2, N4.
P. 523-554. ь-•’ ". о»
"^'Bpmtemaison’L. Particularites de la croissance embryonnaire et dela diapause
nymphale chez le'chematobie (Operophtera brumata'L.) (Lepidoptera, Gebmetridae)
// Bui. Soc. Entomol. France. 1971. Vol. 76-, ’N 5—6/P.-123-E301 " 0
'BonnemaisonL. Action de la photoperiode sur la diapause de la pyrale du mais
(Ostrinia1 niibilalis'Wm'.’, Lepidoptera, Pyralididae)Jy/'C.:i'<Seanc.”Acad. Scii.1, Paris
(D). 1974. Vol. 279, N 11. P. 923-925. ‘ ‘ лдои. :/Л
Braasch Z).eZur Ueberwinterungsmoglichkeit des'WeiSsen Barenspinners
(HyyiAaniriacuneaDrury.Arptiidae, Lepidoptera) inder DeutschenDemokratischen
Republik // Nachrbl. Pflschutz DDR. 1976. Bd.i30,''N>;ll. S.-223-225'. . '
1 Braune 'HJ. Der EinfluB derTemperatur auf Eidiapause undEntwicklung von
Weichwanzen (Heteroptera, Miridae) // Oecologia. 1971. Bd.A.’NA' S. 223-22'5,
261—266.-’ h’”- ' " J : !'"s’
Braune KJ.’The role of temperature in controlling1obligatory‘diapausey/ Ef.
Temp. Ectothermic Organisms. Berlin e.’a. 1973. S. 233—238. ''’ iSl‘
’ Braune H.J. Theinfluence of environmental’fectors onwing’polymorphism in
females o^Leptoptema dolobrata ’(Heteroptera, Miridae) // Oecologia. 1983.‘Vol;60.
P. 340-347. - ” ' ‘
Breeland S. G. Biological studies’ in the army-worm, Pseudaletia'^rilpuncta
(Haw.) in Tennesse (Lepidoptera, Noctuidae) // Ten. Acad. SciR958. Vol.33', N‘4.
j> 263—347. - 71 ю
Brower L.P., Malcolm S.B. Animal migrations: Endangered Phenomena //
Amer? Zool. 1991. Vol. 31. P. 265-276? " " - if ’
" Bues R., Poitout'S., Toubon J.'F. Variabilite du'caractere'diapause estivale'dans
les populatons europennes de Mamestrd brassicae L. (Lep.‘!Noctuidae)'7/’BuL’Groupe
etude rythmes Wol. 1991. '\bl?23,‘N 4. P. IE - ; ’ уч»'
" ' ' Ji'Bues"R.^Poitout S., Robin FC!/ et a/."Etudes eipcondit'ioris controlees des
limites thermiques au developpement de Afy/A(mna uni/>uncto Haw. (Lepidoptera,
Noctuidae) // Acte Oecol. Appl. 1987. Vbl. 8, N LP. 79-89. ;i? ’ ‘ 1
‘nBum A. J., Coaker T.H.'-'Diapause and overwintering of the cariot’Hy, Psila
rosae (F.) (Diptera:Psilidae) // Bui. Entomol. Res, 1981. Vol. 71, N 4. P. 583—590.
' Vanard M. La sensibitite photoperiodique des tarves 'de’ la chrysope Nineta
flava // Entomologia Exp. Appl. 1983. Vol. 34, NT.*P?111— If®. : --1
Canard M. Seasonal change in photoperiodic response of the larvae of the
lacewing Ninetapai/idgy/Entomol.' Exp.’Appl. 1988. Vol" 47',*N 2. P. 153—159.
Canard^M., Grimal A. Insect photoperiodism:’ various ways'’of‘regulating
univoltinism'iri* lacewings (PlanipenniaGhrysopidae)y/cExperientia. 1988. Vol. 22.
P 523-525. "
1 Canard M., ‘Grinial A. The irole ’of photoperiod;in the regulation of the annual
cycle in two’montane conifer-inhabiting 'green lacewings in southwestern Europe
(Insecta: Neuroptera: Chrysopidae) // Advance in Neuropterology (Proc, of HI Int.
Symp? of'Neuroptera). Pretoria: RSA,’4990. Р/39-46: с‘:‘ 3! 41 - 4 *
Chen C.P., Denlinger D.L., Lee R.'E. Cold'shock’injury and "rapid cold-
hardening’ in the fly, Sarcophaga ^crassipalpis /f ’Physiol. ’ Zooli.41987. Vol.' '60? P. 297—
304'. .. ........ \
r‘l A
227
.... ChippendaletGM., Reddy A-S, Temperature and photopejiqfdic regulation of
diapauseiof the southwesfernxorn borer, Jiiatraea grandiosely,//},. Insect.Physiol.
1973. Vol. 19. P. 1397-1408. ' fc>;.
Cfiippeftdale Refldy A.S., Cait C.L. Photoperiodic and .fhexmoperodic
interactions in the regulation of the larval diapauseof Diatraea grandiosely/ / J. Insect
Physiol. 1976. 'VoL.i2,!Ni6.iP...823-828. v-'S •'•Г Г~г ?'-? " '?? ”''7’
fChyzik /Ц 87eml(AL,;flea-Dov.T.Overwinteriiig biology of „the „predatory bug
Orius albidipennis (Hemiptera: Anthoppridacl^n lsxae], // Biocontrpj .Science, and
Technology. 1995^ Vol. 5. P. 287-296. } .. ?: „ ; ;l'< ,
...Coates S.A., Mutchfnor J.A, Tollefson LJ. Regulation of-migratory .flight by
juvenile. hprmof mimic dnb jbito^, ini jthe wes(emtcom rootworm .(Co|eopfera>
Chrysomelidaeh//; An. Entomol? Sop. Am. 1987. Vol. .80, P.: 697—70$. •. j
- t ^.Cooper, \B.A. Ovexwjn^fliig:;o^P/«^<g<unm<>tla^a9<i4^rorkfliim/7uflntomol.
1946. Vol. 79. 17бЛ)(Г ivrf't у ... , ?..« s,.,.r ,-p ,t!.,.
Corbet Ph.S. Environmental factors influencing the induction and termination
ofdiapau?e inflie Emperqr Pragopfly, Jnax щрууаг Leach (Odpnatp: .Aeschnidae)
//J. Exp. Biol. 1956. Vol. 33< N L P.jl-14. ..,,,. >
.. ,ppt<{nrier.M.>.pqn|ribotion>a r.e$uile biqlpgique dCf/’fldiw maculiventris Say
predateur^ameriqu^in^dM Dqryphore // An. dep, Epiphyt-etfle.. phytogen. 19,38.
Vol. 4. P. 95—165. ” ?.................. v ' .
Dpnks.yfl.Yf .Insect .dormancy: an ecolog^cjad perspective,// Biol. Survey of
Capad:. 19$/. Mqnogrpphr ,spr. l^-i. 439. p.. ,.y' ..4. . ..т ..>,...u
Danks H.V. The wider integration of studies on insect cold-hardiness//JEur. J.
Entomol,.^996. Vol.,93.:<P. 383—40,3. f? y, .. у ; ... ii;.
Dean J.M. Control of diapause induction by a change, in photoperiod,in
^elgtyplus. Jzmguin^sy/<J»,Inse^,Physjql,.J982yyol->28, J4 .L2. P. 10j35—1040.
r ... tDenlinger Embryoniq determjijatioii ofjpupal diapause, in the Jlesh (fly;;
Sarcophaga crassipalpis // J. Insect Physiol. 1971. Уо1.«17/1№Т0.;,Р.Л815—1822., •
Denlinger jD.-Pupal.djapause in, tropical flesh.flies: environmental and endocrine
regulation, jnetajydiCj^atq gpd;gpnetic splectjop //, Jtol, ?ub mgr,.biol, Lab.> Wpqds
Hole. 1979. Vol. 156, N l/p. 31-46. ' \ < - r , л >
, , jDe«h7iger.j[)rEjPpxmaocyrifl tropicallipse9ts //,An^ev..^|it9moli!Д986. ybl. 31.
Р.Д39г-264. ;"s> т. -t ' ! r 1 1
, ,„j', Denlinger A. Relaiionship.betwqen cold Hardiness and diapause // Insect at
Low Temperature. 1,Edd li^j5'.;£eeJ /r.> D.L. Denlinger,. New-YoxkrLondon, 1991.
p, 174,-19$.j 3;
Deseo K. V., /Tontermini A, Kortelli И/ .The firpt outbreak of the fall webworm
(ffyphantria^y^q prury; Lepidpp|tera,,Ar:ctiidiie) in North; Italy. Preliminary
observations on the^deveiopment,. diapause,..‘and,derivation //Ji,. Appln,.^ntomql..
1986. Vol. 101, N 3. P. 201-206. ... - .
Dicksop.RrBrB.,.Haggis.M.J„ Rainey.RJC-i^PVK^ budwprm moths,flight and
storms; ca§e; study of a cold frpnj system 7/Л C|im.-anfl Appb-Metpo/oL 1983,,
Vol, 22, N ^.Рт.278уГ28б;;7 ,5. у , у ..n .№ у ,;у. -Л
Dingle H. The relation,between age-and;, flight activity in, Xhe milkweed, .bug,
Opcopeltus // Д. Expfl. Biol.; 1965. Vol. f!2, N 2. P. 269-283? ? ?
r Dingle f/zThoexperimental, analysis of migration and Ufe-history .strategies in
insects // Experimental Analysis of Insect Behaviour (Ed.B.Browne). New-York,
1974a. P. 329-342.
22^
.Dingle И. Diapause in a migrant insect, the milkweed,bug Oncopeltus fasciatus
(Dallas) ( Hemipt era,.Lygaeidae) // Oecojogia. 1974b,.jVqL 17, N Д. P. 1—10. •
Dingle H. (Ed.) Evolution of «Insect (Migration, and', biapause^New- York, 1978.
284 p. ( , ' , . !' j V . : , • / 3 ’» ,5
Dingle fl. Ecologyrapd ,evolution of migration //.Animal Migration,, Orientation
anft*.Navigation (Ed..Gaufltreaux).' New-York, 1980. P. 1—101, ; r; n..
Dingle H. Function of migration in the seasonalsynchronizationpf Insects//
Entomol. Exp. Appl. 1982f Vol. 31, P. 36—48, . , •>. 1f A
Dingle H. Migration // Comprehensive insect Physiology, Biochemistry .and
Pharpjacology. -1985, yol. 9.f?. 375-41.5. ... ... ,7 ’ 7 ;J ,•>
,, flfngle H. .Evolutionatjj^Gcnetics of Animal Migration •// Дтер Zool. 1991.
Vol.(31. P. 253—264. ;.
Dingle H., Arora G. Experimental studies of migration in bugs of, the genus
Dysdercus Ц, Oecologia. 1973. Vol. ;12,,N 2.p, 119—140ч|«7 ••riv. >
, - Dingle H., Alden B.M., BJakley NR., eio/.)Variation‘in photoperiodic response
within and among species ofmilkweedbugsI-Oncope/to)//,Evolution. 1980i. Vol. 34,
N:,?. 5P. ,356—37<Q, 1ШИ ;’i'. (!-• •. "is" ' .'X ’
1 (« Dingle H., Boldwin D. Geographic variation, in-life histories; a ^comparison of,
tropical and temperate milkweed bugs {Oncopeltus) // Diapause and life cycle strategies
in insects (Eds. B. Brown,-fl HddeR). Junk, The Hague,, 1983 (Series' EntOmologica.
Vol, 23). P.(J43-r 165. , <j,.- кг..-г . vi 4 '.’r;-
Dixon A.F.G. Evolutionary aspects of parthenogenetic reproduction in aphids
//jAcfa Phyfopathpl. Entomol. Hung,, 1990. У01/25, Nil.-4, P. 41—56- ,
Dolling )у.Я. Photpperiodically determined phase production and diapause
termination'in, Notostira elongata (Geoffroy)(Hemiptera, Miridaefy/’Entonjiol. Gaz.
1973. Vol. 24,7N l.;P- 75-79.- ... •. -,-77^7 ; w
Douglas M.M., Gru la J.W. Thermoregulatory'adaptations/allowing -ecological
range expansion by the (Pierid butterfly, > Na/halis. to/e.-Boisd. //•'Evolution. 1978.
\ r'l -':Л «/M .-i-to ‘ и. Jr-;
Drake V.A. Methods-for studying adult movement>;in Heliothis //Heliothis:
Res. Meth. an<i Prosp. New York etc. l991/P.109—121.. V » 17 'f >
Drake V.A., Farrow R,A. The influence ofiatmospheric structure' and'motions
on insect'.migration '// An. Rev.. Entomol.’ 1988.i'Mj1."33<iP. 183—2>10- ' i ?
Duman J.G., jfforwath K.L., Tomchaney A., его/. Antifreeze agentsofterrestrial
arthropods // Comp. Biochem. Physiol. (A). 1982. Vol. 73, N 4, P. 545-555. «
:;Duman.J,G., Horwath K.L.. Theijrole of haemolymph proteins in the cold
tolerancetof insects 7/*An. Rev. Physiol. 1983, Vol. 45,« P. -261—27BU' . ; «- : . •
Eghterdar E. Zur Biologie und Okologie der Staphyliniden Philonthusfuscipennis
Mannh. y^flOxytelus rugosus GrgN: f/ Pedobiolpgia. 1970.1 Bad.110, N '3'. -Р. 169-
179.5^0 '5 A , . 7* ;Vb'« -'''A'' -
Endo K., Ito T., Chiba К Photoperiodic control of pupal diapause in the
swallowtail.Papi/mxuriiKsL. (Lepidoptera,.Papilionidae) — (sensitive stage andnumber
of required .cycles //-Zool. Sei. 19867,Vol. 3. P. 35L—356.’. ! . '
Endo K., Maruyama Y., Sasaki KI Environmental factors controlling seasonal
morph determination in-the small, copper butterfly, Lycaena phlaeas daimio- Seitz //
J. Insect Physiol.-"1985. W. 317N-7.P.£25—53271 l>! । ' -'• 'и!. i
Endo K, Murakami Y. Photoperiodic control pf the determination ’of three
different seasonal 'phenomena' Of the; swallowtail butterfly,: Papilio xuthus L. // Zool.
Sci. 1985. Vol. 2, N 5. P. 755-760. ' ’i
229
Endo^K./Ueno S.,'Matsufuji M., eta/.-Photoperiodiccontrolofthe determination
of two. different'seasonal diphenisms’bf the ’Asian comma butterfly, :Polygonid c-
aureum L .// Zool. Sei. 1992/ Vol. 9.'«>'725-731*. ‘ > w f -
Endo K, Yamashita I, Chiba Y. Effect of photoperiodic transfer and brain
surgeTy on the photoperiodic control of pupal diapause and seasonaf morphs in the
swallowtail, Papilio xuthus L. (Lepidoptera: Papilionidae) // Appl. Entomol. Zool.
1985."Vol.:,20,;N 4.TP.1'470—478. 'i ,! -••
Ermi T, Saringer G. A burgonyabogar Leptinotarsa decemlineata Say. Budapest,
Mezogazd Kiodo, 1955. P.4-188.' * :• ’ 1
Ferenz HJ. Two-step photoperiodic and hormonal control of reproduction 'in
the*females beetle, Pterostichufnigrttd1// J. Insect -Physiol. 1977. N 23.’ P. 671—676.
Fisher KJ. The migrations of the silver Y-moth in Great Britain// J. Animal.
Ecol. 1938.:-Vol.!j7.'P.'230—247. -V'-
Fujisaki K. Wing form determination and’sensitivity'of stages to environmental
factors in the oriental! chink: bug, !Cavelerius saccharivorus Okajima (Heteroptera:
Lygaeidae)// ApplJEntomol. Zool. 1989. Vol. 24. P.*287-294.' " ’ '
Fujisaki K. Wing reduction in the autumn generation of the oriental chink bug
CaveleriussaccharivorusOkajima (Heteroptera: Lygaeidae) //‘Appl. Entomol. Zool.
1993. Vol. 28, N lv P. 112-115. "
Fukuda S., Endo K. Hormonal control of thedevelopment of seasonal'forms in
the butterfly, Polygonia c-aureum L. // Proc. Jap. Acad. 1966. Vol. 42. P. 1082—
1087. . i
GangavalliR.R., AliniazeeM.T. Diapause'Induction in the oblique banded
leafroller Choristoneura: rosaceana (Lepidoptera? Tortricidae): role of photoperiod
and temperature //>Ji. Tnsect Physiol'4985. Vbl.'-31’,"N 10. P.’831—835. ’ r 1
Gepp J. Die Generation enzahl von Chrysopa per la am Sudostrand der Alpen
//,Nachrichtenblatt.4975. Jg. 24, N 3>S^60-6A '< - 1 1 ‘ {
! Л ! Goettel M.S., PMlogene.ВJ.R. Effects of photoperiod and temperature on the
development of a univoltine population of the banded woollybear’ Pyrrharctia (Isia)
Isabella // J. Insect Physiol. 1978. Vol. 24, N 6-7. P. 523-527?
Gogala M., Micheli S. Vpliv svetlobe in temperature na sezonsko prebarvanje
pri JJezara . viridukt (L.)ri(Heteroptera)7/ Biol. vestn.>1966. Vol. 14-. P. 8'3-90.
Gomi T Effect of temperature on diapauseresponseto photoperiodin/TypAantria
cawea with special referencesto local divergence//Appl. Entom.’Zool. 1995.Vol. 30,
N 3. P. 490-492. ’ И ‘ . . -F F л ; ->' ,
> ,* ’Gomk T... Geographic -variation in critical photoperiod for diapause induction
and its temperature dependence in Hyphantria сипеа Drury (Lepidoptera: Arctiidae)
//,.Oecologia.V199X VolJ 111. P.460-165,<-k >: '
. < Gomi T., Takeda M. The transition to'a trivoltine life cycle and mechanism that
inforce the voltinism change in Hyphantria сипеа Drury (Lepidoptera, Arctiidae)' in
Kobe // Appl. Entomol. Zool. 1990. Ш. 25, N 4. P. 483-489. < (
, . Gomi T,, Takedat M. Geographic variation’ in photoperiodic responses in'an
introduced insect Hyphantria cuned Drury-' (Lepidoptera, Arctiidae)-in1 Japan //
Appl.sEntomoJ. Zool.: 1991.SVoL 26, N.3лP.<357-363,
•I Gomi T., Takeda M. А-quantitative photoperiodic response terminates summer
diapause in the tailed Zygaenid moth, Elcysma westwoodii // J. Insect Physiol. 1992.
\bl. 38. P. 665-670. C , : . , < ‘ >
Grimal A. Exigences photoperiodiques du cycle de la chrysope Tjederina gracilis
// Entomol. Exp. Appl. 1988. Vol. 47, N 2. P. 189-194.
230
. Grosser N. Hyphantria cunea Drury, der amerikanischeWebbar, . in.derDDR?
//Ent. Nachrichten. 1980. Bd. 24,.N 4. S-49-53. • , „I,,..;,, . It .,
Gruner, C., Sauer- K.P. Aestival dormancy in1 the cabbage moth. Mamestra
brassicae L. (Lepidoptera:Noctuidae). 1 Adaptive significance of variability of two
traits; Day length threshpldatfiggnring^aestival t dorapancy and duratipu .ofiaestival
dormancy//Oecologia.1988., Vol. 74, N 4. P. 515-523.
1 Guppy ZC. Life history and behaviour ofthearmy-worm, Psecda/eda um/mncta
(Haw.) (Lepidoptera, Noctuidae)in Eastern,Ontario // Canad. EptojnoL 1961.
Vol.93, N 12. P. 1141-1153.U . ' :
< Hackett D.S., Gatehouse.AG.: Diapause in Hetiothisarmigera (Hubner) and
H.fletcheri (Hurdwick) (Lepidoptera; Noctuidae) in t)te ^ndap-(j^zirg.// Bui.
Entomol. Res. 1982. Vol. 72, N3- R 409-422. :
Harada T. Photosenejttive stages in photoperiodism for -wing form and
reproduction in»a water strider, Aquarius palludupts// iZpol. ScL ,L9912%yol.tl9„N,6.
p. 1252 — . -- - - ч 'Л ...
Harada T., Numata H. Two criticai daylengths fop the determination,of wing
forms, and-the induction., of adult diapause in the ;water strides,AguariuspaUudum
//Naturwissenschaften. 1993. Vol. 80. P. 80—82. • . ; , '
1 Harada-T., Taneda K. Seasonal changes in alary dimorphism of, a water strider,
Gerris palludum,insularis(Motschulsky) Ц,i. Insect Physiol. 1989.,, VoL 35, N |,2.
P 919-924. •, . >-; . , ' r.
Harcourt D:C., Cass Z,.M,.>Pho,tqperipdis,m and fecundity in Plutella
maculipennis (Curt.) // Nature;<London).1966. Vol. 21,0, N 5032.P.217-218.
Harvey.W.R. Metabolicaspects of Insect diapause//An.Rev,Entomol. 1962.
Vol. 7, РЛ57—80. , -’t t , ; ; • . f-
HayesD.K.,RedfernR.E.,-Schmidtn?am KT.,Diapause in the fape .fly, Musca
autumnalis. //• Chronobiologia.:f1.983, Vol. 10, N 2. P. .1(33г^ -,L, ...
Hayes J.L. Diapause and diapause dynamics of Colias alexandra (Lepidoptera:
Pieridae)// Decplogia. 1982, V»L 53, N 3. P. 317-322;,., . . x
Hazel W.N., West D.A. The effect oft larval-, photoperiod on pupalcolour and
diapause in swallowtail butterflies // Ecol. EntomoL 1983.- Vol. 8.,P. 37—42.
Helfert B. Die regulative Wirkupg von Photoperiode nnd Temperatur. auf den
Lebenszyklus okologisch untershiedlocher terrigoniiden, Arten, (Orthoptera:
Saltatoria). 1. Teil: Larvalentwicklung, Reproduction und Lebensdaver der
Parentalgeneration // Zool. Jb.. Syst. 1^80, Bd. 107, N, 2. S,. ,159-182.
Herman JF.X The endocrine:basis of,reproductive inactivity,in Monarch
tButterflies.overwintering in Central .California // J. Insect Physiol, 1973, Vol. 19,
N.9. R-1883-1887.
Herman W.S. -Studies on the adult reproductive diapause of the-. Monarch
butterfly, iDanaus piexippus //«Bioh,Bui. 1-981. ,Vo). 160, N 1. P. 89—106. >.
. Herman W.S. Endocrinology of the- monarch butterfly.// Sci Sep. Natur, Hist.
Mus. Los. Angeles County. 1993. N 38. P. 143—146. •'
>' Hidaka T. Adaptation and Speciation in, the Fall Webworm, Kodansha LTD,
Tokyo, 1977. 179 p. v- , , .
, Hidaka T., Aida S. Day length as main factor of the.seasonal form determination
in Polygonia c-aureum- (Lepidoptera,! Nymphalidae) // Zool, Mag. .1963, Vol. 72,
N3. P. 77-83. > ,, ,
Hodek I. Zimovani slunecek (6 prispevek к studyin ekologie slunecek) // Cas
cesk. Spol. Entomol., 1960. Roc. 57, N 1. S. 1—70.
231
1 1 Hodek I. Diapause in females1 of Pyrrhocoris apterus L. - (Heteroptera) // Acta
Entomol. Bohemoslov. 1968. Vol; 65, N 6. P.^Z-isS. .» л <•
''/Jodek/.Tennination 'of adult ’diapause1' in1 Pyrrhocoris apterus (Heteroptera:
PyrrhocoridaeyHn the>field!//1-Entomol.!Exp. Appt.- 1971a.;VoLf,l'4i Pj 212-222.
1'v'' Hodek 7.1 Sensitivity of larvae to'photoperiods controlling theadult diapause of
two insects // J. Insect Physiol. fT97lb. Vol. 17. P. 206-216. ” -a’’
Hodek I. Inductionof adult diapause in PyrrAocorir ap/eruj L. by short cold
exposure*’//“Oecologia. •197fc,.’VoEi6,,N 2. PJ709— fl?.'”"' ;f ui: .. ,
Hodek I. Biology of Coccinellidae. Junk, The Hague, IWS'. 260 p. ' ; 1
ib i Hodek 7.*life"teyde‘ strategies;- «diapause and ’migration in- aphidophagous
Coccinellidae'"(Minireview) // Ecol. Aphidophaga. Vol. 2: P-roc.Symp./Zvikovske
Podhradi, Sept. 2—8, 1984. Praha, 1986. P.'155-166. ' * : . i . /
° “HoelUcher C^£:, 'Vinson ‘S.B. The sex -ratio of a 'hymenopterous^arasitoid,
Campoletis perdistinctus, as affected by photoperiod$ mailing and- temperature 7/ An.
Entomol. Soc. Am. 1972. Vol. 64. P. 1373-1376. i ? !
11 HoffmannK.H.Anpassswgenim Stoffwechselvonlnsekten an tagesperiodisch.
Wechseltemperatiiren // Verh. Dtsch. Zool. Ges. 73JJahresversamml. Berlin, 26-31
Mai, 1980. Stuttgart, New-York, 1980. "S.‘214-227. /// ;> <<; s!. ,, / i
' ‘‘Hoffmanti'iC.J. Environmental control of seasonal variation in’the4>utterfly
Colias eurytheme: Effects of photoperiod and temperature on pteridine pigmentation
// J. Insect Physiol. 1974. Vol. 20. P. 1913-1924. < ’>
‘‘‘‘'Hogon T". WCThe onset'and1 duration' of 'diapause in eggs of АсИеКГсоттоЛи
(Walk.)'(Orthoptera)-// Aust. J. Biol.'Sci. 1960. Vol.'13.> P. 14-29. ‘ s >
>,r ' 77o/A'dayjV.ZEffectsoftemperatureonwintermothpupae, Operophterabrumata
(Lepidoptera, Geometridae) // Can. Entomol. 1983. Vol. 115, N 3. ‘P. 243—249. ;’
** HolzaphePC.M., “BradshraW W.£.' Geography- of larval’dormancytin-the tree-
hole mosquito, Aedes triseriatUs Say'111 Can. J.'Zool. l93ll:;'Vol.159,‘ Ni6. РлЮ'14—
1021/ ' а ?" 'ГГь • -а > atv; «ччг!',
Honek A. Relationship of 'colour changes andr diapause'in natural populations
ofChrysopa camea'Stepli. (Neuroptera/Chrysopidae) // Acta-entomol. Bohemoslov.
1973. Vol. 70,‘N 4?P. 254-258.'“ ” -r • -us?-.-
i-:; ‘‘‘Honek A^Vfing polimoTphfcm'in"' f^rrhocoris 'apterus Heteroptera:
Pyrrhocoridae):’influence ‘o^^photoperiod’//'Vest?Cs. 'Spot Zool. -1974>Vol. 38.
P. 241-24’2'. '; ! Si)? si,: . ;£ii-
Honek A: ’Maternal regulation of wing1 polymorphism- in Pyrrhocoris‘apterus:
Efficiency of cold activation1// Experientia. 1980.' Vol. 36. P. 418—419. T
....‘Honek Л/ Temperature and wing polymorphism in natural”populations of
Pyrrhocoris apterus L (Heteroptera, Pyrrhocoridae) // Zool. Jb. Syst1. 1981.;Vbl. 108.
R 487^-501' 1 ' > jo.-w
Honek A: Ecophysiological differences between brachypterous and'macropterous
morphs!!in Pyrrhocoris’apterus (Heteroptera, Pyrrhocoridae)<//lActa Entomol.
Bohemoslov. 1985. Vol. 82. P. *347-354. ' ' ' ' . s/"«/
‘ 1 Honek A. Inheritance of wing tormWPyrrhocoris apterus //-J. Hered. 1986.
Vol. 77. P. 465-467. ; ’ -
' ; "Hdriek''A:“Wing ’polymorphism in 'Pyrrhocoiris apterus (Heterbptera,
Pyrrhocorid'ae): penetrance of‘recessive1, homozygotes and duration of larval
development // Genetica. 1987. Vol. 73. P. 211-215. ’ '• 1
232
Honek A., Kocourek F. Temperature and .development time in insects: a general
relationship between thermal constants //Zool. Jahrb Abt. Syst. Ocol. und Geogr.
Here. 1990. Bd. 117, N 4. S. 401-439. . '"XX
Honek A., Sramkova K..rBehayioral regulation of developmental cycle in
Pyrrhqeoris (tpierus L,t4Heteroptera: JJyrrhocoridae),// Oecologi^’/1'9.76/$^ £.4,
P. 277-281. • ”
Sewrw-4 Д0. The army-worm // U.S. Dept., Age EptompL ,С/г<^. N 4.
P. 1 — 5. i •< • ' Д, ,, ,
Horwath K.L., Duman J.G. Photoperiodic and thermal regulation,of antifreeze
protein levels in the:beetle. Dendroidps canadensis //-J.,Insect Physiol. ,1983г»уо1. 29,
N 12. P..907-917, «, « « . ,X« „/Xt ,• ”.. X" - -/X'1-’/'
Horwath K.L., PoggioliC.M., Myerss.K., SchnorrJ.D- Tracking,the profileofa
specific,«antifreeze pipteinamLits contribution to the,thermal hysteresis, activity in
cold hardy insects // Eur. J. Entomol. 1996. Vol. 93, N 3. P. 419—433. । . . t ,
Hsiao T.ff.Host plant adaptations among geographic populations of. the
Colorado potato beetle // Entomol. Exp. Appl. 1978. \fbl.,2.4. P. 437—447.'
Hsiao T. /A Ecophysiological adaptations among geographic populations .of the
Colorado,, potato beetle -in North .America //Advapcesin Potato pest management,
Proc. Conf. New Brunswick, N.J. 1981. P. 69—85. X, . , . /
Hsiao Т.Н. Host specificity,, seasonality and bionomics.ofjLeptinolarsa^beeties
// Ser. Entomolog.1988. Vol. 42- P. 580—599. ,
Huggins H C. An early Plusid gamma // Entomol. 1958. Vol. ,91. P. 99. /'
Ichinose f., Negishi H. Pupal diapause in some Japanese papilionid butterflies.
IL. The difference ip the,inductioppf diapause.between,the fwosubspecies ,of Papilio^
protenor Cramer // Kontyu. 1979. Vol. 47, N 2. P. 89—98. .. '*/
Ikeda-KikueK., Numata ff.Effectof .low temperatureon the termination of
photoperiodic and food-mediated diapause, in The cabbage bug, JEurydema. rugosa
Motsphulsky (Heteroptera/»,fpntXopiida^,// Appl. Entopiol. Zool/, 1994-"Уо1. .29,1
N?. P. <229-236. , X -‘/"X"-'' XXX/,...SXX
Iperti. G., Hodekd. -Induction alimentaire ,de la dormapce imaginale chez
Semiadalia undecimnotata Schn. (Coleoptera, Coccinellidae) .pour aider a la
conservation dps Coccinelles elevees au laboratore avant une utilization ultprieure
//An. Zool, Ecol.tAnim. 19.74. V>1 6, N. I. P. 45—51. /' / //,. /'/ .
Ishii M., Hidaka T. Seasonal polymorphism of the~ adult rice' plant skipper,
Parnara guttata (Lepidoptera, Hesperiidae) and its consol,// Appl. Entomol, Zool.
1979. Vbl.’‘14, N. 2./R 123^184. . = i’ Г X/'. • '
. , Ishii Mt^ .JIidflka T. Characteristics of Pupal Diapause'-in the/Upivoltine
Papilionid, Luehdorphia japonica (Lepidoptera, Papilionidae) //.Kontyu, 1982.
Vol. 50, N 4. P. 610-620. . ”X, XX' rX/"XX
. . Itp Y., Mfyashita.Jf.^ Yamada H. Biology of Hyphantria сипеа Drury
(Lepidoptera, Arctiidae) ,in .Japan. YI. Effect of temperature on development of
immature stages // Appl. Entomol. ZooL. 1968. \bl.,3. *P. ‘163—175,.'/'"‘X.’*'
Iwao S. Studies of the,phase variation and related- phenomena ,in some
lepidppterouSj insects //Mem.«Col. Agric. Kyoto Univ. 1962. Vol. 84,'P<.Lr|80.
James D.-G. Ovarian dormancy in tya/iaus plexippus (L.) (Lepidoptera:
Nymphalidae) — oligopause not diapause // J?Austral. Entomolog.Soc.1982, Vol,'Jll,
P. 31-35... : .... . ,,(i Xi' '
233
‘ Janie J - D;G.' Induction ’oPRepioductive 'Dormancy iin ‘Australian' -Monarch
Butterflies' ‘Dahaus plexippus (L.1)®// Austral. J. Zool.1983. Vol. 31,'N 4. P. 491—
498. ' 1 to!
” '’'Jasic' J.^MackoW: Some1 results of experimentaFstudy of falP'webworm
(Hyphantria cuneaDrury) (Lepidoptera,1Arctiidae'XV/ Ecology. Bratislava,>1961..
P. 1—127. . ' .m -i
Jernty Ti Experiments onthe factors govering diapause in the' codling‘moth,
Cydia pomonella L.(Lepidoptera, Tortricidae) //Acta Phytopath. Hung. 1967. Vol. 2, ’
Rjl-^49-60?°’d-Ч Сэt
^Johnson'C.G. Migration and dispersal1 oflnsects'by flight;London, 1969. 7>63'p.*‘!
Jordan R.G. Geographic differentiation in the development of Aedes sierrensis
(Dj'pterat'Culicidae) in’nature // 'Cah. Entomoi? 1980? Vol. 112. P.. 205—210.‘-
”' n 'Kaaker'S'.i NorqaardJiVloTp. a. Fauna. 1970. Vol. 76.fFund'af Stormsommerfulge
fraDanmark, 1969. ' ,л '1: -..а® Д ж< -lo
! jCawi/e'/.'ItamigratiowiindS'•expansion1 der' llepidopteren1 in‘Finland in den
Jahren 1869-1960 // Acta'Entomol. Fen1. 1962. Ser/18.' S.41-452. f ; M w-
’' Kane/vo'KUbertias'Massenauftretender’Gammaeuie/P/iy/o/nerra gamma L.
(Lepidoptera,1 Noctuidae),’tnSommer 1946'in’Finland // An.iEntomol. Feni’4947.'
Vol. 13. P. 89—104., ; ;iWf -- «с : .uoi'-i
‘' ’Kappus'' KiD., Venard'C.E. The effecW'of photoperiod^and'temperature on the
induction of diapause in Aedes triseriatus (Shy) // J’. Insect1 Physiol.*1967-.< Vol. 13.'
P. 1007—1019;'’ *'•*' ,,WM I w» v.!.'? u-hae la ..
*'л Raster ‘Showers11 WsS Evidence ’of spring migration hnd’ autumn
reproductive*diapause offybe'adult blackscutwofm'inlowa 7/ Enviroh/'Entomol.'
1982. Vol. 11, P. 306-312? ' * '• - л л' v' 5i ‘ ..
° 1 ‘ !kem subtropical population Ы Еигета
hecabe (Lepidoptera, Tieridae) /7 ‘Jap'- * J?iEntomol.fT987i1 Vol.' 60 , N 2. P. 305—318?
''1 ‘Katd Y,,1 Honda H. Seasonal polyphenism'in a subtropical population'of Eurema
hacabe (Lepidoptera, Pieridae) // Jap. J. Entomol. 1992. Vol. 60, N42-. P. 305—318.
'Kennedy j.£ Al‘turning’ point in the ’study' of insect ‘migration // Nature. 1961.
Vol.;i89’,:N’47167:'R‘'785-791? ' - '." •'' ь'-»-м.кг i ,iv,Z
’Ki/nu>a'T.vGeogiaphic yariation and genetic aspectsWreproductive diapause
in Drosophila triauraria and D.quadraria // Physiol.'"Ehtomol.' 1983? Vol.’ 8, N‘-2.'
P " " *' J’ 41''E'VH'Cj ! i Ь.Ъ-'ЛЛН - ? v?;
’ Kimurat., AfasaA:i5Brachypterismand seasonaladaptationsin Orgyia thyellina
Butler (Lepidoptera, Lymantriidae) // Kontyu. 1977?Vor,'45, N I.1?..97—1'06. ’ !’1
, '"Kihtura' T.,' ‘Tdkatib’ H./ MasakPK. 'Photoperiodic programming of summer
diapauseMernibernation i^Spilarctia’inipari/is Butler(Lepidoptera: Arctiidae) //
AppL^ntomol. Zool. 1982.,M>1. 17, N 2. P. 218-226. ‘ •! ’
Kisimotoit. Studies1 on the 'diapause ih,!the planthoppters'and leafhoppers.
IIL'Sensitivityof variouslarvaf stages tophotoperiodand'the'fbrmsof ensuing1 adults’
in the green rice leafhopper, Nephotettix1 cincticeps Uhler'//"Jap. J. Appl. Entomol.
Zoto*()1959’'',\ftg/3,M 3.P. 200-207. ,7tv 1 ,sii" ‘':г •> ‘
Klun Z4.B*Insect sex pheromones:intraspecific pheromonal variability of Ostrinia
nuMa/is in'North America'and Europe // Environ.1 Ehtomol.' 1975*. M>1’.‘4; N 6.
p 891—894 ujuaws л ' ггцфшищ ‘
Kobayashi S., Numata H. Photoperiodic responses controlling the induction of
adult diapause and the determination of seasonal form in the bean bug, Riptortus
clavatus // Zool. Sci. 1993. Vol. 10, N 6. P. 983—990.
234
‘ - Kobayashi S., Numata' iH. ;Effects’of temperature and photoperiod5 on the
induction of diapause andthe; determination of body, colaration in the.beanbug,
Riptortus clavatus // Zool. Sci. 1995. Vol. 12. P. 343—348/ ’ ' >J “
Jft . - Koch Warum* wandern fteinige)VSchme4terlingsarten?‘// Entomol.
Abhandlungen. 1965. Bd. 32. S. 203-211. s<’< - > ' . s
’ ^'Kowalska^T. Effect of photoperiod and temperature sow the growthof owlet
mothsEuxoaexclamationis L. and Rhyacia1-c-nigrumL.(Lepidoptera,-Noctuidae)
/1 Ecol. Polska. Ser. A. 1964. Vol. 12, N 14. P. 235-241. ' 1 ft<“
- Krysan J.L. Embryonic stage of Diabroticavirgifera (Coleoptera^Chryso-
melidae) at diapuase // An. Entomol. Зое. Ami 11972. Vol. 65, N 3. P? 768—769...’
< .1 Xo'san ZE. Diapaiiseinthe nearctic speciesofthe virgifera group of Diabrotica:
evidence-5 for tropical origin and temperate adaptations- ///An. Entomol. Sac. Amer.
1982. Vol. 75, N 2. P. 136-142. Г ь f f!, ft-/
. •! Krysan J.L., Branson? T.F.,,Castro <G.P. <Diapausa'in Diabrotica 'virgifera
(Coleoptera, Chrysomelidae)c a comparison of eggs from temperate and> subtropical
climates /'/ Entomol. Exp. Appt : 1977.*ЛЫ. 22,’Р.ЗЛ—89. /' 1 • ' >.:н-чгокм..l
Krysan' J.L^i Jackson JJ.< Lew A.C. Field termination of eggs- diapause in
Diabrotico, with new evidence of extended diapause in J).6ar/>«n (Coleoptera,
Chrysomelidae>//. Environ. Entomol. 1984. Vol. 13,ftN; 5.-E. 1237—1240aV
KukaM.,DumanJ. G.ySerianni\A.S;^Co'ld-indaced mitochondnaldegradation
and cryoprotectant synthesis in freeze-tolerant arctic caterpillars // J.iComp. Physiol.
4989ft Vol. 158.- Ri«61-671.';w;!/./ftb« !-/ ft.;/,../ft ims / г/-- ..i
Kurahashi Hi^Ohtakt T. Geographic variation in the incidence' of - pupal
diapause ;in Asian and’ Oceanian species of the flesh fIy Eoe«chemca>(Diptera:
Sarcophagidae) // Physiol. Entomol. 1989. Vol. M, N 4;''<R>^6NL-29&Au--4f
Leather S.R., Walker K.F.A, Bale J.S. The ecologyofinsecf overwintering.
Cambridge, 1993/255'p..: > ,-i,--r.r;n . ZsW.i
• । < ’ '< Lee R.E.,'Jr.,- Denlinger D.L.LColdstolerance in'diapausing^and non-diapausing
stages of the flesh fly, Sarcophaga cpassipalpis Ц Physiol. 'Entomol. 1985ii.VbL;10.
'₽. 309-315//.-- /-'-ft/' /7 .-'«<#' 7 '
ft Lee R.E., Jr., Chen С-P., MeachamM.H.,DenlingerD.L.Ontogenetic patterns
of cold-hardiness and glycerol production in Sarcophaga crassipalpis Ц J. Insect
Physiol. 198.7a.. VolJ33. R’587-592.-м
’ ' LeeiR.Ei, Jr.,-Chen С-P., Denlinger D.L. Anapid cold hardening process an
insects//Science. 1987b. Vol. 238. P, 1415-1417. К ; ft '
; -.''ii'JLees A.D: Environmental.factorsacontrolling'the'evocation and termihation of
diapause iwMetaielranychus ulmi Kodh // An. Appl. jBiol.-1953. Vol. i40, !N’3.
P. 449-486. -<* " fti i.w’ .'ft'ft
::S‘'Lees‘\A.D. The physiology of diapause in arthropods //‘Cambridge/Monogr.
Exper. Biol.; 1955: Vaf 4. P. 1—151. . • “<- - - -v" . . . ..iwm , //
: - Lees A.D. The role of photoperiod and temperature; in the determination of
parthenogenetic* and sexual., forms, inftthe- aphid 'Megoure .vicitee/Buckton.i 1 /The
influence of these factors on apterous virginoparae and their progeny //'> J. Insect
Physiol. 1959. Vol. 3. P. 92-Ц7. - m v, t т.ь'! . ’ • V./Ьа.ЛА
?' ft 'Lees AiDi The location! of thet photoperiodic’receptors; in; the'^aphith Megoura
viciae Bucktttn // I. Exp. Btot 1964i Vol/41, N. 1. -Р.’119^133. ft ft/./ : <• у
ft;-LeenAnD. The control of polymorphism in aphidsv// Adv. insecttlPhysiol.
1966. Vol. 3.-P 207—277. ' /‘ft- sftm'-’r- -ro w
235
Lees.E., Tilley. R. J J). Influence of photoperiod and temperature on larval
development in Pararge ajegeria (L.)(Lepidoptera; Satyridae), //i Entomol. Gaz.
1980. Vol. 31, N.I. P. 3-6. i > ! ? .
Leonard D.E. Diapause,inthe Gypsy moth,// J. Econ. Entomol., 1968. Vol. 61,
N 3. P. 596-598. , ,.
,,•> iLipkowiE. Biologisch-okologisceiUntersuchungen uber,Tachyporus-Arten und
7ac/iieur-ru/}'peX>(Coleoptera, .Staphylinidae) // Pedobiologia«1966. Bd. 6,: N 2.
P. 140-177. ' »’ . нг, ' г-. . r. ' -;i
Mansingh;A. Physiological classification Of dormancies in insects//iCanad.
Entomol. 1971. Vol. 103, N. 7isP. 983-1009.S *.<- =< f
' . MarcovitchS. (The, migration of the Aphididae and the appearance of the sexual
forms as afiected by the relative length*otdaily light exposure // JrAgric. Res. 1924.
Vol. 27. P. 513-522. л (
.sr‘Masaki ;S.*The local variation in the diapause’ patterniof the cabbage moth,
Barathra brassicae Linne.withparticularreference to the aestival diapause
(Lepidoptera, Noctuidae)?// Bul.iFac. Agric. Mie Univ. 1956. Vol. il3. P. 29—46.
>‘tMasaki\<.S. Geographical‘variation of diapause in insects.// Bui. Fac. Agr,
HirosakLUpiv.. 1961. M>l.i7.?P; 66—98, ? гг-а j h
Masaki S. Geographic variation1, in the intrinsic incubation period1: "a
physiological, cline in the emma.field crickets// Bul.sFac. Agric. Hirosaki Univ. 1965.
N.H. P.59-90. *=: -yr-H . ns
Masaki S. Seasonal and latitudinal adaptations in the life? cycle of crickets
// Eyolutiomof insect migration and diapause. New-York,* 1978. P.72—100.
* Masaki & Summer diapause •//>. An, Rev.. Entomol. 1980. Vol. 25. P.v.l—25.
Masaki-S. Climatic speciation in'Japanese ground‘crickets // GeoJournal.
1983.:>M>1.^7,bL64.P. 48;3—490, ... - s .’ 's'Л-7. i. .'
Masaki S. Unity and diversity in insect photoperiodism // (Eds. Poster, Collins).
Photoperiodic iregulation of jinsect andrnolluscan hormones., Ciba Found. Symp.
104; Pitman, London, 1984. P. 7-25.- .* ( . , r c
Masaki S., Ohmachi F, Divergence of photoperiodic response and hybrid
developmenton 7e/eogry//us (Orthoptera: Gryliidae) //.Kontyu, 1967. Vol. 35. P. 83—
405.' \ ". №. u- (* .so'/u . , c
Masaki S., Sakai T. Summer diapause irithe seasonal life cycle of Mamestra
brassicae Linne (Lepidopterat;Noctuidae) // Jap. J. Appl. Entom. Zool.4965. Vol. 9,
N 3. P. 191-205. : '
Masaki S^FSeno.E, Effect of selection on wingdimprphismMn the’.ground
cricket Dianemobius fascipes (.Walker) //. Bol. San. Veg. Plagas (Fuera de serie).
1990. Vol. 20. P. 381-393.
* !i, Masaki S., Shirado F,' Nagase A. Tropical, subtropical* and (temperate life
cycles in ground crickets // Insect Sci. Appl. 1987. Vol. 8. P. 475—481. ** и
Masaki S., SugaharaT. Photoperiodic control of larval-development and wing
form in Modicogryllus rp,(Orthoptera: Gryliidae) // Ecob. Reszl992. Vol. 7, N 1.
P.25—30,. veSji; - A •:,« I, ' «,<
Masaki S., Umeya K., Sekiguchi Y. et al. Biology of Hyphantria сипеа Drury
(Lepidoptera; Arctiidae) in Japan. Ill.Photoperiodicinduction of diapause in relation
to the seasonal life cycle // Appl. EntomoL ZooL 1968. VoL 3. P. 55-66.. *
Matsuura H.i, Miyasbita K. Response tophotoperiod of4grori.s ipsilonin relation
to overwintering // Jap. J. Appl. Entomol. Zool. 1978. Vol..,22.* B 7—11. ' '
236
Matteson J.W., Decker G.C. Development of thei European co'm’borer at
controlled constant and variable temperatures // J. EconAEntotnoL 1965.<M>L58,
N 2. P. 344-349. • s =., < .. ... , ’ v --.AW
Me.Lead E.G. Experimental induction and termination ofadultdiapauseand
autumn coloration in Chrysopa carnea (Neuroptera) //J. Insect PhysioL 1967.
Vol. 13. P. 1343-1349. •.
Me.Mullen R.D. The effects of photoperiod, temperature and food supplyon
rate of development and diapause in Coccinella'novemnotata //.Can.Entomol. 1967.
Vol.,99, N 6. P. 578-586. . . ? - . ”,
Mc.Pherson J.E. Sensitivity of immature Thyanta calceata (Hemiptera:
-Pentatomidae) to photoperiod as reflected by adult colour and.pubescence// Great
Lakes Entomol..1978. tVol. 11, N.1-.-PsTl—76..,o . .-.v . t s
Mc.Pherson J.E. Effectsof various photoperiods on 'morphology, in Euschistus
.tristigmus (Hemiptera,-Pentatomidae).// Great Lakes Entomol. 1979-,:Vol.?12, N 1.
P. 23-261. м.-': <; x-i:
Mc.Pherson G.E. A list of the preyspeciesof Podisus maeuliventris (Hemiptera,
Pentatomidae), //Great Lakes Entomol. 1980. Vol. 1-3,-N I. P.47—24-. ’ ’
Mell R. Wanderungen als Aasgleich meteorologisch-sexueller Spannungen’ bei
Lepidopteren (gezeigt am Windenschwarmer Ostasien) //.Entomol. Zeitschr. 1953.
Bd. 63. S. 17-18. ' --.у '
Meudec M. Influence de la temperature et de la lumiere pendantle development
sur 1’etati ovarien aTeclosion cher-Acrolepia assectella Zeller (Insecta,Lepidoptera)
//C. R. Acad. Sci. 1966. N 263. P,554-557, - • . . i
Mikkola K. The interpretation of longrange migrations of Spodoptera exigua
Hb. (Lepidoptera.’’ Noctuidae) // J. Anim. Ecol.< 1970. Vol, 39, N. 3. P. 593—598.
Mikkola K, Salmensuu P. Migration of Laphygma exigua Hb. (Lepidoptera,
Noctuidae) in^Northwestern Europe in 1964.//An. Zool. Fen.’T965.: Vol. 2, N 2.
P. 124-139. • -'-И
MinamiN., Kimura M. T. Geographical variation of photoperiodic adult diapause
in .Drosophila auraria //. Jap,?J. Genet. 1980.’ Vol. 55s, N 5. P. 819—324.
’ : Miya K. The embryonic developmentsachrysomelid beetle, Atrachya menetriesi
Faldermann (Coleoptera, Chrysomelidae). 1. The stages of development and changes
of external form?.//J. Fac. Agric. Twate Univ. T965. Vol. 7, N 2—3. P. 155-166.
Miyata T. Studies on-diapause in Actias moths (Lepidoptera, Saturniidae). Ш.
Effectsof photoperiod andtemprature on the occurence of seasonal forms I I Kontyu,
1977. Vol. 45, N 2. P. 320-329.
Mogi M. Studies on Aedes togoi (Diptera: Culicidae). 1. Alternative, diapause
in the Nagasaki strain // J. Med. Entomol. 4981 • WL.18, N*6'. P. 477—480.
Moran N.A. Aphid life cycle: two evolutionary steps // Amer. Natur. 1990.
Ш. 136, N L.P. -135-138. ? л .
’ Morgan. T.D.,‘ Chippendale G~M. Free-amino acids of the hemolymph of-the
southwestern com borer-and the‘European com borer" in relation to their diapause
//J. Insect Physiol. 1983. V>1. 29, N 10, P. 735-740. A
Morris R.E. Factors inducing diapause in Hyphantria-сипеа Ц-Can'.’Ent?1967.
Vol. 99. P. 522-529. ’ ’.«=.•
Muller H.J. Die Wirkung exogener Factoren auf die zyklische Formenbildung
der Insekten, insbesondere der Gattung EuscelisBrulle // Zool. Ib. Sy st. 1957.
Bd.85,H. 4-5. S. 317-430. ’ ” - v ь. ?
237
ir . Mutter /L/.Problemeder Insektendiapause Ц Verhandl.Deutsch. Zool. Ges.
г1965лВЛ29. S. 192-222. ' • , r v fe: . Ьп/'.-гм
Mutter H.J. Effects of photoperiod and temperature on leafhopper vector //
Leafhopper’'Vectors’and Plant-Disease-Agents./ Eds: Manamorasch K.,Harris K.F.
London^4<979.P.;29-94... *.->>-/., v»sw,- -r,
Muller H.J. Dormonz bei Arthropoden. Jena, Stuttgart, New-York; 1992.
289®. 1- OH 5<r.-Ai....a<3'. ' .1.7.; ! *Vi. •• >>ЬМ
Muraji M.; Miura T., Nakasuji XPhenological studies on-the wing dimorphism
of a semi-aquatic bug, Microvelia douglasi (Heteroptera: Veliidae) // Res. Pop. Ecol.
1989..’Vol.H31, N'd. P. 129-138. < > - i->v' '< 1 -> > V.
..; :>'Nakai T:/Takeda Af/Temperature andphotoperiodic regulation ofsummer
diapause and reproduction in Pyrrhalta humeralis (ColeopteraiChrysomelidae)//
AppE.EntomoVZooL 1995k Voh'3O,?N'2. P.<295—3O1.> Л -V
/ 'UfakasujiF., Ishii<M,HoJ.-Z: Rice skippersdn Taiwan andtheir* life histories
// Tech. Bui. (Taiwan). 1984. N 82. P. 1-10. > ’.? i
i Nakasuji F./Kimura M: Seasonal polymorphism of egg size in a migrant skipper,
Pamara guttata guttata (Lepidoptera, Hesperiidae) // Kointyu. l984. Vol 52. P. 253—
i259>X '• '.of; !' 11 ..‘-If- in'"-- r A' - •; /
’ . Nechols J.R., Tauber C.A.,.Tauber M. J. Geographical Variability in ecophy-
siological traits controlling dormancy in Chrysopa oculata (Neuroptera, Chrysopidae)
,7/ JyyInsect Physiol. 1987. Vol. 33, N 9. P. 627-633. sr. . ’ ' > .
Neumann D., Heimbach F. Das Wachstunvdes Kohlweiblings bei konstanten
und tagesperiodisch wechselnden Temperaturen // Oecologia. 1975. Bd. 20, N 2.
>S. 135—141. To №' ’.iXinf •' :ч .'Д '
Nielsen F.T. Onthemigratiomofinsects’// Ergebn.Biol.l964.Vol.’27. P./162—
,193!.p.: . id'i ''iH'Г'и’/х’; ‘ .susw. '•
* Nishizuka M:,'Azuma‘A.,.Masaki 3. Diapause! response-tophotoperiod and
temperature in Lepisma saccharina Linnaeus (Thysanura: Lepismatidae) // Entomol.
Sci;.fl998. Vol. l, N4;. P. 7—14'' ' ~ Ч W'*.;-
NorrisM.J. The influence of-constant and-changing photoperiods on imaginal
.diapause. in.the Red Locusi-(Nomadacris septemfasciata Serv)r//:Ji HnsectiPhysiol.
1965.lVol. il, N 8. £1105-1119. ^ '
Novak I. The /hibernation of the. silver Y-moth (Autographa gamma^L.)
(Lepidopterav Noctuidae) Mnderthe climatic conditions , of Central Europe // Труды
ХПЦЧеждународного энтомологического конгрессам:2^9 августа 1968 г. М.,
1968. Т I. С. 533. Ь' 7 ’ '-1-
Novak J.. Spitzer К. The: relationship between migration and diapause during
phylogeny- and ontogeny of some Lepidoptera // J. Res; Lepid. 1972. Vol. 10. T> 1181-
1841.! '.И , . .'W'1.- .-p '?*• -чЛеУ
Numata H., Hidaka T. Photoperiodic control of adult diapause in the bean
pug, -Riptortus clavatus Thunberg (Heteroptera: -Coreidae). Г. Reversible induction
and termination of diapause //lAppl.r.';Entomol. 'Zool.i1982..VoL17. P. 530^-538-.
Numata H., Hidaka T. fPhotoperiodic control,of adult'diapause in'the bean
bug, iRiptortus clavatus Thunbeig-.(Heteioptera:,Coreidae). П. Terminal ion ofdiapause
induced under different photoperiods // Appl. Entomol Zool 1983. Vol 18,1 N 3.
P.4i3,9r'44L ’ /">/:, ;сл.- (. -ни..- 'А' • : *. t. -w.1., -'
* -! Numata H:, Kobayashi .5.'Threshold and Quantitative-photoperiodic Responses
exist in an insect // Experientia. 1994. Vol. 50. P. 969—971.. -.
238
Numata H.. Saulich А.Н., VolkovichT.A. Photoperiodic responses of the linden
bug, Pyrrhocoris >a/rferusi/under.conditions of constant temperatureandunder.
thermoperiodic conditions // Zool. Sci. 1993. Vol. 10. P. 521—527. " ’’ *
Numata 'JL,> Yamamoto К. .Feeding- on seeds;induced diapause in(the cabbage
bug, Eurydema rugosa // Entomologia. 1990. ’Vol. 57. P. 281—284. '
r'Nylin S., Wickman P.-O.WiklundC. Life-cycle:regulation and Ijfe.history
plasticity in the speckled wood butterfly: are- reaction norms predictable?,,//,,Biol. J,-.,.
LinneanSoc. 1995» Vol. 55, N 2. P. 143M57. . ,< • , -4'
Obrycki J. J., Tauber MJ. Seasonal synchrony of the parasite Perilitus cpccinellae
and itsRhost, CdleomegiUanmaculata'// Environ: Entomol,.. 1979,^01:8. P, 400^-405.
- 'Oku T. Aestivationand migration-in noctuid-moths // -Diapause and life cycle,
strategies in insect (Eds. Prawn, Hodek). Junk, The Hague, 1383} (Series Entomolo-
gicaj. Vol.<231 P. -219—231. w..:-' u--.-, -'"t',' .» 4 -,vk
Oku T., Kobayashi ^“Migratory behaviour and life cycle of noctuid moths .
(Insecta, Lepidoptera), with notes ontherecent status ofEmigrant species in, northern
Japan// Bid. Tohoku Nat. Agr. Exp. Stn. 1978, Vol, 58. P. 9/—209. ’ : , >
Paarman W. Der Einfluss von' Temperatur Und ;Lichtwechsel, auf die
Gonadenreifung desLaufkafersRrorcus laevigatus Dej. (Coleoptera, Carabidae) aus
Nordafrica//Oecologia.: 1974. Vol. 15,>-N 1; S. ;8,7-92. 4 ; , »
Paarman W. Die Bedeutung exogener Faktoren fur die Gonadenreifung yon ;
Orthomusbarbarus atlanticus (Coleoptera,, Carabidae) aus Nordafrica // Entomol.
Exp-. Appl.-1976.s Vol. l9, N I S.,23-36. u. <. .
Pease R.W. Factors causing seasonal forms in Ascia monuste //Science. 1962.,-j
Vol. 13. P. 987. v- «,-л
Penner M.P., Orshan L. Reversible reproductive-diapause and intermediate.;
states between diapauseand full reproductive -activity ip- -male Oedipadaminiata
grasshoppers // Physiol. Entomol. 1980. Vol. 5, N 4. P. 417-426.; ' г 'Z .</
• .Penney. M.5Diapauseandrreproduction:in<Afe6r<a brpvicollisll.(Et) (.Coleoptera,
Carabidae) // J. >Anim; 'Ecol. 1969: Vol. 38,tNT»';?P: 219—233». и:. E. - ri;, •v-:'1-? ' ’r,
Perez Y., Verdier M., Pener M.P. The effect of photoperiod on male,'sexual;
behavior in a porth adriatic stain of,the migratory locust,// EntomoE^Exp.sjAppl,
1971. Vol. 14? P. 245-250. 1 < ?c ).< < m л i. . • Jt.,,<7j
- . ’ Pinger R.R.,'Eldridg/e RE, :,The effect oPphotopetiodSon.-diapause induction in
Aedes canadenlsis,and,RmropAorm/ercx (Diptera< Culicidae)s// Ап.)ЕрГоде>1- SopH
Amer. 1977. Vol. 70. P. 737-740. .. y, . 'p-f.j-
; PoitoutiS., PueS’R. Mise en evidence d’onediapause nymphale de type-festival»
dans'’des' populations suropeennes de deux lepidop(eres, Noctuidae, Hadeninae
(Mamestra oleracea L. et Mamestra brassicae L.). Facteurs d’induction et apercu sur
la уапаЫ1йес чп1егрори1а11оп.4е соеаГас1₽ге'//гС, rdAcad. Scj.pD, WAiVol, 276,
NK. R{ 336-2-3370?-А. >..-ч ' n-
Poitout S., Cayrol R., Causse R., Anglade P. Deroulement du programme,
d’etudes sue les migration de lepidopteres Noctuidae realjseen mpntagneet principaux
resultats acquis // An-.; Zool., Ecol.- Anime, 1974 (1973).,W.>6yNMhfP.,'585--r587-.‘H
> Pond.'.DtD. (Life'history, studies?of the army-worm Pseudaletiacunippncta
(Lepidoptera, NoctuidaeJ in New‘Brunswick//An.eEntomoLi Soc. Amer.,1960.
Vol. 53, N 5. P. 661-665. : 51. 1
> PpuvreaitA. Cycle biologique et- interpretation de 1’arret de. development chez
Pyrrhocoris apterus L (Heteroptera,Gymnocerate, Pyrrhocoridae) //.Bull. Soc. Zool..
France. 1963. W. 88. P. 180-196.
239
Pruess Л'./’.' Migration -of the army cutworm, Chorizagrotis auxiliaris
(Lepidoptera: Noctuidae). lv Evidence for migration//:An, Entomol'. Soc. Amer.
1967. Vol. 60. P. 910-920. ’ : A- 7 , -л- ,<
Raina A.K., BellR^A; A'nondiapausing strain of;pink boHvorm'from southern
India // An. Entomol. Soc. Amer. <I974.'\4>1. 67, N*4. P. 685—686. v
‘ Rankin M.A. Hormonal control of insect migratory behaviour // Evolution of
insect migration' and diapause* (Ed.* Dingle). New-York, 1978. P. >5—32. • s
Rankin M.A. Endocrine effects on migration';// Amer. Zool. 1991. 'Vol. 31..».
p. 217-230. >'*•' !<->=,.- .. .-V i:..,,, ,, ‘K v ..3
’ 'Rankin S.M.yxRankin M.A. The hormonal control, of migratory flight behaviour
in the1 convergent-ladybird beetle, Hyppodamia convergens // Physiol.> Entomol.
198O. Vbl. 5, N. 2; R 175-182.
Ratte H.T. Temperature and insect development // Environ. Physiol, and
Biochem. of Insectsfed. K. Hoffmann). -Berlin, 1984. P. 33—66. •/
- Riedl HlAnalysis of codling moth phenology in relation to latitude, climatic
and food availability //-Brown V.K., Hodek T (eds) Diapause and life strategies in
insects. Junk, The Hague, 1983. P. 733—253; •"
*• Riedl W., Croft B.A. The effect ofphotoperiod and effective temperatures on the
seasonal phenology of the codling moth (Lepidoptera: Tortricidae) // Can. Entomol.'
1978. Vol*.ПО?P. 455—470. i
Riley T.J. Effect bflarval photoperiod on mating and reproductive diapause in
seasonal forms of Anaea andria (Nymphalidae) //• J. Lepidopter. Soc. 1988. Vol 42,
N 4. P. 263-268. ' ** .. , - , - '
RingR. Maternal induction of diapause in the larva of Lucilia caesarL. (Diptera,
Calliphoridae) // J/Exp/BioL 1967. Vol. 46,’ N 1. P. 123-Г36.
’RingR.; Tesar Di Adaptations to cold. inCanadian arctic .insects //.Cryobiology.
1981. Vol. 18, N 2. Р/199-211. **.'.- :< 5 -
- •’ /{ocfc G. Thermaland thermoperiodic effects on larval and pupal development
and survival in tufted apple bud moth (Lepidoptera: Tortricidae) //.Environ. Entomol.
1985iW.si4, N & P> 637-640. ''< П - *.Л
''Rodriguez J.;Jordano D.<, Haeger J., Fernandez A. Spatial' heterogenity an a
butterfly-host plant interaction // J. Anim. Ecol. 1994. Vol.'63, N Г. P. 31-38. ’
• ’Roonid R.E/РпраЗ'/diapause in Heliothis armigera (Hubner) (Le-'pidoptera:
Noctuidae^in Botswana: it* regulation by enviriomental'factors //:Bul. Entomol. .Res.
1979. Vol. 69, N 1. P 149-160. К ' ! :
’•** 'Sakai T./AfasakPS. Photoperiod as *a factor causing seasonal -forms in Lycaena
pWfle<w;dm'»»'o!Seitz.''(Lepidoptera: ’Lycaenidae). // Kontyu. -1965J Vol.,33,lfN 3.
R275-283.* ’ ** S’ «4. < «л И ->** Г? .1 * .-a
’ Sakurat'H., Takeda'S. Characteristics of diapause in Coccinella septempunciata
brucki in Japan // Ecol. Aphidophaga. 1986. Vol. 2. Proc. Symp., Praha, Sept. 2—8,
1984cE8173-178. s п-s; S 4 . X s ’ л : ' ? .1.
T’Snringer G.'Diapausetin Hyphantria'cunea Drury (Lepidoptera, Arctiidae); in
Hungary // Acta'Agronom. Acad. Sci. Hung. 1974. V>1. 23,'N; 1—2. P. 17—24.
•» ’ -Saringer'G. Diapause-Versuche niit der ungarischen Population*; von Ostrinia
nubilalis Hbn. (Lepidoptera, Pyraustidae) // Z. Angew. Entomol. 1-976. Bd. 80, N 4*.
P. 426-434. > - s
. Sauer K.P., Gruner’ C. Aestival dormancy in the cabbage moth Mamestra
irossicae L.(Lepidoptera:Noctuidae). 2. Geographical variation in two traits: day.
240
length thresholds triggering aestival, dormancy! and duration'ofaestival>dormancy
//(Oecologia. 1988. АЫ.«76. P. '89—96. : • ri’ »с; г '.4 Л;
Sauer K.P., Spieth H. Zeitliche Orientierung ih' fluktuierender Umwelt durch
dasMessen einer AnzahlvonLangtagen//Zool. Jahrb;\Abt. Syst,, OkoL und Geogr.
Tterft l986. Bd. 113,>N 3. S j 373-388; i .
Sauer K.P., Spieth H., Gruner C. Adaptive ^significance- of ^genetic variability of
photoperiodism in- Mecoptera and Lepidoptera;// The. Evolutionof Insect Life- Cycle
(Eds.F.Taylor,R. -AarAan>. New-York, 1986.P.153—172.; !. 1 ?
Saunders D.S. The temperature-compensated photoperiodic clock «program-
ming» “development and pupal diapause -in'-the flesh-fly,- Sarcophaga torgyrosioma //
L Ihsect Physiol. 197К Vol. 17,’ N 5. P. 801-812. " . r
i5aunderj /).5'. Thermoperiodic control of diapause inaninsect.'theory of
internal coincidence^// Science.P1973.'\bL 181. Р.-Збв-гЗбОЛг'р.Ъ , iUj. - > -
Saunders D.S. Insect clocks. Oxford, 1976. 280 p.
Saunders D.S. Some effects; of constant temperature andphotoperiodon the
diapause response, of the flesh-fly, Sarcophaga argyrostoma // Physiol./.Entomol.
1980. Vol. 5, N 2; P. 191-198- u- : \ М .
Saunders D.S. Insect clocks- Oxford, 4982. .409 р/'м - •* .. •> - :
Saunders D.S. A diapause induction-termination; asymmetry in the photo-
periodic responses , of the Linden bug Pyrrhocoris, apterus and the effectjOf near
critical photoperiods on developme/it // J. Insect Physiol. 1983. Vfl.£9? N 5. P. 399—
405. '•;? . /* 'A .
Schaick Z.P.G.van. Seasonal synchronization of the life-cycle of Ptero-stichus
oblongopunctatus..(Coleoptera: Carabidae)-.// Entomol. Gen., 1985.-JfoL 11, N 1—2.
P/33 — 39.'- .'.'..'"nV '•«" K---’"?. l-VG ’ ' - kir.
Schmidt-Koenig K. Migration strategies of monarch butterflies // Contrib.
Mar. Sci. 1985. Vol. 27,.Suppl. P. 786-79.8. ! -i; ;; ; 3 <
Scott J.A. Hibernal diapause of North American Papilionoidea and Hesperioidea
11 J. Res. Lepid. 1981. 18 (1979), N 3. P. 171-200. s/ r
ScottR.W.,Achtemeier J.L Estimating pathways, of migrating insects carried at
atmospheric winds //Environ .''EntOTnol'.1*1987. V>1. .6a<P. 1244-1254. -.д >,
' '«Shapiro AAf.vMovementsfof Hymphalis cab/omfca-.(Njanphalidae) im 1972 //
J. Lepidopter. ,-Soc.:1974.,Vol./28v N 1. P. 75-78. : j«£ ; ,.}
Shapiro A.M. Evidence for a return migration of Vanessa cardui in northen
California (Lepidoptera; Nymphalidae).// Pan-Pacif. Entomok J980. Vol. 56, N. 4.
P. 319-320, •.’«.»! ,л -я;- ; --a........f^.;
Shapiro A. Redundancy in pierid polyphenisms: pupal chilling induces vernal
phenotype ia Pierisaccidentalis (Pieridae) // J. Lepidopter,,Soc.i l982. Vai. 36, N 3.
P. 174-177. - «Г— ?' -' ?< - .И fe-jV
ShimadaiK. Reduction in the critical number of short days for-pupal diapause
in Papilib machnoncwith precocious metamorphosis //J^Ipspct Physiol. 1985. ЛЫ.- 31,
N 9. P. 683-688.
л л и ShimizuTc, Masaki S. Genetic variabilityro£ithejwingLfor-m'response to
photoperiod in. a subtropical population; of the- gpxind;icnc}№l^Dianemobiustfas(dps
// Zoof. Sen 1993. Vol.10.i P. 935-944. .
Showers (V.B., Chiang H.C., KeasterA.J., et al. Ecotypes of the European corn
borer in North > America, //Environ. Entomol’. 1975. Vol. 4„N 5. P.J753—760.
’ ' ’ tW. rtjfti • . i , т . <ч r,
241
Showers W.B., Keaster A.J., Hendux W.N.,et al. Mechanism of southward
migration of a noctuid moth (Agrotis ipsilon Hafnagel): a complete migrant // Ecology.
1993. Vol, 74, N 8. P. 2303-2314. v ,
Shroyer D. A., .CraigG.B.jJr. Egg hatchability and diapause in Aedes triseriatus.
(Diptera: Culicidae): Temperature- and photoperiod-induced latencies / An. Entomol.
SocuiAmer.-1980. Vol. 73. R. 39-43. K
! -«• SiewY.C. Some physiological aspects of adult reproductive diapausein Galeruca
tanaceti L. (Coleoptera: :Chrysomelidae) //i Trans. Roy Entomol. Soc», London.
1966,.Vol. 118. P. 359-374. - -,.'7' ,
.'Sims S.R. -Diapause dynamics and host plant suitability of Papilio zelicaon
(Lepidoptera: Papilionidae) // Amer. Midi. Nat. 1980, W. 103, N 2. P. 375-384.
* Sims £/L The genetic and environmental- basis of pupal- colour dimorphism in
Papilio zelicaon (Lepidoptera: Papilionidae)-// Heredity.1983. Vol. 50, N,2. P.159—
168. ‘ " - -7 ' . -7--’ - •' , '->
’ 'i< Solbreck C.’ Induction of diapause in a-migratory seed bug, Neacoryphusbicrucis
(Say) (Heteroptpta: Lygaeidae) //Oecologia. 1979. VoL43, N l. P. 41-49. <•
Solbreck C., Pehrson J. Relation between environment, migrationiand
reproduction in a seed bug,- Neacoryphus bicrucis (Say) (Heteroptera: Lygaeidae) //
Oecologia/ 1979. Vol. 43, N I P. 51-62. f
; Somme L. Supercooling and winter survival,in terrestrial arthropods // Comp.
Biochem. Physiol. (A). 1982. N-. 73. P. 519-543. i
Somme. L. Kuldetoleranse hos insekter // Fauna (Nor.) 1985. Vol. 38, N. 4.
P.132—139. • •bv; w . '7 .....
Souty-Grosse't'C.,iBauchon D., Mocquard J.P. et al. Interpopulation variability
of the seasonal reproduction in the terrestrial isopod Armadillidium vulgare Latr.
(Crustacea, Oniscidea): a review 11 Acta Ecologies. 1994. Vol. 15, ?N 1. P. 79-91.
Spark A. A review of the biology of the,fall armyworm // Fla Entomol.-1979.
M>1. 62, N 2. P. 82—87. • «
Spieth H.R. Die Anpassung'.des Entwicklungszyklus an unterschiedlich lange
Vegetationsperioden beim Wanderfalter Pieris brassicae L. (Lepidoptera: Pieridae)
11 Zool. Jahrb.'Abt. Syst. Oekol. Geogr. Tiere.4985. Bd. 112. S. 35-69. '
Stinner^R.E.y Gutierres A.P., Butler GiD. Amalgorithm fo* temperature-
dependent growth rate simulation // Canad. Entomol. 1974. Vol. 106, N 5. P. 519-
524.; -• 1 h- -a 7 -.ы .г. .., - -
, Sugiki T.. Masaki 5,-Photoperiodic control of larvae and- pupal development
in Spilarctia imparilis Butler (Lepidoptera: Arctiidae) 11 Kontyu. 1972. Vbr.‘4O, N 4.
P.- 269—278. ' дак-.Ь,! : .к
r1. . Sylven E. Undersokningar over gammaflyer, Phytometra.gamma L. -/1 Stat.
Vaxtskyddsanst., Medd. N. r. Stockholm. 1947. Vol. 48, N 2. P. 42.
,&TakedacM. An «hourglass-like» feature in the photoperiodic time-measurement
in Diatraea grandiosella (Pyralidae) // J. Insect Physiol. 1985.'Vol. 31,N 5i P. 397—
406. r i i
I'akeda'M. Photoperiodic induction, maintenance and termination of winter
diapause in two-geographic ecotypes of the rice,stem maggot, Chlorops.oryzae
Matsumura (Diptera: Chloropidae) // AppL Entomol.’ Zool. 1996. Vol. 31, N ‘3,
R 379—388. ; 1
Takeda M., Chippendale G.M. Phenological adaptations of a: colonising insect:
the southwestern com borer, Diatraea grandiosella // Oecologia (Berl). 1982. Vol. 53,
N 3. P. 386-393.
242
Tanaka К. Diapause and seasonal- life cycle strategy in the house spider,
Achaearanea lepidariorum (Araneae, Theridiidae) // Physiol. Entomol,'1991.Vol. 16.
P. 249-262. V' .
Tanaka K. Photoperiodic control of diapause, and climatic adaptation of the
house spider, Achaearanea lepidariorum (Araneae, Theridiidae) // Funct. Ecol.
1992. Vol. 6. P. 545-552. > . ’
Tanaka S. Effect of changing photoperiod on nymphal development in
Pteronemobiusnitidus bolivar (Orthoptera, Gryllidae) // Jap. J. Entomol. 1978. \bl. 46,
N 1. P135-151. / • ' - -’.y .
Tanaka S. Seasonal control of nymphal diapause in the. spring ground cricket,
Pteronemobias nitidus (Otthoptera: Gryllidae), P. 35—53 // V.K. Brown and I.Hodek
(Eds.), Diapause and life cycle strategies in insects. Junk, The Hague, 1983 (Series
Entomologies)/Vol. 23. 283*p.: ' ; . t •
TanakaS., Tsubaki Y.Seasonal dimorphism, growth and food consumption in
the swallowtail butterfly Papilio xuthus // Kontyu. 1984. Vol. 52, N 3. P.390—398.
Tauber M. J., Tauber C.A. Adult diapause in Chrysopa camea: stage sensitive to
photoperiodic induction // J.< Insect‘Physiol. 1970. Vol. *16/ N 11. P. 2075—2080.
Tauber M.J., Tauber C. Л/Geographic variation in critical photoperiod and in
diapause intensity of Chrysopa camea (Yiewtopleta) // J. Insect Physiol. 1972. Vol. 18,
N 1. P 25-30. ' ' ' - v ;
Tauber M. J., Tanier C.H. Seasonarregulation of dormancy in Chrysopa camea
(Neuroptera) //J'. Insect Physiol. I973a. Vol. 19, :N 7. P. 1455—1464. ’’
Tauber M.J., Tauber C.A. Quantitative response to daylength during‘diapause
iri insects // Nature, bond. 1973b. Vbl.;!244 (5414)- ?. 296—297. '
Tauber M.J., Tauber C. Nutritional' and photoperiodic control' of theseasonal
reproductive cycle in Chrysopa mohave (Neuroptera) // J. Insect Physiol/ 1973c.
Vol. 19, N 4. P. 729-736.
TauberM.J., Tauber C.A. Natural daylengths regulate inSect ’seasonality by two
mechanisms // Nature, bond. 1975. Vol. 258 (5537). P. 711—712. ’ !
Tauber M.J., Tauber C.A. Developmental requirements of the univoltine species
Chrysopa dawnesi: photoperiodic stimuli‘and sensitive stages // J. Insect Physiol.
1976a. Vol. 22, N 2. P. 331-335. ' < • » ,
Tauber M.J., Tauber C.A. Environmental control of univoltinism audits evolution
in an insect speicies // Can. J. Zool, 1976b. Vol. 54, N 2. P. 260—265. ' •
Tauber M.J', Tauber C.A., Masaki S. Seasonal adaptations of insects’.'New-
York, Oxford, 1986. 411 p. x :
' Tduber C, Tauber M., Nechols J.R. ThermaTrequirements for development in
Chrysopa oculata: a geographically stable trait //Ecology. 1987. Vol. 68, N’5. P. 1479-
1487.
Tauber M.-, Tauber C.A., ObryckiJ.J., etal. Geographical variation in responses
to photoperiod and temperature by Leptinotarsa decemlineata Say (Coleoptera:
Chrysomelidae) during and after dormancy // An. Entomol.'Sbc. Amer. 1988.’Vbl. 81,
N. 5. P. 764-773. ‘ ' ' ' ’.
Tauber M.J., Tauber C.A., Roberson J.R., et al. Dormancy in Lymantria dispar
(Lepidoptera: Lymantriidae); Analysis of Photoperiod and Thermal Responses //
An. Entomol. Soc. Amer. 1990. Vol. 83, N 3. P. 494-503.
Taylor L.R., Trench R.A., Woiwod I.P. et al. Synoptic monitoring for migrant
insect pests in Great Britain and Western Europe. 1. Establishing expected values for
243
species content, .population stability andphenology ofaphids and moths// Rothamsted
Exp.rStatl Repti. 1980. N 2; P.i41-104j ii -гг -.4 . , , . «
Taylor L.R., French R.A., Woiwod I.P. et al. Synoptic monitoring for migrant
insect pestsdn Great Britain->and-Western Europe //Rothamsted Exp. Stat. Rept.
1984. Harpenden.4985. P. 251-275. — ; л
Taylor F., Spalding J.R. Fitness function for alternative developmental pathways
in the-timing of diapause Induction // Amer. Natur. 1988. Vol. 131, .N-51-.R 678-699.
? 'ThieleHU. DieSteuerung der JahresrhythmikvonCarabidendurchexogene
, und endogene Faktoren // Zool. Jb. Syst. 1971. Bd. 98, N. 3. S. 341—371.
, Thiele H.U. Interaction between photoperiodism and temperature, with respect
-to the Control ofdormancyin'-theadujt Stage of PterostichusoblongopunctatusF.
(Coleoptera, Carabidae). l. Experiments on gonad maturation under different climatic
conditions in the laboratory // Oecologia. 1975. Vol 19, N L P. 39-47,. , i
и; Thiele, HJkiMeasurement ofdaylengtlvas; a basic for photoperiodism and
annual periodicity in [the carabid beetle Pterostichusnigrita ,F. J) Oecologia. 1977.
Vol. 30, N 4. P. 331-348, \ К-
\ ToppW. VergleichendeDormanzuntersuchungen an St aphylinidenf Coleoptera)
/AZool.Jb., Abt. Syst. Okol. ,G!eogr.1Tiere. 1979. JBd, 106, N l. P. 1-49. ,
t ? 'Topp W. Imaginal aestivation in, the rove beetle species Omalium^iyulare
(Coleoptera: Staphylinidae) // Entomol. Gen. 1986. Vol. 12. P. 51—55.
. , Tppp, W.- Selection -for an optimal .monovpltine life cycle in an unpredictable
environment.-Studies1 on the beetle Catops nigricans,,Spepcs{Coleopter^, G^tPPldae)
,-//Qppolpgia. 1990ч Vol., 84j. 134—14.1. т -; , . T 1 - -
Topp W., Kirste^E.'- Synchronisation of preimaginal development and
rep reduct ivesuccpss, in thewinter.moth Operophtera brumalaF, //J. Appl, Entomol.
1991. Vbl, 111, NAP 137-146;; - ... .... .
Tsuji H. Studies on the diapause of the Indian-meal moth, Plodia interpunctella
. Hubner. II, Theeffectof population densjty on the induction of diapause,// Jap. J.
Appl. Entomol. Zool. 1959. Vol.,3, N.2. P. 34-40. - , ~ : ; r
, - Urguhaft F,A.„ Urguhart H.R, Autumnalrmigration routs of the eastern
population of the- Monarch butterfly (Danaus plexippus L.;. Danaidae; Lepidoptera)
in North America to the overwintering site in the.neovolcanic plateau of. Mexico //
Can, Д» 2^)01., 1978. Vol. 56. P. 1,759-1764,. ,
Vepsalainen K. The role of gradually changing;day length in determination of
. wing length, alary, dimorphism and diapause in a Gerris odontogaster (iZett.)population
in South Finland // An. Acad. Sci. Fen. A. 1971. N 183. P.i 1—25.
p , pYepsalqinen K-Tlye life cycles and wing length in Finnish Gerris Fabr. species
(Hetjeroptera, Gerridae)-//,Acta Zool.,,Fen. 1974.,VoL 1.41. P. 1-73.,
Vepsalainen K. Wing dimorphism and diapause in Gerris: determination and
adaptive significance //-Evolution of insect migration and diapause/Ed.H. Dingle).
^ew-york./^yg. P/218-i53. ' 7." . - ".‘Л " . {
> - Voigt Ж Der Einflub vonPhotoperiode und Temperatur, auf den Saisondimor-
phismus und die Induktion der reproduktiven Diapause bei Polygonia.c-album L.
/Lepidoptera, Nymphalidae) // Zool. Jahrb. .Abt. Syst. Okol. und Geogr. Tiere.
j99^..Bd~M> 3-4. P- 3Q9—32L< - V - : ‘ ,
Vojnist A. Az «igazi» vandorlepkek // Folia Entomol, Hung. .1966. Vol. 8.
s. 167-175. ,, v,. . j;/',:,;:
244
Warren L.O., Bjegovic P. Uticaj duzine dnevnog osvetljenja na razvoj stenica
Podisus placidus Uhl. i Podisus maculiventris Say (Hemiptera, Pentatomidae) //
Zast. bilja. 1972. Vol. 23. N 117-118, S. 7-10.
Warren L.O., Tadic M. The fall webworm, Hyphontria cunea (Drury) // Ark.
Agric. Expt. Sta. Bui. 1970. \bl. 759. P. 1-106.
Walkev T.J. Butterfly migration from and to peninsular Florida // Ecol.
Entomol. 1991. Vol. 16, N 2. P. 242-252.
Walker T.J., Wineriter S.A. Marking technigues for recognizing individual
insects // Fla Entomol. 1981. Mil. 64, N I. P. 18-29.
Wellington W.G. Biometeorology of dispersal // Bui. Entomol. Soc. Amer.
1983. Vol. 29, N 3. P. 24-29. s ir ' л
West D.A., Snelling W.M., Herbek T.A. Pupal color dimorphism and its
environmental control in Papilio polyxenes asterius Stoll (Lepidoptera: Papilionidae)
// J.N.Y. Entomol. Soc. 1972. Vol. 80, N 4. P. 205-211.
Wigglesworth KS. The physiology of insect metamorphosis. 1954.‘JCambridge
Monogr. exp. biol. Vol. 1. P. 1—152.
Wilde de J. Developpment embryonnaire et postembryonnaire du Doryphere
(Leptinotarsa decemlineata)cn fonction de la temperature// Proc. YIH-th Intern.
Congr. entomol. Stokhobn, 1950?“P. З'Ю-З'г!.1' ' "i* ’ J
Wilde de J., Hsiao Т.Н. Geographical diversity of the Colorado potato beetle
and its infestation in Eurasia// Lashomb'J.,'CasagrandeR. (eds).'Advances in potato
pest management. Stroudsburg,.>1981'. •P^4Jr-68v»''W::‘-<'‘ ' !
Willmer P.G., I6iww> DJlti^Field,analysis of insectiheatbudgetrreflectance, size
and heating rates // Oecologia. 1984.АЫ. 5(bP. 25O-r-255i s
Winning E. Bewahrte Metoden der 'Aufzucht yon 'Kartoffelkafern im
Laboratorium // Dt. Pflanzenschutzdienst..l95I..Bd.!:5,?NS7.’S.4>28-130.
Wolff N. The zoology of Iceland. 1971. Vol. 3, N 45. P. 1—193. -
Wood F.E., Nordin EH. Studies on'the low temperature induced biogenesis "of
glycerol by adult Protophormia terrae-novae*//J.; Insect Physiol?’i976. Vol. 22,
N 12. P.1665-1673. ... r-ь ' <-’V>.•„••'I
Wbude H.A., van der. Verhoef H.A. Reproductive diapause and cold* hardiness
in temperate Collembola Orchesella cincta and' Tomocerus minor // J. Insect Physiol.
1988. Vol. 34. P. 387-392. • • ;
Yan-I-Chu. The migration of Diamondback Moth // Diamondback Moth
Management. Proc. 1-st Intemat. Workshop. Taiwan, 1986. P, 77,-81. -
Yothom S., Rivnay E. Phenology' and distribution"'of Phytometrinae in Israel
// Z. Angew.:Entomol. 1968. M>1. 61,fN 1. Р.Ч*-1б1.«! • “ : < ' " !
ZachariassenJCE. Physiology of’cold tolerance’in insects*//Physiol. Rev.
1985. Vol. 65, N 4. P. 799-832. i v ' ’«'? ” -
Zero A.J. Wing polymorphism-in waterstriders/Gerridae: Hemiptera):
Mechanism of morph determination and fitness‘difference betyveen morphs //
Migration: Mechanisms and’ Adaptive ’Significance. (Ed. Rankin M.A.) Contrib. Mar.
Sci. 1985. P. 661-673.
ОГЛАВЛЕНИЕ
f4< ' ' .'iV .
Предисловие „................................................ 3
ЧАСТЬ I
ЭКОЛОГИЧЕСКАЯ РЕГУЛЯЦИЯ
СЕЗОННОГО ЦИКЛА НАСЕКОМЫХ
БЦ1М 1. Рим климата и сезонке адядгаци^ ,........................ 5
1.1. Активная жизнедеятельность и физиологический покой.........—
, .1.2. Диапаза как наиболее совершенная форма покоя. "
Краткая характеристика и значение...................... 8
1.3. Зимняя диапауза и.холодоустойчивость ...л.......... 10
1.4. Летняя диапауза и ее 'экологическое значение'......... 12
Глава 2. Экологической механизм регуляции сезонного.цикла............ 14
v - ' . * ' ; 1 к
2.1. Количественные фотопериодические реакцин.
Их экологическая роль................................... л___16
- : 2Д. Качественные.фотопериодические реакции...................л..20
2.2.1. Цикломорфозы тлей.................л................... 21
2.2.2. Сезонный полиморфизм............................ 22
. :, , , .2.2Д Индукция и терминация диапаузы ............... >....31
Глава 3. Факториальная изменчивость, фотопериодической реакции.л.......34
-Il S,
3.1. Температурная изменчивость основных параметров ФПРч^....... —
3.2. Влияние суточных колебаний температуры: на ФПР...........41
3.3. Роль изменяющейся длины дня в индукции диапаузы........„.42
3.4. Другие факторы, влияющие, на ФПР ................... 43
Глава 4. Географическая изменчивость сезонных адаптаций ...................................47
4.1. Вольтинизм и диапауза................................... -
4.2. Днапаузирующая и чувствительная к длине дня стадии.......48
4.3. Параметры фотопериодической реакции......................49
246
ЧАСТЬ II
ЭКСПЕРИМЕНТАЛЬНЫЙ АНАЛИЗ
СЕЗОННОГО РАЗВИТИЯ НАСЕКОМЫХ
Глава 5. Методы исследования.........................................59
5.1. Фенологические наблюдения и полевые опыты...............60
5.2. Лабораторные опыты.......................................-
5.3. Методы расчета сроков развития насекомых................64
Глава 6. Гетеродииамное развитие.....................................83
6.1. Поливольтинизм и его регуляция...........................—
6.1.1. Чешуекрылые.......................................84
6.1.2. Жесткокрылые (колорадский жук)...................103
6.1.3. Полужесткокрылые (клоп солдатик).................121
6.2. Моновольтинизм. и механизмы его поддержания............128
Глава 7. Значение абиотических факторов при формировании
вторичных ареалов адвентивных видов ,............................. 150
7.1. Американская белая бабочка, Hyphantria сипеа Drury.....151
7.2. Хищный клоп Podisus maculiventris Say..................156
7.3. Колорадский-жук, Leptinotarsa decemlineata,Say1........161
7.4. Особенности ФПР,‘определяющие возможность
акклиматизации-разных видовнановойтерритории................168
И х:'-?
Глава 8. Гомодинамное развитие..................................... 173
8.1. Возможность постоянного обитания в умеренных широтах
видов с гомодинамным развитием................................—
8.2. Сравнительное изучение реакций на температуру и
длину дня у видов, с гетеро- я гомодинамным типом
развития ...........,........................... 176
8.2.1. Капустная совка, Mamestra brassicae................—
8.2.2. Луговая совка, Mythimna unipuncta................177
8.2.3. Совка гамма, Autographa gamma, и малая наземная
совка, Spodoptera exigua..........................179
8.3. Холодостойкость разных стадий онтогенеза...............181
8.4. Устойчивость к длительному действию низких
положительных температур................................185
8.5. Сезонное развитие..................................... 190
8.5.1. Капустная, совка, Mamestra brassicae>..:.........191
8.5.2. Палевые опыты с гомодинамными видами.............195
' - ".Г< ’• ЗГ '
Глава 9. Миграции насекомых и их .место в системе
сезонио-циклическихадаптаций .................................... 203
Указатель литературы ............................;.....'............211
Научное м-зданме
Аида Хаматовна Саулич
^СЕЗОННОЕ РАЗВИТИЕ ’
/НАСЕКОМЫХ И ВОЗМОЖНОСТИ
ИХ РАССЕЛЕНИЯ
'1 ’Редактор' Т.Н. Пескова
ОбложкаЕ.АСоловьевой <4-- •
Компьютерная верстка А.В. Павлова
1 •* 1 “ J . 5 Ч.« ' . 1 . • 1 Т1 > • ,
Лицензия ЛР.№ 040050 от-, 15.08.96
. К ;-U't '• 1 •’ - -С> '
Подписано в печать 04.11.99. Формат 60x84 1/16.
Бумага офсетная.1*Печать'офсетная. г ’’ и:* ’' ’"
Усл'. печ. л. 14,41. Уч.-изд. л. 15,22. Тираж 450Ъкз. Заказ № 382.
- Издательство СПбГУ. 199034, С.-Петербург, Университетская наб., 7/9.
Типография Изд-ва СПбГУ.
199034, С.-Петербург, Университетская наб., 7/9.